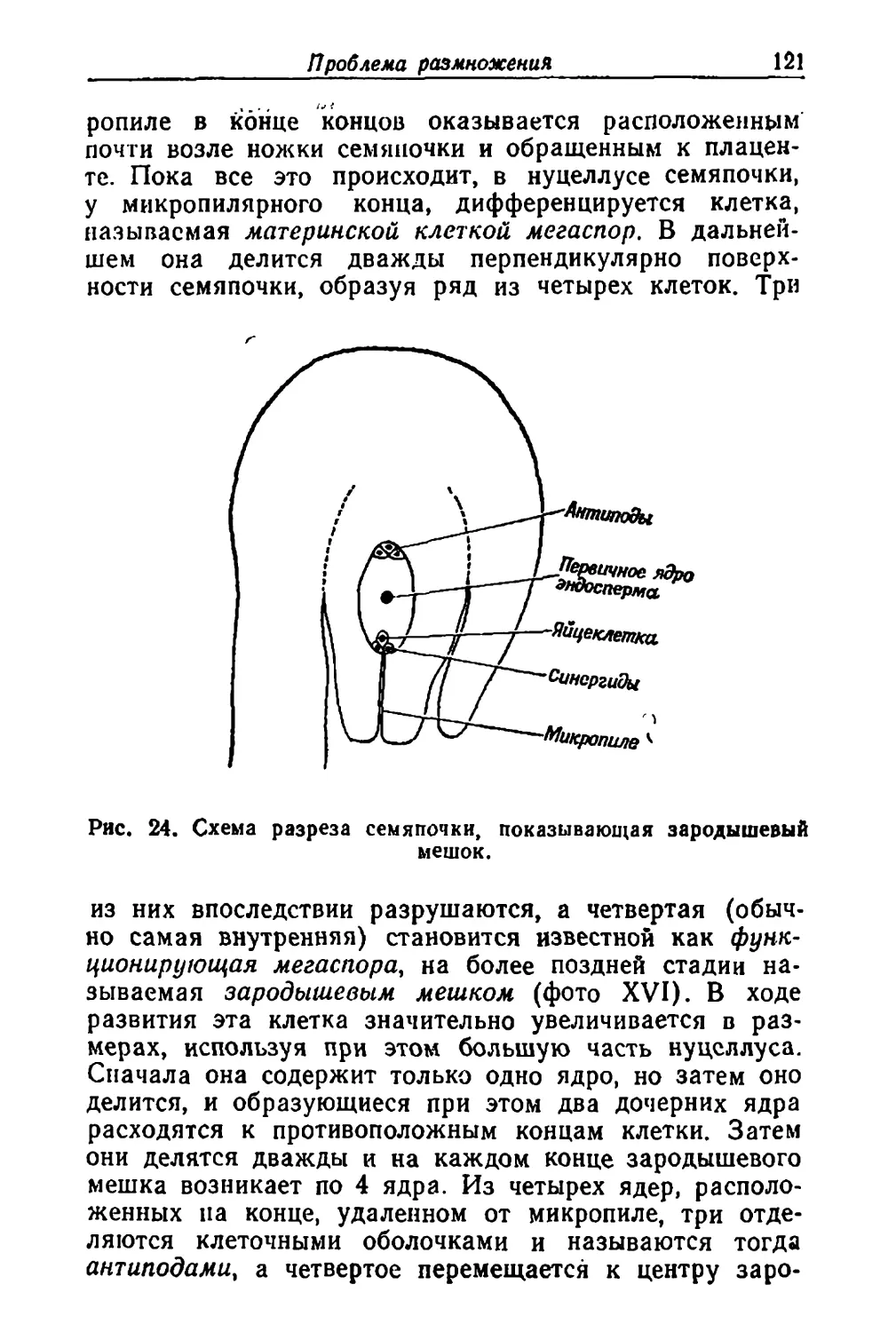
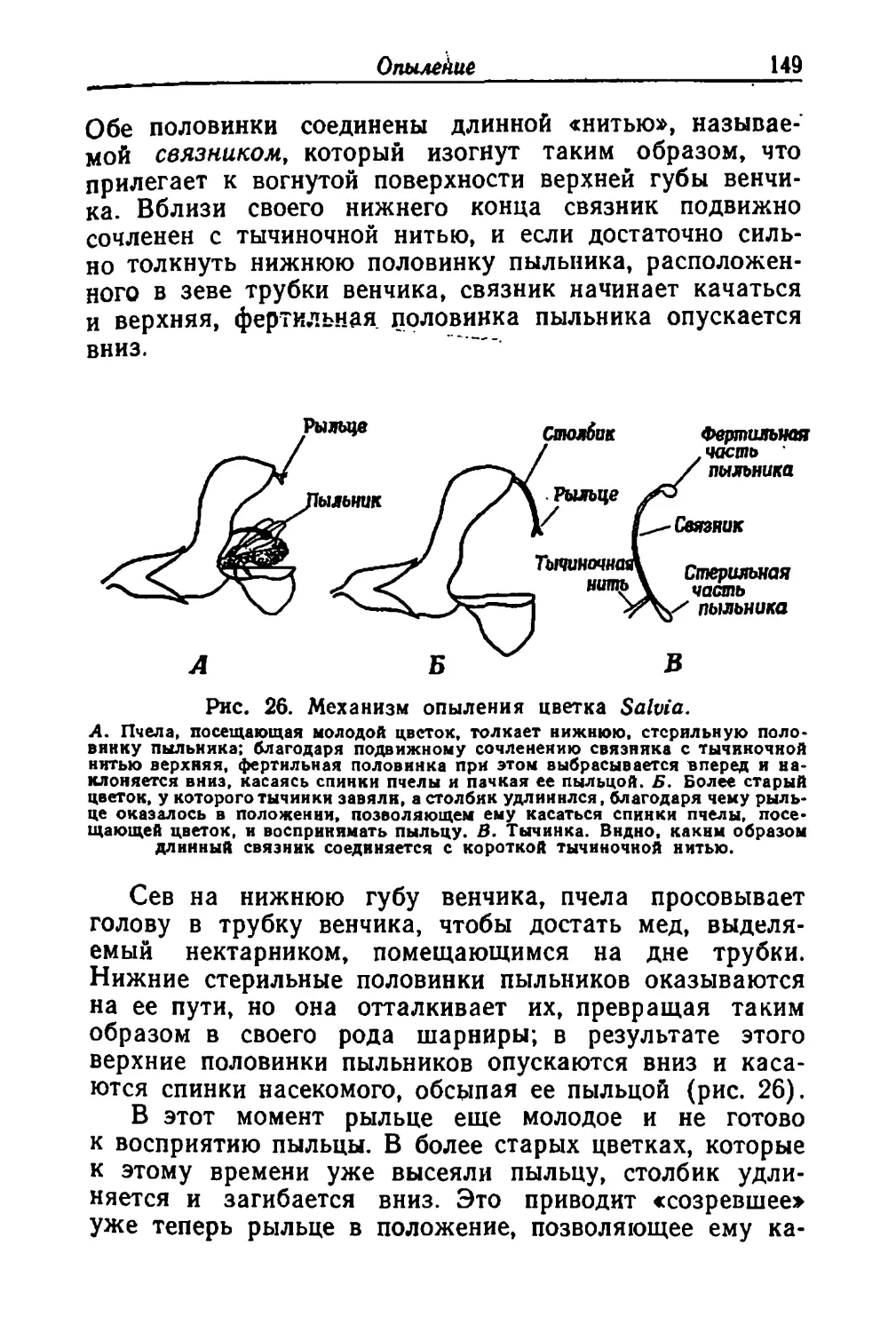
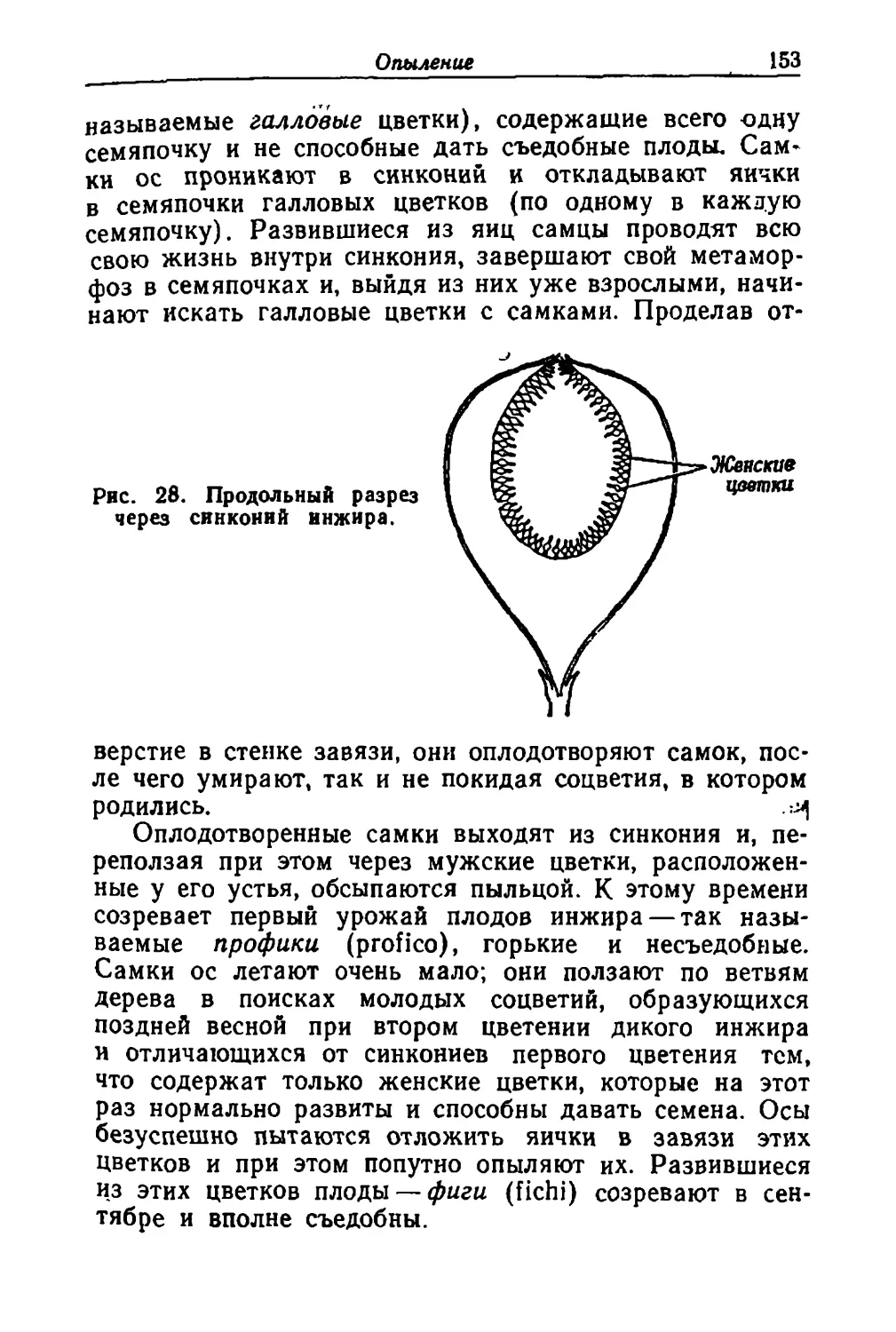
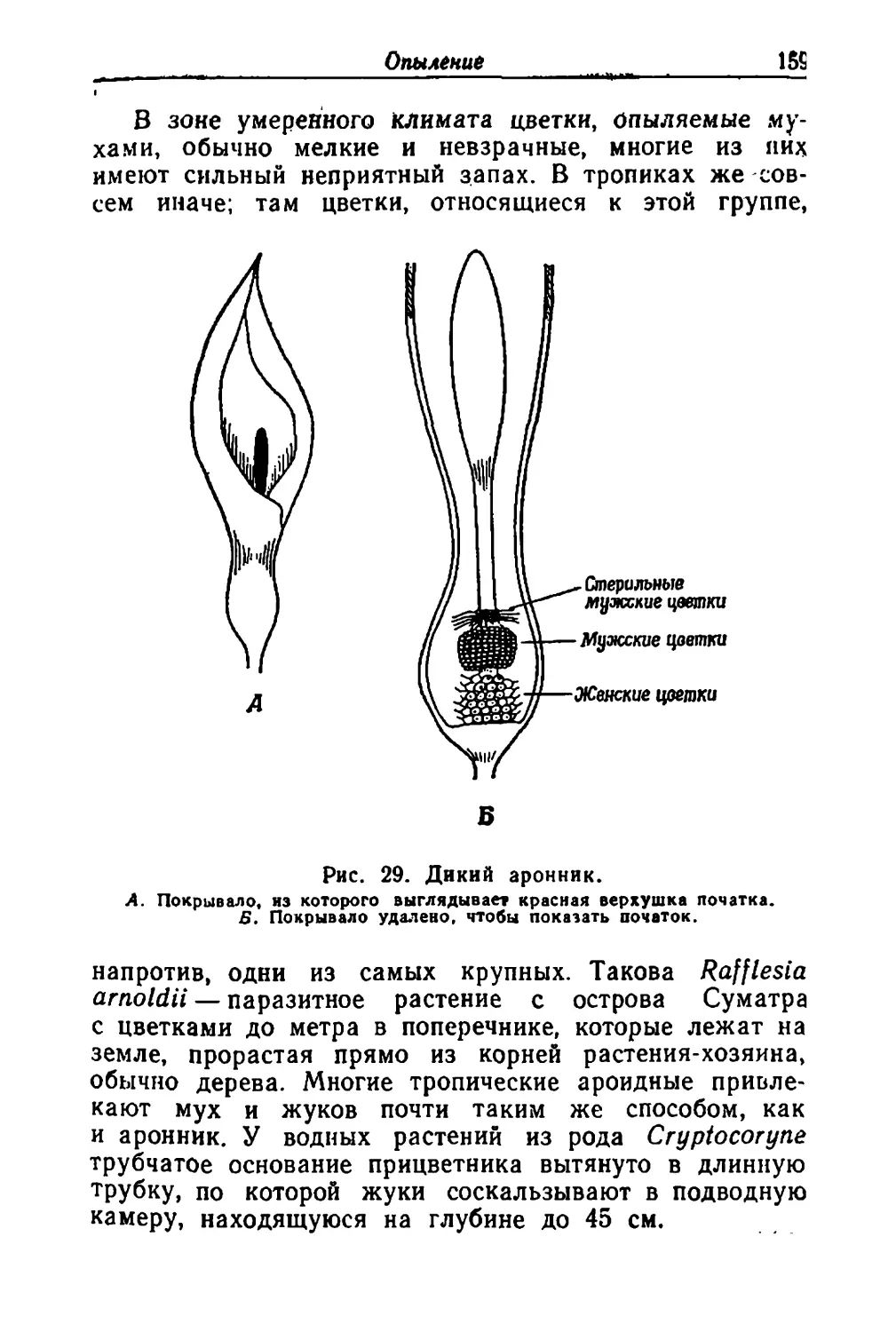
/


























































































































































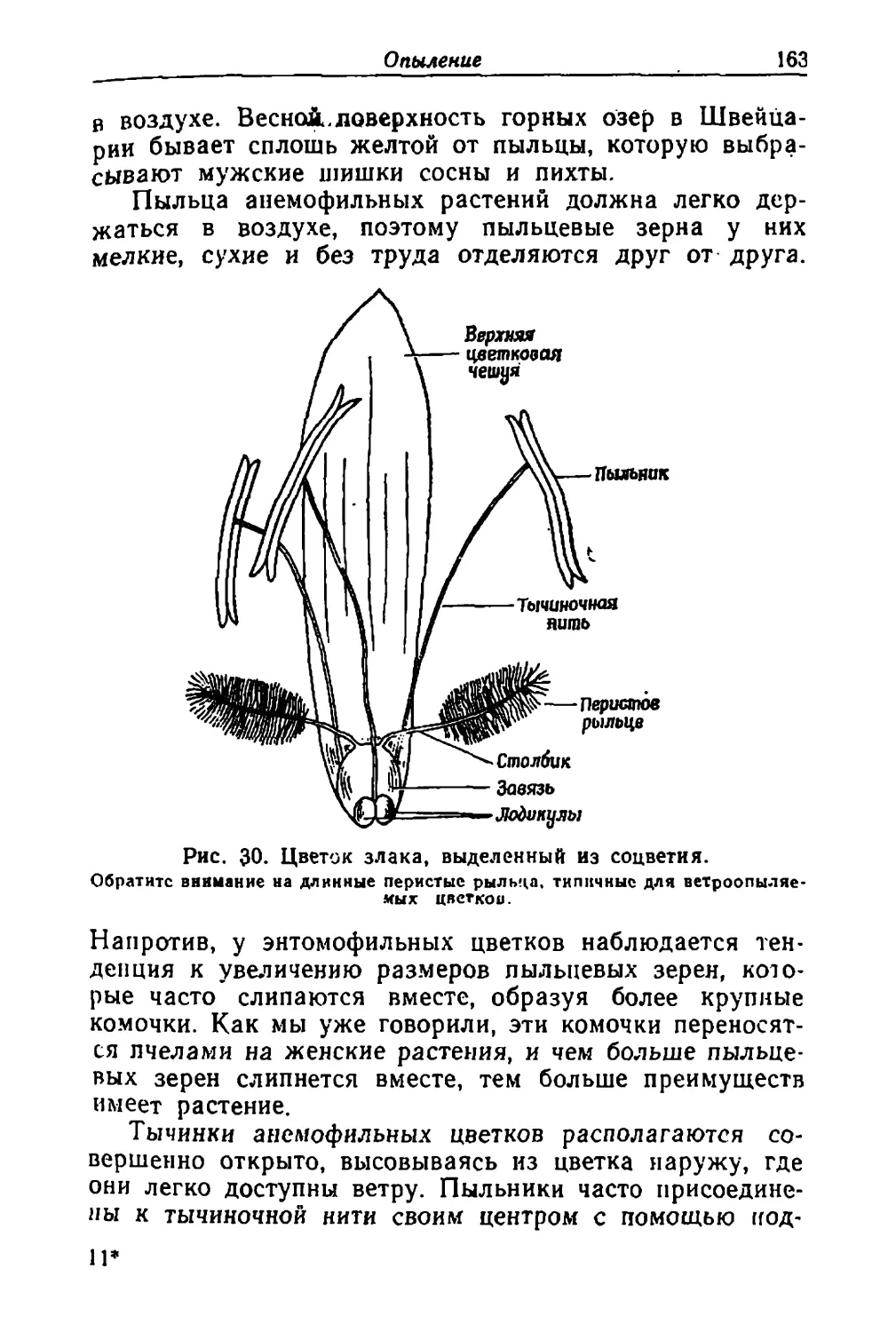









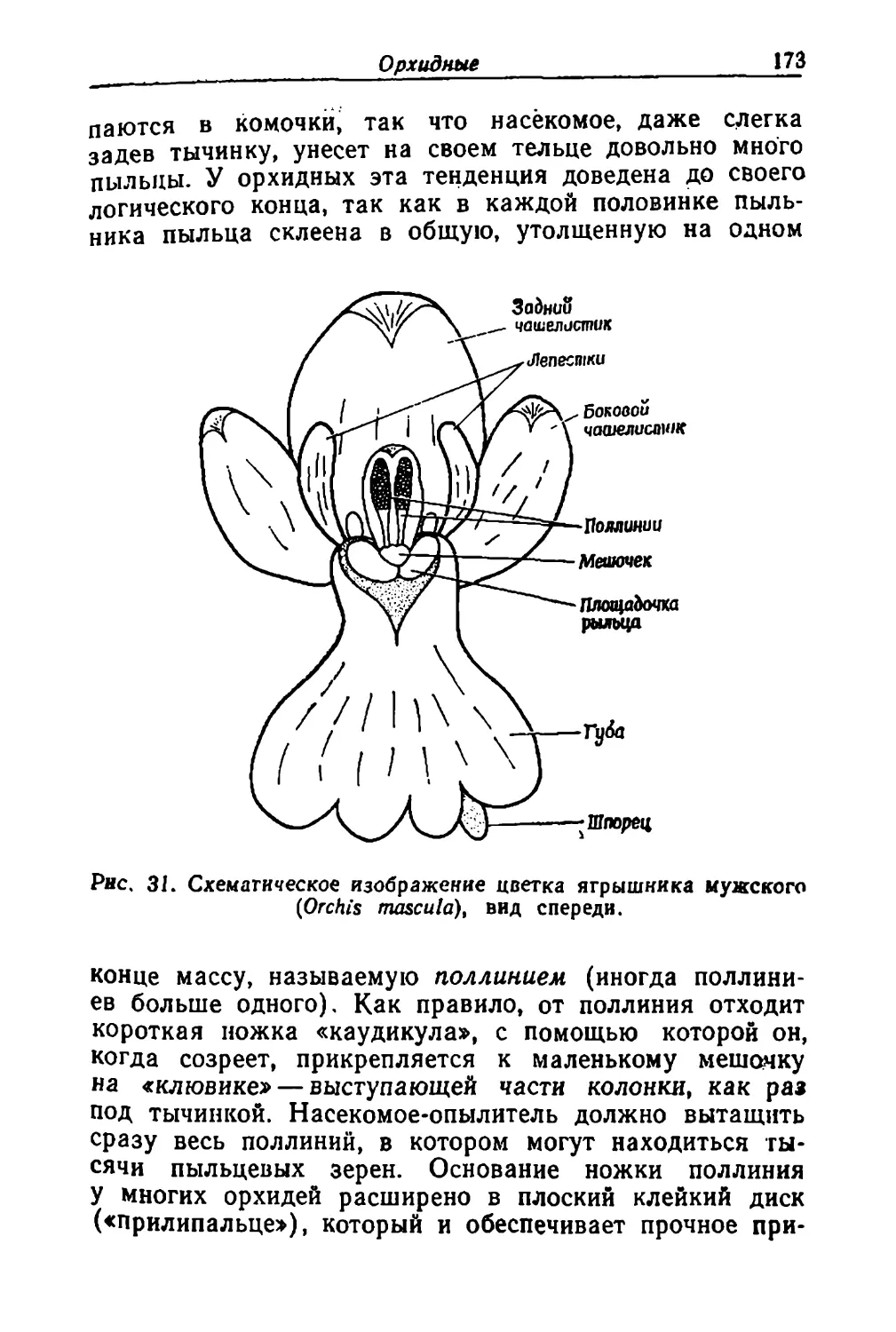




























































































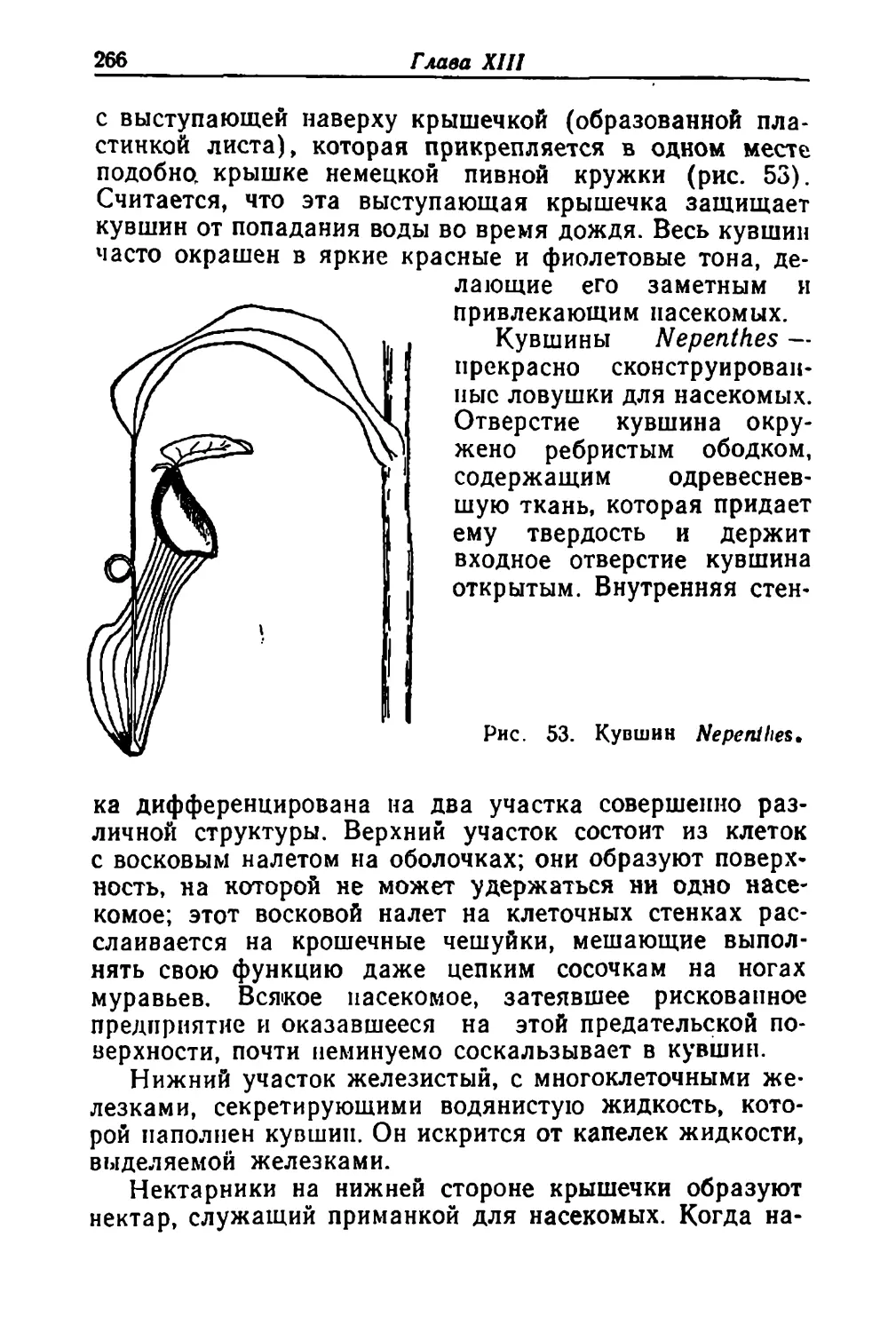



















































Text
Эволюционная
ботаника
К Дядошгтом
К
Evolution
in plant design
C. L. Duddington
Faber and Faber Limited • London 1969
К. Даддингтон
Эволюционная
ботаника
Перевод с английского
Е. Е. БЕРЕНГАРТЕН
Под редакцией к с предисловием
д-ра биол. наук
Т. И. СЕРЕБРЯКОВОЙ
Издательство «Мир»* Москва «1972
УДК 681.4
Популярная книга по ботанике, написанная на
вполне современном уровне н прекрасно иллюстриро-
ванная. Даны основные сведения по анатомии, морфо-
логии, физиологии и экологии цветковых растений,
причем не в стандартной последовательности учебного
пособия, а в виде оригинально построенного и занима-
тельного повествования о жизни растений на фоне их
эволюционного развития. При этом язык и стиль книги
таковы, что автора никак нельзя упрекнуть ни в чрез-
мерном упрощении, ни тем более в вульгаризации.
Предназначена для широкого круга читателей —
учащихся старших классов средней школы и технику-
мов, преподавателей, студентов биологических факуль-
тетов, сельскохозяйственных и педагогических инсти-
тутов и просто для людей, интересующихся природой.
Редакция биологической литературы
2-10-5, 4-3-1
127—72
ПРЕДИСЛОВИЕ К РУССКОМУ ИЗДАНИЮ
Научно-популярная литература — очень трудный
жанр. Человек, взявшийся за этот труд, должен не
только в совершенстве владеть своей отраслью науки,
но и уметь выразить свою мысль ярко, образно и до-
ходчиво и при этом отобрать из обширного арсенала
фактов и теорий только главное и необходимое. К со-
жалению, сочетание таких качеств встречается не
часто.
Две опасности подстерегают автора научно-попу-
лярной книги: это, с одной стороны, чрезмерная «науч-
ность» стиля и языка и перегрузка материалом, делаю-
щие книгу сухой и скучной, а с другой — вульгаризация
научных данных, преднамеренное упрощение и даже
искажение их в угоду занимательности и мнимой «про-
стоте». С точки зрения влияния на читателя эти две
крайности далеко не равноценны. Если первая из них
приводит лишь к тому, что книгу будут читать неохотно
и автор не получит должного признания, то вторая при-
носит громадный вред, создавая в умах широкой публи-
ки неверные представления, путаницу и хаос. К сожа-
лению, в ботанической научно-популярной литературе
наряду с превосходными произведениями можно найти
и упомянутые крайности.
До наших дней непревзойденным образцом популя-
ризации остается «Жизнь растения» К. А. Тимирязева,
написанная почти сто лет назад. Сохраняет свою све-
жесть выдержавшая с тридцатых годов несколько изда-
ний «Занимательная ботаника» А. В. Цингера. Очень
хороши неоднократно переиздававшиеся книги рано
умершего А. В. Кожевникова — «По тундрам, лесам,
степям и пустыням» (первое издание вышло в 1937 го-
ду) и «Весна и осень в жизни растений» (первое изда-
ние— в 1938 году). Заслуженным признанием пользу-
ются книги Н. М. Верзилина — «Путешествие с домаш-
ними растениями», «По следам Робинзона» и другие.
Есть и другие неплохие популярные книги о растениях,
6 Предисловие к русскому изданию
но в общем их в последнее время немного, особенно по
сравнению с популярной литературой в других областях
естествознания.
Предлагаемая читателю книга К- Даддингтона обла-
дает всеми качествами Отличного научно-популярного
сочинения, и издание ее на русском языке представляет-
ся весьма целесообразным. Она охватывает широкий
круг вопросов, касающихся строения растений, а также
деятельности отдельных органов и тканей, причем воз-
никновение всех структур рассмотрено в свете эволю-
ционной теории и очень ярко показано приспособитель-
ное значение каждой из них. По существу это интерес-
ное и популярное изложение основ общей ботаники —
морфологии, анатомии и физиологии растений.
Оригинальное название книги — «Evolution in plant
design»—трудно перевести на русский язык. Англий-
ское слово «design» имеет множество смысловых оттен-
ков. Инженерное, техническое значение этого термина —
эскиз, чертеж, конструкция. Однако специалисты по при-
кладному искусству и технической эстетике употребляют
термин «дизайн» как непереводимый, для обозначения
внешней формы предметов, наилучшнм образом соответ-
ствующей их функции. Вот в этом последнем смысле
автор книги и обсуждает строение растений. Пожалуй,
можно предъявить автору одну претензию: он нередко
увлекается приспособительным значением структур на-
столько, что впадает в чрезмерный «телеологизм», то и
дело ссылаясь на «цель» того или иного приспособления
и пренебрегая причинной стороной явления. Но это
довольно распространенный в учебной и популярной ли-
тературе недостаток.
Очень важно, что К. Даддингтон ведет изложение на
вполне современном уровне ботанических знаний, ни-
когда не вульгаризирует объяснений и не боится пре-
поднести читателю очень трудные вещи. Он приводит
новые факты и теории, умеет показать противоречивые
гипотезы, их положительные и отрицательные стороны,
подчеркнуть еще не понятые явления. Посмотрите, как
он остроумно сравнивает и обсуждает хотя бы теории
поднятия воды по сосудам древесины! С тонким анг-
лийским юмором К- Даддингтон показывает, что в об-
Предисловие к русскому изданию
1
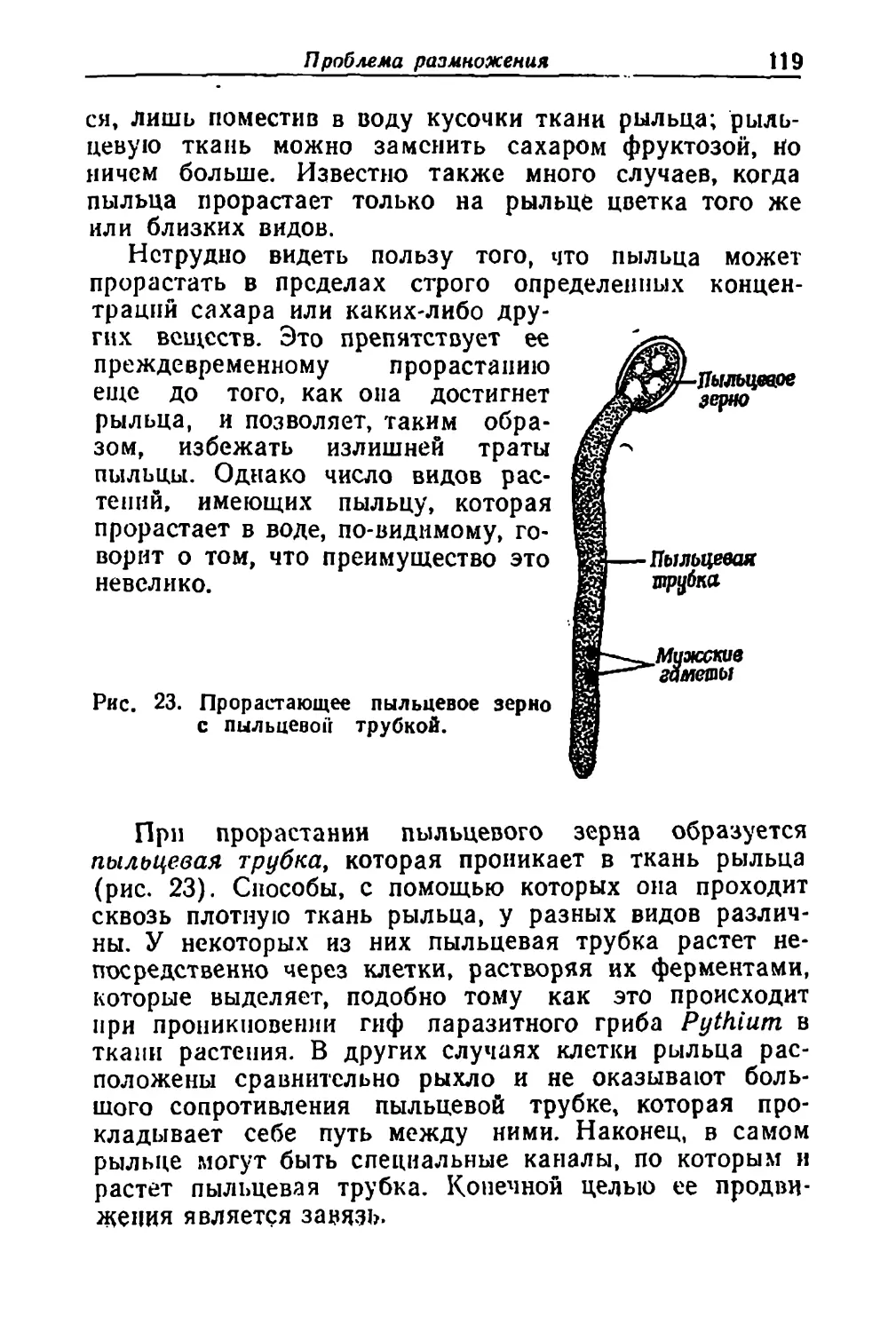
щем-то очень немногие проблемы в ботанике можно
считать до конца решенными, а в большинстве случаев
остается еще непочатый край работы для новых иссле-
дований.
Таким образом, читатель получает не застывшие
истины «в последней инстанции», а ощущает противоре-
чия (разные точки зрения как раз и способствуют про-
грессу науки) и чувствует науку в ее движении.
Не все разделы книги равноценны. Нам кажется бо-
лее удачной первая часть, касающаяся структуры п
функций вегетативных органов и общих приспособлений
растений к жизни на суше. Несколько «стандартнее»
звучат разделы об органах размножения: о цветке,
плодах и семенах, а также об опылении, хотя и здесь
масса интересных фактов. Совсем бегло, даже поверх-
ностно, автор затрагивает интереснейшие вопросы пе-
резимовки, покоя, возобновления растений, их жизнен-
ных форм. Но ни один раздел не страдает существенны-
ми пороками, и везде радует живая, легкая и доходчи-
вая манера изложения.
Мы сочли необходимым снабдить книгу довольно
многочисленными примечаниями. Иногда это вызвано
тем, что автор придерживается теоретических представ-
лений, отличных от принятых у нас, например о роли
естественного отбора, о принадлежности тех или иных
групп растений к низшим и высшим. Но чаще в приме-
чаниях приводятся дополнительные сведения о растени-
ях флоры СССР, .потому что автор, естественно, исполь-
зует в качестве примеров преимущественно растения
Англии, не всегда встречающиеся у нас. Кроме того,
кое-где в связи с обсуждаемыми автором проблемами в
примечаниях упомянуты доступные и достаточно попу-
лярные русские книги по этим вопросам, чтобы заинте-
ресовавшийся читатель при желании мог с ними позна-
комиться.
Книга К- Даддингтона может быть полезной не толь-
ко школьникам, учителям и студентам-биологам, но и
просто широкому кругу людей, любящих природу и ин-
тересующихся ею.
Г. И. Серебрякова
Предисловие Автора
Растения существуют уже очень давно. Ископаемые
остатки растений, которые удалось обнаружить, отно-
сятся к кембрийскому периоду, т. е. их возраст около
500 млн. лет, однако можно с уверенностью сказать,
что некоторые формы растительной жизни, несомненно,
существовали на Земле гораздо раньше. Первые на-
ходки наземных растений, о которых мы знаем, извест-
ны из силура; их ископаемые остатки, которым более
300 млн. лет, позволяют говорить о том, что эти ра-
стения были сравнительно высокоорганизованы, по-
скольку у них имелась уже развитая проводящая ткань,
служащая для поднятия воды вверх по стеблю. По этой
и по ряду других причин логично предположить, что они
не были первыми растениями, вышедшими из моря в
завоевавшими сушу.
В течение 300 млн. лет или около того, прошедших
после силурийского периода, высшие растения далеко
продвинулись эволюционно к создали тот очень разно-
образный и сложный ансамбль, который мы видим теперь
вокруг. Это явилось результатом естественного отбора,
отдававшего предпочтение более приспособленным и
постоянно отбрасывавшего неудачников. Благодаря это-
му отбору, происходившему во все времена, современ-
ные растения оказались превосходно «оснащенными»,
т. е. прекрасно подготовились к той жизни, которую они
должны вести.
Эта книга посвящена в основном высшим растениям
и путям, по которым они шли в своем приспособлении
к жизни в таком перенаселенном мире. Она предназна-
чена не только для ботаников, но скорее для неиску-
шенного читателя, интересующегося растениями, — для
садовода, натуралиста и для всех тех, кто просто любит
растения. Поэтому я старался избегать научных терми-
нов там, где это возможно; там же, где нельзя было
обойтись без ботанической терминологии, я объяснял по
мере их появления значения всех терминов и этим, наде-
Предисловие автора 9
юсь, облегчил задачу читателям, не обладающим специ-
альными знаниями. Мне казалось, что говоря о гидрав-
лике растений (гл. III) и о фотосинтезе (гл. IV), проще
дать уравнения, нежели ограничиться словесным объяс-
нением.
Мне хотелось бы поблагодарить моих коллег за ту
помощь, которую они охотно оказывали мне в процессе
работы над книгой и без которой она никогда не была
бы написана. Я хотел бы также выразить благодар-
ность Лоуренсу Д. Хиллсу как за то, что он подал мне
мысль написать эту книгу, так и за его неоценимую по-
мощь при работе над предварительным ее вариантом.
К ингстон-иа-Темзе,
19Gb г.
К. Даддингтон
ГЛАВА I
ПРОБЛЕМЫ ПРИСПОСОБЛЕНИЯ РАСТЕНИИ
К НАЗЕМНОЙ ЖИЗНИ
Жизнь зародилась в море. Когда и где это произош-
ло, мы не знаем, но большинство биологов согласны
между собой в одном: первые комочки живого вещества
появились именно в океане, в среде, до того бывшей
всецело неорганической. Это замечательное событие
произошло по меньшей мере 2 млрд, лет назад, а воз-
можно, и гораздо раньше.
Первые комочки жизни были очень просты в биоло-
гическом смысле, но очень сложны в сравнении с окру-
жавшими их неорганическими молекулами. Мы не зна-
ем, что представляли собой эти предшественники пер-
вичных простейших организмов, но разумно предполо-
жить, что они состояли из белковых молекул, а это вы-
зывает мысль о сходстве с некоторыми существующими
в настоящее время простейшими вирусами. Из этих
первых предшественников возникли живые клетки, воз-
можно, напоминавшие микроскопические организмы,
стоящие на границе между растениями и животными,
которые мы называем теперь жгугикочыми. Приблизи-
тельно в этот период в природе обособились два царст-
ва— растений и животных, дальнейшее развитие кото-
рых пошло различными путями. У растений сохранился
зеленый пигмент, называемый хлорофиллом., который
поглощает солнечную энергию, а благодаря этому со-
хранилась и способность создавать из простых соедине-
ний, имеющихся в «мастерской» природы, — углекисло-
го газа, воды и некоторых минеральных солей — веще-
ства, необходимые для их жизни. Эгог замечательный
процесс называется фотосинтезом. Животные лишились
хлорофилла и поэтому вынуждены были добывать орга-
ническую пищу, питаясь растениями или охотясь на
своих собратьев.
В обоих царствах природы простейшие одноклеточ-
ные организмы дали начало сложным, многоклеточным
формам. Вначале жизнь растений была связана исклю-
чительно с водой, поскольку суша еще не была заселена.
12
Глава I
Однако эволюционные возможности моря не могли со-
храняться вечно. Растениям необходимо было завоевать
сушу, которая в те времена была совсем иной, чем сей-
час: бесплодные скалы и пустыни, на которых не было
видно не только ни травинки, но отсутствовала даже
почва, если не считать рассеянной то здесь, то там пы-
ли. Тем не менее это была земля; «фигурально выра-
жаясь», земля сильно привлекала к себе первых выход-
Рис. 1. Водоросль Pelvetia сапа-
liculaia.
цев из моря, достаточно смелых для того, чтобы поки-
нуть гостеприимные объятия океана и попытать счастья
на берегу.
Первые растения-колонисты появились на суше в си-
лурийский период, т. е. более чем 300 млн. лет назад;
однако еще до этого растения, по-видимому, начали
проникать то тут, то там на береговую полосу, затоп-
ляемую приливом, хотя их было недостаточно много
для того, чтобы можно было назвать их колонистами.
Сначала это проникновение шло очень медленно. Расте-
ния, растущие вблизи границы прилива, постепенно пе-
редвигались к берегу, пока не оказались выше этой
границы, где на них попадали только брызги — в силь-
ный шторм или при исключительно высоком уровне во-
ды весной. В настоящее время такую же картину мож-
но наблюдать у водоросли Pelvetia canaliculata, которая
растет так высоко на берегу, что является почти сухо-
путным растением (рис. 1). Отсюда был уже только
Проблемы приспособления растений к наземной Исиэни 13
один маленький шаг до полного отрыва от моря (в про-
странстве), хотя предстояло сделать гораздо более зна-
чительный шаг в смысле необходимой адаптации.
Проблемы наземной жизни растений
Как только растения покинули водную среду, они
столкнулись со множеством проблем, требовавших свое-
го разрешения. Прежде всего это была проблема снаб-
жения водой: ведь для того, чтобы в теле растения мог-
ли протекать жизненные процессы, оно должно полу-
чать воду, без которой жизнь вообще невозможна. Для
водорослей не существует такой проблемы, так как они
по крайней мере часть дня бывают погружены в воду.
Существование же наземного растения зависит от поч-
венной воды, и чтобы эта вода стала доступной, расте-
ние должно было развить корневую систему, которая
может поглощать воду из почвы. У водорослей корней
нет, так как они им не нужны; самое большее, что они
имеют, — это «пятки», прикрепляющие их к камням, но
через них ничего не всасывается. Первые наземные ра-
стения не имели корней, но у них были подобные во-
лоскам органы, называмые ризоидами, которые прони-
кали в почву и служили для всасывания воды. Однако
вскоре после того, как растения окончательно обоснова-
лись на суше, у них развились корни.
Корни наземных растений всасывают из почвы не
только воду, но и минеральные соли, необходимые для
питания растений. Перед водорослями и в данном слу-
чае не возникает проблемы, так как источником мине-
ральных веществ для них служит море, и вода, которая
омывает их, представляет собой своего рода питатель-
ный бульон, который обеспечивает все необходимое для
их жизни. Переселившись на сушу, растения должны
были найти какой-то другой способ снабжения мине-
ральной пищей, поэтому их корни стали служить для
поглощения не только воды, но и минеральных веществ.
По мере того как увеличивались размеры растений,
становилась все острее проблема поднятия воды (а вме-
сте с ней и минеральных веществ) от корней к самым
верхним ветвям. Калифорнийское мамонтово дерево, на-
14
Глава I
пример, достигает в высоту 107 м, и вода, поглощаемая
его корнями, должна дойти до каждого его листа. Спе-
циальные водопроводящие клетки развились в ответ на
эту потребность, а сила сцепления между молекулами
воды обеспечила поднятие воды против силы тяжести.
Большая часть органических веществ, необходимых
растению, образуется в листьях при участии хлорофил-
ла и солнечного света. Выработанные в листьях пита-
тельные вещества должны распределиться между всеми
частями растения, вплоть до кончиков корней. Для это-
го нужна налаженная проводящая система, клетки ко-
торой приспособлены к переносу аминокислот, сахаров
и других органических веществ. Подобную систему мож-
но наблюдать в зачаточной форме у некоторых крупных
морских водорослей, где специальные клетки образуют
как бы трубку, по которой передвигается вода с рас-
творенными в ней питательными веществами. У назем-
ных растений, тело которых расчленено на стебель, кор-
ни и листья, потребность в такой проводящей системе
еще больше, и в ответ на это у них развилась ткань, на-
зываемая флоэмой.
Наземное растение сталкивается и со многими ме-
ханическими проблемами, чуждыми водорослям. Первая
из них — это необходимость стоять прямо, тогда как
водоросль поддерживает вода. Теоретически предела,
которого могут достигать размеры этих растений, не
существует, и длина крупной бурой водоросли Масго-
cystis, например, может составлять более 180 м
(рис. 2). Иначе обстоит дело с наземными растениями.
Если такое растение не хочет провести всю свою жизнь
стелясь по земле, оно должно иметь стебель, достаточно
прочный для того, чтобы не только поддерживать его
ветви, но и противостоять порывам ветра, склоняющим
растение- вниз. Это обеспечивается развитием ксилемы,
или древесины, и ее расположением в стебле я корне,
позволяющим стеблю сопротивляться изгибу, а корню,
с помощью которого растение закрепляется в почве,
противостоять «тянущему» усилию, сохраняя в то же
время максимальную гибкость.
У первых наземных растений был простой древесин-
ный цилиндр, проходящий в центре их стебля; он с бес-
Проблемы приспособления растений к наземной жизни 15
печивал 'проведение воды и придавал прочность. По
мере увеличения размеров и усложнения строения ра-
стений эти центральные цилиндры расщепились на бо-
лее мелкие единицы, называемые проводящими пучками
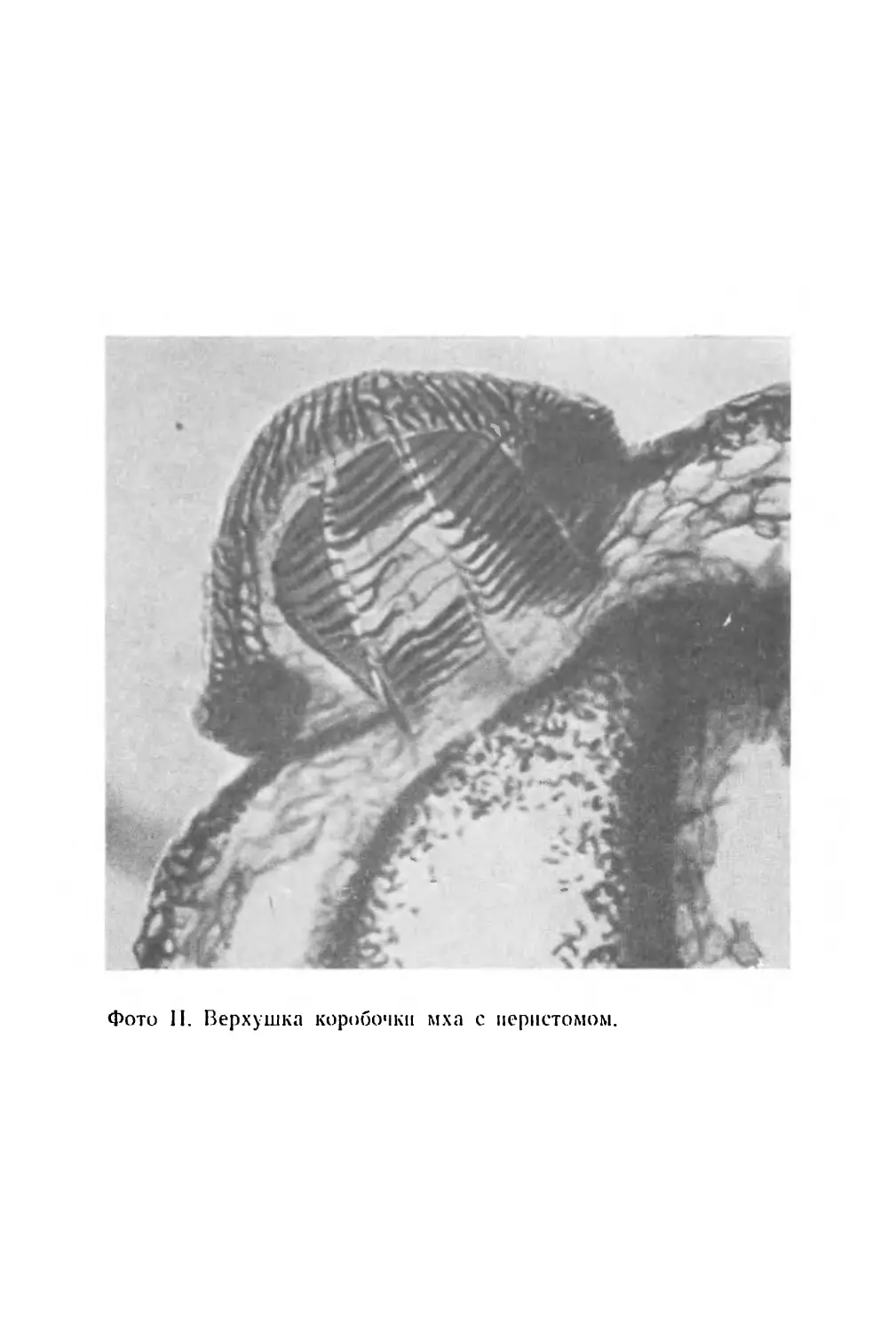
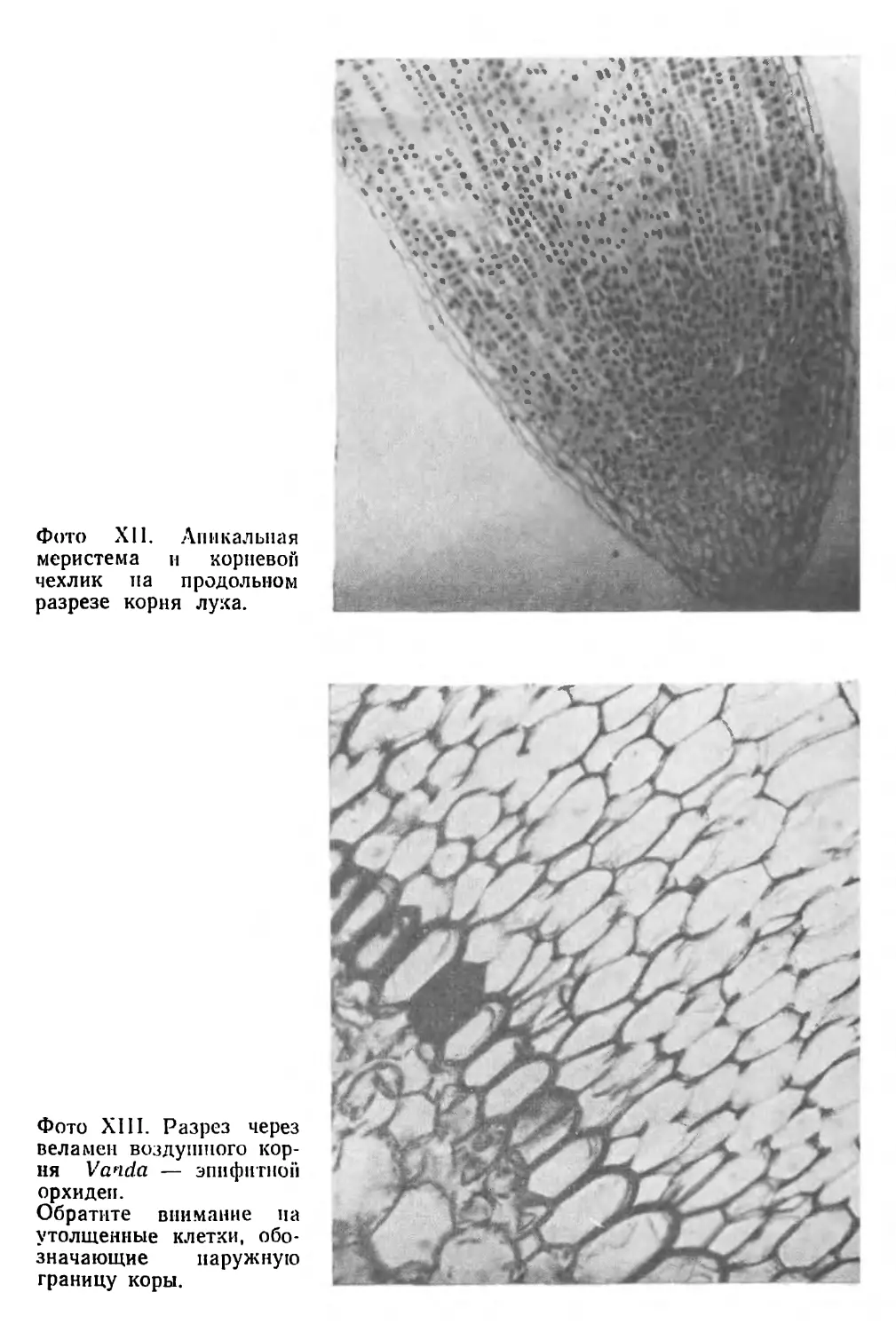
(фото I)1, которые расположились различным образом,
что позволило им наилучшим способом удовлетворять
нужды растения. Характер клеток, составляющих дре-
весину, тоже менялся в связи с изменением этих по-
требностей. Развились ряды трубчатых клеток, называе-
мых сосудами, единственной функцией которых было
проведение воды; другие клетки, называемые волокнами,
Рис. 2. Крупная бурая водоросль
Macrocystis pyrifera:
с сильно утолщенными и упроченными благодаря отло-
жению твердого вещества, называемого лигнином, стен-
ками приняли на себя обязанность укрепления стебля и
придания ему прочности. В наши дни эти волокна ис-
пользуют для изготовления тканей и веревок; полотно,
например, вырабатывают из волокон льна (Linum usita-
tissimum), после того как мягкие наружные ткани стеб-
ля вымачивают в воде и удаляют.
Увеличение размеров и усложнение строения расте-
ний повлекло за собой образование листьев — первичных
органов фотосинтеза. Водоросли не расчленены на сте-
бель и листья, и у крупных водорослей процесс фото-
синтеза осуществляется всеми клетками наружной ча-
сти плоского пластинчатого тела. Первые наземные
растения функционировали таким же образом, т. е. вся
их надземная часть фотосинтезировала. С развитием
1 Все фотографии помещены в конце книги.
16
Глава I
листа возник новый орган; его функцией было образо-
вание крахмала. Этот орган стал плоским и тонким, так
что клетки, в которых образовывался крахмал, получа-
ли весь свет, необходимый им для этого процесса. Раз-
вилась листовая мозаика: каждый лист располагался
таким образом, чтобы занять пространство, оставшееся
открытым благодаря просвету между вышерасположен-
ными листьями, так что ни один лист не затенялся боль-
ше, чем это было абсолютно необходимо. Вместе с тем
в листьях возникла эффективная система внутренней
вентиляции и, таким образом, необходимый для питания
растений углекислый газ мог проникать через отверстия,
называемые устьицами, и быстро доходить до фотосин-
тезирующих клеток.
Однако вентиляция листа таила в себе и опасности
для растения, так как если углекислый газ мог прони-
кать внутрь через устьичные щели, то через них же мог-
ла испаряться вода. Поскольку снабжение наземного
растения водой обычно ограничено, это могло привести
к гибели его от 'высыхания. Эта проблема была частич-
но разрешена благодаря развитию вокруг устьичных ще-
лей специальных клеток, называемых замыкающими
клетками, которые закрывают устьица на ночь, когда из-
за отсутствия света процесс фотосинтеза прекращается.
С выходом растений из водной среды па сушу, по-
жалуй, особенно сильно изменились условия для их
размножения. При половом размножении происходит
слияние женской яйцеклетки с мужским сперматозои-
дом. К услугам водорослей для переноса мужской га-
меты к женской всегда имеется вода, причем спермато-
зоиды обычно, хотя и не всегда, бывают снабжены ма-
ленькими протоплазматическими жгутиками, которые
осуществляют волнообразные движения, а это позволяет
сперматозоидам двигаться. Все, что должна сделать во-
доросль,— это выпустить сперматозоиды в воду, где они
уже найдут необходимый им путь к женским яйцеклет-
кам (этому немало способствуют водные течения). Кро-
ме того, яйцеклетки обычно выделяют в воду вещества,
привлекающие сперматозоиды.
Покинув океан и выбравшись на берег, растения тем
самым лишились той всепроникающей среды, в которой
Проблемы приспособления растений к наземной жизни 17
сперматозоиды легко распространялись, и перед ними
особенно остро встала проблема размножения. Но эта
проблема не могла быть разрешена немедленно, так как
те сотни миллионов лет, когда сперматозоиды были
«плавающими», означали, что растения могут размно-
жаться половым путем только в условиях влажности,
а это было возможно лишь тогда, когда влага, покры-
вающая растения, давала бы крошечным сперматозои-
дам воднхю среду, в которой они могли передвигаться.
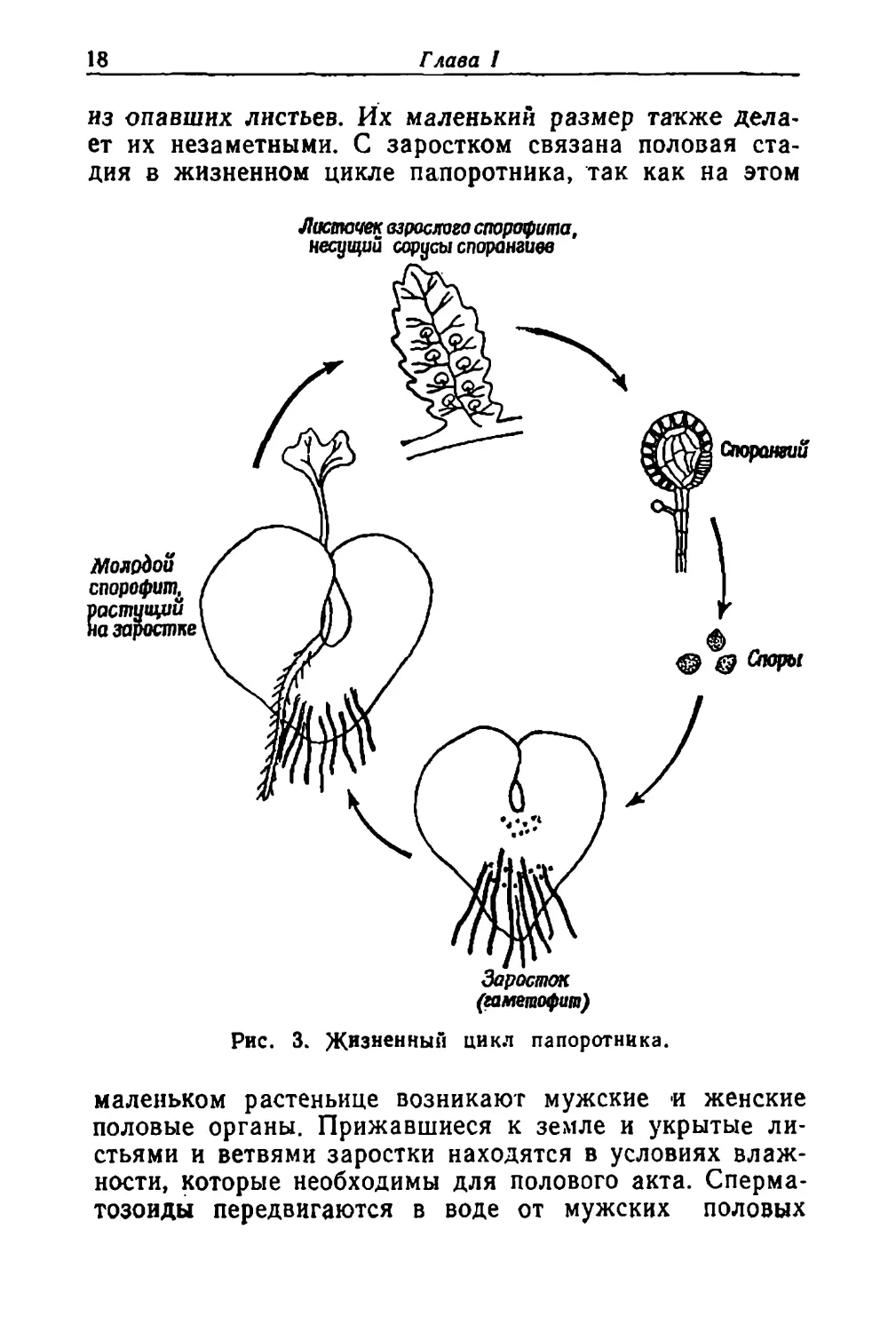
Жизненный цикл папоротников
Некоторые наземные растения так и не эволюциони-
ровали далее этой стадии. Папоротники и родственные
им растения, как, например, хвощи и плауны, а также
мхи и печеночники, до сих пор живут, не умея обходить-
ся без воды, необходимой для передвижения спермато-
зоидов. Однако они весьма любопытным образом обош-
ли связанные с этим трудности и тем самым вступили на
путь, который в конечном счете привел к возникнове-
нию семенных растений, навсегда освободившихся от
этой зависимости.
Само растение папоротника вообще не размножается
половым путем. Вместо этого на нижней поверхности его
листочков или по их краям возникают тысячи крошеч-
ных образований, называемых спорангиями, в которых
развиваются споры. Спора — это мельчайшая структу-
ра, служащая для размножения; она довольно похожа
на семя, но гораздо меньше и проще. Для образования
спор не нужен процесс оплодотворения. Спора папорот-
ника в тех условиях влажности и температуры, которые
ей необходимы, прорастает и развивается далее, однако
при этом не возникает нового растения папоротника.
Вместо этого из споры развивается растение, называе-
мое заростком. Оно представляет собой плоскую пла-
стинку, имеющую приблизительно сердцевидную форму,
и при окончательном своем развитии достигает в попе-
речнике не более сантиметра.
Очень немногие люди наблюдали заростки папорот-
ника, так как они находятся на земле среди подстилки
2—302
18
Г лава /
из опавших листьев. Их маленький размер также дела-
ет их незаметными. С заростком связана половая ста-
дия в жизненном цикле папоротника, так как на этом
Листочек взрослого спорофита,
несущий сорусы спорангиев
Рис. 3. Жизненный цикл папоротника.
маленьком растеньице возникают мужские -и женские
половые органы. Прижавшиеся к земле и укрытые ли-
стьями и ветвями заростки находятся в условиях влаж-
ности, которые необходимы для полового акта. Сперма-
тозоиды передвигаются в воде от мужских половых
Проблемы приспособления растений к назеяЯбй жизни 10
органов к женским и оплодотворяют яйцеклетку. Из
оплодотворенной яйцеклетки возникает маленькое ра-
стеньице-молодой спорофит, который затем растет и
развивается, превращаясь в новое растение папоротника
(рис. 3).
Таким образом, в жизненном цикле папоротника
имеется два отчетливо выраженных поколения — само
растение папоротника, несущее споры, и заросток с
половыми органами. Эти два поколения точно сменяют
друг друга, и растение папоротника дает начало заро-
стку, а заросток — вновь растению папоротника. Такое
чередование поколений наложило неизгладимый отпеча-
ток на жизненный цикл не только папоротников, но и
всех зеленых растений, находящихся на более высокой
ступени эволюционного развития, чем водоросли. Таким
образом растения и разрешили проблему полового раз-
множения на суше.
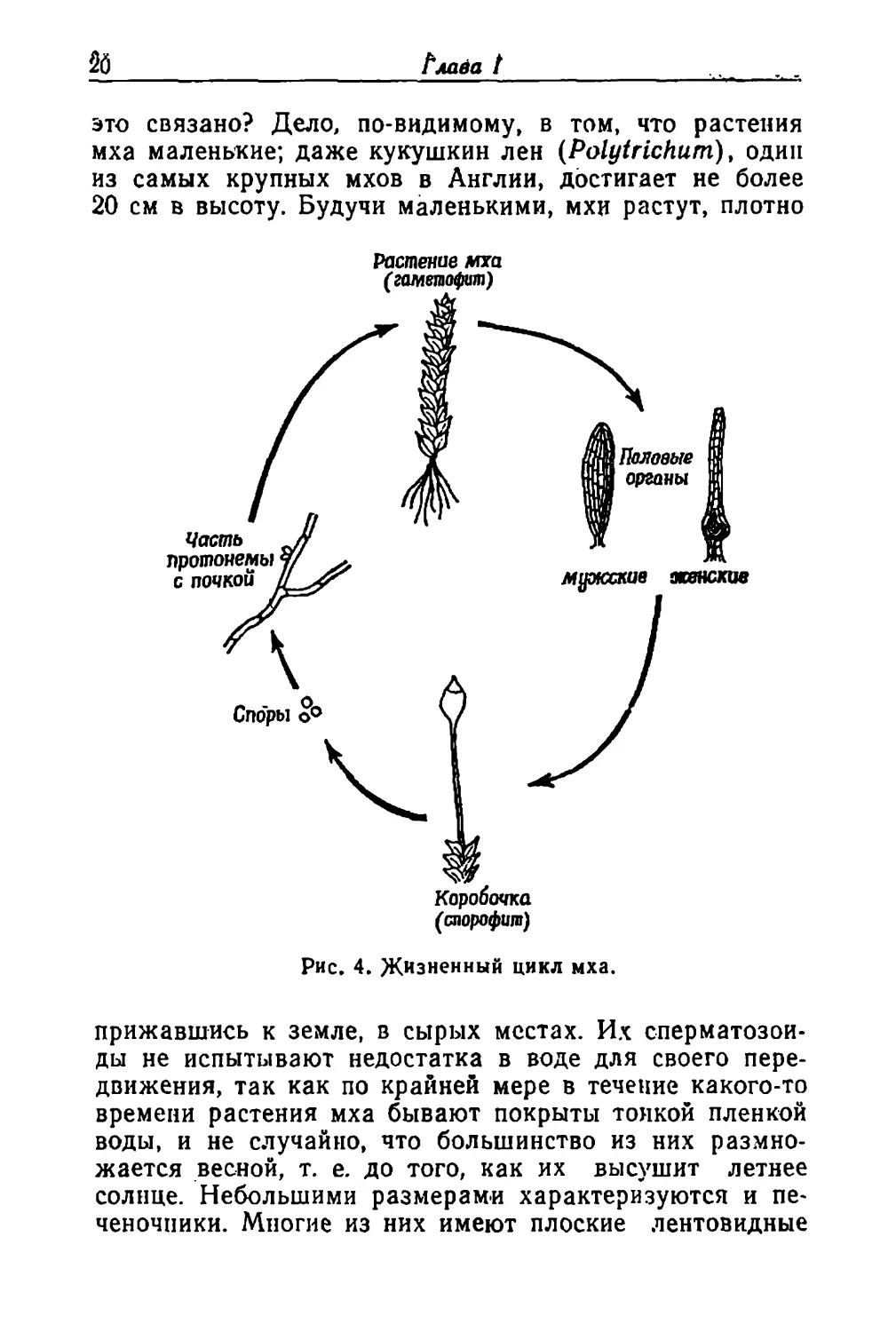
Мхи и печеночники
Смена поколений проявляется различно у разных
групп растений. У папоротников и других сходных с ни-
ми растений заросток чередуется с более крупным обра-
зующим споры поколением. У мхов и печеночников де-
ло обстоит иначе. У них «главное» растение несет поло-
вые органы, а спорообразующее поколение, возникаю-
щее в результате полового размножения, это всего-на-
всего коробочка, прикрепленная к главному растению
ножкой. Весной легко увидеть эти коробочки, возвышаю-
щиеся над растеньицами мха на своих тонких, нежных
стебельках. В легкой коробочке много спор, которые вы-
сыпаются и в подходящих условиях прорастают, давая
начало растению мха или печеночника. В сущности ко-
робочки паразитируют на материнских растениях или
полностью (как у печеночников), или частично (как у
мхов) (рис. 4).
Можно задать вопрос, почему у растений мха глав-
ное поколение несет половые органы, а развитие спор
перенесено в коробочку, тогда как у папоротников все
по-другому: само растение папоротника производит спо-
ры, а половые органы образуются на заростке. С чем
2*
20
Глава t
это связано? Дело, по-видимому, в том, что растения
мха маленькие; даже кукушкин лен (Polytrichum), один
из самых крупных мхов в Англии, достигает не более
20 см в высоту. Будучи маленькими, мхи растут, плотно
Растение мха
Коробочка
(спорофит)
Рис. 4. Жизненный цикл мха.
прижавшись к земле, в сырых местах. Их сперматозои-
ды не испытывают недостатка в воде для своего пере-
движения, так как по крайней мере в течение какого-то
времени растения мха бывают покрыты тонкой пленкой
воды, и не случайно, что большинство из них размно-
жается весной, т. е. до того, как их высушит летнее
солнце. Небольшими размерами характеризуются и пе-
ченочники. Многие из них имеют плоские лентовидные
Проблемы приспособления растений к ноэбЬтоА асизнй *21
талломы и растут во влажных местах, например на
берегах ручьев или старых водостоков. Листостебельные
же печеночники по размерам даже меньше мхов.
Но если для мхов и печеночников проблема воды не
стоит столь остро, то почему у них вообще имеется спо-
рообразующее поколение? Кажется, проще было бы
избежать смены поколений, т. е. чтобы одно половое
поколение непосредственно давало начало следующему
половому поколению. Почему же у этих растений все-
таки наблюдается смена поколений? Здесь две причины.
Прежде всего, хотя из-за отсутствия ископаемых остат-
ков мы не знаем происхождения мхов и печеночников,
весьма вероятно, что их предками были растения с
установившимся чередованием поколений. А то, что бы-
ло с таким трудом завоевано в процессе эволюции,
имеет тенденцию сохраняться, даже когда нужда в
этом отпадает. Однако имеется и более важная причи-
на. Она состоит в том, что наличие спор дает растению
возможность расселяться, которую половое поколение
дать не может.
Для водорослей расселение не составляет труда, так
как оплодотворенные яйцеклетки уносятся от материн-
ского растения водой. Иначе обстоит дело с наземными
растениями. Если у них нет специальных приспособле-
ний, обеспечивающих рассеивание, подобных тем, какие
имеются у плодов и семян цветковых растений, то опло-
дотворенная яйцеклетка должна прорасти в том месте,
где она остается после гибели материнского растения.
Поэтому для растений, обычно проводящих всю свою
жизнь на одном месте, способность к расселению имеет
решающее значение.
Рассеивание спор вообще не проблема. Маленькие и
легкие, они легко переносятся ветром, иногда на сотни
километров, как, например, споры Puccinia graminis —
гриба, вызывающего черную ржавчину пшеницы. Для
наземных растений спора — превосходный агент рас-
селения, тогда как оплодотворенная яйцеклетка — пло-
хой. Кстати, у мхов споры «выставлены» вверх в их ко-
робочках и имеют, таким образом, больше всего шансов
быть подхваченными порывом ветра. Даже сама коро-
бочка имеет устройство, помогающее высыпанию спор
.22
Глава t
наружу в сухую погоду. Камера, в которой находятся
споры, закрывается рядом зубцов — это так называе-
мый перистом,—отходящих от края коробочки внутрь
(фото II). Зубцы образованы клетками, лишенными жи-
вого содержимого, стенки которых утолщены таким об-
разом, что в сухую погоду зубцы отгибаются наружу,
освобождая споры, а если воздух влажный, они воз-
вращаются в прежнее положение, замыкая отверстие
споровой камеры. Следовательно, споры высеиваются
лишь при наличии условий, благоприятствующих их рас-
пространению ветром.
Папоротники тоже «поднимают» свои спорангии со
спорами вверх, на листочки, тогда как заростки, несу-
щие половые органы, прижимаются к влажной земле.
Как и мхи, папоротники имеют специальное устройство,
обеспечивающее рассеивание их спор преимущественно
в сухую, а не в сырую погоду.
Цветковые растения лучше всех других приспособле-
ны к жизни на суше. Это и не удивительно: ведь они —
самое совершенное из всего того, что дала эволюция ра-
стений. Прошло 200 млн. лет после рискованного пере-
хода растений из водной среды на сушу, прежде чем
появились цветковые растения. Выйдя из колыбели эво-
люции, они быстро заняли на Земле господствующее по-
ложение.
Чередование поколений у цветковых растений
Для цветковых растений, как и предшествовавших
им папоротников, характерно чередование полового по-
коления — гаметофита — и бесполого — спорофита, но
адаптация здесь зашла так далеко, что эти два поколе-
ния нелегко распознать. Само цветковое растение — это
спорофит, образующий споры двух типов. Одни из них —
микроспоры — дают гаметофит, несущий мужские поло-
вые органы, тогда как другие — мегаспоры — дают на-
чало женскому гаметофиту. Микроспоры хорошо знако-
мы каждому, потому что это не что иное, как .пыльца.
Но для того, чтобы увидеть мегаспоры, нам необходим
уже микроскоп. Внутри завязи (центральная женская
часть цветка) находятся семяпочки или незрелые семе-
Проблемы приспособления растений к наземной жизни 23
на. Если мы сделаем тонкий срез семяпочки и иссле-
дуем его под микроскопом, то обнаружим в ней одну
крупную клетку, называемую зародышевым мешком;
это женское половое поколение, которое развивается из
мегаспоры внутри семяпочки.
Половое поколение у цветковых растений настолько
сильно редуцировано, что его никак нельзя назвать за-
ростком. При прорастании пыльцевое зерно (микроспо-
ра) образует пыльцевую трубку, которая растет через
ткань завязи к семяпочке. В пыльцевой трубке имеют-
ся два мужских половых ядра, или гаметы1. Одно из
них сливается в зародышевом мешке с ядром яйцеклет-
ки, или женской гаметой; теперь оплодотворенная семя-
почка может развиваться в семя.
Сведя половое поколение почти на нет, цветковые
растения тем самым оказались в состоянии размножать-
ся половым путем без воды. Вместо плавающих спер-
матозоидов появились мужские ядра, доставляемые к
яйцеклетке пыльцевой трубкой. При условии же, что
корни цветковых растений могут доставлять нужное
для жизни количество воды, некоторые из них могут
размножаться и размножаются даже в центре пустыни
Сахара.
Современное цветковое растение — это организм,
прекрасно приспособленный к условиям окружающей
его среды и к своему образу жизни. Так и должно быть.
В борьбе за существование между растением и живот-
ным и растением и растением нет места для слабых.
Чтобы выжить, растения должны были приспособиться,
а те, которым это не удавалось, погибали, уступая ме-
сто более жизнеспособным. Дарвин назвал это «выжи-
ванием наиболее приспособленных».
Основной движущей силой этого процесса — эволю-
ции, начавшейся с появления жизни на Земле, — служит
естественный отбор. Те организмы, которые по какой-
х Хотя часто мужские гаметы представляются под световым
микроскопом как «голые» ядра, более тонкое изучение, как прави-
ло, показывает наличие у них тончайшей плазматической пленки,
содержащей митохондрии и пластиды. Таким образом, мужские
гаметы представляют собой настоящие клетки, а не просто ядра. —
Прим. ред.
24
Глава I
либо причине были лучше приспособлены, чтобы выдер-
жать борьбу за существование, выжили и передали
свои благоприятные качества потомкам, которые в свою
очередь оказались способными пережить других. Од-
нажды возникшие полезные свойства таким образом
закреплялись и со временем усиливались. Наоборот,
особи с неблагоприятными отклонениями не могли кон-
курировать со своими собратьями и погибали, унося с
собой качества, явившиеся причиной их гибели. Приро-
да следит за тем, чтобы оставались жить только при-
способленные.
Естественный отбор действует так же, как и искус-
ственный, часто применяемый человеком при выведении
новых пород скота или создании различных сортов ра-
стений. Встретив новую разновидность гороха с более
крупными стручками или более сочными семенами, чем
обычно, селекционер непременно использует ее для вы-
ведения новых растений гороха, надеясь, что у них эти
свойства закрепятся. Растения же с более низкими каче-
ствами, чем обычно, никогда не используют для селек-
ции. Бывает, что новые разновидности возникают сти-
хийно, но это случается не всегда. Человек же — селек-
ционер животных и растений — может обдуманно со-
здавать новые сорта и расы путем скрещивания особей,
обладающих какими-то желательными качествами. Ког-
да впервые были обнаружены растения пшеницы, устой-
чивые к черной ржавчине, они давали плохой урожай,
но, скрещивая их с высокоурожайными растениями, хотя
и склонными к черной ржавчине, проф. Биффен (Кемб-
ридж) вывел сорт пшеницы Иомен, сочетающий устой-
чивость к ржавчине с высокой урожайностью1.
1 В СССР выведено много сортов пшенвцы, высокоурожайных
и в то же время устойчивых к грибным заболеваниям, в том числе
к ржавчине. Так, например, в нечерноземной полосе Европейской
части СССР районирован сорт Московка, выведенный В. Е. Писа-
ревым, а также пшенично-пырейный гибрид 599 (Шехурдиновка),
полученный Н. А. Цициным и Г. Д. Лапченко. На юге Украины
распространен урожайный и устойчивый к болезням сорт Одес-
ская 12, в Ставропольском и Краснодарском краях — гибрид 481, в
Заволжье— Безенчукская 98 и т, д. Интересно, что мировое призна-
ние, как самый устойчивый против грибных заболеваний вид,
Проблемы приспособления растений к наземной жизни 2S
Возникают ли нужные качества сами по себе, т; е.
спонтанно, или благодаря стараниям селекционера, ре-
зультат всегда одинаков — улучшение сорта. Обладая
специальными знаниями, человек может иногда — но
отнюдь не всегда — быстро достигнуть желаемой цели.
Естественный отбор, действующий, как известно, слепо,
делает то же самое, по медленнее. У природы много
времени, и ей некуда спешить.
Важно отметить, что естественный отбор не может
сам создавать новые виды; он может лишь направлять
вариации, которые уже подготовлены для его деятель-
ности. Здесь выступает огромное значение полового
размножения. Когда два организма размножаются по-
ловым путем, их признаки смешиваются в потомстве:
в одних отношениях оно напоминает отца, а в других —
мать. Но при этом потомки обладают и некоторыми соб-
ственными признаками, свойственными им одним, так
как физические носители наследственности, или, как
мы их называем, гены, обоих родителей могут различ-
ным образом комбинироваться, и в результате возника-
ют особенности, которых не было ни у одного из них.
Если кто-нибудь надеется вырастить сливу сорта
Виктория из семян, его ждет разочарование: может ока-
заться, что из пятидесяти разных слив, которые он по-
лучит, ни одна не будет Викторией. Об этом позаботят-
ся насекомые, производящие перекрестное опыление.
Если вы хотите развести сливу сорта Виктория или ка-
кое-нибудь другое наше обычное плодовое дерево, вы
должны привить частичку материнского дерева на со-
ответствующий подвой. Только в этом случае вы може-
те быть уверены, что получите именно то, что хотите.
Дело в том, что половое размножение ненадежно, когда
речь идет о том, чтобы сохранить неизменными призна-
ки какого-либо сорта или вида.
Именно эта «незапрограммированность» результатов
полового акта и дает материал для деятельности естест-
имеет пшеница «Зандури», принадлежащая к особому виду Triti*
cum Timopheevi и культивируемая только в некоторых районах
западной Грузии. Один из гибридов этой пшеницы, выведенный
П. М. Жуковским, был назван Triticum [ungicidum, что в переводе
значит сгрибобойная пшеница». — Прим, ред.
26
Глава 1
венного отбора; в этом заключается «жизненная сила»
эволюции и в противном случае процесс этот не мог бы
продолжаться, т, е. пришел бы к печальному концу.
Однако гибриды возникают не только прн половом
размножении; гораздо более важно то, что сама приро-
да время от времени дает резкие уклонения, или мута-
ции, которые и подхватывает естественный отбор.
На первый взгляд может показаться странным, что
возникающие в природе мутации почти всегда вредны,
но это вполне объяснимо. Существуя в природе десятки
миллионов лет, растение или животное прекрасно при-
спосабливается к своему образу жизни. Не будь это
так, оно бы не выжило. Другими словами, гены, конт-
ролирующие его наследственность, «работают» гармо-
нично с внешней средой. Мутация — это изменение, про-
исходящее в самом гене. А в хорошо сбалансированном
наборе генов это по большей части нарушает порядок.
Лишь в редких случаях мутации приводят к улучшению
признаков. В таких случаях естественный отбор бывает
направлен на сохранение благоприятной мутации.
Таким образом, мы можем представить себе процесс
эволюции как постепенное изменение путем мутаций.
При этом мутанты проходят через сито естественного
отбора и благоприятные мутации сохраняются, а вред-
ные (их большинство) отбрасываются1. Борьба за суще-
ствование является в этом процессе отсева движущей
силой. Все, что может помочь организму в этой борьбе,
имеет шансы на сохранение, тогда как все вредное отме-
тается. Таким образом в процессе эволюции возникли
цветковые, хвойные, папоротники, мхи и все другие
растения в течение многих веков, прошедших с того мо-
1 Автор несколько упрощенно толкует естественный отбор
лишь как <сито>, просеивающее уже готовые формы. В природе дело
обстоит гораздо сложнее. Естественный отбор проявляется в разных
формах и выступает не только как отсеивающий и распределяющий,
но и как единственный направляющий фактор, от которого зависят
вероятность и частота возникновения новых мутаций и комбина-
ций. В этом смысле эволюционисты всегда подчеркивают творче-
скую роль естественного отбора. Подробнее об этом см. Н. В. Т и-
м о ф е е в-Р есовский, Н. Н. Воронцов, А. В. Я б л о-
ков, Краткий очерк теории эволюции, изд-во <Наука», М.а
1969. — Прим. ред.
Проблемы приспособления растений к наземной жизни 27
мента, когда жизнь впервые попыталась появиться на
нашей планете. Так из примитивных микроскопических
водорослей, плававших в докембрийском океане, т. е.
500 млн. лет назад, возник в своем бесконечном разно-,
образин и сложности растительный мир.
Направляемые естественным отбором растения в про-
цессе эволюции приспособились к свойственному им
образу жизни и окружающей среде. То, что естествен-
ный отбор действует вслепую, никак не сказывается на
его результатах. И нам часто приходится поражаться
гармонии, существующей между строением растения и
условиями его жизни.
ГЛАВА II
РАСТЕНИЯ-ИНЖЕНЕРЫ
Одна из первых проблем, которую должны были
разрешить растения, выбравшись из океана на сушу,—
это необходимость выносить свои стебли вверх, к свету,
и поддерживать их без помощи воды, которой уже не
было вокруг них. Для водорослей этой проблемы не
существует. Омывающая их вода служит им опорой, и
для сохранения плавучести им необходимо лишь некото-
рое количество пузыревидных вздутий, наполненных воз-
духом и разбросанных там и сям по их телу. Благодаря
этим пузырям их ассимилирующие органы выкосятся
ближе к свету и могут осуществлять жизненно важную
функцию фотосинтеза. Даже у гигантской водоросли
Macrocystis ветви, достигающие 180 м, могут держаться
близ поверхности воды, где они грациозно колышутся в
океанических течениях.
Другое дело наземные растения. Ни одно из них по-
ка не способно образовывать пузырьки, наполненные во-
дородом, которые дали бы растению возможность па-
рить в воздухе, подобно аэростату. И весьма сомни-
тельно, чтобы это когда-нибудь произошло, хотя я и
не решаюсь утверждать, что для растения есть что-то
невозможное. Наземное растение должно было быть
построено прочно, чтобы стебель не только стоял прямо
благодаря его собственной жесткой структуре, но и про-
тивостоял порывам ветра, склоняющим его вниз.
Небольшие травянистые растения могут сохранять
прямостоячее положение благодаря гидростатическому
давлению, возникающему при наполнении их клеток во-
дой. Этот же принцип лежит в основе многих других
явлений. Если, например, из внутренней камеры велоси-
педной покрышки выкачать воздух, ей легко можно при-
дать любую форму и она не способна сопротивляться.
Камера же, надутая воздухом, принимает форму круга
и противостоит любой попытке ее деформировать. В на-
дутом виде она становится почти такой же твердой, как
само колесо,
Растения-инженеры
29
Тургор, прочность и проведение воды
С маленькими растениями происходит то же самое.
Когда клетки насыщены водой, т. е. становятся, как го-
ворят ботаники, тургесцентными, растение приобретает
твердость, и всякая попытка согнуть стебель может при-
вести к разрыву тканей. Однако такой тургорный меха-
низм годится только для очейь маленьких растений или
их отдельных частей, например для черешков листьев.
Вы можете видеть действие этой системы — вернее, ее
бездействие, — если заглянете в огород морозным зим-
ним утром. При этом обратите внимание на то, как
безвольно висят листья брюссельской капусты, вместо
того чтобы держаться гордо и стоять прямо. Дело в
том, что в холодную погоду корням растения трудно на-
сасывать воду из почвы, поэтому тургор листовых че-
решков ее может поддерживаться и листья поникают.
Это явление называют завяданием; всем хорошо извест-
но, как вянут срезанные в саду цветы и как они быстро
оживают, если поместить их в воду.
Но растение растет, и наконец наступает момент,
когда оно достигает определенной величины и тургор-
ный механизм оказывается не в состоянии обеспечивать
твердость главному стеблю. Это происходит потому,
что необходимость насасывать большое количество во-
ды для поддержания тургора во всех клетках стебля
создавала бы невыносимое напряжение в корневой си-
стеме. Кроме того, когда растение становится выше, его
вес увеличивается уже не пропорционально высоте и
прочности стебля. Прочность стебля примерно пропор-
циональна его поперечному сечению: при увеличении
площади этого сечения в два раза прочность возрастает
в четыре раза. Но объем растения, определяющий его
вес, находится в кубической зависимости от линейных
размеров; если эти размеры увеличиваются в два раза,
растение становится тяжелее не в четыре, а в восемь
раз. Поэтому, если растение становится больше, оно ско-
ро достигает предела, когда никакой тургорный меха-
низм уже не в состоянии обеспечить твердость.
Разрешение проблемы прочности шло рука об руку
с решением столь же неотложной проблемы проведения
30
Глава II
воды с растворенными в ней минеральными вещества-
ми, необходимыми растению, из корней вверх по стеблю.
У первых наземных растений для удовлетворения обеих
этих потребностей выработался единый механизм.
Вдоль их стебля проходили ряды клеток, называемых
трахеидами (фото III, IV). Они имели веретеновидную
форму, и их длина значительно превышала ши-
рину (рис. 5). Их первичные оболочки, подобно
оболочкам большинства растительных клеток,
состояли из целлюлозы, но с внутренней сторо-
ны откладывались слои вторичного утолщения,
содержащие уже упоминавшееся твердое вещест-
во, называемое лигнином. Эти клетки во взрос-
лом состоянии теряли свое живое содержимое,
но утолщенные клеточные оболочки сохранялись.
Заостренные концы трахеид переплетались друг
с другом, образуя прочную структуру, способную
выдержать сгибание. Благодаря трахеидам стеб-
ли растений оказались в состоянии выдерживать
вес листвы, не прибегая к помощи тургорного
механизма.
Рве. 5. Трахеида с окаймленными порами.
Но это не все. Полые трахеиды, лишенные клеточно-
го содержимого, были хорошо приспособлены для про-
ведения воды. Они образовывали сплошной ряд трубок,
т. е. как бы трубопровод, проходивший от основания
стебля к его верхушке, по которому от корней к верху-
шечным листьям могла подниматься вода с растворен-
ными в ней минеральными солями.
Трахеиды этих спервобытных» растений были пер-
вой стадией в развитии специализированной ткани —
ксилемы, или древесины. В ходе дальнейшей эволюции
растений древесина становилась все более сложной.
Образованию трахеидами эффективной трубчатой си-
стемы мешало то, что каждая трахеида отделялась от
следующей клеточной оболочкой, которая была пропита-
на лигнином, что должно было бы создать непроннцае-
Рас*еНия-инжене0ь1
31
мый барьер для йрохождения воды. Но эти оболочки
одревесневали не полностью: в них оставались тонкие
участки, называемые порами, через которые и могла
проходить вода.
Почти не приходится сомневаться в том, что трахеи-
ды возникли как элемент, служащий для двух целей.
Одревесневшие вторичные оболочки трахеид, их заост-
ренные и переплетенные между собой концы и то, что,
как правило, трахеиды располагаются в стебле группа-
ми, примыкая друг к другу, — все это позволяет им
успешно осуществлять механическую функцию. В то же
время отсутствие в трахеидах живого содержимого де-
лает их вполне подходящими для проведения воды. Это
приспособление работало очень хорошо, и до сих пор
трахеиды верно служат растениям; так, даже гигантская
секвойя, самое большое из современных деревьев, поль-
зуется трахеидами для проведения воды вверх по стволу
на высоту свыше 100 м. Однако, хотя трахеиды и эф-
фективны, их можно еще более усовершенствовать.
Из-за того что трахеиды несут функцию проведения,
они должны сохранять определенный диаметр. Если бы
процесс утолщения стенок продолжался дальше како-
го-то определенного предела, их внутренний диаметр
стал бы слишком маленьким для эффективного прове-
дения воды (как вы помните, утолщение происходит на
внутренней стороне первичной оболочки.благодаря де-
ятельности живого, содержимого клетки). Кроме того,
если бы стенка стала слишком толстой, это затруднило
бы образование пор, через которые вода проходит от
трахеиды к трахеиде. В связи с этим толщина оболочки
трахеиды имеет предел, а значит, и прочность отдель-
ной трахеиды не может возрастать бесконечно.
Волокна и членики сосудов
Предположим, что трахеида перестанет выполнять
проводящую функцию и сосредоточится только на ме-
ханической. В таком случае она могла бы откладывать
На внутренней стороне первичной оболочки столько сло-
ев вторичного утолщения, сколько ей нравится, помня,
однако, о том, что внутри клетки все же должно оста-
S&
Глава II
ваться небольшое пространство для живого содержимо-
го, обеспечивающего одревеснение. В результате полу-
чился бы элемент гораздо более прочный, чем обычная
трахеида, хотя, конечно, он был бы почти непригоден
для проведения воды. Именно это и произошло с ра-
стительными клетками, называемыми волокнами. Во-
локна— длинные и узкие клетки, иногда почти игло-
видной формы (рис. 6). Оболочки волокон силь-
но одревесневают, так что в центре клетки оста-
ется только небольшое пространство или полость,
где находится живая цитоплазма, которая су-
ществует до тех лор, пока не закончит свою ра-
боту— вторичное утолщение, а затем отмирает.
Клетка подобного типа специализирована исклю-
чительно на выполнении опорных функций.
Чтобы увидеть волокна, находящиеся на наи-
высшей ступени своего развития, мы должны
обратиться к цветковым растениям, хотя начали
они свою эволюцию еще у хвойных. Технические
волокита, используемые для изготовления тканей
или веревок, могут достигать фантастической
длины. Одно волокно льна может быть до
Рис. 6. Волокно.
Разрыв в середине означает, что в действительности волокно в
несколько раз длиннее, чем показано на рисунке. Обратите вни-
мание на дегенерацию окаймленный лор.
70 мм длиной, а волокно рами (Boehtneria nivea), расте-
ния из семейства крапивных (Urticaceae), используемо-
го в текстильной промышленности, имеет совершенно не-
правдоподобную длину — 250 мм.
Волокна непригодны для проведения воды, но это не
имеет существенного значения, поскольку другие трахеи-
ды, не подвергшиеся специализации, выполняют эту
функцию. Более важными компонентами древесины
цветковых растений являются клетки, называемые чле-
никами (или элементами) сосудов и служащие только
для проведения воды.
Растения-инженеры
33
Членики сосудов возникли из трахеид, специализи-
ровавшихся в противоположном по сравнению с волок-
нами направлении. Для волокна главное — прочность;
членики же сосудов пожертвовали прочностью ради спо-
собности проводить воду. Они стали важнейшими про-
водящими клетками и встречаются почти исключительно
у цветковых растений.
Основной недостаток трахеид как водопроводящих
клеток заключается в том, что вода вынуждена прохо-
дить из трахеиды в трахеиду через поры в их стенках.
При этом передвижение воды замедляется в связи с не-
обходимостью преодолевать множество очень узких от-
верстий, затянутых поровой мембраной, которая пред-
ставляет собой первичную оболочку трахеиды. Если бы
не было этих конечных стенок! Тогда вода могла бы
свободно проходить из трахеиды в трахеиду, встречая на
своем пути не больше препятствий, чем в водопровод-
ных трубах.
Именно это и было достигнуто в члениках сосудов.
Вообразите ряд трахеид, лежащих друг над другом, ко-
торые соединены между собой концами й сообщаются
через поры в их конечных стенках. Затем представьте
себе, что .поровые мембраны исчезают и несколько пор,
увеличиваясь, сливаются, образуя одно большое отвер-
стие; наконец конечные стенки трахеид разрушаются и
между ними появляется свободный проход. Трахеиды
превратились в членики сосудов (рис. 7).
У цветковых растений мы находим именно такие чле-
ники сосудов, примыкающие вплотную друг к другу по-
добно сегментам водопроводной трубы. Так образуются
водопроводящие трубки, идущие до верхушек самых
высоких стеблей. Ничто не препятствует воде подни-
маться по такой системе сосудов, за исключением силы
тяжести, которая стремится повернуть поток обратно
вниз.
Чем шире членики сосудов, тем свободнее вода мо-
жет проходить по ним; поэтому не удивительно, что в
процессе специализации сосудов отдельные их членики
становились все шире и короче. Мы можем проследить
такую эволюционную линию от примитивных типов дре-
весины к более совершенным через ряд семейств цвет-
»-*й
34
Г лава II
ко вых растений. Например, членики сосудов у ив при-
митивны: они длинные и сравнительно узкие, с заострен-
ными концами. У дуба — усовершенствованный тип дре-
весины: членики сосудов короткие и широкие, с гори-
зонтальными поперечными стенками.
Членики сосудов в значительной сте-
Рис. 7. Членик
пени утратили свою первоначальную
опорную функцию, и их роль свелась к
проведению воды. Но это не страшно,
так как механическую функцию взяли на
себя многочисленные волокна. Наряду с
члениками сосудов и волокнами у многих
цветковых растений имеются и простые
трахеиды. Членики сосудов, однако, со-
храняют свои утолщенные оболочки. Это
необходимо, поскольку отложение в этих
оболочках твердого лигнина предотвра-
щает спадание стенок и таким образом
обеспечивает непрерывность передвиже-
ния воды по сосудам. Имеется и другая
важная причина, почему стенки сосудов
должны быть жесткими, но мы отложим
обсуждение этого вопроса до следующей
сосуда. главы.
Ткани стебля
Стебель растения состоит из многих различных ти-
пов клеток, расположенных определенным образом; эю
расположение хотя и варьирует в зависимости ст вида
растения, но в общем не выходит за рамки некоего об-
щего плана. В качестве иллюстрации расположения тка-
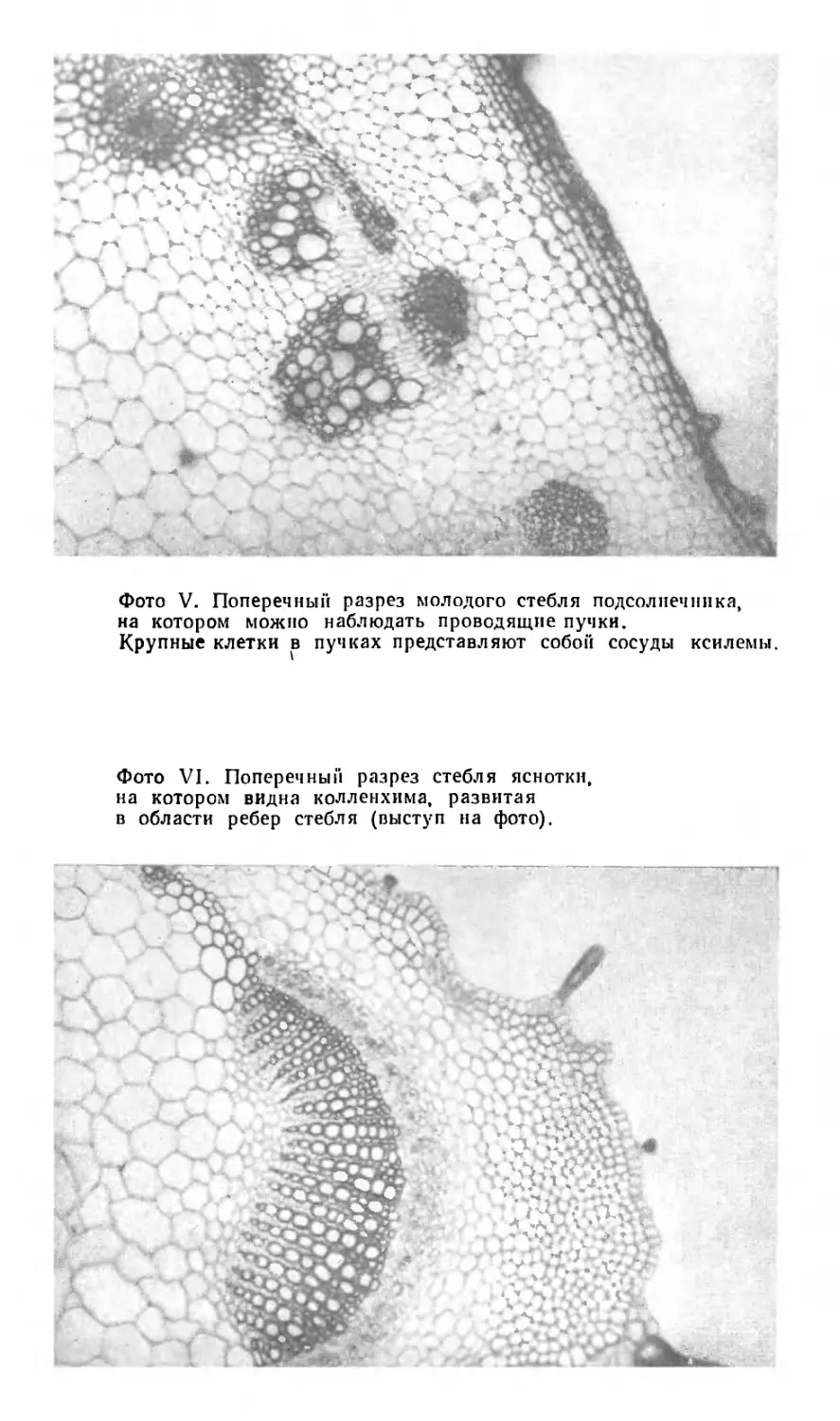
ней нам может служить молодой стебель подсолнечни-
ка. Подсолнечник — однолетнее растение; его стебель
хотя и достигает значительных размеров, но не приспо-
соблен к многолетнему существованию, так что древе-
сины в нем немного.
Если мы рассмотрим тонкий поперечный срез стебля
подсолнечника, то прежде всего увидим, что он состо-
ит в основном из тонкостенных клеток, вместе состав-
ляющих нечто похожее на пчелиные соты, хотя и менее
Растения-инженеры
35
правильные. Эти клетки имеют целлюлозные оболочки
и обозначаются общим термином паренхима. Паренхи-
ма может быть названа основной тканью стебля. Сна-
ружи стебель покрыт кожицей, или эпидермисом, со-
стоящим из одного слоя клеток.
Основная ткань разграничена на две зоны: наруж-
ную — кору и центральную — сердцевину, разделенные
кольцом ясно выраженных групп специализированных
Рис. 8. Схема поперечного разреза молодого стебля подсолнечника.
клеток, которые называются проводящими пучками; они
названы так потому, что содержат клетки, проводящие
воду и питательные вещества. В промежутках между
проводящими пучками кора смыкается с сердцевиной
(рис. 8).
Проводящие пучки содержат клетки различного типа
и построены соответственно одному общему плану
(фото V). Ближе к центру стебля располагается кси-
лема, или древесина. Она состоит преимущественно из
толстостенных клеток, и если мы, прежде чем иссле-
довать срез стебля под микроскопом, погрузим его на
несколько минут в подкисленный раствор флороглюци-
на, то увидим, что оболочки этих клеток окрашиваются
з»
36
Глава Л
в ярко-розовый цвет, что свидетельствует об их силь-
ном одревеснении. В древесине прежде всего обращают
на себя внимание членики сосудов — крупные клетки,
имеющие форму неправильных многоугольников. Эти
членики сосудов расположены радиальными рядами,
а между ними находятся клетки меньшего размера,
с тонкими оболочками, которые не окрашиваются фло-
роглюцином; это клетки древесинной паренхимы.
Кнаружи от древесины в каждом проводящем пучке
имеются два или три слоя уплощенных тонкостенных
клеток, напоминающих ряды прямоугольных коробочек.
Это камбий — ткань, играющая важную роль, когда мо-
лодой стебель начинает расти в толщину. Снаружи
к камбию примыкает флоэма (луб) (стр. 14). Это
ткань, связанная главным образом с проведением пла-
стических веществ в растении; более подробно мы по-
говорим о ней в гл. V. Луб состоит из больших тонко-
стенных и на вид почти пустых клеток, называемых си-
товидными трубками. Между ними разбросаны мень-
шие по размеру клетки с густым содержимым. Это
клетки-спутники. Среди ситовидных трубок и клеток-
спутников имеется также некоторое количество клеток
флоэмной паренхимы. Оболочки клеток камбия и фло-
эмы не становятся розовыми при действии на них фло-
роглюцина; следовательно, они не одревесневают.
С наружной стороны каждого проводящего пучка
подсолнечника имеется группа клеток с очень толстыми
оболочками, как бы шапочка, надетая на пучок. Стенки
этих клеток ярко окрашиваются флороглюцином, что
говорит об их сильном одревеснении. Это волокна, яв-
ляющиеся основными опорными элементами молодого
стебля.
Распределение групп волокон по периферии стебля
не случайно, потому что именно здесь они более всего
нужны. Ведь если вы имеете некоторое количество ме-
талла и хотите соорудить из него нечто устойчивое, вы
не будете отливать его в форме сплошного стержня,
а скорее придадите ему форму трубки большего диа-
метра. Чем шире трубка, тем больше устойчивость (при
условии, что стенки трубки имеют некоторую минималь-
ную прочность). Этот факт хорошо известен инжене-
Растения-инженеры
37
рам1. В расчет принимается лишь материал, располо-
женный снаружи; ведь стальная трубка, подобная тем,
какие используются для велосипедных рам, почти так.
же прочна, как и цельная металлическая полоса тако-
го же диаметра, хотя, конечно, ее абсолютная проч-
ность меньше.
Следовательно, расположение волокон по периферии
стебля, т. е. там, где они могут обеспечить ему макси-
мальную прочность без лишних затрат материала, пра-
вильно отражает принципы, существующие в инженер-
ной практике.
Такой тип распределения опорных элементов мы
встречаем в стеблях многих травянистых растений.
В этом отношении белая яснотка, или глухая крапива,
представляет собой именно то, что нам нужно. Это
растение имеет четырехгранный стебель, и если мы
посмотрим на поперечный срез этого стебля под мик-
роскопом, то увидим, что проводящие пучки тоже об-
разуют четырехгранник, располагаясь близ поверх-
ности стебля. Кроме проводящих пучков, у белой
яснотки есть и другие участки механической ткани, но
уже другого типа (фото VI). В области ребер ее четы-
рехгранного стебля, непосредственно под эпидермисом,
располагаются группы клеток, стенки которых утолще-
ны неравномерно. Утолщение сосредоточено в углах
клеток. Оболочки таких клеток не одревесневают, и их
утолщение происходит в основном за счет целлюлозы;
в этом отношении они резко отличаются от волокон,
которые почти всегда бывают одревесневшими. Другое
отличие этих клеток от волокон и трахеид заключается
в том, что и в зрелом состоянии они сохраняют живые
протопласты.
Этот весьма обычный в растениях тип опорной тка-
ни носит название колленхимы. Особенно часто она
встречается у маленьких растений и в таких небольших
1 Очень интересные исследования были проведены советским
ботаником, анатомом растений и инженером по образованию,
В. Ф. Раздореким. Сформулированные им «строительно-механиче-
ские принципы в конструкции органов растений» популярно изло-
жены в его книге «Архитектоника растений», изд-во «Советская нау-
ка», М., 1955. — Прим. ред.
38
Глава ll
частях крупных растений, где необходима некото-
рая жесткость, например в черешках и пластинках
листьев. Тенденция к образованию колленхимы имеется
у проростков, но впоследствии присущая ей опорная
функция передается древесине.
Стебель глухой крапивы полый — еще одна обыч-
ная для травянистых растений черта строения. Хотя
центральная часть стебля занята сердцевиной, она не
играет заметной роли в жизни растения и поэтому
часто полностью разрушается.
Особенно хорошо эта полая структура заметна
в стеблях многих злаков, включая такой замечатель-
ный пример инженерной конструкции растений, как сте-
бель бамбука. Это одна из самых твердых растительных
структур, используемая даже для изготовления лезвий
ножей.
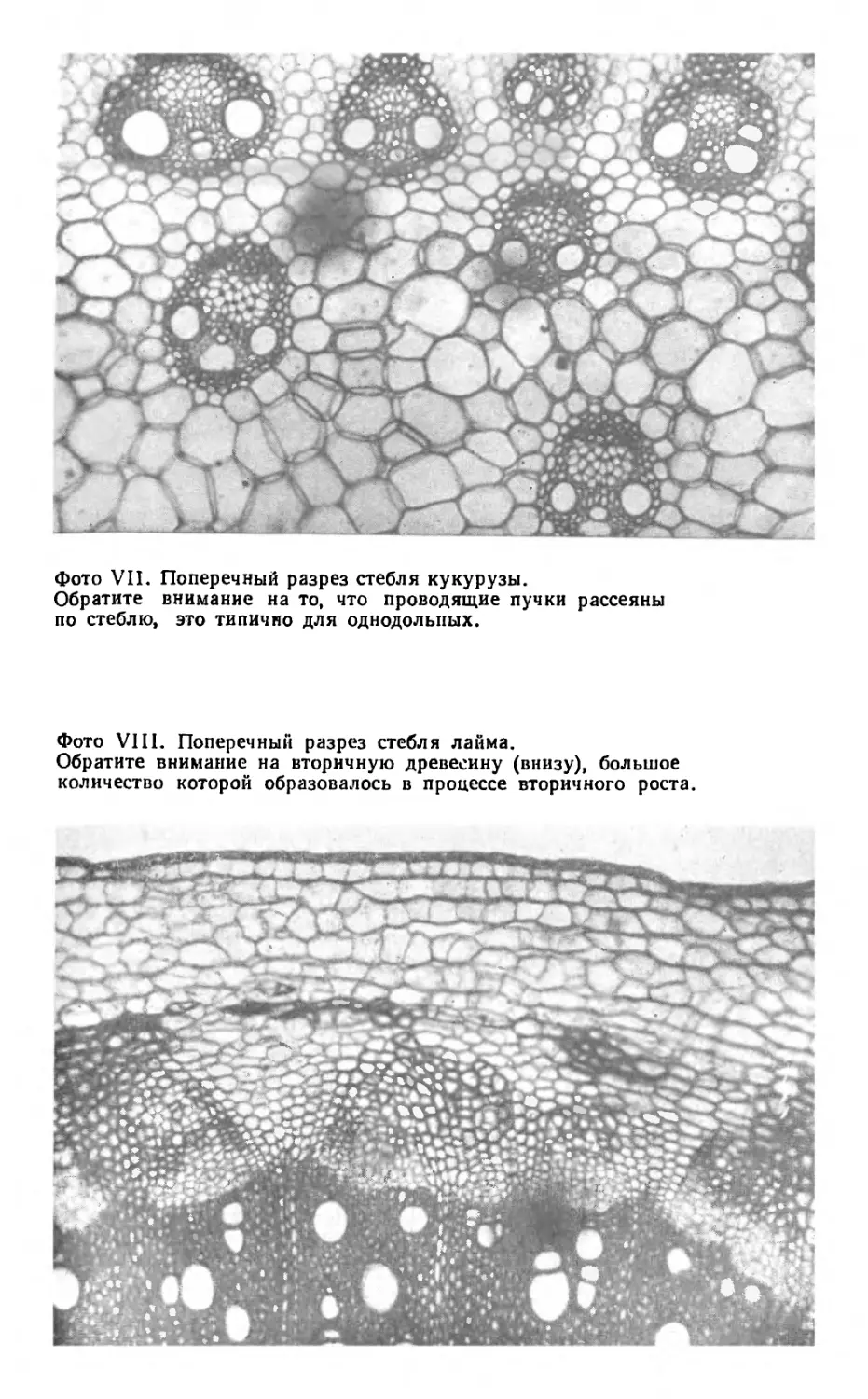
Бамбук и вообще злаки принадлежат к однодоль-
ным— группе цветковых растений, к которым относят-
ся также орхидеи, лилии, нарциссы, ирисы и другие рас-
тения с лентовидными листьями. Расположение прово-
дящих пучков в их стеблях, как правило, отличается от
кольцевого, типичного для двудольных — другой боль-
шой группы цветковых растений, куда относятся под-
солнечник, глухая крапива и большинство наших обыч-
ных трав, кустарников и деревьев.
У однодольных проводящие пучки не образуют на
поперечном срезе кольцо, а рассеяны по всему стеблю
(фото VII). Это можно наблюдать у бамбука. Его сте-
бель полый, за исключением узлов, или «суставов»,
а проводящие пучки сконцентрированы на периферии
стебля. В коре, сразу под эпидермисом, располагается
масса волокон, в которую погружены наиболее удален-
ные от центра проводящие пучки, что придает стеблю
большую прочность. Наружные клетки стебля тоже спо-
собствуют его прочности благодаря тому, что их стен-
ки пропитаны твердым кремнеземом, который расто-
ния-алхимики извлекают из раствора минеральных со-
лей, поглощаемого их корнями из почвы; этот кремне-
зем откладывается там, где он нужнее всего.
Бамбук находит самое разнообразное практическое
применение, особенно в восточных странах. Его стебли
Растения-инженеры
39
сочетают легкость с твердостью и в то же время легко
расщепляются. Из цельных стеблей бамбука строят
дома, а если их расщепить, получается прекрасная «че-
репица» для крыши. Стены домов делают из циновок,'
сплетенных из тонких бамбуковых пластинок. Полые
стебли бамбука могут служить в качестве труб для про-
ведения воды в дом, а расщепленные на две половинки,
они представляют собой идеальные водосточные желоба.
Толстые стебли этого растения служат опорами для
мостов, а из тонких можно изготовить любую вещь на
свете: трости, флейты, мебель, домашнюю утварь, ульи,
сельскохозяйственные орудия и множество других по-
лезных вещей. Расщепленные стебли бамбука с острыми
краями используются даже как ножи для косилок.
Его молодые побеги можно есть, как спаржу, а на стеб-
лях Bambusa arundinacea имеются наросты кремнезе-
ма, которые, как считается на Востоке, исцеляют почти
от всех болезней. Создав бамбук, природа превзошла
самое себя.
Функция камбиального слоя
До сих пор мы рассматривали только травянистые
растения, стебли которых отмирают каждый год при
приближении зимы. Но кустарникам или деревьям нуж-
но что-то более совершенное. С увеличением размеров
растения возрастает и нагрузка на главный стебель,
или ствол, в связи с чем для выполнения всех его функ-
ций возникает необходимость в развитии большой мас-
сы древесины. К тому же многочисленные листья дере-
вьев предъявляют большие требования к снабжению
водой, и ксилема как проводник воды должна их удов-
летворять. Поэтому не удивительно, что главное изме-
нение, которое мы замечаем в ходе превращения сла-
бого стебля проростка в ствол дерева, заключается
в образовании большого количества древесины, спо-
собной справиться с такой исключительной нагрузкой.
Одновременно происходят и другие изменения, но имен-
но прирост древесины лежит в основе этого превра-
щения.
40
Глава И
Вернемся к нашему исходному примеру со стеблем
подсолнечника и посмотрим, что с ним происходит,
когда он становится старше. Достигнув определенного
размера, стебель претерпевает процесс так называемого
вторичного роста. Он начинается в слое камбия — тка-
ни, состоящей из несколько уплощенных клеток, лежа-
щих между ксилемой и флоэмой в проводящем пучке.
Первое, что мы замечаем,— это деление камбиаль-
ных клеток; одна клетка делится на две, которые,
дорастая до размеров исходной материнской клетки,
в свою очередь снова делятся. Таким образом камбий
проводящих пучков начинает образовывать новые
клетки.
В то же время некоторая активность обнаруживает-
ся и в промежутках между проводящими пучками.
Клетки, лежащие на одном уровне с камбием, находя-
щимся в пучках, также начинают делиться. Новый
камбий закладывается в паренхиме — ткани, располо-
женной между проводящими пучками, и смыкается
с пучковым камбием. В результате по окружности стебля
вскоре возникает полное, активно делящееся кольцо
камбия.
Для деятельности камбия характерны две особен-
ности. Во-первых, камбиальные производные обычно
располагаются в стебле в виде радиальных рядов, т. е.
либо по направлению к центру, либо к периферии
и очень редко между двумя камбиальными клетками.
С утолщением стебля длина окружности камбиального
кольца тоже увеличивается благодаря происходящим
время от времени тангентальным делениям клеток кам-
бия1; однако число таких делений всегда ограничено —
не больше, чем это необходимо для приведения в со-
ответствие размеров стебля и камбиального кольца.
1 Автор здесь и в других местах книги, говоря о направлении
деления клеток, имеет в виду направление, в котором откладываются
новые клетки, но не направление возникающих при делении клеточ-
ных перегородок. В данном случае тангентальные ряды клеток
формируются за счет перегородок, образующихся радиально.
В других случаях радиальные ряды клеток образуются благодаря
тангентальным делениям. В анатомической литературе принято
обычно обозначать именно последний признак. — Прим. ред.
Растения-инженеры
41
Во-вторых, судьба двух дочерних клеток, образов
вавшихся при делении камбиальной клетки, различна.
Как правило, наружная из них становится новой клет-
кой камбия и снова делится обычным способом. Внут-
ренняя же клетка перестает быть камбиальной и диф-
ференцируется в какой-нибудь элемент ксилемы—тра-
хеиду, членик сосуда, волокно или клетку древесинной
паренхимы. Следовательно, камбий все время форми-
рует новую ксилему и она располагается всегда
ковнутри от камбиального кольца.
Однако иногда случается обратное. Вместо наруж-
ной дочерней клетки новой клеткой камбия становится
внутренняя. В этом случае наружная клетка превра-
щается не в древесинный, а во флоэмный элемент.
Таким образом, если цилиндр новой древесины форми-
руется по направлению к центру от камбиального коль-
ца, то флоэмный цилиндр подобным же образом откла-
дывается кнаружи от него. Камбий остается в сере-
дине, продолжая делиться на протяжении всей жизни
растения.
Новая древесина, образованная камбием, в общем
похожа на первичную древесину (определенные разли-
чия, конечно, имеются, но мы не будем их касаться);
она известна как вторичная древесина (фото VIII).
Аналогичным образом флоэма, возникшая из камбия,
называется вторичной флоэмой.
Теперь мы представляем себе, как образуется мас-
сивный ствол дерева. Начиная свою жизнь проростком,
не слишком отличающимся в анатомическом отноше-
нии от подсолнечника, дерево в процессе вторичного
роста, обусловленного деятельностью камбия, разви-
вает огромную массу древесины. Этот активный слой
камбия существует в течение всей жизни дерева, по-
стоянно увеличивая количество древесины. Никогда
нельзя сказать, что дерево выросло окончательно; это
одно из отличий многолетних растений от большинства
животных. Ведь животное растет до определенной ве-
личины, после чего его рост прекращается, растение
же продолжает расти всю жизнь.
По мере увеличения внутреннего цилиндра древеси-
ны увеличивается обхват ствола. Камбиальное кольцо
42
Глава ll
тоже увеличивается в диаметре благодаря тангенталь-
ным делениям своих клеток1; если бы не эти происхо-
дящие время от времени деления, камбиальное кольцо
вскоре растянулось бы и разорвалось. Расположенная
кнаружи от камбия вторичная флоэма не может увели-
чивать свой диаметр подобным образом, так как ее
клетки, однажды дифференцировавшись в ситовидные
трубки или другие элементы флоэмы, не способны
к дальнейшим изменениям. Дело в том, что клетка мо-
жет продолжать деление, только пока она остается
простой и неспециализированной, подобно клеткам кам-
бия; становясь же сложной, как, например, ситовидная
трубка, она обычно, хотя и не всегда, теряет способ-
ность к делению. Поэтому древесина, расширяясь, ока-
зывает давление на вторичную флоэму, и ее клетки
разрушаются. Впрочем, они тут же заменяются новыми,
возникающими пз камбия. В результате всего этого
вторичная флоэма имеется кнаружи от камбия всегда,
но лишь в виде тонкого слоя. Вторичная древесина,
составляющая основную массу ствола взрослого дере-
ва, может иметь очень большой диаметр, тогда как слой
вторичной флоэмы очень тонкий.
Пробковый камбий
То, что применимо ко вторичной флоэме, в равной
степени применимо и к более наружным тканям — коре
и эпидермису, представляющим собой защитные ткани
молодого растения. Разрастающаяся кнаружи древеси-
на все больше давит на окружающие ее ткани и разру-
шает их. Поскольку кора взрослого растения несет не
так уж много функций, это было бы не столь важно,
если бы не две вещи. Во-первых, если бы ствол не был
покрыт такой водонепроницаемой оболочкой, вода испа-
рялась бы из него в воздух, возможно, даже быстрее,
чем корни успевали бы пополнять ее запасы. Ни одно
дерево не может позволить себе таким образом терять
воду: ведь вода — это самое ценное и для большого,
и для маленького растения. Во-вторых, совершенно не-
1 См. примечание к стр. 40,
Растения-инженеры
43
защищенный ствол стал бы прекрасным убежищем для
различных паразитных грибов, чьи споры готовы вос-
пользоваться малейшей трещиной в наружной «оборо-
не» дерева, как, например, рана или даже укол насеко-
мого. Всякий хороший садовод знает, что обрезку сли-
вовых деревьев лучше проводить летом, когда меньше
всего спор гриба Stereum purpureum, вызывающего
заболевание слив. Вот почему после обрезки деревьев
поверхности срезов обязательно замазывают варом или
краской.
К счастью, развитие нового защитного покрова
взамен тканей, разрушенных в процессе вторичного
роста, не требует от растения большой изобретатель-
ности, и этот процесс очень сходен с деятельностью
камбия, хотя менее сложен. По существу он заклю-
чается в заложении где-нибудь в поверхностных тканях
стебля нового камбиального слоя, который, следуя
за утолщением стебля, образует новый наружный
покров, замещающий старый. Этот новый камбий на-
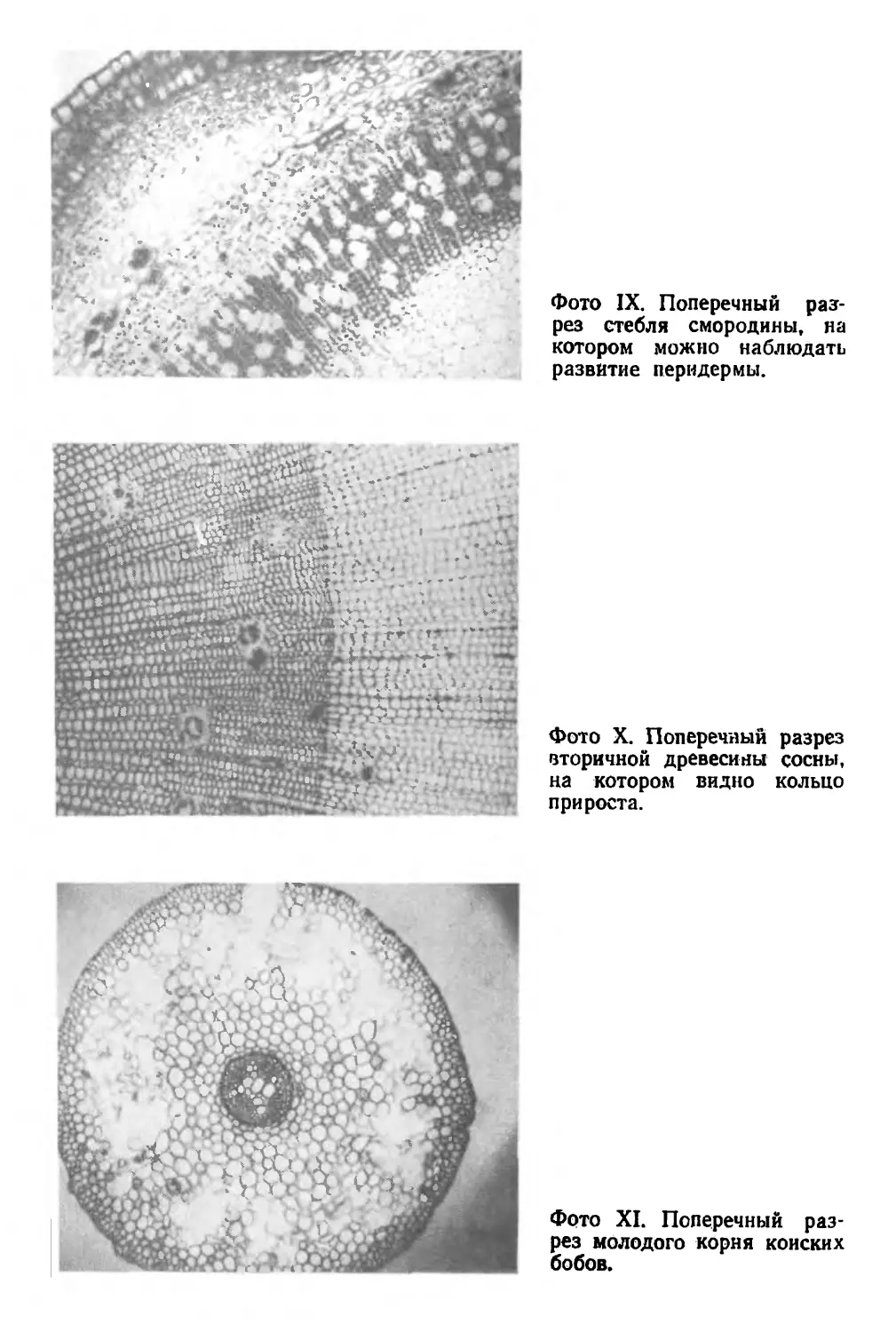
зывается пробковым камбием, или феллогеном, а обра-
зуемая им ткань — перидермой (фото IX).
Местоположение пробкового камбия варьирует у раз-
личных растений. В большинстве случаев он возникает
в коре, а иногда и непосредственно под старым эпи-
дермисом. Клетки феллогена делятся тем же способом,
что и клетки камбия. Деления радиальны по отноше-
нию к стеблю1, причем одна из дочерних клеток остает-
ся частью пробкового камбия и делится дальше, а дру-
гая дифференцируется в клетку перидермы. Ее судьба
зависит от того, как она расположена по отношению
к феллогену. Если эта клетка находится снаружи, она
превращается в клетку феллемы, или пробки. При этом
ее стенки пропитываются водонепроницаемым вещест-
вом, называемым суберином, а затем, достигнув зрело-
го состояния, она лишается живого содержимого и ста-
новится частью грубого наружного покрова ствола.
Если же такая клетка расположена с внутренней сторо-
ны пробкового камбия, то она преобразуется в клетку
феллодермы — ткани, состоящей из довольно однй-
1 См. примечание к стр. 40.
44
Глава П
родных клеток с живым содержимым, оболочки кото-
рых не содержат суберина.
Со временем в результате все продолжающегося
роста стебля пробковый камбий и перидерма, которую
он образует, разрушаются. Тогда в более глубоких
слоях коры обычно закладывается новый феллоген. Это
происходит неоднократно, и в конце концов феллоген
может возникнуть в наружном слое вторичной флоэмы,
а первичная кора полностью исчезает.
Перидерма составляет часть коры дерева; более гру-
бая наружная часть ствола в значительной степени
состоит из слоев пробки. Однако кора включает в себя
не только перидерму; если содрать кору с дерева, под
ней обнаружится обнаженная древесина. Следовательно,
термином «кора» обозначаются все ткани, расположен-
ные кнаружи от вторичной древесины, включая первич-
ную кору (если от нее что-нибудь остается), перидерму
и флоэму. Вот почему так вредно сдирать кору с дере-
ва: ведь в состав ее входит живая флоэма — ткань, от-
ветственная за перенос образующихся в листьях плас-
тических веществ к корням.
Если кора имеет такое важное значение не только
для защиты от проникновения паразитных грибов, но
и из-за жизненно важных функций флоэмы, то не уди-
вительно, что растение имеет свои собственные способы
устранения ее повреждений. Так, если мы удалим узкую
полоску коры со ствола, то вскоре обнаружим, что оба
края раны начинают расти по направлению друг к дру-
гу и в конце концов, если рана не слишком широкая,
смыкаются в центре. Эта ткань, образующаяся вблизи
краев раны, называется каллюсом; сначала клетки ее
не дифференцированы, но впоследствии они способны
превратиться в клетки нового пробкового камбия и да-
же новой флоэмы и возместить таким образом то, что
было утрачено.
Степень повреждения, которое может быть ликвиди-
ровано благодаря такому росту каллюса, ограничена.
Если кора удалена со слишком большого участка, ее
восстановление может оказаться невозможным. Удале-
ние полного кольца коры по окружности ствола приво-
дит иногда к гибели дерева.
Растения-инженеры 48
Так как в стебле имеются живые клетки, которые
все время дышат, то им необходимо постоянное снабже-
ние воздухом, и если бы стебель был сплошь покрыт
непроницаемым слоем пробки, ему грозило бы удуше-
ние. Избежать этой опасности позволяет система пор,
осуществляющих газообмен и называемых чечевичками.
Чечевичка представляет собой небольшой участок на
поверхности стебля, где клетки пробки отсутствуют; они
заменены рыхлой тканью,, что делает возможной сво-
бодную диффузию воздуха. Таким образом, способ по-
дачи воздуха к живым клеткам можно сравнить, по-
жалуй, с вентиляцией внутри корабля, которая осу-
ществляется специальными вентиляторами. Это еще раз
подтверждает, что в структурной организации растения
ничто не забыто и не оставлено на волю случая.
Некоторые деревья имеют очень толстую перидерму.
Например, у пробкового дуба (Quercus suber), дерева,
растущего в Испании, пробка достигает толщины 22 см.
Ее срезают и используют как бутылочную пробку, при-
чем удаление коры производят через определенные про-
межутки времени, с тем чтобы дерево успело опра-
виться. Снимают пробку на границе перидермы с флоэ-
мой, чтобы как можно меньше повредить дереву, и эта
работа требует высокой квалификации. Темные полосы,
проходящие через бутылочную пробку, представляют
собой чечевички.
Старение стволов деревьев
Массивный ствол лесного дерева содержит огромную
массу древесины, состоящей в основном из водопрово-
дящих клеток, которые, однако, не все время проводят
воду. По мере старения ствола проводящая функция
передается более молодым клеткам наружной зоны
древесины, тогда как внутренние клетки теряют эту
способность. Процесс старения сопровождается много-
численными изменениями; в частности, клетки паренхи-
мы, расположенные рядом с сосудами, образуют вырос-
ты, которые прорастают в полости этих древесинных
элементов и закупоривают их; такие выросты парен-
химных клеток называются тиллами (рис. 9). Иногда
46
Глава II
с возрастом древесина пропитывается различными орга-
ническими соединениями, как, например, камеди, смо-
лы, масла, таннины и пигменты, и приобретает более
темную окраску. В конце концов живые паренхимные
клетки древесины отмирают.
Эти изменения не влияют на прочность древесины,
но оказывают влияние на другие ее свойства. Старую
древесину трудно резать, и она более устойчива к гри-
бам и бактериям, которые вызывают гниение древесины.
Она также менее проницаема для различных жидкостей,
в том числе и для искусственных консервирующих жид-
Рис. 9. Продольный разрез части
членика сосуда, закупоренного тил-
лой — выростом соседнего сердце-
винного луча.
костей. Старую древесину принято называть ядровой,
противопоставляя ее древесине, которая еще функцио-
нирует и называется заболонью.
Но нередко у растущих деревьев ядровая древесина
оказывается менее прочной, чем заболонь. Например,
у дуплистых деревьев, встречающихся на лесных про-
галинах или на обочинах дорог, ядровая древесина сгни-
вает, а деревья продолжают жить благодаря тому, что
наружная заболонь остается еще живой и функциони-
рует. Это вполне возможно, потому что заболонь в жи-
вом дереве в отличие от мертвого заполнена водой
и поэтому вследствие плохой аэрации менее подверже-
на нападению грибов, вызывающих гниение древесины.
Степень развития ядровой древесины, а также со-
отношение между ней и заболонью очень изменчивы
у различных деревьев. Некоторые из них не имеют
четко дифференцированного ядра (тополь, ива, пихта,
ель). У других деревьев, например у тиса и шелковицы,
Растения-инженеры
47
заболонь образует тонкий слой, тогда как у бука, ясе-
ня и клена эта зона широкая. Время превращения за-
болони в ядро также варьирует; у одних видов’ это
происходит рано, у других же заболонь более долго-
вечна.
В промышленности лесоматериалов различают
твердую и мягкую древесину в зависимости от того, по-
лучена ли она из хвойных или лиственных деревьев.
Эти термины, впрочем, немного обманчивы, так как
«мягкая» древесина не обязательно мягкая, а «твердая»
Рис. 10. Продольный разрез части
трахеиды тиса.
Видны третичные утолщающие полосы,
придающие древесине тиса прочность и
эластичность,
не всегда бывает твердой. Технически «мягкая» дре-
весина корабельной сосны (Pinus palustris) иа самом
деле — одна из самых твердых древесин, которые из-
вестны, и в то же время каждый, кто имеет дело с из-
готовлением моделей самолетов, знает о мягкости дре-
весины бальзы (Ochroma lagopus), технически «твердо-
древесинного» дерева. Но в общем у хвойных древесина
действительно более мягкая, чем у лиственных. Кроме
того, древесина хвойных обычно имеет более простую
структуру и более гомогенна, чем у цветковых древес-
ных растений (разница между хвойными и цветковыми
растениями будет объяснена ниже, см. стр. 123—124).
Довольно интересна древесина тиса обыкновенного
(Taxus baccata); на стенках его трахеид образуются
утолщения в виде полос (так называемые «третичные
утолщающие полосы»), проходящие спирально по
внутренней поверхности вторичного утолщения кле-
точной стенки (рис. 10), что придает древесине большую
прочность и эластичность. Благодаря этим качествам
древесина тиса — очень подходящий материал для из-
готовления луков.
48
Глава If
Как растения решают проблемы,
связанные с преодолением силы тяжести
Живые растения вынуждены сопротивляться самым
разнообразным нагрузкам. Некоторыми из них растения
бывают обязаны человеку, например определенные сорта
пшеницы имеют слишком тяжелые колосья, а у сливы
сорта Виктория ветки часто ломаются под тяжестью
плодов. Но и помимо этих тягот цивилизации растения
испьиывают многочисленные внутренние трудности,
связанные с поддержанием определенного положения
вопреки собственному весу.
Почему плакучая ива «плачет» или, правильнее ска-
зать, почему не все деревья плачут? Превращение мо-
лодого деревца во взрослое, как правило, не сопро-
вождается существенными изменениями его формы.
Молодые веточки саженца располагаются под опреде-
ленными углами по отношению к стволу, и эти углы
остаются примерно такими же, когда саженец станет
большим взрослым деревом. Но по мере образования
новой древесины ветви, естественно, становятся все
тяжелее и тяжелее. Инженер сравнил бы ветвь дерева
с тяжелым кронштейном. Ведь по мере того, как такая
ветвь становится тяжелее, она должна сгибаться все
больше и больше, пока в конце концов, достигнув опре-
деленной длины, не согнется под собственной тяжестью
и не повиснет вертикально вниз. И все же, за некото-
рыми исключениями, этого не происходит.
Механические напряжения, возникающие в ветви
дерева, можно считать такими же, как в любом крон-
штейне. Ткани, расположенные ближе к концу ветви,
«работают» на растяжение, а расположенные у ее ос-
нования— на сжатие. Чтобы избежать изгибов, древе-
сина должна в равной степени выдерживать и те и дру-
гие напряжения. Каким образом сохраняется это явно
предопределенное положение, мы не знаем.
Если мы изучим древесину на нижней стороне
зетвей хвойных или на верхней стороне ветвей листвен-
ных деревьев, то обнаружим, что она заметно отличает-
ся от остальной древесины в структурном отношении,
^тд древесина называется реактивной и отличается <jt
Растения-инженеры
4$
обычной как анатомически, так и химически. У хвойных,
где ее называют древесиной сжатия, она имеет более
темную по сравнению с нормальной древесиной окраску
и ее клеточные оболочки одревесневают сильнее. Тра-
хеиды здесь короче, чем обычно, и древесина в этой
зоне более компактна. Одревесневшая вторичная обо-
лочка, которая откладывается на внутренней поверх-
ности первичной оболочки, у этих элементов в обычной
древесине состоит из трех слоев; у трахеид же древеси-
ны сжатия она имеет только два слоя, а внутренний
ее слой отсутствует.
У лиственных пород деревьев нет древесины сжа-
тия, но зато древесина, расположенная на верхней сто-
роне ветвей и называемая древесиной растяжения, пре-
терпевает некоторые структурные изменения. Сосуды
в древесине растяжения более узкие, а волокна имеют
толстый внутренний слой во вторичной оболочке — же-
латинозный слой, который очень хорошо отражает свет
и содержит вместо лигнина главным образом целлю-
лозу.
Недавние исследования показали, что развитие
реактивной ткани не является привилегией древесины,
поскольку вторичная флоэма, кора и пробковый кам-
бий тоже обнаруживают модифицированный рост.
Изучение реактивной древесины — сравнительно но-
вая область ботаники, и нам еще многое предстоит
выяснить, прежде чем мы сможем сказать, что вполне
представляем себе ее образование. Ее роль в жизни
растения пока не известна, ио ряд компетентных в дан-
ной области исследователей выражает мнение, что
реактивная древесина некоторым образом ответственна
за то, что ветви дерева сохраняют первоначальное
направление роста и не меняют его под действием
напряжений, обусловленных их собственным весом. Та-
кой взгляд на функцию реактивной древесины объяс-
няет ее положение в растении. Однако, если регуля-
ция роста и в самом деле является функцией реактив-
ной древесины, доказать это будет чрезвычайно трудно.
Стебель хвойного дерева, такого, например, как
сосна, пихта или лиственница, очень сходен со стеблем
лиственных деревьев пр всем основным свойствам,
50
Глава П
Вообще же древесина хвойных имеет более простое
строение, чем древесина лиственных деревьев; в ней
отсутствуют сосуды, а ее волокна весьма примитивного
типа. Вторичный рост происходит уже описанным для
лиственных пород способом, т. е. благодаря деятель-
ности камбиального слоя, в результате чего обхват
ствола увеличивается.
Кольца прироста
Хорошо известно, что возраст дерева можно хотя
бы приблизительно определить, сосчитав ясно различи-
мые на поверхности среза пня годичные кольца; в боль-
шинстве случаев действительно каждый год образуется
по кольцу, но все же это не всегда бывает так, поэтому
будет точнее называть их кольцами прироста (фото X).
В наших климатических условиях кольца прироста
различимы благодаря тому, что клетки весенней дре-
весины и дрёвесины, образованной предыдущим летом,
различаются по размерам. После зимнего периода
жизнь растений весной возрождается с новой силой.
Рост идет быстро, клетки образуются крупные. С приб-
лижением лета происходит общее замедление роста;
формирующиеся в этот период клетки меньше по разме-
ру. Затем наступает зима и начинается новый период
покоя, вновь сменяющийся следующей весной усилен-
ным ростом. Таким образом получается резкая граница
между весенней древесиной и слоем прироста преды-
дущего года, что и выражается в виде кольца, хоро-
шо заметного на пне, который остается после того, как
дерево спилено.
Разница между весенней и летней древесиной в зна-
чительной степени обусловлена различной потребностью
дерева в воде. В период активного весеннего роста
транспирация идет чрезвычайно интенсивно и поэтому
только крупные сосуды или трахеиды могут обеспечить
достаточное количество воды. Образовавшаяся позднее
древесина осуществляет в основном механическую
функцию, скорее способствуя укреплению стебля, чем
участвуя в проведении воды. Вот почему элементы лет-
ней древесины имеют меньший диаметр.
Растения-инженеры 51
Кольца прироста лучше заметны в быстрорастущей
древесине; у сосны, например, они видны очень хоро-
шо. С другой стороны, саговники — любопытные расте-
ния, родственные хвойным и часто называемые «живы-
ми ископаемыми» из-за их древнего происхождения
и примитивного строения, — растут столь медленно, что
слои прироста незаметны.
Таким образом, у растений северных широт кольца
прироста возникают благодаря разнице между весен-
ней и летней древесиной, однако не везде это происхо-
дит именно так. В муссонных лесах Индии не холод
зимой, а жара и засуха летом задерживают рост и дают
начало кольцам прироста. Во влажных тропических ле-
сах, например в Бразилии, Западной Африке и на Су-
матре, круглый год жарко и влажно. Там нет небла-
гоприятного для роста сезона и кольца прироста обычно
отсутствуют1.
На поперечном срезе древесины крупные сосуды
выглядят как отверстия, которые специалисты по дре-
весине называют порами. Различают два типа древе-
сины в зависимости от распределения этих пор по коль-
цу прироста. Если они имеют приблизительно одина-
ковый диаметр и равномерно распределены по всему
кольцу, древесину называют рассеяннососудистой. При-
мером может служить древесина клена, березы, бука,
граба, грецкого ореха и тополя. Напротив, если сосуды
гораздо крупнее в весенней древесине, чем в летней,
что видно очень отчетливо, то древесину называют коль-
цесосудистой. Такого типа древесина встречается у нас-
тоящего каштана, ясеня, дуба и ильма. В целом коль-
цесосудистый тип, по-видимому, является признаком
высокой специализации и встречается у древесин срав-
нительно немногих видов; большинство из них — обита-
тели северной умеренной зоны.
1 В тех случаях, когда у тропических деревьев кольца
прироста все же выражены, они обычно связаны не с временем
года, а с развертыванием новых почек. Периоды активной дея-
тельности камбия могут повторяться несколько раз в год (у
гевеи бразильской — до 7 раз) или, наоборот, длиться больше
года. — Прим. ред.
52
Глава it
Структура и функции корня
В анатомическом отношении корень поразительно
отличается от стебля. Ему приходится иметь дело
с совсем другими нагрузками, чем стеблю. Корень не
только не нуждается в твердости, но даже, наоборот,
она мешала бы ему, потому что на своем пути между
частицами почвы он обычно извивается, обходя такие
препятствия, как крупные камни. В то же время корень
часто вынужден противостоять резким рывкам, кото-
рые испытывает растение при сильном ветре; стебель
при этом может быть вырван из почвы, и корень при-
Рис. 11. Схематическое
изображение клетки эндо-
дермы с полосой Каспари.
зван обеспечить его прочное заякоривание. Верхушка
стебля кончается почкой, но на самом кончике корня
нежная структура невозможна, потому что на пути
сквозь твердую почву она неизбежно повреждалась бы.
Поэтому кончик корня нуждается в защите нежной
растущей части (апикальной меристемы) <ут трения
о почву.
Принимая все это во внимание, мы не удивимся,
обнаружив, что ткани древесины сосредоточены в цент-
ре корня, т. е. там, где они более всего нужны. Если
мы посмотрим на поперечный разрез молодого корня,
то увидим, что его центральная часть занята компакт-
ной массой древесины, которая имеет форму звезды
с лучами, расходящимися к периферии (фото XI). В не-
которых корнях сердцевина отсутствует, тогда как
в центре других она имеется. Флоэма представлена в ви-
де тяжей, чередующихся с лучами древесины. Как
и в стебле, между флоэмой и ксилемой располагается
камбий. Однако в корне мы можем обнаружить и неч-
то такое, чего мы не видели в стебле. Кнаружи от флоэ-
_________________Растения-инженеры S3
мы расположен слой клеток, полностью отграничиваю-
щих центральную массу древесины; эти клетки назы-
ваются эндодермой, и их легко узнать по характерным
полосам на стенках. Эти полосы содержат суберин
и охватывают клетку как поясок, надетый на ко-
робочку (рис. 11). По имени открывшего их ученого они
называются полосами Каспари или пятнами Каспари.
Функция их не совсем ясна, но предположительно они
имеют отношение к передвижению воды в корне, как
мы узнаем из следующей главы.
Кнаружи от эндодермы лежит кора, состоящая обыч-
но, как и в стебле, из тонкостенных клеток. В молодом
корне самый наружный слой клеток образует эпидермис,
но по мере старения корня он исчезает, и тогда наруж-
ным оказывается слой, называемый экзодермой. Оболоч-
ки клеток экзодермы содержат суберин н в достаточной
мере, хотя и не вполне, непроницаемы для воды.
Корень замечательно приспособлен для выполнения
одной из главных своих функций — сопротивления <тя-
нушей силе». Древесина, сконцентрированная в центре,
придает ему максимальную прочность в сочетании с пре-
дельно возможной гибкостью. Структура корня напоми-
нает структуру каната или троса, который не только
удерживает прикрепленный к его концу груз, но и спо-
собен изгибаться. Жесткая структура, наблюдаемая
в стебле, никак не подошла бы для корня. Таким обра-
зом, мы вновь убеждаемся, что растения (с инженерной
точки зрения) сконструированы наиболее целесообразно
для выполнения функций, присущих их органам.
Выше мы уже говорили, что нежная точка роста
корня подвергается опасности и может быть стерта в ре-
зультате трения, возникающего между прокладывающим
себе путь кончиком корня и частицами почвы. Поэтому
корневая меристема защищена специальным образо-
ванием, называемым корневым чехликом (фото XII).
Если мы посмотрим на продольный разрез корня, то
увидим, что корневой чехлик состоит из довольно рых-
ло расположенных тонкостенных клеток, полностью по-
крывающих точку роста и защищающих ее от повреж-
дений. Когда эти клетки стираются при прохождении
корня через почву, они заменяются новыми, возникаю-
64
Глава tl
щими под ними, так что кончик корня всегда остается
защищенным корневым чехликом.
Хотя у корня нет эпидермиса, он может образовы-
вать перидерму точно таким же образом, как и стебель,
особенно если оказывается на открытом воздухе. Это
можно видеть там, где почва со временем вымывается
из-под крупных корней дерева. Такие корни могут быть
покрыты мощно разросшейся корой.
Для корня может быть характерен вторичный рост,
происходящий тем же путем, что и в стебле, хотя не-
которые корни всю свою жизнь сохраняют первичное
строение. Процесс вторичного роста начинается с того,
что по окружности корня, кнаружи от древесины,
появляется кольцо камбия. Сначала камбий имеет из-
вилистые очертания, так как он окружает вытянутые
лучи древесины, но в ходе вторичного роста в вогну-
тых участках камбий работает быстрее и в конце
концов приобретает очертания кольца. Кнаружи камбий
откладывает рторичную флоэму, а внутрь — вторичную
ксилему, подобно тому как это происходит в стебле.
Помимо того что корень твердо закрепляет расте-
ние в почве, он должен всасывать из нее воду и мине-
ральные вещества. Корень хорошо приспособлен для
этой деятельности. Выкапывая молодой проросток,
можно заметить, что к его корню всегда остается при-
соединенным комок почвы, который часто невозможно
отделить. Это происходит потому, что вблизи от кончи-
ка корня имеются крошечные образования, называемые
корневыми волосками. Они играют огромную роль в
присущей этому органу функции всасывания.
Под микроскопом видно, что корневой волосок
представляет собой вырост поверхностной клетки корня,
причем его полость является продолжением полости
клетки. Ядро клетки, образующей корневой волосок,
переходит в него и располагается примерно в середине
корневого волоска или вблизи его верхушки (рис. 12).
Корневые волоски чрезвычайно увеличивают общую
площадь поверхности корня и повышают таким обра-
зом его эффективность как органа поглощения. На
1 мм2 поверхности корня может располагаться от 200
до 400 волосков, благодаря чему, как было подсчитано,
Растения-инженеры______________________55
его всасывающая поверхность увеличивается примерно
в 18 раз.
Корневые волоски появляются только вблизи кончи-
ка корня сразу же за зоной роста и занимают участок
длиной в 2... 3 см. Обычно по прошествии некоторого
времени более старые волоски перестают функциониро-
вать и засыхают, заменяясь другими, возникающими
ближе к растущему кончику корня. После того как эти
Рис. 12. Схема строения корневого волоска.
более старые корневые волоски сморщатся и отпадут,
клетки, образовавшие их, как правило, пропитываются
суберином (опробковевают) и формируют часть экзо-
дермы корня; при этом они утрачивают, но, по-видимо-
му, не полностью способность поглощать воду. Иногда,
однако, корневые волоски могут быть и более долго-
вечными. Так, корни многих травянистых растений из
семейства сложноцветных (Compositae) развивают почти
по всей своей длине корневые волоски, которые могут
функционировать в течение трех лет1.
Когда пересаживают молодой проросток, многие
корневые волоски обламываются и засыхают, а остав-
шиеся неповрежденными могут засохнуть на открытом
воздухе. Вот почему так важно при пересадке оставлять
вокруг корней ком земли; это в какой-то степени за-
щитит корневые волоски и некоторые из них могут вы-
жить. Пересаженный проросток развивает новую сеть
корневых волосков; этот период можно назвать кри-
тическим, потому что поглощение корнями воды в это
1 Так же долго могут сохраняться корневые волоски у инте-
ресного злака белоуса (Nardus stricta), растущего по краям болот
и на пустошах. — Прим, ред.
Б6
Глава И
время очень затруднено и проросток легко может по-
гибнуть, если его как следует не поливать.
Развитие корневых волосков — нечто гораздо боль-
шее, чем способ увеличения водопоглощающей поверх-
ности корня. Корневой волосок имеет очень нежную
оболочку и растет в тесном контакте с частицами почвы,
обвивая их как можно плотнее. Наружные слои его
оболочки слизистые и сливаются с почвенными кол-
лоидами, так что корневой волосок и частица почвы
образуют единую структуру. Это помогает корневому
волоску в его всасывающей деятельности.
Помимо главных функций — закрепления растения
в почве и поглощения воды и растворенных в ней солей,
корни имеют и много других, что можно наблюдать
у некоторых растений. Часто они являются как бы
<складами», где накапливаются питательные вещества,
например у моркови и пастернака. Они могут также
служить и для запасания воды. Особенно хорошо это
видно на примере воздушных корней некоторых тро-
пических орхидей, которые растут высоко на стволах
деревьев (иногда 30 м или более от земли) и их корни
не имеют никакого контакта с почвой. У некоторых
орхидей эти воздушные корни, свисающие вниз как
длинные узкие ленты, имеют специализированную
область клеток в наружной части коры — так назы-
ваемый веламен (фото ХШ). Клетки веламена крупные
и пустые и могут впитывать воду подобно губке. Во
время дождя клетки веламена наполняются водой
и таким образом эти эпифитные (т. е. растущие на дру-
гих растениях) орхидеи могут хранить воду, пока в .ней
не возникнет нужда.
Некоторые лазящие растения, такие, например, как
плющ, взбираются вверх при помощи придаточных
корней на стеблях (придаточным называется любой
корень, возникающий не на своем обычном месте — на
конце стебля). У плюща придаточные корни образуются
на стороне стебля, обращенной к стене или дереву,
служащим опорой, и проникают в любые щели или тре-
щины, цепко удерживаясь там и выдерживая вес всего
растения (рис. 13). Эти лазящие корни, оказывается,
имеют естественную тенденцию избегать срета и растут
Растёния-инжбмры
&
по направлению к любому темному месту, которое они.
могут найти, врастая, например, в трещину кирпичной
стены. Такое поведение необычно для корней, которые
в норме безразличны к свету и темноте.
Придаточные корни могут служить и для размно-
жения растения, например когда они появляются на
срезанном конце черенка. Еще более поразительное
явление можно наблюдать, если надрезать лист бего-
нии и поместить его во влажный песок, прикрыв чем-ли-
бо. Через некоторое время вокруг надреза возникнут
придаточные корни и из листа вырастет новое растение.
Рис. 13. Лазящие корни плюща.
Иногда растения имеют специальные корни, втяги-
вающие их в землю. Крокус вырастает из клубнелуко-
вицы, причем каждый год на верхушке старой клубне-
луковицы образуется новая. Постепенно приближаясь
таким образом к поверхности земли, клубнелуковица
в конце концов вышла бы наружу, если бы не корни,
возвращающие ее в прежнее положение. Дело в том,
что, кроме обычных питающих корней, крокус имеет
специальные, более длинные корни, которые могут
сокращаться, становясь при этом морщинистыми
(рис. 14). При сокращении этих корней клубнелукови-
ца погружается глубже в землю, так что она все время
остается на одной глубине.
Чтобы увидеть корни, достигшие необычайного раз-
вития, мы должны обратиться к тропикам, потому что
тепло и влажность тропического дождевого леса или
мангрового болота как нигде создают условия для
исключительного роста. Во влажном тропическом лесу
очень обычны ходульные корни, большей частью у не-
больших деревьев. Они представляют собой развившие-
Ьй
Рлава И
ся на главном стебле придаточные корни, которые, за-
гибаясь вниз, достигают почвы и внедряются в нее.
Такие корни особенно типичны для мангров (рис. 15),
но их можно видеть и у многих других деревьев влаж-
ного тропического леса. Они могут быть круглыми
в поперечном сечении или сплюснутыми и встречаются
у деревьев многих, совсем не родственных друг другу
семейств.
крокуса с втягивающими
корнями.
Рис. 15. Мангровое расте-
ние (Rhizophora) с ходуль-
ными корнями.
Чаще всего ходульные корни имеются у деревьев,
растущих на неустойчивой болотистой почве, поэтому
очень соблазнительно предположить, что они служат
растениям дополнительной опорой. Однако в биологии
всегда опасно принимать кажущееся очевидным объяс-
нение без дальнейших исследований. Если мы вникнем
в дело поглубже, то обнаружим, что ходульные корни
обычно встречаются лишь у небольших деревьев,
тогда как имеются и более высокие деревья, которые
действительно подвергаются опасности быть согнутыми
ветром. Легко представить себе, что если бы деревья
развивали ходульные корни для опоры, то это были
бы прежде всего именно высокие деревья.
Более вероятная причина возникновения ходульных
корней—теплая, влажная атмосфера нижних ярусов
влажного тропического леса: такие условия способст-
вуют развитию придаточных корней. Это не означает, что
Растения-инженеры
59
ходульные корни бесполезны для растения, потому что
растение, естественно, извлекает пользу из любой опо-
ры, которая оказывается в его распоряжении. Растения
очень хорошо умеют обращать случайные особенности
своей структуры в преимущество; история эволюции
растений изобилует примерами такого рода.
Еще большее впечатление, чем ходульные, произво-
дят досковидные опорные корни, которые также можно
встретить во влажном тропическом лесу. Они более
распространены, чем ходульные, и список семейств
растений, в которых их можно обнаружить,
очень длинен. Эти корни напоминают доски,
прислоненные верхними концами к дереву. £
В то время как ходульные корни являются л
придаточными корнями, которые возникают £
на стволе и растут вниз, образование доско- £
видных корней связано с боковыми корнями, £
растущими непосредственно под поверхностью £
почвы. Боковые корни развивают вертикаль- £
Рис. 16. Схема поперечного разреза досковидпого
корпя, на котором видны кольца прироста.
ные выросты, треугольные в поперечном сечении, кото-
рые и прилегают к стволу дерева.
Древесина досковидных корней часто тверже, чем
остальная древесина того же дерева, и нередко они
бывают покрыты тонким слоем коры. Судя по кольцам
прироста, видным на поперечном разрезе этих корней,
вначале это нормальные, округлые в сечении корни, но
затем они развивают очень сильное утолщение на
верхней стороне (рис. 16). Число досковидных корней
У дерева варьирует от одного до десяти, но в большин-
стве случаев их три или более.
Досковидные корни особенно характерны для влаж-
ного тропического леса, хотя их распространение этим
не ограничивается. Вообще подобного типа корни,
по-видимому, развиваются там, где климат влажный
60
Глава ll
и теплый. Иногда они встречаются н в лесах умеренной
зоны, в частности у ломбардского тополя (Populus ita-
lica) и вяза.
В тропиках образование досковидных корней особен-
но заметно у деревьев, растущих в болотистых лесах
и на плохо дренированных почвах. В равнинных лесах
тропиков почвы обычно недостаточно аэрированы,
главным образом из-за температуры и влажности, и кор-
ни растений конкурируют друг с другом за кислород,
а также за воду и минеральные соли. Вполне возможно,
что образование досковидных корней как-то связано
с этой конкуренцией.
Причина возникновения досковидных корней ясна не
более, чем причина появления ходульных корней1. Рань-
ше их считали приспособлением, помогающим дереву
противостоять ветру, что особенно важно для растений,
растущих на зыбких болотистых почвах. Это мнение
подтверждается тем фактом, что дерево редко имеет
один или два таких корня; большинство, как мы уже
знаем, развивает по крайней мере три — минимальное
число корней, способное создать достаточную опору.
Досковидные корни, бесспорно, дают растению преиму-
щество в отношении устойчивости, но это не значит, что
они развились именно поэтому. В равной мере это мо-
жет быть просто еще одним случаем использования
растением особенностей своего строения, возникших
совсем по другой причине.
Среди различных теорий, объясняющих образование
досковидных корней, нет ни одной вполне удовлетвори-
тельной. Но все же наибольшего доверия заслуживает
теория «натяжения», предполагающая, что развитие
досковидпых корней представляет собой непосредствен-
ную реакцию дерева на натяжение, создаваемое ветром.
В пользу этой теории говорят результаты исследований
образования таких корней у ломбардского тополя
в Швейцарии, стремящегося развить досковидные корни
на стороне, обращенной к ветру, т. е. той стороне, ко-
1 Подробное описание досковидных корней и разные гипотезы,
объясняющие их возникновение, приводятся в превосходной книге
П. Ричардса «Тропический дождевой лес», ИЛ, М., 1961.— Прим. ред.
Растения-инженеры
61
торая должна противостоять тянущей силе, когда дует
ветер.
Какова бы ни была причина образования доско-
видных корней, их наличие — характерная особенность
деревьев тропического дождевого леса. Нередко их ис-
пользуют для промышленных целей; так, досковидные
корни произрастающего в Индонезии дерева Koompassia
excelsa идут на изготовление обеденных столов.
Не менее интересный тип корней представляют со-
бой пневматофоры, или дыхательные корни, многих
мангров. Мангры — это не название растения, а скорее
тип растительности, связанной с такими болотистыми
районами, которые встречаются, например, в южной
Флориде1. Под общим названием мангров объединены
разные роды, из которых наиболее обычен род Rhizopho-
ra. Кроме пневматофоров, мангры имеют еще некото-
рые замечательные особенности; в частности, они жи-
вородящие, т. е. у них зародыш без периода покоя про-
растает в плоде, находящемся еще на дереве (см. гл. XI
и стр. 126, 197, 198).
Обитая в тропических болотах, мангры страдают от
недостатка кислорода, необходимого для дыхания кор-
ней; отсюда и необходимость в специальных дыхательных
корнях, восполняющих этот недостаток. Дыхательные
корни бывают различного рода. У одних видов из ила
выступают коленчатые изгибы корней, тогда как у дру-
гих растут вверх и выходят на поверхность просто концы
ответвлений корней, растущие против силы земного при-
тяжения, а не наоборот, что свойственно большинству
этих органов.
Выходящие из почвы дыхательные корни имеют хо-
рошо развитую систему чечевичек, а их внутренние
ткани изобилуют межклетными полостями, по которым
воздух может проникать к подземной корневой си-
стеме. Это придает вероятность точке зрения, согласно
которой они в самом деле являются дыхательными
корнями, но, как оказалось, эти корни несут еще одну
х Мангровые заросли приурочены в тропиках к побережью
океана и связаны с полосой прилива — отлива. Таким образом,
Речь идет о «болотах> особого рода, ничем не напоминающих болсь
Та» находящиеся в глубине материков. — Прим, ред.
62
Глава If
функцию. Уровень почвы в мангровом болоте постоян-
но повышается за счет растительного отпада и различных
наносов, которые задерживаются корнями растений
при приливах. Главную всасывающую часть корневой
системы мангров составляют тонкие корешки, сосредо-
точенные близ поверхности почвы, и эти корешки обра-
зуются на дыхательных корнях, которые формируют но-
вые их группы в своих более верхних частях при каждом
последовательном повышении уровня почвы. При неко-
торых обстоятельствах эта функция дыхательных корней
имеет даже более важное значение, чем аэрация. Обсуж-
дение особенностей мангров мы продолжим в гл. XI
(стр. 222).
Впрочем, не только у мангров можно наблюдать
дыхательные корни. Так, болотный кипарис (Taxodium
distichum), хвойное дерево, обитающее в болотах суб-
тропической и умеренной зон, имеет хорошо развитые
коленчатые корни.
Поистине фантастического развития достигают при-
даточные корни баньяна (Ficus benghalensis), принадле-
жащего к семейству тутовых (Могасеае). Его мясистые
плоды, похожие на инжир, поедают летучие мыши, ко-
торые и распространяют семена. Вырастающее из се-
мени дерево образует на своих ветвях множество при-
даточных корней, которые растут вниз, к земле, укоре-
няются и развивают собственную корневую систему. Со
временем одно дерево баньяна образует маленькую ро-
щу со многими «стволами», в действительности пред-
ставляющими собой придаточные корни. В тени одного
такого дерева может разместиться небольшая деревня.
Экземпляр, растущий в Ботаническом саду в Калькутте,
кроме основного ствола диаметром более 3,5 м имеет
и более двухсот корневых «стволов», а диаметр всей
его кроны —около 100 м. Э. Дж. X. Корнер в книге
«Живое растение» описывает баньян в долине Андхра,
имеющий 610 м в окружности, с 320 «стволами», под-
держивающими его тяжелую крону.
В Индии баньян считается священным. В его тени
принято проводить собрания жителей деревни, а почву
под ним специально обрабатывают для молодых корней,
которые защищают трубками из бамбука.
ГЛАВА Ш
ГИДРАВЛИКА РАСТЕНИЙ
Листья растений постоянно испаряют воду, особенно
в дневные часы. Поверхность листьев усеяна многочис-
ленными микроскопическими отверстиями, называемыми
устьицами, причем у большинства растений их значи-
тельно больше на нижней поверхности листа, чем на
верхней. Международное название устьиц — stomata —
происходит от греческого слова «стома», что означает
«рот»1. Пока устьица остаются открытыми, водяной пар
диффундирует из листа; ночью же, когда темно, боль-
шинство устьиц закрыто и испарение воды через них
прекращается. Однако даже ночью лист продолжает те-
рять воду, хотя и гораздо медленнее, чем днем, так как
некоторое количество воды может проходить через эпи-
дермис, минуя устьица.
Испарение воды листьями называется транспира-
цией, и просто удивительно, насколько много растения
теряют ее таким путем. Например, одно растение под-
солнечника в течение всего вегетационного периода
испаряет до 50 л воды и более, а средней величины бе-
реза за один жаркий день теряет 400 л воды.
Вся эта потерянная вода должна компенсироваться
водой, поглощаемой корнями из почвы, так как, если
этого не произойдет, растение завянет, а при продолжи-
тельном завядании гибель его неминуема. Ни одно
растение не может долго жить без воды, хотя некото-
рые, как мы увидим, могут обходиться очень малым
ее количеством. То, что теряется, должно быть возме-
щено, и поэтому адекватное поступление воды —одно
из первых условий, которое должно обеспечить себе
растение, чтобы выжить.
1 Русское слово «устьице», вероятно, также связано с древне-
славянским «уста». Интересно, что в толковом словаре русского
языка В. Даля слово «устье», производным от которого является
«устьице», отнесено к смысловой группе слов, имеющих общий
корень со словом «уста» — рот. — Прим. ред.
64
Глава lit
Осмос
Прежде чем приступить к обсуждению вопроса
о поглощении воды корнями, полезно вспомнить очень
простой опыт, который вы наверняка проводили или
наблюдали в школе. Возьмем стеклянную воронку, за-
тянутую на широком конце мембраной, например, из
целлофана, пергамента или коллодия, и, перевернув,
наполним ее сильно концентрированным рас-
> твором сахара. Затем, закрепив воронку зажи-
мом, поместим ее на 24 ч в сосуд с водой.
На следующий день мы обнаружим, что вода
прошла через мембрану в раствор сахара, за-
ставив жидкость подняться по отводной труб-
ке воронки (рис. 17). Это явление называется
осмосом.
Рис. 17. Опыт, демонстрирующий явление осмоса
(см. стр. 64— 66).
Прохождение воды через нашу мембрану
объясняется одним ее необычным свойством:
свободно пропуская сквозь себя молекулы воды, она не
пропускает молекул сахара. Из-за различной проницае-
мости по отношению к сахару и воде такого типа мем-
браны получили название полупроницаемых. Итак, с од-
ной стороны мембраны — чистая вода, молекулы кото-
рой легко проникают через нее, с другой —молекулы
сахара и воды, из которых только молекулы воды могут
проходить сквозь мембрану.
Молекулы жидкости и вещества, растворенного в ней,
находятся в непрерывном движении. Двигаясь, молеку-
лы воды постоянно ударяются о мембрану с обеих сто-
рон и некоторые из них проходят через нее (с одной сто-
роны о мембрану ударяются и молекулы сахара, одна-
ко для них она непроницаема). За секунду большее число
молекул воды будет ударяться о мембрану с наружной
Гидравлика растений
6б
стороны, где находится чистая -вода, чем со стороны
раствора сахара, где концентрация воды ниже. Следо-
вательно, за каждый данный промежуток времени боль-
ше воды будет проникать в раствор сахара, чем выхо-
дить из него. Постепенно в растворе сахара будет на-
капливаться все больше и больше молекул воды, раствор
будет все более разбавляться. Такова сущность явле-
ния осмоса. Надо сказать, что приведенное здесь объяс-
нение крайне упрощено, на самом же деле различные
термодинамические явления усложняют картину; к
счастью, мы можем не принимать их во внимание, хо-
тя они и представляют интерес для теоретиков, стре-
мящихся разобраться в вопросе о поступлении воды в
растение.
Я не попытался объяснить, почему вода проходит
через полупроницаемую мембрану, а сахар нет, и не имею
намерения это делать. Предполагается, что мембрана
действует подобно молекулярному ситу, сквозь которое
проходят сравнительно небольшие молекулы воды, но
не проходят большие молекулы сахара. Это, по-видимо-
му, верно, по крайней мере до некоторой степени, но
почти наверное к этому объяснению можно что-то до-
бавить. Полное же объяснение не входит в наши задачи,
и мы с благодарностью уступаем эту честь специалистам
по физической химии, которые любят такого рода
проблемы.
Предположим, что описанный выше опыт продолжа-
ется несколько дней; что произойдет в этом случае?
Мы увидим, что сначала вода довольно быстро входит
в воронку, заставляя раствор подниматься по отводной
трубке. Но постепенно этот процесс замедляется, и на-
конец жидкость совсем перестанет подниматься. К это-
му времени мы будем вынуждены прикрепить к концу
отводной трубки стеклянную трубку, чтобы поместилась
вся проникшая в воронку вода; желательно даже поста-
вить нашу установку у высокой стены, чтобы удобнее
было наращивать трубку.
Высота, на которую благодаря осмосу может под-
няться вода в трубке, зависит от концентрации раство-
ра и порой бывает поистине фантастической. Если наш
раствор содержит 342 г сахара на 1 л воды (а такую
66 Глава 1И
концентрацию можно легко получить), теоретическая
высота столба превысила бы 230 м. Я говорю теорети-
ческая, поскольку различные факторы будут препятст-
вовать поднятию воды на полную расчетную высоту,
в особенности разрыв мембраны, вследствие громадного
и все возрастающего давления столба воды.
Этот вопрос о давлении исключительно важен. Ведь
как только жидкость начнет подниматься по трубке, она
станет давить на мембрану, и это давление измеряется
разницей в высоте уровня воды внутри и снаружи во-
ронки. Разница в 10 м эквивалентна одной атмосфере,
или примерно 1 кг/см2. Теперь понятно, почему при
большей продолжительности опыта мембрана разры-
вается.
Давление столба воды в трубке служит мерой осмо-
тического давления раствора. Всякий раствор имеет
определенное осмотическое давление, пропорциональное
его молекулярной концентрации, т. е. количеству моле-
кул растворенного вещества в данном объеме раствора.
Одна грамм-молекула вещества (его молекулярная
масса в граммах), растворенная в литре воды, имеет
осмотическое давление, равное 22,4 атм, или примерно
22,4 кг/см2.
Еще одно замечание: я сказал вначале, что воронка
с раствором сахара погружается в чистую воду. Это не
обязательно, и не будет принципиальной разницы в ре-
зультате, если воронка будет погружена в раствор, лишь
бы он имел более низкую молекулярную концентрацию,
чем в воронке. Вода все равно будет проникать в во-
ронку, хотя не так быстро и поднимаясь при этом на
меньшую высоту. С другой стороны, если раствор в со-
суде будет иметь более высокую молекулярную концент-
рацию, чем в воронке, вода будет выходить из воронки,
а не входить в нее, и уровень жидкости в трубке начнет
падать, а не подниматься.
А можно ли все это применить к корням растения?
Корень состоит преимущественно из живых тонкостен-
ных клеток, пористые целлюлозные оболочки которых
свободно пропускают жидкость. Изнутри к клеточной
стенке прилегает живая протоплазма, а внутри нее,
занимая большую часть внутриклеточного пространства,
Гидравлика растений 67
находится так называемая вакуоль, заполненная кле-
точным соком — сложным раствором сахаров, органи-
ческих кислот, минеральных солей и других веществ.
Каждая клетка содержит также и ядро. Оно может
рассматриваться как специализированная часть прото-
пласта, контролирующая по существу все функции
клетки.
Протоплазматические мембраны
и работа клетки
Для проникновения воды в клетку особенно важен
слой протоплазмы, лежащий непосредственно под кле-
точной оболочкой. Именно здесь осуществляются все
реакции, придающие клетке свойства живого, т. с. по
существу протоплазма — важнейшая живая часть клет-
ки. Многие компоненты входят в состав протоплазмы;
главные среди них — сложнейшие органические вещест-
ва, называемые белками. Протоплазма не гомогенна;
в частности, при исследовании в электронном микроско-
пе в пей обнаруживаются мембранные структуры, играю-
щие важную роль в жизнедеятельности клетки.
Мембраны состоят из белков и фосфолипидов — ве-
ществ жировой природы. Две мембраны имеют особое
значение в поглощении воды клеткой. Одна из них —
плазмалемма — целиком окружает содержимое клетки
непосредственно под оболочкой; другая же — тоно-
пласт— находится с внутренней стороны протоплазмы,
отграничивая ее от вакуоли.
Наличие этих мембран, в особенности тонопласта,
придает протоплазме свойство полупроницаемости, как
и у мембран в нашем опыте с осмосом. Таким образом,
живая клетка вполне способна к осмосу и всем сопро-
вождающим его явлениям.
Я спешу добавить, что протоплазму нельзя назвать
полупроницаемой в строгом смысле этого слова, пото-
му что в таком случае ipso facto1 она не могла бы про-
1 В силу самого факта (лат.). — Прим. ред.
68
Глава III
пускать в клетку ничего, кроме воды, а мы знаем, что
это не так. Однако вода проникает в клетку легче, чем
большинство растворенных веществ, и в этом смысле
протоплазма полупроницаема. Некоторые из таких ве-
ществ попадают в клетку гораздо легче, чем другие,
но этот вопрос мы сейчас рассматривать не будем.
Важно, что при осмосе протоплазма может играть роль
полупроницаемой мембраны.
Клеточный сок представляет собой раствор умерен-
ной концентрации, обычно с осмотическим давлением
от 5 до 30 атм, окруженный полупроницаемой про-
топлазмой. Если клетка находится в воде или в раство-
ре с более низким осмотическим давлением, чем в кле-
точном соке, вода будет входить в клетку в соответствии
с законами осмоса. Вот каким образом в общем вода
поступает в корпи растения.
Почвенная вода представляет собой сильно разбав-
ленный раствор разных солей — карбопатов, хлоридов,
сульфатов, нитратов и т. д. различных металлов, напри-
мер кальция, магния, натрия, калия и многих других.
Этот раствор, конечно, обладает осмотическим давле-
нием, но оно незначительно, так как общая концентра-
ция солей мала. Осмотическое давление в корневых
волосках, как правило, гораздо больше, чем в почвен-
ном растворе, поэтому вода входит в эти клетки. Из
корневых волосков вода проходит через кору корня
в трахеиды или сосуды древесины. Эти элементы дре-
весины содержат раствор минеральных солей и часто
органических веществ, который имеет более высокую
концентрацию, чем почвенный раствор, поэтому вода
из почвы может поступить в трахеиды и сосуды.
Однако, прежде чем признать эту простую теорию
поступления воды в растение, необходимо выяснить
множество деталей и значительно ее расширить. Начнем
с того, что для успешного прохождения воды по коре
осмотическое давление должно возрастать от клетки к
клетке, т. е. по пути следования воды должен существо-
вать градиент повышающегося осмотического давления.
Но у нас нет оснований полагать, что так оно и есть.
Чем же в таком случае объясняется прохождение воды
через кору?
Гидравлика растений
69
Осмотическое и тургорное давления
Рассматривая поступление воды в клетку путем осмо-
са, я до сих пор ничего не сказал о клеточной оболочке,
так как она состоит из целлюлозы, свободно пропуска-
ющей воду. Тем не менее оболочка играет важную роль
в данном процессе. Когда вода входит в клетку, пос-
ледняя, естественно, растягивается. Целлюлозная стенка,
хотя она и кажется жесткой, обладает, однако, некото-
рой эластичностью и немного растягивается вместе
с клеткой; но ее способность к растяжению ограничена,
и когда растяжение усиливается, оболочка начинает
оказывать давление на содержимое клетки. Чем боль-
ше растянута клетка, тем такое давление больше. Это
направленное внутрь давление клеточной стенки назы-
вается тургорным давлением.
Следовательно, на проникновение воды в клетку
влияют две противодействующие друг другу силы —
осмотическое давление, заставляющее воду войти в клет-
ку, и тургорное, выталкивающее ее из клетки. Соотно-
шение между осмотическим давлением, тургорным
давлением и поступлением воды можно выразить уравне-
нием
ОД—ТД=ДДД,
где ОД — осмотическое давление клеточного сока,
ТД — тургорное давление, а ДДД — величина, назы-
ваемая дефицитом давления диффузии, которая опре-
деляет тенденцию воды к поступлению в клетку.
Термин «дефицит давления диффузии» нуждается
в некотором разъяснении*. До недавнего времени счи-
талось, что раствор притягивает воду через полупрони-
цаемую мембрану благодаря своему осмотическому
давлению, но это все равно, что утверждать, будто по-
возка тянет лошадь. Раствор не втягивает в себя воду.
Правильнее сказать, что давление диффузии воды
в растворе меньше, чем чистой воды, или что давление
1 В русской ботанической литературе в этом же смысле упо-
требляется термин «сосущая сила», который, как видно из дальней-
шего разъяснения автора, неудачен. — Прим. ред.
70
Глава III
диффузии воды в концентрированном растворе меньше,
чем в более разбавленном. Более концентрированный
раствор обладает дефицитом давления диффузии по
отношению к менее концентрированному. Следовательно,
если эти два раствора разделить полупроницаемой
мембраной, вода будет диффундировать из разбавлен-
ного в концентрированный раствор до тех пор, пока
давление диффузии воды не станет одинаковым по обе
стороны мембраны.
Способность клетки поглощать воду извне опреде-
ляется не абсолютным осмотическим давлением кле-
точного сока, а его ДДД, хотя в определенных случаях
он бывает равен осмотическому давлению. В клетке,
которая из-за потери воды близка к «завядаиию», тур-
горное давление будет, очевидно, равно нулю, поскольку
оболочка вялой клетки спадается до минимума и не
оказывает никакого давления на содержимое клетки.
Из уравнения
ОД-ТД = ДДД
ясно, что, если ТД=0, ДДД=ОД.
С другой стороны, ДДД тоже может быть равен ну-
лю, даже при довольно высоком осмотическом давлении.
По мере насасывания воды клеткой тургорное давление
возрастает до тех пор, пока не станет равным осмоти-
ческому давлению клеточного сока. С этого момента
клетка находится в состоянии полного насыщения н вса-
сывание воды прекращается. ДДД становится равным
нулю. И вновь из уравнения
ОД-ТД = ДДД
очевидно, что, если ТД=ОД, ДДД=0.
Ради полноты изложения я упомяну еще один слу-
чай, который иногда наблюдается, особенно в экспери-
ментальной работе. Предположим, что концентрация на-
ружного раствора выше, чем концентрация клеточного
сока. Тогда вода будет выходить из клетки, а не вхо-
дить в нее. Клеточная оболочка при этом сожмется до
минимума и потом сжатие ее прекратится. Протоплазма,
однако, не обладает жесткостью, и она продолжает
Гидравлика растений
?1
сжиматься и после этого, отставая от оболочки и при-
нимая округлую или овальную форму. Такое состояние
клетки называется плазмолизом.
Из всего сказанного выше ясно, что не абсолютное
осмотическое давление клетки определяет возможность
насасывания ею воды, а ДДД — разность между осмо-
тическим давлением и давлением оболочки. Клетка мо-
жет иметь достаточно высокое осмотическое давление,
но если давление оболочки тоже велико, то ДДД клетки
незначителен. С другой стороны, клетка с более низким
осмотическим давлением может обладать большим ДДД,
что наблюдается в том случае, если давление ее обо-
лочки мало.
Таким образом, градиент повышающегося от клетки
к клетке осмотического давления совсем необязателен
для передвижения воды по коре; решающим фактором
здесь является градиент увеличивающегося ДДД. Если
он существует, вода может проходить сквозь кору.
К сожалению, попытки продемонстрировать этот
градиент ДДД оканчивались неудачей, но это не значит,
что его вовсе нет. К тому же соседние клетки, как,
например, в корне, взаимодействуют друг с другом. Если
клетка в ткани начнет набухать, она будет оказывать
давление на окружающие ее клетки, которые в свою
очередь будут передавать это давление на соседние
клетки. Таким путем в ткани развивается напряжение,
усиливающее действие простого тургориого давления.
В том, что ткани могут находиться в состоянии
напряжения, легко убедиться, разрезав продольно цве-
тонос одуванчика. Обе его половинки тотчас же закру-
тятся наружу. Дело в том, что наружные ткани цве-
тоноса относительно нерастяжимы и поэтому централь-
ные клетки находятся под давлением. При разрезании
же цветоноса это давление снимается и внутренние
клетки расширяются, вызывая закручивание обеих по-
ловинок.
Точные детали передвижения воды по коре пока
неясны, хотя можно без особого риска говорить об
огромной роли осмотического давления в этом процес-
се. Однако есть основания предполагать, что осмоти-
ческое давление не единственный фактор и что есть еще
какая-то сила, хотя, к сожалению, точно не известно, ка-
72
Рлава til
кая именно. Это в какой-то мере связано с тем, что
клетка живая, и поэтому эту силу принято называть
«активным переносом воды», чтобы показать ее связь
с жизненной активностью клетки. Возможно, она зави-
сит от изменений электрических ’ зарядов в клетке по
обеим сторонам клеточной мембраны.
Если этот активный перенос воды в самом деле имеет
место, то он нуждается в дополнительной энергии извне
(в противоположность простому осмосу, который проис-
ходит исключительно за счет кинетической энергии
самих молекул и не требует дополнительных затрат).
Эта энергия, по-видимому, доставляется дыханием,
и интересно, что в некоторых случаях, как это было за-
мечено, скорость движения воды в тканях растения
действительно зависит от интенсивности дыхания.
Я надеюсь, у читателя не сложилось впечатления,
что его водят за нос рассуждениями об осмосе, а потом
сознаются, что точно не известно, каким'образом вода
проходит через кору корня. Доминирующая роль, не-
сомненно, принадлежит осмосу; мы не уверены лишь
в отношении деталей.
Пройдя через кору корня, вода должна поступить
в основную водопроводящую систему растения — тра-
хеиды и сосуды древесины. Теоретически это, кажется,
не составляет труда. Листья растения постоянно отдают
воду при транспирации, а древесина поставляет новую.
По мере испарения воды листьями в древесине возни-
кает состояние натяжения водных нитей, которое пере-
дается вниз, к корням. Вода поэтому втягивается в эле-
менты древесины и восполняет ту воду, которая была
потеряна при транспирации.
Это состояние натяжения воды в древесине можно
продемонстрировать, если стебель увядшего растения
разрезать в воде, содержащей немного красного краси-
теля— эозина. Эозин быстро проникает в древесину,
окрашивая ее в красный цвет на значительном расстоя-
нии выше и ниже среза.
Жидкость в сосудах и трахеидах — это обычно не
чистая вода, а вода, содержащая в растворенном виде
различные вещества. Ее осмотическое давление в норме
выше, чем осмотическое давление почвенного раствора,
Гидравлика растений
73
и она склонна к насасыванию воды из почвы путем
осмоса, а наружные ткани корня работают как По-
лупроницаемая мембрана. Как вы помните, элементы
ксилемы имеют очень жесткие одревесневшие стенки,
в связи с чем давление оболочки в этих клетках мало
или совсем отсутствует. Поэтому, хотя осмотическое
давление содержащегося в них раствора может быть
не очень высоким по абсолютной величине, их ДДД
может оказаться намного больше, чем в клетках коры,
где он снижается тургорным давлением и за счет обще-
го натяжения тканей. Следовательно, осмос немало спо-
собствует поступлению воды в древесину.
Роль эндодермы
Говоря о передвижении воды в корне, я пока что не
упоминал эндодерму. Как вы помните, опа представляет
собой слой клеток с характерными полосками, назы-
ваемыми полосами или пятнами Каспари, на оболочках,
окружает древесину и изолирует ее от коры. Эндодерма
отчетливо дифференцирована в корнях всех растений,
но редко встречается в стеблях наземных растений,
хотя стебель подсолнечника — это один из немногих, ко-
торый обнаруживает признаки недоразвитой эндодермы.
У погруженных же в воду стеблей водных растений
эндодерма четко выражена.
Долгое время функция эндодермы в корнях оста-
валась совершенно неизвестной, да и теперь на этот
счет имеются только всевозможные предположения.
Постоянное наличие эндодермы в корнях и отсутствие
в стеблях наводит, однако, на мысль, что она имеет
какое-то отношение к проведению воды. В состав полос
Каспари входит много жировых веществ, которые
должны сделать клетки водонепроницаемыми, но неко-
торые клетки эндодермы, называемые пропускными,
составляют в этом смысле исключение и не имеют отло-
жений жировых веществ. Может быть, это и есть
ключ к разгадке роли эндодермы.
Мы не знаем точно, проходит ли вода через живые
клетки коры или нет, потому что она может проходить
и между ними, через клеточные стенки. Напоминаю,
74
Глава IH
что клеточные стенки состоят из целлюлозы, которая
свободно пропускает воду. Однако, когда вода дости-
гает эндодермы, ее дальнейший путь лежит через кле-
точные оболочки, «забаррикадированные» жировыми
веществами, которые они содержат, и поэтому она вы-
нуждена проходить через живое содержимое клеток.
Таким образом, вода немедленно попадает под контроль
осмоса.
Можно ли это считать причиной образования эндо-
дермы в корнях и какие преимущества дает растению
такой способ поступления воды, не совсем ясно. Но
поскольку ничего лучшего пока не предложено, эта
гипотеза о роли эндодермы представляется наиболее
вероятной.
Дойдя до древесины, вода поднимается по стеблю
к листьям (каким образом — пока неважно). От основ-
ного массива древесины стебля отходят тяжи ксилемы
и флоэмы, называемые листовыми следами-, они пересе-
кают кору и (Идут по листовым черешкам в листья, где
образуют жилки, хорошо заметные почти на каждом
листе. Жилки ветвятся, становясь все мельче, а их дре-
весина в конце концов уменьшается до одного сосуда
или трахеиды, пока совсем не исчезнет в ткани листа.
Эти пучковые окончания, как их называют, столь мно-
гочисленны, что любая клетка листа оказывается вбли-
зи одного из них. Таким образом драгоценная вода
попадает в каждую клетку листа.
Структура н функция листа
Строение листьев в основном соответствует одному
общему плану, хотя и широко варьирует в деталях.
Сверху лист покрыт обычной кожицей, или эпидермисом,
состоящим из одного слоя довольно тесно примыкающих
друг к другу клеток; если смотреть сверху, они напо-
минают кусочки составной картинки. Наружная по-
верхность эпидермиса покрыта воскоподобным вещест-
вом, образующим мало проницаемую для воды кути-
кулу, которая препятствует чрезмерному испарению во-
ды из листа.
Гидравлика растений
15
Под верхним эпидермисом находится слой крупных
прямоугольных клеток, длинная ось которых перпенди-
кулярна поверхности эпидермиса (фото XIV). Эти
клетки называют палисадными из-за их явного сходства
со столбиками, из которых делают ограду. Палисадные
клетки набиты многочисленными мелкими дисками, на-
зываемыми хлоропластами; это именно те образования,
которые содержат зеленый хлорофилл, обеспечивающий
питание растений. Наличие хлоропластов в клетках и де-
лает растение зеленым.
Под палисадной тканью мы находим несколько слоев
клеток неправильной формы, расположенных рыхло,
с большими межклетными пространствами. Эти клетки
тоже содержат хлоропласты, но не так много, как па-
лисадные клетки; они называются губчатым мезофиллом
за свое рыхлое сложение и положение в середине
листа1.
Наконец, под губчатым мезофиллом располагается
нижний эпидермис. Он в основном сходен с верхним
эпидермисом и тоже покрыт кутикулой. Однако для не-
го характерна одна особенность, которая часто отсутст-
вует в верхнем эпидермисе. Так, нижний эпидермис
повсюду перфорирован мельчайшими отверстиями —
устьицами (фото XV). Устьичные щели расположены
между двумя специальными клетками, называемыми
замыкающими клетками, которые в плане имеют почко-
видную форму. На разрезе замыкающих клеток можно
видеть, что стенки их утолщены очень своеобразно. На-
ружные стенки остаются тонкими, а внутренние нерав-
номерно утолщаются. Вследствие жесткости утолщен-
ных и эластичности тонких частей оболочек замыкаю-
щие клетки, находясь в состоянии тургора (когда они
наполнены водой), изгибаются и устьичная щель откры-
вается. Когда же эти клетки теряют воду, они спадают-
ся и сближаются друг с другом, закрывая щель. Таким
образом, если лист насыщен водой, устьица откры-
ваются, позволяя ей испаряться; при недостатке же во-
ды они стремятся закрыться, тем самым сокращая
1 Греческое слово «мезофилл» происходит от «мезо» — средни!
и «филлои» — лист. — Прим. ред.
76
Глава HI
транспирацию. Все это как будто говорит о том, что
устьица с замыкающими клетками являются почти
идеальным механизмом, регулирующим транспирацию
в зависимости от нужд листа, но на самом деле это не
так. Устьица далеко не совершенные регуляторы водно-
го режима, и мы это сейчас увидим.
Рассматривая водный режим листа, важно помнить,
что функция листа — это образование органической
пищи для растения из углекислого газа воздуха и воды
посредством фотосинтеза. Листья кормят растение, а как
они это делают, мы увидим из следующей главы. Функ-
ция эта — самая важная; без этого растение погибло
бы от голода. Если иметь в виду этот простой факт,
то можно объяснить несколько своеобразное поведение
листьев по отношению к воде.
В клеточном соке клеток мезофилла содержатся
растворы различных веществ, и вода путем осмоса
входит сюда из окончаний пучков. Эти клетки листа
находятся в тургесцентном состоянии, а их клеточные
оболочки сохраняются влажными. Вода испаряется из
таких влажных оболочек в воздушные пространства
между клетками мезофилла. Эти воздушные полости,
или межклетники, сообщаются друг с другом и с по-
лостями, существующими под каждым устьицем. Водя-
ной пар диффундирует по системе межклетников
и в конце концов выделяется из листа через устьица
наружу. Некоторое количество водяного пара проходит
через кутикулу эпидермиса, однако такая кутикулярная
транспирация, как ее называют, играет гораздо мень-
шую роль, чем испарение воды через устьица.
Общая площадь устьичных отверстий обычно неве-
лика по сравнению со всей площадью листа; опа сос-
тавляет около 3% его общей поверхности. Казалось бы,
через устьица может испаряться лишь относительно
небольшое количество воды, однако это не так. Изме-
рения показали, что относительная транспирация листа,
т. е. количество воды, испаряющееся из листа, по сравне-
нию с ее испарением со свободной поверхности воды
той же площади составляет примерно 95%. Иначе го-
воря, вода может выходить через устьица почти так же
легко, как если бы их и эпидермиса вовсе не было и она
Гидравлика растений 77
испарялась бы йэ влажных клеток мезофилла прямо
в окружающий воздух.
Это несколько неожиданное явление становится по-
нятным, если мы займемся изучением прохождения йаг
ра через перегородку со множеством малых отверстий,
как это сделали Браун и Эскомб. Мы обнаружим, что,
дойдя до перегородки, молекулы не двигаются по пря-
мой, а сбиваются в группы около каждого отверстия;
пройдя через них, молекулы снова расходятся (рис. 18).
Рис. 18. Диффузия пара через
перегородку с малыми отвер-
стиями.
Это позволяет им преодолеть препятствие гораздо быст-
рее, чем можно было ожидать.
Здесь можно провести полезную аналогию с толпой,
входящей на стадион за полчаса до начала матча. Как
вы знаете, у входа обычно устанавливают ряд турнике-
тов. Около стадиона образуется толпа желающих по-
пасть на матч, но так как места достаточно, они не
мешают друг другу. Однако близ турникетов они ску-
чиваются, чтобы пройти через них. Те болельщики, ко-
торые идут к турникету по прямой, проходят через
него почти без задержки, но тем, кто подходит к нему
с правой или с левой стороны, приходится протискивать-
ся через массу людей, скопившихся у турникета. Чем
дальше они находятся, тем труднее им протискиваться.
Пройдя через турникет, люди снова расходятся и даль-
ше идут уже сравнительно свободно.
Если наблюдать за толпой с самолета, то у нас
создается несколько иное впечатление от всего происхо-
дящего. Мы не увидим всех деталей толкотни у турни-
кетов, и нам покажется, что люди проходят на стадион,
почти нс задерживаясь у этих барьеров, хотя мы, ко-
78
Глава III
нечно, заметим скопления людей возле них. В резуль-
тате мы запишем в свой блокнот, что турникеты практи-
чески не препятствуют толпе зрителей попасть на ста-
дион, и по крайней мере в одном отношении мы будем
правы.
Подъем сока по стеблям и стволам
Теперь мы знаем, как вода поступает в растение
и как она снова удаляется из него, но мы еще не зада-
вали себе вопроса, каким же образом она поднимается
по стеблю к листьям. Этот вопрос всегда был трудным
для физиологов растений, и хотя в настоящее время
на этот счет существует вполне приемлемая теория, тем
не менее он не может считаться окончательно решенным.
Трудность заключается в большой высоте, на кото-
рую вода должна подняться по стволу дерева. Высота
более 30 м обычна даже для деревьев нашей страны,
а если взять калифорнийское Мамонтове дерево (высо-
та более 90 м)' или почти такой же высокий австралий-
ский эвкалипт, то проблема становится просто устра-
шающей.
Возможны три варианта механизма подъема воды.
Во-первых, она может проталкиваться из корней вверх
под действием осмотического давления, капиллярности
или каких-то других сил. Вторая теория предполагает
присасывание воды листьями, причем осмос и здесь
является движущей силой. В-третьих, вода может нагне-
таться вверх «рывками», а живые клетки древесины
каким-то образом играют при этом роль насосных
станций. Давайте рассмотрим каждую из этих возмож-
ностей.
Теория о присасывающем действии листьев, по-види-
мому, сразу же отбрасывается, во всяком случае в при-
менении к деревьям выше 10 м (высота водяного ба-
рометра). Присасывание зависит от атмосферного давле-
ния, выжимающего столб жидкости вверх. Если всосать
в себя воздух из трубки, то в ее верхней части создается
частичный вакуум, куда тотчас же устремляется воздух.
Если нижний конец трубки находится под водой, давле-
ние воздуха на поверхности воды заставляет воду подни-
Г ид рае лика растений
79
маться по трубке и заполнять свободное пространство.
Но даже если вакуум в верхней части трубки полный,
атмосферное давление может только удерживать столб
воды на определенной высоте. Вес столба воды высотой
примерно Юм уравновешивает атмосферное давление,
после чего устанавливается равновесие и дальнейший
подъем воды становится невозможным. Следовательно,
данная теория не в состоянии объяснить передвижение
воды вверх по стволу даже средней величины дуба, не
говоря уже о мамонтовом дереве.
Рассмотрев гипотезу о толкании воды снизу, мы
обнаруживаем здесь два возможных механизма. Один
из них — капиллярность. Явление капиллярности можно
наблюдать, погрузив конец очень узкой трубки в воду;
вода будет подниматься по трубке, причем чем меньше
диаметр трубки, тем выше она поднимется. Посколь-
ку трахеиды и сосуды древесины представляют собой
очень узкие трубки, то явление капиллярности к ним
вполне применимо. В таком случае мы имеем еще одну
версию механизма подъема воды от корней к листьям.
К сожалению, от нее тоже придется отказаться.
Хотя элементы ксилемы имеют малый диаметр, а поры
в их стенках еще уже, капиллярные силы недостаточно
велики, чтобы обеспечить поднятие воды на значитель-
ную высоту в стволах деревьев.
Другой механизм, который мог бы обусловливать
восходящий ток воды по стеблю,— это явление, назы-
ваемое корневым давлением. Если стебель растущего
растения перерезать непосредственно у поверхности
почвы или вблизи нее, то из оставшегося пенька начи-
нает вытекать вода. Насадив на пенек ртутный мано-
метр, можно убедиться в том, что вода вытекает под
некоторым давлением. По высоте столба ртути в мано-
метре можно судить о величине развившегося давления.
Существование корневого давления было обнаруже-
но Стефаном Гельсом в 1727 году. Измерения, которые
он произвел, показали, что у виноградной лозы (vitis
vinifera) оно достигает 1 атм. Некоторые другие расте-
ния развивают и более высокое корневое давление;
например, у томата (Lycopersicum esculentum) недавно
былр обнаружено, что оно составляет 8... 10 атм.
во
Глава III
Есть основания считать, что корневое давление —
явление осмотическое. Если корпи исследуемого вами
растения поместить в раствор, осмотическое давление
которого равно давлению в сосудах древесины, корне-
вое давление исчезает. Вот почему мы полагаем, что
его движущей силой является осмос. Но, с другой сто-
роны, здесь, вероятно, перед нами нечто большее, чем
простой осмос. Вода, поступая в сосуды, вскоре вымыла
бы из них растворенные вещества, снизив тем самым
довольно быстро корневое давление до нуля; но этого
не происходит. Напрашивается мысль, что живые клетки,
связанные с сосудами, снабжают их этими веществами
по мере их вымывания.
Некоторые ученые высказывали мнение, что корне-
вое давление зависит от активности живых клеток корня
в большей степени, чем от простого осмоса; в подтверж-
дение они приводили тот факт, что недостаток кислоро-
да и введение ядов в корень отрицательно сказываются
на корневом давлении. Этот аргумент, однако, ошибо-
чен. Если поддержание корневого давления зависит от
живых клеток, выделяющих растворенные вещества
в сосуды древесины, то недостаток кислорода или яды
будут скорее влиять именно па эту сторону их деятель-
ности, а не на корневое давление.
На первый взгляд может показаться, что корневое
давление и есть та сила, которую мы ищем. Но, рас-
смотрев этот вопрос внимательнее, мы обнаружим, что
корневого давления, как и капиллярности, недостаточно.
Этим давлением в сочетании с присасывающим дейст-
вием листьев можно легко объяснить йередвижение во-
ды по небольшому растению. Если же мы обратимся
к деревьям, то оказывается, что у них все обстоит по-
иному. Прежде всего корневое давление, по-видимому,
отсутствует у хвойных, а ведь именно они самые высо-
кие. К тому же оно сильно варьирует в зависимости от
состояния растения. Высокое корневое давление наблю-
дается только у активно растущих растений, а иногда
оно имеет даже отрицательную величину. В таких слу-
чаях вода, вместо того чтобы подниматься вверх по
стеблю, будет опускаться вниз. По этим причинам мы
Гидравлика растений
81
не можем считать корневое давление той силрй, кото-
рая определяет подъем воды.
Наш третий вариант состоял в том, что живые клетки
ствола дерева могут играть роль своеобразных насосов,
нагнетающих воду вверх. Такая гипотеза привлекатель-
на, но факты против нее. Страсбургером и другими уче-
ными в начале нашего столетия было установлено, что
растворы различных ядов, например пикриновой кисло-
ты, введенные в ствол через разрезанные сосуды, подни-
мались на очень большую высоту и даже достигали
листьев, хотя они убивали все встречавшиеся на их пу-
ти живые клетки. Примерно тогда же было замечено,
что. если живой ствол дерева положить горизонтально,
вода с равной скоростью движется в обоих направле-
ниях— вверх к листьям и вниз к корням. Таким образом,
стало ясно, что в стебле растения нет ничего похожего
на клапаны, обеспечивающие односторонний ток воды,
наличие которых было бы обязательно в случае нагне-
тания ее живыми клетками.
Кажется, мы зашли в тупик. Подъем воды нельзя
объяснить ни присасывающим действием транспирации,
ни поднятием ее вверх за счет корневого давления (разве
что в небольших растениях). Несостоятельной оказа-
лась и третья теория, предполагающая существование
нагнетающего клеточного механизма. Невольно напра-
шивается мысль, что вода поднимается по воле божьей,
однако ученые не склонны привлекать для объяснения
затруднительных фактов потусторонние силы. Давайте
лучше посмотрим, не упустили ли мы чего-нибудь.
Обратившись вновь к теории присасывания воды
сверху, мы обнаружим сразу множество сил, способных
на это; дело в том, что в результате транспирации в ме-
зофилле листа устанавливается осмотический градиент
с давлениями намного выше 10...12 атм, чего вполне
достаточно для подъема воды даже до верхушки
Sequoia. Но если посмотреть на эту теорию с практи-
ческой точки зрения, возникает множество трудностей.
Присасывание здесь ни при чем, так как мы не можем
обойти тот факт, что барометрическая высота не пре-
вышает 10 м. Необходимо участие дополнительной силы,
способной обеспечить распространение присасывающегд
82
Глава III
действия листьев вниз по стеблю к корням, что дало
бы возможность протягивать воду вверх. По-видимому,
ответ надо искать в теории сцепления, предложенной
Диксоном и Джоли. Компактность твердого тела обус-
ловлена силой сцепления его молекул, которые обра-
зуют жесткую кристаллическую решетку, устойчивую
к разрушению. В жидкости межмолекулярные силы
слабее; молекулы имеют значительную свободу передви-
жения, но все же удерживаются силами сцепления и не
расходятся совсем. Только накопив достаточную кине-
тическую энергию (энергию движения), они преодоле-
вают силу сцепления и отрываются друг от друга —
другими словами, испаряются.
Согласно теории сцепления Диксона, вода в сосу-
дах образует непрерывные нити, проходящие от корней
до самых верхних листьев. Силы сцепления не до-
пускают разрыва водных нитей, поэтому, когда осмо-
тический градиент, созданный транспирацией, тянет
верхний конец ннтн, весь столб воды начинает подни-
маться. Заметьте, что мы теперь избавились от теории
присасывания и трудностей с водяным барометром.
Сила, тянущая воду вверх, имеет осмотическую приро-
ду и передается вниз по стеблю благодаря силе сцепле-
ния между молекулами воды.
Почему же в таком случае мы не обнаруживаем
эту силу сцепления в повседневной жизни? Например,
если трубку малого диаметра вы наполните до краев
водой и перевернете, то почему вода не останется в труб-
ке, а выльется? Не противоречит ли это теории Диксона?
Ответ прост. Никакая трубка, сделанная человеком, не
может быть абсолютно чистой; на ее стенках всегда
остаются какие-то частицы. Теория же Диксона предпо-
лагает, что вода смачивает всю внутреннюю поверхность
трубки и таким образом прилипает к ее стенкам. Части-
цы пыли мешают прилипанию частиц воды к стенкам,
уменьшают силу сцепления ее молекул и тем самым
создают слабое звено в этой цепи, что позволяет воде
выливаться из трубки. Это можно проверить, используя
изящный маленький приборчик, известный без всякой
очевидной на то причины как «водяной молоток». Он
состоит из стеклянной трубки, изогнутой в форме ла-
Гидравлика растений
83
тинской буквы 1 Тщательно промыв трубку, наполним
«е водой и перевернем. Если все сделано достаточно
аккуратно, вода не выльется (рис. 19). Теория. Диксона,
таким образом, подтверждена.
рис. 19. Водяной молоток.
В шарик налита вода (Л). Если осто-
рожно наклонить прибор так, чтобы
вода перелилась в трубку, а затем
перевернуть, вода будет удерживаться
в трубке силой сцепления молекул (Б).
А
Сила сцепления, связывающая молекулы воды, по-
видимому, весьма значительна и в состоянии обеспе-
чить целостность водного столба в сосудах дерева. Бы-
ли проведены, например, измерения силы сцепления,
необходимой для вскрывания спорангиев папоротника,
и получились цифры, превышающие 200 атм для натя-
жения, которое в конце концов разрывает водную плен-
ку. У нас есть основания думать, что эта цифра завы-
шена, но надо полагать, что столб воды наверняка вы-
держит натяжение в 30 атм или около того, прежде чем
разорваться, а этого вполне достаточно для наших це-
лей. Однако одно дело — выдвинуть теорию, объясняю-
щую восходящий ток воды в растении, и совсем дру-
гое— доказать ее правильность. В этом смысле и тео-
рия сцепления Диксона сталкивается с трудностями.
Никому не удалось доказать, что эта теория действует
в природе, и даже имеются факты, ставящие под сомне-
ние ее справедливость. В одном из опытов на стволах
деревьев с двух противоположных сторон и на различ-
ной высоте делали неполные поперечные распилы; оба
84
Елада lit
распила проходили через центр ствола и перекрывали
друг друга. Теоретически это должно было остановить
движение воды вверх по стволу, поскольку нарушалась
непрерывность всех столбов воды в сосудах, и все же
вода продолжала подниматься.
Мы еще не можем объяснить это явление. Трудно
поверить, что вода могла обходить оба распила, под-
нимаясь по диагонали стебля, хотя вообще это возмож-
но. А может быть, непрерывный столб воды в действи-
тельности не необходим для действия теории Диксона
или же вода проходит через субмикроскопические поры
в стенках сосудов вместо того, чтобы идти через их
полости. Мы не узнаем этого, пока нс будут проведены
дополнительные эксперименты.
В настоящее время теория сцепления Диксона счи-
тается одной из условно принятых. Безоговорочному
ее признанию мешают многочисленные возражения, по
пока никто не предложил ничего лучшего.
Транспирация и устьица
Мы должны еще рассмотреть роль устьиц листа
в «контролировании» транспирации. Здесь мнения также
разделяются и последнее слово еще не сказано. На
первый взгляд регулирование транспирации путем от-
крывания и закрывания устьиц кажется столь очевид-
ным, что едва ли нуждается в доказательствах, но,
как часто бывает в науке, явно «очевидные» вещи на
самом деле представляют собой совсем не то, чем ка-
жутся. Действительно, устьица устроены таким образом,
что благодаря специфическим утолщениям стенок за-
мыкающих клеток они открываются при достаточном
тургоре и закрываются при нехватке воды. Однако,
взглянув пристальнее, мы замечаем, что зависимость
степени открытости устьиц от потребности растения в
воде, мягко выражаясь, не слишком строгая. Прежде
считали, что эффективность работы устьиц строго опре-
деляется нуждами транспирации, тогда как в последнее
время эти взгляды были пересмотрены и стало ясно,
что контроль транспирации при участии устьиц —явле-
ние бол^е сложное.
Гидравлика растений
&
На скорость испарения воды из листа влияют два
фактора: во-первых, скорость прохождения водяного па-
ра через устьица, а во-вторых, концентрация паров во-
ды непосредственно над устьичными отверстиями.
Что касается первого фактора, то мы уже выяснили,
что водяной лар может диффундировать через ряд ма-
леньких отверстий в перегородке почти так же быстро,
как если бы этой перегородки вовсе не было. Устьица
очень многочисленны, и логично предположить, что,
если их отверстия станут меньше, это очень Мало или
совсем не повлияет на скорость прохождения через них
водяного пара. Иначе говоря, если даже устьица до не-
которой степени и закроются, это не очень отразится на
скорости транспирации.
Это действительно так. Транспирация не прекра-
щается сразу, как только устьица начинают закрывать-
ся. До какой степени они должны закрыться, чтобы это
ощутимо сказалось на транспирации,— вопрос сложный;
это зависит от типа листа, структуры самих устьиц
и условий, господствующих в данный момент, в частно-
сти от того, ветреный день или тихий. В связи с этим
степень открытости устьиц, обеспечивающая нормаль-
ную транспирацию, варьирует от ’/а до ‘До-
говоря о влиянии устьиц на транспирацию, очень
важно учитывать и условия вне листа, в особенности
скорость ветра. В тихую погоду водяной пар скаплива-
ется над устьичной щелью, потому что кет ничего, что
«сдуло» бы его прочь. В таких условиях пар не выхо-
дит из листа, даже если устьица широко открыты.
По-видимому, при отсутствии ветра степень открытости
устьиц не имеет значения и они практически не контро-
лируют транспирацию.
Когда ветрено, пары воды не могут задерживаться
у устьичных отверстий, потому что они тотчас же уно-
сятся ветром. При таких условиях устьица, вероятно,
способны достаточно тонко регулировать транспирацию.
Обращаясь к вопросу о том, что же заставляет устьи-
ца открываться и закрываться, мы обнаруживаем, что
и здесь все обстоит не так, как предполагалось. Закры-
вание устьиц происходит при потере тургора замыкаю-
щими клетками. Когда тургорное давление в них ста-
86
Глава HI
новится ниже, чем в соседних эпидермальных клетках,
устьица закрываются, а открываются вновь, когда тур-
гор становится выше, чем в окружающих клетках. Не-
трудно догадаться, что при завядании листа устьица
закрываются, потому что замыкающие клетки теряют
способность поддерживать тургорное состояние. Имен-
но так в общих чертах это и происходит, хотя все это
слишком упрощенная схема; впрочем, описывать детали
истинной, более сложной картины мы пока не будем.
Устьица и фотосинтез
Если бы то, что мы выяснили выше, было все, что
можно сказать о функционировании устьиц, то картина
была бы ясна, однако это не так. На простой тургорный
механизм накладываются очень сложные взаимоотноше-
ния между устьицами и светом. Поведение устьиц у раз-
ных растений различно, но обычно они открываются на
свету и закрываются в темноте. Таким образом, сущест-
вует определенный суточный ритм устьичных движений,
которые поддерживаются независимо от водного снаб-
жения и часто даже в том случае, если растение какое-
то время находится в условиях постоянного освещения.
Такое поведение устьиц будет вполне понятно, если
мы усвоим, что их деятельность связана не только
с потерей воды. Они играют важную роль в фотосинте-
зе, так как через устьица входит в лист углекислый газ.
Если устьица закрыты, углекислый газ не может посту-
пать в листья и фотосинтез прекращается. Закрывание
же устьиц на продолжительное время, как это случа-
ется при длительной засухе, грозит растению голодной
смертью. Снижение урожая зерна при сильной засухе
объясняется обезвоживанием в такой же мере, как и го-
лоданием из-за закрытых устьиц.
Таким образом, поведение устьиц под влиянием света
вполне объяснимо; это справедливо и для большинства
функций других органов растений. Днем, когда проис-
ходит фотосинтез и необходим доступ углекислого газа
в лист, устьица открыты, если только какие-нибудь при-
чины тому не препятствуют. Ночью фотосинтез прекра-
Гидравлика растений
87
щается из-за отсутствия света. На это время устьица
обычно закрываются, чтобы сохранить воду.
Природа механизма, обеспечивающего открывание
устьиц на свету и их закрывание в темноте, далеко нё
ясна, хотя многие исследователи сходятся во мнении, что
это связано с кислотностью клеточного сока в замыкаю-
щих клетках. Понижение кислотности каким-то образом
вызывает открывание устьиц, тогда как ее повышение
влечет за собой их закрывание. Изменение кислотности,
по-видимому, связано с количеством имеющегося угле-
кислого газа. В процессе активного фотосинтеза (на яр-
ком дневиом свету) потребляется много углекислоты,
поэтому кислотность в замыкающих клетках падает.
Ночью же, когда фотосинтез прекращается, происходит
накопление углекислого газа, образующегося в результа-
те дыхания, и кислотность возрастает.
Полагают, что влияние изменений кислотности замы-
кающих клеток связано с соотношением между крахма-
лом и сахаром. Понижение кислотности благоприятству-
ет распаду крахмала до сахара под действием фермента
фосфорилазы. Крахмал нерастворим в воде и поэтому
не влияет на осмотическое давление, тогда как раство-
римый в воде сахар осмотически активен. Появление
в замыкающих клетках сахара вместо крахмала повы-
шает осмотическое давление клеточного сока, поэтому
под действием осмотических сил вода из окружающих
клеток переходит в замыкающие клетки и устьица, при-
обретая тургор, открываются. Повышение кислотности,
с другой стороны, способствует превращению сахара
в крахмал, и события получают обратный ход.
Многих ботаников не удовлетворяет такое объяснение
влияния света на открывание устьиц, и не без основания.
Прежде всего устьица некоторых растений (например,
лука), которые никогда не содержат крахмала в замы-
кающих клетках, все же открываются на свету и закры-
ваются в темноте. Есть и другие возражения против этой
теории. Пока же мы можем сказать лишь то, что эта
теория объясняет больше фактов, чем все остальные.
Однако будущие исследования, несомненно, заставят нас
если не отказаться от нее, то по крайней мере внести ка-
кие-то изменения.
88
Глава Ilf
Какой же смысл растению испарять столь громадное
количество воды, используя при этом лишь небольшую
ее часть? К сожалению, мы до сих пор не можем дать
исчерпывающего ответа на этот вопрос. Определенное
количество воды, конечно, необходимо для фотосинтеза;
кроме того, вода непосредственно участвует в передви-
жении минеральных солей к листьям и поддерживает
ток веществ во флоэме (гл. V). Наконец, транспирация
в жаркие дни оказывает охлаждающее действие, которое
может быть значительно; в солнечный день у растений
з теплице могут наблюдаться ожоги листьев, если из-за
избыточной влажности воздуха транспирация затрудне-
на. И все же ни одной из этих причин нельзя объяснить
того, почему такое огромное количество воды расходу-
ется на испарение. Это можно сравнить разве что с ис-
пользованием Ниагары на то, чтобы наполнить ванну.
Растения тратят воду, которую часто так трудно добыть,
с расточительной непринужденностью.
Однако мы забыли о настоятельной потребности рас-
тения фотосинтезировать при любой возможности, что
важнее всего. Для этого необходима достаточная венти-
ляция листьев, чтобы углекислый газ мог свободно до-
ходить до фотосинтезирующих клеток. В воздухе содер-
жится не более 0,03% углекислого газа; это довольно
мало, если учесть, что вся огромная масса Мамонтова
дерева образована из этого газа. Это сразу же про-
ясняет проблему. Если углекислый газ будет проникать
в лист, вода, несомненно, будет из него испаряться, ибо
нельзя гарантировать одностороннее движение через
устьица. И чтобы не погибнуть от голода, растение вы-
нуждено примириться с такой огромной потерей воды.
Вот как все просто.
Поглощение минеральных веществ
До сих пор мы говорили только о поступлении воды
в растение, ее передвижении по растению и испарении
в воздух. Однако корни поглощают из почвы не только
воду, но и минеральные соли. Все необходимые для жиз-
ни неорганические вещества поступают в растение через
корни, поскольку иного пути для них нет.
Гидравлика растений
89
Скажу сразу, что мы не знаем, как поглощаются
минеральные соли. Для школьника, начавшего впервые
изучать биологию, это просто: соли входят и корни «ос-
мотическим путем». В действительности же такое явле-
ние совершенно невозможно. Осмосом можно объяснить
поступление воды, по не растворов, поскольку этот про-
цесс требует наличия полупроницаемых мембран.
Одно время считали, что соли просто диффундируют
в корни; при этом предполагали, что клеточные мембра-
ны проницаемы для одних веществ и непроницаемы для
других. Эта идея проста, но явно неверна. Во-первых,
неверно, что клеточный сок — это просто более концент-
рированный наружный почвенный раствор. Во-вторых,
клетки способны накапливать определенные вещества
против градиента концентрации, а это не может иметь
место при простой диффузии. Некоторые морские водо-
росли, например, накапливают в клетках относительно
большие количества иода, хотя концентрация его в воде
очень мала. Здесь должно действовать что-то более су-
щественное, чем просто диффузия.
На основании современных исследований предполага-
ется, что поглощение минеральных солей растениями
связано с метаболическими процессами в живой клет-
ке, и особенно с дыханием. Как оказалось, поглощение
неорганических веществ кусочками клубней картофеля
возрастает с увеличением интенсивности дыхания. Похо-
же, что прохождение минеральных веществ через клеточ-
ные мембраны, в особенности через мембрану, окружаю-
щую вакуоль — тонопласт (по-видимому, он играет
важную роль в этом процессе), связано с электрически-
ми зарядами поглощаемых ионов. Как именно это про-
исходит, пока не известно.
ГЛАВА IV
КАК РАСТЕНИЕ ПИТАЕТСЯ
Растения — это не только производители органиче-
ского вещества; они — главные поставщики энергии на
Земле. Так, каменный уголь — не что иное, как остатки
растительности каменноугольного периода (270 млн. лет
назад). Происхождение нефти неизвестно, но считается,
что она, по крайней мере частично, образовалась из рас-
тений. Сжигая каменный уголь, мы сжигаем остатки
древнейших лесов, а бензин, используемый для автома-
шин, когда-то был водорослями. Большую часть электро-
энергии дают нам уголь и нефть — продукты фотосинте-
за растений. Лршь в немногих случаях получение энер-
гии не связано с растениями (ветряная мельница, водя-
ное колесо, ядерная энергия). Путешествуя по железной
дороге или на самолете, мы передвигаемся за счет расте-
ний, которые много лет назад связали энергию солнца
и сохранили ее для будущего.
Даже мышечную энергию, которую мы расходуем,
например, при ходьбе, мы получаем в конечном счете от
растений, потому что без растений не было бы пищи.
Вегетарианец обязан жизнью непосредственно растени-
ям, так как он не ест ничего другого, но и те, кто пред-
почитает обычную диету, все равно в таком же долгу
у растений, так как животные, мясо которых мы едим,
сами питаются растениями.
Мы не знаем, когда возникло чудо фотосинтеза, но,
вероятно, оно так же старо, как само растительное цар-
ство. Первые растения содержали хлорофилл или что-то
аналогичное и были способны питаться пе за счет дру-
гих живых существ, а поглощая из воздуха углекислый
газ и строя из него сахар с использованием энергии све-
та; хлорофилл при этом играл роль ловушки энергии,
превращая солнечную энергию в химическую. Можно
поэтому с уверенностью сказать, что фотосинтез продол-
жается уже 2 млрд, лет, а возможно, и дольше,
Как растение питается
91
У большинства современных растений главным орга-
ном фотосинтеза служит лист, хотя эту функцию частич-
но или полностью несет иногда стебель, но так было не
всегда. У первых наземных растений листьев не было.-
Одно из самых древних растений—это Zosterophyllum,
найденный в Австралии в отложениях силурийского пе-
риода (т. е. произраставший 350 мли. лет назад). Это
было маленькое растение, состоявшее из массы перепле-
тенных между собой веточек, которые, по-видимому, на-
ходились под землей, образуя нечто вроде корневища,
подобного тому, которое имеется у мяты или купены
(«соломоновой печати»). От этих подземных корневищ
отходили вертикальные ветви, очевидно надземные, так
как они были покрыты эпидермисом и кутикулой, слу-
жившими для защиты растения от высыхания на возду-
хе. Эти ветви достигали 0,2 см в толщину и не имели
листьев, поэтому можно предположить, что именно они
осуществляли фотосинтез.
Хотя у первых наземных растений не было листьев,
вскоре эти органы появились и стали выполнять основ-
ную работу по фотосинтезу. В уже упомянутых отложе-
ниях в Австралии, где был обнаружен Zosterophyllum,
были найдены и остатки растения, названного Baragwa-
nathia. Оно было крупнее, чем Zosterophyllum, со стебля-
ми около 7 см в диаметре и со множеством спирально
расположенных листьев. Эти листья были длинными и
тонкими (0,05... 0,1 см шириной и до 4,4 см длиной).
Каждый лист имел единственный проводящий пучок
(жилку), проходивший в середине листа и связанный
с проводящим цилиндром стебля, который явно предна-
значался для переноса воды к листу и для оттока из него
продуктов фотосинтеза.
Поскольку остатки Baragwanathia были найдены в
отложениях того же силурийского периода, что и остат-
ки Zosterophyllum, их возраст представляется вполне
сравнимым; следовательно, развитие листьев началось
вскоре после выхода растений на сушу.
Как основной орган фотосинтеза лист прекрасно при-
способлен к выполнению своей функции. Он обычно бы-
вает тонким и плоским, что обеспечивает создание мак-
симальной поверхности не только для поглощения угле-
92
Глава IV
кислого газа, но и для освещения солнцем. Правда, есть
листья и несколько иного строения, однако их модифи-
кации были вызваны каким-то другим фактором, глав-
ным образом необходимостью сохранять воду. Внутрен-
ность листа хорошо вентилируется,-так что углекислый
газ может без задержки доходить до фотосинтезирую-
щих клеток. Вдобавок лист имеет в своих жилках много-
численные проводящие пучки, которые не только достав-
ляют воду, но и обеспечивают отток по флоэме органиче-
ских веществ, образующихся при фотосинтезе. Наконец,
лист надежно покрыт эпидермисом и кутикулой, не до-
пускающими чрезмерного испарения воды, а его устьица'
обеспечивают свободный доступ углекислого газа к внут-
ренним воздушным полостям.
Реакция растений на действие силы тяжести
Стебли растений, как правило, растут против силы
тяжести и по направлению к свету, вынося свои листья
туда, где света больше. Если семя посадить «вниз голо-
вой», то корешок все равно не будет расти вверх, а сте-
бель— вниз. Как только молодой стебель и корень по-
явятся из семенной кожуры, сила тяжести немедленно
заставит их принять нормальное положение. Ростовая
реакция на действие силы тяжести заложена в растении;
оно должно «учиться» этому не больше, чем детеныш
учится сосать мать.
Реакция на силу тяжести обусловлена веществом, на-
зываемым ауксином, которое образуется в точках роста
стебля и корня. Влияние ауксина на точку роста стебля
заставляет вновь образующиеся клетки быстрее удли-
няться, и, таким образом, рост стебля ускоряется. Обыч-
но ауксин перемещается из верхушки к основанию стеб-
ля, распространяя свое влияние равномерно по окружно-
сти. Если же придать стеблю горизонтальное положение,
то концентрация ауксина на нижней его стороне стано-
вится выше, чем на верхней. Мы не знаем точно, как
это происходит, хотя подозреваем, что это связано с раз-
личием в электрических потенциалах. В результате ниж-
Как растение питается
93
ляя сторона стебля, где концентрация ауксина выше, рас-
тет быстрее, и стебель изгибается вверх. В вертикально
растущем стебле равновесие восстанавливается,
Корень реагирует на силу тяжести иначе, чем стебель,
хотя механизм реакции тот же. Как выяснилось, ауксин
тормозит рост корней; следовательно, в горизонтально
расположенном корне повышенная концентрация аукси-
на на нижней стороне приводит к тому, что она растет
медленнее, чем верхняя, что и заставляет корень
изгибаться вниз.
Ботаники были очень заинтересованы этим явным
различием в реакции корней и стеблей на действие одно-
го и того же вещества — ауксина. Исследования показа-
ли, что ауксин, содержащийся в корнях, идентичен аукси-
ну стеблей; различие же состоит в реакции на его дейст-
вие. Вопрос стал ясным, когда обнаружилось, что очень
маленькие количества ауксина не тормозят, а стимулиру-
ют рост корней; в то же время рост стеблей задерживает-
ся под влиянием больших доз ауксина. Таким образом,
удалось установить общий принцип действия этого ве-
щества на стебли и корни: низкие*концентрации ауксина
ускоряют рост, а высокие тормозят. Разница между
корнем и стеблем заключается в том, что корни бо-
лее чувствительны к ауксину, чем стебли, поэтому до-
зы, маленькие для стебля, оказываются большими для
корня.
Когда и как установилась различная реакция корпя
и стебля на ауксин, мы не знаем, но, по-видимому, это
произошло вскоре после того, как определилось функ-
циональное различие этих органов. Ауксин — очень древ-
нее вещество. Оно было обнаружено в яйцеклетках и в
различных других частях некоторых водорослей. Веро-
ятно, выйдя из моря, растения сохранили способность
образовывать ауксин и реагировать на него и исполь-
зовали эту способность в новой жизни на берегу. Вот
еще один пример удивительной экономии, столь харак-
терной для растений. Иногда им это не удается (напри-
мер, громадная потеря пыльцы при опылении ветром),
но в основном растения живут и умирают, затрачивая
на это минимум усилий.
94
Глава IV
Рост по направлению к свету
Стебель растения растет не только против силы тя-
жести, но и по направлению к свету. Это может наблю-
дать каждый, кто выращивает растения в горшках на
окне, потому что стебель всегда изгибается в том на-
правлении, откуда падает свет. Реакция эта опять-таки
зависит от ауксина, который накапливается на затенен-
ной стороне стебля больше, чем на освещенной; зате-
ненная сторона поэтому растет быстрее и стебель пово-
рачивается к свету. Рост стеблей по направлению к све-
ту, так же как и против силы тяжести, дает растению
возможность вынести листья вверх, где они получат
больше света и воздуха, необходимых для фотосинтеза.
За некоторыми исключениями (например, лазящие
корни плюща и подземные корни белой горчицы), на-
правление роста корней не зависит от света.
Когда растение находится в условиях слабого осве-
щения, наприцер если оно окружено и затенено другими
растениями, в росте стебля наблюдаются некоторые из-
менения. Он становится длинным и тонким, а листья
располагаются на необычно большом расстоянии друг
от друга; такие стебли называют этиолированными.
Другое следствие этиолирования — бледно-зеленая
окраска, потому что в отсутствие света хлорофилл не
образуется. Наибольший же эффект этиолирования мож-
но наблюдать на растениях, выращенных в полной тем-
ноте. Стебель при этом становится тонким и слабым,
а все растение приобретает желтоватую окраску, ничего
общего не имеющую с зеленой окраской нормальных
растений. Листья тоже становятся гораздо мельче, рас-
положены реже и имеют тот же нездоровый желтоватый
оттенок, что и стебель. Стоит отметить, что в темноте
удлинение стеблей идет заметно быстрее, чем на свету,
так как свет тормозит рост.
В то же время явления, сопровождающие этиолиро-
вание, весьма полезны для растения, которое затеняется
соседями. Благодаря быстрому вытягиванию стебля оно
вскоре выходит на свет и имеет шансы вернуться к нор-
мальному росту, если этиолирование зашло не слишком
далеко.
Как растение питается
96
Как мы вйделйТ’для осуществления фотосинтеза рас-
тению нужны два простейших химических соединения —
углекислый газ и вода; кроме того, хлорофилл и свет
для его активации. Необходимость других веществ менее
очевидна. Например, в листе должен присутствовать са-
хар рибулоза, а также группы атомов, известные как
фосфатные группы: они названы так потому, что, соеди-
няясь с водородом, образуют фосфорную кислоту.
Хлорофилл и другие пигменты
Хлорофилл листа распределен в протоплазме не рав-
номерно, а сосредоточен в мелких дисковидных тельцах,
называемых хлоропластами. Исследования с помощью
электронного микроскопа, который может давать неиз-
меримо большие увеличения, чем оптический микроскоп,
подтвердили предположение о том, что хлоропласты име-
ют чрезвычайно сложную структуру.
Хлорофилл в хлоропластах заключен в субмикроско-
пических частицах, называемых гранами, которые погру-
жены в бесцветную основу — строму хлоропласта. Грана
не состоит целиком из хлорофилла, а представляет собой
многослойную структуру, напоминающую сандвич, в ко-
торой слои хлорофилла (толщиной в одну молекулу)
расположены между слоями белка и липида (жировое
вещество) в определенной последовательности: белок,
хлорофилл, липид, хлорофилл, белок, хлорофилл, липид
и т. д. Каждая палисадная клетка листа содержит мно-
жество хлоропластов, каждый хлоропласт содержит мно-
жество гран, а каждая грана имеет такую сложную слои-
стую структуру.
Функция хлорофилла заключается в поглощении све-
товой энергии и передаче ее другим молекулам в форме,
удобной для осуществления химических реакций. Свет,
попадая на молекулу хлорофилла, переводит ее в воз-
бужденное состояние (разумеется, не в обычном, а в хи-
мическом смысле). Электроны, вращающиеся вокруг яд-
ра, поглощая эту световую энергию, перескакивают на
орбиту, более удаленную от ядра. Возвращаясь впослед-
ствии на исходные орбиты, они отдают поглощенную
Флава IV
&
энергию. В этом и заключается тайна улавливания
энергии хлорофиллом.
Химическая структура хлорофилла сложна. В сущно-
сти имеется несколько различных форм хлорофилла, из-
вестных как хлорофилл а, Ь, с и т. д.; формы а и b най-
дены в хлоропластах высших растений, другие —у раз-
личных групп морских и пресноводных водорослей. Хло-
рофилл а (синевато-зеленого цвета) и хлорофилл b
(желто-зеленый) содержатся в хлоропластах в соотно-
шении примерно 3:1; при таком сочетании этих пигмен-
тов получается травянисто-зеленая окраска, характерная
для большинства высших растений. Хлорофилл а наибо-
лее важен для процесса фотосинтеза, хотя хлорофилл b
тоже принимает в нем участие, и как это происходит, мы
увидим ниже.
Помимо этих двух форм хлорофилла, хлоропласты
содержат еще два пигмента, или, точнее, две группы
пигментов. Это ксантофиллы и каротины, цвет которых
варьирует от желтого до красного. Подобно хлорофиллу,
эти пигменты фотохимически активны и, хотя непосред-
ственно не участвуют в фотосинтезе, важны для этого
процесса, оказывая на него косвенное влияние.
Белый свет в действительности представляет собой
смесь всех цветов радуги, па которые он и разлагается,
проходя через стеклянную призму. Если пучок белого
света пропустить через раствор хлорофилла в подходя-
щем растворителе, например в эфире, а затем проанали-
зировать его с помощью спектроскопа (прибор, содержа-
щий призму для разложения белого света), нетрудно
заметить, что хлорофилл поглощает свет избирательно.
Зеленый свет хлорофилл пропускает почти полностью,
а красные и сине-фиолетовые лучи весьма интенсивно'
поглощает. Вследствие этого в красной и сине-фиолето-
вой частях спектра хлорофилла появляются темные по-
лосы, называемые полосами поглощения.
Чтобы энергия света могла быть использована при
фотосинтезе, свет должен сначала поглощаться хлоро-
филлом. А поскольку максимум поглощения света хло-
рофиллом приходится на красную область спектра, мож-
но предположить, что в красном свете фотосинтез идет
активнее. И это действительно так. Если мы накроем
Как растение питается
У!
растение красным'"?геклянным колпаком и измерим ин-
тенсивность фотосинтеза при данной интенсивности света
за определенный промежуток времени, то она окажется
несколько выше, чем у растения, фотосинтезирующего
в синем свете, и гораздо выше, чем интенсивность фото-
синтеза в зеленом свете.
Если бы в листьях растения присутствовал только
хлорофилл а, оно могло бы использовать для фотосинте-
за лишь очень узкий участок спектра. Весь остальной
свет пропадал бы зря, конечно, если говорить о «произ-
водстве» сахара. Процесс фотосинтеза был бы крайне
неэффективным, так как большая часть белого света,
падающего на листья, оставалась бы неиспользованной.
Наличие хлорофилла Ь, ксантофилла и каротина улуч-
шает положение. Их спектры поглощения отличаются
от спектра поглощения хлорофилла а: они способны по-
глощать свет других длин волн и при этом их молекулы
переходят в возбужденное состояние. Хотя сами эти ве-
щества не принимают непосредственного участия в фото-
синтезе, возбуждение их молекул может передаваться
на хлорофилл а, который затем использует полученную
энергию на фотосинтез.
В этом, по-видимому, и заключается роль ксантофил-
ла и каротина, а возможно, и хлорофилла Ь. Поглощая
свет тех длин волн, которые недоступны хлорофиллу а,
и передавая ему приобретенную энергию, они делают
возможным фотосинтез в более широком диапазоне
длин волн.
Даже с помощью дополнительных пигментов — каро-
тина и ксантофилла — фотосинтез не идет одинаково в
различных областях спектра. Совершенно бесполезным
остается, например, зеленый свет. Но белый свет в при-
сутствии этих пигментов более эффективен, чем без них.
Так как фотосинтез зависит от поглощения света,
возникает вопрос, насколько эффективно листья растений
поглощают падающий на них свет. Наши наблюдения
говорят о том, что эта эффективность очень велика: один
хлоропласт может поглотить 30... 60% падающего па
него света. А так как хлоропласты расположены в клет-
ках палисадной ткани весьма густо, то можно предполо-
жить (и это будет справедливо), что большая часть све-
&8
Глава IV
та, попадающего на лист, натолкнется на какой-нибудь
хлоропласт. Даже свет, который не попадает на хлоро-
пласты или отражается от них не поглощаясь, не обя-
зательно теряется для фотосинтеза. Многочисленные воз-
душные полости листа вместе с оболочками клеток ме-
зофилла превращаются в отражающие поверхности, ко-
торые отсылают свет назад, прежде чем он пройдет через
лист. Этот отраженный свет еще может быть использован
для фотосинтеза, если будет уловлен хлоропластом.
Разные типы листьев отражают разное количество
света; чтобы убедиться в этом, достаточно взглянуть на
некоторые растения. Растения, листья которых не погло-
щают, а отражают большую часть света, возможно, даже
имеют преимущество, так как слишком яркое освещение
вредно для хлорофилла. Вообще же замечено, что у жи-
вущих на ярком солнечном свету растений листья более
мелкие и толстые, чем у тех, которые любят тень. Неко-
торые растения, особенно бук, образуют близ верхушки,
наиболее подверженной солнечной радиации, световые
листья, а внизу, где света меньше, теневые листья. У бу-
ка световые листья толще, чем теневые, палисадная
ткань у них двухслойная.
В зависимости от окружающих условий количество
света, поглощенного растением, меняется. Это происхо-
дит потому, что отдельные хлоропласты листа могут
изменять свою форму в зависимости от степени освещен-
ности: при слабом свете они становятся округлыми, а при
ярком уплощаются и в то же время поворачиваются
к свету ребром, с тем чтобы он падал на возможно мень-
шую их поверхность. В соответствии с освещением хло-
ропласты меняют также свое положение в клетках, со-
средоточиваясь на освещенных сторонах клеток при сла-
бом свете и распределяясь вдоль вертикальных, менее
освещенных сторон при сильном свете, что хорошо за-
метно у ряски трехдольной (Lemna trisulca) (рис. 20).
У многих тенелюбивых растений, например у адоксы
(Adoxa moschatellina) и у кислицы (Oxalis acetosella),
наружные стенки клеток эпидермиса заметно выпуклы,
что позволяет им действовать подобно выпуклым линзам,
фокусируя свет внутрь клетки, где находятся хлоро-
пласты.
Как растение питается
99
Листья деревьев расположены в виде мозаики, и каж-
дый из них прикреплен так, чтобы по возможности не
затенять листья, находящиеся непосредственно под ним.
Такое расположение дает возможность максимально ис-
пользовать доступный свет. Примером хорошо выражен-
ной листовой мозаики может служить бук; поэтому он и
отбрасывает очень густую тень, которая в сочетании с
поверхностной корневой системой этого дерева практи-
чески не позволяет расти под ним никаким другим рас-
тениям. Вы только взгляните вверх в буковом лесу в
солнечный день и сразу увидите листовую мозаику.
Рис. 20. Распределение хлоропластов в клетке листа ряски.
А — на слабом свету; Б — на сильном свету.
Листья многих растений (например, фасоли) способ-
ны поворачиваться на черешках, подставляя плоскость
листа умеренному свету, тогда как края листа оказыва-
ются обращенными к яркому свету.
Углекислый газ воздуха проникает в лист через устьи-
ца и проходит в межклетники. Соприкасаясь с влажными
оболочками клеток мезофилла, углекислый газ раство-
ряется в воде, образуя угольную кислоту, в виде которой
он и достигает хлоропластов.
Как вы помните, воздух содержит 0,03% СО2, т. с.
три части на десять тысяч. И все же, несмотря на столь
низкую его концентрацию, растения ухитряются получить
в достаточном количестве то, что им необходимо для пи-
тания. Поистине лист должен быть чрезвычайно эффек-
тивным органом извлечения углекислого газа из воз-
духа.
Другой вид сырья для фотосинтеза— это вода. Опа
поглощается из почвы и от корней поднимается по сосу-
100
Глава IV
дам или трахеидам ксилемы вверх по стеблю и к листь-
ям, достигая, наконец, через жилки клеток мезофилла.
В листе вода, углекислый газ и хлорофилл встречаются
и, если светит солнце, образуют сахар.
Химия фотосинтеза
Химизм процесса фотосинтеза крайне сложен, и я
скажу о пем только вскользь. В основном его можно
представить таким химическим уравнением:
со,+н2о*=(сн2о)+о;.
Из этого уравнения следует, что углекислый газ сое-
диняется с водой, образуя углевод, который мы условно
изображаем как (СН2О), а кислород выделяется как
побочный продукт. Однако это уравнение — только об-
щепринятый способ выразить реакцию и в сущности дает
очень мало. Во-первых, «углевода» СН2О не существует;
эта формула:—не более чем условное изображение угле-
вода, образующегося в результате фотосинтеза, будь то
глюкоза СвН120б, сахароза С12Н220ц или что-нибудь еще.
Во-вторых, уравнение предполагает, что весь процесс
осуществляется в одну стадию, тогда как на самом деле
этих стадий несколько.
Мы видим, что процесс фотосинтеза нуждается в
энергии света, но это не значит, что свет нужен для каж-
дой его стадии. В прошлом велась оживленная полемика
относительно стадии, на которой, собственно, и использу-
ется световая энергия, — так называемой «световой реак-
ции» в противоположность «темновым реакциям», не
требующим света. Теперь выяснено, что световая реак-
ция— это, вероятно, фотолиз воды, т. е. ее разложение
под влиянием света. Разложение воды можно изобра-
зить так:
нао=н+он,
но это опять-таки всего лишь условное изображение.
Атомы водорода предположительно объединяются с
каким-то веществом, образуя соединения, названные за
легкость, с которой они отдают приобретенные атомы,
Как растение питается
101
донорами водорода; после отщепления водорода такие
соединения снова приобретают способность присоеди-
нять атомы водорода, образующиеся при фотолизе воды.
В клетках растения присутствует вещество, способное
играть роль донора; это никотинамидадениндинуклео-
тидфосфат (сокращенно НАДФ).
От группы ОН молекулы воды тоже надо избавиться.
Это можно представить так:
4ОН=2Н,О+Ог,
хотя, конечно, и здесь все гораздо сложнее, чем пред-
полагается уравнением. Вполне возможно, что сначала
группа ОН образует перекись водорода:
4ОН=2Н2О2,
которая затем разлагается на воду и кислород:
4Н2О2=2Н2О + О2.
Однако более вероятно, что образуется какая-то орга-
ническая перекись, которая потом разлагается.
Но в одном мы можем быть уверены. Весь кислород,
образующийся в ходе фотосинтеза (что видно из первого
приведенного мною в этом разделе уравнения), получа-
ется из воды, а не из углекислого газа.
Взглянув опять на уравнение
СО2 + Н2О=(СН2О) + О2,
мы видим, что гипотетический углевод (CHjO) содержит
меньше кислорода и больше водорода, чем углекислый
газ (СОг), что означает восстановление углекислого газа
в процессе фотосинтеза. Это темновая реакция, не за
висящая от света. Но перед восстановлением должно
произойти «связывание» углекислого газа, т. е. присоеди-
нение его к какому-нибудь веществу в листе. После мно-
гих экспериментальных исследований и споров мы те-
перь достаточно уверены в том, что знаем это вещество:
им, по-видимому, является рибулозодифосфат.
Рибулоза (см. упоминание на стр. 95) сама пред
ставляет собой углевод с химической формулой С5НюО5;
поскольку ее молекула содержит пять атомов углерода.
102
Глава IV
этот сахар относится к пентозам. При соединении мо-
лекулы рибулозы с углекислым газом возникает соедине-
ние, содержащее шесть углеродных атомов, что соответ-
ствует уже молекуле глюкозы (CeHuOe). Поскольку са-
хар глюкоза широко распространен -в органическом мире
и вполне может быть конечным продуктом фотосинтеза,
напрашивается мысль, что с присоединением углекисло-
го газа к рибулозе процесс закапчивается. Но все не так
просто. Дело в том, что эта шестиуглеродная молекула
немедленно расщепляется, образуя две трехуглеродные
молекулы фосфоглицериновой кислоты.
На первый взгляд образование шестиуглеродной мо-
лекулы только для того, чтобы сейчас же расщепиться,
также кажется излишним осложнением. Но дело здесь
гораздо сложнее. Мы ведь еще ни разу не наблюдали
бессмысленной или чересчур расточительной деятельно-
сти растений, и этот случай не исключение. Фосфоглице-
риновая кислота — промежуточный продукт дыхания, в
процессе которого углевод окисляется до углекислого
газа и воды. Дыхание — процесс, противоположный фо-
тосинтезу, и при образовании фосфоглицериновой кисло-
ты эти два процесса перекрещиваются, что таит в себе
огромные биохимические возможности. Начиная от фос-
фоглицериновой кислоты, все вещества могут синтезиро-
ваться сравнительно просто.
Кроме того, мы должны помнить, что расходуемый
в процессе фотосинтеза рибулозодифосфат должен ка-
ким-то образом восполняться, чтобы обеспечить непре-
рывность фотосинтеза. Экспериментальные данные поз-
воляют полагать, что по мере использования запас его
пополняется за счет превращения части образованных
сахаров вновь в рибулозодифосфат.
Фосфоглицериновая кислота, образованная в ходе
фотосинтеза, превращается затем в фосфоглицернновыи
альдегид. В этой реакции, в которой участвует водород,
получившийся при фотолизе воды, фосфоглицериновая
кислота восстанавливается (восстановление — это любая
химическая реакция, сопровождающаяся удалением из
молекулы кислорода или присоединением водорода). Как
вы помните, этот водород присоединяется к НАДФ, об-
разуя восстановленный НАДФ, или НАДФ-Н, который
Как растение питается
103
в свою очередь отдает водород фосфоглицериновой кис-
лоте (говоря химическим языком, он действует как донор
водорода); образующийся после регенерации НАДФ сно-
ва может присоединять водород воды, образуя НАДФ • Н.
Таким образом, этот процесс циклический и теоретически
может продолжаться бесконечно.
Образование фосфоглицерипового альдегида открыва-
ет почти безграничные возможности. Из этого исходного
вещества теоретически может синтезироваться почти все,
и, по-видимому, большинство, если не все сложные ор-
ганические соединения, присутствующие в растениях, так
или иначе ведут свое происхождение от фосфоглицери-
пового альдегида. Нитраты, поглощенные с почвенной
водой, восстанавливаются и вступают в реакцию с угле-
родсодержащими соединениями, образуя аминокислоты,
т. е. те самые кирпичики, из которых потом строятся
белки. Подобным же образом фосфаты доставляют фос-
фатные группы, используемые при построении сложных
нуклеотидов и нуклеиновых кислот, включая универсаль-
ную ДНК (дезоксирибонуклеиновую кислоту), являю-
щуюся основой жизни. Вот почему растения так неза-
висимы: получая СОг и немногие другие простые химиче-
ские вещества из числа тех, что имеются в любой лабо-
ратории, они способны создавать все, что им нужно для
жизни.
В этом очень кратком описании я поневоле крайне
упростил процесс фотосинтеза. Физиология растений не
является предметом данной книги п изложена здесь
лишь постольку, поскольку это необходимо для объясне-
ния структуры растений. Читатели, которые захотят
узнать о фотосинтезе больше, могут обратиться к обшир-
ной специальной литературе по этому вопросу.
В результате фотосинтеза в листьях растений при
дневном свете появляется сахар, который обычно не
накапливается у большинства из них. Некоторое его
количество тотчас же уносится по флоэме проводящих
пучков жилок (гл. V). Однако только сравнительно не-
большая часть сахара успевает уйти из листьев, основная
же масса сахара остается в листьях-и у большинства
растений превращается в крахмал. Это сравнительно
легкий процесс, сопровождающийся удалением воды из
104 Глава IV
молекул глюкозы, которые при этом связываются в боль-
шие «супермолекулы» крахмала. Таким образом, сахар
превращается в нерастворимое вещество, которое бла-
годаря своей нерастворимости не влияет на осмотические
свойства клеток листа.
Крахмал, накапливающийся в листьях днем, получил
название транзиторного, так как ночью он снова превра-
щается в сахар и удаляется из листьев. Этот процесс
будет темой следующей главы.
ГЛАВА V
ТРАНСПОРТНАЯ СИСТЕМА РАСТЕНИИ
Итак, мы уже узнали из предыдущей главы, что
листья представляют собой центры фотосинтеза, в кото-
рых образуется сахар. У простых водорослей не возника-
ет серьезной проблемы транспортировки органических
веществ: сахар у них в основном используется в тех
местах, где синтезируется; только у крупных морских
водорослей можно наблюдать перемещение сколько-ни-
будь значительных количеств веществ из одной части
растений в другую.
С увеличением размеров и усложнением строения
растений передвижение веществ становилось все более
острой проблемой. У деревьев и минеральные соли и
синтезированные питательные вещества должны переме-
щаться с одного конца дерева па другой — на расстоя-
ние, составляющее у Мамонтова дерева или у эвкалипта
60 100 м. Это требует весьма изощренной системы
тр а нспортиров ки.
Передвижение органических веществ
О передвижении воды по растению мы уже говорили
в гл. III. Минеральные соли, поглощенные корнями, в ос-
новном движутся вместе с водой по сосудам и трахеи-
дам. Перемещение же сахаров и других выработанных
в листьях органических соединений — это совсем другое
дело. Вскоре после того, как ботаника стала самостоя-
тельной наукой, было выяснено, что перемещение орга-
нических веществ в растениях — проблема, не связанная
с передвижением воды. Вода и минеральные соли дви-
жутся по ксилеме проводящих пучков, тогда как с са-
харом и другими органическими соединениями все об-
стоит иначе.
Бывает, что вода в сосудах древесины содержит са-
хар— иногда много сахара, например у сахарного клена
(Acer saccharum). В конце зимы концентрация сахара
в ксилемном соке может быть очень высокой, но он нс-
106
Глава V
чезает весной в период развертывания листьев. Это объ-
ясняется тем, что живые паренхимные клетки древесины
накапливают очень много крахмала. Однако ксилема не
может служить главной системой транспорта органиче-
ских веществ. Прежде всего ток веществ в древесине
восходящий, по направлению к листьям, но летом, как
известно, сахара из листьев направляются вниз, к кор-
ням. Едва ли растворы органических веществ могут
передвигаться по ксилеме против
течения.
Теперь известно, что перемещение
органических веществ осуществляется
по флоэме проводящих пучков (гл. II).
Главная роль в этом принадлежит си-
товидным трубкам и клеткам-спутни-
кам, которые составляют основную
массу флоэмы большинства высших
растений.
Ряс. 21. Ситовидная трубка с клетками-спут-
никами.
Зрелая ситовидная трубка представляет собой удли-
ненную клетку, конечные стенки которой пронизаны мно-
гочисленными мелкими отверстиями и называются поэто-
му ситовидными пластинками. Соседние ситовидные
трубки смыкаются концами и сообщаются с помощью
ситовидных пластинок (рис. 21)‘. Ситовидные трубки
имеют тонкие тяжи протоплазмы, но не имеют ядра —
случай весьма необычный. Прилегающие к ним с бо-
ков клетки-спутники уже и заполнены протоплазмой.
В этих клетках имеется и ядро, которое, очевидно, несет
двойную функцию, контролируя метаболизм ситовидных
трубок и клеток-спутников.
1 В современной литературе по анатомии растений принято
называть ситовидными трубками вертикальные ряды клеток;
отдельные клетки в зависимости от степени специализации назы-
вают «ситовидными клетками» и «члениками ситовидных трубок»
по аналогии с трахеидами и члениками сосудов. — Прим, ред.
Транспортная система растений
id?
Многочисленные экспериментальные данные подтвер-
ждают, что передвижение органических веществ происхо-
дит именно по флоэме. Если удалить с дерева кольцо
коры, включающей в себя флоэму (так называемое
«кольцевание»), то легко видеть, что органические ве-
щества, в том числе углеводы и азотистые соединения,
накапливаются над кольцом, в то время как части расте-
ния, расположенные ниже, обнаруживают признаки го-
лодания. Это говорит о том, что с удалением флоэмы
транспорт питательных веществ прекращается. Кроме
того, замечательные и ставшие классическими опыты
с хлопчатником показали, что увеличение содержания
сахара и органического азота в листьях сопровождается
повышением содержания этих веществ в коре, а сни-
жение количества сахара п белкового азота в листьях
немедленно вызывало соответствующее падение их уров-
ня в коре. В древесине же подобных изменений не от-
мечалось.
В последние годы было предложено несколько изящ-
ных способов определения состава флоэмного сока в жи-
вом растении. Один из них заключается в использовании
способа питания тлей — мелких насекомых из отряда
Hemiptcra, куда относятся обычные зеленые и черные
тли. Эти насекомые живут исключительно за счет выса-
сывания сока растений и могут это делать потому, что
их ротовые части превращены в ряд очень тонких стиле-
тов, образующих сосательную трубку. Насекомое по-
гружает стилеты в ткань листа, и стилеты блуждают
там, пока не попадут в отдельную ситовидную трубку.
Тогда насекомое принимается за дело и насыщается
соком растения.
Если оторвать тельце тли от стилета, так чтобы он
остался неповрежденным, то получится тонкая микро-
пипетка, через которую сок из ситовидной трубки про-
должает вытекать, часто в течение 24 ч. Этот сок можно
собрать и проанализировать и таким образом очень
точно определить состав содержимого ситовидной труб-
ки, практически не повреждая растения. Последнее очень
важно, потому что при исследовании в лаборатории рас-
тение может вести себя совершенно иначе, чем в естест-
венных условиях.
108
Глава V
После подобных опытов исчезли всякие сомнения
в том, что именно флоэма служит главным проводником
органических веществ. Что касается минеральных ве-
ществ, то в случае необходимости они тоже могут пере-
мещаться вниз по флоэме (восходящий их ток, как вы
помните, идет по древесине). Некоторые неорганические
вещества, в частности кальций, по-видимому, не могут
передвигаться вниз; достигнув листьев, они остаются
там навсегда.
Многочисленными опытами установлено, что основ-
ным углеводом, который транспортируется по флоэме,
является сахароза. На образовании именно этого сахара
для проведения по флоэме и специализировались цвет-
ковые растения в противоположность водорослям, у ко-
торых флоэмы нет и сахароза так легко не образуется.
Почему сахарозе отдано предпочтение, мы не знаем, но
у нее относительно высокий молекулярный вес, а зна-
чит— низкая осмотическая активность по сравнению с
моносахаридами, например глюкозой; кроме того, опа
легко расщепляется на глюкозу и фруктозу, когда в этих
сахарах возникает необходимость.
Передвижение веществ по флоэме — это явление, ко-
торое можно использовать при применении определенных
гормональных веществ — гербицидов, в частности 2,4-Д,
для борьбы с сорняками. 2,4-Д, сходный с ауксином, пе-
реносится по флоэме с нисходящим током углеводов; по-
этому если опрыснуть этим веществом листья, то можно
быть уверенным, что оно проникнет в подземные части
растения — корни или корневища, где запасаются угле-
воды, и вызовет гибель этих подземных частей, так же
как и листьев. 2,4-Д чрезвычайно полезен, если прихо-
дится иметь дело с такими сорняками, как вьюнок, ко-
торые нельзя искоренить никаким другим путем, по-
скольку даже небольшая часть корневища или корня,
оставшаяся в почве, дает новое растение.
Азот переносится по флоэме в форме аминокислот,
из которых особенно важна аминокислота аспарагин.
Аминокислоты — это азотсодержащие органические кис-
лоты. Объединяясь друг с другом при помощи так на-
зываемых пептидных связей, они образуют более слож-
ные органические соединения. Простейшей из аминокис-
Транспортная система растений 109
лот является глицйй (аминоуксусная кислота) —
CH2(NH2) -COOH. Группа СООН здесь кислотная,- а
NHz-rpynna — основная; следовательно, две молекулы
глицина могут вступить в реакцию с образованием дп-
глицина:
2CH2(NH2)-COOH=CH2(NH2)-CONHCH2-COOH + H2O.
Диглицин тоже представляет собой аминокислоту, по-
тому что его молекула содержит кислотную группу
СООН и аминогруппу ЫН2; следовательно, он может
присоединяться к другой аминокислоте при помощи пеп-
тидной связи. Это может продолжаться бесконечно;
объединяясь друг с другом, аминокислоты образуют
чрезвычайно сложные молекулы. Таким образом строят-
ся белки — сложные азотистые соединения, составляю-
щие основу жизни.
В числе других органических соединений, передви-
гающихся по флоэме, есть некоторые вещества, контро-
лирующие рост и размножение и действующие подобно
гормонам животных. Их и называют растительными гор-
монами. Важнейший среди них — ауксин, регулирующий
рост стеблей и корней. Позднее мы поговорим и о дру-
гих гормонах растений.
Использование питательных веществ
Мы еще не выяснили, куда поступают все питатель-
ные вещества, передвигающиеся по флоэме. Питатель-
ные вещества, синтезируемые в листьях, используются
растением для трех основных целей. Во-первых, они
нужны растущим органам, например точкам роста стеб-
ля и корня, для построения новых тканей. Во-вторых,
все живые клетки растения расходуют питательные ве-
щества при дыхании — процессе, в результате которого
живые существа, как животные, так и растения, получа-
ют энергию. Наконец, в-третьих, пища, в данный момент
являющаяся избыточной, откладывается про запас, как
правило в нерастворимой форме, до тех пор пока опа не
понадобится.
Давайте рассмотрим прорастающее семя. В его се-
менных листьях (семядолях) или в эндосперме (гл. VI)
Глава V
ИО
содержатся значительные запасы, пищи в виде масла или
крахмала, а также большее или меньшее количество
белка. Эти питательные вещества нерастворимы и поэто-
му при прорастании семени прежде всего переводятся
в растворимую форму и перемещаются по флоэме из
мест хранения в места использования — в данном случае
в зону роста. Распад запасных веществ осуществляется
при участии особого класса веществ, называемых фер-
ментами, которые представляют собой органические ка-
тализаторы и способны ускорять химические процессы,
сами при этом не изменяясь.
Различные ферменты действуют на разные классы
питательных веществ. Диастаза, например, переводит
крахмал в мальтозу, липаза разлагает жиры на глице-
рин и жирные кислоты, а протеазы способствуют распаду
белков до аминокислот. Продукты распада затем транс-
портируются в зоны роста, где происходит образование
новых тканей и дыхание наиболее активно.
При созревании плодов имеет место обратное явле-
ние. В этом случае растворимые вещества, транспорти-
руемые по флоэме из активно фотосинтезирующих листь-
ев, переводятся в нерастворимую форму и откладывают-
ся в еще незрелых семенах молодых плодов.
С приближением осени для запасающих органов
всегда наступает горячая пора (это время можно срав-
нить по активности только с наступлением весны). Фо-
тосинтез при этом замедляется и некоторые минеральные
вещества, находящиеся в листьях, уходят из них и воз-
вращаются в эти хранилища. В листьях папоротника
орляка, например, летом накапливается много углекис-
лого калия, но зимой большая часть его возвращается
в подземное корневище, где и хранится до следующей
весны.
Однако не все минеральные вещества запасаются.
Как я уже упоминал, кальций, попав в листья, остается
в них навсегда. В этом есть свой смысл, поскольку в про-
цессе метаболизма растений образуется некоторое ко-
личество щавелевой кислоты, являющейся сильным ядом.
У животных она немедленно выводится из организма
вместе с другими ненужными веществами, но у растений
выделение, характерное для животных, вряд ли возмож-
Транспортная система растений Ш
но из-за отсутствия выделительной системы. Вредная
щавелевая кислота соединяется с кальцием, образуя
оксалат кальция, который можно часто видеть в клетках
листа в виде кристаллов. Благодаря своей нераствори-
мости это соединение не может участвовать в обмене
веществ и потому его присутствие в клетках не опасно
для растения.
Точно так же двухлористая ртуть (сулема) представ-
ляет собой смертельный яд для животных, тогда как
хлорид ртути (каломель) гораздо менее опасен. Это
происходит потому, что двухлорнстая ртуть растворима
в воде, а хлорид ртути практически нерастворим. Когда
листья растений сбрасываются при листопаде, вместе
с ними удаляются и кристаллы оксалата кальция. Та-
ким образом, опасная щавелевая кислота окончательно
исчезает из растения.
Из этого не следует, что растения вообще не способ-
ны к экскреции; по крайней мере у некоторых из них
происходит очень активное выделение через корни. Иног-
да выделения корней используются различными парази-
тами растений, ожидающими возможности напасть на
них, как, например, в случае с картофельной нематодой,
где эти выделения служат своеобразным сигналом к на-
чалу атаки на растение.
Иногда говорят, что растение «выделяет» углекислоту
через корни, и в качестве доказательства ссылаются на
то, что корни, соприкасающиеся с оказавшимися в почве
кусками мрамора, оставляют на них свои отпечатки. Но
это нельзя назвать выделением в прямом смысле, так
как углекислота — это обычный продукт дыхания.
Механизм передвижения веществ по флоэме пока
недостаточно ясен. Теория массового потока, предложен-
ная в 1930 году Мюнхом, считается принятой, но только
потому, что пока кет ничего лучшего. В основе ее лежит
представление о флоэме как о непрерывной осмотической
системе, проходящей через все растение. В местах синте-
за органических веществ, например в листьях, поступле-
ние асспмилятов в проводящую систему представляет
собой активный процесс, требующий энергии дыхания.
Это значит, что в тех участках флоэмы, где происходит
ее «загрузка», должно существовать повышенное осмоти-
112 Глава V
ческое давление. В других же частях растения, где флоэ-
ма «разгружается:», она отдает питательные вещества
нуждающимся в них клеткам. Здесь осмотическое давле-
ние должно быть пониженным. Вода поступает, туда,
где осмотическое давление высокое (в места «загрузки»),
и выходит там, где оно низкое (в местах «разгрузки»).
В результате этого поступления и выхода воды во флоэ-
ме создается массовый поток от «загрузочных» к «раз-
грузочным» пунктам. Вода, конечно, поступает из ксиле-
мы и потом возвращается в нее, покидая флоэму.
В теории Мюнха много «тонких мест», но пока никто
ее не опроверг. Если бы удалось показать, что какое-то
вещество или два разных вещества могут передвигаться
в противоположных направлениях по одной и той же
ситовидной трубке, эта теория тотчас же оказалась бы
несостоятельной, но до сих пор никто этого не сделал.
ГЛАВА VI
ПРОБЛЕМА РАЗМНОЖЕНИЯ
Высшие растения размножаются половым путем
при помощи семян, или, правильнее сказать, семя яв-
ляется результатом полового процесса. Под высшими
здесь имеются в виду семенные растения — цветковые
и их более скромные сородичи — хвойные. Низшие рас-
тения — папоротники, папоротникообразные, мхи, пече-
ночники, водоросли и грибы1 — размножаются другими
способами, без образования семян. В этой главе речь
пойдет в основном о цветковых растениях;
Части цветка
Цветки чрезвычайно разнообразны по форме, ок-
раске, размерам и степени сложности, но, к счастью
для систематиков растений, это, так сказать, всегда
вариации на одну тему. Типичный цветок состоит из че-
тырех серий элементов, прикрепленных к цветоложу —
обычно несколько вздутому концу цветоносного стебля.
Первое, что мы видим, — это чашелистики, со-
ставляющие наружные покровы цветка. Чашелисти-
ки обычно зеленые, и пока цветок не распустился, они
закрывают его и защищают от повреждений. Впрочем,
иногда они бывают лепестковидными, т. е. окрашены
подобно лепесткам, и тогда их трудно или невозможно
различить. У некоторых цветков чашелистики сраста-
ются вместе, тогда как у других они могут быть раз-
дельными. В совокупности чашелистики образуют ча-
шечку (рис. 22).
Во-вторых, мы видим лепестки. В большинстве слу-
чаев (но не всегда) они очень эффектно и ярко окра-
шены и благодаря этой яркости являются главными
1 Точка зрения автора не разделяется большинством ботаников.
К низшим растениям относят бактерии, водоросли, слизевики,
грибы и лишайники, а мхи, в том числе печеночники,'и все папорот-
никообразные причисляют к высшим. — Прим. ред.
114
Глава VI
органами, привлекающими насекомых-опылителей. По-
добно чашелистикам, лепестки могут срастаться или
быть свободными. Совокупность лепестков называют
венчиком.
Внутри венчика мы видим тычинки. Это мужские
элементы цветка, образующие пыльцевые зерна (мик-
роспоры), с помощью которых цветки опыляются. Ты-
чинка обычно состоит из пыльника, в котором содер-
жится пыльца, прикрепленного к ножке, называемой
тычиночной нитью. В своей совокупности тычинки об-
разуют андроцей.
Рис. 22. Схематическое изображение цвегка, разрезанного попо-
лам, чтобы показать расположение его частей.
Наконец, в центре цветка находится гинецей. Это
женская часть цветка. Гинецей образован одним или
(обычно) несколькими плодолистиками, а каждый из
них состоит из трех частей. Главная часть — это полая
завязь, над которой находится образование, назы-
ваемое рыльцем; оно прикреплено к завязи при помо-
щи длинной или короткой ножки — столбика. Обычно
плодолистики, образующие гинецей, срастаются вме-
сте, составляя сложную структуру, от вершины кото-
рой отходит один или несколько столбиков, но у неко-
торых цветков, например у лютика и магнолии, они
обособлены.
Хотя термины «завязь» и «гинецей» часто употреб-
ляют на равных правах, но это не вполне синонимы.
Строго говоря, завязь — это лишь пижняя полая часть
плодолистика (или плодолистиков), тогда как понятие
Проблема размножения
115
«гинецей» относится ко всему образованию. Гинецей
цветка сначала называли пестиком, и это старое назва-
ние еще существует.
Внутри завязи можно видеть от одного до очень
многих мелких круглых телец — это семяпочки*. Позд-
нее они превратятся в семена, но, прежде чем это слу-
чится, цветок должен быть опылен, а семяпочка опло-
дотворена.
В цветке могут присутствовать и различные второ-
степенные части. Среди них особенно важны нектарни-
ки — участки железистой ткани, выделяющие нектар —
сахаристую жидкость, которая привлекает пчел и дру-
гих насекомых и тем самым способствует опылению.
Цветок без нектарников все же может опыляться насе-
комыми, так как их привлекает не только нектар. Пче-
лы, например, великие собиратели пыльцы, и цветки
без нектара, посещаемые этими насекомыми ради
пыльцы, называют «пыльцевыми цветками».
Часто непосредственно под цветком имеется неболь-
шой лист, называемый прицветником.
Типичный цветок состоит из чашечки, венчика, анд-
роцея и гинецея, но не всегда все эти части имеются
в наличии. Очень часто венчик или чашечка отсутст-
вуют, и в этом случае обычно принято называть остав-
шуюся часть околоцветником. Термин «околоцветник»
употребляется и по отношению к чашечке и венчику
вместе, если они присутствуют.
Цветок, содержащий андроцей и гинецей, т. е.
и мужскую и женскую части, называется обоеполым.
Большинство цветков именно таковы, но некоторые бы-
вают однополыми. Они лишены или апдроцея, или
гинецея. Собственно говоря, такие цветки можно назы-
вать мужскими или женскими. Иногда мужские
и женские цветки находятся на одном и том же расте-
нии, которое в этом случае называется однодомным-,
двудомными называют растения, образующие цветки
только одного типа.
* В последнее время в русской литературе часто употребляется
термин «семезачатки>, что лучше отражает действительное поло-
жение вещей. — Прим. ред.
116
Глава VI
Как я уже упоминал в гл. II, цветковые растения
можно разделить на две группы — двудольные и одно-
дольные, что зависит от числа семядолей. Точность
этого подразделения подтверждается и другими призна-
ками. Двудольные имеют листья с‘ сетчатым жилкова-
нием, одно кольцо проводящих пучков в стебле и, как
правило, способны к вторичному росту посредством
деятельности камбия (гл. II). Однодольные обычно
имеют лентовидные листья с параллельным жилкова-
нием, проводящие пучки разбросаны по всему стеблю,
и, за редким исключением, они не обнаруживают вто-
ричного роста.
Различие между двудольными и однодольными про-
является и в цветках. Мы обычно узнаем цветки дву-
дольных потому, что каждый круг частей цветка — ча-
шелистики, лепестки, тычинки и плодолистики — пред-
ставлен четырьмя или пятью членами или числом, крат-
ным четырем или пяти. В цветках однодольных число
этих членов равно или кратно трем.
Разумеется, из этих общих правил есть немало
исключений; ботаника была бы наполовину забавой,
если бы строение растений подчинялось нескольким
строгим правилам. Я иногда смущаю начинающих сту-
дентов: показываю им травянистое растение вороний
глаз (Paris quadrifolia) и спрашиваю их, двудольное
это или однодольное. Это растение имеет широкие
листья с сетчатым жилкованием, члены частей цветка
у него в числе, кратном четырем, но это превосходный
представитель однодольных, принадлежащих к семей-
ству Trilliaceae, родственному лилейным *. Вещи не
всегда таковы, какими они кажутся.
Опыление и оплодотворение
«
Прежде чем цветок даст семена, должно произойти
его оплодотворение, а еще раньше — опыление; опыле-
ние и оплодотворение отнюдь не одно и то же. Акт
1 Во многих руководствах и определителях растений вороний
глаз относят к семейству лилейных (Liliaceae) в более широком его
понимании. — Прим. ред.
Проблема размножения „117
опыления заключается в переносе пыльцы из тычинок
того же или другого цветка на рыльце. Мы не ошибем-
ся, если скажем, что цветок (как структура) развился
в связи с необходимостью обеспечить опыление (обыч-
но пыльцой с другого цветка — перекрестное опыле-
ние). Некоторые цветки, в частности у пшеницы, вооб-
ще опыляются их собственной пыльцой (самоопыле-
ние), но они составляют исключение.
Пыльник „тычинки состоит из 4 полостей, или пыль-
цевых мешков, в которых лежат пыльцевые зерна. При
созревании он вскрывается двумя продольными щеля-
ми, обнажая пыльцу. Обычно, особенно у опыляемых
насекомыми (энтомофильных) цветков, пыльца остает-
ся в пыльнике, пока не будет удалена насекомыми,
слегка задевающими тычинки; у ветроопыляемых (ане-
мофильных) цветков она высеивается при раскачива-
нии пыльника ветром. Иногда, в частности у цветков
последнего типа, она удаляется при помощи специаль-
ного устройства. Хорошо известным примером расте-
ния, имеющего такое устройство, является постенница
(Parietaria diffusa)1. Это растение, растущее в трещи-
нах старых стен и в тенистых местах, образует муж-
ские и женские цветки. В нераскрывшемся мужском
цветке эластичные тычиночные нити загнуты внутрь
(это обеспечивается имеющими форму лодочек листоч-
ками околоцветника). При раскрывании цветка тычи-
ночные нити выпрямляются и благодаря этому резкому
движению пыльники лопаются и пыльца выбрасывается
в виде маленького облачка.
Внешние особенности пыльцевых зерен варьируют
от вида к виду настолько сильно, что часто можно
легко сказать, какому растению принадлежит пыльца,
исследуемая нами под микроскопом. Таким образом
удалось получить много данных о распределении дре-
весной растительности в прошлые времена, так как
пыльцевые зерна, сохранившиеся в залежах торфа,
можно идентифицировать через многие тысячи лет. По
форме пыльцевые зерна обычно круглые или овальные,
1 У нас несколько видов Parietaria есть на Кавказе. — Прим,
ред.
Ив
Глава Vi
хотя имеются и некоторые исключения. У морской
травы (Zostera), например, они нитевидные и обматы-
ваются вокруг рыльца женского цветка. К роду Zoste-
ra относятся морские цветковые растения, растущие
неглубоко под водой в прибрежной полосе, поэтому
в данном случае столь необычная форма пыльцы, поз-
воляющая ей держаться на поверхности воды, вполне
оправданна.
Размеры пыльцевых зерен в пределах какого-то од-
ного вида растений довольно постоянны, однако у раз-
личных видов очень сильно колеблются. Так, пыльца
незабудки имеет 0,0025 мм в диаметре, тогда как
у огурца она в 100 раз больше. Поверхность пыльце-
вых зерен может быть гладкой или неровной. У ветро-
опыляемых растений она большей частью гладкая,
а у опыляемых насекомыми — обычно неровная. Эта
неровная поверхность помогает пылинкам прикреплять-
ся к насекомым, а также способствует их слипанию.
Иногда, как у некоторых представителей Onagraceae
(кипрейных), на пыльцевых зернах имеются нити из
клейкого вещества, помогающие им соединяться друг
с другом.
Пыльца, перенесенная на рыльце цветка, оседает
там, чему часто способствует неровность воспринимаю-
щей поверхности, которая к тому же обычно бывает
клейкой; липкая жидкость, выделяемая рыльцем, не
только помогает улавливанию и удержанию пыльцевых
зерен, но, возможно, ускоряет н их прорастание.
Пыльца разных растений может прорастать в раз-
ных условиях. У одних видов она очень легко прора-
стает в растворе сахара или в некоторых случаях
в воде. Допустимая концентрация раствора при этом
сильно варьирует: для пыльцы яблони, например, она
составляет 2,5...10%, у подснежника пыльца прораста-
ет лучше при концентрации 1...3%, а у одного из видов
кассии может прорасти в сахарном растворе 70%-ной
концентрации. Пыльца примулы прорастает в дистил-
лированной воде, но не прорастает в водопроводной.
В некоторых случаях для прорастания пыльцы необхо-
димо наличие веществ, выделяемых рыльцем, в част-
ности прорастания пыльцы Mussaenda можно добить-
Проблема размножения
119
ся, лишь поместив в воду кусочки ткани рыльца; рыль-
цевую ткань можно заменить сахаром фруктозой, Но
ничем больше. Известно также много случаев, когда
пыльца прорастает только на рыльце цветка того же
или близких видов.
Нетрудно видеть пользу того, что пыльца может
прорастать в пределах строго определенных концен-
траций сахара или каких-либо дру-
гих веществ. Это препятствует ее
преждевременному прорастанию
еще до того, как она достигнет
рыльца, и позволяет, таким обра-
зом, избежать излишней траты
пыльцы. Однако число видов рас-
тений, имеющих пыльцу, которая
прорастает в воде, по-видимому, го-
ворит о том, что преимущество это
невелико.
Рис. 23. Прорастающее пыльцевое зерно
с пыльцевой трубкой.
Пыльцевое
зерно
Пыльцевая
трубка
Мужские
гаметы
При прорастании пыльцевого зерна образуется
пыльцевая трубка, которая проникает в ткань рыльца
(рис. 23). Способы, с помощью которых она проходит
сквозь плотную ткань рыльца, у разных видов различ-
ны. У некоторых из них пыльцевая трубка растет не-
посредственно через клетки, растворяя их ферментами,
которые выделяет, подобно тому как это происходит
при проникновении гиф паразитного гриба Pythium в
ткани растения. В других случаях клетки рыльца рас-
положены сравнительно рыхло и не оказывают боль-
шого сопротивления пыльцевой трубке, которая про-
кладывает себе путь между ними. Наконец, в самом
рыльце могут быть специальные каналы, по которым и
растет пыльцевая трубка. Конечной целью ее продви-
жения является завязь.
120
Глава VI
Для роста пыльцевой трубки необходимо что-то,
что направляло бы ее путь, который в цветках с длин-
ным рыльцем и столбиком может быть действительно
очень длинным для такого маленького образования.
Как правило, эта направляющая сила носит двоякий
характер: трубка стремится удалиться от воздуха
и приблизиться к некоторым химическим веществам.
Если пыльцевые зерна проращивать на поверхности
желатины, их пыльцевые трубки сразу же изогнутся
прочь от окружающего воздуха, в глубь желатины.
А при наличии кусочков рыльцевой ткани они будут
расти к ним. В большинстве случаев химическим ве-
ществом, привлекающим пыльцевые трубки, является
сахар, но не всегда; у нарцисса (Narcissus tazetta)
и некоторых других растений таким активным вещест-
вом оказался белок.
Достигнув полости завязи, пыльцевая трубка растет
по направлению к семяпочке и проникает в нее обычно
через небольшое отверстие, называемое микропиле.
Вблизи кончика пыльцевой трубки находятся два ядра;
это мужские половые ядра, или гаметы1, которые оп-
лодотворяют семяпочку. Закладывается семяпочка
в виде бугорка ткани, формирующего нуцеллус семя-
почки— ткань, которая впоследствии бывает окружена
наружным покровом, или интегументом (их может
быть один или два). Этот покров возникает потом как
кольцеобразный вырост из основания семяпочки. Инте-
гумент (или интегументы) обрастает семяпочку, окру-
жая весь нуцеллус, за исключением одного места на
вершине семяпочки, где эти покровы не смыкаются,
так что остается отверстие, через которое открывается
поверхность нуцеллуса. Это и есть микропиле, и через
него пыльцевая трубка проникает в семяпочку.
В начале роста семяпочки тот ее конец, где должно
образоваться микропиле, направлен от семяносца, или
плаценты (части завязи, где располагается основание
семяпочки). У некоторых растений семяпочка при сво-
ем развитии удерживает первоначальное положение,
у большинства же она переворачивается, так что мик-
Dl|!lll|! к стр. 23.
Проблема размножения
121
ропиле в конце концов оказывается расположенным
почти возле ножки семяпочки и обращенным к плацен-
те. Пока все это происходит, в нуцеллусе семяпочки,
у микропилярного конца, дифференцируется клетка,
называемая материнской клеткой мегаспор, В дальней-
шем она делится дважды перпендикулярно поверх-
ности семяпочки, образуя ряд из четырех клеток. Три
famunofa
’Яйцеклетка.
Синергий
Микропиле '•
Рис. 24. Схема разреза семяпочки, показывающая зародышевый
мешок.
из них впоследствии разрушаются, а четвертая (обыч-
но самая внутренняя) становится известной как функ-
ционирующая мегаспора, на более поздней стадии на-
зываемая зародышевым мешком (фото XVI). В ходе
развития эта клетка значительно увеличивается в раз-
мерах, используя при этом большую часть нуцеллуса.
Сначала она содержит только одно ядро, но затем оно
делится, и образующиеся при этом два дочерних ядра
расходятся к противоположным концам клетки. Затем
они делятся дважды и на каждом конце зародышевого
мешка возникает по 4 ядра. Из четырех ядер, располо-
женных па конце, удаленном от микропиле, три отде-
ляются клеточными оболочками и называются тогда
антиподами, а четвертое перемещается к центру заро-
1»
Глава VI
дышевого мешка. Из четырех же ядер на микропиляр-
ном конце вокруг трех скопляется протоплазма и они
образуют яйцевой аппарат, состоящий из яйцеклетки
и двух синергид, или вспомогательных клеток. Остав-
шееся ядро направляется к центру зародышевого меш-
ка, где оно встречается с ядром, переместившимся
с другого его конца. Эти два ядра называют полярны-
ми ядрами; в конечном счете они сливаются и образу-
ют первичное ядро эндосперма (рис. 24).
Происходящие в зародышевом мешке события
варьируют в деталях у разных растений (слияние двух
полярных ядер, например, может задержаться), но эти
различия не меняют общей картины.
Бесполое размножение (спорами)
Как вы помните, в жизненном цикле папоротника
имеются два поколения — дающий споры спорофит
и гаметофит, да котором возникают половые органы.
Спорофит (т. е. само растение папоротника) размно-
жается бесполым путем при помощи спор, которые,
прорастая, развиваются в заросток (маленькое ра-
стеньице, не больше 0,6 см в поперечнике). Несущий
половые органы заросток представляет собой гамето-
фит, и в результате его полового размножения вновь
образуется растение папоротника. Таким образом цикл
развития, который мы уже описывали на стр. 17, за-
вершается.
У некоторых растений, родственных папоротникам,
например у селягипеллы (Selaginella), образуются два
типа спор, которые при прорастании дают гаметофиты
двух родов. Более мелкие споры (микроспоры) про-
растают, давая начало мужскому гаметофиту, который
в данном случае настолько редуцирован, что состоит
лишь из нескольких клеток. Более крупные споры
(мегаспоры) образуют при прорастании небольшую
«кучку» зеленой ткани, на которой развиваются женские
половые органы. Сперматозоиды, образуемые мужским
гаметофитом, оплодотворяют яйцеклетки на женском
гаметофите, и таким образом образуется новое расте-
ние Selaginella.
Проблема размножения
123
У цветкового,” или покрытосеменного, растения
гаметофит гораздо более редуцирован, хотя его еще.
можно различить. Подобно Selaginella, цветковое рас-
тение имеет два вида спор. Пыльцевые зерна представ-
ляют собой микроспоры, но редукция мужского гамето-
фита зашла так далеко, что его, по сути дела, не су-
ществует1; просто в кончике пыльцевой трубки находят-
ся две мужские гаметы и еще одно ядро, называемое
ядром трубки. Женский гаметофит представлен зароды-
шевым мешком в семяпочке. Ни женский, ни мужской
гаметофит нельзя даже с натяжкой назвать растением.
Преимущество, полученное благодаря этой предель-
ной редукции гаметофитного поколения, заключается
в том, что половое размножение цветкового растения
оказывается совершенно независимым от воды. В цик-
ле его развития уже нет нежного гаметофита, который
может существовать только во влажных местах, а уст-
ранение из этого цикла плавающих сперматозоидов
приводит к тому, что зависимость от воды полностью
ликвидируется. Благодаря такому шедевру структуры,
как семена, цветковое растение может размножаться
половым путем даже в пустыне.
Размножение голосеменных
Другая большая группа семенных растений, о кото-
рой я до сих пор говорил мало, — это голосеменные;
в эту группу входят такие известные хвойные деревья,
как сосна, пихта, ель, лиственница и кедр, а также
множество интересных, но менее знакомых нам расте-
ний. Голосеменные появились задолго до цветковых рас-
тений. Первые ископаемые остатки голосеменных обна-
ружены в позднем девоне, т. е. они жили почти 300 млн.
лет назад; первые цветковые растения появились лишь
спустя 100 млн. лет.
У голосеменных нет цветков. Вместо этого они фор-
мируют женские и мужские шишки, состоящие из че-
шуек, спирально расположенных вокруг центральной
х Зрелое пыльцевое зерно обычно представляет собой уже п;> >
росшую микроспору, т. е. содержит редуцированный мужской • i
метофит. — Прим. ред.
124
Глава VI
оси, и несущие либо пыльиевые мешки, либо семяпочки
в зависимости от пола шишки. Семяпочки голосемен-
ных не заключены в плодолистики, как у цветковых,
или покрытосеменных, растений, а лежат открыто на
чешуйках, что составляет важнейшее различие между
этими группами. Своим названием голосеменные тоже
обязаны этой особенности развития. При опылении
пыльцевое зерно должно попасть непосредственно на
семяпочку, проникнуть в нее через микропиле и обра-
зовать затем пыльцевую трубку.
Голосеменные являются менее совершенными расте-
ниями, чем цветковые. Хотя преимущество, полученное
благодаря образованию семян, делает их размножение
независимым от воды, все же этот процесс у них не так
хорошо специализирован, как у цветковых. У голосе-
менных семяпочки нс заключены в плодолистики, поэ-
тому они менее защищены, чем семяпочки цветковых
растений, а сам факт отсутствия плодолистиков озна-
чает, что у них нет плодов, благодаря чему растения
лишаются всёх тех возможностей расселения, которые
дает наличие плодов. Голосеменные — более ранняя
«модель», чем цветковые растения, и их «конструкция»
еще имеет недостатки, которые до сих пор не устране-
ны. В частности, длинный путь их эволюции начался
раньше, чем появились насекомые, так что их опыление
еще зависит от ветра.
Тем не менее не следует думать, что голосемен-
ные — это какие-то неудачники. Они широко распро-
странены во всем мире, а дугласова пихта и калифор-
нийское Мамонтове дерево относятся к числу самых
высоких деревьев. В северных широтах они доминируют
над всей остальной растительностью н не обнаружива-
ют никаких признаков вымирания.
Развитие семени
Оплодотворенная семяпочка развивается в семя.
Процесс оплодотворения заключается в том, что одно
из двух мужских ядер, находящихся в пыльцевой труб-
ке, сливается с ядром яйцеклетки. У голосеменных это
единственное слияние ядер, которое имеет место, так
Проблема размножения
125
как второе мужское ядро не нужно и оно обычно раз-:
рушается вскоре после того, как его собрат оплодотво-
рит семяпочку. Однако у цветковых растений оплодо-
творение двойное. Одно -из мужских ядер сливается,
у них с ядром яйцеклетки, а другое — с первичным яд-
ром эндосперма1. В ходе дальнейшего развития семя-
почки в семя образовавшееся в результате последнего
слияния ядро многократно делится, формируя ткань,
называемую эндоспермом. В клетках эндосперма нахо-
дится много питательных веществ, и он служит для пи-
тания зародыша на ранних стадиях развития.
После оплодотворения семяпочка увеличивается
в размерах и превращается в семя. Из оплодотворен-
ной яйцеклетки развивается зародыш. Его верхняя
часть у двудольных состоит из двух семядолей, или
семенных листьев, с почечкой, заключенной между ни-
ми; у однодольных развивается только одна семядоля
и почечка занимает по отношению к ней боковое поло-
жение. В нижней части зародыша у обеих групп расте-
ний появляется зародышевый корешок. Между зароды-
шем и микропиле формируется структура, называемая
подвеском. Подвесок состоит из множества плоских
клеток и, удлиняясь, толкает развивающийся зародыш
по направлению к центру семени, где он окружается эн-
доспермом, снабжающим его питательными веществами.
По мере своего развития зародыш потребляет эндо-
сперм, причем в разной степени у разных растений.
У многих двудольных он расходует весь эндосперм
и семядоли поэтому становятся толстыми и мясистыми.
Набухшие семядоли составляют основную массу зрело-
го семени, и когда оно прорастает, запасные питатель-
ные вещества семядолей используются проростком, да-
вая ему возможность пережить то время, пока листья
его достаточно не разовьются, чтобы питать его за счет
фотосинтеза.
У других двудольных и большинства однодольных
дело обстоит иначе. Семядоли у них остаются тонкими,
а основная масса запасных питательных веществ сох-
1 В ботанической литературе это ядро обычно называют <вто-
ричным ядром зародышевого мешка» (по происхождению). —
Прим. ред.
126
Глава VI
ранястся в эндосперме вплоть до прорастания, когда
они идут на питание проростка. Примером такого типа
растений из двудольных является, в частности, кле-
щевина, а из однодольных — пшеница и кукуруза,
у которых также большое количество запасенной пищи
сохраняется в эндосперме до прорастания.
В то время, когда все это происходит внутри семе-
ни, интегумент превращается в твердый наружный пок-
ров, или семенную кожуру, которая окружает его. На-
конец семя созревает и переходит в состояние покоя.
В этот период все жизненные процессы приостанавли-
ваются, рост прекращается и даже дыхание, присущее
всем живым клеткам, замедляется до такой степени,
что его не удается обнаружить. Семя остается в покое,
пока прорастание снова не пробудит его к жизни.
Продолжительность периода покоя семян очень раз-
лична. У некоторых тропических мангров он совсем
отсутствует. Семя у них прорастает, как только оно об-
разовалось на .родительском растении, и когда осво-
бождается из плода, то стрелой летит вниз, зарываясь
в ил тем концом, где находится молодой корешок. На-
против, семена других растений, прежде чем они про-
растут, нуждаются в длительном периоде покоя.
Прорастание
Для прорастания всех семян необходимы влага,
подходящая температура и кислород для дыхания. Не-
которым семенам нужен еще свет (или темнота). Семе-
на белой омелы (Viscum album), а также некоторых
сортов салата прорастают только на свету. С другой
стороны, семена лука надо держать в темноте, потому
что свет замедляет или даже делает совсем невозмож-
ным их прорастание.
В чем именно заключается влияние света, неизвест-
но, но, во всяком случае, оно никак не связано с фото-
синтезом, так как на этой стадии он еще не начинает-
ся. Можно только предположить, что некоторые тонкие
физиологические процессы, протекающие в семени, ка-
ким-то образом нарушаются, если семя получает слиш-
ком много света в одних случаях или слишком мало-—
Проблема размножения 12?
в других. Некоторые семена, прорастающие в темноте
при низких температурах, при повышении температуры
начинают нуждаться в свете. Если семена того сорта
салата, который обычно прорастает в темноте, обрабо-
тать кумарином (веществом, тормозящим прорастание),
свет становится обязательным условием их прораста-
ния. Это наводит на мысль, что влияние света на «све-
толюбивые» семепа заключается в разрушении какого-
то ингибитора прорастания, присутствующего в семени.
Если при наличии всех необходимых условий, вклю-
чая свет (или темноту), семя не прорастает, мы знаем,
что оно находится в состоянии покоя. Это семя может
прорасти и прорастет, когда придет время, но, значит,
время это еще не наступило. Зародыш остается заклю-
ченным в семенную кожуру и ждет чего-то, что должно
случиться и освободить его из «тюрьмы», чтобы он мог
начать развиваться в новое растение.
Иногда покой может быть обусловлен чисто физи-
ческими причинами, например непроницаемостью се-
менной кожуры, препятствующей проникновению воды.
Именно это наблюдается у части семян душистого го-
рошка. Если перед тем, как сажать их, нанести на се-
менную кожуру (пилочкой) царапины, они прорастают
гораздо быстрее. В других случаях, как, например,
у дурнишника (Xanthium), через семенную кожуру не
может проникнуть кислород. Кроме того, эта кожура
может быть настолько твердой, что, пока она не раз-
мягчится в результате гниения, находящийся в заточе-
нии зародыш не может развиваться. В природе почвен-
ные бактерии и грибы успешно разрушают эти непо-
датливые покровы семян.
Твердая семенная кожура далеко не единственная
причина, обусловливающая период покоя. Бывает, что,
в то время как семя во всех других отношениях вполне
созрело, сам зародыш еще недостаточно развит и поэ-
тому не способен к прорастанию1. Такие семена, оче-
1 О разнообразии семян, содержащих недоразвитые зародыши,
и о процессах,- происходящих перед их прорастанием, можно про-
читать п интересной книге И. В. Грушвицкого «Роль недоразвития
зародыша в эволюции цветковых растений» (из серии «Комаровские
чтения»), Изд-во АН СССР, М. — Л., 1961. — Прим. ред.
128
Глава VI
видно, требуют дополнительного периода развития; для
некоторых представителей семейства орхидных это
обычное явление.
В некоторых случаях семена, содержащие ко време-
ни созревания вполне развитый зародыш, все же не
прорастают, пока не пройдет определенный период
времени (послеуборочное дозревание). У розоцветных,
например, он может длиться около шести месяцев.
Благодаря этой особенности семена, образовавшиеся
осенью, не прорастают до следующей весны, что весь-
ма ценно для растения, так как, если кратковременные
зимние оттепели вызовут прорастание семян, проростки
могут погибнуть при первом же заморозке.
Пока не совсем ясно, что же происходит в проме-
жуток времени между созреванием семени и его про-
растанием. Иногда семя содержит тормозящее рост
вещество, которое должно быть удалено до того, как
может начаться прорастание. Для некоторых растений
пустыни необходим дождь, чтобы вымыть этот ингиби-
тор. Такие семена прорастают только после сильного
дождя; быстро проходящего же ливня для вымывания
ингибитора недостаточно. Это опять-таки, что совер-
шенно очевидно, выгодно для растений, так как, если
семя прорастало бы после первого ливня, проростки
могли бы засохнуть, когда дождь прекратится. Только
сильный дождь, достаточный для того, чтобы обеспе-
чить последующий рост растения, вызывает прораста-
ние семепи.
Для прорастания других семян, как оказалось, нуж-
ны вещества, стимулирующие рост. Образование этих
веществ вызывается различными внешними факторами,
особенно длительным воздействием низких температур
в условиях влажности. К этому типу семян относятся
семена яблони. И в этом случае условия, необходимые
семени для его прорастания, точно совпадают с теми,
в которых оно находится во время обычной зимы.
Жизнеспособность семян
Долговечность семян — это вопрос, который оказал-
ся сильно запутанным благодаря многим преувеличен-
ным и вводящим в заблуждение данным прошлых лет.
Проблема размножения
129
При достаточно долгом хранении (даже в идеальных
условиях) все семена в конце концов теряют способ-
ность к прорастанию, но у разных видов растений семе-
на сохраняют всхожесть в течение весьма различного
времени. Лишь немногие из них прорастают сразу пос-
ле опадения. Семена кислицы (Oxalis aceiosella), на-
пример, не могут противостоять высыханию; семена са-
говников также, как оказалось, крайне недолговечны.
Напротив, семена других растений сохраняют жизне-
способность в течение многих лет; в частности, щавель
курчавый (Rurnex crispus) прорастает через 80 лет.
Наибольшая же продолжительность жизни, о которой
имеются достоверные данные, свойственна семенам
индийского лотоса (Nelumbo nucifera). В 1940 году во
время бомбежки Британского музея семена этого
растения, хранившиеся в гербарии, были подмочены
и проросли, несмотря на то что, судя по дате на гер-
барном листе, их возраст составлял 237 лет.
Проросли также семена индийского лотоса, обна-
руженные в торфяных отложениях Маньчжурии. Их
возраст, возможно, достигает и 400 лет, но об этом
нельзя говорить с уверенностью, так как возраст тор-
фяных отложений точно не установлен. История о се-
менах индийского лотоса, якобы проросших через
4000 лет (публикация этих данных наделала много
шума), точно так же как и широко распространенное ут-
верждение о прорастании семян пшеницы из египетских
гробниц, явно неправдоподобна. Семена пшеницы, как
известно, сравнительно недолговечны и едва ли могли
бы сохранить всхожесть даже по прошествии в 100 раз
меньшего срока.
Недавно появилось сообщение о том, что семена
индийского лотоса, 10000 лет пролежавшие в норах
леммингов, сохранили способность к прорастанию.
Если данные об их возрасте подтвердятся, мы вынуж-
дены будем пересмотреть наши представления о долго-
вечности семян.
Растения размножаются отнюдь не только семена-
ми. Каждый садовод знает, что многие растения раз-
множаются луковицами, клубнелуковицами, клубнями,
отпрысками, черенками и т. д. Все многочисленные
130
Глава Vt
способы размножения растений, исключающие половой
процесс, приводящий к образованию семян, объеди-
няются термином вегетативное размножение.
Вегетативное размножение
Усы земляники —хорошо известный пример эффек-
тивной системы вегетативного размножения. К концу
периода цветения у нее образуются длинные ползучие
стебли (усы), которые стелются по поверхности почвы
и несут немного мелких чешуевидных листьев. Со вре-
менем’ верхушка уса изгибается вертикально вверх,
а сам он продолжает расти дальше, за счет новой, ве-
точки, которая появляется в пазухе листа (так назы-
вают угол между листом и стеблем). Когда новое
растение образует розетку листьев, на стебле развива-
ются корни, называемые придаточными, и внедряются
в почву. После укоренения этого растения часть уса,
связывающая его с родительским растением, отмирает
и молодое растение становится совершенно самостоя-
тельным1.
Особенность, присущая усам земляники, — большая
скорость роста; благодаря этому новые растения ока-
зываются достаточно удаленными от родительского,
что снижает конкуренцию. Сходным способом распро-
страняется и ежевика. Она образует длинные стебли,
растущие сначала вверх, а затем изгибающиеся вниз.
Коснувшись земли, верхушка стебля укореняется, да-
вая начало новому кусту ежевики, который через не-
сколько лет становится независимым. Размножаясь
таким образом, ежевика образует непроходимые за-
росли.
Иногда на корнях возникают придаточные побеги,
выходящие на поверхность и дающие начало новому
растению. Вот почему бодяк полевой (Cirsium arvense)
так трудно уничтожить. Главный корень образует мно-
жество белесоватых боковых корней, которые тянутся
1 Вернее, только после этого его и можно назвать <растенвем>.—
Прим. ред.
Проблема размножения
131
под землей на какое-то расстояние, а затем дают над-
земные побеги.
Весьма обычным органом вегетативного размноже-
ния является корневище. Это подземный стебель, кото-
рый служит растению почти так же, как и усы земля-
ники, но в отличие от них растет под землей и поэтому
защищен от морозов. Корневища пырея ползучего
(Agropyron repens), например, знакомы всякому, кто
хотел искоренить этот сорняк в своем саду. Мята
и ландыш тоже относятся к обычным корневищным
растениям. Купену (Polygonatum) называют еще соло-
моновой печатью из-за круглых рубцов, остающихся
на корневищах после отмирания надземных побегов.
Корневище у купены толстое и мясистое, потому что
оно служит подземным хранилищем запасов, где пита-
тельные вещества откладываются на зиму и исполь-
зуются затем весной возникающими из него молодыми
растениями.
Многие видовые названия растений (repens, replans,
radicans) говорят о наличии усов или корневищ. В ка-
честве примера можно назвать лютик ползучий
(Ranunculus repens) и гораздо более редкий вид R. rep-
tans, растущий у берегов озер в Озерном округе
и в Шотландии.
Сочетание функций запасания пищи и вегетативного
размножения наиболее ярко выражено в стеблевом
клубне. Типичный клубень такого типа у картофеля.
Сразу под поверхностью почвы растение картофеля об-
разует множество боковых побегов, на концах которых
возникают утолщения — клубни. Сверху клубень по-
крыт защитным слоем перидермы («шелуха»), а в пазу-
хах его чешуйчатых листьев находятся почки (глаз-
ки)1. Если разрезать клубень на кусочки, имеющие
глазки, и посадить их в землю, то из каждого такого
кусочка можно получить новое растение. Истинная при-
рода глазков картофеля выявляется при проращивании
1 Автор преувеличивает, говоря о «чешуйчатых листьях» иа
клубне картофеля. Эти листья остаются совсем недоразвитыми и
выглядят как валнкообразные рубцы, называемые по-русски «бров-
ками». В их пазухах н располагаются «глазки». — Прим. ред.
132
Глава VI
клубней перед посадкой, когда каждый глазок разви-
вается в побег, несущий зачаточные листья.
Следует отметить, что картофель отнюдь не корне-
плод, хотя его часто так называют. Ведь на корнях не
бывает, чешуйчатых листьев с пазушными почками;
значит, клубни картофеля представляют собой часть
стебля.
Иногда корень может быть клубнеобразным, как
у ятрышника (Orchis fuchsii) и других представителей
рода Orchis. В этом случае на концах корней образуют-
ся утолщения, служащие хранилищами питательных
веществ, за счет которых растение весной трогается
в рост. Как правило, растение развивает больше одно-
го корневого клубня, поэтому они, как и стеблевые
клубни, могут служить средством вегетативного раз-
множения.
Клубнелуковица крокуса образуется путем утолще-
ния основания стебля. Если выкопать растение крокуса
в апреле, то можно увидеть новую клубнелуковицу,
возникающую среди оснований листьев, на верхушке
клубнелуковицы нынешнего года, только что давшей
цветоносный побег; в свою очередь под ней часто мож-
но обнаружить сморщенные остатки клубнелуковицы
прошлого года. Позднее, но в этом же году, новая
клубнелуковица достигнет своих окончательных разме-
ров. Она покрывается чешуйчатыми листьями, в пазу-
хах которых находятся почки, развивающиеся на сле-
дующий год в новые цветоносные побеги.
Клубнелуковицы очень обычны как у дикорасту-
щих, так и у культивируемых видов. Примерами могут
служить лютик луковичный (Ranunculus bulbosus),
который легко узнать по отогнутым чашелистикам,
и аронник (Arum maculatum)'.
В клубнелуковице пища запасается в утолщенном
основании стебля. В луковицах же, например у тюль-
пана или желтого нарцисса, запасные питательные
вещества откладываются в толстых мясистых листьях,
1 Из широко известных культивируемых растений сюда отно-
сится гладиолус. — Прим. ред.
Проблема размножения
133
тогда как стебель, от которого они отходят, редуциро-.
ван до плоского дисковидного образования.
В луковице тюльпана имеется центральная почка,
окруженная мясистыми листьями. Весной, используя,
отложенные в них запасные вещества, эта верхушечная
почка дает начало цветоносному побегу с настоящими
зелеными листьями. В пазухе одного из мясистых
листьев имеется почка, которая, развиваясь за счет
питательных веществ, доставляемых в результате фото-
синтеза развернувшимися зелеными листьями, увеличи-
вается в размерах и превращается в луковицу следую-
щего года. Иногда таких пазушных почек в луковице
развивается больше, чем одна, и соответственно в на-
ступающем году образуется две или более луковиц.
В этом случае можно сказать, что вегетативное размно-
жение растения обеспечено.
Другого типа луковицы у желтого нарцисса (Nar-
cissus pseudonarcissus) и подснежника (Galanthus
nivalis). Здесь мясистые луковичные чешуи образуются
из оснований листьев предыдущего года или несколь-
ких предыдущих лет, как у нарцисса. Настоящие зеле-
ные листья появляются на старом стебле, который
сохраняется из года в год, тогда как цветок возникает
на пазушном побеге. Главная, или верхушечная, почка
тоже существует из года в год, а основания надземных
зеленых листьев утолщаются, образуя луковицу ново-
го сезона. Дочерние луковицы образуются в пазухах
наружных луковичных чешуй.
Существенное различие между луковицами тюльпа-
на и нарцисса состоит в том, что развивающаяся каж-
дый год луковица тюльпана представляет собой совер-
шенно новое образование, тогда как у нарцисса и под-
снежника по крайней мере часть луковицы постоянна.
Говоря ботаническим языком, луковица тюльпана воз-
никает «симподиально», а луковицы нарцисса и под-
снежника — «моноподиально».
Луковица Эндимиона (Endymion nonscriptus) соче-
тает в себе признаки обоих описанных типов луковиц.
Подобно луковице тюльпана, она возникает симподи-
ально, но образована преимущественно утолщенными
основаниями листьев (как у нарцисса),
134
Г лава VI
Некоторые растения размножаются выводковыми
почками — небольшими почками, которые легко отде-
ляются от материнского растения и, падая на землю,
прорастают в новые растения. Как и у луковиц, их
листья богаты питательными веществами. Такой способ
размножения характерен и для обычного сорного
растения чистяка (Ranunculus ficaria)1. У лука вино-
градного (Allium vineale var. compactum) вместо цвет-
ков в соцветиях развиваются выводковые почки в виде
луковичек.
Для садовода вегетативное размножение представ-
ляет собой чрезвычайно удобный способ получения по-
томства, которое было бы сходно с родительским. При
размножении растений черенками мы знаем, что новое
растение будет совершенно сходно со старым по всем
важным для нас признакам, так как при этом не про-
исходит смешения наследственного материала двух ро-
дителей. Многие наши культурные растения размножа-
ют только таким путем, так как они представляют
собой гибриды, полученные при скрещивании двух
разновидностей, и потому при выращивании из семян
далеко не всегда будут идентичны родительскому
растению.
Каждого, кто посадит семена сливы Виктория в на-
дежде получить дерево того же сорта, почти наверняка
ждет разочарование. Дело в том, что этот сорт сливы
получен в результате скрещивания многих сортов, по-
этому дерево, выросшее из его семян, будет очень мало
напоминать родительское. Правда, таким путем можно
получить новый и даже лучший сорт, но шансов для
этого крайне мало.
1 В СССР чистяк растет массово в сырых лиственных лесах и
в лесных оврагах, но никогда не оказывается в роли «сорняка».
Выводковые почки чистяка бывают двух типов. Одни связаны с
клубневидно утолщенными придаточными корешками, легко отде-
ляющимися от материнского растения, другие образуются на стеб-
ле, в пазухах листьев, благодаря клубневидному же утолщению оси
почки. Их, конечно, нельзя считать по структуре сходными с на-
стоящими луковичками, так как здесь органы запаса не листово-
го происхождения. — Прим. ред.
Проблема размножения I5S
Если вы хотите получить сливу сорта Виктория во
всем ее совершенстве, это надо делать путем прививки.
Только в этом случае вы можете быть уверены, что
потомство будет напоминать материнское растение во
всех отношениях.
Так же обстоит дело с картофелем. Семенной кар-
тофель, который мы сажаем, — это вовсе не семена,
а небольшие клубни, образовавшиеся из стебля друго-
го растения картофеля. Настоящие же семена обра-
зуются из цветков обычным способом, но, если их вы-
сеять, они не дают начало растениям того же сорта,
а иногда образуют совсем другие растения, по-видимо-
му, не имеющие никакого экономического значения.
Изредка, правда, при этом может получиться новый
и ценный сорт, но такой случай бывает один на десят-
ки тысяч.
Преимущества семян
Среди дикорастущих видов вегетативное размноже-
ние очень распространено и, надо сказать, имеет по
сравнению с половым определенное преимущество: оно
просто, надежно и к тому же обеспечивает быстрое
расселение вида на ограниченном пространстве. Боль-
шая часть семян диких растений по той или другой
причине пропадает зря. Они могут упасть на камени-
стую почву или быть склеванными птицами; наконец,
другие растения могут не дать им прорасти. В этом
смысле вегетативное размножение гораздо надежнее,
однако и оно имеет свои недостатки. Хотя такое раз-
множение, «а,к мы видели, очень полезно, делая воз-
можным быстрое расселение растения на какой-то ог-
раниченной площади, оно не может обеспечить рассе-
ление на больших пространствах. Молодое растение
при этом обязательно вырастает в непосредственной
близости к родительскому. Семя же имеет в своем
распоряжении все те остроумные способы распростра-
нения, которые были выработаны в процессе эволюции,
поэтому расстояния здесь измеряются не сантиметрами
и дециметрами, а километрами и даже сотнями кило-
метров.
136
Глава VI
Семена имеют и другое преимущество: свойствен-
ный им период покоя помогает пережить зиму, когда
«вегетативная жизнь» над землей приостанавливается.
Упаковка внутри семени — самый надежный способ для
будущего растения, позволяющий ему охранить себя
от зимних морозов. Для однолетнего растения это фак-
тически единственный способ сохранения вида. Семя —
надежная защита не только от холода, но и от других
неблагоприятных условий. Растениям пустыни, напри-
мер, надо опасаться не льда и снега, а солнечных ожо-
гов. И наконец, семя, образуясь в результате полового
процесса, заключает в себе возможность возникновения
новых признаков у потомства, потому что при слиянии
мужской гаметы из пыльцевого зерна с ядром яйце-
клетки в семяпочке происходит смешивание признаков
родительских особей. Это имеет место не только при
перекрестном опылении, но даже при самоопылении.
В результате проросток несколько отличается от обоих
родителей. »
Все это дает сырье для деятельности естественного
отбора, и только благодаря существующей изменчи-
вости вообще возможна эволюция. Если многообра-
зие— это, так сказать, «приправа» к жизни, то для
эволюции оно составляет самую сущность.
ГЛАВА VII
ОПЫЛЕНИЕ
Как я уже объяснял в предыдущей главе, опылени-
ем называется перенос пыльцы с тычинок на рыльце
того же или другого цветка. Для большинства расге-
ний характерно перекрестное опыление, и многие из
них имеют хитроумные приспособления, гарантирующие
его осуществление. Преимущество перекрестного опы-
ления, как я уже упоминал, состоит в том, что призна-
ки двух родителей при этом смешиваются и, если меж-
ду родителями имеются различия, возможно возникно-
вение у потомства новых признаков.
Опыление может производиться различными аген-
тами, наиболее важные из которых — ветер и насеко-
мые. Прежде считалось, что ветроопыляемые цветки
появились раньше, чем опыляемые насекомыми, кото-
рые возникли в результате изменения механизмов,
первоначально служивших для опыления ветром.
Теперь мы в этом не так уверены. Вспомним, что насе-
комые появились задолго до цветковых растений; в ка-
менноугольный период над лесными болотами летали
гигантские стрекозы с размахом крыльев около 70 см.
Насекомые каменноугольного периода были весьма
примитивны, но позднее эта группа живых существ
претерпела эволюционный взрыв такой силы, что в ре-
зультате насекомые могли бы завоевать весь мир, если
бы этому не помешали некоторые непоправимые недо-
статки в их организации. Так или иначе, процесс эво-
люции насекомых, приведший к возникновению 28 от-
рядов, в которые они объединяются в настоящее время,
шел одновременно с эволюцией цветковых растений
и превращением их в доминирующую группу раститель-
ного мира. Что могло быть более естественным, чем
развитие этих двух групп в содружестве? Насекомые
питались на цветках и в обмен на нектар, который
цветки вскоре начали вырабатывать, переносили пыль-
цу с одного цветка на другой. В настоящее время ги-
потеза о том, что опыление насекомыми является более
138
Глава VII
древним способом, чем опыление ветром, имеет много
приверженцев. Разумеется, оба эти способа опыления
могли возникнуть и бок о бок, но при современном
состоянии наших знаний на этот счет нельзя сказать
ничего определенного* 1.
Первобытные цветки, возможно, походили на цве-
ток магнолии — цветок открытого типа с неопределен-
ным числом частей околоцветника, не разделенного на
чашечку и венчик, с множеством тычинок, дающих
массу пыльцы, и гинецеем, образованным не срастаю-
щимися друг с другом плодолистиками. Такой цветок
позволяет легко представить происхождение цветка
лютика. Современные цветки подобного типа опыляют-
ся самыми различными насекомыми, поскольку они не
специализированы в приспособлениях к опылению толь-
ко одним определенным насекомым, посещающим их,
например пчелой или бабочкой.
Цветок лютика может служить хорошим примером
этого довольно примитивного типа строения цветка.
В обиходе лютиками называют обычно несколько раз-
ных видов этого рода, из которых наиболее распрост-
раненными являются лютик луковичный (Ranunculus
bulbosus), лютик ползучий (/?. repens) и лютик луго-
вой, или едкий (/?. acris)1. Все они имеют цветки сход-
1 Среди наших отечественных ученых одним из наиболее ярких
сторонников первичности насекомоопыления, точнее «кантарофй-
лии», т. е. опыления при помощи жуков, является А. Л. Тахтаджян.
Он очень убедительно пишет об этом в популярной книге «Происхож-
дение покрытосеменных растений» («Наука», М., 1961). С другой
стороны, в книге Н. В. Первухиной «Проблемы морфологии и би-
ологии цветка» («Наука», 1971), написанной тоже очень живо
и достаточно популярно, выдвигаются не менее убедительные дока-
зательства того, что первые цветковые растения имели цветки, мало
специализированные в смысле агентов опыления, и, смотря по об-
стоятельствам, могли пользоваться и ветро- и насекомоопыле-
нием. — Прим. ред.
1 Во флоре Советского Союза имеется несколько десятков ви-
дов лютиков. В лесной зоне наиболее обычны указанные Даддинг-
тоном лютики ползучий и едкий. Довольно широко распространены
на лугах также лютик многоцветковый (R. polya nt he mas), лютик-
прыщинец (R. flammula), лютик золотистый (/?. auricomus), а в
лиственных лесах — лютик кашубский (R. cassubicus). Приводи-
мый Даддингтоном лютик луковичный встречается у нас редкф. —
Прим- ре<?,
OnMAtHUt
13d
ного типа с пятью свободными чашелистиками и пятью
лепестками, широко открытые и доступные для любого
насекомого. Характерно для них множество тычинок
и гинецей, состоящий из м.ногих отдельных плодолисти-
ков, расположенных на возвышенной части цветоло-
жа — утолщенного конца цветоножки, к которому при-
крепляются все части цветка. У основания каждого
лепестка, на внутренней его стороне, имеется нектар-
ник, скрытый маленькой чешуйкой (рис. 25).
Рис. 25. Цветок лютика, разрезанный пополам, чтобы можно было
видеть его части.
Цветок лютика посещают различные насекомые, не-
которые ради пыльцы, как, например, маленькая ноч-
ная бабочка Micropteryx calthella, но большинство ради
нектара. Дневные и ночные бабочки, пчелы, осы, жуки,
клопы и мухи — всех можно увидеть на цветке лютика,
который представляет собой удобную посадочную пло-
щадку для насекомых, как больших, так и маленьких.
Посещают его и трипсы, сосущие из лепестков сок.
Любое из этих насекомых может быть ответственно за
перенос пыльцы с цветка на цветок, и, поскольку таких
визитеров множество, а какие-либо специальные при-
способления, предотвращающие самоопыление, у люти-
ка отсутствуют, можно было бы ожидать, что оно про-
исходит часто. В действительности же это случается
редко и объясняется заметной самостерильностью
лютиков, т. е. физиологической неспособностью к оаы-
140
Глава VII
лению собственной пыльцой. Самоопыление происходит
только у отдельных особей, тогда как для большинства
ЛЮТНКов оно оказывается невозможным. Таким обра-
зом (за редкими исключениями) гарантируется пере-
крестное опыление.
Самостерильность отнюдь не редкость среди цвет-
ковых растений и может быть обусловлена многими
причинами. Обычно пыльца не прорастает на рыльце
того же цветка, а если прорастание все-таки происхо-
дит, рост пыльцевой трубки вскоре прекращается. Для
прорастания пыльцы золотого дождя {Laburnum апа-
gyroides), кустарника из семейства мотыльковых, необ-
ходимо легкое повреждение рыльца; небольшой ранки,
наносимой пчелой-опылителем, вполне достаточно для
стимуляции прорастания. Каждому плодоводу известно,
что самостерильностью обладают некоторые сорта
яблонь и многие сорта груш; будучи предоставлены
самим себе, эти деревья не плодоносят.
Специализация в направлении
обеспечения опыления
По мере эволюционного совершенствования цветки
специализировались для опыления определенными ви-
дами насекомых. Превосходным опылителем является
пчела. Опуская свой длинный хоботок в цветок, она
отыскивает спрятанный в нем нектар. У цветков, опы-
ляемых пчелами, а в еще большей степени у тех, кото-
рые опыляются дневными и ночными бабочками, нек-
тарник скрыт на дне трубочки, образованной лепестка-
ми, обычно срастающимися между собой. Только насе-
комое с длинным хоботком может добраться до такого
нектарника, тогда как более мелкие насекомые часто
не могут вползти в трубку венчика и завладеть некта-
ром из-за волосков, преграждающих им путь, хотя
некоторые из них научились красть нектар, «пробурав-
ливая» трубку вблизи нектарника.
В текущем столетии была проделана большая рабо-
та по изучению органов чувств пчел, которая дала нам
возможность подойти к пониманию того, чем же обу-
словлен интерес пчел только к определенным видам
Опыление
141
цветков. Видимо* здесь важны три фактора: окраска
цветков, их запах и характер узоров, имеющихся на
некоторых из них; обнаруженные в 1793 году Кристиа-
ном Шпренгелем, эти узоры получили в наши дни наз-
вание указателей меда. Долгое время полагали, что
пчелы не имеют цветового зрения и видят все в сером
цвете, а вся гамма цветов представляется им лишь как
различные оттенки серого. В 1914 году Фриш начал
свои классические исследования органов чувств пчел
и вскоре доказал, что они обладают хорошо развитым
чувством цвета, хотя и отличающимся от нашего. Фриш
приучал пчел брать сироп из блюдечек, помещенных
на синие квадраты. Когда пчелы привыкли связывать
синий квадрат с пищей, он предложил им целую серию
квадратов, из которых один был синий, а остальные —
различных оттенков серого, темнее и светлее синего
квадрата. Эти КБПДраты не имели никакой связи с пи-
щей. Пчелы нисколько не растерялись: они направились
прямо к синему квадрату и начали искать сирой. Ясно,
что они без труда могут отличить синий цвет от серого.
Повторяя этот опыт с квадратами разных цветов,
Фриш сумел показать, что пчелы имеют тонкое чувство
цвета, хотя оно и отличается от нашего. Человек спо-
собен различать около 60 цветовых оттенков (худож-
ник больше), а пчела—только 4 цвета. В участке
спектра от оранжевого до желтовато-зеленого мы ви-
дим множество оттенков, пчела же видит только жел-
тый. Такая же история с зеленовато-синим и синим
участками спектра. Таким образом, из всего много-
образия различных цветовых оттенков, делающих
для нас мир таким прекрасным, пчела может видеть
лишь три.
Все же в одном отношении пчела превосходит нас.
Я сказал, что она различает четыре цвета. Четвертый
находится в ультрафиолетовой части спектра, невиди-
мой для нас. Что это за «четвертый цвет», боюсь, мы
никогда не узнаем. Даже если бы можно было спросить
пчелу — а я не оставил бы без внимания возможность
того, что некоторые будущие исследователи окажутся
настолько изобретательными, что найдут средства об-
щения с этими замечательными насекомыми, — она
142
Глава VII
смогла бы рассказать нам об этом не больше, чем мы
можем объяснить слепому от рождения человеку, как
выглядит красный цвет.
Эта особенность зрения пчел ставит их над нами, но
мы имеем преимущество в другом отношении. Пчелы
не видят красного цвета, и все цвета, расположенные
в области между оранжевым и красным, смешивают
с темно-серым. Это в какой-то мере восстанавливает
престиж человека.
Если выводы, к которым пришел Фриш, правильны,
они должны быть применимы к окраске цветков, обыч-
но посещаемых пчелами. Проведенные исследования
подтвердили это. Большинство опыляемых пчелами
цветков синие, желтые или фиолетовые. Даже исключе-
ния подтверждают открытия Фриша. Так, пчелы не
должны замечать ярко-красные цветки мака, однако
они часто их посещают. При дальнейшем изучении
выяснилось, что красные лепестки мака сильно отра-
жают ультрафиолетовый свет, который мы не можем
видеть, а пчелы видят.
Многие другие цветки также отражают ультрафио-
летовые лучи, и это позволяет объяснить, почему неко-
торые из них, неприметные на наш взгляд, очень при-
влекательны для пчел. Вот, например, растение пере-
ступень, или бриония (Bryonia dioica). Его небольшие,
зеленовато-желтые цветки почти не выделяются на
фоне листвы, и тем не менее пчелам они бросаются
в глаза.
Помимо окраски цветка, необходимо принимать во
внимание и его форму. Фриш показал, что пчелы легко
различают форму предмета, например звездчатую или
овальную, но все же главное для них — это цвет. Если
пчелам, приученным прилетать к синей звезде, предло-
жить выбирать между желтой звездой и синим овалом,
они предпочтут синий овал, несмотря на изменение
формы.
Позднее Хертц установил, что пчелы охотнее выби-
рают узор из небольших пятен, чем сплошные пятна
той же площади. Следовательно, они предпочтут разъ-
единенные группы мелких цветков одному большому
цветку.
Опыление
143
Цвет и запах
Окраска цветков бывает обусловлена одним из двух
факторов. Большинство желтых цветков и некоторые
красные содержат хромопласты — структуры, окрашен-
ные пигментами из группы каротиноидов. Другие цвет-
ки (особенно синие) содержат пигменты, называемые
антоцианами, которые растворены в клеточном соке.
Антоцианы меняют окраску в зависимости от реакции
клеточного сока — от красной (в кислой среде) до си-
ней или фиолетовой (в щелочной среде). Этим и объ-
ясняется различная окраска цветков окопника {Sym-
phytum officinale), варьирующая от розовой до фиоле-
товой даже у одного и того же растения.
Окраска других частей растения может зависеть от
тех же факторов. Благодаря присутствию в клетках
хромопластов зрелые томаты имеют красную, а мор-
ковь— оранжевую окраску, тогда как темно-красный
цвет свеклы и роскошная окраска листьев колеуса
создаются антоцианами. Осенняя окраска листьев вы-
звана отчасти влиянием продуктов разрушения хлоро-
филла, а отчасти накоплением в стареющих листьях
антоцианов.
Большинство цветков, опыляемых пчелами, привле-
кает этих насекомых не только своей окраской, но
и запахом. В процессе изучения влияния различных
запахов на пчел Фришу впервые удалось доказать, что
обонятельные рецепторы у них располагаются на уси-
ках; при удалении усиков пчелы полностью утрачива-
ли обоняние. Он пытался с помощью дрессировки за-
ставлять пчел связывать различные запахи с пищей
и показал, что они способны распознавать целый ряд
запахов. Как выяснилось, пчел можно «обучить» при-
летать на запахи искусственного происхождения, напри-
мер лизола и сероуглерода (последнее вещество имеет
запах, напоминающий запах гнилой капусты).
Фриша интересовала также сравнительная роль за-
паха и окраски в привлечении пчел. Выработав у пчел
условный рефлекс на синюю карточку, пахнущую жас-
мином, он предложил им на выбор желтую карточку
с. дапахрм жасмина и синюю карточку без запаха'
’44
Глава VII
Большинство пчел предпочло желтую карточку, у кото-
рой был привычный запах, но незнакомый цвет. Затем
между этими карточками он поместил ширму длиной
около 1 м, так что пчелы, еще находясь на некотором
расстоянии от карточек, вынуждены были решать, к ка-
кой же из них лететь. На этот раз большая часть пчел
направилась к синей карточке. Таким образом Фриш
пришел к заключению, что, пока пчела находится на
некотором расстоянии от предмета, более сильным сти-
мулом для нее является окраска, но вблизи первосте-
пенное значение приобретает запах предметов.
Этот принципиально верный вывод Фриша оказался
правильным лишь до некоторой степени. Дело в том,
что строение глаз у пчел таково, что они, по нашим
понятиям, крайне близоруки. Дальше чем примерно за
метр они не могут видеть цветков, поэтому, когда пче-
лы летят издалека, они должны руководствоваться за-
пахом, а не окраской. Примером может служить такая
культура, как клевер, у которого большая масса цвет-
ков группируется вместе. Едва ли можно сомневаться
в том, что именно запах множества цветков и привле-
кает пчел. У отдельных же цветков запах вряд ли на-
столько сильный, чтобы привлечь пчелу, когда цветки
находятся вне пределов видимости для нее.
По-видимому, в процессе привлечения пчел к цзет-
кам можно выделить три стадии. Со сравнительно боль-
шого расстояния пчелу может привлекать запах, если
он достаточно сильный, и этим определяется то направ-
ление, в котором она должна лететь. Когда цветки на-
ходятся в поле зрения насекомого, т. е. в пределах при-
мерно 1 м, запах уже не имеет значения, зрительные
ощущения берут верх и пчела подлетает к цветку, ру-
ководствуясь лишь зрением. Но в непосредственной
близости от цветка обоняние снова становится домини-
рующим чувством — пчела ни за что не сядет на цве-
ток, если у него «не тот» запах.
Чувствительность пчелы к запаху — вопрос спорный.
Фриш на основании своих работ пришел к заключению,
что обоняние у пчелы такое же тонкое, как и у человека,
но более поздние исследования с применением усовер-
шенствованных методов показали, что пчелы имеют го-
Опыление
145
раздо более чувствительные «носы*. Сейчас считают»
что чувство обоняния у пчел не менее чем в 10, а воз-
можно, даже в 100 раз тоньше, чем у нас.
Запах цветков обычно обусловлен присутствием ле-
тучих эфирных масел, представляющих собой смеси уг-
леводородов (т. е. веществ, состоящих только из водо-
рода и углерода), называемых терпенами, и их кисло-
родных производных. Хорошо известны лавандовое
масло (из Lavandula vera), анисовое масло (из Pimpi-
nella anisum) и мятное масло (из Mentha piperita).
Многие из этих растений являются ценным сырьем для
парфюмерной промышленности. Эфирные масла прида-
ют аромат таким специям, как гвоздика (цветочные бу-
тоны Eugenia caryophyllata), имбирь (корневище Zingi-
ber officinale), черный перец (ягоды Piper nigrum) и ко-
рица (кора Cinnamomum zeylanicum). Хмель (Humulus
lupulus), придающий горький привкус пиву, а также
служащий средством, предохраняющим его от порчи,
содержит эфирные масла лупулины, которые продуци-
руются железками, расположенными на прицветниках
женских соцветий.
То, что я только что рассказал о чувствах пчел, от-
носится главным образом к медоносной пчеле, являю-
щейся объектом многочисленных научных исследова-
ний, но почти не приходится сомневаться в том, что все
это применимо и к пчелам, ведущим одиночный образ
жизни, и к шмелям. Медоносные пчелы имеют одну
важную, присущую только им особенность. Они посы-
лают нескольких пчел-разведчиц на поиски подходящих
мест для кормежки; возвратившись в улей, они «рас-
сказывают* остальным пчелам при помощи сложного
танца, куда следует лететь. Шмели же могут рассчиты-
вать только на собственную изобретательность, хотя,
надо сказать, вполне в этом преуспевают.
Указатели меда и нектар
Теперь мы подошли к вопросу о роли, которую игра-
ют различные метки на цветках, известные как указа-
тели меда. Они очень обычны, особенно у высокоорга-
низованных типов цветков. Часто они состоят из черто-
10—302
1'6
Глава VII
чек, сходящихся к месту расположения нектарника,
как, например, у душистой фиалки (Viola odoraia), на
нижних лепестках которой имеются ярко выраженные
полоски, но могут иметь и форму пятен на трубке вен-
чика, как у пурпуровой наперстянки (Digitalis purpu-
rea)', кроме того, центр цветка может быть иначе окра-
шен, чем остальная его часть, как, например, желтая
середина цветка незабудки (Myosotis).
Предположение Шпренгеля о том, что эти знаки
указывают пчеле путь к нектарнику, скрытому в цветке,
вполне подтвердилось в ходе дальнейших исследований.
Когда пчела садится на цветок, она обычно опускается
на край венчика, привлекаемая, очевидно, прежде всего
его окраской, контрастирующей с окружающим фоном.
Однако, если на цветке есть указатели меда, ориенти-
ром насекомому служит их окраска, отличающаяся от
окраски остальной части венчика, и оно садится не на
край цветка, а ближе к центру, где имеет больше шан-
сов найти нектарник.
Конечно, многие опыляемые пчелами цветки не име-
ют никаких указателей меда, и в этом случае невольно
напрашивается вопрос, как же пчелы без них обходят-
ся. Но, оказывается, пчелы обладают замечательной
способностью учиться на опыте и после нескольких по-
пыток легко находят нектарники и в таких цветках.
К тому же вполне возможно, что на цветках имеются
указатели меда, которых мы не видим. Как уже гово-
рилось, пчелы способны видеть ультрафиолетовый свет.
Кроме того, было обнаружено, что в некоторых цвет-
ках запах неравно.мерно распределен по лепесткам
н бывает, что указатели меда выделяются своим осо-
бым запахом, а не окраской. Это может быть и в том
случае, когда присутствуют видимые для нас указате-
ли меда; было показано, что линии-указатели могут
иметь другой запах, чем остальная часть цветка.
Строение пыльцевых цветков
Пчелы посещают цветки не только ради нектара, но
и для сбора пыльцы, и среди опыляемых ими цветков
Мнргце дйщены нектара. Это и есть т^к называемые
Опыление
147
«пыльцевые» цветки. Примером может служить цветок
ракитника (Sarothamnus scoparius), интересный также
н в смысле приспособлений, обеспечивающих пере-
крестное опыление. Цветок имеет форму бабочки, ха-
рактерную для всех мотыльковых (Papilionaiceae),—
семейства, к которому относятся также горох и бобы.
Верхний лепесток у такого цветка крупный и располо-
жен вертикально; он называется флагом или парусом
и, хотя у ракитника цветки пыльцевые, несет на себе ука-
затели меда, имеющие вид темных линий. Два боковых
лепестка называются веслами или крыльями, а дса
нижних соединены друг с другом, образуя лодочку —
структуру, окружающую андроцей и гинецей. Края ле-
пестков лодочки сочленяются с краями лепестков-
крыльев. Андроцей образован десятью тычинками со
сросшимися тычиночными нитями, пять из которых
длинные, а пять короткие. Гинецей состоит из одного
плодолистика с длинной завязью и длинным столби-
ком, а на вершине находится рыльце. Тычинки и стол-
бик спрятаны внутри лодочки и находятся в состоянии
сильного напряжения.
Когда насекомое (обычно это бывают тяжелые
шмели) садится на крылья цветка, то под его тяжестью
нарушается сочленение между лодочкой и крыльями,
что позволяет лепесткам, образующим лодочку, разъе-
диниться; что разъединение начинается во внутренней
части лодочки и распространяется к ее вершине. По
мере разделения двух лепестков, составляющих лодоч-
ку, первыми освобождаются пять коротких тычинок,
которые ударяют шмеля по брюшку, выбрасывая на
него пыльцу. Затем резко освобождаются столбик
и длинные тычинки, высыпая пыльцу на спинку шмеля.
Столбик находится в состоянии даже большего напря-
жения, чем тычинки, поэтому он освобождается немно-
го быстрее и касается шмеля до того, как тычинки
обсыпят его пыльцой. Если шмель побывал перед этим
на другом цветке ракитника, то рыльце воспримет при-
несенную пыльцу. Коснувшись шмеля, столбик про-
должает изгибаться, что позволяет рыльцу уклонить-
ся от тычинок в тот момент, когда из них высыпается
пыльца.
)48
Глава VU
Этот взрывной механизм необратим, поэтому, если
лодочка открылась, освободившиеся тычинки и столбик
уже не могут снова в нее спрятаться, а края крыльев и
лодочки не могут опять сочлениться. Взглянув на расте-
ние ракитника, можно узнать опыленные цветки по без-
вольно висящим лепесткам; такие цветки впоследствии
дадут семена, тогда как цветки, которые не посещались
шмелями, не дадут их. Описанный нами механизм, по-
видимому, успешно предупреждает самоопыление.
У утесника (Ulex)—вида, близкого ракитнику,—
сходный механизм опыления, но менее стремительный.
Когда насекомое, садясь на цветок, опирается на
крылья, лодочка открывается и столбик, появляясь
первым, касается его брюшка; только после этого по-
казываются тычинки и высыпают на него пыльцу. По-
скольку в цветке утесника столбик первым покидает
лодочку, перекрестное опыление здесь даже более ве-
роятно, чем у ракитника1.
Цветки, образующие нектар
Совершенно другой механизм опыления пчелами
наблюдается у шалфея (Salvia) — растения, принадле-
жащего к семейству губоцветных (Labiatae). Пять ле-
пестков, соединяясь, формируют трубку венчика, но
верхние их концы остаются свободными и образуют ряд
выступающих долей, причем хорошо заметны две губы.
Верхняя губа, состоящая из двух сросшихся лепест-
ков, расположена вертикально, и в ее вогнутой внут-
ренней части скрыты две тычинки и молодой столбик.
Нижняя губа венчика служит посадочной площадкой
для пчел. В основании завязи, спрятанной на дне труб-
ки венчика, помещается нектарник. Наиболее специфи-
ческое строение имеют тычинки. Каждый пыльник
состоит из двух отделенных друг от друга половинок,
нижняя из которых стерильна и не содержит пыльны.
1 Даддингтон описывает опыление у растений, обычных для
флоры Англии. У нас эти виды редки, но подобный же механизм
опыления наблюдается у многих других мотыльковых, в частности
у русского ракитника {Cytisus ruthenicusy, нечто похожее — у лю-
церны (JUedicago [atcata). — Прим. ред.
Опылекие
149
Обе половинки соединены длинной «нитью», называе-
мой связником, который изогнут таким образом, что
прилегает к вогнутой поверхности верхней губы венчи-
ка. Вблизи своего нижнего конца связник подвижно
сочленен с тычиночной нитью, и если достаточно силь-
но толкнуть нижнюю половинку пыльника, расположен-
ного в зеве трубки венчика, связник начинает качаться
и верхняя, фертильная половника пыльника опускается
вниз.
Рис. 26. Механизм опыления цветка Salvia.
А. Пчела, посещающая молодой цветок, толкает нижнюю, стерильную поло-
винку пыльника; благодаря подвижному сочленению связника с тычиночной
нитью верхняя, фертильная половинка при этом выбрасывается вперед и на-
клоняется вниз, касаясь спинки пчелы и пачкая ее пыльцой. Б. Более старый
цветок, у которого тычинки завяли, а столбик удлинился, благодаря чему рыль-
це оказалось в положении, позволяющем ему касаться спинки пчелы, посе-
щающей цветок, и воспринимать пыльцу. В. Тычинка. Видно, каким образом
длинный связник соединяется с короткой тычиночной нитью.
Сев на нижнюю губу венчика, пчела просовывает
голову в трубку венчика, чтобы достать мед, выделя-
емый нектарником, помещающимся на дне трубки.
Нижние стерильные половинки пыльников оказываются
на ее пути, но она отталкивает их, превращая таким
образом в своего рода шарниры; в результате этого
верхние половинки пыльников опускаются вниз и каса-
ются спинки насекомого, обсыпая ее пыльцой (рис. 26).
В этот момент рыльце еще молодое и не готово
к восприятию пыльцы. В более старых цветках, которые
к этому времени уже высеяли пыльцу, столбик удли-
няется и загибается вниз. Это приводит «созревшее»
уже теперь рыльце в положение, позволяющее ему ка-
150
Глава Vll
саться спинки пчелы, посещающей цветок. Если пчела
только что побывала на более молодом цветке, на ее
спинке находится пыльца, часть которой остается на
рыльце.
Кроме замечательных качающихся тычинок, еще
одно приспособление помогает шалфею (Salvia) избе-
жать самоопыления. Это неодновременное созревание
пыльцы и рыльца — так называемая дихогамия, очень
обычная для растений с обоеполыми ( т. е. развиваю-
щими и тычинки, и гинецей) цветками. У Salvia первы-
ми созревают тычинки — явление, известное как про-
тандрия и очень широко распространенное; мы наблю-
даем его, в частности, у иван-чая (Chamaenerion
angustifolium), почти у всех сложноцветных (Сотро-
sitae), зонтичных (Umbelliferae) и многих других. Про-
тогиния (так называется более раннее созревание
рыльца) менее распространена, но все же встречается
у многих видов, в том числе у подорожника (Plantago),
желтого василнстника (Thalictrum flavum) и зимовни-
ка (Helleborus foetidus).
Дихогамия очень обычна у ветроопыляемых цвет-
ков. Это вполне понятно: при таком зависящем от мно-
жества случайностей процессе, как опыление ветром,
трудно найти более надежный способ избежать само-
опыления, чем разделение полов.
Еще одним средством предупреждения самоопыле-
ния является гетеростилия, выражающаяся в том, что
растение имеет цветки двух или более типов, с тычин-
ками и столбиками разной длины. Известнейший при-
мер гетеростилии— примула (Primula vulgaris). Если
рассмотреть цветки примулы, собранные с разных эк-
земпляров, то нетрудно заметить, что они двух типов —
с длинным и с коротким столбиком. У цветков первого
типа рыльце находится в зеве трубки венчика и вы-
глядит маленьким округлым пятнышком, если смотреть
на цветок сверху. Вы не можете видеть тычинки, пото-
му что пыльники расположены близ середины трубки
и загораживаются более длинным столбиком. У цветков
второго типа, наоборот, пыльники тычинок видны в зе-
ве трубки венчика и скрывают рыльце, доходящее
лишь до середины трубки, т. е. занимающее то же по-
Опыление
151
ложение, что и тычинки в длинпостолбчатых цветках
(рис. 27).
Примула опыляется медоносной пчелой. Когда пче-
ла погружает свой длинный хоботок в трубку венчика
короткостолбчатых цветков, чтобы достать нектар, она
пачкается пыльцой как раз на том уровне, где будет
рыльце длинностолбчатого цветка, и наоборот. С гете-
ростилией связаны и дополнительные приспособления
против самоопыления. У цветков с высокорасположен-
Рис. 27. Гетеростилия у при-
мулы.
А. Цветок с длинным столбиком.
5. Цветок с коротким столбиком.
ними пыльниками пыльца крупнее, чем в пизкопри-
крепленных пыльниках, а сосочки рыльца имеют боль-
шие размеры у цветков с длинным столбиком, чем
у короткостолбчатых. В результате более крупные
пыльцевые зерна цветков последнего типа (коротко-
столбчатых) лучше удерживаются на рыльцах длинно-
столбчатых цветков, чем небольшие пыльцевые зерна
цветков противоположного типа.
Как было показано, тип цветка у примулы контро-
лируется одним геном, причем короткостолбчатостъ
является доминантным признаком по отношению
к длинностолбчатости. Примерно у 70% примул цвет-
ки короткостолбчатые и только у 30%—длинностолб-
чатые.
У дербенника иволистного (Lythrum salicaria) гете-
ростилия зашла еще дальше — у него три типа цветков:
с длинными, короткими и средними столбиками и ты-
ЧИНКамц. В цветках с длинным столбиком развиваются
152
Глава VII
как короткие, так и средние тычинки и т. д. Каждый
цветок может быть опылен пыльцой цветков любого из
двух других типов.
Один из наиболее замечательных механизмов опы-
ления насекомыми можно наблюдать у орхидей. Цвет-
ки орхидей крайне специализированы и во многих отно-
шениях совсем непохожи на цветки других растений.
Они имеют всего одну тычинку, а пыльца сливается
у них в общую массу, образуя в каждом пыльнике гак
называемый поллиний, который целиком переносится
пчелой. Созревшие поллинии обычно прикреплены
к маленькому мешочку, как раз над поверхностью
рыльца, и в таком виде готовы прилипнуть к тельцу
пчелы (рис. 31).
Я описал лишь некоторые из многочисленных путей
приспособления цветков к опылению пчелами. Но пче-
ла не единственный опылитель, и некоторые приспособ-
ления цветков к опылению другими насекомыми также
не менее интересны.
Опыление осами, ночными бабочками,
жуками и мухами
Одно из самых необычайных приспособлений к опы-
лению насекомыми мы находим у инжира, который
опыляется осой Blastophaga grossorum. Дикий инжир,
или смоква (Ficus carica) растет в Италии1. Его «цве-
ток» на самом деле представляет собой полое соцветие,
так называемый синконий, грушевидной формы с от-
верстием наверху, внутри которого находятся цветки.
В устье синкония помещаются мужские цветки, а ниже
их — женские (у инжира цветки однополые). Плод
у инжира ложный, состоящий из разросшегося синкония
с настоящими плодами (семянками) внутри (рис. 28).
У дикого инжира первое цветение происходит ран-
ней весной. Тогда в устье синкония образуются много-
численные мужские цветки, а ниже — женские (гак
1 В СССР дикий и одичавший инжир встречается в Закавказье,
Крыму,*в горах Колет-Дага и западного Памиро-Алая. — Прим,
ред.
Опыление
153
называемые галловые цветки), содержащие всего одну
семяпочку и не способные дать съедобные плоды. Сам*
кн ос проникают в синконий и откладывают яички
в семяпочки галловых цветков (по одному в каждую
семяпочку). Развившиеся из яиц самцы проводят всю
свою жизнь внутри синкония, завершают свой метамор-
фоз в семяпочках и, выйдя из них уже взрослыми, начи-
нают искать галловые цветки с самками. Проделав от-
Рнс. 28. Продольный разрез
через синконий инжира.
верстие в стенке завязи, они оплодотворяют самок, пос-
ле чего умирают, так и не покидая соцветия, в котором
родились.
Оплодотворенные самки выходят из синкония и, пе-
реползая при этом через мужские цветки, расположен-
ные у его устья, обсыпаются пыльцой. К этому времени
созревает первый урожай плодов инжира — так назы-
ваемые профики (profico), горькие и несъедобные.
Самки ос летают очень мало; они ползают по ветвям
дерева в поисках молодых соцветий, образующихся
поздней весной при втором цветении дикого инжира
и отличающихся от синкониев первого цветения тем,
что содержат только женские цветки, которые на этот
раз нормально развиты и способны давать семена. Осы
безуспешно пытаются отложить яички в завязи этих
цветков и при этом попутно опыляют их. Развившиеся
из этих цветков плоды — фиги (fichi) созревают в сен-
тябре и вполне съедобны.
154
Г лава VII
Пока эти плоды созревают, начинается третье цве-
тение инжира. На этот раз синконии, или мамме (mam-
me), содержат только галловые цветки, rf осы, которым
не удалось отложить яички во время второго цветения,
откладывают их в эти галловые цветки. Здесь личинки
перезимовывают, а ранней весной выбираются из син-
кониев и начинают жизненный цикл сначала1.
Таким образом, дикий инжир дает плоды три раза
в год, но съедобны только плоды второго урожая (фи-
ги). Растение инжира не может завершить свой жиз-
ненный цикл без осы, а оса полностью зависит от ин-
жира в отношении пищи. Гибель растения неминуемо
влечет за собой гибель насекомого, и наоборот.
От дикого инжира произошли многие разводимые
его сорта, например Ficus carica var. domestica, кото-
рый очень похож на дикий вид Ficus carica var. сар-
rifica. Хотя этот дикий вид не образует съедобных
плодов, его присутствие совершенно необходимо для
развития плодов у культурного инжира.
Культурный инжир опыляется тем же видом ос, что
и дикий. Осы проводят зиму в синкониях (мамме) ди-
кого инжира, а ранней весной покидают их и проника-
ют в синконии (профики) этого инжира и так называе-
мые «fiori di fico» у культурного. У дикого вида (var.
caprifica) в этих синкониях находятся галловые цветки,
в которые насекомые откладывают яички, но у культур-
ного инжира (var. domestica) они содержат только
стерильные женские цветки, и здесь отложение яичек
1 Следует несколько уточнить терминологию, относящуюся к
цветению и плодоношению инжира. Синконии с развитыми мужски-
ми цветками и недоразвитыми женскими (галловыми) принято на-
зывать каприфигами. Длина столбиков в женских цветках здесь
в точности равна длине яйцеклада бластофаги. Синконии с реду-
цированными мужскими цветками и развитыми женскими называют
фигами. Столбики здесь длинные, значительно длиннее яйцеклада
осы, почему она и не может отложить здесь яички в завязь. Урожай
от первого цветения (профики) представлен преимущественно не-
съедобными каприфигами; съедобных фиг очень мало. При втором
цветении (маммони) образуются главным образом фиги, дающие
основной урожай плодов; каприфиг мало. В результате третьего
цветения (мамме) образуются только каприфиги, в которых и зи-
муют осы. — Прим. ред.
Опыление
155
невозможно: за исключением некоторых сортов, эти
синконии опадают с дерева, не образовав плодов.
В начале лета перепачканные пыльцой самки ос
вылетают из синкониев (профик) дикого инжира и ока-
зываются перед выбором, в какие синконии направить-
ся. Те осы, которые проникают в синконии культурной
разновидности (pedagnuoli), обнаруживают там раз-
витые женские цветки и, тщетно пытаясь отложить
в них яички, опыляют их пыльцой дикого инжира. Осы,
проникшие в синконии дикой разновидности, называе-
мые маммони (mammoni), находят галловые цветки,
ожидающие их, и откладывают в них свои яйца.
Синконии культурной разновидности созревают
и дают основной урожай съедобных плодов. Из яиц,
отложенных в галловые цветки дикой разновидности,
появляется новое поколение самок, которые оплодотво-
ряются самцами. Выйдя из синкониев и перепачкав-
шись при этом пыльцой мужских цветков, расположен-
ных вблизи устья, самки ос опять имеют две возмож-
ности. Некоторые из них проникают в синконии
(cimaruoli) культурной разновидности и, найдя там
функционирующие женские цветки, в которые невоз-
можно отложить яйца, опыляют их пыльцой из синко-
ниев маммони. Впоследствии эти цветки могут дать
зимний урожай плодов. Другие осы забираются
в синконии мамме дикой разновидности и откладывают
яички в галловые цветки. Здесь из яичек выводятся
личинки, которые зимуют, а весной появляются взрос-
лые осы, и все повторяется сначала.
Плодоношение культурного инжира, образующего
только женские цветки, всецело зависит от дикого ви-
да, дающего пыльцу, и от ос, которые производят опы-
ление. Это хорошо известно крестьянам Южной Ита-
лии, поэтому среди культурных деревьев инжира они
всегда сажают дикие1. Завезенный в Калифорнию ин-
жир не плодоносил до тех пор, пока туда не завезли
дикий вид и осу Blastophaga,
1 Опыление зависит от наличия каприфиг, поэтому приемы,
связанные с их доставкой, называют каприфнкацвей. — Прим. ред.
156
Глава VII
На севере Италии культурный инжир выращивает-
ся один, причем культивируются те его сорта, которые
дают плоды без оплодотворения {партенокарпические
сорта), поэтому в этом случае в капрификации нет не-
обходимости.
С ботанической точки зрения история инжира и его
опыления чрезвычайно интересна, поскольку это исто-
рия однодомного растения (однополые цветки разви-
ваются на одном и том же растении), утратившего
мужские цветки в процессе окультуривания. Это уни-
кальный случай в растительном царстве. Если семена
Ficus carica var. domestica посеять, они дадут дикий
вид, поэтому культурный инжир можно размножать
только прививкой или черенками.
Многие цветки, хорошо известным:: примерами кото-
рых являются жимолость {Lonicera periclymenum),
ослинник {Oenothera.) и душистый табак {Nicotiana
affinis), опыляются ночными бабочками. Для цветков,
относящихся к этой группе, характерны следующие
особенности: длинная трубка венчика (так как у бабо-
чек очень длинный хоботок), белая или светлая окра-
ска, заметная в темноте, и сильный запах, появляю-
щийся обычно к вечеру, когда летают ночные бабочки.
В отличие от цветков, опыляемых пчелами, у них обыч-
но нет «посадочной площадки», поскольку дневные
и ночные бабочки не садятся на цветок, а сосут нектар,
порхая над ним (фото XVII).
Но одним из самых удивительных случаев взаим-
ной зависимости насекомого и растения являются
взаимоотношения между южноамериканской юккой
{Yucca filamentosa) из семейства лилейных (Liliaceae)
и ее опылителем — особым видом моли Pronuba
yuccasella. У этой моли, как и у других представителей
рода Pronuba, верхняя челюсть снабжена шиповатыми
хватательными отростками. Привлеченная запахом,
который цветки юкки распространяют по ночам, моль
сначала направляется к тычинкам, где собирает пыль-
цу, используя при этом верхнечелюстной отросток, что-
бы слепить ее в твердый комочек размером примерно
в три раза больше собственной головы, и уносит этот
комочек прочь. Затем она летит к завязи другого цвет-
Опыление
157
ка, где длинный яйцеклад позволяет ей отложить яич-
ки среди семяпочек. После этого моль спускается по
свисающему вниз столбику и, достигнув рыльца, запи-
хивает принесенный комочек пыльцы в его углубления,
обеспечивая тем самым опыление цветка.
После оплодотворения начинается развитие плода,
и хотя гусеницы пронубы питаются семяпочками по
мере их развития в семена, у юкки они столь много-
численны, что оставшихся семяпочек вполне достаточ-
но для образования семян. Связь между юккой и про-
нубой настолько тесная, что они стали жизненно
необходимыми друг другу; в Европе, где пронубы не
водятся, юкка никогда не приносит плодов.
Некоторые растения приспособились к опылению
цветочными мухами (Syrphidae), которые тоже являют-
ся хорошими опылителями. Примером таких растений
может служить обычная у нас по сыроватым местам
и обочинам дорог вероника дубравная (Veronica cha-
maedrys). Ее мелкие синие цветки имеют четыре ле-
пестка (верхний образован двумя сросшимися лепест-
ками), две свисающие вниз тычинки и очень длинный
столбик, висящий между ними. На первый взгляд
лепестки кажутся раздельными, но, приглядевшись
внимательнее, нетрудно заметить, что они срослись
у основания, образуя короткую трубку венчика, на дне
которой помещается нектарник.
Когда муха садится на цветок вероники, рыльце
касается ее брюшка и воспринимает пыльцу другого
цветка. Доставая нектар, насекомое обхватывает обе
тычинки передними лапками и притягивает их цруг
к другу, обсыпаясь при этом пыльцой. Затем муха несет
пыльцу на другой цветок, где повторяется та же цепь
событий. Цветки вероники заметно протогиничны, что
снижает вероятность самоопыления.
Цветочная муха принадлежит к отряду двукрылых
(Diptera), куда относится и комнатная муха. Опылять
цветки могут и многие другие двукрылые, но именно
с опылением мухами зачастую связаны такие харак-
терные особенности цветков, как грязноватая коричне-
вая, зеленая или фиолетовая окраска и неприятный
запах.
158
Глава VII
Дикий аронник, или арум кукушечный (Arum
maculatumy, высоко специализирован для опыления
мухами. Крупный кроющий прицветник, или покрывало,
окружает центральную ось, несущую цветки, и сужа-
ется книзу, образуя «перехват». Верхушка оси со-
цветия, или початка, вытянута и образует на конце
придаток — булавовидное утолщение вроде колоса
красноватого цвета. На початке располагаются цвет-
ки, делящиеся на три группы. В самом низу находятся
женские цветки, над ними — мужские, а еще выше —
группа недоразвитых мужских цветков, которые видо-
изменены в кольцо направленных вниз волосков. Эти
волоски находятся как раз в области «перехвата»
и таким образом частично загораживают проход, веду-
щий вниз, к функционирующим цветкам (рис. 29).
Мелкие комарики и мушки, привлеченные неприят-
ным запахом цветков, а также соблазнившись красным
початком, заползают внутрь прицветника, преодолев
зону волосков. Наружу, однако, они выбраться уже не
могут из-за направленных вниз волосков, которые ме-
шают им, и скользких масел, выделяемых клетками
нижней части оси, которые еще более затрудняют путь
наверх.
В это время рыльца женских цветков готовы к при-
ему пыльцы, и если попавшие в плен насекомые при-
несли пыльцу с ранее посещенных цветков, они опыля-
ют рыльца. Через некоторое время созревают мужские
цветки; волоски, преграждавшие путь, вянут, масло
высыхает, и насекомые могут выбраться наружу. Пере-
ползая через мужские цветки, они оказываются обсы-
панными пыльцой. Не удовлетворенные первой попыт-
кой, мухи могут посетить другое соцветие и, попав
в него, опылить пыльцой, которую они переносят на
своем тельце, следующую группу женских цветков.
Ароннику не грозит самоопыление, потому что
рыльца женских цветков вянут до того, как в мужских
цветках созреет пыльца.
1 У нас три вида аронника произрастают в Крыму и на Кавказе;
кроме того, некоторые виды аронника и близкого к нему Eminium
встречаются в Средней Азии. — Прим. ред.
Опыление
15?
В зоне умеренного климата цветки, опыляемые му-
хами, обычно мелкие и невзрачные, многие из них
имеют сильный неприятный запах. В тропиках же сов-
сем иначе; там цветки, относящиеся к этой группе,
Рис. 29. Дикий аронник.
А. Покрывало, из которого выглядывает красная верхушка початка.
Б. Покрывало удалено, чтобы показать початок.
напротив, одни из самых крупных. Такова Rafflesia
arnoldii — паразитное растение с острова Суматра
с цветками до метра в поперечнике, которые лежат на
земле, прорастая прямо из корней растения-хозяина,
обычно дерева. Многие тропические ароидные привле-
кают мух и жуков почти таким же способом, как
и аронник. У водных растений из рода Cryptocoryne
трубчатое основание прицветника вытянуто в длинную
трубку, по которой жуки соскальзывают в подводную
камеру, находящуюся на глубине до 45 см.
160
Глава VII
Цветки, опыляемые жуками, не редкость, особенно
в тропиках. Сюда относятся цветки Magnolia и водя-
ных лилий. Гигантская Victoria amazonica с плаваю-
щими на воде листьями до 2 м в поперечнике и цветка-
ми, достигающими 35 см в поперечнике, опыляется
одним из видов хрущей (Melolontha melolontha), кото-
рые поедают ее тычинки и поэтому попадают в ловуш-
ку, когда гигантские цветки закрываются утром; вече-
ром цветки раскрываются и жуки вылетают. Возмож-
но, слово «ловушка» звучит здесь не совсем удачно:
хрущи, ведущие ночной образ жизни, вполне могут
оценить тепло и «комфорт» закрытых цветков, нахо-
дясь там в течение дня, не говоря уже о защите от
врагов, которую обеспечивает им это «жилище».
Цветки, опыляемые жуками, как правило, наименее
специализированы и относятся к примитивному, откры-
тому типу. Это обстоятельство привело к весьма убе-
дительной теории о том, что жуки были первыми насе-
комыми-опылителями, а способ опыления путем пере-
носа пыльцы более специализированными насекомыми
(например, пчелами) появился позже1. Жуки — очень
древняя группа насекомых, гораздо более древняя, чем
пчелы. Впрочем, жуки очень неловкие опылители: часть
тычинок они обычно поедают, а оставшиеся ломают.
С появлением пчел, имеющих длинный хоботок для
искусного сбора нектара, стала возможной более тон-
кая техника опыления.
Опыление птицами и летучими мышами
В теплых странах, таких, как Бразилия, Южная
Африка, южная Австралия и Патагония, очень распро-
странены цветки, опыляемые птицами. В Америке это
главным образом колибри (Trochilidae) и цветочницы
(Coerebidae); в Австралии — медососы (Meliphagidae)
и попугаи лори (Trichoglossidae); в Африке — множе-
ство различных птиц, в частности нектарницы (Necta-
rinidae) и цветососы (Dicaeidae). Для цветков, опы-
ляемых птицами, характерны яркая окраска, отсутст-
вие запаха, прочная, эластичная структура и обилие
1 См. примечание к стр. 138.
Опыление
161
довольно водянисто.го нектара. Эта последняя особен-
ность свойственна многим из них; так, американскую
Erythrina cristagalli называют «плачущей» из-за нек-
тара, капающего из ее перевернутого соцветия. На
Яве цветки Erythrina во время сухого сезона являются
основным источником влаги для таких крупных птиц,
как голуби. Некоторые австралийские виды Protea об-
разуют нектар в таком количестве, что местные жите-
ли собирают его.
Многие цветки, опыляемые птицами, имеют ярко-
красную окраску, что не удивительно, если знать стро-
ение глаза птицы. Дело в том, что в сетчатке большин-
ства птиц имеется желтоватое масло, играющее роль
своеобразного светофильтра и дающее такой же эф-
фект, что и оранжевые очки. В результате птицы сов-
сем не реагируют на синий цвет, но зеленый и в осо-
бенности красный различают хорошо. В 1915 году был
опубликован список 159 «орнитофильных» (т. е.
опыляемых птицами) цветков, из которых 84% имели
красную окраску.
Обычно птицы «потягивают» нектар, порхая в воз-
духе, поэтому орнитофильные цветки в отличие от цвет-
ков, опыляемых пчелами, как правило, не видоизменя-
ются, чтобы обеспечить опылителям «посадочную
площадку». Южноафриканская Strelitzia reginae, у ко-
торой видоизмененный прицветник образует выступ,
составляет в этом смысле исключение.
Там, где дело касается цветков, птицы еще более
неуклюжи, чем жуки: они рвут цветки и ломают ты-
чинки. Поэтому не удивительно, что у многих «орнито-
фильных» цветков нежная завязь спрятана в цветоложе
(нижняя завязь), а тычинки имеют прочные деревя
нистые тычиночные нити.
Какой же способ опыления более древний — опыле
ние птицами (орнитофилия) или насекомыми (энтомо
филия), — точно мы не знаем, но на этот счет сущест
вует весьма правдоподобная теория, согласно которой
в меловом периоде в тропиках преобладала орнитофи
лия, затем, в условиях более умеренного климата, воз
никла энтомофилия, а еще позднее — анемофилия (опы
ление ветром).
11-302
162
Глава VII
Опыление летучими мышами (хироптерофилия) —
естественное следствие орнитофилии и характерно для
многих тропических ночных цветков, в том числе
Crescentia cujete, Kigelia, Oroxylon. и др. Некоторые
кактусы из рода Cereus опыляются летучими мыша-
ми-вампирами.
Цветки, опыляемые летучими мышами, имеют круп-
ные размеры и прочную структуру, так как эти живот-
ные сильно повреждают их своими когтями. Подобно
орнитофильным цветкам, они образуют большое коли-
чество нектара. Раскрываются такие цветки обычно
вечером, имеют чаще всего зеленовато-желтую, корич-
неватую или фиолетовую окраску и специфический, до-
вольно неприятный запах. Данные об опылении
улитками (малакофилия) имеются лишь в отношении
немногих растений, в частности селезеночника (Chry-
sosplenium alternifolium). Однако доказательства того,
что такое опыление существует, весьма сомнительны;
более вероятно, что улитки просто поедают цветки.
Опыление ветром
Многие цветки опыляются ветром, который, кстати,
является единственным опылителем хвойных и родст-
венных им растений. По своим особенностям ветроопы-
ляемые (анемофильные) цветки обычно резко отлича-
ются от опыляемых насекомыми и птицами. Крупные
яркие лепестки у них отсутствуют, потому что они
только мешали бы сдуванию пыльцы ветром, не прино-
ся никакой пользы. Нектарники и запах также отсут-
ствуют. Цветок выглядит очень строго, без всяких не-
нужных украшений (рис. 30).
Как правило, анемофильные цветки образуют ог-
ромное количество пыльцы, что оказывается очень
кстати, так как анемофилия — дело рискованное, и на
каждое пыльцевое зерно, попавшее в цель, т. е. на
рыльце, приходятся десятки или сотии тысяч (а иногда
и миллионы) потерянных. Злаки относятся к ветроопы-
ляемым растениям, и каждый, кто страдает от сенной
лихорадки, знает, как много пыльцы злаков носится
Опыление
163
в воздухе. Весной.ловерхность горных озер в Швейца-
рии бывает сплошь желтой от пыльцы, которую выбра-
сывают мужские шишки сосны и пихты.
Пыльца анемофильных растений должна легко дер-
жаться в воздухе, поэтому пыльцевые зерна у них
мелкие, сухие и без труда отделяются друг от друга.
Рис. 30. Цветок злака, выделенный из соцветия.
Обратите внимание на длинные перистые рыльца, типичные для ветроопыляе-
мых цветков.
Напротив, у энтомофильных цветков наблюдается тен-
денция к увеличению размеров пыльцевых зерен, кото-
рые часто слипаются вместе, образуя более крупные
комочки. Как мы уже говорили, эти комочки переносят-
ся пчелами на женские растения, и чем больше пыльце-
вых зерен слипнется вместе, тем больше преимуществ
имеет растение.
Тычинки анемофильных цветков располагаются со-
вершенно открыто, высовываясь из цветка наружу, где
они легко доступны ветру. Пыльники часто присоедине-
ны к тычиночной нити своим центром с помощью иод-
11*
164
Глава VII
вижного сочленения, так что пыльца высыпается из них
при малейшем порыве ветра. Выделяются у анемо-
фильных растений также крупные и пушистые рыльца;
созрев, они, как и тычинки, высовываются из цветка.
Обратите внимание на то, как много наших ветрооиы-
ляемых деревьев цветет рано, до того как развернутся
почки и появятся листья, мешающие распространению
пыльцы.
Ветер — слишком неопределенный посредник, раз-
брасывающий пыльцу где попало, и структурные при-
способления, предотвращающие самоопыление, у ане-
мофильных цветков отсутствуют. Вместе с тем дихога-
мия (неодновременное созревание пыльцы и рылец)
встречается у них часто. Так бывает и у однодомных,
и у двудомных растений. Благодаря этим обстоятель-
ствам анемофильным цветкам удается свести самоопы-
ление до минимума, хотя время от времени оно должно
случаться. Раздельнополость цветков — это еще одно
преимущество. В этом случае можно произвести муж-
ских цветков гораздо больше, чем женских, и таким
образом обеспечить избыток пыльцы и меньшую поте-
рю семяпочек из-за отсутствия опыления. Орхидные,
весьма любопытным образом приспособившиеся к опы-
лению насекомыми, могут быть расточительными в про-
изводстве семяпочек, которые развиваются тысячами
в одном цветке, что гарантирует опыление значитель-
ной их части. Другое дело — ветроопыляемые злаки.
В каждом цветке у них развивается всего одна семя-
почка, поэтому все усилия направлены на ее опыление.
Другой агент переноса пыльцы — это вода, хотя
опыляемые таким образом цветки встречаются и не
очень часто. Большинство водных растений образуют
соцветия над поверхностью воды и, таким образом,
могут пользоваться для этой цели услугами ветра или
насекомых; мы только что приводили пример Victoria
amazonica, которая опыляется одним из видов хрущей.
Другие примеры энтомофильных водных растений —
частуха (Alisma) и стрелолист (Sagittaria sagittifolia),
тогда как среди анемофильных следует отметить камыш
озерный (Schoenoplectus lacustris) и рогоз широколист-
ный и узколистный (Typha latifolia и Т. angustifolia).
Опыление
165
Гидрофилия
Некоторые водные растения опыляются водой (гид-
рофилия), даже если их соцветия располагаются на ее
поверхности. У Vallisneria americana ко времени цве-
тения цветоножки женских цветков удлиняются и вы-
косят цветки на поверхность. Крошечные мужские
цветки состоят из двух тычинок и околоцветника,
образованного тремя сегментами. Цветки отрываюгся
от растения под водой и всплывают на поверхность,
где сегменты околоцветника откидываются назад и под-
держивают цветок. Беспорядочно плавая, они прибли-
жаются к женским цветкам, чему немало способствует
поверхностное натяжение воды. При столкновении муж-
ские цветки опрокидываются, касаясь своими тычинка-
ми рылец. Если вода захлестывает женские цветки,
они вместе с мужскими цветками окружаются пузырь-
ком воздуха. Когда опыление произошло, женские
цветки снова погружаются, их цветоножки скручива-
ются спирально и втягивают цветки под воду, где и раз-
виваются плоды.
Морская трава (Zostera marina)—цветковое расте-
ние, вернувшееся в море, откуда вышли его предки.
Оно целиком погружено в воду и обитает в прибреж-
ной полосе от границы малой воды до глубины 3,5 м
или около этого. Пыльца освобождается под воюй
и переносится водными течениями к рыльцам женских
цветков. Она нитевидная и обматывается вокруг ры-
лец; считается, что такая форма пыльцы помогает ей
плавать в воде (стр. 118).
Самоопыление
Хотя большинство растений старается избегать са-
моопыления, все же бывает, что по той или иной при-
чине оно становится желательным. Не говоря уже
о таких культурных растениях, как пшеница, которые
всегда самоопыляются, может случиться, что перекрест-
ное опыление не произойдет. В таком случае, чтобы
все-таки дать семена, растению выгодно, -если оно
166
Глава VII
в состоянии, прибегнуть как к последнему средству
к самоопылению. Нередко у растений имеется в запасе
такая возможность.
У адоксы мускусной (Adoxa moschatellina) ты ihh-
ки располагаются в цветке ниже, чем рыльце, что де-
лает невозможным самоопыление. Однако позднее они
вырастают и соприкасаются с рыльцем. Таким обра-
зом, если перекрестное опыление не удалось, цветок
все же может опылиться собственной пыльцой. Анало-
гичное явление наблюдается у многих камнеломок.
У вербейника (Lysimachia nemorum) и зверобоя
(Hypericum perforatum) тычинки сначала наклонены от
рыльца, но потом загибаются внутрь и касаются его.
Во всех этих случаях у цветка имеется возможность
перекрестного опыления, прежде чем он прибегнет
к самоопылению.
Более специализированный способ обеспечения са-
моопыления на случай отсутствия перекрестного опы-
ления можно наблюдать у клейстогамных цветков,
Которые довольно распространены; наиболее знакомый
нам пример — душистая фиалка (Viola odorata). Это
растение цветет рано весной, но цветки его, хотя и пах-
нут приятно, не бросаются в глаза. Несмотря на то что
их посещают насекомые, они редко дают семена. Одна-
ко позже растение образует клейстогамные цветки,
которые прячутся среди листьев и никогда не раскры-
ваются, но благодаря самоопылению обычно дают мно-
жество семян*.
Хотя можно говорить о том, что назначение клей-
стогамных цветков состоит в обеспечении самоопыления
как крайнего средства на случай отсутствия перекре-
стного опыления, эта точка зрения не доказана, и, бо-
лее того, против нее имеются веские доводы. Можно
предположить, что такие цветки могли развиться и под
влиянием плохих условий роста, например недостатка
1 В нашей флоре имеется довольно много видов фиалок с клей-
стогамными цветками, в том числе фиалка удивительная (Viola
mirabilis) и фиалка пушистая (V. hirta), очень обычные в средней
полосе. Кроме того, это же явление можно наблюдать у кислицы
(Oxalis acetoeel la), — Прим. ред.
Опыление 167
питания. Недотрога (Im pat lens noli-tangere), кстати,.
образует клейстогамные цветки, если растет на бедной
почве. У некоторых растений такие цветки появляются
раньше нормальных, а у шалфея (Salvia horminoides)
они образуются почти всегда, тогда как нормальные
цветки — сравнительно редко.
ГЛАВА Viil
ОРХИДНЫЕ
В орхидее всегда было что-то таинственное и даже
зловещее. Дорогие и экзотичные цветки орхидеи не по-
хожи ни на какой другой цветок. Можно подумать,
будто они специально созданы природой для украше-
ния кинозвезд и принцесс, и кажется невероятным, что
они несут ту же функцию, что и цветки лютика. И все
же части их цветков в основном те же, что и у других
растений, хотя они сильно изменились в процессе эво-
люции, особенно с целью обеспечения перекрестного
опыления насекомыми. Необычная форма цветков и то,
что орхидные (Orchidaceae) — самое большое семейст-
во растений, побудило меня посвятить им отдельную
главу.
Название «орхидея» очень старое, восходящее к вре-
менам Древней Греции. Считается, что первым описал
орхидею Теофраст, ученик Платона1; во всяком случае,
он нашел растение с парными утолщенными корнями,
по форме напоминавшими семенники, и назвал его
orchis, что по-гречески означает «семенник». Мы не
можем быть уверены в том, что это действительно была
орхидея. Сейчас название Orchis применяется к одному
из основных родов семейства орхидных.
Сходство клубневидных корней орхидей с семенни-
ками было замечено и в средние века; в то время тор-
говцы лечебными травами собирали корпи орхидей
и использовали их для приготовления разного рода
«приворотных зелий,» которые очень ценились. В неко-
торых частях света орхидные издавна применяются
1 Теофраст (IV в. до н. э.), которого обычно называют «отцом
ботаники», начинал свое образование действительно в школе Пла-
тона, но очень скоро перешел к Аристотелю. Принято считать Теоф-
раста не только любимым учеником, но и научным наследником
Аристотеля. — Прим. ред.
Орхидные
169
в медицине1. Зулусы используют препараты Habenaria
как рвотное средство; на островах Индонезии против
кожных инфекций применяют экстракт Dendrobium;
в Южной Америке используют Spiranthes в качестве
мочегонного средства, a Epidendrutn — как глистогон-
ное.
Орхидные принадлежат к однодольным; насчиты-
вается в общей сложности от 20000 до 35000 различ-
ных видов орхидей — никто не. может сказать точно,
сколько именно. В настоящее время они объединяются
приблизительно в 600 родов. Все орхидные — травы,
хотя некоторые тропические виды — это лазящие расте-
ния. Распространены они по всему свету, от Арктики
до Антарктики, и растут почти во всех возможных ме-
стообитаниях, от тропических влажных лесов до вер-
шин гор, от болот до пустынь.
Эпифитные и наземные орхидеи
Многие тропические орхидеи являются эпифитами,
т. е. растут на других растениях, но не как паразиты,
а просто поселяясь на них везде, где можно найти
опору для корней. Особенно ярко это выражено во
влажных тропических лесах, где можно увидеть расту-
щие на ветвях деревьев эпифитные орхидеи всюду,
куда бы вы ни бросили взгляд. Эпифитные орхидеи
могут поселяться даже на телеграфных проводах, ка-
ким-то образом удерживаясь на столь ненадежной
опоре и образуя цветки.
Корни орхидных, ведущих эпифитный образ жизни,
конечно, не контактируют с почвой. У большинства та-
ких орхидей имеются длинные зеленоватые или белые
1 Клубневидные корни некоторых орхидных умеренной зоны
(главным образом из рода Orchis) и теперь имеют применение в ме-
дицине и постоянно упоминаются в справочниках по лекарственным
растениям (например, в хорошей книге А. Ф. Гаммерман, М. Д. Шу-
пинской и А. А. Япенко-Хмелевского «Растения-целители», изд-во
«Высшая школа»,'М., 1963). Они служат источником получения сли-
зистых веществ,^'применяемых при желудочных катарах у детей,
при отравлениях и т. п. Эти слизи известны под'названием «салеп»,
в высушенные корни орхидей — «клубни салеп», — Прим. ред.
170
Глава VIII
корни, позволяющие им цепляться за опору, иногда
в самых невероятных положениях, примером чему мо-
жет служить Cattleya citrina, которая предпочитает
расти «вниз головой», т. е. корни ее находятся в возду-
хе, а стебель свисает вниз, к земле. Эпифитные орхидеи
часто имеют любопытное образование в коре корней,
так называемый веламен, состоящий из ряда пуегых
клеток в наружной части коры. Функция веламена по-
ка остается предметом дискуссий между ботаниками,
но большинство из них сходится во мнении, что эти
пустые клетки действуют как своего рода губка, пог-
лощая воду. Это резонно, потому что при отсутствии
контакта корней с почвой снабжение водой должно
представлять в какой-то степени проблему для орхидей
даже во влажном тропическом лесу.
Орхидеи умеренных и холодных зон земного шара
в основном наземные и растут в почве, как и другие
растения1. У наземных орхидных преобладают клубне-
видные корни,, которые и послужили причиной их наз-
вания.
Некоторые из орхидей — сапрофиты; их листья ре-
дуцированы до чешуек, не содержащих хлорофилла,
поэтому в отношении питания они зависят от разлага-
ющихся органических веществ почвы и растут, как
правило, в лесной подстилке, богатой гниющими орга-
ническими остатками. В Англии встречаются три вида
таких орхидных: гнездовка (Neottia nidus-avis), ладьян
(Corallorhiza trifida) и надбородник (Epipogium aphyl-
lum)1 2. Эти сапрофитные орхидеи имеют обильно раз-
витую подземную часть, в которой особенно хорошо
1 Во флоре СССР известно несколько десятков видов наземных
орхидей, растущих преимущественно в лесах, на лугах и болотах.
Большинство их относится к большому роду Orchis, в частности
в средней полосе особенно обычен ятрышник крапчатый, или ку-
кушкин цвет — Orchis maculaia. Одна из наиболее популярных ор-
хидей наших лесов — так называемая «ночная фиалка», или любка
двулистная (Platanthera bifolia), — отличается изящными зелено-
вато-белыми цветками и особенно ароматом. Ее иногда неправиль-
но называют просто «фиалкой». — Прим. ред.
2 У нас они тоже встречаются в лесной зоне. Для средней по-
лосы особенно обычна гнездовка в лиственных и смешанных ле*
сах. — Прим. ред.
Орхидные
171
развиваются микоризные грибы (стр. 283). Гифы
гриба пронизывают окружающую лесную подстилку,
переваривая органические остатки при помощи фермен-
тов, а затем передавая питательные вещества назад
орхидее. У гнездовки сплетение массы корней напоми-
нает птичье гнездо (отсюда ее название). У ладьяна
и надбородника корней совсем нет, а на их месте раз-
вивается подземный стебель, или корневище. Корневи-
ще ладьяна распадается на множество веточек, каждая
из которых кончается несколькими короткими веточка-
ми второго порядка, похожими на шишечки; такой
кораллообразный вид корневища и послужил причиной
латинского названия растения Corallorhiza. Надбород-
ник имеет утолщенное корневище, несущее множество
округлых отростков и также напоминающее коралл.
Гнездовка и надбородник совершенно лишены хло-
рофилла; цветоносный побег — единственная надзем-
ная часть растения — несет только бесцветные чешуй-
чатые листья. Обе эти орхидеи должны добывать всю
необходимую им пищу из лесной подстилки, так как
путем фотосинтеза они не могут выработать даже са-
мое небольшое ее количество. Они во всем зависят от
своих партнеров — грибов. С другой стороны, ладьян
содержит некоторое количество хлорофилла в цветонос-
ном побеге и поэтому может самостоятельно синтези-
ровать некоторые питательные вещества, которых,
правда, недостаточно для жизни.
Некоторые сапрофитные орхидные живут невероят-
но долго. В. С. Саммерхейс в книге «Дикорастущие
орхидные Англии» упоминает один австралийский вид,
у которого все растение целиком, вместе с цветоносным
побегом, находится под землей. Эта орхидея была об-
наружена при вспашке никогда прежде не обрабаты-
вавшейся земли. Опыляется ли она почвенными насеко-
мыми или же образует клейстогамные цветки, неиз-
вестно.
Другая интересная орхидея — это японская Gastro-
dia elata, которая является вторичным паразитом дуба
и других деревьев. С этой орхидеей связан микоризный
гриб опенок (Armillaria mellea), паразитирующий на
Деревьях, в том числе и на дубе. Гриб хорошо известен,
т
Г лава Vi II
так как образует толстые подземные нити, называемые
ризоморфами (иногда многометровой длины),при помо-
щи которых он может распространяться от дерева
к дереву. И эти же ризоморфы, кроме того, оказывают-
ся связанными с клубнями Gastrodia. Опенок погло-
щает из дерева-хозяина питательные вещества, часть
которых достается растению.
Типы цветков орхидных
Цветки орхидных уникальны по своему строению.
Будучи совершенно непохожими на цветки других
растений, они обнаруживают предельную специализа-
цию для опыления насекомыми. Если не среди всех
цветковых растений, то по крайней мере среди одно-
дольных цветки орхидных являются наиболее совер-
шенными. Некоторые из них мелкие, чуть больше I мм
в диаметре, другие достигают 30 см и более. Цветок
орхидей имеет три лепестковидпых, т. е. окрашенных
подобно лепесткам, чашелистика и три лепестка, при-
чем чашелистики и лепестки могут иметь одинаковую
или совершенно различную окраску. Один из чашели-
стиков может отличаться от двух других своими раз-
мерами, или окраской, или же по обоим этим призна-
кам; это всегда верхний чашелистик. Самый нижний
из трех лепестков тоже видоизменяется, иногда очень
сильно, и причудливая форма, которую он приобретает,
так же как и необычная его окраска, в основном и при-
дают цветкам многих орхидей экзотический вид. Обыч-
но нижний лепесток выступает из цветка, образуя так
называемую губу, которая может быть разделена на
несколько долей. Губа часто продолжается за цветок,
образуя трубчатую структуру, называемую шпорцем
(рис. 31).
Но больше всего специализация цветка орхидеи
проявляется в строении андроцея и гинецея. Они объ-
единены в сложное образование — колонку, на вершине
которой располагается единственная тычинка с двумя
половинками пыльника, содержащими пыльцу. Как
уже говорилось (стр. 163), пыльца энтомофильных
цветков часто бывает клейкой, пыльцевые зерна ели-
Орхидные
173
паются в комочкщ так что насекомое, даже слегка
задев тычинку, унесет на своем тельце довольно много
пыльны. У орхидных эта тенденция доведена до своего
логического конца, так как в каждой половинке пыль-
ника пыльца склеена в общую, утолщенную на одном
Рис. 31. Схематическое изображение цветка ятрышника мужского
(Orchis mascula), вид спереди.
конце массу, называемую поллинием (иногда подлинн-
ее больше одного). Как правило, от поллиния отходит
короткая ножка «каудикула», с помощью которой он,
когда созреет, прикрепляется к маленькому мешочку
на «клювике» — выступающей части колонки, как раз
под тычинкой. Насекомое-опылитель должно вытащить
сразу весь поллиний, в котором могут находиться ты-
сячи пыльцевых зерен. Основание ножки поллиния
У многих орхидей расширено в плоский клейкий диск
(«прилипальце»), который и обеспечивает прочное при-
174
Глава Vlll
клеивание сравнительно массивного поллиния к насе-
комому; у некоторых орхидных нет прилипальца, но
липкая жидкость, выделяющаяся из клювика, прочно
приклеивает поллиний к насекомому.
Под клювиком располагаются площадочки рыль-
ца, которых у большинства орхидей две, по одной на
каждой стороне колонки. Они часто бывают в той или
иной степени соединены друг с другом. Под остальной
частью цветка располагается завязь; она очень длин-
ная, и зачастую ее трудно отличить от цветоножки.
Завязь содержит много сотен, а то и тысячи семяпо-
чек.
Другая особенность цветка орхидных, не замечае-
мая случайным наблюдателем, — это явление «пере-
вертывания», т. е. вращение всего цветка по мере его
развития, пока он не окажется «вниз головой». Когда
цветок закладывается в почке, губа находится наверху,
но, развиваясь, цветок поворачивается на 180°, так
что, когда он ,наконец раскрывается, губа оказывается
внизу.
Механизмы опыления
Цветки орхидей энтомофильные и благодаря сво-
ему сложному строению обычно приспособлены к опре-
деленному виду насекомых. Крайняя степень адапта-
ции к перекрестному опылению, наблюдаемая у неко-
торых орхидей, поистине фантастична, и ни один
цветок не может в этом с ними состязаться.
Для начала разберем довольно простой механизм
опыления, наблюдаемый у листеры, или тайника
(Listera ovata), — орхидеи с зеленоватыми цветками,
растущей и в буковых лесах, и на выгонах, и на пес-
чаных дюнах. Механизм этот замечателен тем, что
в нем учитывается психология насекомого. Цветок
имеет очень длинную губу, которая в нижней части
разделена на две доли, а вверху имеет проходящее по
всей ее длине углубление, в которое выделяется нек-
тар. Остальные небольшие по размеру чашелистики
и лепестки образуют «шлем», заключающий в себе
Орхидные
175
довольно короткую толстую колонку. Тычинка содер-
жит два поллиния, которые, созрев, располагаются на
самом краю вогнутого клювика таким образом, что их
основания направлены вперед. Листера опыляется
некоторыми мелкими жуками, а из перепончатокры-
лых — наездниками. Они взбираются вверх по губе,
чтобы достать спрятанный в желобке нектар, и при
этом касаются края клювика. Тотчас же из клювика
выбрасывается капля клейкой жидкости и приклеивает
поллинии к голове или спинке насекомого; через не-
сколько секунд жидкость затвердевает, и поллинии
приклеиваются очень прочно. Такое внезапное выделе-
ние жидкости из клювика пугает насекомое, которое,
надо полагать, в глубоком возмущении отлетает от
цветка, чтобы «обдумать» случившееся. Возможно, че-
рез одну-две секунды, решив, что нектар стоит голо-
вомойки, оно снова попытается проникнуть в цветок,
но может случиться и так, что за время вынужденного
«размышления» насекомое уже перелетит на другое
растение. Если цветки нового растения находятся на
той же стадии развития, что и цветки предыдущего,
с насекомым произойдет та же история, и к его тельцу
приклеятся новые поллинии. Однако тайник — про-
тандричное растение, т. е. его тычинки созревают
прежде, чем рыльце становится готовым к приему
пыльцы. Если же насекомое посетит более старый цве-
ток, оно обнаружит уже загнувшийся кверху клювик
цветка и обнажившуюся часть рыльца, которая
и воспримет поллинии.
Обратите внимание, что в этом механизме внезап-
ное выделение клейкой жидкости из клювика дает два
совершенно различных эффекта. Капля жидкости проч-
но приклеивает поллинии к насекомому и в то же время
сильно пугает его, заставляя покинуть цветок и пере-
лететь на другой, находящийся обычно на другом
растении. Благодаря этому возрастает вероятность
перекрестного опыления.
Более специализированный механизм опыления
У ятрышника мужского (Orchis mascula)—одного из
самых распространенных в Англии видов, встречаю-
щегося в лесах и на открытых пастбищах, главным
176
Глава VIII
образом на известковых почвах1. Колосья его фиолето-
вых цветков образуются в мае и июне и опыляются
пчелами. Верхний чашелистик и два боковых лепестка
образуют шлем, заключающий в себе колонку, которая
располагается как раз перед входом в шпорец, образо-
ванный задней частью губы. На вершине колонки на-
ходится пыльник с двумя поллиниями, основания кото-
рых вытянуты в длинные ножки (каудикулы) с клей-
кими дисками, или прилипальцами. У зрелых тычинок
поллинии располагаются на клювике, имеющем кро-
шечное образование, называемое мешочком, в котором
находятся прилипальца поллиниев. Благодаря находя-
щейся в мешочке клейкой жидкости прилипальца оста-
ются липкими до тех пор, пока цветок не опылится.
Пчела садится на губу цветка и всовывает свой
хоботок в отверстие шпорца, чтобы добраться до жид-
кости, содержащейся в клетках, выстилающих его
стенку. При этом опа надавливает головой на створча-
тую сторону мешочка, обнажая поллинии, один из
которых (или оба) приклеивается к верхней части ее
головы. Покидая цветок, она уносит на голове полли-
нии, торчащие вертикально, как рожки, и прочно при-
клеенные липкими прилипальцами, клейкое вещество
которых застывает и твердеет в несколько секунд.
Во время полета насекомого к другому растению
положение поллиниев меняется. Примерно через пол-
минуты или около этого они загибаются вниз под
прямым углом, так что располагаются параллельно
спинке пчелы и торчат вперед, как копье идущего
в атаку всадника. Когда пчела посещает следующий
цветок, поллинии из-за своего положения не попадают
на клювик, проходя под ним, и оказываются как раз
против нижерасположенных площадочек рыльца. Часть
пыльцы отделяется от поллиниев и приклеивается
к площадочкам рыльца, так что цветок опыляется.
Заметьте, что загибание поллиниев вниз и вперед
занимает около полминуты. Это дает пчеле время закон-
чить свои дела на цветках колоса, где она собирала полли-
нии, и перелететь на следующее растение. Самоопыле-
1 У нас этот вид встречается главным образом на северо-западе
Европейской части СССР. — Прим. р$.
Орхидные
177
ние поэтому очень маловероятно. Если же пчела сядет
на другой цветок, прежде чем поллинии успеют заг-
нуться вниз, они просто ударятся о выступающий край
клювика и опыления не произойдет. С одного соцветия
пчела может собрать дюжину и больше подлиннее
и сможет опылить много цветков, прежде чем ей надо
будет, так сказать, перезарядиться.
Описанный механизм легко наблюдать в действии,
если у вас есть возможность собрать соцветия муж-
ского (Orchis mascula) или пятнистого (О. fuchsii)
ятрышника, которые опыляются одним и тем же спо-
собом. Возьмите остро отточенный карандаш и на
мгновение введите его в отверстие шпорца. Вынув
карандаш, вы увидите поллинии, торчащие вертикаль-
но на кончике карандаша, приклеенные своими прили-
пальцами. Если подождать несколько секунд, то мож-
но увидеть, как они загнутся вперед и окажутся
«в опыляющей позиции».
Чтобы наблюдать некоторые из наиболее замысло-
ватых механизмов опыления орхидных, мы должны
обратиться к тропическим видам. У Coryanthes губа
имеет форму ковша, который наполнен слегка опьяня-
ющей жидкостью. Насекомые-визитеры падают в этот
ковш и не могут из него выбраться, так как стенки
ковша скользкие; положение насекомого ухудшается
еще и потому, что оно слегка пьянеет под действием
жидкости в ковше. Существует, однако, один довольно
доступный выход — через узкое отверстие, лежащее
непосредственно под поллиниями. Через него насекомое
и выбирается наружу, захватывая при этом поллинии
и перенося их на другое растение.
Еще более любопытный механизм наблюдается
у южноамериканской орхидеи Gongora maculata. Пче-
ла, садящаяся на губу в поисках нектара, обнаружи-
вает, что губа скользкая, и не может удержаться на
ногах. Волей-неволей пчела скользит вниз по изогну-
той и свисающей вниз колонке и в конце своего пути,
захватывая поллинии, переносит их на своей спинке
на другое растение. Те же трудности поджидают пчелу
и на новом цветке, но на этот раз она оставляет при-
несенные поллинии на клейком рыльце.
178
Глава VIII
Псевдокопуляция
Цветки некоторых орхидей обнаруживают порази-
тельное сходство с определенными животными, В Анг-
лии, например, встречаются виды Ophrys apifera,
Ophrys insectifera и Aceras anthropophorum, цветки ко-
торых соответственно напоминают пчелу, муху и чело-
века. Несомненно, что в большинстве случаев эта
мимикрия — чисто случайное явление, но иногда она,
привлекая насекомых, способствует опылению. У сре-
диземноморского вида Ophrys speculum и австралийско-
го Cryptostylis leptochila форма и окраска губы цветка
делают ее очень похожей на самку насекомого-опыли-
теля. Цветки посещаются самцами насекомых, которые
пытаются спариться с «самками» и при этом или уно-
сят, или оставляют на рыльце принесенные с другого
цветка поллинии. Во всех этих случаях самцы насеко-
мых выходят из куколок раньше, чем самки, а цвете-
ние орхидей происходит в период, когда самцы не име-
ют возможности спариться с самками. Когда же появля-
ются самки, самцы больше не интересуются орхидеями.
Это явление, получившее название псевдокопуля-
ции, по-видимому, весьма распространено у орхидных,
которые имеют сходство с насекомыми. Чтобы описан-
ный механизм сработал, необходимо, чтобы цветение
орхидеи было точно приурочено к периоду, когда сам-
цы насекомого, служащего моделью, летают в поисках
самок и не находят их. У орхидных такая точность,
по-видимому, строго соблюдается.
Может случиться, что, несмотря на прекрасно отла-
женный механизм опыления насекомыми, перекрестного
опыления не произойдет. На этот случай многие ор-
хидные имеют запасную систему, с помощью которой
они могут самоопылиться. Для растений, которые
столь тщательно приспособились к опылению опреде-
ленным видом насекомых, это особенно важно, так как,
если по случайной прихоти природы численность опы-
лителей внезапно упадет, нормальный механизм опы-
ления может отказать. Способы, обеспечивающие само-
опыление, в случае если перекрестного опыления
почему-либо не произойдет, варьируют в зависимости
Орхидные
i7S
от вида орхидей. У ЪрИгуз apifera, например, по мере
старения цветка колонка сморщивается, поллинии вы-
талкиваются из мешочка и свисают вниз, как раз на-
против рыльца, так что при малейшем сотрясении
цветка они соприкасаются с воспринимающими пло-
щадочками рыльца, производя самоопыление.
У других видов рода Ophrys механизм самоопыле-
ния несколько сложнее. В молодых цветках колонка
прямостоячая, но по мере их старения в тканях колон-
ки начинается автолиз (самопереваривание), так что
она свисает вниз и поллинии касаются рыльца.
Прорастание семян
Изменения, которые следует за опылением, не менее
замечательны, чем сами механизмы опыления. Семена
орхидей очень мелкие и образуются в огромных коли-
чествах. Миллион семян в одном плоде — вполне
обычное явление, а в коробочке Cycnoches ventricosum
их четыре миллиона. Дарвин подсчитал, что если бы
все семена, образованные одним растением пятнистого
ятрышника, развились в новые растения, они заняли
бы площадь 0,4 га, а если бы все семена от этого
и последующих поколений дали в свою очередь новые
растения, то четыре поколения потомков одного расте-
ния пятнистого ятрышника заселили бы всю сушу. Так
что, может быть, и к лучшему, что большая часть се-
мян орхидей гибнет.
В семенах орхидных эндосперм или вовсе не раз-
вивается, или развивается крайне слабо и весь запас
питательных веществ находится в самом зародыше.
Более того, зародыш не дифференцирован на корешок,
почечку и семядолю, как у большинства однодольных,
а состоит из небольшой массы недифференцированных
клеток. Дифференциация зародыша до прорастания
не происходит.
Когда семя орхидеи прорастает, то прежде всего
возникает клубневидное образование, так называемый
протокорм. В таком состоянии растение остается иног-
да до двух лет и не может нормально развиваться
дальше до тех пор, пока не будет заражено микориз-
180
Глава VIII
ным грибом, обычно каким-то видом Rhizoctonia. По-
сле этого оно очень медленно продолжает расти. Из
верхушки протокорма появляются первые листья, по-
том из его основания вырастают корни. Весь этот про-
цесс идет крайне медленно, и проходит много лет,
прежде чем растение станет настолько взрослым, что-
бы образовать цветки, а затем семена. (О микоризе
орхидных см. стр. 283).
Необходимость заражения протокорма микоризным
грибом для дальнейшего его развития — причина того,
что садовники считают очень трудным успешное выра-
щивание орхидей из семян. Стало обычным говорить,
что семена орхидных вообще не могут развиваться без
гриба. Однако Кнудсону удалось доказать ошибочность
этой крайней точки зрения.
Он предположил, что роль гриба не столь уж зна-
чительна и ограничивается тем, что он просто «одал-
живает» растению некоторые ферменты, которых у ор-
хидеи нет, и это дает ей возможность разлагать
сложные органические соединения до более простых,
например до сахара. Он помещал семена орхидей
в специальную стерильную среду, содержащую в числе
других веществ сахар, и успешно доводил развившиеся
из них растения до взрослого состояния и цветения без
участия гриба. Все же роль микоризного гриба выяс-
нена еще не полностью. На первый взгляд кажется,
что Кнудсон доказал правильность своей точки зрения
и что гриб, участвующий в этом союзе, служит лишь
источником ферментов, нужных растению-партнеру.
Однако дальнейшие исследования показали, что это не
совсем так. Некоторые орхидеи нельзя вырастить на
среде Кнудсона, хотя они прекрасно растут при добав-
лении стерильных экстрактов гриба или витаминов.
Детально этот вопрос пока не изучен1.
1 У нас весьма успешно выращиваются из семян разнообраз-
ные орхидеи в оранжерее Главного ботанического сада АН СССР.
Разработка оригинальных методов культуры орхидей — заслуга
В. А. Поддубной-Арнольди и В. А. Селезневой. Есть книга
В. А. Поддубной-Арнольди и В. А. Селезневой «Орхидеи и их куль-
турам, Изд-во АН СССР, М., 1957, а также В. А. Селезневой «Тро-
пические и субтропические орхидеи», изд-во «Наука», М., 1965.
ГЛАВА IX
РАССЕЛЕНИЕ РАСТЕНИЙ
Растения обречены всю жизнь оставаться на одном
месте; впрочем, ползучая пальма может «передвигать-
ся», образуя из кроны растущие вниз и укореняющиеся
стебли, но «движется» она чрезвычайно медленно. Да-
же такие растения, как земляника, имеющая в своем
распоряжении стелющиеся по земле во всех направле-
ниях и укореняющиеся столоны, не могут таким спо-
собом путешествовать на дальние расстояния. Для того
чтобы растение в случае нужды могло попасть в любое
место земного шара, оно должно иметь какие-то более
эффективные средства передвижения, чем те, которые
в состоянии обеспечить вегетативное тело растения.
У цветковых это было достигнуто путем превращения
плодов или семян в агентов расселения.
Семя — это прежде всего покоящаяся структура, обо-
рудованная так, что внутри прочной кожуры зародыш
растения может в безопасности переждать, пока не кон-
чится зима (или в некоторых странах—сухой сезон)
и условия снова станут благоприятными для роста. Се-
мя — компактная структура, идеально приспособленная
для путешествий, так как оно маленькое и легкое и не
нуждается до прибытия на место назначения ни в воде,
ни в пище. Семя — или плод, в котором оно содержит-
ся,— может ехать зайцем в шерсти животного, путешест-
вовать за отворотами брюк, в желудке птицы или даже
в багажнике самолета, причем сам переносчик об этом
и не догадывается. Упав наконец в землю, иногда в ты-
сячах километров от места своего происхождения, семя
готово прорасти в новое растение, если климат и почва
благоприятствуют этому. В случае же неблагоприятной
погоды семя может и подождать; спешить ему незачем.
Семена образуются из семяпочек цветка после опло-
дотворения. Клетки внутри крошечной семяпочки делят-
ся, формируя зародыш растения, состоящий из зачаточ-
182
Глава IX
ного стебелька с парой семенных листьев (семядолей)
по обеим сторонам; если растение однодольное, то вместо
двух образуется одна семядоля (фото XVIII). Как пра-
вило, имеется и зачаточный корешок. По мере развития
семени питательные вещества, состоящие в основном
из масла или крахмала, а также небольшого количества
белка, накапливаются в семядолях или во внезароды-
шевой ткани, называемой эндоспермом-, они пойдут на
питание проростка, пока в его листьях не образуется
хлорофилл и он не сможет питаться самостоятельно.
Наконец, покровы семяпочки утолщаются и твердеют,
превращаясь в семенную кожуру, защищающую семя до
его прорастания.
Образование плодов
Пока семя развивается, завязь цветка превращается
в плод. Она разрастается, иногда очень сильно, претер-
певая в то же время изменения, необходимые для вы-
полнения ее двойной функции — защиты семени и уча-
стия в его распространении. Стенка завязи может стать
мягкой и сочной, как у томата, делая плод привлека-
тельным для птиц, или же грубой, покрытой крючкова-
тыми щетинками, которые позволяют плодам цеплять-
ся за шерсть переносящих их животных, как у подма-
ренника цепкого (Galium aparine). Плод может быть
маленьким и твердым, с парашютиком из волосков, об-
легчающим его парение в воздухе и распространение
ветром, как, например, у одуванчика (Taraxacum offi-
cinale). Поскольку плод подсыхает, находясь на мате-
ринском растении, то может случиться, что вследствие
развивающегося в его тканях натяжения он «взорвет-
ся», разбрасывая семена, как шрапнель, куда попало;
примером такого растения может служить утесник
(Ulex). Таким образом, способы, благодаря которым
семена распространяются с помощью плодов, весьма
разнообразны.
Ботаники различают настоящие плоды, образованные
одной только видоизмененной завязью и ее содержи-
мым, и ложные плоды (псевдокарпии), включающие.
Расселение растений
помимо завязи и другие структуры'. Томат и слива —
настоящие плоды, поскольку их мякоть образована раз-
росшейся стенкой завязи. Земляника же ложный плод,
так как ее нежная мякоть в действительности представ-
ляет собой разросшееся цветоложе, а вовсе не завязь.
Настоящие плоды земляники — это зернышки, сидящие
на поверхности ложного плода; каждое из них — целый
плод, состоящий из затвердевшей завязи с единствен-
ным семенем внутри. Яблоко — еще один пример лож-
ного плода: его мясистая часть тоже развилась из раз-
росшегося цветоложа, а настоящий плод—только «серд-
цевина». Ежевика, малина и логанова ягода—это
в действительности целые собрания плодов, так как
каждый из маленьких шариков, составляющих такой
«плод» — это отдельный плодик. Он имеет такое же
строение, что и слива, и называется костянкой. Ежевика,
таким образом, сложный плод, состоящий из собрания
костянок.
Следовательно, то, что мы обычно называем плодом,
с ботанической точки зрения может быть одним из не-
скольких разных по происхождению образований.
К счастью, чтобы оценить качества плода, вовсе не обя-
зательно знать его ботаническую природу. Когда мы
с наслаждением едим зрелый персик или ломтик дыни,
мы делаем только то, что назначено нам природой, ибо
мясистые, сочные плоды для того и созданы, чтобы их
поедали животные (в Англии главным образом птицы);
пройдя через пищеварительный тракт животного непов-
режденными, заключенные в плодах семена могут по-
пасть в землю вместе с испражнениями. Различают два
основных типа сочных плодов (за исключением ложных,
как у земляники): ягоды и костянки. У ягоды весь около-
плодник (перикарп) сочный, как у крыжовника и тома-
та (может показаться странным называть томат яго-
дой, но для ботаника это типичный пример ягоды).
В околоплоднике различают твердую наружную кожи-
1 Вопросы классификации плодов нельзя считать вполне ре-
шенными. Рекомендуем читателю книгу Р. Е. Левиной «Плоды»
(Саратов, 1967), где изложено современное состояние вопроса. —
Лрим. ред.
184
Глава IX
цу, или эпикарп, и сочную центральную часть, или ме-
зокарп, составляющий основную массу плода. В центре
плода находятся семена — «зернышки». Околоплодник
костянки делится не на два, а на три слоя: эпикарп, или
кожицу, сочный мезокарп и самую внутреннюю твер-
дую часть, или эндокарп, который окружает семя (боль-
шинство костянок односеменные, тогда как в ягодах
может быть одно или несколько семян). Примерами
костянок являются слива и вишня. Косточка сливы —
это не семя, а эндокарп, содержащий семя, в чем мож-
но убедиться, осторожно расколов косточку.
Многие из употребляемых в пищу плодов — или яго-
ды, или костянки, хотя часто имеют необычную структу-
ру, придающую им специфические качества. Апельсин,
лимон, грейпфрут, лайм и мандарин — все это различ-
ные виды рода Citrus (семейство Rutaceae); их плоды —
это ягоды, у которых и эндокарп, и мезокарп жесткие.
Эпикарп образует кожицу и содержит многочисленные
масляные железки в виде полостей, заполненных эфир-
ным маслом, обусловливающим характерный запах
и вкус плода. Мезокарп более толстый и представлен
белой губчатой массой ткани, окружающей мясистую
часть плода. Плодолистики, которые мы едим, богаты
витамином С и содержат большое количество пектина;
получение этих веществ из плодов — немаловажная
отрасль промышленности в районах, где выращивают
цитрусовые. Внутренность плода состоит из сильно уве-
личенных плодолистиков завязи, причем каждый из сег-
ментов, на которые можно разделить плод, представ-
ляет собой отдельный плодолистик. Сок продуци-
руется многочисленными волосками, вырастающими из
стенки плодолистика; за время развития плода они
образуют большое количество сока, а характерное
для данного вида эфирное масло придает ему особый
привкус.
Плод банана — тоже ягода, хотя и не соответствую-
щая обычному представлению об этом типе плодов (фо-
то XIX). Разводимые сорта бананов триплоидны, т. е.
их клеточные ядра содержат тройной набор хромосом
(палочковидных телец, являющихся носителями наслед-
ственных признаков) вместо двойного набора в обычных
Расселение раСГений
185
(диплоидных) ядрах. По этой причине нормальное опло-
дотворение цветков трудноосуществимо, плоды разви-
ваются без оплодотворения и не содержат семян, хотя
можно заметить, что завязь состояла из трех плодолисти-
ков, и даже увидеть семяпочки — маленькие коричневые
образования в центре плода. Дикорастущие бананы
диплоидны (с двойным набором хромосом, характер-
ным для большинства растений); плоды у них разви-
ваются обычным путем, после оплодотворения, и содер-
жат семена.
Плод банана имеет резко выраженный изгиб, кото-
рый отнюдь не случаен. Цветки этого растения собраны
вместе на длинном повислом соцветии: вверху —муж-
ские, посредине — обоеполые, внизу — женские. По мере
развития плоды изгибаются в направлении, противопо-
ложном действию силы* тяжести (отрицательный геотро-
пизм), так что их концы оказываются отогнутыми нару-
жу. Плоды грецкого ореха (Juglans regia) и кокосового
ореха (Cocos nucifera) — костянки, как у сливы и виш-
ни, но, прежде чем их продавать, у них удаляют сочный
мезокарп, оставляя только твердый окружающий семя
эндокарп. В эндокарпе кокосового ореха, как н у мно-
гих других костянок, процесс одревеснения, т. е. отвер-
дения клеточных стенок вследствие отложения на вну-
тренней их стороне твердого вещества лигнина
(гл. 11), распространился за пределы сосудов и волокон
древесины; практически одревесневает все, включая
паренхимные клетки коры. Эндокарп кокосового
ореха — одна из самых твердых структур в раститель-
ном мире.
В скорлупе кокосового ореха мы обнаруживаем
единственное семя. Семенная кожура срастается с эн-
докарпом плода, а внутри находится маленький зародыш
и большая масса эндосперма. Молодой эндосперм состо-
ит из довольно тонкого наружного слоя, окружающего
большую полость, заполненную жидкостью, так назы-
ваемым «кокосовым молоком». В процессе созревания
семени жидкость постепенно заменяется плотным ве-
ществом, хотя полностью молоко никогда не исчезает.
Молоко молодых кокосовых орехов — восхитительный
напиток, но, чтобы его попробовать, надо ехать в тро-
Глава 1Х
пики, где можно достать свежие кокосовые орехи. К то-
му времени, когда орехи попадают к нам, от молока
остается нечто пресное и безвкусное.
В кокосовом молоке содержится несколько неиденти-
фицированных веществ, ускоряющих рост растений. Ес-
ли небольшие кусочки апикальной меристемы растения
(верхушка стебля или корня, где идет активное деление
клеток) поместить в раствор, содержащий все извест-
ные нам необходимые для роста вещества, это почти
не повлияет на процесс деления. Но если подобным же
образом обработать кусочки зрелой ткани, например
флоэмы корня моркови, деление клеток сильно замед-
лится или даже совсем прекратится. Добавление к это-
му раствору небольшого количества кокосового молока
вызывает активное клеточное деление. Совершенно оче-
видно, что молоко содержит какое-то вещество или
вещества, действующие как стимуляторы роста. Анализ
кокосового молока подтвердил присутствие по меньшей
мере шести таких веществ.
Другой приМер стимулирующего рост действия коко-
сового молока — это улучшение состояния маленьких,
еще недоразвитых зародышей дурмана (Datura stramo-
nium), помещенных в питательный раствор, куда добав-
лено кокосовое молоко и ауксин. Без кокосового моло-
ка никакого роста не происходит. Более зрелые зароды-
ши могут обходиться без кокосового молока; очевидно,
что на этой стадии развития они или получают необхо-
димые ростовые вещества из семени, или же (что более
вероятно) уже способны сами их синтезировать.
Теория распространения океаном
(кокосовые орехи)
Способы распространения кокосовых орехов яви-
лись предметом оживленной полемики среди ботаников.
Согласно традиционной точке зрения, они расселяются,
плавая по морям, и то, что кокосовые пальмы встреча-
ются почти на всех островах Тихого океана, придает
правдоподобность этой версии. В большинстве учебни-
ков по основам ботаники кокосовый орех рассматрива-
ется как пример плода, приспособленного к переносу
Расселение растений
187
морскими течениями. Вполне возможно, что это и так,
но в доказательствах этой гипотезы обнаруживаются
некоторые противоречия: рассказы очевидцев не соот-
ветствуют друг другу. Возможно, что теория 0 распро-
странении кокосовых орехов морем — одна из тех по-
спешных теорий, в которые верят только потому, что ни-
кому не пришло в голову в них усомниться.
Когда Тур Хейердал и его товарищи пересекали Ти-
хий океан на плоту «Кон-Тики», кокосовые орехи, на-
ходившиеся в корзинах на палубе, оставались жизнеспо-
собными в течение всего путешествия из Америки в По-
линезию; другие же орехи, которые хранились под
палубой, где они были доступны морской воде, вскоре
испортились или под действием воды, или же по вине
так называемых «сборщиков отбросов», постоянно очи-
щающих океан от любых плавающих органических
остатков. С другой стороны, один ботаник из Гонолулу
ПО дней держал кокосовые орехи в воде залива Пирл-
Харбор, и после этого они ни в коей мере не утратили
жизнеспособности. За столь долгое время кокосовый
орех вполне мог быть отнесен морскими течениями на
расстояние около 5000 км. Таким образом, эти два на-
блюдения прямо противоположны друг другу.
Единственным заслуживающим доверия доказатель-
ством переноса кокосовых орехов океаном служит на-
личие их на некоторых вулканических островах, в част-
ности группы Кракатау, где в 1883 году наблюдалось
сильнейшее в истории извержение. В 1928—1930 годах
в результате вулканической деятельности возник новый
остров Анак Кракатау1У, который начал заселяться
всем, что могло принести море. Д-р ван Лёвен, посетивший
остров спустя почти 18 мес после его появления, обна-
ружил, что берег заселен растениями, принадлежащими
к так называемому сообществу Barringtonia. В это
растительное сообщество, часто встречающееся в тропи-
ках по берегам океана, входит баррингтония (Barring-
tonia), панданус (Pandiinus) и индийский миндаль (Тег-
minalia catappa). В числе прочих растений д-р ван Лё-
вен насчитал 41 прорастающий кокосовый орех;
последние, несомненно, были принесены морем, посколь-
ку остров находился еще в очень опасном состоянии,
188
Глава IX
и ни одно туземное каноэ не отважилось бы пристать
к нему. К несчастью, неизвестно, развились ли бы из
этих орехов кокосовые пальмы, так как в 1932 году
в результате вулканической деятельности остров был
разрушен.
Известно, правда, несколько, но только несколько,
достоверных примеров, когда случайно занесенные оре-
хи образовывали взрослые пальмы. Например, кокосо-
вые пальмы на восточном берегу Тринидада, как гово-
рят, появились после кораблекрушения французской
шхуны, груженной кокосовыми орехами.
Из всего вышесказанного следует, что кокосовые
орехи могут переноситься морем (иногда на большие
расстояния) и прорастать там, где их прибьет к берегу.
Однако это случается очень редко. Не говоря уже
о трудностях и опасностях морского путешествия, су-
ществует множество факторов, препятствующих прора-
станию и развитию ореха во взрослое растение. Здесь
надо принять во внимание и жгучее тропическое солн-
це, и трудность выживания проростка среди массы уже
растущих растений. Даже если проросток чудом попадет
на подходящее место и начнет старательно расти, почти
наверняка найдется множество животных, например ди-
ких свиней, которые позаботятся, чтобы от него ничего
не осталось. Мы должны, следовательно, прийти к вы-
воду, что перенос кокосовых орехов морскими течения-
ми— вещь почти невероятная.
Как же в таком случае кокосовый орех пересекает
море? Ответ очень прост: на каноэ! Человек использует
кокосовые орехи уже тысячи лет и во всякое путешест-
вие в тропиках всегда брал их с собой. Торговые шхуны,
появившиеся гораздо позднее, тоже сыграли в этом свою
роль. Таким образом, продвижение кокосовой пальмы
в глубь материка — целиком.заслуга человека; почти то
же самое можно сказать и о ее распространении по бе-
регам морей. Кокосовый орех — это плод, распростра-
няемый животными, и в данном случае таким живот-
ным является человек.
Удивительным союзником кокосового ореха в зоне
Тихого океана является краб Пальмовый вор (Birgus
lalro), обнаруженный Дарвином на Кокосовых островах
Расселение растений
189
(острова Килинг) в 1836 году во время путешествия на
корабле «Бигль». У этого животного на передней паре
ног имеются обыкновенные клешни, но они необычайно
большие и тяжелые; клешни задней пары ног меньше,
и слабее. Волокно за волокном краб сдирает с кокосо-
вого ореха скорлупу всегда на том конце, где располо-
жены три «глазные ямки». Добравшись до этих ямок,
он несколько раз прокалывает одну из них, пока не про-
делает в твердой скорлупе дырку, а затем вставляет
в скорлупу одну из задних клешней и вытаскивает эн-
досперм. Этот краб питается исключительно кокосовыми
орехами; не будь это так, трудно представить, чтобы
у него могла выработаться столь своеобразная форма
клешней, не говоря уже об инстинкте, побуждающем его
отдавать предпочтение такой внешне непривлекатель-
ной пище. Не только в растительном мире действует
естественный отбор, создающий высокоприспособленные
организмы.
Способ распространения омелы
Белая омела (Viscum album)' расселяется интерес-
ным способом, естественно вытекающим из ее паразити-
ческого образа жизни на широколиственных деревьях,
в особенности на яблоне. Хорошо известные белые «яго-
ды» омелы (на самом деле это не ягоды, а ложные пло-
ды, включающие в себя разросшееся цветоложе цветка)
весьма привлекательны для птиц, особенно для дрозда-
дерябы. Сочная часть плода содержит клейкую массу,
которая употребляется для изготовления птичьего клея.
Когда птица поедает плод, на ее клюве остается это
клейкое вещество, и, чтобы избавиться от такого не-
удобства, она чистит клюв о ветку дерева, на котором
сидит. При этом семена омелы остаются на коре, удер-
живаясь на ней липкими остатками плода. Таким обра-
зом омела переносится с дерева на дерево.
х В СССР омела встречается только в западных и юго-запад-
ных районах; это западноевропейское растение. — Прим, ред,
190
Глава IX
Другие способы расселения
Очень любопытный механизм расселения у Rafflesia
arnoldii, паразитного растения Индонезии, которое, как
говорят, распространяется слонами. Rafflesia паразити-
рует на корнях лесных деревьев. Она совсем не обра-
зует надземных стеблей и всю жизнь проводит под
землей, но во время цветения развивает громадные,
иногда до метра в поперечнике цветки, лежащие на
земле. Плоды у нее мягкие и сочные (см. стр. 159).
Проблема состоит в том, чтобы семена раффлезии
попали в спрятанные под землей корни дерева. Когда
слоны наступают на плоды и раздавливают их, семена
пристают к их широким ступням, а йотом вдавливаются
в корни деревьев под тяжестью слона. Только такое же
тяжелое, как слон, животное, ноги которого при ходьбе
глубоко уходят в мягкую почву, может осуществить
этот необычный способ распространения.
В Англии, когда говорят о сочных плодах, привле-
кающих животнкх, имеют в виду главным образом птиц,
хотя не надо забывать и о поедающих плоды млекопи-
тающих, в том числе и человеке. В тропиках у птиц
гораздо больше конкурентов из числа млекопитающих,
желающих полакомиться легкодоступными плодами.
Слоны, свиньи, различные виды оленей, тапиры, обезья-
ны и множество других животных, а также человек яв-
ляются участниками пира. Насекомые борются за остат-
ки со стола более крупных животных.
Плод не готов к распространению, пока он не со-
зреет и содержащиеся в нем семена полностью не
сформируются. Образование множества плодов, если
все они будут съедены до созревания семян, было бы
для растения напрасной тратой энергии. Поэтому вкус,
окраска и запах появляются у сочных плодов только
после того, как семена созреют. В детстве большинство
из нас на собственном опыте испытало неприятные
последствия налетов на фруктовые сады, когда яблоки
еще не поспели. Для защиты от нетерпеливых потреби-
телей незрелые плоды обычно содержат неприятные на
вкус вещества, в частности органические кислоты (на-
пример, яблочную кислоту). Только после созревания
Расселение растений
191
плода эти кнздоды исчезают и заменяются сахаром,
придающим плоду сладкий вкус. В то же время окрас-
ка плода изменяется от зеленой, незаметной на фоне
листвы, до более яркой, «рекламирующей» его местона-
хождение. Не случайно, что так много сочных плодов,
созрев, имеют красную окраску — красный цвет на фоне
зеленого лучше всего заметен для птиц и млекопитаю-
щих.
Сухие плоды могут распространяться животными,
обычно прикрепляясь к их шерсти. Таковы различные
виды цепких плодов, «репьев», которые, как правило,
Рис. 32. Плоды колдун-травы
с крючочками.
снабжены небольшими крючками, чтобы цепляться за
шерсть животных или за перья или лапки птиц. Эти
крючочки могут быть частью самого плода, как у под-
маренника цепкого (Galium aparine) и колдун-травы
(Circaea lutetiana) (рис. 32), или принадлежать к ка-
кой-нибудь другой части цветка, включенной в плод,
чтобы способствовать его расселению. Так, у гравилата
городского (Geum urbanum) столбик не вянет, а сох-
раняется и загибается на конце в виде крючка (рис. 33).
Цепкий плодик череды (Bidens) имеет модифицирован-
ную чашечку: вместо обычного для большинства слож-
ноцветных (к которым принадлежит череда) хохолка —
«паппуса» из кольца волосков — она формирует две или
три жесткие щетинки, покрытые массой направленных
192
Глава IX
вниз шипиков (рис. 34). У обычного репешка (Agri-
mania eupatoria) чашечка цветка остается при плоде
и покрывается крючочками.
Цепкие плоды прочно прикрепляются к любой шер-
шавой поверхности, в чем легко убедиться, попытав-
шись снять их с шерсти собаки или кошки или, если
на то пошло, с собственных брюк.
Рис. 35. Султанчик волосков
на семени иван-чая.
Многие плоды разносятся ветром, и мы видим
у растений массу приспособлений, способствующих это-
му. Самое простое из них заключается в том, чтобы
образовывать огромное количество мелких и легких се-
мян; когда плод раскрывается, освобождая семена
(иногда резко их выбрасывая), они тут же подхваты-
ваются даже легким дуновением ветерка. Этим способом
пользуется наперстянка (Digitalis purpurea) и многие
орхидные, которые настолько специализировались в этом
отношении, что их семена напоминают пыль — такие они
мелкие.
Бывает, что на самих семенах имеются султанчики
волосков для улавливания ветра, как у иван-чая (Cha-
maenerion angustifolium) (фото XX). Еще будучи при-
крепленным к растению, плод раскрывается, выставляя
пучки волосков (рис. 35), и в ветреный день в воздухе
носится множество семян1.
1 Всем известны также опушенные семена тополя («тополиный
пух»), заполняющие городские" улицы в начале лета. Плоды топо-
ля—двустворчатые коробочки — раскрываются при созревании. —
Прим. ред.
Расселение растений
133
Весьма обычны крылатые плоды, как у вяза, ясеня
и клена; крыло, как правило, образуется из плоского
выроста околоплодника. Форма крыла варьирует у раз-
ных растений. У вяза оно овальное (рис. 36), тогда как
у ясеня — длинное й слегка скрученное (рис. 37). Дву-
семянный плод клена при созревании расщепляется на
две половинки, хотя половинки эти друг от друга не
отделяются; семена остаются скрепленными, с ножкой
Рис. 36. Крылатый плод вяза.
Рис. 37. Крылатые плоды
ясеня.
между ними и двумя длинными плоскими крыльями, вы-
ступающими по обеим сторонам, как пропеллер само-
лета (рис. 38).
Наиболее совершенные приспособления к воздушной
транспортировке имеются у сложноцветных (Composi-
tae). «Цветок» маргаритки, подсолнечника или нивяни-
ка — на самом деле не отдельный цветок, а собрание
цветков. Если мы осторожно вытащим один из лепест-
ков подсолнечника, то обнаружим, что в сущности это
отдельный, очень маленький цветок. При рассмотрении
плоской середины «цветка» (диска) оказывается, что
он тоже состоит из множества крошечных цветков, хо-
тя они не имеют длинных лентовидных лепестков, укра-
шающих краевые цветки. В крупной головке подсолнеч-
194
Глава IX
ника могут быть сотни таких крошечных цветочков. Тес-
ное сближение мелких, но многочисленных цветков
и образование головки (правильнее — корзинки) делает
соцветие в целом гораздо более заметным для насеко-
мых-опылителей, чем отдельный цветок.
Некоторые сложноцветные имеют и срединные цвет-
ки без лентовидных лепестков, и краевые, выделяющиеся
своим венчиком; венчик этот в действительности обра-
зован пятью сросшимися лепестками, хотя часто внешне
Рис. 39. Плод
одуванчика.
это незаметно. У других все цветки язычковые, т. е.
с лентовидными лепестками, как, например, у одуван-
чика.
Отдельный цветок сложноцветных крайне упрощен.
Хотя завязь образована двумя плодолистиками, она
имеет только одно гнездо и содержит единственное се-
мя. Чашечка обычно представлена кольцом волосков,
или хохолком (паппусом), который служит парашюти-
ком, позволяющим маленькому плоду иногда часами
парить в воздухе, так как даже легкий порыв ветра не
дает ему опуститься на землю. Плод сложноцветных мо-
жет пролететь по воздуху многие километры, прежде
чем наконец приземлится.
У некоторых сложноцветных, например у одуванчи-
ка (рис. 39) и козлобородника (Tragopogon pratensis),
мы видим дальнейшее усовершенствование. По мере
развития плода хохолок отделяется от него длинной
Расселение растений
195
ножкой. На пушистой головке одуванчика это отчетливо
видно. У козлобородника волоски хохолка перистые, со
множеством мелких ответвлений, что еще более облег-
чает его роль как парашюта.
Разбрасыванию семян из плодов иногда способству-
ет движение стеблей в ветреную погоду. «Кадильный
механизм» мака — именно такой случай. Зрелые плоды
при этом открываются рядом отверстий, через которые
семена высыпаются при сотрясении плода. Когда по-
являются цветочные почки, цветоножки мака становят-
ся положительно геотропичными (т. е. растут в на-
правлении, совпадающем с действием силы тяжести)
и цветки поникают. Благодаря такому положению —
чашечкой вверх — они в какой-то мерс защищены от
непогоды.
Однако по мере образования плодов цветоножки ме-
няют свое отношение к силе тяжести, становясь отрица-
тельно геотропичными, и растут вверх. Будучи слегка
эластичной, цветоножка под действием ветра совершает
резкие движения, что заставляет семена высыпаться из
плода. Поскольку отверстия находятся в верхней части
плода, семена не сразу падают па землю, а разбрасы-
ваются в стороны, так что по крайней мере часть из них
приземляется на некотором расстоянии от материнского
растения и имеет шансы прорасти, не испытывая затене-
ния. Это, между прочим, одна из целей большинства
приспособлений к распространению семян.
Изменение реакции цветоножки мака на действие
силы тяжести происходит в последний момент, что наи-
лучшим образом способствует разбрасыванию семян.
У некоторых растений изменяется реакция не на силу
тяжести, а на свет. Из их числа отметим льнянку (Cym-
balaria muralis)—очаровательное маленькое растеньи-
це, впервые появившееся в Англии в 1640 году и впо-
следствии ставшее широко распространенным; растет
оно в расщелинах старых стен. Сначала его цветонож-
ки положительно фототропичны (т. е. растут по направ-
лению к свету), так что сиреневые цветки доступны
опыляющим их пчелам, но когда начинают завязывать-
ся плоды, отношение к свету меняется. Цветоножки ста-
новятся отрицательно фототропичными и, избегая света,
196
Глава IX
укрываются в трещинах стен. Здесь семена и освобож-
даются из плодов.
У некоторых растений мы видим более усложненный
вариант «принципа катапульты», характерного для цве-
тоножки мака: внезапно и сильно «взрываясь», плод от-
брасывает семена на некоторое расстояние. Созревшие
бобы утесника обыкновенного (Шех europaeus) вслед-
ствие неравномерного подсыхания различных слоев око-
лоплодника находятся в состоянии натяжения. При ма-
лейшем воздействии зрелый плод лопается и семена раз-
летаются в разные стороны. Если в летний день забрать-
ся в заросли утесника, подальше от шума машин и воя
реактивных самолетов, то можно услышать треск ло-
пающихся бобов* 1.
Однако при распространении семян утесник не по-
лагается только на свой «взрывчатый механизм» — само
семя у него снабжено еще одним приспособлением, воз-
можно даже более эффективным, чем «взрывающиеся»
бобы. Каждое семя имеет сочный оранжевый вырост,
привлекающи'й муравьев. Нередко можно видеть, как
они «возятся» с семенем и с трудом тащат его, а по-
скольку муравьи передвигаются по дорожкам, то счи-
тается, что именно поэтому утесник, как правило, растет
вдоль тропинок.
Органы, привлекающие муравьев, так называемые
элайосомы, весьма обычны в растительном мире и раз-
виваются на семенах или плодах. Как правило, они бы-
вают пропитаны маслом, которое любят муравьи. Хо-
рошо известными примерами таких растений являются
мускатный орех и африканский боб (рис. 40), а из
более близких нам —дымянка (Fumaria), живучка
(Ajuga reptans), пальчатая осока (Carex digitata)
и многие другие2. У растений рода Allium и ряда других
1 У нас обычно такое явление можно наблюдать в посадках
«желтой акации», или караганы (Caragana arborescens). Эффект-
ные «стреляющие» плоды имеют виды недотроги (Impatiens noli tan-
gere, I. parviflora). — Прим. ped.
1 В нашей флоре элайосомы имеют еще такие обычные растения,
как копытень (Asaru/n еигораеит), большинство видов фиалок
(Viola), чистотел' (Chelidonium та]us), хохлатки (Corydalis), ожики
(Luzula) и другие. — Прим. ред.
Расселение растений
197
однодольных маслом пропитана тонкая семенная ко-
жура. Распространение семян муравьями чрезвычайно
важно для лесных растений — ведь муравьи в основном
лесные жители.
Плоды и семена некоторых водных растений распро-
страняются водой, хотя надо помнить, что эти растения
могут использовать и некоторые другие средства (мно-
гие, например, распространяются птицами). Плоды, ко-
торые путешествуют по воде, обычно имеют какое-ни-
будь приспособление, придающее им плавучесть. Так,
плоды частухи — водяного подорожника (Alisma plan-
tago-aquatica) держатся на воде благодаря рыхлой
Рис. 40. Африканский боб с элайосомой.
и губчатой стенке плода, заполненной воздухом; ту же
роль выполняют и опробковевшие воздухоносные ткани
семенной кожуры ириса, или касатика (Iris pseudaco-
rus). Как мы видели, кокосовый орех, о котором обыч-
но говорят как о величайшем морском путешественни-
ке, в действительности нс распространяется морем; но
множество морских растений, включая часто встречаю-
щуюся на побережьях тропическую Barringtonia, рас-
селяются именно таким способом. Плод Barringtonia
имеет заполненные воздухом ткани, помогающие ему
плавать на поверхности воды, но может обходиться
и без них. У берегов Англии растут морская капуста
(Crambe maritima), какиле (Cakile maritima), морская
свекла (Beta vulgaris var maritima) и виды солероса
(Salicornia), которые переносятся с одного места на
другое морской водой.
Мангры часто расселяются с помощью морских те-
чений, но у многих из них путешествуют не плоды и се-
мена, а проростки. Мангры — это не отдельное растение
198
Глава IX
и даже не род растений (более подробно о них говорит-
ся на стр. 58, 120 и 222). Это общее название, приме-
няемое к растениям с определенным обликом, живущим
в определенных местообитаниях. Многие мангровые
живородящи, т. е. их семена прорастают еще в плоде,
прикрепленном к материнскому растению. Направ-
ляемый силой тяжести молодой корень проростка растет
вертикально вниз, и когда наконец проросток выпадает
из плода, он летит подобно стреле, корнем вперед, и за-
рывается в ил мангрового болота. В случае если он упа-
дет не в ил, а в воду, проросток без всякого для себя
вреда может проплыть большое расстояние, пока не
окажется на берегу.
Некоторые плоды и семена не располагают специ-
альными средствами расселения, и тем ие менее одно
поколение у них сменяет другое. Такова ромашка па-
хучая (Matricaria matricarioides) —сорняк, обычный по
обочинам дорог. Ее плоды мелкие и бескрылые, воло-
систый хохолок, характерный для многих сложноцвет-
ных, рудиментарен или вовсе отсутствует, и все же это
растение становится все более распространенным на
Британских островах. Полагают, что этим оно обязано
главным образом плодам, которые вместе с грязью при-
стают к колесам автомобилей. Этим способом могут
распространяться и другие сорняки.
Семена распределяются не только в пространстве,
но у некоторых наиболее процветающих сорняков и во
времени. Другими словами, у них не все семена про-
растают в один и тот же год. Существует старая
крестьянская пословица: «Только год рос сорняк, семь
лет теперь от него не избавишься:», но это буколическое
изречение несколько преуменьшает возможности семян
сорняков: для дикой горчицы, например, этот срок со-
ставляет не 7, а около 40 лет. Отмечались случаи появ-
ления этого вредного сорняка па почве, которая по край-
ней мере в течение 40 лет была занята лесом; то же
можно сказать о щавеле курчавом (Rumex crispus), по-
дорожнике большом (Plantago major) и некоторых дру-
гих сорняках, растущих па пустырях и пахотных землях.
Выше я касался вопроса о долговечности семян, но
необходимо учесть, что цифры, приведенные на стр. 129,
Расселение растений
199
справедливы для семян, хранящихся в более или менее
сухом месте. Однако в почве вовсе не сухо, и гораздо
труднее определить продолжительность покоя семян
в естественных условиях, чем на сухом гербарном листе.
Об этом пока известно мало. Иногда длительный период
покоя можно, не сомневаясь, отнести за счет твердости
семенной кожуры, но этим нельзя объяснить, почему се-
мена, годами находящиеся в почве, остаются в покое.
За это время семенная кожура непременно должна раз-
мягчиться до такой степени, чтобы прорастание стало
возможным.
Была проделана работа по изучению прорастания се-
мян черной горчицы (Sinapls nigra), которые, находясь
в почве, могут долгое время оставаться в состоянии по-
коя. Оказалось, что семена, насыщенные водой, не про-
растают, если содержание углекислого газа в окружаю-
щем воздухе выше определенного уровня. Углекислый
газ освобождается при разложении органических ве-
ществ в почве, и вполне возможно, что именно этим
в значительной степени объясняется наблюдаемая там
и сям задержка прорастания. Но чтобы создать на этот
счет какую-то теорию, нужны дальнейшие исследования.
Преимущество неодновременного прорастания семян
состоит в том, что, растягивая это прорастание на дли-
тельный период времени, растение получает возмож-
ность использовать множество различных климатиче-
ских ситуаций. Если бы все семена прорастали одно-
временно, большинству или даже всем проросткам
угрожала бы опасность погибнуть в случае внезапного
наступления неблагоприятных условии, например за-
сухи.
ГЛАВА X
ВЫЖИВАНИЕ РАСТЕНИЙ
Во влажном тропическом лесу растения могут расти
круглый год, так как постоянные высокая температура
и влажность устраняют необходимость периода покоя.
В большинстве других мест каждый год наступает не-
благоприятное для жизни растений время. В Англии
это зима, когда вследствие промерзания почвы и ледя-
ных ветров прекращается рост почти всех растений, за
исключением морозостойких, составляющих меньшин-
ство. В других странах неблагоприятным может быть не
столько холод, сколько сухой сезон; именно так обстоит
дело в районах, где муссонные леса представляют со-
бой коренной тип растительности, например в некото-
рых частях Индии. Палящее солнце в сухое время года
является там таким же препятствием для развития
растений, как Ъимние холода в Англии.
В это трудное для растений время некоторые из них
могут спастись исключительно благодаря выживанию
семян. Я имею в виду однолетние растения. Они живут,
пока не кончится благоприятный период, а затем пол-
ностью отмирают, оставляя после себя только семена,
обеспечивающие сохранение вида. Иногда за один се-
зон может смениться несколько поколений, как у мокри-
цы, растущей всюду па рыхлых почвах. Этому малень-
кому растению нужно всего несколько недель, чтобы
вырасти, достигнуть зрелости и дать семена.
В некоторых отношениях это может быть идеальным
решением проблемы неблагоприятного сезона, ибо ничто
так хорошо не приспособлено к выживанию в трудных
условиях, как семя. Толстая семенная кожура и состоя-
ние покоя, в которое оно погружено, гарантируют, что
никакие капризы погоды не нанесут ему серьезного вре-
да. Но однолетний образ жизни все же имеет один
серьезный недостаток: поскольку весь цикл должен за-
нимать не больше года, размеры, которых может достиг-
нуть растение, ограничены. Большинство однолетников
маленькие.
Выживание растений
201
Растение, выживающее из года в год, называют-
многолетним, и чтобы оно могло пережить суровое вре-
мя, по крайней мере какие-то его части должны оста-
ваться живыми и вырасти снова на следующий год.
Это достигается с помощью зимующих почек, или, точ-
нее, почек возобновления, так как неблагоприятным пе-
риодом не всегда бывает зима.
Древесные многолетники — деревья и кустарники —
образуют почки на ветвях. В течение зимы растение мо-
жет лишиться листьев, но почки остаются, чтобы
раскрыться весной. Однако не все многолетники дере-
вянистые. У травянистых многолетников, например
у дельфиниума, или живокости, с наступлением зимы
мягкие травянистые стебли отмирают, но корневище
с почками возобновления остается живым, хотя и на-
ходится в состоянии покоя. От зимних морозов почки
защищены подстилкой из мертвых листьев и окружаю-
щей почвой. Когда весеннее солнце снова прогреет зем-
лю, они пробуждаются к жизни и образуют надземные
побеги.
Датский ботаник Раункиер первым предложил раз-
делить растения на несколько жизненных форм1; хотя
это было более 60 лет назад, его идеи не утратили своей
ценности и в наши дни. По Раункиеру, жизненные фор-
мы классифицируются в соответствии с расположением
почек возобновления (в климатических условиях Англии
это то же самое, что и зимующие почки). Так, напри-
мер, растения, у которых зимующие почки остаются на
воздухе (у нас это деревья и большинство кустарников),
называются фанерофитами-, растения, которые, подобно
крокусу, скрывают почки под землей,— геофитами и т. д.
В настоящее время различают семь основных групп
жизненных форм.
1 Хотя заслуги Раункиера в классификации и изучении жиз-
ненных форм действительно велики, автор неправ, приписывая ему
приоритет в разделении растений на «жизненные формы». История
учения о жизненных формах уходит своими истоками еще к Теофра-
сту, а самый термин предложен соотечественником Раункиера
Вармингом лет на 20 раньше. См. в Большой советской энциклопедии
статью «Жизненные формьГрастений», а также книгу^И. Г. Се-
ребрякова «Экологическая морфология растений», изд-во «Высшая
Школа», М-Л 1962. — Прим. ред.
202
Глава X
Жизненная форма фанерофитов
К фанерофитам относятся все деревья и почти все
кустарники. Их почки расположены в воздухе высоко
над поверхностью почвы и испытывают на себе все тя-
готы зимы. Возможно, именно поэтому фанерофиты —
растения преимущественно мягкого климата, где моро-
зы играют небольшую роль или вовсе не относятся
к числу подстерегающих растения опасностей. Это не
означает, что некоторые фанерофиты не приспособились
к более суровому климату; северные хвойные леса —
тип растительности, простирающейся от Северной Аме-
рики через Северную Европу до Сибири, служат дока-
зательством того, что некоторые фанерофиты способны
выдерживать холод. Но число видов этих северных фа-
нерофитов в противоположность числу особей очень ма-
ло по сравнению с числом их видов в тропиках. Если мы
хотим увидеть деревья во всем их великолепии, мы
должны отправиться в экваториальные области земно-
го шара. Во влажном тропическом лесу на сравнительно
небольшом пространстве число разных видов деревьев
может исчисляться сотнями.
В таком лесу почки могут быть совершенно не защи-
щены чешуйками, так как при отсутствии какого бы то
ни было неблагоприятного периода ничто не мешает им
сразу же начать развиваться. В местах с резко выра-
женным сухим периодом различным образом защищен-
ные почки становятся обычным явлением1. Наружные
листочки могут быть покрыты волосками, которые, за-
держивая пары воды, регулируют испарение из почек.
Той же цели служит и растительный клей или смола,
покрывающая почки. У некоторых растений почки за-
щищены прилистниками — листовидными выростами,
расположенными у основания листового черешка.
По мере продвижения в более холодные области, где
зимы могут быть длительными и суровыми, мы обычно
1 Имеются многочисленные данные о том, что и во влажных тро-
пических лесах достаточно много видов с почками, закрытыми че-
шуйками; это явление связано с внутренним ритмом роста. —
Прим, ред.
Выживание растений
203
обнаруживаем закрытые почки, защищенные почечными
чешуями; это можно наблюдать, например, у большин-
ства фанерофитов Англии. Иногда защитные свойства
чешуек могут быть усилены опушением или клейкой
смолой, как у хорошо известных клейких почек конского
каштана.
Другое приспособление фанерофитов к неблагоприят-
ным условиям — это осенний листопад. Если во влаж-
ном тропическом лесу преобладают вечнозеленые де-
ревья, то большинство наших деревьев и кустарников
листопадны, они теряют листья с наступлением зимы.
Благодаря этому предотвращается чрезмерная потеря
воды путем транспирации, что особенно важно в тот
период, когда охлаждение почвы затрудняет поглощение
воды корнями в количестве, достаточном для нужд расте-
ния. Подробнее мы поговорим о листопаде в гл. XI, где
речь пойдет о ксероморфных приспособлениях.
Чем дальше фанерофиты проникают в высокие ши-
роты, тем меньше становятся их размеры. Та же тенден-
ция наблюдается у них и с увеличением высоты над
уровнем моря. Мы можем проследить это на примере
некоторых наших деревьев и кустарников. Например, на
юге Нортамбсрлепда карликовая береза (Beiula папа)
не встречается, а растет главным образом в болотистых
местностях, находящихся на высоте 250...850 м над уров-
нем моря, распространяясь дальше на север, к Ислан-
дии. Ветви этого небольшого, не более метра высотой,
кустарника прижимаются к почве. Чем ближе к земле,
тем лучше защищены зимующие почки1.
Жизненная форма хамефитов
От таких фанерофитов, как карликовая береза, не-
далеко до второй жизненной формы — хамефитов. Почки
возобновления у них расположены близ поверхности
земли или невысоко над пей. Надземные побеги с цвет-
1 В СССР эти закономерности превосходно прослеживаются как
по широтным ботанико-географическим зонам (лее -— лесотундра —
тундра), так и по высотным поясам (см. превосходную книгу
А. В. Кожевникова <По тундрам, лесам, степям и пустыням», Гео-
графгиз, М., 1955).
204
Рлава X
ками свободно растут вверх, в воздух, но у травянистых
хамефитов осенью могут отмирать, и тогда от них оста-
ются лишь участки, несущие расположенные близ по-
верхности земли почки следующего года. Холодной зи-
мой снежный покров может защитить их от высыхания
и от морозов. В более теплых местностях, где им гро-
зит скорее высыхание, чем замерзание, они обыкновен-
но укрываются под подстилкой из мертвых раститель-
ных остатков. Из числа хамефитов, обычных для Англии,
отметим лесную звездчатку (Stellaria holostea), луговой
чай (Lysimachia nummularia) и веронику лекарственную
(Veronica officinalis)1.
Жизненная форма гемикриптофитов
Там, где явно выражен неблагоприятный период,
особенно если этот период холодный, почки нуждаются
в лучшей защите, чем это возможно у хамефитов. Сле-
дующую стадию в смысле защиты почек можно на-
блюдать у гемикриптофитов. Здесь зимующие почки
формируются в поверхностном слое почвы, где они за-
щищены не только самой окружающей почвой, но и за-
сохшими остатками надземных побегов, отмирающих
с наступлением холодов.
Категории гемикриптофитов
По сравнению с хамефитом почки гемикриптофита
защищены лучше, поэтому не удивительно, что боль-
шинство наших трав и почти половина всех растений
Центральной Европы принадлежат к этому типу. Как
и другие жизненные формы, гемикриптофиты подразде-
ляются на категории по степени защищенности почек.
Начиная с протогемикриптофитов, характеризующихся
равномерным распределением листьев по всему цвето-
носному побегу вплоть до соцветия, они прогрессируют
1 Хамефиты чаще всего представляют собой кустарнички и
полукустарники. Они господствуют, с одной стороны, в тундрах,
а с другой — в полупустынях и некоторых типах пустынь (напри-
мер, разные виды полыни). У кустарничков побеги не отмирают на
зиму. — Прим. ред.
Выживание растений
205
через полу розеточное растение, у которого нижние
листья заметно крупнее верхних, к розеточному расте-
нию, где прямостоячий стебель несет только цветки,
а листья собраны в розетку у основания стебля, как
у обычного одуванчика.
Зимующие почки гемикриптофитов нс зарываются
глубоко в почву, а располагаются в поверхностных ее
слоях. Защитный покров для них создается разными
способами, среди которых важнейшую роль играет пе-
ремешивание почвы земляными червями. Корни многих
гемикриптофитов втягивающие (контрактильные)- они
втягивают растение в почву, если его части с течением
времени становятся обнаженными.
Иногда благодаря деятельности кротов или по ка-
ким-либо другим причинам подземные побеги, наобо-
рот, погружаются слишком глубоко в землю. В этом
случае почки возобновления образуются на побеге вы-
ше и снова оказываются расположенными близко от по-
верхности почвы.
У многих гемикриптофитов имеются столоны — пол-
зучие стебли, которые время от времени укореняются
и служат для вегетативного размножения. Они могут
стлаться по земле или же располагаются непосредствен-
но под поверхностью почвы. Земляника — пример геми-
криптофита, столоны которого стелются по земле, тогда
как подбел (Petasites hybridus) имеет подземные сто-
лоны.
Некоторые растения, в частности крапива двудомная
(Vrtica dioica) и чистец лесной (Stachys sylvatica),
образуют столоны обоих типов.
Розеточные растения
По-видимому, форма розеточных растений, таких,
например, как одуванчик, в какой-то степени определя-
ется светом. Розеточный облик создается за счет уко-
рочения междоузлий стебля (промежутков между ли-
стьями). У стебля, растущего под землей, междоузлия
нормальные, но стоит ему выйти на свет, как они уко-
рачиваются и листья сближаются. Если главный сте-
бель одуванчика засыпать землей, чтобы создалось впе-
206
Глава X
чатление, будто он только собирается показаться из
земли, междоузлия будут удлиняться до тех пор, пока
верхушка стебля снова не пробьется к свету; после
этого они укорачиваются и растение принимает свойст-
венный ему розеточный облик. Иное дело — цветоносный
побег. Свет не влияет на его рост, и поэтому он тянет-
ся вверх, в воздух. Листьев он не образует.
Жизненная форма геофитов
От гемикриптофитов, которые характеризуются рас-
положением зимующих почек в верхних слоях почвы,
легко перейти к следующей жизненной форме — геофи-
там. Их зимующие почки скрыты в земле; побеги на-
чинают расти и выходят наружу только тогда, когда
погода становится благоприятной. Все выращиваемые
садовниками луковичные и клубнелуковичныс (нарцисс,
тюльпан, гиацинт, крокус) принадлежат к этой катего-
рии.
Почки возобновления геофитов защищены лучше,
чем у любого другого растения. Укрытые почвой, они
могут не опасаться ни высыхания, ни морозов. Особен-
но хорошо геофиты приспособлены к жизни в областях
с длительным сухим периодом: поэтому естественно,
что в засушливых степях они являются преобладающей
жизненной формой. Тем не менее геофиты могут су-
ществовать и в тех районах, где вегетационный период
длинный, а зима суровая. Доказательством тому служат
мириады крокусов в Швейцарских Альпах, радующих
глаз весной.
В условиях короткого вегетационного периода (в сте-
пях) для геофитов очень важен ускоренный темп роста.
Надземный побег, несущий листья и цветки, должен
развиваться быстро. Следовательно, геофит должен
иметь подземные запасы питательных веществ, за счет
которых надземная часть могла бы начать развиваться
сразу же, как только наступит благоприятный период.
У многих геофитов роль хранилищ выполняют лукови-
цы или клубнелуковицы. У других для этой цели слу-
жит толстый подземный стебель (корневище).
Выживание растений 207
Жизненная форма гелофитов
Растения, растущие на болотистых, насыщенных во-
дой почвах и укрывающие свои зимующие почки под
землей, называются гелофитами. Таков, в частности, ро-
гоз (Typha). Некоторые гелофиты, например стрело-
лист обыкновенный (Sagittaria sagiitifolia), могут расти
и на отмелях, оставляя почки возобновления зимовать
в илистом дне.
Жизненная форма гидрофитов
Последняя категория гелофитов сливается со следу-
ющим классом, называемым гидрофитами, чьи почки
в неблагоприятное время укрыты и защищены водой.
К этой группе относятся многие хорошо знакомые нам
водные растения, в том числе белая кувшинка и желтая
кубышка (N'ymphaea alba и Nyphar lutea), у которых
корневища с зимующими почками располагаются на
дне водоема, где растут растения. У некоторых гидро-
фитов корневища отсутствуют. Их зимующие почки от-
деляются от растения и опускаются на дно водоема, где
и дожидаются прихода весны. Как раз таким гидрофи-
том является рдест (Potamogeton obtusifolius).
Жизненная форма терофитов
Седьмая, последняя жизненная форма представлена
растениями, которые в конце вегетационного периода
полностью отмирают, доверяя свое выживание семенам.
Это терофиты, о которых мы уже говорили. В некото-
рых отношениях они могут рассматриваться как наи-
лучшим образом защищенные, поскольку ничто так не
устойчиво в трудных условиях, как семя.
Подобно семенам, зимующие почки обычно перехо-
дят в состояние покоя и в естественных условиях не
выходят из него, пока не придет время. Это несомнен-
ное биологическое преимущество для растения; ведь бы-
ло бы бесполезно защищать зимующие почки, если бы
они могли начать развиваться преждевременно, подвер-
гая нежные молодые побеги действию холода или за-
сухи. Покой почек — явление- всеобщее, отсутствующее
только в местах с исключительно благоприятными уело-
208
Глава X
виями, например во влажном тропическом лесу’, и осо-
бенно ярко выраженное у фанерофитов и криптофитов,
произрастающих в зонах умеренного климата, где зи-
мы холодные.
Покой почек
В климатических условиях северной и Центральной
Европы, включая Британские острова, холод, оказывает-
ся, выступает в качестве естественного фактора, нару-
шающего покой почек. Зимующие почки у фанерофитов
Англии трогаются в рост только после того, как в те-
чение некоторого времени подвергнутся действию темпе-
ратуры ниже 9 °C. Длительность периода воздействия
варьирует у различных растений; для некоторых доста-
точно 200 ч, чтобы нарушить состояние покоя, а для
других этот период составляет несколько тысяч часов.
Состояние покоя можно снять и искусственным пу-
тем, например обработкой кислотами или гибберелли-
ном (веществе), продуцируемое грибом Fusarium moni-
liforme), воздействием парами эфира или синильной
кислоты, введением в почки спирта или погружением ве-
точек на несколько часов в тепловатую воду с после-
дующим перенесением их в теплицу. Почему эти обра-
ботки эффективны, неизвестно.
Внутренние факторы, обусловливающие состояние
покоя, точно еще не установлены. Можно предположить,
что в этом участвует ауксин (см. гл. IV), поскольку он
тормозит развитие почек, возникающих ниже растущей
верхушки побега, но, по-видимому, он не является при-
чиной покоя. В некоторых случаях похоже на то, что
ингибирующее вещество образуется в самих спящих
почках. В спиртовой вытяжке из почек и листьев клена
{Acer pseudoplatanus) имеется вещество, замедляющее
рост колеоптилей пшеницы и некоторых других частей
растений. В начале зимы содержание в почках этого
пока еще не идентифицированного вещества наи-
большее, а когда растение начинает активно ра-
сти — наименьшее; полностью оно никогда не исчезает.
1 У большинства тропических деревьев покой почек, хотя и
кратковременный, все же выражен, но он не связан с внешними
условиями. — Прим, ред,
Выживание растений
209
Важной особенностью этого довольно таинственного
ингибитора является то, что он образуется (по крайней
мере частично) при выращивании растения в условиях
короткого дня. Длина дня, т. е. период суток, в течение
которого растение освещается, как мы узнаем из сле-
дующей главы, чрезвычайно важна для растений, поэто-
му не было бы удивительным, если бы переход почек
в состояние покоя в какой-то мере зависел и от нее.
Было обнаружено, что в листьях клена, находящихся на
искусственном коротком дне, образуется больше инги-
битора, чем в нормально освещаемых листьях; это наво-
дит на мысль, что в природе более короткие дни, пред-
вещающие наступление осени, вызывают увеличение
количества ингибитора, заставляя почки переходить
в состояние покоя.
Зависимость периода покоя от длины дня обнаружи-
вают и другие растения; если в тот период, когда дни
становятся короче, давать растениям дополнительное
освещение, период покоя наступает позже. Тюльпанное
дерево {Liriodendron tulipifera) продолжает расти всю
зиму в условиях искусственного длинного дня и подхо-
дящей температуры. Более того, тот факт, что допол-
нительное освещение не обязательно должно быть очень
сильным, опровергает гипотезу, согласно которой за-
держка наступления покоя обусловливается увеличе-
нием интенсивности фотосинтеза.
Образуется ли ингибитор, вызывающий переход по-
чек в состояние покоя, и в других растениях, кроме
клена, мы не знаем. Мы несведущи и в том, каким
образом короткий день вызывает состояние покоя, и мо-
жем лишь сказать, что у некоторых растений, которые
были исследованы, это явление отмечается.
В настоящее время покой почек — это широчайшее
поле для исследований, результаты которых, несомнен-
но, будут представлять интерес с научной, а может
быть, и с практической точки зрения1.
1 О перезимовке растений, покое и пробуждении почек и дру-
гих сезонных явлениях можно прочитать в превосходной популяр-
ной книге Л. В. Кожевникова «Весна и осень в жизни растений>,
Изд-во МГУ, М., 1950. — Прим. ред.
ГЛАВА XI
ПРИСПОСОБЛЕНИЕ К СРЕДЕ
Впервые покинув море и поселившись на j суше,
растения вынуждены, были приспосабливаться к/совер-
шенно иным условиям среды; в гл. I мы говорили'о том,
как совершенствовались некоторые из их приспособле-
ний. Для закрепления растений на месте и поглощения
воды и минеральных веществ из той почвы, которая уже
была, развилась корневая система, а для проведения
воды и этих веществ в те части растений, которые боль-
ше всего в них нуждались, сформировались проводящие
ткани. Наземные растения не появились в один день;
чтобы вполне приспособиться к жизни на суше, им по-
требовались миллионы лет непрерывной эволюции.
Начавшийся в те далекие времена процесс адапта-
ции продолжается и сейчас. Растения не могут позво-
лить себе оставаться неизменными, так как это означало
бы вступить на путь вымирания. Прежде всего с го-
дами окружающая среда меняется, и растение, превос-
ходно приспособленное к определенным условиям, мо-
жет оказаться в изменившихся условиях. Климат мо-
жет стать теплее или холоднее, более влажным или
более сухим. Растение, которое не в состоянии приспо-
собиться к меняющейся обстановке, должно или погиб-
нуть, или переселиться в другое место.
Когда на смену позднему палеозою пришел ранний
мезозой, климат резко изменился. Болотистые леса ка-
менноугольного периода после перми уступили место
пустыням триаса. В жизни растений это произвело на-
стоящий переворот. Многие виды отказались от борьбы
и вымерли, оставшиеся же обнаружили, что мест, где
они могли бы жить, становится все меньше. В то же
время усилилась конкуренция со стороны новых видов,
организация которых лучше отвечала новым условиям.
Изменение среды предъявляло к растениям новые
требования, но вместе с тем открывало перед ними но-
вые возможности. Прежние виды в большинстве своем
уже не удовлетворяли этим требованиям, однако пере-
Приспособление к среде
211
мены, приведшие”'» вымиранию одних видов, явились
стимулом к эволюции других. Как всегда, время опас-
ностей было и временем возможностей.
Новая катастрофа разразилась в плейстоцене, когда
на территории Англии наступило внезапное похолодание
и льды распространились от Северного полюса почти
до современного побережья Суссекса. Растения отсту-
пали перед надвигавшимися льдами, и только па сво-
бодных от льда возвышенных участках, называемых ну-
натаками, возможно, выжили немногие выносливые ви-
ды. Когда климат снова потеплел и льды отступили,
назад вернулась лишь часть растений; многие из них
так и не совершили обратного путешествия и навсегда
исчезли из нашей флоры.
Считается, что на протяжении плейстоцена было че-
тыре больших ледниковых эпохи, разделенных тремя
межледниковыми. Первое оледенение началось прибли-
зительно 600 тыс. лет назад, и прошло около 20 тыс. лет
с тех пор, как ледники наконец начали отступать. Жи-
вем ли мы и сейчас в межледниковую эпоху, за кото-
рой последует новое оледенение, никто не знает.
Хотя в настоящее время условия на земном шаре
сравнительно стабильны (в физическом смысле), в при-
роде по-прежнему существует жестокая конкуренция.
Растения ведут нескончаемую борьбу со средой — под
«средой» я подразумеваю все климатические, физико-
географические и почвенные факторы, в совокупности
составляющие местообитание растений, а также влия-
ние соседних животных и растительных организмов
и других особей того же вида, конкурирующих с ними
за место и пищу. Чтобы иметь шансы выжить, растение
должно быть исключительно хорошо приспособлено
к окружающим условиям.
Некоторые растения научились противостоять под-
стерегающим их превратностям судьбы. Обыкновенный
одуванчик, например, является одним из наиболее вы-
носливых созданий природы. Он способен расти почти
везде, где есть почва, и даже там, где никаких внешних
признаков ее пет. Одуванчик может проникнуть в замк-
нутое сообщество и, оказавшись там, готов бороться
с кем угодно и с чем угодно, пока его не вытопят. Да-
212
Глава Xi
же если его целиком выдернуть из земли, он снова вы-
растет из какого-нибудь крошечного кусочка корневой
системы, не замеченного потенциальным врагом.
Не все растения обладают такой живучестью, как
одуванчик. Некоторые избегают вступать в борьбу
с другими видами и предпочитают уйти в какое-нибудь
укромное место, где конкуренция менее жестока и где
им, возможно, удастся спокойно прожить жизнь.
Растения-пионеры
Такие растения — вовсе не трусы или снобы в расти-
тельном царстве. Как раз наоборот. Они — завоеватели,
и их вполне можно сравнить с безрассудно смелыми
людьми, которые, не боясь неизвестности, покидают об-
житые, но перенаселенные места. Этим растениям уда-
лось обратить в свою пользу некоторые отклонения
структуры или физиологии, что позволило им жить там,
где другие погибли бы. Они стали обитателями крайне
малообещающих мест, не пригодных для развития боль-
шинства растений, и буйно там разрослись. Так, какту-
сы захватили негостеприимные пустыни, эдельвейс (Le-
ontopodium alpinum) сделал своей собственностью вы-
сокогорья Альп, а морская трава даже передвинула
часы истории на 300 млн. лет назад и обосновалась
в море.
Ксероморфные признаки
Растение, обитающее в областях с исключительно
сухим климатом, называется ксерофитом, а особенности
структуры, позволяющие снижать чрезмерную потерю
влаги и тем самым дающие ему возможность жить в та-
ких местах, — ксероморфными признаками. Понятия
«ксерофит» и «ксероморфные признаки» не обязательно
совпадают. Растение засушливых местообитаний, кото-
рому удается благодаря быстрому росту завершить
свой жизненный цикл в течение короткого влажного се-
зона, можно справедливо назвать ксерофитом, хотя за
ненадобностью ксероморфные признаки у него могут
и отсутствовать. С другой стороны, большинство солон-
Приспособление к среде „ . ,....... 213
чаковых растений обнаруживает хорошо выраженную
ксероморфную структуру, хотя их корни постоянно омы-
ваются водой, а надземные части могут затопляться
при каждом приливе. Это объясняется тем, что вода
в этом случае соленая и корням трудно поглощать со-
леную воду из-за ее высокого осмотического давления
(или низкого давления диффузии). Подобные растения
постоянно страдают от так называемой физиологиче-
ской сухости; это как раз тот случай, когда «вода, во-
да кругом, но нет ни капли, чтобы напиться».
Ксероморфные растения могут иметь разнообразные
приспособления, ведущие к уменьшению потери воды.
Их листья могут быть толстыми и жесткими, кутикула
у них сильно утолщается и часто покрыта мощным
восковым налетом. Очень распространенный ксероморф-
ный признак — образование на листьях волосков. В ве-
треную погоду волоски задерживают струи воздуха, не
допуская их к устьичным отверстиям, и таким образом
замедляют испарение из устьиц паров воды. Располо-
жение устьиц ниже общей поверхности листа — еще один
способ уменьшить транспирацию; пары воды при этом
собираются в углублениях над устьицами, что затруд-
няет выдувание их ветром. Непосредственно под эпидер-
мисом у листьев ксероморфных растений часто распола-
гается слой толстостенных клеток (гиподермис). Этот
слой, по-видимому, выполняет двойную функцию, ре-
гулируя испарение и в то же время придавая листу
прочность, что затрудняет его сморщивание при потере
воды.
Растения с предельно развитой ксероморфной струк-
турой могут пойти дальше этих простых мер предосто-
рожности против потери воды. Вся морфология листа
у них может быть приспособлена к выполнению одной
этой необходимой функции. В листе олеандра (Nerium
oleander), например, распределение устьиц приурочено
к рядам углублений, которые сами защищены волоска-
ми. У вереска (Calluna vulgaris) устьица заключены
в желобок (устьичную камеру), проходящий по нижней
стороне толстого, сочного листа.
Листья многих злаков могут свертываться таким об-
разом, что устьица оказываются на внутренней поверх-
214
Глава XI
ности получающейся при этом трубочки. Способность
к свертыванию зависит от наличия на этой внутренней
поверхности групп так называемых «шарнирных клеток».
Прекрасным примером может служить песколюб пес-
чаный (Ammophila arenaria), крупные свернутые листья
которого характерны почти для любой песчаной дюны
(фото XXI). Относительно гладкая наружная поверх-
ность листа этого растения покрыта толстой кутикулой
и не песет устьиц. Внутренняя поверхность образует
множество складок, на дне каждой из которых находит-
ся группа «шарнирных клеток». Рядом с каждой груп-
пой шарнирных клеток, в складке листа, имеется участок
фотосинтезирующей ткани, клетки которой богаты хло-
ропластами. Устьица расположены лишь в этой зоне. До-
полнительной защитой от чрезмерной транспирации слу-
жат многочисленные жесткие волоски по наружным
краям желобков.
Преимущество такого расположения состоит в том,
что транспирация может регулироваться в зависимости
от потребностей растения. Когда лист насыщен влагой,
шарнирные клетки становятся тургесцентными и набу-
хают. Это вызывает развертывание листа, так что угле-
кислый газ может свободно входить в устьица. Если
это приводит к чрезмерной потере воды, шарнирные
клетки теряют тургор и спадаются, так что лист опять
свертывается.
Листья кссроморфпых растений часто бывают мел-
кими и могут редуцироваться до чешуек, а функцию
фотосинтеза принимает на себя стебель. Чрезвычайно
ярко это выражено у австралийской казуарины (Casua-
rina\ фото XXII). Здесь листья представляют собой ма-
ленькие чешуйки, а поверхность веток изрезана мно-
жеством глубоких продольных желобков. На дне желоб-
ков, защищенных по краю волосками, находится зеленая
фотосинтезирующая ткань. Всю фотосинтетическую ра-
боту выполняют эти зеленые веточки.
Стебли многих ксерофитов не только выполняют
функции листьев, но во многих случаях они действитель-
но выглядят как листья. В частности, у иглицы (Ruscus
aculeatus) из семейства лилейных, растущей в сухих
лесах и среди скал на юге Англии, имеются мелкие,
Приспособление к среде
215
жесткие, остроконечные образования, которые можно
принять за листья (рис. 41). Однако при тщательном
рассмотрении выясняется, что «листья» являются веточ-
ками стебля. На верхней стороне каждого такого
«листа», в его центре, располагается крошечный чешуй-
чатый лист, а в период цветения на этих своеобразных
плоских побегах появляются цветки.
Стебли, подобным образом выдающие себя за листья,
называются кладодиями'. Иногда ножка (черешок)
листа принимает листовидную форму, а пластинка
Рис. 41.
А. Часть побега иглицы с листовидными кладодиями. Ь’. Кладодий с цветоч-
ной почкой на верхней стороне.
листа утрачивается. Это мы наблюдаем у многих видов
Acacia. Такие образования называются филлодиями
(рис. 42).
Может показаться странным, что растение, утратив-
шее листья ради экономии воды, развивает листовидные
стебли или черешки, очевидно, сводящие на нет приобре-
тенное преимущество. Однако в стеблях и черешках
обычно больше механической ткани, чем в листьях.
В случае недостатка воды они оказываются в состоянии
лучше противостоять сморщиванию и поэтому менее
уязвимы.
1 Кладодий — более общий термин, обычно относимый к лю-
бым плоским стеблевым образованиям, которые принимают на себя
функцию фотосинтеза при редукции листьев. Если они вполне ли-
стовидны, как у иглицы, то нх называют филлокладиями. — Прим.
216
Глава XI
Многие ксероморфные растения несут колючки, кото-
рые представляют собой видоизмененные листья или
ветви. Колючки терна (Prunus spinosa) и боярышника
(Crataegus monogyna), например, стеблевого происхож-
дения, а у барбариса (Berberis vulgaris)—листового.
Утесник (Ulex) имеет и стеблевые и листовые колючки.
Проросток утесника не образует колючек до тех пор,
пока его стебель не достигнет определенной высоты.
Очевидно, причина здесь в том, что у поверхности поч-
вы воздух сравнительно влажный, поэтому надобность
в кссроморфных приспособлениях, расположенных в
нижней части растения, отпадает.
Рис. 42. Часть побега Acacia,
на котором можно наблюдать
переход от листьев к филло-
диям.
Колючки выполняют полезную функцию защиты от
животных, ощипывающих молодые побеги. Интересно
предположить, что признак, первоначально возник-
ший как ксероморфный, может сохраниться для выпол-
нения совершенно иных функций. Это прекрасно гармо-
нировало бы с общей тенденцией растений к экономии.
Сбрасывание листьев с наступлением зимы у листо-
падных деревьев — тоже мера предосторожности про-
тив потери воды. В холодные зимние месяцы корням
трудно насасывать воду из почвы и если бы такие де-
ревья, как дуб или береза, сохраняли листья круглый
год, в холодную погоду их водный баланс оказался бы
род угрозой. Благодаря осеннему сбрасыранию листьев
Приспособление к среде
21?
эта опасность устраняется1. Конечно, это значит, что
с опадением листьев фотосинтез прекратится, но, по-
скольку зимой рост крайне замедлен, это не имеет осо-
бого значения. У вечнозеленых деревьев и кустарников,
сохраняющих листву круглый год, листья обычно ксеро-
морфные. Возьмем, например, сосну с ее длинными, уз-
кими и жесткими иглами. На поперечном срезе такого
листа ясно видны типичные ксероморфные признаки:
толстая кутикула, погруженные устьица, толстостенный
гиподермис и округлая форма, уменьшающая отноше-
ние поверхности к объему (фото XXIII).
Осенний листопад
Хотя сосну называют вечнозеленой, это не значит,
что ее листья сохраняются вечно; прогулка в сосновом
лесу по толстому ковру игл, покрывающему землю, до-
казывает обратное. Вечнозеленое — это такое растение,
которое не теряет все свои листья каждый год. У раз-
ных видов сосны листья живут от двух до двадцати лет.
Осенний листопад — не случайный процесс; подобно
большинству функций растения он тщательно подготов-
лен. Близ основания листового черешка, по его окруж-
ности формируется слой клеток, называемый отдели-
тельным слоем (фото XXIV). Он обыкновенно состоит
из более мелких клеток, чем остальная часть черешка,
и в месте отделения листовой черешок может быть да-
же несколько сужен. Клетки, расположенные ниже от-
делительного слоя, опробковевают и одревесневают, тог-
да как клетки самого этого слоя разъединяются и лист
держится только на эпидермисе и проводящих пучках.
Малейшего дуновения ветра достаточно, чтобы эти тон-
кие тяжи разорвались и лист опал. Рубец, оставшийся
после листа, покрывается защитным слоем пробки, что-
бы предотвратить потерю воды и, что еще более важно,
проникновение спор грибов.
1 Автор правильно подчеркивает приспособительное значение
листопада, однако не следует думать, что оно могло послужить
непосредственной причиной возникновения листопадности. Это лишь
способствовало отбору листопадных форм в соответствующих ус-
ловиях. — Прим. ред.
21S
Глава XI
Прежде чем лист окончательно отомрет, в нем про-
исходят определенные химические изменения. Хлоро-
филл разрушается, так что лист приобретает желтую
окраску, обусловленную ксантофиллом; в то же время
во многих листьях образуются антоцианы. Красные
и желтые краски осени — результат этих процессов.
В окрашивании листьев могут принимать участие и про-
дукты разрушения хлорофилла.
Суккуленты
Ксероморфпыс приспособления, которые мы обсуж-
дали до сих пор, были направлены на снижение транс-
пирации, но ими ни в коей мере не исчерпываются воз-
можности растений приспосабливаться к сухим место-
обитаниям. Еще один путь решения проблемы — это
развитие водозапасающих тканей. Эти ткани имеются
у суккулентных растений, к которым относятся некото-
рые наиболее специализированные ксерофиты, такие,
например, как обитающие в пустынях кактусы, а из
растений нашей флоры — очитки (Sedum) и иглица.
У многих суккулентов часть листа или стебля заня-
та крупными, бесцветными, тонкостенными клетками,
предназначенными для запасания воды впрок. У видов
очитка, солянки — поташника (Salsola kali) и шведки
(Suaeda maritima) — водоносная ткань развита в мя-
систых листьях, тогда как у Cactus, листья которого
превращены в колючки,—в суккулентных стеблях.
Суккулентное растение может выдерживать значи-
тельное высушивание. По мере расходования воды в за-
пасающей ткани растение сморщивается, но, видимо,
это нисколько не вредит ему, так как, получив воду, оно
легко восстанавливает свои прежние размеры.
Листья водных растений
Водные растения вынуждены сталкиваться с усло-
виями, противоположными тем лишениям, которые ис-
пытывают ксерофиты. Их побеги в большей или мень-
шей степени погружены в воду, пе говоря уже о постоян-
но омываемых ею корнях. Во многих отношениях вода
П риспособление к среде 2К
представляет собойблагоприятную для растений среду,
но она ставит и свои собственные специфические про-
блемы.
Условия жизни растений в воде на всем земном ша-
ре гораздо более однообразны, чем па суше. Это нашло
свое отражение в более широком распространении мно-
гих водных растений по сравнению с наземными. Вода
имеет высокую удельную теплоемкость: она медленно
нагревается и так же медленно охлаждается. Поэтому
водное растение не подвергается резким и частым из-
менениям температуры, имеющим место на суше. Это
благоприятствует вегетативному росту растения.
Различны и условия питания водного и наземного
растений. Углекислый газ лучше растворим в воде, чем
кислород, и, поскольку растению нужно больше угле-
кислого газа для фотосинтеза, чем кислорода для ды-
хания, это способствует его вегетативному росту. Вод-
ные растения часто обнаруживают бурный рост в соче-
тании с усиленным вегетативным размножением, что
редко наблюдается у наземных растений (по крайней
мере в зоне умеренного климата).
Помимо углекислого газа, всей погруженной поверх-
ностью своего тела водные растения всасывают и мине-
ральные соли. В связи с этим кутикула на подводных
стеблях и листьях тонкая или вовсе отсутствует. Погру-
женные листья глубоко рассечены и часто напоминают
пучки волосков, что увеличивает поглощающую поверх-
ность. Это составляет резкий контраст с плавающими
листьями, обычно крупными, округлой или овальной
формы, с черешками, прикрепленными вблизи центра
нижней стороны листа, так что черешок не' наклоняет
лист и не тянет его под воду. Такие листья у белой кув-
шинки и желтой кубышки.
У многих водных растений имеются как погружен-
ные, так и плавающие листья, и в этом случае оба типа
листьев совершенно различной формы. Подводные
листья либо длинные и лентовидные, либо тонко рассе-
ченные, тогда как плавающие — более или менее округ-
лые. В качестве примера можно назвать водяной лютик
(Ranunculus heterophyllus) (рис. 43). У стрелолиста
(Sagittaria sagittifolia), который растет по берегам рек,
220
Глава XI
на одном растении могут быть листья трех типов, осо-
бенно в тихих неглубоких местах: 1) погруженные
листья — длинные и лентовидные, 2) плавающие
листья — овальные, с выемчатыми основаниями, и
3) выступающие над водой воздушные листья стрело-
видной формы, из-за которых растение и получило свое
название.
Рис. 43. Водяной лютик (Ranunculus heterophyllus), на примере
которого можно наблюдать различие между воздушными и по-
груженными листьями.
Погруженные листья водных растений не имеют
устьиц, потому что углекислый газ поглощается ими не-
посредственно из раствора, в форме бикарбонатных
ионов. У плавающих листьев на нижней стороне устьиц
нет, по по верхней стороне расположены нормальные
устьица. Верхняя сторона может иметь толстую, покры-
тую воском кутикулу, что затрудняет намокание листа,
а в некоторых случаях края плавающих листьев бывают
загнуты вверх, как у огромных листьев гигантской кув-
Приспособление к среде
221
шинки Victoria arridzonica, растущей в Амазонке. Листья
этого замечательного растения могут достигать 2 м в по-
перечнике. Выше уже говорилось о его опылении одним
из видов майских хрущей (гл. VII, стр. 160).
Стеблям водных растений приходится нести неболь-
шую нагрузку, так как все растение или его часть под-
держивается водой. Не удивительно поэтому, что дре-
весина в них развита слабо; некоторые из наиболее удач-
но приспособившихся водных растений, такие, как
полностью погруженный роголистник {Ceratophyllum de-
mersum), вовсе не имеют ксилемы, и на месте древеси-
ны у них образуется полость, потому что клетки, кото-
рые обычно дают начало ксилеме, разрушаются. Флоэма
же, напротив, развита хорошо. Это тоже вполне естест-
венно: поскольку растение может всасывать воду всей
поверхностью тела, нет надобности в водопроводящей
ткани, но образующиеся в листьях органические веще-
ства должны, как обычно, доставляться ко всем частям
растения.
Если же у водного растения древесина развивается,
она, как правило, располагается не по периферии, как
у большинства растений, а вблизи центра стебля. Это
разумно, так как водное растение вынуждено сопротив-
ляться тянущей силе, создаваемой водными течениями,
а пе силе, пытающейся согнуть растение. Поэтому по
расположению древесины стебель здесь больше похож
на корень.
Другая особенность водных растений — обилие воз-
душных полостей в коре стебля и корня (фото XXV).
Газ в этих полостях представляет собой преимуществен-
но кислород, который расходуется в процессе дыхания.
Полости тянутся вниз по стеблю и в корень, но в них
может быть ряд перегородок, образованных в узлах
и перфорированных маленькими отверстиями, которые
пропускают только газ, а воду нет. Если отламывается
часть стебля, случайно или при вегетативном размноже-
нии, внутренние полости не заполняются водой.
Один из недостатков водной среды состоит в том,
что вода очень быстро снижает интенсивность света; да-
же на глубине 10... 15 см это снижение значительно. Это
означает меньше света для фотосинтеза. Чтобы как
222
Глава XI
можно более эффективно использовать поступающий
свет, содержащие хлорофилл клетки должны распола-
гаться у поверхности растения. У водных растений хло-
ропласты нередко находятся в эпидермисе. У наземных
растений единственные эпидермальные клетки, кото-
рые содержат хлоропласты,— это замыкающие клетки
устьиц.
Размножение водных растений
При общей пышности вегетативного роста, свойст-
венной водным растениям, не удивительно, что вегета-
тивные способы размножения у них хорошо развиты
и весьма продуктивны. Некоторые водные растения ред-
ко размножаются семенами. Яркий пример тому — эло-
дея (Elodea canadensis). Это небольшое, но вездесущее
растеньице было завезено в Англию в 1842 году из
Ирландии, куда шестью годами раньше оно попало из
Канады и распространилось подобно чуме. Кстати, «чу-
ма»— вполне подходящее в этом случае слово1, так как
вскоре элодея начала блокировать каналы Англии, слу-
жившие в те времена важным средством транспорта.
Такого успеха элодее удалось добиться исключительно
благодаря вегетативному размножению, так как расте-
ние это двудомное (мужские и жепские цветки на от-
дельных растениях), а мужские экземпляры в Англии
встречаются редко.
Elodea размножается путем фрагментации, заклю-
чающейся в том, что от главного побега отламываются
веточки и начинают самостоятельную жизнь. Этот спо-
соб размножения настолько эффективен, что отсутствие
семян, видимо, не причиняет растению никакого неудоб-
ства.
Вспомним снова о манграх. Эти полуводные расте-
ния представляют собой вечнозеленые деревья, обитаю-
щие в зоне приливов и отливов на побережье океана
в тропиках, в частности во Флориде, на полуострове
1 В России элодея появилась в шестидесятых годах прошлого
века и также стала самым обычным растением водоемов. По-русски
ее тоже часто называют «водяная чума». — Прим. ред.
Приспособление к среде
223
Малакка и на островах Индонезии. Они принадлежат
к нескольким совершенно не связанным между собой’
семействам, но благодаря сходным условиям местооби-
тания у них появилось много общих признаков, таких,
например, как дыхательные корни и семена, прорастаю-
щие еще на материнском растении (о живородящих
мангровых см. стр. 198). Они составляют не таксоно-
мическую, а экологическую группу и являются хорошим
примером конвергенции, т. е. примером того, как силь-
но различающиеся по происхождению организмы ста-
новятся похожими друг на друга в результате приспо-
собления к одним и тем же условиям среды. Подобное
явление известно и в зоологии; например, внешнее сход-
ство между акулой (рыба), ихтиозавром (рептилия)
и китом (млекопитающее).
Как мы уже видели, мапгровые обитают в болоти-
стых местах, ежедневно затопляемых приливом; их пи-
тающие корни проникают в ил эстуария, или лимана,
а из нижних частей стеблей развивается часто сложная
система ходульных корней, так что кажется, будто расте-
ние стоит па беспорядочно расположенных подпорках.
Ходульные корни задерживают осадки, наносимые при-
ливом, и уровень почвы в мангровом болоте непрерывно
повышается. Мапгры, таким образом, могут явиться
важным фактором в борьбе с береговой эрозией.
Другая интересная особенность мангров, о которой
я уже упоминал,— это доставка воздуха к погруженной
корневой системе при помощи дыхательных корней, или
пневматофоров. Нужда в дыхательных корнях возни-
кает потому, что омывающая корни мангровых вода
бедна кислородом и корням трудно добыть его в нуж-
ном для дыхания количестве. Заболоченная почва всегда
содержит мало кислорода, а в мангровом болоте его
нехватка возрастает из-за массы гниющих органических
остатков, накапливающихся на дне. Так как бактериям,
разлагающим эти остатки, тоже нужен кислород, они
конкурируют за воздух с корнями.
Коленчатые дыхательные корни не являются моно-
полией мангровых; они имеются и у некоторых других
болотных растений, например у болотного кипариса
(Taxodium distichum).
224
Глава Xi
Лазящие и вьющиеся растения
Лазанье — еще одно приспособление, встречающееся
в различных семействах растений. Множество примеров
можно привести и из флоры Англии, но наибольшего
развития это свойство получило в-тропиках, особенно
во влажном тропическом лесу, где лазящие растения,
или, как их часто называют, лианы, являются важным
элементом флоры.
Растения лазают различными способами. Дарвин,
чья книга о лазящих растениях должна быть прочитана
каждым, кто интересуется жизнью растений, разделял
их на четыре основных класса: 1) растения, лазящие
с помощью разнообразных крючков; 2) вьющиеся расте-
ния; 3) растения с чувствительными лазящими органа-
ми, такими, как усики, и 4) растения, лазящие с по-
мощью придаточных корней.
Растения, лазящие с помощью крючков, самые про-
стые и наименее специализированные из этих четырех
групп. Они скорее цепляющиеся, чем лазящие, и к на-
стоящим лазящим растениям имеют примерно такое же
отношение, какое человек, гуляющий по холмам, имеет
к альпинизму. Такие растения снабжены загнутыми
вниз шипами, подобными, например, шипам ежевики,
действующими как своеобразные крюки и позволяющи-
ми им цепляться за другие растения и карабкаться по
ним (рис. 44).
В Англии к этому типу лазящих растений относятся
ежевика (Rubus fruticosus), малина (R. idaeus), неко-
торые виды роз и подмаренников (Galium), но в тропи-
ках растения такого облика широко распространены.
У некоторых тропических видов в крючки модифициро-
ваны целые стебли или листья вместо скромных эпи-
дермальных шипов, характерных для растений нашей
зоны (рис. 45).
Вьющиеся растения лазают, обвиваясь стеблем во-
круг опоры, как это делает, например, декоративная фа-
соль. Они лучше приспособлены к лазанью, чем расте-
ния, снабженные крючками. Способность виться обуслов-
лена явлением нутации, которое свойственно большин-
ству растений, но наиболее резко выражено у вьющих-
Приспособление к среде
225
ся. Нутация — это круговое движение растущей вверх
верхушки стебля. В поисках опоры кончик стебля опи-
сывает широкую спираль. Найдя вертикальную палку
или другой подходящий предмет, стебель крепко обви-
вается вокруг него, удерживая все растение.
Многие растения вьются всегда в одном и том же
направлении. Хмель (Humulus lupulus), гречишка вьюн-
ковая (Polygonum convolvulus) и жимолость (Lonicera
Рис. 44. Шип на стебле еже-
вики.
Рис. 45. Цепляющиеся крючки
Ancistrocladus.
periclymenum), например, всегда обвивают опору по ча-
совой стрелке, тогда как вьюнок заборный (Calystegia
septum) и декоративная фасоль — против часовой стрел-
ки (фото XXVI). Причины такой избирательности не-
ясны. В то же время некоторые вьющиеся растения, по-
видимому, индифферентны в этом отношении.
Рост вьющегося растения вокруг опоры, вероятно,
вызван тенденцией стеблей расти против силы тяжести.
Когда кончик стебля обвисает под собственной тяже-
стью, отрицательный геотропизм заставляет нижнюю
его сторону расти быстрее верхней, в результате чего
он снова загибается вверх. Это постоянное изгибание
верх вместе с резко выраженной нутацией и обусловли-
226
Слава XI
вает спиральный характер роста, в результате чего сте-
бель обвивается вокруг опоры. Доказательств, что здесь
действует какой-либо другой фактор, нет.
Растения с чувствительными лазящими органами,
например усиками,— наиболее специализированные из
лазящих растений. Усик чувствителен к прикосновению.
Когда усик гороха соприкасается с возможной опорой,
например стеблем другого растения, он изгибается по
направлению к той стороне, которая первой коснулась
опоры. При этом в контакт с опорой вступают и дру-
гие части усика, усиливая, таким образом, ответную ре-
акцию, и он плотно обвивается вокруг предмета, за ко-
торый цепляется. Эта реакция на раздражение, вызван-
ное соприкосновением, называется гаптотропизмом.
Когда усик прочно укрепился на опоре, в его нижней
части происходят дальнейшие изменения. Она закручи-
вается в спираль, образуя своеобразную пружину. Это
позволяет усику пружинить и не отрываться от опоры,
когда она сотрясается ветром. Где-то по ходу спирали
направление закручивания меняется; это естественное
следствие попытки свернуть спиралью закрепленный на
обоих концах предмет. По мере старения усика витки
часто становятся деревянистыми.
Изгибание усика к опоре вызвано перераспределени-
ем ауксина — вещества, ответственного за реакцию кор-
ней на действие силы тяжести, а стеблей — на свет
и силу тяжести (гл. IV). При соприкосновении усика
с твердым телом ауксин накапливается на стороне, про-
тивоположной опоре; в результате эта сторона усика
растет быстрее и он изгибается к опоре.
Имеются данные, что стимулом, вызывающим ответ-
ную реакцию усиков, служит не столько соприкоснове-
ние, сколько трение. Если усики переступеня белого
(брионии) слегка погладить по одной стороне, они бу-
дут изгибаться в этом направлении, даже когда мы от-
нимем руку. Если же к ним просто прикоснуться, не
поглаживая, усики никак не прореагируют. Усик, осто-
рожно прикрепленный к опоре таким образом, чтобы он
не мог двигаться, не изгибается. Следовательно, значе-
ние имеет тип раздражения, а не длительность его дей-
ствия.
Приспособление к среде 227
Усики нечувствительны к контакту с жидкостями.
Капли воды, попадающие на них, не вызывают ника-
кого изгиба, и даже ртуть, в 13 с лишним раз более
плотная, чем вода, и самая тяжелая из жидкостей, на
них не действует. Для растения, вынужденного нахо-
диться на открытом воздухе в любую погоду, от-
сутствие чувствительности к жидкостям представляет
собой несомненное преимущество. Усики способны вы-
полнять свою работу постоянно, не прерывая ее в случае
дождя.
Часть растения, видоизмененная в усики, варьирует
в зависимости от вида, но чаще всего к лазанью приспо-
сабливается лист или часть листа. Особенно хорошо это
можно видеть у многих бобовых, куда относится боль-
шое число лазящих с помощью усиков растений. Листья
бобовых, как правило, сложные, т. е. разделены на мно-
жество листочков, расположенных по обеим сторонам
общего черешка. У вики (Vicia) один или несколько ко-
нечных листочков утратили пластинки и превратились
в усики. У душистого горошка (Lathyrus odoratus) все
листочки, кроме одной пары, видоизменены в усики (фо-
то XXVII). Чтобы компенсировать потерю фотосинтези-
рующей поверхности листа, прилистники (два листовид-
ных выроста, возникающих у места присоединения ли-
стового черешка к стеблю) чрезвычайно увеличились
и взяли на себя функции листа. Кроме того, по обеим
сторонам стебля у душистого горошка имеется пара
уплощенных, сбегающих вниз крыльев. Они зеленые
и тоже участвуют в фотосинтезе.
Ломонос лозный (Clematis vitalba) и другие виды
рода Clematis образуют усики иным путем. У них пла-
стинка листа не изменяется и растение лазает с по-
мощью чувствительных листовых черешков. Таким же
образом лазают и садовые настурции (Tropaeolum).
У Gloriosa и Littonia, тропических представителей се-
мейства лилейных, роль усиков выполняют кончики
листьев (рис. 46).
Иногда усиком может быть модифицированная вет-
ка, как у винограда (Vitis vinifera) и некоторых стра-
стоцветных (Passif(oraceae), в частности у Passiflora
edulis и Р. maliformis.
228
Глава Х.1
Истинная природа усиков у переступеня белого точ-
но не выяснена. Сопровождаемые короткими веточками,
они появляются в пазухах листьев; некоторые ботаники
считают их модифицированными первыми листьями бо-
ковых побегов, тогда как другие рассматривают их как
дополнительные ветви.
Некоторые тропические лианы лазают при помощи
загнутых вниз крючков, расположенных на концах ви-
доизмененных соцветий. Они выступают из стебля,
а найдя опору, плотно охватывают ее и одревесневают.
Рис. 46. Листовой усик Gtoriosa.
Эта манера лазанья' наблюдается у растений из самых
различных семейств, в том числе у Hugonia (семейство
лилейных), Landolphia (семейство кутровых) и Оигои-
paria (семейство мареновых).
К лианам, лазящим с помощью придаточных корней,
относится плющ (Hedera helix). На его стебле на неко-
тором расстоянии друг от друга образуются пучки кор-
ней, которые, как мы уже знаем, называют придаточны-
ми, потому что они не являются частью первичной кор-
невой системы (рис. 13). Эти лазящие корни интересны
тем, что избегают света (большинство корней к свету
безразлично) и благодаря этому легко находят щели
в опоре, будь то шершавая кора дерева или старая стена.
Необычны они и в том отношении, что не реагируют на
силу тяжести, тогда как у большинства корней сильно
выражена тенденция расти вниз, к земле. Это тоже бла-
гоприятствует использованию лазящих корней в каче-
стве органов прикрепления, потому что они растут го-
ризонтально и, таким образом, соприкасаются с опорой.
Приспособление к среде 229
Лазящие корни имеются у различных тропических
лиан, в том числе у многих Агасеае и у ряда лазящих
папоротников. У некоторых растущих на ветвях де-
ревьев видов Philodendron (Агасеае) воздушные корни
обвиваются вокруг веток дерева-опоры, спускаясь к
земле. /
Смысл лазанья состоит в том, что ото позволяет
растению достигнуть значительной высоты, не тратя
времени и энергии на укрепление стебля. В совершен-
стве овладели искусством лазанья лианы, в таком изо-
билии произрастающие во влажном тропическом лесу.
Эти громадные растения со стеблями толщиной в чело-
веческое бедро, могут взобраться на вершину высочай-
шего дерева, снова спуститься на землю и подняться на
другое дерево, достигая при этом фантастической дли-
ны. Стебель лазящей пальмы, измеренный Трейбом
в 1883 году, был более 200 м в длину — Vs километра!
Возможно, эго исключительный случай, но тропическая
лиана длиной 60 м — вполне обычное явление.
Поскольку лиану поддерживает дерево, рядом с ко-
торым она растет, она мало нуждается в механической
ткани, служащей для придания стеблю твердости. Во-
локна у лиан развиты слабо или отсутствуют вовсе. Это
выгодно для растения, так как рост волокон идет мед-
ленно, а используя энергию в ином направлении, оно
может расти быстрее. С другой стороны, проведение во-
ды легко может стать проблемой, так как вода у лиан
должна проходить длинный путь по относительно тонко-
му стеблю. Поэтому у таких растений имеется хорошо
развитая ксилема с очень широкими сосудами.
Другая существенная особенность стебля лианы —
это гибкость; чтобы взбираться на вершину дерева, она
должна быть способной перекручиваться и изгибаться.
Традиционное распределение ксилемы в стебле древес-
ного растения в виде непрерывного кольца здесь не го-
дится. Проблему помогают решить разнообразные ано-
малии вторичного роста лианы, направленные на со-
четание гибкости с хорошо развитыми проводящими
элементами ксилемы. Clematis, например, вместо тоге
чтобы образовывать вторичную ксилему и флоэму пс
всей окружности стебля, образует через определенные
230
Глава XI
промежутки тонкостенную ткань, так что ксилема на
протяжении всей жизни растения остается расчлененной
на отдельные пучки (фото XXVIII). Структура сосудов
у некоторых тропических лиан, в частности у многих бо-
бовых (Bauhinia и др.) и кирказоновых (Aristolochia),
чрезвычайно сложна.
Если растение растет на слабом свету, оно обнару-
живает ряд симптомов, в результате сочетания кото-
рых возникает состояние, называемое этиолированием.
К этим симптомам относятся длинный слабый стебель
и длинные междоузлия (междоузлие — это участок
стебля между двумя следующими друг за другом ли-
стьями). В связи с тем что именно эти признаки свойст-
венны большинству лазящих растений, было высказано
предположение, что лазящий образ жизни мог развить-
ся в результате этиолирования: пытаясь пробиться
к свету и воздуху, растение образовывало слабый сте-
бель, нуждающийся в опоре. С этой точки зрения, ве-
роятно не случайно, что лианы обитают главным обра-
зом во влажном тропическом лесу, где густые кроны
деревьев создают условия для этиолирования появив-
шегося из земли растения. Если лиана в самом деле
возникла из этиолированного проростка, это еще раз
иллюстрирует умение растений извлекать выгоду из
собственных несчастий.
Лианы естественно подводят нас к следующей линии
в приспособлении растений к окружающей среде —
к эпифитам. Это растения, которые совсем покинули
землю и проводят жизнь на ветвях дерева или в ка-
ком-нибудь другом труднодоступном месте; как мы уже
знаем, некоторые эпифитные орхидеи поселяются даже
на телеграфных проводах.
Приспособления эпифитов
Некоторые растения могут стать эпифитами волей
случая, если их семена упадут в развилину дерева
и прорастут там. Я видел белую яснотку, вполне ком-
фортабельно устроившуюся на ветке дуба. По-видимо-
му, нанесенной ветром почвы, скопившейся в углубле-
нии коры, оказалось достаточно для удовлетворения
Приспособление к среде
231
скромных нужд растения. На вершинах ветел часто
растут как эпифиты небольшие цветковые растения.
Однако все эти заблудившиеся растения— лишь слу-
чайные эпифиты. Настоящий эпифит, т. е. растение, для
которого жизнь на другом растении является обычной,
морфологически и физиологически приспособился к свое-
му любопытному образу жизни; иначе он не смог бы
выжить.
В прохладном климате эпифиты чувствуют себя не-
важно. В Англии растет только один настоящий эпифит
из числа высших растений; это многоножка (Polypodium,
vulgare) — папоротник, живущий на деревьях и среди
скал. Все остальные наши эпифиты относятся к низ-
шим растениям (мхи1, водоросли и лишайники). Эпи-
фиты из числа цветковых растений процветают там, где
климат более теплый, и поэтому их можно найти в изо-
билии во влажных тропических лесах, а еще больше —
в горных и субтропических дождевых лесах. Жизнь эпи-
фита полна опасностей даже в самых благоприятных
условиях, и надеяться, что они, кроме всех прочих труд-
ностей, вынесут такой климат, как наш, значило бы же-
лать слишком многого.
Чтобы обосноваться па дереве, которое в дождевом
лесу может достигать головокружительной высоты
в шестьдесят и более метров, эпифиту необходима эф-
фективная система распространения семян. Только два
агента — ветер и птицы -г- могут осуществлять его си-
стематически и наверняка, и поэтому надо полагать, что
почти все эпифиты пользуются тем или другим из этих
способов распространения семян.
Эпифиты обычно образуют сочные плоды, многие из
которых содержат клейкую мякоть, так что после еды
птице приходится чистить клюв о кору дерева, попутно
втирая в нее семена. Этот механизм был описан для
нашей омелы, которая является скорее частичным пара-
зитом, чем эпифитом, хотя и растет на деревьях. Он
свойствен также тропическому роду RhipsaUs и многим
эпифитным бромелиевым. Птицы часто заносят семена
1 См. примечание к стр. 113.
232
Глава XI
бромелиевых на телеграфные провода, где растения мо-
гут обосноваться.
Чрезвычайно мелкие и легкие споры папоротников без
труда поднимаются ветром на значительную высоту. То
же можно сказать и о семенах орхидей, которые бывают
настолько мелкими, что похожи на пыль. Семя Stanho-
реа oculata, например, весит всего 0,0000028 г. Как пра-
вило, семена тропических и субтропических эпифитов
снабжены приспособлениями для парения в воздухе.
Рис. 47. Южноамериканский эпифит-
Tillandsia usneoides.
Луизианский «мох» (Tillandsia usneoides) —интерес-
ное цветковое растение, обитающее в тропиках и суб-
тропиках Америки. Внешне он напоминает лишайник
Usnea — отсюда и его латинское название. Обматываясь
основанием стебля вокруг ветви дерева, он свешивается
вниз длинными серыми космами. По мере роста стебля
к земле более старые его части отмирают, оставляя не-
поврежденным только центральный тяж волокнистой
ткани, похожий на конский волос. Для поглощения сте-
кающей сверху воды служат чешуйчатые волоски, по-
крывающие все растение (рис. 47). Цветки образуются
редко, и растение распространяется частично ветром,
отрывающим куски напоминающей бороду массы, а ча-
стично птицами, использующими ее для постройки гнезд.
Приспособление к среде
233
Когда семя эпифита попало в подходящее место
и проросло, встает вопрос, как там закрепиться. Это
почти всегда осуществляется с помощью корневой си-
стемы. В частности, проросток орхидеи Aeschyanthus
имеет уплощенный диск и снабжен многочисленными
корневыми волосками, при помощи которых он при-
крепляется к опоре и удерживается на ней до тех пор,
пока окончательно не оформится. У многих ароидных
(Агасеае), орхидных (Orchidaceae) и представителей
других семейств имеются корни двух типов — питающие
и служащие исключительно для прикрепления. У броме-
лиевых, которые представляют собой высокоспециали-
зированные эпифиты, поглощение воды осуществляется
скорее волосками на листьях, чем корнями, которые
приспособлены исключительно для прикрепления. Жест-
кие и похожие на проволоку, они совсем не насасывают
воду (или поглощают ее в крайне небольшом количестве).
Но вот семя занесено в подходящее место на стволе
или ветви дерева, и растение должным образом там обо-
сновалось; тут для эпифита и начинаются настоящие
трудности. Будучи совершенно оторванным от почвы,
он должен как-то собрать достаточное для поддержа-
ния жизни количество воды и вместе с ней необходимые
для удовлетворения его нужд минеральные соли. Это
совсем не легко, и поэтому наиболее существенные при-
способления к эпифитному образу жизни связаны с вод-
ным режимом.
Ботаник Шимпер, работы которого относятся к кон-
цу XIX века, выделял четыре класса эпифитов с раз-
личной степенью специализации. Это протоэпифиты,
гнездовые и «.кармашковые» эпифиты, цистерновые эпи-
фиты и полуэпифиты. У протоэпифитов нет особых
структур для собирания воды и почвы. Многие из них
обнаруживают ксероморфные признаки, например сук-
кулентные листья; обычны водозапасающие органы.
Многие орхидеи, принадлежащие к этой группе, разви-
вают воздушные корни со специальной водопоглощаю-
щей тканью — веламеном. О функциях этой ткани мы
говорили в начале VIII гл. на стр. 170. Веламен рас-
полагается на внешней поверхности корня и состоит из
крупных пустых клеток, часто со спиральными или сет-
234
Глава XI
чатыми утолщениями на стенках; через имеющиеся в них
отверстия клетки сообщаются друг с другом. Эта ткань
действует подобно промокательной бумаге, поглощая
воду, которая легко входит в пустые клетки. Находящая-
ся под веламеном экзодерма корня видоизменена как
эндодерма: ее клетки имеют утолщенные стенки, за ис-
ключением некоторых, остающихся подобно пропускным
клеткам тонкостенными( гл. II). Едва ли можно сомне-
ваться, что это связано с поглощением воды веламеном.
Протоэпифиты — наименее специализированная груп-
па эпифитов, и поэтому они больше других страдают от
«перемежающейся» засухи.
Строение гнездовых и кармашковых эпифитов тако-
во, что позволяет им накапливать органические остатки,
создавая таким образом собственный запас перегноя,
или гумуса. У гнездовых эпифитов сплетение корней на-
поминает птичье гнездо. Гумус и гниющие остатки ска-
пливаются между корнями, служащими укрытием для
муравьев, которые, строя под ними свои гнезда, попол-
няют тем самым запас перегноя.
У кармашковых эпифитов некоторые или же все
листья, прилегающие к стволу дерева, образуют воронки.
В этих воронках накапливается гумус, чему нередко
'способствуют муравьи.
Хорошим примером кармашковых эпифитов могут
служить виды папоротника олений рог (Platycerium),
произрастающего в лесах полуострова Малакка (фо-
то XXIX). У этого папоротника два типа листьев. Листья
первого типа называют покровными листьями. У Р. gran-
de нижняя часть покровных листьев прилегает к стволу
дерева, на котором растет папоротник, тогда как верхняя
часть оттопыривается и образует кармашек, в котором
собирается перегной. У Р. bifurcatum весь покровный
лист плотно прижат к стволу дерева-опоры; накапли-
вающийся гумус образуется здесь только за счет гние-
ния старых покровных листьев с возможным добавле-
нием коры дерева. У обоих видов листья второго типа,
иногда называемые спорофиллами, свободно свешивают-
ся вниз. Они сильно разветвлены, наподобие оленьих
рогов (отсюда название растения), и несут на своих
концах спорангии,
Приспособление к среде
235
Между муравьями и эпифитами существует тесная
связь, доходящая в иных случаях почти до симбиоза
(см. начало гл. XIV); такой союз, где оба партнера жи-
вут вместе, но в то же время свободны и временами
могут идти каждый своим путем, носит название ком-
менсализма. У эпифита Myrmecodia, произрастающего
Рис. 48. Стеблевой клубень
Myrmecodia, в полостях ко-
торого живут муравьи.
на островах Индонезии, имеются крупные стеблевые
клубни, содержащие связанные ходами полости, в кото-
рых поселяются муравьи (рис. 48). По-видимому, основ-
ной функцией клубней является запасание воды, поэто-
му трудно понять, почему вообще возникли эти полости
и ходы, ибо муравьи, очевидно, не приносят растению
никакой пользы1.
У живущего в лесах Новой Гвинеи и островов Фид-
жи эпифита Hydnophyium тоже развиваются стеблевые
клубни, похожие на клубни Myrmecodia, в которых по-
селяются муравьи. И Hydnophytum, и Myrmecodia при-
надлежат к семейству мареновых (Rubiaceae).
Цистерновые эпифиты
Цистерновые эпифиты встречаются только среди бро-
мелиевых— семейства, обнаруживающего высокую спе-
циализацию к эпифитному образу жизни. Бромелиевые
1 По мнению некоторых исследователей, это явление, называе-
мое мирмекофилией, все же имеет некоторое биологическое значе-
ние, так как эти муравьи защищают растения от других муравьев-
листорезов. —- Прим. ред.
236
Глава X/
растут в тропической Америке и Вест-Индии. Замеча-
тельные цистерны бромелиевых, улавливающие и хра-
нящие воду, образованы розетками сочных листьев, ко-
торые, плотно смыкаясь у основания, вверху образуют
нечто наподобие воронки. В получившихся кувшинчи-
ках скапливается вода вместе с различным мусором,
а также живые растения и животные и мертвые их
остатки. Флора и фауна этих цистерн чрезвычайно раз-
нообразна. Так, некоторые бразильские виды пузыр-
чатки (Utricularia) можно найти только в цистернах
бромелиевых. Фауна цистерн представлена большим ко-
личеством личинок и куколок комаров и другими водя-
ными насекомыми; время от времени в них выводятся
и лягушки.
Воду из этих хранилищ бромелиевые поглощают при
помощи чешуйчатых волосков на листьях. Сами листья
тоже богаты водозапасающей тканью. Вместе с водой
растение получает питательные вещества, извлеченные
из скопившегося в цистернах гумуса. Корни цистерно-
вых бромелиевых не несут всасывающей функции и слу-
жат только для прикрепления.
Приспособления полуэпифитов
Полуэпифиты (гемиэпифиты) —это растения, кото-
рые начинают жизнь как эпифиты, но развивают длин-
ные воздушные корни, растущие вниз до тех пор, пока
не достигнут почвы. Как только установлен контакт
с почвой, проблему добывания воды можно считать ре-
шенной. Полуэпифит Coussapoa fagifolia из семейства
тутовых (Могасеае), растущий в тропиках Южной Аме-
рики,— довольно крупный кустарник, тогда как многие
ароидные (Агасеае) относятся к травянистым членам
этой группы эпифитов.
При изучении эпифитной флоры тропического леса
поражает тот факт, что чем выше деревья, на которых
растут эпифиты, тем ярче у них выражены ксероморфные
признаки. Эпифиты, которые живут на более низких
деревьях, образующих нижний из трех ярусов, обычно
различимых во влажном тропическом лесу, слабо или
почти совсем не приспособлены к экономии воды. Живя
_______________Приспособление к среде ДЗУ
в постоянно влажной атмосфере и будучи защищены от
ветра, эти растения не испытывают особых трудностей
со снабжением водой. С другой стороны, эпифитам, ра;
стущим на верхних ветвях самых высоких деревьев,
чтобы выжить, необходимо иметь хорошо развитую ксе-
роморфную структуру.
Интересно поразмыслить о том, как возник эпифит-
ный образ жизни, хотя в настоящее время ничего опре-
деленного о его происхождении не известно. Некоторые
эпифиты, возможно, начинали свое существование как
лйаны, а кончали полной утратой связи с почвой. Это
можно наблюдать на примере орхидеи Dipodium pictum,
произрастающей на островах Индонезии, которая в на-
чале жизни погружена корнями в почву, но в конце кон-
цов переходит к эпифитному образу жизни. То же про-
исходит и с некоторыми ароидными. В этой связи инте-
ресно отметить, что в семействе ароидных, известном
своими лианами, имеются и настоящие эпифиты, такие,
как Philodendron, которые в течение всей своей жизни
оторваны от почвы. Однако это позволяет правдоподоб-
но объяснить происхождение некоторых эпифитов, но не
эпифитпого образа жизни вообще.
Шимпер выдвинул теорию о том, что сначала эпи-
фиты были наземными растениями — обитателями влаж-
ных тенистых лесов. Но поскольку большинство эпифи-
тов почти совсем не выносит затенения, эта теория едва
ли правильна, разве что для немногих «теневых эпифи-
тов», растущих в нижних ярусах леса. Более правдопо-
добно звучит предположение, что эпифитные бром ел ие-
вые, во всяком случае, произошли от предков, обитав-
ших на границах южноамериканских пустынь; оно пред-
ставляется справедливым и для некоторых других эпи-
фитов, в частности эпифитн-ых Cactaceae. Привлекатель-
ная сторона этой идеи в том, что многие характерные
для эпифитов признаки растения развили еще тогда,
когда корни их находились в почве.
Эпифиты ни в коем случае не паразитируют на тех
деревьях, где они живут, а используют своих «хозяев»
только как опору. Все же, несмотря н-а это, между не-
которыми видами деревьев и обитающими на них эпи-
фитами существует тесная связь. На Филиппинах, на-
238
Глава X/
пример, папоротник Stenochlaena areolaris встречается
исключительно на панданусе (Pandanus utilissimus).
Прежде думали, что специфичность определенных эпи-
фитов по отношению к определенным деревьям вызвана
физическими причинами, например степенью неровности
коры. Вероятно, во многих случаях так оно и есть, но
недавно выяснилось, что эта связь теснее, чем можно
объяснить с точки зрения физических условий, и скорее
всего должна быть отнесена за счет каких-то химических
явлений. Снабжение эпифита водой зависит, по крайней
мере частично, от воды, стекающей по стволу дерева.
В этой воде могут содержаться вещества, выделяемые
хозяином. В воде, стекающей с веток одного из видов
каштана (Castanopsis argentea), например, можно об-
наружить таннин. Для эпифитов, предпочитающих опре-
деленный вид дерева, может иметь значение и химиче-
ский состав наружных слоев коры.
Приспособления галофитов
Химический состав окружающей среды может ока-
зывать сильное влияние на жизнь растения и требует
предельной адаптации. Наилучшим образом это выра-
жено у галофиюв — растений, живущих по берегам мо-
рей, на солончаках и в засоленных степях. В таких ме-
стах в почве имеется избыток солей, что чрезвычайно
щтрудняет поглощение воды корнями.
Как вы помните (гл. III), поступление воды в корни
1роисходит только в том случае, если осмотическое дав-
дение в корнях выше, чем в почвенной воде, или, точнее
жазать, если давление диффузии почвенной воды выше,
дем в корнях. При высоком осмотическом давлении дав-
дение диффузии воды соответственно низкое. Вода, со-
держащая большое количество соли в растворе, имеет
1ысокое осмотическое давление; давление диффузии во-
пя при этом, несомненно, будет небольшим, что явится
[репятствием к ее поступлению в корни растений.
Галофиты могут в какой-то степени устранить это
фепятствие, повысив осмотическое давление клеточного
ока. Измерение осмотического давления в корнях не-
оторых галофитов показало, что именно так они и по-
Приспособление к среде
239
ступают. В клетках нормальных растений оно обычно
составляет 5 10 атм, редко поднимаясь до 20 атм
(за исключением запасающих органов, содержащих
много сахара). Измерения осмотического давления в
клетках Atriplex confertifolia — растения, обитающего
близ Большого Соленого озера в Америке, дали пора-
зительную цифру—153 атм.
Такое высокое осмотическое давление может повлечь
за собой физиологические проблемы, от решения кото-
рых зависит нормальное существование растения. В
частности, высокое осмотическое давление в листьях мо-
жет повлиять на фотосинтез.
Поскольку галофитам трудно насасывать воду, они,
естественно, должны стремиться сохранить уже име-
ющуюся. Вот почему галофиты, как правило, обладают
теми или иными ксероморфными признаками. Хотя они
могут и не быть ксерофитами (растения солончаков
определенно таковыми не являются), галофиты растут в
условиях физиологической сухости, которая бывает не
менее изнуряющей, чем настоящая засуха. Следователь-
но, нет ничего удивительного в том, что у галофитов име-
ется толстая кутикула, погруженные устьица, мясистые
листья и все остальные ксероморфные приспособления.
Кальцифилы и кальцифобы
Растения, произрастающие на известняках, напри-
мер в некоторых районах Америки, подобно галофитам,
страдают от высокого осмотического давления почвен-
ной воды и могут обнаруживать такие же приспособле-
ния. Эти растения сталкиваются и с другой проблемой,
заключающейся в щелочности почвы. Та же проблема,
правда менее остро, стоит перед растениями, растущими
на меловых обнажениях. Большинство плодородных
почв слегка кислые, но в местах, где мел выходит на
поверхность, как в районе так называемых Северных
и Южных меловых холмов в Англии, реакция почвы ста-
новится явно щелочной. В таких местах появляется осо-
бая флора, которую называют меловой флорой. Типич-
ными меловыми растениями являются лен слабитель-
240
Глава XI
ный (Linum catharticum), зеленка (Blackstonia perfolia-
ta), золототысячник (Centaur ium minus) и многие орхид-
ные, произрастающие на Британских островах.
Некоторые растения могут расти только на извест-
ковых почвах и известны как кальцифилы. Другие из-
бегают мела, обитая исключительно на кислых почвах,
например на пустошах или торфяных болотах; они на-
зываются кальцифобами. Между этими двумя крайне
специализированными группами располагаются расте-
ния, по-видимому, безразличные к реакции почвы, кото-
рые одинаково хорошо растут на почвах любого типа.
Это было известно давно, но что же все-таки заставляет
кальцифилов любить мел, а кальцифобов избегать его,
пока не ясно. Возможно, что некоторые кальцифилы ра-
стут на мелу потому, что их устраивают физические усло-
вия, характерные для известняков. Но это объяснение
нельзя отнести ко всем кальцифилам. Сравнительные ис-
следования растительности некоторых участков в пре-
делах распространения песчаников, где задолго до этого
в почву было рнесено большое количество извести, и
обычной для тех мест флоры дали интересные результа-
ты. Участки с обогащенной известью почвой ничем не
отличались от соседних, за исключением повышенного
содержания извести (4,5%); в обычных же здешних
почвах следы извести едва обнаруживаются. Тем не ме-
нее такие ярко выраженные кальцифилы, как крово-
хлебка малая (Poterium sanguisorba) и кульбаба шер-
шаво-волосистая (Leontodon hispidus), встречались
только на опытных участках и нигде больше. Напраши-
вается вывод, что эти растения — химически обусловлен-
ные кальцифилы, которым нужен карбонат кальция как
таковой, а не просто влияние, оказываемое им на почву.
Чем вызвана столь настоятельная потребность этих
растений в извести, пока трудно сказать. Правда, каль-
ций— элемент, играющий существенную роль в жизни
всех растений, но те небольшие его количества, которые
нужны растению, могут быть получены почти из любой
почвы. Следовательно, причина здесь более глубокая.
Не выносящие извести кальцифобы лучше всего
1увствуют себя на кислых почвах. К ним относятся тор-
фяной мох (Sphagnum), вереск обыкновенный (Calluna
Приспособление к среде
241
vulgaris) и вереск болотный (Erica cinerea). Некоторые
растения настолько нетерпимы к извести, что ее при-
сутствие в воде оказывает на них даже отравляющее
действие; следовательно, кальцифобность этих расте-
ний, без всякого сомнения, обусловлена химически.
Однако причины такой нетерпимости остаются пока не-
ясными.
Фотопериодизм
Продолжительность светлого времени суток от рас-
света до сумерек оказывает существенное влияние на
зацветание некоторых растений. Впервые это отметил
Турнуа, обнаруживший, что коноплю, которая в северных
широтах с длинным световым периодом обычно не цве-
тет, можно заставить зацвести, искусственно сократив
длину дня. Приблизительно в то же время Клебс уста-
новил, что цветение одного из видов молодила (Semper-
vivutn funkii) можно вызвать зимой, увеличив продол-
жительность светового периода путем дополнительного
освещения электрическими лампами. Таким образом,
конопле для зацветания требовался короткий день, а
молодилу — длинный.
Вскоре были найдены и другие, столь же чувствитель-
ные к длине дня растения. В 1920 году Гарнер и Аллард,
работавшие с новыми сортами табака, обнаружили, что
сорт Мэриленд Мамот в окрестностях Вашингтона в от-
крытом грунте не цветет, хотя он цвел осенью и зимой
при выращивании в теплице. Это было очень неудачно,
потому что они хотели скрестить Мэриленд Мамот с
другими сортами, но не могли сделать это из-за отсут-
ствия цветков. Когда же исследователи попытались со-
кратить длину дня, помещая растения ежедневно через
определенное число часов в темноту, они были возна-
граждены обилием образовавшихся цветков, что позво-
лило им провести намеченное скрещивание.
Успех эксперимента побудил Гарнера и Алларда ис-
пытать этот метод на других растениях, которые в тех
местах обычно не цвели; результаты показали, что реак-
ция на укороченный день присуща не только табаку
сорта Мэриленд Мамот. Так было открыто общее для
242
Глава XI
всех растений явление, получившее название фотоперио-
дизма.
В зависимости от фотопериодической реакции расте-
ния можно разделить на три группы. К.ороткодневные
растения образуют наибольшее количество цветков, ког-
да светлый период суток составляет 12 ч или менее; к чис-
лу таких растений принадлежат конопля, табак сорта
Мэриленд Мамот, многие тропические цветковые расте-
ния и многие растения умеренной зоны, цветущие обыч-
но весной или осенью. С другой стороны, для цветения
длиннодневных растений оптимальным является фотопе-
риод продолжительностью 12 или более часов. К этой
группе относятся молодило (Sempervivum funkii), белена
(Hyoscyamus niger), картофель, пшеница и все растения
умеренных широт, которые обычно цветут летом. Поми-
мо растений короткого и длинного дня, существует
группа фотопериодически нейтральных растений, цвете-
ние которых, по-видимому, не зависит от длины дня.
В эту группу входят томат и одуванчик.
Надо сказать, что эта физиологическая классифика-
ция, основывающаяся на фотопериодической реакции
растений, не совпадает с обычной ботанической класси-
фикацией. Фотопериодическая реакция двух близкород-
ственных растений может быть совершенно различной.
Все сорта табаков, например, принадлежат к одному и
тому же виду Nicotiana tabacum, и тем не менее Мэри-
ленд Мамот по своей фотопериодической реакции отли-
чается от большинства других сортов. Томат и карто-
фель принадлежат к одному семейству (Solanaceae),
однако картофель — длиинодневное растение, а томат —
нейтральное. По-видимому, степень ботанического род-
ства и фотопериодическая реакция никак не связаны
между собой.
Приведенная выше классификация весьма прибли-
зительна. Каждому отдельному растению свойствен
собственный, вполне определенный фотопериод; в част-
ности, критический фотопериод для табака сорта Мэри-
ленд Мамот лежит между 13 и 14 ч, более продолжи-
тельный световой период полностью подавляет цвете-
ние. Длиннодневное растение белена не будет цвести,
если ежедневная экспозиция ее на свету составляет ме-
Приспособление к среде
249
нее 10...11 ч (критический фотопериод белены). Следо-
вательно, фотопериоды табака сорта Мэриленд Мамот
и белены перекрывают друг друга: при 12-часовом дне
оба растения будут цвести.
Для короткодневных растений решающим фактором,
определяющим время их цветения, по-видимому, явля-
ется длинный темновой период, а не короткий день.
С другой стороны, длиннодневные растения совсем не
требуют темнового периода; они цветут даже при не-
прерывном освещении.
Чтобы растение зацвело, не обязательно всю жизнь
выращивать его в условиях необходимой ему продолжи-
тельности освещения, так как во многих случаях бывает
достаточно поразительно недолгой экспозиции при соот-
ветствующей длине дня. В этом смысле рекорд, вероят-
но, принадлежит Xanthium — короткодневному расте-
нию, которое может долго расти, получая неправильный
фотопериод, и тем не менее зацветает, получая всего
один короткий день. Ускорение зацветания путем корот-
кой экспозиции на правильном фотопериоде называется
фотопериодической индукцией.
Стимулирующее влияние света на зацветание длин-
нодневных растений или тормозящее его влияние на за-
цветание короткодневных растений зависит не столько
от интенсивности освещения, сколько от его продолжи-
тельности. Освещения, которое дает 100-ваттная лампа,
находящаяся на расстоянии 45 см, вполне достаточно
для большинства растений, а некоторые из них гораздо
более чувствительны к свету. Цветение Xanthium, напри-
мер, можно предотвратить, поместив на расстоянии
30,5 см от него лампу в 0,3 свечи, что эквивалентно од-
ной 100-ваттной лампе, удаленной на 150 м!
Отсюда совершенно очевидно, что фотопериодизм в
данном случае не может быть связан с фотосинтезом.
Вместе с тем фотопериодическая индукция неэффектив-
на, если на время индукции из воздуха удалить углекис-
лый газ.
Для обеспечения фотопериодического эффекта не!
необходимости освещать все растение; как показали
опыты, освещать надо листья, а не почки. Шпинат
(Spinacia oleracea) —длиннодневное растение, но он за-
244
Глава XI
цветет и в условиях короткого дня, если его листья бу-
дут получать длинный день; если же, наоборот, освещать
почки, а листья держать в темноте, это не даст никакого
эффекта.
Значение освещения листа убедительно доказывают
опыты с прививкой; так, если лист,- подвергшийся фото-
периодической индукции, привить на растение, выращи-
ваемое при «неправильном» фотопериоде, зацветание
растения ускорится. Даже изолированный лист может
воспринять фотопериодическую индукцию и, будучи при-
витым на растение, окажет свое стимулирующее
влияние.
Результаты описанных опытов с прививкой позволя-
ют предположить, что под действием необходимого для
растения фотопериода в листьях образуется какое-то
вещество, которое диффундирует затем в точки роста,
ускоряя начало цветения. Дальнейшие исследования
подтвердили это предположение, хотя несмотря на мно-
гочисленные попытки, вещество, способное стимулиро-
вать зацветание, из листьев выделить не удалось. Тем
не менее это' гипотетическое вещество было названо
флоригеном*.
Географические эффекты фотопериодизма поистине
поразительны. В тропиках, где дни относительно корот-
кие, произрастают короткодневные растения, а те корот-
кодневные растения, которые мы обнаруживаем в зоне
умеренного климата, цветут весной или осенью. С дру-
гой стороны, длиннодневные растения являются типич-
ными летнецветущими растениями умеренных широт, где
длинные летние дни благоприятствуют этому2.
1 Термин принадлежит советскому физиологу М. X. Чайлахя-
ну (1937\ автору гормональной теории развития растений. —
Прим. ред.
1 Приспособительное значение фотопериодизма, теснейшим об-
разом связанное с географическим распространением видов, впер-
вые было проанализировано в работе советских физиологов
В. Н. Любименко и О. А. Щегловой «О фотопериодической адапта-
ции растений», Журнал Русского ботанического общества, т. 12,
1927. Много нового внесли в изучение этой проблемы Б. С. Мошков,
В. М. Катунский, Г. А. Самыгин, В. В. Скрипчинский и другие. —
Прим. ред.
Приспособление к средб
245
Поселились ли короткодневные растения в тропиках,
потому что предпочитают короткий день, или же такое
отношение к короткому дню выработалось у них под
влиянием жизни в тропиках? На этот вопрос мы не мо-
жем ответить. Как бы то ни было, приспособление к
короткому или длинному дню является еще одним при-
мером пригнанности структуры и функций растений к
условиям их обитания.
Кальцифильность или кальцифобность и фотоперио-
дизм— это лишь некоторые из многочисленных физио-
логических приспособлений к окружающей среде. Фи-
зиологическая специализация не всегда столь очевидна,
как структурные изменения, позволившие растениям
стать лианами или эпифитами (фото XXX), но она не
менее важна. Мир полон растений, настолько полон, что
каждое растение вынуждено бороться за свое существо-
вание, начиная с того момента, как оно вступает в жизнь
многообещающим проростком, и до того времени, пока
не образует плодов, а однолетник — пока не отомрет,
оставив после себя лишь семена. Любая причуда мор-
фологии или физиологии, благодаря которой растение
может опередить своих собратьев, должна быть исполь-
зована. И то, что это действительно происходит, прекрас-
но, потому что именно этому мы обязаны всем тем заме-
чательным многообразием растений, которые мы видим
и о которых слышим.
В наше время незанятых мест для растений в мире
осталось очень мало. Для аркто-альпийских видов, мо-
жет быть, найдется немного свободного места на скло-
нах Килиманджаро и других гор в Экваториальной
Африке, если они смогут туда добраться и «развить»
приспособления, которые дали бы им возможность по-
селиться немного выше своих конкурентов; но такие не-
занятые территории немногочисленны и разбросаны.
Между тем в любой ситуации растения должны опи-
раться на уже имеющиеся у них преимущества, борясь
за каждое небольшое усовершенствование, дающее да-
же самое незначительное преимущество над конкурен-
тами. Отказ от борьбы в природе означает гибель ин-
дивидуума и вымирание вида.
ШВА ХИ
ПАРАЗИТЫ И САПРОФИТЫ
Большинство растений получает весь необходимый
им углерод из углекислого газа воздуха, а минеральные
соли из почвы при помощи корней, однако некоторые из
них приспособились к другим способам питания. Боль-
шая группа грибов с самого начала была лишена хло-
рофилла; весь углерод они добывают из гниющих орга-
нических остатков или высасывая жизненные соки из
другого живого организма-хозяина. Насколько нам из-
вестно, у их предков тоже никогда не было хлорофилла
(если мы не придерживаемся еще бытующей точки зре-
ния, будто грибы произошли от водорослей).
Некоторые из цветковых растений избрали тот же
способ питания, что и грибы; они либо живут на других
растениях как ^паразиты, либо извлекают питательные
вещества из гниющих органических остатков в лесной
подстилке или еще где-нибудь. В отличие от грибов они
произошли от предков, которые имели хлорофилл; в са-
мом деле, в листьях некоторых паразитов (чтобы быть
совершенно точным — частичных паразитов) до сих пор
сохранился хлорофилл, и они способны синтезировать
все или часть необходимых органических соединений,
находясь в зависимости от растения-хозяина лишь в от-
ношении минерального питания. Такова, в частности,
омела (Viscutn album).
Омела знакома каждому1. Она растет как частичный
паразит на ветвях многих листопадных деревьев, в осо-
бенности на яблоне. Иногда она встречается и на хвой-
ных, например на сосне и лиственнице, и очень редко —
на вечнозеленых лиственных деревьях. С омелой связано
множество полурелигиозных мифов. Обычай украшать
омелой жилище (как это принято на рождество) восхо-
дит к древним языческим временам. Друиды высоко це-
1 См. примечание к стр. 189.
Паразиты и сапрофиты
247
нили дубовую омелу, вероятно, потому что она редко по-
селяется на дубе и представляет собой, таким образом,
необычное растение.
Полупаразитнческий образ жизни омелы
Своеобразный механизм распространения семян оме-
лы уже был описан в главе IX. Прикрепившись клейкой
слизью плода к ветви дерева, семя омелы не прорастает
тотчас же. Оно должно подождать до весны, когда более
высокая температура и более длинные дни позволят ему
начать этот процесс; ведь прежде, чем семя омелы смо-
жет прорасти, оно должно получить не только тепло, но
и много света. Когда это происходит, из семенной ко-
журы появляется маленькое белое цилиндрическое обра-
зование, напоминающее молодой корень.' На самом деле
это не корень (у омелы нет настоящих корней), а гипо-
котиль, т. е. часть оси, лежащая ниже семядолей, или
семенных листьев.
Сначала гипокотиль растет вверх, против силы тяже-
сти, но вскоре теряет способность к гравитационной ре-
акции и начинает расти от света. Вследствие этого он
изгибается по направлению к ветви, на которой растет,
и через короткое время соприкасается с ней. После этого
кончик гипокотиля разрастается в круглый клейкий
диск, плотно прижимающийся к поверхности ветви. Из
центра нижней стороны диска вырастает присоска. При-
соска врастает в кору хозяина, причем диск служит
твердым основанием, от которого она может проклады-
вать себе путь через ткани хозяина. Присоска продолжа-
ет расти, пока не достигнет древесины, после чего в ней
развиваются тяжи проводящей ткани, соединяющейся с
древесиной хозяина. На это первичное вторжение оме-
ла тратит первый год жизни.
В начале второго года появляется первая пара листь-
ев; рост, поначалу идущий крайне медленно, с течени-
ем времени ускоряется, и растение приобретает типич-
ную для него форму кустарника.
Пока куст омелы развивается, на наружной поверх-
ности дерева, в его коре, тоже происходят многочислен-
248
Глава XII
ные невидимые изменения (рис. 49). На первичной при-
соске появляются выросты, которые проникают в более
глубокие слои коры, под перидерму. Эти новые присоски
располагаются главным образом вдоль ветви. На при-
сосках здесь и там образуются почки, из которых выра-
стают новые кусты омелы, так что из одного семени в
конечном счете развивается множество растений.
Пытаться освободить дерево от омелы, срезая кусты
там, где они появились, бесполезно, так как присоски
проникают в кору, где их не может достать нож садов-
ника.
Рис. 49. Срез через ветвь яб-
лони, на которой паразити-
рует омела.
Природа присосок омелы неясна. Некоторые рас-
сматривают их как видоизмененные придаточные корни,
но у присосок нет ни одного из морфологических призна-
ков, которые были бы характерны для корней. Видимо,
лучше считать их специфическими органами поглощения,
развившимися в ответ на образ жизни этого своеобраз-
ного растения. Такие органы часто называют гаустория-
ми — термин, заимствованный из морфологии грибов.
У омелы много зеленых листьев, и поэтому она мо-
жет удовлетворять потребности в углероде, не прибегая
к помощи хозяина, однако всецело зависит от него в от-
ношении воды и неорганических веществ. Их она высасы-
вает прямо из ксилемы хозяина, через проводящие эле-
менты в присосках. Омелу поэтому надо считать частич-
ным паразитом (полупаразитом).
Может возникнуть вопрос, является ли она вообще
паразитом. Поскольку омела имеет листья и способна
фотосинтезировать, может быть, это не грабеж, а скорее
обмен, при котором она платит выработанными в про-
Паразиты и сапрофиты
249
цессе фотосинтеза сахарами за взятую у растения:хозя-
ина воду и минеральные соли? То, что это не так, пока-
зали результаты хорошо известного опыта, проведенного
Молишем в 1920 году. С молодой яблони, на ветвях ко-
торой поселилось несколько кустов омелы, Молиш срезал
все листья. Через короткое время дерево погибло от го-
лода, а вместе с ним и омела. Совершенно очевидно, что
омела не снабжает своего хозяина углеводами, а, как и
все паразиты, берет все, ничего не давая взамен. Это
характерный признак паразита.
Семейство Loranthaceae, к которому принадлежит
омела, включает множество кустарников, являющихся
полупаразитами. В Англии растет только один вид —
Viscum album, но более чем 500 других видов живут в
умеренных и тропических областях. Почти все они ведут
полупаразитический образ жизни, однако некоторые
растут в почве. Так, Nuytsia (Западная Австралия) вы-
растает в небольшое дерево высотой около 9 метров.
Arceuthobium minutissimum — интересный вид, который
обитает в Гималаях и растет как полный паразит. Его
хозяин — македонская сосна (Pinus рейсе). Паразит
развивается полностью внутри хозяина, только цветки
появляются над поверхностью коры.
К семейству норичниковых (Scrophulariaceae) при-
надлежат такие известные полупаразиты, как погремок
(Rhinantlius), марьянник (Melampyrum) .мытник болот-
ный и мытник лесной (Pedicularis palustris и Р. sylvati-
са)и различные очанки (виды рода Euphrasia). Все они
живут как частичные паразиты на корнях других расте-
ний.
Другие полупаразиты
Очанки и погремки — не облигатные паразиты, так
как могут существовать самостоятельно, без хозяина.
Что же касается марьянника, то прикрепление к корням
Других растений — необходимое условие его нормально-
го развития. Все эти растения имеют хлорофилл и спо-
собны в достаточной мере обеспечить себя углеводами.
Они растут на пастбищах и лугах, где много корней зла-
250
/’лава ХН
ков, к которым можно присосаться. Семена их прораста-
ют обычным путем. На ранних этапах жизни проросток
образует боковые корни, которые утолщаются, превра-
щаясь в клубеньки. Если один из этих клубеньков сопри-
касается с корнем злака, он выпускает отросток,
внедряющийся в его корень и развивающий проводящую
ткань, через которую паразит «грабит» своего хозяина.
По-видимому, главное, что нужно паразитам от хозяи-
на, это вода, поскольку их слабо развитая корневая си-
стема не в состоянии восполнить ее потерю при транспи-
рации.
Семейство санталовых (Santalaceae) состоит в значи-
тельной степени из полупаразитов, одни из которых по-
селяются, подобно омеле, на ветвях деревьев, а другие,
как, например, очанка, паразитируют на корнях. В Анг-
лии встречается один представитель этого семейства —
ленец (Thesium humifusum), являющийся корневым па-
разитом и растущий на мелах и известняках1.
Настоящие паразитные растения
Весьма любопытны растения рода Tozzia (семейство
норичниковых), которые в молодом возрасте живут как
настоящие паразиты, но впоследствии становятся полу-
паразитами. Этот род представлен всего двумя видами,
один из которых обитает в Альпах, а другой — в Кар-
патах. При прорастании семян обоих видов образуется
подземное корневище, полностью паразитирующее на
корнях хозяина. Позднее появляется цветоносный побег
с бледно-зелеными, бедными хлорофиллом листьями.
Образовав надземный побег, Tozzia переходит к полупа-
разитическому образу жизни и придерживается его
вплоть до периода цветения; цветки Tozzia напоминают
цветки очанки. Образовав семена, паразит отмирает.
Близкое к норичниковым семейство заразиховых
(Orobanchaceae) состоит исключительно из растений-па-
1 В СССР имеется несколько видов этого рода, также приуро-
ченных к известнякам и распространенных в степных и горных райо-
чах. — Прим. ред.
Паразиты и сапрофиты
251
разитов. Наивысшего расцвета оно достигает в теплых
умеренных зонах северного полушария, но и в Англии
растет несколько видов, в том числе петров крест
{Lathraea squamaria)1 и множество видов заразих
(Orobanche).
Петров крест паразитирует на корнях древесных
растений, главным образом на орешнике {Corylus avel-
lana) и ильме (Ulmus). Он- растет в сырых лесах и в
живых изгородях на богатых почвах, в частности обычен
на известняках. Это растение образует толстый цвето-
носный побег, несущий белые или фиолетовые шлемо-
видные цветки. На надземном побеге имеются также че-
шуйчатые листья, лишенные хлорофилла.
Подземная часть растения состоит из толстого
разветвленного корневища, несущего мясистые чешуе-
видные листья. Эти листья перегнуты назад таким обра-
зом, что их верхняя часть соединяется с нижней, обра-
зуя полые мешки. Внутренняя поверхность каждого меш-
ка покрыта железками, функцией которых является, ве-
роятно, выделение воды. По-видимому, эти необычные
листья служат для запасания пищи, но им приписывали
и другие, зачастую совершенно невероятные функции.
Так, высказывалась мысль, что секретирование воды
железками заменяет транспирацию. Вполне возможно,
что так оно и есть, поскольку петров крест связан с ос-
новными водопроводящими элементами хозяина и может
транспирировать только через чешуйчатые листья. Кро-
ме того, было предположено, что железки несут выдели-
тельную функцию, удаляя излишки аммиака, фосфатов
и сульфатов, и что мелкие почвенные организмы, попав-
шие в мешки, перевариваются и поглощаются паразитом.
Нет никаких данных, которые подтверждали бы эти
гипотезы, да и вообще они представляются маловеро-
ятными.
Считается, что семя петрова креста прорастает толь-
ко в контакте с корнями подходящего хозяина; видимо,
сначала оно должно быть вмыто в почву дождем. На
1 В лиственных лесах Европейской части СССР он тоже обы-
чен. — Прим. ред.
/
252
Глава XII
корне проростка возникают присоски; встретив корень
хозяина, они обвивают его и закрепляются на нем при
помощи многочисленных крошечных выростов. Прочно
закрепившись в этом, положении, присоска выпускает
отросток, врастающий в древесину хозяина, и с этого
момента петров крест получает всю свою пищу из корней
растения-хозяина.
Кроме Lathraea squatnaria, в некоторых районах
Англии натурализовались и другие виды Lathraea.
В частности, L. clandestina паразитирует на корнях ивы
и тополя и растет во влажных тенистых местах. Перво-
начально это растение было распространено в Испании,
Италии, Франции и Бельгии.
В Англии произрастают одиннадцать видов заразих1,
и все они — корневые паразиты. Их крошечные семена
вмываются в почву дождем и, наткнувшись на корень
хозяина, прорастают. При этом на корне проростка обра-
зуется присоска, проникающая в корень хозяина. Осо-
бенностью корйевого паразитизма заразих служит тес-
ный контакт между тканями паразита и хозяина. По ме-
ре развития паразита ткани совмещаются: древесина
паразита точно совпадает с древесиной хозяина, кора —
с корой, и даже эпидермис заразихи срастается с эпи-
дермисом растения-хозяина.
Из подземной части заразихи вырастает цветонос-
ный побег. Как и у петрова креста, он состоит из стебля
с лишенными хлорофилла чешуйчатыми листьями, окан-
чивающегося соцветием.
Паразит совсем иного рода — это повилика (Cuscuta),
два вида которой произрастают в Англии. Третий вид
паразитирует на льне и изредка попадает к нам с се-
менами льна. Повилика принадлежит к семейству вьюн-
ковых (Convolvulaceae) 2 и представляет собой вьющееся
растение; обвивая стебель хозяина, она высасывает из
него питательные вещества при помощи присосок.
Повилика европейская (Cuscuta еигораеа) парази-
тирует на жгучей крапиве (Urtica dioica) (фото XXXI),
1 В СССР их несколько десятков. — Прим. ред.
1 В СССР более 30 видов повилик. Их обычно выделяют в осо-
бое семейство Cuscutaceae. — Прим. ред.
Паразиты и сапрофиты
255
тогда как повилика тимьянная (Cuscuta epilhymum) по-
ражает множество различных растений, в том числе
утесник, вереск и клевер (Trifolium). Семя обыкновен-
ной повилики маленькое и очень простое по строению
Прорастая, оно образует крошечный нитевидный про-
росток, у которого нельзя ясно разграничить зароды-
шевый корешок, почечку и семядоли.
Проросток способен некоторое время существовать
самостоятельно, и в этот период его стебель обнаружи-
вает ярко выраженную нутацию, характерную для вью-
щихся растений. Если он встречает стебель хозяина,
Рис. 50. Повилика (Cuscuta
еигораеа), растущая на иве.
остальная часть проростка засыхает и отмирает, а сте-
бель паразита крепко обвивает свою жертву; таким об-
разом, на этой стадии развития повилика никак не свя-
зана с почвой и все питательные вещества должна по-
лучать от хозяина (рис. 50).
Присоски повилики появляются из стебля. Сначала
они распластываются на стебле хозяина, и тогда из цент-
ра уплощенного участка образуется вырост, который
врастает в ткани хозяина, пока в конце концов не до-
берется до проводящей ткани, где сильно разрастается.
Хотя вначале вырост состоит из недифференцированной
ткани, достигнув проводящей системы хозяина, он разви-
вает ксилему и флоэму, которые соединяются с соответ-
ствующими тканями хозяина. Итак, связь между прово-
дящими тканями паразита и хозяина установлена и пи-
тание повилики обеспечено.
Вьющийся стебель взрослого растения повилики име-
ет желтую или красную окраску и кажется безлистным;
254
Глава XU
однако, приглядевшись внимательнее, можно увидеть
на нем то здесь, то там крошечные чешуйчатые
листья. Они лишены хлорофилла и не способны к фото-
синтезу.
Такой же паразитический образ жизни свойствен и
Cassytha — тропическому роду, принадлежащему к се-
мейству лавровых (Lauraceae). Этот род насчитывает
около 15 видов, обитающих в тропических областях Ста-
рого Света.
При паразитизме у цветковых растений наблюдается
явная тенденция к общему упрощению строения. Изба-
вившись от необходимости питаться самостоятельно,
растение может сконцентрировать свою энергию на раз-
множении, в особенности на образовании огромного
количества семян; последнее совершенно необходимо,
поскольку подходящего хозяина не так легко встретить.
То же самое наблюдается и у тех животных, которые
являются эндопаразитами. Так как самостоятельно пере-
двигаться им больше не нужно, конечности у них исче-
зают; часто исчезают и пищеварительные органы, по-
скольку такое животное питается уже переваренной пи-
щей хозяина. Органы размножения, напротив, обычно
хорошо развиты, поскольку риск не найти хозяина, не-
обходимого при паразитическом образе жизни, очень
велик.
Предельная адаптация к паразитическому образу
жизни среди растений наблюдается у тропической Raf-
flesia arnoldii, произрастающей в Индонезии и парази-
тирующей на корнях виноградных. Вегетативное тело
Rafflesia редуцировано до ряда нитей, напоминающих
гифы гриба, которые разветвляются в тканях растения-
хозяина. Нормальны только цветки. Они возникают пря-
мо из корней хозяина, так что кажется, будто они появ-
ляются из почвы. Цветки R. arnoldii самые крупные в ра-
стительном царстве (стр. 190). Они имеют тусклую
красноватую окраску и запах, напоминающий запах
гнилого мяса, в связи с чем раффлезию иногда называют
«вонючей лилией». Судя по запаху, можно предполо-
жить, что цветки этого растения опыляются мухами.
Интересный способ распространения плодов паразита
слонами был описан выше.
Паразиты и сапрофиты
255
Сапрофиты
Сапрофиты — это растения, питающиеся за счет пе-
регнивающих органических остатков. Сапрофитизм ши-
роко распространен среди грибов, но у цветковых расте-
ний встречается редко. Причины этого пока неясны. На
первый взгляд многое говорит в пользу сапрофитного
образа жизни. Лесная подстилка обычно богата органи-
ческими остатками, и, кроме того, сапрофиту не прихо-
дится заботиться ни об освещении, ни о конкуренции за
питательные вещества со стороны других растений.
Тот факт, что сапрофитный образ жизни избрали
лишь немногие цветковые растения, скорее всего объяс-
няется отсутствием у них ферментов, необходимых для
разложения органических остатков до такого состояния,
в котором шеи легко поглощались бы корнями. Хотя до-
казано, что корни большинства растений в некоторой
степени способны поглощать молекулы органических ве-
ществ, все же это происходит не настолько легко, чтобы
они могли удовлетворять свои потребности в естествен-
ных условиях, несмотря на то что в определенных лабо-
раторных условиях это им удается (горох, выращивае-
мый в темноте, на питательной среде, содержащей сахар,
даже образовывал цветочные почки). Видимо, растениям
легче стать паразитами, чем сапрофитами.
Важно отметить, что почти все сапрофитные цветко-
вые растения микотрофны, т. е. на их корнях поселя-
ется гриб-партнер (гл. XIV). Единственным исключени-
ем из этого, как считают, является Wullschlaegelia
aphylla — сапрофитная орхидея, которая по общему
мнению, свободна от гриба, хотя определенно этого
опять-таки сказать нельзя. Вполне возможно, что для
разложения органических остатков сапрофиты полага-
ются на ферменты, продуцируемые грибом, или по край-
ней мере передают грибу функцию поглощения этих
остатков. Если это так, то по меньшей мере сомнитель-
но, что сапрофитное цветковое растение действительно
паразитирует на грибе.
Наиболее известным сапрофитом флоры Англии яв-
ляется гнездовка (Neottia nidus-avis) — орхидея, расту-
щая в буковых лесах на известковых почвах. Своим на-
256 Глава XII
званием растение обязано густому сплетению корней, на-
поминающему птичье гнездо. Надземный побег несет
коричневые чешуйчатые листья, содержащие очень мало
хлорофилла (если он вообще имеется).
Из числа других сапрофитов Англии следует отме-
тить также подъельник (Monotropa hypepithys), встре-
чающийся в буковых и сосновых лесах. Все растение
желтоватое или цвета слоновой кости и выглядит со-
стоящим как бы из воска; хлорофилла крайне мало или
совсем нет. В лесах Америки растет близкий к нему вид
М. uniflord1.
1 В СССР растут и гнездовка (Neotiia nidus-avis) и подъельник
(Monotropa hypopithys); первая обычна в лиственных лесах, вто-
рой — в хвойных. — Прим. ред.
ГЛАВА Х1П
ХИЩНЫЕ РАСТЕНИЯ
Мы уже убедились (гл. VII), насколько важное зна-
чение для растений имеют насекомые как основные аген-
ты при перекрестном опылении. Насекомые и растения,
так сказать, сжились, и приспособления некоторых цвет-
ков к своим опылителям граничат с чудом. Но в природе
насекомых есть и другая сторона, пагубная для расте-
ний. Листоеды, тли и прожорливая саранча — злейшие
враги растений, и ущерб, который они наносят расти-
тельности каждый год, неисчислим.
Поэтому едва ли удивительно, что в некоторых слу-
чаях обычные взаимоотношения хищника и жертвы на-
рушаются и растения, вместо того чтобы пассивно слу-
жить пищей для насекомых, наносят ответный удар и
сами питаются насекомыми. Насекомоядность неодно-
кратно возникала в растительном царстве в процессе эво-
люции, и растения изобрели замечательные приспособле-
ния, направленные на то, чтобы сделать насекомых своей
добычей.
Росянка круглолистная {Drosera rotundifolia)—не-
большое растеньице, встречающееся на болотах и сырых
торфяниках по всей Англии (фото XXXII)*. Оно имеет
розетку длинночерешковых красных листьев с округлыми
пластинками около 1 см в поперечнике. В период цвете-
ния, продолжающегося с июня по август, цветонос, не-
сущий кисть белых цветков, возвышается на 10... 30 см
над розеткой: маленькое, ничем не примечательное рас-
теньице трудно заметить, если не искать его специально.
Росянка как ловушка для насекомых
По краям и на верхней стороне листьев росянки рас-
полагается множество чувствительных щетинок (фото
XXXIII). Каждая щетинка состоит из ножки с шаровид-
1 В СССР это обычное растение сфагновых болот. — Прим. ред.
258
Глава ХШ
ной головкой на конце; головка увенчана блестящей
капелькой жидкости, так что кажется, будто листья
смочены росой. Щетинки, сидящие по краям листьев,
имеют относительно длинные ножки, тогда как ножки
щетинок на верхней стороне листьев более короткие.
На одном листе может насчитываться до 200 щетинок.
При рассмотрении щетинок под микроскопом виден
тяж ксилемы, поднимающийся по ножке и расширяю-
щийся на ее конце. Здесь проводящий пучок окружен
тремя слоями клеток. Самый внутренний слой, клетки
которого имеют утолщенные стенки, несколько напоми-
нает эндодерму, а два наружных слоя содержат красный
пигмент, обусловливающий окраску листа; все эти три
слоя также несут секреторную функцию.
Было высказано предположение, что листья росянки
обладают некоторой привлекательностью для насекомых
благодаря своей окраске, блестящему секрету или запа-
ху, но это пока не доказано. Во всяком случае, любое
насекомое, опрометчиво севшее на лист росянки, вскоре
раскаивается в этом, хотя для напрасных сожалений
у него остается очень мало времени. Коснувшись чувст-
вительной щетинки, насекомое тотчас же увязает в вы-
деляемой ею чрезвычайно липкой жидкости. Для удер-
жания добычи росянка не полагается на один липкий
секрет — как только насекомое поймано, щетинка начл-
нает сгибаться у основания, другие щетинки присоеди-
няются к ней, так что тело жертвы оказывается погре-
бенным под массой клейких щетинок. Сгибанием щети-
нок дело не ограничивается; весь лист начинает склады-
ваться по центру пластинки и накрывает насекомое,
удерживая его и одновременно приводя в действие дру-
гие щетинки.
Поймав насекомое, росянка переваривает его. Ще-
тинки начинают выделять пищеварительный фермент,
похожий на имеющийся в желудке человека пепсин,
и кислоту, создающую необходимую для деятельности
фермента кислую среду. Фермент растворяет мягкие
части жертвы, расщепляя белки до более простых сое-
динений, которые могут поглощаться листьями. Всосав
все удобоваримые части насекомого, лист снова раскры-
вается и готов ловить новую добычу.
Хищные растения
259
Сгибание щетинок росянки, как и гаптотропическая
реакция усика лазящего растения на раздражение при-
косновением (гл. XI), по-видимому, обусловлено нерав-
номерным ростом. Полагают, что в ответ на прикосно-
вение происходит перераспределение ауксина, хотя это
еще не доказано. Сгибание щетинок росянки, наблюдае-
мое при прикосновении к ним каким-нибудь твердым
нейтральным предметом, например куском стекла, го-
ворит о том, что единственным раздражителем, вызы-
вающим реакцию, служит прикосновение. Подобно уси-
кам лазящего растения, щетинки росянки нечувствитель-
ны к жидкостям; капли дождя скатываются с них, не
оказав никакого влияния. Частью щетинки, чувствитель-
ной к прикосновению, является ее шаровидная головка,
тогда как сгибание происходит у основания ее ножки.
Щетинки реагируют не только на прикосновение, но
и на химические раздражители — при обработке раство-
рами некоторых веществ они сгибаются. Дарвин первым
обнаружил, что все вещества, способные вызвать реак-
цию щетинок, содержат азот. Он обрабатывал листья
Drosera различными веществами, не содержащими азота,
включая гуммиарабик, сахар, крахмал и спирт, и ни
в одном из этих случаев ему не удалось вызвать движе-
ния щетинок. Но когда он применял такие содержащие
азот продукты, как молоко, белок яйца или мясной экст-
ракт, щетинки каждый раз реагировали. Большую чувст-
вительность они проявляли, в частности, к растворам
солей аммония.
Росянка, несомненно, выигрывает от ловли и перева-
ривания добычи; впервые это было доказано Фрэнсисом
Дарвином, сыном великого Чарлза Дарвина. Он взял
шесть сосудов с росянками и каждый разделил пере-
городкой на две половины. Росянки, находившиеся по
одну сторону перегородки, получали мясо, а тех, которые
были по другую сторону, не кормили совсем. Через не-
которое время растения, питавшиеся мясом, выглядели
более здоровыми и содержали больше хлорофилла н
крахмала. Однако при сравнении обеих групп растений
оказалось, что наиболее сильно мясная диета повлияла
на количество образованных цветков. В августе при сре-
зании цветоносов обнаружилось, что на каждые 100 цвет-
260
Глава XIII
ков, образованных контрольными растениями, приходит-
ся 165 цветков растений, питавшихся мясом, а соотно-
шение числа семян составляет соответственно 100 : 240.
Следовательно, главная польза от питания насекомыми
заключается, по-видимому, в том, что репродуктивная
способность растений увеличивается.
Растениям росянки, растущим в естественных усло-
виях, насекомоядность, вероятно, столь же выгодна, как
и растениям, выращиваемым в лаборатории. Какие имен-
но элементы питания они получают из насекомых, неиз-
вестно, но, естественно, возникает мысль, что таким эле-
ментом является азот, особенно если учесть реакцию
щетинок на азотистые вещества. С этой точки зрения,
возможно, немаловажен тот факт, что росянка растет на
болотах, где кислая среда снижает активность нитрифи-
цирующих почвенных бактерий. Что могло быть для рас-
тения более естественным, чем воспользоваться случаем
и дополнить свою диету несколькими кусочками богатого
белками мяса насекомых?
В Англии, произрастают три вида росянок, отличаю-
щихся друг от друга формой листьев*. Большой род
Drosera содержит 90 видов растений, многие из которых
тропические. Все они ловят насекомых одним и тем же
способом. Некоторые тропические виды крупнее росянок,
произрастающих в Англии; у D. binata, например, листья
длинные и раздвоенные. Близкий к росянке кустарник
Drosophyllum lusilanicum, растущий в Испании, Порту-
галии и Марокко, ловит насекомых при помощи клейкой
жидкости, выделяемой железками на листьях; листья
при этом остаются неподвижными. Португальские
крестьяне используют листья D. lusitanicum как липкую
бумагу от мух.
Венерина мухоловка (Dionaea muscipula) — другой
представитель семейства росянковых (Droseraceae), для
которой характерен совсем иной способ ловли насеко-
мых. Она растет по сырым местам, главным образом
в Северной и Южной Каролине, и примечательна тем,
что была первым насекомоядным растением, деятель-
1 В СССР тоже встречаются три вида и гибриды между ними. —
Прим. ред.
Хищные растения
261
ность которого стала объектом научного наблюдения;
в 1768 году натуралист из Лондона Эллис послал опи-
сание венериной мухоловки Карлу Линнею, который
отозвался о ней как о «чуде природы» («miraculum
naturae»). Линней и не предполагал, что это растение
н самом деле питается насекомыми, так как думал, что
они захватываются случайно и впоследствии освобож-
даются.
Dionaea ловит насекомых при помощи листьев и рас-
тет в болотистых местах, но на этом сходство с росянкой
Рис. 51. Лист венериной му-
холовки.
Обратите внимание на чувствитель-
ные щетинки в середине листа.
кончается. Более или менее округлые листья мухоловки
состоят из двух половинок, способных захлопываться
подобно книге; края листьев украшены многочисленными
краевыми зубцами. Когда лист закрывается, зубцы за-
ходят друг за друга, как зубцы капкана (рис. 51).
На верхней стороне каждой половинки листа, близ
ее середины,"сидят по три нежных волоска, чувствитель-
ных к прикосновению. Если муха или небольшой червяк,
пробираясь над листом, касаются одного из этих волос-
ков, лист внезапно захлопывается и несчастная жертва
оказывается в ловушке, образованной двумя половинка-
ми листа. Поверхность листа усеяна маленькими крас-
ными железками, которые выделяют жидкость, содер-
жащую муравьиную кислоту и фермент, напоминающий
пепсин. Мягкие части насекомого перевариваются, и об-
разующиеся продукты усваиваются растением. Процесс
переваривания может длиться неделю или дольше, и вес
это время лист остается закрытым. После того как
262
Глава XIII
жертва полностью переварена, лист слова раскрывается
и вновь ждет добычу.
Пищеварительная жидкость выделяется железками
листа в значительном количестве. Дарвин делал надрез
на листе, поймавшем крупную муху, и наблюдал, что
жидкость продолжала вытекать из него в течение всех
девяти дней, которые потребовались растению для пере-
варивания насекомого.
Aldrovanda vesiculosa — близкая родственница Dio-
naea. Это лишенное корней, плавающее растение живет
в освещенных солнцем водоемах южной и Центральной
Европы, Индии и Австралии. Его листья мельче, чем
у Dionaea, и обычно наполовину раскрыты подобно
створкам раковины мидий. На поверхности листьев, за-
крывающихся так же, как и листья Dionaea, сидят чув-
ствительные волоски. Добычей растению служат водя-
ные блохи, личинки насекомых и диатомеи. Хотя на
листьях имеются железки, неизвестно, выделяют ли они
пищеварительную жидкость.
Другие хищные растения
Все описанные выше насекомоядные растения при-
надлежат к семейству росянковых. Тоже хищным, но
не родственным росянковым является семейство пузыр-
чатковых (Lentibulariaceae), насчитывающее во всех
частях света почти 250 видов. Наиболее простые расте-
ния этого семейства — жирянки (Pinguicula), четыре ви-
да которых встречаются в Англии1.
Жирянка обыкновенная (Р. vulgaris) обитает на бо-
лотах и влажных пустошах; она широко распространена
на Британских островах, хотя на юге встречается редко.
Она хорошо растет на заболоченных местах в горах,
поднимаясь на высоту более 900 м над уровнем моря.
Название Pinguicula переводится как «маленькое, жир-
ное» и относится к распластанной на земле розетке мя-
систых листьев с единственным, необычайно красивым
1 В СССР растут 7 видов жирянки; главным'обраэом это оби-
татели тундр, в^том числе горных, сырых лугов и болот. — Прим,
ред.
Хищные растения
263
фиолетовым, цветком, возвышающимся из ее середины.
Насекомое приклеивается к листьям липкими выделения-
ми многочисленных железок, имеющих ножки; другие
железки, сидячие, секретируют пищеварительную жид-"
кость.
Кроме фермента, напоминающего пепсин, пищевари-
тельная жидкость жирянки, видимо, содержит вещество,
сходное с реннином, выделяющимся в желудке теленка.
Поскольку реннин обладает свойством вызывать сверты-
вание молока, в Лапландии жирянку используют при
изготовлении сыра.
Четыре обитающих в Англии вида пузырчатки
(Utricularia)1 имеют ловушку для насекомых гораздо
более сложного типа, нежели клейкие листья жирянки;
пузырчатка — одно из самых специализированных хищ-
ных растений. Это водные растения, без корней, свободно
плавающие в озерах, прудах и канавах, главным обра-
зом в горных местностях. Тело растения, по-видимому,
состоит из множества коротких веточек, несущих изящно
рассеченные листья; впрочем, такая интерпретация его
структуры вызвала сомнения, и вполне возможно, что
сами «веточки» представляют собой видоизмененные
листья. Летом в течение месяца или дольше над водой
возвышаются сидящие на вертикальном цветоносе ярко-
желтые цветки, благодаря чему плавающее в небольшом
горном озере растение становится хорошо заметным.
В остальное время года Utricularia не привлекает к се-
бе внимания, так как растеньице это маленькое и листья
его в основном погружены в воду.
Utricularia ловит добычу при помощи небольших
пузырьков (у наиболее распространенной в Англии
U. vulgaris их длина составляет около 0,25 см), в зна-
чительном количестве сидящих на концах листьев, за-
меняя некоторые крайние дольки листа. Способ развития
пузырька указывает на то, что это видоизмененный лис-
точек. Пузырек имеет яйцевидную или грушевидную
форму и прикрепляется к листу короткой ножкой, возни-
кающей сбоку.
* В СССР обитает 6 видов пузырчатки главным образом на
торфяных болотах, в озерках и канавах. — Прим. ред.
204
Глава XIII
Входное отверстие пузырька прикрыто небольшим
клапаном, или дверкой ловушки, открывающимся внутрь;
когда пузырек закрыт, клапан опирается на утолщенный
валик, окружающий отверстие. На внутренней поверхно-
сти пузырька располагается множество четырехраздель-
ных волосков, которые всасывают воду; в результате
гидравлическое давление внутри пузырька ста-
V ловится ниже наружного давления, о чем свиде-
тельствуют его сдавленные стенки.
На внешней поверхности клапана сидят кро-
шечные чувствительные волоски, реагирующие
на прикосновение. Стоит мелкому водяному су-
ществу случайно задеть один из них, как «пру-
жинка», держащая клапан закрытым, момен-
! тальню расслабляется. Толкаемая давлением во-
ды снаружи дверка ловушки открывается, и во-
! да устремляется в пузырек, увлекая за собой
\ незадачливую жертву, по собственной глупости
J пустившую в ход ловчий механизм. Клапан за-
। крываетЬя, и животное поймано.
V рис. 52. Листовой кувшинчик Genlisea.
Добычей пузырчатке служат главным образом водя-
ные блохи, например Daphnia и Cyclops, но вообще ее
жертвой может стать любое существо, достаточно мел-
кое, чтобы попасть в пузырек, и в то же время достаточ-
но крупное, чтобы вызвать раздражение чувствительных
волосков. Пузырьки не выделяют никаких пищеваритель-
ных соков; растение ждет, когда пленники погибнут и
бактерии разложат их тела. Затем растворимые продук-
ты разложения вместе с некоторым количеством воды,
имеющейся в пузырьке, всасываются водопоглощающими
волосками. После этого пузырек готов к приему новой
порции пищи.
Крупный род Utricularia содержит свыше 200 видов,
большей частью тропических. Многие пузырчатки — на-
Хищные растения
265
земные растения, а йёкоторые даже эпифиты. Все они
хищные.
В тропиках Америки и Африки обитает интересный
род семейства пузырчатковых Genlisea. Это наземное
растение со странными кувшинообразными листьями,
тесно прижатыми к земле. Кувшин образован нижней
частью листа и открывается вильчатой шейкой с двумя
изогнутыми веточками (рис. 52). На этих веточках име-
ется ряд направленных вниз щетинок, которые позволяют
мелким почвенным животным заползать в кувщип, но
не дают им выбраться наружу. На внутренней поверх-
ности кувшина располагаются железки, но выделяют ли
они пищеварительные соки или же растение ждет, пока
бактерии переварят жертву, мы не знаем.
Растения-кувшины
Растения этого типа принадлежат к трем семействам:
непентесовые (Nepenthaceae), саррацениевые (Sarrace-
niaceae) и цефалотовые (Cephalotaceae). Большей частью
это тропические или субтропические виды, но один из
них, Sarracenia purpurea, в 1906 году был завезен в бо-
лота центральной Ирландии, где акклиматизировался и,
по-видимому, хорошо себя чувствует.
Семейство Nepenthaceae представлено единственным
родом Nepenthes, встречающимся главным образом на
эстровах тропической Азии и особенно процветающим
на острове Калимантан, хотя область его распростране-
ния простирается до Мадагаскара, Бенгальского залива,
Южного Китая и северной Австралии. Насчитывается
эколо 60 видов непентеса; в большинстве своем это ла-
зящие кустарники, растущие в болотистых местах и под-
держивающие свое тело с помощью видоизмененных
листьев. Замечательные кувшины, в которые они улав-
ливают насекомых, возникают на концах листовых
усиков.
Черешок листа Nepenthes у основания плоский, затем
переходит в усик, который обвивается один раз вокруг
эпоры, закручивается книзу, а затем вновь поднимается
и образует на своем конце кувшин. Длина кувшинов
зарьирует от 2,5 до 30,5 см; они имеют форму урны
266
Глава ХШ
с выступающей наверху крышечкой (образованной пла-
стинкой листа), которая прикрепляется в одном месте
подобно, крышке немецкой пивной кружки (рис. 53).
Считается, что эта выступающая крышечка защищает
кувшин от попадания воды во время дождя. Весь кувшин
часто окрашен в яркие красные и фиолетовые тона, де-
лающие его заметным и
привлекающим насекомых.
Кувшины Nepenthes —
прекрасно сконструирован-
ные ловушки для насекомых.
Отверстие кувшина окру-
жено ребристым ободком,
содержащим одревеснев-
шую ткань, которая придает
ему твердость и держит
входное отверстие кувшина
открытым. Внутренняя стен-
Рис. 53. Кувшин Nepenthes.
ка дифференцирована на два участка совершенно раз-
личной структуры. Верхний участок состоит из клеток
с восковым налетом на оболочках; они образуют поверх-
ность, на которой не может удержаться ни одно насе-
комое; этот восковой налет на клеточных стенках рас-
слаивается на крошечные чешуйки, мешающие выпол-
нять свою функцию даже цепким сосочкам на ногах
муравьев. Всякое насекомое, затеявшее рискованное
предприятие и оказавшееся на этой предательской по-
верхности, почти неминуемо соскальзывает в кувшин.
Нижний участок железистый, с многоклеточными же-
лезками, секретирующими водянистую жидкость, кото-
рой наполнен кувшин. Он искрится от капелек жидкости,
выделяемой железками.
Нектарники на нижней стороне крышечки образуют
нектар, служащий приманкой для насекомых. Когда на-
Хищные растения
267
сеномые садятся на ободок кувшина, им достаточно
сделать несколько шагов, чтобы попасть на скользкий
слой, откуда нет возврата. Сначала жидкость, выделен-
ная железками кувшина, имеет нейтральную реакцию,
но как только жертва поймана, реакция становится кис-
лой и в жидкости появляется фермент, напоминающий
пепсин, который переваривает мягкие
части ее тела.
Интересно, что в жидкости кувшинов
Nepenthes, если их разрезать, всегда
можно обнаружить разнообразную флору
и фауну. Сюда входят сине-зеленые,
десмидиевые, диатомовые водоросли,
Protozoa, нематоды, клещи, мухи и их
личинки, личинки мотыльков и бабочек.
Все эти жильцы, по-видимому, прекрасно
себя чувствуют и находятся там скорее
добровольно, чем как пленники. Удиви-
тельно, как.они защищаются от действия
переваривающего фермента, содержаще-
гося в жидкости. Вероятно, они инактиви-
Рис. 54. Кувшин Sarracenia.
руют его с помощью какого-то антифермента. Подобные
вещества, как известно, существуют.
Семейство Sarraccniaceae включает три рода расте-
ний с кувшинчатыми листьями, из которых наиболее
важен род Sarracenia. Sarracenia растет на болотах
Атлантического побережья Северной Америки, где кли-
мат теплый. От Nepenthes она отличается тем, что у нее
весь лист преобразован в кувшин, тогда как у Nepenthes
кувшин образован только частью листа. Кувшины Sar-
racenia имеют трубчатую форму (рис. 54) и располага-
ются вертикально. Как и у Nepenthes, отверстие кувши-
на защищено крышечкой. Край кувшина расширен и
образует плоский вырост, который не только увеличивает
фотосинтезирующую поверхность, но и служит удобной
Глава XiП
площадкой, по которой насекомые могут переползти
к отверстию кувшина.
Жесткий ободок по краю отверстия кувшина Sarrace-
nia не дает ему спадаться. Нектарники, которыми изо-
билуют крышечка, ободок и наружная поверхность кув-
шина, представляют большой соблазн для насекомых.
Насекомые, ищущие нектар, завлекаются на внутренний
край ободка, где начинается скользкая зона. Она состо-
ит из клеток с гладкой кутикулой, на которой отклады-
вается воск; эти клетки заходят друг за друга, как че-
репица на крыше. Ниже скользкой зоны располагается
участок с длинными щетинками, каждая из которых
направлена вниз, в глубь кувшина; в этой зоне между
щетинками тоже имеются нектарники, так что насеко-
мые заманиваются еще дальше. Наконец, вблизи дна
кувшина находится участок с гладкой поверхностью,
лишенный щетинок и ясно выраженных железок.
Обычно, хотя и не всегда, кувшины Sarracenia содер-
жат некоторфе количество жидкости, но секретируется
ли опа железками, выделяющими нектар, недифферен-
цированным эпидермисом нижней части кувшина или же
это в основном просто дождевая вода, неизвестно. Преж-
де считалось, что Sarracenia не вырабатывает пищевари-
тельных соков, но теперь установлено, что в жидкости
кувшинов присутствует какой-то фермент. Удобовари-
мые части тела насекомых разлагаются, и образовавшие-
ся продукты всасываются стенками кувшина.
У некоторых видов Sarracenia жидкость в кувшинах
имеет не кислую, а щелочную реакцию. Если перевари-
вающий фермент лучше действует в щелочной среде,
чем в кислой, то он должен отличаться от фермента
Nepenthes, напоминающего пепсин.
Sarracenia, видимо, менее приспособлена к насеко-
моядному образу жизни, чем Nepenthes. У Darlingtonia,
другого представителя саррацениевых, растущего в Ка-
лифорнии, кувшины имеют сложную структуру; фермен-
ты для переваривания насекомых в них отсутствуют,
эта функция осуществляется бактериями в жидкости
кувшинов (рис. 55).
Третий небольшой род саррацениевых — это Heliam-
phora, обитающая в Гайане и Венесуэле.
Хищные растения
269
Единственным представителем цефалотовых (Ccpha-
lotaceae) является Cephalotus follicularis, растение-кув-
шин, произрастающее на банке Кинг-Джордж-Саундс
в Западной Австралии. У него только нижние листья
Рис. 55. Кувшин Darlingionia,
образуют кувшины, верхние же плоские и зеленые. Кув-
шины у Cephalotus действуют почти так же, как и у ЛГе-
penthes.
Хищные грибы
Плотоядность встречается не только у высших расте-
ний, но развита сильно и у некоторых групп грибов.
Она неоднократно возникала в ходе эволюции цветковых
растений, поскольку не все семейства, для которых ха-
рактерен этот образ жизни, родственны друг другу и
способы ловли добычи у них различны. Точно так же
и хищные формы грибов обнаруживаются в далеко от-
стоящих друг от друга порядках, что указывает па их
независимое развитие.
Хищные грибы, как и насекомоядные цветковые рас-
тения, ловят мелких животных живыми и питаются ими.
Добычей служат главным образом простейшие и немато-
ды,хотя иногда попадаются и коловратки. Среди про-
стейших жертвами хищных грибов чаще всего оказыва-
ются мелкие амебы или их родственники, как, например,
Dtfflugia, тело которой заключено в твердую раковинку.
Коловратки хорошо известны тем, кто любит работать
270
Глава Xlll
с микроскопом; вокруг ротового отверстия у них имеется
венчик постоянно колеблющихся волосков, или ресничек,
движение которых создает впечатление вращающегося
колеса. Нематоды — это чрезвычайно широко распрост-
раненные круглые черви микроскопических размеров.
При рассматривании под микроскопом они очень напо-
минают угрей, в особенности из-за быстрых извивающих-
ся движений.
Рис. 56. Три амебы, пой-
манные мицелием одного
из видов Stylopage— хищ-
ного гриба, принадлежа-
щего к Zoopa gales.
Как плотоядная группа среди низших грибов выделя-
ется порядок Zoopagales, все члены которого (за исклю-
чением нескольких видов) хищники. Питаются они в ос-
новном амебами и другими близкими простейшими; эти
грибы имеют простое строение и состоят из необыкно-
венно изящно ветвящихся нитей, или гиф. Коснувшись
гифы одного из Zoopagales, амеба приклеивается к ней.
очевидно, липкой жидкостью, выделяемой гифой. Затем
гриб выпускает гаусторию («присоскуэ), которая про-
никает в тело животного, разветвляется там и высасы-
вает его содержимое (рис. 56).
Сначала амебу, по-видимому, мало беспокоит втор-
жение чужеродного тела, так как ее обычные функции
продолжаются, насколько можно судить, беспрепятствен-
но. Однако, по мере того как содержимое ее тела погло-
щается, движения сократительной вакуоли (пульсирую-
щая вакуоль, которая действует подобно трюмной помпе
Хищные растения
271
корабля, откачивая из животного избыток воды) замед-
ляются и, наконец, прекращаются. По мере высасывания
ее протоплазмы грибом амеба становится все более
прозрачной, и в какой-то момент (точно трудно сказать
когда) ее уже можно считать мертвой. Гриб же продол-
жает поглощение до тех пор, пока от амебы не останется
только прикрепленный к гифе сморщенный наружный
покров, или пелликула.
Рис. 57. Нематода, схва-
ченная клейким мице-
лием Slylopage grandis.
Хотя большинство Zoopagales ловит простейших, не-
которые из них, более сильные, питаются нематодами.
Нематода крупнее амебы и гораздо активнее; гриб, охо-
тящийся за нематодой, можно сравнить с человеком,
ловящим акул удочкой. Гифы таких грибов — охотников
за нематодами— толще, чем у других Zoopagales, доволь-
ствующихся простейшими. Способ ловли, однако, тот же
самый. Нематоды захватываются с помощью клейкого
секрета гифы, который держит их так же надежно, как
птичий клей захватывает в ловушку порхающих птиц
(рис. 57).
Нематода — более предприимчивая жертва, нежели
флегматичная амеба. С того момента, как она поймана,
нематода борется за свободу, как попавшийся на крю-
чок тарпон (большая морская рыба), извиваясь во все
стороны и натягивая держащую ее гифу, пока освобож-
27$
Глава XI it
дение кажется возможным. Однако гриб не менее на-
стойчив и держит нематоду, как бульдог, пока она, из-
нуренная борьбой, не станет неподвижной. Тогда из той
части гифы, которая находится в контакте с жертвой,
появляются новые гифы, врастающие в тело нематоды
и поглощающие его содержимое. Менее чем через 24 ч
от пленницы не остается ничего, кроме наружного по-
крова, заполненного пустыми гифами, содержимое кото-
рых перешло обратно в гриб.
Zoopagales, видимо, являются облигатными хищни-
ками, поскольку никому еще не удалось вырастить их на
искусственной питательной среде, без организмов, кото-
рыми они питаются в естественных условиях, т. е. их
добычи. Очень много их в почве, навозе и разлагающих-
ся растительных остатках; Zoopagales могут быть всюду,
где живут их жертвы. Но, несмотря на столь широкое
распространение, они были открыты сравнительно не-
давно: пока Дречслер не опубликовал в Америке первое
из своих многочисленных описаний этих грибов, никто
и не подозревал об их существовании. Первые Zoopaga-
les были описаны им в 1935 году, и за последние 30 лет
их список сильно увеличился. Тридцать лет назад Zoopa-
gales были маленькой ботанической диковинкой, сегодня
это крупнейшая группа грибов.
Чтобы увидеть охоту на нематод во всем блеске,
лучше всего обратиться к другой группе грибов, весьма
далекой от Zoopagales как по строению, так и по степе-
ни родства. Это хищные Moniliales. У Zoopagales нет
специальных морфологических приспособлений для лов-
ли добычи; большинство грибов этой группы и не нужда-
ется в них, так как Protozoa — легкая добыча и лишь
несколько видов «метят выше». Что касается хищных
Moniliales, то у них только один или два вида доволь-
ствуются столь скромной добычей; у остальных же ловля
нематод стала главной в их жизнедеятельности. Нс уди-
вительно поэтому, что у них имеются настоящие ловушки
для нематод, зачастую не менее искусные, чем пузырьки
Utricularia или кувшины Nepenthes.
В книге «Полезные грибы» («The friendly fungi»,
1957) я подробно описал строение и жизнедеятельность
хищных Moniliales; здесь же я лишь в общих чертах
Хищные растения
273
обрисую их замечательные приспособления к хищному
образу жизни. Способы ловли добычи варьируют у раз-
ных видов. У одного из наиболее распространенных ви-
дов, Arthrobotrys oligospora, гифы снабжены рядом пе-
тель, образующих системы трехмерных сетей (фото
XXXIV). Петли эти липкие, и нематоды, случайно слегка
задевшие их, оказываются пойманными и удерживаются,
как и у Zoopagales. Извиваясь, они окончательно запу-
тываются в сетях. После обычного периода энергичной,
но бесполезной борьбы животные затихают и, очевидно,
погибают, хотя истинная причина их гибели неясна.
Предполагать, что они гибнут в результате борьбы, нет
оснований, поэтому невольно напрашивается мысль о
действии какого-то яда, вырабатываемого грибом. Неко-
торые исследователи утверждали, что они выделили та-
кой яд, но их данные нуждаются в подтверждении,
прежде чем могут быть безоговорочно приняты.
Как только нематода перестает двигаться, сеть гриба
выпускает крошечный сосочек, который проникает сквозь
наружные покровы животного и разрастается в его теле,
образуя инфекционный шарик. Развивающиеся из него
гифы заполняют нематоду, высасывая ее содержимое.
После того как все поглощено, протоплазматическое
содержимое гнф переходит обратно в сети и использу-
ется для питания гриба, а от жертвы остается лишь
наружный покров, прикрепленный к сети и заполнен-
ный пустыми гифами.
Чтобы поверить в прожорливость Arthrobotrijs
oligospora, надо ее видеть. Гриб может полностью истре-
бить нематод в чашке Петри за несколько дней, если
серьезно возьмется за дело.
У многих других видов хищных Moniiiales наблюда-
ются и другие варианты клейких сетей. Некоторые,
в частности Monacrosporium cionopagum, развивают на
гифах клейкие короткие боковые отростки, действующие
подобно сетям Arthrobotrys. У М. ellipsosporum и других
видов к гифам при помощи коротких ножек прикреплены
небольшие липкие шишечки. Детали, касающиеся ловли
нематод, здесь тс же, что и у Arthrobotrys.
Не все хищные Moniiiales при охоте на нематод по-
лагаются на клейкие ловушки. У Dactylaria Candida ло-
274
Глава Xlll
вушка механическая. Гифы снабжены множеством ма-
леньких колец, каждое из которых образовано тремя
изогнутыми клетками, сидящими на коротких ножках.
Если извивающаяся нематода каким-то образом попадет
в кольцо, она основательно застрянет в нем, так как
диаметр кольца немного меньше диаметра ее тела. Не
сознавая того, что надо уйти, пока не поздно, нематода
пытается пролезть через кольцо и в результате неизбеж-
но застревает в нем все сильнее. Затем события разви-
ваются обычным путем, и содержимое тела нематоды
высасывается гифами, вырастающими из кольца.
Кольца Dactylaria Candida не клейкие и не сжима-
ются вокруг тела нематоды, которая оказывается пой-
манной исключительно благодаря собственным усилиям
протиснуться через кольцо. Однако у некоторых хищных
Moniliales кольца гораздо более смертоносны: стоит
только нематоде попасть в кольцо, как оно сжимается,
держа пленницу так же крепко, как силки — кролика.
Dactylaria gracilis — один из хищных грибов, снаб-
женных сжимающимися кольцами, очень похожими на
несжимающиеся кольца D. Candida, но они имеют более
прочную структуру и более короткие и толстые ножки.
Внутренние стороны трех клеток, составляющих кольцо,
чувствительны к прикосновению, и когда попавшая в
кольцо нематода трется о них, она тем самым приводит
ловушку в действие; клетки кольца внезапно набухают
с внутренней стороны, увеличиваясь при этом примерно
в три раза (фото XXXV). В результате просвет кольца
закрывается, и нематода оказывается крепко зажатой,
без всякой надежды на спасение. Поимка животного,
как обычно, сопровождается врастанием в его тело по-
глощающих гиф. Сжимание колец происходит порази-
тельно быстро: начинающееся после нескольких секунд
задержки (лаг-фазы) набухание клеток занимает всего
0,1 с. Если ловчий механизм пущен в ход, у нематоды
уже нет шансов спастись. Проделана большая работа по
изучению устройства сжимающихся колец, но механизм
их действия еще неясен.
В отличие от Zoopagales хищных Moniliales можно
легко изолировать и выращивать в лаборатории, хотя
□ни обычно не образуют ловушек, если в культуре не
Хищные растения
27Е
присутствуют также и нематоды. Легкость выращивания
этих грибов привела многих исследователей в Англии,
Америке, Франции и СССР к попытке использовать- их
как биологическое средство борьбы с нематодами — Па-
разитами растений, являющимися злейшими врагами
урожая. На экспериментальном уровне достигнуты не-
которые успехи, но еще многое предстоит сделать, преж-
де чем появится надежда на спасение хотя бы части из
тех 2 млн. фунтов стерлингов в год, в которые обходится
Англии ежегодно одна только картофельная нематода.
Хищные Moniliales распространены столь же широко,
как и Zoopagales, а может быть, даже шире. Они живут
в почве, экскрементах животных, гниющих растительных
остатках — в сущности везде, где есть нематоды, почти
наверняка можно встретить и хищных Moniliales, пи-
тающихся ими.
Хищные растения представляют собой замечательную
и интересную экологическую группу. В настоящее время
эти растения не имеют экономического значения (если
не считать листья Drosophyllum lusitanicum, используе-
мые в Португалии вместо липкой бумаги от мух), но
это не умаляет интереса к ним. Кроме того, заранее ни-
чего нельзя сказать. Возможно, когда-нибудь будет вы-
ведено растение томата, питающееся собственным вре-
дителем-мухой алейродидой, или же будут найдены
и перенесены из лесов Амазонки в теплицы Англии по-
настоящему эффективные пожиратели тлей и пилиль-
щиков. Вряд ли, однако, мне удастся дожить до этих
времен.
ГЛАВА XIV
СОЖИТЕЛЬСТВО РАСТЕНИЙ
Грибы обычно считают заклятыми врагами высших
растений и, вероятно, не без оснований, так как чаще
всего болезни растений вызывают именно грибы, а не
что-либо иное. Однако их взаимоотношения— это не
всегда взаимоотношения хищника и жертвы; бывает,
что гриб и высшее растение вступают в сожительство
ради взаимной выгоды.
Рис.
58. Микоризные корни бука.
Если вырыть несколько поверхностных питающих кор-
ней бука, то ближе к их концам мы обнаружим мно-
жество коротких,боковых корешков, которые в основном
и несут функцию поглощения воды и минеральных солей
из почвы (рис. 58). На поперечном срезе такого корешка
видно, что он густо оплетен нитевидными гифами гриба,
образующими своеобразный чехол толщиной 20...30 мкм
(1 микрон равен 1/1000 мм).
Здесь и там от чехла отходят гифы, пронизывающие
почву. В какой-то мере гифы внедряются и в корень, про-
ходя между клетками коры и образуя так называемую
сеть Гартига. Слишком глубоко в корень эти внутренние
гифы не проникают; обычно они ограничиваются наруж-
ными клетками коры и никогда не заходят глубже эндо-
дермы.
Симбиотические союзы
Сначала может показаться, что гриб паразитирует
<а буке, но это не так. Дерево нисколько не страдает от
юселившегося на его корнях гриба и даже выигрывает
)т этой связи. Сожительство двух организмов приносит
саждому из них какую-то пользу. Для таких союзов,
;ак этот, где оба партнера соединяются вместе с целью
Сожительство растений
277
обоюдной выгоды, биологи предложили термин симбиоз.
Весьма обычны симбиозы между растением и животным
или растением и растением, а этот частный случай сим-
биоза между грибом и корнями высшего растения назы-
вается микоризой (буквально «грибокорень»).
Подобная связь существует между корнями сосны
и грибом. Как и у бука, гриб оплетает короткие и тол-
стые боковые корешки, расположенные вблизи оконча-
ний крупных корней. У сосны эти микоризные корешки
в большинстве случаев разветвляются, а их концы ха-
рактерно утолщаются, за что их и называют коралло-
видными.
При возникновении микоризы образование корневых
волосков вблизи кончика корпя в значительной степени
или полностью подавляется и их место занимают нити
гриба. Делают они это настолько ловко, что в сущности
являются более эффективными органами поглощения,
чем настоящие корневые волоски. Микоризы этого типа
можно обнаружить на корнях многих лесных деревьев.
Среди хвойных они имеются у сосновых (сосна, ель,
лиственница и др.), встречаются также у березовых
(Betulaceae), буковых (Fagaceae) и некоторых других
семейств цветковых растений. Грибы, образующие такую
микоризу, почти всегда принадлежат к базидиомице-
там — группе грибов, включающей съедобные и не-
съедобные шляпочные грибы. Многие из столь обычных
в лесах поганок представляют собой плодовые тела
грибов, связанных с корнями деревьев. В некоторых
случаях гриб строго специфичен к определенному виду
дерева; Boletus elegans, например, всегда связан с лист-
венницей (Larix decidua). Однако бывает, что один гриб
может вступать в сожительство с множеством различных
деревьев. Рекорд в этом отношении пока принадлежит
Cenococcum graniforme, который образует микоризы с
сосной, елью, пихтой, псевдотсугой, тсугой, лиственницей,
можжевельником, дубом, буком, березой, лещиной, ги-
кори, лаймом, тополем, ивой и ольхой. Лишь для немно-
гих микоризных грибов мы можем привести такой об-
ширный список, большинство же занимает промежуточ-
ное положение между привередливым Boletus elegans
и универсальным Cenococcum,
278
Глава XIV
Остается неясным, в какой степени существование
микоризных грибов зависит от их хозяев. Некоторые
виды микоризообразователей, такие, как Boletus bovinus,
по-видимому, способны жить самостоятельно, не имея
никакой связи с корнями деревьев, но в отношении боль-
шей части грибов это сомнительно. Что касается деревь-
ев, то некоторые виды, обычно образующие микоризу,
могут достаточно хорошо развиваться и без участия
гриба, во всяком случае па богатой почве; на бедной
почве нужда в грибе возрастает. Рейнер н Нелсон-Джонс
в своих опытах показали, что сосны, не зараженные ми-
коризными грибами, ла бедной минеральными вещества-
ми почве растут плохо.
Хотя за последние два десятилетия была проделана
большая работа по изучению проблем, касающихся ми-
коризы, мы еще не знаем, что дает сожительство каждо-
му партнеру. Прежняя, получившая широкую гласность
теория состояла в том, что гриб способен разлагать
сложные органические соединения в лесной подстилке
и таким образЬм делать их доступными для дерева. Не-
смотря на привлекательность этой теории, к ней, за
исключением немногих случаев, надо относиться очень
осторожно. Во-первых, экспериментально доказано, что
известные нам свойства самих грибов делают любое та-
кое явление очень маловероятным. Большинство мико-
ризных грибов не способно добывать углерод из сложных
органических соединений, и в качестве источника угле-
рода им необходим сахар. Гораздо более вероятно, что
микориза повышает поглощающую способность корней
благодаря большей протяженности грибных гиф по срав-
нению с обычными корневыми волосками, а также, воз-
можно, за счет большей эффективности гиф в поглоще-
нии минеральных солей как из почвы, так и из массы
гниющих остатков в лесной подстилке. Как пример легко
поглощаемых гифами солей упоминались, в частности,
фосфаты.
Предполагая, что именно в этом состоит выигрыш
Дерева от сожительства, мы должны еще решить, каким
образом поглощенные грибом вещества переходят к рас-
тению-хозяину. Теоретически нет причин, по которым
вещества, передвигающиеся по гифам, не могли бы про-
Сожительство растений
279
пикать в клетки корня через сеть Гартига, но экспери-
ментально это не доказано. Количество растворимого
вещества, образующегося при разложении органических
остатков, сильно варьирует. Временами гриб может уси-
ленно всасывать фосфаты или другие минеральные соли,
передавая их в случае надобности хозяину.
Если мы не знаем, каким образом дерево извлекает
пользу из своего партнера, мы ничего не можем сказать
и о преимуществах микоризообразования для гриба.
Согласно наиболее приемлемой теории, гриб получает
от дерева выработанные им органические соединения;
скорее всего гриб нуждается в сахаре. В этой связи,
вероятно, важно, что установление микоризных связей
зависит от способа питания дерева; проростки, например,
не поражаются (если это слово здесь применимо) ми-
коризными грибами, пока у них не развернутся первые
листья и молодое деревце не начнет само вырабатывать
углеводы путем фотосинтеза.
Эктотрофная и эндотрофная микоризы
Микориза, образуемая лесными деревьями, называ-
ется эктотрофной, потому что гриб располагается глав-
ным образом на наружной поверхности корня. Для ми-
коризы другого типа — эндотрофной — характерно гораз-
до более глубокое проникновение гриба в ткани хозяина;
благодаря внедрению гиф в клетки корня между обоими
растениями устанавливается более тесная связь. Первый
тип микоризы не столь широко распространен, как вто-
рой; чем больше мы узнаем об эндотрофной микоризе,
тем с большим основанием мы можем назвать ее почти
универсальной. Очень немного найдется таких семейств
растений, где бы какой-нибудь представитель не обра-
зовывал эндотрофной микоризы.
Быть может, стоит пожалеть, что термином «мико-
риза» принято обозначать оба эти типа, так как, за
исключением редких случаев, между ними мало общего.
Кроме того, несомненно, существует несколько различ-
ных видов эндотрофной микоризы, и некоторые из них
сильно отличаются друг от друга. Растения из семейства
280
Глава XIV
вересковых (Ericaceae) образуют эндотрофную микори-
зу, в некоторых отношениях промежуточную между экто-
трофной микоризой лесных деревьев и высокоспециали-
зированной эндотрофной микоризой орхидных. Корни
таких растений, как болотный вереск (Erica) и черника
(V actinium), необычайно тонкие, с небольшого диаметра
корой и центральным цилиндром. Они заключены в рых-
лый чехол из грибных гиф, которые проникают в кору,
причем эти нити активно внедряются в клетки коры,
образуя в них плотную массу. Гифы, проникшие в клетки
коры, впоследствии могут быть переварены хозяином.
Процесс заражения молодого корня развивается в оп-
ределенной последовательности. Весной наблюдается
лишь слабое заражение. В это время корни усиленно
растут и гифы, если им удается проникнуть в кору,
быстро перевариваются. Со временем заражение рас-
пространяется, достигая максимального уровня ранней
осенью.
Как по образованию наружного чехла из гиф, так
и по сезонному изменению степени заражения микориза
Ericaceae напоминает эктотрофную микоризу лесных де-
ревьев. Однако степень внедрения гриба в клетки коры
в микоризах вересковых и микоризах лесных деревьев
различна. В микоризе деревьев проникновение в клетки
коры наблюдается редко, потому что гифы гриба, обра-
зующие сеть Гартига, распределяются между наружны-
ми клетками коры, а не внедряются в них. Для микоризы
вересковых характерно свободное проникновение гриба
в клетки коры; в этом, а также в тщательном перевари-
вании гнф клетками хозяина заключается основное раз-
личие между эндотрофной и эктотрофной микоризой.
Рейнер и другие исследователи, занимающиеся мико-
ризой, сообщили о присутствии гриба, помимо корней,
и в других частях растения — в стеблях, листьях и даже
цветках и плодах. При этом гриб, найденный в надзем-
ных частях растений, как отмечалось, находится в «исто-
щенном» состоянии: его гифы тоньше, чем у гриба, встре-
чающегося в корнях, и окрашиваются менее интенсивно.
Это привело к убеждению, что в микоризе вересковых
заражение системное, т. е. поражается обычно все
растение.
Сожительство растений 281
Позднее это сообщение было проверено многими ис-
следователями, но результаты оказались отрицательны-
ми, и в свете этих недавних работ можно заключить,
что системное заражение нельзя считать нормальным
явлением для микоризы вересковых, где гриб, как прави-
ло, поражает корни и только корни. Откуда взялись
истощенные гифы, обнаруженные Рейнер и другими, не-
известно; возможно, они принадлежали совсем другому
грибу, а может быть, в отдельных аномальных случаях
системное заражение все-таки происходит, например ког-
да устойчивость хозяина к грибу почему-либо понижает-
ся. Между симбиозом и паразитизмом в случае эндотроф-
ной микоризы существует тонкое равновесие, и нет при-
чин полагать, что микоризный гриб при соответствующих
условиях пе может стать настоящим паразитом.
Идентификация гриба — микоризообразователя вере-
сковых тоже явилась предметом широкой дискуссии.
Сначала считали, что это Phoma radicis. Он был выделен
из корней многих видов Ericaceae, в том числе и из бо-
лотного вереска (Erica tetralix) Тернетсом в 1907 году.
Рейнер и Левисопу в 1940 году несколько раз удалось
обнаружить его в семенах вереска и снова выделить из
корней, на этот раз клюквы (Vaccinium oxycoccos). Но
чтобы доказать, что именно этот гриб образует микоризу,
недостаточно выделить его из корней вересковых и опре-
делить; для этого необходимо получить микоризу в ла-
боратории, заражая грибом проростки, выращенные в
стерильных условиях. До сего времени Phoma radicis пе
выдерживал этой проверки. Следовательно, мы не можем
принять гипотезу, согласно которой Phoma radicis явля-
ется микоризообразователем у Ericaceae. Выделение же
гриба из семян в значительной степени теряет свою цен-
ность, поскольку теория системного заражения уже утра-
тила былую убедительность. У лас в руках остается,
таким образом, только факт выделения гриба из корней
Тернетсом 60 лет назад и позднее Рейнер и Девисоном.
Вполне возможно, что гриб, полученный в этих случаях,
не был микоризообразователем, а просто был связан
с растением как обычный паразит.
Но если гриб, о котором идет речь, пе Phoma radicis,
то какой же? Ответ на этот вопрос можно найти в рабо-
282
Глава XIV
тах Доука, который в 1928 году выделил стерильные
гифы какого-то гриба из корней Vaccinium corymbosum
и V. pennsylvanictim. Так как мицелий этого гриба не
давал спороношений, идентифицировать его было невоз-
можно, но Доук сумел доказать, что при заражении сте-
рильных проростков гриб образует типичные микоризы.
Успех опыта с заражением немедленно дал грибу Доука
большие права на то, чтобы считаться микоризообразо-
вателем, чем были у Phoma radicis.
Работа, начатая Доуком, была продолжена другими
с теми же результатами. К несчастью, гифы, выделенные
из корней этими исследователями, также не образовыва-
ли спор, так что определить, кому они принадлежат, так
и не удалось; несомненно было одно: это не Phoma
radicis.
Фиксация азота
Темой еще одцой дискуссии, возникшей вокруг мико-
ризы вересковых, явился вопрос о том, могут ли мико-
ризные грибы Ericaceae фиксировать атмосферный азот.
В некоторых ранних работах высказывалась мысль, что
гриб, заставляя атмосферный азот соединяться с чем-то
содержащимся в его гифах, дает, таким образом, возмож-
ность растению-хозяину использовать его для питания.
Эти гипотезы были приняты без должного критического
разбора и подтверждения, и поэтому появилось что-то
вроде волшебной сказки о «фиксации азота» вересковы-
ми. Современные исследования показали ошибочность по-
добных теорий, и сегодня никто серьезно не верит в спо-
собность Ericaceae усваивать азот с помощью своих
микоризных грибов. Насколько нам известно, пп одни
гриб (независимо от его типа) не может связывать ат-
мосферный азот.
Хотя теперь мы уверены в том, что микоризные гри-
бы вересковых не фиксируют азот, мы еще далеки от
выяснения, какую же пользу они приносят хозяину. На-
блюдали, что ранний рост проростков Ericaceae ускоря-
ется в присутствии грибов, но это относится, по-видимо-
му, почти ко всем грибам, а не только к тем из них,
которые образуют микоризу. Сходство микориз Ericaceae
Сожительство растений
283
и микориз лесных деревьев предполагает и сходные
взаимоотношения партнеров; возможно, микоризные кор-
ни лучше поглощают минеральные соли из почвы, чем
незаряженные, но точные доказательства отсутствуют.
Возможно также, что гриб способен разлагать органи-
ческие соединения в почве, делая их доступными хозяи-
ну. Пока мы не будем знать гораздо больше, чем теперь,
о микоризах и образующих их грибах, мы не сможем бо-
лее определенно ответить на этот вопрос.
Что получает гриб от хозяина, тоже пока остается
неясным. Напрашивается предположение, что хозяин
снабжает его сахарами, но в биологии не всегда бывает
верно то, что кажется очевидным. Еще многое надо сде-
лать, чтобы узнать, почему грибы вступают в этот не-
обычный союз.
Микоризы орхидных
У орхидей образуются микоризы другого типа. Здесь
нет чехла из грибных гиф, облекающего корни; гифы
почти целиком находятся внутри корня, хотя некоторые
могут покидать его и выходить наружу в почву. Как мы
уже видели, орхидные — это высокоспециализированное
семейство растений, одной из особенностей которых яв-
ляется развитие огромного количества крошечных семян.
Семя орхидеи имеет простое строение. Из оплодотворен-
ной яйцеклетки развивается маленький нерасчлененный
зародыш, который иногда может быть образован даже
восемью клетками, а вообще число клеток редко превы-
шает сто. Семядолей нет; зародыш представляет собой
просто яйцевидную или веретеновидную массу клеток
с точкой роста на конце.
При прорастании семя орхидеи набухает и семенная
кожура разрывается, после чего следует период слабого
роста за счет запасных питательных веществ, отложен-
ных в семени. В естественных условиях дальнейшее раз-
витие зависит от заражения подходящим микоризным
грибом, поскольку следующая стадия роста проростка
у всех орхидных сапрофитная. Питаясь органическими
веществами, поглощаемыми извне, зародыш развивается
в протокорм (стр. 179). На нем дифференцируются раз-
284
Глава XIV
личные органы, затем в листьях появляется хлорофилл,
и молодое растение постепенно начинает питаться само-
стоятельно, путем фотосинтеза.
Важная особенность этого способа развития, имею-
щая отношение к микоризе орхидных (и притом очень
большое), состоит в том, что все орхидеи проходят сапро-
фитную стадию. У большинства из них эта стадия ко-
роткая и растения становятся зелеными, как только
в первых листьях появляется хлорофилл, но некоторые
орхидные все время ведут сапрофитный образ жизни.
Вспомним, что гнездовка (Neottia nidus-avis) не имеет
хлорофилла и всю жизнь остается сапрофитом.
Поскольку молодой протокорм — первая часть про-
ростка орхидеи, заражаемая микоризным грибом, можно
предположить, что от него заразятся и корни, однако
этого не происходит. Как и протокорм, корни должны
быть заражены из почвы. Протокорм сильно поражается
грибом, так же как и молодые корни, клубни же остают-
ся свободными от него. Когда в следующем году из
клубней вырастает новые корни, они заражаются гифа-
ми из почвы. Этот процесс повторного заражения обычно
происходит ежегодно.
Проникшие в кору гифы гриба внедряются в ее клет-
ки, образуя клубки, которые сначала выглядят совершен-
но здоровыми (фото XXXVI). Позднее они переварива-
ются клетками хозяина, образуя бесструктурную массу.
Переваривание гиф может осуществляться в любом
участке коры, но часто ограничивается определенным
пищеварительным слоем.
Грибы, образующие микоризы у орхидных, довольно
легко выделить, и многие из них идентифицированы. По
всей вероятности, они разделяются на две группы. Гри-
бы, выделенные из зеленых орхидей, принадлежат в ос-
новном к роду Rhizoctonia, который из-за отсутствия
у него полового размножения отнесен к несовершенным
грибам. С другой стороны, грибы, выделенные из корней
сапрофитных орхидей, — это главным образом базидио-
мицеты, как, например, Xerotus javanicus и Marasmius
coniatus. К микоризным грибам относится и опенок
(Armillaria mellea), вступающий в союз с такими орхи-
деями, как Gastrodia и Gdleola,
Сожительство растений 28?
Было показано, что в отношениях между определен-
ными видами Rhizoctonia и определенными видами ор-
хидных наблюдается некоторая специфичность, хотя не
все виды Rhizoctonia одинаковы в этом смысле. Прежде
существовало мнение, что многие виды и расы Rhizocto-
nia строго специфичны к хозяину, т. е. определенная
раса гриба образует микоризы с определенным видом
или группой родственных видов орхидей. Теперь уста-
новлено, что это не так, хотя зачастую конкретная раса
Rhizoctonia легче заражает какой-то один вид орхидеи,
чем другие виды. Однако мы еще недостаточно знаем об
этом, чтобы делать какие-то обобщения.
Изучение физиологии грибов, образующих микоризы
у орхидей, показало, что в отличие от грибов эктотроф-
ных микориз они способны использовать сложные источ-
ники углерода. Это важно с точки зрения предполагае-
мой роли этих грибов в сапрофитном питании орхидных.
Если бы они, подобно грибам лесных деревьев, не могли
добывать углерод из сложных органических соединений
и нуждались для этого в сахарах, они не играли бы
большой роли в жизни сапрофитных орхидей. Но по-
скольку грибы орхидпых обладают такой способностью,
разумно предположить, что разложение сложных орга-
нических веществ является их основным, если пе единст-
венным, вкладом в микоризный союз.
Предположив, что грибы орхидей снабжают своих
хозяев продуктами разложения сложных органических
соединений, остается выяснить, каким же образом эти
продукты переходят от гриба к орхидее. Конечно, орхи-
дея может получать пищу, переваривая грибные гифы
в клетках корня (и, без сомнения, так оно и есть), но,
вероятно, это не единственный способ использования
гриба для питания орхидеи. Можно по крайней мере
предполагать, что по гифам гриба осуществляется пере-
мещение питательных веществ и что какая-то часть уже
перемещенных веществ проникает через стенки гиф
в клетки коры орхидеи. Это привело бы к сходству
взаимоотношений в микоризе орхидей с предполагае-
мыми взаимоотношениями в эктотрофной микоризе
лесных деревьев. Хотя это и не доказано, но вполне
вероятно.
286
Глава XIV
Что выигрывает гриб от связи с корнями орхидей,
еще не выяснено. Между обоими растениями существуют
довольно сложные отношения; бывает, что гриб начинает
паразитировать на орхидее и губит ее, тогда как в дру-
гих случаях ему не удается произвести даже первичного
заражения проростка. Переваривание грибных гиф в
клетках коры орхидеи —явно защитное приспособление;
если деятельность гриба в ее тканях начинает досаждать
орхидее, она отвечает перевариванием гриба. Видимо,
это случай сбалансированного паразитизма; пока равно-
весие поддерживается, оба партнера процветают, но пре-
успевание одного из них трагически кончается для дру-
гого. На вопрос о том, что же выигрывает гриб от та-
кого союза, можно ответить так: он получает то же, что
и любой внутренний паразит, однако с условием, что его
паразитическая деятельность находится под контролем
хозяина.
Микоризы подобного типа имеются у многих других
растений. В частности, среди Triuridaceae и Burmannia-
сеае встречаются’ сапрофиты, микоризы которых сходны
с микоризами орхидных, и из них тоже был выделен
гриб Rhizoctonia. Такие микоризы обнаружены не толь-
ко у цветковых растений; они наблюдаются, например,
у сапрофитного гаметофита Psilotum — примитивного
родича папоротников, а также у многих печеночников.
Микоризы, образованные низшими грибами
Грибы, участвующие во всех описанных здесь мико-
ризах, имеют септированные гифы, т. е. гифы, разделен-
ные поперечными перегородками, или септами, на от-
дельные клетки. Наличие септированных гиф — признак
высших грибов. У низших грибов таких гиф нет; их гифы
представляют собой длинные трубки, выстланные прото-
плазмой и содержащие непрерывающуюся центральную
вакуоль. Гифы такого типа встречаются также в мико-
ризах высших растений; в сущности, микоризы, обра-
зованные низшими грибами, гораздо более многочислен-
ны, чем любой другой тип микориз. Недавние работы
показали, что они распространены гораздо шире, нежели
полагали ранее. Почти все изученные семейства растений
Сожительство растений 287
содержат по крайней мере несколько родов с такой ми-
коризой.
В пораженных растениях обнаруживаются несептиро-
ванные гифы, часто большого диаметра, проходящие че-
рез кору корня и нередко образующие в клетках вет-
вистые структуры. Эти похожие па кустарнички вет-
вистые системы называют арбускулами. На концах или
в других участках гиф могут появляться также утолще-
ния, называемые везикулами (пузырьками).
О взаимоотношениях между хозяином и грибом в та-
ких микоризах ничего не известно, но поскольку расте-
'ние-хозяин выглядит совершенно здоровым, видимо, они
носят скорее характер симбиоза, чем паразитизма. По-
добные микоризы наблюдаются у многих хлебных зла-
ков, имеющих большое экономическое значение, в том
числе у пшеницы, ячменя, овса, ржи и кукурузы. Встре-
чаются они также у клевера и у многих других бобовых
растений1.
Грибу, участвующему в микоризе этого типа, дали
название Rhizophagus; правда, это было сделано скорее
для удобства, так как не известно, образованы ли все
микоризы данного типа одним и тем же видом гриба
или же (что кажется более вероятным) мы имеем дело
с целой группой грибов. Есть доказательства, что по
крайней мере некоторые из этих микориз вызываются
обычным в почве грибом Endogone. Кроме того, есть ос-
нования полагать, что к микоризообразованию может
иметь отношение и вид Pythium. Мы с нетерпением ожи-
даем новых работ по этим интересным грибам и образуе-
мым ими микоризам.
Микоризы, образованные Rhizophagus, примечатель-
ны своей древностью: гифы с везикулами на концах най-
дены в девонских отложениях, т. е. их возраст превыша-
ет 270 млн. лет. Самая ранняя зарегнстрнрованная ока-
менелость— гриб Palaeomyces asteroxyli — обнаружена
в коре Asteroxylon mackiei, одного из древнейших на-
земных растений.
1 Вообще распространение микоризы очень широко: в нашей
лесной, луговой и болотной флоре очень многие виды, если не
большинство, являются микоризными. В частности, микориза обыч-
на для всех луговых злаков. — Прим. ред.
288
Глава XIV
Водоросли как партнеры
Помимо образования микориз с грибами, многие выс-
шие растения вступают в симбиоз с водорослями. Так,
сине-зеленую водоросль Ahabaena можно обнаружить
в союзе с крошечным водяным папоротником Azolla, в не-
которых корнях саговников и во многих других расте-
ниях. Другая сине-зеленая водоросль, Nostoc, сожитель-
ствует с печеночником Anthoceros, с саговниками и с не-
которыми цветковыми растениями. Есть доказательства,
что по крайней мере в некоторых случаях водоросль
способна приносить пользу своему хозяину, связывая
атмосферный азот.
Среди химических элементов, необходимых для под-
держания жизни, азот особенно важен, так как он тре-
буется в больших количествах, чем почти все другие
элементы, и его часто не хватает. Атмосфера земного
шара почти на 4/s состоит из газообразного азота; это
значит, что над ^каждым гектаром земной поверхности
висит 75000 т азота. К сожалению, очень немногие ор-
ганизмы могут использовать этот неистощимый запас
азота, так как в газообразном состоянии он химически
инертен и плохо соединяется с другими веществами.
Некоторые почвенные микроорганизмы являются фик-
саторами азота. Они способны поглощать этот газ из
воздуха и, соединяя его с другими молекулами, строить
белки и другие азотистые соединения. Некоторые из
этих фиксаторов азота принадлежат к сине-зеленым во-
дорослям, другие — к бактериям. Из последних лучше
всего известны свободно живущие формы — Azotobacter
chroococcum и Clostridium pasteurianum, которые в зна-
чительной степени возмещают теряемый почвой азот;
уменьшение содержания азота в. почве происходит по
многим причинам, в частности, азот удаляется вместе
с убранным урожаем.
Азотфиксирующие бактерии
Азотфиксирующие бактерии, вступающие в симбиоз
с высшими растениями, имеют более важное значение,
чем только что упомянутые свободно живущие фиксато*
Сожительство растений
289
ры азота. Наиболее'йЗВестны среди них виды Rhizobium,
обитающие в корневых клубеньках растений, относящих-
ся к порядку бобовых и особенно семейству мотыльков
вых, к которому принадлежат горох, бобы и клевер.
Живущие в почве бактерии Rhizobium маленькие
и цилиндрические, но в корневых клубеньках своих хо-
зяев они часто образуют необычной формы разветвлен-
ные клетки, известные как бактероиды. Проростки бобо-
вых растений заражаются клубеньковыми бактериями
еще в начале своей жизни. Они проникают в их корни
Рис. 59. Клубеньки на
корешках декоративной
фасоли.
из почвы, через корневые волоски. Внедрившись в кор-
невой волосок, бактерии образуют инфекционную нить,
представляющую собой массу бактериальных клеток,
заключенных в футляр из целлюлозы, гемицеллюлоз и
пектинов, вырабатываемых пораженной клеткой расте-
ния. Инфекционная нить растет и многократно ветвится,
переходя из одной клетки коры корня в другую.
Заражение корня сопровождается образованием кор-
невых клубеньков — небольших шаровидных вздутий на
наружной поверхности корня (рис. 59). У гороха и бо-
бов клубенек развивается из коры корня, тогда как
у клевера и люцерны — из перицикла, т. е. наружной
части центрального цилиндра корня.
Полностью сформировавшийся клубенек состоит из
двух слоев клеток: тонкого наружного слоя, или коры,
и центральной части, или сердцевины. Весь клубенек пок-
рыт тонким слоем клеток, принадлежащих коре корня.
С центральной областью корня, проводящей воду и пи-
тательные вещества, он связан тяжами проводящей тка-
ни (рис. 60).
290
Глава XIV
В сердцевине клубенька находится огромное коли-
чество бактерий; каждая ее клетка содержит несколько
тысяч, а весь клубенек может вмещать от 1 млн до
1 млрд этих крошечных постояльцев. В молодом клубень-
ке большинство бактерий живые и функционируют, но
в старых клубеньках 99% из них или больше погибают.
Связь между бобовым растением и бактериями, не-
сомненно, носит симбиотический характер. Растения, ра-
стущие на бедной азотом почве без соответствующих
бактерий, имеют бледную окраску и плохо растут; они
Рис. 60. Схема разреза через корневой клубенек.
обнаруживают все признаки азотного голодания. С дру-
гой стороны, при наличии нужных бактерий они растут
хорошо, даже если содержание азота в почве низкое.
Что получают бактерии от бобового растения взамен
отданного азота, менее ясно, но, по всей вероятности,
хозяева снабжают их сахарами.
От симбиоза Rhizobium с растениями из семейства
мотыльковых выигрывают не только оба партнера,
по и почва. В нее поступают азотистые ве-
щества, выделяемые живым растением, а когда оно уми-
рает, почва получает соединения азота при разложении
его тела. Даже если растение убирают целиком, то
и в этом случае почва обогащается азотом за счет за-
пахиваемого в нее жнивья. То, что бобовые культуры
повышают плодородие почвы, было известно задолго до
Сожительство растений
291
предположения о существовании этого симбиоза; еще
тогда был разработан четырехпольный севооборот, где
пшеница — культура, нуждающаяся в азоте,— следова-
ла за клевером.
Каждый из многочисленных видов Rhizobiutn может
сожительствовать с определенной группой бобовых ра-
стений. R. legumitiosarum, например, встречается в кор-
нях гороха, конских бобов, викн и чечевицы; R. met Hoti
поселяется в корнях люцерны и некоторых видов клеве-
ра; R. phaseoli связан с обыкновенной и декоративной
фасолью; R. trifolii—с красным, белым и пунцовым кле-
вером; R. lupi.nl вступает в симбиоз с люпинами,
a R. japonicum — с соей. Чтобы установилась симбиоти-
ческая связь, необходимо присутствие в почве именно
того вида бактерий, который необходим данной культуре.
Известна по меньшей мере 21 такая растительно-бакте-
риальная группа. Бактерии из одной группы могут иног-
да встречаться в клубеньках растений из другой группы,
но подобные союзы часто бывают неудовлетворитель-
ными.
Специфичность к хозяину заходит даже дальше, чем
только что описанная: внутри каждого вида Rhizobiutn
имеются различные расы, связанные с определенными
сортами растения-хозяин а. Одна раса R. trifolii, напри-
мер, может усваивать больше азота, находясь в союзе
с красным клевером, чем с белым, и т. д. Такого рода
строгая специфичность вовсе не редкость в тех случаях,
когда два живых организма тесно связаны друг с дру-
гом.
Связь между бобовыми культурами и Rhizobiutn име-
ет большое значение для сельского хозяйства, и об одном
из способов использования этой связи — роли клевера
в севообороте — мы уже говорили. Другой метод — это
использование бобовых растений, например люцерны,
в качестве зеленого удобрения. Бобовая культура может
усваивать в среднем 125 кг азота на I гектар, а при бла-
гоприятных условиях — более 250 кг. Это дает возмож-
ность сократить расходы на удобрения.
Если почва богата нитратами, союз Rhizobiutn и хо-
зяина нарушается. Растение поглощает нитраты обыч-
ным путем, через корни, а клубеньковые бактерии или
292
Глава XIV
вовсе ие связывают азот, или связывают его очень мало,
поскольку они легко могут получить все, что им надо,
из почвы. В этих условиях бактерии перестают быть сим-
бионтами и переходят к обычному паразитированию на
своих хозяевах.
В нормальных условиях бобовые растения могут за-
разиться Rhizobium из почвы, но если нужная раса бак-
терий отсутствует или имеется в недостаточном количе-
стве, может возникнуть необходимость специального за-
ражения почвы. Его можно осуществить различными
путями. Традиционный способ заключается в том, чтобы
взять верхний слой почвы с другого поля, на котором
недавно выращивалась бобовая культура, и разбросать
ее на новом поле из расчета 500 кг на 1 гектар. Можно
также почву с поля, зараженного нужной расой Rhizo-
Ыит, поместить в воду, а затем замочить в этой воде
предназначенные для посева семена; перед посевом се-
мена надо подсушить. Более современным методом яв-
ляется обработка семян чистой культурой Rhizobium.
Лишайники как пример симбиоза
Наконец, ни одно сообщение о симбиозах растений не
будет полным, если не упомянуть о лишайниках. Эти
своеобразные растения известны еще с античных времен,
если не раньше; от греков же перешло к нам и слово
«лишайник», что означает «чешуйчатый». Оно как нель-
зя более подходит для описания их сухих, зеленовазо-
серых талломов.
Лишайник состоит из водоросли и гриба, находящих-
ся в симбиозе. Ббльшая часть таллома лишайника обра-
зована грибом и состоит из переплетенных гиф. Почти
у всех лишайников одноклеточные водоросли, входящие
в их состав, располагаются в определенной зоне внутри
лишайника, сразу под поверхностью, и относятся обычно
к зеленым водорослям (Chlorophyta). У некоторых же
лишайников водоросли равномерно распределены по все-
му телу и в этом случае относятся, как правило, к сине-
зеленым водорослям (Cyanophyta). Гриб-симбионт почти
всегда принадлежит к большой группе грибов, известных
под названием сумчатых ( Ascomycetes). Однако в обра-
Сожительство растений
293
зовании некоторых лишайников участвуют и базидиаль-
ные грибы (Basidiomycetes)—группа, к которой принад-
лежат поганки.
Лишайники растут в самых различных местах, часто
там, где ни один из партнеров не смог бы существовать
самостоятельно, что представляет собой одно из преиму-
ществ совместной жизни. Xanthoria parietina, например,
поселяется на голых прибрежных скалах, образуя на них
большие оранжевые пятна. Многие лишайники живут на
стволах деревьев или на крышах ветхих домов. Единст-
венное, чего ни один лишайник не может вынести, это
загрязнение воздуха фабричным дымом; вы редко встре-
тите лишайники вблизи большого города. Исключение
составляет Lecanora conizaeoides, по-видимому, облада-
ющая неплохой устойчивостью к загрязнению воздуха;
в окрестностях городов это часто единственный лишай-
ник.
Размножение лишайников осложняется тем, что это
двойные организмы. Большинство видов размножается
вегетативно; участки таллома лишайника, содержащие
гриб и водоросль, отделяются от материнского растения
и растут самостоятельно. Будучи одноклеточными, водо-
росли могут размножаться простым делением клетки на
две и таким образом не отставать в росте от лишайника.
Гриб образует споры, которыми, конечно, может размно-
жаться только он сам.
И гриб, и водоросль способны существовать незави-
симо друг от друга, по крайней мере до некоторой сте-
пени. Водоросли в лишайниках часто представлены са-
мыми обычными видами; такие роды, как Chlorella
и Trebouxia, например, чрезвычайно широко распростра-
нены как сами по себе, так и в качестве составной части
лишайников.
Поскольку многие водоросли, входящие в состав ли-
шайников, весьма обычны, можно предположить, что
спора грибного компонента может прорасти, образовав
грибную основу, которая заражается соответствующей
водорослью и таким образом дает начало новому лишай-
нику. Крайне сомнительно, однако, что это происходит
на самом деле. Никому пока не удалось получить лишай-
ник, выращивая гриб в чистой культуре и добавляя к
294
Глава XIV
нему культуру водоросли. Конечно, неудачи лаборатор-
ных опытов еще не доказывают невозможность этого в
природе *.
Что дают друг другу участники этого симбиоза, мы
точно не знаем. Поскольку водоросль способна к фотосин-
тезу, напрашивается мысль, что она вносит свою лепту
в виде сахара или какого-то другого углевода, но опре-
деленно этого сказать нельзя. Может быть, гриб пред-
почитает получать «из вторых рук» синтезируемые во-
дорослью азотистые вещества или какой-то необходи-
мый для роста витамин.
Многие грибы могут создавать все нужные для роста
вещества из соответствующих органических и неоргани-
ческих соединений, но некоторые не способны к этому.
Известно, например, что для нормального развития мно-
гих грибов совершенно необходимы небольшие коли-
чества тиамина (витамин Bi); некоторым грибам тре-
буются и другие витамины. Так как эти витамины вы-
рабатываются зелеными растениями, то для лишайнико-
вого гриба, возможно, важнее получить от водоросли
синтезированные ею витамины, чем сахар.
Исходя из этого, многие исследователи занялись вы-
яснением вопроса о том, насколько выделенные из ли-
шайников грибы нуждаются в витаминах. Результаты
оказались разноречивыми. Для нормального роста одних
грибов требовалось наличие в среде тиамина, тогда как
рост других не зависел от добавления витаминов.
Еще меньше известно о выгодности подобного симби-
оза для водоросли. Гриб, несомненно, дает ей убежище,
и под его защитой она может расти там, где иначе не
смогла бы выжить. Но мы не знаем, ограничиваются ли
этим получаемые ею преимущества. Предполагают, что
продуцируемая грибом аскорбиновая кислота использу-
ется водорослью в процессе дыхания, но это не доказано.
Особенностью всех лишайников является их крайне
медленный рост. Согласно имеющимся данным, макси-
1 Автор, по-видимому, неправ: во многих сводках и учебниках
говорится, что такие «синтезы» экспериментально были получены
многократно (см. Н. А. К о м а р н и ц к и й, Л. В. Кудря-
шов, А. А. У р а и о в, Систематика растений, Учпедгиз, М.,
1962).
Сожительство растений
295
мальная скорость их роста не превышает 4 см в год,
причем у большинства она гораздо меньше. Живут они,
вероятно, очень долго, хотя точными данными о про-
должительности жизни лишайников мы не располагаем.
Считается, что возраст некоторых из них достигает
400 лет, а арктический вид Rhizocarpon geographicum,
как говорят, насчитывает 4500 лет, т. е. почти столько
же, сколько одна из гигантских секвой.
К числу самых интересных особенностей многих ли-
шайников относится образование лишайниковых веществ,
большинство которых свойственно только им. Это раз-
личные органические соединения, составляющие в сред-
нем от 2 до 5% сухого веса лишайника, а иногда и 20%.
Обычно они находятся вне клеток растения, и причина
их образования неизвестна.
Большинство лишайниковых веществ бесцветно, но
некоторые из них ярко окрашены и в прошлом широко
использовались в качестве красителей. Именно им шот-
ландские ткани обязаны своей яркостью, хотя в наши
дни предпочтение отдается анилиновым кра.сителям.
Многие из этих красителей являются производными ор-
сина, которые при обработке раствором аммиака дают
прекрасные цвета. Недавно лишайниковые красители
снова начали применять в небольшом масштабе для
окраски домашней пряжи.
Получение лакмуса, используемого в химических ла-
бораториях в качестве индикатора кислотности или ще-
лочности, является побочной отраслью производства ли-
шайниковых красителей, особенно процветающей в Гол-
ландии.
Лишайниковые вещества используются и для других
целей. В старину их применяли в медицине, хотя
и с сомнительным успехом, но в настоящее время выде-
ленные из лишайников антибиотики кажутся весьма
многообещающими. В частности, усниновая кислота, по-
лучаемая из некоторых видов Usnea, успешно применя-
ется вместе со стрептомицином для лечения туберку-
леза.
В последнее время лишайники снова стали входить
в состав пудры и косметических средств для волос Для
этой цели, а также для закрепления запаха духов и туа-
19*
296
Глава XIV
летного мыла часто используют виды Evernia и Ramu-
Una. Но, что самое важное, лишайники в северных райо-
нах служат пищей для оленей. В Лапландии и других
местах растут в огромных количествах олений мох
(Cladonia rangiferina) (рис. 61) и исландский мох
(Cetraria islandica). Их хватает не только для обеспече-
ния оленей прекрасным кормом, но эскимосы заготовля-
ют его для оленей и впрок, на зимнее время.
Рис. 61. Олений мох
(Cladonia rangiferina).
Человек также иногда употребляет лишайники в пи-
щу. Считают, что библейская «манна» — не что иное,
как лишайник Lecanora esculenta, который растет в го-
рах и часто заносится ветром в пустыни.
Вот мы и подошли к концу нашего рассказа о строе-
нии и жизнедеятельности растений, об их замечательных
приспособлениях к определенному образу жизни. В их
морфологии, физиологии или экологии нет ни одной
черты, которая не была бы в свое время отмечена пе-
чатью естественного отбора. В длительном процессе эво-
люции неудачники отметались, а преуспевающие особи
совершенствовались, так что большинство современных
растений хорошо приспособлено к той жизни, которую
они ведут. Процесс адаптации идет и сейчас, так как
единственное, чего эволюция не может сделать,— это
остановиться.
Пока растения существуют, они будут эволюциони-
ровать; с течением времени старые структуры будут усту-
пать место новым, лучше приспособленным, чтобы вы-
жить в бесконечной борьбе за существование.
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Азотфиксирующие бактерии
288—292
Андроцей 114
Анемофилия 162—164
Антиподы 121
Антоцианы 143, 218
Апикальная меристема 52
Арбускулы 287
Ауксин 92, 93, 109, 226, 259
Бактероид 289
Вакуоль 67
Веламен 56, 170, 233,.234
Волокна 15, 32, 35, 38, 41,
50, 229
Вторичный рост 40, 50, 54
— — аномалии 229, 230
Древесина кольцесосудистая 5К
— рассеян нососудистая^51
— растяжения 49
— реактивная 48, 49
— сжатия 49
— ядровая 46
Желатинозный слой 49
Жизненные формы 201—207
Заболонь 46
Завязь 114
— нижняя 161
Зародышевый^” мешок^ 23, 121
Заросток 17
Интегумент 120, 126
Галофиты 238, 239
Гамета 23, 120
Гаметофит 18, 20, 122
Гаптотропизм 226
Гаустория 248
Гелофиты 207
Гемикриптофиты 204, 205
Геофиты 201, 206
Гетеростилия 150
Гидрофилия 165
Гидрофиты 207
Гипокотиль 247
Годичные кольца 50, 51, 59
Губчатый мезофилл 75
Давление диффузии 69—73
•--дефицит 69, 238
— корневое 79—81
— осмотическое 66, 78, 80, 87,
238, 239
___ тургорное 69
Дихогамия 150, 164
Древесина 14
— вторичная 41
Каллюс 44
Кальцифилы 239—241
Кальцифобы 239—241
Камбий 36
— пробковый 43
— функция 39—42
Капиллярность 79
Каротин 96
Кладодий 215
Клетки замыкающие; см. Усть-
ица
— палисадные 75
Клетки-спутники 36, 106
Клеточный сок 67
Клубень 131
Клубнелуковица 132, 206
Колленхина 37
Комменсализм 235
Кора, образование 42—45
Корневище 131, 206, 251
Корневой чехлик 53
Корневые волоски 54—56
— клубеньки 289
Корни втягивающие (контрак-
тильные) 57, 205
298
Предметный указатель
Корни досковидные 59—61
— дыхательные (пневматофо-
ры) 61, 62, 223
— коралловидные 277
— придаточные 56, 57, 130,
228
— строение и функция 52—62
— ходульные 07, 59, 223
Коробочка 19
Костянка 183
Ксантофилл 96, 218
Ксероморфные признаки 212—
217, 233, 236, 239
Ксерофиты 212
Ксилема 14, 30, 35, 41, 52,
106, 221, 229
— вторичная 41, 54
Лепестки 113
Лигнин 15, 30/34, 49
Листовая мозаика 16, 99
Листовые следы 74
Листопад 217, 218
Листья водных растений 218—
222
— световые 98
— структура и функция 74—
78
— теневые 98
Луковица 133, 206
Малакофилия 162
Материнская клетка мегаспор
121
Мегаспора 22
— функционирующая 121
Междоузлие 205, 230
Микориза 277—287
— эктотрофная 279—282
— эндотрофная 279—282
Микропиле 120
Микроспоры 22, 114
Мирмекофилия 235
Мутация 26
Нутация 224/225, 253
Нуцеллус 120
Опыление 116—122, 137—167
174—177
— перекрестное 117
Орнитофилия 160, 161
Осмос 64—66, 78, 80
Отделительный слой 217
Отрицательный геотропизм
185, 225
Паразиты 246, 250—254
Паренхима 35, 40
— древесинная 36, 41
— флоэмная 36
Первичное ядро эндосперма
122
Перидерма 43
Перикарп’183
Перистом 22
Плазмалемма 67
Плазмолиз 71
Плацента 120
Плоды ложные 182
— настоящие 182
— образование 182—186
— сложные 183
— способы расселения 186—
199
Покровные листья 234
Поллиний 152, 173
Полосы Каспари 53
Полупаразиты 247—250
Полуэпифиты 233, 236—238
Полярные ядра 122
Почки возобновления 201—208
— выводковые 134
— покой 208, 209
Прицветник^! 15
Проводящий . пучок 15, 35—
38
Протандрия 150
Протокорм 179, 180, 284
Протоэпифиты 233, 234
Псевдо копуляция 178
Пыльцевая трубка 119, 123'J
Пыльцевой мешок 117
Пыльцевые зерна 117—119,
123
Размножение бесполое (спо-
рами) 122, 123
Предметный указатель
299
Размножение вегетативное
130—135, 222
Растения, выживание 200—
209
— двудольные 115, 222
— многолетние 201
'— однодольные 115
— однолетние 200
— полурозеточные 205
— розеточные 205, 206
— хищные 257—275
Растительные гормоны 109
Ризоиды 13
Ризо морфы 172
Рыльце 114
Самоопыление 117, 165—167,
178, 179
Самостерильность 139, 140
Сапрофиты 246, 255, 256
Связник 149
Семя (ена) 180, 182
— жизнеспособность 128—130
— покой 126—128
— прорастание 126—128, 179,
180
— развитие 124—126
— распространение 186—199,
231 —233
Семядоли 109, 125
Семяпочка 115, 120, 121, 123—
125
Симбиоз 277
Симбиотические союзы 276—
296
Синергиды 122
Ситовидная пластинка 106
— трубка 36
Сосуды 15, 32—34, 41, 50, 79,
105
— членики (элементы) 32—34
Спорангий 17, 234
Спорофиллы 234
Споры 17, 19, 21, 22
Столоны 181, 205
Суберин 43
Суккуленты 218
ерофиты 207
иллы 45
онопласт 67
Транзиторный крахмал 104
Транспирация 63, 76, 84-7-86,
88, 213, 214
— кутикулярная 76
Трахеиды 30—33, 41, 79, 105
Тургор 29, 75, 84, 85, 87
Тычинки 114
Указатели меда 141, 145, 146
Усики 224, 226—228
Устьица 16, 63, 75, 76, 84—
88, 99, 213, 214, 220
— замыкающие клетки 16, 75,
85—87, 222
Фанерофиты 201—203, 208
Феллема 43
Феллоген 43
Феллодер ма 43
Ферменты 110
Филлодий 215
Филлокладий 215
Флориген 244
Флоэма 14, 44, 52, 106—112,
221
— вторичная 41, 42, 54
Фосфолипиды 67
Фотопериодизм 241—245
Фотопериодическая индукция
243
Фотосинтез 11, 76, 86—88, 90,
91, 95—104, 214
Хамефиты 203, 204
Хироптерофилия 160—162
Хлоропласты 75, 95, 96, 98,
99, 222
— граны 95
— строма 95
Хлорофилл 11, 14, 75, 90, 94—
96, 98, 218, 246, 251, 254,
256
Хромопласты 143
Цветки галловые 153
— клейстогамные 166
— обоеполые 115, 150
— образующие нектар 148—
152
— однополые 115
— пыльца 117, 118
300
Предметный указатель
Цветки пыльца прорастание
118—120
-- пыльцевые, строение 147
— части 113—116
Цветоложе 113
Чашелистики 113
Чечевички 45
Экзодерма 53, 234
Элайосомы 196, 197
Эндодерма 53, 234
— роль 73, 74
Эндокарп 184
Эндосперм 109, 125, 182
Энтомофилия 147—161
Эпидермис 35, 42, 74, 75
Эпикарп 184
Эпифиты 230—238
— гнездовые 233. 234
— «кармашковые» 233 234
— дистерновые 233, 235, 236
Этиолирование 94, 230
Ягода 183
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИИ
Acacia 215, 216
Acer pseudo plat anus 208
— saccharum 105
Aceras anthropophorum 178
Adoxa moschatellina 98
Aeschyanihus 233
Agrimonia eupatoria 192
Agropyron repens 131
Ajuga reptans 196
Aldrovandra vesiculosa 262
Alisma 164
— planiago-aquaiica 197
Allium 196
— vineale var. compactum 134
Ammo phi la arenaria 214
Anabaena 288
A ncistrocladus 225
Anthoceros 288
Araceae 229, 233, 236
Arceuthobium minutissimum249
Aristolochia 230
Ar millaria mellea 171, 284
Arthrobotrys oligospora 273
Arum maculatum 132, 158
Asarum europaeum 196
Ascomycetes 292
A triplex confertifolia 239
A 201 la 288
Azoto batter chroococcum 288
Bambusa arundinacea 39
Вага&ыма/Ыа 91
Barringtonia 187, 197
Basidlomycetes 293
Bauhinia 230
Berber is vulgaris 216
Beta vulgaris var. maritima 197
Betula папа 203
Betulaceae 277
Bidens 191
Blackstonia perfoliata 240
Boehmeria nivea 32
Boletus bovinus 278
— elegans 277
Bryonia dioica 142
Burmanniaceae 286
Cactaceae 237
Cakile maritima 197
Calluna vulgaris 213, 240
Calystegia sepium 225
Caragana arborescens 196
Carex digital a 196
Cassyiha 254
Castanopsis argentea 238
Casuarina 214
Cattleya citrina 170
Cenococcum graniforme 277
Centaurium minus 240
Cephalotaceae 269
Cephalotus follicularis 269
Ceratophyllum demersum 221
Cereus 162
Cetraria islandica 296
Chamaenerion angustifolium
150, 192
Chelidonium ma]us 196
Chlorella 293
Chlorophyta 292
Chrysosplenium alternifolium
162
Cinnamomum zeylanicum 145
Circaea luietiana 191
Cirsium arvense 130
Citrus 184
Cladonia rangiferina 296
Clematis vitalba 227
Clostridium pasteurianum 28&
Cocos nucifera 185
Compositae 150
Convolvulaceae 252
Corallorhiza trifida 170
Coryanthes 177
Corydalis 196
Corylus aveIlana 251
Coussapoa fagifolia 236
Crambe maritima 197
Crataegus monogyna 216
Crescentia cujete 162
Cryptocoryne 159
Cryptostylis leptochlla 178
Cuscuta 252
— e pithy mum 253
— europaea 252
Cyanophyta 292
Cycnoches ventricosum 179
302
Указатель латинских названий
Cymbalaria muralis 195
Cytisus ruthenicus 148
Dactylaria Candida 274
— gracilis 274
Darlingtonia 268, 269
Datura stramonium 186
Dendrobium 169
Digitalis purpurea 146, 192
Dionaea museipula 260, 261,
262
Dipodium pictum 237
Drosera binata 260
— rotundifolia 257—260
Drosophyllum lusitanicum 260,
275
Elodea canadensis 222
Eminium 158
Endymion nonscruptus 133
Epidendrum 169
Epipogiun aphyllum 170
Erica 280
— cinerea 241 \
— tetralix 281
Ericaceae 280—282
Erythrina cristagalli 161
Eugenia caryophyllata 145
Euphrasia 249
Evernia 296
Fagaceae 277
Ficus benghalensis 62
— carica 152
-----var. caprifica 154
-----var. domestica 154
Eumar ia 196
Fusarium moniliforme 208
Dalanthus nivalis 133
Galeola 284
Galium 224
— aparine 182, 191
Gastrodia 172, 284
— elata 171
Genlisea 265
Geum urbanum 191
Gloriosa 227
Gongora maculata 177
Habenaria 169
Hedera helix 228
Heliamphora 268
Helleborus foetidus 150
Hugonia 228
Humulus lupulus 145, 225
Hydnophytum 235
Hyoscyamus niger 242
Hypericum perforatum 166
Impaiiens noli tangere 167
Iris pseudacorus 197
Juglans regia 185
Kigelia 162
Koompassia excelsa 61
Labiatae 148
Laburnum anagyroides 140
Landolphia 228
Larix decidua 277
Lathraea clandestina 252
— squamaria 251, 252
Lathyrus о do rat us 227
Lauraceae 254
Lavandula vera 145
Lecanora conizaeoides 293
— esculenta 296
Lemna trisulca 98
Lentibulariaceae 262
Leontodon hispidus 240
Leontopodium al pi num 212
Linum catharticum 240
— usitatissimum 15
Liriodendron tulipifera 209
Listera о vat a 174
Littonia 227
Lonicera periclymenum 156, 225
Luzula 196
Lycopersicum esculentum 79
Lysimachia nemorum 166
— nummularia 204
Lythrum salicaria 151
Macrocyst is 14
— pyrifera 15
Magnolia 160
Marasmius coniatus 284
Matricaria matricarioides 198
Medicago falcata 148
Melampyrum 249
Указатель латинских названий
303
Mentha piperita 145 7
Monacrosporium cionopagum 273
— ellipsosporum 273
Moniliales 272—275
Monotropa hypo pithys 256
— uniflora 256
Moraceael62, 236
Mussaenda 118
Myosot is 146
Myrmecodia 235
Narcissus pre udonarc issue 133
— tazetta 120
Nardus strict a 55
Nelumbo nucifera 129
Neottia nidus-avis 170, 255,
256 284
Nepenthes 265—269, 272
Nerium oleander 213
Nicotiana tabacum 242
Nostoc 288
Nuytsia 249
Nymphaea alba 207
Nyphar lutea 207
Ochroma lagopus 47
Oenothera 156
Ophrys apifera 178, 179
— insect if era 178
— speculum 178
Orchidaceae 233
Orchis 132, 168
— fuchsii 132, 177
— maculaia 170
— mascula 173, 175, 177
Orobanchaceae 250
Orobanche 251
Oroxylon 162
Ourouparia 228
Oxalis acetosella 98, 129
Palaeomyces asieroxyli 287
Pandanus 187
— utilissimus 238
Parietaria diffusa 117
Paris quadrifolia 116
Passifloraceae 227
Passiflora edulis 227
— maliformis 227
Pedicular is palusiris 249
— sylvatica 249
Pelvetia canaliculate 12
Petasites hybridus 205
Philodendron 229, 237
Phoma radicis 281, 282
Pimpinella anisum 145
Plnguicula 262
— vulgaris 262
Pinus palusiris 47
— рейсе 249
Piper nigrum 145
Plantago 150
— major 198
Platanthera bifolia 170
Plaiycerium 234
— difurcatum 234
— grande 234
Polygonatum 131
Polygonum convolvulus 225
Polypodium vulgare 231
Polytrichum 20
Populus italica 60
Potamogeton obtusifolius 207
Poterium sanguisorba 240
Primula vilgaris 150
Protea 161
Prunus spinosa 216
Psilotum 286
Puccinia graminls 21
Pythium 119, 287
Quercus suber 45
Rafflesia arnoldii 159, 254
Ramulina 296
Ranunculus acris 138
— auricomus 138
— bulbosus 132, 138
— cassubicus 138
— ficaria 134
— flammula 138
— heterophyllus 219, 220
— polyanthemus 138
— repens 131, 138
— reptans 131
Rhinanthus 249
Rhipsalis 231
Rhizobium 289—292
— japonicum 291
— leguminosarum 291
— lupini 291
meliloti 291
304
Указатель латинских названий
Rhizobium phaseoli 291
— trifolii 291
Rhizocarpon geographicum 295
Rhizoctonia 180, 284—286
Rhizophagus 287
Rhizophore 61
Rubus f rut icos us 224
— idaeus 224
Rumex crispus 129, 198
Ruscus aculeatus 214
Rutaceae 184
Sagittaria sagittifolia 164, 207,
219
Salicornia 197
Salsola kali 218
Salvia 148—150
— horminoides 167
Santalaceae 250
Sarothemnus sco paries 147
Sarracenia 267, 268
— purpurea 265
Schoenoplectus locust r is 164
Scrophulariaceae 249
Sedum 218
Sequoia 81
Selaginella 122, 123
Sempervivum funkii 241, 242
S inapis nigra 199
Solanaceae 242
Sphagnum 240
Spinacia oleracea 243
Spiranthes 169
Stachys sylvatica 205
Stanhopea оси I at a 232
Stellaria ho lost e a 204
Stenochlaena areolaris 238
Stereum purpureum 43
Strelitzia reginae 161
St у lopage 270
— grandis 271
Suaeda maritime 218
Symphytum officinale 143
Taraxacum officinale 182
Taxodium distichum 62, 223
Taxes baccata 47
Terminalia catappa 187
Thalictrum flavum 150
Thesium humifusum 250
Tillandsia usneoides 232
Tozzia 250
Tragopogon prate ns is 194
Trebouxia 293
Trifolium 253
Triurldaceae 286
Tropaeolum 227
Typha 207
— angustifolia 164
— latifolia 164
Ulex 148, 182, 216
— europaeus 196
Ulmus 251
Umbelliferae 150
Usnea 232, 295
Urtica dioic a 205, 252
Utricularia 236, 263, 272
— vulgaris 263
Vaccinium 280
— corymbosunfl2S2
— oxycoccos 281
— pensylvanicum 282
Vallisneria americana 165
Veronica chamaedrys 157
— officinalis 204
Vida 227
Victoria amazonica 160, 164r
221
Viola odoraia 146, 166
Viscum album 126, 189, 246,
249
Vitis vinifera 79, 227
Wullschlaegelia aphylla 255
Xanthium 127, 243
Xanihoria parietina 293
Xerotus jaoanicus 284
Yucca filameniosa 156
Zingiber officinale 145
Zoopagales 270—275
Zostera 118
— marina 165
Zositrophyllum 91
ОГЛАВЛЕНИЕ
Предисловие к русскому изданию..................... 5
Предисловие автора..............................» С
ГЛАВА I. ПРОБЛЕМЫ'ПРИСПОСОБЛЕНИЯ РАСТЕНИЙ
К НАЗЕМНОЙ ЖИЗНИ » Н
Проблемы наземной жизни растений......... 13
Жизненный цикл папоротников.............. 17
Мхи и печеночники.................... . 19
Чередование поколений у цветковых растений 22
ГЛАВА Н. РАСТЕНИЯ-ИНЖЕНЕРЫ........................ 28
Тургор, прочность и проведение воды ...» 29
Волокна и членики сосудов........ , . . 31
Ткани стебля............................ 34
Функция камбиального слоя............... 39
Пробковый камбий . •.................... 42
Старение стволов деревьев .............. 45
Как растения решают проблемь^ связанные с
преодолением силы тяжести............... 48
Кольца прироста ........................ 50
Структура и функции корня............... 52
ГЛАВАМИ. ГИДРАВЛИКА РАСТЕНИЙ , ,..................... 63
Осмос..................................... 64
Протоплазматические мембраны и работа
клетки................................ 67
Осмотическое и ту prop ное давления....... 69
Роль эндодермы............................ 73
Структура и функция листа................. 74
Подъем сока по стеблям и стволам.......... 78
Транспирация и устьица.................... 84
Устьица и фотосинтез...................... 86
Поглощение минеральных веществ ..... 88
ГЛАВА IV. КАК РАСТЕНИЕ ПИТАЕТСЯ.......... W
Реакция растений на действие силы тяжести . 92
Рост по направлению к свету................ -94
Хлорофилл и другие пигменты ....... 95
Химия фотосинтеза............................ 100
ГЛАВА V. ТРАНСПОРТНАЯ СИСТЕМА РАСТЕНИЙ . . 105
Передвижение органических веществ . . . . • 105
Использование питательных веществ . . . , . 109
306
Оглавление
ГЛАВА VI. ПРОБЛЕМА РАЗМНОЖЕНИЯ........ИЗ
Части цветка................................... из
Опыление и оплодотворение............< . . 116
Бесполое размножение (спорами) 1........... 122
Размножение голосеменных................... 123
Развитие семени........................... 124
Прорастание................................ 126
Жизнеспособность семян..................... 123
Вегетативное размножение................... 130
Преимущества семян......................... 135
ГЛАВА VII. ОПЫЛЕНИЕ.......................... 137
Специализация в направлении обеспечения
опыления........................... 140
Цвет и запах............................. 143
Указатели меда и нектар.................. 145
Строение пыльцевых цветков............... 146
Цветки, образующие нектар................ 148
Опыление осами, ночными бабочками, жуками
и мухами............................ 152
Опыление птицами и летучими мышами ... 160
Опыление ветром.......................... 162
Гидрофилия........................ . . . 165
СакЛюпыление............................. 165
ГЛАВА V111. ОРХИДНЫЕ................ 168
Эпяфитные и наземные орхидеи.......... 169
Типы цветков орхидных................. 172
Механизмы опыления ................... 174
Псевдокопуляция......................... 178
Прорастание семян ..................... 179
ГЛАВА IX. РАССЕЛЕНИЕ РАСТЕНИЙ...................... 181
Образование плодов ..................... 182
Теория распространения океаном (кокосовые
орехи).................................. 186
Способ распространения омелы........... 189
Другие способы расселения............. 190
ГЛАВА X. ВЫЖИВАНИЕ РАСТЕНИЙ........................ 200
Жизненная форма фанерофитов........... 202
Жизненная форма хамефитов............. 203
Жизненная форма гемикриптофитов....... 204
Категории гемикриптофитов............... 204
Розеточные растения..................... 205
Жизненная форма геофитов.............. 206
Жизненная форма гелофитов............... 207
Оглавление 307
Жизненная форма гидрофитов............... 207
Жизненная форма терофитов................ 207
Покой почек.............................. 208
ГЛАВА XI. ПРИСПОСОБЛЕНИЕ К СРЕДЕ .................\ 210
Растения-пионеры........................ 212.
Ксероморфные признаки................... 212
Осенний листопад........................ 217
Суккуленты.............................. 218
Листья водных растений.................. 218
Размножение водных растений............. 222
Лазящие и вьющиеся растения ............ 224
Приспособления эпифитов............... 230
Цистерновые эпифиты..................... 235
Приспособления полуэпифитов............. 236
Приспособления галофитов................ 238
Кальцифилы и кальцифобы................. 239
Фотопериодизм........................... 241
ГЛАВА XII. ПАРАЗИТЫ И САПРОФИТЫ.................... 248
Полупаразитический образ жизни омелы . . . 247
Другие полупаразиты.................... 240*
Настоящие паразитные растения.......... 250
Сапрофиты.............................. 255-
ГЛАВА XIII. ХИЩНЫЕ РАСТЕНИЯ........................ 257
Росянка как ловушка для насекомых . . . 257
Другие хищные растения............ 262.
Растения-кувшины................... 265
Хищные грибы....................... 269
ГЛАВА XIV. СОЖИТЕЛЬСТВО РАСТЕНИЙ................ 276
Симбиотические союзы............... 276
Эктотрофная и эндотрофная микоризы . . . 279
Фиксация азота.................... 282:
Микоризы орхидных.................. 283
Микоризы, образованные низшими грибами 286
Водоросли как партнеры............ 288-
Азотфиксирующие бактерии........... 288
Лишайники как пример симбиоза...... 292
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ . ..............• . . ♦ . 297
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ.................... 300
УВАЖАЕМЫЙ ЧИТАТЕЛЬ!
Ваши замечания о содержании книги,
ее оформлении, качестве перевода и другие
просим присылать по адресу:
129820, Москва, И-110, ГСП,
1-й Рижский пер., д. 2,
издательство «Мир»
К. Даддингтон
ЭВОЛЮЦИОННАЯ
БОТАНИКА
Редактор Н. А. Сыснна
Художник А. Д. Смеляков
Художественный редактор Ю. Л. Максимов
Технический редактор Е. Н. Лебедева
Корректор Г. А, Секачева
Сдано в набор 24/IV 1972 г.
Подписано к печати 27/VII 1972 г.
Бумага № 2 84Х108‘/м-5,13 бум. л. Усл. печ. л. 17,22.
В т/ч. 1.05 усл. печ. л. иллюстраций. Уч.-изд. л. 15.94
Изд. № 4/6056 Цена 1 р. 33 к. Зак. 302
ИЗДАТЕЛЬСТВО «МИР»
Москва. Ьй Рижский пер.. 2
Московская типография № 11 Главполнграфдрома
Комитета .по печати при Совете Министров СССР.
Москва, 88. Угрешская, 12.
Фото L Проводящий пучок из стебля подсолнечника.
Фото II. Верхушка коробочки мха с перистомом.
Фото 1П. Трахеиды с окай-
мленными порами иа про-
дольном тапгепталыюм сре-
зе древесины сосны.
Фото IV. Трахеиды с окай-
мленными порами па про-
дольном радиальном срезе
Древесины сосны.
Фото V. Поперечный разрез молодого стебля подсолнечника,
на котором можно наблюдать проводящие пучки.
Крупные клетки в пучках представляют собой сосуды ксилемы.
Фото VI. Поперечный разрез стебля яснотки,
на котором видна колленхима, развитая
в области ребер стебля (выступ на фото).
Фото VII. Поперечный разрез стебля кукурузы.
Обратите внимание на то, что проводящие пучки рассеяны
по стеблю, это типично для однодольных.
Фото VIII. Поперечный разрез стебля лайма.
Обратите внимание на вторичную древесину (внизу), большое
количество которой образовалось в процессе вторичного роста.
Фото IX. Поперечный раз-
рез стебля смородины, на
котором можно наблюдать
развитие перидермы.
Фото X. Поперечный разрез
вторичной древесины сосны,
на котором видно кольцо
прироста.
Фото XI. Поперечный раз-
рез молодого корня конских
бобов.
Фото XII. .Апикальная
меристема и корневой
чехлик па продольном
разрезе корня лука.
Фото XIII. Разрез через
веламен воздушного кор-
ня Vanda — эпнфнтнон
орхидеи.
Обратите внимание па
утолщенные клетки, обо-
значающие наружную
границу коры.
Фото XIV. Поперечный разрез через верхний эпидермис
и палисадную ткань листа бирючины. В палисадных клет-
ках можно видеть хлоропласты.
Фото XV Устьица на нижней поверхности листа Trades-
сапй’а.
Фо г о XVI. 3 а родышевы и
мешок в цветочной почке
лллля.
Фото XVII. Жимолость
растение, опыляемое ба-
бочками.
Обратите внимание на блед-
ную окраску цветка и длин-
ный трубчатый венчик.
Фото XVIII. Поперечный сре* плода пастушьей сумки.
На срезе, проходящем через семя и зародыш, который оно
содержит, можно видеть две семядоли и зародышевый ко-
решок. '
Фото XIX. Развитие плода банана.
Фото XX. Развивающиеся
плоды иван-чая.
Фото XXI. Поперечный раз-
рез листа песколюба песча-
ного.
Фото XXI1. Поперечный разрез части фотосинтезирующей
коры веточки Casuarina.
Обратите ш;ил;анне, на подобные палисадным клетки, набитые
хлороиластамп.
Фото ХХ.111. Поперечный разрез листа сосны.
Фото XXIV. Отделительный слой, формирующийся у ос нона и и я
листового черешка бузины незадолго до листопада.
Фото XXV) Поперечный разрез через кору стебля водного
растения (Myriophyllurn), на котором видны большие воз-
ду шные полости.
Фото XXVI. Вьющийся сте-
бель декоративной фасоли,
обвивающийся вокруг палки
в направлении против часо-
вой стрелки.
Фото XXVII. Усик душисто-
го горошка, охватывающий
палку.
Фото XXVIII. Часть поперечного разреза стебля Clematis (лианы).
Обратите внимание на то, что вторичная ксилема разделена тон-
костенными клетками на отдельные участки, что придает стеблю
гибкость.
Фото XXIX. Тропический папоротник олений реи
(Platycerium), растущий как эпифит на ствола?
деревьев.
Фото XXX. Ваниль (Ботанический сад, Кыо)
Фото XXXI. Жгучий во-
лосок крапивы под мик-
роскопом.
Фото XXXII. Росянка.
Обратите внимание на
чу ветви тел ьные щети н к<г
па листьях.
Фото XXXIII. Срез листа росянки, иа котором видна
чувствительная щетинка.
Фото XXXIV. Нематода, пойманная хищным грибом
Arthrobotrys robusta (сильно увеличено).
Справа виден передний конец нематоды, прилипший
к клейкой сетке. В разных местах тела нематоды
можно наблюдать питающие гифы гриба.
Фото XXXV. Нематода, схваченная хищным грибом
Dactylaria gracilis.
Она попала в одно из сокращающихся колец, имею-
щихся у гриба; на микрофотографии видны набух-
шие клетки кольца, окружающие нематоду.
Фото XXXVI. Микоризный слой в коре корня орхид-
ного.