/























































































































































Author: Лаос-Бельтра Р.
Tags: математика биология экология численные методы серия мир математики точные науки
ISBN: 978-5-9774-0723-6
Year: 2014
Similar




Text
Мир
МАТЕМАТИКИ
28
Математика
жизни
Численные модели в биологии и экологии
D^AGOSTINI
Мир математики
Мир математики
Рафаэль Лаос-Бельтра
Математика жизни
Численные модели в биологии и экологии
Москва - 2014
TCAGOSTINI
УДК 51(0.062)
ББК 22.1
М63
М63 Мир математики: в 40 т. Т. 28: Рафаэль Лаос-Бельтра. Математика жизни. Численные
модели в биологии и экологии. / Пер. с исп. — М.: Де Агостини, 2014. — 160 с.
Жизнь — одно из самых прекрасных и сложных явлений на планете, изучением кото-
рого с начала XX века занимается не только одна биология. Физики, а затем и математики
обнаружили, что некоторые биологические явления можно описать с помощью матема-
тического языка. Так родилась новая дисциплина — математическая биология, или био-
математика. Благодаря ей сегодня можно получить ответы на множество важных вопросов,
касающихся биологии и биомедицины. Эта книга представляет собой панорамный обзор
различных явлений, которые изучает биоматематика.
ISBN 978-5-9774-0682-6
ISBN 978-5-9774-0723-6 (т. 28)
УДК 51(0.062)
ББК 22.1
© Rafael Lahoz-Beltra, 2010 (текст)
© RBA Coleccionables S.A., 2011
© ООО «Де Агостини», 2014
Иллюстрации предоставлены: Getty Images, Age Fotostock.
Все права защищены.
Полное или частичное воспроизведение без разрешения издателя запрещено.
Содержание
Предисловие......................................................... 9
Глава 1. Математическая биология в исторической перспективе......... И
Рождение математической биологии.................................... И
Теория эволюции................................................. 14
Развитие математических методов теории эволюции.................. 15
Роль компьютера в математическом анализе жизни..................... 16
Открытия Алана Тьюринга......................................... 17
Зарождение биологии систем.......................................... 23
1970-е — время перемен............................................. 26
Современная математическая биология................................. 27
Математические модели в биологии.................................... 30
Компьютер как пробирка............................................. 31
Программы для символьных вычислений................................. 33
Некоторые примеры использования математики в биологии.............. 36
Изучение популяций оленей, белок и других животных.
Матрица Лесли................................................. 36
Клеточные автоматы.............................................. 39
Модель «хищник — жертва» и клеточные автоматы.................... 43
Глава 2. Жизнь — изменчивое явление................................. 47
Как извлечь игрушку из пасхального яйца............................ 49
Дифференциальное уравнение Парка юрского периода................ 51
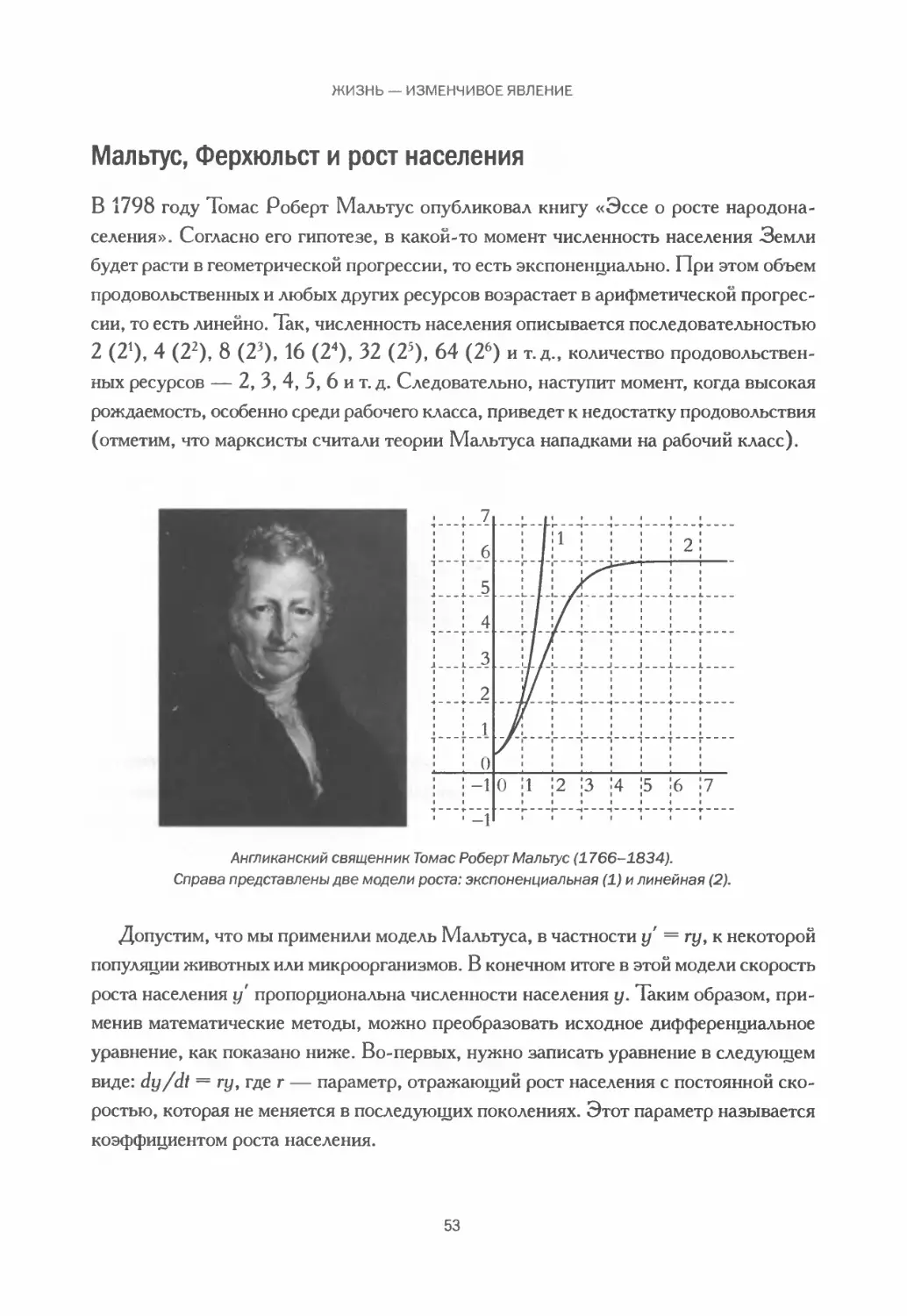
Мальтус, Ферхюльст и рост населения................................. 53
Дифференциальные уравнения в биотехнологии.......................... 56
Математическое изучение рака: опухоли в компьютере.............. 58
спид, свиной грипп и другие заболевания,
которые можно изучить с помощью математики.................... 62
Число е и колония бактерий Escherichia coli..................... 64
Глава 3. Микробиолог, покорившийся хаосу........................... 67
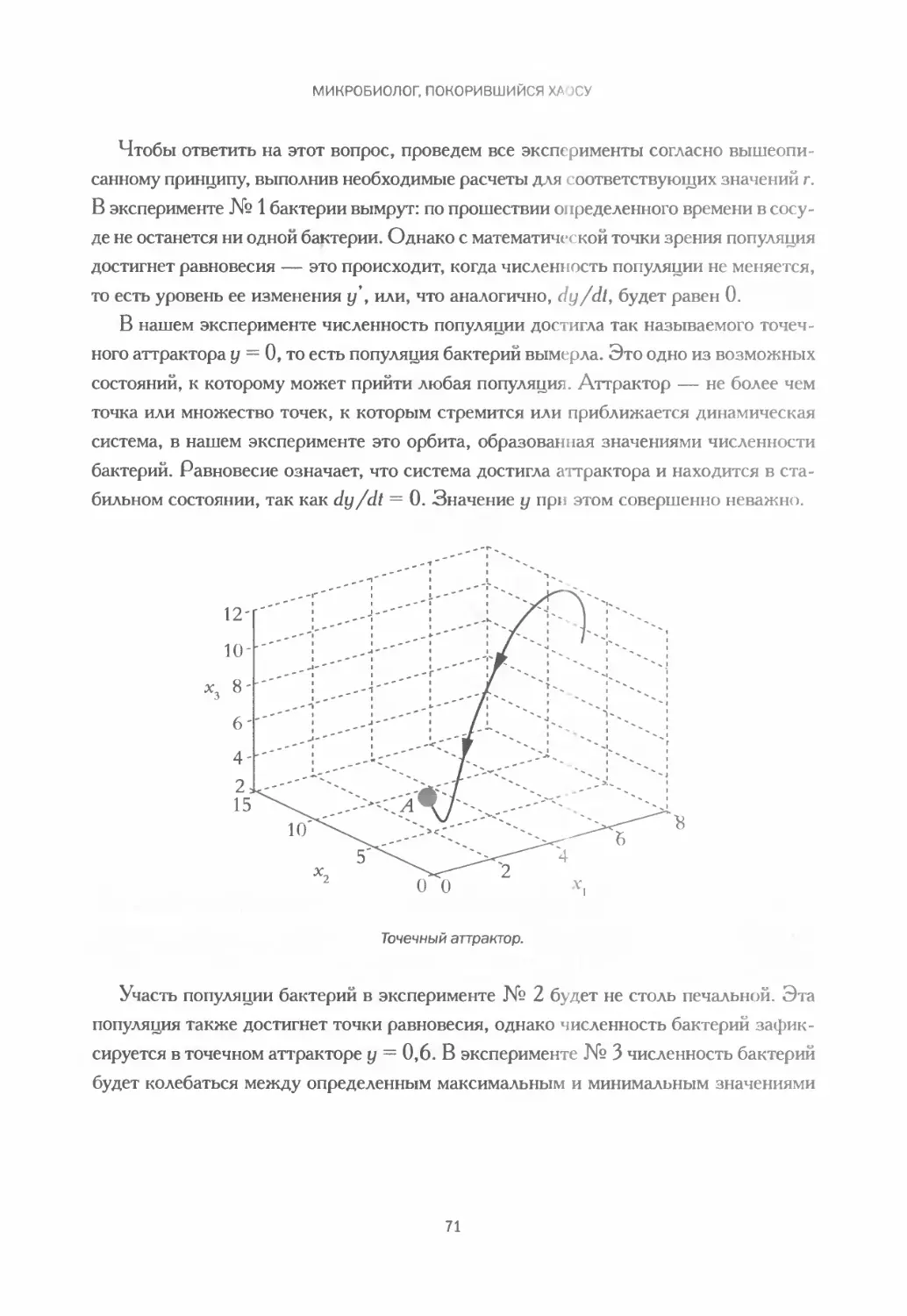
Дифференциальное уравнение у’ = гу и его ограничения............... 67
Четыре эксперимента в качестве примера.......................... 70
5
СОДЕРЖАНИЕ
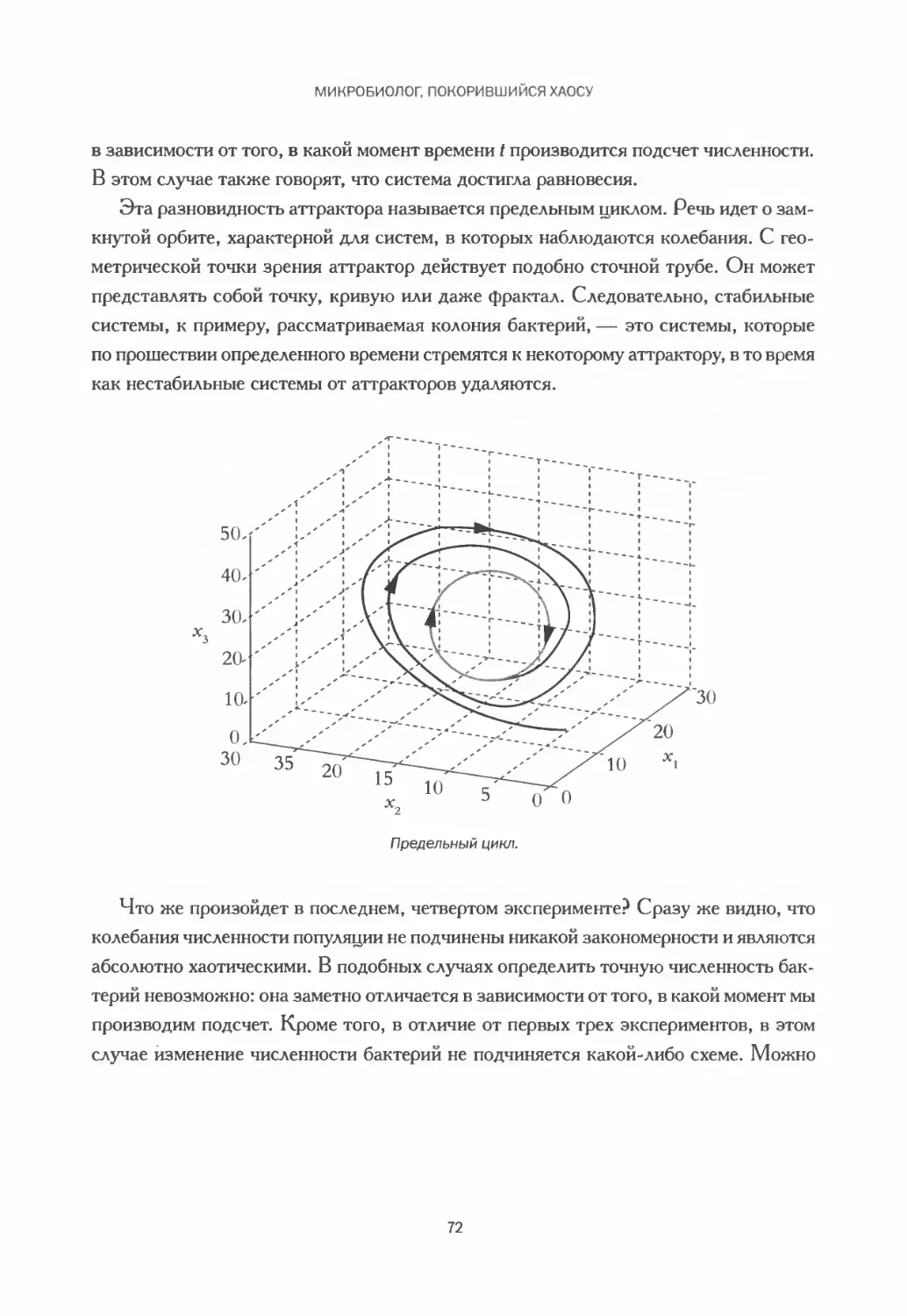
Изучение природы шаг за шагом..................................... 73
Является ли природа фрактальной?.................................. 78
Множества Жюлиа и Мандельброта..................................... 81
Игра в хаос Барнсли................................................ 83
Глава 4. Судоку жизни.............................................. 85
Таблицы, судоку и матрицы.......................................... 85
Операции над матрицами............................................ 89
Сложение....................................................... 89
Вычитание...................................................... 90
Умножение...................................................... 91
Умножение матрицы на вектор и применение этой операции......... 93
Транспонирование матриц........................................ 95
Определители...................................................... 95
Определитель квадратной матрицы размером 3x3.................... 97
Как делить матрицы. Обратные матрицы и их применение
в биологии......................................................100
Матрицы и горошины: законы Менделя............................... 101
Является ли наследование признаков независимым?................105
Марковские матрицы, ДНК и биоинформатика .........................107
Глава 5. Векторные величины. Векторы в биомеханике,
нейронные сети и системы уравнений..............................109
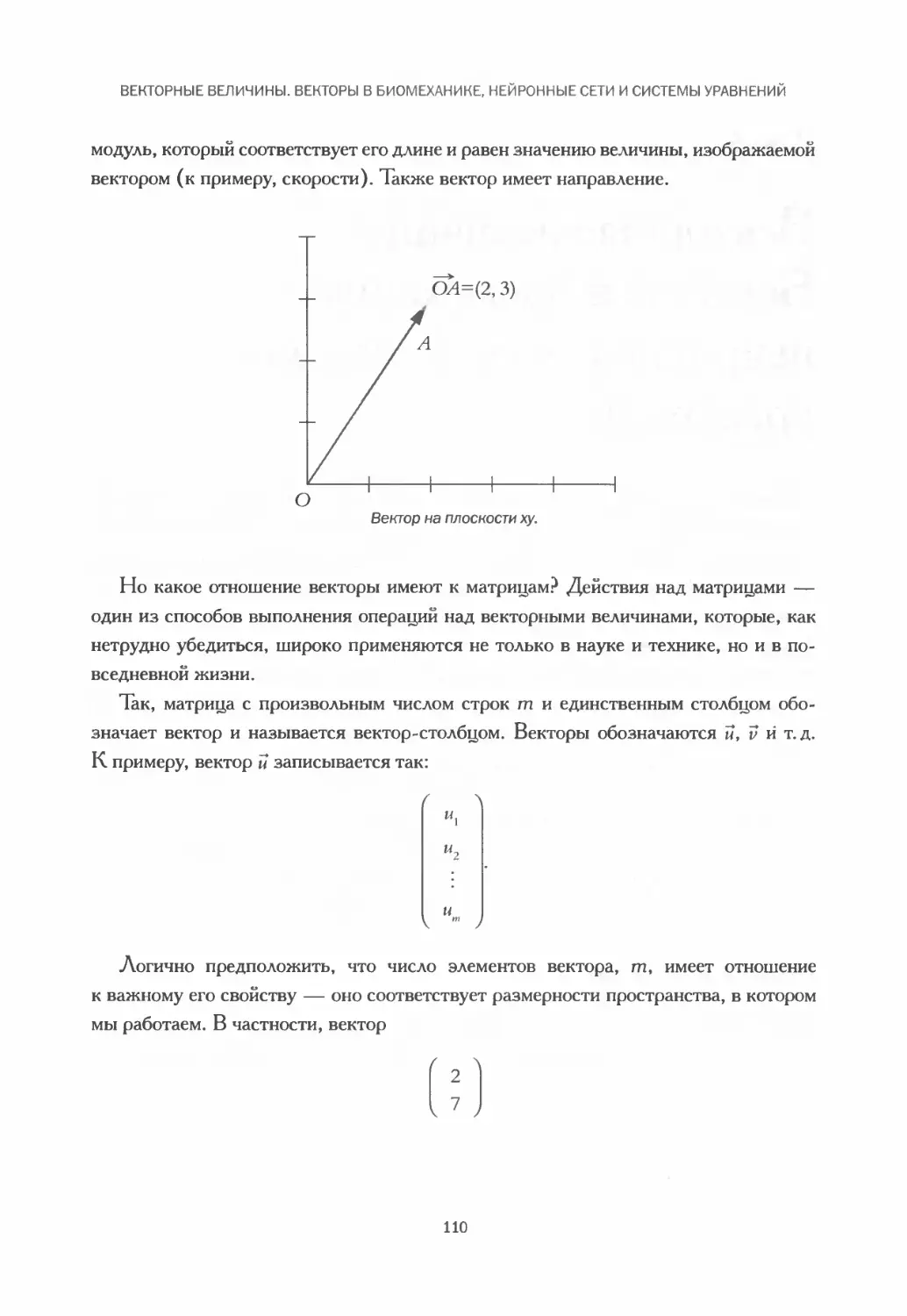
Векторы и матрицы.................................................109
Сложение векторов: сокращение мышц и локомоция................... 112
Умножение векторов и применение этой операции в нейронных сетях.. 115
Обучение. Пример с распознаванием звуков...................... 117
Векторное, или внешнее, произведение...........................120
Модель памяти животных и человека............................. 121
Решение систем уравнений. Эксперимент энтомолога..................124
Имеет ли система уравнений решение?............................126
Сколько молодых и взрослых насекомых поймал энтомолог.
Правило Крамера..............................................127
Глава 6. Экология и математика. Взаимовыгодное сотрудничество.... 131
6
СОДЕРЖАНИЕ
Построение моделей.............................................132
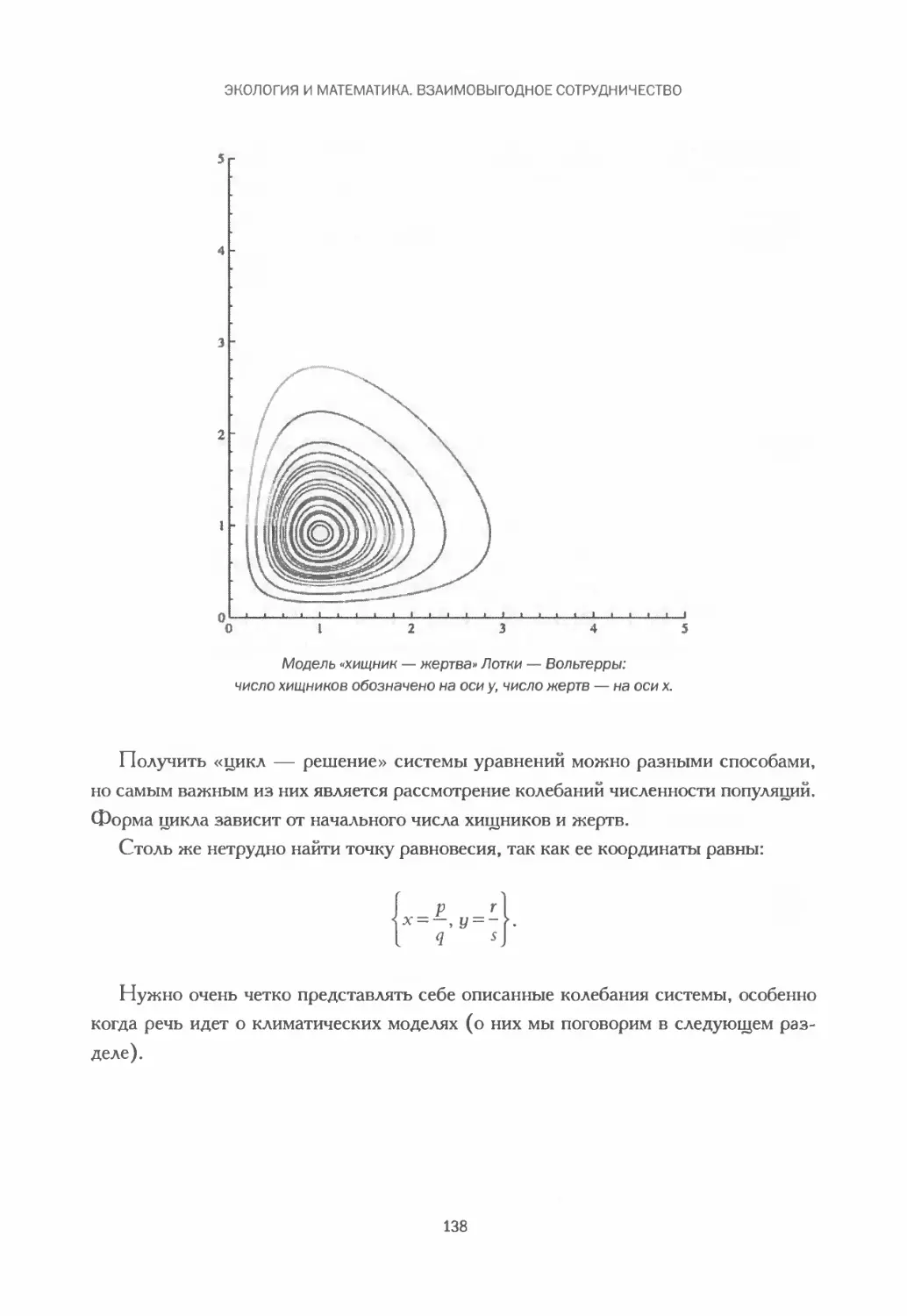
Модель Лотки — Вольтерры: волки и зайцы........................134
Математические модели климата.
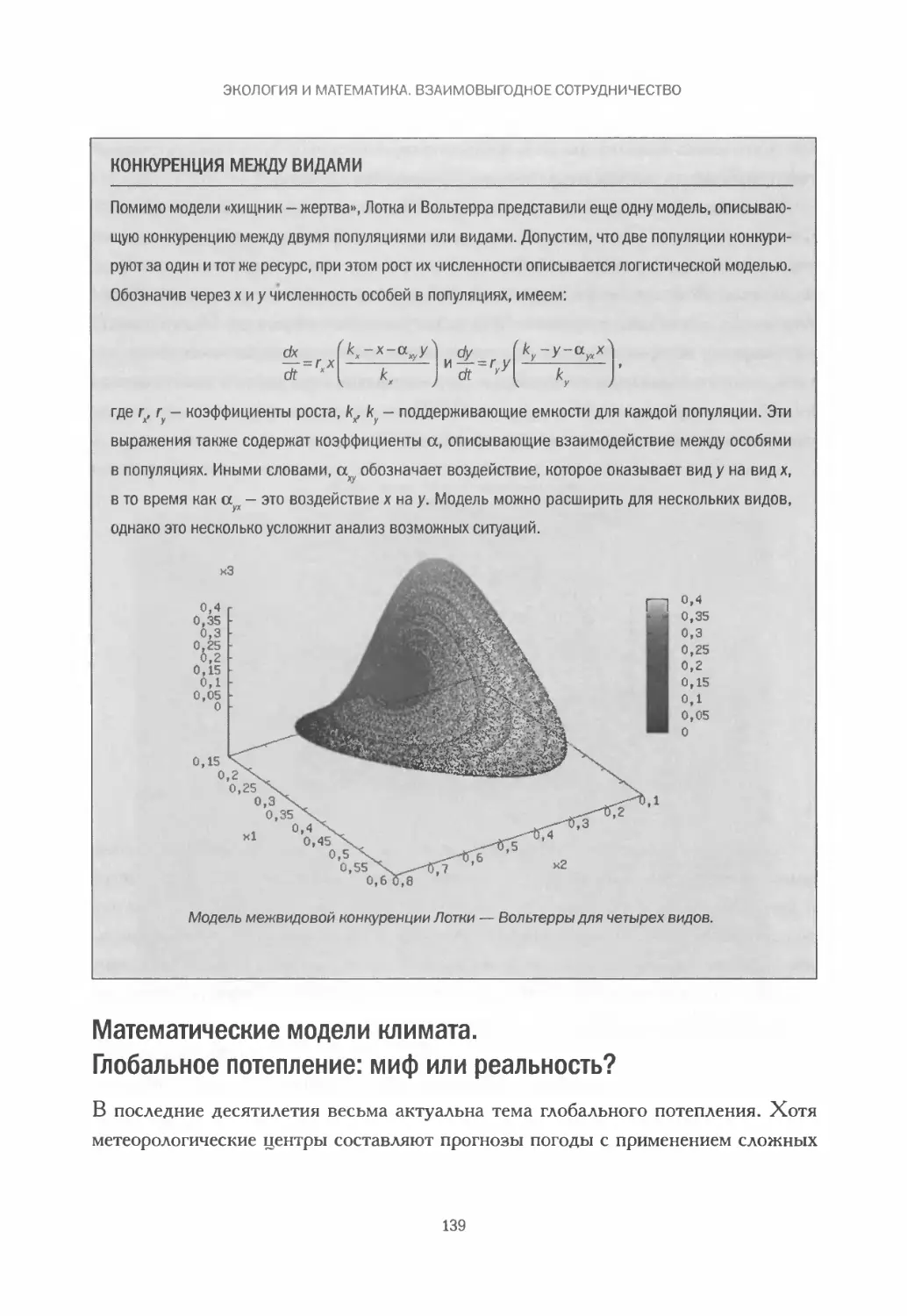
Глобальное потепление: миф или реальность? ..................139
А что, если мы сами меняем атмосферу? Гея и «Маргаритковый мир».146
Приложение. Магия комплексных чисел........................... 151
Библиография.................................................. 155
Алфавитный указатель...........................................157
7
Предисловие
Жизнь — одно из самых прекрасных и сложных явлений на планете, изучением
которого с начала XX века занимается далеко не одна биология. Физики, а затем
и математики обнаружили ряд биологических явлений, которые можно описать на ма-
тематическом языке. Николай Рашевский, Карл Людвиг фон Берталанфи и Алан
Тьюринг положили начало плодотворному союзу математического формализма и нау-
ки о жизни, а компьютеры позволили ученым проводить количественные исследования
биологических явлений. Так родилась новая дисциплина — математическая биология,
или биоматематика. Математическая биология внесла и продолжает вносить свой
вклад в развитие биологии как посредством теоретического изучения динамических
систем (мозга, муравейника или экосистем), так и благодаря решению практических
задач в ходе изучения раковых заболеваний, эпидемий СПИДа или свиного гриппа.
Сегодня ответы на множество вопросов биологии и биомедицины можно дать
с помощью математического анализа. Так, размножение раковых клеток в опухолях
определенного типа описывается функцией Гомпертца. Во многих процессах в сфе-
ре биотехнологий при мониторинге биореакторов используются дифференциальные
уравнения. Даже такие проблемы современности, как возможное изменение клима-
та Земли, изучаются с помощью математических моделей, в частности климатиче-
ской модели Лоренца.
В этой книге представлен панорамный обзор различных аспектов биоматематики.
В первой главе мы говорим об основных достижениях этой науки и ее историческом
развитии. Во второй главе показана возможность использования дифференциальных
уравнений для описания динамики биологических явлений, то есть явлений, благодаря
которым становится возможным сохранение жизни. Эти уравнения очень важны
для человечества, так как позволяют решить бесчисленное множество задач, от де-
мографических проблем, о которых писал еще Мальтус в 1798 году, до определения
возраста ископаемых посредством радиоуглеродного анализа (этот метод предложил
Уиллард Либби в 1950 году).
Математика, конечно же, не смогла остаться в стороне от еще одного притя-
гательного явления. Хаос, о котором мы поговорим в третьей главе, присутствует
повсеместно, будь то рост населения, поведение биржевых индексов или электроэн-
цефалограмма человека. В этой же главе мы рассмотрим еще одну тему, связанную
9
ПРЕДИСЛОВИЕ
с хаосом, — фракталы, их присутствие в природе (в частности, в виде снежинок или
ветвей деревьев), способы графического представления фракталов с помощью ком-
пьютера. Хаос и фракталы нельзя изучить без краткого рассмотрения комплексных
чисел, а не имея представления о комплексных числах, невозможно понять даже самые
яркие и наглядные особенности мира фракталов.
В четвертой главе показано, что математическая биология по большей части ос-
нована на использовании числовых таблиц, или матриц, и основную роль в ней игра-
ют операции над матрицами. В завершение главы мы рассмотрим законы Менделя
и познакомимся с одним из важнейших понятий биологии — полным факторным
экспериментом. В пятой главе освещается еще одно математическое понятие, играю-
щее особую роль благодаря множеству способов применения, — векторы. Мы опи-
шем использование векторов в биомеханике, при моделировании нейронных сетей
и решении систем линейных уравнений.
И в завершение удивительного путешествия вы узнаете о взаимосвязи матема-
тики и экологии. Сегодня ни один проект по охране окружающей среды не обходит-
ся без использования формального математического аппарата. В шестой главе мы
определим понятие экосистемы и представим матричные популяционные модели,
особенно полезные при изучении и сохранении популяций. Отдельно мы рассмотрим
одну из классических моделей математической биологии — модель «хищник —
жертва» Лотки — Вольтерры1. Следующий дискуссионный вопрос, на котором мы
остановимся, звучит так: ждет ли нас глобальное изменение климата? Вы увиди-
те, что проблема изменения климата имеет математическую природу, поэтому ответ
на поставленный вопрос нельзя дать без знания климатических моделей и применяе-
мого в них математического аппарата. Книга завершается анализом «Маргаритко-
вого мира» — математической модели, созданной Джеймсом Лавлоком в 1980-е
годы на основе гипотезы Геи. Эта модель бросает вызов дарвинизму и классическим
представлениям о сохранении жизни на планете. 1
1 Для читателя более привычным может оказаться неправильный вариант — Вольтерра. — Примеч. ред.
10
Глава 1
Математическая биология
в исторической перспективе
В начале XX века Россия напоминала бурлящий котел. Глубокий экономический
кризис и социальное недовольство, возникшие после поражения в русско-японской
войне 1904—1905 годов и начала Первой мировой войны с Германией в 1914 году,
привели к Октябрьской революции. Из-за этих событий физик-теоретик украинско-
го происхождения Николай Рашевский (1899—1972), который сегодня считается
создателем математической биологии, вместе с супругой Эмилией покинул страну.
Сменив несколько государств, в 1924 году Рашевские осели в США.
Рождение математической биологии
Оказавшись на американской земле, Рашевский приступил к работе в исследователь-
ской лаборатории компании Westinghouse, где занялся изучением деления клеток.
Таким образом, деление клеток впервые было рассмотрено с точки зрения физики
и математики — подобный подход в те годы считался невероятно передовым.
В 1934 году Николай Рашевский (или, как его стали называть к этому вре-
мени, Николас Рашевски) получил должность старшего преподавателя кафедры
физиологии Чикагского университета. Вскоре благодаря этому ученому произошли
два события, имевшие большое значение для развития математической биологии.
В 1938 году была опубликована его первая научная статья по биоматематике, зна-
менитая «Биофизическая математика: физико-математические основы биологии».
В 1939 году Рашевски создал первый научный журнал, посвященный исследовани-
ям в математической биологии,— The Bulletin of Mathematical Biology («Вестник
математической биологии»).
С тех пор математическая биология прошла долгий и непростой путь, пока на-
конец не обрела статус полноценной научной дисциплины.
И
МАТЕМАТИЧЕСКАЯ БИОЛОГИЯ В ИСТОРИЧЕСКОЙ ПЕРСПЕКТИВЕ
THE BULLETIN OF
Mathematical
BIOPHYSICS
№t VHIVU 1ТГ <» CHICAGO НШ . CHICAGO . IllIHOn
VOLUME 4
NUMBER 1
SEPT. 1948
Николас Рашевски основал Общество математической биологии, а в 1939 году стал редактором
первого журнала по этой дисциплине. Первоначальное название журнала — The Bulletin of
Mathematical Biophysics («Вестник математической биофизики») — позднее сменилось
на The Bulletin of Mathematical Biology («Вестник математической биологии»).
В последующие годы Рашевски занимался теоретической работой и применил
теорию множеств и логику высказываний в исследованиях биологических систем. Он
изучал различные общества и способы организации живых существ, а также иерар-
хии, которые они образуют. Сегодня ответы на стоявшие перед ним вопросы кажутся
очевидными: рассмотрим, к примеру, последовательность молекулы —> клетки —>
ткани —> органы —> системы —> индивид —> популяция. Рашевски создал теорию био-
логических отношений — реляционную биологию, а также ввел понятие «множество
организмов». Все эти открытия до недавнего времени оставались незамеченными
большинством биологов, которых в основном интересовали полевые исследования или
работа в лаборатории. Подлинный размах и возможности теоретического аппарата,
терпеливо выстроенного Рашевски, стали очевидны лишь с возникновением так на-
зываемой биологии сложных систем. А развитие этой дисциплины, в свою очередь,
было бы невозможным без распространения компьютеров.
Наверное, одной из важнейших особенностей первого этапа развития математиче-
ской биологии, который мы будем дальше называть этапом зарождения биоматема-
тики, стало влияние на нее физики. Это неудивительно, если учесть, что в 1921 году
12
МАТЕМАТИЧЕСКАЯ БИОЛОГИЯ В ИСТОРИЧЕСКОЙ ПЕРСПЕКТИВЕ
Рашевски преподавал теорию относительности в Праге. Подобно Эйнштейну, по
святившему последние годы жизни работе над «единой теорией», в 1960-е Рашевски
пытался создать единую теорию биологии. Он мечтал выразить на языке математики
биологические принципы, описывающие жизнь во всех ее проявлениях, будь то рас-
тения, животные или микроорганизмы.
Подобно другим физикам того времени, например Шрёдингеру, Рашевски также
задавался вопросом: что такое жизнь? К сожалению, полет его фантазии оборвал
сердечный приступ в 1972 году, а ответ на этот вопрос до сих пор не получен, хотя
со смерти ученого прошло уже много лет.
Австрийская банкнота с портретом Эрвина Шрёдингера (1887-1961),
лауреата Нобелевской премии по физике 1933 года и автора книги «Что такое жизнь?»,
опубликованной в 1944 году и оказавшей огромное влияние на развитие биологии.
В отличие от теоретического направления математической биологии, пионером
которого был Рашевски, работы других ученых, например Карла Людвига фон
Берталанфи, носили более прикладной характер. Фон Берталанфи родился в Ав-
стрии в 1901 году, учился в университетах Инсбрука и Вены, работал в Лондонском
университете, различных канадских институтах, а закончил карьеру в Университете
штата Нью-Йорк. Он внезапно умер от сердечного приступа в том же 1972 году,
что и его коллега Рашевски. Хотя основным вкладом фон Берталанфи в науку стала
общая теория систем, о которой мы поговорим позже, ему принадлежат и различные
открытия в математической биологии. Так, в 1938 году он сформулировал знамени-
13
МАТЕМАТИЧЕСКАЯ БИОЛОГИЯ В ИСТОРИЧЕСКОЙ ПЕРСПЕКТИВЕ
тое уравнение роста, которое в наши дни применяется в рыбоводческих хозяйствах.
Фон Берталанфи связал размер рыбы L(t) с ее возрастом / (LK — максимальный
размер, Lq — начальный размер, k — постоянная роста):
L(/) = LK-(LK-L0) е-к‘.
Теория эволюции
Эволюция — одна из важнейших тем биологии, которой уделяется большое внимание
и в математической биологии с момента ее зарождения в 1930-е годы. В целом эволю-
ция — это физиологические и другие изменения, претерпеваемые живыми существа-
ми с течением времени. По прошествии миллионов лет в результате этих изменений,
а также изменений окружающей среды одни виды выживают, другие — вымирают.
Известно, что изменения живых существ вызваны определенными биологическими
механизмами. Среди всех теорий, известных на сегодняшний день, наибольший успех
имела теория естественного отбора Чарльза Дарвина, представленная им в 1859 году,
в расцвет викторианской эпохи, в книге «Происхождение видов».
THE OHIGIN OF SPECIES
ВУ MEtSS OF MiTIKAl. ЯЯ I K A,
l n 4 ,jhv.... • « r THE STBUC'iLE
Ю* СИАПШ* lUKWIK M.A
ЪОИТЮЬ
Л1НЧ Ml ALBEMARLE tfBKKT
rfc f **< Г Л
На этой фотографии Чарльз Дарвин изображен в возрасте 51 года, вскоре
после публикации своего революционного труда «Происхождение видов».
14
МАТЕМАТИЧЕСКАЯ БИОЛОГИЯ В ИСТОРИЧЕСКОЙ ПЕРСПЕКТИВЕ
Согласно теории Дарвина, живые существа, будь то растения, животные или
микроорганизмы, представляют собой различные решения задачи адаптации к окру-
жающей среде. Под окружающей средой понимаются различные условия существо-
вания, начиная от океанов или озер и заканчивая наземно-воздушной средой. При
этом в каждой отдельно взятой среде наблюдается большое разнообразие живых
существ: например, джунгли, дубовый лес или пустыня очень отличаются между
собой. Согласно Дарвину, чем лучше «решение», которое представляет собой живой
организм, точнее биологический вид, тем лучше он приспособлен. А чем выше при-
способленность организма, тем больше его шансы на выживание и, следовательно,
на достижение репродуктивного возраста. Репродукция, по Дарвину, является на-
градой: если организму удалось размножиться, гены счастливчика будут переданы
следующему поколению.
Но как живые организмы находят новые решения в изменяющейся или неблаго-
приятной среде? Ответить на этот вопрос помогает генетика. За поиск новых решений
отвечают механизмы, случайным образом меняющие генетический код, — мутации.
Чем выше изменение генов в определенных пределах, тем лучше для вида: его пред-
ставители получают большой набор возможных «решений», который поможет им
адаптироваться к будущим изменениям окружающей среды. По Дарвину, окружа-
ющая среда отбирает виды, наиболее пригодные для обитания в ней.
Развитие математических методов теории эволюции
В второй половине XIX века, после публикации книги Дарвина, в Великобритании
возникла английская биометрическая школа, к которой принадлежали такие видные
ученые, как Фрэнсис Гальтон и Карл Пирсон. Представители этой школы впервые
применили в биологии методы статистики. Позднее, в 1930 году, Рональд Эйлмер
Фишер, внесший огромный вклад в развитие биоматематики и биостатистики, сфор-
мулировал основную теорему естественного отбора, в которой дарвиновская теория
эволюции путем естественного отбора объясняется на языке математики.
Согласно Фишеру, при определенных условиях и за определенное время / ритм или
скорость, с которой повышается средняя приспособленность конкретного вида, равна
разнообразию возможных значений генов. Обозначим средний рост приспособлен-
ности через ДИ/, среднюю приспособленность — через W, множество возможных
15
МАТЕМАТИЧЕСКАЯ БИОЛОГИЯ В ИСТОРИЧЕСКОЙ ПЕРСПЕКТИВЕ
значений генов (генных вариаций) — через (у^, и получим обычную запись теоремы
Фишера в математической биологии:
W
Эта теорема — прекрасный пример того, сколь важную роль сыграла математи-
ка в последующем развитии биологии. Фишеру всего в одной формуле удалось точ-
но выразить описанные выше идеи. В итоге биологические задачи начиная с 1930-х
годов начали выражаться на языке математики, и развитие количественных методов
биологии было уже не остановить. Еще одним важным событием для математиче-
ской биологии стала модель, известная как модель «хищник — жертва» Дотки —
Вольтерры (ее предложил Альфред Джеймс Лотка в 1925 году и Вито Вольтерра
годом позже). Это одна из самых ярких математических моделей математической
биологии и одна из самых популярных моделей в экологии. Мы подробнее расска-
жем о ней в главе 6.
Роль компьютера в математическом анализе жизни
По окончании Второй мировой войны в Великобритании и США появились пер-
вые компьютеры. Два союзных государства начали борьбу за право называться их
родиной, и толчком к началу этого соперничества стала возможность использования
компьютеров прежде всего в военных целях. Новая техника создавалась для борьбы
с общим врагом — СССР. Напомним, что именно эти годы стали началом эпохи
холодной войны, и изменение политической обстановки повлияло на работы ученых
во всем мире.
Хотя историки науки считают, что первый компьютер, известный как ENIAC
(Electronic Numerical Integrator and Computer — «электронный числовой интегратор
и вычислитель»), был сконструирован в США в 1946 году, сегодня мы знаем, что
до него существовал «Колосс», созданный в Великобритании в 1944 году.
В 1950 году Алан Тьюринг, один из самых плодовитых британских уче-
ных XX столетия, сконструировал компьютер АСЕ (сокр. от англ. Automatic
Computing Engine — «автоматическая вычислительная машина») в Национальной
физической лаборатории. Этот компьютер имел возможности хранения данных и ра-
боты программ, весьма схожие с возможностями первых компьютеров Macintosh,
16
МАТЕМАТИЧЕСКАЯ БИОЛОГИЯ В ИСТОРИЧЕСКОЙ ПЕРСПЕКТИВЕ
«Колосс» — первый компьютер в истории, построенный в Великобритании в 1944 году.
созданных только в 1980-е годы. Компьютер Тьюринга был британским конкурентом
американского EDVAC (от Electronic Discrete Variable Automatic Computer — «уни-
версальный автоматический компьютер с дискретными переменными»), созданного
на базе ENIAC. В конструировании EDVAC участвовал еще один гениальный уче-
ный того времени — Джон фон Нейман.
Британским ответом на EDVAC стал EDSAC (Electronic Delay Storage Automatic
Computer — «электронный автоматический вычислитель с памятью на линиях за-
держки») — еще один компьютер с похожими характеристиками. В это же время
в США был сконструирован UNIVAC (Universal Automatic Computer — «универ-
сальный автоматический компьютер») — прямой потомок ENIAC и EDVAC. Из-
готовившая его компания Remington Road стала первым в мире производителем
коммерческих компьютеров.
Открытия Алана Тьюринга
В 1948 году в Университете Манчестера находился один из самых мощных компью-
теров того времени, а в 1951 году университет получил компьютер Ferranti Mark I,
на котором работал Тьюринг. С 1952 года до своей смерти в 1954 году Тьюринг был
одним из первых ученых, кто использовал компьютер для математического модели-
рования биологических задач.
17
МАТЕМАТИЧЕСКАЯ БИОЛОГИЯ В ИСТОРИЧЕСКОЙ ПЕРСПЕКТИВЕ
Компьютер Ferranti Mark I, на котором работал Алан Тьюринг (на фото справа, стоит).
В то время Тьюринга очень интересовало математическое изучение морфогенеза.
Одна из самых любопытных задач этой дисциплины заключается в том, чтобы объ-
яснить, как живые организмы обретают конечную форму: почему ветви деревьев об-
разуют именно такую структуру, почему членистоногие словно состоят из отдельных
кусочков, а кольчатые черви — из колец. Еще одна классическая задача морфогенеза
заключается в изучении узоров, например на коже некоторых позвоночных — по-
лосок у зебр или круглых пятен у далматинцев.
Тьюринг первым попытался решить биологические задачи с помощью компьюте-
ра, став одним из пионеров вычислительной биоматематики. Таким образом, его ис-
следования придали этой дисциплине более прикладной характер, сблизив ее с при-
вычными биологическими исследованиями в лаборатории. Биологи и другие ученые
под влиянием работ Тьюринга также начали изучать жизнь с математической точки
зрения. Подобные исследования проводились в разные годы XX века; проводятся
они и сейчас. Кроме того, Тьюринг открыл новую область математической биологии,
предложив первую математическую теорию морфогенеза. В одной из своих работ
для анализа формы растений он использовал числа Фибоначчи.
Последовательность Фибоначчи 0, 1, 1, 2, 3, 5, 8, 13, 21, 34, 55, 89, 144 и т.д.
образуется по следующему правилу: если принять первое число Фибоначчи, рав-
ным 0, второе число, а2, равным 1, то каждое последующее число будет определяться
18
МАТЕМАТИЧЕСКАЯ БИОЛОГИЯ В ИСТОРИЧЕСКОЙ ПЕРСПЕКТИВЕ
как сумма двух предыдущих. Иными словами, ап = ап _ + ап _ 2. Любопытно, что
числа Фибоначчи описывают количество лепестков цветов, расположение чешуек
шишек и листьев растений.
Число спиралей на этой шишке в каждом направлении (8 и 13 соответственно)
выражается последовательными числами Фибоначчи.
Эта особенность растений получила название филлотаксиса Фибоначчи. Так, чис-
ла Фибоначчи описывают расположение листьев растений, при котором их освещен-
ность будет оптимальной. Представьте, что лист соперничает с соседними за доступ
к солнечному свету. Каким будет оптимальное расположение листьев, обеспечива-
ющее наибольшую освещенность каждого листа? Ответ дает последовательность
Фибоначчи.
Продолжив исследования, Тьюринг совершил свое самое знаменитое открытие
в этой области — он создал математическую модель «реакция — диффузия». Свои
идеи ученый изложил в статье «Химическая основа морфогенеза», опубликованной
в престижном научном журнале Лондонского королевского общества в 1952 году.
Тьюринг был математиком, а не биологом, поэтому он попытался объяснить интересо-
вавшее его явление с помощью дифференциальных уравнений. Он задался вопросом:
каким образом в однородной ткани клеток, в зачаточном состоянии очень похожих
друг на друга, например клеток кожи позвоночных, образуются полоски или пятна?
С биологической точки зрения эти полоски или пятна — проявление различий между
пигментными и непигментными клетками. Как следствие, полоски на шкуре зебры
будут результатом нарушения изначального единообразия зародышевых клеток кожи.
19
МАТЕМАТИЧЕСКАЯ БИОЛОГИЯ В ИСТОРИЧЕСКОЙ ПЕРСПЕКТИВЕ
Тьюринга интересовал биологический механизм, ведущий к появлению подобных
узоров. Ученый предполагал, что полученный узор представляет собой нестабильное
состояние, поскольку стабильным состоянием является единообразие зародышевых
клеток без характерного узора. С помощью компьютера Ferranti Mark I Тьюринг про-
вел ряд экспериментов по моделированию и доказал, что полученный узор на коже
зависит от значений параметров математической модели.
Полоски на шкуре зебры — один из примеров, описываемых уравнениями
«реакция — диффузия» Тьюринга.
Параметр математической модели — это значение, соответствующее какому-ли-
бо свойству, которое нельзя оценить напрямую, в ходе наблюдений. Тьюринг выявил
несколько закономерностей, очень похожих на те, что описывают распределение
щупалец гидры или расположение лепестков цветка. Предположив, что клетки име-
ют круглую форму, Тьюринг смоделировал многоклеточный зародыш — бластулу.
Бластула — один из этапов развития зародыша, на котором уже можно заметить
появление узоров. Тьюринг изучил зародыши амфибий и ежей, которые сегодня бла-
годаря своим особым свойствам широко используются в качестве моделей при изу-
чении морфогенеза. Ученый предположил, что узоры образуются в результате про-
20
МАТЕМАТИЧЕСКАЯ БИОЛОГИЯ В ИСТОРИЧЕСКОЙ ПЕРСПЕКТИВЕ
цессов реакции — диффузии. Согласно его гипотезе, в зародышевой ткани, то есть
в группе клеток, сгруппированных на плоскости, будут присутствовать пигментные
клетки, продуцирующие вещество морфоген. Как только молекулы этого загадоч-
ного вещества распространятся в результате диффузии по зародышевой ткани, они
вступают между собой в реакцию. Распределение продуктов этой химической реак-
ции определяет так называемое поле концентраций — отпечаток, согласно которому
и формируется узор зародышевых клеток. Следовательно, полоски, пятна и любые
другие узоры, которые мы можем увидеть на шкуре животных, есть не более чем
реплики поля концентраций. Мы не будем рассматривать знаменитые уравнения ре-
акции — диффузии Тьюринга во всех подробностях, а только приведем их:
—= f(M ,М ) + D ^2М ,
ЗА/,
—J-=S(MA,Ml)+Dy2Mr
ot
Эти выражения объясняют, как с течением времени изменяется объем или кон-
центрация двух веществ, предложенных Тьюрингом, которые он назвал морфогеном-
активатором (МА) и морфогеном-ингибитором (М/). Как мы уже отмечали, эти два
вещества производятся только пигментными клетками. В свою очередь, f(MA, М{)
и g(MA, — две функции, обозначающие реакцию между активатором и ингиби-
тором, а выражения DA\72MA и /J; V2A/; указывают, как эти два класса морфогенов
распространяются по ткани. Так, когда морфогены высвобождаются пигментными
клетками, начинается процесс их диффузии, подобный диффузии песчинок сахара
в стакане с водой. По Тьюрингу, морфоген-активатор стимулирует воспроизвод-
ство себя самого и морфогена-ингибитора. Еще одна любопытная особенность этой
реакции заключается в том, что морфоген-ингибитор распространяется на большее
расстояние, чем морфоген-активатор. Расстояния, на которые распространяются мор-
фогены, зависят от Da — коэффициентов диффузии морфогенов — активатора
и ингибитора соответственно.
В 1954 году, в возрасте 41 года, Алан Тьюринг покончил с собой. Так оборвалась
жизнь одного из величайших ученых XX века. Его гениальность доказывает и тот
факт, что химические вещества, существование которых он предсказал математи-
чески (так называемые морфогены), были открыты экспериментально лишь много
21
МАТЕМАТИЧЕСКАЯ БИОЛОГИЯ В ИСТОРИЧЕСКОЙ ПЕРСПЕКТИВЕ
лет спустя, в начале 1990-х. Кроме того, некоторые узоры из изученных Тьюрингом
на компьютере Ferranti Mark I были обнаружены на чешуе рыбы полукруглый ангел,
или Pomacanthus semicirculatus. В настоящее время морфогенез — одна из областей
математической биологии, и удивительным путем, на который первым вступил Алан
Тьюринг, проследовали такие видные ученые, как Мюррей, Мейнхардт и другие.
ЖИЗНЬ - ЭТО ИНФОРМАЦИЯ
За год до кончины Тьюринга, в 1953 году, Уотсон и Крик предложили спиралевидную модель
ДНК. Ранее Джон фон Нейман и Алан Тьюринг, предвосхитив создание этой модели, писали:
«Жизнь - это информация». Тем не менее модель ДНК, которая сегодня принимается всеми
учеными, в свое время произвела фурор. Ее цепочка образована четырьмя азотистыми основа-
ниями, которыми кодируются гены: А - аденин, Т - тимин, Г - гуанин и Ц - цитозин.
Параллельно с этим произошло еще одно важное событие - появилась информатика как наука.
В компьютерах используется двоичная система счисления, и это означает, что вся информация
кодируется последовательностями, состоящими всего из двух цифр, 0 и 1. Как следствие, ком-
пьютер - это машина, с помощью которой можно естественным образом исследовать жизнь, от-
крывать ее элементы, проникать в тайны тончайших
ее механизмов и делать прогнозы. С момента соз-
дания компьютер стал инструментом, позволившим
установить тесную взаимосвязь между математикой
и биологией. Со временем вычислительный подход,
основанный Тьюрингом, не только способствовал
укреплению этой взаимосвязи, но и привел к слия-
нию биологии и математики в новую дисциплину -
математическую биологию.
Молекула ДНК, описанная Уотсоном
и Криком в 1953 году.
22
МАТЕМАТИЧЕСКАЯ БИОЛОГИЯ В ИСТОРИЧЕСКОЙ ПЕРСПЕКТИВЕ
Зарождение биологии систем
Начиная с 1950—1960-х годов в математических исследованиях живых существ
и жизни в целом, проводимых с помощью компьютеров, предполагалось, что рас-
тения, животные и микроорганизмы находятся в так называемом стационарном со-
стоянии, и эта стабильность возможна благодаря механизмам саморегуляции, или
гомеостаза. Чтобы поддерживать саморегуляцию, живым существам требовалось
тратить большое количество энергии. Важность гомеостаза в биологии привлекла вни-
мание ученых уже в 1940-х годах благодаря передовым исследованиям британского
ученого Уильяма Росса Эшби. К примеру, организм человека естественным образом
стремится к содержанию в крови определенного количества глюкозы. При ее избытке
поджелудочная железа вырабатывает инсулин, при недостатке — глюкагон. Иными
словами, для сохранения стабильности телу нужно постоянно работать.
При изучении жизни с математической точки зрения по возможности предпола-
гается, что изучаемое явление имеет так называемое линейное поведение. Линейные
системы изучать проще всего, так как их общее состояние или поведение на мате-
матическом языке описывается как сумма состояний или поведений частей такой
системы. Представим себе примитивное живое существо (назовем его z), настолько
простое, что оно имеет всего два органа — х и у. Если мы обозначим физиологиче-
ские состояния х и у через f(x) и /(у), то жизненное состояние организма /(z) будет
равно сумме состояний его органов: f(x) + /(у). В стационарном состоянии произ-
водная /(z) будет равна 0. Иными словами, математическая функция, описывающая
жизненное состояние организма, не будет ни возрастать, ни убывать.
Линейные системы
проще всего изучать
с математической
точки зрения.
23
МАТЕМАТИЧЕСКАЯ БИОЛОГИЯ В ИСТОРИЧЕСКОЙ ПЕРСПЕКТИВЕ
Математическое изучение линейных систем связано с комплексным и органици-
стическим представлением о жизни Карла Людвига фон Берталанфи. Это пред-
ставление, которое имеет отношение не только к биологии, но и к другим дисци-
плинам, описано в статье, опубликованной в 1968 году под названием «Общая те-
ория систем: основы, развитие, применение» (General System Theory: Foundations,
Development, Applications). По сути, эта теория оказала огромное влияние на то, как
ученые используют компьютер для моделирования, то есть воссоздания, описания
и прогнозирования столь разных явлений, как климат, метаболизм, жизнь клеток или
поведение финансовых рынков. Система — это множество реально существующих
объектов (частей или элементов системы) и абстрактных переменных, атрибутов,
свойств и, что более важно, связей и взаимоотношений между этими элементами.
Важный момент теории систем фон Берталанфи заключается в том, что части
системы взаимодействуют между собой, а сами системы являются незамкнутыми
и взаимодействуют с окружающей средой. При этом из среды в систему поступает
входная информация, результатом обработки или преобразования которой является
ответ системы, или выходная информация, поступающая обратно в среду. Такие по-
нятия, как саморегулирование и обратная связь, баланс и гомеостаз, в этой модели
возникают естественным образом.
Глобальное видение природы, в которой система рассматривается как «всё», из-
вестно как холизм. В XX веке холизм оказал огромное влияние на то, как мы видим
мир. Это влияние проявилось не только в биологии, но и в социологии, экономике,
химии и даже лингвистике. Холизм повлиял и на способы применения математики
для изучения реального мира. В экологии он был введен школой североамерикан-
ских экологов во главе с Говардом Одумом. В 1950-е годы Одум радикально изме-
нил методы изучения всех проблем, связанных с окружающей средой, что вызвало
появление системной биологии. В рамках этой дисциплины ученые рассматривают
любое биологическое явление с холистической точки зрения и описывают событие
посредством математической модели. К примеру, одна из классических моделей этой
дисциплины — первая модель органа, созданная с помощью компьютера, а именно
модель сердца, представленная Денисом Ноблом в 1960 году в журнале Nature. Этот
британский исследователь сыграл важную роль в международном проекте Physiome,
начатом в 1990-е годы, целью которого была расшифровка генома — совокупности
24
МАТЕМАТИЧЕСКАЯ БИОЛОГИЯ В ИСТОРИЧЕСКОЙ ПЕРСПЕКТИВЕ
генов организма. Расшифровка производилась с помощью компьютерного модели-
рования с использованием математических моделей физиологии.
Fatwito* Safeя ’
' Пучком*#m aibro human — Я ’ J l|l ” Pijm»» StinAd’ Hanwnianta* -
PbyslemBjp
| Home
insiiicoML Physiome.jp -toward in silico human
InslllcoIDE Phystomejp has been estebhshed to provide building Mocks useful to devu^p m atitco human The blocks wif include
dowr*>ML matoemabcal models and experimental data representing physwtogicai functions
РПузюте jp is e part of toe WgrtdwiQe Intogrut^ Bmmedicnj Research Cooperation to promote Physome and Systems Butogy
F*Q The building blocks (modules models, biotogicel data) representing biologic functions and structure are detabased and served
I n Bill co D В as elements in toe catalogue of human knowledge They can be reused for deeper understanding of human physwtogy
M aventualy contributing to estaMshment of in sihco medicine end predictive medicine
Platform Overview
Our devetoping ptatfcrm is composed of
three pieces
Ans/teoMl an XML based description
language for mathemeticaf models of
physiological funebons
/лей^оЮЕ an Integrated Development
Environment which can be used as a
viewer, an editor and a simulator of ISML
models
/пайИс oDB a database to store ISML
models
Веб-страница одного из множества учреждений,
связанных с проектом Physiome в сфере системной биологии.
Одной из особенностей проекта была интеграция разных уровней, начиная
от биохимии и отдельных клеток и заканчивая целыми органами. Любопытная черта
системной биологии заключается в том, что в этой дисциплине проекты реализуются
междисциплинарными рабочими группами с участием биологов, физиков, математи-
ков, информатиков и других специалистов. Противоположным подходом является
редукционизм, который довольно долго применялся в биологии под влиянием мно-
гочисленных успехов молекулярной биологии. Прогресс в этой дисциплине привел
к тому, что математическая биология на некоторое время ушла в тень, как и любые
попытки «заняться математикой жизни». И все же накопление экспериментальных
данных молекулярной биологии, а также удивительные успехи в изучении генов,
белков и метаболизма во второй половине XX века привели к появлению геноми-
ки, протеомики и метаболомики — трех новых дисциплин, которые быстро начали
набирать популярность во всем мире. Это заставило вновь вспомнить о системной
25
МАТЕМАТИЧЕСКАЯ БИОЛОГИЯ В ИСТОРИЧЕСКОЙ ПЕРСПЕКТИВЕ
биологии, а вместе с ней — и об изучении жизни количественными методами. Си-
стемная биология вновь вошла в моду лишь в конце XX столетия, и одновременно
с этим вновь пробудился интерес к математической биологии.
1970-е - время перемен
В 1970-х годах ученые начали принципиально иначе рассматривать биологические
явления, изменилась и «математика жизни». Решающее влияние на этот процесс
оказали идеи Ильи Романовича Пригожина, лауреата Нобелевской премии по химии
1977 года. Согласно его теории диссипативных структур, системы, которые непре-
рывно обмениваются материей и энергией с окружающей средой (к ним относятся
сложные химические реакции или ураганы), функционируют благодаря тому, что
далеки от равновесного состояния. Одной из характеристик диссипативных систем
является образование сложных структур, которые порой кажутся хаотичными. Эта
особенность привлекла внимание ученых, вновь пересмотревших решения классиче-
ских задач биологии. Биоматематики вернулись к давно известным проблемам, ин-
терпретировав их в соответствии с теориями Пригожина. В качестве примера можно
привести узоры, изученные Тьюрингом. По мнению ученого, однородная ткань, со-
стоящая из очень похожих друг на друга зародышевых клеток, например клеток кожи
позвоночных, находится в равновесном состоянии. Но как только между клетками
начинают возникать отличия, на шкуре животного проявляется узор из полосок или
пятен. Сохранение этого узора в течение всей жизни животного Тьюринг и Приго-
жин трактовали как ситуацию, далекую от равновесного состояния. Как следствие,
уравнения реакции — диффузии стали одним из основных формальных инструментов,
которые позволили биоматематикам изучить некоторые диссипативные системы, на-
пример уже упомянутые узоры на шкуре некоторых позвоночных.
Бельгийская марка, выпущенная
в честь Ильи Пригожина (1917-
2003) за два года до смерти
этого выдающегося русского
ученого.
26
МАТЕМАТИЧЕСКАЯ БИОЛОГИЯ В ИСТОРИЧЕСКОЙ ПЕРСПЕКТИВЕ
Еще одной характеристикой систем, далеких от равновесного состояния, являются
их колебания. В качестве примера приведем знаменитые уравнения «хищник — жерт-
ва» Лотки — Вольтерры. К сожалению, не существует универсальных принципов,
управляющих формированием описанных узоров в диссипативных системах. Однако
если система находится в равновесии, образования узоров не происходит. К при-
меру, трехмерное представление белка всегда остается неизменным. Почему? Ответ
прост: белок находится в наиболее стабильном состоянии, требующем минимальных
энергозатрат. Еще один пример системы, находящейся в равновесном состоянии, —
химическая реакция:
А + В С + D.
Если вещества А и В преобразуются в С и D с той же скоростью, что С и D пре-
образуются обратно в Л и В, то реакция находится в равновесном состоянии. Пред-
положим, что равновесие оказалось нарушено. Если скорость, с которой вещества
А и В преобразуются в С и D, не равна скорости протекания обратного процесса,
реакция будет находиться в неравновесном состоянии. Общих правил, описывающих
неравновесные, диссипативные системы, не существует, как и общего математиче-
ского метода их изучения, поэтому используется компьютерное моделирование —
особенно полезное с учетом того, что в жизни встречается множество примеров дис-
сипативных систем. Описанные выше идеи постепенно сформировали современное
видение биологии и, как следствие, способствовали ее математической формализации.
Современная математическая биология
Изучение систем, находящихся в неравновесном состоянии, и поиск вычислительных
методов, позволяющих смоделировать подобные системы, стали популярны в 1980-е
и 1990-е годы при изучении нелинейных систем, то есть систем, поведение которых
нельзя представить как сумму поведений их частей. Основная причина этого в том,
что части нелинейных систем взаимодействуют друг с другом. Вновь рассмотрим
примитивный живой организм z и предположим, что он имеет всего два органа — х
и у. Если поведение этого организма нелинейное, то жизненное состояние организма
/(2) будет равно, к примеру, произведению, а не сумме состояний его органов /(х)
и /(у). В качестве примера из повседневной жизни можно привести прием лекарств.
27
МАТЕМАТИЧЕСКАЯ БИОЛОГИЯ В ИСТОРИЧЕСКОЙ ПЕРСПЕКТИВЕ
Если вы примете два лекарства или более, их совокупный терапевтический эффект
не будет равен сумме эффектов отдельных медикаментов. Как правило, они вступают
в реакцию между собой, причем часто во вред организму.
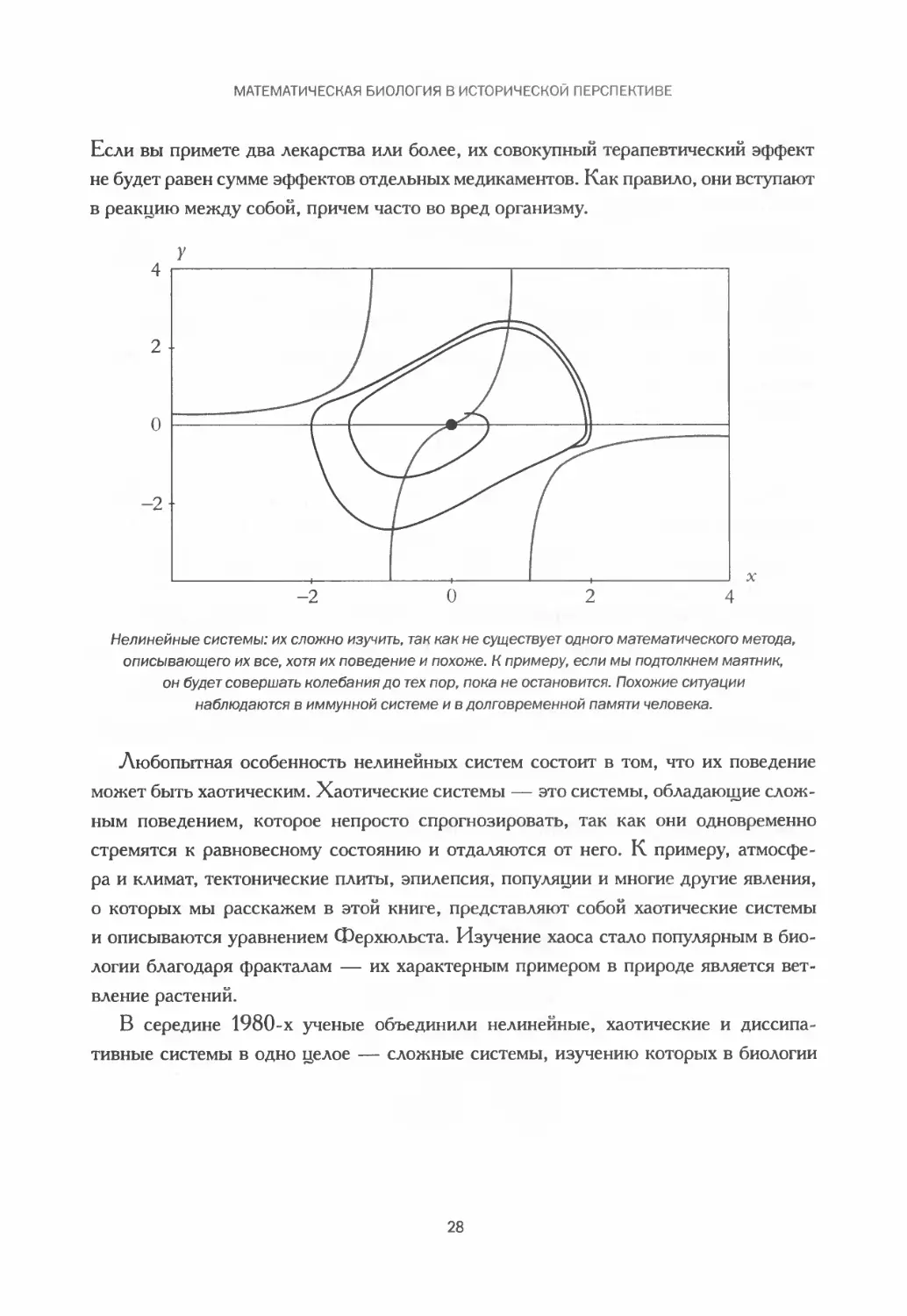
Нелинейные системы: их сложно изучить, так как не существует одного математического метода,
описывающего их все, хотя их поведение и похоже. К примеру, если мы подтолкнем маятник,
он будет совершать колебания до тех пор, пока не остановится. Похожие ситуации
наблюдаются в иммунной системе и в долговременной памяти человека.
Любопытная особенность нелинейных систем состоит в том, что их поведение
может быть хаотическим. Хаотические системы — это системы, обладающие слож-
ным поведением, которое непросто спрогнозировать, так как они одновременно
стремятся к равновесному состоянию и отдаляются от него. К примеру, атмосфе-
ра и климат, тектонические плиты, эпилепсия, популяции и многие другие явления,
о которых мы расскажем в этой книге, представляют собой хаотические системы
и описываются уравнением Ферхюльста. Изучение хаоса стало популярным в био-
логии благодаря фракталам — их характерным примером в природе является вет-
вление растений.
В середине 1980-х ученые объединили нелинейные, хаотические и диссипа-
тивные системы в одно целое — сложные системы, изучению которых в биологии
28
МАТЕМАТИЧЕСКАЯ БИОЛОГИЯ В ИСТОРИЧЕСКОЙ ПЕРСПЕКТИВЕ
уделяется наибольшее внимание. К таким системам относятся, например, муравей-
ники, мозг, иммунная система, клетка, морфогенез или экосистемы. В некоторых
случаях сложные системы изучаются с применением стандартных методов матема-
тической биологии. Однако некоторые системы настолько сложны, что изучить их
можно только альтернативными компьютерными методами, позволяющими найти
лишь приближенные решения. Такие методы называются эвристическими. К при-
меру, в настоящее время метод клеточных автоматов является одной из альтернатив
моделированию сложных систем, для которых неизвестны описывающие их диф-
ференциальные уравнения. Классический пример клеточного автомата — колония
муравьев. В некоторых случаях, несмотря на то что дифференциальные уравнения,
описывающие систему, известны (например, в случае с пятнами на коже позвоноч-
ных), поведение системы быстрее и удобнее смоделировать с помощью клеточных
автоматов. Кроме того, клеточные автоматы позволяют наглядно изобразить узоры,
к примеру полоски зебры, что при использовании дифференциальных уравнений
невозможно. Еще одним примером служит клеточный автомат Ba-Тор, описываю-
щий модель «хищник — жертва» Дотки — Вольтерры.
В этой главе мы коротко обрисовали основные этапы развития математической
биологии. Обратите внимание, что не только зарождение, но и последующее раз-
витие этой дисциплины неизменно находилось под большим влиянием преобладав-
ших на тот момент физических интерпретаций жизни. Более того, математическая
биология — это дисциплина, которая способствовала тщательному анализу биоло-
гических явлений и экспериментальных данных. Сегодня одним из самых важных
достижений математической биологии являются математические модели, позволяю-
щие проводить с помощью компьютера сложные эксперименты.
29
МАТЕМАТИЧЕСКАЯ БИОЛОГИЯ В ИСТОРИЧЕСКОЙ ПЕРСПЕКТИВЕ
Математические модели в биологии
Человек всегда испытывал потребность понимать, контролировать и предсказывать
поведение всего сущего. Для этого ученые всех времен и народов создавали модели
окружающего мира, то есть представления или абстракции некоторой системы или
явления.
Модель обладает несколькими полезными свойствами. С одной стороны, она
позволяет понять и объяснить то или иное явление — в качестве примера можно
привести модель клеточного цикла или метаболизма глюкозы. С другой стороны,
что особенно важно, она позволяет предсказать состояние или поведение изучаемой
системы в будущем: это может быть прогнозирование климата или описание какой-
либо гипотетической ситуации, например воздействия аварии на атомной электро-
станции на флору и фауну региона.
Также компьютерное моделирование позволяет ученым проверить те или иные
гипотезы. К примеру, можно провести эксперимент, опровергающий гипотезу о про-
исхождении жизни или позволяющий рассмотреть механизм эволюции конкретно-
го вида. Модель может использоваться и для того, чтобы вдохновить, например,
группу инженеров на поиски решения задачи. В любом случае построение моделей
очень важно как в силу их практической ценности, так и из-за того, что моделиро-
вание — единственный способ, который позволяет постепенно выстроить картину
окружающего мира.
В биологии, как и в других науках, наиболее полезны математические модели:
они в абстрактной форме представляют систему или явление с использованием языка
и формальных средств математики. К примеру, в модели клетки, сердца или экосисте-
мы составные части объекта и взаимодействие между ними представлены математиче-
скими выражениями. Эти выражения связывают множество входных переменных
/2, / и выходную переменную О. Входные переменные обозначают величины,
которые можно наблюдать (и измерить) в ходе эксперимента. Обычно одна из этих
переменных — время, I. Она обозначает момент времени, в который были получе-
30
МАТЕМАТИЧЕСКАЯ БИОЛОГИЯ В ИСТОРИЧЕСКОЙ ПЕРСПЕКТИВЕ
ны входные значения /t(f), /2(f), •••» ^п(0- Как только эти значения определяются
экспериментально или любым другим способом (например, на основе каких-либо
теоретических предпосылок), они вводятся в модель. Используя математические
выражения модели, ученый определяет значение выходной переменной 0(f). которое
отражает какое-либо свойство системы. Обычно этим свойством является состояние
или поведение системы в определенный момент времени t.
В математических выражениях используются параметры. В отличие от входных
и выходных переменных, они обозначают величины, которые нельзя наблюдать в ходе
эксперимента напрямую, например уровень рождаемости, константа распада, ско-
рость биохимической реакции и т. д. Как следствие, значения параметров устанав-
ливаются в лаборатории или при полевых исследованиях.
Для определения приближенного значения параметра используются сложные
статистические методы. Однако иногда это значение уже известно: его можно найти
в таблицах, опубликованных другими исследователями. В качестве примера можно
привести калорийность продуктов в модели, связанной с диетами. Другие извест-
ные параметры — это сезонный уровень заболеваемости гриппом или время роста
культуры бактерий. Параметры связывают входные переменные /2(f), ..., In(t)
с выходной переменной 0(f) посредством выражений математической модели.
Математическая модель, входные переменные (I) и выходная переменная (О).
Компьютер как пробирка
Моделирование — одно из основных понятий современной науки — заключается
в прогнозировании будущего состояния системы, 0(f + 1), на основе определенной
31
МАТЕМАТИЧЕСКАЯ БИОЛОГИЯ В ИСТОРИЧЕСКОЙ ПЕРСПЕКТИВЕ
вычислительной модели. К примеру, прогноз погоды на ближайшие дни основан
на вычислительной модели климата, прогнозирование численности волков и зайцев
в определенном регионе производится на основе модели «хищник — жертва», а число
людей, которые заболеют сезонным гриппом, можно спрогнозировать с помощью
вычислительной модели эпидемии гриппа. Таким образом, для составления прогнозов
требуется вычислительная модель.
В общем случае такая модель — это компьютерная программа, написанная на од-
ном из языков программирования (Visual Basic, C/C++, Java и т. д.). Моделирование
заключается в том, чтобы заставить математическую модель работать на компьютере
в поисках ответа на вопросы, касающиеся будущего состояния системы: «что про-
изойдет, если...?». Таким образом, компьютер превращается в пробирку, подлинную
лабораторию, где можно исследовать явления, которые нельзя изучить при полевых
исследованиях или в лаборатории.
Существует несколько способов компьютерного моделирования. Во-первых,
оно может заключаться в определении начальных условий и будущего состояния
системы. Начальные условия — это значения входных переменных модели (они
известны), на основе которых выполняется прогноз. Ученые называют отправную
точку модели нулевым моментом времени, поэтому начальные условия записывают-
ся так: //0), /2(0)..., /п(0). К примеру, если на сегодняшний день свиным гриппом
заболели 1247 человек, из которых 1240 выжили, семь — умерли, то начальные
условия таковы: /t(0) = 1247, /2(0) = 1240 и /3(0) = 7. Зная эти начальные условия
и применив вычислительную модель эпидемии, можно задаться вопросом: сколько
человек заболеют гриппом через семь дней?
Во-вторых, моделирование может заключаться в изменении параметров и оценке
воздействия новых значений на будущее состояние системы. Что произойдет в при-
мере со свиным гриппом, если вместо уровня смертности в 0,78 % использовать
значение в 2,96 %? Каким в этом случае будет уровень смертности через месяц?
В-третьих, моделирование может заключаться в определении будущего состояния
системы при заданных начальных условиях и некоторых значениях определенных
параметров.
32
МАТЕМАТИЧЕСКАЯ БИОЛОГИЯ В ИСТОРИЧЕСКОЙ ПЕРСПЕКТИВЕ
СРАВНЕНИЕ МОДЕЛЕЙ
В некоторых ситуациях моделирование может состоять в прогнозировании явления путем сравне-
ния прогнозов, полученных с помощью различных вычислительных моделей. Такая ситуация может
сложиться, когда одно явление описывается несколькими математическими моделями. К при-
меру, можно сравнить различные математические модели климата для одной и той же ситуации,
смоделировать поведение колонии муравьев с помощью разных вычислительных моделей или
определить число хищников и жертв, сравнив данные, полученные с использованием клеточных
автоматов, с данными, полученными по уравнениям Лотки - Вольтерры.
Увеличение объема метана
в земной коре и стратосфере
согласно вычислительной
модели в сравнении с другими
моделями, описывающими
это же явление.
Программы для символьных вычислений
Программы для символьных вычислений позволяют обрабатывать математические
выражения в символьном виде. Подобные программы появились в 1960-е и стали
первым коммерческим продуктом, в котором использовался искусственный интел-
лект. Первыми пользователями этих программ стали физики, со временем к ним
присоединились и другие ученые. На заре эпохи символьных вычислений родились
такие программы, как Schoonschip и MathLab, однако лишь с развитием muMath,
Reduce, Macsyma и Derive программы для символьных вычислений обрели популяр-
ность в научных кругах. Сегодня эти приложения используются в университетах,
учебных центрах, а также при реализации научных и инженерных проектов. Самыми
33
МАТЕМАТИЧЕСКАЯ БИОЛОГИЯ В ИСТОРИЧЕСКОЙ ПЕРСПЕКТИВЕ
популярными коммерческими программами для символьных вычислений являются
Maple и Mathematica, а также бесплатные SciLab и Octave.
SciLab — бесплатная программа для научных расчетов и символьных вычислений.
Эти приложения незаменимы в математической биологии — при изучении ди-
намических систем в экологии, эпидемиологии, фармакологии и т. д. Программы
для символьных вычислений не только позволяют редактировать и исправлять вы-
ражения, но и содержат много других возможностей: с их помощью можно строить
графики в двух и трех измерениях, использовать внешние программы или библи-
отеки процедур, имеющих различное применение в вычислительной химии и т.д.
В них используются методы эволюционных вычислений, методы биоинформатики,
34
МАТЕМАТИЧЕСКАЯ БИОЛОГИЯ В ИСТОРИЧЕСКОЙ ПЕРСПЕКТИВЕ
статистические методы, дифференциальные уравнения и многое другое. Среди за-
дач, решаемых с помощью программ символьных вычислений, выделяется упроще-
ние выражений, разложение в ряд Тейлора, разложение многочленов на множители,
вычисление пределов, производных и интегралов, выполнение операций с матрица-
ми и векторами.
С помощью языков программирования пользователи могут создавать приложе-
ния с собственными «рецептами» вычислений. Использование программ символь-
ных вычислений для решения определенного класса задач биологии привело к тому,
что в математической биологии появился новый раздел — алгебраическая биология.
Эта дисциплина возникла в 2005 году, ее цель — создание моделей, объясняющих
биологические явления, при этом большую важность имеет биологическая задача,
а не математические преобразования символьных выражений. В настоящее время
алгебраическая биология считается частью биологии систем и используется для ана-
лиза и моделирования биологических систем при создании моделей биохимических
реакций, регулировании экспрессии генов, а также для решения различных задач
клеточной и молекулярной биологии.
Также алгебраическая биология применяется в эпидемиологии, при изучении по-
пуляций организмов и решения таких задач, как построение филогенетических де-
ревьев, показывающих эволюционные взаимосвязи между различными видами. Не
вдаваясь в детали, отметим, что с помощью этой дисциплины удалось создать модель
одного из известнейших механизмов молекулярной биологии — лактозного оперона
бактерии Escherichia coli, который был открыт Франсуа Жакобом и Жаком Моно
(Нобелевская премия по медицине 1965 года). Чем может быть полезна вычисли-
тельная модель чего-то, открытого еще в 1960-е? Дело в том, что вычислительная
алгебра — это относительно новый и очень мощный инструмент, позволяющий соз-
давать биологические модели и системно анализировать их. И в этом случае речь
идет не об открытиях, а о методе, позволяющем исследователям яснее понять все
составляющие сложного механизма лактозного оперона и даже поиграть с ними.
35
МАТЕМАТИЧЕСКАЯ БИОЛОГИЯ В ИСТОРИЧЕСКОЙ ПЕРСПЕКТИВЕ
Некоторые примеры использования математики в биологии
Последний раздел этой главы посвящен четырем важным примерам использования
математики в биологии. Мы поговорим о матрице Лесли, клеточных автоматах, мо-
дели «хищник — жертва» и клеточных автоматах, а также о применении теории
множеств в математической модели иммунной системы.
Изучение популяций оленей, белок и других животных.
Матрица Лесли
Патрик Лесли родился в 1900 году. Он был экологом и работал в Оксфорде, в ор-
ганизации, занимавшейся подсчетом численности животных. В 1945 году Лесли
опубликовал модель структуры популяции, которая нашла широкое применение в эко-
логии популяций и демографии. Эта модель позволяет определить рост популяции
с учетом ее возрастной структуры. Сведя воедино две функции (первая описывала
рождаемость, вторая — уровень смертности), ученый определил матрицу популяции,
известную под названием матрицы Лесли. Эта матрица является квадратной, то есть
имеет одинаковое число строк и столбцов, совпадающее с числом составляющих неко-
торого вектора. Также в этой модели предполагается, что популяция является изо-
лированной и не пополняется в результате миграции. Поскольку модель применяется
для животных, которые размножаются половым путем, в ней учитываются только
самки: число самцов на рост популяции не влияет. Составляющие вектора, о котором
мы упоминали выше, обозначают число особей определенного возраста.
Объясним модель Лесли на следующем примере. Предположим, что в природ-
ной среде, например в национальном парке или заповеднике, была зафиксирова-
на следующая численность самок оленей, которые в момент времени t (связанный,
к примеру, с датой выборки) принадлежали к возрастным группам под номерами 0,
1,2,3и4: Nlt, N*, N? и TV?- Обратите внимание, что 0,1, 2, 3 и 4 — всего лишь
обозначения, указывающие возрастные интервалы, к примеру в годах, от меньше-
го возраста к большему. В нашем примере предполагается, что оленей можно раз-
36
МАТЕМАТИЧЕСКАЯ БИОЛОГИЯ В ИСТОРИЧЕСКОЙ ПЕРСПЕКТИВЕ
делить на пять возрастных групп согласно ожидаемой продолжительности жизни.
Также предполагается, что плодовитость всех особей известна, то есть экологи, ра-
ботающие в заповеднике, знают среднее число детенышей у самок определенного
возраста. Если рассмотреть всю популяцию, то число новорожденных оленей, ко-
торые включаются в возрастную группу, образованную самыми молодыми особями
в следующем поколении (то есть в момент времени t + 1), будет равно:
n'’+i = /o-N? + /г№'+/2-М2 + /3- N? + f4-Nr
Теперь будем учитывать смертность оленей, вызванную различными причи-
нами. В этом случае особь не перейдет из текущей возрастной группы в следую-
щую, так как не достигнет нужного возраста. Обозначим через «0, s2 и $3 долю
выживших особей в каждой возрастной группе, которые, таким образом, перей-
дут в следующую возрастную группу. Это число выражается в долях едини-
цы и обозначает вероятность. Как следствие, в рассматриваемой модели число
самок, перешедших в следующую возрастную группу, определяется формулой
Nh-i = 50N?, N?+i = n', = s2HN?+i ~ 53Nr В математической биологии мо-
дель Лесли иллюстрирует очень элегантную и оригинальную формулировку. Все
представленные выше выражения сведены в матрицу перехода L, которая получила
название матрицы Лесли:
Л /1 А А Л
50 о о о о
О S1 О О О
О О S2 О О
О 0 0 $, О
к 3 7
Представим в виде вектора Л для поколения I число самок в каждой возрастной
. О , Т1 „ _2 . т3 „ _4
группе, то есть N,, N,, N, и /V,:
37
МАТЕМАТИЧЕСКАЯ БИОЛОГИЯ В ИСТОРИЧЕСКОЙ ПЕРСПЕКТИВЕ
N?
n!
N«
Nt
Представим в виде вектора N + число самок в каждой возрастной группе для
следующего поколения, t + 1:
ж то
N f+i
n!+i
N?+1
N,3+1
< N'+1 >
В конце концов объединим матрицу L и векторы /V и N + , описанные выше,
в одно выражение в матричной нотации. Сразу же увидим, что для получения воз-
растной структуры популяции, начиная от поколения / и заканчивая следующим по-
колением, t + 1, достаточно найти произведение вектора, соответствующего поколе-
нию t, и матрицы L:
Ж Т0 N г+1 л A A A A N?
nL 50 0 0 0 0 n!
Nt+i = 0 S1 0 0 0 N,
Nt+i 0 0 S2 0 0 N.
Nt+i , 0 0 0 S3 0 7 < N‘
В сокращенном виде это записывается так: /Vf+1 = L • N.
Не описывая пока подробности выполнения операций над матрицами (об этом
мы поговорим в главе 4), предположим, что экспериментальные данные о возрастах
38
МАТЕМАТИЧЕСКАЯ БИОЛОГИЯ В ИСТОРИЧЕСКОЙ ПЕРСПЕКТИВЕ
популяции оленей таковы: 190, 80, 56,18 и 6 (численность особей от меньшего воз-
раста к большему). Матрица L будет выглядеть следующим образом:
0,2 1,0 2,0 0,8 0,2
0,9 0 0 0 0
0 0,8 0 0 0
0 0 0,6 0 0
0 0 0 0,4 0
Если мы умножим вектор с исходными данными на матрицу L, получим следу-
ющий вектор:
( "1 Ni ( 0,2 1,0 2,0 0,8 0,2 190 '
n] 0,9 0 0 0 0 80
n. 0 0,8 0 0 0 56
N? 0 0 0,6 0 0 18
< 0 0 0 0,4 0 > k 6 >
Если мы умножим полученный вектор на матрицу L, получим новый вектор, ко-
торый затем вновь умножим на матрицу L, и т. д. По прошествии 10 единиц вре-
мени, рассчитав последовательные итерации модели, получим, что общая числен-
ность популяции будет разделена по возрастным группам (от меньшего возраста
к большему) в следующей пропорции: 48, 29, 16, 6 и 4 %.
Клеточные автоматы
В 1940-е годы Станислав Улам и Джон фон Нейман впервые описали метод моде-
лирования, известный под названием метод клеточных автоматов. Улам изучал рост
кристаллов, а фон Нейман впервые рассмотрел с теоретической точки зрения бес-
полое размножение, которое он назвал самовоспроизведением. Улам и фон Нейман
встретились в лаборатории Лос-Аламоса, где была создана первая атомная бомба
и где в те годы работал еще один их гениальный современник — Алан Тьюринг.
39
МАТЕМАТИЧЕСКАЯ БИОЛОГИЯ В ИСТОРИЧЕСКОЙ ПЕРСПЕКТИВЕ
Джон фон Нейман вместе со Станиславом Уламом
описал клеточные автоматы.
Клеточные автоматы — это модели, позволяющие описать тот же класс ситуа-
ций, который описывается дифференциальными уравнениями. Однако клеточные
автоматы имеют некоторые преимущества: при их использовании необязательно
знать какое-либо уравнение, описывающее явление или систему, они не требуют
мощного компьютера и позволяют быстро получить результат. При этом прогноз
будущего состояния системы, то есть искомое решение, представляется в графиче-
ском виде. Так, модель «реакция — диффузия», с помощью которой Тьюринг опи-
сал узоры на шкуре позвоночных, можно описать (и наглядно представить на ком-
пьютере) с помощью клеточных автоматов без использования уравнений.
В 1970 году англичанин Джон Хортон Конвей сделал клеточные автоматы по-
пулярными, создав игру «Жизнь». Сегодня эта игра покинула стены лабораторий
и обосновалась на множестве домашних компьютеров. В 2002 году Стивен Вольфрам,
создатель программы Mathematica и один из тех, кто занимался изучением клеточных
автоматов, написал книгу «Новый вид науки», вокруг которой развернулась бурная
полемика. Вольфрам предсказал рождение новой физики, основу которой будут со-
ставлять клеточные автоматы. В настоящее время клеточные автоматы позволяют
моделировать столь непохожие явления, как кольца планет (в частности, кольца Са-
турна), столбы дыма (а следовательно, агрегацию частиц), поведение групп муравьев,
40
МАТЕМАТИЧЕСКАЯ БИОЛОГИЯ В ИСТОРИЧЕСКОЙ ПЕРСПЕКТИВЕ
рост кристаллов, узоры на шкуре позвоночных, форму раковин моллюсков и даже
электоральные предпочтения.
На экране изображены некоторые классические конфигурации из игры «Жизнь»,
созданной Джоном Хортоном Конвеем. Различные формы или узоры,
образованные этими автоматами, — это решения, альтернативные тем,
которые можно получить с помощью дифференциальных уравнений.
Клеточный автомат — это решетка ячеек, находящихся в одном из множества воз-
можных состояний. К примеру, если возможны всего два состояния, то ячейки могут
находиться либо в состоянии 1 (черный цвет; «вкл.»), либо в состоянии 0 (белый цвет;
«выкл.»). Ячейки называются конечными автоматами. К примеру, светофор — это
конечный автомат с тремя возможными состояниями: зеленый, желтый, красный.
Каждая ячейка имеет так называемое соседство, куда обычно входят ячейки, смеж-
ные с ней. Существует множество разновидностей соседства. Одно из возможных
соседств состоит из клеток, расположенных выше, ниже, слева и справа от данной
ячейки, включая ее саму.
В любой модели начальное состояние ячеек (t = 0) решетки определяется соглас-
но некоторому критерию. Затем по заранее установленным правилам определяется
актуальное состояние ячеек (/ + 1). При этом учитывается как текущее состояние
рассматриваемой ячейки, так и состояние ее соседей. Этот процесс повторяется сно-
ва и снова, пока моделирование не будет завершено. На решетке клеточного авто-
мата образуются узоры, порой имеющие удивительную форму. Эти узоры можно
считать решением модели, то есть они представляют собой отпечаток будущего со-
стояния изучаемой системы.
41
МАТЕМАТИЧЕСКАЯ БИОЛОГИЯ В ИСТОРИЧЕСКОЙ ПЕРСПЕКТИВЕ
КЛЕТОЧНЫЕ АВТОМАТЫ В ПРИРОДЕ
В одном из классических примеров математической биологии рассматриваются моллюски родов
Conus и Cymbiola. Их раковины имеют характерные узоры, образованные пигментами - актива-
торами и ингибиторами, которые, по всей видимости, подчиняются так называемому правилу
30 - одному из правил, изученных Стивеном Вольфрамом. Неудивительно, что некоторые считают
этих моллюсков прекрасным примером клеточных
автоматов в природе.
Экземпляр моллюска Conus Textile. Рисунки
на его раковине напоминают узор, получаемый
при рассмотрении клеточного автомата,
который описывается правилом 30.
В игре «Жизнь» каждый конечный автомат имеет восемь соседей, расположен-
ных выше, ниже, справа, слева и по диагоналям от нее. Будем считать, что каждый
конечный автомат имеет всего два возможных состояния — 0, или «мертв», и 1, или
«жив», — которые мы будем обозначать разными цветами. Суть игры в том, чтобы
последовательно определять состояния конечных автоматов по установленным пра-
вилам перехода.
Правило № 1: Будущее состояние конечного автомата равно предыдущему,
если число соседей конечного автомата в состоянии 1 равно 2.
Правило № 2: Конечный автомат переходит из состояния 0 в состояние 1, если
число его соседей в состоянии 1 равно 3.
Правило № 3: Третье правило моделирует соседство с большим или малым чис-
лом «живых» автоматов, то есть автоматов в состоянии 1. Если число соседних авто-
матов в состоянии 1 меньше 2, то есть 1 или 0, либо больше 3, то есть 4, 5, 6, 7 или 8,
то конечный автомат «умирает», то есть переходит из состояния 1 в состояние 0.
Последовательно применяя правила перехода для всех конечных автоматов
клеточного автомата, мы увидим, как в процессе эволюции постепенно появляются
характерные шаблоны и фигуры.
42
МАТЕМАТИЧЕСКАЯ БИОЛОГИЯ В ИСТОРИЧЕСКОЙ ПЕРСПЕКТИВЕ
Модель «хищник - жертва» и клеточные автоматы
Модель «хищник — жертва» Лотки — Вольтерры стала одной из первых матема-
тических моделей в биологии и, возможно, одной из самых важных в математиче-
ской биологии. Как мы уже отмечали, одно из преимуществ клеточных автоматов
заключается в том, что для их использования не требуется знать дифференциальное
уравнение, описывающее явление или систему. Модель «хищник — жертва» Лот-
ки — Вольтерры была представлена в 1984 году Александром Дьюдени в статье
«Акулы и рыбы ведут экологическую войну на тороидальной планете Ва-Tbp» (Shark
and Fish Wage an Ecological War on the Toroidal Planet Wa-lbr). He используя ни одно
из уравнений, представленных Лоткой и Вольтеррой, Дьюдени получил похожие
результаты на компьютере со стандартными для 1980-х годов характеристиками.
Целью Дьюдени было найти подходящие значения параметров модели, допускавшие
сосуществование на небольшой решетке популяции хищников (акул) и жертв (рыб).
Дьюдени рассмотрел следующие параметры:
— число жертв (рыб);
— временной порог размножения рыб: если рыба выживает в течение опреде-
ленного числа циклов (или заранее установленного времени моделирования)
и ячейка остается свободной, в ней рождается рыба;
— число хищников (акул);
— максимальное время голодания хищников: если акула не может поймать рыбу
в течение определенного числа циклов (или заранее установленного времени
моделирования), она умирает;
— временной порог размножения акул: этот параметр определяется аналогично
соответствующему параметру для рыб, однако значения этих параметров не-
обязательно совпадают.
43
МАТЕМАТИЧЕСКАЯ БИОЛОГИЯ В ИСТОРИЧЕСКОЙ ПЕРСПЕКТИВЕ
«ест to depend on the initial random
distribution of a specified number of
sharks and fish. Nor does it seem to d.
pend in an ocukntal v*y on the actual
random movement of sharks and fish,
instead the likelihood of a population
crash appears to follow clouly the fish-
shark geometry that manifests itself
on O'lr screens: the more highly organ-
ized and localized either population be-
come", the likelier it it dirt the ecology
h doomed. Meditating on this theme,
we were led to wonder how we might
choose the five parameters in a way
tending to break up the geometry. Then
came a flash of insight-, if sharks had
congregated at the edges of a school of
fish, one way to break up the resulting
geometry would be to have the sharks
breed less often. The congregation itself,
after all, was less rhe result of motion
than it was of breeding.
Before forming this hypotnest; we
had chosen roughly equal breeding
times for sharks and fish. Balanced re-
production rates, we thought, would re-
juli in balanced populations. This kind
of vague thinking probably accounts
for many woes in today's technological
world In any event, I put 200 fish and 20
sharks in my -2-by 14 ocean and set the
fish to breed every three chronons but
barred any shark .rom reproducing be-
fore 10 chronons had elapsed. Shark
A realistic ne* sharks JMl
Фрагмент статьи Александра Дьюдени, посвященной модели «хищник — жертва»
и опубликованной в декабрьском номере американского журнала Scientific American за 1984 год.
Клеточный автомат модели имеет тороидальную форму, выбранную для того, что-
бы устранить границы решетки и обеспечить схожесть с настоящим морем. Ячейки
имеют всего три состояния: 1) в ячейке находится рыба, 2) в ячейке находится акула,
3) ячейка свободна. Рыбы (цветные ячейки) «плавают» случайным образом в на-
правлении одной из четырех соседних ячеек (на север, юг, запад или восток), если
одна из них или более свободны (не имеют цвета). Акула «съедает» рыбу, если они
находятся в смежных ячейках. Если в соседних ячейках нет рыбы, акула плывет
в свободную ячейку.
44
МАТЕМАТИЧЕСКАЯ БИОЛОГИЯ В ИСТОРИЧЕСКОЙ ПЕРСПЕКТИВЕ
Динамика эксперимента аналогична той, что описывается уравнениями модели
«хищник — жертва» Лотки — Вольтерры. Если акул немного, численность рыб
быстро увеличивается. С увеличением числа рыб численность акул также возрас-
тет, что ведет к постепенному снижению числа рыб. В зависимости от численности
акул и их расположения на тороидальной решетке рыбы могут полностью исчез-
нуть. В этом случае популяция акул в отсутствие пищи, то есть рыб, также быстро
вымрет. Какими должны быть условия сосуществования акул и рыб, необходимые
для сохранения обеих популяций? Приглашаем читателя поиграть с моделью Ва-Тор
и самостоятельно определить наиболее подходящие параметры.
45
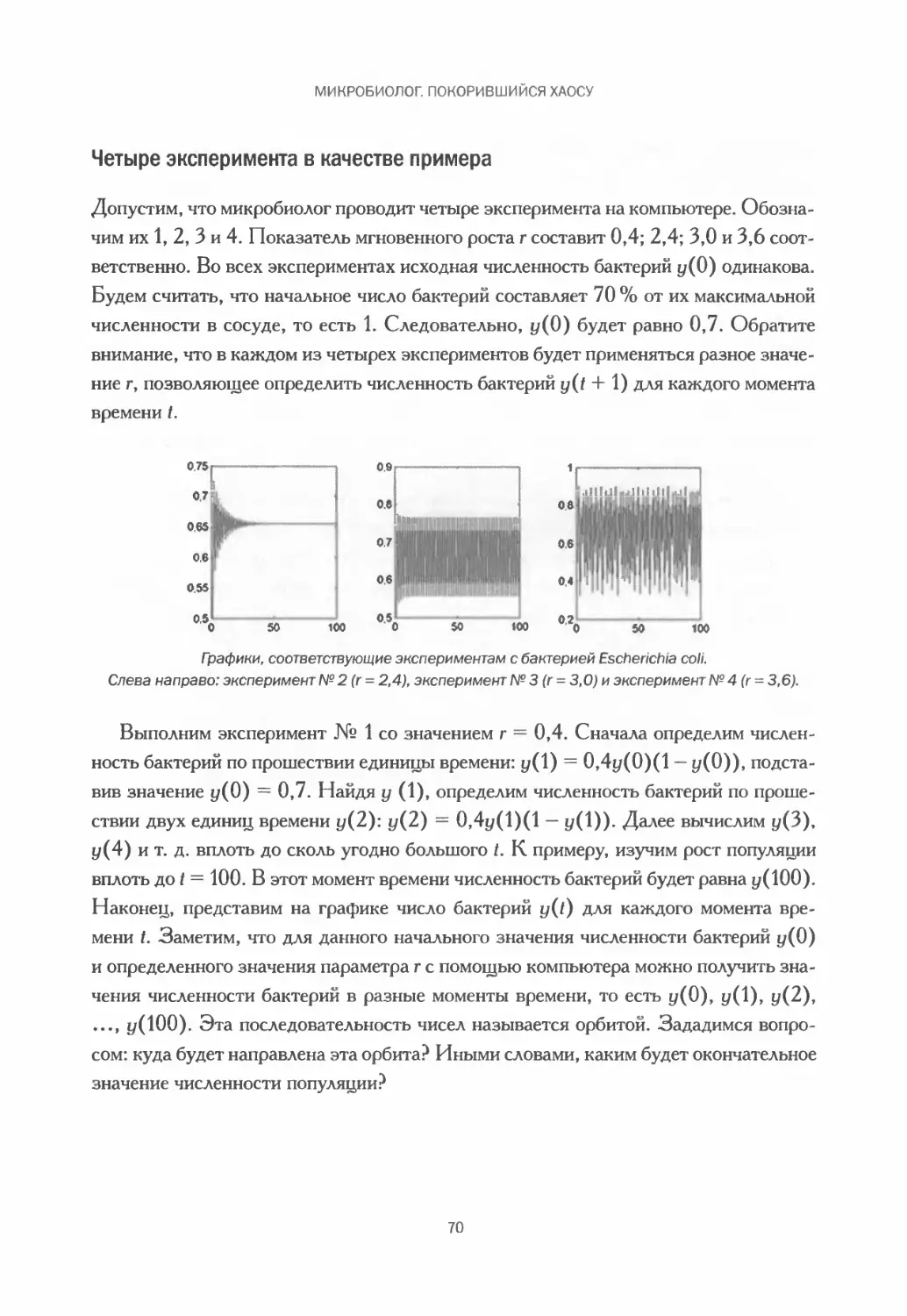
Глава 2
MZ
шизнь
изменчивое явление
Кажется, что живые существа постоянно решают самые разные задачи, в том числе
и для того, чтобы поддерживать такое удивительное и сложное явление, как жизнь.
Постоянная беготня муравьев, переносящих пропитание и различные материалы,
движение красных кровяных телец, образование стай птиц, беспрерывная передача
сигналов между нейронами мозга, преобразование одних веществ в другие в ходе
клеточного метаболизма, сердцебиение, этапы развития эмбриона с момента зачатия
до момента рождения, изменения, происходящие с головастиком, — лишь некоторые
примеры, демонстрирующие динамическое поведение живых существ. Как следствие,
живые организмы представляют собой подвижные системы, состояние и поведение
которых со временем меняются. Если бы мы могли увидеть все, что происходит внутри
простой клетки на протяжении одной секунды, мы бы поразились количеству преоб-
разований за это время. Системы, обладающие подобными свойствами, называются
динамическими.
Жизнь — результат множества динамических явлений,
благодаря которым становится возможным ее поддержание и развитие.
На иллюстрации — жизненный цикл лягушки,
изображенный на немецкой гравюре конца XIX столетия.
47
ЖИЗНЬ — ИЗМЕНЧИВОЕ ЯВЛЕНИЕ
Для изучения живых существ и экосистем могут использоваться те же теории
и методы, что и для изучения любых других динамических систем. Так, биологические
системы образованы множеством элементов, будь то муравьи, нейроны, вещества,
участвующие в метаболизме, или птицы, причем их состояние или поведение (идет ли
речь о муравейнике, мозге или стае птиц) со временем изменяется.
Еще одно важное свойство биологических систем заключается в том, что их со-
стояние или поведение является результатом взаимодействия между их элементами.
К примеру, состояние муравейника в момент времени t будет результатом взаимо-
действия между отдельными муравьями в рассматриваемый период времени В силу
этого свойства кажется очевидным, что математическая модель должна включать
наблюдаемые характеристики, репрезентативные для состояния или поведения изу-
чаемой системы. В случае с муравейником это будет численность рабочих муравьев,
муравьев-солдат и других членов колонии, в примере с метаболизмом — объемы
веществ А, В, С и т. д.
Для математика наблюдаемые характеристики системы, значение которых мож-
но получить экспериментально, являются переменными модели и обозначаются х,
у, ..., z. Если известны значения этих переменных в разные моменты времени х(/),
ПРОГНОЗИРОВАНИЕ И МОДЕЛИРОВАНИЕ
Прогнозирование стало неотъемлемой частью жизни обще-
ства в XX-XXI веках. Определяющее влияние на будущее
людей оказывает возможность контролировать изменения:
социальные, экономические, эпидемические и т.д. С мо-
мента изобретения компьютера прогнозирование будущего
с помощью математического моделирования чрезвычайно
широко используется в науке и технике. Моделирование ох-
ватывает все сферы человеческой деятельности, от изучения
экологических (рост численности населения, экосистемы,
климатические модели и т.д.) и физиологических систем
(обмен веществ, клетки, сердце, мозг, мышцы) в биологии
и медицине до изучения социальных систем (опросы обще-
ственного мнения, анализ безработицы, состояние рынков
и т.д.) в политологии и экономике.
Компьютерное моделирование
поведения белков, связанных
с болезнью Паркинсона.
48
ЖИЗНЬ — ИЗМЕНЧИВОЕ ЯВЛЕНИЕ
у (Г), .z(f), то известно, каким будет состояние или поведение системы (муравей-
ника, мозга, метаболизма или стаи птиц) в момент времени t. Обратите внимание,
что в динамической системе время t является независимой, или входной, переменной.
Выходной переменной, в свою очередь, будет состояние системы, которое определя-
ется множеством зависимых переменных x(t), y(t),..., z(t). Математическая модель
позволяет описать состояние системы в определенный момент времени t, а также
с ее помощью предсказать будущее состояние системы для значения t, достаточно
далеко отстоящего от текущего момента, — именно это происходит при составлении
прогнозов погоды или прогнозировании уровня заболеваемости во время эпидемии.
В последние десятилетия стало актуальным прогнозирование уровня заболеваемо-
сти гриппом, коровьим бешенством или СПИДом.
Как извлечь игрушку из пасхального яйца
Одна из классических математических задач звучит так. Предположим, что мы хотим
найти неизвестную функцию у, для которой известна ее производная у'. Допустим,
производная у' неизвестной функции у равна Зх2 (говоря математическим языком,
у' = З/2). Необходимо определить у. Эта задача допускает несколько возможных
решений (они называются первообразными), но если нам также известно значение
функции в некоторой точке, например у(0) = 0, то решение будет единственным.
Чтобы найти у, нужно вычислить интеграл j Зх2 dx.
Сначала вынесем число 3 за знак интеграла: 3 j х2 dx. Затем, поскольку мы имеем
дело с табличным интегралом, достаточно вспомнить следующую формулу:
где С — константа интегрирования. В нашем случае функция у выглядит так:
3[х2 dx = 3--1-С.
J 3
В нашем случае С = 0, так как у(0) = 0. Упростив выражение, получим искомую
функцию: у = /3. Задача была успешно решена стандартными методами интегрального
исчисления.
49
ЖИЗНЬ — ИЗМЕНЧИВОЕ ЯВЛЕНИЕ
Что произойдет, если производная у' не будет напрямую выражена в виде /(/),
например у' = З/2, как в нашем примере, а будет зависеть от функции у, которую
нужно найти? Именно так выглядят дифференциальные уравнения, в которых значе-
ние у' связано со значением у. Производная функции по времени обозначается у' либо
dy/dt. Эти обозначения эквивалентны. В простейших случаях дифференциальное
уравнение имеет следующий вид:
Расскажем немного подробнее об элементах дифференциального уравнения. Что
означает у', или dy/dt? Производная выражает уровень изменений, скорость или
ритм изменения системы. Напомним, что одной из характеристик динамических си-
стем является зависимость их состояния от взаимодействия между их элементами,
при этом любое изменение произвольного элемента влияет на общее состояние си-
стемы у. Иными словами, если известно состояние системы в момент времени t, на-
пример y(f), и мы подставим это значение в дифференциальное уравнение, то опре-
делим степень изменений системы — она будет характеризоваться значением у'.
Заметьте, что дифференциальные уравнения в силу своих свойств наиболее удобны
для построения математических моделей динамических систем и поэтому играют
важную роль в математической биологии — с их помощью были успешно смоде-
лированы многие биологические и экологические явления, о которых мы расскажем
в этой главе.
Но как найти у в дифференциальном уравнении? Эта задача в общем виде реша-
ется не так просто, как в предыдущем примере, когда, зная производную функции
у' = Эх2, мы смогли вычислить саму функцию. В дифференциальных уравнениях
наряду с у' фигурируют и другие члены.
Если сравнить дифференциальное уравнение с шоколадным пасхальным яйцом,
внутри которого находится игрушка, то решение уравнения будет равносильно тому,
чтобы извлечь игрушку путем последовательных действий. К примеру, сначала нуж-
но снять с яйца обертку, затем съесть шоколад, и только тогда вы увидите игрушку
или, в случае с дифференциальным уравнением, найдете искомую функцию у. Сле-
довательно, решить дифференциальное уравнение означает найти функцию у. Если
выполнить с этим уравнением различные действия, вы получите и, если возможно,
примените аналитические методы решения. На одном из этапов решения мы исполь-
зуем интегральное исчисление, однако не столь явно, как в предыдущем примере.
50
ЖИЗНЬ — ИЗМЕНЧИВОЕ ЯВЛЕНИЕ
Пример решения представлен в следующей главе для уравнения модели Мальтуса,
которое играет основную роль в демографии и при изучении динамики популяций.
Если методы интегрального исчисления заведут нас в тупик, мы также сможем най-
ти приближенное решение с помощью компьютера. Приближенное решение означа-
ет, что мы выберем желаемую точность результата и применим алгоритм, который
позволит найти решение с точностью, превышающей указанную. Теория численного
анализа гарантирует, что определенный алгоритм позволяет найти решение с на-
перед заданной (или более высокой) точностью.
Наиболее известные численные алгоритмы решения дифференциальных уравне-
ний — это метод Эйлера и метод Рунге — Кутты. Эти методы используют не толь-
ко математики, но и экологи, а также сотрудники фармакологических лабораторий.
Метод Рунге — Кутты более известен и обеспечивает прекрасное соотношение
между временем расчетов на компьютере и точностью результата. Метод Эйлера
проще, но менее точен.
Дифференциальное уравнение Парка юрского периода
В 1950-е годы Уиллард Либби разработал интересный метод определения примерного
возраста ископаемых. В основе метода Либби лежало измерение содержания радио-
активного изотопа углерода С-14 в изучаемом объекте, например, в ископаемом или
в Туринской плащанице.
Американский химик
Уиллард Либби (1908-1980)
на обложке журнала Time.
51
ЖИЗНЬ — ИЗМЕНЧИВОЕ ЯВЛЕНИЕ
С-14 — это изотоп углерода, концентрация которого в атмосфере Земли по-
стоянна. Живые организмы в течение жизни накапливают углерод С-14, полу-
чая его с дыханием и при питании другими живыми существами. Каким бы путем
С-14 ни попадал в организм, его содержание также будет неизменным. После смер-
ти накопление С-14 прекращается, и его концентрация в тканях начинает постепен-
но снижаться.
Чтобы получить формулу для определения возраста объектов, используем урав-
нение роста из модели Мальтуса. Обозначим через у содержание С-14 в определен-
ный момент времени t, через у0 — содержание С-14 в ископаемом. Кроме того, ис-
комая формула будет включать г — так называемую константу распада, известную
для всех изотопов: у = уоег/.
Так как известно, что период полураспада С-14 составляет 5600 лет, предыду-
щее выражение примет вид:
После ряда преобразований получим у = у0е~0,00012378/. Выразив время t из этого
выражения, найдем формулу, с помощью которой палеонтологи и археологи опреде-
ляют возраст ископаемых и археологических находок:
W)
0,00012378
Это выражение можно использовать в случаях, когда возраст анализируемого
объекта не превышает 50 тысяч лет.
К примеру, мы обнаружили кость доисторического животного, содержащую
1/100 изотопа С-14. Чему равен возраст находки? По условию задачи, с будет рав-
но 100. Подставив это значение в исходное выражение, имеем:
1п(100) ^4,60517018,372р1
0,00012378 0,00012378 ~
Можно сделать вывод: возраст кости составляет примерно 37 тысяч лет.
52
ЖИЗНЬ — ИЗМЕНЧИВОЕ ЯВЛЕНИЕ
Мальтус, Ферхюльст и рост населения
В 1798 году Томас Роберт Мальтус опубликовал книгу «Эссе о росте народона-
селения». Согласно его гипотезе, в какой-то момент численность населения Земли
будет расти в геометрической прогрессии, то есть экспоненциально. При этом объем
продовольственных и любых других ресурсов возрастает в арифметической прогрес-
сии, то есть линейно. Так, численность населения описывается последовательностью
2 (21), 4 (22), 8 (23), 16 (24), 32 (25), 64 (26) и т. д., количество продовольствен-
ных ресурсов — 2, 3, 4, 5, бит.д. Следовательно, наступит момент, когда высокая
рождаемость, особенно среди рабочего класса, приведет к недостатку продовольствия
(отметим, что марксисты считали теории Мальтуса нападками на рабочий класс).
Англиканский священник Томас Роберт Мальтус (1766-1834).
Справа представлены две модели роста: экспоненциальная (1) и линейная (2).
Допустим, что мы применили модель Мальтуса, в частности у' = ту, к некоторой
популяции животных или микроорганизмов. В конечном итоге в этой модели скорость
роста населения у' пропорциональна численности населения у. Таким образом, при-
менив математические методы, можно преобразовать исходное дифференциальное
уравнение, как показано ниже. Во-первых, нужно записать уравнение в следующем
виде: dy/dt = гу, где г — параметр, отражающий рост населения с постоянной ско-
ростью, которая не меняется в последующих поколениях. Этот параметр называется
коэффициентом роста населения.
53
ЖИЗНЬ — ИЗМЕНЧИВОЕ ЯВЛЕНИЕ
Затем перенесем dt в правую часть так, что dy = rydt. Это уравнение словно
подсказывает, что нужно сгруппировать в одной части все члены, связанные с у.
Следовательно, перенесем у в левую часть. Имеем dy/y = rdt.
Наконец, чтобы решить уравнение, нужно взять интеграл от обеих его частей,
как показано далее:
У
На этом этапе у читателя может создаться впечатление, что мы не решаем зада-
чу, а только усложняем рассуждения. Внимательно рассмотрим выражение. В пра-
вой его части записан простейший табличный интеграл. Так как г — константа, ее
можно вынести за знак интеграла. Имеем:
J^ = rjA.
У
Напомним, что ^dt = t + C- Правая часть равенства будет выглядеть так:
J^=rt+c.
У
В левой части также записан табличный интеграл. Обратите внимание, что, по-
скольку dy записано в числителе, у — в знаменателе, интеграл будет равен логариф-
му у, а именно:
|^/у = 1п(у) + С.
У
Поэтому
ln(y)= rt+C.
Если мы избавимся от логарифма и сгруппируем члены выражения, то найдем
решение дифференциального уравнения у' = гу. Для этого подставим в выражение
54
ЖИЗНЬ — ИЗМЕНЧИВОЕ ЯВЛЕНИЕ
величину, обозначающую исходное число бактерий (ранее мы обозначили его че-
рез у0). Определим функцию у:
у = у(/-
В1838 году математик Пьер Франсуа Ферхюльст видоизменил модель Мальтуса
с учетом того, что размеры окружающей среды ограничены, поэтому должно суще-
ствовать некоторое максимальное значение численности населения k, известное как
поддерживающая емкость среды. Ферхюльст получил следующее дифференциальное
уравнение: у' = ry(k — у).
Бельгийский математик Пьер Франсуа Ферхюльст (1804-1849),
один из величайших специалистоь по теории чисел первой половины XIX века.
Решением уравнения Ферхюльста является знаменитое логистическое уравне-
ние, которое описывает не только рост населения, но и распространение эпидемий
и рост социальных сетей в интернете:
y{)+(k-y0)e~rt
Логистическое уравнение применимо для анализа S-образного роста — экспо-
ненциального, но ограниченного количеством ресурсов, будь то физическое про-
странство, продовольствие, емкость рынка мобильной связи или число пользова-
телей социальной сети. Экспоненциальный рост является неограниченным, то есть
утопичным, возможным только в мире с неисчерпаемыми ресурсами. В логистиче-
ской же модели рассматривается реальный мир, к примеру планета Земля, ресурсы
которой, что очевидно, ограничены.
55
ЖИЗНЬ — ИЗМЕНЧИВОЕ ЯВЛЕНИЕ
Любопытно отметить, что эти модели были предложены в XIX веке, в разгар
промышленной революции. В эту эпоху жили такие ученые, как Чарльз Дарвин,
создатель теории эволюции путем естественного отбора, и Чарльз Бэббидж, изо-
бретатель аналитической и разностной машин — прообразов современных компью-
теров. Эти любопытные совпадения предвосхитили плодотворный союз математики
и компьютерных технологий, который сыграл в XX веке определяющую роль в изу-
чении жизни.
Дифференциальные уравнения в биотехнологии
Биотехнология — это раздел биологии, с помощью которого методы генной инже-
нерии и выращивания клеточных культур находят широкое применение в сельском
хозяйстве, фармакологии, медицине и диетологии. Основной инструмент биотехноло-
гов — хемостат, резервуар или биореактор, в котором посредством культивирования
клеток вырабатываются полезные вещества.
Биореактор в лаборатории. Внутри биореактора находятся клетки.
56
ЖИЗНЬ — ИЗМЕНЧИВОЕ ЯВЛЕНИЕ
Цель подобных исследований — достичь состояния, при котором число микро-
организмов N и объем питательных веществ С были бы практически постоянными,
а рост численности микроорганизмов — экспоненциальным. Именно при таком ро-
сте вырабатываются полезные вещества, например антибиотики. В ходе экспери-
мента необходимо постоянно пополнять запас питательных веществ и одновременно
убирать жидкости, токсины и любые другие продукты метаболизма микробов, при
этом объем среды культивации должен оставаться неизменным. В промышленности
хемостаты используются для выработки этанола, ферментированных продуктов пи-
тания (например, сыров), белков, обладающих лечебными свойствами (в частности,
инсулина), и т. д. Хемостаты также применяются при изучении экологии микроор-
ганизмов, а также для анализа их эволюции.
Как вы уже, наверное, догадались, инженеры-биохимики и другие специалисты
в сфере биотехнологий в своих экспериментах с биореакторами успешно и широ-
ко применяют дифференциальные уравнения, которым посвящен отдельный раздел
математической биологии. К примеру, дифференциальные уравнения, описывающие
процессы, происходящие в хемостате, выглядят так:
— = K(C)N-F—,
dt V
— = -aK(C)N-F—+F—,
dt V V
где N — число микроорганизмов, С — концентрация питательных веществ, F —
поток (при этом Fmoj = Рды о^), V — объем. В дифференциальных уравнениях К(С),
а и Со — параметры модели. Обратите внимание, что первое дифференциальное
уравнение, описывающее изменение N, напоминает логистическое уравнение (о нем
мы поговорим позже). Оно также содержит поток F. На основе этих и других вы-
ражений были разработаны программы, управляющие хемостатами, которые исполь-
зуются, в частности, для компьютерного контроля ферментации.
57
ЖИЗНЬ — ИЗМЕНЧИВОЕ ЯВЛЕНИЕ
Математическое изучение рака: опухоли в компьютере
Рак — это заболевание, при котором наблюдается бесконтр< >льный рост группы кле-
ток, образующих опухоль, разрушающую близлежащие клетки и ткани (это определе-
ние не вполне подходит для лейкемии). В действительности раковая опухоль содержит
и обычные клетки, злокачественная трансформация которых ведет к образованию
раковых. Поскольку рак чрезвычайно распространен, он стал одним из объектов изу-
чения математической биологии. Лечение раковых заболеваний настолько важно, что
сегодня существует целая база данных QCDB (от англ. Quantitative Cancer Modelling
DataBase — «база данных для количественного моделирования рака»), предоставля-
ющая доступ к информации об этом заболевании биоматематикам всего мира.
Математическое изучение раковых заболеваний проводится с использованием
математических моделей и компьютерного моделирования. Математика при этом,
во-первых, помогает выдвигать новые гипотезы о причинах образования опухолей,
а во-вторых, использование математических моделей позволяет лучше проанализи-
ровать огромные объемы накопленных экспериментальных и клинических данных.
Биологи и математики рассматривают опухоли как сложные системы. Раковые
клетки в них взаимодействуют между собой и с другими клетками, при этом их пове-
дение нельзя объяснить, если мы будем рассматривать раковые клетки изолированно
от других. Согласно этому подходу, предполагается, что опухоль образуется не в ре-
зультате сбоя в конкретном гене. Причиной рака является общий сбой взаимодействия
между генами. Проводя параллель с интернетом, рак можно считать результатом
нарушения работы множества компьютеров в сети (DNS-серверов, маршрутизаторов
и т. д.), а не результатом сбоя какого-то конкретного компьютера.
В 1964 году исследователь по фамилии Лэйрд заметил, что рост опухолей в ус-
ловиях ограниченного пространства и питательных веществ описывается функцией
Гомпертца. Классическим примером является рак груди. Скорость роста опухоли
в этих условиях, у’ (размер опухоли может быть выражен через ее объем или число
клеток), описывается следующим дифференциальным уравнением:
58
ЖИЗНЬ — ИЗМЕНЧИВОЕ ЯВЛЕНИЕ
y' = aln — у.
\У J
В этом выражении 01 — параметр, описывающий способность раковых клеток
опухоли к росту, К — максимально возможный размер опухоли (напомним, что объ-
емы ткани, в которой находится опухоль, и количество питательных веществ огра-
ничены). Решением этого дифференциального уравнения будет функция Гомпертца,
предложенная английским математиком Бенджамином Гомпертцем в 1825 году как
уточнение модели Мальтуса. Функцию Гомпертца первыми применили страховые
компании. Основная ее идея заключается в том, что с увеличением возраста уровень
смертности возрастает в геометрической прогрессии.
Гомпертц описал любопытную связь между уровнем смертности, который мы
обозначим через Rm, и возрастом t:
R =R/'+A
Особенность этого выражения заключается в том, что А играет удивительную
роль: эта величина отражает воздействие на уровень смертности факторов, не свя-
занных с возрастом человека. К примеру, рост средней продолжительности жизни
в развитых странах обусловлен именно тем, что удалось значительно снизить значе-
ние А благодаря росту уровня жизни и созданию более здоровой среды. На сниже-
ние А могли повлиять рост городов, появление зданий, защищенных от воздействия
климата, улучшение гигиены, питания и т. д. Тем не менее параметр [3 остается неиз-
менным. Сердечные и раковые заболевания стали причиной того, что, с одной сто-
роны, Rm в результате снижения А уменьшилось, но, с другой стороны, с возрастом
Rm увеличивается. Дастся ли нам когда-нибудь устранить или существенно снизить
воздействие возраста, или t, на R?
59
ЖИЗНЬ — ИЗМЕНЧИВОЕ ЯВЛЕНИЕ
Если мы используем функцию Гомпертца в ином контексте, в частности приме-
нительно к раковым заболеваниям, то размер опухоли у будет описан выражением:
где у(0) — начальный размер опухоли. Если пациент проходит лечение, то у(0) будет
меньше К, в противном случае размер опухоли будет увеличиваться.
Кривая Гэмпертца, описывающая рост раковой опухоли
(N — размер опухоли, t — время).
Эта функция весьма схожа с сигмоидой (логистической функцией): рост опухоли
замедлен в начале и конце процесса. Замедление в конце процесса кажется очевид-
ным, если учесть, что по мере роста опухоли клетки, расположенные внутри нее,
получают меньше кислорода, отмирают и вызывают некроз ядра опухоли. В резуль-
тате ее размер стабилизируется: рост внешней части уравновешивается отмиранием
клеток во внутренней части.
Этому же закону подчиняется и динамика роста некоторых предприятий, в част-
ности тех, где большую роль играют технологии, — фармацевтических компаний
или операторов мобильной связи. Вначале затраты на исследования, патенты и т. д.
превышают доходы от продаж, затем компания переживает период бурного роста
и получает прибыль. На следующем этапе продажи падают, так как рынок постепенно
насыщается. Также функцией Гомпертца описывается рост органов эмбриона или, что
еще любопытнее, регенерация хвоста у ящерицы.
60
ЖИЗНЬ — ИЗМЕНЧИВОЕ ЯВЛЕНИЕ
Хотя приведенное выше выражение может показаться сложным, следует по-
нимать, что благодаря компьютерам вычислить его значение сравнительно легко.
По сути, речь идет о достаточно простом выражении вида у = аеЬе, в котором по-
казатель степени возводится в новую степень.
В 1980-е годы исследователь Уэлдон заметил, что этой функцией не очень точно
описывается рост опухолей малых размеров, поскольку в ней не учтены некоторые
биологические аспекты, в частности роль иммунной системы. В поправке Уэлдона
утверждается, что на первом этапе роста опухоли раковые клетки не сражаются за до-
МАТЕМАТИКА И НОВЫЕ ПУГИ ИССЛЕДОВАНИЯ
В 2005 году исследователь Антонио Бру из мадридского университета Комплутенсе предположил,
что на поздних стадиях раковые заболевания можно излечивать, вызывая сильное и продол-
жительное воспаление тканей вокруг опухоли. Эта гипотеза стала результатом математических
исследований роста раковых клеток. В ходе исследований было отмечено, что рост всех клеток
подчиняется одной схеме, которую Бру назвал схемой универсальной динамики роста опухолей.
В этой модели клетки на границе опухоли играют определяющую роль в методе лечения, пред-
ложенном Бру. Первоначальное скептическое отношение к гипотезе отчасти было вызвано тем,
что использованная математическая модель отличалась от классических моделей раковых за-
болеваний. Во-первых, в ней предполагалось, что рост клеток подчиняется не экспоненциальному,
а линейному закону, а во вторых, считалось, что рост опухоли зависит не от количества пита-
тельных веществ, а от свободного пространства. Это прекрасный пример того, как математика
подсказывает исследователям новые пути лечения рака.
Математическая модель и результат компьютерного моделирования
роста раковой опухоли.
61
ЖИЗНЬ — ИЗМЕНЧИВОЕ ЯВЛЕНИЕ
ступные ресурсы, и их рост описывается экспоненциальным законом, или моделью
Мальтуса. Однако по достижении некоторого критического размера рост опухоли
будет описываться уже не моделью Мальтуса, а функцией Гомпертца.
СПИД, свиной грипп и другие заболевания,
которые можно изучить с помощью математики
В 1983 году французский исследователь Люк Монтанье описал вирус СПИДа,
или ВИЧ (вирус иммунодефицита человека). Он представляет собой сферу диаме-
тром 100 нанометров и имеет внешнюю белковую оболочку. Вирусологи называют
этот вирус ретровирусом, так как его геном образован цепочкой РНК. По данным
Всемирной организации здравоохранения, в 2006 году в мире насчитывалось при-
мерно 39,5 миллиона человек, зараженных вирусом СПИДа, примерно 24 миллиона
из них проживали на Африканском континенте.
В 2009 году средства массовой информации сообщили о начале пандемии сви-
ного гриппа. По данным Всемирной организации здравоохранения, возбудителем
заболевания является вирус H1N1/09. Его геном представляет собой смесь ДНК
птиц, свиней и человека, поэтому вирус способен преодолевать межвидовые барье-
ры. Свиной грипп был самой популярной темой в СМИ летом и осенью 2009 года.
Изначально процент смертельных случаев среди заболевших был высоким, однако
со временем он снизился, что совпало с началом широкого использования противо-
вирусных препаратов.
Подобные заболевания, носящие характер пандемии, становятся источником
напряженности в обществе. Как санитарные службы всего мира прогнозируют
и отслеживают распространение заболеваний? Как определяется момент начала
эпидемии в определенной стране? Когда следует начинать вакцинацию людей, вхо-
дящих в группу риска? Ответы на эти вопросы дает ряд математических моделей,
составляющих формальное ядро эпидемиологии, которая изучает факторы, влияю-
щие на здоровье и заболеваемость населения. Эпидемиология привлекла внимание
математиков еще в начале XX века, а сегодня она стала одной из областей изучения
математической биологии.
62
ЖИЗНЬ — ИЗМЕНЧИВОЕ ЯВЛЕНИЕ
Первыми, кто рассмотрел эпидемии с точки зрения математики, были Уильям
Хаммер и Рональд Росс. Для анализа эти ученые применили закон действующих
масс. Позднее Лоуэлл Рид и Уэйд Фрост разработали модель Рида — Фроста, свя-
зав число здоровых людей, восприимчивых к заболеванию (S), число заболевших (/)
и число людей, невосприимчивых к заболеванию.
Анализировать распространение заболеваний специалистам вновь помогают диф-
ференциальные уравнения. Допустим, что численность населения составляет N чело-
век, из которых I заражены вирусом. Это означает, что число здоровых людей равно
/V — I. Так как люди, зараженные вирусом, живут рядом со здоровыми, последние
подвергаются риску заражения (S). Следовательно, 5 = N — 1.
В одной из классических моделей эпидемиологии утверждается, что изменение
числа зараженных в зависимости от времени описывается дифференциальным урав-
нением: Г — kI(N — /). В упрощенном виде оно выглядит так: Г = kIS. Решением
этого дифференциального уравнения будет знаменитое логистическое уравнение,
описывающее ход любой эпидемии:
Обратите внимание, что в начале эпидемии (то есть при t, стремящемся к нулю)
логистическое уравнение будет приблизительно эквивалентным уравнению экспо-
ненциального роста, то есть уравнению модели Мальтуса. Это отражает тот факт,
что в начале эпидемии число зараженных резко увеличивается. В случае с заболева-
ниями, которые становятся причиной напряжения в обществе, например СПИДом
или свиным гриппом, сообщения о росте эпидемии, распространяемые СМИ, толь-
ко усугубляют панику.
Если предположить, что предельное число заболевших равно числу здоровых
людей, восприимчивых к заболеванию, то есть N, то начиная с определенного мо-
мента рост эпидемии замедлится, как и рост числа новых заболевших, I. Это значе-
ние, столь важное для органов здравоохранения любой страны, достигается, когда
63
ЖИЗНЬ — ИЗМЕНЧИВОЕ ЯВЛЕНИЕ
число заболевших 1 составляет половину численности восприимчивого к вирусу на-
селения, то есть N/2. После этого количество новых случаев заболевания стабили-
зируется вплоть до окончания эпидемии.
В настоящее время благодаря использованию компьютерного моделирования
можно оценить распространение эпидемии (например, сезонного гриппа), что по-
зволяет органам здравоохранения формировать календарь вакцинации населения.
В эпидемиологии используются такие компьютерные программы, как Epigrass,
AnyLogic Model-Builder и STEM (SpatioTemporal Epidemiological Modeler).
Число e и колония бактерий Escherichia coli
Нет такой области науки, где рано или поздно не появилось бы число е, будь то мо-
лекулярная биология или статистика, физика или химия. Это вездесущее число об-
наруживается во множестве природных явлений. Число е — иррациональное. Это
означает, что его десятичная запись никогда не заканчивается и не повторяется, что
роднит его с числом Л. Считается, что число е открыл Якоб Бернулли при изучении
следующего предела:
liml 1 + -
п
получив результат 2,71828. Программа символьных вычислений, подобная Derive,
позволяет мгновенно вычислить этот предел и получить указанный выше результат:
LIM ((1 + 1/пУп, п, INF, 0).
Приближенное значение числа е.
64
ЖИЗНЬ — ИЗМЕНЧИВОЕ ЯВЛЕНИЕ
В 1618 году Непер уже упоминает это число в своих логарифмических табли-
цах как основание натуральных логарифмов Zog(x) или, в сокращенном виде, /п(х).
Обозначение в виде буквы е ввел математик Леонард Эйлер в 1727 году. Среди
любопытных фактов, связанных с этим числом, выделяются те, что относятся к экспо-
ненциальной функции ех. Во-первых, производной функции /(х) = ех является эта же
самая функция, то есть f (х) = ех. Производная в точке х = 0 равна / ‘(х) = 1. Во-
вторых, интерес представляет интеграл этой функции: fex dx = ех + С. Наконец, сумма
бесконечного числа членов ряда 1/0! + 1/1! + 1/2! + ... + 1/п! равна числу е.
Обратите внимание, что знаменателями членов ряда являются факториалы после-
довательных чисел, которые определяются, к примеру, так: 4!= 4 • 3 • 2 • 1. Если бы
ряд имел вид 1 + х + х2/2! + х3/3! + х4/4! + ... + хп/и!, то сумма этого ряда при-
ближалась бы к экспоненциальной функции ех.
Подобные соотношения наблюдаются при изучении множества явлений. В част-
ности, к ним относится экспоненциальный рост населения, проанализированный
Мальтусом. Примером экспоненциального роста является рост колонии бактерий.
Пусть N — начальное число бактерий Escherichia coli в лабораторной чашке Петри,
г — показатель роста численности бактерий. Тогда N — численность бактерий в ко-
лонии в момент времени t — будет определяться следующим выражением:
Так как следующее поколение бактерий образуется каждые 30 минут, по про-
шествии некоторого времени в чашке Петри будет находиться несколько миллионов
бактерий. Другой пример, представляющий практический интерес для математиче-
ской биологии, — это распределение Пуассона, или закон распределения вероят-
ностей, описывающий случайные события, имеющие малую вероятность. Допустим,
мы подвергли чашку Петри с культурой бактерий Escherichia coli ультрафиолетовому
излучению. Вероятность получения определенного числа мутантов х будет равна:
где А, — параметр закона распределения вероятностей. Число е также фигурирует
в гиперболических функциях, широко применяющихся в биологии.
65
Глава 3
Микробиолог,
покорившийся хаосу
Представьте себе микробиолога, который хочет провести ряд экспериментов, требу-
ющих достаточно много бактерий Escherichia coli. Для этого он выращивает культуру
в чашке Петри. Обозначим начальное число бактерий в чашке Петри через yQ. Благо-
даря современным теориям математической биологии известно, что рост популяции
описывается дифференциальным уравнением у’ = гу. Это означает, что скорость
роста численности бактерий у’ равна численности бактерий в момент времени t, ум-
ноженной на г, где г — мгновенный уровень роста.
Важно отметить, что рост численности бактерий будет описываться этим диффе-
ренциальным уравнением при условии, что питательные вещества в среде с бактери-
ями не должны заканчиваться. Похожую ситуацию можно воспроизвести в микро-
биологической лаборатории с помощью хемостата, о чем мы уже рассказывали. Также
рост бактерий не должен быть ограничен физическим пространством. Это означает,
что культуре должно быть предоставлено, по меньшей мере в теории, неограниченное
пространство для все новых и новых поколений. Если бы эти условия были возможны
в природе, мы имели бы дело со сценарием экспоненциального роста, описанным
Томасом Мальтусом в 1798 году.
Дифференциальное уравнение у* = гу и его ограничения
Решив дифференциальное уравнение у' — гу, получим следующее выражение:
у = уУ‘-
67
МИКРОБИОЛОГ, ПОКОРИВШИЙСЯ ХАОСУ
Это выражение означает, что число бактерий у в определенный момент времени t
будет равно начальной численности бактерий у0, умноженной на экспоненту. Здесь
экспонента — это число е, возведенное в степень, равную произведению г на время t.
Значение параметра г нельзя определить в результате наблюдений, но можно найти
путем оценки или по справочнику. Напомним, что приближенное значение числа е
равно 2,71828.
Обратите внимание, что число бактерий будет возрастать экспоненциально при
соблюдении указанных условий. Зная выражение, приведенное в начале этого раз-
дела, микробиолог сможет определить, сколько времени потребуется, чтобы получить
необходимое количество бактерий.
Сканер, показывающий рост группы бактерий Escherichia coli.
Однако он должен обеспечить поступление к культуре Е. coli неограниченного
количества питательных веществ и предоставить им необходимое физическое про-
странство. Для этого микробиолог может подсчитать максимальное число бактерий,
которое уместится в лабораторной посуде. Обозначим это параметр через k (макси-
мальная численность колонии бактерий также называется поддерживающей емкостью
среды). Следовательно, в уравнение мальтусовской модели у’ = ту следует ввести
дополнительный множитель (k — у). Мы получили дифференциальное уравнение,
68
МИКРОБИОЛОГ, ПОКОРИВШИЙСЯ ХАОСУ
отражающее новое ограничение — максимальный размер популяции, способной уме-
ститься в той или иной среде:
у =ry(k-y).
ото выражение, как вы знаете из предыдущей главы, называется уравнени-
ем Ферхюльста и ввел его бельгийский математик Пьер Франсуа Ферхюльст
в 1838 году. Очевидно, что если численность бактерий у растет, значение k — у
уменьшается, и наоборот, с уменьшением численности бактерий у значение k — у
увеличивается, что обеспечивает регулирование размеров колонии. Этот механизм
регуляции подобен тому, который используется в термостате и называется механиз-
мом обратной связи.
Обратите внимание, что в примере с бактериями (и многими другими живыми
существами) их максимальная численность может достигать нескольких миллионов.
Работать со столь большими числами неудобно, но ничто не мешает нам обозначить
максимальный размер популяции через 1. Проведя это изменение масштаба, получим,
что размер популяции будет варьироваться от 0 до 1 и составлять, например, 0,67,
0,16 и т. д. Зная, какой величине соответствует 1, можно определить фактический
размер популяции. С учетом изменения масштаба дифференциальное уравнение будет
выглядеть так:
у' = гу(Д-у).
Перед тем как рассмотреть уравнение подробнее, выполним последнее действие:
заменим у' на у(/ + 1), у — на у(/). Эти величины обозначают численность бакте-
рий в момент времени t + 1 в будущем и в настоящий момент времени t соответствен-
но. Теперь исходное дифференциальное уравнение приняло вид:
у(/+ 1)=гу(0(1-у(0)-
Именно это выражение использовал биолог Роберт Мэй в 1976 году при ком-
пьютерном моделировании роста популяций.
69
МИКРОБИОЛОГ. ПОКОРИВШИЙСЯ ХАОСУ
Четыре эксперимента в качестве примера
Допустим, что микробиолог проводит четыре эксперимента на компьютере. Обозна-
чим их 1, 2, 3 и 4. Показатель мгновенного роста г составит 0,4; 2,4; 3,0 и 3,6 соот-
ветственно. Во всех экспериментах исходная численность бактерий у(0) одинакова.
Будем считать, что начальное число бактерий составляет 70 % от их максимальной
численности в сосуде, то есть 1. Следовательно, у(0) будет равно 0,7. Обратите
внимание, что в каждом из четырех экспериментов будет применяться разное значе-
ние г, позволяющее определить численность бактерий y(t + 1) для каждого момента
времени t.
Графики, соответствующие экспериментам с бактерией Escherichia coli.
Слева направо: эксперимент №2(г = 2,4), эксперимент №3(г = 3,0) и эксперимент №4(г = 3,6).
Выполним эксперимент № 1 со значением г = 0,4. Сначала определим числен-
ность бактерий по прошествии единицы времени: у(1) = 0,4у(0)(1 — у(0)), подста-
вив значение у(0) = 0,7. Найдя у (1), определим численность бактерий по проше-
ствии двух единиц времени у(2): у(2) = 0,4у(1)(1 — у(1)). Далее вычислим у(3),
у(4) и т. д. вплоть до сколь угодно большого /. К примеру, изучим рост популяции
вплоть до t = 100. В этот момент времени численность бактерий будет равна у(100).
Наконец, представим на графике число бактерий у(/) для каждого момента вре-
мени t. Заметим, что для данного начального значения численности бактерий у(0)
и определенного значения параметра г с помощью компьютера можно получить зна-
чения численности бактерий в разные моменты времени, то есть у(0), у(1), у(2),
..., у(100). Эта последовательность чисел называется орбитой. Зададимся вопро-
сом: куда будет направлена эта орбита? Иными словами, каким будет окончательное
значение численности популяции?
70
МИКРОБИОЛОГ, ПОКОРИВШИЙСЯ ХА >СУ
Чтобы ответить на этот вопрос, проведем все эксперименты согласно вышеопи-
санному принципу, выполнив необходимые расчеты для соответствующих значений г.
В эксперименте № 1 бактерии вымрут: по прошествии определенного времени в сосу-
де не останется ни одной бактерии. Однако с математической точки зрения популяция
достигнет равновесия — это происходит, когда численность популяции не меняется,
то есть уровень ее изменения у’, или, что аналогично, dy/dt, будет равен 0.
В нашем эксперименте численность популяции достигла так называемого точеч-
ного аттрактора у = 0, то есть популяция бактерий вымерла. Это одно из возможных
состояний, к которому может прийти любая популяция. Аттрактор — не более чем
точка или множество точек, к которым стремится или приближается динамическая
система, в нашем эксперименте это орбита, образованная значениями численности
бактерий. Равновесие означает, что система достигла аттрактора и находится в ста-
бильном состоянии, так как dy/dt = 0. Значение у при этом совершенно неважно.
Участь популяции бактерий в эксперименте № 2 будет не столь печальной. Эта
популяция также достигнет точки равновесия, однако численность бактерий зафик-
сируется в точечном аттракторе у = 0,6. В эксперименте № 3 численность бактерий
будет колебаться между определенным максимальным и минимальным значениями
71
МИКРОБИОЛОГ, ПОКОРИВШИЙСЯ ХАОСУ
в зависимости от того, в какой момент времени t производится подсчет численности.
В этом случае также говорят, что система достигла равновесия.
Эта разновидность аттрактора называется предельным циклом. Речь идет о зам-
кнутой орбите, характерной для систем, в которых наблюдаются колебания. С гео-
метрической точки зрения аттрактор действует подобно сточной трубе. Он может
представлять собой точку, кривую или даже фрактал. Следовательно, стабильные
системы, к примеру, рассматриваемая колония бактерий, — это системы, которые
по прошествии определенного времени стремятся к некоторому аттрактору, в то время
как нестабильные системы от аттракторов удаляются.
10 5
Предельный цикл.
Что же произойдет в последнем, четвертом эксперименте? Сразу же видно, что
колебания численности популяции не подчинены никакой закономерности и являются
абсолютно хаотическими. В подобных случаях определить точную численность бак-
терий невозможно: она заметно отличается в зависимости от того, в какой момент мы
производим подсчет. Кроме того, в отличие от первых трех экспериментов, в этом
случае изменение численности бактерий не подчиняется какой-либо схеме. Можно
72
МИКРОБИОЛОГ, ПОКОРИВШИЙСЯ ХАОСУ
сделать вывод: хаос есть отсутствие закономерности колебаний. Подобное поведение
наблюдается при достижении критического значения параметра дифференциального
уравнения. В нашем эксперименте этим критическим значением является г = 3,6.
В этом случае орбита значений численности бактерий у(0), у(1), у(2), у(100)
приближается к так называемому странному аттрактору. Он описывает поведение
таких динамических систем, как климат на Земле, поведение биржевых индексов или
электроэнцефалограмма человека.
500
400
300
200 ‘
100
0 ,
-60
Странный аттрактор.
Изучение природы шаг за шагом
В предыдущей главе вы познакомились с тем, как математическая биология изучает
биологические системы и явления с помощью дифференциальных уравнений. Однако
их использование — не единственный метод изучения динамических систем, а сле-
довательно, не единственный метод моделирования биологических явлений. Ввиду
все более широкого использования компьютеров еще одним популярным методом
являются функции, или отображения. Их применение кажется более простым, чем
использование дифференциальных уравнений. Классическим примером является ло-
73
МИКРОБИОЛОГ. ПОКОРИВШИЙСЯ ХАОСУ
СВОЙСТВА ХАОСА
Одно из свойств хаотических систем заключается в том, что они ведут себя так, словно являются
стабильными и нестабильными одновременно. Это означает, что существует апрактор, к кото-
рому система приближается и от которого она в то же время отдаляется. Благодаря подобному
поведению система обладает очень интересными свойствами. К примеру, хаотические системы
очень чувствительны к изменению начальных условий. Небольшие отклонения, составляющие
несколько десятых от величины ошибки измерения, приводят к тому, что будущее состояние си-
стемы значительно изменяется (в качестве примера можно привести климат Земли). Даже при
известном текущем состоянии хаотической системы нельзя точно спрогнозировать ее поведение
в последующие моменты времени. Рассмотрим в качестве примера колебания биржевых индек-
сов: решение небольшой компании о покупке или продаже акций в зависимости от состояния
индексов, дня и часа может оказать значительное воздействие на биржевые котировки спустя
несколько часов. Таким образом, хотя хаотические системы описываются дифференциальными
уравнениями, они характеризуются сложным поведением, которое объясняется существованием
странного апрактора.Так, например, странным аттрактором в простейшей модели земного кли-
мата является апрактор Лоренца. Еще один странный апрактор - апрактор Хенона, связанный
с тем, как происходит запоминание информации в нейронных сетях мозга человека и животных
(за это отвечают связи между нейронами, или синапсы). Странными апракторами являются
множества Жюлиа, представляющие собой фракталы и объясняющие свойства некоторых био-
логических организмов, в частности бактерий и простейших.
Аттрактор Лоренца.
74
МИКРОБИОЛОГ. ПОКОРИВШИЙСЯ ХАОСУ
гистическое отображение. Вместо дифференциального уравнения Ферхюльста у’ =
= ry(k — у) используется выражение у = ryn(k — у ), которое называется уравне-
нием в конечных разностях. Напомним, что последнее выражение мы использовали
в предыдущей главе, говоря об изучении хаоса.
В уравнениях в конечных разностях п обычно обозначает время t. Теперь, что
очень важно, время является дискретным, то есть принимает конкретный набор
фиксированных значений 0, 1, 2, 3, ..., t, как на циферблате цифровых часов. Об-
ратите внимание, что при использовании логистического отображения в компьютер-
ном моделировании мы ограничиваемся тем, что вычисляем одно и то же выражение
для множества значений. Иными словами, после того как выбраны значения k, г
и начальный размер популяции, к примеру, бактерий, у (0), остается лишь вычис-
лить значения у(1), у(2), у(3) и т. д. Для этого используется выражение:
у(/ + 1) = г*у(/)*(/?-у(0),
где * — оператор умножения, используемый в языках программирования. Мате-
матики обычно говорят, что для заданных начальных условий у(0) последователь-
ность значений у(1), у(2), у(3) и т. д. является орбитой точки у(0). Если вы хотя бы
немного знакомы с программированием (на любом языке), то сможете убедиться, что
программа для вычисления орбиты точки сводится к следующему набору операций:
‘начальные условия и параметры логистического отображения
у(0): г: k
‘максимальное время моделирования
tmax
‘уравнение в конечных разностях
For t — 0 ТО tmax
y(t + 1) = r*y(t)*(k - y(t))
print t, y(t + 1)
End
Отличие этого метода от использования дифференциальных уравнений состоит
в том, что интегрирование заменяется итерированием. Иными словами, нам больше
не требуется вычислять интегралы — вместо этого мы снова и снова повторяем одни
и те же математические операции. Повторные вычисления позволяют определить ото-
75
МИКРОБИОЛОГ, ПОКОРИВШИЙСЯ ХАОСУ
бражение. Так, экологи определяют будущее состояние системы y(t + 1), к примеру
озера или пастбища, на основе текущего состояния y(f). Если повторить эти действия
снова и снова, можно предсказать изменения, которые произойдут в системе.
Что произойдет с предыдущей программой, если вместо логистического отобра-
жения использовать уравнение Ферхюльста? В этом случае потребуется уже не ите-
рировать, а интегрировать, о чем мы рассказали в предыдущей главе.
Интегрирование дифференциального уравнения можно выполнить разными ме-
тодами, которые используются не только в экологии, физиологии и фармакологии,
но и в экономике, химии и многих других науках, включая политологию.
Методы интегрирования дифференциальных уравнений с помощью компьютера
называются численными методами интегрирования. Самый простой и популярный
из них метод Эйлера, однако более точным является метод Рунге — Кутты чет-
вертого порядка. Любой из этих методов позволяет найти приближенные значения
у(1), у(2), у(3) и т. д. Чтобы получить точные значения, нам потребовалось бы под-
ставить в полученное выражение значения t = 1, 2, 3 и т.д. При использовании
метода Эйлера программа будет выглядеть так:
‘начальные условия и параметры уравнения Ферхюльста
У(О): г: k
‘максимальное время моделирования
tmax
‘увеличение времени
Inct
‘дифференциальное уравнение
For t = О ТО tmax
y(t + 1) - y(t) + Inct*( r*y(t)*(k ~ y(t)) )
print t, y(t + 1)
End
76
МИКРОБИОЛОГ, ПОКОРИВШИЙСЯ ХАОСУ
Сразу видно, что решение дифференциального уравнения на компьютере по мето-
ду Эйлера заключается в том, чтобы предсказать будущее состояние системы y(t +
+ 1) по текущему состоянию y(t), как если бы мы выполняли итерирование уравнения
МАГИЯ КОМПЛЕКСНЫХ ЧИСЕЛ
С древних времен человек чувствовал непреодолимое желание превзойти самого себя, и это
стремление лежит в основе всевозможных изобретений в самых разных областях, однако осо-
бенно ярко творческие способности человека проявились в математике. В качестве примера
приведем комплексные числа. Существует ли число, квадрат которого является отрицательным?
Существуют ли квадратные корни из отрицательных чисел? Утвердительный ответ на оба этих во-
проса дали итальянские математики Джероламо Кардано и Рафаэль Бомбелли в середине XV века.
Найденные ими числа названы мнимыми. С того времени из соображений удобства мнимая еди-
ница обозначается буквой /. Ее значение равно i=4-i или, что аналогично, /2 - -1. На основе
мнимой единицы были определены так называемые комплексные числа. Ученые и инженеры
используют их в самых разных областях с той же легкостью, с какой мы в повседневной жизни
применяем 10,23, 6 1/2 и т. д. Комплексные числа состоят из двух частей - вещественной а
и мнимой Ь. К примеру, комплексными являются 3 + 2/, 6 + 7/ и т. д. В стандартной форме они
записываются в виде а + Ы (обратите внимание, что а и b - вещественные числа). По-настоящему
удивительно то, что комплексные числа находят широкое применение в биологии, инженерном
деле, прикладной математике и т.д. Подробнее о действиях с ними рассказано в приложении.
Слева — представление комплексных чисел. На оси X
откладывается вещественная часть, на оси Y — мнимая. В центре представлена сумма двух
комплексных чисел (1 + 3i) + (2 + /), справа — произведение (1 + 7i) и мнимой единицы i.
77
МИКРОБИОЛОГ, ПОКОРИВШИЙСЯ ХАОСУ
в конечных разностях. Сделаем оговорку: в итоговые вычисления мы включаем произ-
ведение времени, прошедшего между y(t + 1) и y(t), и выражения, которое является
решением дифференциального уравнения. Можно убедиться, что выражение Эйлера
не более чем уравнение прямой, угол наклона которой описывается дифференциаль-
ным уравнением. С помощью этой касательной прямой в точке можно предсказать
будущее состояние или поведение системы у(/ + 1) с большей точностью, чем при
использовании разностных уравнений.
Помимо дифференциальных уравнений и отображений для моделирования дина-
мических систем используются клеточные автоматы.
Является ли природа фрактальной?
Природа состоит из неодушевленных предметов (облаков, гор, побережий, снежи-
нок), живых организмов (растений, микроорганизмов, водорослей, кораллов) и их
структур (бронхиальной системы, кровеносной системы или нейронных сетей), в ко-
торых снова и снова повторяется один и тот же узор. На каком бы уровне ни произво-
дились наблюдения, в любом масштабе (на расстояниях порядка нескольких метров,
сантиметров и даже миллиметров) существует некая повторяющаяся схема. Подобные
структуры, обладающие самоподобием, называются фракталами.
Основная идея проста: природа выстраивает структуры, следуя принципу наи-
меньших затрат, а затем этот шаблон или образец повторяется в разном масштабе.
Классический пример — ветвление растений. Нетрудно видеть, что отдельная боль-
шая ветвь и отходящие от нее мелкие ветви есть не что иное, как уменьшенная версия
всего растения в целом.
Ветвление растений —
пример фрактальной
структуры в природе.
78
МИКРОБИОЛОГ, ПОКОРИВШИЙСЯ ХАОСУ
Сегодня считается, что температура здорового тела, дыхательный ритм и арте-
риальное давление подобны фракталам. Если мы будем измерять температуру тела
и артериальное давление каждые десять минут, раз в неделю или в месяц, а затем
представим данные графически, построенный график будет обладать самоподобием.
Но во время заболевания самоподобие на графиках не наблюдается. Заболевание —
это утрата системой сложности, когда физиологические переменные перестают опи-
сываться фракталами.
Другой пример — электроэнцефалограмма человека. Ее аттрактор также име-
ет фрактальную природу. Размерность аттрактора D (о понятии размерности по-
говорим чуть позже) будет неким числом. С точки зрения нейрофизиологии по-
настоящему интересно то, что значение D зависит от состояния человека и отлича-
ется у здоровых людей в состоянии бодрствования, здоровых людей, находящихся
под воздействием анестезии, и у эпилептиков.
Какие свойства фракталов делают их столь интересными? Их размерность
не выражается целым числом, а мы к этому привыкли в нашем мире, который ма-
тематики называют евклидовым. Так, лист бумаги имеет длину и ширину, поэтому
его размерность D равна 2. Если мы проведем прямую линию на листе бумаги, ее
размерность D будет равна 1. Комната, в которой стоит стол, на котором лежит лист
бумаги, на котором мы провели линию, имеет длину, ширину и высоту. Это означает,
что комната имеет три измерения, и ее размерность D = 3. Говоря, что размерность
фракталов не выражается целыми числами, мы имеем в виду, что их размерность D
равна 1,8, 2,6 или другим подобным значениям.
Еще фракталы бесконечны: независимо от масштаба наблюдений кажется, что они
никогда не заканчиваются. С помощью компьютерных программ Fractint или Ultra
Fractal можно убедиться, что увеличивать определенный участок фрактала можно
до бесконечности. Во время эксперимента вы также увидите, что фракталы обладают
самоподобием, иными словами, вам будет казаться, что вы постоянно видите один
и тот же узор, который проявляется во всей красоте и сложности и не изменяется
в зависимости от масштаба наблюдений.
79
МИКРОБИОЛОГ, ПОКОРИВШИЙСЯ ХАОСУ
ИЗОБРАЖЕНИЕ ФРАКТАЛОВ НА КОМПЬЮТЕРЕ
Одна из классических компьютерных программ для изображения фракталов называется Fractint
Существуют ее версии для Windows, Linux и Mac OS. Одна из самых интересных особенностей
программы - широкие возможности увеличения изображений: она позволяет просматривать
фракталы с точностью до 1600 знаков после запятой. Несомненно, это одно из лучших приложе-
ний с точки зрения мощности алгоритмов и математической точности.
В подобных программах важен набор доступных инструментов. К примеру, программа Ultra
Fractal позволяет изменять цвета фракталов, использовать слои и маски, генерировать анима-
ции и т д. Используя слои вы сможете объединить несколько фракталов и получить поистине
прекрасный рисунок. Также следует отметить программу Aphysis, предназначенную для изобра-
жения «туманных» фракталов. Она создана Марком Таунсендом и позволяет на основе заранее
определенных фракталов генерировать их вариации. Некоторые приложения отличаются осо-
бым обилием настроек. К примеру, Fractal Explorer содержит 148 классов фракталов и позволя-
ет создавать странные аттракторы (аттракторы в 3D), включает системы итерируемых функций
и 22 класса так называемых и .атернионов. кроме того, в этой программе можно генерировать
спецэффекты с помощью 117 фильтров, она содержит 12 алгоритмов выбора цвета, поддерживает
создание анимаций в формате AVI и т.д. Интересный компонент приложения - FEParser (© Kyle
McCord) - компилятор формул, ориентированный на продвинутых пользователей, с помощью
которого можно создавать собственные фракталы. Другие программы, например XenoDream, вы-
деляются благодаря художественной составляющей. Они позволяют добавлять текстуры и эффекты
освещения, создавать фракталы в 3D, анимации и стереограммы. Мы упомянули лишь некоторые
из множества приложений, предназначенных для решения самых разных задач, от генерации
интересных картинок до математического изучения фракталов или создания на их основе на-
стоящих произведений искусства.
Палитра цветов
программы Fractint.
80
МИКРОБИОЛОГ. ПОКОРИВШИЙСЯ ХАОСУ
Этими свойствами обладают все фракталы, созданные математиками. Иными
словами, фракталы, сгенерированные на компьютере, — это идеальные математи-
ческие объекты, в отличие от неидеальных фракталов конечного размера, созданных
природой. И все же красота и сложность фракталов в природе не перестают удив-
лять нас.
Снежинка (иллюстрация Уилсона Бентли) и соответствующий ей фрактал — снежинка Коха.
Множества Жюлиа и Мандельброта
Французский математик Гастон Жюлиа первым начал исследовать фракталы. Он
сделал ряд важных открытий, однако умер в 1978 году, так и не став известным.
Отчасти это объяснялось тем, что компьютеры в то время еще не могли изобразить
всю красоту и кажущуюся сложность фракталов. Жизнь Жюлиа вообще сложилась
непросто: в Первую мировую войну он был ранен в лицо и вынужден был всю жизнь
скрывать его часть.
Фрактальное множество Жюлиа.
81
МИКРОБИОЛОГ. ПОКОРИВШИЙСЯ ХАОСУ
Рассмотрим функцию вида Z = Z2 +С, где Z — переменная, С — константа.
Значение левой части этого выражения на итерации п + 1 равно квадрату пере-
менной Z на предыдущей итерации, п, увеличенному на константу С. Само по себе
это выражение не представляет особого интереса, если только переменная Z и кон-
станта С не являются комплексными числами. Именно этот случай и рассмотрел
Жюлиа. Несколько лет спустя Бенуа Мандельброт также изучил этот класс итера-
тивных выражений с помощью компьютера. В некотором роде Мандельброт про-
должил исследование фракталов, начатое Гастоном Жюлиа, и внес огромный вклад
в их популяризацию. В 1958 году Мандельброт начал работать в исследовательском
центре IBM в США, где изучал возможности использования фракталов не только
в физике, но и в других дисциплинах: гидродинамике или даже экономике. Именно
он в 1975 году ввел термин «фрактал» для обозначения подобных объектов. Изучив
множества Жюлиа с помощью компьютера, Мандельброт обнаружил множества,
которые теперь носят его имя — множества Мандельброта.
Фрактал Мандельброта.
В 1982 году была опубликована книга Мандельброта под названием «Фракталь-
ная геометрия природы», в которой он объяснил, как природа создает фрактальные
формы (к примеру, линии побережий, горы, растения, кровеносные сосуды и легкие).
На множествах Мандельброта рассматривается орбита точки 0. Ученый предложил
изучить последовательные итерации выражения Z = Z2 + С для частного случая
Zo = 0. Его идея заключалась в том, чтобы определить, каким в этих условиях будет
множество значений С, для которых рассматриваемая орбита точки не будет уходить
в бесконечность. Именно это множество значений и стало называться множеством
Мандельброта. На множествах Жюлиа, напротив, комплексная константа С имеет
82
МИКРОБИОЛОГ. ПОКОРИВШИЙСЯ ХАОСУ
фиксированное значение, для которого изучается поведение функции, орбита и зна-
чения Х(), Z2, ... и т. д. Поскольку компьютер не может работать с комплексными
числами, переменная и константа записываются в следующем виде:
Z = X + iY
С = р + iq,
а функция Z = Z2 + С раскладывается на вещественную часть X и мнимую часть Y.
Компьютер позволяет представить точки орбиты на комплексной плоскости, то есть
в декартовой системе координат, и изобразить точки, для которых выражение
\ X2 + Y2 меньше заданного значения и орбита которых не уходит в бесконечность.
Эта орбита является границей множества точек, орбиты которых уходят в бесконеч-
ность, и это множество называется множеством Жюлиа.
«Игра в хаос» Барнсли
Хаотические системы представляют собой разновидность динамических систем, что
доказал преподаватель Технологического института Джорджии Майкл Барнсли,
объяснявший теорию хаоса на примере математической игры под названием «Игра
в хаос». Барнсли показал любопытную особенность хаотических систем: их беспо-
рядочность была лишь кажущейся, и в конечном итоге в хаотической системе можно
было увидеть одну и ту же фигуру. Эта фигура, очевидно, являлась аттрактором,
в частности фракталом, однако проявлялась она только на поздних этапах модели-
рования. Напомним, что аттрактор — это не более чем точка или множество точек,
к которым стремится или приближается динамическая система. «Игра в хаос» заклю-
чается в графическом представлении точек (х, у), которые описываются следующим
матричным выражением:
Изначально расположение полученных точек будет казаться случайным. Но бли-
же к концу эксперимента, после того как мы изобразим достаточное их количество,
всегда будет виден странный аттрактор — треугольник Серпинского.
Идея Барнсли была простой, элегантной и в то же время удивительной. Любая
точка, которую мы изобразим в будущем, то есть в момент времени t + 1, будет
83
МИКРОБИОЛОГ. ПОКОРИВШИЙСЯ ХАОСУ
иметь координаты (х/ + 1. У,+ 1)- Они будут рассчитаны на основе координат (х, yt)
точки, которую мы изобразим в момент времени t. Обратите внимание, что здесь
речь идет об отображении, схожем с логистическим отображением или уравнением
Ферхюльста. О том, что такое матрицы и как они применяются в математической
биологии, поговорим в следующей главе. Пока лишь укажем, что приведенное выше
выражение можно преобразовать в два более привычных и определить координаты
новой точки на основе следующих преобразований:
х х| — ах +Ьу +е,
Г+1 t ’
У,+1=а,+(1у, + /.
Эксперимент Барнсли заключался в определении трех преобразований.
Преобразование 1 Преобразование 2 Преобразование 3
= 0,5xt+ 1 Ум = °-5УЛ 1 х(+1 = 0,5xf + 50 Ум = °-5yt+ 1 хм = °-Ч+ 1 Ум = °’5У(+ 50
Для произвольной начальной точки с координатами (х, у) путем жеребьевки опре-
делялось число 1, 2 или 3. К примеру, если при броске кубика выпадало 1 или 2 очка,
выбиралось преобразование 1, если выпадало 3 или 4 очка — преобразование 2, если
выпадало 5 или 6 очков — преобразование 3. С помощью соответствующих урав-
нений определялись координаты новой точки (х/ + ;, У1 + 1)- Далее путем жеребьевки
выбиралось новое преобразование, которое применялось к предыдущей точке для
определения координат новой точки, и т. д.
Обратите внимание, что в эксперименте, результатом которого является треуголь-
ник Серпинского, and всегда равны 0,5, b и с — 0. Значения ей/ изменяются для
каждого преобразования. Другие природные фракталы, например листья растений,
ветки папоротника и т. д., можно получить, рассмотрев различные значения а, Ь, с,
d, ей/.
Описанная процедура, названная системой итерируемых функций, представляет
собой один из наиболее интересных методов построения фракталов на компьюте-
ре. Эксперимент привел к удивительному результату: многие биологические формы
и структуры являются фракталами. Если, к примеру, мы применим преобразова-
ния (повороты, переносы, изменение масштаба) к точкам, представляющим клетки,
то получим структуру, которая будет фракталом, изображающим, к примеру, лист
растения.
84
Глава 4
Судоку жизни
Одна из классических научных задач — наблюдение за природой и проведение экс-
периментов. Наблюдение явления подразумевает сбор каких-либо данных. В качестве
примера приведем изучение загрязнения окружающей среды, в котором как индика-
торы используются некоторые виды лишайников, количество мутаций определенной
бактерии или вес мышей из одного помета. На этом этапе исследования ученый под-
считывает, например, количество муравьев, проходящих через определенное место
за минуту, или число красных кровяных телец. Вместо подсчетов в некоторых случаях
могут требоваться измерения, например кислотности среды, веса, роста или любых
других показателей, значение которых, по определению измерения, будет содержать
несколько десятичных знаков.
Таблицы, судоку и матрицы
Данные, собранные в ходе эксперимента, объединяются в таблицы. Допустим, что
в оранжерее на четырех грядках растет по семь растений, при этом на каждую гряд-
ку вносится свое удобрение. Чтобы выяснить, дает ли оно положительный эффект,
по прошествии определенного времени производится подсчет числа листьев на каж-
дом растении. Обозначим это число через х. Подобные данные обычно представлены
в виде таблиц.
1 2 3 4
1 хп Х12 Х13 Х14
2 *21 Х22 Х23 Х24
3 Х31 Х32 хзз Х34
4 Х41 Х42 Х43 Х44
5 Х51 Х52 Х53 Х54
6 Х61 Х62 Х63 Х64
7 Х71 Х72 Х73 Х74
85
СУДОКУ жизни
К примеру, х23 будет обозначать число листьев растения 2 с удобрением 3, х74 —
число листьев растения 7 с удобрением 4. Математики проводили подобные экспе-
рименты с древних времен, используя так называемые латинские квадраты, то есть
таблицы или матрицы, в которых символ (число или сочетание нескольких символов)
встречается в каждой строке и каждом столбце только один раз. Разновидностью
латинского квадрата является популярная сегодня игра судоку.
Аа ву С6 DP
вр Аб Dy Са
су Da Ар В6
D6 Ср Ва Ау
Слева — пример популярной японской головоломки судоку, справа — латинский квадрат.
Теперь предположим, что мы хотим дать общее определение таблице, использо-
ванной в эксперименте с растениями и удобрениями. Можно сказать, что даны m
растений и п видов удобрений, и записать представленную выше таблицу в круглых
скобках:
Такая форма представления данных называется матрицей. Таким образом, ма-
трица размера m х п — это всего лишь множество из тп х и элементов, записанных
в m строк и п столбцов. Матрицы обычно обозначаются заглавными буквами — А,
В, С и т. д. Они позволяют удобно представлять не только данные, но и системы
уравнений. Рассмотрим в качестве примера следующую систему линейных уравне-
ний:
86
СУДОКУ жизни
Зх+2у = 7
5х—у = 4.
В матричном виде эту систему уравнений можно представить так:
3 2
5 -1
7
4
У
С помощью матриц можно проанализировать также химическую структуру мо-
лекулы. К примеру, если мы присвоим произвольные обозначения атомам углеро-
да С в молекуле витамина А, или ретинола, как показано на рисунке
то молекула витамина А будет представлена следующей матрицей.
0 10 0
10 10
0 10 1
0 0 10
0 0 0 1
10 0 0
0 0 0 0
0 0 0 0
0 0 0 0
0 0 0 0
0 0 0 0
0 0 0 0
0 0 0 0
о о о о
0 10 0 о о о о о о
0000000000
0000000000
1 000000000
0100000000
1010000000
0101000000
0010100000
0 0 01010000
0000101000
0000010100
0000001010
0000000101
0000000010
Обратите внимание, что х = 1, если между атомами i и j существует связь, если же
связь между атомами отсутствует, х.. = 0.
87
СУДОКУ жизни
ПРЕЛЕСТЬ МАТРИЦ - В ИХ РАЗНООБРАЗИИ
Всевозможные обозначения, связанные с матрицами, встречаются очень часто. Разъясним неко-
торые популярные термины.
Квадратная матрица - это матрица, в которой число строк и столбцов одинаково:
2 3-1
5 2 8
9 7 6
Симметричная матрица - это квадратная матрица, в которой выполняется соотношение/ = х^:
Единичная матрица - это квадратная матрица, все элементы которой равны 0, и только эле-
менты главной диагонали равны 1. Единичная матрица обозначается буквой Е.
1 О О '
О 1 о
о
J О 0 1 ,
Диагональная матрица - это квадратная матрица, все элементы которой равны 0, за исклю-
чением элементов главной диагонали:
' 2 О О '
0-1 о
о
k 0 0 0 5 >
Нулевая матрица - матрица (необязательно квадратная), все элементы которой равны 0:
' о о о '
ООО
ООО
< 0 0 0 >
Треугольная матрица - это квадратная матрица, в которой все элементы расположенные над
главной диагональю или под ней, равны 0. Слева представлен пример верхнетреугольной матри-
цы, справа - нижнетреугольной.
8
О
О
3
-5
О
-2 Y 2 0 0 '
9 -14 0
1 Л 5 6 7 >
88
СУДОКУ жизни
Соотношение между элементами представимо с помощью графа. К примеру, эле-
менты нейронной сети или клеточного метаболизма могут быть представлены узлами,
связанными между собой дугами. Таким образом, можно сопоставить матрицу графу,
как мы показали в примере с витамином А.
С помощью матрицы можно представить экспериментальные данные, системы
уравнений и графы. И по-настоящему важно, что над матрицами мы можем вы-
полнять различные действия. С середины XIX века известны правила операций над
«данными, расположенными в строках и столбцах», к примеру, сложение и умножение
матриц. С того времени была создана матричная алгебра, составляющая основу мно-
гих количественных методов математической биологии и других дисциплин. К при-
меру, изучение динамических систем в экологии или физиологии, анализ и решение
многочисленных задач генетики, как правило, проводятся с помощью операций над
матрицами.
Операции над матрицами
В этом разделе мы опишем некоторые наиболее частые операции над матрицами,
знание которых поможет понять многочисленные способы применения матриц.
Сложение
Это одна из простейших операций над матрицами. Допустим, в эксперименте рас-
сматриваются две матрицы размером 2x2, которые мы обозначим Л и В:
^11 ^12
^21 ^22
такие, что
,в=
О -1 ) I 3 1 J
Как будет выглядеть матрица С, равная их сумме, то есть А + В? Матрица С
образуется последовательным сложением элементов исходных матриц: си = + Ьп,
С12 — а12 + ^12’ С21 — а21 + ^21 И С22 “ ° 22 ^22'
89
СУДОКУ жизни
с=
I 3 о )
Сложение матриц возможно в том случае, если они имеют одинаковый размер.
Предположим, что в лаборатории при изучении детородной функции человека ис-
пользуется модель, в которой яйцеклетка или сперматозоид, несущие доминантный
ген/4, обозначены 1, рецессивный ген а — 0. Если сперматозоид оплодотворяет яй-
цеклетку, то все возможные эмбрионы для рассматриваемого гена будет описывать
следующая сумма матриц:
1 1
0 0
1 0
1 0
7 к
В этом примере 2, 1 и 0 соответствуют эмбрионам с генами АА, Аа и аа соот-
ветственно.
Вычитание
Разность матриц А — В определяется как сумма Л + (—1)В. Чтобы найти разность
матриц А и В из первого примера:
2 4
0 -1
5 -6
сначала нужно сменить знак элементов матрицы В:
-5 6
I -3 -1
и сложить полученную матрицу с А. Результат будет искомой разностью:
1-3 -2 J
90
СУДОКУ жизни
Умножение
Еще одна простейшая операция над матрицами — это умножение матрицы на число.
Пусть k — произвольное число, к примеру константа или даже функция у(х) (мате-
матики называют подобные величины скалярными), А — матрица.
Произведение k • А:
равно
Допустим, что на земельном участке со сниженной плодородностью почвы испы-
тываются четыре удобрения с разным соотношением азота, фосфора и калия. После
внесения удобрений на четыре разных участка уровень всхожести семян на первом
участке составил 5 %, на втором — 15 %, на третьем — 8 %, на четвертом — 27 %.
Пусть на каждом участке посеяно 80 семян. Обозначим всхожесть на первом, вто-
ром, третьем и четвертом участках через аи, а12, а21 и а2Г Число семян, взошедших
на каждом участке, будет равно:
0,05 0,15 U 4 12
0,08 0,27 J [ 6 22
Умножение матриц очень часто используется в математической биологии и других
дисциплинах. Чтобы матрицы А и В можно было умножить друг на друга, число
столбцов и матрицы Л должно быть равно числу строк п матрицы В. Произведением
матриц Л и В будет матрица С, имеющая т строк и р столбцов. Будем использовать
обозначения, которые обычно применяются в математической литературе, и проил-
люстрируем операцию умножения матриц.
Пусть даны две матрицы А и В:
91
СУДОКУ жизни
«11 «12
«21 ^22
1 а ->
ги! m2
ь., ••• ь,
12 1 р
^22 Ь2р
ь, ••• ь
п2 пр
Их произведением будет матрица С:
С11 С12
С _ С21 С22
С . С -
ml m2
Элемент с., матрицы С в общем виде задается следующим выражением:
... + а. b ..
il 1/ iZ Zj in nj
Таким образом, чтобы вычислить элемент с, нужно взять элементы аи, а12,..., Ьп,
... и найти значение выражения а,,Ь,, + а.3Ь^. + ... + а. b
Проще всего продемонстрировать вычисление произведения матриц на примере.
Пусть даны матрицы А и В:
А= 0 -1
< 7 4
6 ,В= 6 1
5 J ^-3 4,
Найдем их произведение А • В, выполнив следующие действия:
(3)-(2)+(2)-(6)+(1)-(-3)
(0).(2)+(-1).(6)+(6)-(-3)
(7)-(2)+(4)-(6)+(5)-(-3)
(3)-(0)+(2).(1)+(1)-(4)
(0).(0)+(-1).(1)+(6)-(4)
(7)-(0) + (4)-(1) + (5)-(4)
Получим матрицу:
92
СУДОКУ жизни
-24 23 .
< 23 24 >
Важно учесть, что сумма матриц А + В не изменится, если мы поменяем слагае-
мые местами (следовательно, операция сложения матриц является коммутативной),
в то время как произведение матриц не обладает этим свойством: произведение ма-
триц Л • В не равно произведению В • А. С учетом этого одним из самых интерес-
ных действий является умножение матрицы А на матрицу В, состоящую из одного
столбца, то есть умножение Л * В г Матрица, состоящая из одного столбца, на-
зывается вектор-столбцом.
Умножение матрицы на вектор и применение этой операции
Чтобы показать, как применяется произведение матрицы на вектор, представим, что
нейробиолог составляет модель нейронной сети. В этой сети группа нейронов (обо-
значены кругами), которые мы будем называть афферентными, или чувствительны-
ми, получает импульсы из внешнего мира, к примеру посредством органов чувств.
Афферентные нейроны отправляют сигналы другим нейронам, которые называются
эфферентными, или двигательными, через соединения, или синапсы. Задача эффе-
рентных нейронов — отправить ответ на некоторый входной сигнал, поступивший
к афферентным нейронам. Этот ответ поступает к мышцам, железам и т. д. Обозначим
входной сигнал, поступающий к афферентным нейронам, вектором и. Элементами
вектор-столбца хр х2, ..., х. будут, к примеру, характеристики некоторого объекта.
Обозначим через у., у7,... у. составляющие вектора v, который будет обозначать
ответ эфферентных нейронов после обработки входного сигнала.
Модель нейронной сети.
93
СУДОКУ жизни
Наконец, представим синапсы между афферентными и эфферентными нейронами
в виде матрицы М. Назовем ее матрицей памяти. Каждый элемент с., этой матрицы
обозначает связь между входным, или афферентным, нейроном i и выходным, или
эфферентным, нейроном j. Учитывая вышесказанное, модель нейронной сети можно
сформулировать с помощью матричной алгебры. Имеем:
Если нейробиолог, используя эту модель, захочет узнать ответ эфферентного ней-
рона № 2, зная все остальные значения, ему достаточно будет вычислить:
у, = с91х. + с„х9 + ... + с9.х..
^2 21 1 22 2 2/ 1
А если нужно определить выходное значение для первого эфферентного нейрона?
В этом случае достаточно вычислить yt — с11%1 + с12%2 + ... + сгх..
Пусть дана нейронная сеть с тремя входными, или афферентными нейронами,
и тремя выходными, или эфферентными нейронами. Матрица связей, или синапсов,
между нейронами М приведена ниже:
0,2 0,6 0,8
0,3 0,9 0,4
0,1 0,2 0,3
Если к слою входных нейронов поступает извне следующий сигнал:
0
1)
каким будет выходное значение для первого эфферентного нейрона? В соответствии
с описанной моделью имеем:
94
СУДОКУ жизни
0,2
0,3
0,1
0,6
0,9
0,2
0,8
0,4
0,3
У1
у2
Уз
1
0
1
Искомое выходное значение равно у1 = 0,2 • 1 + 0,6 • 0 + 0,8 • 1, и, как следствие,
У,= 1.
Сегодня этот класс математических моделей используется для распознавания об-
разов — букв, чисел, фотографий и т. д. в системах искусственного интеллекта.
Транспонирование матриц
Еще одна привычная операция над матрицами называется транспонированием. Для
того чтобы получить транспонированную матрицу А‘ для матрицы А, достаточно
поменять строки исходной матрицы на столбцы. Пусть дана матрица А:
А= о
7 -1
1 2
4 3 >
транспонированная матрица А' будет выглядеть так:
6 0-7
7 1 4
<-12 3 )
Транспонированная матрица определяется мгновенно. В самом деле, если мы
транспонируем транспонированную матрицу, то есть найдем (Л')\ то вновь получим
матрицу Л. Покажем, где применяется эта операция.
Определители
Предположим, что ученый работает с моделью, в которой определена квадратная
матрица, то есть матрица с равным числом строк и столбцов. В этом случае суще-
95
СУДОКУ жизни
ствует особое число, соответствующее матрице, которое указывает на некоторые ее
любопытные свойства. Это число называется определителем матрицы. Рассмотрим
простейший случай — матрицу 2x2 (две строки и два столбца), элементы которой
обозначим через а, Ь, с и d:
Определитель матрицы А будет равен:
Значение определителя будет равно а • d — b • с. Иными словами, нужно найти про-
изведение элементов на главной диагонали и вычесть из него произведение элементов
на побочной диагонали. Определитель матрицы А будет равен 2, так как:
Швейцарский математик Габриэль Крамер (1704-1752) сформулировал
правило, носящее его имя, которое позволяет решать системы линейных
уравнений с помощью определителей.
96
СУДОКУ жизни
Отметим, что мы заменили круглые скобки, типичные для матрицы, двумя прямы-
ми линиями — именно так обозначаются определители. Любопытно, что определите-
ли были созданы раньше, чем матрицы, и были известны в Древнем Китае за 300 лет
до Рождества Христова. Древние китайцы использовали понятие, схожее с понятием
определителя, располагая неизвестные системы линейных уравнений на шахматной
доске. Европейские математики впервые применили определители для решения систем
линейных уравнений лишь в 1750 году (это сделал Габриэль Крамер). В XIX веке
с определителями работали другие математики, в частности Коши. Матрицы по-
явились позднее, так как нужно было дать какое-то название объекту, для которого
рассчитывались определители.
Определитель квадратной матрицы размером 3x3
Мы уже показали, как вычислить определитель второго порядка. Сделаем еще один
шаг вперед. Пусть дана квадратная матрица третьего порядка:
Будем считать, что каждому ее элементу соответствует знак + или —, как если бы
речь шла о кристалле хлорида натрия, то есть обычной поваренной соли:
97
СУДОКУ жизни
Выберем, к примеру, первую строку матрицы и исключим ее из рассмотрения.
Затем исключим элементы первого столбца матрицы:
Выполним над элементами матрицы следующие операции:
Й11 (й22 а33 а23 а\2^'
Обратите внимание, что ап положительно, так как этому элементу матрицы со-
ответствует знак +.
После того как мы исключили из рассмотрения первую строку и первый столбец
матрицы, оставшиеся элементы образуют новую матрицу. Определитель полученной
матрицы называется минором М.., где i и j — номер строки и столбца, исключенных
из рассмотрения. В нашем примере z = 1, j — 1.
Выполним аналогичные действия для второго столбца матрицы:
Учитывая, что элемент а12 имеет знак —, получим:
й12 (й21 ' й33 й23 ’ Й31 )'
Повторим аналогичные действия для третьего столбца:
С учетом того, что а13 имеет знак +, получим:
98
СУДОКУ жизни
Й21 ^22
Й31 й32
й13 ’ (й21 ’ й32 й22 " Й31 )•
Теперь, чтобы вычислить определитель матрицы А, нужно свести полученные
выше результаты в одно выражение:
Й11 (й22 " й33 й23 ' й32 ) й12 (й21 ’ й33 й23 ' Й31) й13 (й21 ’ й32 й22 ' Й31)’
Пусть дана матрица А:
2 4 1
3 7 5
6 1 4
Ее определитель вычисляется следующим образом:
Предположим, что даны три вектора, исходящие из одной точки. Допустим, их
координаты таковы: м=(2,—1,4), р = (3,3,— 2) ий) = (—3,2,1). Если мы вычислим опре-
делитель:
2-14
3 3-2
-3 2 1
получим 71. Что означает это число? Поскольку в нашем примере векторы исходят
из одной точки, значение определителя равно объему параллелепипеда, построенного
на этих трех векторах.
99
СУДОКУ жизни
Как делить матрицы. Обратные матрицы
и их применение в биологии
Любопытно, что деление матрицы на матрицу невозможно. Однако на помощь придет
математическая смекалка. Допустим, что мы хотим разделить 5 на 2, то есть найти
значение 5/2, при этом использовать операцию деления нельзя. Напомним, что:
5 1 1
— = 5- — и5-~ = 5-2
2 2 2
Следовательно, если мы заменим числа 5 и 2 матрицами А и В, получим:
где В-1 — матрица, обратная В. Обратите внимание, что произведение В • В-1 бу-
дет равно единичной матрице Е. Отметим, что матрица В должна быть квадратной,
то есть иметь одинаковое число строк и столбцов. Кроме этого, матрица В будет иметь
обратную только в том случае, если ее определитель отличен от 0.
Найти обратную матрицу для матрицы 3x3 несложно, хотя для этого потребу-
ются трудоемкие вычисления. Читатель легко найдет всю интересующую информа-
цию по этому вопросу самостоятельно. Обратную матрицу для матрицы 2x2 очень
просто найти следующим способом. Пусть дана матрица Л:
Обратная ей матрица А 1 определяется напрямую. Она имеет вид:
1 d —b
ad — bc\^—c а
Напомним, что 1/(ас/— be) — величина, обратная определителю матрицы. При-
менив программу символьных вычислений Derive, найдем матрицу, обратную матри-
це Л (не будем приводить все промежуточные действия):
100
СУДОКУ жизни
6 9 1
2 7 8
Если мы запишем в программе выражение А (—1), то получим А \ то есть об-
ратную матрицу:
65 19 43
— —— —
223 223 223
46 14 27
_ I .
223 223 223
24 17 15
223 223 223
Обратные матрицы часто используются в трехмерном компьютерном модели-
ровании, а умножение матриц полезно для обсчета поворотов, например при ком-
пьютерном моделировании поворота головы динозавра. Подобные модели широко
применяются в биомедицине, а обратные матрицы — при шифровании сообщений,
а также в некоторых основных статистических методах многовариантного анализа,
который представляет собой совокупность статистических методов, применяющихся
для анализа данных в биологии и медицине. Также операции над матрицами исполь-
зуются для решения систем линейных уравнений, о чем мы расскажем в следующей
главе.
Матрицы и горошины: законы Менделя
С исторической точки зрения законы Менделя, сыгравшие важную роль в зарож-
дении генетики, не только знаменуют одну из важнейших вех в биологии, но и пред-
ставляют собой прекрасный пример полных факторных экспериментов. В этом разделе
мы представим элементарную модель знаменитых законов наследования, в которой
используются матрицы.
Первый закон Менделя, или закон единообразия гибридов (Аа) первого поко-
ления Fp был выведен экспериментально следующим образом. Представьте, что мы
скрестили два растения, относящихся к разным чистым линиям (АА и аа). У расте-
ния АА все горошины желтые, у растения аа — зеленые. Скрещивание выполняет-
ся методом перекрестного опыления: мы отрезаем ножницами тычинку одного рас-
101
СУДОКУ жизни
тения, например аа, чтобы избежать самоопыления. Затем мы собираем пинцетом
пыльцу с другого растения, АА, и переносим ее на первое растение, аа. Изучив по-
томство, которое Мендель называл первым поколением, или F}, можно убедиться,
что все горошины имеют желтый цвет и принадлежат к гибридному типу Аа.
АА
аа
Первый закон Менделя.
После экспериментов, которые помогли Менделю сформулировать первый закон
наследования, ученый захотел узнать, отличаются ли желтые горошины (F^, получен-
ные в ходе эксперимента, от горошин растений чистой линии (АА). Чтобы найти ответ
на этот вопрос, Мендель провел самоопыление растений из поколения F1 и изучил их
потомство, рассмотрев в общей сложности 8023 горошины. После тщательного под-
счета ученый обнаружил, что 3/4 горошин были желтыми, 1/4 — зелеными. Этот
результат привел к открытию второго закона наследования, или закона расщепления
признаков во втором поколении.
Аа
Аа
АА Аа Аа аа
Второй закон Менделя.
102
СУДОКУ жизни
В матричном представлении второй закон Менделя выглядит так:
4
1
Обратите внимание, что этот вектор отражает соотношения, которые генетики
называют расщеплением по фенотипу. Сумма элементов матрицы (которая в этом
случае состоит из одного столбца) равна единице. В биологической математике та-
кая матрица называется стохастической.
Представьте, что мы провели эксперимент и получили 660 горошин, 510 из них
оказались желтыми, 150 — зелеными. Соответствуют ли эти результаты второму
закону Менделя? Чтобы узнать это, необходимо ответить на вопрос: каким должно
быть количество желтых и зеленых горошин в точном соответствии с этим законом?
Умножим общее число горошин, 660, на вектор-столбец, описывающий второй за-
кон Менделя:
660-
3
4
J_
4
Получим вектор-столбец:
195
165
Таким образом, мы должны были получить 495 желтых горошин и 165 зеленых.
Опишем метод, с помощью которого генетики проверяют, соответствуют ли резуль-
таты эксперимента математическому закону, а мы сможем узнать, соответствуют ли
наши результаты второму закону Менделя.
103
СУДОКУ жизни
Схема экспериментального метода применительно к менделевским законам
наследования — столпам математической биологии.
Для этого мы используем один из самых популярных в биологии статистических
критериев — критерий согласия хи-квадрат. Не вдаваясь в детали, вычислим сумму
следующих выражений, которая позволит оценить отклонение фактических данных
от результатов, на 100% соответствующих второму закону Менделя. Обозначим
отклонение греческой буквой % (хи):
2 (510-495)2 (150-165)2
495 165
Затем сравним эту сумму с эталонным значением, которое назовем критическим
значением хи-квадрат и обозначим у2. Значение у2 для экспериментов Менделя со-
ставляет 3,84. Применим следующий критерий: если сумма у2 больше, чем у2, наши
результаты не соответствуют второму закону Менделя. Если же сумма у2 меньше,
чем у2, экспериментальные результаты соответствуют второму закону Менделя.
Так как 1,81 меньше критического значения 3,84, результаты эксперимента соот-
ветствуют этому закону, а отклонение между фактическими значениями (510 и 150)
и теоретическими (495 и 165) обусловлено случайными факторами, не имеющими
значения для эксперимента.
104
СУДОКУ жизни
Является ли наследование признаков независимым?
Напомним, что, согласно второму закону Менделя, признаки А или а передаются
независимо друг от друга. Далее Мендель сделал еще один шаг и задался вопросом:
если индивид обладает двумя признаками одновременно, как они передаются следу-
ющему поколению? Является ли наследование признаков независимым?
Чтобы ответить на этот вопрос, Мендель рассмотрел следующие два признака
горошин: гладкая (Л) или морщинистая (о) поверхность и желтый (В) или зеле-
ный цвет (Ь). После того как Мендель выбрал два анализируемых признака, он
скрестил растения двух чистых линий. Горошины растений первой линии были глад-
кими и желтыми (ААВВ), горошины растений второй линии были морщинистыми
и зелеными (aabb). Растения, полученные в результате скрещивания, представляли
поколение Р, и от их признаков напрямую зависели признаки первого поколения по-
томков F}. Обратите внимание, что все гаметы растений ААВВ имели тип АВ, все
гаметы растений aabb — тип ab. После перекрестного опыления растений из раз-
личных линий все растения в поколении имели гладкие желтые горошины (гено-
тип АаВЬ). Получив этот результат, Мендель задался вопросом: какими должны
быть потомки растений с генотипом АаВЬ, принадлежащих к поколению F}, или
каким будет генотип растений поколения F2? В итоге Мендель сформулировал тре-
тий закон, или закон независимого наследования признаков.
АаВЬ х АаВЬ
г
АВ АЬ аВ ab
АВ ААВВ ААВЬ АаВВ АаВЬ
АЬ ААВЬ ААЬЬ АаВЬ Aabb
аВ АаВВ АаВЬ ааВВ ааВЬ
ab АаВЬ Aabb ааВЬ aabb
Третий закон Менделя.
В самом деле, 16 растений поколения F2 были получены путем комбинирования
четырех классов гамет растений АаВЬ, то есть АВ, аВ, АЬ и ab, на решетке Пен-
нета, которая крайне полезна при определении потомства из поколения F2 согласно
третьему закону Менделя.
105
СУДОКУ жизни
Обратите внимание, что 16 полученных генотипов можно разделить на следую-
щие группы:
• 9 горошин — желтые гладкие (АхВу);
• 3 горошины — желтые морщинистые (ааВу);
• 3 горошины — зеленые гладкие (Axbb);
• 1 горошина — зеленая морщинистая (aabb).
При этом х может иметь значение А или а, у — В или Ь. Если мы представим
этот закон наследования в матричном виде, получим:
16
3
16
3
16
1
<9 * * * * * * 16 ,
В заключение этого раздела опишем еще один похожий эксперимент. На этот раз
по результатам эксперимента было получено 1425 горошин: 807 — желтые гладкие
(АхВу), 270 — желтые морщинистые (оаВу), 265 — зеленые гладкие (Axbb),
93 — зеленые морщинистые (aabb). Соответствуют ли эти результаты третьему
закону Менделя? Чтобы убедиться в этом, вначале нужно определить эталонные
значения:
9
16
3
16
3
16
1
16
1425-
802
267
267
89
106
СУДОКУ жизни
МЕНДЕЛЬ И ЗАКОНЫ НАСЛЕДОВАНИЯ
Ботаник Грегор Мендель родился в Австрии в 1822 году. В 1843 году он постригся в монахи
августинского монастыря в Брюнне (ныне Брно, Чехия). В своем маленьком саду размерами
всего 35 на 7 метров Мендель в 1857 году начал эксперименты по перекрестному опылению
гороха, а позднее на основании своих экспериментов сформулировал знаменитые законы на-
следования признаков.
Гениальность Менделя проявилась в том, что он изучил всего несколько признаков гороха
(Pisum sativum) и рассматривал их по отдельности. Кроме того, ботаник различал доминантные
признаки, которые обозначал заглавными буквами (А, В, С, ...) и рецессивные (их он обозна-
чал строчными буквами а, Ь, с, ...) При исследовании он сосредоточился на форме горошин
(гладкие А или морщинистые а), их цвете (желтый В или зеленый Ь) и некоторых других призна-
ках - положении цветка, форме и цвете стручков и т. д. Одним из залогов успеха Менделя стало
использование так называемых чистых линий (АА, аа) и отсутствие самоопыления во время экс-
периментов.
Основной вывод исследователя заключался в том, что наследование признаков можно описать
простыми математическими законами. Он отправил полученные результаты знаменитому швей-
царскому ученому Карлу Вильгельму фон Негели, однако тот проигнорировал письмо. Мендель
опубликовал свою работу под названием «Опыты над растительными гибридами» (Versuche Ober
Pflanzenhybriden) и в 1865 году представил ее в Обществе естествоиспытателей в Брно, но его труд
вновь остался без внимания. По-видимому, Мендель писал о полученных результатах и Дарвину,
но тот также не прочел его послание. 35 лет труды ботаника оставались незамеченными. Умер
Мендель в 1884 году, а его работы были повторно открыты лишь около 1900 года.
Затем нужно оценить, насколько велико отклонение между эталонными значе-
ниями и результатами эксперимента. Для этого вновь применим критерий согласия
хи-квадрат. Дальнейшие расчеты оставляем заинтересованному читателю.
Марковские матрицы, ДНК и биоинформатика
Одна из самых перспективных областей биоинформатики, дающая ответ на вопрос
об эволюции видов, — это изучение генома отдельных видов, то есть совокупно-
сти генетической информации, записанной в ДНК его клеток. Для анализа гено-
107
СУДОКУ жизни
ма используются матрицы, элементами которых являются вероятности. Отметим,
что вероятность — это величина, характеризующая частоту, с которой наблюдается
определенный результат или исход того или иного события. Вероятность — число,
заключенное в интервале от 0 до 1. Если результат эксперимента невозможен, говорят,
что его вероятность равна 0. Если результат эксперимента наблюдается всегда, его
вероятность равна 1. К примеру, при броске кубика вероятность получения любого
результата {1, 2, 3, 4, 5, 6} равна 1/6. Вероятность того, что выпадет четное число
очков {2, 4, 6}, равна 1/2.
Одна из особенностей подобных экспериментов заключается в том, что кубик
или монета не имеют памяти, то есть исход эксперимента не зависит от предыдущих
результатов. Математик Андрей Марков изучил последовательности случайных со-
бытий, обладающие памятью, которые получили название цепей Маркова. В них
вероятность того или иного исхода зависит от предыдущих результатов. Таким об-
разом, цепи Маркова описывают эксперименты или явления, в которых последний
результат влияет на последующие. К примеру, вероятность того, что в определенный
момент в будущем, t + 1, определенный участок цепочки ДНК будет содержать то или
иное азотистое основание {А, Т, Г, Ц} (аденин, тимин, гуанин или цитозин), зависит
от того, какое основание занимало этот участок цепочки ДНК в момент времени /.
Если учесть все возможные перестановки или мутации определенного азотистого
основания ДНК, получится следующая матрица:
Р р р р
АА AT AG АС
Р Р Р Р
ТА ТТ TG ТС
Р Р Р Р
GA GT GG GC
Р Р Р Р
СА СТ CG СС
В соответствии с этой матрицей для некой молекулы ДНК вероятность того, что
в определенном участке цепочки аденин А сменится цитозином Ц, равна Р .
Цепи Маркова используются при изучении структуры белков, для прогнозиро-
вания областей генома, которыми кодируются белки, а также при изучении эволю-
ции видов путем анализа цепочек ДНК.
108
Глава 5
Векторные величины.
Векторы в биомеханике,
нейронные сети и системы
уравнений
В повседневной жизни мы довольно часто измеряем значения величин, к примеру,
температуру в комнате или вес пакета кофе. В науке измерения также имеют боль-
шое значение. Химики в лаборатории измеряют кислотность вещества, определяя
показатель pH, а биологи оценивают парциальное давление О2. Особенность этих
измерений состоит в том, что значение величин не зависит от направления измерения.
Так, температура окружающей среды не зависит от того, где именно мы поместим
термометр, при измерении pH вещество считается достаточно однородным, поэтому
местоположение электрода также не влияет на результат. Такие величины, значение
которых выражается одним числом, называются скалярными.
Однако существует множество ситуаций, когда направление измерения так же
важно, как и значение величины. К примеру, в физике скорость, ускорение, сила
и электрическое поле характеризуются не только числом, но и направлением. Меха-
низмы сокращения мышц позвоночных устроены так, что мышца сокращается толь-
ко в определенном направлении. Подобные величины, которые характеризуются
не только числом, но и направлением, называются векторными.
Векторы и матрицы
Как следует из названия, для представления векторных величин используются мате-
матические объекты, называемые векторами. Вектор имеет численное значение —
109
ВЕКТОРНЫЕ ВЕЛИЧИНЫ. ВЕКТОРЫ В БИОМЕХАНИКЕ, НЕЙРОННЫЕ СЕТИ И СИСТЕМЫ УРАВНЕНИЙ
модуль, который соответствует его длине и равен значению величины, изображаемой
вектором (к примеру, скорости). Также вектор имеет направление.
Вектор на плоскости ху.
Но какое отношение векторы имеют к матрицам? Действия над матрицами —
один из способов выполнения операций над векторными величинами, которые, как
нетрудно убедиться, широко применяются не только в науке и технике, но и в по-
вседневной жизни.
Так, матрица с произвольным числом строк тп и единственным столбцом обо-
значает вектор и называется вектор-столбцом. Векторы обозначаются u, v й т.д.
К примеру, вектор и записывается так:
( \
и2
и
\ m J
Логично предположить, что число элементов вектора, т, имеет отношение
к важному его свойству — оно соответствует размерности пространства, в котором
мы работаем. В частности, вектор
110
ВЕКТОРНЫЕ ВЕЛИЧИНЫ. ВЕКТОРЫ В БИОМЕХАНИКЕ, НЕЙРОННЫЕ СЕТИ И СИСТЕМЫ УРАВНЕНИЙ
соответствует векторной величине на плоскости. Начало этого вектора находится
в точке (0, 0), конец — в точке (2, 7). Следующий вектор расположен в простран-
стве (очевидно трехмерном):
1
15
Начало этого вектора находится в точке (0, 0, 0), конец — в точке с координатами
(3,1, 5).
Чему равно значение величины, изображаемой этим вектором? Если обозначить
рассматриваемый вектор через и, достаточно будет вычислить:
9 2 7
и~ + и. + .
12 tn
В математике модуль вектора обозначается м . К примеру, модули двух описанных
выше векторов равны:
л/22+72 = 7,28 и \/з2 + 12+52 = 5,91.
ВЕКТОРНОЕ ПРОСТРАНСТВО
В результате изучения матриц и систем линейных уравнений в XVII веке было определено по-
нятие векторного пространства. Не будем останавливаться на нем подробнее, отметим лишь,
что с точки зрения математики возможность сложения векторов (то есть выполнения операции
и+v) и умножения произвольного числа к на вектор и (к - и ) вкупе с соблюдением некоторых
свойств позволяет определить векторное пространство как множество векторов, обладающее
определенными характеристиками. Векторное пространство является одним из основных поня-
тий в математической биологии. Оно используется в изучении филогенеза, при классификации
цепочек ДНК, в экологических моделях, при исследовании метаболизма или восприятия цветов,
а также в других областях.
111
ВЕКТОРНЫЕ ВЕЛИЧИНЫ. ВЕКТОРЫ В БИОМЕХАНИКЕ, НЕЙРОННЫЕ СЕТИ И СИСТЕМЫ УРАВНЕНИЙ
Если возникает необходимость определить вектор-строку, достаточно приме-
нить операцию транспонирования:
(и. и, ... и ).
' 1 Z тх
Далее вы узнаете, как векторы используются в изучении локомоции (перемещения
животных) и при анализе нейронных сетей.
Сложение векторов:
сокращение мышц и локомоция
Один из самых интересных способов применения векторов — изучение локомоции
животных. Кузнечики прыгают, люди могут поднимать тяжести руками, рыбы плава-
ют, птицы летают. Понять механику этих движений помогают операции с векторами.
Если мы рассмотрим движение руки человека, один вектор можно будет сопоста-
вить бицепсу (этот вектор будет обозначать силу сокращения мышц), второй вектор
будет обозначать противодействующую силу, третий вектор — указывать вес объ-
екта, который поднимает рука.
Сложение векторов также помогает понять функцию некоторых мускулов. Один
из методов сложения векторов — это известное правило параллелограмма. Заклю-
чается оно в том, что нужно привести два вектора, сумму которых мы хотим найти,
к общему началу. Затем на этих двух векторах нужно построить параллелограмм.
К примеру, если рассмотреть ногу человека и обозначить боковую часть четырехгла-
вой мышцы бедра вектором р , а среднюю часть этой мышцы — вектором р , сумму
L М
этих векторов можно найти по правилу параллелограмма. Иными словами, сумма
векторов р{ + р^ будет обозначать суммарную силу четырехглавой мышцы F.
112
ВЕКТОРНЫЕ ВЕЛИЧИНЫ. ВЕКТОРЫ В БИОМЕХАНИКЕ, НЕЙРОННЫЕ СЕТИ И СИСТЕМЫ УРАВНЕНИЙ
Сумма векторов, соответствующих мышцам ноги, найденная по правилу параллелограмма.
Другой классический пример — сила F, с которой сокращаются мышцы-сгибатели
предплечья. Если представить эту силу в виде вектора, то она будет равна сумме
двух других векторов, соответствующих другим мышцам. Один из этих векторов,
р , перпендикулярен предплечью, второй вектор, р , параллелен предплечью.
и W
Сумма векторов, соответствующих мышцам руки, найденная по правилу параллелограмма.
Если векторов больше двух, их сумму можно найти по правилу многоугольника.
Заключается оно в том, что конец каждого вектора совмещается с началом следу-
ющего. Суммой исходных векторов будет вектор, начало которого совпадает с на-
113
ВЕКТОРНЫЕ ВЕЛИЧИНЫ. ВЕКТОРЫ В БИОМЕХАНИКЕ, НЕЙРОННЫЕ СЕТИ И СИСТЕМЫ УРАВНЕНИЙ
чалом первого вектора, конец — с концом последнего вектора. Этот метод полезен
при вычислении скорости движения корабля, полета птиц, перемещения пловца или
рыбы.
В примерах с птицей или рыбой результирующая скорость будет равна сумме
всего двух векторов. Но в силу особенностей задачи для сложения векторов исполь-
зуется не правило параллелограмма, а правило многоугольника.
Допустим, что рыба или птица движется в воде или в воздухе со скоростью, обо-
значаемой вектором Vа, — скорость течения воды (или ветра). Как следствие,
вектор результирующей скорости у будет равен сумме векторов JZ и у , определя-
я лг
емой по правилу многоугольника.
Сумма векторов в примере с полетом птицы, найденная по правилу многоугольника.
Достаточно помнить, что во всех подобных примерах, если вы хотите найти ре-
зультат как вектор-столбец, к примеру F, у:
нужно сложить векторы по тому же
F +F nV +V соответственно.
L М А м
правилу, что и матрицы, то есть р +FW.
114
ВЕКТОРНЫЕ ВЕЛИЧИНЫ. ВЕКТОРЫ В БИОМЕХАНИКЕ. НЕЙРОННЫЕ СЕТИ И СИСТЕМЫ УРАВНЕНИЙ
Умножение векторов и применение
этой операции в нейронных сетях
Помимо сложения, существует множество способов применения других операций
над векторами. Так, умножение векторов успешно используется в математических
моделях, описывающих наиболее характерные функции мозга.
Когда мы говорили об операциях над матрицами, мы представили модель ней-
ронной сети, основанную на произведении вектора и матрицы:
Нейронную сеть также можно представить в более простом виде:
М-и = v.
В соответствии с вышесказанным, и — вектор, представляющий слой входных,
или афферентных, нейронов, вектор и — слой выходных, или эфферентных нейро-
нов.
М — матрица связей между нейронами этих двух слоев, также известная как ма-
трица памяти. Это название указывает на то, что именно в связях между нейронами,
синапсах, мозг хранит всю известную нам информацию.
Эту гипотезу выдвинул испанский исследователь Сантьяго Рамон-и-Кахаль,
а позднее развил американский ученый Дональд Хебб. В настоящее время нейро-
биологи считают, что именно в связях между нейронами фиксируются черты лиц
знакомых нам людей, очертания букв, чисел и многие другие образы.
115
ВЕКТОРНЫЕ ВЕЛИЧИНЫ. ВЕКТОРЫ В БИОМЕХАНИКЕ, НЕЙРОННЫЕ СЕТИ И СИСТЕМЫ УРАВНЕНИЙ
Сантьяго Рамон-и-Кахаль (1852-1934)
в лаборатории. Справа изображен один из его
рисунков, описывающих нейронные сети.
Следовательно, если мы рассмотрим произвольную строку матрицы М как век-
тор-строку, описывающий связи между определенным выходным нейроном и всеми
входными нейронами, то состояние этого выходного нейрона можно будет вычис-
лить так, как мы объясняли в прошлой главе. Операция над векторами называется
скалярным произведением. Рассмотрим два вектора: вектор-строку а (исключи-
тельно из формальных соображений дополним это обозначение буквой t, что оз-
начает «транспонированный») и вектор-столбец Ь. Скалярное произведение этих
двух векторов будет равно:
2
6
4
= (1)-(2)+(—3)-(6)+(5)-(4).
Выполнив указанные арифметические действия, получим итоговый результат,
равный 4.
Скалярное произведение, которое также называют внутренним произведением
векторов, — это число, указывающее длину проекции вектора-строки а на вектор Ь.
Если известны длины обоих векторов, |«| и |б|, а также угол (X между ними, то скаляр-
ное произведение векторов а- Ь будет равно |д|-|б|- cos01. Этот результат представляет
для нас особый интерес, если учесть, что |д|- cos 01 — это значение проекции вектора
а на вектор Ь.
116
ВЕКТОРНЫЕ ВЕЛИЧИНЫ. ВЕКТОРЫ В БИОМЕХАНИКЕ, НЕЙРОННЫЕ СЕТИ И СИСТЕМЫ УРАВНЕНИЙ
Скалярное произведение векторов а- b
ЧЕМУ РАВНА РАБОТА, КОГДА МЫ ТЯНЕМ ИЛИ ТОЛКАЕМ ГРУЗ?
Вычисление работы, которую мы совершаем, когда тянем груз по земле, - еще один пример, ког-
да используется скалярное произведение векторов. Согласно законам классической механики,
работа определяется как скалярное произведение действующей на предмет силы F и переме-
щения D. Иными словами, если векторы F и D расположены под углом друг к другу, работа А бу-
дет равна |f| p|-cosa. Обратите внимание, что при неизменной силе F работа будет изменяться
в зависимости от угла между векторами. В самом деле, если векторы F и D имеют одинаковое
направление, угол между ними равен 0, и работа будет максимальной, так как косинус 0 равен 1.
Нетрудно видеть, что при а > 0° работа будет меньше максимума.
Обучение. Пример с распознаванием звуков
С начала XX века благодаря работам Сантьяго Рамон-и-Кахаля нейробиологи знают,
что обучение с точки зрения биологии заключается в видоизменении синапсов. Обу-
чение животного или человека можно смоделировать, изменив одно или несколько
значений связей, представленных вектором-строкой а. В результате этих изменений,
то есть в результате обучения, меняется значение или состояние выходного нейрона,
а следовательно, реакция субъекта на некоторый стимул.
Допустим, что свойства некоторого повторяющегося звука представлены следу-
ющим вектором:
117
ВЕКТОРНЫЕ ВЕЛИЧИНЫ. ВЕКТОРЫ В БИОМЕХАНИКЕ, НЕЙРОННЫЕ СЕТИ И СИСТЕМЫ УРАВНЕНИЙ
О
Единицы обозначают присутствие определенных характеристик звука, нули — отсут-
ствие. Допустим, что первая характеристика обозначает громкость звука: если гром-
кость превышает 30 децибел, эта характеристика равна 1, в противном случае — 0.
Громкость в 30 децибел соответствует шуму на полупустой улице. В качестве примера
звука, громкость которого меньше этого значения, приведем шелест страниц книги.
Вторая характеристика описывает частоту звука: она равна 1, если частота звука за-
ключена в интервале 100—5000 герц (именно в этом интервале лежат частоты всех
звуков или шумов, доставляющих неудобство), в противном случае — 0. Третья
характеристика, которую описывает вектор, — это мощность звука. Ее значение
равно 1, если мощность звука превышает 1 ватт (это сопоставимо с пневматическим
молотом или реактивным самолетом), и 0, если мощность звука меньше 1 ватта (ис-
точником такого звука может быть автомобиль, пианино и т. д.). Теперь рассмотрим
связи между тремя входными нейронами (их функция заключается в том, чтобы
считывать вектор с данными о звуке, воспринимаемом органами слуха) и выходными:
( 0,8 1 0,2 )
Если мы вычислим скалярное произведение этих двух векторов, то увидим, что
состояние выходного нейрона равно 1,0, так как 0,8 • 1 + 1 • 0 + 1 • 0,2.
Предположим, что мы улучшили нейрон выходного слоя, чтобы сделать модель
более реалистичной. Будем считать, что нейрон реагирует активно, если его выходное
значение превышает определенную пороговую величину, выбранную произвольным
образом. Обозначим эту пороговую величину через 0 и примем ее значение рав-
ным 0,6. Если скалярное произведение больше либо равно 0, нейрон станет активным
(обозначим это состояние через 1). Если скалярное произведение меньше 0, нейрон
останется в состоянии покоя (обозначим это состояние через 0). В нашем примере
скалярное произведение равно 1, что превышает пороговое значение 0,6. Следова-
тельно, нейрон возбужден и принимает состояние 1.
118
ВЕКТОРНЫЕ ВЕЛИЧИНЫ. ВЕКТОРЫ В БИОМЕХАНИКЕ, НЕЙРОННЫЕ СЕТИ И СИСТЕМЫ УРАВНЕНИЙ
Пример нейронной сети до обучения.
Но что произойдет, если человек будет сталкиваться с разными звуками? Что
происходит, когда человек или животное обучается? В этом случае значения связей
меняются. Допустим, в результате обучения значение одной из связей изменилось:
( 0,3 1 0,2 ).
Это означает, что важность этой связи снизилась с 0,8 до 0,3 и громкость звука
в децибелах стала менее важной. Если вычислить скалярное произведение, то есть
простимулировать нейронную сеть тем же стимулом, то после обучения состояние
выходного нейрона будет равно 0,5 — нетрудно видеть, что 0,3 • 1 + 1 • 0 + 1 • 0,2.
Если теперь мы сравним состояние эфферентного нейрона с пороговым значением 0,
равным 0,6, то увидим, что нейрон находится в состоянии покоя, то есть 0.
Пример нейронной сети после обучения.
Обратите внимание, что обучение можно интерпретировать как поворот вектора-
строки связей а относительно Ь. Чем больше проекция вектора связей а на вектор
стимула Ь, тем сильнее реакция выходного нейрона.
119
ВЕКТОРНЫЕ ВЕЛИЧИНЫ. ВЕКТОРЫ В БИОМЕХАНИКЕ. НЕЙРОННЫЕ СЕТИ И СИСТЕМЫ УРАВНЕНИЙ
Реакция нейрона максимальна, когда вектор связей имеет то же направление,
что и стимул. Что произойдет, если эти векторы будут перпендикулярны? Реакция
выходного нейрона будет равна 0, так как cos 90° = 0.
Таким образом, мы смоделировали обучение — один из самых удивительных
процессов, протекающих в мозгу человека и животных, и выразили биологическое
значение этого процесса с помощью операции над векторами. Математическая мо-
дель обучения была представлена Мак-Каллоком и Питтсом в 1946 году. Впослед-
ствии она стала основой для моделирования различных аспектов работы мозга с ис-
пользованием элементарных нейронных сетей.
Векторное, или гчешнее, произведение
Еще одной операцией умножения векторов является векторное произведение, которое
также называется внешним произведением.
Объясним вычисление векторного произведения на примере тех же векторов, для
которых мы рассчитывали скалярное произведение.
Даны вектор а и вектор Ь. Их векторное произведение равно:
После необходимых действий результирующий вектор будет равен:
—46 i +6 j + V2k.
Обратите внимание, что мы обозначаем векторное произведение знаком X, чтобы
отличить его от скалярного произведения. Более того, если скалярное произведе-
ние представляет собой число, то векторное произведение — это вектор. Еще одно
различие заключается в том, что скалярное произведение a' -h обозначает проекцию
120
ВЕКТОРНЫЕ ВЕЛИЧИНЫ. ВЕКТОРЫ В БИОМЕХАНИКЕ, НЕЙРОННЫЕ СЕТИ И СИСТЕМЫ УРАВНЕНИЙ
вектор-строки а на вектор-столбец Ь, а векторное произведение «х// представляет
собой вектор (обозначим его F), перпендикулярный плоскости, определяемой двумя
исходными векторами.
Векторное произведение векторов axb.
Модуль нового вектора F будет схож со скалярным произведением, однако его
значение будет равно |д|-|£|- sin ОС. Модуль векторного произведения векторов будет
равен площади построенного на них параллелограмма. Направление вектора f опре-
деляется по известному правилу буравчика, или правилу правой руки.
В биологии векторное произведение используется при изучении молекул, играю-
щих основную роль в поддержании жизни, к примеру таких белков, как миоглобин.
Сюда же относится самая знаменитая из всех известных сегодня молекул — молекула
ДНК. При их изучении биофизики используют классические понятия физики и изме-
ряют величины, рассчитываемые как векторное произведение, к примеру дипольный
момент — электромагнитную силу, действующую на частицу в магнитном поле.
Модель памяти животных и человека
Существует еще одна поистине замечательная операция — тензорное произведе-
ние, которое применяется в математических моделях нейронных сетей, описывающих
память животных и человека. Представим, что вектор v состоит только из единиц
и нулей, то есть является двоичным вектором. Каждый из его элементов обозначает
121
ВЕКТОРНЫЕ ВЕЛИЧИНЫ. ВЕКТОРЫ В БИОМЕХАНИКЕ. НЕЙРОННЫЕ СЕТИ И СИСТЕМЫ УРАВНЕНИЙ
наличие (1) или отсутствие (0) той или иной характеристики некоторого объекта.
Если мы вычислим тензорное произведение v и v, то получим следующую матрицу:
1
0
1
0
0
о
1
о
о
1
10011)=
10 10 0 11
10 10 0 11
0 0 0 0 0 0 0
10 10 0 11
0 0 0 0 0 0 0
0 0 0 0 0 0 0
10 10 0 11
10 10 0 11
Обратите внимание, какие действия мы выполнили, чтобы получить эту матрицу:
(1)-(1) (1)-(0) (1)-(0)
(1)-(1) (1)-(1) (1)(0) (1)-(1) (1)-(0) (1)-(0) (1)-(1) (1)-(1)
(0)(1) (0)(1) (0)-(0) (0)(1) (0)-(0) (0)-(0) (0)-(1) (0)-(1)
(1)-(1) (1)-(1) (1)-(0) (1)-(1) (1)-(0) (1)-(0) (1)-(1) (1)-(1)
(0)(1) (0)-(1) (0)-(0) (0)-(1) (0)-(0) (0)-(0) (0)-(0) (0)-(1)
(0)(1) (0)-(1) (0)-(0) (0)-(1) (0)-(0) (0)-(0) (0)-(0) (0)-(1)
(1)-(1) (1)-(1) (1)-(0) (1)-(1) (1)-(0) (1)(0) (1)-(1) (1)-(1)
(1)-(0) (l)-(l) (1)-(0) (1)-(0) (!)•(!)
Несмотря на кажущуюся сложность, эта операция на самом деле проста. Мы
получили матрицу памяти, обладающую свойством запоминать предмет, показанный
нейронной сети. Она позволяет смоделировать на компьютере способность людей
и животных запоминать различные объекты. Так как элементы матрицы обозначают
связи между нейронами, в модели предполагается, что каждый нейрон связан со всеми
другими нейронами. Как следствие, все элементы главной диагонали матрицы должны
быть равны 0. Исправим значения элементов главной диагонали, равные 1:
122
ВЕКТОРНЫЕ ВЕЛИЧИНЫ. ВЕКТОРЫ В БИОМЕХАНИКЕ, НЕЙРОННЫЕ СЕТИ И СИСТЕМЫ УРАВНЕНИЙ
0 10 10 0 11
1 0 0 1 0 0 1 1
00000000
1 1 0 0 0 0 1 1
оооооооо’
00000000
1 1 0 1 0 0 0 1
к 1 1 0 1 0 0 1 о }
Существуют математические методы, позволяющие восстановить объект, пред-
ставленный матрицей, и смоделировать процесс вспоминания и распознавания об-
разов.
СЕТЬ ХОПФИЛДА
Механизм обучения, запоминания букв, цифр и сигналов светофора можно смоделировать с по-
мощью нейронной сети. Модель памяти, определяемая с помощью тензорного произведения, из-
вестна как сеть Хопфилда. Она названа в честь исследователя Джона Джозефа Хопфилда, который
представил эту модель в 1980-е годы. Сегодня модель Хопфилда используется в самых разных
цифровых системах: не только для решения множества физических задач, но и в электронике,
и при обработке изображений.
Модель памяти Хопфилда из восьми нейронов.
Каждый нейрон в этой модели связан со всеми остальными.
123
ВЕКТОРНЫЕ ВЕЛИЧИНЫ. ВЕКТОРЫ В БИОМЕХАНИКЕ, НЕЙРОННЫЕ СЕТИ И СИСТЕМЫ УРАВНЕНИЙ
Решение систем уравнений.
Эксперимент энтомолога
Обратные матрицы применяются также для решения систем уравнений. Рассмотрим
систему из трех уравнений с тремя неизвестными:
а..х + а.л} + a.^z — b.,
И Iz17 13 г
а21Х + а21У + fl23Z = Ь2’
V + аз1У + аззг = ^з-
Матрицы также используются для представления систем уравнений:
Это равенство равносильно следующему:
А -Х = В.
Если мы найдем матрицу, обратную А, то есть А \ а затем умножим обе части
равенства на эту обратную матрицу:
А~х А •Х = Л"1-В,
то, поскольку произведение А- А~х равно единичной матрице Е, имеем:
Е-Х = А~'-В.
Кроме того, так как произведение любой матрицы на единичную матрицу Е рав-
но исходной матрице, получим:
Х = А~Х-В.
124
ВЕКТОРНЫЕ ВЕЛИЧИНЫ. ВЕКТОРЫ В БИОМЕХАНИКА НЕЙРОННЫЕ СЕТИ И СИСТЕМЫ УРАВНЕНИЙ
Таким образом, решить систему уравнений, то есть определить значения х, у, z,
можно с помощью обратной матрицы коэффициентов: нужно умножить ее на вектор-
столбец свободных членов системы уравнений.
Продемонстрируем этот метод на примере под названием «эксперимент энтомо-
лога». Допустим, что мы отправились в поле в поисках определенного вида насеко-
мых и разместили ловушки там, где эти насекомые водятся. Спустя несколько дней
мы вернулись к ловушкам, чтобы собрать насекомых. В лаборатории мы установили,
что в ловушках оказалось 180 насекомых. Мы разделили их на молодых (обозначим
их через х) и взрослых (у) особей. Имеем первое уравнение системы:
х + у — 180.
На основе результатов аналогичных экспериментов, проведенных ранее, мы зна-
ем, что для насекомых этого вида соотношение молодых и взрослых особей равно
2 к 1. Кроме того, в силу естественных причин 6 взрослых насекомых умерло:
2х = у — 6.
Чтобы определить численность молодых и взрослых особей, нужно решить сле-
дующую систему уравнений:
х + у = 180,
2х = у - 6.
Второе уравнение можно записать в виде: 2х — у — — 6. Система примет вид:
х + у = 180,
2х~ у = -6.
В матричной нотации эта система уравнений записывается так:
1
2
1
-1
У
180
< “6 >
125
ВЕКТОРНЫЕ ВЕЛИЧИНЫ. ВЕКТОРЫ В БИОМЕХАНИКЕ. НЕЙРОННЫЕ СЕТИ И СИСТЕМЫ УРАВНЕНИЙ
Имеет ли система уравнений решение?
Проницательные математики имеют одну достойную привычку — они не тратят
время на бесполезные действия. Одним из наиболее ярких примеров этому является
решение систем уравнений. Рассмотрим все возможные группы систем уравнений.
Во-первых, система может не иметь решений — в этом случае она называется
несовместной. Представим, что система состоит из двух уравнений, описывающих две
параллельные прямые. Поскольку прямые не пересекаются, система не будет иметь
решений. Во-вторых, система может иметь бесконечно много решений, то есть быть
неопределенной. Продолжив аналогию с прямыми, такая система состоит из двух
уравнений, описывающих две совпадающие прямые, имеющие бесконечно много
общих точек. Наконец, если система из двух уравнений описывает прямые, пересе-
кающиеся в одной точке, она называется совместной и определенной. Ее решением
будет единственная точка пересечения прямых (х, у).
Рассмотрим систему из трех уравнений, которая в матричном виде выглядит так:
Система является совместной и определенной, если определитель матрицы А
а32 а33
отличен от нуля. Если определитель А равен 0, система будет либо совместной и нео-
пределенной, либо несовместной. К примеру, система уравнений в эксперименте эн-
томолога в матричном виде будет записываться так:
126
ВЕКТОРНЫЕ ВЕЛИЧИНЫ. ВЕКТОРЫ В БИОМЕХАНИКЕ, НЕЙРОННЫЕ СЕТИ И СИСТЕМЫ УРАВНЕНИЙ
I 2
180
к ~6 >
Поскольку эта система является совместной и определенной, ее можно решить.
И действительно, если мы вычислим определитель
= (1) •(-!)-да)=-з,
он будет отличен от нуля.
Сколько молодых и взрослых насекомых поймал энтомолог.
Правило Крамера
Правило Крамера — это метод решения систем линейных уравнений с помощью
определителей. Он был представлен Габриэлем Крамером в 1750 году.
Промежуточный этап решения системы уравнений по правилу Крамера в программе MathLab.
Значения неизвестных определяются путем вычисления определителя для двух
типов матриц, D. и D. Правило Крамера можно использовать только тогда, когда
127
ВЕКТОРНЫЕ ВЕЛИЧИНЫ. ВЕКТОРЫ В БИОМЕХАНИКЕ, НЕЙРОННЫЕ СЕТИ И СИСТЕМЫ УРАВНЕНИЙ
число уравнений равно числу неизвестных, а определитель матрицы коэффициентов
отличен от нуля (det(D) 0). Объясним правило Крамера на примере эксперимента
энтомолога. Система линейных уравнений выглядит так:
Обозначим через D матрицу коэффициентов системы:
Г 1 1
[ 2 -1
Определитель этой матрицы det(D) равен —3. Так как система имеет две неиз-
вестные, х и у, имеем две матрицы D.: D и Dy. Чтобы составить матрицу D., нуж-
но заменить j-й столбец матрицы D на вектор-столбец, образованный свободными
членами системы:
180
< "6 >
В нашем эксперименте первой неизвестной является х, поэтому j будет равен 1.
Если мы заменим первый столбец матрицы D на вектор-столбец, образованный
свободными членами системы, матрица D* примет вид:
180 1
< -6 -1 ,
det(D ) будет равен —174, так как (180) • (—1) — 1 • (—6) = —174. Рассуждая ана-
логичным образом и учитывая, что второй неизвестной является у, то есть j — 2,
получим, что матрица D имеет вид:
1 180
<2 -6,
Ее определитель равен —366, так как det(D^) равен 1 • (—6) — 180 • 2.
128
ВЕКТОРНЫЕ ВЕЛИЧИНЫ. ВЕКТОРЫ В БИОМЕХАНИКЕ. НЕЙРОННЫЕ СЕТИ И СИСТЕМЫ УРАВНЕНИЙ
Правило Крамера гласит, что решение системы уравнений можно найти, вычис-
лив следующие выражения:
det(D ) det(D) x= , у = det(D) det(D)
Следовательно, в эксперименте энтомолога получим:
-174 -366 x = , y = -3 -3
Энтомолог поймал 58 молодых особей (х) и 122 взрослых (у).
129
Глава 6
Экология и математика.
Взаимовыгодное сотрудничество
Живые существа, будь то растения, животные или микроорганизмы, взаимодейству-
ют между собой и с окружающей средой. Биологические организмы, принадлежащие
к различным видам, образуют общую природную среду — экосистему. В экосистеме
можно выделить некоторые физические факторы, также называемые абиотическими,
поскольку они не имеют биологической природы, и биотические факторы, которые
относятся к живым обитателям экосистемы. Абиотические факторы — это все фак-
торы, связанные с геологией и климатом: свет, вода, температура, атмосфера и состав
почвы. К биотическим факторам относятся растения, травоядные и хищные живот-
ные, грибы, бактерии и т. д.
Эрнст Гэнрих Гэккель (1834-1919)
первым ввел термин «экология».
Справа изображено созданное им
«древо жизни».
131
ЭКОЛОГИЯ И МАТЕМАТИКА. ВЗАИМОВЫГОДНОЕ СОТРУДНИЧЕСТВО
Экосистемы изучает экология, появившаяся в XIX веке как подраздел биологии.
Она преимущественно рассматривает задачи, связанные с многообразием живых
существ (биологическим разнообразием), взаимосвязи между живыми организмами
и окружающей средой. С момента появления экологии в ней использовались инстру-
менты математической биологии для построения моделей, позволяющих описывать
и прогнозировать экологические явления. Это привело к быстрому развитию новой
науки и появлению в ней многих понятий и теорий, имеющих математическую основу.
Построение моделей
Передача энергии между живыми организмами, населяющими экосистему, происхо-
дит в результате питания. Так как одни организмы питаются другими, образуется пи-
щевая цепочка, которая обычно выглядит так: растение —> травоядные —> хищники —>
—> детритофаги. В математической модели каждое звено пищевой цепи характеризу-
ется численностью организмов, описывается циркуляция энергии и другие аспекты.
Как правило, экосистемы достаточно сложны ввиду большого числа населяющих их
видов и множества связей между ними, поэтому экологи стремятся упростить модели.
К примеру, экосистемы грибы и бактерии могут быть объединены в рамках единой
подсистемы детритофагов. Кроме того, модель экосистемы объединяется с други-
ми моделями и становится частью итоговой общей модели. Одной из составляющих
итоговой модели является окружающая среда, будь то суша, океан и т. д. Ну а для
построения моделей широко используется компьютерное моделирование.
Подобным подходом к экологии мы во многом обязаны Говарду Одуму, который
в 1950-е годы впервые применил для описания экосистем схемы, напоминающие
схемы электрических цепей, которые сегодня лежат в основе компьютерного мо-
делирования. Также важным вкладом Одума в науку стало объединение экологии
и теории систем.
Первые математические экологические модели описывали динамику популяций.
Авторы этих моделей стремились описать изменение численности популяции и ее
возрастное распределение в результате взаимодействия с окружающей средой. Эти
исследования берут начало в XVIII веке, когда Томас Мальтус составил модель экс-
132
ЭКОЛОГИЯ И МАТЕМАТИКА. ВЗАИМОВЫГОДНОЕ СОТРУДНИЧЕСТВО
Модель, созданная на основе схем
Гэварда Одума. Его инновационная
методология легла в основу нового
способа изучения экосистем.
поненциального роста населения, а позднее, в 1938 году, Пьер Франсуа Ферхюльст
представил логистическую модель роста.
В 1920 -е годы Вито Вольтерра и Альфред Джеймс Лотка описали две модели,
которые стали настоящими столпами математической экологии. Это были модель
межвидовой конкуренции и модель «хищник — жертва». Обе имели похожую струк-
туру, однако в первой предполагалось, что рост популяций описывается логистическим
уравнением, а во второй рассматривался экспоненциальный рост. Примерно двадцать
лет спустя, в 1940-е годы, Патрик Лесли представил матрицу Лесли — модель струк-
туры популяции, в которой он объединил динамику роста популяции и демографию.
В то время как в основе моделей Мальтуса, Ферхюльста и Лотки — Вольтерры
лежали дифференциальные уравнения, заслуга Лесли состояла в том, что он показал
применимость математических моделей, использующих матрицы.
Матричная алгебра оказалась достойным инструментом экологического и ком-
пьютерного моделирования. Матричные модели получили название BIDE (от ан-
глийского Births, Immigrants, Deaths, Emigrants — «рождение, иммиграция, смерть,
эмиграция»). Очевидное преимущество операций с матрицами заключалось в том,
что их могли выполнять компьютеры.
В модели BIDE популяция в общем виде описывается следующим выражением:
Л , = Л + В + / - D - Е.
»+1 /
133
ЭКОЛОГИЯ И МАТЕМАТИКА. ВЗАИМОВЫГОДНОЕ СОТРУДНИЧЕСТВО
ЭЛЕМЕНТАРНАЯ МОДЕЛЬ BIDE
Опишем одну из простейших экологических моделей. Допустим, что мы хотим изучить популяцию
оленей, для которой известна численность самок NJ, не достигших репродуктивного возраста,
и число выживших самок в этой группе SJ на временном интервале от t до t + 1, а также числен-
ность взрослых самок NA и число выживших самок в этой группе S* за этот же промежуток вре-
мени. Если обозначить через Р7 число выживших молодых самок в пересчете на каждую самку
репродуктивного возраста, то численность популяции будет изменяться по следующей модели:
( . А ( \
N,+1 = sJpJ $ApJ Nt
Ц S' sA Д/V?
На основе этого класса элементарных моделей можно составить более сложные, к примеру
модель с матрицей Лесли.
Численность популяции в момент времени t + 1 в будущем определяется ее численно-
стью в настоящий момент времени t и совокупностью всех факторов, которые ведут
к ее росту или сокращению: В — число родившихся в период времени с t по t + 1,
I — число иммигрантов, присоединившихся к популяции в период времени с t по t +
+ 1, D — число умерших в период с/по/ + 1иЕ — число эмигрантов, покинувших
популяцию в период с t по t + 1.
Модель Лотки - Вольтерры: волки и зайцы
В течение всей Первой мировой войны, с 1914 по 1918 год, на севере Адриатического
моря была приостановлена рыбная ловля. После войны улов рыбы вернулся на преж-
ний уровень. Именно тогда, в 1920-е годы, итальянский биолог Умберто д’Анкона
провел количественный анализ различных видов рыбы, продававшейся на рынках
Венеции, Риеки и Триеста. Ученый обнаружил, что на рынках продавалось намного
больше хищных рыб, чем рыб, которые были естественной добычей для хищников.
Д’Анкона испытывал романтические чувства к дочери знаменитого математика того
времени Вито Вольтерры и предложил ему провести математический анализ ситуации
134
ЭКОЛОГИЯ И МАТЕМАТИКА. ВЗАИМОВЫГОДНОЕ СОТРУДНИЧЕСТВО
и объяснить различие в численности хищников и жертв. Именно так его будущий
тесть в 1926 году предложил систему дифференциальных уравнений. Похожую си-
стему годом ранее, совершенно независимо от Вольтерры, разработал американский
физик и химик Альфред Джеймс Лотка. Результаты их работ стали известны как
уравнения Лотки — Вольтерры.
Система дифференциальных уравнений, описывающая взаимодействие «хищ-
ник — жертва», стала одной из первых и наиболее известных моделей математиче-
ской биологии. Она применяется в экологии при восстановлении численности вида
в регионе или при определении численности рыб, которые будут сосуществовать
с хищниками, например с акулами, в океанариуме. Также модель применима в других
областях: в иммунологии при изучении взаимодействия вируса или раковых клеток
с иммунной системой, в паразитологии — при изучении взаимосвязи между парази-
том и хозяином, в экономике — при изучении соотношения количества потребителей
и ресурсов и так далее.
Обозначим через х численность хищников — акул, волков и т. д., через у — чис-
ленность жертв (рыб, зайцев и т. д.). Будем предполагать, что хищники питаются
только жертвами, а жертвы также имеют достаточно пропитания. Модель, описы-
вающая взаимодействие между акулами и рыбами или между волками и зайцами,
будет корректной для многих тысяч взаимодействий между хищниками и жертвами,
будь то насекомые, простейшие и т. д. Проиллюстрируем модель на примере волков
и зайцев. Допустим, что их популяции изолированы друг от друга. Это означает, что
хищники не могут питаться жертвами, то есть вымрут от голода. Скорость, с кото-
рой будет снижаться популяция хищников, описывается следующим выражением:
dx
— = —рх.
dt
Это простое дифференциальное уравнение, в котором коэффициент р со знаком
минус обозначает уровень смертности хищников. Решение этого уравнения выгля-
дит так:
= хое~р1‘
135
ЭКОЛОГИЯ И МАТЕМАТИКА. ВЗАИМОВЫГОДНОЕ СОТРУДНИЧЕСТВО
Что это означает? В указанном выражении х{) обозначает начальную численность
волков, которая с течением времени /(*(/)) будет уменьшаться экспоненциально.
С другой стороны, численность зайцев, которые живут в изоляции от хищников
и имеют достаточно корма, будет расти, и скорость этого роста описывается следу-
ющим выражением:
dy
~Т = ГУ-
dt
Это простое дифференциальное уравнение, подобное тому, которое описывает
скорость снижения численности хищников, однако здесь положительный коэффи-
циент г обозначает уровень рождаемости жертв.
У(О = Уоег'-
Как вы можете видеть, речь идет о мальтусовской модели экспоненциального
роста. Таким образом, если у0 — начальная численность жертв (зайцев), то чис-
ленность жертв y(f) за время t будет возрастать экспоненциально согласно модели
Мальтуса.
Теперь предположим, что хищники и жертвы обитают в одном регионе. В этом
случае необходимо учесть их взаимодействие в выражениях, описывающих числен-
ность изолированных групп хищников и жертв. Как следствие, в выражение, опи-
сывающее скорость изменения численности хищников, нужно добавить член qxy,
который будет отражать рост численности хищников как результат встречи с жерт-
вами (у — параметр, описывающий подобную встречу). В нашем примере q можно
определить как коэффициент, связанный с употреблением в пищу зайцев. Результи-
рующее выражение выглядит так:
dx
— = ~px + qxy.
136
ЭКОЛОГИЯ И МАТЕМАТИКА. ВЗАИМОВЫГОДНОЕ СОТРУДНИЧЕСТВО
Обратите внимание, что естественная убыль хищников компенсируется ростом
их численности при охоте на жертв (именно поэтому qxy имеет знак плюс). Следо-
вательно, хищники не вымрут, имея достаточно пропитания.
Аналогично хищники компенсируют естественный рост численности жертв, по-
этому в дифференциальное уравнение включается член —sxy со знаком минус, где
s — параметр, описывающий взаимодействие хищников и жертв:
dy
- = ry- sxy.
dt
Одна из предпосылок моделей Вольтерры и Лотки заключалась в том, что от-
дельные особи, к примеру волки и зайцы, ведут себя подобно частицам газа. Про-
ведя параллель с так называемым законом действующих масс, мы обнаружим, что
число взаимодействий между хищниками и жертвами зависит от столь очевидных
параметров, как число хищников х и число жертв у. По этой причине их взаимо-
действие обозначается произведением этих чисел, то есть х ♦ у с соответствующими
коэффициентами: qxy и —sxy.
Объединив два описанных выше дифференциальных уравнения в систему:
dx dy
— = —рх + qxy и — = ry— sxy,
получим знаменитую систему уравнений Лотки — Вольтерры.
После того как мы учли взаимодействие хищников и жертв, решить эту систему
уже не так просто, как раньше. В соответствии с моделью, при высокой численности
жертв возрастет и число хищников, однако по мере поедания жертв популяция хищ-
ников также сократится. В свою очередь, ввиду снижения численности хищников
число жертв вновь возрастет, и это вновь приведет к увеличению числа хищников.
Если представить подобные циклические колебания численности популяций на од-
ном графике, мы получим цикл, имеющий название «цикл — решение», так как он
является решением уравнений Лотки — Вольтерры.
137
ЭКОЛОГИЯ И МАТЕМАТИКА. ВЗАИМОВЫГОДНОЕ СОТРУДНИЧЕСТВО
Модель «хищник — жертва» Лотки — Вольтерры:
число хищников обозначено на оси у, число жертв — на оси х.
Получить «цикл — решение» системы уравнений можно разными способами,
но самым важным из них является рассмотрение колебаний численности популяций.
Форма цикла зависит от начального числа хищников и жертв.
Столь же нетрудно найти точку равновесия, так как ее координаты равны:
Нужно очень четко представлять себе описанные колебания системы, особенно
когда речь идет о климатических моделях (о них мы поговорим в следующем раз-
деле).
138
ЭКОЛОГИЯ И МАТЕМАТИКА. ВЗАИМОВЫГОДНОЕ СОТРУДНИЧЕСТВО
КОНКУРЕНЦИЯ МЕЖДУ ВИДАМИ
Помимо модели «хищник - жертва», Лотка и Вольтерра представили еще одну модель, описываю-
щую конкуренцию между двумя популяциями или видами. Допустим, что две популяции конкури-
руют за один и тот же ресурс, при этом рост их численности описывается логистической моделью.
Ооозначив через х и у численность особей в популяциях, имеем:
dx , (kx-x-avy
dt к
dy [к-у-ах
иагг4 к,
где г, г - коэффициенты роста, к*, ку - поддерживающие емкости для каждой популяции. Эти
выражения также содержат коэффициенты а, описывающие взаимодействие между особями
в популяциях. Иными словами, аху обозначает воздействие, которое оказывает вид у на вид х,
в то время как аух - это воздействие х на у. Модель можно расширить для нескольких видов,
однако это несколько усложнит анализ возможных ситуаций.
Модель межвидовой конкуренции Лотки — Вольтерры для четырех видов.
Математические модели климата.
Глобальное потепление: миф или реальность?
В последние десятилетия весьма актуальна тема глобального потепления. Хотя
метеорологические центры составляют прогнозы погоды с применением сложных
139
ЭКОЛОГИЯ И МАТЕМАТИКА. ВЗАИМОВЫГОДНОЕ СОТРУДНИЧЕСТВО
математических моделей, на их основании довольно трудно дать ответ на вопрос,
действительно ли наблюдается глобальное изменение климата.
Математические модели, используемые в метеорологии, называются климати-
ческими моделями. Они основаны на описаниях атмосферных процессов и компью-
терном моделировании взаимодействия атмосферы и океанов, суши и шапок льда
на полюсах. Эти модели представляют собой дифференциальные уравнения, в осно-
ве которых лежат законы физики. При их составлении поверхность Земли делится
на квадраты, которые описываются уравнениями. Затем вычисляется скорость ве-
тров, относительная влажность воздуха, теплопередача и так далее, а также взаимо-
действие между соседними областями. На основе интерпретации итоговых резуль-
татов моделирования метеорологи и составляют свои прогнозы.
Математические модели атмосферы позволяют предсказывать погоду.
К наиболее надежным метеорологическим данным относятся так называемые
климатические режимы, которые учитывают преобладающее направление ветра
и другие параметры, описывающие ожидаемую погоду — солнечную, облачную или
дождливую. Климатические режимы определяются с помощью статистических мето-
дов на основе данных о погоде прошлых лет в стране или регионе. Однако поскольку
атмосфера представляет собой хаотическую систему, стоит задуматься, насколько
достоверны прогнозы погоды?
Подобные вопросы навели Эдварда Лоренца на мысль изучить атмосферу с точ-
ки зрения теории хаоса. Первые попытки составить прогноз погоды были пред-
140
ЭКОЛОГИЯ И МАТЕМАТИКА. ВЗАИМОВЫГОДНОЕ СОТРУДНИЧЕСТВО
приняты задолго до 1960-х годов, когда работы Лоренца увидели свет, — это по-
пытались сделать Жюль Чарни, Филип Томпсон, Ларри Гейтс, Рагнар Фьюртофт
и гениальный Джон фон Нейман в 1950-х годах с помощью одного из первых ком-
пьютеров в истории — ENIAC.
В то время на вычисление прогноза погоды на следующий день требовалось це-
лых 24 часа, а его точность оставляла желать лучшего. Именно в те годы родилась
фраза «климат — это то, что ожидается, а погода — то, что будет». В модели,
использованной для составления прогноза погоды, атмосфера была представлена
решеткой из 270 точек, расположенных на территории Северной Америки и уда-
ленных друг от друга на расстояние 700 километров. Метод конечных разностей
позволил решить с помощью ENIAC дифференциальные уравнения, а также урав-
нение завихренности, которое использовалось в упрощенной модели атмосферы.
Компьютер ENIAC и Бетти Зайндер (справа) — одна из первых программистов.
141
ЭКОЛОГИЯ И МАТЕМАТИКА. ВЗАИМОВЫГОДНОЕ СОТРУДНИЧЕСТВО
В 1960-е годы Лоренц работал в престижном Массачусетском технологическом
институте (США). Ученый рассматривал атмосферу как турбулентный поток, крайне
чувствительный к малым изменениям. Чтобы описать этот поток математически, он
составил максимально простую модель климата, включив в нее всего три перемен-
ные х, у, z. Это означает, что для Лоренца «погода на сегодня» обозначалась точкой
с координатами (х, у, z) в трехмерном пространстве. Прогноз погоды на несколько
дней в этой модели представлял собой линию, соединяющую эту точку с другими
точками. Как следствие, все возможные ситуации описывались так называемым ат-
трактором Лоренца.
В 1963 году ученый получил три дифференциальных уравнения, объясняющих
конвекцию в атмосфере, то есть движение молекул в потоке, которое и является од-
ним из основных механизмов передачи массы и тепла. По этой причине уравнения
Лоренца очень важны при изучении климата и составлении прогнозов погоды. Эти
уравнения обладают интересной особенностью: хотя поведение атмосферы полно-
стью предсказуемо, оно подвержено резким изменениям, которые кажутся слу-
чайными. Именно поэтому, подобно уравнениям Лотки — Вольтерры, уравнения
Лоренца являются нелинейными. Хаотическое поведение системы, описываемой
ими, для некоторых значений параметров было доказано лишь в 2001 году. Лоренц
предложил следующую математическую модель, ставшую прообразом всех клима-
тических моделей:
dx dy dz D
— = U(y-x), — = x(p-z)-y, — = xy-fiz,
dt dt dt
где G, p и P — параметры модели.
Конвекция в природе возникает при переходе системы в нестабильное состояние
и проявляется в движении масс. К примеру, конвекция будет наблюдаться, если мы
нагреем воду в сосуде: так как горячая вода менее плотная, она будет смещаться бли-
же к поверхности; холодная вода, более плотная, напротив, будет опускаться на дно.
Подобные явления характеризуются числом Рэлея, которое в уравнениях Лоренца
обозначается параметром р. Также этот параметр объясняет поведение атмосферы
в следующей простой модели: когда значение р достигает 28, атмосфера начинает
демонстрировать хаотическое поведение.
142
ЭКОЛОГИЯ И МАТЕМАТИКА. ВЗАИМОВЫГОДНОЕ СОТРУДНИЧЕСТВО
Отсюда можно сделать вывод: аттрактор Лоренца — это осциллятор, представ-
ляющий модель климата в трех измерениях. Как мы отмечали, переменные модели,
х, у, z, описывают эволюцию хаотического потока, в которой отсутствуют какие-либо
закономерности. Хотя модель Лоренца очень проста, два «больших крыла бабочки»
аттрактора Лоренца объясняют особенности климатических режимов.
Любопытно, что известный эффект бабочки описал сам Лоренц в 1972 году. Его
исходная формулировка звучала так: «Может ли взмах крыльев бабочки в Бразилии
вызвать торнадо в Техасе?». На практике это означает, что даже малейшие изменения
начальных условий, исчисляемые несколькими знаками после запятой, оказывают
огромное влияние на погоду. Если сегодня в определенном месте наблюдается по-
143
ЭКОЛОГИЯ И МАТЕМАТИКА. ВЗАИМОВЫГОДНОЕ СОТРУДНИЧЕСТВО
года, которой соответствует точка (х, у, z) на правом «крыле бабочки», то погода
в ближайшие дни будет описываться траекторией, берущей начало в точке (х, у, z).
Погода будет более или менее схожа с сегодняшней, если эта траектория будет при-
надлежать правому «крылу бабочки». Однако может случиться, что она достигнет
левого «крыла бабочки», и прогноз существенно изменится.
Это изменение зависит от метеорологических параметров атмосферы «на сегодня»,
то есть от начальных условий модели: траектория, описывающая погоду в последую-
щие дни, будет иметь тот или иной вид в зависимости от начальных условий в точке
(х, у, z). И если погода представляет собой хаотическую систему, ее прогноз является
прогнозом «индивидуальной траектории» в аттракторе климата. На практике погоду,
то есть «индивидуальную траекторию», можно спрогнозировать на основе начальных
условий, то есть погоды на сегодняшний день. В решении этой задачи метеорологам
помогают мощные компьютеры, в которых используются климатические модели с на-
много большим числом переменных, чем три.
Кроме того, прогнозирование усложняется еще и потому, что климат в разных
частях Земли описывается различными сценариями. В средних широтах и в тропиках
погода существенно отличается. В средних широтах любое изменение погоды вы-
зывается атмосферными явлениями, в то время как погода в тропиках определяется
взаимодействием атмосферы и океана, известным как Эль-Ниньо. Самые известные
его проявления — это ураганы и муссоны. Более того, некоторые явления, наблю-
даемые в Тихом океане, вносят элемент неопределенности в начальное состояние
атмосферы, на основе которого составляется прогноз погоды.
Карта с прогнозом атмосферного давления на пять дней вперед
144
ЭКОЛОГИЯ И МАТЕМАТИКА. ВЗАИМОВЫГОДНОЕ СОТРУДНИЧЕСТВО
В 1963 году благодаря Лоренцу стало известно, насколько сложно составление
точных прогнозов ввиду хаотической природы климата. Поэтому ученые решили
использовать различные математические модели или начальные условия и значения
параметров и составлять на их основе различные прогнозы. Существуют методы,
ДЖЕЙМС ЛАВЛОК И ГИПОТЕЗА ГЕИ
Джеймс Лавлок - английский исследователь (род. в 1919 году), известный как автор гипотезы
Геи. Согласно Лавлоку, живые организмы отвечают за поддержание земной атмосферы. Суша,
океаны, живые существа и атмосфера в совокупности образуют кибернетическую систему. Идею
о кибернетической системе развила американский исследователь Линн Маргулис. Она предпо-
ложила, что планета Земля в действительности представляет собой суперорганизм, способный
регулировать условия, благодаря которым возможны эволюция и поддержание жизни.
Идеи Джеймса Лавлока и Линн Маргулис разделяют не все. Несогласие с ними с самого на-
чала выразили эволюционные биологи Стивен Джей Гулд и Ричард Докинз. Дарвинисты выдви-
нули вопрос: быть может, именно окружающая среда отбирает живых организмов в соответствии
с дарвиновской теорией эволюции путем естественного отбора? И если это так, то как объяснить,
что живые организмы медленно видоизменяют окружающую среду, в которой обитают, делая ее
более благоприятной для себя? Несмотря на эти возражения, существуют примеры, подтвержда-
ющие гипотезу Геи. Так, бактерии и водоросли поддерживают температуру земной поверхности,
регулируют соленость морей и участвуют в накоплении углерода в осадочных горных породах.
Согласно этой экологической гипотезе, вмешательство бактерий и водорослей в окружающую
среду в кибернетике описывается циклами обратной связи. В качестве примера подобного ме-
ханизма можно привести систему кондиционирования воздуха в комнате: повышение или пони-
жение температуры в комнате выявляется термостатом, после чего система кондиционирования
соответственно нагревает или охлаждает воздух.
Согласно I ипотезе Гёи, Земля —
это кибернетическая система планетарного
масштаба, или суперорганизм.
145
ЭКОЛОГИЯ И МАТЕМАТИКА. ВЗАИМОВЫГОДНОЕ СОТРУДНИЧЕСТВО
которые позволяют оценить согласованность или степень совпадения различных про-
гнозов и получить средний прогноз. В этой методике также учитывается, сколько раз
прогноз погоды оказывался верным для каждой модели из множества используемых.
Этот подход, в котором вместо единственной модели, как во времена фон Неймана
и ENIAC, используется множество моделей климата, называется мультимодельным
(англ, multi-model ensemble).
Наконец, открытым остается вопрос о том, существует ли глобальное измене-
ние климата. Прежде чем дать ответ на него, следует пояснить, что имеется в виду.
С точки зрения математики глобальное изменение климата означает изменение фор-
мы и положения аттрактора климата. Это изменение может быть вызвано непод-
контрольными нам силами природы, действиями человека или сочетанием разных
факторов. Допустим, что изменение климата произошло. Является ли оно значимым
или же это всего лишь небольшое колебание, вызванное одной из особенностей ат-
трактора? Вероятно, вскоре мы узнаем ответ на этот вопрос. Изменения климата
также могут быть вызваны действиями живых существ — вспомним о гипотезе Геи,
предложенной Джеймсом Лавлоком в 1980-е годы. Согласно ей, живые организмы
отвечают за поддержание земной атмосферы. На основе гипотезы Лавлока была
создана упрощенная математическая модель под названием «Маргаритковый мир»,
о которой мы расскажем далее.
А что, если мы сами меняем атмосферу?
Гея и «Маргаритковый мир»
«Маргаритковый мир» (англ. Daisyworld) — это математическая модель, предло-
женная Эндрю Уотсоном и Джеймсом Лавлоком в 1983 году. Подобно тому, как
уравнения Лоренца представляют собой упрощенную модель климата, модель Уотсона
и Лавлока представляет упрощенный мир, населяемый исключительно маргаритками
белого и черного цвета. Целью этой модели было объяснить, как живые организмы
способны менять атмосферу, в частности температуру воздуха на планете.
146
ЭКОЛОГИЯ И МАТЕМАТИКА. ВЗАИМОВЫГОДНОЕ СОТРУДНИЧЕСТВО
Джеймс Лавлок, автор гипотезы Гёи. 1993 год.
В модели предполагается существование Солнца с характеристиками, схожими
с нашим Солнцем, и, как следствие, присутствие солнечного света. В модели так-
же учитывается альбедо, то есть доля солнечного света, отражаемого маргаритками
(черными или белыми), а также земной поверхностью, на которой не растут мар-
гаритки. Если выбрать в качестве базового значения альбедо земной поверхности,
не заселенной маргаритками, то альбедо белых маргариток будет выше базового
значения, а их температура — ниже, чем температура земной поверхности. Черные
маргаритки, напротив, будут отражать меньше света, чем незаселенная поверхность,
в результате их температура будет выше. В модели Уотсона и Лавлока энергетиче-
ский баланс планеты рассчитывается с учетом того, что на части суши произрастают
белые маргаритки, на части — черные, а часть суши остается незаселенной. Кро-
ме того, в модели рассматривается показатель снижения численности маргариток,
а также математическая функция температуры. Модель содержит два дифференци-
альных уравнения, позволяющих смоделировать рост числа черных и белых марга-
риток на воображаемой планете:
^ = л-(₽(Т ).S-y), ^ = у(₽(Т)5-т).
dt dt у
Эти уравнения описывают скорость, с которой численность маргариток возрас-
тает или убывает. Согласно первому уравнению, скорость, с которой меняется число
белых маргариток х, зависит от их численности в момент времени t, коэффициента
роста |3(Т*), а также от доли земной поверхности, где не растут маргаритки (5),
и показателя снижения численности у. Обратите внимание, что рост числа белых
147
ЭКОЛОГИЯ И МАТЕМАТИКА. ВЗАИМОВЫГОДНОЕ СОТРУДНИЧЕСТВО
маргариток Р(7*), в свою очередь, зависит от температуры Т* участка земной по-
верхности, заселенной маргаритками.
Второе уравнение описывает динамику численности черных маргариток. В этом
случае у — число черных маргариток в момент времени t, |3(7у) — коэффициент
роста, который зависит от локальной температуры на участке суши, где произрас-
тают черные маргаритки, S — часть суши, где не растут маргаритки, у — показатель
снижения численности. Будем считать, что значение у для обоих видов маргариток
одинаково.
Также будем считать, что поведение коэффициента роста для белых и черных
маргариток одинаково и описывается параболой. В соответствии с динамикой этого
коэффициента предполагается, что температура, оптимальная для роста обоих видов
маргариток, равна 22,5 °C. Таким образом, при оптимальной температуре коэф-
фициенты роста белых и черных маргариток, Р(7) и Р(7у), максимальны и равны
единице. Так как коэффициенты роста описываются параболой, их значение будет
уменьшаться до нуля по мере того, как температура будет приближаться к 5 °C или
40 °C (пороговые значения в модели выбраны произвольно). Следовательно, ко-
эффициенты роста для двух видов маргариток варьируются от 0 до 1 и описаны по-
хожими выражениями, которые приведены ниже:
Р(Т ) = 1 - 0,003265(22,5-7 )2,
0(7*) = 1 - 0,003265(22,5-7* )2.
Важно заметить, что в этих выражениях учитывается локальная температура.
При температуре от 5 °C до 40 °C число маргариток будет возрастать. Так как бе-
лые маргаритки отражают больше солнечного света, область, в которой они произ-
растают, начнет охлаждаться. Черные маргаритки, напротив, поглощают солнечный
свет, и область, в которой они произрастают, будет нагреваться. В результате темпе-
ратуру в областях, где произрастают белые маргаритки Т* и черные маргаритки 7 ,
можно выразить так:
7 = Q(A - А ) + 7 ,
х х' m
7 = Q(A - А ) + 7 ,
У у7 m
где Q — коэффициент поглощения тепла (его значение в модели равно 20), 7 —
средняя температура на планете, А — альбедо планеты, А* — альбедо, вызванное
белыми маргаритками, А^ — альбедо, вызванное черными маргаритками.
148
ЭКОЛОГИЯ И МАТЕМАТИКА. ВЗАИМОВЫГОДНОЕ СОТРУДНИЧЕСТВО
Средняя температура на планете Т рассчитывается согласно закону Стефана —
Больцмана. Этот закон гласит, что энергия, излучаемая телом, пропорциональна тем-
пературе этого объекта, возведенной в четвертую степень. Применив этот закон,
получим следующее выражение:
-273,
где О = 5669 • 10 8 Вт/м2 • К4 — постоянная Стефана — Больцмана, Е — сол-
нечная энергия, получаемая планетой. Обратите внимание, что если бы мы захотели
смоделировать безжизненную планету, то есть планету, не населенную маргаритками,
то значение альбедо было бы равным А = 0,5. Альбедо планеты Л зависит от того,
в какой степени поверхность планеты покрыта белыми и черными маргаритками. Это
означает, что альбедо планеты А определяется следующим выражением:
А = А • S + А • S + А - S ,
S X х у у
где As — альбедо той части суши, где не растут маргаритки (площадь этой части
суши равна S), А* — альбедо, вызванное белыми маргаритками (площадь суши, где
растут белые маргаритки, равна SJ, Ау — альбедо, вызванное черными маргаритками
(доля земной поверхности, где они произрастают, равна SJ). Обратите внимание, что
выражение, по которому рассчитывается альбедо, описывает модель энергетического
баланса планеты. Напомним, что черные маргаритки отражают меньше света, чем
открытая суша, а белые маргаритки — больше, чем суша. Иными словами, выпол-
няется следующее неравенство: А^ < As< А
Цель простейшего эксперимента с моделью «Маргариткового мира» — изучить,
как изменяется доля поверхности планеты, где произрастают белые и черные марга-
ритки. Как правило, в модели используются следующие значения: А = 0,25, А^ =
= 0,50, А* = 0,75, у = 0,30. Планета, где в изобилии растут белые маргаритки, бу-
дет холоднее, чем планета, изобилующая черными цветами. После увеличения числа
белых маргариток, вызванного нагреванием планеты, температура будет снижаться,
и напротив, любое снижение температуры на планете будет компенсироваться ростом
числа черных маргариток. Кроме того, для некоторых значений параметров возмож-
ны два равновесных состояния: планета будет либо совершенно безжизненной, либо
полностью покрытой маргаритками.
149
ЭКОЛОГИЯ И МАТЕМАТИКА. ВЗАИМОВЫГОДНОЕ СОТРУДНИЧЕСТВО
На страницах этой книги мы совершили увлекательную экскурсию, во время
которой показали, что причиной удивительного прогресса биологии и биомедицины
во многом стало плодотворное сотрудничество математики и науки о жизни. Ис-
пользование дифференциальных уравнений, теории хаоса, программ символьных
вычислений, операций над векторами и матрицами (и линейной алгебры) позволяет
биоматематикам описать красоту и сложность жизни с помощью математических
выражений. Благодаря математике ученые могут создавать модели, делать прогно-
зы относительно самых разных проявлений жизни, будь то гены, белки, клетки, ор-
ганизмы, популяции или экосистемы. Мы надеемся, что после прочтения этой книги
читатель захочет подробнее узнать о математической биологии, или биоматемати-
ке — науке с богатыми традициями и большими перспективами.
150
Приложение
Магия комплексных чисел
По какой-то странной причине комплексные числа тесно связаны со многими физиче-
скими явлениями. Они присутствуют в электромагнетизме, используются в электро-
нике, электротехнике, квантовой механике и при изучении волн. В математической
биологии комплексные числа применяются при изучении биологических ритмов, за-
нимают важное место в теории хаоса, без них невозможно представить фракталы
на компьютере.
Графическое представление комплексных чисел очень просто. Если предположить,
что комплексное число — это точка z, то в декартовой системе координат, которую
далее мы будем называть комплексной плоскостью, на горизонтальной оси X будет
откладываться его вещественная часть а, на вертикальной оси Y — мнимая часть Ь.
Если мы также обозначим через г расстояние от начала координат до точки z, нетруд-
но показать, что это расстояние (оно называется модулем комплексного числа и обо-
значается |z|) равно у]а2 +Ь2- Более того, если учесть, что вещественная часть а равна
косинусу угла ОС между осью х и радиус-вектором точки z, умноженному на модуль
комплексного числа, а мнимая часть b — синусу угла ОС, умноженному на модуль
комплексного числа, то в так называемых полярных координатах число z будет за-
писываться следующим образом:
z = r(cos(OC) + i sin(oc)).
Одно из самых любопытных свойств комплексных чисел заключается в том, что
они расширяют возможности моделирования реальности, так как на них не распро-
страняются ограничения, свойственные вещественным числам. Чтобы совершать
с ними действия, можно представить, что, находясь на комплексной плоскости, уче-
ный одной ногой стоит в вещественном мире (ему соответствует часть а), другой —
в мнимом мире (ему соответствует часть Ь). При необходимости он беспрепятствен-
но путешествует из одного мира в другой. Таким образом, к примеру, операции сло-
жения и умножения комплексных чисел расширяют значение этих двух операций,
что вы могли видеть при изучении фракталов Мандельброта и множеств Жюлиа.
151
ПРИЛОЖЕНИЕ
Сложение и вычитание комплексных чисел
С комплексными числами могут выполняться операции сложения и вычитания.
На первый взгляд это кажется сложным, но в действительности это не так. Сумма
двух комплексных чисел а 4- bi и с 4- di рассчитывается следующим образом:
(а + bi) + (с + di) = (а 4- с) 4- (Ь 4- d)i.
К примеру, (2 4- 5z) 4- (3 — i) = (2 4- 3) 4- (5 ~ l)z = 5 4- 4i.
Вычитание — операция, обратная сложению, следовательно, разность ком-
плексных чисел а + Ы ис + di рассчитывается так:
(а 4- bi) — (с 4- di) = (а — с) 4- (Ь — d)i.
К примеру, (1 4- 3i) — (4 4- 2i) = (1 — 4) 4- (3 — 2)z = — 3 4- i.
Умножение и деление комплексных чисел
Также для комплексных чисел определены умножение и деление.
Произведение двух комплексных чисел а + Ыис + di определяется так:
(а 4- bi) ’ (с 4- di) = (ас — bd) 4- (ad 4- bc)i.
Обратите внимание, что результат умножения можно получить следующим, бо-
лее понятным способом:
(а 4- bi) • (с 4- di) = а • с 4- а • d • i 4- b • с • i 4- b • d • i2.
Напомним, что i2 — —1. Имеем:
a'c + a‘d'i + b’i-c — b'd.
Приведем подобные слагаемые:
(ас — bd) 4- (ad 4- bc)i.
152
ПРИЛОЖЕНИЕ
К примеру, (2 + 6i) • (8 + 2i) = (2 • 8 — 6 • 2) 4- (2 • 2 + 6 • 8)i — 4 + 52z.
Частное двух комплексных чисел а + Ыис + di определяется так:
а + Ы
с + di
ac + bd) ( lx — ad 1
c2+</2J+u2+d2)
Например,
3-5»
6 + 2»
S-6-5-2^ f-5-6-3-2
, 62+22 J\ 62+22
Г18 —10^1 f-30-6\
----- + ----- »
4 36 + 4 J 36 + 4 ?
_8__36 .
40 40 ’
И вновь обратите внимание, что частное двух комплексных чисел — это резуль-
тат выполнения следующей последовательности действий:
а + Ы (а + Ы)-(с —di) a-c — a-d-i + b-c-i — b-d-i2
с + di (с — di) (с — di) (с — di) (с — di)
Приведем подобные слагаемые:
ac + bd
2 . j2
с +d
he —ad
c2+d2
Формула Эйлера, одно из прекраснейших
математических выражений
Комплексные числа полезны не только для графического изображения фракталов. Их
постоянно используют инженеры при работе с электрическими цепями. Так, мощность
бытовой техники выражается вещественными числами, мощность промышленных
устройств — комплексными. Изучение биологических циклов, которые переживает
человек, и анализ колебаний (к примеру, колебаний тела, закрепленного на пружине)
в физике отчасти схожи: для решения этих задач используются комплексные чис-
ла. По этой причине те, кто знаком с комплексными числами, обычно используют
формулу, которая считается одной из самых красивых и полезных в математике. Это
153
ПРИЛОЖЕНИЕ
формула Эйлера, связывающая мнимую единицу i, степень числа е и тригонометри-
ческие функции синуса и косинуса:
еа' = cos(a) + i sin(oc) •
Нетрудно показать, что если мы изобразим окружность единичного радиуса
на комплексной плоскости, то на вещественной оси X будет откладываться косинус
угла ОС, на мнимой оси Y — синус угла ОС. По-видимому, эта формула была открыта
несколькими учеными независимо друг от друга в XVIII веке. В 1748 году Эй-
лер получил ее в результате преобразований степенного ряда, однако никто не смог
представить эту формулу на комплексной плоскости. Столь простая идея представ-
ления комплексных чисел в декартовых координатах появилась позднее, примерно
в 1800 году, благодаря Жану Роберу Аргану. Именно тогда математики впервые
смогли увидеть графическое отображение одной из прекраснейших формул матема-
тики.
154
Библиография
BATSCHELET, Е., Introduction to Mathematics for Life Scientists, Berlin-Heidelberg,
Springer-Verlag, 1971.
BaZIN, M. (ed.), Mathematics in Microbiology, Londres, Academic Press, 1983.
BRIGGS, J., Peat F. D., Turbulent Mirror. An Ilustrated Guide to the Chaos Theory and
the Science of Wholeness, Nueva York, Harper & Row Publisher, 1989.
LAHOZ-BELTRA, R., Bioinformdtica. Simulation, vida artificial e inteligencia artificial,
Madrid, Ediciones Diaz de Santos, 2004.
—: cfuega Darwin a los dados? Madrid, Nivola, 2008.
—: Turing. Del primer ordenador a la inteligencia artificial, 2a edition, Madrid, Nivola,
2009.
Martinez Calvo, M.C., Perez de Vargas, A., Metodos matemdticos en biologi'a,
Madrid, Centro de Estudios Ramon Areces, 1993.
Martinez Calvo, M.C., Fernandez Bermejo, E., Gonzalez Manteiga, M.T.,
LAHOZ-BELTRA, R., PERALES Gravan, C., Matemdticas bdsicas para biologos, CD-
ROM. Innovation Educativa (ISBN 84-7491-786-7), Madrid, Editorial Complu-
tense, 2005.
RlETMAN, E., Exploring the Geometry of Nature. Computer Modeling of Chaos, Fractals,
Cellular Automata and Neural Networks, Nueva York, Windcrest, 1989.
155
Алфавитный указатель
Derive 34, 64, 100
ENIAC 16-17,141,146
Escherichia coli 64—65, 67—70
аттрактор
Лоренца 74, 142, 143
предельный цикл 72
странный 73, 74, 80
точечный 71
Хенона 74
Барнсли, Майкл 83—84
биоинформатика 35, 107—108
биология
алгебраическая 35
реляционная 12
систем 23-26
системная 24-26
биореактор 9, 53, 54
вектор
столбец 93, 103, 110, 114, 116, 121,
125,128
стохастический 103
строка 116-117, 120-121
векторное пространство 111-112
величины
векторные 109—110
скалярные 109
витамин А 87, 89
Вольтерра, Вито 16, 42, 133, 134, 137
Геи гипотеза 145—147
гипотеза 30,146
гомеостаз 23
Дарвин, Чарльз 14—15, 57, 107
ДНК 22,107-108,111,121
естественного отбора теория 14—15, 57,
145
Жюлиа, Гастон 81—83
игра в хаос 83-84
ископаемые 51—52
искусственный интеллект 33, 95
клеточные автоматы 29, 39—42, 78
Ва-ТЬр 29, 42, 44
игра «Жизнь» 40, 41
климат 28, 32, 33,139-146
Крамера правило 127—129
Лавлок, Джеймс 145—147
Лесли, Патрик 36, 133
локомоция 112
Лоренц, Эдвард 140, 142—145
Лотка, Альфред Джеймс 16, 133, 135,
137
Лотки — Вольтерры уравнение 42, 43,
133-135,142
модель межвидовой конкуренции
133,139
модель «хищник — жертва» 16, 27,
29, 42-44,138-139
Мак-Каллока — Питтса 120
Мальтус, Томас 54—55, 65, 67,132
Мандельброт, Бенуа 82
«Маргаритковый мир» 146, 149
Маркова цепи 108
матрица
диагональная 88
единичная 88
квадратная 88, 95, 97-99,103
Лесли 36-39,133,134
157
АЛФАВИТНЫЙ УКАЗАТЕЛЬ
Маркова 107—108
нулевая 88
обратная 99—101, 124—125
операций 39, 89—93, 99,110
перехода 37
симметричная 88
стохастическая 103
транспонированная 95
треугольная 88
Менделя законы 101—107
методы
численные 29, 51, 76
минор 98
множества
Жюлиа 74,1-82,151
Мандельброта 81—82
теория 12, 36, 44—46
моделирование 27—33, 41—43, 48
(
модель
BIDE 133-134
климатическая 142, 143
конкуренции 133, 139
математическая 20, 24, 30—32,
44-46, 48-49
реакция — диффузия 19—21, 26, 40
эпидемии 49, 62—64
модуль 110, 111, 121, 151
морфогенез 18—22, 28
начальные условия 32-33, 74-76,
144-145
нейрон 74, 78, 93-94,115-123
нейронная сеть 89, 93—94, 115, 119,
121-123
сеть Хопфилда 123
обратная связь 24, 69
общая теория систем 13
определитель 95-100,126-128
орбита 70—75, 82—83
отображение 73—76, 78, 84
переменные 31—32
подсчет 72, 85,102
популяция 36—44, 54—57, 63—73,
132-139,146-148
порядок 97
правило
многоугольника ИЗ
параллелограмма 112-114
Пригожин, Илья 26
произведение
векторное 120-123
скалярное 116-121
Пуассона распределение 65
равновесие 26-28, 71-72,128
рак 58-62,135
Рашевски, Николас 11—13
ритм 15, 50, 67,135-136
рост
экспоненциальный (см. мальтусов-
ский) 53, 62, 63, 65,133,136
логистический (см. Ферхюльст) 133
мальтусовский 55—56, 62, 67, 68
функция Гомпертца 9, 58, 59
рыбоводческие хозяйства 14
Серпинского треугольник 83—84
символьные вычисления 29, 33-35, 64,
100
системы
динамические 34, 47—50, 73, 78, 83
диссипативные 26—28
изменяющиеся 47
итерируемых функций 80, 84
линейные 23—24, 96
158
АЛФАВИТНЫЙ УКАЗАТЕЛЬ
матричные 83 нелинейные 27—28 сложные 12, 28—29, 58 уравнений 111, 124—129 хаотические 28, 74, 83 скорость 27, 114 теория множеств 12, 36, 44—46 Тьюринг, Алан 16—22, 26, 39, 40 углерод С-14 51—52 уравнение в конечных разностях 75, 78 дифференциальное 50—51, 53—59, 67-69, 75-78 Мальтуса 63, 133, 136 реакция — диффузия 19—21, 26, 40 Ферхюльста 28, 57, 69, 76, 84 уровень изменений 50, 71 Ферхюльст, Пьер Франсуа 54—57, 69, 133 Фибоначчи, числа 18—19 Фишер, Рональд Эйлмер 15—16 фон Берталанфи, Карл Людвиг 13, 24 фон Нейман, Джон 17, 22, 39—40, 141, 146 фрактал 78—84,151,153 хаос 67, 73—75 хи-квадрат, критерий 104, 107 число е 64-65, 68,154 комплексное 151, 153 мнимое 77, 154 эволюция 14—16, 30, 76,107—108,145 эвристические методы 29 экосистема 48, 131—133 эпидемия 57, 62—64 свиного гриппа 32, 62—64 СПИДА 62-64 «эффект бабочки» 143
159
Научно-популярное издание
Выходит в свет отдельными томами с 2014 года
Мир математики
Том 28
Рафаэль Лаос-Бельтра
Математика жизни.
Численные модели в биологии и экологии
РОССИЯ
Издатель, учредитель, редакция:
ООО «Де Агостини», Россия
Юридический адрес: Россия, 105066,
г. Москва, ул. Александра Лукьянова, д. 3, стр. 1
Письма читателей по данному адресу не при-
нимаются.
Генеральный директор: Николаос Скилакис
Главный редактор: Анастасия Жаркова
Выпускающий редактор: Людмила Виноградова
Финансовый директор: Наталия Василенко
Коммерческий директор: Александр Якутов
Менеджер по маркетингу: Михаил Ткачук
Менеджер по продукту: Яна Чухиль
Для заказа пропущенных книг и по всем вопро-
сам, касающимся информации о коллекции, за-
ходите на сайт www.deagostini.ru, по остальным
вопросам обращайтесь по телефону бесплатной
горячей линии в России:
® 8-800-200-02-01
'Телефон горячей линии
для читателей Москвы:
S 8-495-660-02-02
Адрес для писем читателей:
Россия, 600001, г. Владимир, а/я 30,
«Де Агостини», «Мир математики»
Пожалуйста, указывайте в письмах свои кон-
тактные данные для обратной связи (телефон
или e-mail).
Распространение:
ООО «Бурда Дистрибьюшен Сервисиз»
УКРАИНА
Издатель и учредитель:
ООО «Де Агостини Паблишинг» Украина
Юридический адрес: 01032, Украина,
г. Киев, ул. Саксаганского, 119
Генеральный директор: Екатерина Клименко
Для заказа пропущенных книг и по всем вопро-
сам, касающимся информации о коллекции, за-
ходите на сайт www.deagostini.ua, по остальным
вопросам обращайтесь по телефону бесплатной
горячей линии в Украине:
® 0-800-500-8-40
Адрес для писем читателей:
Украина, 01033, г. Киев, a/я «Де Агостини»,
«Мир математики»
Украша, 01033, м. Ки!'в, а/с «Де Агостпп»
БЕЛАРУСЬ
Импортер и дистрибьютор в РБ:
ООО «Росчерк», 220037, г. Минск,
ул. Авангардная, 48а, литер 8/к,
тел./факс: (+375 17) 331-94-41
Телефон «горячей линии» в РБ:
+ 375 17 279-87-87 (пн-пт, 9.00-21.00)
Адрес для писем читателей:
Республика Беларусь, 220040, г. Минск,
а/я 224, ООО «Росчерк», «Де Агостини»,
«Мир математики»
КАЗАХСТАН
Распространение:
ТОО «КГП «Бурда-Алатау Пресс»
Издатель оставляет за собой право увеличить реко-
мендуемую розничную цену книг. Издатель остав-
ляет за собой право изменять последовательность
заявленных тем томов издания и их содержание.
Отпечатано в соответствии с предоставленными
материалами в типографии:
Grafica Veneta S.p.A Via Malcanton 2
35010 Trebaseleghe (PD) Italy
Подписано в печать: 11.06.2014
Дата поступления в продажу на территории
России: 29.07.2014
Формат 70 х 100 / 16. Гарнитура «Academy».
Печать офсетная. Бумага офсетная. Печ. л. 5.
Усл. печ. л. 6,48.
Тираж: 34 000 экз.
© Rafael Lahoz-Beltra, 2010 (текст)
© RBA Collecionables S.A., 2011
© ООО «Де Агостини», 2014
ISBN 978-5-9774-0682-6
ISBN 978-5-9774-0723-6 (т. 28)
@
Данный знак информационной про-
дукции размещен в соответствии с требования-
ми Федерального закона от 29 декабря 2010 г.
№ 436-ФЗ «О защите детей от информации, при-
чиняющей вред их здоровью и развитию».
Издание для взрослых, не подлежит обязатель-
ному подтверждению соответствия единым требо-
ваниям, установленным Техническим регламентом
Таможенного союза «О безопасности продукции,
предназначенной для детей и подростков» ТРТС
007/2011 от 23 сентября 2011 г. № 797.
Математика
жизни
Численные модели в биологии и экологии
Жизнь - одно из самых прекрасных и сложных явлений
на планете, изучением которого с начала XX века занимается
не только одна биология. Физики, а затем и математики
обнаружили, что некоторые биологические явления можно
описать с помощью математического языка. Так родилась новая
дисциплина - математическая биология, или биоматематика.
Благодаря ей сегодня можно получить ответы на множество
важных вопросов, касающихся биологии и биомедицины.
Эта книга представляет собой панорамный обзор различных
явлений, которые изучает биоматематика.
12+