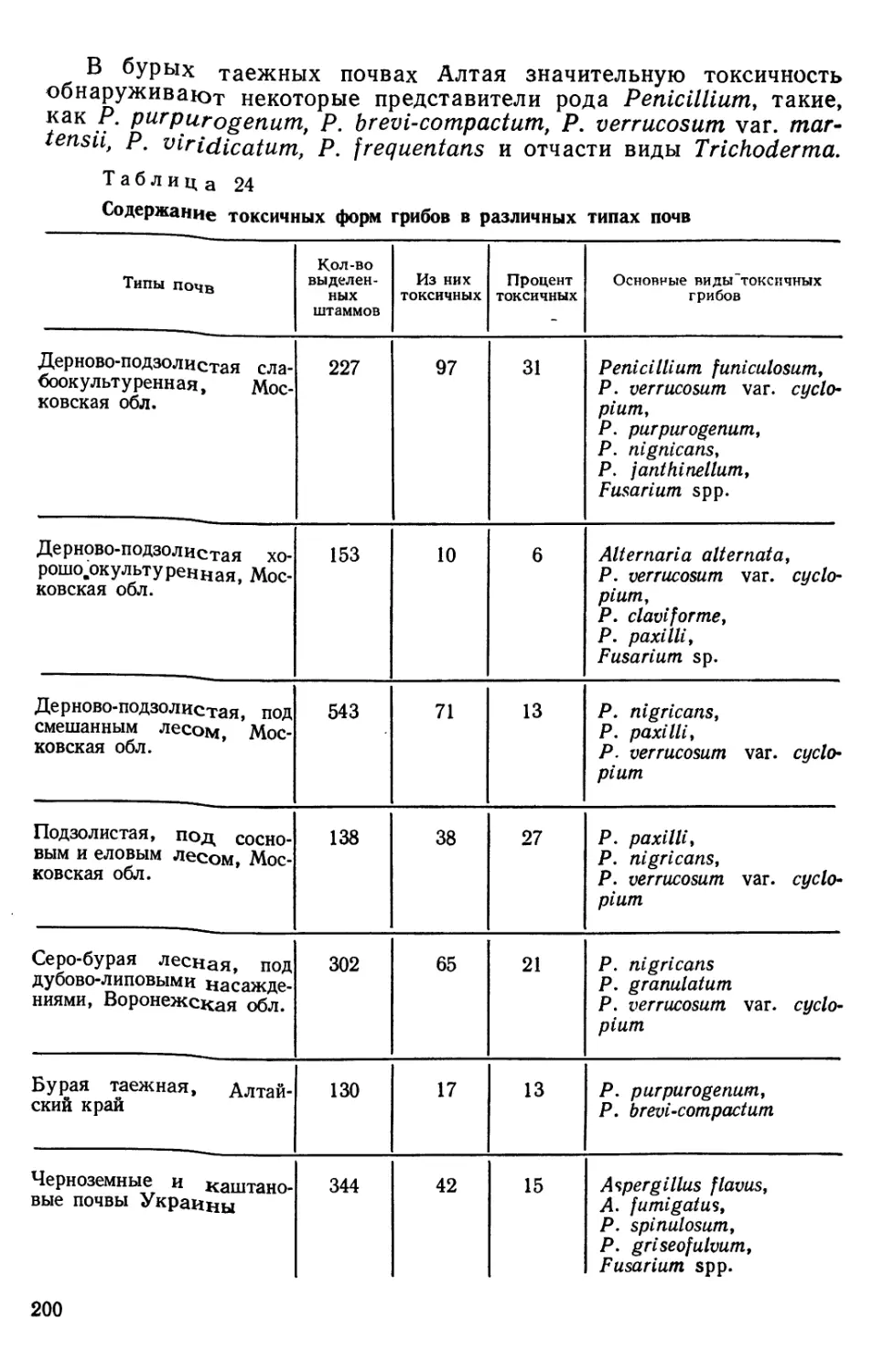
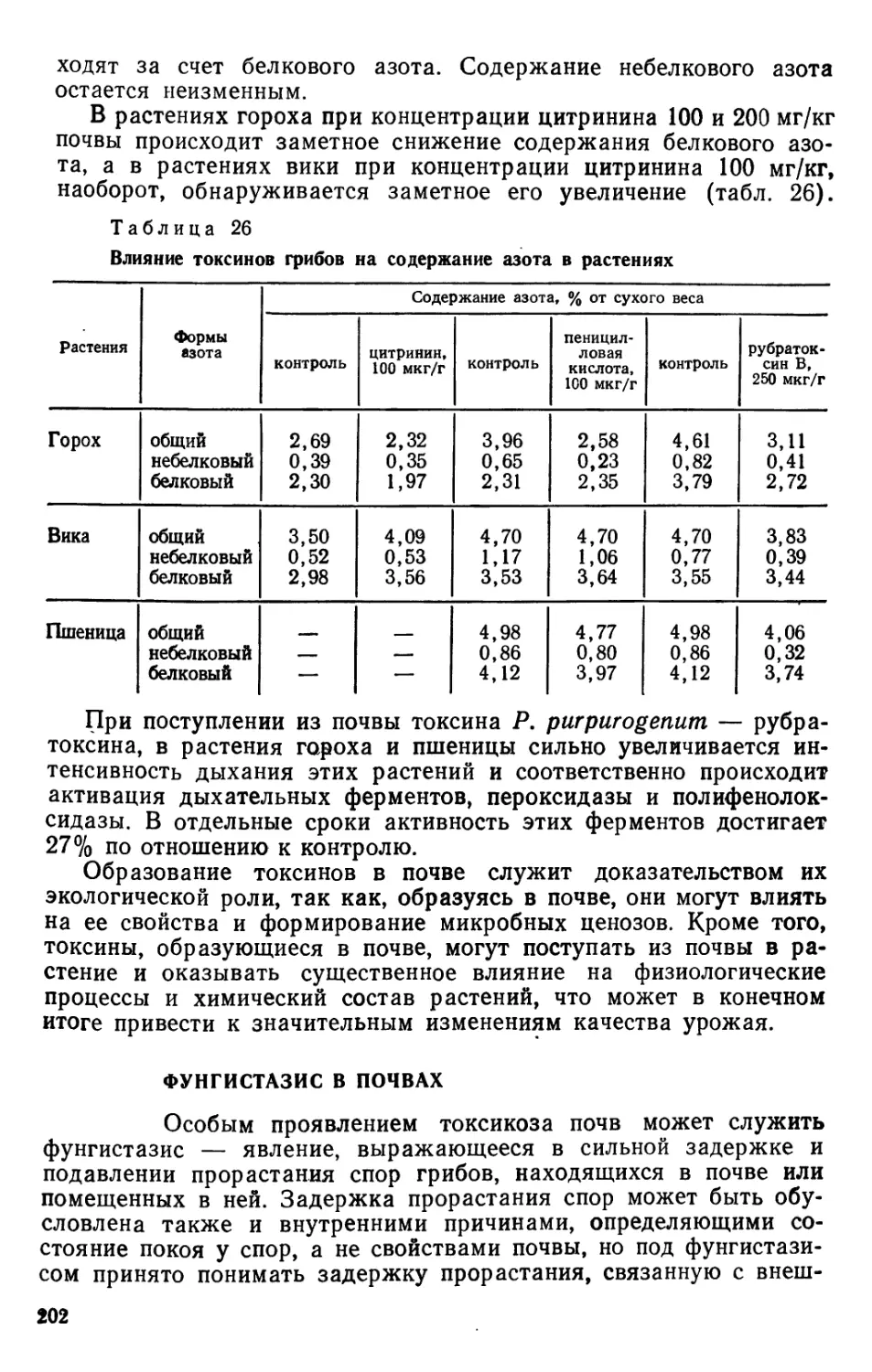
/























































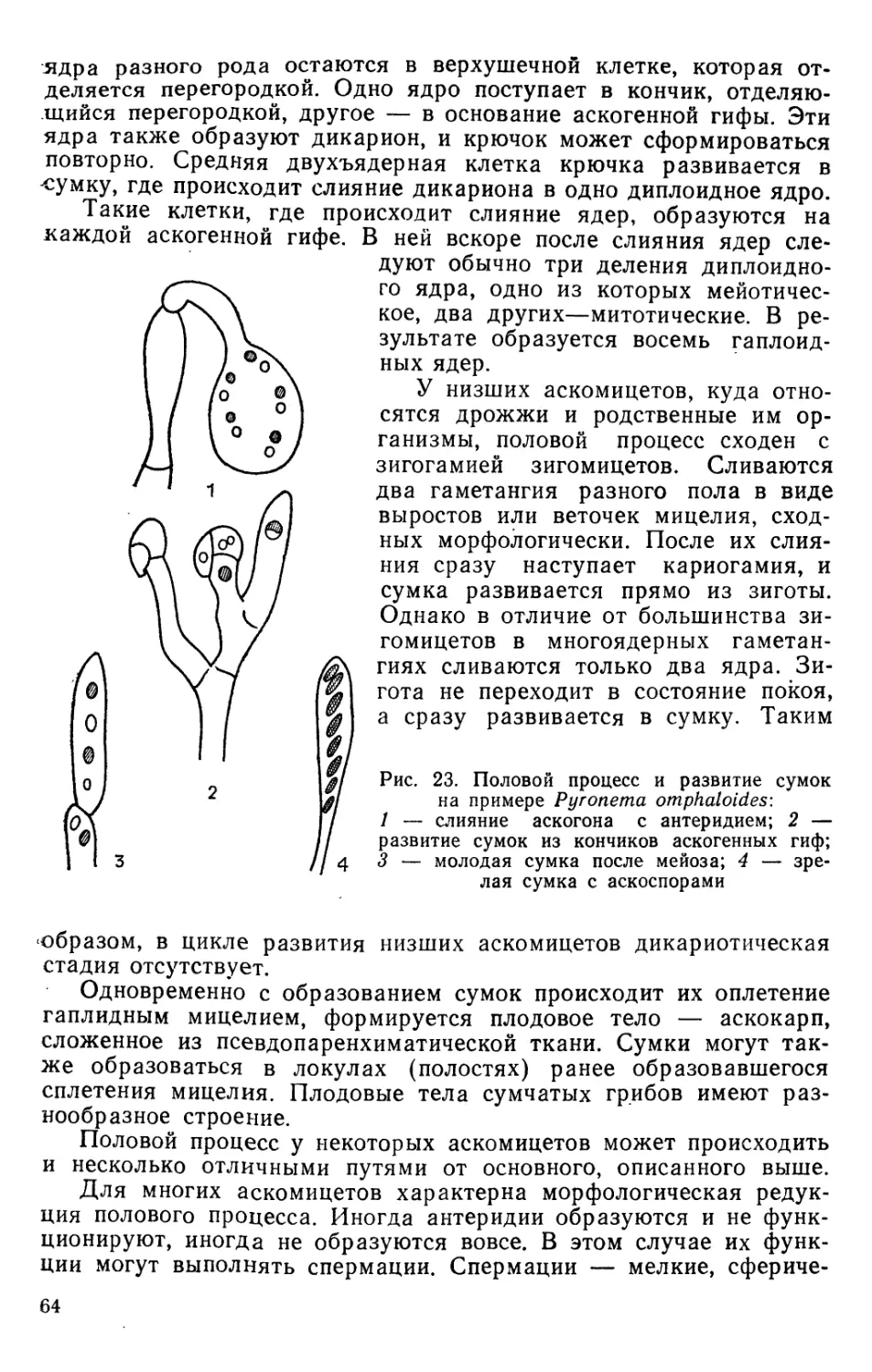

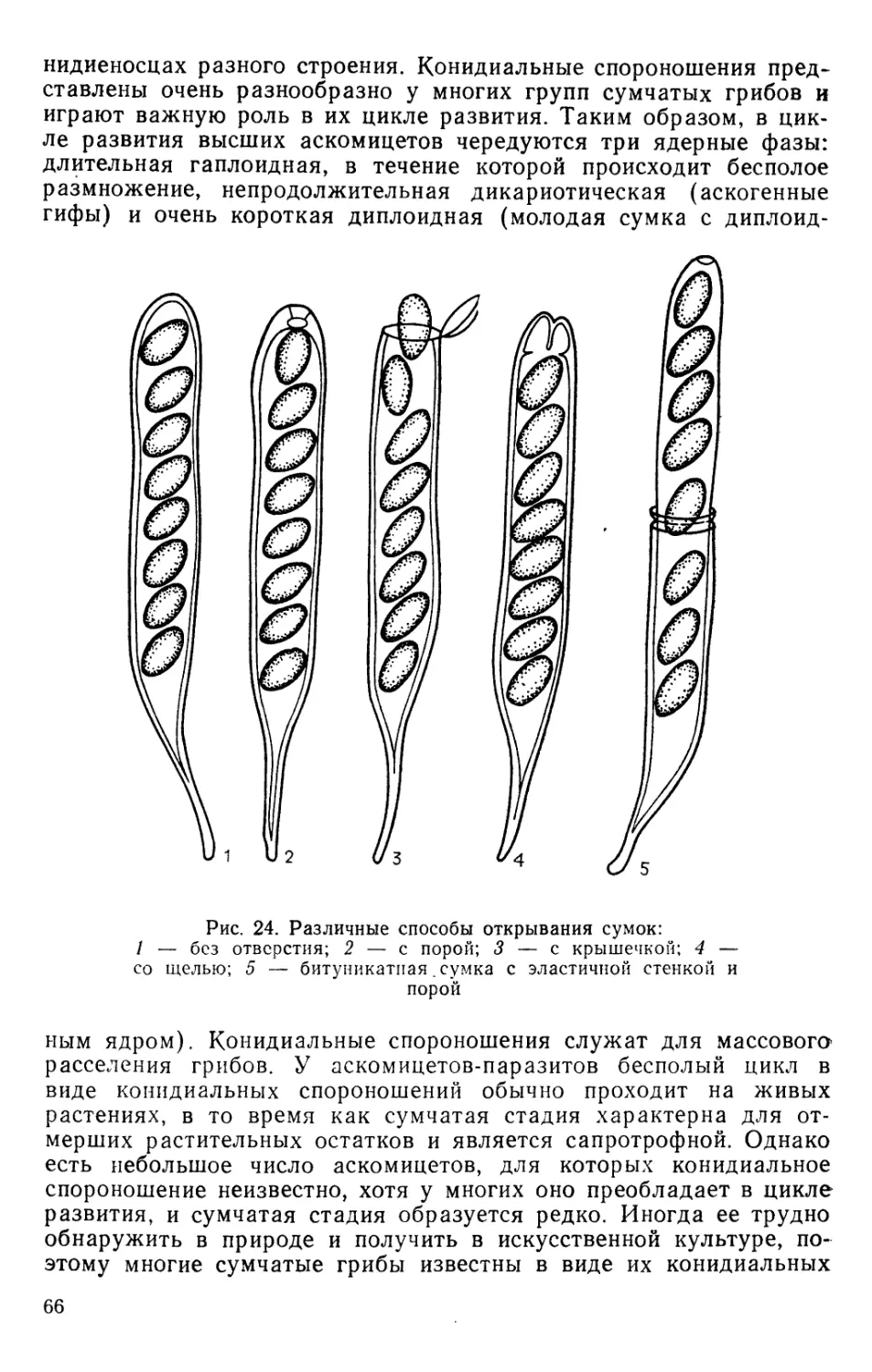




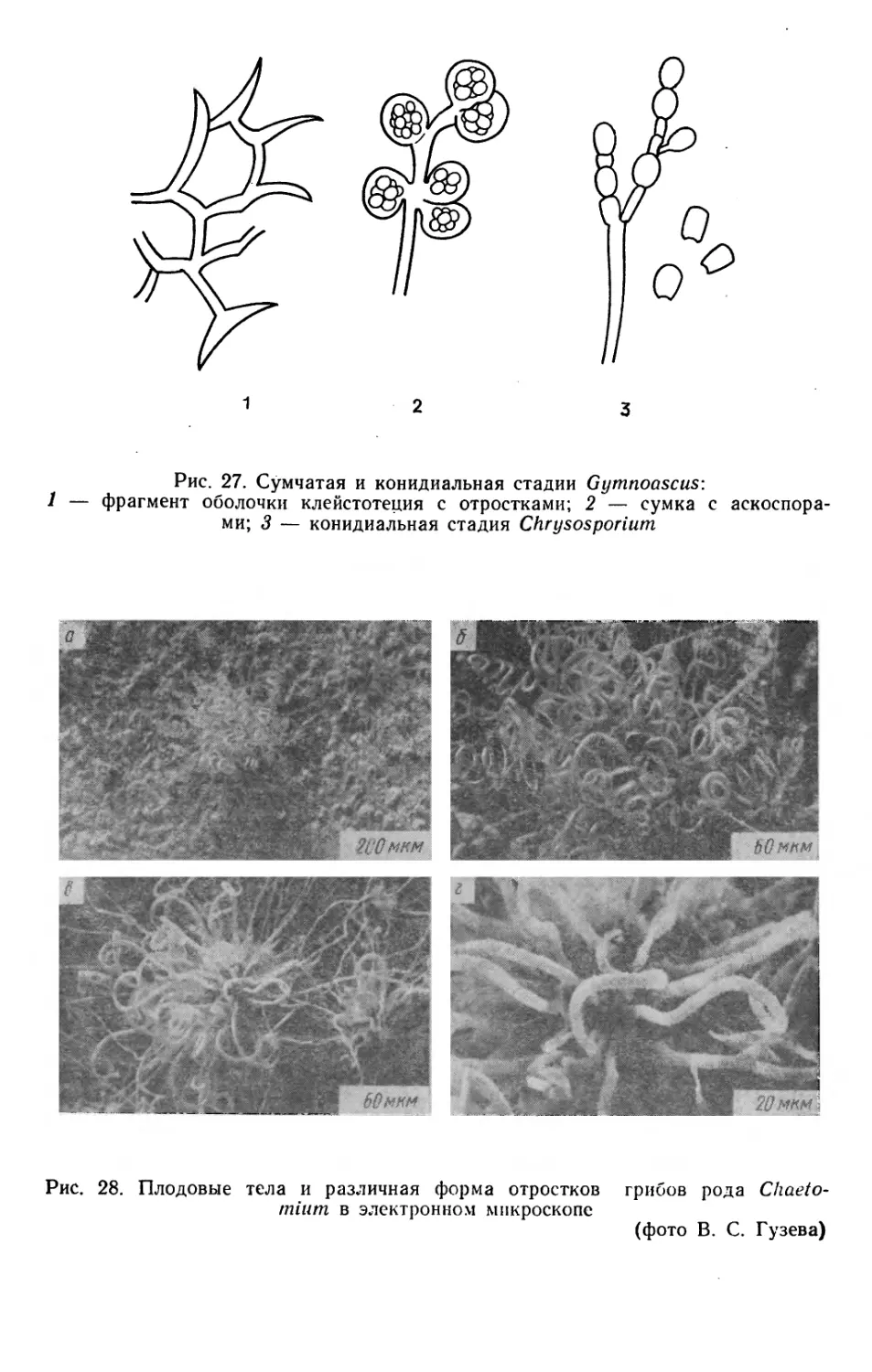
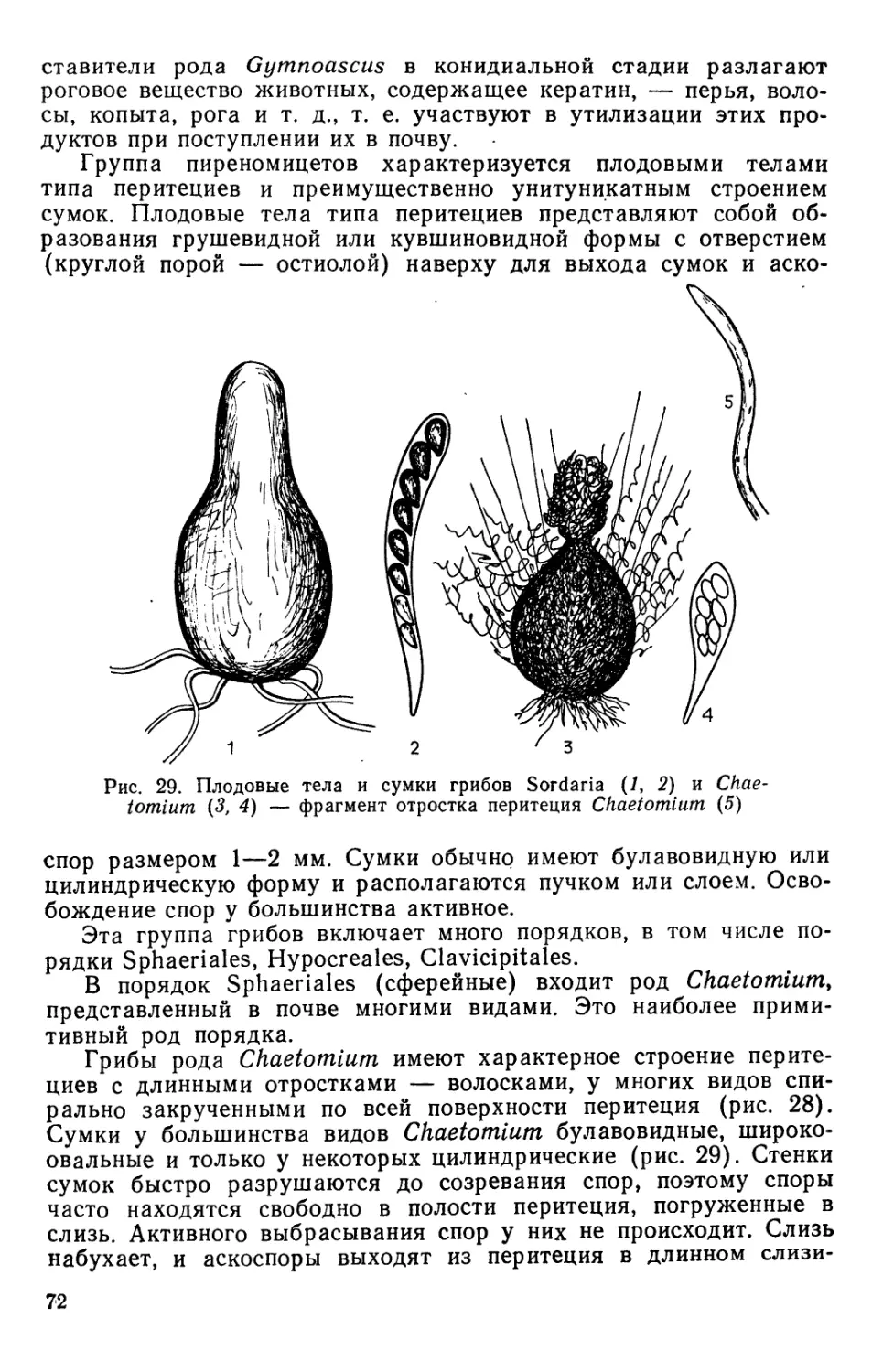






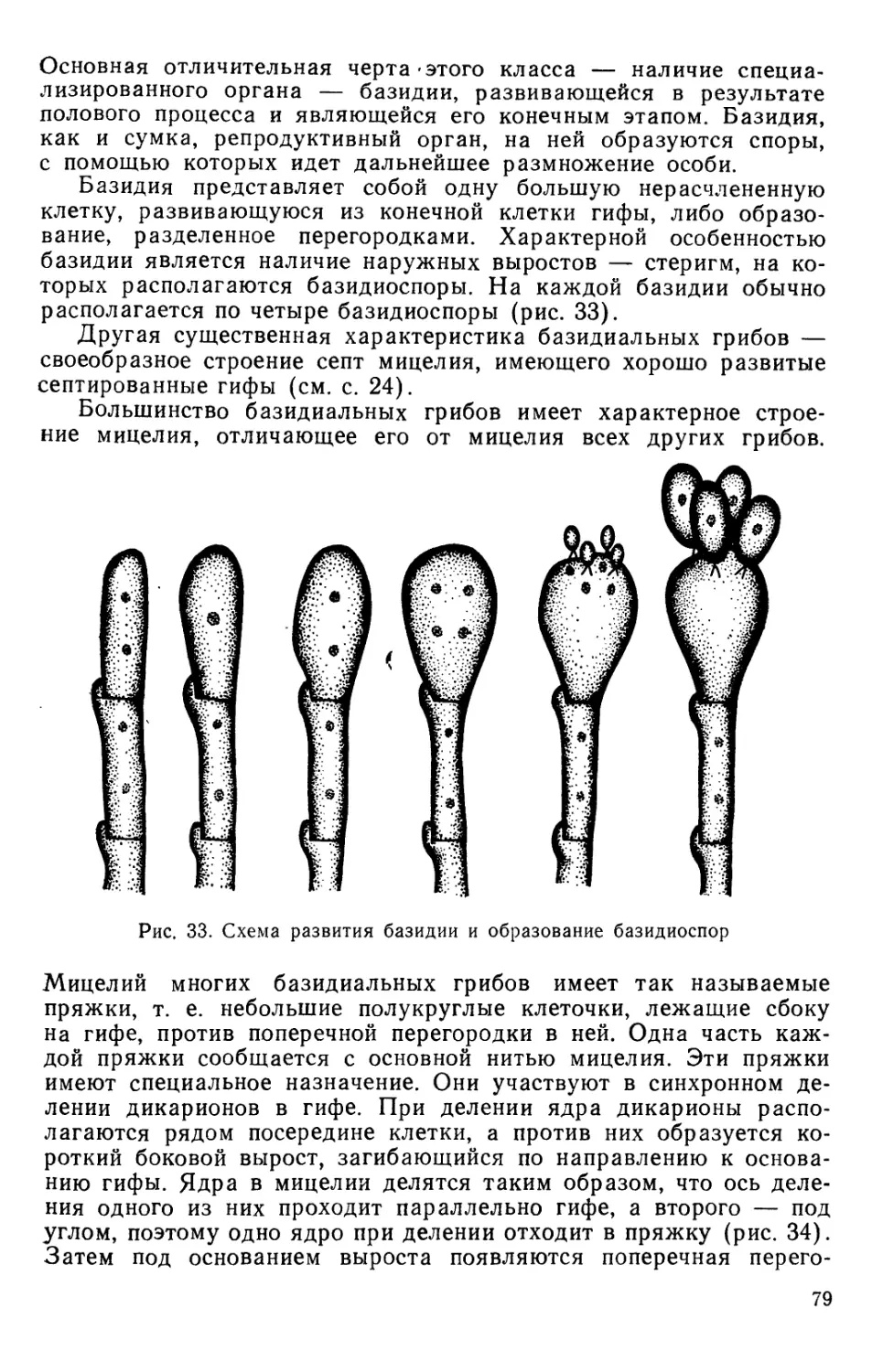















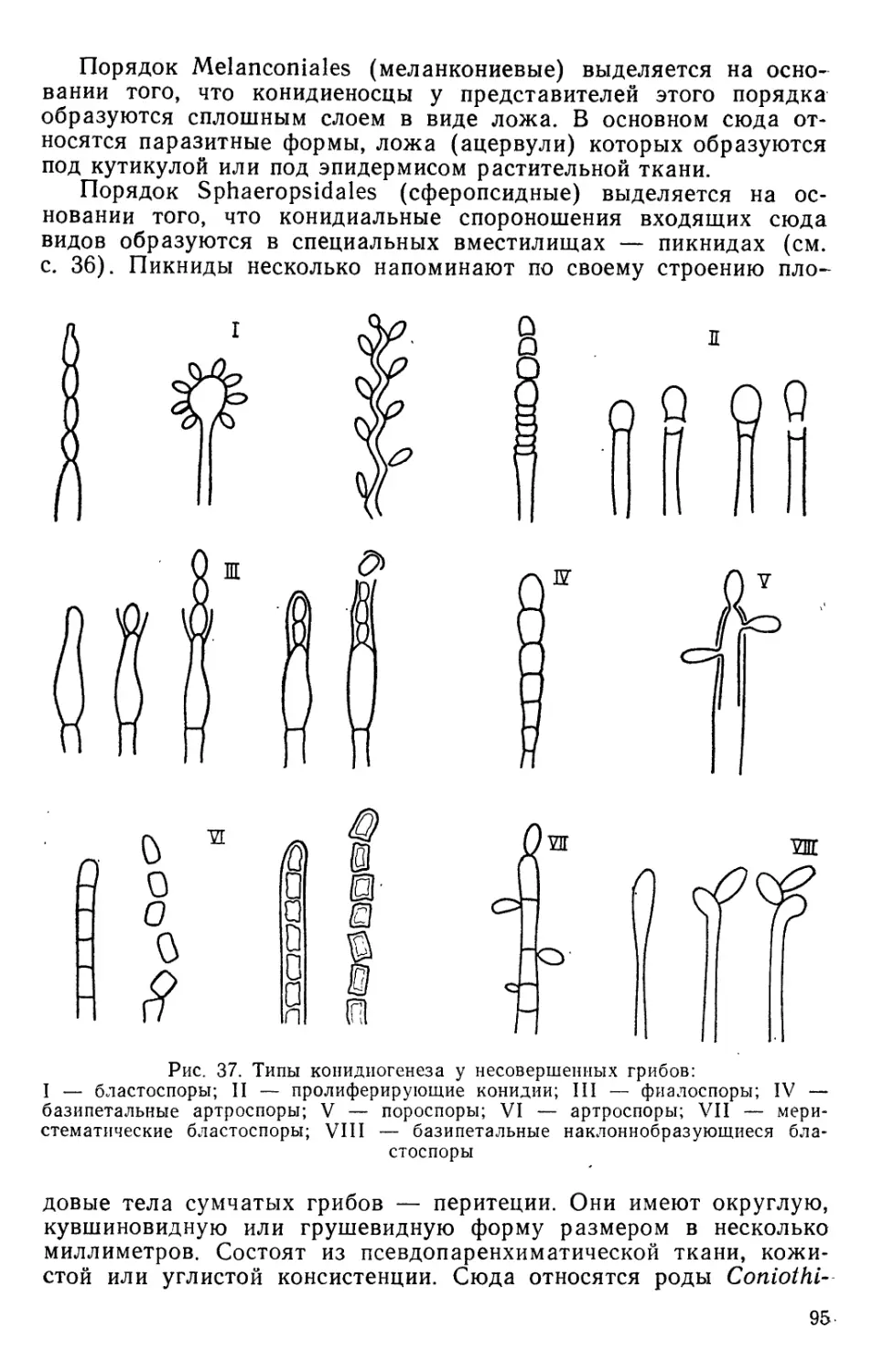
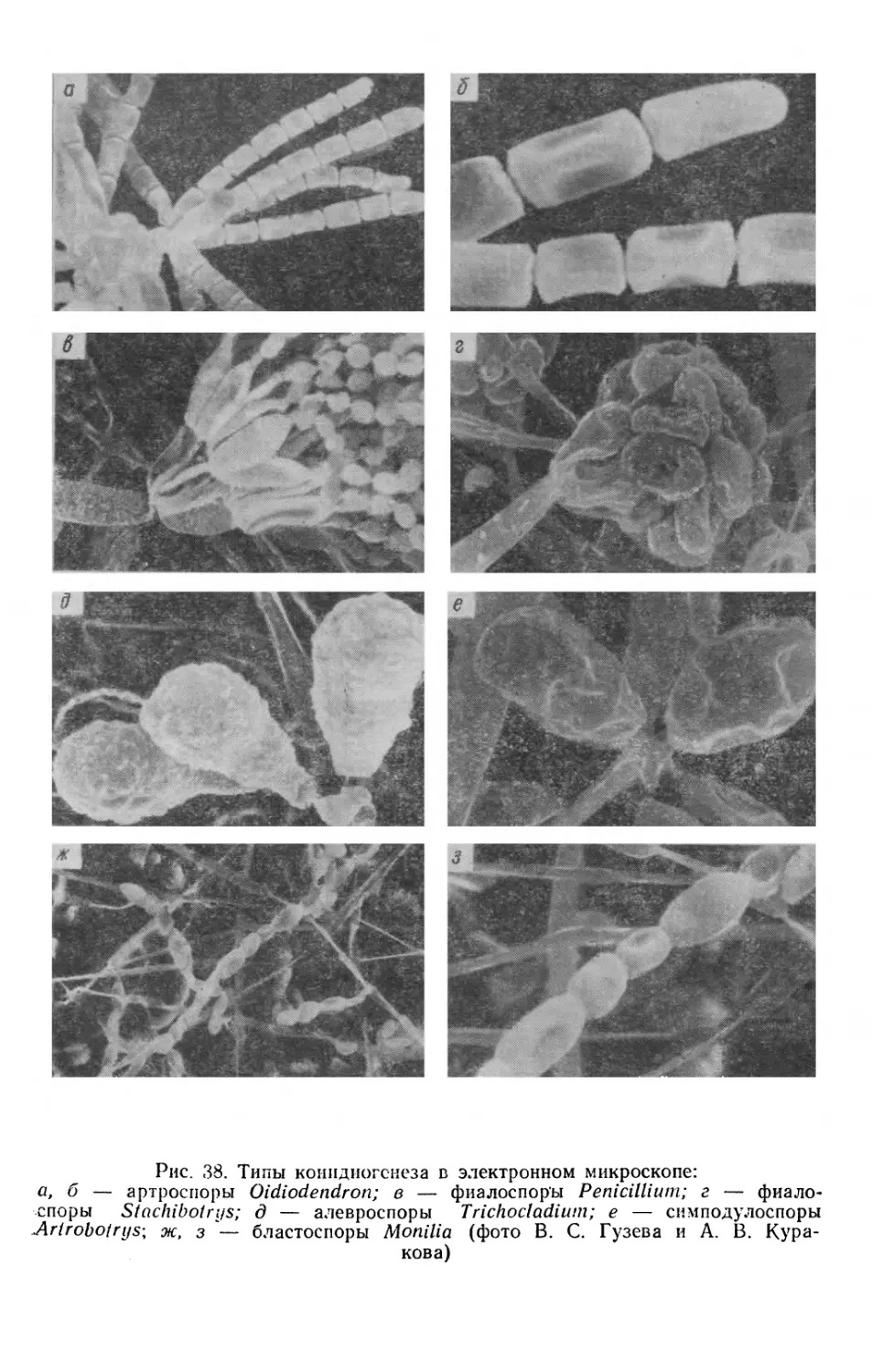








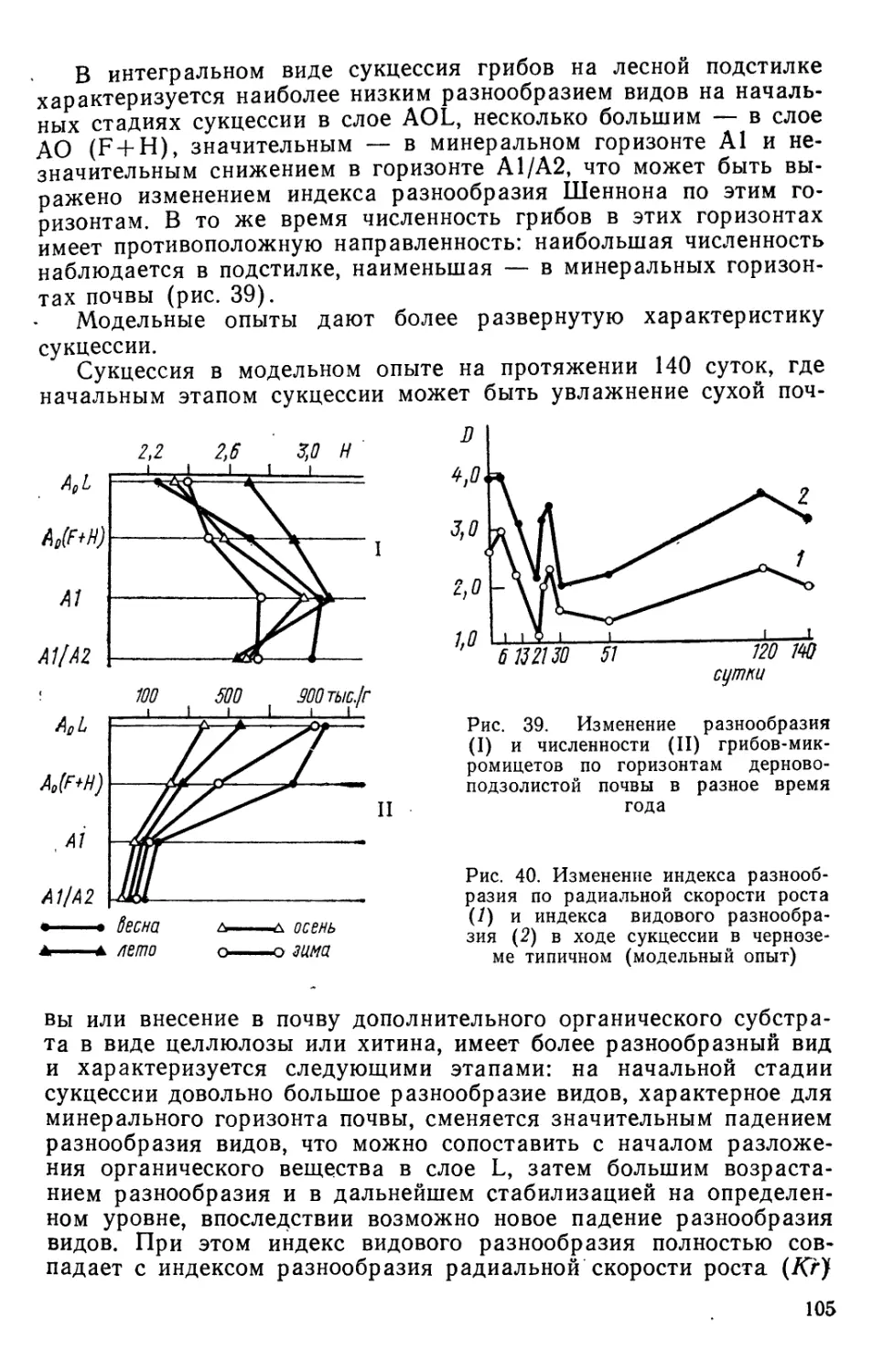




































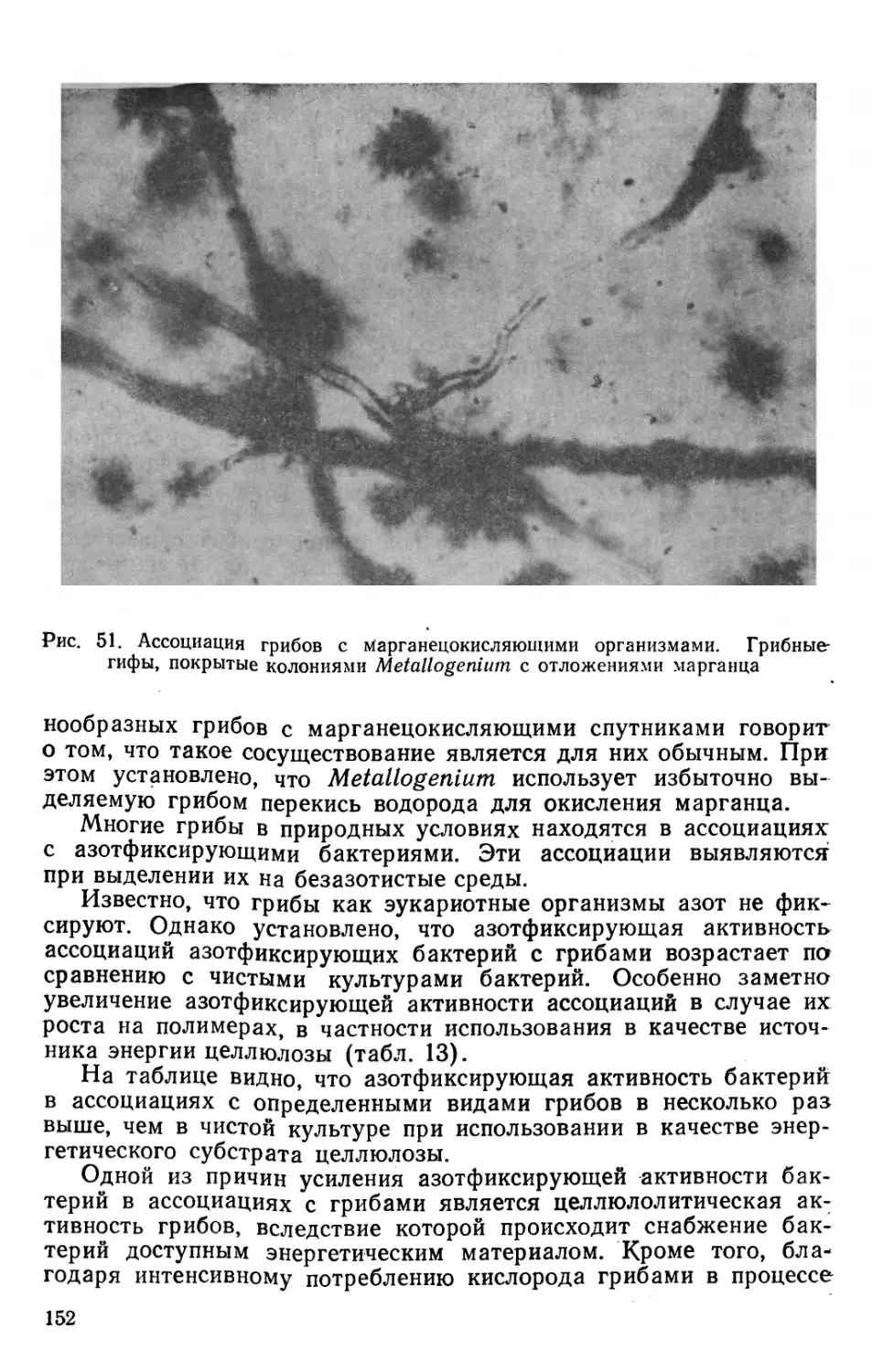




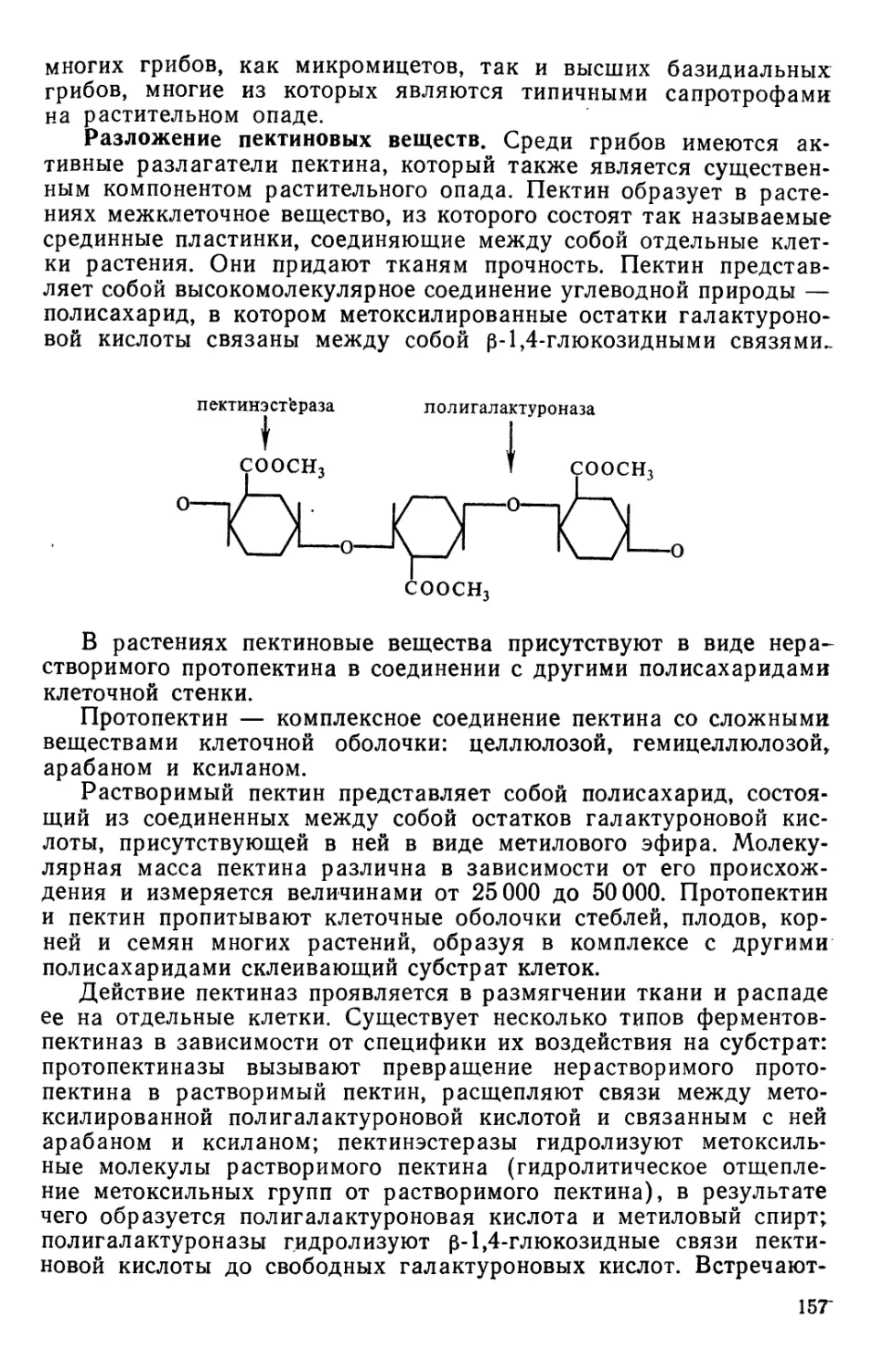

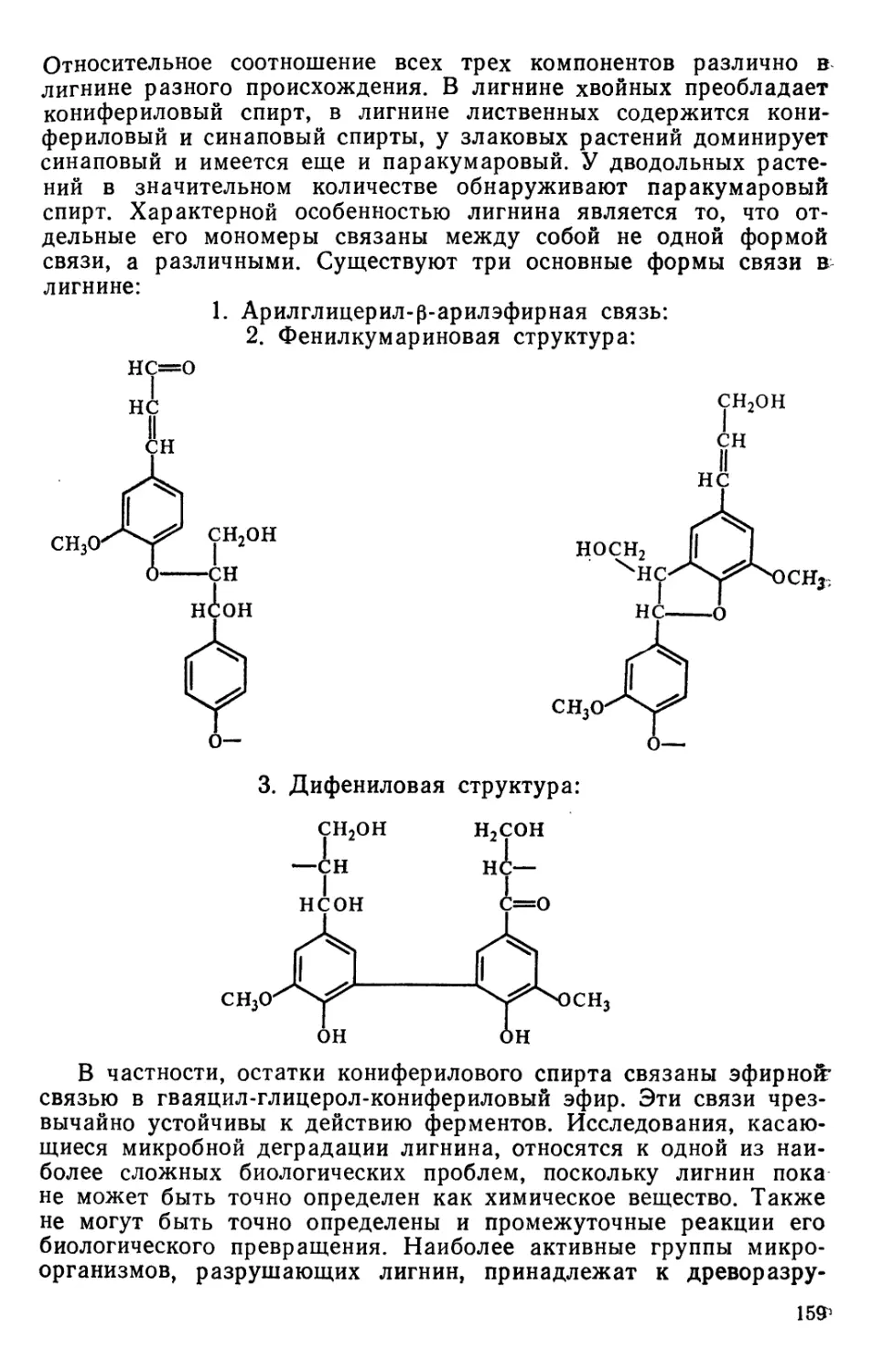

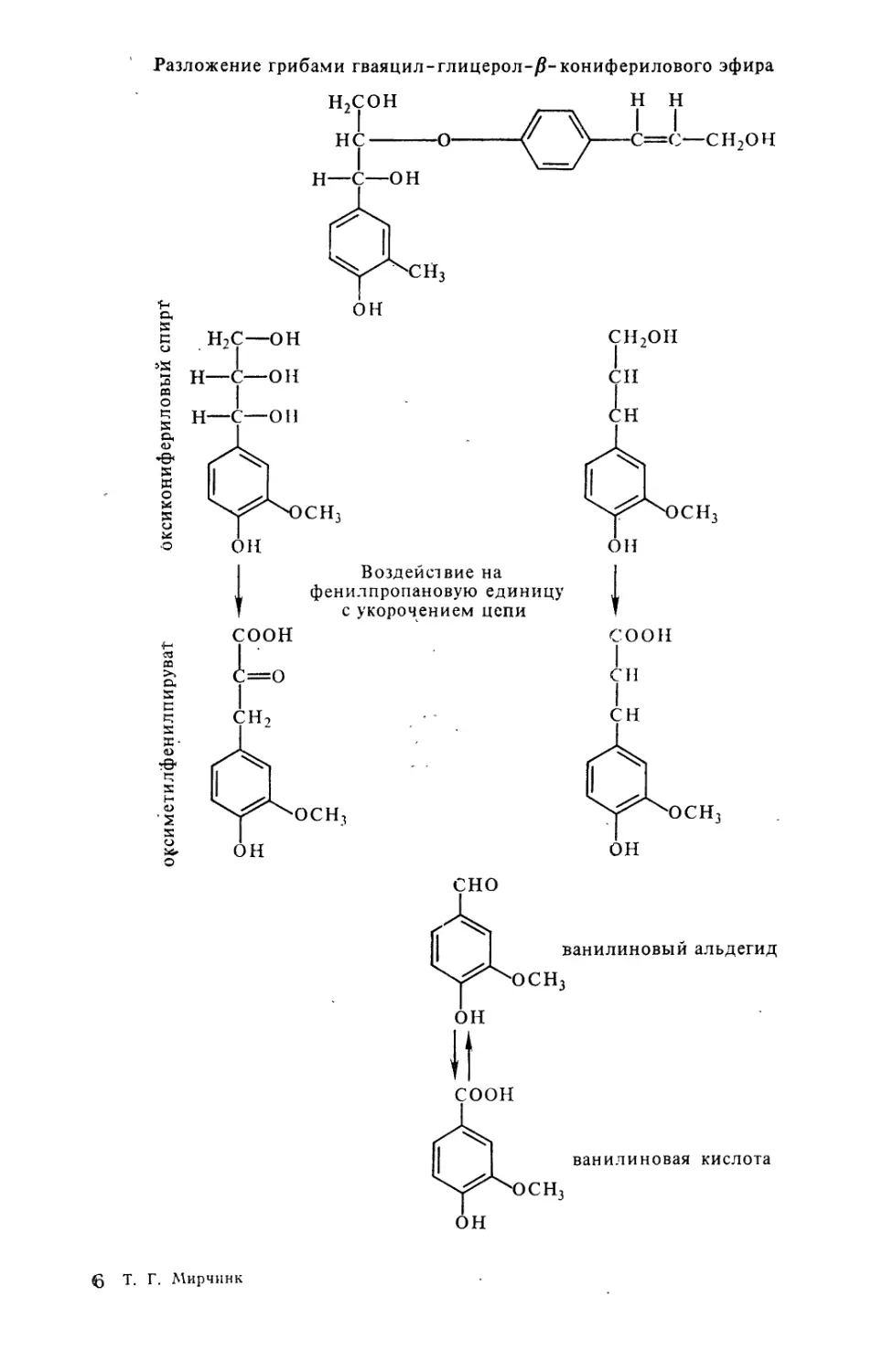















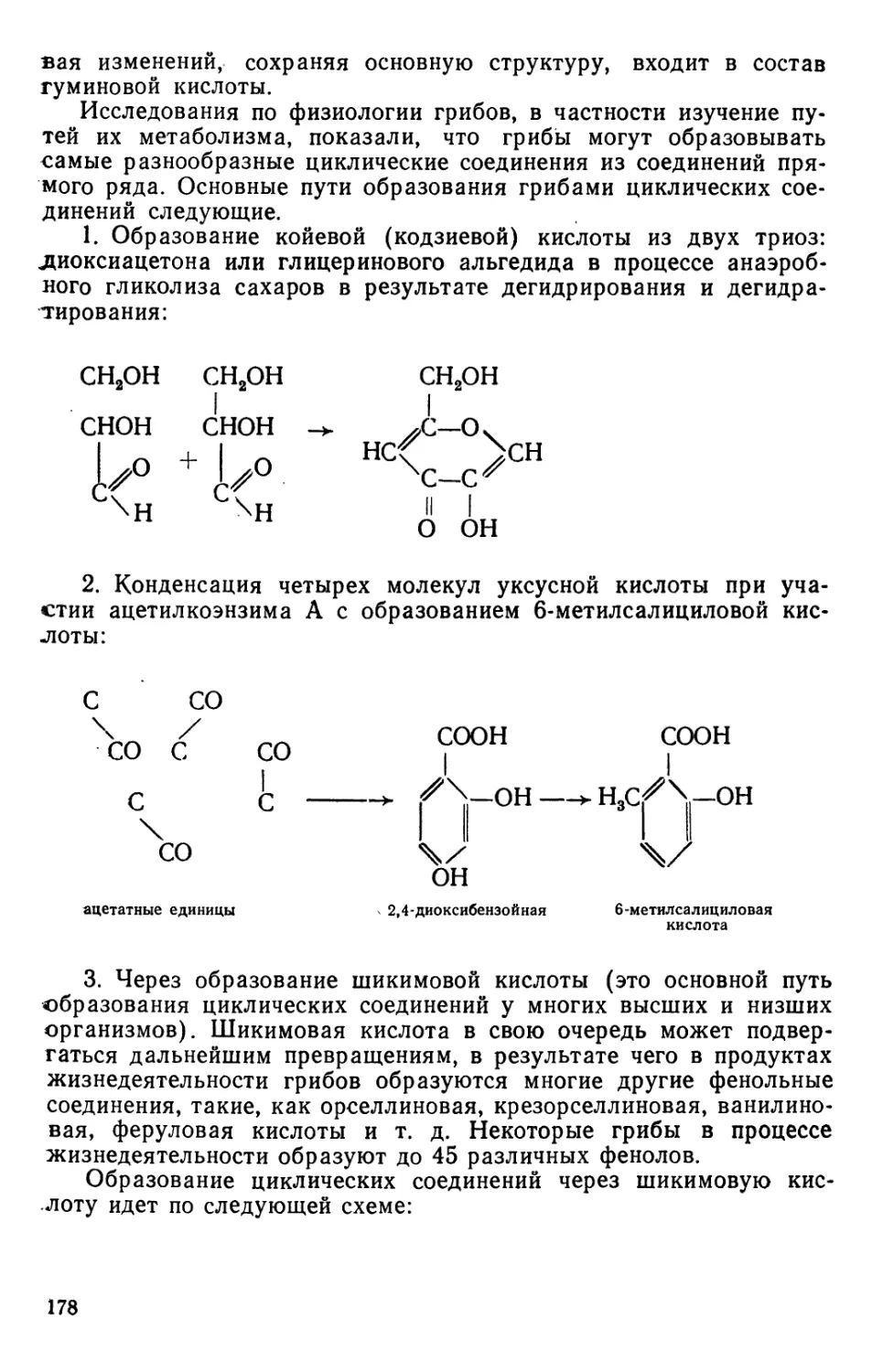
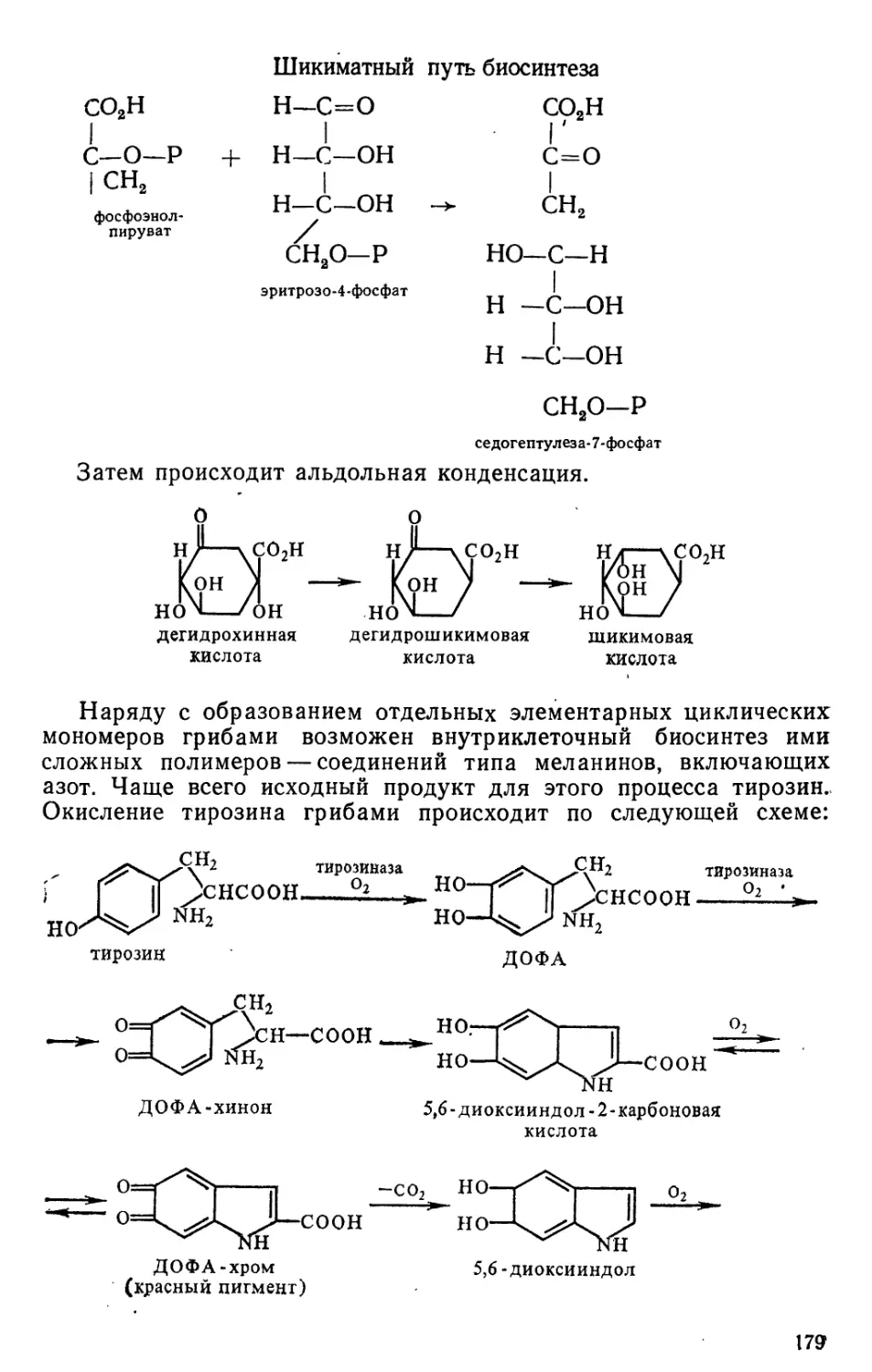








































Text
Т* Г. Мирчинк
микология
Т. Г. МИРЧИНК
ПОЧВЕННАЯ
микология
Допущено Министерством высшего и среднего
специального образования СССР в качестве
учебного пособия для студентов высших учебных
заведений, обучающихся по специальности
«Агрохимия и почвоведение»
ИЗДАТЕЛЬСТВО
МОСКОВСКОГО УНИВЕРСИТЕТА
1988
УДК 631
Мирчинк Т. Г. Почвенная микология: Учебник. — М.: Изд-во МГУ 1988 —
220 с —ISBN 5—211—00157—5
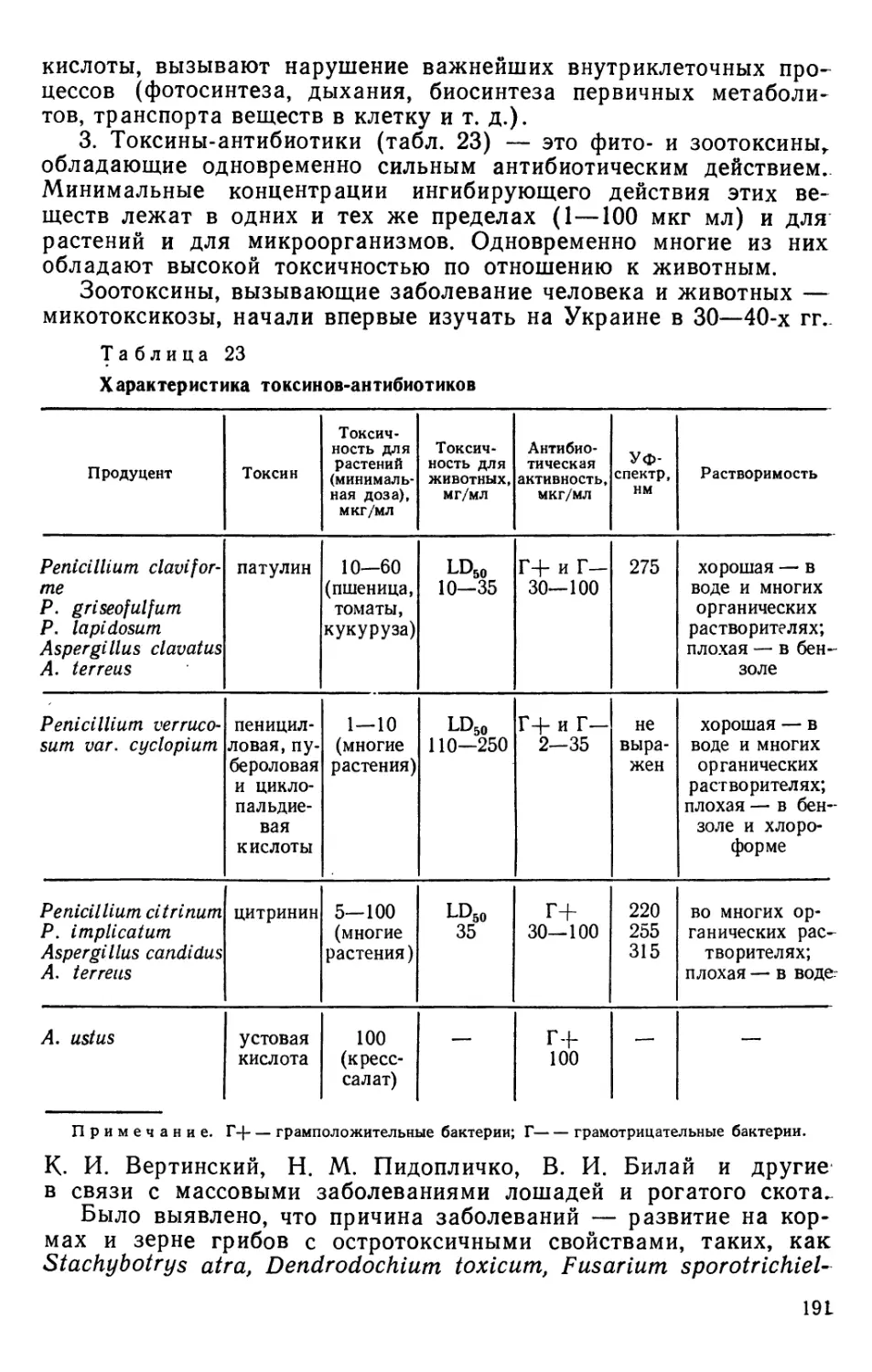
Настоящее издание — первое учебное пособие по почвенной микологии,
который обобщает и систематизирует обширный материал по почвенным грибам,
накопленный к настоящему времени. В нем содержатся общие сведения
о грибах как самостоятельном царстве живого мира, изложенные на основе
современных данных по цитологии, ультраструктуре и химическому строению
грибной клетки и мицелия. Излагаются особенности экологии почвенных
грибов, раскрывается их роль в почвообразовательных процессах. Большое
внимание уделяется роли грибов в гумусообразовании и токсикозе почв.
Учебное пособие не имеет аналогов как у нас в стране, так и за рубежом.
Рецензенты:
кафедра микробиологии ТСХА
(зав. кафедрой профессор В. Т. Емцев),
доктор биологических наук Л. В. Калакуцкий
м 3802020000 (4309000000)—026 157—88
077(02)—88 © Издательство Московского»
ISBN 5—211—00157—5 университета, 1988
ВВЕДЕНИЕ
Основной объект почвенной микологии представляют
грибы, обитающие в почве и принимающие активное участие в
процессах, протекающих в ней.
Почвенными можно считать грибы, которые постоянно и
закономерно выделяются как из почвы, так и из различных
субстратов, находящихся в ней (опавших листьев, подстилки, корней
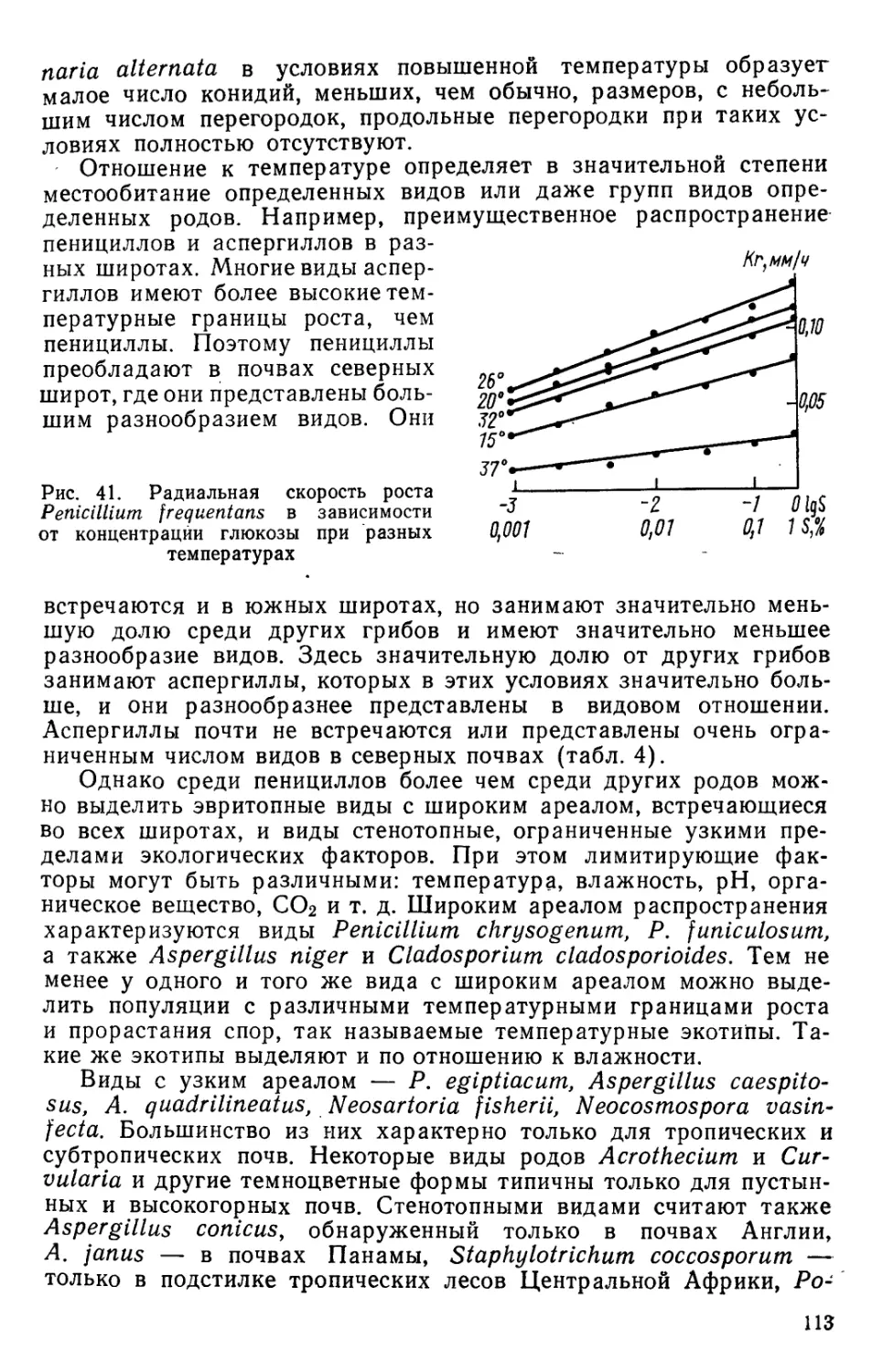
и семян растений, других растительных остатков и животных
субстратов).
Почвенные грибы не являются единой таксономической
группой и представлены самыми разнообразными в систематическом
отношении формами. Они не представляют собой также единой
экологической группы, так как включают разные эколого-трофи-
ческие группы: сапротрофы, патогены растений, микоризообразо-
ватели, грибы-хищники. Их объединяет только общее
местообитание в широком смысле — почва. Однако основную массу
почвенных грибов все же составляет группа сапротрофов, т. е.
организмов, осуществляющих разложение отмерших органических
субстратов, которые могут быть выделены как непосредственно
из почвы, так и из различных животных и растительных остатков.
Несмотря на большое таксономическое разнообразие почво-
обитающих сапротрофных грибов и их функций (в настоящее
время к почвенным грибам можно отнести до 3000 видов), они
представляют собой единое звено в трофической цепи любой
экосистемы, в состав которой входит почва, осуществляя в ней
функцию организмов-редуцентов. Их основная роль проявляется в
разложении органического вещества в разных его формах, в
результате чего происходит новообразование различных соединений,
поступающих в почву и вовлекаемых непосредственно в
дальнейший обмен, таких, как разнообразные органические кислоты,
физиологически активные вещества, высокомолекулярные
полимерные соединения, часть из которых свойственна метаболизму
только грибов.
Почвенная микология как наука о почвенных грибах
естественно возникла на основе собственно микологии, науки о грибных
организмах, их строении, развитии, эволюции, физиологии,
биологии. Тем не менее почвенные грибы и их роль в почве часто
рассматривают как раздел биологии почв об одном из
компонентов почвенной биоты.
Однако своеобразие объектов, их огромное самостоятельное
3
значение в процессах, протекающих в почве, свои проблемы if
задачи определяют обособление почвенной микологии в
самостоятельную науку.
Поскольку объектом исследования почвенной микологии, как
было уже сказано, являются грибы, то естественно, что
почвенная микология основывается на тех знаниях, которыми
располагает собственно микология.
Грибы имеют огромное значение в природе и жизни человека.
Они были известны еще в древности. Однако систематическое
изучение грибов было начато только в конце XVIII в. и связана
с именем голландского ученого X. X. Персона (1761—1836),
который попытался систематизировать известные к тому времени
виды грибов вслед за К. Линнеем и дать им бинарную
номенклатуру. До этого времени Линней, создавая свою систему
животного и растительного мира, системой грибов не занимался.
Одним из первых крупных микологов-систематиков был
шведский ученый Е. М. Фриз (1794—1878). Он положил основу
номенклатуры грибов и дал правильное описание многих видов.
Часть видов описана им совместно с Персоном. Таким
образом, Персона и Фриза заслуженно считают «коллективным»
Линнеем. Описания их настолько ясны и точны, что сохранились и
до настоящего времени.
Далее одним из основополагающих исследователей в
микологии, сделавшим эпоху, следует назвать немецкого миколога А. Де-
Бари (1831—1888). Он известен рядом фундаментальных работ
в микологии. В первую очередь это введение новых принципов»
новых методологических подходов. Ему принадлежит заслуга
разработки классификации грибов на основе эволюционных
принципов. Он первый ввел в микологию использование
онтогенетического метода исследования. Его работы ознаменовали таким
образом новое направление в микологии. Ему принадлежит также
большое число работ по изучению биологии, строения, цикла
развития многих грибов, в том числе паразитных. Выяснение
природы заболеваний растений и открытие их возбудителей, сделанные
Де-Бари, могут явиться образцом микологических исследований.
Таким образом, им были заложены основы фитопатологии.
Из крупных систематиков необходимо назвать Саккардо
(1845—1920), итальянского миколога, который систематизировал
и свел воедино все имеющиеся (описанные) к этому времени
виды грибов (около 80 000), что составило сводку из 25 томов, и
дал всем латинское описание.
Начало онтогенетическому направлению в русской микологии
было положено М. С. Ворониным (1838—1903), учеником
Де-Бари. Большой вклад в развитие отечественной микологии внес
А. А. Ячевский. Основные его труды посвящены систематике
грибов и фитопатологии.
Большую многолетнюю и разностороннюю деятельность в
области микологии вел Л. И. Курсанов (1877—1954), основатель
кафедры низших растений в Московском университете.
4
Крупным советским микологом был Н., А. Наумов (1888—
1959). Он известен и как миколог-систематик, исследователь видо-
и формообразования паразитизма у грибов, а также прикладных
вопросов и фитопатологии.
Почвенная микология начала развиваться естественно, после
того как в микологии был применен метод выделения грибов на
искусственные питательные среды.
До этого микроскопические грибы изучались только на
поверхности субстратов в виде налетов и плесеней, чаще всего на
растениях, как они были найдены в природе, и подвергались микро-
скопированию. Человечеству были известны либо шляпочные
грибы, обитающие в лесу и на лугах, либо плесени и налеты на
растениях или пищевых продуктах. Основным местообитанием
микроскопических грибов считали растения, но не почву.
Первые самые ранние работы по почвенным грибам носили
рекогносцировочный характер и имели целью выяснить, является
ли почва их естественной средой обитания или только местом
резервации грибных зародышей.
Первым изолировал грибы из почвы на питательной среде
И. Адамец (1886) в Германии. Им было изолировано всего 11
видов грибов, и 4 вида дрожжей из глинистой и песчаной почвы,
и он пришел к выводу, что две эти почвы не отличаются "по
составу грибов в них. Более точное исследование состава
почвенных грибов было проведено А. Одеманс и Ц. Конинг в Голландии
в 1902 г. Ими было выделено 45 видов грибов, дана их
классификация и полное описание, а в 1904 году Конинг придал методу
выделения грибов из почвы количественное выражение,
используя определенную навеску почвы и точное разведение этой
навески для посева на питательную среду. Далее Хагем (1907—
1910) выделил и описал значительное число мукоровых грибов
в почвах Норвегии, 8 из которых были новыми для науки. Им
были подмечены также некоторые закономерности
распространения мукоровых грибов в почвах. В частности, большее их
число и разнообразие в окультуренных, чем в лесных почвах, их
способность утилизировать богатые азотом органические
вещества.
Однако систематическое изучение почвенных грибов началось
с 1916—1917 гг., когда С. Ваксман, исследовав состав грибов
многих различных почв, показал наличие в них определенного
и постоянного набора видов. Причем набор видов одних почв
отличался от набора видов в других почвах наряду с некоторыми
общими видами для всех почв. На основании этого Ваксман
заключил, что почва является естественной средой обитания грибов
и в ней существует определенный, только ей присущий состав
видов грибов. С этого времени почвенная микология оформилась
как наука.
На этом этапе ею решались такие задачи, как изучение
распространения грибов в различных почвах, т. е. исследовался
основной набор видов грибов в различных типах почв, в первую
5
очередь в лесных почвах, изучалось отношение выделенных
грибов к некоторым факторам среды, в частности пределы
выносливости к рН среды, температуре, влажности и т. д.
С. Ваксман описал большое число грибов, встречающихся в
почве (около 200 видов), и показал определенные закономерности
в распространении грибов в зависимости от некоторых свойств
почв окультуренных и целинных, величины рН, содержания
органического вещества и т. д., а также привел данные о
способности грибов разлагать клетчатку и лигнин.
Он впервые отметил такие закономерности в распространении
грибов, как преобладание грибов из родов Penicillium и Мисог
в северных почвах и видов рода Aspergillus в южных, что
неоднократно подтверждалось затем рядом авторов.
После того как положение о том, что почва является
естественной средой обитания для грибов, было доказано, было
выполнено большое число работ многими исследователями по
инвентаризации состава видов грибов в разных почвах.
В этих работах рассматривалась также роль грибов в
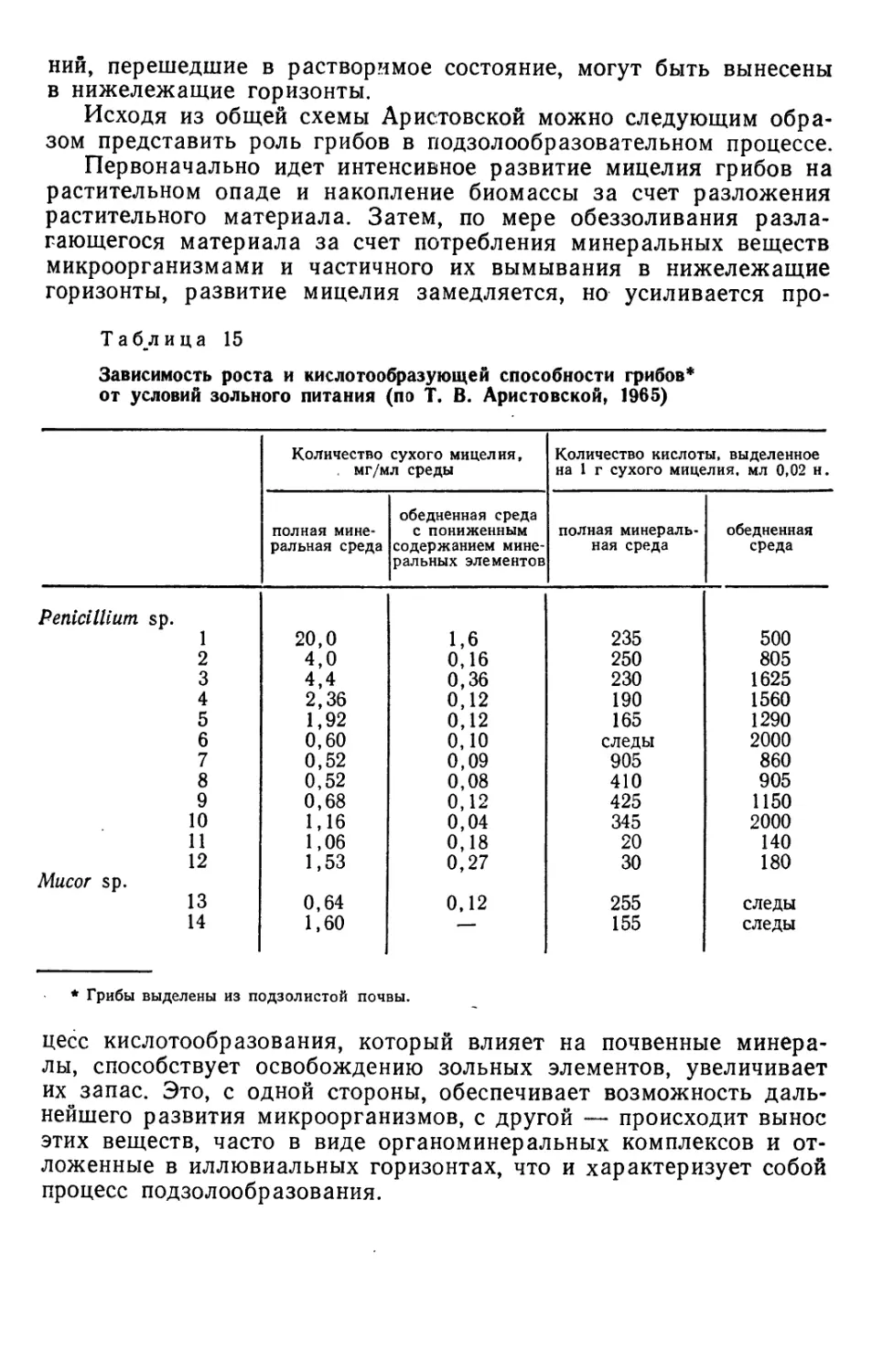
круговороте азота, в частности участие грибов в процессах
аммонификации, где особенно активны мукоровые грибы. В них освещалась
также роль грибов в процессах разложения клетчатки и лигнина.
Что касается русских работ по почвенной микологии, то в
качестве первой из них, выполненной в таком плане, можно назвать
работу М. М. Самуцевич (1927), которая представляет собой
первую попытку охарактеризовать видовой состав грибов целинных
и окультуренных почв в подзолистой зоне Советского Союза по
различным глубинам. Ею выделено 58 видов грибов,
характерных для почв, установлено уменьшение их числа с глубиной.
Однако разницы в видах грибов в исследованных ею почвах она не
установила.
Еще одной работой по почвенным грибам, опубликованной
годом позже, является работа А. И. Райлло (1928), в которой
почвенные грибы изучались в широком географическом аспекте.
Пункты взятия образцов для исследования различались по
географическому положению и по типам почв. Райлло сравнивала
состав грибов в почвах Кольского полуострова, Ленинградской
области и Каменной степи. В результате для каждого типа почв
был выявлен свой набор видов, за исключением четырех общих
для всех исследованных почв форм Verticillium glaucum, Mucor
racemosus, Monosporium sp., Stysanus stemonites. Было
подтверждено также наблюдение Ваксмана о преобладании грибов из
родов Penicillium и Мисог в северных почвах и Aspergillus в
южных.
Продолжением этих исследований является работа А. А.
Рихтера и А. Вернера (1931) по составу и количеству грибов в
почвах нижнего Поволжья, где приводятся материалы по количеству
грибных зачатков и основным видам грибов в черноземах,
каштановых и светло-каштановых почвах. Далее в обстоятельной
работе Л. И. Курсанова и Т. Н. Шкляр (1938) содержатся резуль-
6
таты изучения состава грибов дерново-подзолистых почв
Подмосковья и красноземов ь районе Батуми в связи с их активностью
в разложении целлюлозы. Показано, что почвы Подмосковья и
батумские почвы имеют ряд общих видов, но по большинству
видов различаются между собой. Общие для двух типов почв
виды, такие, как Trichoderma lignorum и некоторые виды Peni-
cillium, обладают, однако, различной активностью к разложению
клетчатки. Грибы из Батумских красноземов значительно более
активны в отношении разложения целлюлозы.
Следует сказать, что все эти работы, выполненные как у нас
в стране, так и за рубежом, можно рассматривать как первую
попытку охарактеризовать состав грибов почвы. Они ставили
своей целью, во-первых, подтвердить существование постоянного
состава грибов и выявить наиболее характерные виды грибов для
тех или иных почв. Было показано, что почва не только место
резервации грибных зародышей, которые попадают туда
случайно и где споры их сохраняются до времени прорастания после
попадания на благоприятный субстрат, но естественная их среда
обитания, где они осуществляют активную деятельность.
Во-вторых, насколько характерен и специфичен состав грибов разных
типов почв, каковы различия в составе грибов в различных
почвенных условиях и растительных ассоциациях.
К сожалению, работы на этих первых этапах проводились в
отрыве от систематических почвенных исследований, поэтому они
не содержали достаточной характеристики почвенных условий и
анализа экологических факторов. Таким образом, экологического
подхода в современном понимании в них не было осуществлено.
Тем не менее эти работы имели определенное положительное
значение, так как дали возможность судить о существовании в
почве постоянного, только ей присущего состава грибов и
составить некоторое представление о наборе видов грибов,
встречающихся в почве, а также о различии в составе видов в разных
почвах.
Следующая, более поздняя серия работ характеризуется
изучением грибов в зависимости от конкретных экологических
условий. Первой из них следует упомянуть работу И. X. Варкупа
(1951), который впервые сделал попытку связать изменение
состава грибов с направленностью почвообразовательного процесса,
в частности выяснить изменение состава грибов при развитии
подзолообразовательного процесса от карбонатных к подзолистым
почвам. Им изучался видовой состав грибов в почвах различной
стадии оподзоливания. Были взяты почвы, сформированные на
меловой гальке и интергляциальном песке с различной глубиной
залегания известняков, следовательно, с различной степенью кар-
бонатности и величиной рН. При расположении известняков
близко к поверхности они имели высокое содержание карбоната
кальция и высокое значение рН. По мере углубления залегания
известняков снижалась насыщенность поглощающего комплекса
основаниями и уменьшалось значение рН. При отсутствии изве-
7
стняков формировался типичный подзол. И. X. Варкуп установил,
что почвы, имеющие крайние значения рН, бедны грибами.
Наибольшее количество грибов было найдено в кислых
дерново-подзолистых почвах, имеющих высокую кислотность на поверхности
и щелочность глубоких слоев. Также богаты грибами почвы с рН,
близким к нейтральному. Было показано, что в щелочных почвах
содержится значительное число грибов, при этом в них
выделяется наибольшее разнообразие видов. Ряд видов был выделен
только из щелочных почв, среди них Fusarium sambucinum, Pe-
nicillium luteum, Thielavia sp. При развитии подзолообразователь-
ного процесса появляются виды, характерные только для этих
почв, такие, как Penicillium janthinellum, P. frequentans, P. ter-
likowskii, Mortierella rumanniana.
Таким образом, с изменением почвообразовательного процесса
изменяется и набор видов грибов, характеризующий процесс.
Такой же подход, как и у И. X. Варкупа, осуществлен в
работе Р. Торнтона (1956). Он проследил изменение состава
почвенных грибов в связи с оподзоливанием бурой почвы в
результате постепенной смены растительности. Бурые почвы под дубом
под влиянием вереска подвергались оподзоливанию, изменялись
опад, кислотность почвы, сумма обменных оснований и другие
признаки, вследствие чего изменился состав почвенных грибов.
Далее эти работы развили Кендрик, Паркинсон, Бургес, Саксен
и многие другие.
Экологию почвенных грибов у нас в стране изучали 3. Э. Бек-
кер, Т. П. Сизова, А. Г. Романкова, Т. П. Супрун, Е. С. Лисина-
Кулик и т. д. По систематике грибов известны работы М. А.
Литвинова, Т. П. Сизовой, 3. Э. Беккер и Т. П. Супрун, где особое
внимание уделяется растительности как экологическому фактору,
формирующему состав грибов, рассматриваются различные типы
растительных ассоциаций и характерный для той или иной
растительной ассоциации состав почвенных грибов. Вскрываются
взаимоотношения почвенных грибов с ризосферой растений.
Вышеперечисленные работы, начиная с работы Варкупа, в
большей или меньшей степени характеризуют экологическое
направление в почвенной микологии.
Современный этап экологических исследований в почвенной
микологии характеризуется изучением почвенных грибов с
позиций системного подхода. С этих позиций почвенные грибы
рассматриваются как одно из звеньев гетеротрофного блока такой
сложной системы, какую представляет собой биогеоценоз, и в
связи с этим оценивается их роль в превращении вещества и
энергии. Такой подход делает необходимым для учета общей
продуктивности БГЦ количественное определение содержания грибной
биомассы в почве, а также строгое и достоверное измерение
скорости новообразования, отмирания и реутилизации грибной
биомассы в основных зональных типах почв, что является одной из
важных задач почвенной микологии.
При изучении грибов как компонента БГЦ ставятся задачи
8
вскрыть трофические и иные связи грибов с другими
составляющими БГЦ, их отношение к разным органическим субстратам,
причины и направление грибных сукцессии, а также определение
скорости разложения и усвоения субстрата.
Большое значение в почвенной микологии приобретает
изучение грибов как показателя почвенных условий. Эти исследования
также характеризуются в настоящий период использованием
количественных критериев для выделения комплекса типичных
видов для каждой почвы и сравнения по этому комплексу как
естественных почв, так и подвергшихся антропогенному воздействию.
В содержание почвенной микологии входит также изучение
формы существования популяции грибов в почве (мицелий,
споры, соотношения мицелия и спор в тех или иных почвах, т. е.
биоморфологическая структура грибов), циклы развития,
представленность сапротрофных стадий, патогенных грибов в почве,
формы переживания ими неблагоприятных условий.
Изучаются морфологические, физиолого-биохимические
свойства индивидов и популяций в зависимости от различных факторов
среды, определяющие возможность их существования в той или
иной среде.
К вопросам почвенной микологии относится изучение
взаимоотношений грибов с другими организмами, обитающими в почве,
растениями, животными, микроорганизмами.
Существенное место в почвенной микологии занимают работы
по роли грибов в круговороте веществ, в частности по
превращению грибами органического вещества, поступающего в почву, в
первую очередь растительного опада как в целом, так и
отдельных его компонентов, главным образом клетчатки, лигнина,
пектиновых веществ.
В последнее время изучаются ферментные комплексы, с
помощью которых осуществляется этот процесс на разных стадиях
разложения. Делаются попытки раскрыть пути биохимического
превращения лигнина, последовательность стадий его
превращения.
Большой вопрос почвенной микологии, представляющий собой
целую проблему, составляет изучение образования грибами
специфических веществ гумуса в связи со способностью грибов
синтезировать в процессе обмена внутриклеточно сложные
полимерные высокомолекулярные соединения, имеющие темную окраску
и азот гетероциклов в своем составе, по основным показателям
сходные с гуминовыми кислотами почв.
Специальный и совершенно новый вопрос почвенной
микологии представляет собой изучение биологически активных веществ
почвенных грибов: антибиотиков, токсинов, стимуляторов роста.
Условия, способствующие или подавляющие их образование
в почве, влияние на свойства почвы и жизнедеятельность
почвенных организмов. Возможность регулирования численности
продуцентов и их активности в почве.
Часть I
ОБЩАЯ ХАРАКТЕРИСТИКА ГРИБОВ
Глава 1
ОСНОВНЫЕ ЧЕРТЫ СТРОЕНИЯ ГРИБОВ
Грибы — большая группа хемоорганотрофных
организмов, включающая около 100 000 видов. Это одноклеточные или
многоклеточные организмы, микроскопические и
макроскопические, не имеющие дифференциации на органы и ткани. По образу
жизни они могут быть сапротрофами, патогенами растений и
животных и симбионтами.
Все грибы, за немногим исключением, имеют мицелиальное
строение, т. е. их вегетативное тело (таллом) состоит из мицелия
или грибницы, которая представляет собой систему ветвящихся
нитей (гиф), пронизывающих субстрат и имеющих с ним большую
поверхность соприкосновения, через которую осуществляется
адсорбция питательных веществ.
Грибы — неподвижные организмы. Подвижными бавают их
отдельные элементы на определенных стадиях развития.
Все грибы лишены хлорофилла и не являются хемоавтотро-
фами, поэтому необходимое условие их существования — наличие
органического углерода.
Грибы относятся к аукариотным организмам и обладают
всеми чертами, присущими этому типу клеток: они имеют
двуслойную ядерную мембрану и сильно развитую систему внутренних
цитоплазматических мембран, эндоплазматический ретикулум,
аппарат Гольджи, лизосомы, михотондрии, наличие белков-гистонов
в хромосомах.
ОСНОВНЫЕ СТРУКТУРЫ ГРИБНОЙ КЛЕТКИ
Мицелий грибов либо одноклеточный (ценоцитный) и
тогда представляет собой единую громадную клетку, либо
разделен на клетки.
Клеточная стенка. Клетка грибного мицелия, за немногими
исключениями, всегда одета ригидной оболочкой, что определяет
ее устойчивую форму благодаря наличию клеточной стенки.
Клеточная стенка представляет собой плотную упругую полимерную
структуру, которая несет опорно-механическую функцию и
защищает клетку от воздействия внешних факторов. Клеточная стенка
10
обладает избирательной проницаемостью для веществ различной
химической природы. Возможно, что в ней происходят
синтетические процессы. Внутрь от клеточной стенки располагается
пристенная цитоплазматическая мембрана — плазмолемма,
окружающая внутреннюю часть клетки, — сферопласт.
Цитоплазматическая мембрана трехслойна и состоит из липо-
протеидов. Поверхность ее может быть ровной или фестончатой.
На цитоплазматической мембране располагаются ферменты,
такие, как инвертаза, кислые фосфатазы, аденозинтрифосфатазы, а
также окислительно-восстановительные ферменты и транслоказы,
контролирующие перемещение различных ионов и молекул в
клетку.
Клеточная стенка у грибов состоит на 80—90% из
полисахаридов, азотсодержащих, каковым является хитин, и безазотистых.
Кроме полисахаридов в клеточной стенке грибов содержатся
белки и липиды, а также имеются полифосфаты и неорганические
ионы. Есть данные о наличии в клеточной стенке грибов
нуклеиновых кислот. У большинства грибов основной полисахарид —
хитин и только у грибов класса оомицетов — преимущественно
целлюлоза. У очень немногих грибов, а именно у представителей
небольшого класса гифохитридиомицетов, хитин и целлюлоза
присутствуют вместе так же, как у некоторых оомицетов наряду с
целлюлозой обнаружен хитин. В качестве дополнительных
безазотистых полисахаридов у грибов представлены различные глю-
каны, полимеры D-глюкозы, отличающиеся от целлюлозы
характером связи между мономерами. В определенных слоях клеточной
стенки глюканы объединены с протеинами. Для внутреннего слоя
клеточной стенки многих грибов характерен хитин, образующий
комплекс с глюканами. Состав полисахаридов клеточной стенки
различен для представителей крупных таксономических групп
грибов.
Для грибов класса оомицетов характерен микрофибриллярный
комплекс целлюлоза—глюкан, для зигомицетов — хитозан с по-
лиуронидами. У хитридиомицетов, аскомицетов и дейтеромицетов
в клеточных стенках содержатся хитин и глюканы. У базидио-
мицетов в качестве мономеров полисахаридов комплекса с
хитином выступают ксилоза, фруктоза и глюкуроновая кислота.
Иногда присутствуют небольшие количества рамнозы, рибозы, ара-
бинозы. У грибов с дрожжеподобным ростом вне зависимости от
их систематического положения количество хитина в клеточных
стенках снижено, но содержатся маннаны — полимеры маннозы.
Хитин и целлюлоза придают клеточной стенке фибриллярное
строение, микрофибриллы этих полисахаридов сцементированы
аморфными веществами, в качестве которых выступают белки и
безазотистые полисахариды. Хитин может быть также связан с
каротиноидами, меланинами.
Таким образом, клеточная стенка грибов представляет собой
систему микрофибрилл, встроенных в аморфный матрикс, и
состоит из четырех слоев: 1) наружный глюкановый слой; 2) сле-
1Г
дующий — гликопротеидный слой; 3) слой, состоящий из белка;
4) внутренний слой — хитин, составляющий арматуру клеточной
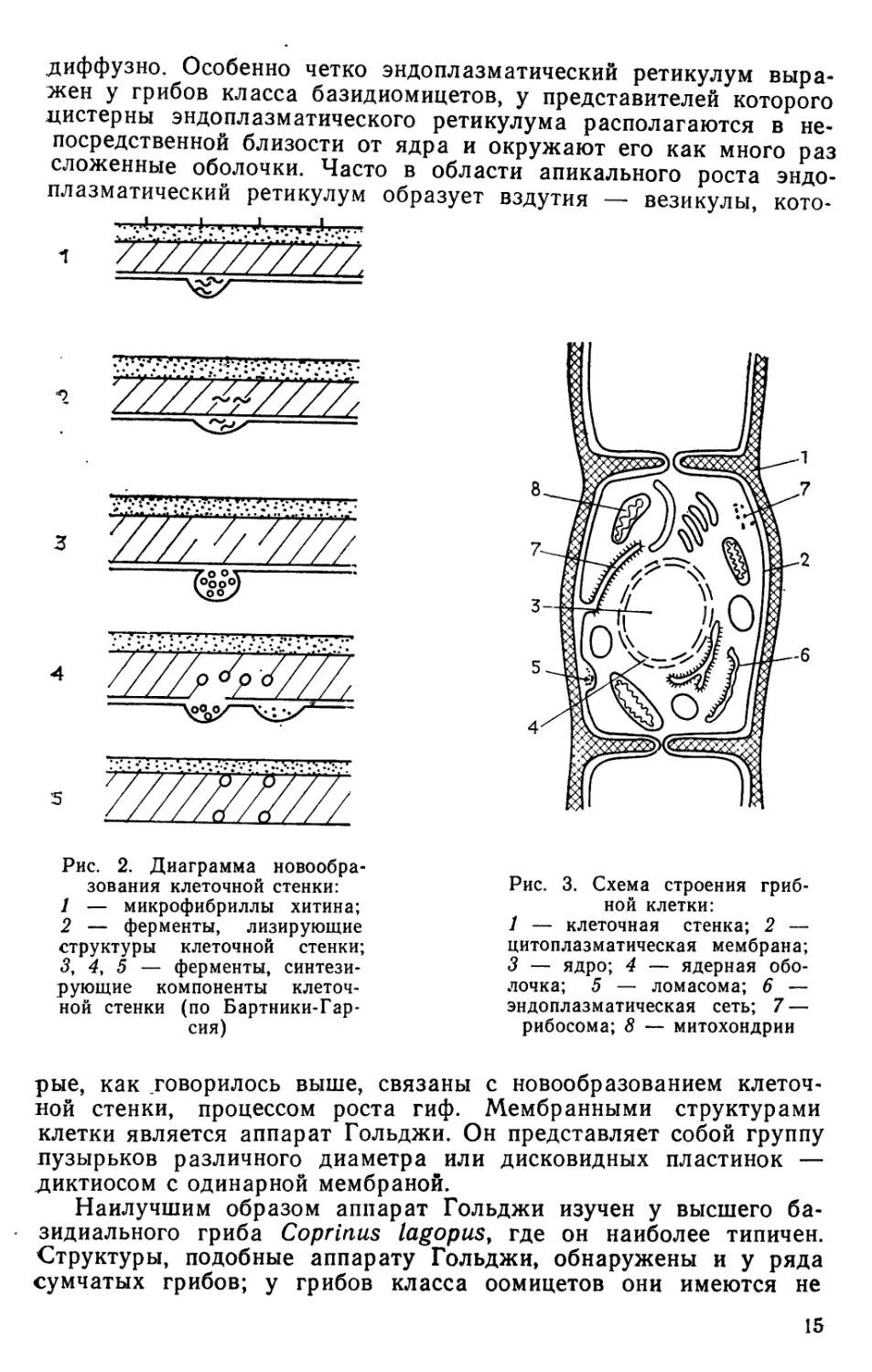
стенки (рис. 1).
Наличие фибриллярного строения установлено методом
дифракции рентгеновских лучей и с помощью электронной
микроскопии. Диаметр микрофибрилл составляет 250 А. Чаще всего
микрофибриллы располагаются в два слоя, идущих в разных
направлениях, что придает клеточной стенке пла-
4 ?^) стинчатый характер. Внутренний слой ориенти-
i^ji рован параллельно главной оси гифы,
з || ^ протеиновой фракции клеточной стенки гри-
0^-—^Ь бов преобладают дикарбоновые кислоты, обуслов-
КС^^Э Рис. 1. Диаграмма расположения основных слоев клеточной
F;:,' '' •';• j стенки Neurospora crassa:
1 ry.. ' \:.;| / — глюканы; 2 — гликопротеидный комплекс; 3 — белок;
?:.". -# •'. .';*] 4 — микрофибриллы хитина, погруженные в слой белка
4*<^Lj->^ (по Хансли и Барнет)
ливающие их кислую реакцию. Характерная для грибов,
аминокислота а-аминоадипиновая. Диаминопимелиновая кислота,
характерная для клеточной стенки бактерий, имеет место только у
оомицетов; у всех других групп грибов она отсутствует.
Постоянные компоненты клеточной стенки грибов — липиды.
В отличие от липидов цитоплазмы в клеточной стенке они
представлены в основном насыщенными жирными кислотами. В состав
клеточной стенки входит также РНК в количестве 1% от сухого
веса стенки и некоторые ферменты, при участии которых идет ее
наращивание в длину. В наружных частях клеточной стенки
нередко откладываются пигменты — меланины, представляющие
собой полимерные соединения с большим молекулярным весом.
Меланины составляют 2—3% от сухого веса клетки. Меланин
распределяется по клеточной стенке, в основном локализируется
во внешних ее частях в виде гранул. Частично он ассоциируется
с ее хитиновой фракцией во внутреннем слое, что придает
большую устойчивость клеточной стенке к микробным ферментам.
Содержание меланина составляет от 16,3 до 18,2% от сухого веса
клеточной стенки.
Состав клеточных стенок спор значительно отличается от
состава их в вегетативном мицелии. Отличия эти проявляются, в
частности, в составе липидов. Так, поверхностные липиды спор
Rhizopus stolonifer представлены преимущественно C2i—Сгз-алка-
нами, в то время как в клеточной стенке мицелия присутствуют
Cis—С32-алканы.
Наличие на поверхности спор липидов необходимо для
сохранения жизнеспособности этих структур, так как споры
приобретают гидрофобные свойства. Кроме того, липидный слой на
поверхности споры препятствует ее быстрому высушиванию.
В поверхностном слое спор некоторых грибов наряду с липи-
12
дами присутствуют соединения необычной химической природы —
споридесмолиды (депсипептиды). У гриба Pitomyces chartarum
эти соединения образуют вместе с липидами игловидные
образования — спикулы. Установлено, что они имеют определенное
функциональное значение. Образование споридесмолидов тесно
связано с синтезом антибиотика валиномицина. По-видимому,
расположенные на поверхности споры спор идесмол иды ингибиру-
ют рост сопутствующих бактерий и создают более благоприятные
условия для прорастания спор.
У некоторых мукоровых грибов на поверхности спорангия
имеются игловидные выросты, которые состоят из дегидрата ок-
салата кальция, как, например, у Mucor plumbeus.
Клеточные стенки спор грибов отличаются от клеточных
стенок вегетативного мицелия и по составу полисахаридов. В
клеточных стенках спор Mucor rouxii характерно наличие глюкозы
(около 42%), в то время как в клеточных стенках гиф она
отсутствует. В гифах грибов в зависимости от стадии развития
содержится различное количество уроновых кислот. В спорах ее
обнаружить не удается. Клеточные стенки спор некоторых мукоровых
грибов содержат больше белка, хитина, хитозана и глюкозы, чем
клеточные стенки мицелия.
Наблюдаются также значительные различия в составе
клеточных стенок, зигот мукоровых грибов и спор. В зиготах
значительно больше глюкозамина, но меньше Сахаров и уроновых
кислот. Основные моносахариды клеточных стенок зигот — фукоза
и ксилоза; в спорах они полностью отсутствуют. В зиготах
большую долю, чем в спорах, составляет липидная фракция.
Клеточная стенка зигот значительно толще, чем в спорах, и содержит
до 10 различных слоев.
Верхний слой зигот состоит из меланина, хитина и хитозана,
а внутренний — из одного хитина. Кроме того, в зиготах и спор-
ангиоспорах некоторых мукоровых грибов найден спорополле-
нин — продукт окислительной полимеризации каротина. Наличие
в зиготе этих соединений обеспечивает ей хорошую сохранность
и устойчивость к ряду ферментов микроорганизмов и другим
внешним воздействиям. На поверхности одиночных клеток
некоторых грибов (споридиях головневых грибов и клетках дрожжей)
обнаружены тонкие нитевидные структуры — фимбрии,
аналогичные фимбриям бактерий. Они имеют длину 0,5—10 мкм и
толщину 6—7 нм. Предполагают, что фимбрии играют существенную
роль в агрегации клеток.
Пути новообразования клеточной стенки. Новообразование
клеточной стенки мицелия происходит следующим образом: в
апикальной области гифы находятся в большом количестве везикулы
сферической или неправильной формы, содержащие литические
ферменты, ферменты синтетазы и предшественники синтеза
биополимеров. Эти везикулы постоянно образуются аппаратом Гольд-
жи, являясь его производными. Везикулы направляются к
первичному району роста клеточной стенки и выбрасывают свое содер-
13
жимое в периплазматическое пространство. Литические ферменты
(хитиназы, глюконазы, эндогидролазы) разрывают молекулу
полимера, затем происходит некоторое расхождение разорванных
частей молекул и последующая их достройка при участии
ферментов синтетаз (хитинсинтетазы, глюкансинтетазы), когда к
разорванным концам присоединяются новые молекулы глюкозамина.
Вышеназванные ферменты также освобождаются из апикальных
везикул (рис. 2). Хитинсинтетазы ассоциированы со
специальными сферическими структурами — хитосомами. Фермент находится
в них в неактивной форме, в то время как активная хитинсинте-
таза располагается в плазмолемме, где и происходит синтез
хитина. Таким образом, рост клеточной стенки осуществляется путем
разрыва биополимеров в результате действия автолизинов (ли-
тических ферментов) и встраивания вновь синтезированного
материала. Наличие строгого баланса между лизисом и синтезом
лежит в основе образования клеточной стенки.
Мембранные структуры. Внутреннее содержимое клетки
составляет цитоплазма, которая представляет собой коллоидную
фазу, имеющую различную степень вязкости. В ней содержатся
структурные и несвязанные с органеллами ферментные белки,
аминокислоты, рибонуклеиновые кислоты, углеводы, липиды и
различные низкомолекулярные вещества, а также гранулы
запасных веществ различной природы. Через цитоплазму
осуществляется непосредственная связь между органеллами клетки.
В цитоплазме располагаются мембранные структуры. В клетке
грибов имеется пристенная мембрана и внутренняя система
мембран. Цитоплазматическая пристенная мембрана (плазмолемма)
составляет единое целое с внутренней системой мембран. Роль
мембранной системы многообразна. Она увеличивает поверхность
клетки, при участии мембран происходит транспорт веществ и
многие метаболические процессы. Мембранная система делит
клетку на отдельные участки жидкой фазы, которые
представляют собой цитоплазматический матрикс. Цитоплазматический мат-
рикс — это среда для клеточных органелл — ядра, митохондрий,
рибосом, лизосом. В цитоплазматическом матриксе находятся
цитоплазматические нити, представляющие собой палочковидные
структуры. Иногда эти нити ориентированы в микротрубочки,
представляющие собой детали механизма движения. Они всегда
присутствуют в двигательных органеллах, в частности в веретене
и кинетосомах, с помощью которых осуществляется расхождение
ядер в клетке.
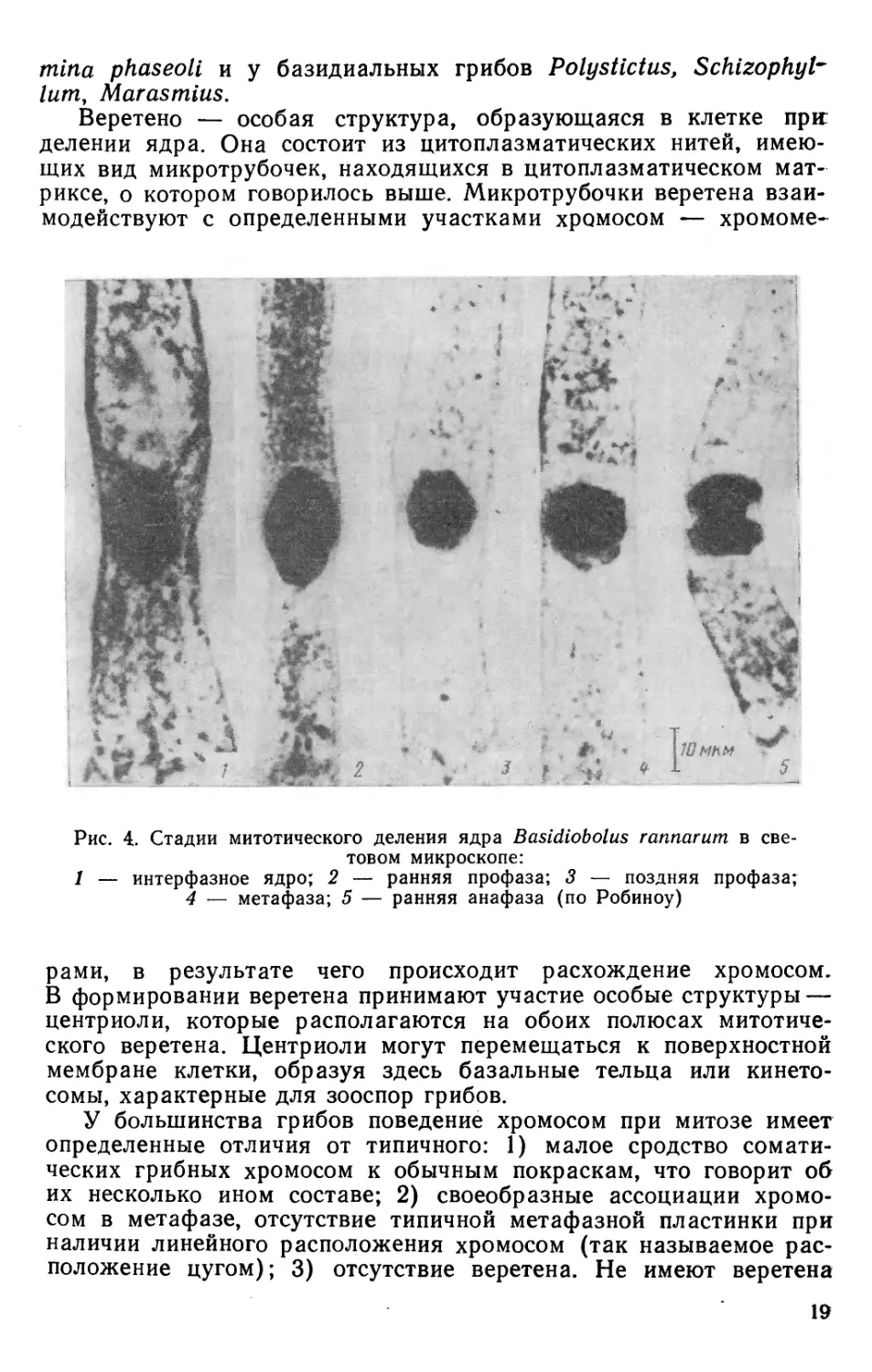
Клетке грибов присущи многие структуры мембранной
системы, характерные для клетки эукариотных организмов (рис. 3).
Грибная клетка имеет сильно развитую систему внутренних
мембран — эндоплазматическую сеть (эндоплазматический ретику-
лум), которая формируется из пристенной мембраны. Она
представляет собой систему канальцев, пузырьков или цистерн.
С внутренней стороны они усажены гранулами-рибосомами, хотя
рибосомы также содержатся в большом количестве в цитоплазме
14
диффузно. Особенно четко эндоплазматический ретикулум
выражен у грибов класса базидиомицетов, у представителей которого
дистерны эндоплазматического ретикулума располагаются в
непосредственной близости от ядра и окружают его как много раз
сложенные оболочки. Часто в области апикального роста
эндоплазматический ретикулум образует вздутия — везикулы, кото-
'*• 9,А> V«':" \9'9' !'¦ '.?*» *!»'.«ч * «v..*1.
~^У
'f'." VJ ^ 'V ¦?>'1 • *&'•'!•'«•| ¦ 'г •."»"Л1 •'; .
.'..у'.'.'.-.".
^
Рис. 2. Диаграмма
новообразования клеточной стенки:
/ — микрофибриллы хитина;
2 — ферменты, лизирующие
структуры клеточной стенки;
3, 4, 5 — ферменты,
синтезирующие компоненты
клеточной стенки (по Бартники-Гар-
сия)
Рис. 3. Схема строения
грибной клетки:
1 — клеточная стенка; 2 —
цитоплазматическая мембрана;
3 — ядро; 4 — ядерная
оболочка; 5 — ломасома; 6 —
эндоплазматическая сеть; 7 —
рибосома; 8 — митохондрии
рые как говорилось выше, связаны с новообразованием
клеточной'стенки, процессом роста гиф. Мембранными структурами
клетки является аппарат Гольджи. Он представляет собой группу
пузырьков различного диаметра ^или дисковидных пластинок —
диктиосом с одинарной мембраной.
Наилучшим образом аппарат Гольджи изучен у высшего ба-
зидиального гриба Coprinus lagopus, где он наиболее типичен.
Структуры, подобные аппарату Гольджи, обнаружены и у ряда
сумчатых грибов; у грибов класса оомицетов они имеются не
15
всегда, но описаны у Pythium debaryanum, Phytophthora spp.y
Peronospora manshurica и некоторых других. В целом у грибов
аппарат Гольджи не так четко выражен, как у других
организмов.
Мембраны, аппарата Гольджи морфологически связаны с
ядерной мембраной и мембранами эндоплазматического ретикулума.
Роль его полифункциональна. Пузырьки аппарата Гольджи — эта
транспортная структура. Известно, что на отдельных его
элементах происходит синтез новых мембран.
Производными аппарата Гольджи являются лизосомы,
представляющие собой разнообразные по форме тельца. Они отшну-
ровываются от аппарата Гольджи в виде пузырьков и
распределяются по клетке. Йизосомы выполняют различные функции.
В них'Содержатся протеолитические ферменты, осуществляющие
расщепление белков. Кроме того, лизосомы осуществляют защиту
клеток от повреждений, в них отлагаются и расщепляются
подлежащие удалению шлаки.
Специфическими элементами грибной клетки являются,
своеобразные мембранные структуры — ломасомы. Они обнаружены
у грибов всех классов, а. также у красных водорослей. Ломасомы
представляют собой образования в виде пузырьков, возникающие
между клеточной стенкой и плазматической мембраной в
результате отслоения мембраны от стенки, и могут быть обнаружены
в любой части стенки. В ломасомы поступают ферменты лизиса
и синтеза клеточной стенки из апикальных везикул, и через них
же идет ее новообразование.
В цитоплазматическом матриксе грибной клетки имеются и
другие мембранные органеллы, характерные для клетки эукари-
отного организма. В нем находятся типичные митохондрии,
которые представляют собой структуры с двойной мембраной.
Наружная поверхность митохондрий гладкая, внутренняя — образует
впячивания, называемые кристами, которые увеличивают
активную поверхность митохондрий.
На митохондриях располагаются многие ферменты, в
частности окислительного фосфорилирования, цепи переноса электронов
и др. Ферменты дыхательной цепи составляют 25% белка
митохондрий и представляют собой не только функциональные, но и
структурные элементы митохондрий. На митохондриях
происходит биосинтез самых разнообразных веществ (нуклеиновых
кислот, белков, углеводов, липидов).
Митохондрии содержат собственную ДНК, локализованную в
определенных зонах, которые можно назвать митохондриальными
«нуклеоидами». Поэтому митохондрии являются
саморепродуцирующимися структурами, а не возникают в клетке de novo. В
основном митохондрии грибов сходны с таковыми у высших
организмов. Однако у грибов они меньших размеров и представлены
иногда сферическими телами, менее регулярными, чем в клетках
высших растений. Они обнаружены у всех изученных грибов.
В грибной клетке содержатся вакуоли, хорошо различимые
16
структуры, которые часто локализуются около клеточной стенки.
Вакуоли отсутствуют или их мало в апикальной части гифы, но
появляются в более старой части и число их увеличивается с
возрастом.
Вакуоли имеют двоякое происхождение. Это или производные
эндоплазматического ретикулума, или аппарата Гольджи. Эти
структуры различаются и в функциональном отношении.
Вакуоли, производные эндоплазматического ретикулума, содержат
запасные питательные вещества клетки. Вакуоли, происшедшие от
аппарата Гольджи, представляют собой структуры,
аккумулирующие чужеродные токсические веществу. Вакуоли связаны с
разделительными мембранами и могут передаваться через поры в
септах. Запасные питательные веществд, которые содержат
вакуоли, в основном волютин, липиды и гликоген (см, рис. 3).
, . Болютин, или метахроматин,, скапливается в вакуолях в виде
коллоидного раствора или в виде гранул./Гранула волютина
представляет собой сложный комплекс рибонуклеиновой кислоты,
белка и липидов; последние покрывают ее с периферии. В центре
гранулы располагаются кислоторастворимые полифосфаты,
связанные с РНК. Гранулы волютина могут быть локализованы
непосредственно в цитоплазме.
Жиры как запасные питательные вещества грибной клетки
представлены в основном ненасыщенными жирными кислотами,,
которые могут находиться как в вакуолях, так и в цитоплазме
мицелия в виде капель, а также в соединении с фосфатами и
белками, образуя фосфолипидные или фосфопротеидные гранулы.
Гликоген также может содержаться как в вакуолях, так и
непосредственно в цитоплазме. Кроме запасных веществ вакуоли
могут содержать пигменты, различные кристаллы и аморфные
вещества, природа которых не установлена. Непосредственно в
цитоплазме содержатся глобулы липидов, включающие пигменты,
такие, как каротиноиды и др., гранулы волютина, метафосфатов,
гликогена. Грибная клетка содержит кристаллы Са и другие
кристаллы, а иногда бывает ими инкрустирована с поверхности.
Ядро. Основная клеточная структура любого эукариотного
организма — ядро. Оно осуществляет генетическую, формативную
и метаболическую функции, контролируя дифференцировку
клетки, синтез белка, липопротеидов и других соединений, обменные
процессы, а также размножение, синтез клеточных структур de
novo.
Обычно, за немногими исключениями, в одной грибной клетке
содержится не одно, а несколько ядер. Ядро у грибов имеет
четкие границы, определяемые двойной мембраной. Оно содержит
ядрышко и хромосомы. Покоящиеся ядра грибов окружены
мембраной обычного вида, которая достаточно отчетливо видна на
электронных снимках (см. рис. 4). Ядерная мембрана имеет поры,
осуществляющие перенос макромолекул из ядра в цитоплазму,
так называемые нуклеопоры. После фиксации хроматин
покоящихся ядер обычно нежно или гру0о гранулирован и имеет нит-
17
-чатое строение. Большую часть времени он не организован в
хромосомы и образует так называемую хроматиновую сеть.
Покоящееся интактное ядро обычно содержит одно относительно
большое ядрышко. У многих грибов ядрышки играют определенную
роль при митозе. Участие ядрышка в митозе известно для грибов
классов оомицетов и зигомицетов родов Saprolegnia, Mucor, Phy-
-comyces, Basidiobolus и др, У грибов этих родов ядрышко
удлиняется при делении и разделяется между дочерними ядрами.
К моменту деления ядра хроматин организуется в хромосомы.
Происходит уплотнение и спирализация хроматиновых нитей.
Хромосомы у грибов, как у всех эукариотных организмов, состоят
из ДНК и гистонов. Число гаплоидных хромосом у грибов
зависит от вида организма, но никогда не бывает меньше двух.
Обычно оно варьирует от 3 до 28 и чаще бывает равно 8. Размеры
ядер грибов находятся в пределах 1—3 мкм, но в отдельных
случаях достигают 20—25 мкм. В вегетативных гифах они меньше,
в репродуктивных органах они несколько больше. В вегетативном
мицелии по мере роста и деления клеток преимущественно
осуществляется митотическое деление ядра, при репродукции
(образовании гамет или прорастании зиготы) — мейоз.
ДЕЛЕНИЕ ЯДРА
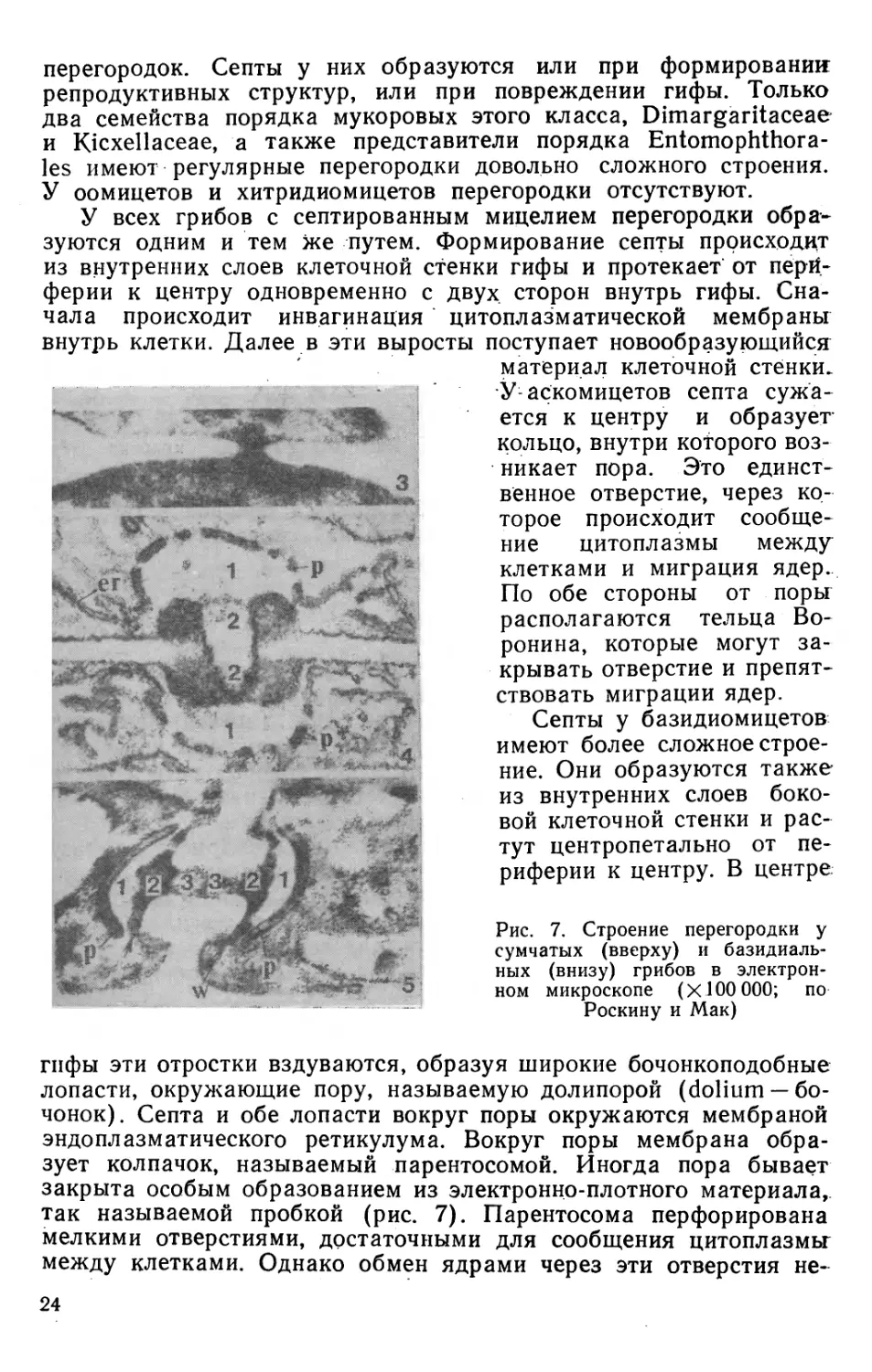
Митотическое деление ядра. У грибов имеются
различные способы митоза, несходные с хорошо известными. Этим грибы
отличаются от высших эукариотических организмов. Наряду с
этим некоторые виды грибов имеют все стадии, характерные для
этого способа деления. В частности, классическим примером
такого деления могут служить грибы рода Basidiobolus. Этот род
относится к классу Zygomycetes, порядку Entomophthorales,
семейству Basidiobolaceae; представители этого рода имеют как
исключение для этого класса грибов постоянные септы и строго
одноядерные клетки. Ядра очень крупные, достигают 25 мкм.
Деление ядра в клетке эукариотных организмов представляет
собой единый непрерывный процесс, но условно принято
разделять его на главные четыре стадии. Все стадии деления ядра
хорошо описаны цитологически, особенно у высших организмов.
Эти стадии, которые наиболее хорошо наблюдаются у
Basidiobolus ranarum, следующие: 1) профаза — обособление хроматино-
вой нити в хромосомы, уплотнение и скручивание нитей; 2) ме-
тафаза — концентрация хромосом в центре ядра с образованием
метафазной пластинки; 3) анафаза — на этой стадии происходит
разделение двойной спирали ДНК хромосом с последующей
саморепродукцией и расхождением таких хромосом к полюсам;
4) телофаза — обособление разделившихся ядер с образованием
собственной оболочки (рис. 4). Расхождение хромосом к
полюсам происходит у Basidiobolus так же, как у высших организмов
при участии веретена. Помимо Basidiobolus все стадии деления
и веретено обнаружены при митозе у сумчатого гриба Macropho-
1®
mina phaseoli и у базидиальных грибов Polystictus, Schizophyl*
lumf Marasmius.
Веретено — особая структура, образующаяся в клетке при:
делении ядра. Она состоит из цитоплазматических нитей,
имеющих вид микротрубочек, находящихся в цитоплазматическом мат-
риксе, о котором говорилось выше. Микротрубочки веретена
взаимодействуют с определенными участками хромосом — хромоме-
Рис. 4. Стадии митотического деления ядра Basidiobolus rannarum в
световом микроскопе:
/ — интерфазное ядро; 2 — ранняя профаза; 3 — поздняя профаза;
4 — метафаза; 5 — ранняя анафаза (по Робиноу)
рами, в результате чего происходит расхождение хромосом.
В формировании веретена принимают участие особые структуры —
центриоли, которые располагаются на обоих полюсах
митотического веретена. Центриоли могут перемещаться к поверхностной
мембране клетки, образуя здесь базальные тельца или кинето-
сомы, характерные для зооспор грибов.
У большинства грибов поведение хромосом при митозе имеет
определенные отличия от типичного: 1) малое сродство
соматических грибных хромосом к обычным покраскам, что говорит об
их несколько ином составе; 2) своеобразные ассоциации
хромосом в метафазе, отсутствие типичной метафазной пластинки при
наличии линейного расположения хромосом (так называемое
расположение цугом); 3) отсутствие веретена. Не имеют веретена
19
лри митотическом делении, например, виды родов Conidiobolus,
Neurospora, Aspergillus, Penicillium, Helminthosporium.
У мукоровых грибов ядерный материал просто разделяется
на две половины, более интенсивно окрашенные. Между ними
образуется тяж, т. е. происходит прямое деление.
У большинства грибов имеет место так называемый эндоми-
тоз, т. е. все стадии ядерного митотического деления проходят
внутри ядерной оболочки без ее разрушения, что является
отличительной чертой митоза этих грибов от классического. Оболочка
исчезает только к концу телофазы. Фрагментация ядерной
оболочки и ее исчезновение в профазе, как и у всех высших эукариот-
ных, имеет место только у базидиомицетов.
Мейотическое деление. Для грибов характерно наличие
мейоза в цикле развития, т. е. деление ядра в этом случае
происходит не с разделением ДНК хромосом и их последующей
достройкой, а с расхождением и, следовательно, с редукцией числа
хромосом, в результате чего восстанавливается гаплоидный
набор, характерный для жизненного цикла многих грибов.
Мейотическое деление ядер у грибов наступает часто вслед за
слиянием ядер и образованием диплоидной зиготы. Образованию
зиготы и слиянию ядер у грибов предшествуют различные формы
полового процесса или парасексуальный процесс (см. с. 37).
В протекании мейоза у грибов нет существенных отличий от
других организмов. Имеются отличия только второстепенного
порядка. У некоторых грибов из-за малых размеров ядер мало данных
о мейозе и не всегда можно установить, на какой стадии
жизненного цикла он происходит, хотя у многих грибов он происходит
при прорастании зиготы.
Мейотическое деление ядер у грибов было очень точно
описано у Neurospora crassa (Мак Клинток, Синглентон, 1953);
электронная микроскопия подтвердила эти наблюдения. Основная
часть жизненного цикла у этого гриба происходит в гаплоидном
состоянии.
Перед мейозом у N. crassa происходит кариогамия, т. е.
слияние ядер в одно ядро, в результате чего образовавшееся после
слияния ядро содержит двойной набор хромосом. Хромосомы в
это время находятся в диффузном состоянии.
Вскоре после слияния ядер два ядрышка сливаются в одно
большое, в то время как хромосомы находятся еще некоторое
время в двух отдельных группах.
В профазе мейоза ядро достигает самых больших размеров
при наличии довольно малых размеров хромосом и веретена.
Ядерная мембрана при мейозе в отличие от митотического
деления обычно дегенерирует к концу профазы. Ядрышки также
исчезают к концу профазы или несколько позднее.
Далее профаза характеризуется тем, что хромосомы
собираются вместе в хорошо красящееся пятно на ядерной мембране,
названное «центральным телом» (central body). В конце профазы
хромосомы обособляются и сильно сокращаются. Затем насту-
20
пает стадия метафазы, в которую происходит наибольшее
сокращение' хромосом.
В метафазе мейоза хромосомы так же, как и при митозе, у
грибов не всегда слагаются в четкую метафазную пластинку, они
могут быть и беспорядочно расположенными, но чаще собираются
в центральной части ядра. На этой стадии происходит спирали-
зация хромосом и имеет место кроссинговер, когда парные
хромосомы спирально закручиваются одна вокруг другой и
осуществляется обмен отдельными гомологичными участками. В
результате происходит перераспределение наследственного материала
во вновь образующихся ядрах.
В метафазе или в поздней профазе ядерная мембрана
разрушается. В это время ядрышко, которое удваивается в размере,
дегенерирует в ядре или позднее в цитоплазме, где оно
отделяется от ядра. Затем происходит расхождение хромосом при
участии веретена, что характеризует собой стадию анафазы. При
этом хромосомы начинают удлиняться, достигая максимума,
который у Neurospora crassa составляет 22,4 мкм.
Ядро вступает в стадию телофазы. В это время и
последующую интерфазу ядрышко снова появляется и дегенерирует
только к следующему делению. В течение телофазы происходит
восстановление ядерной мембраны. Поскольку образование септ в
гифах отстает от деления ядра, то в каждой грибной клетке
содержится по нескольку ядер, как это было сказано выше.
ГЕТЕРОКАРИОЗ У ГРИБОВ
Грибы характеризуются особым свойством — наличием
генетически различных ядер в мицелии одного индивидуума.
Среди них могут находиться ядра как одного и того же генотипа,
так и разных. Часть из них может быть гаплоидной, часть
диплоидной.
Такое явление существования различного рода ядер в
мицелии одного индивидуума носит название гетерокариоза, а
индивидуум, обладающий такими ядрами, называется гетерокарион-
том. Мицелий у гетерокарионтов можно рассматривать как
интегрированный организм, или популяцию ядер.
Гетерокариоз возникает в грибном .талломе следующими
путями.
1. При прорастании многоядерной гетерокариотической споры,
которая затем дает гетерокариотический мицелий.
2. В результате образования анастомозов вегетативных гиф
с различной генетической основой. При этом неродственные
ядра с различной генетической информацией поступают в мицелий,
размножаются там и распределяются по мицелию.
3. При мутациях в многоядерных, гомокариотических
структурах с постепенным размножением и распространением мутант-
ных ядер среди дикого типа ядер. При этом мутируют не все
ядра, а только одно или несколько ядер, поэтому в цитоплазме
21
одной клетки создаются различные отношения между ядрами с
различной наследственной информацией.
4. Как естественное следствие обмена генетическим
материалом при слиянии отдельных ядер и образовании диплоидов при
половом процессе, когда сначала происходит объединение ядер-
с различной наследственной информацией, а затем их
многократное деление.
Таким образом, в гетерокариотичном мицелии возможны пять
типов ядер: два типа гаплоидов, два типа гомозиготных
диплоидов и гетерозиготный диплоид. Эти типы ядер могут находиться
в самых разнообразных соотношениях в клетке грибного мицелия
(рис. 5).
Если у диплоидных организмов, имеющих по одному ядру в
клетке, отношения между генами, несущими доминантные и ре-
Рис. 5. Разные типы ядер в мицелии и распределение их1 в конидиях:
1 — гаплоид 1-го типа; 2 — гаплоид 2-го типа; 3 — гомозиготный диплоид
1-го типа; 4 — гомозиготный диплоид 2-го типа; 5 — гетерозиготный диплоид
цессивные признаки, могут быть только трех вариантов: 2:0;
1:1; 0:2, то у грибов-гетерокарионтов процентное отношение
генов с доминантными и рецессивными признаками может
варьировать от 0 до 100, так как могут быть самые различные
соотношения ядер, несущих те или иные признаки в хромосомах,
например соотношение генов, определяющих гладкую и складчатую
поверхность колонии.
СТРОЕНИЕ ГРИБНОГО МИЦЕЛИЯ
Вегетативное тело грибов, за небольшими
исключениями, представлено мицелием, который состоит из ветвящихся
нитей, называемых гифами, имеющих апикальный рост и боковое
ветвление. Толщина гиф мицелия варьирует от 2 до 20 мкм.
Мицелий грибов образуется при прорастании споры сначала в
короткую ростковую трубку. Обычно спора прорастает двумя или
несколькими ростковыми трубками. Первоначально проростки
22
растут не строго радиально, так как скоро начинают ветвиться,
что способствует быстрому истощению субстрата в
непосредственной близости от растущих гиф. Лимитация по субстрату
приводит к конкуренции между
верхушками гиф. Это и определяет
радиальный рост и в дальнейшем
сферическую форму колонии
|рис. 6). Тем не менее боковые
ветви образуются только на
определенном расстоянии от
вершины. Предполагается, что ну-
-жен определенный минимальный
объем цитоплазмы в гифе,
чтобы могло начаться ветвление.
Пока этот объем не достигнут, ~ с „ (1
J ' Рис. 6. Развитие колонии (/ —
имеет место только верхушеч- Х220, 2 — Х65)
ный рост. Как только
критический объем достигается, может образоваться боковая ветвь.
Частота ветвления пропорциональна линейной скорости роста гифы.
Эту зависимость выражает отношение
общая длина гиф
число апексов
и обозначает как единицу роста гифы, считая эту величину
постоянной для каждого штамма при температуре 20—37°.
Повреждение верхушечной части гифы (апекса) вызывает
рост боковых ветвей раньше обычного, в то время как при
активном верхушечном росте гифы образование боковых ветвей
подавляется. В этом подавлении проявляется механизм апикального
доминирования, свойственный и высшим растениям.
Для характеристики скорости роста мицелия гриба
используется показатель радиальной скорости роста колонии (Кг),
который определяется как увеличение диаметра колонии за
определенный отрезок времени. Радиальная скорость роста для каждого
гриба при всех прочих равных условиях величина постоянная, но
обнаруживает четкую зависимость от концентрации субстрата,
температуры, влажности и т. д.
Мицелий грибов может быть без перегородок и с
перегородками. Мицелий, гифы которого не имеют септ, представляет собой
одну гигантскую клетку и носит название ценоцитного.
Индивидуальная клетка септированной гифы может содержать одно, два
или много ядер, как у большинства грибов.
Наличие большого числа ядер в клетке многоклеточного
мицелия является результатом того, что образование перегородки
между клетками значительно отстает от деления ядер. Сначала
в клетке происходит серия последовательных делений ядер и
только затем образуется перегородка. Перегородки всегда есть у ас-
комицетов и базидиомицетов. Большинство зигомицетов не имеют
2с
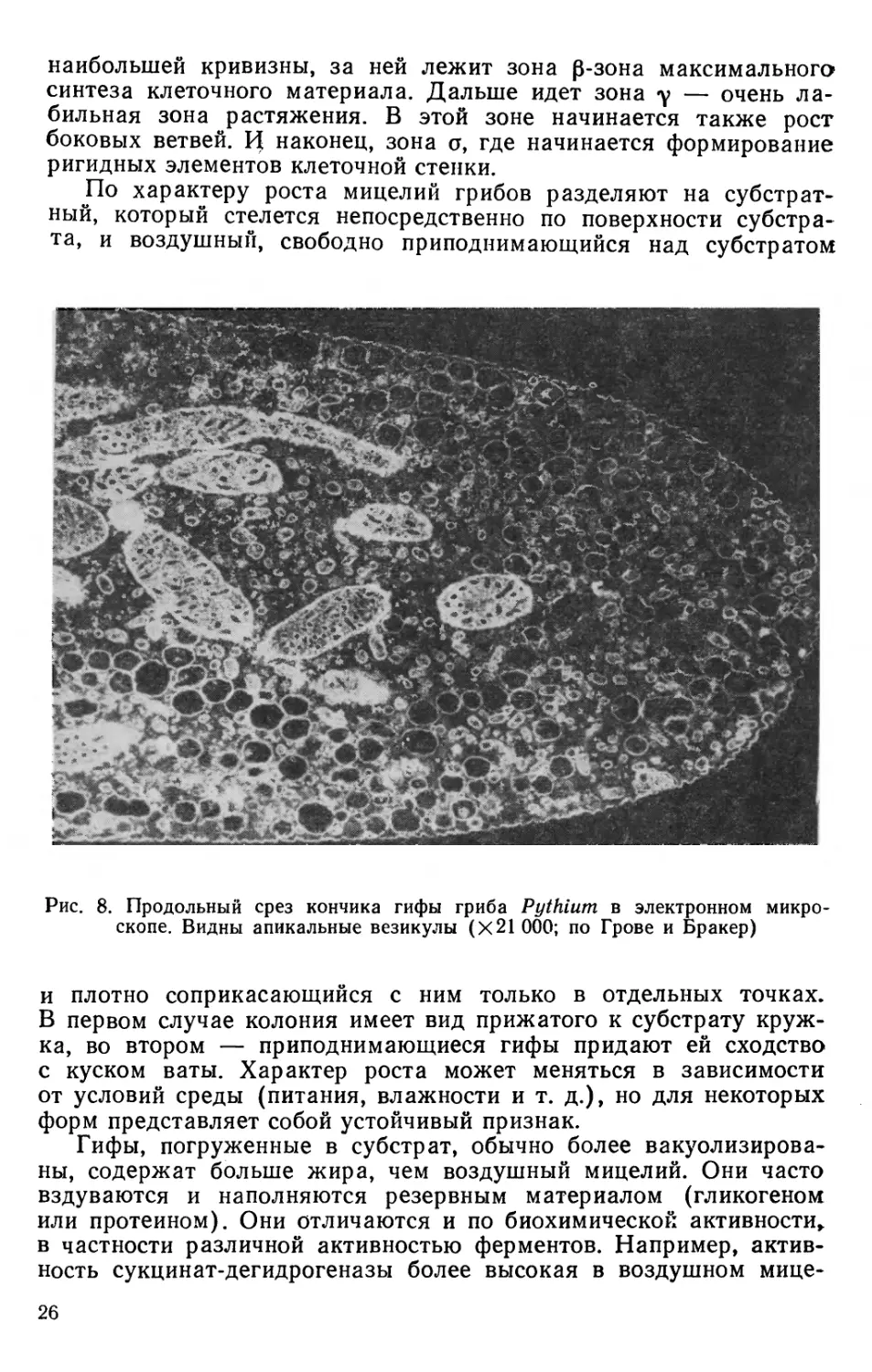
перегородок. Септы у них образуются или при формировании:
репродуктивных структур, или при повреждении гифы. Только
два семейства порядка мукоровых этого класса, Dimargaritaceae
и Kicxellaceae, а также представители порядка Entomophthora-
les имеют регулярные перегородки довольно сложного строения.
У оомицетов и хитридиомицетов перегородки отсутствуют.
У всех грибов с септированным мицелием перегородки
образуются одним и тем же путем. Формирование септы происходит
из внутренних слоев клеточной стенки гифы и протекает от
периферии к центру одновременно с двух сторон внутрь гифы.
Сначала происходит инвагинация цитоплазматической мембраны
внутрь клетки. Далее в эти выросты поступает новообразующийся
материал клеточной стенки.
У-аскомицетов септа
сужается к центру и образует
кольцо, внутри которого
возникает пора. Это
единственное отверстие, через
которое происходит
сообщение цитоплазмы между
клетками и миграция ядер.
По обе стороны от поры
располагаются тельца
Воронина, которые могут
закрывать отверстие и
препятствовать миграции ядер.
Септы у базидиомицетов
имеют более сложное
строение. Они образуются также
из внутренних слоев
боковой клеточной стенки и
растут центропетально от
периферии к центру. В центре.
Рис. 7. Строение перегородки у
сумчатых (вверху) и базидиаль-
ных (внизу) грибов в
электронном микроскопе (X100 000; по
Роскину и Мак)
гифы эти отростки вздуваются, образуя широкие бочонкоподобные
лопасти, окружающие пору, называемую долипорой (dolium
—бочонок). Септа и обе лопасти вокруг поры окружаются мембраной
эндоплазматического ретикулума. Вокруг поры мембрана
образует колпачок, называемый парентосомой. Иногда пора бывает
закрыта особым образованием из электронно-плотного материала,
так называемой пробкой (рис. 7). Парентосома перфорирована
мелкими отверстиями, достаточными для сообщения цитоплазмы
между клетками. Однако обмен ядрами через эти отверстия не-
24
возможен, что определяет стабильное содержание одной пары
ядер в клетке. Предполагалось, что такое строение септ
характерно только для класса базидиомицетов и является наряду с
базидией их отличительной чертой. Однако в настоящее время
сложно устроенные поры обнаружены и у других грибов, не
относящихся к классу базидиомицетов. В частности, грибы Pipto-
cephalis, Dispira, Linderina порядка Mucorales, а также род
сумчатых грибов Ambrosiazymae имеют пробку в порах. Септы с
пробками имеет сумчатый гриб Sporormia australis, где такие
¦септы образуются только между клетками аскогенных гиф и
крючка.
В первичном, монокариотическом мицелии базидиомицетов,
образующемся при прорастании споры, септы напоминают аско-
мицетные, через которые ядро беспрепятственно мигрирует.
Слияние проростковых мицелиев и последующее спаривание ядер
вызывают образование мицелия со сложной септой.
Мицелий всех грибов имеет хорошо выраженный верхушечный
рост, т. е. включение новых пластических веществ в мицелий
происходит только в апикальной области.
Скорость роста гифы мицелия может быть несколько
различной и находится в пределах от 0,1 до 6 мм/ч. Она регулируется
в значительной степени скоростью поступления питания и его
перемещением к вершине.
При благоприятных условиях гифы гриба Neurospora crassa
растут со скоростью 100 мкм/мин (6 мм/ч). Верхушечная часть
гифы размером в 100 мкм представляет собой область наиболее
активных синтетических процессов, куда быстро перемещаются
питательные вещества из более старых частей. Она отделена от
более старой, сильно вакуолизированной области и на расстоянии
100 мкм от вершины лишена органелл.
В апикальной части гифы, находятся цитоплазматические
пузырьки — апикальные везикулы. Среди них есть хитосомы — ор-
ганеллы, участвующие в синтезе хитина (рис. 8).
Везикулы представляют собой образования, производные от
аппарата Гольджи, которые осуществляют транспорт ферментов,
необходимых для образования клеток.
Во-первых, они содержат литические ферменты, глюканазы,
осуществляющие разрыв микрофибрилл клеточной стенки;
во-вторых, ферменты-синтетазы, осуществляющие достраивание
участков фибрилл, образовавшихся в результате их разрыва под
действием литических ферментов. Глюканазы разрезают молекулу
глюкана, к концам которой присоединяются новые молекулы
глюкозы. Таким образом, рост клеточной стенки осуществляется
путем разрыва биополимеров и встраивания вновь синтезированного
материала и регулируется двумя противоположными
механизмами, один из которых контролирует лизис стенки, а другой — ее
синтез.
Верхушечную область гифы в 100 мкм можно разделить еще
на несколько зон. Крайняя верхушка гифы (а-зона) — область
25
наибольшей кривизны, за ней лежит зона р-зона максимального
синтеза клеточного материала. Дальше идет зона у — очень
лабильная зона растяжения. В этой зоне начинается также рост
боковых ветвей. И наконец, зона а, где начинается формирование
ригидных элементов клеточной стенки.
Jlo характеру роста мицелий грибов разделяют на
субстратный, который стелется непосредственно по поверхности
субстрата, и воздушный, свободно приподнимающийся над субстратом
Рис. 8. Продольный срез кончика гифы гриба Pythium в электронном
микроскопе. Видны апикальные везикулы (Х21 000; по Грове и Бракер)
и плотно соприкасающийся с ним только в отдельных точках.
В первом случае колония имеет вид прижатого к субстрату
кружка, во втором — приподнимающиеся гифы придают ей сходство
с куском ваты. Характер роста может меняться в зависимости
от условий среды (питания, влажности и т. д.), но для некоторых
форм представляет собой устойчивый признак.
Гифы, погруженные в субстрат, обычно более вакуолизирова-
ны, содержат больше жира, чем воздушный мицелий. Они часто
вздуваются и наполняются резервным материалом (гликогеном
или протеином). Они отличаются и по биохимической активности,
в частности различной активностью ферментов. Например,
активность сукцинат-дегидрогеназы более высокая в воздушном мице-
26
лии, а активность р-галактозидазы более высокая в субстратном.
У Neurospora crassa чувствительный к кислороду р-каротин
почти не образуется в воздушном мицелии. Основное место его
образования — субстратный мицелий.
Субстратные гифы могут иметь некоторые модификации в
виде сильно разветвленных ризоидов (например, у видов Rhizopus),
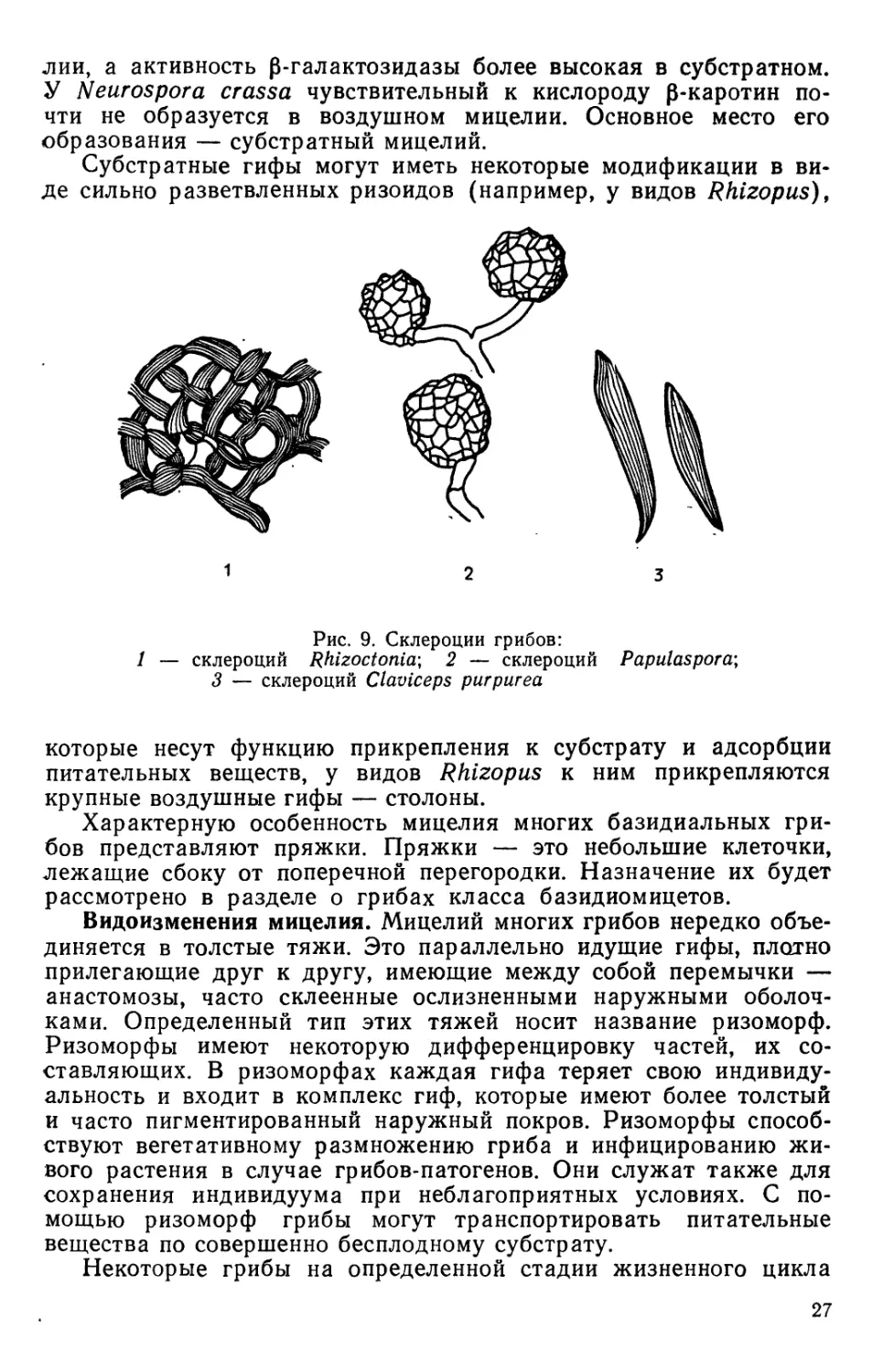
Рис. 9. Склероции грибов:
/ — склероции Rhizoctonia; 2 — склероции Papulaspora;
3 — склероции Claviceps purpurea
которые несут функцию прикрепления к субстрату и адсорбции
питательных веществ, у видов Rhizopus к ним прикрепляются
крупные воздушные гифы — столоны.
Характерную особенность мицелия многих базидиальных
грибов представляют пряжки. Пряжки — это небольшие клеточки,
лежащие сбоку от поперечной перегородки. Назначение их будет
рассмотрено в разделе о грибах класса базидиомицетов.
Видоизменения мицелия. Мицелий многих грибов нередко
объединяется в толстые тяжи. Это параллельно идущие гифы, плотно
прилегающие друг к другу, имеющие между собой перемычки —
анастомозы, часто склеенные ослизненными наружными
оболочками. Определенный тип этих тяжей носит название ризоморф.
Ризоморфы имеют некоторую дифференцировку частей, их
составляющих. В ризоморфах каждая гифа теряет свою
индивидуальность и входит в комплекс гиф, которые имеют более толстый
и часто пигментированный наружный покров. Ризоморфы
способствуют вегетативному размножению гриба и инфицированию
живого растения в случае грибов-патогенов. Они служат также для
сохранения индивидуума при неблагоприятных условиях. С
помощью ризоморф грибы могут транспортировать питательные
вещества по совершенно бесплодному субстрату.
Некоторые грибы на определенной стадии жизненного цикла
27
образуют еще одно видоизменение мицелия — склероции,
представляющие собой тела плотной консистенции, обычно темного-
цвета или ярко окрашенные, размером от одного и нескольких
миллиметров до нескольких десятков сантиметров (рис. 9).
Склероции могут образоваться в результате плотного переплетения
нитей мицелия и активного его ветвления, создающего подобие-
ткани. Такая ткань, имеющая место у грибов, в отличие от
истинной ткани у высших растений носит название параплектен-
химы, или псевдопаренхимы.
Из псевдопаренхимы у грибов образованы различные типы
соматических и репродуктивных структур. Соматические — эта
строма и склероции. Репродуктивные структуры, сложенные из
псевдопаренхимы, — это различные типы плодовых тел грибов.
Наибольшие из известных склероциев — это подземные
склероции Polyporus mylittae, обитающего в Австралии, размером с
человеческую голову, которые весят 15 кг. Их едят аборигены,
и они известны под названием «black fellow's bread». При
образовании склероциев происходит очень значительная отдача воды
мицелием, так что зрелые склероции имеют всего 5—10% воды,
в то время как мицелий грибов содержит до 90% воды, но
характеризуются большим запасом питательных веществ, в
частности содержание жира в них составляет 30%. Склероции также
обнаруживают некоторую «дифференцировку тканей»: снаружи
они покрыты корой из нескольких слоев, обычно
пигментированных, внутри — имеют белую сердцевину. Склероции обычно
служат организму для перенесения неблагоприятных условий. Они
могут оставаться покоящимися долгое время и затем прорастают
мицелием или специализированными структурами со спорами
после наступления благоприятных условий.
Склероции образуются преимущественно на мицелии, у
некоторых патогенных форм внутри пораженного растения или
мумифицируют целые органы, пронизывая мицелием, например,
плоды фруктовых деревьев. В этом случае они представляют собой
псевдосклероции. Таким образом, псевдосклероции — это
образования, состоящие не только из одного мицелия, но и другого
субстрата (растительного материала, почвы и других),
связанного с помощью мицелия в плотную массу.
Глава 2
РАЗМНОЖЕНИЕ И РАЗВИТИЕ ГРИБОВ
Грибы имеют разнообразные способы размножения:
вегетативное, бесполое и половое. Структуры бесполого и
полового размножения грибов чрезвычайно разнообразны по
строению и характеру образования и являются основой для
классификации этих организмов.
28
ВЕГЕТАТИВНОЕ РАЗМНОЖЕНИЕ
Вегетативное размножение — это размножение без
образования каких бы то ни было специализированных структур
размножения. Чаще всего это размножение отдельными
участками мицелия. Это свойство мицелия используется для
поддержания роста грибов на искусственных питательных средах путем
переноса кусочков мицелия с одной среды на другую. Широко
применяемый для размножения высших базидиальных грибов
метод тканевой культуры является также примером вегетативного
размножения. Этот способ заключается в том, что из кусочка
плодового тела, которое представляет собой плотное
переплетение мицелия гриба, стерильно вырезается кусочек и помещается
на питательную среду. Вегетативным является также
размножение, при котором происходит деление одноклеточного организма
на две дочерние клетки или многоклеточного таллома на большое
число фрагментов, каждый из которых вырастает в новый
индивидуум (фрагментация), а также почкование соматических
клеток. При фрагментации гифы предварительно образуют частые
перегородки и затем распадаются по ним на отдельные клетки
овальной или цилиндрической формы, называемые
соответственно оидиями или артроспорами. Если такие клетки образуют тол-
Рис. 10. Хламидоспоры гриба Verticillium nigrescens
стостенную оболочку, то они называются хламидоспорами.
Хламидоспоры могут быть верхушечными и интеркалярными,
образуются одиночно или группами и способны переносить
неблагоприятные условия (рис. 10). В дальнейшем они также могут
подвергаться фрагментации. Некоторые грибы могут
образовывать почкующийся мицелий, который может быть определенной
стадией в цикле развития. Вегетативное размножение у дрожжей
происходит путем почкования клеток.
2$
БЕСПОЛОЕ РАЗМНОЖЕНИЕ
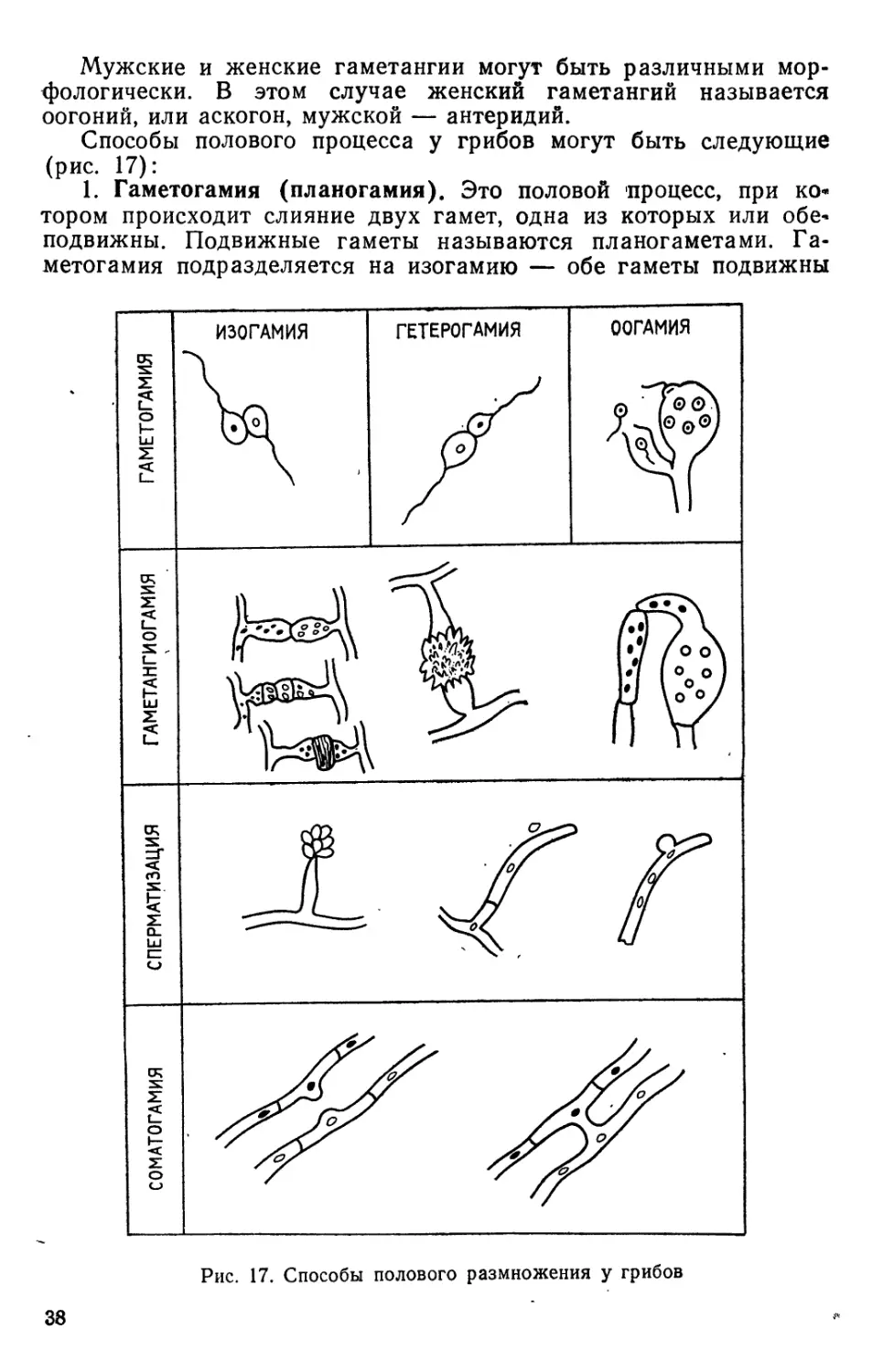
Бесполым размножением называется размножение с
образованием специализированных структур, образованию
которых не предшествует предварительное слияние клеток или
объединение ядер, в противоположность половому размножению,
которое всегда включает предварительное слияние клеток и
слияние или объединение ядер. В образовавшихся
специализированных репродуктивных структурах — спорангиях или конидиеносцах
всегда образуются споры, с помощью которых и осуществляется
размножение. У некоторых грибов весь таллом превращается в
репродуктивные органы; такие формы называются голокарпиче-
-скими. Но у большинства репродуктивные органы возникают как
часть таллома, в то время как оставшаяся часть нормально
соматически активна. Такие грибы называют эукарпическими.
Бесполый цикл у грибов обычно повторяется много раз, в то время
как половой цикл бывает значительно реже.
Бесполые споры у одних грибов образуются в специальных
вместилищах — спорангиях и называются спорангиоспорами, у
других они образуются открыто на специализированных гифах
или на мицелии различным способом и называются конидиями.
Спорангий — это закрытое образование, внутреннее
содержимое которого состоит обычно из многих спор.
У мицелиальных форм грибов спорангий образуется на
специализированной гифе, называемой спорангиеносцем, который
поднимается кверху от субстрата. Такое расположение
спорангиев способствует распространению спор токами воздуха, после
того как они освобождаются из спорангия. Спорангиеносец может
быть неветвящимся, но может ветвиться моноподиально, симпо-
диально или дихотомически. При образовании спорангия вершина
гифы прекращает расти, слегка вздувается (рис. 11). Спорангио-
споры могут быть подвижными или неподвижными. Подвижные
спорангиоспоры называют зооспорами, или планоспорами,
неподвижные — апланоспорами. Подвижность зооспор определяется
наличием у них жгутиков. Зооспоры грибов снабжены одним или
двумя жгутиками. Жгутики у грибов имеют общий план строения
эукариотических организмов, который выражается формулой 9 + 2.
Главный стержень жгутика состоит из двух одинарных
центральных фибрилл (микротрубочек), которые окружены кольцом из
девяти дублетов фибрилл. Такой план строения имеют жгутики
зооспор зеленых водорослей, мхов, папоротников, простейших, но
он отличается от плана строения жгутиков у бактерий.
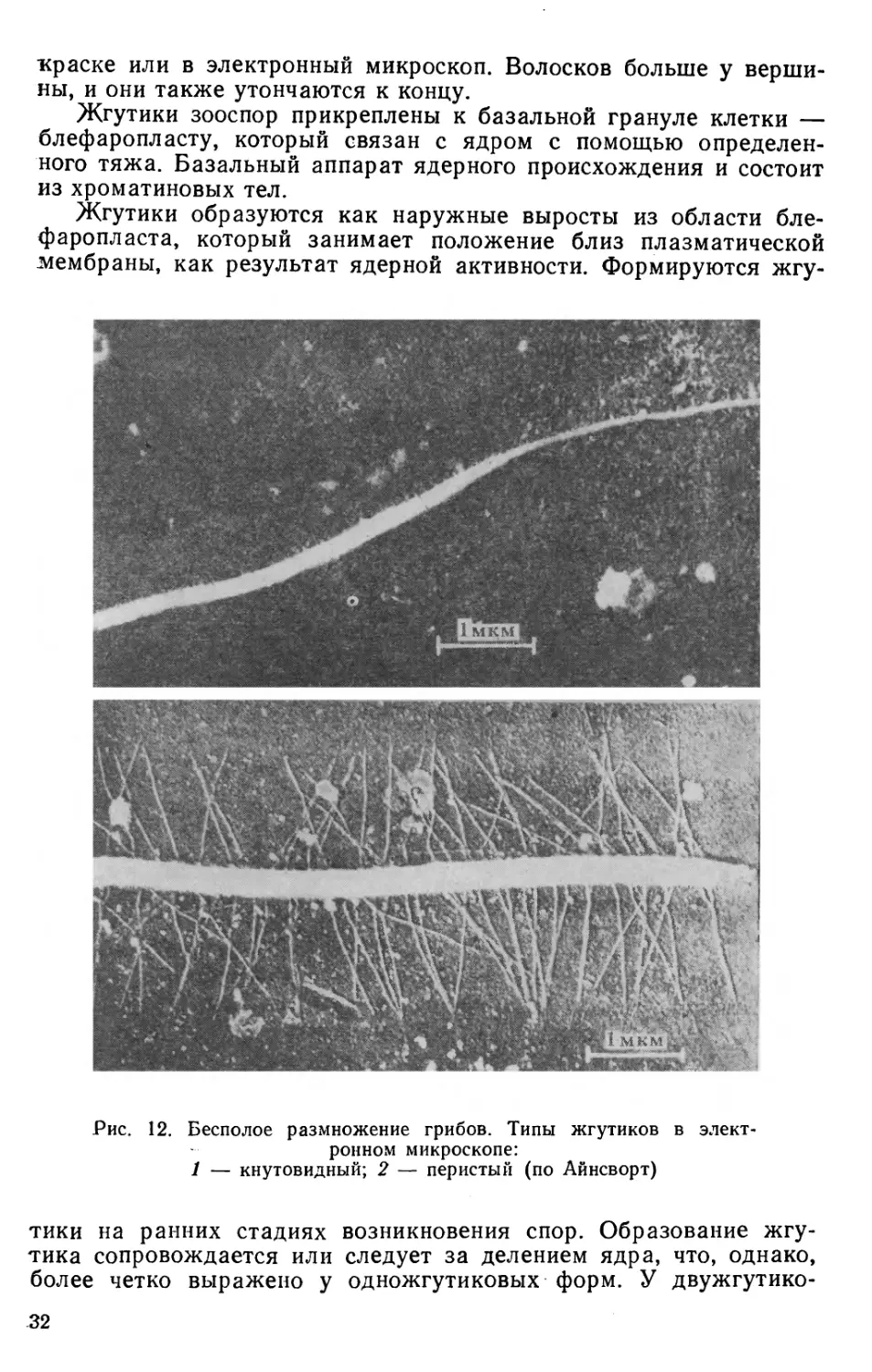
Зооспоры грибов имеют два типа жгутиков: 1) жгутики в виде
кнута, или бичевидные; 2) жгутики, снабженные придатками,
перистые (рис. 12). Бичевидный жгутик гладкий, не имеет
никаких придатков, боковые волоски отсутствуют. Жгутик
постепенно утончается к концу, но утончение происходит за счет
наружных слоев, а не главной оси. Жгутики с придатками имеют
боковые волоски — мастигонемы, которые видны только при по-
30
шШ
f^*^****^*
1 ¦ 50мкм
шшш
mm' 1
ННк.
SOmhm
бОмнм
20мт
е\
иди
%'¦,.:¦ «~ 1Ш
»; |^
Шшш
,--
у^^^Я
бит
^щ
^ЩЗ^^т^ш^^ВЯ^шн^Ш^^^^
шШШШ$ШШт Mm
т
Рис. 11. Бесполое размножение грибов. Различные типы спорангиев в
электронном микроскопе:
а, б, в, г — Actinomucor elagans; д, е — Mortierella ramanniana; ж, з — Ми-
cor plutnbeus (фото В. С. Гузева)
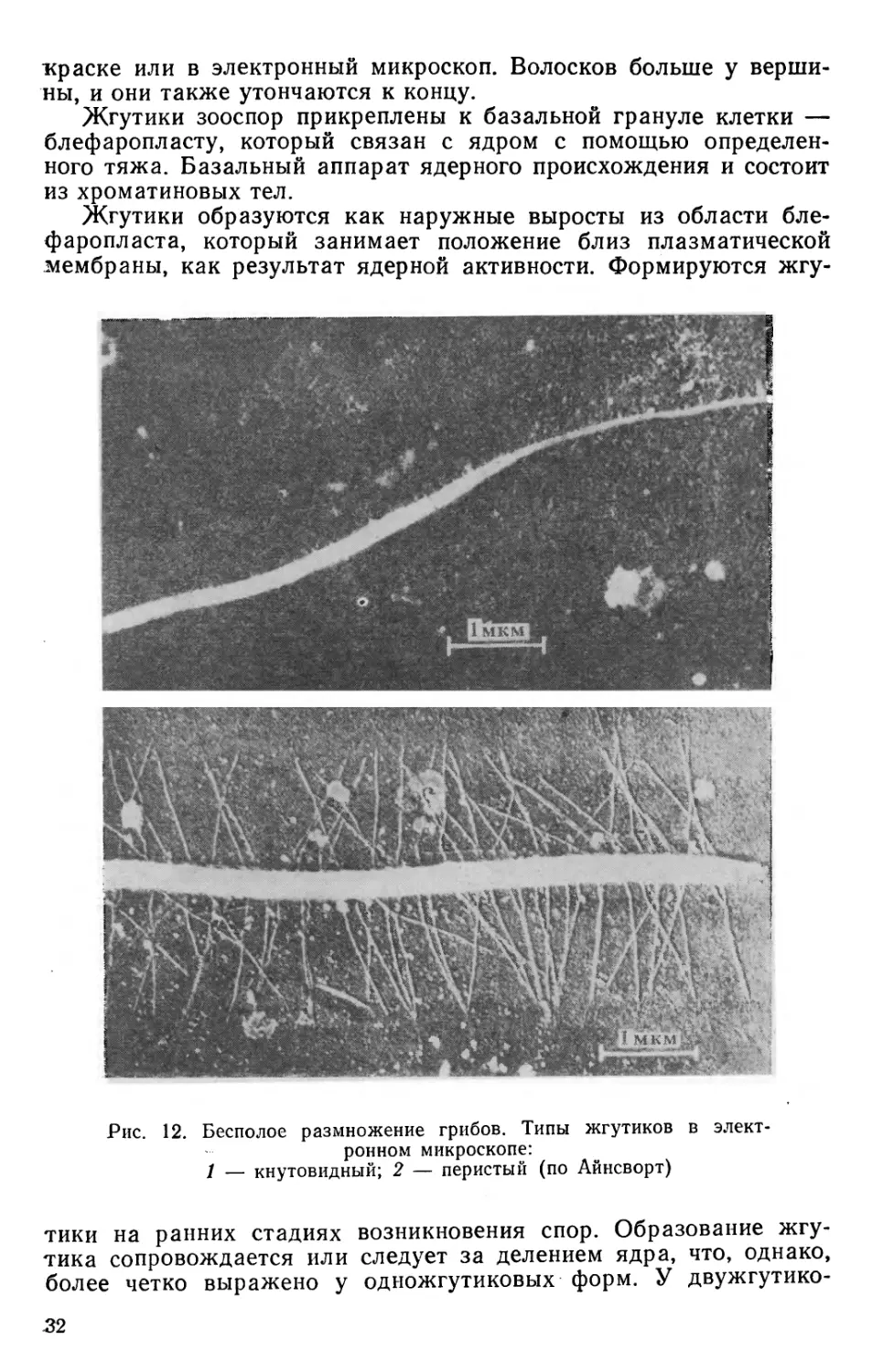
краске или в электронный микроскоп. Волосков больше у
вершины, и они также утончаются к концу.
Жгутики зооспор прикреплены к базальной грануле клетки —
блефаропласту, который связан с ядром с помощью
определенного тяжа. Базальный аппарат ядерного происхождения и состоит
из хроматиновых тел.
Жгутики образуются как наружные выросты из области бле-
фаропласта, который занимает положение близ плазматической
мембраны, как результат ядерной активности. Формируются жгу-
Рис. 12. Бесполое размножение грибов. Типы жгутиков в
электронном микроскопе:
1 — кнутовидный; 2 — перистый (по Айнсворт)
тики на ранних стадиях возникновения спор. Образование
жгутика сопровождается или следует за делением ядра, что, однако,
более четко выражено у одножгутиковых форм. У двужгутико-
32
вых зооспор это не связано так тесно с делением ядра и жгутики
образуются после периода ядерной активности. После окончания
подвижной фазы жгутики исчезают. Жгутик или сбрасывается
или остается, превращаясь в шишку.
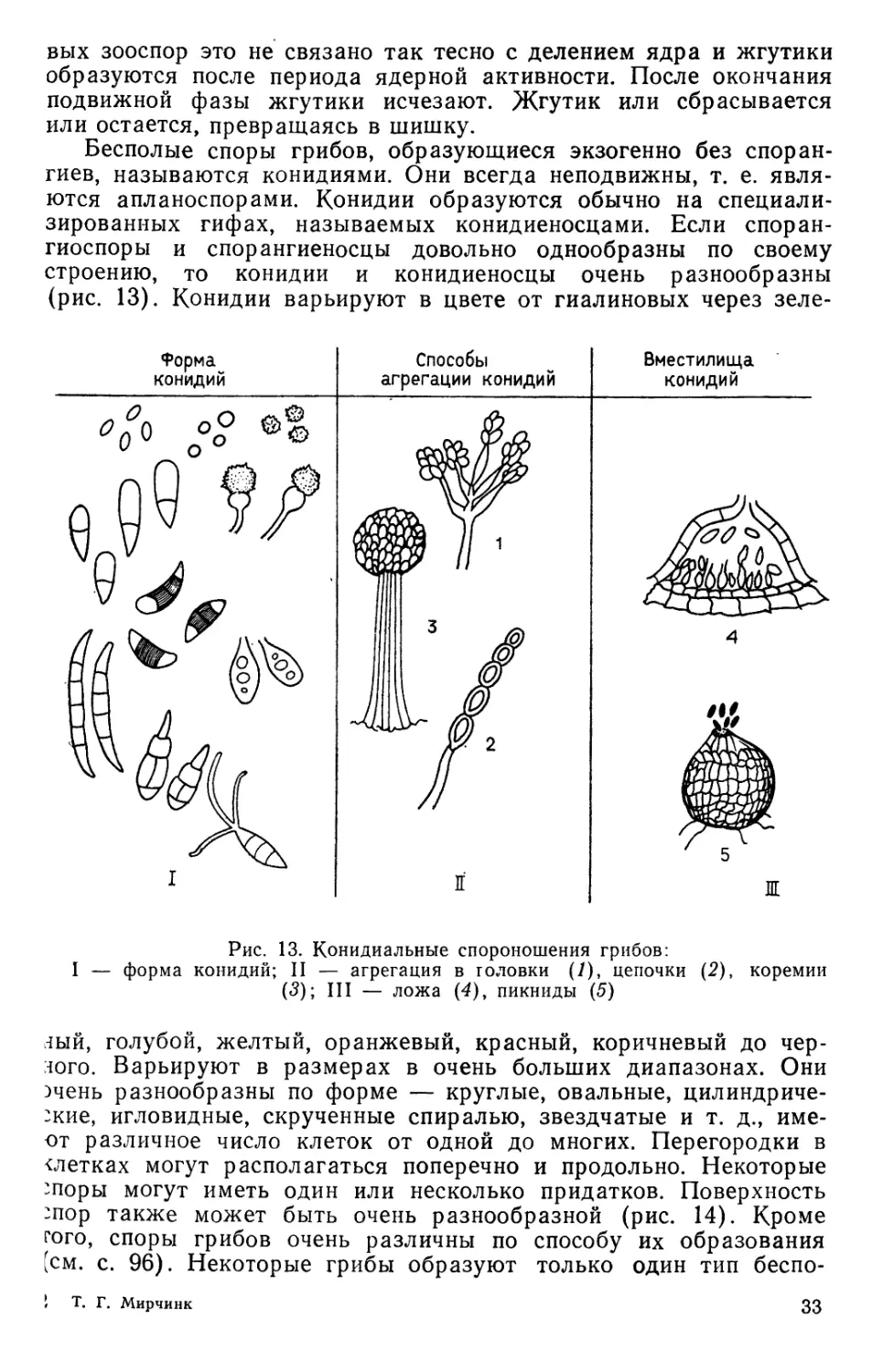
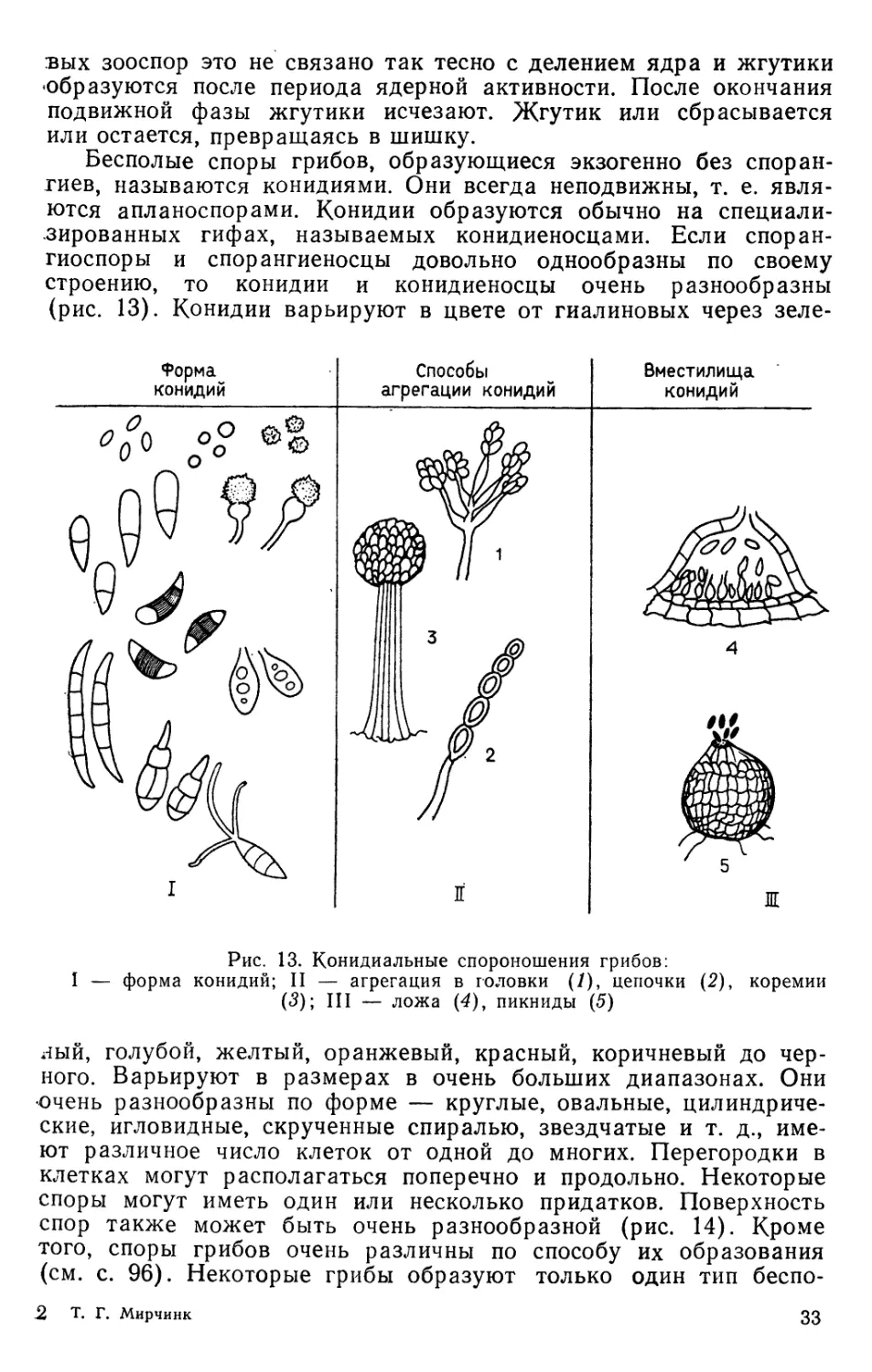
Бесполые споры грибов, образующиеся экзогенно без
спорангиев, называются конидиями. Они всегда неподвижны, т. е.
являются апланоспорами. Конидии образуются обычно на
специализированных гифах, называемых конидиеносцами. Если споран-
гиоспоры и спорангиеносцы довольно однообразны по своему
строению, то конидии и конидиеносцы очень разнообразны
(рис. 13). Конидии варьируют в цвете от гиалиновых через зеле-
Форма
конидий
Способы
агрегации конидий
Вместилища
конидий
Ж
Рис. 13. Конидиальные спороиошения грибов:
форма конидий; II — агрегация в головки (1), цепочки (2), коремии
(3); III — ложа (4)у пикниды (5)
г!ый, голубой, желтый, оранжевый, красный, коричневый до
черного. Варьируют в размерах в очень больших диапазонах. Они
эчень разнообразны по форме — круглые, овальные, цилиндриче-
:кие, игловидные, скрученные спиралью, звездчатые и т. д., име-
от различное число клеток от одной до многих. Перегородки в
клетках могут располагаться поперечно и продольно. Некоторые
'поры могут иметь один или несколько придатков. Поверхность
:пор также может быть очень разнообразной (рис. 14). Кроме
того, споры грибов очень различны по способу их образования
(см. с. 96). Некоторые грибы образуют только один тип беспо-
\ Т. Г. Мирчинк
33
Ш
¦ -
ь-
" ' ?МКМ
С<
4^ *
шшшш^шшяш*?
¦ :' ¦
"
¦fir
;'-¦
f й
8Д№? йкр;
яр
' &'," .'¦¦:
уЙМНМ
: f MAW
Рис. 14. Различная орнаментация конидии в электронном микроскопе:
а, б, в, г — виды рода Penicillium; д, е, ж, з — виды родов Aspergillus
(фото В. С. Гузева)
s"...
<#
.
•:f
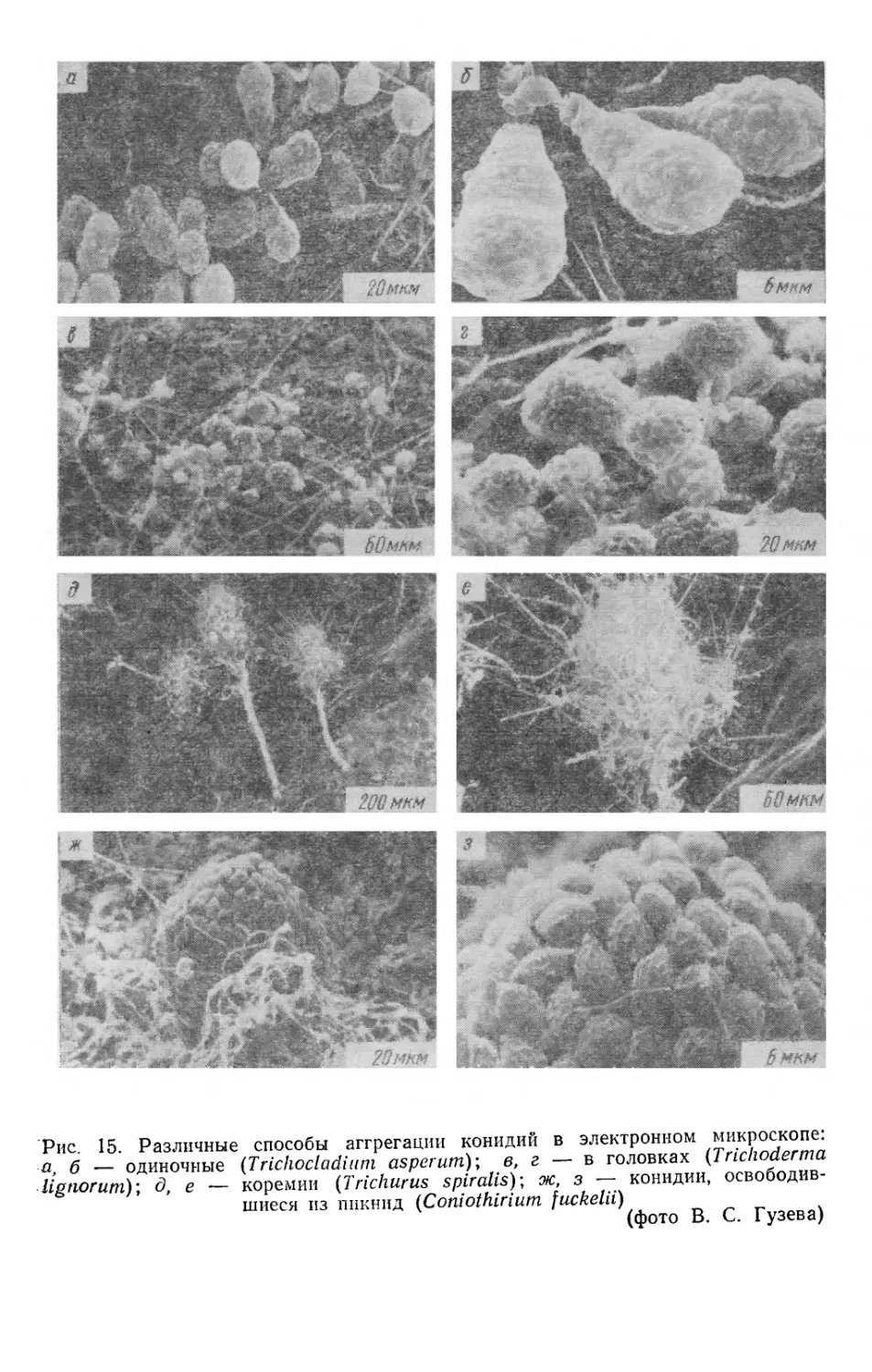
Рис. 15. Различные способы аггрегации конидии в электронном микроскопе.
а, б - одиночные (Trichocladium asperum); в г - в ^овках У"с^"а
lignorum); д. е - коремии (Trichurus spiralis); ж з - конидии,
освободившиеся из шшнид (Coniothirium fuckelu)
(фото В. С. Гузева)
краске или в электронный микроскоп. Волосков больше у
вершины, и они также утончаются к концу.
Жгутики зооспор прикреплены к базальной грануле клетки —
блефаропласту, который связан с ядром с помощью
определенного тяжа. Базальный аппарат ядерного происхождения и состоит
из хроматиновых тел.
Жгутики образуются как наружные выросты из области бле-
фаропласта, который занимает положение близ плазматической
мембраны, как результат ядерной активности. Формируются жгу-
Рис. 12. Бесполое размножение грибов. Типы жгутиков в
электронном микроскопе:
/ — кнутовидный; 2 — перистый (по Айнсворт)
тики на ранних стадиях возникновения спор. Образование
жгутика сопровождается или следует за делением ядра, что, однако,
более четко выражено у одножгутиковых форм. У двужгутико-
32
зых зооспор это не связано так тесно с делением ядра и жгутики
образуются после периода ядерной активности. После окончания
подвижной фазы жгутики исчезают. Жгутик или сбрасывается
или остается, превращаясь в шишку.
Бесполые споры грибов, образующиеся экзогенно без
спорангиев, называются конидиями. Они всегда неподвижны, т. е.
являются апланоспорами. Конидии образуются обычно на
специализированных гифах, называемых конидиеносцами. Если споран-
гиоспоры и спорангиеносцы довольно однообразны по своему
строению, то конидии и конидиеносцы очень разнообразны
(рис. 13). Конидии варьируют в цвете от гиалиновых через зеле-
Форма
конидий
Способы
агрегации конидий
Вместилища
конидий
° ®%
Рис. 13. Конидиальные спороношения грибов:
I — форма конидий; II — агрегация в головки (/), цепочки (2), коремии
(3); III — ложа (4), пикниды (5)
ный, голубой, желтый, оранжевый, красный, коричневый до
черного. Варьируют в размерах в очень больших диапазонах. Они
•очень разнообразны по форме — круглые, овальные,
цилиндрические, игловидные, скрученные спиралью, звездчатые и т. д.,
имеют различное число клеток от одной до многих. Перегородки в
клетках могут располагаться поперечно и продольно. Некоторые
споры могут иметь один или несколько придатков. Поверхность
спор также может быть очень разнообразной (рис. 14). Кроме
того, споры грибов очень различны по способу их образования
(см. с. 96). Некоторые грибы образуют только один тип беспо-
2 Т. Г. Мирчинк
33
лых спор, в то время как другие по крайней мере до четырех
видов различных спор. Кроме того, конидии могут быть
агрегированы на несущих их структурах в головки и цепочки.
Конидиеносцы также имеют более разнообразное строение,
чем спорангиеносцы. Они могут иметь и разнообразное ветвление:
поочередное, супротивное, мутовчатое, симподиальное,
дихотомическое. Кроме того, могут быть различные вздутия на вершине
и на протяжении конидиеносца, короткие разветвления и выросты
фиалиды. Конидиеносцы могут быть объединены тесными
группами, образуя единое целое, так называемую коремию (коремия
представляет собой пучок тесно сближенных конидиеносцев),
располагаться тесным слоем на специальной поверхности из пе-
Рис. 16. Эволюция бесполых спороношений у грибов:
/ — спорангий у Мисог; 2 — спорангий и спорангиоли у 77шт-
nidium; 3,4 — спорангиоли у Helicostylum и Blakeslea; 5 —
конидии у Choanephora
реплетения гиф, образуя ложе, и формироваться в специальных
вместилищах разнообразной формы и строения — пикнидах (рис. 13,
15). У некоторых грибов конидиеносцы образуют слой на
поверхности выпуклого сплетения гиф в виде подушечек, называемых
спородохиями.
Более сложное и разнообразное строение конидиальных спо-
36
роношений по сравнению со спорангиальными свидетельствует о
более высоком уровне их эволюционного развития. Вероятно, что
конидиальные спороношения произошли в результате эволюции
от спорангиальных. Подтверждением этому могут служить
различные стадии редукции спорангиев с образованием спорангио-
лей. Спорангиоли — это спорангии, содержащие очень небольшое
число спор (2—3). Уменьшение числа спор до одной приводит к
образованию фактически конидии. Многие стадии редукции
спорангиев можно проследить на примере различных представителей
мукоровых грибов (рис. 16).
ПОЛОВОЕ РАЗМНОЖЕНИЕ
У грибов половому размножению обязательно
предшествует слияние специализированных клеток и последующее
объединение или слияние ядер. Продукт слияния клеток — зигота или
зигоспора. Конечный результат полового процесса — образование
специальных спор, с помощью которых и осуществляется
дальнейшее размножение особи. Половой процесс состоит из трех
стадий: плазмогамии, кариогамии и мейоза.
Плазмогамия представляет собой слияние клеток и
объединение двух протопластов, которые приносят два различных ядра в
одну клетку. Кариогамия — слияние ядер в одно диплоидное
ядро, ядро зиготы. Мейоз — это деление диплоидного ядра с
редукцией числа хромосом и восстановление в ядрах гаплоидного
набора хромосом. В истинно половом цикле эти процессы протекают
в регулярной последовательности и в фиксированном месте.
У одних грибов мейоз может быть перед образованием половых
клеток, но у большинства он происходит при прорастании зиготы.
У одних грибов кариогамия следует непосредственно за
плазмогамией. У других эти два процесса разделены во времени и
пространстве. Результатом плазмогамии у этих организмов
является образование двуядерной клетки, или клетки, содержащей
несколько пар ядер, ассоциированных строго попарно. Такие
пары ядер называются дикарионами. Эти пары ядер могут не
сливаться неограниченно долгое время и, синхронно делясь строго
попарно, переходят из клетки в клетку или в растущую нить
мицелия — аскогенную гифу без перегородок. В определенный
период жизненного цикла пара таких ядер сливается в одно
диплоидное ядро, обычно это происходит в верхушечной клетке и
рано или поздно сопровождается мейотическим делением.
Другие грибы не образуют дифференцированных половых
клеток или структур, и половая функция принадлежит соматическим
гифам. Талломы таких грибов могут или не могут размножаться
половым путем в зависимости от комбинации с совместимыми
или несовместимыми партнерами.
Половые структуры у грибов называются гаметангиями. Они
могут образовывать половые клетки, называемые гаметами, или
могут содержать вместо них одно или больше гаметных ядер.
37
Мужские и женские гаметангии могут быть различными
морфологически. В этом случае женский гаметангии называется
оогоний, или аскогон, мужской — антеридий.
Способы полового процесса у грибов могут быть следующие
(рис. 17):
1. Гаметогамия (планогамия). Это половой процесс, при
котором происходит слияние двух гамет, одна из которых или обе-
подвижны. Подвижные гаметы называются планогаметами.
Гаметогамия подразделяется на изогамию — обе гаметы подвижны
о
1-
ш
ИЗОГАМИЯ
ГЕТЕРОГАМИЯ
ООГАМИЯ
2
о
S .
1_
и
Z
<с
го
ЗЕ.
о
г:
о
Рис. 17. Способы полового размножения у грибов
38
и морфологически неразличимы, гетерогамию — обе гаметы
подвижны, но различаются размерами или другими признаками и
оогамию — половой процесс, при котором женские гаметы
неподвижны, в то время как мужские гаметы (сперматозоиды)
подвижны. Подвижные гаметы проникают в оогоний и
оплодотворяют яйцеклетку.
При оогамном половом процессе у многих грибов
сперматозоиды не образуются, и яйцеклетки оплодотворяются выростами
антеридиев. Гаметогамия и оогамия характерны для низших
грибов классов хитридиомицетов и оомицетов.
2. Гаметангиогамия. Этот способ полового процесса
характеризуется слиянием двух специализированных половых структур —
гаметангиев, не дифференцированных на гаметы. Гаметангии
обычно многоядерны, и в результате их слияния наряду со
слиянием цитоплазмы происходит множественное слияние ядер. Такой
тип полового процесса характерен для зигомицетов и аскомице-
тов. У зигомицетов сливающиеся гаметангии в большинстве
случаев морфологически хорошо отличимы от мицелия, на котором
они образуются. В результате слияния формируется зигоспора,
которая прорастает после периода покоя в спорангий.
У аскомицетов один из гаметангиев — женский — часто
бывает разделен на две клетки — аскогон и трихогину. Через
последнюю содержимое мужского гаметангия — антеридия —
поступает в аскогон. При этом происходит только плазмогамия, т. е.
сливается цитоплазма клеток без слияния ядер. Неслившиеся
ядра образуют дикарион — ассоциацию двух ядер, которые
затем строго синхронно делятся.
Из оплодотворенного таким образом аскогона сразу же без
периода покоя вырастают гифы, носящие название аскогенных,
в которые переходят синхронно делящиеся дикарионы. Слияние
дикарионов в одно диплоидное ядро, т. е. кариогамия, происходит
на вершине аскогенной гифы. Таким образом, наблюдается
характерный для многих грибов разрыв во времени между
плазмогамией и кариогамией. Из верхней клетки аскогенной гифы, где
произошло слияние дикарионов, формируется сумка. Диплоидное
ядро при этом делится мейотически, затем следуют два митоти-
ческих деления и вокруг восьми образовавшихся гаплоидных ядер
формируются восемь аскоспор, с помощью которых происходит
дальнейшее размножение индивидуума. У некоторых сумчатых
грибов число аскоспор может быть иным (2, 4, 16). Сумки обычно
образуются в плодовых телах различного строения.
3. Сперматизация. Некоторые грибы образуют
многочисленные мелкие одноядерные спороподобные образования наподобие
конидий на конидиеносце, но способные к копуляции,
называемые спермациями, так как они несут функцию мужских половых
структур. Спермации распространяются насекомыми, ветром,
водой или какими-либо другими путями к женскому гаметангию,
который представляет собой специально воспринимающую гифу
(рецептор) или даже соматическую гифу, к которой они при-
39
крепляются. В месте контакта образуется пора, и содержимое
спермация проникает в клетку-рецептор, где происходит слияние
цитоплазмы и ядра спермация с цитоплазмой и ядром клетки, в
результате чего образуется зигота, прорастающая аскогенными
гифами. Сперматизация имеет место у некоторых сумчатых
грибов, например рода Neurospora.
4. Соматогамия. У многих высших грибов не образуется
никаких половых структур и соматические клетки несут половую
функцию, т. е. сливаются обычные соматические клетки мицелия.
Слияние происходит путем образования перемычек (анастомозов)
между гифами мицелия. Такой тип полового процесса
характерен для базидиомицетов и некоторых сумчатых грибов. У бази-
диомицетов, как и у аскомицетов, плазмогамия и кариогамия при
половом процессе разделены во времени. После плазмогамии,
наступающей вскоре после прорастания спор, формируется ди-
кариотический мицелий с синхронно делящимися в нем ядрами.
Дикариотический мицелий у базидиомицетов существует
значительно более продолжительное время, чем у аскомицетов. На нем
образуются базидии, в которых ядра дикариона сливаются в
диплоидное ядро, делящееся сначала мейотически, а затем мито-
тически. В результате на базидии образуются четыре базидио-
споры с гаплоидными ядрами. У многих групп базидиомицетов
базидии возникают на плодовых телах разнообразного строения
и размеров.
На основании пола и половой совместимости грибы делятся
на: 1) обоеполые организмы — женские и мужские структуры
развиваются на одном талломе; 2) раздельнополые, или
диморфные, организмы — мужские и женские талломы разделены.
Обоеполые организмы у грибов обычно называют гомоталличными,
раздельнополые организмы носят название гетероталличных.
У гетероталличных грибов осуществление полового цикла
возможно только при наличии двух разных талломов.
Морфологически неразличимые штаммы обозначаются
условно как ( + )- и (—)-штаммы, что определяет различие полов. Ге-
тероталлические грибы по половой совместимости делятся в свою
очередь на три группы:
1. Биполярные гетероталлические. Грибы этой
категории имеют два половых типа индивидуумов,
различающихся по одному фактору, определяющему пол, А и а, которые
различаются по половой совместимости. Каждое ядро одного
полового типа имеет ген А и каждое ядро другого полового типа
имеет ген а. Только талломы, имеющие ядра с
противоположными генами, совместимы. Образующаяся после слияния зигота
имеет генотип Аа.
2. Тетр апол яр ные гетер от а л л ич н ые. Грибы этой
категории содержат 4 половых типа индивидуумов. Совместимость
здесь определяется двумя парами факторов, определяющих пол.
Например, одни имеют набор половых генов Аа, другие Вв,
тогда набор половых признаков в зиготе будет АаВв. При редук-
40
ционном делении тетраполярные особи должны дать в равном
числе четыре сорта гаплоидных ядер: Ав, аВ, АВ и ав. Слияние
возможно только в тех случаях, если ядра двух мицелиев
отличаются обоими половыми факторами, т. е. между АВ и ав или
Ав и аВ, но невозможно, например, между Аа и Ав. Поэтому при
наличии тетраполярного разделения полов имеют место четыре
группы неодинаковых в половом отношении спор, так что каждая
группа реагирует только с одной из трех остальных.
3. Вторично гомоталличные. У некоторых
биполярных гетероталличных грибов при образовании спор два ядра
противоположного пола регулярно объединяются в каждой споре.
Каждая спора поэтому после прорастания дает таллом, который
содержит А и а ядра и затем ведет себя как гомоталличный.
Такое явление называют вторичным гомоталлизмом.
ПАРАСЕКСУАЛЬНЫЙ ПРОЦЕСС
Некоторые грибы не имеют истинного полового цикла,
который характеризуется образованием специализированных
структур, но тем не менее обмен генетической информацией у них
происходит так же, как при половом процессе. Половой процесс
у этих организмов заменяется так называемым парасексуальным
циклом.
Парасексуальный цикл — это процесс, в котором
плазмогамия, кариогамия и гаплоидизация хромосом происходят, но не
в специфическом определенном месте таллома или определенной
стадии жизненного цикла, а в любой вегетативной гифе мицелия.
Парасексуальный цикл имеет большое значение у несовершенных
грибов — дейтеромицетов, которые и получили свое название
из-за отсутствия полового процесса. У некоторых грибов
парасексуальный цикл имеет место и при наличии полового процесса.
Явление пар асексуальности впервые было открыто Г. Понтекорво
в 1952 г. у Aspergillus nidulans (конидиальная стадия гриба Ете-
ricella nidulans). Он был обнаружен у многих несовершенных
грибов и базидиомицетов. Основные ступени пар асексуального
цикла — это гетерокариоз, слияние несходных ядер в гетерока-
риотической гифе и последующая рекомбинация участков
хромосом при митозе. Парасексуальный цикл имеет большое значение
в изменчивости несовершенных грибов и у всех ценоцитных
грибов при их вегетативном росте. В результате пар асексуального
цикла в гетерокариотическом мицелии происходит слияние
гаплоидных ядер разных генотипов, а также и одинаковых и
соответственно образуется гетерозиготный или гомозиготный диплоид.
При последующем размножении диплоидных ядер имеет место
митотический кроссинговер. Это наиболее важная фаза пара-
сексуального цикла, как и полового процесса, при котором
происходит обмен гомологическими участками хромосом, в результате
чего осуществляется перераспределение признаков во вновь
образующихся ядрах. В ходе этого процесса происходят различные
41
рекомбинации хромосом. Однако, по данным Понтекорво,
рекомбинации при парасексуальном цикле осуществляются в 500 раз
реже, чем при половом. У грибов, образующих одноядерные
конидии на гетерокариозном мицелии, можно разделить диплоидные
и гаплоидные конидии и получить отдельно диплоидный и
гаплоидный мицелий. Диплоидные конидии обычно в 1,3 раза
больше по размеру гаплоидных.
Диплоидные ядра претерпевают гаплоидизацию в мицелии и
отсортировываются; часто они образуют сектор в диплоидной
колонии. Гаплоидные штаммы, полученные от таких секторов,
генетически отличны от их родителей в результате митотических
рекомбинаций, происходящих при делении.
В гетерокариотном мицелии после пар асексуального цикла
появляются следующие варианты ядер: гаплоидные ядра,
подобные обоим родителям; гаплоидные ядра с новыми генетическими
рекомбинациями; гомозиготные диплоиды; гетерозиготные ди-
плоиды.
Все вышесказанное касается морфологического и
генетического контроля полового процесса, но есть еще и физиологический
контроль. У грибов имеются специальные регуляторные
механизмы, действующие через выделение специальных метаболитов,
несущих функцию гормонов.
ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯ ПОЛОВОГО ПРОЦЕССА
У ГРИБОВ
Один из способов, с помощью которого клетки
взаимодействуют друг с другом, — это диффундирующие в среду
метаболиты, структура и состав которых в соответствующих
количествах дают «адрес» клетке, по которому она двигается в нужном
направлении. Такие вещества у грибов называют прогамонами,
или ферромонами. Они выполняют функцию гормонов. Выделение
такие половых «гормонов» гетероталличными штаммами
увеличивает возможность слияния двух штаммов противоположного
знака. Через них осуществляется воздействие на особи
противоположного пола.
Ферромоны индуцируют образование половых структур на
мицелии другого знака или служат аттрактантами для привлечения
гамет или мицелия партнера по спариванию и способствуют
окончательному слиянию половых клеток. Таким образом, эти
соединения являются своеобразными «средствами общения» между
особями отдельных видов.
Естественно, что изучение способа действия этих веществ и
их роли в процессе развития требует знания их химической
структуры. В настоящее время для многих ферромонов установлена
химическая структура. Известна эмпирическая и структурная
формула ферромона рода Allomyces из класса хитридиомицетов,
получившего название «сиренин» (С15Н24О2, молекулярная
масса 236). Биологическая активность сиренина проявляется от
42
10-5 до 10~10 М. Грибы рода Allomyces имеют подвижные
женские и мужские гаметы. Однако женские гаметы раньше теряют
подвижность и привлекают мужские гаметы, выделяя сиренин.
После привлечения мужской гаметы обе гаметы сливаются,
образуя зиготу, которая теряет подвижность и прорастает в
диплоидный мицелий. Клетка зиготы быстро теряет чувствительность
к сиренину и начинает привлекаться в направлении источника
аминокислот. В направлении градиента концентрации этих
веществ движение постепенно подавляется.
У грибов рода Achlya из класса оомицетов, в частности А. Ы-
sexyalis, механизм полового процесса, по Раперу, выглядит
следующим образом: соматическая гифа женского мицелия выделяет
ферромон А, который индуцирует образование антеридиальных
ветвей мужского мицелия. Антеридиальные ветви продуцируют
ферромон В, названный в настоящее время антеридиолом и
относящийся к группе стероидов, который стимулирует образование
оогониального вздутия. Последнее образует ферромон С, или оого-
ниол, который притягивает антеридий мужского мицелия к
женскому. После слияния (плазмогамии и кариогамии)
происходит отделение антеридия и оогония перегородкой и
образуется ооспора, прорастающая после периода покоя.
Предшественником обоих этих соединений является фукосте-
рол, который в свою очередь может стимулировать" образование
ооспор у Phytophthora и Pythium этого же класса оомицетов.
Таким образом, гриб Achlya, несмотря на то что является
примитивным эукариотным организмом, имеет ферромоны
стероидного типа аналогично высшим животным, которые можно
отнести к истинным гормонам.
Среди класса зигомицетов в отношении гормональной
регуляции полового процесса изучены несколько видов Mucor mucedo,
Blakeslea trlspora, Phycomyces blekesleanus. Например, у М.
mucedo образуются половые гифы — зигофоры или гаметангии,
когда мицелии разных половых знаков достигают друг друга.
Зигофоры хорошо отличаются от вегетативных гиф, они растут
навстречу друг другу и после контакта вздуваются, сливаются
и образуют зигоспору. X. Бургеф в 1924 г. показал, что
образование зигоспор контролируется низкомолекулярным веществом,
которое диффундирует через полупроницаемую мембрану.
Установлено, что индуцирование зигофоров осуществляется триспоро-
вой кислотой и ее предшественниками, каковыми являются каро-
тиноиды. Она имеет эмпирическую формулу С20Н25О5,
молекулярную массу от 314 до 374 и представляет собой соединение
неароматической природы с ненасыщенными связями.
У базидиомицетов известен ферромон тремеллоген А-10. Этот
фактор ингибирует ядерное и клеточное деление и индуцирует
образование копуляционных гиф. У базидиомицетных дрожжей
Rhodosporidium вещество родоторуцин — липофильный пептид,
задерживающий почкование и индуцирующий образование
копуляционных трубок.
43
Таким образом, основные черты гормональной регуляции
полового процесса гетероталличных грибов характеризуются тем,
что один из них или оба продуцируют сигнальные метаболиты в
малых количествах. В ответ на это партнер выделяет второе
эффективное вещество, которое вызывает ряд химических
взаимодействий, приводящих к образованию половых структур,
привлечению их друг к другу и окончательному слиянию.
Особенностью триспоровой кислоты, образуемой мукоровыми грибами,
является ее способность индуцировать половые структуры в обоих
половых типах Mucor mucedo без образования другого ферромона.
ОНТОГЕНЕЗ ГРИБОВ (ЦИКЛЫ РАЗВИТИЯ)
И СМЕНА ЯДЕРНЫХ ФАЗ
Грибы имеют сложный цикл индивидуального
развития — онтогенеза, в процессе которого они проходят несколько
морфогенетических и ядерных стадий (ядерных фаз).
Жизненный цикл грибов можно рассматривать как развитие
организма от прорастания споры, через образование мицелия,
различных форм репродуктивных органов, до образования спор.
Большинство грибов характеризуются сложным циклом развития,
состоящим из нескольких, следующих друг за другом
морфогенетических стадий. Они образуют различные, сменяющие друг
друга типы спороношений. Морфогенетические стадии в цикле
развития следующие: 1) прорастание споры; 2) образование
вегетативного мицелия; 3) образование бесполых спороношений с
последующим образованием спор; 4) образование половых
спороношений с последующим образованием спор.
Иногда цикл развития грибов ограничивается образованием
только бесполых спороношений. Эта стадия развития носит
название анаморфы. Цикл может включать и только половую
стадию — телеоморфу. Бесполая и половая стадии, представленные
одновременно, называются голоморфой.
В жизненном цикле грибов может быть ряд следующих друг
за другом бесполых спороношений (анаморф), которые затем
сменяются половым спороношением (например, у ржавчинных).
Наличие разнообразных по морфологии типов спороношения, т. е.
морфогенетических стадий, у одного вида гриба носит название
плеоморфизма.
Бесполые и половые спороношения у разных грибов имеют
очень разнообразное строение. Каждой морфогенетической
стадии развития гриба соответствует ядерная фаза, которая
характеризуется определенным набором хромосом в ядре. На
соответствующих морфогенетических стадиях развития организм
содержит либо одинарный (гаплоидный), либо двойной (диплоидный)
набор хромосом.
Таким образом, у всех организмов существуют две ядерные
фазы. Все высшие организмы основную часть цикла разви-
44
тия проходят в диплоидной фазе, гаплоидны только половые
клетки.
Различные низшие организмы имеют преобладание в цикле
развития либо диплоидной, либо гаплоидной стадии.
Грибы обладают уникальной особенностью: у многих из них
жизненный цикл характеризуется тремя ядерными фазами —
гаплоидной, диплоидной и дикариотической. Гаплоидная фаза
возникает в результате мейоза (редукционного деления ядер),
характеризующегося расхождением хромосом и наличием одинарного
набора хромосом. Диплоидная фаза
начинается с кариогамии, т. е. слияния ядер и ^^^^^/
образования двойного набора хромосом в
ядре, и кончается мейозом. Дикари-
отическая фаза образуется - в резуль-
Рис. 18. Схема Рапера по соотношению
гаплоидной (I), дикариотической (II) и диплоидной (III)
ядерных фаз у грибов:
М — наличие мейоза.
Циклы развития: / — бесполый; 2 — гаплоидный;
3 — гаплоидно-дикариотический; 4 — гаплоидно-
диплоидный; 5 — диплоидный
тате ассоциации ядер в строго параллельные синхронно
делящиеся пары — дикарионы без их слияния. У разных групп
грибов существуют различные соотношения ядерных фаз в цикле
развития, соответствующие морфогенетическим стадиям, на
основании чего можно выделить пять основных типов жизненных
циклов грибов: 1) бесполый; 2) гаплоидный; 3)
гаплоидно-дикариотический; 4) гаплоидно-диплоидный; 5) диплоидный. Эти
циклы развития можно демонстрировать схемой Рапера (рис. 18).
ДИФФЕРЕНЦИАЦИЯ ГРИБОВ В ОНТОГЕНЕЗЕ
В процессе онтогенеза у грибов в связи с образованием
вегетативных и репродуктивных структур происходит
дифференциация клеток мицелия по форме, размерам, химическому строе
нию. В основе дифференциации лежит процесс активации — реп
рессии определенных генов. Активация одних генов и репрессия
других происходят в ответ на сигнал, поступающий из
окружающей среды.
У высших высокодифференцированных организмов источником
сигнала служат в основном физические и химические
взаимодействия между клетками. Сигналы возникают как в результате пря*
мых межклеточных контактов, так и через выделение специфиче
ских гормонов. У грибов как низших эукариотных организмов
без выраженной дифференциации на органы и ткани реализация
45
программы индивидуального развития осуществляется за счет-
сигналов, поступающих из внешней среды.
Примером регуляции дифференциации фактором внешней
среды может служить гриб из класса хитридиомицетов Blastocla-
diella emersonii, открытый Е. Кантино в 1961 г. Гриб В. enter-
sonii характеризуется тем, что он может развиваться двумя
путями. Первый путь — образование активного зооспорангия с
подвижными зооспорами. Спорангий формируется из мешковидной
ценоцитной клетки путем возникновения базальной септы,
отделяющей эту клетку от ризоидов. Второй путь — образование
покоящегося толстостенного спорангия с меланизированной
оболочкой. Сигналом для образования того или иного спорангия
является концентрация бикарбоната или СОг в среде. При низком
уровне содержания бикарбоната образуется зооспорангий с
подвижными зооспорами, которые через некоторое время теряют
подвижность, инцистируются и прорастают в ростковую трубку.
Высокий уровень бикарбоната в среде приводит к образованию
покоящегося спорангия. Установлено, что в ходе этого процесса
подавляется активность целого ряда ферментов-дегидрогеназ три-
карбонового цикла, что приводит к накоплению отдельных кислот
этого цикла и изменению направления синтетических процессов.
Происходит образование меланинов в клеточной стенке и каро-
тиноидов — в цитоплазме, увеличивается синтез хитина, липидов
и растворимых полисахаридов.
Таким образом, концентрация бикарбоната в среде влияет
на пусковую реакцию организма, которая может переключать
развитие гриба с одного пути морфогенетического развития на
другой.
СПОРЫ ГРИБОВ И ИХ ПРОРАСТАНИЕ
Процесс бесполого и полового размножения у грибов^
заканчивается обязательным образованием спор, которые служат
дальнейшему размножению индивидуума.
Споры грибов по своему биологическому значению можно*
разделить на две основные группы. Одни — покоящиеся споры,,
обеспечивают сохранение вида в течение периода,
неблагоприятного для вегетации. Другие — споры размножения, или пропага-
тивные споры, служат для быстрого размножения.
Покоящиеся споры образуются обычно в небольшом числе
нередко внутри субстрата и лишены особых приспособлений для
рассеивания. Они не способны к немедленному прорастанию и
нуждаются в определенном периоде покоя, во время которого
происходит, по-видимому, дозревание. Покоящиеся споры
принадлежат к диплоидной фазе развития гриба и у многих являются
непосредственным продуктом оплодотворения, как ооспоры ооми-
цетов и зигоспоры мукоровых. К покоящимся спорам относятся
и хламидоспоры, телеоспоры и некоторые другие.
46
Пропагативные споры образуются обычно в больших
количествах и имеют целый ряд приспособлений для рассеивания.
Пропагативные споры принадлежат к гаплоидной фазе развития и
являются либо продуктами бесполого размножения, либо хотя
и берут начало от полового воспроизведения, но имеют перед
своим образованием редукционное деление. К пропагативным
спорам относятся спорангиоспоры, конидии, зооспоры, аско- и ба-
зидиоспоры.
Однако не все пропагативные споры способны к немедленному
прорастанию. Пропагативные споры по скорости прорастания
также можно разделить на две группы.
I. Прорастают после латентного периода от 30 мин до
нескольких часов.
II. Прорастают с периодом покоя от одной недели до месяца.
Прорастание — стадия, которая отличается от покоя
морфологическими, цитологическими и биохимическими критериями.
Период прорастания споры можно разделить на три стадии.
Первая стадия — начало прорастания. На этой стадии не
происходит никаких внешних морфологических изменений, но
осуществляется активация ферментов в споре. В первую очередь
активируются гидролитические ферменты — фосфатазы, полифосфат-
киназы, окислительные ферменты — каталаза и осуществляется
активный синтез нуклеиновых кислот.
Вторая стадия — набухание. Цитоплазма на этой стадии
наполняется водой, спора вздувается. Все споры обычно содержат
мало воды, и поэтому гидратация с набуханием споры очень
существенная ступень в прорастании. Сначала изменяется
проницаемость стенки спор. На этой стадии исчезают жировые капли,
увеличивается базофилия.
Третья стадия — на этой стадии происходят определенные
морфологические изменения, намечается полярный центр роста,
появляется ростковая трубка.
Образование ростковой трубки требует синтеза многих
клеточных компонентов: протеина, РНК, полисахаридов, липидов,
используемых в синтезе клеточной стенки, мембран, органелл.
Многие споры грибов прорастают просто в воде, т. е. обладают
достаточными конструктивными и энергетическими резервами.
Другие образуют ростковую трубку только при снабжении
источниками углерода и азота, солями или витаминами. Прорастание
спор обычно сопровождается увеличением синтеза протеинов и
концентрации циклических нуклеотидов.
Состояние покоя у спор может быть конститутивным, т. е.
обусловленным внутренними причинами, морфологическими и
физиологическими особенностями, или экзогенным, зависящим от
внешних условий. Например, аскоспоры Neurospora tetrasperma
представляют собой споры с типичным конститутивным покоем,
который обусловлен наличием у них толстой меланизированной
оболочки, клеточная стенка которой содержит хитин. Другой
причиной конститутивного покоя может быть наличие в спорах внут-
47
ренних ингибиторов. В настоящее время многие из них выделены
и идентифицированы. Например, в уредоспорах ржавчинного
гриба Uromyces phaseoli находится самоингибитор метил-чыс-3,4-ди-
метоксицинамат (МДС), в уредоспорах другого ржавчинного
гриба Puccinia graminis — метил-^ис-ферулат (MF), в конидиях Ре-
ronospora tabacina, вызывающего болезнь мучнистой росы табака,
5-изо-бутирокси р-ионон (киесон), ингибирующий синтез протеина.
Самоингибиторы очень специфичны и активны в очень низких
концентрациях. Однако они не всегда имеют уникальное
химическое строение: у Phomopsis viticola это элементарная сера, у Col-
letotrichum musae — железо.
Для прекращения конститутивного покоя требуется удаление
оболочки, а также необходима активация спор, которая может
быть достигнута нагреванием до 50—60° С, автоклавированием,
обработкой химическими реагентами: кислотой, фурфуролом или
другими гетероциклическими соединениями, мочевиной, гуаниди-
ном, диметилсульфоксидом. Предполагают, что они действуют или
как естественные катализаторы, или инактивируют имеющиеся
в споре естественные ингибиторы. В результате такой активаций
возрастает проницаемость клеточной стенки для анионов и
катионов и прорастание может наступить через час после
помещения в дистиллированную воду. При этом утилизируются
эндогенные резервы аскоспор, жиры на 27% сухого веса споры, углеводы,,
состоящие в основном из трегалезы, на 33%.
В этот период возрастает содержание пируватов, этанола, аце-
тальдегида, что происходит в связи со смещением метаболизма
с гликолитического в сторону окислительного, увеличивается
содержание продуктов окислительного обмена, таких, как а-кето-
глютаровая кислота. При этих условиях становится необходимым
снабжение спор экзогенными гексозами. Меченая глюкоза
непосредственно включается в аминокислоты, и содержание протеинов
увеличивается вдвое за 2 ч активации.
Экзогенный покой определяется внешними факторами:
влажностью, температурой, рН, концентрацией СОг, токсинами во
внешней среде. Спорам многих грибов достаточно для прорастания
быть суспендированными в воде или инкубированными при
высокой влажности. Отсутствие достаточного эндогенного питания
также может быть причиной покоя у спор. Поэтому многие споры
прорастают при добавлении глюкозы, NH4N03, P04~, S04~.
Некоторые виды грибов для прорастания спор требуют наличия
специфических веществ, например споры Memnoniela echinata
нуждаются в биотине, а споры Glomierella в наличии С-, N-, Р- и
S-содержащих соединений. Споры миксомицета Dictiostellum dis-
coideum прорастают при добавлении смеси триптофана, фенила-
ланина, метионина. Возможно, что они активируют
метаболические пути, которые позволяют спорам прорастать.
Глава 3
ОСНОВНЫЕ ТАКСОНОМИЧЕСКИЕ ГРУППЫ
ГРИБОВ
Со времени создания Дарвином эволюционной теории*
в основу классификации живых организмов кладется
филогенетический принцип, имеющий целью классифицировать организмы
нл основе родственных связей, установленных путем раскрытия
основных направлений эволюции живого мира. Для этого
используют следующие основные методы: морфолого-анатомический,
цитологический, онтогенетический, палеонтологический, геогр а-
фический.
Однако для грибов почти невозможно использование целого
ряда вышеперечисленных методов, таких, как
палеонтологический, так как ископаемые остатки грибов почти не сохранились,
ограниченно применение географического метода из-за часто
очень широких, неограниченных ареалов некоторых видов грибов
и т. д. Для построения классификации грибов требуется
привлечение дополнительных новых методов, например физиолого-био-
химического, экологического и др.
Традиционно, следуя представлениям Линнея, грибы всегда
рассматривали среди растительного царства, исходя из
существования двух царств живых организмов. Однако по мере
накопления знаний стало очевидным, что есть много организмов,
которые не являются ни растениями, ни животными, хотя им присущи
некоторые черты и тех и других. Система двух царств перестала
удовлетворять современных биологов. Поэтому весь живой мир
ряд исследователей, в частности Уиттакер и другие, предлагают
делить не на два царства: животных и растений, как это было
предложено Линнеем, а на пять царств.
1. Monera (Procaryota) — его представители четко
отличаются от всех других организмов отсутствием ядерной мембраны,
присущей эукариотным организмам.
2. Protista — одноклеточные подвижные микроскопические
организмы с ядром и ядерной мембраной. Одни из них
пигментированные и автотрофны, другие бесцветные и гетеротрофны.
Эти организмы дали, по-видимому, начало многоклеточным
организмам, которые эволюционировали на ранних этапах независимо
и имеют фундаментальные различия в типе питания и обмене
веществ.
3. Planta (Metaphyta) — растения, организмы с автотрофным
обменом веществ.
4. Animalia (Metazoa) — животные с гетеротрофным обменом,
питание которых осуществляется по типу заглатывания.
5. Mycota (Mycophyta) — грибы, гетеротрофные организмы с
адсорбционным типом питания (рис. 19).
Выделение грибов в самостоятельное царство обусловлено тем,
что они занимают четко обособленное положение в системе жи-
49
вых организмов, которое определяется их самостоятельным путем
эволюции. По типу строения они одноклеточные и многоклеточные
эукариотные организмы, но не имеющие тканевого строения, а
следовательно относящиеся к низшим эукариотам, куда относят
водоросли и простейшие. Однако от первых отличаются
фундаментально иным способом получения энергии, от вторых — иным
адсорбционным типом питания в отличие от голозойного у
простейших. Кроме того, они имеют уникальные, свойственные только
грибам черты — гетерокариоз (разнокачественность ядер в одной
клетке), наличие парасексуального процесса, наличие дикарио-
тической стадии в цикле развития.
Они выделились в самостоятельный ствол еще до разделения
живых организмов на животных и растений и представляют собой
самостоятельную филогенетическую линию. Подтверждением это-
Piaqtae Fungi Animalia
Рис. 19. Схема пяти царств живых организмов (по Вуттакеру)
му служат общие черты и тех и других. Черты животных
организмов, присущие грибам, следующие: гетеротрофный тип обмена
по углероду, потребность в витаминах, наличие гликогена в
клетках, синтез хитина, образование и накопление мочевины,
первичная структура цитохромов и транспортной РНК. Черты, характер-
50
ные для растений, — клеточная полярность, верхушечный рост,,
строение клеточной стенки, образование поперечных перегородок.
Уникальные черты, свойственные только грибам, — гетерокариоз
(разнокачественность ядер в одной клетке), наличие парасексу-
ального процесса, наличие дикариотической стадии в цикле
развития.
В систематике и классификации грибов из всех перечисленных
широко используются морфологический, цитологический и
онтогенетический методы. Последний основывается на изучении всех
стадий жизненного цикла, в котором большое место занимает
характеристика полового воспроизведения, а у более примитивных
грибов — и характер жгутиковой стадии. В качестве
вспомогательных характеристик организмов служат некоторые
биохимические признаки, которые в последнее время приобретают все
большее значение, хотя материала по хемотаксономии грибов еще
очень мало.
Основные таксономические категории, которые приняты в
классификации грибов, следующие: царство, отдел, класс, порядок,
семейство, род, вид. Например: царство Mycota, отдел Eumycota,
класс Ascomycetes, подкласс Euascomycetidae, порядок Eurotiales,
семейство Trichocomataceae, Emericella nidulans. Многие из этих
категорий разделяются еще на подгруппы.
Царство — самая большая категория, включает несколько
отделов, каждый отдел несколько классов и т. д. Виды разбивают
на варианты, биологические штаммы, физиологические расы.
В соответствии с Международным кодексом ботанической
номенклатуры названия отделов грибов имеют окончания — mycota,
подотдела — mycotina, класса — mycetes, подкласса — myceti-
deae. Названия порядков — ales, семейств — асеае. Роды и виды
не имеют стандартных названий.
Для каждого организма принята биноминальная система
названий, предложенная Линнеем, т. е. название организма состоит
из двух слов. Первое — это род, к которому организм
принадлежит, второе — видовой эпитет. Дальше стоит имя ученого,
описавшего вид. Если стоит два имени, то первое в скобках
принадлежит автору, впервые описавшему данный организм, но
отнесшему его к другому роду. Следующее имя без скобок — автора,
отнесшего его к роду, к которому он правильно должен
относиться, на основании более детального исследования. Например: Мог-
tierella ramanniana (Moeller) Lin. Мёллер первый описал этот
организм, но отнес его к роду Mucor, когда он должен быть
отнесен к роду Mortierella, к которому его отнесла Линнеман.
Указание на двух авторов помогает найти первое описание
организма.
Система пяти царств также имеет свои недостатки, так как
критерии выделения каждого царства неравнозначны (наличие
ядерной мембраны, одноклеточность, многоклеточность, тип
питания).
Ежегодно в мире описывают новые виды грибов. В среднем
51
„до 600—700 видов за год. Одновременно осуществляется ревизия
старых видов, ошибочно отнесенных к иному роду, чем они
должны относиться. Такая работа проводится в ряде коллекций мира.
Наиболее известные из них следующие: CBS — Голландия CMJ—
Англия, АТСС - США, ССМ - Чехословакия, ВКМ - СССР
и др.
Царство грибов делится на несколько отделов, которые в свою
очередь включают ряд классов. Подразделение грибов на классы
основано на использовании комплекса признаков. Главными
признаками, которые кладутся в основу деления грибов на классы,
являются характер развития спор полового процесса, типы
полового процесса и бесполого размножения, количество, строение и
расположение жгутиков грибов, имеющих подвижные стадии в
цикле развития, состав клеточной стенки.
По современным представлениям система грибов выглядит
следующим образом.
Царство Mycota
отдел Oomycota
класс Hyphochytridiomycetes
класс Oomycetes
отдел Chytridiomycota
класс Chytridiomycetes
отдел Eumycota
класс Zygomycetes
класс Ascomycetes
класс Basidiomycetes
класс Deuteromycetes (Fungi imperfecti)
В настоящее время наиболее обосновано представление о ди-
фелитическом происхождении грибов от двух групп бесцветных
Flagellata (жгутиконосцев). От двужгутиковых флагеллят
произошли грибы класса оомицетов, имеющие два жгутика у зооспор.
Все остальные грибы — от одножгутиковых флагеллят, часто
способных терять жгутик в процессе вегетативного роста. Эти грибы
можно расположить в следующий эволюционный ряд: от самых
примитивных хитридиомицетов до более высокоразвитых бази-
диальных грибов: хитридиомицеты — зигомицеты — сумчатые —
базидиальные.
Ниже приводятся классы и порядки грибов, которые имеют
своих представителей в почве.
КЛАСС CHYTRIDIOMYCETES
Этот класс включает примерно 110 родов и 575 видов.
Представители его характеризуются либо полным отсутствием
мицелия (вегетативное тело представлено плазменной массой),
либо зачаточным ризомицелием в виде тонких неветвящихся
нитей, отходящих от основной округлой или удлиненной клетки
таллома, либо более развитым ветвящимся ризомицелием без
перегородок.
52
Если вегетативное тело немицелиального строения, то
клеточная стенка у такой плазменной массы отсутствует. Хитридиевые
грибы такого строения обычно являются внутриклеточными
паразитами растений; всей поверхностью они всасывают питательные
вещества из клетки хозяина. Если есть мицелий, то имеется и
клеточная оболочка, а основной ее компонент представлен
комплексом хитин — глюкан, В состав протеинового комплекса
входит аминоадипиновая кислота (AAA). Процентное содержание
ГЦ-оснований в ядерной ДНК колеблется от 34 до 64%.
Характерной особенностью этого класса, отличающего его от
всех других грибов, является то, что зооспоры и планогаметы
{элементы размножения) имеют один задний жгутик,
построенный по типу кнута.
Бесполое размножение хитридиомицетов осуществляется с
помощью зооспор, образующихся в зооспорангиях. В молодой
стадии спорангии наполнены цитоплазмой, содержащей много ядер.
По мере развития спорангия протопласт распадается на большое
число участков, каждый из которых развивается в одноядерную
зооспору различного строения и формы, но всегда с одним биче-
видным гладким жгутиком, прикрепленным сзади. Зооспоры
плавают некоторое время в воде или почвенном растворе, инцисти-
руются, теряют жгутик, затем прорастают, после короткого
периода покоя. Иногда эти клетки, потерявшие жгутик, снова
прорастают зооспорой. У некоторых хитридиевых грибов все
вегетативное тело превращается в спорангий, и жизнь данной особи
прекращается (голокарпические формы), у других только какая-
то часть становится зооспорангием (эукарпические формы).
Половое размножение осуществляется как гаметогамия
следующими способами.
1. Копуляцией планогамет, которая включает слияние изога-
мет и слияние гетерогамет. При этом способе конъюгации две
гаметы, копулирующие между собой, морфологически сходны, но
физиологически могут быть различны. При втором способе
конъюгации одна гамета значительно больше другой.
2. Оплодотворением неподвижной женской гаметы (оогония)
подвижной мужской гаметой. Подвижная мужская гамета
выходит из мужского гаметангия (антеридия) в воду или почвенный
раствор и плавает некоторое время. После встречи такой гаметы
с оогонием происходит их копуляция. Зигота, возникающая в
результате копуляции половых клеток, превращается в покоящуюся
спору, одетую толстой оболочкой, содержащей хитин, которая и
прорастает у некоторых видов в диплоидный ценоцитный
(неклеточный) таллом. Бесполая стадия развития у большинства
представителей протекает в гаплоидной фазе, половая — часто в
диплоидной (рис. 20).
Многие Chytridomycetes — типичные водные организмы, но
некоторые из них обитают в почве, а также в тканях наземных
растений как паразиты или на отмерших остатках органического
материала, на котором они растут. Некоторые водные предста-
53
вители класса хитридиевых — паразиты на водорослях,
вызывающие нередко их массовую гибель, или сапротрофы, разрушающие
отмершие остатки водорослей, являясь звеном в цепи питания
водных животных.
В составе класса имеются три порядка, различающихся
степенью развития таллома и формой полового процесса: Chytridia-
les (хитридиевые), Blastocla-
2 з diales (бластокладиевые), Мо-
noblepharidales (моноблефа-
ридиевые).
Порядок Chytridiales
характеризуется отсутствием
истинного мицелия, но у некоторых
есть ризомицелий и ризоиды.
В вегетативном состоянии
эти организмы не имеют
клеточной оболочки, оболочку
имеет только зооспорангий и
зигота. Половое размножение
их осуществляется
подвижными изо- и гетеропланогамета-
ми. Представители порядка
Рис. 20. Цикл развития Olpidium
viciae:
1 — зооспоры; 2 — заражения-
клетки хозяина; 3 — протопласт
паразита в клетке хозяина; 4 —
зооспорангий паразита в клетке
хозяина; 5 — прорастание зооспоран-
гиев; 6 — слияние гамет; 7 — пла-
нозигота; 8 — поступление
содержимого планозиготы в клетку
хозяина; 9 — цисты в клетке
хозяина; 10 — мейоз в цисте
Chytridiales роды Synchytrium и Olpidium — паразиты важных
сельскохозяйственных растений. В частности, гриб Synchytrium endo-
bioticum — возбудитель распространенного в Европе заболевания
картофеля, носящего название «рак картофеля». Olpidium brassi-
сае — возбудитель гнили капустной рассады — болезни,
известной как «черная ножка».
Порядок Blastocladiales характеризуется истинным мицелием,,
без перегородок. Бесполое размножение осуществляется
зооспорами, половое — с помощью подвижных изо- и гетерогамет.
Представители этого порядка роды Allomyces и Blastocladiella —
важные объекты для изучения морфогенеза грибов. Виды рода
Allomyces обитают в воде и почве на остатках растений и трупах
животных.
На представителях рода Allomyces показана физиологическая,,
54
гормональная регуляция полового процесса у грибов. Мужские
гаметы привлекаются к женским, даже если они отделены друг
от друга коллодиевой пленкой. Установлено, что причиной этого
феномена является растворимый зиготропный гормон сиренин,
выделяемый женскими гаметами.
Виды рода Allomyces — одни из первых грибов, у которых
были обнаружены каротиноидные пигменты в виде у-каротина,
который преимущественно содержится в мужских гаметах. На
видах рода Blastocladiella, в частности В. emersonii, показана
возможность регуляции развития воздействием определенных
внешних факторов среды.
Порядок Monoblepharidales имеет истинный мицелий.
Бесполое размножение осуществляется зооспорами, половой процесс
оогамный. Половое размножение происходит с помощью
неподвижных оогониев и антеридиев, в которых образуются подвижные
гаметы. Грибы этого порядка развиваются как сапротрофы в
пресной воде, на сучьях, ветках, плодах и трупах насекомых.
Представитель этого порядка — род Monoblepharis.
КЛАСС OOMYCETES
Класс объединяет около 70 родов и 550 видов. Класс
оомицетов в современном понимании представляет собой хорошо
очерченную группу, объединяемую на основании характерного
полового процесса — оогамии, строения элементов бесполого
размножения — зооспор, своеобразного, отличного от других грибов
химического состава клеточной стенки. Вегетативное тело
варьирует от одноклеточного образования до хорошо развитого цено-
цитного (неклеточного) мицелия. Основной полисахаридный
комплекс клеточной стенки этих грибов представлен комплексом
целлюлоза — глюкан со связями р-1,3; (3-1,4; р-1,6 в отличие от
многих других грибов, где основу клеточной стенки представляет
комплекс хитин-глюкан, а протеин клеточной стенки в качестве
основной аминокислоты содержит диаминопимелиновую кислоту
(ДАП) в противоположность другим грибам, содержащим а-ами-
ноадипиновую (AAA). Процентное содержание ГЦ-оснований в
ядерной ДНК колеблется в пределах от 45 до 62%.
К классу оомицетов относят грибы, зооспоры которых имеют
два жгутика, один из которых перистый и направлен вперед,
другой — кнутовидный и направлен назад.
Этот класс характеризуется оогамным половым процессом. Он
выражается в оплодотворении женской яйцеклетки, находящейся
в специальном образовании — оогонии, недифференцированным
содержимым мужской клетки через посредство оплодотворяющего
отростка — антеридия (рис. 21).
Оогоний имеет шаровидную форму и одет довольно толстой
оболочкой, в которой заметны более тонкие места — поры. Через
них происходит впоследствии оплодотворение. Первоначально
сплошное многоядерное содержимое оогония распадается затем
55
на несколько одноядерных яйцеклеток, свободно лежащих в виде
шаров. Внутри оболочки иногда остается только одна
яйцеклетка, на образование которой идет центральная часть плазмы.
Мужская клетка — антеридий — представляет собой округлую
или цилиндрическую клетку, развивающуюся на том же мицелии
или на другом со многими клеточными ядрами, но лишенную
дифференцировки на
обособленные гаметы. Антеридий к
моменту созревания плотно
соприкасается с оогонием и
пускает через поры отростки,
после чего содержимое антеридия
поступает внутрь яйцеклетки и
затем осуществляется
плазмогамия и кариогамия, которые
не разделены во времени.
После оплодотворения
яйцеклетки оогоний превращает-
Рис. 21. Цикл развития Phytophtho-
ra infestans:
1, 2 — зооспорангий; 3 — выход,
зооспор; 4 — прорастание
ростковой трубкой; 5 — внедрение гифы
или зооспоры в ткань растения; 6 —
слияние оогония и антеридия; 7 —
оплодотворенный оогоний; 8 —
ооспора; 9 — прорастание ооспоры
ся в покоящуюся клетку — ооспору с толстой, часто скульптуриро-
ванной оболочкой.
Большинство оомицетов имеют бесполый и половой циклы
развития. Часто наблюдается несколько генераций бесполого
цикла и затем только одна — полового. У многих оомицетов в
качестве элементов бесполого размножения образуются зооспоры, на
иногда спорангий сам выполняет функцию споры и прорастает
непосредственно ростковой трубкой, которая дает мицелий.
Основная часть жизненного цикла оомицетов проходит в
диплоидном состоянии. Мейоз наступает при заложении половых органов.
Гаплоидны только яйцеклетки и ядра антеридия. После их
слияния снова восстанавливается диплоидность. Многие оомицеты —
наземные облигатные паразиты, проводящие полный жизненный
цикл на растении-хозяине, однако образование зооспор говорит
об их водном происхождении.
Класс Oomycetes делится на несколько порядков: из них
почвенные обитатели — представители порядков Saprolegniales (ca-
пролегниевые) и Peronosporales (пероноспоровые). Представители
порядка Saprolegniales в большинстве своем гомоталличные
формы и образуют совместимые оогоний и антеридии на одном
талломе. Но есть среди них и двудомные, гетероталличные, имеющие
- ^%
56
двух индивидуумов для половой репродукции — женский и
мужской. Такие виды, как Achlya bisexualis и Achlya ambisexualis,
использованы Рапером в исследованиях полового механизма. Ра-
пер продемонстрировал, что если потенциально мужские и
женские талломы растут рядом, то система имеет несколько
гормонов, которые инициируют начало полового процесса (см. с. 43).
Почвенные обитатели порядка Saprolegniales представлены
родом Aphanomyces. Среди них есть паразиты корней сосудистых
растений, вызывающие заболевание сахарной свеклы, гороха и
других растений, которое носит название «корнееда».
Порядок Peronosporales содержит водные, амфибоидные и
наземные организмы, многие из них высоко специализированные
облигатные паразиты. Основные представители почвенных грибов
относятся к семейству Pythiaceae и Peronosporaceae. Бесполое
размножение грибов семейства Pythiaceae осуществляется через
образование спорангиев, которые могут прорастать зооспорами
или прямо в ростковую трубку в зависимости от внешних
условий, главным образом влажности и температуры. Основные роды
этого семейства — Pythium и Phytophthora. Род Pythium
включает как водные формы, паразитирующие на водорослях, так и
почвенных обитателей, представляющих собой паразитов на
высших растениях, но способных расти и сапротрофно в почве.
Например, P. debaryanum, будучи патогеном, вызывает гниль
сеянцев, но может жить и как сапротроф.
Род Phytophthora включает как сапротрофные формы,
могущие жить длительно в почве или на растительных остатках, так
и специализированные паразиты на наземных органах растений.
К числу последних относится известный картофельный гриб Ph.
infestans, поражающий как клубни, так и ботву картофеля.
Мицелий этого гриба полностью эндофитный, т. е. находится в тканях
растения, проходит по межклетникам и дает отростки —
гаустории — внутрь клеток. На эндофитном мицелии образуются спо-
рангиеносцы, которые высовываются через устьица наружу и на
концах образуют спорангии в виде лимонообразных тел.
Характерная черта спорангиеносцев фитофторы — симподиальный тип
ветвления, при котором рост спорангиеносца не заканчивается
образованием конечного спорангия, а дает боковую ветвь,
сдвигающую в сторону первый спорангий, и т. д. Спорангий отпадает
целиком, и прорастание его чаще всего происходит зооспорами,
но при определенных условиях, в частности низкой влажности и
высокой температуре, спорангий прорастает как конидия,
развиваясь прямо в гифу, внедряющуюся в ткань растения.
Половые структуры Phytophthora infestans в виде типичных
оогониев и антеридиев образуются не в ткани питающего
растения, а в сапротрофных условиях, хотя образование ооспор
наблюдается редко, так как гриб гетероталличен и для их образования
необходимо присутствие обоих типов спаривания.
Ph. infestans завезен в Европу из Америки в 30-х гг.
прошлого столетия. Первое массовое развитие его в Европе было в
57
1849 г., которое вызвало гибель большой части урожая
картофеля, особенно в Ирландии, что вызвало усиленную иммиграцию
населения Ирландии в Америку.
Семейство Peronosporaceae включает много родов,
различающихся ветвлением спорангиеносцев, в отличие от представителей
семейства Pythiaceae, которые не имеют дифференцированных
спорангиеносцев. Сюда входят роды Plasmopara, Peronospora,
Bremia и др. Многие роды Peronosporaceae имеют спорангии,
прорастающие зооспорами, только представители родов
Peronospora и Bremia прорастают ростковой трубкой и таким образом пред-
ставляют собой фактически конидию. Микологи предпочитают
называть эти образования спорангиями по аналогии с другими
Peronosporales, фитопатологи — конидиями.
Образующиеся в результате полового процесса ооспоры у
Peronospora прорастают после периода покоя спорангием. Ооспоры
образуются в ткани пораженного растения и перезимовывают на
опавших листьях. Все виды семейства Peronosporaceae — обли-
гатные паразиты растений, вызывающие болезнь мильдью на
различных растениях. Например, Plasmopara viticola вызывает
мильдью на винограде, Peronospora destructor — на луке, Bremia
lactucae — на салате. Все названные паразиты наносят большой
вред сельскому хозяйству.
КЛАСС ZYGOMYCETES
Этот класс насчитывает до 85 родов и более 500 видов.
Класс Zygomycetes представляет собой группу грибов, полностью
утративших подвижные стадии в цикле развития.
Главными характеристиками этого класса являются: 1)
половое размножение — гаметангиогамия или зигогамия, откуда и
произошло название класса; 2) бесполое размножение
осуществляется неподвижными спорангиоспорами или конидиями.
Гаметангиогамия (зигогамия) выражается в слиянии двух
клеток мицелия, содержимое которых лишено какой бы то ни было
дифференцировки на особые гаметы. Эти клетки носят название
гаметангиев. Большей частью они неразличимы внешне, но могут
различаться размером. Гаметангии образуются как вздутия на
вершине двух совместимых гиф или ветвей гиф, которые
направляются друг к другу и сливаются (рис. 22). Стенка между ними
разрушается и содержимое смешивается. Образуется одна
клетка, отделяющаяся перегородками от несущих гиф, в которой
происходит кариогамия. Образующаяся зигоспора (зигота)
покрывается толстой оболочкой и прорастает после периода покоя. На
зиготе остаются участки гиф, называемые суспензорами. Иногда
зигота приподнимается на них.
Плазмогамия и кариогамия у этих организмов не разделены
во времени и следуют непосредственно друг за другом, в
результате чего образуется диплоидная зигота. При прорастании зиготы
имеет место мейоз и весь основной жизненный цикл этих грибов
58
протекает в гаплоидной фазе. Бесполое размножение
осуществляется только неподвижными элементами, чаще спорангиоспо-
рами, реже конидиями (рис. 22).
При образовании спорангия кончик спорангиеносца
шаровидно вздувается и отделяется перегородкой. Затем первоначально
сплошная цитоплазма спорангия распадается на несколько тысяч
спорангиоспор, каждая из которых может прорастать в мицелий.
Спорангиоспоры различны по раз- ^
меру, объему, форме, цвету. У
большинства они круглые и овальные, у
некоторых видов — цилиндрические,
иногда имеют поперечную исчер-
ченность. Спорангиоспоры
образуются путем расслоения протопла-
та спорангия вокруг ядер; споры
освобождаются в результате
растворения оболочки спорангия. У
большинства они сухие, имеют вид
порошка и распространяются
ветром и токами воздуха, но у
некоторых споры
лей слизистой
рая высыхает
споры.
окружены кап-
жидкости, кото-
и освобождает
Рис. 22. Цикл развития Rhizopus stolonifer:
1 — мицелий; 2 -— спорангий; 3 — спо-
рангиоспора; 4 — прорастание
спорангиоспоры; 5 — мицелий двух разных
половых знаков; 6 — слияние двух мицелиев;
7 — начало формирования зиготы; 8 —
зигота; 9 — прорастание зиготы; 10 —
спора; // — прорастание споры
У грибов этого класса мицелий хорошо развит, у
большинства — ценоцитный, неклеточный, за исключением двух семейств,
выделенных Бенжамином в 1959 г., Dimargaritaceae и Kickxella-
сеае порядка Mucorales, имеющих регулярные перегородки
довольно сложного строения, и представителей порядка Entomoph-
thorales очень рано приобретают перегородки и имеют
многоклеточный мицелий.
Полисахаридный комплекс клеточной стенки представлен
сочетаниями: хитин—глюкан, хитозан—глюкан, хитин—полиурони-
ды. В состав протеинов входит а-аминоадипиновая кислота.
Процентное содержание ГЦ-оснований в ядерной ДНК для
всего класса колеблется в очень широких пределах от 27,5 до
60% и не может служить характеристикой для всего класса в
целом, но значительно более определенно оно в порядке
Mucorales, где содержание ГЦ-оснований в геноме колеблется от 35 до
48%, а у рода Мисог составляет от 37 до 39 мол.%;.
59
Класс Zygomycetes включает несколько порядков. Почвенные
грибы входят в порядки Mucorales (мукоровые) и Endogonales
(эндогоновые).
Большинство мукоровых грибов — широко распространенные
почвенные сапротрофы; эндогоновые грибы представляют
основную массу симбионтов эндотрофных микориз.
Порядок мукоровых включает несколько семейств. Различия
между семействами проявляются в строении органов бесполого
размножения, в частности в строении спорангиеносцев и
спорангиев. У рода Mucor семейства Mucoraceae спорангиеносцы или
неветвящиеся, или имеют моноподиальный или симподиальный
тип ветвления.
У рода Thamnidium семейства Thamnidiaceae они мутовчатые;;
очень сложные закругленные ветвления у Spirodactylon семейства
Kickxellaceae.
На примере развития спорангиев у мукоровых грибов можно
проследить эволюцию бесполых спороношений от спорангиев к
конидиям. Наиболее примитивные — крупные спорангии
семейства Mucoraceae. Они образуются на вершине спорангиеносца как
круглое вздутие, внутри которого отграничены колонка или
столбик как продолжение спорангиеносца внутри спорангия.
Типичный спорангий содержит несколько тысяч спор. В
противоположность такому спорангию представители порядка из семейства
Thamnidiaceae и Choanephoraceae образуют маленькие спорангии,
содержащие всего несколько спор, или даже моноспоровые. Они
называются спорангиоли (см. рис. 16). Иногда и спорангии и спо-
рангиоли образуются вместе на одной особи. Это имеет места,,
например, у Thamnidium; только спорангиоли образуются у
родов Chaetocladium, Helicostylum. Спорангиоли с одной спорой^
как это имеет место у семейства Cunninghamellaceae,
представляют собой фактически одну конидию. В этом случае часто
стенка спорангия и споры сливаются воедино. Семейство Mortierel-
laceae имеет многоспоровые спорангии без колонки, простые или
разветвленные. Кроме того, это семейство характеризуется
образованием зачаточного плодового тела у некоторых его видов.
Большинство представителей порядка Mucorales —
сапротрофы, живущие на таких субстратах, как навоз, разлагающиеся
остатки растений. Большое число мукоровых грибов — типичные
почвенные обитатели, развивающиеся или на растительных
остатках, поступающих в почву, или в самой почве, обычно богатой
органическим веществом, — это различные виды родов Mucor,
Rhizopus, Zygorhinchus. Однако некоторые виды обитают в
почвах при низком содержании доступных органических веществ —
Mortierella, Mucor, Absidia и др. Есть виды, энергично
расщепляющие пектин (Mucor hiemalis), виды рода Mortierella
обладают хитиназами и разлагают хитин и развиваются в природе на
соответствующих субстратах. Многие из них синтезируют
важные промышленные продукты. Rhizopus stolonifer (R.
nigricans) — промышленный продуцент фумаровой кислоты, с его по-
60
мощью осуществляется производство кортизона. R. oryzae
используется для производства спирта. Различные виды Rhizopus
способны образовывать молочную кислоту. Другие мукоровые
образуют лимонную, щавелевую и а-кетоглютаровую кислоты. Многие
мукоровые образуют каротиноидные пигменты. Гриб Choanephora
trispora (Blakeslea trispora) — промышленный продуцент р-каро-
тина.
Некоторые Mucorales — слабые паразиты на фруктах,
вызывают болезни при их хранении. Rhizopus siolonifer — возбудитель
заболевания клубники, Choanophora cucurbitaerum является
причиной заболевания тыквы, дыни, порчи фруктовых соков. Absidia
corymbifera и некоторые представители Мисог и Rhizopus
вызывают заболевания, связанные с поражением нервной системы
человека. Грибы из семейства Piptocephalidaceae — облигатные
паразиты на других грибах, главным образом на грибах порядка
Mucorales.
На мукоровых грибах американский генетик А. Блексли в
1904 г. открыл явление гетероталлизма, т. е. среди грибов были
обнаружены раздельнополые виды. Блексли установил, что у
многих мукоровых грибов, выращенных из одной споры, не
образуется зигот. Они развиваются только при встрече двух мицелиев,
при этом не всяких, а только некоторых, хотя и одинаковых по
виду, но разных в половом отношении.
Поскольку мужские и женские особи не различались между
собой морфологически, было предложено обозначать их условно
знаками ( + ) и (—), а само явление раздельнополости было
названо гетероталлизмом в отличие от обоеполости, названной го-
моталлизмом. При этом обозначения ( + )- и (—)-штаммы не
являлись случайными для каждого отдельного вида, но
устанавливались по сравнению с первоначальными штаммами, которые
получили эти обозначения в первых исследованиях Блексли.
В случае, когда штаммы имели некоторые морфологические
различия, женские обозначались как ( + )-штаммы, а мужские как
(—)-штаммы.
Бургеф в 1924 г. показал, что у мукоровых грибов имеется
вещество, ответственное за начало полового воспроизведения гете-
роталличных штаммов.
Гифы мицелиев двух различных знаков направляются друг к
другу, и одни из них индуцируют образование гаметангиев у
других, даже если мицелии разделены коллодиевой пленкой. Это
было первое обнаружение гормональной природы полового
механизма у грибов.
Работами М. Племпеля 60-х гг. и его последователей эти
гормоны были выделены в виде кристаллического вещества и
установлена их химическая природа (см. с. 43).
Порядок Endogonales представлен одним семейством Endo-
gonaceae. Для многих эндогоновых грибов характерны особые,
находящиеся в почве плодовые тела или спорокарпы. Они
представляют собой округлые тельца желтоватого цвета, образован-
61
яые из переплетенного мицелия величиной от нескольких
миллиметров до 2—3 см. Внутри спорокарпа находятся либо спорангии,
либо зигоспоры, либо хламидоспоры. Зиготы образуются в
результате слияния двух одноядерных или многоядерных клеток,
отделившихся перегородками от гиф мицелия. Есть гомо- и гетеро-
талличные виды. В почве распространены виды родов Endogone,
Glomus, Gigaspora и др. Всего в настоящее время известно 7
родов и более 30 видов эндогоновых грибов. Грибы рода Endogone
образуют спорокарп с зигоспорами внутри. Большинство видов
Glomus характеризуется отсутствием спорокарпа. Грибы
образуют круглые хламидоспоры по одной на вершине гифы размером
100—300 мкм с толстой оболочкой, представляющие собой
покоящиеся споры. При созревании спора отделяется от гифы септой.
Хламидоспоры образуются либо в почве, либо в корневой
паренхиме корня.
Грибы рода Gigaspora образуют в почве азигоспоры —
крупные клетки размером более 250 мкм на вздутых подвесках,
сходных с теми, которые образуются при половом процессе, но,
слияние клеток перед этим не наблюдается. Поэтому такие структуры
в отличие от зигоспор называют азигоспорами. Азигоспоры
образуются только в почве. Род Sclerocystis образует хламидоспоры
в спорокарпах, хламидоспоры располагаются сплошным слоем в
верхней части спорокарпа. Центральная часть остается
свободной и представлена плотным переплетением гиф. Спорокарпы,
имеющие размеры до 2—3 см, образуются либо свободно в
почве, либо на различных органических остатках, во мху и листьях.
Все роды содержат виды, образующие эндотрофную микоризу.
Это — основные симбионты травянистых растений. Наиболее
обильно эти грибы представлены в южных географических
зонах — пустынных и полупустынных почвах.
КЛАСС ASCOMYCETES
Аскомицеты, или сумчатые грибы, — один из
обширнейших классов грибов, включающий более 2000 родов и 15 000
видов, разнообразных как по строению, так и по образу жизни.
Основная характеристика класса сумчатых грибов, отличающая
их от всех других, — это наличие у них сумки, или аска,
благодаря которой этот класс и получил свое название. Сумка
представляет собой специализированную клетку, формирующуюся в
результате полового процесса и являющуюся его конечным
этапом, которому предшествуют плазмогамия, кариогамия и мейоз.
Сумка — это репродуктивный орган, в ней образуются споры, с
помощью которых идет дальнейшее размножение особи.
У большинства сумчатых грибов в сумке содержится строго
определенное число спор (чаще всего восемь), но у некоторых
видов оно может варьировать в разных пределах.
Другие характеристики класса Ascomycetes — наличие строго
септированного мицелия и полное отсутствие подвижных форм.
62
Образование септ у сумчатых грибов происходит в результате-
инвагинации пристенной цитоплазматической мембраны от
периферии к центру и заполнения ее материалом клеточной стенки.
В результате такого процесса остается отверстие в центре септы,
через которое цитоплазма сообщается между клетками, а также
могут мигрировать органеллы клетки и даже ядра. Поры в
септах играют существенную роль в переносе питательных веществ
по гифам в зону роста. Клетки сумчатых грибов обычно
многоядерны, хотя имеется небольшое число видов с одноядерным
мицелием. У некоторых низших аскомицетов, к которым, в
частности, относятся дрожжи, настоящего мицелия нет, а вегетативное
тело представлено одиночными почкующимися или делящимися
клетками, иногда образующими псевдомицелий.
Основной полисахаридный комплекс клеточной стенки
сумчатых грибов хитин и глюканы, которые представляют собой
полимеры D-глюкозы, отличающиеся от целлюлозы характером связи
между мономерами. Содержание хитина у аскомицетов ниже, чем
у других грибов, и составляет 20—25% полисахаридов клеточной
стенки. Глюканы составляют до 80—90%. У дрожжей вместо
глюканов преобладают маннаны — полимеры маннозы. В состав
протеинового комплекса входит, как и у многих других грибов,
сс-аминоадипиновая кислота.
Процентное содержание ГЦ-оснований в ядерной ДНК
варьирует в пределах от 48 до 60 мол.% ГЦ, но основная масса — от
50 до 55% мол.% ГЦ.
Существенной особенностью сумчатых грибов является то, что
у большинства из них плазмогамия и кариогамия при половом
процессе разделены во времени.
Половой процесс у сумчатых грибов в типе представляет
собой гаметангиогамию. Он протекает следующим образом.
Большинство сумчатых грибов — гомоталличные формы. На их
мицелии закладываются специализированные половые структуры:
женский гаметангий обычно состоит из двух клеток — нижней
вздутой, называемой аскогоном, и верхней цилиндрической,
называемой трихогиной, или же он представлен одной клеткой.
Мужской антеридий всегда одноклеточный. Как аскогон, так ш
антеридий многоядерны, но содержимое их лишено какой-либо
дифференцировки на отдельные гаметы. При созревании половых
структур антеридий соприкасается с трихогиной или аскогоном,
и мужские ядра поступают через пору в этих образованиях в
аскогон. Однако ядра после плазмогамии не сливаются, а
ассоциируются строго попарно в так называемые дикарионы.
Из продукта слияния двух половых клеток — зиготы —
начинают развиваться короткие ветвящиеся нити мицелия, носящие
название аскогенных гиф, в которые, строго синхронно делясь,
поступают дикарионы (рис. 23).
На аскогенных гифах образуются сумки. Верхняя часть аско-
генной гифы загибается в виде крючка, и в ней происходит
последнее деление пары ядер. Из образовавшихся четырех ядер два;
63,
ядра разного рода остаются в верхушечной клетке, которая
отделяется перегородкой. Одно ядро поступает в кончик,
отделяющийся перегородкой, другое — в основание аскогенной гифы. Эти
ядра также образуют дикарион, и крючок может сформироваться
повторно. Средняя двухъядерная клетка крючка развивается в
^сумку, где происходит слияние дикариона в одно диплоидное ядро.
Такие клетки, где происходит слияние ядер, образуются на
каждой аскогенной гифе. В ней вскоре после слияния ядер
следуют обычно три деления
диплоидного ядра, одно из которых мейотичес-
кое, два других—митотические. В
результате образуется восемь
гаплоидных ядер.
У низших аскомицетов, куда
относятся дрожжи и родственные им
организмы, половой процесс сходен с
зигогамией зигомицетов. Сливаются
два гаметангия разного пола в виде
выростов или веточек мицелия,
сходных морфологически. После их
слияния сразу наступает кариогамия, и
сумка развивается прямо из зиготы.
Однако в отличие от большинства зи-
гомицетов в многоядерных гаметан-
гиях сливаются только два ядра.
Зигота не переходит в состояние покоя,
а сразу развивается в сумку. Таким
Рис. 23. Половой процесс и развитие сумок
на примере Pyronema omphaloides:
1 — слияние аскогона с антеридием; 2 —
развитие сумок из кончиков аскогенных гиф;
3 — молодая сумка после мейоза; 4 —
зрелая сумка с аскоспорами
образом, в цикле развития низших аскомицетов дикариотическая
стадия отсутствует.
Одновременно с образованием сумок происходит их оплетение
гаплидным мицелием, формируется плодовое тело — аскокарп,
сложенное из псевдопаренхиматической ткани. Сумки могут
также образоваться в локулах (полостях) ранее образовавшегося
сплетения мицелия. Плодовые тела сумчатых грибов имеют
разнообразное строение.
Половой процесс у некоторых аскомицетов может происходить
и несколько отличными путями от основного, описанного выше.
Для многих аскомицетов характерна морфологическая
редукция полового процесса. Иногда антеридии образуются и не
функционируют, иногда не образуются вовсе. В этом случае их
функции могут выполнять спермации. Спермации — мелкие, сфериче-
64
ские или бациллярные мужские половые клетки, образуются
часто подобно конидиям, на конидиеносце, соединяются с
женской клеткой — аскогоном и освобождают в нее свое содержимое.
Ядро спермация мигрирует через пору в септе аскогона. Сперма-
ции отделяются от материнской гифы и переносятся насекомыми,
ветром, водой к рецепторному органу.
У некоторых аскомицетов имеет место слияние соматических
гиф двух совместимых мицелиев. Ядра при этом мигрируют из
одного мицелия в другой через перфорированные септы.
Среди сумчатых грибов есть гомоталличные и гетероталлич-
ные виды. Большинство гетероталличных видов имеют
биполярное распределение полов (см. с. 40), однако есть и тетраполяр-
ные виды.
Аскоспоры обычно одноядерны. Как правило, они такими
остаются, но у некоторых видов ядра еще делятся в аскоспоре и
они становятся двухъядерными. У других видов аскоспоры
становятся многоядерными, между отдельными ядрами образуются
септы. Форма аскоспор очень разнообразна — от шаровидных или
эллипсоидальных до нитевидных.
Сумки также имеют различную форму, строение оболочки и
способы ее раскрывания. Наиболее важная черта морфологии
сумки — строение ее оболочки. По строению оболочки и
функциям сумки выделяют три типа сумок: прототуникатные, униту-
никатные и битуникатные. Прототуникатные сумки имеют
тонкую недифференцированную оболочку из одного слоя, которая
разрушается, пассивно освобождая споры. Оболочка унитуникат-
ных сумок состоит из двух тонких слоев, образуя
морфологически единое целое, и выглядит однослойной. На вершине таких
сумок обычно имеется апикальный аппарат различного строения,
служащий для их вскрывания. Споры из сумок могут
освобождаться через пору на вершине, крышечку или щель (рис. 24).
У битуникатных сумок оболочка состоит из двух морфологически
различимых слоев — жесткого наружного и растягивающегося
эластичного внутреннего. При созревании аскоспор наружный
слой растрескивается ниже вершины, а внутренний слой под
действием увеличивающегося тургорного давления растягивается, и
аскоспоры активно выбрасываются.
Сумки с активным выбрасыванием спор имеют удлиненную
булавовидную или цилиндрическую форму.
Для грибов с замкнутыми плодовыми телами без активного
распространения спор характерны круглые или овальные сумки.
Сумки могут располагаться в плодовом теле на разных уровнях,
иметь ножку, их прикрепляющую. Сплошной ряд сумок
называется гимением. У некоторых аскомицетов сумки образуются на
аскогенных гифах цепочками по ходу аскогенных гиф из их
конечных клеток или боковых выростов.
Бесполое размножение у сумчатых грибов осуществляется с
помощью конидиальных спороношений. Споры бесполого
размножения — конидии — образуются на гаплоидном мицелии — ко-
3 Т. Г. Мирчинк
65
нидиеносцах разного строения. Конидиальные спороношения
представлены очень разнообразно у многих групп сумчатых грибов и
играют важную роль в их цикле развития. Таким образом, в
цикле развития высших аскомицетов чередуются три ядерные фазы:
длительная гаплоидная, в течение которой происходит бесполое
размножение, непродолжительная дикариотическая (аскогенные
гифы) и очень короткая диплоидная (молодая сумка с диплоид-
Рис. 24. Различные способы открывания сумок:
/ — без отверстия; 2 — с порой; 3 — с крышечкой; 4 —
со щелью; 5 — битуникатная .сумка с эластичной стенкой и
порой
иым ядром). Конидиальные спороношения служат для массового-
расселения грибов. У аскомицетов-паразитов бесполый цикл в
виде конидиальных спороношений обычно проходит на живых
растениях, в то время как сумчатая стадия характерна для
отмерших растительных остатков и является сапротрофной. Однако
есть небольшое число аскомицетов, для которых конидиальное
спороношение неизвестно, хотя у многих оно преобладает в цикле
развития, и сумчатая стадия образуется редко. Иногда ее трудно
обнаружить в природе и получить в искусственной культуре,
поэтому многие сумчатые грибы известны в виде их конидиальных
66
стадий (анаморф) и имеют соответственно самостоятельные
видовые наименования, нередко более распространенные, чем
названия сумчатых стадий (телеоморф). Например, многие пени-
циллы и аспергиллы, образующие сумчатые стадии, более
известны под названием их конидиальных стадий. Так, часто
обнаруживаемый в почве аскомицет Talaromyces flavus обычно
называют Penicillium vermiculatum. Для большого числа грибов,
встречающихся в природе в гаплоидной конидиальной стадии, половые
стадии неизвестны. Такие грибы относят к классу дейтеромице-
тов, или несовершенных грибов (см. с. 92).
Класс Ascomycetes включает несколько тысяч видов. На
основании наличия или отсутствия плодовых тел и их строения он
делится на три подкласса.
Подкласс Hemiascomycetidae — голосумчатые, включает
наиболее примитивные грибы из класса сумчатых, которые не
образуют ни аскокарпов, ни аскогенных гиф, но имеют сумки с про-
тотуникатным строением оболочки, образующиеся прямо на
мицелии. В этот подкласс включают также сумчатые дрожжи,
объединяя их в порядок Endomycetales. Некоторые микологи
выделяют их в самостоятельный класс Endomycetes, объединяя в
нем не только примитивные сумчатые грибы, но и примитивные
базидиальные грибы, имеющие дрожжевые стадии, в частности
порядок Ustilaginales — головневые грибы.
Подкласс Euascomycetidae — собственно сумчатые, включает
грибы с хорошо развитыми аскокарпами различного строения,
имеющие прототуникатное и унитуникатное строение оболочки
сумок.
К подклассу Loculoascomycetidae — локулоаскомицетные,
относят грибы, сумки которых образуются в локулах (полостях)
ранее образовавшейся аскостромы (переплетения гиф) и
представляют собой битуникатные образования. К этой группе
преимущественно относятся паразитные грибы.
Подкласс Hemiascomycetidae включает порядки
Endomycetales и Taphrinales. В первом значительное место занимают
дрожжи. Второй порядок представлен исключительно паразитными
грибами, вследствие чего эти группы как и подкласс Loculoasco-
mycetideae здесь не рассматриваются.
Подкласс Euascomycetidae — обширный подкласс собственно
сумчатых грибов, для удобства подразделяют на группы
порядков соответственно типу плодовых тел и строению сумок, которым
обычно не присваивают ранг таксонов. Это плектомицеты, пире-
номицеты, дискомицеты.
В группе плектомицетов наиболее важный и обширный
порядок Eurotiales, в группе пиреномицетов — Sphaeriales, Нуросгеа-
les, Clavicipitales, а в группе дискомицетов — Helotiales, Peziza-
les, Tuberales.
Плектомицеты характеризуются плодовыми телами,
называемыми клейстотециями, и имеют прототуникатное строение
оболочки сумок. Плодовые тела типа клейстотециев представляют
3*
67
собой округлые образования размером от десятых долей
миллиметра до нескольких миллиметров, не имеющие специальных
отверстий для выхода спор. Клейстотеции могут быть образованы
как рыхлым паутинистым сплетением гиф, так и плотным псев-
допаренхиматическим. Споры освобождаются только после
разрушения плодового тела. Наиболее примитивные формы имеют
зачаточные плодовые тела в виде рыхлого переплетения гиф.
Сумки, беспорядочно расположенные в таком плодовом теле,
образуются на разных уровнях аскогенных гиф или цепочками,
имеют округлую или овальную форму.
В распространении большинства грибов этой группы большую
роль играет конидиальное спороношение, немногие в цикле
развития имеют- только сумчатую стадию, а большое число видов ее
утратили вовсе.
Одну из наиболее важных групп основного порядка (эвроцие-
вых) составляют грибы, конидиальные стадии (анаморфы)
которых относятся к родам Penicillium и Aspergillus. Каждый из
этих двух родов содержит по несколько сотен видов, причем у
многих из них известны только конидиальные стадии. Такие виды
относят к несовершенным грибам.
У ряда видов пенициллов и аспергиллов, по традиции также
относимых к несовершенным грибам, известны сумчатые стадии,
относящиеся к разным родам эвроциевых. Поскольку согласно
Международному кодексу ботанической номенклатуры основным
наименованием плеоморфных грибов является наименование их
совершенной стадии, эти виды должны рассматриваться среди
аскомицетов.
Конидиальные спороношения рода Penicillium имеют вид
кисточки, откуда этот гриб получил свое русское название гриб-кис-
тевик. Конидиеносец прямостоящий, имеет разветвления только
наверху, в виде одного, двух или трех ярусов мутовок. Верхний
ярус мутовок обычно состоит из коротких бутыльчатых
образований, носящих название фиалид. Следующий ярус составлен из-
таких же, но несколько более длинных образований, называемых
метулами, и третий самый нижний представлен веточками. От
последнего, самого верхнего яруса мутовок, отходят цепочки
конидий. Такое строение придает спороношению сходство с кисточкой.
Соответственно конидиеносцы с одним ярусом мутовчатых
разветвлений носят название моновертициллятных (Monoverticillata),
с двумя ярусами — бивертициллятных (Biverticillata),
разветвления с тремя ярусами обычно бывают асимметричными и носят
название асимметричных (Asymmetrica).
Примером эвроциевых с этой конидиальной стадией может
служить род Talaromyces, образующий мелкие шаровидные или
неправильной формы клейстотеции, обычно ярко-желтой или
оранжевой окраски, придающей окраску всей колонии. Кониди-
альная стадия Т. flavus — Penicillium vermiculatum относится к
секции Biverticillata (рис. 25).
Конидиальные спороношения Aspergillus представлены невет-
68
вящимися прямостоячими конидиеносцами, имеющими на
вершине округлое или булавовидное вздутие. От поверхности вздутия
отходят фиалиды, которые могут располагаться или
непосредственно на вздутии конидиеносца, или на профиалидах.
Соответственно конидиальные спороношения рода Aspergillus могут быть
одно- и двухъярусными.
На фиалидах образуются цепочки конидий. Сумчатая стадия
их представлена родами Eurotium, Emiricella, Neosartoria
Рис. 25. Сумчатая и конидиальная стадия Talaromyces flavus:
клейстотеций; 2 — сумка; 3 — аскоспоры; 4 — конидиальная
стадия Penicillium vermiculatum
(рис. 26). Они имеют клейстотеций разной окраски, содержащие
круглые сумки с аскоспорами различной формы и строения.
Представители родов Penicillium и Aspergillus повсеместно
распространены во многих почвах земного шара от Арктики до
тропиков и являются типичными почвенными обитателями. Кроме
того, они могут быть обнаружены на различных субстратах
растительного происхождения. На поверхности плодов и овощей
они развиваются как слабые паразиты, вызывая их порчу при
хранении. Некоторые из них вызывают порчу пищевых
продуктов, повреждают изделия из пластмасс, металлов и других
материалов. Некоторые представители рода Aspergillus могут быть
причиной тяжелых заболеваний внутренних органов человека и
животных — аспергиллезов.
Представители родов Penicillium, Aspergillus, Acremonium
69
(последний имеет сумчатую стадию, относящуюся к роду Emiri-
cilopsls) являются основными продуцентами антибиотиков —
пенициллина, цефалоспорина, гризеофульвина, цитринина, аспергил-
лина, фумагиллина и др. Некоторые образуют сильно
действующие токсины группы афлатоксинов (A. flavus), рубротоксинов
(P. rubrum, P. purpurogenum) и др.
Виды Aspergillus — активные продуценты органических
кислот: лимонной, коевой, итаконовой и др. Они используются также
для получения ряда ферментов (протеиназ, амилаз и др.).
Многие представители рода Eurotium — ксерофилы, т. е.
развиваются при пониженной влажности, неблагоприятной для роста
Рис. 26. Сумчатая и конидиальная стадии Neosartoria fi-
sheri:
1 — клейстотеций; 2 — сумка; 3 — аскоспоры; 4 —
конидиальная стадия Aspergillus fisheri
других грибов. Например, Е. repens вызывает плесневение зерна
и многих других продуктов при влажности 13—15%. Этот же вид
может служить примером осмофильного организма, т. е. он
развивается на средах с высоким осмотическим давлением, например
с повышенным содержанием сахара до 20% и более, и часто
образует плесень на джемах и вареньях.
Грибы рода Gymnoascus имеют плодовые тела —
клейстотеций, состоящие из рыхлого сплетения мицелия с характерными
придатками в виде крючков (рис. 27). В плодовых телах
содержатся сумки с восемью аскоспорами. Некоторые грибы этого рода,
существующие только в конидиальной стадии, являются дерма-
тофитами и вызывают кожные заболевания человека и животных,
например виды Chrysosporium, Microsporum и др. Многие пред-
70
Рис. 27. Сумчатая и конидиальная стадии Gymnoascus:
фрагмент оболочки клейстотеция с отростками; 2 — сумка с аскоспора-
ми; 3 — конидиальная стадия Chrysosporium
** -: '
;
. у
-¦*.***
ВОмнм
6§мШ
^Щшм » : *"^ ^Н
Щ^/^^Шмш':
Рис. 28. Плодовые тела и различная форма отростков грибов рода Chaeto-
mium в электронном микроскопе
(фото В. С. Гузева)
ставители рода Gymnoascus в конидиальной стадии разлагают
роговое вещество животных, содержащее кератин, — перья,
волосы, копыта, рога и т. д., т. е. участвуют в утилизации этих
продуктов при поступлении их в почву.
Группа пиреномицетов характеризуется плодовыми телами
типа перитециев и преимущественно унитуникатным строением
сумок. Плодовые тела типа перитециев представляют собой
образования грушевидной или кувшиновидной формы с отверстием
(круглой порой — остиолой) наверху для выхода сумок и аско-
Рис. 29. Плодовые тела и сумки грибов Sordaria (/, 2) и Chae-
tomium (3, 4) — фрагмент отростка перитеция Chaetomium (5)
спор размером 1—2 мм. Сумки обычно имеют булавовидную или
цилиндрическую форму и располагаются пучком или слоем.
Освобождение спор у большинства активное.
Эта группа грибов включает много порядков, в том числе
порядки Sphaeriales, Hypocreales, Clavicipitales.
В порядок Sphaeriales (сферейные) входит род Chaetomium,
представленный в почве многими видами. Это наиболее
примитивный род порядка.
Грибы рода Chaetomium имеют характерное строение
перитециев с длинными отростками — волосками, у многих видов
спирально закрученными по всей поверхности перитеция (рис. 28).
Сумки у большинства видов Chaetomium булавовидные,
широкоовальные и только у некоторых цилиндрические (рис. 29). Стенки
сумок быстро разрушаются до созревания спор, поэтому споры
часто находятся свободно в полости перитеция, погруженные в
слизь. Активного выбрасывания спор у них не происходит. Слизь
набухает, и аскоспоры выходят из перитеция в длинном слизи-
72
стом шнуре. Аскоспоры обычно темные и сильно варьируют по
форме у разных видов. Из нескольких десятков видов Chaetomium
только некоторые образуют конидии. Виды Chaetomium,
активные разрушители целлюлозы, играют существенную роль в
разложении растительных остатков. Наряду с этим приносят
большой вред, разрушая бумагу и книги. Они постоянные обитатели
почв. Солома, навоз и сходные материалы также обычные
субстраты для этих грибов.
В порядок Sphaeriales входят еще грибы родов Neurospora,
Sordaria, Pleurage и др., характеризующиеся темными,
кожистыми или углистыми перитециями, обитающие как в почве, так и
на навозе и пищевых продуктах. Neurospora в настоящее время
представляет собой для микологов то же, что Drosophila для
генетиков, так как на ней сделано очень много генетических
работ. Основные из них сделаны на N. sitophila и N. tetrasperma
и особенно на N. crassa. N. sitophila — один из наиболее
известных видов этого рода, известен как «пекарская плесень», или
«красная плесень хлеба», так как обычно приносит вред в
хлебопечении. Этот организм очень быстро заражает лаборатории и
чистые культуры.
Мицелий N. sitophila содержит обильные ветвящиеся гифы.
Воздушный мицелий образует массу розовых овальных конидий,
развивающихся в ветвящихся цепочках на разветвленных кони-
диеносцах. Наряду с образованием сумок гриб может
размножаться только с помощью конидий. Гриб в этой стадии был
описан в 1843 г. под названием Monilia sitophila. Сумчатая
стадия его стала известна только в 1927 г. благодаря работам
Доджа. Neurospora, как и большинство представителей этого порядка,
отличается быстрым ростом. N. sitophila и N. crassa восьмиспо-
ровые, гетероталличные. Женские элементы представлены прото-
перитециями с многоядерными аскогонами. Аскогоны образуют
длинную ветвящуюся гифу, которая функционирует как трихо-
гина; антеридиев нет. Мужские элементы представлены
микроконидиями, спермациями, образующимися в цепочках на микро-
конидиеносцах и прорастающими ростковой трубкой. У этих видов
наблюдается дегенерация мужской половой функции и половой
процесс осуществляется как сперматизация.
Другой широко известный род Sordaria (см. рис. 29) имеет
типичные перитеции с отверстием наверху, цилиндрические
сумки, расположенные пучком у основания перитеция. В сумках
находятся темно-коричневые аскоспоры, окруженные слизистой
капсулой. Иногда она довольно толстая и отчетливая, иногда
трудно различимая.
Sordaria fimicola — наиболее типичный представитель копро-
фильных грибов. Гриб имеет приспособление к активному
разбрасыванию спор. Выброшенные из перитеция аскоспоры
благодаря слизистым обверткам прилипают к траве, попадают с ней
в кишечный тракт травоядных животных, а затем, попав на
навоз, прорастают.
73
У представителей порядка Clavicipitales (клавицепсовые)
перитеции образуются в хорошо развитых стромах, состоящих из
гиф гриба. Строма обычно развивается на пораженных органах
растения-хозяина или из склероциев. К порядку клавицепсовых
относится хорошо известный паразитный гриб Claviceps
purpurea — возбудитель болезней многих хлебных злаков, носящий
название спорыньи. Гриб образует темные твердые склероции в
завязи растения-хозяина. Аскоспоры этого гриба
распространяются ветром. Весной, попадая на цветущую рожь в соцветие,
прорастают ростовыми трубками, вызывая инфекцию.
Развивающийся мицелий разрушает завязь. Вместо нее формируется
плотный слой конидиеносцев, образующий мелкие овальные конидии.
Эти конидии находятся в сладких нектароподобных выделениях,
привлекающих насекомых, быстро разносящих конидии на неин-
фицированные растения. Тем временем мицелий в завязи
сплетается, уплотняется, твердеет и образует черно-фиолетовый
склероции, имеющий вид рожка. Завязь полностью разрушается, и зерно
не образуется. Многие склероции к осени падают на землю,
зимуют на ней и весной прорастают плотным сплетением — стромой.
Строма у С. purpurea имеет вид головки на ножке, сходной с
плодовым телом базидиального гриба, и также образована псев-
допаренхиматической тканью. Внутри головки стромы, близко к
поверхности, возникают многочисленные полости, окруженные
псевдопаренхиматической тканью. В этих полостях
осуществляется половой процесс с образованием аскогона, антеридия,
осуществлением плазмогамии, который приводит к образованию
сумок и нитевидных аскоспор. В полостях вокруг половых клеток
образуются перитеции, которые открываются длинным шееподоб-
ным отверстием.
Склероции гриба содержат ядовитые алкалоиды, вызывающие
заболевание животных и человека, называемое эрготизмом.
Болезнь проявляется в поражении нервной системы. Под действием
алкалоидов происходит сокращение гладкой мускулатуры и
сосудов, в результате чего возникают конвульсии. Сейчас токсикоз
у людей, вызванный алкалоидами спорыньи, попавшими в зерно,
наблюдается очень редко.
Алкалоиды спорыньи широко применяются в современной
медицине как лекарственные препараты для лечения
сердечно-сосудистых и нервных заболеваний, используются в акушерской и
гинекологической практике. Склероции спорыньи содержат две
группы алкалоидов. Первая группа — производные лизергиновой
кислоты — алкалоиды эрготамин, эргозин, эргокристин и др.
Некоторые производные лизергиновой кислоты обладают
галлюциногенными свойствами. Вторая группа — клавиновые
алкалоиды — агроклавин, элимоклавин и др. Используют два метода
получения алкалоидов спорыньи: из склероциев, выращенных на
растениях, и путем биосинтеза в сапротрофной культуре спорыньи.
В порядок Hypocreales (гипокрейные) входит большое число
родов, многие из которых представлены почвенными грибами. Для
74
этого порядка характерны мягкие или мясистые перитеции яркой
или светлой окраски. Перитеции могут образоваться на мицелии
или на стромах, т. е. плотном сплетении мицелия такого же
характера, как и перитеций. В цикле развития гипокрейных часто
есть и конидиальная стадия, имеющая большое значение в
распространении вида, а иногда полностью замещающая сумчатую
стадию.
Среди гипокрейных известно много паразитных форм
растений, грибов и насекомых, а также сапротрофных организмов,
обитающих в почве. Нередко их анаморфные (конидиальные) стадии
бывают патогенными, а телеоморфы сапротрофны. Род Nectria
может быть паразитом в той и другой стадии.
Вид Gibberella fujikuroi поражает корни, стебли, семена
растений многих семейств. Этот вид распространен преимущественно
в конидиальной стадии, известной как Fusarium moniliforme.
Перитеции гриба образуются только на отмерших частях растений.
Особенно часто этот гриб вызывает заболевание злаков — риса,
сахарного тростника, сорго. Вызываемая им болезнь риса носит
название «баканэ», или «болезнь дурных побегов». При
поражении грибом у растений вытягиваются междоузлия, листья
становятся более узкими. Эти симптомы вызывают специфические
ростовые вещества — гиббереллины, образуемые этим грибом.
Другой вид Gibberella zeae, развиваясь на злаках, вызывает
их серьезное заболевание, приводящее к сильному снижению
урожая. Пораженное зерно содержит токсин, который вызывает
отравление. Причиной заболевания также является конидиальная
стадия гриба — Fusarium graminearum (рис. 30).
Еще один гриб порядка Micronectriella nivalis имеет кониди-
альную стадию Fusarium nivale, вызывающую заболевание
«снежная плесень». Болезнь развивается ранней весной, сразу после
таяния снега. Конидиальные спороношения образуются у
основания стеблей и на остатках погибших растений в течение всего
периода вегетации, перитеции — преимущественно на отмерших
растениях. К этому порядку относятся также грибы рода Hypo-
crea. H. rufa представляет собой сумчатую стадию широко
распространенного в почвах, на древесине и растительных остатках
целлюлолитического гриба Trichoderma viride — активного
продуцента целлюлаз, на котором впервые была показана
множественность ферментов, участвующих в процессе разложения
целлюлозы и ступенчатость этого процесса.
Группа, относимая к дискомицетам, включает грибы,
имеющие преимущественно крупные плодовые тела макроскопических
размеров, дисковидной или чашевидной формы, называемые апо-
тециями. Плодовые тела некоторых из них могут иметь также
форму губки или колокольчика с сильно извитой поверхностью.
Однако их объединяет общий признак: они открыты и сумки
образуются на поверхности плодовых тел в больших открытых
полостях тесным слоем. Сплошной слой сумок, чередующийся со
стерильными гифами, носит название гимения. Сумки у дискоми-
75
цетов обычно булавовидные или цилиндрические унитуникатного
строения. При созревании сумок и аскоспор последние с силой
выбрасываются наружу, образуя облако спор. Плодовые тела
этих грибов обычно ярко окрашены в красный, желтый или
оранжевый цвет. Некоторые из них коричневой окраски и очень
немногие — черной. Большинство этих грибов сапротрофы и растут
на земле, гниющих ветках, листьях или навозе животных; но есть
и паразитные формы.
В группу дискомицетов входит несколько порядков,
наибольшее значение из которых имеют порядки Helotiales, Pezizales,
Tuberales.
Грибы порядка Helotiales (гелоциевые) имеют сумки,
вскрывающиеся в виде трещины или поры. Представители этого поряд-
Рис. 30. Gibberella zeae: Рис. 31. Monilinia fructigena:
1 — перитеции на колосе пшеницы; / — пораженный плод яблони с ко-
2 — разрез перитеция; 3 —¦ сумка с ас- нидиальными спороношениями; 2 —
коспорами; 4 — аскоспоры конидиеносцы
ка или строгие сапротрофы, или паразиты, но имеющие сапро-
трофную стадию в цикле развития. Паразитная у них
преимущественно конидиальная стадия. Сумчатая стадия обычно сапро-
трофная и развивается на опавших листьях и отмирающих
растительных остатках. Апотеции образуются на строме или скле-
роциях. Они средних размеров, обычно коричневые, на ножке.
Сапротрофные гелоциевые обитают на древесине,
растительных остатках и на почве. Примером могут служить Calycella
citrina, ярко-желтые апотеции которой развиваются на
поваленных стволах деревьев и крупных ветвях, и Spathularia flavida,
имеющая крупные шпателевидные апотеции, образующиеся прямо
на почве в хвойных лесах.
Важнейшие роды паразитических гелоциевых — Monilinia и
Sclerotinia. M. fructigena образуют склероции в плодах растений
из семейств розоцветных и брусничных. В цикле ее развития
76
всегда имеется конидиальная стадия типа монилия (МопШа).
Конидиальная стадия представлена подушечками коротких кони-
диеносцев, на которых образуются длинные, часто ветвящиеся
цепочки конидий. М. fructlgena вызывает плодовую гниль яблок
и груш. Гриб заражает плоды с поврежденной кожицей.
Инфекция часто переносится жуком-казаркой. На пораженных плодах
образуются пятна отмершей ткани, а на них концентрическими
кольцами развиваются подушечки конидиального спороношения
гриба (рис. 31). Зараженные плоды опадают с дерева и служат
источником инфекции. Плоды, оставшиеся на дереве,
превращаются в склероции. Склероции зимуют, а весной на них снова
развивается конидиальное спороношение. Апотеции у этого вида
образуются редко.
Sclerotinia sclerotiorum вызывает белую гниль различных
растений. Он поражает как вегетирующие растения, так и овощи
при хранении. Как следует из названия, для склеротинии
характерно образование склероциев, являющихся обязательной
стадией в их цикле развития. На поверхности пораженных частей
растения, часто на моркови, свекле, образуется белый войлочный
мицелий, на котором формируются склероции. Конидиальное
спороношение у видов этого рода отсутствует. Апотеции образуются
на склероциях весной. Грибы порядка Peziziales — пецициевых
имеют сумки, открывающиеся на вершине крышечкой. Они
характеризуются большим разнообразием в форме, размерах и окраске
плодовых тел. Размеры аскокарпов варьируют у них в пределах
от нескольких миллиметров до 10—12 см. Некоторые имеют
красивую яркую окраску, другие — черные и коричневые. В
умеренном климате известны Sarcoscypha coccinea и Aulevria auran-
tiaca с ярко-красным гимениальным слоем, Peziza и Ascobolus
имеют темную окраску плодовых тел (рис. 32). Peziza,
Sarcoscypha, Aleuria, Ascobolus имеют блюдцевидные или чашевидные
апотеции. У Peziza, Sarcoscypha, Aleuria они довольно крупных
размеров 1—6 см, у Ascobolus — мелкие, несколько миллиметров.
Большинство существует в сумчатой стадии, конидиальное
спороношение у них отсутствует. Среди них есть съедобные и
ядовитые формы. Многие из них сапротрофы, растут в лесах на
растительном опаде, мертвой древесине, почве, а также на навозе
животных. Типичными копрофилами являются представители
рода Ascobolus. Небольшое число этих видов пецициевых
паразитирует на растениях.
К грибам с колокольчатым аскокарпом, иначе их называют
еще моршелловидным или гельвелловидным апотецием по
названию основных родов, относятся разнообразные сморчки. Это са-
протрофные грибы, растущие на почве. Они имеют крупные
плодовые тела (6—10 см), состоящие из ножки и шляпки, которые
имитируют плодовые тела базидиальных грибов. Шляпка имеет
складчатую поверхность. Складки образуют ячейки, выстланные
гимением на наружной стороне. Конидиальная стадия у этих
грибов отсутствует.
77
Сюда относятся ценные съедобные грибы, появляющиеся в
лесу весной. Некоторые образуют микоризу с хвойными деревьями-
Основные роды Helvetia, Verpa, Morchella — сморчки, Gyromyt-
ra — строчок (рис. 32), М. esculenta и М. conica — съедобные
грибы. Однако их следует употреблять в пищу после кипячения
для удаления гельвелловой кислоты. Gyromytra — условно съе-
Рис. 32. Апотеции:
1 — Peziza\ 2 — Morchella (сморчка); 3 — Gyromitra (строчка)
добный, некоторые его экземпляры образуют токсин гиромитрин„
не удаляющийся и кипячением. У сморчков гиромитрин не
обнаружен.
Грибы порядка Tuberales (трюфелевых) имеют замкнутые в
зрелом состоянии плодовые тела клубневидной формы размером
от 1 до 10 см с плотной кожистой поверхностью. Сумки
трюфелевых располагаются в плодовых телах гимениальным слоем или
гнездообразно. Освобождение аскоспор пассивное после
разрушения плодового тела или поедания его животными.
Трюфелевые — обязательные микоризообразователи.
Наиболее ценный представитель этого порядка — черный французский
трюфель Tuber melanosporum, формирует микоризу с дубом,
буком и грабом. Этот вид распространен на юге Франции. В
некоторых районах нашей страны встречается другой вид этого
рода — летний трюфель Г. aestivum, образующий микоризу с теми
деревьями, что и первый вид. Он имеет плодовые тела довольно
крупного размера. Мякоть плодового тела желтовато-белая с
многочисленными плотными жилками. Гриб съедобен, но по
качеству уступает черному французскому трюфелю.
КЛАСС BASIDIOMYCETES
Это обширный класс, содержащий больше тысячи
родов и более 30 000 видов разнообразного строения и размеров.
78
Основная отличительная черта этого класса — наличие
специализированного органа — базидии, развивающейся в результате
полового процесса и являющейся его конечным этапом. Базидия,
как и сумка, репродуктивный орган, на ней образуются споры,
с помощью которых идет дальнейшее размножение особи.
Базидия представляет собой одну большую нерасчлененную
клетку, развивающуюся из конечной клетки гифы, либо
образование, разделенное перегородками. Характерной особенностью
базидии является наличие наружных выростов — стеригм, на
которых располагаются базидиоспоры. На каждой базидии обычно
располагается по четыре базидиоспоры (рис. 33).
Другая существенная характеристика базидиальных грибов —
своеобразное строение септ мицелия, имеющего хорошо развитые
септированные гифы (см. с. 24).
Большинство базидиальных грибов имеет характерное
строение мицелия, отличающее его от мицелия всех других грибов.
Рис. 33. Схема развития базидии и образование базидиоспор
Мицелий многих базидиальных грибов имеет так называемые
пряжки, т. е. небольшие полукруглые клеточки, лежащие сбоку
на гифе, против поперечной перегородки в ней. Одна часть
каждой пряжки сообщается с основной нитью мицелия. Эти пряжки
имеют специальное назначение. Они участвуют в синхронном
делении дикарионов в гифе. При делении ядра дикарионы
располагаются рядом посередине клетки, а против них образуется
короткий боковой вырост, загибающийся по направлению к
основанию гифы. Ядра в мицелии делятся таким образом, что ось
деления одного из них проходит параллельно гифе, а второго — под
углом, поэтому одно ядро при делении отходит в пряжку (рис. 34).
Затем под основанием выроста появляются поперечная перего-
79
родка и перегородка в основании пряжки. Другая часть пряжки
соприкасается с гифой, между ними образуется отверстие. Ядро
из пряжки переходит в клетку гифы и восстанавливает ее дву-
ядерность.
Отличительный биохимический признак класса базидиомице-
тов — состав полисахаридного комплекса клеточной стенки.
Основу клеточной стенки составляют комплексы хитин—ксилан и
хитин—фруктозан. Кроме того, в клеточной стенке базидиальных
грибов присутствуют уроновые кислоты.
Грибы класса базидиомицетов имеют наивысший процент ГЦ-
оснований в геноме (от 50 до 68%), таким образом относятся к
ясно выраженному ГЦ-типу. Половой процесс у грибов этого
класса также состоит из трех стадий и включает плазмогамию,
ГХ
±D
3
Рис. 34. Схема деления дикариотических ядер у
базидиомицетов
кариогамию и мейоз. Однако базидиомицеты не имеют
специальных половых структур. Половой процесс представлен у них сома-
тогамией и осуществляется в результате слияния мицелиев
разного полового знака. Слияние мицелиев происходит на ранних
стадиях прорастания базидиоспор. Базидиоспоры обычно
одноядерны и имеют гаплоидный набор хромосом. После прорастания
базидиоспоры в ростковую трубку происходит слияние ростковых
трубок мицелиев разных половых знаков, так как большинство
базидиомицетов гетероталличны. Так же, как у сумчатых грибов,
плазмогамия и кариогамия у базидиомицетов разделены во
времени. Однако этот разрыв гораздо более значителен, чем у аско-
мицетов. В результате слияния двух гаплоидных мицелиев вскоре
после прорастания базидиоспор происходит слияние их
цитоплазмы, но кариогамии при этом не происходит, а ядра ассоциируются
строго попарно, образуя так называемые дикарионы, так же, как
это имеет место в. аскогенных гифах аскомицетов. Гаплоидный
мицелий очень недолговечен, он представлен только ростковыми
80
трубками прорастающих спор, и основное развитие мицелия
осуществляется в дикариотическом состоянии. Из дикариотического
мицелия слагается плодовое тело — базидиокарп,
представляющее собой образование из псевдопаренхиматической ткани, на
котором далее формируются базидии. Таким образом, мицелий
большинства базидиомицетов проходит три хорошо выраженные
стадии: гаплоидную, дикариотическую и диплоидную.
Почти все базидиомицеты образуют плодовые тела, за
исключением грибов из порядка Uredinales, Ustilaginales и Exobasidia-
les, являющихся высокоспециализированными паразитами.
Плодовые тела имеют самую разнообразную форму и размеры от
микроскопических до полутора метров. Образование базидий
происходит или на поверхности плодового тела, или внутри его из
кончика гифы дикариотического мицелия. Базидии часто
располагаются тесным слоем, называемым, как и у сумчатых грибов, ги-
мением. Развитию базидии предшествует слияние ядер дикариона.
Участок гифы, в котором происходит это слияние, отделяется
перегородкой и образует материнскую клетку базидии. Она
несколько увеличивается в размерах и в ней происходят два
последовательных деления ядра, одно из которых мейотическое, в
результате чего образуются четыре ядра. В это же время на
поверхности базидии образуются четыре выроста, вздувающиеся на
концах, в них поступают по одному ядру. Эти выросты,
содержащие ядра, образуют базидиоспоры. В деталях строение и развитие
базидии может быть несколько различным. Иногда споры
образуются не на верхнем конце ее, а сбоку, число базидиоспор в
большинстве случаев равно четырем, но известны и односпоровые
базидии, двух-, шести- и восьмиспоровые. Есть базидии, у
которых число базидиоспор достигает нескольких десятков.
Сама базидия у одних базидиомицетов остается
одноклеточной, у других вслед за делением ядер она также делится
поперечными или продольными перегородками на четыре клетки.
Есть базидии, которые развиваются из толстостенной клетки,
представляющей собой покоящуюся спору. Эти покоящиеся
клетки называются пробазидиями, или телиоспорами. Они типичны
для порядков паразитных грибов Ustilaginales и Uredinales,
относящихся к подклассу Teliomycetidae.
Половое размножение у базидиальных грибов, как было
сказано выше, представлено преимущественно соматогамией. Однако
есть и некоторые другие пути полового процесса. В частности,
у навозного гриба Coprinus lagopus, образующего оидии,
плазмогамия осуществляется слиянием оидии с соматической гифой.
Оидии обычно обнаруживаются в слизи и разносятся водой или
насекомыми. Ржавчинные грибы образуют спермации, которые
сливаются с мицелием, и таким образом осуществляется плазмогамия.
Среди базидиомицетов есть гомоталличные виды, но
преобладают гетероталличные, составляющие около 90% всех известных
видов с тетраполярным разделением полов.
Иногда бывает, что уже на одной базидии образуются все
81
четыре типа спор, иногда же споры одной базидии попарно
одинаковы, а на другой базидии развиваются две другие пары,
несходные с первыми.
Базидиоспоры разбрасываются активно благодаря повышению
тургорного давления, которое происходит в результате
ферментации гликогена. Но тургорное давление передается через узкий
канал стеригмы, поэтому базидиоспора получает очень
незначительный толчок и отлетает на несколько миллиметров.
Бесполое спороношение есть у очень немногих базидиомице-
тов. Более распространено у них вегетативное размножение
путем образования почек, фрагментации мицелия с образованием
артроспор. Бесполое размножение, если оно есть, представлено
конидиальными спороношениями. У гриба Fomitopsis annosa оно
представлено спороношениями типа Oedocephalum, есть кониди-
альные спороношения у головневых грибов. У ржавчинных
грибов конидиальные спороношения — оидии — образуются в
специальных вместилищах.
Таким образом, в цикле базидиальных грибов прослеживается
большая гомология с циклом развития сумчатых грибов.
Гомология имеется в развитии сумки и базидии. У базидиальных грибов
так же, как и у сумчатых, плазмогамия и кариогамия разделены
во времени. В результате плазмогамии и у тех и у других
образуются дикарионы. Кариогамия с образованием диплоидного ядра
происходит в клетке, которая затем становится материнской
клеткой сумки или базидии. Как в сумке, так и в базидии
происходит редукционное деление с последующим образованием аско-
и базидиоспор. Только у аскомицетов они образуются эндогенно,
внутри сумки, а у базидиомицетов — экзогенно. Кроме.того, ба-
зидиомицеты имеют более растянутую дикариотическую стадию.
Класс Basidiomycetes на основании строения и формирования
базидии разделяется на три подкласса: Holobasidiomycetidae, He-
terobasidiomycetidae и Teliomycetidae.
ПОДКЛАСС HOLOBASIDIOMYCETIDAE
Этот подкласс характеризуется нерасчлененной бази-
дией, обычно булавовидной формы. В него входят две более или
менее естественные группы, но которые не рассматриваются как
строго установленные таксономические категории. Это группы
гименомицетов и гастеромицетов.
Гименомицеты
Сюда относятся грибы, известные в быту. Наиболее
характерным признаком этой группы грибов является
расположение базидии плотным слоем — гимением — на поверхности
плодового тела. Поверхность плодового тела, несущего гимений,
называется гименофором. Гименофор может быть гладким или
зубчатым, трубчатым, пластинчатым и т. д. Гимений открыт уже
82
с ранних стадий развития. Только у немногих простейших
представителей гимений зачаточный или совсем не выражен, так же
как и плодовые тела.
В типичном случае гимений состоит из базидий и
чередующихся с ними стерильных образований парафиз. У некоторых
гименомицетов развиваются еще особые цистиды в виде более
крупных, чем парафизы, образований, часто причудливой формы,
возвышающихся своими концами над общим уровнем гимения.
Большинство гименомицетов относится к почвенным
обитателям. Значительное количество их также живет в качестве сапро-
трофов на отмершей древесине или паразитов деревьев. Они
включают основную массу грибов-микоризообразователей древесных
пород. Среди них много съедобных, некоторые поддаются
культивированию в условиях теплиц. Есть и ядовитые грибы.
В группу гименомицетов входят два порядка: Aphyllophorales
(непластинчатые) и Agaricales (пластинчатые). Первый порядок
объединяет грибы, плодовые тела которых разного типа,
преимущественно деревянистой или кожистой консистенции. Они очень
разнообразны по строению. Наиболее примитивные имеют
плодовые тела в виде пленки или корочки с гладкой поверхностью.
Более сложные — коралловидной, воронковидной формы или
состоят из шляпки и ножки, имеют складчатый, шиповатый или
трубчатый гименофор. На основании строения и консистенции
плодовых тел и гименофора, формы и окраски спор в этом
порядке выделяют несколько семейств.
Семейство Thelephoraceae — телефоровые. Грибы имеют
хорошо различимые плодовые тела кожистой или деревянистой
консистенции с гладким шероховатым или складчатым гименофором.
Часто располагаются на субстрате черепитчато. Систематическим
признаком телефоровых грибов служит присутствие телефоровой
кислоты. Грибы этого семейства растут как сапротрофы на
опавших листьях, древесине и на почве. Например, Thelephora ter-
restris растет на почве обычно в сухих сосновых лесах.
Семейство Clavariaceae — рогатиковые. Характерная черта
этого семейства — коралловидное ветвление плодового тела, но
есть булавовидные и цилиндрические. Гимений у многих
представителей покрывает все плодовое тело и отсутствует только у его
основания. Консистенция плодовых тел мясистая, кожистая,
иногда хрящеватая. Основные представители — сапротрофные грибы
на растительном опаде. Sparassis crispa и Ramaria flava —
съедобные грибы. Виды рода Typhula паразитируют на диких и
культурных растениях.
Семейство Cantharellaceae — лисичковые. Виды этого
семейства имеют воронковидные или трубковидные плодовые тела.
Образуют гимений на наружной стороне плодового тела. Основные
представители Cantharellus (лисички) и Cratarellus имеют
мясистые плодовые тела со складками, воронковидными шляпками
соответственно желтого и дымчато-серого цвета. Сапротрофные
съедобные грибы образуют микоризу с сосной.
83
Семейство Hydnaceae — ежовиковые. Плодовые тела имеют
тименофор в виде шипиков или зубчиков. Они могут быть в виде
корочек, кораллов или иметь шляпку и ножку. Коралловидные
ллодовые тела гораздо более мягкие, чем в семействе Clavaria-
сеае.
Представитель этого семейства Hericium coralloides имеет
большие белые коралловидные плодовые тела, которые
образуются на пнях и в дуплах деревьев. Другой представитель Hydnum
repandum имеет типичную шляпку и ножку с зубчатым гименофо-
ром и обитает на почве, в хвойных и лиственных лесах, съедобен.
Семейство Coniophoraceae — кониофоровые. Плодовые тела
распростертые. Гименофор гладкий, складчатый, лабиринтосетчатый
или пористый. Наиболее известный представитель Serputa lacry-
mans — настоящий домовый гриб, разрушающий древесину в
постройках. Этот гриб сапротроф, использующий целлюлозу клеток
древесины, вызывает гниение деревянных конструкций. Плодовые
тела его распростертые, неопределенных очертаний, до 0,5 м в
диаметре. Обильно выделяет капли жидкости, откуда и получил
свое название (lacrymans — плачущий). Другие виды этого
семейства обитают в лесу и разрушают древесный опад.
Семейство Polyporaceae — трутовиковые. Плодовые тела
грибов этого семейства могут иметь вид корочек, копытообразную
форму деревянистой консистенции или образовывать шляпку и
ножку. К первым относится род Porta, виды которого разрушают
древесину или вызывают корневую гниль. Другая группа,
известная под названием трутовиков, имеет копытообразные плодовые
тела, являющиеся также возбудителями различных болезней
лесных деревьев. Эти организмы ответственны и за разложение
древесного опада, так как активно разрушают лигнин и целлюлозу
благодаря наличию в них соответствующего комплекса
ферментов. Трутовиковые грибы являются почти монопольными
разрушителями лигнина. Можно назвать следующие основные виды
этого семейства. Большое экономическое значение имеет Laetipo-
rus sylfureus или серо-желтый трутовик, который вызывает гниль
дубов и других деревьев. Forties fomentarius — березовый
трутовик, имеет серый цвет и обитает на ослабленных березах. Во
многих лесах широко распространен Ganoderma applanatum, который
лоражает мертвую древесину березы и другие виды твердой
древесины. Его копытообразные плодовые тела могут жить 80 лет и
более. Fomes annosa вызывает болезнь ели, называемую корневой
губкой. Плодовые тела F. annosa достигают больших размеров.
Это один из немногих представителей порядка, имеющий кони-
диальную стадию в цикле развития.
Порядок Agaricales включает грибы, плодовые тела которых
могут быть либо съедобными, либо ядовитыми. Среди последних
есть сильно ядовитые формы. Характерная черта этих грибов —
мясистые плодовые тела, обычно состоящие из шляпки и ножки.
Базидии образуются у них на поверхности пластинчатого или
трубчатого гименофора, на нижней стороне плодового тела. Их
34
трубчатый гименофор отличается от гименофора трутовых грибов
тем, что легко отделяется от мякоти и своим происхождением
связан с пластинчатым.
Грибы порядка имеют двуядерный многолетний мицелий, на
котором из года в год образуются однолетние плодовые тела.
Мицелий растет от центра во всех направлениях, образуя
концентрическую колонию. Плодовые тела формируются на концах гиф
и образуют кольцо. Особенно хорошо образует так называемое
«ведьмино кольцо» Marasmius oreadus — луговой опенок. В
центре такого кольца трава обычно бывает особенно зеленая, что
обусловлено накоплением азотистых веществ, которые образуются
по мере отмирания мицелия. Такой мицелий у М. oreadus может
существовать много лет.
Базидиокарпы (плодовые тела) у грибов порядка Agaricales
начинают развиваться на мицелии из небольшого клубка гиф в
маленькое круглое или овальное тело. В это время уже
закладываются пластинки. У некоторых видов пластинки прикрыты так
называемым частным покрывалом — пленочкой из бесплодных
гиф, прижимающих края шляпки к ножке, например у рода Aga-
ricus — шампиньона. По мере роста гриба наружная часть
шляпки растет быстрее покрывала и оно разрывается; остатки его
видны в виде бахромы или кольца на ножке. У других родов,
например Amanita (мухомор, бледная поганка), плодовое тело
целиком покрыто покрывалом (общее покрывало). Остатки этого
покрывала при вытягивании плодового тела остаются в виде воль-
вы или влагалища в основании и в виде белых пятен и чешуи
на шляпке. У них есть и частное покрывало.
Для порядка Agaricales почти неизвестно бесполое
размножение. Только немногие виды, такие, как Coprinus fimentarius,
С. lagopus, Coliybia conigena, образуют оидии. Оидии выполняют
двойную функцию: они могут прорастать в мицелий и могут
осуществлять функцию спермациев, сливаясь с гифой
противоположного пола. Некоторые виды Agaricus образуют хламидоспоры
(Agaricus campestris).
Представители этого порядка распространены по всему
земному шару. Многие из них растут в лесах, но такие, как Agaricus
campestris (шампиньон) — на открытых лугах, a A. rodmani,
Marasmius oreadus — на лужайках даже в городах. Некоторые из
них образуют микоризу главным образом с древесными
породами; другие известны как разрушители древесины и растительного
спада, немногие из них паразиты. Многие из Agaricales образуют
разнообразные антибиотики, в частности полиацетиленовые
антибиотики с тремя ненасыщенными связями, такие, как агроцибин
и немотиновая кислота, свойственные только этой группе грибов.
Гриб Psilocybe mexicana известен как галлюциноген, который
считался священным грибом у древних ацтеков. Они использовали
этот гриб в своих ритуальных обрядах. Psilocybe mexicana
образует галлюциногенное вещество — псилоцибин, вызывающее
галлюцинации. Псилоцибин близок к алкалоидам, содержащим
85
индольное кольцо, имеет в своем составе азот и фосфор,
действует на нервную систему. Имеются попытки использования era
для лечения шизофрении.
Гриб Atmiliar'iella mellea (опенок) — активный паразит,
вызывает гниль яблонь и многих лесных деревьев. A mellea еще
известен тем, что его мицелий способен к биолюминесценции, в
результате чего он светится в темноте. Он содержит субстрат —
люциферин, который подвергается действию имеющегося у него
фермента люциферазы. Под действием этого фермента и при
использовании энергии АТФ происходит свечение.
В качестве съедобных грибов специально культивируется Aga-
ricus bisporus, Pleurotus ostreatus — вешенка, а в Японии — Cor-
tinellus berkelyanus и Volvariella esculent. Вкусовые качества этих
грибов определяются набором специфических белков. Эти грибы
содержат и некоторые витамины. В качестве основных семейств
можно назвать следующие.
Семейство Boletaceae — болетовые. Представители этого
семейства хорошо отличаются от других Agaricales трубчатым ги-
менофором. Все трубочки равной длины и легко отделяются от
основной части плодового тела. Виды этого семейства
распространены по всему земному шару и являются исключительно
обитателями лесов. Плодовые тела мясистые и мягкие, легко и быстро
загнивают. Образование плодовых тел в большой степени зависит
от влажности и температуры окружающей среды. Многие
образуют микоризу с сосной, лиственницей, березой и другими
деревьями. Большинство из них — съедобны. Например, Boletus edulis —
белый гриб считается в некоторых странах лучшим съедобным
грибом.
К этому семейству относятся также Leccinum aurantiacum
(подосиновик), Leccinum scabrum (подберезовик), Ixocomus luteus
(масленок), Suillus grevillei (масленок лиственничный).
Семейство Amanitaceae — мухоморовые. Имеют типичный
пластинчатый гименофор. У большинства представителей имеется
общее покрывало, оставляющее влагалище в основании ножки.
Наиболее известный род Amanita — мухоморы. Большинство
видов этого рода — ядовитые грибы. A. phalloides — бледная
поганка и A. virosa — белая поганка смертельно ядовиты, образуют
токсины фаллоидин и аманитин полипептидной природы,
состоящий из а-аминокислот. A. muscaria — мухомор также сильно
ядовит. Образуемый им токсин действует не только как яд, но и как
галлюциногенное вещество. Род Pluteus включает съедобный гриб
P. cervinus, растущий на гниющих пнях и валежнике.
Семейство Agaricaceae — шампиньоновые. Базидии
образуются на пластинчатом гименофоре. Всегда имеется частное
покрывало, оставляющее кольцо на ножке. Представители семейства
различаются по цвету спор: белые, розовые, коричневые,
пурпурные, черные. Основные широко распространенные роды Agaricus
(шампиньон), Lepiota (зонтик), Macrolepiota (гриб-зонтик) —
съедобные грибы. A. bisporus — один из немногих грибов, ко-
86
торые успешно культивируют в специальных помещениях более
чем в 40 странах. Этот вид легко выращивается и в
лабораторных условиях, его плодовые тела можно получать даже в
чашках Петри.
Семейство Russulaceae — сыроежковые. Выделяются на
основании строения плодового тела и пластинок. Грибы этого
семейства имеют толстые пластинки. Мякоть плодовых тел хрупкая,
состоит из двух типов гиф: тонких и толстых. В это семейство
входят два рода: Russula — сыроежка и Lactarius — млечник,
включает грузди, рыжики, волнушки и др. Род Russula имеет
ломкие плодовые тела и не имеет млечного сока. Существуют
съедобные и ядовитые формы. Грибы рода Lactarius выделяют
млечный сок. Род большой, есть съедобные формы.
Представители обоих родов широко распространены в разных типах леса.
Гастеромицеты
К гастеромицетам относятся голобазидиальные грибы,
т. е. грибы с нерасчлененной базидией, в которых гимений
закладывается внутри замкнутого плодового тела и базидиоспоры
освобождаются только либо вследствие разрыва, либо общего
разрушения оболочки. Обычно базидии несут по четыре споры, но
бывает и больше. У многих гастеромицетов гимениальный слой
неразличим к моменту созревания спор, так как базидии быстро
разрушаются.
Гастеромицеты имеют довольно разнообразную форму
плодовых тел. Чаще всего она шаровидная и остается такой до
полного созревания, у некоторых видов она звездчатая, в виде ножки
с головкой или корзиночки и т. д. Плодовые тела имеют хорошо
выраженную оболочку, называемую перидием, которая или
разрывается после созревания спор, или споры остаются внутри до
освобождения внешними агентами. Перидий, часто темно
окрашенный снаружи, представляет собой плотную обвертку из
бесплодных гиф, у одних она бывает однородной, у других
дифференцирована на несколько анатомически различимых слоев.
Репродуктивная часть базидиокарпа называется глебой. Она
состоит из полостей и камер различной формы и прослоек
бесплодной ткани между ними. На внутренней поверхности камер
расположены базидии с базидиоспорами. При созревании плодового
тела базидии разрушаются и споры свободно лежат внутри
перидия. Почти все гастеромицеты — почвенные сапротрофы, лишь
немногие встречаются на гнилой древесине. Для некоторых видов
известна способность к микоризообразованию.
Основные порядки гастеромицетов следующие: Lycoperdales,
Sclerodermales, Nidulariales и Phallales.
Большой порядок Lycoperdales — дождевиковые, включает
дождевики (Lycoperdon) и звездовики (Geastrum) (рис. 35). При
созревании глеба этих грибов становится пылящей массой от
огромного количества образующихся в ней спор.
87
Большинство представителей — почвенные сапротрофы,
растут на пнях, гниющей древесине или на земле в лесу, на
пастбищах и лужайках. Некоторые съедобны в молодом возрасте, до
созревания глебы. Большой род этого порядка — Lycoperdon.
L. perlatum — дождевик шиповатый, один из самых широко
распространенных видов,
встречающийся на всех континентах, ра-
n^a.-i*1 \ Г Л стущий на лугах и выгонах.
В центре его плодового тела по
направлению к периферии
закладываются лабиринтообраз-
ные полости, на которых обра-
Рис. 35. Плодовые тела гастеромицетов:
1 — Lycoperdon perlatum (дождевик
1 2 шиповатый); 2 — Geastrum pectina-
turn (звездовик гребенчатый)
3УЮтся базидии. Они располагаются на дикариотическом мицелии
и заполняют всю полость. К моменту созревания базидиоспоры
заполняют всю полость базидиокарпа, а базидии разрушаются.
Род Geastrum — звездовик, или земляная звездочка, имеет-
характерное строение плодового тела. Его перидий состоит из
Нескольких слоев. Наружный слой к моменту созревания
растрескивается и образует звезду. Лопасти звезды отгибаются вниз. На
Вершине внутреннего слоя перидия возникает отверстие, через-
второе 'рассеиваются споры. Лопасти наружного слоя перидия
отгибаются вниз, приподнимая внутреннюю часть перидия как бы
на ножках во влажную погоду, и загибаются кверху — в сухую.
angermannia gigantea имеет самые крупные плодовые тела
среди грибов.
Представители порядка Sclerodermales — ложнодождевико-
вЫе, имеют толстый твердый перидий и темную глебу; плодовые
тела ложнодождевиковых шаровидной и клубневидной формы.
Сличаются от дождевиковых тем, что не имеют четко
выраженного гимения, особенно на ранних стадиях развития, а глеба —
жестко-плотная в отличие от рыхлой глебы дождевиковых.
Основной род Scleroderma — ложнодождевик. S. aurantium —
обычный гриб в лесах разного типа с плотным наземным плодовым
телом и бородавчатой оболочкой, ядовит.
Порядок Nidulariales — гнездовиковые. Названы они так
потому, что их плодовые тела содержат маленькие твердые
структуры, похожие на яйца в птичьих гнездышках. Они представляют
собой образования, называемые перидиолями, залагающиеся в
глебе и обособляющиеся вместе с окружающей их оболочкой
внутреннего перидия. Внутренняя щелевидная полость их выстлана
слоем- базидии. При созревании общий перидий плодового тела
Раскрывается на вершине и перидиоли обнажаются в виде
маленьких белых яичек, лежащих как бы в гнезде (в общем пери-
38
дии). В роде Nidularia перидиоли лежат в перидии свободно, а в
родах Crucibullum и Cyathus они прикреплены к стенке
плодового тела каждая особым жгутом гиф, достигающим в
расправленном состоянии нескольких сантиметров длины. После
созревания перидиоли выпадают из перидия. Содержащиеся в них
базидиоспоры освобождаются благодаря разрушению оболочки
перидиолей. Виды выше перечисленных родов преимущественно
встречаются на гнилой древесине и принимают участие в ее
разложении. Основные роды Cyathus и Nidularia встречаются
повсюду.
Порядок Phallales — веселковые. Представители этого
порядка в основном распространены в тропических областях земного
шара. Имеет наиболее сложное строение плодового тела среди
гастеромицетов и всех грибов вообще. На территории СССР
наиболее обычен гриб Phallus impudicus — веселка обыкновенная,
встречающийся как сапротроф в лиственных лесах. Плодовое тело
у этого гриба, как и у большинства гастеромицетов,
закладывается подземно, но затем выходит на поверхность в виде
яйцевидных тел, одетых белой плотной оболочкой — вольвой, внутренний
слой которой значительно ослизнен. Во внутренней части
плодового тела дифференцируется бесплодное цилиндрическое полое
тело со стенками губчатого строения, занимающее осевое
положение в яйце. Вокруг его верхней части в виде колокола
дифференцируется плодущая глеба. При созревании вольва
разрывается, стерильная часть глебы сильно вытягивается и выносит
кверху плодущую часть с базидиями в виде ячеистой шляпки, быстро
ослизняющейся. Растяжение может происходить в течение 5 ч
после разрыва перидия при благоприятных условиях. Затем глеба
очень быстро расплывается и споры погружаются в
желеобразную неприятно пахнущую слизь. Этот запах привлекает мух,
которые затем разносят споры.
ПОДКЛАСС HETEROBASIDIOMYCETIDAE
Грибы этого подкласса имеют сложную базидию,
состоящую из двух частей: нижней, называемой гипобазидией, и
верхней — эпибазидии, на которой образуются базидиоспоры.
Зпибазидия состоит из двух, чаще из четырех клеток и отделена
от нижней части перегородкой. Такая базидия называется гетеро-
*базидией.
Плодовые тела гетеробазидиомицетов студенистой
консистенции, реже сухие, напоминающие корочку или подушечки. В сухую
погоду они быстро теряют воду и становятся незаметными, но при
повышенной влажности набухают и приобретают характерные
черты. Большинство этих грибов сапротрофы, развивающиеся на
гниющей древесине, но есть и паразиты насекомых и зеленых
растений. Подкласс объединяет три небольших порядка.
Порядок Auriculares — аурикуляриевые, характеризуется
поперечными перегородками у базидии. Стеригмы клеток базидии
89
неодинаковы по длине. У некоторых видов имеется толстостенный
пробазидий, проходящий перед образованием базидии период
покоя. У видов рода Helicobasidium широко распростертые
плодовые тела. Несовершенная стадия Н. purpureum — Rhizoctonia
crocorum; паразитирует на корнях многих сельскохозяйственных
культур. Для видов рода Auricularia характерны своеобразные
студенистые плодовые тела. A. auricula напоминает ушную
раковину человека. Развивается на отмерших ветвях бузины. Это один
из первых базидиомицетов, у которого были открыты долипоры.
Порядок Tremellales — дрожжалковые, объединяет грибы с
базидиями, разделенными взаимно перпендикулярными
продольными перегородками. Представители этого порядка имеют
разнообразные по форме и цвету плодовые тела, которые могут быть
тонкими распростертыми и крупными студенистыми. Иногда весь
базидиокарп представлен тонким слоем желеобразных гиф,
образующих базидии. Цвет плодовых тел может быть серым,
пурпурным, коричневым. Многие представители содержат каротиноиды.
У представителей порядка тремеловых базидиоспоры прорастают
вторичными спорами или конидиями, впоследствии почкующимися,
в результате чего формируются дрожжевые колонии. Эту стадию
некоторые авторы по ряду признаков сближают с аспорогенными
дрожжами рода Cryptococcus, считая их несовершенной стадией
отдельных родов порядка Tremellales. Представителем может
служить Tremella mesenterica, имеющая студенистое плодовое
тело оранжевой окраски. Большинство видов этого рода — сапро-
трофы на древесине.
Представители порядка Dacryomycetales имеют
неразделенные базидии цилиндрической формы, на конце вильчато
разветвленные на два отростка, на каждом из которых развивается по
одной базидиоспоре. Плодовые тела разнообразны:
распростертые, булавовидные, кустовидные. Все они сапротрофы на
древесных остатках. Dacryomyces ellisii, растущая на агаре, не образует
базидии, но образует четыре типа бесполых спор: оидии, хлами-
доспоры, эллипсоидные споры, растущие в пучках (конидии), к
баллистоспоры. Последние представляют интерес, так как дают
доказательство родства баллистоспоровых дрожжей Sporobolomy-
cetaceae с Dacryomycetales.
ПОДКЛАСС TELIOMYCETIDAE
В этот подкласс входят организмы, у которых базидия
развивается из толстостенной клетки — пробазидий, или телио-
споры, которая рассматривается как предшественник базидии и
прорастает в промицелий. На промицелии формируется сложная
базидия, состоящая из гипобазидии и эпибазидии, сходная с ба-
зидией гетеробазидиальных грибов. Однако отсутствие плодовых
тел и ряд других признаков существенно отличают подкласс от
гетеробазидиомицетов (рис. 36).
Телиоспоры (телейтоспоры) представляют собой стадию, в ко-
190
торой гриб переносит неблагоприятные условия. Плодовых тел в
подклассе нет. Они утрачены вследствие паразитического образа
жизни его представителей. В подкласс входят порядки Ustilagi-
nales (головневые) и Uredinales (ржавчинные), представленные
исключительно паразитными формами. Головневые грибы в
природе паразитируют на растениях, но они не являются строго об-
ъ
Off
=ф=^
1
Рис, 36. Цикл развития Rhodosporidium toruloides:
1 — телиоспора; 2 — прорастание телиоспоры в промицелий
и базидию; 3 — базидиоспоры; 4 — копуляция базидиоспор;
5 — образование дикариотического мицелия
лигатными паразитами. Их можно культивировать на
искусственной питательной среде, сохраняя полный жизненный цикл —
от телиоспоры до телиоспоры.
В 1968 г. был обнаружен новый сапротрофный род базидиаль-
лых грибов, также не имеющий плодовых тел, названный Filoba-
sidium и выделенный в специальное семейство Filobasidiaceae,
которое на основании строения базидии и цикла развития
сближается с порядком Ustilaginales, хотя его цикл развития
несколько отличается от типичного цикла этих грибов. Его базидиоспоры
прорастают в бластоспоры и образуют дрожжеподобные колонии.
Затем в результате слияния бластоспор образуется пряжковый
мицелий, образующий пробазидии, из которых развиваются
базидии и базидиоспоры. Базидии обычно несут восемь базидиоспор.
Наличие у этого гриба дрожжевой стадии говорит о близости его
к дрожжевым организмам. Кроме того, по циклу развития к
порядку головневых близки дрожжи родов Rhodosporidium и Leuco-
sporidium.
91
Представители порядка Uredinales до последнего времени
рассматривались как строгие облигатные паразиты и
культивирование их считалось невозможным. Однако в 1966 г. группой
австралийских ученых Сиднейского университета впервые была
получена культура грибов, а затем получение таких культур было
осуществлено еще в ряде стран. Тем не менее получение
культуры ржавчинных грибов по-прежнему представляет большие
трудности. Все ржавчинные грибы в природе не способны к сапро-
трофному существованию.
КЛАСС DEUTEROMYCETES (FUNGI IMPERFECTI)
Дейтеромицеты — один из крупнейших классов грибов.
Он объединяет формы с хорошо развитым септированным
мицелием, не имеющие половых спороношений. Реже и конидиальные
спороношения у них отсутствуют, а образуются только склероции
или же они представлены стерильным мицелием. Их цикл
развития проходит в гаплоидной конидиальной стадии с
относительно небольшим числом диплоидных ядер в мицелии и без смены
ядерных фаз.
Некоторые несовершенные грибы представляют собой кониди-
альную (анаморфную) стадию аскомицетов, реже базидиомице-
тов. Подтверждением этому служит то, что многие из них
действительно удалось связать с совершенными стадиями аскомицетов
или базидиомицетов, доказать, что они представляют собой
конидиальные стадии высших грибов. Например, сумчатые стадии
были найдены у многих аспергиллов, пенициллов, фузариумов
и др.
По мере обнаружения высших спороношений у несовершенных
грибов они распределяются в соответствующие классы и порядки
высших грибов, занимают определенное место в естественной
системе и получают название по совершенной стадии. Совершенные
стадии многих грибов были обнаружены в природе или получены
на питательных средах в культуре часто много лет спустя после
того как гриб был описан впервые. В последнем случае при
получении совершенных стадий в культуре невозможность их
образования ранее объяснялась гетероталличностью вида, при котором
отсутствие партнера по спариванию делает невозможным
образование половых спороношений. Поэтому долгое время
дейтеромицеты рассматривали как временную, искусственную группу,,
объединяющую высшие грибы, у которых известны только
конидиальные стадии, а половые стадии (сумчатые или базидиальные)
пока не обнаружены. Предполагали, что у всех дейтеромицетов
со временем по мере их изучения будут найдены совершенные
стадии и они займут соответствующее место в системе среди
аскомицетов и базидимицетов.
В настоящее время эта точка зрения на положение
дейтеромицетов в общей системе грибов существенно изменилась. Есть
убедительные доказательства того, что эволюция этой группы
92
идет в направлении совершенствования конидиального аппарата,
а также, вероятно, и механизмов, заменяющих у этой группы
отсутствующий половой процесс — гетерокариоз и парасексуаль-
ный процесс.
Такое усложнение конидиального аппарата с постепенной
потерей полового спороношения можно наблюдать у родов Penicil-
lum и Aspergillus порядка Eurotiales — сумчатых грибов, на что
впервые обратила внимание Т. П. Сизова. У рода Penicillium
такое усложнение конидиального спороношения происходит в
направлении от секции Monoverticillata, имеющей наименее
развитые одномутовчатые конидиеносцы, через секцию Biverticillata-
symmetrica с двухъярусными конидиеносцами, к секции Asymmet-
rica — с мощными трехъярусными асимметричными кисточками.
Одновременно наибольшее число видов, имеющих половую
стадию, приходится на секцию Monoverticillata, значительно меньше
их в секции Biverticillata-symmetrica. И наконец, в очень
обширной секции Asymmetrica они представлены единичными видами.
У рода Aspergillus также большинство видов с сумчатой
стадией имеют менее развитые конидиеносцы с одним ярусом фиа-
лид; у большинства видов, имеющих конидиеносцы с двумя
ярусами фиалид, половая стадия утрачена.
Утрата половых спороношений и увеличение роли бесполого
спороношения могли происходить в самых отдаленных группах
аскомицетов и приводить к образованию сходных конвергентных
форм конидиальных спороношений у разных родов сумчатых
грибов. В результате разные роды по сумчатой стадии можно
объединить в один род по конидиальной. Так, различные роды
сумчатых стадий Eurotium, Emiricella, Neosartoria можно объединить
в один род конидиальной стадии Aspergillus, роды Talaromyces,
Carpenteles — в род Penicillium. Нередко в один род дейтероми-
цетов входят виды, связанные по происхождению не только с
представителями разных родов, как в приведенном выше случае,
но и порядков и даже классов высших грибов. Род Acremonium,
например, связан с совершенными стадиями Emiricillopsis (эвро-
циевые), Chaetomium (сферейные), Nectria (гипокрейные), Сог-
diceps (спорыньевые), а конидиальные стадии типа Oedocepha-
lum известны как у сумчатых дискомицетов, так и у некоторых
базидиомицетов, и наоборот, у одного рода сумчатых грибов,
например, у рода Hypomyces конидиальное спороношение может
относиться к разным родам Fusarium, Tubercularia, Dactylium,
Diplosporium.
Это свидетельствует о том, что в процессе эволюции
изменения конидиальных спороношений и сумчатых стадий происходили
разными темпами и независимо. Класс дейтеромицетов
представляет собой гетерогенный по происхождению класс, поэтому его
и называют формальным классом. Хотя таксономические
категории у несовершенных грибов те же, что и у других групп
грибов, — это искусственные формальные группы, выделяемые на
основе чисто внешнего сходства, а не их родства. Например, род
9а
в этом классе часто не соответствует истинному роду в обычном
понимании — группе близкородственных видов. Он представляет
собой искусственную группу, объединяющую виды со сходными
конидиальными спороношениями, как было показано выше,
называемую формальным родом.
Большинство микологов для классификации несовершенных
грибов пользуются до настоящего времени системой Саккардо.
Эта система искусственна и служит только для каталогизации
форм и создания схемы определения. Однако она удобна для
практических целей и в настоящее время наиболее полно отражает
многообразие форм в группе несовершенных грибов.
На основании строения и расположения конидиального
аппарата несовершенных грибов Саккардо выделил три порядка: Ну-
phomycetales, Melanconiales., Sphaeropsidales. Некоторые микологи
рассматривают порядок Hyphomycetales на уровне класса и
выделяют еще один класс Coelomycetes, объединяющий порядки
Melanconiales и Spaeropsidales.
Порядок Hyphomycetales (гифомицетовые) — наиболее
многочисленный порядок несовершенных грибов, включающий до
10 000 видов.
Среди представителей этого порядка много типичных
почвенных обитателей — сапротрофов, играющих в почве большую роль.
Многие представители — патогены растений, некоторые
патогенны для человека. К этому порядку относятся дрожжи и дрожже-
подобные грибы, не образующие сумок и базидий.
К порядку Hyphomycetales относятся формы, имеющие кони-
диеносцы, свободно образующиеся прямо на мицелии или
соединенные в коремии или спородохии (см. с. 36), но без
специальных вместилищ.
Порядок Hyphomycetales Саккардо разделяет на четыре
семейства: Moniliaceae и Dematiaceae — на основании отсутствия
или наличия темного пигмента; Stilbaceae — на основании
объединения конидиеносцев в коремии; Tuberculariaceae — на
основании расположения конидиеносцев тесным слоем в виде
спородохии. К семейству Moniliaceae, имеющему светлоокрашенный
или бесцветный мицелий и спороношения, относятся широко
распространенные в почве роды Aspergillus, Penicillium, Verticillium,
Acremonium, Trichothecium и др. К семейству Dematiaceae
относятся грибы, имеющие темные гифы и конидии; иногда темными
бывают либо гифы, либо конидии. Основные представители —
роды Cladosporium, Alternaria, Stemphylium, Ulocladium и др.
Большинство из них почвенные сапротрофы или факультативные
паразиты растений, некоторые — эпифиты на растениях.
К семейству Stilbaceae относятся роды Doratomyces, Graphium,
Trichurus и другие — сапротрофы или факультативные паразиты;
к семейству Tuberculariaceae — роды Tubercularia, Volutella, Fu-
sarium и другие преимущественно фитопатогенные грибы.
Некоторые виды перечисленных родов имеют сумчатую стадию. Такие
виды относятся к классу аскомицетов.
94
Порядок Melanconiales (меланкониевые) выделяется на
основании того, что конидиеносцы у представителей этого порядка
образуются сплошным слоем в виде ложа. В основном сюда
относятся паразитные формы, ложа (ацервули) которых образуются
под кутикулой или под эпидермисом растительной ткани.
Порядок Sphaeropsidales (сферопсидные) выделяется на
основании того, что конидиальные спороношения входящих сюда
видов образуются в специальных вместилищах — пикнидах (см.
с. 36). Пикниды несколько напоминают по своему строению пло-
о9 99
Ш
п
о
о
й Ш
о о
ч ш
И В
И и
In! U
к
W
Ж
Рис. 37. Типы конидиогенеза у несовершенных грибов:
I — бластоспоры; II — пролиферирующие конидии; III — фиалоспоры; IV —
базипетальные артроспоры; V — пороспоры; VI — артроспоры; VII — мери-
стематические бластоспоры; VIII — базипетальные наклоннобразующиеся
бластоспоры
довые тела сумчатых грибов — перитеции. Они имеют округлую,
кувшиновидную или грушевидную форму размером в несколько
миллиметров. Состоят из псевдопаренхиматической ткани,
кожистой или углистой консистенции. Сюда относятся роды Coniothi-
95 •
MS ^^^ШШЯШк.
У**
r <r
|
з ||и р^:
.';-г ¦ , ..
Рис. 38. Типы коиидиогснеза в электронном микроскопе:
а, б — артроспоры Oidiodendron; в — фиалоспоры Penicillium; г —
фиалоспоры Stachibotrys; д — алевроспоры Trichocladium; е — симподулоспоры
Artrobolrys; ж, з — бластоспоры МопШа (фото В. С. Гузева и А. В. Кура-
кова)
rium, Aposphaeria, Phoma, Dendrophoma — почвенные сапротро-
фы и факультативные паразиты на растениях.
Еще выделяется искусственный порядок — Mycelia Sterilia,
объединяющий грибы, вообще не образующие спороношений.
Поэтому выделение видов в этом порядке еще более условно.
Наиболее известен род Rhizoctonia, имеющий половую стадию Heli-
cobasidium, относящийся к базидиомицетам.
В последние десятилетия сделаны попытки создания системы
несовершенных грибов на иных принципах нежели система Сак-
кардо, основанной на способах конидиогенеза. По способу кони-
диогенеза несовершенных грибов разные авторы выделяют их в
разное число групп (рис. 37). Наибольшее распространение
получило деление Хьюза. Основные группы, предлагаемые им
следующие.
Бластоспоровые — конидии развиваются как вздутие конца
конидиогенной клетки, затем отшнуровываются от нее и
отделяются перегородкой, но сохраняются в цепочках, как например,
у видов Cladosporium, Alternaria (этот процесс напоминает
почкование у дрожжей).
Алевроспоровые — конидии формируются из отделившейся
перегородкой части конидиогенной клетки, разрастающейся и
дифференцирующейся в зрелую конидию (виды Humicola, Mycogone).
Фиалоспоровые — клеточная стенка конидии формируется
заново и стенка конидиогенной клетки не участвует в ее
образовании. При образовании конидий по типу фиалоспор на вершине
конидиогенной клетки — фиалиды образуется расширение в виде
воротничка (виды Phialophora, Penicillium).
Пороспоровые — конидии образуются из внутреннего слоя
клеточной стенки конидиогенной клетки через поры в стенках кони-
диеносцев (виды Helminthosporium).
Артроспоровые — артроконидии образуются в результате
фрагментации конидиеносца или гифы сохраняя цепочки (виды Geot-
richum, Oidiodendron) (рис. 38). Цепочки конидий могут
образовываться в акропетальном (снизу вверх) или базипетальном
(сверху вниз) порядке.
Однако новая система несовершенных грибов находится пока
в стадии разработки и охватывает далеко не все известные
сейчас несовершенные грибы. Морфогенез конидий хорошо изучен
только у гифомицетов, а из двух других порядков он известен
только лишь у отдельных представителей. Кроме того, при
построении системы нельзя использовать только один даже
достаточно стабильный признак. В основу должен быть положен
комплекс признаков.
Часть II
ЭКОЛОГИЯ ПОЧВЕННЫХ ГРИБОВ
Глава 4
ОБЩИЕ СВЕДЕНИЯ ПО ЭКОЛОГИИ ГРИБОВ
МЕСТО И РОЛЬ ГРИБОВ В БИОГЕОЦЕНОЗАХ
Экология исследует структуру и закономерности
функционирования экологических систем. Основным ее содержанием
является изучение закономерностей жизнедеятельности
популяций и сообществ организмов в экосистеме, которая
осуществляется во взаимодействии их друг с другом и окружающей средой.
Все организмы представляют собой взаимодействующие и
взаимозависимые части более крупных единиц, составляющих единую,
нормально функционирующую систему (экосистему).
Экосистема — это совокупность биотических (живых организмов) и
абиотических элементов (неживой части), связанных пространственно
и функционально, в результате взаимодействия которых
создается достаточно стабильная система, где имеет место круговорот
веществ и осуществляется процесс трансформации энергии и
органического вещества. Конкретным примером наземных экосистем
служат биогеоценозы, которые включают основные элементы
экосистемы: живую часть — биоценоз и костное тело, которое в
биогеоценозе представляет органоминеральная часть почвы. Биоценоз
включает организмы — продуценты (зеленые растения), конеу-
менты первого порядка (травоядные животные макро- и
микроскопические), консументы второго порядка (хищники макро- и
микроскопические), редуценты (разлагатели органических
веществ). Отличие редуцентов от консументов (и те и другие гете-
ротрофы, органотрофы) заключается в том, что консументы
потребляют большую часть (более 50%) превращенного ими
органического вещества и закрепляют в собственном теле, редуценты
значительно меньшую (10—20 редко 40%), возвращая большую
часть в круговорот в виде неорганических соединений.
Живые организмы в биоценозе экосистемы представлены
популяциями, группами популяций и сообществами. Популяция в
экологии — это совокупность близких особей часто одного вида,
выполняющих одну функцию в экосистеме на ограниченном
пространстве. Группы популяций разных видов с разными функциями
составляют сообщество. Сообщество включает функционально
зависимые друг от друга популяции организмов.
98
Часто популяции микроорганизмов в почве биогеоценозов
бывают разобщены в пространстве и не связаны функционально, хотя
их совокупность характерна для тех или иных почв. В этом
случае для такой совокупности употребляют понятие комплекса
микроорганизмов. Понятие комплекс применяется и к грибам-микро-
мицетам, так как отдельные популяции грибов-микромицетов
разобщены в почве и более тесно связаны с другими
компонентами биогеоценоза.
Существует понятие общей экологии (синэкологии) и частной
(аутоэкологии). Предмет общей экологии — изучение баланса
энергетических и продукционных процессов, происходящих в
экосистеме и биосфере во всех ее звеньях и уровнях, распределение
вещества и энергии по пищевым цепям в каждой конкретной
экосистеме. Однако функционирование экосистемы в целом
невозможно без понимания процессов, протекающих в отдельных ее
блоках. Необходимо изучение места и роли отдельных групп
организмов в экосистеме, в частности грибов.
Грибы-редуценты входят таким образом наряду с
бактериями в гетеротрофный блок экосистемы. В качестве редуцентов
грибы выполняют в экосистеме ряд функций.
Предметом изучения частной экологии, или аутоэкологии,
является исследование взаимоотношений организмов внутри вида
и приспособление к среде отдельных видов на уровне популяций,
изучение закономерностей изменения численности структуры
популяций и влияния факторов внешней среды на развитие той или
иной популяции и организмов, ее составляющих, и их обратное
влияние на среду. Изучение особенностей организмов,
позволяющих им развиваться в тех или иных условиях внешней среды.
Грибы имеют ряд морфологических, физиологических и
генетических особенностей, которые определяют специфику их
взаимоотношений со средой. Они характеризуются мицелиальным
строением, обеспечивающим им большую величину отношения
поверхности гиф к их объему. Такое строение дает высокую
степень контакта со средой. Грибы обладают высокой скоростью
роста и размножения, позволяющей им в короткие сроки заселять
обширные массы субстрата и накапливать большое число про-
пагативных структур. Высокая активность метаболизма
проявляется у них в широком интервале действия различных
экологических факторов: температуры, влажности, света, кислотности
среды, аэрации и т. п. Многим грибам свойственна способность
образовывать токсические продукты, в том числе и антибиотики,
что еще больше повышает их конкурентоспособность в освоении
субстрата. Они обладают способностью быстро реагировать на
действие неблагоприятных факторов среды переходом к анабиозу,
возможностью длительного пребывания в этом состоянии, не
теряя своей жизнеспособности, и также переходить к активной
жизнедеятельности при наступлении благоприятных условий.
Наличие гетерокариоза у грибов делает их весьма лабильными
в генетическом отношении и обусловливает их высокую адаптив-
4*
99
ность к условиям окружающей среды. Многократная смена кони-
диальных спороношений и смена половых и бесполых циклов
способствуют увеличению жизненности вида у грибов, так как
каждое спороношение приводит к образованию большого числа
спор, что в свою очередь способствует быстрому заселению
субстрата и закреплению популяции на субстрате.
Наличие изоферментов у грибов, т. е. разных ферментов с
одинаковой субстратной специфичностью, но работающих в
различных диапазонах условий, позволяет грибам функционировать в
больших интервалах температуры, влажности, значения рН и
других факторов среды.
В экологических исследованиях все грибы условно делят на
микромицеты и макромицеты. К первой группе относят формы с
микроскопическими плодовыми телами, не превышающим 1 мм.
К ним относятся хитридиомицеты, оомицеты, зигомицеты, дей-
теромицеты и большинство сумчатых грибов. Ко второй группе —
организмы с крупными плодовыми телами от нескольких
сантиметров до 1,5 м. Это главным образом представители класса ба-
зидиомицетов и некоторые сумчатые грибы.
Наиболее общие вопросы изучения экологии грибов связаны
с изучением положения грибов в экосистеме в целом. Положение
организмов в любой экосистеме определяется в первую очередь
их пищевыми трофическими связями, т. е. местом, которое они
занимают в цепи превращений вещества и энергии.
По взаимоотношению со средой и другими организмами на
основе пищевых связей почвенные грибы можно разделить на
шесть трофических групп:
сапротрофы
патогены растений факультативные
патогены растений облигатные
симбиотрофы-микоризообразователи
грибы-хищники
грибы — паразиты животных.
В свою очередь патогены и микоризообразователи входят в-
группу биотрофов, так как связаны в своей жизнедеятельности с
живым растением.
Только облигатные патогены имеют ограниченную связь с
почвой, так как для них почва служит только средой резервации
их покоящихся структур, остальные группы играют в ней
значительную роль.
Сапротрофы и факультативные патогены (последние могут
проходить в почве цикл развития как сапротрофы в отсутствие
растения-хозяина) являются основными разлагателями
органического вещества и играют большую роль в круговороте углерода
в природе. Грибы-микоризообразователи и грибы-хищники играют
значительно меньшую роль в этом процессе. Их положение в
экосистеме основывается на взаимоотношениях с живыми
организмами.
100
СУКЦЕССИИ ГРИБОВ В БИОГЕОЦЕНОЗАХ
Несмотря на то что все грибы являются гетеротрофами,
они очень разнообразны по своим пищевым потребностям и
возможностям в освоении субстрата, различаются по своим
функциям и поэтому составляют еще несколько функциональных
подгрупп. Их различия в функциях объясняются различием в наборе
ферментов, которыми они обладают. Эти подгруппы следующие:
1) грибы, использующие простые углеводы («сахарные грибы»);
2) гидролитики, осуществляющие гидролитическое расщепление
гемицеллюлозы, пектина, целлюлозы, хитина, кератина; 3) грибы,
разлагающие лигнин. Эти группы хорошо выявляются при
разложении грибами какого-либо животного или растительного
субстрата в определенной последовательности, представляющей
собой сукцессию (смену видов на субстрате).
По мере разложения субстрата создается определенная смена
(сукцессия) видов. Эту смену на корнях растений можно
представить следующим образом: слабые корневые патогены (грибы,
внедряющиеся в корни отмирающих растений) сменяются сапро-
трофными грибами, усваивающими легкодоступные вещества —
моно- и олигосахара, гемицеллюлозу, их в свою очередь сменяют
целлюлолитические, а затем и лигнинразрушающие грибы.
Группа корневых паразитов представляет собой слабых
паразитов, которые продолжают существовать некоторое время на
поверхности корня после его отмирания. Эта группа существенно
отличается от специализированных паразитов. Паразитизм у них
почти на грани сапротрофного существования, так как они
быстро умерщвляют растение-хозяина и развиваются в отмерших
тканях, они локально ограничены, представлены в основном
грибами родов Fusarium, Pythium, Corticium, некоторыми видами
Cladosporium. Такая экологическая группа относится к некро-
трофам.
Мицелий у этих грибов обычно растет поверхностно на
корнях, и внедрение его внутрь клеток встречает сопротивление
растения-хозяина. Они открывают дорогу для инвазии быстро
захватывающим пространство «сахарным грибам». Не выдерживая
конкуренции с сапротрофными грибами, заселяющими мертвые
растительные остатки, они переходят в состояние покоя.
В группу «сахарных грибов» входят грибы, быстро
использующие доступные углеводы (сахара, крахмал). Несмотря на то
что сахар доступен всем организмам, группа «сахарных грибов»
немногочисленна, так как ее составляют только формы,
способные быстро захватывать пространство. Основная особенность
грибов этой группы — быстрый рост мицелия, быстрое прорастание
спор и покоящихся клеток при наличии подходящего субстрата.
К «сахарным грибам» с такими свойствами относятся в первую
очередь грибы из класса зигомицетов — мукоровые грибы;
частично к ним примыкают несовершенные (дейтеромицеты),
главным образом из родов Penicillium, Aspergillus и небольшое чис-
101
ло других форм. Многие из них характеризуются способностью
образовывать антибиотики, что еще больше способствует захвату
субстрата и закреплению своих позиций.
В группу целлюлозоразрушающих грибов входят более
медленно растущие грибы, не выдерживающие конкуренции с
«сахарными грибами» за легкодоступные субстраты. Она
представлена сумчатыми и несовершенными грибами (видами Chaetomi-
ит spp., Humicola grisea, Mycogone nigra). Последующая группа
сукцессии — грибы, разрушающие лигнин. В эту группу входят
главным образом базидиомицеты. Они растут медленно,
начинают развиваться тогда, когда все легкодоступные углеводы уже
использованы первичными колонизаторами. Таким образом,
смена видов на субстрате определяется наличием тех или иных
ферментов, что и определяет их распределение в пространстве, в
частности по слоям лесной подстилки.
Однако возможно и несколько иное направление сукцессии,
а именно: на растительном опаде еще некоторое время
продолжают развиваться медленно растущие грибы-эпифиты, часто
обладающие пектинолитической активностью. Их могут сменять
целлюлозоразлагающие грибы, такие, как Trichoderma, Fusarium,
которые могут развиваться параллельно с быстро растущими
«сахарными грибами» или опережать их. Последние снова
сменяются медленно растущими целлюлолитиками, грибами родов Chae-
tomium, Humicola grisea, Mycogone nigra, а затем и лигнинраз-
рушающими подстилочными сапротрофами. Кроме того, на
каждой стадии сукцессии к основным разлагателям того или иного
субстрата могут подключаться грибы с высокой экономичностью
обмена, довольствующиеся малым содержанием доступных
Сахаров, так называемая «микрофлора рассеяния».
Сукцессия видов грибов при использовании какого-либо
субстрата, например только Сахаров или только целлюлозы,
определяется уже не наличием тех или иных ферментов, а различными
скоростями роста при разных концентрациях данного субстрата,
а также антагонистическими взаимоотношениями между
организмами.
В результате временной смены видов происходит и различное
распределение грибов в пространстве. Смену видов на лесной
подстилке лиственных пород в соответствии с делением ее на слои
можно представить следующим образом: сразу после опадения
листьев и до 6—8 месяцев, т. е. в слое L, преобладающими
формами являются Aureobasidium pullulans, Cladosporium herbarum,
Epicoccum nigrum, Coniosporium sp., грибы рода Мисог, часть из
которых наряду с разложением Сахаров обладает высокой
пектинолитической активностью (Aureobasidium pullulans).
Через 6 месяцев, т. е. в слое F (ферментационном слое), где
наиболее активно идут процессы разложения опада, появляются
в большом количестве Mycogone nigra, Humicola grisea, Chaeto-
mium sp., грибы рода Penicillium (P. frequentans, P. thomii),
типичным для этого слоя подстилки хвойного опада являются грибы
L02
Phialophora, Thysanophora, грибы родов Mucor, Trichoderma.
Большинство из них обладают высокой целлюлолитической
активностью. Постоянные компоненты слоя Н (слоя гумификации) Мог-
tierella ramanniana, Mucor hiemalis представляют собой
«микрофлору рассеяния» наряду с целлюлозоразлагающими грибами
Chaetomium, Mycogone, Humicola и лигнинразлагающими бази-
диальными грибами, представленными в этом слое обильным
мицелием.
Горизонт гумификации имеет сравнительно более низкую
биологическую активность. Однако добавление к нему растительных
остатков сильно стимулирует развитие популяции грибов. Сначала
очень обильно развиваются виды Mucor и другие «сахарные
грибы», что сопровождается исчезновением сахара и крахмала, а
затем появляются виды родов Penicillium, Aspergillus, а также
грибы, разрушающие целлюлозу: Chaetomium spp., Humicola gri-
seay виды Stachybotrys, Doratomycesf Oidiodendron в зависимости
от типа почвы. Все эти виды являются обычными компонентами
горизонта А1 в почвах, к которым добавляется еще целый ряд
видов в зависимости от типа почвы. Так, в дерново-подзолистых
почвах под хвойной растительностью добавляются виды Chryso-
sporium sulfureum, Verticillium terrestre и некоторые другие
формы. В результате в верхних горизонтах почвы состав видов
грибов оказывается наиболее разнообразным, хотя многие виды
находятся там в покоящемся состоянии в виде спор и могут начать
развиваться при поступлении дополнительного органического
вещества и увлажнения.
Таким образом, происходит постепенное увеличение видового
разнообразия в ряду поверхность растений — подстилка — почва.
Максимальная численность наблюдается в подстилке и
значительно снижается в почве, где видовое разнообразие максимально. На
поверхности растений численность грибов минимальна.
Сукцессия на травянистых растениях может начинаться либо
с эпифитов, либо с патогенных форм, представленных дейтеро-
мицетами. Первые поселенцы обычно имеют быстро
прорастающие споры при довольно низкой влажности, это дает им
преимущество первичного заселения субстрата (Cladosporium herbarum,
tpicoccum nigrum, Alternaria alternata). Вторая группа,
представленная формами, споры которых могут прорастать только при
высокой влажности. При недостаточной влажности она выпадает.
Третья группа состоит из сумчатых и несовершенных грибов.
Последняя стадия, представленная базидиальными грибами, на
этом субстрате может отсутствовать. Несмотря на то что
конкретный состав видов на каждой стадии сукцессии может быть
различен в разных природных зонах, последовательность смены
определенных трофических групп одна и та же.
Сукцессия на древесине выглядит следующим образом:
начальный этап сукцессии определяется паразитными базидиомицета-
ми — трутовиками, мицелий которых пронизывает живое дерево,
что приводит к его быстрой гибели. На отмершей древесине их
103
сменяют несовершенные и сумчатые деревоокрашивающие грибы,
использующие внутреннее содержимое клеток. Затем появляется
группа базидиальных грибов-сапротрофов, преимущественных раз-
лагателей клетчатки, которая сменяется также сапротрофными
базидиальными грибами, разлагающими лигнин.
Смена экологических групп в зависимости от пищевых
потребностей очень хорошо прослеживается на таком субстрате, как
навоз травоядных животных. Эта смена видов грибов-копрофилов
изучена давно и является классической. Первые поселенцы на
навозе мукоровые грибы, т. е. Pilobolus spp., Pilaira, Mucor и
другие — грибы с быстро прорастающими спорами, активно
растущим мицелием, поэтому имеющие преимущество в захвате
легкодоступного субстрата. Затем они сменяются сумчатыми
грибами (виды родов Ascobolus, Rhyporohius, Humaria и др.)>
растущими более медленно, не успевающими захватить легкодоступные
углеводы, но приспособленные к использованию целлюлозы и
имеющие соответствующий набор ферментов. После них
развиваются медленно растущие базидиомицеты, разлагающие лигнин
(виды родов Coprinus, Stropharia).
На субстратах животного происхождения прослеживается своя
смена видов. Важный и весьма специфический субстрат
животного происхождения представляет собой кератин (роговое
вещество) в виде остатков волос, рогов, копыт, птичьих перьев и т. д.
На нем можно выделить особую экологическую группу кератин-
разрушающих грибов.
Кератинофильные грибы содержатся в почвах многих стран
всех континентов и являются постоянными обитателями почв.
Наиболее часто встречаются Keratinomyces ajelloi (Microsporum
ajelloi), Trichophyton terrestre и Chrysosporium. Большинство
из них представляют собой конидиальную стадию сумчатых
грибов семейства Gymnoascaceae — эвроциевых грибов.
Кератинофильные грибы чаще встречаются в почвах, богатых
органическим веществом, в местах, посещаемых животными, в
садовых почвах и на фермах. Торфянистые, болотные и другие
преимущественно кислые почвы их почти не содержат.
Другим распространенным, но трудноразлагающимся
субстратом, содержащимся в животных, является хитин. Он содержится
в клеточных стенках грибов у животных, в покровных тканях
насекомых, моллюсков, нематод и простейших.
На субстратах животного происхождения первыми
поселенцами так же, как и на растительных субстратах, могут быть
паразитные формы, часто из зигомицетов порядка энтомофторовых
грибов, их сменяют виды, использующие белки и углеводы,
представленные мукоровыми грибами и дейтеромицетами. Затем
поселяются кератинофильные грибы, большинство из которых
представляют конидиальную стадию сумчатых грибов, и хитинразла-
гающие, представленные в почвах умеренного климата видами
Мисог и Mortierella, а в тропических сумчатыми грибами Euro-
Hum, Emlricella, Chaetomium, Thielavia.
104
В интегральном виде сукцессия грибов на лесной подстилке
характеризуется наиболее низким разнообразием видов на
начальных стадиях сукцессии в слое AOL, несколько большим — в слое
ДО (р + н), значительным — в минеральном горизонте А1 и
незначительным снижением в горизонте А1/А2, что может быть
выражено изменением индекса разнообразия Шеннона по этим
горизонтам. В то же время численность грибов в этих горизонтах
имеет противоположную направленность: наибольшая численность
наблюдается в подстилке, наименьшая — в минеральных
горизонтах почвы (рис. 39).
Модельные опыты дают более развернутую характеристику
сукцессии.
Сукцессия в модельном опыте на протяжении 140 суток, где
начальным этапом сукцессии может быть увлажнение сухой поч-
2,6 2,0 И
J L
А1/А2
61321J0
100 500 900 тысJr
\ i i i i \'
120 W
сутки
Рис. 39. Изменение разнообразия
(I) и численности (II) грибов-мик-
ромицетов по горизонтам дерново-
подзолистой почвы в разное время
года
Рис. 40. Изменение индекса
разнообразия по радиальной скорости роста
(/) и индекса видового
разнообразия (2) в ходе сукцессии в
черноземе типичном (модельный опыт)
вы или внесение в почву дополнительного органического
субстрата в виде целлюлозы или хитина, имеет более разнообразный вид
и характеризуется следующими этапами: на начальной стадии
сукцессии довольно большое разнообразие видов, характерное для
минерального горизонта почвы, сменяется значительным падением
разнообразия видов, что можно сопоставить с началом
разложения органического вещества в слое L, затем большим
возрастанием разнообразия и в дальнейшем стабилизацией на
определенном уровне, впоследствии возможно новое падение разнообразия
видов. При этом индекс видового разнообразия полностью
совпадает с индексом разнообразия радиальной скорости роста (Лг)
105
лых почвах грибы подвержены меньшей конкуренции со стороны
бактерий, они развиваются там в большем числе, но большее
разнообразие видов наблюдается в щелочных почвах. В видовом
составе грибов кислых и щелочных почв существуют заметные
различия, хотя ряд видов может быть общим. Только в кислых
песчаных подзолах Англии выделены Penicillium janthinellum,
Таблица 2
Численность грибов по генетическим горизонтам различных почв,
тыс./г
Дерново-подзолистая
суглинистая почва
под ельником,
Московская обл.
горизонт
А1
А1/А2
А2
В1
В/С
глубина, см
0-8
8—30
30—45
45—73
73—84
число грибных
зародышей
52,6
17,7
4,3
10,2
2,1
Подзолистая песчаная почва
под вереском (по Thornton, 1956)
горизонт
А2
В1
В2
С
глубина, см
4—20
20—30
30—50
органическое
вещество, %
0,70
3,24
0,71
0,09
X
Р.
5,9
4,8
5,1
число грибных
зародышей
85
112
61
30
Бурая лесная песчаная почва
под дубом (по Thornton, 1956)
горизонт
А
В
С
глубина, см
4—10
10—20
20—40
органическое
вещество, %
0,89
0,74
0,04
X
сх
5,1
5,0
5,2
число грибных
зародышей
46
18
4
P. frequentans, Mortierella ramanniana, только в щелочных
карбонатных почвах выделены Fusarium sambucinum, P. luteum, Thie-
lavia sp. Одни виды грибов рода Mortierella встречаются только
в кислых почвах, другие только в щелочных. Только в кислых
почвах обнаруживается М. ramanniana, только в щелочных —
М. alpina, M. minutissima. Грибы рода Mortierella (кроме М.
ramanniana) характерны для субстратов, богатых CaS04, SiC^, CaO.
В .известняковых пещерах на стенках сталактитов обнаружен
мицелий, принадлежащий грибам рода Acremonium. В некоторых
случаях гифы этого гриба связывают вновь образующиеся
кристаллы кальция с телом сталактитов. Гриб существует за счет
органического вещества, выщелачиваемого из почвы
карбонатами кальция.
Прямые методы учета наряду с высевом на питательные среды
позволяют констатировать, что кислые почвы с рН 4,0—5,0 более
богаты грибным мицелием, чем нейтральные и щелочные с рН
6,0—7,5. Минимальное количество грибных зачатков в щелочных
почвах составляет 24 тыс/г, максимальное в кислых почвах —
460 тыс/г. Особенно много грибного мицелия в кислых, богатых
органикой почвах и торфах.
При искусственной нейтрализации почв численность грибов
меняется незначительно, но при этом всегда очень существенно
повышается численность бактерий и актиномицетов, что сильно
изменяет отношение грибов к сумме бактерий и актиномицетов.
108
Однако видовой состав грибов при этом меняется значительно.
Так, в кислых дерново-подзолистых окультуренных почвах
Московской области с рН 4,8—5,1 постоянно с высокой частотой (до
100%) встречающимися видами являются Penicillium funiculosum,
P. janthinellum, P. verrucosum, var. cyclopium, Talaromyces flavus,
Trichoderma viride.
При известковании этих почв и повышении вследствие этого
рН до 6,5—6,8 доля этих видов сильно сокращается при общем
увеличении разнообразия видов. Видовой состав заметно
изменяется. Увеличивается представленность несовершенных грибов
семейства Dematiaceae, в частности Aureobasidium pullulans, Му-
cogone nigra, а также таких грибов, как Monilia geophila, P.
nigricans, P. simplicissimum, Rhizopus nigricans. Пределы значений
рН, в которых могут расти грибы, демонстрирует табл. 3
(интенсивность роста дана в 5-балльной системе).
Влажность почвы. Вода необходима всем формам жизни, в
том числе и грибам, так как любые жизненные процессы могут
Таблица 3
Рост грибов при различных значениях рН
Виды грибов
Mucor hiemalis
Mortierella ramanniana
.Zigorhinchus vuielemini
Absidia cylindrospora
Trichoderma lignorum
Aspergillus fumigatus
Sclerotium agricola
Mycogone nigra
JFusarium orthoceras
1.4
о
0
0
0
1
0
0
0
0
1
2,0 |
0
о
0
1
3
1
0
0
1
1
2>51
2
1
0
5
3
1
0
0
1
3,1
3
2
3
5
4
4
1
1
2
рН среды
3,7
3
—
3
5
5
5
—
—
2
4,3
4
3
3
1 5
5
5
3
3
3
i i
5,5 1 6,0 '
4
—
3
5
4
5
—
—
—
5
5
5
4
3
5
5
5
4
6,5
5
3
3
3
3
5
5
5
5
7,2
5
2
2
3
3
5
4
4
3
7,8
4
1
2
3
2
5
3
3
5
8,8
4
0
1
3
1
4
2
2
3
протекать только при наличии, свободной воды. Во-первых, вода
необходима для непосредственного роста организма; во-вторых,
элементы питания становятся доступными благодаря наличию
воды; действие токсинов и антибиотиков большей частью
осуществляется в водной среде.
Наличие влаги и ее дефицит сильно влияют на морфогенез
грибов, степень ветвления, интенсивность споруляции,
репродукцию и т. д.
Для получения воды из почвы организм должен обладать
энергией, преодолевающей водоудерживающие силы почвы. Наиболее
важными из них являются капиллярные силы и осмотическое
давление почвенного раствора.
Многие грибы обладают достаточно высокой энергией для
преодоления водоудерживающей силы почвы.
Доступная влага в почве выражается величиной aw, при
которой данный организм может существовать. Процентное
содержание влаги в почве не отражает количества реальна доступной
109
влаги. Так, в почвах глинистых и песчаных одно и то же
процентное содержание влаги обладает различной доступностью для
микроорганизмов. В глинистой почве большая часть ее находится
в связанном состоянии и малодоступна для живых клеток. В
песке она находится в свободном состоянии и легко поглощается
микроорганизмами. Поэтому в настоящее время используется
понятие активности воды (aw), или потенциала влаги:
_ Р
где Р — давление пара над раствором, Р0 — давление пара над
чистой водой.
Активность свободной воды равна единице, при
взаимодействии воды с какими-либо веществами она становится меньше
единицы. Таким образом, активность воды (aw) характеризует
степень связанности ее молекул и тем самым доступность для
микроорганизмов. Чем меньше aw, тем больше водоудерживающие
силы, которые требуется преодолеть организму для извлечения
воды из субстрата. Наряду с показателем aw для характеристики
водоудерживающей силы водного потенциала в субстрате
используют и другие единицы, в частности бары и паскали.
Способность микроорганизмов существовать при различных
значениях водного потенциала говорит об их различной
способности преодолевать силы, которыми вода связана с субстратом,
в частности водоудерживающие силы почвы.
Границы прорастания спор и активного роста мицелия обычна
различны. Для роста мицелия и образования репродуктивных
органов требуются более высокие значения aw, чем для
прорастания спор. Причем более высокие для формирования
репродуктивных органов, чем для роста мицелия. Способность грибов к
прорастанию можно рассматривать как показатель их
жизнеспособности, а способность к активному росту как показатель
жизнедеятельности. Границы водного потенциала жизнеспособности,
и жизнедеятельности грибов различны.
Рост мицелия обычно происходит наиболее быстро при
потенциале влаги ниже нуля, т. е. при aw ниже единицы. Это
объясняется тем, что для его роста необходима определенная
концентрация органического субстрата в среде, которая несколько
снижает потенциал влаги.
Микроорганизмы могут развиваться на средах со значением
активности воды от 1,00 до 0,60. При этом пределы активности
воды у грибов гораздо ниже, чем у бактерий. У бактерий — от
1,00 до 0,93, у грибов — от 1,00 до 0,60 и ниже.
По отношению к влаге грибы разделяют на следующие группы:
1) гигрофилы — минимум для роста aw 0,95 (—80 бар),
максимум — 1,00, т. е. они развиваются при наличии высокой
активности воды; 2) мезофилы — минимум aw 0,90 (—150 бар),
максимум aw 97—98 (—50—30 бар); 3) ксерофилы — минимум aw ниже
80 (ниже —300 бар).
и о:
К гигрофилам относятся некоторые мукоровые грибы и
многие фитопатогенные грибы, такие, как Ophiobolus graminis, Ver-
ticillium albo-atrum. Грибы родов Fusarium, Rhizopus, Phycomy-
ces — мезофиллы, к ксерофилам относятся виды рода Aspergillus.
Особо высокой ксерофильностью обладают грибы Alternaria
alternata, Ulocladium botryosum, Penicillium funiculosum,
выделенные из пустынных почв Туркмении. Пределы прорастания спор
этих видов лежат в диапазоне аю от 0,75—0,55 (—400 до —750 бар).
Прорастание спор у типичного для пустынных почв гриба Асго-
thecium apicale наблюдается даже при aw —0,45, т. е. —1000 бар.
Аналоги вышеупомянутых видов Alternaria alternata,
Ulocladium botryosum, Penicillium funiculosum, выделенные из
дерново-подзолистых почв умеренной зоны, требуют несколько более
высоких значений потенциала влаги с нижней границей aw от 0,86
до 0,75 (—200 400 бар), т. е. вышеперечисленные виды имеют
экологические расы (экотипы) с различными пределами
выживания в условиях влажности.
Частный случай ксерофилии — осмофилия и галофилия. Ос-
мофильные организмы способны жить в условиях пониженной
влажности, определяемой высокой концентрацией Сахаров, гало-
фил ьные — высокой концентрацией солей. Осмофильные и гало-
фильные организмы имеют высокое осмотическое давление
внутри клеток, поэтому приспособлены к существованию в условиях
пониженной влажности окружающей среды.
Примерами осмофильных и галофильных грибов могут
служить представители рода Aspergillus. Виды Л. repens и Л. сап-
didus растут на средах с концентрацией сахара 20%, Л. glau-
?us — 30—70%. Для этих же видов оптимальной концентрацией
NaCl является ее содержание от 10 до 20%.
Температура. Температура и температурный режим в
поверхностных слоях почвы представляют собой один из наиболее
важных факторов окружающей среды, от которых зависит развитие
грибов. Она оказывает прямое и непосредственное влияние на
«формирование комплекса микромицетов. Преимущество всегда
имеют грибы с температурным оптимумом, близким к
окружающим условиям.
По отношению к температуре так же, как и к влажности,
грибы можно разделить на три группы, которые выделяются по
температурным границам роста:
психрофилы — минимум ниже 0°, оптимум +5 ЬЮ°,
максимум Ь20°,
мезофилы — минимум +5 1-15°, оптимум — +25 1-30°,
максимум Ь37°,
термофилы — минимум +20°, оптимум Ь35 Ь40°,
максимум + 45 Ь55°.
Границы-прорастания спор обычно несколько шире, чем границы
активного роста, хотя не для всех грибов. Кроме того, существуют
лсихротолерантные организмы, у которых оптимум лежит при
низких температурах, но они могут существовать и за пределами
ill
максимальных для истинных психрофилов температур, и
термотолерантные виды, которые растут при высокой температуре, но
имеют нижние границы, выходящие за пределы минимальных для
термофилов температур, т. е. имеют гораздо более широкие
температурные границы роста, чем для истинных психрофилов или
соответственно термофилов. В целом температурные границы
роста грибов (от 0 до 55°) значительно уже, чем таковые у
бактерий (от 0 до 70°).
Облигатных психрофилов среди грибов почти нет, за
исключением некоторых видов рода Acremonium.
Большинство грибов мезофилы, однако некоторые имеют
сдвиги температурных границ в сторону психротолерантности или
термотолерантности. Психротолерантные виды обитают
преимущественно в северных почвах тундры, хотя небольшое их числа
можно выделить из почв умеренной зоны. Это виды Geomyces рап-
погит и некоторые виды Мисог и Cladosporium.
К мезофилам относится гриб Penicillium frequenetans,
обитающий в умеренной зоне (оптимум 26°, минимум 15°, максимум 37°),
Cladosporium cladosporioides, обитающий как в умеренной зоне,,
так и далеко за ее пределами. Однако популяции этого вида^
обитающие в разных зонах, имеют разные температурные
границы роста и прорастания спор. Мезофилами также можно
считать многие виды Penicillium, Fusarium, Mucor и многие другие.
К термофилам относятся многие организмы, выделяющиеся
из саморазогревающихся торфов, такие, как Chaetomium thermo-
phila, Sporotrichum thermophila, Talaromyces emersonii, Thermo-
ascus aurantiacus, виды Humicola.
К термофилам и термотолерантным формам принадлежат
многие виды Aspergillus. Так, популяция A. terreus, выделенная из
пустынных почв Туркмении, имеет границы роста от 20 до 47а
при оптимуме 37°.
При оптимальных условиях температуры наблюдается
отчетливая зависимость роста грибов от концентрации субстрата в
среде, однако при экстремальных условиях такая зависимость
почти отсутствует и температура становится решающим фактором,
определяющим рост грибов (рис. 41).
Экстремальная для определенных видов температура
оказывает существенное влияние на морфологию организма и
прохождение им цикла развития. Так, у Penicillium frequentans при
повышении температуры до 37° спорообразование полностью
подавляется. А у Л. terreus при температуре 42° и выше в случае
высоких концентраций субстрата формируются некомпактные,
деформированные головки с укороченными цепочками конидий.
При той же температуре, но при низких концентрациях субстрата
спороношения представлены кисточками, напоминающими
кисточки пенициллов. Характерные для аспергиллов вздутия почти
полностью редуцируются. При более высокой температуре +47°
образуются цепочки с очень малым количеством конидий, а
вздутая конидиеносцез становятся конусовидными. Также гриб Alter-
Ш
naria alternata в условиях повышенной температуры образует
малое число конидий, меньших, чем обычно, размеров, с
небольшим числом перегородок, продольные перегородки при таких
условиях полностью отсутствуют.
Отношение к температуре определяет в значительной степени
местообитание определенных видов или даже групп видов
определенных родов. Например, преимущественное распространение
пенициллов и аспергиллов в
разных широтах. Многие виды
аспергиллов имеют более высокие
температурные границы роста, чем
пенициллы. Поэтому пенициллы
преобладают в почвах северных
широт, где они представлены
большим разнообразием видов. Они
Рис. 41. Радиальная скорость роста
Penicillium frequentans в зависимости
от концентрации глюкозы при разных
температурах
Кг, мм/ч
0,1 1 S,%
встречаются и в южных широтах, но занимают значительно
меньшую долю среди других грибов и имеют значительно меньшее
разнообразие видов. Здесь значительную долю от других грибов
занимают аспергиллы, которых в этих условиях значительно
больше, и они разнообразнее представлены в видовом отношении.
Аспергиллы почти не встречаются или представлены очень
ограниченным числом видов в северных почвах (табл. 4).
Однако среди пенициллов более чем среди других родов
можно выделить эвритопные виды с широким ареалом, встречающиеся
во всех широтах, и виды стенотопные, ограниченные узкими
пределами экологических факторов. При этом лимитирующие
факторы могут быть различными: температура, влажность, рН,
органическое вещество, С02 и т. д. Широким ареалом распространения
характеризуются виды Penicillium chrysogenum, P. funiculosum,
а также Aspergillus niger и Cladosporium cladosporioides. Тем не
менее у одного и того же вида с широким ареалом можно
выделить популяции с различными температурными границами роста
и прорастания спор, так называемые температурные экотипы.
Такие же экотипы выделяют и по отношению к влажности.
Виды с узким ареалом — P. egiptiacum, Aspergillus caespito-
susf A. quadrilineatus, Neosartoria fisherii, Neocosmospora vasin-
fecta. Большинство из них характерно только для тропических и
субтропических почв. Некоторые виды родов Acrothecium и Сиг-
vularia и другие темноцветные формы типичны только для
пустынных и высокогорных почв. Стенотопными видами считают также
Aspergillus conicus, обнаруженный только в почвах Англии,
Л. janus — в почвах Панамы, Staphylotrichum coccosporum —
только в подстилке тропических лесов Центральной Африки,
Polls
lyscitalum pustulant — отмечен в высокогорных и пустынных
почвах.
Солнечная радиация. Она представляет собой также
экологический фактор, который может определять существование одних
видов в каких-то определенных условиях и исключать другие.
Особенно большое значение этот фактор имеет для высокогорных
и пустынных почв.
В высокогорных и пустынных почвах Памира,
характеризующихся высокой солнечной радиацией, содержатся специфические
•виды грибов, представленные в основном темноокрашенными фор-
Таблица 4
Соотношение процентного содержания и числа видов родов
Penicillium и Aspergillus в различных почвах
Почвы
"Подзолистые
Дерново-подзолистые
Серые лесные
Серые лесостепные
Черноземы
Каштановые
Сероземы
Красноземы тропиков
.Аллювиальные тропические почвы
Penicillium
% от
общего числа
70
77
70
62
40-80
| 37
24
20—40
15—20
число
видов
34
38
30
22
42—55
24
19
10-15
10—15
Aspergillus
% от
общего числа
1
2,5
5
5,2
7—20
56
38
50—60
I 50—60
число
видов
3
5
6
4
10—11
8
12
20
20
Автор
Сизова, Супрун,
1962
Мирчинк, 1963
Мирчинк, 1963
Лисина-Кулик,
1969
Коваль, Элан-
ская, 1984
[Лисина-Кулик,
1969
Беккер, Супрун,
1963
Farrow, 1954
Nour, 1956
мами. Темноцветные грибы, имеющие соответственно
черно-коричневые пигменты типа меланинов, доминируют среди грибов в
высокогорных почвах Памира, где на высоте 5000 м они могут
быть единственными представителями грибов-микромицетов.
Наиболее часто встречаются там виды родов Cladosporium,
Stemphylium, Ulocladium, Alternaria, Polyscitalum. Большой процент тем-
ноокрашенных грибов обнаружен также в пустынных почвах
Тянь-Шаня и пустынях Туркмении, где они представлены в
основном родами Alternaria, Stemphylium, Ulocladium, Acrothecium,
Curvularia. Много их содержится в почвах южных широт.
Наличие темноцветных грибов в большом числе отмечают в
пустыне Сахара. Для пустынных почв Невады характерны
следующие виды темноокрашенных грибов: Aternaria alternata,
Cladosporium herbarum, Stachybotrys atra, Stemphylium illicis, Dora-
tomyces medius, виды родов Phoma, Aureriumbasidium, Tetracocco-
jsporium.
114
Наоборот, в умеренной зоне они встречаются в почвах в
небольших количествах и спорадически. Отдельные представители
их есть в почвах всех зон в количестве 1—10%. Несколько
больше их содержится в северных районах в тундровых и подзолистых
почвах, где большое место среди них занимают Aureobasidium
pullulans и виды Cladosporum.
В подзолистой зоне темноокрашенных грибов больше в
почвах низменных и заболоченных мест. Однако значительно больше
их в ризосфере растений, а также на разлагающемся
растительном опаде и растительных остатках. Там они представлены в
первую очередь родами Cladosporium, Alternaria, Ulocladium,
Stemphylium, Aureobasidium и некоторыми другими. Высокое
содержание темноцветных грибов отмечается в. составе эпифитной
микофлоры. Обилие их на растительном опаде и пожнивных
остатках связано с их предшествующим развитием на поверхности
растений, что подтверждается наличием одних и тех же форм
Таблица 5
Распространение темноокрашенных грибов в различных почвах
Район
Памир
Тянь-Шань
Пустыня Мары
Туркмения
Египет
Московская обл.
Московская обл.
Почва и субстрат
примитивно-рухляковая, 4800 над у р. моря
то же
то же
примитивно-рухляковая, 5200 над у р. моря
горно-луговая степная
то же
пустынный серозем
то же
мелкозем в расщелинах скал
пустынная почва под саксаулом
пустынная почва под тамариксом
лугово-карбонатная дельта Нила
то же
песчаная пустынная
дерново-подзолистая слабо окультуренная
дерново-подзолистая хорошо окультуренная
дерново-подзолистая под лесом у .
дерново-подзолистая (пожнивные остатки
ячменя)
пожнивные остатки пшеницы
пожнивные остатки ржи
пожнивные остатки овса
опад (лесная подстилка)
торф
Число
грибов в 1 г
1600
800
1000
9 800
10 200
1800
1200
1430
4 080
10 000
9 500
4 000
17 000
32 500
33 600
25 000
40 000
174 000
44 000
103 000
71000
19 900
26 000
%
темноцветных
66
43
50
100
100
81
57
23
35
65
42
27
17
66
0—20
0—3
0—3
65
92
73
89
73
20
как на поверхности листьев, так и на опаде (табл. 5). Однако*
не все темноокрашенные формы попадают в подстилку с опадом,
так как есть виды, для которых подстилка их естественное
местообитание.
Темноокрашенные грибы, являющиеся многочисленной и раз-
115
зюобразной в систематическом отношении группой, содержат в
мицелии и спорах черный пигмент сложной полимерной
структуры, который на основании ряда характеристик можно отнести к
меланинам. Наличие черного пигмента типа меланинов определяет
защитные свойства грибов против облучения, и дает им
возможность существовать в условиях повышенной электромагнитной
радиации, в том числе УФ- и у-излучения, какие имеют место в
высокогорных и пустынных почвах, а также на поверхности
растений. Известно, что чем выше местность над уровнем моря, тем
больше возрастает доля УФ- и у-лучеи в солнечном излучении.
Экспериментально показано, что споры многих темноцветных
грибов устойчивы к ультрафиолетовым лучам. Они выдерживают
облучение УФ-лучами при длине волны от 230 до 370 нм, при
неизменяющемся высоком проценте выживания (до 50%) в
течение 1—3 и более часов, в то время как непигментированные и
светлопигментированные формы полностью погибают через
несколько минут.
Имеются экспериментальные данные о большой устойчивости
темноцветных грибов к v-облучению. При сравнении ряда видов
темноокрашенных грибов с беспигментными и светлоокрашенными
по устойчивости к ^-облучению обнаружено, что наименее
устойчивы беспигментные виды, такие, как Gliocladium fimbriatum и
Acremonium charticola. Они почти полностью погибают при дозе
80—100 кР. Такой же чувствительностью обладают грибы,
имеющие желто-оранжевые пигменты некаротиноидного типа.
Несколько большую устойчивость обнаруживают грибы с
красно-фиолетовыми пигментами, и значительно большей устойчивостью
обладают меланизированные грибы. Самой высокой устойчивостью
отличаются виды родов Stemphylium, Ulocladium и Alternaria.
Они выдерживают дозу в 625 кР при проценте выживаемости 1,2—
1,7% (рис. 42).
На примере пигментных грибов (их популяций и видов)
можно демонстрировать значение физико-химических факторов среды
и соответствующих защитных свойств организмов в определении
их ареалов, что является одной из сторон аутоэкологии.
Как было отмечено выше (см. с. 101), все сапротрофные
грибы, занимающие в экосистеме место редуцентов (к ним можно
также отнести факультативных паразитов), составляют еще
несколько функциональных групп по способности утилизировать
тот или иной субстрат. Они занимают соответствующие
экологические ниши в природе, в результате чего возможна сукцессия
видов на субстрате.
Грибы в разных географических зонах, находящиеся на одной
стадии сукцессии, различны по видовому составу, хотя и сходны
но своим пищевым потребностям. Эти различия определяются уже
не субстратом, а физико-химическими факторами среды.
Особенное значение имеет приспособленность организма к воздействию
экстремальных факторов. Например, пигментация грибов как
приспособление к солнечной радиации. Наличие или отсутствие пиг-
116
10 20
80 100
ПО 180
Доза, кР
10 20
100
ПО 180
Доза, кР
Рис. 42. Выживание различно пигментированных грибов при разных
дозах облучения:
I. / — Acremonium charticola\ 2 — Talaromyces flavus; 3, 4 — Fu-
sarium moniliforme.
II. 1,3 — Ulocladium botrytis; 2, 4 — Alternaria alternata
ментов будет дифференцирующим фактором, разделяющим
ареалы сходных по пищевым потребностям организмов.
В связи с этим в группе грибов определенной стадии
сукцессии, сходных по пищевым потребностям, беспигментные формы
обитают в почвах, в ризосфере растений, частично на опаде,
преимущественно в равнинных условиях и умеренном климате, а
пигментированные — на поверхности растений, на пожнивных
остатках и опаде, в высокогорных и пустынных почвах близко к
поверхности. Существование последних в этих условиях, с одной
стороны, обусловлено конкуренцией с беспигментными
формами, которую они часто не выдерживают, так как представители
беспигментных форм часто высококонкурентоспособны, быстра
растут, быстро осваивают субстрат, обладают большим набором
ферментов и антибиотиков, а с другой стороны, наличием
пигмента, который дает темноцветным формам возможность
существовать в условиях, недоступных или труднодоступных для других
грибов. Такая же дифференциация может идти и по другим
признакам (дефицит влаги, высокая температура, высокое или низкое
значение рН и т. д.).
Растительный покров. Для формирования грибных комплексов
очень важны характер и степень развития высшей растительности.
Изменение растительного покрова влечет за собой изменения в
составе грибов. Так, например, по данным Торнтона, в результате
внедрения вереска в травянистый покров на песчаной каменистой
бурой лесной почве Англии идет изменение почвообразовательного
процесса в сторону оподзоливания, которое сопровождается
обеднением травянистого покрова. Одновременно происходит
обеднение почвы видами грибов, хотя общая их численность в
подзолистых почвах выше, чем в бурых. В подзолистых почвах под
вереском много грибов рода Penicillium, представленного
несколькими видами, среди которых преобладает. P. frequentans. Только
в подзолистых почвах встречается Mortierella ramanniana,
которая полностью отсутствует в бурых почвах. В то же время для
бурых почв характерны другие виды рода Mortierella, такие, как
М. gracilis, M. humilis, M. папа, а также виды родов Sporotrichum,
Paecilomyces, Fusidium. По нашим данным, четко
обнаруживаются различия в составе и структуре комплексов микромицетов
(см. с. 119) в различных растительных ассоциациях,
сформированных на сходных дерново-подзолистых почвах, — ельнике и
березняке волосисто-осоковом Малинского стационара
Московской области. Так доминирующими в верхнем горизонте (А1) почв
ельника являются виды Chrysosporium sulfureum, M. ramanniana,
Verticillium terrestre. Доминантами в этом же горизонте почв
березняка оказались виды P. daleae, Trichoderma koningii.
Также различными видами представлены доминанты лесных
подстилок (горизонт A0L) в этих растительных ассоциациях,
В подстилке ельника — это Aureobasidium pullulans, в подстилке
березняка — Aposphaeria pulviscola и Cladosporium herbarum.
118
ПОЧВЕННЫЕ ГРИБЫ ОСНОВНЫХ ПРИРОДНЫХ ЗОН
Каждая природная зона, в которой формируется
определенный тип почвы, характеризуется определенным типом
растительности и специфическими, присущими ей климатическими
условиями, включающими режим влажности, температуры,
интенсивность солнечной радиации, диапазон значений рН почвы. Все
это определяет также своеобразие состава почвенных
микроорганизмов, в том числе специфику почвенных грибов каждой зоны,
т. е. подчинение их распространения в почвах общему закону
зональности. Закон зональности микроорганизмов в целом был
сформулирован Е. Н. Мишустиным в 1954 г. Его конкретное
содержание очень четко проявилось на примере грибов-микроми-
цетов. Для характеристики своеобразия состава видов грибов
разных типов почв в настоящее время используется понятие
комплекса типичных видов. Комплекс типичных видов выделяется
на основе пространственной и временной частоты встречаемости
вида.
Пространственная частота встречаемости характеризуется
числом образцов, в которых вид обнаружен, к общему числу
исследованных образцов, а временная — отношением числа сроков в
тоду, когда вид обнаружен, к общему числу сроков отбора
образцов.
Виды с высокой пространственной и временной
встречаемостью (выше 60%) относят к доминирующим, виды с
пространственной и временной встречаемостью выше 30% — к часто
встречающимся, виды с низкой пространственной (ниже 10%) и
высокой временной (более 30%) — к редким, но типичным видам,
виды с низкой пространственной и временной (ниже 10%) — к
случайным видам.
Первые три категории видов представляют собой комплекс
типичных видов, четвертая категория — виды случайные и в
комплекс не входят. Комплекс типичных видов невелик и
представлен 15—20 видами.
Распределение видов в комплексе с учетом их встречаемости
характеризует структуру комплекса. Комплексы типичных видов
основных зональных почв приведены в табл. 6.
В качестве примера дифференцированный комплекс типичных
видов грибов-микромицетов дерново-подзолистой почвы, в
котором отражена его структура, приведен в табл. 7, где выделение
основных групп видов основано на частотах их встречаемости.
Такой дифференцированный комплекс может служить
количественной мерой для сравнения разных почв по микромицетам.
Для сравнения степени сходства и различия комплексов мик-
ромицетов почв в количественном выражении используют
коэффициент Сьеренсена — Чекановского, который рассчитывают не
только по количеству видов, но и по их встречаемости.
Коэффициент Сьеренсена — Чекановского (S) имеет
следующий вид:
119
Таблица 6
Типичные виды микромицетов разных почвенных зон
Почвы
Тундровые
Подзолистые
и дерново-
подзолистые
Серые лесные
Черноземы
Виды грибов
Geomyces pannorum
Penicillium nigricans
Mucor plumbeus
Mortierella ramanniana
Chrysosporium sulfureum
Penicillium daleae
P. frequentans
P. raciborskii
P. spinulosum
P. verrucosum var. cyclo-
pium
P. thomii
P. nigricans
Verticillium t err est re ,
Trichoderma alba
Mucor hiemalis
Paecilomyces sp.
Cladosporium herbarum
Gliocladium penicilloides
Oidiodendron gracile
Penicillium brevi-compacA
turn
P. cinereo-airum
P. paxilli
Paecilomyces sp.
Chrysosporium sulfureum
Verticillium album
Trichoderma viride
Mortierella ramanniana
Aspergillus alUaceus
A. wentH
A. ustus
Gliocladium penicilloides
Acremonium charticola
P. janthinellum
P. lapidosum
P. funiculosum
P. raciborskii
P. purpurogenum
P. roseo-purpureum
P. tardum
P. verrucosum var. cycloA
pium
Почвы
Черноземы
Каштановые
Сероземы
Красноземы
Виды грибов
P. simplicissimum
Humicola grisea
Mycogone nigra
Fusarium nivale
Aposphaeria pulviscola
Acremonium charticola
Aspergillus ochraceus
A. ustus
A. terreus
A. wentii
Chrysosporium merdatium
Cladosporium herbarum
Fusarium solani
Paecilomyces lilacinus
Penicillium raciborskii
P. chrysogenum
P. lividum
P. simplicissimum
P. restrict urn
Aspergillus fumigatus
A. nidulans
A. terreus
A. flavus
Alternaria aliernaia
Cladosporium cladosporioi-
des
Penicillium chrysogenum
P. claviforme
P. frequent ans
P. funiculosum
Stachibotrys atra
Chaetomium olivaceum
Penicillium daleae
P. chrysogenum
P. verrucosum var. cyclo-
pium
P. simplicissimum
P. janthinellum
P. digit at um
Trichoderma koningii
Acremonium charticola
Broomella acuta
Mortierella ramanniana
120
А + В '
где С — сумма частот встречаемости общих для двух вариантов
видов по минимальным значениям встречаемости, А и В— сумма
всех частот встречаемости видов первого (А) и второго (В) ва-
Та блица 7
Дифференцированный комплекс грибов-микромицетов
дерново-подзолистой почвы
Преобладающие
Частые
Редкие
Случайные
Mortierella гатап-
niana
Chrysosporium sul-
fureum
Penicillium daleae
Trichoderma alba
Thysanophora peni-
cilloides
Vert ici Ilium t err est-
re
Phialophora sp.
Penicillium simpli-
cissimum
P. frequentans
Penicillium verruco-
sum var. cyclopi-
um
Cladosporium herba-
rum
Paecilomyces sp.
P. thomii
Aureobasidium puU
lulans
Aposphaeria pulvis-
cola
риантов. Комплексы микромицетов можно считать неразличаю-
щимися при коэффициенте сходства более 80%.
Сходство комплексов микромицетов разных типов почв
представлено на рис. 43. Чем меньше расстояние между
сравниваемыми почвами, тем больше
между ними сходство, и наоборот,
чем расстояние больше, тем
меньше сходство.
Группу сходных по микромице-
там почв составляют почвы степ-
Сходстдо
1,0 0,90,8 0,7 0,6 0,5 0,4 0,3 0,1 0,7 0
Рпггтпянир
0 0,1 0,2 0,3 0Л 05 0,6 0,10,8 0,9 1,0
I—п—i—I—гп—I—п—г
Рис. 43. Степень сходства комплексов
микромицетов между различными
типами зональных почв:
/ — дерново-подзолистая; 2 —
чернозем типичный; 3 — чернозем
выщелоченный; 4 — каштановая; 5 —
краснозем; 6 — серозем типичный светлый;
7 — серая лесная
1
3
1
5
7
6
ной зоны: черноземы (типичный и выщелоченный) и каштановая
почва. Другая группа сходных почв — дерново-подзолистые, серые
лесные, и красноземы. Наибольшее отличие от первой и второй
групп почв имеют светлые сероземы.
Большее сходство и большее различие комплексов
грибов-микромицетов различных типов почв в значительной степени
соответствует степени сходства и различия между этими почвами по
морфологии, физико-химическим свойствам, общему направлению
121
почвообразовательного процесса. Таким образом, формирование
комплекса типичных видов микромицетов подчиняется
определенным закономерностям, согласующимся с общими
закономерностями происхождения и свойств почв, и комплекс микромицетов
может служить дополнительной характеристикой почв, которая
может быть использована в решении спорных вопросов генезиса
почв.
ИСПОЛЬЗОВАНИЕ КОМПЛЕКСА ГРИБОВ-МИКРОМИЦЕТОВ
ДЛЯ ОЦЕНКИ АНТРОПОГЕННОГО ВОЗДЕЙСТВИЯ
Окультуривание почвы, представляющее собой один из
примеров антропогенного воздействия, которое включает
распашку, внесение удобрений, полив и другие агротехнические
мероприятия, вызывает существенные изменения в комплексе
микромицетов.
Во-первых, эти изменения можно характеризовать
коэффициентом сходства. Чем меньше коэффициент сходства между
целинной и окультуренной почвой, тем значительнее степень
воздействия окультуривания.
Наиболее отчетливо выраженные изменения происходят при:
окультуривании дерново-подзолистых почв, особенно при
внесении азотных удобрений (коэффициент сходства целинных и
окультуренных почв от 0,09 до 0,20; табл. 8). Большие изменения про-
Та бли ца 8
Коэффициенты сходства Сьеренсена целинных и
окультуренных почв
Почва
Коэффициент
сходства
Дерново-подзолистая целинная
Дерново-подзолистая окультуренная
Чернозем типичный целинный
Чернозем типичный окультуренный
Каштановая целинная
Каштановая окультуренная
Светлый серозем целинный
Светлый серозем окультуренный
Чернозем выщелоченный целинный
Чернозем выщелоченный окультуренный
0,09—0,20
0,25
0,40
0,28
0,50
являются также при окультуривании черноземов и сероземов
(коэффициент сходства 0,25—0,28 соответственно). Несколько
меньше реагирует комплекс микромицетов при окультуривании
каштановых почв (коэффициент сходства 0,40) и наиболее
устойчив комплекс микромицетов выщелоченных черноземов
(коэффициент сходства 0,50).
Различия в комплексе микромицетов, которые проявляются
между основными типами почв, несколько сглаживаются между
122
этими почвами при окультуривании (рис. 44). Так, значительно
меньше проявляются различия между окультуренными дерново-
подзолистыми почвами и выщелоченными черноземами, между
выщелоченными черноземами и каштановыми почвами, между
каштановыми почвами и сероземами, между выщелоченными
черноземами и сероземами. На том же уровне, что и между
основными типами целинных почв,
остаются различия между окультурен- Сходстбо
ными дерново-подзолистыми поч- W М G*s 0>7 °>6 р^с^0^ие0,2
вами и черноземами, между дер- 0 а1 да 0t3 до 0)5 0t6 Д7 о&
ново-подзолистыми почвами и каш- i i i ' i i i i г
2
Рис. 44. Степень сходства комплексов мик- * *"
ромицетов между различными типами 5 •-
Ъ
7 -
окультуренных почв:
/ — дерново-подзолистая; 2 — чернозем
типичный; 3 — чернозем выщелоченный; J • ¦
4 — каштановая; 5 — серозем типичный
тановыми. Низкая степень различия наблюдается между
окультуренными дерново-подзолистыми почвами и окультуренными
сероземами.
Во-вторых, антропогенное воздействие можно характеризовать
по индексу видового разнообразия, рассчитанного по формуле
Шеннона, и, в-третьих, по ранговому распределению видов на
основе их частот встречаемости.
Индекс видового разнообразия (Н) выражается следующим
образом: Н = — S/Vlog2Pi,
где Pi — вероятность попадания встречаемости вида в
определенный ранг значений.
Для расчета сначала производится распределение видов
комплекса по рангам их встречаемости (число видов с
встречаемостью от 1 до 10, от 11 до 20 и т. д.), а затем рассчитывается по
вышеприведенной формуле.
Окультуривание почвы с применением комплекса мероприятий
сопровождается заметным обогащением видами, увеличением
индекса разнообразия, т. е. усложнением комплекса микромицетов.
Такие результаты получены, в частности, на выщелоченных
черноземах и на типичных сероземах при использовании комплекса
мероприятий по окультуриванию: орошения, внесения
минеральных удобрений, противовилтовых препаратов. При этом в почвах
сероземов, которые характеризуются отсутствием доминирующих
видов в результате этих мероприятий, происходит увеличение
числа видов и появляются доминирующие виды.
Однако простая распашка целины и использование ее под
посевы сельскохозяйственных культур приводят к значительному
упрощению комплекса микромицетов, снижению его
разнообразия. Особенно наглядно это проявляется на типичных черноземах,
каштановых почвах, красноземах. Такое же явление наблюдается
123
и на дерново-подзолистых почвах при внесении азотных
удобрений без известкования (табл. 9).
В этом случае окультуривание почв, т. е. использование их
под сельскохозяйственные культуры, отрицательно сказывается
Таблица 9
Индексы видового разнообразия основных зональных типов почв
Почва
Индексы
целинная
2,69
2,61
2,54
2,23
2,68
2,58
1,15
окультуренная
2,15
—
2,34
2,99
2,25
2,П
2,47
Дерново-подзолистая . .
Серая лесная .....
Чернозем типичный . .
Чернозем выщелоченный
Каштановая
Краснозем типичный . .
Светлый серозем . . . .
100]
* 40
I 20
ах
. * 100]
"* 60\
*> щ
20
ЕЧ5н
J » i i
Г I I
J I
2 4 6 8 10 12 14 16 18
IZ?!±
Й&Й
I 1 I I I Г~Т^Г^Т 7
2 4 6 8 10 12 14 16 18 20
Чи с/1 о бидов
х х окультуренная почва
*-—-# це/шнная почда
**г**х
на некоторых свойствах почв,
что отражается и в комплексе
микромицетов.
Графическое изображение
распределения видов по частотам
их встречаемости дает
представление о том, за счет каких
структурных единиц идет
изменение комплекса (рис. 45)
В дерново-подзолистых
почвах в вариантах с длительным
внесением минерального азота
оно происходит за счет сокра-
Рис. 45. Ранговое распределение
типичных видов комплекса по частотам их
встречаемости:
I — дерново-подзолистая почва; II —
серозем типичный;
щения общего числа видов, снижения частых и редких видов,
увеличения степени доминирования отдельных немногих видов.
В целинных почвах сероземов, характеризующихся полным
отсутствием доминирующих видов при окультуривании (внесение
удобрений, орошение), происходит увеличение числа видов и
появляются доминирующие виды. В черноземах и каштановых
почвах происходит снижение всех структурных единиц комплекса.
Коэффициенты сходства Сьеренсена, индексы видового
разнообразия и распределение видов в комплексе по частотам их
встречаемости могут служить количественной мерой любого
антропогенного воздействия на микромицеты почв, такого, как удоб-
124
рения, загрязнения тяжелыми металлами, пестицидами
рекреационных нагрузок, пастбищной дигрессии и т. д.
КОЛИЧЕСТВЕННОЕ СОДЕРЖАНИЕ ГРИБОВ
В ПОЧВАХ И ИХ БИОМОРФОЛОГИЧЕСКАЯ СТРУКТУРА
Для того чтобы оценить роль грибов как деструкторов-
растительного опада в почве того или иного биогеоценоза,
необходимо иметь представление об их количественном содержании,
выраженном в суммарной биомассе мицелия и спор. Наряду с
этим грибной мицелий — это реальные запасы пищи для других
организмов, в первую очередь для почвенных животных, и
иммобилизованные питательные элементы для растений.
Количественное содержание грибов в почве нельзя
характеризовать по числу колоний, вырастающих на чашках с
питательной средой, потому что для огромной гетерогенной как в
таксономическом, так и в физиологическом отношении группы организмов
не может быть одной универсальной среды, кроме того, не все
грибы вырастают на питательных средах.
Для количественного определения грибов наиболее объективно-
определение биомассы мицелия и спор. Для этих целей наиболее
часто используют метод прямого микроскопирования на
мембранных фильтрах, который заключается в том, что навеску почвы,
разведенную в определенном объеме воды, фильтруют через
мембранный фильтр, который окрашивают красителем и осветляют
иммерсионным маслом, а затем подсчитывают длину
находящегося на фильтре мицелия с помощью окулярной линейки,
используя формулу
_ b.x<\0-*.S-n-\0-2
50-P.V-C-10-8 *
где а — суммарная длина гиф в 1 г почвы (см), b — суммарная-
длина гиф на фильтре в 50 полях зрения в единицах окуляра-
микрометра, х — цена деления окуляра-микрометра (мкм), 5 —
площадь фильтра (мм2), п — разведение почвенной суспензии,.
Р — площадь поля зрения (мкм2), V — объем наносимой
суспензии (см3), С — навеска почвы (г). Расчет биомассы мицелия
производят, учитывая, что вес одного метра мицелия равен 0,02 мг..
Число спор в 1 г почвы рассчитывают по формуле
SO-P-V-C-Wr*
где М — число спор в 1 г почвы, т — число спор в 50 полях
зрения, остальные обозначения те же, что и для расчета длины гиф.
Биомассу спор определяют по формуле
g=V.dt
где V — объем клетки, d — удельный вес, равный 1,2.
В зональном ряду почв наблюдаются различия в содержании
125
мицелия в разных типах почв, т. е. каждый тип почвы
характеризуется определенным уровнем биомассы мицелия и общей
биомассой в целом.
Наиболее высоко содержание мицелия и соответственно его
биомассы на 1 г почвы в некоторых почвах тундры, где длина
мицелия составляет более тысячи метров, а его сухая биомасса
несколько миллиграммов на 1 г почвы (табл. 10).
Много грибного мицелия содержится в подзолистой и
дерново-подзолистой почвах, в лесных биогеоценозах, где мицелий
измеряется сотнями метров на 1 г почвы, а сухая биомасса
соответственно десятыми долями миллиграммов. В этих почвах
жизнедеятельность грибов протекает не только в подстилке, но и в
минеральных горизонтах почвы, где они осуществляют процессы,
связанные преимуЩественН0 с превращением органического
вещества.
Сотни метров и более достигает содержание мицелия в серых
лесных почвах лесостепной растительной формации, т. е. такого
же уровня, что и в подзолистых и дерново-подзолистых почвах
таежно-лесной зоны.
На порядок меньше, чем в основной массе подзолистых почв,
грибная биомасса в черноземах. Но тем не менее и в этих почвах
содержится грибной мицелий, измеряемый десятками метров на
1 г почвы. Соответственно и биомасса его в этих почвах
составляет доли миллиграмма.
Длина мицелия и его биомасса в красноземах в буково-гра-
бово-каштановой растительной ассоциации приблизительно такого
же порядка что и в черноземах, измеряется несколькими
десятками метров и долями миллиграммов соответственно.
В период весенней вегетации можно обнаружить мицелий и в
пустынно-песчаных почвах илаково-белосаксаульной растительной
ассоциации, измеряемый десятками метров в грамме.
Соотношение мицелия и спор характеризует
биоморфологическую структуру грибов.
Относительное содержание спор (в процентном отношении от
общей биомассы грибов) имеет в зональном ряду почв
соответствующих биогеоценозов обратное соотношение с содержанием
мицелия. По мере продвижения с севера на юг заметно
изменяется соотношение спор и мицелия по крайней мере в весенний
период. Несмотря на динамичный характер взаимопревращения
споры — мицелий —споры, различия в биоморфологической
структуре между разными типами почв сохраняются. Наименьшую
долю споры по отношению к мицелию занимают в почвах тундры.
Также меньшую долю по отношению к мицелию составляют споры
в подзолистых почвах лесной зоны. По мере продвижения на юг
к серым лесным почвам, черноземам и каштановым почвам
лесостепной и степной зоны доля спор по отношению к мицелию
возрастает, что свидетельствует о возрастании неактивного запаса
грибных зачатков (рис- 46).
Однако несмотря на высокое содержание мицелия в почвах
126
о
я
а*
о
3~
s
О rt
о
5 *
к s
Ч ев
рибов
и,
Биомасса
спор, %
«7:
1 о *
1 я
Биом
К
3
о.
СПО
(О
<->
обща
иомас
Биомасса
мицелия,
м/г 1
пина мицелия,
м/г
tt
онт, 1
а, см
со х
О >,
to
CQ
О
С
ция
СЗ
ассоци
ительная
Раст
Зона
О
17,3
^
о"
СЛ
4,0
3,38 1
05
Г-
СО"
СО
СМ
СО
1
о
о
<
овая
о-
п
1 тун
R
икова
X
устар]
,
х
R X
m 5
CQ S
CJ &
Ч
u
соково-к
я АССР
0-01
ш
О
X
ска
н
>>
«
°гс
s>
R
[дрова
| 33,9
ю
СО
о"
см
о
0,67 |
ю
cd"
-н
CD
CD
CM
1
ю
CM
<
-ПОД-
CJ
X
1 сред
3
сличи
as
тая
U
S
1 зол
бл.).
о
чернично-к
(Ярославская
X
X
X
CU
1=5
л о
ч ©
CJ
ес-
жно-л
Тае
ная
74,2
ст>
ю
о"
о
0,8
99*0
СМ
cd"
+1
о
оо"
—ч
ст>
1
о
^
<
r"
;есна
ч
1 серая
»Х
3
осист
ч
;овово
вно-осок
тра
о
X
СП
со
0В-
Моск
лес
убовый
»=С Ч
А ю
2 о
CQ
О
X
S
О.СЗ
яст-
CQ
8
О)
рОКОЛ!
НЫХ Л
я
3
вен
R
СО
и:
о
73,0
^
<м
о"
СО
е'о
0,08
7,5±2,4 1
^-н
о
1
ю
fcl
<
& 3
о
X
1 чер
6
X
л
ч
0-КОВЬ
шотравн
со
СХ
Л
с
CU
н
CJ
пная
Сте
X
а"
о
ур-
R
екосим*
ровая, н
g
«
о
н
CJ
о
3
X
СТ
К
1 тип
1
R
со
*
CJ
85,0
см
CD
о"
СО
z*o
0,11
3,3±5,5
СМ
О
1
ю
^
<
озем
X
CJ
1 КР3
нный
CD
)ЛИСТВ
ныи
сг
X
| тип
вый,
о
аштан
э-широкс
рабово-к
X
X
со
в
о>
S
6
CQ
О
*3
>>
о VO
шные
ч
CQ
S
тропи
ю
>>
CJ
R
СО
жара
«=t
ее (А
вянои л
со
О,
о(Х
«U
0,12 |
см
^"
+1
со
CD
^^
^
ю
1
о
_н
<
IH0-
14
CU
с
рк-
я(Ту
со
аульн
шная
л
пуст
белосакс
ССР)
ВО-1
о
W
СО
ч
я
:тыни
С
R
со
Ы
и
X
CU
S
северных широт и большую его долю от имеющихся спор, он
обладает меньшей активностью, чем в почвах южных широт.
В почвах южных широт, в черноземах и красноземах
создаются благоприятные условия для роста вегетативного мицелия.
Благодаря высокой напряженности биологических процессов в
этих почвах, что было отмечено в трудах Е. Н. Мишустина, до-
0 0,2 ОЛ 0,6
О 0,2 ОЛ 0,6 08
j
О 0,2 ОЛ
^мг/г
28.1
О 0S2 0{Л 0,6 0{8 1,0 1,2 1f4 1,6 0 0,2 ОЛ 0,6 0 0,2 ОЛ 0,6
А] А 2
МГ/Г
О 0,2 ОЛ 0,6 0,8 34 36 38 40 42
О 0,2 ОЛ
31. Ш
ш
Рис. 46. Распределение грибного мицелия (/) и спор (2) в профиле почв:
I — дерново-карбонатная; II — серая лесная; III — тундровая глеевая
перегнойная; IV — чернозем типичный
128
стутшый субстрат в этих зонах быстро исчерпывается, что
приводит к массовому образованию спор, запас которых может быть
в любое время мобилизован при поступлении новых порций
органического вещества.
В подзолистых почвах и тундре имеется избыток субстрата
в виде мощной подстилки и неразложившейся органики, поэтому
в этих почвах процессы спорообразования замедлены. Кроме того,
большее абсолютное содержание вегетативного мицелия в
северных почвах может быть также обусловлено значительным
вкладом мертвого мицелия, который не успевает полностью
разложиться за короткий вегетативный период.
Распределение биомассы грибов на всю глубину почвенного
профиля (г/м2) также обнаруживает определенные
закономерности по почвенным зонам. Биомасса мицелия на весь небольшой
профиль тундровой почвы (37 см) больше, чем на профиль в
68 см подзолистой почвы. В то же время и биомасса мицелия в
Таблица 11
Содержание грибной биомассы по горизонтам
профиля разных типов почв
Почва
Тундровая глеевая
перегнойная
Дерново-среднепод-
золистая
Чернозем типичный
Краснозем
типичный
Горизонт, мощность,
см
А0/А1 0—3
А1 3—12
В 12—37
0—37
А1 0—6
А1/А2 6—15
А2 15—38
А2/В 38—68
0—68
А1 0—4
А1 4—64
АВ 64—130
Вк 130—165
С 165—180
0—180
А1 0—15
ВС 15—28
ВС 43—59
С 59—71
С 71—85
С 85—100
0—100
1 Биомасса, г/м2
мицелия
34,5
33,1
5,9
73,5
13,9
16,5
13,0
4,7
48,1
1,9
8,1
2,6
1.0
0,0
12,7 i
13,5
1,5
1,4
1,3
0,7
0,3
19,6
спор
7,3
10,3
7,0
24,6
6,0
59,1
84,2
5,2
154,5
6,7
105,7
25,8
5,4 |
1.4 !
145,0
79,5
3,0
2,5 !
2,4
0,2
0,2
91,4
общая
41,8
43,4
12,9
98,1
19,9
75,6
97,2
9,9
202,6
8,6
113,8
28,4
5,5
1,4
157,7
93,0
4,5
4,0
4,0
3,0
2,5
111,0
Процент
биомассы спор
от общей
биомассы
17,5
23,7
54,3
25,0
30,1
78,1
86,6
52,5
76,3
77,9
92,9
90,8
98,2
100,0
91,3
85,0
64,0
63,2
72,0
75,0
88,0
83,3
5 Т. Г. Мирчинк 129
профиле тундровой почвы и особенно в профиле подзолистой
почвы значительно больше, чем во всем профиле чернозема
(180 см; табл. 11).
Максимальные запасы общей биомассы (мицелий + споры)
приходятся на подзолистые и дерново-подзолистые почвы под
хвойными лесами (более 200 г/м2). Тундровые почвы содержат около
100 г/м2 грибной биомассы, включающей мицелий и споры, т. е.
приблизительно столько же, сколько в профиле краснозема
(111 г/м2), и немногим меньше, чем в большом профиле чернозема
(157 г/м2).
Относительная доля спор по отношению к мицелию возрастает
с глубиной по профилю почвы, причем в дерново-подзолистой
почве, вплоть до верхней границы иллювиального горизонта,
увеличивается не только
относительное, но и абсолютное
содержание спор, т. е. также
изменяется биоморфологическая
структура грибов по профилю почвы.
Соотношени грибов и бактерий:
в почвах разных зон можно
демонстрировать рис. 47.
Биомасса грибов во многих:
Рис. 47. Соотношение грибов (а) и
бактерий (б) в почвах разных зон:
/ — дерново-подзолистая; 2,3 —
чернозем типичный; 3 — краснозем
типичный; 4 — песчано-пустынная
1
т
ш
1
о
!
¦
Шгл Ш
почвах значительно превышает биомассу бактерий. Особенно
заметно это превышение в подзолистых почвах и красноземах,
наблюдается оно также и в почвах пустынь.
Приблизительно в равных соотношениях находятся биомассы
грибов и бактерий в черноземах. Причиной доминирования
грибов в почве в первую очередь является их способность разлагать
высокополимерные субстраты лигноцеллюлозного комплекса,
кроме того, мицелиальная организация дает грибам целый ряд
преимуществ.
Грибной мицелий обладает на 1—2 порядка большей
линейной скоростью роста (50—1000 мкм/ч) по сравнению с бактериями
(1—10 мкм/ч), и поэтому грибной мицелий более эффективно
колонизирует поступающий в почву органический субстрат. Также
легкорастворимые вещества, вносимые в почву, потребляются:
преимущественно грибами, о чем свидетельствует вспышка роста
мицелия в первые сутки после обогащения почвы глюкозой, тогда
как бактерии развиваются лишь спустя 12—18 дней на продуктах:
грибного ресинтеза.
130
Г л ава 5
ВЗАИМООТНОШЕНИЯ ПОЧВЕННЫХ ГРИБОВ
С ДРУГИМИ ОРГАНИЗМАМИ
Между грибами и другими членами биотического
сообщества (биоценоза) в экосистеме складываются разнообразные
связи как трофические (пищевые), так и метабиотические
(посредством продуктов обмена веществ, выделяемых во внешнюю
среду), строгое разделение между которыми у микроорганизмов,
в том числе грибов, трудно установить. Такие связи существуют
у грибов с высшими растениями, животными и другими
микроорганизмами (бактериями, актиномицетами и грибами).
Связи грибов с высшими растениями главным образом кон-
сортивные, т. е. связи, в которых растение представляет собой
вид-эдификатор, являясь автотрофным организмом. Оно образует
ядро консорции, от которого зависят связанные с ним популяции
грибов как гетеротрофных организмов. Функциональные группы
грибов, связанные с растениями консорционными связями, могут
быть биотрофы и сапротрофы. К биотрофам относятся
патогенные грибы, питающиеся содержимым клеток живого растения,
эккрисотрофы, использующие выделения живого растения, через
листья, хвою (эпифиты или грибы филосферы) и через корни
(грибы прикорневой зоны), и симбиотрофы. К сапротрофам —
грибы, разлагающие отмершие ткани растений.
Связи грибов с беспозвоночными животными в основном
являются трофическими: симбиотрофными и паразитическими.
Причем грибы могут поедать животных (нематофаговые и амебофа-
говые грибы) и могут сами служить для них пищей.
Взаимоотношения грибов с бактериями, актиномицетами и
грибами мог^т складываться как симбиотические, паразитические,
антагонистические, а также грибы могут быть просто субстратом
для этих микроорганизмов.
ВЗАИМООТНОШЕНИЯ ГРИБОВ С РАСТЕНИЯМИ
Фитопатогенные почвенные грибы
Среди грибов, обитающих в почве, имеется ряд
патогенных видов, способных вызывать болезни растений и оказывать
влияние на их продуктивность. Эти грибы вызывают разные по
симптомам проявления болезней. Это могут быть выпревание
растений, гнили молодых растений, корневые гнили, увядание
растений и др. Для них почва является основным резервуаром и
источником инфекции. Наряду с этим у многих фитопатогенных
грибов часть жизненного цикла может проходить в почве: в нее
попадают споры, которые там накапливаются и сохраняются, там
происходит их прорастание, поиск мицелием восприимчивого
хозяина и его инфицирование.
.5*
131
Так, например, ежегодно потери урожая только от корневых
болезней, вызванных грибами, составляют 5—15%, однако они
могут достигать и значительно больших величин 40—70%.
Фитопатогенные грибы в почвах очень разнообразны в
таксономическом и физиолого-биохимическом отношении. Среди них
есть представители почти всех таксономических групп. Так,
например, Olpidium brassicae, Synchytrium endobioticum и другие —
из хитридиомицетов, Pythium и Phytophthora — из класса ооми-
цетов, Cochliobolus, Sclerotinia, Thielaviopsis — из аскомицетов,
Corticium, Fomitopsis — из базидиомицетов и многие виды, такие,
как Fusarium Drechslera, Bipolaris, Verticillium — из дейтероми-
цетов.
Самое распространенное заболевание, которое вызывают
фитопатогенные грибы, обитающие в почве, — корневые гнили.
Заболевание встречается во всех зонах земледелия и поражает все
культурные растения, а также и дикорастущие. Возбудителями
обыкновенной корневой гнили являются грибы из рода Fusarium,
Bipolaris — В. sorokiniana, F. graminearum, F. avenae, F. oxyspo-
rum, F. solani. Они вызывают заболевание яровых злаков.
Корневые гнили озимых злаков — офиоболезы, вызывают гриб^
Ophiobolus graminis, относящийся к локулоаскомицетам.
Корневые гнили бобовых вызывают грибы Pythium debarianum, P. ul~
morumy а также F. oxysporum, в частности его
специализированные формы.
Гнили молодых растений вызывают виды Bipolaris, Drechslera,
Rhizoctonia, Thielaviopsis. Они могут обусловливать гибель семян:
и проростков в результате почти полного разрушения их
вегетативных тканей. На ослабленных проростках в качестве вторичных
паразитов появляются виды Alternaria, Cladosporium, Botrytis и
даже сапротрофные Penicillium.
Главные возбудители увядания растений, носящего название
вилта, — грибы из родов Fusarium и Verticillium, повсеместно-
распространенные во всех почвенно-климатических 30Hiax. Грибы
проникают в растение из почвы и разрастаются в тканях корней
и стеблей. Разрастание мицелия происходит в основном по
проводящим тканям, в результате чего происходит их закупорка кг
разрушение. Вследствие этого нарушается обмен веществ
растений и появляются симптомы увядания.
Серьезным источником инфекции являются также
растительные остатки, находящиеся в почве. На них фитопатогенные грибы
могут существовать не только в виде спор, но и активного мице-
лия, способного заражать корни растений. Инфекция может
передаваться и с помощью почвенных животных, в первую очередь
беспозвоночных.
Грибы-эпифиты
Мицелиальные грибы-эпифиты, обитающие на
поверхности листьев, представлены небольшим разнообразием видов и:
.132
по численности значительно уступают дрожжевым организмам и
некоторым бактериям. Наиболее типичными среди эпифитов на
многих растениях грибы семейства Dematiaceae, родов Alternaria,
Cladosporium, Peyronella, Papularia.
Преимущественно это темнопигментированные формы,
содержащие в мицелии меланины, служащие защитой от интенсивного
облучения грибов на поверхности растений.
После отмирания растений эпифитные грибы поступают с опа-
дом в почву, но постепенно вытесняются там типичными сапро-
трофными грибами, и их численность в почве обычно на порядок
ниже, чем сапротрофных грибов.
Грибы прикорневой зоны
Грибы наряду с другими микроорганизмами —
постоянные обитатели прикорневой зоны растений, представляют собой
либо типичных эккрисотрофов корней, использующих корневые
выделения растений, либо патогенных и потенциально
патогенных организмов.
Прикорневую зону растения разделяют на область,
находящуюся вокруг корней и прилегающую к корням, — ризосферу и
область непосредственно на поверхности корней — ризоплану.
Ризосфера представляет собой слой почвы, испытывающий
воздействие корневых выделений, значительно отличающийся по
своим физико-химическим свойствам от остальной почвы.
Почвенные частицы здесь более оструктурены. В связи с этим
улучшается процесс движения корней и микроорганизмов, лучше
сохраняется влага, поддерживается более постоянная
температура. Более высокая влажность почвы в зоне ризосферы связана
как с изменением структуры почвы, так и со способностью
корневых систем растений активно изменять влажность окружающей
среды.
Прикорневая почва также заметно отличается величиной рН.
Растения изменяют также восстановительный потенциал корней.
Чаще эти изменения происходят в сторону понижения гН почвы.
Ризосфера — область, богатая питанием, обусловленным
выделениями корней (экзоосмосом), в которых обнаружены сахара,
аминокислоты, витамины, ауксины, фосфатиды и различные
ароматические вещества. Корни выделяют по меньшей мере 10
разных Сахаров, но больше всего глюкозы и фруктозы. Наблюдаются
определенные различия в составе корневых выделений разных
растений. Корневые выделения кукурузы, кормовых и овощных
культур богаты сахарами. Злаковые растения выделяют много
органических кислот, таких, как уксусная, щавелевая, лимонная,
яблочная, которые сильно подкисляют почвенный раствор. В
частности, в подзолистых почвах рН в зоне корней злаков
изменяется от 4,5 до 5,5. Корневые выделения бобовых содержат много
нейтральных аминокислот, вследствие чего почва становится
менее кислой. Некоторые растения выделяют антибиотики и другие
133
физиологически активные вещества. В этой зоне создается
повышенная концентрация продуктов метаболизма микроорганизмов.
В ризосфере растений более энергично протекают многие
химические и биохимические процессы. Например, быстрее
разрушаются различные минералы, горные породы — известняк,
мрамор и др. Процесс этот вызывается не только корневыми
выделениями, но и микроорганизмами ризосферы.
Под влиянием микроорганизмов в ризосфере растений
создаются условия более высокой растворимости железа и марганца,
что обусловлено изменением окислительно-восстановительного
потенциала. Железо и марганец в ризосфере находятся в
соединениях с органическими веществами — аминокислотами,
органическими кислотами и другими соединениями, обладающими
сильными хелатирующими свойствами и образующими прочные
комплексы, сохраняющиеся в почве длительное время. Они
используются растениями как источники железа, марганца и других
минеральных элементов. В ризосфере такие органометаллические
комплексы находятся в больших количествах, чем вне ее,
вследствие более высокого содержания органических веществ, а
растение лучше усваивает железо и марганец в виде комплексов,
которые образуются при участии микроорганизмов. Таким образом,
вокруг корней создается особая зона с благоприятными условиями
не только для микроорганизмов, но и для самих растений.
Общее число грибов так же, как и других микроорганизмов,
в ризосфере больше, чем вне ризосферы.
Однако видовое разнообразие их заметно убывает по мере
приближения к поверхности корня, вследствие чего видовой
состав в ризосфере значительно более однообразный, чем вне
ризосферы.
Если в почве, гетерогенной и мозаичной среде, мицелиальная
организация грибов может дать ряд важных преимуществ для
развития, в частности высокая линейная скорость роста, на один-
два порядка превышающая скорость роста бактерий, дает
возможность более эффективно колонизировать поступающий в почву
органический субстракт, то в зоне ризосферы, относительно
гомогенной по многим свойствам, преимущества мицелиальной
организации исчезают, подвижные бактерии в этой среде, насыщенной
влагой, передвигаются намного экономичнее и быстрее.
В зависимости от того, бедна почва гумусом или сильно гуму-
сирована, численность грибов в ризосфере изменяется различно.
В почвах, бедных органикой, увеличение численности грибов в
ризосфере более значительно, чем в сильно гумусированных, так
как в первых более заметна добавка органического вещества за
счет корневых выделений.
Выделяемые корнями вещества играют значительную
селективную роль в формировании сообществ микроорганизмов, в том
числе влияют на состав грибов ризосферы.
Специфика корневых выделений определяет различный состав
видов грибов в корневой зоне растений. Так, в ризосфере злако-
134
вых преобладают грибы родов Fusarium и Alternaria. В ризосфере
бобовых, лютиковых и крестоцветных много пенициллов.
Особенно существенны различия в наличии или отсутствии
патогенных форм грибов, например рода Fusarium, Verticillium,
Helminthosporium и других, в корневой зоне устойчивых и
поражаемых сортов растений в связи с значительными различиями в
составе их корневых выделений, в частности большое значение
имеет состав аминокислот. Так, эксудат корней бананов содержит
набор аминокислот, 13 из которых являются общими как для
устойчивых сортов, так и для поражаемых патогенным грибом
Fusarium oxysporum f. cubense в корневых выделениях
поражаемых сортов отсутствуют цистеин и треонин, а у устойчивых —
лейцин, серии, тирозин.
Состав грибных ценозов может также регулироваться
действием физиологически активных веществ, выделяемых корнями
растений. При этом возможно прямое стимулирующее действие
корневых выделений, прямое их ингибирующее действие. Кроме
этого возможна стимуляция роста микробов-антагонистов в
ризосфере к определенным видам грибов, в частности к фитопатоген-
ным грибам. Тогда последние угнетаются не корневыми
выделениями, а микробами-антагонистами. Наконец, возможно угнетение
роста микробов-антагонистов. Тогда могут усиленно размножаться
многие грибы ризосферы, в том числе фитопатогенные и
некоторые симбионты. Действие корневых выделений на грибы
представлено в табл. 12.
Микроорганизмы ризосферы в свою очередь могут оказывать
влияние на растение через корневую систему. Некоторые из них
снабжают растение витаминами, ростовыми веществами,
антибиотиками. Работами Н. А. Красильникова показана возможность
поступления антибиотиков в растение, в результате чего может
повышаться устойчивость растений к заболеваниям.
Ризоплана растений, как указывалось выше, — это область
непосредственно поверхности корней. Распространение грибов на
поверхности корней не является только результатом
специфичности субстрата, а зависит также от конкурентной способности
грибов. На поверхности корней обитают сапротрофные грибы и
факультативные паразиты. Сапротрофные грибы, обитающие на
поверхности корней, менее конкурентоспособны, чем истинные
обитатели почв. Грибы этой зоны в основном представлены тем-
ноокрашенными стерильными формами. Некоторые из них
относятся к базидиомицетам. Факультативные паразиты на
поверхности корней в основном представлены грибами-возбудителями
корневых гнилей. Это специфические виды, присущие данной
культуре или данному типу заболеваний. Например, грибы родов Olpi-
dium, Pythium, Phytophthora, Aphanomyces, Ophiobolus, некоторые
грибы Fusarium и др. Они активно образуют экстрацеллюлярные
ферменты, разрушающие стенки растения-хозяина и проникают
внутрь клеток, но могут быстро переходить к сапротрофному
образу жизни.
135
В отличие от грибов корневой зоны возбудители заболеваний
семян и проростков не специфичны (мало специализированы), и
один и тот же вид способен поражать большой круг растений —
зерновые, бобовые, технические культуры, древесные породы и пр.
Таблица 12
Характер действия корневых выделений высших растений на грибы
Действие
Стимуляция роста
Ингибирование роста
Стимуляция прорастания
Игнибирование прорастания
Высшие
растения
редис
салат
земляника
овес
картофель
томаты
горох
лук
томаты
редис
салат
бобы
бананы
горох
Грибы
Rhizoctonia solani
то же
Rhizoctonia sp.
Bissoclamys fulva
Spongospora subterranea
Colletotrichum atramentarium
Aphanomyces lutiches
Sclerotium cepivorum
то же
Fusarium spp.
Fusarium spp.
F. solani f. phaseoli
F. oxysporum f. cubense
F. oxysporum f. pi si
Они рассматриваются как почвенные грибы, хотя многие из них
находятся в почве в состоянии покоя и прорастают только тогда,
когда стимулируются эксудатами корней.
Грибы-микоризообразователи
Микоризообразователи представляют собой одну из
эколого-трофических групп, выделяемую по типу
взаимоотношений в группу симбиотрофов. Грибы-микоризообразователи,
вступая в симбиоз с высшими растениями, образуют микоризу.
Микориза, что означает в переводе с греческого грибокорень
(термин, введенный Франком в 1885 г.), представляет собой
симбиоз мицелия гриба с корнями высших растений. В результате
такого симбиоза корень претерпевает существенные
морфологические и анатомические изменения и приобретает специфические
черты.
Первые работы, посвященные изучению микориз, были сделаны
русским ученым Каменским в 1881 г. на микоризах бесхлоро-
фильного растения подъельника — Hypopitis monotropa.
В природе образование микориз у растений является
правилом, их отсутствие — исключением. Различают микоризу
эндотрофную (везикулярно-арбускулярную) и эктотрофную. Везику-
лярно-арбускулярные микоризы (ВАМ) характеризуются тем, что
в коровой паренхиме корневых окончаний растения мицелий
гриба, проникая через эпидермис, образует характерные древовид-
13Б
но-разветвленные гифы (арбускулы) (рис. 48) в сочетании с
отходящими от корня и распространяющимися в почве гифами.
Снаружи корень не претерпевает никаких изменений в
морфологии, его ветвление не нарушается, на поверхности корня
сохраняется корневые волоски. По мере воздействия клетки растения —
хозяина на находящийся в ней мицелий гриба образуются клубки
гиф со сферическими или
овальными вздутиями на
концах или утолщенными
посередине везикулами.
Рис. 48. Везикулярно-арбускулярная
(эндотрофная) микориза:
1 — арбускулы; 2 — везикулы
Более обильное развитие арбускул указывает на менее
уравновешенный характер взаимоотношений с растением и на
наличие более интенсивных защитных реакций со стороны растения.
При более уравновешенных взаимоотношениях гриба и
растения происходит образование клубков из мицелия. -'
Эктотрофные микоризы (эктомикоризы или чехольчатые
микоризы) характеризуются образованием чехла из плотного
сплетения гиф на поверхности инфицированного корня из псевдош;•
ренхиматической ткани с многочисленными отходящими от него
свободными гифами. Отдельные гифы проникают в межклетники
коровой паренхимы корня, образуя так называемую сеть Гартига.
Корневые волоски при этом исчезают (микоризный корень не
имеет корневых волосков). Иногда отдельные гифы проникают
внутрь клеток корня; через них осуществляется питание гриба.
При образовании эктотрофных микориз происходят и
значительные морфологические изменения корней. Под влиянием гриба
они усиленно ветвятся. Особенно типичны такие изменения,у
корневых окончаний сосны, имеющих характерное вильчатое или
коралловидное ветвление (рис. 49). Усиленное ветвление
корневых окончаний стимулируется выделением ауксинов, в частности
Р-индолилуксусной кислоты (ИУК), образование которой
показано для грибов-микоризообразователей.
Существенные морфологические изменения корней в
микоризах вызваны также образованием кинетинов, в частности цито-
кинина.
Из чистых культур микоризных грибов Suillus conthurnatus,
S. punetipes и Amanita rubescens цитокинины получены в
кристаллически чистом виде. Эти соединения, ранее обнаруженные у
кукурузы, представляют собой зеатин и рибонуклеозид зеатина.
Микоризы всех травянистых растений, в том числе и
возделываемых человеком, относятся к везикулярно-арбускулярному типу.
К нему относятся также микоризы многих древесных растений
137
тропиков и субтропиков, а также некоторых древесных растений
умеренной зоны — яблони, клена, вяза, ольхи. Грибной компонент
этих микориз относится к различным родам семействам Endogo-
naceae порядка эндогоновых класса зигомицетов, таких, как_ G/o-
Рис. 49. Ветвление корневых
окончаний сосны с микоризой:
I — вильчатая микориза; II —
коралловидная микориза; III —
черная микориза, образованная
грибом Сепососсит graniforme
mus, Gygaspora, Acaulospora, их споры (зиго- или азигоспоры, а
также хламидоспоры), имеющие крупные размеры 100—300 мкм,
находятся в почве и могут быть извлечены из нее либо последо-
138
нательным просеиванием через ряд сит с отверстиями различного
диаметра, либо центрифугированием в градиенте плотности или
флоатационным методом. Однако плотность их в почве очень
невелика, она варьирует от нескольких спор до нескольких тысяч
на 100 г почвы.
Везикулярно-арбускулярные микоризы преимущественно
представлены в пустынных и полупустынных почвах. Они редки в
подзолистых и болотных почвах. В обрабатываемых почвах их
содержание, как правило, выше. В луговых, степных, пустынных,
горных биогеоценозах эндогоновые грибы играют ведущую роль
в микоризообразовании. Большинство представителей флоры
лесов, тундры и болот также вступают в симбиоз с эндогоновыми
грибами. Однако у растений доминантов и субдоминантов этих
зон микоризы преимущественно образованы высшими грибами.
Крупные покоящиеся споры эндогоновых грибов обеспечивают
им длительную выживаемость в почве при неблагоприятных
условиях и в отсутствие растения-хозяина. Однако способ их
культивирования на питательных средах пока не найден.
Одну из разновидностей эндотрофных микориз представляют
собой микоризы растений из родов Erica, Calluna, Vaccinium,
Rhododendron. Отличие от типичных эндотрофных микориз
заключается в том, что наряду с внутриклеточным мицелием в виде
клубков на поверхности корня образуется небольшой чехол. Грибной
симбионт этих микориз относится к дискомицетам порядка пеци-
циевых.
Эндотрофные микоризы орхидных образует гриб Rhizoctonia.
У орхидных существует наиболее тесная связь с грибом, так как
эти растения в обычных условиях не могут развиваться без
микориз, а семена не прорастают в отсутствие гриба. Обычно
мицелий гриба уже находится в семени растений. У некоторых
древесных пород, в частности у клена, в результате внедрения
мицелия в клетки растения-хозяина происходит сильная деформация
клеточного ядра. Ядро увеличивается в размерах и деформируется
в своих очертаниях, принимая амебоидную форму. Одновременно
увеличивается число и размеры ядрышек. Эти изменения
начинаются еще в клетках, содержащих неизмененные гифы, но своего
максимального выражения достигают в клетках, где происходит
усвоение мицелия.
Увеличение линейных размеров ядра, по данным Н. П.
Горбуновой, составляет 56%, ядрышка — 105%. В клетках, где идет
переваривание гриба, увеличение линейных размеров ядра и
ядрышка еще большее и составляет 80 и 120%.
В отдельных случаях гриб вызывает образование в растении
фитоалексинов — веществ, аналогичных антителам животных, т. е.
оказывает влияние на растение, подобное патогенным грибам.
Большинство травянистых растений уже изначально имеют
везикулярно-арбускулярную микоризу, грибной симбионт которых
не является строго специфичным, поэтому искусственная мико-
ризация (инокуляция) грибами ВАМ дает наиболее ощутимые
139
результаты на почвах с низким содержанием фосфора. Кроме
того, хорошие результаты дает применение грибов ВАМ в
искусственном грунте, на почвах, подвергшихся фумигации и на
рекультивируемых территориях.
Положительное влияние ВАМ в первую очередь сказывается
в снабжении растений фосфором. Растения с такой микоризой
активнее поглощают фосфор из почвы: его содержание в тканях
микоризных растений выше, чем у немикоризных.
Микоризные растения утилизируют труднодоступные
соединения фосфора, такие, как фосфатиты, фосфориты, трикальцийфос-
фат. Более интенсивная адсорбция фосфора микоризными
растениями была доказана опытами с 32Р.
Микоризные растения по сравнению с немикоризными
выносят из почвы намного больше азота и калия, а также улучшают
снабжение растений микроэлементами — цинком, медью, серой.
Элементы, минерального питания из почвы поглощаются гифами
эндофита и транспортируются к внутренним грибным структурам,
а затем используются растением.
Эндомикориза повышает водообеспечение растений в
экстремальных засушливых условиях благодаря тому, что грибы
устойчивее высших растений к низким значениям водного потенциала.
Возможно, что эндофиты вырабатывают ростовые фитогормоны
типа цитокининов. Виды рода Glomus, образующие микоризу,
стимулируют фиксацию азота атмосферы клубеньковыми
бактериями. Грибы-микоризообразователи задерживают корневую
инфекцию фитопатогенами (бактерий, грибов, нематод).
На микоризах орхидей показано, что между эндофитными
грибами Rhizoctonia и растениями существует тесная взаимосвязь
по синтезу и потреблению витаминов. Гриб синтезирует пирими-
диновую, а растение тиазоловую часть витамина Bi.
Эктотрофная микориза имеется у большинства древесных и
кустарниковых пород, таких, как береза, дуб, осина, сосна,
лиственница и многие другие.
Грибные симбионты этих растений, главным образом высшие
базидиальные грибы, представители родов Boletus, Suillus, Rus-
sula, Lactarius, Amanita, Cortinarius и других из порядка Agari-
cales, гастеромицеты и небольшое число сумчатых грибов из
родов Elaphomyces и Tuber.
Некоторые из грибов-микоризообразователей высокоспециали-
зированы, например Suillus greillei — лиственничный масленок,
образующий микоризу только с лиственницей. В большой степени
специализированы грибы Suillus granulatus — масленок
зернистый, S. luteus — масленок желтый, Leccinum scab rum —
подберезовик обыкновенный. Наоборот, менее специализированы грибы
рода Amanita, Russula и т. д. Пример наименее
специализированного микоризообразователя представляет гриб Сепососсит grani-
forme, образующий так называемую черную микоризу более чем
со 130 породами кустарников и деревьев. Этот гриб
распространен по всему земному шару во многих типах почв, таких, как
140
подзолистые, дерново-подзолистые, горные лесные, пойменные и
т. д. Он не образует никаких спороношений, а существует в почве
в виде мицелия и склероциев. На этом основании его долгое
время относили к несовершенным грибам. В настоящее время на
основании строения септ, имеющих простые поры и строения
псевдопаренхимы микоризного чехла, он отнесен к сумчатым грибам.
Поскольку грибы-микоризообразователи находятся в тесном
контакте с растением, выделение их в чистую культуру на
искусственные питательные среды очень затруднено. Выделение
возможно либо непосредственно из микоризных окончаний корней с
предварительной обработкой антисептиками, для устранения сап-
ротрофных микроорганизмов, находящихся на поверхности
корней, либо из плодовых тел грибов, предполагаемых микоризооб-
разователей. В состав среды обязательно должен входить набор
аминокислот и витамин Вь
В том и другом случае выделенная культура проверяется на
способность ее образовывать микоризу, т. е. должен быть
осуществлен синтез микоризы из чистой культуры гриба и стерильно
выращенного высшего растения. Для этого сеянцы, например
сосны, предварительно выращивают в стерильных условиях. Семена
стерилизуют и помещают в стерильные колбы Эрленмейера
емкостью 500 мл, в которые насыпают песок, промытый крепкой
соляной кислотой и водой. Песок увлажняют раствором
питательных солей. Спустя четыре месяца после прорастания семени колбу
заражают мицелием гриба, предполагаемого микоризообразова-
теля. Через два месяца определяется наличие микоризы по
морфологии и анатомии корня, строению чехла и сети Гартига.
Отличительная особенность базидиомицетных грибов-микори-
зообразователей — низкая активность гидролитических
ферментов, в частности у грибов рода Boletus полностью отсутствуют
инвертаза (р-глюкозидаза) и р-амилаза. Они не содержат также
полифенолоксидаз, за исключением Boletus subtomentosus и рода
Lactarius, представляют собой хорошо очерченную
физиологическую группу, способную к утилизации только простых Сахаров.
Вследствие отсутствия или слабой активности гидролитических
ферментов они не выдерживают конкуренции с грибами —
подстилочными сапротрофами, активно разлагающими растительный
спад, что обусловило их совместное существование с корневыми
окончаниями деревьев, где они черпают необходимые им
легкодоступные углеводы, в большом количестве отлагающиеся в
корневых окончаниях растений.
Обильные микоризы образуются при наличии оптимального
освещения, когда идет активный синтез углеводов, и при
некотором дефиците азота или фосфора как элементов, лимитирующих
синтез белков и приводящих к накоплению углеводов в. корнях.
Зто доказано экспериментально с помощью изотопа 14С.
Грибы-микоризообразователи витаминозависимы по витамину
Вь который они получают от растения. Возможно также
использование ими некоторых аминокислот растений.
Hi
В свою очередь гриб оказывает определенное положительное
влияние на растение. Доказано, что микоризный чехол с
отходящими от его поверхности многочисленными грибными гифами
увеличивает поверхность всасывания и снабжения растения водой и
минеральными элементами. Особенно большое значение это имеет
на малоплодородных почвах. У микоризных растений существует
более интенсивный обмен питательными элементами с почвой, чем
у немикоризных. Это доказано с помощью изотопов. Микоризные
корни более активно поглощают радиоактивные азот, фосфор,
кальций и магний, а также частично железо и калий и
аккумулируют их в грибном чехле в больших количествах, чем ткани
растений.
Микоризная корневая система активнее адсорбирует
органические соединения азота, чем немикоризная. Экспериментально
показано, что микоризные грибы могут воздействовать на
минералы посредством различных органических кислот, таких, как
яблочная, щавелевая, гликолевая, шикимовая, ^мс-аконитовая и
другие, активно образуемые этими грибами. Под действием
органических кислот идет извлечение калия из силикатов: биотита,
мусковита, вермикулита, а также фосфора и калия из ортоклаза
и апатита.
В настоящее время существует мнение, что любые симбиоти-
ческие отношения представляют собой сильно
эволюционировавшие отношения паразитизма. То же самое касается и микориз.
Их можно рассматривать как уравновешенное сожительство гриба
и высшего растения в течение длительного периода времени.
Эволюция паразитизма идет в направлении все большего
взаимного приспособления гриба и хозяина и специализации
паразитизма. Одна из точек зрения на происхождение паразитизма у
грибов — это возникновение его из сапротрофного способа
питания.
В процессе конкуренции между сапротрофными грибами
многие виды вытеснялись из экосистем более сильными
конкурентами. Однако возможны случаи, когда эти сапротрофы попадал»
внутрь тканей растения через раневые каналы. Большинство из
них погибали под действием защитных реакций растения. Но
некоторые выживали и оказались способными размножаться внутри
хозяина. Способность существовать внутри хозяина все более
закреплялась естественным отбором и оказалась выгодным
приспособлением для таких, ставших паразитными, грибов, так как
защищала их от конкурентов и антагонистов. Патогенные грибы
обрели экологическую нишу. Это привело к серии биохимических
адаптации к такому образу жизни: выработке «ферментов
нападения», дающих возможность проникать не через раны
поврежденных растений, а непосредственно через кутикулу, появлению
«средств защиты» от таких метаболитов хозяина, как фитоалек-
сины, ферменты, и других, адаптации к условиям ограниченного
доступа кислорода внутри тканей хозяина, появлению веществ,
регулирующих рост хозяина и позволяющих ему использовать
142
питательные вещества растения (токсины, регуляторы роста типа
ауксинов, гиббереллинов, цитокининов).
Факультативные паразиты имеют несбалансированный
паразитизм, т. е. вызывают очень быстрое отмирание тканей
питающего растения и вскоре начинают развиваться в мертвых тканях.
Примером такого рода могут быть древоразрушающие грибы.
Наоборот, высокоприспособленные патогены долго не
обнаруживают своего губительного действия на клетки растения-хозяина
и сначала даже вызывают повышение его жизнедеятельности.
В таких случаях нередко растение, пронизанное мицелием гриба,
развивается почти так или лучше, чем здоровое, и даже
плодоносит. Число ферментов нападения сокращается, а ферменты сап-
ротрофного питания исчезают полностью, примером такого
наиболее приспособленного паразита может служить ржавчинный
гриб Chrysomyxa pirolae, мицелий которого сохраняется из года в
год в корневище грушанки (Pirola rotundifolia). Мицелий
проникает в заложившиеся в почве зачатки листьев, и на следующий
год все листья оказываются пронизанными мицелием, хотя
выглядят совсем здоровыми. Только на второй год на листьях
появляются спороношения гриба и листья отмирают. Следовательно,
здесь наблюдается длительный период уравновешенного
сожительства паразита и растения-хозяина. Токсины патогена перестают
быть факторами поражения, а становятся облигатными ростовыми
и морфогенетическими факторами. В частности, они стимулируют
образование в растении специфических веществ — микобоэтинов,
от которых в дальнейшем зависит их существование. В
отсутствие таких веществ паразитные грибы не могут пройти полный
цикл развития.
Такая эволюция взаимоотношений между патогенными
грибами и растениями-хозяевами достигает наивысшей ступени в
явлениях симбиоза (мутуализма), в частности в микоризах. В
микоризах гриб извлекает из сожительства с растением такую же
пользу, как и при любой форме паразитизма. Что же касается
высшего растения, то "оно также извлекает определенную пользу,
изложенную выше.
Трудности культивирования многих микоризных грибов
объясняются, по-видимому, также отсутствием в среде веществ типа
микобоэтинов.
ВЗАИМООТНОШЕНИЯ ГРИБОВ С ПОЧВЕННОЙ
МИКРОФАУНОЙ
Грибы-хищники
Особое место среди почвенных грибов занимают
хищные нематофаговые грибы с особым типом сапротрофного образа
жизни. Нематофаговые хищные грибы, которые имеют пищевые
связи с нематодами, по образу жизни нельзя причислять ни к
паразитам, существующим на живых животных и растениях, ни
143
к обычным сапротрофам, поселяющимся на мертвых
растительных или животных остатках. Хищничество у них, по мнению
Н. А. Мехтиевой, — это особая форма сапротрофного образа
жизни, при котором гриб улавливает здоровые живые особи
нематод, быстро их умерщвляет и питается за счет убитых им
животных. Таким образом хищные грибы отличаются от обычных
сапротрофов по способу добывания источников питания. Это
почвенные грибы, занимающие особую экологическую нишу.
Первое наблюдение хищных грибов было сделано М. С.
Ворониным в 1864 г., обратившим внимание на своеобразные петли
гриба Arthrobotrys oligospora. В. Цорф в 1888 г. установил их
значение в улавливании и умерщвлении нематод. Углубленное
изучение хищных грибов и хищничества началось в 1933 г. за
рубежом с работ Ц. Дречлера и Ц. Даддингтона, а в Советском
Союзе — Ф. Ф. Сопрунова и Н. А. Мехтиевой.
В связи с особым образом жизни хищные грибы имеют
специфические морфологические черты — специальные ловчие
приспособления для улавливания микроскопических животных,
главным образом нематод (рис. 50).
Они могут быть следующие: загнутый на конце и внешне
ничем не отличимый от обыкновенных гиф отросток, дифференци-
Рис. 50. Ловчие приспособления хищных грибов:
1 — петля; 2 — цепочковидные соединения липких почек; 3 — липкие
почки
рованные, изогнутые или кручковидные отростки. Одиночные
липкие почки, или их цепочковидные соединения; шаровидные или
широкоэллиптические липкие головки, расположенные на гифо-
подобных ножках, липкие петли и их сложные соединения в виде
сети; округлые двух-, трехклетные сжимающиеся кольца,
расположенные на толстых ножках, гантелевидные головки,
окруженные шаровидной липкой каплей и расположенные на почковидных
выростах. В случае наличия петель нематода проникает в петлю,
в случае липких головок последние прилипают к ее поверхности.
В обоих случаях постепенно движение нематоды прекращается,
мицелий гриба пронизывает все тело и полностью ассимилирует
ее содержимое.
Умерщвление нематоды происходит в результате как
механического поражения животного, так и токсического действия про-
144
дуктов метаболизма гриба. Животное попадает в ловчее кольцо,,
которое сжимается, и сильное сжатие губит его. При улавливание
нематод гриб выделяет токсическое вещество «нематотоксин»,
парализующее нематоды, близкое к производному терпенов три-
хотецину. В результате такого воздействия нематоды гибнут
быстро. Только после, умерщвления нематоды или после
прекращения ее движения в теле нематоды образуется инфекционная
луковица гриба, которая дает ростки, во всех направлениях
пронизывающие тело нематоды. Они служат ассимилятивными гифами,
которые используют вещества уже мертвой нематоды. Таким
образом, гриб активно улавливает здоровые живые особи животных,
хотя питается за счет тканей убитых им нематод; таким образом,,
источником органического азотного питания хищных грибов
являются животные белки. Попадание и улавливание нематод
ловчими аппаратами не случайно, так как происходит активное
привлечение нематод к ловчим аппаратам благодаря выделению ими
летучих пахучих продуктов, имеющих аттрактирующее значение.
Они же могут обладать одновременно нематоцидными свойствами.
Это липкие нерастворимые в воде вещества относятся к группе
ненасыщенных соединений, близких к терпенам. Таким образом,
пахучие вещества имеют двоякое значение: привлечение нематод
и их отравление.
Хищные грибы способны к обычному сапротрофному питанию^
и могут быть выделены в чистую культуру.
В отличие от обычных сапротрофных грибов хищные грибы
предпочитают среды с повышенным содержанием азота (C:N =
= 20:1). Необходима также добавка витаминов и
микроэлементов. Среды с повышенным содержанием белка стимулируют
образование ловчих аппаратов.
.Хищные нематофаговые грибы принадлежат к порядку Hypho-
mycetales несовершенных грибов семейства Moniliaceae и
представлены ограниченным числом систематически обособленных
родов гифомицетов. В настоящее время известно 12 родов немато-
фаговых грибов. Наибольшим числом видов представлены роды
Arthrobotrys и Golovinia. Многие описаны как новые (роды и
виды) Н. А. Мехтиевой только в 70-е гг. (например, род Nematopha-
gus — в 1975 г., род Kafiaddinia — в 1978 г.).
Хищные грибы широко распространены на земном шаре. Они
обнаружены во всех странах, где была сделана попытка их
изучения. Хищные грибы обитают на разнообразных субстратах,
богатых органическими веществами, на разлагающихся
растительных остатках, подстилке, мхах, экскрементах животных, а также
в водоемах, иле и т. д. Основное местообитание хищных грибов —
почва. Нахождение хищных грибов на различных других
субстратах представляет результат соприкосновения этих субстратов с
почвой или перенесения почвенных частиц разными путями на
эти субстраты.
Необходимые условия для развития хищных грибов —
наличие органического вещества, доступного для сапротрофных
микрона
организмов, оптимальная влажность, слабокислая или
нейтральная реакция среды, достаточная обеспеченность среды кислородом,
защищенность от прямого действия солнечных лучей, осеменен-
ность почвы нематодами.
В почве имеется большое количество микроорганизмов,
способных подавлять развитие хищных грибов. Главным образом это
актиномицеты и споровые бактерии, такие, как Вас. subtilis, Вас.
mesentericus, Вас, megatherium. Среди грибов ярко выраженный
антагонизм по отношению к хищным грибам обнаруживается
главным образом у пенициллов и аспергиллов.
Хищные грибы, по-видимому, произошли от сапротрофных
грибов, обитавших на животных остатках. О приспособленности к
.животным субстратам свидетельствует их более интенсивный
белковый обмен и в связи с этим наличие активных протеаз в
отличие от грибов сапротрофов, обитающих на растительных остатках,
для которых характерны потребность в углеводах и наличие
гидролитических ферментов. Ловчие аппараты их образуются в
основном в присутствии животных субстратов, но не растительных.
По мере приспособления к захвату организмов возникали и
совершенствовались их специальные ловчие аппараты,
обеспечивающие им связь с субстратом, улавливание и умерщвление
жертвы. Приспособление их к захвату животного субстрата в виде
активно существующих организмов обеспечило им большую
конкурентоспособность.
Есть попытка практического использования хищных грибов в
борьбе с нематодозами человека, животных и растений как у нас
в стране, так и за рубежом. При применении хищных грибов в
случаях, когда условия внешней среды благоприятствуют
развитию гриба, инокуляция ими почвы приводит к значительному
снижению числа нематод и процента зараженности ими растений.
Наилучший эффект от применения хищных грибов получают на
огурцах, пораженных галловой нематодой в условиях
вегетационного опыта и закрытого грунта. Однако для эффективного
использования хищных грибов в практических целях необходимо
соблюдение целого ряда условий, выбор активных штаммов и
установление степени и продолжительности их активности в
почве, создание оптимальных условий для их развития.
Грибы-паразиты
Среди грибов-паразитов беспозвоночных животных
представляют интерес широко распространенные в почвах амебофа-
говые грибы. Амебофаговые грибы представляют собой облигат-
ных эндо- и эктопаразитов почвенных амеб и таким образом
занимают особую экологическую нишу.
Амебофаговые грибы относятся преимущественно к зигомице-
там, семейству Zoopagaceae порядка Zoopagales. Наиболее часто
встречаются представители родов Zoopage, Stylopage, Cochlenema.
Амебофаговые грибы можно выделить на газоне бактерий Azo-
146
tobactcr chroococcum, который служит пищей для почвенных амеб.
Газон культуры Azotobacter инокулируют исследуемой почвой. На
нем развиваются амебы, многие из которых содержат амебофаго-
вые грибы.
Эндопаразитные амебофаговые грибы инфицируют своих
хозяев конидиями, которые либо поглощаются ими, либо
приклеиваются к их наружным покровам.
Конидия прорастает ростковой трубкой, которая продолжает
свой рост внутри эндоплазмы хозяина. Позднее мицелий гриба
спирально закручивается.
Эктопаразиты улавливают амеб липкими гифами либо также
заражают их мелкими и многочисленными экзогенными
конидиями. Мицелий их образует гаустории, проникающие в амебу.
Амебофаговые грибы распространены в почве многих стран
и являются ее постоянными обитателями или могут быть
обнаружены на субстратах, имеющих контакт с почвой.
Грибы как пищевые субстраты
беспозвоночных животных
Для многих беспозвоночных грибы служат пищевым
субстратом. Как было сказано выше, нематоды служат основной
пищей для сапротрофных грибов. Наряду с этим есть нематоды,
для которых грибы служат пищевым субстратом, в частности
микофаговые нематоды, питающиеся грибами родов Alternaria,
Fusarium и др. Многие другие беспозвоночные также питаются
мицелием грибов.
Энхитреиды (щетинковые черви) обитатели подстилок
используют определенные виды грибов в качестве пищевого субстрата
(виды Cladosporium и Mortierella).
Дождевые черви преимущественно потребляют грибы-пеницил-
лы. Среди клещей есть микофаги, относящиеся к орибатидам,
использующие многие несовершенные грибы разных родов, но не
пенициллы. Клещи рода Tyroglyphus охотно поедают споры
хищных грибов. Для панцирных клещей грибы являются их
основными объектами питания. Их связи с грибами складываются как
симбиотические. Они используют ферменты грибов, которых они
заглатывают с пищей, для дальнейшего разложения
растительной ткани.
Для многоножек (кивсяков), ногохвосток (коллембол) грибы
не являются регулярными пищевыми объектами. В подстилках
лесов обитают брюхоногие моллюски — микофаги.
Между грибами и насекомыми существуют самые
разнообразные взаимоотношения, начиная от случайных (непрочных)
ассоциаций, до вполне сложившихся симбиотических отношений и
отношений паразитизма.
Жесткокрылые насекомые и их личинки являются микофагами
и питаются преимущественно грибами. Среди них есть формы,
культивирующие грибы.
147
Очень интересны примеры сожительства грибов с жуками —
древоточцами и короедами. Последние прогрызают
многочисленные ходы в древесине и коре, эти ходы заполняет грибной
мицелий, который они постоянно переносят в своем теле. Мицелий
служит пищей для личинок насекомых. Частично он сохраняется
в их теле, а потом снова попадает наружу с экскрементами и
таким образом может перемещаться на значительные
пространства вместе с насекомыми. Грибы в таком сожительстве принято
называть «амброзией».
Аналогичная ситуация имеет место с древесной осой. Ее
внутренние органы бывают заражены древоразрушающим базидиаль-
ным грибом Stereum sanguinolentum, так что яйца, которые
откладывает насекомое в древесину, оказываются уже
зараженными. Древесина инфицируется и постепенно разрушается грибом,
а личинка осы питается при этом продуктами разложения
древесины.
Большинство грибов «амброзия» не растут на сусло-агаре, но
растут на гидролизате казеина и пептоне. Многие обнаруживают
потребность в витаминах группы В и не способны утилизировать
неорганический азот, но активно используют жиры. Видимо, они
восполняют свои потребности в витаминах, органическом азоте
и жирах за счет партнера. Жуки в свою очередь поедают
мицелий и споры грибов.
Амброзиевые жуки, таким образом, являются
распространителями грибной инфекции в древесине и стимулируют тем самым
процессы ее разложения. Многие личинки жуков могут
потреблять не только грибные гифы, но и продукты их
жизнедеятельности, а также растительные ткани, предварительно
переработанные грибами.
Двукрылые насекомые откладывают личинки в плодовые тела
.высших базидиальных грибов и питаются ими, вызывая
червивость этих грибов.
Совершенно уникальны взаимоотношения с грибами
некоторых муравьев. Наиболее хорошо изучены в этом отношении
муравьи родов Atta и АсготугтеХу распространенные в Южной
Америке. Эти муравьи питаются только грибами, которые они сами
культивируют, создавая так называемые грибные плантации или
сады в своих гнездах.
Сначала муравьи приготовляют субстрат для разведения
грибницы в виде листьев и цветов, которые они обгрызают с веток и
измельчают. Затем они спрессовывают их челюстями, превращая
в бесформенную массу, смоченную слюной, кроме того,
смачивают жидкими фекалиями. Этот субстрат муравьи переносят в
гнезда с помощью передних ног и челюстей и заражают мицелием
грибов, перенесенным из старого гнезда.
Муравьи родов Atta и Асготугтех используют в качестве
субстрата очень многие растения. Другие виды муравьев
выбирают маленькие фрагменты гниющей древесины или остовы
насекомых.
148
Первым описал грибы, которые выращивают в своих «садах»
муравьи рода Atta, Меллер в 1893 г. Эти грибы относятся к ба-
зидиальным грибам Leucocoprinus gongylophora, Leucoagaricus
gongylophora и Attomyces bromaticus. Видовое название гриб
получил по наличию на концах специфических колбовидных
образований — бромат. Другие роды муравьев культивируют еще
грибы из родов Daldinia или Hylaria и Auricularia, но чаще всего из
рода Lepiota. Плодовые тела последнего удалось получить в
чистой культуре.
Грибы растут в «садах» в виде вегетативного мицелия и не
образуют спороношений. Благодаря постоянному обгрызанию
муравьями гиф мицелия, на нем образуются головки из вздутых
клеток, которые и составляют пищу муравьев.
Грибные «сады» — это источник пищи для муравьев, это
место, где они выводят потомство, здесь располагается самка. Когда
самка кладет яйца, рабочие муравьи берут и расселяют их по
саду.
Самые крупные «сады» у наиболее крупных муравьев Atta и
Асготугтех. Первые имеют размеры 10—13 см, вторые — 8—
12 см. Зрелая колония использует от 1 до 2 кг свежих листьев
для «сада».
Муравьиные гнезда содержат много органического вещества
в виде грибных садов. Это специфическое органическое вещество,
переработанное муравьями и снабженное продуктами их
жизнедеятельности — слюной и фекалиями. Кроме того, оно
подвергается активному воздействию растущих на нем грибов и дает
возможность для размножения бактерий, нематод, насекомых и
других организмов, которые также активно изменяют исходный
субстрат. Переработанный субстрат поступает в почву, оказывая
существенные изменения в составе органического вещества почв.
Таким образом, муравьи через свои сады снабжают почву
специфическими органическими веществами. Им принадлежит
значительная роль в образовании органического вещества почв
тропических лесов.
Своеобразный характер взаимоотношений складывается между
грибами и насекомыми, когда грибы поселяются внутри тела
насекомых, в кишечнике или самих клетках. Наиболее яркий
пример такого внутриклеточного сожительства — так называемые
мицетомы или мицетоциты, известные у многих тлей, щитовок и
некоторых муравьев. Это группы довольно крупных клеток
насекомых, в протоплазме которых находится масса дрожжеподоб-
ных грибов. Постоянное присутствие таких мицетом в теле
некоторых видов насекомых, их определенное положение в теле
насекомого, а также передача грибов при размножении через яйца
позволяют заключить, что здесь имеет место не случайная
инфекция, а урегулированный симбиоз. Однако сущность этих
взаимоотношений не раскрыта.
Инфекция мицетомы может появиться при заглатывании гриба
насекомым или как результат заражения поверхности яиц гри-
149
бами. В некоторых случаях удается культивировать эти грибы
на искусственных средах при добавлении в среду витаминов
группы В и дрожжевых стеролов. Видимо, в мицетомах эти вещества
гриб получает от насекомого. Каждое насекомое имеет своего
определенного дрожжевого партнера, но пока их не удается
идентифицировать. Многие из них очень плеоморфны.
В 1948 г. в кишечнике артропод обнаружены виды новой
группы грибов — трихомицетов, характеризующихся настолько
своеобразным строением, что систематическое положение их до конца
не ясно. Эти организмы представлены простыми септированными
нитями, размножаются амебоидными или неподвижными спорами.
В некоторых случаях они образуют структуры, напоминающие
зигоспоры. Взаимоотношения их с насекомыми и другими
членистоногими также недостаточно ясны. Некоторые из них удается
культивировать на питательных средах.
ВЗАИМООТНОШЕНИЯ ПОЧВЕННЫХ ГРИБОВ
С ДРУГИМИ МИКРООРГАНИЗМАМИ
Грибы в почве в большинстве случаев существуют не
изолированно, а в ассоциациях с другими микроорганизмами.
Заселение поверхности грибных гиф бактериями и актиноми-
цетами часто можно наблюдать на стеклах обрастания и педоско-
пах, на что впервые обратил внимание Н. Г. Холодный. На
стеклах обрастания сначала появляются грибные гифы, затем на их:
поверхности поселяются бактерии. Они располагаются
небольшими агрегатами или покрывают гифу рядами толщиной в один или:
больше слоев клеток, а иногда образуют мощные чехлы.
Виды бактерий, которые заселяют грибные гифы, относятся к
Pseudomonas fluorescens, Achromobacter denitrificans, Proteus
vulgaris, Вас. mesentericus, Вас. cereus.
Более активно заселяют грибные гифы актиномицеты, чем
бактерии. Возможно, что заселение гиф грибов микроорганизмами
объясняется выделением грибами определенных продуктов
метаболизма.
Летом и осенью все микробное население почвы особенно
интенсивно концентрируется около грибных гиф. Являясь
организмами очень активными в биохимическом отношении, грибы
создают вокруг себя среду, насыщенную продуктами обмена, что и
привлекает другие микроорганизмы. Установлено, что мицелий
грибов выделяет такие вещества, как аминокислоты, белки, пептиды,
мочевину, моносахара, органические кислоты, витамины и
минеральные вещества, главным образом фосфор и серу. Таким
образом, грибы представляют собой организмы-эдификаторы. Такая
связь грибов с другими микроорганизмами проявляется не только
в случае хорошо развитого мицелия, но и на стадии прорастания
спор. Однако иногда колонизация мицелия микроорганизмами
заканчивается лизисом его, тогда отношения между ними
становятся паразитическими. Тем не менее никакой специфики в лизи-
.150
рующих организмах не наблюдается. Самые различные бактерии
и актиномицеты могут вступать в такие отношения.
Среди микроорганизмов, лизирующих мицелий грибов, можно
назвать Streptomyces spp., Вас. cereus, Вас. macerans, Вас.
megatherium, Pseudomonas spp., а также некоторые актиномицеты.
Лизису могут подвергаться грибы родов Rhizoctonia, Phyco-
myces, а также разнообразные представители несовершенных
грибов, многие из которых являются фитопатогенными.
Иногда одни грибы могут лизировать мицелий других.
Например, Penicillium purpurogenum лизирует Aspergillus niger.
Бактерии и актиномицеты лизируют грибной мицелий, так как
обладают соответствующим набором ферментов, в том числе глю-
козидазами, хитиназами, липазами. Эти ферменты внеклеточные,
и поэтому лизис мицелия в почве может осуществляться и без его
колонизации, за счет выделения экзоферментов. Есть наблюдения
о проникновении бактериальных клеток и гиф актиномицетов
внутри грибной споры (конидии). Это хорошо показано на примере
Cochliobolus sativus.
Наличие меланинов в клеточной стенке грибов существенно
препятствует ее лизису. В связи с этим лизис мицелия темноокра-
шенных грибов, принадлежащих к семейству Dematiaceae, почти
не имеет места.
Отмерший грибной мицелий может служить питательным
субстратом для других микроорганизмов. При внесении отмершего
мицелия в почву происходит заметное увеличение числа
почвенных бактерий в 10—70 раз (через 7 дней).
В свою очередь микроорганизмы могут оказывать влияние на
жизнедеятельность грибов. Бактерии могут развиваться на гифах
грибов как эпифиты, используя грибные метаболиты и не
причиняя им никакого вреда. Они могут вызывать также явление фун-
гистазиса за счет выделения специфических веществ,
задерживающих прорастание грибных спор. Бактерии и актиномицеты могут
вступать в конкурентные взаимоотношения с грибами за
питательный субстрат, причем их конкурентоспособность повышается
за счет выделения антибиотиков и токсинов, свойственных
многим микроорганизмам.
Особые отношения складываются у грибов с марганецокис-
ляющим организмом Metallogenium. При использовании
специальных сред, содержащих в своем составе марганец, на них
выделяются грибы, имеющие постоянным спутником Metallogenium
{рис. 51).
Грибы со спутником обнаружены в различных горизонтах
почвенного профиля, преимущественно в почвах подзолистого типа.
Такие ассоциации есть в подстилке и растительном опаде и даже
в составе эпифитных микроорганизмов на поверхности растений.
В числе грибов, входящих в такие ассоциации, находятся самые
разнообразные виды, типичные как для почв, так и для растений,
например Coniothyrium fuckelii, Mycogone nigra, виды Alternaria,
Cladosporium, Acremonium и др. Постоянная встречаемость раз-
151
Рис. 51. Ассоциация грибов с марганецокисляющими организмами. Грибные-
гифы, покрытые колониями Metallogenium с отложениями марганца
нообразных грибов с марганецокисляющими спутниками говорит
о том, что такое сосуществование является для них обычным. При
этом установлено, что Metallogenium использует избыточно
выделяемую грибом перекись водорода для окисления марганца.
Многие грибы в природных условиях находятся в ассоциациях
с азотфиксирующими бактериями. Эти ассоциации выявляются1
при выделении их на безазотистые среды.
Известно, что грибы как эукариотные организмы азот не
фиксируют. Однако установлено, что азотфиксирующая активность
ассоциаций азотфиксирующих бактерий с грибами возрастает па
сравнению с чистыми культурами бактерий. Особенно заметно
увеличение азотфиксирующей активности ассоциаций в случае их
роста на полимерах, в частности использования в качестве
источника энергии целлюлозы (табл. 13).
На таблице видно, что азотфиксирующая активность бактерий
в ассоциациях с определенными видами грибов в несколько раз
выше, чем в чистой культуре при использовании в качестве
энергетического субстрата целлюлозы.
Одной из причин усиления азотфиксирующей активности
бактерий в ассоциациях с грибами является целлюлолитическая
активность грибов, вследствие которой происходит снабжение
бактерий доступным энергетическим материалом. Кроме того,
благодаря интенсивному потреблению кислорода грибами в процессе
152
Таблица 13
Азотфиксирующая активность бактерий в ассоциации с грибами
Виды грибов в ассоциации
JPenicillium claviforme
P. fellutanum
Trichoderma hamatum
T. harzianum
T. koningii
P. verrucosum var. cyclopium
T. koningii
Aspergillus niger
T. koningii
T. pseudokoningii
JP. notatum
Азотфиксирующая активность,
10~3 мг N 10 мл/ч
в составе
ассоциации
19,9
22,6
45,8
87,2
37,2
39,8
74,5
в чистой
культуре бактерий
3,4
13,5
12,6
29,7
10,3
18,5
33,4
Целлюлоти-
ческая
активность грибов,
% от контроля
10
10—20
30—40
30—35
25—30
20—35
25—60
разложения целлюлозы, снижается
окислительно-восстановительный потенциал среды, что также способствует процессу азотфик-
сации. В свою очередь грибы могут пополнить дефицит азота за
счет своего бактериального спутника.
Ч а сть III
РОЛЬ ГРИБОВ В ПОЧВЕННЫХ
ПРОЦЕССАХ
Гл ава 6
РОЛЬ ГРИБОВ В ПРЕВРАЩЕНИИ
ПРИРОДНЫХ СУБСТРАТОВ
РАЗЛОЖЕНИЕ ГРИБАМИ УГЛЕРОД СОДЕРЖАЩИХ
СОЕДИНЕНИЙ РАСТИТЕЛЬНОГО ПРОИСХОЖДЕНИЯ
Процессы распада сложных органических веществ
биологического происхождения, таких, как клетчатка и лигнин,
представляют собой одну из важных проблем микробиологии и
почвоведения. Эти вещества — главные составные части
растительного опада и древесины и от их разложения зависит круговорот
соединений углерода в природе.
Общая продукция органического вещества на земном шаре
ежегодно определяется в 50—100 млрд т. Основную массу его
составляют соединения растительного происхождения. Общие запа
сы фитомассы суши — 3,1 -1012—1,2-1013 т. Ежегодное количество
растительного опада составляет для таежной зоны 2—7 т га, для
лиственных широколиственных лесов — 5—13 т га, опад остепнен-
ных лугов колеблется в пределах от 5,2 до 8,5 т га.
В состав растительных клеток входят разнообразные
химические вещества, такие, как простые углеводы, крахмал,
органические кислоты, спирты, белки, аминокислоты, жиры воска и многие
другие, но преобладают по массе такие сложные полимерные
соединения, как целлюлоза, гемицеллюлозы, пектин и лигнин.
Если низкомолекулярные органические соединения минерала
зуются многими микроорганизмами, то разложение сложных
полимерных соединений, особенно в составе древесных растений,
осуществляется грибами и только в аэробных условиях.
Разложение целлюлозы
Содержание целлюлозы в древесном опаде составляв!
34—59%, несколько меньше содержание гемицеллюлоз и пектина.
Грибы — активные агенты разрушения целлюлозы. Установлено,
что разложение целлюлозы — это многоступенчатый процесс,
обусловленный ее сложным строением, который идет в несколько
стадий. Он осуществляется комплексом ферментов — целлюлаз.
154
Целлюлоза — это полисахарид, нерастворимое вещество,
которое в природных растительных субстратах всегда связано со
многими другими соединениями, гемицеллюлозами, лигнином, тан-
нином, а также восками, смолами, терпенами. Молекулы
целлюлозы представляют собой мицеллы или микрофибриллы, на
поверхности которых прочно адсорбируются вышеперечисленные
соединения или они могут быть включены в молекулу.
Микрофибриллы сложены из линейных макромолекул,
агрегированных в пучки. Элементарные единицы линейных
полимерных молекул представляют собой ангидроглюкозу. Молекулы
клетчатки содержат до 14 000 элементарных единиц с молекулярной
массой до 500 000. Они не имеют определенной длины, и число
глюкозных остатков может варьировать.
Линейные молекулы целлюлозы образуют участки с
параллельным расположением цепей и соединены между собой
водородными связями, которые обусловливают такие свойства, как
скорость растворения, гигроскопичность, реакционную способность.
Из-за высокой прочности водородных связей молекулярные
ассоциации целлюлозы в воде нерастворимы.
В образовании водородных связей участвуют все свободные
тидроксилы, имеющие различную прочность связи. Поэтому в
микрофибриллах чередуются участки с более высокой степенью
упорядоченности, образующие кристаллиты, с участками, в которых
нет упорядоченности, — аморфными.
Высокая степень упорядоченности служит причиной их
устойчивости к ферментативному гидролизу. Истинными целлюлозо-
разлагающими организмами являются только организмы, способ*
ные воздействовать на нативную целлюлозу, но не на ее
производные. Разложение ее осуществляется ферментами Ci (целлюло-
>биогидролаза), Сх (р-1-4 глюконазы) и р-глюкозидазой через
следующие стадии:
нативная целлюлоза с* целлюлодекстрины сх целлобиоза
(возможно образование целлотриоз и целлотетроз)
р-глюкозидаза ^ гЛЮКОЗа.
Таким образом, целлюлаза представляет собой комплекс фер-
гментов, действующих на разных стадиях разложения целлюлозы.
Все ферменты, участвующие в разложении целлюлозы, являются
гидролитическими ферментами. Фермент Q необходим для
гидролиза высокоорганизованной кристаллической целлюлозы. На
следующей стадии группа ферментов Сх гидролизует более
короткие полиангидроглюкозные цепи до растворимых небольших
молекул, способных к диффузии в клетку, т. е. гидролизует глю-
козидные связи, последовательно отщепляя глюкозные остатки
зсонца целлюлозной цепи с образованием восстановленных
Сахаров (целлобиоз, целлотриоз, целлотетроз). Следующая стадия
заключается в воздействии р-глюкозидазы на глюкозидные связи
целлобиозы и образование глюкозы — конечного продукта
разложения целлюлозы.
155
Фермент Сь экстрацеллюлярный фермент, представляет собой
гомогенную фракцию и является гликопротеином с соотношением
углеводов и белков 1:1. Его молекулярный вес равен 61 000, он
термолабилен и теряет при нагревании 50% активности. Фермент
Сх представляет собой целую группу неоднородных белков; 2
фермента этой группы отличаются механизмом гидролиза, так как
представляют собой эндо- и экзоглюконазы.
Целлюлазная активность всего комплекса ферментов
обнаружена у некоторых сумчатых и несовершенных грибов, хотя они:
различаются по содержанию ферментов Ci и Сх.
Chaetomium globosum, Aspergillus niger, A. fumigatus, Mem-
noniella echinulata, Penicillium verrucosum var. verrucosum, Tri-
choderma virtue, Fusarium solani и некоторые другие образуют
индуцированные Сг и Слгферменты. Высокой целлюлолитической
активностью обладают Penicillium daliae, Cladosporium herbarum,
Alternaria alternata.
У Paecilomyces varioty обнаружены неиндуцированные d- и
Qc-целлюлазы. Виды Fusarium и Alternaria alternata образуют в
основном неиндуцированную Сх-целлюлазу.
Наличие всего комплекса целлюлолитических ферментов
особенно характерно для высших базидиальных грибов. С этим
свойством связано их активное участие в разложении нативной
молекулы целлюлозы растительного опада.
Активные целлюлазы могут быть обнаружены в фильтратах
грибов. Для большинства грибов наибольшая их продуктивность
характерна для молодого мицелия и верхушек гиф. Это имеет
большое экологическое значение, так как дает возможность
молодому растущему мицелию активно внедряться в клеточную
стенку растения.
Для определенных видов грибов необходимы субстраты —
индукторы образования целлюлаз.
Наряду с этим среди продуктов метаболизма грибов есть к
репрессоры их образования. Репрессорами могут служить сахара,
т. е. продукты распада целлюлозы. Репрессорами — ингибитора- N
ми целлюлаз — могут быть также метилцеллюлоза, глюконолак-
тоны, полиолы.
Наличие репрессоров может иметь большое биологическое
значение, препятствуя проникновению патогенных грибов в растение*
а также регулируя внедрение в корни растений грибов-микоризо-
образователей.
Разложение гемицеллюлоз. Гемицеллюлозы — полисахариды*
сахара и уроновые кислоты — присутствуют во всех растительных
тканях. Ксилан, относящийся к гемицеллюлозам, — полимер
ксилозы, занимает по количеству в растениях второе место после
целлюлозы. В древесине хвойных его 12%, лиственных
деревьев — до 25%.
Разложение гемицеллюлоз — процесс неспецифический и
осуществляется многими микроорганизмами. Большое место в этом
процессе занимают грибы, в частности фермент ксиланаза есть у
156
многих грибов, как микромицетов, так и высших базидиальных
грибов, многие из которых являются типичными сапротрофами
на растительном опаде.
Разложение пектиновых веществ. Среди грибов имеются
активные разлагатели пектина, который также является
существенным компонентом растительного опада. Пектин образует в
растениях межклеточное вещество, из которого состоят так называемые
срединные пластинки, соединяющие между собой отдельные
клетки растения. Они придают тканям прочность. Пектин
представляет собой высокомолекулярное соединение углеводной природы —
полисахарид, в котором метоксилированные остатки галактуроно-
вой кислоты связаны между собой р-1,4-глюкозидными связями-
пектинэст'ераза полигалактуроназа
¦ 1
СООСН3 * СООСНз
Чхлргхь
СООСНз
В растениях пектиновые вещества присутствуют в виде
нерастворимого протопектина в соединении с другими полисахаридами
клеточной стенки.
Протопектин — комплексное соединение пектина со сложными
веществами клеточной оболочки: целлюлозой, гемицеллюлозой,
арабаном и ксиланом.
Растворимый пектин представляет собой полисахарид,
состоящий из соединенных между собой остатков галактуроновой
кислоты, присутствующей в ней в виде метилового эфира.
Молекулярная масса пектина различна в зависимости от его
происхождения и измеряется величинами от 25 000 до 50 000. Протопектин
и пектин пропитывают клеточные оболочки стеблей, плодов,
корней и семян многих растений, образуя в комплексе с другими
полисахаридами склеивающий субстрат клеток.
Действие пектиназ проявляется в размягчении ткани и распаде
ее на отдельные клетки. Существует несколько типов ферментов-
пектиназ в зависимости от специфики их воздействия на субстрат:
протопектиназы вызывают превращение нерастворимого
протопектина в растворимый пектин, расщепляют связи между мето-
ксилированной полигалактуроновой кислотой и связанным с ней
арабаном и ксиланом; пектинэстеразы гидролизуют метоксиль-
ные молекулы растворимого пектина (гидролитическое
отщепление метоксильных групп от растворимого пектина), в результате
чего образуется полигалактуроновая кислота и метиловый спирт;
полигалактуроназы гидролизуют р-1,4-глюкозидные связи
пектиновой кислоты до свободных галактуроновых кислот. Встречают-
15Г
ся главным образом у различных видов бактерий и грибов.
Многие грибы образуют пектинолитические ферменты.
Высокая пектинолитическая активность обнаружена у некоторых эпи-
фитных грибов, главным образом Aureobasidium pullulans и видов
Cladosporium. Пектинолитические грибы занимают значительное
место среди типичных представителей лесной подстилки — это
виды родов Cladosporium, Alternaria, Aposphaeria, Penicillium, фи-
топатогенные грибы родов Fusarium, Verticillium, Botrytis cinerea,
Sclerotinia sclerotiorum и др.
Ферментативное разрушение пектиновых веществ в растениях
имеет значение в патогенезе некоторых заболеваний. Фитопато-
генные грибы разрушают пектин срединной пластинки и пектаты
в первичных клеточных оболочках, что приводит к изменению их
физико-химических свойств и создает условия для внедрения
паразита, а также в результате действия пектинэстеразы
образуются вещества — полигалактурониды, способные закупоривать
сосуды, что в конечном итоге приводит к увяданию растений.
Есть сведения о наличии пектиназ в верхушечной части гиф
трибов-микоризообразователей, что обеспечивает им внедрение в
корень растения.
Существенное значение разрушение пектиновых веществ
грибами имеет при разложении растительного опада. Практическое
использование пектиназ грибов — применение в пищевой
промышленности при приготовлении фруктовых соков для их осветления,
а также при мочке льна.
Разложение лигнина. Грибы — почти единственные
разрушители лигнина. Способность грибов осуществлять глубокое
разрушение лигнина представляет собой уникальное явление.
Лигнин — наиболее распространенное в природе полимерное
циклическое соединение. В наибольшем количестве лигнин
содержится в древесине и древесном опаде. Содержание его в опаде
хвойных пород составляет 28—34%, лиственных пород — 18—28%.
В химическом отношении лигнин не является индивидуальным
веществом с вполне определенными свойствами и составом.
Известно, что лигнин — высокомолекулярный разветвленный
полимер, состоящий из фенилпропановых единиц. Основными мо-
зномерами его являются три спирта — производные фенилпропана.
сн2оы сн2он
L
осн
н он он
паракумаровый конифериловый синаповый
158
Относительное соотношение всех трех компонентов различно в
лигнине разного происхождения. В лигнине хвойных преобладает
конифериловый спирт, в лигнине лиственных содержится кони-
фериловый и синаповый спирты, у злаковых растений доминирует
синаповый и имеется еще и паракумаровый. У дводольных
растений в значительном количестве обнаруживают паракумаровый
спирт. Характерной особенностью лигнина является то, что
отдельные его мономеры связаны между собой не одной формой
связи, а различными. Существуют три основные формы связи в-
лигнине:
1. Арилглицерил-р-арилэфирная связь:
2. Фенилкумариновая структура:
нс=о
не
сн3о
сн,он
СН2ОН
сн
НС
носн2
сн3о
о—
3. Дифениловая структура:
сн3о
В частности, остатки кониферилового спирта связаны эфирной?
связью в гваяцил-глицерол-конифериловый эфир. Эти связи
чрезвычайно устойчивы к действию ферментов. Исследования,
касающиеся микробной деградации лигнина, относятся к одной из
наиболее сложных биологических проблем, поскольку лигнин пока
не может быть точно определен как химическое вещество. Также
не могут быть точно определены и промежуточные реакции его
биологического превращения. Наиболее активные группы
микроорганизмов, разрушающих лигнин, принадлежат к древоразру-
15^
шающим базидиомицетам, вызывающим белую гниль. Однако до
настоящего времени неизвестны полностью все стадии
ферментативных реакций в процессе разложения лигнина, т. е. известны
далеко не все ферменты, осуществляющие этот процесс.
Отличия деградации лигнина от деградации других полимеров
заключается в том, что такие полимеры, как протеины,
полисахариды, нуклеиновые кислоты, состоят из регулярно
повторяющихся единиц, в то время как лигнин состоит из различных
мономеров, имеющих различные типы связей. Большинство
микроорганизмов, воздействующих на лигнин, вызывают в нем очень
незначительные изменения, которые проявляются в основном в
уменьшении числа метоксильных групп и очень слабой потере в
весе. Некоторые сумчатые и несовершенные грибы могут расти
на средах, содержащих препарат лигнина в качестве
единственного источника углерода, такие, как Fusarium lactis, F. nivale и
некоторые другие, но они не вызывают существенных изменений
в молекуле лигнина. Вещества, представляющие собой
производные лигнина, — ванилин, сиреневый альдегид и другие,
используются грибами родов Chaetomella, Coniothyrium, Cylindrocarpon,
Torula, Hormiscium и др.
Исследования последних лет показывают, что полное
разложение лигнина с разрушением ароматического кольца могут
осуществлять только базидиальные древоразрушающие грибы,
вызывающие белую гниль: Coriolus versicolor, Forties fomentarius,
и некоторые подстилочные базидиомицеты, такие, как виды
родов Collybia, Marasmius, Mycena и др.
Высокая молекулярная масса и низкая растворимость лигнина
препятствуют его прямой ассимиляции микроорганизмами.
Предварительно происходит его расщепление экзоферментами во
внешней среде. Своеобразное химическое строение лигнина делает его
труднодоступным для ферментных систем микроорганизмов.
Многие микроорганизмы обладают ферментными системами,
способными к превращению простых ароматических соединений. Однако
полизамещенные ароматические соединения — мономеры
лигнина — доступны лишь немногим представителям отдельных родов,
так как для их разложения требуются весьма редкие у
микроорганизмов ферменты деметилирования и декарбоксилирования
ароматических структур. Необычен для микроорганизмов и характер
связей между мономерами, некоторые из них не имеют аналогий
среди соединений, типичных для живой клетки. Они совершенно
недоступны для ферментов — деполимераз микроорганизмов,
легко разрушающих белковые, полисахаридные, полинуклеиновые
молекулы. Неясно, какие ферменты могут расщеплять связь ди-
фениловой структуры. Трудности для расщепления представляет
собой связь фенилкумаровой структуры. Обычные для микробного
разложения эфирные связи становятся труднодоступными из-за
соседних полизамещенных ароматических структур. Грибы-бази-
диомицеты образуют ферменты, воздействующие на арилглице-
рол-р-ариловые эфирные связи в лигнине. В результате образу-
,160
Разложение грибами гваяцил-глицерол-/?-кониферилового эфира
НХОН Н Н
/~\ 1 I
о— (' х) с=с—сн,оч
он
он
соон
сн.
Воздействие на
фенилпропановую единицу
с укорочением цепи
осн3
он
сно
сн,
осн
он
ванилиновый альдегид
ОСН,
ванилиновая кислота
ОСН
он
% Т. Г. Мирчннк
ются продукты распада лигнина в виде гваяцил-глицерола, что
можно обнаружить хроматографически. Процесс осуществляется
экзоферментами. Затем идет дальнейшее разложение молекулы
лигнина, сопровождающееся укорачиванием боковых цепей,
окислением и карбоксилированием их, в результате чего образуются
ванилиновая кислота и ванилиновый альдегид.
Далее возможно деметилирование и окисление фенолов в хи-
ноны с образованием темноокрашенных продуктов. Эти процессы
идут при участии фенолокисляющих систем — полифенолоксидаз.
Такие ферментативные системы могут быть обнаружены в
культурах грибов, вызывающих белую гниль.
Деградировавший при белой гнили лигнин содержит меньше
углерода, метоксилов, водорода, но значительно больше
кислорода, а также характеризуется более высоким содержанием карбо-
нилов, карбоксилов, гидроксилов.
Способность многих грибов разлагать клетчатку и лигнин
определяет их активное участие в разложении растительного опада.
В растительном опаде в древесине лигнин и целлюлоза образуют
природный комплекс, в котором его составляющие структурно и
химически связаны между собой. Разложение лигноцеллюлозного
комплекса грибами установлено серией экспериментов по
разложению растительного опада, проведенных В. Я. Частухиным. Он:
разработал специальную методику для лабораторного изучения;
процессов распада, которая позволила создать модель процессов,,
происходящих в природе, в условиях точно контролируемых
опытов.
Такая методика давала возможность использовать в опыте
естественный растительный субстрат в виде лесной подстилки,,
а также обеспечивала длительное проведение опытов в течение
3—6 месяцев, при поддержании
определенного режима
влажности, близкого к природному, и.
удалении продуктов распада.
Для этой цели Частухин
сконструировал специальные
колбы со сливными трубками для
выращивания культур грибов на
растительном опаде (рис. 52).
Колбы такой конструкции мо-
__\sJI Рис. 52. Колбы со сливными трубкам»
В. Я. Частухина:
2 1 — общий вид колбы; 2 — колпачки:
для стерильной смены жидкости
гут неограниченно долго сохранять стерильность, обеспечивая
периодическую подачу воды со дна колбы через соединение
колокольчатых запоров одних колб с каучуковыми трубками других.
В колбах создается определенный режим: поступление влаги:
162
происходит снизу, а подсыхание подстилки начинается, как в
лрироде, с верхних слоев.
Колбы для выращивания загружали растительным опадом
(4—5 г листьев на колбу) и в таком виде стерилизовали. Затем
каждую из них заражали испытуемой культурой гриба.
Периодически, раз в месяц, через сливные трубки брали образцы
продуктов распада и осуществляли стерильную подачу воды.
В опыте использовали микроскопические грибы, выделенные
из различных слоев подстилки: виды Alternaria, Cladosporium —
из верхнего слоя, Penicillium, Trichoderma — из нижнего, Мисог
и Mycogone — из гумусированного, а также мезоскопические
грибы Mycosphaerella punktiformis, Venturia tremulae и
макроскопические базидиальные — Collybia dryophila, Clitocybe (несколько
видов), Marasmius androsaceus, Phallus impudicus, Lepiota pro-
cera. Все виды типичны для подстилок дубрав.
Микроскопические грибы дают очень скудный рост на
растительном опаде. К концу опыта существенных изменений в
строении листьев не наблюдалось. Листья несколько темнели, но не
теряли механической прочности.
Мезоскопические грибы не росли вовсе. Видимо, эти грибы
ведут паразитический образ жизни на растениях. Базидиальные
грибы росли очень активно на растительном опаде, вызывали
поселение листьев и потерю их механической прочности. Сперва
исчезали наименее стойкие части ткани, в основном содержимое
клеток, межклеточные пластинки и др. Сосудисто-волокнистые
лучки разрушались в последнюю очередь.
Под влиянием высшего базидиального гриба Collybia driophila
листья дуба почти полностью разложились за 8 месяцев.
Наиболее энергично разлагают растительный опад высшие
базидиальные грибы. Разложение ими растительных остатков
осуществляется более чем на 80% (табл. 14). Значительно слабее
несовершенные грибы, и очень слабо осуществляется этот процесс
бактериями. Остаток веществ после воздействия грибов
представляет сложную массу, состоящую из неразложившихся элементов
листа, нерастворимых продуктов жизнедеятельности гриба и
грибного мицелия. При воздействии на опад микромицетов в остатке
превалируют ткани листа, при воздействии базидиомицетов
значительная масса оставшегося вещества приходится на долю
грибного мицелия. Таким образом, базидиальные грибы —
подстилочные сапротрофы — вызывают почти полную минерализацию
листьев и представляют собой поэтому основных агентов
разложения опада в лесах, разлагая его лигноцеллюлозный
комплекс.
По характеру воздействия на опад выделяют три группы
подстилочных сапротрофов.
I группа — грибы, преимущественно разлагающие клетчатку,—
Lepiota procera.
II группа — грибы, преимущественно разлагающие лигнин,—
Phallus impudicus.
<6*
163
Ill группа — грибы, осуществляющие смешанное
воздействие, — Coilybia, Marasmius, Clitocybe.
Хвойный опад разлагается медленнее, чем лиственный, так как
он значительно беднее основаниями и легкоподвижными
органическими соединениями и содержит смолы. Особенно большое
значение в процессе разложения имеет содержание азота в опаде.
Чем выше содержание азота и ниже соотношение С: N, тем
интенсивнее идет процесс разложения.
Мицелий подстилочных сапротрофов из базидиальных грибов,
осуществляющих процесс разложения растительного опада, мо-
Та блица 14
Интенсивность разложения опада разными культурами
грибов на листьях дуба и клена
Группы
Базидиальные грибы
Несовершенные грибы
Бактерии
Виды грибов
Ctytocybe flaccida
С. nebularis
Phallus impudicus
Lepiota procera
Coilybia dryophila
Marasmius androsaceus
Cladosporium herbarum
Mycogone nigra
Trichoderma lignorum
PeniciIlium sp.
Cytophaga hutchinsonii
% потери в весе
62,39
53,23
73,84
76,10
82,20
55,04
11,63
15,35
10,58
18,28
9,50
КОНТРОЛЬ 20,0
жет переносить длительное высыхание в течение 9—10 месяцев,,
не теряя ферментативной активности. Эта биологическая
особенность обеспечивает им устойчивость к недостатку влаги и
сохранение при этом ферментативной активности, что очень важно в*
природных условиях и увеличивает их значение в процессах
распада.
Г. Линдбергом были проведены опыты по определению
интенсивности разложения клетчатки и лигнина подстилочными сапро-
трофами, принадлежащими к различным видам Marasmius,
непосредственно в растительном опаде с точным учетом исходного и
конечного содержания этих продуктов в субстрате. Почти все
виды Marasmius оказались способными разлагать лигнин, но
интенсивность разложения лигнина у разных видов различна. Также
было различным отношение количеств разложенных лигнина и
клетчатки. Таким образом, внутри рода Marasmius можно выде-
164
лить группы, преимущественно разлагающие лигнин, клетчатку
или оба компонента вместе.
В целом большинство видов Marasmius разлагает 48—59%
лигнина. Очевидно, виды Marasmius, часто и в большом
количестве встречающиеся в хвойных и лиственных лесах, занимают
значительное место в разложении клетчатки и лигнина.
Грибы осуществляют также активное разложение древесины,
разложение древесины начинается еще в растущем дереве под
влиянием паразитных грибов. Сначала происходит изменение
окраски, вызываемое деревоокрашивающими грибами, —
сумчатыми и различными несовершенными, таким, как Ceratostomella,
Cladosporium, и некоторыми другими (серо-синее и коричневое
окрашивание). Розовую окраску вызывают Fusarium и Penicil-
lium. Истинными разрушителями древесины, ее лигноцеллюлоз-
ного комплекса являются базидиальные грибы — возбудители
бурой и белой гнили древесины. Возбудители бурой гнили
превращают древесину в красновато-коричневую массу, разрушая
преимущественно ее целлюлозные и гемицеллюлозные
компоненты, почти не действуя на лигнин (деструктивное разложение). Это
грибы рода Coniophora, Daedalia quercina, Polyporus squamosus.
Возбудители белой гнили разрушают древесину, превращая
ее в белую массу. Они действуют в первую очередь на лигнин и
почти не затрагивают целлюлозу (коррозионное разложение).
Это грибы Pleurotus ostreatus, Poria vaporaria, Forties
fomentarius. Разложение лигнина грибами белой гнили — это
окислительный процесс, включающий деметилирование, образование
дифенолов, расщепление их под действием диоксигеназ. Кроме
того, грибы белой гнили способны расщеплять ароматические
кольца лигнина, связанные в лигниновый полимер.
Грибы, разлагающие и целлюлозу и лигнин, в равной степени
объединены в третью группу грибов мягкой гнили.
Лигноцеллюлоза обычно быстрее разлагается, чем чистый
лигнин. Это объясняется тем, что целлюлоза ускоряет деградацию
лигнина, являясь дополнительным источником энергии. Кроме
того, у грибов белой гнили обнаружен комплексный фермент —
целлобиозо: хинон-оксидоредуктаза, который, с одной стороны,
восстанавливает хиноны в фенолы, так как хиноны сильно
препятствуют расщеплению ароматического кольца, таким образом
это препятствие устраняется, с другой стороны, окисляют целло-
биозу — продукт разложения клетчатки.
Грибы, разрушающие твердую древесину, характеризуются
очень эффективным использованием связанного азота, которого
в древесине очень мало. Отношение С : N в древесине может быть
350: 1 и даже 1250: 1.
Участвуя в разложении многих углеродсодержащих веществ
растительного опада и древесины в первую очередь труднораз-
лагаемых полимерных соединений, где грибам принадлежит
ведущая роль, они занимают значительное место в круговороте
углерода, являясь поставщиками С02 в атмосферу.
165
Среди грибов есть организмы, разлагающие жиры и воска,
входящие в состав растительных и животных тканей. Это
определяется наличием у них ферментов липаз. Наибольшей
активностью липолитических ферментов обладают виды Mucor lipoly-
ticus, Rhizopus nigricans, Aspergillus niger, Penicillium verrucosum
var, cyrfopium, P. roquefortii. Многие выделены с поверхности
растений, являются эпифитами и способны разлагать также
восковые налеты на поверхности растений.
Известна также способность грибов разлагать как
алифатические, так и ароматические углеводороды. В этом
отношении наибольшей активностью характеризуются грибы рода As-
pergillus.
РОЛЬ ГРИБОВ В ПОДЗОЛООБРАЗОВАТЕЛЬНОМ
ПРОЦЕССЕ
При разложении растительного опада грибы
утилизируют все основные компоненты растительной клетки, начиная от
простых углеводов и белков и кончая трудноусвояемыми
клетчаткой и лигнином. Разложение этих веществ приводит к
образованию разнообразных органических кислот цикла Кребса —
щавелевая, лимонная, яблочная, фумаровая и др.
В результате разложения лигнина образуются органические
кислоты — производные фенолов, такие, как ванилиновая, сирин-
говая, параоксибензойная и др.
Характерной особенностью обмена веществ у грибов является
то, что эти кислоты часто не вовлекаются далее полностью в
обмен, а в значительном количестве могут накапливаться в
окружающей среде. При определенных условиях органические кислоты
могут составлять 40—50% от исходного органического вещества,
используемого грибом.
Со способностью грибов образовывать и накапливать
органические кислоты при разложении растительного опада связывают
их роль в процессах подзолообразования. Эти представления
нашли свое развитие в работах Т. В. Аристовской. Она исходит из
того, что подзолистые почвы, формируясь под хвойной
растительностью, опад которой беден минеральными элементами,
характеризуются слабой обеспеченностью необходимыми питательными
веществами, высоким содержанием в опаде трудноразлагаемых
органических соединений и наличием бактерицидных веществ, а
также высокой кислотностью. Они характеризуются
относительно бедным набором микроорганизмов, в составе которого большое
место занимают грибы.
В подзолистых почвах представлены как микроскопические
мукоровые, сумчатые, несовершенные грибы, так и базидиальные,
мицелий которых обильно пронизывает лесную подстилку.
Соотношение грибов и бактерий в подзолистых почвах по их биомассе
выражается как 90: 10.
166
Характерная особенность микроорганизмов подзолистых почв —
способность использовать источники углерода, присутствующие в
очень низких концентрациях: например, Mortierella ramanniana
имеет максимальную радиальную скорость роста при
концентрации глюкозы 0,005%, a Penicillium daliae — в пределах от 0,01
до 0,1%. Им свойственна высокая эффективность использования
субстрата и, возможно, способность потреблять специфические
вещества гумуса. Все эти особенности есть результат
приспособления к тем специфическим условиям, которые создаются в
подзолистых почвах.
Эти черты в значительной мере присущи грибам, обитающим
в них. Они могут развиваться на чрезвычайно бедных
питательных субстратах, и, следовательно, усваивать элементы питания
из рассеянного состояния, хотя и не проявляют заметной олиго-
трофности, т. е. высокие концентрации не действуют на них
угнетающе, так как они могут существовать в широком диапазоне
концентрации субстрата и осмотического давления.
Вторая особенность грибов, обитающих в подзолистых почвах,
это то, что особенно активное образование ими органических
кислот (лимонной, щавелевой, фумаровой) происходит при
наибольшем дефиците минеральных элементов. Образование кислоты
в этих условиях в 5—8 раз превышает ее образование в
нормальных условиях обеспеченности минеральными элементами.
Особенно отчетливо эта закономерность проявляется у грибов рода
Penicillium, составляющих многочисленную группу грибов-микроми-
цетов в подзолистых почвах (табл. 15). Это имеет адаптивное
значение и связано с постоянной конкуренцией за минеральное
питание в условиях подзолистых почв. Образующаяся кислота
воздействует на минералы породы, что приводит к переходу
отдельных зольных элементов породы в растворимое состояние.
Тем самым грибы осуществляют процессы выветривания
минералов, что имеет большое значение в процессах
почвообразования.
Процессы выветривания минералов под действием
органических кислот и других подобных продуктов обмена грибов и
бактерий, таких, как полисахариды и другие, осуществляются
интенсивно, так как эти соединения способны образовывать хелаты с
элементами, входящими в состав минеральной породы, и тем
способствовать быстрейшему выветриванию. В виде таких
комплексных соединений происходит перемещение минеральных элементов
в нижележащие слои почвы, где формируется горизонт вмыва-
ния — иллювиальный горизонт. Воздействие грибов на минералы
нашло экспериментальное подтверждение в работах Т. В. Ари-
стовской и Р. С. Кутузовой по разложению нефелина культурой
Penicillium notatum. В результате разложения нефелина
продуктами метаболизма этого гриба соотношение K20/Si02 в
растворившихся продуктах распада возросло в 8. раз по сравнению с
соотношением в исходном минерале. Na20/Si02 увеличилось в
4 раза, a Al203/Si02 — в 5 раз. Следовательно, калий и алюми-
167
ний, перешедшие в растворимое состояние, могут быть вынесены
в нижележащие горизонты.
Исходя из общей схемы Аристовской можно следующим
образом представить роль грибов в подзолообразовательном процессе.
Первоначально идет интенсивное развитие мицелия грибов на
растительном опаде и накопление биомассы за счет разложения
растительного материала. Затем, по мере обеззоливания
разлагающегося материала за счет потребления минеральных веществ
микроорганизмами и частичного их вымывания в нижележащие
горизонты, развитие мицелия замедляется, но усиливается про-
Т а белица 15
Зависимость роста и кислотообразующей способности грибов*
от условий зольного питания (по Т. В. Аристовской, 1965)
Penicilliutn sp.
1
2
3
4
5
6
7
8
9
10
И
12
Mucor sp.
13
14
Количество сухого мицелия,
... мг/мл среды
полная
минеральная среда
20,0
4,0
4,4
2,36
1,92
0,60
0,52
0,52
0,68
1,16
1,06
1,53
0,64
1,60
обедненная среда
с пониженным
содержанием
минеральных элементов
1,6
0,16
0,36
0,12
0,12
0,10
0,09
0,08
0,12
0,04
0,18
0,27
0,12
Количество кислоты, выделенное
[ на 1 г сухого мицелия, мл 0,02 н.
полная
минеральная среда
235
250
230
190
165
следы
905
410
425
345
20
30
255
155
обедненная
среда
500
805
1625
1560
1290
2000
860
905
1150
2000
140
180
следы
следы
* Грибы выделены из подзолистой почвы.
цесс кислотообразования, который влияет на почвенные
минералы, способствует освобождению зольных элементов, увеличивает
их запас. Это, с одной стороны, обеспечивает возможность
дальнейшего развития микроорганизмов, с другой — происходит вынос
этих веществ, часто в виде органоминеральных комплексов и
отложенные в иллювиальных горизонтах, что и характеризует собой
процесс подзолообразования.
Глава 7
РОЛЬ ГРИБОВ В ПРОЦЕССАХ ПРЕВРАЩЕНИЯ
АЗОТА
Роль грибов в превращении азота в природе менее
многообразна, чем бактерий, хотя они используют, и превращают
в процессе жизнедеятельности различные соединения азота.
Таковыми являются различные формы органического азота и
неорганический азот в виде солей аммония и нитратов. Существовавшие
некоторое время представления о способности грибов, в первую
очередь эндотрофных микоризообразователей, фиксировать
атмосферный азот в настоящее время не подтвердились. Более того,
установлено,' что способностью фиксировать азот обладают
только прокариотные организмы. Эукариотные организмы лишены этой
способности.
Все грибы без исключения используют и превращают
органические формы азота, почти все, за небольшими исключениями,
ассимилируют аммонийный азот и несколько меньший набор
видов используют нитратный азот.
Ассимиляция нитратного азота. Нитратный азот ассимилируют
некоторые представители из класса Chytridiomycetes, сумчатые
грибы, среди них представители родов Chaetomium, Gymnoascus,
небольшое число базидиальных грибов — Armiliariella, Lentinus,
многие несовершенные грибы. Не используют нитратный азот
многие мукоровые грибы, большинство базидиальных грибов,
некоторые немногие сумчатые грибы, такие, как Ceratostomella и др.
Усвоение нитратного азота грибами представляет собой
универсальный процесс, характерный для растений и
микроорганизмов. Он проходит через восстановление нитратного азота по
следующей схеме при участии соответствующих ферментов: нитрат-
редуктазы, нитритредуктазы, гидроксиламинредуктазы.
HNO нитРатРедуктазы > HNO нитРитРедуктазы >nh ОН >
гидроксиламинредуктаэы^ын^аминоксилоты^белок<
Таким образом, способность к ассимиляции нитратов
объясняется наличием у грибов так же, как и у растений,
соответствующих ферментов на каждом этапе восстановления нитратов. В
качестве нитратредуктазы выступает никотинамидадениндинуклео-
тидфосфат (НАДФН), являющийся донором водорода,
передающий водород через флавиновые ферменты, содержащие молибден,
железо и медь.
Вследствие наличия этих ферментов некоторые сумчатые и
несовершенные грибы могут расти на чистой азотной кислоте.
В частности, Aspergillus при концентрации НЫОз 0,015 моля
использует ее на 90%. Процесс поглощения нитратов и их *
восстановления в организме носит название ассимиляторной дени-
трификации. К дисимиляторной денитрификации во внешней сре-
169
де, приводящей к высвобождению молекулярного азота в отличие
от бактерий грибы не способны.
Усвоение аммонийного азота. Азот аммонийных солей
усваивают почти все грибы за небольшими исключениями,
относящимися к некоторым видам сумчатых и базидиальных грибов.
Обычно грибы, использующие N03~, всегда усваивают NH4+, но
не наоборот. При наличии в среде соли NH4N03 сначала
потребляется ион NH4+ и только после его исчерпания начинается
потребление N03~. Это объясняется тем, что ион аммония подавляет
активность нитратредуктазы и тем самым возможность усвоения
иона N03~. Аммиак, поглощенный грибом в виде ионов аммония,
вступает в реакцию с кетокислотами, образует аминокислоты.
Такой путь синтеза аминокислот является основным как для
грибов, так и для растений. Например, реагируя под действием
фермента аланиндегидрогеназы с пировиноградной кислотой, аммиак
образует аминокислоту — аланин NH3+CH2CO—СООН2 + 2Н^:
^CH3CHNH2—СООН + Н20. Очень легко аммиак реагирует с
а-кетоглютаровой кислотой при участии фермента глютаматдегид-
рогеназы, в результате чего образуется глютаминовая кислота:
НООС-СН2—СН2—СО—СООН + NH3 + 2Н ^ НООС—СН2—
—СН2—CHNH2—СООН + Н20.
Превращение органического азота
Процессы аммонификации. Все грибы без исключения
используют органический азот или в виде белков и протеидов,
или отдельных аминокислот, тем самым активно участвуя в
разложении белков и входящих в их состав соединений. Они
обладают активными протеолитическими ферментами, действующими
на пептидные связи в молекуле белка, в результате чего
освобождаются отдельные аминокислоты. Под действием ферментов
грибов этот процесс может идти и дальше до образования конечных
продуктов распада белка, каковыми являются аммиак, С02,
сульфаты, вода и др. Процесс минерализации органических
соединений с образованием аммиака носит название
аммонификации.
Аминокислоты могут ассимилироваться грибами, включаясь
в их конструктивный обмен, или могут быть разрушены до
конечных продуктов.
Есть виды грибов, которые нуждаются в определенных
аминокислотах. К ним относятся некоторые сумчатые грибы и многие
базидиальные, особенно микоризообразователи. Например, гриб
Сепососсит graniforme нуждается в гистидине, Trichophyton
spp. — в* гистидине и аргинине. Есть аминокислоты, которые
включаются в обмен в неизменном состоянии, это так называемые
«первичные» аминокислоты. Другие первоначально подвергаются
дезаминированию.
Дезаминирование у грибов может идти теми же путями, что
и у других организмов:
170
а) дезаминирование путем гидроксилирования с образованием
оксикислот и аммиака:
R-CH (NHJ-COOH^-^R-CH (OH)-COOH + NH3;
б) окислительное дезаминирование с образованием кетокислот
и аммиака:
2R—СН (NH2)-COOH-^2R— CO-COOH + 2NH3;
в) дезаминирование с одновременным декарбоксилированием:
R-CH (NH2)-COOH-^-^R-CH2OH + С02 + NH3.
Грибам свойственна также универсальная способность к пёреами-
нированию аминокислот. Этот процесс установлен для Neurospora
crassa и других грибов. Переаминирование происходит в
результате взаимодействия между аминокислотами и кетокислотами по
следующей схеме:
R'—CNH2—СООН + R"—СО-СООН =
= R'—СО—СООН + R"—CHNH2—COOH.
Например, между а-кетоглютаровой кислотой и глютаминовой,
между пировиноградной и аланином.
Использование органического азота, в частности аминокислот,
идет в двух направлениях: с одной стороны, он непосредственно
используется как конструктивный материал" в синтетических
процессах при наличии дополнительного источника углерода. С
другой стороны, в качестве конструктивного и энергетического
материала, когда он представляет собой единственный органический
субстрат. В этом случае идет дезаминирование аминокислот и
грибы проявляют себя как организмы-аммонификаторы,
осуществляя процесс аммонификации, так как при этом идет окисление
субстрата с образованием большого количества аммиака, который
выделяется во внешнюю среду.
Процесс аммонификации осуществляют многие грибы, но
наиболее активны в этом отношении грибы порядка Mucorales, а
также представители родов Aspergillus, Trichoderma и некоторые
другие.
Избыточный аммиак, образующийся в процессе дезаминиро-
вания аминокислот в мицелии, обезвреживается либо путем
связывания, либо путем образования мочевины. В последнем
случае мочевина играет такую же роль в грибах, как амиды —
аспарагин и глютамин в растениях. Она сохраняется в мицелии
в качестве резервного продукта в случае недостатка углеводов и
снова вовлекается в обмен при участии уреазы в результате
дополнительного поступления углеводов.
Мочевина в грибах может образовываться при
гидролитическом распаде аргинина под действием фермента аргиназы. При
этом образуются орнитин и мочевина:
171
V:hn(ch2)3—chnh,cooh+ ?е=^
nh/
24>c0 + nh2 + (ch2)3—chnh2-cooh
nh/
мочевина орнитин
Особенно много мочевины содержится в высших базидиальных
грибах. По данным Н. Н. Иванова, ее содержание в мицелии
шампиньона составляет 13,2%, дождевика — 10,7%.
Многие грибы активно разлагают мочевину, так как они
обладают ферментом уреазой. Разложение мочевины приводит к
образованию аммиака и углекислоты:
мн
2\С0 =—™^-а-^2Ш3 + С02
NH2/ н*°
Образовавшийся аммиак используется при синтезе белка или
выделяется в свободном виде, характеризуя участие грибов в
процессе аммонификации.
Участие грибов в процессах нитрификации. До последнего
времени считалось, что окисление аммиака до нитратов и
нитритов осуществляется только хемоавтотрофными микроорганизмами.
В настоящее время установлено, что многие гетеротрофные
микроорганизмы могут окислять восстановленные соединения азота
(аммиак, аминный азот, гидроксиламин и т. д.) и большое
количество органических соединений. Среди этих организмов
значительное место занимают грибы, осуществляющие процесс
нитрификации: Наиболее хорошо изучены в этом отношении некоторые
виды рода Aspergillus, такие, как Aspergillus flavus и Л. wentii
и некоторые другие. Этот процесс проявляется в окислении
азотистых соединений, в том числе солей аммония до нитритов и
нитратов при участии ферментов — дегидрогеназ НАД, ФАД и
цитохромов. Весь процесс осуществляется одним грибом с начала
и до конца без разделения на две стадии, как это имеет место у
бактерий, по следующей схеме:
NH4OH^NH2OH-^HN02->HN03
Однако этот процесс не используется грибами как
энергетический и наступает обычно после периода активного размножения
культуры. Он протекает как побочный процесс параллельно с
окислением органического вещества при наличии дополнительного
энергетического субстрата и носит название гетеротрофной
нитрификации. Процесс гетеротрофной нитрификации может
осуществляться в среде, содержащей пептон, аминокислоты,
аммонийные соли.
172
В природе он происходит везде, где аммиак образуется в
условиях высокого содержания органического вещества.
Возможно окисление грибами этих соединений и не до
конечных продуктов нитратов и нитритов. Процесс окисления может
заканчиваться образованием гидроксиламина, гидроксимовых
соединений, аминооксидов, нитрозо- и нитросоединений. Хотя ни одно
из этих соединений не образуется в больших количествах, но
такие химически активные вещества, как гидроксиламины и нитро-
зосоединения, обладают высокой биологической активностью и
токсичностью в очень малых концентрациях. Поэтому значение
гетеротрофной нитрификации следует оценивать не по
количественной значимости в валовом превращении азота в природе, а по
потенциальной роли этого процесса в экологии микроорганизмов
л его влиянию на общее состояние внешней среды.
Некоторые продукты гетеротрофной нитрификации играют
роль факторов роста, например гидроксамовые кислоты.
Некоторые токсические и даже мутагенные ее продукты, например
нитрозосоединения, возможно играют роль в конкуренции
микроорганизмов.
Глава 8
ПИГМЕНТЫ ГРИБОВ И ИХ РОЛЬ
В ПРОЦЕССАХ ГУМУСООБРАЗОВАНИЯ
Основные группы пигментов как классов химических
соединений, обнаруженных у грибов, представлены каротиноида-
лли, хинонами и меланинами. Все грибы лишены хлорофилла, и у
лих отсутствуют также пигменты-флавоноиды (антоцианы, фла-
воны, флавонолы и др.).
ПИГМЕНТЫ-КАРОТИНОИДЫ
Каротиноидные пигменты представляют собой
ненасыщенные алифатические соединения терпенового ряда,
производные изопрена. Характерный признак каротиноидов — большое
число сопряженных двойных связей, что объединяет их в группу
полиенов и обусловливает характерные спектры поглощения в
видимой области. Большое число двойных связей придает им
неустойчивый характер.
Известны две основные группы каротиноидов: каротиноиды-
углеводороды или собственно каротины и кислородсодержащие
соединения. Примерами каротиноидов-углеводородов являются а-,
р-, ^-каР0ТИНы и ликопин. Примерами кислородсодержащих
каротиноидов могут быть торулородин и нейроспорин.
У грибов преимущественно содержатся каротиноиды-углеводо-
роды. Есть каротиноиды с карбоксильными группами, которые
встречаются только у дрожжей, но совершенно отсутствуют каро-
173
тиноиды, содержащие метоксильные и эпоксигруппы. В этом
смысле каротиноиды имеют определенное систематическое
значение и служат характеристикой таксонов микроорганизмов. Каро-
тиноидные пигменты образуют целый ряд грибов, принадлежащих
к различным систематическим группам. В частности, они
образуются низшими грибами классов Chytridiomycetes, Zygomycetes, a
также у высших грибов классов Ascomycetes, Basidiomycetes,
Fungi imperfecti.
Так, они есть в классе хитриодиомицетов в порядке Chytridi-
ales у Rhizophlyctes rosea, в порядке Blastocladiales того же
класса каротиноидные пигменты образуют многочисленные
представители рода Allomyces. У вида A. javanicus каротиноидные
пигменты были обнаружены впервые у грибов. Очень многообразны
каротиноидные пигменты у представителей класса зигомицетов,
В порядке Mucorales этого класса они присутствуют у Phycomy-
ces blakesleanus преимущественно в виде р-каротина. Этот гриб
давно стал излюбленным объектом для изучения каротиноидов,
так как в виде чистого соединения из гриба р-каротин впервые
был выделен из этого объекта. Каротиноидные пигменты есть у
Mucor hiemalis различных видов Choanephora, в частности С. tri-
spora (Blakeslea trispora), которая используется для
промышленного получения каротина, а также целого ряда других
представителей этого класса.
Среди высших грибов каротиноиды есть в классе аскомицетов
у представителей порядков Taphrinales и Protomycetales в
группах пиреномицетов и дискомицетов у родов Protomyces, Neuro-
spora, Sarcoscypha и др. В классе базидиомицетов каротиноиды
имеются у ржавчинных грибов и у некоторых гименомицетов.
Каротиноидные пигменты широко представлены у многих
несовершенных грибов, у светлоокрашенных родов Acremonium, Tricho-
thecium, Arthrobotrys, Fusarium, Verticillium, семейства Moniliaceae
и некоторых других. У темноокрашенных несовершенных грибов
в семействе Dematiaceae они обнаружены у родов Epicoccum, Cla-
dosporium, Stemphylium, Curvularia, Bipolaris, Helminthosporiutn
и присутствуют как дополнительная фракция к темным пигментам.
Функции каротиноидов у грибов разнообразны, они принимают
участие в фотореакциях, связанных с фототропизмом грибов и
фототаксисом подвижных клеток, в процессах репродукции
некоторых грибов, в защите клеток от повреждения, вызываемого
поглощением видимого света, вследствие фотоокисления.
Каротиноиды в связи с их высокой реакционной способностью могут
блокировать сильные окислители.
ПИГМЕНТЫ-ХИНОНЫ
Пигменты-хиноны — многочисленная группа
соединений, разнообразно окрашенных веществ, включающих бензохино-
ны, нафтахиноны, антрахиноны, фенантрохиноны и др. Пигменты-
хиноны широко представлены у грибов. К ним, в частности, отно-
174
сятся многие желтые пигменты некаротиноидной природы.
Окраска хинонов разнообразна. Они образуют широкую гамму цветных
оттенков от бледно-желтого (тектохинон) до почти черного (те-
лефоровая кислота), что связано со структурными изменениями
хинонной молекулы по числу колец от простых бензохинонов до
полициклических соединений. Однако присутствие хинонов не
всегда бывает заметно у грибов, так как эти соединения так же, как
и каротиноиды, обычно бывают замаскированы такими
соединениями, как меланины. ^
Многие хиноны часто меняют окраску в зависимости от рН,
являясь таким образом индикаторами. Наибольшее число
исследований по хинонам грибов посвящено родам Penicillium,
Aspergillus и Fusarium. У этих родов найдены три типа хинонов (бен-
зо-, нафта- и антрахиноны). Бензохиноны в большинстве случаев
метаболиты Penicillium и Aspergillus, представляют собой
желтые, фиолетовые и красные пигменты. Типичный пример
бензохинонов — фумигатин, образуемый A. fumigatus.
Нафтахиноны реже встречаются в грибах. Они в небольшом
количестве образуются грибами рода Penicillium, Aspergillus и
наиболее типичны для Fusarium, например нафтахиноны фуза-
рубин и яваницин у F. solanL
В грибах найдены разнообразные антрахиноны с различным
числом гидроксильных групп и других заместителей, чаще всего
производные эмодина и хризофановой кислоты. Эти два вещества
свойственны как высшим растениям, так и грибам. Некоторые
хиноны, например исландицин и скирин, встречаются в грибах,
но не найдены в высших растениях. Антрахиноны обнаружены
главным образом у представителей порядков Hyphomycetales,
Sphaeropsidales у их темноокрашенных форм, таких, как Alterna-
ria solani, Cladosporium fulvum, родов Curvularia, Helminthospo-
rium и других представителей класса несовершенных грибов.
Хиноны обладают антибиотическим действием (бактериостати-
стическим и бактерицидным). Они участвуют в процессе переноса
электронов. Витамины К и Е также представляют собой хиноны,
убихиноны и пластохиноны участвуют в процессах дыхания.
ТЕМНЫЕ ПИГМЕНТЫ-МЕЛАНИНЫ
Наибольшее значение в почвенных процессах имеют
темные пигменты грибов типа меланинов, которые занимают
особое положение среди прочих пигментов грибов. Это черные или
коричневые полимерные соединения, которые присущи
организмам разного филогенетического уровня и разнообразным
таксономическим группам грибов. Пигмент отлагается в клеточной
стенке вегетативного мицелия и в репродуктивных органах. На
основании наличия темных пигментов в мицелии или
репродуктивных органах выделяется целое семейство несовершенных
грибов — семейство Dematiaceae порядка Hyphomycetales. Кроме
видов этого семейства темным пигментом обладают представители
175
семейства Stilbaceae и других порядков несовершенных грибов:
Melanconiales и Sphaeropsidales, многие сумчатые грибы,
особенно их плодовые тела и споры, а также базидиальные. Темные
пигменты содержатся в зиготах многих мукоровых грибов и
относятся к истинным меланинам.
К истинным меланинам относят высокополимерные
соединения с молекулярным весом несколько тысяч и десятков тысяч,,
образующиеся при ферментативном окислении фенолов, главным
образом пирокатехина и тирозина, и последующей окислительной
полимеризации этих соединений. Таким образом, меланины могут
быть азотсодержащими и безазотистыми соединениями.
Поскольку в образовании меланинов участвуют разнородные
по строению исходные мономеры без строгой закономерности в
их чередовании, строение меланинов весьма сложно и
разнообразно. Элементный состав вещества в какой-то степени
характеризует его химическое строение. Однако для меланинов эта
характеристика довольно условна, так как элементный состав
отдельных меланинов сильно варьирует из-за большой неоднородности
данной группы веществ. Тем не менее при изучении элементного
состава меланинов разных групп организмов выявилась
определенная закономерность: меланины животных всегда
азотсодержащие соединения, а растительные меланины, или фитомеланины,
либо полностью лишены азота, либо содержат его в
незначительных количествах. Так, если в меланинах животных содержание
азота колеблется от 5 до 8%, то в фитомеланинах оно составляет
0,3—0,7% или азот полностью отсутствует.
Меланины грибов занимают в этом отношении промежуточное
положение. Они содержат в своем составе азот, в том числе и
негидролизуемый, но содержание общего азота составляет от I
до 4,5%; в редких случаях оно достигает 8%. Они характеризу-
Та блиц а 16
Элементный состав меланинов различного происхождения
Животные
Растения
Грибы
Объекты
пигмент глаза
пигмент шерсти
Sepia officinalis
Helianthus annuus
Citiullus vulgaris
Ustilago maydis-
Rhizoctonia solani
Nadsoniella nigra
Cladosporiurn cladospori6idesr
Uloclodium botrytis *
C, %
60,4
61,2
60,8
65,5
63,5
62,7
62,0
54,2
60,7
43,9
H, %
4,6
4,0
3,4
4,1
4,1
3,4
4,8
5,6
4,0
5,1
N. %
8,7
5,1
8,5
0,7
0,7
1,0
3,0
2,9
2,2
з,а
176
ются также большими колебаниями водорода (3-— 7%) и
особенно углерода (45—63%).
Табл. 16 характеризует элементный состав меланинов.
В грибной клетке меланины обычно присутствуют в связанном
с белками состоянии и представляют собой хромопротеиды (ме-
ланопротеиды). Кроме того, они прочно связаны со структурными
элементами клетки, в частности образуют устойчивый комплекс:
с хитином.
Меланины в клетке микроорганизмов, в том числе и грибной,,
играют защитную роль. Они позволяют меланизированным
организмам существовать в экстремальных условиях, повышенной
инсоляции, сухости и высокой температуры, а также препятствуют
лизису мицелия этих организмов.
РОЛЬ ГРИБОВ В ГУМУСООБРАЗОВАНИИ
Изучение гуминовых кислот почвы развивалось
независимым путем от изучения меланинов. Лишь после накопления
достаточного количества экспериментальных данных постепенна
становилось ясным, что между этими двумя группами соединений
существует большое сходство.
На возможность участия темноокрашенных продуктов
метаболизма микробов в образовании почвенного гумуса указывали
еще П. А. Костычев, М. Бейеринк, В. Л. Омелянский, С. Н. Вино-
градский. Дальнейшие работы подтвердили это положение.
Почвенный гумус представляет собой смесь многих
органических соединений, как специфической, так и неспецифической
природы. Неспецифические вещества гумуса представлены
углеводами, белками, жирами, органическими кислотами и др.
Специфические вещества составляют 90% гумуса и представлены двумя
типами полимеров: гуминовыми и фульвокислотами.
Гуминовые кислоты, составляющие 50—80% почвенного
гумуса, представляют собой полимерные высокомолекулярные
соединения, состоящие из фенольных и индольных единиц, связанные
с аминокислотами, пептидами и другими органическими
соединениями.
Химическое строение их, как и многих полимеров, до конца
не расшифровано, так как последовательность этих
циклических соединений может быть различна в каждом отдельном
случае.
Долгое время существовало мнение о том, что гуминовые
кислоты образуются только в результате участия лигнина,
содержащего фенольные мономеры, которые могут быть окислены в хи-
ноны, образуя темноокрашенные продукты. Основоположником
этой теории был С, Ваксман. Предполагалось, что эти фенольные
мономеры и даже полимеры целиком входят в состав гумуса,
образуя лигнопротеиновый комплекс с аминокислотами и
пептидами, т. е. обязательным для образования гумуса был лигнин.
Предполагалось, что ароматическое ядро лигнина, не претерпе-
17Г
вая изменений, сохраняя основную структуру, входит в состав
гуминовой кислоты.
Исследования по физиологии грибов, в частности изучение
путей их метаболизма, показали, что грибы могут образовывать
самые разнообразные циклические соединения из соединений
прямого ряда. Основные пути образования грибами циклических
соединений следующие.
1. Образование койевой (кодзиевой) кислоты из двух триоз:
диоксиацетона или глицеринового альгедида в процессе
анаэробного гликолиза Сахаров в результате дегидрирования и
дегидратирования:
СН2ОН СН2ОН СН2ОН
I I
снон снон -> j:—оч
А
п + I о нс\ ^сн
\Н 4i
О ОН
2. Конденсация четырех молекул уксусной кислоты при
участии ацетилкоэнзима А с образованием 6-метилсалициловой
кислоты:
С СО
Чсо { со у°°н ^оон
с с >• j^V-oh—>н,с/\-он
\ I ..
со ч/ \/
он
ацетатные единицы у 2,4-диоксибензойная 6-метилсалициловая
кислота
3. Через образование шикимовой кислоты (это основной путь
образования циклических соединений у многих высших и низших
организмов). Шикимовая кислота в свою очередь может
подвергаться дальнейшим превращениям, в результате чего в продуктах
жизнедеятельности грибов образуются многие другие фенольные
соединения, такие, как орселлиновая, крезорселлиновая,
ванилиновая, феруловая кислоты и т. д. Некоторые грибы в процессе
жизнедеятельности образуют до 45 различных фенолов.
Образование циклических соединений через шикимовую
кислоту идет по следующей схеме:
178
со2н
1
С—0—Р
|СН2
фосфоэнол-
пируват
Шикиматный
Н—С=0
|
+ Н—С—ОН
1
Н—С—ОН
СН20-Р
эритрозо-4-фосфат
путь
-
биосинтеза
С02Н
С=0
1
сн2
но-с—н
н —с—он
1
н —с—он
СН20-Р
седогептулеза-7-фосфат
Затем происходит альдольная конденсация.
HON /ОН
дегидрохинная
кислота
Н/ ЧС02Н
и/"
дегидрошикимовая
кислота
Ш
)Н
. м
НО
шикимовая
кислота
С02Н
Наряду с образованием отдельных элементарных циклических
мономеров грибами возможен внутриклеточный биосинтез ими
сложных полимеров — соединений типа меланинов, включающих
азот. Чаще всего исходный продукт для этого процесса тирозин..
Окисление тирозина грибами происходит по следующей схеме:
НО
СН2
II Зснсоон-
11 NH2
тирозиназа
02
тирозин
.*. Сш
Н—СООН,
ДОФА-хинон
снсоон
тирозиназа
°2 '
ДОФА
НО
НО
СООН
Н
5,6-диоксииндол-2-карбоновая
кислота
0=
0=
^
-со2
СООН
га
ДОФА-хром
(красный пигмент)
5,6-диоксииндол
179
NH
меланопротеид
индол- 5,6 -хинон
Физико-химические свойства продуктов жизнедеятельности
грибов типа меланинов, образовавшиеся по вышеприведенному
пути, и гуминовых кислот имеют большое сходство по целому
ряду признаков.
1. Черно-коричневая окраска.
2. Не растворяются в воде, кислотах и обычных органических
растворителях, но растворяются в щелочи, откуда могут быть
осаждены кислотами.
3. Обесцвечиваются сильными окислителями, такими, как
перекись водорода, бромная вода и др.
4. Спектры поглощения щелочных растворов в
ультрафиолетовой и видимой области имеют вид пологих кривых с
постепенным уменьшением оптических плотностей по мере возрастания от
250 до 750 нм.
5. Величины оптической плотности при Е°'°^% укладываются
в пределы от 0,01 до 0,1.
6. Имеют сходный элементный состав и процентное
содержание негидролизуемого азота.
7. Обнаруживают парамагнитные свойства, определяемые с
помощью электронного парамагнитного резонанса,
свидетельствующие о присутствии высоких концентраций свободных
радикалов порядка 1018—1019 спинов на 1 г сухого вещества.
8. Общность ИК-спектров.
. 9. Сходная молекулярная масса.
10. Однотипный набор аминокислот.
Пигменты с такими свойствами обнаружены и исследованы по
этим признакам у многих видов преимущественно темнопигмен-
тированных грибов, в том числе Aspergillus nidulans, Cladospo-
rium herbarum, С. cladosporioides, Nadsoniella nigra, Stachybo-
trys atra, Alternaria alternata, Stemphylium botryosum, видов
Aureobasidium, Cladosporium и др. Характеристика гуминовых
кислот и продуктов жизнедеятельности грибов приведена в
табл. 17.
Сходство пигментов грибов с гуминовыми кислотами
подтверждается и сходством их отдельных фракций. При
фракционировании на сефадексах как гуминовых кислот, так и пигментов,
выделяются две основные фракции: бурая с молекулярной массой
100000 и коричневая с молекулярной массой 20 000. Обе фракции
не имеют выраженных максимумов поглощения в видимой обла-
1180
сти спектра. Кроме
того, как в гуминовых
кислотах некоторых
типов почв, так и в
пигментах многих
видов темноокрашенных
хрибов присутствует
фракция зеленого
пигмента с молекулярной
массой порядка 500 и
.характерными
максимумами поглощения в
области 430, 450, 570 и
620 нм (рис. 53). Эта
фракция была
обнаружена японским
исследователем К. Кумада
в 1965 г., а затем и
рядом других ученых в
дерново - подзолитых,
<бурых лесных,
пойменных, горно-луговых
почвах многих стран,
в том числе и СССР.
Фракция зеленого
лигмента имеет в
щелочи четко
выраженную зеленую окраску.
Она была выделена в
кристаллическом виде.
Пигмент сходен по
спектру поглощения с
синтетическим 4,9-ди-
гидрооксиперилен, 3,10-
хиноном. Подобно
синтетическому
веществу пигмент обладает
индикаторными
свойствами. Точка
плавления пигмента 360° С,
эмпирическая
формула его — C3oHi808. В
водном спирте
пигмент изумрудно-зеле-
лого цвета.
Эта фракция
присутствует также у
многих видов темно-
окрашенных грибов.
!>
S
я
Ос
ISSSS-
3 5 «j p.— w
8**
Я О) Я
<и К н
с о <j
sb-s
О О
~ Я
н
=f S.
H X
о о о о
ОО СО О СО
ОО
I
о"
о
о
о
о
ооооо
о" о" о" о*4 о"
oggo
о* о
OOCOWO
о о" о" о" о"
+ I+++
со о><м
о о" о" о"
+ 1 +
2 Я
goo 5 | со
Ю Tf< CO t^-
COCO ОЮСО
СО CD t^. "^ О)
CS —« CMCN —«
оо t^ ^co оо
сч~.-Г^сч~с<Г
со со со со со
О ч* СМ СЧ Ю
(М СО Tf ^ СО
<м т^ t^. coco
TjTt^ of сГсч
тр ^ <N CO Tf
20,3
19,9
8,1
21,8
Tt Tf 00 h-
ofcsfc^oT
00 <M —<т*<
05 LQ О СО
СО СО ч* СО
ЮЮО—|
t^cvT оъся
СО rf CO Tt4
<М
см
«л СМ О
сч 5
О о
<М с
•g со JfcO О g
3 О 3 3 4>
3 СО
н о
о о
§2
° й
о«2
181
Кумада выделил ее из склероциев широко
распространенного микоризообразующего гриба Сепососсит gram-
forme, а затем она была обнаружена и в мицелии многих других
грибов, таких, как Alternaria alternata, Aureobasidium pullulatis,
Diplococcium resinae, виды Cladosporium и Ulocladium.
Зеленый пигмент грибов по всей вероятности является одним
из непосредственных источников фракции в гуминовых кислотах
почв, где содержание этих грибов значительно.
Таким образом, не отрицая существенной роли лигнина и era
отдельных структурных единиц в образовании гумуса, необходима
учитывать активное участие гри-
jj бов в синтезе сложных
полимерных соединений типа гуминовых
кислот. Грибы могут синтезиро-
Рис. 53. Спектры поглощения зеленой
фракции грибных пигментов и гумино-
вой кислоты Р-типа:
1 — гуминовая кислота
дерново-подзолистой почвы; 2 — пигмент
Сепососсит graniforme; 3 — пигмент
Alternaria alternata
SO О Л,НМ
вать внутриклеточно не только фенолы, но и сложные
полимерные соединения меланопротеиды, почти полностью идентичные
гуминовым кислотам. Меланопротеиды, синтезируемые грибами
внутриклеточно, после отмирания мицелия поступают в почву, и
тогда участие грибов в гумусообразовании может быть оценена
исходя из количества пигмента, которое образуется грибами в той
или иной почве, и может быть рассчитано по количеству темно-
окрашенного мицелия, учитываемого прямым микроскопическим
методом.
Так, в дерново-подзолистой почве количество пигмента,
поступающего с темноцветным мицелием, единовременно составляет
0,1—0,3% от гуминовой кислоты, в серой лесной почве — 0,02—
0,06, в черноземе — 0,001—0,03%.
Учитывая многократное поступление отмершего мицелия в
почву в течение года и трудность разложения таких
высокомолекулярных соединений, как меланины, можно считать, что последние
занимают значительное место в образовании гумуса.
РОЛЬ ГРИБОВ В РАЗЛОЖЕНИИ ГУМУСА
Почвенный гумус устойчив к разложению. Сохранность
гумуса, определяемая методом радиоактивного углерода,
составляет от 250 до 3000 лет. Однако некоторые почвенные грибы
способны разлагать гумус и использовать его в качестве
единственного источника азота или углерода, или того и другого вместе.
Существует несколько методов определения разложения
гумуса микроорганизмами. Наиболее распространенный — определе-
182
иие интенсивности роста микроорганизмов на гуматных средах.
Гуминовая кислота добавляется в синтетическую питательную
среду в виде гумата натрия в качестве единственного источника
углерода и азота.
К росту на гуматных средах способны грибы Trichothecium ro-
seum, Trichocladium as per ит, некоторые виды Aspergillus, Pent-
-cillium, Helminthosporium и др. Однако этот метод не дает
представления о действительном разложении гумуса, так как
организмы могут развиваться за счет примесей в гуматных средах.
Для более точной оценки разложения гумуса определяется:
элементный состав гуматов до и после роста на них микроорганизмов;
изменение молекулярного веса отдельной фракции при
фракционировании на сефадексах; разрушение ароматического ядра по
изменению отношений зкстинкций при различных длинах волн
Е465: Ебб5. Чем меньше эта величина, тем более высокое
содержание ароматических соединений в составе полимера. Увеличение
этого показателя свидетельствует об уменьшении содержания
ароматических соединений, т. е. их разрушении.
Процесс разложения гумуса могут проводить неспецифические
микроорганизмы. Потеря основных элементов гумуса (в
процентах от исходного содержания) под воздействием различных
микроорганизмов характеризуется данными, приведенными в табл. 18,
Таблица 18
Разложение гуминовой кислоты микроорганизмами
(по С. Мазуру, 1967)
Роды и виды бактерий и грибов
% потери
С
1 5-9
11
13
19
7,7
3,0
9,6
% потери
N
5,7
0
9,6
17,3
—
2,5
% потери
С+ N
Agrobacterium sp. . . .
Pseudomonas sp. . . .
Arthrobacter sp
Penicillium frequentans
Acremonium sp
Fusarium sp
Aspergillus versicolor .
7—11,
15
11
17,3
7,7
5,7
5,7
19, 20, из которых следует, что высокой активностью в
разложении гумуса обладает гриб Penicillium frequentans.
Определение изменения молекулярной массы отдельных
фракций под влиянием гриба P. frequentans дало следующие
результаты (табл. 19).
Величины экстинкций гуматов до и после разложения гумуса
грибом характеризуется следующими показателями (табл. 20).
Эти данные говорят о значительном разрушении
ароматического ядра микроорганизмами и особенно под влиянием P.
frequentans. Это подтверждается и изменением цветности гуматов, что
можно определять колориметрически.
По данным А. Дубовской, количество фульвокислоты в модель-
183
ных опытах убывало за месяц до 54% под действием грибов**
Aspergillus versicolor, Penicillium decumbens, а гуминовой
кислоты за этот же срок — на 64%. Количество углерода снижалось
соответственно в фульвокислоте на 10—45%, в гуминовой
кислоте — на 12—38%.
Чистые меланины, полученные в виде препаратов и внесенные
в почву, так же, как гуминовые кислоты, отличаются большой
устойчивостью к микробному разрушению. Основной причиной в.
Таблица 19 Таблица 20
Изменение молекулярной массы
отдельных фракций
гуминовой кислоты
под влиянием гриба
P. frequentans
Отношение величин
экстинкций гуматов до
и после воздействия
Р, frequentans
Фракции
Потери MB, %
Образцы
<5000
5 000—10 000
10 000—50 000
>50 000
22,0
34,0
23,0
53,0
Гумат исходный
Гумат после роста
P. frequentans
Е4вб/Евв5
4,71
5,33
сохранении гуминовых кислот и меланинов является
пространственная их разобщенность с ферментами и адсорбция их на
твердых поверхностях, которая препятствует доступу ферментов к тем
местам полимера, на которые они должны воздействовать. Более
быстро гумус разрушается в присутствии доступных
микроорганизмам водорастворимых органических соединений, т. е. в
условиях кометаболизма (соокисления).
Процесс разложения гумуса микроорганизмами приводит к
освобождению азота, который в гумусе находится в закрепленном
состоянии и недоступен для растений.
Соотношение углерода к азоту в гумусе в среднем равно 10: 1,.
а потребление этих элементов микробами, в том числе и грибами,,
обычно осуществляется в отношении 25:1. Поэтому при воздейт
ствии микроорганизмов на гуминовые кислоты освобождается да
60% азота, который далее может быть потреблен растением.
РОЛЬ ГРИБОВ В СТРУКТУРООБРАЗОВАНИИ
Первые высказывания о роли микроорганизмов в
формировании почвенной структуры были сделаны П. А. Костычевым
в 1886 г. Он и предположил, что продукты жизнедеятельности,
микроорганизмов могут быть цементирующими веществами,
склеивающими почвенные частицы в структурные элементы. Такую же
роль могут играть продукты распада органических веществ,
образующихся в результате жизнедеятельности микробов.
В дальнейшем экспериментально было показано, что важная
роль в этом процессе принадлежит грибам. Грибы еще при жизни,.
184
развиваясь около живых корней или разлагая растительные
остатки, вызывают цементирование почвы, пронизывая ее мицелием
и пропитывая различными органическими веществами. После
отмирания цементирующую роль могут играть продукты их
автолиза.
Структура — существенное свойство почвы, от которого
зависит ее плодородие. Особенно большое значение она имеет на
глинистых и суглинистых почвах. В структурной почве много
промежутков, она легко впитывает влагу. Средние и крупные поры
быстро освобождаются от воды и в них образуется запас воздуха,
поэтому в таких почвах устанавливается благоприятное
соотношение воды и воздуха. Хорошо оструктуренная почва
характеризуется высоким содержанием (70—80%) макроструктурных
водопрочных элементов, которые имеют размеры от 0,25 до 10 мм.
В оструктуривании почвы принимают участие многие виды
грибов. Такие данные были получены при внесении в
бесструктурные почвы чистых культур разных видов грибов или смеси
культур, а также для сравнения чистых культур бактерий и ак-
тиномицетов.
В частности, грибы Mucor hiemalis, Alternaria alternata, Sta-
chybotys atra, Aspergillus niger, Penicillium chrysogenum создают
стабильные агрегаты с достаточно высокой водопрочностью в
образцах пылеватой глины с добавлением небольшого количества
органического вещества, устойчивость которых не изменяется в
течение длительного времени и начинает уменьшаться только
через 36 дней.
Гриб Trichoderma lignorum значительно повышает
структурность некоторых черноземных почв. Хорошей агрегирующей
способностью в образовании крупных водопрочных структур —
макроагрегатов обладают грибы родов Choanephora, Aspergillus и
особенно Acremonium, агрегирующие свойства которых были
проверены на бесструктурной сероземной почве Голодной степи,
при внесении незначительного количества органического
вещества.
На окультуренных почвах Ротамстедской опытной станции был
произведен подсчет микроагрегатов, сформировавшихся под
влиянием гиф грибов и слизеобразующих бактерий. Культуры грибов
Absidia glauca и Entericella nidulans, выросшие в стерильной
почве, обогащенной глюкозой, образовали 242 и 374 м гиф на 1 г
почвы и связывали 96,5 и 80,3% соответственно почвенных частиц
в стабильные агрегаты.
Агрегирующая роль грибов была установлена в песчаных
почвах Австралии и в песчаных дюнах Англии. К. Д. Бонд показал,
что эти почвы содержат в большом числе гифы грибов, которые
участвуют в агрегировании почвы. Гифы могут быть двух типов:
одни из них имеют клейкую поверхность, благодаря которой они.
легко прилипают к поверхности отдельных песчаных зерен,
другие — имеют восковидную сухую поверхность; поэтому в
агрегации почвенных частиц имеют значение только гифы первого типа.
185
Наблюдалась прямая связь между степенью оструктуренноста
почвы и развитием мицелиальных грибов и актиномицетов.
При определении устойчивости агрегатов, сцементированных
различными органическими веществами, было установлено, что
наиболее прочные агрегаты образовывались при добавлении
грибного мицелия.
Большинство экспериментов, проведенных по оструктуриванию
почв с помощью микроорганизмов, подтвердили важную роль
грибов в этом процессе. Особенно хорошей агрегирующей
способностью обладают грибы из родов Absidia, Mucor, Rhizopus, Chaeto-
mium, Fusarium, Aspergillus, Acremonium. Все организмы
распределяются в порядке убывания их агрегирующей способности
следующим образом: грибы и слизеобразующие бактерии>акти-
номицеты>дрожжи>большинство бактерий. При использовании
смеси культур их агрегирующая способность располагается
следующим образом: грибы>грибы + актиномицеты + бактерии> ак-
тиномицеты> бактерии.
Роль грибов в структурообразовании может быть
многообразной. Во-первых, грибы осуществляют механическое оплетение
мицелием почвенных частиц, тем самым скрепляя их. Во-вторых,
грибы, адсорбируя на гифах мелкие частицы почвы, образуют
пористые, не разрушающиеся водой агрегаты. В-третьих, грибы
выделяют разнообразные продукты обмена, которые
адсорбируются на почвенных частицах и способствуют их цементированию.
Многие грибы и дрожжи образуют полисахариды, содержащие
в своем составе уроновые кислоты.
Микробные полисахариды более устойчивы, чем растительные.
Еще более устойчивыми они становятся, когда вступают в
соединения с другими веществами, например Na-бентонитом, Na-каоли-
нитом и гидроокисью железа.
Образование комплексов между металлами (Са, Си, Zn, A1,
Fe) и полисахаридами, содержащими уроновые кислоты, может
резко уменьшать степень разрушения полисахаридов в почве.
Полисахариды, связанные с глинистыми минералами, более
устойчивы к микробному разрушению, что приводит к образованию
стабильных агрегатов.
Кроме полисахаридов, большое значение в агрегации
почвенных частиц имеют протеины. Некоторые протеины адсорбируются
внутри кристаллической решетки глин монтмориллонитового типа,
причем адсорбция делает их более устойчивыми к действию
ферментов. Другими агрегирующими веществами могут быть жиры,
масла, воска и смолы, но их влияние на агрегацию значительна
меньше.
Липидоподобные вещества, образуемые некоторыми
почвенными грибами, могут значительно увеличить стабильность
агрегатов, покрывая поверхность их пленкой и предохраняя от
разрушающего действия воды. Например, гидрофобность песчаных почв
некоторых районов Австралии частично обязана липидоподобным
продуктам метаболизма базидиомицетов. Почвенные частицы об-
186
волакиваются органической, непроницаемой для воды пленкой,
и вода проникает только по узким каналам, оставляя большие
участки сухими.
Таким образом, грибы играют очень существенную роль в
формировании макроагрегатов почвы. Опутывая гифами
минеральные частицы почвы, они механически скрепляют их. Гифы грибов
адсорбируют частицы почвы, их метаболиты склеивают частицы.
Образование основного цемента почвенной структуры — гумино-
вых веществ — происходит при существенном участии грибов, как
в разложении растительных остатков, так и в синтезе полимеров.
Кроме того, косвенная роль грибов в структурообразовании
проявляется в том, что разрушение грибных гиф бактериями
приводит к образованию слизистых веществ — активных агрегирующих
агентов.
Глава 9
СПЕЦИФИЧЕСКИЕ ФИЗИОЛОГИЧЕСКИ
АКТИВНЫЕ ВЕЩЕСТВА ГРИБОВ
АНТИБИОТИКИ И ТОКСИНЫ ПОЧВЕННЫХ ГРИБОВ
В процессе жизнедеятельности грибы образуют
разнообразные вторичные метаболиты, которые могут оказывать
существенное влияние как на развитие самого организма-продуцента,
так и на процессы, происходящие во внешней среде, в первую
очередь в почве.
Вторичные метаболиты — соединения, синтез которых не
связан непосредственно с ростом и накоплением биомассы
синтезирующих их организмов, т. е. трофофазой. Образование их
происходит в период торможения или прекращения роста в идиофазе.
Как уже говорилось выше (см. с. 100), взаимоотношения
организмов в любой экосистеме формируются на основе
трофических связей и конкуренции между ними. Поэтому большое
значение в формировании микробных сообществ в экосистеме имеют
характер субстрата и антагонизм между микроорганизмами,
которые усиливают конкурентную способность организмов.
Антагонизм микроорганизмов в первую очередь определяется
образованием антибиотиков и токсинов.
Антибиотики и токсины не единый класс химических
соединений, а представляют собой разнообразную группу химических
веществ, объединяемую по их биологическому действию. Это
могут быть белки, полипептиды, полисахариды, органические
кислоты, в том числе производные фенолов и хинонов, глюкозиды,
стеролы, изопреноиды, разнообразные гетероциклические
соединения, включающие различные неорганические элементы, и т. д.
Антибиотики, по определению Н. С. Егорова, — это
высокоспецифические продукты жизнедеятельности, обладающие высокой фи-
1.87
зиологической активностью по отношению к определенным
группам микроорганизмов, избирательно задерживая их рост или
полностью подавляя развитие. Механизм действия этих веществ^
различен. Классификация антибиотиков и токсинов строится на
нескольких принципах: 1) по их биологическому происхождению
(бактерии, актиномицеты, грибы и т. д.); 2) по механизму
биологического действия (ингибиторы синтеза клеточной стенки,
нарушения функции мембран, синтеза нуклеиновых кислот и т. д.);
3) по химическому строению; 4) по объекту действия
(микроорганизмы, растения, животные).
Биологическая роль этих соединений для самого организма-
продуцента многообразна. Во-первых, они могут играть защитную
роль и подавлять конкурентов за источник питания; во-вторых,,
установлено их влияние на отдельные звенья процесса
метаболизма через активацию или ингибирование ферментных систем;
в-третьих, некоторые антибиотики играют роль в
дифференциации организма в процессе онтогенеза, принимая участие в
образовании спор бактерий-продуцентов, либо задерживают
прорастание спор.
Есть мнение, что общая высокая синтетическая активность
организма, в том числе и активность биосинтеза антибиотиков^
дает ему возможность легче выжить.
Токсины — вещества биологического происхождения,
вызывающие любые нарушения функции организма, в первую очередь
животных и растений, интегральное действие которых
выражается в ингибировании роста и развития организма. Обычно токсины
противопоставляют антибиотикам или проводят границу между
ними. Принято считать, что токсины.— это вещества, вызывающие
нарушение жизнедеятельности высших организмов — макроорга-
«измов, антибиотики — ингибиторы микроорганизмов. Безусловно,
такое деление на определенном этапе было оправдано, так как
действие тех и других рассматривалось как
взаимопротивоположное. Токсины — причина болезни, антибиотики — средство
подавления продуцента токсинов, следовательно, подавления источника
заболевания.
Однако по мере изучения спектра действия этих веществ
становилось ясно, что такое деление в значительной степени условно,,
так как обнаружилось, что действие некоторых из них совпадает
в отношении одних и тех же объектов (например, антибиотик**
патулин, пеницилловая кислота и некоторые другие являются
одновременно высокотоксичными для животных и растений, а
такие токсины, как дендродохин, роридин, афлатоксин, обладают
антимикробным действием).
Если в действии вещества проявляется значительное
преобладание активности по отношению к тем или иным объектам, то их
можно считать соответственно либо истинными антибиотиками,
либо истинными токсинами. В общебиологическом смысле
истинные антибиотики можно рассматривать как токсины, действие
которых направлено на микроорганизмы, т. е. как-бактериотоксиньь
188
Если действие антибиотиков проявляется и по отношению к
растениям и животным, их можно считать токсинами-антибиотиками..
Условность деления продуктов жизнедеятельности на антибиотики,
и токсины определяет и сходство методов их изучения.
Изучение токсинов проводится в различных аспектах. Начала
изучения токсинов связано с медицинской микробиологией, с
обнаружением их у патогенных микробов, возбудителей инфекцион-"
ных заболеваний человека и животных. Действие этих токсинов
определяло симптомы заболеваний. Тогда же было показано, что-
токсины, образуемые патогенными бактериями, представляют
собой высокоспецифические вещества, действующие в очень малых
дозах и являющиеся протеинами с высокой молекулярной массой,,
обладающие антигенными свойствами. Поэтому надолго
укоренилось мнение, что токсины — это только белки. Однако с
развитием фитопатологии токсины были обнаружены у многих
микроорганизмов, в том числе грибов, вызывающих заболевания
растений. Изучение их строения и свойств обнаружило их
разнообразную химическую природу. Токсины иной, чем белки,
химической природы были обнаружены и у патогенов животных. Токсины
разнообразного химического строения были выявлены у сапро-
трофных организмов, обитающих в почве.
Среди токсинов почвенных сапротрофных грибов обнаружено
много таких, которые одновременно с токсическим и фитотокси-
ческим действием обладают также антимикробной активностью.
Таким образом, действие токсинов почвенных грибов может быть
направлено на самые разнообразные макро- и микрообъекты.
Наибольшее распространение получила классификация
токсинов по организмам, на которые направлено действие. По
направленности действия токсические вещества грибов можно разделить
на три группы.
1. Токсины, являющиеся ядами для животных, — зоотоксины.
Они представляют собой преимущественно вещества
общеплазматического действия. Поскольку эти токсины широко
представлены у грибов, их часто называют микотоксинами. Помимо
действия на животные организмы они часто обладают специфической
антимикробной и фитотоксической активностью (табл. 21). Эти
токсины могут быть причиной серьезных микотоксикозов человека
и животных, многие из которых приводят к смертельному исходу.
2. Специфические фитотоксины — токсины, действие которых
направлено только на растения и антибиотическим действием они
не обладают. Образуются преимущественно фитопатогенными
грибами (табл. 22). Специфичность их проявляется в том, что они
угнетают только строго определенные виды и даже сорта
растений, а высокая физиологическая активность выражается в очень
малых концентрациях их активности в пределах десятых и даже
сотых долей микрограмма на миллилитр. Наиболее высокой
специфичностью обладают фитотоксины — викторин и перикулярин,
представляющие собой соединения белковой природы, а также
высоко специфичный гликопротеин гриба Cladosporium fulvum.
189>
Если высокомолекулярные фитотоксины белковой природы
Действуют либо как экзоферменты, вызывающие деградацию
углеводов клеточной стенки хозяина, что часто приводит к заку-
Таблица 21
Физиологическая активность зоотоксинов
Продуцент
bendrodochium toxicum
Aspergillus flavus
Stachybotrys atra
Penicillium rubrum
^. purpurogenum
Fusarium sporotrichiella
Токсин
дендродохин А
афлатоксин Bi
афлатоксин В2
афлатоксин Gx
афлатоксин G2
стахиботриоток-
син
рубратоксин
то же
спорофу зарин
Токсичность для
растений,
мкг/мл
+
25
—
—
—
+
5—55
5—55
+
Токсичность для
животных, мг/кг
LD60 — 20
LD50 —55
LD50 — 60
LD60 — 60
LD50-10
LD50- 0,2-0,4
LD60 —0,3-1,5
Антибиотическая
активность
5—20 мкг/мл
(дрожжи и
грибы)
неактивен
неактивен
неактивен
Примечание, -j— токсичность обнаружена; нет данных.
порке сосудов, либо как структурные элементы, способные
связываться со специфическими белками, то низкомолекулярные
токсины различной природы, такие, как фузариевая, альтернариевая
Таблица 22
Щ f: Физиологическая активность фитотоксинов
Продуцент
Helminthosporium victo-
riae
Periconia circinata
Fusarium oxysporum
F. moniliforme
F. orthoceras
Alternaria alternata
A. solani
Rhizoctonia solani
Alternaria kukuchiana
Cochelobotus miabeanus
Colletotrichum fuscum
Токсин
викторин
периконин
фузариевая
кислота
альтернариевая кислота
фенилуксус-
ная кислота
альтеин
кохлиоболин
коллетоксин
Токсичность для растений;
минимальная доза, мкг/мл
0,02 (на чувствительные
сорта овса)
0,1 (на чувствительные
сорта сорго)
1—10 (хлопчатник,
томаты)
1—10 (на проростки
редиса, капусты, томатов)
5—50 (корни свеклы,
риса, семена люцерны,
хлопчатника)]
0,02 (листья груши)
1—10 (проростки риса)
30—60 (томаты)
Антибиотическая
активность, мкг/мл
неактивен
неактивен
0,1—1,0
(грибы,
прорастание спор)
неактивна
неактивен
—
1
• 190
кислоты, вызывают нарушение важнейших внутриклеточных про-
цессов (фотосинтеза, дыхания, биосинтеза первичных
метаболитов, транспорта веществ в клетку и т. д.).
3. Токсины-антибиотики (табл. 23) — это фито- и зоотоксины,
обладающие одновременно сильным антибиотическим действием.
Минимальные концентрации ингибирующего действия этих
веществ лежат в одних и тех же пределах (1—100 мкг мл) и для
растений и для микроорганизмов. Одновременно многие из них
обладают высокой токсичностью по отношению к животным.
Зоотоксины, вызывающие заболевание человека и животных —
микотоксикозы, начали впервые изучать на Украине в 30—40-х nv
Та блица 23
Характеристика токсинов-антибиотиков
Продуцент
Penicillium clavifor-
те
P. griseofulfum
P. lapidosum
Aspergillus clavatus
A. terreus
Penicillium verruco-
sum var. cyclopium
Penicillium citrinum
P. implicatum
Aspergillus candidus
A. terreus
A. ustus
Токсин
патулин
пеницил-
ловая, пу-
бероловая
и цикло-
пальдие-
вая
кислоты
цитринин
устовая
кислота
Токсичность для
растений
(минимальная доза),
мкг/мл
10—60
(пшеница,
томаты,
кукуруза)
1—10
(многие
растения)
5—100
(многие
растения)
100
(кресс-
салат)
Токсичность для
животных,
мг/мл
LD50
10—35
LD50
110—250
LD50
35
—
Антибиотическая
активность,
мкг/мл
Г+иГ-
30—100
Г+иГ—
2—35
г+
30—100
г+
100
Уф-
спектр,
нм
275
не
выражен
220
255
315
—
Растворимость
хорошая — в
воде и многих
органических
растворителях;
плохая — в
бензоле
хорошая — в
воде и многих
органических
растворителях;
плохая — в
бензоле и
хлороформе
во многих
органических
растворителях;
плохая — в воде:
—
Примечание. Г-| грамположительные бактерии; Г грамотрицательные бактерии.
К. И. Вертинский, Н. М. Пидопличко, В. И. Билай и другие
в связи с массовыми заболеваниями лошадей и рогатого скота,-
Было выявлено, что причина заболеваний — развитие на
кормах и зерне грибов с остротоксичными свойствами, таких, как
Stachybotrys atra, Dendrodochium toxicum, Fusarium sporotrichiel-
19L
la, образующие соответственно токсины стахиботриотоксин, ден-
дродохин, спорофузарин. Усиленно изучается в настоящее время
во многих зарубежных странах группа токсинов, получившая
название афлатоксины. Появление интереса к этой группе токсинов
-связано с массовыми заболеваниями домашней птицы и мелкого
скота, вызванных отравлениями такими токсинами. Позднее были
обнаружены токсины такого же действия у грибов рода Penicil-
lium исландиотоксин, рубратоксин и др.
Характерно, что у сапротрофных грибов среди образуемых им#
физиологически активных веществ преобладают
токсины-антибиотики. У фитопатогенных грибов преобладают более
высокоспецифичные фитотоксины. Это объяснимо с биологической точки
зрения. По представлениям М. В. Горленко, токсины
сапротрофных грибов обеспечивают им преимущества в борьбе за
питательный субстрат, являются их основным оружием в конкурентной
борьбе. Фитотоксины служат в основном для обеспечения
организму паразитического существования. Вещества всех
перечисленных выше групп, образуясь в почве и поступающем в почву
органическом материале — опаде, лесной подстилке, соломе, сене,
зерне, могут придавать ей токсические свойства и влиять на
почвенное плодородие и далее на развитие произрастающих на ней
растений. Тем не менее для почвы основное значение имеют
токсины сапротрофных грибов, так как патогены в почве либо
погибают, либо находятся в неактивном состоянии в виде покоящихся
форм. Сапротрофные формы могут вызывать токсикоз почв по
отношению к микроорганизмам и растениям.
РОЛЬ ГРИБОВ В ТОКСИКОЗЕ ПОЧВ
ч
При образовании в почве токсинов они могут отравлят/
ее, делать токсичной, а через почву оказывать действие на
растение и почвенные микроорганизмы, вызывая перестройку
сообщества. В результате складываются сложные взаимоотношение
между микроорганизмами, почвой и растением, где почва стако-
\вится посредником между двумя другими компонентами в био
ценозе. Тогда влияние микроорганизмов на растение сказывается
через почву, что в свою очередь может иметь значение для
раскрытия взаимоотношений отдельных компонентов в биоценоза*
При образовании токсинов в почве возникает явление токсикоза
почв и как частный случай его — почвоутомление. Почвоутомле
ние известно в практике сельского хозяйства давно, значительно
раньше, чем токсикоз почв. Оно проявляется в резком угнетении
растений и снижении их урожая, которое возникает при бессме"
ном культивировании какой-то одной культуры (монокультуре;;
Широко известно льноутомление, клевероутомление, утомление
лочв под плодовыми деревьями. Токсикоз почв — более широкое
понятие. Оно включает проявление угнетения роста растений на
целинных и окультуренных почвах не только в монокультуре, но
и там, где происходит смена культур.
192
Проявление токсикоза почв, не связанного с явлением
почвоутомления и обнаруживаемого в угнетении развития сосны в
почвах Узрхемского лесничества, отмечают в своих работах
П. 3. Бриан, М. Рейнер и В. Нельсон-Джонс. Прорастание
желудей подавляли почвы из-под дубовых насаждений, а также почвы
из-под степной растительности. Сильное проявление токсикоза
почв по отношению к ячменю, клеверу, травам и другим
растениям наблюдается на дерново-подзолистых почвах, особенно при
внесении кислых азотных удобрений без серьезных сдвигов в
агрохимических свойствах этих почв.
Наиболее четко определить токсикоз почв можно с помощью
метода инициированного микробного сообщества (ИМС),
предложенного В. С. Гузевым.
Еще С. Н. Виноградским была высказана мысль о том, что
активную деятельность микробного населения почвы можно
наблюдать после внесения в нее органического вещества. Эту идею
применительно к грибам реализовал С. Гаррет, который
наблюдал активное заселение свежих растительных остатков,
внесенных в почву, сначала мицелием грибов, а затем актиномицетами
и бактериями.
Метод ИМС заключается в том, что на поверхность
исследуемой почвы в чашках Петри наносят тонкий слой крахмала в виде
полоски с четкими границами в качестве инициирующего
органического вещества в концентрации 0,05—0,1% относительно всей
массы почвы. Этот метод дает возможность благодаря инициации
микроорганизмов почвы с помощью крахмала как органического
субстрата с малым селективным действием вычленить из всего
микробного населения почвы активно функционирующую ее часть,
а благодаря использованию сканирующего электронного
микроскопа, дающего возможность работать в широком диапазоне
увеличений, наблюдать всех представителей микробного сообщества
^гочвы одновременно и установить, какую долю в нем занимают
активно функционирующие грибы.
Благодаря локальному внесению крахмала удается
сконцентрировать микроорганизмы и продукты их жизнедеятельности, в
частности токсины, в одном месте. О токсикозе при
использовании этого метода можно судить по задержке прорастания семян,
наносимых непосредственно на зону развития инициированного
микробного сообщества после месячной инкубации.
^ Метод дает возможность определить микробную токсичность
Непосредственно в почве не только по отношению к растениям,
-но и по отношению к почвенным животным.
^ О микробном токсикозе можно судить по степени поедаемости
биомассы грибов инициированного микробного сообщества
почтенными беспозвоночными животными. Хорошая поедаемость
клещами мицелия грибов свидетельствует об отсутствии у грибов
токсичных продуктов. Грибы, не поедаемые клещами, содержат
токсичные метаболиты.
Многолетними исследованиями Н. С. Авдонина и его сотруд-
7 Т. Г. Мирчинк
193
ников было установлено, что длительное применение азотных
удобрений отдельно и совместно с калийными на кислых дерново-
подзолистых почвах приводит к резкому ухудшению плодородия
этих почв и падению урожая без заметного изменения их
агрохимических свойств. Это явление было названо Н. С. Авдониным
скрытым отрицательным действием минеральных удобрений на
кислых почвах.
Однако микробиологические исследования показали, что при
почти неизменяющихся агрохимических свойствах происходит
резкое изменение комплекса грибов-микромицетов в этих почвах и
активно функционирующей его части в сторону преобладания
токсинообразующих форм.
Если в контрольной почве доминантные грибы представлены
видами Chaetomium, Penicillium janthinellum, часто
встречающимися Oideodendron griseum и Monila geophila и рядом других
видов, то при внесении азотных и калийных удобрений идет
резкое сокращение разнообразия видов, причем пенициллы
составляют большую часть комплекса микромицетов. Единственными
доминантами сообщества становятся токсинообразующие грибы
P. funiculosum, Talaromyces flavus, P. verrucosum var. cyclopium,
а токсинообразующий гриб Р. purpurogenum попадает в число
часто встречающихся видов (рис. 54). При этом численность
грибных зачатков увеличивается в 2—3 раза по сравнению с почвами
без минеральных удобрений. Коэффициент сходства Сьеренсена
между микромицетами почв без минеральных удобрений и с
азотными и калийными удобрениями становится равным 0,40—0,45,
что подтверждает большие изменения, произошедшие в составе
комплекса микромицетов.
Анализ влияния возрастающих концентраций азотных и
калийных удобрений на структуру и состав микробного сообщества
почвы в модельных опытах позволяет выделить четыре зоны
концентрации и типа реакции микробной системы почвы на действие
антропогенного фактора, каковым является в первую очередь
внесение азотных и калийных минеральных удобрений. Небольшие
дозы этих удобрений для дерново-подзолистой почвы не вызывают
изменений в структуре и составе ИМС. Эта зона концентрации
названа В. С. Гузевым зоной гомеостаза (отсутствие изменений).
Следующая зона, соответствующая интервалу концентраций от
100 до Ю00 кг/га для этой же почвы, — зона стресса, по типу
реакции характеризуется существенным изменением структуры,
сообщества. Доминирующие виды зоны гомеостаза постепенно
исчезают, а некоторые редко встречающиеся виды переходят в
разряд доминирующих и часто встречающихся организмов. В этой
зоне изменение структуры микробного сообщества приводит к
увеличению доли фитотоксичных микромицетов, которые
обусловливают микробный токсикоз почвы. В дерново-подзолистой почве
резкое возрастание фитотоксических свойств микробного
сообщества соответствует дозе азота и калия 400—600 кг/га. Такое же
проявление микробного токсикоза может происходить в полевых:
194
условиях при значительно меньших дозах этих удобрений, но при
длительном и постоянном внесении, т. е. длительное внесение
минеральных удобрений в малых дозах может вызвать тот же
эффект, что и одноразовое внесение высоких доз удобрений. Таким
образом, скрытое отрицательное действие минеральных
удобрений на кислых почвах можно объяснить коренной перестройкой
Дозы минерального азота v кг/га
30
100
зоо
1000'
5000
—I 1 1,
10000 30000 100001)
зона
гамеостаза
зона
стресса
титотоксичность
не проявляется
микробный
токсикоз
зона
резистентности
зона
репрес-
солебой
токсикоз
Ctiaetomium sp.
Streptomyces sp.
гр. griseus
Monllla geophila
Penicillium janthinellum
—^JjjiatenulaTum
1. Didlodendron
griseum
Рис. 54. Влияние минерального азота на структуру инициированного
микробного сообщества дерново-подзолистой почвы
195
микробного населения почвы в сторону преобладания токсичных
форм.
Третья зона еще более высоких концентраций минеральных
удобрений (зона резистентности, которая используется только в
модельных опытах) по типу реакции проявляется в смене видов
микробного сообщества. В этой зоне концентраций развиваются
только микроорганизмы, устойчивые к высоким концентрациям
соли. При еще больших концентрациях минеральных удобрений
в почве полностью подавляется развитие микроорганизмов, что
определяет зону репрессии микробной системы почв.
Поэтому отрицательное действие высоких доз минеральных
удобрений на микробную систему почвы можно охарактеризовать
величиной зоны гомеостаза и дозой, вызывающей микробный
токсикоз. Таким образом, одной из причин проявления микробного*
токсикоза может быть доза вносимых минеральных удобрений,
вызывающая перестройку микробного сообщества.
Пока микробная система находится в состоянии гомеостаза,
токсикоз не проявляется. Он начинает проявляться при переходе
в зону стресса.
Аналогичная реакция почв с последующим проявлением
микробного токсикоза обнаруживается и по отношению к другим
факторам антропогенного воздействия, таким, как тяжелые металлы,,
пестициды, нефтяные загрязнения, вещества, подавляющие
процессы нитрификации. Среди них минеральные удобрения
занимают место нестойких загрязнителей.
Однако устойчивость разных почв к воздействию разного рода
загрязнителей, в том числе и минеральным удобрениям, различна.
Наименее устойчивы к действию минеральных удобрений и
других загрязнителей дерново-подзолистые почвы. Это значит, что
небольшие дозы азотных и калийных удобрений выводят
микробное сообщество из зоны гомеостаза в зону стресса и вызывают
микробный токсикоз. Более устойчив к воздействию химических:
реагентов типичный серозем, зона гомеостаза его равна 300—
400 кг/га, а микробный токсикоз проявляется при концентрации:
1000—1200 кг/га.
Наиболее устойчив чернозем, для него зоны гомеостаза равны
так же, как и для серозема, 300—400 кг/га, но микробный
токсикоз, обусловленный преобладанием в сообществе грибов Peni-
cillium funiculosum, P. purpurogenum, Aspergillus ustus, наступает
только при дозах удобрения 2500—3000 кг/га (рис. 55).
Таким образом, микробные системы почв по способности
выдерживать нагрузку минеральных удобрений располагаются в
обычный ряд, обусловленный увеличением их буферности к
химическим воздействиям: дерново-подзолистая почва, типичный
серозем, чернозем.
Следующие микроорганизмы вызывают микробную токсичность
при внесении химических реагентов. В удобряемой
дерново-подзолистой почве микробный токсикоз связан с возрастанием
количества грибов P. funiculosum, Talaromyces vermiculatum, P. jan-
196
thinellum, в выщелоченном черноземе — с P. funicutosum, P. pur-
purogenum, P. velutinum, P. janthinellum, Aspergillus ustus, в
типичном сероземе — со Stachybotrys atra, Aspergillus ustus.
Внесение извести в дерново-подзолистые почвы сильно
повышает устойчивость ее микробной системы к внесению
минеральных удобрений и приближает ее тем самым к черноземам.
Под влиянием извести сильно меняется состав микробного
сообщества. Вместо доминирования видов Penicillium, Gliocladium,
Величина зон
гомеостаза ,
кг/га
Азот
400
а
I И III
то
Доза, вызывающая
микробный токсикоз,
кг/ггг
600
1200
I II III
Калий
80
а
/ // ш
2500
400
а
п
Не ВышВает
I II III
Полное
минеральное удобрение
600
300
Ш
I II III
4500
Р
10001
и
то
-пгш
Рис. 55. Устойчивость микробных систем дерново-подзолистой почвы (I),
выщелоченного чернозема (II), типичного серозема (III) к воздействию
минеральных удобрений
Coniothyrium появляются и становятся преобладающими Мусо-
gone nigra, Monilia geophila бактерии рода Bacillus и Strepto-
myces. При этом увеличивается устойчивость микробной системы
к внесению высоких доз азотных и калийных удобрений,
возрастает величина гомеостаза, т. е. почва выдерживает значительно
более высокие дозы удобрений без заметных изменений
микробной системы и микробный токсикоз начинает проявляться при
значительно более высоких дозах.
Многие выявленные при высоких дозах загрязнения микроми-
цеты синтезируют фитотоксичные метаболиты при росте на
питательных средах, природных субстратах и в почве.
197
P. verrucosum var. cyclopium образует несколько токсинов,
такие, как пеницилловая, пубероловая, циклопальдиевая кислоты.
P. funiculosum и P. purpurogenum продуцируют рубратоксин,
обладающий как высокой фитотоксической активностью, так и
токсичностью для животных.
P. janthinellum — образует фитотоксичный антибиотик хода-
Дидин. Asp. ustus известен как продуцент токсичной устовой
кислоты, a Stachybotrys atra образует стахиботриотоксин,
обладающий высокой токсичностью как для растений, так и для
животных. Он проявляет также токсическое действие на
растительные ткани, вызывая их некрозы и отмирание.
Токсины некоторых видов грибов могут быть обнаружены в
почве при наличии в ней достаточного содержания органического
вещества, особенно при поступлении туда свежих растительных
остатков, а в окультуренных почвах при внесении мульчи.
Патулин образуется грибом Penicillium griseofulvum и на
соломе, применяемой для мульчирования почвы. Мак Калла
с сотрудниками извлекли его из соломы органическими
растворителями и идентифицировали хроматографически и по
характерному спектру поглощения в УФ-свете. Заметное образование
патулина грибом P. griseofulvum было обнаружено О. А. Бере-
стецким в почвах под яблоневыми садами, проявлявшими
сильный токсикоз по отношению к саженцам яблонь.
В наших работах была показана возможность образования
токсинов в почве при заражении ее культурами грибов P.
verrucosum var. cyclopium, P. citrinum и P. purpurogenum.
Предварительно у этих культур была определена способность
образовывать токсины соответственно пеницилловую кислоту, цитринин и
рубратоксин на питательной среде. Два вида грибов P.
verrucosum var. cyclopium и P. citrinum образуют метаболиты,
обладающие наряду с фитотоксической активностью широким
антимикробным спектром, т. е. типичные токсины-антибиотики. P.
purpurogenum продуцирует метаболит, являющийся как фито-, так и
зоотоксином. Благодаря этому для идентификации первых двух
можно использовать метод суммарной хроматографии с
проявлением по микробному биотесту. Токсин P. purpurogenum
определяется по растительному тесту (семенам). Метод суммарной
хроматографии основан на различной скорости прохождения
антибиотика-токсина в разных системах растворителей. На основании
данных по скорости прохождения вещества в разных системах
растворителей строится график. Сходные вещества имеют
сходные графики распределения. Для идентификации этих веществ
используется также метод спектрофотометрии. Например, цитри-
ЦИн и рубратоксин имеют характерные хорошо выраженные
спектры поглощения в УФ-свете (рис. 56).
Помимо установления возможности образования токсинов в
почве очень важен вопрос о длительности их сохранения,
адсорбции и инактивации. Длительность их сохранения зависит как от
свойств токсина, так и от свойств почвы.
198
Ю. А. Худякова установила, что токсин, образуемый Penicil-
lium verrucosum, var. cyclopium, плохо адсорбируется почвой,
слабо инактивируется и может находиться в почвах в активном
состоянии. Однако длительность сохранения его в почвах невелика,
так как он легко вымывается, будучи непрочно связанным с
почвой. Токсин P. purpurogenum — рубротоксин, по данным Ф. Г. Бон-
даревской, при внесении в почву легко вымывается из нее водой
*f
220 250
WO
90
70
50
50
10
-
-
1-1 1
и
I Т
Г*
I I
2 J 4 5 6 7 8
Рис. 56. УФ-спектр (I) и схема хроматограммы (И) цитринина в культуре и
почве:
/ — чистый цитринин; 2 — вытяжка из почвы
и в большой концентрации содержится в промывных водах.
Максимальная длительность сохранения токсина в почве без
промывания — 8 суток, сильное ингибирующее действие токсина
проявляется до 5 суток, на 10-е сутки токсин полностью исчезает.
Токсин P. citrinum — цитринин, по данным Р. А. Скоробога-
товой, также недолго сохраняется в почве. Уже на 6-е сутки его
действие сильно ослабляется, а на 12-е сутки он исчезает вовсе.
В связи с образованием токсинов в почве важное значение
имеет распространение грибов-токсинообразователей в различных
почвах. При исследовании различных типов почв Советского
Союза и зарубежных стран выяснилось, что наиболее токсичные
почвы с большим числом потенциально токсичных видов —
подзолистые, дерново-подзолистые, серо-бурые лесные и бурые таежные
почвы. Это можно иллюстрировать табл. 14. .
В подзолистых почвах среди токсинообразователей наиболее
часто встречаются виды Penicillium janthinellum, P. funiculosum,
P. verrucosum var. cyclopium; несколько меньше — P. nigicans,
P. verrucosum var. martensii. Наиболее токсичны, но менее
распространены виды P. purpurogenum и P. claviforme.
Некоторой токсичностью обладают также Alternaria alternate.
и виды Fusarium spp. В серо-бурых лесных почвах — P. granu-
latum, P. nigricans, P. verrucosum, var. cyclopium.
199
В бурых таежных почвах Алтая значительную токсичность
обнаруживают некоторые представители рода Penicillium, такие,
как P. purpurogenum, P. brevi-compactum, P. verrucosum var. mar-
tensii, P. viridicatum, P. frequentans и отчасти виды Trichoderma.
Таблица 24
Содержание токсичных форм грибов в различных типах почв
Типы почв
Дерново-подзолистая сла-
боокульту ренная,
Московская обл.
Дерново-подзолистая хо-
рошо.окультуренная,
Московская обл.
Дерново-подзолистая, под
смешанным лесом
Московская обл.
Подзолистая, под
сосновым и еловым лесом,
Московская обл.
Серо-бурая лесная, под
дубово-липовыми
насаждениями, Воронежская обл.
Бурая таежная,
Алтайский край
Черноземные и
каштановые почвы Украины
Кол-во
выделенных
штаммов
227
153
543
138
302
130
344
Из них
токсичных
97
10
71
38
65
17
42
Процент
токсичных
31
6
13
27
21
13
15
Основные виды~токсичных
грибов
Penicillium funiculosum,
P. verrucosum var, cyclo-
pium,
P. purpurogenum,
P. nigricans,
P. janthinellum,
Fusarium spp.
Alternaria alternata,
P. verrucosum var. cyclo-
pi urn,
P. claviforme,
P. paxilli,
Fusarium sp.
P. nigricans,
P. paxilli,
P. verrucosum var. cyclo-
pium
P. paxilli,
P. nigricans,
P. verrucosum var. cyclo-
pium
P. nigricans
P. granulatum
P. verrucosum var. cyclo-
pium
P. purpurogenum,
P. brevi-compactum
Aspergillus flavus,
A. f и mi gat us,
P. spinulosum,
P. griseofulvum,
Fusarium spp.
200
В черноземных и темно-каштановых почвах Украины
Aspergillus flavus, Л. fumigatus, P. spinulosum, P. funiculosum, P. verm-
cosum var. cyclopium, Fusarium spp. и др. Распределение
токсичных форм среди основных групп грибов можно представить
следующим образом (табл. 25).
Наибольшее число токсичных штаммов, обитающих в почвах,
приходится на род Penicillium. Число фитотоксичных штаммов в
Таблица 25
Распределение токсичных штаммов среди почвенных грибов
Группа грибов
Число
выделенных
штаммов
Из них
токсичных
Количество
токсичных, %
Penicillium
Aspergillus
Fungi imperfecti
Всего
674
246
206
1126
198
54
24
276
30
22
12
24
этой группе составляет около 30%, несколько меньше их в группе
Aspergillus (22%) и содержание токсичных штаммов среди
других форм несовершенных грибов составляет 12%.
Род Penicillium в целом и его токсические формы наиболее
широко представлены в подзолистых и дерново-подзолистых
почвах, которые характеризуются одновременно и наибольшей
токсичностью. Представителям этого рода принадлежит основная
роль в образовании токсических веществ в почве.
Образуясь в почве, токсины поступают в растение, вызывая
серьезные изменения в процессах обмена веществ в нем и влияют
на урожай. Н. А. Красильниковым была показана возможность
поступления антибиотиков в растение из растворов и почвы.
Р. А. Скоробогатова применила флюоресцентный метод для
определения в растении флюоресцирующего токсина — цитринина, и
установила, что токсин поступает в растение и распределяется в
подземных и наземных органах. У гороха большее содержание
токсина в листьях, у пшеницы — в корнях.
Влияние токсинов пеницилловой кислоты и рубратоксина на
растение проявляется в сильном изменении содержания азота в
надземных частях гороха и вики, причем снижается содержание
как белкового, так и небелкового азота. Заметно снижается и
количество свободных аминокислот в этих растениях, однако
изменений в качественном составе их не происходит. Содержание
азота в корнях остается почти без изменений.
Влияние цитринина, внесенного в почву, проявляется в
изменении содержания азота в надземных частях гороха и вики.
Причем, в отличие от действия вышеназванных токсинов, изменения
в содержании азота под действием цитринина в основном проис-
201
ходят за счет белкового азота. Содержание небелкового азота
остается неизменным.
В растениях гороха при концентрации цитринина 100 и 200 мг/кг
почвы происходит заметное снижение содержания белкового
азота, а в растениях вики при концентрации цитринина 100 мг/кг,
наоборот, обнаруживается заметное его увеличение (табл. 26).
Таблица 26
Влияние токсинов грибов на содержание азота в растениях
Растения
Горох
Вика
Пшеница
Формы
азота
общий
небелковый
белковый
общий
небелковый
белковый
общий
небелковый
белковый
Содержание азота, % от сухого веса
контроль
2,69
0,39
2,30
3,50
0,52
2,98
—
цитринин,
100 мкг/г
2,32
0,35
1,97
4,09
0,53
3,56
—
контроль
3,96
0,65
2,31
4,70
1Д7
3,53
4,98
0,86
4,12
пеницил-
ловая
кислота,
| 100 мкг/г
2,58
0,23
2,35
4,70
1,06
3,64
4,77
0,80
3,97
контроль
4,61
0,82
3,79
4,70
0,77
3,55
4,98
0,86
4,12
рубраток-
син В,
250 мкг/г
3,11
0,41
2,72
3,83
0,39
3,44
4,06
0,32
3,74
При поступлении из почвы токсина P. purpurogenum — рубра-
токсина, в растения гороха и пшеницы сильно увеличивается
интенсивность дыхания этих растений и соответственно происходит
активация дыхательных ферментов, пероксидазы и полифенолок-
сидазы. В отдельные сроки активность этих ферментов достигает
27% по отношению к контролю.
Образование токсинов в почве служит доказательством их
экологической роли, так как, образуясь в почве, они могут влиять
на ее свойства и формирование микробных ценозов. Кроме того,
токсины, образующиеся в почве, могут поступать из почвы в
растение и оказывать существенное влияние на физиологические
процессы и химический состав растений, что может в конечном
итоге привести к значительным изменениям качества урожая.
ФУНГИСТАЗИС В ПОЧВАХ
Особым проявлением токсикоза почв может служить
фунгистазис — явление, выражающееся в сильной задержке и
подавлении прорастания спор грибов, находящихся в почве или
помещенных в ней. Задержка прорастания спор может быть
обусловлена также и внутренними причинами, определяющими
состояние покоя у спор, а не свойствами почвы, но под фунгистази-
сом принято понимать задержку прорастания, связанную с внеш-
202
ними факторами, свойственными почве. Наиболее сильно
фунгистазис проявляется в почвах с большим количеством
микроорганизмов и дефицитом питательных веществ.
Один из методов определения фунгистазиса — метод
агаровых блоков. Агаровые блоки помещают на поверхность почвы и
выдерживают определенное время. После экспозиции на почве
на поверхность агаровых блоков помещают грибные споры, а
затем определяют процент проросших спор по сравнению с
контролем.
Причиной фунгистазиса может быть дефицит элементов
питания в почвах или недоступность их для спор, прорастание которых
зависит от экзогенного питания, а также наличие специфических
ингибиторов биотической природы — физиологически активных
веществ, и абиотической — ионов металлов СаСОз, алюминия,
величиной рН.
Споры грибов, выделяющие питательные и ростактивирующие
вещества, обусловливают усиленное развитие бактерий на их
поверхности. Последние выделяют антибиотические вещества,
задерживающие прорастание спор грибов. Есть и летучие фунги-
статические вещества, образуемые в первую очередь актиномице-
тами. Часто образование таких веществ индуцируется внесением
хитина, который стимулирует развитие актиномицетов.
Фунгистазис могут вызвать и продукты разложения лигнина и этилен.
В некоторых случаях фунгистазис может определяться
дефицитом влаги, который имеет значение на ранних этапах
прорастания спор. Возможно прямое поражение спор грибов
паразитными бактериями и актиномицетами.
Одной из причин фунгистазиса может быть известкование
почвы. Внесение извести ингибирует прорастание спор определенных
видов грибов.
Исследованиями, проведенными совместно с О. Е. Марфени-
ной, было установлено, что более подвержены фунгистазису
мелкие споры, чем крупные с толстой меланизированной оболочкой
и большим запасом питательных веществ. В результате
известкования подавляется прорастание мелких спор, зависящих от
экзогенного питания, в частности грибов рода Penicillium, многие
представители которого сильные токсинообразователи.
Возможно, что подавление известью прорастания спор токси-
нообразователей — одна из причин смены доминирующих форм
и снятия с помощью извести токсикоза почв.
Фунгистазис как одно из свойств почвы может способствовать
длительному сохранению спор грибов в покоящемся состоянии,
даже если они не имеют специальных покоящихся структур, и,
в частности, тормозит прорастание спор патогенных грибов, в чем
проявляется отрицательная роль этого явления. Внесение
органического вещества стимулирует «провокационное» прорастание
спор патогенов, способствуя очищению почвы от инфекции.
Аналогичное действие оказывают корневые выделения.
203
ФИТОГОРМОНЫ, ОБРАЗУЕМЫЕ ГРИБАМИ
Фитогормоны — это физиологически активные
вещества, осуществляющие регуляцию физиологических процессов в
организме, протекающих на основе целой цепи физических и
химических изменений. Обычно они действуют не в тех частях
организма, где они вырабатываются.
Фитогормоны отличаются исключительно высокой
физиологической активностью, так как проявляют свою регуляторную
функцию в очень низких концентрациях, отличающихся на несколько
порядков от тех концентраций, в которых организму нужны
вещества субстрата. Таким образом, фитогормоны — это
высокоспециализированные регуляторы ростовых и формообразовательных
процессов на всех этапах онтогенеза организма.
В первую очередь фитогормоны вызывают активное деление
клеток или растяжение меристематической ткани, являясь
регуляторами процессов дифференциации. Наряду с этим
фитогормоны регулируют интенсивность и направленность обмена веществ
клеток и органов, уже закончивших свой рост, и влияют на их
функциональную активность. В настоящее время к фитогормонам
относят ауксины, гиббереллины, цитокинины, абсцизовую кислоту
и этилен. Первые три относят к природным стимуляторам роста,
а два вторых — к ингибиторам этого процесса. Однако все эти
фитогормоны обладают широким спектром физиологического
действия, которое включает в себя индукцию или активацию одних
процессов и подавление или ослабление других.
Все эти фитогормоны имеются и у грибов. Наиболее изучены
у этих организмов гиб>береллины. Это наиболее обширная группа
фитогормонов. К 1981 г. выделено всего 62 гиббереллина,
которые принято обозначать буквой А с разными индексами — Аь Аг,
А3 и т. д., 11 из них только грибного происхождения, 37 находятся
только в растениях и 14 являются общими для растений и грибов.
Гиббереллины, впервые обнаруженные у грибов, оказались
нормальными фитогормонами цветковых растений, открытыми у них
значительно позднее. Они представляют собой группу весьма
близких по строению тетрациклических карбоновых кислот,
относящихся к классу дитерпенов. Гиббереллины продуцируются
как вторичные метаболиты в результате конденсации ацетильных
остатков и образования изопренов. Изопрены в свою очередь
являются предшественниками каротиноидов, а также геранилгера-
ниола, из которого затем путем циклизации возникает гибберел-
ловая кислота.
Существуют эндогенные и экзогенные гиббереллины.
Эндогенные гиббереллины — это истинные фитогормоны широкого
физиологического действия, экзогенные — действуют как стимуляторы
роста растений. Они действуют на интактные растения и на
изолированные части при наличии активной меристематической зоны.
В результате происходит вытягивание междоузлий, усиление роста
боковых побегов, возрастание числа цветоносов, увеличение дли-
204
иы соцветий. Это вызвано как усилением растяжения клеток, так
и повышением митотической активности меристематической ткани.
Предполагается, что эндогенные гиббереллины участвуют в тро-
пизмах растений и в явлении апикального доминирования.
Согласно современным представлениям геотропические и
фототропические реакции растений определяются соотношением стимуляторов
и ингибиторов роста. В явлениях апикального доминирования,
которое проявляется в том, что при верхушечном росте растения
подавляется развитие боковых почек, основную роль играют
ауксины и гиббереллины.
Большое разнообразие гиббереллинов в растении
предполагает, что менее активные из них, т. е. гиббереллины с более узким
спектром действия, осуществляют функции регуляторов роста в
различные периоды жизни растения. Возможно, что в процессе
онтогенеза происходит смена гиббереллинов узконаправленного
действия, тем более что химические взаимопревращения этих
соединений легко осуществимы.
Наиболее известный и активный продуцент экзогенного гиб-
береллина — конидиальная (анаморфная) стадия фитопатоген-
ного гриба Gibberella fujikuroi, которая носит название Fusarium
moniliforme. Он образует преимущественно гиббереллин А3,
называемый часто гибберелловой кислотой. Гиббереллина А3 в
цветковых растениях немного. Гиббереллин А3, образуемый грибом,
сильно стимулирует гидролитическую активность растения,
прорастание семян, вегетативный рост и т. д. Поэтому образование
или действие А3 на растение в значительных количествах может
вызывать серьезные нарушения физиологических процессов.
В этом случае его правильнее рассматривать не как фитогормон,
а как образуемый грибом фитотоксин.
Гиббереллины у F. moniliforme были обнаружены японским
исследователем Куросава в связи с изучением болезни риса,
называемой «баканэ» или «болезнью дурных побегов»,
распространенной в Японии, Китае, Индии и ряде других южных стран.
Болезнь проявляется в сильном вытягивании междоузлий, они
начинают усиленно ветвиться; в местах междоузлий образуются
дополнительные побеги. Впервые гиббереллин А3 (гибберелловая
кислота) был выделен в кристаллическом виде из культуры
F. moniliforme, в Японии Ябута и Сумики и его сотрудниками
в 1939 г.
Экзогенные гиббереллины сильно стимулируют рост побегов,
листьев, плодов и в меньшей степени корней у растений. Они
влияют не только на рост побегов, но и на их образование. Под
влиянием гибберелловой кислоты трогаются в рост и интенсивно
растут боковые побеги, ускоряется цветение и плодоношение.
Сеянцы двулетних культур, таких, как морковь, свекла и другие,
цветут и плодоносят на первом году жизни.
Очень существенный эффект гиббереллина, полученного в
виде препарата культуры — гриба F. moniliforme — угнетение
развития семян в плодах и получение бессемянных растений. Осо-
205
бенно большое значение он имеет для получения бессемянного*
винограда в производственных условиях.
Многие сорта винограда, в первую очередь бессемянные,
реагируют на обработку гиббереллином увеличением размеров ягод.
Увеличиваются размеры плодов и у некоторых плодовых
растений.
Гиббереллин действует на активность ферментов после
обработки им растения, особенно на активность оксидазы индолил-
уксусной кислоты и инвертазы, а также на амилазную активность
семян злаков. Гиббереллин, локализованный в зародыше семени,
поступает в эндосперм и индуцирует синтез а-амилазы и других
гидролитических ферментов, способствующих мобилизации
веществ эндосперма.
Другие фитогормоны, играющие роль регуляторов роста
растений, — ауксины, которые также образуются некоторыми
грибами, в частности грибы-микоризообразователи синтезируют
ауксин.
Природный активный ауксин как растений, так и грибов
представляет собой (3-индолил-уксусную кислоту (ИУК). Ауксины в
растении индуцируют или активируют широкий спектр ростовых
и формообразовательных процессов. Передвигаясь вниз по стеблю
из его растущей верхушки, они могут ингибировать рост боковых
почек, т. е. участвуют в явлении апикального доминирования.
Ауксин грибов-микоризообразователей вызывает усиленное
ветвление корневых окончаний у растений. Его образование
установлено и у факультативно патогенных грибов Alternaria dauci,
Fusarium solani /. faseoli и Sclerotium rolfsii. Первые два
образуют ИУК в питательной среде с добавлением триптофана,
последний '•— как в присутствии триптофана, так и без него.
У некоторых микоризообразующих грибов открыты цитокини-
ны — производные аминопурина. Цитокинины грибов
представлены зеатином, его аналогами и производными. Они также
регулируют ростовые процессы, функциональную активность клеток,,
задерживают их старение.
К фитогормонам относят абсцизовую кислоту и этилен,
которые также образуются некоторыми грибами. Абсцизовая
кислота — оптически активный сесквитерпеноид, этилен —
газообразное вещество, выполняющее роль регулятора физиологических
процессов у растений, широко представлен у грибов. На основе
взаимодействия стимуляторов и ингибиторов осуществляется
процесс дифференциации в растении.
Некоторые из ростстимулирующих веществ образуются
грибами в почве. Работами Ю. А. Худяковой показана возможность
образования в почве гиббереллинов.
При внесении в нестерильную дерново-подзолистую почву
культуры продуцента гиббереллина Fusarium moniliforme,
предварительно размноженной на увлажненном сене, в почве образуется
гиббереллин, который может быть обнаружен как по биотесту
(специфическое вытягивание междоузлий гороха), так и методом
206
хроматографии. Гиббереллин может сохраняться в нестерильной
почве и оставаться физиологически активным 40—45 дней.
Значительно быстрее он инактивируется в почвах с высоким
содержанием органического вещества.
Гиббереллин адсорбируется почвой и может сохраняться в ней
в адсорбированном состоянии. Продуцируемый в почве грибами,
он всасывается растением, что можно определить по
стимулирующему эффекту растительных вытяжек на биотесты и хромато-
графически.
Таким образом, грибы представляют собой очень обширную и
многообразную группу организмов. Они разнообразны по
строению, способам размножения и циклам развития, хотя и
объединяются рядом общих черт.
В определенных экологических условиях они образуют
характерные комплексы, являющиеся неотъемлемой частью биоценоза
в целом, и вступают в различные взаимоотношения с другими
организмами. Комплекс типичных видов и изменение его
структуры могут быть индикаторами на определенный тип
почвообразовательного процесса и степень антропогенного воздействия.
Роль грибов в почвенных процессах очень разнообразна.
Обладая большим набором ферментов, они являются активными
агентами в разложении растительного опада и минерализации
органического вещества, тем самым принимая активное участие
в круговороте углерода в природе. В этом процессе наибольшее
значение их проявляется в разложении клетчатки, лигнина,
пектиновых веществ. Образующиеся кислоты влияют на
почвообразовательные процессы.
Грибы наряду с бактериями принимают активное участие в
аммонификации белковых веществ. В последнее время
установлена нитрифицирующая роль почвенных грибов.
Грибы характеризуются выраженным полихимизмом,
проявляющимся в продуцировании разнообразных вторичных
метаболитов. Им свойственно образование разнообразных пигментов.
Основные из них — каротиноиды, хиноны, меланины. Пигменты,
хиноны и особенно меланины играют значительную роль в гуму-
сообразовании. Как высокополимерные соединения, сходные по
строению с гуминовыми кислотами, они могут быть основой
образования гуминовых веществ. Некоторые грибы могут разлагать
гумус, тем самым способствуя освобождению закрепленного азота,
потребляемого растением.
Благодаря мицелиальному строению и образованию слизистых
веществ, в первую очередь полисахаридов, грибы принимают
участие в структурообразовании.
Очень значительна и разнообразна роль грибов в образовании
физиологически активных веществ — антибиотиков и токсинов,
фитогормонов. Являясь активными продуцентами этих веществ,
грибы оказывают существенное влияние на свойства почвы, ее
плодородие.
ЛИТЕРАТУРА
Аристовская Т. В. Микробиология подзолистых почв. М.—Л., 1965.
Аристовская Т. В. Микробиология процессов почвообразования. Л., 1980.
Бабьева И. П. Биология почв. М., 1983.
Берестецкий О. А. Фитотоксины почвенных микроорганизмов и их
экологическая роль. Л., 1978.
Билай В. И. Основы общей микологии. Киев, 1984.
Билай В. И., Пидопличко Н. М. Токсинообразующие
микроскопические грибы. Киев, 1970.
Великанов Л. Л., Сидорова И. И. Некоторые биохимические
аспекты в экологии грибов//Успехи микробиологии. Т. 18, 1983.
Виноградский С. Н. Микробиология почвы. М., 1953.
Горленко М. В. Миграции фитопатогенных микроорганизмов. М., 1975.
Дурынина Е. П., Великанов Л. Л. Почвенные фитопатогенные грибы-
М., 1984
Жданова Н. Н., Василевская А. И. Экстремальная экология грибо*
в природе и эксперименте. Киев, 1982.
Егоров Н. С. Основы учения об антибиотиках. М., 1979.
Звягинцев Д. Г. Некоторые концепции строения и функционирования
комплекса почвенных микроорганизмов // Вестн. Моск. ун-та. Сер. почвоведение,.
№ 17, 1978.
Кононова М. М. Формирование гумуса в почве и его разложение//Ус-
пехи микробиологии. Т. 11. 1976.
Красильников Н. А. Микроорганизмы почв и высшие растения. М.,
Кулаева О. Н. Гормональная регуляция физиологических процессов у
растений. М., 1982.
Кураков А. В. Некоторые аспекты экологии везикулярно-арбускулярной
микоризы//Сельскохозяйственная биология. 1985. № 10.
Курс низших растений/М. В. Горленко. М., 1981.
Курсанов Л. И. Микология. М., 1940.
Лобанок А. Г., Бабицкая В. Г. Микробиологический синтез белка на
целлюлозе. Минск, 1976.
Лях С. П. Микробный меланиногенез. М., 1981.
Мехтиева Н. А. Хищные нематофаговые грибы — гифомицеты. Баку
1979.
Микромицеты почв/В. И. Билай. К., 1984.
М и ш у с т и н Е. Н., Е м ц е в В. Т. Микробиология. М., 1978.
Муромцев Г. С. Гиббереллины. М., 1979.
О дум Ю. Основы экологии. М., 1975.
Орлов Д. С. Гуминовые кислоты почв. М., 1974.
Почвенные организмы как компонент биогеоценоза/Мишустин Е. Н. М.,,
1984.
Родин Л. Е., Базилевич Н. И. Динамика органического вещества и
биологический круговорот в основных типах растительности. М.—Л., 1965.
Селиванов И. А. Микосимбиотрофизм как форма консортивных связей
в растительном покрове. М., 1981.
Стриганова Б. Р. Питание почвенных сапрофагов. М., 1980.
Федоров В. Д., Гильманов Т. Г. Экология. М., 1980.
Феофилова Е. П. Клеточная стенка грибов. М., 1983.
208
Частухин В. Я., Николаевская М. А. Биологический распад и ре-
синтез органических веществ в природе. Л., 1969.
Тахтаджан А. Л. Четыре царства органического мира//Природа. 1973.
№ 2,
Агх J. A. von Pilzkunde. Ein kurzer Abriss der Mycologie unter beson—Be-
rucksichtigung der Pilze in Reinkultur//J. Cramer. Lehre. 1967.
Ainsworth a. Bisby Dictionery of fungi//7th edition. Commonwealth My-
cological institute. L., 1984.
Bartnicki-Garca S. Cell wall chemistry morphogenesis a. taxonomy of
fungi//Ann. Rev. of Microbiol. 1968. Vol. 22.
Bartniki-Garcia S. Fundamental aspects of hyphal morphogenesis//
Symp. Soc. Gen. Microbiol. 1973. N 23.
Burnett J. H. Fundamental of mycology. London, 1976.
Domsch К. Н., Gams W., Anderson Т. Н. Compendium of soil fungi.
Vol. 1. N. J. — Sydney, 1980.
Garret S. D. Root pathogenic fungi. Cambridge, 1970.
Mycology series/P. A. Lemke. Vol. 4. N. J. Basel, 1984.
Pirt Fungal grouwth and hyphal walls. 1982.
Talbot P. H. B. Principals of Fungal Taxoriomy//Glasgow, 1971.
Wittaker R. H. New concepts of kindoms of organisms//Science. 1969.
Vol. 163.
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
1би?ос„ОоВрааЯ|2ИСЛОТа204.206
Активность воды 110
Алевроспора 96
Альтеин 190
^аЖнРИ8ебВаЯ КИСЛ°Та 19°
«Амброзия> 1(48
Анае5о?Я6 ГРИбЫ U6' Ш
Анастомоз 27
Анафаза 1$
Антеридиол 43
Антеридий 39
^^т™**1™23> 205
Апотеций 73
Арбускула 137
Артроспора 29, 96
Аск (сумка) 62, 63
Аскогенная гифа 39, 63
Аскогон 39, 63
Аскокарп 64
Аскострома 67
Ауксины 204, 206
Аутоэкология 99;
Афлатоксины 188, 190, 192
Базидиокарп 81
Базидиоспора 79, 93
Базидия 79
«Баканэ» 205
Биогеоценоз 98
БГотро^ы^Гш351 СТРУКТУРа 126
Биоценоз 98 '
210
Бластоспора 9!6
Броматы 1(49
Вакуоль 16
Веретено 19'
Везикулярно-арбускулярная
микориза 136
Викторин 189, 190
Вольва 85
Временная частота встречаемости 119
Галофилия 11|
Гамета 38
Гаметангий 39
Гаметангиогамия 319
Гаметогамия 38
Гаустория 57
Гетероталлизм 61 ¦ '»
Гетеробазидия 108
Гиббереллины 204, 205, 206
Гигрофилы ПО
Гимений 75, 81, 82
Гименофор 82
Гипобазидия 89
Гиромитрин 78
Гифа 10
Глеба 87
Голоморфа 44
Голокарпические формы 30, 62
Гомоталлизм 61
Дендродохин 188, 100
Дикарион 39, 63, 64
Диктиосома 1Э
Долипора 24
Зеатин 206
Зигоспора 37, 5,8
Зигота 37, 58
Зона гомеостаза 194
Зона репрессии 196
Зона ресистентности 196
Зона стресса 194
Зооспора 30, 53, 55
Изогамия 38
Индекс видового разнообразия 123
Инициированное микробное
сообщество (ИМС) 193, 194
Исландиотоксин 192
Кариогамия 37
Кинетосома 19
Клейстотеций 67
Коллетоксин 190
Комплекс микроорганизмов 99
Конидия 33
Консорция 13(1
Консументы 98
Копрофилы 104
Коремия 36
Кохлиоболин 190
Коэффициент Съеренсена — Чеканов-
ского 11;9
Ксерофилы 80
Лизосомы 16
Ложе 36, 95
Локула 671
Ломасома 16
Люцифераза 86
Люцеферин 86
Макромицеты 100
Мастигонема 30
Мейоз 20, 37
Метафаза 18
Микромицеты 100
Микобоэтины 143
Митоз 18
Митохондрия 16
Мицелий L0
Некротрофы 101
Нематотоксины 145
Нематофаговые грибы 145
Оидия 29
Оогамия 39
Оогониол 43
Оогоний 38
Осмофилия 111
Остиола 84
Параплектенхима 28
Парафиза 83
Парентосома 24
Перидий 87
Перидиоль 88
Пеницилловая кислота 188, 191
Периконин 190
Перитеций 72
Пикнида 36, 95
Плазмогамия 37
Плазмолемма 11, 14
Планогамета 38
Плеоморфизм 44
Популяция 98
Пороспора 96
Почвоутомление 192, 193
Пробазидия 90
Прогамон 42
Пропагативные споры 46
Пространственная частота
встречаемости 119
Прототуникатные сумки 65
Профаза 18
Пряжка 79
Псевдосклероций 28
. Псилоцибин 85
Психротолерантные организмы 111
Психрофилы 1111
Пубероловая кислота 191
Редуценты 98
Ризоиды 27
Ризомицелий 52
Ризоморфа 27
Ризоплана 133
Ризосфера 133
Родоторуцин 43
Рубратоксин 190
Синэкология 99
Сиренин 43, 55
Склероций 27
Соматогамия 40
Сообщество 98
Сперматизация 39
Спермаций 39
Спикула 13
Спорангий 30
Спорангиоль 37
Споридесмолиды 13
Спородохия 36
Спорополленин 13
Спорофузарин 190
211
Стахиботриотоксин ШО, 192
Стенотопные виды ИЗ
Столон 27
Строма 74
Сукцессия 101
Сферопласт 11
Телеоморфа 44
Телиоспора 90
Телофаза 18
Тельца Воронина 24
Термотолерантные организмы 112
Термофилы 1'1Д
ТетРаПолярное распределение полов
Тремеллоген 43
Триспоровая кислота 43
Трихогина 39, 63
Унитуцикатные СуМКИ 65
Устовая кислота 191
4>аллоИДин 86
Ферромон 43)
^иалида 36, 68
<Риалоспора 96
Фимбрия 13
Фитоалексины li42
Фитотоксины 189
Фузариевая кислота 190
Хитосома 14
Хламидоспора 29
Ходацидин Ю8
Ценоцитный мицелий 10, 23
Центриоль 1,9'
Циклопальдиевая кислота 198
Цистида 83
Цитокинины 204, 206
Цитоплазматический матрикс 14
Эвритопные виды 113
Экосистема 98
Эккрисотрофы 131
Эктотрофная микориза 136
Эндомитоз 20
Эндоплазматический ретикулум 14
Эпибазидия 89
Эрготизм 74
Эукариотические организмы 10
Эукарпические формы 30
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
Absidia 60, 186
Absidia glauca 185
A corymbifera 61
A. cylindrospora 109
A. caulospora 138
Achlia 43
Achlia ambisecsualis 57
A bisecsualis 57
Achromobacter 150
Acremonium 69, 93, 94, 108, 112, 151,
174, 183, 185, 186
A charticola 116, 117, 120
Acrothecium 113, 114
A apicale 111
Actinomucor elegans 31
Acromyrmex 148, 149
Agaricaceae 86
Agaricales 83, 84, 85, 86, 140
Agaricus 85, 86
A bisporus 86
A cam pest ris 85
A rodmiani 85
Agrobacterium 183
Allomyces 42, 43, 54, 55, 174
Allomyces javanicus 174
Alternaria 94, 96, 114, 115, 116, 132,
133, 135, 147, 151, 163
A alternata 103, 111, 113, 114, 117,
120, 156, 180, 181, 182, 185, 190,
199, 200
A. dauci 206
A kuckuchiana 206
Amanitaceae 86
Amanita 85, 86, 140
A muscaria 86
A phalloides 86
A rubescens 137
A virosa 86
Ambrosiazymae 25
Anlmalia 49
Aphanomyces 57, 135
A lutiches 136
Aphillophorales 83
Aposphaeria 96, 158
A pulviscola 118, 120, 121
Armillariella 169
A mellea 86
Arthrobacter 183
Arthrobotrys 97, 145, 174
A oligospora 144
Ascobolus 77, 104
Ascomycetes 51, 52, 62, 67, 174
Aspergillus 20, 34, 68, 70, 93, 101,
103, 111, 166, 169, 171, 175, 183,
185, 186
A alliaceus 120
A caespitosus 113
A candidus 111, 191
A clavatus 191
A conicus 113
A //auws 70, 120, 172, 190, 200, 201
A fumigatus 109, 120, 156, 175, 200,
201
A glaucus 111
A /anws 113
A nidulans 41, 120, 180
A niger 113, 151, 153, 156, 166, 185,
195
A ochraceus 120
A quadrilineatus 113
A repens 111
A terras 112, 120, 191
A versicolor 183, 184
A ws/as 120, 195, 196, 197, 198
A. wentii 120, 172
Asymmetrica 68, 93
Atta 148, 149
Attomyces bpomaticus 149
Aulevria 77
A aurantiaca 77
213
Aureobasidium 114, 115
A. pullulans 102, 109, 115, 118, 121,
158, 182
Auriculares 89
Auricularia 90, 149
A. auricula 90
Azotobacter chroococcum 147
bacillus 197
Вас. cereus 150, 151
Вас. macerans 151
Вас. meghaterium 146, 151
Вас. mesentericus 146, 150
Вас. subtilis 146
Basidibolaceae 18
Basidiobolus 18
B. ranarum 18, 19
Basidomycetes 52, 174
Bipolar is sorokiniana 132
Bissochlamus fulva 136
Biverticillata 68
Biverticillata symmetrica 93
Blakeslea 36
J5. trispora 43, 61, 174
Blastocladiales 54, 174
Blastocladiella 54, 55
fi. emersonii 46, 55
Boletaceae 85
Яо/e/tts 140, 141
B. edulis 85
B. subtomentosus 141
Botrytis 132
Л. cinerea 158
Bremia 58
5. lactucae 58
Broomella acuta 120
Calycella citrina 76
Cantharellaceae 83
Cantharellus 83
Cenococcum grant] or me 140, 170, 182
Ceratostomella 165, 169
Chaetocladium 60
Chaetomella 160
Chaetomium 70, 72, 93, 102, 104, 169,
186, 194, 195
С globosum 156
С olivaceum 120
С thermophila 112
Choanephoraceae 60
Choanephora 36, 174, 185
С cucurbit arum 61
Chrisomyxa pirola 143
Chrysosporium 70, 71, 104
C. merdarium 120
C. sulfureum 103, 118, 120, 121
Chytridiomycota 52
Chytridiales 54, 174
Chytridiomycetes 52, 53, 169, 174
214
Citiulus vulgaris 176
Cladosporium 94, 96, 101, 112, 114,.
115, 132, 147, 151, 158, 163, 165,
174, 180, 182
С cladosporioides 112, 113, 120, 176,
180, 181
С fulvum 175
С herbarum 102, 103, 114, 118, 120,
121, 156, 180
Clavariaceae 83
Claviceps purpurea 74
Clavicipitales 67, 72, 74
Clitocibe 163, 164
С flaccida 164
Cochlenema 146
Cochliobolus 132
C. miaveanus 190
C. sativus 151
Colletotrichum atramentarium 13d
С fuscum
С musae 48
Со/Ша 160, 164
С. conigena 85
С. driophila 163
Conidiobolus 20
Coniophora 165
Coniophoraceae 84
Coniosporium 102
Coniothirium 95, 160, 197, 195
С. /ыс/геШ 35, 151
Coprinus 104
С lagopus 15, 81, 85
C. fimentarius 85
С or dice ps 93
Coriolus versicolor 160, 165
Corticium 101, 132
Cortinarius 140
Cortinellus berkeleanus 86
Cratarellus 83
Crucibulum 89
Cryptococcus 90
Cunninghamelaceae 60
Curvularia 113, 114, 174, 175
Cyathus 89
Cylindrocarpon 160
Dacryomycetales 90
Dacryomyces ellisii 90
Dactylum 93
Daedalia 149
Z). quercina 165
Dematiaceae 94, 133, 151, 174, 175
Dendrodochium toxicum 190, 191
Dendrophoma 96
Deuteromycetes 52
Dictiostellum discoideum 48
Dimargaritaceae 24, 59
Diplococcium resinae 182
Diplosporium 93
Dispira 25
Doratomyces 103
D. medius 114
Drechslera 132
Elaphomyces 104
Emiricella 69, 93, 104
E. nidulans 41, 51, 185
Emiricillopsis 70, 93
Endogonaceae 61, 138
Endogone 62
Endogonales 60, 61
Endomycetales 67
Endomycetes 67
Enthomophtorales 18, 24, 59
Epicoccutn 174
E. nigrum 102, 103
Euascomycetidae 51, 67
Eumycota 51, 52
Eupenicillium 93
Eurotiales 51, 67, 93
Eurotlum 69, 70, 93, 104
E. repens 70
Exobasidiales 81
Filobasidiaceae 91
Filobasidium 91
Forties annosa 84
F. fomentarius 160, 165
Fungi imperfecti 52, 174, 201
Fusarium 93, 101, 111, 132, 135, 147,
156, 158, 165, 174, 186, 199, 100,
201
F. graminearum 75, 132
F. lactis 160
F. moniliforme 117, 190, 205, 206
F. nivale 75, 120, 132, 160
F. orthoceras 109, 190
F. oxysporum 132, 190
F. oxysporum var. cubense 135, 136
F. oxysporum var. pisi 136
F. sambucinum 108
F. solani 106, 120, 132, 156, 175, 206
F. solani var. phaseoli 136
F. sporotrichiella 190, 191
Fusidium 118
Qanoderma applanatum 84
Oeastrum 87, 88
G. pectinatum 88
Oeomyces pannorum 112, 120
Geotrichum 96
Oibberella fujicuroi 205
О. геае 75, 76
Gigaspora 62, 138
Cliocladium 197
С fimbriatum 116
С penicilloides 120
Glomierella 48
Glomus 62, 138, 140
Golovinia 145
Graphium 94
Gymnoascaceae 104
Gymnoascus 70, 71, 169
Gyromytra 78
Relianthus anuus 176
Helicobasidium 90, 96
#. purpureum 90
Helicostylum 36, 60
Helminthosporium 20, 96, 135, 174,
175, 183
H. victoriae 190
Helotiales 67, 76
Яе/?/е//а 78
Hemiascomycetidae 67
Heteroascomycetidae 82
Hericium coralloides 84
Holobasidiomycetidae 82
Hormiscium 160
Humaria 104
Humicola 96, 103, 112
#. grisea 102, 103, 120
Hydnaceae 84
Hylaria 149
Hyphochitridiomycetes 52
Hyphomycetales 145, 175
Hypocrea 75
#. ru/a 75
Hypocreales 67, 72, 74
Hypomyces 93
Hypopitis monotropa 136
ixocomus luteus 85
Kaffiadinia 145
Keratinimyces ajelloi 104
Kicxellaceae 24, 59, 60
Lactarius 87, 140, 141
Laetiporus sulfureus 84
Langermania gigantea 88
L. scabrum 85
Lentinus 169
Lepiota 163
Lepiota procera 163
Leucoagaricus gongulospora 149
Leucocoprinus gongulospora 149
Leucosporidium 85
Linderina 25
Loculoascomycetidae 67
Lycoperdales 87
Lycoperdon 87, 88
L. perlatum 88
215
Macrolepiota 85
Macrophomina phaseoli 19
Marasmius 19, 160, 164, 165
Af. androsaceus 163
Af. oreadus 85
Melanconiales 94, 95, 176
Memnoniella echinata 48, 156
Metallogenium 151, 152
Metaphyta 49
Metazoa 49
Micronectriella nivales 75
Microsporium 70
Af. ajelloi 104
Monera 49
Monilia 77, 97
Af. geophila 109, 194, 195, 197
Af. sitophila 78
Moniliaceae 94, 145, 174
Monilia 76
Af. fructigena 76, 77
Monoblepharidales 54, 55
Monoblepharis 55
Monoverticillata 68, 93
Morchella 78
Morchella conica 78
Af. esculenta 78
Мог tier ella 60, 104, 107, 108, 118, 147
Af. alpina 108
Af. angulispora 106
Af. gracilis 118
Af. humilis 118
M. minutissima 108
Al лапа 118
Af. ramanniana 31, 51, 103, 108, 109,
118, 120, 121, 167
Mortierellaceae 60
Mucor 18, 36, 51, 60, 61, 102, 103,
104, 112, 163, 168, 186
M. hiemalis 60, 103, 109, 120, 174,
185
Af. lypoliticus 166
Af. mucedo 43, 44
Af. plumbeus 13, 31, 120
Af. гошш 13
Mucoraceae 60
Mucorales 25, 59, 60, 171, 174
Mycelia sterilia 96
Mycena 160
My со gone 96, 103, 163
Af. nigra 102, 109, 120, 151, 195, 197
Mycophyta 49
Mycospherella punktiformis 163
Mycota 49, 51, 52
Myrothecium striatosporum 106
Nadsoniella nigra 176, 180
Nectria 93
Nematophagus 145
Neocosmospora vasinfecta 113
Neosartoria 69, 93
N. fisheri 113
N euros pora 20, 40, 73, 174
N. crassa 12, 20, 21, 25, 27, 73, 171
N. sitophila 73
N. tetrasperma 47, 73
Nidulariales 88
Nidularia 89
Oedocephalum 93
Oidiodendron 96, 97, 103
O. gracile 120
O. griseum 194
Olpidium 54, 135
O. brassicae 54, 132
О. ша'ае 54
Oomycetes 52, 55, 56
Oomycota 52
Ophiobolus 135
O. graminis 111, 132
Paecilomyces 118, 120, 121
P. lilacinus 106, 120
P. varioty 156
Papularia 133
Penicillium 20, 34, 68, 93, 96, 101, 103,
112, 117, 132, 158, 163, 165, 167,
175, 183, 192, 197, 200, 201, 203
P. brevi-compactum 120, 200
P. citrinum 191, 198, 199
P. chrysogenum 113, 120, 185
P. cinereo—atrum 120
P. claviforme 120, 153, 191, 199, 200
P. daleae 118, 120, 121, 156, 167
P. decumbens 184
P. digitatum 120
P. egyptiacum 113
P. fellutanum 153
P. frequentans 102, 108, 112, 113, 118,
120, 121, 183, 184, 200
P. funiculosum 109, 111, 113, 120, 194r
196, 198, 200, 201
P. granulatum 199
P. griseofulvum 191, 198, 200
P. janthinellum 106, 108, 109, 120, 192,
195, 196, 197, 198, 199, 200
P. implicatum 191
P. lanosum 195
P. lapidosum 120, 191
P. lividum 120
P. Шешп 108
P. nigricans 106, 109, 120, 199, 200
P. notatum 153, 167
P. рах/Ш 120, 200
P. purpurogenum 70, 120, 151, 190,
194, 196, 198, 200, 202
P. raciborskii 120
P. roquefortii 166
P. restrict um 120
P. roseo-purpureum 120
216
P. rub rum 70, 190
P. simplicissimum 109, 120, 121
P. spinulosum 120, 200, 201
P. tardum 120
P. thomii 102, 120, 121
P. velutinum 197
P. vermiculatum 67, 68, 69
P. verrucosum var. cyclopium 109,
120, 121, 153, 166, 191, 194, 198
P. verrucosum var. verrucosum 156
Periconia circinata 190
Peronospora 58
P. destructor 58
P. manshurica 16
P. tabacina 48
Peronosporaceae 57
Peronosporales 56, 57
Peyronella 133
Peziza 77
Pezizales 67, 76, 77
Phallales 87, 89
Phallus impudicus 89, 163
Phialophora 96, 103, 107, 121
Phoma 96, 114
Phomopsis viticola 48
Phycomyces 18, 111, 151
P. blakesleanus 43, 174
Phytophthora 16, 43, 57, 132, 135
P. infestans 56, 57
Pt/ana 104
Pilobolus 104
Piptocephalidaceae 61
Piptocephalis 25 i
Pitomyces char tar urn 13
Planta 49
Plasmopara 58
jP. viticola 58
Pleurage 73
Pleurotus ostreatus 86, 165
Pluteus 86
jP. cer virtus 86
Polyporaceae 84
Polyporus mylittae 28
P. squamosus 165
Polyscitalum 114
P. pustulant 114
Polystictus 19
Porm vapor aria 165
Proteus vulgaris 150
Pona 84
Pseudomonas 151, 183
Ps. fluorescens 150
Protomyces 174
Protomycetales 174
Psylocybe mexicana 85
Puccinia graminis 47
Pyronema omphaloides 64
Pythiaceae 57
Pythium 26, 43, 57, 101, 132, 135
P. debarianum 16, 57, 132
P. ulmorum 132
Ramaria flava 83
Rhizoctonia 96, 132, 139, 140, 151,
136
P. crocorum 90
P. so/am 136, 176, 190
Rhizophlyctes rosea 174
Rhisopus 27, 60, 61, 111, 186
P. nigricans 60, 109, 166
P. oryzae 61
P. stolonifer 59, 60, 61
Rhodosporidium 43, 91
P. toruloides 91
Rhyporobius 104
Russula 87, 140
Russulaceae 87
Saprolegnia 18
Saprolegniales 56
Sarcoscypha 77, 174
S. coccinea 77
Schizophillum 19
Sclerocystis 62
Scleroderma 88
S. aurantium 88
Sclerodermales 87
Sclerotinia 76, 132
S. sclerotiorum 77, 158
Sclerotium agricola 109
5. cepivorum 136
5. ro//s*7 206
Sep/a officinalis 176
Serpula lacrimans 84
Sordaria 73
S. fimicola 73
Sparassis crispa 83
Spathularia flavida 76
Sphaeriales 67, 72, 73
Spharopsidales 94, 95, 175, 176
Spirodactylon 60
Spongospora subterranea 136
Sporobolomycetaceae 90
Sporormia australes 25
Sporotrichum 118
5. thermophila 112
Stachybotrys 97, 103
5. afra 114, 120, 180, 185, 190
197, 198
Staphylotrichum coccosporium 11
StemphyIlium 94, 114, 115, 116,
«S. botryosum 180, 181
5. i7/i«s 114
Stereum sanguinolentum 146
Stilbaceae 94, 176
Streptomyces 150, 195, 197
Stropharia 104
Stylopage 146
Sw7/«s 140
5. conturnatus 137
S. granulatus 140
5. grevilei 86
S. luteus 140
S. punctipes 137
Synchytrium 54
S. endobioticum 54, 132
Talaromyces 68, 93
7. enterssonii 112
7. /7ay«s 67, 68, 109, 117, 194,
196
Taphrinales 67, 174
Teliomycetidae 81, 82y 90
Tetracoccosporium 114
Thamnidiaceae 60
Thamnidium 36, 60
Thelephora terrestris 83
Thelephoraceae 83
Thermoascus aurantiacus 112
Thielavia 104, 108
Thielaviopsis 132
Thysanophora 103
7. penicilloides 121
7orw/a 160
Tremella mesenterica 90
Trmellales 90
Trichocladium 97
7. asperum 35, 183
Trichocomataceae 51
Trichoderma 102, 103, 163, 171, 2i
7. a/fca 120, 121
7. hamatum 153
7. harzianum 153
7. koningii 118, 120, 153
7. lignorum 35, 109, 185
7. pseudokoningii 153
7. u/rute 75, 109, 120, 156
Trichophyton 170
7. terrestre 104
Trichothecium 94, 174
7. roseum 183
Trichurus 94
7. spirales 35
7w6er 140
7. aestivum 78
7. melanosporum 78
Tuberales 67, 76, 78
Tubercularia 93, 94
Tuberculariaceae 94
Typhula 83
Venturia tremulae 163
1/erpa 78
Verticillium 94, 132, 135, 158, 174
F. a/6o—a/rwm 111
V. <Шшт 120
V. nigrescens 29
1Л terrestre 103, 118, 120, 121
Volutella 94
V. esculenta 86
Zoopagaceae 146
Zoopagales 146
Zoopage 146
Zygomycetes 18, 52, 58, 60, 174
Zygorhinchus 60, 107
Z. vuielemini 109
ОГЛАВЛЕНИЕ
Введение « • 3
Часть I. ОБЩАЯ ХАРАКТЕРИСТИКА ГРИБОВ 10
Глава 1. Основные черты строения грибов Ю
Основные структуры грибной клетки Ю
Деление ядра IS
Гетерокариоз у грибов 21
Строение грибного мицелия 22
Глава 2. Размножение и развитие грибов 28
Вегетативное размножение 29
Бесполое размножение 30
Половое размножение 37
Пар асексуальный процесс 41
Гормональная регуляция полового процесса грибов .... 42
Онтогенез грибов (циклы развития) и смена ядерных фаз . 44
Дифференциация грибов в онтогенезе 45
Споры грибов и их прорастание 46
Глава 3. Основные таксономические группы грибов 49
Класс Chytridiomycetes 52
Класс Oomycetes 55
Класс Zygomycetes 58
Класс Ascomycetes 62
Класс Basidiomycetes 78
Подкласс Holobasidiomycetidae 82
Подкласс Heterobasidiomycetidae 89
Подкласс Teliomycetidae 90
Класс Deuteromycetes (Fungi imperfecti) 92
Часть II. ЭКОЛОГИЯ ПОЧВЕННЫХ ГРИБОВ 98
Г,л а в а 4. Общие сведения по экологии грибов 98
Место и роль грибов в биогеоценозах 98
Сукцессии грибов в биогеоценозах 101
Зависимость видовых популяций почвенных грибов от
факторов внешней среды 106
Почвенные грибы основных природных зон 119
Использование комплекса грибов-микромицетов для оценки
антропогенного воздействия 122
Количественное содержание грибов в почвах и их
биоморфологическая структура 125
Глава 5. Взаимоотношения почвенных грибов с другими организмами . 131
Взаимоотношения грибов с растениями 131
Взаимоотношения грибов с почвенной микрофауной . . 143
Взаимоотношения почвенных грибов с другими
микроорганизмами ... 150
219
Часть III. РОЛЬ ГРИБОВ В ПОЧВЕННЫХ ПРОЦЕССАХ
154
Глава 6. Роль грибов в превращении природных субстратов 154
Разложение грибами углеродсодержащих соединений
растительного происхождения .154
Роль грибов в подзолообразовательном процессе .... 166
Глава 7. Роль грибов в процессах превращения азота 169
Глава 8. Пигменты грибов и их роль в процессах гумусообразования . 173
Пигменты-каротиноиды 173
Пигменты-хиноны 174
Темные пигменты—меланины 175
Роль грибов в гумусообразовании 177
Роль грибов в разложении гумуса 182
Роль грибов в структурообразовании 184
Г л а в а 9. Специфические физиологически активные вещества грибов . 187
Антибиотики и токсины почвенных грибов 187
Роль грибов в токсикозе почв 192
Фунгистазис в почвах 202
Фитогормоны растений, образуемые грибами . . . . . 204
Литература 208
Предметныйуказатель . . . . , „ > . . . , . 210
Указатель латинских названий ,..»,.... 213
УЧЕБНОЕ ПОСОБИЕ
Мирчинк Татьяна Георгиевна
ПОЧВЕННАЯ МИКОЛОГИЯ
Зав. редакцией Н. М. Глазкова
Редактор Н. А. Жук
Художественный редактор М. Ф. Евстафиева
Переплет художника Б. С. Казакова
Технический редактор Л. Р. Черемискина
Корректоры М. И. Эльму с, JVL А. Мерецкова
ИБ № 3023
Сдано в набор 23.06.87.
Подписано в печать 17.12.87.
Л-62401. Формат 60X90/16. Бумага типогр. № 1.
Гарнитура литературная. Высокая печать
Усл. печ. л. 14,0 Уч.-изд. л. 16,73.
Тираж 2940 экз. Заказ 131 Цена 90 коп.
Изд. № 4948
Ордена «Знак Почета»
издательство Московского университета.
103009, Москва, ул. Герцена, 5/7.
Типография ордена «Знак Почета» изд-ва МГУ.
119899, Москва, Ленинские горы.