/








































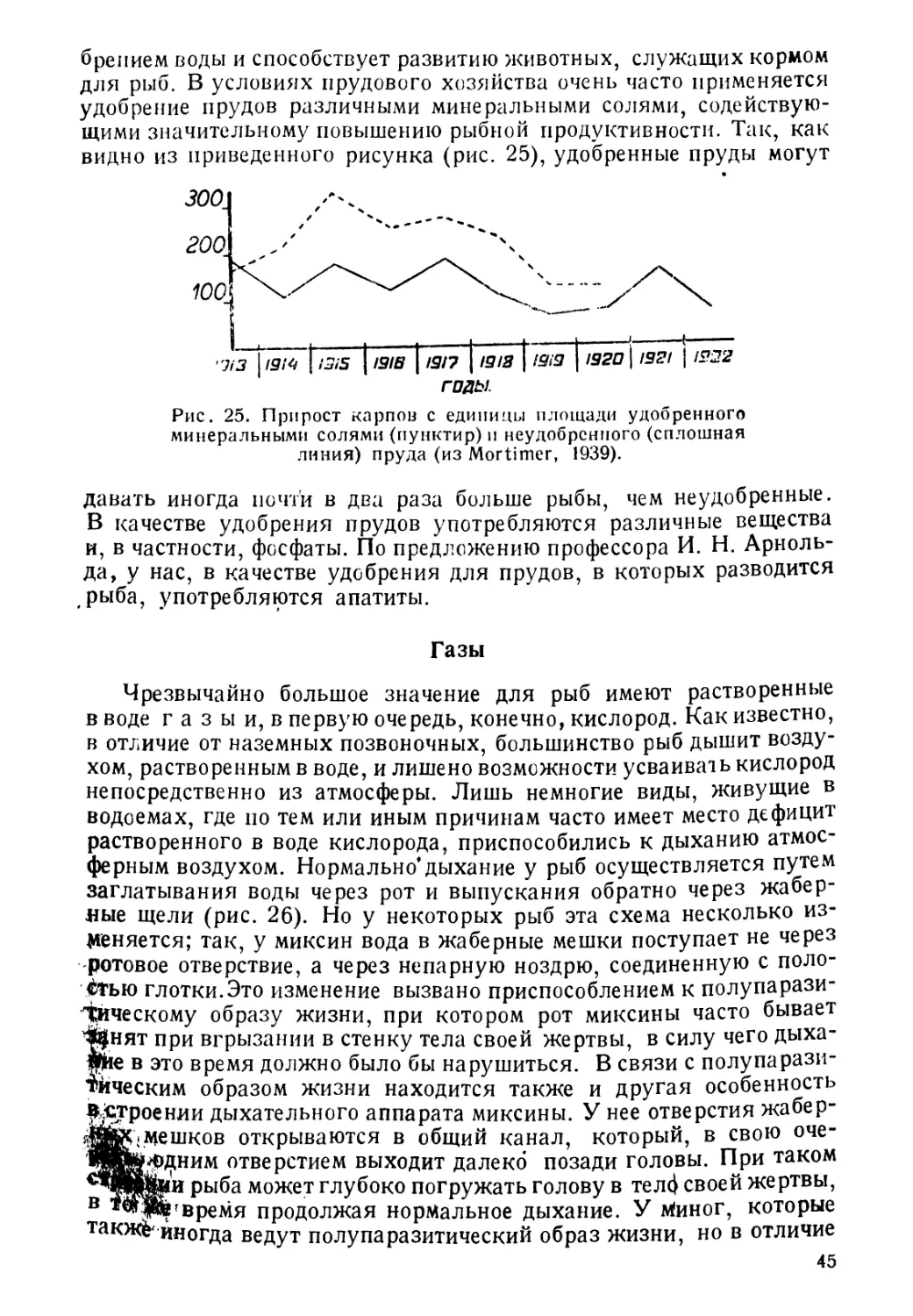



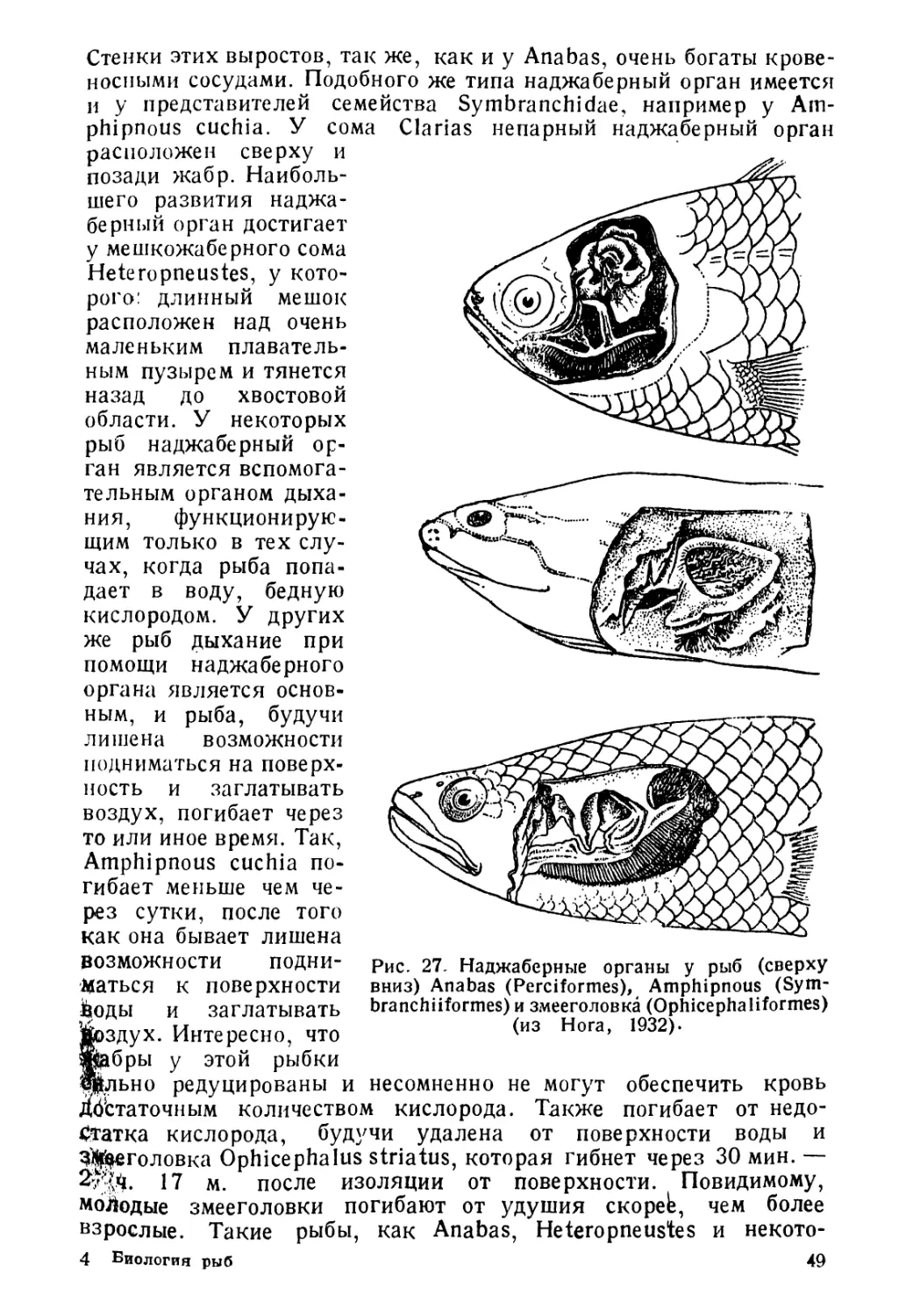





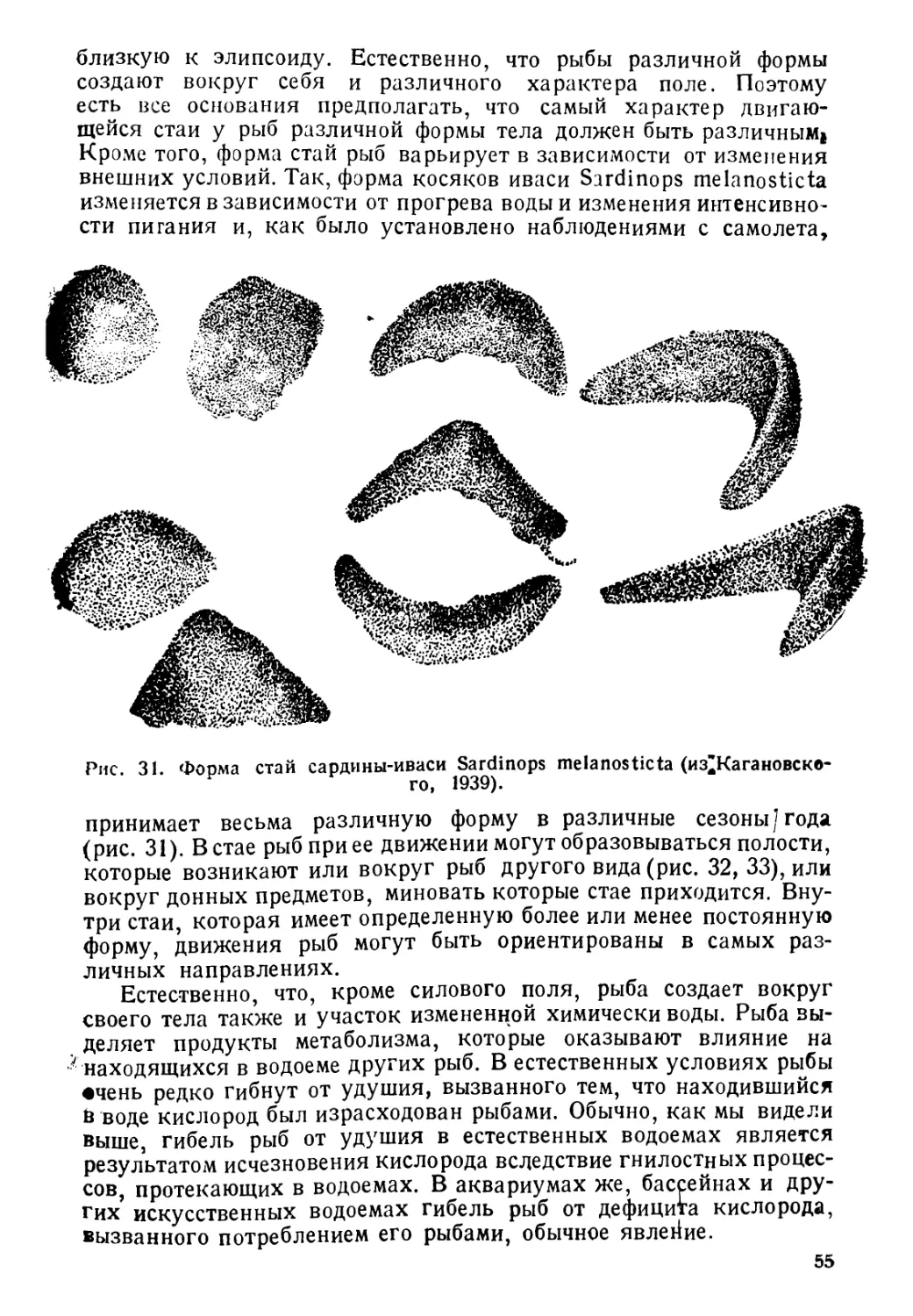





























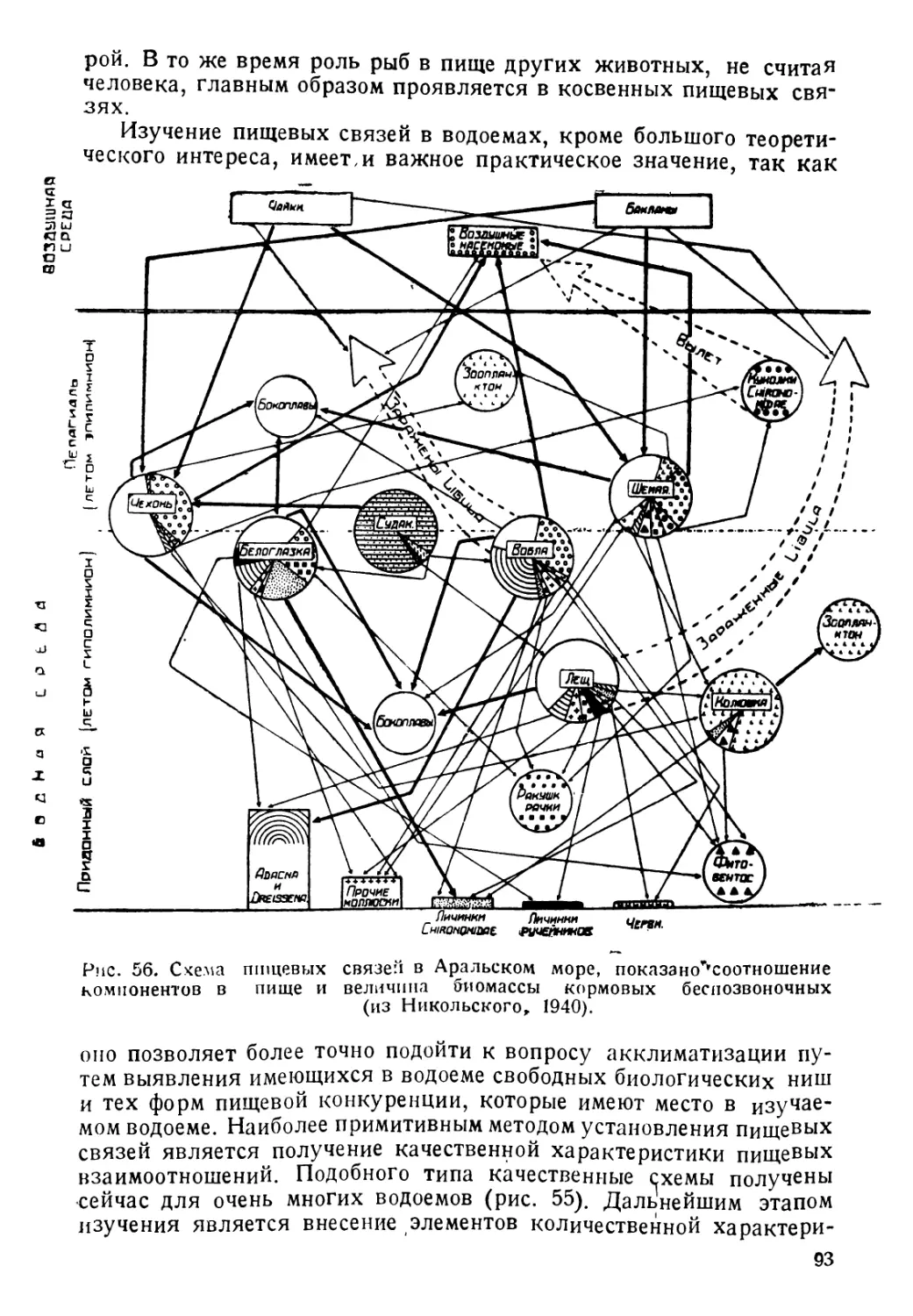
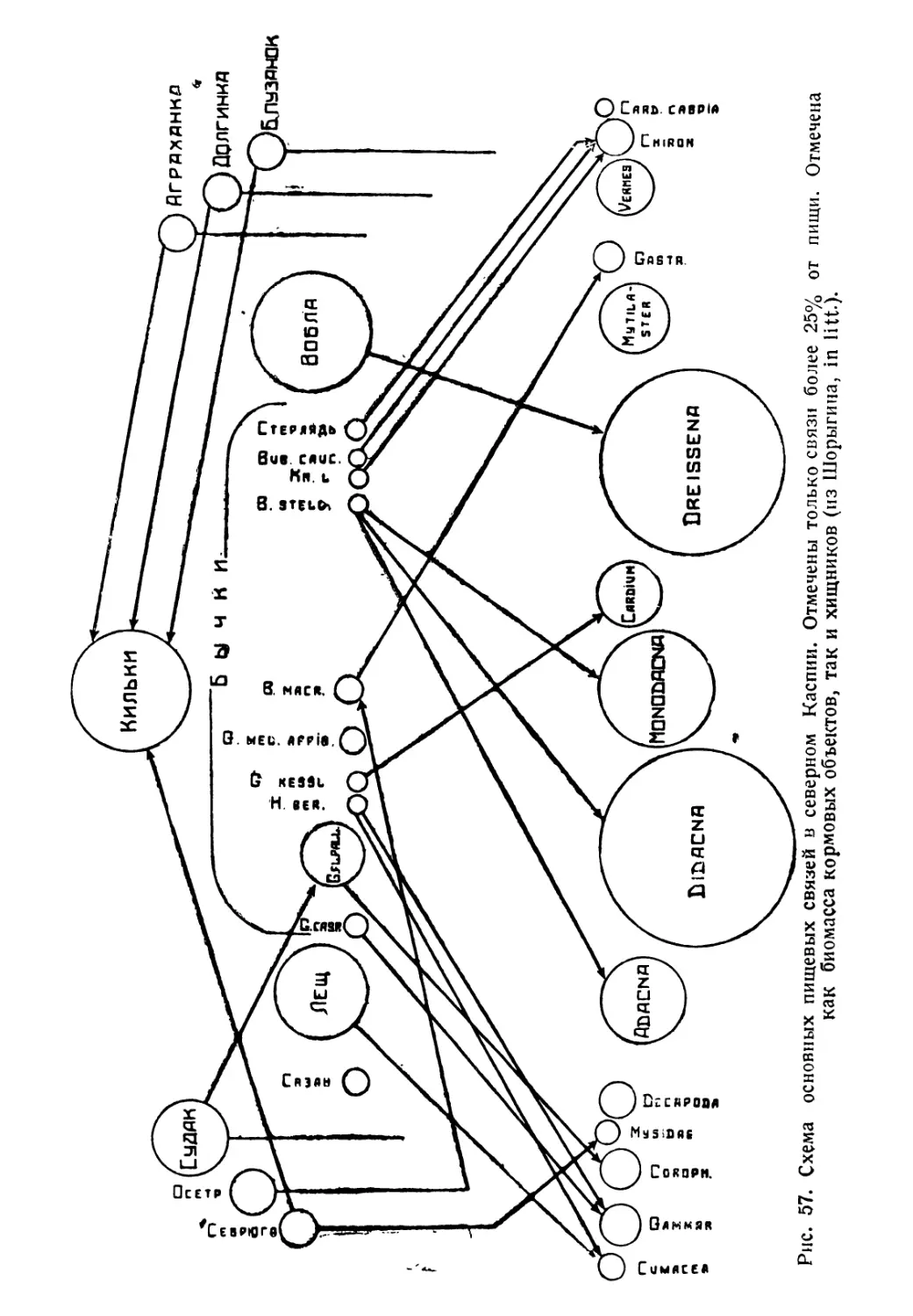



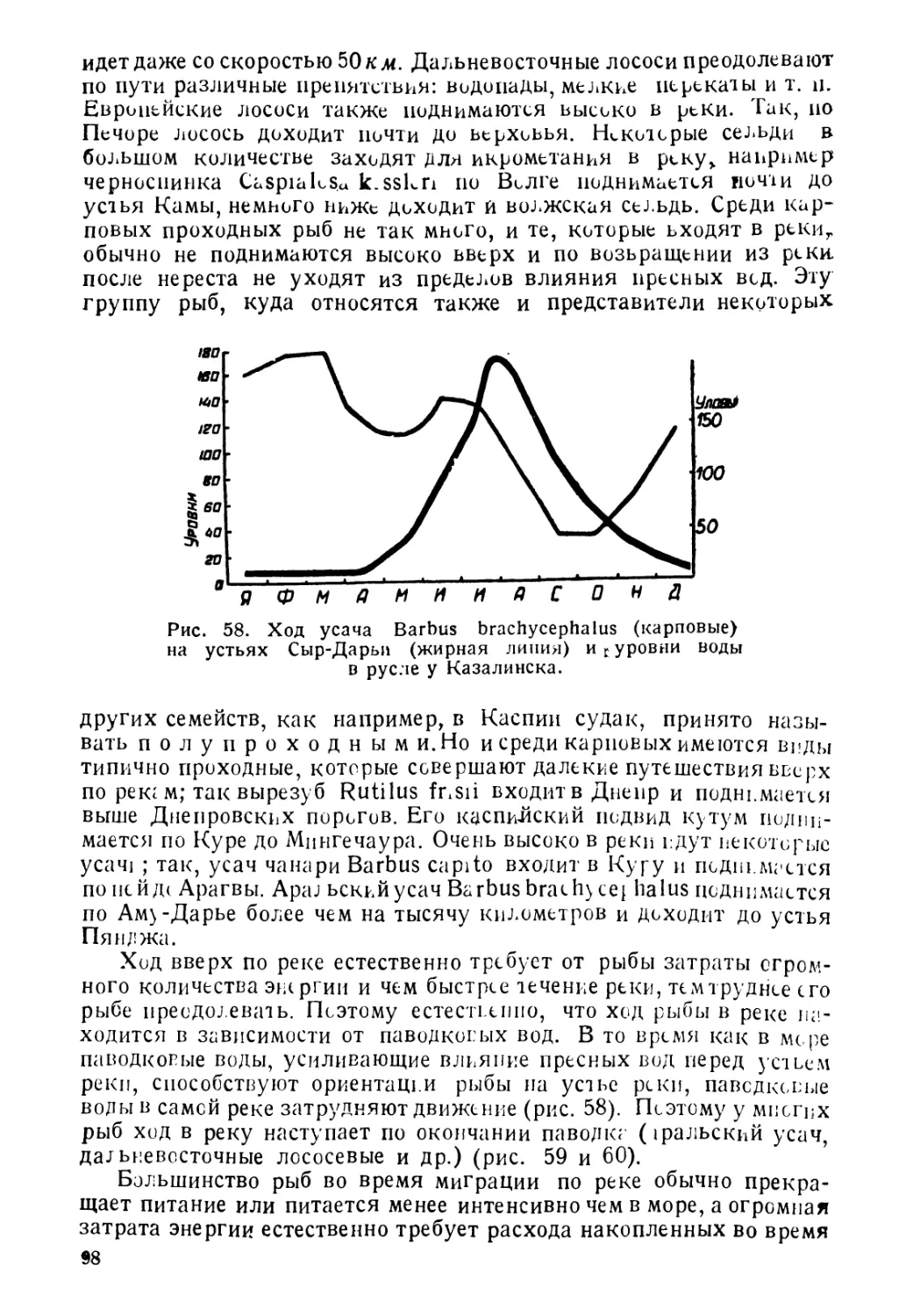
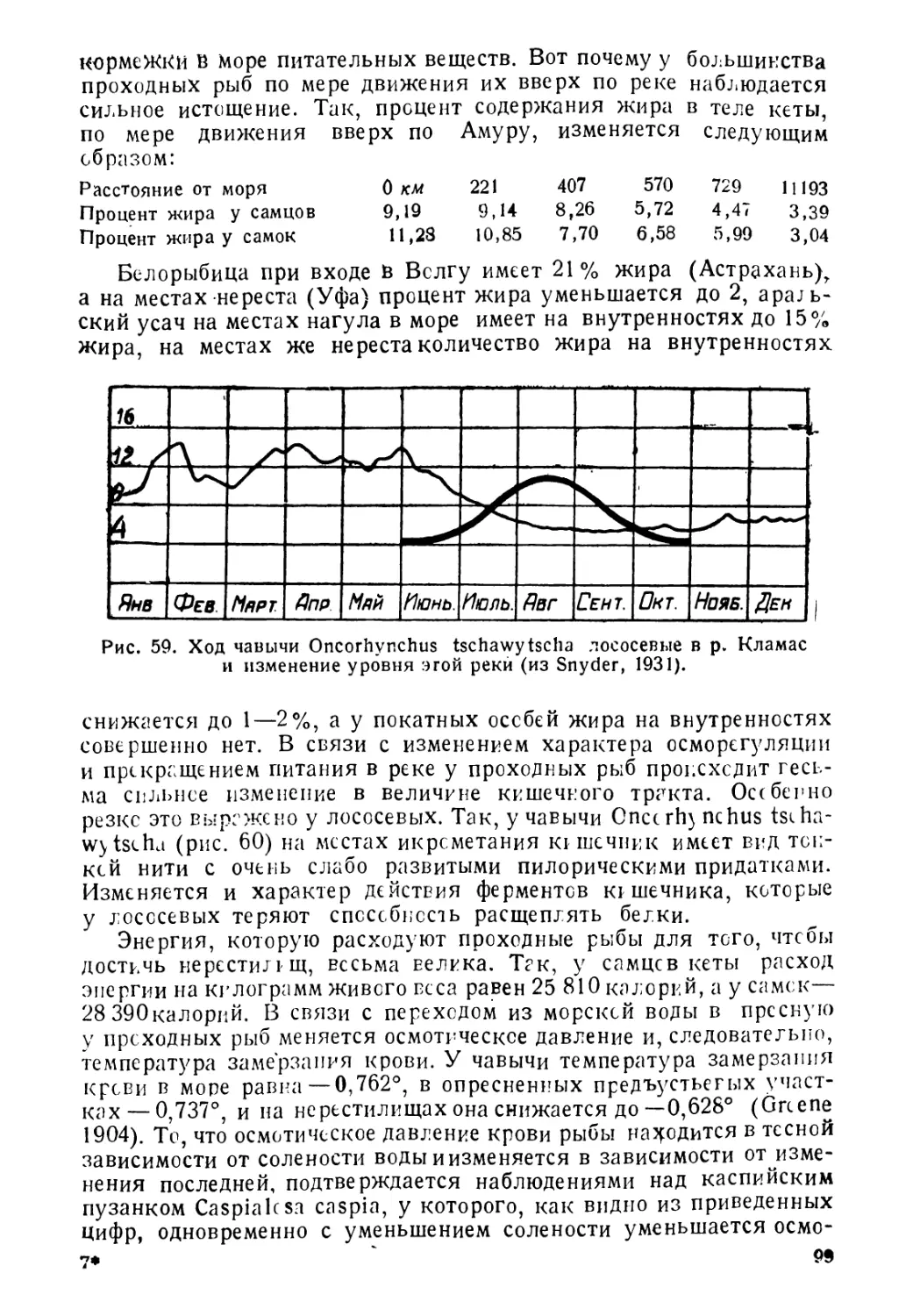


















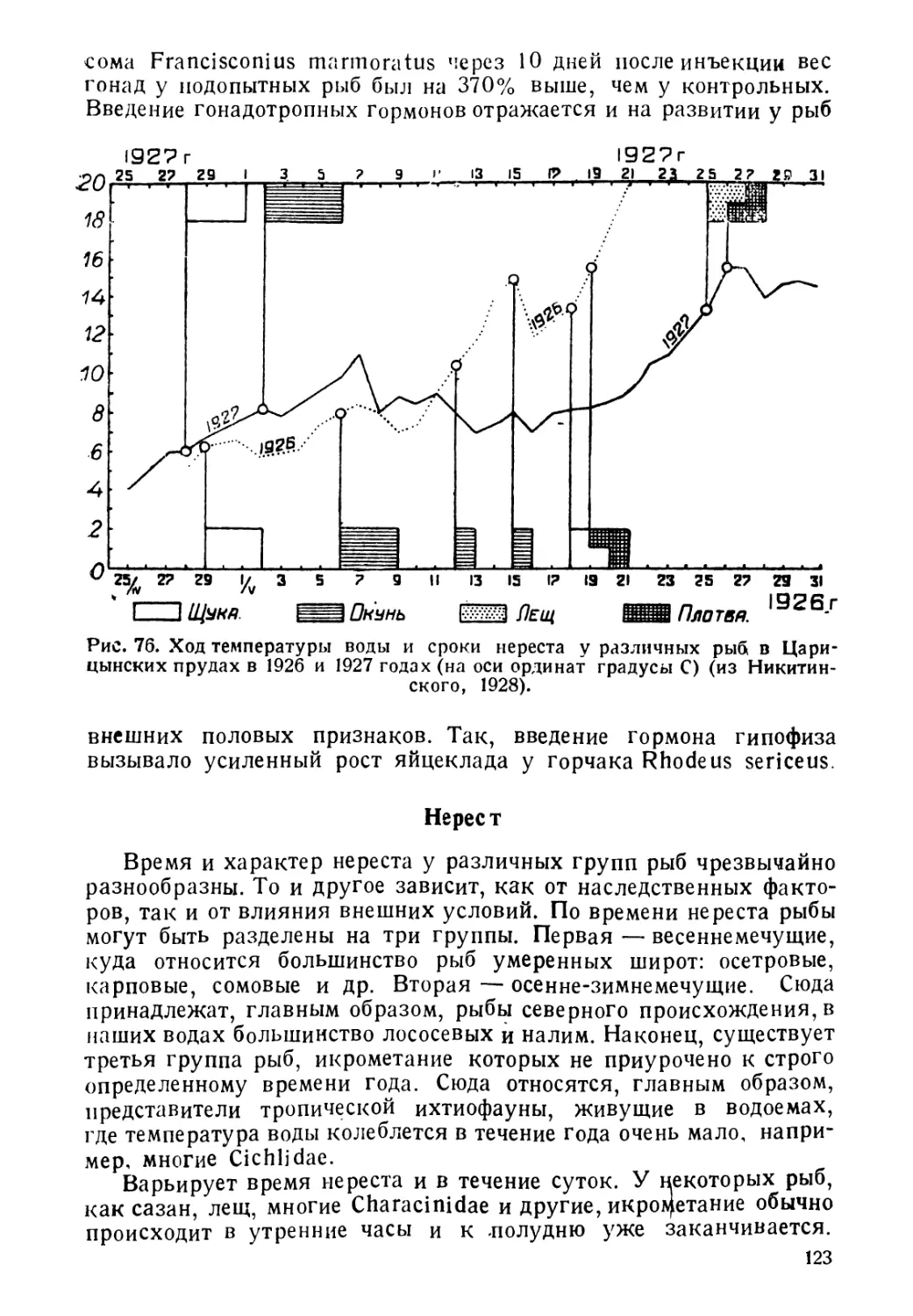

































































































Author: Никольский Г.В.
Tags: биология ихтиология рыбы пособие для студентов рыбная промышленность зоология позвоночных
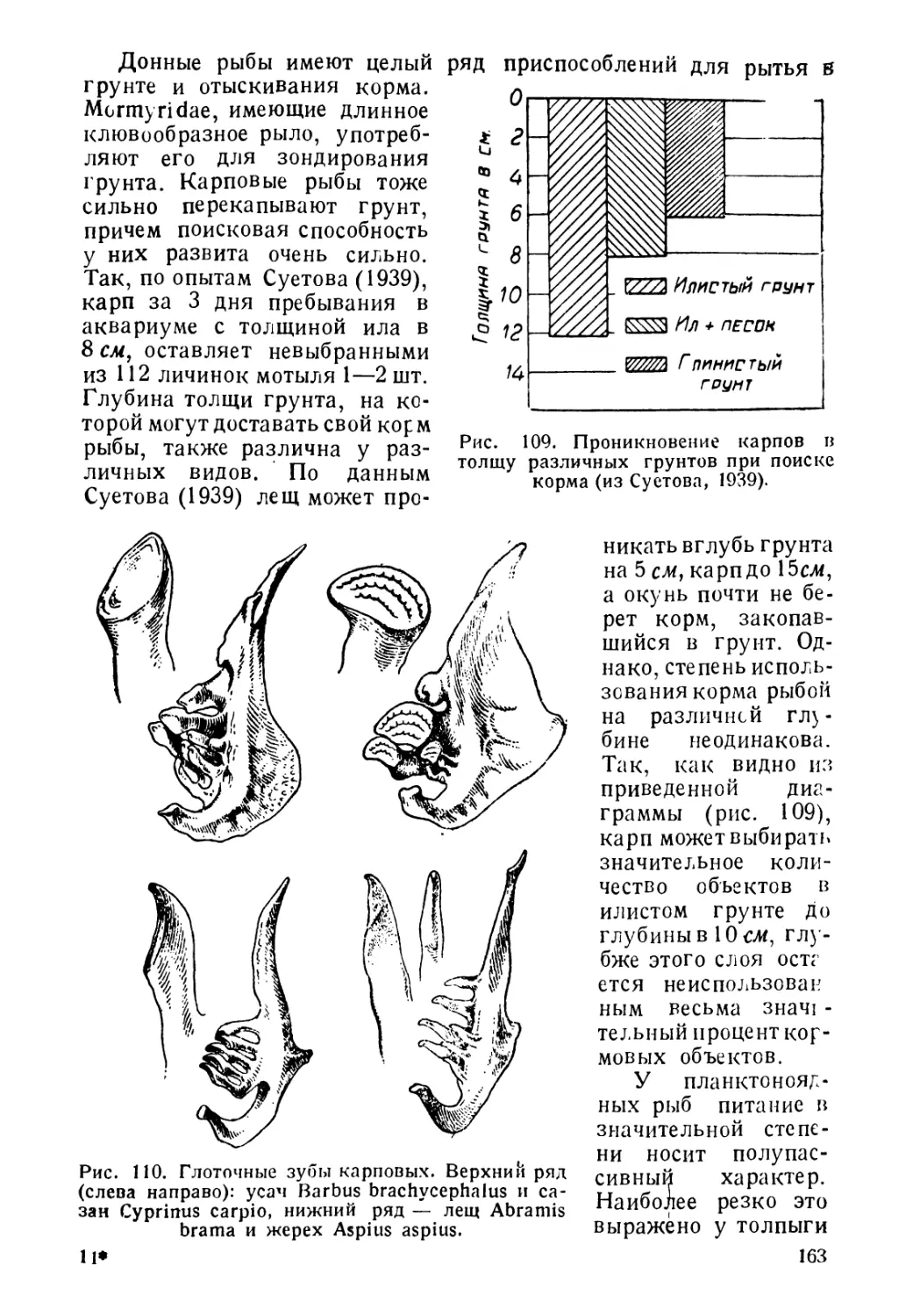
Year: 1944




Text
Проф. Г. В-НИКОЛЬСКИЙ
БИОЛОГИЯ
РЫБ
СОВЕТСКАЯ НАУКА-
Проф. Г В. НИКОЛЬСКИЙ
БИОЛОГИЯ РЫБ
Допущено Всесоюзным Комитетом
по делам высшей шко^ы п[ и СИ К
СССР в качестве учебника для био-
логических факультетов и институ-
тов рыбной промышленности
I
ГОСУДАРСТВЕННОЕ ИЗДАТЕЛЬСТВО «СОВЕТСКАЯ НАУКА*
МОСКВА • 1944
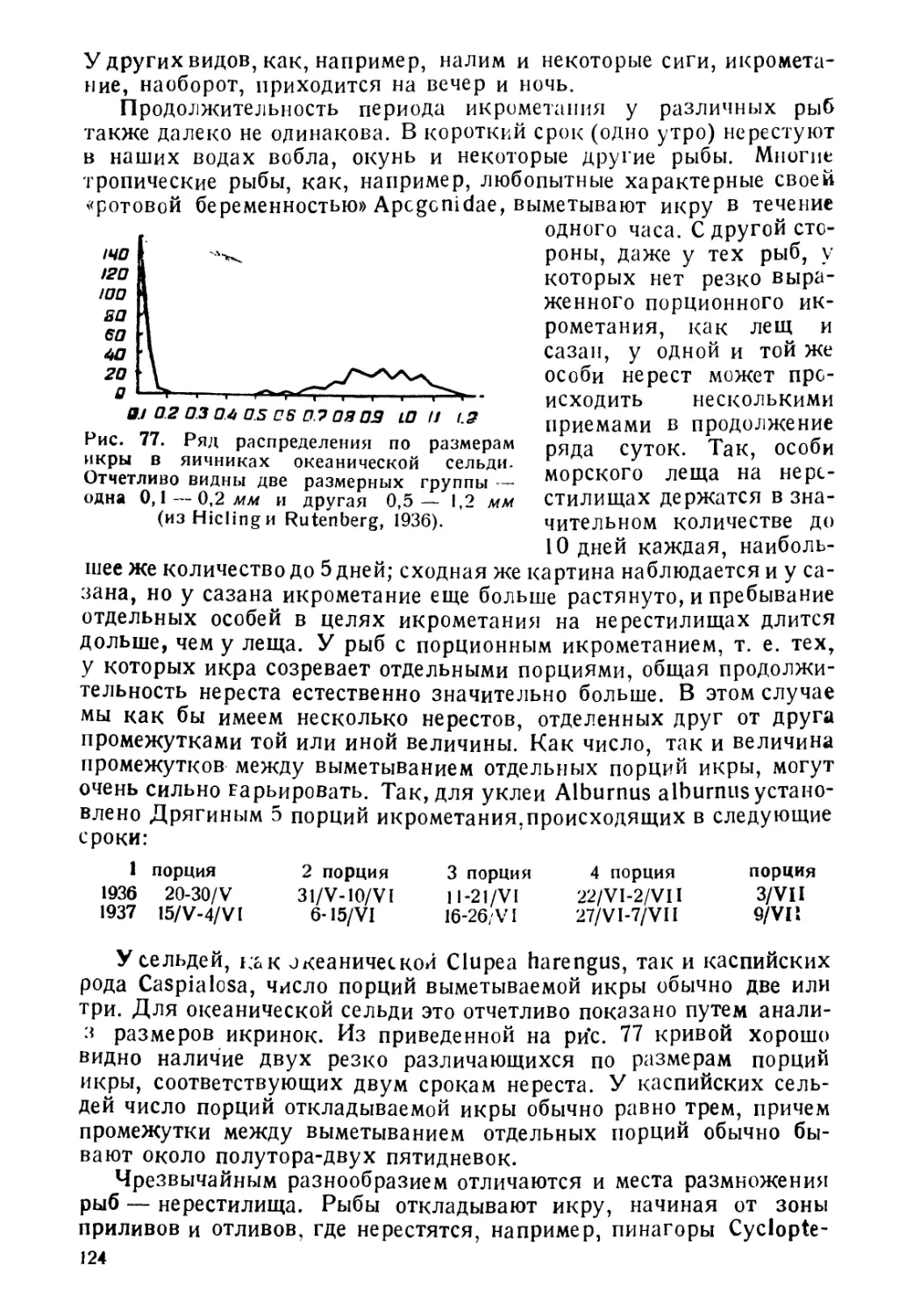
Книга «Биология рыб» предназначается для сту-
дентов Биологических факультетов университетов и
институтов рыбной, промышленности, для которых
ихтиология не является основной специальностью.
В основу книги положен курс, который автор в
течение ряда лет читал для студентов-биологов Мо-
сковского университета, специализирующихся по
зоологии позвоночных.
В отличие от иностранных руководств подобного
типа, предлагаемая книга построена несколько по
иному плану: первая часть посвящена рассмотрению
взаимоотношений рыб с факторами внешней среды
неорганической и органической. Вторая часть содер-
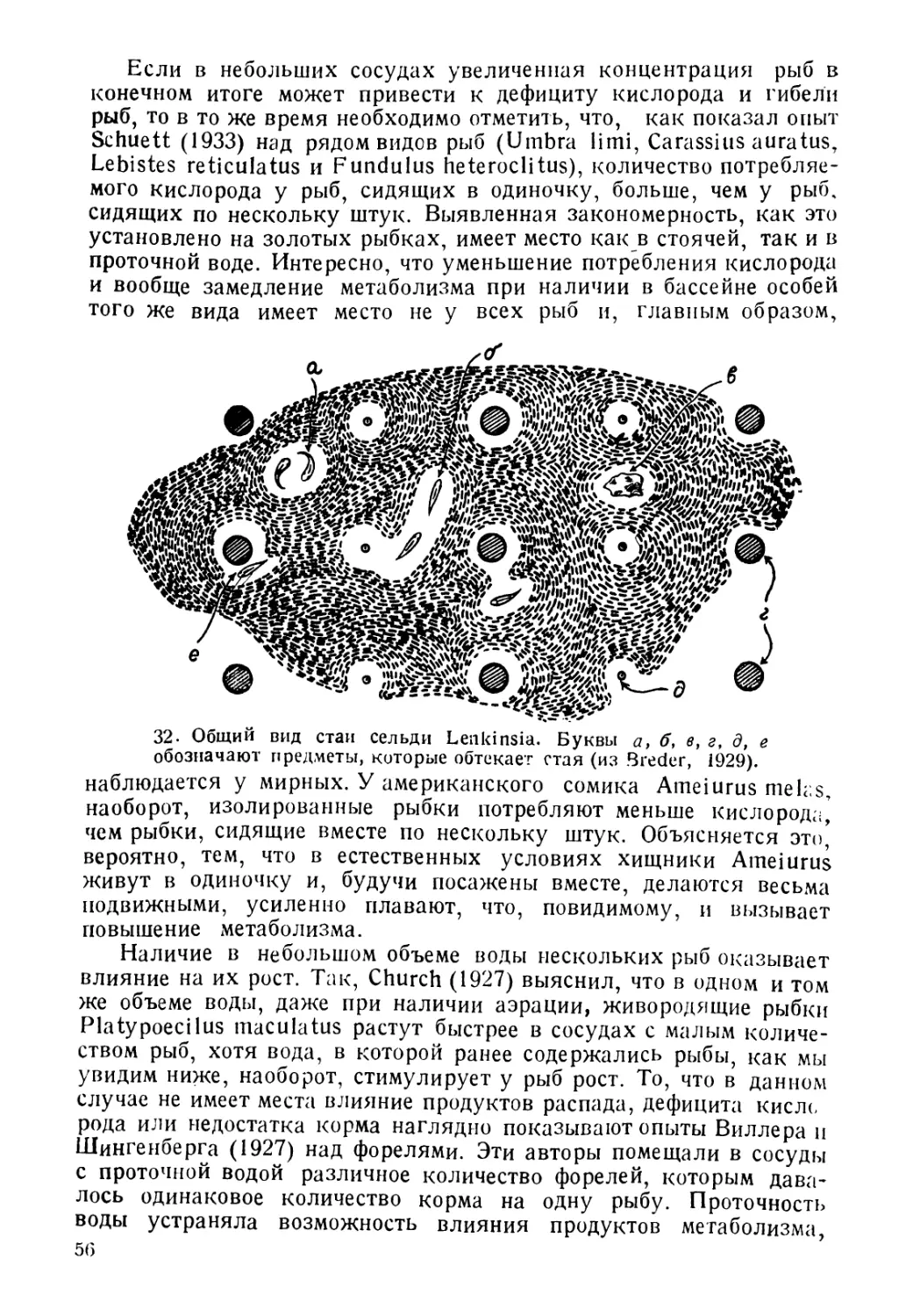
жит обзор основных звеньев жизненного цикла рыбьи
Наконец, третья представляет краткий обзор образа
жизни основных промысловых групп.
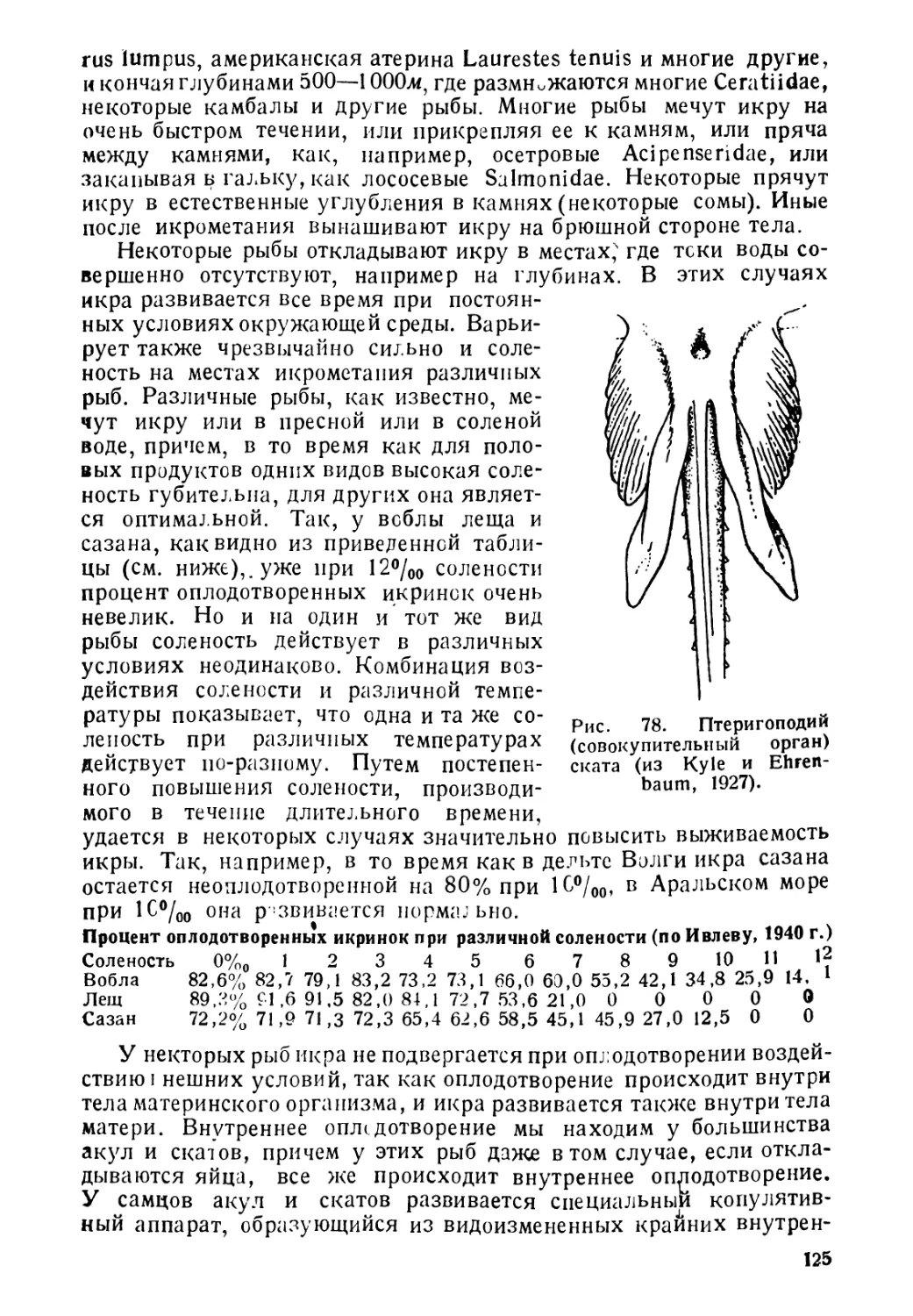
ВВЕДЕНИЕ
Из общей поверхности земного шара, определяющейся прибли-
зительно в 510 миллионов кле2, около 361 миллиона км2, т. е.
71% всей площади занято поверхностью океанов и морей. Кроме
того, около 2,5 миллионов км2, т. е. около % % площади земного
шара занято внутренними водоемами. Максимальная известная
глубина океана достигает около 11 000 м. Площадь океанов с
глубинами свыше 3 000 м занимает 51—58% (по различным
авторам) всей площади морских вед. Таковы размеры той жизнен-
ной арены, на которой распространены рыбы. Совершенно естествен-
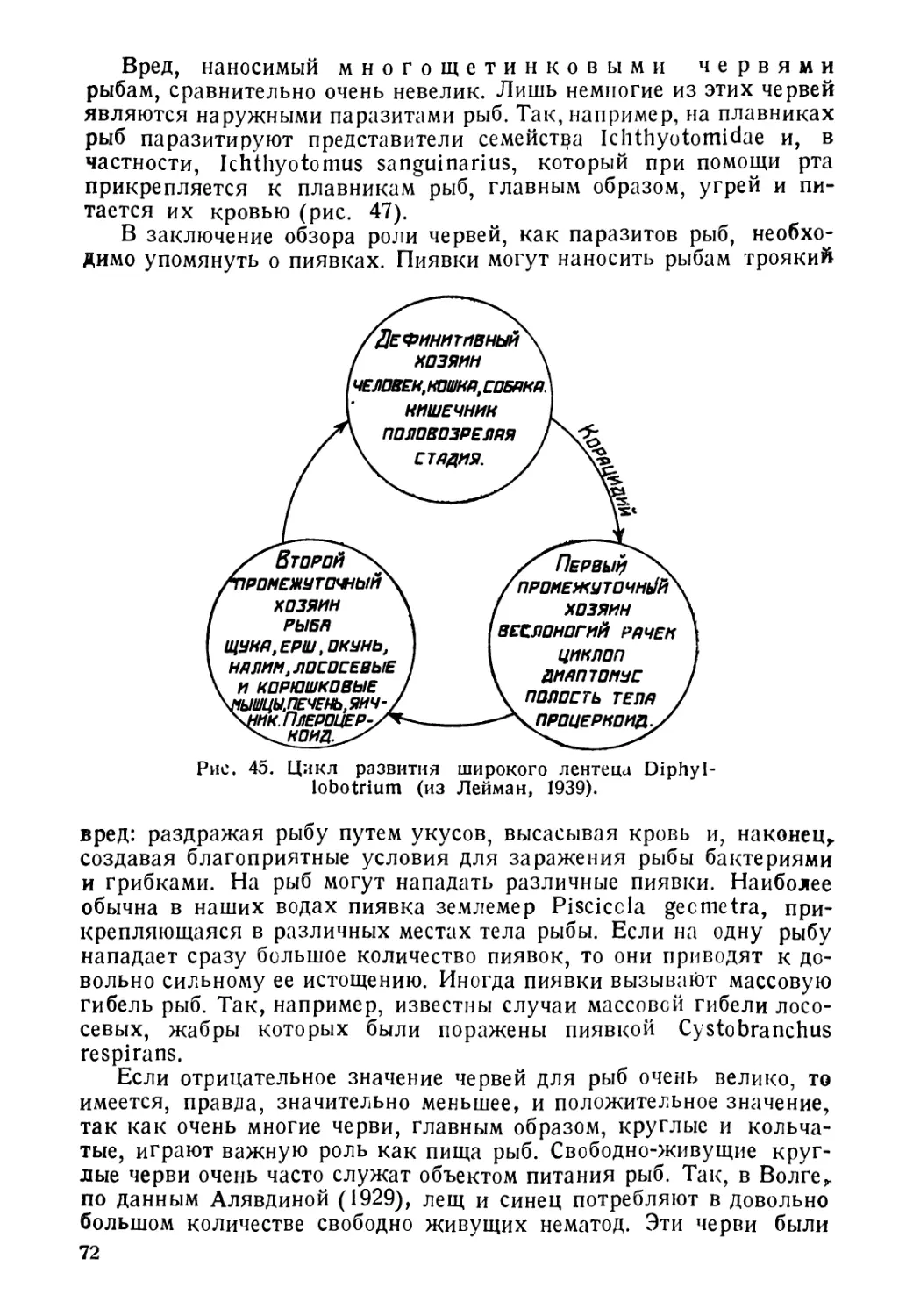
но, что водная среда, охватывающая земной шар от полюсов до
экватора и встречающаяся в пределах высот около 6 000 м над
уровнем океана и глубин свыше 10 000 м, чрезвычайно разнообраз-
на по условиям существования для рыб. Поскольку рыбы, следуя
в своем распространении за водной средой, также распространены,
начиная от высокогорных водоемов и кончая океаническими глу-
бинами, и встречаются, как в полярных водах, так и под тропи-
ками, постольку естественно, что для жизни в столь разнообразных
условиях они должны были выработать у себя целый ряд весьма
различных приспособлений, позволяющих им существовать и в
горных потоках, со скоростью движения воды до 2 и более метров
в секунду, и на огромных глубинах свыше 10 000 м, где давление
достигает до 1000 атмосфер. Столь же велика и амплитуда
приспособлений к такому фактору, как температура. Рыбы жи-
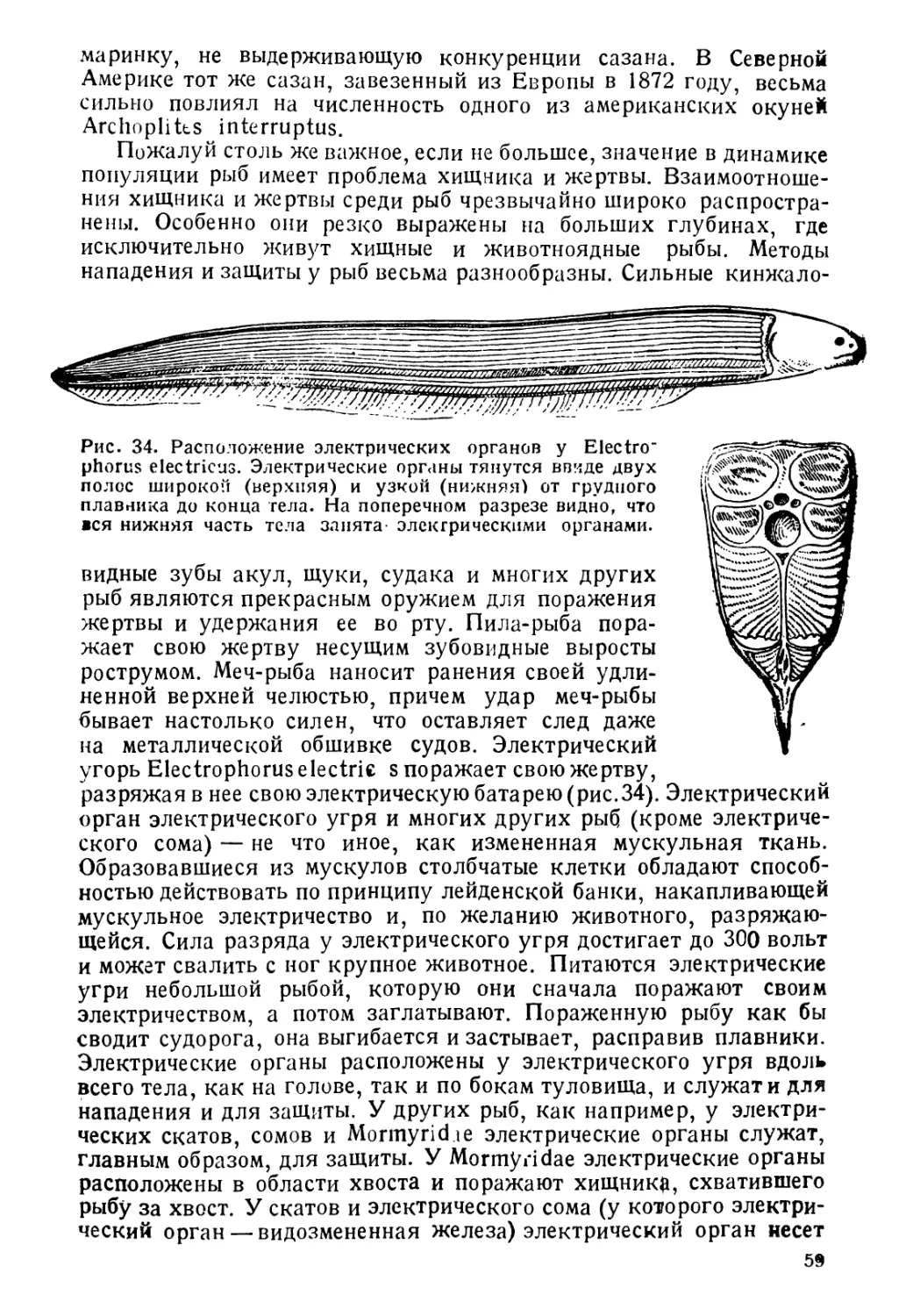
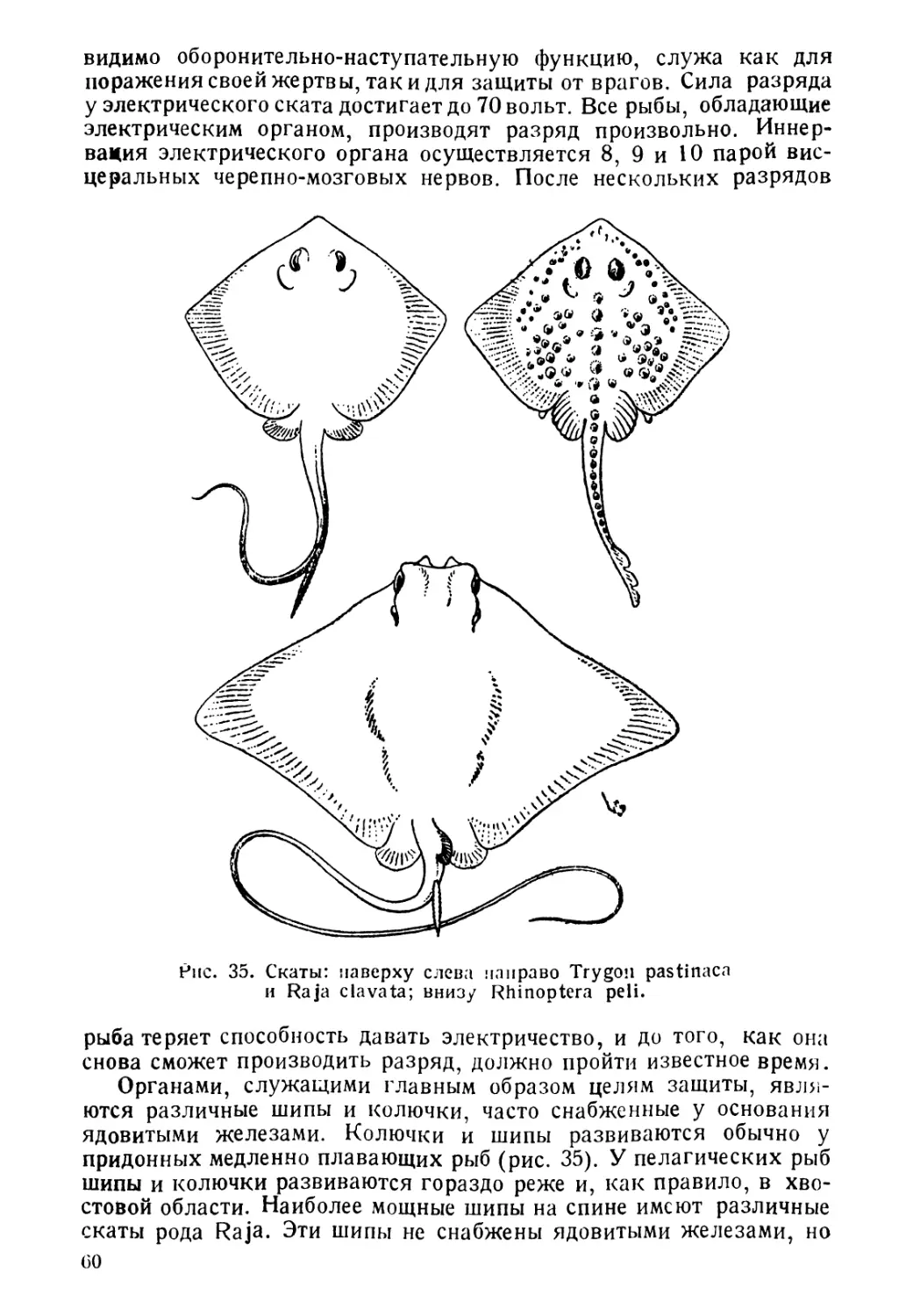
вут и при температурах, близких к температурам замерзания
соленой воды (—2),—3°) и при + 50°С. Точно так же ве-
лика и амплитуда колебаний солености, при которой могут
жить рыбы.
Столь разнообразные условия существования естественно спо-
собствуют адаптивной радиации организмов и несомненно, что
этим объясняется тот факт, что средй позвоночных рыбы являются
классом, наиболее богатым видами. Таким образом, морфологи-
ческое разнообразие рыб неразрывно связано с разнообразием их
образа жизни. Но все же, несмотря на все разнообразие условий
существевания, рыбы в соответствии с их образом жизни могут
быть разбиты на определенные группы. Сейчас обычно принимается
следующая классификация рыб, согласно их образу жизни. Прав-
да, и эта классифеКения далеко не идеальна и несомненно нуж-
дается в значительней доработке и уточнении. Придерживаясь ее,
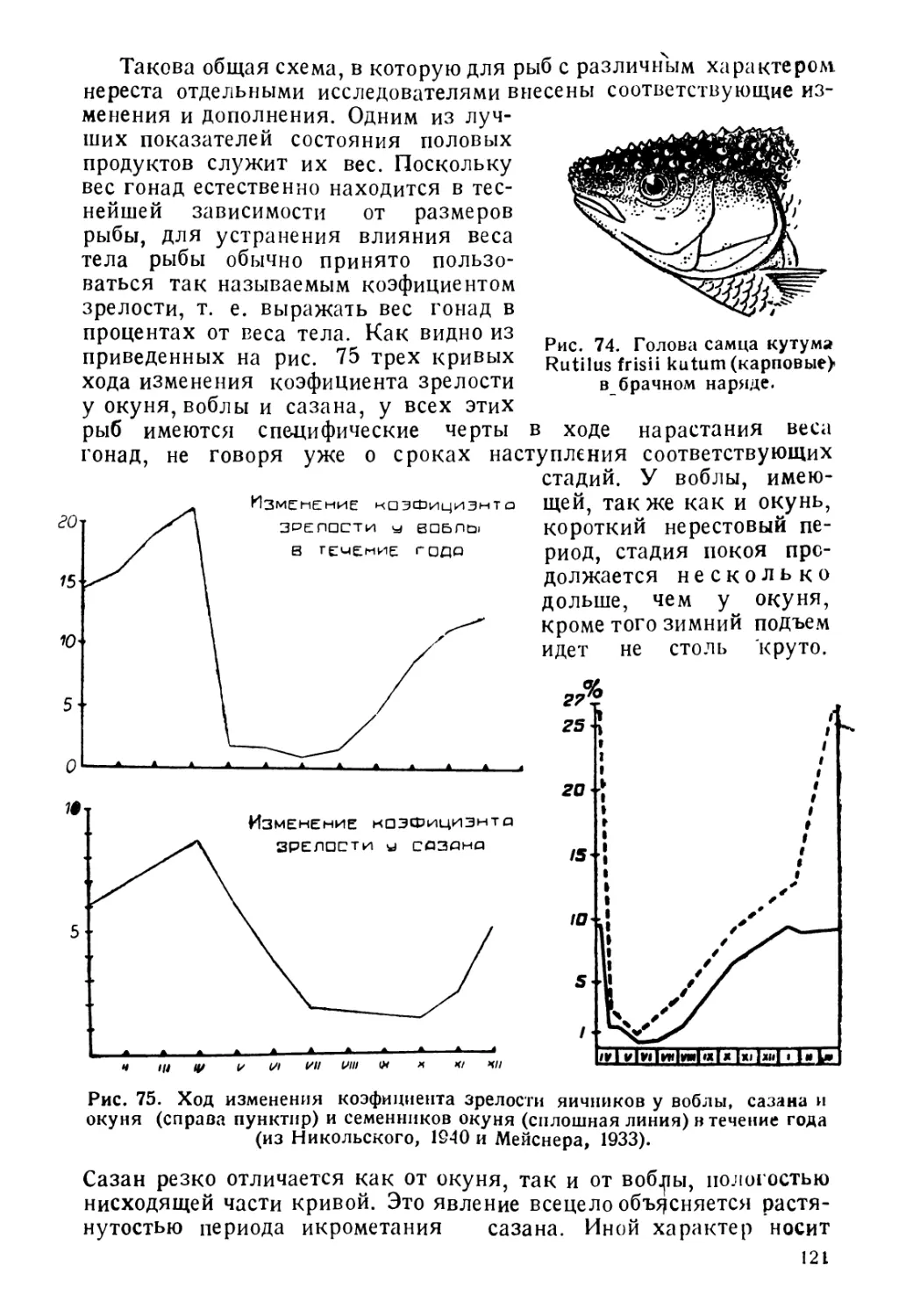
рЫ5 можно разделить на: морских, пресноводных, 'проходных и
солоноватоводных.
3
1. Морские рыбы живут и размножаются в морской соленой
воде, избегают пресных и Даже опресненных вод и, будучи по-
мещены в пресную воду, как правило, погибают. Морские рыбы
подразделяются на пелагических и донных.
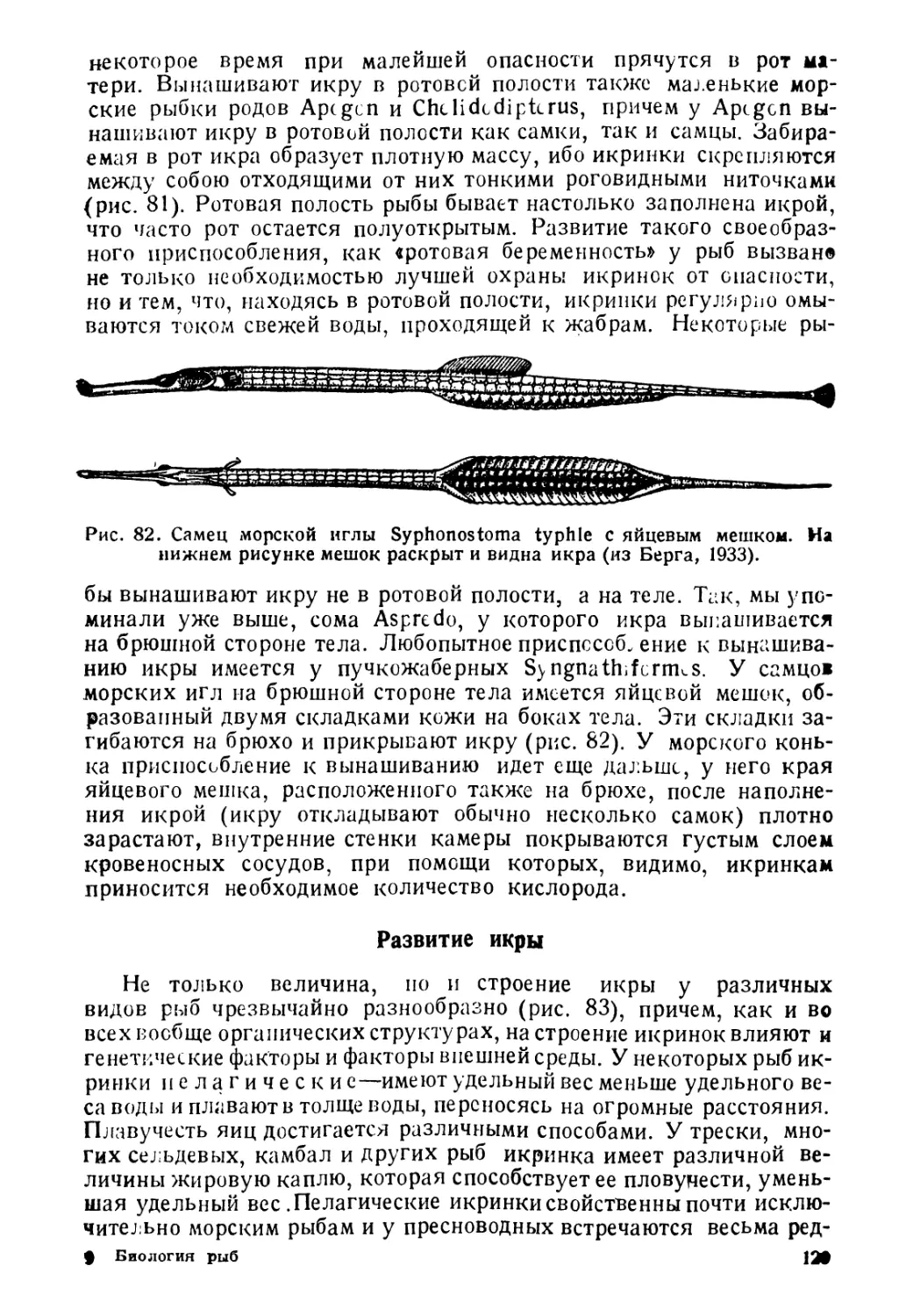
А. Пелагические рыбы живут в толще воды, начиная
от поверхности вплоть до больших глубин (батипелагические).
Размножаются они или в толще воды, или для размножения под-
ходят к берегу.
Сюда относятся, например, сельдевая акула, хамса, скумбрия
и Др.
Б. Донные рыбы живут в непосредственной близости
от дна, обычно пользуясь дном как опорой. Распространены, начи-
ная от прибойной зоны и до наибольших глубин. К этой группе
принадлежат многие камбалы, бычки и др.
Некоторые авторы считают необходимым каждое из указанных
подразделений в свою очередь разбить еще на рыб мигрирующих,
совершающих более или менее значительные периодические переме-
щения, и рыб оседлых, постоянно держащихся в одном и том же
месте.
II. Пресноводные рыбы постоянно живут в пресной воде и,
как правило, вморскей и даже ссолоненнсй воде не встречаются.
Эта группа распадается в свою очередь на 3 подгруппы: речных,
рыб стоячих вод и общепресновсдных.
А. Р еч н ы е рыбы — реофильные. Почти всю жизнь прово-
дят в текучих водах. Так же, как и морские, они распадаются
на: 1) пелагических, как например, уклея, амурская вост-
робрюшка и др. и 2) донных, держащихся обычно между
предметами, лежащими на дне и, как правило, в отличие от
речных пелагических рыб, плохо плавающих, например, пескари,
гольцы, подкаменщики.
В свою очередь большинство из'этих групп речных рыб может
быть подразделено на рыб, размножающихся в реке, там же, где они и
живут все время, и рыб, уходящих для размножения из русла реки
в придаточные водоемы поймы.
Б. Рыбы стоячих вод — лимнофильные. Населяют
стоячие водоемы — озера и пруды, текучих вод избегают. Под-
разделяются на: 1) пелагических, как например, многие
европейские и американские озерные сиги, американская озерная
кумжа и др. и 2) придонных, как например,карась,четырех-
рогий бычок.
В. Общепресноводные рыбы — водящиеся как в
стоячих, так и в текучих водах; почти исключительно обитатели
толщи воды, держащиеся обычно в одиночку или только периоди-
чески собирающиеся в стаи.
Две последующие группы рыб являются промежуточными между
морскими и пресноводными и несомненно произошли как от тех,
так и от других.
III. Проходные рыбы, совершающие для икрометания переход
или из моря в реки, или из рек в море, делятся на:
4
А. Питающиеся в море, которые, в свою очередь,
разделяются на:
а) пелагические, как например, сельдь менхеден и
I) донные, как русский осетр.
Б. П и тающиеся в реках. Например, угорь.
IV Солоноватоводные рыбы населяют опресненные участки
окаенов и морей перед устьями рек и внутренние неполносоленые
моря. В свою очередь могут быть разделены на полупроходных и
жилых.
А. По л»у и ро ходные — заходящие для размножения в
реки, но высоко по рекам не поднимающиеся. Таковы вобла, сом
и др.
Б. Ж и л ы е рыбы постоянно населяют солоноватые воды,
где кормятся и размножаются. В свою очередь делятся на пелаги-
ческих и донных.
Естественно, что наряду с таким биологическим разнообразием,
связанным со столь же большим разнообразием морфологическим,
у рыб имеется ряд черт строения, вызванных вообще жизнью в
водной среде и поэтому присущих всей группе. Такими признаками
являются в первую очередь жабры, обеспечивающие дыхание в
водной среде, конечности в виде плавников, способствующие пере-
движению в воде, хвост, который служит органом поступательного
движения и горизонтальным рулем, и кожа, богатая железами,
которые выделяют слизь, способствующую уменьшению трения
тела с окружающей средой. Правда, у незначительного количества
рыб три последних признака могут быть несколько видоизменены,
но для всей группы рыб в целом приведенные признаки являются
наиболее характерными.
Рыба, живя в водной среде, подвергается воздействию целого
ряда различных факторов, как неорганической, так и органической
среды. Эти факторы служат сильнейшим орудием отбора, который
позволяет существовать в том или ином участке гидросферы только
определенным, к данным условиям приспособленным формам.
В свою очередь и рыбы оказывают значительное влияние на окружа-
ющую их неорганическую и органическую среду, подвергая ее
зачастую весьма сильным изменениям.
На фоне взаимоотношений организма рыбы с окружающей сре-
дой проходят основные звенья ее жизненного цикла: размножение,
рост и т. д. Знание образа жизни рыб имеет огромнее значение
для человека. Используя большое количество рыбы, человек стре-
мится к наиболее полному освоению рыбных запасов, однако, к
такому освоению и использованию, которое, давая максимальный
эффект, в то же время не приводило бы к подрыву запасов объектов
промысла. Подобного рода рациональное рыбное хозяйство, есте-
ственно, невозможно без знания основных черт образа жизни про-
мысловых рыб. I
ЧАСТЬ I
ВЗАИМООТНОШЕНИЯ РЫБ С ФАКТОРАМИ ВНЕШНЕЙ СРЕДЫ
А* ВЛИЯНИЕ ФИЗИЧЕСКИХ СВОЙСТВ ВОДЫ НА РЫБ
Плотность и давление воды
Плотность воды, как известно, зависит от давления, под которым
находится вода, от содержания в воде солей и от температуры.
С повышением количества растворенных в воде солей плотность воды
повышается. Наоборот, с повышением температуры плотность
уменьшается.
Живое вещество, как правило, тяжелее воды, — его удельный
вес от 1,02 до 1,06. Следовательно, рыба, чтобы держаться в толще
воды, должна обладать какими-то специальными приспособлениями,
которые могут быть, как мы увидим ниже, довольно разнообразны.
Основным органом, при помощи которого рыбы могут регули-
ровать свой удельный вес, а следовательно, и приуроченность к
определенным слоям воды, является плавательный пузырь. Лишь
немногие рыбы, обитатели толщи воды, не имеют плавательного
пузыря. Нет плавательного пузыря у акул и некоторых скумбрие-
вых. Эти рыбы регулируют свое положение в том или ином слое воды
при помощи движения своих плавников.
У примитивных костистых рыб, как мягкоперые и костнопузыр-
ные, а также и у всех нижестоящих групп рыб, как двоякодышащие,
многоперовые, костные и хрящевые ганоиды, плавательный пузырь
соединен с кишечником при помощи протока.
У более специализированных костистых рыб, как колючеперые,
трескообразные и др., плавательный пузырь у взрослых особей с
кишечником не соединяется. Как показали наблюдения Ледебура
(Ledebur) и ряда других авторов,, наполнение газом плавательного
пузыря происходит у открытопузырных рыб и у рыб с замкнутым
пузырем не сразу по выходе из икринки. Пока личинки проходят
стадию покоя, подвесившись к стеблям растений или лежа на дне,
газа в плавательном пузыре у них нет. Наполнение плавательного
пузыря газом происходит из атмосферы. У рыб, у которых во взрос-
лом состоянии ductus pneumaticus (проток, соединяющий кишечник с
пузырем) отсутствует, наполнение пузыря газом у личинок проис-
ходит также через этот впоследствии редуцирующийся проток. Это
наблюдение подтверждается следующим опытом. Из икры колюче-
перых рыб выводились личинки в таком сосуде, поверхность воды
в котором была отделена от дна тонкой сеткой, непроницаемой для
личинок. В естественных условиях наполнение пузыря газом про-
6
исходит у колючеперых на 2-й — 3-й день по выходе из икры. В
опытном же сосуде рыбы выдерживались до 5—8-дневного возраста,
после чего преграда, отделявшая их от поверхности, удалялась.
Однако, к этому времени связь между плавательным пузырем и
кишечником прерывалась, и пузырь оставался ненаполненным
газом. Таким образом, первоначальное наполнение плавательного
пузыря газом и у открытопузырных, и у рыб с замкнутым плаватель-
ным пузырем происходит одинаково. Дальнейшее же регулирование
количества газа в плавательном пузыре идет уже различными спо-
собами. У тех рыб, у которых плавательный пузырь соединен с
кишечником, поступление и выделение газа из плавательного пу-
зыря происходит в значительной степени через кишечник. У рыб
с замкнутым плавательным пузырем, после первоначального напол-
нения газом из атмосферы, дальнейшие изменения количества и
состава газа происходят путем абсорбции газа кровью. У рыб с замк-
нутым пузырем на внутренней стенке пузыря имеется так называемое
красное тело, чрезвычайно густо пронизанное кровеносными
капиллярами образование. Так, в двух красных телах, находящихся
з плавательном пузыре угря, имеется 88 000 венозных и 116 000
зртериальных капилляров общей длиной 352 и 464 м. В то же время
збъем всех капилляров в красных телах у угря всего лишь 64 льи3,
г. е. не больше капли средней величины. Красное тело варьирует
/ различных рыб от небольшого пятна, имеющегося даже у некоторых
зткрытопузырных рыб, до мощной газоотделительной железы,
юстоящей из цилиндрического железистого эпителия.
Состав газа в плавательном пузыре рыб различен как у различ-
<ых видов, так и у разных особей одного и того же вида. Так, у
линя количество кислорода обычно около 8%, у окуня 19—25%,
! щуки около 19%, у плотвы 5—6%. Так как из кровеносной системы
«огут проникать в плавательный пузырь почти исключительно
сислород и углекислота, то при наполнении газом пузыря в нем
Преобладают именно эти газы, азот же составляет весьма малый
фоцент. Наоборот, при удалении газа из плавательного пузыря
крез кровеносную систему, в пузыре резко возрастает процентное
«держание азота. Как правило, у морских рыб количество кисло-
юда в плавательном пузыре больше, чем у пресноводных. Объяс-
няется это, видимо, главным образом, тем, что в море преобладают
вйбы с замкнутым плавательным пузырем. Особенно велико коли-
истро кислорода в плавательном пузыре у вторично глубоководных
ШВеличина давления газа в плавательном пузыре у рыб обычно
'ЧМ’ИЛи иным путем передается к слуховому лабиринту. Так, у
ещэдевых, тресковых и некоторых других рыб передняя часть
[лабательного пузыря имеет парные выросты, которые доходят
.□затянутых перепонкой отверстий слуховых капсул (как у треско-
вых) или даже входят внутрь последних (например, у сельдевых). У
Юетнопузырных передача давления плавательного пузыря к лаби-
ринту осуществляется при помощи так называемого Вебе рова аппара-
а ряда косточек (по три с каждой стороны), соединяющих плава-
7
плавательного пузыря
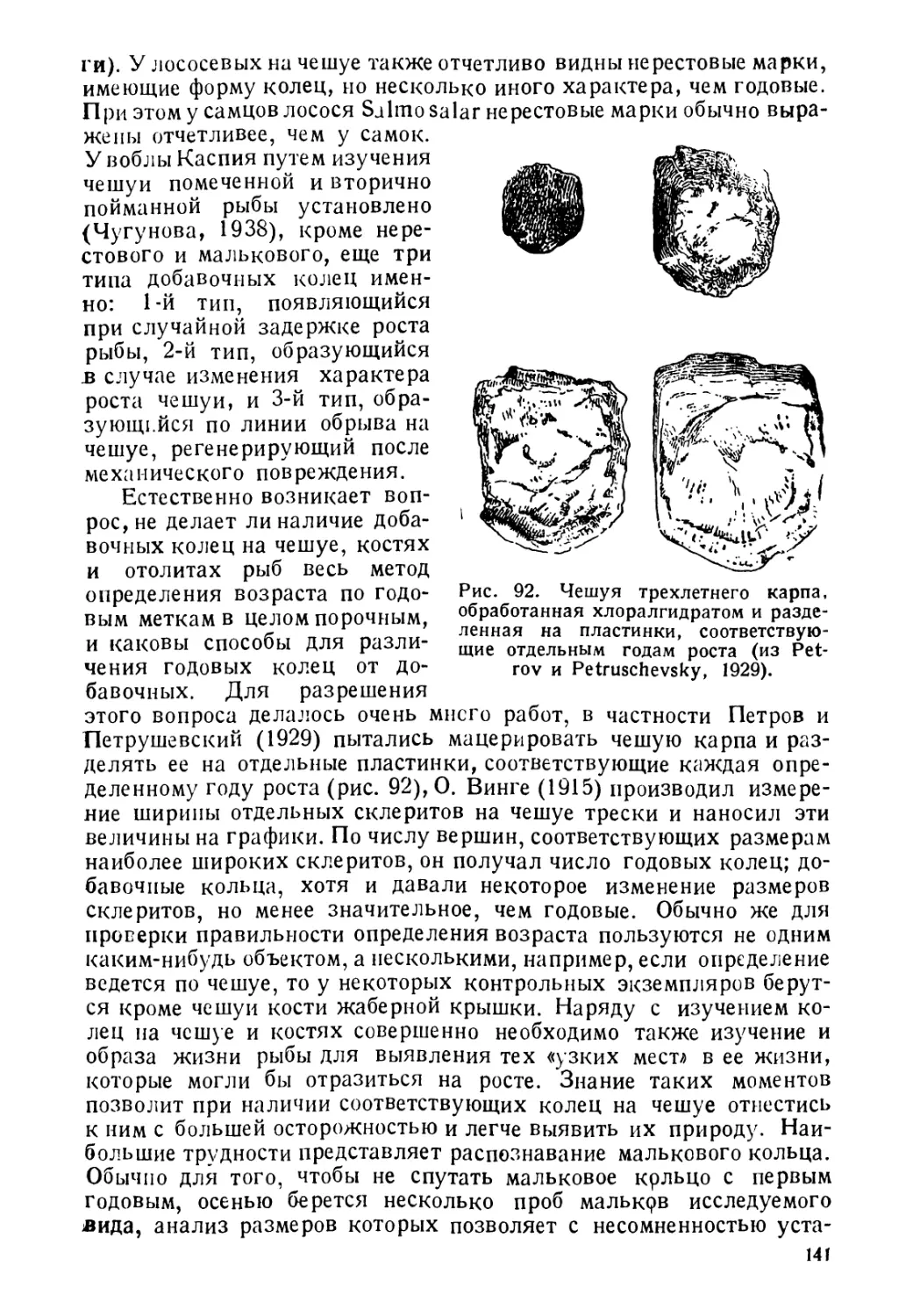
Рис. 1 Схема соединения
с органом слуха (слева направо) у сельди Clup’ea
harengus (сельдеобразные), карпа Cyprinus carpio
(карпообразные) и Physiculus japonicus (треско-
образные) (из Kyle и Ehrenbaum, 1926, Wunder,
1936 и Световидов, 1937).
тельный пузырь с лабиринтом. Эти косточки, служащие также, ве-
роятно, и подсобным органом звукоулавливающего аппарата, яв-
ляются видоизмененными частями передних позвонков (рис. 1).
Как уже отмечалось, рыба не может быстро изменять давление
газа в плавательном пузыре, и поэтому при подъеме на поверхность
происходит выдавливание плавательного пузыря через рот или
раздувание тела. Это делает понятным, почему у рыб, которые
совершают быстрые переходы с больших глубин на поверхность и
обратно, плаватель-
ный пузырь отсут-
ствует, и погруже-
ние в глубину и
поднятие на поверх-
ность происходит
при помощи мус-
кульных движений.
Редуцируется плава-
тельный пузырь так-
же у многих при-
донных рыб, напри-
мер, у многих быч-
ков, некоторых мор-
ских собачек В1ея-
nius и др. Редукция
пузыря у этих рыб
вызвана тем, что они
держатся все время,
касаясь дна, и для
них биологически
целесообразнее иметь
удельный вес тела
больший, чем вес
воды. Сокращается
также плавательный
потоков. Так, напри-
свободная от костной
пузырь и у донных рыб быстротекущих
мер, у различных речных вьюновых задняя
капсулы часть пузыря редуцируется, и сохраняется лишь передняя
часть, заключенная в костную капсулу и играющая, видимо,
главным образом роль звукоулавливающего аппарата. У некото-
рых рыб, в том числе у отдельных видов бычков Gobiidae плава-
тельный пузырь сохраняется всю жизнь, как например, у Aphya.
У других плавательный пузырь имеется только у личинок, веду-
щих пелагический образ жизни, как например, у Gcbius niger.
У третьих и личинки и взрослые особи совершенно лишены
плавательного пузыря. К этой группе относится Gobius mela-
nostomus.
Как указывалось выше, давление воды очень быстро возрастает
с глубиной, и естественно, что рыбы, живущие на глубинах, должны
обладать рядом специальных приспособлений для того, чтобы не
быть уничтоженными тем огромным давлением, которое существует
на глубине. Все глубоководные рыбы характеризуются слабым
развитием скелета и мускулатуры. Давление внутри тела рыбы,
влагодаря проницаемости ее тканей, устанавливается столь же
высокое, как и в наружной среде, и это делает возможным жизнь
рыбы на больших глубинах (рис. 2).
Большинство глубоководных рыб обладает плавательным пу-
зырем, не соединенным с кишечником при помощи протока ductus
pneumaticus. Те глубоководные рыбы, которые принадлежат к
группам, имеющим соединение плавательного пузыря с кишечни-
ком, приспособившись к глубоководной жизни, теряют вообще
плавательный пузырь. Биологический смысл это редукции вполне
понятен, так как естественно, что при тех больших давлениях, ко-
торые существуют на глубинах, воздух из плавательного пузыря,
соединенного с кишечником, неминуемо должен был бы быть вы-
давлен.
По своему происхождению глубоководные рыбы неоднородны.
Они ргзко распадаются на две группы. Первая из них — истинно
глубоководные, которые принадлежат к таким семействам, все
представители которых живут на глубинах. Поэтому адаптивные
к глубоководности признаки обычно свойственны всему семейству
в целом. Большинство истинно глубоководных семейств—космо-
политы. И< глубоководность носит весьма древний характер.
К этой группе относятся Myctophiidae, Chauliodcntidae, Sto-
i'atidae, MelampHaenidae и др.
Ко вторей группе глубоководных рыб принадлежат виды, глу-
ководность которых исторически более поздняя. Это так называ-
ые вторично глубоководные рыбы. Сюда принадлежат формы
логенетически более молодые, главным образом представители
пючеперых. Обычно семейства, к которым относятся вторично
/боководные рыбы, в основной массе включают обитателей кон-
нентальной ступени. Приспособления к глубинной жизни у
)рично глубоководных рыб менее специфичны, чем у представи-
тей перво группы. Обычно они сводятся к студенистости и про-
чности тела, увеличению слизевых каналов, изменению окраски
’. д. Глубоководные представители этой группы имеются в семей-
>ах Cottidae, Liparidae, Blenniidae, Zoarcidae и др.
Большие давления воды оказывают влияние, естественно, не
пько на взрослых рыб, но и на икру. Так, как показали опыты
арег и Edwards (1934), до тех пор, пока давление ниже 13С0 фун-
i на 1 кв. дюйм, развитие Fundulus heteroclitus идет нормально.
( ри повышении давления с 1300 до 15С0 фунтов на 1 кв. дюйм ско-
рость деления клеток замедляется (вместо 30 минут—3 часа). Пуль-
сация сердца у эмбрионов Fundulus также становится под большим
давлением замедленной.
8^, Если высокие давления при жизни рыб на глубинах вызы-
перестройку всего их организма, то приспособление к жизни
» MtHiee глубоких слоях воды также требует известных гидростати-
ческих приспособлений, позволяющих удерживаться в том слое
9
10
юды, в каком это наиболее выгодно организму. Наиболее свое-
образны различные приспособления, развивающиеся у пелаги-
зеских личинок рыб и способствующие их парению в толще воды.
<ак известно, чем больше площадь по отношению к весу, тем боль-
шее сопротивление оказывает такое тело при погружении и тем
легче ему держаться в том или ином слое воды. Подобного рода
приспособления в виде различных шипов и выростов, увеличиваю-
щие поверхность тела и способствующие удержанию в толще воды,
развиваются у многих пелагических животных и в том числе у ли-
чинок рыб. Так, например, у пелагической личинки донной рыбы
морского чорта Lophius piscatorius (рис. 3) имеются огромные вы-
росты спинного и брюшных плавников, помогающие ее парению
в толще воды; подобные же изменения плавников наблюдаются и
у личинки Trachypterus. На теле личинок луны-рыбы Mola mola
имеются огромные шипы, так что эти личинки немного напоминают
увеличенную планктонную водоросль Ceratium. У некоторых
пелагических личинок рыб увеличение их поверхности идет путем
сильного уплощения тела. Подобного рода изменения наблюда-
ются, например, у личинок речного угря, у которых тело значитель-
но более высоко и плоско, чем у взрослых особей.
Чрезвычайно важным фактором, обусловливающим возмож-
ность приспособления рыб к жизни в водной среде различной плот-
ности, является более или менее полное постоянство внутренней
среды. Это постоянство достигается развитием различных осморе-
Еуляторных приспособлений, позволяющих регулировать осмоти-
ческое давление жидкостей тела. Чем совершеннее осморегулятор-
Йые приспособления, тем больше независимость состава и давления
внутренней жидкости животного от внешней среды. У рыб, в отли-
тие от большинства морских беспозвоночных, имеется разница
(ического‘давления жидкостей тела и внешней среды. По ха-
ру осмотического давления внутренней среды рыбы разде-
:я на три группы: 1) хрящевые рыбы, имеющие давление боль-
чем давление внешней среды, 2) морские костистые рыбы, у
ых осмотическое давление меньше, чем давление внешней
[ и 3) пресноводные рыбы, давление соков тела которых больше,
жружающей среды.
►став веществ, содержащихся во внутренних жидкостях рыб
i различен. У акул, кроме хлористого натрия, составляющего
12 до 1,77% веса жидкости, значительную роль играет еще и
;ина, которой в крови акул содержится от 2 до 2,6%, а у ска-
г 1,42 до 2%. У акул и скатов, держащихся в пресной воде,
ина также содержится в большом количестве в полостных
Рис. 2 Глубоководные рыбы.
fflgy^yptopsarus couesii (Lophiiformes)* 2—Nemichthys ayocetta (Anguil-
j _ Chauliodus sloani (Clupeiformes); 4 — Ypnops murrayi (Sco-
pwfoSiies); 5— Gastrostomus bairdi (Anguilliformes); 6 — Argyropelecus olfersii
(Scop£liformes); 7 — Neoliparis mucosus (Perciformes); 8 — Coelorhynchus car-
minatus (Macruriformes); 9 — Ceratoscopelus maderensis (Scopcliformes).
11
Рис. 3. Приспособления к парению у пелагических личинок рыб. Слева
снизу вверх— Trachypterus cristatus, Lophius piscatorius, Physiculus dalwij-
kii. Справа—Hymenocephalus italicus
12
жидкостях, причем осмотическое давление у пресноводных акул
и скатов выше, чем у пресноводных костистых рыб.
Морские костистые рыбы имеют в жидкостях внутренней среды
меньше солей, чем хрящевые, но, поскольку давление их жидкостей
меньше, чем окружающей среды, то и этих солей оказывается почти
достаточно, и роль органических соединений, как мочевина у
акул, у костистых рыб очень невелика. Органические соединения у
морских костистых рыб обусловливают лишь 7—8% осмотического
давления. У пресноводных рыб количество органического вещества
в крови примерно то же, что и у морских, но количество минераль-
ных солей довольно сильно понижается.
Круглоротые в отношении своего осмотического давления пред-
ставляют разнородную'группу. Миксина Myxine glutlncsa имеет в
полостных жидкостях большее количество мочевины и по этому
признаку приближается к хрящевым рыбам. Наоборот, у Polis-
totlfemi stouti осмотическсе давление в основном обусловлено солями.
Большинство рыб приспособлено к жизни в растворах более или
менее определенного осмотического давления, и перенесение их в
.среду с иным осмотическим давлением довольно скоро приводит к
!х гибели. Хрящевые рыбы, выработавшие у себя в организме
юханизм, позволяющий накапливать большее количество моче-
ины, достаточное, для поддержания гипертонии полостных жид-
:остей по сравнению с морской водой, при перенесении в пресную
оду 'очень быстро понижают осмотическое давление внутренних
жидкостей и погибают. Те немногие виды хрящевых рыб, которые
[риспособились к жизни в пресной воде, для сохранения высокого
биотического давления в жидкостях тела выработали у себя спо-
обность усиленного выделения мочи. При помощи этого приспособле-
ия из тела удаляется огромное количество воды (см. табл. 1). Почки
рыб задерживают мочевину, и основная экскреция мочевины у
Iостистых рыб (до 70%) происходит через жабры, у хрящевых же
ыб жабры плохо проницаемы для мочевины, что и дает этим рыбам
озможность накапливать мочевину в полостных жидкостях.
количество выделяемой мочи морскими и пресноводными хрящевыми рыбами
(на 1 кг веса рыбы в день)
Морские
Scyllium stellare . 5,25 см3
1 Scyllium catulus . .2—4 см3
Acanthias acanthias .4,7—12,2 см3
Пресноводные
£;• Pristis microdon . 250 см3
Морские костистые рыбы в значительной массе с т е н о Га-
лин н ы, т. е. приуроченные к определенной солености живот-
ные и, будучи перемещены в пресную веду, довольно быстро поги-
№При значительных колебаниях солености осмотическсе да-
J внутренних жидкостей меняется у них дсвсльно мало. Так,
ки при изменении температуры замерзания во|ы от 1,09 де
S крови изменяется всего с 0,73 до 0,76° (т. е. при изменении
внеШней среды на 74 % Д крови меняется на 4%). У морских кости-
13
стых рыб, в связи с тем, что внутренняя среда гипотонична по
отношению к внешней, имеется необходимость удержания воды в
организме. Следовательно, морские костистые рыбы должны полу-
чать из окружающей гипертонической внешней среды воду. Эта
необходимость усугубляется еще тем, что моча костистых рыб по
сравнению с морской водой или гипотонична, или в лучшем случае
изотонична. Пополнение водой организма морской костистой рыбы
происходит через кишечник. Все морские рыбы «пьют» воду, причем,
как установлено опытами над Myoxocephalus, рыбки заглатывают
ежедневно от 50 до 200 см3 воды на 1 кг веса. Прекращение доступа
воды в кишечник приводит к сильному исхуданию рыбы и к гибели
на 3—4-й день. Удаление солей из крови костистых рыб происходит
двояким путем: или через почки (главным образом двувалентные
ионы), или через жабры (одновалентные ионы).
Пресноводные рыбы имеют, как отмечалось выше, гипертони-
ческую, по сравнению с внешней средой, внутреннюю жидкость,
Быстрое перенесение пресноводных рыб в морскую воду вызывает
их гибель. Однако, если приучение к морской воде производить
постепенно, то рыба может выжить, но к концу опыта наступает
полная изотония между внешней и внутренней средой. В отличие
от морских рыб у пресноводных костистых основная задача организ-
ма сводится не к удержанию воды, а к удалению избыточного количе-
ства воды, поступающего осмотическим путем. Поэтому по сравне-
нию с морскими рыбами пресноводные костистые рыбы так же, как
это мы наблюдали у хрящевых рыб, как правило, выделяют значи-
тельно больше мочи. Проходные рыбы, как перемещающиеся для
икрометания из рекв моря и обратно, имеют осмотическое давление,
претерпевающее, правда, незначительные изменения. У проходных
рыб при переходе из морской воды в пресную, в результате деге-
нерации слизистых оболочек кишечника, происходит почти полное
прекращение поступления воды через кишечник в организм живот-
ного (см. ниже в главе о миграциях).
Таким образом, как видно из изложенного, приспособления
рыб к различному давлению воды и изменению ее плотности чрезвы-
чайно разнообразны, что естественно, способствует весьма широ-
кому расселению представителей этой группы.
Температура воды
Как известно, различные рыбы могут жить при самой разно-
образной температуре. Наиболее высокую температуру переносит
рыбка из семейства Cyprincdontidae, Cyprinodon macularius, которая
живет в небольших горячих источниках Калифе ры и и при темпе ра-
туре выше 52°С. С дгугей стороны, такие рыбы, как карась Carassius
carassius и далия Dalia pectoralis, выдерживают даже обмерзание,
правда, при условии, если соки тела остаются позамерзп г ми.
Наряду с приспособленностью к особенно высока м и сссбенно
низким температурам весьма важное значение для возможности
расселения и жизни в различных условиях имеет и амплитуда
14
температур, при которых могут жить рыбы. Амплитуда темпера-
тур, при которой живут различные виды рыб, далеко не-
одинакова, некоторые из них выдерживают колебания температуры
в несколько десятков градусов, другие же не переносят разницы
температуры в 5—7°. На основании этого признака рыб можно
разделить на эвритермных, выдерживающих широкие изме-
нения температуры, и стенотермных, не переносящих резких
изменений температуры.
Основное различие между Животными, обладающими «теплой
кровью», и животными холоднокровными заключается в количе-
ственной стороне процесса теплообразования. У холоднокровных
животных этот процесс идет значительно медленнее, чем у тепло-
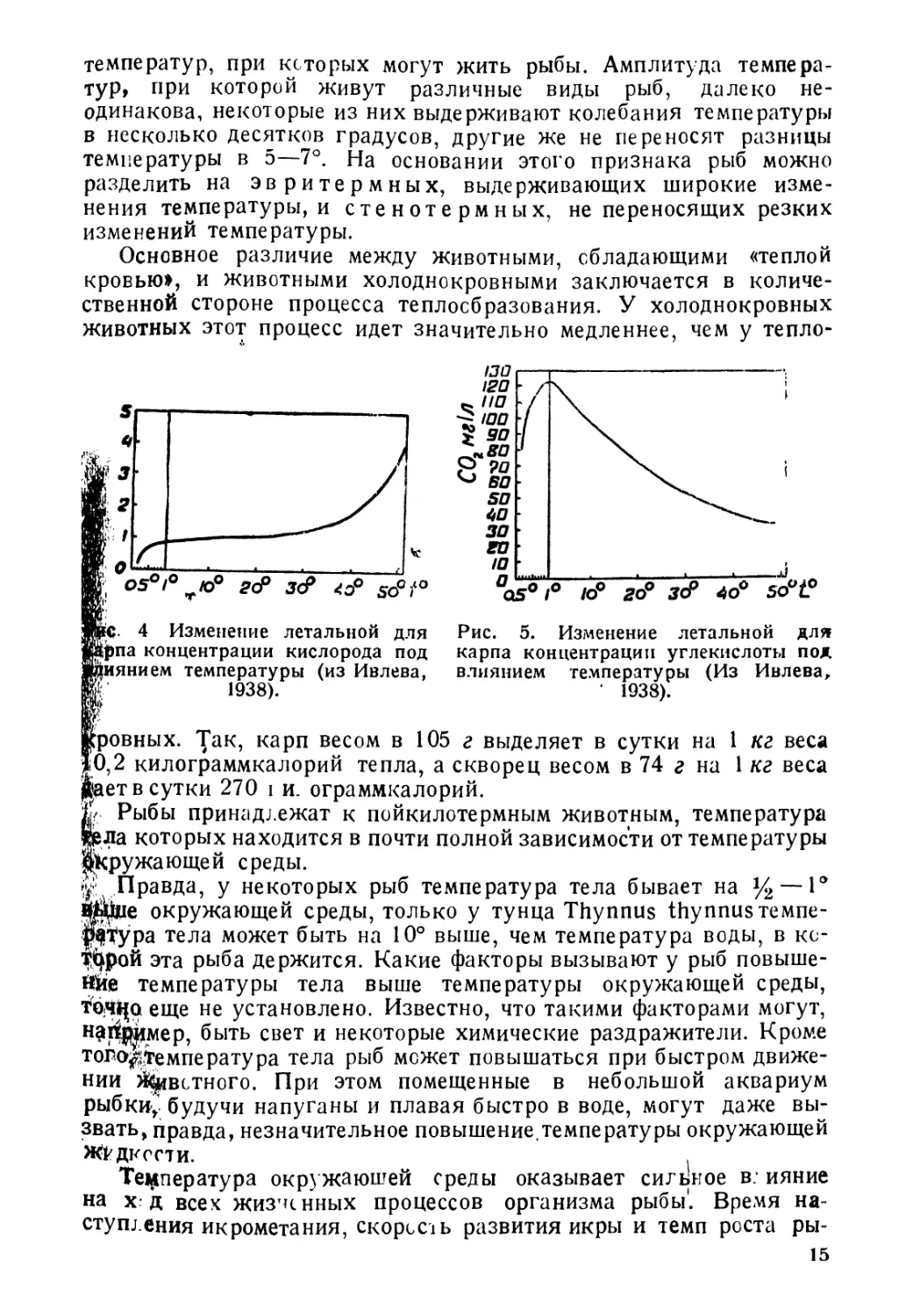
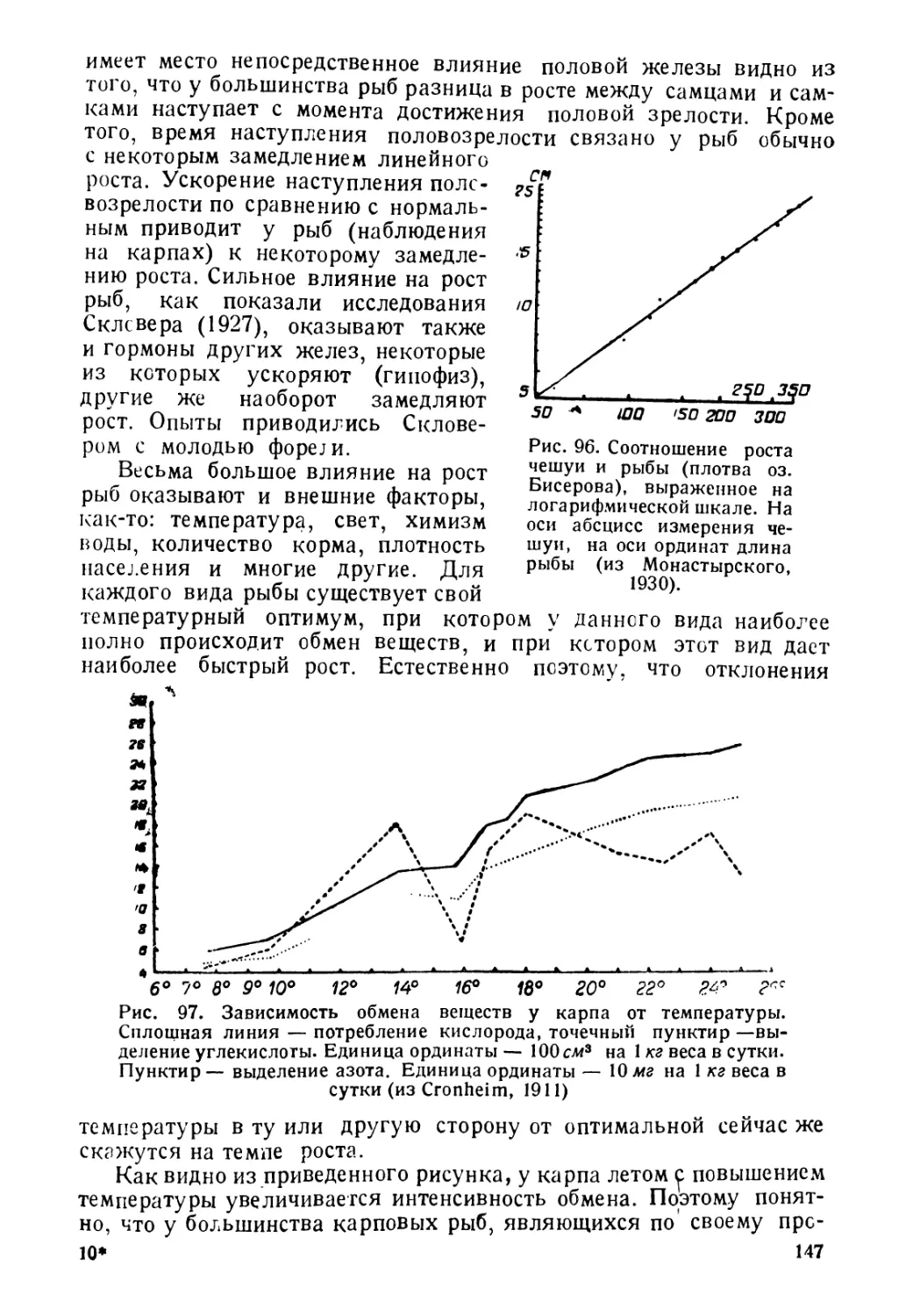
№С. 4 Изменение летальной для
арпа концентрации кислорода под
шиянием температуры (из Ивлева,
Рис. 5. Изменение летальной для
карпа концентрации углекислоты под
влиянием температуры (Из Ивлева,
Г 1938). 1938).
кровных. Так, карп весом в 105 г выделяет в сутки на 1 кг веса
10,2 килограммкалорий тепла, а скворец весом в 74 г на 1 кг веса
|Ваетвсутки 270 i и. ограммкалорий.
Рыбы принадлежат к пойкилотермным животным, температура
тела которых находится в почти полной зависимости от температуры
Окружающей среды.
^Правда, у некоторых рыб температура тела бывает на %— 1®
ИЭДще окружающей среды, только у тунца Thynnus thynnusTeMne-
wfypa тела может быть на 10° выше, чем температура воды, в ко-
ЗДрой эта рыба держится. Какие факторы вызывают у рыб повыше-
ние температуры тела выше температуры окружающей среды,
таяцц еще не установлено. Известно, что такими факторами могут,
нэод^мер, быть свет и некоторые химические раздражители. Кроме
того^:Температура тела рыб может повышаться при быстром движе-
нии рвотного. При этом помещенные в небольшой аквариум
рыбки, будучи напуганы и плавая быстро в воде, могут даже вы-
звать, правда, незначительное повышение,температуры окружающей
Жидкости. .
Температура окружающей среды оказывает сильное в: ияние
на х д всех жизненных процессов организма рыбы'. Время на-
ступления икрометания, скорсезь развития икры и темп роста ры-
15
xt
A
20
42 .
►4?
t
бы находятся в непосредственной зависимости от температуры
•кружающей среды (см. ниже в главах, посвященных вопросу
размножения и роста).
Чрезвычайно сильно температура влияет на газообмен рыб.
При различных температурах ко. ичестьо потребляемого рыСой
кис? орода изменяется дово, ьно
сильно, увеличиваясь обычно при
повышении темьератхры и пони-
жаясь при се понижении. В связи
с этим изменяется и минимальная
концентрация кислорода, при ко-
торой рыбы погибают. Так, для
карпа при температуре в 1° мини-
мальная концентрацг я кис? орода
будет около 0, 8 мг л, а при темпера-
туре в 30° она возрастет до 1,3
мг л. Столь же сильное влияние
(рис. 4 и 5) оказывает температура
и на изменение? етальнойконцентра-
ции сьсбодной углекис. оты, но ход
изменения здесь будет естественно
почти обратным наблюдавшемуся
нами для кислорода. Правда, при
место некоторое увеличение леталь-
температуры, но при темпе-
АО
0\-------------------—
Аму-дяръя Lmp-дяръя Чу
Рис. б. Изменение числа чешуй
в боковой линии (И) и числа лу-
чей в анальном плавнике (А) у
€ыстрянки Alburnoides taeniatus,
в связи с температурой (из Ни-
кольского, 1937).
температурах от Одо
ного количества СО2
ратурах выше 1°
наблюдается резкое
понижение леталь-
ного количества СО2,
снижающегося с 120
мг л при 1° до 55-60
мг л при 30°
Естественно, что
зависимость от тем-
пературы и других
процессов, проте-
кающих в организме
рыбы, очень велика.
Так, например, ско-
рость переваривания
Г имеет
при повышении
>1
hl
ГЦ
tJ ex
ki
&
ID
0
AO
30
ско-
Г___________Г--- - ___, воб-
лой при разных температурах (из Боковой, 1940).
Температуря
Рис. 7. Суточное потребление (пунктир) и
рость переваривания корма (сплошная линия)
Ul
£
£
пищи стоит в непосредственной зависимости
от температуры. У воблы скорость переваривания пещи при 15—20*
в три раза больше, чем при температуре 1—5°. В связи с увели-
чением скорости переваривания возрастает и интенсивность по-
требления корма. С повышением температуры, разумеется, до
известного предела, интенсивность потребления корма у рыбы
илавно возрастает (см. главу о питании)(рис. 6 и 7).
Сильное понижение температуры может вызвать у многих
видов рыб впадение в своего рода спячку. Многие карповые ив част-
ности, например, карась, сазан и некоторые другие, а также многие
осетровые собираются зимой на ямы, где лежат неподвижно на дне,
скопляясь иногда в огромных количествах. Поверхность тела зи-
мующих таким образом рыб обычно покрывается толстым слоем сли-
зи который изолирует тело рыбы от влияния внешней среды. Об-
мен веществ у рыбы, находящейся в спячке, чрезвычайно сильн»
снижается. Подобное состояние спячки наблюдается как у пресно-
водных, так и у морских рыб. У нас в массовом масштабе залегание
на ямы в зимнее время можно наблюдать в дельте Волги, где в одну
и ту же яму залегают часто различные виды рыб. Осетровые ямы
хорошо известны рыбакам, как по Волге, так и по Уралу. Из мор-
ских рыб нечто сходное с зимней спячкой наблюдается у некоторых
дальневосточных камбал. Последние в зимнее время скопляются
в огромном количестве в сравнительно мелководных участках и
лежат, совершенно не питаясь и почти не совершая никаких дви-
жений.
Некоторые виды, как например, серебряный и золотой караси,
а также далия, могут перезимовывать вмерзшими в ил, но толькв
в том случае, если соки их тела не промерзают. Как показали опы-
ты, лед может окружать все тело рыбы, но внутренние соки остают-
ся незамерзшими и имеют температуру до —0,2, —0,3°. Дальней-
шее охлаждение приводит, при условии замораживания рыбы в во-
де, к плавному снижению температуры тела рыбы, замерзанию жид-
кости внутренней среды и смерти. Если рыба замерзает вне воды,
то обычно промерзание ее связано с предварительным переохлажде-
нием и падением температуры тела на короткое время даже до —4,8*,
после чего наступает замерзание жидкостей тела и некоторое повыше-
ние температуры в результате выделения скрытой теплоты замер-
зания. Если промерзают внутренние органы и жабры, то гибель
рыбы неизбежна. 1
Температура оказывает влияние и на формообразование у рыб.
Так, как показали Jordan (1883) и Берг (1922), число позвонков,
чешуй в боковой линии и лучей в непарных плавниках у рыб на-
ходится в тесной зависимости от температуры окружающей среды.
У рыб, живущих при низких температурах в высоких широтах или
Л верховьях горных рек с ледниковым питанием, число позвонков,
$ешуй в боковой линии и лучей плавников, как правило, больше,
у близких видов, живущих при более высоких температурах,
^вершенно естественно, что колебания числа позвонков и других
Меристических признаков у одного и того же вида в зависимости
температуры ограничены известными очень жесткими рамками,
3 пределах которых только и могут происходить изменения. Эта же
закономерность сохраняется и у представителей различных видов
или даже близких родов одного семейства (рис. 6).
Причины подобного рода изменений числа позвонков, чешуй и
лучей в плавниках в зависимости от температуры еще недостаточно
Ясны. Как показали опыты над некоторыми морскими и пресновод-
ными рыбами, число позвонков зависит у них от температуры среды
на ранних стадиях развития и, естес геню, сохранится лишь в
том случае, если это изменение оказывается адаптивным.
2 Биология рыб 17
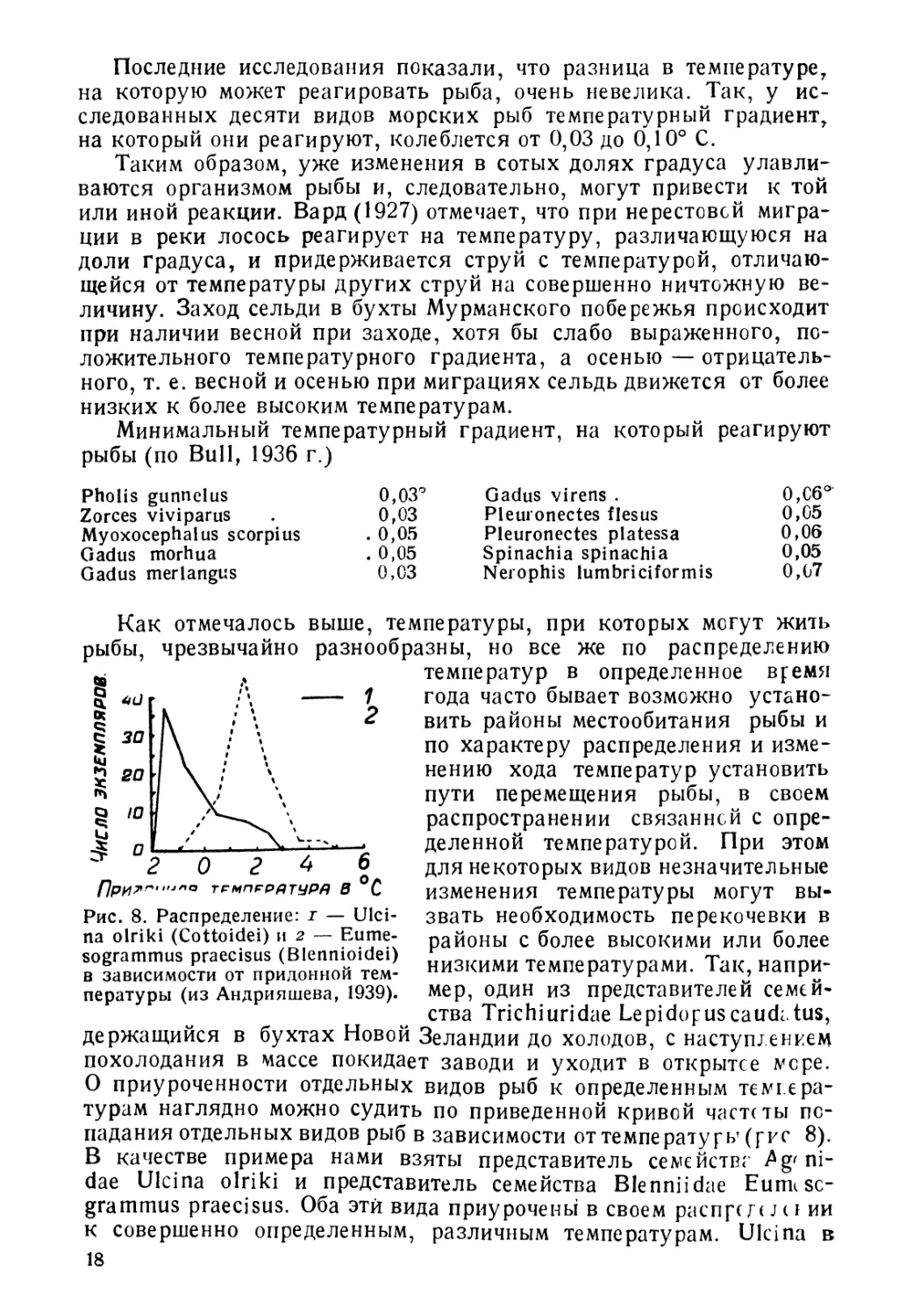
Последние исследования показали, что разница в температуре,
на которую может реагировать рыба, очень невелика. Так, у ис-
следованных десяти видов морских рыб температурный градиент,
на который они реагируют, колеблется от 0,03 до 0,10° С.
Таким образом, уже изменения в сотых долях градуса улавли-
ваются организмом рыбы и, следовательно, могут привести к той
или иной реакции. Вард (1927) отмечает, что при нерестовсй мигра-
ции в реки лосось реагирует на температуру, различающуюся на
доли градуса, и придерживается струй с температурой, отличаю-
щейся от температуры других струй на совершенно ничтожную ве-
личину. Заход сельди в бухты Мурманского побережья происходит
при наличии весной при заходе, хотя бы слабо выраженного, по-
ложительного температурного градиента, а осенью — отрицатель-
ного, т. е. весной и осенью при миграциях сельдь движется от более
низких к более высоким температурам.
Минимальный температурный градиент, на который реагируют
рыбы (по Bull, 1936 г.)
Pholis gunnelus 0,03°
Zorces viviparus . 0,03
Myoxocephalus scorpius . 0,05
Gadus morhua . 0,05
Gadus merlangus 0,03
Gadus virens . 0,C6°
Pleuronectes flesus 0,05
Pleuronectes platessa 0,06
Spinachia spinachia 0,05
Nerophis lumbriciformis 0,07
----- 1
2
4 б
_ О л
riPH^n,,,jna ГРМПРРЙТУРЙ в С
Рис. 8. Распределение: г — CJlci-
na olriki (Cottoidei) и 2 — Eume-
sogrammus praecisus (Blennioidei)
в зависимости от придонной тем-
пературы (из Андрияшева, 1939).
Как отмечалось выше, температуры, при которых могут жить
рыбы, чрезвычайно разнообразны, но все же по распределению
температур в определенное время
года часто бывает возможно устано-
вить районы местообитания рыбы и
по характеру распределения и изме-
нению хода температур установить
пути перемещения рыбы, в своем
распространении связанней с опре-
деленной температурой. При этом
для некоторых видов незначительные
изменения температуры могут вы-
звать необходимость перекочевки в
районы с более высокими или более
низкими температурами. Так, напри-
мер, один из представителей семей-
ства Trichiuridae Lepidopuscaudatus,
держащийся в бухтах Новой Зеландии до холодов, с наступлением
похолодания в массе покидает заводи и уходит в открытсе мере.
О приуроченности отдельных видов рыб к определенным темгера-
турам наглядно можно судить по приведенной кривой чаете ты по-
падания отдельных видов рыб в зависимости от температуры (рис 8).
В качестве примера нами взяты представитель семейства ^g' ni-
dae Ulcina olriki и представитель семейства Blenniidae Eumisc-
grammus praecisus. Оба эти вида приурочены в своем распрей jci ии
к совершенно определенным, различным температурам. Ulcina в
18
максимуме встречается при температуре —1,0, —1,5°, a Eumeso-
graminus— при температуре +1,4-2°.
Примером рыб, напротив, мало связанных с водой определенной
температуры, может служить хотя бы обыкновенная маринка Schi-
zothorax intermedins, которая распространена от ледниковых ру-
чьев с температурой, близкой к 0°, и до равнинных озер, где темпе-
ратура воды поднимается иногда до 4-25°С.
Стенотермные рыбы, приуроченные в своем распространении
и. определенным температурам, при изменении температуры бывают
вынуждены или переместиться в другое место, или погибнуть. При-
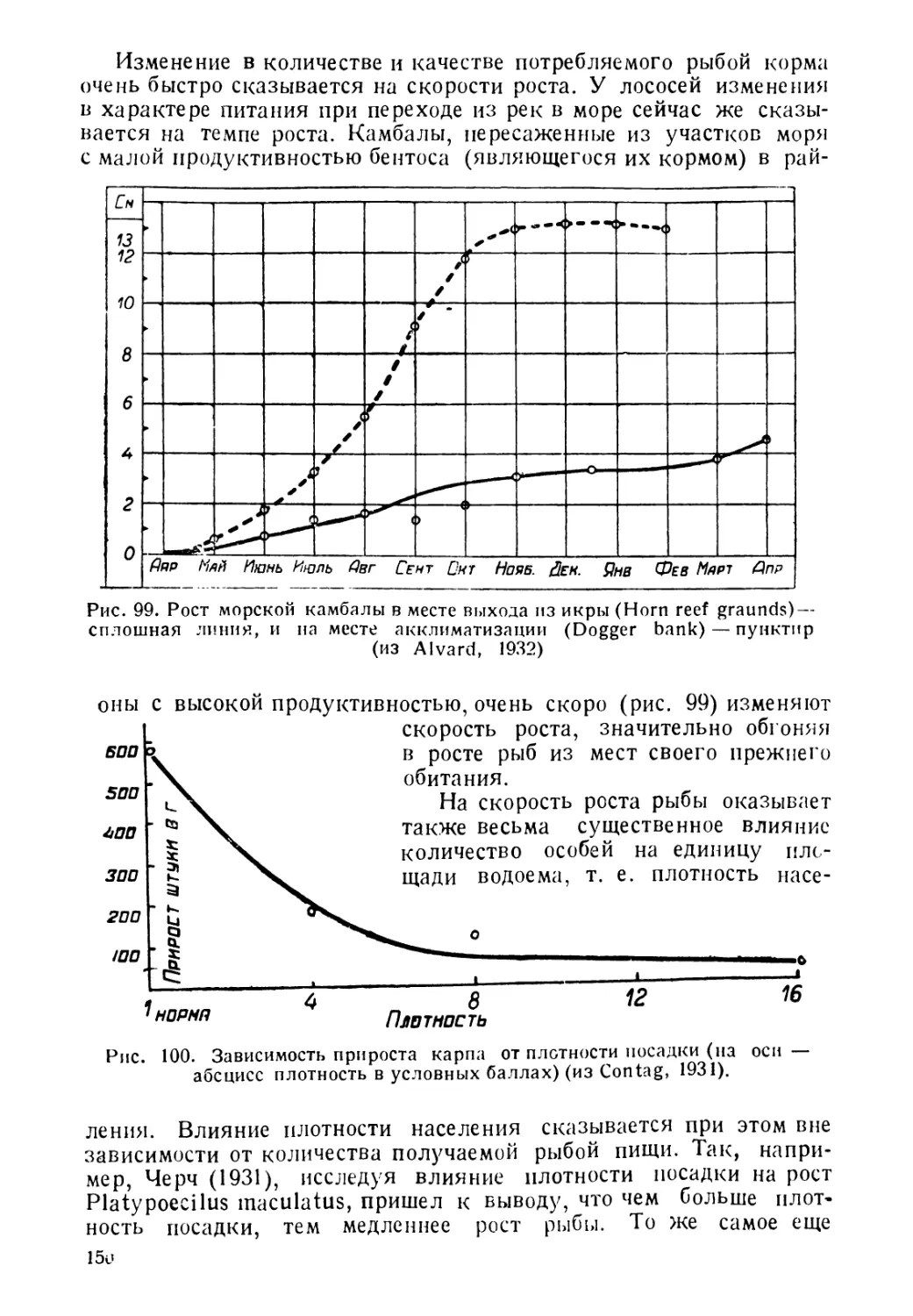
Рис. 9. Хамелеоноголов Lopholatilus hamaeleonticeps (из Boulenger, 1904)
мерой массовой гибели рыб, в результате изменения температуры
воды, может служить случай с хамелеоноголовсм Lcpholatiluscha-
maekcnticeps (рис. 9). Этот вид был неизвестен до 1879 года у юж-
ных берегов Новой Англии, но в последующие годы, в связи с по-
теплением, он появился здесь в большом количестве и стал объектом
промысла. В результате резкого похолодания, наступившего в мар-
те 1882 года, масса особей этого вида погибла, и они покрыли сво-
ими трупами поверхность моря на многие мили. После этого случая
на долгое время Lopholatilus chamaeleonticeps совершенно исчез
из этого района и лишь в последние годы снова опять появился в
Довольно значительном количестве. Периодическое потепление вод,
имеющее место в северных морях, позволяет бореальным видам в
более теплые периоды проникать в высокие широты, а в периоды
похолодания, наоборот, арктическим видам проникать в более юж-
ные районы. Периодические колебания температуры вызывают так-
же изменение и самого состояния рыбы. Так, как показали наблю-
дения Иорта (1912), количество трескового жира находится в не-
посредственной зависимости от температуры вод Гольфштрема,
oni еделяющей ебщую продуктивность водоема.
В связи с наблюдавшимся в последние годы потеплением в Ар-
ктике удалось отметить проникновение на север целого ряда более
теплсвсдных форм. Так, например, в 1820 году у западных берегов
1 Ренландии было очень много трески, затем она исчезла и появи-
лась снова в 1845 г. В последнее время, начиная с 1925 года, треска
2* 19
снова стала ловиться в этом районе. Периодические подходы трески
к Гренландии, а у нас заходы в Карское море связаны в значитель-
ной мере с периодичностью температурного режима Гольфштрема.
Периодичность температурного режима гидросферы, связанная в
значительной степени с интенсивностью солнечной радиации и в част-
ности с количеством солнечных пятен, сильно отражается не только
на распространении и состоянии рыб, но также и на величине попу-
ляции— урожае отдельных видов. Хорошо известные колебания
уловов семги Salmo salar, сельди Clupea harengus, трески G..dus
mcrhua являются в значительной степени результатом этих перио-
дических колебаний количества тепла в гидросфере (см. ниже).
Наряду с такими периодическими изменениями в распространении
и численности отдельных видов рыб в зависимости от колебаний
температуры мы знаем много примеров непериодических изменений,
связанных с изменениями ‘термического режима. Так, в связи с
общим потеплением, наблюдающимся со второй половины прошлого
столетия в северном и южном полушарии, ряд рыб, так же как и
других животных, в своем распространении несколько продвинул-
ся в более высокие широты, чем те, в которых он был распростра-
нен раньше. Так, у побережья Камчатки стали попадаться в послед-
нее время иваси Sarctnops melancsticta и были констатированы слу-
чаи захода в камчатские воды японского анчоуса EngrauLs japcnicus
и брамы Brama sp. В Белое мере за последнее время начали прони-
кать некоторые более тепловодные рыбы, например, пикша Gadus
aegkfinus, морской окунь Seb..stes marinus и некоторые другие.
Изменения в распространении рыб в зависимости от изменения
температуры окружающей среды имели место и в предыдущие гео-
логические эпохи. Установлено, например, что в водоемах, распо-
ложенных на месте бассейна современного Иртыша, в неогене встре-
чались рыбы, значительно более тепловодные, чем те, которые на-
селяют бассейн Оби теперь. Так, в состав неогеновой иртышской
фауны входили представители родов Chondrcstoma, Alburnoides,
Blicca, которые сейчас в бассейне Ледовитого океана на территории
Сибири не встречаются, а распространены, главным образом, в
Понто-Арало-Каспийской провинции и, повидимому, были вытес-
нены из бассейна Ледовитого океана в результате изменения кли-
мата в сторону похолодания. И в более позднее время мы находим
примеры изменения ареала распространения и численности видов
под влиянием изменений температуры окружающей среды. Так,
похолодание, вызванное наступанием ледников, привело к тому,,
-что представители семейства лососевых, в своем распространении
приуроченные к холодным водам, смогли продвинуться значитель-
но на юг и попасть в бассейн Средиземного моря, включая и реки
Малой Азии и Северной Африки. В это время лососи были значитель-
но более обильны и в Черном море, на что указывает большое коли-
чество костей этой рыбы в остатках пищи палеолитического чело-
века.
В послеледниковое время колебания климата также приводили
к изменениям в составе ихтиофауны. Так, например, во время кли-
20
матического оптимума (около 5000 лет тому назад), когда имело место
некоторое потепление климата, фауна рыб бассейна Белого моря
содержала до 40% более тепловодных видов, таких, как жерех
Aspius aspius, красноперка Scardinius erythrophthJmus и синец
Abranrs balkrus, которые сейчас в бассейне Белого моря не встре-
чаются и несомненно были вытеснены отсюда наступившим похо-
лоданием, происшедшим еще до начала нашей эры.
Как видно из изложенного, значение температурного фактора
для распространения отдельных видов чрезвычайно велико. Привя-
занность отдельных фаунистических комплексов к определенным
термическим условиям обусловливает тот факт, что границы между
отдельными зоогеографическими областями в море очень часто сов-
падают с определенными изотермами. Так, например, Чукотская
умеренно арктическая провинция характеризуется весьма низкими
температурами и соответственно преобладанием арктической фау-
ны. Большинство бореальных элементов проникает лишь в восточ-
ную часть Чукотского моря вместе со струями теплого течения.
Фауна Белого моря, выделенная в особый зоогеографический уча-
сток, по своему составу значительно белее хслсдсвсдна, чем распо-
ложенная севернее его фауна южней части Баренцева меря. Обо-
собление беломорской фауны стало возможным лишь благодаря
специфическому термическому режиму, затруднившему обмен фау-
нами.
В заключение необходимо остановиться на влиянии льда на рыб.
Это влияние очень разнообразно, но главным образом оно выража-
ется не во влиянии температурном, а в других фермах воздействия.
Температурное влияние льда сказывается в повышении температу-
ры воды при образовании льда и в понижении температуры при тая-
нии. Значительно важнее иные формы влияния льда на рыб. Особен-
но важно значение ледового покрова, как изолятора воды от атмосфе-
ры. Во время ледостава почти полностью прекращается влияние
ветров на воду, поступление кислорода из воздуха ит. п. (см. ниже).
Изолируя воду от воздуха, лед затрудняет также проник-
новение в воду света. Наконец, лед сказывает иногда на рыб, и меха-
ническое влияние. Известны случаи, когда в прибрежной полосе
выносимым на берег льдом раздавливались державшиеся у берега
рыбы. Играет некоторую роль лед и в изменении химического
состава воды и величины солености. Солевой состав льда отличен
от солевого состава морской воды, и при массовом льдообразовании
изменяется не только соленость воды, при этом повышающаяся,
но и соотношение солей. Таяние льда, наоборот, вызывает понижение
солености и изменение солевого состава в обратную сторону.
Свет
Свет оказывает огромное влияние на рыб, как и i^a других жи-
вотных, в течение всей их жизни. Сравнительно оч^нь небольшой
процент видов рыб приспособился к жизни в полной темноте больших
глубин или водоемов пещер.
21
Условия освещения в воде чрезвычайно разнообразны и зависят
от многих причин, основной из которых является прозрачность воды.
Прозрачность воды в различных водоемах чрезвычайно разнообраз-
на, начиная от мутных кофейного цвета рек Индии, Китая и Сред-
ней Азии, где предмет, погружаемый в воду, делается невидимым
как только покроется водой, и« кончая прозрачными водами Саргас-
сова моря (прозрачность 66,5 к), центральной части Тихого океана
(59 м) и ряда других мест, где белый круг становится невидимым
для глаза только после погружения на глубину более 50 м.
Естественно, что условия освещения в различных водоемах,
дажз расположенных в одинаковых широтах на одной и той же глу-
бине, будут весьма различными, не говоря уже о различных глуби-
нах, ибо, как известно, с глубиной степень освещенности быстро
понижается. Как показали опыты Бииба (1936), человеческий глаз
еще может различать следы света на, глубине около 500 м. На глу-
бине 1000 м фотографическая пластинка чернеет после экспозиции
в течение 1 часа 10 минут, на глубине же 1700 м фотографическая
пластинка даже после 2-часовой экспозиции не обнаруживает
никаких изменений. Таким образом животные, в том числе и рыбы,
живущие, начиная с глубин порядка 1500 м и кончая максимальными
глубинами мирового океана свыше 10000 лг, совершенно не подвер-
жены влиянию дневного света и живут в полной темноте, нарушае-
мой только светом, исходящим от органов свечения различных глу-
боководных животных. *
Интенсивность света,'на которую реагируют рыбы, у различных
видов, повидимому, неодинакова. У американской рыбки Lepomis
из семейства Centrarchidae глаз еще улавливает свет интенсивностью
к 10—10 от нормального дневного. Подобная сила освещения на-
блюдается в наиболее прозрачной воде Саргассова моря на глубине
430 м от поверхности. Lepomis — пресноводная рыбка, обитатель-
ница сравнительно неглубоких водоемов. Поэтому, весьма вероят-
но, что глубоководные рыбы, особенно имеющие телескопическое
зрение, способны реагировать на значительно более слабое осве-
щение.
Влияние света на рыбу идет по целому ряду направлений. У
рыб, живущих при сильном освещении, вырабатывается различ-
ного типа покровительственная окраска, делающая их менее замет-
ными на фоне грунта, поверхности воды или среди зарослей подвод-
ной растительности и коралловых рифов. У рыб, живущих на боль-
ших глубинах, где количество света чрезвычайно мало, органы зре-
ния или развиваются очень сильно, или деградируют. У глубоко-
водных рыб очень часто на теле образуются особые органы свече-
ния, иногда, как показали наблюдения Бииба, излучающие доволь-
но сильный свет. Рыбы—обитатели темных пещер очень часто
слепы.
Свет оказывает влияние на ход созревания половых продуктов
У рыб, на скорость развития икры, на ход обмена веществ и т. д.
Суточная периодичность освещения вызывает выработку у рыб су-
точной периодичности их деятельности (питания, миграций и т. п.),
22
з сезонные изменения в инсоляции оказывают серьезное влияние
на сезонные ритмы жизнедеятельности рыб.
Кроме видимой части спектра на рыб оказывают влияние как
инфракрасные, так и ультрафиолетовые лучи, а также лучи Рент-
гена. Некоторые формы влияния этих лучей весьма резки и вносят
серьезные изменения в нормальную деятельность организма рыбы,
вызывая иногда сильные нарушения созревания половых продуктов,
обмена веществ и т. д.
Из сказанного видно, что формы влияния видимого света и дру-
гих видов лучистой энергии на организм рыбы чрезвычайно разно-
образны, и связанные с этим IDa\
приспособления могут вно-
сить весьма серьезные изме-
нения в организм рыбы.
Условия освещения в
воде отличаются от таковых
в воздушной среде не только
ио интенсивности освещения,
но и по степени проникнове-
ния в глубину воды отдель-
ных лучей спектра. Как из-
вестно, коэфициент поглоще-
ния водой лучей с различ-
ной длиной волны далеко
не одинаков. Наиболее силь-
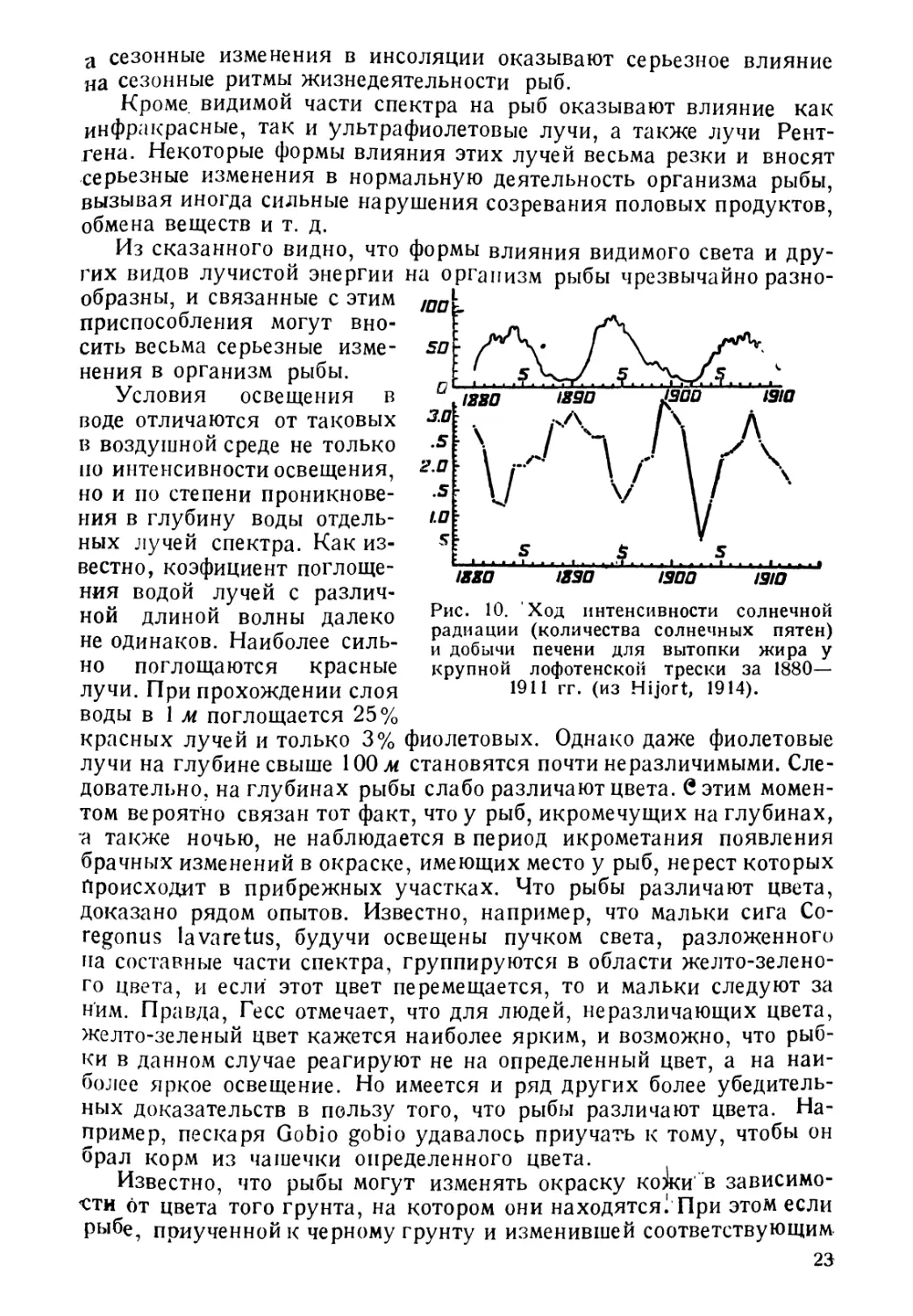
Рис. 10. Ход интенсивности солнечной
радиации (количества солнечных пятен)
и добычи печени для вытопки жира у
но поглощаются красные
лучи. При прохождении слоя
воды в 1 м поглощается 25%
крупной лофотенской трески за 1880—
1911 гг. (из Hijort, 1914).
красных лучей и только 3% фиолетовых. Однако даже фиолетовые
лучи на глубине свыше 100 м становятся почти неразличимыми. Сле-
довательно, на глубинах рыбы слабо различают цвета. С этим момен-
том вероятно связан тот факт, что у рыб, икромечущих на глубинах,
а также ночью, не наблюдается в период икрометания появления
брачных изменений в окраске, имеющих место у рыб, нерест которых
Происходит в прибрежных участках. Что рыбы различают цвета,
доказано рядом опытов. Известно, например, что мальки сига Со-
regonus lavaretus, будучи освещены пучком света, разложенного
на составные части спектра, группируются в области желто-зелено-
го цвета, и если этот цвет перемещается, то и мальки следуют за
ним. Правда, Гесс отмечает, что для людей, неразличающих цвета,
желто-зеленый цвет кажется наиболее ярким, и возможно, что рыб-
ки в данном случае реагируют не на определенный цвет, а на наи-
более яркое освещение. Но имеется и ряд других более убедитель-
ных доказательств в пользу того, что рыбы различают цвета. На-
пример, пескаря Gobio gobio удавалось приучать к тому, чтобы он
брал корм из чашечки определенного цвета.
Известно, что рыбы могут изменять окраску кожи в зависимо-
сти от цвета того грунта, на котором они находятся?При этом если
рыбе, приученнойк черному грунту и изменившей соответствующим
23
подтверждает изучение строения
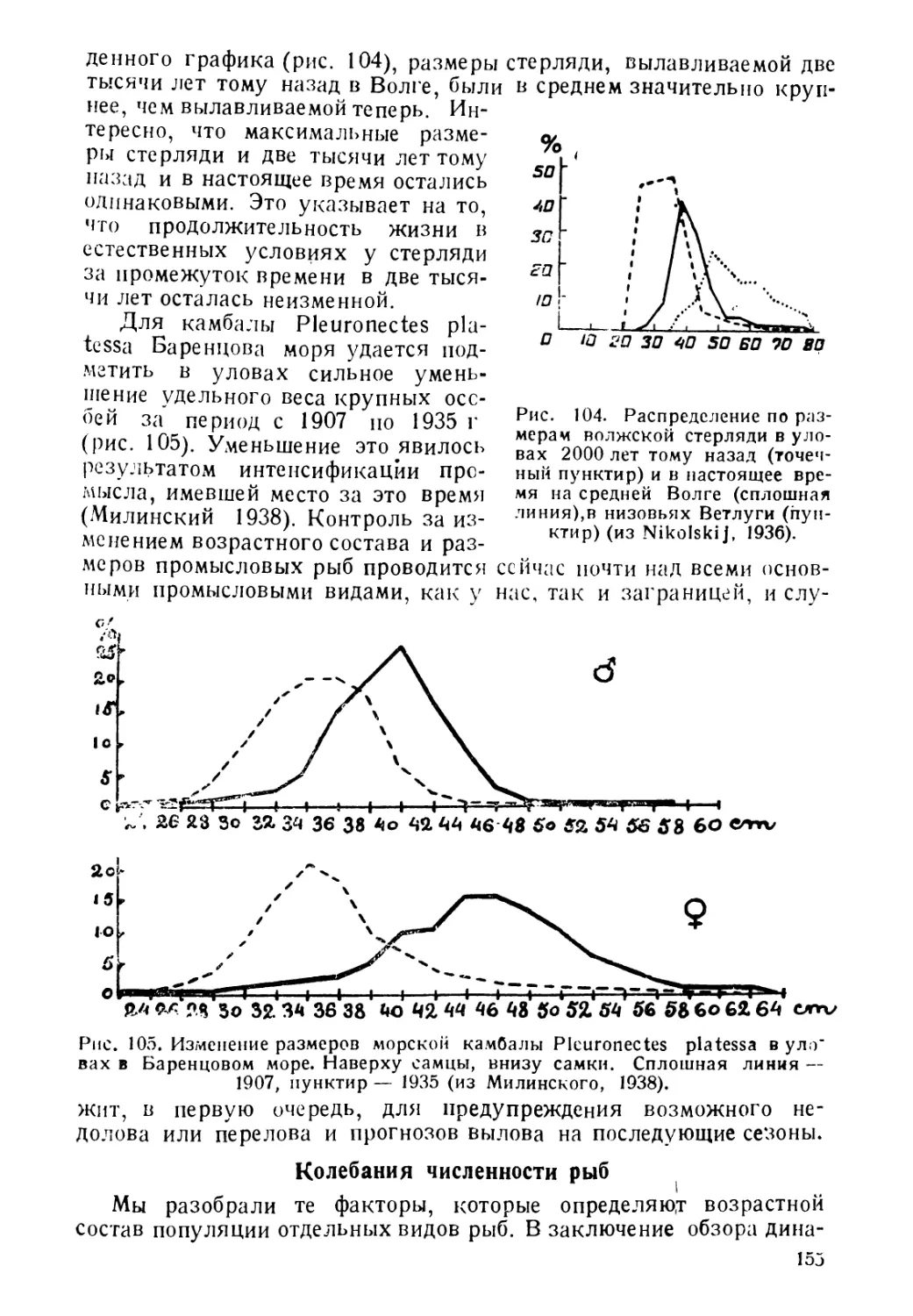
Рис. 11. Зависимость окраски камбалы
•т цвета грунта, на котором находится
ее голова (из Книповича, 1933).
образом свою окраску, давали на выбор ряд грунтов различной
расцветки, то рыба выбирала обычно тот грунт, к которому она
выла приучена и окраске которого соответствует окраска ее кожи.
Специфика освещения в водной среде отражается и на строении
глаза как рыб, так и других водных животных. Хрусталик глаза
рыб в отличие от хрусталика глаза наземных позвоночных имеет
шаровидную форму. Вундер (1936) связывает это с особенностями
преломления света в водной среде. Высказанное предположение
глаза американской рыбки из се-
мейства An: blepidae—An; bleps
t(trorhthalmus. Anableps tetra-
phtalmus живет в мелких водо-
емах и глаза ее приспособлены
для зрения, как в воздушной,
так и в водной среде. Каждый
глаз этой рыбки разделен на
две половины горизонтальной
полоской (рис. 12). Поделена
надвое, как роговая, так и ра-
дужная оболочки. Верхняя,
приспособленная для зрения в
воздушной среде, часть хруст; -
лика имеет более плоскую фор-
му, чем нижняя, служащая для
зрения в воде. Плавая близ
поверхности воды, эти рыбки
одновременно наблюдают за
тем, что происходит на воздухе
и в воде. Как мы отмечали вы-
1 ше, рыбы могут несколько из-
менять окраску в связи с тем
грунтом, на котором они нахо-
дятся. Особенно резкие измене-
ния в цвете тела на различных грунтах наблюдаются у камбал, при-
чем меняется не только тон, но и рисунок, в зависимости от характера
грунта, на котором находится рыба. Каков механизм этого явления,
еще точно не выяснено. Известно только, что изменение окраски
происходит в результате соответствующего раздражения глаза.
Семнер (1933), надевая на глаза рыбы прозрачные цветные колпач-
ки, вызывал у рыб изменение окраски в тон с тем цветом, в который
были окрашены колпачки. Камбала, тело которой находится на
грунте одного цвета, а голова — на грунте другого цвета, приобре-
тает окраску тела под тон к цвету того грунта, на фоне которого
находится голова (рис. 11).
Слабое освещение на глубинах заставляет вырабатываться у
глубоководных рыб целому ряду приспособлений для зрения при
слабом свете (рис. 13). У многих глубоководных рыб глаз достигает
вгромных размеров. Так, например, у Bathymacrops macrolepis
из семейства Microstomidae диаметр глаза составляет около 40% от
24
длины головы. У Polyipnus из семейства Sternoptychidae диаметр
глаза 25—32% от длины головы, а у Myctophium rissci из семейства
Myctophiidae даже до 50%. Очень часто изменяется у глубоководных
рыб и форма зрачка, он становится продолговатым, и концы его за-
ходят за хрусталик, благодаря чему так же, как и путем общего
увеличения размеров глаза, увеличивается его светопоглотительная
Рис. 12. «Четырехглазая» рыба — Anableps tetrophthalmus (из Bowlee-
ger, 1904).
способность. У Argyropelecua (рис. 13) из семейства Sternoptychi-
dae в глазу имеется специальный светящийся орган, поддерживаю-
щий сетчатку в состоянии постоянного
раздражения и тем самым повышающий ее
чувствительность к попадающим в глаз
извне световым лучам. У многих глубоко-
водных рыб для увеличения чувствитель-
ности зрения глаза становятся телескопиче-
скими, что расширяет поле зрения. Наиболее
любопытные изменения органа зрения имеют
место у лич1 нки глубоководной рыбы Idiacan-
thus (рис. 14). У нее глаза расположены на
длинных стебельках, что позволяет сильно
увеличить поле зрения. У взрослых рыб сте-
бельчатость глаз теряется. Наряду с сильным
развитием органа зрения у одних глубоко-
водных рыб, у других, как уже отмечалось,
орган зрения или становится очень малым
(Benthcsaurus и др.), или совсем, исчезает
(Ipncps). Наряду с редукцией органа зрения
у этих рыб обычно развиваются различные
выросты на теле: или удлиняются лучи парных
и непарных плавников, или усики. Все эти
выросты служат органами осязания и в из-
вестной степени компенсируют недостаток ор-
Рис. 13. Головы Аг-
gyropelecus affinis
(сверху) и Myctophi-
um rissoi (внизу) (из
Fowler, 1936).
ганов зрения.
Окраска глубоководных рыб также весьма специфична. Рыбы,
населяющие большие глубины, обычно скрашены или в темные,
иногда почти черные, или в красные тона. Объясняемся это тем, что
даже при сравнительно небольших глубинах, красный цвет под
водой кажется черным и плохо заметен для хищников.
25
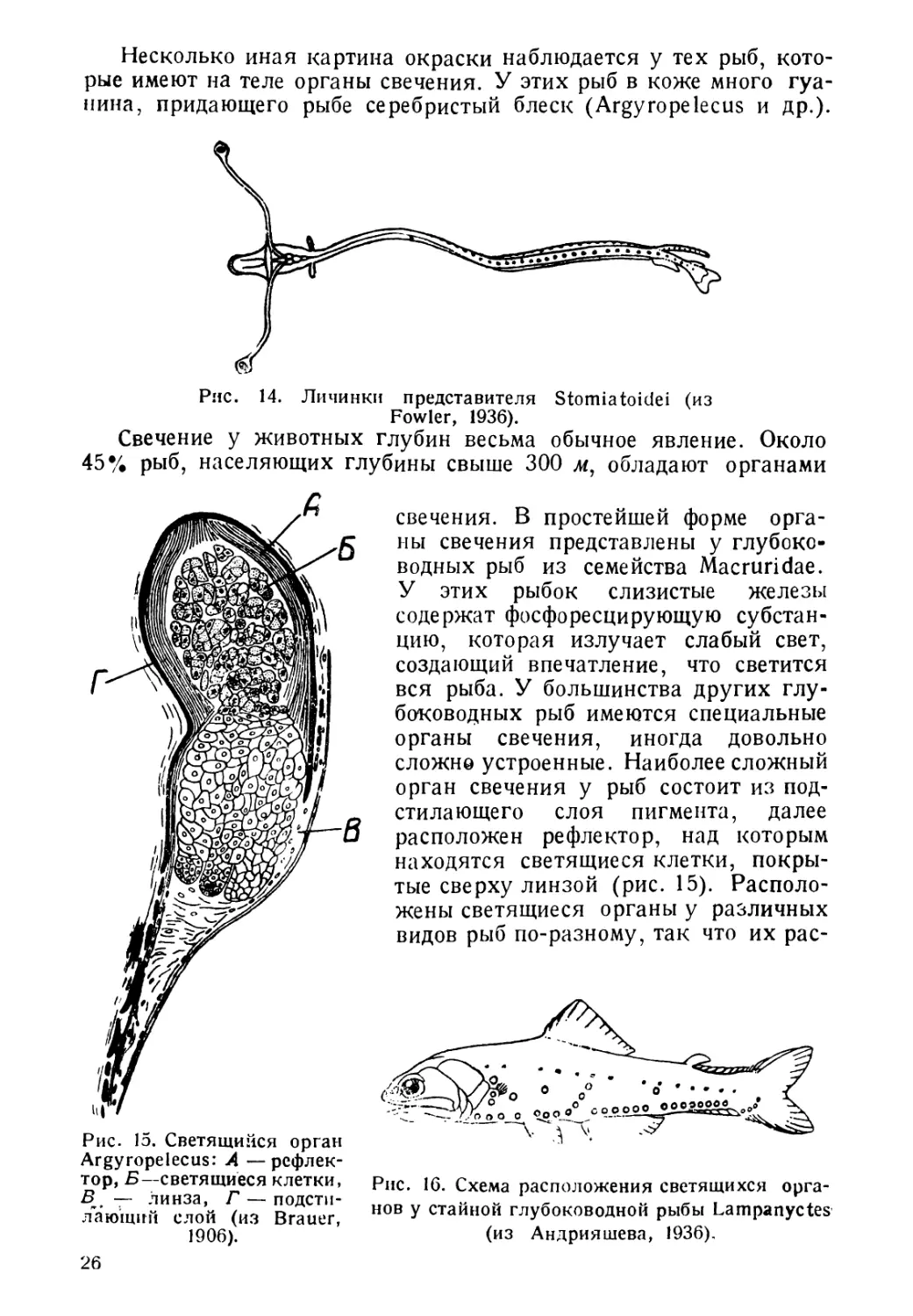
Несколько иная картина окраски наблюдается у тех рыб, кото-
рые имеют на теле органы свечения. У этих рыб в коже много гуа-
нина, придающего рыбе серебристый блеск (Argyropelecus и др.).
Рис. 14. Личинки представителя Stomiatoidei (из
Fowler, 1936).
Свечение у животных глубин весьма обычное явление. Около
45% рыб, населяющих глубины свыше 300 л/, обладают органами
Рис. 15. Светящийся орган
Argyropelecus: А — рефлек-
тор, Б—светящиеся клетки,
В — линза, Г — подсти-
лающий слой (из Brauer,
1906).
свечения. В простейшей форме орга-
ны свечения представлены у глубоко-
водных рыб из семейства Macruridae.
У этих рыбок слизистые железы
содержат фосфоресцирующую субстан-
цию, которая излучает слабый свет,
создающий впечатление, что светится
вся рыба. У большинства других глу-
боководных рыб имеются специальные
органы свечения, иногда довольно
сложно устроенные. Наиболее сложный
орган свечения у рыб состоит из под-
стилающего слоя пигмента, далее
расположен рефлектор, над которым
находятся светящиеся клетки, покры-
тые сверху линзой (рис. 15). Располо-
жены светящиеся органы у различных
видов рыб по-разному, так что их рас-
Рис. 16. Схема расположения светящихся орга-
нов у стайной глубоководной рыбы Lampanyctes
(из Андрияшева, 1936).
26
положение может служить во многих случаях систематическим при-
знаком (рис. 16).
Свечение происходит обычно в результате соприкосновения сек-
рета светящихся клеток с водой, но у рыбки Acropoma japonicum
свечение вызывается находящимися в железе микроорганизмами.
У одной и той же рыбы интенсивность свечения варьирует и зави-
Рис. 17. Пещерные рыбы (сверху вниз): Chologaster,
Typhlichthys и Amblyopsis (Cyprinodontiformes) (из
Jordan, 1925).
сит от целого ряда факторов. Особенно интенсивно у многих рыб
свечение бывает в период размножения.
Каково биологическое значение свечения глубоководных рыб,
полностью еще не выяснено, но несомненно, что у разных рыб роль
светящихся органов различна. У Ceratiidae светящийся орган на
конце первого луча спинного плавника видимо служит для примани-
вания добычи. Ту же функцию вероятно выполняет светящийся
орган на конце хвоста Saccopharynx. Расположенные по бокам тела
светящиеся органы Argyropelecus, Lampanyctes, Myctophium, Vin-
ciguerria и многих других рыб позволЯют’им в темноте на больших
глубинах находить особей того жз вида. Особенно), повидимому,
это имеет значение для рыб, держащихся стаями. <
Также в полном мраке, даже ненарушаемом светящимися орга-
низмами, живут пещерные рыбы. По тому, насколько плотно живот-
27
ные связаны с жизнью в пещерах, их принято подразделять на:
1) троглобионтов — постоянных обитателей пешер, 2) тро-
г л о ф и лов — преимущественных обитателей пещер, но встре-
чающихся и в других местах, 3) т р о г л о к с е н о в — широк®
распространенные формы, заходящие и в пещеры. Так же, как
и у рыб глубин, и у пещерных форм наиболее сильные изменения
в организации связаны с изменением освещения. Среди пещерных
рыб можно найти всю цепь переходов от рыб с хорошо развитыми
глазами до совершенно слепых. Так, у Chclcgistcr cornutus (сем.
Amblyopsidae) глаза развиты нормально и фу» кциснируют как ор-
ган зрения. У близкого вида Chckgaster papilliferus, хотя все эле-
менты глаза имеются налицо, но ретина дегенерирует. У Typh-
lichthys зрачок еще не закрыт, и нервная связь с головным мозгом
глаза сохранена, но колбочки и палочки отсутствуют. У Amblyopsis
и зрачок уже закрыт, и, наконец, у Trcglichthy s глаза ред) цврсваны
очень сильно (рис. 17). Интересно, что у молодых Trcglichthys гла-
за развиты лучше, чем у взрослых.
В качестве компенсации дегенерирующего органа зрения у пе-
щерных рыб обычно очень сильно развиты органы боковой линии,
особенно сильно представленные, как правило, на голове. У пещер-
ных сомов Бразилии из семейства Pimelcdidae, Typhkbrgrus kro-
ne!, Coecoramdia urichi и Coeccramdella brasiliensis очень сильно
развиты усы, служащие органом осязания.
Рыбы, населяющие пещеры, весьма разнообразны. В настоящее
время в пещерах известны представители ряда групп. Тут мы нахо-
дим представителей отряда Cypriniformes (Aulopyge, Paraphoxinus,
Chondrcstoma, американские сомы и др.), Cyprincdontiformes (Chc-
logaster, Amblyopsis), бычков и других групп.
Кроме влияния на органы зрения и окраску рыб свет оказывает
воздействие также на ход созревания половых продуктов рыб. По-
ставленные над американской палией Salvelinus fcntinaLs опыты
показали, что рыбы, подвергавшиеся усиленному освещению, со-
зревали раньше, чем контрольные, находившиеся при нормальном
свете. Повидимому, у рыб в высокогорных условиях так же, как у
некоторых млекопитающих в условиях искусственного освещения,
свет, после стимулирования усиленного развития половых желез,
может вызвать резкое падение их деятельности. В связи с этим у
древних высокогорных форм развивается интенсивно окрашенная
брюшина, предохраняющая гонады от чрезмерного воздействия
света. Динамика интенсивости освещения в течение года в значи-
тельной степени определяет ход полового цикла у рыб. То, что у
тропических рыб размножение происходит в течение всего года,
а у рыб умеренных широт только в определенное время, в значи-
тельной степени связано с интенсивностью инсоляции.
Свет оказывает весьма сильное влияние и на развивающуюся
икру (см. ниже, стр. 132), причем на икру некоторых рыб свет дей-
ствует, ускоряя развитие, у других воздействие света сказывается
на замедлении развития.
Любопытное приспособление к защите от чрезмерного освещения
28
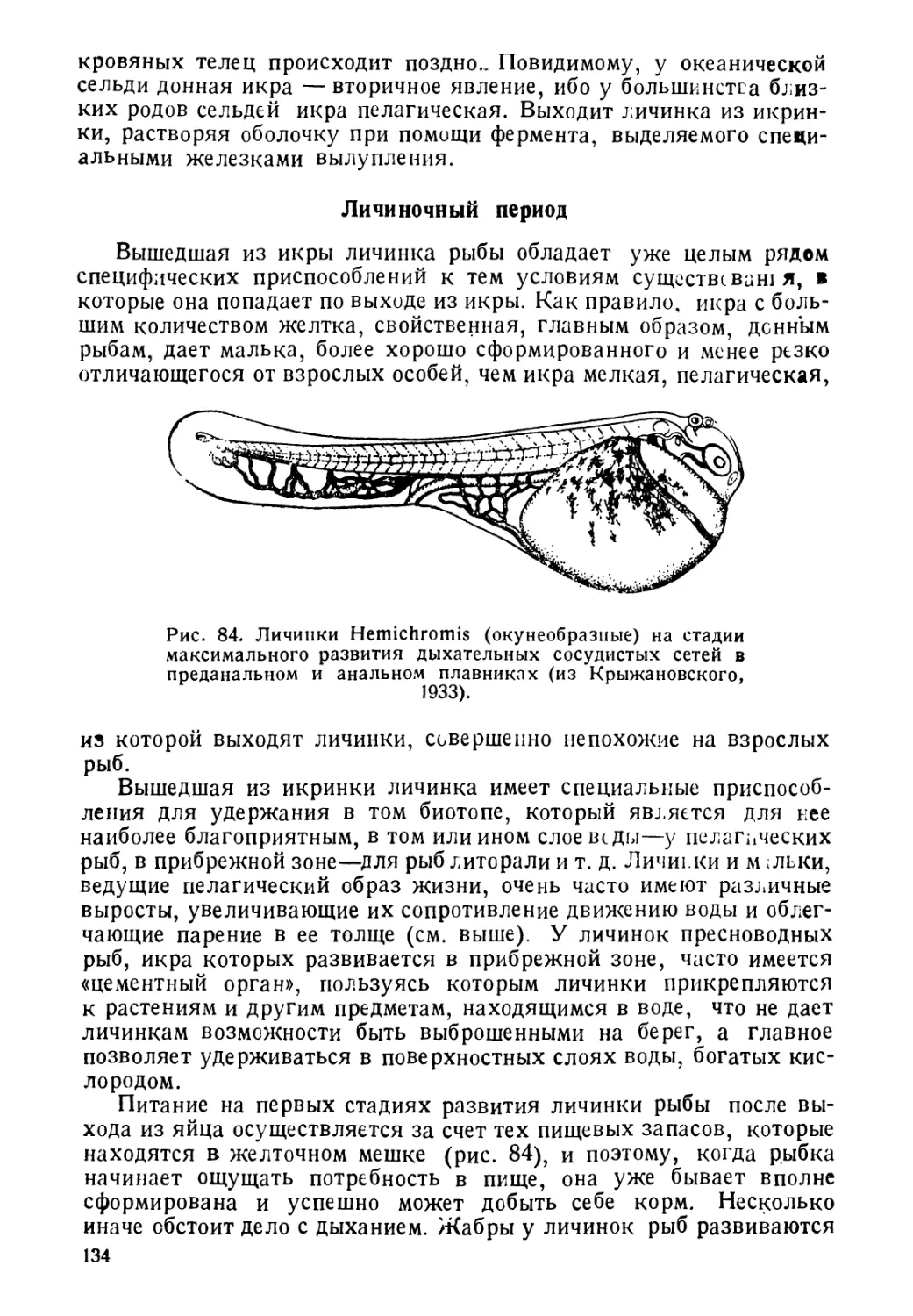
наблюдается у личинок многих пелагических рыб. Так, у личинок
сельдей родов SpratJla и S. rcLna над нервней трубкей развивается
черный пигмент, предохраняющий нервную систему и лежащие
ниже органы от чрезмерного воздействия света. Интересно, что у
близких видов, имеющих донную икру, и личинок, держащихся в
придонных слоях, подобного пигмента нет. С резорбцией
желт чного пузыря, пигмент над нервной трубкой у мальков
исчезает.
Весьма серьезное влияние оказывают солнечные лучи на ход
обмена веществ у рыб. Опыты, проведенные на гамбузии (Gambusia,
affinis), показали, что у гамбузий, лишенных света, развивается
довольно быстро авитаминоз, вызывающий, в первую очередь,
потерю способности к размножению.
Здесь же необходимо также остановиться на влиянии на орга-
низм рыбы других форм лучистой энергии и, в частности, лучей
Рентгена. Как показали опыты Самохваловой (1930 г.) над Lobistes
reticuLtus, доза в 6ОН является летальной для рыбок. Меньшие
доз>| при воздействии на половую железу Lebistos rcticulatus вы-
зывают уменьшение помета и дегенерацию железы. Облучение мо-
лодых неполовозрелых самцов вызывает у них недоразвитие вторич-
нополовых признаков.
Грунт и взвешенные в воде частицы
Рыбы, меньше чем какие-либо другие группы позвоночных жи-
вотных, связаны с твердым субстратом, как опорой. Многие виды
рыб никогда за всю свою жизнь не прикасаются к дну, незначитель-
ная, пожалуй, ббльшая часть рыб, находится в той или иной связи с
грунтом водоема, который они населяют. Наиболее часто зависи-
мость между грунтом и рыбой — не непосредственная, а через по-
средство кормовых объектов, приуроченных к определенному виду
субстрата. Например, приуроченность леща в Аральском море, в
определенное время года, к серым илистым грунтам целиком объ-
ясняется высокей биомассой бентоса этого грунта (бентос служит
пищей леща). Но имеет лесто и непосредственная зависимость ры-
бы от характера грунта, вызываемая приспособлением рыбы к опре-
деленному типу субстрёта. Так, например, закапывающиеся рыбы
всегда приурочены в своем распространении к мягким грунтам;
рыбы, приуроченные в своем распространении к каменистым грун-
там, часто обладают присоской для прикрепления к донным пред-
метам и т. п. Многие рыбы выработали у себя ряд иногда довольно
сложных приспособлений для ползания по грунту. Некоторые ры-
бы, вынужденные иногда передвигаться по суше, также имеют ряд
особенностей в строении своих конечностей и хвоста, приспособлен-
ных к движению по твердому субстрату. Наконец, окраска рыб
как мы видели выше, в значительней степени определяется цветом
и рисунком того грунта, на котором рыба находился. Не только
взрослая рыба, но денная—демерсальная икра (см. ниже) и ли-
чинки также находятся в весьма тесной зависимости от характера
29
грунта водоема, на который откладыается икра или у которсго
держатся личинки.
Рыб, проводящих значительную часть своей жизни зарывшись
в грунт, сраьнительно немного. Значительную часть времени про-
водят в грунте, например, личинки миног, которые могут в течение
нескольких дней находиться в грунте и не подниматься на поверх-
ность. Значительное время проводит в грунте среднеевропейская
шиповка Ccbitis taenia, которая так же, как и пескоройка, может
даже питаться, закопавшись в грунт. Большинство же рыб зака-
пывается в грунт или во время опасности или в период пересыха-
ния водоемов. Почти все рыбы, которые закапываются в грунт,
имеют змееобразно вытянутое тело и ряд других приспособлений,
связанных с закапыванием. Так, у роющей ходы в жидком иле
индийской рыбки PhisoodonophiS boro ноздри имеют вид трубочек
и расположены на вентральной стороне головы. Это приспособление
позволяет рыбке успешно проделывать свои ходы заостренной го-
ловой, и ноздри ее при этом не забиваются илом. Процесс закапы-
вания производится путем ундулирующих движений тела, сходных
с теми движениями, которые рыба проделывает при плавании. Рыба,
становясь под углом к грунту головой вниз, как бы ввинчивается
в грунт. Другая группа закапывающихся рыб имет плоское тело,
как например, камбалы и скаты. Эти рыбы обычно закапываются
значительно хуже представителей первой.группы и никогда не за-
рываются столь глубоко. У них процесс закапывания происходит
несколько иным способом: рыбы как бы накидывают на себя грунт
и обычно целиком не зарываются, а выставляют наружу голову и
часть тела.
Закапывающиеся в грунт рыбы — главным образом обитатели
мелководных внутренних водоемов или прибрежных участков мо-
рей. Среди рыб глубинных частей моря и внутренних водоемов мы
этого приспособления не наблюдаем. Из пресноводных рыб, приспо-
собившихся к закапыванию в грунт, можно указать африканского
представителя двоякодышащих — Protopterus, закапывающегося в
грунт водоема и впадающего в своего рода летнюю спячку во время
засухи. Из пресноводных рыб умеренных широт закапываются в
грунт вьюн Misgurnus fcssilis, зарывающийся главным образом во
время пересыхания водоемов; шиповка Ccbitis taenia, для которой
закапывание в грунт служит главным образом средством защиты.
Многие рыбы, хотя сами и не закапываются в грунт, но в поисках
пищи, могут относительно довольно глубоко проникать в грунт,,
выкапывая объекты своего питания. Перекапывают грунт в большей
или меньшей степени почти все бентосоядные рыбы. Раскапывание
грунта производится обычно струей воды, выпускаемой из ротового
отверстия и относящей в сторону мелкие иловые частицы. Непосред-
ственно роющие движения у бентосоядных рыб наблюдаются реже.
В качестве примеров зарывающихся морских рыб можно приве-
сти песчанку Ammodytes, которая также закапывается в песок,
главным образом спасаясь от преследования. Некоторые бычки
Oobiidae выкапывают себе неглубокие норки, в. которые прячутся
зо
Рис. 18. Грудные плавники донных рыб
(сверху вниз): многопера (Polypterl),
морского петуха — триглы (Percifor-
mes) и uOgcocephaIus (Lophiiformes).
от опасности. Закапывающиеся в грунт камбалы и скаты прибегают
к этому также, главным образом, чтобы быть менее заметными.
Очень часто перекапывание грунта у рыб связано с постройкой
гнезда. Так, например, гнезда в виде ямки строят некоторые пред-
ставители семейства Cichlidae, в частности, Geoplugus brasiliensis,
который выкапывает в грунте ямку, куда откладывает икру. Для
защиты от врагов многие рыбы закапывают икру в грунт, так что
она проходит свое развитие внутри грунта. Развивающаяся в грунте
икра обладает целым рядом
специфических приспособлений
и вне грунта развивается ху-
же (см. ниже, стр. 130), чем
внутри него. В качестве при-
мера морских рыб, закапы-
вающих икру, может быть
указана атерина Laurcstes te-
nuis, а из пресноводных —
большинство лососей, у кото-
рых как икра, так и личинки
на ранних стадиях, разви-
ваются закопанными в гальку,
будучи таким образом защи-
щены от многочисленных вра-
гов. У рыб, зарывающих икру в
грунт, инкубационный период
бывает обычно весьма длите-
лен от 10 до 100 и более дней.
У многих рыб оболочка икрин-
ки, попадая в воду, становится
клейкой, благодаря чему икриг
ка прикрепляется к субстрату.
Рыбы, держащиеся на твер-
дом грунте, особенно в при-
брежной зоне или на быстром
течении, очень часто имеют раз-
личные органы прикрепления
к субстрату (см. ниже, стр. 30,40)
или в виде присоски, образо-
ванной путем видоизменения
ижнейгубы, грудных или брюшных плавников, или в виде шипов
и зацепок, обычно развивающихся на окостенениях плечевого и
брюшного поясов и плавников, а также жаберной крышки.
Как мы уже указывали выше, распространение многих рыб при-
урочено к определенным грунтам, причем часто близкие виды од-
ного и того же рода встречаются на разных грунтах. Так, например,
бычок Icelus spatula приурочен в своем распространении к камени-
стогалечниковым грунтам, а близкий вид Icelus spiniger к песча-
ным и илистопесчаным грунтам. Причины, вызывающие приурочен-
ность рыб к определенного типа грунту, как указывалось выше,
31
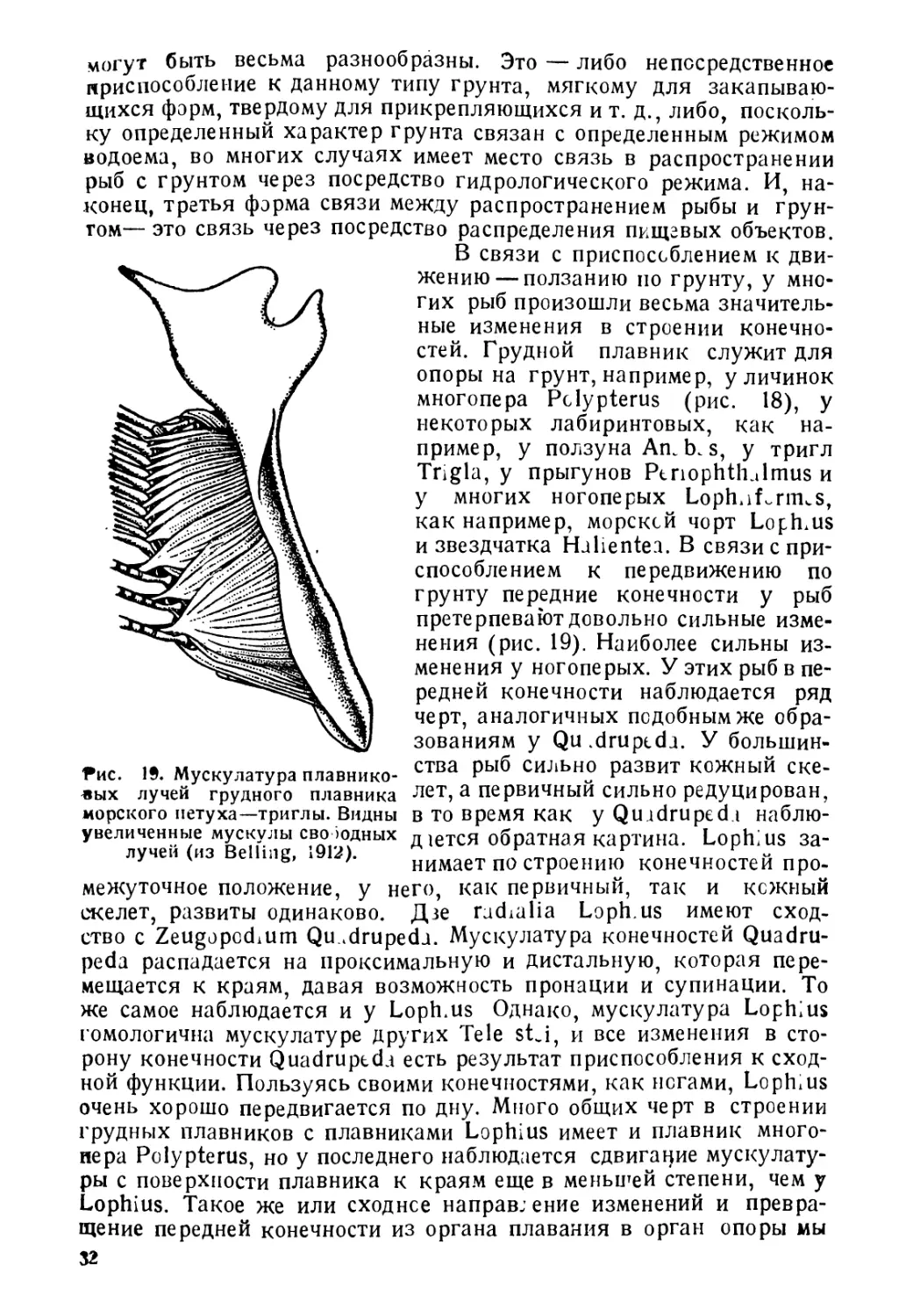
Рис. 19. Мускулатура плавнико-
вых лучей грудного плавника
морского петуха—триглы. Видны
увеличенные мускулы свободных
лучей (из Belling, 1912).
могут быть весьма разнообразны. Это — либо непосредственное
приспособление к данному типу грунта, мягкому для закапываю-
щихся форм, твердому для прикрепляющихся и т. д., либо, посколь-
ку определенный характер грунта связан с определенным режимом
водоема, во многих случаях имеет место связь в распространении
рыб с грунтом через посредство гидрологического режима. И, на-
конец, третья форма связи между распространением рыбы и грун-
том— это связь через посредство распределения пищевых объектов.
В связи с приспособлением к дви-
жению— ползанию по грунту, у мно-
гих рыб произошли весьма значитель-
ные изменения в строении конечно-
стей. Грудной плавник служит для
опоры на грунт, например, у личинок
многопера Pclypterus (рис. 18), у
некоторых лабиринтовых, как на-
пример, у ползуна An. b. s, у тригл
Trigla, у прыгунов PtnophthJmus и
у многих ногоперых Loph.if-rniuS,
как например, морской чорт Lophius
и звездчатка Halientea. В связи с при-
способлением к передвижению по
грунту передние конечности у рыб
претерпевают довольно сильные изме-
нения (рис. 19). Наиболее сильны из-
менения у ногоперых. У этих рыб в пе-
редней конечности наблюдается ряд
черт, аналогичных подобным же обра-
зованиям у Qu.drupeda. У большин-
ства рыб сильно развит кожный ске-
лет, а первичный сильно редуцирован,
в то время как у Qu idruped.i наблю-
дщтся обратная картина. Lophius за-
нимает по строению конечностей про-
межуточное положение, у него, как первичный, так и кожный
скелет, развиты одинаково. Дзе radialia Loph.us имеют сход-
ство с Zeugopodium Qtcdrupeda. Мускулатура конечностей Quadru-
peda распадается на проксимальную и дистальную, которая пере-
мещается к краям, давая возможность пронации и супинации. То
же самое наблюдается и у Loph.us Однако, мускулатура Lophius
гомологична мускулатуре других Tele stj, и все изменения в сто-
рону конечности Quadrupeda есть результат приспособления к сход-
ной функции. Пользуясь своими конечностями, как ногами, LophxUs
очень хорошо передвигается по дну. Много общих черт в строении
грудных плавников с плавниками Lophius имеет и плавник много-
пера Polypterus, но у последнего наблюдается сдвигание мускулату-
ры с поверхности плавника к краям еще в меньшей степени, чем у
Lophius. Такое же или сходнее направл ение изменений и превра-
щение передней конечности из органа плавания в орган опоры мы
32
наблюдаем у прыгуна Periophthalmus. Последний живет в мангро-
вых зарослях и значительную часть времени проводит на суше. На
берегу он гоняется за наземными насекомыми, которыми питается.
Двигается эта рыбка по .суше прыжками, которые совершает при
помощи хвоста и грудных плавников.
Своеобразное приспособление для ползания по грунту имеется
у триглы, у которой три первые луча грудного плавника обособлены
и приобрели подвижность. Пользуясь этими лучами, тригла ползает
по грунту; кроме того, эти лучи служат рыбе также органом осяза-
ния. В связи с особой функцией первых трех лучей происходят и
некоторые анатомические изменения; в частности мускулы, приво-
дящие в движение свободные лучи, развиты значительно сильнее,
чем все остальные (рис. 19). Представитель лабиринтовых—ползун
Anabas, перемещаясь по суше, пользуется для движения грудными
плавниками, а также иногда жаберными крышками.
В жизни рыб огромную роль играет не только грунт, но и взве-
шенные в воде твердые частицы. Выше отмечалось, какое значение
в жизни рыб имеет прозрачность воды, в значительной степени опре-
деляющаяся в небольших внутренних водоемах и прибрежных участ-
ках морей примесью взвешенных минеральных частиц. Но кроме
влияния через посредство изменения прозрачности, взвешенные ча-
стицы в воде оказывают воздействие на рыбу в самых различных
направлениях. Наиболее сильное влияние оказывают на рыб взве-
шенные в воде частицы мутных речных вод, где содержание твердых
частиц часто достигает до 4% по объему. Здесь в первую очередь
сказывается непосредственное механическое влияние несущихся
в воде минеральных частиц различной величины, от нескольких
микронов до гальки в 2—3 см в диаметре. В связи с этим у рыб
мутных рек вырабатывается ряд приспособлений, как например,
резкое уменьшение размера глаз. Малоглазость характерна для
лопатоносов, живущих в мутных водах гольцов и различных со-
мов. Сокращение размеров глаза объясняется необходимостью умень-
шения незащищенной площади, которая может быть повреждена
несомой потоком взвесью.
Наличие большого количества взвеси в воде естественно должно
затруднять также и дыхание рыбы. Повидимому, в связи с этим у
рыб, живущих в мутных водах, выделяемая кожей слизь обладает
способностью очень быстро осаждать взвешенные в воде частицы.
Наиболее детально это явление изучено для американского чешуй-
чатника Lepidcsiren paradoxa, коагулирующие свойства слизи ко-
торого помогают ему жить в тонком илу водоемов Чако. Для Phi-
soodonophis boro установлено, что его слизь в сильной степени об-
ладает способностью осаждать взвесь. Прибавление одной—двух
капель слизи, выделяемой кожей рыбки, к 500 см3 мутной воды вы-
зывает осаждение взвеси в 20—30 секунд. Такое быстрое осаждение
..приводит к тому, что даже в очень мутной воде рыба живет как бы
Окруженная футляром чистой воды. Сама слизь, выделяемая кожей,
при соприкосновении с мутной водой, меняет сво|Ю химическую
реакцию. Так, установлено, что pH слизи при соприкосновении с
3 Биология рыб 33
водой резко снижается, падая с 7,5 до 5,0. Естественно, что коагули-
рующее свойство слизи важно, как предохранение жабр от забивания
взвешенными частицами. Но несмотря на то, что у рыб, живущих
в мутных водах, имеется ряд приспособлений для защиты от воздей-
ствия взвешенных частиц, все же, если количество мути превысит
определенную величину, может наступить гибель рыб. При этом
смерть, невидимому, происходит от удушия в результате забивания
жабр наносами. Так, известны случаи, когда во время силей, когда
мутность потоков возрастала в десятки раз, имела место массовая
гибель рыб. Подобное явление зарегистрировано в горных районах
Афганистана и Индии, причем погибали даже столь приспособлен-
ные к жизни в мутной воде рыбы, как туркестанский сомик Glyptc-
sternum reticulatum и некоторые другие.
Движение воды
Разнообразные формы движения воды, как поступательные (те-
чения, вертикальная циркуляция), так и колебательные (волнения,
приливы),оказывают на рыб чрезвычайно сильное влияние, как не-
посредственно, так и косвенно.
Особенно быстрое течение мы наблюдаем в реках, где иногда
скорость движения воды достигает скорости падающего тела. В
реках, берущих начало с гор, скорость движения воды является
основном фактором, определяющим распределение животных, в
том числе и рыб на протяжении русла потока. Основной формой
влияния речного течения, несомненно, является непосредственное
механическое воздействие на организм животного движущегося
потока, причем при больших скоростях лишь специально приспо-
собившиеся формы могут сопротивляться воздействию этого факто-
ра. Приспособление к сопротивлению быстроте течения у различных
Представителей ихтиофауны идет различными путями. По характеру
местообитания в быстром потоке и связанным с этим приспособле-
нием индусский исследователь Нега (1930) делит всех рыб, населяю-
щих быстрые потоки, на четыре группы: 1) Мелкие еиды, живущие
в застойных местах, в бочагах под водопадами, заводях и т. п. Эти
рыбы по своему строению наименее приспособлены к жизни в быст-
ром потоке. Примером представителей этой группы могут с; у жить:
быстрянка Alburnoidcs bipunctatus, дамский чулочек D< то ге-
rio и др. 2) Хорошие пловцы с сильным вальковатым телом, пре-
восходно преодолевающие быстрое течение. Сюда относятся многие
речные виды: лосось S. Imo solar, маринки Sch.zcthorax, некоторые,
как азиатские (Barbus brachycephalus В. ter), так и африканские
(B rbus radcl.ffi) виды усачей, и многие другие. 3) Мелкие донные
рыбы, обычно живущие между камнями дна потока й плавающие
от камня к камню. Эти рыбки, как правило, имеют веретенообраз-
ную слегка вытянутую форму. Сюда относятся многие гольцы Ne-
machJus, пескарь G.bio g-bio и др. 4) В эту группу входят такие
формы, которые выработали у себя специальные органы прикреп-
ления (присоски, шипы), при помощи которых они прикрепляются
34
к донным предметам. Обычно относящиеся к этой группе рыбы име-
ют уплощенную дорзовентрально форму тела (рис. 20). Присоска
образуется или на губе (Garra и Др.), или между грудными плавни-
ками (Glyptotharax), или путем слияния брюшных плавников. В
эту группу входят D.scognathichthys, многие виды семейства Si-
soridae, большинство пр.дставител-й своеобразного тропического
семейства Homaloptaridue и др.
Как видно, механическое воздействие текущего потока отби-
рает рыб, имеющих форму тела, приспособленную к преодолению
Рис. 20. Присоски для прикрепления к грунту у речных рыб,
сомика Glyptothorax (слева) и Garra из карповых (справа)
(из Нога, 1923 и Annandale, 1919).
сопротивления движущейся воды. Естественно, что в реках с быст-
рым течением мы не находим рыб с уплсщенным с боков телом или
со слабо развитым хвостом, — основным органом поступательного
движения. По мере замедления течения при движении от верховья
к низовью реки постепенно в русле начинают появляться рыбы,
менее приспособленные к большим скоростям — худшие пловцы.
У рыб — обитателей вод с медленным течением тело более уплощено
и, если это не хищники, то они обычно не столь хорошие пловцы,
как обитатели быстрых рек (рис. 21). Постепенное изменение формы
тела рыбы от верховья к низогыо реки, связанное с постепенным
изменением скорости течения, носит вполне закономерный характер.
В тех местах реки, где течение замедляется, держатся рыбы, менее
приспособленные к жизни в быстром потоке, в местах с чрезвычай-
но быстрым движением воды сохраняются только наиболее приспо-
собленные формы, — типичные обитатели быстрого потока — рео-
филы. Ван-дем Борне, пользуясь распределением рыб в реке, раз-
деляет реку на отдельные участки (для рек западной Европы).
1) Участок форели—горная часть потока с быстрым течением и ка-
менистым грунтом характеризуется рыбами с вальковатым телом
(форель, голец, гольян, подкаменщик). 2) Участок усача — рав-
нинное течение, где скорость потока все же еще! значительна.
Появляются уже рыбы с более высоким телом. Для этого
участка характерны усач, елец и др. 3) Участок леща — течение
з* 35
замедлено, грунт — частью ил, частью песок, в русле появляется
подводная растительность, преобладают рыбы с уплощенным с
боков телом, как лещ, плотва, красноперка и др. Конечно, границу
между этими отдельными экологическими участками провести весь-
ма трудно, и смена одних рыб другими происходит обычно весьма
постепенно, но в общем намеченные Борне участки выделяются
в большинстве рек с горным питанием весьма отчетливо, и уста-
новленные им закономерности для рек Европы сохраняются как в
реках Америки, так Азии и Африки. Отбирающее влияние скорости
течения сказывается не только на отборе различных видов, но и
Рис. 21. Поперечный раз-
рез различных рыб быстро-
текущих вод. Верхний ряд
слева направо: форель,
гольян, голец, подкамен-
щик в стоячих и медленно
G00W
среди особей одного и того же вида реч-
ные формы имеют, как правило, строение
тела, более приспособленное к жизни в
движущемся потоке, чем у особей из
стоячих вод. Так, ха'риус Thymallus
arcticus из Байкала имеет более высокое
тело и более длинный хвостовой стебель,
в то время как представители того же
вида из Ангары более низкотелы и корот-
кохвосты, что свойственно хорошим плов-
цам. Более слабые молодые особи речных
рыб (усачи, гольцы), как правило, имеют
более низкое вальковатое тело и более
текущих водах. Нижний
ряд слева направо: лещ,
карась, карп, плотва, крас-
ноперка. Рыбы взяты оди-
наковой высоты (из Hesse,
короткий хвостовой стебель, чем взрос-
лые. Кроме того, обычно в горных реках
взрослые, более крупные и сильные осо-
би держатся выше по течению, чем моло-
1924). дые. Если двигаться вверх по тече-
нию реки, то средние размеры особей
одного и того же вида, например, гребенчатохвостого и
тибетского гольцов все увеличиваются, и наиболее крупные
особи наблюдаются близ верхней границы распространения вида
(Турдаков, 1939). Речные течения влияют на организм рыбы
не только непосредственно механически, но и косвенно, через
посредство других факторов. Как правило, водоемы с быстрым
течением характеризуются.|перенасыщением кислородом. Поэтому
реофильные рыбы одновременно являются и оксифильными, т. е.
кислородолюбивыми, и, наоборот, рыбы, населяющие медленно
текущие или стоячие воды, обычно лучше приспособлены к различ-
ному кислородному режиму и более хорошо переносят дефицит
кислорода. Течение, влияя на характер грунта потока, а тем самым
на характер донной жизни, естественно оказывает влияние и на пи-
тание рыб. Так, в верховьях рек, где грунт образует неподвижные
глыбы, обычно может развиваться богатый перифитон, служащий
основной пищей для большинства рыб этого участка реки. В силу
этого рыбы верховий характеризуются, как правило, очень длин-
ным кишечным трактом, приспособленным для переваривания рас-
тительности, а также развитием рогового чехлика на нижней губе.
По мере движения вниз по течению грунты становятся мелкими,
36
и под влиянием течения приобретают подвижность. Естественно, что
на подвижных грунтах богатая донная фауна развиваться ье может,
и рыбы переходят к питанию рыбами же или пищей, падающей с
суши. По мере замедления течения постепенно начинается заиле-
ние грунта, развитие денной фауны, и в русле опять появляются
растительноядные виды рыб с длинным кишечным трактом.
Течение в реках влияет изменяющим образом не только на стро-
ение организма рыбы, но и на ее поведение, ив первую очередь из-
меняет характер размножения. Многие обитатели рек с быстрым
течением имеют прилипающую икру. Некоторые виды откладывают
икру, закапывая ее в песок. Амер» каискг е сомы из рода Plecosto-
mus откладывают икру в специальные пещерки, другие роды(см. раз-
множение) вынашивают икру на своей брюшной стороне. Изме-
няется и строение внешних половых органов. У некоторых видов
развивается менее длительная живучесть спермы и т. д.
Таким образом мы видим, что формы влияния на рыб течения в
реках весьма разнообразны, и в некоторых случаях неожиданные
перемещения больших масс воды, например сили или прорывы
плотин горных озер, могут приводить к массовой гибели ихтиофау-
ны, как это, например, имело место в Читрале в 1929 году. Скорость
течения иногда является прекрасным изолирующим фактором, при-
водящим к разобщению фауны отдельных водоемов и содействую-
щим ее обособлению. Так, например, пороги и водопады между круп-
ными озерами восточной Африки не являются преградой для силь-
ных крупных рыб, но непроходимы для мелких и приводят к изо-
ляции отдельных локальных форм в разъединенных таким образом
бассейнах (рис. 22—см. вклейку в конце книги). Естественно, что те
своеобразные приспособления, которые мы наблюдаем у рыб в связи
с жизнью на быстром течении, как видно из изложенного, в усло-
виях горного быстрого потока, характеризующегося, как правило,
низкой температурой, огромными скоростями движения воды и вы-
сокой насыщенностью кислородом, не могли выработаться сразу, а
являются результатом весьма длительного эволюционного процесса.
По современным воззрениям фауны горных быстрых рек умеренных
и высоких широт северного полушария—реликты ледниковой эпохи.
В то же время фауна горных потоков тропических, а частично и
умеренных широт, развилась в результате постепенного приспособ-
ления к своеобразным условиям и постепенного переселения орга-
низмов в высокогорные водоемы из равнин. Для ряда групп пути
приспособления к жизни в горных потеках прослеживаются доста-
точно ясно и могут быть восстановлены (рис. 23). Как в реках, так
и в стоячих водоемах, течения оказывают на рыб весьма сильное
влияние, но в то время как в реках основная форма влияния прояв-
ляется в непосредственном механическом воздействии движущегося
потока, влияние течения в морях и озерах сказывается больше через
посредство изменений, вызываемых 'течением в распределении
Других факторов среды (температура, соленость и д)р.). Непосред-
ственное механическое воздействие движущегося 'потока также
отражается на биологии рыб в стоячих водоемах. Механическое
37
влияние течений в первую очередь сказывается на переносе рыб,
личинок и икры инсгда на огромное расстояние. Так, например, ли-
чинки сельди Clupea harengus, выведшиеся у берегсв Норвегии, те-
чением выносятся далеко на север. Расстояние от Лофотен, — места
нереста сельди, идо Кольского меридиана мальки сельди проходят
GLyPTtJSTERNUM
(HPCROPTER4M)
GLyPTOSTERNUn
GlyPTDSTERNUM
(srOLiCZHRE И ДР.)
Ere tristes
Npnorr
----► Насыщение воды кислородом.
Рис. 23. Предполагаемый ход приспособления сомиков се-
мейства Sisoridae по отношению к двум основным факторам
горных потоков. На диаграмме видно, что ряд видов вто-
рично стал менее реофильным (из Нога, 1930).
примерно за 3 месяца, в июле (нерест в марте) появляясь у бере-
гов Мурмана .Пелагические яйца многих рыб также переносятся тече-
ниями иногда на очень значительные расстояния. Так, например,
яйца камбал, отложенные у берегов Франции, относятся к берегам
Дании, где происходит выход молоди. Продвижение личинок угря
от нерестилищ до устьев рек Европы в значительной своей части
приурочено к Гольфштрему, которым личинки почти пассивно пере-
носятся к берегам Европы. Взрослые рыбы, особенно плохие плов-
цы, также часто переносятся течениями на значительные расстоя-
ния. Так, известно несколько случаев заноса луны-рыбы в район
Владивостока теплым течением, идущим с юга на север.
38
Многие взрослые рыбы при своих миграциях ориентируются
иа течения, придерживаясь их при передвижении. Например, даль-
невосточные лососи идут на струи воды, выходящие из устьев рек.
Акула Cetcrhinus в своем распространении почти целиком привя-
зана к северо-атлантическому течению.
Скорости течения, на которые рыба начинает реагировать, для
большинства рыб неизвестны, но для тех рыб, для которых они опре-
делены, они не менее 5 см в секунду (для хамсы).
Органом, воспринимающим движение воды, служат клетки бо-
ковой линии. В простейшем виде у акул они представляют собою
чувствующие клетки, расположенные в эпидермисе. В процессе
эволюции (например у химеры) эти клетки погружаются в кагал,
который постепенно (у костистых рыб) замыкается и соединяется с
окружающей средой только посредством трубочек, прободающих
чешую и образующих боковую линию. У различных рыб боковая
линия развита далеко не одинаково. У сельдевых каналы боковой
линии имеются только на голове. У многих пресноводных рыб бо-
ковая линия бывает неполная (например у верховки и некоторых
гольянов). Иннервируют органы боковой линии Nervus facialis и
N. vagus. Органы боковой линии позволяют рыбе ощущать движение
и колебания воды, причем многие морские рыбы почти не реагируют
на течения при помощи органов боковой линии. Боковая линия им
служит только для ощущения колебательных движений воды. У
речных же рыб боковая линия позволяет ориентироваться на те-
чение. Вундер (1936) так определяет функцию органов боковой ли-
нии: 1) помощь в определении положения подвижных кормовых
объектов (у хищников обычно органы боковой линии развиты лучше,
чем у мирны'х рыб), 2) ощущение приближения врага, 3) ощущение
предметов, встреченных на пути движения рыбы или приближающих-
ся к нему, 4) ориентировка на течение или на колебания воды.
Значительно больше чем непосредственное влияние сказывается
косвенное влияние течения на рыбу, главным образом путем изме-
нения режима вод. Хслсдные те'ения, идущие с севера на юг, по-
зволяют проникать арктическим ф рмам далеко в умеренную область.
Так, например, холоднее лабрадорское течение далеко к югу отодви-
гает распространение ряда теплсвсдных форм, которые по побережью
Европы, где сильно сказывается теплое течение Гольфштрема, про-
двигаются далеко на север. В Баренцевом море распространение
отдельных высоко арктических видов семейства Zoarcidae приуро-
чено к промежуткам с холодной водой между струями теплог»
течения. В ветвях же этого течения держатся более тепловодные
рыбы, как, например, скумбрия и другие.
Течения коренным образом могут изменять и химический режим
водоема и, в частности, влиять на его соленость, внося или более
соленую, или более пресную воду. Так, Гольфштрем вносит в Ба-
ренцево море более соленую воду, и к его струям приурочены бо-
лее соленоводные организмы. К течениям, образованным пресными
водами, вынесенными сибирскими реками, в значительной степени
приурочены в своем распространении сиги и сибирский осетр. В
Зв
Рис. 24. Брюшные плавники морских рыб
видоизмененные в присоску: слева—-бычка
Gobius и справа—колючего пинагора Еп-
microtremus (из Берга, 1933 и Перминова,
1936).
местах стыка холодных и теплых течений образуется обычно зона
весьма высокой продуктивности, так как в таких областях происхо-
дит массовое отмирание беспозвоночных и растений планктона,
дающих огромную продукцию органического вещества, которое
позволяет развиваться немногим эвритермным фермам в массовом
количестве. Примеры подобного рода стыков холодных и теплых
вод довольно обычны, например, близ западного побережья Южной
Америки у Чили, на Ньюфаундлендских банках и др.
Значительную р. ль в жизни рыб играют вертикальные
токи воды. Непосредственное механическое воздействие это-
го фактора удается наблюдать редко. Обычно влияние вертикальней
циркуляции вызывает перемешивание нижних и верхних слоев во-
ды; а тем самым выравнивание распределения температуры, солено-
сти и других факторов, что,
в свою очередь, создает бла-
гоприятные условия для
вертикальных миграций рыб.
Так,'например, в Аральском
море воблд вдали от берегов
весной и осенью ночью под-
нимается за пищей в поверх-
ностные слои, а днем опус-
кается в придонные. Летом
же, когда устанавливается
резко выраженная стратифи-
кация, вобла все время дер-
жится в придонных слоях.
Перейдем теперь к рассмотрению влияния на рыб колеба-
тельных движений воды. Основной формой колеба
тельных движений воды являются волнения, воспринимаемые,
как мы говорили1 выше, органами боковой линии рыбы. Волны
производят на рыбу огромное влияние. Как правило, во время
сильных волнений в море пелагические рыбы опускаются в более
глубокие слои воды, где волнение на них ге влияет. Особенно
сильное влияние оказывает на рыб волнение в прибрежных
участках. В этих местах сила удара волны достигает до полутора
тонн на один кв. метр и, естественно, рыбы должны выработать
специальные приспособления для предохранения себя и своей
икры от влияния прибоя. Большинство рыб, живущих в при-
брежной зоне, имеют специальные приспособления, помогаю-
щие им держаться на месте и не быть оторванными волнением и
разбитыми о камни. Так, например, типичные обитатели прибреж-
ных вод различные бычки Gcbiidae имеют брюшные плавники,
измененные в виде присоски, при помощи которой рыбы удержи-
ваются на камнях. Несколько иного характера присоски име-
ются у пинагоров Cyclopteridae, также живущих в прибрежной
зоне (рис. 24).
Кроме огромного непосредственного механического воздей-
ствия волнения оказывают на рыбу и большое косвенное влия-
40
ние, способствуя перемешиванию воды и утопанию слоя скачка.
Так, например, в последние годы в связи с понижением Каспий-
ского моря в результате увеличения зоны перемешивания
(эпилимнион) верхняя граница придонного слоя, где происходит
накопление биогенных веществ, также понизилась. Тем самым
часть биогенных вещств вошла в круговорот органического
вещества в водоеме, вызвав увеличение количества планктона,
а тем самым, следовательно, и кормсвой базы для каспийских
планктоноядных рыб.
Другим видом колебательных движений морских вод яв-
ляются приливно-отливные движения, достигающие в некото-
рых участках моря весьма значительной амплитуды. Так, у
берегов Северной Америки разность уровней прилива и отлива
достигает 16 м. Естественно, что у тех рыб, которые живут в
приливно-отливной, периодически обсыхающей зоне, или в
прибрежных участках моря, над которыми четыре раза в сутки
проносятся огромные массы воды, должны выработаться
специальные приспособления к жизни в небольших лужицах,
остающихся после отлива. Все обитатели приливно-отливной зоны—
литорали имеют или дорзовентрально уплощенную или змеевидную
или вальковатую форму тела. Высокотелых рыб мы на литорали
не находим. Так, на Мурмане обычно остаются на литорали
бельдюга Zoarces viviparus и маслюк Pholis gunnelus, виды е
удлиненной формой тела, а также большеголовые подкаменщики,
главным образом, Myoxocephalus scorpius.
Своеобразные изменения происходят у рыб приливно-отливной
зовы в биологии размножения. Многие из рыб на время икромета-
ния просто отходят из литоральной полосы (подкаменщики); не-
которые приобретают способность живорождения, как, например,
бельдюга, икра которой проходит инкубационный период в теле
матери. Пинагор, который обычно откладывает икру ниже уровня
отлива, в тех случаях, когда икра его все же обсохнет, поливает
ее водой изо рта и плещет на нее хвостом. Наиболее любопытное
приспособление к размножению в приливно-отливной зоне наблю-
дается у американской рыбки Laurestes tenuis, которая отклады-
вает икру в сизигийные приливы в той части приливно-отливной
зоны, которая не покрывается квадратурными приливами, так что
икра развивается вне воды во влажной атмосфере. Инкубационный
период длится до следующего сизигия, когда молодь выходит из
икры и уходит в воду. Приливно-отливные течения, так же, как и
вертикальная циркуляция, оказывают влияние на рыб, перемешивая
донные отложения и вызывая, таким образом, лучшее освоение их
органического вещества, а тем самым повышение продуктивности
водоема. Так, в горле одной из бухт залива Фунди, где наблюдается
весьма большая амплитуда приливно-отливных течений (четыре
в сутки проносится масса воды около двух биллионов тонн)
нМ^?Йается и очень богатое развитие планктона 1и совершенно
исключительные уловы £ыбы. '
Несколько особняком стоит влияние такой формы движения
41
воды как смерчи. Смерчи захватывают из моря или внутренних
водоемов большое количество воды и переносят ее вместе со всеми
животными, в том числе и рыбой, на огромные расстояния. В Индии
во время муссонов довольно часто имеют место рыбные дожди,
когда обычно вместе с ливнем на землю падает и живая рыба.
Иногда эти дожди захватывают довольно значительные площади.
Подобные рыбные дожди бывают в самых различных частях света,
они описаны для Норвегии, Испании, Индии и ряда других мест
Биологическое значение рыбных дождей несомненно в первую
очередь выражается в способствовании расселению рыб, причем
при помощи рыбных дождей могут преодолеваться преграды, в
обычных условиях для рыб непреодолимые.
Таким образом, как видно из изложенного, формы влияния на
рыбу движения воды чрезвычайно разнообразны и накладывают
на организм рыбы неизгладимый отпечаток, заставляя вырабаты-
вать специфические приспособления, обеспечивающие рыбе суще-
ствование в различных условиях.
Б. ВЛИЯНИЕ ХИМИЧЕСКИХ СВОЙСТВ ВОДЫ НА РЫБ
Соленость
Влияние на рыб растворенных в воде веществ чрезвычайно ве-
лико. Как известно, амплитуда солености воды, при которой могут
жить рыбы, весьма значительна, начиная от совершенно пресной
воды до солености в 70°/^ и выше. По отношению к солености рыб
принято разделять на эвригалинных, выдерживающих
значительные колебания солености, и стеногалинных, ко-
торые не могут переносить сильных изменений в количестве раство-
ренных в воде солей. Примером рыб, относящихся к первой группе,
могут служить многие бычки, как Bubyr caucasicus, выдерживающий
колебания солености от десятых долей °/00 до 6С°/00. Каспийская
игла Svngnathus nigrol.neatus caspius живит в воде с соленостью
от О,27°/оо до 38°/00 и выше. Примерами стеногалинных форм могут
служить многие колючеперые, обитающие среди коралловых рифов.
Эги рыбки выдерживают колебания солености всего в несколько */ог
Также, повидимому, не выносят сильных колебаний солености и
морские глубоководные рыбы. Из пресноводных рыб, которые не
переносят осолонения воды, могут быть упомянуты хотя бы лопа-
тоносы, живущие в пресной воде и при попадании в воду с соле-
ностью в 2—3°/00 уже погибающие. Основной причиной привязан-
ности рыб к воде с определенной соленостью является, как уже ука-
зывалось выше, приспособление к жизни в воде с постоянней плот-
ностью и зависимость от плотности воды, в значительной мере об-
условливаемой ее соленостью, характера осморегуляторных про-
цессов, протекающих в организме рыбы. Химический состав солей,
растворенных в воде, также оказывает известное влияние на рыб^.
В то время как некоторые виды рыб хорошо живут в разбавленной
морской воде, они очень скоро погибают, будучи перенесены в во-
42
ду с тем же количеством солей, но с составом солей озерного типа,
т. е. с преобладанием сернокислых солей над хлоридами.
Взсьма важное значение, особенно за последнее время, при-
обретают для рыб растворимые в вс^де вещества, которые не входят
нормально в состав морской и пресной воды. Это те вещества, которые
понадают в воду в результате различных форм деятельности чело-
века — лесосплава, спуска сточных вод предприятий, разработки
различных горных пород, попадания в воду нефти и т. п. Список
различных веществ, попадающих в воду в результате деятельности
человека и оказывающих влияние на водную фауну и в том числе
и на рыб, очень велик. Ellis (1937) указывает до ста названий по-
добного рода веществ. Влияние на рыбу попадающих в воду кислот
очень велико, но исследование их токсического действия затруднено
тем, что кислота неодинаково влияет на рыбу при различной жест-
кости воды и различной концентрации водородных ионов. Так,
например, серная кислота при концентрации 1 7450 в мягкой
воде,вызывает гибель золотой рыбки через 6—9 часов, в жесткой
же воде рыба .продолжает жить в течение многих дней. Винная
кислота при концентрации 1 1000 в мягкой воде убивает рыбку
через 20—25 минут, а в жесткой воде при той же концентрации
кислоты рыбка.выживает 3 часа—3 ч. 20 м. Это, повидимому, объ-
ясняется тем, что при низких pH кислота менее диссоциирована и
легче проникает через кожные покровы и жабры. По силе токси-
ческого действия кислоты на основании опытов над золотой рыбкой
могут быть распределены в следующий ряд (цифры показывают
количество вещества в миллиграммах, на литр, при котором насту-
пает гибель рыбы):
Дубильная кислота 10
Хромовая кислота 1С0
Серная кислота 130
Соляная кислота 159
Азотная, щавелевая, бензойная 200
Уксусная кислота 348
Молочная и лимонная 625
Механизм действия различных кислот на рыбу неодинаков.
Некоторые кислоты, обжигая жабры, приводят к нарушению дыха-
ния и тем самым вызывают гибель рыбы (например уксусная кис-
лота), другие влияют через посредство кишечного тракта, проникая
через стенки кишечника в соки тела рыбы и нарушая нормальный
,ход обмена веществ. Обычно же токсическое действие кислот на
организм рыбы бывает комбинированным, т. е. поражаются не
только жабры, или действие осуществляетсЯ';не только через кишеч-
ник, а имеют место одновременно оба пути влияния.
Соединения различных металлов, особенно тяжелых, оказы-
вают весьма сильное влияние на рыб. Так, одна часть свинца, раз-
веденная на 3 миллиона частей воды, уже вызывает гибель рыбы
(Carpenter, 1927). Железо и алюминий также при высоких концентра-
циях вызывают гибель рыбы. Доза, свыше 0,2 мг железа на литр
уже приводит к гибели рыбы. Алюминий смертелен для рыб
Ц^^Й^ентрациях свыше 0,5 мг на литр.
Как показали опыты Минкиной, концентрация железа в 0,2 мг
на литр воды вызывает резкое снижение газообмена у рыб
43
и замедление их роста. При кислой среде и высоких концентрациях
железа также наблюдается депрессия роста рыбы. Наоборот, не-
большие дозы железа (менее 0,1 мг на литр) особенно в щелочной
среде вызывают усиленное по сравнению с нормальным потребление
кислорода и ускорение )роста рыбы.
Токсическое действие металлов сводится в первую очередь к
свертыванию слизистых выделений жабр и протоплазмы живых
клеток, что приводит к нарушению газообмена. Жабры при отра-
влении железом покрываются плотным бурым налетом, естественно
затрудняющим дыхание.
Влияние других попадающих в воду соединений, как органи-
ческих, так и неорганических, также довольно разнообразно.
Правда, большинство из растворенных в воде веществ влияет на
ход газообмена, и поэтому чем медленнее идет у рыбы газообмен,
тем медленнее сказываются токсические свойства того или иного
растворенного в воде вещества. Поэтому понятно, что с понижением
температуры, вызывающим замедление газообмена, действие токси-
ческих веществ, как правило, понижается. Другие вещества, как
например, фенолы, вызывают у рыб паралич нервно-мускульного
аппарата и гемолиз крови. Амиловый спирт нарушает координацию
движений у рыбы и также вызывает полупараличное состояние.
Влияние растворенных в воде веществ сказывается, естественно,
не только на взрослых рыбах, но и на икре. Так, например, сточ-
ные воды бумажного производства, содержащие сернистскислый
натрий, меркаптан и другие вещества, вызывают в незначительных
концентрациях нарушение нормального хода дробления икры и
развития эмбрионов, а при более высоких концентрациях и довольно
быструю их гибель. Интересно, что на разных стадиях развития
эмбрионы рыб не одинаково чувствительны к воздействию различных
веществ. Так, как показали опыты, воздействие динитрсфенолом
на ранних стадиях развития рыбы вызывает значительно больший
процент гибели и большее количество уродов, чем воздействие на
стадии появления глазных пузырей. Токсическое действие различ-
ных растворенных в воде веществ меняется и в течение года. Наи-
большая восприимчивость к действию различных ядов у рыб при-
ходится на время усиленного питания и обычно совпадает с наиболее
теплым временем года; очень возможно, что это связано с увеличе-
нием скорости взаимодействия ядов и белков протоплазмы при
повышении температуры.
Некоторые соли, в частности фосфорные и.азотные, попадая в
воду, способствуют развитию органической жизни в воде и благо-
приятно отражаются на рыбах. Мы видели, что в районах постоян-
ного взмучивания донных отложений, способствующего вымыва-
нию солей из грунта, обычно развивается богатый планктон и бентос,
и скопляется большое количество рыбы. Отмечается, что количество
молоди некоторых из морских промысловых рыб находится в тесной
зависимости от количества в воде фосфатов, способствующих богатому
развитию планктона. В местах гнездовья птиц, где в воду падает
огромное количество помета, последний служит прекрасным удо-
44
бреиием воды и способствует развитию животных, служащих кормом
для рыб. В условиях прудового хозяйства очень часто применяется
удобрение прудов различными минеральными солями, содействую-
щими значительному повышению рыбной продуктивности. Так, как
видно из приведенного рисунка (рис. 25), удобренные пруды могут
300J
,ГЛЗ \l9№ | I9IB | 1917 | 1918 | I9i9 | 1920 j 1921 \ 1922
годы.
Рис. 25. Прирост карпов с единицы площади удобренного
минеральными солями (пунктир) и неудобренного (сплошная
линия) пруда (из Mortimer, 1939).
давать иногда почти в два раза больше рыбы, чем неудобренные,
качестве удобрения прудов употребляются различные вещества
и, в частности, фосфаты. По предложению профессора И. Н. Арноль-
ntrfio НаС’ В к^честве Удобрения для прудов, в которых разводится
.рыба, употребляются апатиты.
Газы
Чрезвычайно большое значение для рыб имеют растворенные
в воде г а з ы и, в первую очередь, конечно, кислород. Как известно,
отличие от наземных позвоночных, большинство рыб дышит возду-
растворенным в воде, и лишено возможности усваивать кислород
п°СРеАСТВеИИ0 из атмосФеры. Лишь немногие виды, живущие в
Доемах, где по тем или иным причинам часто имеет место дефицит
растворенного в воде кислорода, приспособились к дыханию атмос-
фер IM воздухом. Нормально*дыхание у рыб осуществляется путем
uui/1 а*ывания воды через рот и выпускания обратно через жабер-
мрм (рис. 26). Но у некоторых рыб эта схема несколько из-
яется> Та1^’ миксин в°Да в жаберные мешки поступает не через
0ТВеРствие> а через непарную ноздрю, соединенную с поло-
Л0ТКИ-/т° изменение вызвано приспособлением к полупарази-
образу жизни, при котором рот миксины часто бывает
при bi рызании в стенку тела своей жертвы в силу чего дыха-
а BJT0 ВР?МЯ Д°лжн° было бы нарушиться. В связи с полупарази-
образом жизни находится также и другая особенность
^^роении дыхательного аппарата миксины. У нее отверстия жабер-
етйчлМешков открываются в общий канал, который, в свою оче-
^Я^^ВДним отверстием выходит далеко позади головы. При таком
в теПию Может глубоко погружать голову в тел4 своей жертвы,
такж&^лреМЯ пР°ДШ1Жая нормальное дыхание. У Миног, которые
гда ведут полупаразитический образ жизни, но в отличие
45
Рис. 26. Схема дыха-
ния рыбы. Наверху
вдох, внизу выдох
(из Воскобойникова,
1936).
©т миксин не имеют ноздри, соединенной с полостью глотки, дыхание
происходит путем всасывания и удаления воды через жаберные
отверстия. У скатов в то время, когда они лежат на дне, и рот их
обращен к грунту, поступление воды к жабрам осуществляется
через брызгальце. У рыб, у которых голова и тело покрыты плотным
панцирем; а жаберные отверстия малы и крышки неподвижны,
смена воды у жабр осуществляется при помо-
щи токов воды, создаваемых постоянным дви-
жением грудных плавников. Так дышат мно-
гие кузовки, луна-рыба и некоторые другие.
Наряду с жабрами, а иногда и вместо жабр,
у рыб функционируют некоторые другие орга-
ны, служащие для улавливания кислорода,
растворенного в воде. У личинок рыб, до того
момента, пока у них окончательно не сформи-
руются жабры, имеется ряд дополнительных
органов дыхания, как-то: наружные жабры,
псевдобранхия, кровеносная система плавни-
ков и т. д. (см. ниже стр. 134), У взрослых рыб
в качестве дополнительного органа дыхания
служит также кожа, причем роль кожи как
органа выделения СО2 у рыб особенно велика.
Так, у вьюна через кожу выделяется до 92%
всего количества СО2, а через жабры только
8%. Нормально у вьюна 63% кислорода по-
требляется через кожу и 37% —через жабры.
В случае, если жабры выключаются (путем
перевязки зева и жабр), процент потребления
кислорода через кожу возрастает до 85%. Ко-
личество кислорода, потребляемого различны-
ми видами рыб, далеко неодинаково. Лососе-
вые могут жить при содержании 7—8 см3 кис-
многие карповые легко переносят количество
лерода в воде, а
khcj орода в 3 см3 на литр.
Пи количеству кислорода, необходимого для нормального дыхания
рыбам, последних обычно разделяют (Вундер, 1936) на четыре
группы: 1) рыбы, требующие очень большего количества кислорода;
нормальные условия для них 7—11 см3 на литр, и при 5 см3 на
литр некоторые уже чувствуют себя плохо. Сюда откосятся кумжа
S;:lmo trutta, гольян Phoxinus phoxinus, гольцы Nennahilus bar-
batulus, подкаменщик Ccttusgobio и многие другие, главным образом,
рыбы холодных быстрых рек. 2) Рыбы, требующие большего коли-
чества кислорода и хорошо живущие при 5—7 см3. Сюда относятся
xapiyc Thjm.Jlus thymallus, голавть Leucist us ccphalus, подуст
Chondrt stoma nasus, пескарь Q. bio gobjc, налом Lota lota и др.
3) Рыбы, потребляющие сравните/ьно небс/ыгое количество кисло-
рода и могущие свободно жить даже при 4 см3 на литр. Сюда отно-
сятся такие виды, как плотва Rutilus rutilus, окунь Perea fluviatilis,
ерш Acerina cernua и др. 4) Рыбы, выдерживающие очень слабое
46
насыщение воды кислородом и зимой живущие даже, когда в воде
остается всего у2 см3 на литр. Таковы сазан, линь, караси и др.
Вышеприведенное деление применимо, главным образом, к пресно-
водным рыбам, так как морские подвержены очень слабым коле-
баниям кислорода. Морская вода обычно бывает или полностью
насыщена, или часто даже перенасыщена кислородом. В море дефи-
цит кислорода может наблюдаться только изредка, главным образом,
в отшнуровавшихся заливах и бухтах, или ночью в местах мас-
сового развития планктонных водорослей. Соверши нноцная картина
имеет место в пресных водах. Целый ряд водоемов в зимнее или,
наоборот, в Летнее.время, в результате гнилостных процессов со-
вершенно теряет кислород, растворенный в воде. В зимнее время
заморы бывают особенно сильны, так как наличие ледового покрова
делает невозможным пополнение кислородом из атмосферного
воздуха. При заморах рыбы стремятся, если есть возможность,
уйти из заморного водоема, входят в впадающие в озеро реки, скоп-
ляются у устьев ручьев и поднимаются к прорубям. Заморы в озерах
и прудовых хозяйствах, вызывая массовую гибель рыб, иногда
наносят весьма серьезный ущерб рыбной промышленности. Для
борьбы с зимними заморами обычно делают проруби для допуска
атмосферного воздуха к воде. Однако, при сильных заморах эта
мера оказывается часто неэффективней. В тех случаях, когда можно
увеличить проточность озера или пруда, необходимо это делать.
Последняя мера обычно приводит к прекращению или, в худшем
случае, к ослаблению замора.
В наиболее сильной степени заморы наблюдаются у нас в Союзе
на реке Оби. Они захватывают в зимнее время огромный район от
Васюгана, а иногда и выше, до низовьев этой реки. Причина замора
на Оби —это вынос из болотистых местностей бассейна Васюгана
и других рек гуминовых кислот и закисных соединений железа,
которые, окисляясь, отнимают из воды весь растворенный в ней
кислород. Так как ледовый покров делает невозможным поступле-
ние кислорода из воздуха, то замор распространяется очень быстро и,
начавшись в конце декабря или начале января в районе Васюгана,
к февралю и началу марта достигает Обсксй губы. От замера в
Оби рыба уходит в придаточную систему, а частично спускается в
Обскую губу. Небольшая часть обских рыб, однако, ежегодно
^огибает от замора.
£ Летние заморы возникают в водоемах только в ночное время.
причиной служит богато развитая водная растительность,
Йртенсивно потребляющая ночью кислород, что межет вызвать его
Дефицит и привести к гибели рыб. Если зимние замеры имеют место
Только в умеренном и холодном климате и для своего развития
Требуют наличия ледового покрова, то летние заморы возникают
также и в тропиках, где они бывают, пожалуй, деже чаще, чем r
умеренных широтах. По своему масштабу, однако, летние ночные
заморы естественно значительно меньше, чем зимний.
Недостаток растворенного в воде кислорода, необходимого для
дыхания рыб, привел к тому, что у рыб выработался целый ряд
47
различных приспособлений, позволяющих им дышать атмосферным
воздухом. При дефиците кислорода в воде у рыб довольно быстро
расходуется кислород, содержащийся в плавательном пузыре. Это
наиболее примитивная форма использования плавательного пузыря
для целей дыхания. Но у некоторых рыб, и в частности, у двояко-
дышащих, многопера и некоторых других плавательный пузырь
действительно превратился в орган дыхания атмосферным воз-
духом. Структура плавательного пузыря при этом сильно изменилась,
появилась губчатость, напоминающая легкие высших позвоночных.
У двоякодышащих превращение плавательного пузыря в «легкие»
позволило этим рыбам жить в сильно заболоченных водоемах, где
кислород в воде очень часто совершенно отсутствует. В этом случае
рыба и может перейти на дыхание атмосферным воздухом. Proto-
pterus переносит полное высыхание водоема и на период засухи
закапывается в ил, где находится в состоянии, напоминающем
спячку.
Другой способ дыхания атмосферным воздухом — это кишеч-
ное дыхание. Кишечное дыхание развито у различных групп рыб.
например, среди костноиузырных (сомы, вьюновые), или средн
Symbranchidae. Кишечное дыхание также является дополнением к
жаберному дыханию и имеет место при недостатке кислорода в
воде. Кишечник у рыб, имеющих кишечное дыхание, претерпе-
вает некоторые изменения. Кровеносные капилляры подходят близ-
ко к внутренней поверхности, в некоторых местах переваривающая
функция выпадает, ворсинки отсутствуют, и кишечник выполняет
в этих местах только дыхательную функцию. У рыб, дышащих
кишечником, он обычно бывает длиннее, чем у близких видов, у
которых кишечник этой функции не несет. У некоторых рыб для
дыхания приспособлена часть желудка. Так, у сомика Otocinclus
имеется специальный слепой мешок желудка, который обычно
наполнен воздухом и несет дыхательную функцию. Заглатываемый
рыбами при дыхании воздух теряет в кишечнике около 5% кисло-
рода и получает около 3 % углекислого газа. Выходит воздух обратно
или через рот, как у многих сомов, или через анальное отверстие,
как, например, у вьюна.
У некоторых рыб, дышащих частично и за счет атмосферного
воздуха, иногда развиваются различного типа наджаберные камеры,
которые служат резерваторами атмосферного воздуха (рис. 27).
Кислород из заглатываемого атмосферного воздуха поглощается
или стенками наджаберного органа, как, например, у Anabantidae
и Ophicephalidae, или расположенными ниже жабрами. Строение
наджаберного органа у различных групп рыб весьма разнообразно.
У Anabas это парные выросты жаберной полости, расположенные
симметрично над жабрами. Внутри каждой полости имеются склад-
ки. стенки которых пронизаны мельчайшими кровеносными со-
судами. Заглоченный воздух попадает в эти полости, которые функ-
ционируют как легкие, при помощи богатой сети капилляров усваи-
вая кислород из воздуха и отдавая СО2. У змееголовок Ophicepha-
lidae наджаберный орган представлен парными выростами в глотке.
48
Стенки этих выростов, так же, как и у Anabas, очень богаты крове-
носными сосудами. Подобного же типа наджаберный орган имеется
и у представителей семейства Symbranchidae, например у Ат-
phipnous cuchia. У сома Clarias непарный наджаберный орган
расположен сверху и
позади жабр. Наиболь-
шего развития наджа-
берный орган достигает
у мешкожаберного сома
Heteropneustes, у кото-
рого: длинный мешок
расположен над очень
маленьким плаватель-
ным пузырем и тянется
назад до хвостовой
области. У некоторых
рыб наджаберный ор-
ган является вспомога-
тельным органом дыха-
ния, функционирую-
щим только в тех слу-
чах, когда рыба попа-
дает в воду, бедную
кислородом. У других
же рыб дыхание при
помощи наджаберного
органа является основ-
ным, и рыба, будучи
лишена возможности
подниматься на поверх-
ность и заглатывать
воздух, погибает через
то или иное время. Так,
Amphipnous cuchia по-
гибает меньше чем че-
рез сутки, после того
как она бывает лишена
Рис. 27. Наджаберные органы у рыб (сверху
вниз) Anabas (Perciformes), Amphipnous (Sytn-
branchiiformes) и змееголовка (Ophicephaliformes)
(из Hora, 1932)-
Возможности подни-
маться к поверхности
Лоды и заглатывать
воздух. Интересно, что
ЖЗбры у этой рыбки
Шльно редуцированы и несомненно не могут обеспечить кровь
Достаточным количеством кислорода. Также погибает от недо-
статка кислорода, будучи удалена от поверхности воды и
Змееголовка Ophicephalus striatus, которая гибнет через 30 мин. —
17 м. после изоляции от поверхности. Повидимому,
моЛодые змееголовки погибают от удушия скорей, чем более
взрослые. Такие рыбы, как Anabas, Heteropneustes и некото-
4 Биология рыб
49
рые другие, могут свободно жить в хорошо аэрированной воде
и не подниматься к поверхности для заглатывания атмосферного
воздуха. Наджаберный орган у этих рыб является добавочным ор-
ганом дыхания и функционирует как основной лишь в исключитель-
ных случаях, позволяя таким рыбам, как, например, Anabas, при
пересыхании одних водоемов уходить из них и по суше переползать
в другие.
Кроме кислорода весьма существенное значение в жизни рыб
имеют и другие газы. Как указывалось выше, СО2, даже сравни-
тельно в небольших дозах, вызывает гибель рыбы. Механизм
действия углекислоты на рыб, как и на других животных, сводится
к понижению способности крови усваивать кислород. Правда,
количество свободной углекислоты в воде, особенно в морской, очень
невелико, ибо магний, кальций и другие катионы ее очень быстро
связывают. Количество СО2 в воде находится в обратной зависимо-
сти от концентрации водородных ионов, изменение которой также
оказывает на рыб серьезное влияние. С повышением кислотности
воды уменьшается интенсивность газообмена, хотя дыхательный
ритм учащается в более кислой среде по сравнению с нейтральной
и щелочной.
Сероводород оказывает на рыб весьма отрицательное влияние.
Вообще он может образоваться и накопиться в водоеме лишь при
условии бедности воды кислородом, ибо при наличии в воде кисло-
рода происходит очень быстрое окисление серы. Действие серо-
водорода на различных рыб неодинаково. Такие виды, как линь
Tinea tinea и карась Carcssius auratus погибают через 8—10 дней
после выдерживания в'течение 3 часов в воде с содержанием серо-
водорода 1 : 10000, форель же погибает через 15 мин. при концент-
рации сероводорода 1 : 100000. Американский ушастый окунь при
этой же концентрации сероводорода погибает через 1 час. Сила
действия сероводорода, так же как и других газов, меняется доволь-
но сильно в зависимости от температуры. При высокой температуре
гибель наступает скорее, чем при низкой.
В. ВЛИЯНИЕ НА РЫБ ДРУГИХ ОРГАНИЗМОВ
Наряду с рассмотренными выше взаимоотношениями рыб с
факторами абиотической среды колоссальную роль, как в жизни
отдельных индивидуумов, так и развитии всей группы рыб в целом,
имеют взаимоотношения рыб с другими организмами, как животны-
ми, так и растительными. Эти взаимоотношения идут по большому
количеству путей: взаимоотношения хищника и жертвы, конкурен-
ции из-за пищи, влияния на режим биотопа и т. д. Все эти формы
связи имеют существенное значение в жизни рыб. Естественно,
что в первую очередь имеют значение взаимоотношения между
рыбами, как между особями одного и того же вида, так и между
представителями различных видов.
50
Взаимоотношения рыб друг с другом
По характеру поведения рыбы резко различны друг от друга.
Некоторые держатся почти всю жизнь стаями, и единичные особи,
если и попадаются иногда изолированно от стаи, то это имеет места
обычно лишь в том случае, если стар или была
напугана, или разбита, например, сильным вол-
нением. Подобного типа постоянные скопления
мы наблюдаем, например, у некоторых сель-
девых Clupeidae, анчоусов Engraulidae, Scopeli-
dae и многих других рыб. Полная противопо-
ложность им — рыбы, ведущие одиночный образ
жизни и даже к моменту размножения не собираю-
щиеся стаями. Таковы многие акулы, некоторые
сомы, пинагоры и т. д. Наиболее же обычны
рыбы, которые периодически в определенную
фазу годового цикла собираются в скопления,
в другое же время держатся разреженно. Спай-
ный образ жизни есть приспс соб; ент е или для
поиска пищи, или для н хождения миграцион-
ного пути г ли для защиты от хищнике в. Во
всех эр х с. уч: ях стая бьстрее чем один(Ч-
ная рыба ориентируется, находит корм или за-
мечав! хищника.
У многих стайных и периодически стайных
рыб имеет место образование группировок —
стад, обособленных биологически, а иногда и
морфологически. Группировкам этим свойстве-
нен свой ареал распространения, отличный от
других стад во времени или пространстве, места
размножения и места нагула, а также своеобра-
зие динамики биологических показателей, ха-
рактеризующих ход годового цикла. Предста-
вители одного стада, как правило, ни на ме-
стах нагула, ни, особенно, на местах икромета-
ния, не смешиваются с представителями другого
стада. При дальнейшем обособлении расхождение
в биологии и морфологии может привести к
образованию самостоятельных рас, уже отли-
чающихся друг от друга четко выраженными
морфол' гическими особенностями. Примером
Рис, 28. Места на-
гула (горизонталь-
ная штриховка) к
размножения стад
(крестообр азная
штриховка) у ти-
хоокеанской сель-
ди Clupea harengus
pallasi близ юго-
восточной Аляски
(из RounsefelJ и
Dahlgren, 1935).
таких стад могут быть хотя бы группировки сельди, выявленные
R unsefellnD hlgren (рис. 28) Эти группировки сельди имеют обо-
собленные места нереста и нагула. Отдельные стада известны также,
например, у аральского леща. Так, лещ. ккрсмечущий в восточней
части дельты Сыр-Дарьи и кормящийся в районе Левушкиной
горы, является несомненно обоссбленным стадом (рис. 29).
В пределах каждого стада у стайных рыб, согласно Н. В. Лебе-
деву (1943), особи группируются в биологически обособленные
4* 51
популяции, которые возникают после нереста из выведшихся
одновременно особей, обладают одинаковым темпом жизненного
цикла, постоянным соотношением полов и у многих рыб, пови-
димому, являются пожизненным образованием.
Биологическое значение этих популяций, надо думать, следует
рассматривать, как приспособление к миграциям.
Различные формы скоплений рыб могут иметь весьма сложную
структуру и генезис и несомненно являются продуктом длительной
30’ 61° 30'
Рис. 29. Места нагула (горизонтальная штриховка) и
размножения (крестообразная штриховка) восточно-
сыр-дарьинекого стада леща.
эволюции образуясь в результате приспособления к определенным
воздействиям внешней среды (Рагг, 1931). Причины, вызывающие
скопления рыб, весьма разнообразны. Обычно принято различать
П нерестовые скопления, возникающие в целях раз-
множения и состоящие почти исключительно из половозрёлых
особей Молодые особи попадаются в них лишь случайно. При-
меоом' подобных скоплений может служить скопление трески
на нерестилищах у Лофотенских островов, скопления воблы в
полоях и т п- 2) с к о п л е н и я м и г р а ц и о н н ы е; возни-
кают на путях движения рыбы. Следует иметь в виду, что очень
часто этот вид скопления переходит в другой, например, в нересто-
вое скопление. Примером подобного рода скопления могут служить
скопления лососей, подымающихся на нерестилища по рекам Даль-
него Востока, или скопления хамсы, проходящей через Керченский
пролив на зимовку в Черное море; 3) с ко п л е н и я н а г у л ь-
н ы е образующиеся на местах кормежки рыбы и вызывающиеся
главным образом скоплениями пищевых объектов. Эта группа ско-
плений наиболее разнообразна. Первые две группы скоплений,
52
как правило, образуются из особей одного вида, и лишь в небольшом
количестве к ним примешиваются особи других видов, главным
образом хищники, питающиеся за счет стайных рыб. Кроме того,
обычно скопления первых двух типов имеют специфический воз-
растной состав. Нерестовые скопления всегда состоят из взрослых
рыб, ходовые — или из взрослых, или из молодых, смешанные же
стаи ходовых рыб — явление более редкое. Наоборот, нагуль-
ные часто могут состоять как из рыб различных видов, так и из
рыб различного возраста, взрослых и молодых, хотя обычно
рыбы одного возраста кормятся вместе, именно отдельно взрослые и
отдельно молодые, не говоря уже о молоди; 4) с к о п л е н и я зи-
мовальные. В наиболее резкой форме скопления эти выраже-
ны в Каспийском море, в устье Волги и на глубоких местах Волги
и Урала, где на зиму залегает огромное количество различных
видов рыб, причем, как показали водолазные работы, при залега-
нии на ямы имеет место все же достаточно четко выраженное видо-
вое обособление. Как уже отмечалось, стайный инстинкт рыб об-
разуется в результате длительного развития. Как правило, наи-
более сильно развита стайность у рыб пелагических, которые со-
вершают длительные перемещения. Обычно хищные рыбы не обра-
зуют стай, что всецело объясняется условиями питания. Донные
рыбы обычно не образуют таких больших стай, как пелагические.
Наиболее резко стайность выражена среди представителей семейств
Clupeidae, Gadidae, Atherinidae, Mugilidae, Belonidae, Scom-
beresocidae, Hemiramphidae, Exocoetidae, Carangidae и Gerridae
в меньшей степени, но также достаточно резко у отдельных пред-
ставителей семейств Cyprinidae, Salmonidae u Sciaenidae (Parr, 1931).
Образование скоплений у рыб стимулируется весьма различными
факторами. Некоторые из них заложены в самой рыбе, другие во
внешней среде. Причина нерестовых скоплений заключается в
стремлении к размножению, вызываемом развитием половых желез.
Нагульные скопления обусловливаются наличием объектов питания
и т. п.
Как мы увидим ниже, стая рыб не представляет собою обычно
какого-то бесструктурного образования, в особенности ходовые
стаи имеют часто совершенно определенную организацию. Основны-
ми органами, при помощи которых рыбы реагируют на других рыб
и, которые служат для ориентации при построении в стаи, являются
органы зрения и органы боковой линии. Как показали опыты Bowen
(1932) над американскими сомами, рыбки скорее собираются при
освещении в стаи, чем в темноте. Спунер (1931) показал, что Могопе
labrax движется за другими особями того же вида, ориентируясь,
в первую очередь, при помощи органов зрения. Спунер употреблял
при своих опытах систему зеркал, комбинируя которые он получал,
что «стая рыб» образовывалась изображением самой подопытной
рыбы. Весьма существенное значение ‘при ориентации рыбы на
рыб,у.имеют и органы боковой линии. Как показали о^ыты с ослеп-
ленными рыбами, последние хорошо реагируют на проплывающих
мимо других особей и могут следовать за ними.
53
У некоторых рыб для образования стай имеют значение и органы
•сязания: усы, удлиненные лучи плавников и т. п. Так, американ-
ские сомы, будучи ослеплены, следовали за другими рыбами, лишь
слегка касаясь их усами. При этом близкие виды рыб (например,
различные виды того же рода Ameiurus) сомы не различали, но
Рис 30. Силовое поле вокруг
двигающихся рыб: А — ры-
бы взаимно притягиваются;
Б — рыбы не влияют друг на
друга; В — рыбы взаимна
отталкиваются (из Шулей-
кина, 1940).
хорошо отличали восковой
(весьма тщательно сделанный)
макет от живых особей.
Всякая рыба, двигаясь в
воде, создает вокруг себя груп-
пу силовых линий, весьма
близких по расположению к ли-
ниям магнитного диполя. Попадание одной рыбы в поле другой и
наоборот естественно окажет влияние как на ту, так и на другую.
Как видно из приведенных рисунков, при различном расположении
рыб относительно друг друга, силовое поле будет иметь весьма раз-
личный характер. В первом случае(рис. 30) имеет место наличие
между рыбами сильных притягательных струй. Во втором слу-
чае, повидимому, действие как притягивающих, так и оттал-
кивающих сил взаимно уравновешивается, и механическое
воздействие одной рыбы на другую при подобном располо-
жении равно нулю, и, наконец, в третьем случае имеет место
преобладание отталкивающих сил над притягивающими. Есте-
ственно, поэтому, что при передвижении в стаи рыбы вынуждены
считаться с этими силами и точно так же, как и птицы при пере-
летах, принимать известный определенный порядок, который
создавал бы наиболее благоприятные гидрофизические условия для
движения всех особей стаи. Формы стаи рыб еще почти не изучены,
но, повидимому, стая движущихся рыб очень часто принимает форму,
54
близкую к элипсоиду. Естественно, что рыбы различной формы
создают вокруг себя и различного характера поле. Поэтому
есть все основания предполагать, что самый характер двигаю-
щейся стаи у рыб различной формы тела должен быть различным^
Кроме того, форма стай рыб варьирует в зависимости от изменения
внешних условий. Так, форма косяков иваси Sardinops melanosticta
изменяется в зависимости от прогрева воды и изменения интенсивно-
сти питания и, как было установлено наблюдениями с самолета,
Рис 31. Форма стай сардины-иваси Sardinops melanosticta (изТКагановско-
го, 1939).
принимает весьма различную форму в различные сезоны’года
(рис. 31). Встае рыб при ее движении могут образовываться полости,
которые возникают или вокруг рыб другого вида (рис. 32, 33), или
вокруг донных предметов, миновать которые стае приходится. Вну-
три стаи, которая имеет определенную более или менее постоянную
форму, движения рыб могут быть ориентированы в самых раз-
личных направлениях.
Естественно, что, кроме силового поля, рыба создает вокруг
своего тела также и участок измененной химически воды. Рыба вы-
деляет продукты метаболизма, которые оказывают влияние на
-находящихся в водоеме других рыб. В естественных условиях рыбы
•чень редко гибнут от удушия, вызванного тем, что находившийся
в воде кислород был израсходован рыбами. Обычно, как мы видели
выше, гибель рыб от удушия в естественных водоемах является
результатом исчезновения кислорода вследствие гнилостных процес-
сов, протекающих в водоемах. В аквариумах же, бассейнах и дру-
гих искусственных водоемах гибель рыб от дефицита кислорода,
вызванного потреблением его рыбами, обычное явлейие.
55
Если в небольших сосудах увеличенная концентрация рыб в
конечном итоге может привести к дефициту кислорода и гибели
рыб, то в то же время необходимо отметить, что, как показал опыт
Schuett (1933) над рядом видов рыб (Umbra limi, Carassius auratus,
Lebistes reticulatus и Fundulus heteroclitus), количество потребляе-
мого кислорода у рыб, сидящих в одиночку, больше, чем у рыб,
сидящих по нескольку штук. Выявленная закономерность, как это
установлено на золотых рыбках, имеет место как в стоячей, так и в
проточной воде. Интересно, что уменьшение потребления кислорода
и вообще замедление метаболизма при наличии в бассейне особей
того же вида имеет место не у всех рыб и, главным образом,
32. Общий вид стаи сельди Lenkinsia. Буквы а, б, в, г, д, е
обозначают предметы, которые обтекает стая (из Breder, 1929).
наблюдается у мирных. У американского сомика Ameiurus mek.s,
наоборот, изолированные рыбки потребляют меньше кислород;;,
чем рыбки, сидящие вместе по нескольку штук. Объясняется это,
вероятно, тем, что в естественных условиях хищники Ameiurus
живут в одиночку и, будучи посажены вместе, делаются весьма
подвижными, усиленно плавают, что, повидимому, и вызывает
повышение метаболизма.
Наличие в небольшом объеме воды нескольких рыб оказывает
влияние на их рост. Так, Church (1927) выяснил, что в одном и том
же объеме воды, даже при наличии аэрации, живородящие рыбки
Platypoecilus maculatus растут быстрее в сосудах с малым количе-
ством рыб, хотя вода, в которой ранее содержались рыбы, как мы
увидим ниже, наоборот, стимулирует у рыб рост. То, что в данном
случае не имеет места влияние продуктов распада, дефицита кисло
рода или недостатка корма наглядно показывают опыты Виллера и
Шингенберга (1927) над форелями. Эти авторы помещали в сосуды
с проточной водой различное количество форелей, которым дава-
лось одинаковое количество корма на одну рыбу. Проточность
воды устраняла возможность влияния продуктов метаболизма,
56
и все же в тех сосудах, где находилось большое количество рыб,
рост последних был более медленный, чем у рыб, сидящих в одиноч-
ку. В естественных водоемах, где к фактору объема водоема еще
присоединяется и конкуренция из-за пищи, размеры рыб находятся
в весьма тесной зависимости от плотности населения и, в частности,
вылов, вызывающий разрежение стада, оказывает на рост рыб очень
сильное влияние (стр. 151). Как показали наблюдения ряда авторов,
размеры рыб тесно связаны с величиной водоема, в котором рыба
живет, и густотой рыбного населения. В маленьких водоемах, с
густым ^населением,
рыбы, как правило,
мельче, чем в боль-
ших с разреженным
населением,
венно, однако,
разрежение
оказывает
на рост рыб
известного
ла, после
уменьшение
сти населения
отражается на росте
рыбы. Если в данном
случае мы имеем при-
Естест-
что
стада
влияние
лишь до
предо
которого
плотно-
не
I
‘</7
)
Рис. Структура стаи сельди Jenkinsia
(из Breder, 1929).
мер отрицательного
влияния рыбы на рыбу, выражающегося в замедлении роста, то
имеет место и обратное, т. е. положительное воздействие одной
рыбы на другую. Как установлено опытами Allee, Clyde и В<-
wen (1932), повышение плотности рыбного населения понижает
вредное действие на рыб отравляющих веществ. Упомянутые ав-
торы показали, что рыба, посаженная в коллоидальный раствор
серебра, погибает скорее, когда она находится в сосуде одна,
чем в том случае, если в сосуде сидит несколько рыб, хотя объем
раствора на каждую рыбу остается один и тот же. Каков механизм
этого действия, еще точно не установлено, но во время опытов по
влиянию ядовитых веществ на рыб рыбы собираются, как правило,
вместе и держатся все время одна около другой.
Необходимо отметить, что вода, в которой некоторое время на-
ходилась рыба, претерпевает целый ряд изменений, как своих хи-
мических, так и физических свойств, становясь тем, что английские
и американские исследователи называют fish conditioned water,
т. е. водой, измененной наличием рыб. Эта вода обладает, например,
способностью довольно сильно флуоресцировать под влиянием
освещения ультрафиолетовыми лучами. В то же время дестиллирс-
ванная вода совершенно не флуоресцирует, а вода естественных
водоемов флуоресцирует в весьма слабой степени (Merker, 1932).
Химические изменения такой! воды, в основном сводятся к абсорби-
рованию продуктов выделения рыбы. Это приводит к тому,что
57
«биологически кондицированиая вода» является благоприятной сре-
дой для роста беспозвоночных и во многих случаях благотворно
действует и на рыб. Так, например, Shaw (1932) содержал в одинако-
вом объеме воды и при одинаковых условиях кормления рыбок
Platypoecilus miculatus, но в одном сосуде предварительно содер-
жались пресноводные моллюски, а в другом была налита обычная
вода. Действие же моллюсков на воду сходно с действием рыб. В ре-
зультате за 3 дней пребывания в воде, в которой ранее содержа-
лись моллюски, рыбки прибавили в длину на 6,3%, в чистой же воде
прирост был всего на 2,7%. В воде, в которой ранее содержались
рыбы или какие-либо другие водные животные, заживание ран у
рыб идет значительно быстрее, чем в обычной воде.
Оказывает влияние рыба на рыбу и в форме выработки у рыб
условных рефлексов. Так, например, золотые рыбки и Fundulus
heteroclitus скорее выучивались проникать в изолированную камеру
за кормом, если к ним подсаживалась уже ранее выученная рыбка,
которая им, так сказать, «показывала дорогу». Но иногда имеет
место и обратное явление, когда рефлекс, выработанный у рыбы,
сидящей в одиночку, при помещении ее в аквариум, где сидят еще
другие, также тренированные рыбы, затухает. Так, например,
собачья рыба Umbra limi приучилась при освещении красным све-
том выпрыгивать за кормом из воды. Если рыбка сидела одна, то
этот рефлекс у нее сохранялся очень долгое время, но если к такой
дрессированной рыбке подсаживалась другая, тоже дрессированная
рыба, то число прыжков сильно сокращалось, а когда вместе по-
мещались три рыбки, то прыжки совершенно прекращались. Оказы-
вается, что когда одна рыба устремлялась к поверхности воды за
кормом, другая в это время пыталась кусать ее в живот, что очень
скоро отучило рыб выскакивать из воды. В данном случае мы уже
имеем пример перехода к другой группе взаимоотношений между
рыбами, — к взаимоотношениям пищевым.
Конкуренция из-за пищи — это пожалуй наиболее острая форма
борьбы за существование. Сильное развитие одного какого-либо
вида Может привести к тому, что другбй вид, конкурентом в пище с
которым является первый, очень часто должен сильно уменьшиться
в числе. Так, например, общее потепление вод в северной Атлантике
привело к тому, что гигантская акула Cetorhinus maximus получила
возможность продвинуться на север и, в частности, в массе появи-
лась у западных берегов Великобритании. Как известно, эта акула
питается мелкими планктонными организмами и, продвинувшись
.на север, становится весьма серьезным конкурентом океанической
'сельди Clupea harengus. Каждая акула съедает примерно столько же
планктона, сколько съедает миллион сельдей. Вот почему появление
массы акул у берегов Великобритании вызвало сильное уменьшение
сельди, то, : в ою очередь, привело к снижению промысла. Конку-
ренция в пище между отдельными видами очень часто приводит к
постепенному вымиранию одного вида и замене его другим. Подоб-
ную роль, например, в водоемах Средней Азии играет сазан, который
очень часто из многих озер вытесняет менее выносливую форму—
58
маринку, не выдерживающую конкуренции сазана. В Северной
Америке тот же сазан, завезенный из Европы в 1872 году, весьма
сильно повлиял на численность одного из американских окуней
Archoplitts interruptus.
Пожалуй столь же важное, если не большее, значение в динамике
популяции рыб имеет проблема хищника и жертвы. Взаимоотноше-
ния хищника и жертвы среди рыб чрезвычайно широко распростра-
нены. Особенно они резко выражены на больших глубинах, где
исключительно живут хищные и животноядные рыбы. Методы
нападения и защиты у рыб весьма разнообразны. Сильные кинжало-
Рис. 34. Расположение электрических органов у Electro"
phorus electricus. Электрические органы тянутся ввчде двух
полос широкой (верхняя) и узкой (нижняя) от грудного
плавника до конца тела. На поперечном разрезе видно, что
вся нижняя часть тела занята- электрическими органами.
видные зубы акул, щуки, судака и многих других
рыб являются прекрасным оружием для поражения
жертвы и удержания ее во рту. Пила-рыба пора-
жает свою жертву несущим зубовидные выросты
рострумом. Меч-рыба наносит ранения своей удли-
ненной верхней челюстью, причем удар меч-рыбы
бывает настолько силен, что оставляет след даже
на металлической обшивке судов. Электрический
угорь Electrophorus electric s поражает свою жертву,
разряжая в нее свою электрическую батарею (рис.34). Электрический
орган электрического угря и многих других рыб (кроме электриче-
ского сома) — не что иное, как измененная мускульная ткань.
Образовавшиеся из мускулов столбчатые клетки обладают способ-
ностью действовать по принципу лейденской банки, накапливающей
мускульное электричество и, по желанию животного, разряжаю-
щейся. Сила разряда у электрического угря достигает до 300 вольт
и может свалить с ног крупное животное. Питаются электрические
угри небольшой рыбой, которую они сначала поражают своим
электричеством, а потом заглатывают. Пораженную рыбу как бы
сводит судорога, она выгибается и застывает, расправив плавники.
Электрические органы расположены у электрического угря вдоль
всего тела, как на голове, так и по бокам туловища, и служат и для
нападения и для защиты. У других рыб, как например, у электри-
ческих скатов, сомов и Mormyrid.ie электрические органы служат,
главным образом, для защиты. У Mormyridae электрические органы
расположены в области хвоста и поражают хищнику, схватившего
рыбу за хвост. У скатов и электрического сома (у которого электри-
ческий орган — видозмененная железа) электрический орган несет
59
видимо оборонительно-наступательную функцию, служа как для
поражения своей жертвы, так и для защиты от врагов. Сила разряда
у электрического ската достигает до 70 вольт. Все рыбы, обладающие
электрическим органом, производят разряд произвольно. Иннер-
вация электрического органа осуществляется 8, 9 и 10 парой вис-
церальных черепно-мозговых нервов. После нескольких разрядов
Рис. 35. Скаты: наверху слева направо Trygon pastinaca
и Raja clavata; внизу Rhinoptera peli.
рыба теряет способность давать электричество, и до того, как она
снова сможет производить разряд, должно пройти известное время.
Органами, служащими главным образом целям зашиты, явля-
ются различные шипы и колючки, часто снабженные у основания
ядовитыми железами. Колючки и шипы развиваются обычно у
придонных медленно плавающих рыб (рис. 35). У пелагических рыб
шипы и колючки развиваются гораздо реже и, как правило, в хво-
стовой области. Наиболее мощные шипы на спине имеют различные
скаты рода Raja. Эти шипы не снабжены ядовитыми железами, но
60
попадающая в ранку выделяемая ими слизь вызывает также бо-
лезнетворные явления. Скаты семейств Trygonidae и Myliobatidae
имеют у основания хвоста одну или две колючки, которыми эти
рыбы могут наносить серьезные, долго незаживающие поражения.
На теле у многих костистых рыб также имеются мощные шипы.
Так, у некоторых камбал, как например, у черноморского кал-
Рис/36. Рыбы, покрытые панцирем (сверху вниз): морская игла (Syngnathi-
formes), кузовок (Tetrodontiformes), каймановая щука (Lepidosteiformes) и
панцирный сом (Cypriniformes).
кана, глазная сторона покрыта мощными шипами. У многих рыб
защитную функцию выполняют лучи плавников, иногда, так же,
как колючки, снабженные ядовитыми железами. Так, например,
у водящейся у берегов Ост-индского архипелага бородавчатки
Synanceia horrida у основания лучей спинного плавника имеются
ядовитые железы. Сила действия яда этих желез достаточно велика
и обычно вызывает смерть даже человека. Последняя наступает
через различные промежутки времени—от 22 часов до нескольких
суток.
Для защиты от хищников у многих рыб служит также иногда
Достигающий довольно мощного развития панцыр^. У различных
групп рыб панцырь развивается конвергентно (рис. 36). В неко-
торых случаях он образуется из разросшихся чешуй, в других
61
случаях это — костные пластинки, 'но функция у всех этих об-
разований одна и та же — защитная.
Паразитизм рыб на рыбах же распространен сравнитель-
но слабо. Myxine glutinosa вгрызается в тело живых рыб, постепен-
но поедая внутренности и приводя к гибели рыб, на которых она
напала. Подобный же образ жизни ведут и глубоководные угри
семейства Simenchehdae, вгрызающиеся в крупных рыб и питаю-
щиеся их внутренностями. Полупаразитический образ жизни на-
кладывает как на миксину, так и на угря Simenchelys своеобразный
отпечаток. Об изменении дыхательного аппарата миксины в связи
Рис. 37. Прилипало Echeneis remora.
с полупаразитическим образом жизни мы говорили выше. У Si-
menchtlys жаберное отверстие очень мало и плотно закрывается.
Как у миксины, так, правда, в несколько меньшей степени, и у
Simenchelys кожа содержит большое количество слизеотделитель-
ных желез, очевидно понижающих своими выделениями силу тре-
ния рыбы о стенки тела хозяина. Среди рыб мы наблюдаем и при-
меры другого рода паразитизма. Так, сомик Steg jh.lus паразити-
рует в жаберной полости более крупного сома Platystcma и пи-
тается, высасывая кровь из жабр своего хозяина. Самцы некоторых
Ceratiidae питаются за счет соков тела самок.
Довольно широко среди рыб развит коменсализм, т. е.
такая форма взаимоотношения между животными, когда одно жи-
вотное извлекает из этого взаимоотношения пользу, а для другого
оно безразлично. Наиболее характерным примером ксменсализма
являются взаимоотношения между акулой и рыбкой прилипало
Echcneisremara и др. (рис. 37). У прилипало первый спинной плавник
видоизменен в расположенную на верхней стороне головы и начале
спины присоску. При помощи этой присоски прилипало прикре-
пляется к акулам и путешествует вместе с ними, питаясь объедками
с их стола. Нисколько прочно прилипало присасывается к рыбам,
показывает тот факт, что туземцы используют прилипало для лсвли
черепах. Они привязывают к хвосту прилипало тонкий шпагат и,
увидев в воде черепаху, бросают прилипало рядом с ней. Рыбка
сейчас же направляется к черепахе и присасывается к ее панцырю.
Осторожно подтягивая за бичевку, рыбак подтаскивает к себе
черепаху.
Примеры симбиоза между двумя видами рыб весьма
редки. Видимо подобного рода отношения существует между не-
которыми акулами и рыбой лоцманом Naucrates. Эти небольшие
62
рыбки следуют все время за акулами и бросаются на остатки пищи,
выбрасываемые с кораблей, как бы указывая акуле на дсбычу.
Акула никогда не ест держащихся около нее лоцманов. В чем вы-
ражаются симбиотические отношения между акулой и лоцманом,
точно не установлено. Возможно, что лоцман, действительна
обладая более острым, чем акула, зрением, указывает ей добычу.
Некоторые авторы отмечают, что лоцманы питаются паразитическими
ракообразными, прикрепляющимися к коже акул.
Влияние растений на рыб
Взаимоотношения рыб и други xj орган и’з м о в
еще более разнообразны, чем существующие внутри группы рыб.
Представители всех главнейших групп органического мира
•т бактерий до человека играют в жизни рыб весьма существенную
роль.
Бактерии, кроме той колоссальной роли, какую они играют
в круговороте органического вещества в природе, без деятельности
которых жизнь рыб, как и других животных, была бы невозможна,
имеют важнее значение и специально для рыб, вызывая, главным
образом, различные заболевания. Так, например, злокачественную
краснуху карпов вызывает бактерия Pseudomonas pkhniae. Бакте-
риального происхождения также водянка, некоторые злокачествен-
ные опухоли и многие иные заболевания у рыб. Имеют значение
для некоторых рыб бактерии, живущие у них в органах свечения и
вызывающие излучение света. Имеют ли бактерии какое-либо зна-
чение как пища рыб, неизвестно.
Высшие расте н и я также имеют колоссальное значение в жизни
рыб. Выделяя при свете кислород и поглощая углекислоту, растение
способствует созданию более благоприятных условий для жизни
рыб. Наоборот, при сильном затемнении, когда фотосинтеза не
происходит, и растения выделяют углекислоту, этот процесс может
иногда достигнуть таких размеров, что приводит к гибели рыб в
водоеме.
Растения имеют важнее значение и как субстрат для икры
многих рыб. Откладывая икру на растения, рыбы, с одной стороны,
несколько защищают ее от хищников, а с другой стороны, помещают
икру в благоприятные для ее развития условия гидрохимического
режима. Так, например, сазан часто откладывает икру в заросли
рдестов, где насыщение воды кислородом и днем и печью значитель-
но выше, чем в окружающих открытых участках водоема. Многие
рыбы строят из растений специальные гнезда, в которых содержат
икру. П. добного рода гнездо устраивают различные колюшки
G; sten st us, Pung t. us. Нечто вроде гнезда делает ильная рыба
Amia calva’ и другие. Растительность, как элемент ландшафта,
имеет значение и для взрослых рыб.’ Многие рыбы специально
приспособились к жизни в зарослях, обладают соответствующей
покровительственней окраской или формой, напоминающей те
заросли, среди которых рыба обитает. Так, например, поперечная
волосатость окуня и многих других рыб, напоминающая тень от
стеблей растений, делает этих рыб очень плохо заметными среди
зарослей. Длинные выросты плавников у морского конька тряпич-
ника Phyllopteryx eques в сочетании с соответствующей окраской
делают его совершенно незаметным среди подводных зарослей
(рис. 38).
Но наиболее серьезное значение для рыб имеет растительность,
как пища. Начиная от низших планктических водорослей и кончая
макрофитами, вся водная растительность служит объектом питания
рыб. В связи с питанием растительной пищей у рыб выработался
Рис. 38. Тряпичник Phyllopteryx eques.
ряд весьма своеобразных приспособлений. Такие замечательные
изменения жаберного аппарата, какие, например, мы наблюдаем у
толстолобика (см. ниже), целиком приспособившегося к питанию
планктоном, или чрезвычайное удлинение кишечного тракта у за-
каспийской храмули, питающейся перифитоном, являются при-
способлениями к тому или иному виду растительной пищи. Есте-
ственно, что удельный вес растительной пищи в питании различных
рыб чрезвычайно различен. Многие рыбы совершенно не питаются
растениями, у других рыб они встречаются в пище лишь случайно,
j' третьих растения составляют всегда определенный процент пище-
вого рациона и, наконец, четвертые целиком питаются только рас-
тительными организмами.
Растения, наряду с большой положительной ролью, которую они
играют в жизни рыб, имеют и отрицательное значение.
Некоторые растения, как, например, пузырчатка Utricularia,
специально приспособились к питанию водными животными и в
том числе личинками рыб. На тонких стеблях у этого растения
сидят вильчатые листья, снабженные пузырьками. Эти пузырьки
имеют входное отверстие, закрывающееся клапаном, который может
пропускать животных только внутрь пузырька, но не наружу
.04
(рис. 39). В эти пузырьки очень часто попадают различные животные
и в том числе только что вышедшие из икры личинки рыб. Попавшее
в пузырек животное, лишенное возможности выйти, погибает,
разлагается, и продукты его распада идут на построение организма
пузырчатки. Личинок рыб иногда потребляют также Aldrovandia,
но способ лова рыбок у этого растения — иной, чем у пузырчатки.
Рис. 39. Рыбоядные растения: слева Aldrovandia, справа
пузырчатка Utricularia (изТалиева, 1922 и Липина, 1941).
Листья Aldrovandia складываются вместе и образуют раскрываю-
щуюся камеру, внутренняя сторона которой покрыта волосками..
При прикосновении к этим волоскам какого-нибудь животного,
листок захлопывается, и животное оказывается пойманным. Вред,
наносимый насекомоядными растениями рыбам, сравнительно все
же очень невелик. Значительно более серьезный вред приносят
рыбам некоторые грибки, вызывающие различные заболевания.
В первую очередь необходимо упомянуть грибок Saprolegnia, по-
ражающий обычно раны у рыб. Постепенно разрушая ткани рыбы,
этот грибок при успешном развитии может привести к гибели особи.
Взаимоотношения рыб и других животных
Большую роль в жизни рыб играют простейшие. Многие
рыбы на ранних стадиях развития питаются различными одно-
клеточными, главным образом, инфузориями. Так, например, как в
естественных условиях, так и в аквариуме, молодь!лабиринтовых
Биология рыб 1 СБ
первое время по выходе из икры питается исключительно простей-
шими. Простейшими питается также и молодь многих других рыб.
Для взрослых рыб простейшие, как объект питания, не имеют су-
щественного значения.
Отрицательная роль простейших также очень велика, особенно
как возбудителей различных заболеваний. Такая болезнь, как
костиазис, вызываемая жгутиковым
Рис. 40. Жгутиковые (Flagellata) паразиты рыб.
Верхний ряд слева направо Trypanosoma, Octo-
mytus, циста Octomitus; внизу Costia на коже
рыбы, циста Costia (из Догеля, 1932)
Costia nacatrix, приносит
огромный вред рыбному
хозяйству, уничтожая
большое количество фо-
релей, карпа и других
рыб. Этот паразит по-
ражает кожу и приво-
дит к разрушению эпи-
дермиса. Некоторые
жгутиковые паразити-
руют в кишечнике рыб.
Таковы Octomitus, най-
денный у форели, и
Cryptcbia, обнаружен-
ный у пинагора. Пара-
зитируют жгутиковые
и в крови рыб. Так,
некоторые Trypanosoma
найдены в крови ска-
тов, акул, лещей, оку-
ней, камбал и других
рыб (рис. 40). Парази-
тируют на рыбах и кор-
неножки, которые по-
ражают, главным обра-
зом, органы пищеваре-
ния и выделения. Из
простейших наиболее
серьезное патогенное
значение имеют раз-
личные споровики. Так,
на коже паразитируют
Myxobolus, которые вызывают различной величины вздутия и
язвы. Хрящевые части скелета и мозг поражает споровик Lentospo-
га, часто приводящ! й к массовой гибели рыб в форелевых пру-
довых хозяйствах. Паразитами рыб являются также многие другие
споровики (рис. 41).
На рыбах паразитируют также иинфузории. Так, на жаб-
рах и коже живут Chilodcnn Ichthyophthirius, в кишечнике встре-
чается инфузория Bakntd.um.
Кишечнополостные также связаны с рыбами целым
рядом различных форм взаимоотношений. Среди кишечнополостных
мы находим как экто-, так и эндопаразитов рыб. Polypodium
66
hydriforme паразитируют на половых продуктах волжской стерля-
ди. Из колониальных форм Hydrichthys паразитирует на рыбах
Южной Африки. Но основной вред, причиняемый кишечнополостны-
ми рыбам, — это, конечно, непосредственное хищничание — пита-
ние рыбой, причем питаются рыбой как сидячие, так и свободно
плавающие формы. Гидра приносит часто очень значительный вред,
уничтожая личинок рыб, которых она захватывает своими щупаль-
цами. Многие морские колониальные формы также захватывают
щупальцами молодь рыб и ею питаются. Большое значение, как
пожиратели рыб, имеют и медузы. Из семисот с лишком известных
видов морских медуз для двадцати семи установлено, что они пита-
Рис. 41. Карп, пораженный споровиком Myxobolus
(из Лейман, 1939).
ются рыбами, но несомненно, что и многие другие медузы также
потребляют в пищу рыб, причем вред, приносимый медузами, види-
мо иногда может быть очень велик. Так Бииб (Beebe) наблюдал у
берегов Гаити тысячи медуз Linuche unguiqulata, многие из которых
захватили молодь рыб. Добыча сначала поражается стрекатель-
ными клетками, затем захватывается щупальцами и препровождается
в рот. Объектами пищи медуз чаще становятся мальки и личинки
рыб. Медузы едят личинок камбал, песчанок, трески, взрослых
морских игл и многих других рыб. При этом очень часто малень-
кие медузы нападают на рыб, значительно больших по разме-
рам, чем они сами. Питаются молодью рыб и некоторые
гребневики. С другой стороны, среди кишечнополостных и рыб
мы находим примеры симбиоза. Так, некоторые рыбы из семейства
Pomacentridae живут среди щупалец гигантских морских анемон
Stoichactis и Disccsoma, широко распространенных в Индопацифике.
Живущие среди щупалец морских анемон рыбки: Amp hi р ri с n percula,
A. frenatus, A. perederaion, Premnas biaculeatus и др. имеют малень-
кие размеры и весьма ярко окрашены. Что мы имеем здесь дело с
симбиозом, совершенно несомненно, так как. в то время как всякое
Другое животное, попадающее в район расположения щупалец
анемоны, немедленно поражается, Amphiprion плавает среди них
совершенно безнаказанно, питается объедками со стола анемоны и
Даже иногда объедает у нее со щупалец отбросы пиши. Анемона в
свою очередь получает остатки пищи, добытой Amphiprion. Инте-
ресно, что Amphiprion, схватив свою добычу, приносит ее в свой
5* 67
«дом», где и съедает, а объедки остаются анемоне. Голландские ис-
следователи указываюттакже, что Amphiprion содействует движению
воды перед щупальцами анемоны. Каждый вид Amphiprion связан
с определенным видом анемон и, будучи посажен в аквариум, где
Рис. 42. Медуза, поймавшая рыб.
находятся особи другого вида, к ним не приближается (Негге, 1936).
Около одной морской анемоны держится иногда до десятка рыбок.
Другой пример симбиоза кишечнополостных и рыб мы можем
наблюдать и в наших водах. Под колоколом крупной арктической
медузы Cyanea arctica очень часто держится большое количество
молоди пикши, трески и других рыб. При этом иногда мальки, так
же, как и Amphiprion у актинии, объедают мусор, налипший на
68
щупальце медуз, но в то же время медузы мальков не трогают. Здесь,
так же, как и в первом случае, мы имеем пример симбиоза. Подобное
сожительство медуз и молоди рыб известно и в тропических водах.
Так, Бииб и Ти Ван (Beebe and Тсс Van, 1928) указывают, что под
колпаком медуз близ Гаити держится иногда свыше 300 мальков
рыб, одновременно и медузы им никакого вреда не причиняют.
Пищей рыб кишечнополостные служат довольно редко. Лишь
у немногих видов рыб они составляют основную пищу, обычно же
кишечнополостные попадаются в их пище лишь случайно.
Черви наносят рыбам вред главным образом как паразиты.
У большинства промысловых рыб мы почти не можем найти особей,
которые не были бы заражены тем или иным видом паразитических
червей. Среди сосальщиков имеется огромнее количество парази-
тов рыб, поражающих, как наружные, так и внутренние органы.
Многие сосальщики паразитируют на жабрах у рыб. Так, например,
на жабрах осетровых паразитируют Nitzchia sturionis, приводящая
иногда к массовой гибели этих рыб от истощения, вызванного боль-
шой потерей крови. У некоторых рыб на жабрах часто скопляется
более 50Э экземпляров этого паразита, потребляющих огромное
количество крови и нарушающих нормальный ход кровообращения.
На жабрах у карпа паразитирует сосальщик Dactylogyrus vastator.
Заражение происходит через свободноплавающих личинок, которые
первое время передвигаются в воде при помощи ресничек. После
периода свободней жизци личинка прикрепляется к рыбе, сбрасы-
вает ресничный покров и держится на теле рыбы при помощи
своих крючочков. Этот червь поражает главным образом молодых
особей карпа и у рыб старших возрастов встречается реже. Объя-
сняется это, видимо, такими изменениями в строении жабр рыбы с
возрастом, которые затрудняют условия прикрепления паразита.
Большинство других сосальщиков из Moncgenea также локализи-
руется или на жабрах или на коже рыб. В отличие от м о н о г е-
нетических сосальщиков, поражающих, главным
образом, кожные покровы и жабры рыб, дигенетические
сосальщики, развитие которых происходит с одним или двумя про-
межуточными хозяевами, поражают, главным образом, внутренние
органы рыб. Дигенетические сосальщики, как например, Bucepha-
lus polymorphus, локализуются в кишечнике, откуда их яйца по-
падают в воду и заражают различных моллюсков, в которых и про-
ходит развитие до стадии церкарии. Вышедшие в воду церкарии
попадают под кожу мирным рыбам, /Где инкапсулируются. Хищ-
ные рыбы, поедая мирных, получают этого сосальщика в виде ки-
шечного паразита. У некоторых форм окончательным хозяином
являются водоплавающие птицы, заражающиеся при поедании
рыб. Некоторые дигенетические сосальщики локализуются в почках
и мочевом пузыре. В качестве примера рассмотрим Catopteroides
macrccotyle. Заражение этим червем происходит следующим пу-
тем: яйца паразита гопадают с водой в промежуточного хозяина—
моллюска Dreissena. Из моллюска церкарии выходят уже в виде
цисты с редуцированным хвостом и всплывают в поверхностные
69
слои воды, где случайно вместе с водой заглатываются рыбами и,
попадая в кишечник, добираются до мочеточников, по которым до-
ходят до почек. Поражают дигенетические сосальщики и нервную
систему рыб. Так, в мозговой жидкости локализуются многие черви
из рода Tolyclelphus. Встречаются дигенетические сосальщики в
желчном пузыре, кровеносной системе и других органах рыб.
j Так же весьма большое значение, как паразиты рыб имеют л е н-
точные черви. Наибольший вред рыбному хозяйству при-
носит ленточный червь Ligula intestinalis. Окончательный хозяин
Рис. 43. Цикл развития ремнеца Ligula (из Леймап,
1939).
лигу ли — рыбоядные птицы, у которых паразит локализуется
в кишечнике. В птицу лигуля попадает из рыбы, у которой она жи-
вет в полости тела. Первым хозяином этого паразита являются ве-
слоногие рачки (рис. 43). Лигуля наносит значительный ущерб рыб-
ному хозяйству, часто вызывая массовую гибель рыб. ЛигулеЗ вы-
зывает у рыбы замедление роста, нарушение обмена веществ и, при
сильном развитии паразита, смерть рыбы. Когда находящийся в
полости тела паразит достигнет больших размеров, рыба теряет
способность плавать (рис. 44), поднимается в поверхностные слои
воды, где легко становится доступной для рыбоядных птиц. Боль-
шинство других ленточных червей локализуется в кишечнике ры-
бы, причем иногда взрослые особи находятся в кишечнике, а личин-
ки паразитируют в каком-нибудь другом органе. Так, у Triaeno-
phorus nodulosus взрослые особи живут в кишечнике у рыбы, а
личинки поражают печень.
Многие рыбы являются промежуточными хозяевами ленточных
червей, половозрелая стадия которых паразитирует в кишечнике
70
человека. Так, Diphyllobotrium latus во взрослом состоянии живет
в кишечнике человека, а финны находятся в мускульных тканях
рыбы. Заражение человека происходит в том случае, если будет
съедена недостаточно проваренная или прожаренная рыба (рис. 45).
Большое значение, как паразиты рыб, имеют и круглые
черви Nematodes. Локализуются круглые черви в самых раз-
Рис. 44. Плотва, пораженная’гремнецом’1Л§и1а; внизу
вскрытый экземпляр.
нообразных органах рыб, они поражают кишечник, плавательный
пузырь, печень, брюшину, мускулы, кожу и т. д. Некоторые из них
развиваются с промежуточным хозяином. Так, например, Camal-
lanus lacustris, локализующийся в взрослом состоянии в кишечни-
ке рыбы, имеет промежуточным хозяином водяного ослика Asel-
lus aquaticus. Другие круглые черви еще не выработали у себя спо-
соба размножения с промежуточным хозяином. У некоторых, как
например, у Raphidascaris acus ход развития протекает без про-
межуточного хозяина, однако, хищные рыбы заражаются, поедая
мирных рыб, в печени и брюшине которых находятся личинки это-
го червя (рис. 47). Подобный «полупромежуточный хозяин» назы-
вается резервуарным хозяином.
Паразитами многих рыб являются также скребни Асап-
thocephala. Эти черви локализуются главным образрм вкишеничке.
Они обычно имеют одного, а иногда и двух (Промежуточных
хозяев.
71
Вред, наносимый многощетинковыми червями
рыбам, сравнительно очень невелик. Лишь немногие из этих червей
являются наружными паразитами рыб. Так, например, на плавниках
рыб паразитируют представители семейства Ichthyotomidae и, в
частности, Ichthyotomus sanguinarius, который при помощи рта
прикрепляется к плавникам рыб, главным образом, угрей и пи-
тается их кровью (рис. 47).
В заключение обзора роли червей, как паразитов рыб, необхо-
димо упомянуть о пиявках. Пиявки могут наносить рыбам троякий
широкого лентеца Diphyl-
Лейман, 1939).
хозяин
ЧЕЛОВС^КОШНЯГОБЛКР.
КИШЕЧНИК
ПОЛОВОЗРЕЛЙЯ
СТнДИЯ.
проиежуточнь1й
хозяин
веслоногий рдчек
циклоп
диятомнс
ПОЛОСТЬ ТЕПЛ
ПРОЦЕРКОИД.
Второй
РОМЕЖУТОЧНЫИ
хозяин
РЫБЯ
ЩУНЛ, ЕРШ .ОКУНЬ
НДЛИМЛ ЛОСОСЕВЫЕ
И КОРЮШКОВЫЕ
коид.
Рис. 45. Цикл развития
lobotrium (из
Дефинитивный
ШШЦЫ,ПЕЧЕНЬгЯИЧ-
нк. Плероцер-
Первые
вред: раздражая рыбу путем укусов, высасывая кровь и, наконец,,
создавая благоприятные условия для заражения рыбы бактериями
и грибками. На рыб могут нападать различные пиявки. Наиболее
обычна в наших водах пиявка землемер Pisciccla gecmetra, при-
крепляющаяся в различных местах тела рыбы. Если на одну рыбу
нападает сразу большое количество пиявок, то они приводят к до-
вольно сильному ее истощению. Иногда пиявки вызывают массовую
гибель рыб. Так, например, известны случаи массовой гибели лосо-
севых, жабры которых были поражены пиявкой Cystobranchus
respirans.
Если отрицательное значение червей для рыб очень велико, то
имеется, правда, значительно меньшее, и положительное значение,
так как очень многие черви, главным образом, круглые и кольча-
тые, играют важную роль как пища рыб. Свободно-живущие круг-
лые черви очень часто служат объектом питания рыб. Так, в Волге
по данным Алявдиной (1929), лещ и синец потребляют в довольно
большом количестве свободно живущих нематод. Эти черви были
72
зарегистрированы в пище 17% всех исследованных синцов и 8% ле-
щей. В пище волжских рыб преобладают Dcrylaimus stagnalis.
Значительно большую роль, чем нематоды, в пище рыб играют
малощетинковые и многощетинковые черви. В пище рыб Варенцо-
ва моря сидячие многощетинковые черви составляют весьма солид-
ную часть. Так, пр Зацепину (1939), в пище пикши Gadus aeglefinus
многощетинковые черви составляют в различных районах от 10 до
Рис. 46. Схема* цикла разви-
тия Raphidascaris (с наличием
резервуарного хозяина) (из
Лейман, 1939).
Рис. 47. Паразитически#
многощетинковый червь
Ichthyotomus sanguina-
rius на плавнике рыбы
(из Fauvel, 1923).
20%. Из малощетинковых червей основным представителем, по-
требляемым рыбами в пресных водах умеренных широт, является
Tublfex.
Положительное значение пиявок, как пищи рыб, очень невели-
ко. Обычно пиявки в пище рыб попадаются случайно и в незначи-
тельном количестве.
Так же, как среди рассмотренных выше групп, и среди игло-
кожих имеются самые разнообразные взаимоотношения с ры-
бами. Многие иглоксжие питаются рыбой. Морские звезды и, в
частности, Asterias rubens нападают на рыб, схватывая их своими
лучами. При этом, как показали наблюдения leningls (1907), морские
звезды нападают на рыб, обычно не превышающих'по размерам са-
мой морской звезды (расстояния от конца одного луча до противо-
73
положного). Исключение составляют морские иглы, которые ста-
новятся жертвами морских звезд, даже будучи много больше их
по размерам (рис. 48). Иногда одну рыбу захватывают две звезды
и начинают ее пожирать с разных концов. Захватывание рыбы про-
исходит следующим способом: рыба, проплывая, з: д.вает морскую
звезду ? в тот же момент морская звезда схватывает свою жертву педи-
Цилляриями и постепенно перемещает ее хвостом вперед к ротово-
му отверстию. Захваченная рыба переваривается по частям, и в то
время, как задняя часть рыбы уже переварилась, передняя нахо-
Рис. 48. Морская звезда, поедающая рыбу (из Goree, 1939).
дится еще снаружи. Основными жертвами морских звезд являются
те рыбы, которые плавают в, придонных слоях воды. Интересно,
что рыбы, живущие непосредственно на дне среди камней близко
от морских звезд, при прикосновении к последним не вызывают у
них хватательной реакции. Известны случаи поедания морскими
звездами различных Scarus, Nerophis,. Syngnathus, Syphonostoma,
Fundulus и некоторых других рыб. К сожалению, до настоящего
времени не выяснено, какой процент в пище морских звезд
составляют рыбы. Морские ежи также потребляют в пин1у рыб,
захватывая их при помощи своих педициллярий. Как объект пи-
щи ежей зарегистрированы следующие рыбы: Gobius paganellus,
Labrus mixtus, Blennius, Pleuronectes и многие другие. Так же,
как это имело место в отношении морских звезд, и у морских ежей
наблюдаются случаи нападения двух животных на одну и ту же ры-
бу. В то время, как один еж переваривает переднюю часть рыбы,
другой переваривает заднюю.
Если для иглокожих рыбы являются видимо все же случайным
объектом питания, то для многих рыб иглокожие составляют основной
объект пищи, и у некоторых рыб вырабатываются специальные при-
способления в связи с питанием иглокожими. Так, зубатки Апаг-
74
rhichas, питающиеся главным образом иглокожими, имеют мощные
конусовидные зубы, прекрасно приспособленные для раздробления
панцирей морских ежей и звезд. Большую роль иглокожие играют
в пище у многих скатов.
Нечто вр де коменсализма между иглокожими и рыбами мы встре-
чаем на примере взаимоотношений между рыбкой Fierasfer и голо-
туриями. Эта рыбка живет в клоакальной полости голотурии. По-
видимому, это странное жилище служит ей убежищем от врагов.
Питается Fierasfer, главным образом, теми мелкими животными,
которые поп ’Дчют вместе с водой внутрь голотурии. При отсутствии
голотурий Fierasfer поселяется иногда внутри морских звезд и в
мантийной пол сти крупных двустворчатых моллюсков. Один из
близких к Fierasfer родов описан Семмнером уже как паразит го-
лотурии. Проникая внутрь своей жертвы, эта рыбка постепенно
поедает ее внутренности.
Весьма большую роль в жизни рыб играют водные м о л л ю с-
к и.
Роль моллюсков, как паразитов рыб совершенно ничтожна. На
рыбах гагааитнруют только некст< рые личгньи —глохидии двух-
створчатых, р кушки Unio и беззубки An: donta. Глохидии, будучи
выкинуты в наружную воду из материнского организма, при прикос-
новении к телу рыбы захлопывают створки и таким образом прикреп-
ляются к своему хозяину. В результате раздражения эпителий
рыбы в месте пр' крепления глох, дия начинает довольно быстро
разраст; ться вокруг него. Обросшие эпителием глохидии имеют
вид белых крупинок или бугорков, питаются они за счет тканей и
соков хозяина. Локализуются глохидии, главным образом, на жаб-
рах и плавниках у рыб. Продолжительность пребывания в таком
состоянии у различных видов мол; юсков ва рьирует от 10 до 75 дней.
Даже у одного и того же вида сроки пребывания изменяются в за-
висимости от температуры и ряда других факторов. По окончании
периода развития ткань рыбы, окружающая моллюска, лопается,
и он выходит на свободу. Глохидии оказывают на рыбу истощающее
влияние лишь в том случае, если они поражают ее в большом ко-
личестве. Случаи гибели рыб от поражения глохидиями в естест-
венных условиях неизвестны. Интересно, что многие виды рыб яв-
ляются имунными по отношению к глохидиям, и последние, будучи
окружены тканями хозяина, постепенно уничтожаются его лейко-
цитами.
Из моллюсков питаются рыбами только некоторые головоногие.
Но вред, наносимый этими животными рыбам, видимо, очень неве-
лик.
Если вредная роль моллюсков для рыб очень незначительна,
то польза от них, как кормовых объектов, огромна. Так, например,
в Северном Каспии у основных промысловых видов рыб, питающих-
ся донными организмами — бентосом, различные моллюски состав-
ляют в пище по весу весьма значительный процент. В других вну-
тренних морях роль моллюсков в пище рыб также очень велика.
75
Многие виды, как например, вобла в Каспии, почти целиком пи-
таются за счет этой группы животных.
Потребление моллюсков рыбами в Северном Каспии
Вобла Лещ Русский осетр Gobius caspiuS Gobius f 1 uvi ati lis pallasi Gobius melanosto- mus Bentophi- lus macrc- 1 cephalus
Monodacna . . 16,0 1,0 0,6 3.4 29,6
Adacna minima . . 14,0 8,0 1 I ч А Л 1 A
A. plicata и A. laeviuscula . 1,5 — ’к 4,0 I — 0,3
Didacna trigonoides 5,5 0,7 4,0 1,0 3,7 3,0
Cardium edule 1,3 0,003 — 10,0 2,0 20,5 5,0
Bee Cardidae . . 38,3 9,7 1,3 18,0 4,6 27,6 44,1
Dreissena polymorpha . 42,1 2,1 0,2 — 9,0 13,6 0,2
Mytilaster lineatus — —— — — — 5,0 —
Gastropoda 2,0 0,1 — — 0,7 8,0 35,0
Все моллюски 82,4 11,8 1,5 18,0 14,3 54,2 79,3
В пресных ведах роль моллюсксв в пище рыб несколько снижа-
ется. Правда, для некоторых видев, например в Великих озерах
Северной Америки Sphaeriidae служат основным объектом пита-
ния. Пищей рыб являются не только донные моллюски, но также
и моллюски-обитатели толщи веды. Так, крылоногими моллюсками
Clio borealis питаются в значительном количестве треска и пик-
ша.
Между моллюсками и рыбами возможен и коменсализм. В ка-
честве примера такового можно указать на использование моллюс-
ков горчаком для откладки им в мантийную полость икры, разви-
вающейся затем внутри моллюска.
Ракообразные, так же как и другие группы водных беспо-
звоночных, имеют важное значение для рыб. На рыбах паразитируют
представители трех отрядов ракообразных, именно: веслоногие
рачки, жаброхвостые и равноногие. Все ракообразные, парази-
тирующие на рыбах, являются нар жными паразитами, поражаю-
щими кожу или жабры рыб. Паразитируют ракообразные на ры-
бах, как в пресной, так и в соленой воде. Из пресноводных весло-
ногих следует указать Ergasilus sicbcldi, выдерживающего и неболь-
шое осолонение воды. В Невской губе, по данным В. А. Догеля,
этим рачком заражено около 13% щук и 6,5% плотвы. На одной
рыбе зарегистрировано от 1 до 213 паразитов. Прикрепляется Erga-
silus к жабрам рыб при помощи второй пары антенн, которые из-
менены у него в крючки. Паразит этот может наносить серьезный
вред, вызывая иногда массовую гибель рыб в прудовых хозяйствах.
Главным образом на морских рыбах паразитируют другие предста-
вители веслоногих рода Caligus (рис. 49). Прикрепляются эти рачки
к рыбам при помощи присосок и видоизмененных в крючочки вто-
рых антенн. Представители этого рода распространены преимуще-
79
ственно в северных морях, они встречаются на треске, пикше и
других рыбах Варенцова моря. Повышение температуры выше 18—
20° действует на этих паразитов губительно. Некоторые веслоногие
в результате паразитического образа жизни столь сильно изменяют-
ся, что совершенно теряют облик ракообразного. Так, например,
чрезвычайно сильные изменения претерпели рачки Lernaea branchi-
alis, паразитирующие на жабрах трески. Цикл развития этого ра-
Рис. 49. Ракообразные паразиты рыб (слева направо): Argulus
(Branchiura), Caligus (Copepoda), Aega (Isopoda).
koooразного довольно сложен, личиночная стадия развивается на
жабрах камбал. Достигнув половозрелости, эти раки некоторое
время живут в толще воды, где самцы оплодотворяют самок и сами
погибают. Самки же прикрепляются к жабрам тресковых, причем
тело самок принимает мешкообразную форму и сильно увеличи-
вается в размерах по сравнению с размерами во время свободной
жизни. Увеличение идет главным образом за счет первого поло-
вого членика брюшка. Паразит глубоко внедряется в ткань жабр
и питается за счет крови хозяина, вызывая замедление роста и
истощение рыбы.
Одним из наиболее серьезных вредителей прудового рыбного
хозяйства является представитель жаброхвостых—карпоед Argu-
lus foliaceus (рис. 49). Особенно сильный вред паразит этот наносит
в карповых хозяйствах, иногда приводя к гибели значительной ча-
сти поголовья молоди. Карпоед присасывается к коже хозяина,
Питается соками его тела и вызывает обычно сильное изъязвление
Кожи и истощение рыбы, в конечном итоге приводящее к ее гибели.
Паразитируют на рыбак также некоторые равноногие ракообраз-
ные.; Равноногие встречаются на рыбах, главным образом, в морских
воДах. Крупные Ega (рис. 49) очень часто попадаются на жабрах,
7Т
в ротовой полости и на теле, обычно у основания плавников трески’
и пикши. Некоторые представители этой группы паразитируют на
жабрах сельдевых и некоторых других рыб. Поселяясь на теле своей
жертвы, эти крупные ракообразные (Ega достигают в длину до 4 и
более сантиметров) иногда выедают глубокие раны, часто приводя-
щие к гибели рыбы. Так же, как и представители рассмотренных
выше групп, равноногие раки питаются, главным образом, высасы-
вая кровь из своих хозяев. Кроме вреда, npi носимого некоторыми
ракообразными,являющимися паразитами рыб, ракообразные влия-
ют на рыб и как переносчики глистных инвазий, являясь проме-
жуточными хозяевами некоторых червей. Выше мы указывали, что
переносчиками лигулеза являются веслоногие рачки. Широкий
лентец также имеет промежуточным хозяином веслоногого рака.
Форма взаимоотношения хищника и жертвы между ракообраз-
ными и рыбами выражена почти полностью односторонне, а имен-
но в роли хищника выступает рыба. Обратное сочетание встречает-
ся значительно реже. Так, некоторые крабы нападают на больных
и умирающих рыб и поедают их. Личинки рыб иногда являются
объектом питания некоторых циклопов (Дзюбан, 1939); обычно же
ракообразные питаются уже уснушей и начин; кщой разлагаться
рыбой и на живую сильную рыбу не нападают. Исключение состав-
ляют лишь некоторые представители селейства Portunidae, которые
могут свободно плавать и иногда ловят д<же столь быстрых рыб,
как например макрели. Ракообразные в некоторых случаях явля-
ются и конкурентами рыб в пище. Так, например, китайский краб
Ereiocaeir sinensis питается теми же объектами, что и многие ос-
новные промысловые рыбы, именно моллюсками и червями. Раз-
множаясь в огромном количестве, китайский краб может нанести
серьезный вред состоянию запасов рыб, с которыми он конкурирует
в пище.
Несомненно, однако, что вред, причиняемый ракообразными,
совершенно ничтожен по сравнению с тем положительным значе-
нием, которое имеют ракообразные для рыб, поскольку они являют-
ся одним из основных объектов их питания. Во всех морях те или
иные промысловые рыбы являются ракоедами. В пище сельди ра-
кообразные, главным образом веслоногие рачки, занимают ведущее
место. Большинство рыб, питающихся планктоном, имеют своим
основным кормом планктических ракообразных. Мальки почти всех
как бентосоядных, так хищных и растительноядных рыб, проходят
стадию, во время которой они питаются планктоном и, главным об-
разом,планктонными ракообразными. В той или иной стегени рако-
образные встречаются в пище большинства рыб Черного,Азовского,
Каспийского и Аральского морей. Так, в Касгии (Желтенкгва,
1939) у воблы ракообразные составляют 6,8 % ri ши, у л о ша — 54,0%,
у севрюги—45,9%, у осетра—19,8%, у Gcbius fluviatilis palla-
si—70,5 % и т. д. В Баренцевом море основными потребителями
ракообразных являются сельдь и мойра, питающиеся ракообраз-
ными планктона (главным образом Calanus), а также треска, пикша,
морской окунь, малая зубатка, у которых раки играют весьма суще-
78
ственнуюрольв пище. В пресныхводах у взрослых рыб ракообразны
в пище встречаются реже, что объясняется их меньшим количеством
здесь. Большую роль играют ракообразные в пище рыб (главным,
образом форели) в горных реках и озерах. Так, например, в озере
Севан основную пищу местных предствителей рода Salmo состав-
ляют бокоплавы.
Следуя за своей пищей — ракообразными, рыбы совершают
иногда значительные перемещения. Некоторые перемещения имеют
суточный ритм, другие повторяются в одни и те же сезоны года.
Так, например, в Аральском море бокоплавы ночью поднимаются
к поверхности воды, а днем опускаются на дно. Следуя за бокопла-
вами, также перемещаются чехонь и шемая, которые днем кормятся
в придонных слоях, а ночью поднимаются в поверхностные. Приме-
ром сезонных миграций рыб, связанных с перемещениями ракооб-
разных, могут служить перемещения рыбки люме Harpodon nehereus
из семейства Scopelidae.
В период дождей в Индии, приходящихся на время с июня по
октябрь, огромные массы веды устремляются в реки, снося туда
большое количество биогенных веществ. Эти биогенные вещества,
будучи вынесены в море, позволяют в непосредственной близости
от устьев рек и, в частности, Ганга развиваться громадным массам
планктонных водорослей, которые привлекают из отдаленных от
устьев рек районов моря ракообразных, собирающихся в местах
массового развития планктелных водорослей огромными скопле-
ниями. За ракообразными сюда же прикочевывает и люме. Туземцы
прекрасно знают время прикочевки рыб и, как только наступает
период дождей, сейчас же начинают готовиться к промыслу этой
рыбы.
Также огромное значение в жизни рыб имеют инасекомые.
Следует указать, что в отличие от ракообразных насекомые играют
значительно большую роль для рыб в пресных водах, чем в морских.
Многие насекомые и их личинки являются хищниками, уничтожаю-
щими иногда большое количество икры, молоди, а реже и взрослых
рыб. Из жуков многие представители семейства Dityscidae нападают
на мальков рыб и поедают их. Вред, приносимый эи ми жуками,
особенно широко распространенным окаймленным плавунцом Di-
tyscus marginalis, может быть весьма велик. Так, известны слу-
чаи, когда плавунцы уничтожали все потомство сигов и карпов
в выростных прудах.
По данным Петрович (1939) различные водные насекомые по-
едают следующее количество мал ьков рыб:
Вид насекомого В опыте Сколько съедено в день
Число съеденных мальков За сколько дней
Окаймленный плавунец взрослый 102 22 1 4,5
Окаймленный плавунец личинка . 91 12 7,6
Плавунчик взрослый 18 8 2,2
79
Вид насекомого
Плавунчик личинка
Гладыш взрослый
Гладыш личинка.
Водяной скорпион .
Corisa
Продолжение
В опыте Сколько съедено в день
Число съеденных мальков За сколько дней
131 24 5,5
45 9 5
103 15 7
17 8 2,1
36 17 2,1
Также весьма серьезный вред наносят рыбам водяные клопы и
из них особенно водяной скорпион и гладыш. Водянсй скорпион
Nepa нападает главным образом на молодь рыб и взрослым рыбам
вреда не приносит, но, попадая в большом количестве в нерестовые
пруды, он может, уничтожая молодь, нанести существенный вред.
В противоположность водяному скорпиону гладыш NoLnecta уни-
чтожает и небольших взрослых рыб, которых он убивает яд см, вво-
димым в ранку при помощи хоботка. Некоторый вред наносят ры-
бам и клопы Corisa, которые поедают личинок рыб. Мальками рыб
питаются и личинки крупных стрекоз, как например, коромысла.
Так же, как и ракообразные, некоторые насекомые, главным об-
разом жуки и клопы, конкурируют с рыбами из-за пищи, но пови-
димому эта конкуренция сравнительно очень невелика. Значение
насекомых, как пищевых объектов для рыб, в пресных водах и внут-
ренних морях весьма велико. Личинки многих воздушных насе-
комых, живущие в воде, являются основным кормом рыб. Таковы
личинки комаров, ручейников, веснянок, стрекоз, мух и др. Так,
например, в Северном Каспии у осетра личинки комаров толкун-
цов Chircnomidae составляют в среднем около 7% пищи. В реках
с быстрым течением и песчаным грунтом, где биомасса бентоса чрез-
вычайно мала и не может удовлетворить всю потребность рыб в
корме, большую роль в пище рыб играют личинки насекомых, глав-
ным образом, ручейников, которые массами скопляются на упавших
в воду корягах. В Урале за счет личинок, сидящих на корягах,
питаются главным образом осетровые. Та же картина наблюдается
в реках Средней Азии. Большую роль в пище рыб играют также и
насекомые, падающие в воду с берега. В летние месяцы живущая
в мелких горных речках форель преимущественно питается за счет
воздушных насекомых. Так, у форели в Англии воздушные насе-
комые составляют следующий процент в пище по месяцам:
Январь Февраль Март Апрель Май Июнь Июль Август Сентябрь
О 0,7 1,7 21,5 52,5 32,2 25,0 93,0 73,4
Октябрь Ноябрь Декабрь!
1,7 1,5 О
Во время массового вылета насекомых, как, например, веснянок
или комаров толкунцов, на питание воздушными насекомыми пе-
80
реходят даже те рыбы, которые обычно ими не питаются. Роль на-
секомых в пище рыб внутренних водоемов очень велика, как
у экватора, так и за полярным кругом. Разница заклю-
чается только в том, что в водоемах умеренных или высоких широт
потребление насекомых и их личинок носит резко выраженный се-
зонный характер, в то время как в водоемах тропиков оно в течение
всего года подвержено сравнительно незначительным колебаниям.
Позвоночные других классов играют в жизни рыб также весьма
значительную роль. Амфибии имеют известное значение для
рыб внутренних водоемов.Отрицательная роль амфибий может быть
иногда весьма велика. Очень многие лягушки питаются молодью
рыб, причем уничтожают ее иногда огромное количество. Идельсон
(1938) показал, что в дельте Волги зеленые лягушки Rana ridibun-
da уничтожают очень большое количество мальков промысловых
рыб. Так, на Ильмене «Лощина» лягушки уничтожили за полойный
период (2—3 месяца) в среднем 110тысяч штук молоди с одного га,
из них 75,4% воблы, 6,3% сазана, 13,3% уклеи, 1,3% леща и 3,7%
густеры. Процент съеденных лягушками мальков из числа скатив-
шихся из Ильменя выражается солидной цифрой 58,7%. Подобная
же картина наблюдается и в других водоемах. Так, в Северной
Америке рыбу очень в большом количестве уничтожает лягушка
бык Rana catesbyana. Рыбою питается также японская саламандра
Megalobatrachus japonicus и ряд других крупных земноводных.
Имеются указания, что не только лягушки, но и тритоны поедают
икру и личинок рыб.
Положительная роль афмибий для рыб определяется их значе-
нием, как объектов питания. Многие рыбы питаются как личинками,
так и взрослыми амфибиями. В наших водах в очень большом ко-
личестве потребляет лягушек сом, который в Волге, Днепре и мно-
гих других реках специально подходит к берегу для охоты за ля-
гушками. Лягушка считается одной из наиболее хороших наживок
на сомовые крючья. Существует даже особый способ лова сома, так
называемое «клочение»: в том месте, где выставлены крючки, на-
живленные лягушками, производят путем ударов опрокинутой
деревянной чашкой по воде своеобразное бульканье, имитируя из-
даваемые лягушкой звуки, на которые сом приближается и хватает
наживку.
Взаимоотношения между рыбами и рептилиями осуще-
ствляется главным образом в форме взаимоотношения хищника и
жертвы, причем в роли хищника выступает пресмыкающееся. В
умеренных широтах почти исключительно рыбой питается водяной
уж Natrix tesselata и некоторые другие ужи. В некоторых местах,
особенно там, где рыбу разводят на рисовых полях, вред, прино-
симый ужами рыбному хозяйству, может быть весьма серьезен.
В тропических широтах в пресных водах серьезный вред рыбному
населению приносят также некоторые змеи. Так, в озерах Восточ-
ной Африки, питается рыбой водяной уж Natrix oliyaceus. В мор-
ских водах нападают на рыб представители семейству Hydrophidae,
распространенные в тропиках. В отличие от пресноводных рыбояд-
6 Биология рыб 81
ных змей морские рыбоядные змеи ядовиты. Рыбы составляют так-
же значительную часть пищи крокодилов. Так, в озере Виктория-
Нианца нильский крокодил в основном питается рыбой. Правда,
у некоторых особей в пище в большом количестве попадаются и
беспозвоночные. По наблюдениям Грахэм (1929) нильские кроко-
дилы легко ловят плывущих рыб. Крокодилы наносят довольно
значительный вред и рыбному промыслу, вырывая рыб, попавшихся
в сети.
Питаются рыбой черепахи как морские, так и пресноводные.
Из морских необходимо указать каретту Chelone imbricata и зеле-
ную морскую черепаху Chelonia mydas, а из пресноводных пале-
арктических черепах — представителей подсемейства Emydinae(B
СССР Emys orbicularis и Clemmys caspica) и тропических и субтро-
пических Trionyx, которые даже ловятся местными жителями в Ки-
тае и Африке на удочки, наживленные рыбой.
Каково количество рыбы, уничтожаемой различными репти-
лиями, точно неизвестно, и все проводившиеся до последнего вре-
мени исследования по питанию водных рептилий носят чисто ка-
чественный характер.
Если для многих рептилий рыбы являются основным объектом
питания, то в пище рыб рептилии не играют почти никакой роли.
Только у некоторых пресноводных рыб изредка попадаются в пище
(например у сома) упавшие в воду при обвалах берега ящерицы, но
это носит совершенно случайный характер.
Значительно более сложны и разнообразны взаимоотношения,
существующие между рыбами и птицами. Основной пищей
многих птиц является рыба, причем питаются рыбой представители
самых различных отрядов птиц. Среди соколообразных Falconifcr-
ines некоторые представители почти целиком перешли на пищу ры-
бой. Таковы например: скопа Pandion haliaetos, орланы-белохвост
Heliaetos albicilla и долгохвост Н. leucoryphos, камчатский орлан
Talassaetos pelagicus и некоторые другие. Все эти крупные хищники
уничтожают большое количество взрослой, часто довольно большой
по размерам рыбы. Питается рыбой и большинство крупных пред-
ставителей отряда Colymbiformes, как например, гагары, поганки
и некоторые другие. Потребляют рыбу и некоторые ракшеобраз-
ные, главным образом различные зимородки, особенно широко рас-
пространенные в тропических широтах. Но особенно важное зна-
чение имеют такие рыбоядные птицы, как цапли, бакланы, пелика-
ны, чайки и утки, причем в зависимости от способа лова у птиц,
держащихся даже вблизи одного и того же водоема, состав уничто-
жаемых рыб бывает весьма различен. Так, например, у цапель пе-
лагические рыбы в пище, как правило, отсутствуют, у чаек же они
составляют довольно солидный процент содержимого их желудка.
Количество рыбы, уничтожаемое рыбоядными птицами, весьма ве-
лико, и поэтому в тех местах, где рыбоядные птицы встречаются в
большом количестве, они могут наносить серьезнейший вред рыб-
ному хозяйству. Известно, что состав пищи баклана в дельте Волги:
судак 24,4%, сельди 22,6%, окунь 11,6%, бычки 10,4%, густера
82
9.1%, красноперка 6,7%, лещ 6,7%, щука 6,1%, вобла 3%, т. е.
основную пищу баклана составляют главнейшие промысловые ры-
бы. В день взрослый баклан поедает около 700 г рыбы, молодой —
(коло 500 г. Кроме того, во время охоты за рыбой бакланы ранят
огромное количество рыб, которые впоследствии погибают. Для
района дельты Волги подсчитано, что бакланы ежегодно уни-
чтожают около 50 тысяч центнеров рыбы, т. е. около 1% % от всего
улова по северному Каспию. На птичьих базарах в Перу птицами
ежегодно съедается 5,5 млн. килограмм рыбы. Многие рыбоядные
птицы в своей биологии настолько тесно связаны с определенными
видами рыб, что по появлению этих птиц можно судить о подходе
рыбы, которой эти птицы питаются.
Кроме непосредственного вреда, приносимого птицами путем
уничтожения рыб, они оказывают иногда прямее влияние на рыб-
ный промысел. В частности, на некоторых озерах западной Сибири
лов ставными сетями временами делается почти совершенно невоз-
можным, так как гнездящиеся в изобилии на этих озерах чайки
сейчас же спутывают сетку, пытаясь выдрать попавшуюся в сети
рыбу. Также отрицательное влияние птиц на рыб сказывается и в
форме переноса некоторых глистных заболеваний и в первую оче-
редь лигулеза, причем заражение рыб лигулезом через птиц в том
или ином водоеме бывает иногда настолько сильно, что приводит
к массовой гибели рыбы.
Наряду с довольно серьезным отрицательным значением, которое
имеют для рыб птицы, в некоторых случаях последние выступают
и как положительный фактор в жизни рыб. Так, иногда птицы спо-
собствуют расселению рыб, перенося из водоема в водоем на лапах
и перьях икру. Повидимому, таким образом была, например, за-
несена южная колюшка из бассейна Аральского моря в замкнутые
водоемы, расположенные по древному «Тургайскому проливу»,
служащему пролетным путем для многих птиц. Роль птиц, как объ-
ектов питания для рыб, сравнительно невелика. Правда, вблизи
птичьих базаров скопляются иногда в большом количестве хищные
рыбы, главным образом акулы, которые питаются плавающими в
воде птицами. Из пресноводных рыб поедают, преимущественно
птенцов, сом и щука, но ни у одного вида рыб птицы не являются
основным объектом питания.
. Серьезное значение имеют также для рыб птицы, как удобрите-
ли водоемов. В местах гнездовий водяных птиц в воду падает еже-
дневно огромное количество птичьего помета, за счет которого раз-
вивается фитопланктон и водные беспозвоночные, служащие кор-
мом рыб. Поэтому вблизи птичьих базаров часто наблюдаются
скопления рыбы, питающейся за счет развившегося здесь богатого
планктона. В настоящее время роль птиц, как удобрителей во-
доемов, используется и человеком. Так, например, сейчас уже вхо-
дят в практику комбинированные рыбно-птичные прудовые хозяй-
ства, где одновременно в прудах разводятся карпы, и|на этих же
прудах держатся утки, которые своим пометом удобряют карповые
пруды, значительно повышая их продуктивность.
б»
Млекопитающие, так же, как и птицы, играют в жизни
рыб немаловажную роль. Большинство водных млекопитающих в
той или иной степени питается рыбой. Многие киты поедают огром-
ное количество стайных рыб: анчоусов, сельдей, сардин и других
рыб, стаи которых они сопровождают. Значительное количество
рыбы, главным образом, мойвы и сельдей, поедает сельдяной кит
Balaenoptera physalus, также главным образом питаются рыбой Ва-
laenoptera acutirostrata и Hyperoodon rostratus. Значительную долю
пищи составляет рыба и у дельфинов. Пищею белухи Delphinapte-
rus leucas служат лососевые (кета, горбуша и др.), тресковые (глав-
ным образом сайка, треска и навага), а также и представители дру-
гих семейств рыб. Нарвал Monodon monoceros тоже поедает рыбу —
камбал, тресковых и скатов. И в пищу настоящих дельфинов (под-
семейство Delphininae) рыбы входят в значительном количестве.
Представители рода Phocaena — морские свиньи главным образом
поедают мелких рыб, держащихся стаями. В Баренцовом море их
пищей служат: мойва и сельдь, в южных водоемах анчуосы, бычки
и др. Пищей обыкновенного дельфина Delphinus delphis в Черном
море служат хамса, шпрот, южная пикша, ставридка, барбулька,
луфарь и др. В значительной части рыбой питается также и офали-
на Tursiops tursio, в ее пище в Черном море преобладают: хамса,
скаты, камбалы и южная пикша. Касатка Orca orca наиболее хищ-
ный дельфин — питается самой разнообразной рыбой от сельди
до акул включительно.
Тюлени также в значительном количестве употребляют в пищу
рыбу. Так, хохлач питается главным образом рыбой глубинных
слоев воды (морской окунь и треска). Рыба служит основной пищей
для тевяка Halichoerus gryphus, Histiophoca fasciata, Phoca caspica
и других тюленей.
Из млекопитающих, связанных с водой меньше, чем ластоно-
гие и китообразные, многие питаются рыбой. Много рыбной пищи
потребляет выдра Lutra lutra. Медведи, как белый, так и бурый, в
значительной части своей пищи зависят от рыбы; правда, для буро-
го медведя сказанное выше правильно не для всех районов его оби-
тания. Особенно много рыбы бурый медведь поедает на Камчатке
и по побережью Охотского моря, где во время хода лососевых в
реки этот зверь почти целиком переходит на рыбную пищу.
Довольно часто питаются рыбой лисы и шакалы, причем первые
иногда совершают набеги за рыбой даже в прудовые хозяйства, где
разводятся карпы. Из насекомоядных в большом количестве поедает
рыбукутора Neomysfodiens. Этот маленький зверек часто напада-
ет на рыбу, значительно превышающую кутору по размерам. В неко-
торых районах кутора может принести довольно серьезный вред
рыбному хозяйству. Известное место в пище занимают рыбы и у
выхухоли. Как на врага рыб можно указать еще на ондатру Ondatra
zibethica, которая также питается рыбой. Проникая в питомники,
где зимуют карпы, этот зверь может нанести весьма серьезный вред,
вылавливая сеголеток. Вредит рыбному хозяйству ондатра, также
разрушая плотины и другие земляные сооружения.
84
Если место рыбы в пище многих млекопитающих сравнительно
велико, особенно у представителей китообразных и ластоногих, то
роль млекопитающих, как объекта питания рыбы совершенно ни-
чтожна. Некоторые акулы нападают на молодых тюленей, бельки
каспийского тюленя изредка попадаются и в пище белуги. Мыши,
землеройки и другие мелкие млекопитающие, попадая в воду, ча-
сто достаются в пищу щуке, сому и большинству других хищных
рыб, как умеренных широт, так и тропиков. Из пресноводных рыб
Рис. 50. Пиранья Serrasalmo piraya (из[Брэма, 1895);2
активно нападает на млекопитающих сом Silurus glanis, который
часто топит переплывающих реки собак и более мелких животных.
На крупных млекопитающих нападают некоторые представители
подотряда харациновых(рис. 50) Characinoidei: так, амазонская пи-
ранья Serasalmo piraya держится стаями и стаями же нападает на
переплывающих реку крупных млекопитающих. Несмотря на свой
маленький размер (до 30 см) эти рыбки сильно ранят своими острыми
зубами попавшее в воду животное, выхватывая даже большие кус-
ки мяса. Если зверь, даже такой крупный как бык или тапир, бы-
вает застигнут этими хищниками на середине реки, то очень часто
он не успевает достигнуть берега и, истощенный от потери крови,
тонет и окончательно становится добычей маленьких разбойников.
Млекопитающие связаны с рыбами и как окончательные хозяе-
ва некоторых паразитов, преимущественно ленточных червей, про-
межуточными хозяевами которых являются рыбы (см. выше).
Некоторые звери, как например, бобры, производят сильные
изменения в режиме реки, строя плотины и создавая новые водоемы,
и этим несомненно оказывают некоторое влияние на ихтиофауну
(рис. 51). Поселяясь на заморных водоемах, как показали наблю-
дения С. Е. Клейненберга в Хоперском заповеднике бобры, про-
делывая во льду продухи, часто приводят к тому, что водоем
становится незаморным.
Для некоторых водоемов Северной Америки отмечены случаи,
когда бобровые плотины перегораживали нерестовы1е речки и от-
резали нерестилища от мест нагула рыб (Brown, 1938).
85
Огромную роль в жизни рыб играет человек. Формы его
влияния чрезвычайно разнообразны. Влияние может быть, как
косвенное, через воздействие на водоем, в котором живет рыба, так
и прямое, путем вылова, акклиматизации и т. д. Мы не можем оста-
навливаться здесь подробно на всех формах влияния человека на
рыб и вынуждены ограничиться лишь кратким обзором.
Формы косвенного влияния человека на ихтиофауну чрезвычайно
разнообразны,причем некоторые из этих форм отражаются на ихтио-
Рис. 51. Бобровая плотина и измененный ею водоем.
фауне отрицательно, вызывая или сокращение численности погс-
ловья, или гибель рыб, или сокращение области распространения.
Вырубание лесов, как известно, приводит к сильному изменению
режима стока. С обезлесением водосборной площади увеличивается
быстрота таяния снега, что приводит к увеличению мощности весен-
него паводка; последний становится зато более кратковременным и по-
сле его окончания реки быстро мелеют. Обмеление рек приводит к то-
му,что ряд рыб, особенно те, ход которых приходится на летние и осен-
ние месяцы (главным образом лососевые), перестают заходить в обме?
левшие реки для размножения, и область их распространения ста-
новится более ограниченной, или они совершенно покидают те реки,
в которых произошли указанные изменения. Повидимому, причиной
’сокращения поголовья каспийского лосося в Волге именно и яви-
лось изменение режима стока<ее вод.
86
Развитие на реках лесосплава также может вредно отразиться
iia рыбе. Так, в некоторых реках Дальнего Востока и Тихоокеанг
ского побережья Северной Америки лесосплав сильно повлиял на
условия нереста лососевых. Сплавляемый лес скопляется на местах
икрометания лососей; при этом кора, попадая на каменистое дно,
образует местами толстый слой, куда лососевые вместо гальки
и закапывали свою икру. Последняя, будучи окружена гниющей
корой, не могла развиваться и погибала. Существенное отрицатель-
ное влияние на рыб оказывает также спуск в воду сточных вод раз-
личных предприятий, что в некоторых случаях может привести к
полному вымиранию всей ихтиофауны, населяющей тот или иной
водоем. Попадание в воду большого количества нефтепродуктов,
образующих на поверхности воды тонкую пленку, вредно отражает-
ся как на ихтиофауне, так и на кормовых объектах, понижая.возмож-
ность насыщения воды кислородом за счет атмосферного воз-
духа.
И положительное и отрицательное влияние на ихтиофауну ока-
зывает гидростроительство. Плотины, перегораживая русло рек,
делают для проходных рыб зачастую невозможным проход к нере-
стилищам, что при неумелом хозяйствовании может привести к
подрыву запасов этих рыб1. Для устранения перечисленных форм
вредного влияния человека на ихтиофауну разработан ряд меро-
приятий, устраняющих или понижающих вредное влияние той или
иной формы воздействия. Регулирование вырубки лесов, создание
лесоохранных зон и т. п. способствует снегозадержанию, а тем са-
мым равномерности стока. Установление запретных рек для лесо-
сплава и сплав леса, очищенного от коры, содействует сохранению
нерестилищ. Ограничение спуска сточных вод, установление со-
ответствующих фильтров и очистительных установок предохраня-
ют от отравления ядовитыми веществами водоемов, в которые про-
изводится спуск сточных вод. Для предотвращения вредного дей-
ствия плотин гидростанций устраиваются специальные рыбоходы
в виде расположенных обычно в форме лестницы, один над другим,
бассейнов, через которые рыба может обходить плотины (рис. 52—
См, вклейку в конце книги). Рыбоходы иногда делаются и виде
шлюзов. К сожалению, в отношении далеко не всех видов рыб, рыбо-
ходы оказываются эффективными, и даже те рыбы, которые могут
подниматься через рыбоходы, проходят часто в слишком незначи-
тельном количестве и не обеспечивают восстановление поголовья
стада. Проведенный на Боневильской плотине (США) учет прохода
рыбы через рыбоход показал, что с 7 мая 1938 г,-по 31 января
1939 г. через рыбоходы (рыбные лестницы) прошло: чавычи 271.085,
колумбийского лосося 107.026, нерки 75.040, кижуча 15.188, кеты
"2.119, сигов 3.122, прочих лососевых 2.227, Catcstomus 281.873,
миног 227.626, карповых 92.196, сельдевых 5.274, осетровых 44.
Итого за 9 месяцев через рыбоход прршло 1.082.820 рыб. В тех
случаях, когда рыбоходы не могут быть использованы, приходится
итти путем создания ниже плотины условий благоприятных для
нереста, причем для того, чтобы производители отдавали икру,
87
им делается инъекция гонадотропных гормонов, которые стимули-
руют отдачу икры самками (см. ниже).
Положительные формы косвенного влияния человека на ихтио-
фауну также весьма разнообразны. Это влияние в первую очередь
выражается в форме создания новых водоемов. Образование огром-
ных водохранилищ в результате перегораживания плотинами элек-
тростанций многих рек способствует увеличению поголовья озер-
ных рыб, условия жизни которых в значительной степени улучша-
ются. Развитие ирригационного строительства приводит к созда-
нию целых систем новых водоемов, заселяемых своей ихтиофауной.
Сбросовые озера, оросительные каналы, рисовые поля и т. п. — все
эти водоемы вполне пригодны для жизни рыб и очень быстро ими
заселяются.
Строительство судоходных каналов, соединяющих различные
бассейны, способствует обмену фаунами и расселению отдельных
видов животных, в том числе и рыб. Постройка Панамского канала
способствовала обмену фаунами между Атлантическим и Тихоокеан-
ским побережьями Америки. Такую же роль, правда, в меньшей сте-
пени, сыграл и Суэцкий канал, по которому в Средиземное море
проникло 12 новых видов рыб (Стейниц, 1927). Сток в водоемы вод
с пахотных полей приносит огромное количество биогенных веществ,
которые служат базой для развития организмов, являющихся пи-
щей рыб. Прорубание прорубей для получения воды в некоторых
случаях может способствовать предотвращению или ослаблению
действия заморов.
Но наиболее существенно прямое влияние, оказываемое чело-
веком на рыбу и выражающееся в первую очередь в форме вылова,
а также в виде рыборазведения, акклиматизации и т. п. Человек
начал употреблять рыбу в пищу очень давно. Уже остатки чело-
века древнего каменного века (палеолита) находят вместе с остат-
ками рыбы, которая несомненно служила ему пищей. С тех пор роль
человека, как потребителя рыбы, все возрастает. Сначала люди
используют только рыбу внутренних водоемов, лишь постепенно
переходя на море. Дальнейшим этапом было появление у человека
навыков, позволяющих ему сохранять рыбу впрок. Это дало воз-
можность резко увеличить вылов рыбы во время ее подхода к бе-
регам. Следующей ступенью явилось развитие лова рыбы в удален-
ных от берега районах как в толще воды (лов пелагических рыб),
так и в придонных слоях. Правда, и до настоящего времени промыш-
ленное рыболовство придонных рыб еще не вышло за пределы
глубин в 200 м и лишь в незначительной степени использует мате-
риковый свал.
Как показал печальный опыт мирового рыболовства, запасы
промысловых рыб, как пресноводных, так даже и морских, далеко
не безграничны, и интенсивный бесконтрольный лов в более или ме-
нее короткое время приводит к подрыву запасов и прекращению
добычи, которая становится нерентабельной. Наоборот, умеренный
вылов с хозяйственной точки зрения оказывается полезным, так как
разрежая стадо, он создает более благоприятные условия для на-
88
гула и способствует ускорению темпа роста рыб. Задачей рацио-
нального рыбного хозяйства и является установление такой вели-
чины вылова, при которой сохраняющееся число производителей
полностью обеспечивало бы воспроизводство запасов, и в то же вре-
мя в водоеме не происходило бы накопления рыб старших возра-
стов, которые в экономике водоема являются отрицательным фак-
тором, потребляя большое количество корма и одновременно да-
вая малый прирост веса. Для сохранения запасов промысловых рыб
применяется ряд мер, выражающихся или в мероприятиях охран-
ного порядка, или в улучшении условий жизни рыб в во-
доеме. Охранные мероприятия сводятся к запрещению ло-
ва маломерной рыбы, лова вредными орудиями и способами (приме-
нение мелкоячейных сетей, глушение и отравление рыбы и т. п.),
а также к установлению запретных для лова районов, главным об-
разом, на местах икрометания. Вторая группа мероприятий, улуч-
шающих условия жизни рыб в водоемах, сводится к мелиорации
нерестилищ, разведению кормовых объектов, уничтожению живот-
ных, наносящих вред рыбам и т. п. Если все перечисленные меро-
приятия разумно выполняются, то вылов рыбы человеком из факто-
ра отрицательного превращается в фактор положительный в жизни
ихтисфауны.
Другой путь активного влияния человека на ихтиофауну, путь,
получивший наиболее широкое распространение в последнее вре-
мя, — это акклиматизационные мероприятия. Введение видов рыб,
которые раньше в данном водоеме не встречались, применяется сей-
час в очень широких масштабах; от пересадки из одного озера в
другое, расположенное в бассейне той же реки (например, форель)
до перевозки рыб из Америки в Европу (черный окунь Huro salmoi-
des) или из Европы в Австралию (кумжа). Умелый выбор акклимати-
зируемого объекта в случае успешной акклиматизации позволяет
полнее использовать кормовые ресурсы водоема и тем самым спо-
собствует увеличению его ихтиомассы. Однако, проведение аккли-
матизационных мероприятий, выполняемое без надлежащих иссле-
дований, может способствовать заносу паразитов, которые вызы-
вают иногда гибель даже части местной ихтиофауны. Иногда новый
вид рыбы, конкурируя с местными видами, приводит к их постепен-
ному вымиранию. Акклиматизируются не только рыбы, но и их
кормовые объекты.
Влияние человека на ихтиофауну выражается и в форме одо-
машнивания некоторых видов рыб, выведения путем искусственного
отбора домашних рас у рыб, обладающих теми признаками, ко-
торые человеку нужны (рис. 53). Выводя различные расы рыб, че-
ловек может преследовать две цели, с одной стороны, получение
более товарно-ценных пород, или скороспелых, или обладающих
быстрым ростом, или более жирных и т. п. С другой стороны,
человек выводит различные расы рыб в декоративных целях, созда-
вая такие формы, как телескопы, вуалехвосты, кометы и т. д.
(рис. 54).
Таковы основные формы влияния человека на рыб. Но, как уже
89
видно из изложенного, и рыбы в жизни человека имеют весьма боль-
шое и в первую очередь положительное значение. Вылов рыбы за
1933 год по всему земному шару достиг весьма солидной цифры в
12 247 тысяч тонн. Сказанное делает понятным, что значение рыбы,
как объекта питания человека, чрезвычайно велико. Используются
рыбу человеком не только как пищевой продукт, но так же как ко-
жевенное сырье; отходы идут на удобрение, из рыб изготовляется
клей, чешуя используется для приготовления жемчуга и т. д. Но
положительное значение рыбы для человека этим не ограничивается.
Рис. 53. Различные расы карпа. Наверху слева направо” кожистый карп,
зеркальный карп, внизу дикий сазан.
Насекомоядные рыбы, поедая водных личинок насекомых, служа-
щих переносчиками ряда заболеваний и, в первую очередь, маля-
рии, способствуют понижению численности этих насекомых и тем
самым сокращению заболевания. Некоторые виды рыб, как напри-
мер, гамбузия Gambusia affinis, американский окунь Eupomptis
gibbosus и другие, специально разводятся в водоемах, богатых лг:
чинками комаров, причем в некоторых случаях массовое заселение
гамбузией водоемов, где встречались личинки малярийного комара,
быстро приводило к уничтожению личинок и постепенному пониже-
нию заболеваемости малярией.
Отрицательное значение рыб для человека во много раз меньше
положительного. Лишь немногие рыбы используют человека в пи-
щу. прйчец во всех случаях он является для рыб случайной пищей.
Правда, у берегов Индии и в других местах тропиков, во времялов-
ли жемчуга или при промысле губок довольно большое количество
людей становится жертвой акул. В некоторых местах акулы делают
невозможным даже купание в море. Из пресноводных рыб известны
•90
Рис. 54.. Различные вариации золотой рыбки, сверху вниз: комета,
телескоп, вуалехвост, львиноголовца, золотая рыбка.
91
случаи нападения на человека крупных сомов. Так, в 1934 году в
Аму-Дарье близ города Керки сом утопил 14-летнего мальчика.
Как мы уже отмечали выше, гибель человека может быть вы-
звана поражением ядом бородовчатки Synanceia horrida. Ежегодно
у побережья Индомалайского архипелага несколько человек по-
гибают, наколовшись на шипы "этой рыбы. Вредное влияние рыб
на человека сказывается и в форме передачи глистных заболеваний,
так как рыбы являются промежуточными хозяевами многих червей,
взрослая стадия которых живет у человека.
Нрыпные\
ХИЩНИКИ \
LfiTJESHiJDROCHDH.
8PBHUS ИДР
Планк ТОНОЯДНЫЕ
РЫВЫ
EHOMUUCUPRiB hihutus
Stqlqthrissp.
НЯСЕКОКОЯДНЫЕ
РЫБЫ,
Сьялтаз,
ENBRRULlCUPRiS CONDtCVS.
CiiPRiNODnHTi^X
Растения
и пл.
\Нясеконые\
I /7j7/w*hw.|
[Моллюски]
Беитосоядные
РЫБЫ.
HoRHKRUife
CtfCNOBLRNIS,
E. Личинки
HiROHORlDOE.
Открытия область Прибреьнняя оклпсть.
Рис. 55. Схема пищевых связей в озере Танганайка, отражающая только
качественную сторону пищевых взаимоотношений (из Worthington и
Ricardo, 1936).
Примером косвенного вредного влияния может служить вред,
наносимый небольшой рыбкой из семейства Ophichthydae Phiso-
donophis boro. Эта рыбка ведет роющий образ жизни и, попадая на
рисовые поля, перекапывает грунт и выкапывает стебли риса, чем
иногда довольно сильно снижает урожай.
В заключение необходимо вкратце остановиться на той роли,
какую играют рыбы в круговороте органического вещества в водо-
еме и в жизни гидросферы вообще. Взаимоотношения между рыбой
и остальным органическим миром идут, в первую очередь, о ли-
нии пищевых связей. Эти связи могут быть весьма различны, обла-
дать большей или меньшей сложностью и в различных водоемах
рыбы в этих связях играют большую или меньшую роль. Обычно
принято различать два типа связей: 1) прямые связи, когда живот-
ное или растение питается другим животным или растением, потреб-
ляя его в живом виде, 2) косвенные, когда животное и растение по-
требляет уже не сам живой организм, а продукты его распада. В
питании рыб первая форма связи значительно преобладает над вто-
92
рой. В то же время роль рыб в пище других животных, не считая
человека, главным образом проявляется в косвенных пищевых свя-
зях.
Изучение пищевых связей в водоемах, кроме большого теорети-
ческого интереса, имеет,и важное практическое значение, так как
с
Рнс. 56. Схема пищевых связей в Аральском море, показано^соотношение
компонентов в пище и величина биомассы кормовых беспозвоночных
(из Никольского, 1940).
оно позволяет более точно подойти к вопросу акклиматизации пу-
тем выявления имеющихся в водоеме свободных биологических ниш
и тех форм пищевой конкуренции, которые имеют место в изучае-
мом водоеме. Наиболее примитивным методом установления пищевых
связей является получение качественной характеристики пищевых
взаимоотношений. Подобного типа качественные схемы получены
сейчас для очень многих водоемов (рис. 55), Дальнейшим этапом
изучения является внесение элементов количественной характери-
93
F
ДГРА*рнкР
tit
Прлгинкд
БЛУЗДНПК
Кильки
Б Ы 4 к и‘
ВОбПА
ЛЕЩ
Ordiuh
toRCNO
bifcACNA
пищи. Отмечена
MQNOBflCW
DREISSENP
связи более 25% от
связей^северном Каспии. О^ХвТз Ш°РЫГИПа’ *
Рис. 57. Схема основных^пиШевь«х RopMOBblX объектов.
стики процесса, именно: установление соотношения пищевых объ-
ектов в водоеме и в пище отдельных рыб, определение биомассы
кормовых объектов и т. п. (рис. 56). Следующая ступень изучения
пищевых взаимоотношений в водоеме —полный переход на количест-
венную методику —осуществляется тогда, когда удается определить
с той или иной степенью точности величину биомассы не только кор-
мовых объектов, но и рыб (рис. 57). Эта фаза исследования позво-
ляет подойти уже вплотную к определению биомассы водоема и,
в частности, его ихтиомассы. Наконец, дальнейшее изучение дина-
мики органического вещества в водоеме, одним из звеньев в кото-
рой являются рыбы, должно пойти по пути учета сезонной и суточ-
ной динамики всех форм органического вещества. При этом учет
этих изменений должен проводиться количественным методом, но
со строгим учетом специфики каждого отдельного звена процесса.
К сожалению, подобного рода работы еще только начинаются и пока
почти не коснулись рыб. Примером же подобного рода исследования,
проведенного на кормсвых объектах для рыб, может служить работа
Боруцксго о динамике бентоса ’профундали.
Естественно, что не все органическое вещество рыб снова возвра-
щается обратно в круговорот органического вещества в водоеме,
часть остатков рыб, иногда довольно значительная, отлагается в
виде осадков и вместе с остатками других, как животных, так и
растительных организмов, идет на образование грунтов. Скопления
значительных масс органического вещества, как животного, так и
растительного происхождения, в донных отложениях, претерпе-
вали затем изменения, которые в пресных водоемах приво-
дили к образованию бокхедов или углей смешанного происхожде-
ния, а в условиях морских глубин, особенно при наличии сероводо-
родного брожения — к образованию первичных нефтей, возникно-
вение которых стало возможным на земле только после появления
органической жизни (Стадников 1933)., Наконец, последнее время
значительное количество органического вещества извлекается из
морских и пресных вод человеком и идет уже главным образом на
построение континентальных отложений, лишь в незначительной
части в виде растворимых соединений, возвращающихся обратно в
водоемы.
ЧАСТЬ II
ОСНОВНЫЕ ЧЕРТЫ ЖИЗНЕННОГО ЦИКЛА РЫБ
МИГРАЦИИ РЫБ
Очень немногие рыбы живут постоянно на одном и том же месте,
не совершая сколько-либо значительных перемещений. Почти все
рыбы ведут более или менее подвижный образ жизни. Некоторые
в течение своей жизни совершают периодические путешествия —
миграции, другие менее подвижны, и их передвижения не носят
закономерного характера. У многих рыб пелагические икра и личин-
ки течениями переносятся иногда на огромные расстояния, совер-
шая пассивные миграции. Взрослые особи, преодоле-
вая сопротивление течения, возвращаются для размножения обрат-
но на те места, где была отложена икра, совершая активные
миграции. Ряд примеров пассивных миграций под действием
морских течений был приведен выше при разборе влияния движе-
ния воды на рыб. Подобных примеров можно привести очень много;
почти у всех морских рыб с пелагической икрой, последняя как и
только что выведшиеся из нее личинки, относятся течениями от
нерестилищ. Так же в реках, у тех рыб, икра которых не прикре-
пляется к субстрату (например у чехони Pelecus cultratus), она
постепенно сносится вниз по течению. Как пассивные миграции
можно рассматривать также и скат от места икрометания вниз по
течению рек личинок тех рыб, икра которых или приклеивается
к донным предметам, или закапывается в грунт. Скат личинок
осетровых, личинок некоторых лососевых и ряда других рыб в
значительной степени происходит пассивно.
Значительно более разнообразны у рыб активные миграции.
По характеру активные миграции можно разделить на: 1) нересто-
вые миграции, 2) кормовые, 3) зимовальные и 4) связанные с изме-
нениями внешних факторов. Первые три вида активных миграций
являются звеньями одного цикла ч неразрывно связаны одна с
другой.
Нерестовые миграции
Нерестовые миграции, связанные с переходом на места раз-
множения, наиболее сложны и разнообразны. Многие рыбы, так
называемые проходи ые, кормятся в море, а для размножения
входят в реки, совершая а на дромн ые миграции.
Анадромные миграции свойственны миногам, осетровым, лососе-
вым, некоторым сельдям, карповым и др. Другая часть проходных
96
рыб кормится в реках, а для икрометания уходит в море, совершая
катадромную миграцию. Таков угорь и др. Мигра-
ция может происходить против течения — контранатант-
н а я миграция, или по течению —д енатантная ми-
грация. У некоторых рыб нерестовая миграция бывает частью
контранатггнтной, частью денатантной. Так, например, угорь,
спускаясь по течению реки, совершает денатантную миграцию, а
двигаясь в Атлантическом океане против течения,—контранатант-
ную. Чисто морские рыбы также совершают нерестовые миграции,
или подходя с глубин к берегам, или наоборот от берегов отходя на
глубины. Многие из них совершают довольно значительные пере-
ходы на места икрометания, иногда за тысячу и более километров.
Некоторые рыбы, обычно живущие на глубинах, поднимаются
для размножения в поверхностные слои воды, другие же наоборот,
из поверхностных слоев опускаются в более глубокие. Проходные
рыбы с анадромной миграцией приурочены в основной массе к
умеренным и холодным частям северного полушария. Ни в умерен-
ных и высоких широтах Евразии, ни в Северной Америке почти
нет впадающей в море реки, куда бы не заходили для размножения
проходные рыбы.
Начало захода в реку приходится у различных рыб на разное
время года и зависит в первую очередь от времени икрометания
и от того, будет ли рыба метать икру в том же году, или будет зимо-
вать в реке и нереститься уже на следующий год. У многих рыб,
как например, некоторые осетровые, благородный лосось Salmosslar
и др. имеются озимые и яровые расы, отличающиеся по срокам
захода в реки; так, например, лосось, входящий в реки Белого
моря, представлен двумя формами: одна входит в реки осенью,
имеет слабо развитые половые продукты и мечет икру на следую-
щий год, а другая входит в реку летом, имеет хорошо развитые
половые продукты и мечет икру в том же году. Некоторые рыбы
входят в реку или только весной, как например, вобла, жерех и
др., или только летом (аральский шип), или же, наконец, только
осенью (некоторые сиги) Наблюдается также большое разнообразие
в отношении высоты подъема вверх по реке. Последняя варьирует
от нескольких километров до нескольких тысяч километров. Миноги,
входящие в реку для икрометания, как осенью, так и весной (Ивано-
ва-Берг 1936) поднимаются на различные расстояния. Невская
минога Lampetn fluviatilis высоко по реке не идет, каспийская же
минога Caspicmvz n wagneri, наоборот, поднимается по Волге
иногда выше устья Камы. Осетровые, как шип, осетр, севрюга и
белуга, могут подниматься по реке на тысячу и более километров,
доходя до мест с галечниковым дном, где они откладывают икру,
но тот же осетр в Сефидруде и Тереке не поднимается и сотни
километров от устья. Высота поднятия осетра по реке зависит от
наличия удобных для икрометания мест. Проходные лососевые
также поднимаются по рекам очень высоко. Так нерка Oncorhynchus
легка в реке Юкон поднимается вверх до 3600км, идя, как установле-
но путем поимки меченых рыб, со скоростью 30—40/си в сутки, а кета
7 Биология рыб С7
идет даже со скоростью 50кл<. Дальневосточные лососи преодолевают
по пути различные препятствия: водопады, мелкие перекаты и т. и.
Европейские лососи также поднимаются высоко в реки. Так, но
Печоре лосось доходит почти до верховья. Некоторые сельди в
большом количестве заходят для икрометания в реку,, на пример
черноспинка CaspialcSu k.sskn по Волге поднимается почти до
устья Камы, немного ниже доходит й волжская сельдь. Среди кар-
повых проходных рыб не так много, и те, которые ьходят в реки,,
обычно не поднимаются высоко вверх и по возвращении из реки
после нереста не уходят из пределов влияния пресных вед. Эту
группу рыб, куда относятся также и представители некоторых
Рис. 58. Ход усача Barbus brachycephalus (карповые)
на устьях Сыр-Дарьи (жирная линия) и с уровни воды
в русле у Казалинска.
других семейств, как например, в Каспии судак, принято назы-
вать п о л у п р о х о д н ы м и. Но и среди карповых имеются виды
типично проходные, которые совершают далекие путешествия вверх
по рек: м; так вырезуб Rutilus fr.sii входитв Днепр и поднимается
выше Днепровских порогов. Его каспийский подвид кутум подни-
мается по Куре до Мпнгечаура. Очень высоко в реки вдут некоторые
усач) ; так, усач чанари Barbus capito входит в Куру и поднимается
по пейд( Арагвы. Apaj ьскийусач Barbus brach^cej halus поднимается
по Ам)-Дарье более чем на тысячу километров и доходит до устья
Пянд жа.
Ход вверх по реке естественно требует от рыбы затраты огром-
ного количества эщргии и чем быстрее течение реки, темтруднее его
рыбе преодолевать. Поэтому естественно, что ход рыбы в реке на-
ходится в зависимости от паводковых вод. В то время как в море
паводковые воды, усиливающие влияние пресных вод перед устьем
реки, способствуют ориентации рыбы на устье реки, паводковые
воды в самой реке затрудняют движение (рис. 58). Поэтому у многих
рыб ход в реку наступает по окончании паводка (фальский усач,
дальневосточные лососевые и др.) (рис. 59 и 60).
Большинство рыб во время миграции по реке обычно прекра-
щает питание или питается менее интенсивно чем в море, а огромная
затрата энергии естественно требует расхода накопленных во время
98
кормежки В Море питательных веществ. Вот почему у
проходных рыб по мере движения их вверх по реке
сильное истощение. Так, процент содержания жира
по мере движения вверх по Амуру, изменяется
образом:
большинства
наблюдается
в теле кеты,
следующим
Расстояние от моря 0 км 221 407 570 729 11193
Процент жира у самцов 9,19 9,14 8,26 5,72 4,47 3,39
Процент жира у самок 11,23 10,85 7,70 6,58 5,99 3,04
Белорыбица при входе В Волгу имеет 21% жира (Астрахань),
а на местах нереста (Уфа) процент жира уменьшается до 2, араль-
ский усач на местах нагула в море имеет на внутренностях до 15%
жира, на местах же нереста количество жира на внутренностях
16
/
4
Янв Фев. Пярт Дпр Мдй Июнь. Июль. Явг Свит. Окт. Нояб. Ден
Рис. 59. Ход чавычи Oncorhynchus tschawytscha лососевые в р. Кламас
и изменение уровня этой реки (из Snyder, 1931).
снижается до 1—2%, а у покатных оссбей жира на внутренностях
совершенно нет. В связи с изменением характера осморегуляции
и прекращением питания в реке у проходных рыб происходит тесь-
ма сильнее изменение в величине кишечного тракта. Осебенно
резко это выражено у лососевых. Так, у чавычи Onccrhj nchus tsiha-
wytseha (рис. 60) на местах икрометания кишечник имеет вид топ-
кой нити с очень слабо развитыми пилорическими придатками.
Изменяется и характер действия ферментов кгшечника, которые
у лососевых теряют способность расщеплять белки.
Энергия, которую расходуют проходные рыбы для того, чтобы
достичь нерсстилиц, весьма велика. Так, у самцов кеты расход
энергии на килограмм живого веса равен 25 810 калорий, а у самок—
28 ЗЭОкалорий. В связи с переходом из морской воды в пресную
у проходных рыб меняется осмотическое давление и, следовательно,
температура замерзания крови. У чавычи температура замерзания
крови в море равна—0,762°, в опресненных предъустьегых участ-
ках— 0,737°, и на нерестилищах она снижается до—0,628° (Greene
1904). То, что осмотическое давление крови рыбы находится в тесной
зависимости от солености воды и изменяется в зависимости от^изме-
нения последней, подтверждается наблюдениями над каспийским
пузанком Caspiaksa caspia, у которого, как видно из приведенных
Цифр, одновременно с уменьшением солености уменьшается осмо-
7* ' 99
тическое давление
ния).
Открытое
море
крови (цифры
Опресненный
район
Вода —0,121 —0,040
Кровь —0,792 —0,747
При переходе из соленой воды
выражают понижение /° замерза-
Устье реки Нерестилища
—0,018 —0,015 —0,000
—0,731 —0,697 —0,666
у приходных рыб меняется и ха-
рактер содержащихся в тканях витаминов. Так, по данным W. Id
(1938), содержащаяся в тканях глаза проходных рыб в море форма
Рис. 60. Изменение кишечника чавычи
Oncorhynchus tschawytscha по мере хода
в реку. Наверху — на устье, посредине—
в начале хода, внизу — на нерестилище. Раз-
меры рыбы одинаковые (из Scheuring, 1929).
витамина А—Ах при пе-
реходе из соленой воды
в пресную заменяется
формой А4.
Как видно из изло-
женного, изменения, про-
исходящие в организме
проходной рыбы во время
нерестовой миграции,
чрезвычайно велики. Ор-
ганизм рыбы, как бы пре-
терпевает перестройку,
переходя от одних внеш-
них условий к другим, от
жизни в стоячей воде к
преодолению движения
текучей воды. Эти изме-
нения у многих видов
рыб столь значительны и
истощение столь сильно,
что после миграции и
нереста рыба не может
уже снова перестроиться
к морской жизни и гиб-
нет.
Какие же факторы
вызывают миграцию к
нерестилищам у проходных рыб? Непосредственным стиму-
лом нерестовой миграции является начало созревания половых
продуктов, но, как показывают наблюдения, этот фактор
связан с началом миграций далеко не у всех рыб. Многие
рыбы начинают нерестовую миграцию, когда половые железы
у них развиты еще очень слабо. Так, у самца кеты при входе
в Амур гонады составляют-всего 0,6 — 1,2% от веса тела,
в то время как на нерестилищix они достигают 8—10%. Такую же
картину мы наблюдаем у многих осетровых, у озимой расы лосося,
у аральского усача и у некоторых других проходных рыб, для ко-
торых стимулом начала нерестовой миграции несомненно не служит
созревание половых желез. Весьма возможно, что у тех рыб, кото-
рые входят в реку осенью и перезимовывают в реке перед икроме-
100
танием, стимулом миграции является достижение определенной
упитанности. Миграция у зимующих в реке рыб в своей первой
части (до мест зимовки) скорее зимовальная, чем нерестовая.
Если время начала нерестовой миграции несомненно опреде-
ляется изменениями во внутреннем состоянии рыбы (созревание
половых продуктов, достижение высокой упитанности и т. п.), то
направление миграции несомненно зависит от целого ряда гпешьих
Рис. 61. Миграционные пути сельдей, осетра и севрюги в сев.-зап. части
Черного моря (из Великохатько, 1938).
факторов и вырабатывается у рыбы из поколения в поколение,
лишь постепенно закрепляясь наследственно. Большинство лосо-
севых, как показали опыты меченья, идет метать икру в ту реку,
где они вывелись. Основным ориентиром направления движения
рыбы в море к устью реки является ток опресненных вод, идущих
от устья, и у многих рек распространяющийся очень далеко в море.
Если первоначально ориентиром служит течение речных вод, то
впоследствии направление течения может изменитьёя, ориентир ме-
няется, а выработавшийся у рыбы миграционный путь сохраняется,
иногда даже позволяя установить направление движения струй реч-
101
ной воды в предыдущие времена. Так, например, миграционный путь
основных проходных рыб, входящих в Днепр (осетр, сельдь),полнос-
тью следует тем изменениям в положении русла Днепра, какие имели
место в четвертичное время (рис. 61). Раньше зимовка проходных
рыб была расположена перед устьем Днепра, но с начала четвертич-
ного периода устье Днепра стало перемещаться вправо, образовав
ряд кос. Рыбы не изменили своего миграционного пути и сначала
заходят в Джарлыгачский залив к устью старого Днепра, а затем
уже вдоль берега движутся к современному Днепру. В то время
как устье Днепра совершило значительные перемещения, положе-
ние устья Буга осталось за четвертичное время без изменений.
Входящая в Буг севрюга не заходит в расположенные на ее пути
заливы, а прямо идет в Днепробугский лиман к устью Буга, совер-
шая значительно более короткий иугь, чем заходящие в Днепр
осетр и сельдь. Если в некоторых случаях, несмотря на значительные
изменения внешних условий, миграционные пути у проходных
рыб сохраняют прежнее направление и характер, то при более рез-
ких изменениях характер и длина миграционного пути может из-
мениться. Так, севрюга раньше поднималась по Дону значительно
выше чем теперь; лосось и белорыбица заходили в ряд притоков
Волги, куда они сейчас не заходят в результате изменения характера
паводка, последовавшего за вырубкой лесов и приведшего к унич-
тожению нерестилищ. Таким образом, здесь мы имеем примеры из-
менения миграционного путц под влиянием изменившихся внеш-
них условий.
Происхождение сложного процесса миграций у проходных рыб,
совершающих анадремную миграцию, до настоящего времени
не ясно, и имеется ряд гипотез, по-разному объясняющих их воз-
никновение. То, что почти все проходные рыбы, в своем распро-
странении приурочены к северному полушарию, делает весьма
правдоподобной связь возникновения миграций с ледниковым пе-
риодом. Пэ одной гипотезе значительное опреснение, имевшее место
в результате таяния ледников, позволило речным рыбам в целях
поиска пищи выходить из рек в более кормные опресненные районы
мора, постепенно приспособляться к жизни в соленой воде и воз-
вращаться в реку только в целях икрометания. Другая гипотеза
рассматривает проходных рыб, как генеративно морских, ставших
входить в реку постепенно и выработавших размножение в реке в
результате приспособления к пресным водам, что стало возможным
благодаря сильному опреснению морей, имевшему место по оконча-
нии ледникового периода (см. стр. 108).
Значительно реже среди проходных рыб наблюдаются примеры
кат' Доомных миграций, когда река служит рыбе местом кор-
межки, а море — местом икрометания. Наиболее сложны нере-
стовые катадромные миграции у американского и особенно
европейского угрей. Европейский угорь по достижении полово-
зрелости (на 9—12 году жизни, а иногда и позже) начинает ухо-
дить из рек, где он кормился и рос, в море. Ход угря вниз по реке
мало заметен, так как совершается обычно ночью. В море угорь
102
движется от берегов Европы к берегам Центральной Америки, г
на больших глубинах находятся места его размножения. Так ж
как и другие проходные рыбы, угорь за время путешествия от ме<
кормежки в реках Европы до мест размножения у берегов Америк
претерпевает весьма сильные изменения, которые, к сожалении
еще недостаточно хорошо изучены. К моменту выхода в море у угр
начинают увеличиваться глаза, становящиеся иногда в четыре ра?
больше чем нормальные. Одновременно заостряется рыло и изменяв'
ся окраска: спина становится темнее, а брюхо из желтого делаете
серебристым. Начинает угорь миграцию упитанным, но за врем
пути, в результате расхода огромного количества энергии, он силь
но худеет. Мигрирующий угорь не питается, и кишечник его сильн
дегенерирует. При переходе из реки в море у угря повышаете:
осмотическое давление крови. Изменяется также величина плава
тельного пузыря, который становится меньше. Стимуломнерестово!
миграции угря, повидимому, является созревание половых про
дуктов, начинающееся Уже в реке. В реке в декабре икринки угр$
имеют диаметп 0 03—Э.09 мм. в сентябре следующего года 0,\мм.
в октябре 0,16, а в ноябре 0,18—0,23, т. е. имеет место созревание,
хотя и медленное.
Причины возникновения столь сложного миграционного пути
угря до настоящего времени остаются окончательно невыясненными.
Сторонники теории Вегенера о движении материков объясняют
пути миграции угря тем, что якобы в мезозое угри нерестились во
внутреннем озере, расположенном в люм месте, где находятся сейчас
нерестилища угря. Здтем, в результате раздвигания материков,
миграционный путь все увеличивался и, наконец, достиг тех раз-
меров, которые он имепт сейчас. По мнению Экмана (1932), угорь
иД'т в направлении повышения солености и температуры. Места
икрометания угря расположены в наиболее теплых участках глу-
бинной области. Шмидт (1936) предполагает, что современная ми-
грация угря есть результат похолодания, имевшего место в ледни-
ковое время. В то время как в третичный период, по предположению
Шмидта, места нереста угря благодаря более высоким температурам
на глубинах были расположены у берегов Европы, с началом насту-
we ия ледника теплые волы и приуроченные к ним места икроме-
тания угрей были отодвинуты на запад и заняли современное положе-
ние, совместившись с местами икрометания американского угря.
Таким образом, современные места икрометания угря являются
ненормально удаленными от мест кормежки; в связи с этим сильне
растягивается и время развития европейского угря, продолжаю-
щееся значительно дольше, чем у американского, места икроме-
тания которого лежат ближе к рекам, где он кормится.
Мы рассмотрели нерестовые миграции проходных рыб,^ т. е.
тех, которые при нерестовых миграциях переходят из соленой воды
в пресную или наоборот. Но нерестовые миграции имеют место не
только у проходных рыб, они достаточно отчетливо выражены и у
морских и у пресноводных рыб. 1
Некоторые морские рыбы, как например треска, в поисках
103
пищи далеко отходят от мест икрометания, которые у основной
массы трески Варенцова моря находятся у берегов Норвегии, глав-
ным образом, в районе Лсфотенских островов. Треска же в своих
пищевых миграциях доходит на восток до Новый земли и проникает
даже в Карское море. Отсюда треска, нагулявшись, начинает дви-
гаться на запад и совершает переход в несколько тысяч километров
от мест нагула до мест икрометания (рис.62). Океаническая сельдь
Clupea harengus в целях поисков пищи также отходит далеко от
мест икрометания
и ко времени не-
реста должна, что-
бы достигнуть не-
рестилищ, совер-
шать значитель-
ный перехс д почти
от Шпицбергена
до южных берегов
Норвегии. Не-
сколько иной ха-
рактер имеет ми-
грация у камбал.
Морская камбала
Pleurcnectes pla-
tessa обычно отхо-
дит для икромета-
Рис. 62. Схема миграций трески Gadus morhua в ния от берега на
Варенцовой море (из Мейснера, 1933). значительные глу-
бины. Так, камба-
ла в Северном море размножается, главным образом, в Глубоком же-
лобе против входа в Ламанш. Сюда подходят камбалы, как от бе-
регов Англии, так и от Ламанша. В Советских водах нерестилища
камбал расположены еще на больших глубинах чемв Северном море.
Правда, расстояние нерестилищ от берега не особенно велико (Те-
риберская и Кильдинская банки). Иной характер носят миграции
некоторых дальневосточных камбал. Здесь камбалы зимуют на
больших глубинах, а для икрометания подходят в прибрежную
зону. Также различны и нерестовые миграции пресноводных рыб.
Многие пресноводные рыбы для икрометания входят из озер в реки.
Так, например, сиг Coreg<.nus lavuretus живет в Ладожском озере,
а для икрометания входит в реки. Некоторые рыбы из открытого
озера для икрометания выходят в прибрежные заросли и заболо-
ченные места. Так, щука Escx lucius во время разлива рек выходит
для икрометания на залитые луга и часто мечет икру на затопленной
растительности. Африканский чешуйчатник также уходит для
икрометания из открытых частей озер в прибрежные заросли и
болота.
Таковы основные типы нерестовых миграций пресноводных рыб.
Нерестовые миграции у пресноводных рыб значительно проще,
чем у проходных и морских. В первую очередь это объясняется,
104
конечно, сокращением самой области обитания; даже представители
одного и того же вида, живущие в меньшем водоеме, часто не совер-
шают миграций, в то время как живущие в более крупном водоеме
мигрируют. Так, например, вобла в Каспии является типичной
полупроходной рыбой, в то же время плотва, как правило, значи-
тельных нерестовых миграций не совершает. Незначительное коли-
чество пресноводных рыб с пелагической икрой объясняет мало
развитие пассивных миграций, а, следовательно, и обратных ми-
граций на места икрометания взрослых особей.
Кормовые миграции
Вторая группа миграций—кормовые миграции или миграции,
связанные с добычей корма. У многих, как проходных, так морских
и пресноводных рыб, кормовые миграции бывают выражены весьма
отчетливо. У тех проходных рыб, которые не погибают после икре
метания, обратный скат в море можно рассматривать, как миграцию
к местам кормежки. Но и по выходе в море, проходные рь.бы
продолжают передвижение, и иногда места их нагула находятся на
весьма значительном расстоянии от устьев рок, в которые они вхо-
дят для икрометания. Так, например, каспийская севрюга Acipenser
stellatus, входящая для размножения в Куру, кормгтся преимуще-
ственно близ восточного побережья Каспия. Как указывалось,
дальневосточные лососи по выходе в молодом возрасте из Амура
отходят на кормежку к островам Японии. Отчетливо выражены
кормовые миграции у морских рыб. Некоторые рыбы совершают
правильные путешествия, переходя от одних мест, богатых кормом,
к другим. Так, треска, после икрометания у берегов Норвегии,
начинает, усиленно питаясь, двигаться на восток. По окончании
нереста, треска чрезвычайно худа, и печень ее содержит ничтожное
количество жира. По мере же движения на восток треска стано-
вится, в результате усиленного питания, все упитаннее, и к концу
периода нагула она достигает максимальной упитанности и почти
прекращает питаться.
Океаническая сельдь Clupea harengus после нереста совершает
также значительную кормовую миграцию, отходя от берега и дер-
жась в местах скопления планктонных организмов, служащих ей
пищей. Миграции многих морских хищных рыб в значительной
степени связаны с перемещением объектов их питания—других
рыб. Так, сельдевая акула L. mna ccrnublca в своих перемещениях
в значительной мере связана с распространением и перемещениями
сельди, и при подходах сельди к берегу обычно сразу появляется
и большое количество акул.
Кроме горизонтальных кормовых мр граций, ряд морских рыб
совершает и регулярные вертикальные перемещения, связанные с
поисками пищи. Так, скумбрия поднимается в поверхностные слои
воды тогда, когда здесь наблюдается наиболее богатое развитие
планктона. При опускании планктона в более глубокие слои скум-
брия опускается туда же. Меч-рыба во время откочёвки основных
105
объектов своего питания (главным образом сардин и Coryphaena)
для кормежки опускается на большие глубины, где переходит на
питание держащимися здесь представите; ями Scopclidae. Интересно
отметить, что меч-рыба в отличие от многих других рыб обладает
способн >стью чрезвычайно быстро перемещаться с больших глубин
на поверхность. Многие пелагические личинки морских рыб совер-
шают суточные вертикальные кормовые миграции, следуя в своих
Рис. 63. Динамика упитанности и зрелости белоглаз-
ки с мая по ноябрь в Аральском йоре (из Николь-
ского, 1940)
перемещениях за
суточными пере-
мещениями своего
объекта питания—
планктонных ор-
ганизмов, которые
днем опускаются
глубже, а ночью
поднимаются в по-
верхностные с; ои
воды.
У рыб внутрен-
них водоемов так-
же отчетливо вы-
ражены кормовые
миграции, особен-
но хорошо замет-
ные в больших
озерах. Так, в
Аральском море
лещ и белоглазка
после икромета-
ния отходят от
берега на боль-
шие глубины, где
держатся до окон-
чания периода на-
гула. По мере
отхсда от берега и
усиленного пита-
ния упитанность этих рыб все возрастает, до тех пор пока
не достигнет своей максимальной величины (рис. 63). Тогда
лещ и белоглазка прекращают питание и начинают обрат-
ное продвижение к берегу. У многих пресноводных рыб очень
отчетливо выражены суточные перемещения. Так, многие
рыбы, держащиеся днем в русле реки и слабо питающиеся,
ночью заходят в затоны, где имеется, как правило, богатая
фауна беспозвоночных, служащих объектом и с питания. Подобные
миграции совершают ельцы, иногда пескарь, быстрянки и т. д. Сле-
дуя за мирными рыбами в заводи, на кормежку заходят и хищники:
сом, жерех и др. Так же, как и у морских рыб, и у пресноводных
бывают отчетливо выражены вертикальные кормовые миграции,
главным образом, носящие суточный характер. Так, некоторые
бычки Байкала, следующие за своим объектом питания — рако-
образными, перемещаются из одних слоев воды в другие. Чехонь в
Аральском море ночью перемещается из придонных слоев в поверх-
ностные вслед за поднимающимися к поверхности куколками дву-
крылых и бокоплавами, которыми она питается. Подобные же ми-
грации совершают в Аральском море также шемая и весной и осенью
вобла. В озерах Восточной Африки живущий на глубинах сомик
Synod mts также по ночам поднимается в поверхностные слои
воды в погоне за личинками двукрылых.
Зимовальные миграции
Тргтьей группой миграций являются миграции зимовальные.
Как мы указывали выше, осенний ход в реку озимых рас проходных
рыб можно рассматривать скорее как зимовальную миграцию, чем
как нерестовую. Многие полупроходные рыбы, не уходящие далек»
из опресненного предъустьевого района и не поднимающиеся высок»
по рекам, по окончании периода нагула подходят к устьям рек и
залегают на зиму на ямы. На таких ямах иногда скопляются многие
тысячи особей.
У морских рыб часто также бывают резко выражены зимоваль-
ные миграции. Так например, азовская хамса Engrajulis encr; sicholue
но окончании нагула на зиму выхеднт из Азсвского моря в Черное.
Стимулом начала зимовальной мгграции у хамсы, повидимому,
является достижение определенной величины упитанности, при
которой рыба прекращает i итание и начинает миграцию. У рыв
внутренних водоемов зимовальные миграции выражены обычн»
менее резко, но иногда все же бывают отчетливо заметны. Так, как
мы указывали выше, арагьский лещ, по окончании периода нагула,
начинает подхсдить к берегам, вблизи которых и зимует. Так же
как для хамсы, так и для леща стимулом начала зимовальной ми-
грации является достижение определенней величины упитанности.
Внешние факторы не служат непосредственной причиной начал»
зимовальной миграции. Даже сильное похолодание не вынуждает
особей леща, еще не достигнувших высокой упитанности и продол-
жающих питаться, начать мигрировать к берегу. В то же время
лещ, достигнувший высокой упитанности, еще до начала осениег»
похолодания начинает продвигаться к берегу. Некоторые бычки
в Б. йкале на зиму уходят в более глубокие слои воды, где держатся
до начала нерестсвсй миграции.
В:е рассмотренные нами миграции — нерестовые, кормовые
и зимовальные являются, как уже было сказано, звеньями одног»
и того же общего миграционного цикла, повторяющегося в жизни
различных рыб от одного до десяти и более раз. Как видно из из-
ложенного выше, стимулами рассмотренных миграций в перву»
очередь являются внутренние факторы, т. е. различные фазы в
состоянии рыбы — созревание половых продуктов, достижение вы-
ЮТ
сокой упитанности, исхудание после икрометания. Внешние фак-
торы могут изменять срок миграции, их направление и интенсив-
ность, но значительно реже внешние факторы являются поводом
к миграции.
Миграции под влиянием внешних факторов
Обычно изменение внешних факторов вызывает неперио-
дические перемещения. Случаи же, когда псд влиянием внеш-
них абиотических факторов рыба совершает закономерные
периодические перемещения, весьма редки. Примером миграции,
невидимому вызываемой внешними факторами, является зимняя
миграция полярной тресочки Borecg dus saida. Эта рьбха летом
у нас на Мурмане ге встречается, зимою же с понижением темпера-
туры она появляется в значите; ьнсм количестве. Привязанная к
атлантическому течению американская сельдь Менхеден Brtve-
ortia tyrannus приближается к берегам периодически в те годы,
когда струи течения подходят ближе к берегу и, когда струи отходят
от бгрега, удаляется от него.
Причины возникновения миграций
Каковы же ‘те причины, которые привели у различных групп
рыб к выработке стооь сложного биологического явления, какое
представляет миграция? Несомненно, что миграция есть приспособле-
ние, обеспечивающее в одних случаях существование особи, в дру-
гих случаях — вида. Допустим, например, что все лососи,
которые входят для икрометания в ту или иную реку, остались бы
в ней на кормежку, а не ушли бы для нагула в море; несомненно,
что очень скоро кормовые ресурсы реки были бы исчерпаны, и по-
головье стада должно было бы сократиться. Естественно, что самки,
расходующие большое количество энергии на развитие икры и обыч-
но бывающие у проходных рыб бб. ыиих размеров, чем самцы, для
обеспечения как собственного существования, так и развития икры
больше чем самцы нуждаются в усиленном питании. Это питание в
достаточном количестве они могут получить только в море. Действи-
тельно, в природе мы наблюдаем, что у многих рыб самцы или часть
самцов постоянно живут в реке, а самки ведут проходной образ
жизни. Это явление зарегистрировано у лосося,у усача и некоторых
других рыб.
Но у нас, естественно, возникает вопрос, почему тогда проход-
ные рыбы, находящие в море более благоприятные условия для
питания, не переселились полностью в море, как это сделал ряд
ныне живущих в море генеративно-пресноводных рыб, а сохранили
свои места размножения в пресной воде.
Как известно, большинство морских рыб, откладывающих донную
икру, ее охраняет. В противном случае икра очень быстро нацело
108
съедалась бы разными хищниками. Следовательно, при переносе
нерестилщ в море проходным рыбам пришлось бы охранять икру,
так как иначе она была бы съедена. Почти все проходные рыбы
<кроме миног и сельдей), как крупные животные, имеющие значи-
тельный охотничий район и совершающие большие кормовые пере-
мещения, вынуждены были бы при охране икры, в течение длитель-
ного инкубационного периода, часто голодать, а то и гибнуть от
голода. В реках же и без активной охраны развивающаяся икра
подвергается значительно меньшим опасностям, чем в море. Зака-
пывать в море икру в грунт, как это делают лососевые в реках,
очевидно нельзя из-за неблагоприятного кислородного режима
грунтов значительных глубин и постоянной подвижности гальки
прибрежной зоны.
Таким образом, переход на кормежку в море позволяет виду
значительно увеличить свое поголовье за счет часто огромных кор-
мовых ресурсов морских водоемов. Прирост поголовья стада огра-
ничивает, однако, и величина нерестовых площадей. Как указывает
Кузнецов, с увеличением концентрации гнезд на нерестилищах
лососевых свыше одного гнезда на 2 кв. м, резко возрастает процент
гибнущей икры в гнездах, что автоматически лимитирует величину
поголовья стада. Площадь, пригодная для нерестилищ в реках,
далеко небезгранична, а часто бывает и очень ограниченной.
Следовательно, у проходных генеративно-пресноводных рыб
основной причиной возникновения миграционного цикла является
недостаточная обеспеченность кормом в реках.
Для генеративно-морских рыб, ведущих проходной образ жизни,
возникновение миграции также объясняется сходными причинами:
поскольку условия развития икры в реках более благоприятны чем
в море, постольку рыбы приспособились к нересту в текучих водах.
Перейти же полностью к жизни в реке сельди преимущественно
планктоноядные рыбы не смогли из-за недостаточности в реках
планктона, хотя размеры проходных генеративно-морских рыб
обычно меньше, чем генеративно-пресноводных. Такие же генера-
тивно-морские рыбы, как налим, питающиеся рыбой, нашли для
себя в реках достаточно корма и смогли полностью переселиться в
текучую воду.
У морских и пресноводных рыб причина возникновения мигра-
ционного цикла та же, что и у проходных, так как ни одно стадо
мигрирующих рыб не смогло бы прокормиться в районе своих не-
рестилищ и вынуждено было бы или сократить поголовье, или рас-
ширить свой кормовой ареал. Последнее обстоятельство приводит
к возникновению миграции от нерестилищ к местам нагула и об-
ратно.
Если причины возникновения миграционного'цикла,как мы видим,
у большинства рыб более или менее сходны, то обстоятельства, при ко-
торых они возникают, могут очень сильно разниться. У многих
рыб (лососи, сиги) переход к проходному образу жиз|ни оказался
облегченным в результате ледникового опреснения, у других же
переход к проходному образу жизни произошел при иных условиях.
10»
Знание миграционных путей, характера и сроков миграций
имеет огромное значение для рыбного промысла, обычно добываю-
щего рыбу на ее миграционных путях. Поэтому понятно, что при
научно-промысловых исследованиях огромное внимание уделяется
изучению миграций. Наиболее простым способом изучения мигра-
ций является анализ динамики промысловых уловов. Повышение
улова на одно орудие указывает на подход рыбы к берегу и, на-
Иорот, понижение уловов — на отход. К сожалению, этот метод
Рис. 64. Различные метки для мечения рыб.
Наверху метка Гильберта и щипцы для мече-
ния. Внизу метки для мечения камбал (из Ка-
раваева, 1938 и Меек, 1916).
может дать в лучшем случае лишь общую схему миграций, вы-
яснение же путей перемещения отдельных косяков становится воз-
можным в результате мечения и обратного выпуска рыб с последую-
щим учетом случаев поимки. Кроме анализа миграционного пути,
путем мечения рыбы удается разрешить ряд других вопросов, как-
например, процент вылова, темп роста и т. д.
Д: я мечения обычно употребляются различного типа метки
(рис. 64), прикрепляемые или к челюсти или на жаберную крышку
или к плавникам. У нас в СССР наиболее распространена метка
Гильберта, прикрепляющаяся обычно при помощи ос(бых щипцов
кжаберной крышке. На метке, делающейся из нержавеющего металла,
имеется номер и название учреждения, проигзе д> вшегр меченгс.
При мечении рыба обычно измеряется, взвешивается, и от нее бе-
рется несколько чешуй для определения возраста. Мечение рыб
производится сейчас почти на всех морях. У нас в Союзе оно ведется
на севере, на Черном, Каспийском и Аральском морях, а также на
Дальнем Востоке.
11»
Форма тела * движение рыб
Как видно из изложенного, многие рыбы могут совершать пере-
мещения на 1000 км и преодолевать значительные скорости течения.
Естественно поэтому, что
у рыб должны были вы-
работаться прекрасные
приспособления к плава-
нию. Как правило, рыбы,
совершающие далекие
странствования, имеют
торпедообразное хорошо
обтекаемое тело, приспо-
собленное к рассеканию
воды. Но далеко не у
всех рыб форма тела оди-
наково приспособлена к
преодолению сопротивле-
ния при движении в
водной среде. Лучшими
мореходными качествами
обладают такие рыбы, как
пеламида PcLmys s„rda,
ставр| да Traehurus tra-
churus, кефали Mug 1, боль-
шинство акул и многие
другие, наихудшими мо-
реходными качествами об-
ладают скаты, камбалы
и т. п. (1 ис. 65.)
По форме тела рыб
Пелйиидл\
Сгявридд
Кефрль______!
Дкаля_______
Зъленухл
Мор, ерш
Мор. петух
Бычек_______
Mop. OHBHtf
Мор кярясь
Султяння
Зеленушхя
Мор. нялии
Мор. собячкр
Кямеяля_____
Севрюгя
Морской язык
Рис. 65. Мореходные качества некоторых
черноморских рыб (из Шулейкина, 1940).
можно разделить на следующие типы:
1) Торпедовидный — наиболее хорошие пловцы, обита-
тели тот щи воды. К этой группе относятся скумбрия, кефаль, сель-
девая акула, лосось и др.
2) Стреловидный, близок к предыдущему, но тело более
вытянуто и непарные плавники отодвинуты наза Хорошие пловцы,
обитатели тот щи воды: сарган, щу ка.
3) Сплющенный с боков — этот тип наиболее сильно
Bapi ирует. Обычно его принято разделять на: а) лещевидный, б) тип
рыбы луны ив) тип камбалы. По условиям обитания принадлеж; -
щие сюда рыбы также очень разнообразны; от жителей толщи воды,
как луна рыба, до придонных, как камбала.
4) Змеевидный. Тело сит ьно вытянуто, поперечный раз-
рез тела почти круглый, обычно обитатели зарослей — угри, мор-
ские иглы и др.
5) Лентовидный. Тело сильно вытянуто и (сплющено с
боков. Плохие пловцы: сельдяной король Regalecus, Trachypterus
и др.
иг
6) Шаровидный — тело почти шарообразное, хвостовой
плавник развит обычно слабо: кузовки Tetrodon, некоторые пина-
горы и др.
7) Плоский — тело сплющено дорзовентрально: различные
скаты, морской чорт.
Рис. Go. Рыбы различной формы: 1 — сарган, 2 -- скумбрия, 3—лещ, 4 —
луна, 5 — камбала, 6—угорь, 7—сельдяной король, ^--кузавок, д—скат.
Таковы основные формы тела, которые встречаются у рыб
(рис. 66), но естественно, что между этими крайними формами
имеется ряд переходов, так что иногда бывает трудно отнести
некоторых рыб к тому или другому типу. Так, например, обыкновен-
ная шиповка Cobits taenia занимает по форме тела промежуточное
место между змеевидными и ленточными типами.
112
8 Биология рыб
Рис. 67. Различные типы~ хвостов
у рыб. Сверху вниз: эпибатиче-
ский, изобатический, гипобати-
— ческий.
w
Рис. 68. Движение электрического угря при помощи анального плавника и
ската при помощи грудных плавников (из Norman. 1930).
Рис. 69. Движение трески и угря. Видно как по телу рыбы идет волна
(из Gray, 1933).
Основную роль в плавании играет все тело рыбы, совершающее
волнообразные движения. Хвостовой плавник главным образом
парализует тормозящее движение конца тела и ослабляет обратные
токи. По характеру действия хвосты рыб принято разделять на:
1) изобатический, где верхняя и нижняя лопасти
равновелики; подобный тип хвоста имеется у скумбрии, тунца, мино-
ги и многих дру-
гих; 2) э п и б а ти-
че с к и й, у кото-
рого верхняя лопасть
развита лучше, чем
Рис. 70. Схема движения
летучей рыбы при взле-
те, вид сбоку и сверху
(из Шулейкина, 1940).
нижняя; этот хвост
облегчает движение вверх; подобного рода
хвост характерен для акул и осетровых;
3) гипобатический, когда нижняя
лопасть хвоста развита больше верхней и
способствует движению вниз; гипобатиче-
ский хвост имеется у летучей рыбы, леща
и некоторых других рыб (рис. 67). У
скатов движение осуществляется путем вол-
нообразных колебаний грудных плавников
(рис. 68). У морского конька и некоторых
других рыб органом плавания служит спин-
ной плавник. У электрического угря дви-
жение осуществляется путем ундулирующих
колебаний анального плавника.
Основную функцию рулей глубины вы-
полняют у рыб грудные, но также и брюш-
ные плавники. При помсщи их же осу-
ществляется отчасти и поворачивание рыбы
в горизонтальной плоскости. Роль непар-
ных плавников (спинного и анального) у тех
рыб, у которых они не несут функции
двржетеля, сводятся (Васнецов, 1941) к содей-
ствию пов( ротам вверх и вниз и лишь от-
части к роли килей-стабилизаторов.
Для преодоления сопротивления воды
чрезвычайно важно сведение до минимума трения тела о воду. Это
достигается путем максимального сглаживания поверхности и
смазки ее соответствующими понижающими трения веществами.
У всех рыб, как правило, кожа имеет большое количество бока-
ловидных желез, которые выделяют слизь, смазывающую поверх-
ность тела и сильно понижающую трение.
Некоторые рыбы приспособились к передвижению вне воды, к
ползанию по берегу и к полету. Тропические бычки прыгуны Peri-
ophthalmus вылезают на отмель берега, где охотятся за своей
добычей. Передвигаются они при помсщи прыжков, осуществляе-
мых путем комбинированного действия парных плавников и хвоста.
Обыкновенный угорь (рис. 69) передвигается по суше путем змее-
114
образных изгибаний тела. Ползуны Anabas переползают из водоема
в водоем при помощи грудных плавников. Наиболее замечательным
приспособлением к передвижению по воздуху является приспособле-
ние к полету у летучей рыбы. У этой рыбы грудные плавники разви-
ты чрезвычайно сильно и выполняют ту же функцию, что крылья
самолета или скорее планера (рис. 70). Основным движетелем, даю-
щим начальную скорость при полете, является хвост и, в первую
очередь, его нижняя лопасть. Выскочив на поверхность воды,
летучая рыба еще некоторое время скользит по водной поверхности,
оставляя на воде кольцевые волны, расходящиеся в стороны. Уже в
то время, когда вся летучая рыба находится над водой, а в воде
остается только ее хвост, она все еще продолжает увеличивать
скорость, и прекращение нарастания скорости движения наступает
только после полного отрыва от поверхности воды. Держаться в
воздухе летучая рыба может около 10 секунд и пролетать при этом
расст(Я1пе свыше 100 м.
Как видно из изложенного, способы передвижения рыб чрезвы-
чайно разнообразны, и приспособления к различным видам передви-
жений могут вносить коренные изменения в строении отдельных
органов.
РАЗМНОЖЕНИЕ И РАЗВИТИЕ РЫБ
Размножение рыб естественно носит ряд черт, специфичных для
водных животных, черт, обусловленных жи31 ью в водной среде.
У большинства рыб оплодотворение икры происходит вне материн-
ского организма, следовательно в отличие от половых клеток боль-
шинства н; земных Ж1 вс тных неоплодотворенные яйца и спермин
должны пробыть до оплодотворения известное время вне тела роди-
тельского организма. В связи с тем, что вне организма матери опас-
ности, которым подвергается развивающийся эмбрион, значительно
увеличиваются, гибель эмбрионов также сильно возрастает. Для
обеспечения сохранения вида у рыб обычно развивается огромная
плодовитость, и;и вырабатывается забота о потомстве. В некоторых
же случаях наблюдается появление внутреннего оплодотворения и
живорождения.
Плодовитость и созревание половых продуктов
Как правило, чем больше плс довитость рыб, тем меньше у них раз-
вита забота о потомстве. К( личество икры, откладываемое раз-
личными рыбами, варьирует чрезвычайно сильно, от нескольких
штук у акул до трехсот миллионов у луны-рыбы. Наибольшей
плод витсстью отличаются морские пелагические рыбы (луна-
рыба, откладывающая до 300 миллионов икринок, морская щука
и др.). Также более плодовиты те рыбы, которые имеют плавающую
икру. Последний момент объясняется очевидно тем, что плавающая
икра подвергается большей возможности быть съеденной другими7
рыбами или выброшенной на берег, в'то время как1 донная под-
вержена меньшим случайностям. Из морских рыб наименее плодо-
8* 115
виты донные малоподвижные, откладывающие донную же икру,
которую они, как правило, сами охраняют (например пинагоры,
бычки и др.). Количество икры у пресноводных и проходных рыб
в среднем ниже, чем у морских.
У одного и того же вида число откладываемых икринок под-
вержено также чрезвычайно сильным изменениям и в первую очередь
варьирует в зависимости от размеров и возраста рыбы. У большин-
ства рыб количество икринок с возрастом сначала постепенно
увеличивается, а затем с началом старения особи начинает обычно
снова уменьшаться (см. табл.) Правда, большинство наших про-
мысловых рыб не доживает до такого возраста, чтобы начался про-
цесс старения особи, а погибает или бывает выловлено.
Возрастные изменения числа икринок у различных рыб
Севрюга Acipenser stellatus Pall, (по Державину, 1922)
Длина в см................ 100-110—120—130—140—150—160—170—180—190
Число икринок (в тысячах) 40 56 78 107 126 157 179 220 254 258
Семга Salmo salar L. (по Бергу, 1935)
Длина в см . . . . . 50 — 60 — 70 — 80 — 90 — 100
Число икринок (в тысячах) 5,9 9,4 12,2 14,0 19,4
Сазан Cyprinus carpio L. (по Никольскому, 1934)
Длина в см............ . 35 — 40 — 45 — 50 — 55 — 60 — 65 — 70
Число икринок (в тысячах) 181 229 375 428,5 550 525 525
Судак Lucioperca luciopeica (L.) (по Чугуновой, 1931)
Длина в см . . . . .... 30 — 40 — 50 — 60 — 70 — 80
Число икринок (в тысячах) 203 331 487 685 851
Из приведенных примеров видно, что у всех рыб по мере роста
наблюдается увеличение числа икринок, причем момент, когда
начинается старение особи, заметен только у сазана, у которого,
начиная с размеров в 60 слг, число икринок становится меньше.
Определение плодовитости производится обычно следующим
образом: после того, когда рыба измерена и взвешена, у нее выни-
маются ястыки (яичники), взвешиваются, и от них берется навеска
в 1 или 5 г (в зависимости от размеров икры). Просчет числа икри-
нок в навеске ведется под лупой. Полученное число умножается
на вес ястыкив. Если икра в различных частях ястыков не одина-
кова по размерам, то надо брать несколько навесок из различных
участков ястыка и выводить из них среднее. Если у рыбы икра
различных размеров, то просчет надо вести отдельно крупной и
отдельно мелкой икры (см. инструкцию ВНИРО).
Размеры зрелых икринок, так же как и плодовитость, подвер-
жены у рыб сильным изменениям: от долей миллиметра у некоторых
сельдей и камбал до 100 и более миллиметров у акул и химер.
Размеры зрелой икры у некоторых рыб
Миксина Myxine glutinosa . . . 17—30 мм
Пилохвост Pristiurus melanostomus 65 мм (без рог)
Морской пес ScyHium canicula ... 65 мм f> *
Полярная акула Somniosus microcephalus 80 мм
116
Стерлядь Acipenser rufhenus ! ,3—2 мм
Калуга Huso dauricus 3,2—4 мм
Сельдь Clupea harengus 0,9—1,5 мм
Горбуша Oncorhynchus gorbuscha 4,5—6,5 мм
Сазан Cyprinus carpio 0,9—1,2 мм
Песчанка Ammodytes tobianus 0,7—1,0 мм
Окунь Регса fluviatilis 2,0—2,5 мм
Обычно у рыб, имеющих плавающую икру, она меньших раз-
меров, чем у рыб, у которых икра прикрепляется к донным пред-
метам или закапывается в грунт.
У многих рыб икра, находящаяся в ястыках, неоднородна по
размерам. В некоторых случаях это объясняется неодновременно-
стью и растянутостью сроков икрометания. При растянутом нересте
более.мелкая икра обычно располагается или в головной части
яичника или вблизи его стенок. Но не всегда более мелкие
икринки, находимые в яичнике рыбы, указывают на порционность
икрометания. У многих рыб мелкая икра остается после икромета-
ния в яичниках и постепенно рассасывается. Интересно, что процент
мелких рассасывающихся после нереста икринок подвержен у не-
которых рыб довольно сильным возрастным изменениям. Так, на-
пример, для сазана, как видно из ниже приводимой таблицы, у
более молодых и у более старых особей процент мелкой «остаточной»
икры больше, чем у рыб среднего возраста.
Изменение процента мелкой икры (от общего числа икринок)
в зависимости от размеров сазана
Длина рыбы 35 — 40 — 45 — 50 — 55 — 60 — 66
Процент мелкой икры 24,20/ 21,5% 16,1% 15,8% 12,9% 34,7%
Соотношение полову рыб довольно разнообразно.
У большинства оно обычно близко один к одному, но, например, у
обыкновенного окуня наблюдается резкое преобладание числа самок
над числом самцов, и в некоторых водоемах самцы составляют
только 10% от всего стада окуня. Наоборот, у многих лососевых
самцы несколько преобладают над самками. Вообще соотношение
полов у рыб не является чем-то совершенно постоянным и меняется
в зависимости от ряда факторов. Так, например, при спаривании
молодых самок с более старыми самцами у некоторых живородящих
рыб из семейства Poecilidae в потомстве получалось резкое пре-
обладание самок, и наоборот, при оплодотворении старой самки
молодым самцом, наблюдалось сильное преобладание самцов. По-
видимому подобное явление не специфично для указанных рыб, но
свойственно и другим группам.
Как правило, большинство рыб раздельнополы, и гермафроди-
тизм у них встречается лишь как аномалия у отдельных особей.
Однако, у некоторых видов гермафродитизм—постоянное явление:
морской окунь Serranus scriba, Pagellus mormyrus и некоторые
другие рыбы—гермафродиты. Обычно половая железа'у рыб, являю-
щихся гермафродитами, выделяет попеременно то мужские, то
женские половые продукты. Таким путем устраняется возможность
117
самооплодотворения. Исключение составляет морской окунь Ser-
ranus scribu, у которого разные части половой железы одновременно
могут выделять и мужские и женские половые продукты.
Время наступления половой зрелости у рыб весьма различно.
Некоторые бычки Aphya, Crystallogobius, карпозубые Cypri-
nodontidue, некоторые Scopelidae и др. становятся половозрелыми,
еще не достигнув годовалого возраста. С другой стороны, некоторые
осетровые: ;белуга Huso huso, калуга Huso dauricus созревают в
Рис. 71. Спинные плавники самца и самки Labeo dero (карповые)
и грудные плавники тибетского гольца Nemachilus stoliczkai
(вьюновые) (из Нога, 1936 и Берга, 1933).
возрасте 15—20 лет. Обычно средние размеры рыбы при,достижении
половсзрелости в два раза меньше средней максимальной длины
(Fulton,1906 г., Дрягин, 1934). Время наступления половой зрелости
у одного и того же вида подвержено очень сильным колебаниям.
Время созревания зависит как от внешних, так и от внутренних
факторов. Сильно сказывается на времени наступления полово-
зрелости температура окружающей среды. У большинства широко
распространенных видов половозрелость наступает в высоких ши-
ротах позднее, чем в низких. Так, например, плотва в Финляндии
становится половозрелой по достижении 5—6 лет, в средней части
Европы — 4—5 лет, а в южной части Европы по достижении 3 лет.
Оказывает влияние на срок наступления половозрелости и наличие
достаточного количества корма. Например, аквариумная рыбка
Barbus conchonius при интенсивном кормлении уже на следующий
год по выходе из икры становится половозрелой. При недостаточ-
ном же кормлении этотвид созревает к 2, а иногда и 3 годам. Прежде-
временное наступление половозрелое™ может быть вызвано так-
же путем инъекции различных гонадотропных веществ (гормоны
некоторых желез внутренней секреции; см. ниже).
С началом созревания половых продуктов у многих рыб про-
является половой диморфизм, который наиболее часто выражается
в различной величине самцов и самок. Характер вторичнополовых
признаков у различных групп рыб весьма разнообразен. У карпо-
вых самцы обычно несколько меньше самок, обладают относительно
большими, как парными, так и непарными плавниками (рис. 71).
118
У многих вьюновых Cobitidae на грудных Плавниках самцов,между
лучами, образуются вздутия, также несколько вздуваются и щеки
у этих рыб. У некоторых бычков, например у Cotticomephorus
самцы значительно крупнее самок и имеют увеличенные грудные
плавники. У самцов меченосца Xiphophcrus из Poecilidae хвосто-
вой плавник имеет длинный вырост, отсутствующий у самок. Но
наиболее резкая раз-
ница между самцами и
самками наблюдается
у глубоководных рыб
из семейства Ceratiidae.
У некоторых из этих
рыб самцы во много раз
меньше самок и пара-
зитируют на их теле,
питаясь их соками
{рис. 72). Вторично-
половые признаки, раз
появившись у рыбы,
сохраняются в течение
всей ее жизни, оста-
ваясь обычно почти не-
изменными.
Наряду с вторично-
половыми признаками
у многих рыб ко вре-
мени размножения по-
является «брачный на-
ряд». Особенно резко
«брачный наряд» выра-
жен у лососевых. Про-
исходящие у этих рыб
Рис. 72. Паразитические самцы у Ceratioidei*
Наверху Edriolychnus schmidtii. У самки ниж"
няя часть жаберной крышки отвернута и видно’
что самец прирос к телу самки. Внизу самцы
Ceratias, приросшие к брюху самки.
в период размножения изменения выражаются не только в окраске,
которая становится значительно ярче, причем в ней появляются
отсутствовавшие ранее цвета, но одновременно изменяется даже
скелет челюстей. Последние выгибаются и на них вырастают большие
зубы (рис. 73). У карповых брачный наряд обычно выражается в
появлении более яркой окраски и своеобразных бугорков из рого-
вого эпителия на голове и на теле (рис. 74). Весьма сильные изме-
нения в окраске наблюдаются у самцов некоторых бычков Gobius.
Созревание половых продуктов после нереста или у молодых
особей различных групп рыб идет весьма неодинаково и зависит
от целого ряда причин. Знание характера созревания половых
продуктов у рыб имеет, кроме теоретического, и весьма важное
практическое значение, позволяя ориентироваться в сроках и осо-
бенностях размножения, а также в некоторых случаях выяснять
расовую принадлежность отдельных стад рыб. Для удобства анали-
за процесса созревания половых продуктов и разграничения от-
дельных стадий полового цикла в настоящее вреМя разработана
119
целая серия шкал зрелости применительно к различным группам
рыб (для тресковых по Паульсену, для камбал по Гейнке-Майеру,
для каспийских сельдей по Киселевичу, для осетровых по Недоши-
вину, для карповых и окуневых по Кулаеву и Мейену и многие
другие) (см. инструкцию ВНИРО, 1938). Эти шкалы стремятся
подчеркнуть специфические черты в ходе созревания половых про-
дуктов у данной группы рыб, но все они в основу кладут уни-
Рис. 73. Брачные изменения у горбуши Oncorhynchus gor-
buscha (лососевые), внизу лошалый самец, наверху гор- ।
буша без ^брачных изменений.
версальную шкалу, являющуюся простейшей из применяемых
сейчас шкал и дающую только в общих чертах представление о ходе
созревания половых прсдуктсв. Эта шкала имеет следующий видх
I стадия — молодые особи еще ни разу не размножавшиеся.
II стадия — покоя. Половые продукты или еще не начали развиваться или
уже выметаны, воспалительный процесс у полового отвер-
стия окончился, половые железы очень малого размера,
икра простым глазом не заметна.
HI стадия — созревания. Икра заметна простым глазом, наблюдается чрез-
вычайно быстрое увеличение веса половых желез.
Молоки из прозрачных становятся бледно-розовыми.
IV стадия— зрелости. Икра и молоки созрели. Половые железы достигают
максимального веса, но при легком надавливании
половые продукты еще не вытекают.
V стадия — размножения. Половые продукты вытекают уже при самом легком
поглаживании брюшка. Вес гонад от начала икромета-
ния до его конца быстро уменьшается.
VI стадия—выбоя. Половые продукты выметаны и половое отверстие воспале-
но. Половые железы в виде спавшихся мешков, обычно у
самок с единичными оставшимися икринками, а у самцов
с остатками спермы.
12*
Такова общая схема, в которую для рыб с различным характером
нереста отдельными исследователями внесены соответствующие из-
менения и дополнения. Одним из луч-
ших показателей состояния половых
продуктов служит их вес. Поскольку
вес гонад естественно находится в тес-
нейшей зависимости от размеров
рыбы, для устранения влияния веса
тела рыбы обычно принято пользо-
ваться так называемым коэфициентом
зрелости, т. е. выражать вес гонад в
процентах от веса тела. Как видно из
приведенных на рис. 75 трех кривых
хода изменения коэфициента зрелости
у окуня, воблы и сазана, у всех этих
рыб имеются специфические черты
Рис. 74. Голова самца кутума
Rutilus frisii kutum (карповые}
в брачном наряде.
в ходе нарастания веса
гонад, не говоря уже о сроках наступления соответствующих
стадий. У воблы, имею-
щей, также как и окунь,
короткий нерестовый пе-
риод, стадия покоя про-
должается несколько
дольше, чем у окуня,
кроме того зимний подъем
идет не столь круто.
Рис. 75. Ход изменения коэфициента зрелости яичников у воблы, сазана и
окуня (справа пунктир) и семенников окуня (сплошная линия) в течение года
(из Никольского, 1940 и Мейснера, 1933).
Сазан резко отличается как от окуня, так и от воб^ы, пологостью
нисходящей части кривой. Это явление всецело объясняется растя-
нутостью периода икрометания сазана. Иной характер носит
121
динамика коэфициента зрелости у тропических рыб, размножаю-
щихся в течение почти всего года. У них икрометание — обычно
порционное, и колебание размеров гонад (например, у Tilapia escu-
knta из озера Виктории) в течение года сравнительно невелико.
У большинства рыб северного полушария максимум веса гонад
приходится на весенние месяцы. Летом коэфициент зрелости имеет
наименьшую величину, с осени же начинается подъем.Иная картина
наблюдается у рыб с осенним и зимним икрометанием (большинство
лососей и налим). У них максимальная велична коэфициента зре-
лости приходится на осенние месяцы.
Процесс созревания половых продуктов у различных рыб идет
также весьма разно. У карповых, как мы указывали, вторая стадия
продолжается довольно долго (у плотвы май— август). В это время
в половых железах наблюдаются у самок только овогонии, а у
самцов—сперматогонии. С начала осени у карповых наступает
быстрое развитие гонад и появление сперматоцит первого и второго
порядка. На этой стадии обычно карповые и зимуют. Этот момент
на кривых динамики коэфициента зрелости соответствует участку
медленного подъема. Окончательное созревание происходит в очень
короткий срок за две—три пятидневки до начала нереста.
Иная картина наблюдается у окуневых.У обыкновенного окуня
овогонии и сперматогонии очень скоро переходят в сперматоциты.
Начало появления сперматоцит у окуня приходится на июль.
В отличие от карповых у окуня заканчивается в основном сперма-
тогенез еще осенью, и зимовка уже происходит при наличии в семен-
никах сперматид, а иногда даже в значительном количестве и спер-
матозоидов. Женские половые продукты, как мы видели по весу,
несколько запаздывают в развитии, и основное нарастание веса
происходит перед нерестом, когда появляются первые генерации
новых овогоний.
Так же, как это отмечалось в отношении наступления половозре-
лости, и на созревание половых продуктов оказывают сильное
влияние различные факторы: температура, свет, интенсивность
потребления и характер корма и т. д. Почти у всех наших пресно-
водных рыб время начала икрометания находится в непосредствен-
ной зависимости от температуры воды (рис. 76). Как видно из при-
веденного на рис. 76 графика, наступление нереста у ряда рыб при-
ходится на время подъема температуры воды, и, напротив, падение
температуры приводит к понижению интенсивности размножения.
Сильно ускоряется созревание половых продуктов путем введе-
ния различных гонадотропных веществ, например гравидана—гор-
мона гипофиза и др. В настоящее время метод внутричерепных
инъекций вытяжки из гипофиза, детально разработанный у нас
Гербильским, применяется при рыбоводных мероприятиях, главным
образом, для стимулирования текучести икры осетровых рыб и су-
дака.
Насколько велико стимулирующее действие экстракта гипофиза
(гипофиз брался тоже от рыб, применение гипофиза от других по-
звоночных эффекта не дало), указывает хотя бы то, что у бразильского
122
сома Francisconius marmoratus через 10 дней после инъекции вес
гонад у подопытных рыб был на 370% выше, чем у контрольных.
Введение гонадотропных гормонов отражается и на развитии у рыб
Рис. 76. Ход температуры воды и сроки нереста у различных рыб в Цари-
цынских прудах в 1926 и 1927 годах (на оси ординат градусы С) (из Никитин-
ского, 1928).
внешних половых признаков. Так, введение гормона гипофиза
вызывало усиленный рост яйцеклада у горчака Rhode us sericeus.
Нерес т
Время и характер нереста у различных групп рыб чрезвычайно
разнообразны. То и другое зависит, как от наследственных факто-
ров, так и от влияния внешних условий. По времени нереста рыбы
могут быть разделены на три группы. Первая — весеннемечущие,
куда относится большинство рыб умеренных широт: осетровые,
карповые, сомовые и др. Вторая — осенне-зимнемечущие. Сюда
принадлежат, главным образом, рыбы северного происхождения, в
наших водах большинство лососевых и налим. Наконец, существует
третья группа рыб, икрометание которых не приурочено к строго
определенному времени года. Сюда относятся, главным образом,
представители тропической ихтиофауны, живущие в водоемах,
где температура воды колеблется в течение года очень мало, напри-
мер, многие Cichlidae.
Варьирует время нереста и в течение суток. У некоторых рыб,
как сазан, лещ, многие Characinidae и другие, икрометание обычно
происходит в утренние часы и к полудню уже заканчивается.
123
140
120
IUD
за
ва
40
20
о
OJ 0.2 0.3 0.4 OS OS 0.7 03 03 4/7 // 1.3
Рис. 77. Ряд распределения по размерам
икры в яичниках океанической сельди.
Отчетливо видны две размерных группы —
одна 0,1 - 0,2 лш и другая 0,5— 1,2 мм
(из Hiding и Rutenberg, 1936).
У других видов, как, например, налим и некоторые сиги, икромета-
ние, наоборот, приходится на вечер и ночь.
Продолжительность периода икрометания у различных рыб
также далеко не одинакова. В короткий срок (одно утро) нерестуют
в наших водах вобла, окунь и некоторые другие рыбы. Многие
тропические рыбы, как, например, любопытные характерные своей
«ротовой беременностью» Apcgcnidae, выметывают икру в течение
одного часа. С другой сто-
роны, даже у тех рыб, у
которых нет резко выра-
женного порционного ик-
рометания, как лещ и
сазан, у одной и той же
особи нерест может про-
исходить несколькими
приемами в продолжение
ряда суток. Так, особи
морского леща на нере-
стилищах держатся в зна-
чительном количестве до
10 дней каждая, наиболь-
шее же количество до 5 дней; сходная же картина наблюдается и у са-
зана, но у сазана икрометание еще больше растянуто, и пребывание
отдельных особей в целях икрометания на нерестилищах длится
дольше, чем у леща. У рыб с порционным икрометанием, т. е. тех,
у которых икра созревает отдельными порциями, общая продолжи-
тельность нереста естественно значительно больше. В этом случае
мы как бы имеем несколько нерестов, отделенных друг от друга
промежутками той или иной величины. Как число, так и величина
промежутков между выметыванием отдельных порций икры, могут
очень сильно гарьировать. Так, для уклеи Alburnus alburnus устано-
влено Дрягиным 5 порций икрометания,происходящих в следующие
сроки:
1 порция 2 порция 3 порция 4 порция порция
1936 20-30/V 31/V-10/VI 11-21/VI 22/V1-2/VII 3/VII
1937 15/V-4/VI 6- 15/VI 16-26/VI 27/V1-7/VII 9/VI’
У сельдей, как океанической Clupea harengus, так и каспийских
рода Caspialosa, число порций выметываемой икры обычно две или
три. Для океанической сельди это отчетливо показано путем анали-
з размеров икринок. Из приведенной на рис. 77 кривой хорошо
видно наличие двух резко различающихся по размерам порций
икры, соответствующих двум срокам нереста. У каспийских сель-
дей число порций откладываемой икры обычно равно трем, причем
промежутки между выметыванием отдельных порций обычно бы-
вают около полутора-двух пятидневок.
Чрезвычайным разнообразием отличаются и места размножения
рыб — нерестилища. Рыбы откладывают икру, начиная от зоны
приливов и отливов, где нерестятся, например, пинагоры Cyclopte-
124
Рис. 78. Птеригоподий
(совокупительный орган)
ската (из Kyle и Ehren-
baum, 1927).
повысить выживаемость
rus lumpus, американская атерина Laurestes tenuis и многие другие,
и кончая глубинами 500—1000л/5 где размножаются многие Ceratiidae,
некоторые камбалы и другие рыбы. Многие рыбы мечут икру на
очень быстром течении, или прикрепляя ее к камням, или пряча
между камнями, как, например, осетровые Acipenseridae, или
закапывая в гальку,как лососевые Salmonidae. Некоторые прячут
икру в естественные углубления в камнях (некоторые сомы). Иные
после икрометания вынашивают икру на брюшной стороне тела.
Некоторые рыбы откладывают икру в местах,' где теки воды со-
вершенно отсутствуют, например на глубинах. В этих случаях
икра развивается все время при постоян-
ных условиях окружающей среды. Варьи-
рует также чрезвычайно сильно и соле-
ность на местах икрометания различных
рыб. Различные рыбы, как известно, ме-
чут икру или в пресной или в соленой
воде, причем, в то время как для поло-
вых продуктов одних видов высокая соле-
ность губительна, для других она являет-
ся оптимальной. Так, у воблы леща и
сазана, как видно из приведенной табли-
цы (см. ниже),, уже при 12°/00 солености
процент оплодотворенных икринок очень
невелик. Но и на один и тот же вид
рыбы соленость действует в различных
условиях неодинаково. Комбинация воз-
действия солености и различной темпе-
ратуры показывает, что одна и та же со-
леность при различных температурах
действует по-разному. Путем постепен-
ного повышения солености, производи-
мого в течение длительного времени,
удается в некоторых случаях значительно
икры. Так, например, в то время как в дельте Волги икра сазана
остается неоплодотвореиной на 80% при 1С°/00, в Аральском море
при 1 С°/оо она развивается норма; ьно.
Процент оплодотворенных икринок при различной солености (по Ивлеву, 1940 г.)
Соленость 0%о I 2 3 4 5 6 7 8 9 10 П 1?
Вобла 82,6% 82,7 79,1 83,2 73,2 73,1 66,0 63,0 55,2 42,1 34,8 25,9 14, 1
Лещ 89,3% 91,6 91,5 82,0 84,1 72,7 53,6 21,0 О О О О О
Сазан 72,2% 71,9 71,3 72,3 65,4 62,6 58,5 45,1 45,9 27,0 12,5 О О У
У некторых рыб икра не подвергается при оплодотворении воздей-
ствию! нешних условий, так как оплодотворение происходит внутри
тела материнского организма, и икра развивается также внутри тела
матери. Внутреннее оплодотворение мы находим у большинства
акул и скатов, причем у этих рыб даже в том случае, если откла-
дываются яйца, все же происходит внутреннее оплодотворение.
У самцов акул и скатов развивается специальный копулятив-
ный аппарат, образующийся из видоизмененных крайних внутрен-
125
них лучей брюшных плавников, эти лучи увеличиваются в размерах,
ебразуя на каждом брюшном плавнике массивный вырост, на внут-
ренней стороне которого имеется желобок (рис. 78). При спаривании
самец складывает оба выроста вместе и вводит их в клоаку самки.
Интересно, что у наиболее древних ископаемых акул — совокупи-
тельного аппарата, птеригоподиев, видимо, не было, и, вероятно,
оплодотворение было наружным. Нет птеригоподиев и у современ-
ной полярной акулы Somniosus niicrocephalus, но это несомненно
явление уже вторичное. Среди костистых рыб также имеется боль-
шое число живородящих форм. В богатом видами отряде Cyprino-
dontiformes значительная часть видов производит на свет живых
детенышей. Копулятивным органом служат у этих рыб видоизме-
ненные лучи анального плавника (рис. 79) (обычно третий и четвер-
Рис. 79. Гоноподий (совокупительный орган) гамбузии (из Линд-
берга, 1934).
тый),которые превращены в весьма сложно устроенный копулятивный
орган—гоноподий. Интересно, что у живородящих рыб этот орган
приобретает подвижность и может поворачиваться вперед и назад.
Живорождение наблюдается также у некоторых других кости-
стых рыб. Так, из представителей нашей фауны живородящим яв-
ляется морской окунь Sebastes mannus, причем интереснр, что у
него в яичниках сразу может находиться огромное количество яиц,
достигающее по Эренбауму (1936) до 150 000, но одновременно ока-
зывается оплодотворенной и развивается лишь незначительная их
часть (до 1000 и редко более). Как происходит оплодотворение у
морского окуня,точно неизвестно, ио всяком случае внешних по-
ловых органов у него нет. Также живородяща наделяющая литораль
наших северных морей бельдюга Zoarccs v.Viparus. Число эмбрионов
у этой рыбки редко превышает ЗОЭ штук. Встречаются примеры жи-
ворождения и у некоторыхдругих рыб (С) m;.tcg; ster. D. tri ma и др.).
Сам процесс оплодотворения икры связан у большинства рыб с
брачными играми и иногда бывает весьма слсжным. У дальнево-
сточных лососей имеют место драки между самцами из-за самки.
У этих рыб самка выкапывает движением хвоста в гальке ямку и
откладывает туда икру, которая в это же время поливается моло-
ками одним из самцов, отгоняющим обычно других, более слабых,
держащихся невдалеке и все время стремящихся принять участие
в нересте. Весьма сильные бои за обладание самке й происходят у
лабиринтовых рыб; у бойцовой рыбки Betta spkndins драки ме-
жду самцами достигают такой силы, что часто один самец убивает
1261
другого. В юго-восточной Азии, где водятся эти рыбки, их держат
в аквариумах и устраивают между самцами бои, так же, как эта
делается с бойцовыми петухами. Очень многие представители се-
мейства Cichlidae также устраивают во время икрометания бои за
обладание самкой, причем у этих рыбок бои часто кончаются ги-
белью одного из дерущихся самцов. Сперматозоид проникает в
икринку через особое отверстие —микропиле. Обычно у большин-
ства рыб микропиле бывает одно, но для осетровых указывается на-
личие нескольких микропиле, следовательно, у осетровых возмож-
но проникновение в икринку сразу нескольких спермиев.
После оплодотворения происходит отставание оболочки от ик-
ринки, что делает невозможным дальнейшее проникновение в яйце
спермиев. Неоплодотворенная икра, пробыв сравнительно непро-
должительное время в воде, также теряет способность к оплодотво-
рению из-за отставания оболочки. Теряют оплодотворяющую спо-
собность и спермин. Так, у волжской сельди Caspialcsa voljensifc
через минуту после помещения спермиев в воду, сходную с водой
нерестилища, сохраняло подвижность 10% спермиев, а через 3*
минут двигались лишь единичные спермии (Строганов, 1938 г.).
Партенсгенетически в естественных условиях рыбы не размно-
жаются, но искусственно партеногенез иногда удается вызвать.
Правда, развитие идет обычно очень недолго (опыты ставились на
леще, окуне, ерше и других рыбах) и лишь в редких случаях до-
ходит до выхода малька. Во всех случаях, даже когда удается по-
лучить малька, развитие проходит резко отлично от нормы, причем
основные паталогические изменения имеют место в ходе дробления.
Причины, которые могут вызвать партеногенетическое разви-
тие, еще недостаточно ясны. Повидимому, они кроются во внешних
факторах, возможно в изменении количества кислорода или актив-
ной реакции среды.
В связи с наружным оплодотворением рыб у них довольно лег-
ко производится искусственное осеменение икры. Из всех способов
искусственного осеменения наиболее распространен сейчас в прак-
тике так называемый сухой способ, разработанный русским рыбо-
водом Врасским в 1854 г Заключается этот способ в том, что икра
из самки выдавливается не в воду, а на сухую тарелку и, лишь бу-
дучи полита молоками, погружается в воду. Значительно труднее
искусственное осеменение живородящих рыб. В настоящее время
метод искусственного осеменения разработан только для живородя-
щих представителей сем. Poecilidae. При помощи капиллярной
пипетки самке этих рыбок вводится в половое отверстие сперма,
получаемая от самца, так же как и у икрометных рыб, путем лег-
кого надавливания на брюшко. Сперма вводится самке вместе с
раствором Рингера.
У некоторых рыб, как мы указывали, например у лососевых,,
процесс икрометания связан с псстрсйксй гнезда. Гнездо у этих рыб
устроено наиболее просто и имеет вид ямки. Иногда же |оно представ-
ляет собою весьма сложную конструкцию. Так, макропод Масго-
pcdus opercularis откладывает икру в устроенное на поверхности
127
воды гнездо из пены; в этом гнезде икринки охраняются самцом.
Такое же гнездо строят и некоторые другие лабиринтовые. Инте-
ресно, что пузырьки воздуха, из которых строится гнездо, обвола-
киваются более прочной чем вода пленкой, выделяемой рыбой изо
рта. Пленка предохраняет пузырьки от лопанья.
Весьма сложное гнездо устраивают колюшки Gasterosteus и
Pungitius.
У этих рыб самец строит из обрывков растений шарообразное
полое гнездо, куда самки откладывают яйца, после чего самец
остается у гнезда
и его охраняет.
Среди многих отрядов рыб имеются пред-
ставители, вынашивающие на своем теле
икру во время ее развития после оплодотво-
рения. Довольно распространено среди рыб
вынашивание икры в ротовой полости. Это
приспособление конвергентно развивается в
ряде групп, например, у сомовых и колю-
чеперых.
Рис. 80. Брюшной плав-
ник сома Galeichthys fe-
lis, слева самца, справа
самки (из Lee, 1937).
У американского морского сома Ga-
leichthys felis самец вынашивает в ротовой
полости до 50 яиц. Во время вынашива-
ния икры самец, видимо, совершенно не пи-
тается. Интересно, что у
самок многих сомов, у ко-
торых самцы вынашивают
икру в ротовой полости,
происходят своеобразные
изменения в строении брюш-
ных плавников, превра-
щающихся в своеобразный
ковш, приспособленный к
сохранению яиц в одной
кучке в момент оплодо-
творения и до того времени,
покуда они не будут подо-
браны самцом (рис. 80). От-
метавшие икру самки поеда-
ют те яйца, которые не
успеет подобрать самец.
У видов рода Tilapia, ко-
торые также вынашивают
свою икру в ротовой поло-
Рис. 81. Самец Apogon semilineatus (оку-
необразные) с икрой в ротовой полости.
Наверху отдельные икринки на разных
стадиях развития (из Шмидта, 1935).
сти, этим занимается самка.
Самки некоторых видов метут забирать в рот более 100 икри-
нок, причем приводят их в ротовой полости в движение (оче-
видно для лучшей аэрации) до момента выхода малька из
икринки. Инкубационный период у некоторых видов, по наблю-
дениям в аквариуме, длится от 10 до 15 дней. В это время
самка почти не питается. По выходе из икры мальки также
128
некоторое время при малейшей опасности прячутся в рот ма-
тери. Вынашивают икру в ротовсй полости также маленькие мор-
ские рыбки родов Apcgcn и Che li de di pte rus, причем у Aptgcn вы-
нашивают икру в ротовой полости как самки, так и самцы. Забира-
емая в рот икра образует плотную массу, ибо икринки скрепляются
между собою отходящими от них тонкими роговидными ниточками
(рис. 81). Ротовая полость рыбы бывает настолько заполнена икрой,
что часто рот остается полуоткрытым. Развитие такого своеобраз-
ного приспособления, как сротовая беременность» у рыб вызван»
не только необходимостью лучшей охраны икринок от опасности,
но и тем, что, находясь в ротовой полости, икринки регулярно омы-
ваются током свежей воды, проходящей к жабрам. Некоторые ры-
Рис. 82. Самец морской иглы Syphonostoma typhle с яйцевым мешком. На
нижнем рисунке мешок раскрыт и видна икра (из Берга, 1933).
бы вынашивают икру не в ротовой полости, а на теле. Так, мы упо-
минали уже выше, сома Aspredo, у которого икра вынашивается
на брюшной стороне тела. Любопытное приспособ, ение к вынашива-
нию икры имеется у пучкожаберных SyngnathifcrnKS. У самцов
морских игл на брюшной стороне тела имеется яйцевой мешок, об-
разованный двумя складками кожи на боках тела. Эти складки за-
гибаются на брюхо и прикрывают икру (рис. 82). У морского конь-
ка приспособление к вынашиванию идет еще дальше, у него края
яйцевого мешка, расположенного также на брюхе, после наполне-
ния икрой (икру откладывают обычно несколько самок) плотно
зарастают, внутренние стенки камеры покрываются густым слоем
кровеносных сосудов, при помощи которых, видимо, икринкам
приносится необходимое количество кислорода.
Развитие икры
Не только величина, по и строение икры у различных
видов рыб чрезвычайно разнообразно (рис. 83), причем, как и во
всех вообще органических структурах, настроение икринок влияют и
генетические факторы и факторы внешней среды. У некоторых рыб ик-
ринки пелагически е—имеют удельный вес меньше удельного ве-
са воды и плавают в толще воды, переносясь на огромные расстояния.
Плавучесть яиц достигается различными способами. У трески, мно-
гих сельдевых, камбал и других рыб икринка имеет различной ве-
личины жировую каплю, которая способствует ее пловунести, умень-
шая удельный вес .Пелагические икринки свойственны почти исклю-
чительно морским рыбам и у пресноводных встречаются весьма ред-
9 Биология рыб 129
ке и то, главным образом, у тех рыб, которые являются^генератив-
но морскгми. В нашей пресноводной фауне пелагические икринки
имеют только чархальская полька Clupecnclla de licatula tscharc halen-
sis, чехонь Pelccus cultratus и амур Ctcnophary ngedon idcllus, да и
то, собственно, у двух последних форм икра скорее батипелагиче-
ская, т. е. хотя и плавающая, но держащаяся в придонных слоях ве-
ды; тюлька же генеративно морская форма. Среди близких видов,
имеющих пелагическую икру, виды высоких широт имеют икру бо-
лее крупную, чем обитатели более низких. Так, более теплсводные
тресксвые G< dus eismi.rki, G. luscus, G virens, G. mcrlangus имеют
икринки 1,0—1,3 мм в диаметре, более северные —треска G. morhua
и пикша G. aegkfinus — 1,2—1,5 мм и высокоарктические сайка
Boricgadus saida и навага Eleginus navaga 1,5—1,9 мм. Такая ж
Рис. 83. Икра различных рыб (слева направо):' миксина\ Myxine glu-
tinosa, бычок Hyrcanogobius bergi, сарган Belone acus, иваси Sardi-
nops melanosticta, пузанок Caspialosa caspia, хамса Engraulis encra-
sicholus.
картина подмечена и для некоторых других групп рыб (Blenniidae,
Scorpaenidae, Pleuronectformcs. Имеются различия и в характере
и времени появления пигментации эмбрионов. Пигментация у арк-
тических видов наступает позднее, чем у более южных.
Другая группа икринок — икринки д е м е р с а л ь н ы е, имею-
щие обычно уде; ьный вес больше удельного веса воды. Они, как мы
указывали, как правило, почти всегда крупнее, чем пелагические и
часто имеют различные приспособления для прикрепления. У яиц
миксин на концах развивается пучок нитей с зацепками, при помощи
которых яйца и прицепляются к субстрату и сцепляются друг с
другом. У многих акул роговидная оболочка яиц имеет по углам
или на конце нитеобразные выросты, служащие также для прикреп-
ления. Ряд рыб из разных систематических групп имеет прилипаю-
щую икру. Оболочка икринки, попавшей в веду, становится липкой
и приклеивается к субстрату. Интересно, что неприклеившиеся
почему-либо икринки через опреде;енное время пребывания в воде
теряют клейкость. Несколько отличен способ прилипания к субстра-
ту у икры некоторых бычков Gebiidae. У их икры после оплодот-
ворения верхний к; ейкв й слой оболочки расположен лишь на неболь-
шом участке псверхности икринки. Этим-то участком икринка и
пргкреш яется к донным предметам.
В тех случаях, когда оплодотворение происходит вне тела ма-
териисксго организма, количество оплодотворенных икринок весь-
ма си; ьно зависит кроме внешних факторов и от продолжительности
жизни в наружной среде спермин и икринки до момента оплодотво-
130
рения. Для разных рыб эта продолжительность жизни весьма раз-
лична. Так, спермин наших обычных пресноводных рыб живут в
воде от 20 сек. (некоторые лососевые) до 5 и более минут (сазан),
причем у рыб быстротекучих вод спермги, как правило, живут бо-
лее короткий срок, чем у рыб, населяющих стоячие и медленно те-
кущие воды. Икринки также мсгут жить в воде до оплодотворения
сравнительно непродолжительное время, после чего оболочка от-
стает и оплодотворение становится невозможным. Длительность
возможности оплодотворения икринки, так же как и выживаемость
у различных рыб снермиев, разнится дово/ьно сильно, но обычно
первая всегда несколько выше, чем вторая. У пресноводных рыб спо-
собность к оплодотворению у икринки сохраняется от 5 до 30 ми-
нут, но она варьирует не только у различных видов, в различных
условиях, даже и у одного и того же вида в одинаковых условиях
различные икринки обладают различной выживаемостью. Так, у
американской рыбки Fundulus hotcroclitus икра может быть опло-
дотворена в срок от 6 до 20 минут по выметывании. В связи с тем,
что далеко не все икринки могут быть после выметывания найдены
спермиями и оплодотворены, стоит тот факт, что количество опло-
дотворенных икринок сказывается неско/ько меньшим, чем число
отложенных. У лососей Дальнего Воетека процент неоплсдство-
ренней икры варьирует обычно от 14 до 20%. Вообще, как правило,
чем больше икры откладывает рыба, тем больший процент неоплс-
дотворенной икры у нее имеется.
Как только икринка оплодотворена, в ней сейчас же начинаются
изменения, приводящие в конечном итоге к выходу малька.
Начало редукционных делений ядра у икринок наступает лишь
после того, как произойдет их оплодотворение. Оно начинается
через несколько минут, а то и неско/ько часов (у форели 5—7) по-
сле самого акта оплодотворения. Продолжите/ьнссть инкубацион-
ного периода у различных рыб чрезвычайно варьирует: от 12 часов
у Djnio rerio и многих других тропических рыб, до 100 и более дней
у некоторых лососевых и других рыб высоких широт. П род о/жите яв-
ность инкубационного периода может варьировать у одного и того
же вида в зависимости от изменения различных факторов среды и,
в первую очередь, от температуры. На ряде рыб установлено, что
число градусодпей при условиях, близкгх к оптимальным, потреб-
ных для развития икры остается для данного вида более или менее
постоянным. Так, для форели оно будет равно 410 при различных
температурах.
Температура Продолжительность Число градусодней
инкубации
2° 205 дней 2x205=410
5° 82 » 5х 82=410
10° 41 >> 10х 41=410
Конечно из приведенных закономерностей отнюдь йе следует,
что икринку форели межно держать при температуре 20,5° и она при
этом разовьется в 20 дней. Это правило, как мы уже сказали, оста-
вь 131
ется в силе только для условий, близких к оптимальным. Известно,
что в морской воде, температура замерзания которой ниже нуля,
многие икринки, в частности у трески, наваги, сайки, некоторых
камбал и других рыб, развиваются часто при отрицательных темпе-
ратурах. Минимальную температуру, при которой может итги разви-
тие данной рыбы, Рейбиш(1902) назвал «порогом». Для трески такой
теоретический вычисленный порог равен—3,6°, для наваги—2,3е, для
морской камбалы—2,4° и для речной—1,8° Установлено, что изме-
нение величины инкубационного периода при изменении темпера-
туры на один градус при низких температурах значительней, чем
при высоких. Характер зависимости между ’температурой и скоро-
стью развития икры у рыб более или менее соответствует установлен-
ному Вантгоффом правилу зависимости хода химических реакций
от температуры. Как известно, по правилу Вантгоффа, при повыше-
нии температуры, начиная от 10е и выше, на 10° скорость химических
процессов увеличивается в 2—3 раза. Полученная цифра и ж сит
название коэфициента Вантгоффа и обычно обозначается Q10. Для
более низких температур от Одо 10е эта величина возрастает до 5—7
раз. Вычисленный для рыб температурный коэфициент развития
икры довольно близко подходит к величине коэфициента Вант-
гоффа.
Температурный коэфициент развития икры у рыб (Державин, 1922).
Вид Температурные границы @10 Вид Температурные границы ^10
Стерлядь 7,2—18,8 3,36 Чавыча 5,6-10,6 3,32
Русский осетр . 12,1—25,6 2,48 Речная камбала 6—12 2,59
Немецкий » 10 —22 2,82 Морская » 5-12 2,51
Севрюга . 20 —28,6 2,09 Треска 3-14 2,47
Пузанок 18 —24 3,24 Мерланг 5—14 2,97
Черноспинка 16,5-22,7 2,63 Пикша 3—14 2,47
Средняя величина <2К, для рыб равна 2,73.
На развитие икры огромное влияние оказывает и свет, а именно
для ряда рыб усиление освещения вызывает значительное уско-
рение развития икры. Так, икра камбалы развивается в тени на пол-
тора — два дня медленнее, чем на свету (Jjhansen и Krogh 1914).
Этот же факт—влияние света указан для американской сельди шед;
у нее та :же имеет место ускорение развития икры на свету и замед-
ление в темноте. У севрюги развитие икры на свету тоже идет быстрее,
чем в темноте (Кулинченко 1939). Обратная картина наблюдается
у тех рыб, у которых и в естественных условиях икра развивается
в темноте. Так, у форели, развитие икры которой проходит в грунте,
икра развивается при освещении на 4—5 дней медленнее, чем в
темноте (Wilier 1927).
Большое влияние на скорость развития икры оказывает и ко-
личество растворенных в воде газов и, в первую очередь, кислорода.
Как установлено Wilier для форели, и другими авторами для иных
видов рыб, понижение в воде количества кислорода, против нормаль-
132
ного для данного вида, приводит к замедлению развития икры и удли-
нению инкубационного периода. При этом при понижении количе-
ства кислорсда в воде ниже критической точки, различной для раз-
ных вгд( в рыб, происходит гибель икры.
Также сильно влияет на развитие икры и соленость воды. У воб-
лы и леща в Каспии икра не развивается, как в дестиллированной
воде, так и в воде свыше 10%о солености (Ивлев, 1940г.). У хам-
сы Engraulis encrassichtlus maeoticus икра может развиваться в
строго определенных рамках солености, не ниже 3%о. Оптимальной
для этой рыбки является соленость в 11,7—11,9%в (Морозова и
Каракаш 1939). Перед тем как соленость достигнет концентрации,
прй которой икринка погибает, имеется значительная амплитуда
солености, при которой развитие икры хотя и происходит, но по-
лучается значительный процент уродливых мальков.
Осмотическое давление в икринке — величина более или менее
постоянная, не меняющаяся в зависимости от изменения осмотиче-
ского давления окружающей среды. Оболочка икринки проницае-
ма для газов (кислород проникает в икринку, а углекислый газ из
икринки проникает наружу), но слабо проницаема для воды и рас-
творенных в ней солей. Это делает непонятным указанный выше факт
сильного влияния солености на развитие икры, но, повидимому, с
одной стороны, слишком большая разница в парциальном давлении
наружной среды и внутри икринки является для нее губительной
(Светлов 1930) и, с другой, у многих рыб пелагические икринки при
низких соленостях, сохраняя свой прежний удельный вес, опуска-
ются на дно и попадают в условия, где их развитие происходить
не может. Это явление отмечено для трески и камбалы (Dakin 1910 г.).
Наоборот, икра бывшая донной в опресненных участках, при по-
вышении солености может всплыть, как это имеет место, например,
у чехони.
Не toj ько продолжительность, но и характер развития у рыб,
живущих в различных экологических условиях, идет далеко не оди-
наково.
После оплодотворения увеличение размеров икринки, про-
исходящее в результате проникновения между яйцом и оболочкой
воды и отде;ения оболочки от желточного мешка, у одних видов
идет быстрее, и размеры икринки возрастают относительно больше,
чем у других видов, у которых разбухание приводит к меньшему
увеличению размеров икринки. Резко различается также время и
последовательность закладки отдельных органов, но эти особенности
в значительной степени связаны с систематической принадлежностью
вида. Физиологическая сторона процессов, происходящих в орга-
низме эмбриона, главным образом зависит от его образа жизни.
Так, например, появление кровяных телец у эмбрионов в донных
икринках происходит гораздо раньше, чем у эмбрионов рыб с пела-
гической икрой, у которых образование красных кровяных телец
приурочено к выходу личинки из икринки, а у некоторых даже еще
позже. Исключение составляет океаническая сельдь Clupea haren-
gus, у которой икринки, хотя и донные, но образование красных
133
кровяных телец происходит поздно.. Повидимому, у океанической
сельди донная икра — вторичное явление, ибо у большинства близ-
ких родов сельдей икра пелагическая. Выходит личинка из икрин-
ки, растворяя оболочку при помощи фермента, выделяемого специ-
альными железками вылупления.
Личиночный период
Вышедшая из икры личинка рыбы обладает уже целым рядом
специфических приспособлений к тем условиям существе ваш я, в
которые она попадает по выходе из икры. Как правило, икра с боль-
шим количеством желтка, свойственная, главным образом, денным
рыбам, дает малька, более хорошо сформированного и менее резко
отличающегося от взрослых особей, чем икра мелкая, пелагическая,
Рис. 84. Личинки Hemichromis (окунеобразные) на стадии
максимального развития дыхательных сосудистых сетей в
преданальном и анальном плавниках (из Крыжановского,
1933).
из которой выходят личинки, совершенно непохожие на взрослых
рыб.
Вышедшая из икринки личинка имеет специальные приспособ-
ления для удержания в том биотопе, который является для нее
наиболее благоприятным, в том или ином слое веды—у пелагических
рыб, в прибрежной зоне—для рыб литорали и т. д. Личинки и м ;льки,
ведущие пелагический образ жизни, очень часто имеют различные
выросты, увеличивающие их сопротивление движению воды и облег-
чающие парение в ее толще (см. выше). У личинок пресноводных
рыб, икра которых развивается в прибрежной зоне, часто имеется
«цементный орган», пользуясь которым личинки прикрепляются
к растениям и другим предметам, находящимся в воде, что не дает
личинкам возможности быть выброшенными на берег, а главное
позволяет удерживаться в поверхностных слоях воды, богатых кис-
лородом.
Питание на первых стадиях развития личинки рыбы после вы-
хода из яйца осуществляется за счет тех пищевых запасов, которые
находятся в желточном мешке (рис. 84), и поэтому, когда рыбка
начинает ощущать потребность в пище, она уже бывает вполне
сформирована и успешно может добыть себе корм. Несколько
иначе обстоит дело с дыханием. Жабры у личинок рыб развиваются
134
довольно поздно (у окуня через две недели] по выходе из икры), по-
этому на ранних личиночных стадиях у большинства рыб должны
были выработаться дополнительные органы дыхания. У личинок
двоякодышащих Dipnoi и многоперовых Polypteri развиваются
наружные жабры (рис. 85). У костистых рыб личиночным органом
Дыхания служит сосудистая система желточного мешка, плавников,
жаберной крышки псевдобранхия (псевдорбанхия—жабра, рас-
положенная в. области жаберной крышки). Все эти части тела про-
Рис. 85. Личинки Protopterus (наверху) и L.epidosiren (внизу) с
наружными жабрами (из Budget, 1902 и Kerr, 1899).
низаны у личинок богатой сетью кровеносных сосудов, в которых
и нроисходит окисление крови до тех пор, пока не разовьются дефи-
нитивные жабры.
Стадия икринки и личиночный период жизни для рыбы является,
пожалуй, наиболее узким местом в ее сущее.вовании. В это время
рыба гкдвергается наибольшим опасностям, как со стороны живых
организмов, так и со стороны факторов окружающей абиотической
среды. Поэтому совершенно естественно, что на личиночный период
жизни рыбы приходится ее наибольшая смертность. Для промысло-
вых рыб северного Каспия определено, что из всех выведшихся из
икры личинок не больше 10% скатываются в море в виде сформиро-
вавшихся рыбок, остальные же 90% погибают.
Знание величины урожая молоди имеет огромное хозяйственное
значение. Год, характеризующийся высоким урожаем молоди, в
течение ряда последующих лет дает повышенные уловы взрослых
особей, наоборот неурожайный год дает пониженные уловы. Каковы
же те условия, которые определяют урожай молоди рыб?
В основном они сводятся к следующим четырем факторам:
1) количество производителей на местах икрометания и коли-
чество отложенной икры, 2) благоприятные внешние условия для
развития икры и выхода молоди (температура, соленость, содержа-
ние кислорода и др.), 3) наличие достаточного количества корма для
молоди, 4) количество животных, поедающих икру и мальков.
136
Знание указанных четырех условий позволяет с большей или мень-
шей точностью давать прогнозы уловов на последующие года.
Влияние отдельных факторов из числа упомянутых выше на
урожай молоди чрезвычайно велико. Так, при запуске рыболовства,
имевшем место в бассейне Волги в 1918 году, когда почти все про-
изводители осетровых были пропущены к нерестилищам, улов ска-
тывающихся мальков за час траления в море перед устьем Волги
возрос почти в три раза. Второй фактор — благоприятные внешние
условия — имеет также немаловажное значение. Известно, что в
1904 г. трески на нерестилищах у Лофотен было мало, а количество
выведшихся мальков больше, чем обычно. Это целиком объясняется
теми благоприятными условиями, которые имели место в данном
году.
Промерзание нерестилищ в суровые зимы может привести к.
массовой гибели икры лососевых, откладывающих икру часто в
мелких речках.
То, что икра и личиночные стадии рыб подвержены столь силь-
ному влиянию внешних условий заставило за последние годы пе-
ресмотреть и методику рыбоводных мероприятий. Раньше обычно
выпуск выведшихся из икры мальков (это касается главным обра-
зом лососевых) производился очень скоро по выходе их из икры и,
как показали исследования ряда американских ученых, давал очень
низкие результаты, так как по выпуске в реку большинство рыбе-
иек погибало. Поэтому сейчас признано более целесообразным вы-
держивать выведшуюся молодь некоторое время в искусственных
водоемах и выпускать уже подросших и окрепших рыбок.
РАЗМЕРЫ, РОСТ И ВОЗРАСТ РЫБ
Рыбы, насчитывающие около 30000 видов, чрезвычайно разно-
образны по величине. Самая малая по размерам из известных нам
рыб, а пожалуй и самое маленькое позвоночное вообще — это бы-
чок Mystichthys lusonensis, водящийся в водах Филиппинских
островов. Размеры этой рыбки всего 12—14 мм. Немного больше
бычки европейских вод Aphya и Cr\ stallogcb.us, а также многие
представители отряда Cyprincdontifcrnus, населяющие пресные воды
тропических широт. С другой стороны, среди рыб встречаются и
такие гиганты, как полярная акула Cetcrhinus niaximus, достигаю-
щая до 15 м в длину, а возможно даже и более. Некоторые ископа-
емые акулы несомненно были еще более крупных размеров (рис. 86).
Из пресноводных рыб наиболее крупные — это белуга Huso
huso, достигающая до тонны весом, южно-американская арапайма
Arapaima gigas — до 160 кг и обыкновенный сом Silurus glanis —
до 200 кг.
Продолжительность жизни различных видов рыб также весьма
различна. Некоторые представители семейств Gobiidae и Scopelidae
живут не более года, такие же виды, как белуга, доживают до ста
и более лет. Такие старые особи зарегистрированы далеко не для
всех промысловых рыб, кроме белуги они известны у карпа, щуки
13в
Рис. 86. Челюсть гигантской ископаемой
акулы.
и некоторых других. Нормальная же продолжительность жизни
основных морских промысловых рыб, как тресковые, камбаловые и
другие, вне влияния промысла не превышает тридцати лет, но обыч-
но, как мы увидим ниже, под влиянием усиленного промысла рыбы
не достигают своего предельного возраста и не умирают естествен-
ной смертью, а бывают выловлены. Основным признаком старости
является потеря способности к размножению. У некоторых видов
рыб старение связано и с морфологическими изменениями, так,
например, у старых особей трески Gadus morhua нижняя челюсть
крючкообразно изгибается, что затрудняет питание и приводит
постепенно к смерти от истощения. Весьма многие виды рыб поги-
бают от истощения после первого или одного из последующих не-
рестов. Для угря Anjui 11а
anguilla дальневосточных
лососевых рода Oncorhyn-
chus и многих других
видов рыб смерть после
первого икрометания —
постоянное явление, и
случаи вторичного икро-
метания, например, у
угря, неизвестны.
Рыбы в отличие от
позвоночных с постоянной
температурой тела, как
правило, обладают пос-
тоянным ростом. В то
время как у теплокров-
ных позвоночных рост
организма прекращается
по достижении половозре-
лости, у рыб, хотя на-
ступление половозрелости
и отражается на темпе
роста, но рост не прекра-
щается, а обычно только
несколько замедляется.
Полное прекращение ро-
ста наблюдается лишь с
началом старения особи.
Второй характерной чертой роста рыб является, как у всех жи-
вотных с непостоянной температурой тела, его периодичность. В
отдельные определенные сезоны года рыба растет быстрее, в другие—
медленнее. Эта неравномерность роста в течение года хорошо за-
метна на отдельных костях скелета рыб и на чешуе. Периоды за-
медленного роста отпечатываются на сцелете в виде светлых полос
или колец (в падающем свете) (рис. 88). Эти полосы состоят из более
мелких уплотненных клеток и в проходящем свете Скажутся более
темными. 1
I
Периоды усиленного роста характеризуются на скелете широ-
кими темными (в проходящем свете, наоборот, светлыми) полями
или кольцами.
Пзрвое указание на то, что кольца на чешуе и костях рыб соот-
ветствуют периодам замедленного и ускореннсго роста рыбы и что
по числу этих колец можно определить возраст рыб, прингд/ежат
известному голландскому естеавсиспытмтелю Л.вснгуку (1684 г.).
Несмотря на то, что первые указания на возможность определения
возраста рыб по чешуе и костям были сделаны много вр.мени назад,
практическая разработка этого метода, имеющего огромное хозяй-
ственное значение, является достижением последних пятидесяти
лет.
Значительную ро: ь в разработке методики иезлед вания воз-
раста сы рали работы русских ученых. Рааоты В. К. С.л;атова—
по изучению i оз астного состава о.евровых Амура го к< стям.
Исследования И. Н. Арнольда, Н. Л. Чугунова, Е. К. Суворова,
Г Н. Монастырского, Н. И. 4} гуновои и дру! их очень мнего
дали к к для теоретической разработки вопроса, так и в деле
пр ктическэго применения метода для решения практических
задач.
Сильное напряжение промысла ряда рыб, приведшее к снижению
их уловов (морская камбала, лосось и другие), вызвало необходи-
мость постоянного контроля возрастного состава вылавливаемых
особей для того, чтобы предотвратить нежелательный вылсв молодых
неполовозрелых экземпляров. Провидимый из г<да в год анализ
возрастного состава вылавливаемых рыб позволяет во-время под-
метить увеличение примеси младших возрастов и уменьшение стар-
ших, указывающее на чрезвычайное напряжение промысла, могу-
щее в конечном итоге привести к подрыву запасов; с другой стороны,
наличие в^уловах большого количества рыб старших возрастов,
наоборот, указывает на возможность дальнейшего увеличения про-
мысла. Необходимо, конечно, учитывать величину урожая отдель-
ных годов, а также время наступления половозрелости у исследуе-
мого объекта.
Так, например, если в улове воблы Rutilus rutilus caspicus
большое количество семи- и восьмилетних особей будет указывать,
как правило, на недолов (вобла обычно становится половозрелой
по достижении трехлетнего возраста), то наличие в улове осетра
особей Acipenscr gulcknstadti, главным образом, в возрасте 7—8
лет будет указывать на катастрофическое положение промысла
(осетр становится половозрелым обычно не раньше 8—10-летнего
возраста), так как в исследованном улове осетра преобладают
неполовозрелые особи.
Методика исследования возраста и роста
Анализ темпа роста рыб очень часто позволяет различать от-
дельные стада промысловых рыб, которые морфологически или сов-
сем неразличимы, или плохо различимы (например у трески, сель-
13*
ди и других рыб). Сравнение темпа роста представителей одноге и
того же вида, но взятых из различных водоемов, имеет важное зна-
чение и при таксации рыбохозяйственных угодий, позволяя вы-
явить их ценность для нагула рыб.
Большая важность определения возраста рыб для разрешения
вопросов рыбного хозяйства привела к тому, что за последнее время
в этой области, как в заграничной, так и в советской литературе
накопилось очень много материалов. Основным объектом для оп-
ределения возраста у большинства рыб служит чешуя. Сбор чешуи
прост. Чешуя легко обрабатывается и у многих видов рыб позволяет
надежно определять возраст. Определение возраста по чешуе про-
изводится почти у всех лососевых, сельдевых, карповых, трески и
многих других рыб. Чешуя, собираемая для определения возраста
рыб, берется обычно под основанием первого спинного плавника'
и складывается между листками блокнота из непроклеенной бума-
ги, и на той его странице, на которую была положена чешуя, запи-
сываются данные об экземпляре рыбы, чья чешуя была собрана
(длина, вес, пол, место поимки, орудие лова ит. д.). Для определения
возраста чешуя, предварительно промытая в слабом растворе на-
шатырного спирта, закладывается между двумя предметными стек-
лами и просматривается или под лупой, или под микроскопом в за-
висимости от ее размеров. Нежную чешую, как, например, у трес-
ки, иногда лучше помещать в глицерин-жслатину (в канадский
бальзам чешую для определения возраста класть нежелательно,
так как она сильно просветляется, и годовые кольца становятся
плохо заметны). У окуневых, налима и некоторых других рыб хо-
рошим объектом для определения возраста служат плоские кости,
как, например, жаберная крышка и Cleithrum. Видимость на костях
бывает часто не менее отчетлива, чем на чешуе, и у тех видев рыб,
у которых возможно определение возраста и по чешуе можно опре-
делить возраст и по костям, которые служат хорошим контрольным
материалом. Приготовление костей для определения возраста очень
просто: кость очищается от мяса, обезжиривается и высушивается.
По костям, собранным с тухлой рыбы или с рыбы, лежащей в фор-
малине, определять возраст нельзя, так как такие кости обычно мут-
неют.
У тресковых и камбал определение возраста часто производится
по отолитам—ушным камням (рис. 87—см. вклейку в конце книги),
которые извлекаются из черепа путем вскрытия : абиринта. Для оп-
ределения возраста отолиты должны быть обезжирены и в некоторых
случаях, для придания им большей прозрачности, несколько подшли-
фованы. У осетровых, сома и некоторых акул (Acanthias) определение
возраста производится по лучу плавника; у акул непарного,у осетро-
вых и сома—грудного. При этом у акул кольца видны на плавникогом
луче уже непосредственно после очистки луча. У осетровых же и
сома для определения возраста получу из него приходится изготов-
лять поперечный срез, который затем путем шлифсвания доводится
до прозрачности, после чего приклеивается на предметное стекло
канадским бальзамом и может служить для определения возраста
11$
Рис. 88. Схема годовых колец на плоских
костях и шлифе плавникового- луча рыб. Три
верхних рисунка—кости плечевого пояса,
слева внизу жаберная крышка, справа шлиф
луча грудного плавника (из Чугунова, 1926).
(рис. 88). Для приготовления шлифов сконструирована специаль-
ная машинка, состоящая из двух параллельно закрепленных круг-
лых пилок, которые приводятся в движение электромотором. Регу-
лируя расстояние между
пилками, можно изготов-
лять шлифы различной
толщины. Естественно,
что поскольку образова-
ние годовой отметки
происходит в результате
изменения внешних усло-
вий, вызывающих изме-
нения в скорости роста,
постольку возникает во-
прос, не могут ли обра-
зовываться «годовые коль-
ца» несколько раз в году
не только в результате
прекращения питания в
зимнее время, но и в ре-
зультате, например, голо-
довки во время икромета-
ния, перехода от одного
менее питательного вида
пищи к другому, более пи-
тательному и т. п. Основ-
ным способом ответа на
поставленный вопрос яв-
ляется изучение рыб, воз-
раст которых известен или
которые прожили опреде-
ленное время, после того
как у них была взята че-
шуя. Как показало изуче-
ние рыб, живших определенное количество лет в прудах и аквариу-
мах, а также поимки помеченных ранее рыб в открытых водоемах
(треска, вобла)число колец на чешуе и слоев на костях соответствует,
как правило, числу лет,прожитых рыбой. Правда, на чешуе
и костях действительно попадаются иногда добавочные кольца, пс
явление которых часто бывает закономерным и связано с определен-
ным моментом в жизни рыбы. Так, например, у некоторых рыб име-
ется кольцо внутри кольца первого года (елец, вобла и др.), так на-
зываемое мальковое, которое связано с моментом перехода от пи-
тания планктонными организмами к питанию донными организмами.
У очень многих рыб на чешуе имеются нерестовые кольца и марки,
являющиеся результатом прекращения питания во время икромета-
ния. Так, например, у каспийских сельдей во время нереста чешуя
начинает разрушаться,что приводит к образованию на ней определен-
ного типа отметки (Здмахаев 1940) (рис. 91—см. вклейку в конце кни-
140
Рис. 92. Чешуя трехлетнего карпа,
обработанная хлоралгидратом и разде-
ленная на пластинки, соответствую-
щие отдельным годам роста (из Pet-
rov и Petruschevsky, 1929).
ги). У лососевых на чешуе также отчетливо видны нерестовые марки,
имеющие форму колец, но несколько иного характера, чем годовые.
При этом у самцов лосося Salmo salar нерестовые марки обычно выра-
жены отчетливее, чем у самок.
У воблы Каспия путем изучения
чешуи помеченной и вторично
пойманной рыбы установлено
(Чугунова, 1938), кроме нере-
стового и малькового, еще три
типа добавочных колец имен-
но: 1-й тип, появляющийся
при случайной задержке роста
рыбы, 2-й тип, образующийся
в случае изменения характера
роста чешуи, и 3-й тип, обра-
зующийся по линии обрыва на
чешуе, регенерирующий после
механического повреждения.
Естественно возникает воп-
рос, не делает ли наличие доба-
вочных колец на чешуе, костях
и отолитах рыб весь метод
определения возраста по годо-
вым меткам в целом порочным,
и каковы способы для разли-
чения годовых колец от до-
бавочных. Для разрешения
этого вопроса делалось очень много работ, в частности Петров и
Петрушевский (1929) пытались мацерировать чешую карпа и раз-
делять ее на отдельные пластинки, соответствующие каждая опре-
деленному году роста (рис. 92), О. Винге (1915) производил измере-
ние ширины отдельных склеритов на чешуе трески и наносил эти
величины на графики. По числу вершин, соответствующих размерам
наиболее широких склеритов, он получал число годовых колец; до-
бавочные кольца, хотя и давали некоторое изменение размеров
склеритов, но менее значительное, чем годовые. Обычно же для
проверки правильности определения возраста пользуются не одним
каким-нибудь объектом, а несколькими, например, если определение
ведется по чешуе, то у некоторых контрольных экземпляров берут-
ся кроме чешуи кости жаберной крышки. Наряду с изучением ко-
лец на чешуе и костях совершенно необходимо также изучение и
образа жизни рыбы для выявления тех «узких местл в ее жизни,
которые могли бы отразиться на росте. Знание таких моментов
позволит при наличии соответствующих колец на чешуе отнестись
к ним с большей осторожностью и легче выявить их природу. Наи-
большие трудности представляет распознавание малькового кольца.
Обычно для того, чтобы не спутать мальковое крльцо с первым
годовым, осенью берется несколько проб малькрв исследуемого
вида, анализ размеров которых позволяет с несомненностью уста-
141
навить, что мы имели дело с годовым или мальковым кольцом. В об-
щем же, как правило, добавочные кольца и марки всех типов бывают
выражены менее резко, чем годовые, и при некотором навыке обычно
могут быть выявлены без особых трудностей. Мы разобрали случаи,
когда число зон на чешуе и костях больше, чем число лет, прожитых
рыбой, но может быть и так, что число колец на чешуе и меток на
костях наоборот меньше, чем число прожитых рыбой лет. Этот воп-
рос имеет особое значение при определении возраста по чешуе.
Регенерировавшая чешуя имеет меньшее количество годовых ко-
лец, чем нормальная. У некоторых рыб, как например у угря, у
которого чешуя образуется на третьем или четвертом году жизни,
число годовых ко;ец па чешуе заведомо меньше, чем число прожи-
тых рыбой лет и чем количество марок на костях.
Как видно из всего изложенного, перечисленные отклонения
0т нормы ни в коей мере не снижают общей ценности метода опре-
деления возраста рыб по чешуе и костям. При пользовании этим ме-
тедсм необходим лишь внимательный анализ природы колец и ин-
дивидуальный педхед к каждому изучаемому объекту, а главное
учет специфических черт экологии рыбы.
Весьма существенным при изучении возраста рыб является во-
прос о времени закладки годового кольца. Если годовое кольцо
на чешуе является непосредственным результатом замедления ро-
ста в зимний период жизни рыб, то время появления кольца оче-
видно должно падать на весну. Кроме того, казалось бы, у рыб*
тропиков, где режим вед в летнее и зимнее время почти одинаков,
годовые кольца не должны были бы образовываться. Между тем
этого не наблюдается: как у морских, так и у пресноводных рыб
тропиков имеются на чешуе и костях годичные отметки, следова-
тельно рост идет периодически. Из морских рыб годичные кольца,
напоимер, наблюдались у сельди Ethan losa fimbriata Шнейдером
(1910 г.), а между тем эта рыба живет в тропиках вдоль Атлантиче-
ского побережья тропической Африки. Из пресноводных рыб мож-
но указать, например, на недавно изученного представителя семей-
ства харациновых Curimatius ekg ns из Бразилии и Парагвая,
у которого бразильские исследователи Azevedo, Dias и Vieira (1938)
также наблюдали наличие годовых колец на чешуе. Наличие
периодического роста как у рыб тропиков, так и рыб умеренных
широт указывает на то, что образование годового коаьца не яв-
ляется непосредственным результатом смены холодного и теплого
времен года. Это подтверждается также тем, что у рыб умеренных
широт время образования годичного кольца сильно варьирует
как у различных видов, так и у особей одного и того же вида. Обыч-
но начало быстрого роста у рыб умеренных широт происходит
ран! ей весной, когда и появляется граница между годовыми коль-
цами, но у ряда видов время появления коаьца на чешуе имеет
место позднее, а следоватеаьно и перехсд от замедленного к более
быстрому росту наступает позже. Так, например, Щ\ ка в Псковских
озерах не имеет еще новего прироста в мае; то же отмечено и для
осетровых Азовского моря. У различных возрастных групп момент
142
перехода от замедленного роста в зимний период к быстрому росту
в летний происходит в различное время. Так, для судака из Азов-
ского моря Чугунова (1931) указывает, что прирост второго года
начинается ранней весной, прирост четвертого года—в апреле—
мае, а прирост пятого — в июне и иногда даже позднее.
Весьма существенен вопрос об обозначении отдельных возраст-
ных групп. В ихтиологической литературе по этому вопросу нет
единогласия, и имеется довольно значительная путаница. Обычно
сейчас принята следующая терминология:
Название возрастной группы Число колец Обозначение
Сеголеток нет 0+
Годовик одно 1
Двухлеток одно 1 +
Двухгодовик два 2
Трехлеток два 2-F
Трехгодовик три 3
Знаком плюс обозначается начало прироста следующего года.
Также весьма важным методическим вопросом, имеющим огром-
нее, как теоретическое, так и хозяйственное значение, являются
вопросы методики исследования темпа роста рыб и, в частности,
определение размеров рыбы за предшествующие периоды ее жизни.
Основной вопрос, который при этом требует разрешения, это вопрос
о характере зависимости между приростами на костях и чешуе
и приростами тела.
Норвежский ученый Эйнар Леа (Einar Lea, 1910) принял, что
длина рыбы и длина чешуи с возрастом изменяются пропорционально
Друг др^гу и что эта пропорциона;ьнссть носит прямолинейный
характер, т. е. выражается уравнением первой степени. В резуль-
тате проделанных Леа над сельдями исследований им была пред-
ложена формула:
L — —- • L
у ч
где L измеренная длина рыбы, V — длина чешуи от центра до края,
измеренная по средней линии, Ln—вычисленная дтина рыбы в
возрасте п лет, Vn — расстояние от годовего котьца до центра
чешуи в возрасте п лет. Выведенная Леа зависимость хорошо ил-
люстрируется приведенным рисунком (рис. 93), где мы имеем ряд
подобных треугольников, у которых пропорциона14>ные стороны
выражают размер рыбы и размер чешуи за данный год. Поа ьзуясь
этой зависимостью, Эйнар Леа сконструировал специальную доску,
143
п® которой производил расчисление темпа роста рыб. Позднее
эта доска была значительно усовершенствована Г. Н. Монастыр-
ским (1924) (рис. 94), который предложил непосредственно проеци-
ровать чешую при помощи рисовального аппарата на доску и,
двигая движком по линейке с сантиметровой шкалой, при помощи
нитки, соответствующей на рисунке линии АВ, отмечать длину
рыбы во время поимки. Эта же линия совмещается с краем чешуи,
в то время как центр чешуи совпадает с вершиной прямого угла,
Рис. 93. Соотношение между скоростью роста рыбы и ее чешуи (принцип
линейной зависимости, положенный в основу формулы Эйнар Леа).
•бразованного стороной треугольника, проходящей через нуль
икалы, на которой поставлена длина рыбы, и линией, соединяющей
центр чешуи с линией АВ. Передвигая движок по сантиметровой
линейке, совмещают нить с годовыми кольцами и на сантиметровой
линейке читают размеры рыбы, соответствующие данному году
роста. Сейчас методом Эйнар Леа пользуются очень многие ихтио-
логи, так как для практических целей он дает результаты вполне
достаточной точности.
В 1920 году английская исследовательница Роза Ли на осно-
вании накопившихся к этому времени материалов, предложила
внести некоторые уточнения в положение, выдвинутое Эйнар Леа.
По мнению Розы Ли, размеры чешуи и длины рыбы не пропорцио-
нальны друг другу, пропорциональны же только приросты длины
и чешуи. Основной причиной, нарушающей пропорциональность
между длиной рыбы и чешуи, является то, что закладка чешуи
нроисходит не в момент рождения рыбы, а несколько позднее,
когда рыба уже достигает определенных размеров. Поэтому в фор-
мулу Леа Роза Ли предлагает включить дополнительную величину
144
а, соответствующую длине рыбы в момент закладки у нее
чешуи.
После введения поправки Розы Ли формула Эйнар Леа при-
нимает следующий вид:
= у (L а) + а.
Последующие исследования и, в частности, работы Г Н. Ме-
настырского (1926, 1930) показали, что для некоторых рыб зави-
Рнс. 94. Доска системы Г. Н. Монастырского для расчисле-
ния темпа роста по методу Einar Lea (из Монастырского,
1924).
симость между линейным ростом чешуи и костей и ростом тела рыбы
имеет криволинейный характер, т. е. выражается уравнением не
первой степени. В результате своих тщательных многолетних ис-
следований Монастырский пришел к выводу, что приращения
логарифмов длины рыбы пропорциональны приращениям логариф-
мов длины чешуи, и уравнение, характеризующее эту зависимость,
имеет следующий вид:
lgy= IgA: +«lgx,
где у —длина рыбы, х— соответствующая длина *^ешуи, IgK—
отрезок, отсекаемый прямой уравнения на оси ординат^ мп — угло-
вой коэфициент.
Ю Биология рыб 145
Рис. 95. Доска для расчисления темпа роста рыв по логарифмической шка-
ле. Л— логарифмическая линейка, В — движок логарифмической линейки,
С — вертикальный движок (из Монастырского, 1934).
Факторы, определяющие рост
Каковы же те факторы, которые определяют характер и темп
роста рыбы? Несомненно, что общий характер роста является на-
следственным признаком, весьма трудно поддающимся изменениям
и общим для целых групп рыб, как это было, например, устано-
влено для карповых Васнецовым (1934). С другой стороны, на
рост рыбы оказывают чрезвычайно сильное влияние как внешние,
так и внутренние факторы. Из внутренних факторов в первую
очередь на росте рыбы сказывается влияние различных гормонов
и особенно гормонов половых желез. Как указывалось вын.е (см.
раздел «Размножение», стр. 127), у большинства рыб самцы и самки
бывают не одинаковых размеров, следовательно, гормон мужской
и женской половой железы различно влияет на рост. То, что здесь
146
имеет место непосредственное влияние половой железы видно из
того, что у большинства рыб разница в росте между самцами и сам-
ками наступает с момента достижения половой зрелости. Кроме
того, время наступления половозрелости связано у рыб обычно
с некоторым замедлением линейного
роста. Ускорение наступления полс-
возрелости по сравнению с нормаль-
ным приводит у рыб (наблюдения
на карпах) к некоторому замедле-
нию роста. Сильное влияние на рост
рыб, как показали исследования
Склсвера (1927), оказывают также
и гормоны других желез, некоторые
из которых ускоряют (гипофиз),
другие же наоборот замедляют
рост. Опыты приводились Склове-
ром с молодью форе; и.
Весьма большое влияние на рост
рыб оказывают и внешние факторы,
как-то: температура, свет, химизм
Рис. 96. Соотношение роста
чешуи и рыбы (плотва оз.
Бисерова), выраженное на
логарифмической шкале. На
оси абсцисс измерения че-
воды, количество корма, плотность Шуи, на оси ординат длина
населения и многие другие. Для Рыбы (из Монастырского,
каждого вида рыбы существует свой
температурный оптимум, при котором у данного вида наиболее
полно происходит обмен веществ, и при котором этот вид даст
наиболее быстрый рост. Естественно поэтому, что отклонения
Рис. 97. Зависимость обмена веществ у карпа от температуры.
Сплошная линия — потребление кислорода, точечный пунктир—вы-
деление углекислоты. Единица ординаты — 100 см3 на 1 кг веса в сутки.
Пунктир — выделение азота. Единица ординаты — 10 мг на 1 кг веса в
сутки (из Cronheim, 1911)
температуры в ту или другую сторону от оптимальной сейчас же
скажутся на темпе роста.
Как видно из приведенного рисунка, у карпа летом f повышением
температуры увеличивается интенсивность обмена. Поэтому понят-
но, что у большинства карповых рыб, являющихся по' своему прс-
Ю* И7
исхождению обитателями субтропиков и умеренных широт, при
одинаковой кормовой базе в низких широтах наблюдается обычно
ускорение темпа роста по сравнению с высокими широтами (см.
таблицу).
Рост леща в различных водоемах СССР
Возраст /1 /2 /з Ц
Псковское озеро 6,2 И,2 16,7 21,6
Ильмень 6,5 12,0 16,8 21,3
Аму-Дарья 8,7 14,6 19,6 23,4
Каспий 7,3 16,2 25.3 30,7
Азовское морс 9,5 18,9 25.5 30,7
Темп роста рыб, живших в бассейне Ледовитого океана около
пяти тысяч лет тому назад, когда условия здесь были более благо-
Рис. 98. Темп роста океанической сельди с
декабря 1909 по апрель 1911г. (из HJort,
1914).
приятны и температура
была выше, чем теперь,
был значительно более
быстрым, несмотря на
имевшее тогда место
сильное перенаселение в
результате отсутствия вы-
лова. Изменение темпера-
туры не только вызывает
общее замедление или
ускорение роста, но от-
дельные более холодные
или более теплые года
также вызывают реакцию
в организме рыбы, отве-
чающем или замедлением
или ускорением роста.
Как видно из приведенной таблицы роста корюшки Osmerus eper-
lanus из Кандалакшского залива (по Кирпичникову), в годы, с
отклонением температуры от средней в сторону похолодания, у
рыб наблюдалось замедление темпа роста.
Приросты1 Годы 1924 1925 1926 1927 1928
/1 4,3 4,5 3,9 4,5 4,2
/г 8,3 7,0 8,2 6,3
/з 5,3 6,9 6,3
/ч 4,5 4,0
/5 2,7
Отклонения от средней
годовой температуры+ 0,13 -1-0,34 — 0,72 Н-0,68 — 0,44
Температура является одним из тех факторов, которые несомнен-
но влияют на периодичность роста у рыбы. Как правило, в умерен-
ном климате наиболее быстрый рост у рыб приходится па месяцы
1 В ихтиологии, как правило, буквой t обозначаются расчисленные при-
росты, например tlt значит прирост первого года, t2 = (Z2 — Zx) (где /—длина
рыбы) означает прирост второго года и т. д.
148
с температурой выше средней годовой. Примером сказанному может
служить кривая роста сельди в течение года (рис. 98). Интенсивность
и продолжительность освещения различными частями спектра
оказывают также непосредственное влияние на рост рыб, причем
влияние света на рост различных видов рыб сказывается неодина-
ксвым, и, как правило, свет наиболее сильно влияет на рыб в моло-
дом возрасте. Обычно свет оказывает положительное влияние,
ускоряя рост рыб. Лишь у тех форм, у которых икра развивается в
темноте, как например лососевые, сильное освещение может вы-
звать замедление роста. Химический состав воды, ее соленость и
насыщение растворенными в воде газами также естественно отра-
жаются на росте рыбы, влияя на ход обмена веществ.
Чрезвычайно сильное влияние на рост рыбы естественно оказы-
вает корм, его количество, состав и размеры кормовых объектов.
Рыбы, живущие в более или менее сходных экологических условиях,
где сильная разница наблюдается только в количестве корма,
резко различаются по темпу роста. Это наглядно видно из приво-
димых ниже данных потемну роста сазана из озер Средней Азии,
отличающихся по величине биомассы бентоса.
Связь темпа роста сазана и величины биомассы бентоса в некоторых озерах
Водоем । Воз /, рас т Число грамм J биомассы бентоса на 1 Кв. м
1з
Ильджик . 6,9 i 12,6 17.0 i ; 0,26 г
Каракуль 8,0 14,2 18,1 20,0 ! 1,43 »
Джолдыбай 8,6 1 14’7 1 i !8,2 23,2 | 1,83»
Аральское море 12,6 i 21,5 29,3 36^0 1 18,0 » i
Естественно, что на рост рыбы влияет не только количество,
но также качество и размеры кормовых объектов. Опыты по влия-
нию на рост карпов различного рода кормов, как и следовало ожи-
дать, показывают, что более калорийные корма дают, как правило,
более быстрый рост. На рост рыб также оказывает влияние и наличие
в пище витаминов. В наибольшей степени замедление роста рыбы
вызывается отсутствием в пище витамина В. Витамины А и D
резервируются в жирах, и отсутствие их в пище продолжительное
время не позволяет подметить какого-либо изменения в росте, по-
скольку их дефицит в корме пополняется за счет запасов в жирах
тела рыбы.
Кроме пищевой ценности корма на рост рыбы оказывает также
влияние и размер кормовых объектов. Как показали опыты с кар-
пами, чем крупнее до известных пределов куски, заглатываемые
рыбой, тем быстрее она растет. Естественно, что последнее наблю-
дение не может быть распространено на всех рыб, 'но для очень
многих оно является верным.
149
Изменение в количестве и качестве потребляемого рыбой корма
очень быстро сказывается на скорости роста. У лососей изменения
в характере питания при переходе из рек в море сейчас же сказы-
вается на темпе роста. Камбалы, пересаженные из участков моря
с малой продуктивностью бентоса (являющегося их кормом) в рай-
Рис. 99. Рост морской камбалы в месте выхода из икры (Horn reef graunds) —
сплошная линия, и на месте акклиматизации (Dogger bank) — пунктир
(из Alvard, 1932)
оны с высокой продуктивностью, очень скоро (рис. 99) изменяют
скорость роста, значительно обгоняя
в росте рыб из мест своего прежнего
обитания.
На скорость роста рыбы оказывает
также весьма существенное влияние
количество особей на единицу пло-
щади водоема, т. е. плотность насе-
ООО
500
400
300
200
/00
4
f НОРмп
8
Плотность
Рис. 100. Зависимость прироста карпа от плотности посадки (па оси —
абсцисс плотность в условных баллах) (из Contag, 1931).
12
16
ления. Влияние плотности населения сказывается при этом вне
зависимости от количества получаемой рыбой пищи. Так, напри-
мер, Черч (1931), исследуя влияние плотности посадки на рост
Platypoecilus maculatus, пришел к выводу, что чем больше плот-
ность посадки, тем медленнее рост рыбы. То же самое еще
15и
раньше установлено Гофбауером (1902) для карпов (рис. 100), а
также Виллером и Шнигенбергом (1927) для молоди форели.
Отрицательное влияние плотности посадки на рост рыбы
начинается, однако, лишь после того, как эта плотность достигнет
известного предела, различного для каждого вида рыбы. Так,
для карпов (по Contag 1931) нормальной плотностью является
посадка 122 рыб весом 26,4 кг на гектар площади пруда. Превыше-
ние этой посадки, как видно из приводимого рисунка, сейчас же
сказывается на темпе роста рыб, который начинает снижаться.
Естественно, что в природных условиях плотность населения в
первую очередь сказывается на интенсивности конкуренции за
пищу. Конкуренция эта весьма сильно увеличивается при увели-
чении плотности населения и сильно снижается при уменьшении
таковой. Поэтому увеличение интенсивности промысла, особенно в
водоемах, не подвергавшихся ранее облову, сейчас же отражается
па росте рыб, вызывая его некоторое
ускорение. Так, например, бражни-
ковская сельдь Caspialosa brashnikovi
Рост бражниковской сельди на
юге и севере Каспия
Возраст
Водоем
Юг
Север.
Л Z2 'з '4
11,3 11,8 19,2 24,2 25,7 33,1| 29,4 35,1 33,6 37,4
в Южном Каспии растет хуже чем в
Северном, видимо потому, что в Южном
Каспии бражниковская сельдь исполь-
зуется промыслом и сейчас еще не-
достаточно, на севере же Каспия ее
промысел весьма сильно напряжен, и
лов производится уже давно.
Возрастной состав стада
Возрастной состав популяции различных видов рыб есть про-
изводное от действия целого ряда как внешних, так и внутренних
факторов. Возрастной состав зависит, как уже указывалось выше,
от естественной продолжительности жизни особей. Этим же факто-
ром определяется частично и соотношение возрастных групп.
Основные факторы, которые определяют количество особей
отдельных возрастов в популяции, это урожайность отдель-
ных годов, смертность рыб отдельных возрастных групп под влия-
нием различных факторов и интенсивность вылова. Характер
распределения отдельных возрастных групп в тех случаях, когда
влияние промысла отсутствует, зависит исключительно от естествен-
ной смертности и пополнения. Размерный состав в популяции ти-
бетского гольца рыбы, неиспользуемой промыслом, имеет следующий
вид (рис. .101). Растянутость или компактность этой кривой раз-
меров и возрастного состава у различных видов рыб зависит почти
исключительно от продолжительности жизни и роста особей. В
промысловых уловах, в которых младшие возрастные группы обыч-
но отсутствуют, кривая распределения рыб по возрастам носит
несколько иной характер, чем в популяции. Но, поскольку при
практической работе обычно приходится оперировать с возрастным
составом промысловых уловов, и для оценки состояния запасов
151
стада наиболее важное значение имеют старшие возрастные группы,
поэтому в рыбохозяйственной литературе обычно приводится воз-
растной состав уловов, который, как указано, отличается от воз-
Длина в сантинетРах
Рис. 101. Кривая распределения по длине популяции
тибетского гольца Nemachilus stoliczkai (из Никольского,
1938).
растного состава популяции отсутствием или незначительным коли-
чеством молодых, обычно неполовозрелых особей.
Важным моментом, определяющим возрастной состав стада
рыбы, является урожайность поколений отдельных годов. Она
зависит, как мы видели выше, от ряда факторов. В зависимости
от влияния отдельных внешних факторов урожайность отдельных
годов очень сильно варьирует. Как видно из диаграммы, для воблы
северного Каспия наиболее урожайным был 1931 год (рис. 102)
(Монастырский, 1938). Этот урожайный год отразился на промысло-
вых уловах, начиная с 1933 года, дав в 1933 г. значительный
процент двухлеток, а в 1934 г трехлеток (рис. 103). Урожайные
по молоди рыб годы сказываются естественно не только на соотно-
шении возрастных групп в промысловых уловах, но и на величине
этих уловов. Подобные примеры зависимости величины уловов от
урожая молоди очень многочислены, и на основании величины
урожая молоди можно с большой долей вероятности делать прогнозы
величины улова в последующие годы. Поэтому на учет урожай-
ности молоди промысловых рыб и у нас и заграницей сейчас обра-
щается большое внимание.
Вторым фактором, определяющим соотношение возрастов в по-
пуляции, является гибель отдельных возрастных групп от тех или
иных причин. Довольно обычной причиной гибели служат эпизо-
отии, уничтожающие часто значительную долю стада и, в зависимо-
152
сти от характера заражения, поражающие или взрослых, или мо-
лодых рыб (конечно преимущественно в том случае, если заражение
происходит через пищу). Оказывают влияние на соотношение воз-
Рис. 102. Среднее количество мальков воблы на час градирования в север-
ном Каспии. Вертикальная штриховка — Волжский район, горизонтальная
Урало-Эмбенский, без штриховки — среднее по всему северному Каспию (пз
Монастырского, 1938)
растов также и хищники, потребляющие, как правило, рыб опреде-
ленных размеров; поэтому урожай того или иного хищника, скажем,
в Каспии, сома или судака, приводит к более значительному уничто-
жению то одних, то других возрастных групп таких промысловых
рыб, как вобла и сазан. Из факторов неорганической среды необ-
ходимо указать влияние заморов. Более сильные заморы, уничто-
жающие большое количество рыбы, приводят в последующие годы
к выпаданию из популяции старших возрастных групп, почему в
уловах возрастные группы рыб годов рождения, предшествующих
замору, характеризуются очень малым удельным весом. Но несо-
мненно основным фактором, определяющим соотношение отдел! ных
возрастных групп в популяции рыб, имеющих хозяйственное значе-
ние, является промысел. Интенсивный промысел может привесы
к тому, что рыба будет вылавливаться в основной массе на не] вых
годах жизни и не доживать до старших возрастов. Поскольку вылов
будет базироваться главным образом на рыбах, еще не достигших
ноловозрелости и ни разу не размножавшихся, постольку небольшое
количество особей, избежавших орудий лова и давших приплод,
не сможет покрыть вылов, в результате чего в более или менее не-
продолжительный срок должен будет произойти подрыв запасов и
прекращение промысла. Подобные примеры значительного под-
рыва запасов весьма обычны.
Так, например, в результате слишком интенсивного промысла
сейчас совершенно подорваны запасы и прекращена добыча амери-
канского пресноводного осетра Acipenser fulvescens и немецкого
осетра Acipenser sturio в реках Западной Европы. ^Отражаясь на
сокращении в популяции рыб более старых возрастов, промысел
естественно влияет и на величину вылавливаемых особей. Интенсиь-
153
ный промысел вызывает постепенное измельчание рыб и, наоборот,
некоторое (правда, до известных пределов) ослабление промысла
%г
60
50.
40
30
20
10
О
60
50’
40
30
20
10
о
60 •
50
40
30
20
10
О»
60
50
40
30 •
20
10
0 =
60-
50
40 •
30
20’
10-
qSSI
60-
50’
40’
30
1927 г
1928 г
1932г
1929г
1931г
Д3_.гтп______
1930 г
Рис. 103. Изменение возраст-
ного состава воблы Северного
Каспия в зависимости от уро-
жайности отдельных поколе-
ний (из Монастырского, 1938).
йозмстЗ 3 4 5 6 7 8 9
вызывает увеличение размеров. Поэтому показательными для суж-
дения о состоянии запасов рыб являются и те изменения в размерах,
которые происходят в результате интенсификации вылова. Две
тысячи лет тому назад на Волге промысел рыб был почти не развит, и,
естественно, что он не мог вносить изменения в возрастной состав
и размеры популяции отдельных видов. В дальнейшем с интенси-
фикацией вылова начинается сильное уменьшение размеров вылав-
ливаемых особей и снижение их возраста. Так, как видно из приве-
154
денного графика (рис. 104), размеры стерляди, вылавливаемой две
тысячи лет тому назад в Волге, были в среднем значительно круп-
нее, чем вылавливаемой теперь. Ин-
тересно, что максимальные разме-
ры стерляди и две тысячи лет тому
назад и в настоящее время остались
одинаковыми. Это указывает на то,
что продолжительность жизни в
естественных условиях у стерляди
за промежуток времени в две тыся-
чи лет осталась неизменной.
Для камбалы Pleuronectes pla-
te ssa Варенцова моря удается под-
метить в уловах сильное умень-
шение удельного веса крупных осе-
ней за период с 1907 по 1935 г
(рис. 105). Уменьшение это явилось
результатом интенсификации про-
мысла, имевшей место за это время
(Милинский 1938). Контроль за из-
менением возрастного состава и раз-
меров промысловых рыб проводится
ными промысловыми видами, как у
%.
Рис. 104. Распределение по раз-
мерам волжской стерляди в уло-
вах 2000 лет тому назад (точеч-
ный пунктир) и в настоящее вре-
мя на средней Волге (сплошная
линия),в низовьях Ветлуги (пун-
ктир) (из Nikolski j, 1936).
сейчас почти над всеми основ-
ное, так и заграницей, и слу-
Рис. 105. Изменение размеров морской камбалы Pleuronectes platessa в уло’
вах в Баренцевом море. Наверху самцы, внизу самки. Сплошная линия —
1907, пунктир — 1935 (из Милинского, 1938).
жит, в первую очередь, для предупреждения возможного не-
долова или перелова и прогнозов вылова на последующие сезоны.
Колебания численности рыб (
Мы разобрали те факторы, которые определяют возрастной
состав популяции отдельных видов рыб. В заключение обзора дина-
155
мики возрастного состава необходимо остановиться на тех причи-
нах, которые вызывают колебания численности отдельных видов и
практически выражаются в колебаниях уловов.
То, что величина уловов из года в год претерпевает изменения и
что у многих видов рыб удается установить известную периодич-
ность уловов, было известно еще давно. Впервые детально на этом
вопросе остановились норвежские исследователи Гелланд Гансен и
Нансен (1909), которые проследили, что периодические колебания
уловов трески находятся в зависимости от температуры Гольфштро-
ма, причем, как было установлено впоследствии исследованиями Дер-
жавина (1922), Йенсена (1927), Берга (1935) и др., для большинства
промысловых рыб колебания уловов имеют наибольшие максимумы
через 9—11 лет. Десяти-одиннадцатилетний период максимумов уло-
вов установлен для самых различных по образу жизни рыб: для осет-
ровых, сельдей, лосося, трески и др., причем, как это устаноь-
лено для лосося, периоды максимальных уловов не всегда сов-
падают по всей области распространения. Так, по сравнению с
Норвегией, в наших водах уловы лосося часто опаздывают на
год, но периодичность уловов и в Канаде и в СССР одна и та же,
и разница во времени наступления максимумов уловов редко бы-
вает больше года. Обычно же максимумы и минимумы совпадают
по всей области распространения объекта промысла. Так, 1928 г
дал минима; иные уловы лосося во всех промысловых районах.
Приведенные примеры периодических колебаний уловов связаны
с общими климатическими причинами. Формы влияния этих общих
климатических причин могут быть весьма различными, или через
посредство сокращения кормовой базы, или ухудшения условий
нереста, изменения зим( вки и т. д. Но имеются, как периодиче-
ские, так и не периодические колебания уловов, причины которых
носят местный характер. Как мы отмечали выше, урожайность
молоди, колебания численности хищников или конкурентов из-за
пищи, эпизоотии и многие другие причины оказывают также весь-
ма сильное влияние на динамику величины того или иного ст; да
рыб и определяются не только общими климатическими прич! -
нами. Так, например, величина уловов судака Lucioperca lucioperca
в Азовском море находится в непосредственной зависимости от
\ рожая его основного объекта питания— тюльки Clupeontlla
delicatula (Майский 1938). Влияние эпизоотий хорошо иллюстри-
руется примером шипа Acipenser nudiventris, который в конце
1935 г. был заражен в Аральском море паразитическим червем
Nitzschia sturionis; в результате весной 1936 г. имела место мас-
совая гибель рыб, после чего уловы резко упали и находились на
низком уровне длительное время.
Изложенное показывает, что для решения рыбохозяйственных
вопросов и, в частности, выяснения причин изменения темпа роста
рыб, динамики возрастного состава и динамики численности по-
пуляции отдельных видов рыб, т. е. запасов объектов промысла,
требуется знание, как всего комплекса окружающих абиотических
и биотических факторов среды, так и самого организма рыбы.
1Г6
Без этих знаний решение основных рыбохозяйственных задач
окажется невозможным.
ЭКСТЕРЬЕР И УПИТАННОСТЬ РЫБ
Как уже указывалось выше, упитанность разных рыб под-
вержена весьма различным изменениям. У некоторых рыб в течение
года происходят весьма сильные изменения упитанности и жир-
ности тела (например лососевые), у других эти изменения совер-
шенно ничтожны (например Cichlidac).
При сравнении упитанности одного и того же вида в ихтиологии
принято обычно пользоваться отношением веса к длине рыбы. Чем
эга величина больше, тем рыба считается упитаннее. Естественно,
что определяемая, таким образом, упитанность очень сильно зависит
от экстерьера рыбы. У рыб высокотелых и в то же время имеющих
широкую спину отношение веса к длине больше, чем у рыб про-
гоннстых.
Для математического выражения величины упитанности обычно
в настоящее время применяется предложенная Фультоном (1902)
формула:
1У-Ю0
где W —вес тела рыбы, L — се дл^на и Q — коэфициент упитан-
ности. Длина берется в кубе, потому что приращение веса идет
пропорционально приращению объема рыбы. Пользуясь коэфиНиен-
том упитанности Фультона, удается выявить сезонные изменения
упитанности рыбы, изменения упитанности в зависимости от воз-
раста и пола рыбы и разницу в упитанности одного и того же вида
в различных водоемах, что может служить одним из показателей
продуктивности водоема. К сожалению, пользуясь общим весом
тела при вычислении коэфициента упитанности,.не удается устранить
искажающего влияния веса гонад, достигающих иногда более 15%
веса тела рыбы, а также величины содержимого кишечника, дости-
гающего у многих рыб значительной величины до 30 и более про-
центов от веса тела. Обе эти величины могут очень сильно изменить
величину коэфициента упитанности и часто скрыть действи-
тельную картину изменения упитанности рыбы. Для устранения
искажающего влияния веса гонад и пищи, содержащейся в кишечни-
ке, Кларком (1928) было предложено вычислять коэфициент упитан-
ности, пользуясь весом рыбы без внутренностей. Применение этого
способа позволяет устранить влияние веса гонад и съеденной пищи,
но зато при удалении внутренностей с кишечником и другими ор-
ганами удаляется одновременно внутренний жир, количество кото-
рого подвержено также сильным изменениям и в значительной мере
связано с упитанностью рыбы. Поэтому при изучении упитанности
рыбы лучше пользоваться параллельно обоими показателями, вы-
числяя коэфициент упитанности, как По весу всей piii6bi, так и рыбы
без внутренностей.
157
У различных рыб величина коэфициента упитанности варьирует
очень сильно от 0,30—0,40 у длинных змеевидных или лентовидных
рыб и до 4,00 и более у рыб, приближающихся по форме к шару.
Как известно, с возрастом рыбы претерпевают весьма существенные
изменения формы тела, что, естественно, приводит к изменению
коэфициента упитанности. У некоторых рыб величина коэфициента
упитанности уменьшается с возрастом (например у Trachypterus),
у других—увеличивается (вобла), у третьих же до достижения
определенного размера коэфициент упитанности увеличивается,
а затем начинает уменьшаться (сазан). Самцы от самок у большин-
ства рыб также обычно довольно сильно отличаются по величине
коэфициента упитанности, но при сравнении коэфициента упитан-
ности у разных полов необходимо определять упитанность не толь-
ко по общему весу, но и по весу рыбы без внутренностей, так как в
противном случае можно легко впасть в ошибку, получив 66; ьшую
«упитанность,) самок за счет большего веса половых продуктов.
Наиболее важно изучение сезонной динамики коэфициента упи-
танности, позволяющее установить, с одной стороны, время, когда
рыба представляет наибольшую ценность, как объект промысла
и с другой —связь величины коэфициента упитанности с поведе-
нием рыбы (интенсивность питания, миграции и т. д.).
Так же как и упитанность, весьма важным показателем состоя-
ния рыбы и ее ценности, как пищевого продукта, является жир-
ность. Жирность рыбы естественно различна у различных видов
и подвержена так же, как и упитанность, весьма сильным сезон-
ным и возрастным изменениям, связана к тому же с полом и зависит
от условий обитания. Наиболее точным методом учета величины
и динамики жиров в теле рыбы является, конечно, химический
анализ. Путем химического анализа определено количество жира
в тканях очень многих рыб, и выявлен целый ряд весьма интересных
и практически важных закономерностей в-динамике жирности,
как по сезонам, так и в отдельных водоемах. К сожалению, этот
метод очень сложен, требует громоздкой аппаратуры и специальных
знаний, почему далеко не всегда может быть применен при мас-
совых исследованиях в полевой обстановке. Поэтому при ихтиоло-
гических исследованиях часто приходится пользоваться более
простыми способами, позволяющими с большей или меньшей точ-
ностью судить о жирности рыбы. Мы остановимся лишь на неко-
торых из них. У тех рыб, у которых жир во время нагула отла-
гается в значительном количестве на внутренностях и сравнг-
тсльно легко может быть удален (например у леща) для определения
степени жирности рыбы, вес «нутреного» жира выражают в про-
центах от веса всего тела. Изменения этого показателя в значи-
тельной степени отражают изменения общей жирности рыбы.
Для сельдей Портом (1909 г.) предложено оценивать жирность в
баллах, основываясь только на вскрытии рыбы. Пользуясь шкалой
Иорта, можно давать лишь в первом, однако, достаточном для хо-
зяйственных целей приближении характеристику жирности
сельди.
158
Как упитанность, так и жирность различных рыб подвержена
весьма сильным колебаниям. Как видно из таблицы, процент
жира в теле различных промысловых рыб может варьировать от
0,2% у пикши до 22,0% у угря.
Средний процент жира в теле различных промысловых рыб
Осетр 8,0% Вобла 2,5 Бычки 1,2
Севрюга 11,0 Лещ 5,5 Судак 0^5
Салака 4,5 Сазан 1,5 Окунь 0,7
Лосось . 11,0 Сом 4,0 Палтус 5,0
Кета 7,0 Щука 0,5 Камбала 2,0
Белорыбица 18,0 Угорь речи. 22,0 Треска 0,3
Сиги 1,5 Кефаль 1,5 Пикша 0,2
Снеток . 3,0 Скумбрия 8,0 Навага . 0,5
Жирность также изменяется в зависимости от возраста и пола
рыбы. Так, например, у черноморских сельдей процент жира с
возрастом меняется: от 12% у мелкой сельди (промысловой но-
менклатуры от 300 до 1000
шт. в 16 кг) до 20% у круп-
ной (около 150 шт. на 16 кг).
То же наблюдается и у мнг-
гих других рыб. Так, у
леща в северном Каспии у
рыб трехлетнего возраста
жир составляет в среднем
4,85% от веса тела, а у се-
милетних 8,2%.
Сезонные изменения жир-
106. Схема соотношения икры
жира у сельди в течение года.
схема соотношения размеров
ности также очень велики _
Рис.
и весьма тесно связаны со
сроками питания и размно-
жения. На рисунке представлена
половых продуктов и жира в теле океанической сельди в течение
года. Как это видно из рисунка, рост величины гонад и коли-
чества жира сначала идет параллельно, но, начиная с момента
прекращения питания, количество жира быстро уменьшается, рост
же гонад, продолжается (рис. 106).
ПИТАНИЕ РЫБ
По характеру потребляемой пищи рыбы чрезвычайно разно-
образны. Как величина, так и характер пищи у рыб варьирует
больше, чем в каком-либо другом классе позвоночных. По харак-
теру питания рыб принято разделять на: 1) растительноядных,
как например, закаспийская хромуля Varicorhinus heratensis; 2)жи-
вотноядных, питающихся мелкими беспозвоночными, как на-
пример, лещ, сиги и др., 3) хищных, питающихся рыбой, как на-
пример, сельдевая акула, щука, сом и др. Это деление, естественно,
весьма условно, так как большинство рыб питаются смешанной
пищей, и часто животноядные рыбы могут становиться хищными,
15»
или наоборот. Разнообразие в характере пищи естественно связано
с чрезвычайным разнообразием в приспособлениях как к
добыванию пищи, так и к ее усвоению.
Приспособления к добыванию пищи
Строение ротового аппарата, служащего для захватывания,
размельчения и проглатывания пищи, изменяется от воронки у
Рис. 107. Различное по-
ложение рта у карповых
рыб, сверху вниз: верх-
ний рот — чехонь Ге1е-
cus cultratiis, конечный
рот -сазан Cyprinus саг-
pio, полунижний рот —
вобла Rutilus rutilus,
нижний рот — осгро-
лучка Capoetobrama
kuschakevitschi.
миноги до «клюва» у сростночелюстных.
Но все же все разнообразие в строении ро-
тового аппарата может быть объединено в
несколько типов: 1) рот х в а т а т е л ь-
н ы й, широкий с острыми зубами как на
челюстных костях, так часто на сошнике и
небных костях. Жаберные тычинки корот-
кие, редкие, служащие только для предо-
хранения жаберных лепестков от повреж-
дения пищей, но не для отцеживания
пищевых объектов; кишечник, как правило,
короткий, желудок обычно обособлен; к
этой группе относятся хищные рыбы: су-
дак, сом, щука и многие другие; 2) р о т в
виде присоска, челюстей нет, зубы
в виде ороговевших бугорков; на языке то-
же есть «зуб», который и служит основным
органом вгрызания; к этой группе отно-
сятся полупаразиты—миксины и миноги;
3) рот всасывательный в виде
более или менее длинной трубки, иногда
выдвижной, как правило без зубов, слу-
жащий обычно для питания донными бес-
позвоночными, реже беспозвоночными тол-
щи воды; к этой группе относятся, например,
Mormyridae, лещ, морские иглы и ряд дру-
гих; 4) р о т дробящий иногда в виде
клюва (кузовки) с мощными зубами, иногда
в виде пластин (скаты орляки), или шипов
(зубатки), служит, как правило, для раз-
дгабливания твердых панпырей беспозво-
ночных: моллюсков, иглокожих, корал-
лов; 5) рот планктоноед а, обычно
большой или средней величины, как пра-
вило, не выдвижной, зубы мелкие или
вовсе отсутствуют. Жаберные тычинки
длинные, действующие как сито (рис. 108),
к этой группе относятся сельди, сиги, не-
которые карповые и др., 6) рот п е р и-
ф и т о н о е д а — питающегося раститель-
ными обрастаниями. Расположен на ниж-
ней стороне головы, имеет вид поперечной
160
Рис. 108. Жаберный аппарат сига
Coregonus lavaretus (из Берга, 1932).
щели, нижняя губа несет обычно острый режущий край иногда
одетый роговым чехликом. Зубов обычно нет. К этой’ группе
относятся подуст, храмуля, некоторые маринки и т. п.
Таковы основные типы строения рта, естественно связанные
между собою рядом переходов. Как исключение у некоторых рыб
в связи с переходом к парази-
тическому образу питания рото-
вое отверстие редуцируется; так,
самцы многих Ceratiidae, как на-
пример, глубоководной рыбы
Edriolichnus schmi dti, срастаются
кожей ротового отверстия и язы-
ка с телом самки и питаются за
счет тех соков, которые посту-
пают из кровеносных сосудов
самки. Кишечник у этих рыб
дегенерирует. Самки же питаются
нормально, как и другие рыбы,
и органы пищеварения у них не
претерпевают изменений.
В дополнение ко всему вышеуказан-
ному следует упомянуть, что в 1909 г.
немецким ученым Пюттером было вы-
двинуто положение, согласно которому
большинство водных животных, в том числе и рыб, питается не только путем
заглатывания оформленных кусков пищи, но также и путем усвоения раство-
ренных в воде питательных веществ, проникающих внутрь тела, через его стен-
ки; иначе говоря, питание водных животных происходит также, как у гете-
ротрофных растений и у эндопаразитов, живущих все время в окружении
растворенной пищи. Свою теорию Пюттер распространил и на рыб, указывая,
что и у этих животных имеет место всасывание питательных веществ из воды
через жабры и кишечный тракт. Последующие работы, и в частности работы
Крога показали, что теория Пюттера, во всяком случае в том объеме, как
это предполагал ее автор, едва ли соответствует действительности. Крог до-
казывает, что у рыб поглощение может итти только через кишечный тракт,
что, кроме того, органические вещества, растворенные в воде водоемов
земного шара, представляют собой соединения, недоступные для использо-
вания животными. Поэтому, пожалуй, будет правильным сказать, что вв
всяком случае в отношении рыб теория Пюттера представляет сейчас, пови-
ли мому, уже главным образом исторический интерес.
Кроме разнообразия в характере строения ротового аппарата,
в основном зависящего от характера поедаемой пищи, сильно из-
менчиво и положение рта, связанное прежде всего со способом
добывания пищи. Рот может быть верхний, т. е. лежать выше
осп тела, как например, у чехони, коне ч н ы й, расположен-
ный па оси тела, как / ельца, сельдей и многих других рыб, и
нижи и й, расположенный с нижней стороны тела, как напри-
мер, у скатов, осетровых и многих других.
Для осязания пищи у рыбы, как правило, имеется вокруг рта на
рубах очень большое количество чувствующих клеток, иногда распо-
ложенных на специальных выростах—усах. Число усов и их величина
11 Биология рыо 161
очень сильно варьируют; так, у тресковых Gadidae имеется
всего один непарный усик, расположенный на нижней челюсти.
При этом у рыб этого семейства, питающихся преимущественно
донными беспозвоночными, этот усик длиннее,а у хищников—короче.
У некоторых сомов число усиков достигает до 16 шт., и иногда
некоторые усики бывают длиннее тела рыбы, как например, у
индомалайского сома Siluredcs eugeniatus. У Mcrmyndae на конце
хобота имеется вырост, иногда доволвно значительных размерив,
богатый чувствительными окончаниями. Роясь своим длинным ры-
лом в донных отложениях, Mcrmyridae, благодаря наличию на кон-
це хобота большого количества чувствующих окончаний, легко
нащупывают зарывшихся беспозвоночных, которыми они питаются.
У некоторых, главным образом, глубоководных рыб лучи парных
или спинного плавников сильно удлиняются и превращаются в ор-
ганы осязания. Так, у глубоководной рыбы Bathypterois ater лучи
грудных плавников вытянуты в длинные нити, в полтора раза
превышающие длину рыбы и богатые чувствительными окончаниями.
У многих представителей лабиринтсвых AnabantJdei брюшные
плавники вытянуты в длинные нити и служат органами осязания.
У многих ногоперых Lophnformes сильные изменения претерпе-
вают первые лучи спинного плавника, они очен > сильно удлиняются,
часто превышая длину тела рыбы, и на конце их возникают сьое-
образные особые образования, част о снабженные светящимся органом,
с помощью которых рыбы подманивают cloio добычу.
Жесточайшая борьоа за существование, которая ведется в приро-
де, приводит к тому, что объекты питания рыб всячески приспо-
сабливаются к тому, чтобы не быть съеденными, а рыбы приспо-
сабливаются к тому, чтобы их сьесть. Отсюда понятно, что разно-
образие в способах добывания пищи у рыб чрезвычайно велико.
Многие хищники непосредственно преследуют свою добычу (жерех,
судак), догоняя ее, другие, как например сом, подкарауливают и
хватают свою жертву, когда та приближается. Брызгун Toxotcs peu-
lat.r, питающийся воздушными насекомыми, сбрызгивает их струей
изо рта с прибрежной растительности. Высота струи часто бывает до
полуметра и больше. У некоторых рыб для поражения своих объектов
питаш я вырабатываются специальные приспособления: так, у меч
рыбы Xiphias gL.d.us верхняя челюсть вытягивается в мечевидный
отросток, при помощи которого рыба поражает своих жертв. У iiiij.bi
рыбы PriStiS и пилоноса Pnstiophcrus рострум выт$ н\ т в пилообраз-
ный вырост, при помощи которого эти рыбы поражают мелких пела-
гическг х рыб, служащих им лицей. Электрический угорь Elcetn-
pherus elec tri с us, прежде чем схватить свою жертву, поражает ее
электрическим разрядом при помощи специального органа, являю-
щегося видоизмененной мускул ьной тканью и расположенного вдол ь
всего тела рыбы. Электрический разряд, достигавший у крупных
особей, как указывалось выше, до 300 вольт, уторь может произво-
дить произвольно, но после нескольких разрядов нуждается в неко-
тором перерыве, так как на время теряет способность давать разряд.
162
Донные рыбы имеют целый
грунте и отыскивания корма.
Mormyridae, имеющие длинное
клювообразное рыло, употреб-
ляют его для зондирования
грунта. Карповые рыбы тоже
сильно перекапывают грунт,
причем поисковая способность
у них развита очень сильно.
Так, по опытам Суетова (1939),
карп за 3 дня пребывания в
аквариуме с толщиной ила в
8 см, оставляет невыбранными
из 112 личинок мотыля 1—2 шт.
Глубина толщи грунта, на ко-
торой могут доставать свой корм
рыбы, также различна у раз-
личных видов. По данным
Суетова (1939) лещ может про-
ряд приспособлений для рытья в
Рис. 109. Проникновение карпов в
толщу различных грунтов при поиске
корма (из Суетова, 1939).
Рис. ПО. Глоточные зубы карповых. Верхний ряд
(слева направо): усач Barbus brachycephalus и са-
зан Cyprinus carpio, нижний ряд — лещ Abramis
brama и жерех Aspius aspius.
11*
никать вглубь грунта
на 5 с л/, карпдо 1 Ьсм,
а окунь почти не бе-
рет корм, закопав-
шийся в грунт. Од-
нако, степень исполь-
зования корма рыбой
на различней глу-
бине неодинакова.
Так, как видно из
приведенной диа-
граммы (рис. 109),
карп может выбирать
значительное коли-
чество объектов в
илистом грунте до
глубины в 10 см, глу-
бже этого слоя оста
ется неиспользован
ним весьма знач! -
тельный процент кор-
мовых объектов.
У планктонояд-
ных рыб питание в
значительной степе-
ни носит полупас-
сив ный характер.
Наиболее резко это
выражено у толпыги
163
Hypophthalmichthys molitrix, у которой основной кормовой объ-
ект — планктонные водоросли — вместе с водой для дыхания по-
ступает в ротовую полость, отфильтровывается сеткой жаберных
тычинок и перемещается дальше в кишечник. Активное преследо-
вание или поиски добычи здесь отсутствуют.
Зубы рыб, как мы видели выше, чрезвычайно разнообразны по
своему строению, начиная от кинжаловидных у щуки и сельдевой
акулы и кончая трущими пластинками у многих скатов. У тех рыб,
у которых зубы на челюстях отсутствуют или развиты слабо, зубы
обычно развиваются на глоточных костях. Так, у карповых на
нижнеглоточных костях развивается от одного до трех рядов зубов,
Весьма сильно варьирующих, как в отношении формы, так и числа.
В зависимости от рода пищи Геккель (1843) разделяет глоточные
зубы на четыре типа: 1) выдолбленные —excavati тип усач, 2) же-
вательные m sticatorii тип сазан, 3) крючковидные с жевательной
площадкой unc.nato subm lares тип лещ, 4) крючковидные без жева-
тельной площадки — uncinato conici тип голавл и жерех (рис. 110).
В свою очередь эти типы подразделяются еще на ряд более дробных.
Над нижнеглоточными зубами, на нижней стороне мозгового черепа
у карповых рыб имеется образование из рогового эпителия в виде
небольшой мозоли, так называемый жерновок. В паре с жерновком
и работают нижнеглоточные зубы. У таких рыб, как например,
губаны Labridae, у которых имеются и верхне-и нижнеглоточные
зубы, жерновок отсутствует.
Усвоение пищи и пищевой рацион
Столь большое разнообразие В строении ротового аппарата
находится в непосредственной связи с разнообразием в строении
кишечного тракта у рыб. У хищников, как прййило,- имеется ко-
роткий кишечник и хорошо развитый Желудок. У ж'и'вотноядных и
растительнрядцых рыб желудоХ бывает сильно редуцирован, или
как например, у карповых, мож^Т совсем отсутствовать. Даже у
представителей одного и того же семейства характер пищи нахо-
дится в непосредственной связи с1 характером строения ки-
шечного тракта. У различных видов се мейства CicMidae величина
Желудка и длина кишечника непосредственно бвя-заны с образом
питания, свойственным тому или другому хчиду (рис. 111). У расти-
тельноядных форм, нуждающихся в большей всасывающей поверх-
ности, кишечник, как правило, длиннее чем у хищников. Наиболее
длинный кишечный тракт имеют рыбы, питающиеся детритом,
растительными обрастаниями и фитопланктоном'. Так, кишечный
тракт закаспийской храмули Varicorhinus her.it.nsis> основным объ-
ектом питания которой является Детрит и перифитон, составляет
около 700% от длины тела. У толпыги HypophthalmicMhys molitrix,
питающейся почти исключительно фитопланктоном,- кишечный
тракт в тринадцать раз превышает длину рыбы. У древних прими-
тивных рыб, как акулы, двоякодышащие и осетровые, увеличение
всасывательной поверхности кишечника идет путем образования
164
спиральной складки, у различных рыб имеющей различную кру
тизну и размеры.
В трсной зависимости от
характера питания находится
и количество пилорических
придатков у рыб. Так, хари-
ус Ангары Thymallus arcticus
baicalensis ваicalensis, питаю-
щийся главным образом Amphi-
poda и воздушными насекомы-
ми, имеет в среднем 15,3 пило-
рических придатка. Хариус из
Байкала Th. arct. baical. brevi-
pinsis, питающийся главным об-
разом рыбой, имеет в среднем
19,1 пилоричеких придатка. Та
же картина наблюдается у кас-
пийских сельдей,у которых чис-
ло пилорических придатков и
число и характер тычинок на
жаберных дугах связаны с
планктонеядным или хищным
•бразом питания. Как прави-
ло, у хищников число пилори-
ческих придатков больше, а >
их меньше.
Рис. 111, Кишечники различных пред-
ставителей сем. Cichlidae (Perciformes).
Сверху вниз — хищник Cichla temen-
sis, животноед — Geophagus brasilien-
sis, растительноядный — Tilapia heu-
deloti (из Pellegrin, 1921).
:аберные тычинки короче и числе
Связь между характером питания и числом тычинок и пилорических
придатков у каспийских сельдей (по Световидову, 1932)
Вид сельди Число тычинок Среднее число пило- рических придатков Характер пищи
Долги нка
Caspialosa brashnikovi 31-32 51,16 Рыба
Большеглазый пузанок
Caspialosa saposhnikovi 30-31 45,94 Рыба
Бешен ка
Caspialosa kessleri 7и—80 48,16 Рыба и планктон
Волжская сельдь
Caspialosa volgensis НО (средн.) 44,28 Рыба и планктон
Каспийский пузанок
Caspialosa caspia 85—138 40,84 Планктонные ракообразные
У некоторых планктоноядных рыб (многие кефали, сельди и
др.) пилорическая часть кишечника утолщается и, видимо, служит
165
для перетирания панцирей пелагических ракообразных (Свето-
видов, 1932).
Естественно, что разница в характере пищи и в строении кишеч-
ника приводит к различию и в процессе пищеварения. Уже срав-
нение действия ферментов в кишечнике хищной и мирной рыбы
показывает значительные отличия.
Ферменты кишечного тракта хищных и мирных рыб
(по Вундер, 1936)
Ферменты переваривающие
Характер питания Орган Углеводы Жиры Белки
Хищные рыбы желудок1 — — Пепсин, соляная
кислота
(с желудком) Пищеварительные Амилаза Липаза Трипсин
железы
Кишечник — — Энтерокиназа
Мирные рыбы эрепсип
(без желудка) Пищеварительные Амилаза Липаза Трипсин
железы мальтаза
Кишечник — — Энтерокиназа
эрепсин
Не только у различных групп, но и среди рыб, принадлежащих
к одной и той же группе, может нисколько изменяться как характер
распределения фермента, так и его свойства. У близких форм
Арктики и умеренных широт изменяется холодоустойчивость не-
которых ферментов (трипсин), которые у холодноводных рыб, как
правило, более холодостойки, чем у рыб, живущих в более теплых
водах (Коштоянц и Коржуев 1937). У различных групп рыб
меняется и сила действия различных ферментов; так, у карпа панкре-
атическая амилаза действует во мнего раз (около 150) сильнее,
чем у щуки (Банк 1927). У колючей акулы действие амилазы
близко по силе к таковому у щуки.
В связи с отсутствием желудка (карповые) или его малой вели-
чиной (некоторые Cichlidae) мирные рыбы обычно лишены возмож-
ности потреблять одновременно значительное количество пищи;
для нормального питания ni ща должна у них поступать часто не-
значительными порциями. П этому у карповых и других безжелу-
дочных рыб обычно не бывает резкой разницы в интенсивности
питания в различное время суток. Так, например, вобла, имеющая
в достаточном количестве корм, потребляет пищу, примерно, через
каждые четыре часа (Бокова 1938). Хищные рыбы питаются значи-
тельно реже, и иногда промежуток между отдельными кормежками
бывает более суток.
У мирных и хищных рыб различен не только химизм пищева-
рительного процесса, но неодинакова и механика разрушения пи-
щ.вого комка. В то время как у хищных рыб разрушение пищево-
го комка происходит то; ько в желудке (треска, бычки Cot-
tide), у мирных рыб разрушение пищевого комка даже при наличии
желудка, как например, у речной камбалы Pkuronectes flesus, про-
исходит и в кишечнике (Карпевич и Бокова 1937).
166
Величина кусков пищи, заглатываемых хищными рыбами, очень
сильно варьирует Иногда, особенно у глубоководных рыб, как
напоимер, Chiosm^dus, жертва может в два и белее раза превышать
размеры хищника. Отношение величины кормового объекта и хищ-
ника лаже у одного и того же вида бывает подвержено сильным
изменениям. Обычно у большинства хищников по мере их рос-
та число штук кормовых объектов, находящихся одновременно в
же гудке, уменьшается, а величина отдельного съеденного орга-
низма увеличивается.
Меняется величина пищевого объекта и в зависимости от време-
ни гнпл: та1<, треска зимой открывает рот менее широко и потреб-
ляет более мелкую пищу, чем летом (Маккензи 1939).
Большое хозяйственное значение имеет знание суточного раци-
она пыбы и интенсивности потребления рыбой корма в течение
гота Д^я определения величины наполнения кишечника, а тем
самым, по известной степени, и интенсивности питания, сбычно
попкзуются индексом наполнения кишечника, выражающимся от-
ношением веса содержимого кишечного тракта к весу рыбы, причем
некптопые яртопы выражают это отношение в процентах веса тела
(н^ппммер,, Никитинский, 1929), другие же в десятитысячных долях
(Зенкевич 1933, Шорыгин 1939 и ДР ).
Суточный рацион, естественно, у различных рыб весьма раз-
личен, но даже у одного итого же вида, он подвержен чрезвычайно
сильным изменениям и находится в зависимости от целого ряда
факторов. Так, суточный рацион стоит в непосредственной зависи-
мости от возраста рыбы. Годовалая щука потребляет в день мотыля
Chi голоmus в количестве 15—25% своего веса. В то же время
двухгодовалая щука потребляет того же корма только 9% своего
веса (Шольц 1932). ^Вообще количество потребляемого" корма, от-
несенное к единице веса рыбы, находится в обратной зависимости
от ее размеров. Весьма сильно зависит пищевой рацион и от харак-
тера потребляемого копма; так, годовалая щука съедает в день цикло-
пов в количестве 16—17,5% своего веса, энхитрей—15—33%, моты-
ля—15—25%, бокоплавов—11—12%, рыбы—3—5% (Шольц 1932).
Каспийская вобла ппи 20° воды потребляет моллюсков Dreissena
в количестве—28.43% веса своего тела, а ракообразных Mysidae —
17 % (Ежова 1938). Различное потребление корма находится в не-
посредственной связи с его калорийностью; чем больше калорий-
ность, тем меньше количество потребленного корма. Интенсив-
ность потребления корма весьма сильно зависит также от упитан-
ности рыбы. Так, наппимер, в Апальском море осенью лещ, коэфи-
циент упитанности которого—2,3, имеет индекс наполнения кишеч-
ника в среднем 197°/000, а упитанный лещ, коэф’Щиент упитанности
которого 2,5, имеет индекс наполнения в среднем всего 86°/000,
т. е. питается вдвое менее интенсивно. Многие рыбы ьре-
кращают питание или питаются менее интенсивно во время
процесса размножения или даже во время не!рестовых ми-
граций.
167
Весьма сильное влияние оказывают на интенсивность потреб’
ления корма и внешние факторы, в первую очередь температура.
Так, оптимальной температурой для питания щуки является 19®
Вобла потребляет наибольшее количество корма в Каспийском
море при температуре 20—26°. Интенсивность потребления корма
находится также в прямой зависимости от плотности населения
кормовых объектов. До известных пределов интенсивность потреб-
Рис. 112. Интенсивность питания леща в связи с его} концентрацией на
местах питания. Сплошная линия — взрослые рыбы, пунктир — молодь,
(из Воробьева, 1937).
ления корма идет прямо пропорционально нарастанию плотности
кормовых объектов (Суетов 1939).
Интенсивность питания зависит также от концентрации кормя-
щихся рыб/, причем для некоторых рыб, как например для леща,
отмечено (Воробьев 1937), что молодь при больших концентрациях
питается более интенсивно, чем при малых, взрослая же рыба,
наоборот, снижает интенсивность питания при значительных кон-
центрациях (рис. 112). Так же как для взрослых особей леща, так
и для трески указывается, что с увеличением концентрации особей
уменьшается интенсивность потребления пищи рыбой (Бродская
1933). Все сказанное делает понятным, что количество потребля-
емого рыбой корма в течение года весьма сильно варьирует. Как
правило, зимой рыба питается менее интенсивно, чем летом. Так,
каспийская вобла наиболее интенсивно потребляет пищу в мае
И июне, к осени же интенсивность питания сильно снижается..
Сходная с воблой картина наблюдается для многих других рыб,
как лещ, сазан, судак и др.
168
Пищевые объекты и конкуренция из-за пищи
Выше мы видели, что пища различных рыб может быть весьма
разнородной: растительность, как например у храмули и красно-
перки; мелкие животные толщи воды, как у многих сигов; донные
беспозвоночные, как у воблы, леща, зубатки и многих других;
живая рыба, как например у акулы, щуки, сома и мертвая рыбаг
как у миног и миксин и, наконец, соки тела представителей того
же вида, как например, у самцов многих глубоководных Ceratiidae.
По отношению рыбы к пище последнюю можно разделить на
несколько категорий: 1) основная пища та, которой рыба обычно
кормится и которая составляет основное содержимое кишечника,
2) второстепенная пища, которая встречается постоянно в кишеч-
нике рыбы, но в менее значительном количестве, 3) случайная
пища, которая лишь изредка попадается в кишечнике рыбы. Кроме
вышеуказанных видов пищи необходимо выделить вынужденную
пищу, ту, на которую рыба переходит в результате отсутствия ос-
новной. Для определения избирательной способности рыбы но
отношению к тому или иному виду пищи вычисляется индекс из-
бирательной способности, получаемый путем деления процента
объекта питания в кишечнике рыбы на процент этого объекта в
том сообществе, где рыбы кормятся (для планктоноядных рыб—
в улове планктонной сетки, для бентосоядных — в пробе, взятой
дночерпателем-, и для хищников — в улове тралом). Если индекс
выбора больше единицы, —это значит, что кормовой объект рыбой
выбирается, если же меньше единицы, то избегается (Шорыгин
1939, Желтенкова 1939 и др.). Но при пользовании индексом из-
бирательной способности к нему нельзя подходить механически,
и необходимо учитывать ряд факторов, которые могут исказить
истинную картину. В первую очередь необходимо обратить внима-
ние на качество орудий лова, которыми производится сбор. Так,
например, медленная .тяга трала позволит более быстрым рыбам
уйти из него, что исказит соотношение видов. Дночерпатель, опус-
каемый на дно, очень часто разгоняет током воды придонных
животных, главным образом ракообразных, что также искажает
действительное соотношение кормовых объектов. Все же во многих
случаях, пользуясь индексом избирательной способности, удается
получить весьма интересные результаты. Так, например, для судака,
в открытых частях Аральского моря, основным кормом яв-
ляется чехонь, индекс избирательной способности которой дает
•громную величину 24,2; в то же время для леща в пище су-
дака индекс избирательной способности получается равным 0,37,
т. е. лещ судаком в качестве пищи явно избегается. У каспийской
воблы излюбленными объектами питания являются Dreiss^na
(индекс—13,5), Adacna minima (7,3) и Corophiidae (39,3).
Как интенсивность потребления корма, так и состав пищи,,
подвержены даже у одного и того Же вида рыб чрезвычайно силь-
ным изменениям. Основные изменения прежде всего .связаны, как
уже указывалось выше, с изменением размеров рыб. Так, например,
1б©
как видно, из приведенной таблицы, у бычка Gobius fluviatilis
pall *si количество в пище различных ракообразных чрезвычайно
сильно изменяется с возрастом рыбы. Количество бокоплавов по
мере роста рыбы сильно возрастает, а удельный вес Mysidae умень-
шается (Киналев 1937).
Изменение удельного веса различных компонентов пищи с ростом
Gobius fluviatilis pallasi
Размеры рыбы 5 см —7 —9 —11 — 13 — 15
Бокоплавы 16 ,8% 32,0 35,1 67,1 74,4
Mysidae 24,3 11,8 3,0 5,0 0,0
Состав пищи сильно варьирует и в течение года. Основными
причинами, вызывающими изменение состава пищи в зависимости
от времени года, являются динамика биомассы кормовых объектов,
а также перемещения рыб в течение времени нагула из одного
района в другой, отличающийся от первого кормовыми объектами.
Так, в Аральском море лещ весной и осенью потребляет больше
моллюсков, держащихся ближе к берегу, а летом вдали от берега
переходит на питание бокоплавами. Плотва вдали от берега летом
воздушных насекомых почти не потребляет, так как наличие верти-
кальной стратификации температуры лишает ее возможности под-
ниматься к поверхности воды. Осенью и весной, когда температура
воды у дна и у поверхности одинакова, плотва ночью поднимается
к поверхности, где питается падающими в воду воздушными насе-
комыми', главным образом, комарами-толкунцами.
Сезонные изменения удельного веса воздушных насекомых
в пище плотвы в Аральском море
Май Июнь Июль Август Сентябрь
33,08 0,08 1,64 8,3 53,0%
Сильные изменения наблюдаются и в составе пищи у рыб в
различное время суток. Вылет двукрылых, который приходится
обычно на вечерние и ночные часы, приводит к тому, что у таких
рыб, которые питаются воздушными насекомыми, как например,
чехонь, количество воздушных насекомых ночью в кишечнике будет
больше, чем днем. У леща, в связи с тем, что личинки комара-тол-
кунца поднимаются ночью в поверхностные слои грунта и стано-
вятся для этой рыбы доступными1, количество их в пище в ночное
время увеличивается за счет бокоплавов, которые ночью поднима-
ются из придонных слоев, где держится лещ, к поверхности воды и
выпадают из зоны его кормежки.
Несомненно, однако, что главнейшим фактором, вызывающим
разницу в составе пищи одного и того же вида в различных водо-
емах, является состав кормовых объектов. Так, окунь в озерах,
где есть бокоплавы, питается главным образом ими. В тех же во-
доемах, где бокоплавов нет, он поедает рыбу. Сазан в водоемах,
где есть ракушковые рачки, личинки комара-толкунца — мотыль
и моллюски, питается, главным образом, ими и растительности
не ест. При недостатке же этих кормовых объектов он переходит
на питание растительностью.
170
Как мы видели, интенсивность потребления рыбой пищи на-
ходится в зависимости от количества кормовых объектов и, с;едо-
вате;ьно, количество рыб, которое может прокормиться в водоеме,
в значительной степени зависит ст количества объектов корма.
Некоторые авторы делали даже попытку, пользуясь величг ной
биомассы кормовых объектов рыб, вычислить величину рыбной
продукции водоема. В частности шведской исс; сдовате; ь Alm
(1922, 1924) ввел так называемый F/B коэфициент F.schirci Benthos
production koefficiont, где В—биомасса бентоса, a F — рыбная про-
дукция. Но, как показали последующие исследования, подобная
зависимость значительно сложнее, и связать цифры по биомассе
живого не рыбного населения с величиной массы рыбного мяса не
представляется возможным (Ксрзгнюн 1935 г.), хотя, конечно,
величина биомассы кормовых объектов несомненно является одним
из показателей рыбной продукт! внести водоема.
Весьма важным моментом в жизни рыб является конкуренция
из-за пищи. При ограниченности запасов корма разрежение стада
рыб всегда приводит к ускорению их темпа роста. В водоемах, где
совместно встречаются лещ и ерш, ;ещ, как правило, растет хуже,
чем в тех водоемах, где ерша нет. Почти нет водоемов, где бы не
было межвидовой конкуренции из-за пищи. Так, среди донных
рыб северного Каспия, как видно из приведенной таблицы, с вов-
лой в пище конкурируют пуголовка и некоторые другие рыбы,
с лещем —два вида бычков, среди хищных рыб между собою кон-
курируют осетр, севрюга и судак.
Группы рыб
Основные пищевые организмы и их
значение в нище рыб
I. Моллюскоеды
Вобла Rutilus rutilus caspicus
Пуголовка Bentophilus macroce-
phalus
Бычок Gobius melanostomus
II. Ракоеды
Лещ Abramis brama
Бычок Gobius caspius
Бычок Gobius ftuviatilis
III. Хищники
Осетр Acipenser giildenstadtii
Севрюга Acipenser stellatus
Судак Lucioperca lucioperca
Monodaena (16%) Adaena* minima (14%)
Dreissena polymorpha (42,1%)
Monodaena (29,6%) и Gastropoda
dium edule (20,5%) Gammaridae (19%
питается как моллюсками, так и рач-
ами)
Cumacea (30%) Corophiidae (15%)
Cumacea (17%) Gammaridae (36%) Myei-
dae (16%)
Corophiidae (19%) Cumacea (19%) Gamraa-
ridae (25%)
Килька (14,1%) Пуголовка (44,2%) Core-
phiidae (7,1 %)’Gammaridae '7,^%)
Килька (36,8%) Mysidae (30,3%)
Бычки и пуголовки (55%), килька (21%)
Mysidae (10%)
Напряжение конкуренции между отдельными видами рыб мож-
но определить до известной степени путем сравнения их спектров
171
Вис. 113. Совпадение спектров
витания Gobius fluviatilis и
Benthophilus macrocephalus (из
Шорыгина, 1939).
питания, т. е. процентов по весу отдельных компонентов пищи.
Так, два наиболее распространенных каспийских бычка Gobius
fluviatilis и Benthophilus macrocephalus имеют наиболее значитель-
ную конкуренцию из-за ракообразных (рис. ИЗ), из-за моллюсков
же конкуренция ничтожна.
Шорыгин (1939) для выяснения совпадения состава пищи у раз-
личных видов рыб предлагает определять ксэфициент пищевого
сходства, который равняется сумме
меньших величин в спектрах пита-
ния. Так, если у одного вида рыб
в спектре питания нитчатые водо-
росли составляют 50%, ручейни-
ки — 10% и моллюски — 40%, а у
другого вида рыбы нитчатые водо-
росли — 75%, ручейники — 20% и
моллюски — 5%, то коэфкциент
пищевого сходства будет равен
50%-Н0%^5%, т. е. будет равен
65%. Для бычков северного Каспия
наибольший ксэфициент пищевого
сходства наблюдается между Gcbius
fluviatilis и Gobius caspius, для
которых он равен 60%, а наимень-
ший ксэфициент пищевого сходства
между Gobius kessleri и Benthophilus
stellatus, у которых он равен нулю.
Естественно, что сходство в пище
рыб подвержено, как видно из выше-
изложенного, чрезвычайно сильным
изменениям, как в зависимости ст
сезонов, так и от роста рыбы. Так,
например, если лещ и вобла в глу-
бинной зоне Аральского моря летом
конкурируют из-за пищи довольно
сильно, то в прибрежной зоне у
особей этих же видов конкуренция
)евает. Колюшка, питающаяся план-
ктоном, является конкурентом молоди большинства других рыб,
в то же время с взрослыми особями она совершенно в пише не
конкурируст.
Ксэфициент пищевого сходства дает представление об относи-
тельном значении объектов питания, из-за которых рыбы конкури-
руют. Однако он дает лишь весьма слабое представление об интен-
сивности конкуренции (Шорыгин 1939). Для определения силы
пищевой конкуренции между отдельными видами рыб необходимо
знание, кроме состава пищи и интенсивности ее потребления, еще
целого ряда дополнительных факторов. Поэтому все попытки циф-
рового определения пищевой конкуренции могут носить пока еще
лишь сугубо ориентировочный характер.
172
весной и осенью сильно
Вопросы, связанные с питанием рыб,имеют огромной важности
хозяйственное значение. С ними приходится сталкиваться при
разрешении почти всех главнейших проблем, связанных с вопросами
сырьевой базы рыбной промышленности.
Рациональное прудовое хозяйство немыслимо без знания основ-
ных вопросов питания рыб. Нормы посадки рыбы в пруд, необходи-
мость и величина подкорма, изменение величины подкорма в тече-
ние сезона нагула рыбы и целый ряд других вопросов неразрывно
связаны с биологией питания рыбы.
Рыбохозяйственная таксация водоемов не может быть успешно
проведена, если не будут учтены кормовые ресурсы водоемов, их
распределение в нем и сезонная динамика, а также и использова-
ние их рыбами.
Акклиматизационные работы, как по заселению водоема рыбами,
так и кормовыми объектами, должны проводиться только после
детального изучения пищевых взаимоотношений и запасов кормо-
вых объектов в водоеме.
Поисковые работы, проводимые для выяснения распределения
в море промысловых рыб, сейчас в значительной степени базиру-
ются на знании распределения кормовых объектов. У нас на Мур-
мане тральщики снабжены специальными альбомами пищевых ор-
ганизмов, скопления которых являются местами кормежки рыбы.
В Англии для определения концентрации сельди судами, произ-
водящими дрифтерный лов, сконструирован специальный прибор—
лланктониндикатор Гарди, позволяющий в полевых условиях опре-
делять количество планктона—• кормового объекта сельди, в мес-
тах значительной концентрации которого и держится сельдь в»
>ремя нагула.
Таковы основные вопросы рыбного хозяйства,связанные с пита-
нием рыб.
ЧАСТЬ III
ОБЗОР БИОЛОГИИ ОСНОВНЫХ ПРОМЫСЛОВЫХ РЫБ СССР
Перейдем к обзору биологии основных промысловых рыб СССР
Изложение мы ведсм в систематическом порядке.
Миноговые Petrcmj zcnidae—морские, проходные и пресно-
водные рыбообразные животные. Семейство заключает семь родов,
распространенных, как в северном, так и в южном полушарии,
именно: Petromyzon, Ichthyomyzcn, Caspiomyzon, Entosphenes,
Lampetra—в северном, Geotria и Mcrdacia — в южном.
В СССР встречаются представители родов Petromyzon, Cas-
piomyzon, Lampetra, содержащие первые два по одному виду, а
последний 9—10 видов, из которых в пределах Союза встречается
шесть.
Морская минога Petromyzon merinos распространена от Мурмана
на запад вдоль берегов Европы до Адриатического моря и по Атлан-
тическому побережью Северной Америки на юг доФлориды.Морская
минога живет в море и лишь иногда заходит в реки. В море известна
до глубин около 500 м. В некоторых озерах Северной
Америки образует местные пресноводные формы, никогда
не выходящие в море. Размножение у морской миноги
происходит весной на песчаных местах, обычно в устьях
рек. В большие стаи для икрометания, видимо, не собирается,
а совершает нерестовые миграции и икрометание по 2—4 особи
вместе. Морская минога для размножения выкапывает в грунте
неглубокую ямку, иногда длиной до 2 м и шириной до 50сл/, в ко-
торой и происходит откладка икры и ее оплодотворение. У самцов
миног имеется своеобразное образование, выполняющее в извест-
ной степени функцию penis, хотя настоящее внутреннее оплодо-
творение у миног отсутствует. Личинки выводятся при температуре
18—23° на седьмой-восьмой день после откладки икры. Продолжи
тельность личиночного периода для морской миноги точно не уста-
новлена. В отличие от большинства других миног морская минога
после икрометания не всегда погибает. Взрослая морская минога
имеет размеры от 50 до 100 см. Озерные формы морской миноги
ме; ьче, чем морские.
Пгщу морской миноги составляют, главным образом, рыбы,
но иногда в ее кишечнике оказываются и придонные беспозвоноч-
ные. Ко времени икрометания морская минога прекращает питаться.
Промысловое значение морской миноги очень невелико, несмотря
на то, что мясо ее вкусно и весьма жирно. Встречаясь, даже во время
174
размножения, единичными экземплярами, она нигде не дает значи-
тельных уловов, хотя промышляется во время ее размножения,
как у берегов Германии, так Франции и Англии.
Каспийская минога Caspiomyzon wagncri проходная форма,
распространенная в Каспийском море и входящая для размножения
в впадающие в Каспий реки, главным образом, в Волгу и Куру.
Нерестовый ход в Волгу у каспийской миноги начинается с сентября
и длится до конца декабря. В отличие от предыдущего вида ход
носит массовый характер. Размножение происходит с марта по
июнь, количество икринок, откладываемых одной особью, колеб-
лется от 20 до 32 тыс. Икра откладывается на песчаных или каме-
нистых грунтах,видимо так же, как и у предыдущего вида в специаль-
но построенные примитивные гнезда. Во время хода каспийская
минога сильно худеет и становится менее жирной; так, на устье
Волги процент жира у миноги равен 34%, у Сталинграда он падает
уже до 20%, а на местах икрометания количество жира в теле
миноги сходит на нет. После икрометания каспийская минога по-
гибает. Во время хода в реку каспийская минога претерпевает ряд
брачных изменений. Особенно резкие изменения наблюдаются в
строении зубов, которые становятся тупыми и не позволяют ми-
ноге вгрызаться в тело своих жертв, и, если, например, в Куре
во время хода минога часто присасывается к идущему в одно время
с ней лососю, то это, видимо, делается, главным образом, с целью
передвижения вверх по течению. Состав пищи каспийской миноги
изучен очень плохо. В Волге у миноги в кишечнике находили остат-
ки водорослей. Каспийская минога представ; ена двумя формами:
крупной до 50 и более см длиной (обычно 37—41 см) и мелкой
(forma ргаесох) от 19 до 35 см длиной. Продолжите; ьпость жизни
и время превращения для каспийской миноги точно не уста-
нов; ены.
В Волге и Куре каспийская минога имеет важное промысловое
значение. В 1913 году в низсгье Волги было выловлено 33,4 мил
Лиона штук миног весом 22 768 центнеров. Добывается минога во
время ее входа в реки, главным образом, при помсщи различных
морд, верш и других прутяных ср^дгй лова.
Речная минога L; mpetra fluviatilis проходная, распростра-
ненная в бассейнах Балтийского, Немецкого и западной части
Средиземного морей, в Сибири заменена подвидом. Морской период
жизни речной миноги совершенно не изучен. Вероятно в море ми-
нога находится одно —два лета. Ход в Неву начинается весной в
первой половине мая, кончается в конце июня. Идущая в реку
речная минога претерпевает, так же как каспийская, ряд измене-
ний, именно: зубы, в море острые, при входе миноги в реку тупеют,
кишечник дегенер! рует, желчный пузырь атрофируется, в связи
с чем питание естественно прекращается. Интересно, что действие
имеющихся в ротовой воронке миноги железок, секрет которых
препятствует свертыванию крови, в реке видимо ослаб; ястся.
Тело миноги во время хода в реку укорач! вается, причем укоро-
чение во времени нереста достигает 23—27% первоначальной
176
Рис. 114. Промысловые рыбы СССР
-4 (сверху вниз и слева направо) —невская минога, хамса, осетр, черноспинка,
мурманская сельдь, невский сиг, белуга, хариус, севрюга, семга, белорыбица;
В (сверху вниз и слева направо)—вобла, усач, окунь, лещ, судак, сом, бычок
песочник, бычок жаба, щука, морской окунь, пикша, треска, навага.
2 Биология рыб 177
длины. Спинные плавники делаются выше и соприкасаются друг с
другом; у самок появляется анальный плавник.
Кроме весеннего хода у невской миноги имеет место и осенний
ход, который происходит с первых чисел августа (единичные особи
начинают итти уже в июне) и длится до ноября. У идущей в реку
осенью миноги брачных изменений не наблюдается. Икрометание
происходит в июне. Входящая осенью в реки минога икромечет
на следующий год. Плодовитость речной миноги колеблется от 4
до 39 тысяч икринок. После икрометания производители погибают.
Личиночный периоду речной миноги длится не менее четырех лет.
Превратившиеся миноги имеют длину 8,6—15 см.
Длина ходовой речной миноги 30—35 (редко 40) см. У речной
миноги, как у европейской, так и у сибирской, так же как и у кас-
пийской миноги имеется мелкая форма (forma ргаесох), которая
становится половозрелой, уже достигнув 12,5 см длины. Иногда
мелкая речная минога образует в озерах оседлую жилую расу. Реч-
ная минога для питания присасывается к рыбам. В кишечнике
миноги находили чешую и икру корюшки и других рыб. Промы-
словое значение речной миноги довольно велико. У нас в СССР
она добывается в Финском заливе и Неве.
Восточная речная минога Lampetra japonica распространена
от Кореи на север по Тихоокеанскому побережью Сибири и Север-
ной Америки. По образу жизни близка к обыкновенной речней
миноге. В массовом количестве входит в реки. Икрометание в июне.
Икра мельче, чем у предыдущего вида, 80—100 тысяч икринок
в одной самке. После икрометания производители погибают. Про-
мысловое значение пока невелико, но возможности для развития
промысла имеются и освоение запасов миног Дальнего Востока
уже начато.
Украинская минога Lampetra mariae распространена в бассейне
Черного моря от Прута на западе до речек Закавказья на востоке.
Пресноводная форма, икромечущая в апреле и мае. Плодовитость
около 2—3 тысяч икринок. Икра крупная. Зубы во время и после
нереста у одних особей бывают острые, у других тупые. Биология
этого вида изучена очень плохо. Промысловое значение невелико.
Ручьевая минога Lampetra planeri распространена в бассейнах
Балтийского и Немецкого морей, проникает в некоторые реки
Черного и Каспийского морей (Дунай, Волга). В море никогда не
выходит, всю жизнь проводя в реках. Икрометание в мае, плодо-
витость около 1,5 тысяч икринок. Икра крупная. Личинки также
значительно крупнее, чем у речной миноги (до 20 см). Продолжи-
тельность личиночного периода около 5 лет. Метаморфоз происходит
зимою. Взрослая ручьевая минога по окончании метаморфоза не
питается, имеет тупые зубы и дегенерированный кишечник. В ту
же весну после метаморфоза нерестится и по окончании икромета-
ния погибает. Брачные изменения ручьевой миноги сходны с тако-
выми речной.
Промыслового значения ручьевая минога почти не имеет, упо-
требляясь лишь в качестве наживки на крючки.
178
Осетровые Acipenseridae. Проходные и пресноводные р
бы, распространенные в Северном полушарии. Семейство заклю-
чает четыре рода: белуги Huso, осетры Acipenser, лжелопатоносы
Pseudoscaphirhynchus и лопатоносы Scaphirhynchus. Осетры рас-
пространены, как в Старом, так и в Новом свете', белуги и лжело-
патоносы только в Старом и лопатоносы—только в Новом.
Белуги— два вида; белуга Huso huso, распространенная в
бассейнах Адриатического, Черного и Каспийского морей, и
калуга Huso dauricus, водящаяся в бассейне Амура. Белуга —
проходная рыба, для размножения поднимающаяся высоко вверх
по рекам. По Волге белуга поднимается в .значительном количестве
не выше Куйбышева, но единичные особи доходят значительно
выше. В Урале доходит до Чкалова. Калуга не совершает столь
значительных миграций.
Ход белуги в реки, как и многих других осетровых, происхо-
дит дважды: весной, когда в реку идет белуга с зрелыми половы-
ми продуктами, икромечущая той же весной, и осенью, когда
в реку заходит белуга, которая в реке зимует и мечет икру уже
на следующий год. Калуга, в отличие от белуги, более привязана
к пресной воде и в море почти не выходит. Нерестилища белуги
и калуги находятся в равнинном течении рек. Белуга мечет икру
на каменистом грунте и быстром течении. Икра липкая. Нерест
происходит при температуре 12—i4° Калуга нерестится обычно
на песчаных косах в конце мая, в июне.
Время наступления половозрелости и плодовитость осетровых
Первый нерест Плодовитость (в тыся- чах)
Белуга 15—18 лет 500—1400
Калуга 18-20 500—4500
Русский осетр 8-15 70-400
Немецкий осетр 10—15 800—2400
Шип 10—15 200—400
Стерлядь . 4-6 11—67
Сибирский осетр — 50-500
Севрюга . 5-20 80—500
Большой лопатонос . 6-7 Неизвестна
Выведшиеся мальки белуги скатываются в предъустьевые
пространства, где и кормятся, по мере роста отходя от устья реки.
Мальки калуги держатся в реке и часто вообще в море не выходят.
Питается белуга в море почти исключительно рыбой: сельдями,
воблой, бычками и др.; иногда даже заглатывает бельков каспий-
ского тюленя. Калуга также хищнйк, питается кетой, горбушей,
толстолобиком и другими рыбами. Во-время икрометания, как у
белуги, так и у калуги, питание прекращается. Белуга и калуга
наиболее крупные пресноводные рыбы северного полушария. Они
иногда достигают веса свыше тонны и возраста более 100 лет.
12* 179
Бглуга, как в Каспийском, так и в Азовском морях, растет быстрее
калуги, обгоняя ее в росте иногда на 20 и более сантиметров в
девяти-, десятилетнем возрасте.
Белуга и калуга являются весьма ценными объектами промысла.
Только в одном Волго-Каспийском районе вылавливается ежегод-
но свыше 75 тысяч голов белуги. Значение калуги, как объекта
промысла, много меньше. Как указывает В. К. Солдатов, в довоен-
ное время ее в Амуре добывалось всего около двух тысяч цент-
неров.
Осетры Acipenser, или пресноводные (Acipenser fulvescens амери-
канский озерный осетр и Acipenser ruthenus стерлядь), или проход-
ные рыбы. В пределах СССР восемь видов: шип Acipenser nudi ventris,
распространенный в бассейнах Арала, Каспия и Черного моря,
русский осетр A. gtildenstadtii, водящийся в бассейнах Каспия
и Черного моря, сибирский осетр A. baeri, распространенный в ре-
ках севера от Печоры до Колымы и предъустьевых пространствах
Полярного моря. Амурский осетр A. schrenki населяет Амур и Амур-
ский лиман, сахалинский осетр A. medirostris водится по Американ-
скому и Азиатскому побережьям Тихого океана в северном полу-
шарии, севрюга A. stellatus встречается в бассейнах Черного, Азов-
ского и Каспийского морей, немецкий осетр A. sturio распространен
в северной части Атлантического океана по американскому и ев-
ропейскому побережью от западного Мурмана до Черного моря,
стерлядь A. ruthenus водится в реках, впадающих в Черное, Кас-
пийское моря и в Ледовитый океан, на восток до Енисея включи-
тельно.
Икрометание у всех весной, как правило, на каменистом, ^еже
песчаном грунте. Икра или прикрепляется к субстрату (стерлядь),
или закапывается в грунт (амурский осетр).-Инкубационный период
несколько дней варьирует в зависимости от температуры. Мальки
обычно скатываются в предъустьевые пространства, где кормятся
до достижения половозрелости. Половозрелость, как видно из при-
веденной таблицы, у большинства представителей рода Acipenser
наступает довольно поздно. Многие осетровые мечут икру не каждый
год. Ход в реки у многих видов происходит летом и осенью, затем
имеет место зимовка в реке (осетр, шип) и икрометание на следую-
щий год. У некоторых, как севрюга, русский осетр и другие, суще-
ствуют, как озимые, так и яровые расы, т. е. рыбь^, мечущие икру
как в тот же год (яровые), так и на следующий после захода в реку
(озимые). После окончания икрометания, которое у большинства
осетров приходится на конец апреля и май, проходные рыбы спу-
скаются из рек и приступают к нагулу. Места кормежки осетров
или расположены во внутренних морях, как Каспий и Арал, или
приурочены к опресненным пространствам перед устьями рек, впа-
дающих в океан. Пища осетров довольно разнообразна. Шипи амери-
канский осетр питаются, главным образом, моллюсками и в меньшей
степени другими беспозвоночными. Севрюга преимущественно по-
требляет рыбу: тюльку, бычков и др. Преимущественной пищей
стерляди служат насекомые. Наиболее интенсивное [питание имеет
180
место после икрометания. Вовремя начала хода в реку большин-
ство осетровых прекращает питание и вообще, за исключением
стерляди, в реке питается менее интенсивно.
Наиболее быстрым ростом среди видов рода Acipenser обладает
азовская севрюга, далее следует русский осетр из Азовского моря
и затем уже осетр и севрюга Каспия. Значительно медленнее растут
непроходные, постоянно живущие в реках осетровые — в Старом
свете—стерлядь и в Новом — озерный осетр. Продолжительность
жизни осетровых очень различна, но обычно средний максималь-
ный возраст большинства осетров редко превышает 35—40 чет.
Позднее созревание, икрометание у многих видов через год и
значительная ценность как объектов промысла привели к тому
что запасы большинства видов осетровых были подорваны. Так,
например, совершенно прекратился лов немецкого осетра в Эльбе
и Рейне, промысел американского озерного осетра и многих других.
И у нас хищнический лов в царской России в значительной степени
подорвал запасы осетровых Азовского моря и Каспия, и только
запуск рыболовства в период империалистической и гражданской
войны позволили стаду осетровых несколько восстановиться. В
настоящее время по добыче осетровых СССР занимает первое место
среди других стран, являясь почти единственным поставщиком
зернистей и паюсной икры на мировой рынок. Но не только чрезмер-
ный вылов, но и изменение режима рек оказывает влияние на со-
стояние запасов осетровых. В частности строительство на ряде
рек плотин, которые преграждают осетровым доступ к их нерести-
лищам, несомненно должно будет привести к подрыву их запасов,
если своевременно не будут приняты соответствующие меры. В на-
стоящее время уже делаются попытки компенсации естественных
нерестилищ, доступ к которым прегражден плотинами — искус-
ственным рыборазведением ниже плотины, причем отдача рыбой
икры стимулируется при помощи метода Гербильского (см. выше),
а мальки по выходе из икры выдерживаются некоторое время в
садках, пока достаточно не окрепнут, после чего выпускаются в
реку. Подобного рода работы ведутся сейчас у нас на Куре, Волге
и Дону.
Третьим родом осетровых, встречающихся в пределах СССР,
являются лжелопатоносы Pseudoscaphirhynchus— три вида в бас-
сейне Аральского моря. Лопатонос Федченко Pseudoscaphirhynchus
fedtscherikoi водится в Сыр-Дарье, большой амударьинский Р.
kaufmanni и малый амударьинский Р. hermannf лопатоносы в
Аму-Дарье. Лопатоносы — пресноводные рыбы, живущие в русле
рек на быстром течении. Икрометание в апреле месяце. Пищу боль-
шого амударьинского лопатоноса составляют, главным образом ,
рыбы: гольцы, молодь усач1, чехонь и др. Малый амударьинский
лопатонос и лопатонос Федченко питаются беспозвоночными,
главным образом, личинками комаров и других насекомых.
Наиболее крупный большой амударьинский лопотрнос достигает
максимум размеров до 50 см и веса до 1 кг. Два^ другие вида
среднеазиатских лопатоносов еще мельче.
181
Промысловое значение в СССР имеет только большой лопато-
нос, который добывается, главным образе м, главной сетью и
идет на местный рынок. Американские лопатоносы, водящиеся в
Миссисипи, имеют гораздо большее промысловые значение.
Сельдевые Clupeidae —чрезвычайно широко распростра-
ненные во всех морях земного шара рыбы. Некоторые для размно-
жения входят в реки, другие постоянно живут в пресной воде.
В пределах СССР 11 родов: 1) океанические сельди Clupea, 2) кас-
пийско-черноморские сельди Caspialosa, 3) финты или шэд Alosa,
4) Ilisha, 5) уруме-иваси Etrumeus, 6) пятнистые сельди Clupa-
nedon, 7) кильки Spratella, 8) тюльки Clupeonella, 9) сардинки
Sardinella, 10) иваси Sardinops, 11) Har^ngula. Промысловое значе-
ние в пределах Союза имеют: океаническая сельдь, каспийско-
черноморские сельди, кильки, тюльки и иваси. Представители
остальных редев встречаются в пределах СССР редко и как объект
промысла иснильз/ются лишь случайно.
Океанические сельди представлены у нас типичной формой
Clupea harengus harengus на Мурмане; в Дальневосточных водах
водится малопозвонковая сельдь С. harengus pallasi, являющаяся
подвидом океанической. Близкая к да; ьневосточной сельдь рас-
пространена в Белом море, С. harengus maris albi. В Балтийском
море водится салака С. harengus membras. Из представителей этсго
вида наибольшее промысловое значение в наших водах имеет океа-
ническая сельдь. Эта сельдь распространена в северной части Ат-
лантического океана по американскому побережью на юг до мыса
Код и по европейскому до Бискайского залива. На север идет в
Баренцево море до'77° северной широты и на восток до Н<,воземель-
ского мелководья. У нас на Мурмане океаническая сельдь пред-
ставлена двумя группами: фиордовой, живущей в прибрежной
зоне, и морской, лишь периодически подходящей к берегу. Сроки
размножения океанических сельдей очень сильно варьируют: есть
формы, которые мечут икру весной, другие летом и некоторые
расы осенью. Наша мурманская морская сельдь икромечет с марта
по май в районе Лофотенских островов и на север до Нордкапа.
Фиордовая сельдь мечет икру в заливах Кольском и Мотовском.
У сельдей рода Clupea, в отличие от всех других родов этого семей-
ства, за исключением Pomalobus, икра донная, откладывающаяся
на подводные предметы. Плодовитость мурманской сельди от 37
до 280 тысяч икринок. Диаметр зрелых икринок 1,2—1,5 мм.
Инкубационный период при температуре воды 15° длится 7—8 дней.
После выхода из икры личинки сельди, имеющие 5—8 мм длины,
начинают подниматься в верхние слои воды и одновременно дрей-
фовать с Атлантическим течением на восток. К Мурману мальки
сельди подходят через 4,5—5 месяцев по выходе из икры, т. е. в кон-
це июля—августа, достигнув уже размеров 35—50 мм. К осени
мальки мурманской сельди опускаются в придонные слои, где зи-
муют4, а со следующей весны уже активно начинают расселяться по
Варенцову морю, доходя далеко на север. Значительная часть мур-
манской сельди становится половозрелой на пятом году жизни.
182
Сельди старше 7 лет встречаются в Баренцевом море крайне редко
и составляют в уловах ничтожный процент. Мурманская сельдь ред-
ко бывает крупнее 27см. Как видно из приводимых цифр, мурманская
сельдь растет по сравнению с мелкой беломорской много быстрее.
Рост различных подвидов океанической сельди
Возраст | 11 1 1* ) 13 11 ^5
Мурманская сельдь 9,0 13,5 17,5 20,5 22,2 23,7
Беломорская сельдь, мелкая раса 6,3 11,5 14,7 17,6 — —
Беломорская сельдь, крупная раса . 6,2 13,5 19,8 24,8 — —
Чешская сельдь 8,0 13,8 17,8 21,0 — —
Дальневосточная сельдь 12,9 23,3 28,5 31,0 32,7 34,1
Пищу мурманской сельди составляют преимущественно планк-
тонные ракообразные, главным образом, Calanus и Euphasiidae.
Наиболее интенсивно сельдь питается летом, к осени питание
ослабляется, зимою сельдь питается очень слабо. Весной перед
икрометанием интенсивность питания несколько возрастает.
Беломорская сельдь представлена рядом рас, различающихся
как временем нереста, так размерами и плодовитостью. В каждом
заливе имеется несколько местных стад сельдей. Но вся беломор-
ская сельдь распадается на две биологические группы: на мелких
сельдей, подходящих для икрометания весной и характеризующихся
малой плодовитостью—б—7 тысяч икринок (так паз. егорьевская
сельдь), и крупных сельдей, икромечущих летом и обладающих
большей плодовитостью (27—30 тысяч икринок).
Тихоокеанская сельдь, подобно беломорской, принадлежит к
уппе малопозвенковых сельдей, имеющих 53—54 позвонка
вместо 57—58 у океанических сельдей. Так же, как и беломорская
сельдь, дальневосточная больше заходит в пресную веду. Нерест—
с марта по май, плодовитость 10—14 тысяч икринок. Растет дальне-
восточная сельдь много быстрее северной. Продолжите.) ьность жизни
также бо.)ьшс: до 13—14 лет.
Каспийско-черноморские сельди Caspialosa. Около б—1 видов
в бассейнах Черного и Каспийского морей. Распадаются на две
биологические группы: прогонистых рыб обычно с меньшим коли-
чеством жаберных тычинок и отчетливо развитыми зубами—преи-
мущественно хищников сельдей — и рыб с высоким телом, обычно
частыми длинными жаберными тычинками — планктоноядных пузан-
ков. В Каспии насчитываются следующие виды сельдей и пузанков:
сельдь черноспинка Caspialosa kessleri: волжская сельдь С. volgen-
sis, долгинская сельдь C.brashnikovi, пузанок каспийский С. caspia,
пузанок большеглазый С. saposchnikovi, круглоголовый пузанок
С. sphaerocephala, пузанок полосчатый С. curensis.'В свою очередь
почти каждый из этих видов распадается на несколько рас, так,
183
например, каспийский пузанок образует форму, мечущую икру при
повышенной солености, и форму, размножающуюся в пресно
воде.
Некоторые сс;ьди всю жизнь проводят в морской воде, как на-
пример, некоторые расы долгинской сельди. Другие, как например^
черноспинка — проходные рыбы, поднимающиеся высоко вверх
по Волге и другим рекам. Так же как и проходные лососевые, черно-
спинка во время миграций очень сильно истощается и в массе поги-
бает после нереста. Весною все сельди подходят к берегам и вдоль
берегов двигаются к своим нерестилищам, которые расположены
или в реках или на мелких местах у берега. Икра у всех сельдей
рода Caspialosa пелагическая, проходящая свое развитие в толще
воды. Плодовитость у различных видов весьма различна. Как пра-
вило, сельди более плодовиты, чем пузанки. Средняя плодовитость
каспийского пузанка 24 тысячи икринок, долгинской сельди 66
тысяч икринок1, во/жсксй сельди—54 тысячи икринок. Полово-
зрелыми пузанки стм овятся обычно на третьем — четвертом году
жизни, а сельди — на третьем (волжская), четвертом (долгинская)
и пятом (чернеет н» г) году. Икрометание происходит весной и ле-
том. Инкубационный период икры колеблется не только в зависимости
от внешних условий, он различен и у различных видов. У черно-
спинки при 19,6° инкубационный период равен 49 часам, а при
20,4°—42 часам. Выведшиеся из икры личинки держатся в толще
воды и питаются планктоном. Продолжительность жизни различных
сельдей различна, так, для пузанка указывается максимальный
возраст—6 лет, для долгинской сельди—8 лет, для черноспинки—
5—6 лет.
Отнерестовавшие сельди, если они после икрометания не
погибли (как например, погибает в массах черноспинка), выходят
в открытое море,где кормятся. Пузанки, кроме большеглазого, пита-
ются, главным образом, планктонными ракообразными, а сельди —
рыбой: килькой, бычками и др. К осени сельди продвигаются на
юг и зимуют в южной части Каспия, откуда весной начинают свою
нерестовую миграцию. Основной промысел сельдей приурочен к их
нерестовому ходу, когда сельди в огромном количестве добываю-
тся вдоль Дагестанского побережья Каспия и в дельте Волги
береговыми неводами.
Очень близка по биологии к проходным сельдям рода Caspialosa,
водящаяся у нас в Балтийском море Alosa finta. Эта сельдь, так же
как и черноспинка, входит для икрометания в реки, по которым
поднимается весьма высоко. Икра батипелагическая. Инкубационный
период 3—4 дня. Пищу этой сельди составляют, главным обра-
зом, ракообразные. Промысловое значение в пределах СССР
невелико, в Западной Европе Alosa является важным объектом
промысла.
Близка по биологии к Alosa и Caspialosa, распространенная в
Индии, Китае и встречающаяся у нас на Дальнем Востоке Ilisha.
Это также проходная сельдь, совершающая довольно значительные
миграции вверх по рекам к местам икрометания. В Индии и Китае
Ilisha является важным объектом промысла.
184
Сардинки Sardinops населяют северную часть Тихого океана:
иваси S. melanosticta по Азиатскому побережью и Калифорнийская
сардинка S. in. cerulea, являющаяся подвидом иваси, по американ-
скому побережью. В пределы СССР иваси входит весной после
икрометания и движется на север, примерно, до мыса Лазарева.
Эта миграция иваси является пищевой. Осенью с наступлением
похолодания воды иваси начинает обратный ход к берегам Японии,
где она зимует, и в конце зимы и ранней весной (с декабря по март
в различных районах) мечет икру, после чего начинает миграцию
на север. Икра иваси пелагическая, проходящая развитие в толще
воды. Плодовитость в среднем около 50 тысяч икринок. Инкубацион-
ный период длится от 2 до 4 дней. Мальки иваси совершают весьма
далекие миграции и уже на первом году жизни попадаются в При-
морьи. Иваси становится половозрелой на втором году жизни,
достигнув 16—18 см длины. Наиболее старые особи, попадающиеся
в уловах, имеют восьмилетний возраст и 25 см длины. В уловах
преобладают рыбы от трехлетнего до восьмилетнего возраста. Как
иваси, так и калифорнийская сардинка, имеют важное хозяйственш е
значение.
Тюльки ClupecneПа—4—5 видов в бассейнах Черного и Каспий-
ска го морей, встречаются и в реках. Наиболее широко распростра-
нена Clupeciiella delicatula, встречающаяся как в Каспийском,
так ив Черном морях, и довольно высоко поднимающаяся по рекам.
В затонах Волги и в озере Чархал образует местную форму, не вы-
ходящую в море. В Каспии весной тюлька (называемая на Каспии
килькой), подходит к берегам. Нерест происходит на мелководьи у
берегов и лишь незначительная часть стада входит в Волгу. Плодо-
витость 35—40 тысяч икринок. Половозрелой тюлька становится на
втором году жизни, имея размеры 79—80 мм. В уловах редко попа-
даются особи старше пятилетнего возраста. Инкубационный период
икры продолжается 2—4 дня. Выведшиеся из икры мальки держатся
в толще воды и на первом году жизни уже достигают 57—60 мм
длины. Пищей тюльки служат планктонные организмы, главным
образом, различные ракообразные. Как в Каспии, так и в Азовском
море, тюлька встречается в огромном количестве, но промыслом
используется еще совершенно недостаточно.
Кильки Spratella в наших водах представлены двумя подви-
дами одного вида: в Балтийском море S. sprattus baltica и в Черном
море S. sprattus phalerica. По биологии несколько напоминают
тюльку, но икромечут осенью. У Новороссийска икра попадается
в конце осени и начале зимы. Инкубационный период 4—5 дней.
Промысловое значение в пределах СССР невелико. Добывается в
Балтийском море и в западной части Черного, но в незначительном
количестве.
Семейство анчоусовых Engraulidae — широко распространен-
ные рыбы в умеренных и тропических широтах. В пре-
делах СССР — два вида Engraulis encrassicholus в бассейнах
Черного и Азовского морей и Е. japonicus в рбдах Дальнего
Востока.
185
Анчоус Е. encrassicholus распространен в северной части Атлан-
тического океана, в Балтийском, Немецком морях, на юг до Канар-
ских островов, встречается в Черном и Азовском морях, в которых
представлен особыми подвидами — хамсой Е. encrassicholus рсп-
ticus в Черном и Е. encrassicholus nieoticus в Азовском море.
Азовский анчоус или хамса размножается в Азовском море с
мая по сентябрь, разгар нереста приходится на весну, икра пела-
гическая, инкубационный период длится 2—4 дня. После икро-
метания взрослые особи нагуливаются в Азовском море и по оконча-
нии нагула выходят через Керченский пролив в Черное море, где
зимуют. Пищу анчоусов составляют планктонные организмы, глав-
ным образом, различные ракообразные.
В СССР промысел хамсы сконцентрирован почти полностью в
Азовском море, где эта рыба ловится в огромном количестве при
выходе из Керченского пролива, когда она идет после нагула на
зимовку.
Семейство лососевые Saline, nidae. Проходные и пресновод-
ные рыбы умеренных и высоких широт северного полушария. Де-
вять родов: Oncorhynchus, Salmo, Salvelinu$, Hucho, Brachymystax,
St.nodus, Coregonus, Salmothy mus, Crlstivonur. В пределах СССР
встречается семь первых.
Род Oncorhynchus —дальневосточные лососи (шесть видев)
распространены по тихоокеанскому побережью Северной Америки
и Азии. В Северной Америке от мыса Барроу до Калифорнии, в
Азии от Лены до побережья Кореи. Из шести видов только сима
О. masu не встречается по американскому побережью. Проходные
рыбы, кормящиеся в море, а для икрометания входящие в реки. Ме-
ста нагула у некоторых видов (кета, горбуша) отстоят иногда очень
далеко от нерестовых рек: так, кета, нерестующая в Амуре, кормится
у острова Иэзо. Некоторые лососи, как например, кижуч, почти не
выходят во время нагула из опресненной зоны перед устьями рек,
в которых они нерестятся. Различные виды становятся половозре-
лыми в различном возрасте (см. таблицу). Нерестятся один раз в
жизни. После икрометания погибают. Плодовитость невелика,
наименее плодовита горбуша О. gorbuscha. Во время хода к нерести-
лищам у дальневосточных лососей появляются весьма резкие брач-
ные изменения. Окраска становится яркой, челюсти вытягиваются
и загибаются, на них развиваются большие зубы, на спине появляет-
ся горб, тело сильно уплощается. Наиболее резки брачные измене-
ния у горбуши, наименее резки у чавычи.
Входящая в реку рыба, как правило, имеет слабо развитые
половые продукты: так, у самцов кеты О. keta гонады составляют
на устьях 0,6—1,2% веса тела, в то время как на нерестилищах —
8—10%. Развитие гонад происходит в основном во время хода по
pc J'
Нерестилища располагаются в речках на каменистом грунте.
Только нерка О. nerka мечет икру и в озерах. Икра закапывается
в грунт. Нерест происходит осенью. Продолжительность инкуба-
ционного периода несколько месяцев, у кеты 69—108 дней. Мальки
186
по выходе из икры скатываются в моря, где нагуливаются до до-
стижения половозрелости. В реках во время миграции и нереста
взрослые особи не питаются, кишечник сильно дегенерирует, и ки-
шечные соки теряют способность расщеплять белки. В море взрос-
лые рыбы питаются мелкой рыбой и ракообразными. Дальневосточ-
ные лососевые имеют огромное промысловое значение, давая ежегод-
но свыше 2700000 центнеров.
Виды Время на- ступления половэзре- лости Плодовитость Число нерестов Время нереста
Кета . 1 З-б 2—4000 Один Осень
Горбуша 2 1090—1629 Один Осень
Чавыча 4-7 4600—14300 Один Осень
Нерка 3-6 1600—6500 Один Осень
Кижуч.. 3-4 4900 (средн.) Один Осень
Сима . 4—6 3200 (средн.) Один Осень
Лосось 4—6 ' 8000 (средн.) 3—4 Осень
Каспийский лосось 3—8 i 16-3 ЮОО Несколько Осень
Палия 4-6 1 1500-9000 Несколько Осень
Ленок . . 4-5 4700-7500 | Несколько Веспа
Белорыбица 6-7 1 240000 (средн.) 2 Осень
Нельма 13 50009 (средн.) Несколько Зима
Ряпущка 2—3 9000 (средн.) Несколько Осень
Омуль 5-6 89600 (средн.) Несколько Осень
Байкальский омуль 3-5 6—40000 Несколько Осень
Лососи Salmo около восьми видов. В пределах СССР пять. Лосось
или семга S. salar распространен по атлантическому побережью
Америки к северу от мыса Код. Встречается в Европе от Франции
на север до реки Кары. В СССР водится в бассейнах Балтийского»
Варенцова и Белого морей. Размеры, до 39 кг, как редкость, до
63 кг. Проходная рыба входит в реки в различное время, часть осо-
бей (озимая раса) в реке зимует. Нерест происходит осенью. В от-
личие от дальневосточных лососевых не все особи погибают после
икрометания, и некоторые мечут икру до 4 раз в жизни. Икра, так
же как у дальневосточных лососей, закапывается в грунт. Мальки
выводятся весной и обычно постепенно скатываются в море, но
некоторые остаются всю жизнь в реке и даже достигают здесь поло-
возрелое™ (самцы). В реке лосось растет очень медленно, по выходе
же. в море рост его резко усиливается. Здесь лосось усиленно питает-
ся, поедая, главным образом, рыбу, особенно песчанку. В море
лососи совершают огромные переходы; так, известно несколько
случаев миграции лососей из Балтийского моря кругом Скандинав-
ского полуострова в Белое море. Один лосось прошел от Сканди-
навского полуострова до Печоры, где был добыт в среднем течении.
В озерах Ладожском и Онежском имеются местные реликтовые
формы лосося. В реке ходовой лосось не питается. Хотя количество
187
вылавливаемого лосося и значительно уступает уловам дальне-
восточных лососей, но все же оно довольно велико, а высокая цен-
ность получаемого продукта делает понятным, что на добычу и
воспроизводство запасов лосося обращено особое внимание.
Кумжа Salmotrutta распространена от Пиренейского полуострова
до Чешской губы. В бассейнах Черного, Каспийского и Аральского
морей заменена подвидами. Кумжа, так же как и лосось, мечет
икру в реках, но для нагула делеко в море не уходит, а держится
обычно в прибрежной зоне, некоторые же особи становятся поло-
возрелыми не выходя в море. В море кумжа растет хуже лосося,
в реке же, наоборот, быстрее. В реке ходовая кумжа, хотя и слабо,
но потребляет пищу. Кумжа мечет икру осенью. Наиболее крупна
каспийская кумжа, не особенно редки особи до 51 о весом. В Араль-
ском и Черном морях кумжа встречается сейчас редко. По указанию
старых авторов кумжа в Черном море была раньше обыкновенна.
В ледниковый период кумжа была распространена дальше на юг,
чем теперь, и встречалась и в Средиземном море. Особи, попавшие
в верхоЕЬя рек и лишенные возможности выйти обратно в море,
образуют карликовые формы — форелей Salmo trutta m. fario.
Подобные карликовые речные формы имеют более широкое рас-
пространение, чем морская кумжа, и встречаются даже в бассейнах
тех морей (Средиземное), где кумжи сейчас нет. Получая доступ
к морю, форель может снова превратиться в кумжу, как то имело
место, например, в Новой Зеландии, где акклиматизированная
форель, попав в море, превратилась в проходную кумжу. В озерах
кумжа, так же как и лосось, образует туводные формы.
Гокчинская форель Salmo ischchan населяет озеро Севан, где
представлена несколькими экологическими расами, отличающимися
по времени икрометания и размерами: ишхан крупный мечет
икру в озере осенью и зимой, боджак мелкий мечет икру в самом
озере в октябре; летний бахтак мечет икру в устьях речек весной
и летом, гагаркуни мечет икру в речках с октября по январь. Наи-
большие размеры до 16 кг. Питаются в озере почти исключительно
бокоплавами. Составляют важный объект промысла.
Камчатская семга Salmo penshinensis — проходная рыба по-
бережья Камчатки, вес до 16 кг, мечет икру весной в мае — июне,
довольно редка. Есть настоящие лососи S. gairdneri и др. и по тихо-
океанскому побережью Америки.
Salmo irrideus и S. mykiss, повидимому, такие же производные
от тихоокеанских видов рода Salmo, как форель от кумжи.
Палии или гольцы Salvelinus, много видов в северных широтах.
Проходные и озерные формы, распространенные в северной Европе
(Salvelinus alpinus, S. salvelinus и др. всего 2—4 вида), Сибири и
на Дальнем Востоке (S. jacuticus, S. boganidae, S. neiva и др. всего
8—10 видов) и в Северной Америке (S. fontinalis и др. 5—6 видов).
Есть и на островах Арктики в Гренландии, Шпицбергене, Медве-
жьем острове, Новой Земле и других.
Проходные формы, как распространенный кругополярно S. al-
pinus, зимуют в глубоких озерах, в которые входят из моря по
188
рекам. Ход в июле, августе и начале сентября. Те особи, которые
будут метать в этом же году, идут раньше, те же, которые идут
только на зимовку, заходят позднее. Ранней весной часто еще под
льдом голец выходит из озер в море, где кормится. В море голец
держится вблизи тех рек, в которые он входит, и, поводимому, дале-
ких миграций не совершает. Икрометание в озерах в октябре —
ноябре. Мальки гольца держатся в реках и в море не выходят. В
пресной воде молодь гольца проводит от двух (Шпицберген) до
четырех лет (Новая Земля). Во время входа в реку питается слабо.
Мальки в реке едят воздушных насекомы^, донных беспозвоночных
и, в меньшей степени, планктон. В море голец питается преимуще-
ственно рыбой: молодью трески, сайки, мойвой, некоторыми рако-
образными (Spirontocaris), червями (Новая Земля). Во многих озерах
голец образует локальные формы, систематика которых плохо
разработана (Байкал, озера Карелии и др.). Длина до 750мм', ма-
териал из Крестовой Губы, Новой Земли дает средний размер
61 см. Имеет промысловое значение. На Новой Земле в 1933 году
было добыто 80 тонн.
Таймени Hucho—четыре вида в Европе и северной Азии, в преде-
лах СССР — три.
Дунайский лосось Hucho hucho распространен в бассейнах Ду-
п in и Прута, в море никогда не уходит, мечет икру осенью, молодь
выклевывается через 3—5 дней. Размеры до метра, изредка больше.
Таймень Hucho taimen распространена от бассейна Волги на
западе до Лены на востоке, есть в Амуре, размеры до метра, плодо-
витость 34 тысячи икринок, нерест весной. Хищник.
Японский таймень Hucho perryi—бассейн Японского моря, про-
ходная рыба, икрометание весной.
Ленки — Brachymystax: один вид—Brachymystax lenok встре-
чается во всех ^>еках Сибири и Дальнего Востока, в море не выхо-
дит. Икрометание в мае. Размеры обычно до 3,5 кг, иногда бывает
крупнее. Питается личинками насекомых, мелкой рыбой, лягуш-
ками, икрой лососевых. Промысловое значение малое.
Белорыбицы—Stenodus. Один вид: Stenodus ieucichthys—нельма,
берорыбица. Распространена в бассейне Ледовитого океана от.Белого
моря на восток до реки. Меккензи, проникла в Каспийское море,
где образовала особый подвид. Проходная форма, в реки вхо-
дит летом, икромечет в октябре, мальки выходят с марта по май.
Вес до 40 кг, в море держится близ устьев рек. В Каспии спускается
довольно далеко на юг. Питается главным образом мелкой рыбой.
Теми роста каспийской белорыбицы гораздо более быстрый, чем у
сибирской нельмы. Имеет важное промысловое значение. В 1926
году на Каспии добыто 40 тысяч штук.
Сиги Coregonus, много видов в Европе, северной Азии и Америке.
Проходные и пресноводные формы.
Обыкновенный сиг Coregonus lavaretus. Широко распространен-
ный, чрезвычайно пластичный вид, образующий массу форм. В основ-
ном эти формы распадаются по образу жизни на д1ве группы: на
морских—проходных и озерно-речных. Проходные кормятся в море,
190
озерно-речные — в озерах. Все сиги, за редким исключением, пи-
таются беспозвоночными. Как и каспийские сельди они распадаются
на две группы: на рыб с длинными частыми жаберными тычинками
и высоким телом — планктоноедов, как например, Coregenus lava-
retus maraenoides, и рыб с короткими редкими тычинками и низким
телом, питающихся донными беспозвоночными и рыбой, как напри-
мер, Coregonus la'varetus mcngclicus. Проходные входят в реки в
различное время обычно летом. Икрометание осенью, у озерных —
иногда зимою.
Муксун Coregonus muksun, проходной сиг, входящий в реки
Ледовитого океана от Печоры до Колымы. Ход в реки осенью, икро-
метание в октябре — ноябре. Нагуливается в предъустьевых про-
странствах. В озерах бассейна Пясины образует озерную форму, не
выходящую в море. В Лапландии имеется особый подвид.
Омуль Coi\ginus autumnalis. Входит во все реки Ледовитого
океана от Мезени до Чаунской Губы (кроме Оби), размеры до 2 кг.
Так же как и другие сиги, нагуливается в опресненных предъусть-
евых пространствах. В Бай кале особый подвид. Входит в реки летом,
икрометание— во второй половине сентября. После нереста скаты-
вается обратно в морс.
Большинство сигов имеет важное промысловое значение и явля-
ется объектом искусственного разведения. Некоторые сиги аккли-
матизированы в водоемах, где они раньше не встречались. Так,
чудской сиг перевезен в Японию, а также выпущен в озеро Севан
на Кавказе.
Как видно из изложенного, образ жизни лососевых чрезвычайно
разнообразен, но среди этого разнообразия могут быть подмечены и
некоторые общие черты, именно: лососевые — проходные рыбы,
размножающиеся в реках и часто образующие здесь жилые —т\-
водные формы. Нерест у большинства лососевых осенний. По своему
происхождению лососи, видимо, пресноводные рыбы, так как они
известны уже из миоценовых пресноводных отложений. К про-
ходному образу жизни,, видимо, они приспособились позднее.
Однако точно генезис лососевых до последнего времени остается
неизученным.
К семейству карповых Суprinidae относятся, главным
( бразом пресноводные, реже проходные и солоноватоводные рыбы,
распространенные в Северной Америке, в Азии до линии Уол;еса,
в Европе и в Африке. Нет карповых в Австралии и Южной Аме-
рике. Это — одно из наиболее богатых видами семейств пресногодных
рыб. В отличие от лососевых карповые приурочены, главным обра-
зом, к умеренным и тропическим областям, и в арктических водах
встречается небольшое число видов. В пределах Союза ССР на-
иболее богаты видами Понтс-Арало-Каспийская и Манджурская
подобласти.
Карповые живут при самых разнообразных условиях, от горных
рек с холодной водой и быстрым течением, где держатся османы и
маринки, до заболоченных луж и прудов с недостатком кислорода,,
в которых живут карась и линь. Столь же велика и амплитуда со-
190
лености. Правда, болыиинстг о Карцевых живет в пресней воде
но мн( гие населяют, например, наши внутренние солоноватоводные
моря (сазан, лещ, шемая и др.) и размножаются при солености
10%о. При наиболее высокой соленности живет дальневосточная
красноперка Leuciscus brandti, которая встречается и при океани-
чески й солености.
Д 'лско нс у всех карповых развиты нерестовые миграции.
Лишь немногие гиды могут бьпь названы проходными рыбами.
Таковы вырезуб Rutilus frisii и его каспийский подвид кутум R. fris-
si kutum, некоторые формы рыбца Vimba vimba и шемаи Cha leal-
burnus chalcoides, а также аральею й усач Barbus brachycephalus,
причем вырезуб и аральский усач поднимаются иногда на многие
сотни километров вверх по рекам. Ряд карповых принадлежит к
группе полупроходных рыб, кормящихся в опресненных участках
морей перед устьями рек, а для размножения входящих в низоеья
рек. К этой группе принадлежат, например в Каст Иском море,
сазан, леш, вобла. Эти рыбы кормятся, главным образом', на взморье
и лишь в сравнительно небольшом количестве уходят в ссс-
лоненную часть моря, размножаются же в разливах и полсях
дельт рек.
Большинство пресноводных карповых для икрометания заходит
в заросли подводной растительности (карась, сазан, лещ), некоторые
карповые мечут икру на камнях и песке на быстром течении реки,
таковы: подъуст, пескарь и др. Во время размножения у карповых
обычно развивается брачный наряд, чаще всего выражающийся в
виде эпителиальных бугорков, покрывающих, как бисером, голову,
а иногда и тело половозрелых самцов, а у некоторых видов (например
лещ) и самок. Изменяется ко времени икрометания у некоторых
видов и окраска. Так, у гольяна Phoxinus phoxinus самцы имеют
весьма яркий брачный наряде красными, черными и темнозелеными
тонами. Соотношение полов у большинства карповых близко один
к одному, но наблюдаются и исклочения. Так, например, у чехони
самки несколько преобладают над самцами. У некоторых других
видов наб; юдается обратная карп на.
Плодовитость карповых также весьма различна, от нескольких
сотен икринок у горчака до миллиона у сазана. Икра у большинства
карповых донная, приклеивающаяся к подводным предметам, но
у некоторых видов икра бывает батипелагическая. Батипслаги-
ческая икра наблюдается, например, у Amj pa Ctenophoryngcdon
idellus и у чехони Pclecus cultratus. Пелагическая икра у карповых
встречается чрезвычайно редко. Достоверно она известна только
Для маленькой африканской рыбки Engraulicypris, водящейся в
больших озерах восточной Афргки.
Нерест у всех карповых, водящихся в умеренных широтах,
происходит в теплее время года, весною или летом. У представите-
лей же этого семейства, водящихся в тропиках, размножение часто
мсж:ет продолжаться в течение всего года, лиш^ с короткими
промежутками, потребными для созревания очередной порции
половых продуктов.
191
Плодовитость некоторых карповых
Тарань.
Лещ
Сазан
Шемая
Рыбец
22—202000
26—282000
350—1000000
9800—50000
16000—112000
Карась
Пескарь
Герчак
Гольян амурский
Краснопер
160—390000
2720-8670
200—300
740—1744
23370—45126
Как upai ило, карповые умеренных широт не проявляют заботы
о потомстве и часто даже поедают свою икру. Исключение со ста
вляют горчаки Rhodens, Acanthorhodeus и др., которые, как известно,
откладывают икру в мантийную полость двухстворчатых моллюс-
ков. Инкубационный период у карповых, как правило, очень не-
продолжителен: он длится от нескольких часов (Danio и др.) до
нескольких дней.
По возрасту и размерам карповые также чрезвычайно разно-
образны, от 5—7 см (верховка) и возраста 2 лет, и до метра и более
длины и возраста до 30 и свыше лет, как например, сазан.
Пищей различных карповых могут служить самые разнообразные
животные и растительные организмы, как живые, так и мертвые.
Среди карповых мы находим как детритоядные и растительно
ядные формы, как например, храмули Varicorhinus, так и рыб, пи-
тающихся беспозвоночными, живущими на дне (лещ, сазан, вобла),
так, наконец, живущих в толще воды—чехонь, шемая. Многие карпе
вые питаются растительной пищей, как донными растениями, как
например, красноперка, так и планктонными водорослями, которые
составляют пищу амурского толстолоба. Есть среди карповых и хищ-
ники, питающиеся другими рыбами. Таковы различные жерехи, уже
с годовалого возраста переходящие на рыбную пищу. Рыбою во взрос-
лом состоянии питается и голавль.Есть хищники и среди маринок,
как например, илийская маринка, достигающая солидных размеров
до 12 кг.
Всего на территории СССР водится около 130 видов карповых
рыб, но лишь очень немногие из них имеют промысловое значение.
Остановимся на биологии главнейших промысловых карповых
несколько подробнее.
Плотва Rutilus rutilus, типичная форма, распространенная в
пресных водах Европы, имеет малое промысловое значение, но ее
полупроходные подвиды, особенно каспийская вобла и, в несколь-
ко меньшей степени, азовско-черноморская тарань и аральская
вобла являются важными объектами промысла. Каспийская вобла
составляет гщ СССР около 14% от улова всей рыбы и 42% от улова
рыбы по северному Каспию. Вобла — полупроходная рыба, весною
с половины марта по май заходящая в реки и здесь на полоях при
температуре не ниже 12° мечущая икру. Наиболее интенсивный
нерест воблы происходит при температуре 14—17°. Средняя плодо-
витость воблы—35000 икринок. Молодь выходит из икры на 4—5
день после оплодотворения.
Взрослые отнерестовавшие особи скатываются в море, где кор-
мятся. Основную пищу воблы составляют моллюски, достигающие
в отдельных участках Каспия свыше 90% всей пищи. Следующее
192
место занимают ракообразные. Наиболее интенсивно вобла питается
весной и летом, к осени интенсивность потребления пищи снижает-
ся, по прекращении нагула вобла подходит к устьям рек, где зи-
мует. Растет вобла в отдельных участках Каспия различно. Наиболее
быстрым ростом обладает куринская и атрекская воблы. Северо-
каспийская вобла растет несколько медленнее.
Рост Rutilus rutilus в различных водоемах
11 1 1з
<ев. Каспий 7,9 12,3 15,9
Кура . 10,2 18,6 24,4
Гассан-кули 15,8 18,0 20,1
зовское море 7,4 12,1 15,5
Арал 7,7 11,8 15,9
В уловах в Каспии преобладают рыбы в возрасте от 3 до 5 лет.
Аральская вобла Rutilus rutilus aralensis по образу жизни близ-
ка к каспийской. Икрометание происходит с начала апреля по май.
После нереста взрослые особи уходят на глубины, где кормятся
до осени, когда снова подходят в береговую зону.
Тарань Rutilus rutilus haeckeli населяет опресненные участки
Черного и Азовского морей. Да>я размножения входит в реки. По-
ловозрелой тарань становится, так же как вобла и аральская плот-
ва, впервые на четвертом году жизни. Икрометание в озерах и
разливах в средине апреля. Выведшиеся из икры мальки скаты-
ваются в опресненные участки моря, где кормятся- Пищу тарани,
как и других подвидов плотвы, составляют донные беспозвоночные.
По окончании нагула тарань подходит к устьям рек и входит в
лоследние.
Лещ Abramis brama после воблы является второй по важности
промысловой рыбой из семейства карповых. Распространен в Европе
В бассейнах Немецкого, Черного, Каспийского и Аральского морей;
изредка в бассейне Белого моря. Нет леща в Италии, Испании,
Сибири и на Дальнем Востоке. Живет лещ, как в солоноватой до
13%о, так, главным образом, в пресной воде. Миграции резко вы-
ражены лишь в крупных водоемах. В Каспии и Арале лещ дважды
подходит в прибрежную зону—весною для икрометания и осенью
по окончании нагула. Половозрелости лещ достигает на 4—5 году
жизни. Нерест леща происходит в Азовском, северной части Каспий-
ского и Аральском морях во второй половине апреля — начале
мая при температуре воды не ниже 15° В южном Каспии нерест
происходит раньше. На севере же в Финляндии и Швеции нерест
вает и в начале июня. Откладывает икру лещ на растительность,
к которой икра прилипает. Инкубационный период длится обычно
3—4 дня, в зависимости от температуры. Выведшиеся мальки дер-
жатся первое время на местах икрометания, позднее откочевывают
на более глубокие места или в заросли подводной растительности.
13 Биология рыб *93
Взрослый лещ, выметавший икру, отходит от берегов на места
кормежки, где держится обычно все лето, пока не достигнет опре-
деленной упитанности, после чего прекращает питаться и начинает
мигрировать к берегам. Пищу леща составляют, главным образом,
ракообразные: в Каспии Corophiidae и Cumacea, в Арале Gamma-
ridae. В реках и озерах лещ питается преимущественно личинками
насекомых. Наиболее интенсивно лещ питается сейчас же после
окончания икрометания. К концу августа, началу сентября интен-
сивность питания обычно сильно снижается. Зимою лещ питается
очень слабо или даже прекращает питание совсем. Растет лещ в
различных водоемах весьма неодинаково. Как видно из приведен-
ной таблицы, наиболее быстрым ростом лещ обладает в наших юж-
ных морях (см. стр. 148).
Предельный возраст леща больше 20 лет, но обычно в промысло-
вых уловах редко попадаются лещи старше 8-летнего возраста.
Преобладают в уловах рыбыввозрасте 3—5 лет. Наибольшее значе-
ние, как объект промысла, лещ имеет на Каспии, Азовском море и
Арале, где он добывается, главным образом, ставными сетями и бе-
реговыми неводами.
Сазан Cyprinus carpio распространен очень широко в бассейнах
Аральского, Каспийского и Черного морей, на Дальнем Востоке—
от Амура на юг до Бирмы. В настоящее время в результате рыбе-
водных мероприятий распространился еще шире. Завезен сазан в
Америку и на острова Индо-Малайского архипелага. В СССР искус-
ственно пущен в Балхаш, озера Барабинской степи и в бассейн
Балтийского моря. Живет сазан как в солоноватой, так и в прес-
ной воде. В отличие от леща и воблы, на время кормежки в наших
южных морях далеко от берега не отходит, а обычно все время
проводит в береговых зарослях. Половозрелым сазан становится на
4—5-м году жизни. Икрометание происходит в береговых заро-
слях с конца апреля по июнь. Нерест очень растянут.Мальки выводят-
ся обычно на 4—5-е сутки. Первое время они держатся вблизи не-
рестилищ, позже откочевывают в другие места. Пищу сазана со-
ставляют, главным образом, водные беспозвоночные: личинки на-
секомых1, ракообразные, моллюски, но питается сазан также и расти-
тельностью, как макрофитами, так и водорослями. Наиболее ин-
тенсивно кормится сазан летом, но продолжает, правда в меньшей
степени, питаться и зимой. Полностью прекращает сазан питанье
зимою только в северной части своего ареала распространения.
Растет сазан, так же как и лещ, в различных водоемах весьма раз-
лично. Наиболее быстрым ростом обладает сазан из южного Каспия.
Рост сазана в различных водоемах________________
1 ‘‘ ' 1
Арал . . 12,6 21,3 29,3 36,0 41,6 46,5
Азовское море 13,0 25,5 34,9 41,0 45,5 49,0
Южный Каспий 15,0 26,0 36,0 47,0 50,0 58,
Балхаш 8,8 16,0 21,0 25,5 28,0 —
194
Из всех южных сазанов наиболее замедленным ростом
обладает сазан из озера Балхаш. Промысловое значение ди-
кого сазана особенно велико в пределах Аральского бассейна,
где он дает в отдельные годы свыше 100 000 центнеров улова. Второе
место занимает северный Каспий и третье — южный Каспий.
Удельный вес сазана в общей добыче по Аралу наибольший.
В результате длительной селекционной работы получены различ-
ные домашние расы сазана, так называемые культурные карпы,
обладающие более высоким телом, большей упитанностью и рядом
других хозяйственно важных признаков. Число различных рас
карпа сейчас очень велико. Некоторые совершенно лишены чешуи,
у других чешуя сохранилась только вдоль боковой линии и т. д.
Как объект сельскохозяйственного рыбоводства карп представляет
наибольшую ценность и карповое прудовое хозяйство, как у нас,
так и заграницей, получило весьма широкий размах.
Сомовые — Silurldae. Восемь родов в Европе и Азии. В
пределах СССР два рода Silurus и Parasilurus.
Обыкновенный сом—Silurus glanis населяет пресные и солоно-
ватые воды Европы от Рейна на восток. В бассейне Ледовитого
океана сейчас отсутствует. Есть в бассейне Черного, Каспийского и
Аральского морей. Крупный хищник, длиной до 5 м и весом до 300
кг. Самцы крупнее самок. Растет сом довольно быстро. Полутора-
метровые рыбы имеют возраст 9—Юлет(Сев. Каспий). Половозрелым
сом становится в возрасте 3—5 лет. Плодовитость от 135 до 470 тысяч
икринок. Икрометание происходит при температуре около 20°
Икра около 3 мм в диаметре, липкая. Нерестится сом парами, при-
чем устраивает нечто вроде гнезда из растений. Самец охраняет
икру. Инкубационный период 2,5—3 суток. Через несколько (4—5)
дней по выходе из икринок (после всасывания желточного пузыря)
сомята переходят к активному питанию. Молодь питается сначала
беспозвоночными, но скоро переходит на питание рыбой. Взрослый
сом большой хищник, питается крупной рыбой, лягушками, напа-
дает на водоплавающую птицу. Известны случаи нападения сома и
на человека. Далеких миграций сом не совершает и в наших южных
водоемах (Каспий, Арал) держится недалеко от берега.
Амурский сом Parasilurus asotus (L) по образу жизни сходен с
обыкновенным, имеет несколько меньшие размеры и вес (до 50 яг).
Обыкновенный сом в наших южных водоемах имеет существенное
промысловое значение. Ловится, главным образом, различными крюч-
ными орудиями.
Щу ко в ы е — Esocidae. Пресноводные рыбы северного по-
лушария. Пять видов, из которых в Старом свете два: обыкновенная
щука — Esox lucius L, распространенная по Европе, Северной
Азии и Америке, и амурская щука Esox reichertii Dyb., населяющая
бассейн Амура, Сахалин и реки Приморья.
Обыкновенная щука населяет как стоячие, так и текучие водо-
емы, живет как в пресной, так и в солоноватой воде наших южных
морей. Держится в равнинном течении рек обычно среди зарослей.
На быстром течении; как правило, не встречается. Щука достигает
13* 195
размеров до 1,5 м и веса до 35 кг (есть указания, что вес щуки может
достигать в единичных случаях до 65 кг). Растет щука быстро. В
некоторых водоемах двухлетняя щука достигает 26—30 см длины,
а пятилетняя—50—60 см. Самки растут несколько быстрее самцов.
Впервые половозрелой щука становится иногда даже в двухлетнем
возрасте, обычно же трех-четырех лет. Икрометание происходит во
время весеннего половодья от марта на юге (Днепр) до июня (ни-
зовье Енисея) при температуре 3—15° (обычно 5—8°). Инкубацион-
ный период от 7 до 14 дней. Плодовитость щуки колеблется от
100 тысяч до 1 миллиона икринок. Икринки диаметром 2,5—3 мм
слабо липкие. Молодь первое время по выходе из икры питается
планктоном, но очень скоро переходит на питание рыбой. Взрослая
щука питается также, главным образом, мелкой рыбой — плот-
вой, красноперкой, уклейкой и другими. Лещ, имеющий высокое
тело, в пище щуки встречается очень редко.
Амурская щука — Esox reichcrtii Dyb. несколько меньших
размеров, чем обыкновенная, —до 1 м длины и 18 кг веса. По образу
жизни очень близка к обыкновенной. Щуки имеют важное хозяй-
ственное значение. Во всей области своего распространения они
являются объектом промысла. Обладая быстрым ростом и питаясь,
главным образом, малоценными видами рыб, щука в ряде случаев
может явиться хорошим объектом для заселения озер, фауна которых
состоит из малоценных видов карповых рыб.
Окуневые — Percjdae. Обитатели Северного полушария.
Водятся в пресных и солоноватых водах как Старого, так и Нового
света. В наших водах пять родов, именно — окуни Регса, судаки
Lucioperca, ерши Acerina, чопы Aspro, перкарины Percarina. Наи-
большее промысловое значение имеют судаки и окуни.
Судаки представлены в нашей фауне тремя видами: обыкновен-
ным судаком — Lucioperca lucioperca, бершом—L. volgensis и мор-
ским судаком — L. marina. Обыкновенный судак'распространен от
Эльбы на западе до бассейна Арала на востоке. В бассейне Балтий-
ского моря редок. Обилен в бассейнах Черного и Каспийского
морей. В бассейне Ледовитого океана отсутствует. Живет судак как
в пресной, так и в солоноватой воде. Имеется жилой и полупроход-
ной судаки. Жилой судак живет постоянно в реках и озерах, полу-
проходной же кормится в солоноватой воде наших южных морей.
Судак предпочитает чистые водоемы и в заболоченных водоемах
не встречается. Длина судака до 120 см и вес до 12кг. Растет полу-
проходной судак быстрее жилого. Нерестится судак впервые на 4—5-м
годах жизни, в Дону с конца апреля по июнь месяц. Судак, кормиь
шийся в море, на нерест идет весной в реки. Часть особей заходит
в реки еще с осени. Плодовитость судака о? 180 до 900 тысяч икри
нок. Нерестится на мелких местах парами, икра откладывается в
ямку и охраняется самцом. Инкубационный период 72—80 часов..
Выведщаяся молодь первое время питается планктоном, со второго
же месяца переходит на питание рыбой. Взрослый судак — хищник,
питающийся почти исключительно мелкой рыбой. В Азовском море
он в огромном количестве поедает кильку и хамсу. Наиболее интен-
196
сивно взрослый судак питается летом и осенью. Судак имеет важное
промысловое значение. Особенно много его добывается в Азовском
море. Здесь уловы его по отдельным районам (Кубань) составляют
более 50% от всей добываемой рыбы.
Берш Lucioperca volgcnsis водится в бассейнах Черного и Кас-
пийске го морей; по образу жизни близок к жилому судаку, в соло-
новатую воду почти не выходит.Половозрелым становится на 3—4-м
году жизни. Питается рыбой. Промысловое значение невелико.
Морской судак Lucioperca marina водится в северо-западной
части Черного моря и в южной части Каспийского, в пресную воду
не заходит. Нерестится в апреле—мае у берегов, часто на каменистом
грунте. Икра крупнее, чем у обыкновенного судака. Охраняет свою
икру. Имеет некоторое промысловое значение.
Обыкновенный окунь Perea fluviatilis населяет Европу, Север-
ную Азию и Северную Америку, где представлен особым подвидом.
Пресноводная озерно-речная рыба, но встречающаяся и в солоно-
ватой воде. Размеры обычно до 30, редко до 50 см. Растет окунь в
разных водоемах различно. Даже в одном и том же водоеме часто
встречаются две формы: одна — прибрежная медленно растущая,
питающаяся, главным образом, беспозвоночными, и другая —бы-
строрастущая, более крупная, держащаяся в открытой части водо-
ема, питающаяся рыбой. Самки растут быстрее самцов. У окуня,
как правило, наблюдается значительнее (до 90%) преобладание
числа самок над самцами. Плодовитость окуня 12—300 тысяч икри-
нок. Нерест происходит с февраля (на юге) по июнь (на севере)
в затишных местах, обычно среди растительности. Икра отклады-
вается в виде лент. Инкубационный период 2—3 недели. Пищу
молоди составляет планктон. Взрослый окунь в малокормных озе-
рах питается рыбой. В водоемах же, где много бокоплавов, питается
исключительно беспозвоночными. Является важным объектом про-
мысла в районах малого рыболовства (Западная Сибирь, Северный
край), добывается, главным образом, ставными сетями.
Балхашский окунь — Perea schrenki населяет бассейны Бал-
хаша и Ала-куля. Весьма эврибионтная, т. е. повсеместно встре-
чающаяся форма.
Живет как в быстрых реках горного типа, так и в сильно заболо-
ченных озерах. Длина до 35 см. Растет в Балхаше очень медленно.
Икрометание у берегов весной. Питается преимущественно рыбой.
Ловится, главным образом, неводами. На Балхаше составляет до
30% от всего улова.
Остальные окуневые не имеют серьезного промыслового значе-
ния.
Скумбриевые Scombridae. Богатое видами семейство мор-
ских рыб —обитателей то; щи воды, главным образом, умеренных
и тропических широт. В наших водах имеют промысловое значение
три—четыре вида.
Обыкновенная скумбрия — Scomber scombrus, распространена
в морях, омывающих воды Европы, от Черного моря до берегов
Норвегии. Есть в западной части Варенцова моря. Встречается и
197
по Американскому побережью Атлантики от Лабрадора до мыса
Гаттерас. Размеры скумбрии До 50—60 см. Половозрелой скумбрия
становится на 3—4-м году жизни. Плодовитость 350—450 тысяч икри-
нок. Нерест происходит в поверхностных слоях воды с марта (Среди-
земное море) по июнь (Северное море). Икра пелагическая, размеры
ее 0,86—1,38 мм с жировой каплей 0,20—0,22 мм. В Северном море
после нереста рассредотачивается по всей площади моря и интен-
сивно питается, главным образом, планктоном и молодью рыб. К
осени скумбрия опускается на глубины и постепенно перемещается
на юг. Зимует скумбрия в глубинных слоях воды.
Скумбрия, зимовавшая в Мраморном море, в районе Батуми и у
Турецкого побережья движется весной вдоль западного и восточ-
ного побережья Черного моря на север и кормится в северной части
его. Осенью после окончания нагула откочевывает назад к южным
берегам. Скумбрия является важнейшим объектом «мирового рыбо-
ловства, ловится она различными кошельковыми неводами, а также
береговыми неводами и различными объячеивающими орудиями.
Малая скумбрия—Scomber colias и близкая к ней Scomber japo-
nicus по образу жизни близки к обыкновенной скумбрии. Основное
отличие заключается в наличии у этих видов плавательного пузыря,
который у обыкновенной скумбрии отсутствует. Эта особенность
приводит к тому, что малая и японская скумбрия не могут столь
быстро перемещаться из поверхностных слоев в глубинные и обратно.
Оба вида являются важными объектами промысла.
Бычковые —Gobiidae небольшие прибрежные иногда прес-
новодные рыбы, распространенные во всех умеренных и тропических
морях. Множество родов. У бычков имеет место ярко выраженный
половой диморфизм. Во время нереста у самцов обычно темнеет
окраска, изменяется форма головы и удлиняются плавники. Самцы
у многих видов устраивают гнезда обычно в виде пещерок и охра-
няют отложенную икру. Питаются во взрослом состоянии преиму-
щественно донными беспозвоночными. Объектом промысла бычки
служат у нас в Черном, Азовском и Каспийском морях. Промышля-
ются преимущественно бычок песочник Gobius mehnostomus,
бабка Gobius fluviatilis, травяник Gobius ophiocephalis, бычок
жаба Mesogobius batrachocephalus.
Бычок-песочник населяет Черное и Каспийское моря, из кото-
рых входит в реки. Эта небольшая рыбка, размеры которой дости-
гают максимум до 250 мм. Самки мельче самцов. Растет бычок
песочник в Каспии следующим образом:
Самцы
Самки
fl /2 fs fj
6,9 8,8 11,1 13,8
6,3 6,7 8,1 8,5
В Черном море растет хуже, чем в Каспийском.
Половозрелыми самки бычка песочника становятся на втором,
самцы— на третьем году жизни. Плодовитость, по данным Кули-
ченко, 250—400 икринок. Соотношение полов приблизительно один
198
к одному. Икрометание в Каспии с конца апреля. Икринки приклеи-
ваются к донным предметам.
Имеет важное промысловое значение, идет, главным образом,
па приготовление консервов.
Бабка Gobius fluviatilis населяет бассейны Черного и Каспий-
ского морей, преимущественно держится в реках и в опресненных
предъустьевых пространствах. По размерам мельче предыдущего,
максимум до 195 мм.
Бычок травяник Gobius ophiocephalus населяет Черное и Среди-
земное моря. Размеры до 250мм. Держится в соленой воде, в пресную
воду обычно не заходит. Встречается обычно среди зарослей морской
травы.
Бычок жаба Mesogobius batrachocephalus населяет Черное море,
изредка входит и в пресную воду. Самый крупный из всех каспий-
ско-черноморских бычков, достигает до 34 см длины. Ранней весной
(в конце марта) подходит к берегам. Нерестится в соленой воде в
конце марта и апреле.
Так же, как и предыдущие виды, является важным объектом
промысла в Черном море и идет, главным образом, для производ-
ства консервов.
Морские ерши Scorpaenidae. Морские рыбы, главным
образом, обитатели умеренных широт, но отдельные виды имеются
как в тропиках, так и в субарктике. Максимум видов в умеренных
широтах Тихого океана. В пределах нашего Союза основной объект
промысла — морской или красный окунь Sebastes marinus. Морской
окунь распространен в Северной Атлантике до Новой Земли и Шпиц-
бергена на севере и востоке и до Шотландии и мыса Код на юге и
западе. Это—относительно глубоководная рыба, редко держится на
глубинах менее 100 м. Размеры морского окуня до 89 ел/, обычно
до 64 см. Растет морской окунь очень медленно. К десятилетнему
возрасту, когда он впервые становится половозрелым, он достигает
в среднем 32 см длины. Продолжительность жизни морского окуня
максимум 27 лет. Морской окунь принадлежит к той мало-
численной группе рыб, которые родят живых детенышей. Пло-
довитость морского окуня очень велика—до 197 тысяч штук
у крупных особей. Правда, одновременно созревает и выме-
тывается только небольшая часть. Выметывание личинок проис-
ходит весной.
У морского окуня так же, как и у обыкновенного окуня, самки
численно преобладают над самцами.
В восточной Атлантике морской окунь совершает миграцию,
напоминающую миграции трески. Весной косяки морского окуня
появляются в западной части Варенцова моря и двигаются, интен-
сивно питаясь, на восток. Более крупные особи идут позднее. От-
мет личинок происходит в западной части Варенцова моря с апреля
по июнь. Поэтому понятно, почему сперва движение на восток
начинают неполовозрелые особи. Миграция на восток является
кормоЬой. Закончив нагул к осени, окунь отходит на запад и зимует
в западной части Варенцова моря и еще юго-западнее.
190
Пищу морского окуня составляют, главным образом, пелагические
ракообразные, рыба (главным образом мойва), головоногие моллюс-
ки и некоторые другие животные.
Морской окунь является весьма важным объектом промысла.
В 1933 году в Европейских водах всеми государствами выловлено
31 250 тонн.
В уловах СССР по Мурманскому побережью окунь составляет
за отдельные годы до 10%.
Добывается морской окунь, главным образом, тралом.
Тресковые —Gadidae преимущественно морские рыбы
(в пресных водах один род) умеренных и арктических вод. В тропиках
живут только на глубинах. Двадцать два рода, из них в пределах
СССР—девять: Gaidropsarus, Loti, Molva, Gadus, Eleginus, Bg-
reogadus, Arctogadus, Theragra, Brosme.
Тресковые — преимущественно придонные рыбы, питающиеся
или бентосом или рыбой. Многие виды выносят сильное опреснение
воды и заходят в реки и предъустьевые пространства (навага, сайка,
полярная треска). Постоянно же живет в пресной воде только на-
лим Lota lota. Почти все тресковые—обитатели холодных и умерен-
ных вод. В тропиках число видов этого семейства чрезвычайно
мало. Некоторые виды приурочены к высоким широтам Арктики и
живут только при температурах, близких к нулю (Arctogadus,
Boreogadus). Икра у всех морских тресковых, за исключением на-
ваги, пелагическая, плавающая в толще воды, у наваги же икра
донная.
Треска Gadus morhua распространена у берегов Европы от Кар-
ского моря до Бискайского залива.'Имеется она и по Атлантическому
побережью Америки от Гренландии на юг до Виргинии. Треска
является основным промысловым видом тралового промысла в
Баренцовом море. Основные нерестилища трески расположены у
Лофотенских островов. Меньшие нерестилища имеются и в других
местах: в Мотовском, Кольском заливах у Териберки и в ряде мест
восточного Мурмана, а также у острова Медвежьего. Икрометание
с февраля по июнь. Наиболее интенсивный нерест происходит в
апреле при температуре -f- 1°—0° Инкубационный период в среднем
около месяца, продолжительность его сильно зависит от температу-
ры. Плодовитость трески очень велика от 2,5 до 10 млн. и более
икринок. Половозрелой треска становится на 5—6-м году жизни.
После окончания икрометания у Лофотенских островов сильно
исхудавшая норвежская треска, усиленно питаясь, двигается на
восток и постепенно доходит до Новой Земли, а часть особей в по-
следние годы даже входит в Карское море. Треска, нерестующая в
бухтах восточного Мурмана, видимо, представляет собою прибреж-
ную малопозвонковую расу, не уходящую далеко в море.
Пища взрослой нагульной трески весьма разнообразна; из рыб
преобладают сельдь, мойва и другие тресковые (главным образом
молодь), а также бентические беспозвоночные. Наиболее интенсив-
но треска питается весной при начале нагула и осенью после начала
обратного движения с востока на запад к нерестилищам. Входящая
2ое
в Баренцево море после икрометания треска очень низкой упитан-
ности, и количество жира, содержащегося в ее печени, незначитель-
но. По мере же движения на восток к осени вес тресковой печени
быстро возрастает, увеличивается количество жира и в тканях
тела. Молодь трески сначала питается планктоном, но очень скоро
переходит на пищу бентосом. Предельный возраст трески в Барен-
цевом море 17 лет. В уловах преобладают особи семи,-двенадцати-
летнего возраста. Трехлетняя океаническая треска достигает в сред-
нем 35 см длины, четырехлетняя—45 см, восьмилетняя—85 см и
десятилетняя—103 см. Треска из Белого моря, являющаяся особым
подвидом, растет медленнее, чем мурманская.
Треска — основной объект тралового промысла советских траль-
щиков в водах Варенцова моря. За отдельные годы она составляет
от 47 до 85% общего тралового улова. В европейских уловах треска
занимает второе место (первое — сельдь). Кроме тралового промыс-
ла, в прибрежной зоне треска в значительном количестве добы-
вается на яр\са и на поддев. Особенно сильно развит прибрежный
промысел в Норвегии, где он дает основную массу трески.
Пикша Gadus aeglefinus распространена от восточной части
Варенцова моря до Бискайскогозалива. В Белом море редка. Имеется
и у Атлантического побережья Северной Америки на север до Грен-
ландии. Так же как и треска, пикша весной входит с запада в Барен-
цево море. Нерестилища пикши расположены в Северном море, у
берегов Норвегии и в западной части Варенцова моря (Мотовский
залив и др.). Икрометание пикши приходится на период времени
с апреля по июнь. В Мотовском заливе наиболее интенсивный нерест
наблюдается в конце мая. Икрометание пикши происходит при
более высокой температуре, чем трески (около 4-5°). Икра пелаги-
ческая. Выведшиеся из икры мальки первое время ведут пелагический
образ жизни, но уже достигнув 20 мм длины, опускаются в придон-
ные слои веды. Половозрелой пикша становится на 4—5-м году жизни.
Плодовитость пикши меньше, чем у трески: она колеблется от 100
тькяч икринок до двух миллионов. Пикша, вошедшая в Баренцево
море, усиленно питается. С марта по июнь в ее пище наряду с бенто-
сом важное значение имеет мойва. Во второй половине лета пикша
почти целиком переходит на питание бентическими беспозвоноч-
ными. Наиболее интенсивно пикша питается весной. В июле ин-
тенсивность питания несколько снижается и снова возрастает в
августе — сентябре. В отличие от трески в пище пикши совершенно
ничтожную роль играют пелагические ракообразные, составляющие
в пище трески в некоторых районах до 27%.
Растет пикша следующим образом.
Возраст /1 /2 /з /4 Z5 Ze /7
Длина 17,6 24,7 33,2 41,4 40,5 51,3 56,0
В уловах попадаются рыбы до 14-летнего возраста. Наиболь-
шие известные размеры пикши в Баренцевом море —^7 см. Пикша
является вторым по важности промысловым представителем трес-
ковых. В европейских уловах она занимает третье место. Удельный
201
вес пикши в траловых уловах колеблется от 6,4 до 37%. Так же как
и треска,пикша является объектом и прибрежного ярусного и под-
девного лова.
Сайда Gadus virens распространена от Канинских банок в Ба-
ренцевом море до Бискайского залива и по американскому побережью
до мыса Код. Места икрометания сайды расположены у берегов
Норвегии не севернее Тронгейм-фиорда и Лофотенских островов.
Молодь сайды появляется на Мурмане в июне, уже достигнув раз-
меров 13—35 мм. В это же время появляются на Мурмане и взрослые
особи сайды. Сайда — пелагическая рыба, держащаяся в толще
воды. Пищу ее составляют, главным образом, ракообразные и рыбы.
Растет сайда быстрее пикши. Двух лет она имеет в среднем 32 см
длины, трех лет—44 см, четырех лет—50 см, пяти лет—57 см, ше-
сти—65 см. Наиболее крупная сайда редко достигает размеров
свыше метра и возраста более 18 лет. В Европе сайда занимает
следующее место в уловах после пикши. В настоящее время в пре-
делах СССР сайды вылавливается немного. Объясняется это не
незначительностью запасов, а тем, что сайда, как пелагическая
рыба, должна добываться другими орудиями лова, чем треска и
пикша, а в настоящее время орудия лова, которыми можно было бы
с успехом добывать сайду на Мурмане, не применяются.
Сайка Boreogadus saida распространена циркумполярно. К югу
далеко не спускается, придерживаясь в своем распространении
кромки льдов. При положительных температурах сайка встречается
редко. Максимальная температура, при которой наблюдалась эта
рыба, — около 5° Зимою сайка опускается из высоких широт
южнее и доходит до восточного Мурмана. Плодовитость сайки от
9 до 18 тысяч икринок. Нерест происходит зимой. Условия его точ-
но неизвестны. Места икрометания сайки отмечены близ Новой
Земли и у Канина. Сайка—обитательница толщи воды ив придонные
слои обычно не опускается. Пищей ей служат пелагические живот-
ные, главным образом, ракообразные и в частности Calanus, а также
фитопланктон. Размеры, сайки не более 36 см, обычно 24 см.
Промысловое значение сайки до последнего времени было очень
невелико. Последнее время добыча ее быстро развивается особенно
в Белом море, Чешской губе и далее на восток. Сайка представляет
интерес также и как объект питания ценных промысловых рыб и
млекопитающих.
Навага Eleginus navaga распространена от восточного Мурмана
на западе до Карского моря на востоке. Обильна в Белом море.
На Дальнем Востоке близкий вид Е. gracilis. Навага больше других
тресковых, исключая налима, связана с пресной водой, заходя
зимой в устья рек. Икрометание наваги происходит в декабре —
январе, икра, в отличие от других тресковых, донная. Количество
откладываемой икры варьирует от 20 до 62 тысяч. Инкубационный
период около 20—40 дней. Значительных миграций навага не совер-
шает. Пищу наваги составляют беспозвоночные и рыбы. Средние
размеры беломорской наваги около 25 см, вес около 100 г. Новага
имеет важное хозяйственное значение. Основной промысел наваги
202
производится в зимнее время и носит еще, главным образом, кустар-
ный характер (удочки). Заготовляется навага, главным образом,
в свежемороженном виде.
Налим Lota lot i. Обыкновенный налим распространен в Европе
и Северной Азии. В Анадыре, Пенжине, Западной и части Северной
Америки заменен подвидом Lota lota leptura. Населяет реки и озера
с холодной водой и благоприятным кислородным режимом. Будучи
рыбой холодноводной, налим на севере достигает значительно
больших размеров (до 24, а редко и до 35 кг), чем на юге. Так, в
бассейне рек Черного моря обычный размер налима около 1 кг.
Налим ведет донный образ жизни, держась или между камней,
или под корягами. Нерест у налима происходит зимой с декабря
по февраль. Плодовитость очень сильно варьирует и колеблется от
33 тысяч до 5 миллионов икринок. Инкубационный период тянется
около одного месяца. Икра, видимо, батипелагическая. Пищу налима
составляют главным образом рыбы. В Каме налим питается преиму-
щественно пескарем и ершом. В Колыме основную пищу налима
составляет молодь самого же налима. Молодь налима питается, глав-
ным образом, донными беспозвоночными, в том числе ручейниками и
личинками хирономид. Растет налим в разных водоемах далеко
неодинаково.
Возраст 1. ^3 /4
Печора . 16,9 23,9 34,2 41,9 44,4
Ильмень 13,3 23,1 31,8 38,0 —
Кама 17,2 21,4 25,6 31,2 41,2
Наиболее быстрым ростом обладают налимы в самых северных
из исследованных водоемов. Промысел налима нигде не носит мас-
сового характера, но в очень многих местах он составляет довольно
значительный прилов. Добывается налим удочками на живца, в
верши и некоторыми другими орудиями лова.
Камбаловые Pleuronectidae. Сорок три рода чрезвы-
чайно широко распространенных в морских водах земного шара
рыб. В водах СССР более сорока видов, из них тридцать два в водах
Дальнего Востока. Семейство распадается на 5 подсемейств. Соб-
ственно камбалы относятся к подсемейству Pleuronectinae, к кото-
рому принадлежат все основные промысловые камбалы СССР. Это
подсемейство распространено в Северном полушарии на юг до Сре-
диземного моря и параллели Нью-Йорка в Атлантическом океане
и в северной части Тихого—на юг до южной Японии и Калифорнии.
Камбалы — придонные рыбы, не совершающие во взрослом состоя-
нии далеких миграций и держащиеся все время примерно в одних и
тех же участках моря. Почти все из них морские рыбы, но некоторые
живут и в пресной воде. Таковы: речная камбала, некоторые
дальневосточные камбалы и отдельные представители семейства
Bothidae. Икра у камбал пелагическая — плавающая в толще воды.
Большинство камбал обладает весьма высокой плодовитостью. У
некоторых видов число икринок превышает несколько миллионов.
Выведшиеся из икры мальки некоторое время имеют симметричную
203
форму тела, затем глаза перемещаются на одну сторону, рыбы
опускаются в придонные слои и ложатся на один бок, который
остается непигментированным, другая же сторона приобретает
окраску, которая у камбал обладает способностью очень сильно
варьировать в зависимости от окраски грунта, на котором лежит
камбала.
Плодовитость камбал
Тюрбо от 1 000 000 до 9 000 000
Палтус . . » 2 000 000 >> 3 500 000
Морская камбала » 90 000 » 500 000
Речная >> •> 400 000 » 1 500 000
Полярная » » 52 000 » 200 000
Пищу камбал составляют, главным образом, донные беспозвоноч-
ные: моллюски, ракообразные, черви. Палтусы частично переходят
на питание рыбой. В Баренцевом море из 11 распространенных
здесь камбал имеют промысловое значение 4 вида. В водах Дальнего
Востока также далеко не все виды камбал используются промыслом.
Морская камбала Pleuronectes platessa распространена в Барен-
цевом море, в Северном море и в северной части Атлантического
океана на юг до южной Франции. Как и другие камбалы, мало
подвижная придонная рыба, не совершающая далеких перемеще-
ний. Наибольшее зарегистрированное расстояние, на которое пе-
редвинулась камбала, составляет менее 500 км. Икрометание про-
исходите прибрежных участках. У нас на Мурмане места икромета-
ния камбалы известны в Мотовском заливе, у острова Кильдина, в
Териберской губе и близ Харловки. Икрометание происходит с
февраля по апрель. Развитие икры длится обычно больше месяца, но
продолжительность инкубационного периода очень сильно варьи-
рует в зависимости от температуры. В Баренцевом море существуют
видимо две расы морской камбалы: одна раса икромечет на глубинах
от Одо 80 л/, а другая раса нерестится на глубине около 200м. Пищу
морской камбалы составляют почти исключительно беспозвоноч-
ные, мелкие моллюски, черви и ракообразные. Рыба имеет в пище
морской камбалы совершенно подчиненное значение. Растет в Барен-
цевом море морская камбала довольно медленно, лишь на седьмом
году жизни достигая размеров в 31 см. Двадцатилетняя камбала
имеет около 50 см длины. Возрастной состав уловов камбалы в
Баренцевом море очень растянут, в трал попадают особи от б до 20
и даже более лет. Преобладающее значение имеют в уловах 8—10-
летние особи. Половозрелыми самцы морской камбалы становятся
на 8—9-м году жизни, самки — на 10—12-м году. Соотношение
полов близко к единице. У старших возрастов самки несколько
преобладают над самцами.
Камбалы являются очень важным объектом тралового промысла.
В Северном море вылавливается ежегодно около 54 тысяч тонн
камбалы, в Баренцевом море ежегодно добывается 6—7 тысяч тонн
камбалы. Большое количество морской камбалы добывается также
у берегов Исландии и в Скагераке, Каттегате и Бельтах. Запасы
204
морской камбалы в Северном море из-за интенсивности промысла
и медленной возобновляемости стада находятся в очень напряжен-
ном состоянии и близки к почти полному подрыву. В Баренцевом
море запасы камбалы, хотя и находятся в лучшем состоянии, но
также нуждаются в очень бережном использовании, так как размеры
камбал по сравнению с предвоенными очень сильно снизились.
Речная камбала Pleuronectes flesus широко распространена от
Енисея на запад вдоль всего побережья Европы до Черного и Азов-
ского морей. Образует несколько географических рас. Придержи-
вается прибрежных участков морей и входит в ки, по которым
поднимается иногда довольно высоко вверх. Миграции в море изу-
чены еще очень плохо. Икрометание у нас на севере с конца апреля
по конец июня и позже. Выведшиеся из икры мальки держатся,
главным образом, в опресненных районах, постепенно по мере роста
отходя в более осолоненные участки моря. Пища речной камбалы
состоит из ракообразных, моллюсков и червей. В меньшей степени
речная камбала питается мелкой рыбой. Половозрелой речная
камбала становится раньше морской, обычно на пятом году жизни.
Предельный возраст речной камбалы—12 лет. Промысловое значе-
ние речной камбалы сравнительно невелико. В СССР она промы-
шляется на Мурмане, у полуострова Канина, в Чешской губе, а
также в Черном море.
Желтобрюхая камбала Pleuronectes quadrituberculatus распро-
странена в Японском, Охотском и Беринговом морях. Придержи-
вается в своем распространении прибрежных участков, не уходя
летом глубже 80 м. Икрометание происходит на глубинах свыше
100 м (зарегистрировано на глубине 156 м при температуре-|-0,4о)
в конце марта — апреле. Питается эта камбала преимущественно
донными беспозвоночными. Длина до 60 см, вес до 3 кг, самцы мень-
ше самок. В настоящее время' промысловое значение невелико, но
с развитием тралового промысла в Татарском проливе и Охотском
море оно может сильно возрасти.
Полярная камбала Liopsetta glacialis. Циркумполярно рас-
пространенный вид. Встречается по всему побережью Сибири, в
Беринговом и Охотском морях, у Аляски, в Баренцевом и Белом
морях. Летом, как и большинство других камбал, держится у бере-
гов. Подход к берегу начинается с мая. У берегов полярная камбала
обычно держится до сентября, когда откочевывает на глубины.
Икрометание у полярной камбалы в Белом море происходит в мае
месяце. Питается полярная камбала, главным образом, моллюсками.
Максимальная длина полярной камбалы редко превышает 20 см и
максимальный возраст—10 лет. Промысловое значение полярной
камбалы очень невелико, и несомненно, что вылов ее особенно в водах
Дальнего Востока может быть значительно увеличен.
Остроголовая камбала Cleisthcnes herzenst-ini. Распространена
в Японском и Желтом морях. Летом держится у берега до глубин
50 м. Зимой уходит на большие глубины. Икрометание в заливе
Петра Великого происходит в июне—июле месяце. Пищей остро-
головой камбалы служат беспозвоночные — офиуры, ракообразные,
2С5
моллюски и рыбы. Иногда в желудке этой камбалы попадается даже
иваси. Средний размер промысловой остроголовой камбалы около
34 см, максимальные размеры до 46 см. Держится эта камбала боль-
шими скоплениями и является важным промысловым объектом в
заливе Петра Великого, где она добывается в ставные невода и
датским неводом.
Желтоперая камбала Limanda aspera распространена в Беринго-
вом, Охотском и Японском морях. Весною подходит с глубин к
берегам, где держится все лето. Время икрометания очень растя-
нуто — с мая по август. Нерест происходит в прибрежной зоне.
Питается, как и другие камбалы, главным образом, беспозвоноч-
ными. Желтоперая камбала из залива Петра Великого растет зна-
чительно быстрее, чем этот же вид камбалы из Татарского пролива.
Так, в заливе Петра Великого самки этой камбалы имеют при длине
23 см — три года, а в Татарском проливе — 6 лет. Половозрелость
наступает в возрасте 4—5 лет, у самцов и 6—7 лет у самок. Средняя
длина промысловой камбалы — около 30 см. Желтоперая камбала
является основной промысловой камбалой в водах Советского Даль-
него Востока. Она вылавливается как датскими, так и ставными
неводами.
Палтус Hippoglcssus hippoglcssus распространен в северной
части Атлантического и Тихого океана. В Баренцевом море встре-
чается только на западе. На юг идет до берегов Франции. Есть у
берегов Гренландии. Иногда попадается в Балтийском море. Весной
палтус вместе со стаями трески входит в Баренцево море и двигается
на восток, держась в воде с температурой от 3 до 8°. В летнее время
палтус приближается к берегам, в зимнее же уходит на более значи-
тельные глубины—до 350 м. Нерест палтуса, добывающегося в
Баренцевом море, происходит у берегов Норвегии в марте — апреле.
Питается палтус, главным образом, рыбами: треской, пикшей и
другими, а также крупными ракообразными. Палтус наиболее
крупная и быстро растущая камбала наших морей. Она достигает
4,7 м длины и до 300 кг весу. В пределах СССР добываются палтусы
до 2 м, 25 см длины. Возраст, которого достигает палтус, определяет-
ся во всяком случае более чем 36 годами. Рыбы в возрасте 7 лет имеют
длину 65 см, Ю лет — 93 см, 12 лет—108 см и 15 лет—123 см.
Палтус — одна из наиболее ценных морских промысловых рыб.
Добывается палтус преимущественно тралом при лове другой рыбы.
За 1933 год мировой улов палтуса составил 14 319 т, наибольшее
количество палтуса дает сейчас Гренландия, Норвежское море и
Исландия. В некоторых районах палтус сильно перепромышлен*
и запасы его подорваны.
206
ОСНОВНЫЕ ПОСОБИЯ ПО БИОЛОГИИ РЫБ
Берг Л. С., Рыбы пресных вод СССР, т. I, 1932 и т. II, 1933.
Книпович Н. М., Гидрология морей и солоноватых вод, 1938.
Мейснер В. И., Промысловая ихтиология, 1933.
Солдатов В. К., Рыбы и рыбный промысел, 1928.
Солдатов В. К., Промысловая ихтиология: ч. I, Общая ихтиоло-
гия, 1934; ч. II, Рыбы промысловых районов СССР, 1938.
Суворов Е. К., Основы общей ихтиологии, 1940.
Пучков Н. В., Физиология рыб, 1941.
Шмидт П. Ю., Миграции рыб, 1936.
УКАЗАТЕЛЬ РУССКИХ НАЗВАНИЙ, ИМЕН СОБСТВЕННЫХ
И ТЕРМИНОВ
А
Абиотическая среда 50, 135
Абиотические факторы, 108, 156
Абсорбция газа кровью 7
Авитаминоз 29
Адаптивная радиация 3
Акклиматизация 86, 88, 89, 93, 173
190
Актинии 68
Акула гигантская 58
колючая 166
полярная 116, 126, 136
сельдевая 4, 105, 111, 159,
164
Акулы 6, 11, 13, 39, 51, 58, 59, 62
66, 83, 84, 85, 90, 111, 114,
115, 116, 125, 130, 139, 164, 169
— ископаемые 126, 136, рис. 86
Алявдина 72
Амилаза 166
Амур 130, 191
Амфибии 81
Анальное отверстие 48
Андрияшев рис. 8, 16
Антенны 76
Анчоус азовский 186
— японский 20
Анчоусовые 185
Анчоусы 51, 84 186
Арапайма 136
Ареал распространения 20, 50, 194
Арктическая фауна 21
Арктические виды 19, 39, 130
Арнольд 45
Атерина 31
— американская 125
Б
Бабка 198, 199
Бакланы 82, 83
Бактерии 63, 72
Барбулька 84
Батипелагические рыбы 4
Бахтак 188
Безжелудочные рыбы 166
Беззубка 75
Белки 166
Белоглазка 106, рис. 63
Белорыбица 99, 102, 159, 187 189
|рис. 114.
208
Белуга 85, 97, 118, 136, 179, 180
рис. 114
Белуха 84
Бельдюга 41, 126
Бентические беспозвоночные см
Донные беспозвоночные
Бентос 44, 75, 200, 201
Бентосоядные рыбы 30, 78, 169
Берг 17, 116, 156, рис.: 24, 82, 108
Берш 196, 197
Беспозвоночные 159, 160, 162, 180,
181, 190, 194, 197, 202, 204, 205,
206
Бешенка 165
Бииб 22, 67, 69
Биогенные вещества 41, 79, 88
Биологически кондицированная во-
да 58
Биологические ниши 93
Биомасса бентоса 29, 80, 149, 171
— кормовых объектов 95, 170,
171 рис.: 56, 57
Биотические факторы 156
Биотоп 50
Бобры 85
Боджак 188
Бойцовая рыбка 126
Бокова 166, 167, рис. 7
Боковая линия 39
Бокоплавы 79, 107, 167, 170, 188 197
Бореальные виды 19
Бородавчатка 61, 92
Боруцкий 95
Брама 20
Брачные игры 126
— изменения 175, 178, 186,
рис. 73.
Брачный наряд 119, 191, рис. 74
Броцкая 168
Брызгальце 46
Брызгун 162
Брэм рис. 50
Брюшина 28, 71
Брюшной пояс 31
Быстряпка 106, рис. 6
Быстрянки 34, 106
Бычки 4, 8, 28, 30, 31, 40, 42, 84
107, 116, 118, 119, 130, 159, 166
170, 171, 172, 179, 180, 184, рис.
24, 83
Бычки-прыгуны см. Прыгуны
Бычковые 198
Бычок жаба 198, 199 рис. 114
песочник 198, рис. 114
четырехрогий 4
В
Ван дем Борне 35, 36
Банк 166
Вард 18
Васнецов 114, 146
Веберов аппарат 7
Вегенера теория 103
Великохатько рис. 61
Верховна 39, 192
Вес гонад 121, 157, 159
— пиши 157
— тела 121, 159, 167, 195, 196,206
Весепнемечущие рыбы 123
Веслоногие рачки 70, 76, 77, 78
Веснянки (личинки 80)
Виллер 56, 151
Винге 141
Витамины в пище 149
— в тканях 100
Внутренние жидкости 13
Вобла 5, 16 40, 52, 76, 81/83, 97,
105, 107, 121, 124, 125, 133, 138,
140, 153, 159, 166, 171, 172, 179,
191, 194, рис.: 7, 75, 107, 114
— аральская 192, 193
— каспийская 167, 168, 169,
192, рис.: 102, 103
Водоплавающие птицы 69, 195
Водянка 63
Водяной ослик 71
скорпион 80
уж 81
Водяные клопы 80
Возбудители заболеваний 66
Возраст 53, ПО, 116, 118, 136, 159
167, 170, 179, 183, 192, 194, 195,
201, 202, 203, 205, 206
— определение 137, 139, 141,
142
Возрастной состав 53, 138, 151,
152,154 , рис. 103
Возрастные группы 143, 151, 152, 153
Воробьев 168, рис. 112
Воскобойников рис. 26
Воспроизводство запасов 89, 188
Востробрюшка амурская 4
Врасский 127
Вторично глубоководные рыбы 7, 9
Вторичнополовые признаки 29, 118,
119
Вуалехвосты 89, рис. 54
Вундер 24, 39, 46, 166
Выделение азота рис. 97
— углекислоты рис. 97
Выдра 84
Вылет насекомых 80, 170
Выпуск мальков 136, 181
- 4 Биология рыб.
Вырезуб 98, 191
Выростные пруды 79
Выхухоль 84
Высокоарктические виды 130
Высшие растения 63
Вьюн 30, 46,48
Вьюновые 8, 48, 119, рис. 71
Г
Гагары 82
Газ плавательного пузыря 6, 8
Газообмен 16, 43, 44, 50
Газоотделительная железа 7
Гамбузия 29,90, рис. 79
Ганоиды костные 6
— хрящевые 6
Гегаркуни 188
Гейнеке-Майер 120
Геккель 164
ГелландТансси 156
Гемолиз крови 44
Генеративно морские рыбы 109, 130
— пресноводные рыбы 108, 10t
Гербильский 122, 181
Гермафродитизм 117
Гесс 23
Гибель возрастных групп 153
— икры 132
— рыб 19, 34, 37 43, 44, 47,
50, 55, 62, 63, 65, 66, 69, 70, 72,
75, 76, 77, 86, 156
Гидра 67
Гидростатические приспособления 9
Гипертоническая среда 14
Гипертония 13
Гипотоничный 14
Гипофиз вытяжка 122
— гормон 122, 147
Гладыш 80
Глаз светопоглотительная способ-
ность 25
— содержание витамина 100
— строение 24, 28, 33, 103
Глаза стебельчатые 25
— телескопические 25
Глазные пузыри 44
Глистные инвазии. 92
Глотка 45, 46, 48
Глоточные зубы
— выдолбленные 164
жевательные 164
крючковидные 164
Глохидии 75
Глубоководные рыбы 9, 22, 24, 42,
119, 162, 167, 199, рис. 2, 16
— животные 22
Гнездо 31, 63, 109, 127, 175, 195, 198
Годовик 143
Годовой4 цикл 51
Годовые кольца 138, 139, 140, 141,
142, 144, рис. 88
Голавль 46, 164, 192
Голец гребенчатохвостый 36
209
— тибетск 36, 151, рис.: 71,
101
Голова 30, 39, 45, 59, 62, 198, рис.:
11, 13, 74
Головной мозг 28
Головоногие 75, 200
Голотурии 75
Гольцы 4, 33, 34, 35, 36 46, 181, 188,
189, рис. 21
Гольян 35, 39, 46, 191, рис. 21
— амурский 192
Гонадотооппые гормоны 87, 118, 122
Гонады 28, 100, 121, 122, 159, 187
Гоноподий 126, рис. 79
Горбуша 84, 117, 179, 186, 187
рис. 73
Гормоны половых желез 146
Горчак 76, 123, 191, 192
Гофбауер 151
Гравидап 122
Гребневики 67
Грибки 65, 72
Гуанин 26
Губа нижняя 35, 36, 160
Губаны 164
Губки 90
Густера 82
д
Давление воды 6, 8, 9
Далия 14, 17
Дамский чулочек 34
Движение рис.: 68, 69, 70
Двоякодышащие 6, 30, 48, 134, 164
Двукрылые (куколки) 107
— (личинки) 107
Двухгодовик 143
Двухлеток 143, 152
Дельфины 84
Демерсальная икра см. Икра
Державин 116, 132, 156
Детрит 164
Детритоядные рыбы 192
Динамика возрастного состава 155,
156
— жирности 158
— ксэфициент упитанности 159,
рис. 63
— органического вещества 95
— промысловых уловпз ПО
— численности популяции 156
Длина головы 25
— рыбы Ь7, 201, 205, 206
рис. 101
Дночерпатель 169
Добавочные кольца 141
Догель 76, рис. 40
Донная икра см. Икра
— фауна 37
Донные беспозвоночные 160, 162,
169, 189, 190, 192, 193, 198, 200,
201, 203, 204, 205
— организмы 140
растения 192
— рыбы 3, 4, 34, 53, 116, 134г
163, 171, рис. 18
Доска Монастырского 144, рис. 94
— Эйнар Леа 143
Дрифтерные сети 185
Дробление икры см. Икра
Дрягии 118, 124
Дыхание 33, 43, 44, 45, 46, 47, 48
49, 134, 164, рис. 26
— жаберное 48
— кишечное 48
Дыхательная сосудистая сеть рис, 84
Дыхательный аппарат 45, 62
— ритм 50
Е
Елец 35, 106, 140, 161
Ерш 46, 127, 171, 196, 203
Ж
Жаберная крышка 31, 33, 46, 110,
135, 139, 141, рис. 72, 88
— полость 48
Жаберные лепестки 160
— мешки 45
— отверстия 46
—’ тычинки 160, 164, 165, 183
190
~ щели 45
Жаберный аппарат 64, рис. 108
Жаброхвостые раки 76, 77
Жабры 5, 13, 14, 34, 43, 46, 48, 49,
62,66,69,72, 75, 76,77, 134, 161
— наружные 46, рис. 85
Жгутиковые 66, рис. 40
Железки вылупления 134
Железы бокаловидные 114
Желтёнкова 78, 169
Желток 134
Желточный мешок 133, 135
— пузырь 29
Желудок 48, 160, 164, 166
Желчный пузырь 70, 175, 195
Жемчуг 90
Жерех 21, 97, 106, 162, 164, 192,
рис. 110
Жерновок 164
Жертва 59, 162, 167, 175
Живородящие рыбы 56, 117, 126,
127, 199
Живорождение 41, 126
Животноядпые рыбы 159, 164, рис.
111
Жидкость тела (замерзание) 17
Жилые расы 178
— рыбы 5, 190, 196
Жир процент содержания 99, 158,
175, рис. 106
— содержание в печени 105 201
Жирность рыб 157, 158
— сезонные изменения 158, 159
210
Жиры 166
Жуки 79, 80
3
Заболевания рыб 63, 65
Забота о потомстве 115, 192
Закапывание 30
Закапывающиеся рыбы 29, 30, 31, 32
Закладка органов 133
Залегание на ямы 17, 53, 107
Замахаев 140, рис. 90, 91
Замораживание рыб 17
Заморы 47, 88, 153
Запасы корма 171, 173
— промысловых рыб 5, 87, 88,
89, 138, 152, 153, 154, 181,
204, 205, 206
Запуск рыболовства 136, 181
Зацепин 73
Защита 30, 61
Звездчатка 32
Звукоулавливающий аппарат 8
Зев 46
Зеленая морская черепаха 82
Землеройки 85
Земноводные 81
Зенкевич 167
Зимовальные скопления 53
Зимовка 180, 186
Зимородки 82
Змеевидный тип формы тела 111, 159
Змееголовки 48, 49, рис. 27
Змеи 81
— рыбоядные морские 82
— рыбоядные пресноводные 81
Золотая рыбка 43, 56, 58 рис. 54
Зоогеографическая область 21
Зрачок 25, 28
Зубатка малая 78
Зубатки 75, 160, 169
Зубы 85, 119, 160, 164, 175, 178,
183, 186
— верхнеглоточпые 164
глоточные 164, рис. НО
кипжаловндпые 164
— нижнеглоточные 164
И
Иванова-Берг 97
Иваси 20, 55, 182, 185, 206, рис.
31, 83
Ивлев 125, 133, рис.: 4, 5
Игла каспийская 42
Иглокожие 73, 74, 75, 160
Идельсон 81
Иенсеп 156
Избирательная способность 169
Изотоничный 14
Изотония 14
Икра 9, 15, 28, 31, 38, 40, 41, 44,
63, 65, 76, 79, 81, 83, 87, 96, 97,
116, 117, 120, 125, 126, 127, 128,
129, 130, 136, 149, 174, 178, 179,
*4*
192’ '96- 197- Рис.: 81,
о2, оэ, 1ио
— батипелагическая 130, 184,
— выживаемость 125, 131
— вынашивание 128 ’
— демерсальная 29, 130
— донная 29, 108, 116 134
182, 191, 200, 202
дробление 44
— зрелая (размеры) 116, 117
количество 115, 116, 175
неоплодотворенная 127, 131
остаточная 117
пелагическая 96, 105 115
117, 129, 133, 134, 184, 185,’
186, 191, 198, 200, 201, 203
— плавающая см. Икра
— пелагическая
порции 124
— прилипающая 37, 130, 179,
193, 195
— размеры 116, 130
— текучесть 122
Икринка см. Икра
Икрометание 15, 41, 51, 87, 89/97,
99, 102, 104, 10S, 117, 122, 123,’
124, 125, 127, 137, 140, 141, 174,
175, 178, 179, 180, 181, 183, 184,
185, 186, 188, 189, 190, 191, 193,
194, 196, 199, 200, 201, 204, 205
— порционное 124
Ильная рыба 63
Индекс избирательной способности
169
— наполнения кишечника 167
Инкубационный период 31, 41, 109,
128, 131, 133, 180, 182, 184, 185,
186, 187, 192, 193, 195, 196, 197,
200, 203, 204
Интенсивность потребления корма
168, 169 171, 172, 193, 194
— промысла 151, 153
— солнечной радиации рис. 10
Инфузории 65, 66
Иорт 19, 158
Искусственное осеменение икры 127
— разведение рыб см. Рыбо-
разведение
Истинно глубоководные рыбы 9
Ихтиомасса 89
Ишхаи 188
К
Кагановскпй рис. 31
Каймановая щука рис. 36
Калорийность корма 167
Калуга 117, 118, 179, 180
Камбала желтобрюхая 205
— желтоперая 206(
— морская 104, 132, 138, 204,
205, рис.: 87, 99, 105, 114
— остроголовая 205
211
— полярная 204, 205
— речная 132, 166, 203, 204, 205
Камбаловые 136, 203
Камбалы 4, 24, 30, 31, 38, 41, 61, 67,
77,84, 104,111, 116, 120, 125,132,
133, 139 150, 155, 159, 203,
204, рис.: 11, 66
— дальневосточные 17, 104
Каналы боковой линии 39
Капилляры артериальные 7
— венозные 7
Караваев рис. 64
Каракаш 133
Карась 4, 14, 16, 47, 50, 190, 191,
192, рис. 21
золотой 17
серебряный 17
Каретта 82
Карзинкин 171
Карликовые формы 188
Карп 15, 16, 66, 69, 79, 83, 84, 136,
141, 147, 149, 151, 163, 166, рис.: 1,
21, 41, 53, 92, 97, 100, 109
— зеркальный 39
-- кожистый 39
Карпевич 166
Карповые 16, 87, 96, 98 119, 120,
122, 123, 139, 146, 147, 160, 163,
164, 190, 191, 192, 196, рис.: 20,
58, 71, 74, 107, 110
— хозяйства 77, 195
Карпоед 77
Карпозубые 118, 127
Карпообразные рис. 1
Карты культурные 195, рис. 25
Касатка 84
Кета 84, 87, 97, 99, 100, 159, 179,
186, 187
Кефаль 111, 159, 165
Кижуч 86, 186, 187
Кили-стабилизаторы 114
Килька 171, 182, 184, 185, 196
Киналев 170
Кирпичников 148
Киселевич 120
Кислоты токсическое действие 43
Китайский краб 78
Китообразные 84, 85
Киты 84
Кишечник 6, 9, 36, 37 43, 64, 66,
69, 70, 71, 99, 103, 160, 161, 164,
166, 175,178, 192, рис: 60, 111
— всасывающая поверхность
164
— пилорическая часть 165
— содержимое 169
Кишечнополостные 66, 69
Кишечный тракт см. Кишечник
Кларк 157
Климатический оптимум 20
Клоака 126
Клопы 80
Клочение 81
«Клюв» 160
Книпович рис. 11
Кожа 5, 26, 33, 46, 62, 63, 66, 69, 71,
76,77, 114
Кожный скелет см. Скелет
Колбочки 28
Колебания уловов см. Уловы
— численности видов 156
Кольца на чешуе см. Годовые кольца
Кольчатые черви 72
Колючеперые 6, 9, 42, 128
Колючки 60, 61
Колюшки 63, 83, 128, 172
Комары (личинки) S0; 90, 181
Комары-толкупцы (imago) 170
— (личинки) 80, 170
Кометы 89, рис. 54
Комменсализм 62, 75, 76
Компоненты пищи см. Пища состав
Конечности 29, 32
— передние 32
Конкуренция из-за пищи 50, 56, 58,
80, 93, 151, 171, 172
— межвидовая 171
— напряжение 171
Консервы 199
Концентрация кислорода 16
— кормящихся рыб 168, 173,
рис. 112
Координация движений 44
Копулятивный аппарат 125, рис.:
78, 79
Коралловые рифы 22, 42
Кораллы 160
Коржуев 166
Корм 45, 56, 58, 78, 80, 83, 89, 118,
134, 149, 150, 163, 166, 167, 169,
рис. 109
Кормовая база 41, 148
Кормовой ареал 109
Кормовые объекты 29, 39, 75, 89, 95,
149, 163, 167, 169, 170, 173,
ресурсы 108, 109, 173
Корненожки 66
Коромысло 80
Корюшка 148, 178
Космополиты 9
Кости 20, 137
глоточные 164
небные 160
нижнеглоточные 164
— плоские 139, рис. 88
челюстные 160
Костиазис 66
Костистые рыбы 6, 14, 39, 61, 126,
135
Костная капсула 8
Костнопузырные 6, 7, 48
Костные ганоиды см. Ганоиды
— пластинки 62
Косяк 55, 110, 199
Коштоянц 166
Коэфициент Вантгоффа 132,
2,2
— зрелости, 121, 122, рис. 75
— пищевого сходства 172
— упитанности 157, 159, 167
Крабы 78
Красное тело 7
Краснопер 192
Красноперка 21, 36, 83,. 169, 192,
196, рис. 21.
— дальневосточная 191
Краснуха карпов 63
Кровеносная система 46, 70
Кровеносные капилляры 48
— сосуды 48, 49, 129, 135, 161
Кровообращение 69
Кровь 13, 14, 49, 50, 62, 66, 69, 77,
78, 100, 135
— температура замерзания 99
Кровяные тельца 133
Крог 161
Крокодил нильский 82
Крокодилы 82
Круглоротые 13
Круглые черви 71, 72
— свободно живущие 72
Круговорот органического вещества
41, 63, 92
Крыжановский рис. 84
Крылоногие моллюски 76
Кузнецов 109
Кузовки 46, 112, 160, рис. 36
Кулаев 120
Кулинченко 132
Куличенко 198
Кумжа 46, 89, 188
— американская озерная 4
Кутора 84
Кутум 98, 191, рис. 74
Л
Лабиринтовые 32, 33, 65, 126, 128,
162
Ластоногие 84, 85
Леа Эйнар 143
Лебедев 51
Левенгук 138
Ледебур 6
Ленок 187, 189
Лентовидный тип формы тела 111,
159
Ленточные черви 70
Летальная концентрация кислорода
рис. 4
— углекислоты 16, рис. 5
Летучая рыба 114, 115, рис. 70
Лещ 29, 35, 36, 66, 72, 78, 81, 83,
106, 114, 123, 124, 125, 127, 133,
148, 158, 159, 160, 163, 164, 168,
169, 171, 172, 191, 192, 193, 194,
196, рис. 21, 29, 66, 110, 112, 114
— аральский 51, 107, 167, 170
— морской 124
Лжелопатоносы 179 181
Ли Роза 144
Лигулез 70, 78, 83
Лигуля 70
Лимнофильные рыбы 4
Линдберг рис. 79
Линь 7, 47, 50, 190
Липаза 166
Липин рнс. 39.
Лиса 84
Литораль 41
Личинки мотыля 163, 167, 170, 203
— насекомых 194
— пелагические 96, рис. 3
рыб 8, 11, 29, 30, 31, 32, 38,
40, 64, 65, 67, 69, 80, 81, 133,
134, 174, 178, 182, 184, 199,
рис.: 14, 84, 85
Личиночный период жизни 135, 174,
178
Логарифмическая шкала Монастыр-
ского 146, рис.: 95, 96
Локальные формы 189
Лопатонос большой амударьинский
179, 181, 182
— малый амударьинский 181
— Федченко 181
Лопатоносы 33, 42, 179
американские 182
Лососевые 20, 46, 84, 86, 87, 96, 97,
99, 101 117, 119, 123, 125, 127,131,
136, 139, 141, 149, 157, 184, 186,
190, рис.: 59, 73
Лососи дальневосточные 39, 98, 105,
126, 131, 137, 186, 187
— европейские 98
Лосось 18, 31, 34, 52, 86, 87, 100,
102, 108, 109, 111, 122, 138, 141,
156, 159, 186, 187, 188, рис. 22
благородный 97
дунайский 189
каспийский 187
колумбийский 87
Лоцман 62
Луна рыба 11, 38, 46, 111, 115,
рнс. 66
Луфарь 84
Лучи анального плавника 126, рис. 6
— брюшных плавников 126
— грудных плавников 139, 162
рис. 19, 88
— непарных плавников 17, 139
парных плавников 162
плавников 54, 61, 119, рис. 88
спинного плавника 27, 61, 162
Львиноголовка рис. 54
Люме 79
Лягушка бык 81
— зеленая 81
Лягушки 81, 195
Ляйман рис.: 41, 43^ 45, 46
М
Майский 156
Маккензи 167
21
Макрель 78
Макропод 127
Макрофиты 64, 194
Малощетинковые черви 73
Мальки 23, 38, 67, 79, 81, 127, 128,
131, 133, 135, 179, 180, 182, 187,
189, 193, 201, 203, 205, рис. 102
Мальковое кольцо 140, 141
Мальтаза 166
Малярия 90
Мантийная полость 76, 192
Маринка илийская 192
Маринки 19, 34, 59, 161, 190, 192
Маслюк 41
Медведь белый 84
— бурый 84
Медузы 67, 68, 69, рис. 42
Мейен 120
Мейснер рис.: 62, 75
Меристические признаки 17
Мерланг 132
Места икрометания 103
Метаболизм 56
Метаморфоз 178
Метка Гильберта 110, рис. 64
Метки 110, рис. 64
Меч рыба 59, 105, 162
Мечение рыб ПО, рис. 64
Меченосец 119
Миграции 14, 22, 39, 96, 98 108,
109, ПО, 159, 179, 185, 187, 189,
193, 195, 199, 202, 203, 205, рис. 62
активные 96
анадромальные 96, 102
— вертикальные 40, 105, 106
горизонтальные 105
— денатантные 97
— зимовальные 96, 101, 107
— катадромальные 97, 102
контранатантные 97
кормовые 96, 104, 105, 106,
107, 185, 199
— нерестовые 18, 96, 100, 103,
104, 107 167, 174, 184,
187, 191,
пассивные 96, 105
пищевые см. М. кормовые
— связанные с изменением внеш-
них факторов 96, 108
— сезонные 79
— суточные вертикальные 106
Миграционный путь 101, 103, НО,
рис. 61
Миграционные скопления 52
Миграционный цикл 107, 109
Мигрирующие рыбы 4
Микропиле 127
Миксина 13, 45, 46, 62, 116, 130,
160, 169, рис. 83
Милинский 155, рис. 105
Минкина 43
Минога каспийская 97, 175, 178
— морская 174
— невская 97, 178, рис. 114
— речная 175, 178
— восточная 178
— европейская 178
— — сибирская 178
ручьевая 178
— украинская 178
Миноги 30, 45, 87, 96, 97, 109, 114,
160, 169, 174
Миноговые 174
Мирные рыбы 69, 71, 106, 166
Млекопитающие 28, 84, 85, 202
Многопер 32, 48, рис. 18
Многоперовые 6, 134
Многощетинковые черви 72, 73, рис.
47
Мозг 28, 66
Мозговая жидкость 70
Мойва 78, 84, 189, 200, 201
Моллюски 69, 75, 76, 78, 160, 167,
170, 172, 180, 192, 194, 204, 205
— двустворчатые 75, 192
— крылоногие см. Крылоногие
пресноводные 58
Моллюскоеды 171
Молодь 38, 41, 44, 65, 67, 79, 81,
136, 152, 168, 189, 192, 196, 198,
201, 202, 203
Молоки 120, 126, 127
Монастырский 144, 152, рис.: 94,
95, 96, 102, 103
Мореходные качества рыб рис. 65
Морозова 133
Морская щука 115
Морские анемоны 67
— беспозвоночные 11
ежи 74, 75
ерши 199
— звезды 73, 74, 75, рис. 48
— иглы 67, 74, 111, 160, рис.
36, 82
костистые рыбы 11, 13, 14
— рыбы 3, 17, 30, 31, 39, 47,
76, 96, 103, 106, 107, 108,
115, 129, 142, 174, 197, 199,
200, 203, рис. 24
— свиньи 84
собачки 8
Морской конек 114, 129
— тряпичник 64, рис. 38
— окунь 20, 78, 84, 117, 118,
126, 199, 200, рис. 114
— пес 116
петух см. Тригла
чорт 11, 32, 112
Моча 13, 14
Мочевина 11, 13
Мочевой пузырь 69
Мочеточники 70
Муксун 190
Мускулатура конечностей 32, рис. 19
Мускульное электричество 59
Мухи (личинки) 80
214
Мыши 85
Мягко пер ые б
Н
Навага 84, 130, 132, 159, 200, 202,
рис. 114
Нагул 51, 88, 99, 104, 105, 106, 107,
108, 109, 139, 170, 173, 180, 186,
188, 193, 198, 199, 200
— места рис.: 28, 29
Нагульные скопления 52, 53
Наджаберные камеры 48
Наджаберный орган 48, 49, 50,
рис. 27
Наживка 178
Налим 109, 122, 123, 124, 139, 200,
203
Нансен 156
Нарвал 84
Насекомоядные 84
— растения 65
— рыбы 90
Насекомые 79, 80, 90, 162, 165, 170,
180, 181, 189
Научно-промысловые исследования
110
Невода 185, 194, 197, 206
Невод кошельковый 198
Недолов 155
Недошивин 120
Нельма 187, 189
Нематоды 72
Неоген 20
Неогеновая фауна 20
Нервная система 29, 70,
Нервно-мускульный аппарат 44
Нервы висцеральные черепно-мозго-
вые 60
Нерест 23, 38, 51, 52, 87, 98, 99,
104, 105, 109, 117 119, 121, 126,
137, 141, 175, 179, 183, 184, 185,
187, 190, 192, 193, 194, 196, 198,
201, 202, 206
— сроки 124, 179, 183, 187, 190,
191, 197, 200, 203, рис. 76
Нерестилища 38, 52, 87, 89, 96, 99,
100, 104, 109, 124, 127, 179, 181,
184, 187, 200, 201, рис. 60
Нерестовые марки 141, рис.: 90, 91
— миграции см. Миграции
площади 109
— труды 80
— скопления 52, 53
Нерестовый ход 175, 184, 186
Нерка 87, 97, 187
Никитинский 167, рис. 76
Никольский 116, рис.: 6, 56, 63, 75,
101
Ногоперые б 32, 134, 162
Ноздри 30
Ноздря непарная 45, 46
Нормы посадки 173
О
Обмен веществ 17, 22 43 70 147
149, рис. 97 ’ ’
Оболочка икринки 127, 131, 133
Общепресноводные рыбы 4’
Объекты пищи см. Пищевые объекты
Ооъект прсмысла 19, 156, 159 180
181, 182, 184, 188, 194, 196* 197*
198, 199, 200, 204
Объячсивающие орудия лова 198
Овогонии 122
Одомашнивание рыб 89
Окончательный хозяин 69, 70, 85
Окраска 9, 64, 119, 186, 191, 204,
рис. 11
— брачные изменения 23
Оксифильные рыбы 36
Окуневые 120, 122, 139, 196, 197
Окунеобразные рис. 81, 84
Окунь 7, 46, 64, 66, 82, 117, 121,
122, 124, 127, 134, 159, 163, 170,
196, рис. 75, 114
— американский 59, 90
— болхашский 197
— обыкновенный 197
— ушастый американский 50
Омуль 187, 190
— байкальский 187
Ондатра 84
Оплодотворение 115, 117, 125, 126,
128, 130, 131, 174, 192
— внутреннее 125, 174
наружное 126, 127
Опухоли злокачественные 63
Органы боковой линии 28, 40, 53
— выделения 66
дыхания 46, 49, 50
— дыхания личиночные 135
— зрения 22, 53
— опоры 32
осязания 28, 33, 54, 162
пищеварения 66, 161
плавания 32, 114
— прикрепления 31, 34
— свечения 22, 26, 63, рис.
15, 16
— слуха рис. 1
Орлан-белохвост 82
— долгохвост 82
— камчатский 82
Оседлые рыбы 4
Осенне-зимнемечуЩие рыбы 123
Осетр 80, 97, 102, 159, 171, 179,
рис. 61, 114
— американский пресноводный
154, 180, 181
— амурский 180
— немецкий 132, 154, 179, 180,
181
— русский 4, 132, 179, 180
— сахалинский 180
— сибирский 39, 179, 180
215
Осетровые 17, 80, 96, 97, 114, 118,
120, 122, 123, 125, 127, 135, 139,
140, 143, 156, 161, 164, 179, 181
Осетровые ямы 17
Османы 190
Ослюрегуляторные приспособления
— процессы 42
Осморегуляция 99
Осмотическое давление 11, 13, 14,
99, 103, 133
Остролучка рис. 107
Открытопузырные рыбы 6
Отолиты 139, 141, рис. 87
Офалина 84
Офиуры 205
П
Палеолит 88
Палеолитический человек (остатки
пищи) 20
Палия 187, 188
— американская 28
Палочки 28
Палтус 159, 204, 206
Панкреатическая амилаза 166
Панцырный сом рис. 36
Панцырь 46, 61, рис. 36
Паразитизм 62
Паразитические ракообразные 63,
рис. 49.
Паразитический образ питания 161
Паразиты 69, 70, 77, 78, 89, рис. 40
— наружные 72, 76
Парение 11, 134, рис. 3
Партеногенез 127
Паульсен 120
Педицеллярии 74
Пелагические личинки рыб И, 106
— рыбы 3, 4, 29, 40, 53, 60, 82,
88, 115, 134, 262, 202
— яйца 38
Пеламида 111
Пеликаны 82
Пепсин 166
Первичный скелет см. Скелет
Переваривание пищи 16, 36, рис. 7
Перелов 155
Перенаселение 148
Переносчики 78
Переохлаждение 17
Периодические подходы рыбы 20
Периодичность роста 137, 148
— уловов 156
Перифитон .36, 64, 164
Перкарина 196
Перминов рис. 24
Пескари 4, 23, 34, 46, 106, 191,
192, 203
Пескоройка 30
Песчанка 30, 67, 117, 187
Петров 141
Петрович 79
Петрушевский 141
Печень 70, 71, 75, 105, 201, рис. 10
Пещерные рыбы 27, рис. 17
— сомы см. Сомы
Пигмент 26
Пигментация эмбрионов 130
Пикша 20, 68, 73, 76, 77, 78, 130,
132, 159, 201, 206, рис. 114
Пила-рыба 59, 162
Пилонос 162
Пилорические придатки кишечника
99, 165
Пилохвост 116
Пинагор колючий рис. 24
Пинагоры 40,41, 51, 66, 112, 116, 124
Пиранья рис. 50
— амазонская 85
Питание 22, 36, 37, 44, 53, 64, 67,
74, 92, 98, 106, 107, 108, 134, 159,
167, 170, 173, 196
активное 195
интенсивность 166, 167, 168,
180, 183, 194, 201, рис. 112
полупассивное 163
прекращение 140, 141, 175,
179, 194
характер 165
Питающиеся в море (рыбы) 4
— в реках 5
Пища (рыб) 29, 37, 40, 64, 67, 69, 72,
75, 79, 80, 82, 88, 95, 102,
104, 105, 150, 153, 159, 160, 161,
162, 164, 165, 180, 181, 183, 184,
186, 192, 193, 194, 197, 199, 200,
201, 202, 204
второстепенная 169
вынужденная 169
основная 169, 180
растительная 64
случайная 169
— состав 169, 170, 172, 175,
рис. 56
Пищеварение 166
Пишевари гельные железы 166
Пищеварительный процесс 166
Пищевая конкуренция см. Конку-
ренция из-за пиши
Пищевой комок 166
— рацион 64
Пищевые взаимоотношения в водо-
еме 173, рис. 55, 56,57
— объекты 32, 52, 64, 66, 67,
72, 74, 78, 80, 81, 83, 85, 95,
105, 106, 160, 162, 163, 164,
169, 170, 172, 202
Пищевые организмы 171, 173
— связи 92, рис.: 55, 56, 57,
Пиявки 72, 73
Плавательный пузырь 6, 9, 48, 49,
71, 103, 198, рис. 1
Плавники 5, 6, 31, 46, 59, 64, 72,
75, 78, НО, 134, 198,
216
— анальный 114, 178, пис.:
68, 84
брюшные 11, 31, 35, 40,
114, 128, 162, рис. 80
грудные 31, 32, 33, 35, 46,
114, 115, 119, рис.: 18, 34,
68, 71
непарные 111, 114, 118
парные 114, 118
— преданальный рис. 84
~ спинной 11, 62, 114, 139,
рис. 71
— хвостовой 112, 114
Плавные сети 182
Плавунец окаймленный 79
Плавунцы 79
Плавунчик 79, 80
Планктические водоросли 64, 79, 192
— ракообразные 78, 165,
183, 184, 200, 202,
Планктон 40, 41, 44, 58, 64, 78, 79
83, 105, 109,165, 172, 173, 184,
189, 196, 197, 198, 201
Планктон индикатор Гарди 173
Планктонная сетка 169
Планктонные организмы 58, 105, 106,
140, 185, 186
Планктоноядные рыбы 41, 109, 163,
165, 169, 183, 190
Плечевой пояс 31, рис. 88
Плодовитость 115, 178, 179, 182, 1S3,
184, 185, 186, 187, 189, 191, 192,
195, 196, 197, 198, 199, 200, 201,
202, 203, 204
Плоский тип формы тела 112
Плотва 7, 36, 46, 105, 118, 122, 170,
192, 196, рис.: 21, 44
Плотность воды 6
населения 56, 57, 150, 151
кормовых объектов 168
— посадки 150, рис. 100
Поведение рыб 37, 51
Поганки 82
Поддев 201
Подкаменщики 4, 35,41, 46, рис. 21
Подкаменщик большеголовый 41
Подкорм 173
Подуст 46, 161, 191
Позвонки передние
— число 17
Позвоночные 81
Поисковые работы 173
Пойкилотермные животные 15
Покатные особи 99
Покровительственная окраска 22, 63
Ползание 33
Ползун 32, 33, 115
Половое отверстие 120
Половозрелость 102, 118, 122, 137,
138, 147, 153, 179, 180, 187, 193
время наступления 118, 187,
206, рис. 63
Половой деморфизм 1 18, 198
цикл 28, 1 19
Половые железы 28, 53, 100 117
120, 122
органы внешние 37, 126
— признаки внешние 123
продукты 67, 97, 117, 120
125, 179, 187
— созревания 22, 100,
101, 103, 107, 119, 120, 122, 191
Полостная жидкость 13
Полость тела 70
Полупаразнтическпп образ жизни
45, 62
Полупаразиты 160
Полупроходпые рыбы 5, 98, 105, 107
189, 191,192,196
Понто -Арало-Каспийская провин-
ция 20
Популяция 20, 52, 59
Послеледниковое время 20
Потребление кислорода рис. 97
— корма 16, 167, рис. 7
Почки 13, 14, 69, 70
Пресноводные рыбы 3, 4, II, 14, 17
31, 39, 47, 83, 90, 103, 104j 106,
116, 122, 129, 131, 136, 174, 179,
180, 186, 189, 190, 195, 197, 198-
— костистые рыбы 13, 14
Прибрежные рыбы 198
Придонная температура рис. 8
Придонные беспозвоночные 174
— рыбы 4, 8, 60, 200, 203,
204
— животные 169
Приливы квадратурные 41
— сизигийные 41
Прилипало 62, рис. 37
Прирост 143, 148, рис.: 25, 100
— поголовья стада 109
Присоска 29, 31, 34, 35, 40, 62 г
рис.: 20, 24
Прогнозы уловов 135, 152, 155
Продолжительность жизни 136, 151,
152, 175, 181, 183, 184, 199
Продуктивность 41, 45, 83
— бентоса 150
— водоема 157
Продукты метаболизма 55, 56
Продукция 40
— водоема рыбная см. Рыб-
ная продукция
Производители 87, 89, 135, 178
Промежуточный хозяин 69, 70 71
78, 85, 92
Промысловое значение 174, 178, 182,
185, 187, 189, 190, 192, 195—197,
199, 202, 204, 205, 206|
Промысловые рыбы 5, 44, 69, 75, 78,
81, 83,116, 135, 136, 1Й6 174, 193
202, 206, рис. 14
Пронация 32
217
Простейшие 65, 66
Профундаль 95
Проходные рыбы 3, 4, 14, 87, 96, 98,
99, 100, 102, 107, 109, 116, 174,
175, 179, 189, 184, 186, 187, 188,
189, 190, 191
Прудовое хозяйство 45, 66, 76, 77,
83, 84, 173
Прутяные орудия шва 175
Прыгуны 32, 33, 114
Псевдобропхия 46, 135
Птеригоподии 126, рис. 78
Птицы 82, 83
Птичьи базары 83
Пуголовка 171
Пузанок 132 , 183, 184, рис. 83
- большеглазый 165, 183
каспийский 99, 165, 183,
184
круглоголовый 183
— полосчатый 183
Пузырчатка 64, рис. 39
Пульсация сердца 9
Пучкожаберные 129
Пюттер 161
Р
Равноногие раки 76, 77, 78
Развитие 115
— икры 22, 31, 63, 108 109,
128, 131, 132, 133, 135, 184,
204
— личинки 134
— эмбрионов 44
Размеры рыб 136, 143, 154, 170, 183,
187, 188, 189, 190, 192, 196, 197 —
199, 202, 203, 206, рис. 104, 105
Размножение 5, 27, 37, 41, 51, 52,
53, 96, 108, 119, 167, 174, 175, 179,
182, 191, 193
— места 108, рис.: 28, 29
— потеря способности 136
— сроки 119, 182, 191
Район местообитания 18
Районы промысла 156
Ракоеды 78, 171
Ракообразные 76, 78, 79, 80. 107,
167, 169, 170, 172, 184—186, 187,
189, 193, 194, 202, 204, 205, 206
Ракушковые рачки 170
Ракшеобразные 82
Распределение кормовых объектов
173
— промысловых рыб 173
Распространение 18, 31, 86, 105,
156, 202
Расселение 14, 42, 83, 88
Растительноядные рыбы 37, 78, 159,
164, 192, рис. 111
Растительные обрастания 160, 164
Расчисление темпа роста 144
Расы 51, 89, 182, 204, рис.' 53
218
—- географические 205
— домашние 195
озимые 97, 100, 107. 180, 187
экологические 188
— яровые 97, 180
Рдесты 63
Редукционное деление ядра у ик-
ринок 131
Редукция пузыря 8
Резервуарный хозяин 71, рис. 46
Резорбция желточного пузыря 29
Рейбиш 132
Реликты 37, 187
Ремнец рис.: 43, 44
Реофильные рыбы 4, 35, 36, рис. 23
Рептилии 81
Ретина 28
Речные рыбы 4, 36, 39, 102, рис. 20
— пелагические рыбы 4
Роговой эпителий 119
Рост беспозвоночных 58
периодический 137, 142,
рыб 5, 44, 56, 70, 89, 116, 136,
137, 141, 142, 145, 148, 151,
170, 172, 179, 181, 187, 193,
194, 195, 201, 203, рис.: 93,
96, 99
— чешуи 145, рис.: 93, 96
Ростру и 59, 162
Рот 41, 45, 46, 48, 59, 72, 128, 129,
рис. 107
— верхний 161, рис. 107
— всасывательный 160
— дробящий 160
— конечный 161, рис. 107
— нижний 161, рис. 107
перифитоноеда 160
планктоноеда 160
— нолунижний рис. 107
— в виде присоски 160
типы строения 160
— хватательный 160
«Ротовая беременность» 124, 129,
рис. 81
воронка 175
— полость 78, 128. 164, рис. 81
Ротовое отверстие см. Рот
Ротовой аппарат 160, 161, 164
Рули глубины 114
Ручейники (личинки) 80, 203
Рыбец 191, 192
Рыбная продукция 171
Рыбные дожди 42
— запасы см. Запасы про-
мысловых рыб
Рыбный промысел ПО
Рыбоводство 195
Рыбообразные 174
Рыборазведение 88, 181, 190
Рыбоходы 87, рис. 52
Рыбохозяйственная таксация см.
Таксация рыбохозяйственных уго-
дий
Рыбоядные змеи см. Змеи
— птицы 70, 82, 83
— растения рис. 39
Рыбный промысел 110
Рыло 103, 162, 163
Ряпушка 187
С
Сазан 16,47,58,63, 81, 116, 117,
121, 123, 124, 125, 131, 149, 153,
159, 164, 169, 170, 191, 192, 194,
195, рис.: 53, 75, 107, ПО
Сайда 202
Сайка 84, 108, 130, 132,189, 200,202*
Салака 159, 182
Самки преобладание 117, 197, 199,
204
Самооплодотворение 118
Самохвалова 29
Самцы неполовозре ibie 29
— паразитические рис. 72
— преобладание 117
Сарган 111, рнс. 66, 83
Сардинка калифорнийская 185
Сардины 84, 106, 182, 185, рис. 31
Светлов 133
Светловидов 165, рис. 1
Светящиеся клетки 26, рис. 15
— организмы 27
— органы см. Органы све-
чения
Свечение 26, 27
Севрюга 78, 97, 102, 116, 132, 159,
171, 179—181, рис.: 61, 114
— азовская 181
— каспийская 105, 181
Сеголетки 84, 143
Сельдевая акула см. Акула
Сельдевые 7, 39, 51, 78, 87, 139, 182
Сельдеобразные рис. 1
Сельди каспийско-черноморские 182,
183
— каспийские 120, 141, 159, 165,
190
— пятнистые 182
— черноморские 159
Сельдь 18, 20, 29, 38, 51, 78, 82, 84,
96, 98, 102, 109, 116,117, 124, 13Р,
142, 149, 156, 158, 160, 161, 165,
173 179, 184, 200, 201, рис.: 1,
32, 33,61, 106
— беломорская рис. 89
— крупная раса 183
—- мелкая раса 183
— бражниковская см. Сельдь
долгинская
— волжская 98, 127, 165, 183,
184
— дальневосточная 182, 183,
рис. 28
— долгинская 151, 165, 183, 184
менхеден 4, 108
— мурманская 182, 183, рис. 114
— океаническая 58, 104, 105,
— 124, 133, 159, 182, рис.: 77, 98
тихоокеанская см. Сельдь
дальневосточная
— фиордовая 182
— тешская 183
Сельдяной кит 84
Король 111, рис. 66
Семга 20, 116, 187, рис. 114
камчатская 188
Семенники 122, рис. 75
Семнер 24, 75
Сероводород 50
Сетчатка 25
Сигневский рис. 114
— обыкновенный 189, рис. 108
— чудекий 190
Сиги 23, 39, 79, 87, 97, 104, 1М
124, 159, 160, 169, 189
— американские озерные 4
— европейские озерные 4
— морские проходные 180
озерно-речные 189
Сила пищевой конкуренции 172
Силовое поле 54, рис. 30
Сима 186, 187
Симбиоз 62, 67
Синец 21, 72
Скат (личинок и мальков) 96
— (обратный после икрометания)
105
Скаты И, 13, 30, 31,46,60,61,66,75,
84, 111, 112, 114, 125, 161,
164 , рис.: 35, 66, 68, 78
— орляки 160
Скворец 15
Скелет кожный 32
— первичный 32
хрящевые части 66
Склериты чешуи 141
Скловер 147
Скопа 82
Скребни 71
Скрытая теплота замерзания 17
Скумбриевые 6, 197
Скумбрия 4, 39, 105, 111, 114, 159,
198 рис. 66
•— малая 198
— обыкновенная 197, 198
Слепые рыбы 28
Слизевые каналы 9
Слизистая оболочка кишечника 14
Слизистые железы 26, 62
Слизь 17, 33, 61, 114
Слуховой лабиринт 7
Слуховые капсулы 7
Смертность 151 ।
Снеток 159
Собачья рыба 58
Совокупительный аппарат см. Ко-
пулятивный аппарат
219
Созревание половых продуктов см.
Половые продукты
Соки тела 14, 43, 77, 119, 169
Соколообразные 82
Солдатов 180
Соленоводные организмы 39
Солоноватоводные рыбы 3, 190
Соляная кислота 166
Сом 5, 33, 48,49 51, 62, 81, 82, 83
85, 92, 106, 125, 128, 129, 136,
139, 140, 153, 159, 160, 162, 169,
рис. 114
амурский 195
бразильский 122
индомалайский 162
мешкожаберный 49
обыкновенный 195
Сомик 62, 107 рис. 20, 23
американский 56
— туркестанский 34
Сомовые 123, 128, 195
Сомы американские 28, 37, 53, 54
— пещерные 28
Соотношение возрастов в популя-
ции 153
— полов 52, 117, 191, 198,
204
Сосальщики 69
— дигенетические 69, 70
— моногенетические 69
Состав газа в плавательном пу ыре 7
Сошник 160
Спаривание 117, 126
Спектр питания 171, рис. 113
Сперма 37
Сперматиды 122
Сперматогенез 122
Сперматогонпи 122
Сперматозоиды 122, 127, 130
Сперматоциты второго порядка 122
- первого порядка 122
Спермин см. Сперматозоиды
Спиральная складка 165
Сплющенный с боков тип формы тела
111
Споровики 66, рис. 41
Способы добывания пищи 162
Спунер 53
Спячка 16, 17, 48
Сростночелюстныс 160
Ставные сети 83, 194, 197
Ставридка 84, 111
Стадии развития 17
Стадия выбоя 120
— зрелости 120
покоя 120, 121
размножения 120
— созревания 120
Стадников 95
Стадо 51, 56, 87, 88, 109, 117, 119,
139, 153, 156, 171, 181, рис. 29,
— поголовье 108
Стаи рыб 27, 51, 53, 54, 55, 84, 85,
22о
174, 206, рис.: 31, 32, 33
Стайность 53
Стайный инстинкт 53
Стайные рыбы 51, 53, 84, рис. 1S
Старение особи 116, 137
Стейниц 88
Стеногалинный 13, 42
Стенотермный 15, 19
Стерлядь 117, 132, 155, 179, 180, 181
— волжская 67, рис. 104
Стимулирование текучести икры 122,
181
Стрекательные клетки 67
Стрекозы (личинки) 80
Стреловидный тип формы тела 111
Строганов 127
Судак 59, 82, 98, 116, 122, 142, 153,
156, 159, 160, 162, 169, 171,
196, 197, рис. 114
морской 196, 197
— обыкновенный 196, 197
Суетов 163, 168, рис. 109
Супинация 32
Суточный рацион 167
Сырьевая база рыбной промышлен-
ности 173
Т
Таймени 189
Таймень 189
— японский 189
Таксация рыбохозяйственных уго
дий 139, 173
Талиев рис. 39
Тапир 85
Тарань 81, 192, 193
Тевяк 84
Телескопическое зрение
Телескопы 98, рис. 54
Темп роста 15, 89, 1Ю,
143, 148, 149, 150,
рис. 94, 98
Температурный градиент
— коэфицпент развития
икры 132
— оптимум 147
— «порог» развития 132
Тепловодные формы 19, 20, 39
Теплокровные животные 15
Ти Ван 69
Токсическое действие 43, 44
Толпыга 163, 164
Толстолоб амурский 192
Толстолобик 64, 179
Торпедовидный тип формы тела 1
Травяник 198
Трал 169, 20(У, 204, 206
Траловый промысел 200—201, 2U4,
205
Треска 13, 19, 20, 52, 68, 76, 77/ 78,
84, 103, 105, 129, 130, 132, 133,
136, 137, 139, 140, 141 156, 159,
22
137-13
171, 189,
18
166, 168, 189, 199, 200, 201, 202,
206, рис.: 62, 69, 114
Треска беломорская 201
— мурманская 201
— лофотенская рис. 10
норвежская 200
— прибрежная 200
Тресковые 7, 77, 84, 120, 130, 136,
139, 161, 200, 202
Тресковый жир 19
Трескообразные 6, рис. 1
Тресочка полярная см. Сайка
Трехгодовик 143
Трехлетка 143, 152
Триглы 32, 33, рис. 18, 19
Трипсин 16(5
Тритоны 81
Троглобионты 28
Троглокссны 28
Троглофилы 28
Тропические рыбы 28, 131
Туводные формы 188, 190
Тунец 15, 114
Турдаков 36
Тюлени 8?, 85, 179
Тюлька 156, 180, 182, 185
— чархальская 130
Тюрбо 204
У
Углеводы 166
Углекислота 63
Угорь 5, 11, 38, 72, 97, 103, 111, 114,
137, 142, 159, рис.: 66, 69
— американский 102, 103
— европейский 102, 103
Угри глубоководные 62
Удельный вес рыб 6
— икры 130, 133
Удобрение прудов 45
Ужи 81
Уклея 4, 81 , 124, 196
Уловы 20,41, 136, 152, 153, 156, 175,
193, 195, 197, 200 -202, 204,
рис.: 104, 105
— снижение 138
Упдулирующие движения тела
Универсальная шкала зрелости 120
Упитанность 101, 105, 107, 157, 167,
194, 195, 201
Урожай молоди 136, 156
Урожайность по годам 138, 151,
152, рис. 103
Уруме-иваси 182
Усачи 34, 35, 36, 98, 108, 164, 181,
рис.: 110, 114
Усач аральский 98, 99, 100, 191,
рис. 58
— чанари 98
Условные рефлексы 58
Усы 28, 54
Утки 82, 83
Ухо внутреннее 139
Ушные камни см. Отолиты
Ф
Фауна беспозвоночных 106
Фаунистический комплекс 21
Ферменты кишечника 99, 166
Финна 71
Финты 182
Фитопланктон 83, 164, 202
Форель 35, 50, 56, 66, 79, 80, 89
132, 147, 151, 188, рис. 21
— гохчинская 188
Форма тела рыб 111, 159, рис. 66
Формообразование 17
Формула Розы Ли 145
— Эйнар Леа 143, рие. 93
Фосфоресцирующая субстанция 26
Фотосинтез 63
Фультон 157
X
Хамелеоноголов 19, рис. 9
Хамса 4, 39, 52, 84, 133, 186, 196,
рис.: 83, 114
азовская 107
Харациновые 85, 142
Хариус 36, 46, рис. 114
— Ангары 165
— байкальский 165
Хвост 5, 33, 35, 41, 59, 61, 62, 114,
126, рпс. 67
— гипобатический 114, рис. 67
— изобатический 114, рис. 67
— эпибатический 114, рис. 67
Хвостовой стебель 36
Химеры 39, 116
Хищники см. Хищные рыбы
Хищные рыбы 53, 59, 63, 69, 71, 78
105, 106, 153, 159, 160, 162 164—
167, 169, 171,179, 183, 189, 192, 195,
196, 197 рис.: 57, 111
Хлористый натрий 11
Хобот 162
Ход рыбы 98, 179, 180, 184, 185, 187,
189, рис.: 58, 59, 60
— весенний 178
— осенний 178
Ходовые рыбы 53, 187, 188
Хозяйственное значение см. Про-
мысловое значение
Холодноводная фауна 21
Холодноводные рыбы 166, 203
Холоднокровные животные 15
Хохлач 84
Храмули 192
Храмуля закаспийская 64, 159, 164,
169
Хрусталик глаза 24 ।
Хрящевые ганоиды см. Ганоиды
— рыбы 11, 13, 14
— морские 13
пресноводные 13
221
ц
Цапли 82
Цементный орган 134
Церкарии 69
Цикл развития рис.: 43, 45, 46
Циклопы 167
Циркумполярное распространение
Ч
Чавыча 87, 99, 132, 187, рис.: 59, 60
Чайки 82, 83
Челюсти ПО, 119, 164, 186, рис. 86
Челюсть верхняя 59, 162
— нижняя 137, 162
Черви 69, 72, 78, 92, 204
Череп 139
— мозговой 164
Черепахи 62, 82
Черноморский калкан 61
Черноспинка 98, 132, 183—184,
рис.: 90, 114
Черный окунь 89
Черч 150
Четырехглазая рыба рис. 12
Чехонь 79, 107, 130, 133, 161, 169
170, 181, 191, 192, рис. 107
Чешуйчатник американский 33
— африканский 104
Чешуя 17 39, 61, 90, ПО, 137, 139,
140 141, 178, 195, рис.: 89, 90,
91, 92
— боковой линии 17, 195, рис. 6
— закладка 144
Численность вида 20
Число икринок 116
— позвонков см. Позвонки
Чоп 196
Чувствующие клетки 39
Чугунов рис. 88
Чугунова 116, 141, 143
Ш
Шакал 84
Шаровидный тип формы тела 112,
159
Шед 132, 182
Шемая 79, 107, 191, 192
Шиповка среднеевропейская 30, 112
Шип 97, 156, 179, 180
— аральский 97
Шипы (на коже рыб) 11, 31, 34, 60
61, 92
Широкий лентец 78, рис. 45
Шкала жирности 158
— зрелости 120
Шмидт 103, рис. 81
Шнейдер 142
Шпигенберг 56, 151
Шольц 167
Шорыгин 167, 169, 172, рис.: 57, 113
Шпрот 84
Шулейкин рис.: 30, 65, 70
щ
Щука 7, 59, 76, 83, 85, 104, 111, 136,
143, 159, 160, 164, 166, 167, 168^
169, рис. 114
— амурская 195, 196
— обыкновенная 196
Щуковые 195
Щупальца 67
Э
Эврибионтные формы 197
Эвригалинный 42
Эвритермный 15, 40
Экман 103
Экологические участки реки 36
Экскреция 13
Экстерьер рыб 157
Эктопаразиты 66
Электрический орган 59, рис. 34
— скат 59, 60
— сом 59
— угорь 59, 114, 162,
рис. 68
Эмбрион 9, 115, 126, 133
Эндопаразиты 66
Энтерокиназа 166
Энхитреи 167
Эрепсип 166
Эпидермис 39, 66
Эпизоотии 153
Эпи ними он 41
Эренбаум 126
Ю
Южная пикш! 84
Я
Ядовитая железа 60
Язык 160
Яичники 116, 117, рис.: 75, 77
Яйца 38, 130
Яйцевой мешок 129, рис. 82
Яйцеклад 123
Японская саламандра 81
Ярус 201
Ястыки 116, 117
Ящерицы 82
222
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ И ТЕРМИНОВ
И ИНОСТРАННЫХ ИМЕН СОБСТВЕННЫХ
А
Abram is brama 171, 193, рис. 110
— ba Herns 21
Acanthias acanthias 13, 139
Acanthocephala 71
Acanthorhodeus 192
Acerina cernua 46
Acipenscr 179, 180 181
— baeri 180
fulvcsccns 154, 180
— giildcnstadti 138, 171, 180
— medirostris 180
nudiventris 156, 180
-- ruthenus 117, 180
— schrenki 180
— stellatus 105, 116, 171, 180
— sturio 154, 180
Acipenseridae 125, 179
Acropoma japonicum 27
Adacna laeviuscula 76
— minima 76, 169, 171
— plicata 76
Aega 77, рис. 49
Agonidae 18
Alburnoides 20
— bipunctatus 34
— taeniatus рис. 6
Alburnus alburnus 124
Aldrovandia 65, рис. 39
Allee 57
Alm 171
Alosa 182
— finta 184
Alvard рис.: 87, 99
Amblyopsidae 28
Amblyopsis 28, рис. 17
Ameiurus 54, 56
— me las 56
Amia calva 63
Ammodytes 30, 117
Amphipnous рис. 27
— cuchia 49
Amphipoda 165
Amphiprion 67, 68
— frenatus 67
— percula 67
— perederaion 67
Anabantoidei 162
Anabantidae 48
Anabas 32, 33, 48, 49, 50, 115
Anablepidae 24
Anableps tetraphthalmus 24,
рис. 12
Anarrhichas 74, 75
Anguilla anguilla 137
Anguilliformes рис. 2
A n n a n d a 1 e рис. 20
Anodonta 75
Aphya 8, 118, 136
Apogon 129, рис. 81
Apogonidae 124
Arapaima gigas 136
Archoplites interruptus 59
Arctogadus 200
Argulus foliaceus 77, рис. 49
Argyropelccus 25, 26, 27, рис. 15
— affinis рис. 13
— olfersii рис. 2
Asellus aquaticus 71
Aspitis aspius 21, рис. 110
Aspredo 1£9
Aspro 196
Asterias rubens 73
Atherinidae 53
Aulopyge 28
A z e v e d о 142
В
Balaenoptera acutorostrata 84
— physa 1 us 84
Balantidium 66
Barbus brachycephalus 34, 98, 191r
— рис.: 58, 110
— capito 98
— conchonius 118
— radcliffi 34
— tor 34
Bathymacrops macrolepis 24
Bathypterois a ter 162
Beebe 67, 69
Belling рис. 19
Be lone acus рис. 83
Belonidae 53
Benthophilus macrocephalus 76, 17f
172, рис. 113
— stellatus 172
Benthosaurus 25
Betca splendens 126
Blenniidac 9, 18, 130
Blennioidei рис. 8
Blennius 8, 74
223
Blicca 20
Boreogadus 200
— saida 108, 130, 202
Bothidae 203
В о u long er рис.: 9, 12
Bowen 53, 57
Brachymystax 186, 189
— lenok 189
Brama 20
Branch iura рис. 49
В r e d e г рис.: 32, 33
Brevoortia tyrannus 108
Brosme 200
Bubyr cauacasicus 42
Bucephalus polymorhus 69
Budget рис. 85
Bull 18
C
Calanus 78, 183, 202
Caligus 76, рис. 49
Camallanus lacustris 71
Capoetobrama Kuschakevitschi
рис. 107
Carangidae 53
Carassius auratus 50, 56
— carassius 14
Cardidae 76
Cardium edule 76, 171
Carpenter 44
Caspialosa 124, 182, 183, 184
— brashnikovi 151, 165, 183
caspia 99, 165, 183,
— рис. 83
— curensis 183
— kessleri 98, 165, 183
— рис. 90, 91
saposhnikovi 165, 183
sphaeroccphala 183
— volgensis 127, 165, 183
Caspiomyzon wagneri 97, 174, 175
— wagneri forma praccox 175.
Catoptero ides macrocotyle 69
Catostomus 87
Centrarchidae 22
Ceratias рис. 72
Ceratudae 27, 62, 119, 125, 161, 169
Ccratioidei рис. 72
Ceratium 11
Ccratoscopelus maderensis рис. 2
Cetorhinus maximus 39, 58, 136
Chalcalburnus chaleoides 191
Characinoidei 85, 123
Chauliodontidac 9
Chauliodus sloani рис. 2
Chelidodipterus 129
Chelone imbricata 82
— mydas 82
Chiasmodus 167
Chilodon 66
Chironomidae 80
Chironomus 167
Chologaster 28, рис. 17
— Cornutus 28
— papilliferus 28
Chondrostoma 20, 28, 46
— nasus 46
Church 56
Cichla temensis рис. Ill
Cichlidae 31, 127, 157, 164, 166 ,
рис. Ill
Clarias 49
Cleithrum 139
Cleisthenes herzensteini 205
Clemmys caspica 82
Clio borealis 76
Clupanodon 182
Clupea harengus 20, 38, 58, 104,
105, 117, 124, 133
— harengus harengus 182
— harengus maris albi 182
— рис. 89
— harengus membras 182
— harengus pallasi 182 рис. 28
Clupcidae 51, 53, 182
Clupeiformes рис. 2
Clupeonella delicatula 156, 185
— delicatula tcharchalcn-
sis 130
Clyde 57
Cobitidae 119
Cobitis taenia 30, 112
Coecoramdella brasiliensis 28
Coecoramdia urichiu 28
Coelorhynchus carminatus рис. 2
Colymbiformes 82
С о n t a g 151, рис. 10
Copepoda рис. 49
Coregonus 186, 189
— autumnalis 190
— lavaretus 23, 104, 188,
рис. 108
lavaretus maraenoides 190
lavaretus mongo licus 190
muksun 190
Corisa 80
Curophiidae 169, 171, 194
Coryphaena 106
Costia nacatrix 66, рис. 40
Cottidae 9, 166
Cottocomephorus 119
Cottoidei рис. 8
Cottus gobio 46
Cristivomer 186
С г о n h e i m рис. 97
Crysta llogobius 118, 136
Cryptobia 66
Cryptopsaras couesii рис. 2
Ctenopharyngodon idellus 130 191
Cumacea 171, 194
Curimatius elegans 142
Cyanea arctica 68
Cycloptcridae 40
224
Cyclopterus lumpus 124, 125
Cymatogaster 126
Cyprinidae 53, 190
Cypriniformes 28, рис. 36
Cyprinodon macularius 14
Cyprinodontidae 14, 118
Cyprinodontiformes 28, 126, 136,
рис. 17
Cyprinus carpio 116, 117, 194
Cystobranchus respirans 72
D
Dactylogyrus vastator 69
Dahlgren 51, рис. 28
Dakin 133
Dalia pectoral is 14
Danio rerio 34, 131
Delphininae 84
Delphinapterus leucas 84
Delphinus delphis 84
Dias 142
Didacna trigonoides 76
Diphyllobothrium latum 71,
рис. 45
Dipnoi 135
Discognathichthys 35
Discosoma 67
Ditrema 126
Dorylaimus stagnalis 73
Draper 9
Dreissena 69, 167, 169
— polymorpha 76, 171
Ductus । njumaticus 6, 9
Dytiscidae 79
Dytiscus marginalis 79
E
Echeneis remora 62, рис. 37
Edriolichnus schmidti 161, рис. 72
Edwards 9
Ehrenbaum рис.: 1, 78
Electrophorus electricus 59, 162,
рис. 34
Eleginus 200 a
— gracilis 202
— navaga 130, 202
Ellis 43
Emys orbicularis 82
Emydinae 82
Engraulicypris 191
Engraulidae 51, 185
Engraulis encrasicholus 107, 185,
186, рис. 83
— encrasicholus maeoticus 133
— 186
— encrasicholus ponticus 186
— japonicus 20, 185
Entosphenes 174
Ereiocheir sinensis 78
Ergasilus sieboldi 76
Esocidae 195
15 Биология рыб
Esox lucius 104, 195
— reichertii 195, 196
Ethmalosa fimbriata 142
Etrumeus 182
Eumesogrammus praecisus 18.
19, рис. 8
Eumicrotremus рис. 24
Euphausiidae 183
Eupomotis gibbosus 90
Exocoetidae 53
F
Falconiformes 82
F a u v e 1 рис. 47
Fierasfer 75
Flagellata рис. 40
F о w 1 e г рис.: 13, 14
Francisconius marmoratus 123
Fulton 118
Fundulus 9, 74
— heteroclitus 9, 56, 58, 131
G
Gadidae 53, 162
Gadus 200
— aeglefinus 20, 73, 130, t01
eismarki 130
-- luscqs 130
— merlangus 18, 130
— morhua 18, 20, 130, 137
200
Virens 18, 130, 202
Gaidropsarus 200
Galeichthys felis 128, рис. 80
Gambusia affinis 29, 90
Gammaridae 171, 194
Garra 35, рис. 20
Gasterosteus 63, 128
Gastropoda 76, 171
Gastrostomus bairdi рис. 2
Geophagus brasiliensis 31, рис. Ill
Geotria 174
Gerridae 53
Glyptosternum reticulatum 34
Glyptothorax 35, рис. 20
Gobiidae 8, 30, 40, 130, 136, 19S
Gobio gobio 23, 34, 46
Gobius 119, рис. 24
— caspius 76, 171, 172
— fluviatilis 76, 78, 170, 171,
172, 198, 199
— kessleri 172
— melanostomus 8, 76, 171,
198
niger 8
— ophiocepha lus 198, । 199
paganellus 74
Goree рис.ч 48
Gray рис. 69
Greene 99
225
н
Haliaetos albicilla 82
— leucoryphos 82
Halichoerus gryphus 84
Ha lien tea 32
Harengula 182
Harpodon ncherus 79
Hemiramphidae 53
H e r r e 68
Hesse рис. 21
Heteropneustes 49
H i с к I i n g рис. 77
Hippoglossus hippoglossus 206
H j о r t рис.: 10, 98
Homalopteridae 35
H о r a 34, рис.:- 20, 23 27, 71
Hucho 186, 189
— hucho 189
— perryi 189
— taimen 189
Huro salmoides 89
Huso 179
— dauricus 117, 118, 179
— huso 118, 136, 179
Hydrichthys 67
Hydrophidae 81
Hymenocephalus italicus рис. 3
Hyperoodon rostratus 84
Hypophthalmichthys molitrix 164
Hyrcanogobius bergi рис. 83
Hystiophoca fasciata 84
I
Icelus spatula 31
— spiniger 31
Ichthyomyzon 174
Ichthyophthirius 66
Ichthyotomidae 72
Ichthyotomus sanguinarius 72.
рис. 47
Idiacanthus 25
Ilisha 182, 184
Ipnops 25
— murrayi рис. 2
Isopoda рис. 49
J e n i n g s 73
Jenkinsia рис.: 32,33
Johansen 132
Jordan 17, рис. 17
К
Kerr рис. 85
Krogh 132
Kyle рис.: 1, 22,^78
226
L
Labeo dero рис. 71
Labridae 164
Labrus mixtus 74
Lamna cornubica 105
Lanipanyctes 27, рис. 16
Lampctra 174
— fluviatilis 97, 175
fluviatilis forma praecox 17$
japonica 178
— mariae 178
— planeri 178
Laurcstes tenuis 31, 41, 125
Lea Einar 143, рис. 94
Lebistcs reticulatus 29, 56
Ledebur 6
Lee рис. 80
Lepidopus caudatus 18
Lepido siren paradoxa 33, рис.
Lepidosteiformes рис. 36
Lepomis 22
Lernaea branchialis 77
Leuciscus brandti 191
— cephalus 46
Ligula intestinalis 70 рис.: 43, 44
Limanda aspera 206
Linuche unguiqulata 67
Liopsctta glacialis 205
Liparidae 9
Lophiiformes 32 162, рис.: 2, 18
Lophius 32
piscatorius 11, рис. 3
Lopholatilus hamaeleonticeps 19,
рис. 9
Lota lota 46, 200, 203
— lota leptura 203
Lucioperca 196
— lucioperca 116, 156, 171
— marina 196, 197
— volgensis 196, 197
Lutra lutra 84
M
Macropodus opercularis 127
Macruridae 26
Macruriformes рис. 2
Meek рис. 64
Megalobatrachus japonicus 81
Melamphaenidae 9
M e г к e r 57
Mesogobius batrachocephalus 198,
199
Microstomidae 24
Misgurus fossilis 30
Mola mola 11
Molva 200
Monodaena 76, 171
Monodon monoceros 84
Monogenea 69
Mordacia 174
Mormyridae 59, 160 162, 163
Morone labrax 53
Mortimer рис. 25
Mugil 111
Mugilidae 53
Myctophiidae 9, 25
Myctophium 27
— rissoi 25, рис. 13
Myliobatidae 61
Myoxocephalus 14
— scorpius 18, 41
Mysidae 167, 170, 171
Mystichthys lusonensis 136
Mytilaster lineatus 76
Myxine glutinosa 13, 62, 116, рис. 83
Myxobolus 66, рис. 41
N
Natrix olivaceus 81
— tcsselata 81
Naucrates G2
Nemachilus 34,
barbatulus 46
stoliczkai рис.: 71, 101
Nematodes 71
Nemichthys ovocetta рис. 2
Neoliparis mucosus рис. 2
Neomys fodiens 84
Nepa 80
Nerophis 74
— lumbriciformis 18
Nervus facia lis 39
Nervus vagus 39
N i к о 1 s к i j рис. 104
Nitzchia sturionis 69, 156
N о r m an рис. 68
Notonecta 80
О
Octomytus 66, рис. 40
Ogcocephalus рис. 18
Oncorhynchus 137, 186
— gorbusha 117, 186, рис. 73
— keta 187
— masu 186
— nerka 97, 187
— tschwytscha 99, рис.: 59,
60
Ondatra zibethica 84
Ophicepha lidae 48
Ophicephaliformes рис. 27
Ophicephalus striatus 49
Ophichthydae 92
Orca orca 84
Osmerus eperlanus 148
Otocinclus 48
P
Fagellus mormyrus 117
Pandion haliaetos 82
Paraphoxinus 28
Parasilurus 195
— asotus 195
Parr 53
Pelamys sarda 111
15*
Pelecus cultratus 96, 130. 191 nur
107 ’ рис.
Pellegrin рис. 111
Penis 174
Perea 196
— fluviatilis 46, 117, 197
— schrenki 197
Percarina 196
Percidae 196
Perciformes рис.: 2, 18, 27, ill
Periophthalmus 32, 33, 114
Petromyzon 174
— mar inns 174
Petromyzonidae 174
Petrov рис. 92
Petruschevsky рис. 92
Phisoodonophis boro 30, 33, 92
Phoca caspica 84
Phocaena 84
Fholis gunnelus 18, 41
Phoxinus phoxinus 46, 191
Phyllopteryx eques 64, рис. 38
Physiculus dalwigkii рис. 3
japonicus рис. 1
Pimelodidae 28
Piscicola gcometra 72
Platypoecilus maculatus 56, 58, 150
Platy stoma 62
Plecostomus 37
Pleuronectes 74
— flesus 18, 166, 205
— platessa 18, 104, 155, 204
рис.: 87, 105
— quadrituberculatus 205
Pleuronectidae 203
Pleuronectiformes 130
Poecilidae 117, 119
Polyipnus 25
Polypodium hydriforme 66
Polypteri рис.: 18, 135
Polypterus 32
Polystotrema stouti 13
Pomacentridae 67
Pomalobus 182
Portunidae 78
Premnas biaculeitus 67
Pristiophorus 162
Pristis 162
— microdon 13
Pristiurus melanostomus 116
Protopterus 30, 48, рис. 85
Pseudomonas plehniae 63
Pseudoscaphirhynchus 179, 181
— fed tsc hen koi 181
— hermanni 181
— kaufmanni 181
Pungitius 63, 128
Q
Quadrupeda 32
227
R
Radialia 32
Raja 60
— clavata рис. 35
Rana catcsbyana 81
— ridibunda 81
Raphidascaris 71, рис. 46
Regalecus 111
Rhinoptera peli рис. 35
Rhodeus 192
— scriceus 123
Ricardo рис. 55
Rounsfell 51, рис. 28
Rutenberg рис. 77
Rutilus frisii 98, 191
— frisii kutum 191, рис. 74
— rutilus 46, 192, 193, рис. 107
— rutilus aralensis 193
— rutilus caspicus 138, 171, 193
— rutilus haeckeli 193
S
Saccopharynx 27
Sa Imo 78, 186
— gairdneri 188
— irridens 188
— ischchan 188
— mykiss 188
— penshinensis 188
— salar 20, 34, 116, 141, 187
— trutta 46, 188
— trutta m. fario 188
Salmonidae 53, 125, 186
Salmothymus 186
Salvelinus 186
— alpinus 188
— beganidae 188
— fontina lis 28
— jacuticus 188
— neiva 188
— salvelinus 188
Saprolegnia 65
Sardina 29
Sardinella 182
Sardinops melanosticta 20, 55, 185,
рис.: 31, 83
— melanosticta caerulea 185
Scaphirhynchus 179
Scardinius erythrophthalmus 21
Scarus 74
Scheuring рис. 60
Schizothorax 34,
intermedius 19
Schuett 56
Sciaenidae 53
Scomber colias 198
— japonicus 198
— scombrus 197
Scomberesocidae 53
Scombridae 197
Scopelidae 51, 79, 106, 118, 136
Scopeliformes рис. 2
228
Scorpaenidae 130, 199
Scyllium canicula 116,
— catulus 13
_stellare 13
Sebastes marinus 20, 126, 199
Serranus scriba 117, 118
Serrasalmo piraya 85, рис. 50
Shaw 58
Siluridae 195
Silurodes eugeneiatus 162
Silurus glanis 85, 136, 195
Simenchelidae 62
Simenchelys 62
Sisoridae 35, рис. 23
Snyder рис. 59
Somniosus microcephalus 116, 12
Sphaeriidae 76
Spinachia spinachia 18
Spirontocaris 189
Spratella 29, 182
— sprattus baltica 185
— sprattus phalerica 185
Stegophilus 62
Stenodus 186, 189
— leucichthys 189
Sternoptychidae 25
Stoichactis 67
Stomiatidae 9
Stomiatoidei рис. 14
Symbranchidae 48, 49
Symbranchiiformes рис. 27
Synanceia horrida 61, 92
Syngnathiformes 129, рис. 36
Syngnathus 74
— nigrolineatus caspius 42
Synodontis 107
Syphonostoma 74, рис. 82
T
Talassaetos pelagicus 82
Tee Van 69
Teleostei 32
Tetrodon 112
Tetrodontiformes рис. 36
Thera gra 200
Thymallus arcticus 36
— arcticus baichalensis baicale*-
sis 165
arcticus baicalensis brevi-
pinnis 165
— thymallus 46
Thynnus thynnus 15
Tilapia 128
— esculenta 122
— heudeloti рис. Ill
Tinea tinea 50
Toxotes jacubator 162
Trachurus trachurus 111
Trachypterus 11, 111, 159
— cristatus рис. 3
Triaenophorus nodulosus 70
Triehiuridae 18
Trigli 32
Trionyx 82
Troglichthys 28
Trygon pastinaca рис. 35
Trygonidae 61
Trypanosoma 66 рис. 40
Tub-Tex 73
Turiiops tursio 84
Tyloclelphus 70
Typhlichthys 28, рис. 17
Typhlobagrus kronei 28
V i e i r r a 142
Vimba v mba 191
Vinciguerria 27
W
Wald
Wilier 132
Worthington рис. 55
Wunder рис. 1
Ulcina olriki 18, рис. 8
Umbra limi 56, 58
Unio 75
Utricularia 64, рис. 39
V
Varicorhinus heratensis 159, 164,
192,
X
Xiphias gladius 162
Xiphophorus 119
Z
Zeugopodium 32
Zoarces viviaparus 18, 41, 126
Zoarcidae 9, 39
ОГЛАВЛЕНИЕ
Введение 3
Часть I
ВЗАИМООТНОШЕНИЯ РЫБ С ФАКТОРАМ ВНЕШНЕЙ СРЕДЫ
А. Влияние физических свойств воды на рыб
Плотность и давление воды б
Температура воды И
Свет 21
Грунт н взвешенные в воде частицы 29
Движение роды 34
Б. Влияние химических свойств воды hi рыб
Соленость 42
Газы 45
В. Влияние нт рыб других организмов
Взаимоотношения рыб друг с другом 51
Влияние растений на рыо 63
Взаимоотношения рыб и других животных 65
Часть п.
ОСНОВНЫЕ ЧЕРТЫ ЖИЗНЕННОГО ЦИКЛА РЫБ
Миграции рыб
Нерестовые миграции 96
Кормовые миграции 105
Зимовальные миграции 107
Миграции под влиянием внешних факторов 108
Причины возникновения миграций 108
Форма тела и движение рыб 111
Размножение и развитие рыб
Плодовитость и созревание половых продуктов 115
Иерест 123
Развитие икры 129
Личиночный период 134
230
Размеры, рост и возраст рыб
Методика исследования возраста и роста
Факторы, определяющие рост.
Возрастной состав стада
Колебания численности рыб
Экстерьер и упитанность рыб 157
Питание рыб
Приспособления к добыванию пищи 1^0
Усвоение пищи и пищевой рацион
Пищевые объекты и конкуренция из-за пищи 16g.
Часть III.
ОБЗОР БИОЛОГИИ ОСНОВНЫХ ПРОМЫСЛ ВЫХ РЫБ СССР 174
Основные пособия по биологии рыб 207
Указатель русски* названий, имен собственных и терминов
Указатель латинских названий и терминов и иностранных имен соб-
ственных 223
Редактор Ю. А. Яризер
Сдано в набор 26ZVI 1944 р. Подписа-
но к печати 1 1 /1 1946 г. Тиран: 5000-
ЕООЮ4, 14,5 п.л. 17 авт. л. Цена 16 pyi-
Переплет 2 руб.
9-я тип. им. Мяги трестач«Полнграфкннга»
Огиза при СНК РСФСР, г. Куйбышев,
уж. Венцека, 60. Закаа М 904.
Рис. 22. Лососи, преодолевающие водопад (из Kyle, 1926).
Рис. 52, Рыбоход бонневильской плотины (из Holmes, 19Ю).
Рис. 87. Отолиты кам-
балы Pleuronectes plates-
sa (из Alvard, 1932)
Рис. 89. Чешуя беломорской ^сельди Clupea harengus wa-
Рис. 90. Чешуя черноспинки
Caspialosa kessliri, край которой
разрушен во время нереста
(фот. Замахаева).
Рис. 91. Чешуя черноспинки Caspialosa
kessleri с регенерировавшими после нере-
стового разрушения краем (фот. Зама-
хаева)