/







































































































































Text
Е.Н. КУРОЧКИН
птицы
ЦЕНТРАЛЬНОЙ
АЗИИ
В ПЛИОЦЕНЕ
АКАДЕМИЯ НАУК СССР АКАДЕМИЯ НАУК МНР
ПАЛЕОНТОЛОГИЧЕСКИЙ ИНСТИТУТ ГЕОЛОГИЧЕСКИЙ ИНСТИТУТ
ЗСБНХУ-ЫН шинжлэх УХААНЫ АКАДЕМИ БНМАУ-ЫН ШИНЖЛЭХ УХААНЫ АКАДЕМИ
ПАЛЕОНТОЛОГИЙН ХУРЭЭЛЭН ГЕОЛОГИЙН ХУРЭЭЛЭН
ACADEMY OF SCIENCES OF THE USSR ACADEMY OF SCIENCES OF THE MPR
PALEONTOLOGICAL INSTITUTE GEOLOGICAL INSTITUTE
Е. Н. КУРОЧКИН
ТОВ АЗИЙН ПЛИОЦЕНЫ
ШУВУУД
монгол -зевлелтиин
ПАЛЕОНТОЛОГИЙН ХАМТАРСАН ЭКСПЕДИЦИ
(Бутээл цуврул 2 6)
эрлэх зевлел:
Р. Барсболд,
академии Б. Лувсанданзан,
академии Л. П. Татаринов (хариуцлагатай эрхлэгч),
Б. А. Трофимов, В. Ю. Решетов, М. А. Шишкин
МОСКВА
«НАУКА» ХЭВЛЭЛИЙН ГАЗАР
1985
Е. Н. КУРОЧКИН
ПТИЦЫ ЦЕНТРАЛЬНОЙ
АЗИИ В ПЛИОЦЕНЕ
СОВМЕСТНАЯ СОВЕТСКО-МОНГОЛЬСКАЯ
ПАЛЕОНТОЛОГИЧЕСКАЯ ЭКСПЕДИЦИЯ
(Труды, в ы п. 26)
РЕДАКЦИОННАЯ КОЛЛЕГИЯ:
Р. Барсболд,
академик Б. Лувсанданзан,
академик Л. П. Татаринов (ответственный редактор),
Б. А. Трофимов, В. Ю. Решетов, М. А. Шишкин
в
МОСКВА
«НАУК А»
198 5
УДК 568.2 : 551.782.2 (5-925)
Е. N. Kurochkin
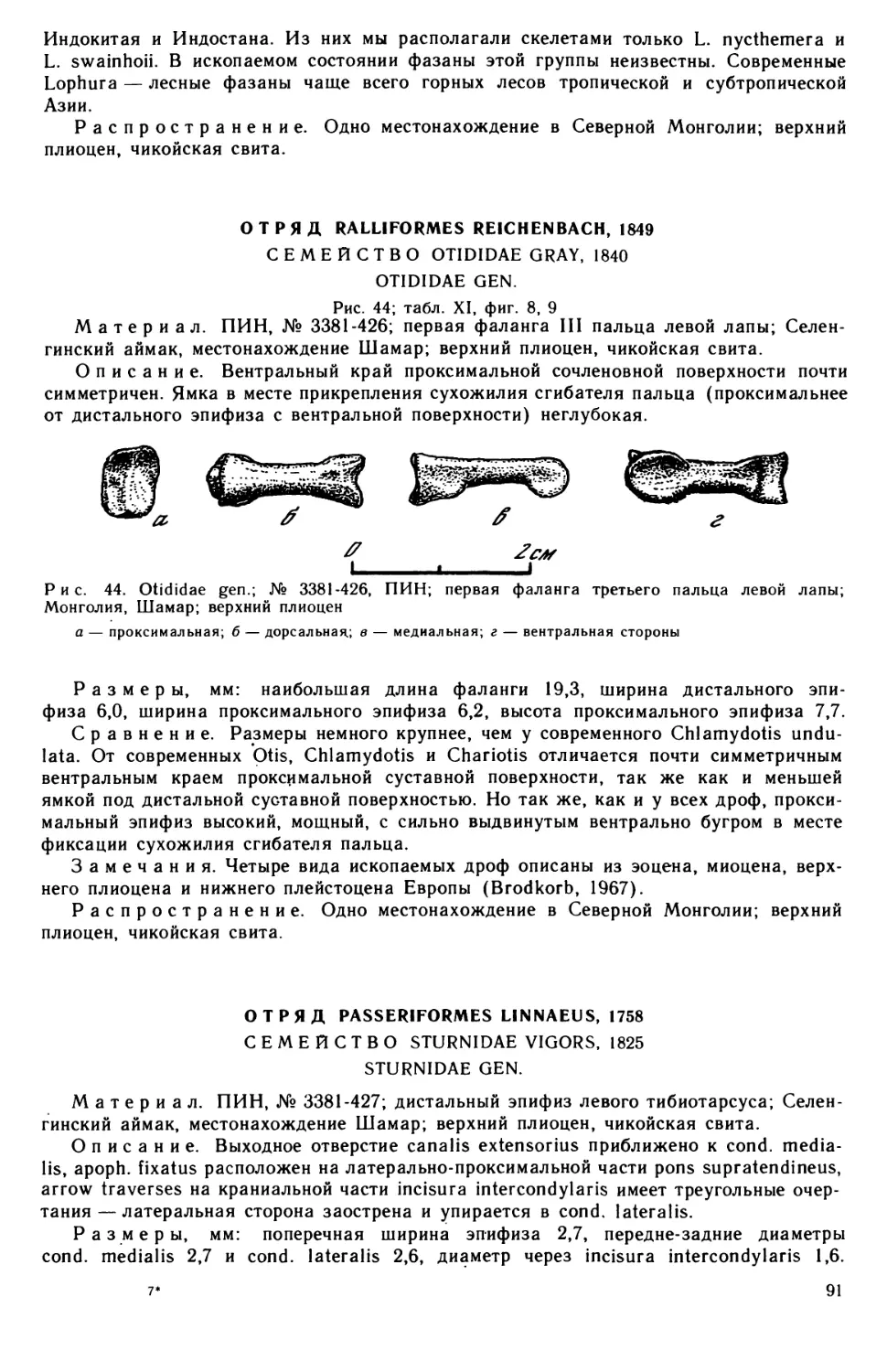
BIRDS OF THE CENTRAL ASIA
IN PLIOCENE
THE JOINT SOVIET-MONGOLIAN
PALEONTOLOGICAL EXPEDITION
(Transactions, vol. 26)
EDITORIAL BOARD:
R. Barsbold,
academician MPR B. Luwsandanzan,
academician L. P. Tatarinov (editor-in-chief),
B. A. Trofimov, V. Yu. Reshetov, M. A. Shishkin
E. H. Курочкин. Птицы Центральной Азии в плиоцене. М/. Наука, 1985, 120 с.
Первая сводка по Азии обобщает многолетние исследования плиоценовых птиц Монго-
лии, Забайкалья и Восточного Казахстана. Описано 56 вымерших форм из 23 семейств птиц,
часть из которых была непосредственными предками современных видов. Многие роды и не-
которые семейства впервые найдены в ископаемом состоянии. Дан систематический, экологичес-
кий, зоогеографический и стратиграфический анализы описанных ископаемых видов и ави-
комплексов, определены их связи с современной авифауной и исторические связи с авифауной
Европы, Северной Америки и Южной Азии. Показаны морфофункциональные особенности не-
которых из изученных видов.
Для зоологов, геологов, палеонтологов и лиц, интересующихся птицами.
Табл. 10, ил. 49, фототабл. 16, библиогр. 109 назв.
Рецензенты:
И. А. ДУБРОВО, М. Ф. ИВАХНЕНКО
Евгений Николаевич Курочкин
ПТИЦЫ ЦЕНТРАЛЬНОЙ АЗИИ В ПЛИОЦЕНЕ
Совм. Сов .-Монгол, палеонтол. экспедиция
(Труды, вып. 26)
Утверждено к печати Палеонтологическим институтом Академии неук СССР
Редактор издательства Р.Л. Цыбульская. Художественный редактор И.Ю. Нестерова
Технические редакторы Н.М. Бурова, И.И. Джиоева. Корректор О А. Разуменко
ИБ № 28883
Подписано к печати 20.04.85. Т - 03425. Формат 70 X 100 1/16. Бумага офсетная № 1
Гарнитура литературная (фотонабор). Печать офсетная
Усл.печ.л. 9,8 + 1,3 вкл. Усл.кр.-отт. 11,3. Уч.-иэд.л. 13,5
Тираж 1000 экз. Тип.зак. 2098. Цена 2 руб.
Ордена Трудового Красного Знамени издательство ’’Наука*’
117864 ГСП-7, Москва В-485, Профсоюзная ул., д. 90
Ордена Трудового Красного Знамени 1-я типография издательства ’’Наука”
199034, Ленинград В-34, 9-я линия, 12
2002000000-261
042 (02)-85
248-85-П
© Издательство «Наука»,
1985 г.
ВВЕДЕНИЕ
История изучения птиц палеогена и неогена Европы, Северной и Южной Америки
насчитывает уже больше 100 лет и оттуда описаны сотни видов ископаемых птиц.
В противоположность этому третичные птицы Азии известны по очень фрагментарным
данным, что может показаться странным на фоне репутации палеонтологического
Эльдорадо, каковой в действительности и является Центральная Азия. По существу же
это объясняется долгим отсутствием конкретного интереса к птицам как к палеонто-
логическому объекту в прошлых экспедициях в Азию для сборов и раскопок ископае-
мых животных. Определенное значение в отсутствии материалов по ископаемым птицам
йз Азии имели невыразительность, фрагментарность и редкость птичьих остатков, на
которых непосредственно при работе на местонахождениях обращают мало внима-
ния, особенно когда палеонтологи и геологи проводят работы на местонахождениях,
отложения которых буквально переполнены скелетами млекопитающих или динозавров,
и когда довлеет гипноз высокой ценности их зубов, челюстей и черепов. Часто, если кости
птиц и попадали в выборки материалов, привозимых из поля, они оседали безвестными
в многочисленных коробочках с неопределимыми фрагментами посткраниального
скелета млекопитающих, черепах или амфибий.
Таким путем из суммы знаний, собираемых палеонтологией о развитии жизни на
Земле, подчас выпадает отдельный большой класс позвоночных, среди наземных —
сейчас самый многочисленный, данные по которому интересны не только сами по себе,
но и для обобщения представлений об эволюции всей биоты в целом, а если помнить и
о задачах геологии, то и для биостратиграфии континентальных (преимущественно)
отложений по всему комплексу ископаемых организмов.
Что же недавно было известно о птицах неогена Азии? В миоцене и плиоцене из
сиваликской серии Индии и Пакистана описаны по костным остаткам Struthio asiaticus,
два вида пеликанов, марабу (L. falconeri), какие-то баклан и крохаль. Из среднего
плиоцена Китая известен таз страуса под названием St. wimani и два вида страусов —
по яйцам и скорлупе, а из местонахождения Эртемтэ сообщалось о находках гуся, уткй,
кулика, фазановой птицы, голубя, ястреба и воробьиных. Из нижнего плиоцена
Павлодара Тугариновым (1935) описаны позвонок страуса и сушкиния (сокол).
Из верхнего миоцена Ирана был описан амфипеларгус ( = Urmiornis maraghanus) и
фаланга страуса. Благодаря трудам Н. И. Бурчака-Абрамовича (1966, 1978) имеются
кое-какие сведения о неогеновых птицах Закавказья. Из сармата им описаны два
вымерших вида Anser, руставиорнис (Phasianinae), скорлупа яиц страусов, из верхнего
плиоцена — таз Struthio transcaucasicus, Anas apscheronica, скорлупа яиц страусов,
а также сообщалось об остатках 16 видов современных или близких к ним видов птиц
из двух местонахождений апшеронского возраста. Из раннего плиоцена Азербайджана
была еще описана крачка — Sterna milne-edwardsii Rjabinin. Фаланга страуса была
найдена в верхнем миоцене Турции. Для павлодарской фауны П. А. Аубекерова
(1965) сообщала о находках Falco cf. tinnunculus, Milvus cf. korschun и Passeres (эти
материалы исследованы нами в данной работе). Опубликовано предварительное опре-
деление М. А. Воинственским остатков птиц из верхнемиоценовых отложений о-ва Оль-
хон на Байкале (Логачев и др., 1964), где приводятся Ardea sp., Crex sp., Porzana sp.,
Anser sp. (2 вида), Anas sp. (4 вида), Branta sp. (?), Nyroca, Striges. Понятно, что
к указаниям на находки современных родов в позднем миоцене, а современных видов
птиц — в раннем плиоцене, надо относиться с осторожностью.
Вот и все сведения, которыми располагал автор, когда начал изучение птиц йз
плиоцена Западной Монголии и Забайкалья.
С началом работ в 1967 г. в Монгольской Народной Республике Совместной
Советско-Монгольской научно-исследовательской геологической экспедиции (СМГЭ)
появились первые птицы из неогеновых местонахождений Западной Монголии, собран-
5
ные Е. В. Девяткиным, И. Г. Лискун и В. С. Зажигиным. Первой коллекции ископае-
мых птиц из плиоцена Монголии была посвящена небольшая статья (Курочкин, 1971).
Находкам птиц значительно способствовал сбор ископаемых материалов по микро-
маммалиям, проводившийся путем промывки В. С. Зажигиным, труд и внимание кото-
рого к птицам принесли материалы по многим мелким формам. С 1969 г. в Монголии
начала работать Совместная Советско-Монгольская палеонтологическая экспедиция
(СМПЭ). В составе этой экспедиции уже квалифицированные палеонтологи В. И. Же-
галло, М. Б. Борисоглебская, Е. Л. Дмитриева и Е. К. Сычевская собрали для нас
множество остатков птиц из плиоцена на западе МНР. Всего, таким образом, оттуда
было накоплено около 200 фрагментов костей птиц из 9 местонахождений нижнего и
среднего плиоцена. Сам автор за время работы экспедиции занимался главным обра-
зом раскопками палеогеновых местонахождений на юге и юго-востоке Монголии.
После обработки плиоценовых монгольских материалов было решено разобраться
в сборах по птицам из плиоценовых местонахождений Забайкалья и Восточного Казах-
стана. Материалы из местонахождения Береговая на р. Чикой в Бурятской АССР были
собраны в 1967 г. В. И. Жегалло и в 1963 и 1967 гг. М. А. Ербаевой, они переданы
в Палеонтологический институт АН СССР (ПИН) для хранения и изучения. А. Г. По-
катилов собрал немного интересных птиц на Береговой и в Тологое в 1964 г., они были
переданы в ПИН временно для исследования Л. Н. Иваньевым и принадлежат Инсти-
туту земной коры (ИЗК) Сибирского отделения АН СССР (Иркутск). Коллекцию из
местонахождения Павлодар составили старые сборы Ю. А. Орлова и М. Ф. Прохо-
рова (1930 г.) и новые сборы В. С. Зажигина (1963—1965 гг.), хранящиеся в коллек-
ции ПИН, а также сборы П. Ф. Савинова 1960—1961 гг. из Института зоологии АН Ка-
захской ССР (ИЗКаз). Последние принадлежат коллекциям этого института и были
предоставлены нам для исследования П. А. Телеубердиной. Многочисленные
остатки амфипеларгусов из местонахождения Калмакпай в районе оз. Зайсан сборов
экспедиции ПИН были уже описаны (Курочкин, 1981).
Изучение ископаемых птиц невозможно без использования современных сравнитель-
ных остеологических материалов. На первых этапах использовались остеологические
коллекции Отдела орнитологии Зоологического музея Московского государственного
университета и Института зоологии АН Украинской ССР в Киеве, руководителям
которых автор хотел бы выразить теперь свою признательность — А. М. Судиловской,
С. В. Луцкой, М. А. Воинственскому и А. С. Уманской. Позднее работа уже велась на
сравнительной коллекции ПИН, созданной нами за эти годы. В пополнении этой кол-
лекции большое участие приняли многие коллеги и товарищи автора — В. Н. Катаев-
ский, В. М. Гудков, В. Н. Калякин, С. М. Успенский, В. Е. Флинт, Ф. Я. Дзержинский,
П. С. Томкович, В. К. Рахилин, Ю. В. Шибаев, Б. К. Шибнев, В. В. Бианки, А. Н. Го-
ловкин, Н. С. Гордиенко, А. И. Кошелев, В. П. Шунтов, С. М. Кудрявцев, А. И. Амелин,
Н. Н. Дроздов, К. Ф. Елкин. Сейчас в коллекции имеется более 900 видов современной
мировой фауны, однако даже при таком (вроде бы большом) общем объеме иногда
отсутствие 1—2 видов из современных родов делает невозможным достоверное опреде-
ление и описание ископаемого вида. В такой ситуации очень плодотворной оказалась
работа с коллекциями Американского музея естественной истории в Нью-Йорке,
Национального музея естественной истории Смитсониановского института в Вашинг-
тоне и проф. П. Бродкорба в университете Флориды, в сумме хранящими наиболее
богатые коллекции скелетов птиц мира. Эта поездка была поддержана Орнитологи-
ческим фондом Ф. Чапмэна, фондом Смитсониановского института и Академией
наук СССР и осуществлена благодаря участию моих американских коллег— У. Бока,
Л. Шорта и С. Олсона.
В процессе подготовки настоящей работы автор постоянно пользовался советами,
консультациями и поддержкой К. К. Флерова, Л. П. Татаринова, Е. В. Девяткина,
В. И. Жегалло, Б. А. Трофимова.
Рисунки выполнены в разное время художниками Ю. П. Алексеевым, Т. Д. Раковой
и К. Н. Улановой, фотографии — М. Н. Бочаровым в фотолаборатории ПИН.
Всем перечисленным здесь лицам и организациям автор выражает свои искренние
признательность и благодарность.
6
МАТЕРИАЛ И МЕТОДИКА
Ископаемые материалы, на которых основано это исследование, представляют собой
типичные материалы такого рода, обычно получаемые и изучаемые палеорнитологами,
это — преимущественно эпифизы костей конечностей. Редко попадались отдельные пол-
ные кости, части таза или грудин. На получение большого числа лучших остатков птиц
рассчитывать часто просто не приходится, и поэтому палеорнитолог удовлетворяется
фрагментами костей. Естественно, отсюда возникают специфические сложности в изуче-
нии ископаемых птиц, возможны ошибки и описание разных видов по разным костям
скелета, особенно разными исследователями. Непременное условие — использование
сравнительных коллекций современных видов. Известны попытки создания руководств
с рисунками для определения костей птиц, но они или имели ограниченное значение,
или закончились неудачно.
Отсутствие в скелете птиц какой-то одной системы, которая бы хорошо сохранялась
в ископаемом состоянии, с одной стороны, а с другой — хорошо бы отражала пути
адаптаций и эволюции, каковыми служат, например, зубы у млекопитающих или черепа
у рептилий и древних амфибий, делает изучение ископаемых птиц исключительно
трудоемкой работой. Она проводится на фоне общей значительной морфологической
монолитности всех птиц, поскольку требования необходимости сохранения способности
к полету накладывают на возможности специализации скелетно-мышечной системы
птиц одинаковые ограничения. Значителен и таксономический объем класса. Однако
вследствие тех же механических ограничений, накладываемых на птиц аэродинамикой,
индивидуальная изменчивость скелета (как и мускулатуры) оказывается у них мини-
мальной, поэтому изменчивость не затушевывает таксономических различий и умень-
шает возможность ошибок при описании ископаемых птиц на основе часто единичных
и фрагментарных объектов.
В палеонтологическом отношении кости разных частей скелета птиц неравноценны
как для диагностики, так и для понимания морфо-функциональных особенностей и
ведущих адаптаций. Разнообразие биологических типов птиц (причем есть веские,
подкрепленные фактами, основания считать, что оно оформилось еще в меловом
периоде) обусловлено иными, чем у остальных позвоночных, особенностями организа-
ции. Ведущие коммуникационные системы у птиц — зрение и слух. В меж- и внутривидо-
вых отношениях они ориентируются в основном на голос и окраску. Ведущие
трофические системы — челюстной аппарат и задние конечности. Аэродинамические ха-
рактеристики близких видов, а следовательно, и скелетно-мышечные структуры
крыла и плечевого пояса довольно сходны. Отсюда проистекает трудность в определе-
нии ископаемых остатков этих частей скелета, по ним же происходит наибольшее число
ошибок при описании ископаемых форм. В узком смысле образ жизни и специфика
биологического облика отдельных, особенно близких, видов птиц определяются в ос-
новном функциональной организацией челюстного аппарата и тазовых конечностей,
которые отражаются в «тонкой» морфологии этих систем. Но черепной и челюстной
материалы встречаются в ископаемом состоянии очень редко. Много более распро-
странены кости задних конечностей, которые в то же время оказываются наиболее
интересными и «понятными» в функциональном и таксономическом планах. Задние конеч-
ности выполняют не только локомоторные функции, часто определяя среду обитания,
но также — функции чесания и оправления оперения, устройства гнезда, умерщвления,
7
переноса и расчленения добычи. Морфология задних конечностей, особенно их дисталь-
ных отделов, отражает многие направления узкой экологической специализации у птиц.
Тщательные сравнительные исследования ископаемых костей позволяют находить раз-
личия между видами и родами, выражающиеся в изменении наклонов суставных
поверхностей, смещениях плоскостей ориентации мыщелков, незначительных переносах
точек и участков прикрепления и отхождения мускулов и т. п. Особенности функцио-
нальной организации скелета, о которых только что говорилось выше, дают у птиц
очень стабильные пропорциональные отношения между размерами отдельных частей
одной и той же кости и разных костей. Этот метод в последнее время стал широко
использоваться в палеонтологической диагностике, главным образом при изучении
четвертичных птиц.
Процедура исследования ископаемых остатков начинается в палеорнитологии
с определения отряда или семейства на современном сравнительном материале. Затем
с использованием сравнительных же коллекций и опубликованных данных по ископае-
мым формам выявленного семейства проводится определение родового и видового
статусов. Таксономический ранг ископаемой формы (родовой или видовой), если она
оказывается новой, а не уже известным вымершим или современным видом, устанавли-
вается на основании размаха структурных различий в отдельных деталях строения
костей современных видов или родов данного семейства. Индивидуальная изменчивость
скелета у птиц, как уже показывалось, довольно незначительна, так что, если мы
располагаем отдельными полными костями или эпифизами с неразрушенными поверх-
ностями, вероятность правильного результата определения в общем объеме материа-
лов, по нашему опыту, превышает 75 %. Известную сложность представляет сопостав-
ление остатков от разных частей скелета, потому что ископаемые кости у птиц чаще
всего собираются изолированными фрагментами. Поэтому синонимия в палеорнитологии
распространена, наверное, шире, чем в других областях палеонтологии позвоночных.
По мере исследования плиоценовых птиц Монголии было опубликовано 4 статьи и
несколько кратких сообщений, в которых описано около 20 новых таксонов птиц и
давался краткий общий анализ этой авифауны (Курочкин, 1971, 1976, 1977, 1982;
Kurochkin, 1970, 1978, 1980). При подготовке работы все описания новых видов и
родов были дополнены, развиты и уточнены. Все диагнозы фактически написаны
заново. В трех случаях было изменено систематическое положение ранее описанных
нами форм. Macrorhamphus finitimus Kurochkin, 1971 переведен в Calidris sp.; Rallus
risillus Kurochkin, 1980 переведен в Porzana risilla (Kurochkin, 1980); род. Bantamyx
Kurochkin, 1982 перемещен из Odontophorinae в Phasianinae. He подтвердилось, таким
образом, нахождение подсемейства американских зубчатоклювых куропаток в Азии.
Не подтвердилось предварительное сообщение о находке в плиоценовой фауне
Монголии Psittacidae. Фрагмент плеча не вполне взрослого экземпляра, на котором оно
основывалось, принадлежал, скорее всего, какому-то представителю Coraciidae. Нао-
борот, во многих случаях дополнительные материалы подтвердили первые описания,
часто выполненные на единичных фрагментах, что позволило шире характеризовать
такие виды, как Cygnus pristinus Kur., 1971, Anser devjatkini Kur., 1971, Phalacrocorax
reliquus Kur., 1976, Ciconia lucida Kur., 1982, Syrmaticus kozlovae sp. nov. и др.
Дополнены и уточнены также описания А. Я- Тугариновым (1935) сокола Sushkinia
и Struthio sp. из павлодарской фауны, что подтвердило правильность оригинальных
определений. Это тем более важно, что у некоторых палеорнитологов из-за плохого
качества иллюстраций и сжатого описания самостоятельность Sushkinia и правиль-
ность ее помещения в Falconidae вызывали сомнения. Остатки птиц из этой фауны,
описанные П. А. Аубекеровой (1965), оказались принадлежащими Sushkinia, а один
фрагмент отнесен к Palaeoperdix sp.
Хотя основа настоящей работы — плиоценовые птицы из Западной Монголии —
представляют довольно значительный объем — 37 видов из 17 семейств — называть его
авифауной, даже в ограниченном понимании такого термина, не представляется
возможным. Лучше всего здесь подойдет определение ископаемый комплекс. В нем пред-
ставлены главным образом водные и околоводные птицы, также наземные виды
открытых пространств, которые регулярно бывают на водопоях. Совершенно отсут-
8
ствуют лесные птицы, такие обитатели открытых ландшафтов, как, например, дрофы,
мало воробьиных птиц, которые занимают большое место в современных зоогеогра-
фических схемах и являются хорошим материалом для ландшафтных реконструкций.
Не совсем ясны в этом комплексе отношения между мигрирующими, гнездящимися и
оседлыми птицами. Все это заставляет осторожно подходить к зоогеографической
трактовке полученных данных.
Большинство описываемых плиоценовых птиц Центральной Азии относятся к совре-
менным родам, но к вымершим видам, другие представляли собой исчезнувшие ныне
роды птиц, которые, однако, оказываются близкими к тем или иным современным
родам, так что их место в семействах становится довольно ясным. Из плиоценовых
центральноазиатских птиц можно сейчас назвать только одного амфипеларгуса
(= урмиорнис), который не имеет прямых родственных связей с ныне живущими пти-
цами. Плиоценовые птицы умеренных широт представляют собой тот комплекс, из
которого сформировалась позже современная авифауна.
Остеологическая номенклатура в палеорнитологических работах имеет особенно
большое значение, потому что сравнение ведется по мельчайшим деталям структуры
костей и для последующего сопоставления описаний важно правильно ориентировать
будущих исследователей. Между тем, как это ни покажется удивительным, в остеоло-
гии птиц нет общепринятой и исчерпывающей терминологии. Удачными последними
попытками создать таковые являются «Nomina Anatomica Avium» (1979), представ-
ляющая собой труд большого международного коллектива ученых, работавшего под
руководством Д. Баумеля, А. Кинга и др., и опубликованный в Чехословакии В. Кома-
реком «Атлас анатомии домашних птиц» (Komarek, 1979). Однако эти солидные руко-
водства не исчерпывают всего спектра остеологических структур у птиц. Мы исполь-
зовали терминологию «Nomina anatomica avium», введя во многих случаях новые
названия отдельных элементов структуры костей птиц, созданные на принципах образо-
вания названий в этом издании. Нами применялись из него также миологическая и син-
десмологическая терминологии.
В палеорнитологической литературе США используется энглизированная остеологи-
ческая номенклатура, разработанная еще в 1929 г. X. Говард (Howard, 1929).
Несмотря на всю ценность этой методической разработки, одобрять полностью ее
использование в современных исследованиях не представляется возможным из-за
вариаций в трактовке и приложимости отдельных терминов и из-за неполного охвата
всех структур рельефа костей птиц. Мы использовали почти исключительно латинские
остеологические термины, чтобы избежать двусмысленности при чтении описаний.
Безусловно морфологический язык должен быть таким же точным и определенным, как
и научная номенклатура животных, но, к сожалению, об этом приходится писать
лишний раз из-за существующего различия позиций в подходах к данной проблеме.
Не менее важной при описании костей ископаемых птиц является правильная их ориен-
тация по отношению к плоскостям сечения и сторонам тела. Впервые внимание на этом
вопросе заострил Бальман (Ballmann, 1969). По его же предложению определение
сторон костей скелета было осуществлено в «Nomina anatomica avium» (1979).
Важность единообразия в промерах отдельных костей так же не требует доказательств.
Общепринятых приемов здесь нет. Нами была предпринята попытка привести в систему
эту методику в морфологии птиц (Курочкин, 1979). Приемы, там описанные, исполь-
зованы в данной работе. Наконец, в орнитологии не установлен приоритет многих
названий таксонов, особенно крупных рангов. Многие из этих вопросов были раз-
решены в последние годы Г. Н. Кашиным. Ориентируясь на одну из его основных
статей (Кашин, 1981) и личные сообщения, за которые крайне ему признательны,
в работе утверждается новый приоритет многих названий таксонов групп отрядов,
подотрядов и семейств.
Здесь автор хотел бы еще раз обратить внимание всех коллег на большое значение
сравнительной остеологической коллекции для всех работ, связанных с изучением
ископаемых птиц, а также — морфологии птиц. Эта коллекция Палеонтологического ин-
ститута АН СССР постоянно и широко используется многими отечественными и зарубеж-
ными орнитологами. Мы просили бы всех орнитологов помочь дальнейшему ее росту.
9
ГЕОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА МЕСТОНАХОЖДЕНИЙ
Основные материалы по птицам происходят из серии плиоценовых местонахожде-
ний, расположенных в Котловине Больших озер на западе МНР с территории Кобдос-
ского и Убсунурского аймаков в районе оз. Хиргис-Нур, нижнего течения р. Дзабхан
и протоки Чоно-Хариах (рис. 1). Они являются сейчас, пожалуй, наиболее полно и
детально изученными и охарактеризованными фауной позвоночных и беспозвоночных
среди местонахождений второй половины неогена во всей Азии. Район избран страто-
типическим для неогена Центральной Азии (Девяткин, Жегалло, 1974). Богатейшая
фауна млекопитающих, рыб, птиц, моллюсков отсюда в значительной мере уже обрабо-
тана и описана.
Можно назвать сводки Е. Л. Дмитриевой по антилопам (1977), В. И. Же-
галло по гиппарионам (1978), И. А. Вислобоковой по оленям (1983), обзор Е. К- Сы-
чевской по рыбам (1983). Детально изучена геология этих местонахождений, их ли-
тология, стратиграфия всего района, спорово-пыльцевые комплексы, наиболее
подробные и важные сведения по которым опубликованы Е. В. Девяткиным (1981),
Е. В. Девяткиным и В. И. Жегалло (1974), И. Г. Лискун и Д. Бадамгарав (1977),
М. А. Певзнер, Э. А. Вангенгейм и др. (1982). Закономерно, что по мере изучения
фауны и геологии местонахождений и корреляции их между собой и с местонахожде-
ниями близкого возраста в других регионах менялись представления об их геологиче-
ском возрасте. Смена взглядов на возраст отложений основных местонахождений
плиоценовых позвоночных Западной Монголии представлена на схеме (рис. 2). Перво-
начально возраст отложений местонахождений Хиргис-Нур 1 и 2, Чоно-Хариах, Дзабхан
и др. определялся средним—поздним плиоценом. Именно в этот интервал неогена вписы-
валась свита Хиргис-Нур (Девяткин, 1970; Девяткин, Жегалло, 1974). Позднее отложе-
ния свиты Хиргис-Нур были отнесены к раннему и среднему плиоцену (Жегалло, 1978;
Девяткин, 1981). Свита Хиргис-Нур была разделена на верхнюю и нижнюю подсвиты,
граница между которыми отвечала определенной фаунистической смене, а вся свита
в целом сопоставлялась с киммерием Восточного Паратетиса. В новейшей работе
нижняя подсвита свиты Хиргис-Нур опущена уже в верхний миоцен (Певзнер и др.,
1982). Последняя позиция основана на сопоставлении эпох магнитной полярности,
абсолютных датировок, времени появления и исчезновения отдельных видов и подвидов
гиппарионов, периодичности смены комплексов зайцеобразных и грызунов в Централь-
ной Азии и в Европе. Граница между верхней и нижней подсвитами свиты Хиргис-Нур
определяется при этом на уровне 5,2—5,3 млн. лет, а нижняя граница свиты — в преде-
лах 6,5—6,7 млн. лет. Коррелируя таким образом отложения этих двух отдаленных
территорий, М. А. Певзнер, Э. А. Вангенгейм и др. (1982) сопоставляют нижнюю под-
свиту свиты Хиргис-Нур с мессинием Средиземноморья, понтом и низами киммерия Во-
сточного Паратетиса, что по европейской стратиграфической шкале соответствует теперь
верхнему миоцену и низам нижнего плиоцена. Верхняя подсвита с верхней границей
в 4 млн. лет оказывается при этом соответствующей нижней половине занклия или
средней части киммерия. В настоящей работе мы придерживаемся рекомендаций
Межведомственного стратиграфического комитета, относя нижнюю подсвиту свиты
Хиргис-Нур (уровни 10—37 местонахождения Хиргис-Нур 2) к нижнему плиоцену, что
отвечает понтическому ярусу, а верхнюю подсвиту (уровни 37—62) — к среднему плио-
цену, сопоставляя ее со средней частью киммерийского яруса. Такая позиция
соответствует в общем виде прежней точке зрения В. И. Жегалло (1978) и принята
в монографии Е. В. Девяткина (1981).
Несмотря на то, что местонахождения неогеновых позвоночных интересующего нас
региона неоднократно описывались ранее, целесообразно, тем не менее, дать общую
характеристику тех из них, где были найдены остатки птиц.
МНР. Мощные толщи неогена распространены на западе Монголии в обширной
депрессии Котловины Больших озер. Наиболее широко здесь развиты плиоценовые
отложения, сформированные озерными и озерно-аллювиальными осадками. Озерная
серия выделена в свиту Хиргис-Нур (Девяткин, 1970). Остатки птиц собраны в место-
10
Рис. 1. Местонахождения птиц в плиоценовых отложениях Центральной Азии
Ранний плиоцен: 1 — Павлодар, 2 — Ошин-Боро; средний плиоцен: 3 — Чоно-Хариах 1 и 2; 4 — Дзагсо-
Хаирхан 1; 5 — Дзагсо-Хаирхан 4; 6 — Дзабхан; 7 — Явор I; 8 — Таталы; 9 — «точка 1080 м» в Шаргаин-
Гобц; 10 — Хунг-Хурэ; 11 — Калмакпай; ранний и средний плиоцен: 12 — Хиргис-Нур 1; 13 — Хиргис-Нур 2;
поздний плиоцен: 14 — Шамар; 15 — Береговая; 16 — Тологой. Условные обозначения возраста: а — ранний;
б — средний; в — ранний и средний, г — поздний плиоцен
нахождениях Хиргис-Нур 1, 2, Чоно-Хариах 1, 2, «точка 1080 м» в Шаргаин-Гоби,
Дзабхан, Дзагсо-Хаирхан 1, 4, Явор 1, Таталы, Ошин-Боро (рис. 1).
Хиргис-Нур 1. Расположено на северном берегу оз. Хиргис-Нур в нескольких кило-
метрах западнее местонахождения Хиргис-Нур 2 у подножия хребта Хан-Хухей.
Основная часть разреза сложена свитой Хиргис-Нур. В этом местонахождении в самом
верхнем уровне свиты Хиргис-Нур найдена единственная фаланга, относимая предполо-
жительно к Cygnus pristinus.
Хиргис-Нур 2. Стратотипическое местонахождение позвоночных свиты Хиргис-Нур
расположено на северном берегу оз. Хиргис-Нур западнее родника Могой-Булак.
Е. В. Девяткин и В. И. Жегалло разделили его отложения на уровни от 0 до 62 м, соот-
ветствующие мощностям слоев в метрах от подошвы разреза до верхнего слоя свиты
Хиргис-Нур. Сначала отложения местонахождения Хиргис-Нур 2 были разделены на
две разнокачественные литологические пачки — А и Б, позднее была выделена базаль-
ная пачка отложений. Детальная их литологическая характеристика приводится
у И. Г. Лискун и Д. Бадамгарав (1977). Уровни 0—10 формируют базальную пролю-
виальную пачку, в которой встречено большое скопление остатков среднего и крупного
размера млекопитающих. Эта пачка относится к свите Алтан-Тэли (Певзнер, Ванген-
гейм и др., 1982) нижнеплиоценового возраста. Основная толща местонахождения от
уровня 10 до уровня 62 сложена разного рода алевритами, глинами и песками озерного
и аллювиального происхождения, почти целиком представляя свиту Хиргис-Нур. Самая
верхняя часть (от уровня 62 и выше) сформирована алевритами и мергелями озерного
происхождения, она формирует пачку Б. В. И. Жегалло (1978) разделил свиту Хиргис-
Нур на верхнюю и нижнюю подсвиты, установив границу между ними, соответствующую
также определенной фаунистической смене, на уровне 37. Нижняя подсвита была
отнесена к нижнему плиоцену, верхняя — к среднему. Нижняя подсвита характери-
зуется остатками раннего Hipparion elegans (тип А), Н. theobaldi mogoicum, Н. t. pla-
tyodus, H. (Neohipparion) houfenense А; оленей: Metacervulus mongoliensis, Eostyloce-
ros blainvillei, Cervavitus sp., Praesinomegaceros asiaticus, Procapreolus latifrons;
грызунов (до уровня 22): Occitanomys cf. adroveri, Lophocricetus cf. vinogradovi,
Scirtodipus cf. kazakhstanica, Microtoscoptes mongolicus и др. (Жегалло, 1978; Висло-
1
0,7
Р и с. 2. Схема стратиграфического положения местонахождений птиц в плиоцене Центральной Азии
бокова, 1983; Певзнер, Вангенгейм и др., 1982). Пачка А местонахождения Хиргис-
Нур 2 характеризуется также определенным комплексом рыб (Сычевская, 1983).
В верхней подсвите указанные виды и подвиды гиппарионов сменяются на Н. раг-
vum, Н. sefvei, Н. (N.) houfenense В; среди оленей появляются Platycemas infans,
Paracervulus bidens; из грызунов — Sinocastor cf. zdanskyi, Lophocricetus cf. grabaui,
Microtodon atavus и др. Следует заметить, что, по данным В. С. Зажигина, самая
12
древняя ассоциация грызунов и зайцеобразных происходит с уровней 18—22 м, более
молодая — с уровня 24 м, а поздняя — с уровней 29—58 м (Певзнер, Вангенгейм и др.,
1982). Остатков птиц в базальной пачке и в пачке Б местонахождения Хиргис-Нур 2 не
найдено, много их собрано на уровнях 37—40 и 50—55, некоторое количество происхо-
дит из отложений нижней подсвиты (ур. 20—21).
Чоно-Хариах 1 и 2. Расположены на северном берегу протоки Чоно-Хариах, между
озерами Хара-Ус-Нур и Хара-Нур, в 10—12 км друг от друга. Отложения их сложены
аллювиальными (в основании) и озерными (в более высоких горизонтах) сериями.
В местонахождении Чоно-Хариах 1 (в тексте далее — просто Чоно-Хариах) представ-
лены отложения верхних горизонтов пачки А и пачка Б, отвечающие второй половине
верхней подсвиты свиты Хиргис-Нур; в местонахождении Чоно-Хариах 2 представлены
в основном отложения пачки Б (Жегалло, 1978; Девяткин, 1981). Большое количество
остатков птиц собрано в желтовато-серых алевритах с прослоями песков местонахож-
дения Чоно-Хариах. В местонахождении Чоно-Хариах 2 из птиц имеются только
Amphipelargus dzabghanensis, Alectoris sp., скорлупа Struthio sp. и неопределимые
ближе крупные Anatidae.
Дзабхан. Расположено на левом берегу р. Дзабхан-Гол, недалеко от ее впадения
в оз. Айрак-Нур. Отложения этого местонахождения, давшие некоторое количество
остатков мелких птиц, составлены осадками верхов пачки А и пачкой Б, относясь
к верхней подсвите свиты Хиргис-Нур.
Явор 1. Расположено недалеко от местонахождения Дзабхан. Отложения этого
местонахождения приурочены к тем же литологическим и стратиграфическим горизон-
там. Здесь найдены только Anas soporata и скорлупа яиц Struthio sp.
Таталы. Расположено южнее Явор 1, с которым сходен и возраст отложений.
Отсюда происходит только одна кость крупного Anatidae.
Дзагсо-Хаирхан 1 и 4. Местонахождения расположены на правом берегу р. Дзаб-
хан-Гол в нижнем ее течении. Из опубликованных данных можно пока сослаться
только на материалы И. А. Вислобоковой (1983), установившей из Дзагсо-Хаирхан 1
Eostyloceros blainvillei и Procapreolus latifrons, что указывает на раннеплиоценовый
возраст отложений (нижняя подсвита свиты Хиргис-Нур). В. И. Жегалло (личное
сообщение) считает отложения Дзагсо-Хаирхан 1 и 4 соответствующими верхам
пачки А местонахождения Хиргис-Нур 2, т. е. верхней подсвите свиты Хиргис-Нур.
В Дзагсо-Хаирхан 1 найдены кости Cygnus pristinus, а в Дзагсо-Хаирхан 4 — не-
сколько остатков Amphipelargus dzabghanensis.
Ошин-Боро-Удзюр-Ула. Местонахождение расположено в Дзергенской впадине
в юго-западных предгорьях хр. Дзун-Джиргаланту-Нуру. В отложениях нижней и
верхней подсвит свиты Ошин, относимых к среднему и верхнему миоцену, встречена
скорлупа яиц Struthio sp.
«Точка 1080 м» в Шаргаин-Гоби. Это местонахождение расположено на северной
стороне Шаргаин-Гоби в Гоби-Алтайском аймаке. В. С. Зажигин в 1967 г. собрал
некоторое количество остатков мелких птиц. Отложения местонахождения, по-види-
мому, можно сопоставить с верхней подсвитой свиты Хиргис-Нур.
Хунг-Хуре. Местонахождение расположено еще дальше к юго-востоку от плиоцено-
вых местонахождений Западной Монголии в депрессии Долины Озер, на юго-восток
от оз. Тациин-Цаган, на территории Убурхангайского аймака. Отложения его отно-
сятся к верхней подсвите свиты Хиргис-Нур. Здесь в 1967 г. была найдена лучевая
кость, принятая позднее голотипом, Cygnus pristinus и несколько других неопредели-
мых остатков птиц.
Шамар. Расположено в Северной Монголии в Селенгинском аймаке на левом берегу
р. Орхон недалеко от пос. Шамар. Богатая фауна млекопитающих этого местонахожде-
ния (Н. (N.) houfenense С, Н. tchikoicum, Orchonoceros gromovi, Presbytis eohanuman,
Mimomys ex. gr. hintoni, Villanyia ex. gr. chinensis и др.) позволяет относить эти отложе-
ния к верхнему плиоцену (нижнему виллафранку) в составе чикойской свиты, коррелируя
их с отложениями около фермы Береговой на р. Чикой, севернее на территории СССР
(Девяткин, Зажигин, 1974; Певзнер, Вангенгейм и др., 1982; Вислобокова, 1983).
СССР. В азиатской части СССР, в районах в значительной мере прилегающих
13
к территории Монголии, плиоценовые местонахождения позвоночных имеют широкое
распространение. Среди них наиболее интересные сборы по птицам были сделаны
в местонахождениях Павлодар, Калмакпай, Береговая и Тологой.
Павлодар («Гусиный перелет») —открыто в 20-х годах Ю. А. Орловым, сейчас
находится фактически в пределах г. Павлодар на левом обрывистом берегу р. Иртыш.
Серо-зеленые глины, вмещающие костеносную линзу, входят в павлодарскую свиту,
относимую к нижнему плиоцену. Эти отложения коррелируются с понтом Восточного
Паратетиса (Жегалло, 1978; Певзнер, Вангенгейм и др., 1982).
Калмакпай. Расположено в Восточно-Казахстанской области, в 60 км на юго-восток
от г. Зайсан, на правом берегу р. Калмакпай. Отложения с массовым захоронением мле-
копитающих относятся к карабулакской свите, возраст которой устанавливается как ран-
ний—средний плиоцен (Дмитриева, 1977) или как средний плиоцен (Жегалло, 1978).
В Калмакпае найдено большое количество остатков Amphipelargus orientals, прежнее
родовое название которого Urmiornis, как недавно выяснилось, имеет старший синоним.
Береговая. Известное местонахождение, расположенное на правом берегу р. Чикой,
притоке р. Селенги, в Бичурском районе Бурятской АССР, у фермы Береговая. Фауна и
геология этого местонахождения детально описывались в ряде работ (Базаров, Ер-
баева и др., 1976; Вангенгейм, 1977; Агаджанян, Ербаева, 1983). В красноцветных
глинах и песках Береговой, датируемых чикойско.й свитой верхнего плиоцена (Певзнер,
Вангенгейм и др., 1982), собрано большое количество остатков различных групп млеко-
питающих, наиболее многочисленные и характерные среди них — Hipparion ex gr.
houfenense, H. tchikoicum, Gazella sinensis, Nyctereutes cf. sinensis, Euryboas lunensis,
Pliolagus, Mimomys cf. reidei, M. minor и др. Здесь же в разные годы были собраны
птицы, особенно много мелких воробьиных, но имеются и остатки представителей
других отрядов, среди них — часть тарсометатарсуса Struthio asiaticus и скорлупа его
яиц. Часть этих материалов описана в настоящей работе.
Тологой — другое позднекайнозойское местонахождение в Забайкалье. Оно распо-
ложено на левом берегу р. Селенги у подножия горы Тологой, в 18 км выше по течению
реки от столицы Бурятской АССР г. Улан-Удэ. В Тологое устанавливаются три толщи
осадков (Александрова, Вангенгейм, 1963), красноцветы которых, охарактеризованные
фауной млекопитающих, по-видимому, близки по возрасту к красноцветам Береговой.
Из Тологоя у нас имелся только один фрагмент кости орлана (Haliaeetus sp.), но
отсюда известна и скорлупа яиц страусов.
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
ПТИЦЫ МЕСТОНАХОЖДЕНИЙ ЗАПАДНОЙ МОНГОЛИИ
ОТРЯД STRUTHIONIFORMES LATHAM, 1790
СЕМЕЙСТВО STRUTHIONIDAE VIGORS, 1825
Род Struthio L., 1758
Struthio sp.
Табл. I; фиг. 1, 2
В основных местонахождениях среднеплиоценовой фауны в Котловине Больших
озер было собрано некоторое количество скорлупы яиц бескилевых птиц. Здесь, как и
во всех других районах Палеарктики, где находят такую скорлупу, этот материал не
сопровождается костными остатками самих птиц, а там, где встречены их редкие кости,
к сожалению, отсутствует скорлупа их яиц. Поэтому скорлуповые и костные материалы
по бескилевым птицам из неогена и плейстоцена многих районов Азии, Африки и
Европы в большинстве своем несопоставимы. Так называемая сиваликская скорлупа
из Индии не может быть привязана к описанному оттуда Struthio asiaticus Milne-
Edwards, 1871, так как при большом возрастном диапазоне этих отложений неизвестна
точная геологическая привязка скорлупы и костных остатков страусов старых сборов
14
из этого района. Отправной точкой для исследования древних материалов по страусо-
подобным птицам могут послужить сборы из верхнеплиоценовых отложений Одесских
катакомб, где наряду с большим количеством костей страусов известна и скорлупа,
а также тщательное исследование скорлупы яиц современного Struthio из разных
районов Африки и Аравийского полуострова.
До появления работ по ископаемой скорлупе бескилевых Ф. Зауера (Sauer, 1972,
1976) с этим материалом все казалось более-менее ясным. Представлялось, что на
огромных степных и саванных пространствах средних широт Старого Света в неогене и
плейстоцене жили настоящие Struthioniformes, потомком которых является современ-
ный Struthio camelus L., 1758, и надо найти только детали, по которым удастся раз-
личать скорлупу яиц разных видов страусов, распространенных не только на обширных
территориях, но и в большом диапазоне времени — от верхнего миоцена до нижнего
плейстоцена. Это и пытались сделать некоторые исследователи, ориентируясь на тол-
щину скорлупы и структуру ее на радиальных шлифах. Однако Ф. Зауер неожиданно
установил, что скорлупа бескилевых из верхнего миоцена Турции, Марокко и с Канар-
ских островов практически неотличима по строению поровых каналов от скорлупы яиц
недавно вымерших эпиорнисов с Мадагаскара (Sauer F., 1976, 1979; Sauer F.,
Sauer E., 1978). Между прочим, эти работы принадлежали перу того же Ф. Зауера,
который в середине 50-х годов XX столетия выполнил пионерские работы по изу-
чению ориентации птиц в планетарии, открывших новую эпоху в изучении навигации
птиц. После работ Зауера, в которых описывалась «эпиорнитоидная» скорлупа из неоге-
новых отложений Малой Азии, Африки и Канарских островов, сразу возникло много во-
просов, которые остались нерешенными. Внести ясность в них смогут только исследова-
ния большого материала по ископаемой скорлупе, которые будут отталкиваться от не
менее обширных исследований скорлупы яиц современных бескилевых.
В Котловине Больших озер скорлупа была собрана в местонахождениях Хиргис-
Нур 2, Чоно-Хариах 2, Дзабхан, Ошин-Боро и Явор 1. Отложения местонахождений
Хиргис-Нур 2, Чоно-Хариах 2, Дзабхан и Явор 1 одновозрастны, они составлены
среднеплиоценовой свитой Хиргис-Нур; верхние горизонты местонахождения Ошин-
Боро относятся к нижнеплиоценовой свите Алтан-Тэли (Девяткин, 1981).
В Палеонтологическом институте АН СССР проводится детальное изучение этих и
других материалов по ископаемой скорлупе и подробный анализ этих данных — обшир-
ное поле специальных исследований. Остановимся здесь только на общих выводах
предварительных публикаций К. Е. Михайлова и данных его личного сообщения.
Условно, по признакам распределения пор и их формы на поверхности скорлупы,
выделяются так называемый «эпиорнитоидный» и «струтиоидный» типы строения.
Замечательно, что большую часть скорлупы из плиоцена Монголии нельзя «чисто»
отнести ни к одному из них, а она имеет смешанный «эпиорнитоидно-струтиоидный»
тип. Видимо, для различения скорлупы определенное внимание следует также при-
давать ее толщине. Полученные данные сведены в табл. 1. Из таблицы следует, что
во всем диапазоне плиоцена встречаются два условных типа скорлупы, а в среднем
плиоцене преобладал смешанный тип строения—«эпиорнитоидно-струтиоидный».
В чистом выражении «эпиорнитоидная» скорлупа не обнаружена в этих местонахожде-
ниях, так же как и не ясно, существовала ли самостоятельно «струтиоидная» скорлупа.
Намечается, что от раннего плиоцена (св. Алтан-Тэли) к среднему плиоцену (пач-
ка «Б» свиты Хиргис-Нур) толщина скорлупы несколько увеличилась. Кому же при-
надлежала эта скорлупа? Как считает К. Е. Михайлов, если отталкиваться от данных
по скорлупе четвертичных Struthioniformes из Африки и Азии, а также учитывать
характер изменчивости в строении скорлупы современных африканских страусов и
вообще полиморфизм всей ископаемой скорлупы, плиоценовая скорлупа из Монголии
принадлежала настоящим страусам — Struthionidae, причем, скорее всего, даже
одному виду Struthio. Как мы пытались показать ранее, в плиоцене и антропогене
во всех подходящих ландшафтах Азии, Африки и Европы был широко распространен
Struthio asiaticus Milne-Edwards, 1871 (Курочкин, Лунгу, 1970; Kurochkin, 1970).
«Эпиорнитоидность» в строении скорлупы может не отвечать ее принадлежности
мадагаскарским ныне Aepyornithiformes, а быть составляющей общего для эпиорнисов
15
Таблица 1
Стратиграфическое распределение и типы структуры скорлупы яиц страусов в плиоцене
Монголии
Местона- хождение Свита Пачка Горизонт Слои Число образцов Тип структуры Толщина, мм
Ошин-Боро Алтан-Тэли — Верхний — Большое количе- ство «Струтиоидный» 1,9—2,2
Хиргис- Нур-2 Хиргис-Нур (нижняя под- свита) А Нижний 18,21 5 «Струтиоид- ный» ? и «эпиор- нитоидно-стру- тиоидный» 1,9—2,4
Хиргис- Нур-2 Хиргис-Нур (верхняя под- свита) А Средний 37—40 1 «Эпиорнитоидно- струтиоидный» 2,3
Хиргис- Нур-2 Хиргис-Нур (верхняя под- свита) А Верхний 51—55 11 «Струтиоид- ный» ? и «эпиор- нитоидно-сгру- тиоидный» 2,0—2,6
Чоно-Ха- риах 2 Хиргис-Нур (верхняя под- свита) А « — 7 «Эпиорнитоидно- струтиоидный» 2,1—2,5
Дзабхан Хиргис-Нур (верхняя под- свита) Б Нижний 50 «Эпиорнитоидно- струтиоидный» 2,0—2,5 (большин- ство 2,3—2,5)
Явор 1 Хиргис-Нур (верхняя под- свита) Б Верхний — 2 «Эпиорнитоидно- струтиоидный» 1,9, 2,1
и настоящих страусов исходного полиморфного морфотипа строения пор. Временами
«эпиорнитоидность» широко распространялась среди позднетретичных страусов в Ста-
ром Свете. Вместе с этим представляется, что изменения в структуре поверхности
в местах выхода поровых каналов и толщине скорлупы скорее всего отвечают смене
климатических условий обитания Struthio и различным условиям, в которых насижива-
лись кладки яиц. В то же время, если учитывать принципиальную обособленность по
строению скорлупы между Aepyornithiformes и Struthioniformes — с одной стороны,
и всеми разными другими бескилевыми — с другой, то можно говорить о более близких
родственных связях двух этих групп бескилевых птиц Старого Света по отношению
ко всем остальным бескилевым.
Следует заметить и подчеркнуть определенную стратиграфическую приуроченность
скорлупы Struthio разного микроструктурного облика и толщины, что определяет
возможность ее использования для целей стратиграфии.
ОТРЯД PODICIPEDIFORMES FURBRINGER, 1888
СЕМЕЙСТВО PODICIPEDIDAE BONAPARTE, 1831
Род Podiceps Latham, 1787
Podiceps solidus Kurochkin, sp. nov.
Рис. 3; табл. I, фиг. 3, 4
Видовое название solidus (лат.) — крепкий, сильный.
Голотип — ПИН, № 3222-37; дистальный эпифиз правого плеча; МНР, север-
ный берег оз. Хиргис-Нур, местонахождение Хиргис-Нур 2; средний плиоцен, свита
Хиргис-Нур.
Материал. Только голотип.
Описание. Дистальная часть дифиза довольно широкая. Impressio lig. epilate-
rale ventrale маленькое. Fossa m. brachialis небольшой глубины, но занимает большую
16
Рис. 3. Podiceps solidus sp. nov.; голотип № 3222-37, ПИН;
правое плечо; Монголия, Хиргис-Нур 2; средний плиоцен
а — дорсальная; б — латеральная; в — вентральная; г — ди-
стальная стороны
площадь как по ширине, так и по длине. Продоль-
ная ось cond. ventralis идет в дистально-краниаль-
ном направлении. Дорсальный участок fossa olec-
rani врезан неглубоко.
Размеры, мм: наибольшая ширина эпифиза
7,2, передне-задний диаметр cond. dorsalis 4,1 (он
частично разрушен, поэтому не подлежит точному из-
мерению), наименьшая передне-задняя толщина
диафиза 3,0, его ширина 4,1.
Сравнение. Среди диагностических призна-
ков Р. solidus по сравнению с современными вида-
ми поганок особенно характерным оказывается рас-
ширенность дистальной части диафиза перед выходом его на эпифиз (дорсальный и вен-
тральный края диафиза, постепенно расширяясь, переходят в эпифиз); дистальная часть
диафиза выглядит поэтому мощнее и шире, хотя абсолютные размеры ее по толщине и
ширине соответствуют таковым одноразмерных экземпляров Р. nigricollis. У современ-
ных Podiceps на уровне середины fossa brachial is происходит резкое расширение дорсаль-
ного и вентрального краев диафиза по направлению к надмыщелкам, особенно по дорсаль-
ному краю, поэтому хорошо выражена головчатость эпифиза по отношению к посадке
его на диафизе. Продольная ось cond. ventralis, будучи направлена в краниально-
дистальном направлении у ископаемой поганки, образует с продольной осью диафиза
широкий тупой угол. У большинства современных видов поганок эти две условные оси
сходятся под прямым углом. Только у Р. auritus этот угол больше раскрыт, т. е. он ту-
пой, хотя и меньшего размера, чем у Р. solidus. Маленькие размеры impres. lig. collate-
rale ventrale у P. solidus характерны по сравнению почти co всеми современными Podi-
ceps, даже с крошечной Р. dominicus. Но вновь у Р. auritus размеры этого impressio
сравнимы с таковыми ископаемого вида. Fossa m. brachialis у Р. solidus в сравнении
с Р. cristatus, Р. grisegena, Р. nigricollis сильно удлинена в проксимальном направлении,
у Р. auritus она уже в основании и несколько короче, у Р. dominicus сильно сужена,
хотя и имеет относительно большую длину. Для Р. solidus специфична и общая незна-
чительная глубина fossa olecrani. Особенно характерна небольшая углубленность дор-
сального заостренного язычка этой ямки, что также отмечается и для Р. nigricollis,
хотя у нее по сравнению с ископаемым видом сильнее углублен вентральный участок
ямки. У Р. cristatus, Р. grisegena и Р. auritus fossa olecrani умеренно углублена по всей
площади, но у Р. dominicus fossa olecrani еще более мелкая, чем у Р. solidus.
Из неогена США и Западной Европы описано 11 видов поганок рода Podiceps.
Большинство их описано по костям задних конечностей или другим костям крыла.
Из среднего плиоцена штата Канзас, США, указана современная Р. nigricollis (Wetmore,
1937), но как считают П. Бродкорб (Brodkorb, 1963) и Б. Муррей (Murray, 1970) этот
экземпляр требует переизучения. Среди остальных ископаемых поганок плечевая кость
известна у Р. pisanus (Portis, 1891). Этот вид был детально переизучен Regalia
(1902), он приводит и достаточно хорошие иллюстрации его остатков из среднеплиоце-
новых морских аргилитов местонахождения Орсиано Пизано в Италии. Этот вид схо-
ден с Р. solidus как общими размерами (ширина эпифиза плеча у него 7,9 мм), так и
деталями строения: маленькими размерами impres. lig. collateral ventrale, незначи-
тельной глубиной и большой шириной fossa m. brachialis, наклоном продольной оси
cond. ventralis, робустностью дистальной части диафиза. Было бы желательно непо-
средственно сравнить оригинальные материалы по этим видам.
Замечания. В современной фауне, кроме рода Podiceps, поганки представлены
двумя видами в родах Podilymbus и Rollandia, монотипическими Aechmophorus и Та-
chybaptus, а в неогеновой — монотипически известными вымершими Pliolymbus, Mio-
2 Заказ 2098
17
baptus, Pliodytes и вымершим видом современного рода Podilymbus majausculus Murray,
1967 из штата Айдахо, США. Целесообразно здесь показать основные отличительные
признаки в строении плечевой кости у этих родов в сравнении с Podiceps. Наиболее
специфично устроено плечо у Aechmophorus. Здесь вентральная половина эпифиза
выдвинута дистально, так что pr. flexorius заходит дистально за уровень cond. dorsalis;
epicond. ventralis у него широкий и далеко простирается проксимально. У Tachybaptus
в отличие от Podiceps и Aechmophorus impres. lig. collaterale ventrale небольшое, оваль-
ное, с закругленным проксимальным концом, который отведен вентрально. Также на-
правлено impressio и у Podilymbus, но общая его площадь относительно намного боль-
шая, кроме того, этот род имеет специфический широкий и короткий треугольной формы
epicond. ventralis. У Podiceps proc, flexorius заканчивается дистально на уровне cond.
dorsalis, impres. lig. collaterale vetrale имеет заостренный проксимальный конец, epicond.
ventralis укороченный у всех видов, но узкий — у мелких и немного расширенный —
у крупных видов. Б. Муррей (Murray, 1970) приводит в качестве отличительного приз-
нака Achmophorus и Podilymbus изогнутость диафиза плеча в дистальной */5 его длины
у этих родов и в дистальной трети у Podiceps, что нам представляется довольно субъек-
тивным.
Среднеплиоценовая Pliolymbus, описанная по фрагменту грудины (голотип) и дру-
гим костям, в том числе и по плечевой, из формации рексроад в штате Канзас, США,
представляет из себя маленькую поганку (ширина дистального эпифиза плеча 5,6 мм,
заметно меньше, чем у Р. solidus), с робустными чертами в строении отдельных костей
посткраниального скелета (Murray, 1970). Поскольку, так сказать, робустность, выра-
женная в расширенности диафиза, характерна и для описываемого нами ископаемого
вида, кажется интересным было бы сравнить эти две формы. Pliodytes lanquisti опи-
сана из нижнего плиоцена штата Флорида по единственному коракоиду (Brodkorb,
1953). Это тоже была маленькая поганка. Нижнемиоценовая (Дольнице, ЧССР) Mio-
baptus walteri представляет собой мелкую поганку, характеризующуюся специфиче-
скими деталями строения известных от нее тарсометатарсуса, коракоида и проксималь-
ной части плечевой кости (Svec, 1982).
Таким образом, ископаемые неогеновые поганки известны не так уж плохо. Ха-
рактерно, что в плиоцене мы не находим современных видов этого отряда (Р. nigricol-
lis из среднего плиоцена Канзаса нуждается в переизучении). Из плейстоцена известны
два вымерших вида: Podiceps parvus (средний плейстоцен, шт. Орегон) и Р. dixi (сред-
ний плейстоцен, штат Флорида). Все известные неогеновые вымершие виды и роды
представлены мелкими формами, а из палеогена поганки неизвестны вовсе. Наряду
со специфическими морфологическими особенностями современных представителей
отряда эти косвенные доводы заставляют считать его эволюционно молодой группой
птиц, специализация которой произошла сравнительно недавно. Поганки не имеют,
конечно, никаких родственных связей с гагарами (Штегман, 1974), но заслуживает
внимания рассмотрение их родства с журавлеобразными (Stolpe, 1953; Storer, 1963;
Zusi, Storer, 1969).
Распространение. Местонахождение Хиргис-Нур 2; средний плиоцен, свита
Хиргис-Нур, верхняя подсвита; уровень 51—55.
ОТРЯД PELECANIFORMES SHARPE, 1891
СЕМЕЙСТВО PHALACROCORACIDAE BONAPARTE, 1853
Род Phalacrocorax Brisson, 1760
Phalacrocorax reliquus Kurochkin, 1976
Рис. 4; табл. II, 1—4
Phalacrocorax reliquus: Курочкин, 1976, с. 52
Голотип —ПИН, № 2614-74; дистальный эпифиз правой плечевой кости; Коб-
досский аймак, местонахождение Чоно-Хариах; средний плиоцен, свита Хиргис-Нур.
Паратип. В дополнение к голотипу из местонахождения Хиргис-Нур 2 имеется
дистальная половина левой плечевой кости, ПИН, № 3222—36.
18
Рис. 4. Phalacrocorax reliquus Kurochkin, 1976; голотип № 2614-74, ПИН; правое плечо;
Монголия, Чоно-Хариах; средний плиоцен
а — дорсальная; б — латеральная; в — вентральная; г — медиальная; д — дистальная стороны
Материал. Фрагмент проксимального эпифиза правой плечевой кости, ПИН,
№ 3222-35, из Хиргис-Нур 2.
Описание. Вершина cond. dorsalis утоньшена и сильно завернута вентрально,
так что при взгляде на эпифиз с краниальной поверхности оказывается открытой зна-
чительная часть epicond. dorsalis. На внутренней части эпифиза вершина cond. dor-
salis продолжается в виде хорошо выраженного поперечного валика. Cond. ventralis
при взгляде с дистального конца значительно (приблизительно на 1 /5 его диаметра)
выдается краниально за уровень cond. dorsalis. Fossa olecrani неглубокая. Pr. flexorius
имеет форму почти прямоугольной площадки, дистальная его вершина укорочена и
слабо выделена. Проксимальный край impres. lig. collateral ventrale имеет заострен-
ную форму и impressio вытянуто строго вдоль продольной оси кости.
Размеры, мм: максимальная ширина дистального эпифиза 16,3 (паратип —
15,2), передне-задний диаметр cond. dorsalis 9,4 (9,7), передне-задний диаметр cond.
ventralis 5,6 (5,5), расстояние от проксимального конца impres. lig. collateral ventrale
12,0 (10,7), передне-задняя толщина диафаза на уровне середины fossa m. brachialis
6,0 (5,3), ширина диафаза на этом же уровне 9,0 (8,5); общая длина голотипа —
46,0, паратипа — 85,5.
Сравнение. Из этого же местонахождения по бедру был описан Phalacroco-
rax mongoliensis Kur., 1971. Учитывая возможную принадлежность фрагментов плеча
к этому виду, были сопоставлены их абсолютные и относительные размеры с таковыми
нескольких современных видов (табл. 2). Из этой таблицы с очевидностью следует, что
отношения между размерами двух ископаемых костей выходят из общего ряда отноше-
ний, характерных для современных бакланов, поэтому можно определенно считать, что
здесь мы имеем дело с разными остатками двух разных видов.
Род Phalacrocorax — один из наилучше известных в ископаемом состоянии среди
отряда веслоногих, потому что бакланы в богатой рыбой водоемах прошлого были,
очевидно, довольно широко распространены, имели высокую численность и многие из
них были связаны с пресными водами. Из неогена всего мира описаны 17 различных
видов бакланов. Почти все они, за двумя исключениями, отнесены к современному же
роду Phalacrocorax и только Pliocarbo из плиоцена Одессы и Valenticarbo из плиоцена
Сивалик, похоже, без достаточных на то оснований выделены в особые роды. Боль-
2*
19
Таблица 2
Абсолютные и относительные размеры плечевой и бедренной костей у ископаемых и некоторых
современных бакланов
Показатель Ph. carbo Ph. pela- gicus Ph. pyg- maeus Ph. atri- ceps Ph. auri- tus Плечо Ph. reliquus и бедро Ph. mon- goliensis
1. Максимальная ширина 17,4 16,3 10,0 17,2 16,6 16,3
дистального эпифиза
плеча
2. Передне-задний диа- 10,5 10,0 6,7 11,4 10,5 9,4
метр cond. dorsalis
3. Максимальная ширина 15,7 14,4 9,7 18,4 15,4 13,0
дистального эпифиза
бедра
4. Передне-задняя тол- 5,7 5,3 4,0 6,0 5,5 4,5
щина дистального эпи-
физа бедра (через
sulcus intercondylari)
Пропорции
1:3 1J1 1,13 1,03 0,94 1,08 1,25
1:2 1,65 1,63 1,49 1,51 1,57 1,73
3:4 2,73 2,72 2,42 3,06 2,80 2,89
3:2 1,49 1,44 1,45 1,61 1,47 1,38
шинство ископаемых бакланов описано по тарсометатарсусам и коракоидам, однако
шесть видов описаны по плечевым костям или таковые от них известны из отпечатков
скелетов. Ph. littoralis (Milne-Edwards, 1863) из нижнего миоцена Франции и ФРГ
отличается меньшими размерами, структурой мышелков и fossa m. brachialis. Гораздо
меньше нового вида и Ph. miocaenus (Milne-Edwards, 1867) из нижнего миоцена
Франции. У Ph. femoralis L. Miller, 1929 из верхнего миоцена Калифорнии дистальная
часть плеча скрыта на плитке другими костями, а проксимальный эпифиз Ph. reliquus
меньшего размера и имеет другие детали строения краниальной поверхности. Близок
к Ph. reliquus по размерам Ph. destefani Regalia, 1902 из среднего плиоцена Италии, но
у него передняя часть cond. dorsalis имеет очень небольшой крючковидный загиб, а ос-
нование этого мыщелка не продолжается в виде поперечного валика по передней по-
верхности эпифиза. Fossa m. brachialis у Ph. destefani не разделяется на дорсальную
и вентральную половины, что свойственно Ph. reliquus. От Ph. anatolicus (Турция,
нижний миоцен) отличается более крупными размерами (приблизительно на 1 /4 круп-
нее) и наличием перерыва на сплошном гребне продольного ребра margo caudalis
в месте перехода его в сг. capiti caudalis. Ph. marinavis Shufeldt, 1915 из нижнего мио-
цена Орегона, хотя и имеет близкие размеры, но отличается отсутствием хорошо раз-
витого крючковидного отростка у cond. dorsalis, острыми очертаниями вершинки
impres. lig. collateral ventrale и неразделенной fossa m. brachialis.
Из 28 видов современных бакланов Ph. reliquus сравнивался с 18 видами рода.
В сравнительной серии, к сожалению, отсутствовал Ph. fuscicollis из Юго-Восточной
Азии. Общий вывод из этого сравнения: Ph. reliquus сходен с отдельными видами по
некоторым деталям строения дистального эпифиза плеча, так же как по другим дета-
лям от них отличается. Полного сходства или полного отличия ни от одного из совре-
менных видов бакланов установить не удается. Impressio lig. collaterale ventrale за-
остренный и вытянутый вдоль оси кости у Ph. reliquus, имеет округлую проксимальную
вершину у Ph. carbo и отведен у него проксимальным концом вентрально. Рг. flexorius
у Ph. reliquus практически прямоугольный и сплошной неразделенный, у Ph. carbo —
укороченный и разделенный на две половины. Во всех остальных деталях строения
эти два вида сходны, хотя Ph. carbo отличается от ископаемого вида относительно бо-
лее крупными размерами всего эпифиза в целом.
20
От Ph. aristotelis ископаемый вид отличается вытянутым строением impres. lig.
collaterale ventrale и более тупым и расширенным cond. dorsalis с краниальной поверх-
ности. Иными пропорциями cond. dorsalis Ph. reliquus хорошо отличается от Ph. urile
и от Ph. bougainvillie. В отличие от Ph. olivaceus и Ph. auritus у Ph. reliquus fossa olecrani
заметно более мелкая, a sulcus m. scapulotricipitis придвинута более дорсально к дорсаль-
ному же краю epicon. dorsalis. Кроме того, у Ph. auritus proc, flexorius короче и толще.
Более глубокая fossa olecrani также у Ph. varius и Ph. penicillatus. Структурой этой ямки
Ph. reliquus отличается также от Ph. punctatus, у которого она расширенных треуголь-
ных очертаний, а у Ph. reliquus — суженная. У ископаемого вида по сравнению с Ph.
punctatus рг. flexorius выдается дистально меньше, хотя общая длина дистально-кра-
ниальной площадки этого отростка больше.
От мелкого Ph. melanoleucos дистальный эпифиз плечевой кости Ph. reliquus отли-
чается более сдвинутой проксимально fossa olecrani, суженной sulcus m. scapulotrici-
pitis и разделенностью двух ямок на дорсальной поверхности epicond. dorsalis. От дру-
гого мелкого вида Ph. pygmacus отличается очень незначительно, у ископаемого вида
fossa olecrani более мелкая, а вершина impres. lig. collaterale ventrale более округлая;
у Ph. pygmaeus последняя сильно заострена, а площадка impressio в целом выглядит
не овалом, а небольшим равносторонним треугольником.
От двух других видов современных бакланов отличия обратного плана. При таких же
размерах Ph. capensis, у Ph. reliquus sulcus m. scapulotricipitis шире, impres. lig.
collaterale ventrale расположена по продольной оси кости, у Ph. capensis последняя
отклонена вершиной вентрально. У мелкого Ph. niger fossa olecrani отодвинута про-
ксимальнее и занимает меньшую площадь.
Велико структурное сходство Ph. reliquus с современным Ph. pelagicus. При одина-
ковых по существу размерах дистального эпифиза плеча их различает по настоящему
только меньший относительный и абсолютный размеры диаметра cond. dorsalis
(табл. 2). Общими у них оказались устройство рг. flexorius, impres. lig. collaterale
ventrale, impres. m. brachialis, передней части cond. dorsalis и краниальной стороны
внутренней площадки эпифиза, рельеф вентральной и дорсальной поверхностей обоих
epicondylorum.
Замечания. Паратип отличается от голотипа немного меньшими размерами.
Отличий в строении между ними нет.
Кроме этих двух фрагментов к Ph. reliquus мы относим также фрагмент прокси-
мального эпифиза плеча № 3222-35 из Хиргис-Нура 2. Основаниями для этого слу-
жат подходящие абсолютные размеры и наибольшее структурное сходство с современ-
ным Ph. pelagicus, с которым также более всего сходен и голотип.
В проксимальном эпифизе плечевой кости у Ph. reliquus в отличие от тех же раз-
меров Ph. capensis нет изгиба sulcus lig. transversus при выходе его на вентральный
край; на краниальной поверхности эпифиза incisura capitis имеет большую абсолютную
длину. По сравнению с мелким Ph. pygmaeus, incisura capitis относительно уже, т. е.
она имеет одинаковые абсолютные размеры у Ph. reliquus и Ph. pygmaeus, но большую
абсолютную и относительную длину. У другого мелкого вида у Ph. niger, как и
у Ph. olivaceus, incisura capitis абсолютно и относительно уже, a caput humeri укорочена
по своей продольной длине. Значительно отличается строение incisura capitis у Ph. urile,
она у него приблизительно в два раза короче, чем у Ph. reliquus. Во многом сходен
с Ph. carbo, но хорошо отличается от него выравненностью участка эпифиза между
incisura capitis и tuberculum dorsale, который у Ph. carbo заглублен. Этими же чер-
тами строения отличается и от Ph. punctatus. В этом отношении сходен с Ph. bougain-
villie, но у последнего более закрытая и глубокая sulcus lig. transversus с обратной
стороны эпифиза. От Ph. carunculatus, Ph. varius, Ph. auritus, Ph. atriceps и Ph. aris-
totelis отличается удлиненной, суженной и неуглубленной incisura capitis, имеются
также различия в строении участка между этой incisura и tuberculum dorsale. Во всех
перечислявшихся особенностях строения Ph. reliquus в значительной мере сходен
с Ph. pelagicus, но отличается от него утолщенностью caput humeri в передне-заднем на-
правлении.
21
Размеры, мм: наибольшая толщина caput humeri 7,1, расстояние от края tuber-
culum dorsale до вентрального края caput humeri 15,2.
Распространение. Местонахождения Хиргис-Нур 2 и Чоно-Хариах; верхняя
подсвита свиты Хиргис-Нур; верхние уровни пачки А.
Phalacrocorax mongoliensis Kurochkin, 1971
Рис. 5; табл. II, фиг. 5, 6
Phalacrocorax mongoliensis: Курочкин, 1971, с. 59.
Голотип —ПИН, № 2614-29, дистальный эпифиз левого бедра; Кобдосский
аймак, местонахождение Чоно-Хариах; средний плиоцен, свита Хиргис-Нур.
Материал. Только голотип.
Описание. Cond. medialis выглядит как бы сжатым спереди назад, дистальная
часть медиального ребра диафиза, непосредственно над медиальным надмыщелком,
имеет прямоугольные очертания, потому что переднее и заднее его ребра идут парал-
лельно. Плантарная часть сочленовной поверхности cond. medialis уплощена. Impres.
m. pubo-ischiofemoralis представлено слабо выраженной шероховатостью, развиваю-
щейся по медиальному краю каудальной части диафиза. Sulcus intercondylaris в своей
каудально-дистальной части довольно узкая и имеет здесь хорошо выраженные сближен-
ные ямки в местах отхождения lig. cruciati cranialis. Надмыщелковая область в ди-
стальной части диафиза на латерально-каудальном его ребре в месте фиксации сухо-
жильной петли (ansa m. iliofibularis) имеет слабо выраженное углубление с покатыми
краями. Fovea tendineus m. tibialis cranialis довольно слабо развита и расположена на
краниально-дистальном участке cond. lateralis. В целом cond. lateralis не на много
мощнее cond. medialis.
Размеры, мм: наибольшая ширина дистального эпифиза 13,2, передне-задний
диаметр cond. medialis 6,9, наименьшая ширина через sulcus intercondylaris 4,6.
Сравнение. Среди 17 известных неогеновых видов бакланов бедро известно
только для трех из них. Кроме того, из плейстоцена Австралии описаны Ph. gregorii
и Ph. vetustus (De Vis, 1906), представленные также бедренными костями. Ph. femora-
lis из верхнего миоцена Калифорнии, США (Miller, 1929) описан по двум отпечаткам
скелетов, на приводимых рисунках которых детали строения бедренных костей неразли-
чимы, не приводятся они и в тексте. Бедро Ph. miocaenus из нижнего миоцена Фран-
ции (Milne-Edwards, 1867—1868), судя по иллюстрациям и описаниям, сходно с бедром
современного Ph. carbo, отличия от которого у ископаемого Ph. mongoliensis будут
приведены дальше. Ph. destefani из среднего плиоцена Италии (Regalia, 1902) под-
ходит к Ph. mongoliensis по абсолютным размерам, но, к сожалению, от него известна
только проксимальная часть бедра. Среди двух австралийских плейстоценовых бакла-
нов бедро у Ph. mongoliensis совершенно отлично от бедра Ph. gregorii, зато имеет ряд
общих черт в строении с Ph. vetustus. У Ph. mongoliensis, как и у Ph. vetustus, ха-
рактерно уплощена плантарная часть сочленовной поверхности cond. medialis, слабо
углублена надмыщелковая зона в месте фиксации ansa m. iliofibularis, слабо развита
fovea tendineus m. tib. cranialis. Различают их значительная разница в размерах между
cond. lateralis и cond. medialis у Ph. vetustus, более расширенная в каудально-дис-
тальной области sulcus intercondylaris, выпукло обозначенное impres. m. pubo-ischio-
femoralis, хорошо отведенная внутрь плоскость cond. medialis.
Сравнение Ph. mongoliensis с современными видами бакланов проводилось
с тем же набором их видов, как и при изучении Ph. reliquus. Ph. mongoliensis отли-
чается в целом от большинства современных видов бакланов меньшими относитель-
ными размерами всего эпифиза по отношению к диафизу, проявляющимися за счет бо-
лее слабо развитого cond. lateralis и меньшей разницы между размерами обоих мыщел-
ков. Сходны в этом плане с Ph. mongoliensis только Ph. carbo и Ph. pelagicus.
Но у Ph. mongoliensis в отличие от Ph. pelagicus очень мелкая ямка в месте фиксации
ansa m. iliofibularis, слабо развита impres. m. pubo-ischiofemoralis, более широкая
и мелкая sulcus intercondylaris в каудально-дистальной части, прямоугольных очерта-
ний дистальная часть внутренней стороны диафиза (у Ph. pelagicus она расширяется
22
Рис. 5. Phalacrocorax mongoliensis Kurochkin, 1971; голотип
№ 2614-29, ПИН; левое бедро; Монголия, Чоно-Хариах; средний
плиоцен
а — каудальная; б — медиальная стороны
в проксимальном направлении), более мелкая и краниально отодвинутая fovea ten-
dineus m. tib. cranialis. По этим структурным деталям Ph. mongoliensis близок
к Ph. carbo. Эти два вида в отличие от всех других бакланов сближает устройство cond.
medialis, вытянутого дистально вдоль продольной оси кости, не отведенного каудально,
не имеющего скошенного среза мыщелка с краниально-дистального угла. По существу,
надежных отличительных признаков между Ph. mongoliensis и Ph. carbo, кроме
в 1,2 раза больших размеров последнего, установить не удается (табл. 2).
В общих чертах отличия Ph. mongoliensis от других видов бакланов сводятся к осо-
бенностям строения плантарной поверхности надмыщелковой зоны у cond. lateralis,
месту расположения fovea tendineus m. tib. cranialis (она расположена более крани-
ально у крупных видов и на дистальной поверхности мыщелка у мелких), разнице
в размерах между cond. lateralis (более крупном) и cond. medialis, строению cond. me-
dialis (срезанному каудально у большинства современных видов бакланов), различиям
в структуре каудально-дистального участка sulcus intercondylaris, который может быть
шире или наоборот уже, с раздвинутыми ямками в местах отхождения lig. cruciati
cranialis (как у мелкого Ph. pygmaeus) или наоборот — с одноямковой структурой
этого участка sulcus (как у другого мелкого вида Ph. melanoleucos).
Распространение. Местонахождение Чоно-Хариах; средний плиоцен, свита
Хиргис-Нур, верхняя подсвита, пачка Б.
ОТРЯД ARDEIFORMES TREVIRANUS, 1802
ПОДОТРЯД ARDEAE TREVIRANUS, 1802
СЕМЕЙСТВО ARDEIDAE VIGORS, 1825
Род Nyctanassa Steineger, 1887
Nyctanassa kobdoena Kurochkin, 1976
Рис. 6; табл. II, фиг. 5, 6.
Nyctanassa kobdoena: Курочкин, 1976, с. 54.
Голотип — ПИН, № 2614-86; дистальный эпифиз правого тарсометатарсуса;
МНР, Кобдосский аймак, северный берег протоки Чоно-Хариах, местонахождение Чоно-
Хариах; средний плиоцен, свита Хиргис-Нур.
Материал. Кроме голотипа, имеется эпифиз левой лопатки, ПИН, № 3222-29,
из местонахождения Хиргис-Нур 2.
Описание. Блоки пальцев широко раздвинуты. Сагиттальная плоскость trochlea
dig. II расположена под небольшим углом к сагиттальной плоскости trochlea dig. III.
В дистальном направлении trochlea dig. II немного не доходит до уровня trochlea
dig. III. Сагиттальные плоскости блоков III и IV пальцев параллельны. С наружной сто-
роны trochlea dig. IV имеет круглую форму. С плантарной стороны блок II пальца и блок
IV пальца соответственно с внутреннего края и с наружного края продолжаются в прок-
симальном направлении невысокими гребнями по краям уплощенной части диафиза.
Foramen vasculare distale имеет очень маленький размер и щелевидную форму как с пе-
редней, так и с задней поверхности. Facies artic. metatarsi I выражена слабо и располо-
23
/7 2см & 2см
1 । ।
Рис. 6. Nyctanassa kobdoena Kurochkin, 1976
Д—№ 3222-29, ПИН; левая лопатка; Монголия, Хиргис-Нур 2; средний плиоцен; а — наружная;
б — краниальная стороны. Б — голотип № 2614-86, ПИН; правый тарсометатарсус; Монголия, Чоно-
Хариах; средний плиоцен; в — краниальная; г — каудальная; д — дистальная стороны
жена от блока II пальца на расстоянии равном поперечной ширине диафиза на уровне
facies.
Размеры, мм: наибольшая поперечная ширина эпифиза 9,4, передне-задний диа-
метр блока II пальца 4,8, то же — для блока IV пальца 4,7.
Сравнение. Nyctanassa kobdoena отличается от современной N. violacea более
параллельно поставленными блоками пальцев и большими расстояниями между ними
при несколько меньших абсолютных размерах ископаемого вида (табл. 3). Диаметр
foramen vase, distale у современного вида (по нескольким экземплярам) всегда больше
по сравнению с N. kobdoena и сечение этого отверстия круглое, в отличие от щелевид-
ного у ископаемого вида. Facies artic. metatarsi I отставлено от блока II пальца на оди-
наковое расстояние у N. kobdoena и N. violacea. Невысокие гребни, идущие по краям эпи-
физа от блоков II и IV пальцев, развиты у обоих видов в равной мере.
Замечания. Сравнение тарсометатарсуса, а также эпифиза лопатки ПИН
№ 3222-29 из Хиргис-Нура 2 (рис. 6,4) от Nyctanassa kobdoena с большой серией
современных Nyctanassa и Nycticorax подтвердило принадлежность ископаемого вида
к ныне американскому роду Nyctanassa. При этом лопатка относится нами к этому же
вымершему виду на основании структурных деталей и сравнительных размеров.
У Nyctanassa акромион не имеет выделенного facies artic. clavicularis, выступаю-
щего дорзально и наружу, как у Nycticorax. Акромион лопатки у N. violacea и N. kobdo-
ena укорочен и закруглен. Facies artic. humeralis у Nyctanassa меньше и более узкая, так
же как и у ископаемого экземпляра, чем у Nycticorax. По относительным размерам тар-
сометатарсуса и лопатки, ископаемый экземпляр заметно ближе к Nyctanassa, чем к Nyc-
ticorax (табл. 3). Ширина дистального эпифиза тарсометатарсуса у Nyctanassa —
меньше или равна высоте эпифиза лопатки, так же как и у ископаемой формы, но она
больше или равна эпифизу лопатки у Nycticorax. Отличия лопатки N. kobdoena от
N. violacea заключаются в большей выдвинутости краниально всего акромиона лопатки
у ископаемого вида.
В дистальном эпифизе тарсометатарсуса Nycticorax отличается от Nyctanassa парал-
лельностью сагиттальных плоскостей блоков пальцев друг к другу (у Nyctanassa плос-
кости блоков II и III пальца расположены под небольшим углом), расширенной сочле-
новной поверхностью блока II пальца с передней стороны, большим диаметром блока
IV пальца (при меньших общих абсолютных размерах всей кости) и отставленностью
facies artic. metatarsi I от блока II пальца проксимально дальше, на расстояние превыша-
ющее ширину диафиза на уровне facies.
Целесообразно указать также и некоторые отличия Nyctanassa от нескольких других
родов современных цаплевых. У Egretta и Ardeola блоки пальцев тесно сближены.
У Cochlearius блоки пальцев спереди расставлены так же широко, но плантарно они на-
правлены под углом друг к другу. У Botaurus блок III пальца склонен наружу. У Ardea
блоки пальцев приближены друг к другу, они относительно большего диаметра и сильно
выдаются плантарно.
В ископаемом состоянии Ardeidae представлены сравнительно небольшим числом
24
Таблица 3
Размеры тарсометатарсуса и лопатки ископаемой и современных ночных цапель
Показатель Nyctanassa kobdoena Nyctanassa violacea Nycticorax nycticorax
крайние среднее n = 4 крайние среднее n = 4
Наибольшая поперечная ширина 9,4 9,7—10,5 10,0 10,6—11,7 11,2
дистального эпифиза Передне-задний диаметр блока 4,5 4,5—5,7 5,5 5,9—6,7 6,2
IV пальца Высота головки лопатки 10,0 9,2—10,2 9,9 10,1 — 11,4 10,8
форм. Недавно их ревизия была проведена П. Бродкорбом (Brodkorb, 1980). Из нижнего
миоцена Флориды известна очень крупная Ardea polkensis Brodkorb, 1955, описанная по
тарсометатарсусу. Из плиоцен—плейстоцена Эфиопии по коракоиду известна еще более
крупная Ardea howardae Brodkorb, 1980. Сравнимая по размерам с Nyctanassa kobdo-
ena ночная цапля Nycticorax fidens Brodkorb, 1963 описана по полной бедренной кости
из нижнего плиоцена Флориды.
Распространение. Средний плиоцен, верхняя подсвита свиты Хиргис-Нур
в местонахождениях Чоно-Хариах и Хиргис-Нур 2, пачка Б и верхние уровни пачки А.
ПОДОТРЯД CICONIAE BONAPARTE, 1854
СЕМЕЙСТВО CICONIIDAE GRAY, 1841
Род Ciconia Brisson, 1760
Ciconia lucida Kurochkin, 1982
Рис. 7; табл. Ill, фиг. 2—7, табл. IV, фиг. 1—3.
Ciconia lucida: Курочкин, 1982, с. 150.
Голотип — ПИН, № 3222-15; дистальная часть правого карпометакарпуса;
МНР, Убсунурский аймак, местонахождение Хиргис-Нур 2; нижний плиоцен, свита Хир-
гис-Нур.
Паратип — ПИН, № 3222-666; полный правый карпометакарпус; местонахож-
дение Хиргис-Нур 2; нижний плиоцен, нижняя подсвита (уровень 24) свиты Хиргис-Нур.
Материал. В дополнение к голотипу и паратипу из нижнеплиоценовых отложе-
ний имеется фрагментарное правое бедро (ПИН, № 3222-56) из нижней подсвиты
(уровни 20—21) свиты Хиргис-Нур местонахождения Хиргис-Нур 2; из среднего плио-
цена (верхняя подсвита свиты Хиргис-Нур) — проксимальная половина первой фаланги
III пальца лапы (№ 3222-600) из местонахождения Хиргис-Нур 1, дистальная половина
первой фаланги III пальца лапы (№ 3222-679) из местонахождения Хиргис-Нур 2
(уровни — 37—43) и головка правой лопатки (№ 2614-72) из местонахождения Чоно-
Хариах.
Описание. Дистальная часть карпометакарпуса массивная и широкая за счет ши-
рокого симфиза между метакарпалиями. Бороздки sulcus extensoris dig. communis и sul-
cus ext. indicus longi далеко проходят дистально — до уровня середины симфиза между
метакарпалиями. Общие размеры птицы крупные.
Размеры: см. табл. 4.
Сравнение. В роде Ciconia известно три ископаемых вида аистов. Из нижнего
миоцена Кении по дистальному эпифизу бедра описан мелкий С. minor Harrison, 1980.
В среднеплиоценовой пикермийской фауне Греции найдена плечевая кость аиста, раз-
мером с современного С. ciconia, описанного как С. gaudri Lambrecht, 1933. Наконец,
очень крупный ябируобразный С. maltha L. Miller, 1910 известен по полным скелетам и
многим отдельным костям в среднем (формация бруней, штат Айдахо) и главным обра-
зом в верхнем плейстоцене США (Ранчо-ла-Бреа в Калифорнии, озерные отложения аме-
рикен фоллз в Айдахо, отложения р. Итчукни и многие местонахождения формации пам-
лико во Флориде) и Кубы (Сьего-Монтеро). Первые два вида явно мельче С. lucida, да и
25
/7
I i -J
Рис. 7. Ciconia lucida Kurochkin, 1982
A — голотип № 3222-15, ПИН; правый карпометакарпус; Монголия, Хиргис-Нур 2; нижний плиоцен;
а — латеральная, б — дистальная стороны. Б — № 2614-72, ПИН; правая лопатка; Монголия, Чоно-Хариах;
средний плиоцен; а — латеральная, б — краниальная стороны
сравнение с ними невозможно ввиду описания их по другим костям скелета. Описания
деталей строения карпометакарпуса и бедра С. maltha мы не нашли ни в одной из много-
численных работ, посвященных этому виду, но общая длина карпометакарпуса у него оп-
ределенно больше — 138—150 мм, средняя — 139 (Miller L., 1932), чем у С. lucida, что
позволяет в известной мере говорить об отличиях между этими видами.
С. lucida был сравнен со всеми современными видами рода: nigra, ciconia, boyciana,
abdimii, episcopus, maguari. Сравнение co всеми современными родами семейства Cico-
niidae показало безусловную принадлежность lucida к роду Ciconia. Основное, и по суще-
ству единственное, отличие С. lucida от современных видов аистов — более широкая сим-
физарная часть между большой и малой метакарпалиями. В результате этого следы про-
хождения сухожилий сгибателей пальцев оканчиваются у С. lucida дистальнее за уров-
нем проксимального края симфизарной перемычки, а у большинства современных аистов
отпечатки сухожилий доходят только до края симфизарной перемычки. У С. maguari
наблюдается промежуточное состояние, у С. abdimii (мелкий вид) симфизарная пере-
мычка относительно широкая, но следы прохождения сухожилий доходят только до
проксимального ее края. По структуре сочленовных фасеток дистального эпифиза
С. lucida не отличим от современных аистов.
В проксимальном эпифизе карпометакарпуса С. lucida отличается от всех других
Ciconia наличием неглубокой канавки на вентральной поверхности эпифиза между ос-
нованиями proc, pisiformis и os metacarpale minus. У современных аистов этот участок
ровный, или даже наоборот на нем имеется приподнятая шероховатость.
Таблица 4
Размеры карпометакарпуса и бедренной кости Ciconia lucida sp. nov. и современных аистов
Показатель С. lucida, № 3222—666 С. lucida, № 3222—15 С. ciconia С. ciconia
Кар пометакар пус
Общая длина 133,2 — 116,2 120,0
Высота проксимального эпифиза 28,6 — 23,3 23,7
Высота сочленовной поверхности дистального 15,4 15,2 14,2 14,3
эпифиза
Ширина fac. artic. digiti major -11,0 10,0 9,9 9,5
Ширина дистального симфиза между метакар- 12,2 10,6 9,2 9,3
палиями
Бедренная кость Ширина проксимального эпифиза № 3222-56 28,2 24,9 24,6
Диаметр caput femoris 11,1 9,8 10,0
Передне-задняя ширина через sulcus intercon- 12,2 9,7 11,0
dylaris
Передне-задний диаметр cond. lateralis -24,8 21,1 21,8
Ширина дистального эпифиза 25,4 23,8 24,5
26
Заметно более крупными размерами С. lucida превосходит почти всех современных
аистов, к нему приближается по размерам только С. boyciana (табл. 4).
Замечания. Было проведено сравнение карпометакарпуса С. lucida со всеми
современными родами Ciconiidae (Ciconia, Mycteria, Ephippiorhynchus, Jabiru, Leptopti-
los, Anastomus). Принадлежность ископаемого вида к роду Ciconia представляется при
этом безусловной. Расширенная симфизарная перемычка дистального эпифиза харак-
терна для Anastomus, но последний имеет мелкие размеры и отличается от С. lucida дру-
гими деталями строения, в том числе и в проксимальном эпифизе карпометакарпуса.
Кроме двух карпометакарпусов от С. lucida были найдены бедро с разрушенным диа-
физом и удовлетворительной сохранности эпифизами, головка лопатки и проксималь-
ная и дистальная половины двух разных первых фаланг III пальца лапы (табл. IV,
фиг. 1—3; табл. III, фиг. 2, 3, 6, 7).
Бедро С. lucida имеет характерно суженную в передне-заднем направлении вертлуж-
ную часть проксимального эпифиза. Близко к нему по строению вертлуга стоит С. epis-
copus, но у последнего сильно наклонена каудально fac. artic. antitrochantericus. Кроме
того, у С. lucida продольная линия trochanter femoris расположена под тупым углом
к продольной оси собственно проксимального эпифиза; в результате этого латерально-
каудальный угол эпифиза при взгляде с проксимального конца выглядит срезанным
наискось. У всех современных аистов продольные оси вертлуга и эпифиза располагаются
под прямым углом, а латерально-каудальный угол прямой. Дистальный эпифиз бедра
С. lucida отличается от всех других Ciconia отсутствием сг. supracondyl. medialis, при-
ближенной к cond. medialis и мелкой sulcus patellaris. У современных Ciconia crista supra-
condylaris начинается непосредственно от condylus, у С. lucida crista отодвинута за-
метно проксимальнее. Размерами (табл. 4) бедро С. lucida заметно превосходит всех
современных аистов, кроме С. boyciana, укладываясь в пределы размеров между самцом
и самкой этого вида.
Фрагмент головки лопатки имеет отогнутый латерально лопастевидный акромион,
слабо выделяющуюся tuberculum coracoideum и небольшой площади fac. artic. hume-
ralis; высота caput scapulae 23 мм.
Фрагменты фаланг среднего пальца лапы относятся к С. lucida, поскольку по строе-
нию они определенно принадлежали представителю рода Ciconia, а по размерам превос-
ходят соответствующие фаланги современных видов рода. Других же представителей
рода Ciconia в этих местонахождениях не найдено, встреча второго столь же крупного
аиста в них маловероятна.
Распространение. Верхняя и нижняя подсвиты свиты Хиргис-Нур местона-
хождения Хиргис-Нур 2, верхняя подсвита свиты Хиргис-Нур местонахождений Хиргис-
Нур 1 и Чоно-Хариах.
С. ciconia С. boyciana 6 С. boyciana ? С. abdimii С. episcopus С. maguari С. nigra С. nigra
116,2 24,2 126,0 26,7 119,5 25,2 77,4 17,2 94,6 24,2 115,0 20,6 111,0 22,0 104,2 21,3
14,9 15,7 15,2 10,7 14,2 12,5 13,3 13,0
10,2 10,5 10,2 7,8 9,4 8,4 8,9 9,0
10,0 9,5 9,2 7,3 9,5 6,9 8,5 8,2
24,4 29,6 27,7 16,8 18,2 24,0 22,6
10,2 12,0 11,0 7,0 7,6 —. 9,7 9,0
10,2 13,0 12,0 7,5 8,1 — 10,2 10,0
21,9 27,0 24,2 15,0 16,8 —. 21,6 19,5
23,1 27,0 25,0 15,9 16,3 — 21,3 21,0
27
ОТРЯД ANSERIFORMES BECHSTEIN, 1804
СЕМЕЙСТВО ANATIDAE VIGORS, 1825
ПОДСЕМЕЙСТВО CYGNINAE VIGORS, 1825
Род Cygnus Bechstein, 1803
Cygnus pristinus Kurochkin, 1971
Рис. 8, 9; табл. IV, фиг. 4—12
Cygnus pristinus: Курочкин, 1971, стр. 60; 1976, с. 55.
Г олотип — ПИН, № 2614-25; дистальный эпифиз левой лучевой кости с частью
стержня; МНР, Убур-Хангайский аймак, Долина Озер, в 3—4 км на юго-восток от оз.
Тациин-Цаган-Нур, северо-западный склон горы Боро-Нур, местонахождение Хунг-Хурэ;
средний плиоцен, свита Хиргис-Нур (табл. IV, фиг. 6).
Паратипы — ПИН, № 2614-30, дистальный эпифиз левой лучевой кости, место-
нахождение Хиргис-Нур 2, верхняя часть пачки А; дистальные эпифизы правой лучевой
кости из местонахождения Хиргис-Нур 2, № 2614-115, пачка А, уровень 37 и № 3222-25,
пачка А, уровень 50—60.
Материал. Кроме голотипа и паратипов, имеется множество фрагментов различ-
ных костей посткраниального скелета из местонахождений Хиргис-Нур 2, Дзагсо-Хаир-
хан 1 и Чоно-Хариах. Плечевая кость. Дистальные эпифизы правого плеча № 2614-108,
Хиргис-Нур 2, пачка Б, уровень 63—65, и левого плеча № 3222-19, Хиргис-Нур 2,
пачка А, уровень 50—55. Лучевая кость. Проксимальный эпифиз правой № 2614-23,
Хиргис-Нур 2, уровень 20—21. Локтевая кость. Дистальные эпифизы № 2614-109,
Хиргис-Нур 2, пачка А, уровень 37 и № 2737-391, Дзагсо-Хаирхан 1. Карпометакарпус,
4 проксимальных и 2 дистальных фрагмента из Хиргис-Нура 2. Проксимальные эпифизы
правых карпометакарпусов № 3222-23 и № 3222-38, пачка А, уровень 50—55;
№ 3222-457, пачка А, уровень 21; левого карпометакарпуса № 3222-671, пачка А,
уровень 40—43. Дистальные эпифизы правых карпометакарпусов № 3222-40 и
№ 3222-41, пачка А, уровень 50—55. Коракоид, 4 дорсальных и один вентральный
фрагмент. Дорсальные эпифизы правых коракоидов из Хиргис-Нура 2, № 3222-32,
пачка А, уровень 50—55, и № 3222-456, пачка А, уровень 21; № 3222-667, пачка А,
уровень 24; из местонахождения Дзагсо-Хаирхан 1 № 2737-390; вентральный фрагмент
правого коракоида № 3222-53, Хиргис-Нур 2, уровень 51—55. Лопатка. 3 фрагмента го-
ловок лопаток из Хиргис-Нура 2, № 2614-27, пачка А; № 2614-111, пачка А, уровень
37; № 3222-6, пачка А, уровень 21. Первая фаланга большого пальца левого крыла
№ 2614-26, Хиргис-Нур 2, пачка A. Ulnare левого крыла № 2614-116, Хиргис-Нур 2,
пачка А. Тибиотарсус, дистальный эпифиз правой кости № 2614-70, Чоно-Хариах.
Тарсометатарсус, средний блок правой кости № 3222-54, Хиргис-Нур 2, уровень 20—21.
Фаланги пальцев лапы, 3 фаланги из пачки А Хиргис-Нура 2; третья фаланга IV пальца
левой лапы № 2614-32, третья фаланга III пальца левой лапы № 2614-113, четвертая
фаланга IV пальца правой лапы № 2614-117; первая фаланга IV пальца левой лапы
№ 3222-599, Хиргис-Нур 1, уровень 73—78, вторая фаланга III пальца левой лапы
№ 2614-73, Чоно-Хариах.
Описание. Дистальная часть стержня лучевой кости уплощена и расширена. Дор-
сальное ребро дистальной части стержня и эпифиза утолщено и притуплено. Tuberculum
aponeurosis занимает большую площадь и сильно поднято над каудальной поверхностью
эпифиза. Facies articularis radiocarpalis заметно выступает над дорсальным ребром
стержня. Наиболее дистальная вершина эпифиза притуплена.
Размеры, мм: наибольшая ширина эпифиза 16,7, ширина стержня на уровне de-
pressio dorsalis 13,5, толщина эпифиза через tuberculum aponeurosis 7,6, общая длина
фрагмента 48,0.
Изменчивость. С. pristinus стоит в ряду самых крупных лебедей по своим
абсолютным размерам. Но на имеющихся у нас экземплярах отчетливо выделяются две
размерные группировки — крупных и мелких экземпляров. Есть все основания с боль-
шой долей вероятия предполагать, что они отвечают самцам и самкам. Причем, по
сравнению с современными лебедями, половой диморфизм в размерах у С. pristinus был
выражен более сильно.
28
Таблица 5
Абсолютные размеры (мм) и индекс расширения дистального отдела лучевой кости
у Cygnus pristinus и современных лебедей
Вид Ширина эпифиза на уровне depressio dorsalis Ширина диафиза в 4 см от дистального конца эпифиза Ин- декс Вид Ширина эпифиза на уровне depressio dorsalis Ширина диафиза в 4 см от дистального конца эпифиза Ин- декс
Cygnus pristi- nus № 2614-25 13,5 10,0 0,74 С. bewickii С. olor 12,3 13,6 7,2 9,5 0,58 0,69
С. pristinus № 3222-25 12,4 — — С. olor С. olor 12,6 12,0 8,8 8,1 0,69 0,67
С. pristinus № 2614-30 11,2 — — С. melano- corhyphus 10,0 6,6 0,66
С. pristinus № 2614-115 13,7 — — Chenopis at- ratus 11,4 7,1 0,62
С. cygnus С. cygnus С. columbianus 13,2 12,0 12,1 8,0 7,5 6,2 0,60 0,62 0,51 Cygnus bucci- nator 14,5 9,6 0,66
Размеры некоторых костей С. pristinus приведены в табл. 5 и 6 и здесь же даны и со-
ответствующие размеры современных форм. Внимание обращают отношения между от-
дельными промерами эпифизов. По многим из них (плечо, локтевая кость, лучевая кость)
С. pristinus отчетливо выходит из ряда современных лебедей. Это является одним из до-
казательств его видоспецифичности. Другие отношения несколько перекрываются тако-
выми у современных видов (табл. 6).
Сравнение. Отдельные черты в строении дистального эпифиза лучевой кости
С. pristinus специфичны для этого вида, другие — общи или с лебедями из группы
кликунов (С. cygnus, С. columbianus, С. buccinator), или с лебедями из группы шипу-
нов (С. olor, С. melanocorhyphus и Chenopis atratus; последнего мы включаем в эту
группу, поскольку по особенностям морфологии он явно ближе к С. olor, чем к С. cygnus).
С. pristinus имеет характерно расширенный дистальный отдел диафиза, постепенно су-
жающийся от эпифиза в проксимальном направлении. У С. cygnus, С. buccinator и других
видов этой группы после эпифиза имеется резкое сужение к диафизу, у С. olor это су-
жение не столь резкое, как у кликунов, но заметно более сильное, чем у С. pristinus. Ука-
занная особенность хорошо иллюстрируется пропорциональными отношениями ширины
эпифиза и диафиза (табл. 5). Наиболее дистальная вершина эпифиза по вентральному
краю притуплена, как у С. bewickii. У остальных современных лебедей она округлая, не
имеет плоскосрезанной вершины, как у С. pristinus, хотя у С. olor — более округла,
а не заострена, как у С. cygnus, С. columbianus и С. buccinator. Дорсальное ребро
стержня перед эпифизами округлое и притупленное у С. pristinus, у всех современных ле-
бедей заостренное, особенно в области depressio dorsalis. Facies artic. radiocarpalis
выступает дорсально у С. pristinus в такой же мере, как и у кликунов, но у С. olor и С. me-
lanocorhyphus почти не выдается над краем стержня.
Замечания. Как видно из образцов, перечисленных в материалах, еще 26 фраг-
ментов различных костей скелета из всех местонахождений свиты хиргис-нур относятся
к Cygnus pristinus. Этот вид в ископаемой авифауне среднего плиоцена Монголии пред-
ставлен наибольшим количеством остатков, что в определенной мере отвечает и его бы-
лой численности. Так же как и структурные детали морфологии голотипа и паратипов,
строение других костей скелета С. pristinus сочетает детали структур с одной стороны —
общие с группой лебедей-кликунов, с другой — общие с группой лебедей-шипунов.
Вместе с преимущественно крупными размерами это был основной критерий отнесения
ископаемых остатков к С. pristinus. Ранее имелись сомнения о возможной близости
этого лебедя из среднего плиоцена Монголии к крупному современному американскому
виду Cygnus buccinator, но после сравнительного исследования, проведенного по скеле-
29
Таблица 6
Абсолютные (мм) и относительные размеры костей крыла и плечевого пояса у лебедей
Вид Плечевая кость Локтевая кость
Наибольшая ши- рина дистального эпифиза Передне-задний диаметр cond. dorsalis Индекс Наибольшая ши- рина дистального эпифиза Высота cond. dorsalis Индекс Ширина на уровне depressio dorsalis
Cygnus pristinus № 2614-108 № 2614-109
36,2 20,2 1,79 23,3 19,6 1,18 13,7
С. pristinus № 3222-19 № 2737-391
19,3 — 22,6 19,0 1,18 13,5
С. olor 1074-2 35,3 19,5 1,81 23,3 18,3 1,27 12,6
С. olor 1074-3 37,5 19,6 1,91 24,0 18,3 1,31 13,6
С. cygnus 1071-1 36,7 20,7 1,77 22,3 17,5 1,27 13,2
С. cygnus 1071-2 37,2 21,0 1,77 23,5 19,0 1,23 13,0
С. columbianus 1073-1 32,3 17,5 1,84 20,5 16,4 1,25 12,1
С. bewickii 1072-1 32,8 17,4 1,88 21,3 16,8 1,26 12,3
С. buccinator 1148-1 37,3 22,0 1,69 24,0 18,2 1,31 14,5
там, присланным проф. Р. Сторером (R. Storer) из Музея зоологии Мичиганского универ-
ситета в Энн-Арборе, США, эти сомнения отпали. При этом сравнение скелета С. bucci-
nator с другими лебедями еще раз показало, что он относится к группе С. cygnuS—
С. columbianus. Покажем специфичность морфологии С. pristinus последовательно по
всем имеющимся костям посткраниума.
Плечевая кость (табл. IV, фиг. 4, 5). В дистальном эпифизе плеча fossa m. brachia-
lis имеет отчетливо выраженный рельеф и четко выраженные границы, как у кликунов, но
не у С. olor. Impressio lig. collaterale ventrale укороченной и округлой формы, как
у С. olor. Fossa olecrani глубокая, как у С. olor и у С. buccinator, но sulcus anconaeus vent-
rale, будучи неглубокой у С. olor, у ископаемого близка по глубине к С. buccinator.
Переход от proc, flexorius к cond. ventralis вытянут и уплощен, как у С. cygnus и С. colum-
bianus, еще более вытянут у С. buccinator, но у С. olor утолщен и укорочен.
Локтевая кость (рис. 8, В). Дистальные эпифизы локтевых костей у современных ле-
бедей практически не различимы по своей структуре. С. pristinus характеризуется утол-
щенной и овальной формой основания и всего tuberculum carpale, который у современ-
ных лебедей — относительно меньше и тоньше.
Лучевая кость (табл. IV, фиг. 6). Проксимальный эпифиз лучевой кости во многом
ближе к С. cygnus, чем к С. olor. Tuberculum capiti большой, равномерно выпуклый
и плавно переходит на диафиз; у С. olor при относительно меньших размерах tubercu-
lum он переходит на диафиз через заметную выемку, которая также выражена у С. cyg-
nus, С. buccinator, Ch. atratus, но при относительно больших размерах tuberculum.
У С. olor дистальнее от tuberculum диафиз сужен, представляя в этом месте как бы
шейку головки. Краниальный край сочленовной поверхности эпифиза при взгляде сбоку
у С. pristinus заметно выше каудального, что свойственно группе кликунов; у С. olor
разница в высоте краев сочленовной поверхности выражена слабо. Приближенно очерта-
ния сочленовной поверхности у С. pristinus, так же как и у кликунов, можно свести к пя-
тиугольнику, а у С. olor и Ch. atratus — к четырехугольнику.
Карпометакарпус (рис. 9, А, В). С. pristinus по строению карпометакарпуса вновь
стоит ближе к С. cygnus и С. columbianus, чем к С. olor и С. buccinator. С первыми его
сближают структура задней поверхности trochlea carpalis, более суженная в средней
части, ориентировка плоскости facies art. dig. alulae, образующей с осью metacarpale
majus тупой угол (у С. olor он — ближе к прямому). Дистальный же эпифиз карпо-
Лучевая кость Лопатка Коракоид
1 Наибольшая ши- рина проксималь- ного эпифиза Индекс Ширина головки от вершины акромиона до вентрального края facies artic. humeralis Наибольшая ши- рина между дор- сальным краем головки и facies artic. humeralis Индекс Высота эпифиза от вершины акрокора- коида до вентраль- ного края cotyla scapularis Ширина шейки эпи- физа между margo cranialis и каудаль- ным краем facies artic. humeralis Индекс
№ 2614-115 № 2614-111 № 3222-456
15,7 0,87 30,5 20,6 1,48 38,3 18,0 2,12
№ 2614-25 № 2614-27 № 3222-32
16,5 0,81 31 21,0 1,47 39 18,3 2,13
15,0 0,84 16,0 0,85 16,4 0,80 17,1 0,76 14,8 0,81 15,4 0,79 17,4 0,83 27,0 17,7 1,52 28,7 18,4 1,55 29,4 19,4 1,51 28,3 19,4 1,45 23,7 17,0 1,39 28,0 17,7 1,58 31,0 20,5 1,51 38,6 20,0 1,93 40,6 20,6 1,97 42,7 19,5 2,18 41,7 19,6 2,12 36,0 16,5 2,18 37,0 15,7 2,35 43,5 19,6 2,21
метакарпуса целиком сходен по своему строению с С. cygnus, где характерна сужен-
ная суставная фасетка metacarpale minus и расширенный симфиз между metacarpalia
majus et minus.
Первая фаланга большого пальца крыла. По длине эта фаланга (47,0 мм) оказы-
вается короткой, такую же длину она имеет у сравнительно небольших С. bewickii
и С. columbianus, но выглядит она у С. pristinus мощнее за счет относительно большей
толщины корпуса. Характерна отчетливо выраженная скульптура поверхности. Переход
от корпуса фаланги к ее уплощенной части обозначен также резко, как у С. cygnus
и Ch. atratus, и он пологий у С. olor. Из-за большей массивности фаланги дорсальные
отделы обоих суставных поверхностей выглядят более расширенными, как у С. cygnus
и С. buccinator, по сравнению с С. olor и Ch. atratus. Корпус фаланги с краниальной по-
верхности представляется равномерно прямым, а у современных лебедей дистальный ко-
нец отведен вентрально.
По карпометакарпусу и первой фаланге большого пальца крыла из нижнеплиоце-
нового местонахождения Чаквар в Венгрии описан Cygnanser csakvarensis (Lamb-
recht, 1933; Kretzoi, 1957). На фаланге Cygnanser отчетливо выражена скульптура по-
верхности и другие особенности, более характерные для Anserinae, чем для Cygninae.
Коракоид (рис. 8, Б). У С. pristinus края collum acrocoracoidei идут параллельно на
всем протяжении шейки, как у С. bewickii и С. columbianus. У С. cygnus, С. buccinator
и С. olor шейка коракоида постепенно расширяется в проксимальном направлении. Пе-
редний край шейки коракоида у С. pristinus утолщен, как и у С. olor; у других лебедей
он заострен, особенно у С. columbianus и у С. buccinator. Форма fac. artic. clavicularis —
угловатая и усеченная с дорсальной и вентральной сторон, сходна у С. pristinus
с С. olor, но отличается по строению у кликунов, имеющих ровную поверхность facies.
В грудинной части коракоида, при общем сходстве ее у всех лебедей, привлекают вни-
мание некоторые различия в структуре надангулярной части передней поверхности
кости. У С. olor она ровная, кликуны в этом месте имеют незначительное углубление,
а у С. pristinus имеет значительную впадину.
Лопатка (рис. 9, Б). Область перехода от уплощенного корпуса лопатки к эпифизу
у С. pristinus плавная и постепенная, более сходная с С. olor и Ch. atratus, чем с С. cyg-
nus—С. buccinator, у которых шейка лопатки относительно сильнее сужена. Tuberculum
coracoideum мощный и сильно выступает краниально, будучи сравним в этом плане
31
Рис. 8. Cygnus pristinus Kurochkin, 1971
A —№ 2614-70, ПИН; правый тибиотарсус; Монголия, Чоно-Хариах; средний плиоцен; а — краниаль-
ная, б — медиальная, в — дистальная стороны. Б — № 3222-456, ПИН; правый коракоид; Монголия,
Хиргис-Нур 2; нижний плиоцен; г — латеральная; д — каудальная стороны. В — № 2614-109, ПИН; правая
локтевая кость; Монголия, Хиргис-Нур 2; средний плиоцен; е — вентральная; ж — дорсальная; з — дисталь-
ная стороны
Рис. 9. Cygnus pristinus Kurochkin, 1971
А — № 3222-38, ПИН; правый карпометакарпус; Монголия, Хиргис-Нур 2; средний плиоцен;
а — медиальная; б — латеральная; в — каудальная стороны. Б — № 2614-111, ПИН; левая лопатка; Мон-
голия, Хиргис-Нур 2; средний плиоцен; г — краниальная; д — латеральная стороны. В — № 3222-40, ПИН;
правый карпометакарпус; Монголия, Хиргис-Нур 2; средний плиоцен; е — медиальная; ж — дистальная
стороны
с С. olor и Ch. atratus, но он маломощный и слабо выделяющийся у С. cygnus—С. buc-
cinator, так же как и у С. melanocorhyphus. Шероховатость, расположенная на латераль-
ной поверхности шейки лопатки, каудальнее и дорсальнее от facies artic. humeralis,
у С. pristinus обширная и отчетливая, хорошо развитая и по дорсальной стороне;
в этом отношении больше сходства с С. olor и Ch. atratus, чем с С. cygnus—С. bucci-
nator. Акромион лопатки направлен в равной степени краниально и вверх. В этом плане
С. pristinus более сходен с С. olor, чем с кликунами, у которых он вверх выставлен силь-
нее, чем краниально. От всех современных лебедей лопатку С. pristinus отличает косая
усеченность краниально-вентрального угла facies artic. humeralis.
Тибиотарсус (рис. 8, А; табл. IV, фиг. 9, 10). По особенностям его структуры С. pris-
tinus стоит гораздо ближе к группе кликунов, чем к С. olor. Незначительная разница
между размерами латерального и медиального мыщелков и более вытянутые в попереч-
ном направлении очертания выходного отверстия canalis extensorius особенно сближают
С. pristinus с кликунами. Расширенная incisura intercondylaris и отставленность cond.
medialis в дистальном направлении за уровень cond. lateralis (при взгляде на эпифиз
сзади), отличают С. pristinus от всех современных лебедей. По размерам данный экзем-
пляр С. pristinus приближается к самым крупным экземплярам С. cygnus.
32
Тарсометатарсус. Средний блок дистального эпифиза тарсометатарсуса из Хиргис-
Нура 2, несмотря на фрагментарность, на основании крупных размеров (передне-зад-
ний диаметр блока 15,3 мм) мы относим к С. pristinus. Он во многом сходен по строению
с С. cygnus — удлиненный овальный боковой профиль блока, слабая расширенность его
в дистальном направлении, если смотреть на эпифиз спереди.
У С. olor, С. buccinator, С. columbianus и С. bewickii средний блок сбоку имеет круг-
лый профиль и расширен в дистальном направлении спереди.
Фаланги пальцев лап. Несколько фаланг пальцев на основании крупных размеров
и специфики строения относятся к С. pristinus. Длина первой фаланги IV пальца левой
лапы (№ 3222-599) —42 мм, длина третьей фаланги III пальца левой лапы (№ 2614-
113) —34,7 мм, длина четвертой фаланги IV пальца правой лапы (№ 2614-117) —
31 мм, длина третьей фаланги IV пальца левой лапы (№ 2614-32) — 31,3 мм. Характерно,
что при тех же размерах сочленовных поверхностей и толщины их тела, эти фаланги ока-
зываются короче, чем соответствующие фаланги крупных экземпляров С. olor, С. cyg-
nus и С. columbianus. Кроме того, дистальные эпифизы фаланг более сильно уплощены
дорсо-вентрально, чем у современных лебедей.
Замечания по экологии и систематике. По фрагментарным остат-
кам Cygnus pristinus трудно воссоздать в деталях внешний облик этого лебедя, хотя в об-
щем он не сильно отличался в этом плане от современных лебедей. Тем не менее строение
отдельных костей, особенно дистальных отделов крыла и ноги, заставляют считать, что
С. pristinus имел более короткие крылья, чем современные лебеди, а пальцы лап у него
были относительно короче. При общей робустности других элементов скелета это рисует
нам более тяжеловесного лебедя, чем современные виды рода Cygnus, задние конечности
которого были больше приспособленными к передвижению по суше, чем у современных
лебедей. Морфологически С. pristinus по одним и тем же костям сочетает детали строения
лебедей из группы кликунов и лебедя-шипуна.
Обращает на себя внимание такой негативный факт, как отсутствие до этого по суще-
ству ископаемых остатков рода Cygnus в третичных местонахождениях, несмотря на не-
плохую палеорнитологическую изученность на сегодняшний день миоцена и плиоцена
Северной Америки, да и Европы тоже. К тому же такие крупные птицы водного образа
жизни должны были сохраниться в ископаемом состоянии в первую очередь. Но в Европе
единственное описание лебедя известно из среднего плейстоцена о-ва Мальты (Cygnus
falconeri), в Северной Америке только из нижнего плейстоцена озерных отложений Хэ-
гермэн в штате Айдахо описан С. hibbardi и из среднего плейстоцена формации Фоссил
Лейк в штате Орегон и Фромэнс Ферри в штате Айдахо описан С. paloregonus, в Австра-
лии в верхнеплейстоценовых отложениях близ оз. Эйр описан С. lacistris и из четвертич-
ных отложений Новой Зеландии — С. sumnerensis (Brodkorb, 1964). Как мы уже указы-
вали, Cygnanser csakvarensis из нижнего плиоцена Венгрии относится не к подсемей-
ству Cygninae, а к подсемейству Anserinae. Другая неогеновая находка Cygnus в Европе
основана на описании по фаланге пальца лапы из верхнего миоцена Бельгии С. herren-
thalsi Van Beneden, 1871, и к ней нельзя относиться серьезно.
Описание С. pristinus является, по сути дела, первым прямым доказательством"
появления и присутствия рода Cygnus в плиоцене. Причем появляется этот вид в Цент-
ральной Азии. Он сочетает в своей морфологии особенности строения, присущие двум
основным линиям современных лебедей. На основании этого можно уверенно считать, что
род Cygnus в его сегодняшнем образе возник на озерах Центральной Азии не позже
раннего плиоцена, а уже потом, в плейстоцене, распространился отсюда по всему миру.
В плейстоцене Западной Европы из многих точек известны находки Cygnus olor,
С. cygnus, С. bewickii, на всей территории Северной Америки от Аляски до Флориды уже
встречаются в этот период в ископаемом состоянии С. buccinator и С. columbianus (Brod-
korb, 1964). Одна из находок С. columbianus достоверно приурочена к отложениям ран-
него плейстоцена оз. Хэгермэн в штате Айдахо (A. Miller, 1948).
Следовательно, можно поэтому принять Cygnus pristinus за предка обеих основных
линий современных лебедей.
Распространение. Местонахождения Хунг-Хурэ в Долиноозерской депрес-
сии в Центральной Монголии, Хиргис-Нур 1, Хиргис-Нур 2, Дзагсо-Хаирхан 1 и Чоно-
3 Заказ 2098 s
33
Хариах в Котловине Больших Озер на западе Монголии. Остатки С. pristinus приурочены
к обеим подсвитам свиты Хиргис-Нур. В местонахождении Хиргис-Нур 2 они встречаются
в основной части пачки А от уровня 21 до уровня 60, будучи наиболее многочисленными
на уровне 50—55. Из основания пачки Б Хиргис-Нура 2 происходит только одна находка
С. pristinus, но много остатков этого лебедя имеется из отложений пачки Б местонахожде-
ния Чоно-Хариах.
Cygnus sp.
Из среднеплиоценовых местонахождений Котловины Больших Озер имеется также
серия фрагментов костей от какого-то небольшого лебедя. От определения его как Cygnus
pristinus останавливают небольшие размеры, сопоставимые по разным костям скелета
с С. columbianus. По структурной же морфологии кости этого мелкого лебедя трудно
отличимы от С. pristinus, но из-за фрагментарности материалов или малой значимости
имеющихся от него костей, мы воздерживаемся от точного указания его систематиче-
ской принадлежности.
Рис. 10. Cygnus sp.
А — № 3222-678, ПИН; правая лопатка; Монголия, Хиргис-Нур 2; нижний плиоцен; а — латеральная;
б — краниальная стороны. Б —№ 2614-101, ПИН; левый карпометакарпус; Монголия, Дзабхан; средний
плиоцен; в — каудальная; г — медиальная стороны
Материал. Лопатка.*! фрагмента правых лопаток из Хиргис-Нура 2, № 3222-598,
пачка А, уровень 26 и № 3222-678, пачка А, уровень 34 (рис. 10, А); размеры, мм: ширина
головки от вершины акромиона до вентрального края facies artic. humeralis 26,2 и 27,4,
ширина между дорсальным краем головки и facies artic. humeralis ~18 и ~19. Карпоме-
такарпус-. 4 фрагмента проксимальных эпифизов — левого № 2614-101, местонахожде-
ние Дзабхан (рис. 10Б), правых и левого из местонахождения Хиргис-Нур 2 с уровня
50—55, № 3222-24, 3222-39, 3222-13; размеры, мм: ширина trochlea carpalis (со-
ответственно в порядке перечисленных фрагментов карпометакарпуса) — 10,5, 10,5,
10,4, 10,2; длина metacarpale alulare — 22,2, —, —, высота проксимального эпифиза —
24,5, —, —, —. Ulnare, 4 экз. из верхних уровней пачки А местонахождения Хиргис-Нур 2,
№ 2614-22, 2614-102, 3222-44, 3222-45. Тибиотарсус, фрагменты дистального эпифиза
правого № 2614-112, Хиргис-Нур 2, уровень 37, и левого № 3222-27, Хиргис-Нур 2,
уровень 50—55. Тарсометатарсус, фрагмент проксимального эпифиза правого
№ 3778-45, Чоно-Хариах. 8-й шейный позвонок № 2737-410, Хиргис-Нур 3, верхняя
подсвита свиты Хиргис-Нур.
34
ПОДСЕМЕЙСТВО ANSERINAE VIGORS, 1825
Род Anser Brisson, 1760
Anser devjatkini Kurochkin, 1971
Рис. И; табл. Ill, фиг. 1
Anser devjatkini: Курочкин, 1971, c. 64; 1976, c. 58.
Г олотип — ПИН, № 2614-28; дистальный эпифиз правого тибиотарсуса; МНР,
местонахождение Хиргис-Нур 2; средний плиоцен, верхняя подсвита свиты Хиргис-Нур.
Паратип — ПИН, № 3222-42; дистальная половина левого тибиотарсуса; МНР,
местонахождение Хиргис-Нур 2; средний плиоцен, верхняя подсвита свиты Хиргис-Нур.
Материал. Голотип и паратип.
Описание. Оба мыщелка дистального эпифиза тибиотарсуса тонкие и сжатые с бо-
ков, значительно выдаются в краниальном направлении. Срединная часть тела эпифиза
при взгляде с дистального конца сравнительно узкая. Дистальная поверхность межмы-
щелковой области мелкая с пологими выходами в обе стороны на мышелки. Гребень
epicond. lateralis усечен слабо с каудально-дистального угла. Выходное отверстие сапа-
lis extensorius большое, расположенное в средней части эпифиза. Медиальная полоска
эпифиза внутрь от pons supratendineus суженная.
Размеры, мм: ширина эпифиза ~' 15,5 (паратипа 15,7), передне-задний диаметр
cond. medialis 16,7 (паратипа 15,6), передне-задний диаметр cond. lateralis — (паратипа
13,5), передне-задняя толщина через incisura intercondylaris 9,3 (паратипа 9,5), наи-
меньшая поперечная ширина диафиза— (паратипа 7,8), наименьшая передне-задняя
толщина диафиза— (паратипа 6,0).
Сравнение. Мыщелки дистального эпифиза тибиотарсуса у A. devjatkini при
взгляде с дистального конца утоньшены и сжаты с боков относительно так же, как у Anser
coerulescens, A. indicus в отличие от A. anser, A. albifrons, A. cygnoides, A. fabalis. Сре-
динная часть тела эпифиза сужена так же, как у A. cygnoides в отличие от других Anser,
у которых она относительно толще. Небольшая глубина межмыщелковой зоны такая же
у A. coerulescens, A. indicus, но у других видов рода, в том числе и у A. cygnoides, она
более глубокая. Epicond. lateralis, не будучи усечен с каудально-дистального угла
у A. devjatkini, так же выглядит у всех Anser, кроме A. anser и A. cygnoides. Выходное
отверстие canalis extensorius дорсо-вентрально сжато у A. devjatkini в отличие от боль-
шинства современных гусей, кроме A. cygnoides и A. coerulescens; у A. indicus оно совер-
шенно круглое. Дистальное окончание диафиза медиальнее от pons supratendineus
сужено так же, как у A. coerulescens, а у других гусей эта полоска относительно и абсо-
лютно шире. В то же время край диафиза латеральнее от pons supratendineus у A. dev-
jatkini расширен и близок по структуре к A. anser, A. cygnoides, а у других Anser он отно-
сительно уже. Incisura intercondylaris спереди довольно широкая, как у A. anser, A. cyg-
noides и A. indicus в отличие от других гусей. При тех же абсолютных размерах, что
и A. fabalis, crista fibularis заканчивается у A. devjatkini заметно проксимальнее.
В ископаемом состоянии из неогена и плейстоцена известно несколько видов рода
Anser, но ни для одного из них тибиотарсус неизвестен.
Замечания. Anser devjatkini относился к группе настоящих гусей и едва ли
можно думать, что он в чем-либо существенно отличался по экологии и образу жизни
Рис. И. Anser devjatkini Kurochkin, 1971; голотип № 2614-28, ПИН; правый тибиотарсус;
Монголия, Хиргис-Нур 2; средний плиоцен
а — краниальная; б — медиальная; в — дистальная стороны
3*
35
от них. Как можно видеть из сравнения, морфологически он имеет больше всего общего
с A. coerulescens, будучи заметно крупнее него по размерам.
От Branta отличается отсутствием выемки на дистальном крае cond. medialis и по-
перечной расширенностью всего дистального эпифиза. От Tadorna отделяется на основа-
нии прямого угла с краниальной стороны между cond. medialis и линией диафиза, этот
угол у Tadorninae — раскрытий, тупой.
Из близких к Anser ископаемых Anserinae по тибиотарсусу из среднего миоцена Ка-
лифорнии (США) был описан Presbychen abavus. От Anser он отличается прямой ориен-
тацией дистального эпифиза вдоль продольной оси кости, выемкой на дистальном ребре
cond. medialis, равновыступающими на краниальную сторону мыщелками. Эти признаки
сближают Presbychen с Branta (Wetmore, 1930).
Распространение. Местонахождение Хиргис-Нур 2; верхняя подсвита свиты
Хиргис-Нур; уровень 50—55.
Anser liskunae Kurochkin, 1976
Рис. 12; табл. Ill, фиг. 10
Anser liskunae: Курочкин, 1976, с. 59.
Голотип — ПИН, № 2614-105; дистальный эпифиз левого плеча; МНР, место-
нахождение Дзабхан; средний плиоцен, свита Хиргис-Нур.
Паратип — ПИН, № 3222-18; дистальный эпифиз левого плеча; МНР, место-
нахождение Хиргис-Нур 2; средний плиоцен, свита Хиргис-Нур.
Материал. Голотип и паратип.
Описание. Дорсальный и вентральный мыщелки с краниальной стороны тесно
сближены. Impressio lig. collateral ventrale имеет округлую, немного вытянутую вдоль
продольной оси форму. Дорсальный край impressio продольно проецируется в дисталь-
ном направлении на вентральный край cond. ventralis. Eminentia m. ext. metacarpi radia-
lis представлена двумя отчетливыми ямками, помещенными непосредственно каудаль-
нее от proc, supracondylaris dorsalis. Fossa olecrani глубокая и широкая, ориентиро-
ванная своей плоскостью больше в каудальном направлении, чем в дистальном.
Размеры, мм: ширина эпифиза 27,4 (паратип ~27), передне-задний диаметр
cond. dorsalis 16,2 (паратип ~15,0), передне-задний диаметр cond. ventralis 10,2 (па-
ратип 9,5), общая длина фрагмента 34,5 (паратип 37,0).
Сравнение. Как отметил Улфинден (Woolfenden, 1961), плечевые кости, осо-
бенно их дистальные эпифизы, Anser, Branta и Chen практически неотличимы. Признаки
различий отдельных видов перекрывают родовые различия. Разделение отдельных видов
современных гусей возможно только по комплексу признаков и по размерам. Диагностич-
ным для A. liskunae в сравнении с современными Anser служат тесно сближенные мы-
щелки эпифиза, приближенность impressio lig. collateral ventrale к cond. ventralis,
углубленная, открывающаяся больше на краниальную сторону, fossa olecrani и круп-
ные размеры. У всех Anser, кроме A. cygnoides, мыщелки спереди расставлены шире. Im-
pressio ligamenti у большинства видов рода Anser, за исключением A. coerulescens и
A. domesticus, далеко отставлено от cond. ventralis; у двух названных видов, как и
у A. liskunae, проекция края impressio перекрывает в дистальном направлении край
cond. ventralis. Форма impressio у A. liskunae округлая, близкая к таковой у A. coerules-
cens, у других Anser она овальная, вытянутая вдоль продольной оси. Отпечатки сухо-
жилия отхождения m. ext. metacarpi radialis у ископаемого A. liskunae более отчетливы
и отставлены на 1 —1,5 мм от вершины proc, supracondylaris dorsalis, а у современных
Anser они неглубокие и приближенные вплотную к processus. Глубокая fossa olecrani
сравнима в общем плане по своему строению с современными Anser, но в отличие от
них она у A. liskunae заметно шире, будучи сходной в этом только с A. cygnoides. Вместе
с тем общая плоскость fossa смотрит у A. liskunae больше на краниальную сторону,
а у всех современных Anser она сильнее развернута в дистальном направлении.
Замечания. Среди известных ископаемых гусей, описанных по плечевым костям,
Anser scaldii (верхний миоцен, Бельгия), A. oeningensis (верхний миоцен, Швейцария),
Eremochen russelli (нижний плиоцен, Орегон, США) и Branta propinqua (средний плей-
36
Рис. 12. Anser liskunae Kurochkin, 1976; голотип № 2614-105, ПИН; левое плечо; Монголия,
Дзабхан; средний плиоцен
а — вентральная; б — латеральная; в — дистальная стороны
стоцен, Орегон, США) представляют более мелких птиц (Brodkorb, 1964). Из ископае-
мых Anserinae, описанных по другим элементам скелета, есть виды, которые по размерам
близки к A. liskunae (Anser cygniformis, верхний миоцен, ФРГ; Anser eldaricus, верхний
миоцен, Азербайджан; Heterochen vicinus, средний плиоцен, МНР; Branta howardae,
нижний плиоцен, Калифорния, США), но сравнение с ними невозможно из-за находок
разных частей скелета.
От Tadorninae отличия A. liskunae заключаются в меньшем выступании дистально
epicond. dorsalis, и особенно proc, flexoris, который заходит дистально за уровень cond.
ventralis, и в удлиненных пропорциях всего epicond. ventralis; у Tadorinae epicond. vent-
ralis в целом укорочен по длине и расширен поперек. На основании различий по этим су-
щественным признакам Anser liskunae никак не может сближаться с Tadorninae, не-
смотря на сходство в устройстве impressio lig. collaterale ventrale и локализации его
по отношению к cond. ventralis.
От видов рода Branta Anser liskunae отличает общее строение epicond. ventralis,
который у казарок продольно короче и мощнее, но в то же время proc, flexorius дальше
идет в дистальном направлении, почти достигая уровня дистального окончания cond.
ventralis. Вместе с этим дистальная часть стержня перед эпифизом у A. liskunae отно-
сительно много шире, чем у казарок.
У паратипа на месте fossa m. brachialis имеется патологическое образование в форме
кратера с наружным диаметром около 8 мм, в центре которого проходит сквозное отвер-
стие.
Распространение. Местонахождения Дзабхан и Хиргис-Нур 2; верхняя под-
свита свиты Хиргис-Нур.
Род Heterochen Short, 1970
Heterochen vicinus Kurochkin, 1976
Рис. 13; табл. II, фиг. 7, 8
Heterochen vicinus: Курочкин, 1976, с. 60.
Голотип — ПИН, №2614-110; проксимальный эпифиз правого тарсометатар-
суса с частью стержня; МНР, местонахождение Хиргис-Нур 2; средний плиоцен, свита
Хиргис-Нур.
Материал Только голотип.
Описание. Латеральные гребни гипотарсуса удлинены, а медиальные — укоро-
чены. Весь эпифиз сильно сдвинут медиально от продольной оси кости. Сочленовная по-
верхность eminentia interglenoidalis расположена почти в горизонтальной плоскости,
cotyla lateralis далеко заходит на переднюю сторону эпифиза. Проксимальная часть
диафиза имеет очень робустные очертания.
Размеры, мм: максимальная поперечная ширина эпифиза 17,4, передне-задняя
ширина эпифиза (от переднего края eminentia interglenoidalis до заднего края crista me-
37
Рис. 13. Heterochen vicinus Kurochkin,
1976; голотип № 2614-110, ПИН; правый
тарсометатарсус; Монголия, Хиргис-
Нур 2; средний плиоцен
а — проксимальная; б — краниальная;
в — каудальная; г — латеральная стороны
dialis hypotarsi) 16,6, передне-задний диаметр cotyla lateralis 10,0, ширина гипотарсуса
по проксимальному краю 10,4, наименьшая поперечная ширина диафиза 7,8, общая длина
фрагмента 54,6.
Сравнение. Тарсометатарсус из Хиргис-Нура 2 поразительно сходен с Н. рга-
tensis Short, 1970 из раннего плиоцена Дэвилс Голч в штате Небраска, США. Сходство
касается характерной робустности диафиза, укороченного гипотарсуса и формы его
внутреннего гребня, ориентировки проксимальной поверхности межсуставного выступа.
Н. vicinus отличается от Н. pratensis более скошенным медиально нижним краем
гипотарсуса и глубже опущенным на переднюю сторону эпифиза краем наружной
сочленовной фасетки. Примечательно, что и некоторые современные виды гусей раз-
деляются по этим же признакам.
Замечания. Сравнение Heterochen со всеми современными и вымершими фор-
мами семейства Anatidae исчерпывающе провел Шорт (Short, 1970), описавший этот
род. При сходстве по отдельным признакам со всеми родами современных Anserinae,
Heterochen не имеет полного сходства ни с одним из современных родов. Шорт
детально сравнил этот род с представителями отдельных триб отряда гусеобразных,
установив наибольшее сходство с Cairinini и Tadornini. Нам представляется, что ни
с теми, ни с другими Heterochen сближать нельзя. Cairinini имеют совершенно специфиче-
ские пропорции тарсометатарсуса, который у них непропорционально короткий и широ-
кий, a eminentia interglenoidalis — округлый спереди, ограниченный заметной выемкой
с наружной стороны. У Heterochen пропорции тарсометатарсуса, несмотря на характер-
ную робустность диафиза, удлиненные, a eminentia interglenoidalis спереди — бобовид-
ный, как у настоящих Anserinae. Также отдален Heterochen и от Tadornini. Определяю-
щими признаками здесь являются общая топография проксимального эпифиза тарсо-
метатарсуса и широкая разведенность в стороны блоков пальцев на дистальном эпифизе.
У Heterochen весь в целом проксимальный эпифиз сильно отведен медиально. У Alopo-
chen, Tadorna и Chloephaga он расположен более-менее симметрично вдоль продольной
оси кости; impressio lig. collateral laterale сильно выдается латерально, питательные
отверстия расположены в проксимальной части fossa infracotylaris dorsalis, impressio
laterale сдвинуто дистальнее уровня питательных отверстий, eminentia interglenoidalis
спереди в общем бобовидная. У Tadornini impressio laterale приподнято слабо и распо-
ложено проксимальнее, питательные отверстия расположены на уровне impressio
в,средней части fossa infracotylaris, eminentia interglenoidalis — в общем шаровидная.
Единственная черта в строении тарсометатарсуса, не позволившая Шорту (1970)
сблизить Heterochen с Anserinae, — широкая расставленность блоков пальцев у них и
кривизна дистальной части стержня с латеральной стороны перед выходом на блок
IV пальца. Но у такого крупного представителя гусиных, как Brauta canadensis, блоки
пальцев, в отличие от многих других представителей подсемейства оказались тесно
сближенными, а наружная грань стержня перед выходом на блок IV пальца — спрям-
ленной. Таким образом, этот признак так же, как и ряд других, не оказался абсолют-
ным среди Anserinae. Эти особенности строения тарсометатарсуса отличают и Hetero-
38
chen. Следовательно, этот род все-таки ближе вего к гусям и прав был Бродкорб
(Brodrorb, 1971а), поместив его в Anserinae.
С трудностью сопоставления отдельных костей посткраниального скелета у птиц
в ископаемом состоянии, найденных разрозненно, хотя и в одних горизонтах, мы сталки-
ваемся вплотную в данном конкретном случае. Положение усложняется невероятным
единообразием строения костей посткраниального скелета у всего отряда гусеобразных,
подтвержденном во всеобъемлющем исследовании Улфиндена (Woolfenden, 1961).
Описанные здесь Anser liskinae sp. nov. и Heterochen vicinus sp. nov., найденные
к тому же в близких горизонтах — в верхах пачки. А местонахождения Хиргис-Нур 2,
вполне сопоставимы по размерам. Плечевые кости Anser liskunae и тарсометатарсус
Heterochen vicinus представляют очень крупных гусей. И только невозможность прямых
сопоставлений и выяснения связи их друг с другом заставляют нас придавать им различ-
ный систематический статус. Таким образом, не исключается возможность происхожде-
ния описанных ископаемых остатков от одного вида крупного необычного гуся.
Распространение. Местонахождение Хиргис-Нур 2; верхняя подсвита свиты
Хиргис-Нур; уровень 37.
ПОДСЕМЕЙСТВО TADORNINAE REICHENBACH, 1852
Род Bonibernicla Kurochkin, gen. nov.
Название рода от bonus (лат.) — хороший и bernicla (лат.) — один из видов
казарок, жен. род.
Типовой вид — Bonibernicla ponderosa sp. nov., средний плиоцен, свита Хир-
гис-Нур; Котловина Больших Озер на западе МНР, местонахождение Хиргис-Нур 2.
Описание. Fossa olecrani плечевой кости глубокая и дистальный ее край обра-
зует ступеньку перед выходом на cond. ventralis. Каудально-дистальный угол proc, flexo-
rius заостренный. Поверхность impressio lig. collaterale ventrale обращена немного ди-
стально от общей плоскости кости, форма impressio округлая, вытянутая незначитель-
ным овалом вдоль длины кости. Вентральный край impressio идет параллельно вент-
рально-каудальному ребру epicond. ventralis. Фасетка в месте отхождения сухожилия
m. pronator superficial is на вентральной плоскости эпифиза располагается на !/3 ши-
рины этой плоскости ближе к краниальному краю. На краниальной стороне эпифиза
выемка между cond. dorsalis и epicond. dorsalis глубоко-выемчатая и дистально дости-
гает уровня ямок в местах фиксации сухожилий на дорсальном надмыщелке (fossae
lig. collaterale dorsale). Грань, образуемая краем этой выемки и краниальной границей
надмыщелка, угловатая и представлена прямой линией.
Видовой состав. Типовой вид.
Сравнение. Bonibernicla относится к подсемейству Tadorninae, поскольку
имеет утонченный изящный epicond. ventralis с заостренной гранью proc, flexorius,
удлиненную глубоко-выемчатую fossa olecrani, округлую impressio lig. collaterale
ventrale и далеко заходящую дистально вырезку между cond. dorsalis и epicond. dorsa-
lis (по краниальной стороне).
Bonibernicla не имеет полного сходства или полного отличия ни от каких современ-
ных и ископаемых пеганок и куриных гусей. С Tadorna сходна структурой всего вент-
рального надмыщелка, округлой формой дистальной проекции cond. ventralis, в том
числе частично дистальной ориентацией impressio lig. collaterale ventrale, заострен-
ностью proc, flexorius. Impres. m. pronator superficialis расположен ближе к краниаль-
ному углу, чем у Tadorna. Fossa olecrani у Bonibernicla глубже, чем у Tadorna, хотя и
имеет у последней небольшую ступеньку перед выходом на вентральный мыщелок.
Из двух ямок, расположенных проксимальнее дорсального надмыщелка на его
дорсальной стороне, проксимальная ямка у Bonibernicla развита лучше дистальной,
у Tadorna они приблизительно одинаково слабо выражены. Весь epicond. dorsalis у Во*
nibernicla не выдается так заметно в дистальном направлении, как у Tadorna, а с дор-
сальной поверхности надмыщелка площадка с двумя отпечатками ligamentorum col-
laterale dorsale у Bonibernicla округлая, а у Tadorna — вытянутая.
39
Bonibernicla сходна c Alopochen заостренностью и утоньшенностью proc, flexorius,
глубокой fossa olecrani, структурой дорсального надмыщелка, который не выступает
сильно дистально, а площадка с отпечатками связок имеет одинаково округлую
форму, идентичностью строения и расположения impressio lig. collaterale ventrale,
fossa m. brachialis. Отличия Bonibernicla от Alopochen касаются строения вентрального
надмыщелка, относительно менее мощного у Bonibernicla; расположением под неболь-
шим острым углом линии вентрально-каудального угла надмыщелка к линии вент-
рально-краниального угла impress, lig. collaterale, формы дистальной проекции cond.
ventralis — немного овальной у Alopochen. Выемка между дорсальными мыщелком и
надмыщелком с краниальной стороны у Alopochen не доходит дистально до уровня
отпечатков связок на дорсальном надмыщелке, а у Bonibernicla, как указывалось
выше, эта выемка доходит дистально почти до середины площадки с отпечатками
связок.
Современный род Chloephaga (сравнено с С. picta и С. melanoptera) сходен с Boni-
bernicla округлыми очертаниями площадки impressio lig. collaterale ventrale, одинако-
вой отставленностью дистально дорсального надмыщелка и одинаковой округлостью
дорсальной поверхности этого надмыщелка с глубокими отпечатками мест отхождений
сухожилий, круглой формой дистальной проекции cond. ventralis, одинаково удлинен-
ной выемкой между дорсальными мыщелком и надмыщелком (дистально до уровня
отпечатков сухожилий на дорсальной поверхности надмыщелка). Отличается Boni-
bernicla от Chloephaga меньшей массивностью всего epicondylus ventralis при большей
его вытянутости вдоль оси кости, более глубокой fossa olecrani и более расширенной
дорсо-вентрально дистальной частью диафиза. Из всех современных Tadorninae (нам
не удалось сравнить, к сожалению, с южноамериканскими Tachyeres и Neochen)
Bonibernocla ближе всего к Chloephaga.
В плейстоцене Северной Америки был широко распространен род Anabernicula,
в котором из многих местонахождений описано 4 вида. Сводки данных по ископаемым
Anabernicula и сравнение их с современными Tadorninae опубликованы Говард (Ho-
ward, 1964) и Шортом (Short, 1970). Bonibernicla отличается от Anabernicula корот-
ким proc, flexorius, который при взгляде с каудальной стороны не достигает дистально
уровня cond. ventralis, значительно более глубокой fossa olecrani, уплощенностью
дистальной части краниальной стороны диафиза (у Anabernicula она — выпуклая),
большей отставленностью fossa m. brachialis от impressio lig. collaterale ventrale.
Одинаково у них строение заостренного proc, flexorius, округлость cond. ventralis,
форма impressio lig. collaterale ventrale, удлиненность выемки между cond. dorsalis и
epicond. dorsalis, расширенность дистальной части диафиза. Anabernicula, по описанию
Шорта (1970), очень сходна с Neochen. Сравнение их по стереофотографиям, приведен-
ным в этой же статье, с Bonibernicla показывает, что Neochen и Anabernicula сходны и
отличны от Bonibernicla по одним и тем же признакам.
Из среднего плейстоцена запада США описан еще один род этого подсемейства —
Brantadorna (Howard, 1963), — известный только по проксимальному эпифизу плече-
вой кости. Сравнение с ним Bonibernicla поэтому невозможно.
Видовой состав. Только типовой вид.
Замечания. Среди гусеобразных средних размеров из авифауны среднего
плиоцена Западной Монголии нами описываются и другие роды и виды гусей. Возни-
кает вопрос, насколько правомочно в данном случае отнести дистальный эпифиз плеча
из Хиргис-Нура 2 к новому роду подсемейства Tadorninae. Не принадлежат ли все они
одним и тем же формам?
Anser liskunae Kur., 1976 описан по дистальным эпифизам той же плечевой кости.
Это был очень крупный гусь, даже несколько крупнее самых крупных подвидов канад-
ской казарки, и много крупнее Bonibernicla. Anser devjatkini Kur., 1971 известен по
дистальным частям тибиотарсуса, от птиц несколько меньших размеров, чем A. lis-
kunae, но все-таки заметно более крупных, чем Bonibernicla ponderosa. Остается еще
Heterochen vicinus Kur., 1976, который по относительным размерам тарсометатарсуса
был раза в полтора крупнее, чем Bonibernicla ponderosa.
40
Bonibernicla ponderosa Kurochkin, sp. nov.
Рис. 14; табл. Ill, фиг. 8, 9
Видовое название ponderosa (лат.) —тяжеловесная.
Г олотип — ПИН, № 3378-50; дистальный эпифиз левого плеча; МНР, местона-
хождение Хиргис-Нур 2; средний плиоцен, свита Хиргис-Нур.
Материал. Только голотип.
Описание. Fossa olecrani глубокая, образующая хорошо выраженную сту-
пеньку перед выходом на cond. ventralis. С краниальной стороны выемка между cond.
ventralis и cond. dorsalis узкая. Proc, flexorius заострен не только сверху и снизу, но и
по каудальному, и по дистальному ребру. Из двух ямок eminentia m. metacarpi radialis,
Рис. 14. Bonibernicla ponderosa gen. et sp.
nov.; голотип № 3378-50, ПИН; левое
плечо; Монголия, Хиргис-Нур 2; нижний
плиоцен
а—медиальная; б — латеральная; в —
дорсальная; г — дистальная стороны
проксимальная заметно глубже. Impressio m. pronator superficialis дистальным краем
находится на уровне проксимальной вершины impressio lig. coHaterale ventrale.
Размеры, мм: наибольшая ширина дистального эпифиза 21,2 передне-задний
диаметр cond. dorsalis 12,4, передне-задний диаметр cond. ventralis 7,2.
Сравнение. В описании морфологии дистального эпифиза плеча. В. ponderosa
приведены особенности строения, которые могли бы отличать данный вид от других
в этом роде. Выведены они из сопоставления Tadorna tadorna с Т. ferruginea, Chloe-
phaga picta с С. melanoptera и Anabernicula robusta с A. oregonensis.
Распространение. Уровень 37 местонахождения Хиргис-Нур 2; основание
верхней подсвиты свиты Хиргис-Нур.
TADORNINAE GEN.
В сборах из местонахождения Хиргис-Нур 2 и Чоно-Хариах имеются фрагментар-
ные остатки двух разных по размерам форм подсемейства Tadorninae. Фрагментар-
ность не позволяет более точно дать их систематическое положение, хотя признаки
принадлежности к Tadorninae представляются довольно отчетливыми.
Материал. Проксимальный эпифиз правой плечевой кости, ПИН, № 2614-34,
Хиргис-Нур 2, верхние уровни пачки А. Два фрагмента дистальных эпифизов левых
тарсометатарсусов № 3222-5, Хиргис-Нур 2, уровень 50—55, и № 2614-71, Чоно-
Хариах.
Сравнение и замечания. Эпифиз плечевой кости № 2614-34 (рис. 15)
относится к Tadorninae, потому что crista capiti caudalis подходит к tuberculum dor-
sale, а не к caput humeri; impressio m. pectoralis имеет небольшую длину и округлые
очертания, tuberculum dorsale угловатый, выдвинут проксимально и при взгляде
с проксимального конца ориентирован на середину головки плеча, головка плеча
округлая и укороченная в дорсо-вентральном направлении. Большая часть этих при-
знаков, как полагал Улфинден (Woolfenden, 1961), специфична для Tadorninae.
Из плейстоцена США известны немного более крупные Brantadorna downsi и несколько
видов рода Anabernicula. В данном случае важно, что на западе Монголии в среднем
плиоцене обитала маленькая пеганковая утка.
41
Z7
i. i— I
Рис. 15. Tadorninae gen.; № 2614-34, ПИН; правое плечо; Монголия, Хиргис-Нур 2; средний
плиоцен
а — проксимальная; б — медиальная стороны
Рис. 16. Tadorninae gen.; № 3222-5, ПИН; левый тарсометатарсус; Монголия, Хиргис-Нур 2;
средний плиоцен
а — медиальная; б — краниальная; в — каудальная стороны
Размеры, мм: ширина проксимального эпифиза 15,0, толщина caput
humeri 5,3.
Дистальные эпифизы тарсометатарсусов № 3222-5 и № 2614-71 принадлежали дру-
гому представителю Tadorninae (рис. 16). Размеры его были крупные — с современ-
ного Anser coerulescens. Ранее оба этих фрагмента были отнесены нами к Anserinae
gen. (Курочкин, 1976). Более тщательный анализ их строения показывает, что по сбли-
женности tr. metatarsi secundii и tr. m. tertii, по большей сдвинутости tr. m. secundii
на заднюю сторону и одновременно проксимально, суженности (почти заостренности)
плантарной части сочленовной поверхности tr. m. tertii и суженности всего основания
этого блока с плантарной же стороны между медиальной и латеральной межблоковой
вырезками эти остатки тарсометатарсусов принадлежали какому-то крупному Tadorni-
nae. В составе среднеплиоценовой авифауны Западной Монголии описана выше
Bonibernicla ponderosa. По абсолютным размерам тарсометатарсусы вполне могли бы
происходить от этого вида.
Размеры, мм: № 3222-5 — передне-задний диаметр tr. m. tertii 10,2; ширина
tr. m. tertii с дистального конца 7,2; передне-задний диаметр tr. m. secundii 9,4;
№ 2614-71 —передне-задний диаметр tr. m. tertii 9.
ANATIDAE SUBFAM.
Из местонахождений среднего плиоцена Котловины Больших Озер имеется не-
сколько других остатков гусеобразных птиц средних размеров, диагностичность и
фрагментарность которых на фоне большого сходства остеологических структур
во всем отряде гусеобразных и описания в этой фауне нескольких представителей
подсемейства Anserinae и Tadorninae не позволяют достоверно говорить даже об их
подсемейственной принадлежности. Тем не менее считаем целесообразным привести
здесь материалы, поскольку они впоследствии могут дополнить какие-то новые находки.
Материал. Коракоид. Дорсальная часть эпифиза левого коракоида № 3222-668,
Хиргис-Нур 2, уровень 24; несколько более крупный, чем у Anser anser, отличающийся
обширной округлой facies artic. clavicularis; размер, мм: ширина акрокоракоида 16,5.
Медиальный фрагмент вентрального эпифиза правого коракоида № 3378-53, Чоно-Ха-
риах 2, основание пачки Б; специфичен заостренным angulus medialis и полого про-
ходящим крылом грудинной фасетки на краниальной стороне медиального угла.
Лопатка. Фрагмент левой лопатки № 2614-17, Чоно-Хариах; по размерам близка
к Branta canadensis. Лучевая кость. Проксимальный эпифиз левой кости № 3376-74,
Таталы, пачка Б, свита Хиргис-Нур; размер, как у A. fabalis. Первая фаланга боль-
шого пальца крыла. Дистальный и проксимальный фрагменты одной и той же фаланги
42
L. I____________I
Рис. 17. Anatidae subfam.; № 2614-21 и № 3222-597, ПИН; первая фаланга большого пальца
правого крыла; Монголия, Хиргис-Нур 2; средний плиоцен
а — проксимальная; б-в — латеральная; г — дистальная стороны
правого крыла № 2614-21 и № 3222-597 (рис. 17) из местонахождения Хиргис-Нур 2,
верхние уровни пачки А. Первый из них был найден в 1965 г. И. Г. Лискун при работах
отряда ССМГЭ, а второй — через 14 лет, в 1979 г., членами отряда В. И. Жегалло
при работах ССМПЭ. Установить это удалось еще три года спустя при ревизии всех
собранных материалов. На поперечном изломе эти два фрагмента имеют точный
до мельчайших щербинок контакт, так что сомнений в принадлежности их одному
экземпляру не остается совсем. По строению и размерам эта фаланга ближе всего
к таковой современного Anser caerulescens.
Размеры, мм: длина 36, ширина проксимального эпифиза 10,5. Тарсометатар-
сус. Средний блок дистального эпифиза левого тарсометатарсуса № 3222-669, Хиргис-
Нур 2, уровень 24. По расширенности основания блока, скорее всего, происходит
от Anserinae средних размеров.
Размеры, мм: передне-задний диаметр 10,1; ширина с дистального конца 7,2.
Фаланга пальца лапы. Первая фаланга третьего пальца задней конечности
№ 2614-106, Хиргис-Нур 2, уровень 62—65. Описана ранее (Курочкин, 1976), как
происходящая от Anserinae.
Позвонки. Имеются три фрагментарных шейных позвонка из местонахождения
Чоно-Хариах 2, № 3378-51, № 3378-52 и 3378-66. Принадлежали каким-то Anserinae
или Tadorninae.
Размеры, мм: длина тела позвонка 25 (№ 51) и 28,5 (№ 66).
ПОДСЕМЕЙСТВО ANATINAE VIGORS, 1825
Род Anas Linnaeus, 1758
Anas soporata Kurochkin, 1976
Рис. 18; табл. V, фиг. 1—6
Anas soporata: Курочкин, 1976, с. 61.
Голотип — ПИН, № 2614-95; дорсальная половина правого коракоида; МНР,
Гоби-Алтайский аймак, местонахождение «точка 1080 м» в Шаргаин-Гоби; средний плио-
цен, свита Хиргис-Нур.
Паратипы — ПИН, № 3379-2; дорсальный эпифиз левого коракоида; МНР,Коб-
досский аймак, местонахождение Явор 1; средний плиоцен, верхняя подсвита свиты
Хиргис-Нур. ПИН № 3222-11; фрагмент дорсальной половины правого коракоида,
МНР, местонахождение Хиргис-Нур 2; нижний плиоцен, нижняя подсвита свиты
Хиргис-Нур.
Материал. В дополнение к голотипу и паратипам из местонахождений Хиргис-
Нур 2, «точка 1080 м» и Чоно-Хариах имеются несколько фрагментов других костей,
по абсолютным размерам относимых к A. soporata. Местонахождение Хиргис-Нур 2
верхние уровни пачки А: ПИН, № 3222-22, дистальная половина левой плечевой кости;
ПИН, № 3222-20, дистальный эпифиз правой плечевой кости; ПИН, № 3222-50, прокси-
мальный эпифиз левой плечевой кости; ПИН, № 3222-673, дистальный фрагмент
правой плечевой кости; ПИН, № 3222-674, дистальный эпифиз правой плечевой кости;
ПИН, № 3222-46, дистальный эпифиз левой локтевой кости. ПИН, № 2614-119, фраг-
мент проксимальной половины правой локтевой кости, местонахождение «точка 1080»
в Шаргаин-Гоби, средний плиоцен, свита Хиргис-Нур. Две лопатки: ПИН, № 3222-8,
43
A — № 3222-46, ПИН; левая локтевая кость; Монголия, Хиргис-Нур 2; средний плиоцен; а — вентраль-
ная, б — дорсо-латеральная стороны. Б — голотип № 2614-95, ПИН; правый коракоид; Монголия,
«высота 1080 м» в Шаргаин-Гоби; средний плиоцен; в — медиальная; г — каудальная; д — латеральная
стороны. В — № 3222-22, ПИН; левое плечо; Монголия, Хиргис-Нур 2; средний плиоцен; е — вентральная;
ж— дистальная; з — латеральная стороны. Г — № 3222-50, ПИН; левое плечо; Монголия, Хиргис-Нур 2;
средний плиоцен; и — медиальная; к — проксимальная стороны
местонахождение Хиргис-Нур 2, уровень 21, нижний плиоцен, нижняя подсвита свиты
Хиргис-Нур; ПИН, № 2614-88, местонахождение Чоно-Хариах, верхняя подсвита свиты
Хиргис-Нур.
Описание. Каудально-медиальная фасетка акрокоракоида в месте расположе-
ния facies artic. clavicularis не разделена на две части и не имеет углубленной facies.
Краниально-медиальный угол акрокоракоида утолщен. Фасетка impressio lig. acrocora-
cohumeralis имеет форму пологой дуги с вершиной, обращенной медиально. Margo
medialis эпифиза заострен.
Размеры, мм: длина дорсального эпифиза 10,7 (голотип), 11,0 (№ 3379-2);
поперечная ширина collum acrocoracoidei на уровне середины facies artic. humeralis 4,6
(голотип), 5,3 (№ 3379-2); длина акрокоракоида от его вершины до дорсального
края facies artic. humeralis 6,7 (голотип); ширина facies artic. humeralis 3,4 (голотип),
3,6 (№ 3379-2), 3,6 (№ 3222-11); наименьшая поперечная ширина диафиза 3,4 (голо-
тип), 3,6 (№ 3222-11); общая длина фрагмента 21,0 (голотип), 19,7 (№ 3379-2).
Сравнение. Из неогена всего мира описано 18 различных видов рода Anas,
из них коракоиды известны у 8 видов (Brodkorb, 1964). A. luederitzensis Lambrecht,
1929, из нижнего миоцена Западной Африки, A. blanchardi Milne-Edwards, 1863 из
нижнего миоцена Франции, Германии и Чехословакии, A. risgoviensis von Ammon,
1918 из верхнего миоцена Западной Германии, A. apscheronica Burchak-Abramovich,
1958 из верхнего плиоцена Азербайджана и Anas integra A. Miller, 1944 из нижнего
миоцена Северной Америки имеют более крупные, чем у A. soporata, размеры кора-
коида и поэтому непосредственно сопоставляться не могут, хотя последний вид сходен
с A. soporata отсутствием выемки по вентральному краю facies artic. clavicularis. Среди
мелких ископаемых уток коракоиды известны от четырех видов, вполне близких по
размерам к A. soporata. Среди них Anas velox Milne-Edwards, 1868 из верхнего мио-
цена Франции и нижнего миоцена Чехословакии (Svec, 1981) отличается более мощ-
ным диафизом коракоида (наиболее узкая ширина диафиза 4,3, по Svec, 1981), под
углом идущими краями collum acrocoracoidei и большим размером дорсального эпи-
44
физа (13,35, по Svec, 1981). A. natator Milne-Edwards, 1868 из нижнего миоцена Фран-
ции и Чехословакии (Svec, 1981) имеет выемчатое строение вентрального края fac.
artic. clavicularis, округлую форму cotyla scapularis и более крупные размеры (длина
дорсального эпифиза 12,0, по Svec, 1981). Сравнение с A. eppelsheimense Lambrecht,
1933 из нижнего плиоцена Германии затруднено из-за плохого качества иллюстра-
ций, хотя размеры их практически одинаковы с A. soporata.
Из шести местонахождений среднего и верхнего плиоцена и из нижнего плейстоцена
США и Мексики описан небольшой чирок Anas bunkeri Wetmore, 1944. От этого вида
известны главным образом карпометакарпусы, но также — фрагменты локтевой кости,
коракоида и тибиотарсуса. В описаниях отмечается, что этот вид структурно очень
близок к современному A. crecca carolinensis, т. е. к американскому аналогу палеаркти-
ческого А. сгесса сгесса, но несколько крупнее его. Можно считать, что A. soporata
отличается от A. bunkeri только меньшими размерами, а в целом эти два вида пред-
ставляются очень близкими.
В сравнении с современными представителями Anas soporata характеризуется
мелкими размерами, вполне сравнимыми с таковыми А. сгесса. Наибольшее сходство
в морфологии дорсальной половины коракоида, выражающееся в общей удлиненности
эпифиза, структуре медиальной поверхности субакрокоракоидальной части, снабжен-
ной небольшой ямкой, утоньшенности и уплощенности субэпифизарной части диафиза,
прослеживается также с этим видом. Но утолщенность и округлость краниально-
медиального угла акрокоракоида отличают A. soporata от А. сгесса и от всех других
видов рода. Это происходит за счет отсутствия углубленной facies artic. clavicularis
и как следствие этого отсутствия видимого разделения каудально-медиальной поверх-
ности акрокоракоида на две половины, выраженные нормально у всех Anas, хотя
развитого в меньшей степени — у маленькой А. сгесса и особенно у A. discors. Изогну-
тость фасетки дугой внутрь в месте прикрепления lig. acrocoracohumeralis выражена
у разных видов рода Anas в разной степени, у А. сгесса и A. discors дугообразность
фасетки почти не выражена и хорошо заметна у A. querquedula и A. formosa. Так же
различна у современных Anas степень заостренности margo medialis эпифиза, нависа-
ние крыла facies artic. clavicularis над sulcus m. supracoracoidei и как следствие разной
степени развития этих двух черт — большее или меньшее развитие глубокой ямки на
медиальной поверхности субакрокоракоидальной части. У A. soporata неглубокая ямка
на субакрокоракоидальной поверхности эпифиза небольшая, но отчетливо выраженная,
накрывается сверху небольшим крылом facies artic. clavicularis, margo medialis
эпифиза на уровне ямки заострен. Сходно устроена эта часть у A. discors, но margo
medialis более притупленный. У А. сгесса субакрокоракоидальная ямка развита плохо,
facies artic. clavicularis почти не нависает крылом. У A. querquedula субакрокоракои-
дальная ямка занимает латеральную половину, а не середину sulcus m. supracoracoidei.
Замечания. Фрагменты плечевых костей, три дистальных эпифиза, один прок-
симальный и один диафиз, а также по одному фрагменту дистального и проксималь-
ного эпифизов локтевых костей относятся к A. soporata, так как они вполне подходят
к этому виду по размерам, а по строению ближе всего среди современных уток
к А. сгесса и A. discors.
Дистальный эпифиз плеча (рис. 18,В) A. soporata имеет характерно сдавленный
проксимально-дистально cond. ventralis, раскрытый тупой угол при взгляде с дисталь-
ного конца между cond. ventralis и proc, flexorius; дорсальный край impressio lig. colla-
terale ventrale на уровне вентрального края cond. ventrale. Это impressio имеет срав-
нительно небольшой размер и не расширено дорсально-вентрально.
У A. discors и A. querquedula cond. ventralis более округлый, как и у остальных
просмотренных нами современных Anas, и только у А. сгесса он немного утолщен
проксимально-дистально. Угол между этим мыщелком и proc, flexorius может быть
прямым и тупым у разных видов рода Anas.
У A. discors и A. querquedula он раскрытый, тупой, как и у A. soporata; у А. сгесса —
прямой. Выемка между cond. ventralis и proc, flexorius с дистального конца у A. sopo-
rata, A. discors и A. querquedula глубже, чем у А. сгесса. Impressio lig. collaterale
ventrale у разных видов рода Anas также может быть сдвинуто по отношению к cond.
45
ventralis дорсальнее или вентральнее. У А. сгесса impressio занимает более дорсальное
положение, вследствие чего вентральный край condylus и дорсальный край impressio
перекрываются. У A. discors и A. querquedula, а также у A. platyrhynchos, A. americana,
A. strepera, как и у ископаемой A. soporata, край мыщелка и край impressio не пере-
крываются, а находятся на одной линии. Форма этого impressio — уменьшенная и
суженная дорсо-вентрально, одинакова у A. soporata с A. querquedula, у других Anas
impressio — расширено дорсо-вентрально.
Проксимальный эпифиз плеча (рис. 18, Г) при близких размерах очень схож
у A. soporata и у мелких Anas, отличаясь только пропорциями головки плеча и строе-
нием каудальной поверхности участка между дорсальным краем головки плеча и пнев-
матической ямкой. A. soporata отличается от мелких современных Anas утолщенной
краниально-каудально и укороченной дорсо-вентрально caput humeri, поэтому caput
при взгляде с проксимального конца выглядит более округлой. Участок каудальной
поверхности эпифиза между caput humeri и дорсальным краем fossa pneumotricipitalis
у A. soporata узкий, имеет меньшую площадь, дорсо-каудальная граница caput и дорсо-
каудальная граница fossa идут параллельно друг другу. У А. сгесса, A. discors и A. quer-
quedula эта площадка расширена, а края caput и fossa раскрываются в дорсо-
каудальном направлении под небольшим острым углом. У более крупных видов рода
Anas данная площадка еще более широкая.
Таким образом, представляется, что среднеплиоценовая монгольская Anas soporata
происходит из группы чирков, в которой находятся современные А. сгесса сгесса и
А. сгесса carolinensis и ископаемый A. bunkeri. Можно даже считать, что A. soporata и
A. bunkeri представляют общий элемент в среднеплиоценовой авифауне Восточной Азии
и Северной Америке.
Размеры, мм: дистальный эпифиз плеча — ширина 9,4 (№ 3222-22), 9,6
(№ 3222-674); передне-задний диаметр cond. dorsalis 6,3 (№ 3222-22), 6,5
(№ 3222-674); передне-задний диаметр cond. ventralis 2,8 (№ 3222-674); дорсо-вент-
ральная ширина диафиза 4,5 (№ 3222-673); передне-задняя ширина диафиза
3,3 (№ 3222-673), 3,4 (№ 3222-22); проксимальный эпифиз плеча (№ 3222-50) —ши-
рина 14, передне-задняя толщина caput humeri 4,7.
Локтевая кость A. soporata в дистальном эпифизе имеет специфически утоньшенный
tuberculum carpale, во всех остальных структурах она не отличается от современных
Anas (рис. 18, А).
Размеры, мм: ширина дистального эпифиза 6,2, диаметр cond. dorsalis 4,6.
Распространение. Местонахождения «точка 1080 м» в Шаргаин-Гоби,
Явор 1, Хиргис-Нур 2, Чоно-Хариах; нижняя и верхняя подсвиты свиты Хиргис-Нур.
Anas molesta sp. nov.
Рис. 19.
Видовое название molesta (лат.) —досадная.
Голотип — ПИН, № 3222-16; дорсальная половина правого коракоида;
местонахождение Хиргис-Нур 2; средний плиоцен, свита Хиргис-Нур.
Паратип — ПИН, № 3378-54; дорсальный эпифиз правого коракоида; МНР,
местонахождение Чоно-Хариах; средний плиоцен, свита Хиргис-Нур.
Материал. Кроме голотипа и паратипа имеются дистальный фрагмент левого
тарсометатарсуса № 3222-677 из местонахождения Хиргис-Нур 2; верхняя подсвита
свиты Хиргис-Нур; и проксимальная часть правой лопатки № 2737-389 из местонахож-
дения Дзагсо-Хаирхан; верхи среднего плиоцена.
Описание. Каудально-медиальная фасетка акрокоракоида резко разделена на
две равные разные части — выпуклую и выемчатую. Facies artic. clavicularis глубокая
и поэтому краниально-медиальный угол акрокоракоида утолщен и заострен. Эпифизар-
ная поверхность sulcus, m. supracoracoidei разделена на две части, на латеральной ее
половине развита глубокая ямка. Margo medialis утолщен и заострен.
Размеры, мм: длина дорсального эпифиза 12,7 (голотип); поперечная ширина
collum acrocoracoidei на уровне середины facies artic. humeralis 6,0 (голотип), 5,9 (па-
46
/7 Sw
I_________I_________I
/7 Лиг
Рис. 19. Anas molesta sp. nov.
Д —№ 2737-389, ПИН; правая лопатка; Монголия, Дзагсо-Хаирхан; средний плиоцен; а — латераль-
ная, б — медиальная стороны. Б — голотип № 3222-16, ПИН; правый коракоид; Монголия, Хиргис-Нур 2;
средний плиоцен; в — медиальная; г— каудальная; д— латеральная стороны
ратип); длина коракоида от его вершины до дорсального края facies artic. humera-
lis 7,5 (голотип), 7,8 (паратип); общая длина фрагмента 19,3 (голотип).
Сравнение. У современных Anas строение внутренней поверхности sulcus
m. supracoracoidei очень видоспецифично, у одних она ровная (A. clypeata), у дру-
гих — немного выемчатая по латеральной половине (A. poecilorhyncha, А. сгесса,
A. acuta, A. discors, A. penelope, A. falcata, A. platyrhynchos), у третьих — очень глубокая
выемка занимает всю поверхность эпифиза в области sulcus (A. formosa). У A. querque-
dula неглубокая выемка занимает только латеральную половину эпифизарной поверх-
ности sulcus. У A. molesta строение этой области эпифиза более всего сходно с таковой
у A. querquedula, но у ископаемого вида ямка — отчетливо глубже. Краниально-ме-
диальный угол акрокоракоида у A. molesta утоньшен и уплощен, как у A. acuta, A. cly-
peata, A. formosa. Margo medialis заострен подобно A. acuta, A. formosa, A. penelope,
A. americana. Этот угол не нависает сильно над margo medialis и сходен в данном
плане у A. molesta и у A. formosa.
По абсолютным размерам эпифиз коракоида A. molesta немного крупнее A.querque-
dula, A. formosa и близок к A. clypeata.
Коракоиды известны от нескольких ископаемых видов рода Anas. A. molesta отли-
чается от A. soporata большими размерами, утоньшенностью краниально-медиального
угла акрокоракоида и ямкой на латеральной половине внутренней поверхности эпи-
физа. От A. blanchardi Milne-Edwards, 1863, A. natator Milne-Edwards, 1868^ и A. velox
Milne-Edwards, 1868 из миоцена Западной Европы (Milne-Edwards, 1868; Svec, 1981)
отличаются более крупными размерами, утоньшенностью краниально-медиального угла
акрокоракоида, но сходны с ними присутствием ямки на латеральной половине внутрен-
ней поверхности эпифиза. A. integra A. Miller, 1944 из нижнего миоцена США — мень-
шего размера и с иными очертаниями акрокоракоида. Сравнение с A. luederitzensis
Lambrecht, 1929 и A.eppelsheimense Lambrecht, 1933 невозможно из-за плохого
качества иллюстраций или фрагментарности материала. От A. apscheronica Burchak-
Abramovich, 1958 из верхнего плиоцена Азербайджана отличается меньшими разме-
рами и уплощенностью акрокоракоида, хотя сходна с ней по наличию ямки на внутрен-
ней поверхности эпифиза.
Замечания. Кроме двух фрагментов коракоидов в материалах из Хиргис-
Нура 2 имеется фрагментарная половина тарсометатарсуса типичной Anas, которая
по абсолютным размерам подходит к утке размером с A. molesta. При общих размерах
близких к A. clypeata этот фрагмент тарсометатарсуса специфичен маленькими разме-
рами блока третьего пальца, которые сходны с размерами блока у А. сгесса.
Проксимальный фрагмент лопатки № 2737-389 (рис. 19, А) из Дзагсо-Хаирхан,
более крупный, чем у A. formosa, и близкий по размерам (немножко крупнее) к A. cly-
peata. Только на основании соответствия в размерах и определенной принадлежности
к уткам рода Anas, мы относим этот экземпляр к A. molesta.
Размеры, мм: высота эпифиза 10,0, длина fac. artic. humeralis 4,8 (у A. cly-
47
peata соответственно 9,4 и 4,5). Имеет заостренный acromion, как у А. сгесса, A. quer-
quedula, а не притупленный, как у A. clypeata и не округлый, как у A. formosa.
Распространение. Верхняя подсвита свиты Хиргис-Нур в местонахождении
Хиргис-Нур 2 (уровни 37—40); местонахождение Чоно-Хариах, пачка, Б; местонахож-
дение Дзагсо-Хаирхан, верхи среднего плиоцена.
Anas sp.
Рис. 20
Кроме двух новых мелких видов рода Anas, описанных из нижнего и среднего плио-
цена Котловины Больших Озер, в этом комплексе определенно был еще один крупный
вид рода, представленный шестью различными фрагментами костей крыла и плечевого
пояса в трех местонахождениях. Материалы по нему такие же фрагментарные, как и по
двум другим ископаемым видам Anas, но в данном случае мы не можем найти замет-
ных диагностических отличий от трех современных видов: Anas platyrhynchos, A. poeci-
lorhyncha и A. strepera. Несколько более мелкими размерами и некоторыми деталями
А —№ 3222-34, ПИН; левый коракоид; Монголия, Хиргис-Нур 2; средний плиоцен; а — латеральная;
б — медиальная стороны. Б — № 2614-118, ПИН; правый карпометакарпус; Монголия, Хиргис-Нур 2;
средний плиоцен; в — латеральная; г — каудальная; д — медиальная стороны
строения остатки этой ископаемой формы кажутся ближе к A. strepera, чем к двум дру-
гим видам. Ограничимся здесь только перечислением материалов с их размерами и
в двух случаях с рисунками.
Материал. Дорсальный эпифиз левого коракоида, ПИН, № 3222-34, Хиргис-
Нур 2, верхние уровни пачки А; свита Хиргис-Нур (рис. 20, А); размеры, мм: длина
эпифиза ~ 16, ширина facies artic. humeralis 6,2. Фрагмент дорсального эпифиза пра-
вого коракоида, ПИН, № 3222-21, Хиргис-Нур 2, верхние уровни пачки А; свита Хир-
гис-Нур; размеры, мм: ширина fac. artic. humeralis 6,2. Проксимальная половина пра-
вого карпометакарпуса, ПИН, № 2614-118, Хиргис-Нур 2, верхние уровни пачки А;
свита Хиргис-Нур (рис. 20, Б); размеры, мм: длина metacarpale alulare 11,3, высота
проксимального эпифиза 12,0, ширина trochlea carpalis 5,3. Проксимальный эпифиз
левой локтевой кости молодого экземпляра, ПИН, № 3378-47, Чоно-Хариах; верхняя
подсвита свиты Хиргис-Нур. Проксимальная половина правой лучевой кости, ПИН,
№ 3222-26, Хиргис-Нур 2, верхние уровни пачки А; свита Хиргис-Нур; размеры, мм:
ширина проксимального эпифиза 5,6 X 4,5. Симфизарная часть вилочки, ПИН,
№ 2614-103, Дзабхан, верхняя подсвита свиты Хиргис-Нур.
48
ПОДСЕМЕЙСТВО AYTHYINAE DELACOUR ET MAYR, 1945
Род Aythya Boie, 1822
Aythya spatiosa Kurochkin, 1976
Рис. 21; табл. VI, фиг. 1, 2
Aythya spatiosa: Курочкин, 1976, с. 62.
Голотип — ПИН, № 2614-81; дистальный эпифиз правого бедра с частью
стержня; МНР, Кобдосский аймак, местонахождение Чоно-Хариах; средний плиоцен,
свита Хиргис-Нур.
Материал. Только голотип.
Описание. Fossa poplitea с глубокой овальной ямкой. Cond. medialis и cond.
lateralis широко развернуты в стороны. След прикрепления сухожильной петли m. biceps
femoris отчетливо развит в виде короткого продольного валика. Проксимальнее его
развита полоска шероховатости немного меньшей длины, чем след петли m. biceps.
Рис. 21. Aythya spatiosa Kurochkin,
1976; голотип № 2614-81, ПИН; правое
бедро; Монголия, Чоно-Хариах; сред-
ний плиоцен
а — медиальная; б — каудальная; в —
латеральная; г — дистальная стороны
/он
I_____।_____I
Причем она идет не как ее продолжение, а сдвинута краниальнее. На середине план-
тарной поверхности дистальной части стержня хорошо выражен след отхождения сухо-
жилия m. perforatus d. IV. Он также существует как отдельная форма рельефа, а не
как продолжение медиальной границы fossa poplitea. Наружная поверхность cr. tibio-
fibularis и плантарная сторона cond. fibularis, ограничивающие с боков sulcus fibula-
ris, при взгляде с дистального конца образуют друг с другом прямой угол.
Размеры, мм: максимальная ширина эпифиза 9,0, передне-задняя толщина
межмыщелковой зоны 3,1, передне-задний диаметр cond. medialis 6,7, передне-задняя
толщина дистальной части стержня 4,3.
Сравнение. Из неогена Европы описано четыре вида рода Aythya (Brodkorb,
1964). Бедренная кость у них неизвестна. Среди современных представителей рода
Aythya spatiosa оказывается по особенностям устройства бедра ближе всего к A. ferina
и A. americana, хотя по размерам она одинакова с A. fuligula. С A. ferina ее сближают
рисунок шероховатостей на наружной поверхности дистальной части диафиза. Струк-
тура sulcus fibularis с прямым .углом между поверхностями ограничивающих ее
поверхностей мыщелков одинакова с A. ferina и A. collaris. У других Aythya этот угол
отчетливо острый. Расположение следа отхождения сухожилия m. perforatus d. IV
у A. spatiosa напоминает таковое у A. ferina.
Распространение. Свита Хиргис-Нур в местонахождении Чоно-Хариах.
Aythya magna Kurochkin, sp. nov.
Рис. 22; табл. VII, фиг. 3, 4
Видовое название magna (лат.) —большая.
Голотип — ПИН, № 3222-28; проксимальный эпифиз левого бедра; МНР,
Убсунурский аймак, местонахождение Хиргис-Нур 2; средний плиоцен, свита Хир-
гис-Нур.
4 Заказ 2098
49
Рис. 22. Aythya magna sp. nov.; голотип № 3222-28, ПИН; левое бедро; Монголия, Хиргис-Нур 2;
средний плиоцен
а — проксимальная; б — латеральная; в — каудальная; г — краниальная стороны
Материал. Только голотип.
Описание. Проксимальная суставная поверхность трохантера широкая, ромбо-
видная профиля, при взгляде с проксимального конца collum femoris утолщенная.
След прикрепления m. iliotrochantericus posterior опущен на краниально-латеральный
угол trochanter femoris.
Размеры, мм: передне-задний диаметр caput femoris 6,1, передне-задняя ши-
рина проксимального эпифиза 8,3, передне-задний диаметр collum femoris 5,0, попереч-
ная ширина проксимального эпифиза 12,0.
Сравнение. A. magna характеризуется крупными размерами и робустными
формами проксимального эпифиза бедра. У наиболее крупной современной чернети
A. marila по 8 экземплярам из коллекции Национального музея естественной истории
в Вашингтоне диаметр caput femoris варьирует от 5,6 до 6,1 (т = 6,0), поперечная
ширина проксимального эпифиза 10,0—11,4 (т=10,6), передне-задняя ширина про-
ксимального эпифиза 7,3—8,8 (т = 7,7). Следовательно, A. magna отличается от
A. marila иными пропорциями проксимального эпифиза — латеральная часть его
крупнее при тех же размерах caput femoris.
Проксимальная суставная поверхность facies artic. antitrochanterica у A. magna
широкая и имеет приближенно ромбовидную форму, у большинства других Aythya
она узкая овальная, только у A. ferina и A, valisineria она расширяется и становится
приближенно треугольной. Для всех современных Aythya характерна сравнительно
узкая collum femoris, а у ископаемой A. magna она широкая.
По-разному у них устроен передне-латеральный угол трохантера в зависимости
от места прикрепления m. iliotrochantericus posterior. У A. magna он как бы усечен
наискось с передне-латеральной стороны, в результате чего след прикрепления m. ilio-
troch. posterior как бы сдвинут проксимальнее и расположен на этой усеченной пло-
щадке. Сходно этот участок устроен у A. collaris; у других видов этого рода след при-
крепления указанного мускула расположен в основном на латеральной стороне по краю
эпифиза.
Распространение. Верхние уровни пачки А, свита Хиргис-Нур; место-
нахождение Хиргис-Нур 2.
ОТРЯД ACCIPITRIFORMES SAVIGNY, 1809
СЕМЕЙСТВО ACCIPITRIDAE SAVIGNY, 1809
Род Haliaeetus Savigny, 1809
Haliaeetus fortis Kurochkin, sp. nov.
Рис. 23; табл. II, фиг. 9, 10
Видовое название fortis (лат.) — крепкий.
Голотип—ПИН, № 3222-31; дорсальный эпифиз левого коракоида; МНР,
Убсунурский аймак, местонахождение Хиргис-Нур 2; средний плиоцен, свита Хир-
гис-Нур.
Материал. Кроме голотипа, имеются две фаланги пальцев правой лапы: первая
фаланга первого пальца № 3222-48 и четвертая фаланга IV пальца № 3222-47 из
слоев 51—55 пачки А местонахождения Хиргис-Нур 2.
50
/7 7см
I______I______I
Рис. 23. Haliaeetus fortis sp. nov.
A — голотип № 3222-31, ПИН; левый коракоид; Монголия, Хиргис-Нур 2; средний плиоцен; а — лате-
ральная; б — каудальная стороны. Б — № 3222-48, ПИН; первая фаланга первого пальца правой лапы;
Монголия, Хиргис-Нур 2; средний плиоцен; в—вентральная; г—латеральная; д—дорсальная стороны
Описание. Акрокоракоид в передне-заднем направлении сужен. Посредине
внутренней поверхности sulcus m. supracoracoidei проходит невысокий пологий, но
хорошо заметный, продольный гребень. Между этим гребнем и fac. artic. clavicularis
имеется неглубокая продольная впадинка. Вход в foramen pneumaticum расположен на
выравненном участке внутренней поверхности sulcus m. supracoracoidei и ориентирован
прямо поперек с внутренней стороны. Условная арка, образуемая последовательно
внутренними краями cotyla scapularis, fac. artic. humeralis и impres. lig. acrocora-
cohumeralis, проходит по пологой дуге большой окружности.
Размеры, мм: ширина fac. articularis humeralis 8,8, длина дорсального эпи-
физа ~29,0, ширина collum acrocoracoidei 17,5.
Сравнение. По карпометакарпусу из верхнего миоцена Франции описан един-
ственный ископаемый вид орланов Haliaeetus piscator Milne-Edwards, 1871. Мы срав-
нили Н. fortis со всеми современными видами рода (albicilla, vocifer, leucogaster,
leucocephalus, pelagicus, leucoryphus), за исключением двух островных видов с Соломо-
новых островов и с Мадагаскара. От всех современных видов отличается суженным
акрокоракоидом, но сходен в этом отношении с Н. leucoryphus и наличием пологого
гребня и продольной впадинки — каудальнее от fac. artic. clavicularis, — на поверх-
ности sulcus m. supracoracoideus. Такая впадинка едва заметно намечается только
у Н. albicilla. Область, окружающая foramen pneumaticum, и ориентировка плоскости
входного отверстия у Н. fortis не имеет себе подобных среди современных Haliaeetus,
у которых плоскость входного отверстия расположена всегда под углом к плоскости
эпифиза, т. е. вход в foramen идет не с медиальной, а больше с вентральной стороны.
Условная арка, ограничивающая с каудальной стороны sulcus m. supracoracoideus,
имеет у Н. fortis приближенно такую же пологую дугу окружности, как у Н. leucory-
phus. У остальных Haliaeetus радиус этой окружности меньше, несмотря на обычно
более крупные размеры коракоида.
Основные другие отличия Н. fortis от отдельных видов современных орланов
сводятся к следующему. От Н. leucogaster ископаемый вид отличается относительно
узкой collum acrocoracoidei при больших абсолютных размерах всего коракоида, чем
у самки этого вида. Также различается collum у Н. fortis и Н. leucocephalus при близ-
ких размерах ископаемого экземпляра и современного самца последнего вида. Н. pela-
gicus имеет очень расширенный в передне-заднем направлении акрокоракоид. У Н. leu-
coryphus дуга каудальной арки sulcus m. supracoracoidei имеет заметно меньшую
окружность, чем у Н. fortis, a foramen pneumaticum придвинуто вплотную к ргос. асго-
coracoideus. У Н. vocifer на наружной поверхности collum acrocoracoidei имеется
высокий бугорок, отсутствующий у Н. fortis и других орланов.
Замечания. Две фаланги пальцев лапы могут быть достаточно уверенно
отнесены к этому виду, потому что они определенно принадлежат, прежде всего,
4*
51
Haliaeetus, будучи весьма специфично устроены у представителей этого рода в отличие
от остальных хищных птиц (рис. 23). Они найдены в тех же слоях местонахождения
Хиргис-Нур 2, что и коракоид.
Размеры, мм: первая фаланга I пальца — длина 31,7, ширина проксимальногс
конца 15,8, ширина дистального конца 7,7; те же для четвертой фаланги IV пальца,
соответственно, 25,3, 6,6, 7,0.
Распространение. Верхние горизонты пачки А свиты Хиргис-Нур в местона-
хождении Хиргис-Нур 2.
ОТРЯД GALLIFORMES TEMMINCK, 1820
СЕМЕЙСТВО GALLIDAE ILLIGER, 1811
ПОДСЕМЕЙСТВО PHASIANINAE VIGORS, 1825
Род Alectoris Каир, 1829
Alectoris sp.
Рис. 24; табл. VI, фиг. 5—7
Alectoris sp.: Курочкин, 1976, с. 63.
Материал. ПИН, № 2614-83 (изображен), дистальный эпифиз правого тибио-
тарсуса; местонахождение Чоно-Хариах; средний плиоцен, свита Хиргис-Нур. ПИН,
№ 3378-67 (не изображен), фрагмент дистального эпифиза левого тарсометатарсуса;
местонахождение Чоно-Хариах 2; средний плиоцен, свита Хиргис-Нур.
Размеры, мм: тибиотарсус — наибольшая поперечная ширина эпифиза 8,8,
наибольшая ширина эпифиза 7,4, передне-задний диаметр cond. lateralis 7,7, передне-
задняя ширина эпифиза через incisura intercondylaris 5,0; тарсометатарсус — передне-
задний диаметр trochlea metatarsi tertii 4,5, поперечная ширина tr. metatarsi tertii 3,7,
передне-задний диаметр tr. metatarsi secundii 4,5.
Zav
-J
Рис. 24. Alectoris sp.; № 2614-83, ПИН; правый
тибиотарсус; Монголия, Чоно-Хариах; средний
плиоцен
а — латеральная; б — краниальная; в — дистальная
стороны
Сравнение. В морфологии данных фрагментов тибиотарсуса и тарсометатар-
суса фактически нет отличий от современного A. graeca. Принадлежность ископаемого
фрагмента тибиотарсуса к роду Alectoris и значительная его близость к современному
A. graeca подтверждается размерными отношениями (табл. 7), по которым Alecto-
ris sp. из Чоно-Хариаха оказывается гораздо ближе к Alectoris, чем к отдельным видам
других Phasianinae. Фрагментарность ископаемых экземпляров не позволяет провести
достоверное сравнение для их видовой идентификации.
В дистальном эпифизе тибиотарсуса сходство ископаемого Alectoris и современ-
ного A. graeca устанавливается следующими особенностями их морфологии, диагности-
чески важными в подсемействе Phasianinae. Форма обоих мыщелков почти одинакова
с краниальной стороны. Центральный бугорок на медиальной поверхности cond. media-
lis сдвинут от центра краниально. Одинаковы рельеф и строение выходного отверстия
canalis extensorius, pons supratendineus и incisura intercondylaris. Три небольших
отличительных черты в строении могут быть отнесены за счет индивидуальной изменчи-
вости, хотя она оказалась по ним довольно мала у 8 экз. A. graeca с Кавказа и Тянь-
Шаня. У ископаемого Alectoris sp. apophysis externa lig. obliqii имеет заостренную
вершинку, на латеральной стороне pons supratendineus отсутствует маленькое пита-
тельное отверстие, краниально-дистальный край окружности cond. lateralis заметно
скошен. У A. graeca apophysis притуплен у вершинки и расширен в основании, пита-
тельное отверстие имеется, а краниально-дистальный край cond. lateralis более округл,
52
Таблица 7
Абсолютные и относительные размеры тибиотарсуса Alectoris sp.
Вид Линейные размеры Пропорции
Наибольшая поперечная ширина эпифиза Передне-задний диаметр cond. lateralis Наименьшая ширина через incisura intercondylaris
а б в б/а в/а а/б • в
Alectoris sp. (№ 2614-83) 8,8 7,7 5,0 0,87 1 1 0,56 5,7
Alectoris graeca 9,0 7,9 5,2 0,87 0,57 5,9
Alectoris graeca 8,0 7,2 4,5 0,90 0,56 4,9
Syrmaticus reevesii 10,5 9,8 6,5 0,93 0,62 6,9
Lophura nycthy- mera 11,6 9,8 5,0 0,84 0,43 5,9
Phasianus colchicus 9,1 8,9 6,2 0,97 0,68 6,3
хотя также немного как бы срезан по хорде. В дистальном эпифизе тарсометатарсуса
для Alectoris характерны укороченная и округлая с боков форма tr. metatarsi tertii,
заостренный и направленный каудально processus на tr. metatarsi secundii.
Ранее по другим костям скелета были описаны только два неогеновых вида кек-
ликов — A. pliocaenica Tug. — из позднеплиоценовых отложений одесских катакомб и
A. bavarica Ball, из среднего миоцена Винтерсхофа в ФРГ.
Распространение. Местонахождения Чоно-Хариах и Чоно-Хариах 2; сред-
ний плиоцен, верхняя подсвита свиты Хиргис-Нур.
Род Bantamyx Kurochkin, 1982
Типовой вид — Bantamyx georgicus Kurochkin, 1982; средний плиоцен Запад-
ной Монголии.
Описание. Tuber brachialis коракоида немного раздвоен неглубокой выемкой
с медиальной стороны и почти не нависает над sulcus m. supracoracoidei. Sulcus m.
supracoracoidei широко раскрыта в медио-каудальном направлении. Facies art. humera-
lis имеет почти одинаковую ширину по всей длине фасетки. Плоскость impressio lig.
coracohumeralis обращена дорсально и латерально. На передней поверхности эпифиза
на уровне вентрального края facies art. humeralis имеется невысокий бугорок с тупой
вершиной. Labrum internum дорсально поднимается к proc, procoracoideus, а в начале
дорсальной трети диафиза образует небольшой гребень.
Видовой состав. Типовой вид.
Сравнение. Коракоид из Хиргис-Нура принадлежал мелкому представителю
подсемейства фазановых. Он относится к отдельному новому роду, поскольку особенности
строения дорсального конца его коракоида не согласуются в полном объеме ни с одним из
просмотренных современных Phasianinae. Из 47 существующих ныне родов нами были
исследованы скелеты 26 родов фазановых (Catreus, Hauppifer, Hierophasis, Lophura,
Crossoptilon, Gennaeus, Caloperdix, Alectoris, Ammoperdix, Melanoperdix, Francolinus,
Pternistis, Perdix, Perdicula, Arborophila, Galloperdix, Tetraogallus, Ithaginis, Chryso-
lophus, Lophophorus, Polyplectron, Pavo, Coturnix, Rollulus, Gallus, Phasianus, Syr-
maticus).
Среди них Bantamyx более всего сходен с Rollulus и Coturnix. С Rollulus его объ-
единяют одинаковая ширина fac. art. humeralis на всем протяжении фасетки, широкое
раскрытие sulcus supracoracoideus и обращенность ее поверхности также каудально,
а не только медиально, сходное прохождение labrum internum, резкий профиль sulcus
supracoracoideus с внутренней стороны эпифиза и диафиза, близкие очертания cotyla
scapularis. Отличается Bantamyx от Rollulus меньшими размерами фасетки акрокора-
коида в месте отхождения сухожилия m. biceps, относительной удаленностью этой
фасетки от краниального крыла fac. articularis humeralis (при вдвое меньших размерах
53
Рис. 25. Bantamyx georgicus Kurochkin, 1982;
голотип № 3222-52, ПИН; правый коракоид;
Монголия, Хиргис-Нур 2; средний плиоцен
а — медиальная; б — латеральная; в — каудальная
стороны
фасетка отставлена от крыла вдвое дальше
у Bantamyx, чем у Rollulus), профилем fac. аг-
ticularis clavicularis (у Bantamyx дорсальный
угол facies сдвинут от середины каудально,
а у Rollulus, наоборот, краниально). Coturnix
сходен с Bantamyx по общим очертаниям fac.
articularis clavicularis (дорсальный угол сдви-
нут от середины facies каудально, посере-
дине facies имеется маленькая ямка), неболь-
шим размером фасетки в месте отхождения сухожилия m. biceps, формой impressio lig.
coracohumeralis (незначительно расширяется в медиальном направлении). Отличия Ban-
tamyx от Coturnix наблюдаются в строении fac. art. humeralis (у Coturnix она расширя-
ется дорсально, а у Bantamyx — нет), impressio lig. coracohumeralis (глубоко-выемчатая
у Coturnix и плоская у Bantamyx), sulcus supracoracoideus, которая у Coturnix раскрыта
почти полностью внутрь, а у Bantamyx заметно развернута также каудально; labrum
internum у Bantamyx подходит к proc, procoracoideus, а у Coturnix ориентирована на
середину cotyla scapularis.
В ископаемом состоянии в неогене ни Coturnix, ни Rollulus неизвестны. Но из
нижнего плиоцена юга Украины (Одесские катакомбы) и верхнего миоцена Франции
(Ла-Грив-Сент-Альбан) описан род Plioperdix, а из верхнего миоцена того же Ла-
Грив-Сент-Альбан во Франции известна еще одна мелкая фазановая птица Palaeo-
cryptonyx. Полный коракоид Plioperdix неизвестен, но, насколько можно ориентиро-
ваться на его размеры, немного больше, чем у современного Coturnix, и частичное
структурное сходство с С. coturnix по другим костям скелета, не исключена возмож-
ность конгенеричности Plioperdix и Bantamyx. Неогеновый Palaeocryptonyx, который
объединил согласно ревизии Бальмана (Ballmann, 1969) некоторых Palaeortyx, Ра-
laeoperdix и Proalector, из миоцена Западной Европы, согласно первоописанию Депере
(Deperet, 1890), сходен в строении коракоида с современными Rollulus и Perdix. К со-
жалению, описание коракоида выполнено очень поверхностно у Депере и совсем не
дано у Бальмана. Однако, насколько можно сравнить изображение коракоида Palaeo-
cryptonyx donnezani (Deperet, 1890, табл. 13, фиг. 6, 6а) с таковым Bantamyx, они по
строению акрокоракоида и sulcus supracoracoideus представляются очень близкими.
Сходны у них и размеры.
Замечания. Первоначально Bantamyx был помещен нами в подсемейство
американских зубчатоклювых куропаток Odontophorinae (Курочкин, 1982). Позднее
появилась возможность сравнить Bantamyx со всем спектром современных родов и
с некоторыми ископаемыми представителями Odontophorinae в больших коллекциях
Национального музея естественной истории в Вашингтоне и Университета Флориды
в г. Гейнсвилл у профессора П. Бродкорба. Эта работа привела нас к заключению, что
коракоид из Хиргис-Нура 2 принадлежал все-таки представителю подсемейства Phasia-
ninae. При этом можно указать только единственный признак, различающий уверенно
коракоиды этих двух подсемейств. У Phasianinae плоскость impressio lig. coracohume-
ralis обращена вверх только дорсально, а его поверхность часто имеет глубокую
выемку, даже у самых мелких родов подсемейства. У Odontophorinae плоскость этой
impressio заметно наклонена немного в краниальном направлении и ее поверхность
у большинства представителей почти плоская.
54
Bantamyx georgicus Kurochkin, 1982
Рис. 25; табл. VII, фиг. 1—3
Голотип — ПИН, № 3222-52; дорсальная половина правого коракоида; место-
нахождение Хиргис-Нур 2; средний плиоцен, свита Хиргис-Нур.
Материал. Только голотип.
Описание. Поверхность facies lig. coracohumeralis плоская и facies незначи-
тельно расширяется в медиальную сторону. Фасетка в месте отхождения сухожилия
длинной головки гл. biceps на краниально-дорсальной поверхности акрокоракоида
Таблица 8
Размеры коракоида некоторых фазановых птиц
Показатель Bantamyx georgicus Palaeocryptonyx donnezani (с ри- сунка у Deperet, 1890) Coturnix coturnix Coturnix japonica Coturnix coroman- delica Coturnix delegorguei Rollulus roul. roul
n-1 n-1 n-2 n-2 n-2 n-5 n-1
Длина
дорсального эпифиза 0,73 0,85 0,71 0,65 0,63 0,71 0,57 0,61 0,63—0,65 1,00
акрокора- коида 0,46 0,48 0,42 0,40 0,39 0,49 0,39 0,42 0,42—0,47 0,62
Поперечная ши- рина диафиза 0,22 0,23 0,21 0,18 0,17 0,23 0,17 0,18 0,18—0,25 0,33
маленькая. На передней поверхности эпифиза на уровне вентрального края fac. arti-
cularis humeralis лежит небольшой бугорок с тупой вершинкой и широким основанием.
Размеры, мм: длина дорсального эпифиза 7,3, ширина fac. artic. humera-
lis 1,7, длина акрокоракоида (от вершины fac. artic. humeralis) 4,6, меньшая попереч-
ная ширина диафиза 2,2, общая длина фрагмента 15,5.
Замечания. Для сравнения целесообразно привести размеры некоторых
современных Phasianinae и Palaeocryptonyx donnezani (табл. 8).
Распространение. Свита Хиргис-Нур, верхние уровни пачки А местонахож-
дения Хиргис-Нур 2.
Род Syrmaticus Wagler, 1832
Syrmaticus kozlovae Kurochkin, sp. nov.
Рис. 26; табл. VII, фиг. 4
Видовое название — в честь Е. В. Козловой, выдающегося отечественного
орнитолога, много сил отдавшей изучению птиц Центральной Азии.
Г олотип — ПИН, № 3222-51; дистальный эпифиз левого плеча; местонахожде-
ние Хиргис-Нур 2; средний плиоцен, свита Хиргис-Нур.
Материал. Кроме голотипа, имеется проксимальный фрагмент правого кора-
коида ПИН № 2614-85 из местонахождения Чоно-Хариах. В одной из статей по этой
фауне (Курочкин, 1976) он был описан в составе подсемейства Phasianinae.
Описание. Ямки на вентральном надмыщелке плеча в местах отхождения про-
наторов лучевой кости (impressio'm. pronator superficialis и impres. m. pro. profun-
dus) тесно сближены друг с другом и с треугольной площадкой tuberculum supracon-
dylare ventrale (в месте отхождения ligamentum collaterale ventrale). Профиль pr. fle-
xoris с передней стороны представлен в виде узкого короткого треугольника. Плоскость
pr. flexoris расположена под прямым углом к плоскости эпифиза. Дистальный углуб-
ленный край fossa m. brachialis подходит вплотную к tuberculum sppracondylare
ventrale.
Размеры, мм: наибольшая ширина дистального эпифиза плеча 12,2, передне-
задний диаметр condylus radialis 7,6, передне-задний диаметр cond. ulnaris 4,8.
55
Рис. 26. Syrmaticus kozlovae sp. nov.
A —№ 2614-85, ПИН; правый коракоид; Монголия, Чоно-Хариах; средний плиоцен; а — медиальная;
б — краниальная; в — каудальная стороны. Б — голотип № 3222-51, ПИН; левое плечо; Хиргис-Нур 2;
средний плиоцен; г — вентральная; д — латеральная; е — медиальная; ж — дистальная стороны
Сравнение. В ископаемом состоянии род Syrmaticus неизвестен. В современной
фауне имеется пять видов, населяющих Японию, о-в Тайвань, Китай и Бирму. Все
они обитают в холмистых или слабогористых местностях, покрытых разреженной
растительностью. У нас имелась возможность сравнения только с тремя видами:
S. reevesii (Северный и Центральный Китай), S. soemmerringii (Япония), S. ellioti
(горы юго-восточного Китая). Отсутствовали в сравнении S. humiae (Южный Китай
и Бирма) и S. mikado (о-в Тайвань). S. kozlovae по структуре вентрального надмы-
щелка ближе всего оказался к S. soemmerringii, у них сходен профиль рг. flexoris
(укороченный и утолщенный) и одинаково ориентированы и сгруппированы ямки
в местах отхождения сухожилий пронаторов (поверхности их больше обращены вент-
рально). У S. reevesii профиль рг. flexoris удлиненный; у S. ellioti, наоборот, — укоро-
ченный и расширенный. Плоскость рг. flexoris образует у S. kozlovae с плоскостью
эпифиза прямой угол, как у S. soemmerringii, у двух других Syrmaticus угол между
рг. flexoris и поверхностью эпифиза тупой, если смотреть на эпифиз с дистального
конца. Ямки от сухожилий пронаторов больше открыты на переднюю сторону.
Замечания. Описанный ранее из Phasianinae gen. (Курочкин, 1976) фрагмент
коракоида № 2614-85 (рис. 26, А; табл. VII, фиг. 4) объединяется теперь вместе с типо-
вым фрагментом плеча и относится к одному виду S. kozlovae. Прежнее неполное опре-
деление было вызвано во многом недостатком современного сравнительного материала.
Теперь нам удалось провести сравнение по основным родам семейства Phasianidae,
перечень которых был дан при описании Bantamyx. Оба остатка при широком сравнении
показали удивительную близость к Syrmaticus и частично к Phasianus. Основанием для
отнесения обоих остатков к одному виду послужили также пропорции между линей-
ными размерами и между разными размерами каждого из фрагментов, сопоставленные
с таковыми некоторых современных Phasianinae (табл. 9). Из таблицы видно, что
пропорции между размерами плеча и коракоида S. kozlovae укладываются в пределы,
полученные для Syrmaticus, Lophura и Phasianus, и отличаются от пропорций для
Alectoris. Отношения между отдельными размерами плеча и между отдельными разме-
рами коракоида подтверждают видовую самостоятельность S. kozlovae.
Дистальные эпифизы плеча S. kozlovae и современных Syrmaticus сходны друг
с другом и отличаются от Phasianus суженной выемкой между cond. ulnaris и cond.
radialis, если смотреть на кость спереди, у Phasianus мыщелки спереди дальше отодви-
нуты один от другого. Impressio lig. collaterale ventrale имеет у Syramaticus меньшую
площадь, чем у Phasianus, и в отличие от последнего площадка impressio обращена
не только вперед, но и немного дистально. У Phasianus площадка impressio расположена
в плоскости эпифиза.
Дорсальный эпифиз коракоида Syrmaticus отличается от Phasianus большей мощ-
ностью наружного крыла facies art. humeralis, меньшей длиной этого крыла вдоль оси
56
Таблица 9
Абсолютные и относительные размеры плечевой кости и коракоида у Syrmaticus kozlovae
Вид
Линейные размеры
Пропорции
Плечевая кость Коракоид то
Наибольшая ширина дистального эпифиза Передне-задний диа- метр cond. radialis Передне-задний диа- метр cond. ulnaris Ширина fac. artic. humeralis Длина дорзального эпифиза Между размерами плечевой кости и коракоида Между разме- рами плечевой кости 1 Между размерами ко коида
ы б 4-М Ь- | а/д б/г в/г б/а в/а 1 г/д
Syrmaticus kozlovae 12,2 7,6 4,8 4,0 13,0 0,94 1,90 1,20 0,62 0,39 0,31
(№ 3222-51)
Syrmaticus reevesii 16,3 8,3 5,7 4,6 18„0 0,91 1,80 1,24 0,51 0,35 0,26
Lophura nycthymera 15,7 8,4 5,7 4,6 16,5 0,95 1,82 1,24 0,53 0,36 0,28
Phasianus colchicus 13,0 7,0 5,1 4,5 15,0 0,88 1,55 1,13 0,54 0,39 0,30
Alectoris graeca 12,1 6,3 4,5 5,3 14,5 0,83 1,19 0,85 0,52 0,37 0,36
Alectoris graeca 10,6 5,0 3,8 4,7 12,7 0,83 1,06 0,81 0,47 0,36 0,37
эпифиза. Сочленовная поверхность facies сужена в средней части у Syrmaticus и
расширена у Phasianus. При первом изучении этого фрагмента коракоида (Курочкин,
1976), было подчеркнуто сходство его с таковыми двух видов рода Lophura. Сейчас
можно считать это заключение ошибочным.
Размеры, мм: длина дорзального эпифиза 13,0, ширина fac. art. humeralis 4,0,
расстояние от вентрального края fac. art. humeralis до дорзального края impres. lig.
acrocoracohumeralis 10,0, наибольшая ширина collum acrocoracoidei 5,4.
Распространение. Средний плиоцен, свита Хиргис-Нур, слой с Unio и
рыбами (уровень 37) местонахождения Хиргис-Нур 2 и пачка Б местонахождения
Чоно-Хариах.
ОТРЯД RALLIFORMES REICHENBACH, 1849
ПОДОТРЯД GRUES BONAPARTE, 1854
СЕМЕЙСТВО GRUIDAE VIGORS, 1852
ПОДСЕМЕЙСТВО BALEARICINAE W. SCLATER, 1924
Род Probalearica Lambrecht, 1933
Probalearica: Lambrecht, 1933, с. 519; Brodkorb, 1967, с. 149;
Cracraft, 1973, с. 87
Типовой, вид — Grus problematicus Milne-Edwards, 1871; нижний миоцен
(аквитан): Франция, местонахождения Сан-Жеран-Ле-Пьи и Жанне.
Диагноз. Tuberositas epicondylaris lateralis развита довольно сильно. Apophysis
externa lig. obliqui короткий и тупой. Задняя поверхность межмыщелковой выемки при
взгляде с дистального конца слабо наклонена внутрь.
Видовой состав. В составе рода описано 4 вида. Типовой вид Р. problema-
tica — из аквитана двух местонахождений Франции. Р. crataegensis Brodkorb, 1963 —
из нижнего миоцена штата Флорида, США. Из верхнего миоцена Молдавии описан
Р. moldavica Kurochkin et Ganea, 1972. P. mongolica sp. nov. происходит из двух место-
нахождений среднего плиоцена Западной Монголии.
Сравнение. В подсемейство Balearicinae П. Бродкорб (Brodkorb, 1967) вклю-
чил вымершие роды Paragrus, Aletornis, Geranopsis, Pliogrus, Eobalearica и два других
рода — Paleogrus и Probalearica. Д. Крэкрафт (Cracraft, 1973) перевел Paragrus
в Geranoididae, Aletornis в Rallidae, a Geranopsis исключил из отряда вообще.
57
Pliogrus из нижнего плиоцена Европы отличается от Palaeogrus и Probalearica
передне-задней сжатостью дистального блока тибиотарсуса. Eobalearica, по нашему
мнению, при сравнении оригинала описания Е. tugarinovi конгенеричен с Palaeogrus
на основании отсутствия apophysis externa lig. obliqui и мелких размеров межблоковой
вырезки с дистальной и задней поверхностями.
Probalearica отличается от Palaeogrus наличием apophysis ext. lig. obliqui, который
у них в отличие от Grus — короткий и расширенный в основании, лучше развитой
tuberositas epicond. lateralis, суженной, углубленной и несколько наклоненной внутрь
задней поверхностью межблоковой зоны при взгляде с дистального конца. От совре-
менного Balearica отличия Probalearica заключаются в сведенности передних обоих
концов мыщелков к средней оси кости. В результате этого поперечная и передне-задняя
ширина эпифиза у Probalearica почти одинакова, а у Balearica поперечная больше
передне-задней. Короткий и широкий apophysis ext. lig. obliqui у Probalearica отличается
от такового Balearica удлиненным и узким в основании apophysis. Выходное отверстие
canalis extensorius у Probalearica более округлое, чем у Balearica.
Замечания. Род Probalearica был создан К. Ламбрехтом (Lambrecht, 1933)
для вида problematicus, описанного А. Мильн-Эдвардсом (Milne-Edwards, 1869—1871)
по верхней челюсти и грудине в составе рода Grus. На основании сходства с современ-
ными Balearica П. Бродкорб (Brodkorb, 1963) описал в составе Probalearica новый вид
балеарикоподобного журавля Р. crataegensis из нижнего миоцена США. Ревизуя всех
журавлеобразных, Д. Крэкрафт (Cracraft, 1973) оставил оба вида в роде Probalearica,
указав на настолько сильнее сходство этого рода с Palaeogrus, что он не мог определенно
отнести crataegensis к одному из них. Р. problematica и Р. crataegensis описаны по
несравнимым костям скелета. Поэтому в дальнейшем, если появятся дополнительные
более полные материалы по четырем видам Probalearica или возникнет возможность
сопоставления их через более полный экземпляр другого вида Probalearica, возможно
подтверждение их конгенеричности или создание нового родового названия для видов
crataegensis, moldavica и mongolica. Пока мы констатируем положение в составе
единого рода двух последних видов в подсемействе Balearicinae. Таким образом,
систематическое разнообразие (роды Palaeogrus, Probalearica и Pliogrus), широкое
географическое (Северная Америка, Европа, Азия) и геологическое (от среднего
эоцена до среднего плиоцена) распространение в прошлом различных Balearicinae
становится очевидным. Современный род Balearica выглядит на этом фоне реликтом
этого древнего подсемейства.
Probalearica mongolica Kurochkin, sp. nov.
Рис. 27; табл. VII, фиг. 5—8
Видовое название mongolica — монгольская.
Голотип — ПИН, № 3378-45; дистальный эпифиз левого тибиотарсуса; МНР,
местонахождение Чоно-Хариах; средний плиоцен, свита Хиргис-Нур.
Материал. В дополнение к голотипу имеется фрагментарный дистальный
эпифиз левого тарсометатарсуса, ПИН, № 3222-43, и левая квадратная кость, ПИН,
№ 3222-670, из местонахождения Хиргис-Нур 2.
Описание. Cond. medialis с боков сильно уплощен и сильно выдается кра-
ниально. Apophysis ext. lig. obliqui короткий. Весь эпифиз на уровне epicond. lateralis
и epicond. medialis в каудальной части заметно сжат с боков. Размеры крупные.
Размеры, мм: передне-задняя ширина cond. lateralis 27,4, передне-задняя
ширина cond. medialis 22,9, наибольшая поперечная ширина на уровне передних кон-
цов мыщелков 21,4, наименьшая поперечная ширина эпифиза с дистального конца 16,5.
Сравнение. Во всех основных характеристиках Р. mongolica сходна с Р. cra-
taegensis и Р. moldavica. Одинаковы у них общие пропорции дистального эпифиза
тибиотарсуса, одинаковы очертания межмыщелковой выемки при взглядах на эпифиз
спереди, с дистального конца и сзади. Tuberositas epicondylaris medialis довольно
сильно развита в виде продольного выступа, отклоненного дистальным концом кау-
дально. Tuberositas epicond. lateralis также хорошо развита и выступает из-за края
58
0 Zr/tf
& Zrtf
I----------------------------------------------------------------I
Рис. 27. Probalearica mongolica sp. nov.
4 — голотип № 3378-45, ПИН; левый тибиотарсус; Монголия, Чоно-Хариах; средний плиоцен; а— ме-
диальная; б—краниальная; в — латеральная; г — дистальная стороны. Б — № 3222-43, ПИН; левый
тарсометатарсус; Монголия, Хиргис-Нур 2; средний плиоцен; д— медиальная; е — краниальная; ж — кау-
дальная стороны
мыщелка при взгляде с дистального конца. Дистальный край cond. lateralis прямой,
а дистальный край cond. medialis с неглубокой выемкой.
Отличия Р. mongolica от двух других видов рода касаются особенностей строения,
указанных при описании нового вида. Cond. medialis латеро-медиально уплощен
у Р. mongolica сильнее, чем у Р. moldavica, и он толще в поперечном направлении
у Р. crataegensis, чем у двух предыдущих видов. Профиль incisura intercondylaris
со стороны cond. medialis при взгляде с дистального конца прямоугольный у Р. mongo-
lica и Р. moldavica и тупоугольный у Р. crataegensis. Apoph. externa lig. obliqui.
специфически укороченная у всех Probalearica, у отдельных видов рода развита в раз-
ной степени. У Р. mongolica apophysis короче, чем у двух других видов. Плантарная
часть эпифиза при взгляде с дистального конца у Р. mongolica и Р. moldavica выгля-
дит значительно суженной по сравнению с передней его кондилярной частью, но
у Р. crataegensis плантарная часть эпифиза шире. Размеры Р. mongolica заметно
крупнее двух других видов.
Замечания. Очень фрагментарный дистальный эпифиз тарсометатарсуса
№ 3222-43 из Хиргис-Нура 2, не имеющий даже блоков пальцев, был трудно диагносци-
руем. После того, как стало ясно, что он принадлежит Gruoidea, удалось установить,
что среди них он явно сближается с современным Balearica. Главные признаки, сбли-
жающие Probalearica и Balearica, в дистальном эпифизе тарсометатарсуса относятся
к блоку II пальца (рис. 27, Б). У Balearica в отличие от всех современных Grus и
Anthropoides tr. metatarsi secundii располагается на эпифизе более дистально и
вынесен на краниальную сторону так, что перекрывает плантарную половину tr. meta-
tarsi tertii и в значительной мере проксимальную половину этого блока с медиальной
стороны. Плантарное ребро основания trochlea dig. II у Balearica острое, а у Grus
округлое. Обе эти особенности, несмотря на фрагментарность, ясно видны у образца из
Хиргис-Нура 2. Кроме того, у Balearica расширена дистальная часть sulcus m. exten-
soris brevis dig. IV на передней поверхности диафиза перед входом в соответствующий
foramen, а у Grus и Anthropoides дистальное окончание sulcus сужено и углублено.
У ископаемого экземпляра мы видим строение sulcus, подобное Balearica. К сожале-
нию, дистальные части тарсометатарсуса неизвестны ни от ранее описанных Probalea-
rica, ни от Palaeogrus. На основании сходства ископаемого экземпляра № 3222-43
59
с современным Balearica и его подходящих относительных размеров с типовым тибиотар-
сусом Р. mongolica мы относим его к этому же виду.
Но фрагмент дистального эпифиза тарсометатарсуса Р. mongolica отличается от
такового Balearica немного большей видвинутостью trochlea dig. II вперед, большей
заостренностью основания этого блока с плантарной стороны, большей расширен-
ностью дистальной части sulcus m. ext. brevis dig. IV.
Квадратную кость № 3222-670 мы также относим к этому виду и роду, поскольку,
будучи построена несколько специфично, по особенностям строения суставной пло-
щадки для нижней челюсти она ближе к Balearica, чем к Grus (табл. VII, фиг. 5, 6).
Balearica pavonina, единственный современный вид подсемейства Balearicinae,
распространен ныне в тропической Африке от средней части долины Нила до Капской
провинции ЮАР. Этот журавль живет оседло и экологически связан с пойменными
лугами крупных рек и приозерных котловин, придерживаясь открытых ландшафтов и
не заходя в леса. Морфология его задних конечностей отвечает приспособлению к пере-
движению по подтопленным лугам и по спловинам, так как дистальная отодвинутость
блока II пальца и выдвинутость его вперед расширяют площадь опоры лапы; к тому же
пальцы у Balearica относительно длиннее, чем у Grus при абсолютно и относительно
более короткой цевке. Аналогичность строения дистального эпифиза тарсометатарсуса
у Probalearica могла иметь такой же адаптивный смысл, что в общем виде согласуется
с большой обводненностью Котловины Больших Озер в Монголии в среднем плиоцене.
Вопрос о филогенетических связах Probalearica и Balearica остается открытым, так
как пока для его решения явно недостаточно материалов. Важно при этом считать, что
в целом подсемейство Balearicinae представляется сейчас наиболее древним из всех
существующих подсемейств подотряда Grues, так как Palaeogrus известен по крайней
мере со среднего эоцена.
Распространение. Верхние горизонты (уровень 51—55) пачки А место-
нахождения Хиргис-Нур 2 и основание пачки Б местонахождения Чоно-Хариах, свита
Хиргис-Нур.
СЕМЕЙСТВО ERGILORNITHIDAE KOZLOVA, 1960
Род Amphipelargus Lydekker, 1891
Amphipelargus: Lydekker, 1891, с. 68; Brodkorb, 1963, с. 288.
Amphipelagus: Harrison, 1981, c. 111.
Urmiornis: Mecquenem, 1908, c. 87; Lambrecht, 1933, c. 521; Бурчак-Абрамович, 1951, c. 83;
Brodkorb, 1967, c. 154; Бендукидзе, 1972, c. 741; Cracraft, 1973, c. 74; Уманская, 1973, c. 31; Ку-
рочкин, 1981, c. 77; Harrison, Walker, 1982, c. 63.
Типовой вид — Urmiornis maraghanus Mecquenem, 1925; верхний миоцен;
Северный Иран.
Описание. Дистальный эпифиз тибиотарсуса повернут относительно диафиза
вокруг продольной оси медиально краниальной стороной, а каудальной — латерально.
Epicond. medialis расположен дистальнее по отношению к epicond. lateralis. Apoph.
fixatus очень мощный. Диафиз постепенно расширяется и плавно по бокам переходит
в более широкий эпифиз. Диафиз почти прямой, только немного выгнут в средней части
краниально. Проксимальный эпифиз тарсометатарсуса мощный, с широким и высоким
одноканальным гипотарсусом; латеральный край гипотарсуса у основания входного
канала выступает проксимально; cotyla medialis и cotyla lateralis расположены почти
на одном поперечном уровне. В дистальном эпифизе практически редуцирован tr. meta-
tarsi secundii, он сохраняется в виде крошечного бугорка; tr. metatarsi quartii укорочен,
доходит только до середины tr. metatarsi tertii, оба сохраняющихся блока пальцев
довольно тесно сближены. Фаланги третьего пальца лапы сильно уплощены дорсо-
вентрально, плантарная сторона их эпифизов в два раза шире дорсальной.
Видовой состав. A. maraghanus (Mecquenem, 1925, верхний миоцен Ирана;
A. majori Lydekker, 1891, верхний миоцен о-ва Самос, Греция; A. ukrainus (Kurochkin,
1981), верхний миоцен Северного Кавказа, Южной Украины и Молдавии, СССР;
A. cracrafti (Harrison et Walker, 1982), верхний миоцен Пакистана; A. oriental is (Kuroch-
kin, 1981), нижний—средний плиоцен Восточного Казахстана, СССР; A. dzabghanensis
sp. nov., средний плиоцен Монголии.
60
Сравнение. В семействе, кроме Amphipelargus, известен еще род Ergilornis
из раннего олигоцена Монголии. Тибиотарсус Amphipelargus отличается от такового
Ergilornis более плавным переходом от диафиза к эпифизу, более мощным apoph. fixatus,
расположенным посередине эпифиза, у Ergilornis apophysis сдвинут латеральнее; раз-
ворот эпифиза вокруг продольной оси диафиза у Ergilornis не выражен. Epicond.
medialis у Amphipelargus сдвинут дистальнее от уровня epicond. lateralis, а каудальные
ребра condylorum также разнесены на разные уровни — у cond. medialis ребро идет
проксимальнее, для Ergilornis характерно расположение обоих этих ребер на одном попе-
речном уровне. В тарсометатарсусе у Amphipelargus характерна прямизна и большая
мощность диафиза, который у Ergilornis выгнут заметной дугой краниально и относи-
тельно более тонок. Проксимальный эпифиз его у Amphipelargus более мощный, переходы
от диафиза к эпифизам по боковым сторонам без заметных углов, плавные; у основания
гипотарсуса с проксимального конца у латерального края Amphipelargus имеет возвы-
шение, которого совсем нет у Ergilornis. Cotyla medialis и cotyla lateralis, расположенные
у Amphipelargus почти на одном поперечном уровне, у Ergilornis сдвинуты друг отно-
сительно друга, плоскость cotyla lateralis располагается дистальнее. В тарсометатарсусе
tr. metatarsi quartii у Amphipelargus отодвинут заметно проксимальнее, чем у Ergilornis,
третий и четвертый блоки у первого более тесно сближены; tr. metatarsi secundii
у Ergilornis иногда сохраняется, хотя и сильно редуцирован. Фаланги третьего пальца
у Amphipelargus много сильнее уплощены дорсоветрально, чем у Ergilornis, и дорсальная
часть их эпифизов у последнего не намного уже, чем плантарная.
Замечания. Амфипеларгус был описан Меквенемом (Mecquenem, 1908, 1925) по
двум находкам из местонахождения Марата в Северном Иране. В дальнейшем в этот
род вошло несколько видов крупных двупалых птиц, как оказалось, достаточно широко
распространенных в Евразии от позднего миоцена до среднего плиоцена (см. видовой
состав). Возможно, Amphipelargus имеется и в Сиваликской фауне в Индии, так как там
найдены плоские крупные фаланги пальцев лапы, которые, скорее всего, принадлежали
этим птицам (Davies, 1880). Недавно Ц. Харрисон (Harrison, 1981) установил, что,
описанный по дистальному эпифизу тибиотарсуса, Amphipelargus Lydekker, 1891 с о-ва
Самос относится к урмиорнисам и, таким образом, становится старшим синонимом
Urmiornis Mecquenem, 1908. Сам Харрисон оставил окончательное решение вопроса
о приемлемости этого старшего синонима открытым, поскольку голотипом Urmiornis
взят тарсометатарсус, а позднее отнесенный Меквенемом к нему дистальный эпифиз
тибиотарсуса принадлежал другому, не типовому экземпляру (Mecquenem, 1925).
Поэтому требовались доказательства конгенеричности этих двух экземпляров. На
основе сравнения оригинальных их описаний и слепков с этих двух костей марагинской
птицы с нашими материалами по Amphipelargus из СССР и Монголии (Калмакпай и
Дзагсо-Хаирхан 4), в которых одновременно были собраны тарсометатарсусы и тибио-
тарсусы, можно уверенно говорить о правильности решения Р. Меквенема и о старшин-
стве названия Amphipelargus. Такая точка зрения уже принята в обзоре С. Олсона
(Olson, 1984) по ископаемым птицам.
Родственные связи Amphipelargus долгое время оставались неясными, несмотря
на то, что автор первоописания правильно отнес его к журавлеобразным. Позднее его
поочередно сближали с фламинго, аистовыми, журавлиными и двупалыми страусами
(Lambrecht, 1921, 1933; Бурчак-Абрамович, 1951; Дементьев, 1964; Уманская, 1973;
Brodkorb, 1971а). Некоторые не исключали возможности своеобразного уродства. Если
об его отношениях с первыми двумя группами вопрос совершенно ясен, сближать их
можно разве только на основании длинноногости тех и других, то сложнее вопрос стоит
об отношениях с настоящими страусами — Struthionidae. В последнее время уже на ос-
новании новых фактов по олигоценовым эргильорнисам и по развитию нелетности у птиц,
а также на основе логических доводов А. Федуччия (Feduccia, 1980) и С. Олсон (1984)
вновь подняли вопрос о родственных связях этих двупалых третичных птиц и страусооб-
разных, считая последних их неотеническими потомками.
До появления материалов по Ergilornis выявить родственные отношения Amphipe-
largus с другими птицами оказывалось трудно. Amphipelargus представлялся специали-
зированным неогеновым родом, стоящим как-то изолированно среди остальных птиц, и
61
действительно, единственное возможное сходство усматривалось у него (правда, на
основании общих черт морфологии его тарсометатарсуса, без рассмотрения деталей)
только с двупалым Struthio. Точно так же и позднее, когда Е. В. Козлова изучала остатки
двупалых раннеолигоценовых птиц из Монголии, она, вероятно, упустила из виду извест-
ных тогда урмиорнисов, посчитала эргильорнисов совершенно оригинальной группой и
создала для них отдельное семейство Ergilornithidae с неясным местом в системе
птиц (Козлова, 1960). После появления дополнительных материалов по эргильорнисам
была установлена их преемственность с урмиорнисами (Brodkorb, 1967; Курочкин,
1971), доказаны место семейства Ergilornithidae в отряде Gruiformes и вероятное
происхождение от эоцен-олигоценовых эогруид (Eogruidae) (Cracraft, 1973; Курочкин,
1974, 1981; Kurochkin, 1976). Eogruidae таксономически близки к Bathornithidae и
Geranoididae, эти три семейства древних палеогеновых журавлеобразных имеют много
общего, связаны общим происхождением. В палеогене они были широко распространены
в Америке и Азии, затем баторнисы и гераноиды вымерли, а эогрусы к началу олигоцена
дали эргильорнисов. Их потомки — амфипеларгусы — просуществовали до конца тре-
тичной эпохи.
Независимость и обособленность этой линии древних журавлеобразных никак не
связывается с происхождением от них Struthioniformes, одним из косвенных логических
доказательств которого А. Федуччия и С. Олсон считают внезапное появление в палеонто-
логической летописи двупалых страусовых в миоцене, отсутствие других возможных
предков страусовых в палеогене, сходные направления модификаций скелета у всех
птиц, утрачивающих способность к полету и возможность сохранения у страусовых при
происхождении их путем неотении палеогнатического типа строения неба, который якобы
имеют в эмбриональном состоянии все килевые птицы.
Прежде всего остатки бескилевых птиц найдены в отложениях Старого Света более
древних, чем миоцен. Из низов среднего эоцена Швейцарии давно описан по тазу
Eleutherornis helveticus Schaub, 1940, а из отложений этого же возраста из ГДР по
проксимальному концу тибиотарсуса недавно стал известен Saurornis matthesi Fischer,
1967. Этих двух птиц относят к Struthioniformes. Из позднего эоцена Египта (Эль-
Файюм) по дистальному эпифизу тибиотарсуса описан Eremopezus eocaenus Andrews,
1904, а из раннеолигоценовых слоев этого же местонахождения по дистальному трехбло-
ковому эпифизу тарсометатарсуса известна Stromeria fajumensis Lambrecht, 1929; их
включают в отряд Aepyornithiformes. Но эпиорнисы среди всех бескилевых по особен-
ностям строения скелета и скорлупы яиц ближе всего оказывается к Struthiornithiformes.
Следовательно, бескилевые птицы из отряда Struthiornithiformes и другие бескилевые,
близкие к ним, существовали в Старом Свете еще до появления в раннем олигоцене
Ergilornithidae. Отсутствие остатков страусообразных в отложениях среднего олиго-
цена—среднего миоцена, так же как и отсутствие в этом же интервале времени остатков
эргильорнитид, вполне объяснимо неполнотой палеонтологической летописи.
Если обратиться к скелету задних конечностей, то, несмотря на внешне сходную
объединяющую их двупалость, можно констатировать несколько принципиально разных
черт в их морфологии, разделяющих Ergilornithidae и Struthionidae, которые не могут
быть связаны с неотеническим недоразвитием последних. Так, у Ergilornithidae: 1) план-
тарный гребень диафиза тарсометатарсуса образован латеральным ребром диафиза, вне
связи с гипотарсусом; 2) этот гребень подходит к проксимальному эпифизу латеральнее
от гипотарсуса; 3) гипотарсус в дистальном направлении ни по медиальной стороне, ни
в середине гребня на диафиз не идет; 4) блок среднего пальца симметричен; 5) в прокси-
мальной суставной поверхности тарсометатарсуса eminentia intercondylaris значительно
выступает проксимально, выходя за уровень основания гипотарсуса; 6) в дисталь-
ном эпифизе тибиотарсуса с краниальной стороны имеется глубокая incisura intercon-
dylaris.
У Struthionidae: 1) плантарный гребень диафиза тарсометатарсуса образован ди-
стальным продолжением гипотарсуса и проходит больше по медиальной стороне; 2) по
латеральной стороне диафиза гребень к гипотарсусу не подходит, в средней части диа-
физа на латеральном его крае имеется короткий невысокий гребень; 3) в дистальном на-
правлении гипотарсус продолжается в виде гребня; 4) блок среднего пальца асимметри-
62
чен; 5) основание гипотарсуса выдается проксимальнее дальше, чем eminentia interglenoi-
dalis; 6) дистальный эпифиз тибиотарсуса не имеет глубокой incisura intercondylaris.
Палеогнатизм в строении черепа и принципиально иная конструкция таза отделяют
Struthioniformes от настоящих птиц (Carinatae). Череп и таз у Ergilornithidae неизве-
стны, но можно не сомневаться, что они были неогнатами, потому что, как показал
П. Хойд (Р. Houde, личное сообщение), не только череп, но и кости посткраниального
скелета в существенных деталях тоже отличаются у палеогнат и неогнат. Де Бир (De
Beer, 1956) указал, и на него ссылается Федуччия (1980), что все неогнатические птицы
проходят в эмбриогенезе палеогнатическую стадию в развитии черепа. Из эмбрио-
логических данных можно, наоборот, сослаться на работу Брума (Broom, 1906), где
показано, что задняя конечность Struthio проходит в эмбриогенезе пятипалую стадию, и,
таким образом, едва ли он происходит неотеническим путем от двупалых Ergilornithidae.
Amphipelargus произошел от Ergilornis, они связаны напрямую по линии предок—
потомок, хотя документированные доказательства их связи в интервале времени от
среднего олигоцена до среднего миоцена отсутствуют. Тем не менее детальная преем-
ственность в морфологии их задних конечностей не оставляет сомнений в прямом филоге-
незе по этой линии. Этот пример — один из немногих в эволюции птиц, когда на протя-
жении многих миллионов лет можно проследить конкретную филогенетическую линию.
Если принять, что предками эргильорнитид были эогруиды, то эта линия протягивается
приблизительно на 40 млн. лет. Нет оснований сомневаться и в принадлежности всех
Ergilornithidae к Gruiformes. На это указывают принципиальное сходство в строении
надмыщелковой зоны дистального эпифиза тибиотарсуса, специфическая конструкция
apoph. fixatus, структура краниальной поверхности проксимального эпифиза тарсомета-
тарсуса, локализация плантарного гребня диафиза тарсометатарсуса на его латеральной
стороне, удлиненная форма блоков дистального эпифиза тарсометатарсуса, не повторя-
ющаяся у каких-либо других птиц.
В позднем миоцене начался второй этап аридизации климата и ландшафта в Евразии.
Первый происходил, как известно, в раннем и среднем олигоцене. В конце миоцена по
всей южной части Палеарктики широко распространилась гиппарионовая фауна, освоив-
шая огромные пространства редколесий, степей, полупустынь и саванн (Дмитриева,
1977; Жегалло, 1978). Амфипеларгусы стали обычным компонентом этой фауны, входя
в ее состав вплоть до среднего плиоцена. Из отложений позднего миоцена (мэотис) они
известны с Северного Кавказа, Южной Украины, Молдавии, Ирана, Пакистана и с о-ва
Самос. В раннем плиоцене амфипеларгус найден в Восточном Казахстане (Павлодар,
Гусиный перелет), в среднем — в Восточном Казахстане (Калмакпай) и в Западной
Монголии (Чоно-Хариах, Дзагсо-Хаирхан 4 и верхние уровни Хиргис-Нур 2). Отчетливо
видно, что от позднего миоцена к среднему плиоцену размеры амфипеларгусов значи-
тельно увеличились. Наиболее поздние амфипеларгусы были крупными птицами, высо-
той в 1,5 м, с совершенно двупалыми задними конечностями, очевидно, совсем лишенные
способности летать, но хорошо бегающими. Надо полагать, что они были при этом
в значительной мере оседлы, совершая при необходимости локальные миграции. Амфи-
пеларгусы обитали одновременно и, возможно, совместно с двупалыми азиатскими
страусами. Можно предполагать, что экологические ниши их перекрывались, как в пита-
нии, так и в предпочитаемых местообитаниях, что явилось причиной вытеснения страу-
сами амфипеларгусов в среднем плиоцене, еще задолго до ледниковых похолоданий.
Причем сначала это произошло в западных частях ареала, так как в многочисленных и
хорошо изученных местонахождениях раннего и среднего плиоцена в Европе совсем не
встречаются остатки этих крупных птиц, а дольше всего они просуществовали в Цент-
ральной Азии (Kurochkin, 1982).
Лапа амфипеларгусов эволюционировала по направлению к однопалости. Об этом
говорит заметная редукция блока наружного IV пальца у среднеплиоценовых амфипе-
ларгусов по сравнению с позднемиоценовыми, маленькие размеры самого пальца и сим-
метричность блока среднего III пальца, сочетающаяся с увеличением его длины у пли-
оценовых амфипеларгусов. Это особенно интересно отметить в сравнении со Struthio,
у которых сохраняется полная опора на наружный палец, имеющий большую длину,
и асимметрия блока среднего пальца.
63
Amphipelargus dzabghanensis Kurochkin, sp. nov.
Рис. 28, 29; табл. VIII, фиг. 1—8
Amphipelargus sp.: Курочкин, 1981, с. 80.
Видовое название — от названия р. Дзабхан в Западной Монголии, в ни-
зовьях которой сосредоточены несколько местонахождений со среднеплиоценовыми пти-
цами.
Голотип — ПИН, № 2737-396; дистальная половина правого тибиотарсуса;
Кобдосский аймак, левый берег протоки Чоно-Хариах, местонахождение Чоно-Хариах 2;
средний плиоцен, свита Хиргис-Нур.
Паратип — ПИН, № 2737-395; дистальный эпифиз левого тибиотарсуса; Коб-
досский аймак, правобережье р. Дзабхан, местонахождение Дзагсо-Хаирхан 4; средний
плиоцен, свита Хиргис-Нур.
Материал. ПИН, № 2737-392, проксимальный эпифиз правого тарсометатарсуса
с частью диафиза; № 2737-393 и 394, два дистальных эпифиза соответственно левого и
правого тарсометатарсусов; все три из местонахождения Дзагсо-Хаирхан 4; № 3222-14,
третья фаланга III пальца лапы, местонахождение Хиргис-Нур 2 (описана Курочкиным,
1981, с. 80).
Описание. Epicond. medialis и apoph. fixatus очень мощные, каудальная половина
cond. medialis сильно выдается в дистальном направлении; передне-задний диаметр
эпифиза, измеренный через incisura intercondylaris, равен половине передне-заднего
диаметра cond. medialis.
Размеры, мм: поперечная ширина дистального эпифиза 33,6, передне-задний
диаметр cond. medialis 31,0, передне-задний диаметр cond. lateralis 28,2, диаметр через
incisura intercondylaris 15,2, общая длина фрагмента 225,0.
Сравнение. Хорошо отличается от A. maraghanus, A. majori, A. ukrainus и
A. orientalis мощно развитыми apoph. fixatus и особенно epicond. medialis. Но у мелкого
A. cracrafti оба этих бугра относительно такие же мощные, как и у A. dzabghanensis.
Также в отличие от первых четырех видов каудальная половина cond. medialis сильно
отставлена в проксимальном направлении. Диаметр через incisura intercondylaris
равен половине передне-заднего диаметра cond. medialis, как у A. ukrainus, это отноше-
ние меньше ’/г У A. orientalis и больше 1/2 у A. maraghanus. В отличие от A. orientalis
краниальный выход из sulcus intercondylaris имеет раскрытую краниально форму
вырезки, у A. orientalis внутренние выступы мыщелков замыкают вырезку с краниальной
стороны, так что ширина sulcus на выходе из нее меньше ширины вырезки в средней
части; A. maraghanus, A. cracrafti и A. ukrainus имеют такую же раскрытую краниально
форму sulcus intercondylaris.
По размерам A. dzabghanensis заметно больше всех других известных амфипеларгу-
сов, ширина дистального эпифиза тибиотарсуса у него 33,6 мм, диаметр cond. medialis
31,0. Соответствующие размеры у A. maraghanus — 26,7 и 24,0, у A. ukrainus — 23,4
(и 26,5) и 25,5, у A. orientalis — 25,5 и 26,0, у A. cracrafti — 18,9 и 17,6. Из такого сопо-
ставления выводится также, что у A. dzabghanensis, A. cracrafti и A. maraghanus попе-
речная ширина дистального эпифиза больше диаметра его внутреннего мыщелка,
а у A. ukrainus и A. orientalis, наоборот, меньше.
Замечания. Паратип № 2737-395 представляет собой сильно разрушенный
дистальный эпифиз левого тибиотарсуса из местонахождения Дзагсо-Хаирхан с диамет-
ром cond. medialis 30,5 мм и диаметром через incisura intercondylaris 16,3 мм, таким
образом, он очень близок по размеру к голотипу. Морфологических отличий между
ними нет.
Как указано в рубрике о материалах, в коллекции имеются еще три фрагмента
тарсометатарсусов и одна фаланга среднего пальца лапы. Для проксимального эпифиза
тарсометатарсуса можно отметить следующие отличия A. dzabghanensis от трех других
видов этого pojja, где эти кости известны. При более крупных размерах переход по бо-
ковым сторонам диафиза на проксимальный эпифиз у A. dzabghanensis более плавный,
чем у A. orientalis и A. ukrainus; у A. maraghanus эти линии представляют собой хорошо
выраженный излом, образующий тупой угол как с медиальной, так и с латеральной
64
Рис. 28. Amphipelargus dzabghanensis sp. nov.; голотип № 2737-396, ПИН; правый тибиотарсус;
Монголия, Чоно-Хариах 2; средний плиоцен
а — латеральная; б — краниальная; в — медиальная; г — дистальная стороны
сторон. Различается у амфипеларгусов и форма сочленовных впадин этого эпифиза.
A. dzabghanensis имеет приближенно округлые cotyla medialis и с. lateralis, как и
A. orientals и A. ukrainus, но у A. maraghanus cotyla lateralis шире по поперечному
диаметру, чем по краниально-каудальному. Еще более существенны и важны различия
в устройстве дистального эпифиза тарсометатарсуса, понятные и с функциональных
позиций. У A. dzabghanensis tr. metatarsi tertii удлиненный, с широким основанием,
с параллельно идущими боковыми поверхностями. У A. maraghanus и A. orientals
блок II пальца короткий, с широким основанием и несколько расширяющийся дистально.
У A. ukrainus блок удлиненный, имеет суженное основание, немного расширяется
дистально. Tr. metatarsi quartii у A. dzabghanensis приближен к tr. metatarsi tertii, они
идут параллельно друг другу, дистальный край tr. metatarsi quartii не доходит до услов-
5 Заказ 2098
65
/7 Zoff
U 1 I
Рис. 29. Amphipeiargus dzabghanensis sp. nov.; № 2737-393, ПИН; левый тарсометатарсус;
Монголия, Дзагсо-Хаирхан 4; средний плиоцен
а — каудальная; б — краниальная; в — латеральная; г — дистальная стороны
ного центра tr. metatarsi tertii по латеральной стороне. У трех других видов амфипелар-
гусов продольные оси блоков III и IV пальцев сходятся подострым углом. У A. maragha-
nus и A. ukrainus блоки сближены, а у A. orientalis tr. metatarsi quartii заметно отставлен
латерально от tr. metatarsi tertii, причем у A. maraghanus и A. ukrainus дистальный край
tr. metatarsi quartii достигает условный центр tr. metatarsi tertii, а у A. orientalis
заходит даже дистально дальше этого центра. Особенности строения блоков пальцев
показывают, следовательно, что у A. dzabghanensis по сравнению с другими амфипелар-
гусами произошло дальнейшее обособление среднего пальца, усиление на него опоры,
а вместе с этим — некоторая редукция наружного IV пальца с уменьшением его опорного
значения, выразившееся в приближении блока этого пальца к блоку среднего пальца
и в отодвинутости блока IV пальца проксимально.
Распространение. Известен из трех местонахождений Котловины Больших
озер: Чоно-Хариах, Дзагсо-Хаирхан 4 и Хиргис-Нур 2; средний плиоцен, верхняя под-
свита свиты Хиргис-Нур.
ПОДОТРЯД RALLI REICHENBACH, 1849
СЕМЕЙСТВО RALLIDAE VIGORS, 1852
ПОДСЕМЕЙСТВО RALLINAE VIGORS, 1852
Palaeoaramides Lambrecht, 1933
Palaeoaramides tugarinovi Kurochkin, 1980
Рис. 30; табл. IX, фиг. 3—7
Голотип — ПИН, № 2614-121; дистальный фрагмент правой плечевой кости;
МНР, местонахождение «точка 1080 м> в Шаргаин-Гоби; средний плиоцен, свита Хир-
гис-Нур.
Материалы. Кроме голотипа имеются дорсальная половина правого коракоида
(ПИН, № 3222-55) и дистальный эпифиз левого тибиотарсуса (ПИН, № 3222-672) соот-
ветственно из нижнеплиоценовых и среднеплиоценовых отложений местонахождения
Хиргис-Нур 2.
Описание. Epicond. ventralis сильно выдается в дистальном направлении, ргос.
flexorius этого epicondylus вытянут и вершина его относительно острая. Proc, supracon-
dylaris dorsalis обычных для близких видов Rallinae размеров, но хорошо выделен
в рельефе передней поверхности эпифиза в виде поперечной ступеньки. Sulcus m. scapu-
lotricipitis имеет небольшую глубину, определяемую степенью развития epicond. dorsalis,
который здесь слабо развит. С дистальной стороны cond. ventralis и proc, flexorius
разделены проксимально углубленной выемкой.
Размеры, мм: наибольшая глубина дистального эпифиза 5,5, передне-задний
диаметр cond. dorsalis 3,3, передне-задний диаметр cond. ventralis 1,9, расстояние от
вершины impres. ligamenti collaterale ventrale до конца proc, flexorius 3,2, наименьшая
передне-задняя толщина дистальной части диафиза 2,1.
66
Рис. 30. Palaeoaramides tugarinovi Kur., 1980
A — № 3222-55, ПИН; правый коракоид; Монголия. Хиргис-Нур 2; нижний плиоцен; а— медиальная;
б — краниальная, « — каудальная стороны; 1 — ioranivii и. supracoracoidei; 2—рг. procoracoideus, 3 —
fac. artic. humeralis, 4 — cotyla scapularis. Б — голотип № 2614-121, ПИН; правое плечо; Монголия,
«точка 1080 м» в Шаргаин-Гоби; г — дорсальная; д — краниальная; е — вентральная; ж — дистальная
стороны; / — incisura intercondylaris; 2 — tuberculum supracondylare ventrale; 3 — рг. supracondylaris
dorsalis
Сравнение. Четыре вида рода Palaeoaramides известны из аквитана и позднего
миоцена Европы (Olson, 1977). Три из них описаны и сравнены по тибиотарсусам и
тарсометатарсусам. Только от Р. beaumontii (Milne-Edwards, 1869—1871) из верхнего
миоцена (гельвет) Франции (местонахождение Сансан в департаменте Жер) известна
плечевая кость. Ее рисунки приведены у Мильн-Эдвардса (Milne-Edwards, 1869—1871)
и стереофотографии у Крэкрафта (Cracraft, 1973). Стереофотографии существенно
помогли определению и сравнению плечевого фрагмента из Шаргаин-Гоби. Р. tugarinivi
и Р. beaumontii очень сходны по форме и взаимному расположению обоих мыщел-
ков дистального эпифиза плеча, по структуре proc, flexoris, характерно вытянутому
дистально и выступающему на внутреннюю поверхность. Одинакова у них impressio
lig. collaterale ventrale — овальная по форме, высоко поднимающаяся наружу и ориенти-
рованная своей плоскостью наружу и дорзально. Одинаковы форма и размер fossa
m. brachialis; cond. ventralis и proc, flexorius у обоих разделены хорошо заметным углуб-
лением, эта деталь структуры очень характерна для всего рода.
Различия выделены в диагнозе. Sulcus m. scapulotricipitis у Р. tugarinovi заметно
мельче, чем у Р. beaumontii. Сильнее развит у Р. tugarinovi по сравнению с Р. beaumontii,
рг. supracondylaris dorsalis, но, наоборот, epicond. dorsalis у нового вида слабее. Размеры
Р. tugarinovi мельче. Для Р. beaumontii наибольшая ширина дистального эпифиза 6,2,
передне-задний диаметр cond. radialis 3,6, передне-задний диаметр cond. ulnaris 1,9
(по Cracraft, 1973).
Замечания. Из нижних уровней (20—21) местонахождения Хиргис-Нур 2
имеются дорсальный фрагмент правого коракоида № 3222-55 (рис. 30, табл. IX) и
дистальный фрагмент левого тибиотарсуса № 3222-672 с уровня 40—43 (табл. IX).
По абсолютным размерам они могут принадлежать птице размером с Р. tugarinovi,
а структурные особенности строения заставляют отнести их в дополнительные материалы
по этому виду.
Коракоид. У четырех известных ископаемых видов Palaeoaramides коракоид неиз-
вестен. Размеры ископаемого экземпляра № 3222-55 в мм: поперечная ширина диафиза
2,4, длина дорзального эпифиза (от вентрального края cotyla scapularis) 5,6, ширина
fac. articularis humeralis 2,4. Размеры коракоида, так же как и размеры плеча лежат
между таковыми Rallus aquaticus и R. longirostris, будучи немного ближе к R. aquaticus.
Среди современных пастушковых сравнение проведено с видами родов Aramides
(4 вида), Rallus (7 видов), Limnocorax, Sarothrura, Micropygia, Coturnicops, Porzana
(4 вида), Neocrex Amaurornis, Pardirallus, Poliolimnas, Gallinula. Среди них наиболь-
шее сходство прослеживается с Aramides и частично с Rallus. Palaeoaramides сходен
с Aramides заостренной формой вершины акрокоракоида, уплощенной формой facies
72 5* 67
artic. clavicularis, которая имеет посередине выступающий дистально выступ, расширен-
ным строением уплощенного медиального края диафиза, подходящего к ргос. ргосога-
coideus, щелевидным строением foramen supracoracoideus и выемчатой задней поверх-
ностью диафиза. Различия между ними вызваны, как нам кажется, прежде всего более
крупными размерами Aramides, отчего у него proc, procoracoideus заметно более мощный,
a fac. artic. clavicularis выставлена медиально при одинаковых у них деталях строения
этих структур. Строение fac. artic. clavicularis, proc, procoracoideus, fac. artic. humeralis
передней стороны акрокоракоида хотя и отличается в мелких деталях у Palaeoaramides
от Rallus, но все-таки более схоже у этих двух родов по сравнению с другими современ-
ными Rallinae.
Тибиотарсус описан для трех из четырех видов рода Palaeoaramides. Экземпляр
№ 3222-672 из Хиргис-Нура 2 (табл. IX), подходя по размерам к Р. tugarinovi (передне-
задний диаметр cond. lateralis 4,8, передне-задний диаметр cond. medialis 4,9, поперечная
ширина между передними концами мыщелков 4,8, передне-задний диаметр через
incisura intercondylaris 3,2), согласуется по всем деталям строения, приводимым
Крэкрафтом (Cracraft, 1973) в родовом диагнозе Palaeoaramides. Cond. lateralis
округлый и приподнятый сзади. Cond. medialis немного заходит вперед за уровень cond.
lateralis. Бугорок на pons supratendineus слабо развит или отсутствует. Впрочем, эти
признаки подходят к дистальному эпифизу тибиотарсуса таких современных родов,
как Rallus и Gallinula, но Aramides, например, имеет cond. lateralis, наоборот, припод-
нятый спереди и хороший бугорок на pons supratendineus. У Crex cond. lateralis также
приподнят спереди и в проекции с дистального конца оба мыщелка почти параллельны
друг другу.
От других видов ископаемых Palaeoaramides отличается меньшими размерами.
Поперечная ширина через передние концы мыщелков у Р. chrystyi — 6,7 и 7,8, Р. beau-
montii — 5,9 и 4,9, Р. lungi — 6,3, Р. tugarinovi — 4,8.
Из верхнего миоцена Молдавии (среднесарматское местонахождение у с. Бужоры)
был описан пастушок Tertiariaporphyrula lungi (Курочкин, Ганя, 1972). При этом ука-
зывалось большое сходство с Rallus beaumonti из верхнего миоцена Франции, на основа-
нии чего beaumonti был включен в род Tertiariaporphyrula. Но Крэкрафт (Cracraft, 1973)
отнес beaumonti в род Palaeoaramides, а вслед за ним на этом основании Олсон (Olson,
1977) включил и lungi в род Palaeoaramides.
Крэкрафт (Cracraft, 1973) указал на общее структурное сходство Palaeoaramides
с современным Rallus и, как мы показали по строению плечевой кости и коракоида, он
очень сходен также с современным Aramides. Действительно, Palaeoaramides ближе
всего к этим двум родам среди современных пастушков по общим пропорциям и деталям
структуры дорсальной части коракоида и дистального эпифиза плеча. Но по строению
дистальной части тибиотарсуса Palaeoaramides, сохраняя много общего с Rallus, сильно
отличается от Aramides.
Экологический облик Р. tugarinovi точно установить трудно. Если ориентироваться на
Aramides, обитающих в болотистых лесах Южной Америки, и на Rallus, связанных во
многом с затопленными кустарниковыми и древесными зарослями в Америке и в Евразии,
то можно было бы приписать и Palaeoaramides подобие такого образа жизни, хотя это
будет откровенная спекуляция на структурном сходстве при неясных родственных
связях. Во всяком случае наличие Palaeoaramides в среднем плиоцене Западной Монго-
лии вместе с двумя другими видами Rallinae в этой фауне указывает на большее разно-
образие и обводненность биотопов в этом регионе в то время, и на более богатое таксоно-
мическое разнообразие Rallinae, по сравнению с сегодняшним состоянием здесь ланд-
шафта и фауны пастушковых.
Распространение. Нижний плиоцен, нижняя подсвита свиты Хиргис-Нур
в местонахождении Хиргис-Нур 2 (уровень 20—21) ; средний плиоцен местонахождения
Хиргис-Нур 2 (уровень 40—43) и местонахождения «точка 1080 м> в Шаргаин-Гоби.
68
Род Crex Linnaeus, 1758
Crex zazhigini Kurochkin, 1980
Рис. 31; табл. IX, фиг. 1, 2
Crex zazhigini: Kurochkin, 1980.
Голотип — ПИН, № 2614-90; дистальный эпифиз левого плеча; МНР, Кобдоский
аймак, местонахождение Чоно-Хариах; средний плиоцен, свита Хиргис-Нур.
Материал. Только голотип.
Описание. Fossa m. brachialis глубокая и широкая. Epicond. dorsalis укорочен.
Epicond. ventralis расширен и выдвинут наружу. Рг. supracondylaris dorsalis тупой и
широкий, структурная полоска вентральнее этого processus занимает около ]/3 ширины
дистальной части диафиза.
Рис. 31. Crex zazhigini Kurochkin, 1980;
голотип № 2614-90, ПИН; левое плечо;
Монголия, Чоно-Хариах; средний плиоцен
а — вентральная; б — краниальная; в —
дорсальная; г — дистальная стороны; 1 — рг.
flexorius; 2 — epicond. dorsalis; 3 — рг. epicon-
dylaris ventralis; 4 — pr. epicondylaris dorsalis
Размеры, мм: наибольшая поперечная ширина эпифиза 6,2, передне-задний
диаметр cond. dorsalis 3,1, то же для cond. ventralis 1,8.
Сравнение. Виды рода Сгех из неогеновых отложений неизвестны. Мы сравнили
ископаемого представителя с современным С. сгех, от которого он отличается несколько
большими размерами. Наибольшая поперечная ширина дистального эпифиза плеча
у С. сгех 5,3—5,5 мм. Отличия в морфологии дистального эпифиза плеча между ними
незначительны. Fossa m. brachialis у С. zazhigini несколько глубже и имеет более отчет-
ливые границы, epicond. dorsalis короче и шире, epicond. ventralis, наоборот, длиннее.
Структурная полоска проксимальнее на краниальной стороне эпифиза вентральнее от
основания pr. supracond. dorsalis у ископаемого вида заметно более отчетливая и длин-
ная, чем у С. сгех. Отчетливы различия в строении pr. supracond. dorsalis, который у С. za-
zhigini расширен в основании и притуплен у вершины, а у С. сгех заострен у вершины.
Замечания. Сравнение с большим набором современных пастушковых подтвер-
дило принадлежность С. zazhigini к роду Сгех. Кроме родов, указанных при описании
Р. risilla, этот ископаемый экземпляр сравнивался с Limnocorax, Neocrex, Atlantisia,
Micropygia. Среди них некоторое сходство прослеживается с Neocrex.
Распространение. Местонахождение Чоно-Хариах; верхняя подсвита свиты
Хиргис-Нур.
Род Porzana Vieillot, 1816
Porzana risilla (Kurochkin, 1980)
Рис. 32; табл. IX, фиг. 8, 9
Rallus risilius: Kurochkin, 1980, c. 72.
Голотип — ПИН, № 2614-100; проксимальная часть левого карпометакарпуса;
МНР, Кобдосский аймак, местонахождение «высота 1080 м» в Шаргаин-Гоби; средний
плиоцен, свита Хиргис-Нур.
69
Z7
I----------------------------------1
Рис. 32. Porzana risilla (Kurochkin, 1980); голотип № 2614-100, ПИН; левый карпометакарпус;
Монголия, «точка 1080 м» в Шаргаин-Гоби; средний плиоцен
а — проксимальная; б — медиальная; в — каудальная стороны; 1 — facies artic. dig. alulae;
2— fossa infratrochlearis; 3 — fovea carpalis caudalis
Материал. Только голотип.
Описание. Proc, pisiformis маленький, узкий и под углом 45° выступает вент-
рально. Этот processus расположен на одной продольной оси с кранио-проксимальным
углом вентральной половины trochlea carpalis. Передняя сторона проксимального
участка os metacarpale majus полого поднимается к facies artic. dig. alulae. Дорсо-вент-
ральный профиль дорсальной половины trochlea carpalis удлиненный и суженный в про-
дольном направлении. Размеры мелкие.
Размеры, мм: поперечная ширина trochlea carpalis 1,5, кранио-каудальная ши-
рина tr. carpalis вместе с proc, extensorius 4,0.
Сравнение. 5 известных ископаемых видов рода Porzana (Р. guti, Р. auffen-
bergi, Р. lacustris, Р. insignis, Р. avita) описаны из верхнего плиоцена, нижнего и сред-
него плейстоцена Северной Америки преимущественно по плечевым костям и тибиотар-
сусам. Однако все они крупнее Р. risilla. Среди 13 современных видов рода мы сравни-
вали новый плиоценовый вид с Р. porzana, Р. pusilia, Р. Carolina, Р. flaviventer. Среди
них Р. risilla более всего сходна с Р. pusilia, но отличается от последней тем, что pr. pi-
siformis сильнее отставлен в сторону, т. е. угол между отростком и os metacarpale majus
у Р. pusilia острый, меньше 45°. Также у Р. pusilia нет плавного приподнятого подъема от
os metacarpale majus к fac. artic. dig. alulae. Pr. pisiformis широкий в основании и за-
остренный у вершины у Р. porzana, он весь расширен и направлен вершиной немного
проксимально у Р. flaviventer и Р. Carolina. В отличие от всех сравниваемых современных
Porzana, у Р. risilla pr. pisiformis расположен на одной продольной оси с кранио-прок-
симальным углом вентральной половины tr. carpalis. У современных Porzana он сдвинут
от продольной оси с углом trochlea каудально, или можно сказать вернее, что угол troch-
lea перемещен краниально. Также Р. risilla отличается от современных Porzana удлинен-
ным профилем дорсальной половины tr. carpalis, который у них заметно более расширен-
ный, т. е. каудальная дуга окружности trochlea более выпуклая.
Замечания. В первоописании этого вида (Kurochkin, 1980) он был помещен
в роде Rallus, но после сравнения оригинала с широким спектром современных родов
[были просмотрены Coturnicops, Amaurornis, Rallus, Porzana (сходство с этими первыми
четырьмя родами наибольшее), Canirallus, Nesophylax, Poliolimnas, Sarothrura, Lateral-
lus, Rallina, Gallicrex, Gallirallus, Himantornis, Porphyriops, Tribonyx, Gallinula, уда-
лось установить более вероятную принадлежность risilla к роду Porzana. У Porzana
и Amaurornis pr. pisiformis заострен и имеет узкое основание, профиль вентральной
половины tr. carpalis более вытянут. У Rallus pr. pisiformis расширен в основании
и у вершины, а вентральная половина tr. carpalis округлая. У Coturnicops в отличие от
этих трех родов проксимальный конец дорсальной половины tr. carpalis сильно отве-
ден дорсально. У Amaurornis в отличие от Porzana fossa supratrochlearis двухямковая.
Распространение. Верхняя подсвита свиты Хиргис-Нур в местонахождении
«точка 1080 м> в Шаргаин-Гоби.
70
ОТРЯД CHARADRIIFORMES HUXLEY, 1867
ПОДОТРЯД CHARADRII HUXLEY, 1867
СЕМЕЙСТВО SCOLOPACIDAE VIGORS, 1825
Род Limosa Brisson, 1760
Limosa lacrimosa sp. nov.
Рис. 33; табл. X, фиг. 1, 2
Видовое название от lacrimosus (лат.) — жалобный.
Г олотип — ПИН, № 2614-82; дистальная часть правого плеча; Кобдосский аймак,
местонахождение Чоно-Хариах; средний плиоцен, свита Хиргис-Нур.
Материал. Только голотип.
Описание. Proc, supracondylaris dorsalis утоньшенный, заостренный и образует
с диафизом острый угол. Impres. lig. collaterale ventrale вплотную подходит к cond.
ventralis. Каудально-дистальный угол proc, flexorius закруглен. Перемычка с дисталь-
ного конца между proc, flexorius и cond. ventralis узкая и заостренная. Боковые ребра
sulcus m. scapulotricipitis острые и высокие, переходящие на диафиз с заметным
изломом.
Рис. 33. Limosa lacrimosa sp. nov.; голо-
тип № 2614-82, ПИН; правое плечо; Монго-
лия, Чоно-Хариах; средний плиоцен
а — латеральная; б — вентральная; в —
дорсальная стороны
Размеры, мм: наибольшая поперечная ширина эпифиза 8,7, передне-задний
диаметр cond. dorsalis 5,6, передне-задний диаметр cond. ventralis 2,7, наименьшая
передне-задняя толщина дистальной части диафиза 3,2.
Сравнение. В роде Limosa было описано три ископаемых вида. Среди них
Numenius gypsorum Gervais, 1844 был сначала помещен Бродкорбом (1967) в род
Limosa, а недавно Мликовский (Mlikovsky, 1981) на основании переизучения слепка
мозга перенес его в Rallidae, описав новый род Montirallus. Сравнение по деталям строе-
ния с L. vanrossemi L. Miller, 1925 из среднего миоцена Калифорнии и L. ossivallis
Brodkorb, 1967 из нижнего плиоцена Флориды невозможно ввиду описания их по другим
костям скелета, но оба эти вида крупнее L. lacrimosa. От современных L. limosa и
L. lapponica новый вид также отличается меньшими размерами (наибольшая попереч-
ная ширина у меньшего L. lapponica 9,3 и 9,6 мм) и структурными особенностями, указы-
ваемыми в диагностическом описании. Proc, supracondylaris у L. limosa такого же
строения, как и у L. lacrimosa, но у L. lapponica он притуплен у вершины и расширен
в основании. У обоих современных видов угол между проксимальным краем этого
processus и диафизом острее, чем у ископаемого вида. Impres. lig. collaterale ventrale
у обоих современных видов немного дальше отставлено от cond. ventralis, каудально-
дистальный угол proc, flexorius у них спрямлен, как бы срезан наискось, перемычка
между proc, flexorius и cond. ventralis расширена у L. limosa, но так же заострена
у L. lapponica, как у L. lacrimosa, ребра sulcus m. scapulotricipitis у современных видов
притуплены и без перелома постепенно переходят на диафиз.
Замечания. Из нижнего миоцена Франции, местонахождение Сан-Жеран-ле-Пьи,
описан по тарсометатарсусу, коракоиду и плечевой кости Elorius paludicola Milne-
Edwards, 1868, подсемейство Scolopacinae. Строение всех трех костей этого вымершего
71
монотипического рода удивительно близко к таковому современного рода Limosa. При
возможности сравнения оригинальных материалов по Elorius нельзя исключить вероят-
ность перемещения его в род Limosa.
Распространение. Верхние горизонты пачки А и нижние горизонты пачки
Б свиты Хиргис-Нур.
Род Tringa L., 1758
Tringa sp.
Рис. 34; табл. X, фиг. 3, 4
Материал. ПИН, № 2614-98; дистальный эпифиз левого тибиотарсуса; МНР,
Шаргаин-Гоби, местонахождение «точка 1080 м»; средний плиоцен, свита Хиргис-Нур.
Описание. Эпифиз в медиально-латеральном направлении уже, чем в крани-
ально-каудальном. Epicond. medialis и impres. lig. medialis слабо развиты. Incisura
intercondylaris спереди глубоко выемчатая с почти вертикальными стенками condylorum,
с одноямковым impres. lig. cruciati cranialis, линия которого наклонена медиально.
Ребра condylorum на каудальной стороне эпифиза заметно сближены.
Размеры, мм: наибольшая поперечная ширина эпифиза 3,0, передне-задний
диаметр cond. medialis 3,5, передне-задний диаметр cond. lateralis 3,2, наименьший
диаметр через incis. intercondylaris (с дистальной стороны) 2,2.
Рис. 34. Tringa sp.; № 2614-98, ПИН;
левый тибиотарсус; Монголия, «точка
1080 м» в Шаргаин-Гоби; средний плио-
цен
а — медиальная; б — краниальная; в —
дистальная стороны
Замечания. Описываемый фрагмент тибиотарсуса сравнивался с современными
представителями всех семейств подотряда Charadrii, кроме Rostratulidae. По общим
пропорциям эпифиза он находится среди представителей семейства Scolopacidae,
а между ними в роде Tringa. Об этом свидетельствуют латерально-медиальная сжатость
эпифиза, отсутствие отведения cond. medialis внутрь, резкий переход от диафиза к эпи-
физу при округлом или почти квадратном профиле дистальной части диафиза. Несмотря
на то, что из 12 видов современной фауны (включая hypoleuca и macularia) в нашем
распоряжении были скелеты всех видов этого рода, кроме Т. melanoleuca, от описания
нового вида останавливает фрагментарность ископаемого экземпляра. Специфические
особенности его строения указаны в описании, по каждой из них он сходен или отличен от
одного из современных видов рода. Ближе всего он находится по деталям структур
к Т. stagnatilis, будучи заметно мельче его. По размерам почти соответствует (немного
мельче) Т. hypoleucos.
В ископаемом состоянии в синонимическом роде Totanus было описано шесть видов
из неогена Европы. Два из них — Т. lartetianus Milne-Edwards, 1863 из нижнего миоцена
(аквитан) Франции и Т. scarabelli Portis, 1888 из нижнего плиоцена (мессиний) Италии
представлены также тибиотарсусами. При сложности решения вопроса о системати-
ческом положении ископаемого Tringa sp. из Шаргаин-Гоби в ряду современных улитов
тем более трудно решить его по отношению к ископаемым формам без сравнения с ори-
72
гинальными материалами. Однако в данном случае важно, что среди прибрежных птиц
в среднем плиоцене в Западной Монголии обитал один мелкий улит.
Распространение. Средний плиоцен, верхняя подсвита свиты Хиргис-Нур
в местонахождении «точка 1080 м» в Шаргаин-Гоби.
Род Calidris Меггеп, 1804
Calidris sp.
Рис. 35; табл. XI, фиг. 5, 6
Macrorhamphus finitimus: Курочкин, 1971, с. 66.
Материал. ПИН, № 2614-31; дорсальная половина правого коракоида;
Убсунурский аймак, местонахождение Хиргис-Нур 2; средний плиоцен, свита
Хиргис-Нур.
Замечания. Данный фрагмент коракоида был описан нами под названием
Macrorhamphus (= Limnodromus) finitimus в первой статье по птицам среднего плио-
цена Западной Монголии. Однако его переизучение показывает, что он принадлежал
одному из видов рода Calidris. Общее строение дорсальной половины коракоида очень
сходно у Calidris, Limnodromus, Lymnocryptes и Micropalama. По общим пропорциям
Рис. 35. Calidris sp.; № 2614-31, ПИН;
правый коракоид; Монголия, Хиргис-Нур 2;
средний плиоцен
а — каудальная; б — медиальная; в —
краниальная стороны
/7 /ГЛ/
I I । 1
proc, acrocoracoideus, структуре facies articularis clavicularis, довольно округлым
очертаниям cotyla scapularis, округлости, а не некоторой уплощенности дорсальной
части диафиза ископаемый экземпляр относится к роду Calidris, но не к другим выше-
перечисленным родам из подсемейств Scolopacinae и Erolinae. Общее же сходство
этой части коракоида у различных видов Calidris и фрагментарность ископаемого
экземпляра на фоне отсутствия в сравнительных материалах по современным куликам
части видов этого рода позволяют указать систематическую его принадлежность только
как Calidris sp.
Размеры, мм: длина дорсального эпифиза (от вентрального края cotyla scapu-
laris) 5,3, ширина fac. artic. clavicularis 3,5, ширина fac. artic. humeralis 2,0, наимень-
шая поперечная ширина диафиза 2,1.
Распространение. Верхняя подсвита свиты Хиргис-Нур в местонахождении
Хиргис-Нур 2, верхние уровни пачки А.
73
ОТРЯД COLUMBIFORMES LATHAM, 1790
СЕМЕЙСТВО PTEROCLIDAE BONAPARTE, 1831
Род Syrrhaptes liliger, 1811
Syrrhaptes kashini Kurochkin, sp. nov.
Табл. XI, фиг. 4, 5
Видовое название дано в честь памяти Г. Н. Кашина, советского орнитолога,
известного своими успешными изысканиями в номенклатуре птиц.
Голотип — ПИН, № 3222-675; synsacrum; МНР, местонахождение Хиргис-Нур
2; средний плиоцен, свита Хиргис-Нур.
Материал. Только голотип.
Описание. Типовой экземпляр синсакрума представляет из себя достаточно
хорошо сохранившуюся его центральную часть, немного разрушенную с краниального
конца и сильнее — с каудального. Crista dorsalis synsacri высокий, узкий. Extremitas
cranialis synsacri краниально суживается до сильно уплощенного с боков гребня.
Sulcus ventralis synsacri в средней части занимает половину общей ширины синсак-
рума с вентральной поверхности. Каудально от последней передней пары больших
pr. costalis синсакрум сначала несколько расширяется, а затем сужается.
Размеры, мм: наибольшая ширина вентрального основания синсакрума 6,2,
общая длина фрагмента 31,1.
Сравнение. Cr. dorsalis у S. kashini более узкий, чем у четырех экземпляров
современной S. paradoxus. Особенно хорошо у них отличается синсакрум с вентраль-
ной стороны. Extrem. cranialis у S. kashini сужается краниально почти до острого
гребня; эта часть синсакрума у современной саджи остается достаточно широкой
вплоть до первого свободного поясничного позвонка, будучи в самой краниальной
части равна половине наибольшей ширины синсакрума в его середине. Отметим, что по
этим структурам у четырех экземпляров S. paradoxus изменчивость оказывается
практически не выраженной. Sulcus ventralis у S. kashini — широкая, состав-
ляя около половины ширины синсакрума с вентральной стороны в средней части,
у S. paradoxus занимает только около */з его ширины. Наиболее широкое место
в синсакруме у S. kashini приходится на некотором расстоянии каудальнее от послед-
ней пары передних рг. costalis, а у S. paradoxus — на уровень расположения этой
пары отростков. Поэтому синсакрум каудальнее от этой пары отростков у S. kashini
еще расширяется, а у S. paradoxus от этого места уже начинается каудальное сужение
синсакрума. Общие размеры и картина отверстий для нервов и сосудов у синсакрумов
S. kashini и S. paradoxus одинаковы.
Замечания. При специфических принципиальных особенностях морфологии
синсакрума, отличающих Pteroclidae от всех других семейств современных птиц,
в диагнозе нового вида приведены детали строения, устанавливающие отчетливую
видоспецифичность этой плиоценовой саджи. Синсакрум рябков (Pterocles) хорошо
отличается от синсакрума садж отсутствием sulcus ventralis synsacri.
В третичных отложениях из фосфоритов Керси во Франции (поздний эоцен—
ранний олигоцен) по тарсометатарсусу и коракоиду были описаны Pterocles validus
и Р. larvatus, еще один вид Р. sepultus описан по тарсометатарсусам из местонахож-
дения Ланги (ранний миоцен) во Франции (Milne-Edwards, 1869—1871). Правиль-
ность отнесения этих трех видов к рябкам указывает то, что ранее саджи в ис-
копаемом состоянии известны не были, если не считать двух находок S. paradoxus
в плейстоцене Европы.
Ниже из Чоно-Хариаха и Хиргис-Нура 2 описываются еще две кости садж, но их
оказывается невозможно включить в состав одного вида, так как эти два дополни-
тельных остатка практически не отличаются морфологически от таких же костей
современной S. paradoxus. В то же самое время синсакрум S. kashini отличается от
этого современного вида совершенно отчетливо, имея те же абсолютные размеры.
К сожалению, в сравнении отсутствовал скелет второго современного вида садж —
S. tibetanus, но скелета этой птицы вообще нет в мировых коллекциях.
Распространение. Местонахождение Хиргис-Нур 2; средний плиоцен,
свита Хиргис-Нур, пачка А, уровни 37—40.
74
Syrrhaptes sp.
Рис. 36; табл. X, фиг. 7, 8
Материал. ПИН, № 3222-33, дорсальная половина левого коракоида; место-
нахождение Хиргис-Нур 2; свита Хиргис-Нур. ПИН, № 2614-75, проксимальная часть
левого карпометакарпуса; местонахождение Чоно-Хариах; свита Хиргис-Нур.
Описание. В строении этих двух фрагментов не удается найти признаки, по
которым их можно было бы отличить от соответствующих костей современной S. para-
doxus.
Размеры, мм: коракоид — поперечная ширина диафиза 3,3, ширина facies
cuticularis humeralis 3,3, длина дорсального эпифиза (от вентрального края cotyla
scapularis) 11,2, общая длина фрагмента 18,4; карпометакарпус — длина os metacar-
pale alulare (от сочленовной поверхности pr. alularis до проксимального края tr.
carpalis) 7,7, ширина tr. carpalis 3,6, наибольшая высота всего проксимального эпи-
физа 8,6, общая длина фрагмента 11,5.
Замечания. Описание этого фрагмента было опубликовано (Курочкин, 1982).
Здесь целесообразно привести признаки, отличающие Syrrhaptes от Pterocles по кора-
коиду и карпометакарпусу, так же как и диагностические особенности в строении
этих костей для семейства Pteroclidae. Некоторые характерные особенности строения
дают возможность уверенно отнести их к семейству рябковых. В коракоиде такими явля-
ются удлиненная и нависающая внутрь форма акрокоракоида, чечевицеобразная форма
cotyla scapularis, отчетливый след прохождения сухожилия m. supracoracoideus с пе-
редне-внутренней стороны sulcus m. supracoracoidei, который создает впечатление об
утолщении основания pr. procoracoideus с внутренней стороны. Карпометакарпус
рябковых имеет специфическое ребро, ограничивающее с проксимальной стороны
fovea carpalis cranialis, и характерный желобок, спускающийся наискось от pr. pisi-
formis к вырезке между двумя метакарпалиями.
Сравнение с современными Syrrhaptes и Pterocles указывает на принадлежность
ископаемых остатков к роду Syrrhaptes. У ископаемого коракоида проксимальная
часть диафиза, прилежащая к cotyla scapularis, сравнительно узкая, передняя поверх-
ность диафиза имеет сглаженные очертания, внутренний край collum acrocoracoidei
заостренный. Эти черты в строении коракоида имеются у современной S. paradoxus.
У Pterocles диафиз, подходя к cotyla scapularis, расширяется, на его передней поверх-
ности проходят два отчетливых продольных ребра, а внутренний край collum acroco-
racoidei притуплен.
В проксимальном эпифизе карпометакарпуса у современной S. paradoxus, так же
как и у ископаемой Syrrhaptes sp., trochlea carpalis незначительно расширяется
Z7 Z/ж Zzm
L„ , ____I -------1 L. I---------------1
Рис. 36. Syrrhaptes sp.
A— № 2614-75, ПИН; левый карпометакарпус; Монголия, Чоно-Хариах; средний плиоцен; а — лате-
ральная; б — краниальная; в — медиальная; г — каудальная стороны. Б — № 3222-33, ПИН; левый
коракоид; Монголия, Хиргис-Нур 2; средний плиоцен; а — медиальная; б — краниальная; в — каудальная
стороны
6*
75
дистально, если смотреть на эпифиз с дистального конца; facies ligamenti lateralis
образует на краю tr. carpalis отчетливый подъем; ребрышко, ограничивающее с про-
ксимальной стороны fovea carpalis cranialis, идет под углом к продольной оси кости.
Pterocles имеют заметно расширяющийся tr. carpalis, у них нет подъема на trochlea,
как у fac. lig. lateralis, и ребрышко с края fovea carpalis cranialis расположено перпен-
дикулярно к продольной оси кости. Таким образом, по всем приведенным здесь деталям
морфологии коракоида и карпометакарпуса ископаемая форма относится к роду
Syrrhaptes.
Семейство рябковых в современном объеме включает два вида рода Syrrhaptes и
многовидовой род Pterocles. Во Внутренней Азии распространены только саджи
(Syrrhaptes), причем один вид — S. paradoxus — населяет равнинные полупустыни
и пустыни, а другой — S. tibetanus — преимущественно высокие нагорья Тибета,
Куньлуня, Каракорума и Памира. Саджи служат хорошим индикатором полупустынного
и пустынного ландшафта в современных условиях. Находки S. kashini и двух костей
Syrrhaptes sp. в двух местонахождениях среднеплиоценового возраста в Котловине
Больших озер могли бы определенно свидетельствовать о наличии и в среднем плиоцене
таких ландшафтов в данном регионе. И хотя этот вывод не противоречит целиком
выводам о ландшафтно-климатических условиях в среднем плиоцене на западе Мон-
голии, полученным на основании изучения фауны млекопитающих, все-таки ясно, что
климат тогда здесь был заметно мягче, чем в наши дни, а для садж очень характерны
периодические дальние инвазии, которые существовали, по крайней мере, уже в сред-
нем плейстоцене, о чем говорят находки S. paradoxus в Европе (Lambrecht, 1933). По-
этому к палеореконструкциям на основании находок садж, несмотря на определенный
экологический облик этих птиц, следует относиться сдержанно.
Распространение. Местонахождения Хиргис-Нур 2 и Чоно-Хариах; средний
плиоцен, свита Хиргис-Нур, верхние уровни пачки А и отложения пачки Б.
ОТРЯД STRIGIFORMES WAGLER, 1830
СЕМЕЙСТВО STRIGIDAE VIGORS, 1825
Род Asio Brisson, 1760
Asio aff. otus L., 1758
Рис. 37
Материал. ПИН, № 2614-89; ростральная часть грудины; МНР, местонахожде-
ние Чоно-Хариах, средний плиоцен.
Описание Labra dorsalia сочленовой вырезки коракоидов образуют глубокую
выемку, почти достигающую основания spina externa. Внутренние края labra схо-
дятся под тупым углом. Spina externa был образован тремя невысокими и тонкими
/7 /ttf
Рис. 37. Asio aff. otus L.; № 2614-89, ПИН;
ростральная часть грудины; Монголия, Чоно-Хариах;
средний плиоцен; вид с краниальной стороны
лепестками. Передняя поверхность основания pila carinae образует с боковыми
поверхностями острые ребра. Основание pila carinae спереди уплощено и spina externa
продолжается на ней только в виде тонкой линии.
Размеры, мм: наибольшая ширина фрагмента 9,1, наибольшая высота 9,4.
Сравнение. Род Asio имеет характерную морфологию ростральной части
грудины—особенно специфичен рисунок вырезки между labra dorsalia и основанием
pila carinae. По глубокой вырезке, уплощенным спереди labra и небольшим размерам
76
spina externa ископаемый экземпляр стоит гораздо ближе к A. otus, чем к A. flammeus.
У A. flammeus вырезка менее пологая, а вертикальный лепесток spina externa
толстый и спускается дальше вертикально. В то же время ископаемый экземпляр
несколько отличается и от A. otus: вырезка у него более глубокая, a spina externa еще
более изящен. Поэтому фрагментарность материала приостанавливает от описания
нового вида.
Распространение. Местонахождение Чоно-Хариах, МНР; средний плиоцен,
свита Хиргис-Нур, верхняя подсвита; пачка Б.
ОТРЯД PASSERIFORMES LINNAEUS, 1758
СЕМЕЙСТВО CORVIDAE VIGORS, 1825
Род Corvus L., 1758
Corvus solitus Kurochkin, sp. nov.
Рис. 38; табл. XI, фиг. 1—3
Видовое название solitus (лат.) —обыкновенный.
Голотип — ПИН, № 3222-458; дистальный эпифиз левого тибиотарсуса; МНР,
местонахождение Хиргис-Нур 2; средний плиоцен, свита Хиргис-Нур.
Материал. Только голотип.
Описание. Дистальная часть диафиза прямая и не расширяется заметно перед
переходом в эпифиз. Выходное отверстие canalis extensorius немного сдвинуто от сред-
ней линии латерально. Наружный профиль окружности cond. lateralis правильно круго-
вой. Внутренний край impres. lig. cruciati medialis чуть приподнят проксимально.
Размеры, мм: наибольшая поперечная ширина эпифиза 7,7, передне-задний
диаметр cond. medialis ~7,5, передне-задний диаметр cond. lateralis ~7,3, диаметр через
incisura intercondylaris с дистальной стороны 4,3. Поскольку каудальные ребра мыщел-
ков немного разрушены, их размер точно указать невозможно.
Сравнение. В современном космополитическом роде Corvus разные авторы
насчитывают 30—35 видов. Несмотря на то, что нам удалось провести сравнение
ископаемого С. solitus только с семью видами палеарктических и неарктических
врановых (С. corax, С. corone, С. frugilegus, С. macrorhinchos, С. ossifragus, С. Ьга-
chyrhynchos, С. nasicus), видоспецифичность новой формы проявляется совершенно
ясно.
По размерам С. solitus мельче всех палеарктических ворон и сравнима с С. nasicus.
Прямой дистальной частью диафиза ископаемый вид отличается от С. frugilegus,
С. macrorhynchos, С. nasicus, С. ossifragus (у них дистальная часть диафиза заметно
отклонена медиально) и сходен в этом плане с С. corax, С. corone, С. brachyrhynchos.
Но у С. solitus дистальная часть диафиза, так же как у С. ossifragus и С. nasicus,
не имеет некоторого расширения в месте перехода в эпифиз, в то время как у осталь-
ных ворон он постепенно равномерно расширяется в дистальном направлении, начиная
от apoph. med. lig. obliqui. Особенно характерна эта черта для С. frugilegus. Неболь-
шой отодвинутостью выходного отверстия canalis extensorius в латеральном направ-
лении, отмечаемому по разной ширине костных участков с боков от выхода канала
(с латеральной стороны от отверстия костный участок более узок, чем с медиальной),
С. solitus хорошо отличается от остальных Corvus. У большинства видов canalis
расположен\юсередине, но у С. corax, С. brachyrhynchos и части экземпляров С. co-
rone он, наоборот, сдвинут медиально. Если смотреть на латеральный мыщелок сбоку,
то его полуокружность у ископаемой вороны идет по дуге правильного круга. Такую же
картину мы наблюдаем у С. nasicus, С. ossifragus и у части экземпляров С. corone.
У всех остальных Corvus профиль мыщелка с латеральной стороны имеет форму овала,
за счет «срезанности» дистально-краниального угла. Особенно заметно эта особенность
проявляется у части экземпляров С. frugilegus и С. согах. Предполагается, что эта
скошенность мыщелка связана с локомоторными адаптациями задних конечностей
ворона и грача, больше приспособленных к наземному, чем к древесному образу жизни.
Ископаемый С. solitus относится, следовательно, к группе преимущественно древесных
77
Рис. 38. Corvus solitus sp. nov.; голотип № 3222-458, ПИН; левый тибиотарсус; Монголия,
Хиргис-Нур 2; средний плиоцен
а — медиальная; б — краниальная; в — латеральная; г — дистальная стороны
ворон. Поперечный след отхождения lig. cruciati medialis на краниальной поверхности
межмыщелковой вырезки у С. solitus своим медиальным краем приподнят немного
проксимально. В некоторой мере эта черта выражена у С. согах и С. brachyrhynchos.
У остальных ворон этот отпечаток сухожилия расположен по всей своей длине перпен-
дикулярно к продольной оси кости.
В роде Corvus из плиоцена и нижнего плейстоцена Европы описано несколько
ископаемых видов. Все они значительно крупнее монгольского С. solitus. От большин-
ства этих видов известны тибиотарсусы. С. pliocaenus (Portis, 1891) был распространен
в верхнем плиоцене Италии, Франции и Румынии, ^еейи полагаться на Бродкорба
(Brodkorb, 1978), сведшего в синонимы этого вида С. betfianus Kretzoi, 1962 и С. рга-
есогах Deperet, 1890. По всем признакам С. pliocaenus был крупный ворон и безуслов-
ный представитель рода Corvus. То же можно сказать и о С. antecorax Mourer-Chau-
vire, 1975 из нижнего плейстоцена Франции и С. hungaricus Lambrecht, 1916 из ниж-
него плейстоцена Венгрии. Нам представляется сомнительным место в роде Corvus,
описанного С. Морер-Шавире (Mourer-Chauvire, 1975) по большому количеству остат-
ков из нижнего плейстоцена Франции, С. pliocaenus janossyi Mourer-Chauvire, 1975.
На дистальном эпифизе тибиотарсуса этой формы cond. lateralis заметно крупнее
cond. medialis, а выходное отверстие canalis extensorius сильно сдвинуто медиально.
Это характерно для рода Pyrrhocorax.
Замечания. Дистальный эпифиз тибиотарсуса представителей рода Corvus
хорошо отличается от других Corvidae одинаковыми размерами мыщелков, протягива-
нием каудального ребра у медиального мыщелка проксимальнее по диафизу, чем
у латерального мыщелка, иной структурой pons supratendineus и апофизов наружной
и внутренней косых связок в районе костного мостика. Ближе всего к Corvus стоит род
Pyrrhocorax, два современных вида которого отличаются от Corvus неравными разме-
рами мыщелков (латеральный больше медиального), и сдвинутостью выходного отвер-
стия canalis extensorius к медиальному краю. Как показано выше, именно такими
чертами строения С. pliocaenus janossyi отличается от других Corvus, поэтому считаем
возможным перевести эту форму в род Pyrrhocorax и называть, следовательно, Pyrrho-
corax janossyi (Mourer-Chauvire, 1975).
Распространение. Верхняя подсвита свиты Хиргис-Нур в местонахождении
Хиргис-Нур 2, верхние уровни пачки А.
78
СЕМЕЙСТВО MOTACILLIDAE VIGORS, 1825
Род Anthus Bechstein, 1805
Anthus sp.
Табл. XII, фиг. 1, 2
Материал. ПИН, № 2614-97; дистальный эпифиз левой плечевой кости; место-
нахождение «точка 1080 м> в Шаргаин-Гоби; средний плиоцен, свита Хиргис-Нур.
Описание. Устроен типично по коньковому образу, но имеет специфически
заостренный рг. epicondylaris dorsalis и мелкую fossa m. brachialis.
Размеры, мм: наибольшая ширина дистального эпифиза 4,5, передне-задний
диаметр cond. dorsalis 2,0, передне-задний диаметр cond. ventralis 1,3, толщина
диафиза 1,3, общая длина фрагмента 8,6.
Сравнение. В ископаемом состоянии в третичных отложениях коньки неиз-
вестны. Из 34 видов этого рода современной фауны имелась возможность сравнения
только с 8 (A. novaeseelandiae, A. campestris, A. trivialis, A. hodgsoni, A. gustavi,
A. pratensis, A. cervinus, A. spinoletta), причем в сравнительном ряду не было большой
отдельной группы коньков, распространенных в Азии и Африке, — виды A. nilghiriensis,
A. sylvanus, A. berthelotii и др. (Morony, Bock, Farrand, 1975). Учитывая значительный
недостаток сравнительного материала и специфические отличия ископаемой формы от
первой группы видов в сравнительном ряду, приходится воздержаться от обозначения
ископаемого Anthus в ранге нового вида. Какой характер следует придавать специфи-
ческим признакам этой формы, пока остается неясным.
В отличие от всех видов сравниваемого ряда рг. epicondylaris dorsalis у ископае-
мой формы укорочен и имеет заостренную вершину; у первых вершина отростка всегда
более округла, и он длиннее у A. pratensis и A. hodgsoni, такой же укороченный, но
с широким основанием у A. novaeseelandiae, A. campestris, A. trivialis, A. gustavi,
A. cervinus и A. spinoletta. Среди этих современных видов fossa m. brachialis самая
глубокая у A. spinoletta, A. campestris и A. hodgsoni, несколько мельче у других видов,
но несравнимо более мелкая данная ямка у Anthus sp. из Шаргаин-Гоби. Некоторые
отличия имеются у этого экземпляра также в структуре рг. flexorius. Общие размеры
его дистального эпифиза (ширина 4,5, диаметр cond. dorsalis 2,0) вполне сопоставимы
с таковыми современных коньков (ширина эпифиза / диаметр cond. dorsalis): trivialis
4,3/2,0 и 4,7/2,2, gustavi 4,1/1,7, hodgsoni 4,4/2,0 и 4,4/2,0, pratensis 4,6/2,2, campestris
4,5/2,2 и 5,0/2,4, novaeseelandiae 4,7/2,2, spinoletta 4,6/2,1 и 4,6/2,2, cervinus 4,0/2,2.
Замечания. Принадлежность данного экземпляра к одному из видов рода
Anthus кажется несомненной. Этот род и все семейство в целом ранее были совсем
неизвестны в ископаемом состоянии из неогена. Недостаток в коллекции всего состава
современных видов удерживают пока от описания нового вида, что, вероятно, окажется
возможным при наличии части видов остальных двух групп этого рода из Азии —
Африки и из Америки.
Большинство современных видов коньков экологически связано с обширными про-
странствами открытых местообитаний, хотя некоторые из них обитают на полянах,
опушках и т. п. местах среди сплошных лесов. Неясность экологического облика нашей
ископаемой формы не позволяет прогнозировать характер ландшафта, в котором мог
жить этот конек.
Аналогичный фрагмент плеча от конька такого же вида найден в местонахождении
Павлодар, нижний плиоцен, в Восточном Казахстане.
Распространение. Известен из двух местонахождений: «точка 1080 м»
в Шаргаин-Гоби и Павлодар; средний плиоцен, свита Хиргис-Нур, верхняя подсвита и
нижний плиоцен, низы павлодарской свиты.
79
СЕМЕЙСТВО MUSCICAPIDAE VIGORS, 1825
ПОДСЕМЕЙСТВО TURDINAE BONAPARTE, 1831
Род Oenanthe Vieillot, 1816
Oenanthe infima Kurochkin, sp. nov.
Табл. XIII, фиг. 1, 2
Видовое название infima (лат.) — самая нижняя.
Голотип — ПИН, № 2614-99; дорсальный эпифиз левого коракоида; местонахожде-
ние «точка 1080 м» в Шаргаин-Гоби; средний плиоцен, свита Хиргис-Нур.
Описание. Pr. lig. sternocoracoidei longi, расширенный в основании, укорочен-
ный, с тупым вентральным ребром. Impres. lig. coracohumerale dorsale расположено
вплотную к дорсо-краниальному краю дорсо-краниального ребра головки кора-
коида. Латеральный валик facies articularis humeralis удлиненный и невысокий.
С внутренней стороны профиль выемки дорсальнее от facies artic. humeralis удлинен-
ный и неглубокий. Impres. lig. acrocoracoacromiale в виде хорошо выраженной удли-
ненной ступеньки, ямка под impressio имеется, но она неглубокая. Cotyla scapularis
хорошо развита. Pr. procoracoideus укороченный с притупленной вершиной.
Размеры, мм: ширина facies artic. humeralis 0,9, длина дорсального эпи-
физа 4,4, расстояние от конца pr. lig. sternocoracoidei longi до вершины акрокора-
коида 2,7, общая длина фрагмента 6,9.
Сравнение. Из 18 современных видов рода Oenanthe для исследования ископае-
мой каменки из Шаргаин-Гоби было использовано только 6 (О. oenanthe, О. pleschanka,
О. hispanica, О. picata, О. deserti, О. isabellina). Однако отличие ископаемой О. infima
от всех этих видов по строению pr. procoracoideus, который у всех современных форм
имеет сжатую с боков заостренную вершину и в виде гребня несколько продолжается
на диафиз, дает возможность обособить ее в качестве самостоятельного вида, не-
смотря на фрагментарность материала. У О. infima pr. procoracoideus имеет расширен-
ную притупленную вершину и в сторону стернального эпифиза не продолжается в виде
узкого гребня, а полого опускается на диафиз вблизи вершины же отростка. О. infima
имеет больше всего общего с О. isabellina. С этим видом она сходна по рисунку impres.
lig. acrocoracoacromiale и общим очертаниям акрокоракоида, отличаясь укороченным
и расширенным в основании, без тонкого вентрального ребра pr. lig. sternocoracoidei
longi («brachial tuberosity», no Hamon, 1964), придвинутой вплотную к дорсо-кра-
ниальному краю impres. lig. coracohumerale dorsale, низким латеральным валиком facies
artic. humeralis, неглубокой выемкой фасетки между этой facies и вершиной акроко-
ракоида, мелкой ямкой под impres. lig. acrocoracoacromiale, хорошо развитой cotyla
scapularis. С О. oenanthe ископаемый вид сходен рисунком impres. lig. acrocoracoacro-
miale, мелковыемчатым участком между facies artic. humeralis и вершиной акрокора-
коида, отчетливо выделенной cotyla scapularis, придвинутостью к краю impres. lig. cora-
cohumerale dorsale и небольшой высотой удлиненного латерального валика facies artic.
humeralis. О. infima отличается от О. oenanthe большой выдвинутостью дорсально
pr. acrocoracoideus, более мощным и утолщенным pr. lig. sternocoracoidei longi, наличием
небольшой ямки под impres. lig. acrocoracoacromiale (у О. oenanthe ямки в этом месте
нет совершенно). Отличия нового вида от О. deserti сводятся к иным очертаниям ргос.
lig. sternocoracoidei longi (у последней тонкий и длинный), размерам латерального
валика facies artic. humeralis, форме impres. lig. acrocoracoacromiale, который у со-
временного вида развит в виде одновершинной шероховатости, более утолщенной
collum acrocoracoidei, различием в степени оформленности cotyla scapularis, которая
у О. deserti уплощена и плохо обособлена. С О. deserti по строению дорсального эпифиза
коракоида очень сходна О. pleschanka, поэтому она отличается от О. infima тем же
рядом структурных характеристик. Довольно сходна с О. hispanica, но отличается от
нее с краниальной стороны менее завернутым на краниальную поверхность impres.
lig. acrocoracohumerale, а с каудальной стороны более отчетливой cotyla scapularis
и укороченным pr. lig. sternocoracoidei longi. По сравнению с О. picata характеризуется
более робустными формами всего эпифиза, меньшей высотой латерального валика
80
Фиг. 1, 2. Скорлупа яиц Struthio sp.
1 — Хиргис-Нур 2; средний плиоцен, верхняя подсвита свиты Хиргис-Нур; уровень 51—55;
струтиоидный тип; 2 — Чоно-Хариах; средний плиоцен, верхняя подсвита свиты Хиргис-
Нур; эпиорнитоидно-струтиоидный тип, Х2,4
Фиг. 3, 4. Podiceps solidus sp. nov.; голотип № 2614-37, ПИН; правое плечо; Хиргис-Нур 2;
средний плиоцен, Х1,8
Фиг. 5, 6. Phalacrocorax mongoliensis Kurochkin, 1971; голотип № 2614-29, ПИН; левое бедро;
Чоно-Хариах; средний плиоцен, Х1,8
Фиг. 1—4. Phalacrocorax reliquus Kurochkin, 1976; Хиргис-Нур 2; средний плиоцен
1,2 — № 3222-36, ПИН; левое плечо, Х0,9; 3,4— № 3222-35, ПИН; правое плечо, Х1.8
Фиг. 5, 6. Nyctanassa kobdoena Kurochkin, 1976; голотип № 2614-86, ПИН; правый тарсометатарсус;
Чоно-Хариарх; средний плиоцен, Х2,7
Фиг. 7,8. Heterochen vicinus Kurochkin, 1976; голотип № 2614-100, ПИН; правый тарсометатарсус;
Хиргис-Нур 2; средний плиоцен; Х0,9
Фиг. 9,10. Haliaeetus fortis sp. nov.; голотип № 3222-31, ПИН; левый коракоид; Хиргис-Нур 2;
средний плиоцен; Х0,9
Фиг. 1. Anser devjatkini Kurochkin, 1971; паратип № 3222-42, ПИН; левый тибиотарсус; Хиргис-
Нур 2; средний плиоцен, Х0,9
Фиг. 2—7. Ciconia lucida Kurochkin, 1982
2, 3 — паратип № 3222-666, ПИН; правый карпометакарпус; Хиргис-Нур 2; нижний плио-
цен, Х0,9; 4,5—голотип № 3222-15, ПИН; правый карпометакарпус; Хиргис-Нур 2;
нижний плиоцен, Х0,9; 6—7 — № 2614-72, ПИН; правая лопатка; Чоно-Хариах; средний
плиоцен; Х1,8
Фиг. 8, 9. Bonibernicla ponderosa gen. et sp. nov.; голотип № 3378-50, ПИН; левое плечо; Хир-
гис-Нур 2; средний плиоцен, Х1.35
Фиг. 10. Anser liskunae Kurochkin, 1976; паратип № 3222-18, ПИН; левое плечо; Хиргис-Нур 2,
средний плиоцен, Х0,9
Фиг. 1, 2, 3. Ciconia lucida Kurochkin, 1982 № 3222-56, ПИН; правое бедро; Хиргис-Нур 2; ниж-
ний плиоцен; Х0,9
Фиг. 4—12. Cygnus pristinus Kurochkin, 1971
4, 5 — № 2614-108, ПИН; правое плечо; Хиргис-Нур 2; средний плиоцен; 6—голотип
№ 2614-25, ПИН; левая лучевая кость; Хунг-Хурэ; средний плиоцен; 7, 8 —
№ 2614-109, ПИН; левая локтевая кость; Хиргис-Нур 2; средний плиоцен; 9, 10 —
№ 2614-70, ПИН; правый тибиотарсус; Чоно-Хариах; средний плиоцен; И, 12 —
№ 3222-456, ПИН; правый коракоид; Хиргис-Нур 2; нижний плиоцен, Х0,9
Фиг. 1—6. Anas soporata Kurochkin, 1976
1,2 — голотип № 2614-95, ПИН; правый коракоид; «точка 1080 м» в Шаргаин-Гоби; сред-
ний плиоцен, Х2,7; 3,4 — № 3222-50, ПИН; левое плечо; Хиргис-Нур 2; средний плиоцен,
X 1,8; 5, 6 — № 3222-22, ПИН; левое плечо; Хиргис-Нур 2; средний плиоцен, Х1,8
Фиг. 1, 2. Aythya spatiosa Kurochkin, 1976; голотип № 2614-81, ПИН; правое бедро; Чоно-Хариах;
средний плиоцен, Х2,7
Фиг. 3, 4. Aythya magna sp. nov.; голотип № 3222-28, ПИН; левое бедро; Хиргис-Нур 2; средний
плиоцен; Х1,8
Фиг. 5—7. Alectoris sp.;
5, 6 — № 2614-83, ПИН; правый тибиотарсус; Чоно-Хариах; средний плиоцен, Х2,7;
7 — то же, Х0,9
Фиг. 1—8. Amphipelargus dzabghanensis sp. nov.
/—3 — голотип № 2737-396, ПИН; правый тибиотарсус; Чоно-Хариах 2; средний плиоцен;
4—6 — № 2737-392, ПИН; правый тарсометатарсус; Дзагсо-Хаирхан 4; средний плиоцен;
7,8 — № 2737-393, ПИН; левый тарсометатарсус; Дзагсо-Хаирхан 4; средний плиоцен, Х0,9
Фиг. 1, 2. Crex zazhigini Kurochkin, 1980; голотип № 2614-90, ПИН; левое плечо; Чоно-Хариах;
средний плиоцен, Х3,6
Фиг. 3—7. Palaeoaramides tugarinovi Kurochkin, 1980; 3, 4 — голотип № 2614-121, ПИН; правое
плечо; «точка 1080 м» в Шаргаин-Гоби; средний плиоцен, Х3,6; 5 — № 3222-56, ПИН;
правый коракоид; Хиргис-Нур 2; нижний,плиоцен; 6, 7 — № 3222-672, ПИН; левый тибиотар-
сус; Хиргис-Нур 2; средний плиоцен, Х2,7
Фиг. 8, 9. Porzana risilla (Kurochkin, 1980); голотип № 2614-100, ПИН; левый карпометакарпус;
«точка 1080 м» в Шаргаин-Гоби; средний плиоцен, Х7,2
Фиг. 1, 2. Limosa lacrimosa sp. nov.; rojioiiin № 2614-82, ПИН; правое плечо; Чоно-Хариах; сред-
ний плиоцен, Х1.8
Фиг. 3, 4. Tringa sp.; № 2614-98, ПИН; левый тибиотарсус; «точка 1080 м» в Шаргаин-Гоби;
средний плиоцен; Х3,6
Фиг. 5, 6. Calidris sp.; № 2614-31, ПИН; правый коракоид; Хиргис-Нур 2; средний плиоцен, Х3,6
Фиг. 7, 8. Syrrhaptes sp.; № 3222-33, ПИН; левый коракоид; Хиргис-Нур 2; средний плиоцен, Х1.8
Фиг. 1, 2, 3. Corvus solitus sp. nov.; голотип № 3222-458, ПИН; левый тибиотарсус; Хиргис-Нур 2;
средний плиоцен, Х1,8
Фиг. 4, 5. Syrrhaptes kashini sp. nov.; голотип № 3222-675, ПИН; синсакрум; Хиргис-Нур 2; сред-
ний плиоцен; 4 — правая сторона, 5 — вентральная сторона), Х1,8
Фиг. 6, 7. Lophura interna sp. nov.; голотип № 3381-425; левый тибиотарсус; Шамар; верхний
плиоцен, Х1,8
Фиг. 8, 9. Otididae gen.; № 3381-426; первая фаланга третьего пальца левой лапы; Шамар; верх-
ний плиоцен, Х1,8
Фиг. 1, 2. Anthus sp.; № 2614-97, ПИН; левое плечо; «точка 1080 м» в Шаргаин-Гоби; средний
плиоцен, Х2,7
Фиг. 3—6. Anthus sp.; Павлодар; нижний плиоцен
3, 4 — № 2614-11; ПИН; левое плечо; 5, 6 — № 2614-10, ПИН; левое плечо, Х2,7
Фиг. 7—12. Anthus seductus sp. nov.; Павлодар; нижний плиоцен
7, 8—голотип № 2614-126, ПИН; левое плечо; 9, 10 — № 2614-12, ПИН; левое плечо
молодого экземпляра; 11, 12 — № 2614-13, ПИН; левый тарсометатарсус, Х2,7
Фиг. 1, 2. Oenanthe infima sp. nov.; голотип № 2614-99, ПИН; левый коракоид; «точка 1080 м»
в Шаргаин-Гоби; средний плиоцен, Х2,7
Фиг. 3, 4. Emberizidae gen.; № 2614-14, ПИН; левый тарсометатарсус; Павлодар; нижний плио-
цен, Х2,7
Фиг. 5—lO.Sushkinia pliocaena Tugarinov, 1935; Павлодар; нижний плиоцен
5, 6 — голотип № 2413/205, ПИН; левый тибиотарсус; 7, 8 — № 3243/61-П, ИЗКаз; пра-
вый тарсометатарсус; 9—11 — № 3239/61-П, ИЗКаз; левый карпометакарпус, Х2,7
Фиг. 11, 12. Palaeoperdix sp.; № 3245/61-П, ИЗКаз; левый тарсометатарсус; Павлодар; нижний
плиоцен, Х2,7
Фиг. 1—4. Limnodromus sp.; Береговая; верхний плиоцен
1,2 — № 2975-3, ПИН; левое плечо; 3,4 — № 2975-6, ПИН; правое плечо, X 1,8
Фиг. 5, 6. Phalaropus eleonorae sp. nov.; голотип № 2975-3, ПИН; левое плечо; Береговая; верхний
плиоцен, Х1.8
Фиг. 7—10. Limnodromus sp.; Береговая, верхний плиоцен
7, 8 — № 2975-121, ПИН; правый коракоид; 9, 10 — № 2975-115, ПИН; левый тибиотарсус,
Х1.8
Фиг. 11, 12. Haliaeetus sp.; № 2483, ИЗК; правый карпометакарпус; Тологой; верхний плиоцен,
Х1.35
Фиг.
Фиг.
Фиг.
1 2 Tadorna petrina sp. nov.; голотип № 2975-2, ПИН; грудина; Береговая; верхний плиоцен,
Х0,9
3—6. Perdix margaritae sp. nov.; Береговая; верхний плиоцен
3 4—голотип № 2975-101, ПИН; правое плечо; 5, 6 — № 2975-98, ПИН; правый кора-
коид, X 1,8
7—13. Plioperdix ponticus (Tugarinov, 1940); Береговая; верхний плиоцен
7f 8 — № 2975-96, ПИН; левое плечо; 9—11 — № 2975-106, ПИН; левый карпометакарпус;
12, 13 —№ 2975-119, ПИН; левый тибиотарсус, Х1,8
Фиг. 1, 2. Struthio sp.; № 2346-7787, ПИН; поясничный позвонок; Павлодар; нижний плиоцен,
Х0.9
Фиг. 3, 4. Struthio asiaticus Milne-Edwards, 1871; № 2975-1, ПИН; блок третьего пальца левого
тарсометатарсуса; Береговая; верхний плиоцен, Х0,9
Фиг. 5, 6. Struthio sp.; № 2413-7472, ПИН; третья фаланга четвертого пальца левой лапы;
Павлодар; нижний плиоцен, Х0,9
facies artic. humeralis, утолщенностью и укороченностью pr. lig. sternocoracoidei longi,
большей длиной impres. lig. acrocoracoacromiale, меньшей глубиной и площадью ямки
под этим impressio.
Замечания. По существу в ископаемом состоянии большое современное семей-
ство Muscicapidae совсем неизвестно. Из третичных отложений по отпечатку скелета
описан только один вид в этом семействе — Palaeoscinis turdirostris Howard, 1957
(средний миоцен, штат Калифорния, США), отнесенный в Myadestinae. Наша находка
является первым указанием на присутствие каменок и всех собственно дроздиных (Tur-
dinae) в неогене. При общем сходстве по деталям строения и общим пропорциям
дорсального эпифиза коракоида с современными Oenanthe, по одному признаку —
особому типу строения рг. procoracoideus — новый вид хорошо от них отличен. Мы
считаем этот признак довольно существенным, и если бы не фрагментарность материала,
то, возможно, следовало бы отнести эту птицу к новому роду.
Современный экологический облик каменок отвечает всегда условиям обитания
в открытых, часто аридных ландшафтах, что вполне укладывается в обстановку
среднего плиоцена на западе Монголии. Характерно, что кроме группы центрально-
азиатских и малоазиатских видов, по которым у нас имелся сравнительный материал,
пять видов этого рода распространены ныне в аридных ландшафтах Южной Африки,
два вида в Северной Африке являются чисто сахарскими и к ним здесь добавляется
несколько других, ареал которых идет в открытые ландшафты Европы и Западной
Азии. Таким образом, сегодня это главным образом центрально-переднеазиатская
группа очень близких видов, довольно монолитная морфологически, сходная эколо-
гически, разделяемая тонкими механизмами этологической изоляции, редко дающая
межвидовые гибриды. По всем канонам микроэволюционных построений каменок
можно было бы считать сравнительно молодой недавно радиировавшей группой птиц.
Однако факт находки даже одного вида этого рода в среднеплиоценовых отложениях
Центральной Азии заставляет полагать во всяком случае, что каменки существовали
еще задолго до плейстоценовых климатических катастроф.
Распространение. Одно местонахождение в Шаргаин-Гоби; средний плио-
цен, верхняя подсвита свиты Хиргис-Нур.
ПТИЦЫ МЕСТОНАХОЖДЕНИЯ ПАВЛОДАР
ОТРЯД STRUTHIONIFORMES LATHAM, 1790
СЕМЕЙСТВО STRUTHIONIDAE VIGORS, 1825
Род Struthio L., 1758
Struthio sp.
Рис. 39; табл. XVI, фиг. 1, 2, 5, 6
Struthio chersonensis: Тугаринов, 1935, с. 82.
Материал: ПИН, № 2346-7787, последний свободный поясничный позвонок;
№ 2413-7472, третья фаланга IV пальца левой лапы; местонахождение Павлодар;
нижний плиоцен, низы павлодарской свиты. Сборы Ю. А. Орлова, 1930 г.
Описание. Facies artic. caudalis позвонка сильно вытянута в поперечном
направлении. Боковые ее поверхности ровные, не вогнутые внутрь. Foramen vertebrale
с краниальной стороны имеет широко открытую входную воронку, занимающую поло-
вину длины мозгового канала. Pr. transversus расположен под углом 35° к горизон-
тальной плоскости. Фаланга пальца лапы широкая и высокая.
Размеры, мм: длина corpus vertebrae с вентральной стороны 49,7, длина тела
позвонка между сочленовными поверхностями 48,0, наименьшая поперечная его ши-
рина 31,2, наименьшая высота fac. artic cranialis 21,5, наименьшая высота fac. artic.
caudalis 19,7, наибольшая ширина fac. artic. caudalis 46,2. Длина фаланги между со-
членовными поверхностями 20,0, по средней линии с плантарной стороны 25,4, наиболь-
шая ширина в средней части 21,8, наибольшая высота проксимальной сочленовной по-
верхности 20,3, наибольшая ширина проксимальной сочленовной поверхности 19,7.
Сравнение. Кроме более крупных размеров, позвонок Struthio sp. из Павло-
81
дара отличается от современного S. camel us также некоторыми чертами строения.
Fac. artic. caudalis у ископаемого Struthio sp. заметно сильнее вытянута в поперечном
направлении. Отношение ширины к высоте fac. artic. caudalis у ископаемого 2,34,
у современного 2,06 и 2,11. Не ясно, каким образом А. Я- Тугаринов (1935) вычислил
соответствующее же отношение, как 1,8, так как 46,2 : 19,7 дает 2,34. Еще более
существенно, что fac. artic. caudalis имеет у ископаемого позвонка ровные боковые
стороны, а у современного страуса они вогнуты внутрь. С краниальной стороны fora-
men vertebrate имеет широко раскрытую входную воронку в начале канала, резко
Рис. 39. Struthio sp., № 2346-7787, ПИН;
третья фаланга четвертого пальца левой лапы;
СССР, Восточный Казахстан, Павлодар; нижний
плиоцен
а — дистальная; б — медиальная стороны
сужающуюся на половине длины отверстия. У современного страуса эта воронка
раскрыта незначительно и сужается в каудальном направлении постепенно. Pr. trans-
versus отходит под углом 35° к горизонтальной плоскости, в то время как у S. camelus
отростки располагаются, как правило, более горизонтально, под углом 15—20°.
Фаланга IV пальца характерна своей большей относительной шириной при боль-
шей же высоте проксимальной сочленовной поверхности. Поэтому по сравнению с та-
кой же фалангой S. camelus она выглядит более уплощенной дорсовентрально и одно-
временно более мощной.
Замечания. Следовательно, в морфологии Struthio sp. из Павлодара имеются
признаки, специфически характеризующие эту форму страуса. Однако обозначать ее
номенклатурно мы считаем преждевременным, потому что все накопившиеся к настоя-
щему времени материалы по ископаемым страусам требуют переосмысления и ревизии.
Ни по одной группе плиоценовых птиц Палеарктики сейчас нет такого большого
количества остатков, какое накопилось по ископаемым Struthioniformes. По ним
описано не только множество находок скорлупы яиц, но и достаточно большое коли-
чество костных остатков, которые, правда, часто относятся к разным частям скелета.
Эти обширные материалы требуют в настоящее время серьезной ревизии как в номен-
клатурном, так и в морфологическом отношении, поскольку нам в действительности
неизвестно, сколько и какие страусы обитали в неогене Палеарктики и как они связаны
с современным страусом? Ограниченную попытку решить этот вопрос мы предприни-
мали ранее (Kurochkin, 1970; Курочкин, Лунгу, 1970), сведя всех описанных плиоце-
новых, плейстоценовых и частично миоценовых ископаемых страусов в один вид —
S. asiaticus Milne-Edwards, 1871. После этих работ прошло достаточно много времени,
появились новые материалы, и стало ясно, что решать проблему с ископаемыми страу-
сами необходимо во всей полноте, причем с учетом данных по ископаемой скорлупе.
Распространение. Местонахождение Павлодар; нижний плиоцен, низы
павлодарской свиты.
ОТРЯД ACCIPITRIFORMES SAVIGNY, 1809
СЕМЕЙСТВО FALCONIDAE ANONYMOUS, 1820
Род Sushkinia Tugarinov, 1935
Sushkinia: Тугаринов, 1935, с. 84; Brodkorb, 1964, с. 291; Дементьев, 1964, с. 692
Типовой вид — Sushkinia pliocaena Tugarinov, 1935; нижний плиоцен; Восточ-
ный Казахстан.
Диагноз. Краниально-латеральное ребро диафиза тибиотарсуса узкое, след
сухожилия m. peroneus longus выражен слабо. Отпечатки мускулов сгибателей паль-
цев, расположенные латеральнее и медиальнее от сг. hypotarsi протягиваются по
82
каудальной поверхности тарсометатарсуса более дистально. В проксимальном эпифизе
локтевой кости tuberculum lig. collat. ventrale занимают большую площадь, а его
поверхность располагается в одной плоскости с cotyla ventralis; дорсо-вентральная
ширина cotyla dorsalis больше таковой cotyla ventralis; impres. m. scapulotricipitis
протягивается дистально почти до уровня основания отростка, на котором находится
cotyla dorsalis. В проксимальном эпифизе лучевой кости tuberculum bicipitale развит
очень сильно, выпячиваясь на вентральную сторону. В проксимальном эпифизе карпо-
метакарпуса pr. extensorius — мощный и уплощенный в дистально-проксимальной
плоскости; pr. pisiformis сдвинут краниально, поэтому ямка, расположенная краниаль-
нее от этого отростка, имеет небольшую площадь, в дистальном эпифизе вентральное
крыло краниальной части fac. artic. dig. major выпукло дистально. Первая фаланга
большого пальца крыла имеет значительную поперечную ширину, вентральное крыло
fac. artic. osi metacarpi majus направлено почти вентрально. На каудальной поверхности
вентральной половины коракоида impres. m. sternocoracoidei протягивается дорсально,
образуя острую вершинку на середине поверхности диафиза.
Видовой состав. Один вид — Sushkinia pliocaena Tug., 1935.
Сравнение. Тибиотарсус. Уплощенное краниально-латеральное ребро дисталь-
ной трети диафиза в месте отхождения m. peroneus longus у Sushkinia очень узкое,
его ширина равна ’/4—’/5 ширины диафиза в этом месте, в то время как у мелких
современных Falco—1/3—’/2 общей ширины дистальной части диафиза. Дистальнее
от этого места располагается след сухожилия m. peroneus longus. У Sushkinia он почти
не обозначен, краниально-латеральное ребро эпифиза в этом месте узкое; у Falco по
краниально-латеральному ребру, дистальнее от латеральной ветви pons supratendineus
проходит хорошо обозначенный след в месте прохождения сухожилия этого мускула,
ограниченный с боков продольными выступами. У Sushkinia очень слабо развит также
apoph. epicondylaris medialis; этот бугорок в разной степени выражен у современных
Falco, лучше всего у F. tinninculus, хуже всего у F. subbuteo и F. columbarius. Apoph.
epicondylaris lateralis очень слабый у Sushkinia, мощно развит у мелких современных
Falco, однако по незначительной отодвинутости apophysis в каудальном направлении
линии по продолжению краниально-латерального ребра дистальной части диафиза
Sushkinia более сходна с F. columbarius, F. tinnunculus и F. subbuteo, чем с другими
мелкими соколами. Слабое развитие apophysis можно частично отнести за счет
неполной взрослости ископаемого экземпляра.
Тарсометатарсус. Отличия Sushkinia от Falco в проксимальной половине тарсомета-
тарсуса очень незначительны. От всех современных просмотренных видов этого рода
Sushkinia отличается более дистальным простиранием отпечатков коротких сгибателей
пальцев, расположенных по каудальной поверхности кости по бокам от сг. hypotarsi;
у Sushkinia они занимают около 1 /4 длины диафиза, у всех Falco — только около ’/з- На
краниальной поверхности эпифиза под eminentia interglenoidalis у Sushkinia имеется
глубокая ассиметричная ямка, локализованная на латеральной стороне участка дис-
тальнее От eminentia. Такую же отчетливую ямку подобного же расположения мы
видим у F. columbarius, несколько более мелкую у F. subbuteo и констатируем полное
отсутствие таковой у других мелких соколов.
Локтевая кость. Необычайно большой по площади tuberculum lig. collat. ventrale
отличает Sushkinia от Falco, кроме того, этот tuberculum, расположенный у Sushkinia
параллельно cot. ventralis, у Falco образует с краем cotyla острый угол, будучи откло-
нен более или менее дистально. Cot. dorsalis у Sushkinia несколько шире, чем cot.
ventralis в дорсо-вентральном направлении; у всех Falco (мелких и крупных) эти две
сочленовные впадины имеют одинаковую дорсо-вентральную ширину. С обратной сто-
роны проксимального эпифиза локтевой кости impres. m. scapulotricipitis протяги-
ваются в дистальном направлении дальше, почти до уровня основания pr. cotylaris
dorsalis, чем у различных Falco.
Лучевая кость. Ее проксимальный эпифиз у Sushkinia отличается от всех Falco
сильно развитыми ямкой и бугром (tuberculum bicipitis) в месте прикрепления одного
из сухожилий m. bicipitis, площадка в месте крепления сухожилия сильно выступает
на вентральную сторону, за уровень края диафиза.
83
Карпометакарпус. Вершинная часть рг. extensorius у Sushkinia в значительной мере
развернута своей плоскостью поперек продольной оси карпометакарпуса, основание
отростка расширенное и мощное; близкое к этому состояние отмечается только
у F. rusticolus, у остальных Falco плоскость отростка более параллельна продольной
оси кости, сам отросток более плоский и слабый. Рг. pisiformis у Sushkinia отодвинут от
основания os metacarpale minus краниально, поэтому площадь ямки, находящейся
краниальнее от этого отростка и ограниченной с противоположной стороны основанием
рг. extensorius, уменьшается. У современных Falco рг. extensorius сидит фактически на
основании os metacarpale minus, а указанная ямка относительно более широкая, лишь
у F. rusticolus наблюдаются некоторый сдвиг processus в том же направлении и
соответственное сужение ямки. Дистальный эпифиз карпометакарпуса Sushkinia харак-
теризуется выпуклостью вентрального крыла краниальной фасетки fac. artic. dig. major;
у Falco оно вогнутое (у большинства) или плоское (F. tinnunculus, F. rusticolus).
Первая фаланга большого пальца крыла. Соответственно краниальной сочленовой
фасетке дистального эпифиза карпометакарпуса устроено вентральное крыло fac.
artic. os metacarpi majus, которое не заворачивается каудально, как у всех Falco,
а направлено почти прямо вентрально. От всех Falco (кроме F. rusticolus) эта фаланга
отличается также большей абсолютно или относительно (по сравнению с крупными
Falco) шириной.
Коракоид. Sushkinia отличается от всех Falco очертаниями impres. m. sternocora-
coidei, которое в дорсальной части образует острую вершину, расположенную в медиаль-
ной половине средней части диафиза. У современных Falco латеральный край impres-
sio подходит вплотную к медиальному краю диафиза.
Замечания. Сравнение показывает ясные отличия Sushkinia от современного
Falco, которые на фоне общего морфологического единства различных представителей
этого рода, подтверждают родовой ранг ископаемого сокола, установленный А. Я. Ту-
гариновым. Следует, однако, учесть, что в нашем сравнении не было современного
Polihierax, два вида которого распространены ныне в Южной и Юго-Восточной Азии
и в Африке, отсутствовал также и мелкий Microhierax, а из 37 видов современных
Falco использованы скелеты только F. cherrug, F. peregrinus, F. rusticolus, F. subbuteo,
F. columbarius, F. tinnunculus, F. sparverius, F. naumanni, F. vespertinus, F. amurensis.
Последнее, однако, не кажется нам значительной погрешностью, поскольку остеология
этого рода довольно единообразна.
Палеогеновая история соколов совсем неизвестна. Из неогена описаны обособленный
Badiostes (средний миоцен Аргентины), два настоящих Falco (средний миоцен США и
средний плиоцен Италии), и Falco sp. (ранний плиоцен США), для плейстоцена США
установлены три вымерших вида Falco (Brodkorb, 1964).
Sushkinia pliocaena Tugarinov, 1935
Рис. 40; табл. XIII, фиг. 5—10
S. pliocaena: Тугаринов, 1935, с. 84.
S. pliocaenica: Brodkorb, 1964, c. 291.
Голотип —ПИН, № 2413/205; дистальный эпифиз левого тибиотарсуса; место-
нахождение Павлодар; нижний плиоцен, низы павлодарской свиты. Сборы Ю. А. Ор-
лова, 1930 г.
Материал. ИЗКаз, № 3236/61 П, полная левая локтевая кость; ИЗКаз,
№ 3237/61 П, проксимальная часть левой лучевой кости, и ИЗКаз, № 3238/61 П —
правой лучевой; ИЗКаз, № 3239/61 П, полный левый карпометакарпус; ИЗКаз,
№ 3240/61 П, первая фаланга большого пальца левого крыла; ИЗКаз, № 3241/61 П,
вентральная половина правого коракоида; ИЗКаз, № 3242/61 П, проксимальная поло-
вина правого тарсометатарсуса; ИЗКаз, № 3243/61 П и № 3244/61 П, две когтевых
фаланги пальцев лапы. Перечисленные кости скорее всего принадлежали одной особи.
Сборы П. Ф. Савинова (Институт зоологии АН Казахской ССР) 1960 и 1961 гг.,
оригиналы хранятся в Лаборатории палеобиологии этого же института, слепки имеются
в коллекции ПИН. Собраны в костеносной линзе местонахождения «Гусиный перелет»
84
A — № 3236/61 П, ИЗКаз; левая локтевая кость; а — вентральная; б — медиальная стороны. Б —
№ 3239/61 П, ИЗКаз; левый карпометакарпус; в—каудальная; г—медиальная; д—латеральная
стороны. В —№ 3240/61 П, ИЗКаз; первая фаланга большого пальца левого крыла; е — краниальная;
ж — латеральная; з — проксимальная стороны. Г — голотип № 2413/205, ПИН; левый тибиотарсус;
и—медиальная; к — краниальная; л — дистальная стороны. Д— № 3241/61 П, ИЗКаз; правый тарсо-
метатарсус; м — проксимальная; н — краниальная; о — медиальная стороны. Все кости из местонахождения
Павлодар («Гусиный перелет>), Восточный Казахстан, СССР; нижний плиоцен
85
в г. Павлодаре, на берегу р. Иртыш; возраст — ранний плиоцен (низы павлодарской
свиты).
Описание. Основные морфологические особенности скелета Sushkinia pliocaena
даны в диагнозе рода. Две когтевые фаланги показывают вооруженность пальцев
сильными когтями, которые не только довольно крупные, но и имеют более высокие
суставные основания с удлиненными бугорками в местах фиксации сгибателей пальцев.
Размеры, мм: тибиотарсус — поперечная ширина дистального эпифиза 6,5,
передне-задний диаметр cond. medialis 5,3, диаметр через incisura intercondylaris 3,7,
общая длина фрагмента 11,2; локтевая кость — общая длина 65,3, ширина проксималь-
ного эпифиза ~7,3, диагональная ширина проксимального эпифиза ~8,3, ширина cot.
ventralis 3,4, ширина cot. dorsalis ~3,7, наименьшая ширина диафиза 3,6, наибольшая
ширина дистального эпифиза 6,2, передне-задний диаметр cond. dorsalis 5,7; лучевая
кость — наибольший диаметр проксимального эпифиза 4,2 и 4,2, наименьший диаметр
его 2,5 и 2,6; карпометакарпус — общая длина 37,4, длина os metacarpale alulare 6,7,
ширина проксимального эпифиза 10,0, ширина краниальной фасетки fac. artic. dig.
major 4,7, длина дистально симфиза 3,4; первая фаланга большого пальца крыла —
общая длина (между суставными поверхностями) 13,5, наибольшая ширина 6,2;
коракоид — ширина вентрального эпифиза 12,3; тарсометатарсус — поперечная ширина
проксимального эпифиза 7,6, передне-задние диаметры cot. lateralis 4,4 и cot. media-
lis 3,8.
Сравнение. В роде Sushkinia известен единственный вид.
Замечания. При сравнении Sushkinia с современными Falco не удалось избе-
жать описания отличий ее от отдельных видов этого современного рода, поскольку
в некоторых случаях требовалось показать специфику этого ископаемого сокола по
отдельным элементам структуры, изменяющихся у разных видов Falco. Если принимать
принадлежность всех дополнительных материалов по Sushkinia одной особи, то выри-
совывается сокол несколько отличного облика от типа современного Falco — с более
длинными крыльями, большей подвижностью в пальцевых суставах крыла (сустав
между первой фалангой большого пальца крыла и карпометакарпусом позволяет боль-
шую свободу движений; сустав пальца крылышка (fac. artic. dig. alulae) имеет две
широко расставленные фасетки, ориентированные в разных плоскостях и определяю-
щие большую подвижность крылышка), но с более короткими ногами, при лапах снаб-
женных сильной мускулатурой коротких сгибателей (отпечатки их мускульных брюшек
занимают значительную длину диафиза тарсометатарсуса) и мощных пальцах (когти
относительно крупнее, а основания когтевых фаланг выше). Лапа Sushkinia была
больше приспособлена к схватыванию подвижной добычи, чем мы это знаем для мел-
ких современных Falco. Вероятно, Sushkinia питалась главным образом грызунами.
Распространение. Местонахождение Павлодар; нижний плиоцен, низы
павлодарской свиты.
ОТРЯД GALLIFORMES TEMMINCK, 1820
СЕМЕЙСТВО GALLIDAE ILLIGER, 1811
ПОДСЕМЕЙСТВО PHASIANINAE VIGORS, 1825
Род Palaeoperdix Milne-Edwards, 1871
Palaeoperdix: Lydekker, 1891, с. 139; Lambrecht, 1933, с. 431; Brodkorb, 1964, с. 313.
Типовой вид—Palaeoperdix longipes Milne-Edwards, 1871; верхний миоцен,
гельвет; Франция, Сансан.
Описание. А. Мильн-Эдвардс дал полный и исчерпывающий диагноз этого
рода, у которого «тарсометатарсус такой же, как у Palaeortyx, но без выступающего
наружного гребня гипотарсуса, существующего у Perdix, Cryptonyx и др.; межмускуль-
ные линии отчетливы и поверхность прикрепления сгибателя большого пальца большая
и глубокая; передняя поверхность тарсометатарсуса уплощенная, отпечаток прикреп-
ления сухожилия m. tibialis anterior очень высокий и сильно сдвинут внутрь. Верхние
отверстия маленькие и размещены приблизительно на одинаковом уровне. Кости зад-
них конечностей расположены почти так же, как у Perdix. Гипотарсус имеет одно боль-
86
шое отверстие и наружные борозды на задней поверхности. Palaeoperdix отличается от
Palaeortyx менее уплощенным тарсометатарсусом; гипотарсус заметно сдвинут наружу
и наружная бороздка задней его поверхности менее выделена» (Milne-Edwards, 1869—
1871, t. 2, s. 245). Можно добавить, что самцы имеют шпору, tuberositas m. tibialis
cranialis придвинуто близко к foramen vascularis proximalis medialis, краниально-
латеральный угол краниальной поверхности диафиза выступает заметно краниальнее,
чем его краниально-медиальный угол.
Видовой состав. Три вида из позднего миоцена Франции и один вид из ран-
него миоцена Чехословакии.
Сравнение. Хорошо отличается от близких к нему миоценового Palaeortyx
и современного Perdix отсутствием выступающего сг. lateralis hypotarsi, приближен-
ностью tuberositas m. tib. cranialis к for. vase. prox. medialis, скошенной, из-за выдви-
нутого вперед краниально-латерального угла, краниальной стороной диафиза.
Palaeoperdix sp.
Рис. 41; табл. XIII, фиг. 11, 12
Материал. ИЗКаз, № 3245/61-П; проксимальная половина левого тарсометатар-
суса; Павлодар; нижний плиоцен, низы павлодарской свиты.
Описание. Хорошо сохранившийся проксимальный фрагмент тарсометатарсуса
самца (сохранилось основание шпоры) с частично разрушенным гипотарсусом.
Несет характерные признаки рода Palaeoperdix.
Рис. 41. Palaeoperdix sp.; № 3245/61 П, ИЗКаз; левый тарсоме-
татарсус; СССР, Восточный Казахстан, Павлодар; нижний плио-
цен
а — проксимальная; б — краниальная; в — латеральная стороны
7? 7/w
I I - J
Размеры, мм: поперечная ширина эпифиза 8,6, передне-задний диаметр cot.
lateralis 3,2, передне-задний диаметр cot. medialis 4,4, общая длина фрагмента 26,5.
Сравнение. Практически не отличается от Р. longipes по изображениям и
описанию. Абсолютные размеры сходны. От Р. priscus отличается более крупными
размерами.
Замечания. Тарсометатарсус фазановых недостаточно диагностичен не только
на видовом, но даже и на родовом уровнях. Почти уверенно относя наш экземпляр
к Palaeoperdix, мы затрудняемся установить его видовой ранг, ориентируясь только
на иллюстрации и описания.
Из миоцену и плиоцена Европы описано большое количество видов куриных в родах
Palaeoperdix, (Palaeortyx, Palaeocryptonyx и Plioperdix. Бальман (Ballman, 1969)
предпринял попытку частичной их ревизии из местонахождения Ла-Грив. Общая
картина прояснилась при этом в малой степени и группа требует общей исчерпываю-
щей ревизии, основанной на современных данных.
Распространение. Павлодар; нижний плиоцен, низы павлодарской свиты.
87
ОТРЯД RALLIFORMES REICHENBACH, 1849
ПОДОТРЯД GRUES BONAPARTE, 1854
СЕМЕЙСТВО ERGILORNITHIDAE KOZLOVA, 1960
Род Amphipelargus Lydekker, 1891
Amphipelargus sp.
Рис. 42
Материал. ПИН, № 2413-7471; дистальная половина первой фаланги III пальца
левой лапы; местонахождение Павлодар; нижний плиоцен, низы павлодарской
свиты.
Описание. Дистальная сочленовная поверхность фаланги довольно высокая,
плантарный гребень ее медиального мыщелка направлен почти вертикально.
/7
I- -I - I
Рис. 42. Amphipelargus sp.; № 2413-7471, ПИН; первая фаланга третьего пальца левой лапы;
СССР, Восточный Казахстан, Павлодар; нижний плиоцен
a — дорсальная; б — медиальная; в — дистальная стороны
Размеры, мм: наименьшая высота дистальной сочленовной поверхности 7,2,
наибольшая ее ширина 13,1; наименьшая ширина диафиза фаланги 9,4, наименьшая
его высота 7,0.
Сравнение. По сравнению с аналогичными фалангами A. orientalis из место-
нахождения Калмакпай, отложения которого имеют более молодой — среднеплиоцено-
вый возраст, Amphipelargus sp. из Павлодара характеризуется более высокой первой
фалангой III пальца лапы. Это касается не только диафиза фаланги, который имеет
большую высоту и меньшую ширину, но также и дистальной суставной поверх-
ности, которая отчетливо более высокая при той же абсолютной ширине. Плантарные
ребра дистальной суставной поверхности фаланги, особенно медиальной ее стороны,
направлены более вертикально, в то время как у A. orientalis — под углом в стороны.
Абсолютные размеры одинаковы с A. orientalis (по нескольким фалангам).
Распространение. Местонахождение^Павлодар; нижний плиоцен, павлодар-
ская свита.
ОТРЯД PASSERIFORMES LINNAEUS, 1758
СЕМЕЙСТВО MOTACILLIDAE VIGORS, 1825
Род Anthus Bechstein, 1805
Anthus seductus Kurochkin, sp. nov.
Табл. XII, фиг. 7—12
Видовое название seductus (лат.) — отдаленный.
Голотип — ПИН, № 2614-126; фрагментарная левая плечевая кость; место-
нахождение Павлодар; нижний плиоцен, низы павлодарской свиты. Сборы В. С. Зажи-
гина 1963—1965 гг.
Материал. ПИН, № 2614-12, дистальный фрагмент левого плеча молодой
особи, ПИН, № 2614-13, дистальная половина левого тарсометатарсуса.
Описание. Fossa m. brachialis обширная, глубокая, близко подходит своим
дорсальным углом к tuberculum m. ext. metacarpi radialis. Impres. lig. collaterale
ventrale проксимально вытягивается узким носиком по вентральной границе fossa* m.
88
brachialis. Pr. epicondylaris dorsalis укороченный с широким основанием, отросточек на
проксимальном ребре этого processus практически отсутствует.
Размеры, мм: наибольшая ширина дистального эпифиза 4,7, передне-задний
диаметр cond. dorsalis 2,1, передне-задний диаметр cond. ventralis 1,4, наименьшая
толщина диафиза 1,5, длина фрагмента 19,3.
Сравнение. Отличается от Anthus sp. из местонахождения «точка 1080 м»
в Шаргаин-Гоби и из Павлодара глубокой и широкой fossa m. brachialis; вытянутой и
заостренной impres. lig. collaterale ventrale, укороченным pr. epicondylaris dorsalis,
имеющим закругленную вершину, широкое основание и лишенным дополнительного
отросточка на проксимальном ребре. Крупными размерами и строением pr. epicond.
dorsalis сходен с A. novaeseelandiae, но у этого вида fossa m. brachialis при достаточ-
ной глубине не занимает такой большой площадки, a impres. lig. collaterale ventrale
в проксимальном направлении не вытянуто узким носиком. От всех остальных совре-
менных Anthus (перечислены при описании Anthus sp. из Шаргаин-Гоби) отличается
расширенным, коротким, с закругленной вершиной и без дополнительного отросточка
pr. epicond. dorsalis; широкой fossa m. brachialis, дорсальная сторона которой подсту-
пает близко к tuberculum m. ext. metacarpi radialis, удлиненным проксимально impres.
lig. collaterale ventrale. Строение pr. epicond. dorsalis у сравниваемого ряда современ-
ных Anthus изменяется от заостренного до округлого, но с более узким основанием и
маленьким дополнительным отросточком; fossa m. brachialis у них может быть доста-
точно глубокой, но никогда не подходит вплотную к tuberculum m. ext. metacarpi radia-
lis; impres. lig. collaterale ventrale в проксимальном направлении короче и не протя-
гивается вершиной по вентральному краю fossa m. brachialis.
Замечания. Дистальный эпифиз плеча молодой особи A. seductus не отличается
по размерам от голотипа (ширина эпифиза 4,6, передне-задний диаметр cond. dorsalis
2,1), но имеет характерную для молодых птиц выщербленную поверхность и не сформи-
рованный вентральный борт fossa m. brachialis. Кроме плечевых костей, мы относим
к этому виду и дистальный эпифиз тарсометатарсуса, основанием для чего служат
довольно крупные его размеры и наибольшее сходство с A. novaeseelandiae. Попереч-
ная ширина эпифиза 2,6, поперечная ширина диафиза 1,1, передне-задняя толщина
диафиза 0,9, передне-задний диаметр tr. metatarsi secundi 1,2, то же tr. m. tertii 1,4,
то же tr. m. quartii 1,1, длина эпифиза (от for. vasculare distale до конца tr. m. ter-
tii) 2,4. Отличается от всех коньков сравниваемого ряда более проксимально отстав-
ленным for. vasculare distale от вырезки между trochleae metatarsi quartii и tertii.
Сходен с A. novaeseelandiae широкими trochleae, но с узкими щелями между ними
(trochleae тесно сближены), как и у этого вида tr. m. qfiartii хорошо сформирован и
выступает до уровня tr. m. tertii. У A. pratensis trochleae тоже тесно сближены,
tr. m. quartii хорошо сформирован, но все trochleae узкие, поэтому общая ширина
эпифиза относительно небольшая.
Распространение. Местонахождение Павлодар; нижний плиоцен, низы
павлодарской свиты.
Anthus sp.
Материал. Два дистальных эпифиза левых плечевых костей ПИН № 2614-10
и № 2614-11 из местонахождения Павлодар (табл. XII, фиг. 3—6).
Описание. Морфологически совершенно неотличимы от описываемого в данной
работе по плечевой кости Anthus sp. из местонахождения «точка 1080 м» в Шаргаин-
Гоби, МНР.
Размеры, мм: ширина эпифиза 5,1 и 4,7, передне-задний диаметр cond. dorsa-
lis 2,3 и 1,9.
Сравнение. Отличается от Anthus seductus sp. nov. строением pr. epicondy-
laris dorsalis, fossa m. brachialis, impres. lig. collaterale ventrale, описанными
выше.
Распространение. Местонахождение Павлодар в Восточном Казахстане и
«точка 1080 м» в Шаргаин-Гоби, МНР; нижний (низы павлодарской свиты) и средний
(верхняя подсвита свиты Хиргис-Нур) плиоцен.
7 Заказ 2098
89
ДРУГИЕ ВОРОБЬИНЫЕ
В сборах В. С. Зажигина из Павлодара имеются еще два фрагмента костей воробь-
иных птиц, систематический статус которых ввиду фрагментарности и недостатка срав-
нительных материалов можно указать только до уровня семейства. Дистальный эпифиз
левого тарсометатарсуса ПИН, № 2614-14, принадлежал какой-то мелкой овсянке,
Emberizidae gen. (табл. XIII, фиг. 3, 4). Ширина дистального эпифиза 1,9 мм. передне-
задние диаметры tr. metatarsi tertii 1,2 и quartii 0,9 мм, длина эпифиза (от for. vascu-
lare до конца tr. m. tertii) 1,7. Дистальный эпифиз левой локтевой кости, ПИН,
№ 2614-127, относился к какому-то средних размеров жаворонку, Alaudidae gen.
Ширина эпифиза 3,4 мм, диаметр cond. dorsalis 2,4.
ПТИЦЫ МЕСТОНАХОЖДЕНИЯ ШАМАР
ОТРЯД GALLIFORMES TEMMINCK, 1820
СЕМЕЙСТВО GALLIDAE ILLIGER, 1811
Род Lophura Fleming, 1822
Lophura inferna Kurochkin, sp. nov.
Рис. 43; табл. XI, фиг. 6, 7.
Видовое название inferna (лат.) —подземная, из подземного царства.
Голотип — ПИН, № 3381-425; дистальный эпифиз левого тибиотарсуса; Селен-
гинский аймак, местонахождение Шамар; верхний плиоцен, чикойская свита. Сборы
В. П. Чичагова 1969 г.
Описание. Apoph. fixatus выглядит невысоким укороченным зубчиком. Выход-
ное отверстие canalis extensorius касается своим медиально-дистальным краем cond.
medialis. Костная перемычка между медиальным краем выходного отверстия canalis
I______________________________I —_________________________I
Рис. 43. Lophura inferna sp. nov.; голотип
№ 3381-425, ПИН; левый тибиотарсус; Монголия,
Шамар; верхний плиоцен
а — краниальная; б — латеральная; в — дисталь-
ная стороны
extensorius и медиальным краем кости узкая. След сухожилия m. peroneus longus
плохо выражен. Медиальный профиль краниальной окружности cond. medialis сжат
в дистально-проксимальном направлении.
Размеры, мм: ширина эпифиза 7,5, передне-задние диаметры cond. media-
lis 7,3 и cond. lateralis 6,8, передне-задний диаметр через incisura intercondylaris 4,4.
Сравнение. Сходен с L. swainhoii укороченными apoph. fixatus, который длин-
нее у L. nycthemera; но у L. inferna sp. n. apophysis выходит на край отверстия canalis
extensorius, как и у L. nycthemera. Так же сходно устроен у L. inferna sp. п. и L. nyct-
hemera дистально-медиальный край отверстия canalis extensorius, касающийся cond.
medialis; у L. swainhoii эти две структуры раздвинуты. Костная перемычка между
медиальным краем отверстия canalis extensorius и медиальной стороной кости у всех
трех узкая. След сухожилия m. peroneus longus выражен у L. inferna очень слабо, но
хорошо отпечатан и ограничен костными стенками у двух современных видов. Сужен-
ность краниальной части медиального профиля cond. medialis у L. inferna отличается
как от L. nycthemera, так и от L. swainhoii, у которых этот профиль ближе к форме
круга.
Замечания. В род Lophura согласно Морони и др. (Могопу and oths., 1975)
входит 10 видов фазанов, распространенных в тропических лесах Южного Китая,
90
Индокитая и Индостана. Из них мы располагали скелетами только L. nycthemera и
L. swainhoii. В ископаемом состоянии фазаны этой группы неизвестны. Современные
Lophura — лесные фазаны чаще всего горных лесов тропической и субтропической
Азии.
Распространение. Одно местонахождение в Северной Монголии; верхний
плиоцен, чикойская свита.
ОТРЯД RALLIFORMES REICHENBACH, 1849
СЕМЕЙСТВО OTIDIDAE GRAY, 1840
OTIDIDAE GEN.
Рис. 44; табл. XI, фиг. 8, 9
Материал. ПИН, № 3381-426; первая фаланга III пальца левой лапы; Селен-
гинский аймак, местонахождение Шамар; верхний плиоцен, чикойская свита.
Описание. Вентральный край проксимальной сочленовной поверхности почти
симметричен. Ямка в месте прикрепления сухожилия сгибателя пальца (проксимальнее
от дистального эпифиза с вентральной поверхности) неглубокая.
I________।- I
Рис. 44. Otididae gen.; № 3381-426, ПИН; первая фаланга третьего пальца левой лапы;
Монголия, Шамар; верхний плиоцен
а — проксимальная; б — дорсальная; в — медиальная; г — вентральная стороны
Размеры, мм: наибольшая длина фаланги 19,3, ширина дистального эпи-
физа 6,0, ширина проксимального эпифиза 6,2, высота проксимального эпифиза 7,7.
Сравнение. Размеры немного крупнее, чем у современного Chlamydotis undu-
lata. От современных Otis, Chlamydotis и Chariotis отличается почти симметричным
вентральным краем проксимальной суставной поверхности, так же как и меньшей
ямкой под дистальной суставной поверхностью. Но так же, как и у всех дроф, прокси-
мальный эпифиз высокий, мощный, с сильно выдвинутым вентрально бугром в месте
фиксации сухожилия сгибателя пальца.
Замечания. Четыре вида ископаемых дроф описаны из эоцена, миоцена, верх-
него плиоцена и нижнего плейстоцена Европы (Brodkorb, 1967).
Распространение. Одно местонахождение в Северной Монголии; верхний
плиоцен, чикойская свита.
ОТРЯД PASSERIFORMES LINNAEUS, 1758
СЕМЕЙСТВО STURNIDAE VIGORS, 1825
STURNIDAE GEN.
Материал. ПИН, № 3381-427; дистальный эпифиз левого тибиотарсуса; Селен-
гинский аймак, местонахождение Шамар; верхний плиоцен, чикойская свита.
Описание. Выходное отверстие canalis extensorius приближено к cond. media-
lis, apoph. fixatus расположен на латерально-проксимальной части pons supratendineus,
arrow traverses на краниальной части incisura intercondylaris имеет треугольные очер-
тания— латеральная сторона заострена и упирается в cond. lateralis.
Размеры, мм: поперечная ширина эпифиза 2,7, передне-задние диаметры
cond. medialis 2,7 и cond. lateralis 2,6, диаметр через incisura intercondylaris 1,6.
7*
91
Сравнение. Недостаток современного сравнительного материала не дает воз-
можности определить таксономический статус этого скворца. Принадлежность его
к Sturnidae кажется несомненной, но эта форма не относится к современным родам
Sturnus, Pastor, Gracula и Acridotheres.
Распространение. Одно местонахождение в Северной Монголии; верхний
плиоцен, чикойская свита.
ПТИЦЫ МЕСТОНАХОЖДЕНИЙ ЗАБАЙКАЛЬЯ
ОТРЯД STRUTHIONIFORMES LATHAM, 1790
Род Struthio L., 1758
Struthio asiaticus Milne-Edwards, 1871
Табл. XVI, фиг. 3, 4
Материал. ПИН, № 2975-1; изолированный блок III пальца левого тарсомета-
тарсуса; Бурятская АССР, р. Чикой, местонахождение Береговая; верхний плиоцен,
чикойская свита. Сборы В. И. Жегалло 1967 г.
Описание. Центральная ямка на латеральной поверхности блока расположена
в центре его, но место срастания оснований trochleae tertii и quartii дистально при-
двинуто вплотную к этой ямке. Сосудистые отверстия на боковых поверхностях внеш-
них периметров блока расположены в канавках. Боковая асимметрия блока с кра-
ниальной поверхности слабо выражена.
Размеры, мм: ширина блока с краниальной стороны 52,0, передне-задний
диаметр по медиальной стороне 52,0.
Сравнение. Характеризуется по сравнению с современным S. camel us заметно
большими размерами. У нескольких экземпляров S. camelus из коллекций Зоологиче-
ского музея МГУ и ПИН размеры блока среднего пальца тарсометатарсуса в среднем
на 20 % меньше S. asiaticus из Береговой характеризуется также иными пропорциями
этого блока, который более округл и передняя ширина его к наибольшему (медиаль-
ному) диаметру относится как 1 : 1. У S. camelus блок более удлинен и относительно уже
с краниальной стороны, указанное отношение в среднем по 5 экз. составляет 1 : 1,2.
Trochleae metatarsi tertii и quartii срастаются у S. asiaticus близко к центральной
поверхности, а у S. camelus место срастания блоков отодвинуто проксимальнее от
ямки. По наружным периметрам боковых поверхностей блока у S. asiaticus имеется
много сосудистых отверстий, расположенных в 2—5 мм длины канавках. У S. camelus
сосудистые отверстия представлены как таковые, он они не имеют канавок.
Чикойский экземпляр характеризуется также менее выраженной асимметрией ме-
диальной и латеральной половины блока, у S. camelus латеральная половина заметно
уже, чем медиальная.
Распространение. Местонахождение Береговая; верхний плиоцен, чикой-
ская свита.
ОТРЯД ANSER1FORMES BECHSTEIN, 1804
ПОДОТРЯД ANSERES BECHSTEIN, 1804
СЕМЕЙСТВО ANATIDAE VIGORS, 1825
ПОДСЕМЕЙСТВО ANSERINAE VIGORS, 1825
Род Anser Brisson, 1760
Anser tchikoicus Kurochkin, sp. nov.
Рис. 45
Видовое название tchikoicus — от p. Чикой, в Забайкалье, на которой рас-
положено местонахождение Береговая.
Г олотип — ИЗК № 2476; полный левый карпометакарпус; Бурятская АССР,
Бичурский район, р. Чикой, местонахождение Береговая; верхний плиоцен, чикойская
свита. Сборы А. Г. Покатилова 1964 г. (рис. 45, А).
92
A — голотип № 2476, ИЗК; левый карпометакарпус; а — медиальная; б — каудальная; в — латеральная
стороны. Б — № 2479, ИЗК; левая локтевая кость; г—вентральная; д — латеральная; е — дистальная
стороны. В — № 2477, ИЗК; первая фаланга большого пальца левого крыла; ж — проксимальная;
з — латеральная; и — медиальная; к — краниальная стороны. Все кости — из местонахождения Береговая
в Забайкалье; верхний плиоцен
Материал. В дополнение к голотипу имеются дистальный эпифиз левой локте-
вой кости ИЗК № 2479 (рис. 45, Б), первая фаланга большого пальца левого крыла
ИЗК № 2477 (рис. 45, В), дистальная половина левой лучевой кости ИЗК № 2482,
фрагмент дистального эпифиза левой плечевой кости ИЗК № 2480, фаланга левого
крылышка (1-го пальца крыла) ИЗК № 2481, фрагмент грудины с коракоидной
фасеткой ИЗК № 2478. Все кости происходят, очевидно, от одной особи гуся.
Описание. Окружность каудального ребра facies artic. ulnocarpalis имеет поло-
гую дугу. Fac. artic. dig. minor «утоплена» проксимально и в дистальном направлении
не доходит до уровня fac. artic. dig. major. Краниальное ребро fac. artic. dig. major
поднято дорсально.
Размеры, мм: общая длина 94,1, длина metacarpale alulare 20,3, ширина
tr. carpalis 9,4, ширина fac. artic. dig. major 9,2, наибольшая ширина проксимального
эпифиза 25,0, длина дистального симфиза 11,6.
Сравнение. Среди всех современных Anser хорошо отличается от A. cygnoides
дорсальным разворотом ребра fac. artic. dig. major, которое у A. tchikoicus, как и у дру-
гих Anser, развернуто род углом 60—70 к плоскости карпометакарпуса в дорсальном
направлении; у A. cygnoides это ребро лежит почти в плоскости краниальной стороны
metacarpale majus. От всех Anser отличается пологой дугой окружности ребра fac.
artic. ulnocarpale, которое у современных представителей имеет более крутую (с корот-
93
ким радиусом) дугу. Две фасетки дистального эпифиза находятся на разных уров-
нях — fac. artic. dig. minor в дистальном направлении не достигает уровня fac. artic.
dig. major. У всех современных Anser они находятся на одном уровне. Подобная
A. tchikoicus картина наблюдается у некоторых Branta и Tadorninae (например,
у В. leucopsis и Т. tadorna).
Замечания. Обзор всех ископаемых находок Anserinae имеется у Шорта
(Short, 1969) при описании Paracygnus plattensis из верхнего плиоцена штата Неб-
раска, США. После выхода этой работы было описано два новых гуся: Anserobranta
Kurochkin et Ganja, 1972 и крупный Anser/Branta из верхнего миоцена Пакистана
(Harrison, Walker, 1982). Anser tchikoicus ясно отличается от ископаемых A. oeningen-
sis (Meyer, 1865) из верхнего миоцена Швейцарии, A. azerbaidzhanicus Serebrovsky,
1940 из Бинагадов, от которых известны те же кости, и от ископаемых Branta и Anse-
robranta. Сравнить с пакистанским верхнемиоценовым Anser/Branta мы не могли
из-за отсутствия его опубликованных изображений. Карпометакарпус tchikoicus более
сходен с различными современными Anser, чем с Branta. Отличается от Branta струк-
турой дорсальной поверхности проксимального эпифиза и выдвинутостью в дистальном
направлении фасетки для фаланги крылышка. Дистальный эпифиз локтевой кости
A. tchikoicus представляет собой явного Anser по строению tuberculum carpale, выпук-
лого по проксимальному краю с округлой вершиной. Немного отличается от других
современных Anser более проксимально протягивающимися sulci tendini на дорсальной
поверхности эпифиза. Наибольшая ширина эпифиза 17,4, передне-задний диаметр cond.
dorsalis 14,6, ширина tr. carpalis 9,5. Первая фаланга большого пальца крыла в дета-
лях строения не отличается от современных Anser, но имеет по сравнению с ними
относительно меньшую длину и большую ширину в дистальной трети. Общая ее
длина 39,2, наибольшая ширина 12,2, наибольшая ширина проксимального эпи-
физа 11,3, наибольшая высота дистального эпифиза 10,7. Имеющиеся от A. tchikoicus
фрагменты лучевой и плечевой костей, грудины и фаланга крылышка фактически не
отличаются структурно от таковых современных Anser, хотя тело фаланги и лучевой
кости более утолщено по сравнению, например, с A. fabalis, имеющим такие же абсо-
лютные размеры.
В целом A. tchikoicus представлял собой, вероятно, типичного гуся, размером
с современного A. fabalis, но отличался от современных гусей деталями строения,
указанными в описании, и пропорциями крыла. Так, карпометакарпус был относи-
тельно более длинным, а первая фаланга большого пальца крыла, наоборот, относи-
тельно короче, чем у A. fabalis, имевшего в общем такие же размеры. Все кости крыла
A. tchikoicus выглядят по сравнению с тем же A. fabalis более мощными и толстыми, как,
например, у более абсолютно крупного A. anser.
Распространение. Местонахождение Береговая в Забайкалье, верхний
плиоцен, чикойская свита.
ПОДСЕМЕЙСТВО TADORNINAE REICHENBACH, 1849
Род Tadorna Boie, 1822
Tadorna petrina Kurochkin, sp. nov.
Табл. XV, фиг. 1, 2
Видовое название petrina (лат.) — каменная.
Голотип — ПИН, № 2975-2; фрагментарная грудина; Бурятская АССР, Би-
чурский район, р. Чикой, местонахождение Береговая; верхний плиоцен, чикойская
свита. Сборы В. И. Жегалло, 1967 г.
Материал. Только голотип.
Описание. Spina interna приподнятая, с выемкой посередине. Между боковым
краем этой spina и началом labrum dorsale имеется достаточно широкое углубление.
Incisurae intercostalia глубокие, processi costalia между ними высокие, замкнутые
сверху, в целом ряду реберных вырезок довольно длинный.
Размеры, мм: длина грудины от spina interna до каудального края 95,0, длина
реберного ряда от первого до последнего pr. costalis 32,1.
Сравнение. Более сходна с огарями (Т. ferruginea, Т. tadornoides), чем с пе-
94
ганками (Т. tadorna). У огарей намечается выемка между боковым краем spina interna
и началом labrum dorsale, которая довольно широка у Т. petrina. Выемка в средней
части spina interna у Т. petrina более узкая и глубокая, чем у Т. ferruginea и Т. tador-
noides. Межреберные вырезки у Т. petrina более глубокие, чем у современных видов,
соответственно реберные отростки этого ряда более высокие, они замкнуты сверху,
а внутри первые четыре из них имеют костную мембрану. При меньших, чем у Т. ferru-
ginea и Т. tadornoides, размерах грудины (длина от spina interna до каудального края
у обоих видов 98 мм), длина реберного ряда у ископаемого вида на 1—4 мм больше.
Широкий, выемчатый с дорсальной стороны, выход от конца реберных вырезок на
trabecula lateralis и уплощенное треугольное основание середины каудального края
с вентральной стороны, очень сходны с таковыми у современных огарей.
Замечания. Род Tadorna в ископаемом состоянии в третичных отложениях
ранее был неизвестен. В этом подсемействе несколько видов крупных Anabernicula
и Brantadorna описаны из плейстоцена Северной Америки, в настоящей работе из сред-
него плиоцена Западной Монголии описаны крупная Bonibernicla и другие, неопреде-
лимые ближе, Tadorninae. По всем накопившимся к настоящему времени данным под-
семейство Tadorninae в плиоцене и плейстоцене в Северном полушарии было представ-
лено большим разнообразием форм, чем в современной фауне.
Распространение. Бурятская АССР, местонахождение Береговая; верхний
плиоцен, чикойская свита.
ОТРЯД ACCIPITRIFORMES SAVIGNY, 1809
СЕМЕЙСТВО ACCIPITRIDAE SAVIGNY, 1809
Род Haliaeetus Savigny, 1809
Haliaeetus sp.
Табл. XIV, фиг. 11, 12
Материал. Дистальный эпифиз правого карпометакарпуса, ИЗК, № 2483,
из местонахождения Тологой на р. Селенге в 18 км от Улан-Удэ, Бурятской АССР;
палевые супеси верхнего плиоцена, Чикойская свита. Сборы А. Г. Покатилова, 1964 г.
Описание. В деталях строения практически не отличается от Н. albicil 1а,
за исключением более обширной fovea carpalis cranialis и утоньшенного дистального
ребра pr. extensorius osi metacarp, alulare, что может быть вызвано несколько более
крупными размерами ископаемого экземпляра.
Размеры, мм: ширина tr. carpalis 12,1, длина metacarpale alulare 26,3, высота
эпифиза 30,0.
Замечания. Из верхнего миоцена Франции (Сансан) описан по карпометакар-
пусу Н. piscator Milne-Edwards, 1871. Место этого вида в роде Haliaeetus, насколько
можно судить по иллюстрациям, кажется сомнительным. Из среднего плиоцена Запад-
ной Монголии по нескольким фрагментам выше описан Н. fortis. Проксимальный фраг-
мент карпометакарпуса из Тологоя по существу ничем, кроме более крупных размеров,
не отличается от современных Н. albicilla и Н. leucocephalus. Фрагментарность ис-
копаемого остатка и отсутствие в сравнении скелетов двух других современных органов
не позволяют диагносцировать этот материал на видовом уровне.
Распространение. Местонахождение Тологой в р-не Улан-Удэ, верх, плиоцен.
ОТРЯД GALLIFORMES TEMMINCK, 1820
СЕМЕЙСТВО GALLIDAE ILLIGER, 1811
Род Perdix Brisson, 1760
Perdix margaritae Kurochkin, sp. nov.
Рис. 46; табл. XV, фиг. 3, 4
Видовое название margaritae — в честь палеонтолога М. А. Ербаевой.
Г олотип — ПИН, № 2975-101, проксимальная половина правой плечевой кости;
Бурятская АССР, Бичурский район, р. Чикой, местонахождение Береговая; верхний
плиоцен, чикойская свита. Сборы М. А. Ербаевой, 1963 г.
95
Рис. 46. Perdix margaritae sp. nov.
A — голотип № 2975-101, ПИН; правое
плечо; Забайкалье, Береговая; верхний
плиоцен; а — латеральная; б — медиаль-
ная; в — проксимальная стороны. Б —
№ 2975-98, ПИН; правый коракоид;
г — медиальная; д — каудальная; е — кра-
ниальная стороны
Материал. Кроме голотипа, имеются дорсальный эпифиз правого коракоида,
ПИН, № 2975-98, рис. 46, табл. XV, фиг. 5, 6; дистальный эпифиз правого бедра,
ПИН, № 2975-137; головка правой лопатки, ПИН, № 2975-119. Все они происходят из
местонахождения Береговая. Сборы М. А. Ербаевой 1963 г. и В. И. Жегалло, 1967 г.
Описание. Fossa tricipitalis занимает около половины пространства между
crus dorsale fossae и дорсальным краем эпифиза, вентральной стороной fossa прибли-
жена к этому гребню. Костная перемычка между fossa tricipitalis и incis. capitis
сравнительно тонкая. Eminentia m. latissimi dorsi posterioris расположена на вообра-
жаемом продолжении crus dorsale fossae в дистальном направлении. Проксимальная
половина fossa pneumaticum в глубине закрыта костной мембраной.
Размеры, мм: ширина проксимального эпифиза 13,7, толщина caput humeri 5,2.
Сравнение. В роде Perdix описан только один вымерший вид Р. jurcsaki
Kretzoi, 1962 из нижнего плейстоцена Румынии, охарактеризованный при описании
как типичный Perdix, только более крупного размера. Р. margaritae по размерам схо-
ден с крупными экземплярами современной Р. dauuricae, крупнее Р. perdix, но отли-
чается от них мелкими деталями структуры проксимальной части плеча, будучи в ос-
новных чертах типичной Perdix. У Р. margaritae fossa tricipitalis занимает меньшую
площадь внутренней поверхности эпифиза, чем у Р. perdix и Р. dauuricae, у которых
она простирается более чем на половину участка между пневматическим отверстием
и дорсальным краем. Вследствие этого данная fossa выглядит у Р. margaritae прибли-
женной к crus dorsale fossae, а у других современных видов как бы сдвинута дор-
сальнее. Костная перемычка (или выступ) в треугольнике между incis. capitis, fossa
tricipitalis и fossa pneumaticum у P. margaritae — утоньшена и имеет маленький раз-
мер, у современных Perdix этот участок расширен в виде небольшой треугольной
площадочки с фасеткой. Место фиксации m. latis. dorsi posterioris у Р. margaritae распо-
ложено так же, как у Р. dauuricae — на линии продолжения в дистальном направлении
crus dorsale fossae, у Р. perdix оно сдвинуто от этой линии дорсальнее. Проксимальная
половина fossa pneumaticum у Р. margaritae на некотором углублении от входа затя-
нута костной мембраной, которая отсутствует у современных куропаток.
Замечания. В материалах с Береговой имеется также по одному фрагменту
коракоида лопатки и бедра, которые по относительным размерам, если их сопоставить
с современными Perdix, уверенно подходят к Р. margaritae, а общее морфологическое
сходство их с современным родом подтверждает возможность включения этих остатков
в число материалов по новому ископаемому виду.
Дорсальный эпифиз правого коракоида (№ 2975-98) имеет немного более глубоко
вырезанный вентральный край fac. artic. clavicularis с вершиной вырезки, расположен-
ной посередине фасетки. У современных Perdix указанная вырезка мельче, а вершина
96
ее немного сдвинута от середины каудально. Медиальный угол fac. scapularis
у Р. margaritae так же заострен, как у Р. perdix, но у Р. dauuricae он — закругленный.
Во всех остальных отношениях коракоиды ископаемой и современных куропаток суще-
ственно сходны, особенно по элементам строения дорсальной поверхности акрокора-
коида и краниальной поверхности эпифиза, являющимися диагностичными для различ-
ных родов Phasianidae.
Головка лопатки Р. margaritae (№ 2975-119), почти идентичная по строению акро-
миона и fac. artic. humeralis с современными Perdix, хорошо отличается от них большей
утоньшенностью участка эпифиза между акромионом и fac. artic. humeralis, особенно
хорошо видной с краниальной стороны. Поэтому углубление на латеральной поверх-
ности эпифиза, проходящее от этого участка в каудальном направлении, выглядит
у Р. margaritae более глубоко выемчатым.
Дистальный эпифиз бедра (№ 2975-137), являясь в основных своих структурах
принципиально сходным с бедром современных Perdix, в то же время подходит по
абсолютным размерам к размерам Р. margaritae. Это позволяет включить его в число
остатков данного ископаемого вида. В то же время он имеет ряд важных отличитель-
ных деталей в строении. Прежде всего обращает внимание округлость с каудально-
дистального края tr. fibularis, который у современных Perdix прямо срезан с каудаль-
ной стороны. Fossa poplitea глубже врезана в медиальную сторону к каудальному углу
cond. medialis. Краниальное ребро этого condylus относительно выше, чем у современ-
ных видов рода.
Размеры дополнительных материалов, мм: коракоид — ширина
fac. artic. humeralis 3,4, длина дорсального эпифиза (от вентрального края fac.
scapularis) 12,0, длина акрокоракоида 7,7; лопатка — высота caput scapulae 9,1,
длина fac. artic. humeralis 4,8; бедро — ширина дистального эпифиза 9,9, передне-
задний диаметр cond. lateralis 7,3, передне-задний диаметр cond. medialis 8,3, передне-
задняя толщина через sulcus intercondylaris 4,6.
Из верхнего миоцена Европы описано четыре вида вымершего рода Palaeoperdix.
Неустановленный вид этих палеокуропаток найден и описан выше из нижнего плиоцена
Павлодара. Европейские Palaeoperdix известны по разнообразным и многочисленным
остаткам из местонахождений Сансан и Ла-Грив-Сант-Альбан во Франции. У нас име-
лась возможность сравнить забайкальские материалы не только с опубликованными
изображениями, но и со слепками плечевых костей Palaeoperdix edwardsi (Deperet,
1887), любезно присланными из Университета Клауда Бернарда в Лионе, Франция
благодаря вниманию и стараниям д-ра С. Морер-Шавире. Palaeoperdix представля-
ется отчетливо близким к современному Perdix, но отличается от него в плечевой кости
большей площадью fossa tricipitalis, относительно меньшим диаметром входного отвер-
стия foramen pneumaticum и более глубокой sulcus lig. transversus. Другой палеогено-
вый и миоценовый род европейских фазановых — Palaeortyx (описано семь видов по
большим материалам, слепок плечевой кости Р. phasianoides также имелся у нас),
существенно отличается от Palaeoperdix и Perdix строением fossa tricipitalis, cr.
pectoralis, места фиксации m. latissimus dorsi и др.
Распространение. Бурятская АССР, р. Чикой, местонахождение Береговая;
верхний плиоцен, чикойская свита.
Род Plioperdix Kretzoi, 1955
Ammoperdix: Тугаринов, 1940, с. 311.
Pliogallus: Тугаринов, 1940, с. 312.
Plioperdix: Kretzoi, 1955, с. 367.
Ammoperdix: Brodkorb, 1964, с. 318.
Типовой вид — Ammoperdix ponticus (Тугаринов, 1940); верхний плиоцен;
юго-запад Украины, юг Молдавии, юг Бурятской АССР в Забайкалье.
Описание. Диафиз тарсосметатарсуса почти квадратный в поперечном сече-
нии, с заметным сужением боковых его сторон к средней линии каудально. Блоки паль-
цев тесно сближены, особенно приближен к среднему блоку tr. met. secundii, латераль-
ная сторона которого при взгляде с плантарной стороны перекрывает медиальную сто-
97
рону tr. met. tertii. Tr. met. secundii сильно отставлен плантарно, почти на всю высоту
блока. Cr. plantaris medius гипотарсуса имеет очень короткое и невысокое продолже-
ние на диафизе. В проксимальном эпифизе cr. lat. hypotarsi вздут плантарно, выходное
отверстие канала гипотарсуса открывается в середине гипотарсуса на невысоком
гребне cr. plant, medius. Дистальный эпифиз тибиотарсуса расширен в поперечном
направлении, cond. medialis с краниальной стороны сжат с боков. Проксимальный эпи-
физ бедра не имеет поднятого дорсо-краниально крыла tr. femoris, латеральная
поверхность эпифиза не расширена краниально-каудально, диафиз бедра довольно
прямой. В дорсальном эпифизе коракоида fac. artic. clavicularis имеет ровный вент-
ральный край и каудальный угол facies не нависает крючком над sulcus, фасетка на
медиально-вентральном углу краниальной поверхности акрокоракоида — плоская,
маленькая. На краниальной поверхности эпифиза нет выемчатого углубления. В прок-
симальном эпифизе плечевой кости cr. pectoralis опускается на диафиз крутой ступень-
кой, вентрально-каудальный край входного отверстия fossa pneumatica — тонкий и
узкий, дорсальная граница глубокой fossa tricipitalis спускается до emin. m. latis.
dorsi posterioris. В дистальном эпифизе плеча epicond. ventralis заострен в дистальном
направлении, impres. lig. collat. ventrale и emin. m. metac. radialis не выступают в виде
бугорков дорсально и вентрально. В карпометакарпусе pr. intermetacarpalis развит
слабо, os metac. minus — тонкая, не выступающая углом каудально, на вентральной
поверхности карпометакарпуса у основания и в конце os metac. minus имеется по
хорошо заметной ямке.
Видовой состав. Один вид — Р. ponticus, широко распространенный в верх-
неплиоценовых отложениях юга Украины, Молдавии и Забайкалья. Очевидно, все
ссылки на Р. coturinoides во многих плиоценовых местонахождениях Украины (Дуб-
рово, Капелист, 1979) относятся к Р. ponticus. К этому же виду следует приписывать
и материалы из «молдавского руссильона», перечисленные у Хозацкого и Курочкина
(1977).
Сравнение. Plioperdix сравнен с современными Ammoperdix, Tetraogallus,
Alectoris, Francolinus, Perdix, Coturnix, Excalfactoria, Arborophila, Rollulus, Bambu-
sicola, Galloperdix, Ithaginis, Crossoptilon, Hierophasis, Lophura, Gallus, Phasianus,
Syrmaticus, т. e. c 18 из 46 родов современных Phasianinae. Среди них он более всего
сходен с современными Coturnix и Excalfactoria, и это сходство настолько заметно
отделяет их от остальных фазановых, что можно пренебречь отсутствием в сравни-
тельном ряду многих родов крупных азиатских фазанов, хотя было бы желательно все-
таки сравнить Plioperdix с мелкими азиатскими и африканскими Pternistes, Perdicula,
Crytoplectron и Tropicoperdix. С неогеновыми Palaeortyx, Palaeocryptonyx, Palaeoperdix
и c Bantamyx сравнение проведено по слепкам, оригиналу и описаниям.
В тарсометатарсусе у Plioperdix блоки пальцев тесно сближены, особенно это
относится к tr. met. secundii. Медиальная и латеральная стороны соответственно
tr. met. tertii и tr. met. secundii при плантарной проекции перекрываются, что можно
видеть только у Excalfactoria, Rollulus и Galloperdix; у Coturnix они находятся на
одной линии, у других фазановых не достигают одна другой. Tr. met. secundii отставлен
у Plioperdix так сильно плантарно, что дорсальный его край только немного перекры-
вает плантарный край tr. met. tertii при медиальной проекции, этот признак свойствен
только Rollulus, у остальных блоки перекрываются с медиальной стороны на ’/з или
’/г диаметра. Но Rollulus и Galloperdix имеют отличия общего характера, отделяющие
их от Plioperdix — дистальная часть диафиза тарсометатарсуса у них сильно уплощена
краниально-плантарно, tr. met. tertii заметно сдвинут медиально, tr. met. quartii
укорочен, сильно развит cr. plant, mediana. В проксимальном эпифизе Plioperdix имеет
специфически вздутый плантарно cr. lat. hypotarsi. Эта особенность строения не встре-
чается у остальных фазановых. Выход единственного канала гипотарсуса у ископае-
мого рода происходит на середине его — на cr. hyp. mediana, как это в известной
степени имеется у Perdix, Alectoris, Tetraogallus и Galloperdix; у остальных канал
открывается с медиальной стороны завершения гипотарсуса.
Тибиотарсус. Имелся только один дистальный эпифиз из Береговой, относимый
к Plioperdix. Эпифиз расширен в поперечном направлении, как у Alectoris и Ammoper-
98
dix, а не сжат с боков, как у Coturnix, Excalfactoria, Bambusicola, Galloperdix и
Rollulus. Perdix имеет промежуточную характеристику. Очень сходен с Alectoris,
отличаясь от него, кроме меньших размеров, относительно более узким с краниальной
стороны cond. medialis. От Ammoperdix хорошо отличается менее сдвинутыми меди-
ально sulcus и canalis extensorius.
Бедро. Характеризуется у Plioperdix отсутствием сильно поднятого дорсо-кра-
ниально крыла tr. femoris, поэтому поверхность проксимального эпифиза сбоку выгля-
дит суженной в отличие от всех фазановых, кроме Rollulus. У последнего структура
этой области эпифиза такая же, но он хорошо отличается от Plioperdix сильно расши-
ренной fac. artic. distalis. В то же время рисунок отпечатков mm. obturatoriae и ilio-
trochanterici у Plioperdix и Rollulus также сходен. Один из трех фрагментов бедра
сохранил значительную часть диафиза, которая так же слабо выгнута дугой кра-
ниально, как у Coturnix, Excalfactoria, Rollulus в отличие от Perdix, Alectoris и Ammo-
perdix, имеющих сильный краниальный выгиб в середине его.
Коракоид. Fac. artic. clavicularis ассиметрична, каудальная ее ’/3 — шире кра-
ниальной, но каудальный угол facies не нависает крючком над sulcus m. supraco-
racoidei, вентральный ее край ровный. Фасетка на краниальной поверхности меди-
ально-вентрального угла рг. acrocoracoideus — маленькая, не поднимающаяся бугор-
ком над остальной поверхностью. Краниально-дорсальный угол fac. artic. humeralis
закругленный, он не выступает поднятым гребнем краниально. По всем этим признакам
Plioperdix очень сходен с Coturnix, Excalfactoria и с Bantamyx из среднего плиоцена
Хиргис-Нура 2, отличаясь от других фазановых, особенно от близкого по размерам
Ammoperdix. Незначительная особенность строения коракоида разделяет Plioperdix и
Coturnix с Excalfactoria — отсутствие у ископаемого рода выемчатого углубления на
краниальной стороне эпифиза между ребром fac. artic. humeralis и медиальным его
краем, которого нет также и у Bantamyx.
Плечевая кость. Проксимальный эпифиз имеет высокий cr. pectoralis, опускаю-
щийся крутой ступенькой на диафиз. Такая картина имеется только у Coturnix и
Excalfactoria, но ни у одного из современных или неогеновых (Palaeortyx, Palaeocryp-
tonyx, Palaeoperdix) фазановых. Вентрально-каудальный край входного отверстия
fossa pneumatica — тонкий и узкий, что также свойственно лишь Coturnix и Excalfac-
toria. Fossa tricipitalis глубокая и дорсальная ее граница проходит наискось заметным
выступом до emin. m. latis. dorsi posterioris, что отмечается и для Coturnix, Excalfactoria
и Palaeortyx. Дистальный эпифиз плеча в общем у всех фазановых построен довольно
единообразно. Plioperdix имеет заостренный дистально (если смотреть с вентральной
стороны) epicond. ventralis, подобным же образом устроенный у Alectoris и Francoli-
nus, но у них между epicondylus и cond. ventralis на краниальной поверхности суще-
ствует также заметная ямка, развитая и у большинства фазановых, кроме Plioperdix
и Coturnix. Impres. lig. collat. ventrale и emin. m. metac. radialis умеренно развитые,
невыдающиеся заметно соответственно вентрально и дорсально, как у Coturnix, но не
у остальных фазановых.
Карпометакарпус у Plioperdix устроен довольно специфично. Тугариновым (1940)
правильно отмечено, что рг. intermetacarpalis довольно слабо развит, os metac.
minus — тонкая, не вздымается каудально, а щель между метакарпалиями поэтому
узкая. Эти особенности строения характерны и для Coturnix, в меньшей мере — для
Excalfactoria, несмотря на его меньшие размеры. Их также сближает не выступающий
дистально за уровень fac. artic. dig. major сочленовный выступ для dig. minor. Но от
этих двух родов Plioperdix хорошо отличен наличием двух заметных ямок в начале и
в конце os metac. minus на вентральной стороне кости. Подобная небольшая дисталь-
ная ямка намечена у Ammoperdix, проксимальная — у Perdix, однако обе они, хотя и
меньшей глубины, присутствуют у Odontophorinae (Colinus, Lophortyx).
Замечания. Плиопердикс — широко распространенная форма в позднеплиоце-
новых отложениях. Однако оригинальное описание и изображение его не позволяют
достоверно исследовать все другие материалы. С этим связан детальный разбор его
таксономии, родовой и видовой характеристик в настоящей работе. Мы располагали
достаточно обширным материалом по плиопердиксу, чтобы попытаться разобраться
99
в уже описанных и новых данных по этому виду. Тугаринов (1940, с. 312) описал по
карпометакарпусу из верхнемиоценовых отложений катакомб Одессы новый род и вид
Pliogallus coturinoides. Кретцой (Kretzoi, 1955) установил, что название Pliogallus
преоккупировано Gaillard, 1939 и предложил новое — Plioperdix, что было принято
в сводке Бродкорба (Brodkorb, 1964). В той же работе на с. 311 Тугаринов описал
новый вид в современном роде Ammoperdix — A. ponticus, голотипом и единственным
материалом по которому явился тарсометатарсус с разрушенным проксимальным
эпифизом. Оригиналы описания coturinoides, по всей видимости, утеряны. Мы пытались
их разыскать в ЗИН АН СССР, ПИН АН СССР и в Институте зоологии АН УССР.
Оригинал ponticus был обнаружен нами в ЗИН АН СССР и в настоящее время хра-
нится в ПИНе.
Бродкорб (Brodkorb, 1964) в известном каталоге включил в род Plioperdix еще
три вида фазановых из верхнемиоценового местонахождения Ла-Грив-Сант-Альбан
во Франции — Р. grivensis (Lydekker, 1893), Р. depereti (Ennouchi, 1930) и Р. joleaudi
(Ennouchi, 1930). Бальман (Ballmann, 1969), ревизуя массовые материалы по птицам
из этого местонахождения, восстановил эти виды в роде Palaeortyx, сведя grivensis
в синоним depereti, и возвратил этот род в семейство Phasianidae в отличие от Брод-
корба, помещавшего его в Cracidae. В этой же работе Бальман подтвердил самостоя-
тельность Plioperdix. Нами установлено, что ponticus и coturinoides, основанные на
разных костях скелета, представляют один вид.
Plioperdix ponticus (Tugarinov, 1940)
Рис. 47; табл. XV, фиг. 7—13
Ammoperdix ponticus: Тугаринов, 1940, с. 311.
Pliogallus coturinoides: Тугаринов, 1940, с. 312.
Plioperdix coturinoides: Brodkorb, 1964, с. 317.
Ammoperdix ponticus: Brodkorb, 1964, c. 318.
Голотип — Институт зоологии АН УССР, № 6489; правый тарсометатарсус;
Украинская ССР, Одесса, катакомбы; верхний плиоцен.
Материал. Дистальный эпифиз левого плеча № 2975-96, ПИН (рис. 47, табл. XV,
фиг. 7—8); дистальный эпифиз правого плеча № 2975-132; левый карпометакарпус
№ 2975-106 (табл. XV, фиг. 9—11); правый карпометакарпус без проксимального
эпифиза № 2975-107; дорсальные эпифизы левого № 2975-120 и правого № 2975-122
коракоидов: проксимальные эпифизы левых № 2975-124 и № 2975-126 и правого
№ 2975-125 бедер; дистальный эпифиз левого тибиотарсуса № 2975-119 (табл. XV,
фиг. 12, 13) ; дистальный эпифиз правого тарсометатарсуса № 2975-112. Все кости про-
исходят из местонахождения Береговая на р. Чикой; верхний плиоцен, чикойская
свита; сборы М. А. Ербаевой 1963 и 1967 гг.
Кроме материалов из Забайкалья, в нашем распоряжении находились сборы
1963—1966 гг. Р. ponticus из Молдавии и Юго-Западной Украины, переданные в ПИН
благодаря вниманию и любезности Л. И. Хозацкого (Ленинградский университет),
Н. А. Константиновой (ГИН АН СССР) и И. М. Гани (Институт зоологии и физиоло-
гии АН МССР).
Плечевые кости: дистальные эпифизы левые № 2614-51, 62, 66 (местонахождение
Этулия, Вулканештский район, МССР; верхний плиоцен, «молдавский руссильон»),
№ 2614-145 (Войничево, Одесская область, УССР, долина р. Кучурган; верхний плио-
цен); проксимальный эпифиз правого плеча № 2614-59 (Этулия; верхний плиоцен).
Карпометакарпусы: проксимальный эпифиз правого № 2614-137 (Котловина, Одесская
область УССР, западный берег оз. Ялпуг; верхний плиоцен, «молдавский руссильон»),
дистальная часть правого № 2614-50 (Этулия; верхний плиоцен). Тарсометатарсусы:
левые, почти полный № 2614-64 (Котловина, верхний плиоцен), с разрушенным прокси-
мальным эпифизом № 2614-63 (Чишмекиой, Вулканештский район, МССР; верхний
плиоцен, «молдавский руссильон»), правые, с разрушенным проксимальным эпифизом
№ 2614-146 (Войничево; верхний плиоцен), дистальный эпифиз № 2614-138 (Котло-
вина; верхний плиоцен).
Описание. То же, что и при описании рода.
100
Размеры голотипа, мм: наименьшая поперечная ширина диафиза 2,3,
наименьшая передне-задняя ширина диафиза 1,5, поперечная ширина дистального
эпифиза 5,2, передне-задний диаметр 2,7 и поперечная ширина tr. met. tertii 2,1,
расстояние от for. vase, distale до конца tr. met. tertii 5,1.
Сравнение. В роде Plioperdix достоверно известен только один вид.
Замечания. Исследование материалов по мелкому виду плиоценовых фазано-
вых из Береговой, руссильонских местонахождений Молдавии и юга Украины, ориги-
нала Ammoperdix ponticus и описания Р. coturinoides у А. Я. Тугаринова (1940) пока-
зывают, что разные кости, по которым были описаны эти два вида, принадлежали
Рис. 47. Plioperdix ponticus (Tugarinov,
1940).; № 2975-96, ПИН; левое плечо
а—медиальная; б — латеральная; в — ди-
стальная стороны. Забайкалье, Береговая; верхний
плиоцен
L- I I
одной и той же мелкой фазановой птице, относившейся к отдельному, хорошо обособ-
ленному роду, отличающемуся от всех современных и неогеновых ископаемых фазано-
вых. Род сохраняет название, данное ему Кретцоем (Kretzoi, 1955), а старшим сино-
нимом вида оказывается имя ponticus, так как оно опубликовано на предыдущей
странице — перед coturinoides (Тугаринов, 1940). Тарсометатарсус ponticus совер-
шенно подходит по абсолютным размерам к coturinoides. Определенное морфологиче-
ское сходство обоих с современным Coturnix, подчеркнутое Тугариновым для coturi-
noides, также ставит их рядом, так как тарсометатарсус ponticus имеет наибольшее
число черт сходства с этим же родом. Но нельзя поэтому согласиться с Тугариновым
о включении ponticus в род Ammoperdix, от которого он хорошо обособлен сближен-
ными блоками пальцев, утолщенным в дорсо-плантарном направлении диафизом и
отсутствием острого гребневидного продолжения на диафизе cr. plant, medius. Одно-
именные кости Р. ponticus из разных местонахождений полностью сходны, а разные
кости хорошо подходят по относительным размерам друг к другу, представляя фаза-
новую птицу немного более крупного размера, чем современный перепел.
Морфологическое своеобразие Plioperdix хорошо видно из сравнения при описании
рода. Прямых филогенетических связей Plioperdix с современными фазановыми уста-
новить не удается. Можно считать, что этот род вымер в плиоцене, не оставив потом-
ков. Однако намечается линия от среднеплиоценового Bantamyx к Plioperdix.
Размеры дополнительных материалов’ (мм): тарсометатарсус —
общая длина 28,4, поперечная ширина эпифизов, проксимального 5,2, дистальных
5,3, 5,3, 5,5, передне-задние диаметры cond. lateralis — 2,4, cond. medialis 2,5, передне-
задний диаметр tr. met. tertii 2,6, 2,4, 2,6, 2,4, расстояние от for. vase, distale до
конца tr. met. tertii 4,1, 4,0, 4,0, 4,1, поперечная ширина диафиза 2,3, 2,3, 2,4, передне-
задняя ширина диафиза 1,4, 1,5, 1,6; тибиотарсус — поперечная ширина дистального
эпифиза 5,0, передне-задние диаметры cond. medialis 5,0, cond. lateralis 4,4, диаметр
через incis. intercondylaris 3,0; бедро—поперечная ширина проксимального эпифиза
7,2, 7,2, 6,5, диаметр cap. femoris 3,0, 2,6, диаметр col. femoris 2,5, 2,7, 2,7, передне-
задняя ширина проксимального эпифиза 5,4. Коракоид: ширина fac. artic. humeralis
2,2, 2,3, длина дорсального эпифиза (от вентрального края cot. scapularis) 9,0, 9,0;
плечо—ширина дистального эпифиза 6,7, 6,9, 7,0, 7,1, передне-задние диаметры cond.-
epicond. dorsalis 3,8, 3,9, 3,9, 4,0, 4,2, cond. ventralis — 2,2, 2,5, 2,5, 2,4, 2,6. Карпо-
метакарпус—общая длина 2,4, ширина диафиза (наибольшая между метакарпалиями)
101
4,0, 3,6, ширина tr. carpalis 2,5, 2,3, высота проксимального эпифиза 5,7, 5,6, длина
дистального симфиза 3,7, 4,0.
Данные по функциональной морфологии. Р. ponticus представлял
небольших размеров куропаткоподобную птицу, приближаясь размерами к Ammoper-
dix griseogularis. Если сопоставить относительные размеры костей его крыла и задней
конечности с подобными у обыкновенного современного перепела и пустынной куро-
патки, то оказывается, что при относительно такой же длине задней конечности крылья
у Р. ponticus были относительно длиннее, чем у этих двух современных видов. Это,
очевидно, указывает, что плиопердикс летал лучше и пользовался полетом чаще, чем
это свойственно современным перепелу и пустынной куропатке, которые в основном на
местах гнездования передвигаются пешком, редко используя полет, главным образом
только в случае опасности, перелетая при этом на короткие расстояния. Своеобразие
морфологии костей задних конечностей Р. ponticus сводится к более сближенным бло-
кам пальцев тарсометатарсуса при сильно отставленном плантарно блоке II пальца,
расширенному дистальному эпифизу тибиотарсуса с узким cond. medialis. Особенно
обращает на себя внимание устройство проксимального эпифиза бедра, имеющего
небольшой трохантер и узкую дорсальную сочленовную поверхность на шейке бедра,
а также довольно прямой диафиз бедра. Эти особенности морфологии говорят о том,
что плиопердикс имел лапу со сближенными пальцами, тарзальный сустав, способный
к множественным сгибательным движениям при ротации цевки внутрь во время сги-
бания, большие степени свободы движений в тазобедренном суставе. При этом можно
видеть прямые аналогии с морфологией и функционированием задней конечности
у мелких и среднего размера пастушковых. Эти морфофункциональные особенности
прямо указывают на приспособление задней конечности плиопердикса к быстрому и
постоянному бегу по земле и, вероятно, на преимущественное обитание в совершенно
открытых местах, где эта птица, с одной стороны, много и быстро передвигалась по
земле, а с другой — часто и далеко летала.
Распространение. Местонахождение Береговая в Забайкалье и многие
местонахождения на юге Украины и в Молдавии; верхний плиоцен, чикойская свита
и «молдавский руссильон».
ОТРЯД CHARADRIIFORMES HUXLEY, 1867
ПОДОТРЯД CHARADRII HUXLEY, 1867
СЕМЕЙСТВО SCOLOPACIDAE VIGORS, 1825
Род Limnodromus Wied, 1833
Limnodromus sp.
Рис. 48, 49; табл. XIV, фиг. 1—4, 7—10
Материалы. Плечевая кость: проксимальная половина левой № 2975-3, ПИН;
проксимальные эпифизы левой № 2975-102, 133, 134; дистальные эпифизы левой
№ 2975-6, 131 и правой № 2975-103, 127, 128, 129, 130. Карпометакарпус: проксималь-
ный эпифиз правого № 2975-136. Коракоид: дорсальные эпифизы левого <№ 2975-99 и пра-
вого № 2975-121. Лопатки: эпифизы правых № 2975-9, 104, 105. Краниаль-
ные фрагменты грудины № 2975-8, 100, 123. Тибиотарсус: дистальные эпифизы
правого № 2975-97, ИЗ, 115 и левого № 2975-114 и 116. Все кости — из местонахожде-
ния Береговая в Забайкалье; верхний плиоцен, чикойская свита. Сборы М. А. Ербаевой
1963 и 1967 гг.
Описание и сравнение. Род Limnodromus в ископаемом состоянии неиз-
вестен. В современной фауне он представлен двумя своеобразными видами, более
крупный из которых локально распространен в Азии, а другой в Северной Америке,
откуда заходит на Чукотку. Отсутствие в сравнительной серии азиатского вида —
L. semipalmatus — не позволяет установить видовой ранг ископаемого Limnodromus
из Береговой. Последний отличается от L. griseus более крупными размерами и тон-
кими особенностями в строении костей. Безусловно, было бы важно определить,
к какому из двух современных видов стоит ближе этот ископаемый бекасовидный
веретенник, но пока это не представляется вообще возможным, так как скелеты
102
A —№ 2975-3, ПИН; левое плечо; а — медиальная; б — латеральная; в — проксимальная стороны.
Б — № 2975-6, ПИН; правое плечо; г — дорсальная; д — латеральная; е — вентральная; ж — дистальная
стороны. Забайкалье, Береговая; верхний плиоцен
Рис. 49. Limnodromus sp.
/7
А — № 2975-100, ПИН; краниальная часть грудины; а — спереди; б — сбоку. Б — № 2975-115, ПИН;-
левый тибиотарсус; в — медиальная; г — краниальная; д — латеральная; е — дистальная стороны. В —
№ 2975-99, ПИН; левый коракоид; ж — краниальная; з латеральная; и — каудальная стороны.
Забайкалье, Береговая; верхний плиоцен
азиатского вида в мировых остеологических коллекциях отсутствуют. Здесь важна, тем
не менее, находка многочисленных ископаемых остатков Limnodromus в фауне Береговой.
Размеры, мм: плечевая кость — поперечная ширина проксимального эпифиза
~ 12, ~ 11, ~11, толщина cap. humeri 2,7, ширина дистального эпифиза 7,2, 7,1, 7,1,
7,1, передне-задний диаметр cond.-epicond. dorsalis 4,3, 4,2, 4,4, 4,2, 4,3; карпомета-
карпус: высота проксимального эпифиза 7,1, длина os metac. alulare 6,2; коракоид:
высота дорсального эпифиза 7,7, 8,1, длина акрокоракоида 6,3, 6,5; лопатка: высота
эпифиза — 6,5, 6,5, 6,8; тибиотарсус ширина дистального эпифиза 5,0, 4,7, 4,8, 4,7,
диаметр cond. medialis 4,4, 4,3, 4,3.
Распространение. Местонахождение Береговая; верхний плиоцен, чикой-
ская свита.
103
СЕМЕЙСТВО PHALAROPODIDAE BONAPARTE, 1831
Род Phalaropus Brisson, 1760
Phalaropus eleonorae Kurochkin,sp. nov.
Табл. XIV, фиг. 5, 6
Видовое название eleonorae — в честь Элеоноры Алексеевны Вангенгейм,
внесшей заметный вклад в изучение стратиграфии и палеонтологии позднего кайнозоя
Сибири.
Голотип — ПИН, № 2975-135; полная левая плечевая кость; Бурятская АССР,
Бичурский район, местонахождение Береговая; верхний плиоцен, чикойская свита.
Сборы М. А. Ербаевой 1963 и 1967 гг.
Материал. Только голотип.
Описание. Cap. humeri удлинена дорсо-вентрально, tuberculum dorsale только
доходит до cap. humeri, fossa tricipitalis неглубокая, pr. supracond. dorsalis небольшой,
impres. lig. collat. ventrale маленькое, гребень от него в проксимальном направлении
тонкий, pr. flexorius притуплен с дистальной стороны.
Размеры, мм: общая длина 26,7, ширина эпифизов — проксимального 5,9
и дистального 4,1, диаметры cond.-epicond. dorsalis 2,6 и cond. ventralis 1,1.
Сравнение. Ископаемые Phalaropodidae неизвестны. Размеры Ph. eleonorae
соответствуют размерам самца Ph. lobatus, но мелкие морфологические отличия
в строении плеча, выделенные в описании, указывают на видовую самостоятельность
ископаемого плавунчика. В современной фауне их имеется два вида — Ph. lobatus
и Ph. fulcarius, третий вид этого семейства относится к отдельному роду Zonifer
tricolor и, как можно видеть, он действительно остеологически заметно отличается от
Phalaropus. Cap. humeri у Ph. eleonorae более вытянута дорсо-вентрально, чем у совре-
менных видов, у них она — круглее и толще. Tub. dorsale в месте фиксации одной из
порций т. pectoralis у Ph. eleonorae только доходит до границы cap. humeri, как это
наблюдается у Ph. fulcarius, а у Ph. lobatus tuberculum немного «наползает» на
головку плеча. Более мелкая fossa tricipitalis отличает ископаемый вид от двух совре-
менных, так же как и маленькие размеры невысокого и тонкого pr. supracond. dorsalis,
impres. lig. collat. ventrale и притупленность pr. flexorius. Последний у обоих современ-
ных видов оканчивается дистально заметным округлым бугорком.
Распространение. Местонахождение Береговая; верхний плиоцен, чикой-
ская свита.
ОБЗОР ИСКОПАЕМЫХ КОМПЛЕКСОВ
Из плиоцена Центральной Азии теперь установлено 56 форм вымерших птиц, из
которых 32 представлены новыми видами, а 4 относятся к ранее известным — Struthio
asiaticus, Sushkinia pliocaena, Plioperdix ponticus и Asio aff. otus. Из них лишь послед-
ний относился, вероятно, к ныне живущему виду, все остальные — вымершие. Все
вместе эти птицы представляют 23 семейства 12 отрядов. 26 родов птиц встречается
в современной фауне, только 9 относятся к вымершим.
К сожалению, из нижних уровней нижней подсвиты Хиргис-Нур (отложения ран-
него плиоцена) данных по птицам для анализа состава вымершей авифауны ни-
чтожно мало. Несколько лучше представлены раннеплиоценовые птицы из Павлодара,
где имеется восемь различных форм, из которых три относятся к вымершим родам
(Sushkinia, Palaeoperdix и Amphipelargus). Все они представляют птиц открытых
ландшафтов степного или саванного типа. По-настоящему всесторонне можно рас-
смотреть только ископаемый авикомплекс среднеплиоценовых местонахождений Запад-
ной Монголии, откуда описана большая часть плиоценовых птиц Центральной Азии
(39 форм). Данные о позднеплиоценовых птицах по трем местонахождениям Северной
Монголии и Забайкалья — сравнительно полнее, чем для раннего плиоцена, но в целом
также ограничены. Полностью все наши материалы сведены в табл. 10.
104
Таблица 10
Распределение ископаемых птиц по местонахождениям плиоцена, Центральной Азии
Систематический состав Нижний плиоцен Средний плиоцен Верхний плиоцен
Хиргис-Нур 2, уровни 10—36 Павлодар Хиргис-Нур 2, уровни 37—62 Хиргис-Нур 1 Чоно-Хариах 1,2 «Точка 1080 м> Дзабхан Явор Таталы Дзагсо-Хаир- хан 1 и 4 Хунг-Хурэ Шамар Береговая Тологой
Struthionidae
Struthio sp. (скорлупа) + + 4- 4- 4-
Struthio sp. (кости) +
Struthio asiaticus 4-
Podicepedidae
Podiceps solidus +
Phalacrocoracidae
Phalacrocorax reliquus + +
Phalacrocorax mongo- +
liensis
Ardeidae
Nyctanassa kobdoena + +
Ciconiidae
Ciconia lucida + + + +
Anatidae
Cygnus pristinus + + + + 4- 4-
Cygnus sp. + + + 4-
Anser devjatkini +
Anser liskunae + 4-
Anser tchikoicus 4-
Heterochen vicinus +
Bonibernicla ponderosa +
Tadorna petrina 4-
Tadorninae gen. 1 4-
Tadorninae gen. 2 4-
Anatidae subfam. 1 +
Anatidae subfam. 2 +
Anatidae subfam. 3 4-
Anatidae subfam. 4 +
Anas soporata + + 4- + 4-
Anas molesta + +
Anas sp. + + 4-
Aythya spatiosa +
Aythya magna +
Falconidae
Sushkinia pliocaena +
Accipitridae
Haliaeetus fortis +
Haliaeetus sp. 4-
Phasianidae
Alectoris sp. +
Bantamyx georgicus +
Syrmaticus kozlovae + +
Lophura inferna 4-
Perdix margaritae 4-
Plioperdix ponticus 4-
Palaeoperdix sp. 4-
Ergilornithidae
Amphipelargus dzab- + 4- 4-
ghanensis
8 Заказ 2098
105
Таблица 10 (окончание)
Систематический состав Нижний плиоцен Средний плиоцен Верхний плиоцен
Хиргис-Нур 2, уровни 10—36 Павлодар Хиргис-Нур 2, уровни 37—62 Хиргис-Нур 1 Чоно-Хариах 1,2 «Точка 1080 м> Дзабхан Явор Таталы Дзагсо-Хаир- хан 1 и 4 Хунг-Хурэ Шамар Береговая Тологой
Amphipelargus sp. Gruidae +
Probalearica mongolica 4- +
Rallidae
Palaeoaramides tuga- + + +
rinovi
Crex zazhigini +
Porzana risilla +
Otididae
gen.
Scolopacidae +
Limosa lacrimosa +
Tringa sp. +
Calidris sp. +
Limnodromus sp. +
Phalaropodidae
Phalaropus eleonorae +
Pteroclidae
Syrrhaptes kashini +
Syrrhaptes sp. + +
Strigidae
Asio aff. otus +
Corvidae
Corvus solitus +
Motacillidae
Anthus seductus +
Anthus sp. -1- +
Turdidae
Oenanthe infima +
Emberizidae gen. +
Alaudidae gen. +
Sturnidae gen. +
РАННИЙ ПЛИОЦЕН
Из отложений этого возраста птицы были найдены только в двух местонахожде-
ниях: Хиргис-Нур 2 — уровни 10—36 и Павлодар. Систематический состав их сущест-
венно различен — нет ни одной общей формы. Так же разнятся они и экологически.
С нижних уровней Хиргис-Нура 2 имеются исключительно водные и околоводные
птицы, а из Павлодара — только наземные птицы открытого ландшафта. Раннеплиоце-
новые птицы из Хиргис-Нура 2 фактически не отличаются от среднеплиценовых этого
и других соседних местонахождений Западной Монголии, все четыре идентифициро-
ванные до вида формы проходят в средний плиоцен, причем лебедь (С. pristinus),
крошечная утка (Anas soporata) и большой аист (Ciconia lucida) найдены выше
в нескольких местонахождениях среднего плиоцена, так^ же как и неопределенные
ближе многочисленные остатки Cygnus sp.; палеоарамид (Р. tugarinovi) встречен
106
также выше в двух среднеплиоценовых местонахождениях. Ясно, что этот ограничен-
ный авикомплекс слишком мал и случаен для каких-либо сравнений и заключений. Тем
не менее можно заметить, что остатки С. pristinus и A. soporata в нем были не еди-
ничны. В целом, тут не прослеживается никаких связей с птицами позднего миоцена
или раннего плиоцена других территорий, но имеется прямая преемственность по всем
линиям в систематическом и экологическом отношениях с птицами среднего плиоцена
этого же района.
Удаленный почти на тысячу километров к северо-западу от Хиргис-Нура ави-
комплекс Павлодара составлен в то же самое раннеплиоценовое время крупным страу-
сом (Struthio sp.), крупным амфипеларгусом (Amphipelargus sp.), древней куропаткой
(Palaeoperdix sp.), воробьиными открытых местообитаний (Anthus seductus, Anthus
sp., Alaudidae gen.), древним соколом Sushkinia, обитавшим скорее всего в открытых
местах. Sushkinia и Palaeoperdix (несколько видов которого описано из верхнего
миоцена Европы) подчеркивают древность этого комплекса. Крупные страус и амфипе-
ларгус, наоборот, сближают его с более молодыми комплексами птиц Центральной
Азии, так же как и Anthus sp., который найден и в среднем плиоцене «точки 1080 м»
в Западной Монголии.
СРЕДНИЙ ПЛИОЦЕН
В девяти местонахождениях этого возраста на территории Западной Монголии
имеются представители четырех экологических групп птиц — водные, околоводные,
наземные птицы открытых пространств и хищники. Большинство ископаемых птиц
относится к группе водных видов, это — поганка (Р. solidus), два вида бакланов (Ph.
reliquus и Ph. mongoliensis), лебеди (вероятно, все-таки два вида — Cygnus pristinus
и меньшего размера Cygnus sp.), по крайней мере три вида гусей: Anser devjatkini,
A. liskunae и Heterochen vicinus (вероятно, там существовали еще два—четыре вида
крупных и среднего размера гусей, от которых найдены только фрагментарные, ближе
не определимые, остатки), три вида пеганковых пластинчатоклювых (Bonibernicla
ponderosa, мелкая и крупная Tadorninae), три вида речных уток (Anas soporata, А.
molesta и Anas sp.) и два вида нырковых уток (Aythya spatiosa и A. magna). Всего,
таким образом, около 20 форм настоящих водных птиц, в основном обитателей
водоемов озерного типа. Если сопоставить этот список с современным населением
озерных птиц, например Центрального Казахстана, то можно видеть, что он факти-
чески исчерпывающе представляет полный набор видов, которые могли бы обитать на
крупных водоемах Западной Монголии в плиоцене. Ограниченно здесь представлены
только нырковые утки — единичные остатки двух видов. На фоне большого числа
остатков лебедей, гусей и речных уток этот факт не выглядит случайным, а в опреде-
ленной мере отражает естественную структуру авикомплекса. Очевидно, нырковые
утки были действительно бедно представлены в количественном и видовом отношениях
на плиоценовых озерах Западной Монголии, что можно объяснить только бедностью
в них бентоса, основного корма этих птиц. Это подтверждается данными по рыбам и
моллюскам, которые показывают, что от раннего к среднему плиоцену в озерных
бассейнах Западной Монголии произошло исчезновение многих групп теплолюбивых
рыб и моллюсков и общее обеднение водной фауны (Сычевская, 1983). Это было
связано с постепенным похолоданием климата, утерей связи западномонгольских во-
доемов с западносибирскими и китайскими и, как следствие этого, с некоторым засо-
лением до этого чисто пресноводных, хорошо аэрируемых и даже слабопроточных,
хотя и обширных, водоемов. Большая часть остатков птиц как раз и происходит из
отложений верхней части верхней подсвиты свиты Хиргис-Нур (верхов пачки А),
когда в полной мере проявились эти изменения. Разнообразие же растительноядных
водных птиц — лебедей, гусей, речных уток, наоборот, в этих условиях могло даже
увеличиться.
Околоводные птицы представлены ночной цаплей (Nyctanassa kobdoena), крупным
аистом (Ciconia lucida), монгольским журавлем (Probalearica mongolica), тремя
пастушковыми (Palaeoaramides, Crex zazhigini, Porzana risilla) и куликами (Limosa
lacrimosa, Tringa sp., Calidris sp.). Все данное разнообразие околоводных птиц скорее
8*
107
всего было связано с открытыми лугоподобными биотопами приозерных котловин.
В противоположность водным птицам большинство представителей данного комплекса,
за исключением куликов, в Западной Монголии в настоящее время не встречается. Так
же как и в ситуации с наземными птицами открытых ландшафтов, это указывает на
значительные отличия биотопических и климатических условий данной территории
в среднем плиоцене и в современности. При более теплом климате аридность и контра-
стность между водоемами и берегами в среднем плиоцене не была так резко выражена,
как сегодня, а существовала, очевидно, значительная переходная зона мелководий,
покрытых богатой травянистой растительностью, между открытой водой и настоящим
берегом, в которой могли обитать разнообразные околоводные птицы, очень немного-
численные в этих местах ныне, где сухая степь непосредственно граничит с водой озер
и рек. Причем в среднем плиоцене такая обстановка характеризовала весь этот обшир-
ный район в целом — от современного северного берега оз. Хиргис-Нур до центра
Шаргаин-Гоби, где найдены остатки нескольких видов пастушковых, аиста, журавля,
цапли и куликов.
Птицы наземных открытых местообитаний представлены страусом и амфипеларгу-
сом. И хотя костных остатков страусов в среднеплиоценовых местонахождениях
не найдено, широкая встречаемость скорлупы их яиц указывает на распространенность
страусов в открытых местообитаниях. Вместе с амфипеларгусами страусы в плиоцене
были обычным компонентом гиппарионовых фаун степных пространств Азии и Европы.
Эти крупные двупалые бегающие и совершенно нелетающие птицы могли жить только
в условиях отсутствия зимой устойчивого снежного покрова и всюду — от Восточной
Европы до Западной Монголии — сопутствуют друг другу от позднего миоцена до
среднего плиоцена. В Западной Монголии скорлупа яиц страусов появляется с позд-
него миоцена (Ошин-Боро), не встречается в раннем плиоцене, но в отложениях этого
времени из Павлодара имеются костные остатки крупных страусов. Широко распростра-
нена скорлупа яиц в местонахождениях среднего плиоцена, а в позднем плиоцене
в Забайкалье найдена как многочисленная скорлупа, так и одна кость очень крупного
страуса. Предварительный анализ материалов по скорлупе и по костным остаткам
страусов ведет сейчас к тому, что обширная степная зона Евразии на протяжении всего
плиоцена была населена одним видом страуса.
Амфипеларгус в Центральной Азии впервые становится известным с раннего плио-
цена Павлодара, где найдена фаланга крупного представителя этого рода. В Запад-
ной Монголии несколько хороших остатков задних конечностей самого крупного из
всех известных амфипеларгусов, выделенного в новый вид — A. dzabghanensis, име-
ются из трех среднеплиоценовых местонахождений.
В эволюции этого рода явственно прослеживается тенденция увеличения абсолют-
ных размеров от позднего миоцена к среднему плиоцену. Поэтому небольшие размеры
A. orientalis, описанного по многочисленным остаткам из местонахождения Калмакпай
в Восточном Казахстане (Курочкин, 1981), возраст которого принимается за средний
плиоцен, выглядят довольно странно, если к тому же учитывать отсутствие географи-
ческих преград в виде горных систем между Восточным Казахстаном и Западной
Монголией в среднем плиоцене.
Еще одна группа наземных птиц — фазановые, которых имеется три формы:
какой-то кеклик (Alectoris sp.), своеобразный бантамикс (Bantamyx georgicus),
структурно ближе всего стоящий к некоторым перепелам, и фазан Козловой (Syrmati-
cus kozlovae). Их остатки единичны, но они все-таки указывают на большее разнообра-
зие фазановых птиц в среднем плиоцене в Западной Монголии, чем сейчас. Кеклик
был обитателем каменистых или глинистых склонов, бантамикс скорее всего был связан
с равнинными степными участками, а для фазана Козловой требовались кустарнико-
вые заросли. К этому же комплексу можно отнести и имеющихся в этих местонахожде-
ниях воробьиных птиц — конька (Anthus sp.) и каменку (Oenanthe infima); плиоце-
новая ворона (Corvus solitus), если ориентироваться на нынешнюю эврибионтность
этого рода, могла жить в самых разнообразных условиях. Особое место занимают
находки трех остатков садж. Они относятся, похоже, к двум различным видам, если
параллелизовать степень их структурного сходства, отправляясь от современных видов
108
этого семейства. Все современные саджи связаны в своей жизни с открытыми арид-
ными ландшафтами, хотя во время миграций и на водопои могут залетать очень
далеко от мест своего нормального распространения.
Хищные птицы в силу тафономических причин составляют обычно очень небольшой
процент в ископаемых авикомплексах. Местонахождения среднего плиоцена Западной
Монголии в данном случае — не исключение. Здесь встречен только один фрагмент
совы (Asio aff. otus) и две кости орлана (Haliaeetus fortis).
ПОЗДНИЙ ПЛИОЦЕН
Остатки позднеплиоценовых птиц происходят из трех местонахождений Северной
Монголии и соседствующего с ней Забайкалья, поэтому рассматривать их можно как
один комплекс. Ныне и в прошлом эта территория существенно отличается в ланд-
шафтном и климатическом отношениях от Котловины Больших Озер на Западе Мон-
голии. Преемственность между этими позднеплиоценовым и среднеплиоценовым
западномонгольскими комплексами весьма относительна. Их связывает наличие страу-
сов (хотя и разных форм по строению скорлупы), разных видов родов Limosa и
Haliaeetus, но в общем между ними больше отличий, особенно по фазановым и гусе-
образным.
В общем виде в этом небольшом позднеплиоценовом авикомплексе видим те же
четыре экологические группы: водных (Anser techikoicus и Tadorna petrina), околовод-
ных (Limnodromus sp. и Phalaropus eleonorae) и наземных птиц (страус, дрофа,
Perdix margaritae и Plioperdix ponticus), к которым добавлен один хищник (Haliaeetus
sp.). Экологический облик фазана Lophura указать трудно, потому что в современной
обстановке все виды этого рода придерживаются тропических лесов.
ОБСУЖДЕНИЕ СТАТУСА ПЛИОЦЕНОВЫХ ПТИЦ
Большинство видов ископаемых птиц, исследованных в работе, относится к совре-
менным родам, но прямые родственные связи их с современными видами определяются
лишь в немногих случаях.
Проблема систематического состава и филогении настоящих двупалых страусов
в неогене, как было указано выше, требует сейчас специального рассмотрения с прив-
лечением всех накопленных материалов по костным остаткам и скорлупе. Однако,
очевидно, уже сейчас можно говорить об одном виде крупного настоящего Struthio,
обитавшего на протяжении всего плиоцена в Центральной Азии, вплоть до Забайкалья
на север в позднем плиоцене, и сохранявшегося на юге современной территории Мон-
голии и в Северном Китае до неолита или даже дольше. Этот страус хорошо система-
тически отличается по особенностям строения костей скелета от современного африкан-
ского страуса. Строение скорлупы плиоценового азиатского страуса на протяжении
нескольких миллионов лет жизни этого вида неоднократно менялось, что, очевидно,
было связано со сменами климата и экологических условий гнездования. Этот замеча-
тельный феномен установлен и доказан в самое последнее время (Курочкин, Михайлов,
1986).
Место маленькой Р. solidus в системе рода Podiceps из Хиргис-Нура 2 неясно.
Ископаемых поганок описано довольно много из неогена Западной Европы и Северной
Америки, но они представляют веер несвязываемых друг с другом форм, известных
по единичным фрагментам. Р. solidus — первая находка ископаемой поганки в Азии, не
проясняющая общей картины развития этой группы. В общем виде такой же вывод
можно сделать и о находке двух новых видов бакланов в среднем плиоцене Западной
Монголии, которых еще больше описано из неогена Европы и Северной Америки, один
из миоцена Японии. Общая картина с ними — большой спектр близких видов, почти
все — хорошие Phalacrocorax, родственные отношения между которыми не ясны ни
в неогене, ни в современности. Однако выглядит правдоподобным, что два среднеплио-
ценовых монгольских баклана связаны какими-то отношениями с двумя разными
видами современных бакланов: Ph. reliquus — с Ph. pelagicus, a Ph. mongoliensis —
c Ph. carbo. Интересно, что бакланы и поганки в палеонтологической истории птиц
109
появляются только в миоцене в виде отчетливых родов Phalacrocorax и Podiceps,
широко радиируют в плиоцене и сохраняют это разнообразие близких видов в совре-
менности, но из палеогена их остатки совсем неизвестны.
Nyctanassa kobdoena представляет вторую находку квакв в ископаемом состоянии,
первая была описана из нижнего плиоцена Флориды в США, причем она отнесена
к роду Nycticorax. Монгольская плиоценовая кваква достоверно ближе к Nyctanassa
violacea, чем к Nycticorax nycticorax, другой вопрос, насколько действительно эти
кваквы относятся к двум разным родам. По мнению некоторых систематиков, несколько
видов их в современной мировой фауне составляют только один род. Ископаемые
плиоценовые кваквы остеологически очень близки к современным видам.
Монгольский Ciconia lucida найден в четырех местонахождениях нижнего и сред-
него плиоцена. Это был один из самых крупных аистов среди современных и ископае-
мых видов рода. Морфологически все они очень сходны, но неплохо отличаются от
Mycteria, Jabiru и других родов семейства. Между разными видами рода Ciconia
имеются тонкие различия в деталях строения отдельных костей и с этих позиций
С. lucida, будучи близок по размерам к современному дальневосточному белому аисту
С. boyciana, отличается от последнего структурно. Два вида других вымерших Ciconia
известны сейчас из неогена Африки и Европы, а один — из плейстоцена Северной
Америки. В миоцене и плиоцене аисты имели тот же экологический тип и морфологиче-
ский облик, что и сейчас, и были не более разнообразны в позднетретичное время,
чем ныне.
Ископаемых гусеобразных описано к настоящему времени очень большое число
видов и родов как из палеогена, так и из неогена всех континентов, за исключением
Азии. До наших работ они были отсюда фактически неизвестны. Современные их роды
(такие, например, как Anas и Anser) появляются в палеонтологической летописи
сравнительно рано — с олигоцена. На этом фоне выглядело довольно странным полное
отсутствие находок собственно лебедей в неогеновых отложениях, хотя близкие к ним
роды Cygnanser и Cygnavus были известны из олигоцена Европы и Азии. Поэтому
многочисленные остатки Cygnus pristinus из шести местонахождений нижнего и сред-
него плиоцена Монголии представляют значительный интерес, показывая, что в первой
половине плиоцена этот вид был одной из обычных и многочисленных птиц на озерах
Центральной Азии. Будучи сравним по размерам с современными лебедями, он ясно
сочетает мелкие особенности в строении костей конечностей и плечевого пояса, свой-
ственные двум основным довольно различным морфологически, экологически и геогра-
фически группам современных лебедей — шипунов и кликунов. Мозаика их признаков
у С. pristinus такова, что не позволяет ни формально, ни статистически сблизить его
ни с одной из них. Все это вместе взятое заставляет считать, что этот вид был предком
двух основных линий современных лебедей: С. olor и С. cygnus — С. columbianus —
С. bewickii — С. buccinator во всей Голарктике. Очень возможно, что весь род Cygnus
возник в Центральной Азии, а отсюда уже только в плейстоцене распространился,
разделившись на несколько видов, по всему Северному полушарию. Ситуация с радиа-
цией к австралийскому С. atratus и южноамериканскому С. melanocoryphus за отсут-
ствием фактов выглядит неясной. На озерах Центральной Азии в плиоцене, возможно,
существовал второй более мелкий вид Cygnus.
Данные по настоящим гусям рода Anser из среднего плиоцена Западной Монголии
таковы, что показывают наличие в это время, с одной стороны, настоящих гусей
анзероидного облика, с другой — продолжающийся процесс адаптивной радиации
в этом роде, что подтверждается комплексным сочетанием отдельных деталей строения
многих современных видов рода у двух видов среднеплиоценовых гусей. Похоже, что
здесь были и другие их виды, от которых найдено несколько точно неопределенных
остатков. В несколько более позднее время — в позднем плиоцене из Забайкалья —
мы имеем A. tchicoicus, который уже определенно становится рядом только с одним
современным видом — с A. fabalis. Вместе с ними в плиоцене существовали и позднее
вымерли гуси другого рода — Heterochen. Они были распространены как в Северной
Америке, так и в Азии, где были представлены по крайней мере двумя видами.
Фауна крупных гусеобразных умеренных широт Северного полушария в прошлом
ПО
отличалась от современной более широким представительством крупного размера
видов подсемейства пеганковых Tadorninae. Это особенно ярко проявляется в Север-
ной Америке, где сейчас нет ни одного вида из этого подсемейства, но два их рода
описано из плейстоцена (четыре вида Anabemicula и монотипическая Brantadorna).
Все крупные гусеподобные представители пеганковых обитают сегодня в основном
в Южной Америке и на окружающих ее островах (Cyanochen, пять видов Chloephaga,
Neochen). Только в Африке распространен крупный Alopochen. А в Северном полу-
шарии Старого Света обитают сейчас только два вида средних размеров из рода Ta-
dorna. В среднем плиоцене Западной Монголии встречалась крупная Bonibernicla,
еще какая-то крупная Tadorninae и маленький представитель этого подсемейства. Эта
картина приближается к ситуации, существовавшей с этой группой птиц в неогене
Северной Америки.
Настоящие Anas представлены в плиоцене семи местонахождений Западной Мон-
голии тремя формами — очень маленькой A. soporata, от которой найдено самое боль-
шое число остатков, причем не только в среднем, но и в нижнем плиоцене; довольно
крупной A. molesta — из двух местонахождений и крупной Anas sp. — в трех место-
нахождениях. Это были настоящие Anas, от которых, однако, не протягивается связей
к каким-то конкретным современным видам. Род Anas известен со среднего олигоцена.
Из неогена всего мира описано 20 различных его видов, в общем довольно единообраз-
ных морфологически, что характерно и для современных Anas, которых насчитывается
в мировой фауне 37 видов. В неогене, начиная с раннего миоцена, настоящие речные
утки были уже широко распространены по всему миру.
На фоне большого разнообразия в современной фауне различных видов и родов
нырковых уток трибы Aythini, если к тому же учитывать обычно значительную числен-
ность и стайность некоторых видов, широкое представительство как в морских водах,
так и на пресных внутриматериковых водоемах, довольно бедное их представительство
в ископаемом состоянии не выглядит случайным. Так, в Северной Америке на обоих
побережьях известно несколько богатых местонахождений с морскими птицами и там
не найдено ни одной нырковой утки. Единичные их находки имелись из неогена кон-
тинентальных отложений Европы и Северной Америки, теперь к ним добавились еще
две встречи из Азии — Aythya magna и A. spatiosa. Эти косвенные факты и отсутствие
таковых позволяют предполагать, что нырковые утки в целом — сравнительно очень
молодая группа, бурно радиировавшая в самое недавнее время.
Остатки хищных птиц попадаются в ископаемом состоянии в силу тафономических
причин сравнительно нечасто. Наши материалы подтверждают это правило. Ранне-
плиоценовая павлодарская Sushkinia представлена по существу двумя особями,
так как, кроме голотипа, все остальные кости происходят почти наверняка от одного
экземпляра. Этот материал позволил подтвердить родовую специфичность Sushkinia,
которая выпукло проявляется на фоне остеологической однотипности современного
рода Falco, и показать, что, во всяком случае, в неогене существовали еще какие-то
другие сокола, кроме Falco. Морфология скелета Sushkinia не позволяет, на наш
взгляд, говорить о близости этого рода к каким-то конкретным видам современных
соколов, как об этом писал А. Я- Тугаринов (1935).
Единичная находка Haliaeetus fortis из среднего плиоцена, а затем Haliaeetus sp.
из позднего плиоцена Тологоя показывает только то, что в плиоцене в Азии уже были
настоящие орланы.
Фазановые птицы известны в ископаемом состоянии очень обширно. Их описано
около 10 видов из палеогена и около 40 видов из неогена Европы и Америки. Они
хорошо представлены в неогене Европы, откуда описано около 30 их видов. Но боль-
шинство этих птиц относится к родам Palaeocryptonyx, Palaeortyx, Miophasianus,
Palaeoperdix, генеалогические связи которых с современными фазановыми неясны,
систематика и описания запутаны и в настоящее время весь этот материал нуждается
в полной и серьезной ревизии. Из миоцена и плиоцена Европы имеются также остатки
фазановых, близких к современным их представителям, по нескольку видов в родах
Alectoris, Plioperdix, Ammoperdix, Gallus, Francolinus, которые свидетельствуют
о достаточной древности этих современных родов. В целом же создается впечатление,
111
что в позднем миоцене—раннем плиоцене фауна фазановых птиц Европы была гораздо
богаче и разнообразнее современной, но позднее большая группа каких-то необычных
куропаток вымерла, освободив место в открытых биоценозах куропаткам и кекликам,
близким к современным формам, а в лесах, может быть, тетеревиным.
В Северной Америке настоящих фазановых (Phasianinae) не было. Начиная, во
всяком случае, с раннего миоцена, там появились зубчатоклювые куропатки (Odonto-
phorinae), которые вместе с разнообразными Cracidae представляли фауну куриных
в неогене. Но одному виду каждого из этих подсемейств известно из олигоцена Север-
ной Америки и Европы, что на фоне богатства фауны краксовых (Cracidae) на обоих
континентах в палеогене не кажется случайной игрой тафономии, а заставляет счи-
тать, что фазановые и зубчатоклювые куропатки сменили на переходе от палео-
гена к неогену фауну краксовых как в Европе, так и в Америке. Краксы же до совре-
менности в достаточном разнообразии сохранились в тропических лесах Южной и
Центральной Америки. В современной фауне Азия, и в особенности Юго-Восточная
Азия, имеют наибольшее разнообразие фазановых птиц, представленных здесь малень-
кими и очень крупными птицами, лесными, пустынными и высокогорными формами.
Но именно из Азии палеонтологических остатков фазановых до недавних пор не было
совершенно.
Хотя и немногочисленные, но довольно разнообразные материалы по фазановым
из среднего плиоцена Западной Монголии (Alectoris sp., Bantamyx georgicus, Syrma-
ticus kozlovae и из верхнего плиоцена Монголии и Забайкалья (Lophura inferna,
Perdix margaritae, Plioperdix ponticus) показывают, что в плиоцене фауна фазановых
птиц была здесь, так же как и в Европе, разнообразнее, чем сейчас (по одному виду
родов Coturnix, Phasianus, Alectoris, Perdix и высокогорные Tetraogallus); она явст-
венно ближе к европейскому типу, чем к североамериканскому, но в то же время при-
сутствие родов Lophura, Syrmaticus и, вероятно, Bantamyx, характеризует ее как
специфически азиатскую. Находка единственной кости Palaeoperdix в нижнем плио-
цене Павлодара, с другой стороны, указывает, что связи фазановых птиц Европы и
Центральной Азии, хотя и не очень устойчивые, имеют, очевидно, довольно давний
характер. Не будет удивительно, если в миоцене Центральной Азии вдруг найдут
отдельных представителей сейчас специфически европейских родов Palaeortyx и
Miophasianus, как из плиоцена теперь стал известен Plioperdix.
Журавлеобразные в плиоцене Монголии представлены четырьмя семействами:
Ergilornithidae, Gruidae, Rallidae и Otididae. Все виды, описанные в их составе из
плиоцена Монголии, естественно и логично ложатся в канву представлений о большем
богатстве птичьего населения исследуемой территории в то время, на фоне более разно-
образных ландшафтных условий и более мягкого климата.
Находки остатков двупалых Amphipelargus в составе среднеплибценовой гиппарио-
новой фауны Западной Монголии, после того, как они были найдены в Калмакпае
в Восточном Казахстане, не вызывают удивления. Amphipelargus dzabghaensis был из
известных неогеновых амфипеларгусов самым крупным видом и, похоже, — одним из
самых последних в филогенетическом ряду этих птиц: Появились они в Центральной
Азии еще в раннем плиоцене, о чем говорит фаланга, достоверно относящаяся к пред-
ставителю этого рода, из местонахождения Павлодар. Более древних находок амфипе-
ларгусов отсюда неизвестно, все позднемиоценовые встречи привязаны к далеко
отстоящим на запад территориям Пакистана, Ирана, Украины, Молдавии и Греции.
Однако центром происхождения этой группы можно почти наверняка считать Цент-
ральную Азию, откуда из раннего олигоцена известны эргильорнисы и их вероятные
позднеэоценовые предки эогруиды.
Подробно история возникновения, филогения и морфология эргильорнетид рас-
смотрены в родовой характеристике Amphipelargus. Там же подчеркнуто, что нет
никаких оснований, кроме номинальной двупалости, производить род Struthio от
эргильорнитид. Значительные сомнения вызывает и теория неотенического происхожде-
ния всех Ratitae. Доказательство принципиальной разобщенности Ratitae и Carinatae,
основанное на онтогенезе тарзального сустава, было недавно хорошо обосновано
Макгоуном (McGowan, 1984]. Он установил, что на ранних этапах эмбрионального
Н2
развития внешне сходные отросток претарзальной кости у килевых птиц и восходящий
отросток бескилевых сливаются с совсем разными элементами тарзалий — соответ-
ственно с пяточной костью и с астрагалом. Причем у бескилевых это происходит по
гомологии с тероподными динозаврами.
Находка в среднеплиоценовых отложениях Западной Монголии нового вида рода
Probalearica подкрепляют формировавшуюся в последнее время точку зрения о глу-
бокой древности подсемейства Balearicinae, их прошлом разнообразии и о реликтовости
единственного современного вида африканского Balearica. Известные от среднего
эоцена до среднего плиоцена несколько видов родов Ornitocnemus, Probalearica и
Pliogrus, найденные в Северной Америке, Европе, а теперь и в Азии, показывают на
широкое разнообразие и длительное процветание древних Balearicinae. При этом
Eobalearica Gureev, 1949 по своей морфологии хорошо вписывается в характеристики
Ornitocnemus Zigno, 1876, становясь его синонимом. Родовое название Palaeogrus
Portis, 1884, как показал Бродкорб (Brodkorb, 1967), является младшим синонимом
Ornitocnemus Zigno, 1876, потому что они основаны на разных типовых видах, для
описания которых послужил один и тот же экземпляр.
Три вида трех разных родов пастушковых (Rallidae) представляют различных
мелких пастушков, два из которых относятся к современным родам (Сгех zazhigini
и Porzana risilla), а один — к вымершему роду Palaeoaramides. При этом последний
найден в нижнем и в среднем плиоцене, а представители современных родов — только
в среднем. Р. tugarinovi является самым маленьким по сравнению с четырьмя другими
ископаемыми видами этого рода, известными из верхнего олигоцена и нижнего миоцена
Европы. Теперь, таким образом, эти древние пастушки, сходные одновременно с совре-
менными Aramides и Rallus, найдены и в Азии, причем в более молодых отложениях
среднего плиоцена. Новый вид рода Сгех представляет первую находку коростелей
в третичных отложениях. Несколько вымерших видов Porzana до этого были найдены
только в плиоцене и плейстоцене Северной Америки. Может быть, не случайно пого-
ныши совсем неизвестны из хорошо изученного в отношении птиц неогена Европы.
Происхождение и адаптивная радиация этого рода была связана с другими конти-
нентами, но в Азии, как показывает наша находка, они были уже в среднем плиоцене.
Большого разнообразия их здесь быть не могло. Так же как и сегодня, эти широты
находились в плиоцене на периферии ареала рода, зоной развития которого могли быть
тропики Старого Света или американских континентов. В целом, по сравнению
с сегодняшним днем фауна пастушковых озерных котловин Западной Монголии была
разнообразнее, но объясняется это, по-видимому, в основном иной ландшафтной
обстановкой — мягким климатом, обводненностью территории, наличием обширных
мелководий, покрытых травянистой растительностью.
Дрофы (Otididae) известны по нескольким находкам в эоцене, миоцене и плиоцене
Европы. Слабая их представленность в ископаемом состоянии объясняется только
плохой изученностью в палеорнитологическом отношении Азии и Африки. Находка
единственной фаланги какой-то среднего размера дрофы в верхнем плиоцене Шамара
естественным образом расширяет их распространение в прошлом до Центральной
Азии. Нормально ожидать их находок в дальнейшем из других возрастов и других
районов азиатского континента.
Различные кулики (Charadrii), будучи околоводными птицами, неплохо известны
в ископаемом состоянии. Исчезнувшие семейства этого подотряда появляются уже
в отложениях верхнего мела. В эоцене и олигоцене появляются уже некоторые совре-
менные роды, например Limosa, Vanellus, Charadrius. В плиоцене в Северной Америке
и Европе они представлены главным образом вымершими видами современных родов.
Очевидно, фауна куликов плиоцена немногим отличалась в общих чертах от современ-
ной, характеризуясь таким же разнообразием экологических форм. Довольно редкие
остатки этой группы птиц из среднего и верхнего плиоцена Монголии и Забайкалья
показывают только, что здесь были распространены в то время Limosa, Tringa, Calid-
ris, Limnodromus и Phalaropus, близкие к современным видам.
Интересна находка садж. Их остатки из двух основных местонахождений среднего
плиоцена, вероятно, относятся к двух разным видам. Саджи (Syrrhaptes) ранее
ИЗ
фактически были неизвестны в ископаемом состоянии, хотя три вида рябков (Ptero-
cles) описаны из эоцен—олигоцена и миоцена Франции. Наши находки подтверждают
древние неогеновые связи рода Syrrhaptes с Центральной Азией, где сейчас распро-
странены два вида этого рода, обитающие исключительно в открытых аридных ланд-
шафтах. Из двух ископаемых форм одна очень близка к современной S. paradoxus,
а вторая заметно от нее отличалась.
Единственная находка фрагмента грудины совы (Asio), сходной с современной
A. otus, указывает на присутствие такого вида на западе Монголии в среднеплиоце-
новое время.
Немногие описанные в настоящей работе воробьиные (Passeriformes) представ-
ляют, тем не менее, существенный интерес при недостатке сведений по палеонтологи-
ческой истории этого отряда, что связано во многом не только с редкостью их ископае-
мых остатков, но и с трудностями, вызванными изучением уже имеющихся материалов
из-за огромного разнообразия мелких современных форм и отсутствия достаточных
сравнительных коллекций (Janossy, 1983). Большое количество ископаемых остатков
неогеновых воробьиных птиц накопилось, как известно, во многих музеях мира, но
они лежат неизученными. Среди наших материалов большое их число осталось пока
неописанными из верхнеплиоценового местонахождения Береговая, где они составляют
подавляющее большинство материалов по птицам.
В целом, на фоне немногих известных сейчас данных все-таки можно сказать, что
воробьиные — не такая уж молодая группа, как принято считать. Они появляются
в палеонтологической летописи с палеогена, а в миоцене и плиоцене представлены уже
современными родами, виды которых не отличаются кардинально от современных.
Поэтому можно предполагать, что их адаптивная радиация приходилась также на
палеоген, как и в большинстве других современных отрядов птиц.
В дополнение к трем или четырем вымершим видам рода Corvus, известным до
этого только из плиоцена и плейстоцена Европы, наше описание С. solitus показывает
присутствие еще одного вида в среднем плиоцене Азии. Находки ископаемых Anthus
и Oenanthe представляют первые данные по этим родам из третичных отложений.
Они показывают виды, мало отличавшиеся от современных. Среди них какой-то
конек (Anthus sp.) встречался как в раннем (Павлодар), так и в среднем («точка
1080 м») плиоцене. Вымерший Anthus seductus, найденный в нижнем плиоцене (Павло-
дар), ближе всего оказался к современному A. novaeseelandiae, имеющему огромный
ареал сейчас в Азии, Австралии и в Африке. Неопределимые ближе остатки овсянки
(Emberizidae gen.) и крупного жаворонка (Alaudidae gen.) найдены в нижнем плио-
цене Павлодара, а какой-то скворец (Sturnidae gen.) в верхнем плиоцене Шамара.
СТРАТИГРАФИЧЕСКИЕ ДАННЫЕ
Вопрос о возрасте фаунистических горизонтов Хиргис-Нура, Чоно-Хариаха и дру-
гих местонахождений этого ряда в Западной Монголии, естественно, не может быть
решен на палеорнитологических материалах, но данные по птицам все-таки позволяют
провести некоторые параллели между хорошо известными авикомплексами неогена
Европы и Азии. В Европе известен ряд местонахождений с достаточным количеством
верхнемиоценовых птиц — Ла-Грив-Сант-Альбан, Сансан, Штейнхайм, Регенсбург,
Варница и др. Их фауна птиц очень специфична почти полным отсутствием совре-
менных родов (кроме Anas и Phalacrocorax). Там представлены разнообразные
Palaeortyx, Palaeocryptonyx, Palaeoperdix, Miophasianus, Miofulica, Miorallus и др.
В отчетливо понтических местонахождениях Европы появляются в большом количе-
стве Pliogrus, Amphipelargus, Gallus, Struthio, Anser, Rallus, Pelecanus, Ciconia,
Podiceps, Falco, Alauda и др. В плиоценовых местонахождениях Западной Монголии
также преобладают современные роды, которые вместе с немногими вымершими
(Bonibernicla, крупный Amphipelargus, Probalearica) относят время формирования их
отложений к этапу более позднему, чем ранний плиоцен, т. е. к среднему плиоцену.
114
О более молодом возрасте западномонгольского плиоцена по сравнению, например,
с плиоценом Восточного Казахстана (Калмакпай) говорят и находки очень крупного
Amphipelargus. В общем этот авикомплекс имеет облик немного моложе понтического.
Данные по птицам самых древних горизонтов Хиргис-Нура совсем незначительны,
но они не показывают абсолютно никакой разницы между птицами этих и вышележа-
щих отложений. В то же время отложения Павлодара представляются древнее запад-
номонгольских, так как содержат своеобразных Sushkinia, Palaeoperdix, Anthus и
крупного Struthio. Авикомплекс чикойской свиты носит отчетливо верхнеплиоценовый
характер, подчеркнутый присутствием Plioperdix, Anser — очень близкого к современ-
ному виду, очень крупного Struthio, дрофы, современного рода Perdix, не встречав-
шегося никогда до этого древнее, близких к современным Limnodromus и Phalaropus.
Прослеживается некоторая связь этого комплекса с авифауной «молдавского рус-
сильона» Европы.
* * *
В плиоцене Центральной Азии установлено 56 форм птиц 12 отрядов, из которых
36 описаны до видового ранга, а остальные — до уровня родов и семейств. В раннем
плиоцене найдено 12 форм, в среднем — 41 и в позднем — 13. Ряд видов птиц проходят
без изменений из раннего плиоцена Западной Монголии в средний. Раннеплиоценовые
птиц Павлодара заметно отличаются от плиоценовых западномонгольских.
В целом среднеплиоценовый авикомплекс Западной Монголии кажется довольно
сходным с современным, но полностью от него отличается на видовом уровне, специ-
фичен более богато представленными баклановыми, пеганковыми, фазановыми,
пастушковыми, наличием вымерших страуса, амфипеларгуса, пробалеарики, палеора-
мида, бантамикса. Все данное разнообразие ископаемых видов и родов характеризуют
авифауну этого региона в среднем плиоцене как палеарктическую, свидетельствуя
о возникновении и формировании ее с этого времени, поскольку, если брать хорошо
известную верхнемиоценовую авифауну Западной Европы, то в ней тогда было значи-
тельное число индомалайских и эфиопских элементов. Отдельные же роды (Heterochen,
Nyctanassa, Probalearica, Syrmaticus) указывают на наличие связей между птицами
Центральной Азии с Южной Азией и Америкой.
Позднеплиоценовые птицы Забайкалья и Северной Монголии демонстрируют сла-
бую преемственность их от среднеплиоценовой авифауны, их объединяет только нали-
чие Struthio, Anser и, вероятно, линии Bantamyx—Plioperdix. В то же время этот
комплекс слабо связан и современным, который сформировался в основных своих
чертах, очевидно, позже. Это подтверждают и свежие идеи в изучении географического
распределения современных животных и формирования типов фаун Палеарктики
(Назаренко, 1979; Матюшкин, 1982). На основе наших данных представляется, что
явление дизъюнкции ареалов имеет исторически древнее происхождение, связанное
с особенностями географии и распределения растительности в Старом Свете. Примеры
по амфипеларгусу, палеопердиксу, плиопердиксу, страусу указывают на вероятное
существование дизъюнкций и кольцевых ареалов еще в среднем плиоцене.
По сравнению с современным среднеплиоценовый западномонгольский ави-
комплекс очень сходен по составу водных птиц, хотя и отличается от него добавлением
некоторых специфических гусеобразных. Возможно, эти птицы тогда, как и сейчас,
широко мигрировали, но в целом, если рассматривать всю изученную ископаемую
авифауну, черта оседлости здесь в то время была выражена значительно сильнее.
Мы встречаем разнообразных фазановых, страуса, амфипеларгуса, которые, наверняка
не совершали перелетов, к ним могли относиться и некоторые пастушковые.
115
ЗАКЛЮЧЕНИЕ
Из 11 местонахождений МНР, 2 местонахождений в Восточном Казахстане
и 2 местонахождений в Забайкалье раннего, среднего и позднего плиоцена описана
коллекция ископаемых остатков птиц 56 форм, относящихся к 23 семействам. Большин-
ство видов птиц относится к вымершим, большинство родов птиц представлены
современными таксонами; все семейства, за исключением Ergilornithidae, существуют
и поныне. Вымершие роды птиц в этих фаунах Heterochen, Bonibernicla, Bantamyx,
Probalearica, Palaeoramides, Sushkinia, Plioperdix и Palaeoperdix представлены един-
ственными видами. Они не формировали основы авифаунистических комплексов сред-
него и позднего плиоцена, общий облик которых определялся широким набором ныне
живущих родов птиц, представленных в плиоценовых комплексах Центральной Азии
двумя или большим числом видов, например, у Phalacrocorax, Anser, Aythya, Anthus.
От других найдены только по одному виду в каждом, таких много — Podiceps,
Nyctanassa, Ciconia, Haliaeetus, Alectoris, Syrmaticus, Lophura, Crex, Limosa, Syrrhap-
tes, Corvus и др. Некоторые из них, а также вымершие моноспецифические роды отра-
жали в плиоценовых фаунах Центральной Азии наследие более древних авифаунистиче-
ских комплексов этих территорий. Исчезли они в позднем плиоцене или даже
в плейстоцене. Вместе с этим в плиоцене в Центральной Азии в комплексе водных
птиц была более распространена и разнообразна группа пеганковых гусей. В авифау-
нах плиоцена Центральной Азии, так же как и Северной Америки, это подсемейство
гусеобразных определенно имело больший удельный вес, чем ныне. Не только пеган-
ковые, но и некоторые другие птицы — Nyctanassa, Heterochen, Probalearica, Palaeo-
aramides — указывают на сохранение в плиоцене Центральной Азии более древних
связей с авифауной Северной Америки. О взаимовлиянии авифаун этих континентов
во второй половине неогена говорить не приходится. У отдельных плиоценовых видов
птиц прослеживается тесное сходство и, вероятно, филогенетическая близость к опре-
деленным современным видам. Особенно интересен в этом отношении Cygnus pristinus,
которого можно принять за предка двух современных линий лебедей — кликунов и
шипунов. За предков конкретных современных видов птиц можно принять Podiceps
solidus, Ciconia lucida, Anser devjatkini, Anser tchikoicus, Anas soporata, Lophura
interna, Syrmaticus kozlovae, Crex zazhigini, Anthus seductus, Oenanthe intima.
В целом от раннего до позднего плиоцена в авифауне Центральной Азии на имеющихся
материалах каких-то заметных изменений не улавливается. Общий ее облик был в зна-
чительной степени сходен с современным, но экологически это была авифауна более
гумидных, теплых и покрытых более богатой растительностью территорий, обитавшая
Определенно в условиях отсутствия снежной зимы. Стратиграфическое значение птиц
в этих условиях для расчленения континентальных отложений оказывается невелико.
Из местонахождений раннего плиоцена Котловины Больших Озер в среднеплиоценовые
местонахождения этого региона проходит ряд одних и тех же видов птиц.
116
ЛИТЕРАТУРА
Агаджанян А. К., Ербаева М. А. Позднекайно-
зойские грызуны и зайцеобразные террито-
рии СССР. М.: Наука, 1983. 189 с.
Александрова Л. П., Вангенгейм Э. А. и др.
Новые данные о разрезе антропогеновых
отложений горы Тологой (Западное Забай-
калье) . — Бюл. Комис, по изуч. четвертичн.
периода, АН СССР, 1963, № 28, с. 65—73.
Аубекерова П. А. Новые данные об ископаемых
птицах Казахстана. — В кн.: Новости орни-
тологии. Алма-Ата, 1965, с. 18, 19.
Базаров Д. Б., Ербаева М. А. и др. Геология и
фауна опорных разрезов антропогена Запад-
ного Забайкалья. М.: Наука, 1976. 124 с.
Бендукидзе О. Г. О находке Urmiornis mara-
ghanus Mecq. на Северном Кавказе. — Сообщ.
АН ГрузССР, 1972, т. 66, № 3, с. 741—743.
Бурчак-Абрамович И. И. Урмиорнис (Urmior-
nis maraghanus Mecq.) страусоподобная пти-
ца гиппарионовой фауны Закавказья и Юж-
ной Украины — Изв. АН АзССР, 1951, № 6,
с. 83—94.
Бурчак-Абрамович Н. И. Верхнетретичные пти-
цы Кавказа. — Сообщ. АН ГрузССР, 1966,
т. 44, № 3, с. 651—656.
Бурчак-Абрамович Н. И. История развития
орнитофауны Грузии и прилегающих мест-
ностей (третичный период). — Изв. АН
ГрузССР. Сер. биол., 1978, т. 4, № 6, с. 529—
535.
Вангенгейм Э. А. Палеонтологическое обосно-
вание стратиграфии антропогена Северной
Евразии. М.: Наука, 1977. 170 с.
Вислобокова И. А. Ископаемые олени Монго-
лии. — Тр. Совм. Сов.-Монгол, палеонтол.
экспедиции, 1983, вып. 23, с. 78.
Девяткин Е. В. Геология кайнозоя Западной
Монголии. — В кн.: Геология мезозоя и кай-
нозоя Западной Монголии. М.: Наука, 1970,
с. 44—102.
Девяткин Е. В. Кайнозой Внутренней Азии.
М.: Наука, 1981. 196 с.
Девяткин Е. В., Жегалло В. И. Новые данные
о местонахождениях неогеновых фаун се-
веро-западной Монголии. — Тр. Совм. Сов.-
Монгол. палеонтол. экспедиции, 1974, вып. 1,
с. 320—356.
Девяткин Е. В., Зажигин В. С. Эоплейстоцено-
вые отложения и новые местонахождения
фауны млекопитающих Северной Монго-
лии. — Тр. Совм. Сов.-Монгол, палеонтол.
экспедиции, 1974, вып. 1, с. 357—363.
Дементьев Г. П. Класс Aves. Птицы. — В кн.:
Основы палеонтологии. Земноводные, пре-
смыкающиеся, птицы. М.: Наука, 1964,
с. 660—699.
Дмитриева Е. Л. Антилопы неогена Монголии
и сопредельных территорий. — Тр. Совм.
Сов. Монгол, палеонтол. экспедиции, 1977,
вып. 6. 120 с.
Дуброво И. А., Капе лист К. В. Каталог место-
нахождений третичных позвоночных УССР.
М.: Наука, 1979. 159 с.
Жегалло В. И. Гиппарионы Центральной
Азии. — Тр. Совм. Сов.-Монгол, палеонтол.
экспедиции, 1978, вып. 7, 156 с.
Кашин Г. Н. О названиях некоторых таксонов
класса птиц. — В кн.: Компоненты биоты и
их роль в народном хозяйстве: Докл. МОИП,
зоология и ботаника, 1978 г. М.: Наука,
1981, с. 12—14.
Козлова Е. В. Новые ископаемые птицы из
Юго-Восточной Гоби. — В кн.: Тр. проблемн.
и тематич. совещ. ЗИН АН СССР, 1960,
вып. 9, с. 323—329.
Курочкин Е. Н. Основные вопросы изучения
ископаемых птиц. — В кн.: Итоги науки.
Зоология позвоночных. Вопросы орнитоло-
гии. М.: ВИНИТИ, 1971, с. 116—151.
Курочкин Е. Н. К авифауне плиоцена Монго-
лии. — Тр. Совм. Сов.-Монгол. научно-исслед.
геол, экспедиции, 1971, вып. 3, с. 58—67.
Курочкин Е. Н. Обзор палеогеновых птиц
Азии. — В кн.: Орнитология. М.: МГУ, 1974,
вып. 11, с. 317—328.
Курочкин Е. Н. Новые данные о птицах плио-
цена Западной Монголии. — Тр. Совм. Сов.
Монгол, палеонтол. экспедиции, 1976, вып. 3,
с. 51—67.
Курочкин Е. Н. Птицы среднего плиоцена За-
падной Монголии. — В кн.: VII Всесоюз.
орнитол. конф.: тез. докл. Киев: Наук, думка,
1977, ч. 1, с. 12, 13.
Курочкин Е. Н. Методы изучения ископаемых
птиц. — В кн.: Частные методы изучения
современных экосистем. М.: Наука, 1979,
с. 152—163.
Курочкин Е. Н. Новые представители и эволю-
ция двух семейств архаичных журавлеобраз-
ных в Евразии. — Тр. Совм. Сов.-Монгол.
палеонтол. экспедиции, 1981, вып. 15, с. 59—
85.
Курочкин Е. Н. Новые птицы из среднего плио-
цена Монголии. — В кн.: Орнитология. М.:
МГУ, 1982, вып. 17, с. 150—154.
Курочкин Е. Н., Ганя И. М. Птицы среднего
сармата Молдавии. — В кн.: Позвоночные
неогена и плейстоцена Молдавии. Кишинев:
Штиинца, 1972, с. 45—70.
Курочкин Е. И., Лунгу А. Н. Новый страус из
среднего сармата Молдавии. — Палеонтол.
журн., 1970, № 1, с. 118—126.
117
Курочкин Е. И., Михайлов К Е. Скорлупа
ископаемых Struthioniformes Палеарктики и
ее место в системе представлений об эволю-
ции Ratitae. М., 1986.
Лискун И. Г. Бадамгарав Д. Литология кайно-
зоя Монголии. — Тр. Совм. Сов.-Монгол.
научно-исслед. геол, экспедиции, 1977,
вып. 20, 159 с.
Логачев Н. А., Ломоносова Т. К., Клима-
нова В. М. Кайнозойские отложения Иркут-
ского амфитеатра. М.: Наука, 1964. 169 с.
Матюшкин Е. Н. Региональная дифференциа-
ция лесной фауны Палеарктики в прошлом и
настоящем. — В кн.: Теоретические и при-
кладные аспекты биогеографии. М.: Наука,
1982, с. 59—80.
Назаренко А. А. К истории орнитофауны суб-
альпийского ландшафта гор. Сибири и
Дальнего Востока. — Зоол. журн., 1979,
т. 68, вып. 11, с. 1680—1691.
Певзнер М. А., Вангенгейм Э. А. и др. Корре-
ляция отложений позднего неогена Цент-
ральной Азии и Европы по палеомагнитным
и биостратиграфическим данным. — Изв.
АН СССР. Сер. геол., 1982, № 6, с. 5—16.
Сычевская Е. К. История формирования их-
тиофауны Монголии и проблема фаунисти-
ческих комплексов. — В кн.: Рыбы Монголь-
ской Народной Республики. М.: Наука, 1983,
с. 225—249.
Тугаринов А. Я. Некоторые данные для плиоце-
новой орнитофауны Сибири. — Тр. Палео-
зоол. ин-та, 1935, т. 4, с. 79—89.
Тугаринов А. Я. Новые находки плиоценовой
орнитофауны Одессы. — Докл. АН СССР,
1940, т. 26, № 3, с. 311—313.
Уманская А. С. Новая находка остатков ур-
миорниса из неогеновых отложений Укра-
ины. — Вести, зоол., 1973, вып. 4, с. 30—33.
ХозацкийЛ. И., Курочкин Е. Н. Материалы к по-
знанию плиоценовой орнитофауны Молда-
вии. — В кн.: VII Всесоюз. орнитол. конф.: тез
докл. Киев: Наук, думка, 1977, ч. 1, с. 31—33.
Штегман Б. К. О филогенетических взаимоот-
ношениях между семействами Gaviidae и Ро-
dicipedidae. — В кн.: Орнитология. М.:
Изд-во МГУ, 1974, вып. И, с. 6—19.
Ballmann Р. Die Vogel aus der altburdigalen
Spaltenfiillung von Eichstatt in Bayern. —
Zitteliana, 1969a, Bd. 1, S. 5—60.
Ballmann P. Les oiseaux miocenes de la Grive-
Saint-Alban (Isere). — Geobios, 1969b, N 2,
p. 157—204.
Beer G. de.The evolution of ratites. — Bull. Brit.
Mus. (Natur. Hist.), 1956, vol. 4, p. 59—70.
Brodkorb P. A pliocene grebe from Florida. —
Ann. and Mag. Natur. Hist., Ser. 12, 1953, vol.
6, p. 953.
Brodkorb P. Catalogue of fossil birds. — Bull.
Fla St. Mus., 1963a, vol. 7, N 4, p. 180—293.
Brodkorb P. Miocene birds from the Hawthorne
formation. — Quart. J. Fla Acad. Sci., 1963b,
vol. 26, N 2, p. 159—167.
Brodkorb P. Catalogue of fossil birds. Pt 2. —
Bull. Fla St. Mus. Biol. Sci., 1964, vol. 8,
N 3, p. 195—335.
Brodkorb P. Catalogue of fossil birds. Pt 3. —
Bull. Fla St. Mus. Biol. Sci., 1967, vol. 11,
N 3, p. 99—220.
Brodkorb P. Catalogue of fossil birds. Pt 4. —
Bull. Fla St. Mus. Biol. Sci., 1971a, vol. 15,
N 4, p. 163—266.
Brodkorb P. Origin and evolution of birds. —
In: Avian biology. N. Y.; L.: Acad, press,
1971b, vol. 1, p. 19—55.
Brodkorb P. Catalogue of fossil birds. Pt 5. —
Bull. Fla St. Mus. Biol. Sci., 1978, vol. 23,
N 3, p. 139—228.
Brodkorb P. A new fossil heron (Aves: Ardei-
dae) from the Omo Basin of Ethiopia with
remarks on the position of some other species
assigned to the Ardeidae. — Natur. Hist. Mus.
Los Angeles County. Contribs Sci., 1980,
N 330, p. 87—92.
Broom R. On the early development of the
appendicular skeleton of the ostrich with re-
marks on the origin of birds. — Trans. S. Afr.
Philos. Soc., 1906, vol. 16, pt 4, p. 355—368.
Cracraft J. Systematics and evolution of the
Gruiformes (Class Aves). 3. Phylogeny of the
suborder Grues. — Bull. Amer. Mus. Natur.
Hist., 1973, vol. 151, N 1, p. 1 — 127.
Davies W. On some fossil bird remains from
the Siwalik Hills in the British Museum. —
Geol. Mag. N. S., 1880, dec. 2, vol. 7, N 1,
p. 18—27.
Deperet Ch. Les animaux pliocenes du Roussil-
lon. — Mem. Soc. geol. France. Paleontol.,
1890, N 3, p. 127—139.
Feduccia A. The age of birds. Cambridge
(Mass.); London: Harvard Univ, press, 1980.
196 p.
Hamon J. H. Osteology and paleontology of the
passerine birds of the Reddick, Florida, Pleis-
tocene. — Fla Geol. Surv. Geol. Bull., 1964,
N 44, VII 4- 210 p.
Harrison C. J. O. A re-assignment of Amphipe-
largus majori from Ciconiidae (Ciconiiformes)
to Ergilornithidae (Gruiformes). — Tertiary
Res., 1981, vol. 3, N 3, p. 111 — 112.
Harrison C. J. O., Walker C. A. Fossil birds
from the Upper Miocene of Northern Pakis-
tan. — Tertiary Res., 1982, vol. 4, N 2, p. 53—
59.
Howard H. The avifauna of the Emeryville
shellmound. — Univ. Cal. Pubis Zool., 1929,
vol. 32, N 2, p. 301—394.
Howard N. Fossil birds from the Anza-Borrego
desert. — Natur. Hist. Mus. Los Angeles
County. Contribs Sci., 1963, N 73, p. 1—33.
Howard H. A new species of the ’’Pigmy goose’’
Anabernicula, from the Oregon Pleistocene,
with a discussion of the genus. — Amer. Mus.
Novit., 1964, N 2200, dec. 15, p. 1 —14.
Janossy D. Humeri of Central European smaller
passeriformes. — Fragm. miner, et palaeon-
tol., 1983, vol. 11, p. 85—112.
Komarek V. Anatomia avium domesticarum.
Pt 1. Br.: Priroda, 1979. 16.2 s.
Kretzoi M. Pliogallus Gaillard, 1939 es Plio-
gallus Tugarinov, 1940. — Aquila, 1955,
vol. 59/62, p. 367.
Kretzoi M. Bird-remains from the Hipparion-
fauna of Csakvar. — Aquila, 1957, vol. 63/64,
p. 239—248.
Kurotchkin E. N. New data on Pliocene birds
of the Soviet Union. — In: XV congr. intern.,
ornithol. Abstrs. Hague, 1970, p. 146—147.
Kurochkin E. N. A survey of the Paleogene
118
birds of Asia. — Smithsonian Contribs Paleo-
bioL, 1976, N 27, p. 75—86.
Kurochkin E. N. The birds of the middle Plio-
cene of the West Mongolia. — In: XVII Congr.
intern, ornithol. Abstrs. Berlin (West), 1978,
p. 4—5.
Kurochkin E. N. Middle Pliocene rails from
Western Mongolia. — Natur. Hist. Mus. Los
Angeles County. Contribs Sci., 1980, N 330,
p. 69—73.
Kurockin E. N. On the evolutionary pathways of
didactylous tertiary gruids under increasing
aridization. — In: Evolution and environ-
ment. Pr.: CSAV, 1982, p. 731—736.
Lambrecht K. Aves: Fossilium Catalogus. I.
Animalia, Ps 12. B., 1921.
Lambrecht K. Handbuch der Palaeornitholo-
gie. B.: Verl. G. Borntrager, 1933. XIX +
+ 1024 S.
Lydekker R. Catalogue of the fossil birds in the
British Museum. L.: Taylor and Fransis,
1891. XXVII+368 p.
Mecquenem R. Contribution a 1’etude du gise-
ment des vertebres de Maragha et de ses
environs. — Bull. Mus. nat. hist, natur. Pale-
ontol., 1908, t. 1. p. 27—79.
Mecquenem R. Contribution a 1’etude des fossi-
les de Maragha. — Ann. paleontoi. vertebr.,
1925, t. 14, p. 55, 56.
McGowan C. Evolutionary relationships of rati-
tes and carinates: Evidence from ontogeny of
the tarsus. — Nature, 1984, vol. 307, N 5953,
p. 733—735.
Miller A. H. The whistling swan in the Upper
Pliocene of Idaho. — Condor, 1948, vol. 50,
N 3, p. 132.
Miller L. A new cormorant from the Miocene of
California. — Condor, 1929, vol. 31, N 4,
p. 167—172.
Miller L. The Pleistocene storks of California. —
Condor, 1932, vol. 34, p. 212—216.
Milne-Edwards A. M. Recherches anatomiques et
paleontologiques pour servir a I’histoire des
Oiseaux fossiles de la France. P.: Victor
Masson et Fils, 1867, 1868. T. I. 474 p.
Milne-Edwards A. M. Recherches anatomiques
et paleontologiques pour servir a I’histoire des
Oiseaux fossiles de la France. P.: Libr. de
G. Masson, 1869—1871. T. II. 632 p.
Mlikowsky J. Relatioships of the Eocene bird
„Numenius” gypsorum Gervais. — Bull. Mus.
Natur. Hist. Ser. 4, 1981, vol. 3, sect. C, N 4,
p. 341—343.
Morony J. J., Jr., Bock W. J., Farrand J., Jr.
Reference list the birds of the World. N. Y.:
Amer. Mus. Natur. Hist., 1975. 207 p.
Mourer-Chauvire C. Les oiseaux du pleistocene
moyen et superieur de France. — Doc. Lab.
geol. Fac. sci. Lyon, 1975, N 75, fasc. 1,
p. 261. .
Murray B. A. redescription of two Pliocene cor-
morants. — Condor, <1970, vol. 72, N 3
p. 293—298.
Nomina anatomica avium / Ed. J. J. Baumel et
al.,L.; N. Y.: Acad, press, 1979. XXV + 637 p.
Olson S. A synopsis of the fossil Rallidae. — In:
Rails of the World. Boston: David R. Godine
Publ., 1977, p. 339—373.
Olson S. A selective synopsis of the prequater-
nary fossil record of birds. — In: Avian bio-
logy. N. Y.; L.: Acad, press, 1984, vol. 8,
o. 117—138.
Portis A. Gli ornitoliti del Valdarno superiore e
di alcune altre localita plioceniche di Toscana.
Firenze: Tip. Successor! Le Monnier, 1891.
20 p.
Regalia E. Sette uccelli pliocenici del Pisano e
del Valdarno superiore. — Paleontogr. Ital.,
1902, vol. 8, p. 219—238.
Sauer F. G. E. Ratite eggshells and phylogene-
tic questions. — Bonn. zoll. Beitr., 1972,
Bd. 23, S. 3—48.
Sauer F. E. G. Aepyornithoide eierschalen aus
dem Miozan und Pliozan von Anatolien,
Tiirkei. — Palaeontographica A, 1976, Bd.
153, N 1/3, S. 62—115.
Sauer F. E. G. A Miocene ostrich from Anato-
lia.— Palaeontographica A, 1979, Bd. 121,
N 4, S. 494—501.
Sauer F. E. G., Sauer E. M. Ratite eggshell
fragments from Mio-Pliocene continental sedi-
ment in the district of Quarzazate Morocco. —
Palaeontographica A, 1978, Bd. 161, N 1/3,
S. 1—54.
Short L. L., Jr. A new genus and species of
gooselike swan from the Pliocene of Neb-
raska.— Amer. Mus. Novit., 1969, N 2369,
p. 1—7.
Short L. A new anseriform genus and species
from the Nebraska Pliocene. — Auk, 1970a,
vol. 87, N 3, p. 537—543.
Short L. Mid-Pleistocene birds from western
Nebraska, including a new species of sheld-
goose. — Condor, 1970b, vol. 72, N 2, p. 147—
152.
Stolpe M. Colymbus, Hesperornis, Podiceps:
Ein fergleich ihrer hinteren Extremitat. —
J. Ornithol., 1935, Bd. 83, S. 115—128.
Storer R. W. Courtship and mating behavior and
the phylogeny of the grebes. — In: Proc. XIII in-
tern. ornithol. congr., Ithaca. N. Y., 1963,
p. 562—569.
ivec P. Lower Miocene birds from Dolnice
(Cheb basin), western Bohemia. Pt II. — Cas.
miner, a geol., 1981, roc. 26, c. 1, S 45—56.
Suec P. Two new species of diving birds from
the Lower Miocene of Czechoslovakia. — Cas.
miner, a geol., 1982, roc. 27, c. 3, s. 243—259.
Wetmore A. Fossil bird remains from the Temb-
lor formation near Bakersfield, California. —
Proc. Cal. Acad. Sci. Ser. 4, 1930, vol. 19,
N 8, p. 85—93.
Wetmore A. The eared grebe and other birds
from the Pliocene of Kansas. — Condor, 1937,
vol. 39, N 1, p. 40.
Woolfenden G. E. Postcranial osteology of the
waterfowl. — Bull. Fla St. Mus. Biol. Sci.,
1961, vol. 6, N 1, p. 1 — 129.
Zusi R. L., Storer R. W. Osteology and myology
of the head and neck of the Pied-billed grebes
(Podilymbus). — Mus. Zool. Univ. Mich.,
1969, N 139, p. 49.
ОГЛАВЛЕНИЕ
ВВЕДЕНИЕ........................................................... 5
МАТЕРИАЛ И МЕТОДИКА................................................ 7
ГЕОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА МЕСТОНАХОЖДЕНИЙ...................... 10
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ............................................. 14
Птицы местонахождений Западной Монголии......................... 14
Птицы местонахождения Павлодар................................. 81
Птицы местонахождения Шамар..................................... 90
Птицы местонахождений Забайкалья................................ 92
ОБЗОР ИСКОПАЕМЫХ КОМПЛЕКСОВ...................................... 104
СТРАТИГРАФИЧЕСКИЕ ДАННЫЕ......................................... 114
ЗАКЛЮЧЕНИЕ....................................................... 116
ЛИТЕРАТУРА....................................................... 117
ТАБЛИЦЫ 1—XVI
ГАРЧИГ
УДИРТГАЛ........................................................... 5
МАТЕРИАЛ, АРГАЧЛАЛ................................................. 7
ОЛДВОРТ ГАЗРУУДЫН ГЕОЛОГИЙН ШИНЖ.................................. 10
АНГИЛАЛ........................................................... 14
Баруун монголын олдворт газрын шувууд........................... 14
Павлодарын олдворт газрын шувууд................................ 81
Шаамарын олдворт газрын шувууд.................................. 90
Байгалын чанад олдворт газрын шувууд............................ 92
ЭРМЭЛ ЦОГЦОЛБОРУУДЫН ТОЛОВ....................................... 104
ДАВХРАГА ЗУЙН УЗУУЛЭЛТ......................................... 114
ДУГНЭЛТ.......................................................... 116
АШИГЛАСАН НОМ ЗОХИОЛ........................................... 117
ТАЙЛБАР I—XVI
CONTENTS
INTRODUCTION....................................................... 5
DATA AND METODS.................................................... 7
GEOLOGICAL IDEX OF LOCALITIES..................................... 10
SYSTEMATIC PART................................................... 14
Birds of Western Mongolia localities............................ 14
Birds of Pavlodar locality...................................... 81
Birds of Shamar locality........................................ 90
Birds of Transbaikalia locality................................. 92
SURVEY OF THE FOSSIL COMPLEXES OF BIRDS.......................... 104
STRATIGRAFIC POSITIONS........................................... 114
CONCLUSION....................................................... 116
REFERENCES....................................................... 117
PLATES I—XVI
120