/



























































































Author: Шмидт Р.
Tags: физиология животных биофизика, биохимия и физиология животных и человека физиология
Year: 1984



Text
основы СЕНСОРНОЙ ФИЗИОЛОГИИ
..ЯРЕЙ1». * .шт! i.umi W.WI»|»I II' III W..1 .... w
Fundamentals of Sensory
Physiology
Edited by Robert F. Schmidt
With Contributions by Helmut Altner, Josef Dudel, Otto-Joachim Grusser, Ursula Grusser-Cornehls, Rainer Klinke, Robert F. Schmidt, Manfred Zimmermann
Translated by Marguerite A. Biederman-Thorson
Second, Corrected Edition
Springer-Verlag
New York Heidelberg Berlin 1981
основы СЕНСОРНОЙ ФИЗИОЛОГИИ
Под редакцией Р. Шмидта
Перевод с английского
канд. физ.-мат. наук г. и. рожковой
под редакцией
чл.-корр. АН СССР А. Л. БЫЗОВА
Москва «Мир» 1984
ББК 28.903 075
УДК 591.1
Основы сенсорной физиологии: Пер. с англ./Под ред. 075 Р. Шмидта-М.: Мир, 1984.-287 с., ил.
Книга специалистов из ФРГ, выдержавшая три издания на немецком языке и два на английском, посвяшена одному нз самых интересных разделов нейрофизиологии. Рассмотрены общие вопросы сенсорной физиологии, строение и функции соматовисцеральной, зрительной, слуховой, вкусовой, обонятельной систем, механизмы, лежащие в основе чувств равиовесия. голода и жажды. Материал изложен на современном уровне, компактно и четко.
Предназначена для студентов и аспирантов университетов н медицинских институтов, а также для специалистов в области нейрофизиологии.
2001040000-140 0---------------133-84, ч. 1
041 (01)-84
ББК 28.903 5А2.2
Редакция литературы по биологии
© by Springer-Verlag Berlin Heidelberg 1978, 1981.
All Rights Reserved. Authorized translation from English language edition published by Springer-Verlag Berlin-Heidelberg-New York.
© Перевод на русский язык, «Мир», 1984
Предисловие редактора перевода
Предлагаемая вниманию читателя книга представляет собой учебное пособие по сенсорной физиологии, в котором материал излагается, начиная с азов; предполагается лишь знание читателем самых основ нейрофизиологии. Именно в этом главное отличие этой книги от большинства других переводных и отечественных изданий подобного рода, рассчитывающих на солидный исходный уровень знаний и стремящихся отразить новейшие достижения в рассматриваемой области физиологии. Книга написана коллективом авторов, каждый из которых, будучи специалистом в определенной области, подготовил 1-2 главы. Это обеспечило достаточно высокий общий уровень изложения, хотя главы написаны не равноценно. Кроме конкретных сведений о морфологии и физиологии различных сенсорных систем, даются некоторые общие представления о свойствах рецепторов, принципах обработки сенсорных сигналов, соотношении нейрофизиологического и психофизического подходов. Включены также сведения о чувствах голода и жажды, обычно выпадающих из поля зрения сенсорной физиологии. Все это создает представление о сенсорной физиологии как о составной части нейрофизиологии и физиологии вообще, а также как об основе психофизики восприятия. Жаль только, что в книге полностью отсутствуют упоминания об электрорецепторной системе рыб, интенсивно и интересно изучаемой в течение последнего десятилетия. Вероятно, это связано с ориентацией авторов в первую очередь на физиологию человека и соответственно ее медицинские аспекты.
Книга не заменяет курса лекций. В каких-то отношениях она содержит значительно больше материала, чем любые лекции. В то же время она не составляет им конкуренции хотя бы уже в силу слишком «учебного» стиля изложения (усиленного школьной системой контрольных вопросов и ответов на них). Специальные курсы лекций, более подробно и динамично излагающие те или иные вопросы сенсорной физиологии, явились бы хорошим дополнением пособию, компенсируя его неизбежную лаконичность, недостаток новейших сведений и подходов, а также некоторую статичность изложения. Следует подчеркнуть, что пособие будет полезно не только студентам, которые должны знать «все», но и преподавателям, а также и научным работникам, узкая специализация которых оставляет широкие бреши в системе их представлений о сенсорной физиологии.
А. Л. Бызов
Предисловие ко второму изданию
Первое английское издание этой книги вышло три года назад; за это время авторы получили много полезных отзывов читателей. При подготовке этого исправленного издания мы тщательно проанализировали каждую главу, улучшив и расширив текст там, где это было необходимо; в этом нам очень помогли присланные замечания. Дальнейшие отзывы об этом издании будут для нас весьма ценными.
Мне снова хотелось бы от имени всех авторов выразить благодарность сотрудникам издательства „Шпрингер” за быструю публикацию книги.
Киль, июль 1981 г. Роберт Ф. Шмидт
Предисловие к первому изданию
В области сенсорной физиологии мы имеем дело с вопросом о том, каковы функции наших органов чувств и связанных с ними центральных нервных структур и каким образом эти функции осуществляются. Исследования здесь не ограничиваются описанием физико-химических реакций, протекающих в этих структурах; фундаментальный интерес представляют также те условия, в которых возникают ощущения и восприятия, и законы, которые управляют ими. Таким образом, сенсорная физиология требует, чтобы пытливая мысль того, кто хочет-или должен-это делать, доискивалась до потенциальных возможностей и ограничений человеческого опыта.
Нашей целью является введение студентов, имеющих минимальные предварительные знания по биологии и другим естественным наукам, в сферу сенсорной физиологии.
Первое английское издание было переведено с третьего немецкого издания, появившегося в 1977 г. Два первых немецких издания были исключительно хорошо приняты студентами - физиологами, медиками, биологами и психологами. При подготовке третьего издания мы тщательно переработали текст, в некоторых местах дополнили его и снабдили новыми иллюстрациями; во всем этом нам постоянно помогали г-н Вольф-Рюдигер Гэй и миссис Барбара Гэй из Штутгарта. Благодаря этому книга была приближена к современному уровню и сделана более удобочитаемой. От имени всех авторов я выражаю искреннюю благодарность за ту помощь и поддержку, которую нам оказывали со всех сторон, особенно сотрудничавшие с нами секретари.
Обращаясь к этому изданию на английском языке, я считаю своим долгом выразить большую благодарность д-ру Маргарите Бидерман-Торсон из Оксфорда (Великобритания) за ее прекрасный перевод. Наконец, я особенно благодарен издателям, руководству и персоналу „Шпрингер-Ферлаг” за тесное сотрудничество и тщательность, с которой они подошли к подготовке книги.
Киль, июнь 1978 г.
Роберт Ф. Шмидт
1. ОБЩАЯ СЕНСОРНАЯ ФИЗИОЛОГИЯ, ПСИХОФИЗИКА
Дж. Дудел
В качестве введения в частные аспекты сенсорной физиологии, обсуждаемые в этой книге, мы рассмотрим предмет «общей сенсорной физиологии»-принципы, лежащие в основе любого сенсорного восприятия. Такое обобщение и возможно, и полезно, хотя бы потому, что различные органы чувств весьма сходны между собой как по своей организации и работе, так и по своим связям с центральной нервной системой (ЦНС). Вместе с тем, изучая сенсорное восприятие человека, мы сталкиваемся с проблемой субъективности. Здесь имеется в виду, что внешние стимулы и соответствующие реакции наших органов чувств сопоставляются с отчетами испытуемого о своих ощущениях и представлениях. Так, например, даже физиология мышц имеет некоторую субъективную «психологическую» сторону. Субъект связывает свое «я» с определенными движениями своих конечностей; он «желает» совершить эти движения или «выражает себя» с их помощью. Но по сравнению с этим разумные аспекты сенсорной физиологии высокого уровня кажутся намного богаче и удивительнее. Мы переживаем наши ощущения как сугубо личные события, от которых зависит наше настроение; в среде, из которой исключены сенсорные стимулы-в условиях «сенсорной депривации»,-мы становимся неуравновешенными и больными. Значит, человеческое существо-это «не что иное, как сумма своих переживаний» (D. Hume). Действительно, некоторые философские школы находились под столь большим впечатлением от наличия сильного субъективного компонента во всем сенсорном опыте, что признавали существующим только субъекта, а «окружающую среду» считали продуктом мозга. Эта общая психофизическая проблема, с которой мы так вплотную сталкиваемся в сенсорной физиологии, не может быть разрешена естествоиспытателями, по крайней мере в настоящее время. Психофизические вопросы встают в очень сходной форме для всех органов чувств. Следовательно, наряду с обсуждением основных механизмов функционирования сенсорных систем глава по общей сенсорной физиологии должна содержать введение в трудные вопросы субъективного чувственного опыта.
1.1. Основные понятия общей сенсорной физиологии
Органы чувств. Мы воспринимаем окружающий мир и события, происходящие внутри нас, не непосредственно и не суммарно, а при помощи специализированных органов чувств. Наиболее известные из этих органов-глаз, ухо, кожа как орган осязания, язык как орган вкуса и нос
8
Дж. Дудел
как орган обоняния. Каждый из этих органов устроен таким образом, что он реагирует на определенный диапазон влияний окружающей среды и передает соответствующую информацию в ЦНС. Диапазон стимулов, к восприятию которых приспособлены наши органы чувств, можно объяснить в свете филогенеза. Мы замечаем только такие события в окружающей среде, которые были существенны для выживания в данной среде тех приматов, от которых мы произошли. Возьмем для примера электромагнитные волны, действующие на поверхность нашего тела. Мы не ощущаем действия гамма-излучения, рентгеновских или ультрафиолетовых лучей. Наш глаз воспринимает только излучения с длинами волн между 350 и 800 нм, для которых атмосфера Земли относительно прозрачна. С другой стороны, мы не видим инфракрасного света, но ощущаем длинноволновые тепловые излучения посредством тепловых рецепторов кожи. Во всем своем спектре радиоволны не вызывают ощущений у человека. Но другие животные приспособились к внешней среде, очень отличающейся от нашей, развив иные органы чувств. Так, например, некоторые рыбы, живущие в очень мутной воде, имеют сенсорные органы, которые чрезвычайно чувствительны к изменениям силы электрического поля. С их помощью рыбы обнаруживают изменения электрического поля, связанные с импульсами тока, который они сами генерируют. Эта информация используется для ориентации: данный механизм подобен эхолокации или радиолокации.
Модальность, качество, специфические сенсорные стимулы. Каждый орган чувств обеспечивает сенсорные впечатления, которые могут варьировать по интенсивности, но сходны по качеству. Совокупность сходных сенсорных впечатлений, обеспечиваемых определенным органом, называется чувством или формально более точным термином-модальностью. В число модальностей входят классические «пять чувств»: зрение, слух, осязание, вкус и обоняние. Однако легко добавить сюда и другие модальности. Так, например, кожа воспринимает не только давление и прикосновение, но также холод и тепло, вибрацию и боль. Вдобавок к этим модальностям, охватывающим сенсорные впечатления, порождаемые внешней средой, действующей на поверхность тела, имеются и другие модальности, обеспечиваемые сенсорными органами внутри тела и отражающие его собственное состояние. Примерами таких модальностей могут служить чувство равновесия и наше представление относительно положения конечностей или мышечной нагрузки. Кроме того, есть модальности, связанные с информацией о таких состояниях нашего организма, которые мы не осознаем или осознаем лишь косвенно. Среди прочих сюда входят осмотическое давление крови (жажда) или напряжение СО2 в крови (одышка), а также степень растяжения легких и желудка. Определение «модальность» для этих интероцептивных «чувств» также годится. В каждом случае они объединяют группу сенсорных впечатлений, которые сходны между собой и обеспечиваются специальным органом чувств. Таким образом, число модальностей значительно больше пяти.
1. Общая сенсорная физиология, психофизика
9
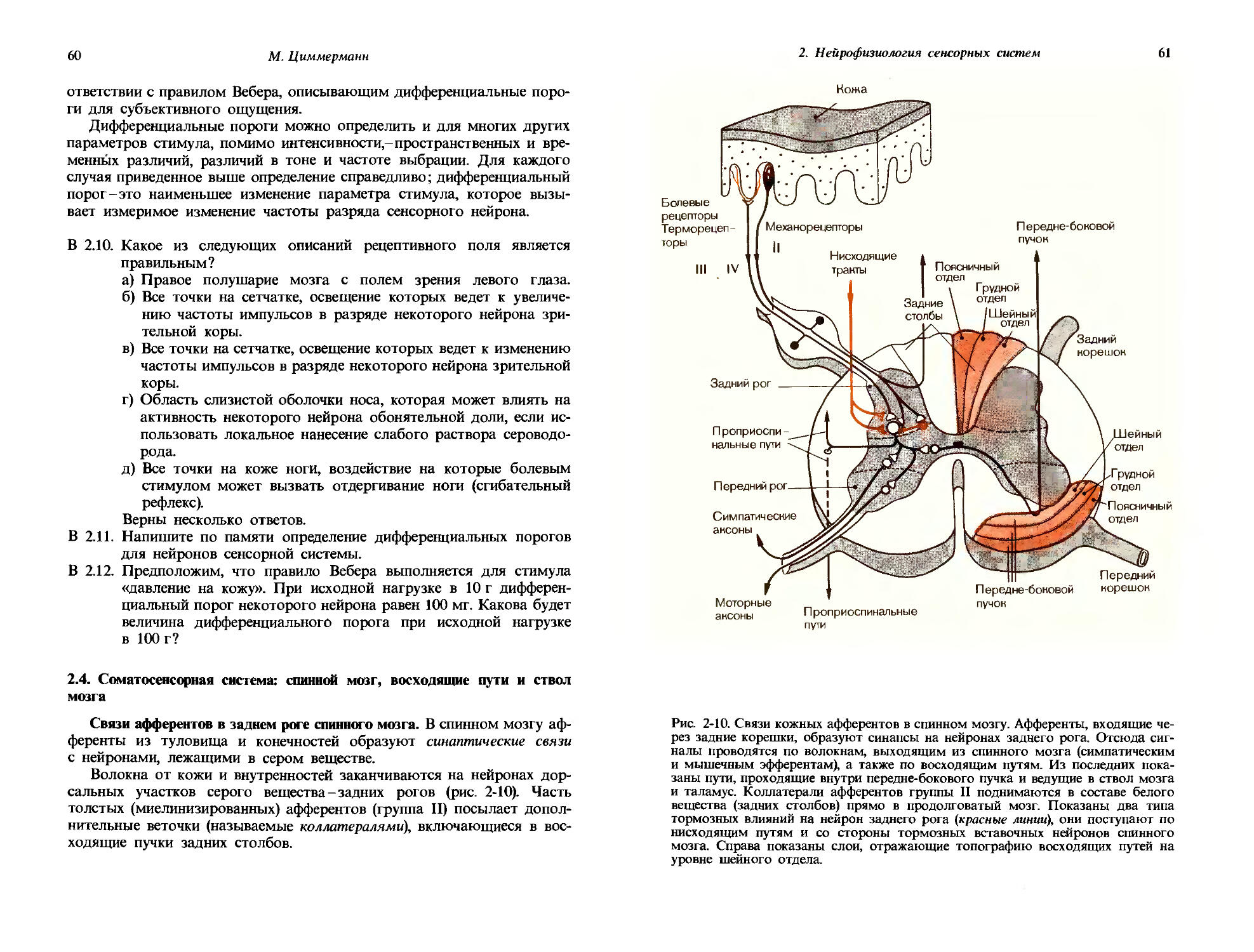
Внутри каждой отдельной модальности оказывается возможным провести дальнейшее разделение в соответствии с видом сенсорного впечатления, или его качеством. Так, например, модальность «зрение» может быть охарактеризована качествами: светлота (позиция на серой шкале), краснота, зелень и синева. Соответствующими качествами для слуха являются высоты различных тонов; качествами вкуса являются сладкое, кислое, соленое и горькое.
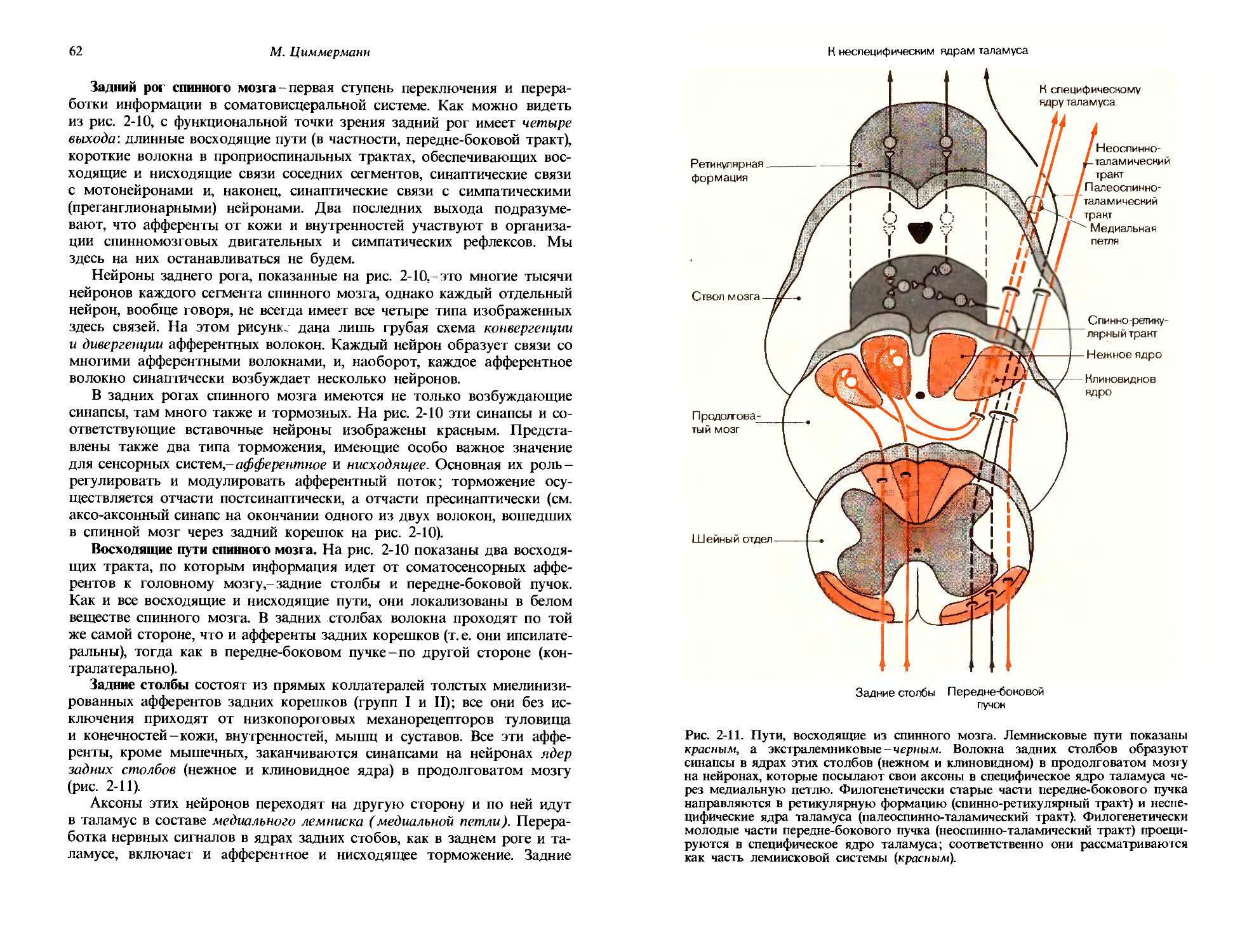
Сенсорное впечатление определенного качества возбуждается тогда, когда на данный сенсорный орган воздействует соответствующий фактор внешней среды. Качество «кислое» ощущается, когда на язык попадает кислота. Факторы, которые возбуждают сенсорные впечатления определенного качества, называются специфическими сенсорными стимулами, или просто стимулами. Стимулы обретают свое качество, вступая в реакцию с обнаруживающими их клетками органов чувств -рецепторами. Эти клетки приспособлены реагировать так сильно и избирательно, как только это возможно, на стимулы определенного специфического качества, что достигается как их местоположением, так и присутствием в них специализированных клеточных органелл. Органы чувств располагаются в местах воздействия их специфических стимулов: вкусовые рецепторы-на языке, световые рецепторы-в сетчатке глаза. Различные типы рецепторов имеют особые свойства, которые гарантируют максимальную возможную чувствительность к качеству специфического стимула. Приведем снова как пример зрительные клетки сетчатки. Каждая из них содержит пигмент, который поглощает свет «своего» специфического качества. Детали этой специализации рецепторов для специфических качеств описаны в следующей главе. В общем специфические стимулы вызывают изменения потенциала рецепторных клеток (рис. 1-1); они в свою очередь генерируют потенциалы действия, которые направляются к центрам по афферентным волокнам. Эти потенциалы действия одинаковы для всех сенсорных качеств. Качество информации, которую они содержат, полностью определяется типом рецепторов, от которых отходят данные волокна. Даже когда такие рецепторы стимулируются сильным «неспецифическим» стимулом, он интерпретируется как специфический: при ударе по глазу мы видим свет -«искры». Еще в прошлом веке для описания этого феномена Иоганн Мюллер сформулировал «закон специфических нервных энергий».
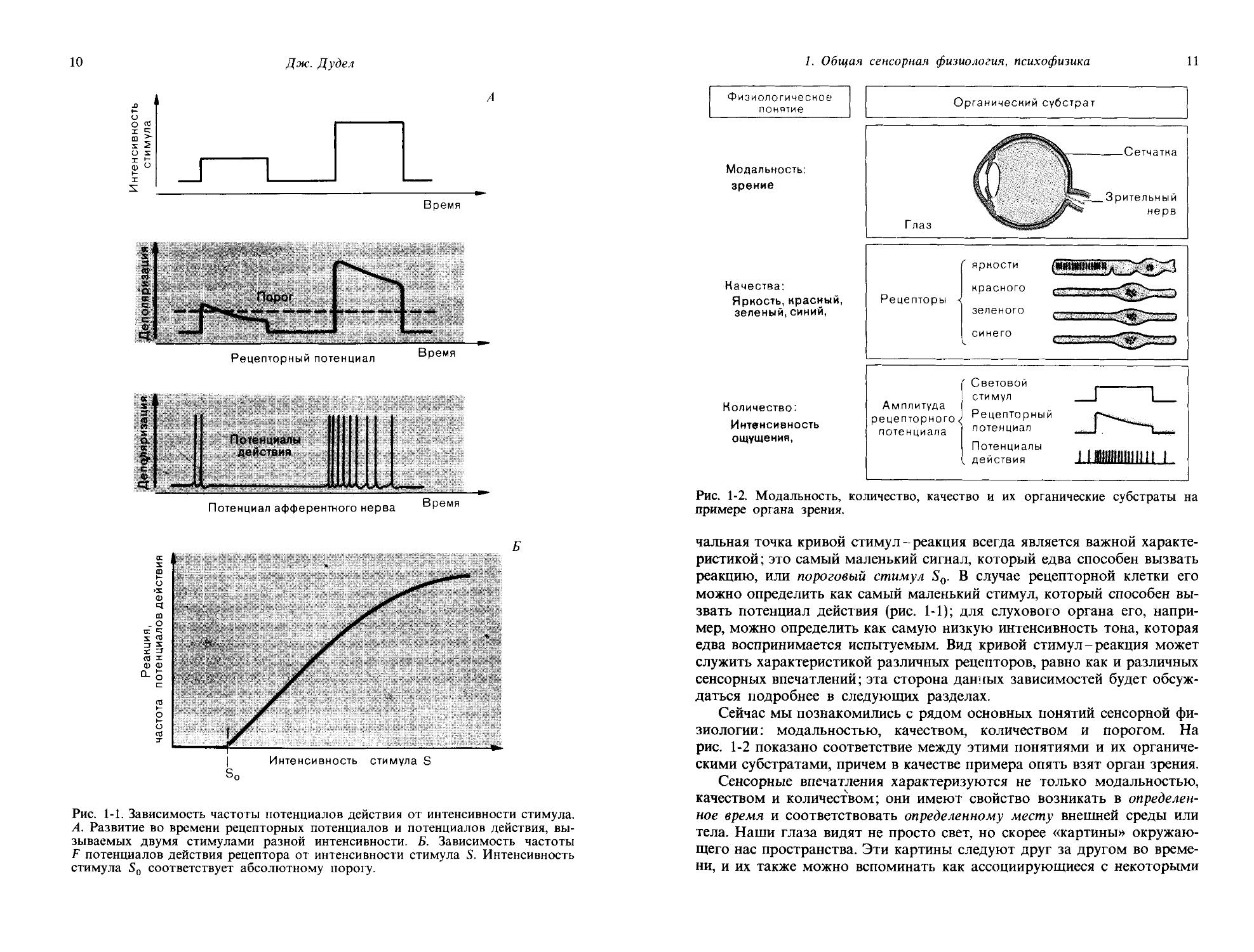
Количество, порог. В то время как вид сенсорного впечатления определяется его модальностью и качеством, его интенсивность можно назвать количеством. Количественная характеристика сенсорного впечатления соответствует силе стимула. На рис. 1-1 показан общий характер реакций рецептора на стимулы нарастающей интенсивности. Рецепторный потенциал становится все больше, и частота генерируемых потенциалов действия увеличивается. На рис. 1-1, Б показано соотношение между интенсивностью стимула и реакцией, выдаваемой рецептором. Такие зависимости можно определять для разных уровней нервной системы, равно как и для субъективного впечатления и восприятия. На-
10
Дж. Дудел
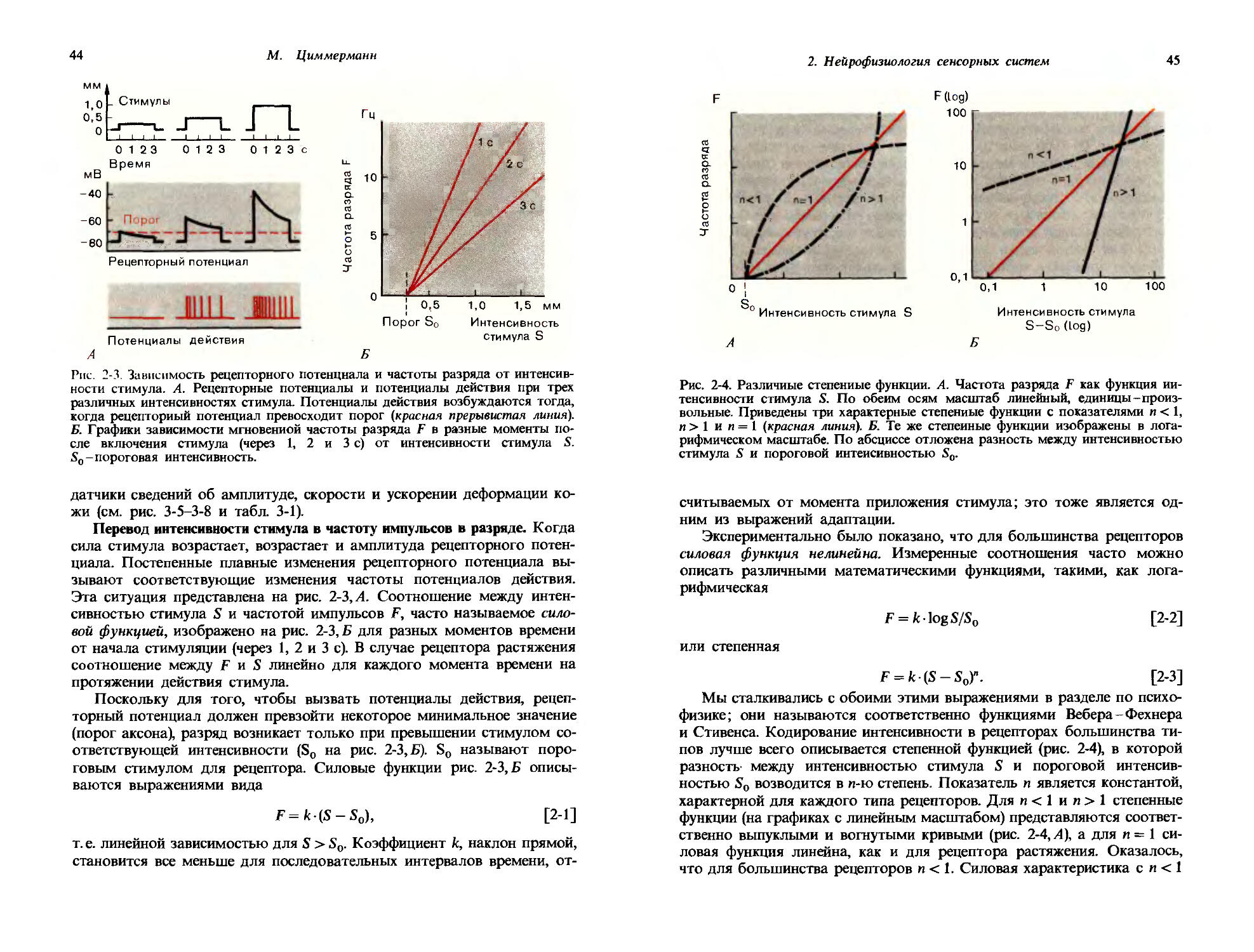
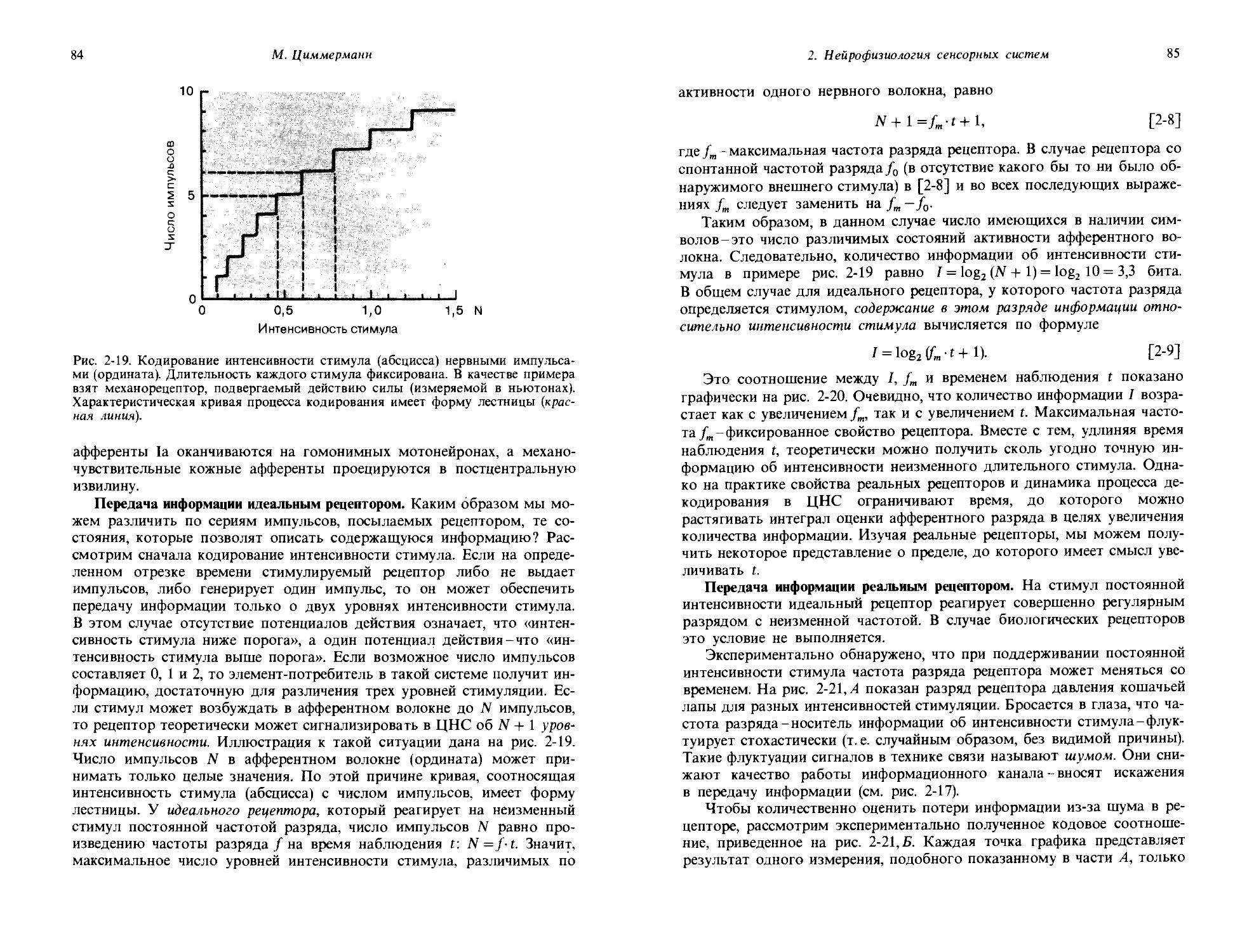
Рис. 1-1. Зависимость частоты потенциалов действия от интенсивности стимула. А. Развитие во времени рецепторных потенциалов и потенциалов действия, вызываемых двумя стимулами разной интенсивности. Б. Зависимость частоты F потенциалов действия рецептора от интенсивности стимула S. Интенсивность стимула So соответствует абсолютному порогу.
1. Общая сенсорная физиология, психофизика
11
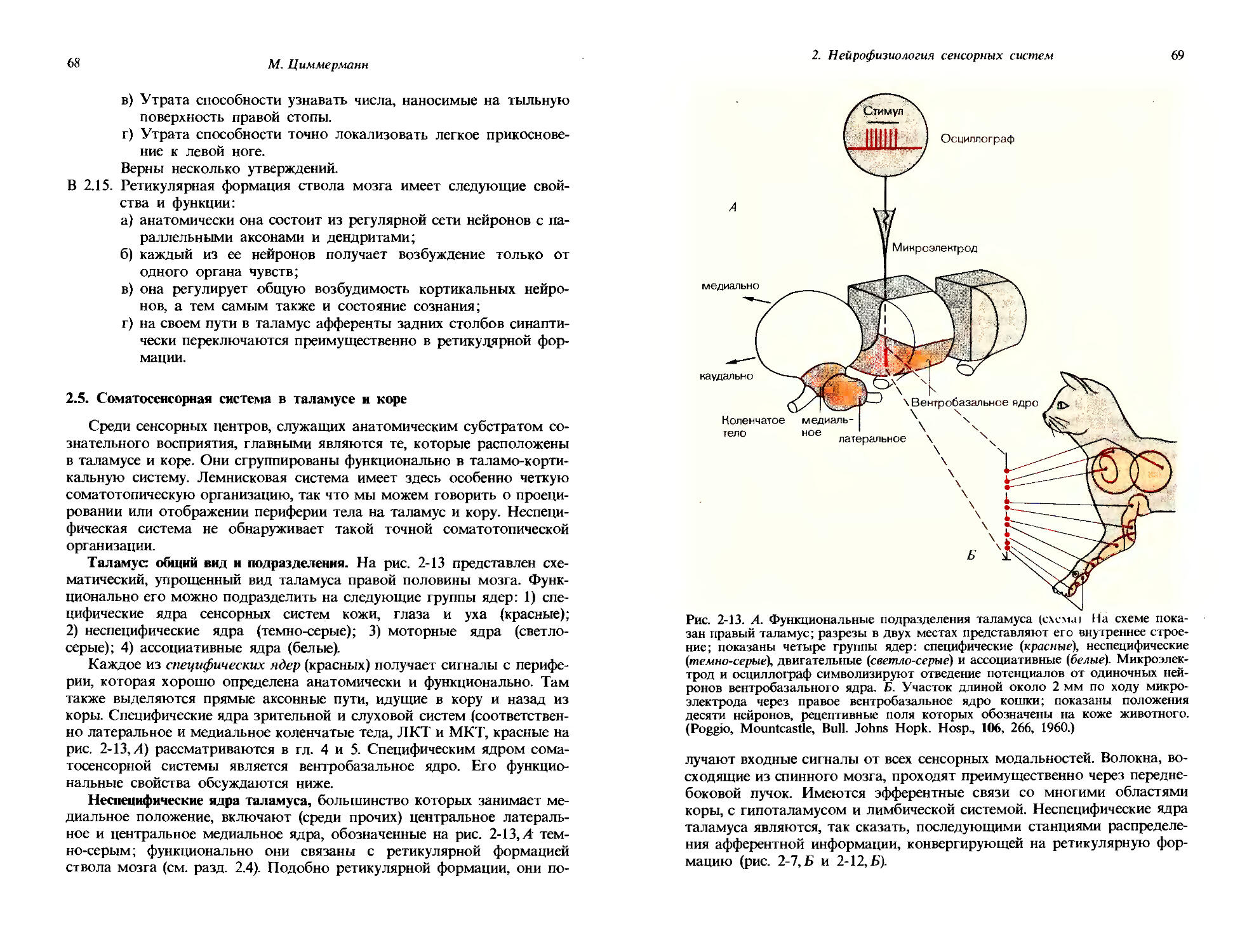
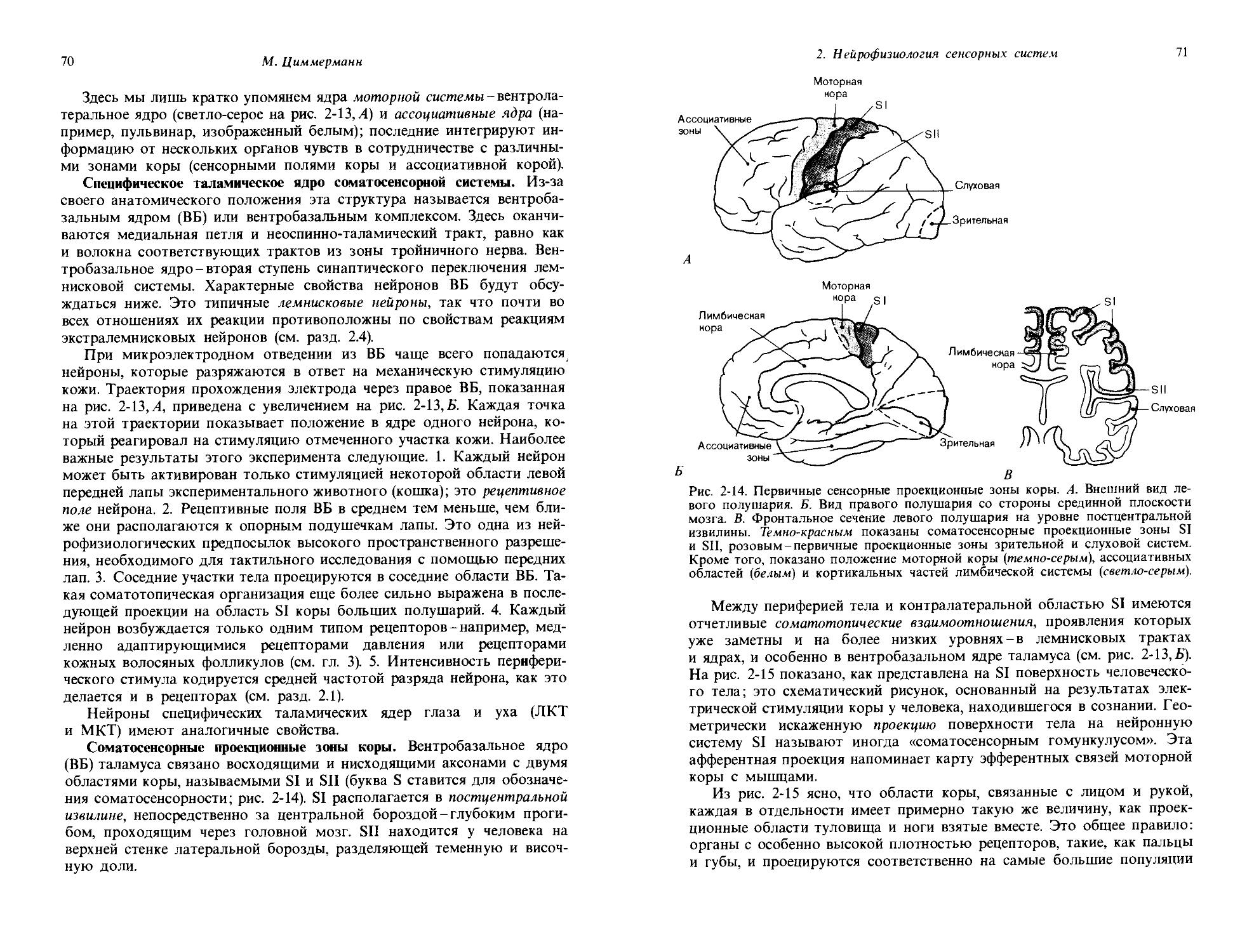
Физиологическое понятие
Качества:
Яркость, красный, зеленый, синий,
Органический субстрат
' Световой стимул Амплитуда
рецепторного^ Рецепторный потенциала | потенциал
Потенциалы
I действия
Модальность: зрение
Количество:
Интенсивность ощущения,
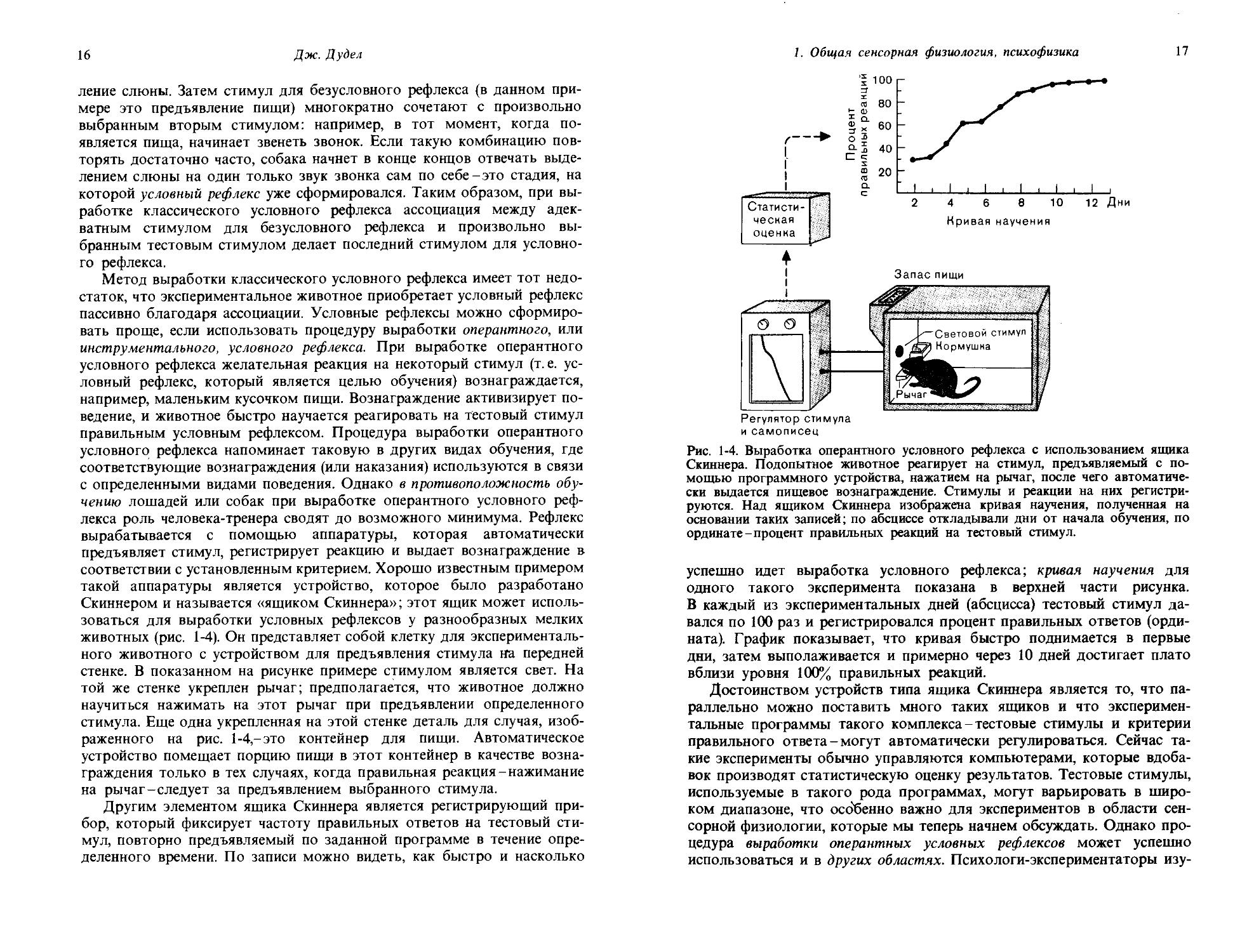
Рис. 1-2. Модальность, количество, качество и их органические субстраты на примере органа зрения.
чальная точка кривой стимул-реакция всегда является важной характеристикой ; это самый маленький сигнал, который едва способен вызвать реакцию, или пороговый стимул So. В случае рецепторной клетки его можно определить как самый маленький стимул, который способен вызвать потенциал действия (рис. 1-1); для слухового органа его, например, можно определить как самую низкую интенсивность тона, которая едва воспринимается испытуемым. Вид кривой стимул-реакция может служить характеристикой различных рецепторов, равно как и различных сенсорных впечатлений; эта сторона данных зависимостей будет обсуждаться подробнее в следующих разделах.
Сейчас мы познакомились с рядом основных понятий сенсорной физиологии: модальностью, качеством, количеством и порогом. На рис. 1-2 показано соответствие между этими понятиями и их органическими субстратами, причем в качестве примера опять взят орган зрения.
Сенсорные впечатления характеризуются не только модальностью, качеством и количеством; они имеют свойство возникать в определенное время и соответствовать определенному месту внешней среды или тела. Наши глаза видят не просто свет, но скорее «картины» окружающего нас пространства. Эти картины следуют друг за другом во времени, и их также можно вспоминать как ассоциирующиеся с некоторыми
12
Дж. Дудел
конкретными моментами времени. Пространственные и временные аспекты сенсорного восприятия, естественно, соответствуют пространственным и временным свойствам стимулов.
Сенсорное впечатление, восприятие. Термин сенсорное впечатление, который мы до сих пор использовали мимоходом, теперь нужно определить более четко. Он применяется для обозначения простейших единиц, элементов сенсорного опыта. Так, например, воспринимаемый цвет «синий» или вкус «сладкий» будут сенсорными впечатлениями. Редко случается так, что мы получаем такие впечатления изолированно; комбинации сенсорных впечатлений такого рода называют ощущением. Как правило, чистое ощущение сопровождается некоторой интерпретацией, учитывающей то, что встречалось и было выучено ранее, и результат называется восприятием. Мы выражаем наше восприятие, когда говорим: «Там стоит стул».
Отображение внешнего феномена в восприятии; объективная и субъективная сенсорная физиология. Цепь взаимосвязей между явлениями во внешней среде и их восприятием, рассмотренная в последних разделах, суммирована на рис. 1-3. Прямоугольники заключают в себе основные явления сенсорной физиологии на последовательных уровнях; рядом изображены стрелки, которые указывают только на соответствие, но не на причинную связь. Стрелки поставлены для выражения отношения, которое называется «отображением». Это значит, что возбуждение нерва может рассматриваться как отображение сенсорного стимула, а восприятие-как отображение сенсорных впечатлений. Понятие «отображение» предполагает, что имеется определенное и единственное правило, посредством которого точки одного объекта ассоциируются с точками другого (или «отображаются» на него), или более абстрактно в математических терминах-однозначное соответствие между элементами двух множеств (хеА->уеВ). Сам по себе объект не является причиной своего представления; отображение можно осуществить с помощью подходящего устройства, например аэрофотокамеры, которая проецирует образ объекта на фотобумагу. Таким образом, представление определяется не только объектом, но также и специфическими условиями построения отображения, вспомогательными инструментами, шкалой и законом проецирования и символами для представления специфических деталей.
В связи с этим под прямоугольниками, которыми на рис. 1-3 обозначены различные уровни, перечислены условия, которые должны выполняться при отображении в каждом случае. Внешние события (слева на рис. 1-3) являются сенсорными стимулами только в том случае, если они взаимодействуют с соответствующим органом чувств. Аналогично на рис. 1-3 указано, что возбуждение, посылаемое от органа чувств в ЦНС и перерабатываемое там, становится сенсорным впечатлением или ощущением только в том случае, если это ЦНС субъекта, находящегося в полном сознании. Закономерности отображения, указанные на рис. 1-3 стрелками, ведущими от событий во внешней среде к интегративным процессам в сенсорных отделах ЦНС, в принципе могут быть
1. Общая сенсорная физиология, психофизика
13
Объективная сенсорная физиология
Субъективная сенсорная физиология
Явления во внешней среде
Сенсорные стимулы
Возбуждение сенсорных нервов
Интеграция в сенсорных областях центральной нервной системы
Сенсорные впечатления, ощущения
Восприятие
Отобра- ।w жение
Взаимодействие с сенсорным органом
I-----►
Соответствующие рецепторы, надпороговый рецепторный потенциал
I-------
Функционирующие мозговые центры надпороговое возбуждение
1
Рис. 1-3. Отображение в сенсорной физиологии. Слова внутри прямоугольников соответствуют основным явлениям сенсорной физиологии, а стрелки указывают на связь в форме «отображения». Внизу перечислены условия, необходимые для отображения.
представлены в виде физических и химических процессов в структурах организма. В связи с этим соответствующая сфера сенсорной физиологии называется объективной сенсорной физиологией. Напротив, взаимосвязь между этими объективными феноменами-сенсорным стимулом вместе с последующими реакциями в нервной системе-и осознанным ощущением не может быть описана в терминах физических и химических процессов. Поэтому сфера ощущений и восприятий в ее отношении к сенсорным стимулам называется субъективной сенсорной физиологией.
Утверждение, что субъективное ощущение и восприятие не поддаются описанию в физических и химических терминах, требует пояснений. С точки зрения естественных наук такое утверждение есть просто констатация современного состояния наших знаний. Естествоиспытатель будет пытаться применить свои методы также и к субъективным явлениям, и тут уже имеются впечатляющие примеры успеха, как это показывает обсуждение психофизики в разд. 1.3 и 1.4. Однако многие специалисты из других академических областей полагают, что реакция субъекта, сфера психического, принципиально непредставима в рамках естественных наук.
В связи с этим сенсорная физиология делится на две части: описание реакций нервной системы на стимул - объективную сенсорную физиологию и анализ высказываний, которые субъект делает относительно своих ощущений и восприятий,-субъективную сенсорную физиологию. Термины объективный и субъективный в этом контексте ни в коем случае не должны рассматриваться как оценки значимости в смысле корректности утверждения. Предложение «красный-это теплый цвет» может быть столь же корректным, сколь и предложение «частота разряда нервного волокна растет с увеличением интенсивности сенсорного сти
14
Дж. Дудел
мула». Как биологи, и в особенности как ученые, которые исследуют людей, мы должны обращаться с субъективными высказываниями об ощущениях и восприятиях так же непредвзято, как это мы делаем с записями клеточных потенциалов. В разд. 1.3 будет показано, что используя подходящие термины, можно высказывать очень точные утверждения, касающиеся субъективной сенсорной физиологии; в этой области вполне могут быть установлены количественные математические соотношения. Несмотря на то что отношение между стимулом и ощущением может быть описано только термином «отображение», а качественные различия между физическим стимулом и субъективным сенсорным впечатлением кажутся непреодолимыми, чтобы выявить некоторую промежуточную функцию между физикой и субъективностью, можно пользоваться методами физиологии поведения, кратко рассмотренными в следующем разделе.
Отвечая на следующие вопросы (здесь и далее везде в книге обозначаемые буквой «В»), вы можете проверить свои знания о предмете данного раздела. Насколько это возможно, при подготовке ответа старайтесь не пользоваться текстом. Запишите свои ответы на листе бумаги и затем сравните их с правильными ответами, помещенными в конце книги.
В 1.1. Совокупность сенсорных впечатлений, сходных между собой и порождаемых некоторым определенным органом чувств, называется ...........Когда сенсорные впечатления разных видов могут
быть различены в пределах одной модальности, они называются
В 1.2. Расклассифицируйте слова из следующего списка, поставив рядом с теми, что означают модальность, букву М, с теми, что означают качество,-букву Q, а с теми, что означают количество-букву I. а) Слух ( ).
б) Интенсивность звука ( ).
в) Красное ( ).
г) Вкус ( ).
д) Растяжение легких ( ).
е) Кислое ( ).
ж) Высота тона ( ).
з) Интенсивность красного цвета ( ).
и) Кожное чувство холода ( ).
В 1.3. Выберите термин, который лучше всего описывает отношение между сенсорным стимулом и ощущением.
а) Причина и следствие.
б) Объективность.
в) Отображение.
г) Увеличение специфичности.
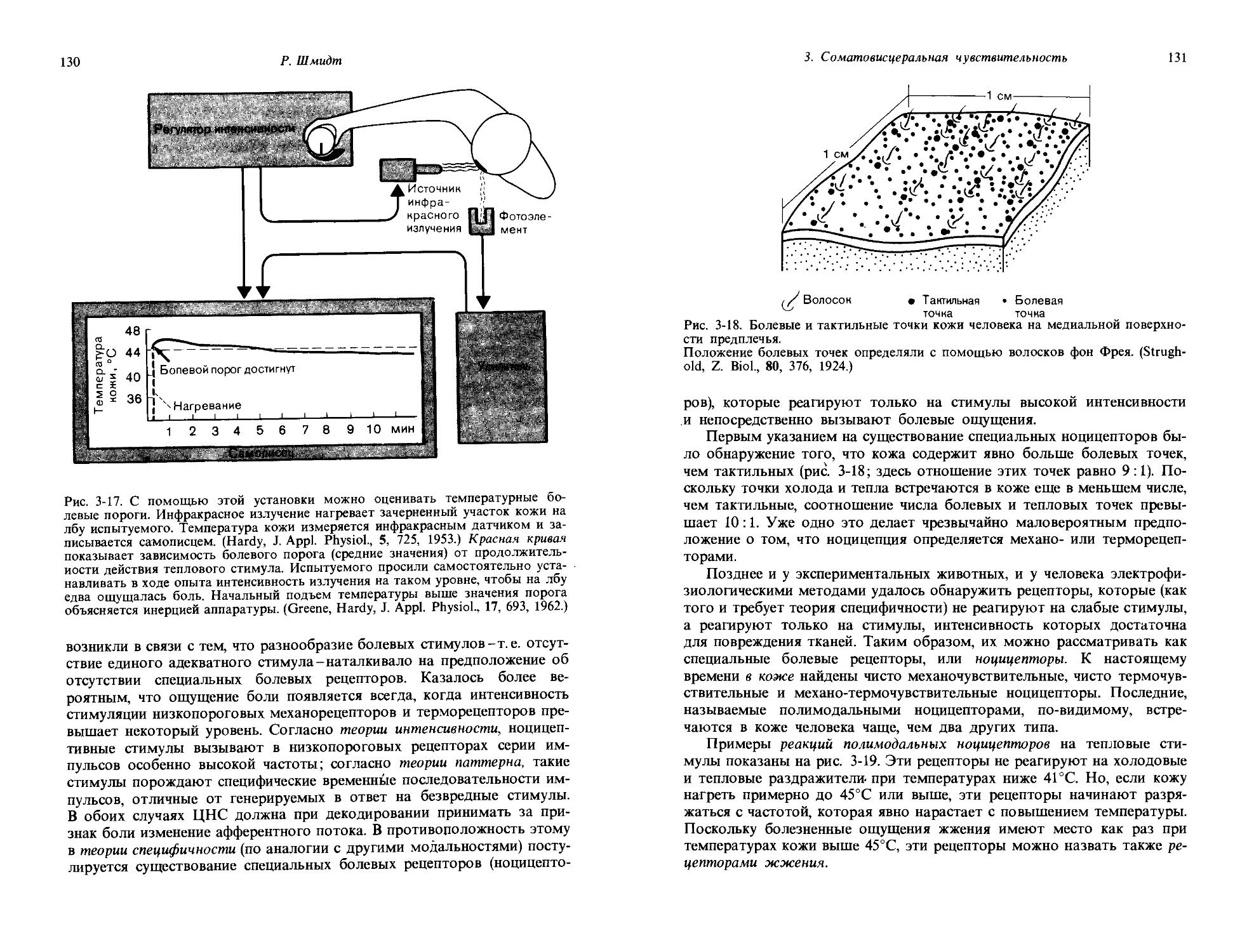
д) Качество.
1. Общая сенсорная физиология, психофизика
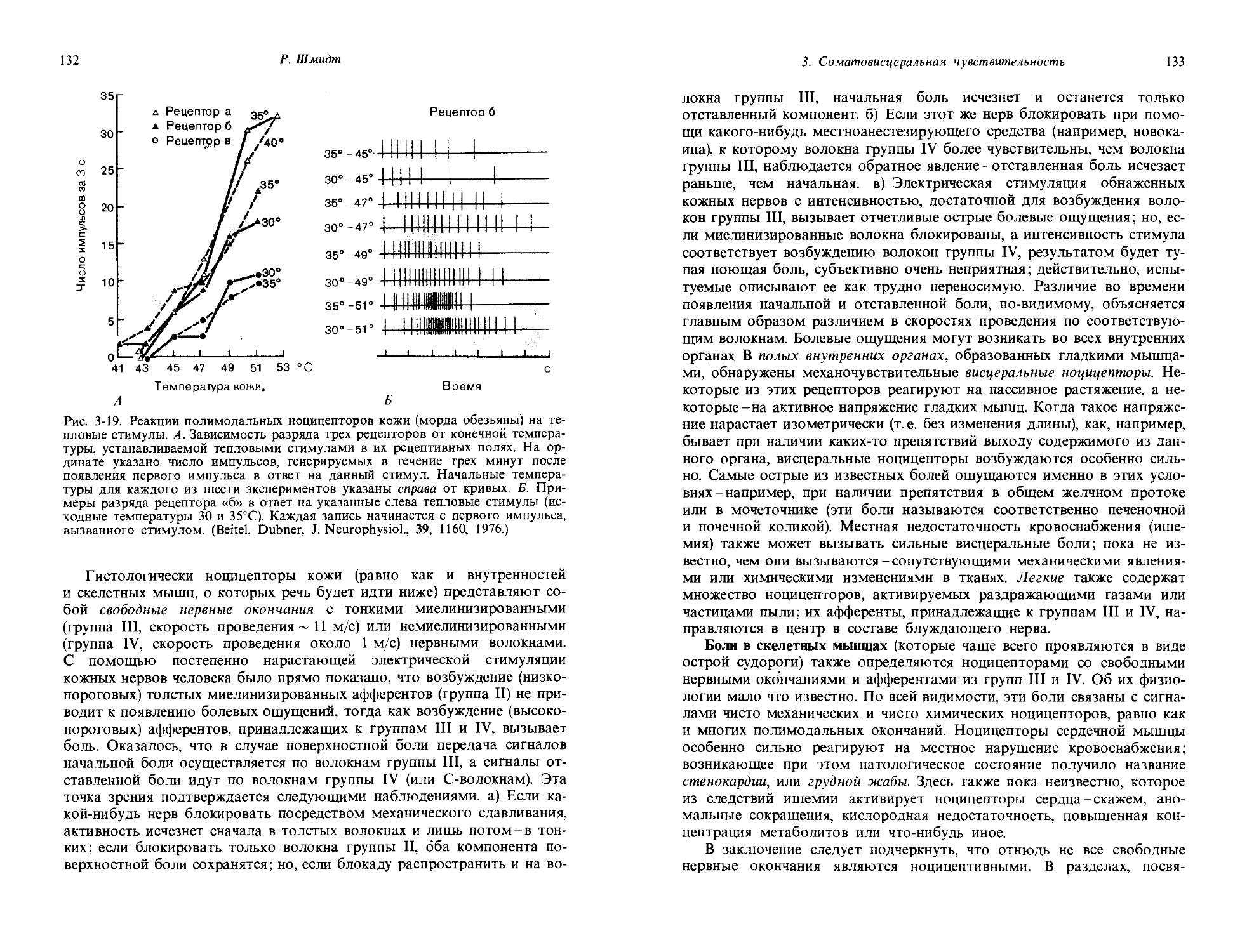
15
1.2. Связь между стимулом и поведением; условный рефлекс
Реакции центральной нервной системы, вызываемые сенсорным стимулом, могут приводить к реакциям всего организма, которые могут быть направлены вовне или внутрь. Если мы слышим сбоку какой-то неожиданный шум, мы поворачиваем голову в ту сторону; олень в лесу поступает точно таким же образом. Когда обезьяны в зоопарке видят своего служителя во время кормления, они становятся беспокойными и шумными. Когда мы ведем машину, появление какого-нибудь препятствия заставляет нас не только затормозить и свернуть в сторону-у нас также возрастает тонус мышц и сердце начинает биться чаще. Во всех этих примерах специфические сенсорные стимулы возбуждают более или менее сложные изменения активности человека или животного. Такую активность, которую можно обычно интерпретировать как целенаправленную, обобщенно называют поведением. Изменения поведения, явившиеся результатом действия сенсорного стимула, могут быть описаны наблюдателем, но могут также регистрироваться и подходящими измерительными приборами. Мы рассматриваем такие изменения поведения как объяснимые, в принципе, реакциями нервной системы животного на стимул даже тогда, когда мы , возможно, еще не понимаем эти реакции во всех деталях. Таким образом, с точки зрения сенсорной физиологии, изучение поведения составляет часть объективной сенсорной физиологии. Некоторые основные понятия и методы из этой области, относящейся также к разделу психологии животных и психологии обучения, будут представлены ниже.
Стимул, поведение, рефлекс. Если на заднюю лапу кошки подействовать болевым стимулом (например, прищемить ее), кошка будет отдергивать лапу, сгибая ее в суставах. Это поведение кошки называют в нейрофизиологии сгибательным рефлексом. Следовательно, часть поведения животных состоит из стереотипных реакций на определенные стимулы-рефлексов. Некоторые из рефлексов врожденные, или безусловные (среди них и сгибательный рефлекс); они базируются на фиксированных нервных связях между рецепторами и эффекторами. Но в контексте сенсорной физиологии особый интерес представляют приобретенные рефлексы. В случае таких рефлексов функциональная связь между рецепторами и эффекторами формируется посредством процесса обучения. Животное научается систематически реагировать на определенный стимул определенной активностью. Такие рефлексы называют также условными, поскольку они обнаруживаются только в условиях предварительного обучения. Примером может служить «автоматическое» нажимание водителем на тормоз при встрече с препятствием.
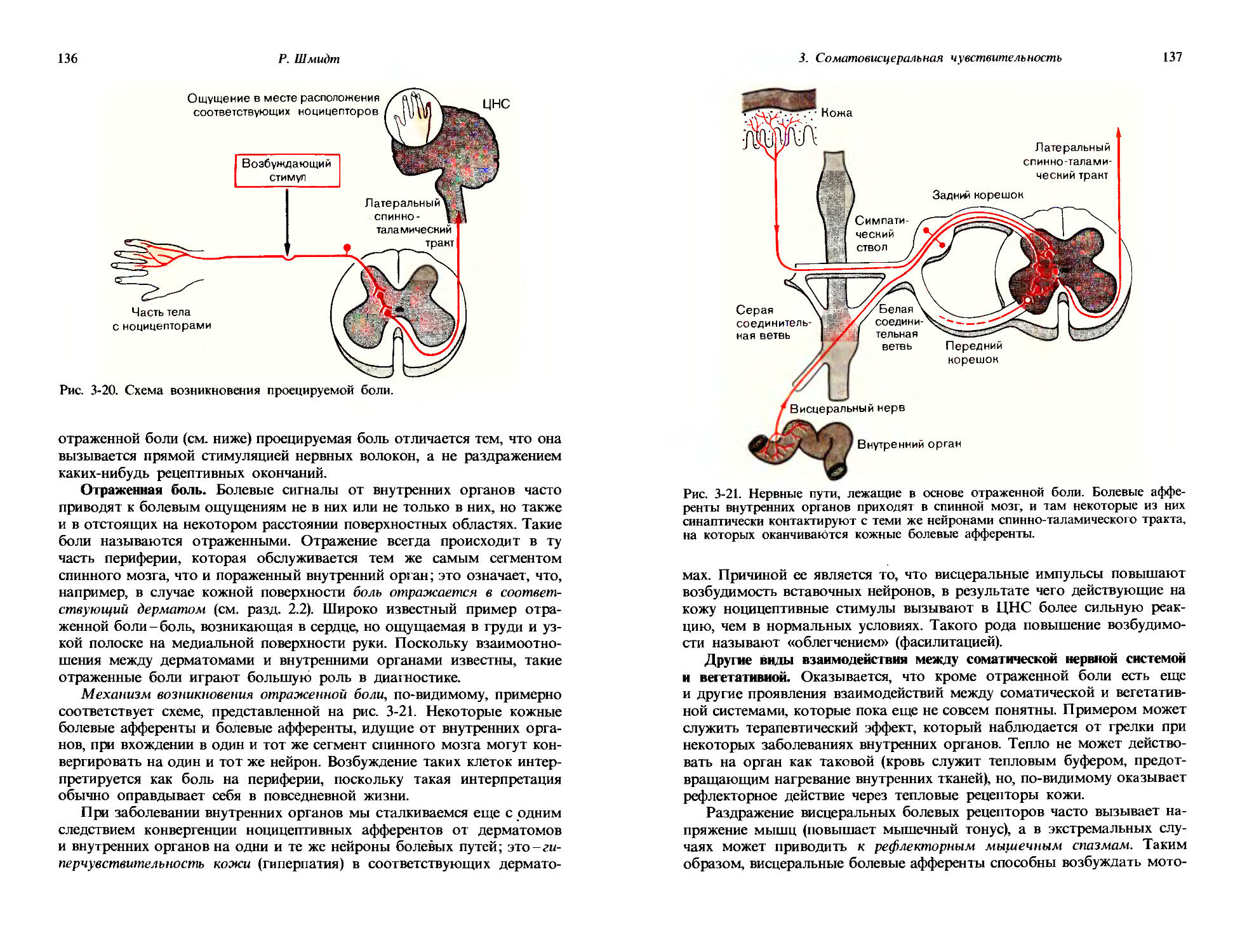
Выработка условного рефлекса. Процесс приобретения условных рефлексов многими животными можно легко исследовать в лаборатории. Первая процедура для достижения этого была разработана И. П. Павловым и называется процедурой выработки классического условного рефлекса. В этой методике сначала запускается безусловный рефлекс — например, собаке предлагается пища, в связи с чем возбуждается выде
16
Дж. Дудел
ление слюны. Затем стимул для безусловного рефлекса (в данном примере это предъявление пищи) многократно сочетают с произвольно выбранным вторым стимулом: например, в тот момент, когда появляется пища, начинает звенеть звонок. Если такую комбинацию повторять достаточно часто, собака начнет в конце концов отвечать выделением слюны на один только звук звонка сам по себе-это стадия, на которой условный рефлекс уже сформировался. Таким образом, при выработке классического условного рефлекса ассоциация между адекватным стимулом для безусловного рефлекса и произвольно выбранным тестовым стимулом делает последний стимулом для условного рефлекса.
Метод выработки классического условного рефлекса имеет тот недостаток, что экспериментальное животное приобретает условный рефлекс пассивно благодаря ассоциации. Условные рефлексы можно сформировать проще, если использовать процедуру выработки оперантного, или инструментального, условного рефлекса. При выработке оперантного условного рефлекса желательная реакция на некоторый стимул (т. е. условный рефлекс, который является целью обучения) вознаграждается, например, маленьким кусочком пищи. Вознаграждение активизирует поведение, и животное быстро научается реагировать на тестовый стимул правильным условным рефлексом. Процедура выработки оперантного условного рефлекса напоминает таковую в других видах обучения, где соответствующие вознаграждения (или наказания) используются в связи с определенными видами поведения. Однако в противоположность обучению лошадей или собак при выработке оперантного условного рефлекса роль человека-тренера сводят до возможного минимума. Рефлекс вырабатывается с помощью аппаратуры, которая автоматически предъявляет стимул, регистрирует реакцию и выдает вознаграждение в соответствии с установленным критерием. Хорошо известным примером такой аппаратуры является устройство, которое было разработано Скиннером и называется «ящиком Скиннера»; этот ящик может использоваться для выработки условных рефлексов у разнообразных мелких животных (рис. 1-4). Он представляет собой клетку для экспериментального животного с устройством для предъявления стимула на передней стенке. В показанном на рисунке примере стимулом является свет. На той же стенке укреплен рычаг; предполагается, что животное должно научиться нажимать на этот рычаг при предъявлении определенного стимула. Еще одна укрепленная на этой стенке деталь для случая, изображенного на рис. 1-4,-это контейнер для пищи. Автоматическое устройство помещает порцию пищи в этот контейнер в качестве вознаграждения только в тех случаях, когда правильная реакция-нажимание на рычаг-следует за предъявлением выбранного стимула.
Другим элементом ящика Скиннера является регистрирующий прибор, который фиксирует частоту правильных ответов на тестовый стимул, повторно предъявляемый по заданной программе в течение определенного времени. По записи можно видеть, как быстро и насколько
1. Общая сенсорная физиология, психофизика
17
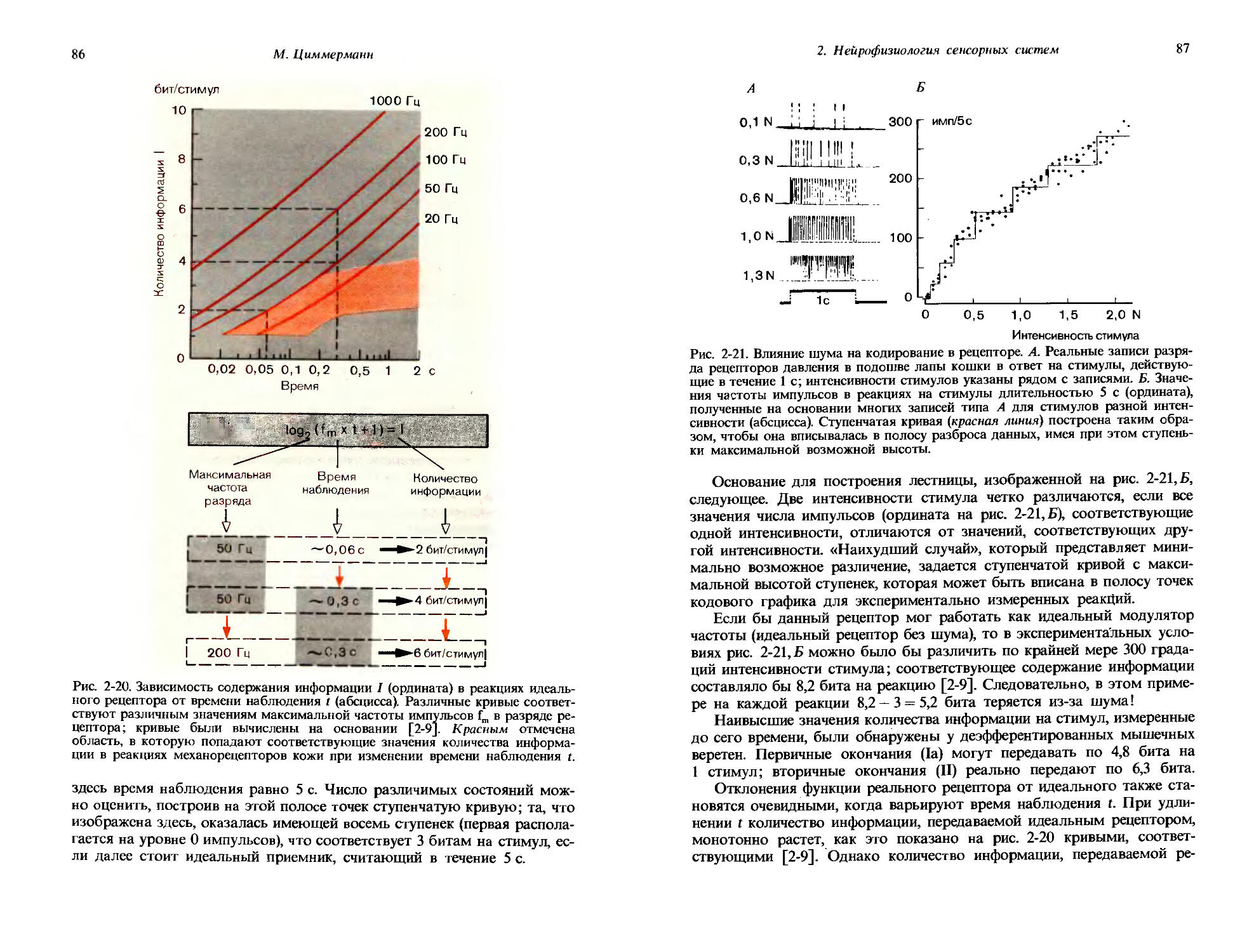
Регулятор стимула и самописец
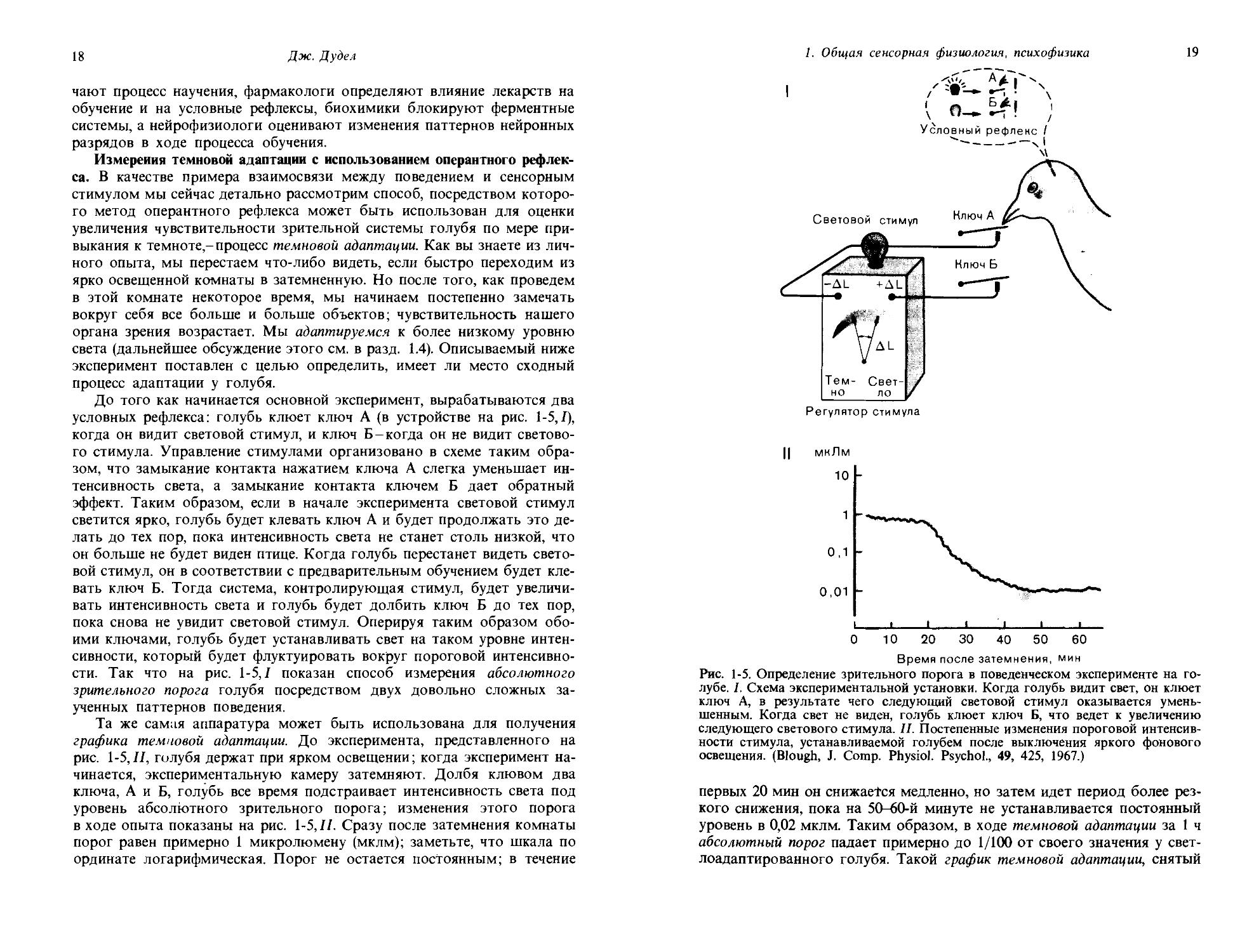
Рис. 1-4. Выработка оперантного условного рефлекса с использованием ящика Скиннера. Подопытное животное реагирует на стимул, предъявляемый с помощью программного устройства, нажатием на рычаг, после чего автоматически выдается пищевое вознаграждение. Стимулы и реакции на них регистрируются. Над ящиком Скиннера изображена кривая научения, полученная на основании таких записей; по абсциссе откладывали дни от начала обучения, по ординате-процент правильных реакций на тестовый стимул.
успешно идет выработка условного рефлекса; кривая научения для одного такого эксперимента показана в верхней части рисунка. В каждый из экспериментальных дней (абсцисса) тестовый стимул давался по 100 раз и регистрировался процент правильных ответов (ордината). График показывает, что кривая быстро поднимается в первые дни, затем выполаживается и примерно через 10 дней достигает плато вблизи уровня 100% правильных реакций.
Достоинством устройств типа ящика Скиннера является то, что параллельно можно поставить много таких ящиков и что экспериментальные программы такого комплекса-тестовые стимулы и критерии правильного ответа-могут автоматически регулироваться. Сейчас такие эксперименты обычно управляются компьютерами, которые вдобавок производят статистическую оценку результатов. Тестовые стимулы, используемые в такого рода программах, могут варьировать в широком диапазоне, что особенно важно для экспериментов в области сенсорной физиологии, которые мы теперь начнем обсуждать. Однако процедура выработки оперантных условных рефлексов может успешно использоваться и в других областях. Психологи-экспериментаторы изу
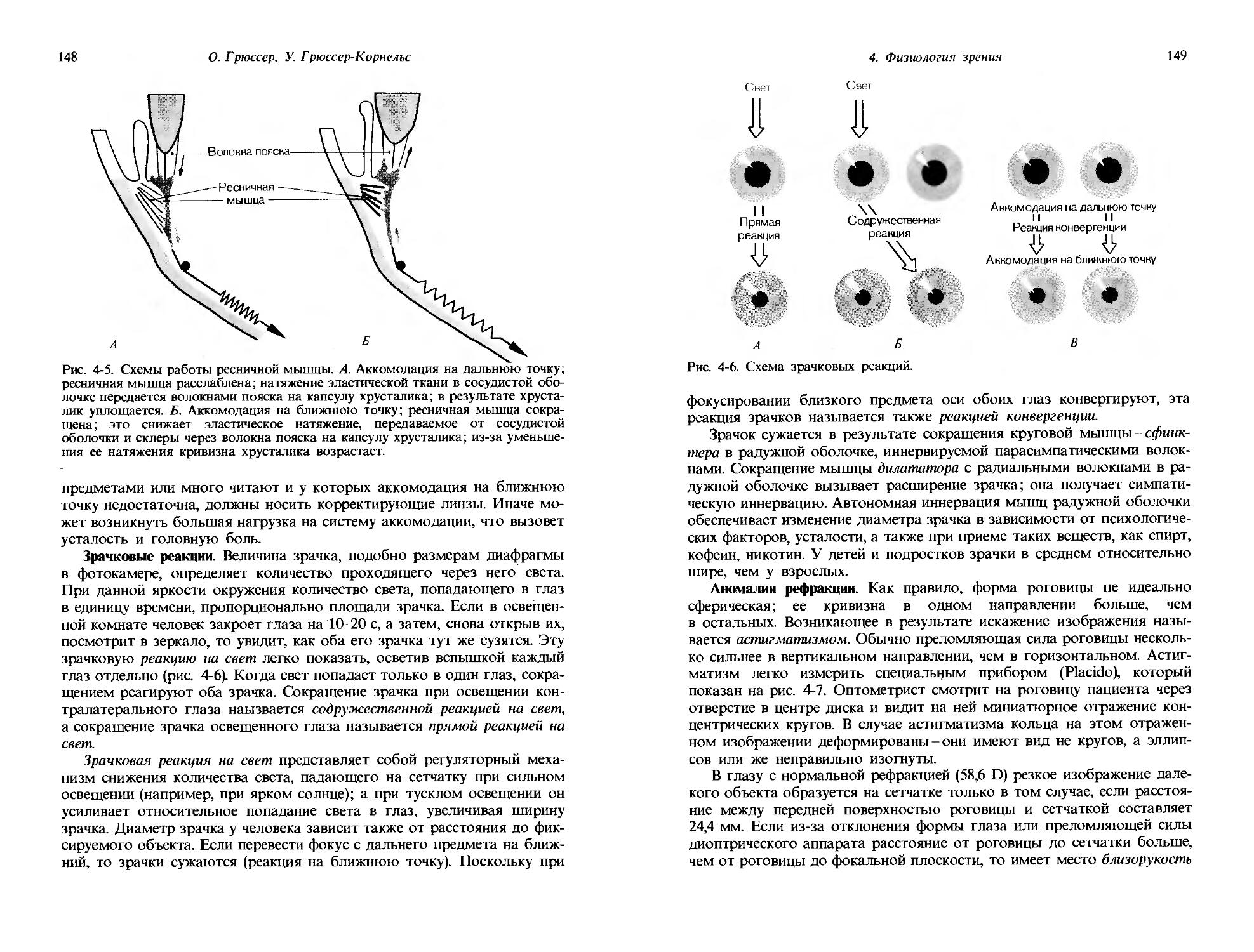
18
Дж. Дудел
чают процесс научения, фармакологи определяют влияние лекарств на обучение и на условные рефлексы, биохимики блокируют ферментные системы, а нейрофизиологи оценивают изменения паттернов нейронных разрядов в ходе процесса обучения.
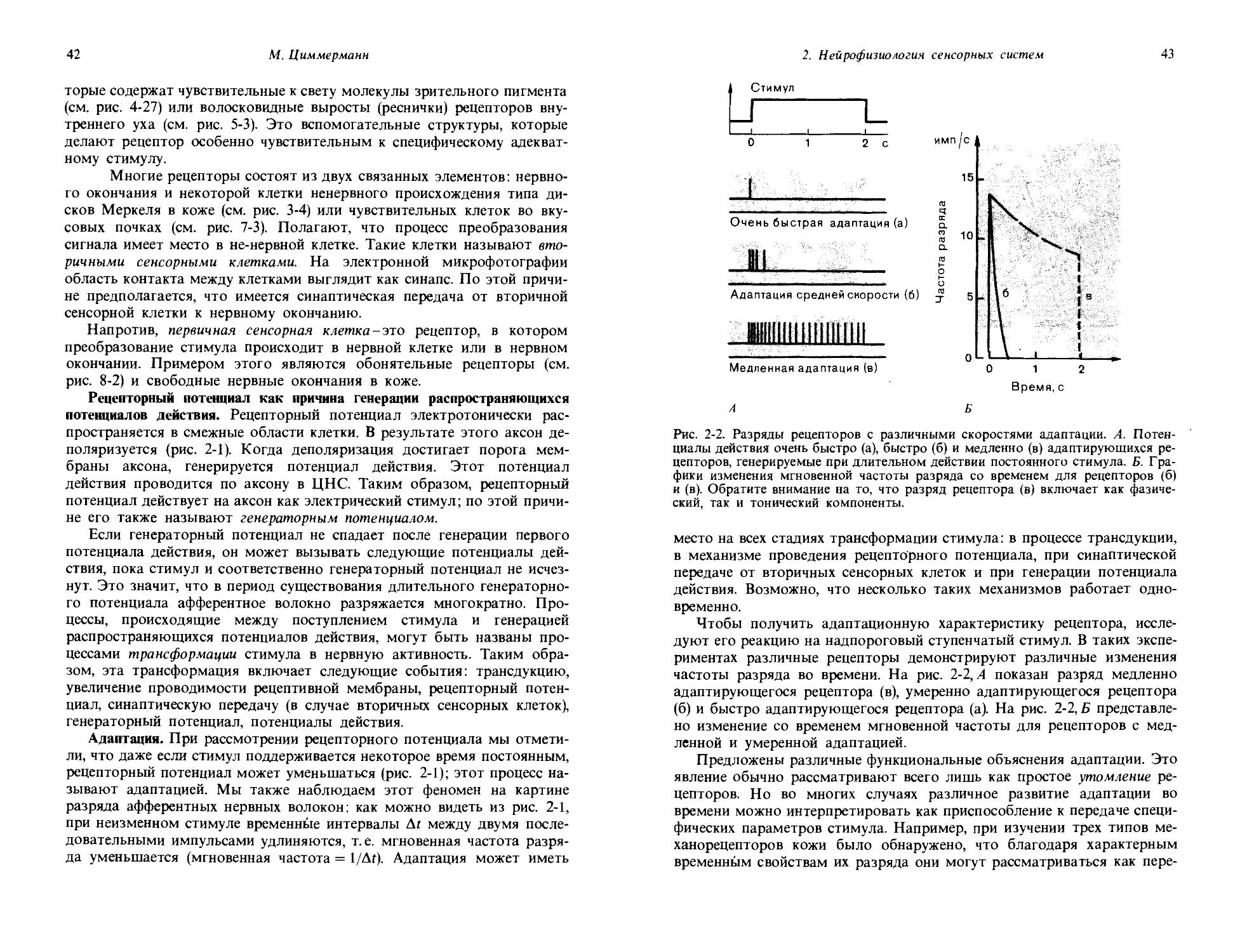
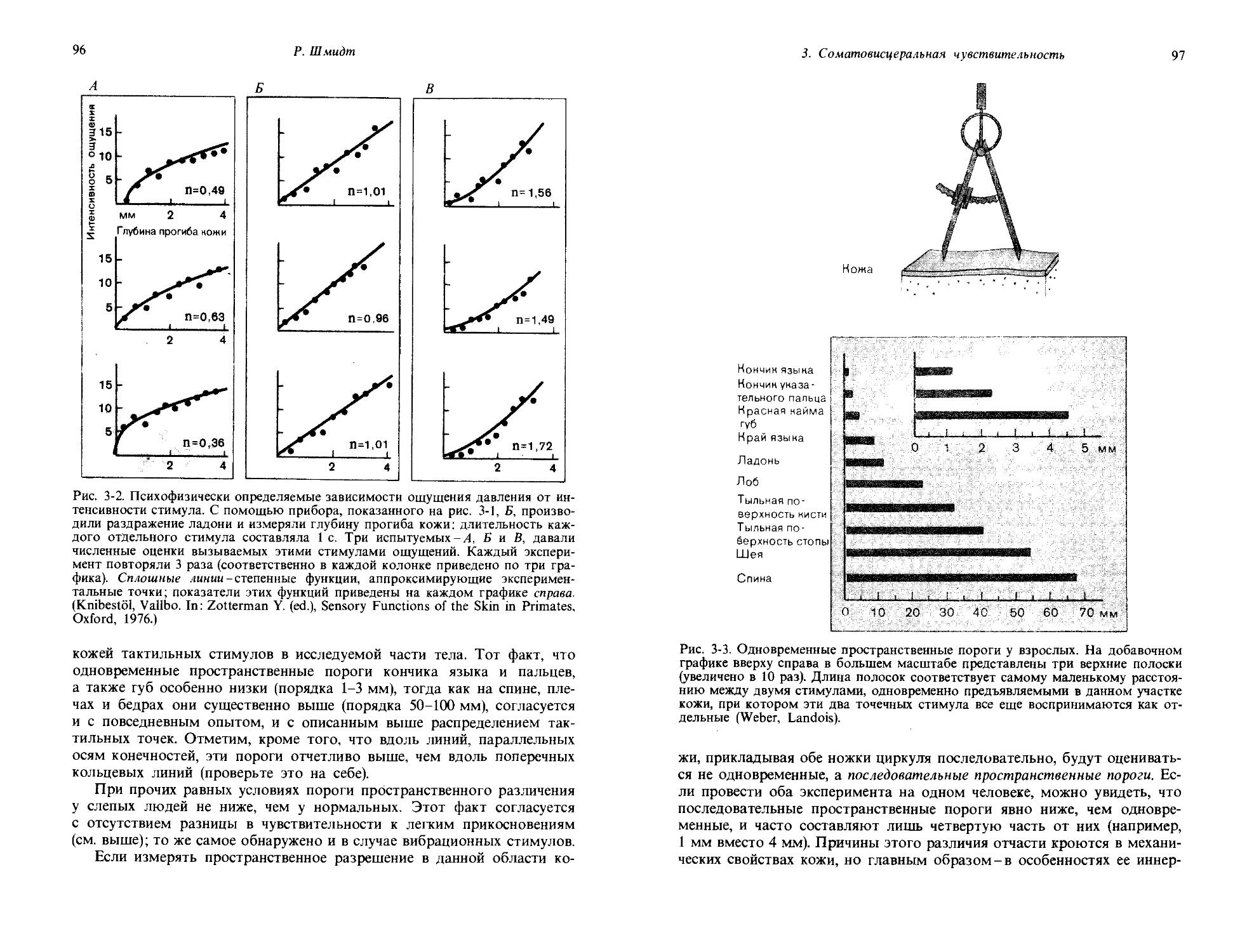
Измерения темновой адаптации с использованием оперантного рефлекса. В качестве примера взаимосвязи между поведением и сенсорным стимулом мы сейчас детально рассмотрим способ, посредством которого метод оперантного рефлекса может быть использован для оценки увеличения чувствительности зрительной системы голубя по мере привыкания к темноте- процесс темновой адаптации. Как вы знаете из личного опыта, мы перестаем что-либо видеть, если быстро переходим из ярко освещенной комнаты в затемненную. Но после того, как проведем в этой комнате некоторое время, мы начинаем постепенно замечать вокруг себя все больше и больше объектов; чувствительность нашего органа зрения возрастает. Мы адаптируемся к более низкому уровню света (дальнейшее обсуждение этого см. в разд. 1.4). Описываемый ниже эксперимент поставлен с целью определить, имеет ли место сходный процесс адаптации у голубя.
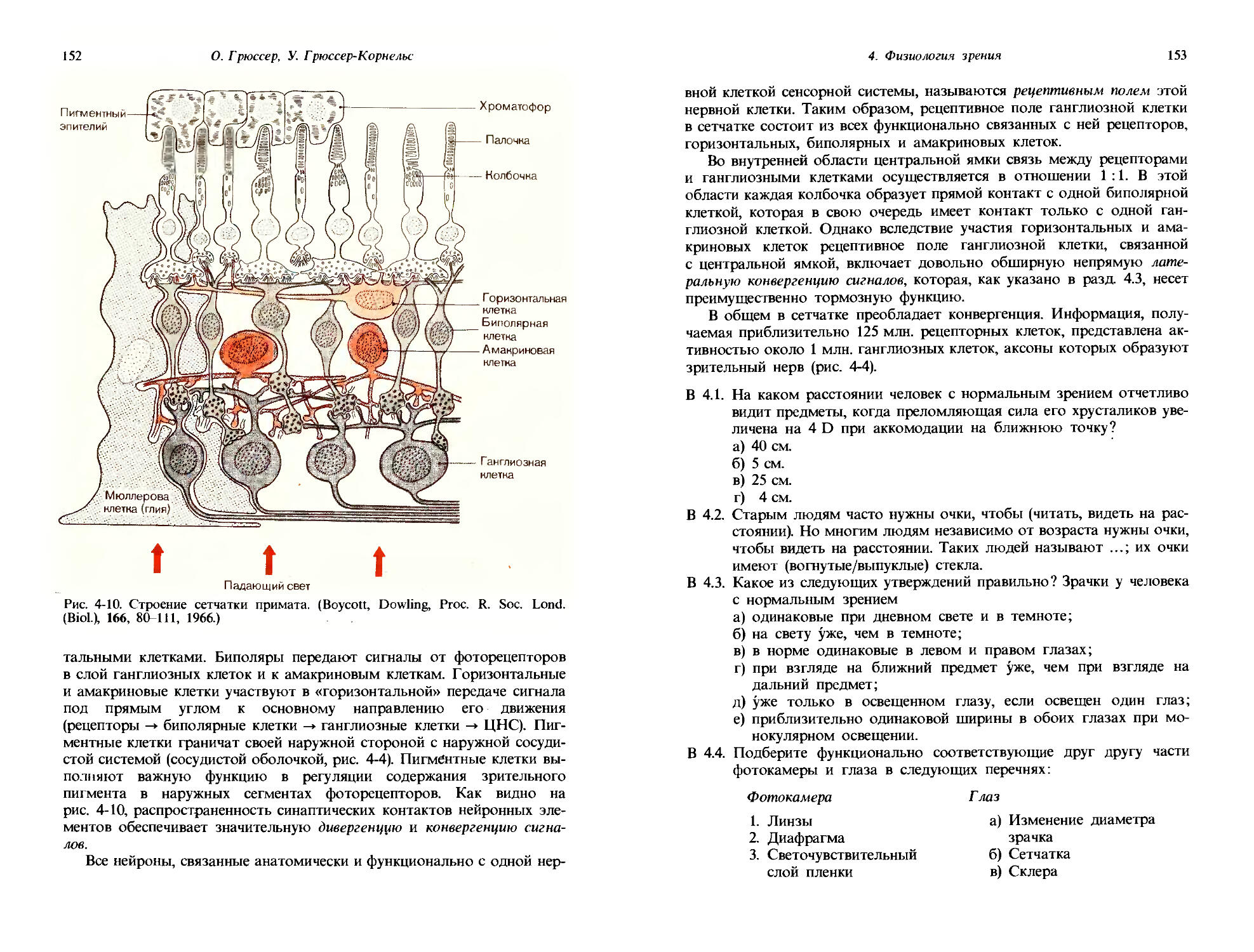
До того как начинается основной эксперимент, вырабатываются два условных рефлекса: голубь клюет ключ А (в устройстве на рис. 1-5,/), когда он видит световой стимул, и ключ Б-когда он не видит светового стимула. Управление стимулами организовано в схеме таким образом, что замыкание контакта нажатием ключа А слегка уменьшает интенсивность света, а замыкание контакта ключем Б дает обратный эффект. Таким образом, если в начале эксперимента световой стимул светится ярко, голубь будет клевать ключ А и будет продолжать это делать до тех пор, пока интенсивность света не станет столь низкой, что он больше не будет виден птице. Когда голубь перестанет видеть световой стимул, он в соответствии с предварительным обучением будет клевать ключ Б. Тогда система, контролирующая стимул, будет увеличивать интенсивность света и голубь будет долбить ключ Б до тех пор, пока снова не увидит световой стимул. Оперируя таким образом обоими ключами, голубь будет устанавливать свет на таком уровне интенсивности, который будет флуктуировать вокруг пороговой интенсивности. Так что на рис. 1-5,1 показан способ измерения абсолютного зрительного порога голубя посредством двух довольно сложных заученных паттернов поведения.
Та же самая аппаратура может быть использована для получения графика темновой адаптации. До эксперимента, представленного на рис. 1-5,II, голубя держат при ярком освещении; когда эксперимент начинается, экспериментальную камеру затемняют. Долбя клювом два ключа, А и Б, голубь все время подстраивает интенсивность света под уровень абсолютного зрительного порога; изменения этого порога в ходе опыта показаны на рис. 1-5,11. Сразу после затемнения комнаты порог равен примерно 1 микролюмену (мклм); заметьте, что шкала по ординате логарифмическая. Порог не остается постоянным; в течение
1. Общая сенсорная физиология, психофизика
19
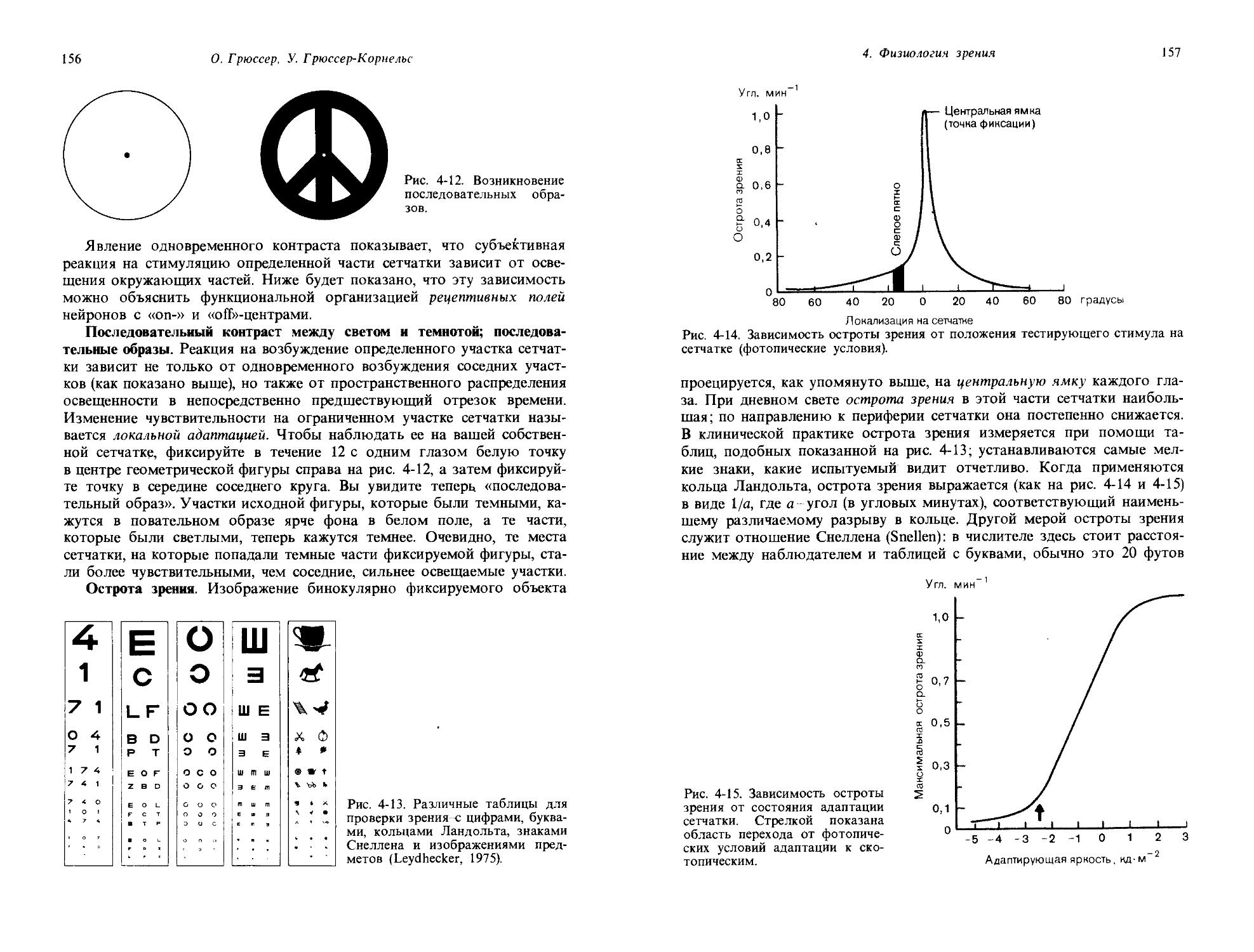
Время после затемнения, мин
Рис. 1-5. Определение зрительного порога в поведенческом эксперименте на голубе. I. Схема экспериментальной установки. Когда голубь видит свет, он клюет ключ А, в результате чего следующий световой стимул оказывается уменьшенным. Когда свет не виден, голубь клюет ключ Б, что ведет к увеличению следующего светового стимула. II. Постепенные изменения пороговой интенсивности стимула, устанавливаемой голубем после выключения яркого фонового освещения. (Blough, J. Comp. Physiol. Psychol., 49, 425, 1967.)
первых 20 мин он снижается медленно, но затем идет период более резкого снижения, пока на 50-60-й минуте не устанавливается постоянный уровень в 0,02 мклм. Таким образом, в ходе темновой адаптации за 1 ч абсолютный порог падает примерно до 1/100 от своего значения у светлоадаптированного голубя. Такой график темновой адаптации, снятый
20
Дж. Дудел
у голубя посредством условных рефлексов, сильно напоминает, и по форме, и по временному ходу соответствующие кривые, снятые у людей методами субъективной сенсорной физиологии (ср. с рис. 4-17).
Аналогичные поведенческие эксперименты могут использоваться для определения и других пороговых значений путем выработки подходящих условных рефлексов. Так, например, эта методика была использована для измерения зрительных порогов на различных длинах световых волн у млекопитающих, лягушек, рыб и даже осьминога, в результате чего оказалось возможным охарактеризовать кривые поглощения зрительных пигментов, имеющихся у этих животных. Соответственно после выработки подходящих условных рефлексов можно определить и слуховые пороги, или пороги различения высоты тона. Порог различения тона-это наименьшая разница в высоте (частоте) двух тонов, при которой эти тоны еще воспринимаются как различные. Таким образом, он характеризует способность отличать тоны друг от друга. Процедура измерения порогов различения тона по аналогии с процедурой получения кривой темновой адаптации на рис. 1-5, И, грубо говоря, должна быть следующей: животное обучают выполнять действие А, когда оно слышит два одинаковых тона один за другим, и действие Б-когда два следующих друг за другом тона различны. Стимул состоит из эталонного тона и последующего варьируемого и сравниваемого с первым тона. Система управления стимулом работает таким образом, что после действия А различие тонов увеличивается, а после действия Б-уменьшается. Таким образом, порог различения тона в области данного эталонного тона будет устанавливаться автоматически.
Итак, вырабатывая подходящие условные рефлексы, можно осуществлять очень детальные исследования в области сенсорной физиологии животных. Многие из действующих в сенсорной физиологии правил, которые будут представлены в следующем разделе как относящиеся к «субъективной сенсорной физиологии» человека, таким путем могут быть распространены на связь между стимулом и поведением у животных; мы не имеем возможности упоминать здесь другие примеры. В области субъективной сенсорной физиологии обычно устанавливают связь между сенсорными стимулами и устными высказываниями об ощущениях и восприятии (рис. 1-3). Но речь не что иное, как форма поведения. Таким образом, метод определения порогов различения тона, описанный в предыдущем абзаце, может быть одинаковым путем применен и к поведенческим экспериментам на животных, и к субъективной сенсорной физиологии человека. Действие А будет просто означать утверждение человека: «Я слышу один тон», а действие Б будет соответствовать заявлению: «Я слышу два тона». Так что формально естествоиспытатели могут рассматривать субъективную сенсорную физиологию как частную главу сенсорной физиологии поведения животных.
Наши ощущения и восприятия сопровождаются осознанием нашей субъективности и личностности, и многие думают, что это составляет
1. Общая сенсорная физиология, психофизика
21
специфически человеческий аспект наших реакций на явления окружающей среды и на сенсорные стимулы (рис. 1-3). Мне как естествоиспытателю представляется очень трудным решить, свойственно ли и животным также некоторое осознание своей субъективности и личностно-сти. Но, если это имеет место, различие между предметом данного раздела и следующего за ним можно считать скорее количественным, чем качественным.
В 1.4. Определите своими словами, что такое условный рефлекс.
В 1.5. Какое из следующих утверждений можно приложить к процедуре выработки оперантного условного рефлекса?' а) Условный рефлекс подкрепляется вознаграждением правильных реакций.
б) Тестовый стимул сочетается с адекватным стимулом для врожденного (безусловного) рефлекса и благодаря повторению этой процедуры становится стимулом для условного рефлекса.
в) При частом повторении условный рефлекс становится безусловным рефлексом.
г) Условный рефлекс вырабатывается в процессе активной деятельности животного.
В 1.6. Кривая научения для случая условного рефлекса
а) представляет частоту ошибок как функцию трудности задачи; б) представляет частоту правильных реакций как функцию времени, в течение которого приобретается навык;
в) сначала плоская, а затем круто поднимается;
г) сначала поднимается круто, но в конечном итоге выходит на плато;
д) может быть определена только для высших животных.
В 1.7. Какая (ие) из следующих процедур подходит (ят) для оценки разрешающей способности зрения кошек (минимального расстояния между двумя точками, при котором они могут восприниматься как отдельные)? (Кошку научили нажимать на рычаг А, когда она видит одну точку, и на рычаг Б, когда она видит две точки.) а) Программа работы управляющей стимулом аппаратуры такова, что пары точек с расстоянием, варьируемым в диапазоне от 0 до 5 мм, предъявляются в случайном порядке. Расстояние между точками, при котором кошка нажимает на рычаг А так же часто, как и на рычаг Б, и соответствует разрешающей способности для точек.
б) Управляющая стимулом аппаратура работает таким образом, что при нажимании на рычаг А расстояние между точками, увеличивается, а когда кошка нажимает на рычаг Б, это расстояние уменьшается. После большого числа испытаний разрыв между точками устанавливается на уровне, соответствующем разрешающей способности для точек.
22
Дж. Дудел
1.3. Измерение интенсивности ощущений. Психофизика
В субъективной сенсорной физиологии мы имеем дело с высказываниями, которые человек делает относительно внешних событий. Как экспериментаторы мы предъявляем ему специфические сенсорные стимулы и регистрируем, что он говорит. В тех пределах, в которых гипотезы, выдвигаемые на основе этих высказываний, проверяемы результатами следующих экспериментов, т. е. до тех пор, пока эти гипотезы имеют возможность быть подтвержденными или отвергнутыми экспериментально (см. принцип опровержимости К. Р. Поппера), субъективная сенсорная физиология представляет собой область естественных наук; как таковая она часто называется психофизикой. Сенсорный опыт также можно трактовать способом более ориентированным на человеческую природу, как это делается в гештальт-психологии, эстетике или гносеологии. Здесь мы будем рассматривать только психофизическую сторону субъективной сенсорной физиологии.
С точки зрения методологического подхода к субъективной сенсорной физиологии мы можем в принципе игнорировать все, что мы знаем об органах чувств, рецепторах и мозговых центрах,-наш интерес направлен целиком на реакцию человека, испытуемого на сенсорный стимул. Тем не менее, субъективная и объективная сенсорная физиология взаимно обогащают друг друга в чрезвычайной степени. Большинство вопросов, поставленных в объективной сенсорной физиологии, первоначально были сформулированы в фазе открытий в субъективной ветви исследований; во многих областях эти два подхода дают результаты, которые могут быть сопоставлены, и часто новые данные, полученные в объективных экспериментах, могут быть проверены путем постановки субъективных опытов.
Оставшаяся часть этой главы может служить лишь кратким введением в общую субъективную сенсорную физиологию. В главах, касающихся отдельных органов чувств, будут представлены конкретные результаты из области субъективной сенсорной физиологии. Здесь мы рассмотрим лишь несколько принципов и проиллюстрируем их примерами. В этом разделе мы будем обращать особое внимание на измерение интенсивности ощущений, а в следующем вернемся к пространственным и временным аспектам.
Субъективные измерения. В объективной сенсорной физиологии как сила стимула, так и амплитуда реакции могут быть измерены физическими или химическими способами. Так, например, интенсивность вкусового стимула может быть задана в виде концентрации стимулирующего вещества в растворе, в миллимолях на литр, а реакция на стимул может быть определена как частота потенциалов действия в нерве, идущем от языка. В противоположность этому в субъективных экспериментах невозможно использовать физические или химические измерительные системы для установления интенсивности ощущения, порождаемого стимулом. Следовательно, нужно ввести новую систему субъективных измерений.
1. Общая сенсорная физиология, психофизика
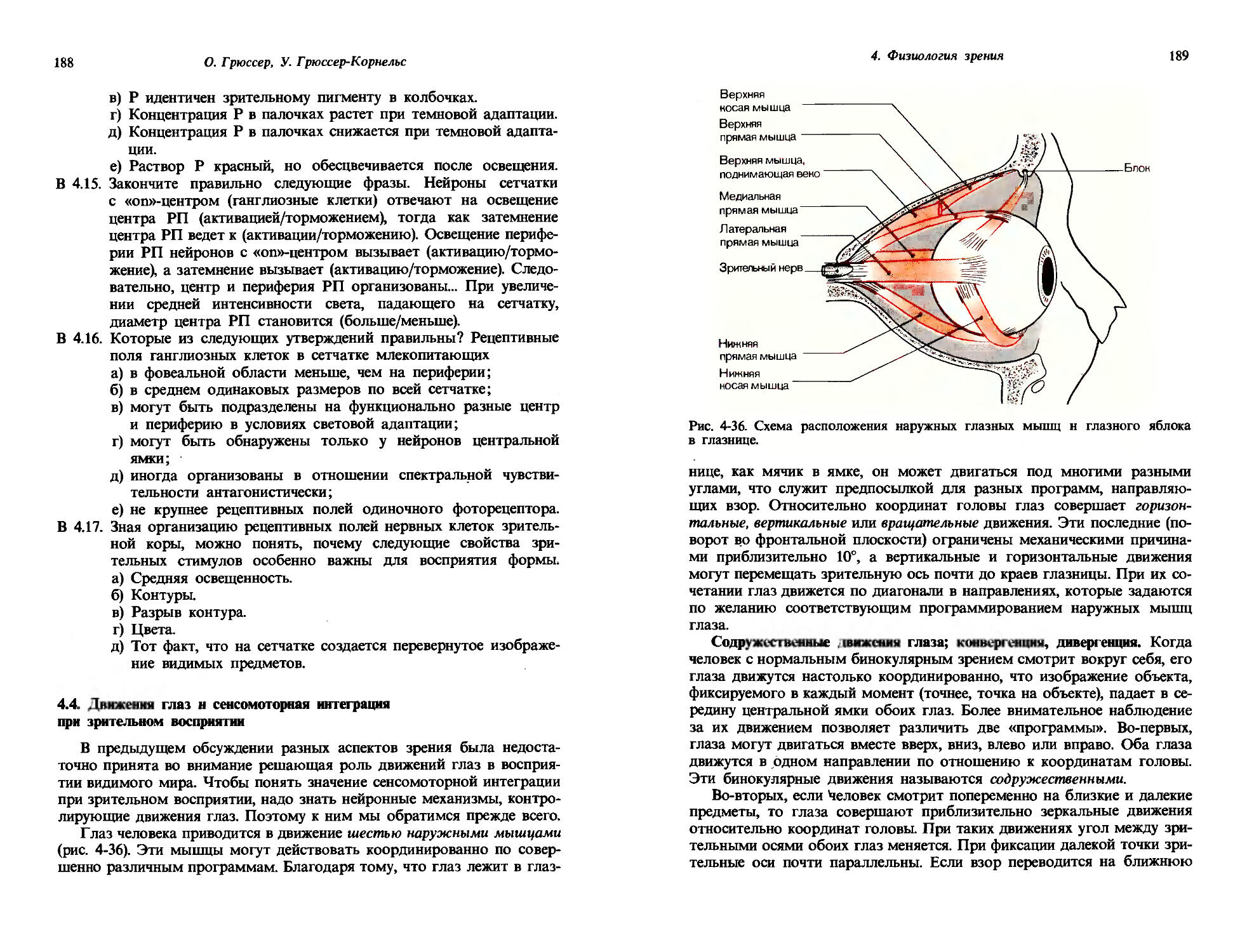
23
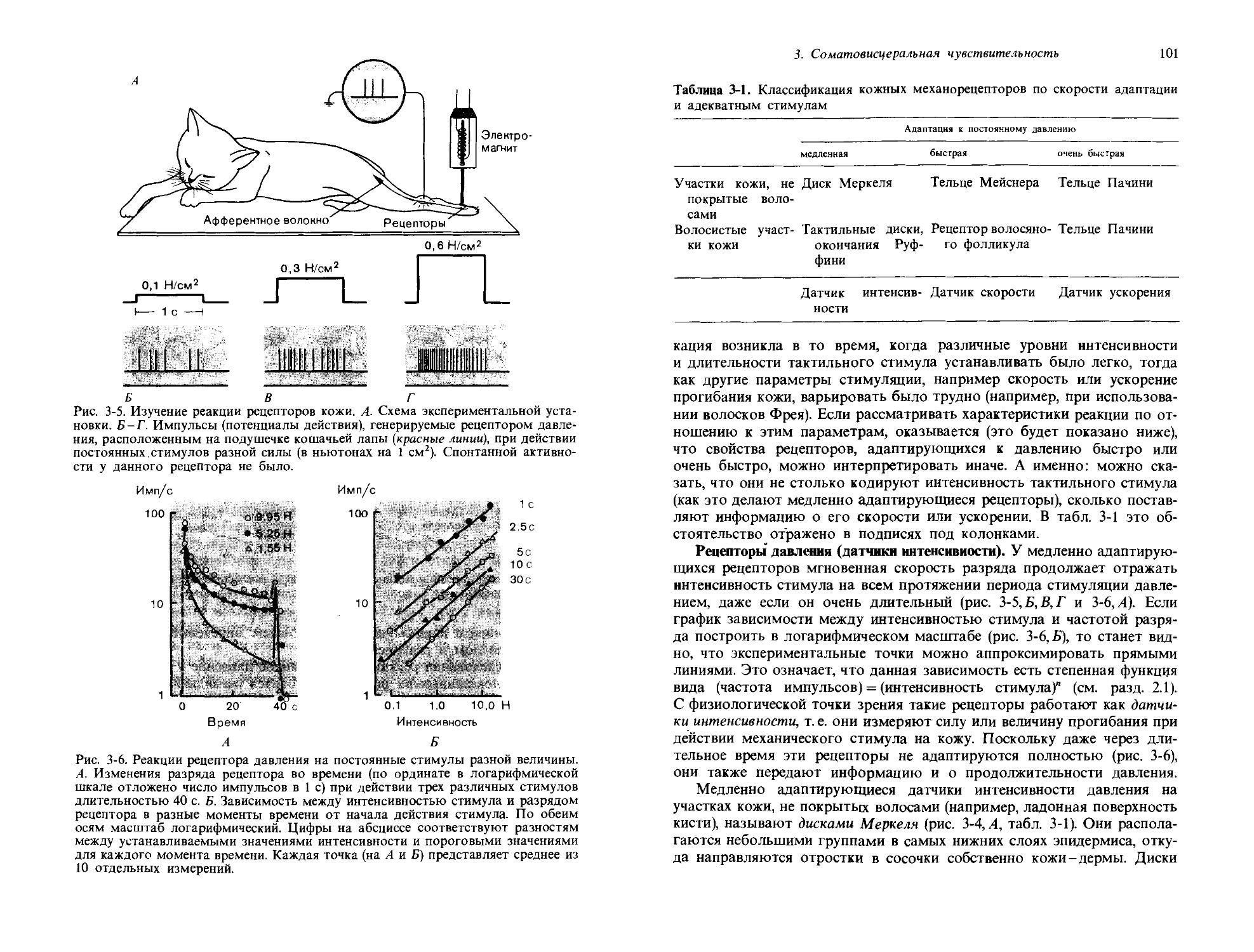
Любая система измерений должна иметь определенную элементарную единицу и фиксированную процедуру, посредством которой определенное число таких единиц может быть приписано величине, которая должна быть измерена. Следовательно, для различных ощущений должны быть установлены подходящие единицы. Одна из таких субъективных единиц, часто используемая, это абсолютный порог ощущения; она берется в качестве эталонной интенсивности ощущения, и другие степени ощущения выражаются числами, показывающими, во сколько раз они превосходят это пороговое ощущение. Но эталонное ощущение можно также установить, взяв в качестве единицы ощущения тот уровень, который порождается каким-то определенным стимулом. Теперь мы опишем три важных метода, используемых для количественного описания интенсивности ощущения.
Оценка отношения интенсивности ощущения к эталонному. Один из возможных способов определения интенсивности ощущения-это определение по оценке испытуемого, во сколько раз данное измеряемое ощущение больше, чем эталонная единица ощущения. Поскольку эта процедура включает определение отношения между ощущением и единицей ощущения, она также называется установлением шкалы отношений. На рис. 1-6 приведен пример такой процедуры. Испытуемым предлагали пробовать растворы лимонной кислоты или сахара в концентрациях, указанных на абсциссе. Затем испытуемые говорили, во сколько раз тестовый раствор кажется им сильнее, чем некоторый стандартный раствор, предложенный для сравнения. Полученные таким путем значения субъективных интенсивностей ощущения отложены (по ординате) красными крестиками. Эти наборы точек могут быть довольно хорошо аппроксимированы прямыми линиями. И по ординате, и по абсциссе масштаб здесь логарифмический, так что прямые линии соответствуют степенным функциям; интенсивность ощущения I пропорциональна п-й степени надпороговой части стимула (S — So):
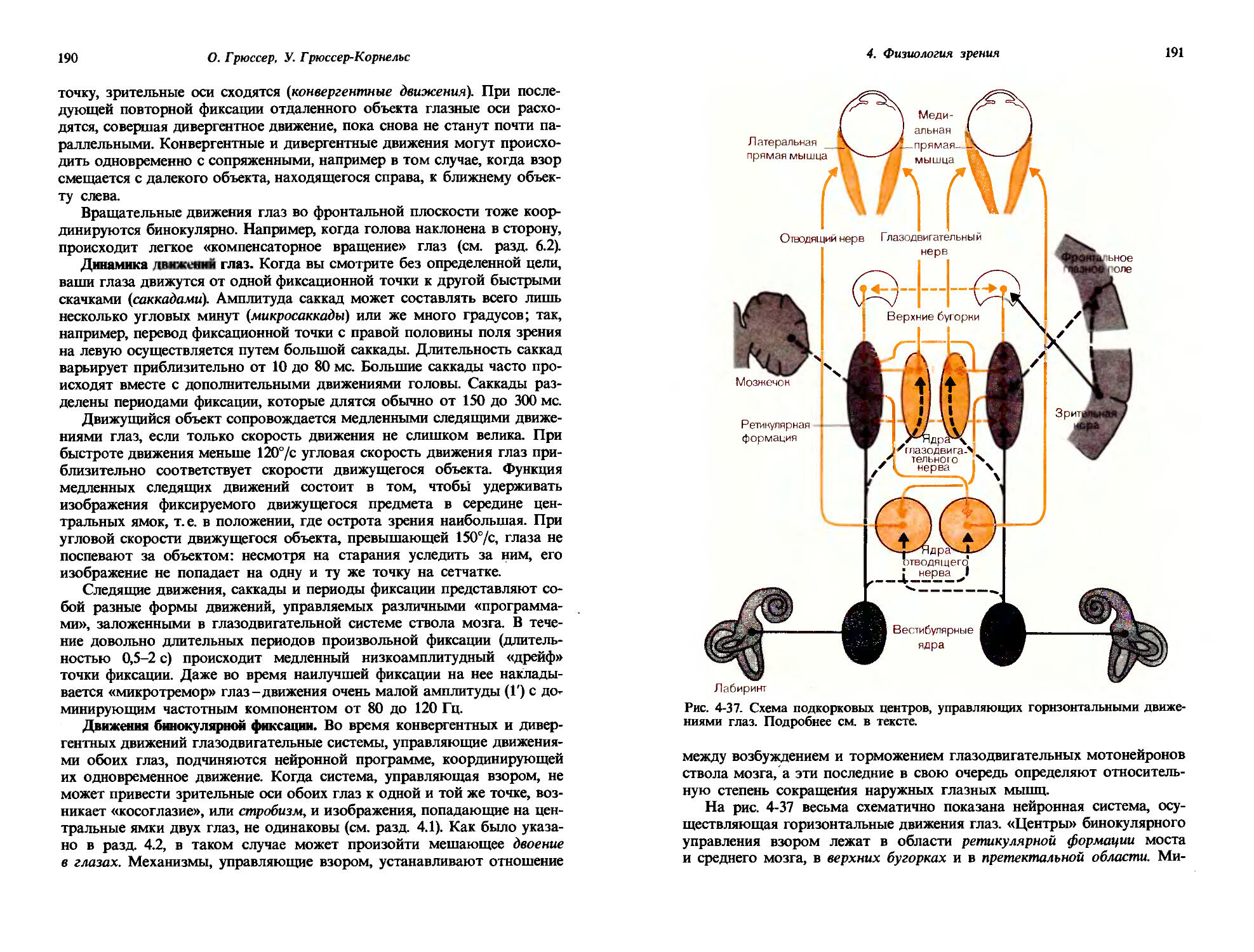
I = k(S-Soy, [1-1]
где /с-константа, a So-пороговая интенсивность стимула. Чтобы понять, почему эта формула представляет прямую линию, заметим, что если взять логарифм от обеих частей [1-1], то получится
log/= nlog(S - So) +/с', [1-2]
где /с'-(равное log/с)-другая константа; таким образом log! является линейной функцией log(S — So). Наклон прямой, получающейся при откладывании интенсивности как стимула, так и ощущения по логарифмическим шкалам (как на рис. 1-6), задается показателем степени п. В случае лимонной кислоты и = 0,85, а для сахарного раствора п= 1,1; эта разница обнаруживается систематически даже на разных испытуемых.
Степенные функции типа [1-1] могут описывать соотношения между стимулом и интенсивностью ощущения в весьма широких диапазонах;
24
Дж. Дудел
Сила стимула-нонцентрация раствора
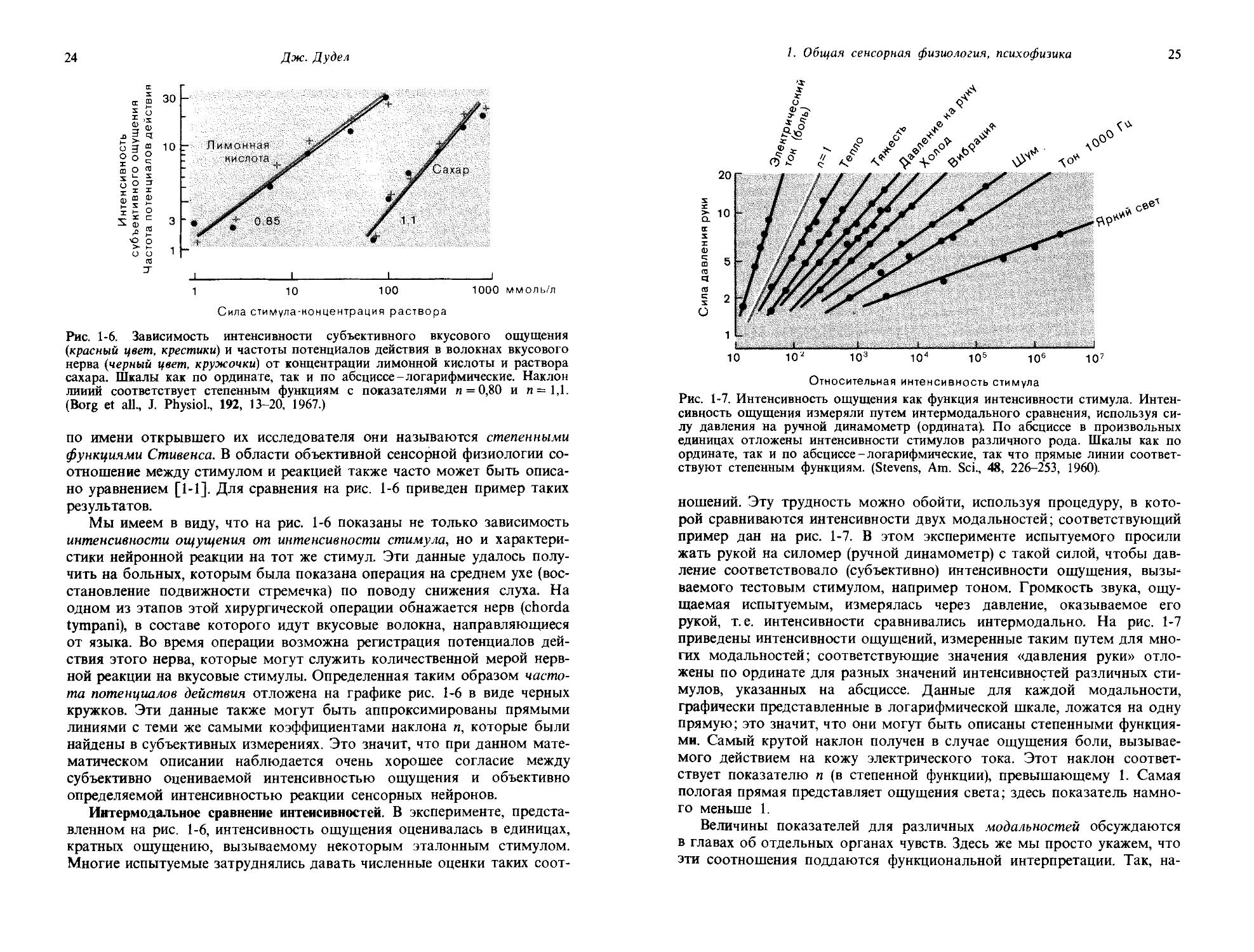
Рис. 1-6. Зависимость интенсивности субъективного вкусового ощущения (красный цвет, крестики) и частоты потенциалов действия в волокнах вкусового нерва (черный цвет, кружочки) от концентрации лимонной кислоты и раствора сахара. Шкалы как по ординате, так и по абсциссе-логарифмические. Наклон линий соответствует степенным функциям с показателями и = 0,80 и и =1,1. (Borg et all., J. Physiol., 192, 13-20, 1967.)
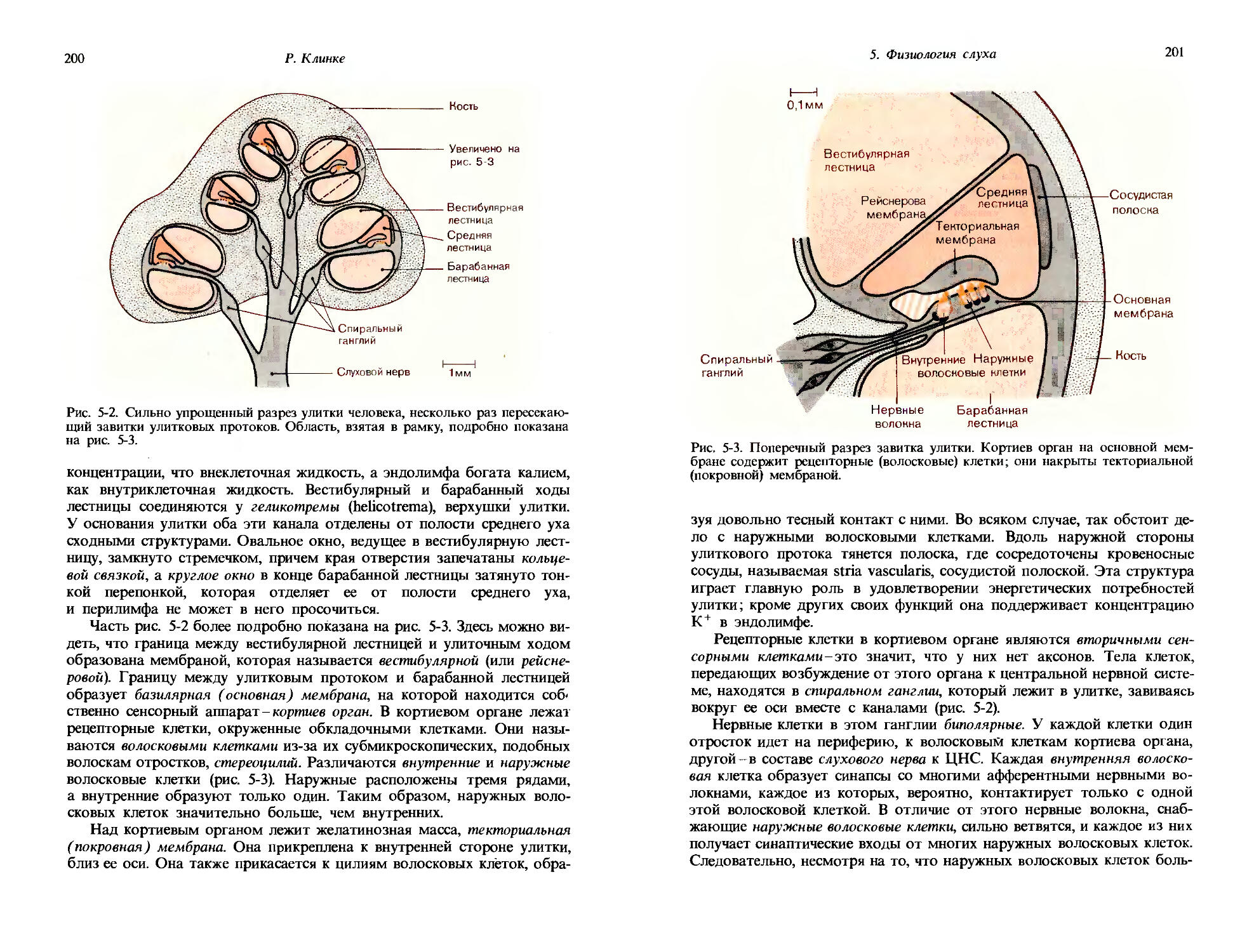
по имени открывшего их исследователя они называются степенными функциями Стивенса. В области объективной сенсорной физиологии соотношение между стимулом и реакцией также часто может быть описано уравнением [1-1]. Для сравнения на рис. 1-6 приведен пример таких результатов.
Мы имеем в виду, что на рис. 1-6 показаны не только зависимость интенсивности ощущения от интенсивности стимула, но и характеристики нейронной реакции на тот же стимул. Эти данные удалось получить на больных, которым была показана операция на среднем ухе (восстановление подвижности стремечка) по поводу снижения слуха. На одном из этапов этой хирургической операции обнажается нерв (chorda tympani), в составе которого идут вкусовые волокна, направляющиеся от языка. Во время операции возможна регистрация потенциалов действия этого нерва, которые могут служить количественной мерой нервной реакции на вкусовые стимулы. Определенная таким образом частота потенциалов действия отложена на графике рис. 1-6 в виде черных кружков. Эти данные также могут быть аппроксимированы прямыми линиями с теми же самыми коэффициентами наклона п, которые были найдены в субъективных измерениях. Это значит, что при данном математическом описании наблюдается очень хорошее согласие между субъективно оцениваемой интенсивностью ощущения и объективно определяемой интенсивностью реакции сенсорных нейронов.
Интермодальное сравнение интенсивностей. В эксперименте, представленном на рис. 1-6, интенсивность ощущения оценивалась в единицах, кратных ощущению, вызываемому некоторым эталонным стимулом. Многие испытуемые затруднялись давать численные оценки таких соот-
1. Общая сенсорная физиология, психофизика
25
Относительная интенсивность стимула
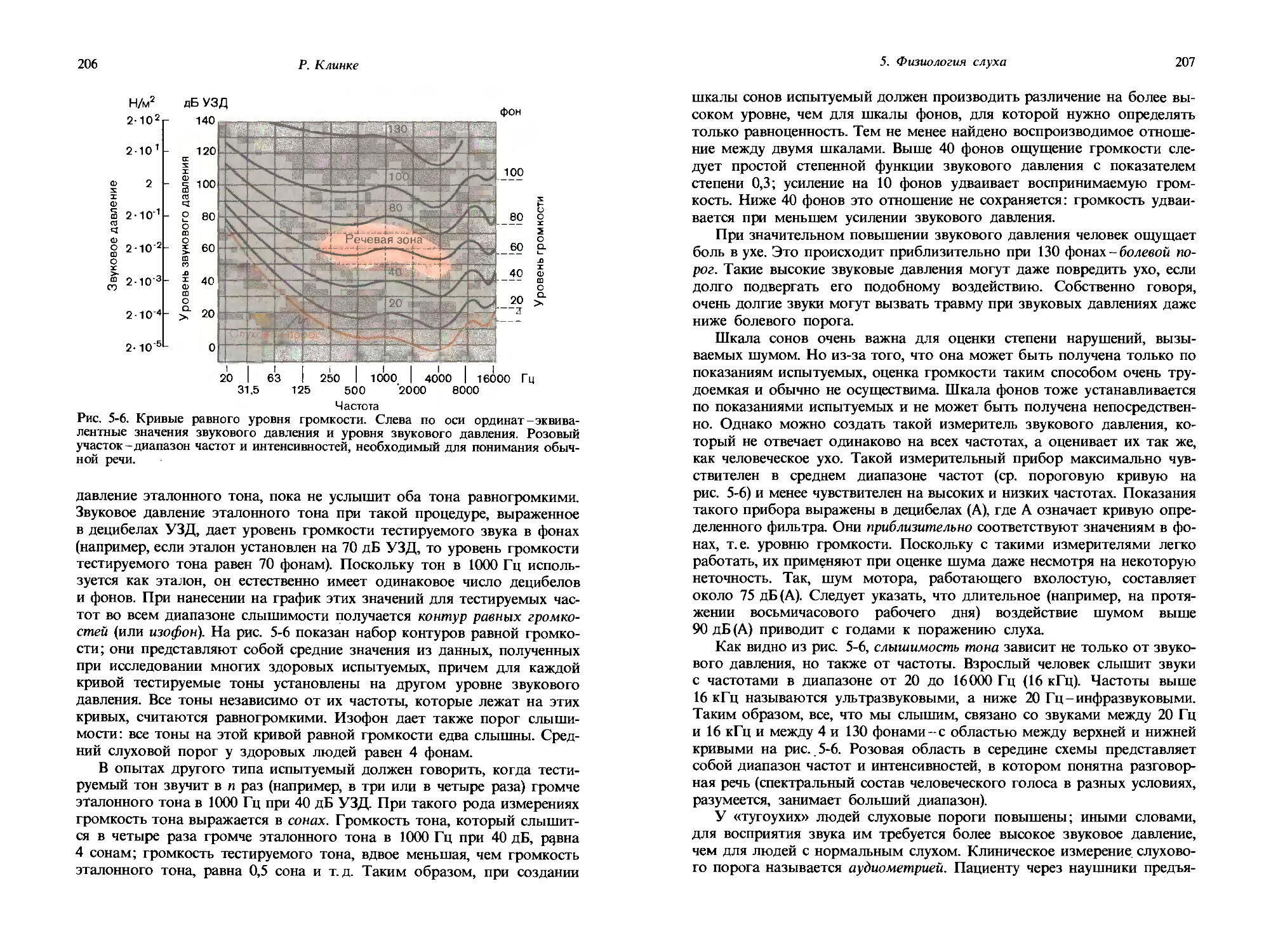
Рис. 1-7. Интенсивность ощущения как функция интенсивности стимула. Интенсивность ощущения измеряли путем интермодального сравнения, используя силу давления на ручной динамометр (ордината). По абсциссе в произвольных единицах отложены интенсивности стимулов различного рода. Шкалы как по ординате, так и по абсциссе-логарифмические, так что прямые линии соответствуют степенным функциям. (Stevens, Am. Sci., 48, 226-253, 1960).
ношений. Эту трудность можно обойти, используя процедуру, в которой сравниваются интенсивности двух модальностей; соответствующий пример дан на рис. 1-7. В этом эксперименте испытуемого просили жать рукой на силомер (ручной динамометр) с такой силой, чтобы давление соответствовало (субъективно) интенсивности ощущения, вызываемого тестовым стимулом, например тоном. Громкость звука, ощущаемая испытуемым, измерялась через давление, оказываемое его рукой, т. е. интенсивности сравнивались интермодально. На рис. 1-7 приведены интенсивности ощущений, измеренные таким путем для многих модальностей; соответствующие значения «давления руки» отложены по ординате для разных значений интенсивностей различных стимулов, указанных на абсциссе. Данные для каждой модальности, графически представленные в логарифмической шкале, ложатся на одну прямую; это значит, что они могут быть описаны степенными функциями. Самый крутой наклон получен в случае ощущения боли, вызываемого действием на кожу электрического тока. Этот наклон соответствует показателю п (в степенной функции), превышающему 1. Самая пологая прямая представляет ощущения света; здесь показатель намного меньше 1.
Величины показателей для различных модальностей обсуждаются в главах об отдельных органах чувств. Здесь же мы просто укажем, что эти соотношения поддаются функциональной интерпретации. Так, на
26
Дж. Дудел
пример, когда возрастают болевой или тепловой стимулы, интенсивность ощущения нарастает заметным образом (и больше или равно 1); такие ощущения имеют характер предупреждающих от более сильного повреждения. Вместе с тем с функциональной точки зрения выгодно, чтобы в случае световых стимулов, диапазон интенсивности которых составляет 5-6 порядков, интенсивность ощущения нарастала сравнительно медленно и благодаря этому позволяла данному диапазону ощущений соответствовать более широкому диапазону стимулов.
Таким образом, интермодальные сравнения интенсивностей позволяют осуществить точные и в широком диапазоне измерения интенсивности ощущения для различных модальностей. Как видно из рис. 1-7, эта процедура дает для степенной функции показатели п, очень хорошо согласующиеся с теми, которые получаются при процедуре, описанной выше,-оценке отношений к стандартному стимулу. Другим примечательным свойством является то, что соотношение между нейронными реакциями и стимулами, измеренное методами объективной сенсорной физиологии, выявляет показатели, очень похожие на те, которые получают при субъективных процедурах, представленных как на рис. 1-6, так и на рис. 1-7.
Измерение посредством дифференциально-пороговых шагов. Третья процедура, используемая в субъективной сенсорной физиологии, состоит в том, что интенсивность ощущения выражается числом дифференциально-пороговых шагов, требующимся для того, чтобы пройти от абсолютного порога (или иного стандартного уровня) к той интенсивности, при которой должно быть измерено ощущение. Дифференциальные пороги (или пороговые различия)-это наименьшие изменения стимула, которые еще могут быть обнаружены. Их часто называют «едва заметными различиями». Измерения интенсивности на этой основе производятся следующим образом. Сначала оценивается пороговая интенсивность того вида стимула, который будет использоваться для данного конкретного испытуемого. Ощущению при этом абсолютном пороге приписывается значение 1. Затем интенсивность стимула увеличивают, пока испытуемый не обнаружит изменение. Ощущению при новой интенсивности приписывают значение 2. Эту процедуру повторяют N раз до тех пор, пока не будет достигнута заданная интенсивность стимула /; ощущению при этой интенсивности стимула приписывается значение Nt. Если таким способом определить для разных интенсивностей ощущения, можно построить график зависимости интенсивности ощущения от интенсивности стимула. Здесь опять получается степенная функция с показателем, близким к найденным с помощью двух других психофизических процедур и путем объективных измерений.
Таким, образом, в отношении определения интенсивности психофизические измерения, выполненные путем подсчета числа дифференциально-пороговых шагов, дают результаты, эквивалентные тем, которые получаются при рассмотренных выше процедурах. Но эту процедуру можно также применить и к другим параметрам стимула-к
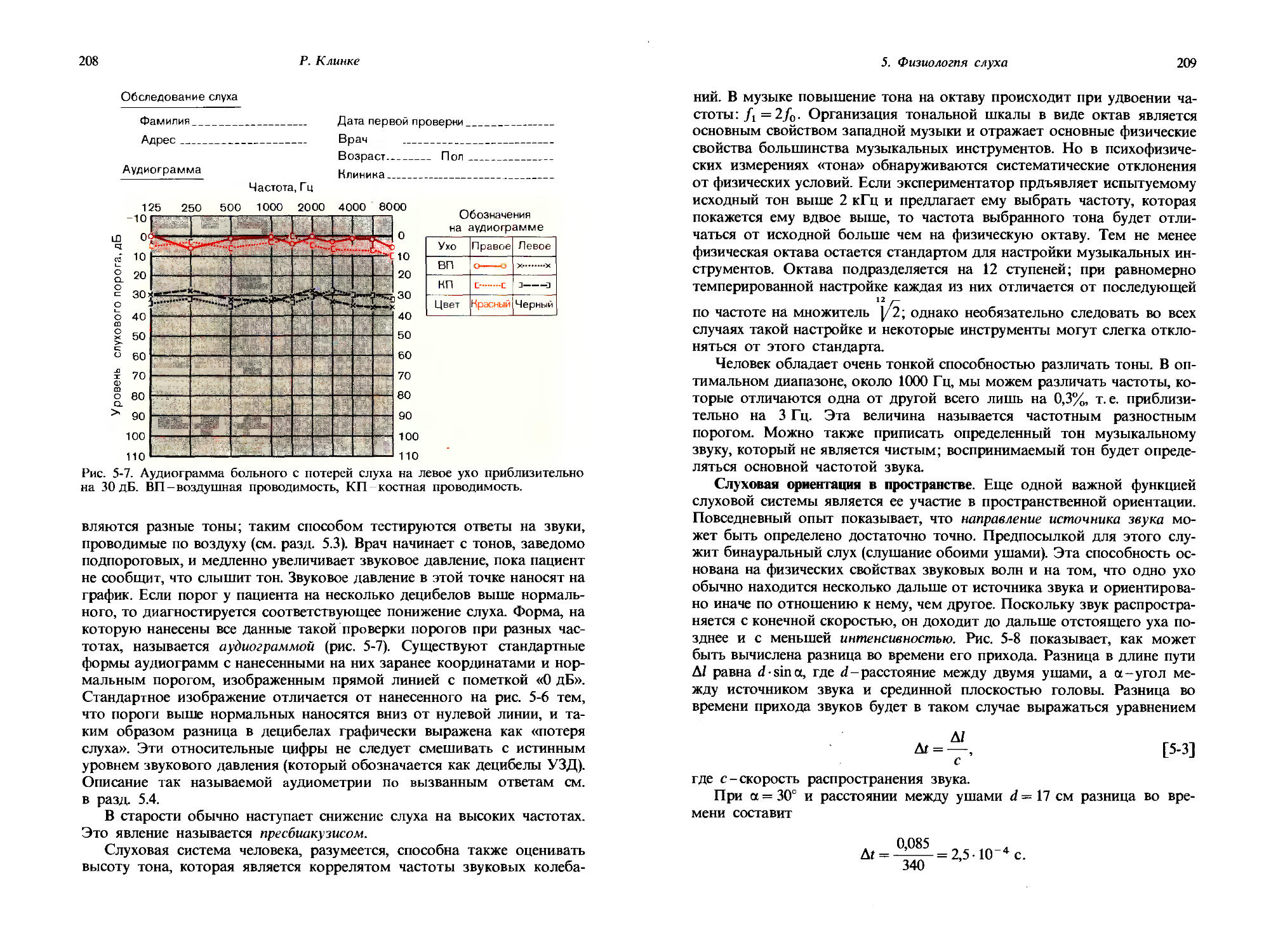
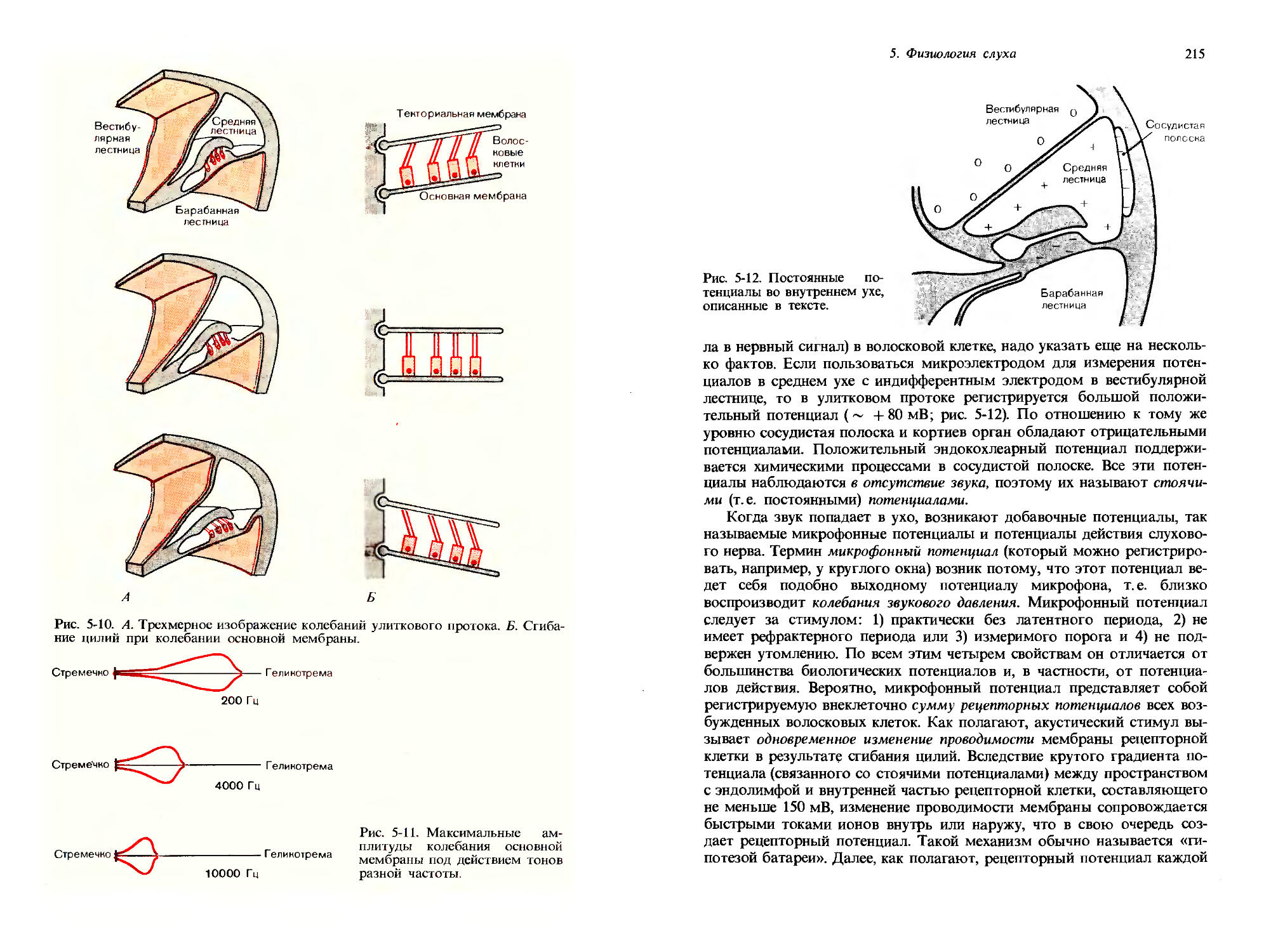
1. Общая сенсорная физиология, психофизика
27
I । < । iii । ।___।___।
О 10 20 30 40 50 60 70 80 90 100
Площадь контакта а
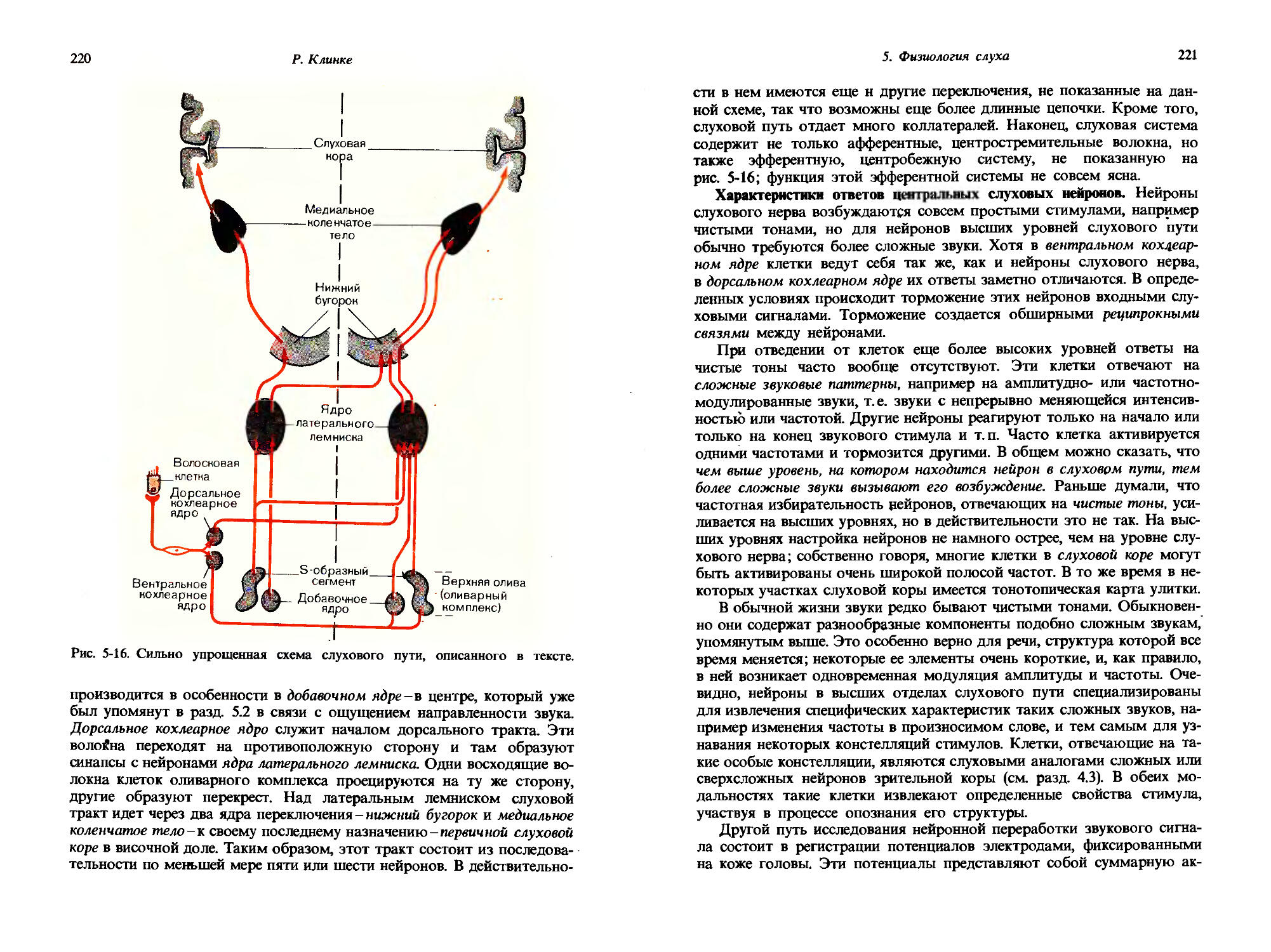
Рис. 1-8. Определение числа N дифференциально-пороговых шагов для стимула «давление на кожу», характеризующегося площадью а и интенсивностью i; данные получены на двух испытуемых (черные и красные стрелки). По ординате откладывали давление, а по абсциссе-площадь, на которую действует стимул, то и другое-в произвольных единицах. Длина стрелок в каждом случае отражает величину дифференциально-порогового шага. Дальнейшие объяснения см. в тексте. (Bergstrom, Lindfors, Acta Physiol., Scand., 44, 170-183, 1958.)
размерностям ощущения, отличным от интенсивности. Дифференциальные пороги могут быть определены не только для интенсивности, но и для протяженности во времени, смещения в пространстве, протяженности по площади и т.д. Для каждой из этих размерностей дифференциальный порог должен рассматриваться как единица, специфическая для каждого субъекта, так что могут быть даны количественные соотношения между величинами различных размерностей.
Таким образом, определяя ряд дифференциальных порогов, можно количественно изучать комбинированные изменения различных параметров или размерностей стимула. Такой эксперимент представлен на рис. 1-8. На графике приведены результаты, полученные на двух испытуемых (черные и красные стрелки); варьировали площадь а, и интенсивность i некоторого стимула (состоящего в давлении на область возвышения большого пальца кисти-thenar); эти параметры откладывались в произвольных единицах по ординате и абсциссе соответственно. Начиная с давления 6,7 и площади 1 площадь контакта увеличивали и шаги стимула, равные дифференциальным порогам, получаемым во время этого увеличения, изображали горизонтальными стрелками. «Черному» испытуемому понадобилось Na = 10 шагов, чтобы добраться до площади стимула 97. Затем, сохраняя площадь стимула постоянной на значении 97, увеличивали интенсивность i и дифференциальные пороги изображали вертикальными стрелками. «Черному» испытуемому понадобилось Nt = 10 пороговых шагов, чтобы достичь давления 10,5.
Теперь начинается интересующая нас часть эксперимента: давление и площадь стимула можно также увеличивать одновременно, что позво
28
Дж. Дудел
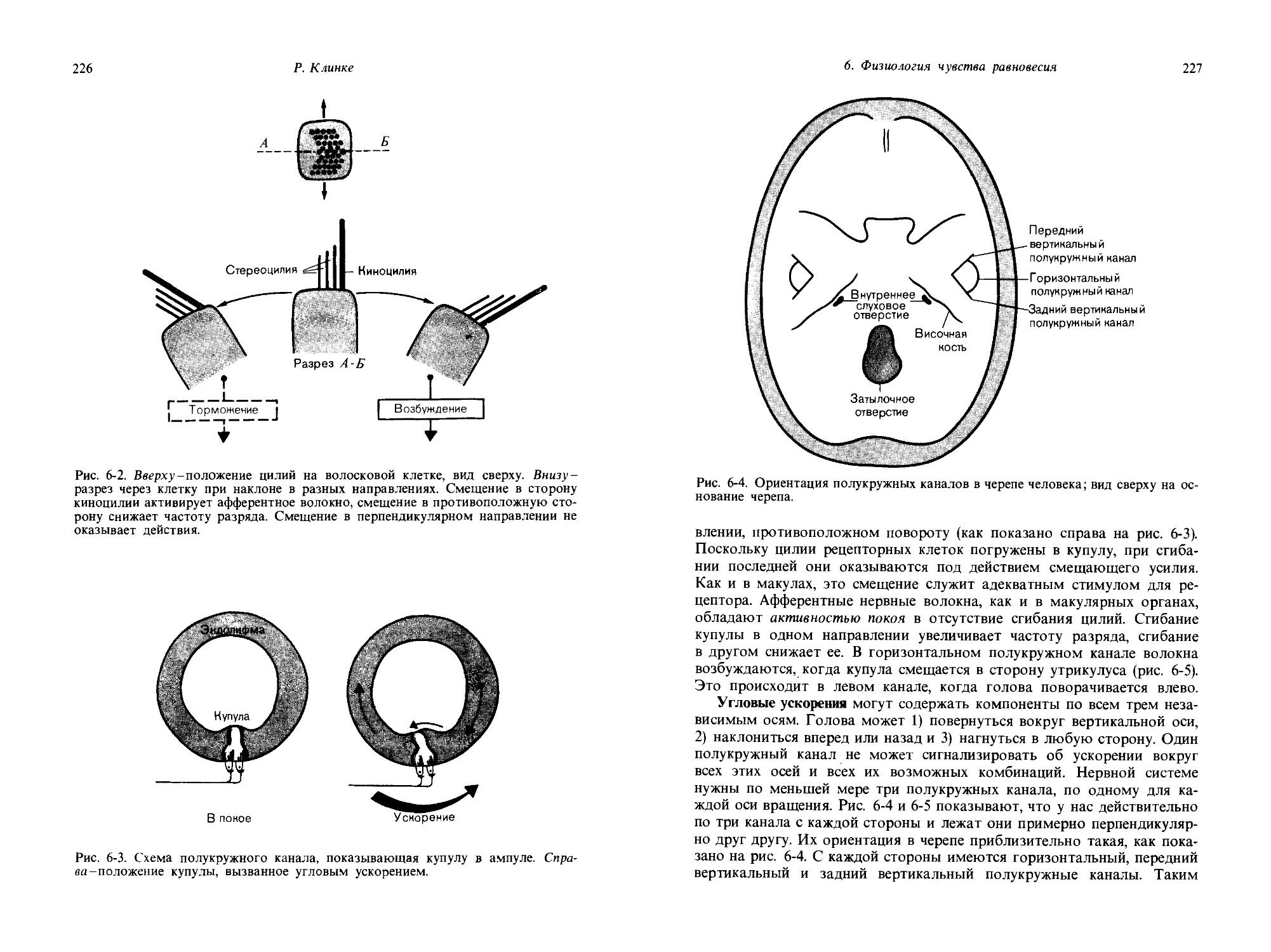
ляет определять комбинированные дифференциальные пороги для одновременного изменения интенсивности и площади. Такие одновременные добавки в эксперименте рис. 1-8 начинаются от значений: площадь контакта = 1, интенсивность давления = 6,7. Последовательные дифференциальные пороги изображены диагональными стрелками. В конечном итоге в результате одновременного изменения давления и площади кривая приближается к финишу, достигнутому в первой части эксперимента,-давлению 10,5 и площади 97. Чтобы достичь этой точки, потребовалось Nai = 14 шагов, равных комбинированным дифференциальным порогам.
Итак, в этом эксперименте мы получили результаты = 10, N/ = 10 и Nai = 14. Можно видеть, что эти цифры приблизительно удовлетворяют следующему соотношению:
Nai = VNa + = /100+ 100 = 14,1. [1-3]
Но это-формула для гипотенузы прямоугольного треугольника, имеющего катеты Na и Nt. Числа дифференциальных порогов математически соответствуют длинам сторон треугольника, образованного стрелками на рис. 1-8.
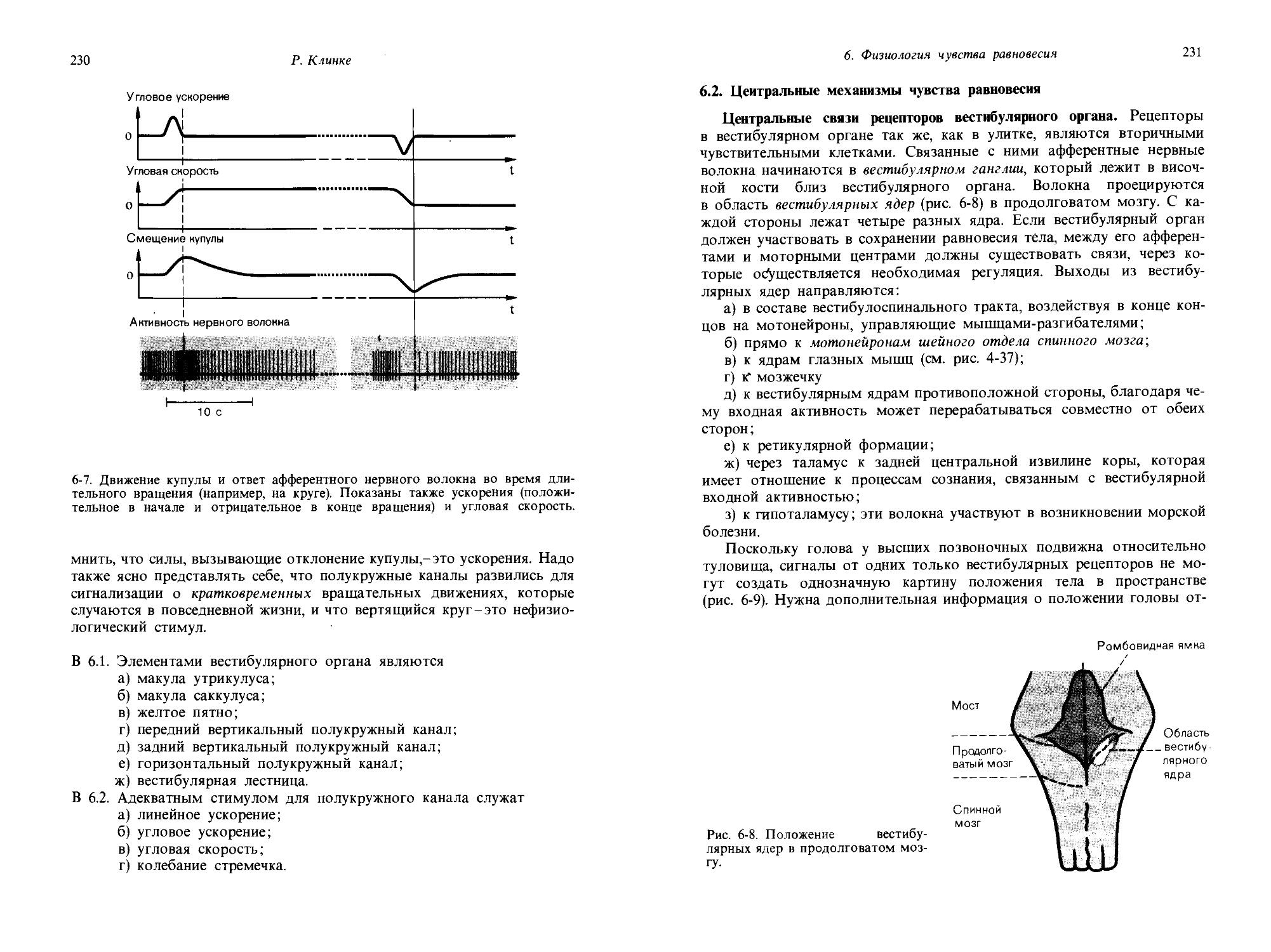
«Красный» испытуемый оказался менее чувствительным, чем «черный»: у него такие же увеличения площади и интенсивности стимула были достигнуты всего за 4 и 5 дифференциально-пороговых шагов соответственно. В согласии с этим и значение Nai = 6 для этого испытуемого оказалось меньше, чем для первого. Причем Nal = 6 опять же достаточно хорошо соответствует длине гипотенузы прямоугольного треугольника со сторонами 4 и 5: )/42 + 52 = |/41 = 6,4.
Таким образом, оказывается, что можно описывать одновременные изменения в различных размерностях ощущения посредством прямоугольной системы координат. Далее, можно сказать, что эти размерности ортогональны друг другу и для них справедлива теорема Пифагора. Пригодность ортогональных соотношений такого рода в определенном диапазоне интенсивностей была продемонстрирована для разных размерностей таких модальностей как зрение, ощущение давления и слух. Тот факт, что одновременные изменения по разным размерностям ощущения могут быть описаны соотношениями, справедливыми для ортогональных величин, делает возможным приложение субъективной сенсорной физиологии к изучению отношений между ощущениями и сложными видами стимулов, приближающимися по свойствам к естественным стимулам из окружающей нас среды. Так, например, когда мы прикасаемся к какому-нибудь объекту, давление и площадь контакта меняются одновременно и непрерывно.
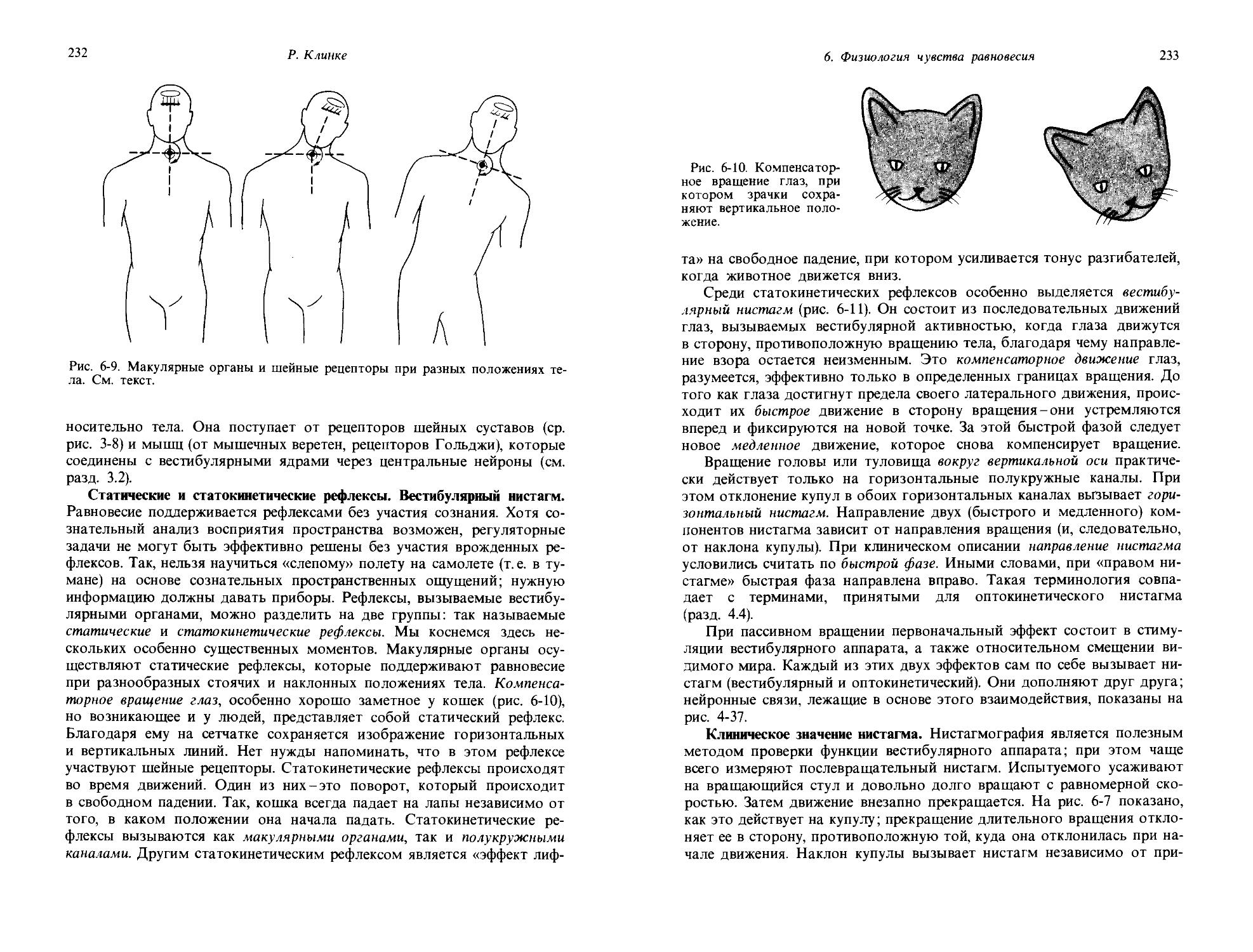
Анализ дифференциальных порогов по рис. 1-8 открывает нам еще одну закономерность, имеющую большое значение в общей сенсорной физиологии. Длины отдельных стрелок указывают величину изменения стимула AS, которая соответствует дифференциальному порогу. Видно, что по мере того, как амплитуда стимула возрастает, стрелки становят
1. Общая сенсорная физиология, психофизика
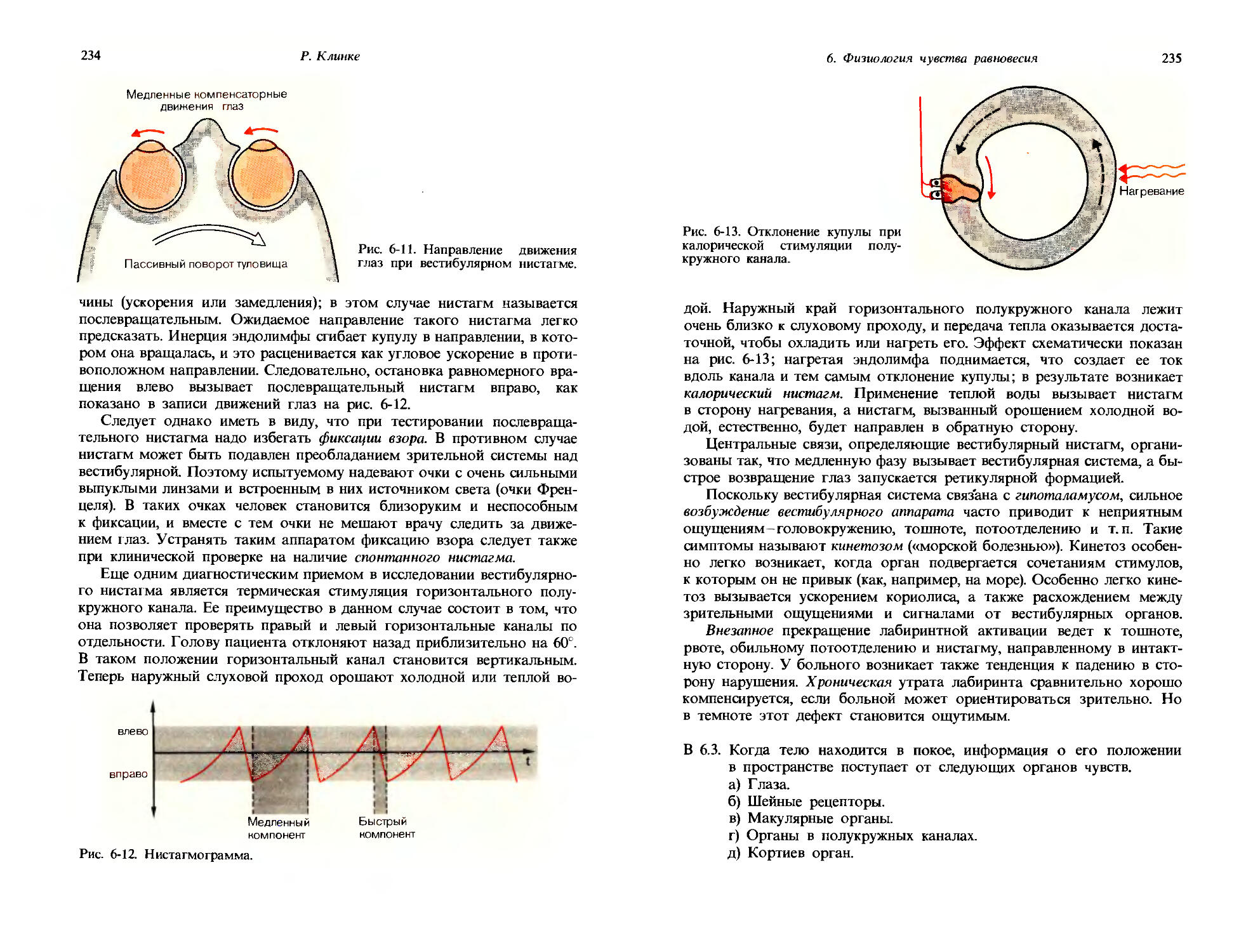
29
ся все длиннее; в среднем, они примерно пропорциональны амплитуде стимула. В прошлом веке на основании подобных наблюдений Вебер сформулировал правило, которое получило название правила Вебера:
AS ----= const.
S
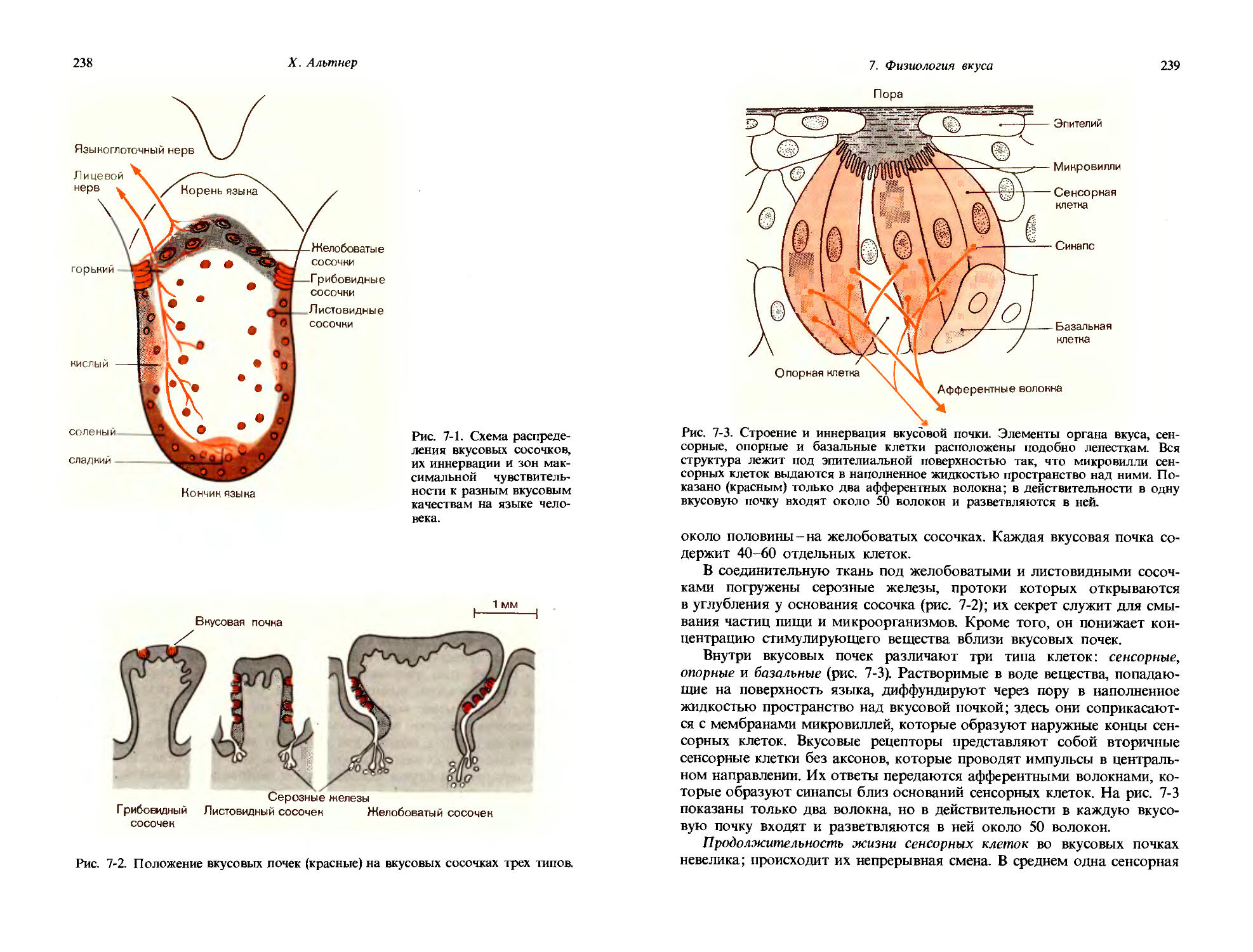
[1-4]
Если применить его к случаю давления на кожу, это правило будет означать, что как бы ни менялась величина стимула, дифференциальный порог всегда будет составлять 3% от исходного уровня.
Если принять, что правило Вебера справедливо всегда, можно сделать заключение, что реакция на стимул пропорциональна логарифму амплитуды стимула. Это соотношение называют законом Вебера-Фех-нера и часто ссылаются на него как на «основной закон психофизики». Однако этот «закон» справедлив только в ограниченном диапазоне интенсивности и применим не ко всем модальностям. Другое соотношение-уже упоминавшаяся степенная функция Стивенса F = = k(S — So)" - имеет гораздо более широкую область применения.
Давайте проиллюстрируем эту разницу примером из области зрения. Реакции клеток зрительной коры могут быть описаны функцией Стивенса в диапазоне яркостей 1 :10000 (см. рис. 1-7), тогда как закон Вебера-Фехнера выдерживается с хорошим приближением лишь для среднего диапазона яркостей, соответствующего отношению интенсивностей 1:100.
В 1.8. Перечислите основные размерности восприятия.
В 1.9. Какая из следующих процедур может быть использована для измерения интенсивности ощущения?
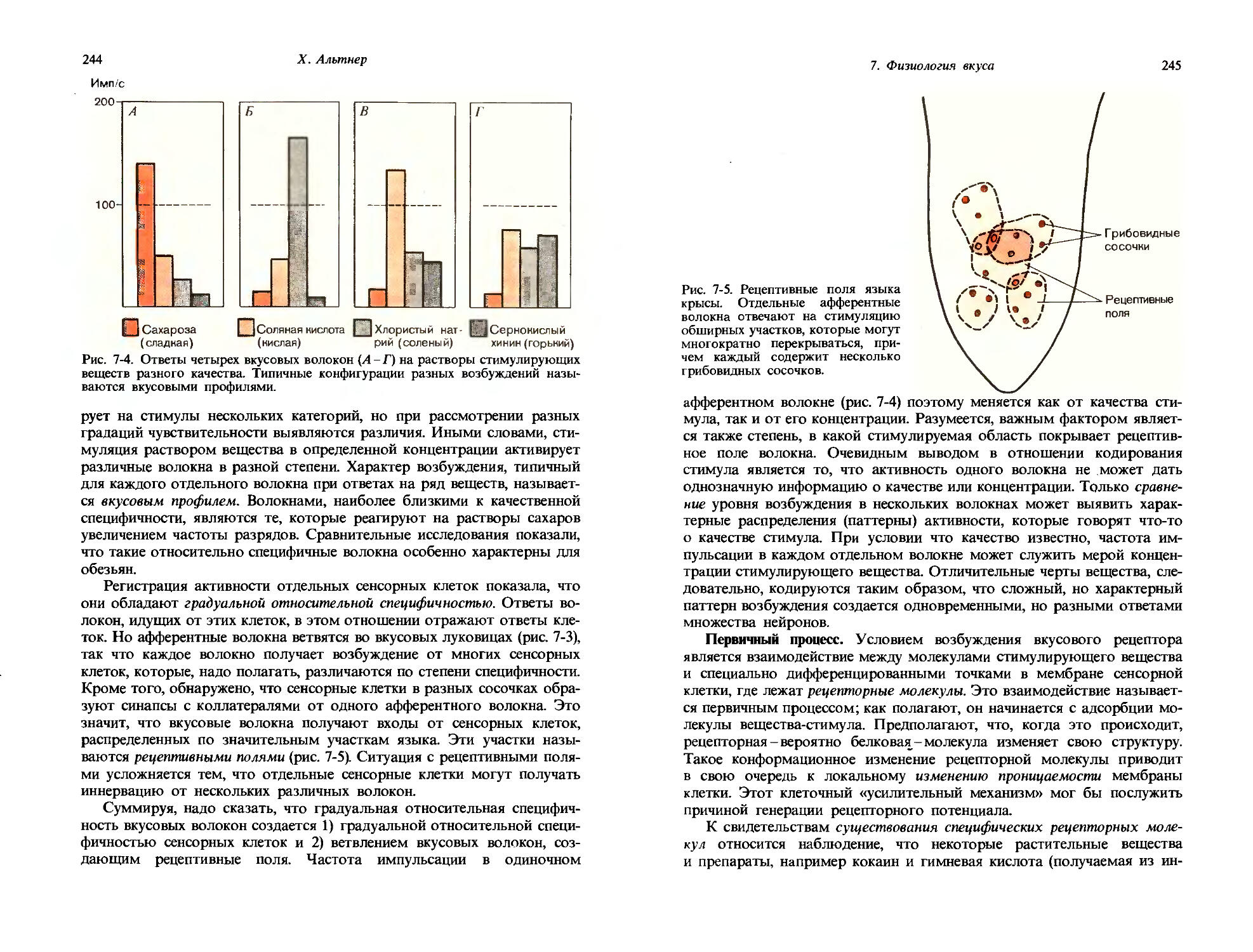
а) Специальное психофизическое определение субъективной интенсивности.
б) Измерение времени, протекающего до исчезновения ощущения.
в) Определение числа дифференциально-пороговых шагов, необходимых для увеличения стимула от абсолютного порога до той интенсивности, при которой оценивается ощущение.
г) Оценка отношения интенсивностей эталонного ощущения и того, которое должно быть измерено.
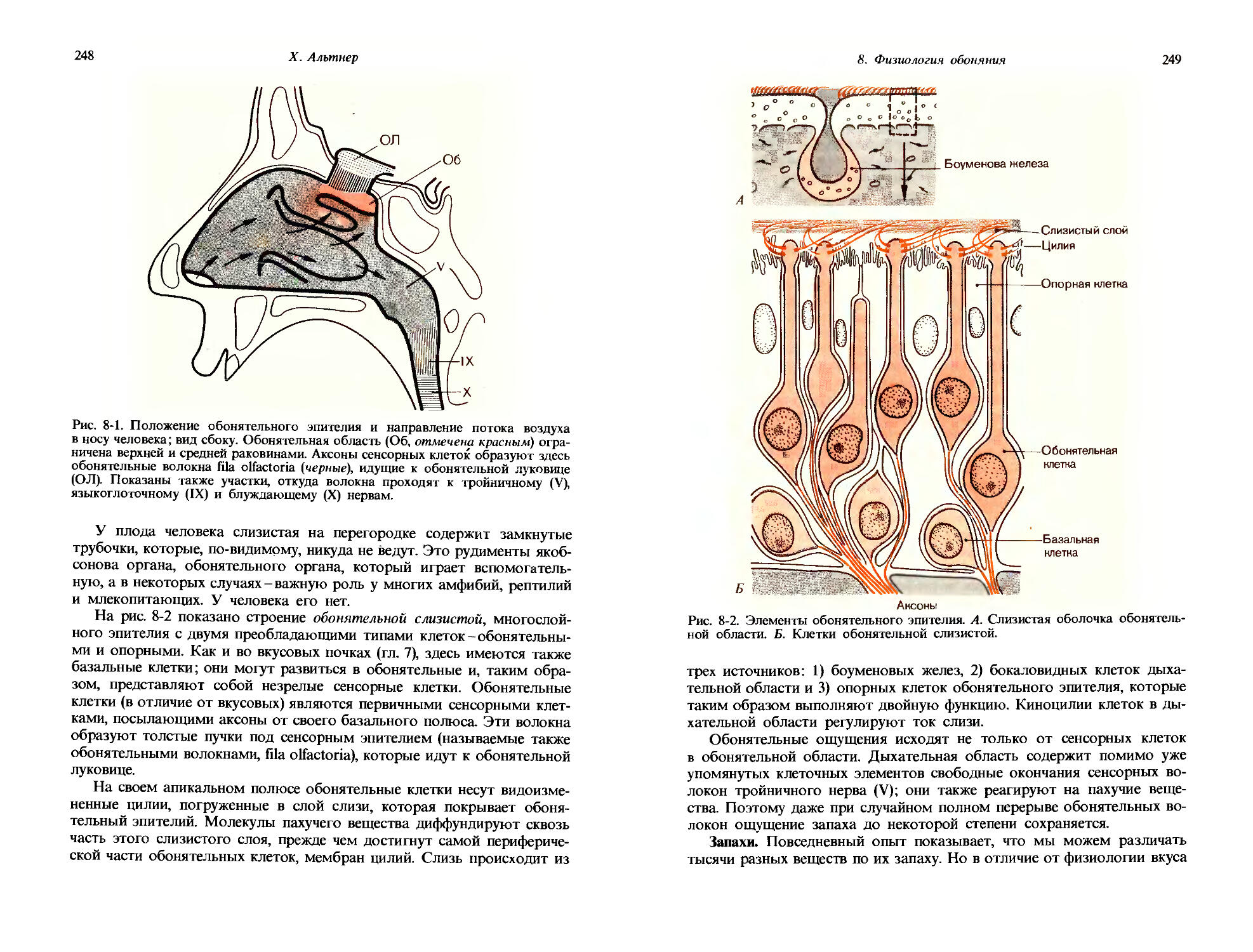
В 1.10. Для измерения светового ощущения в ответ на предъявление яркого пятна могла быть применена следующая процедура: сначала интенсивность точечного источника света увеличили за N. = 3 дифференциально-пороговых шага, а затем площадь источника света увеличили за Na = 4 дифференциально-пороговых шага. Сколько дифференциально-пороговых шагов потребуется для достижения той же интенсивности ощущения, если яркость и площадь увеличить одновременно начиная от абсолютного порога?
30
Дж. Дудел
1.4. Пространственные, временные и эмоциональные аспекты ощущения
Пространственная размерность ощущения. Ощущения имеют не только размерности качества и количества, которые обсуждались до сих пор, но также и размерности пространства и времени. Рассмотрим сначала пространственную размерность. Наши ощущения относятся к окружающему нас пространству; мы воспринимаем стимулы, идущие из определенных мест, как объекты, имеющие разную протяженность и находящиеся на разных расстояниях от нас. Эти параметры ощущений можно определить путем измерения. Так, например, порог пространственного разрешения измеряется как минимальное расстояние между двумя источниками стимула, при котором они еще могут восприниматься как отдельные. На рис. 3-3 показаны результаты такого эксперимента, в котором на кожу помещали два острия иглы. Другим примером может служить эксперимент, результаты которого представлены на рис. 1-8 (абсцисса; здесь переменной является площадь, на которую оказывали давление) и в котором измерение размера или протяженности стимула производили путем подсчета дифференциально-пороговых шагов.
Из этого эксперимента уже стало ясно, что впечатление интенсивности сенсорного стимула усиливается при увеличении площади, на которую данный стимул действует. Это подразумевает, что ощущения, вызываемые различными элементами внутри стимулируемой площади, хотя бы частично суммируются. В определенных диапазонах некоторых модальностей эта суммация подчиняется теореме Пифагора [1-3], и, таким образом, она не является линейной суммацией для надпороговых стимулов-удвоение площади стимула не увеличивает ощущение вдвое по сравнению с исходной интенсивностью. Вместе с тем, в случае пороговых стимулов суммация может быть линейной. Когда стимулом является источник света, произведение пороговой интенсивности стимула на освещаемую площадь есть величина постоянная:
Is А = const. [1-5]
Так, если пороговая интенсивность света равняется 20 при площади освещаемой поверхности 1 (в произвольных единицах), то увеличение площади до 10 будет сопровождаться снижением пороговой интенсивности света до 2. Заметим, что [1-5] справедливо только для малых участков поля зрения; после превышения некоторой критической площади пороговая интенсивность становится не зависящей от площади стимула. Соотношения, подобные [1-5], равно справедливы и для других модальностей. Приблизительно линейная пространственная суммация пороговых стимулов составляет основу для обнаружения крупных объектов при слабом освещении. В сильно затемненной комнате положение стрелок еще удается разглядеть на будильнике с большим циферблатом, но не на ручных часах.
1. Общая сенсорная физиология, психофизика
31
До сих пор мы занимались зависимостью величины ощущения от пространственной протяженности одного стимула. Более интересным аспектом является то, как на восприятие влияет второй стимул, пространственно отделенный от первого. Тут появляются эффекты облегчения и торможения, которые удобнее всего описать на примере явлений, связанных с контрастом.
Контраст. Термин «контраст» в применении к зрительному восприятию означает отношение яркостей смежных частей рассматриваемой картины-например, отношение яркости темного буфета к яркости светлой стены, у которой он стоит. Аналогичным образом контраст можно определить и для других ощущений (например, громкость речи оратора по отношению к фоновому шуму). Только при наличии достаточного контраста объекты настолько выделяются на фоне, что их можно обнаружить. Когда различия в яркости между частями зрительной картины слишком малы, образы становятся туманными и нечеткими, и в случае экрана телевизора приходится использовать электронные усилители, чтобы усилить контраст.
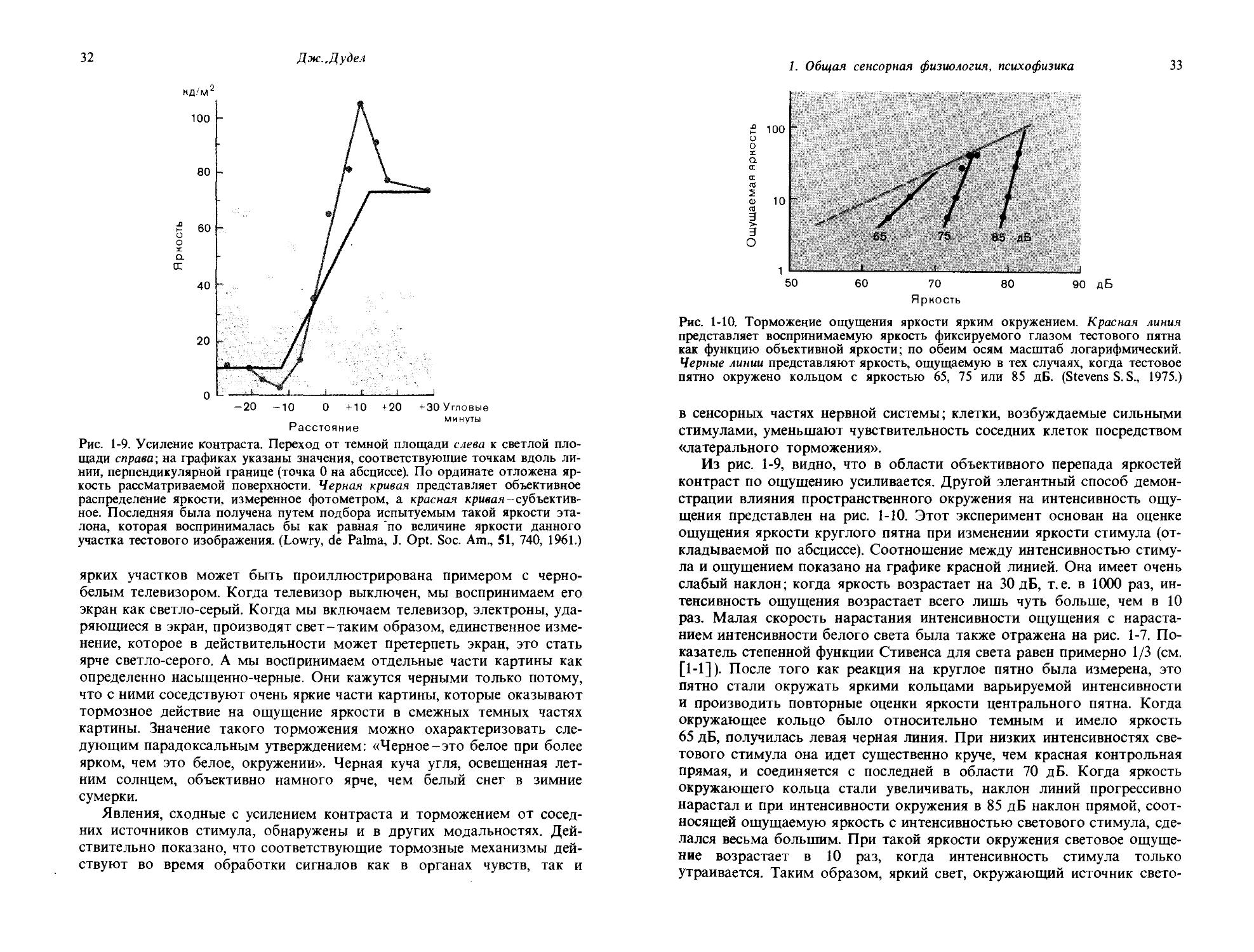
Общим свойством сенсорного восприятия является то, что имеет место усиление контраста. Когда мы смотрим на любую сравнительно большую темную область на светлом фоне, края темной зоны кажутся темнее, чем ее середина, а вокруг темного объекта имеется узкая полоска фона, которая кажется ярче окружения (см. также рис. 4-11). Усиление контраста такого рода можно продемонстрировать количественно. В эксперименте, к которому относится рис. 1-9, испытуемый рассматривал поверхность, где темная область слева соседствовала со светлой областью справа; объективное распределение яркостей, измеренное физическим устройством, показано черной линией. Красной линией изображено субъективно ощущаемое распределение яркостей. Для его определения испытуемого инструктировали устанавливать яркость эталона на уровне, субъективно ощущаемом как равный по яркости различным областям переходной зоны в рассматриваемом образце. Субъективные изменения яркости намного резче, чем объективно измеряемые перепады. Кроме того, субъективно край темной зоны кажется темнее. Таким образом, субъективное усиление контраста легко измерить.
Тот факт, что сразу слева от края на рис. 1-9 ощущаемая яркость намного меньше, чем внутри темной площади, указывает на существование торможения; светлая зона справа уменьшает субъективную яркость смежной темной области. Таким образом, интенсивный световой стимул не только ведет к ощущению большой интенсивности, но и снижает или тормозит ощущение света в соседних частях картины. Увеличенная яркость сразу справа от границы на рис. 1-9 также определяется этим механизмом торможения в смежных областях. Внутри однородной яркой зоны соседние яркие части картины взаимно тормозят друг друга, тогда как на границе яркой зоны часть торможения от смежных (более темных) частей исключается; по этой причине ощущение оказывается относительно «расторможенным». Заметная тормозная функция
32
Дж..Дудел
Рис. 1-9. Усиление контраста. Переход от темной площади слева к светлой площади справа; на графиках указаны значения, соответствующие точкам вдоль линии, перпендикулярной границе (точка 0 на абсциссе). По ординате отложена яркость рассматриваемой поверхности. Черная кривая представляет объективное распределение яркости, измеренное фотометром, а красная кривая-субъективное. Последняя была получена путем подбора испытуемым такой яркости эталона, которая воспринималась бы как равная по величине яркости данного участка тестового изображения. (Lowry, de Palma, J. Opt. Soc. Am., 51, 740, 1961.)
ярких участков может быть проиллюстрирована примером с чернобелым телевизором. Когда телевизор выключен, мы воспринимаем его экран как светло-серый. Когда мы включаем телевизор, электроны, ударяющиеся в экран, производят свет-таким образом, единственное изменение, которое в действительности может претерпеть экран, это стать ярче светло-серого. А мы воспринимаем отдельные части картины как определенно насыщенно-черные. Они кажутся черными только потому, что с ними соседствуют очень яркие части картины, которые оказывают тормозное действие на ощущение яркости в смежных темных частях картины. Значение такого торможения можно охарактеризовать следующим парадоксальным утверждением: «Черное-это белое при более ярком, чем это белое, окружении». Черная куча угля, освещенная летним солнцем, объективно намного ярче, чем белый снег в зимние сумерки.
Явления, сходные с усилением контраста и торможением от соседних источников стимула, обнаружены и в других модальностях. Действительно показано, что соответствующие тормозные механизмы действуют во время обработки сигналов как в органах чувств, так и
1. Общая сенсорная физиология, психофизика
33
Яркость
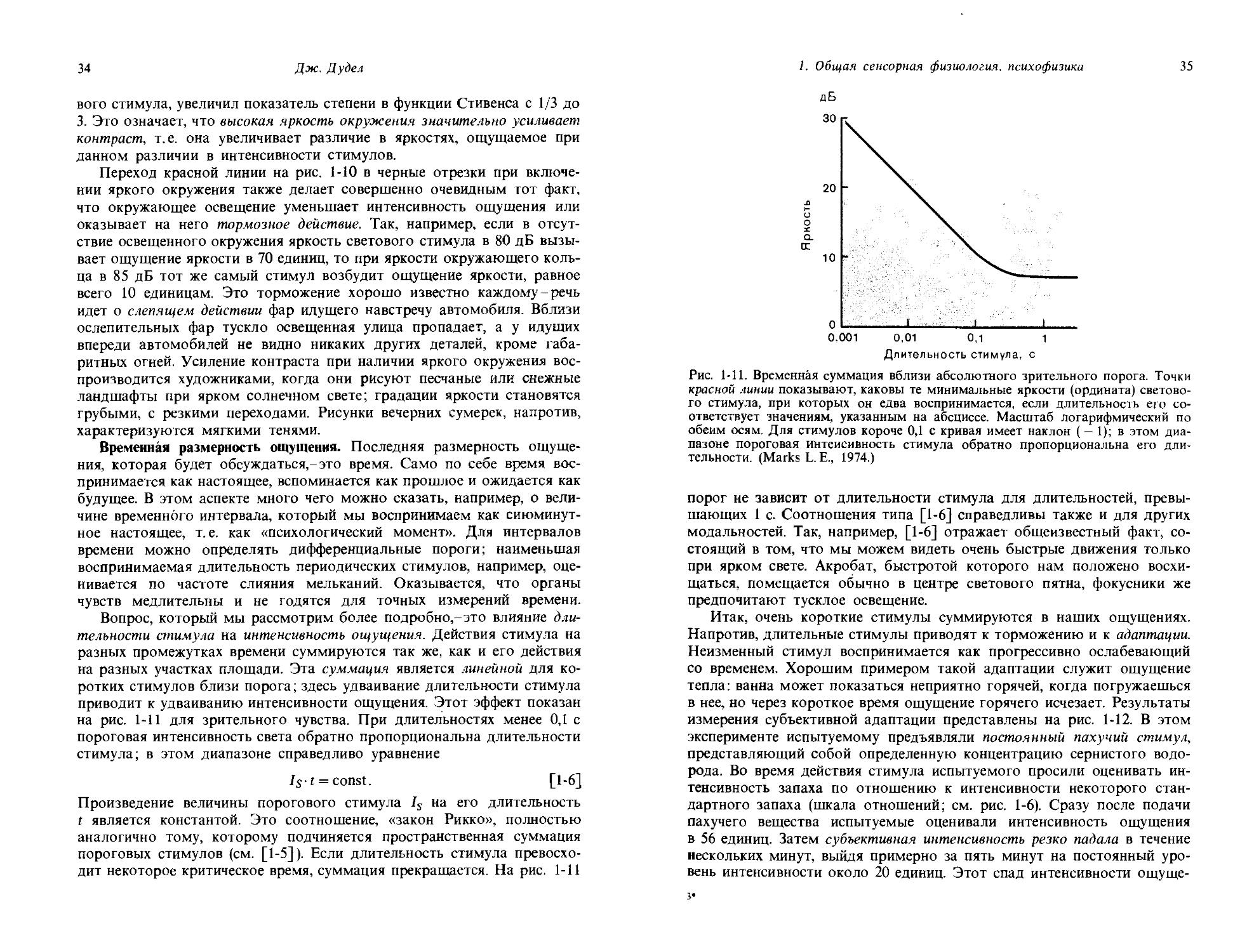
Рис. 1-10. Торможение ощущения яркости ярким окружением. Красная линия представляет воспринимаемую яркость фиксируемого глазом тестового пятна как функцию объективной яркости; по обеим осям масштаб логарифмический. Черные линии представляют яркость, ощущаемую в тех случаях, когда тестовое пятно окружено кольцом с яркостью 65, 75 или 85 дБ. (Stevens S. S., 1975.)
в сенсорных частях нервной системы; клетки, возбуждаемые сильными стимулами, уменьшают чувствительность соседних клеток посредством «латерального торможения».
Из рис. 1-9, видно, что в области объективного перепада яркостей контраст по ощущению усиливается. Другой элегантный способ демонстрации влияния пространственного окружения на интенсивность ощущения представлен на рис. 1-10. Этот эксперимент основан на оценке ощущения яркости круглого пятна при изменении яркости стимула (откладываемой по абсциссе). Соотношение между интенсивностью стимула и ощущением показано на графике красной линией. Она имеет очень слабый наклон; когда яркость возрастает на 30 дБ, т.е. в 1000 раз, интенсивность ощущения возрастает всего лишь чуть больше, чем в 10 раз. Малая скорость нарастания интенсивности ощущения с нарастанием интенсивности белого света была также отражена на рис. 1-7. Показатель степенной функции Стивенса для света равен примерно 1/3 (см. [1-1]). После того как реакция на круглое пятно была измерена, это пятно стали окружать яркими кольцами варьируемой интенсивности и производить повторные оценки яркости центрального пятна. Когда окружающее кольцо было относительно темным и имело яркость 65 дБ, получилась левая черная линия. При низких интенсивностях светового стимула она идет существенно круче, чем красная контрольная прямая, и соединяется с последней в области 70 дБ. Когда яркость окружающего кольца стали увеличивать, наклон линий прогрессивно нарастал и при интенсивности окружения в 85 дБ наклон прямой, соотносящей ощущаемую яркость с интенсивностью светового стимула, сделался весьма большим. При такой яркости окружения световое ощущение возрастает в 10 раз, когда интенсивность стимула только утраивается. Таким образом, яркий свет, окружающий источник свето
34
Дж. Дудел
вого стимула, увеличил показатель степени в функции Стивенса с 1/3 до 3. Это означает, что высокая яркость окружения значительно усиливает контраст, т.е. она увеличивает различие в яркостях, ощущаемое при данном различии в интенсивности стимулов.
Переход красной линии на рис. 1-10 в черные отрезки при включении яркого окружения также делает совершенно очевидным тот факт, что окружающее освещение уменьшает интенсивность ощущения или оказывает на него тормозное действие. Так, например, если в отсутствие освещенного окружения яркость светового стимула в 80 дБ вызывает ощущение яркости в 70 единиц, то при яркости окружающего кольца в 85 дБ тот же самый стимул возбудит ощущение яркости, равное всего 10 единицам. Это торможение хорошо известно каждому-речь идет о слепящем действии фар идущего навстречу автомобиля. Вблизи ослепительных фар тускло освещенная улица пропадает, а у идущих впереди автомобилей не видно никаких других деталей, кроме габаритных огней. Усиление контраста при наличии яркого окружения воспроизводится художниками, когда они рисуют песчаные или снежные ландшафты при ярком солнечном свете; градации яркости становятся грубыми, с резкими переходами. Рисунки вечерних сумерек, напротив, характеризуются мягкими тенями.
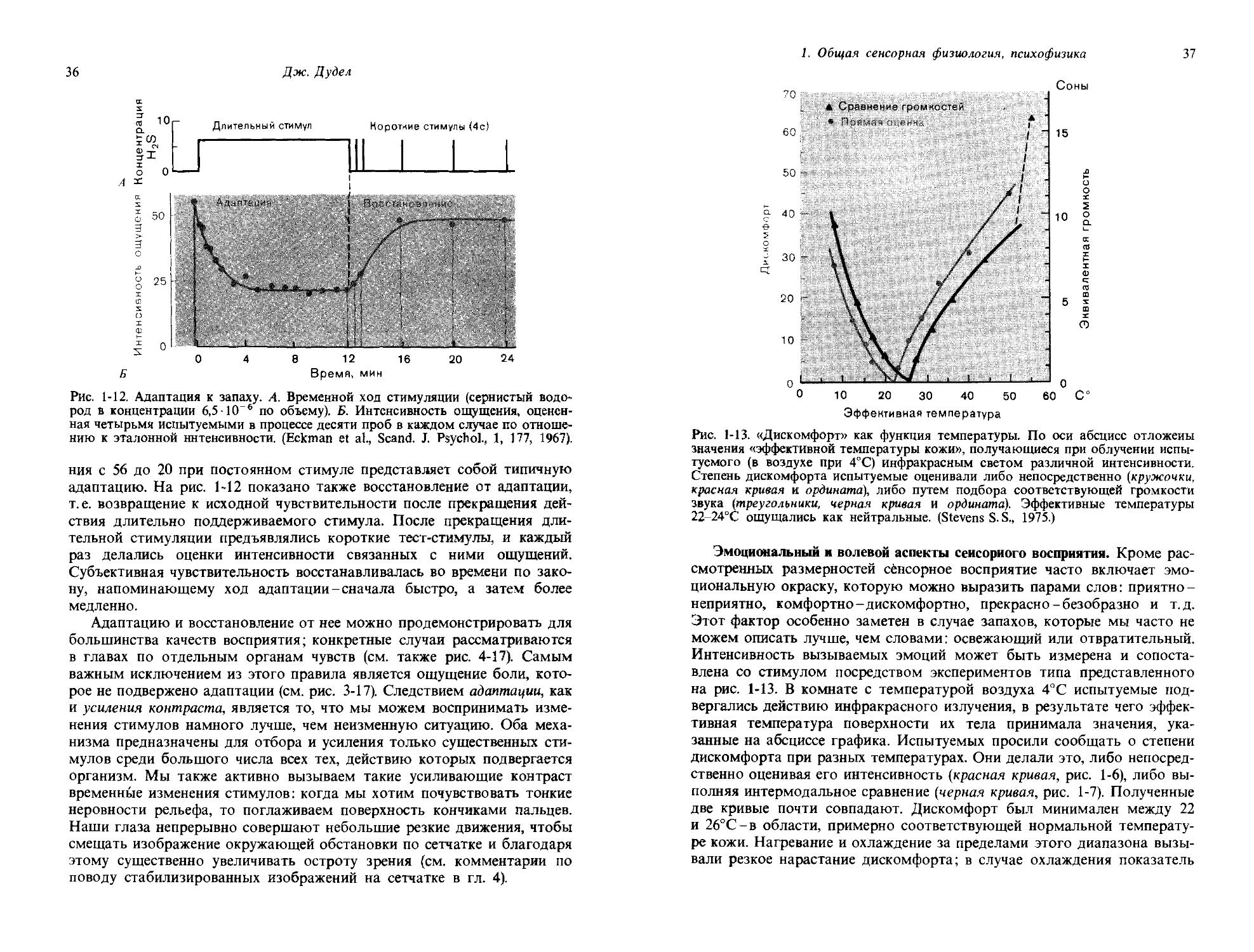
Временная размерность ощущения. Последняя размерность ощущения, которая будет обсуждаться,-это время. Само по себе время воспринимается как настоящее, вспоминается как прошлое и ожидается как будущее. В этом аспекте много чего можно сказать, например, о величине временного интервала, который мы воспринимаем как сиюминутное настоящее, т.е. как «психологический момент». Для интервалов времени можно определять дифференциальные пороги; наименьшая воспринимаемая длительность периодических стимулов, например, оценивается по частоте слияния мельканий. Оказывается, что органы чувств медлительны и не годятся для точных измерений времени.
Вопрос, который мы рассмотрим более подробно,-это влияние длительности стимула на интенсивность ощущения. Действия стимула на разных промежутках времени суммируются так же, как и его действия на разных участках площади. Эта суммация является линейной для коротких стимулов близи порога; здесь удваивание длительности стимула приводит к удваиванию интенсивности ощущения. Этот эффект показан на рис. 1-11 для зрительного чувства. При длительностях менее 0,1 с пороговая интенсивность света обратно пропорциональна длительности стимула; в этом диапазоне справедливо уравнение
Is-t = const. [1'6]
Произведение величины порогового стимула Is на его длительность t является константой. Это соотношение, «закон Рикко», полностью аналогично тому, которому подчиняется пространственная суммация пороговых стимулов (см. [1-5]). Если длительность стимула превосходит некоторое критическое время, суммация прекращается. На рис. 1-11
1. Общая сенсорная физиология, психофизика
35
Длительность стимула, с
Рис. 1-11. Временная суммация вблизи абсолютного зрительного порога. Точки красной линии показывают, каковы те минимальные яркости (ордината) светового стимула, при которых он едва воспринимается, если длительность его соответствует значениям, указанным на абсциссе. Масштаб логарифмический по обеим осям. Для стимулов короче 0,1 с кривая имеет наклон ( — 1); в этом диапазоне пороговая интенсивность стимула обратно пропорциональна его длительности. (Marks L. Е., 1974.)
порог не зависит от длительности стимула для длительностей, превышающих 1 с. Соотношения типа [1-6] справедливы также и для других модальностей. Так, например, [1-6] отражает общеизвестный факт, состоящий в том, что мы можем видеть очень быстрые движения только при ярком свете. Акробат, быстротой которого нам положено восхищаться, помещается обычно в центре светового пятна, фокусники же предпочитают тусклое освещение.
Итак, очень короткие стимулы суммируются в наших ощущениях. Напротив, длительные стимулы приводят к торможению и к адаптации. Неизменный стимул воспринимается как прогрессивно ослабевающий со временем. Хорошим примером такой адаптации служит ощущение тепла: ванна может показаться неприятно горячей, когда погружаешься в нее, но через короткое время ощущение горячего исчезает. Результаты измерения субъективной адаптации представлены на рис. 1-12. В этом эксперименте испытуемому предъявляли постоянный пахучий стимул, представляющий собой определенную концентрацию сернистого водорода. Во время действия стимула испытуемого просили оценивать интенсивность запаха по отношению к интенсивности некоторого стандартного запаха (шкала отношений; см. рис. 1-6). Сразу после подачи пахучего вещества испытуемые оценивали интенсивность ощущения в 56 единиц. Затем субъективная интенсивность резко падала в течение нескольких минут, выйдя примерно за пять минут на постоянный уровень интенсивности около 20 единиц. Этот спад интенсивности ощуще-
36
Дж. Дудел
Рис. 1-12. Адаптация к запаху. А. Временной ход стимуляции (сернистый водород в концентрации 6,5 -10 6 по объему). Б. Интенсивность ощущения, оцененная четырьмя испытуемыми в процессе десяти проб в каждом случае по отношению к эталонной интенсивности. (Eckman et al., Scand. J. Psychol., 1, 177, 1967).
ния c 56 до 20 при постоянном стимуле представляет собой типичную адаптацию. На рис. 1-12 показано также восстановление от адаптации, т. е. возвращение к исходной чувствительности после прекращения действия длительно поддерживаемого стимула. После прекращения длительной стимуляции предъявлялись короткие тест-стимулы, и каждый раз делались оценки интенсивности связанных с ними ощущений. Субъективная чувствительность восстанавливалась во времени по закону, напоминающему ход адаптации-сначала быстро, а затем более медленно.
Адаптацию и восстановление от нее можно продемонстрировать для большинства качеств восприятия; конкретные случаи рассматриваются в главах по отдельным органам чувств (см. также рис. 4-17). Самым важным исключением из этого правила является ощущение боли, которое не подвержено адаптации (см. рис. 3-17). Следствием адаптации, как и усиления контраста, является то, что мы можем воспринимать изменения стимулов намного лучше, чем неизменную ситуацию. Оба механизма предназначены для отбора и усиления только существенных стимулов среди большого числа всех тех, действию которых подвергается организм. Мы также активно вызываем такие усиливающие контраст временные изменения стимулов: когда мы хотим почувствовать тонкие неровности рельефа, то поглаживаем поверхность кончиками пальцев. Наши глаза непрерывно совершают небольшие резкие движения, чтобы смещать изображение окружающей обстановки по сетчатке и благодаря этому существенно увеличивать остроту зрения (см. комментарии по поводу стабилизированных изображений на сетчатке в гл. 4).
1. Общая сенсорная физиология, психофизика
37
Рис. 1-13. «Дискомфорт» как функция температуры. По оси абсцисс отложены значения «эффективной температуры кожи», получающиеся при облучении испытуемого (в воздухе при 4°С) инфракрасным светом различной интенсивности. Степень дискомфорта испытуемые оценивали либо непосредственно (кружочки, красная кривая и ордината), либо путем подбора соответствующей громкости звука (треугольники, черная кривая и ордината). Эффективные температуры 22 24°С ощущались как нейтральные. (Stevens S. S., 1975.)
Эмоциональный н волевой аспекты сенсорного восприятия. Кроме рассмотренных размерностей сенсорное восприятие часто включает эмоциональную окраску, которую можно выразить парами слов: приятно -неприятно, комфортно-дискомфортно, прекрасно - безобразно и т.д. Этот фактор особенно заметен в случае запахов, которые мы часто не можем описать лучше, чем словами: освежающий или отвратительный. Интенсивность вызываемых эмоций может быть измерена и сопоставлена со стимулом посредством экспериментов типа представленного на рис. 1-13. В комнате с температурой воздуха 4°С испытуемые подвергались действию инфракрасного излучения, в результате чего эффективная температура поверхности их тела принимала значения, указанные на абсциссе графика. Испытуемых просили сообщать о степени дискомфорта при разных температурах. Они делали это, либо непосредственно оценивая его интенсивность (красная кривая, рис. 1-6), либо выполняя интермодальное сравнение (черная кривая, рис. 1-7). Полученные две кривые почти совпадают. Дискомфорт был минимален между 22 и 26°С-в области, примерно соответствующей нормальной температуре кожи. Нагревание и охлаждение за пределами этого диапазона вызывали резкое нарастание дискомфорта; в случае охлаждения показатель
38
Дж. Дудел
степени равнялся 1,7, т. е. был почти таким же высоким, как показатель, полученный для боли (рис. 1-7). Боль можно в сущности рассматривать как предельную форму дискомфорта. Дискомфорт и комфорт, как правило, оказывают влияние на поведение. Мы избегаем крайностей-как холода, так и жары-и ищем приятного тепла. Очень резкое нарастание дискомфорта при охлаждении, возможно, защищает нас от опасности переохлаждения.
Чтобы закончить этот неполный очерк субъективной сенсорной физиологии, мы должны снова напомнить, что обычно мы не просто пассивно испытываем ощущения: с одной стороны, они побуждают нас действовать, а с другой-мы часто активно вызываем их своими направленными движениями. Именно это желание ощущения делает его моим личным опытом, заставляет его занимать мое личное время, превращает его в событие, относящееся к моему «я», и, поскольку этот опыт повторяем-и, значит, предсказуем,-он может стать основой для моих действий.
В 1.11. Какие из следующих утверждений относительно пороговой интенсивности света ложны?
а) Она уменьшается с увеличением площади светового пятна, б) Она возрастает с усилением освещенности окружения.
в) Она увеличивается пропорционально квадратному корню из длительности стимула.
г) Она уменьшается в течение длительного периода в темноте, д) Она может быть снижена за счет пространственной суммации.
В 1.12. Адаптация к сенсорному стимулу-это термин, описывающий следующее наблюдение.
а) Долго действующие стимулы воспринимаются как более слабые, чем короткий стимул той же интенсивности.
б) Слабый стимул воспринимается как более интенсивный, если он часто повторяется.
в) После длительного интенсивного стимула чувствительность к коротким тестовым стимулам медленно повышается.
г) Сильный источник стимула снижает интенсивность ощущения, вызываемого стимулами, действующими в области, пространственно смежной с сильным источником.
д) Во время действия продолжительного стимула интенсивность ощущения, вызываемого дополнительными малыми тест-сти-мулами того же качества, уменьшается.
2. НЕЙРОФИЗИОЛОГИЯ сенсорных систем
М. Циммерманн
Термин «сенсорные системы» применяется к тем частям нервной системы, которые получают сигналы из внешней среды и из самого организма, проводят эти сигналы и обрабатывают их. Данная глава дает общее представление о принципах работы этих сенсорных систем и опирается преимущественно на примеры, взятые из исследований соматосенсорной системы (кожные чувства). Наибольшее внимание уделяется объективным, нейрофизиологически измеримым процессам; субъективное восприятие и его психофизические корреляты обсуждались в гл. 1.
2.1. Преобразование стимулов в рецепторах
Адекватный стимул, классификация рецепторов. Рецептор-это специализированная нервная клетка. Она сигнализирует в ЦНС о состоянии и (или) изменениях состояния среды, в которой находится. Те факторы окружающей среды и те их изменения, которые оказывают действие на рецепторы, называются стимулами. Стимулы-это величины, которые можно измерять объективными методами, например деформация кожи, температура и электромагнитное излучение (свет).
Физиологические исследования обнаружили, что каждый рецептор с особой готовностью реагирует на стимулы какого-то одного типа. Это свойство обычно называют специфичностью рецепторов; тот стимул, который является эффективным в каждом случае, иногда называют адекватным стимулом для данного рецептора. Для некоторых рецепторов адекватный стимул можно установить из повседневного опыта. Так, например, мы можем легко определить, что адекватным стимулом для рецепторов наших глаз является свет, тогда как тепловые или механические стимулы с помощью этих рецепторов мы в обычных условиях не обнаруживаем.
Следовательно, каждый рецептор способен передавать в ЦНС информацию только об одном определенном аспекте или размерности окружающей среды. Это значит, что рецептор играет роль фильтра. В связи с этим представляется полезным классифицировать рецепторы на основе их адекватных стимулов. Рецепторы млекопитающих распадаются на следующий четыре группы: механо-, термо-, хемо-и фоторецепторы.
Внутри каждой из этих четырех групп мы можем усмотреть значительную степень специализации. Например, есть различающиеся типы фоторецепторов, по-разному отвечающие на излучения с различными длинами волн; эти рецепторы называют чувствительными к длине во-
40
М. Циммерман»
Мышечные — волокна
Тело
1ООмкм
Аксон (возбудимый)
Дендриты (рецептивная зона)
Рецеп-
Потенциалы действия
Потенциалы действия
Г енераторный —__-|
потенциал
Стимул----
потенциал
мВ
-50 - 60 -70 -80
мВ
0
Д!_ мм
2
- 1
-0
Я
-50
-80
мВ
+ 50 0
-80
Рис. 2-1. Схема электрофизиологических измерений, которые можно провести на рецепторе растяжения ракообразных. Во время стимуляции (изменение длины соответствующей мышцы на AL) микроэлектрод, введенный в тело клетки, регистрирует рецепторный потенциал, который возникает в рецептивной зоне (на дендритах). Являясь генераторным потенциалом, он возбуждает распространяющиеся потенциалы действия в возбудимом аксоне.
лны или цветочувствительными. Терморецепторы можно подразделить на рецепторы тепла и холода в соответствии с тем, что их возбуждает -увеличение или уменьшение температуры. У механорецепторов также можно наблюдать специализацию к различным параметрам стимулов: в коже, например, одни рецепторы чувствительны к вибрации, другие-к давлению.
Рецепторный потенциал. В следующих параграфах мы рассмотрим процессы в рецепторе (подвергаемом действию адекватного стимула), результатом которых является потенциал действия в афферентном волокне. В качестве примера взят рецептор растяжения ракообразных-механорецептор, расположенный между определенными мышечными волокнами животного; он чувствителен к растяжению (рис. 2-1).
Когда микроэлектрод проникает в рецепторную клетку, то потенциал, который регистрируется исходно, как и во всех других нервных клетках,-это потенциал покоя ( — 80 мВ на рис. 2-1). Под действием адекватной стимуляции когда окружающая мышца растягивается-
2. Нейрофизиология сенсорных систем
41
мембранный потенциал меняется в направлении деполяризации. Эта вызываемая стимулом деполяризация-отклонение от потенциала покоя-называется рецепторным потенциалом. Рецепторный потенциал длится столько же, сколько и стимул (изменение ДЕ на рис. 2-1). Однако даже тогда, когда стимул сохраняется постоянным, рецепторный потенциал снижается от своего исходного максимального значения до более низкого уровня. Такое снижение влияния стимула, поддерживаемого в течение некоторого времени на постоянном уровне, можно наблюдать практически у всех рецепторов; оно называется адаптацией.
Рецепторный потенциал в этом случае является результатом повышения проводимости мембраны, которое неспецифично и касается всех мелких ионов (Na+, К+, Са2+, С1“). В нормальных условиях единственный из этих ионов, который может обеспечить деполяризацию, это ион Na+, потому что только этот ион имеет потенциал равновесия, сдвинутый в деполяризационном направлении по отношению к потенциалу покоя. Следовательно, ионы Na+ должны быть главным источником возникновения рецепторного потенциала. Повышение проводимости мембраны, описанное для рецепторов, сходно с тем, что имеет место в субсинаптической мембране возбуждающих синапсов-например в концевой пластинке мотонейрона.
Повышение проводимости мембраны, которое вызывает рецепторный потенциал, локализовано в специализированном участке рецепторной клетки-рецептивной мембране (дендриты на рис. 2-1). Рецепторный потенциал возникает только тогда, когда стимул действует на этот специализированный участок. Далее, воспринимающий участок мембраны пространственно отделен от других частей клетки-например от электрически возбудимой (проводящей импульсы) мембраны аксона. Его отличие можно также выявить и фармакологически: например, возбудимую мебрану можно избирательно «отравить» нанесением тетро-дотоксина (ТТК), так что потенциалы действия больше там возникать не будут. В то же время рецептивная мембрана, а значит, и рецепторный потенциал, как правило, не чувствительны к действию ТТК.
Фоторецепторы позвоночных представляют исключение в том смысле, что при стимуляции светом в них возникает гиперполяризационный потенциал. Однако при синаптической передаче на нейроны высших уровней в сетчатке индуцируется деполяризация-возбуждающий постсинаптический потенциал (ВПСП), который ведет к генерации распространяющихся потенциалов действия.
Процесс трансдукции; первичные и вторичные сенсорные клетки. Промежуточные события между поступлением стимула и появлением рецепторного потенциала пока еще мало изучены; они объединяются под общим названием процессы трансдукции. Этот термин подразумевает главным образом молекулярно-биологические процессы, имеющие место на рецептивной мембране и (или) во внутриклеточных структурах (называемых органеллами). В воспринимающей части рецептора имеется много таких органелл, как мембранные диски фоторецепторов, ко
42
М. Циммерманн
торые содержат чувствительные к свету молекулы зрительного пигмента (см. рис. 4-27) или волосковидные выросты (реснички) рецепторов внутреннего уха (см. рис. 5-3). Это вспомогательные структуры, которые делают рецептор особенно чувствительным к специфическому адекватному стимулу.
Многие рецепторы состоят из двух связанных элементов: нервного окончания и некоторой клетки ненервного происхождения типа дисков Меркеля в коже (см. рис. 3-4) или чувствительных клеток во вкусовых почках (см. рис. 7-3). Полагают, что процесс преобразования сигнала имеет место в не-нервной клетке. Такие клетки называют вторичными сенсорными клетками. На электронной микрофотографии область контакта между клетками выглядит как синапс. По этой причине предполагается, что имеется синаптическая передача от вторичной сенсорной клетки к нервному окончанию.
Напротив, первичная сенсорная клетка -это рецептор, в котором преобразование стимула происходит в нервной клетке или в нервном окончании. Примером этого являются обонятельные рецепторы (см. рис. 8-2) и свободные нервные окончания в коже.
Рецепторный потенциал как причина генерации распространяющихся потенциалов действия. Рецепторный потенциал электротонически распространяется в смежные области клетки. В результате этого аксон деполяризуется (рис. 2-1). Когда деполяризация достигает порога мембраны аксона, генерируется потенциал действия. Этот потенциал действия проводится по аксону в ЦНС. Таким образом, рецепторный потенциал действует на аксон как электрический стимул; по этой причине его также называют генераторным потенциалом.
Если генераторный потенциал не спадает после генерации первого потенциала действия, он может вызывать следующие потенциалы действия, пока стимул и соответственно генераторный потенциал не исчезнут. Это значит, что в период существования длительного генераторного потенциала афферентное волокно разряжается многократно. Процессы, происходящие между поступлением стимула и генерацией распространяющихся потенциалов действия, могут быть названы процессами трансформации стимула в нервную активность. Таким образом, эта трансформация включает следующие события: трансдукцию, увеличение проводимости рецептивной мембраны, рецепторный потенциал, синаптическую передачу (в случае вторичных сенсорных клеток), генераторный потенциал, потенциалы действия.
Адаптация. При рассмотрении рецепторного потенциала мы отметили, что даже если стимул поддерживается некоторое время постоянным, рецепторный потенциал может уменьшаться (рис. 2-1); этот процесс называют адаптацией. Мы также наблюдаем этот феномен на картине разряда афферентных нервных волокон: как можно видеть из рис. 2-1, при неизменном стимуле временные интервалы Аг между двумя последовательными импульсами удлиняются, т.е. мгновенная частота разряда уменьшается (мгновенная частота = 1/Аг). Адаптация может иметь
2. Нейрофизиология сенсорных систем
43
Очень быстрая адаптация (а)
Л
Адаптация средней скорости (б)
MIIIIIIIIIIIHI
Время, с
Б
Медленная адаптация (в)
Рис. 2-2. Разряды рецепторов с различными скоростями адаптации. А. Потенциалы действия очень быстро (а), быстро (б) и медленно (в) адаптирующихся рецепторов, генерируемые при длительном действии постоянного стимула. Б. Графики изменения мгновенной частоты разряда со временем для рецепторов (б) и (в). Обратите внимание на то, что разряд рецептора (в) включает как фазический, так и тонический компоненты.
место на всех стадиях трансформации стимула: в процессе трансдукции, в механизме проведения рецепторного потенциала, при синаптической передаче от вторичных сенсорных клеток и при генерации потенциала действия. Возможно, что несколько таких механизмов работает одновременно.
Чтобы получить адаптационную характеристику рецептора, исследуют его реакцию на надпороговый ступенчатый стимул. В таких экспериментах различные рецепторы демонстрируют различные изменения частоты разряда во времени. На рис. 2-2, А показан разряд медленно адаптирующегося рецептора (в), умеренно адаптирующегося рецептора (б) и быстро адаптирующегося рецептора (а). На рис. 2-2, Б представлено изменение со временем мгновенной частоты для рецепторов с медленной и умеренной адаптацией.
Предложены различные функциональные объяснения адаптации. Это явление обычно рассматривают всего лишь как простое утомление рецепторов. Но во многих случаях различное развитие адаптации во времени можно интерпретировать как приспособление к передаче специфических параметров стимула. Например, при изучении трех типов механорецепторов кожи было обнаружено, что благодаря характерным временным свойствам их разряда они могут рассматриваться как пере-
44
М. Циммерманн
мм
0123 0123
Потенциалы действия
Рис. 2-3. Зависимость рецепторного потенциала и частоты разряда от интенсивности стимула. А. Рецепторные потенциалы и потенциалы действия при трех различных интенсивностях стимула. Потенциалы действия возбуждаются тогда, когда рецепторный потенциал превосходит порог (красная прерывистая линия). Б. Графики зависимости мгновенной частоты разряда F в разные моменты после включения стимула (через 1, 2 и 3 с) от интенсивности стимула X. 80 -пороговая интенсивность.
датчики сведений об амплитуде, скорости и ускорении деформации кожи (см. рис. 3-5-3-8 и табл. 3-1).
Перевод интенсивности стимула в частоту импульсов в разряде. Когда сила стимула возрастает, возрастает и амплитуда рецепторного потенциала. Постепенные плавные изменения рецепторного потенциала вызывают соответствующие изменения частоты потенциалов действия. Эта ситуация представлена на рис. 2-3, Л. Соотношение между интенсивностью стимула Б и частотой импульсов F, часто называемое силовой функцией, изображено на рис. 2-3, Б для разных моментов времени от начала стимуляции (через 1, 2 и 3 с). В случае рецептора растяжения соотношение между F и S линейно для каждого момента времени на протяжении действия стимула.
Поскольку для того, чтобы вызвать потенциалы действия, рецепторный потенциал должен превзойти некоторое минимальное значение (порог аксона), разряд возникает только при превышении стимулом соответствующей интенсивности (So на рис. 2-3, Б). So называют пороговым стимулом для рецептора. Силовые функции рис. 2-3, Б описываются выражениями вида
F=k(S — S0), [2-1]
т. е. линейной зависимостью для Б > Бо. Коэффициент к, наклон прямой, становится все меньше для последовательных интервалов времени, от-
2. Нейрофизиология сенсорных систем
45
Рис. 2-4. Различные степенные функции. А. Частота разряда F как функция интенсивности стимула 5. По обеим осям масштаб линейный, единицы-произвольные. Приведены три характерные степенные функции с показателями и < 1, и > 1 и п = 1 (красная линия). Б. Те же степенные функции изображены в логарифмическом масштабе. По абсциссе отложена разность между интенсивностью стимула S и пороговой интенсивностью So.
Интенсивность стимула S-So (log)
Б
считываемых от момента приложения стимула; это тоже является одним из выражений адаптации.
Экспериментально было показано, что для большинства рецепторов силовая функция нелинейна. Измеренные соотношения часто можно описать различными математическими функциями, такими, как логарифмическая
F = klogS/So
[2-2]
или степенная
F = k(S-Soy. [2-3]
Мы сталкивались с обоими этими выражениями в разделе по психофизике; они называются соответственно функциями Вебера-Фехнера и Стивенса. Кодирование интенсивности в рецепторах большинства типов лучше всего описывается степенной функцией (рис. 2-4), в которой разность между интенсивностью стимула S и пороговой интенсивностью So возводится в n-ю степень. Показатель п является константой, характерной для каждого типа рецепторов. Для п < 1 и и > 1 степенные функции (на графиках с линейным масштабом) представляются соответственно выпуклыми и вогнутыми кривыми (рис. 2-4, А), а для и = 1 силовая функция линейна, как и для рецептора растяжения. Оказалось, что для большинства рецепторов п < 1. Силовая характеристика с и < 1
46
М. Циммерманн
позволяет рецептору кодировать весьма значительный диапазон интенсивностей стимула частотой разряда, ограниченной сверху (максимальная частота разряда толстого миелинизированного нервного волокна составляет 500-1000 имп./с, а у тонких волокон она еще меньше).
Степенные функции часто используются при описании психофизических результатов. Сравнение нейрофизиологически полученной силовой характеристики какого-нибудь рецептора с измерениями субъективного ощущения интенсивности в психофизическом эксперименте (см. рис. 1-6) часто обнаруживает поразительное соответствие; такие сопоставления важны для исследования нейрофизиологической основы восприятия.
Чтобы проверить, можно ли установленную в эксперименте силовую функцию описать степенной функцией, интенсивность стимула и частоту разряда представляют на графике в логарифмической системе координат (рис. 2-4, Б). В такой системе степенная зависимость выглядит как прямая линия, поскольку при логарифмировании обеих частей [2-3J получается
logF = logic + nlog(5 — 50). [2-4]
Это уравнение прямой линии с угловым коэффициентом п для переменной logF при аргументе log (5 — So). Если точки на таком графике могут быть аппроксимированы прямой линией, то они соответствуют степенной функции. Показатель п может быть определен непосредственно по наклону этой линии.
В 2.1. На какие четыре группы можно подразделить рецепторы млекопитающих по их адекватным стимулам?
В 2.2. Рецепторный потенциал
а) это реакция рецепторной клетки типа «все или ничего», которая не возникает до тех пор, пока стимул не превысит некоторого порога;
б) обычно представляет собой деполяризацию рецептивной мембраны, амплитуда которой нарастает с увеличением силы стимула;
в) электротонически распространяется до мембраны аксона, где действует как возбудитель проводимых далее потенциалов действия;
г) возникает благодаря увеличению проводимости, связанной с ионами Н+ (значением pH);
д) медленно нарастает при действии постоянного стимула и длится в течение всего времени стимуляции.
Верны несколько ответов.
В 2.3. В афферентных аксонах многих рецепторов частота импульсов в разряде
а) увеличивается с увеличением интенсивности стимула;
б) увеличивается со временем при действии стимула постоянной интенсивности;
2. Нейрофизиология сенсорных систем
47
в) уменьшается со временем при действии стимула постоянной интенсивности;
г) обычно равна нулю при действии стимулов подпороговой интенсивности;
д) не зависит от величины рецепторного потенциала. Верны несколько ответов.
В 2.4. В случае многих рецепторов степенная функция описывает а) временной ход адаптации;
б) временной ход рецепторного потенциала при увеличении силы стимула;
в) зависимость частоты импульсации F от величины превышения стимулом порога So;
г) связь между амплитудой потенциала действия и величиной рецепторного потенциала.
2.2. Сенсорные функции ЦНС. Общее рассмотрение
Периферические нервы, спинальные нервы, задние корешки и их зоны иннервации. Все рецепторы любой сенсорной системы (например, органа чувств или проприоцептивной системы) вместе взятые составляют периферическую сенсорную поверхность, или просто - периферию. С периферии информация передается в ЦНС в закодированной форме, в виде потенциалов действия, или нервных импульсов. Таким образом афферентные нервные волокна выполняют функцию передачи информации. Пучки этих волокон направляются в ЦНС в составе периферических нервов (идущих от кожи, мышц, суставов и внутренних органов), которые также содержат эфферентные нервные волокна. От всех частей тела, за исключением головы, периферические нервы идут к спинному мозгу, которого они достигают как спинальные нервы. В спинной мозг афферентные нервные волокна входят затем в составе задних корешков.
Если какой-нибудь кожный нерв в результате несчастного случая окажется перерезанным, обозначится относительно резко ограниченный участок кожи, в пределах которого стимулы больше не будут обнаруживаться. Та область кожи, где возникло такое нарушение,-это область иннервации перерезанного нерва или его ветви. Резкость границы обусловлена малой степенью перекрывания областей иннервации соседних нервов (рис. 2-5).
Другим результатом перерезки кожного нерва вдобавок к нечувствительности будет то, что кожа в затронутой области станет сухой и жесткой-нарушится иннервация потовых желез симпатическими эфферентами, которые также идут в составе кожного нерва.
Афферентные волокна каждого заднего корешка передают информацию от определенной периферической зоны. Но эти зоны образуются не путем простого сложения областей иннервации периферических нервов. Между спинным мозгом и периферией нервные волокна перегруппировываются в новые пучки, это касается главным образом сплетения во-
48
М. Циммерманн
Рис. 2-5. Области иннервации кожных нервов и задних корешков (схема). Области иннервации ветвей кожного нерва имеют четкие границы и перекрываются незначительно. Из-за перераспределения пучков периферических волокон при формировании спинальных нервов области иннервации задних корешков определены менее четко и значительно перекрываются. Зоны перекрывания показаны красным.
локон, идущих от конечностей (например, плечевого сплетения на уровне плечевого сустава). Смысл этой перегруппировки показан на рис. 2-5: каждый периферический нерв содержит волокна, расходящиеся по нескольким соседним спинальным нервам, и наоборот, в каждом спинальном нерве имеются волокна из разных периферических нервов. Благодаря такой перестройке зона иннервации каждого спинального нерва или его заднего корешка оказывается менее резко очерченной, чем у периферического нерва, и области иннервации соседних спинальных нервов существенно перекрываются (рис. 2-5). В то время как результатом перерезки самого периферического нерва является четко ограниченная потеря чувствительности (рис. 2-5) в том органе, который он обслуживает (в коже, мышце или во внутреннем органе), перерезка заднего корешка или спинального нерва ослабляет иннервацию всех трех типов органов одновременно, приводя лишь к незначительной потере чувствительности.
Сенсорная зона иннервации каждого спинального нерва в коже называется дерматомом. Дерматомы распределены по поверхности тела в порядке, соответствующем порядку связанных с ними сегментов спинного мозга. Так, например, иннервация тазовой области обеспечивается сегментами спинного мозга, отстоящими от головного мозга дальше, чем те, которые иннервируют плечи. В соответствии с таким отображе-
2. Нейрофизиология сенсорных систем
49
Ядра тройничного нерва
Шейный отдел
Г рудной отдел
Поясничный отдел
Крестцовый отдел
Рис. 2-6. Упорядоченное расположение на поверхности тела областей, иннервируемых разными участками спинного мозга. Жирными линиями показаны границы крупных областей тела, связанных с четырьмя главными отделами спинного мозга и тройничной областью ствола головного мозга. Показано несколько дерматомов отдельных спинальных нервов.
нием поверхности тела на спинном мозге последний подразделяется на четыре основных отдела: шейный, грудной, поясничный и крестцовый (рис. 2-6). Примерные границы связанных с ними дерматомов показаны на рис. 2-6, который предназначен для того, чтобы облегчить читателю их запоминание.
Черепные нервы. Область головы обслуживают 12 пар нервов, входящих в собственно головной мозг на уровне ствола мозга и диэнцефалона. Данные об этих нервах суммированы в табл. 2-1 с указанием их важнейших функций и органов, которые они иннервируют. Некоторые из черепных нервов исключительно сенсорные (I, II, VIII), тогда как другие имеют как сенсорные, так и моторные функции, а в некоторых случаях включают и эфферентные парасимпатические волокна. Нервы IX и
50
М. Циммерманн
Таблица 2-1. Перечень основных черепных нервов с указанием их важнейших сенсорных и моторных функций
н азвание Иннервируемый орган Пример сенсорной функции Пример моторной функции
I Обонятельный Слизистая оболочка носа Ощущение запаха —
II Зрительный Сетчатка Зрение —
III Глазодвигательный Мышцы глаза — Движения глаз
IV Блоковый » » — » »
V Тройничный Лицо и полость рта Соматосенсорная функция Жевание
VI Отводящий Мышцы глаза — Движения глаз
VII Лицевой Мышцы лица, язык Вкусовые ощущения Мимика лица
VIII Слуховой Внутреннее ухо, вестибулярный аппарат Слух, ориентация в пространстве —
IX Языко-глоточный Язык, глотка Вкусовые ощущения Глотание
X Блуждающий Внутренние органы Висцеральные афференты Регуляция сердечной деятельности
XI Добавочный Шея — Движения головы
XII Подъязычный Мышцы языка — Движения языка
X содержат висцеральные афференты от рецепторов, которые действуют как датчики в контрольных цепях, регулирующих внутреннюю среду (например, в сердечно-сосудистой и дыхательной системах).
В составе тройничного нерва (V) идут афференты от лица и полости рта; он иннервирует кожу, а также зубы, слизистую оболочку рта и язык. Таким образом, функционально он принадлежит к соматосенсорной системе. Тройничный нерв занимает особое место в том отношении, что его афференты работают у новорожденных, возбуждая пищевое поведение; это нерв, который обеспечивает младенцу первый опыт сенсорного познания окружающей среды.
Сенсорные системы кожи и внутренних органов вместе называются соматовисцералъной системой. К этой системе относятся афференты спинальных нервов и черепных нервов V, VII, IX и X.
Центральные переключения сенсорных путей. Войдя в спинной мозг через задние корешки, сигналы рецепторов перерабатываются на нескольких уровнях ЦНС. Для соматосенсорной системы такие уровни показаны на рис. 2-7 (сравните с центральными зрительными путями рис. 4-32 и центральными слуховыми путями рис. 5-16). Связи между этими уровнями не следует рассматривать как улицы с односторонним движением: кроме восходящих потоков информации имеется несколько нисходящих, центрифугальных, потоков (не показаны на рис. 2-7), например из коры больших полушарий в спинной мозг. Благодаря их
2. Нейрофизиология сенсорных систем
51
влиянию афферентный поток может модулироваться на всех уровнях (см. разд. 2.3).
При изучении сенсорных систем-и особенно соматосенсорной - оказалось полезным подразделять афферентные центростремительные пути и соответствующие центральные отделы на филогенетически молодую специфическую систему и филогенетически старую неспецифическую систему. Здесь прилагательные «специфический» и «неспецифический» характеризуют ряд противоположных функциональных свойств, характерных для первой и второй систем. В качестве предварительного мы предлагаем следующее определение: термин «специфический» применяется к тем центральным нервным компонентам сенсорной системы, которые имеют только один анатомически и нейрофизиологически идентифицируемый главный вход от одной периферической сенсорной поверхности. К настоящему времени детально изучены специфические системы, связанные с механорецепторами кожи, рецепторами глаза и внутреннего уха. Напротив, в любой неспецифической системе афферентные входы не так четко определены, система возбуждается сигналами, сходящимися от всех сенсорных поверхностей (полимодальная, или полйсенсорная, конвергенция).
Теперь мы возьмем для примера соматосенсорную систему, чтобы проиллюстрировать особенности анатомии двух указанных подсистем; функциональные детали будут обсуждаться в последующих разделах.
Анатомия специфической соматосенсорной системы. Специфическая часть соматосенсорной системы называется лемнисковой системой, поскольку одним из ее важнейших трактов является медиальный лемниск (идущий от продолговатого мозга в таламус). Лемнисковая система (рис. 2-7, Л) состоит из путей, отчетливо различимых анатомически и нейрофизиологически; посредством этих путей кожные афференты спинальных нервов и тройничного нерва проецируются в две кортикальные зоны теменной доли-соответственно первую и вторую соматосенсорные области (SI и SII).
SI получает свой афферентный вход от кожи противоположной (контралатеральной) стороны, тогда как в SII афференты приходят от обеих половин тела (билатерально). Восходящий путь (рис. 2-7, Л) проходит главным образом через задний столб спинного мозга и его ядра в продолговатом мозгу (первый синапс); далее он идет через медиальный лемниск к вентробазальным ядрам таламуса (второй синапс) и затем к областям SI и SII коры больших полушарий (третий синапс). По этому пути сигналы передаются быстро, в оптимальном случае-всего с тремя синаптическими задержками. Характерной особенностью этой системы является ее соматотопическая организация: здесь имеет место упорядоченное (топографическое) отображение кожи, периферической сенсорной поверхности, на всех «станциях» переключения. Мы можем также сказать, что кожа проецируется на все эти центры. Дальнейшие подробности этого проецирования будут обсуждаться ниже (см. разд. 2.4 и 2.5).
Кора
Вентробазальное ядро
Медиальная петля
Тройничный нерв
Ядра заднего столба
Задний столб
Спинальные нервы
Неспецифические ядра таламуса
Ретикулярная формация
Тройничный нерв
Передне-боковой пучок
Спинальные нервы
Рис. 2-7. А. Общая схема анатомической организации специфической части соматосенсорной системы; показаны главные связи афферентов спинальных и тройничного нервов и проекционные поля коры. Б. Общая схема неспецифической соматосенсорной системы; показаны наиболее важные из известных связей и блоков переработки информации.
2. Нейрофизиология сенсорных систем
53
Лемнисковая система особенно высоко развита у приматов-обезьян, человекообразных обезьян и человека. Она является анатомической основой осязания и всех сенсорных способностей (осознаваемых и неосознаваемых), которые включают определение пространственных параметров стимула.
Анатомия неспецифической системы. Эта соматосенсорная подсистема называется также экстралемнисковой, или нелемнисковой, системой. Она менее четко определена анатомически (рис. 2-7, Б), чем лемнисковая система. Ее наиболее важными компонентами являются ретикулярная формация ствола мозга и определенные ядра в медиальном таламусе, называемые неспецифическими ядрами таламуса. Главные спинномозговые восходящие пути неспецифической системы-спинно-ретикулярный и палеоспинно-таламический; оба они локализованы в переднебоковом пучке спинного мозга.
Экстралемнисковая система связана практически со всеми областями коры больших полушарий. Эти связи диффузны, т.е. не строго ограничены в пространстве и, кроме того, в них почти совсем или совсем отсутствует соматотопическая организация. В добавление к этому имеются связи с центрами автономной регуляции в стволе мозга и гипоталамусе, с лимбической системой и подкорковыми центрами моторной системы.
Полисенсорная конвергенция и малая степень выраженности сомато-топики являются характерной особенностью данной неспецифической системы; эти свойства резко отличают ее от специфических систем.
Если обратиться к поведению и восприятию, то среди функций, приписываемых неспецифической системе, можно назвать следующие: эмоциональная окраска восприятия (удовольствие, от вращение), восприятие боли, контроль состояния сознания (сон, бодрствование), ориентировочные реакции (поворот в сторону новых стимулов). Нейрофункцио-нальная организация неспецифической системы изучена очень слабо по сравнению с лемнисковой системой.
Подразделение на «лемнисковую» и «экстралемнисковую» подсистемы не должно создавать у читателя ложного впечатления, что эти две подсистемы могут быть активированы независимо или что каждой из них могут быть приписаны четко различающиеся перцептуальные феномены. Напротив, разнообразные сенсорные стимулы, встречающиеся в повседневной жизни, возбуждают обе системы, и взаимодействия этих двух подсистем многочисленны и разнотипны.
В 2.5. Перерезка кожного нерва на самой крайней периферии ведет к а) исчезновению только температурной и болевой чувствительности;
б) сухости кожи в соответствующей области иннервации из-за денервации потовых желез;
в) вялому параличу из-за рассечения эфферентных моторных аксонов;
54
М. Циммерманн
г) прерыванию сигналов, идущих от высоко- и низкопороговых механорецепторов и терморецепторов.
Верны несколько ответов.
В 2.6. Когда разрушается один из задних корешков, прерываются следующие нервные волокна:
а) эфферентные моторные и симпатические волокна;
б) афферентные волокна от кожи, мышц, суставов и внутренних органов;
в) только афферентные волокна от кожи.
При повреждении спинального нерва прерываются:
г) только афферентные волокна;
д) все афферентные и эфферентные волокна соответствующего полусегмента;
Верны несколько ответов.
В 2.7. Следующие утверждения применимы к понятию «дерматом»: а) дерматом-это патологическое изменение кожи (жесткость, сухость), развивающееся при перерезке соответствующего нерва;
б) так называют зону снижения чувствительности, появляющуюся при перерезке кожного нерва;
в) он определяется как участок кожи, соответствующий зоне перекрывания областей иннервации двух соседних спинальных нервов;
г) это область иннервации кожных нервных волокон спинального нерва.
В 2.8. Изобразите контуры ЦНС в продольном разрезе. Наметьте границы семи главных отделов ЦНС и назовите эти отделы.
В 2.9. Подразделение центральной части сенсорной системы на специфическую и неспецифическую подсистемы основывается на следующих фактах:
а) в кору больших полушарий афферентный поток импульсов поступает как по быстропроводящим и отчетливо выявляемым путям, так и по медленнопроводящим и диффузно распределенным путям;
б) черепные нервы обеспечивают ясное, отчетливое (специфическое) восприятие, тогда как восприятие, определяемое спинальными афферентами, смутное, неотчетливое (неспецифическое);
в) имеет место как сознательное (специфическое), так и бессознательное (неспецифическое) восприятие периферических стимулов;
г) в ЦНС имеются сенсорные области, в которые афференты приходят только от одной специфической сенсорной поверхности, но есть и такие, которые могут возбуждаться неспецифически от нескольких сенсорных поверхностей.
Верны несколько ответов.
2. Нейрофизиология сенсорных систем
55
2.3. Свойства и функционирование сенсорных нейронов и нейронных ансамблей
На каждом уровне сенсорной системы поступление афферентных сигналов вызывает возбуждение и торможение отдельных нейронов, а также взаимодействие между нейронами и группами нейронов. Некоторые виды взаимодействий являются особенно характерными для ряда сенсорных систем. Они будут описаны в следующих разделах.
Латеральное торможение. На рис. 2-8, А показана нейронная сеть, состоящая из двух рецепторов и связанных с ними элементов на двух последующих синаптических уровнях. Обнаруженные здесь конвергентные и дивергентные нервные связи, как и везде в нервной системе, казалось бы, должны были приводить к лавинообразному распространению информации во все стороны на все более высоких уровнях ЦНС. Следовало бы ожидать, что представление точечного стимула в ЦНС будет с каждым синаптическим переключением все увеличиваться и становиться более размытым и менее точным. Однако с такими ситуациями мы встречаемся только в патологических условиях, например в случае отравления стрихнином. Стрихнин блокирует тормозные синапсы в ЦНС. Когда такое случается, малейший периферический стимул действительно порождает в нервных сетях цепные реакции, захватывающие обширные очаги. По-видимому, в норме торможение предотвращает такое катастрофическое распространение возбуждения.
Данный тормозный эффект базируется на характерной пространственной организации тормозных связей, нашедшей выражение в терминах латеральное торможение или торможение со стороны окружения; соответствующая иллюстрация дана на рис. 2-8, Б. На уровне первого синаптического переключения в афферентном пути каждый нейрон оказывает тормозное влияние на ближайшие к нему клетки посредством вставочных нейронов (на схеме они красные). Тот нейрон, который передает самый сильный афферентный сигнал (здесь это средний нейрон), оказывает и самое сильное тормозное влияние на соседей. Это ведет к снижению возбуждения в точках, удаленных от центра возбуждения. Таким образом, по сравнению с ситуацией, изображенной на рис. 2-8, Л, пространственное растекание возбуждения будет снижено.
Было показано, что латеральное торможение существует на всех уровнях сенсорных систем; в соматосенсорной системе оно обнаружено в сером веществе спинного мозга, в ядрах дорсального столба, в таламусе и коре. Роль латерального торможения схематически представляют графики на рис. 2-8, Б. Латеральное торможение-свойство всех сенсорных систем, и особенно специфических. Его можно рассматривать как функциональную компенсацию растекания возбуждения, обусловленного нейроанатомией. Следствием этого является более четкая локализация информации о стимуле. Специфическое субъективное проявление латерального торможения-феномен усиления пространственного контраста в зрении (см. гл. 1).
56
М. Циммерманн
А Стимул
Кожа--------------
Рецепторы -
1.1 IIIIII LLLLLLL
Первое переключение
Рис. 2-8. Латеральное торможение. А. Пространственное распространение возбуждения в результате конвергенции и дивергенции в гипотетической ЦНС без латерального торможения. Вертикальные черточки (красные) символизируют импульсацию аксонов на различных уровнях. Кривая показывает соответствующее пространственное распространение возбуждения в ЦНС; эту кривую можно считать отражающей центральное представление стимула малой площади, показанного вверху.
Нисходящее торможение. Почти во всех сенсорных системах наблюдаются тормозные эффекты, порождаемые вышестоящими центрами. Такое нисходящее (центробежное) торможение может проявляться даже в таких периферических звеньях системы, как рецепторы (см. рис. 8-4) или афферентные окончания в спинном мозгу (рис. 2-10). Существование торможения такого рода подчеркивает важность сделанного выше замечания о том, что сенсорные системы не следует рассматривать как «улицы с односторонним движением».
Аналогично латеральному торможению нисходящее торможение можно считать средством регуляции чувствительности афферентных каналов передачи', вышестоящие части мозга «открывают клапан», управляющий потоком афферентной информации.
Рецептивное поле. Частота потенциалов действия сенсорных нейро-
2. Нейрофизиология сенсорных систем
57
Б. Латеральное торможение (тормозное влияние каждого нейрона на соседние нейроны через тормозные нейроны; показано красным) на каждом синаптическом уровне противодействует распространению возбуждения в результате дивергенции. Как показывают кривые распределения возбуждения, следствием этого является уменьшение области представления в ЦНС пространственно ограниченного периферического стимула.
нов обычно зависит от стимулов, действующих на периферии. Например, частота потенциалов действия какой-нибудь клетки зрительной коры может меняться, когда в определенной области поля зрения появляется определенный световой стимул. Совокупность всех точек на периферии тела, из области которых на данную клетку можно оказывать влияние действием специфических стимулов, называют рецептивным полем этой клетки. На рис. 2-9 приведен пример одного такого поля. На коже предплечья отмечены все точки, легкое прикосновение к которым может менять частоту разряда некоторой определенной клетки таламуса. Зона, внутрь которой попали все эти точки, обведена красным, она называется рецептивным полем таламической клетки для стимула «легкое прикосновение».
Некоторые клетки сенсорных центров имеют очень маленькие рецеп-
58
М. Циммерманн
Рецептивное поле
«Стимуляция!
прикосновением в рецептивном поле
11 1
|Н111111111111111Ш11Н111И
Потенциалы
действия нейрона
Рис. 2-9. Рецептивное поле центрального нейрона. Регистрируется активность центрального сенсорного нейрона; он генерирует потенциалы действия в ответ на прикосновение к коже руки в пределах области, показанной слева. Отметка раздражения и развитие реакции во времени показаны справа.
тивные поля. В качестве примера можно привести нейроны зрительной коры, которые реагируют на световые стимулы, действующие только в области желтого пятна глаза, имеющего площадь 0,02 мм2. Другие клетки, особенно принадлежащие к неспецифическим системам, представляют противоположный крайний случай: например, на такую клетку могут оказывать влияние стимулы, действующие на кожу обширной области тела, скажем всей ноги; причем эта клетка может реагировать и на прикосновение, и на охлаждение, и на вибрацию. В этом случае рецептивное поле имеет большую протяженность и охватывает различные модальности (полимодальная конвергенция).
Все, что отдельный сенсорный нейрон «знает» о положении специфического стимула,-это то, что стимул действует где-то внутри его рецептивного поля. Однако когда рецептивные поля нескольких нейронов одного уровня, которые во многих случаях значительно перекрываются, сравниваются на следующем, более высоком уровне, оказывается возможной более точная локализация стимула. В частности, такого рода переработка информации в нейронных сетях позволяет зрительной системе достичь высокой степени пространственного разрешения, несмотря на то что рецептивные поля отдельных нейронов в зрительных путях сравнительно велики (см. гл. 4).
До сих пор единственным критерием, по которому определялось ре
2. Нейрофизиология сенсорных систем
59
цептивное поле, было «влияние» на данный центральный нейрон стимуляции определенной точки на периферии. Но такие влияния могут быть как возбуждающими, так и тормозными, т.е. имеются возбуждающие и тормозные рецептивные поля или части полей. Для примера на рис. 4-28 показаны рецептивные поля сетчатки; в одном из случаев возбуждающий центр окружен тормозной областью, в другом - ситуация обратная. Такое подразделение рецептивных полей может отражать нейронные связи, включающие латеральное торможение (см. выше).
Свойства рецептивного поля-его положение и размер, модальности и качества специфических стимулов и подразделение на возбуждающую и тормозную части-в принципе могут быть определены для каждой клетки сенсорной системы. Соседние клетки различных центров, как правило, имеют сходные рецептивные поля. Изучая синаптически связанные сенсорные нейроны на различных уровнях, можно выяснить, как происходит видоизменение рецептивных полей, и тем самым описать функциональные операции различных центров. От одного синаптического переключения к другому размер и форма рецептивных полей могут меняться весьма характерным образом, а часто наблюдаются также и изменения вида стимулов, эффективных для разных уровней. Так, например, в зрительной системе для нейронов первого порядка в число существенных параметров стимулов входят такие, как яркость и цвет, а для зрительной коры стимулы могут быть более специализированными-скажем, нейрон может реагировать только на изменение цвета.
Соотношения интенсивность-реакция, дифференциальные пороги. Реакции центральных сенсорных нейронов можно охарактеризовать не только зависимостью от типа и локализации стимулов, но и зависимостью от интенсивности стимулов. Соотношение между частотой разряда нейрона F и величиной превышения стимулом пороговой интенсивности (5 — So) часто может быть описано степенной функцией
F = k(s-soy;
как это было обнаружено для многих рецепторов. У центральных нейронов также обнаружены различные значения п; встречаются и линейные соотношения (п = 1), но у многих нейронов наклон функции уменьшается при возрастании интенсивности стимула, т.е. показатель п меньше 1. Для центральных клеток также можно определять пороговые стимулы. Значение абсолютного порога So было включено в приведенную выше формулу степенной функции. Он измеряется путем нахождения наименьшей интенсивности стимула, на которую клетка реагирует изменением частоты потенциалов действия.
Порог другого типа можно определить, измеряя величину, на которую нужно изменить интенсивность надпорогового стимула, чтобы вызвать изменение частоты потенциалов действия нейрона. Такой порог называется дифференциальным порогом. Обнаружено, что он регулярным образом изменяется с изменением интенсивности стимула в со
60
М. Циммерманн
ответствии с правилом Вебера, описывающим дифференциальные пороги для субъективного ощущения.
Дифференциальные пороги можно определить и для многих других параметров стимула, помимо интенсивности,-пространственных и временных различий, различий в тоне и частоте выбрации. Для каждого случая приведенное выше определение справедливо; дифференциальный порог-это наименьшее изменение параметра стимула, которое вызывает измеримое изменение частоты разряда сенсорного нейрона.
В 2.10. Какое из следующих описаний рецептивного поля является правильным?
а) Правое полушарие мозга с полем зрения левого глаза, б) Все точки на сетчатке, освещение которых ведет к увеличению частоты импульсов в разряде некоторого нейрона зрительной коры.
в) Все точки на сетчатке, освещение которых ведет к изменению частоты импульсов в разряде некоторого нейрона зрительной коры.
г) Область слизистой оболочки носа, которая может влиять на активность некоторого нейрона обонятельной доли, если использовать локальное нанесение слабого раствора сероводорода.
д) Все точки на коже ноги, воздействие на которые болевым стимулом может вызвать отдергивание ноги (сгибательный рефлекс).
Верны несколько ответов.
В 2.11. Напишите по памяти определение дифференциальных порогов для нейронов сенсорной системы.
В 2.12. Предположим, что правило Вебера выполняется для стимула «давление на кожу». При исходной нагрузке в 10 г дифференциальный порог некоторого нейрона равен 100 мг. Какова будет величина дифференциального порога при исходной нагрузке в 100 г?
2.4. Соматосенсорная система: спинной мозг, восходящие пути и ствол мозга
Связи афферентов в заднем роге спинного мозга. В спинном мозгу афференты из туловища и конечностей образуют синаптические связи с нейронами, лежащими в сером веществе.
Волокна от кожи и внутренностей заканчиваются на нейронах дорсальных участков серого вещества-задних рогов (рис. 2-10). Часть толстых (миелинизированных) афферентов (группа И) посылает дополнительные веточки (называемые коллатералями), включающиеся в восходящие пучки задних столбов.
2. Нейрофизиология сенсорных систем
61
Кожа
пути
Рис. 2-10. Связи кожных афферентов в спинном мозгу. Афференты, входящие через задние корешки, образуют синапсы на нейронах заднего рога. Отсюда сигналы проводятся по волокнам, выходящим из спинного мозга (симпатическим и мышечным эфферентам), а также по восходящим путям. Из последних показаны пути, проходящие внутри передне-бокового пучка и ведущие в ствол мозга и таламус. Коллатерали афферентов группы II поднимаются в составе белого вещества (задних столбов) прямо в продолговатый мозг. Показаны два типа тормозных влияний на нейрон заднего рога (красные линии), они поступают по нисходящим путям и со стороны тормозных вставочных нейронов спинного мозга. Справа показаны слои, отражающие топографию восходящих путей на уровне шейного отдела.
62
М. Циммерманн
Задний рог спинного мозга-первая ступень переключения и переработки информации в соматовисцеральной системе. Как можно видеть из рис. 2-10, с функциональной точки зрения задний рог имеет четыре выхода: длинные восходящие пути (в частности, передне-боковой тракт), короткие волокна в проприоспинальных трактах, обеспечивающих восходящие и нисходящие связи соседних сегментов, синаптические связи с мотонейронами и, наконец, синаптические связи с симпатическими (преганглионарными) нейронами. Два последних выхода подразумевают, что афференты от кожи и внутренностей участвуют в организации спинномозговых двигательных и симпатических рефлексов. Мы здесь на них останавливаться не будем.
Нейроны заднего рога, показанные на рис. 2-10,-это многие тысячи нейронов каждого сегмента спинного мозга, однако каждый отдельный нейрон, вообще говоря, не всегда имеет все четыре типа изображенных здесь связей. На этом рисунк дана лишь грубая схема конвергенции и дивергенции афферентных волокон. Каждый нейрон образует связи со многими афферентными волокнами, и, наоборот, каждое афферентное волокно синаптически возбуждает несколько нейронов.
В задних рогах спинного мозга имеются не только возбуждающие синапсы, там много также и тормозных. На рис. 2-10 эти синапсы и соответствующие вставочные нейроны изображены красным. Представлены также два типа торможения, имеющие особо важное значение для сенсорных систем,- афферентное и нисходящее. Основная их роль-регулировать и модулировать афферентный поток; торможение осуществляется отчасти постсинаптически, а отчасти пресинаптически (см. аксо-аксонный синапс на окончании одного из двух волокон, вошедших в спинной мозг через задний корешок на рис. 2-10).
Восходящие пути спинного мозга. На рис. 2-10 показаны два восходящих тракта, по которым информация идет от соматосенсорных афферентов к головному мозгу,-задние столбы и передне-боковой пучок. Как и все восходящие и нисходящие пути, они локализованы в белом веществе спинного мозга. В задних столбах волокна проходят по той же самой стороне, что и афференты задних корешков (т. е. они ипсилате-ральны), тогда как в передне-боковом пучке-по другой стороне (кон-тралатерально).
Задние столбы состоят из прямых коллатералей толстых миелинизированных афферентов задних корешков (групп I и II); все они без исключения приходят от низкопороговых механорецепторов туловища и конечностей-кожи, внутренностей, мышц и суставов. Все эти афференты, кроме мышечных, заканчиваются синапсами на нейронах ядер задних столбов (нежное и клиновидное ядра) в продолговатом мозгу (рис. 2-11).
Аксоны этих нейронов переходят на другую сторону и по ней идут в таламус в составе медиального лемниска (медиальной петли). Переработка нервных сигналов в ядрах задних стобов, как в заднем роге и таламусе, включает и афферентное и нисходящее торможение. Задние
Продолговатый мозг
К неспецифическим ядрам таламуса
Шейный
Ретикулярная формация
Ствол мозга
Неоспинно-таламический тракт Палеоспинно-таламический ракт Медиальная петля
Спинно-ретикулярный тракт
Нежное ядро
Клиновиднов ядро
Задние столбы Передне-боковой пучок
Рис. 2-11. Пути, восходящие из спинного мозга. Лемнисковые пути показаны красным, а экстралемниковые-черным. Волокна задних столбов образуют синапсы в ядрах этих столбов (нежном и клиновидном) в продолговатом мозгу на нейронах, которые посылают свои аксоны в специфическое ядро таламуса через медиальную петлю. Филогенетически старые части передне-бокового пучка направляются в ретикулярную формацию (спинно-ретикулярный тракт) и неспецифические ядра таламуса (палеоспинно-таламический тракт). Филогенетически молодые части передне-бокового пучка (неоспинно-таламический тракт) проецируются в специфическое ядро таламуса; соответственно они рассматриваются как часть лемнисковой системы (красным).
К специфическому ядру таламуса
64
М. Циммермане
столбы и петля-наиболее заметные тракты специфической (лемнисковой) части соматосенсорной системы.
Передне-боковой пучок (рис. 2-11) содержит аксоны, которые идут преимущественно от нейронов заднего рога контралатеральной стороны (рис. 2-10). Этот путь заканчивается в ретикулярной формации ствола мозга и в таламусе; соответственно различают спинно-ретикулярный и спинно-таламический тракты (рис. 2-11). Спинно-таламический тракт сам по себе имеет два компонента, различающиеся своим филогенетическим возрастом: палеоспинно-таламический тракт (старый) и неоспинно-таламический тракт (новый). Последний особенно хорошо развит у приматов; он рассматривается как часть лемнисковой системы. В то же время спинно-ретикулярный и палеоспинно-таламический тракты - важные экстралемнисковые спинномозговые пути.
Задние столбы передают сигналы исключительно от низкопороговых механорецепторов, тогда как информация, проходящая по передне-боковому пучку, поступает от механорецепторов, терморецепторов и болевых рецепторов. В оба тракта афферентные волокна приходят не только от кожи и внутренних органов, но также от мышц и суставов.
Когда один из этих путей перерезается, наблюдаемая сенсорная недостаточность соответствует только что описанной организации связей рецепторов разных типов. Повреждение заднего столба вызывает ипсилатеральное (на той же стороне тела) нарушение способностей, имеющих отношение к локализации тактильных стимулов, в частности способности отличать два точечных прикосновения от одного (см. разд. 3.1) и способности совершать исследование путем ощупывания, которая обеспечивается цепями обратной связи, включающими афференты от кожи, мышц и суставов.
После перерезки передне-бокового пучка в контралатеральных месту повреждения областях тела отмечается потеря болевой и температурной чувствительности. При заболеваниях, сопровождающихся непереносимой болью, которую не удается снять фармакологическими препаратами, можно прибегнуть к операции перерезки передне-бокового пучка (хордотомия). Эта операция прерывает также передачу температурных стимулов, тогда как тактильная чувствительность сохраняется (она передается по задним столбам).
Кроме задних столбов и передне-боковых пучков в спинном мозгу имеются и другие восходящие пути. Спинно-мозжечковые тракты передают в мозжечок информацию от механочувствительных кожных и мышечных афферентов; функционально они принадлежат к моторной системе. Спинно-шейный тракт, ведущий в таламус, несет преимущественно кожные афференты от конечностей. Его функция пока не ясна. Кроме длинных трактов, упоминавшихся до сих пор, существуют восходящие к головному мозгу полисинаптические пути, состоящие из последовательно соединенных нейронов спинного мозга с короткими аксонами.
Аксоны, выходящие из того или иного сегмента, идут внутри восхо-
2. Нейрофизиология сенсорных систем
65
дящих трактов бок о бок. Вследствие этого тракты имеют слоистую организацию, которая схематически представлена на рис. 2-10 для задних столбов и передне-бокового пучка на уровне шейного отдела. Когда аксоны входят в тракт, они поворачивают, чтобы идти параллельно волокнам из нижних отделов в части тракта, находящейся на стороне серого вещества. Границы отдельных пучков не так резко выражены, как это показано на рис. 2-10; в действительности имеют место диффузные переходы и перекрытия зон. В связи с такой топографической организацией при поверхностном повреждении передне-бокового пучка в шейной области (которое может быть вызвано травмой или опухолью) в первую очередь следует ожидать симптомов выпадения в нижней половине тела.
Ядра и восходящие тракты тройничного нерва. Тройничный нерв-со-мато-висцеральный сенсорный нерв лицевой области - входит в головной мозг на уровне моста. Его афференты образуют синапсы в двух ядрах внутри серого вещества моста-спинальном ядре и главном сенсорном ядре. Спинальное ядро тройничного нерва функционально соответствует заднему рогу спинного мозга, тогда как главное сенсорное ядро соответствует ядрам заднего столба спинальных афферентов. Эту аналогию можно распространить и на пути, выходящие из этих ядер. В спинальном ядре афференты от механорецепторов, терморецепторов и болевых рецепторов синаптически переключаются на нейроны, посылающие свои аксоны в ретикулярную формацию и (или) в таламус, куда идут и волокна передне-бокового пучка спинного мозга. В главном сенсорном ядре заканчиваются только афференты от низкопороговых механорецепторов, а аксоны постсинаптических нейронов присоединяются к медиальной петле.
Кроме передачи в таламус, информация, поставляемая афферентами тройничного нерва, используется для двигательных рефлексов мышц области головы. Система тройничного нерва осуществляет жизненно важные функции, особенно у младенцев: например, обеспечивает тактильное узнавание, пищевое поведение и издавание звуков.
Ретикулярная формация. Ретикулярная формация, которая занимает значительную часть ствола мозга, является важным отделом восходящей неспецифической системы (рис. 2-7, Б). Она имеет огромное количество разнообразных афферентных и эфферентных связей, схематически показанных на рис. 2-12, А. Сомато-висцеральные афференты идут по спинно-ретикулярному тракту (передне-бокового пучка) и, возможно, также по проприоспинальным (полисинаптическим) путям, равно как и по соответствующим трактам, выходящим из спинального тройничного ядра. Кроме того, ретикулярная формация получает входные сигналы из всех других афферентных черепных нервов, т. е. от всех органов чувств. Афферентные волокна приходят также из многих других областей головного мозга, например из моторной и сенсорной областей коры больших полушарий, таламуса и гипоталамуса. Эфферентные пути тоже разнообразны: есть спускающиеся в спинной мозг, есть подни-
66
М. Циммерманн
Состояние сознания
Б
Эмоции
Рис. 2-12. Ретикулярная формация: связи и функции. А. Схема афферентных и эфферентных связей; на рисунке представлена комбинация анатомических и физиологических данных; его назначение-дать представление о большом числе входов и выходов. Б. Наиболее важные функции, приписываемые ретикулярной формации.
2. Нейрофизиология сенсорных систем
67
мающиеся через неспецифические ядра таламуса к коре, к гипоталамусу и к лимбической системе.
Многообразие афферентных связей проявляется также и на уровне отдельных нейронов. Микроэлектродные отведения из области ретикулярной формации показали, что обычно там имеется конвергенция на один нейрон двух или трех афферентов из числа представленных на рис. 2-12, А, т.е. преобладает полисенсорная конвергенция. Это отличительная черта ретикулярных нейронов. Другие характерные свойства данных нейронов - обширные рецептивные поля, часто представленные билатерально на поверхности тела, большой латентный период реакций на периферическую стимуляцию (связанный с большим числом синаптических переключений), вариабельность реакций (случайные флуктуации числа потенциалов действия, генерируемых в ответ на повторные предъявления стимула, или усиление реакций со временем при действии серии стимулов). Все эти свойства резко отличаются от свойств нейронов специфических ядер, называемых лемнисковыми нейронами соматосенсорной системы. Нейроны ретикулярной формации представляют собой пример неспецифических, экстралемнисковых нейронов.
В наших знаниях относительно многообразных функций ретикулярной формации пока еще имеются существенные пробелы. Как схематически представлено на рис. 2-12, Б, полагают, что она вносит вклад в осуществление ряда функций, которые можно суммировать следующим образом: а) контроль состояния сознания последством влияния на активность кортикальных нейронов и тем самым участие в ритме сон/ бодрствование (ключевой структурой в этом отношении является восходящая ретикулярная активирующая система); б) посредничество в обеспечении эмоционального влияния сенсорных стимулов, в частности болевых сигналов, передаваемых по передне-боковому пучку путем посылки афферентной информации в лимбическую систему; в) вегетативная регуляция, особенно касающаяся жизненно важных рефлексов (сердечно-сосудистые, дыхательные, глотательные рефлексы, кашель, чихание), когда многие афферентные и эфферентные системы должны работать координированно; г) участие в двигательных механизмах регуляции позы и направленных движений в качестве важного элемента моторных центров ствола мозга.
В 2.13. Нарисуйте схему поперечного среза спинного мозга, указав афферентные волокна и их связи с сегментарными эфферентами и восходящими трактами, напишите на ней названия.
В 2.14. Предположим, что спинной мозг поврежден: его левая половина полностью перерезана на уровне груди. Какие из перечисляемых ниже симптомов будут наблюдаться?
а) Исчезновение температурной и болевой чувствительности на левой половине тела.
б) Исчезновение температурной и болевой чувствительности в области правой ягодицы.
68
М. Циммерманн
в) Утрата способности узнавать числа, наносимые на тыльную поверхность правой стопы.
г) Утрата способности точно локализовать легкое прикосновение к левой ноте.
Верны несколько утверждений.
В 2.15. Ретикулярная формация ствола мозга имеет следующие свойства и функции:
а) анатомически она состоит из регулярной сети нейронов с параллельными аксонами и дендритами;
б) каждый из ее нейронов получает возбуждение только от одного органа чувств;
в) она регулирует общую возбудимость кортикальных нейронов, а тем самым также и состояние сознания;
г) на своем пути в таламус афференты задних столбов синапти-чески переключаются преимущественно в ретикулярной формации.
2.5. Соматосенсорная система в таламусе и коре
Среди сенсорных центров, служащих анатомическим субстратом сознательного восприятия, главными являются те, которые расположены в таламусе и коре. Они сгруппированы функционально в таламо-корти-кальную систему. Лемнисковая система имеет здесь особенно четкую соматотопическую организацию, так что мы можем говорить о проецировании или отображении периферии тела на таламус и кору. Неспецифическая система не обнаруживает такой точной соматотопической организации.
Таламус общий вад и подразделения. На рис. 2-13 представлен схематический, упрощенный вид таламуса правой половины мозга. Функционально его можно подразделить на следующие группы ядер: 1) специфические ядра сенсорных систем кожи, глаза и уха (красные); 2) неспецифические ядра (темно-серые); 3) моторные ядра (светлосерые); 4) ассоциативные ядра (белые).
Каждое из специфических ядер (красных) получает сигналы с периферии, которая хорошо определена анатомически и функционально. Там также выделяются прямые аксонные пути, идущие в кору и назад из коры. Специфические ядра зрительной и слуховой систем (соответственно латеральное и медиальное коленчатые тела, ЛКТ и МКТ, красные на рис. 2-13, Л) рассматриваются в гл. 4 и 5. Специфическим ядром соматосенсорной системы является вентробазальное ядро. Его функциональные свойства обсуждаются ниже.
Неспецифические ядра таламуса, большинство которых занимает медиальное положение, включают (среди прочих) центральное латеральное и центральное медиальное ядра, обозначенные на рис. 2-13, Л темно-серым; функционально они связаны с ретикулярной формацией ствола мозга (см. разд. 2.4). Подобно ретикулярной формации, они по-
2. Нейрофизиология сенсорных систем
69
Осциллограф
Рис. 2-13. А. Функциональные подразделения таламуса (схема) На схеме показан правый таламус; разрезы в двух местах представляю) его внутреннее строение; показаны четыре группы ядер: специфические (красные), неспецифические (темно-серые), двигательные (светло-серые) и ассоциативные (белые). Микроэлектрод и осциллограф символизируют отведение потенциалов от одиночных нейронов вентробазалыюго ядра. Б. Участок длиной около 2 мм по ходу микроэлектрода через правое вентробазалыюе ядро кошки; показаны положения десяти нейронов, рецептивные поля которых обозначены на коже животного. (Poggio, Mountcastle, Bull. Johns Hopk. Hosp., 106, 266, 1960.)
лучают входные сигналы от всех сенсорных модальностей. Волокна, восходящие из спинного мозга, проходят преимущественно через переднебоковой пучок. Имеются эфферентные связи со многими областями коры, с гипоталамусом и лимбической системой. Неспецифические ядра таламуса являются, так сказать, последующими станциями распределения афферентной информации, конвергирующей на ретикулярную формацию (рис. 2-7, Б и 2-12, Б).
70
М. Циммерманн
Здесь мы лишь кратко упомянем ядра моторной системы - вентролатеральное ядро (светло-серое на рис. 2-13, А) и ассоциативные ядра (например, пульвинар, изображенный белым); последние интегрируют информацию от нескольких органов чувств в сотрудничестве с различными зонами коры (сенсорными полями коры и ассоциативной корой).
Специфическое таламическое ядро соматосенсорной системы. Из-за своего анатомического положения эта структура называется вентроба-зальным ядром (ВБ) или вентробазальным комплексом. Здесь оканчиваются медиальная петля и неоспинно-таламический тракт, равно как и волокна соответствующих трактов из зоны тройничного нерва. Вен-тробазальное ядро-вторая ступень синаптического переключения лемнисковой системы. Характерные свойства нейронов ВБ будут обсуждаться ниже. Это типичные лемнисковые нейроны, так что почти во всех отношениях их реакции противоположны по свойствам реакциям экстралемнисковых нейронов (см. разд. 2.4).
При микроэлектродном отведении из ВБ чаще всего попадаются нейроны, которые разряжаются в ответ на механическую стимуляцию кожи. Траектория прохождения электрода через правое ВБ, показанная на рис. 2-13, А, приведена с увеличением на рис. 2-13, Б. Каждая точка на этой траектории показывает положение в ядре одного нейрона, который реагировал на стимуляцию отмеченного участка кожи. Наиболее важные результаты этого эксперимента следующие. 1. Каждый нейрон может быть активирован только стимуляцией некоторой области левой передней лапы экспериментального животного (кошка); это рецептивное поле нейрона. 2. Рецептивные поля ВБ в среднем тем меньше, чем ближе они располагаются к опорным подушечкам лапы. Это одна из нейрофизиологических предпосылок высокого пространственного разрешения, необходимого для тактильного исследования с помощью передних лап. 3. Соседние участки тела проецируются в соседние области ВБ. Такая соматотопическая организация еще более сильно выражена в последующей проекции на область SI коры больших полушарий. 4. Каждый нейрон возбуждается только одним типом рецепторов-например, медленно адаптирующимися рецепторами давления или рецепторами кожных волосяных фолликулов (см. гл. 3). 5. Интенсивность периферического стимула кодируется средней частотой разряда нейрона, как это делается и в рецепторах (см. разд. 2.1).
Нейроны специфических таламических ядер глаза и уха (ЛКТ и МКТ) имеют аналогичные свойства.
Соматосенсорные проекционные зоны коры. Вентробазальное ядро (ВБ) таламуса связано восходящими и нисходящими аксонами с двумя областями коры, называемыми SI и SII (буква S ставится для обозначения соматосенсорности; рис. 2-14). SI располагается в постцентральной извилине, непосредственно за центральной бороздой-глубоким прогибом, проходящим через головной мозг. SII находится у человека на верхней стенке латеральной борозды, разделяющей теменную и височную доли.
2. Нейрофизиология сенсорных систем
71
Рис. 2-14. Первичные сенсорные проекционные зоны коры. А. Внешний вид левого полушария. Б. Вид правого полушария со стороны срединной плоскости мозга. В. Фронтальное сечение левого полушария на уровне постцентральной извилины. Темно-красным показаны соматосенсорные проекционные зоны SI и SII, розовым-первичные проекционные зоны зрительной и слуховой систем. Кроме того, показано положение моторной коры (темно-серым), ассоциативных областей (белым) и кортикальных частей лимбической системы (светло-серым).
Между периферией тела и контралатеральной областью SI имеются отчетливые соматотопические взаимоотношения, проявления которых уже заметны и на более низких уровнях-в лемнисковых трактах и ядрах, и особенно в вентробазальном ядре таламуса (см. рис. 2-13, Б). На рис. 2-15 показано, как представлена на SI поверхность человеческого тела; это схематический рисунок, основанный на результатах электрической стимуляции коры у человека, находившегося в сознании. Геометрически искаженную проекцию поверхности тела на нейронную систему SI называют иногда «соматосенсорным гомункулусом». Эта афферентная проекция напоминает карту эфферентных связей моторной коры с мышцами.
Из рис. 2-15 ясно, что области коры, связанные с лицом и рукой, каждая в отдельности имеет примерно такую же величину, как проекционные области туловища и ноги взятые вместе. Это общее правило: органы с особенно высокой плотностью рецепторов, такие, как пальцы и губы, и проецируются соответственно на самые большие популяции
72
М. Циммерманы
Рис. 2-15. Соматотопическая организация зоны SI. Символически представленное отображение поверхности человеческого тела на постцентральную извилину, выявляющееся при локальном раздражении коры во время нейрохирургических операций (Penfield W., Rasmussen Т., 1950.)
нейронов SI. Следствия этого, касающиеся восприятия, будут обсуждаться ниже.
Как у животных, так и у человека соматотопическое отображение периферии тела на сенсорной коре можно изучать путем стимуляции периферии и регистрации в коре больших полушарий возникающих при этом вызванных потенциалов-, латентные периоды между стимулами и вызванными потенциалами дают указание на число переключений в цепи. Таким способом были построены «карты» соматосенсорных проекционных зон для многих видов животных. У высших приматов проекция SI похожа на соответствующую проекцию у человека, показанную на рис. 2-15. У кошки и крысы непропорционально большое представительство имеют области рта и носа (в особенности тактильные вибриссы), тогда как передние конечности занимают относи
2. Нейрофизиология сенсорных систем
73
тельно малую площадь по сравнению с тем, что характерно для приматов.
В SII также выявляется соматотопическая организация, хотя и хуже выраженная, чем в SI. Особенностью SII является то, что обе половины тела представлены в каждом полушарии, т. е. проекции являются билатеральными.
Регистрация вызванных потенциалов может быть также использована для составления карт первичных проекционных зон в зрительной и слуховой системах. Сетчатка проецируется в ограниченную область затылочной доли, а рецепторы внутреннего уха имеют проекционные поля в височных долях. И те и другие показаны на рис. 2-14 (см. также рис. 4-32 и 5-16). Здесь также имеется поточечное представление периферической сенсорной поверхности на поверхность коры.
Переработка информации нейронами соматосенсорной коры. У нейронов, располагающихся прямо один над другим в 6 слоях SI, рецептивные поля сильно перекрываются, тогда как у нейронов, лежащих рядом по горизонтали, они обычно четко различаются. Этот факт привел к заключению, что кора построена из функциональных единиц, представляющих собой нейронные колонки, ориентированные перпендикулярно поверхности, причем каждая колонка имеет диаметр 0,2-0,5 мм. При этом на нейроны каждой колонки могут влиять только рецепторы одного типа, т. е. колонки специализированы не только в отношении локализации, но и в отношении типа рецепторов. Однако в коре пока еще точно не установлено существование колонок, которые бы обрабатывали исключительно температурные стимулы, и специфических ноцицептивных нейронов в кортикальных проекционных полях пока еще тоже не обнаружено.
Теоретически каждая колонка коры благодаря большому числу содержащихся в ней нейронов (до 105) обладает значительными потенциями в смысле обработки информации с периферии. Было выдвинуто предположение, что колонки функционируют (на основе разнообразных возбуждающих и тормозных взаимодействий между нейронами) по иерархическому принципу; в этом процессе в результате соответствующих связей и операций разные свойства периферических стимулов кодируются разными нейронами. К этой функции ансамблей корковых нейронов применяется термин выделение признаков. Примеры, поясняющие это понятие, даны в разд. 4.3 для случая зрительной коры. После такой обработки информация направляется в другие области коры-например в моторную кору (прецентральная извилина) или в ассоциативные зоны.
Специфическая таламокортикальная система и сознательное восприятие. У человека и других приматов рука и область рта представлены в SI в особо крупном масштабе (рис. 2.15). Вместе с тем это именно те участки кожи, которые выделяются по своим тактильным способностям; пороги пространственного разрешения двух точек в этих областях особенно низки (см. рис. 3-3). Кортикальные проекции кожи, равно как и других органов чувств, принимают участие в процессе различительно
74
М. Циммерманы
го, сознательного восприятия. В подтверждение этого свидетельствует целый ряд фактов, два из которых мы здесь опишем.
При хирургической операции на мозге, производимой под местной анестезией, больной способен сообщить о характере ощущений, возникающих при раздражении коры тонкими электродами. Например, если раздражению подвергается область кисти в зоне SI, больной ощущает прикосновение к кисти. Соматотопическая организация, представленная на рис. 2-15, была установлена именно таким способом. Значит, путем прямой стимуляции сенсорной коры можно вызвать сознательное ощущение, не активируя афферентные пути, включающие периферические нервы, спинной мозг и специфические ядра таламуса. Аналогичным образом стимуляция проекционных зон глаза производит ощущение вспышки света. Однако ощущение боли получить путем стимуляции коры не удается.
Экспериментальное удаление ограниченного участка SI у обезьян вызывает выпадение чувствительности в соответствующем периферическом участке. В одном опыте животное перед операцией научили отличать куб от шара на ощупь, не глядя на предмет. После удаления соответствующей руке сенсорной области коры эта способность оказалась утраченной восстановить ее не удалось.
Аналогичные наблюдения были сделаны и на людях с повреждениями отдельных участков SI (например, в результате огнестрельного ранения или опухоли). Тактильные стимулы еще воспринимаются, но способность локализовать их нарушена; ощущения прикосновения имеют смутный, неопределенный характер. Боль чувствуется, но локализация болевых стимулов менее точна. Таким образом, интактность SI является предпосылкой пространственно организованного сознательного восприятия.
Электрическая стимуляция SII у человека также вызывает ощущения, которые кажутся связанными с соответствующими участками периферии. Однако удаление только участков SII не вызывает отмеченных выше сенсорных выпадений. Роль SII в сенсорном восприятии остается пока не выясненной.
Ассоциативные зоны коры. Все сенсорные проекционные зоны вместе с моторной корой занимают менее 20% поверхности коры (см. рис. 2-14). Оставшиеся области называются ассоциативными зонами. С первичными проекционными зонами их соединяют мощные тракты нервных волокон; обычно каждая ассоциативная область связана таким образом с несколькими проекционными зонами.
Термин «ассоциативная зона» происходит от функции, приписываемой этим участкам; полагают, что здесь осуществляются процессы ассоциации разнообразной, но специфической информации от органов чувств и формируются более сложные элементы сознания. В теменной ассоциативной области, расположенной между соматосенсорной и зрительной проекционными зонами, предположительно возникают наши субъективные представления об окружающем пространстве и нашем со
2. Нейрофизиология сенсорных систем 75
бственном теле («образ тела»). Предполагается, что критическим моментом в формировании этих представлений является процесс научения в ходе развития в раннем детстве, во время которого оценивается и сопоставляется соматосенсорная, проприоцептивная и зрительная информация.
Гипотезы о функции ассоциативных полей у человека сформировались главным образом на основании наблюдения неврологических нарушений, возникающих в результате повреждения коры в ограниченных участках. Примером нарушения вследствие повреждения теменной ассоциативной области является зрительная агнозия. У больного с таким синдромом зрение сохраняется в том смысле, что он может обходить препятствия или подходить к предметам и хватать их. Но он не может определять значение предметов. Когда больного, страдающего зрительной агнозией, просят назвать какую-нибудь вещь, на которую он смотрит, он не может этого сделать, но, взяв ее в руки, узнает ее с помощью осязания.
Относительная площадь, занимаемая ассоциативными зонами у приматов, и особенно у человека, намного больше, чем у других млекопитающих. Специфически человеческие ассоциативные области, расположенные в височной и лобной долях, принимают участие в восприятии смысла речи и в формировании речи (речевые центры). Ассоциативным зонам посвящено очень мало нейрофизиологических исследований.
Экстралемнисковая система и сознательное восприятие. Возбуждение сенсорной коры-необходимое, но не достаточное условие появления сознательного восприятия. Многие клинические наблюдения и результаты экспериментов на животных свидетельствуют в пользу этого утверждения. Например, даже в условиях наркоза или глубокого сна можно вызвать первичные корковые потенциалы, но это не приведет к сознательному восприятию. Кроме того, повреждения верхней части ствола мозга, включающей ретикулярную формацию, приводят к стойкой потере сознания; электрическая стимуляция этой области у экспериментальных и&вотных вызывает пробуждение, настораживание и переключение внимания. Два последних изменения поведения объединяют под названием «активация» (arousal). На основе этих и многих аналогичных данных возникло представление о том, что постоянный возбуждающий приток из ретикулярной формации в кору участвует в определении состояния сознания в диапазоне от глубокого сна через дремотное состояние до активного бодрствования; появился термин «восходящая ретикулярная активирующая система».
Как схематически показано на рис. 2-16, сознательное восприятие, скажем, тактильного стимула, требует наличия двух входов в неокор-текс-специфической проекции по лемнисковому пути (через вентроба-зальное ядро таламуса) и активации посредством неспецифической системы ретикулярной формации (через неспецифические ядра таламуса). Лемнисковая система обеспечивает информацию об объекте восприятия (например, его положении, размере и времени действия), этот вклад
76
М. Циммермане
Поведение
Рис. 2-16. Взаимодействие специфической и неспецифической систем (очень упрощенная схема) в случае трех категорий поведения: сознательного восприятия, двигательного поведения и автономных реакций.
в сознательное восприятие обозначен на рис. 2-16 термином «различение». Вместе с тем неспецифическая активация, в которую могут внести вклад все сенсорные каналы, определяет уровень бодрствования как условие восприятия.
2. Нейрофизиология сенсорных систем
77
Но мы знаем из личного опыта (интроспекции), что каждое восприятие имеет еще один аспект-эмоциональный, или, как его обычно называют, чувственный: наше отношение к объекту восприятия. Эта размерность восприятия приписывается лимбической системе. Например, путем электрического раздражения области лимбической системы можно вызвать у кошки ярость и агрессивное поведение. Лимбическая система состоит из ряда подкорковых областей, грубо говоря располагающихся кольцом вокруг диэнцефалона.
Такая интерпретация ее функции предполагает, что лимбическая система вносит вклад в эмоциональное содержание восприятия. Как схематически показано на рис. 2-16, соматосенсорные входные сигналы поступают в лимбическую систему преимущественно по неспецифическому (экстралемнисковому) пути.
Лимбическая система имеет тесные реципрокные взаимоотношения с гипоталамусом-верховным управляющим центром для многих регуляторных цепей в автономной нервной системе (например, цепей регуляции сердечно-сосудистой активности и температуры тела) и эндокринной системе (например, цепи регуляции водного баланса). Возможность того, что сенсорные стимулы действуют на эффекторы автономных систем через гипоталамус, также учтены на схеме (рис. 2-16).
До сих пор, используя рис. 2-16, мы обсуждали только взаимодействия специфической и неспецифической систем. На том же рисунке сильно упрощенно показаны взаимоотношения между тремя типами поведенческих реакций на внешние стимулы. Это-сознательное восприятие, моторное поведение и бессознательные автоматические реакции тела.
Сознательное восприятие и двигательное поведение имеют по крайней мере две размерности, которые в каждом случае могут быть приписаны соответственно неокортексу и лимбической системе. Для случая восприятия мы уже обсуждали эти размерности - различение и эмоции. В двигательном поведении мы имеем, с одной стороны, когнитивные факторы (неокортикальные); они определяются конкретными сигналами от органов чувств. Вторая размерность в данном случае-это мотивация (drives); считается, что здесь участвует лимбическая система. Двигательные акты можно грубо подразделить на категории «приближение» и «избегание» в соответствии с тем, какая имеется тенденция-приближаться или удаляться.
Автономные реакции тела в значительной степени бессознательны. Их интенсивность, оцениваемая, например, по изменению кровяного давления или электрического сопротивления кожи, связана с интенсивностью эмоционального компонента восприятия. Сила мотивации двигательного поведения также коррелирует с интенсивностью эмоций. «Общим знаменателем» здесь является возбуждение лимбической системы.
Графическое обобщение, представленное на рис. 2-16, следует рассматривать как попытку изобразить то, что мы знаем о центральных
78
М. Циммерманн
частях сенсорных систем, в виде наглядной схемы. Она обеспечивает читателю самую предварительную ориентацию в многочисленных сложных данных.
В 2. 16. Термин «соматосенсорная проекция» означает, что
а) тело нейрона посылает отростки в определенных направлениях;
б) периферическая сенсорная поверхность поточечно отображается на сенсорной коре;
в) существует определенное представительство периферии на сенсорной коре;
г) нейроны соматосенсорной области коры SI связаны волокнами с нейронами ассоциативной коры.
Верны несколько ответов.
В 2.17. Нейроны специфических ядер таламуса получают свои афферентные сигналы
а) от органов чувств, по крайней мере через семь синапсов; б) от нескольких органов чувств одновременно, по крайней мере через три синапса;
в) от терморецепторов и болевых афферентов кожи через ядра задних столбов;
г) от одного органа чувств в каждом случае, хотя бы через два синапса.
В 2.18. Какое из следующих утверждений приложимо к человеку, у которого разрушены области, соответствующие кисти и лицу в правой соматосенсорной коре SI?
а) Никакие объекты не могут быть распознаны на ощупь при исследовании правой рукой.
б) Болевые раздражения левой руки ощущаются, но точность локализации снижена.
в) Отсутствует болевая чувствительность левой руки.
г) Благодаря билатеральной проекции в SII никаких заметных нарушений не появляется.
В 2.19. Латеральное торможение в ЦНС ответственно за
а) компенсацию пространственного распространения возбуждения, обусловленного дивергенцией;
б) уменьшение всех импульсов, вызываемых стимуляцией;
в) нейронный механизм усиления контраста;
г) подавление контралатеральных афферентных сенсорных влияний.
Верны несколько ответов.
2.6. Сенсорная система в свете теории информации
С точки зрения его функции нервное волокно можно сравнить с кабелем, передающим информацию. Подход инженеров связи теорию ин-
2. Нейрофизиология сенсорных систем
79
Источник информации
Передатчик (кодирование)
Канал передачи
Приемник Потребитель
(декодирование)
Источник искажений
Рис. 2-17. Схема, иллюстрирующая понятия и
термины теории
информации.
формации-можно применить также и к нервной системе. Эту область знаний в совокупности с теорией управления часто называют кибернетикой; ее применение к биологии называют биокибернетикой, особенно в Европе, в США такие общие термины используются реже.
Элементы теории информации. Фундаментальная идея, из которой развилась теория информации, представлена в виде блок-схемы на рис. 2-17. Здесь выделены те функциональные компоненты, которые принимают участие в передаче информации: источник информации, передатчик, канал передачи, приемник, потребитель и источник помех, который в данном примере действует на канал передачи. Это простое расчленение можно применить ко всем видам передачи информации как в технике, так и в биологии. Теория информации может дать количественную меру информации, что позволяет выводить утверждения, помогающие описывать и сравнивать системы передачи информации.
Чтобы передавать информацию, нужны символы (например, буквы или цифры); в определенной степени они задаются или определяются источником информации. Эти знаки кодируются в передатчике, обычно преобразуясь в другие виды сигналов, более подходящие для передачи (например, частотно-модулированные электромагнитные волны в случае ЧМ-радиосвязи). Кодирование -это общий термин, используемый для такого преобразования, при котором сохраняется однозначная связь между элементами двух наборов знаков; примером кодирования является перевод букв алфавита в знаки азбуки Морзе. В приемнике переданная информация декодируется и направляется потребителю.
В теории информации особое внимание уделяется влиянию помех. Помехи могут возникать в передатчике (в процессе кодирования), в канале передачи и в приемнике (при декодировании). Такие источники помех представлены на рис. 2-17 как один элемент, действующий на канал передачи. Одной из задач теории информации является нахождение таких процедур кодирования, которые защищают информацию от разрушающего действия помех.
Как можно измерять информацию? В теории информации термин
80
М. Циммерманы
«информация» применяется только к тем характеристикам сообщений, которые поддаются численной оценке и могут описываться математическими формулами. Следующий пример показывает, откуда возникает такое использование этого термина.
Если вы бросите одну игральную кость из пары, то с одинаковой вероятностью может выпасть одно из шести чисел. До бросания мы знаем только то, что результатом будет одно из шести определенных чисел. Акт бросания устраняет эту неопределенность измеримым образом, что мы можем использовать для определения информационного содержания броска. Заметим, что каждое такое событие, т.е. каждое бросание кости с шестью гранями, будет иметь одинаковое информационное содержание. Событие другого типа-подбрасывание монеты (где есть только два возможных результата) будет иметь другое информационное содержание. В общем случае можно сказать, что: информационное содержание события-это количественная мера уменьшения неопределенности наших знаний о ситуации благодаря знанию исхода события. Заметьте, что передача символов от источника к потребителю имеет точную аналогию с вышеприведенными примерами-тут тоже имеется измеримое уменьшение априорной неопределенности.
Легко видеть, что это уменьшение неопределенности сведений об исходе события тем больше, чем меньше вероятность р получения каждого исхода; бросание кости (р= 1/6) устраняет большую неопределенность, чем подбрасывание монеты (р = 1/2). В связи с этим количество информации I имеет смысл определить через величину, обратную р. т. е. через 1/р. Разумным дополнительным условием для измерения информации является требование, чтобы в случае события с одним возможным исходом, который обязательно наступит и, значит, имеет вероятность 1, количество информации равнялось бы нулю. Это условие можно выполнить, используя логарифмическую функцию от 1/р, поскольку log 1 = 0.
Эти рассуждения поясняют, почему количество информации в одном событии (или сообщении, состоящем из одного символа, выбранного из нескольких равновероятных) было определено как
/ = log (1/р). [2-5]
В этом определении используют логарифм по основанию 2 (записывая его в виде log2), поскольку тогда ситуация с р = 1/2 будет давать 1 ед. информации. Таким образом, [2-5] можно следующим образом переписать для определения количества информации I в событии, имеющем равновозможные исходы с вероятностью р:
1 = log2 (1/р). [2-6]
Для игры в кости информационное содержание каждого броска одной игральной кости теперь можно оценить численно: I = = log2 [1 (1/6)] = 2,58 бит. Что такое «бит», мы узнаем позже. Поскольку логарифмические линейки и карманные калькуляторы обычно не
2. Нейрофизиология сенсорных систем
81
дают возможности непосредственно определять функцию log2, полезно знать формулу перевода: log2 п = log10 и/log] 0 2 = 3,32 -log10 п.
Вводя меру информации, мы начали с предположения, что все состояния источника информации могут иметь место с одинаковой вероятностью р, как, например, в случае игральных костей. Это предположение является упрощением; обычно отдельные знаки, или состояния, источника информации не равновероятны.
Примером случая неравной вероятности могут служить буквы алфавита, которые встречаются в тексте с разными частотами (вероятностями). Читатель может легко убедиться в этом самостоятельно; в англий-ском тексте, например, самая распространенная буква-е. Общий случай неравных вероятностей выходит за рамки данного вводного рассмотрения меры информации; интересующиеся читатели могут найти дополнительную литературу по приведенному списку.
Остановившись на частном случае равных вероятностей, заметим, однако, что [2-6] можно записать в другом виде; если « общее число возможных символов, или состояний источника, справедливо равенство р= l/п. Для игральной кости п = 6. Выражая через п количество информации при передаче одного символа, получим
I = log2 п. [2-7]
Это значит, что содержание информации в таком сообщении равно логарифму по основанию 2 числа п всех возможных символов, которые могут быть переданы. Мы применим это соотношение в нейрофизиологии (см. далее).
Конечно, субъективная ценность единицы информации для получателя зависит от многих факторов, не включенных в рассмотренную выше количественную меру информации. Например, при игре в кости субъективная цена, которую игрок назначает разным числам, может варьировать в зависимости от бессознательной мотивации, его отношений с другими партнерами, того, последний ли доллар он сейчас ставит, и т.д. Изучение таких аспектов символьной информации часто называют семантикой.
Двоичные и недвоичные знаки; бит. Как подчеркивалось выше, передача информации требует набора символов, из которых передающий информацию может делать выбор. В простейшем случае набор состоит всего из двух символов, называемых двоичными, или бинарными (например, двух цифр: 0 и 1). Используя их, передающий информацию может посылать сообщения о выборе из двух возможных вариантов (например, «да» или «нет»). Двоичные системы особенно удобны с технической точки зрения; примерами их реализации являются свет-темнота, смена положений тумблера, наличие и отсутствие отверстия в перфокарте. Это одна из причин, по которой в качестве единицы информации было выбрано информационное содержание акта, определяющего один двоичный символ. Этот выбор уже подразумевался в [2-6] и [2-7], так
82
М. Циммерманн
как log22=l. Количество информации, которое передается в сообщении, содержащем один из двух равновероятных двоичных символов, называется одним битом.
Один бит-это относительно небольшое количество информации; если с помощью двоичных символов нужно передавать более пространные сообщения, следует объединять вместе несколько знаков-из двоичных символов нужно составлять слова. Алфавит, из которого составляются такие слова, состоит из «букв» 0 и 1. Длина слов, т.е. число двоичных знаков, используемых на одно слово, есть прямая мера (в битах) количества информации, передаваемой в одном слове. Словом, состоящим из двух двоичных знаков, можно передать два бита, из трех-три и т.д. Число различных слов, которые могут быть составлены из двух двоичных знаков, равно 22 = 4. Это 00, 01, 10 и И. С помощью трех знаков можно составить 23 = 8 слов следующего вида: ООО, 001, 010, ОН, 100, 101, ПО, 111. Очевидно, что используя т двоичных знаков на слово, можно составить п = 2т различных сообщений; процесс, конкретизирующий любое из них, передает т бит информации.
Обсуждавшееся до сих пор определение количества информации можно также применить и к случаям, в которых в качестве носителей информации служат любые произвольные символы. Любой желаемый набор символов можно представить двоичными последовательностями достаточной длины. Чтобы кодировать символы таким образом, т.е. чтобы получить однозначное соответствие между набором из п символов и двоичными словами, длина последних должна составлять в среднем т = log2 п двоичных знаков.
Принцип такого кодирования представлен на рис. 2-18. Это дерево выбора решений, в котором в результате последовательного раздвоения ветвей получается в точности по одной ветви на каждый символ (здесь нужно было представить 8 букв от А до Н). Каждое раздвоение символизирует выбор из двух возможностей. Если выбор левой ветви мы обозначим цифрой 0, а правой 1, то мы сможем построить соответствующие двоичные кодовые слова путем простого прохождения от «ствола» дерева к символам на конечных веточках. Получившиеся двоичные слова написаны под буквами, к которым они относятся.
Если такое кодовое дерево, или дерево выбора, строить для всего английского алфавита, то число выборов (точек раздвоения) будет равно первому целому числу, превышающему log2 26, или 5, так как 25 = = 32. Шесть двоичных слов, не понадобившихся для букв, можно использовать для кодирования знаков препинания и интервалов.
Но если произвольный знак можно заменить двоичным словом, то можно также сказать, что он содержит столько же информации (в битах), сколько и двоичное слово, которым он закодирован. Следовательно, среднее содержание информации I в акте определения одного символа из набора в п символов равно I = log2 п. Это [2-7], которое мы получили выше при определении количества информации.
Настоящее введение в основные концепции теории информации
2. Нейрофизиология сенсорных систем
83
Исходная точна
Рис. 2-18. Кодовое дерево, или дерево выбора. Путем последовательных выборов из двух возможностей буквам от А до Н можно присвоить разные коды. В каждой точке раздвоения повороту влево ставится в соответствие знак 0, а повороту вправо-знак 1. Если начать с исходной точки и последовательно зафиксировать все знаки вдоль какой-то траектории, получится запись в двоичной системе той буквы, которая стоит у конечной точки (от А до Н). Эти двоичные слова приведены под соответствующими буквами.
сильно упрощено, в частности по причине допущения равной вероятности всех символов в наборе. Ссылки на более фундаментальные изложения теории информации приведены в конце книги.
Теория информации в сенсорной физиологии. Теперь мы применим идею, описанную выше в общих терминах, к передаче информации в нервной системе. Подходящим для обсуждения примером является рецептор с афферентным нервным волокном. Здесь информация кодируется и передается дальше в виде нервных импульсов. Источниками информации служат раздражители окружающей среды, передатчиком-рецепторная клетка, каналом передачи-нервное волокно, приемником-синапс некоторого центрального нейрона, а сам этот нейрон может рассматриваться как потребитель.
Физически измеримые параметры стимулов (например, интенсивность давления на кожу, положение стимула на периферической сенсорной поверхности, длины волн светового и звукового стимулов)-это передаваемые величины. На примере рецептора растяжения (разд. 2.1) мы видели, как параметры стимула определяют реакцию рецептора: частота импульсного разряда в афферентном волокне меняется в ту же сторону, что и интенсивность стимула. В этом случае переменная «интенсивность стимула» кодируется переменной «средняя частота нервных импульсов». Такой способ кодирования, сопоставимый с частотной модуляцией в технических системах связи, обнаружен в рецепторах всех модальностей: в мышечных веретенах, в кожных рецепторах давления, в хеморецепторах языка и фоторецепторах сетчатки носителем информации является частота нервных импульсов.
Поскольку афферентные волокна определенных рецепторов связаны с определенными клетками ЦНС, информация, содержащаяся в потоке импульсов, расшифровывается соответствующим образом. Например,
84
М. Циммерманн
Рис. 2-19. Кодирование интенсивности стимула (абсцисса) нервными импульсами (ордината). Длительность каждого стимула фиксирована. В качестве примера взят механорецептор, подвергаемый действию силы (измеряемой в ньютонах). Характеристическая кривая процесса кодирования имеет форму лестницы (красная линия).
афференты 1а оканчиваются на гомонимных мотонейронах, а механо-чувствительные кожные афференты проецируются в постцентральную извилину.
Передача информации идеальным рецептором. Каким образом мы можем различить по сериям импульсов, посылаемых рецептором, те состояния, которые позволят описать содержащуюся информацию? Рассмотрим сначала кодирование интенсивности стимула. Если на определенном отрезке времени стимулируемый рецептор либо не выдает импульсов, либо генерирует один импульс, то он может обеспечить передачу информации только о двух уровнях интенсивности стимула. В этом случае отсутствие потенциалов действия означает, что «интенсивность стимула ниже порога», а один потенциал действия-что «интенсивность стимула выше порога». Если возможное число импульсов составляет 0, 1 и 2, то элемент-потребитель в такой системе получит информацию, достаточную для различения трех уровней стимуляции. Если стимул может возбуждать в афферентном волокне до N импульсов, то рецептор теоретически может сигнализировать в ЦНС об N + 1 уровнях интенсивности. Иллюстрация к такой ситуации дана на рис. 2-19. Число импульсов N в афферентном волокне (ордината) может принимать только целые значения. По этой причине кривая, соотносящая интенсивность стимула (абсцисса) с числом импульсов, имеет форму лестницы. У идеального рецептора, который реагирует на неизменный стимул постоянной частотой разряда, число импульсов N равно произведению частоты разряда/ на время наблюдения t: N =f-t. Значит, максимальное число уровней интенсивности стимула, различимых по
2. Нейрофизиология сенсорных систем
85
активности одного нервного волокна, равно
N+i=fm.t+l, [2-8]
где fm - максимальная частота разряда рецептора. В случае рецептора со спонтанной частотой разряда/0 (в отсутствие какого бы то ни было об-наружимого внешнего стимула) в [2-8] и во всех последующих выражениях fm следует заменить на fm —f0.
Таким образом, в данном случае число имеющихся в наличии символов это число различимых состояний активности афферентного волокна. Следовательно, количество информации об интенсивности стимула в примере рис. 2-19 равно I = log2 (N + 1) = log2 10 = 3,3 бита. В общем случае для идеального рецептора, у которого частота разряда определяется стимулом, содержание в этом разряде информации относительно интенсивности стимула вычисляется по формуле
/ = log2(fm-t+l). [2-9]
Это соотношение между I, fm и временем наблюдения t показано графически на рис. 2-20. Очевидно, что количество информации I возрастает как с увеличением fm, так и с увеличением t. Максимальная частота /„ фиксированное свойство рецептора. Вместе с тем, удлиняя время наблюдения г, теоретически можно получить сколь угодно точную информацию об интенсивности неизменного длительного стимула. Однако на практике свойства реальных рецепторов и динамика процесса декодирования в ЦНС ограничивают время, до которого можно растягивать интеграл оценки афферентного разряда в целях увеличения количества информации. Изучая реальные рецепторы, мы можем получить некоторое представление о пределе, до которого имеет смысл увеличивать г.
Передача информации реальным рецептором. На стимул постоянной интенсивности идеальный рецептор реагирует совершенно регулярным разрядом с неизменной частотой. В случае биологических рецепторов это условие не выполняется.
Экспериментально обнаружено, что при поддерживании постоянной интенсивности стимула частота разряда рецептора может меняться со временем. На рис. 2-21, Л показан разряд рецептора давления кошачьей лапы для разных интенсивностей стимуляции. Бросается в глаза, что частота разряда-носитель информации об интенсивности стимула-флуктуирует стохастически (т. е. случайным образом, без видимой причины). Такие флуктуации сигналов в технике связи называют шумом. Они снижают качество работы информационного канала - вносят искажения в передачу информации (см. рис. 2-17).
Чтобы количественно оценить потери информации из-за шума в рецепторе, рассмотрим экспериментально полученное кодовое соотношение, приведенное на рис. 2-21, Б. Каждая точка графика представляет результат одного измерения, подобного показанному в части А, только
86
М. Циммермана
—
Iog,(fm х t + 1)
_I____
Количество информации
Максимальная частота разряда
Время наблюдения
Г 50 Гц
— 0,06 с —►2 бит/стимул|
50 Гц
200 Гц
0,3с
С,3с
Рис. 2-20. Зависимость содержания информации I (ордината) в реакциях идеального рецептора от времени наблюдения t (абсцисса). Различные кривые соответствуют различным значениям максимальной частоты импульсов fm в разряде рецептора; кривые были вычислены на основании [2-9]. Красным отмечена область, в которую попадают соответствующие значения количества информации в реакциях механорецепторов кожи при изменении времени наблюдения t.
здесь время наблюдения равно 5 с. Число различимых состояний можно оценить, построив на этой полосе точек ступенчатую кривую; та, что изображена здесь, оказалась имеющей восемь ступенек (первая располагается на уровне 0 импульсов), что соответствует 3 битам на стимул, если далее стоит идеальный приемник, считающий в течение 5 с.
2. Нейрофизиология сенсорных систем
87
А Б
Интенсивность стимула
Рис. 2-21. Влияние шума на кодирование в рецепторе. А. Реальные записи разряда рецепторов давления в подошве лапы кошки в ответ на стимулы, действующие в течение 1 с; интенсивности стимулов указаны рядом с записями. Б. Значения частоты импульсов в реакциях на стимулы длительностью 5 с (ордината), полученные на основании многих записей типа А для стимулов разной интенсивности (абсцисса). Ступенчатая кривая {красная линия) построена таким образом, чтобы она вписывалась в полосу разброса данных, имея при этом ступеньки максимальной возможной высоты.
Основание для построения лестницы, изображенной на рис. 2-21, Б, следующее. Две интенсивности стимула четко различаются, если все значения числа импульсов (ордината на рис. 2-21, Б), соответствующие одной интенсивности, отличаются от значений, соответствующих другой интенсивности. «Наихудший случай», который представляет минимально возможное различение, задается ступенчатой кривой с максимальной высотой ступенек, которая может быть вписана в полосу точек кодового графика для экспериментально измеренных реакций.
Если бы данный рецептор мог работать как идеальный модулятор частоты (идеальный рецептор без шума), то в экспериментальных условиях рис. 2-21,5 можно было бы различить по крайней мере 300 градаций интенсивности стимула; соответствующее содержание информации составляло бы 8,2 бита на реакцию [2-9]. Следовательно, в этом примере на каждой реакции 8,2 —3 = 5,2 бита теряется из-за шума!
Наивысшие значения количества информации на стимул, измеренные до сего времени, были обнаружены у деэфферентированных мышечных веретен. Первичные окончания (1а) могут передавать по 4,8 бита на 1 стимул; вторичные окончания (II) реально передают по 6,3 бита.
Отклонения функции реального рецептора от идеального также становятся очевидными, когда варьируют время наблюдения г. При удлинении t количество информации, передаваемой идеальным рецептором, монотонно растет, как это показано на рис. 2-20 кривыми, соответствующими [2-9]. Однако количество информации, передаваемой ре
88
М. Циммерманы-
альным рецептором, не может нарастать беспредельно. На рис. 2-20 красным указана область, в пределах которой изменяются экспериментально полученные значения количества информации на стимул в зависимости от времени наблюдения t у кожных механорецепторов определенного типа. Например, рецепторы давления на кошачьей лапе, по существу, уже достигают своих максимальных возможностей (3 бита на стимул) всего лишь через 1 с.
Хотя частота разряда многих рецепторов может достигать нескольких сотен в секунду, качество их работы с точки зрения передачи информации об интенсивности стимула явно немногим лучше или даже не лучше, чем у идеального рецептора с максимальной частотой разряда 20 с-1.
Избыточность. Неточное кодирование в реальном рецепторе (из-за шума)-это один из видов искажения информации при передаче. Можно ли найти в нервной системе примеры компенсации таких искажений? Методы, разработанные для предотвращения потерь информации в технике связи, используют введение избыточности. Чтобы пояснить это понятие, рассмотрим сначала пример из области лингвистики. Попробуйте расшифровать следующую последовательность букв:
Код.р.в...е в р.цеп...ах ис.аж... и.-з. ш.ма
Информацию, содержащуюся в этом предложении, можно восстановить, хотя около 40% букв отсутствует; письменный текст содержит больше символов, чем это необходимо для точного понимания его смысла; этот излишек и есть избыточность. Она измеряется в битах.
Исследования текстов, из которых систематическим образом изымались отдельные части, показали, что, например, письменный немецкий содержит в среднем лишь по 1,3 бита на букву. Теоретически среднее содержание информации в каждой из 26 букв алфавита равно log2 26 = = 4,7 бита, так что средняя избыточность в этом случае составляет 4,7 — 1,3 = 3,4 бита на букву.
С первого взгляда это может показаться расточительной тратой символов, но это только одна сторона избыточности. Ее достоинства становятся очевидными, когда в канале передачи возможны искажения -например, при плохой телефонной связи, радиоприеме в условиях помех или в случае неразборчивого почерка. Здесь избыточность языка обеспечивает узнавание текста, когда удается идентифицировать всего лишь часть знаков. В теории информации показано в общем виде, что передача информации тем надежнее застрахована от искажений, чем больше избыточность, введенная при кодировании.
Избыточность в нервной системе. Один из эффективных способов использования избыточности для защиты от шума состоит в передаче информации по двум или более каналам параллельно. Такая возможность реализуется в нервной системе. На периферии плотность рецепторов обычно так высока, что даже точечные стимулы возбуждают несколько
2. Нейрофизиология сенсорных систем
89
волокон. При такой параллельной передаче степень фактического использования периферической информации зависит от характера ее обработки в центральной нервной системе. В предположении, что оценка включает суммирование импульсов во всех возбужденных волокнах, получаемую наблюдателем информацию об интенсивности стимула можно вычислить на основании вариабельности суммарного разряда-например, способом, представленным на рис. 2-21, Б. При таком процессе суммации общее число импульсов в интервале наблюдения растет с увеличением числа активированных афферентных волокон приблизительно линейно. Вариабельность разряда (шум), т.е. ширина полосы неопределенности на рис. 2-21, Б, увеличивается в меньшей степени. В связи с этим, по мере того как число афферентных волокон нарастает, число шагов, которые могут быть изображены на экспериментально полученном кодовом графике (типа показанного на рис. 2-21, Б), увеличивается. Отсюда следует, что в суммарном разряде содержится больше информации, чем в разряде отдельного рецептора. Таким образом, обобщая, можно сказать, что избыточность, создаваемая передачей по параллельным каналам, компенсирует искажения, возникающие при кодировании в отдельном рецепторе.
Избыточность, создаваемая параллельной передачей информации, используется и внутри ЦНС. Но тут вступает в действие новый фактор: из-за конвергенции и дивергенции в сетях синаптических связей параллельные каналы могут действовать как весьма сложные цепи, которые (как показано в теории информации) могут в принципе обеспечить дополнительную избыточность.
Но если для сохранения информации об интенсивности стимула столь широко используется передача по параллельным каналам, то неизбежно должно наблюдаться обширное «растекание» возбуждения внутри ЦНС, о котором уже говорилось выше (см. рис. 2-8, Л). Однако, как мы уже видели, такому растеканию может противодействовать латеральное торможение (см. рис. 2-8, Б). В зависимости от силы такого торможения в центральных представительствах периферии может подчеркиваться либо информация об интенсивности стимула, либо информация о его местоположении.
Информация в психологии. В экспериментальной психологии, и в особенности в психофизике (см. гл. 1), также используется количественное определение информации. Здесь мы приведем несколько примеров, подчеркивающих взаимосвязь между нейрофизиологическим и психофизическим измерением информации.
В процессе экспериментов люди могут субъективно оценивать интенсивность какого-нибудь стимула (например, яркость или давление на кожу) и выражать эти оценки каким-нибудь способом (например, численно, см. разд. 1.3). Если такие субъективные оценки изобразить точками против соответствующих значений интенсивности стимула, получится график типа изображенного на рис. 2-21, Б. На нем вместо числа импульсов, генерируемых рецептором (на ординате рис. 2-21, Б), мы те
90
М. Циммерманн
перь имеем численные выражения субъективной оценки интенсивности. Учитывая разброс данных, мы можем оценить соответствующее количество информации, имеющееся в наличии на уровне сознательного восприятия, с помощью уже описанной ступенчатой кривой. В экспериментах, связанных с субъективными оценками интенсивности давления на кожу, были получены значения порядка 3 бит/стимул; это примерно то же самое, что найдено для одиночного рецептора давления. Значит, в этом эксперименте для субъективных оценок интенсивности в восприятии достаточно информации, передаваемой отдельным нервным волокном, и, следовательно, вклад всех других возбужденных волокон -по крайней мере для этого очень частного психофизического вопроса об интенсивности стимула-может рассматриваться как избыточность.
В другом эксперименте испытуемого просили сравнивать два последовательных стимула и определять различия в их интенсивности. Сравнение количеств психофизической и рецепторной информации обнаружило, что, по крайней мере в случае психофизического различения холодовых воздействий на кожу, в решение испытуемого вносит вклад информация, идущая по всем холодовым волокнам стимулируемой области кожи (см. рис. 3-15), в этом случае вся имеющаяся в наличии нейронная информация используется для снижения дифференциального порога.
В заключение мы сравним общее количество сенсорной нейрофизиологической информации, которая может поступать в распоряжение субъекта, с соответствующим количеством психофизической информации. Для этого мы воспользуемся понятием максимального потока информации (бит/с); эту меру называют также емкостью (пропускной способностью) канала.
Емкость канала, например механорецептора (см. рис. 2-21), экспериментально определяется путем прогрессивного учащения последовательности стимулов при одновременном укорочении каждого из них. При снижении длительности стимула до значений менее 1 с содержание информации в реакции на отдельный стимул уменьшается (см. рис. 2-20), но из-за возрастания числа стимулов поток информации обычно возрастает. В табл. 2-2 суммированы данные о емкости каналов всех рецепторов в ряде органов чувств, полученные на основании общего числа афферентных волокон и емкости отдельных каналов. Они сравниваются с измерениями емкости психофизических каналов; это максимальный поток информации на уровне сознательного восприятия. В случае зрительной системы приводимое значение емкости относится к случаю чтения и определено в психофизических экспериментах. Для слуховой системы это емкость, связанная с пониманием речи.
Как мы знаем из нашего собственного опыта, в каждый данный момент времени мы можем полностью направить свое сознательное внимание только на одну сенсорную модальность. По этой причине маловероятно, чтобы человек мог одновременно обрабатывать более одного из тех максимальных психофизических потоков, которые приведены
2. Нейрофизиология сенсорных систем
91
Таблица 2-2. Сравнение потоков информации (емкости каналов) на уровне периферических нервов и сознательного восприятия. Слева указаны общее число рецепторов и афферентных волокон для пяти сенсорных систем и максимальный поток информации в каждой из них. Справа приведены данные по максимальным потокам информации на уровне сознательного восприятия-оценки емкости каналов, полученные в психофизических экспериментах
Сенсорная система Число рецепторов Число афферентов Общая емкость каналов, бит/с Психофизическая оценка емкости каналов, бит/с
Зрительная 2 108 2- 10е 107 40
Слуховая ЗЮ4 2-Ю4 105 30
Тактильная 107 10е 106 5
Вкусовая 3 • 107 103 103 1 (?)
Обонятельная 7 • 107 105 105 1 (?)
в табл. 2-2 справа. Следовательно, мы можем заключить, что максимальный поток информации в процессе сознательного сенсорного восприятия составляет около 40 бит/с т. е. на много порядков ниже, чем передаваемый рецепторами (левая часть табл. 2-2). Таким образом, получается, что наше восприятие, по-видимому, ограничивается ничтожной частью всего обилия информации, имеющейся в наличии на сенсорных входах.
В 2.20. Для случая кодирования рецептором интенсивности стимула количество информации может быть определено
а) как число различимых значений частоты разряда рецептора; б) как число потенциалов действия, генерируемых в единицу времени;
в) как логарифм по основанию 2 числа уровней интенсивности стимула, различаемых по разряду рецептора;
г) только в том случае, если отсутствуют статистические флуктуации частоты разряда (нет шума).
В 2.21. Какие из следующих утверждений относительно избыточности верны?
а) Избыточность в принципе связана с увеличением мощности, затрачиваемой на передачу информации, по сравнению и необходимым минимумом.
б) Избыточность-это увеличение количества нформации, передаваемой по нервному волокну, за счет укорочения рефрактерного периода.
в) Избыточность реализуется в нервной системе путем передачи одной и той же информации по нескольким параллельным каналам.
г) Как правило, подверженность информации искажениям увеличивается с увеличением избыточности.
92
М. Циммерманн
В 2.22. Следующие утверждения применимы к понятию «количество информации».
а) Словам английского языка нельзя приписать никаких значений количества информации, поскольку эти слова имеют смысл, который нельзя измерить (семантическая информация).
б) Количество информации в каждом отдельном броске игральной кости с шестью гранями равно I = log10 (6) бит.
в) Когда произвольный символ кодируется двоичными знаками, количество информации в нем всегда неизбежно становится меньше (потеря информации).
г) Количество информации в акте выбора некоторого символа зависит от вероятности его появления.
3. СОМАТОВИСЦЕРАЛЬНАЯ ЧУВСТВИТЕЛЬНОСТЬ
Р. Шмидт
Кожа не является однородной чувствительной поверхностью. Внутри нее, варьируя по плотности, располагаются рецепторы трех независимых модальностей: давления/прикосновения (механорецепция), тепла и холода (терморецепция) и боли (ноцицепция). В пяти последующих разделах сначала будут обсуждаться психофизика, физиология и гистология этих кожных сенсорных модальностей. Затем мы рассмотрим болевую чувствительность более глубоких слоев тела (мышц, костей, суставов и соединительной ткани), а также внутренностей. Вместе взятые все эти модальности объединяют под названием «соматовисцеральная чувствительность». Общим свойством этих модальностей является то, что их рецепторы не сгруппированы и не образуют органов чувств (таких, как глаз или ухо), а, как правило, широко разбросаны по телу; кроме того, их афферентные волокна не сходятся в специализированные нервы (такие, как зрительный или слуховой), а включаются в многочисленные нервы тела и многие центральные тракты (см. гл. 2).
Вместе с тем работа соматовисцеральной системы вполне сопоставима с работой систем, обслуживающих любую из других модальностей. Термин «низшие чувства», применявшийся ранее к соматовисцеральной чувствительности (возможно, чтобы подчеркнуть отсутствие сложного специального органа чувств), является, следовательно, неподходящим и, более того, вводит в заблуждение.
3.1. Механорецепция
Этот раздел посвящен процессам обнаружения механических стимулов, действующих на кожу, и центральной обработке соответствующих сигналов, т.е. механорецепции (иногда называемой тактильной чувствительностью, или осязанием). Эта модальность объединяет четыре качества: ощущения давления, прикосновения, вибрации и щекотания. В дальнейшем, после обсуждения вопроса о субъективном восприятии механических стимулов, будет показано, что имеющиеся в коже различные механорецепторы обладают свойствами (специфика адекватных стимулов, адаптационное поведение), которые делают их особенно пригодными для восприятия того или иного из этих качеств.
Пороги ощущения и функция интенсивности в случае механической стимуляции кожи. В экспериментах, имеющих целью измерение порога для ощущения механического раздражения кожи, результаты сильно зависят от используемого метода стимуляции. При использовании самой старой процедуры, в которой пороги и распределение механочувстви-
94
Р. Шмидт
Рис. 3-1. Пороги восприятия механического раздражения в различных участках кожи. А. Калибровка «волоска фон Фрея»; измеряется сила (в миллиньютонах, мН), при которой волосок или щетинка (нейлоновая) начинают изгибаться. Б. Пример механического стимулятора с электрически управляемым стержнем. В. Пороги ощущения в разных точках, выраженные через глубину прогиба кожи (в микрометрах, мкм); форма использованных стимулов показана на рис. 3-1, Б. (Lindblom, Lindstrom. In: Zotterman Y. (ed.), Sensory Functions of the Skin in Primates, Oxford, 1976.)
тельности определяются посредством волосков или щетинок, выясняется, что с помощью легкого надавливания (в диапазоне 0,1-0,5 г; метод калибровки представлен на рис. 3-1, Л) можно вызвать ощущение давления или прикосновения (т.е. тактильные ощущения) только в определенных точках кожи. Эти точки называются тактильными точками. Результаты одного такого эксперимента показаны на рис. 3-18. Участки кожи с большим количеством тактильных точек-это кончики пальцев, и в особенности губы; на плечах, бедрах и спине тактильные точки особенно редки.
Эксперименты по определению тактильных точек кожи и порогового
3. Соматовисцеральная чувствительность
95
давления в них информативны, но трудны и утомительны. Кроме того, у них есть тот недостаток, что из-за точечного характера стимуляции нелегко производить прямые сопоставления с обычно встречающимися стимулами, которые занимают ощутимую площадь. В современных стимуляторах, с помощью которых можно в широком диапазоне менять форму, длительность и интенсивность стимулов, используются сменные стержни, кончики которых контактируют с разными площадками на поверхности кожи. Посредством одного такого прибора при использовании формы стимула, показанной на рис. 3-1, Б, были получены значения порогов тактильной чувствительности на внутренней поверхности правой руки, представленные на рис. 3-1, В. Здесь пороги выражены через минимальную глубину прогибания кожи (в микронах), необходимую для появления едва заметного ощущения прикосновения.
Для появления тактильного ощущения на внутренней стороне руки достаточно вдавливания кожи на глубину порядка 0,01 мм (10 мкм). Пороги на кончиках пальцев отчетливо ниже, чем на остальной поверхности кисти. Этот результат согласуется с повседневным опытом. Однако тот факт, что пороги указательного и среднего пальцев не отличаются заметным образом от порогов других пальцев, кажется весьма удивительным. В том же самом исследовании-опять же довольно неожиданно-было обнаружено, что пороги тактильного ощущения у слепых не отличаются заметным образом от порогов у нормальных испытуемых.
Используя надпороговую механическую стимуляцию кожи, можно оценивать зависимость интенсивности ощущения от интенсивности стимула на основе психофизических методов, описанных в разд. 1.3. Пример данных, получаемых в таких исследованиях, приведен на рис. 3-2. В виде графиков, соотносящих величину действующего давления (абсцисса) с субъективно воспринимаемой интенсивностью стимула (ординаты), здесь представлены результаты для трех испытуемых (А, Б и В). Функции интенсивности были получены по три раза для каждого испытуемого; результаты ясно показывают, что эти функции хорошо воспроизводятся от раза к разу для каждого испытуемого, но сильно различаются у разных испытуемых. Этот факт отчетливо выражается в значениях показателей степенных функций, аппроксимирующих полученные данные. Следовательно, при оценке функций интенсивности такого рода (другие примеры см. на рис. 1-6 и 1-7) необходимо принимать во внимание не только условия эксперимента, но и индивидуальные различия в субъективных ощущениях.
Пространственные пороги различения. Используя пару ножек циркуля (с тупыми концами, чтобы избежать болевого раздражения), можно легко определять пространственные пороги различения, т.е. минимальные расстояния между двумя тактильными стимулами, при которых их удается воспринять как раздельные. Если оба кончика касаются кожи одновременно, т.е. исследуются пороги одновременного пространственного различения, на взрослых получаются результаты типа представленных на рис. 3-3. Они являются мерой пространственного разрешения
96
Р. Шмидт
Рис. 3-2. Психофизически определяемые зависимости ощущения давления от интенсивности стимула. С помощью прибора, показанного на рис. 3-1, Б, производили раздражение ладони и измеряли глубину прогиба кожи: длительность каждого отдельного стимула составляла 1 с. Три испытуемых-Л, В и В, давали численные оценки вызываемых этими стимулами ощущений. Каждый эксперимент повторяли 3 раза (соответственно в каждой колонке приведено по три графика). Сплошные линии-степенные функции, аппроксимирующие экспериментальные точки; показатели этих функций приведены на каждом графике справа. (Knibestol, Vallbo. In: Zotterman Y. (ed.), Sensory Functions of the Skin in Primates, Oxford, 1976.)
кожей тактильных стимулов в исследуемой части тела. Тот факт, что одновременные пространственные пороги кончика языка и пальцев, а также губ особенно низки (порядка 1-3 мм), тогда как на спине, плечах и бедрах они существенно выше (порядка 50-100 мм), согласуется и с повседневным опытом, и с описанным выше распределением тактильных точек. Отметим, кроме того, что вдоль линий, параллельных осям конечностей, эти пороги отчетливо выше, чем вдоль поперечных кольцевых линий (проверьте это на себе).
При прочих равных условиях пороги пространственного различения у слепых людей не ниже, чем у нормальных. Этот факт согласуется с отсутствием разницы в чувствительности к легким прикосновениям (см. выше); то же самое обнаружено и в случае вибрационных стимулов.
Если измерять пространственное разрешение в данной области ко-
3. Соматовисцералъная чувствительность
97
Кожа
Кончин языка Кончин указательного пальца Красная кайма губ
Край языка
Ладонь
Лоб
Тыльная поверхность кисти Тыльная поверхность стопы Шея
Спина
Рис. 3-3. Одновременные пространственные пороги у взрослых. На добавочном графике вверху справа в большем масштабе представлены три верхние полоски (увеличено в 10 раз). Длина полосок соответствует самому маленькому расстоянию между двумя стимулами, одновременно предъявляемыми в данном участке кожи, при котором эти два точечных стимула все еще воспринимаются как отдельные (Weber, Landois).
жи, прикладывая обе ножки циркуля последовательно, будут оцениваться не одновременные, а последовательные пространственные пороги. Если провести оба эксперимента на одном человеке, можно увидеть, что последовательные пространственные пороги явно ниже, чем одновременные, и часто составляют лишь четвертую часть от них (например, 1 мм вместо 4 мм). Причины этого различия отчасти кроются в механических свойствах кожи, но главным образом-в особенностях ее иннер
98
Р. Шмидт
вации и центральных связях афферентных нервных волокон (см. разд. 2.3).
Проверка механорецепции. При клиническом обследовании чувствительность к прикосновению обычно оценивают, прикладывая к коже клочок ваты или что-либо похожее и спрашивая пациента, что он чувствует и где ему кажется расположенным место стимуляции. Способность различать острое и тупое проверяют, воздействуя в случайной последовательности то острым кончиком, то головкой булавки со стеклянным шариком. Такне обследования также обычно включают выяснение у пациента того, может ли он узнавать цифры, выводимые на его коже каким-нибудь закругленным предметом, например пальцем или головкой булавки. При этом сначала изображаются крупные цифры, а затем-все меньше и меньше.
Чувствительность к вибрации проверяется в клинике прикладыванием камертона к костным выступам (например, локтю или голени). В экспериментальных условиях или когда желательно более детальное обследование, лучше использовать электрический вибратор или громкоговоритель, приводимый в действие генератором синусоидальных колебаний. Целью, как правило, является нахождение абсолютного порога осознанного ощущения вибрации. Его минимум приходится на частоты около 150-300 Гц. Минимальные амплитуды вибрации, требующиеся на этих частотах, порядка 1 мкм.
Где только возможно, клинические измерения должны включать билатеральное сравнение-определение того же самого параметра на корреспондирующем месте другой стороны тела, причем следует обращать внимание даже на слабые различия. Обычное обследование во врачебном кабинете дает лишь относительно грубое представление о состоянии механорецепторной системы; слабые, но диагностически важные нарушения механочувствительности не всегда обнаруживаются.
Общие сведения о гистологической структуре, афферентной иннервации и адаптации кожных механорецепторов. На рис. 3-4 представлены виды механорецепторов, которые можно обнаружить у человека и других млекопитающих на не покрытых волосами (Л) и на волосистых (Б) участках кожи. Все эти рецепторы иннервируются миелинизированными быстро проводящими афферентными волокнами группы II (диаметр 5-12 мкм, скорость проведения 30-70 м/с). При таких высоких скоростях проведения импульс, возникший на периферии, достигает спинного мозга за несколько миллисекунд.
В экспериментах на животных соотношение стимул-реакция для кожных рецепторов определяется путем регистрации потенциалов действия в одиночных афферентных нервных волокнах (рис. 3-5, Л). Аналогичные отведения можно осуществить и на человеке, погружая через кожу в исследуемый нерв металлические электроды до тех пор, пока не будет уловлена активность одиночных нервных волокон. Путем стимуляции соответствующими приборами (см. рис. 3-1, А, Б) или прикладыванием грузов экспериментатор пытается вызвать раздражение рецепто-
3. Соматовисцеральная чувствительность
99
Роговой слой
Эпидермис
Кориум
Подножная клетчатка
Тельце Диски Тельце
Мейснера Меркеля Пачини
Рис. 3-4. Схема строения механорецепторов и их расположения в коже на не покрытых волосами (А) и волосистых (Б) участках кожи (см. также табл. 3-1).
Рецептор волосяного фолликула
Окончание Руффини
ра, связанного с данным волокном, и затем определить зависимость между стимулом и реакцией.
При исследовании реакции на постоянное давление у различных рецепторов, показанных на рис. 3-4, становится очевидным, что по способности адаптации рецепторы могут быть разделены на три типа: адаптирующиеся очень быстро и реагирующие на каждый стимул только одним или двумя импульсами (тельца Пачини, см. рис. 3-8); адаптирующиеся быстро -разряд в них прекращается спустя 50-500 мс после включения стимула (рецепторы волосяных фолликулов, тельца Мейснера; см. рис. 3-7), и такие, которые адаптируются медленно, продолжая генерировать вызываемые стимулом потенциалы даже тогда, когда давление поддерживается долгое время (тактильные диски, окончания Руффини, диски Меркеля; рис. 3-5,Б,Г и 3-6).
В табл. 3-1 представлена классификация рецепторов, основанная на этом критерии (адаптации к постоянному давлению). Данная классифи-
till и min।ни। •. iiiiiiii
Б В Г
Рис. 3-5. Изучение реакции рецепторов кожи. А. Схема экспериментальной установки. Б-Г. Импульсы (потенциалы действия), генерируемые рецептором давления, расположенным на подушечке кошачьей лапы (красные линии), при действии постоянных ,стимулов разной силы (в ньютонах на 1 см2). Спонтанной активности у данного рецептора не было.
Рис. 3-6. Реакции рецептора давления на постоянные стимулы разной величины. А. Изменения разряда рецептора во времени (по ординате в логарифмической шкале отложено число импульсов в 1 с) при действии трех различных стимулов длительностью 40 с. Б. Зависимость между интенсивностью стимула и разрядом рецептора в разные моменты времени от начала действия стимула. По обеим осям масштаб логарифмический. Цифры на абсциссе соответствуют разностям между устанавливаемыми значениями интенсивности и пороговыми значениями для каждого момента времени. Каждая точка (на А и Б) представляет среднее из 10 отдельных измерений.
3. Соматовисцералъная чувствительность
101
Таблица 3-1. Классификация кожных механорецепторов по скорости адаптации и адекватным стимулам
Адаптация к постоянному давлению
медленная быстрая очень быстрая
Участки кожи, не Диск Меркеля Тельце Мейснера покрытые волосами Волосистые участ- Тактильные диски, Рецептор волосяно-ки кожи окончания Руф- го фолликула финн Тельце Пачини Тельце Пачини
Датчик интенсив- Датчик скорости ности Датчик ускорения
кация возникла в то время, когда различные уровни интенсивности и длительности тактильного стимула устанавливать было легко, тогда как другие параметры стимуляции, например скорость или ускорение прогибания кожи, варьировать было трудно (например, при использовании волосков Фрея). Если рассматривать характеристики реакции по отношению к этим параметрам, оказывается (это будет показано ниже), что свойства рецепторов, адаптирующихся к давлению быстро или очень быстро, можно интерпретировать иначе. А именно: можно сказать, что они не столько кодируют интенсивность тактильного стимула (как это делают медленно адаптирующиеся рецепторы), сколько поставляют информацию о его скорости или ускорении. В табл. 3-1 это обстоятельство отражено в подписях под колонками.
Рецепторы давления (датчики интенсивности). У медленно адаптирующихся рецепторов мгновенная скорость разряда продолжает отражать интенсивность стимула на всем протяжении периода стимуляции давлением, даже если он очень длительный (рис. 3-5, Б, В, Г и 3-6,4). Если график зависимости между интенсивностью стимула и частотой разряда построить в логарифмическом масштабе (рис. 3-6, Б), то станет видно, что экспериментальные точки можно аппроксимировать прямыми линиями. Это означает, что данная зависимость есть степенная функцця вида (частота импульсов) = (интенсивность стимула)" (см. разд. 2.1). С физиологической точки зрения такие рецепторы работают как датчики интенсивности, т. е. они измеряют силу или величину прогибания при действии механического стимула на кожу. Поскольку даже через длительное время эти рецепторы не адаптируются полностью (рис. 3-6), они также передают информацию и о продолжительности давления.
Медленно адаптирующиеся датчики интенсивности давления на участках кожи, не покрытых волосами (например, ладонная поверхность кисти), называют дисками Меркеля (рис. 3-4,4, табл. 3-1). Они располагаются небольшими группами в самых нижних слоях эпидермиса, откуда направляются отростки в сосочки собственно кожи-дермы. Диски
102
Р. Шмидт
Скорость прогибания, мкм/мс
Рис. 3-7. Свойства реакций маханорецептора с быстрой адаптацией (датчик скорости). А. Реальные записи реакций (красные линии) на три механических стимула, различающихся скоростью прогибания кожи стержнем (отметки раздражения-черные линии), но действовавших в течение одного и того же времени. Б и В. Зависимость величины реакций рецептора от скорости прогибания кожи в линейном (Б) и логарнфмичеком (В) масштабе. При пересчете в графике В из значений интенсивности вычитали величину порогового стимула (1,6 мкм/мс; Zimmermann).
Число
Скорость прогибания, мкм/мс
Меркеля можно также обнаружить и на волосистых участках кожи, но там они располагаются в специальных тактильных дисках-небольших возвышениях кожи (рис. 3-4, Б). Кроме тактильных дисков в коже имеются также другие медленно адаптирующиеся рецепторы -окончания Руффини, располагающиеся в собственно коже (рис. 3-4, Б).
Рецепторы прикосновения (датчики скорости). Если на тыльной поверхности кисти отогнуть несколько волосков, не касаясь самой кожи, и оставить их в новом положении, что-то будет ощущаться только во время движения волосков. Как только они «замрут» в новом положении, это ощущение исчезнет. Данный факт указывает на то, что рецепторы волосяных фолликулов регистрируют не столько степень смеще
3. Соматовисцералъная чувствительность
103
ния волосков, сколько само их движение или, возможно, скорость движения.
В коже, не покрытой волосами, также имеются рецепторы, реагирующие сходным образом. Соответствующий пример приведен на рис. 3-7. Рецептор разряжается только во время непрерывного движения (наклонный участок на отметке) стимулирующего стержня; после его остановки разрядов нет, хотя стимуляция (прогиб) сохраняется. Частота импульсов явно зависит от скорости, с которой стержень погружается в кожу, что видно как по реальным записям (рис. 3-7, А), так и по графику (рис. 3-7, Б). Если эти данные представить в логарифмическом масштабе (В), то окажется, что график зависимости частоты разряда от скорости стимула (т.е. от первой производной по времени)-почти прямая линия; такая зависимость описывается степенной функцией. Следовательно, эти рецепторы можно назвать датчиками скорости (табл. 3-1).
Описанные выше датчики скорости-это тельца Мейснера, рецепторы с умеренной адаптацией, обнаруженные на не покрытых волосами участках кожи в сосочках дермы (рис. 3-4, Л). На волосистых участках кожи их роль выполняют рецепторы волосяных фолликулов, располагающиеся на погруженных в кожу частях волосков (рис. 3-4, В).
«Тонические» рецепторы, например рецепторы давления, называемые так из-за своего продолжительного разряда, сигнализируют преимущественно об интенсивности стимула. Соответственно «фазные» рецепторы ведут себя как датчики скорости; кроме того, имеются промежуточные (фазно-тонические) виды рецепторов. Иногда по аналогии с терминами, используемыми для технических датчиков, перечисленные рецепторы называют соответственно пропорциональными (П), дифференциальными (Д) и пропорционально-дифференциальными (ПД) датчиками.
Виброрецепторы (датчики ускорения). На рис. 3-8, А, Б показаны реакции рецепторов третьего типа при действии прямоугольных стимулов. Назависимо от силы стимула (стимул А - околопороговый, а стимул Б в несколько раз превышает пороговую интенсивность) рецептор генерирует только по одному импульсу на каждый стимул. Это значит, что он адаптируется очень быстро (табл. 3-1). Такой рецептор не может передавать информации ни о глубине прогиба кожи, ни о скорости прогибания.
Однако при синусоидальной стимуляции потенциалы действия вызывает каждая волна (рис. 3-8, В, Г). Если амплитуду синусоидальных колебаний установить на таком уровне, чтобы в каждый период возникало в точности по одному импульсу, станет ясно, что необходимая для этого минимальная амплитуда определенным образом зависит от частоты колебаний. При 110 Гц (Г) она меньше, чем при 55 Гц (В). На рис. 3-8, Д показана зависимость минимальной амплитуды колебаний, требующейся для возникновения на каждом периоде по одному импульсу (ордината), от частоты колебаний (абсцисса). Возрастание частоты стимуляции
104
Р. Шмидт
Рис. 3-8. Реакции телец Пачини (датчики ускорения). А-Г. Реакции на механические ступенчатые (А, Б) и синусоидальные (В, Г) стимулы; амплитуда синусоидальных стимулов находится вблизи порога генерации одного импульса на каждую волну; калибровка единая для всех записей. Д. Зависимость пороговой амплитуды от частоты при механических синусоидальных стимулах для трех телец Пачини. Стимулами воздействовали на места максимальной чувствительности каждого рецептора (подушечка кошачьей лапы). Обе шкалы-логарифмические.
с 30 до 200 Гц сопровождается резким снижением порога; на приведенном здесь графике в логарифмическом масштабе тангенс угла наклона кривой в диапазоне 30-200 Гц примерно равен — 2. Следовательно, зависимость пороговой амплитуды стимула So от частоты f можно записать так: So = const-f2, откуда можно заключить, что адекватным стимулом для этих рецепторов является вторая производная по времени от глубины прогибания, т.е. ускорение смещения кожи. В связи с этим мы можем назвать данные рецепторы датчиками ускорения (табл. 3-1). С увеличением частоты выше 200 Гц порог рецепторов этого типа возрастает, и при относительно высоких частотах (400-1000 Гц) реакцию типа 1 :1 получить уже не удается.
Датчики ускорения способны возбуждаться при стимуляции как волосистых, так и не покрытых волосами участков кожи. Как показано в табл. 3-1, на тех и других участках роль датчиков ускорения играют тельца Пачини, располагающиеся в жировой ткани подкожных слоев (рис. 3-4, А, Б). Это относительно крупные терминальные нервные образования, заключенные в слоистые оболочки из соединительной ткани, наподобие лука. Кроме подкожного жира они также обнаружены в разном числе на сухожилиях и фасциях мышц, на надкостнице и в суставных сумках.
Механочувствительные свободные окончания в коже. Кроме миелинизированных афферентов каждый кожный нерв содержит большое число
3. Соматовисцеральная чувствительность
105
(50% и более) немиелинизированных аксонов (группа IV, или С). Некоторые из них-это эфферентные постганглионарные симпатические волокна, обслуживающие такие структуры, как гладкие мышцы сосудов кожи и волосяные фолликулы. Но в данную категорию входят также и афферентные волокна; эти нервные волокна имеют свободные окончания-на них нет корпускулярных структур. Рецепторные функции свободных нервных окончаний в ряде случаев пока неясны. Однако известно, что некоторые из них представляют собой терморецепторы, а многие -болевые рецепторы (см. разд. 3.3 и 3.4). Немногие из них чувствительны к слабым прикосновениям. Такие механорецепторы с немие-линизированными афферентными волокнами встречаются на волосистых участках кожи и изредка-на не покрытых волосами.
Из-за малой скорости проведения у волокон группы IV (порядка 1 м/с) между моментом стимуляции свободных нервных окончаний и прибытием афферентных импульсов в ЦНС проходит значительное время. Так, чтобы дойти по волокну группы IV от большого пальца ноги взрослого человека до спинного мозга (расстояние около 1 м), импульсу понадобится 1 с; тогда как импульсу, идущему по волокну группы II (скорость проведения 50 м/с), чтобы покрыть то же расстояние, потребуется всего 20 мс. Многие рефлексы, вызываемые механическими стимулами, и большая часть наших сознательных ощущений имеют более короткие латентные периоды, чем те, которые способны обеспечить волокна группы IV. Уже по одной этой причине данные волокна не могут играть роли в быстрых рефлексах, по крайней мере на их ранних стадиях.
Исследование особенностей реакций механочувствительных рецепторов, связанных с волокнами группы IV, обнаруживает еще одно важное их отличие от рассмотренных выше корпускулярных механорецепторов: кожные стимулы одинаковой интенсивности вызывают у них разные реакции. Это означает, что рецепторы данного типа не могут поставлять точных сведений об изменениях интенсивности стимула, т.е. в отношении оценки интенсивности стимула возможности одиночных механорецепторов, иннервируемых немиелинизированными нервными волокнами, очень малы. Они позволяют различать не более двух уровней интенсивности. Такая малая точность заставляет предполагать, что эти рецепторы являются пороговыми датчиками, которые сигнализируют только о наличии стимула в данном месте кожи. Кроме того, они могут принимать участие в передаче информации о слабых механических стимулах, движущихся по коже (например, о ползущем насекомом). Обсуждалось также предположение, что активация этих рецепторов, одних или в сочетании с другими, вызывает ощущение щекотки.
Рецептивные поля механорецепторов и плотность иннервации. Рецептивное поле механорецептора определяется как площадь, в пределах которой можно вызвать его возбуждение действием стимула определенной интенсивности; обычно используют интенсивность, в несколько раз превышающую интенсивность порогового стимула. Определяемое та
106
Р. Шмидт
ким образом рецептивное поле часто приблизительно соответствует анатомическим границам рецептора (это, например, справедливо для тактильных дисков). Однако в других случаях возбуждение рецептора могут вызывать и стимулы, действующие на некотором расстоянии (это, например, имеет место в случае телец Пачини). Сходным образом дело обстоит и с другими соматовисцеральными рецепторами.
Оценок плотности иннервации в различных участках кожи проведено немного, и те, как правило, косвенные (например, на основании измерений порогов различения). Кроме того, мы располагаем лишь разрозненными данными, касающимися дивергенции и конвергенции афферентных нервных волокон внутри рецепторных ансамблей. Например, известно, что на одно нервное волокно может приходиться по два-три тактильных диска, а внутри тактильных дисков все диски Меркеля (30-50 штук) обслуживаются одной нервной ветвью. У рецепторов волосяных фолликулов обнаружена более высокая степень дивергенции и конвергенции; тут одно афферентное волокно может обслуживать несколько сотен фолликулов, а каждый фолликул может иннервироваться множеством афферентных волокон. Общая картина усложняется значительными видовыми различиями в иннервации соответствующих участков кожи.
Данные об относительной встречаемости в коже различных механорецепторов также довольно скудны. Проведенные исследования указывают на то, что на волосистых и не покрытых волосами участках кожи обезьян и человека явное большинство составляют датчики скорости. С функциональной точки зрения это разумно, поскольку изменения ситуации для организма, как правило, важнее, чем абсолютные значения интенсивности и длительности стимулов. В этом смысле рецепторы скорости (фазные) иногда также называют детекторами новизны.
Свойства рецепторов и механорецепция. Разумно спросить, не является ли каждый из трех основных типов механорецепторов (подразумеваются датчики интенсивности, скорости и ускорения) ответственным за одно из трех различных ощущений-давления, прикосновения и вибрации соответственно. Тельца Пачини удовлетворяют такому предположению: при частотах стимуляции выше 60 Гц они, по-видимому, несут полную ответственность за ощущение вибрации. Но обычно встречающиеся механические стимулы содержат частоты ниже 60 Гц и поэтому раздражают одновременно несколько типов механорецепторов. На разные стимулы рецепторы этих типов реагируют по-разному, но результирующее ощущение нельзя связать с рецепторами одного типа. Соответственно в повседневной практике нам трудно определить различие между ощущениями давления и прикосновения.
Несколько лет назад, когда был разработан метод регистрации активности одиночных афферентных нервных волокон кожи человека с помощью металлических микроэлектродов, появилась возможность одновременно производить психофизические и нейрофизиологические измерения. Таким методом удалось показать, что очень слабые так
3. Соматовисцеральная чувствительность
107
тильные стимулы (с амплитудой порядка 4 мкм), которые при действии на кончик пальца возбуждают всего один рецептор с умеренной скоростью адаптации, вызывают некоторое ощущение. Однако на ладони пороги ощущения отчетливо выше, чем пороги самих механорецепторов (см. рис. 3-1, В). Это означает, что ЦНС «обращает больше внимания» на импульсы механорецепторов, если они поступают от кожных зон, более важных с точки зрения осязания.
В 3.1. Расположите следующие кожные зоны в порядке нарастания величины одновременного пространственного порога.
а) Край языка.
б) Красная кайма губ
в) Кончик указательного пальца.
г) Ладонь.
д) Спина.
е) Тыльная поверхность кисти.
В 3.2. Который из следующих терминов применим к тельцам Пачини? а) Датчик интенсивности.
б) Датчик ускорения.
в) Пороговый датчик.
г) Датчик скорости.
В 3.3. Которое из следующих структурных образований имеет свойства датчика интенсивности?
а) Тельце Пачини.
б) Тельце Мейснера.
в) Диск Меркеля.
г) Рецептор волосяного фолликула.
В 3.4. Какого типа афферентные нервные волокна обслуживают рецепторы волосяных фолликулов?
а) Группа 1а.
б) Группа 16.
в) Группа II.
г) Группа III.
д) Группа IV.
В 3.5. Какие рецепторы из следующего списка являются датчиками скорости?
а) Тельца Пачини.
б) Тактильные диски.
в) Рецепторы волосяных фолликулов.
г) Тельца Мейснера.
д) Диски Меркеля.
В 3.6. Которое из следующих утверждений применимо к механочув-ствительным элементам с немиелинизированными афферентными волокнами (группы IV)?
а) Эти элементы имеют очень стабильную характеристику стимул-реакция.
108
Р. Шмидт
б) Скорость проведения у этих афферентных волокон выше 2 м/с. в) Число различимых градаций интенсивности больше трех, г) Верны только утверждения бив.
д) Все утверждения (а-г) ложны.
3.2. Проприоцепция
В состоянии бодрствования мы отдаем себе отчет в ориентации наших конечностей по отношению друг к другу. Мы также воспринимаем движения наших суставов и способны довольно точно оценивать сопротивление любому движению, которое мы совершаем. Все эти способности вместе взятые называются проприоцепцией, поскольку источником эффективных стимулов во всех этих случаях служит само тело. Эта модальность имеет три качества-ощущения положения, движения и усилия; они будут рассмотрены ниже. Затем мы обратимся к соответствующим рецепторам-проприоцепторам, которые располагаются преимущественно в мышцах, сухожилиях и суставах. В конце будет показано, что структура осязаемого мира, равно как и нашего субъективного ощущения общего положения тела в пространстве,-это результат интегративной деятельности нервной системы, в которой важнейшую роль играют и другие сенсорные модальности, помимо проприоцепции, а также двигательная система.
Качества проприоцепции. В темноте или при закрытых глазах мы представляем положение наших конечностей и ориентацию их частей относительно друг друга. Это качество проприоцепции называется ощущением положения. Строго говоря, это чувство информирует нас об углах в каждом суставе и тем самым - об относительном положении конечностей. Если мы не двигали конечностями долгое время или только что проснулись после долгого сна, наше ощущение положения конечностей обычно оказывается хорошо сохранившимся. Так что, по всей видимости, ощущение положения подвержено адаптации в слабой степени или совсем не подвержено.
Степень сгибания в суставе невозможно сколько-нибудь точно описать ни в градусах, ни в любой другой словесной форме без специальной тренировки. Однако с помощью следующих двух экспериментов мы можем легко продемонстрировать высокую точность, с которой мы воспринимаем относительное положение различных частей наших конечностей. Во-первых, любое положение одной конечности, в которое она была приведена экспериментатором или же произвольно самим испытуемым, последний может воспроизвести без зрительного контроля с помощью симметричной конечности. Во-вторых, мы можем, опять же без зрительного контроля, очень точно указать любую точку одной из конечностей (по собственному выбору или по желанию экспериментатора) при помощи пальцев кисти другой стороны.
Когда мы без зрительного контроля меняем суставной угол, например сгибаем или разгибаем руку в локте, мы воспринимаем и направле
3. Соматовисцеральная чувствительность
109
ние, и скорость движения. Это качество проприоцепции называется ощущением движения. Здесь порог восприятия зависит от величины и скорости изменения угла. Для проксимальных суставов (например, плечевых) этот порог заметно ниже, чем для дистальных (например, межфаланговых суставов пальцев).
Наша способность оценивать величину мышечного усилия, требующегося, чтобы совершить определенное движение или сохранить определенное положение сустава при действии какого-то сопротивления, также является одним из качеств проприоцепции. Мы называем его ощущением усилия. Поскольку сила, которую должна развивать мышца, зависит от препятствующего движению сопротивления, мы могли бы использовать в качестве синонима выражение «ощущение сопротивления», но этот термин не получил широкого распространения.
При экспериментальной оценке нашей способности определять усилие мы неизбежно наталкиваемся на трудности, связанные с исключением или учетом вклада кожных механорецепторов. Однако легко показать, что в случае ощущения усилия различительная способность явно лучше, чем в случае кожного ощущения давления. Вес какого-нибудь предмета намного легче оценить, если его приподнять, а не просто положить на кожу,-это факт, который знаком каждому из повседневной жизни.
Проприоцепторы. Как отмечалось в начале этого раздела, первыми кандидатами на роль датчиков, обслуживающих данную модальность, являются рецепторы подкожных структур, называемые проприоцепторами. Главные из этих подкожных структур-мышцы, сухожилия и суставные сумки; все они содержат много механочувствительных элементов. В мышцах это в первую очередь мышечные веретена, а в сухожилиях -сухожильные органы Гольджи. Строение и реакции рецепторов этих двух типов подробно рассматривались в руководствах по нейрофизиологии; здесь они обсуждаться не будут. Однако в суставных сумках имеются многочисленные суставные рецепторы, которые мы сейчас опишем.
Поскольку при изменении положения сустава суставная сумка сжимается или растягивается, механорецепторы с подходящими свойствами могут сигнализировать как о положении сустава, так и о направлении и скорости движения, т.е. они могут определять ощущения положения и движения, но не усилия. Например, рецептор, схематически изображенный на рис. 3-9, А, расположен внутри суставной сумки таким образом, что сгибание растягивает его, а разгибание-сжимает. Графики реакций этого рецептора на сгибание сустава от 180 до 112,5° при трех различных- угловых скоростях приведены на рис. 3-9, Б. Во всех трех случаях начальная фаза реакции рецептора состоит из серии учащающихся импульсов, причем частота их тем выше, чем больше скорость движения (сравните на 3-9, Б красные кривые для частоты разряда с черными, представляющими движение). После того как движение завершилось, рецептор начинает разряжаться с постоянной частотой, не
по
Р. Шмидт
А. Механорецептор (схема)
^.Одинаковая скорость, разные углы
Б.Одинаковый угол, разные скорости
Г. Реакция рецептора на сгибание и разгибание сустава
Рис. 3-9. Примеры реакций суставных рецепторов (упрощенные графики, основанные на данных Boyd, Roberts). А-Г. Объяснения в тексте.
3. Соматовисцеральная чувствительность
111
зависящей от скорости предшествующего движения. Однако если данный сустав перевести с одной и той же скоростью в три различные конечные позиции, как это показано на рис. 3-9, В, то во всех трех случаях во время начальной фазы реакции частота разряда будет нарастать с одинаковой быстротой, а конечные частоты будут различными. Иными словами, частота разряда отражает как положение сустава по окончании движения, так и скорость движения (т.е. первую производную от координаты по времени). Другой примечательной особенностью, явствующей из рис. 3-9, Б, В, является то, что при определенном положении сустава разряд рецептора устанавливается на определенной частоте. Это означает, что после фиксации сустава в некотором положении рецептор еще долго посылает правильные сведения о том, какой угол образует сустав.
Фазно-тонические свойства суставных рецепторов проявляются не только тогда, когда сустав движется так, что вызывает возрастание активности рецептора (как на рис. 3-9, Б, В), но и тогда, когда движение производится в обратном направлении. Это иллюстрирует рис. 3-9, Г на примере другого такого же рецептора. В исходном положении этот рецептор разряжался с частотой около 20 имп./с, что показано в левой части графика. Сгибание в суставе вызвало фазно-тоническую реакцию, подобную только что’ описанной. Обратное движение-разгибание за пределы исходного положения (средняя часть графика)-вызвало отрицательную реакцию со столь сильным опусканием ниже начального уровня, что рецептор на время «замолчал». Возвращение к исходному углу (правая часть графика), сопровождавшееся второй положительной реакцией с «перехлестом» за начальный уровень, восстановило исходную частоту разряда.
Рецепторы, представленные на рис. 3-9, реагируют на сгибание увеличением частоты разряда, а на разгибание-уменьшением. Однако можно найти много других рецепторов, поведение которых будет зеркальным отражением этого. Сверх того, в очень подвижных суставах-таких, которые имеют много степеней свободы-можно также обнаружить рецепторы, избирательно чувствительные к вращению, отведению, приведению и т. д. Наконец, следует упомянуть, что в суставных сумках наряду с фазно-тоническими рецепторами имеются и фазные (быстро адаптирующиеся); последние разряжаются только во время движения.
У отдельных суставных рецепторов рабочий диапазон углов обычно меньше, чем это показано на рис. 3-9. Как правило, для перехода от нулевой активности к максимальной достаточно всего нескольких градусов. Достоинством такой особенности является то, что у отдельного рецептора на малые изменения угла приходится большая часть рабочей характеристики; это означает, что в пределах своего характеристического диапазона каждый отдельный рецептор будет регистрировать положение сустава с большой точностью. Такое подразделение всего диапазона движения сустава на перекрывающиеся зоны активности отдельных нейронов из некоторой популяции (называемое «дробление
112
Р. Шмидт
диапазона») дает гарантию того, что в ЦНС будет поступать точная информация о каждом положении сустава. Информация, посылаемая в мозг от многих рецепторов различных суставов, затем интегрируется, чтобы составилось общее представление об относительном положении суставов. Этот процесс интеграции, как и в случае других сенсорных сигналов, начинается в подкорковых сенсорных ядрах. Например, в таламусе обнаружены нейроны, активность которых точно отражает положение сустава в диапазоне углов, превышающем 90°. Для этого там должна иметь место весьма точная и сложная конвергенция на такие нейроны волокон многих рецепторов из одного сустава, каждый из которых обслуживает малую часть общего диапазона движения.
Гистологическая структура суставных рецепторов не вполне ясна. В суставной сумке имеются рецепторы типа окончаний Руффини (см. учебник гистологии), которые являются главными кандидатами на эту роль. Кроме того, там имеются рецепторы типа сухожильных органов Гольджи со сходным расположением внутри связок, и наконец, в меньшем числе там встречаются рецепторы типа телец Пачини. Несколько настоящих телец Пачини, как правило, обнаруживается в соединительной ткани, плотно облегающей сустав, но не в самой сумке. Все эти рецепторы обслуживаются миелинизированными афферентными нервными волокнами из групп II и III. Кроме того, там имеются свободные нервные окончания с немиелинизированными афферентами (волокна группы IV, или С), функция которых неизвестна. Похоже, что они принимают участие в передаче сигналов о болях в суставах.
Ощущения положения и движения могут также обеспечиваться кожными рецепторами в области суставов, так как при движениях суставов кожа сжимается или растягивается. Однако в опытах с применением местной анестезии таких участков кожи было показано, что кожные рецепторы играют в проприоцепции лишь второстепенную роль.
Центральная интеграция. По совокупности свойств суставных рецепторов можно сделать вывод о большой вероятности того, что в первую очередь именно они ответственны за ощущение положения и движения. Об этом свидетельствуют также и другие данные. Если вследствие патологии или местной анестезии передача по нервам суставной сумки прерывается, ощущения положения и движения сильно нарушаются.
Однако в восприятии положения и движения участвуют также и мышечные веретена; если у человека избирательно стимулировать эти рецепторы, воздействуя на сухожилия низкоамплитудной вибрацией (например, частотой 100 Гц и амплитудой в несколько микрометров), суждение испытуемого об истинном положении сустава искажается удивительным образом. Возникающие при этом ложные представления о положении можно выявить, если предложить испытуемому без зрительного контроля придать соответствующему суставу другой половины тела такое же положение, которое установил экспериментатор.
Наиболее вероятными датчиками для ощущения усилия являются рецепторы растяжения мышц-.мышечные веретена и сухожильные ор-
3. Соматовисцеральная чувствительность
113
Суставные рецепторы
Мышечные веретена
Сухожильные органы
Другие рецепторы (?)
Восприятие ' Восприятие положения положения
голоеы тела
Проприоцепция: восприятия положения, движения, усилия
Рис. 3-10. Схема возникновения проприоцептивных ощущений. Чтобы обеспечить восприятие положения, движения и усилия, при обработке сигналов проприоцепторов в нервной системе учитываются моторные эфферентные копии. В восприятие положения тела в пространстве наряду с проприоцептивной информацией вносит свой вклад информация, поступающая от рецепторов вестибулярного органа.
ганы. Следует, однако, помнить, что частота разряда афферентов мышечных веретен зависит не только от мгновенного значения длины мышцы, но также и от активности (а значит, и от напряжения) интрафу-зальных волокон, возбуждаемых моторными у-аксонами. Было также показано, что у животных не удается вырабатывать условные рефлексы, используя избирательную стимуляцию волокон группы I. Исходя из этого следует предположить, что ощущение усилия определяется также и другими рецепторами, кроме уже упомянутых.
Таким образом, ни одна из рассмотренных рецепторных систем не способна независимо от других обеспечить информацию, необходимую для того или иного качества проприоцепции. Следовательно, в случае этой модальности восприятие требует одновременной активации различных рецепторных систем в специфических комбинациях и центральной интеграции таких афферентных сигналов (рис. 3-10).
114
Р. Шмидт
Центральная интеграция включает также и использование сигналов несенсорного происхождения. Один из источников таких сигналов, составляющих важный, но еще мало изученный аспект переработки информации в ЦНС,-центральные отделы моторной системы. Оказывается, что они посылают копии своих «приказов мышцам» (эфферентные копии) в ЦНС для взаимодействия с сенсорными сигналами от проприоцепторов. Эти эфферентные копии поставляют информацию о планируемой активности мышцы и о том движении, которое последует. Значит, их можно использовать для сортировки афферентной информации о движении в смысле выделения той ее части, которая связана с запрограммированной активностью, и той, которая определяется внешними силами и ограничениями.
Перцептивная схема тела и представление о положении тела. Таким образом, сигналы от периферических проприоцепторов и центральные эфферентные копии являются главными переменными, на основании которых строится внутренняя картина, или схема тела в данный момент с учетом движений головы, туловища и конечностей. Проприоцептивные сигналы и параллельно поступающие сигналы от органа равновесия (от рецепторов лабиринта; см. разд. 6.1), которые извещают нас о положении головы в поле земного тяготения, в конечном итоге совместно обрабатываются таким образом, что позволяют нам судить о положении тела в пространстве (рис. 3-10, слева).
Сознательное представление о собственном теле удивительно крепко сидит в человеке и оказывается отделенным от афферентных сигналов проприоцепторов. Приведем в качестве примера тот факт, что после частичной и даже полной ампутации конечности подавляющее большинство пациентов продолжают ощущать отсутствующую конечность еще долгое время, часто до конца жизни. Эта иллюзия во многих случаях бывает столь убедительной, что пациент ощущает свою фантомную конечность более отчетливо, чем оставшуюся. Иногда пациент может по своему желанию двигать фантомной конечностью (влияние эфферентной копии?); в других случаях он ощущает ее постоянно находящейся в неизменном положении. Часто случается, что с фантомной конечностью связываются соматосенсорные ощущения. К несчастью, многие из них неприятны, иногда-настолько болезненны, что синдром фантомных болей, которые не удается устранить никакими терапевтическими средствами, становится невыносимым для больного.
Структура осязаемого мира. Наше представление о пространстве формируется главным образом на основе зрительного восприятия. Но многие свойства нашего окружения лучше познаются, если провести исследование рукой-«потрогать и пощупать». Представьте, например, такие свойства, как жидкое, липкое, твердое, мягкое, гладкое, грубое, рифленое и многие другие, распознаваемые по комбинациям сигналов проприоцепторов, механорецепторов и кожных терморецепторов. Существенным моментом здесь является то, что при простом прикосновении (т. е. если мы просто положим предмет на руку или положим руку
3. Соматовисцеральная чувствительность
115
на предмет и будем держать ее в одном положении) эти свойства плохо распознаются, если вообще распознаются. Однако, когда мы двигаем рукой, мы не испытываем затруднения в определении структуры и формы. Превосходство «ощупывающей» руки над неподвижной частично определяется активацией при движении значительно большего числа кожных рецепторов: адаптироваться рецепторы не успевают, и в ЦНС направляется более детальная информация о раздражителях кожи. Кроме того, при движениях руки в распознавание формы и упругих свойств ощупываемых предметов вносит свою долю проприоцепция.
В
В
3.7. Назовите три качества проприоцепции, о которых вы узнали
в этом разделе.
3.8. Какой (ие) из следующих типов рецепторов в первую очередь ответствен (ны) за ощущение движения?
а) Рецепторы волосяных фолликулов.
б) Тельца Пачини.
в) Суставные рецепторы (типа рецепторов Руффини).
г) Рецепторы мышечных веретен.
д) Сухожильные органы Гольджи.
е) а, г и д совместно.
ж) в, г и д совместно.
3.9. Какие из следующих свойств тела легче определяются на ощупь, а не с помощью зрения?
а) Угловатый. г) Темный. ж) Жидкий.
б) Липкий. д) Мягкий. з) Яркий.
в) Круглый. е) Рифленый. и) Гибкий.
В
в
3.10. Какие из следующих типов рецепторов принимают участие
в формировании представления о силе сопротивления?
(Выберите три типа, которые вы считаете самыми важными.) а) Диски Меркеля.
б) Рецепторы волосяных фолликулов.
в) Суставные рецепторы.
г) Тельца Пачини.
д) Рецепторы мышечных веретен, е) Сухожильные органы Гольджи.
ж) Тактильные диски.
3.3. Терморецепция
В этом разделе мы будем рассматривать температурное чувство (синонимы: терморецепция, восприятие температуры). На основании как объективных, так и субъективных наблюдений с данной сенсорной модальностью можно связать два качества. Это ощущения холода и тепла. Свойства данной системы у человека включают следующее. В коже имеются специальные точки холода и тепла, в которых можно возбудить только рщущения холода и тепла соответственно. Измерения ла
116
Р. Шмидт
тентных периодов реакций показали, что для ощущения холода скорости проведения выше, чем для ощущения тепла. Путем избирательного блокирования нервов можно исключить по отдельности либо только ощущение тепла, либо только ощущение холода. Наконец, в коже имеются специальные рецепторы холода и тепла. Они не только служат датчиками для сознательного ощущения температуры, но и принимают участие в регуляции температуры тела. При выполнении второй функции их работу дополняют и облегчают датчики температуры внутри ЦНС (например, в гипоталамусе и в спинном мозгу). Как правило, активность этих центральных терморецепторов до нашего сознания не доходит; далее они здесь обсуждаться не будут.
При анализе терморецепции оказалось полезным отдельно изучать те физиологические процессы, которые можно зарегистрировать, когда температура кожи постоянна и когда она изменяется. Соответственно в последующем обсуждении психофизических результатов сначала будут рассматриваться статические, а затем динамические ощущения температуры; та же последовательность будет соблюдаться и при обсуждении самих терморецепторов.
Статические ощущения температуры (при постоянной температуре кожи). Когда вы погружаетесь в теплую (~ 33°С) ванну, у вас вначале появляется отчетливое ощущение тепла. Через некоторое время это ощущение исчезает, даже если температура воды поддерживается постоянной. Без сомнения, вам знакомо и противоположное явление: когда вы в жаркий летний день прыгаете в бассейн с температурой воды около 28°С, вам сначала кажется, что вода холодная. Однако спустя некоторое время ощущение холода начинает пропадать. Значит, по крайней мере в диапазоне умеренных температур, нагревание и охлаждение только временно вызывают соответствующие ощущения тепла и холода. Для этого диапазона температур характерна полная адаптация к новой температуре кожи.
Температурный диапазон, в пределах которого происходит полная адаптация и исчезновение температурных ощущений, считается нейтральным диапазоном (иногда он называется «зоной комфорта»). Выше и ниже этого нейтрального диапазона постоянные ощущения тепла и холода сохраняются даже тогда, когда температура кожи длительно поддерживается на одном уровне (вероятно, тут самый простой пример-можно часами ощущать, что ноги замерзли). Для участка кожи в 15 см2 верхняя и нижняя границы нейтральной зоны-это 36 и ЗО°С соответственно. Когда исследуются меньшие участки кожи, эта зона расширяется, а при увеличении площади она становится уже (это указание на то, что в центральных отделах происходит суммация импульсов, приходящих от терморецепторов). В экспериментах, в которых раздетых до гола людей помещали в условия регулируемого климата, эта зона сужалась до 2°: 33-35°С.
Сохраняющееся ощущение тепла, которое не исчезает при постоянных температурах кожи, превышающих 36°С, тем сильнее, чем вы
Соматовисцеральная чувствительность
117
ше температура кожи. При температурах, превышающих 43Ч4 С, ощущение тепла переходит в болезненное ощущение жжения (боль от перегрева). Аналогично этому, при температурах ниже ЗО°С сохраняющееся ощущение холода тем сильнее, чем холоднее кожа. Настоящие боли от холода возникают при температурах кожи около 17°С и ниже, однако уже при температуре 25°С ощущение холода имеет неприятный компонент, особенно когда действию холода подвергаются большие участки кожи. (Человек-создание «тропическое»; он плохо переносит температуры кожи, которые существенно ниже ЗО°С.)
Измерения временного хода адаптации при ступенчатом изменении температуры кожи в пределах нейтральной зоны показали, что переход возникающего температурного ощущения в нейтральное занимает много минут. Как мы хорошо знаем из повседневного опыта, за пределами нейтральной зоны также имеет место некоторая (неполная) адаптация к новой температуре кожи. Погружение руки в теплую воду (42°С) в первый момент вызывает сильное ощущение тепла; затем это ощущение спадает - вначале быстро, потом медленнее-и, наконец, переходит в постоянное слабое ощущение тепла.
Недавние исследования статических температурных ощущений на внутренней поверхности кисти обнаружили, что здесь даже при очень длительной адаптации (30 мин и более) в диапазоне температур 25-40°С возможны воспроизводимые оценки температуры, т. е. здесь нет полной адаптации. На рис. 3-11 представлена зависимость средних оценок температуры, сделанных 18 испытуемыми, от величины той постоянной температуры, которую в действительности поддерживали на поверхности ладони. Испытуемых просили выражать свои оценки двумя способами: давать значение ощущаемой температуры в градусах Цельсия и указывать положение этой температуры между наибольшим (40°С) и наименьшим (25°С) значениями в относительных единицах. Оценки, выражаемые в градусах, были наиболее точными, когда реальная температура была около 37°С; при более высоких и более низких температурах оценки достоверно отклонялись от правильных значений либо в ту, либо в другую сторону. Температуре кожи 25-27°С давалась оценка 10 ! В этих экспериментах температуры около 34°С не вызывали ни ощущения тепла, ни ощущения холода, так что эту область можно рассматривать как нейтральную зону (очень узкую). При 37°С у испытуемых возникало приятное сохраняющееся ощущение тепла.
Динамические температурные ощущения. Можно считать, что ощущения, появляющиеся при изменении температуры кожи, определяются тремя параметрами: исходной температурой кожи, скоростью изменения температуры и площадью кожи, на которую действует стимул.
Влияние исходной температуры на пороги ощущения тепла и холода показано на рис. 3-12. При низких температурах, например при 28°С, порог для появления ощущения тепла - высокий, а для появления ощущения холода-низкий. Если исходную температуру (абсцисса) повысить, тепловые пороги уменьшатся, а холодовые - увеличатся. Приведем
118
Р. Шмидт
Температура поверхности кисти
Рис. 3-11. Психофизические оценки температуры (сплошная красная линия) при стимуляции поверхности ладони после адаптации к каждому значению температуры в течение 30 мин. Штрихами показана линия, на которую ложились бы экспериментальные результаты, если бы оценки температуры совпадали с реальными температурами. Среднее для 18 испытуемых. (Hensel et. al. In: Zotterman Y. (ed.), Sensory Functions of the Skin in Primates, Oxford, 1976.)
конкретные цифры. Чтобы на фоне сохраняющегося ощущения холода возникло ощущение «холоднее», холодную кожу (скажем, при 28°С) достаточно охладить менее чем на 0,2°С; чтобы при такой же исходной температуре кожи появилось ощущение тепла, ее нужно нагреть почти на 1°С. Однако, если исходная температура равна 38°С, уже слабое нагревание ( < 0,2°С) вызовет ощущение «теплее», а для получения ощущения «холоднее» кожу придется охладить примерно на 0,8°С.
Следует также отметить одну особенность данных реакций, которая не полностью отражена на рис. 3-12. Когда изменение температуры таково, что она выходит из нейтральной зоны (31-36°С на рис. 3-12), охлаждение и нагревание приводят к замене нейтрального ощущения на ощущения тепла и холода. В то же время, если температуру кожи повышать от уровня 28°С (для примера), то, прежде чем будет достигнут порог ощущения тепла, указанный на рис. 3-12, испытуемый скажет, что ощущение холода уменьшилось, а затем-перешло в нейтральное. И наоборот, если кожу охлаждают при высокой исходной температуре, то сначала возникают ощущения уменьшения тепла и нейтральности и только потом появляется ощущение холода. Эти ощущения снижения интенсивности существовавших до того ощущений холода или тепла не следует путать с порогами появления ощущений тепла и холода.
Окончательное заключение, которое можно вывести из рис. 3-12, состоит в том, что в зависимости от условий стимуляции одна и та же температура кожи может вызывать и ощущение тепла, и ощущение хо-
3. Соматовисцеральная чувствительность
119
28 30 32 34 36 38 40 С
Температура адаптации
Рис. 3-12. Зависимость порогов ощущения тепла и холода от исходной температуры кожи. Чтобы вызвать ощущение холода или тепла при указанных на абсциссе начальных температурах, к которым кожа адаптировалась определенное время, температуру кожи нужно изменить на величину, указанную на ординате (в градусах Цельсия). Эта схема годится для любых изменений температуры, производимых со скоростью больше 6°С в 1 мин. (Kenshalo. In: Zotterman Y. (ed.), Sensory Functions of the Skin in Primates, Oxford, 1976.)
лода. Например, если исходная температура равна 32°С, нагревание на 0,5е вызовет ощущение тепла, а если исходная температура равна 33°С, охлаждение на 0,5е вызовет очетливое ощущение холода; в то же время конечные температуры кожи в этих двух случаях будут одинаковыми. В правильности вышеизложенного вы легко можете убедиться сами, повторив опыт Вебера с тремя сосудами. Наполните один сосуд холодной водой, второй-тепловатой, а третий-теплой и опустите одну руку в холодную воду, а вторую-в теплую. Если вы теперь перенесете обе руки в сосуд с тепловатой водой, то в одной руке вы получите ощущение тепла, а в другой-холода.
Как показано на рис. 3-13, скорость изменения температуры почти не оказывает влияния на пороги ощущения тепла и холода, если она превышает 0,1°С/с (6°С/мин). При снижении скорости изменения температуры оба порога монотонно нарастают. Это справедливо и для тех скоростей, которые меньше включенных в рис. 3-13. Например, охлаждение кожи со скоростью 0,4°С/мин (0,0067°С/с) от исходной температуры
120
Р. Шмидт
Скорость изменения стимула, °C с
Рис. 3-13. Зависимость порогов ощущения тепла и холода от скорости изменения температуры. Исходная температура во всех случаях 32°С. (Kenshalo. In: Zotterman Y. (ed.), Sensory Functions of the Skin in Primates, Oxford, 1976.)
33,5°C вызовет ощущение холода только после того, как температура упадет на 4,4°С, через И мин после начала охлаждения. Когда процесс охлаждения идет очень медленно, человек может не заметить, что большие участки его кожи стали весьма холодными (и соответственно имели место большие потери тепла), особенно если его внимание отвлечено другими вещами. Возможно, этот фактор способствует простуде.
Что касается площади кожи, на которой меняется температура, то тут пороги ощущений при охлаждении и нагревании в случае малых областей больше, чем в случае больших; кроме того, при данном надпороговом изменении температуры кожи интенсивность ощущения растет с увеличением стимулируемой площади. Это означает, что и в околопороговом, и в надпороговом диапазонах имеет место пространственная суммация импульсных реакций терморецепторов в нервных центрах. Особенно ярко это проявляется в экспериментах с билатеральным предъявлением стимулов. Так, если тепловыми стимулами воздействовать одновременно на кисти обеих рук, пороги будут ниже, чем при действии на каждую кисть в отдельности.
Точки тепла и холода; пространственные пороги. Чувствительные к холоду и теплу элементы человеческой кожи локализованы в разных точках, т. е. имеются точки холода и точки тепла. Их плотность различна для разных участков кожи, а общее их число меньше, чем число тактильных точек в механорецепторной системе. Сравнение плотностей точек холода и тепла на коже показало, что первых заметно больше, чем вторых. Например, на поверхности кисти имеется по 1-5 точек холода на каждом квадратном сантиметре и только по 0,4 точки тепла. Максимальная плотность тех и других точек обнаружена в области кожи, наиболее чувствительной к температуре,-на лице. Точек холода здесь имеется по 16-19 на см2, а чувствительные к теплу элементы этой обла-
Соматовисцеральная чувствительность
121
Рис. 3-14. Активность холодовых (слева) и тепловых (справа) рецепторов кожи обезьяны при постоянной температуре. Потенциалы действия отводились от тонких нервных веточек, идущих к соответствующим рецепторам, как на схеме рис. 3-5. Представлены средние частоты разряда для популяций холодовых и тепловых рецепторов; число рецепторов указано рядом с кривыми. (Kenshalo. In: Zotterman Y. (ed.), Sensory Functions of the Skin in Primates, Oxford, 1976.) сти не удается связать с отдельными точками - они образуют сенсорный континуум.
Низкая по сравнению с плотностью механорецепторов плотность точек холода, и особенно тепла, согласуется с тем фактом, что одновременные пространственные пороги для температурных стимулов относительно велики. Пространственные пороги для холодовых стимулов ниже, чем для тепловых. Кроме того, имеются значительные различия между продольным и поперечным направлениями. Например, на бедре одновременный пространственный порог для тепловых стимулов в продольном направлении составляет 26 см, а в поперечном-9 см; соответствующие значения для холодовых стимулов-16,5 и 2,9 см.
Рецепторы холода и тепла. У приматов (включая человека), также как и у многих других животных, достоверно установлено существование специфических терморецепторов. Они имеют следующие общие характеристики.
- При постоянной температуре кожи они тонически разряжаются с частотой, зависящей от температуры (статическая реакция, рис. 3-14).
- При изменении температуры кожи они увеличивают (или уменьшают) частоту разряда (динамическая реакция, рис. 3-15).
- Они нечувствительны к нетепловым стимулам.
- Пороги их реакций сравнимы с порогами появления ощущений при тепловой стимуляции кожи.
- Они имеют малые рецептивные поля (1 мм2 и менее), и каждое афферентное волокно обслуживает только одну точку тепла или холода или несколько.
- У этих афферентных нервных волокон скорости проведения ниже 20 м/с, а у некоторых видов доходят до 0,4 м/с.
122
Р. Шмидт
Пример результатов, получаемых в случае исследования разряда терморецепторов при постоянной температуре кожи, приведен на рис. 3-14. Средние частоты разряда для двух популяций рецепторов (рецепторов холода и тепла) при разных температурах ложатся на «колоколообразные» кривые, причем у рецепторов холода активность максимальна в области 30°С, а у рецепторов тепла-около 43°С. (Индивидуальные максимумы активности у одиночных рецепторов холода лежат в диапазоне от 17°С до 36°С, а у рецепторов тепла-между 41°С и 47°С.)
Поведение терморецепторов при изменении температуры кожи иллюстрируется данными опыта, в котором регистрировалась активность рецептора холода при охлаждении кожи и возвращении к исходной температуре (рис. 3-15). Охлаждение вызывало динамическую реакцию, зависящую от перепада температур: через несколько секунд устанавливался новый уровень статической активности. Когда же кожу снова нагревали до исходной температуры, сначала наблюдалось временное прекращение активности, а затем восстанавливалась исходная частота разряда (показанная в начале каждой записи и на нижней записи). Поведение тепловых рецепторов при изменении температуры соответствует зеркальному отражению описанного выше. Нагревание приводит к увеличению частоты их разряда, а при охлаждении до исходной температуры частота сначала падает ниже исходного уровня, но потом к нему возвращается.
Гистологическая структура терморецепторов пока не вполне выяснена. Возможно, они представляют собой свободные нервные окончания, причем у человека рецепторы холода, по-видимому, располагаются в эпидермисе и непосредственно под ним, а рецепторы тепла-преимущественно в верхнем и среднем слоях собственно кожи. Рецепторы холода обслуживаются тонкими миелинизированными нервными волокнами (группа III), а рецепторы тепла-немиелинизированными волокнами (группа IV).
Свойства рецепторов и терморецепция. Нет никаких сомнений в том, что за тепловые ощущения ответственна активация терморецепторов тепловыми стимулами. Однако, как это будет ясно из приводимого ниже сравнения реакций рецепторов и психофизических данных, активность рецепторов отражается в сознании в такой форме, которая подразумевает центральную интеграцию потока идущих с периферии сигналов. Мы уже упоминали о пространственной суммации температурных стимулов: это один из аспектов такой переработки.
Как видно из рис. 3-14, когда температура кожи поддерживается постоянной в пределах субъективной нейтральной зоны (31-36°С), импульсы генерируют и рецепторы тепла, и рецепторы холода. Так что сама по себе активность терморецепторов, по крайней мере при низких частотах импульсации, не обязательно вызывает появление субъективных ощущений. Можно думать, что ощущения возникают только тогда, когда частота поступающих в ЦНС импульсов становится доста
3. Соматовисцералъная чувствительность
123
точно большой. Появление сохраняющихся ощущений тепла при температурах выше 36°С можно тогда объяснить тем обстоятельством, что с ростом температуры частота разряда рецепторов тепла растет: в области выше 43°С из-за дополнительного возбуждения тепловых рецепторов ощущение тепла переходит в болезненное ощущение жжения. Однако появление сохраняющихся ощущений холода при температурах ниже 31 °C не удается так просто объяснить возрастанием частот статического разряда холодовых рецепторов. Действительно, нижняя граница нейтральной зоны и положение максимума на графике средней частоты разряда рецепторов холода практически совпадают. Кроме того, например, при 25 и 33°С средние частоты разряда примерно одинаковы, но при одной из этих температур мы испытываем сохраняющееся ощущение холода, а при другой-нейтральное ощущение. Следовательно, в этом случае для суждения о том, какой стороне «колокола» соответствует реальная температура кожи, требуется дополнительная информация. Один из возможных способов ее получения-учесть наличие или отсутствие сопутствующей активности у рецепторов тепла: кроме того, ЦНС может воспользоваться тем фактом, что многие холодовые рецепторы имеют тенденцию в своем среднем диапазоне разряжаться залпами (рис. 3-15). Заканчивая перечисление свидетельств того, что периферические сигналы сильно модифицируются в центрах, прежде чем дойдут до сознания, напомним, что процесс адаптации к новой температуре кожи на уровне субъективных ощущений растягивается на много минут, а у терморецепторов он занимает несколько секунд (рис. 3-15). По-видимому, возбуждение центральных отделов, вызванное афферентным потоком при изменении температуры, угасает довольно медленно.
Особые виды температурных ощущений. Общеизвестным свойством терморецепции является существование следовых ощущений. Например, если вы прижмете ко лбу холодный металлический стержень и подержите его около 30 с, а затем уберете, то отчетливое ощущение холода сохранится и тогда, когда кожа снова нагреется, вопреки нашему ожиданию появления ощущения тепла. Вебер, изучавший это любопытное явление, полагал, что стойкость ощущения холода связана с распространением охлаждения на соседние участки кожи. Однако непосредственные отведения от терморецепторов показали, что рецепторы холода, будучи очень сильно охлажденными, при последующем нагревании не прекращают разряжаться, а вначале даже увеличивают частоту разряда. Таким образом, рассматриваемое следовое ощущение-это «нормальное» ощущение холода. Соответствующие следовые ощущения тепла также были описаны. Очень сильные тепловые стимулы (слишком горячая вода в ванне) часто вызывают парадоксальное ощущение холода. По-видимому, это связано с тем обстоятельством, что холодовые рецепторы, в норме «молчащие» при температурах выше 40°С, в течение какого-то времени разряжаются, если их быстро нагреть до температуры выше 45°С.
124
Р. Шмидт
Охлаждение (скачок температуры)
------- 34' С
Исходная температура
j _______j.......................—циннии81111 и 1111
10
b I I______J............................................................ 1
_____Lbb...____L _-L J. I ..................................1111111111 11111 I
_j_____________j_____l. 11 Til" '1,1I4IIHII HillII. Illi I__________________________________________________________ 8
„I_________\ L ± \__________1____ ill^—llllll II IIIIII 11 i I I 11 I III i I 11 i I
___________I_________\___________lllllllllllllllllllllllllllllltlllllllllllllllllllllllll_Illllll llllll I___________
_A____I____I___I____________I....U1IIIIIMIIIIIII.............................................................. 6
_L J_______I_________Li. .1 IIIIIIIHIIIIIII______________________________________________________________________ Скачок
j_J____________i_____LA_____ i iiiiiiiMiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiia^ ________________I______________ температуры
.J_________L1_J_____________-L 1 iilILililillllIlliIllllalilllibliliilllliillllllllllllllliUlllllll llilllllllllll I_
4
_____LJ______________I___________I llllillllllllllllHIIIIIHIIHIIIIIIIIIIIilllllllllillllllllllll_____________________
I I I I llllll lllllllllli llllll 11111 III 11II11II111111 111 1111 I 111 I
_J_____________L^.J_________I I IIIIIIIIIIIIIIIIIIIIIHIIIIIIIIIIIIIIIIIIIIIIIIIIIIII_________________________________ 2
I____I_____L_________I I I 11111111111 i I 111 I i 11 111 111 I I I I I I I 11 I_____________________________________I
.. _1__________L._L— I___________UJbbLLL-LLI I I I I III__________________________________U_L1_ U I_I__I_____________I
I _ L ..J-------------------L--LL_J--------------LA„A....L^_J------------------LJ-------------------LI. 1 I 0
I__________1_________I------1__________1_________I__________I_________I ।___________________________I_I______________
Секунды
Рис. 3-15. Поведение холодового рецептора при кратковременном снижении температуры. Начальная температура для всех записей 34°С. Перепады температуры указаны справа в градусах Цельсия. Хорошо видны фазно-тонические свойства, а также появление залповой активности при сильном охлаждении. Отведение от кожной ветви срединного нерва (обезьяна) по методике, показанной на рис. 3-5. (Darian-Smith et al., J. Neurophysiol., 36, 325, 1973.)
Для ощущения жжения, которое, как правило, возникает при температурах кожи выше 45°С, нейрофизиологические корреляты пока точно не определены. Однако оказалось, что имеются специальные рецепторы жжения (см. разд. 3.4). Поскольку ощещение жжения болезненно и поскольку сильное нагревание может причинить повреждение, ощущение жжения имеет смысл рассматривать как качество боли, а не как качество терморецепции. Сильный отрицательный эмоциональный компонент характерен также для ощущений типа «бросило в жар» и «бьет озноб». Оба они сопровождаются вегетативными рефлексами: потоотделением и расширением кровеносных сосудов в первом случае, дрожью и сужением сосудов-во втором. Обычно они вызываются либо внешними стимулами, либо психологическими причинами: в редких случаях можно подозревать наличие патологических процессов в ЦНС.
Клинические оценки терморецепции обычно ограничиваются проверкой ощущений тепла и холода с помощью двух пробирок, наполненных горячей и ледяной водой. Нарушение терморецепции в пространственно ограниченной области может служить диагностическим признаком повреждения или блокирования спинномозгового тракта. Обычно при этом затрагивается и ощущение боли, поскольку болевые афференты
3. Соматовисцералъная чувствительность
125
идут в мозг по тем же путям (см. разд. 2.4). Механорецепция и проприоцепция могут при этом сохраняться.
В 3.11. Какое (ие) из следующих утверждений правильно (ны)?
а) Если температуру кожи поддерживать на уровне 10°С, через короткое время температурные ощущения исчезнут.
б) При постоянной температуре кожи 25°С сохраняется постоянное ощущение тепла.
в) При постоянной температуре кожи 33°С сохраняется постоянное ощущение тепла.
г) При постоянной температуре кожи 20°С поддерживается постоянное ощущение холода.
д) Все предыдущие утверждения ложны.
В 3.12. Какие из следующих утверждений ложны?
а) Порог температурного чувства при равномерном изменении температуры не зависит от исходной температуры.
б) При фиксированной исходной температуре появление температурных ощущений не зависит от скорости изменения температуры.
в) В зависимости от исходного уровня и направления изменения температуры в среднем диапазоне температур кожи могут возникать либо ощущения тепла, либо ощущения холода.
г) В нейтральной зоне адаптация на уровне субъективного ощущения длится в точности столько же, сколько и динамическая реакция терморецепторов на изменение температуры.
В 3.13. Следующие утверждения применимы к терморецепции.
а) Не существует ни точек тепла, ни точек холода; все участки кожи имеют одинаковую чувствительность к температуре.
б) Разрешающая способность (одновременные пространственные пороги) для тепловых стимулов ниже, чем для холодовых.
в) Отношение чувствительностей к холоду и теплу одинаково для всех участков человеческой кожи.
г) Область лица с самой высокой чувствительностью к температуре-это кончик носа.
д) Разрешающая способность для термических стимулов (одновременные пространственные пороги) ниже, чем для механических.
В 3.14. Тепловые рецепторы
а) не разряжаются, если температура кожи поддерживается на постоянном уровне ниже 25°С;
б) ведут себя как чисто тонические элементы;
в) обслуживаются преимущественно волокнами группы II;
г) обеспечивают ощущение жжения при большой интенсивности стимула;
д) гистологически идентичны дискам Меркеля.
126
Р. Шмидт
В 3.15. Холодовые рецепторы
а) обслуживаются преимущественно волокнами групп I и II; б) гистологически идентичны тельцам Мейснера;
в) «молчат» при температурах выше 45°С;
г) разряжаются с частотой, в точности пропорциональной мгновенной температуре кожи;
д) имеют общие с механорецепторами проводящие пути в спинном мозгу.
3.4. Соматические и висцеральные боли
Боль радикально отличается от других сенсорных модальностей с точки зрения той информации, которую она поставляет. Она информирует нас о грозящей опасности, так как вызывается вредными (повреждающими ткани) стимулами. Такая специфическая защитная функция не обеспечивается другими модальностями, так что значение чувства боли для нормальной жизни трудно переоценить. Боль также чрезвычайно важна в медицине, так как именно болевое проявление действия вредных факторов приводит пациента к врачу. И боль является столь широко распространенным и общим ощущением-хотя и субъективным,- что простое ее словесное описание часто оказывается решающим для диагноза заболевания.
В этом разделе мы сначала опишем различные качества боли, возникающей как в коже, так и в более глубоких соматических и висцеральных структурах. Затем рассмотрим характеристики ощущения боли и под конец-ее периферическую физиологическую основу (о центральных нейрофизиологических процессах можно прочитать в гл. 2 и разд. 3.5).
Качества боли. Ощущения боли можно расклассифицировать по качествам, исходя как из места их происхождения, так и из их свойств. На рис. 3-16 эти качества даны в розовых прямоугольниках. Модальность «боль» объединяет два типа соматических болей и висцеральные боли.
Если соматическая боль имеет кожное происхождение, ее называют поверхностной болью; если же ее источник находится в мышцах, костях, суставах и соединительной ткани, она называется глубокой болью. Поверхностные и глубокие боли-это два подкласса соматической боли.
Если поверхностную боль вызвать уколом иглы, испытуемый сначала почувствует острую «вспышку» боли-легко локализуемое ощущение, которое быстро исчезнет с прекращением стимуляции. За этой резкой и легко локализуемой начальной болью (первой болью) часто следует, особенно при высоких интенсивностях стимуляции, отставленная боль (вторая боль), имеющая тупой (или ноющий) характер, с латентным периодом 0,5-1 с. Последняя ощущается пространственно как более диффузная и угасает медленно; хорошим примером такой боли является боль, ощущаемая после того, как прищемишь палец дверью.
Боли, возникающие в мышцах, костях, суставах и соединительной
3. Соматовисцеральная чувствительность
127
Рис. 3-16. Качества'боли (розовый фон). Указана также локализация каждого качества (серый фон); даны примеры особых видов боли.
ткани, называются глубокими болями. Как и поверхностная боль, глубокая боль-это разновидность соматической боли. Всем нам известен такой пример болей, как головные боли-возможно, это самые общераспространенные виды болей у людей. Глубокая боль по своему характеру тупая; как правило, она плохо локализуется и имеет тенденцию иррадиировать в окружающие структуры.
Муки и страдания, связанные с болью, а также автономные и моторные реакции на боль зависят от ее качества. Отставленные, и особенно глубокие, боли сопровождаются неприятными ощущениями, часто они вызывают автономные рефлексы, такие, как тошнота, сильное потоотделение и падение артериального давления. Напротив, начальная боль вызывает защитные рефлексы-примером может служить отдергивание ноги, когда ойа наталкивается на острый предмет.
Кроме соматических болей разного качества важный и отдельный вид болей-это висцеральные боли. Висцеральная боль также имеет тенденцию быть по своему характеру тупой и диффузной; она похожа на глубокую боль в том отношении, что сопровождается сходными автономными реакциями (вспомните, например, боли при желчно-каменной болезни, аппендиците и т. д.). Примечательно, что, обнажив внутренности (например, при вскрытии брюшной полости под местной анестезией), их можно сжимать и резать, не вызывая ощущения боли, если
128
Р. Шмидт
только не задевать париетальную брюшину и корень брыжейки. Однако резкое или сильное растяжение полых органов вызывает острую боль. Кроме того, болями сопровождаются спазмы или сильные сокращения гладких мышц, особенно если этому сопутствует нарушение кровообращения (ишемия). Другие особенности висцеральных болей будут обсуждаться в разд. 3.5.
Измерение интенсивности боли. Эксперименты по изучению боли у человека и животных наталкиваются на многочисленные особые трудности. Начнем с того, что не так-то легко найти адекватный стимул, вызывающий боль. Кроме того, из-за повреждающего действия болевых стимулов не всегда удается воспроизводить в эксперименте необходимую постоянную ситуацию. Трудно также бывает сравнивать субъективные ощущения, возникающие у человека, с поддающимися измерению физиологическими коррелятами, наблюдаемыми в опытах на животных; как у человека, так и у животных вызывать боль в эксперименте допустимо лишь в сравнительно узком диапазоне. Наконец, следует помнить о том, что эмоциональный и мотивационный компоненты боли часто оказываются более важными для больного и врача, чем физиологические аспекты, с которыми мы здесь имеем дело. Например, субъективно интенсивность боли зависит не только от интенсивности стимула, но и от того, в какой степени на нем фиксировано внимание испытуемого: если отвлечь внимание, то ощущение боли можно ослабить, а в экстремальных ситуациях (стресс, связанный с несчастным случаем, ранение в бою, гипноз) человек может и вовсе не почувствовать боли.
Для измерения интенсивности поверхностной боли пробовали использовать разные методы. Один из примеров был приведен на рис. 1-7: в психофизических экспериментах с помощью интермодального сравнения там количественно оценивалась интенсивность боли, вызываемой раздражением кожи электрическим током. Измерялись также абсолютные и дифференциальные пороги тупой поверхностной боли, вызываемой надавливанием (т.е. механическими стимулами). Когда такими стимулами воздействуют на лоб, порог составляет около 600 г/см2, а до ощущения максимальной боли насчитывается около 15 дифференциально-пороговых шагов. Для измерения болевых порогов также широко использовали температурные стимулы-в особенности тепловое излучение, которым можно действовать без сопутствующей механической стимуляции. Первые болевые ощущения под действием тепла возникают при температурах между 43 и 47°С, обычно около 45°С. Если температуру кожи повышать дальше, до выхода на уровень максимального болевого ощущения можно различить порядка 20 уровней интенсивности. Химические раздражители, как правило, бывают неэффективными, если просто подействовать на поверхность кожи. Чтобы изучить их действие, сначала с помощью специального раздражителя вызывают образование волдыря. Затем кожу с волДыря срезают, а его основание (базальный слой) орошают тестовыми растворами. Эта про-
3. Соматовисцеральная чувствительность
129
цедура вызвала большой интерес в особенности в связи с тем обстоятельством, что казалось возможным найти таким путем «болевую субстанцию» - общее для всех видов боли вещество, выделяемое из тканей под действием повреждающего стимула. К настоящему времени с помощью таких экспериментов найден целый ряд веществ, в норме имеющихся в организме и в подходящей концентрации вызывающих боль. Совокупность всех полученных данных свидетельствует против гипотезы о существовании единой болевой субстанции.
Адаптация к боли. Кроме характерных особенностей и интенсивности боли с клинической точки зрения важно знать, существует ли адаптация к боли. Субъективный опыт как будто указывает на отсутствие такой адаптации: вспомните, например, длительные (часами не проходящие) ощущения головной и зубной боли. Один из экспериментов, имевших целью оценить адаптацию к боли, представлен на рис. 3-17; на этой установке оценивали боль, вызываемую перегревом кожи. Зачерненную поверхность лба испытуемого облучали с помощью регулируемого источника (лампы инфракрасного излучения). Для измерения температуры кожи использовали инфракрасный датчик, выход которого вместе с сигналом об интенсивности излучения регистрировали с помощью самописца. Такая процедура позволяет воздействовать болевыми стимулами (тепловыми) без сопутствующих механических. В этой ситуации вопрос об адаптации к боли исследовали следующим образом. Испытуемому разрешали самостоятельно регулировать интенсивность облучения, но не давали никакой информации о положении регулятора интенсивности. Нужно было устанавливать такую интенсивность облучения, чтобы она все время находилась на уровне болевого порога. Мерой болевого порога служила температура кожи, так что в данных условиях прогрессивное увеличение температуры кожи подразумевало бы увеличение порога, т.е. наличие адаптации.
Средние результаты таких измерений представлены на рис. 3-17. После того как в течение первой минуты опыта был установлен некоторый порог, температура кожи не претерпевала существенных изменений; следовательно, болевой порог изменялся незначительно. В действительности в ходе эксперимента регистрировалось даже небольшое снижение температуры кожи, указывающее на то, что для появления болевого ощущения испытуемому требовалось все меньшее облучение. Этот факт говорит о том, что имела место не адаптация, а скорее прогрессивная сенситизация стимулируемого участка кожи. Таким образом, мы можем констатировать, что ни повседневный опыт, ни результаты экспериментов с тепловой болью не дают никаких указаний на существование адаптации к боли.
Болевые рецепторы. На протяжении последнего столетия относительно механизма периферического кодирования болевых стимулов были выдвинуты три основные гипотезы. Это теории интенсивности, паттерна и специфичности; только последняя из них получила достаточное экспериментальное подтверждение. Теории паттерна и интенсивности
130
Р. Шмидт
Рис. 3-17. С помощью этой установки можно оценивать температурные болевые пороги. Инфракрасное излучение нагревает зачерненный участок кожи на лбу испытуемого. Температура кожи измеряется инфракрасным датчиком и записывается самописцем. (Hardy, J. Appl. Physiol., 5, 725, 1953.) Красная кривая показывает зависимость болевого порога (средние значения) от продолжительности действия теплового стимула. Испытуемого просили самостоятельно устанавливать в ходе опыта интенсивность излучения на таком уровне, чтобы на лбу едва ощущалась боль. Начальный подъем температуры выше значения порога объясняется инерцией аппаратуры. (Greene, Hardy, J. Appl. Physiol., 17, 693, 1962.)
возникли в связи с тем, что разнообразие болевых стимулов-т.е. отсутствие единого адекватного стимула-наталкивало на предположение об отсутствии специальных болевых рецепторов. Казалось более вероятным, что ощущение боли появляется всегда, когда интенсивность стимуляции низкопороговых механорецепторов и терморецепторов превышает некоторый уровень. Согласно теории интенсивности, ноцицептивные стимулы вызывают в низкопороговых рецепторах серии импульсов особенно высокой частоты; согласно теории паттерна, такие стимулы порождают специфические временные последовательности импульсов, отличные от генерируемых в ответ на безвредные стимулы. В обоих случаях ЦНС должна при декодировании принимать за признак боли изменение афферентного потока. В противоположность этому в теории специфичности (по аналогии с другими модальностями) постулируется существование специальных болевых рецепторов (ноцицепто-
3. Соматовисцеральная чувствительность
131
Волосок • Тактильная • Болевая
точка точка
Рис. 3-18. Болевые и тактильные точки кожи человека на медиальной поверхности предплечья.
Положение болевых точек определяли с помощью волосков фон Фрея. (Strugh-old, Z. Biol, 80, 376, 1924.)
ров), которые реагируют только на стимулы высокой интенсивности и непосредственно вызывают болевые ощущения.
Первым указанием на существование специальных ноцицепторов было обнаружение того, что кожа содержит явно больше болевых точек, чем тактильных (рис. 3-18; здесь отношение этих точек равно 9:1). Поскольку точки холода и тепла встречаются в коже еще в меньшем числе, чем тактильные, соотношение числа болевых и тепловых точек превышает 10:1. Уже одно это делает чрезвычайно маловероятным предположение о том, что ноцицепция определяется механо- или терморецепторами.
Позднее и у экспериментальных животных, и у человека электрофизиологическими методами удалось обнаружить рецепторы, которые (как того и требует теория специфичности) не реагируют на слабые стимулы, а реагируют только на стимулы, интенсивность которых достаточна для повреждения тканей. Таким образом, их можно рассматривать как специальные болевые рецепторы, или ноцицепторы. К настоящему времени в коже найдены чисто механочувствительные, чисто термочувствительные и механо-термочувствительные ноцицепторы. Последние, называемые полимодальными ноцицепторами, по-видимому, встречаются в коже человека чаще, чем два других типа.
Примеры реакций полимодальных ноцицепторов на тепловые стимулы показаны на рис. 3-19. Эти рецепторы не реагируют на холодовые и тепловые раздражители- при температурах ниже 41 °C. Но, если кожу нагреть примерно до 45°С или выше, эти рецепторы начинают разряжаться с частотой, которая явно нарастает с повышением температуры. Поскольку болезненные ощущения жжения имеют место как раз при температурах кожи выше 45°С, эти рецепторы можно назвать также рецепторами жжения.
132
Р. Шмидт
Рецептор б
35° - 45° I I I I I I I |-1----------------
30° -45° | | | | |--1------1---------------
35° -47° |- I 11 | | | | | | | | |---------
30°-47° | IIIH1IIII Illi I I...............
35о_49о Ш1ИШЯ------------------------------
30°-49° IIIIIIIIIIIIIIIIII I I I-----------
35° -51° HIIIII—III------------------------
30°-51 ° -I--1 | IIIHIIIIII I II-I I I I I-
-I---1___I___1__I___I__1_____I
с
Время Б
Рис. 3-19. Реакции полимодальных ноцицепторов кожи (морда обезьяны) на тепловые стимулы. А. Зависимость разряда трех рецепторов от конечной температуры, устанавливаемой тепловыми стимулами в их рецептивных полях. На ординате указано число импульсов, генерируемых в течение трех минут после появления первого импульса в ответ на данный стимул. Начальные температуры для каждого из шести экспериментов указаны справа от кривых. Б. Примеры разряда рецептора «б» в ответ на указанные слева тепловые стимулы (исходные температуры 30 и 35°С). Каждая запись начинается с первого импульса, вызванного стимулом. (Beitel, Dubner, J. Neurophysiol., 39, 1160, 1976.)
Гистологически ноцицепторы кожи (равно как и внутренностей и скелетных мышц, о которых речь будет идти ниже) представляют собой свободные нервные окончания с тонкими миелинизированными (группа III, скорость проведения ~ 11 м/с) или демиелинизированными (группа IV, скорость проведения около 1 м/с) нервными волокнами. С помощью постепенно нарастающей электрической стимуляции кожных нервов человека было прямо показано, что возбуждение (низкопороговых) толстых миелинизированных афферентов (группа II) не приводит к появлению болевых ощущений, тогда как возбуждение (высокопороговых) афферентов, принадлежащих к группам III и IV, вызывает боль. Оказалось, что в случае поверхностной боли передача сигналов начальной боли осуществляется по волокнам группы III, а сигналы отставленной боли идут по волокнам группы IV (или С-волокнам). Эта точка зрения подтверждается следующими наблюдениями, а) Если какой-нибудь нерв блокировать посредством механического сдавливания, активность исчезнет сначала в толстых волокнах и лишь потом-в тонких; если блокировать только волокна группы II, оба компонента поверхностной боли сохранятся; но, если блокаду распространить и на во
3. Соматовисцеральная чувствительность
133
локна группы III, начальная боль исчезнет и останется только отставленный компонент, б) Если этот же нерв блокировать при помощи какого-нибудь местноанестезирующего средства (например, новокаина), к которому волокна группы IV более чувствительны, чем волокна группы III, наблюдается обратное явление-отставленная боль исчезает раньше, чем начальная, в) Электрическая стимуляция обнаженных кожных нервов с интенсивностью, достаточной для возбуждения волокон группы III, вызывает отчетливые острые болевые ощущения; но, если миелинизированные волокна блокированы, а интенсивность стимула соответствует возбуждению волокон группы IV, результатом будет тупая ноющая боль, субъективно очень неприятная; действительно, испытуемые описывают ее как трудно переносимую. Различие во времени появления начальной и отставленной боли, по-видимому, объясняется главным образом различием в скоростях проведения по соответствующим волокнам. Болевые ощущения могут возникать во всех внутренних органах В полых внутренних органах, образованных гладкими мышцами, обнаружены механочувствительные висцеральные ноцицепторы. Некоторые из этих рецепторов реагируют на пассивное растяжение, а некоторые-на активное напряжение гладких мышц. Когда такое напряжение нарастает изометрически (т. е. без изменения длины), как, например, бывает при наличии каких-то препятствий выходу содержимого из данного органа, висцеральные ноцицепторы возбуждаются особенно сильно. Самые острые из известных болей ощущаются именно в этих условиях-например, при наличии препятствия в общем желчном протоке или в мочеточнике (эти боли называются соответственно печеночной и почечной коликой). Местная недостаточность кровоснабжения (ишемия) также может вызывать сильные висцеральные боли; пока не известно, чем они вызываются - сопутствующими механическими явлениями или химическими изменениями в тканях. Легкие также содержат множество ноцицепторов, активируемых раздражающими газами или частицами пыли; их афференты, принадлежащие к группам III и IV, направляются в центр в составе блуждающего нерва.
Боли в скелетных мышцах (которые чаще всего проявляются в виде острой судороги) также определяются ноцицепторами со свободными нервными окончаниями и афферентами из групп III и IV. Об их физиологии мало что известно. По всей видимости, эти боли связаны с сигналами чисто механических и чисто химических ноцицепторов, равно как и многих полимодальных окончаний. Ноцицепторы сердечной мышцы особенно сильно реагируют на местное нарушение кровоснабжения; возникающее при этом патологическое состояние получило название стенокардии, или грудной жабы. Здесь также пока неизвестно, которое из следствий ишемии активирует ноцицепторы сердца - скажем, аномальные сокращения, кислородная недостаточность, повышенная концентрация метаболитов или что-нибудь иное.
В заключение следует подчеркнуть, что отнюдь не все свободные нервные окончания являются ноцицептивными. В разделах, посвя
134
Р. Шмидт
щенных механорецепции и терморецепции, указывалось, что многие из рецепторных элементов с афферентными волокнами из группы III (миелинизированными) и из группы IV (немиелинизированными) обладают специфической чувствительностью к механическим и термическим стимулам низкой интенсивности; возможно, все они также имеют свободные нервные окончания. Таким образом, разным свободным нервным окончаниям могут соответствовать различные адекватные стимулы. Отсутствие гистологической дифференциации не обязательно подразумевает отсутствие функциональной специфичности. Такая специфичность может быть связана с определенными молекулярными свойствами рецепторной мембраны, которые недоступны исследованию с помощью световой и электронной микроскопии.
В 3.16. Какое (ие) из следующих утверждений правильно (ны)? На коже, как правило, а) болевых точек больше, чем тепловых;
б) тепловых точек больше, чем тактильных;
в) холодовых точек меньше, чем тепловых;
г) тактильных точек больше, чем болевых;
д) отсутствуют специальные сенсорные точки; чувствительность всех участков кожи одинакова.
В 3.17. Гистологически ноцицепторы, как правило, представляют собой а) тельца Пачини;
б) диски Меркеля;
в) тельца Мейснера;
г) свободные нервные окончания;
д) рецепторы волосяных фолликулов.
В 3.18. Которая из следующих гипотез относительно периферического механизма ноцицепции лучше всего объясняет известные факты?
а) Теория интенсивности.
б) Теория паттерна.
в) Теория специфичности.
В 3.19. Афферентные нервные волокна, обслуживающие ноцицепцию, принадлежат к группам а) 1а.
б) 16.
в) II.
г) III.
д) IV.
В 3.20. Какие три из следующих описаний лучше всего подходят для глубокой боли?
а) Легко локализуемая.
б) Неадаптирующаяся.
в) Сопровождающаяся вегетативными рефлексами.
г) Острая по своему характеру.
3. Соматовисцеральная чувствительность
135
д) Часто иррадиирует в окружающие структуры.
е) Исходит преимущественно из полых органов брюшной полости.
3.5. Особые и аномальные виды болей; терапия при болях
Физиологию ноцицепции, представленную в предыдущем разделе, мы теперь дополним рассмотрением некоторых специальных вопросов, имеющих клиническое и физиологическое значение, например механизмов особых видов боли - проецируемой и отраженной. Мы также рассмотрим периферические нарушения восприятия боли и приведем несколько примеров нарушения центральных механизмов обработки болевых сигналов. В заключение мы кратко остановимся на физиологических основах терапии при болях.
Проецируемая боль. Резкий удар по локтевому нерву, проходящему в области локтя у самой поверхности, вызывает очень неприятное ощущение, которое трудно описать (смесь покалывания, щекотания и чего-то еще); оно распространяется на те части руки, которые обслуживаются этим нервом, т.е. на участок от локтя до кисти и на саму кисть. Очевидно, что ЦНС (т.е. наше сознание) проецирует активность, вызванную в афферентных нервных волокнах в области локтя, в те части руки, которые обслуживаются этими афферентными волокнами. Мы приучены к тому, что в норме такие сенсорные импульсы поступают от рецепторов этих областей. Нам трудно интерпретировать возникающие при этом ощущения (покалывание и т. д.), так как паттерны импульсной активности, вызываемой прямой механической стимуляцией нерва, отличны от возникающих в обычных условиях. Сходные нетипичные ощущения возникают и тогда, когда по методическим или терапевтическим причинам стимулируют кожные нервы с помощью электродов. В этих случаях интенсивность и характер ощущения зависят от условий стимуляции (см. разд. 3.4).
В принципе проецируемые ощущения могут возникать в пределах любой сенсорной модальности. В примере с локтевым нервом эффекты кратковременны, но многие длительные проецируемые боли имеют клиническое значение. Механизм их возникновения схематически представлен на рис. 3-20. Сигналы, вызываемые аномальной стимуляцией афферентного волокна, обычно передаются в мозг по латеральному спинно-таламическому тракту (см. разд. 2.4); там они порождают ощущения, относимые к области, обслуживаемой данным афферентным волокном. Одной из распространенных причин проецируемых болей является пережатие спинальных нервов в местах их вхождения в спинной мозг, происходящее вследствие повреждения межпозвонковых хрящевых дисков. Афферентные импульсы, вызываемые в ноцицептивных волокнах при такой патологии, порождают болевые ощущения, которые проецируются в область, обслуживаемую стимулируемым спинальным нервом. (Тут, конечно, возможны боли и в области самого диска.) От
136
Р. Шмидт
отраженной боли (см. ниже) проецируемая боль отличается тем, что она вызывается прямой стимуляцией нервных волокон, а не раздражением каких-нибудь рецептивных окончаний.
Отраженная боль. Болевые сигналы от внутренних органов часто приводят к болевым ощущениям не в них или не только в них, но также и в отстоящих на некотором расстоянии поверхностных областях. Такие боли называются отраженными. Отражение всегда происходит в ту часть периферии, которая обслуживается тем же самым сегментом спинного мозга, что и пораженный внутренний орган; это означает, что, например, в случае кожной поверхности боль отражается в соответствующий дерматом (см. разд. 2.2). Широко известный пример отраженной боли-боль, возникающая в сердце, но ощущаемая в груди и узкой полоске на медиальной поверхности руки. Поскольку взаимоотношения между дерматомами и внутренними органами известны, такие отраженные боли играют большую роль в диагностике.
Механизм возникновения отраженной боли, по-видимому, примерно соответствует схеме, представленной на рис. 3-21. Некоторые кожные болевые афференты и болевые афференты, идущие от внутренних органов, при вхождении в один и тот же сегмент спинного мозга могут конвергировать на один и тот же нейрон. Возбуждение таких клеток интерпретируется как боль на периферии, поскольку такая интерпретация обычно оправдывает себя в повседневной жизни.
При заболевании внутренних органов мы сталкиваемся еще с одним следствием конвергенции ноцицептивных афферентов от дерматомов и внутренних органов на одни и те же нейроны болевых путей; это-гп-перчувствительность кожи (гиперпатия) в соответствующих дермато-
3. Соматовисцеральная чувствительность
137
Рис. 3-21. Нервные пути, лежащие в основе отраженной боли. Болевые афференты внутренних органов приходят в спинной мозг, и там некоторые из них синаптически контактируют с теми же нейронами спинно-таламического тракта, на которых оканчиваются кожные болевые афференты.
мах. Причиной ее является то, что висцеральные импульсы повышают возбудимость вставочных нейронов, в результате чего действующие на кожу ноцицептивные стимулы вызывают в ЦНС более сильную реакцию, чем в нормальных условиях. Такого рода повышение возбудимости называют «облегчением» (фасилитацией).
Другие виды взаимодействия между соматической нервной системой и вегетативной. Оказывается, что кроме отраженной боли есть еще и другие проявления взаимодействий между соматической и вегетативной системами, которые пока еще не совсем понятны. Примером может служить терапевтический эффект, который наблюдается от грелки при некоторых заболеваниях внутренних органов. Тепло не может действовать на орган как таковой (кровь служит тепловым буфером, предотвращающим нагревание внутренних тканей), но, по-видимому оказывает рефлекторное действие через тепловые рецепторы кожи.
Раздражение висцеральных болевых рецепторов часто вызывает напряжение мышц (повышает мышечный тонус), а в экстремальных случаях может приводить к рефлекторным мышечным спазмам. Таким образом, висцеральные болевые афференты способны возбуждать мото
138
Р. Шмидт
нейроны (через полисинаптические рефлекторные дуги). В этих рефлексах принимают участие не только мотонейроны данного сегмента, но и мотонейроны функционально связанных мышц. При болях в брюшной полости (например, при аппендиците) отмечается напряжение мышц брюшной стенки. В то же время такие больные часто лежат, согнув ноги в коленях, так как мышцы-сгибатели ног возбуждаются теми же самыми афферентами. Имеющее такое происхождение значительное и длительное повышение мышечного тонуса вызывает мышечные боли и напряжение мышц. Боли такого рода наблюдаются не только при органических заболеваниях, при которых служат важным диагностическим признаком, но и в случае психологического стресса. Типичным примером может служить головная боль, сопровождающаяся болезненным напряжением задних мышц шеи; боль исчезает, как только причина психологического стресса устраняется или под влиянием эффективной психотерапии. Механизм возникновения такой мышечной боли неизвестен. Повышение мышечного тонуса снижает кровоток в мышце, что, по всей вероятности, играет некоторую роль в возникновении боли (возможно, способствуя накоплению в тканях продуктов метаболизма с ноцицептивным действием).
Зуд. Одно из кожных ощущений, о которых мы мало что знаем,-это зуд. Возможно, зуд является особым видом болевого ощущения, возникающим в определенных условиях стимуляции. Подобная интерпретация подтверждается тем фактом, что серия вызывающих зуд стимулов высокой интенсивности приводит к ощущению боли; кроме того, известно, что блокада проведения болевых сигналов по передне-боковому пучку спинного мозга сопровождается исчезновением зуда, тогда как при нарушениях восприятия давления и прикосновения (при повреждении задних столбов) зуд не устраняется. Можно также показать, что кожа чувствительна к вызывающим зуд стимулам только в определенных точках и что эти точки зуда соответствуют болевым точкам.
Однако другие данные указывают на то, что зуд бывает независим от боли и, возможно, определяется особыми рецепторами. Так, зуд можно вызвать только в самых верхних слоях эпидермиса, тогда как боль можно возбудить и в более глубоких слоях кожи. Используя подходящие методы, можно вызывать все степени зуда, не вызывая боли, и наоборот. В заключение следует заметить, что предпосылкой возникновения зуда, по-видимому, служит выделение какого-то химического вещества, возможно гистамина. Внутрикожная инъекция гистамина вызывает острый зуд, а при кожных заболеваниях, сопровождающихся зудом, гистамин обнаруживается в коже в свободном состоянии.
Периферические нарушения ноцицепции. Через несколько часов после повреждения кожи интенсивным ультрафиолетовым излучением возникает «солнечный ожог»: кожа краснеет и ее чувствительность к механическим стимулам повышается. Такие явления наблюдаются также и после повреждения кожи нагреванием, охлаждением, рентгеновскими лучами и механической травмы. Эту сверхчувствительность называют
3. Соматовисцеральная чувствительность
139
гипералгезией. Болевые пороги снижаются, и теперь даже самые обычные нейтральные стимулы (такие, как трение одежды о кожу) могут восприниматься как неприятные или болевые. Гипералгезия и покраснение (последнее связано с местным расширением кровеносных сосудов - вазодилатацией) могут сохраняться в течение нескольких дней. Кажется весьма правдоподобным предположение, что они определяются локальным выделением в ткани из поврежденных клеток какого-то химического вещества типа гистамина, однако относящиеся к этому случаю данные чрезвычайно разноречивы. Повышение болевого порога, называемое гипалгезией, а также полная потеря болевой чувствительности, называемая аналгезией, обычно обнаруживаются только в комбинации с нарушением или ослаблением других модальностей кожной рецепции. Например, в простейшем случае перерезки какого-нибудь кожного нерва, конечно же, одновременно наблюдаются и локальная аналгезия, и исчезновение ощущений других модальностей.
Нарушения механизма центральной переработки болевых сигналов. Когда нарушается процесс переработки болевых сигналов в ЦНС, результатом, как правило, бывает не потеря чувствительности, а видоизменение болевых ощущений; нормальные болевые ощущения возможны только при «правильной» активации корковых и подкорковых отделов ЦНС, относящихся к болевой системе (см. разд. 2.5). Например, если повреждение затрагивает таламические структуры, участвующие в проведении болевых сигналов, болевые стимулы начинают вызывать чрезвычайно неприятные ощущения, которые субъективно объясняются сверхчувствительностью. Кроме того, у пациентов при этом часто возникают невыносимые спонтанные боли в контралатеральной половине тела, с трудом поддающиеся терапии. Такие таламические боли -это лишь один из многих возможных примеров нарушения центральной переработки болевых сигналов. Следует также отметить, что при центральных повреждениях может затрагиваться и эмоционально-мотивационный компонент боли. Например, больные с тяжелым повреждением лобной доли могут едва замечать боль, если чем-то отвлечены или поглощены. Такое наблюдается, несмотря на то что у этих пациентов совсем не регистрируются изменения болевых порогов, если их оценивать по методике, описанной в разд. 3.4.
В связи с этим становится понятным, почему в тех случаях, когда от болей не удавалось избавиться никакими средствами, предпринимались попытки их устранения хирургическим путем-перерезкой трактов, ведущих в лобную долю (эта операция называется лоботомией). Однако этот хирургический метод лечения при болях снискал повсюду дурную славу из-за многих своих недостатков-в частности, из-за того, что приводил к глубоким изменениям личности. В случаях стойких болей, не поддающихся терапевтическим воздействиям, иногда делают другую операцию, называемую цингу лото мией,- перерезку опоясывающей извилины, т.е. складки коры мозга, которая составляет часть лимбической системы, тесно связанной с эмоциями (см. разд. 2.5). Эту операцию, как
140
Р. Шмидт
и распространенную ранее лоботомию, назначали также и при некоторых психических заболеваниях, например при депрессии. Такие операции относятся к психохирургии-области медицины, которая эмпирически прогрессировала до весьма высокого уровня. Своей целью она имеет получение устойчивых изменений в ощущениях и поведении человека путем разрушения или удаления мозговой ткани; к хирургии прибегают тогда, когда никакие другие методы уже не помогают.
В заключение следует заметить, что многие целесообразные и эмоционально нормальные реакции на болевые стимулы, по-видимому, не являются врожденными, а должны вырабатываться у молодого организма на ранних стадиях развития. Если в раннем детстве животные лишены соответствующего опыта, нужные реакции очень трудно выработать позднее. Например, молодые собаки, которых оберегали от всех вредных стимулов на протяжении первых 8 месяцев жизни, оказались неспособными правильно реагировать на болевые стимулы и позднее обучались этому медленно, так и не достигнув нормы. Они снова и снова нюхали открытый огонь и позволяли иглам проникать глубоко в кожу, реагируя лишь локальным рефлексом отдергивания. Аналогичные наблюдения были сделаны и на молодых шимпанзе.
Терапевтическое воздействие на боль. Терапия при болях включает физические, фармакологические и нейрохирургические меры. Меры по устранению боли всегда оправданны, так как боль уже выполнила свою функцию индикатора повреждения; обезболивание, если его проводить правильно, часто вносит решающий вклад в быстрое выздоровление. Но даже в случаях неизлечимых болезней, вернее - особенно в этих случаях, облегчение или снятие боли имеет большое значение. Физические меры, которые могут быть назначены в разных ситуациях- это иммобилизация, холодные или теплые обертывания, диатермия (прогревание глубоких тканей коротковолновым излучением), массаж, упражнения для разрабатывания суставов и многие другие. Химические воздействия, которые могут повлиять на боль, т. е. фармакологические меры, могут проводиться на разных уровнях. На периферии можно предотвращать возникновение импульсов в болевых волокнах и их проведение (местная анестезия). Можно также блокировать передачу по восходящим путям спинного мозга (например, спинномозговая анестезия). Кроме того, можно Снижать возбудимость центральных нейронов, участвующих в проведении и переработке болевых сигналов. Наконец, есть лекарственные средства, которые таким образом действуют на структуры, регулирующие эмоциональное состояние больного, что вызывают у него более нейтральное эмоциональное отношение к боли и делают ее легче переносимой. Среди нейрохирургических мер мы уже отмечали выше цингулотомию и упоминали о хордотомии (см. разд. 2.4).
В терапии хронических болей имеется ряд новых многообещающих достижений; некоторые из них уже были проверены на людях. Эти процедуры базируются на том обстоятельстве, что ноцицептивный афферентный поток, как и все другие сенсорные потоки, на своем пути через
3. Соматовисцералъная чувствительность
141
спинной мозг и высшие отделы ЦНС может подвергаться тормозным влияниям. Идея заключается в том, чтобы вызвать или усилить эти тормозные процессы, как правило, с помощью электрической стимуляции определенных структур, в том числе-периферических нервов. Приемы иглоукалывания (акупунктура), используемые для снижения боли, возможно, как раз связаны с такой активацией и поддержанием тормозных процессов. На такое объяснение наталкивает тот факт, что для получения и поддержания эффекта обезболивания, иглы нужно все время поворачивать или использовать как электроды для периодической электрической стимуляции.
В 3.21. В случае солнечного ожога расширение сосудов (краснота) со прово ждается а) аналгезией;
б) люмбальной анестезией;
в) гипалгезией;
г) местной анестезией;
д) гипералгезией.
В 3.22. Какое из следующих веществ, в норме присутствующих в организме, предполагается вызывающим ощущение зуда?
а) Ацетилхолин.
б) Брадикинин.
в) Гистамин.
г) Норадреналин.
д) Серотонин.
В 3.23. При заболеваниях мочевыводящих путей у больного наряду с другими симптомами часто отмечаются боли в паху. Этот симптом является примером а) гипалгезии;
б) отраженной боли;
в) проецируемой боли;
г) начальной боли;
д) отставленной боли.
В 3.24. Во многих случаях рака молочной железы у женщин надавливание на плечевое нервное сплетение в области подмышечной впадины вызывает острую боль в зонах, иннервируемых этими нервами. Этот симптом является примером а) проецируемой боли;
б) аналгезии;
в) иррадиации по симпатическому стволу;
г) отраженной боли;
д) распространяющейся боли.
В 3.25. Назовите три физические процедуры, используемые для воздействия на боль, и три уровня, на которые могут влиять фармакологические препараты.
4. ФИЗИОЛОГИЯ ЗРЕНИЯ
О. Грюссер, У. Грюссер-Корнельс
Вокруг себя мы видим множество трехмерных предметов. Их можно описать как движущиеся, или неподвижные, или же определенным образом расположенные в пространстве. Они различаются по яркости, цвету, величине и форме. Зрительно воспринимаемые предметы могут иметь для нас особое значение или же быть безразличными, создавать эмоциональное впечатление или же не вызывать никакой субъективной реакции. Физиологи привыкли говорить, что зрительное восприятие возникает потому, что на сетчатку падает изображение окружающего мира. Это изображение вызывает определенные процессы обнаружения сигнала и его переработки в рецепторах и в нервных клетках высшего порядка, что в конечном счете на уровне «сознания» приводит к восприятию (см. гл. 1). Но каждый знает по своим снам, что восприятие возможно и без изображений на сетчатке.
В этой главе мы сознательно ограничим себя «простыми» механизмами зрения, которые можно объяснить в понятиях современных физиологических знаний. Мы не будем подробно рассматривать такие более сложные явления, как восприятие формы, пространства и зрительную обработку знаков, букв и слов. Мы не коснемся также повседневного опыта, который говорит нам, что всякое восприятие связано с «ожиданием» со стороны наблюдателя, которое до какой-то степени зависит от эмоционального состояния. Взгляните на рис. 4-1 и вы увидите, как ожидание может повлиять на восприятие.
В качестве введения в физиологию зрения вам здесь предлагаются несколько простых наблюдений, которые вы можете сделать сами. Закройте рукой левый глаз, фиксируйте предмет правым глазом и обратите внимание на то, что вы видите вокруг себя при фиксации. Вы обнаружите, что только часть из того, что вас окружает, включена в монокулярное поле зрения вашего правого глаза. Когда после этого вы будете фиксировать предмет двумя глазами, бинокулярно, то при этом не только улучшится ваше восприятие глубины, но вы также увидите больше из того, что вас окружает. Бинокулярное поле зрения по очевидным причинам больше монокулярного. Оно состоит из центральной области перекрывания (области, видимой обоими глазами) и двух участков по сторонам, каждый из которых виден только одним глазом. На рис. 4-2 схематично показан результат измерения границ полей зрения, произведенного периметром-аппаратом, которым офтальмологи пользуются для диагностики.
Снова фиксируйте какой-нибудь предмет и теперь постарайтесь решить, насколько отчетливо вы видите другие предметы в бинокулярном
4. Физиология зрения
143
Рис. 4-1. «Старая матушка Хаббард пошла полгядеть в буфете, нет ли там косточки для ее бедной собачки». (Заметили ли вы, какое слово напечатано неправильно?)
поле зрения. Вы заметите, что, чем дальше предмет от фиксационной точки, тем он менее ясно виден. Четкость зрительного восприятия можно определить количественно, измерив остроту зрения (см. разд. 4.2). Фиксируемый участок составляет ту часть поля зрения, где острота последнего наибольшая, если Смотреть нормально при дневном свете. Если вы хотите увидеть ясно предмет, находящийся в нефиксируемой части поля зрения, то обычно переводите взор в направлении этого предмета, из-за чего ранее фиксируемый предмет попадает на периферию поля зрения.
Глаза могут отклоняться приблизительно на 60° в обе стороны от их центрального положения и на 40° вверх и вниз. Поэтому при неподвижной голове все поле зрения (на рис. 4-2) больше поля зрения неподвижного глаза на 120° в горизонтальном направлении и на 80° в вертикальном. Но совершенно очевидно, что человек может разглядывать все вокруг себя, двигая головой и/или телом. Когда он смотрит таким образом, движения головы и глаз координируются центральными нервными механизмами.
Но движения глаз имеют значение не только для их функции перемещения направления взора. Смещение изображения по сетчатке, происходящее при движении глаз, необходимо для нормального зрительного восприятия. Если при помощи надлежащей оптической системы «стабилизировать» ртимул в поле зрения так, чтобы он совсем не смещался по сетчатке даже при движении глаз, то контуры и окраска изображения бледнеют и через несколько секунд исчезают. (Очень мелкие движения, которые трудно подавить даже при усиленной фиксации, предотвращают это исчезновение, которое поэтому трудно продемонстрировать без специального аппарата.) Ясно, что в зрительном восприятии так же, как и в некоторых других модальностях, органы чувств и связанные с ними центральные системы восприятия не просто пассивные «приемники»; напротив, активные двигательные компоненты играют в восприятии важную роль. Можно сказать, что мы «видим» пассивно, но
144
О. Грюссер, У. Грюссер-Корнелъс
Рис. 4-2. Поля зрения левого (сплошная линия) и правого (прерывистая линия) глаз. Бинокулярное поле зрения, где эти два поля наложены друг на друга, показано темно-красным. ЛСП -слепое пятно для поля зрения левого глаза; ПСП-то же для правого глаза. Кругами очерчены углы в 10, 30, 50 и 90° от точки фиксации.
мы также «смотрим», «рассматриваем», «разглядываем»-все эти слова подчеркивают активный компонент зрения. Даже в состоянии, казалось бы, пассивного видения мы «отбираем» наш зрительный мир произвольными и непроизвольными движениями глаз; их амплитуды и направления зависят не только от внутреннего состояния ЦНС (внимания, интереса), но также от характера зрительного стимула.
Ради простоты изложения мы рассмотрим отдельно сенсорный и моторный аспекты зрительного аппарата. Разд. 4.1 посвящен оптической системе глаза, формированию изображений на сетчатке и ее строению. В разд. 4.2 будут описаны элементарные наблюдения и некоторые «законы» из области зрительной психофизики, а лежащие в их основе нейрофизиологические явления и некоторые теоретические следствия изложены в разд. 4.3. В разд. 4.4 рассмотрена физиологическая основа глазодвигательной активности и координация сенсорных и моторных компонентов зрения.
4.1. Глаз
Когда вы смотрите на свои глаза в зеркало, то видите конъюнктиву -белую соединительную ткань, пронизанную мелкими кровеносными сосудами, которая у переднего полюса глазного яблока соединяется с прозрачной роговицей (рис. 4-3). Позади роговицы лежит радужная оболочка-синяя, серая или каряя в зависимости от числа и распределения пигментных клеток. В центре радужки находится зрачок-отверстие, обычно круглое; его диаметр меняется, как у диафрагмы в фотокамере. Между роговицей и радужной оболочкой находится передняя камера
4. Физиология зрения
145
Рис. 4-3. Правый глаз, видимый в зеркало.
глаза, наполненная прозрачной жидкостью, называемой водянистой влагой.
Роговица и конъюнктива покрыты тонкой пленкой слезной жидкости. Слезы образуются в слезных железах, расположенных в наружной (височной) части глазницы над глазным яблоком. Протоки слезных желез оканчиваются в конъюнктиве позади век, над наружным «углом» глаза. Движения век (мигание) равномерно распределяют слезы по роговице и конъюнктиве, и эта тонкая пленка жидкости улучшает оптические свойства поверхности роговицы. В небольших количествах слезная жидкость образуется непрерывно. Часть воды испаряется, остальная стекает в носовую полость через слезный проток.
Слезы на вкус соленые, так как их состав приблизительно такой же, как у ультрафильтрата плазмы крови. Слезы защищают роговицу и конъюнктиву от высыхания и действуют как «смазка» между глазом и веками. Когда между веком и глазом попадает какая-нибудь чужеродная частица, например песчинка, происходит рефлекторное усиление образования слез и мигания. В таком случае слезы действуют как смазывающая жидкость. Они содержат ферменты, которые разрушают бактерии и защищают глаз от инфекции. Наконец, когда человек «плачет», слезы служат средством выражения эмоций.
Оптическая система глаза. На рис. 4-4 дана схема поперечного сечения правого глаза человека. Оптическая система-диоптрический аппарат-представляет собой сложную, неточно центрированную систему линз, которая отбрасывает перевернутое, сильно уменьшенное изображение окружающего мира на сетчатку. Роговица, передняя камера и радужная оболочка составляют самую переднюю часть диоптрического аппарата. Непосредственно за радужкой расположена задняя камера глаза и двояковыпуклая линза-хрусталик. Эластичный хрусталик окружен сумкой; от нее веером расходятся волокна ресничного пояска. Эти волокна соединены с ресничными мышцами и наружным сосудистым слоем сетчатки; таким образом они косвенно связаны с наружной стенкой глазного яблока.
Пространство внутри глаза позади хрусталика заполнено стекло-
146
О. Грюссер, У. Грюссер-Корнельс
видным телом. Эта желатинозная субстанция, прозрачная, как вода, образована коллоидным раствором гиалуроновой кислоты во внеклеточной жидкости. Задняя внутренняя поверхность глаза выстлана сетчаткой, которая состоит из слоев пигментных клеток, рецепторов и нервных клеток (см. рис. 4-10). Зрительная ось (рис. 4-4) пересекает сетчатку в центральной ямке, небольшом углублении в сетчатке. Центральная ямка составляет область наибольшей остроты зрения.
Между сетчаткой и склерой лежит сосудистый слой -сеть кровеносных сосудов, питающих слои сетчатки, ближайшие к склере. Части сетчатки, примыкающие к стекловидному телу, снабжаются сосудистой системой центральной артерии сетчатки, которая проникает в глаз через сосок зрительного нерва.
Регуляторные процессы в диоптрическом аппарате. В этом вводном тексте мы не будем касаться принципов физической оптики и создания изображения в глазу (подробнее см. у Ruch, Patton). Большинство читателей помнят из школьного курса физики, что преломляющая сила линзы измеряется ее фокусным расстоянием f Это то расстояние позади линзы, на котором параллельные пучки света сходятся в одной точке. Преломляющая сила (рефракция) выражается в диоптриях (D) и связана с / следующим образом:
Преломляющая сила = — [D],
[4-1]
где /-фокусное расстояние в метрах. В нормальном глазу общая преломляющая сила диоптрического аппарата составляет около 58,6 D, когда
4. Физиология зрения
147
глаз сфокусирован на дальней точке. Большая часть преломления происходит при переходе из воздуха в роговицу; эта поверхность действует как сильная линза в 42 D. В нормальном глазу с рефракцией в 58,6 D на сетчатке возникает четкое изображение бесконечно далеких предметов (например, звезд). Для получения четкого изображения предмета на каком-то определенном расстоянии оптическая система должна быть перефокусирована. Для этого существуют два «простых» способа смещение хрусталика относительно сетчатки или увеличение его преломляющей силы. В глазу человека используется второй способ, а у лягушки, как и в фотокамере, хрусталик перемещается относительйо чувствительной поверхности.
В человеческом глазу с нормальным зрением при увеличении рефракции хрусталика на X диоптрий плоскость наилучшего фокуса движется от бесконечности на расстояние около 1/Х от глаза. Если человек с нормальным зрением (фокус в бесконечности) хочет ясно видеть страницу книги, которую он держит вЗО см от глаз, рефракция должна возрасти на 1/0,3 м = 3,3 D. Усиление рефракции при фокусировании более близкой точки называется аккомодацией на ближнюю точку, а ее снижение при фокусировании более далекой точки называется аккомодацией на дальнюю точку. Усиление рефракции хрусталика при аккомодации на ближнюю точку достигается увеличением кривизны его поверхности. Иными словами, при аккомодации на ближнюю точку хрусталик становится «более круглым».
Чтобы понять изменение формы хрусталика при аккомодации, нужно знать некоторые анатомические детали. В состоянии аккомодации на дальнюю точку ресничная мышца (рис. 4-4 и 4-5) не находится в состоянии сокращения; волокна ресничного пояска пассивно передают натяжение сосудистой оболочки и стенки глаза сумке хрусталика, и благодаря этому эластичный хрусталик становится относительно плоским. При аккомодации на ближнюю точку ресничная мышца сокращается и вследствие своей связи с волокнами ресничного пояска противодействует эластическим силам сосудистой оболочки. В результате натяжение сумки хрусталика снижается (рис. 4-5, Б). Внутреннее строение хрусталика таково, что при ослабленном натяжении его кривизна увеличивается, в особенности на передней поверхности; фокусное расстояние сокращается. Сокращение ресничной мышцы контролируется автономными нервными волокнами, главным образом парасимпатическими, идущими в составе глазодвигательного нерва (см. разд. 4.4).
С возрастом эластичность хрусталика падает и, по мере того как человек становится старше, способность хрусталика к аккомодации снижается. 10-летний ребенок может аккомодировать в среднем на 10 D, но к 50 годам аккомодация часто снижается до 2 D, а к 70 она составляет всего лишь около 0,5 D. Поэтому пожилым людям, у которых в остальном зрение нормальное, чтобы хорошо видеть вблизи и читать, нужны очки с выпуклыми линзами (такое состояние называется пресбиопией). Люди любого возраста, которые систематически работают с мелкими
148
О. Грюссер, У. Грюссер-Корнельс
Рис. 4-5. Схемы работы ресничной мышцы. А. Аккомодация на дальнюю точку; ресничная мышца расслаблена; натяжение эластической ткани в сосудистой оболочке передается волокнами пояска на капсулу хрусталика; в результате хрусталик уплощается. Б. Аккомодация на ближнюю точку; ресничная мышца сокращена; это снижает эластическое натяжение, передаваемое от сосудистой оболочки и склеры через волокна пояска на капсулу хрусталика; из-за уменьшения ее натяжения кривизна хрусталика возрастает.
предметами или много читают и у которых аккомодация на ближнюю точку недостаточна, должны носить корректирующие линзы. Иначе может возникнуть большая нагрузка на систему аккомодации, что вызовет усталость и головную боль.
Зрачковые реакции. Величина зрачка, подобно размерам диафрагмы в фотокамере, определяет количество проходящего через него света. При данной яркости окружения количество света, попадающего в глаз в единицу времени, пропорционально площади зрачка. Если в освещенной комнате человек закроет глаза на 10-20 с, а затем, снова открыв их, посмотрит в зеркало, то увидит, как оба его зрачка тут же сузятся. Эту зрачковую реакцию на свет легко показать, осветив вспышкой каждый глаз отдельно (рис. 4-6). Когда свет попадает только в один глаз, сокращением реагируют оба зрачка. Сокращение зрачка при освещении контралатерального глаза наызвается содружественной реакцией на свет, а сокращение зрачка освещенного глаза называется прямой реакцией на свет.
Зрачковая реакция на свет представляет собой регуляторный механизм снижения количества света, падающего на сетчатку при сильном освещении (например, при ярком солнце); а при тусклом освещении он усиливает относительное попадание света в глаз, увеличивая ширину зрачка. Диаметр зрачка у человека зависит также от расстояния до фиксируемого объекта. Если перевести фокус с дальнего предмета на ближний, то зрачки сужаются (реакция на ближнюю точку). Поскольку при
4. Физиология зрения
149
Рис. 4-6. Схема зрачковых реакций.
Аккомодация на дальнюю точку II II
Реакция конвергенции
Аккомодация на ближнюю точку
фокусировании близкого предмета оси обоих глаз конвергируют, эта реакция зрачков называется также реакцией конвергенции.
Зрачок сужается в результате сокращения круговой мышцы-сфинктера в радужной оболочке, иннервируемой парасимпатическими волокнами. Сокращение мышцы дилататора с радиальными волокнами в радужной оболочке вызывает расширение зрачка; она получает симпатическую иннервацию. Автономная иннервация мышц радужной оболочки обеспечивает изменение диаметра зрачка в зависимости от психологических факторов, усталости, а также при приеме таких веществ, как спирт, кофеин, никотин. У детей и подростков зрачки в среднем относительно шире, чем у взрослых.
Аномалии рефракции. Как правило, форма роговицы не идеально сферическая; ее кривизна в одном направлении больше, чем в остальных. Возникающее в результате искажение изображения называется астигматизмом. Обычно преломляющая сила роговицы несколько сильнее в вертикальном направлении, чем в горизонтальном. Астигматизм легко измерить специальным прибором (Placido), который показан на рис. 4-7. Оптометрист смотрит на роговицу пациента через отверстие в центре диска и видит на ней миниатюрное отражение концентрических кругов. В случае астигматизма кольца на этом отраженном изображении деформированы - они имеют вид не кругов, а эллипсов или же неправильно изогнуты.
В глазу с нормальной рефракцией (58,6 D) резкое изображение далекого объекта образуется на сетчатке только в том случае, если расстояние между передней поверхностью роговицы и сетчаткой составляет 24,4 мм. Если из-за отклонения формы глаза или преломляющей силы диоптрического аппарата расстояние от роговицы до сетчатки больше, чем от роговицы до фокальной плоскости, то имеет место близорукость
150
О. Грюссер, У. Грюссер-Корнелъс
Рис. 4-7. Диск Плацидо для определения астигматизма роговицы. Врач смотрит через отверстие в диске на зеркальное изображение кругов, создаваемое роговицей больного. Диаметр диска около 20 см.
(миопия)-, если же это расстояние меньше дальнозоркость (гиперметропия). При миопии отдаленные предметы не фокусируются резко на сетчатке, потому что даже в состоянии аккомодации на дальнюю точку преломляющая сила диоптрического аппарата слишком велика (рис. 4-8). Поэтому при миопии, чтобы ясно различать удаленные предметы, следует пользоваться очками с вогнутыми линзами, которые уменьшают общую рефракцию диоптрического аппарата. Преломляющая сила вогнутых линз дается в диоптриях со знаком минус.
В то же время дальнозоркий человек может аккомодировать так, чтобы четко видеть предметы в бесконечности и на больших расстояниях (рис. 4-9). Но его пределы аккомодации обычно недостаточны, чтобы резко фокусировать близлежащие предметы. Такому человеку для компенсации гиперметропии нужны выпуклые линзы. Когда дальнозоркий ребенок старается осуществить аккомодацию на ближнюю точку, чтобы ясно видеть на близком расстоянии, обычно происходит сильная конвергенция одного глаза, создающая конвергентное «косоглазие». Хорошо подобранные очки часто предупреждают развитие косоглазия у ребенка с гиперметропией.
Сетчатка. Рис. 4-10 представляет собой упрощенную схему сетчатки примата. Сетчатка состоит из фоторецепторов, четырех разных классов нервных клеток, глиальных клеток (мюллеровские опорные клетки) и пигментных клеток. Светочувствительная часть сетчатки представлена слоем фоторецепторов. В глазу человека различаются морфологически два класса фоторецепторов -палочки (около 120 млн.) и колбочки (около 6 млн.). В области наиболее ясного видения (в центральной ямке) сетчатка содержит только колбочки. Каждый фоторецептор обладает узким наружным сегментом, который состоит приблизительно из 1000 мембранных дисков (в палочках) или внутрь идущих складок (в колбочках). Молекулы зрительных пигментов (которые поглощают свет и инициируют рецепторную реакцию) погружены в эти мембранные структуры и расположены в них упорядоченно. Палочки и колбочки соединены синаптическими контактами с биполярными и горизон-
Нечеткое изображение
Аккомодация на дальнюю точку
Четное изображение
Аккомодация на ближнюю точку
Аккомодация на дальнюю точку с помощью корректирующей линзы
Рис. 4-8. Миопия (близорукость) и ее коррекция вогнутой линзой.
Нечеткое изображение близкого объекта
Четкое изображение дальнего объекта
Аккомодация на ближнюю точку
с помощью корректирующей линзы
Четкое изображение близкого объекта
Рис. 4-9. Гиперметропия (дальнозоркость) и ее коррекция выпуклой линзой.
152
О. Грюссер, У. Грюссер-Корнельс
Пигментный эпителий
Хроматофор
Палочка
Ганглиозная клетка
Г оризонтальная клетка Биполярная клетка Амакриновая клетка
t t t
Падающий свет
Рис. 4-10. Строение сетчатки примата. (Boycott, Dowling, Proc. R. Soc. Lond. (Biol.), 166, 80-111, 1966.)
---- Колбочка
Мюллерова • клетка (глия)
тальными клетками. Биполяры передают сигналы от фоторецепторов в слой ганглиозных клеток и к амакриновым клеткам. Горизонтальные и амакриновые клетки участвуют в «горизонтальной» передаче сигнала под прямым углом к основному направлению его движения (рецепторы -» биполярные клетки -» ганглиозные клетки -» ЦНС). Пигментные клетки граничат своей наружной стороной с наружной сосудистой системой (сосудистой оболочкой, рис. 4-4). Пигментные клетки выполняют важную функцию в регуляции содержания зрительного пигмента в наружных сегментах фоторецепторов. Как видно на рис. 4-10, распространенность синаптических контактов нейронных элементов обеспечивает значительную дивергенцию и конвергенцию сигналов.
Все нейроны, связанные анатомически и функционально с одной нер
4. Физиология зрения
153
вной клеткой сенсорной системы, называются рецептивным полем этой нервной клетки. Таким образом, рецептивное поле ганглиозной клетки в сетчатке состоит из всех функционально связанных с ней рецепторов, горизонтальных, биполярных и амакриновых клеток.
Во внутренней области центральной ямки связь между рецепторами и ганглиозными клетками осуществляется в отношении 1 :1. В этой области каждая колбочка образует прямой контакт с одной биполярной клеткой, которая в свою очередь имеет контакт только с одной ганглиозной клеткой. Однако вследствие участия горизонтальных и амакриновых клеток рецептивное поле ганглиозной клетки, связанной с центральной ямкой, включает довольно обширную непрямую латеральную конвергенцию сигналов, которая, как указано в разд. 4.3, несет преимущественно тормозную функцию.
В общем в сетчатке преобладает конвергенция. Информация, получаемая приблизительно 125 млн. рецепторных клеток, представлена активностью около 1 млн. ганглиозных клеток, аксоны которых образуют зрительный нерв (рис. 4-4).
В 4.1. На каком расстоянии человек с нормальным зрением отчетливо видит предметы, когда преломляющая сила его хрусталиков увеличена на 4 D при аккомодации на ближнюю точку?
а) 40 см.
б) 5 см.
в) 25 см.
г) 4 см.
В 4.2. Старым людям часто нужны очки, чтобы (читать, видеть на расстоянии). Но многим людям независимо от возраста нужны очки, чтобы видеть на расстоянии. Таких людей называют ...; их очки имеют (вогнутые/выпуклые) стекла.
В 4.3. Какое из следующих утверждений правильно? Зрачки у человека с нормальным зрением
а) одинаковые при дневном свете и в темноте;
б) на свету уже, чем в темноте;
в) в норме одинаковые в левом и правом глазах;
г) при взгляде на ближний предмет уже, чем при взгляде на дальний предмет;
д) уже только в освещенном глазу, если освещен один глаз; е) приблизительно одинаковой ширины в обоих глазах при монокулярном освещении.
В 4.4. Подберите функционально соответствующие друг другу части фотокамеры и глаза в следующих перечнях:
Фотокамера
1. Линзы
2. Диафрагма
3. Светочувствительный
Глаз
а) Изменение диаметра
слой пленки
зрачка
б) Сетчатка
в) Склера
154
О. Грюссер, У. Грюссер-Корнельс
4. Фокусировка путем перемещения объектива
5. Камера
6. Снижение количества падающего света уменьшением отверстия диафрагмы
г) Радужка
д) Аккомодация
е) Роговица
ж) Хрусталик
з) Сужение зрачка
В 4.5. Какое из следующих утверждений верно для рецептивного поля ганглиозной клетки сетчатки, связанной с областью центральной ямки?
а) В центральной ямке имеются только палочки.
б) В центральной ямке имеются только колбочки.
в) Для такого нейрона множество рецепторов конвергируют на одну биполярную клетку.
г) Для такого нейрона связь между рецепторами, биполярными и ганглиозными клетками осуществляется в отношении 1 :1.
д) В фовеальной области нет горизонтальных или амакриновых клеток.
4.2. Психофизика зрительного восприятия
В разделе, посвященном общей субъективной сенсорной физиологии, время, место, качество и интенсивность были представлены как основные размерности при описании любого восприятия. Далее эти основные размерности будут рассмотрены в приложении к некоторым примерам. Рассматривается влияние пространственных и временных свойств зрительных стимулов на видение; обсуждаются такие качества зрительного восприятия, как яркость, цвет и глубина.
«Собственный свет сетчатки» («Eigengrau»). Если испытуемый долгое время находится в совершенно темной комнате, а затем пытается описать свое зрительное восприятие, то он говорит, что видит не «черное» или «темное» поле, а скорее сероватое с более светлыми «туманными пятнами», светящимися точками и даже некоторыми упорядоченными структурами, изменяющимися в пространстве и времени. Разные испытуемые неизменно говорят о сероватом поле (иногда называемом собственным светом сетчатки), тогда как дополнительные зрительные восприятия сильно варьируют. Предполагается, что ощущение серого является субъективным коррелятом спонтанной активности нервных клеток сетчатки и центрального отдела зрительной системы.
Серая шкала. Представим себе, что перед человеком, который сидит в темной комнате и описывает свое восприятие «собственного света сетчатки», находится шахматная доска. Если ее внезапно осветить источником света, достаточно слабым, чтобы не ослепить человека, он немедленно увидит доску «правильно»: черные квадраты будут казаться ему темнее, чем серое поле, которое он видел перед этим, а белые квадраты светлее, хотя теперь на все части сетчатки света падает больше, чем
4. Физиология зрения
155
Рис. 4-11. Одновременный контраст (см. текст).
в предыдущий период полной темноты. Это простое наблюдение показывает, что восприятие черного, серого или белого зависит не только от количества света, падающего на каждый рецептор в единицу времени, но также от относительной освещенности соседних участков сетчатки. При условиях освещения, преобладающих при нормальном дневном свете, можно различить 30-40 разных градаций серого-от совершенно черного до ярчайшего белого.
Поскольку восприятие «внутреннего света» очевидным образом характеризует «состояние покоя» зрительной системы, Геринг (Е. Hering) более 100 лет назад предложил интерпретировать восприятия черного и белого (в примере с шахматной доской) в понятиях двух противоположно действующих нейронных систем; эти «антагонистические» системы могли находиться в сетчатке или же в высших зрительных центрах. С этой точки зрения субъективное восприятие яркости в любом участке поля зрения определяется относительной степенью активации темновой (черной) системы и Светловой (белой) системы. Нейронные системы, постулированные Герингом, действительно были обнаружены как в слое ганглиозных клеток сетчатки, так и в высших зрительных центрах. Эти элементы упоминаются в разд. 4.3; это нейроны с «оп»-центром (светловая система) и нейроны с «о!Т»-центром (темновая система).
Одновременный контраст. На рис. 4-11 показан одновременный зрительный контраст. Серый участок на белом фоне кажется темнее, чем такой же серый участок на черном фоне. Иными словами, свойства фона определяют субъективно воспринимаемую яркость серых полей даже при том, что они одинаковые. Вдоль гранцы между светом и темнотой светлая часть кажется особенно яркой, а более темная часть кажется темнее, чем на расстоянии от границы. Этот эффект называется «усилением краевого контраста», а участки измененного восприятия известны под названием «полос Маха». Увидеть полосы Маха на рис. 4-11 можно лучше всего, фиксируя середину одного из'серых дисков с расстояния около 50 см. При фиксации левого диска его край кажется ярче центральной части, а темная зона сразу же за краем кажется темнее остального фона.
156
О. Грюссер, У. Грюссер-Корнельс
ф
последовательных обра-
Явление одновременного контраста показывает, что субъективная реакция на стимуляцию определенной части сетчатки зависит от освещения окружающих частей. Ниже будет показано, что эту зависимость можно объяснить функциональной организацией рецептивных полей нейронов с «оп-» и «о!Т»-центрами.
Последовательный контраст между светом и темнотой; последовательные образы. Реакция на возбуждение определенного участка сетчатки зависит не только от одновременного возбуждения соседних участков (как показано выше), но также от пространственного распределения освещенности в непосредственно предшествующий отрезок времени. Изменение чувствительности на ограниченном участке сетчатки называется локальной адаптацией. Чтобы наблюдать ее на вашей собственной сетчатке, фиксируйте в течение 12 с одним глазом белую точку в центре геометрической фигуры справа на рис. 4-12, а затем фиксируйте точку в середине соседнего круга. Вы увидите теперь «последовательный образ». Участки исходной фигуры, которые были темными, кажутся в повательном образе ярче фона в белом поле, а те части, которые были светлыми, теперь кажутся темнее. Очевидно, те места сетчатки, на которые попадали темные части фиксируемой фигуры, стали более чувствительными, чем соседние, сильнее освещаемые участки.
Острота зрения. Изображение бинокулярно фиксируемого объекта
Рис. 4-13. Различные таблицы для проверки зрения с цифрами, буквами, кольцами Ландольта, знаками Снеллена и изображениями предметов (Leydhecker, 1975).
4. Физиология зрения
157
Локализация на сетчатке
Рис. 4-14. Зависимость остроты зрения от положения тестирующего стимула на сетчатке (фотопические условия).
проецируется, как упомянуто выше, на центральную ямку каждого глаза. При дневном свете острота зрения в этой части сетчатки наибольшая; по направлению к периферии сетчатки она постепенно снижается. В клинической практике острота зрения измеряется при помощи таблиц, подобных показанной на рис. 4-13; устанавливаются самые мелкие знаки, какие испытуемый видит отчетливо. Когда применяются кольца Ландольта, острота зрения выражается (как на рис. 4-14 и 4-15) в виде 1/а, где а-угол (в угловых минутах), соответствующий наименьшему различаемому разрыву в кольце. Другой мерой остроты зрения служит отношение Снеллена (Snellen): в числителе здесь стоит расстояние между наблюдателем и таблицей с буквами, обычно это 20 футов
Рис. 4-15. Зависимость остроты зрения от состояния адаптации сетчатки. Стрелкой показана область перехода от фотопиче-ских условий адаптации к ско-топическим.
158
О. Грюссер, У. Грюссер-Корнельс
Рис. 4-16. Нахождение слепого пятна правого глаза (см. текст).
(6,1 м). Знаменатель вычисляется после определения величины наименьших читаемых букв и представляет собой расстояние, на котором черные полоски, образующие эти буквы, составляют угол Г. Для людей с «нормальным» зрением эти расстояния равны; отсюда острота их зрения выражается отношением 20/20.
Остроту зрения можно измерять для разных частей сетчатки, если наблюдатель фиксирует одним глазом точку вне таблицы. На рис. 4-14 показано измеренное отношение между остротой зрения и положением на сетчатке. Следует также указать, что на остроту зрения влияет яркость рассматриваемого знака. На рис. 4-15 показана зависимость остроты зрения в фовеальной области (близ точки фиксации) от средней освещенности предъявляемой фигуры. Все знают из собственного опыта, что острота зрения зависит от условий освещения:
Вы не могли бы прочесть этот мелкий шрифт без досгаточного освещения.
Правильный выбор освещения важен не только для чтения, но и на производстве и при домашней работе.
Слепое пятно. При внимательном изучении рис. 4-4 видно, что часть внутренней задней поверхности глаза не покрыта сетчаткой; это сосок, или место выхода зрительного нерва. Часть поля зрения (рис. 4-2), проецирующаяся на сосок,-это слепое пятно. В области слепого пятна не видно «ничего». Чтобы убедиться в существовании у вас слепого пятна, посмотрите на рис. 4-16 и фиксируйте одним правым глазом верхний крест с расстояния около 25 см. Вы заметите, что черный диск справа «исчез»; его изображение попадает на сосок вашего правого глаза.
Но нижняя часть рис. 4-16 показывает, что, хотя стимул на слепом пятне не виден, но область слепого пятна «заполнена», на нее экстраполируется изображение с окружающего участка. Когды вы фиксируете нижний крест на этом рисунке (одним правым глазом на расстоянии около 25 см), то изображение мыши исчезает в слепом пятне, но в вер
4. Физиология зрения
159
тикальной решетке не будет дырки. Как можно нейрофизиологически объяснить такое «перцептивное заполнение», будет показано в разд. 4.3.
Фотопическое и скотопическое зрение. Большинство читателей помнят из школьных курсов биологии, что колбочки-это рецепторы, которыми мы видим преимущественно при дневном свете (фотопическое зрение), а палочки служат главным образом в условиях освещения, соответствующих ясной ночи (скотопическое зрение). В сумерках не преобладает ни та, ни другая рецепторная система (мезопическое зрение). Острота зрения наилучшая в фотопических условиях, в которых хорошо видны также и цвета; максимальная острота зрения приходится на центральную ямку. Временное разрешение быстро меняющихся фигур тоже лучше при фотопическом, чем при скотопическом зрении. В скотопиче-ских условиях время зрительных реакций удлинено.
При скотопическом зрении, осуществляемом системой палочек, преобладает функциональная цветовая слепота («ночью все кошки серы»), а острота зрения в области центральной ямки значительно хуже, чем при фотопическом зрении. Кроме того, положение наибольшей остроты зрения и максимальной чувствительности в скотопических условиях приходится не на середину центральной ямки, а на ее край. Это смещение точки максимальной чувствительности с фиксационной точки легко обнаружить, глядя ночью на звездное небо.
Если фиксировать взором очень бледную звезду, можно заметить, что она исчезает и появляется вновь, если избрать фиксируемую точку слегка в сторону от нее.
Процессы световой и темновой адаптации во времени. Если выйти ночью из ярко освещенной комнаты и пройти в сад, освещенный только звездами, то сначала вы практически не увидите ничего. Но постепенно чувствительность вашей зрительной системы приспособится к низкой интенсивности света в окружающей среде (темновая адаптация). По мере развития темновой адаптации острота зрения постепенно увеличивается и объекты в саду становятся видными хотя бы приблизительно.
Развитие темновой адаптации во времени можно выразить количественно, измеряя пороговую интенсивность стимулов, предъявляемых в разное время после перехода от света к темноте (рис. 4-17). Полученная таким образом кривая темновой адаптации показывает, что глаз не достигает максимальной чувствительности, пока не пробудет в темноте более 30 мин. Нормальная кривая темновой адаптации состоит из двух частей - первая считается колбочковой, вторая-палочковой. Колбочковый компонент можно измерить отдельно при помощи небольших пятен красного света, сфокусированных в фовеальной области. Хотя начальная адаптация (колбочковая) происходит быстрее, чем в палочках, конечная колбочковая чувствительность значительно ниже (т.е. пороги выше), чем палочковая (измеряемая белым или синим светом вне центральной ямки).
Процессом, противоположным темновой адаптации, является световая адаптация. Быстрое течение световой адаптации легко наблюдать,
160
О. Грюссер, У. Грюссер-Корнелъс
Рис. 4-17. Кривые темновой адаптации. Прерывистая красная линия соответствует данным, полученным у полностью цветнослепого наблюдателя (палочковый монохромат); прерывистая черная линия получена при освещении красным светом центральной ямки, а черная сплошная -при освещении белым светом экстрафовеальной области у нормального испытуемого.
если, проведя много времени в темноте, войти затем в ярко освещенное помещение. При очень большой разнице в интенсивности вы будете на время ослеплены; затем за 15-60 с зрительная система приспосабливается к новой интенсивности света. Такого рода слепота связана с пониженной чувствительностью и нарушением восприятия формы. Подобное состояние может наступить, если, едучи ночью по шоссе, встретить машину с зажженными фарами.
Временные характеристики зрительного восприятия. Если смотреть на спицы колес быстро движущегося велосипеда, видна «прозрачная» серая поверхность. Смены света и темноты в движущихся изображениях спиц на сетчатке слишком часты, чтобы их заметить. Это обыденное наблюдение показывает, что существует верхний предел временного зрительного разрешения, называемый частотой слияния мельканий. Прерывистый световой стимул, мелькающий с частотой выше частоты слияния мельканий, неотличим от непрерывного света той же средней интенсивности. Частоту слияния мельканий легко измерить при помощи диска, разделенного на черные и белые секторы, как показано на рис. 4-18. Наблюдатель смотрит на часть такого диска, вращающегося все быстрее, пока не увидит его равномерно серым. Частота слияния мельканий возрастает с увеличением средней освещенности и размеров мелькающего светового стимула (рис. 4-18). Кривая на рис. 4-18 состоит из двух разных отрезков; часть ее, возрастающая более медленно, связана с палочковым зрением, а часть с более крутым наклоном-с колбочковым. Мелькающий свет воспринимается как таковой при частотах ниже 22 Гц в скотопических условиях адаптации и стимуляции. Более высокая частота слияния мельканий колбочковой системы говорит о том, что передача сигнала колбочками происходит значительно быстрее, чем в палочках.
Обычно при измерении частоты слияния мельканий при каждом промере проверяется одна определенная часть поля зрения. Но если
4. Физиология зрения
161
Средняя яркость мелькающего света
Рис. 4-18. Зависимость частоты слияния мельканий от средней интенсивности светового стимула. Измерение при помощи синусоидально модулированного света; амплитуда модуляции-70%.
проецировать последовательные световые стимулы попеременно на разные части сетчатки, то при этом вовлекаются временные и пространственные свойства переработки сигнала в сетчатке и в зрительной системе. Предположим, что в момент времени t0 световой стимул появляется в точке А поля зрения, а в момент t0 + tr он выключается; затем стимул вновь возникает (в момент времени t0 + tr + At) в точке Б. В этих условиях наблюдатель воспринимает мнимое движение света от А к Б, неотличимое от истинного движения, если At < 120 мс. Попеременное освещение участков А и Б создает впечатление, будто световой стимул движется взад и вперед между А и Б. Описанное здесь явление мнимого движения для случая двух простых стимулов используется в кино. Проецируемая пленка состоит из последовательности неподвижных изображений разной конфигурации; они возникают друг за другом на экране с частотой от 18 до 24 кадров в 1 с. Такой частоты достаточно, чтобы вызвать в зрительной системе впечатление мнимого движения, при котором смена образов неотличима от истинного движения.
Цветовое зрение. Для человека с нормальным зрением предметы вокруг него обладают большим разнообразием цветовых качеств, или хроматическим рядом. Удобно рассматривать ступени серого как эквивалентный ряд, называемый также ахроматическим рядом. Он идет от ярчайшего белого до самого глубокого черного, создаваемого одновременным контрастом. Воспринимаемая окраска поверхностей объектов характеризуется тремя объективными величинами: цветовым тоном, насыщенностью и светлотой. Цвета располагаются в естественной замкнутой последовательности («цветовой круг» на рис. 4-19): красный, оранжевый, желтый, зеленый, синий, фиолетовый, пурпурный, красный.
162 О. Грюссер, У. Грюссер-Корнелъс
Рис. 4-19. Положение цветов на цветовом круге. Цвета между (Л) и (Б) не спектральные, а получены смешением красного и синего.
Источник света
Рис. 4-20. Схема аддитивного (Л) и субтрактивного (Б) смешения цветов.
Источник света
4. Физиология зрения
163
Насыщенность цвета зависит от относительной величины хроматических и ахроматических компонентов. (Хроматичность, или окраска, определяется цветовым тоном и насыщенностью. Например, смесь спектрального красного с черным создает коричневый, а розовый создается смешением красного с белым.) Светлота указывает на относительное положение ахроматического компонента на шкале серого, между белым и черным.
Все цветовые тона в цветовом круге или соответствуют определенным цветам в спектре видимого солнечного света, или создаются аддитивным смешением двух спектральных цветов. Такое смешение происходит. когда свет разных длин волн падает на одну и ту же часть сетчатки (рис. 4-20). Для человека с нормальным цветовым зрением любой цвет, создаваемый освещенными предметами, может быть воспроизведен определенным аддитивным смешением трех надлежащим образом выбранных монохроматических стимулов Сь С2 и С3. Такая ситуация полностью и однозначно описывается уравнением ощущения, или цветовым уравнением:
a[Cj + b[C2] + c[C3]^d[Cx].
[4-2]
В этом выражении символ ~ означает «эквивалентность ощущения» или «подравнивание» цветов.
По международному соглашению, три определенные длины волны, основные цвета, обозначают С2, С2 и С3. Это спектральные цвета с длиной волны 700 нм (красный), 546 нм (зеленый) и 435 нм (синий); определенными их соотношениями можно характеризовать любой цветовой оттенок.
Цвета можно также создавать процессом, противоположным аддитивному сочетанию. Такое субтрактивное (вычитательное) сочетание происходит, когда белый свет проходит от источника через один (или более) цветовой фильтр (рис. 4-20). Когда художник смешивает краски (например, синюю и желтую, чтобы получить зеленую), он до некоторой степени использует субтрактивное смешение цветов, поскольку отдельные гранулы синей и желтой красок действуют как цветовые фильтры. Но при разглядывании красок на мозаиках или цветных изображениях известную роль может играть также аддитивное смешение, если две разные маленькие цветные точки попадают на одну группу колбочек. В «точечной манере» некоторых импрессионистов и в работах пуантилистской школы систематически использовался этот эффект. Слияние цветов происходит только при достаточно большом расстоянии между картиной и наблюдателем.
Белое или черное как цвета. Для каждого цвета С2 в цветовом круге имеется второй цвет С2, который при аддитивном смешении с С2 дает белый цвет, т.е.
a [Ci] + b [С2] = W (белый).
[4-3]
164 О. Грюссер, У. Грюссер-Корнелъс
В этом цветовом уравнении смешения цветов коэффициенты а и b зависят от определения «белого». Цвета С( и С2 во всех случах являются дополнительными. Белый свет может исходить от светящихся источников, но это невозможно для черного и разных оттенков серого-для их возникновения нужен механизм контраста. Термин цветовое тело применяется для представления воспринимаемых цветов в трехмерном пространстве. В простейшем цветовом теле его ось (черное/бе-лое) «перпендикулярна» плоскости цветового круга, представляющего чистые цветовые тона.
Результаты опытов по сенсорной психологии с аддитивным смешением цветов [4-2] и [4-3] представлены геометрически двумерным «цветовым треугольником» (или диаграммой хроматичности, рис. 4-21). Если два цвета, входящие в треугольник, сочетаются аддитивно, то возникающий при этом цвет лежит на линии между этими двумя цветами. Цвет, дополнительный к любому другому, лежит на линии, проходящей через белую точку (Е на рис. 4-21). Для человека с нормальным цветовым зрением все цветовые тона излучений (выраженные уравнением [4-2]) удовлетворительно описываются не более чем тремя величинами. Это наблюдение породило термин «трихроматическое зрение». Значения коэффициентов а, b и с в уравнении [4-2], которые требуются для получения любого из цветов в цветовом треугольнике, практически одинаковы для преобладающей части населения; такие люди называются нормальными трихроматами. Для небольшого процента людей коэффициенты иные, и такие люди называются аномальными трихроматами. Около 1% людей -дихроматы, они воспринимают все цвета таким образом, что это можно описать уравнением с двумя членами:
a^J+b^JScHCj. [4-4]
Диаграмма цветов, описывающая восприятие у этих людей, значительно менее дифференцирована, чем у трихроматов. Дихроматические отклонения от нормального цветового зрения, как и случаи аномальных трихроматов, обусловлены генетически. Самая обычная форма дихро-мазии-это «красно-зеленая слепота» (подробнее см. Ruch, Patton). Менее 0,01% людей полностью цветнослепые. Они различают только разные оттенки серого и называются монохроматами. Гены большинства полностью цветнослепых обусловливают существование у них только одной пигментной системы: их палочки и колбочки содержат только один пигмент-родопсин (см. разд. 4-3).
Здесь следует сказать о двух главных теориях цветового зрения. Трихроматическая теория Юнга и Гельмгольца (R. Young, Н. Helmholtz) постулирует существование трех типов колбочек, которые действуют как независимые системы приемников в фотопическом зрении; их сигналы анализируются совместно нейронными системами восприятия яркости и цвета. Эту теорию подтверждают результаты по смешению цветов (уравнение [4-2]), открытие трех разных колбочковых пигментов
4. Физиология зрения
165
Рис. 4-21. Цветовой треугольник, основанный на немецком стандарте DIN 5033. Белая зона окружает точку Е. Основание «цветового треугольника» образовано пурпурными тонами, которых нет в спектре и которые созданы аддитивным смешением спектральных цветов синего и красного.
(см. разд. 4.3) и различная спектральная чувствительность рецепторных потенциалов разных колбочек (см. рис. 4-26).
Оппонентная теория цветов Э. Геринга (Е. Hering) постулирует существование антагонистических нейронных процессов для оппонентных цветов зеленый/красный и желтый/синий в дополнение к системе чер-ный/белый, которая также организована антагонистически. Известная организация (см. разд. 4.3) рецептивных полей цветоспецифических нейронов сетчатки и латерального коленчатого тела, а также многие данные, полученные в опытах по сенсорной психологии, служат подтверждением этой теории. Таким образом, и трихроматическая теория цветового зрения и теория оппонентных цветов-обе «правильны» на разных уровнях афферентной зрительной системы. На возможность синтеза этих двух главных теорий цветового зрения указывал еще в начале века в своей зональной теории фрейбургский физиолог Крис.
166 О.Грюссер, У. Грюссер-Корнелъс
Бинокулярное зрение. Видимый объект представляется нам одним и тем же независимо от того, смотрим ли мы одним правым, одним левым или же обоими глазами. Объединение обоих монокулярных изображений объекта называется бинокулярным слиянием. По сравнению с монокулярным при бинокулярном зрении усилено впечатление пространственной глубины. Поскольку глаза расположены на голове не в одной точке, в изображениях окружающего мира, попадающих на обе сетчатки, имеются небольшие геометрические различия. Эти различия между изображениями, создаваемыми одним фиксируемым объектом на правой и левой сетчатках, тем сильнее, чем объект ближе. Различие (диспаратность) двух изображений служит предпосылкой восприятия глубины (стереоскопии). Чтобы достичь известной степени восприятия глубины при зрении только одним глазом, можно использовать различия в величине, степень перекрывания, параллакс, вызванный движением головы, но результат будет менее информативным, чем при полном бинокулярном зрении.
Разницу в изображениях видимого мира на обеих сетчатках можно продемонстрировать в следующем опыте: протяните правую руку во всю длину, поднимите большой палец и фиксируйте его сначала бино-кулярно, затем монокулярно попеременно правым и левым глазом. При смене глаз палец как бы перемещается по отношению к фону. Отсюда следует, что изображение окружающего мира на левой и правой сетчатке заметно различаются.
Бинокулярное слияние легко нарушается, если при фиксации объекта изменить положение одного глаза в глазнице, слегка нажав пальцем на веко. При этом объект проецируется на смещенную сетчатку «неправильно»: изображение «двоится», так как нарушено бинокулярное слияние.
Простое бинокулярное зрение (т.е. рассматривание одновременно только одного объекта) осуществляется лишь в том случае, когда изображения бинокулярно фиксируемого объекта падают на совершенно определенные части сетчатки в обоих глазах, на корреспондирующие участки сетчатки. При длительной бинокулярной фиксации данной точки (например, точки, отмеченной на рис. 4-22) можно определить эти корреспондирующие участки, передвигая тест-объект и находя те положения в пространстве, в которых он виден как один. Оказывается (как можно убедиться, начертив более подробные геометрические схемы), что все такие положения в пространстве падают на трехмерную изогнутую поверхность, включающую фиксационную точку. Эта поверхность называется гороптером, и, разумеется, для каждой фиксационной точки имеется свой гороптер. Для одной только горизонтальной плоскости ситуация проще-получаемая часть поверхности гороптера это «гороп-терный круг», показанный на рис. 4-22.
На заметном расстоянии от гороптера предметы «двоятся». Например, на рис. 4-22 при бинокулярной фиксации указанной точки обозначенный предмет лежит вне гороптера. Его изображения в обоих глазах
4. Физиология зрения
167
Циклопический глаз
Циклопический глаз
Рис. 4-22. Схема бинокулярного зрения и построение циклопического глаза. Видимый объект лежит здесь снаружи от поверхности гороптера. Его изображение проецируется справа от центральной ямки в левом глазу. При бинокулярном зрении создаются неперекрещивающиеся двойные изображения объекта; их положение можно определить, проецируя правую и левую сетчатки на воображаемую сетчатку циклопического глаза.
явно падают на некорреспондирующие участки, а именно на противоположные стороны центральных ямок (углы а и 0 на рис. 4-22).
Один из способов рассматривать проблему «глаза два, а мир один» состоит при таких ситуациях в том, чтобы представить себе «циклопический глаз», расположенный на продолжении гороптера посередине между глазами. Как показано на рис. 4-22, когда оба глаза, фиксирующие точку, отмеченную на гороптере, смотрят на объект, отдаленный от нее, то циклопический глаз увидел бы два мнимых предмета по обе стороны от реального. Когда же циклопический глаз видит только одно изображение, оно должно быть на гороптере, т.е. пары корреспондирующих участков в двух глазах по определению приходятся на одни и те же участки циклопического глаза.
Существенное различие между изображениями двух объектов, лежа
168
О. Грюссер, У. Грюссер-Корнельс
щих внутри и вне гороптера, легко продемонстрировать в «опыте с двумя пальцами». Держите перед собой правый указательный палец на уровне глаз на расстоянии около 20 см, а левый палец-за ним, на расстоянии около 50 см от глаз. Сначала фиксируйте ближний палец, тогда дальний будет двоиться. Затем закрывайте попеременно правый и левый глаз, и вы увидите, что двойные изображения не перекрещиваются. Это значит, что левый глаз видит левое изображение, а правый-правое. Дальний палец лежит снаружи от гороптера, создаваемого фиксацией ближнего пальца. Затем фиксируйте дальний палец. Теперь двоится ближний, и, попеременно закрывая то один, то другой глаз, вы увидите, что двойные изображения перекрещиваются: правый глаз видит левое изображение, и наоборот. Теперь ближний палец находится внутри гороптера, определяемого фиксацией дальнего пальца. Попробуйте зарисовать эту ситуацию и построить положения возникающих изображений по аналогии с рис. 4-22.
На основании описанных наблюдений можно было бы заключить, что все видимые предметы, изображения которых не прпадают на корреспондирующие участки сетчатки, потому что они лежат внутри или снаружи от гороптера, должны вызывать двойные образы. Но если вы глядите вокруг себя в хорошо структурированном видимом мире, то мешающее двоение не возникает. По-видимому, существуют механизмы, которые препятствуют этому. Один из таких механизмов состоит в низкой остроте зрения на периферии сетчатки. Второй, как полагают, представляет собой бинокулярный тормозный механизм в центральных отделах зрительной системы, подавляющий восприятие того или другого из двух диспаратных изображений. Глядя на разные объекты центральными ямками левого и правого глаза, можно наблюдать этот бинокулярный тормозный механизм. Для этого надо сделать две трубки длиной около 30 см и диаметром 3-4 см. В одну из них смотрите одним глазом на марку, в другую (другим глазом)-на монету. С этими резко различными стимулами бинокулярного слияния не происходит-вы не увидите объединенную «марко-монету». Благодаря бинокулярному тормозному механизму возникает «бинокулярная конкуренция», попеременное восприятие то марки, то монеты. Отдельные части обоих стимулов могут быть видны одновременно, но только рядом, а не наложенные одна на другую. В этой бинокулярной конкуренции контуры, как правило, эффективнее равномерно затемненных поверхностей. Это можно объяснить известной из опытов более выраженной активацией нейронов афферентной зрительной системы при действии контрастных границ (подробнее см. разд. 4.3) по сравнению с активацией однородными поверхностями.
Стереоскопическое зрение. Несмотря на то что от любого достаточно крупного трехмерного объекта, видимого бинокулярно, на центральные ямки обоих газ падают несколько разные изображения из-за рассмотренной выше геометрической ситуации, обычно такой объект не «двоится» в нашем зрении, мы воспринимаем его как расположенное
4. Физиология зрения
169
Рис. 4-23. Стереоскопическое восприятие, основанное на горизонтальной диспаратности.
в пространстве целое. Расхождения в горизонтальной плоскости между изображениями на двух сетчатках (когда голова в нормальном положении), т.е. отклонения от точно корреспондирующих участков сетчаток, называются горизонтальной диспаратностью рецептивных полей. Она служит важным условием бинокулярного трехмерного зрения. Степень диспаратности определяет силу восприятия глубины. Поэтому приведенное выше описание функционального значения корреспондирующих участков сетчаток следует видоизменить: только в случае, когда определенная горизонтальная диспаратность превышена, восприятие биноку-лярно видимого объекта как единого целого уступает место двоящемуся образу. Иными словами, восприятие объекта как единого при стереоскопическом зрении требует функционального анализа не корреспондирующих точек в сетчатках обоих глаз, а связанных между собой изображений, падающих на участки в несколько угловых градусов. Горизонтальная диспаратность двух изображений предмета уменьшается по геометрическим причинам с увеличением расстояния от него до глаз. Поэтому на больших расстояниях точное восприятие глубины становится невозможным. Оптический дальномер увеличивает функционально расстояние между глазами и, кроме того, увеличивает размеры видимых предметов, улучшая тем самым восприятие глубины. Влияние горизонтальной диспаратности на восприятие глубины можно изучать при помощи рис. 4-23. Фиксируйте воображаемую точку за книгой так, чтобы оба больших круга бинокулярно слились. Тогда вы увидите три больших круга, средний из них-обоими глазами. При этом вам будет казаться, что маленький круг парит в пространстве перед плоскостью рисунка. Его изображения в глазах горизонтально диспаратны из-за асимметрии рисунка.
Восприятие формы. Если смотреть на хорошо структурированное окружающее пространство, то видны не изолированные «стимулы», а предметы в их соотношениях между собой. «Расчленение» механизмов зрительного восприятия на простые процессы («сенсорные впечатления», см. с. 5), как это сделано в предыдущих разделах, оказалось полезным для сенсорной физиологии; это очевидный подход к количественному определению и пониманию функции периферических органов чувств. Гипотеза, связанная с таким подходом, а именно что восприятие сложной формы осуществляется элементарными физиологическими механизмами в сетчатке и афферентных путях зрительной системы, под
170
О. Грюссер, У. Грюссер-Корнельс
тверждена прямыми нейрофизиологическими измерениями в некоторых системах (см. разд. 4.3).
В то же время еще неизвестно, каким образом из обнаруженных до настоящего времени элементарных механизмов возникает восприятие вещественного объекта (Gestalt). Нет также объяснения «феноменологической инвариантности» воспринимаемых предметов. Этим термином обозначается тот хорошо известный факт, что чайник воспринимается как один и тот же чайник независимо от условий освещения, его положения и ориентации, а также от того, виден он одним или двумя глазами. Чайник воспринимается как целое, даже если видна только его часть.
В предметном восприятии так же, как в элементарных механизмах восприятия цвета и света или темноты, участвуют многочисленные процессы оценки контраста. При определенных условиях они создают иллюзии. Некоторые из таких иллюзий показаны на рис. 4-24. Воспринимаемая длина и кривизна линии определяются не только ее «объективными» характеристиками, но также длиной и углом линий рядом с ней (рис. 4-24, а, б). Контуры простых форм заполняются, и можно видеть, как край их продолжается через объективно пустое пространство (рис. 4-24, в). Воспринимаемые размеры фигуры зависят от размеров окружающих фигур (рис. 4-24, г). Систематические исследования в области гештальт-психологии, в особенности в первой четверти нашего века, выявили множество правил, управляющих зрительным восприятием сложных форм. Поскольку последние современные нейрофизиологические данные не дают достаточного объяснения этим правилам или вовсе не объясняют их, они не будут здесь приведены. Впрочем, в разд. 4.3 описаны некоторые нейрофизиологические факты, которые позволяют в первом приближении подойти к пониманию восприятия сложных зрительных форм.
В 4.6. Одновременный контраст между светлым и темным является а) ошибкой суждения, которую легко избежать, сосредоточив внимание;
б) следствием нарушения диоптрического аппарата глаза;
в) механизмом, который приобретается обучением;
г) эффектом, вызываемым латеральным торможением в сетчатке и в центральной нервной системе.
В 4.7. Которое из следующих утверждений приложимо к остроте зрения у человека с нормальным зрением?
а) В условиях световой адаптации наибольшей остротой обладает центральная ямка.
б) Острота зрения при световой адаптации выше, чем при темновой.
в) Участок сетчатки с самым острым разрешением варьирует в зависимости от состояния адаптации.
г) Острота зрения в области слепого пятна равна 1,0'.
4. Физиология зрения
171
Рис. 4-24. Примеры иллюзий при восприятии формы, а-Расстояния АБ объективно одинаковые в изображениях двойной стрелки и двойной вилки, но они кажутся разными; б-две толстые вертикальные линии в действительности прямые и параллельные, в чем можно убедиться при помощи линейки; в-белый квадрат посередине создан субъективным «заполнением»; г-круги в центре одинаковой величины на обеих фигурах.
(О
д) Остроту зрения можно измерить кольцами Ландольта.
е) Когда глаза ослеплены ярким светом, острота зрения пони-
жена.
В 4.8. Какова острота зрения у человека, который правильно опознает кольцо Ландольта с промежутком а = 0,8'?
В 4.9. Какие механизмы нужны для нормального стереоскопического зрения (двумя глазами)?
а) Бинокулярное слияние.
б) Световая адаптация.
в) Темновая адаптация.
г) Бинокулярное торможение мешающих двойных изображений (бинокулярная конкуренция).
д) Горизонтальная диспаратность.
е) Одинаковое цветовое восприятие в обоих глазах.
В 4.10. Корреспондирующими участками сетчатки мы называем
а) отношения между центром и периферией рецептивного поля;
б) участки в обеих сетчатках, которые при нормальном бинокулярном зрении связаны между собой;
в) участки одной сетчатки, которые находятся на одинаковых расстояниях от центральной ямки;
г) участки сетчатки, где имеются только колбочки.
В 4.11. Гороптер-это
а) область поля зрения, проецирующаяся на центральную ямку;
б) область бинокулярного поля зрения, проецирующаяся на обе центральные ямки;
в) воображаемая поверхность в пространстве, каждая точка которой проецируется на геометрически соответствующие корреспондирующие участки на двух сетчатках;
г) область окружающей среды, лежащая вне бинокулярного поля зрения;
д) ни одно из приведенных утверждений не правильно.
172
О. Грюссер, У. Грюссер-Корнелъс
В 4.12. Которое из следующих утверждений правильно?
а) Если три разных монохроматических стимула падают на один фоторецептор в глазу, происходит аддитивное смешение цветов.
б) Если два разных монохроматических световых стимула падают на один фоторецептор в глазу, происходит субтрактивное смешение цветов.
в) Когда художник смешивает красную и желтую краски, чтобы получить оранжевую, он производит субстрактивное смешение цветов.
г) Когда художник смешивает желтую и синюю краски, чтобы получить зеленую, он производит аддитивное смешение цветов.
д) Набор всех цветов от световых источников, воспринимаемых человеком с нормальным зрением, описывается правильно и однозначно уравнением смешения цветов, включающим три спектральных цвета (первичных цвета).
4.3. Нейрофизиология зрения
Первичный процесс; фотохимическая реакция. Диоптрический аппарат глаза фокусирует на сетчатке перевернутое и уменьшенное изображение окружающего мира. Если это изображение хорошо структурировано, то в каждый интервал времени на разные участки рецепторной мозаики падает разное количество световых квантов (фотонов) определенной длины волны. Некоторые фотоны поглощаются молекулами зрительного пигмента, погруженными в мембранозные диски (рис. 4-10) фоторецепторов. Каждая молекула пигмента может поглотить только один фотон, вследствие чего она переходит на более высокий энергетический уровень. При этом возникает вероятность (равная около 0,5) перехода в конфигурационное изменение, которое ведет к многоступенчатому процессу распада молекулы пигмента (эта вероятность называется «квантовой эффективностью»). Зрительный пигмент в палочках (родопсин) состоит из белка (опсина) и ретиналя, альдегида витамина АР Максимум поглощения молекулы родопсина лежит при длине волны около 500 нм. При распаде родопсина образуются бесцветные опсин и витамин АР Состав колбочковых пигментов еще не полностью выяснен. Однако полагают, что они обладают структурой, сходной с родопсином. В колбочках сетчатки человека обнаружены три разных пигмента. Их максимумы поглощения составляют около 425, 535 и 570 нм. В сетчатке человека каждая палочка, очевидно, содержит только один пигмент.
Распад молекул пигмента, запущенный поглощением фотона, составляет первую ступень в зрительном процессе трансдукции (см. разд. 2.1), от которого зависят все остальные нейрофизиологические процессы возбуждения. Конфигурационные изменения молекулы пигмента, вероятно, приводят к повышению кальциевой проводимости мембраны диска, так
4. Физиология зрения
173
что ионы кальция (или другого, еще неизвестного внутриклеточного медиатора) диффундируют из мембранных дисков во внутриклеточное пространство наружного сегмента фоторецептора. Там взаимодействие с наружной мембраной наружного сегмента приводит к снижению проницаемости для мелких ионов, особенно для натрия. Это изменение проводимости вызывает рецепторный потенциал фоторецепторов, который будет рассмотрен ниже.
Распавшиеся молекулы пигмента снова восстанавливаются в результате цепи промежуточных стадий химического процесса, протекающего с поглощением энергии. «Мембранный насос» возвращает кальций внутрь мембранных дисков. При постоянном освещении достигается равновесие между фотохимическим распадом пигментов и их химическим ресинтезом. Достижение этого баланса представляет собой физико-химический компонент световой/темновой адаптации; при снижении адаптирующей освещенности концентрация зрительных пигментов в фоторецепторах возрастает.
Рецепторный потенциал. Если микроэлектрод расположен на поверхности или внутри одиночного фоторецептора при полной темноте, то он регистрирует относительно большой электрический ток, протекающий через клеточную мембрану («темновой ток»). В темноте потенциал покоя составляет лишь от — 20 до — 40 мВ. При освещении фоторецептора возникает гиперполяризация, т.е. увеличение электроотрицательности внутри клетки по отношению к внеклеточному пространству. В то же время электрический ток через мембрану слабеет. Таким образом, сопротивление мембраны фоторецептора повышается при освещении. Как уже было упомянуто, это повышение электрического сопротивления объясняется взаимодействием между ионами кальция или медиаторного вещества и клеточной мембраной. Гиперполяризационный ответ на адекватный стимул является особенностью фоторецепторов сетчатки позвоночных. Во всех других исследованных до сих пор рецепторах возбуждение связано с деполяризацией мембраны; потенциал покоя в других рецепторах лежит в пределах между — 60 и — 80 мВ (см. разд. 2.1).
Амплитуда (А) гиперполяризационного рецепторного потенциала, возникающего в ответ на освещение, тем выше, чем больше интенсивность света I относительно предыдущего состояния адаптации (рис. 4-25). Отношение между А и I в известных пределах (двух-трех порядков I) выражается уравнением
А =/с log —мВ, [4-5]
I s
в котором к выбирается таким, чтобы выразить А в милливольтах. Спектральная чувствительность рецепторных потенциалов одиночных фоторецепторов в сетчатке позвоночных животных была определена путем стимуляции монохроматическим светом разных длин волн, урав-
174
О. Грюссер, У. Грюссер-Корнелъс
Вспышка
Рис. 4-25. Рецепторный потенциал одиночной колбочки в сетчатке черепахи. Ответ на короткие световые вспышки прн трех разных интенсивностях. (Baylor, Fuortes, J. Physiol. (Lend.), 207, 72-92, 1970.)
ненных по энергии. При таких измерениях применяются микроэлектроды для внутриклеточного отведения от отдельных фоторецепторов. На рис. 4-26 показано, как амплитуда рецепторного потенциала зависит от длины волны в большом числе рецепторов, исследованных в таких опытах. В этих данных опять-таки выделяются колбочки трех разных классов. Максимум ответа палочек лежит примерно при 500 нм-длине волны максимального поглощения родопсина. Эти нейрофизиологические данные согласуются с упомянутыми выше результатами о наличии разных зрительных пигментов в палочках и колбочках и о присутствии трех разных типов колбочек в сетчатке позвоночных. Вместе взятые они подтверждают изложенную выше трихроматическую теорию цветового зрения.
Переработка сигнала в горизонтальных, биполярных и амакриновых клетках. В опытах на животных удается проникать в нервные клетки сетчатки микроэлектродами и измерять мембранный потенциал внутриклеточно. С помощью разных синаптических контактов нейроны сетчатки образуют тесно связанную нейронную сеть (см. рис. 4-10). В этой сети различаются два направления потока сигналов: главный-от рецепторов к биполярным, а затем к ганглиозным клеткам и латеральная передача сигнала-по слоям горизонтальных и амакриновых клеток. В горизонтальных, биполярных и амакриновых клетках переработка сигнала происходит путем медленных изменений мембранных потенциалов; потенциалы действия здесь не генерируются. Количество освобождаемого синаптического медиатора в этих клетках, очевидно, зависит от мембранного потенциала в синаптических терминальных структурах. На рис. 4-27 показана схема «переработки» сигналов в нервной сети сетчатки. Окончательный результат этой переработки находит отражение в функциональной организации рецептивных полей сетчаточных ганглиозных клеток-«выходных элементов» сетчатки.
4. Физиология зрения
175
Рис. 4-26. Кривые спектральной чувствительности трех типов колбочек в сетчатке рыбы (карп). Несмотря на значительный разброс индивидуальных данных, можно различить три разных типа колбочек; пределы разброса показаны затененными участками. (Tomita et al., Vision Res., 7, 519-531, 1967.)
Нейрофизиология ганглиозных клеток сетчатки. Термин «рецептивная единица» относится ко всей совокупности афферентных нейронных элементов, которые воздействуют на данную нервную клетку сенсорной системы. Таким образом, рецептивная единица ганглиозной клетки сетчатки состоит из всех тех рецепторов, биполярных, горизонтальных и амакриновых клеток, которые прямо или косвенно связаны с этой ганглиозной клеткой. Из рис. 4-10 видно, что рецептивная единица ганглиозной клетки охватывает большую область сетчатки. Рецептивная единица служит анатомической основой определяемых посредством стимуляции рецептивных полей ганглиозных клеток. В то время как мембрана рецепторов горизонтальных и биполярных клеток генерирует только медленные потенциалы, мембрана ганглиозных клеток способна генерировать потенциалы действия. Они проводятся к ЦНС по аксонам ганглиозных клеток. Эти аксоны составляют зрительный нерв, который проходит от соска глаза через стенку черепа в его полость; нерв входит в область промежуточного мозга, и аксоны ганглиозных клеток образуют синапсы с клетками латерального коленчатого тела, верхних бугорков и претектальной области.
Функциональную организацию рецептивных полей ганглиозных клеток сетчатки можно проанализировать, регистрируя микроэлектродами потенциалы действия их аксонов в зрительном нерве. У каждой ганглиозной клетки имеется небольшая область общего поля зрения (см. рис. 4-2), в пределах которой соответствующие световые стимулы могут вызывать возбудительные и тормозные ответы. Такая область называется рецептивным полем (РП). Рецептивные поля сосоедних ганглиозных клеток могут значительно перекрываться. При тестировании ответов многих ганглиозных клеток сетчатки стимулами белого света, проецируемыми на разные части их рецептивных полей, становится очевидным наличие двух больших классов нейронов-нейронов с «оп»-цен-тром и нейронов с «offw-центром. В светлоадаптированной сетчатке
176
О. Грюссер, У. Грюссер-Корнельс
Освещенная
Неосвещенная
Г оризонтальная клетка
Биполярная _ клетка
Амакриновая клетка
Г англиозная клетка
Зрительный нерв
к ЦНС
Рис. 4-27. Схема расположения нервных элементов в сетчатке и потенциалы, отводимые от них внутриклеточными микроэлектродами при освещении (прямом или косвенном) связанных с ними рецепторов. Трансформация медленных мембранных потенциалов в серию потенциалов действия типа «все или ничего» происходит только в ганглиозных клетках. (Dowling, Invest. Opthalmol., 9, 655-680, 1970, с изменениями).
4. Физиология зрения
т
I—1-Ю°—|
Свет падает на
центр РП
периферию РП
центр и периферию
Нейроне ”оп”-центром
центр РП
периферию РП
центр и периферию
Рис. 4-28. Функциональная организация рецептивных полей ганглиозных клеток в сетчатке млекопитающих. Для анализа рецептивных полей белые пятнышки света проецируются либо на центр, либо на периферию поля, или же и туда н сюда.
млекопитающих нейроны обоих классов обладают рецептивными полями, в которых стимулы в центре вызывают иные ответы, чем на периферии (рис. 4-28). В случае нейрона с «оп»-центром освещение центра РП вызывает возбуждение (т.е. повышение частоты потенциалов действия), тогда как освещение периферии РП вызывает торможение-понижение частоты потенциалов действия (рис. 4-28). При одновременном освещении центра и периферии обычно преобладает ответ, связанный с центром. При выключении маленького светового стимула возникает торможение, если он нанесен на центр, и активация, если на периферию, т.е. возникают ответы, обратные ответам на «оп».
Рецептивные поля нейронов с «offw-центром тоже имеют «антагонистическую» организацию, приблизительно обратную рецептивным полям нейронов с «оп»-центром. Освещение центра РП вызывает торможение, а выключение света в центре-возбуждение ганглиозной клетки; освещение периферии РП вызывает активацию, а выключение света на периферии-торможение (рис. 4-28).
Нейрофизиология одновременного контраста. На основе функциональной организации рецептивных полей сетчатки можно представить себе, каким образом возникает одновременный контраст (см. разд. 4.2). На рис. 4-29 в виде схемы показано, как меняется ответ нейрона сетчат-
178
О. Грюссер, У. Грюссер-Корнельс
' ЛНОСИТъЛ'АЙн икмааиии найдена
Рис. 4-29. Схема активации нейрона с «оп»-центром, объясняющая явление одновременного контраста; объяснение см. в тексте.
Б В
8 О
-3 -1
ки с «оп»-центром, когда граница «или край между светом и темнотой проецируется на разные части рецептивного поля. Числа на рисунке означают относительные уровни возбуждения или торможения нейрона. Если край занимает положение А, то и периферия, и центр равномерно освещены светлой частью стимула. В нейроне с «оп»-центром преобладает нейронная активация от центра поля (в итоге возбуждение-4). Но если край находится в положении Б, то освещена только часть тормозной периферии. Поэтому торможение будет слабее, чем при стимуле в положении А, и возбуждение ганглиозной клетки в итоге будет больше (= 5). Если край занимает положение В, освещается только малая часть тормозной периферии, а центр РП остается темным. В этом случае результатом явится торможение спонтанной активности клетки. Таким образом, максимальная активация возникает тогда, когда край находится как раз на границе между центром и периферией рецептивного поля (положение Б). Если принять обоснованное допущение, что субъективно воспринимаемая степень «яркости» положительно коррелирует с частотой импульсации в нейронах с «оп»-центром, то такие ответы ганглиозных клеток должны предсказывать явление одновременного контраста (см. рис. 4-11).
Изменение организации рецептивного поля при темновой адаптации. Антагонистическую организацию РП можно объяснить наложением возбудительного и тормозного процессов в рецептивной единице ганглиозной клетки сетчатки. Рассуждение основано здесь на допущении, что процессы латерального возбуждения передаются преимущественно конвергенцией сигналов от нескольких биполярных клеток на одну ганглиозную. У тормозных процессов порог выше, чем у возбудительных, причем латеральные тормозные процессы передаются горизонтальными
4. Физиология зрения
179
Средняя яркость
Темновая адаптация Световая адаптация
Рис. 4-30. Изменение величины центра РП нейрона с «оп»-центром в сетчатке млекопитающего при разных состояниях световой и темновой адаптации. При темновой адаптации периферия РП (серая зона) исчезает.
и амакриновыми клетками. Относительная сила возбуждения и торможения в рецептивной единице ганглиозной клетки сетчатки зависит от состояния адаптации, т.е. от среднего количества света, падающего на довольно большую область сетчатки.
Рис. 4-30 иллюстрирует тот факт, что функциональный центр РП становится тем меньше, чем выше средняя освещенность предъявляемого паттерна, тогда как общая площадь РП остается постоянной. По мере развития темновой адаптации центр РП расширяется, и, когда она становится полной, антагонистическая организация рецептивных полей полностью исчезает. В этих условиях нейрон с «оп»-центром отвечает на освещение любой части своего рецептивного поля активацией при включении света, а нейрон с «о(Г»-центром отвечает по всему своему рецептивному полю активацией при выключении света.
Плотность рецепторов и размеры РП как функция эксцентриситета на сетчатке. Эксцентриситет на сетчатке означает расстояние участка сетчатки от центральной ямки. Плотность рецепторов (их число на 1 мм2 поверхности сетчатки) меняется в зависимости от эксцентриситета. В колбочковой системе плотность рецепторов наибольшая в центральной ямке. Плотность же палочек наибольшая в парафовеальной области, а фовеа, центральная ямка, не содержит палочек. Плотность палочек также снижается по направлению к периферии сетчатки. На самом краю сетчатки рецепторный слой состоит почти полностью из палочек. Поскольку палочки не различают цветов, крайняя периферия функционально цветно-слепая.
Величина рецептивных полей растет от области центральной ямки к самому краю сетчатки. Данные, полученные на ганглиозных клетках светлоадаптированной сетчатки обезьяны, показывают, что центры рецептивных полей ганглиозных клеток в фовеальной области имеют в диаметре несколько угловых минут и увеличиваются по мере возрастания эксцентриситета. На периферии сетчатки центры РП ганглиозных клеток доходят до нескольких угловых градусов. Сравнение
180
О. Грюссер, У. Грюссер-Корнельс
истинной плотности рецепторов, полученной путем анатомических измерений, с размерами рецептивных полей позволяет думать, что конвергенция сигналов от рецепторов на ганглиозные клетки тоже растет с эксцентриситетом сетчатки. На основании тесной корреляции между диаметрами центров РП и остротой зрения в разных частях сетчатки можно заключить, что, как и следовало ожидать, острота зрения зависит не только от плотности рецепторов, но и от свойств РП. Это заключение подтверждается тем, что острота зрения изменяется при изменении средней освещенности; в этом случае диаметр центров рецептивных полей очень тесно коррелирует с остротой зрения, между тем как плотность рецепторов, разумеется, на зависит от состояния адаптации.
Ответы ганглиозных клеток на цветные стимулы. Количественная оценка поведенческих реакций говорит о том, что цветовое зрение у таких животных, как макак-резус, и у человека сходно. Однако функциональная организация рецептивных полей ганглиозных клеток сет.чатки у этих животных сложнее, чем описанное выше деление на клетки с «оп»- и «о(Т»-центрами. В светлоадаптированной сетчатке, а также в латеральном коленчатом теле и зрительной коре у них можно найти нервные клетки, дающие частично цвето-специфические ответы. Несколько упрощая, можно различить три класса таких клеток (рис. 4-31).
Ганглиозные клетки свето-темновой системы реагируют качественно одинаково (свойства «оп»- и «о(Г»-центров) при разных длинах волны монохроматического светового стимула в пределах видимого спектра (~ 400-700 нм). Спектральная чувствительность в центре и на периферии РП ганглиозных клеток этого класса одинакова.
Ганглиозные клетки красно-зеленой системы обладают частично цвето-специфической антагонистической организацией РП. Монохроматические световые стимулы из красной области спектра вызывают активацию в центре РП и торможение на периферии, а монохроматические стимулы из зеленой области спектра вызывают ответы противоположного характера.
Ганглиозные клетки желто-синей системы при монохроматической стимуляции центра РП активируются желтым светом и тормозятся синим; наоборот, на периферии желтый свет тормозит, а синий возбуждает.
Таким образом, нейроны сетчатки и коленчатого тела осуществляют преобразование сигналов, имеющих отношение к цветовому зрению. Через биполярные, горизонтальные и амакриновые клетки сигналы от трех разных типов колбочек направляются таким образом, что создают в слое ганглиозных клеток одну нейронную систему для «ахроматического зрения» и две цвето-специфические антагонистические нейронные системы. Эти последние составляют четырехцветную оппонентную систему с оппонентными парами цветов желто-синей и красно-зеленой.
Проекции сетчатки на ЦНС. Зрительные нервы обоих глаз соединяются у основания черепа и образуют зрительную хиазму (перекрест)
4. Физиология зрения
181
Свет падает на
Световой стимул “белый»
центр РП
периферию РП
Рис. 4-3 L Схема пространственной организации трех рецептивных полей в слое
центр РП
периферию РП
центр РП
периферию РП
ганглиозных клеток сетчатки и в латеральном коленчатом теле млекопитающего с цветовым зрением. А. Нервная клетка в системе свет- темнота. Б. Нервная клетка в системе красный-зеленый. В. Нервная клетка в системе желтый-синий. В цветоспецифических рецептивных полях (Б н В) центр и периферия организованы антагонистически.
(рис. 4-32). В хиазме человека около полумиллиона волокон зрительного нерва переходят на противоположную сторону; вторая половина остается ипсилатеральной и вместе с перекрещенными аксонами второго зрительного нерва образует зрительный тракт. Следовательно, на каждой стороне около миллиона аксонов ганглиозных клеток сетчатки проходят в зрительном тракте к первым центральным переключательным станциям зрительного пути-к латеральному коленчатому телу, верхнему бугорку и претектальной области ствола мозга. Перекрест нервных волокон в хиазме следует строгому правилу: аксоны ганглиозных клеток в височной части левой сетчатки и назальной части правой сетчатки входят в левый зрительный тракт, а аксоны от ганглиозных клеток назальной части левого глаза и височной части правого глаза образуют правый зрительный тракт (рис. 4-32). Аксоны большей части нервных клеток латерального коленчатого тела проходят по
182
О. Грюссер, У. Грюссер-Корнельс
назальная часть
темпоральная часть
Рис. 4-32. Схема зрительных путей в головном мозгу человека. Справа показаны также некоторые эфферентные связи между зрительной корой н подкорковыми структурами. Зрительные зоны в левом н правом полушариях соединены между собой аксонами, идущими через мозолистое тело. Разделение на поля 17, 18 и 19 сильно упрощено.
зрительной радиации к первичной зрительной коре (поле 17) в затылочной доле большого мозга. Эта область коры связана со вторичными и третичными зрительными центрами, а также с высшими зрительными ассоциативными областями, тоже лежащими в затылочной области коры больших полушарий или в теменной коре. Из этих зрительных зон идут пути к корковым станциям переключения афферентной зрительной системы, показанным на рис. 4-32, к участкам ствола мозга, контролирующим движения глаз (см. разд. 4.4), и к ассоциативным зонам в теменной и височной долях, функция которых связана с речью. Зрительные центры левого и правого полушарий сообщаются через мозолистое тело (рис. 4-32).
Центральный зрительный путь характеризуется топографической организацией. Это значит, что подобно тому, как пространственные отношения в определенной географической области сохраняются на ее карте,
4. Физиология зрения
183
так и пространственное распределение возбуждения в слое ганглиозных клеток сетчатки «картируется» пространственным распределением возбуждения нейронов в латеральном коленчатом теле, верхнем бугорке и зрительной коре. Но детали такого картирования несколько отличны от географического стандарта. Карта чертится в определенном масштабе (например, уменьшение 1:100 000), так что каждый километр горизонтального расстояния естественной местности соответствует фиксированному расстоянию на бумаге, тогда как топографическая проекция сетчатки нелинейна. Маленькая область центральной ямки проецируется на гораздо большее число центральных нейронов, чем область такой же величины на периферии сетчатки. Эта нелинейная проекция отражает гораздо большее функциональное значение центральной ямки по сравнению с периферией, а также снижение остроты зрения по мере удаления от ямки (см. разд. 4.2).
Нейронная переработка сигналов в латеральном коленчатом теле. Нервные клетки латерального коленчатого тела, первой станции переключения на пути между сетчаткой и корой большого мозга, обычно обладают простыми концентрическими РП (судя по измерениям при стимуляции сетчатки светом), подобно РП ганлиозных клеток сетчатки. Бинокулярное взаимодействие между сигналами от левого и правого глаза выявляется здесь только как слабое реципрокное торможение. В трех из шести слоев нервных клеток коленчатого тела «доминирует» один глаз, а в остальных трех слоях-второй. Таким образом, в каждой из двух трехслойных групп происходит переработка сигналов преимущественно одного глаза. Истинная бинокулярная интеграция зрительной информации от обоих глаз происходит впервые в зрительной коре. Функция латерального коленчатого тела до сих пор далеко не ясна, несмотря на обширные исследования. Имеются данные о том, что процессы латерального торможения, столь важные при различении контраста (см. разд. 4.2), особенно выражены в некоторых клетках коленчатого тела. И, как было указано выше, информация о цвете представлена здесь по меньшей мере в трех разных классах нервных клеток (светло-темной, желто-синей и красно-зеленой системах). Возбудимость нервных клеток в латеральном коленчатом теле сильно меняется с переходом от бодрствования ко сну. В состоянии глубокого сна передача сигнала в коленчатом теле, вероятно, резко снижена.
Нейроны зрительной коры. В нервных клетках первичной, вторичной и третичной зрительных зон затылочной коры появляются такие особенности организации РП, которые неизвестны на нижних уровнях. Строение, типичное для нейронов сетчатки и коленчатого тела с концентрическими возбудительными и тормозными зонами, обнаружено лишь для некоторых нейронов первичной зрительной коры. В рецептивных полях остальных корковых зрительных нейронов «оп»- и «ой»-зоны расположены параллельно друг другу (рис. 4-33). Диффузное освещение всего РП обычно мало меняет спонтанную активность этих клеток. Но если на РП проецируется «полоска» света в правильных ориентации
184
О. Грюссер, У. Грюссер-Корнелъс
| On доча | рэрчона | f 'Светлая i ! полоска i
Рис. 4-33. Организация «простого» рецептивного поля нервной клетки в первичной зрительной коре. (Упрощенный рисунок из Hubei, Wiesel, J. Physiol. (Lond.), 160, 106-154, 1962.)
и положении, то наступает сильная активация. Так происходит, например, когда полоска ориентирована, как на рис. 4-33, В. РП с такими параллельными «оп»- и «off»-зонами называются «простыми», потому что их функциональная организация легко устанавливается путем проецирования маленьких пятнышек света на разные части РП; иначе говоря, ответ на полоски можно понять на основании ответов на пятнышки.
Другие же корковые зрительные нейроны обладают «сложными» РП. Для активации таких нейронов нужно проецировать на РП светлотемные контуры со специфической пространственной ориентацией и размерами, перерывами линии, углами и т. п. Все участки РП, раздражение которых «правильным» стимулом вызывает активацию нейрона, называют возбудительным рецептивным полем (ВРП) Обычно ВРП окружено областью, в которой светло-темные стимулы вызывают только торможение. Эта область называется тормозным рецептивным полем (ТРП). Нейроны со сложными РП обычно не отвечают на проецируемые на их РП диффузные неструктурированные световые стимулы. На рис. 4-34 показаны ответы двух нейронов зрительной коры со сложными РП.
Многие корковые нейроны со сложными РП активируются гораздо сильнее не неподвижными, а движущимися стимулами. Кроме того, некоторые из этих нейронов, чувствительных к движению, требуют определенного направления движения стимула. Например, стимул с оптимальной пространственной структурой окажет возбуждающий эффект, только если он движется по ВРП слева направо, а движение в обратном направлении не будет активировать клетку. Чувствительность корковых зрительных нейронов к движению и направлению, несомненно, отра-
4. Физиология зрения
185
Тормозное рецептивное поле Движение
Возбудительное Направление движения Конфигурация стимула рецептивное поле
Нейрон поля 18
Рис. 4-34. Схема ответов двух нейронов в зрительной коре (поля 18 н 19) со сложными рецептивными полями. Стимул обозначен серой штриховкой, стрелка показывает направление движения. (Упрощенный рисунок из Hubei, Wiesl, J. Neurophysiol., 28, 229-289, 1965.)
жает приспособление к тому, что изображение неподвижного окружения всегда смещается по сетчатке из-за непрерывного движения глаз и тела (см. разд. 4.4). «Мозговой образ» неподвижного зримого мира должен быть создан во время коротких периодов фиксации из картин на сетчатке, которые меняются при каждом движении глаза.
Нейроны зрительной коры со сложными РП, как правило, обладают своими РП для каждого глаза. Поэтому эти клетки могут быть возбуждены монокулярно, т.е. раздражением одного глаза. Бинокулярная стимуляция одинаковыми стимулами приводит к более сильной актива
186
О. Грюссер, У. Грюссер-Корнельс
ции (бинокулярная суммация). РП бинокулярно активируемой корковой нервной клетки, приходящиеся на участки левой и правой сетчатки, обладают только приблизительным геометрическим соответствием (см. разд. 4.2). Отклонение этих участков от точного соответствия областей сетчатки варьирует у разных нейронов. Эта диспаратность такова, что стимул, наносимый бинокулярно, вызывает оптимальное возбуждение, когда он лежит несколько снаружи от гороптера. Такой нейронный ответ рассматривается как нейрофизиологический коррелят бинокулярного восприятия глубины, о чем более подробно речь шла выше.
Нейронная основа восприятия формы. Корковые зрительные нейроны со сложными РП отвечают только на узко специфические зрительные стимулы. Эти оптимально активирующие свойства различны для разных классов нейронов. В общей сложности в первичной, вторичной и третичной зрительной коре обнаружено не менее 15 классов нейронов; в каждом случае они распределены по всей корковой проекционной зоне поля зрения. Путем отведения активности по очереди от множества разных нервных клеток в зрительной коре можно реконструировать пространственную картину возбуждения, которая существует в данный момент в корковых нейронах одного класса под воздействием определенного зрительного стимула. На рис. 4-35 дана попытка такой реконструкции в упрощенном виде. Пространственное распределение возбуждения в нейронах разных классов отражает разные свойства стимула. Поскольку проекция сетчатки на зрительной коре нелинейна, это распределение во многих тысячах нейронов данного класса не такое простое, как показано на рис. 4-35. Данная схема имеет целью лишь показать одновременные многочисленные представительства одного изображения, которые проявляются в особом распределении возбуждения в нейронах зрительной коры разных классов. Рисунок помогает также дать читателю представление о сложности переработки сигналов, с которой приходится иметь дело в нейрофизиологических исследованиях отдельных нервных клеток зрительной коры млекопитающих. Нейронные механизмы, воссоздающие форму из множественных предъявлений зрительной информации в виде разных конфигураций возбуждения в зрительной коре, пока еще совершенно неясны. Но из наблюдений над больными, у которых нарушено зрительное восприятие формы и предметов (зрительная агнозия), известно, что для этих функций необходимы не только зрительные проекционные зоны затылочной коры больших полушарий, но также высшие ассоциативные зоны теменной коры.
В 4.13. Которое из следующих утверждений приложимо к рецепторным потенциалам отдельных колбочек в сетчатке позвоночных? а) При освещении возникает деполяризационный рецепторный потенциал.
б) При освещении возникает гиперполяризационный рецепторный потенциал.
4. Физиология зрения
187
Рис. 4-35. Схема процессов возбуждения, вызываемых светящейся буквой К в разных нейронных слоях сетчатки н центральной нервной системы, а. Изображение буквы на сетчатке и пространственное распределение возбуждения в рецепторном слое сетчатки, б, в. Распределение возбуждения на выходе сетчатки в слое ганглиозных клеток. Возбуждение обозначено красными полосами, б. Нейроны с «оп»-центром. в. Нейроны с «оП»-центром. г. Распределение возбуждения в нейронном слое латерального коленчатого тела и некоторых нейронах зрительной коры. Контуры светящейся буквы вызывают возбуждение в нервных клетках..д-и. Распределение возбуждения в разных слоях нервных клеток различных классов в первичной, вторичной и третичной зрительной коре головного мозга. Здесь нервные клетки возбуждаются только контурами определенной ориентации, углами или разрывами линий. На рисунке нейробиологическне отношения сильно упрощены; пространственное распространение возбуждения в разных слоях не связано линейно с размерами стимула.
в) При освещении мембранный потенциал рецептора не меняется.
г) Рецепторный потенциал совершенно не связан с движением ионов через мембрану.
д) Имеется линейная зависимость между амплитудой рецепторного потенциала и интенсивностью стимула.
е) Имеется приблизительная логарифмическая зависимость между амплитудой рецепторного потенциала и интенсивностью стимула.
В 4.14. Которое из следующих утверждений приложимо к родопсину (Р)?
а) Р состоит из опсина и ретиналя.
б) Р состоит из гамма-глобулина и витамина А.
188
О. Грюссер, У. Грюссер-Корнелъс
в) Р идентичен зрительному пигменту в колбочках.
г) Концентрация Р в палочках растет при темновой адаптации, д) Концентрация Р в палочках снижается при темновой адаптации.
е) Раствор Р красный, но обесцвечивается после освещения.
В 4.15. Закончите правильно следующие фразы. Нейроны сетчатки с «оп»-центром (ганглиозные клетки) отвечают на освещение центра РП (активацией/торможением), тогда как затемнение центра РП ведет к (активации/торможению). Освещение периферии РП нейронов с «оп»-центром вызывает (активацию/тормо-жение), а затемнение вызывает (активацию/торможение). Следовательно, центр и периферия РП организованы... При увеличении средней интенсивности света, падающего на сетчатку, диаметр центра РП становится (болыпе/меньше).
В 4.16. Которые из следующих утверждений правильны? Рецептивные поля ганглиозных клеток в сетчатке млекопитающих а) в фовеальной области меньше, чем на периферии;
б) в среднем одинаковых размеров по всей сетчатке;
в) могут быть подразделены на функционально разные центр и периферию в условиях световой адаптации;
г) могут быть обнаружены только у нейронов центральной ямки;
д) иногда организованы в отношении спектральной чувствительности антагонистически;
е) не крупнее рецептивных полей одиночного фоторецептора.
В 4.17. Зная организацию рецептивных полей нервных клеток зрительной коры, можно понять, почему следующие свойства зрительных стимулов особенно важны для восприятия формы, а) Средняя освещенность.
б) Контуры.
в) Разрыв контура.
г) Цвета.
д) Тот факт, что на сетчатке создается перевернутое изображение видимых предметов.
4.4. Движения глаз н сенсомоторная интеграция при зрительном восприятии
В предыдущем обсуждении разных аспектов зрения была недостаточно принята во внимание решающая роль движений глаз в восприятии видимого мира. Чтобы понять значение сенсомоторной интеграции при зрительном восприятии, надо знать нейронные механизмы, контролирующие движения глаз. Поэтому к ним мы обратимся прежде всего.
Глаз человека приводится в движение шестью наружными мышцами (рис. 4-36). Эти мышцы могут действовать координированно по совершенно различным программам. Благодаря тому, что глаз лежит в глаз-
4. Физиология зрения
189
Верхняя
косая мышца Верхняя прямая мышца
Верхняя мышца, поднимающая веко
Медиальная прямая мышца Латеральная _____
прямая мышца
Зрительный нерв__
Нижняя
прямая мышца
Нижняя косая мышца -
Рис. 4-36. Схема расположения наружных глазных мышц н глазного яблока в глазнице.
нице, как мячик в ямке, он может двигаться под многими разными углами, что служит предпосылкой для разных программ, направляющих взор. Относительно координат головы глаз совершает горизонтальные, вертикальные или вращательные движения. Эти последние (поворот во фронтальной плоскости) ограничены механическими причинами приблизительно 10°, а вертикальные и горизонтальные движения могут перемещать зрительную ось почти до краев глазницы. При их сочетании глаз движется по диагонали в направлениях, которые задаются по желанию соответствующим программированием наружных мышц глаза.
Содружествешгые .(важенмя глаза; конвертенцнм, дивергенция. Когда человек с нормальным бинокулярным зрением смотрит вокруг себя, его глаза движутся настолько координированно, что изображение объекта, фиксируемого в каждый момент (точнее, точка на объекте), падает в середину центральной ямки обоих глаз. Более внимательное наблюдение за их движением позволяет различить две «программы». Во-первых, глаза могут двигаться вместе вверх, вниз, влево или вправо. Оба глаза движутся в одном направлении по отношению к координатам головы. Эти бинокулярные движения называются содружественными.
Во-вторых, если Человек смотрит попеременно на близкие и далекие предметы, то глаза совершают приблизительно зеркальные движения относительно координат головы. При таких движениях угол между зрительными осями обоих глаз меняется. При фиксации далекой точки зрительные оси почти параллельны. Если взор переводится на ближнюю
190
О. Грюссер, У. Грюссер-Корнельс
точку, зрительные оси сходятся (конвергентные движения). При последующей повторной фиксации отдаленного объекта глазные оси расходятся, совершая дивергентное движение, пока снова не станут почти параллельными. Конвергентные и дивергентные движения могут происходить одновременно с сопряженными, например в том случае, когда взор смещается с далекого объекта, находящегося справа, к ближнему объекту слева.
Вращательные движения глаз во фронтальной плоскости тоже координируются бинокулярно. Например, когда голова наклонена в сторону, происходит легкое «компенсаторное вращение» глаз (см. разд. 6.2).
Динамика двкжешм глаз. Когда вы смотрите без определенной цели, ваши глаза движутся от одной фиксационной точки к другой быстрыми скачками (саккадами). Амплитуда саккад может составлять всего лишь несколько угловых минут (микросаккады) или же много градусов; так, например, перевод фиксационной точки с правой половины поля зрения на левую осуществляется путем большой саккады. Длительность саккад варьирует приблизительно от 10 до 80 мс. Большие саккады часто происходят вместе с дополнительными движениями головы. Саккады разделены периодами фиксации, которые длятся обычно от 150 до 300 мс.
Движущийся объект сопровождается медленными следящими движениями глаз, если только скорость движения не слишком велика. При быстроте движения меньше 120°/с угловая скорость движения глаз приблизительно соответствует скорости движущегося объекта. Функция медленных следящих движений состоит в том, чтобы удерживать изображения фиксируемого движущегося предмета в середине центральных ямок, т.е. в положении, где острота зрения наибольшая. При угловой скорости движущегося объекта, превышающей 150°/с, глаза не поспевают за объектом: несмотря на старания уследить за ним, его изображение не попадает на одну и ту же точку на сетчатке.
Следящие движения, саккады и периоды фиксации представляют собой разные формы движений, управляемых различными «программами», заложенными в глазодвигательной системе ствола мозга. В течение довольно длительных периодов произвольной фиксации (длительностью 0,5-2 с) происходит медленный низкоамплитудный «дрейф» точки фиксации. Даже во время наилучшей фиксации на нее накладывается «микротремор» глаз-движения очень малой амплитуды (Г) с доминирующим частотным компонентом от 80 до 120 Гц.
Движения бинокулярной фиксации. Во время конвергентных и дивергентных движений глазодвигательные системы, управляющие движениями обоих глаз, подчиняются нейронной программе, координирующей их одновременное движение. Когда система, управляющая взором, не может привести зрительные оси обоих глаз к одной и той же точке, возникает «косоглазие», или стробизм, и изображения, попадающие на центральные ямки двух глаз, не одинаковы (см. разд. 4.1). Как было указано в разд. 4.2, в таком случае может произойти мешающее двоение в глазах. Механизмы, управляющие взором, устанавливают отношение
4. Физиология зрения
191
Рис. 4-37. Схема подкорковых центров, управляющих горизонтальными движениями глаз. Подробнее см. в тексте.
между возбуждением и торможением глазодвигательных мотонейронов ствола мозга, а эти последние в свою очередь определяют относительную степень сокращения наружных глазных мышц.
На рис. 4-37 весьма схематично показана нейронная система, осуществляющая горизонтальные движения глаз. «Центры» бинокулярного управления взором лежат в области ретикулярной формации моста и среднего мозга, в верхних бугорках и в претектальной области. Ми-
192
О. Грюссер, У. Грюссер-Корнелъс
кроэлектродные отведения от ретикулярной формации ствола мозга показали, что саккадические движения глаз регулируются ретикулярной нейронной системой, отличной от системы, управляющей медленными следящими движениями.
Центры ствола мозга, управляющие бинокулярными движениями, сами управляются входами от зрительной коры и «фронтального глазного поля» коры больших полушарий. Эти нейронные связи имеют значение для корреляций между конфигурацией зрительных стимулов и сканирующим движением глаз (см. ниже) и для управления следящими движениями. Нервные клетки в верхних бугорках и претектальной области получают зрительные афференты через кору, а также прямо от глаз по коллатералям некоторых аксонов зрительного нерва (см. рис. 4-32).
На рис. 4-37 показаны также связи органов равновесия (лабиринтов) со стволовыми центрами монокулярных и бинокулярных движений. Эти связи имеют непосредственное отношение к рефлекторным изменениям положения глаз, вызываемым изменениями положения головы. Но как правило, у бодрствующего человека глазодвигательные рефлексы, возникающие при возбуждении лабиринтов, маскируются другими нервными командами, управляющими движениями глаз. Лабиринтные глазодвигательные рефлексы имеют значение главным образом для «удержания» точки фиксации при внезапном движении головы. Но рецепторы в лабиринте могут быть возбуждены искусственно или патологически настолько, что вестибулярное возбуждение становится единственным фактором, определяющим движение глаз. В таких условиях возникают вестибулярный нистагм и связанное с ним головокружение, о котором см. в разд. 6.2.
Точная регуляция движений глаз-в особенности при медленных следящих движениях, хотя в известной степени это касается и точности саккад-возможна при нормальном состоянии мозжечка. В мозжечке вестибулярные и зрительные сигналы перерабатываются в сочетании с сигналами, отражающими положение глаз и головы. Результаты этого анализа передаются по мозжечковым эфферентным путям к стволовым центрам, управляющим бинокулярным движением.
Повседневный опыт показывает, что движения глаз могут быть показателем «внутреннего психологического состояния». Внимание, усталость, интерес, страх и безразличие-все это меняет частоту и амплитуду саккад. Влияние эмоций на движение глаз, вероятно, осуществляется по «неспецифическим» системам нейронов ретикулярной формации, на которые в свою очередь влияет «лимбическая система».
Простым и часто применяемым методом измерения движения глаз, полезным для диагностики, является электроокулография. Она использует то обстоятельство, что между роговицей и сетчаткой каждого глаза имеется разность потенциалов -корнео-ретинальный постоянный потенциал. Это значит, что глаз является диполем, в котором роговица электроположительна по отношению к сетчатке (рис. 4-38). Регистрирующие электроды укрепляются пастой на коже над костным краем
4. Физиология зрения
193
и электроокулограмма оптокинетического нистагма.
глазницы над и под глазом, а также у носа и виска. При движении переднего полюса глаза в сторону одного из электродов между соответствующей парой электродов создается разность потенциала, потому что это движение меняет направление оси электрического диполя. Напряжение, отводимое каждой парой электродов, приблизительно пропорционально смещению глаза в глазнице. Таким образом, электроокулограмма позволяет проследить за положением глаза относительно координат головы.
Движения глаз и зрительное восприятие; нистагм. Сначала рассмотрим только горизонтальные движения. На рис. 4-39 дана электроокулограмма, зарегистрированная при чтении: испытуемый прочел в книге три строчки. В этой записи движение глаз влево вызывает отклонение кривой вверх, а движение вправо-отклонение вниз. Видно, что глаза совершают при чтении быстрые саккады с короткими периодами фиксации между саккадами. Когда точка фиксации достигает конца строчки, она перепрыгивает к началу следующей строчки одной саккадой. В зависимости от амплитуды саккады длятся от 10 до 80 мс; средняя угловая скорость движения глаз составляет от 200°/с до 600°/с.
При рассматривании движущегося стимула возможно циклическое чередование саккад и медленных следящих движений. Если смотреть на
194
О. Грюссер, У. Грюссер-Корнелъс
А А А
1 с
Рис. 4 39. Движения глаз при чтении трех строк текста. Каждая строка сканируется пятью или шестью саккадами. В точках А более крупная саккада возвращает глаз к началу следующей строки.
ландшафт через боковое окно автомобиля или поезда, то глаза попеременно совершают сопряженные медленные горизонтальные движения и быстрые саккады. Во время медленных следящих движений изображение фиксируемого объекта удерживается вблизи центральной ямки. Иными словами, это движение следует за относительным движением объекта. Чередование саккад и медленных следящих движений называется нистагмом. Описанный выше пример-это оптокинетический нистагм, называемый так, потому что он вызывается движущимися оптическими стимулами.
Оптокинетический нистагм легко вызвать у человека, фиксирующего деления на сантиметровой ленте, которую экспериментатор передвигает в горизонтальном или вертикальном направлении. Условились обозначать направление нистагма по направлению быстрой фазы (по саккадам). Таким образом, когда лента движется вправо, если смотреть от наблюдателя, возникает «левый оптокинетический нистагм» - медленные следящие движения вправо сменяются саккадами влево, которые, меняя положение глаз, позволяют установить новую точку фиксации. Стимулом, обычно применяемым для более точного анализа оптокинетического нистагма, служит изображение движущихся горизонтально или вертикально светлых и темных полос, проецируемых на внутреннюю поверхность полуцилиндра.
Движения глаз при обычном рассматривании предметов. Если мы рассматриваем хорошо структурированное изображение, то попеременно возникают саккады и периоды фиксации. Глаз может двигаться в любом направлении. На рис. 4-40 показана двумерная запись последовательных фиксаций у человека, рассматривающего сложную картину. Совершенно очевидно, что движения глаз в известной степени определяются контурами, перерывами контуров или точками, где два контура встречаются. Если изображение представляет собой человеческое лицо, то тогда особенно часто фиксируются рот и глаза. Следовательно, бинокулярная фиксация управляется не только структурой стимула, но и значением его компонентов. Если стимул рассматривается в течение некоторого времени, он приблизительно воспроизводится в виде образа, отражаемого записью сканирующих движений глаз. Частота фиксаций соответствует «важности» некоторых субструктур стимула. Психологи, работающие в области рекламы, использовали это обстоятельство для
4. Физиология зрения
195
Рис. 4-40. Двумерное картирование движений глаз при рассматривании лица. Испытуемый в течение нескольких минут смотрел на помещенную слева фотографию (Yarbus, 1965).
повышения эффективности рекламы: измеряли движения глаз, чтобы определить, какие структуры рекламных изображений больше всего «привлекают взор».
Саккады смещают точку фиксации в пределах поля зрения: во время коротких периодов фиксации зрительный сигнал видим. Во время быстрого саккадического движения изображения по сетчатке оно не воспринимается. Это объясняется разными причинами. Если точка фиксации движется во время саккады по сильно структурированному пространству, то временные флуктуации интенсивности в каждой точке сетчатки обычно намного превышают частоту слияния мельканий (см. разд. 4.2). Тем самым сигнал на сетчатке во время саккады очень близок краткому «серому стимулу». Вторым фактором, по-видимому, является активный центральный тормозный механизм, подавляющий восприятие движений во время саккад. При объяснении этого явления надо принять, что высшие зрительные центры получают по обратным связям некоторые сигналы от стволовых или корковых центров, управляющих движением глаз. Вы можете сами у себя наблюдать торможение восприятия движения в следующем простом опыте. Смотрите на свой глаз в зеркало и переводите взгляд от внутреннего к наружному краю глазницы и обратно. Вероятно, вы не увидите движения вашего глаза. Теперь повторите этот опыт на другом человеке и наблюдайте в зеркало за движением одного из его глаз, когда он также двигает его между краями глазницы. Вы совершенно отчетливо увидите это движение.
Координация движений глаз и зрительного восприятия осуществляется главным образом по связям между зрительной корой и участком ствола мозга, управляющим бинокулярной фиксацией. В разд. 4.3 было указано, что некоторые нервные клетки в зрительной коре обладают сложными рецептивными полями. Для нервных клеток во вторичной и третичной зрительных зонах самыми эффективными стимулами являются контуры, перерывы контуров и соединения контуров. Весьма вероятно поэтому, что эти нервные клетки управляют сканирующими
196
О. Грюссер, У. Грюссер-Корнелъс
движениями глаз по своим связям с подкорковыми центрами, управляющими взором, в особенности в верхних бугорках. О зависимости сканирующих движений также и от высших ассоциативных зон головного мозга свидетельствует связь между движениями глаз и текстом во время чтения. При чтении зрительная информация получается только в периоды фиксации. Фиксационные точки располагаются предпочтительно в начале строчки и в начале слова, в особенности если оно начинается с прописной буквы. Длинное слово может фиксироваться в нескольких точках. Впрочем, вероятность фиксации определяется не только физической структурой текста, но и значением слов. В длинном тексте на одной и той же последовательности слов точки фиксации могут быть в разных местах, когда значение этих слов меняется в зависимости от контекста. Это наблюдение показывает, что во время чтения сенсорная речевая зона в верхней височной извилине коры (зона Вернике) тоже влияет на движение глаз.
Движения глаз, вызываемые стимулами, движущимися на периферии поля зрения. Когда движущийся объект внезапно появляется на периферии поля зрения, он вызывает рефлекторную саккаду, которую может сопровождать движение головы. Движения эти координированы таким образом, что изображение «нового» объекта перемещается на центральную ямку. Этим рефлексом управляют чувствительные к движению нейроны зрительной коры и верхних бугорков. В целом движущиеся объекты на периферии поля зрения эффективнее, чем в центральной ямке. Эти рефлекторные саккады связаны с переключением внимания на новый объект. Описанный выше механизм, очевидно, представляет собой биологическое приспособление, важное для древних млекопитающих и доисторического человека. Повышенная чувствительность к движению на периферии поля зрения и рефлекторное перемещение внимания и направления взора облегчают опознание внезапно возникающего движущегося зрительного стимула, например жертвы и хищника. Повышенная периферическая чувствительность чрезвычайно полезна и в условиях современного транспорта; она позволяет водителю или пешеходу правильно реагировать на неожиданное появление машины на краю его поля зрения. Таким образом, даже в наше время филогенетически древний механизм продолжает повышать вероятность выживания.
В 4.18. Саккады представляют собой а) скачки глаз;
б) медленные следящие движения глаз;
в) вращательные осцилляции глаз;
г) ни одно из этих утверждений не правильно.
В 4.19. Которая из следующих записей электрической активности служит мерой движения глаз?
а) Электрокардиограмма.
б) Электроретинограмма.
4. Физиология зрения
197
в) Электроокулограмма.
г) Электроэнцефалограмма.
д) Электронистагмограмма.
В 4.20. Когда человек смотрит на картину, движения его глаз
а) статистически случайны;
б) зависят от красок на картине;
в) зависят от контуров и их разрывов; .
г) обычно координированы для обоих глаз;
д) независимы у каждого глаза, но зависят от определенных свойств картины.
В 4.21. Когда вы смотрите на пейзаж через боковое окно поезда, идущего с постоянной скоростью, ваши глаза совершают а) скачкообразные движения конвергенции;
б) оптокинетический нистагм;
в) движения, вызываемые из лабиринтов;
г) неупорядоченные движения, независимые от направления движения поезда;
д) ни одно из этих утверждений не правильно.
5. ФИЗИОЛОГИЯ СЛУХА
Р. Клинке
Чувство слуха одно из главных в человеческой жизни; слух и речь составляют вместе весьма важное средство общения между людьми и служат основой сложных социальных взаимоотношений. Неудивительно поэтому, что потеря слуха может привести к тяжелым нарушениям поведения.
С самого начала своей истории люди знали, что слухом ведает ухо. Но до XVII в. никто не подозревал, что слух связан с системой полостей, лежащих в толще кости у основания черепа. То, что эта система служит органом слуха, было окончательно доказано в начале XIX в. Во второй его половине внимание ученых привлекали физиология слуха и проблемы физической акустики. Особенно большой вклад в изучение этих проблем внес Гельмгольц, и его имя часто звучит и сегодня. Тем не менее многие проблемы оставались нерешенными; некоторые из них решены в нашем веке, но многие важные аспекты физиологии слуха до сих пор остаются неясными.
5.1. Анатомия органа слуха
Понимание физиологии слуха требует некоторого знакомства со строением рецепторного аппарата. Ниже дается его краткое не претендующее на исчерпывающее описание. Предполагается, что читатель может обратиться дополнительно к руководствам по анатомии.
Орган слуха состоит из наружного, среднего и внутреннего уха (рис. 5-1). Слуховой проход, соединяющий первые два из них, перегорожен на своем внутреннем конце барабанной перепонкой. Эта тонкая мембрана в здоровом состоянии отсвечивает перламутром, что дает в руки врача ценный диагностический критерий. За барабанной перепонкой лежит наполненная воздухом полость среднего уха. Полость эта соединена с глоткой узким проходом-евстахиевой трубой; при глотании происходит некоторый обмен воздухом между глоткой и средним ухом. Изменение давления наружного воздуха, как, например, в самолете, вызывает неприятное ощущение-«закладывает» уши. Оно объясняется натяжением барабанной перепонки из-за разницы между атмосферным давлением и давлением в полости среднего уха. При глотании евстахиева труба открывается, и таким образом давление по обе стороны барабанной перепонки выравнивается.
В среднем ухе расположены три маленькие косточки -молоточек, наковальня и стремечко (или по-латыни соответственно malleus, incus, stapes). Они гибко связаны между собой и образуют своего рода цепоч-
5. Физиология слуха
199
Орган равновесия
Орган слуха
Наружное ухо • Среднее' Внутреннее ухо ухо |
Рис. 5-1. Схема наружного, среднего и внутреннего уха. М-молоточек, Н-наковальня, С-стремечко. Пунктирные линии близ М, Н и С показывают крайние положения, в которые косточки могут быть приведены колебаниями барабанной перепонки.
проход
ку. Один из отростков молоточка слит с барабанной перепонкой. Когда воздушные колебания приводят в движение барабанную перепонку, оно передается костной цепочке. Стремечко действительно похоже на стремя, основание которого входит в отверстие в кости, называемое овальным окном. Эта пластинка образует границу между полостью среднего уха и третьим отделом органа слуха, внутренним ухом. Таким образом, цепочка из косточек служит мостиком между барабанной перепонкой и овальным окном, между атмосферой и внутренним ухом. По этому пути звуковая энергия достигает внутреннего уха, где, как мы увидим, заложены сенсорные клетки.
Внутреннее ухо находится в височной кости; оно непосредственно сообщается с органом равновесия (см. рис. 5-1). Вместе оба органа называются лабиринтом. Из-за своей формы внутреннее ухо называется также улиткой (по-латыни cochlea; на рисунке она частично развернута). К органу равновесия мы вернемся в следующей главе. Улитка состоит из трех параллельных, свернутых в катушку трубчатых каналов. На рис. 5-2 показан разрез поперек оси улитки, и таким образом каналы, обвивающие ось, перерезаны в нескольких местах. Эти каналы называются scala vestibuli (лестница преддверия, или вестибулярная), scala media (средняя лестница, или улитковый проток) и scala tympani (барабанная лестница). У человека улитка образует два с половиной витка. Общее их расположение показано на рис. 5-1. Пластинка стремечка в овальном окне примыкает к вестибулярной лестнице, которая (как и остальные каналы) заполнена жидкостью. Вестибулярная и барабанная лестницы содержат так называемую перилимфу, а улитковый проток заполнен эндолимфой. Эти жидкости различаются по химическому составу. Перилимфа содержит много натрия, приблизительно в той же
200
Р. Клинке
Рис. 5-2. Сильно упрощенный разрез улитки человека, несколько раз пересекающий завитки улитковых протоков. Область, взятая в рамку, подробно показана на рис. 5-3.
концентрации, что внеклеточная жидкость, а эндолимфа богата калием, как внутриклеточная жидкость. Вестибулярный и барабанный ходы лестницы соединяются у геликотремы (helicotrema), верхушки улитки. У основания улитки оба эти канала отделены от полости среднего уха сходными структурами. Овальное окно, ведущее в вестибулярную лестницу, замкнуто стремечком, причем края отверстия запечатаны кольцевой связкой, а круглое окно в конце барабанной лестницы затянуто тонкой перепонкой, которая отделяет ее от полости среднего уха, и перилимфа не может в него просочиться.
Часть рис. 5-2 более подробно показана на рис. 5-3. Здесь можно видеть, что граница между вестибулярной лестницей и улиточным ходом образована мембраной, которая называется вестибулярной (или рейсне-ровой). Границу между улитковым протоком и барабанной лестницей образует базилярная (основная) мембрана, на которой находится соб< ственно сенсорный аппарат -кортиев орган. В кортиевом органе лежат рецепторные клетки, окруженные обкладочными клетками. Они называются волосковыми клетками из-за их субмикроскопических, подобных волоскам отростков, стереоцилий. Различаются внутренние и наружные волосковые клетки (рис. 5-3). Наружные расположены тремя рядами, а внутренние образуют только один. Таким образом, наружных волосковых клеток значительно больше, чем внутренних.
Над кортиевым органом лежит желатинозная масса, текториальная (покровная) мембрана. Она прикреплена к внутренней стороне улитки, близ ее оси. Она также прикасается к цилиям волосковых клеток, обра-
5. Физиология слуха
201
Спиральный ганглий
0,1 мм
Рейснерова мембрана
Вестибулярная лестница
Кость
Нервные
Барабанная
Основная мембрана
Сосудистая полоска
Внутренние Наружные волосковые клетки
Средняя
лестница
Тенториальная
мембрана
волокна
лестница
Рис. 5-3. Поперечный разрез завитка улитки. Кортиев орган на основной мембране содержит рецепторные (волосковые) клетки; они накрыты текториальной (покровной) мембраной.
зуя довольно тесный контакт с ними. Во всяком случае, так обстоит дело с наружными волосковыми клетками. Вдоль наружной стороны улиткового протока тянется полоска, где сосредоточены кровеносные сосуды, называемая stria vascularis, сосудистой полоской. Эта структура играет главную роль в удовлетворении энергетических потребностей улитки; кроме других своих функций она поддерживает концентрацию К+ в эндолимфе.
Рецепторные клетки в кортиевом органе являются вторичными сенсорными клетками -это значит, что у них нет аксонов. Тела клеток, передающих возбуждение от этого органа к центральной нервной системе, находятся в спиральном ганглии, который лежит в улитке, завиваясь вокруг ее оси вместе с каналами (рис. 5-2).
Нервные клетки в этом ганглии биполярные. У каждой клетки один отросток идет на периферию, к волосковый клеткам кортиева органа, другой-в составе слухового нерва к ЦНС. Каждая внутренняя волосковая клетка образует синапсы со многими афферентными нервными волокнами, каждое из которых, вероятно, контактирует только с одной этой волосковой клеткой. В отличие от этого нервные волокна, снабжающие наружные волосковые клетки, сильно ветвятся, и каждое из них получает синаптические входы от многих наружных волосковых клеток. Следовательно, несмотря на то, что наружных волосковых клеток боль
202
Р. Клинке
ше, основная часть волокон в слуховом нерве идет от внутренних волосковых клеток.
В 5.1. Расположите следующие структуры в порядке следования от наружного воздуха к внутреннему уху.
а) Молоточек
б) Барабанная Перепонка.
в) Стремечко.
г) Наковальня.
В 5.2. Вестибулярная лестница и улитковый проток
а) соединены у геликотремы;
б) соединены у овального окна;
в) соединены у круглого окна;
г) не соединены.
В 5.3. Кортиев орган лежит на
а) вестибулярной (рейснеровой) мембране;
б) покровной мембране;
в) основной мембране;
г) барабанной мембране;
д) сосудистой полоске.
В 5.4. Сосудистая полоска играет основную роль в
а) снабжении внутреннего уха энергией;
б) компенсации движений стремечка;
в) поддержании ионной среды в эндолимфе;
г) образовании перилимфы;
д) всасывании перилимфы.
В 5.5. Эндолимфа богата
а) К+;
б) Na +
и поэтому сходна с
в) внутриклеточной жидкостью;
г) внеклеточной жидкостью.
Положение в перилимфе
д) такое же;
е) противоположное.
5.2. Слух человека
Физические свойства звуковых стимулов. Колеблющееся тело, например камертон или мембрана громкоговорителя, вызывают колебания окружающего воздуха, ускоряя движение молекул воздуха в непосредственной близости от себя. Эти последние в свою очередь передают энергию дальше, так что и там молекулы воздуха начинают колебаться. Эти возмущения поэтому распространяются от источника волнами со скоростью (в воздухе) около 340 м/с. Такой феномен называется звуком, хотя точнее было бы назвать его «звуком, передаваемым воздухом».
5. Физиология слуха
203
Рис. 5-4. Состояние части звукового поля вокруг колеблющегося камертона.
В воде, например, звук распросраняется в четыре раза быстрее, чем в воздухе. Благодаря таким же физическим взаимодействиям колебания распространяются в твердых телах (например, при землетрясениях), но большинство биологов отличают многие из таких «вибраций» от возникающего в воздухе «звука». На рис. 5-4 показано пространственное распределение давления в простой звуковой волне в данный момент времени; зоны повышенного давления (большей плотности молекул воздуха) чередуются с зонами пониженного давления. Через короткое время можно было бы видеть, как этот ряд волн несколько переместился вправо. Таким образом, на большие расстояния перемещаются не молекулы Воздуха, а волна и связанная с ней энергия. А молекулы в действительности движутся взад и вперед (в соответствии с локальным изменением градиентов давления вдоль направления распространения волны); поэтому звуковые волны называют продольными (в отличие от поперечных волн, которые распространяются по струне или поверхности воды).
Амплитуда периодических колебаний давления (рис. 5-4) называется звуковым давлением', его можно измерять при помощи микрофона и применять для описания звука. Как и любое другое, звуковое давление измеряется в ньютонах на квадратный метр. Но диапазон звуковых давлений, действующих на слуховую систему, так велик, что удобнее-и это действительно повсеместно принято в акустике - пользоваться логарифмической шкалой, так называемым уровнем звукового давления. Его установили приняв как исходный произвольно выбранный уровень р0 = = 2-10~5 Н/м2 (который близок к порогу слышимости). Уровень звукового давления (L) данного звукового давления р описывается уравнением
L= 201og10 — Ро
[5-1]
204
Р. Клинке
и получаемые единицы L называются децибелами (дБ). Так, для уровня давления р, равного р0, L= 0 дБ. «Таинственные 20» объясняются просто: логарифм отношения амплитуд давления первоначально назывался «бел» (в честь Александра Грэма Белла, Bell), который, естественно, равен 10 дБ; но децибельная шкала, отражающая мощность (пропорциональную квадрату амплитуды), удобнее, a logp2 = 21ogp; отсюда 2 • 10 = 20.
Например, чтобы выразить в децибелах уровень звукового давления тона со звуковым давлением р = 2-10 2 Н/м2, мы выводим:
р 2-10“2
— =——г=Ю3, Ро 2-10-5
L = 20 loglо (103) = 20 • 3 = 60 дБ.
То есть звуковое давление, равное 2-10-2 Н/м2, соответствует уровню звукового давления в 60 дБ. По ординате на левой стороне рис. 5-6 даны значения звукового давления и соответствующие уровни звукового давления в децибелах.
Поскольку другие величины, например электрический потенциал, иногда тоже выражаются в такой же децибельной шкале, уровни звукового давления (УЗД) часто даются как децибелы УЗД. Такое указание подчеркивает, что значения получены по приведенной формуле, где уровень отсчета ро = 2-10-5 Н/м2.
Второй параметр звука, частота, выражается в циклах в секунду, или герцах (в честь немецкого физика XIX в.), сокращенно Гц. У высокочастотных звуков длины волн короче, чем у низкочастотных (рис. 5-4). Частота /, скорость звука с и длина волны к (лямбда) связаны между собой следующим образом:
с=/-Х. [5-2]
Звук, характеризующийся только одной частотой (например, 2000 Гц), называется тоном (рис. 5-5,А). Но в повседневной жизни чистых тонов практически не бывает. Обычные звуки, от самых музыкальных до самых шумных, почти всегда содержат много частот. Звуки, которые мы считаем музыкальными, состоят из ограниченного числа частот, обычно из основного тона с несколькими гармониками (рис. 5-5, Б). Основной тон определяет «период повторения» сложных флуктуаций звукового давления (Т на рис. 5-5, Б). Гармоники-это обертоны с частотами, кратными основной частоте. С помощью различных приборов можно получить почти чистые тоны, но «тоны», производимые музыкальными инструментами, содержат гармоники. Разные инструменты различаются по числу и относительной интенсивности обертонов, сопутствующих основному тону. Некоторые инструменты не могут издавать определенные обертоны; например, звуки, издаваемые
5. Физиология слуха
205
Рис. 5-5. Кривые изменений звукового давления во времени для чистого тона (Л), музыкального звука (Б) и шума (В).
замкнутыми трубами органа, содержат только нечетные гармоники, частоты /0, 3/0, 5/0 и т.д. Именно эти особенности частотного спектра создают разнообразие звуков в оркестре. Если звук включает очень много частот, то тогда это «шум», а если все частоты в таком звуке обладают равными интенсивностями, то он называется белым шумом. У других шумов иные частотные спектры, но для всех таких звуков характерно, что в записях изменений во времени их уровня звукового давления отсутствует очевидная периодичность (рис. 5-5, В).
Наличие звука и субъективное слуховое ощущение. Повседневный опыт показывает, что наши слуховые способности ограничены. Иными словами, для того чтобы человеческое ухо могло воспринять звук, звуковое давление должно превысить определенный уровень. Звуковое давление, при котором тон едва слышен, называется слуховым порогом. Его давление, зависит от частоты тестируемого тона. Нижняя кривая на рис. 5-6 показывает, как меняется слуховой порог в зависимости от частоты; видно, что ухо наиболее чувствительно в пределах от 2000 до 4000 Гц. На этих частотах порог оказывается превышенным при очень низких звуковых давлениях. При более высоких и более низких частотах требуются большие звуковые давления.
По мере возрастания звукового давления над порогом тон слышится все громче независимо от его частоты. Отношение между (физическим) звуковым давлением и (субъективно воспринимаемой) громкостью описывается количественно. Испытуемый может не только сказать, когда тон стал слышимым (превысил порог слышимости), но и сообщить, когда два тона одинаковой частоты различаются по громкости. В пределах низких интенсивностей такие тоны звучат по-разному, если их звуковые давления различаются всего лишь на 1 дБ. При больших интенсивностях этот дифференциальный порог еще меньше.
Два тона разной частоты обычно воспринимаются с различной громкостью даже при равных звуковых давлениях. Это свойство слуховой системы можно определить количественно в разного вида опытах. Например, испытуемому предъявляют в качестве эталона тон в 1000 Гц и для сравнения с ним тестируемый тон, частоту которого изменяют в последующих применениях. При каждой паре тонов испытуемый устанавливает новое положение потенциометра, меняя тем самым звуковое
206
Р. Клинке
Рис. 5-6. Кривые равного уровня громкости. Слева по оси ординат-эквивалентные значения звукового давления и уровня звукового давления. Розовый участок-диапазон частот и интенсивностей, необходимый для понимания обычной речи.
давление эталонного тона, пока не услышит оба тона равногромкими. Звуковое давление эталонного тона при такой процедуре, выраженное в децибелах УЗД, дает уровень громкости тестируемого звука в фонах (например, если эталон установлен на 70 дБ УЗД, то уровень громкости тестируемого тона равен 70 фонам). Поскольку тон в 1000 Гц используется как эталон, он естественно имеет одинаковое число децибелов и фонов. При нанесении на график этих значений для тестируемых частот во всем диапазоне слышимости получается контур равных громкостей (или изофон). На рис. 5-6 показан набор контуров равной громкости; они представляют собой средние значения из данных, полученных при исследовании многих здоровых испытуемых, причем для каждой кривой тестируемые тоны установлены на другом уровне звукового давления. Все тоны независимо от их частоты, которые лежат на этих кривых, считаются равногромкими. Изофон дает также порог слышимости: все тоны на этой кривой равной громкости едва слышны. Средний слуховой порог у здоровых людей равен 4 фонам.
В опытах другого типа испытуемый должен говорить, когда тестируемый тон звучит в п раз (например, в три или в четыре раза) громче эталонного тона в 1000 Гц при 40 дБ УЗД. При такого рода измерениях громкость тона выражается в сонах. Громкость тона, который слышится в четыре раза громче эталонного тона в 1000 Гц при 40 дБ, р^вна 4 сонам; громкость тестируемого тона, вдвое меньшая, чем громкость эталонного тона, равна 0,5 сона и т. д. Таким образом, при создании
5. Физиология слуха
207
шкалы сонов испытуемый должен производить различение на более высоком уровне, чем для шкалы фонов, для которой нужно определять только равноценность. Тем не менее найдено воспроизводимое отношение между двумя шкалами. Выше 40 фонов ощущение громкости следует простой степенной функции звукового давления с показателем степени 0,3; усиление на 10 фонов удваивает воспринимаемую громкость. Ниже 40 фонов это отношение не сохраняется: громкость удваивается при меньшем усилении звукового давления.
При значительном повышении звукового давления человек ощущает боль в ухе. Это происходит приблизительно при 130 фонах -болевой порог. Такие высокие звуковые давления могут даже повредить ухо, если долго подвергать его подобному воздействию. Собственно говоря, очень долгие звуки могут вызвать травму при звуковых давлениях даже ниже болевого порога.
Шкала сонов очень важна для оценки степени нарушений, вызываемых шумом. Но из-за того, что она может быть получена только по показаниям испытуемых, оценка громкости таким способом очень трудоемкая и обычно не осуществима. Шкала фонов тоже устанавливается по показаниями испытуемых и не может быть получена непосредственно. Однако можно создать такой измеритель звукового давления, который не отвечает одинаково на всех частотах, а оценивает их так же, как человеческое ухо. Такой измерительный прибор максимально чувствителен в среднем диапазоне частот (ср. пороговую кривую на рис. 5-6) и менее чувствителен на высоких и низких частотах. Показания такого прибора выражены в децибелах (А), где А означает кривую определенного фильтра. Они приблизительно соответствуют значениям в фонах, т.е. уровню громкости. Поскольку с такими измерителями легко работать, их применяют при оценке шума даже несмотря на некоторую неточность. Так, шум мотора, работающего вхолостую, составляет около 75 дБ (А). Следует указать, что длительное (например, на протяжении восьмичасового рабочего дня) воздействие шумом выше 90 дБ (А) приводит с годами к поражению слуха.
Как видно из рис. 5-6, слышимость тона зависит не только от звукового давления, но также от частоты. Взрослый человек слышит звуки с частотами в диапазоне от 20 до 16000 Гц (16 кГц). Частоты выше 16 кГц называются ультразвуковыми, а ниже 20 Гц-инфразвуковыми. Таким образом, все, что мы слышим, связано со звуками между 20 Гц и 16 кГц и между 4 и 130 фонами-с областью между верхней и нижней кривыми на рис. 5-6. Розовая область в середине схемы представляет собой диапазон частот и интенсивностей, в котором понятна разговорная речь (спектральный состав человеческого голоса в разных условиях, разумеется, занимает больший диапазон).
У «тугоухих» людей слуховые пороги повышены; иными словами, для восприятия звука им требуется более высокое звуковое давление, чем для людей с нормальным слухом. Клиническое измерение слухового порога называется аудиометрией. Пациенту через наушники предъя-
208
Р. Клинке
Обследование слуха
Фамилия_________________
Адрес __________________
Аудиограмма
Частота, Гц
Дата первой проверни_____________
Врач ____________________________
Возраст_______ Пол_______________
Клиника__________________________
Рис. 5-7. Аудиограмма больного с потерей слуха на левое ухо приблизительно на 30 дБ. ВП-воздушная проводимость, КП-костная проводимость.
Ухо Правое Левое
ВП о— х- -X
КП II——-с □ □
Цвет Красный Черный
вляются разные тоны; таким способом тестируются ответы на звуки, проводимые по воздуху (см. разд. 5.3). Врач начинает с тонов, заведомо подпороговых, и медленно увеличивает звуковое давление, пока пациент не сообщит, что слышит тон. Звуковое давление в этой точке наносят на график. Если порог у пациента на несколько децибелов выше нормального, то диагностируется соответствующее понижение слуха. Форма, на которую нанесены все данные такой проверки порогов при разных частотах, называется аудиограммой (рис. 5-7). Существуют стандартные формы аудиограмм с нанесенными на них заранее координатами и нормальным порогом, изображенным прямой линией с пометкой «0 дБ». Стандартное изображение отличается от нанесенного на рис. 5-6 тем, что пороги выше нормальных наносятся вниз от нулевой линии, и таким образом разница в децибелах графически выражена как «потеря слуха». Эти относительные цифры не следует смешивать с истинным уровнем звукового давления (который обозначается как децибелы УЗД). Описание так называемой аудиометрии по вызванным ответам см. в разд. 5.4.
В старости обычно наступает снижение слуха на высоких частотах. Это явление называется пресбиакузисом.
Слуховая система человека, разумеется, способна также оценивать высоту тона, которая является коррелятом частоты звуковых колеба
5. Физиология слуха
209
ний. В музыке повышение тона на октаву происходит при удвоении частоты: /1=2/0. Организация тональной шкалы в виде октав является основным свойством западной музыки и отражает основные физические свойства большинства музыкальных инструментов. Но в психофизических измерениях «тона» обнаруживаются систематические отклонения от физических условий. Если экспериментатор прдъявляет испытуемому исходный тон выше 2 кГц и предлагает ему выбрать частоту, которая покажется ему вдвое выше, то частота выбранного тона будет отличаться от исходной больше чем на физическую октаву. Тем не менее физическая октава остается стандартом для настройки музыкальных инструментов. Октава подразделяется на 12 ступеней; при равномерно темперированной настройке каждая из них отличается от последующей по частоте на множитель \/2; однако необязательно следовать во всех случаях такой настройке и некоторые инструменты могут слегка отклоняться от этого стандарта.
Человек обладает очень тонкой способностью различать тоны. В оптимальном диапазоне, около 1000 Гц, мы можем различать частоты, которые отличаются одна от другой всего лишь на 0,3%, т.е. приблизительно на 3 Гц. Эта величина называется частотным разностным порогом. Можно также приписать определенный тон музыкальному звуку, который не является чистым; воспринимаемый тон будет определяться основной частотой звука.
Слуховая ориентация в пространстве. Еще одной важной функцией слуховой системы является ее участие в пространственной ориентации. Повседневный опыт показывает, что направление источника звука может быть определено достаточно точно. Предпосылкой для этого служит бинауральный слух (слушание обоими ушами). Эта способность основана на физических свойствах звуковых волн и на том, что одно ухо обычно находится несколько дальше от источника звука и ориентировано иначе по отношению к нему, чем другое. Поскольку звук распространяется с конечной скоростью, он доходит до дальше отстоящего уха позднее и с меньшей интенсивностью. Рис. 5-8 показывает, как может быть вычислена разница во времени его прихода. Разница в длине пути А/ равна d-sina, где d- расстояние между двумя ушами, а а-угол между источником звука и срединной плоскостью головы. Разница во времени прихода звуков будет в таком случае выражаться уравнением
At = —, [5-3]
с
где с-скорость распространения звука.
При a = 30° и расстоянии между ушами d = 17 см разница во времени составит
At =
0,085 340
= 2,5 10“4с.
210
Р. Клинке
Рис. 5-8. Геометрические отношения, применяемые для расчета запаздывания в попадании звука в одно ухо. См. текст.
Однако слуховая система выявляет гораздо меньшую интераураль-ную разницу. Безусловно может быть обнаружена разница всего лишь в ЗЮ5 с (а при оптимальных условиях и вдвое меньшая); такая величина соответствует углу 3° между источником и средней линией.
Что различия во времени прихода звука и в его интенсивности действительно влияют на воспринимаемое направление звуков, можно показать экспериментально. В таком психофизическом опыте на уши надеты телефоны, что позволяет независимо менять в них время стимуляции и ее интенсивность. Оказывается, что задержка в предъявлении звука в одно ухо может быть компенсирована усилением звука в этом ухе, и наоборот. Сходные результаты получены при регистрации активности отдельных нервных клеток в добавочном ядре верхней оливы. Это ядро служит основным местом нейронной переработки сигналов от двух ушей для пространственной ориентации. Результаты такого анализа передаются в высшие центры; на самом высшем уровне, в слуховой коре, имеются клетки, которые активируются только звуком, приходящим под определенным углом относительно головы (см. разд. 5.4).
Однако описанные механизмы не объясняют, почему направление различается независимо от того, находится ли источник звука впереди слушателя или позади него. Рис. 5-8 можно было бы продолжить и показать, что разница во времени прихода звука к обоим ушам и в его интенсивности остается неизменной, когда источник звука расположен сзади. Требуется некоторый дополнительный механизм, и им служит ушная раковина, наружное ухо. Благодаря ее форме звуки, возникающие впереди слушающего, воспринимаются совершенно иными, чем те, которые приходят сзади. Этот факт можно использовать для магнитофонной регистрации музыки. При помощи модели головы, в которую на месте барабанных перепонок вмонтированы микрофоны, получают прекрасные стереофонические записи.
5. Физиология слуха
211
В 5.6. Верхнее «до» певца представляет собой в физическом смысле а) чистый тон;
б) ноту с основным тоном и гармониками;
в) шум.
В 5.7. Звуковое давление, равное 20 дБ УЗД, удваивается. Получаемый при этом уровень звукового давления равен: а) 22 дБ; в) 40 дБ;
б) 26 дБ; г) все указанные значения неверны.
В 5.8. Изофоны-это кривые
а) равного звукового давления;
б) равных уровней звукового давления;
в) равных уровней громкости;
г) равной громкости.
В 5.9. Точки ниже нулевой линии в клинической аудиограмме представляют собой:
а) порог звукового давления, при котором пациент впервые сообщает, что слышит звук;
б) уровень звукового давления слухового порога пациента; в) потерю слуха в дБ.
В 5.10. Локализация источника звука возможна, если он
а) впереди головы или немного сбоку, но не в другом положении;
б) впереди, сзади или сбоку головы;
в) меньше чем на 30° в сторону от линии, направленной от головы прямо вперед.
г) Необходимо участие и глаз, и ушей.
5.3. Функции среднего и внутреннего уха
В последнем разделе речь шла о некоторых возможностях слуховой системы как целого. Настоящий и следующий разделы посвящены тому, каким образом они реализуются.
Роль среднего уха. Как было сказано в разд. 5.1, барабанная перепонка отвечает на звук и передает энергию своих колебаний по цепи косточек внутреннему уху или, точнее говоря, перилимфе вестибулярной лестницы. Колебания, проводимые таким путем, называются воздушными. Но ощущение звука создается также в том случае, если колеблющееся тело, например камертон, прямо соприкасается с черепом. При этом колебание передается через кости черепа-костное проведение звука. В повседневной жизни костное проведение играет роль только при слушании собственного голоса. Во всех остальных случаях возникший в воздухе звук достигает внутреннего уха через барабанную перепонку и косточки. Тем не менее клиническая проверка костного проведения, как мы увидим, имеет большое диагностическое значение. Проводимый по воздуху звук должен быть передан из воздуха жидкости, наполняющей внутреннее ухо. В норме, когда звуковые волны переходят из воз
212
Р. Клинке
духа в жидкость, большая часть звуковой энергии отражается от этой границы. Но совершенно очевидно, что для уха такое отражение было бы непродуктивным. Сложный механизм, состоящий из барабанной перепонки и аппарата косточек, природа «изобрела» для уменьшения потерь из-за отражения. Если пользоваться техническими терминами, то аппарат подстраивает акустический импеданс воздуха к импедансу внутреннего уха. В результате потери из-за отражения значительно уменьшаются, и большее количество звуковой энергии достигает внутреннего уха. Этот механизм в принципе аналогичен применению покрытия на линзах фотографических объективов, которое тоже служит для уменьшения отражения на границе между воздухом и стеклом. Подравнивание импеданса в среднем ухе производится главным образом двумя факторами. Во-первых, площадь барабанной перепонки значительно больше площади основания стремечка. При передаче данной силы по косточкам сама разница этих площадей создает усиление давления у овального окна по сравнению с давлением у барабанной перепонки. Во-вторых, отростки косточек так расположены в цепочке, что действуют как рычаги и еще больше усиливают давление.
Таким образом, по своему действию эта система сходна с трансформатором, хотя здесь участвуют другие факторы. Весь этот механизм улучшает слух приблизительно на 15-20 дБ.
К молоточку и стремечку прикреплены тонкие мышцы, так называемые мышцы слуховых косточек. Они отвечают на звуковые стимулы рефлекторными сокращениями, нарушающими передачу звука.
Прием звука внутренним ухом. Теория места. Когда звук попадает в ухо, стремечко передает энергию перилимфе вестибулярной лестницы (см. рис. 5-1). Стремечко колеблется взад и вперед, увлекая за собой лимфу. Поскольку жидкость во внутреннем ухе почти несжимаема, соответствующее движение должно происходить также и в другом месте, а именно у круглого окна. Когда стремечко перемещается внутрь, мембрана круглого окна выпячивается наружу, и наоборот. При этих движениях стремечка происходят одновременные смещения близлежащих базальных частей улиткового протока, а именно основной и вестибулярной мембран. Эта область колеблется вместе со стремечком, попеременно то в сторону барабанной, то в сторону вестибулярной лестницы. Ради простоты изложения в дальнейшем улитковый проток будет рассмотрен как единое целое-это средняя лестница, наполненная эндолимфой, и ограничивающие ее вестибулярная и основная мембраны. Описанное выше начальное смещение основания улиткового протока вызывает волну, бегущую вдоль него от базального конца к геликотре-ме подобно тому, как можно послать в горизонтальном направлении волну по веревке. Форма этой волны в два момента времени показана на рис. 5-9, на котором улитковый проток изображен в виде одной линии. Длительные тоны приводят стремечко в длительные колебания, и такие бегущие волны перемещаются вдоль наполненного эндолимфой хода.
5. Физиология слуха
213
Направление движения волны.
Рис. 5-9. Бегущие волны в улитковом протоке, который изображен линией. Конец улитки, где лежит стремечко, показан слева, геликотрема-справа. Две показанные волны (сплошная и штриховая линии) возникли в разные моменты времени. Красным дана огибающая волн, которая показывает крайние смещения в каждой точке улитки.
Не пытаясь физически точно описать здесь механику этих волн, мы обратимся к упрощенному описанию ситуации, такой, какой ее выявили опыты Бекеши (Bekesy) несколько десятилетий назад. Под действием звука основная мембрана колеблется вверх и вниз, как показано на увеличенных трехмерных изображениях (рис. 5-10). В действительности амплитуды колебаний очень малы-около 10“11 м в пороговом диапазоне. Поскольку жесткость основной мембраны снижается от стремечка к геликотреме, скорость распространения волн постепенно уменьшается в этом направлении, а амплитуда волн сначала возрастает. Но дальше вследствие некоторых физических особенностей наполненных жидкостью каналов волны затухают и в конце концов исчезают совсем обычно до того, как достигнут геликотремы. Поэтому где-то между местом возникновения волны у стремечка и точкой, где она кончается, лежит точка максимального отклонения. Этот максимум для разных частот приходится на разные места, сдвигаясь в направлении стремечка при повышении частоты и помещаясь ближе к геликотреме для низких частот (рис. 5-11). Таким образом, наличие этих точек максимального смещения помещает каждую частоту на определенное место улиткового протока. Сенсорные клетки возбуждаются главным образом в местах максимумов, и тем самым каждая частота возбуждает «свои» сенсорные клетки. Такова вкратце «теория места».
Рецепция стимула волосковыми клетками. Как показано на рис. 5-10, звук вызывает колебания улиткового протока попеременно в сторону вестибулярной и в сторону барабанной лестницы. Одним из эффектов этого сложного движения является смещение основной и покровной мембран относительно друг друга. Это показано на весьма схематичном рис. 5-10, Б. Из-за того что цилии волосковых клеток плотно соприкасаются с покровной мембраной, это смещение вызывает их отклонение. Сгибание цилий составляет для волосковых клеток адекватный стимул.
Прежде чем описывать процесс трансдукции (преобразования стиму-
Т екториальная мембрана
Рис. 5-10. А. Трехмерное изображение колебаний улиткового протока. Б. Сгибание цилий при колебании основной мембраны.
Стремечко
Стремечко
---------Геликотрема
10000 Гц
Рис. 5-11. Максимальные амплитуды колебания основной мембраны под действием тонов разной частоты.
5. Физиология слуха
215
Рис. 5-12. Постоянные потенциалы во внутреннем ухе, описанные в тексте.
ла в нервный сигнал) в волосковой клетке, надо указать еще на несколько фактов. Если пользоваться микроэлектродом для измерения потенциалов в среднем ухе с индифферентным электродом в вестибулярной лестнице, то в улитковом протоке регистрируется большой положительный потенциал (~ +80 мВ; рис. 5-12). По отношению к тому же уровню сосудистая полоска и кортиев орган обладают отрицательными потенциалами. Положительный эндокохлеарный потенциал поддерживается химическими процессами в сосудистой полоске. Все эти потенциалы наблюдаются в отсутствие звука, поэтому их называют стоячими (т.е. постоянными) потенциалами.
Когда звук попадает в ухо, возникают добавочные потенциалы, так называемые микрофонные потенциалы и потенциалы действия слухового нерва. Термин микрофонный потенциал (который можно регистрировать, например, у круглого окна) возник потому, что этот потенциал ведет себя подобно выходному потенциалу микрофона, т. е. близко воспроизводит колебания звукового давления. Микрофонный потенциал следует за стимулом: 1) практически без латентного периода, 2) не имеет рефрактерного периода или 3) измеримого порога и 4) не подвержен утомлению. По всем этим четырем свойствам он отличается от большинства биологических потенциалов и, в частности, от потенциалов действия. Вероятно, микрофонный потенциал представляет собой регистрируемую внеклеточно сумму рецепторных потенциалов всех возбужденных волосковых клеток. Как полагают, акустический стимул вызывает одновременное изменение проводимости мембраны рецепторной клетки в результате сгибания цилий. Вследствие крутого градиента потенциала (связанного со стоячими потенциалами) между пространством с эндолимфой и внутренней частью рецепторной клетки, составляющего не меньше 150 мВ, изменение проводимости мембраны сопровождается быстрыми токами ионов внутрь или наружу, что в свою очередь создает рецепторный потенциал. Такой механизм обычно называется «гипотезой батареи». Далее, как полагают, рецепторный потенциал каждой
216
Р. Клинке
“Щелчок»
Рис. 5-13. Потенциалы, записанные у круглого окна при действии на ухо щелчка. МП-микрофонный потенциал; КПД-кохлеарный потенциал действия.
волосковой клетки вызывает высвобождение медиатора на ее базальном полюсе. А это вещество возбуждает афферентные нервные волокна.
Потенциалы действия в волокнах слухового нерва сигнализируют центральной нервной системе о возбуждении волосковых клеток. Эти потенциалы действия можно зарегистрировать микроэлектродами. Их свойства будут описаны в следующем разделе.
Между функциями внутренних и наружных волосковых клеток не установлено четких различий.
Когда на ухо действует щелчок (но не тон), волокна в слуховом нерве возбуждаются синхронно и регистрируется суммарный потенциал действия. На рис. 5-13 показаны как кохлеарный микрофонный потенциал, так и суммарный потенциал действия слухового нерва (кохлеарный потенциал действия) в ответ на щелчок. При действии длительного звука отдельные волокна в слуховом нерве не разряжаются синхронно, и суммарный кохлеарный потенциал действия не наблюдается.
Глухота при поражении среднего или внутреннего уха. Выше было сказано, что в костях черепа можно вызывать колебания непосредственно, например соприкосновением с колеблющимся камертоном. При таких сильных стимулах вибрация кости может вызвать возбуждение внутреннего уха. Костная проводимость имеет значение для клинических обледований по следующей причине. Нарушения слуха можно грубо разделить на два вида: 1) нарушение проведения, при котором среднее ухо по разным причинам не передает достаточную звуковую энергию к внутреннему уху, и 2) поражение внутреннего уха, при котором глухота вызвана повреждением улитки-волосковых клеток или афферентных нервных волокон. При аудиометрическом обследовании следует проверять не только ответы на звуки, возникшие в воздухе, но
5. Физиология слуха
217
и эффективность костного проведения. При этом вибратор помещается на сосцевидный отросток, костный выступ за ушной раковиной, с той стороны, которую надо обследовать. Очевидно, что если нарушение слуха вызвано повреждением внутреннего уха, слуховой порог возрастет и при воздушном, и при костном проведении, так как затронут сам процесс рецепции. Если же глухота связана с поражением среднего уха, проведение возникших в воздухе звуков нарушается, а внутреннее ухо продолжает функционировать и порог костного проведения остается неизменным. Аудиограмма на рис. 5-7 свидетельствует о поражении внутреннего уха; слуховой порог больного повышен при обоих видах проведения.
Существует очень простой способ определить у больного с нарушением слуха только с одной стороны, поражено у него среднее или же внутреннее ухо. Этот способ называется пробой Вебера. Ножку колеблющегося камертона (лучше всего с частотой 256 Гц) прижимают к средней линии черепа. Если поражено внутреннее ухо, больной слышит тон на непораженной стороне. Если же нарушено проведение через среднее ухо, он слышит тон на пораженной стороне; пользуясь клиническими терминами, говорят, что он «латерализуется в пораженную сторону».
В 5.11. Аппарат барабанной перепонки и косточек
а) служит исключительно в качестве мостика через пространство, отделяющее барабанную перепонку от овального окна;
б) снижает потери, вызываемые отражением при переходе звука из воздуха во внутреннее ухо;
в) благодаря своему строению из рычагов и звеньев защищает внутреннее ухо от повреждения звуком;
г) является филогенетическим реликтом, который развился из жаберных дуг и лишен какого-либо специального значения у млекопитающих.
В 5.12. Выражение «бегущая волна», применяемое в отношении рецепции звука, относится к тому, что а) звук создает в улитке стоячие волны;
б) волна проходит от геликотремы к стремечку, образуя между ними зависимый от определенной частоты максимум;
в) волна проходит от стремечка к геликотреме, образуя между ними зависимый от определенной частоты максимум;
г) звук распространяется в воздухе в форме волн.
В 5.13. Теория места гласит, что
а) звуковой стимул возбуждает только волосковые клетки на одном месте основной мембраны;
б) каждый частотный компонент звука возбуждает волосковые клетки на своем участке основной мембраны;
в) все волокна слухового нерва начинаются в одном месте-в спиральном ганглии.
218
Р. Клинке
5.4. Слуховой нерв и высшие уровни слухового пути
Условия, при которых стимулируются слуховые нервные волокна. Каждое волокно в слуховом нерве начинается от узкой ограниченной области улитки, в некоторых случаях от одной внутренней волосковой клетки. Поскольку отдельные участки улитки связаны с отдельными частотами, каждое нервное волокно оптимально возбуждается звуком определенной частоты. Эта частота называется характеристической частотой данного волокна. Таким образом, волокно в слуховом нерве подвергается возбуждению легче всего, когда ухо стимулируется звуком частоты, характеристичной для данного волокна (рис. 5-14). Чистые тоны очень низкой интенсивности возбуждают специфические одиночные волокна нерва. Если же ухо стимулируется тонами, частоты которых не являются характеристическими, то оно может быть активировано только при достаточном увеличении интенсивности. Это свойство «настройки» показано на рис. 5-15, где на графике для каждой частоты дано звуковое давление, которое вызывает околопороговый ответ в каждом из двух волокон, различающихся по характеристической частоте (~ 3000 и 4500 Гц). Затененный участок над каждой кривой показывает диапазон частот и интенсивностей, в котором волокно реагирует. Такие кривые называются кривыми настройки. Если звуковой стимул содержит несколько частот, то активируются все соответствующие группы нервных волокон. Длительность стимула кодируется длительностью активации, а интенсивность-степенью активации. При повышении уровня звукового давления не только усиливается возбуждение данных волокон (увеличивается частота их разряда), но в активность вовлекаются также дополнительные волокна (с частотами, близкими к характеристической). На рис. 5-15 показано также, что в ответ на звуковой стимул, параметры которого лежат в области перекрывания, возбуждаются оба волокна.
Итак, можно сказать, что на уровне первичных афферентных волокон звуковой стимул разлагается на свои частотные компоненты. На более высоких уровнях слухового пути переработка информации носит совсем иной характер.
Анатомия слухового пути. На рис. 5-16 приведена сильно упрощенная схема основных частей слухового пути. Для большей ясности показаны пути только от левого уха. Кончиками стрелок обозначены точки переключения, где нейроны образуют синапсы с нейронами более высокого порядка; на схеме не различаются тормозные и возбудительные синапсы. Первичные афферентные волокна идут сначала к кохлеарному ядру, подразделенному на вентральную и дорсальную части. От вентральной части вентральный тракт идет к ипси- и контралатеральным оливарным комплексам. Таким образом нервные клетки в каждом оли-варном комплексе получают входы от обоих ушей. Здесь расположен самый нижний из тех уровней головного мозга, на которых возможно сравнение акустических входов от обоих ушей. Такого рода сравнение
5. Физиология слуха
219
Потенциалы действия нерва А Характеристическая частота, низкое звуковое давление
Б Близкая частота, то же давление, что на А
Тон 1200 Гц
Тон 1400 Гц
JIIIHIIgilllill III
Спонтанная активность
В Характеристическая частота, звуковое давление выше, чем на А
IIIIIIIIHIIIIIIIII III
Г Близкая частота, звуковое давление то же, что на В
Рис. 5-14. Разряд в одиночном волокне слухового нерва в ответ на стимуляцию его характеристической и близкой частотами при тух разных интенсивностях.
Рис. 5-15. Кривые настройки двух волокон слухового нерва. См. текст.
220
Р. Клинке
Рис. 5-16. Сильно упрощенная схема слухового пути, описанного в тексте.
производится в особенности в добавочном ядре-в центре, который уже был упомянут в разд. 5.2 в связи с ощущением направленности звука. Дорсальное кохлеарное ядро служит началом дорсального тракта. Эти волойна переходят на противоположную сторону и там образуют синапсы с нейронами ядра латерального лемниска. Одни восходящие волокна клеток оливарного комплекса проецируются на ту же сторону, другие образуют перекрест. Над латеральным лемниском слуховой тракт идет через два ядра переключения -нижний бугорок и медиальное коленчатое тело-к своему последнему назначению -первичной слуховой коре в височной доле. Таким образом, этот тракт состоит из последовательности по меньшей мере пяти или шести нейронов. В действительно
5. Физиология слуха
221
сти в нем имеются еще н другие переключения, не показанные на данной схеме, так что возможны еще более длинные цепочки. Кроме того, слуховой путь отдает много коллатералей. Наконец, слуховая система содержит не только афферентные, центростремительные волокна, но также эфферентную, центробежную систему, не показанную на рис. 5-16; функция этой эфферентной системы не совсем ясна.
Характеристики ответов центральных слуховых нейронов. Нейроны слухового нерва возбуждаются совсем простыми стимулами, например чистыми тонами, но для нейронов высших уровней слухового пути обычно требуются более сложные звуки. Хотя в вентральном кохлеарном ядре клетки ведут себя так же, как и нейроны слухового нерва, в дорсальном кохлеарном ядре их ответы заметно отличаются. В определенных условиях происходит торможение этих нейронов входными слуховыми сигналами. Торможение создается обширными реципрокными связями между нейронами.
При отведении от клеток еще более высоких уровней ответы на чистые тоны часто вообще отсутствуют. Эти клетки отвечают на сложные звуковые паттерны, например на амплитудно- или частотно-модулированные звуки, т. е. звуки с непрерывно меняющейся интенсивностью или частотой. Другие нейроны реагируют только на начало или только на конец звукового стимула и т.п. Часто клетка активируется одними частотами и тормозится другими. В общем можно сказать, что чем выше уровень, на котором находится нейрон в слуховом пути, тем более сложные звуки вызывают его возбуждение. Раньше думали, что частотная избирательность нейронов, отвечающих на чистые тоны, усиливается на высших уровнях, но в действительности это не так. На высших уровнях настройка нейронов не намного острее, чем на уровне слухового нерва; собственно говоря, многие клетки в слуховой коре могут быть активированы очень широкой полосой частот. В то же время в некоторых участках слуховой коры имеется тонотопическая карта улитки.
В обычной жизни звуки редко бывают чистыми тонами. Обыкновенно они содержат разнообразные компоненты подобно сложным звукам, упомянутым выше. Это особенно верно для речи, структура которой все время меняется; некоторые ее элементы очень короткие, и, как правило, в ней возникает одновременная модуляция амплитуды и частоты. Очевидно, нейроны в высших отделах слухового пути специализированы для извлечения специфических характеристик таких сложных звуков, например изменения частоты в произносимом слове, и тем самым для узнавания некоторых констелляций стимулов. Клетки, отвечающие на такие особые констелляции, являются слуховыми аналогами сложных или сверхсложных нейронов зрительной коры (см. разд. 4.3). В обеих модальностях такие клетки извлекают определенные свойства стимула, участвуя в процессе опознания его структуры.
Другой путь исследования нейронной переработки звукового сигнала состоит в регистрации потенциалов электродами, фиксированными на коже головы. Эти потенциалы представляют собой суммарную ак
222
Р. Клинке
тивность многих нейронов, но они сильно ослаблены и для их анализа нужны ЭВМ. Такие измерения образуют основу так называемой аудиометрии по вызванным потенциалам (АВП); этот метод недавно стали применять как способ диагностики локализации поражения в случаях потери слуха-находится ли оно на уровне улитки или выше.
Адаптация в слуховой системе. Явление адаптации присуще всем сенсорным системам, и слуховая система не составляет исключения. Это свойство как периферических рецепторов, так и центральных нейронов. Адаптация к данному уровню звука приводит к повышению слухового порога, но из этого не следует, что она нежелательна или бесполезна. Адаптация к некоторым интенсивностям и частотам может в действительности понизить дифференциальный порог, усилив различение разных уровней слухового давления. Так, например, тон в 4000 Гц на рис. 5-6 при 33 дБ УЗД имеет уровень громкости 40 фонов, а тот же тон при 52 дБ УЗД-60 фонов. Следовательно, в этой части графика, чтобы получить разницу уровня громкости в 20 фонов, нужно усиление звукового давления на 19 дБ. Но если ухо адаптировано к долгому воздействию 4000 Гц, то изофоны в этой области смещаются вверх; в частности, для низких уровней громкости изофоны сместятся дальше, чем для высоких. В результате разница в уровне громкости в 20 фонов потребует изменения меньше чем на 19 дБ УЗД, например на 14 дБ. Иными словами, в адаптированном состоянии данная разница в уровне звукового давления вызовет большую разницу в уровне громкости. Субъективная разница становится больше, и, следовательно, порог различения понижается.
Предыдущее изложение могло создать впечатление, что ощущение тона основано полностью на локализации смещения в улитке, зависимой от частоты, как это постулирует теория места, или на настройке волокон слухового нерва на их характеристические частоты. Однако не все психофизические наблюдения над тональными ощущениями можно объяснить одной только теорией места. Так, например, при стимуляции уха последовательностью коротких импульсов давления (щелчков) с интервалами в 5 мс, как и следовало ожидать, слышится тон в 200 Гц. Но если предварительно профильтровать звук так, чтобы пропускались частоты только выше определенного уровня (скажем, 600 Гц), то в стимуле уже не будет частоты 200 Гц. Однако субъективно воспринимаемый тон по-прежнему будет 200 Гц. Это явление еще не получило убедительного объяснения, но его нельзя объяснить одной только теорией места. По-видимому, мозг «воссоздает» недостающую основную частоту информации, которая содержится в сохранившихся обертонах.
В 5.14. Предположим, что звук активирует волокна слухового нерва, обладающие несколькими разными характеристическими частотами-около 800, 1600 и 3200 Гц. Волокна с характеристическими частотами, лежащими между этими величинами, не активируются. Причиной этого может быть
5. Физиология слуха
223
а) то, что звук содержит несколько частот;
б) то, что интенсивность звука очень велика;
в) то, что нейроны слухового нерва обладают спонтанной активностью.
В 5.15. Слуховой путь содержит среди других следующие ядра переключения.
а) Кохлеарное ядро.
б) Ретикулярная формация.
в) Верхний бугорок.
г) Медиальное коленчатое тело.
д) Латеральное коленчатое тело.
е) Слуховая кора.
ж) Верхняя олива.
з) Чечевицеобразное ядро.
В 5.16. Возбуждение первичных афферентных волокон в слуховом нерве может быть вызвано
а) любым звуковым стимулом, лишь бы он был выше порога;
б) только чистыми тонами;
в) только сложными звуками, например частотно- или ампли-тудно-модулированными тонами;
г) только началом или концом звука («оп»- и «offw-нейроны).
6. ФИЗИОЛОГИЯ ЧУВСТВА РАВНОВЕСИЯ
Р. Клинке
6.1. Анатомия и физиология периферического органа
Рецепторы органа равновесия и стимулы, их возбуждающие. В височной кости близ улитки лежит вестибулярный орган-преобразователь, тесно связанный с поддержанием равновесия. Относительная ориентация этих частей лабиринта была показана на рис. 5-1 (см. также рис. 6-5). Вестибулярный орган филогенетически связан с ухом, в особенности в том, что касается рецепторов обеих систем. В том и другом случае рецепторами являются волосковые клетки. Адекватным стимулом для волосковых клеток, как было сказано в гл. 5, служит сгибание цилий. То же самое относится к вестибулярному органу, но здесь смещающее усилие возникает иначе. На рис. 6-1 показано действие этого механизма в одном из сенсорных образований вестибулярного органа-макуле. Цилии волосковых клеток вдаются в желатинозную массу, в которую погружены мелкие гранулы с высоким удельным весом. Масса вместе с ее содержимым называется отолитовой мембраной. Цилии разной длины образуют лесенку, причем самая длинная из них (называемая киноцилиеи) построена иначе, чем остальные. Такая дифференциация отличает эти волосковые клетки от клеток в улитке, которые лишены ки-нойилии и обладают только стереоцилиями.
В верхней части рис. 6-1 видна макула в покое. При наклоне головы в ту или другую сторону сила тяжести слегка смещает отолитовую мембрану, как показано на двух нижних рисунках. Смещаясь на это небольшое расстояние относительно сенсорного эпителия, она сгибает цилии. Сгибание служит стимулом для рецепторов. Афферентный нерв передает сигнал об этом состоянии в ЦНС следующим образом. Волокна активны (активность покоя) даже в нейтральном положении. При сгибании цилий в одном направлении частота разряда возрастает, при сгибании в другом-снижается. Усиление активности афферентного нервного волокна происходит, когда смещающее усилие в пучке цилий направлено в сторону киноцилий; торможение наступает при смещении в противоположную сторону (рис. 6-2). Таким образом, главным эффективным стимулом макулы служит сила тяжести. Она соответствует специальному случаю линейного ускорения. Макулы отвечают также на все другие формы линейного ускорения, например такого, какое возникает при пуске в ход или при торможении автомобиля. В таких случаях отолитовая мембрана смещается по отношению к лежащему под ней сенсорному эпителию точно так же, как подвижный предмет скользит вперед при внезапном включении тормозов автомобиля.
Структура и функция статолитовых органов и полукружных каналов. С каждой стороны головы имеются по две макулы (называемые также
6. Физиология чувства равновесия
225
Макула в покое
Рис. 6-1. Изменение положения макулярного органа при наклоне головы (адекватный для этого органа стимул) в двух направлениях.
статолитовыми органами)-макула утрикулуса и макула саккулуса (рис. 6-5). Обе они фиксированы относительно черепа. При прямом положении тела макула в утрикулусе лежит приблизительно горизонтально (как на верхнем изображении рис. 6-1), а макула в саккулусе расположена вертикально. При наклоне головы эти органы смещаются на угол между горизонтальным и вертикальным положением, как на нижних изображениях рис. 6-1. При любой возможной ориентации черепа действие силы тяжести состоит в перемещении отолитовых мембран в определенные положения относительно сенсорного эпителия. В результате в связанных с ним афферентных нервных волокнах создается специфическое соотношение возбуждений. Центральная нейронная оценка этой совокупности возбуждений от вестибулярной системы дает информацию о положении черепа в пространстве.
Каждый вестибулярный орган включает еще одну особую сенсорную систему - полукружные каналы. Рецепторы в полукружных каналах отвечают не на линейное, а на угловое ускорение. Принцип их строения показан на рис. 6-3. В определенном месте в каждом из замкнутых, кольцеобразных, наполненных жидкостью каналов имеется расширение - ампула. В ней находится желатинозная мембрана, купула, которая вдается в жидкость; ее удельная масса точно такая же, как у окружающей эндолимфы, и благодаря этому купула (в отличие от отолитовой мембраны) не перемещается относительно канала во время линейного ускорения. Но угловое ускорение влияет на нее в силу инерции эндолимфы. Если череп, первоначально неподвижный, поворачивается, эндолимфа стремится сохранить прежнее положение; поскольку купула одним концом прикреплена к стенке канала, ее свободный конец отклоняется в напра-
226
Р. Клинке
Рис. 6-2. Вверху-положение цилий на волосковой клетке, вид сверху. Внизу -разрез через клетку при наклоне в разных направлениях. Смещение в сторону киноцилии активирует афферентное волокно, смещение в противоположную сторону снижает частоту разряда. Смещение в перпендикулярном направлении не оказывает действия.
Рис. 6-3. Схема полукружного канала, показывающая купулу в ампуле. Справа -положение купулы, вызванное угловым ускорением.
6. Физиология чувства равновесия
227
Рис. 6-4. Ориентация полукружных каналов в черепе человека; вид сверху на основание черепа.
влении, противоположном повороту (как показано справа на рис. 6-3). Поскольку цилии рецепторных клеток погружены в купулу, при сгибании последней они оказываются под действием смещающего усилия. Как и в макулах, это смещение служит адекватным стимулом для рецептора. Афферентные нервные волокна, как и в макулярных органах, обладают активностью покоя в отсутствие сгибания цилий. Сгибание купулы в одном направлении увеличивает частоту разряда, сгибание в другом снижает ее. В горизонтальном полукружном канале волокна возбуждаются, когда купула смещается в сторону утрикулуса (рис. 6-5). Это происходит в левом канале, когда голова поворачивается влево.
Угловые ускорения могут содержать компоненты по всем трем независимым осям. Голова может 1) повернуться вокруг вертикальной оси, 2) наклониться вперед или назад и 3) нагнуться в любую сторону. Один полукружный канал не может сигнализировать об ускорении вокруг всех этих осей и всех их возможных комбинаций. Нервной системе нужны по меньшей мере три полукружных канала, по одному для каждой оси вращения. Рис. 6-4 и 6-5 показывают, что у нас действительно по три канала с каждой стороны и лежат они примерно перпендикулярно друг другу. Их ориентация в черепе приблизительно такая, как показано на рис. 6-4. С каждой стороны имеются горизонтальный, передний вертикальный и задний вертикальный полукружные каналы. Таким
Перилимфа Эндолимфа
непрерывность пространств, запол-в улитке.
Рис. 6-5. Схема лабиринта, показывающая ненных лимфой в вестибулярном органе и Угловое ускорение
I I
--------1-----------------------------------*
Угловая скорость t
I
Рис. 6-6. Движение купулы и ответ афферентного нервного волокна при кратком вращении (например, при повороте головы). Показаны также эффективное ускорение и угловая скорость.
6. Физиология чувства равновесия
229
образом, вестибулярный орган состоит с каждой стороны из пяти частей: двух макул (в саккулусе и в утрикулусе) и трех полукружных каналов. По своей форме каналы не являются такими идеальными кольцами, какие показаны на рис. 6-3; кроме того, как видно на рис. 6-4 и 6-5, некоторыми своими частями они соединены между собой. Наконец, надо указать, что так называемый горизонтальный полукружный канал при нормальном положении головы расположен не точно горизонтально; его передний край слегка приподнят, и плоскость, в которой он лежит, образует с горизонтальной плоскостью угол в 30°.
Полукружные каналы, утрикулус и саккулус состоят из тонких перепонок, которые образуют замкнутые трубки (отсюда название перепончатый лабиринт). Эта система наполнена эндолимфой, которая сообщается с эндолимфой улитки. Кроме того, перепончатый лабиринт окружен перилимфой, которая опять-таки переходит в перилимфу слухового органа. Система полостей в кости, в которых заключены улитка и вестибулярные органы (рис. 5-1), называется костным лабиринтом.
Поредение купулы при кратковременном и длительном вращении. Некоторые свойства механики купулы представляют особый интерес. На рис. 6-6 графически показаны угловое ускорение, сопровождающее краткое вращение головы при ее повороте из одного положения в другое, а также угловая скорость, вычисленная степень отклонения купулы в горизонтальном полукружном канале и экспериментально установленная активность одного из первичных афферентов, идущих от этой ампулы. Ясно видно, что отклонение купулы и афферентная активность отражают угловую скорость, а не угловое ускорение, несмотря на то что силами, вызывающими отклонение купулы, являются силы ускорения. Роль ускорения более выражена в опыте с длительным равномерным вращением на круге (рис. 6-7). Сначала животному сообщают ускорение, пока не достигается определенная угловая скорость. Эта скорость поддерживается в течение значительного времени. Рисунок показывает, что в фазе ускорения купула отклоняется от положения покоя, а во время постоянного вращения медленно возвращается к нему. Происходит зто потому, что из-за трения между эндолимфой и стенкой канала лимфа постепенно усваивает вращательное движение тела и в конце концов полностью следует за ним; когда зто произошло, разница между вращательным движением тела и эндолимфы исчезает. В этих условиях больше нет сил, которые отклоняли бы эластичную купулу. Поэтому нейронная активность возвращается к уровню покоя. Когда движение прекращается, все явления идут в обратном порядке. Купула отклоняется в противоположную сторону, а после того как тело приходит в состояние покоя, она медленно (за 10-30 с) возвращается в исходное, состояние, поскольку вращение лимфы тоже замедляется.
Различия в поведении купулы, связанные с длительностью вращения (после кратковременного вращения она немедленно возвращается в состояние покоя), объясняются механическими свойствами системы купула-эндолимфа, которых мы здесь не будем касаться. Но важно по-
230
Р. Клинке
6-7. Движение купулы и ответ афферентного нервного волокна во время длительного вращения (например, на круге). Показаны также ускорения (положительное в начале и отрицательное в конце вращения) и угловая скорость.
мнить, что силы, вызывающие отклонение купулы,-это ускорения. Надо также ясно представлять себе, что полукружные каналы развились для сигнализации о кратковременных вращательных движениях, которые случаются в повседневной жизни, и что вертящийся круг-это нефизиологический стимул.
В 6.1. Элементами вестибулярного органа являются
а) макула утрикулуса;
б) макула саккулуса;
в) желтое пятно;
г) передний вертикальный полукружный канал;
д) задний вертикальный полукружный канал;
е) горизонтальный полукружный канал;
ж) вестибулярная лестница.
В 6.2. Адекватным стимулом для полукружного канала служат
а) линейное ускорение;
б) угловое ускорение;
в) угловая скорость;
г) колебание стремечка.
6. Физиология чувства равновесия
231
6.2. Центральные механизмы чувства равновесия
Центральные связи рецепторов вестибулярного органа. Рецепторы в вестибулярном органе так же, как в улитке, являются вторичными чувствительными клетками. Связанные с ними афферентные нервные волокна начинаются в вестибулярном ганглии, который лежит в височной кости близ вестибулярного органа. Волокна проецируются в область вестибулярных ядер (рис. 6-8) в продолговатом мозгу. С каждой стороны лежат четыре разных ядра. Если вестибулярный орган должен участвовать в сохранении равновесия тела, между его афферентами и моторными центрами должны существовать связи, через которые осуществляется необходимая регуляция. Выходы из вестибулярных ядер направляются:
а) в составе вестибулоспинального тракта, воздействуя в конце концов на мотонейроны, управляющие мышцами-разгибателями;
б) прямо к мотонейронам шейного отдела спинного мозга;
в) к ядрам глазных мышц (см. рис. 4-37);
г) г мозжечку
д) к вестибулярным ядрам противоположной стороны, благодаря чему входная активность может перерабатываться совместно от обеих сторон;
е) к ретикулярной формации;
ж) через таламус к задней центральной извилине коры, которая имеет отношение к процессам сознания, связанным с вестибулярной входной активностью;
з) к гипоталамусу; эти волокна участвуют в возникновении морской болезни.
Поскольку голова у высших позвоночных подвижна относительно туловища, сигналы от одних только вестибулярных рецепторов не могут создать однозначную картину положения тела в пространстве (рис. 6-9). Нужна дополнительная информация о положении головы от-
Рис. 6-8. Положение вестибулярных ядер в продолговатом мозгу.
232
Р. Клинке
Рис. 6-9. Макулярные органы и шейные рецепторы при разных положениях тела. См. текст.
носительно тела. Она поступает от рецепторов шейных суставов (ср. рис. 3-8) и мышц (от мышечных веретен, рецепторов Гольджи), которые соединены с вестибулярными ядрами через центральные нейроны (см. разд. 3.2).
Статические и статокинетические рефлексы. Вестибулярный нистагм. Равновесие поддерживается рефлексами без участия сознания. Хотя сознательный анализ восприятия пространства возможен, регуляторные задачи не могут быть эффективно решены без участия врожденных рефлексов. Так, нельзя научиться «слепому» полету на самолете (т. е. в тумане) на основе сознательных пространственных ощущений; нужную информацию должны давать приборы. Рефлексы, вызываемые вестибулярными органами, можно разделить на две группы: так называемые статические и статокинетические рефлексы. Мы коснемся здесь нескольких особенно существенных моментов. Макулярные органы осуществляют статические рефлексы, которые поддерживают равновесие при разнообразных стоячих и наклонных положениях тела. Компенсаторное вращение глаз, особенно хорошо заметное у кошек (рис. 6-10), но возникающее и у людей, представляет собой статический рефлекс. Благодаря ему на сетчатке сохраняется изображение горизонтальных и вертикальных линий. Нет нужды напоминать, что в этом рефлексе участвуют шейные рецепторы. Статокинетические рефлексы происходят во время движений. Один из них-это поворот, который происходит в свободном падении. Так, кошка всегда падает на лапы независимо от того, в каком положении она начала падать. Статокинетические рефлексы вызываются как макулярными органами, так и полукружными каналами. Другим статокинетическим рефлексом является «эффект лиф-
6. Физиология чувства равновесия
233
Рис. 6-10. Компенсаторное вращение глаз, при котором зрачки сохраняют вертикальное положение.
та» на свободное падение, при котором усиливается тонус разгибателей, когда животное движется вниз.
Среди статокинетических рефлексов особенно выделяется вестибулярный нистагм (рис. 6-11). Он состоит из последовательных движений глаз, вызываемых вестибулярной активностью, когда глаза движутся в сторону, противоположную вращению тела, благодаря чему направление взора остается неизменным. Это компенсаторное движение глаз, разумеется, эффективно только в определенных границах вращения. До того как глаза достигнут предела своего латерального движения, происходит их быстрое движение в сторону вращения-они устремляются вперед и фиксируются на новой точке. За этой быстрой фазой следует новое медленное движение, которое снова компенсирует вращение.
Вращение головы или туловища вокруг вертикальной оси практически действует только на горизонтальные полукружные каналы. При этом отклонение купул в обоих горизонтальных каналах вызывает горизонтальный нистагм. Направление двух (быстрого и медленного) компонентов нистагма зависит от направления вращения (и, следовательно, от наклона купулы). При клиническом описании направление нистагма условились считать по быстрой фазе. Иными словами, при «правом нистагме» быстрая фаза направлена вправо. Такая терминология совпадает с терминами, принятыми для оптокинетического нистагма (разд. 4.4).
При пассивном вращении первоначальный эффект состоит в стимуляции вестибулярного аппарата, а также относительном смещении видимого мира. Каждый из этих двух эффектов сам по себе вызывает нистагм (вестибулярный и оптокинетический). Они дополняют друг друга; нейронные связи, лежащие в основе этого взаимодействия, показаны на рис. 4-37.
Клиническое значение нистагма. Нистагмография является полезным методом проверки функции вестибулярного аппарата; при этом чаще всего измеряют послевращательный нистагм. Испытуемого усаживают на вращающийся стул и довольно долго вращают с равномерной скоростью. Затем движение внезапно прекращается. На рис. 6-7 показано, как это действует на купулу; прекращение длительного вращения отклоняет ее в сторону, противоположную той, куда она отклонилась при начале движения. Наклон купулы вызывает нистагм независимо от при-
234
Р. Клинке
Медленные компенсаторные движения глаз
Рис. 6-11. Направление движения глаз при вестибулярном нистагме.
чины (ускорения или замедления); в этом случае нистагм называется послевращательным. Ожидаемое направление такого нистагма легко предсказать. Инерция эндолимфы сгибает купулу в направлении, в котором она вращалась, и это расценивается как угловое ускорение в противоположном направлении. Следовательно, остановка равномерного вращения влево вызывает послевращательный нистагм вправо, как показано в записи движений глаз на рис. 6-12.
Следует однако иметь в виду, что при тестировании послевраща-тельного нистагма надо избегать фиксации взора. В противном случае нистагм может быть подавлен преобладанием зрительной системы над вестибулярной. Поэтому испытуемому надевают очки с очень сильными выпуклыми линзами и встроенным в них источником света (очки Френ-целя). В таких очках человек становится близоруким и неспособным к фиксации, и вместе с тем очки не мешают врачу следить за движением глаз. Устранять таким аппаратом фиксацию взора следует также при клинической проверке на наличие спонтанного нистагма.
Еще одним диагностическим приемом в исследовании вестибулярного нистагма является термическая стимуляция горизонтального полукружного канала. Ее преимущество в данном случае состоит в том, что она позволяет проверять правый и левый горизонтальные каналы по отдельности. Голову пациента отклоняют назад приблизительно на 60е. В таком положении горизонтальный канал становится вертикальным. Теперь наружный слуховой проход орошают холодной или теплой во-
зле во
вправо
Рис. 6-12. Нистагмограмма.
6. Физиология чувства равновесия
235
Рис. 6-13. Отклонение купулы при калорической стимуляции полукружного канала
дой. Наружный край горизонтального полукружного канала лежит очень близко к слуховому проходу, и передача тепла оказывается достаточной, чтобы охладить или нагреть его. Эффект схематически показан на рис. 6-13; нагретая эндолимфа поднимается, что создает ее ток вдоль канала и тем самым отклонение купулы; в результате возникает калорический нистагм. Применение теплой воды вызывает нистагм в сторону нагревания, а нистагм, вызванный орошением холодной водой, естественно, будет направлен в обратную сторону.
Центральные связи, определяющие вестибулярный нистагм, организованы так, что медленную фазу вызывает вестибулярная система, а быстрое возвращение глаз запускается ретикулярной формацией.
Поскольку вестибулярная система связана с гипоталамусом, сильное возбуждение вестибулярного аппарата часто приводит к неприятным ощущениям-головокружению, тошноте, потоотделению и т.п. Такие симптомы называют кинетозом («морской болезнью»). Кинетоз особенно легко возникает, когда орган подвергается сочетаниям стимулов, к которым он не привык (как, например, на море). Особенно легко кинетоз вызывается ускорением кориолиса, а также расхождением между зрительными ощущениями и сигналами от вестибулярных органов.
Внезапное прекращение лабиринтной активации ведет к тошноте, рвоте, обильному потоотделению и нистагму, направленному в интактную сторону. У больного возникает также тенденция к падению в сторону нарушения. Хроническая утрата лабиринта сравнительно хорошо компенсируется, если больной может ориентироваться зрительно. Но в темноте этот дефект становится ощутимым.
В 6.3. Когда тело находится в покое, информация о его положении в пространстве поступает от следующих органов чувств.
а) Глаза.
б) Шейные рецепторы.
в) Макулярные органы.
г) Органы в полукружных каналах.
д) Кортиев орган.
236 Р. Клинке
В 6.4. Сколько времени требуется купулам, чтобы вернуться в состояние покоя после остановки длительного равномерного вращения ?
а) 100 мс.
б) Самое большее 1 с.
в) 10-30 с.
г) Обычно больше 1 мин.
В 6.5. Острое поражение лабиринта вызывает следующие симптомы: а) тошноту;
б) рвоту;
в) потоотделение;
г) нистагм в интактную сторону;
д) нистагм в пораженную сторону;
е) состояние тревоги.
ж) Ни один из этих симптомов, которые появляются только при хронических нарушениях.
7. ФИЗИОЛОГИЯ ВКУСА
X. Альтнер
Органы, обслуживающие вкусовую чувствительность, лежат в ротовой полости, преимущественно на языке. При рассмотрении вкусовой чувствительности полезно сначала обратиться к строению этих органов и их связям с ЦНС. Мы увидим, что рецепторные органы, вкусовые почки, сидят на сосочках (складках кожи языка) трех разных форм. Однако вкусовые почки и находящиеся в них чувствительные клетки нельзя сгруппировать в отдельные морфологические типы. Из основных размерностей вкусового ощущения два-качество и интенсивность - будут рассмотрены детально (разд. 7.1). Имеются четыре основных ощущения'. сладкое, кислое, соленое и горькое. В разд. 7.2 показано, что способность различать эти категории можно вывести из специфичности рецепторных молекул. Но типы рецепторов, специфические для данного качества, не обнаружены; качество и концентрация, очевидно, кодируются градуальными ответами большого числа рецепторов.
7.1. Морфология органов вкуса; субъективная физиология вкуса
Ориентация и строение вкусовых почек. Язык у человека покрыт слизистой оболочкой, складки которой во многих местах образуют маленькие выпуклости в форме колышков, называемые сосочками. На рис. 7-1 показано распределение трех типов сосочков-желобоватых, листовидных и грибовидных - по поверхности языка.
Эти три типа распределены по-разному. Только грибовидные сосочки рассеяны по всей поверхности. Желобоватые сосочки, которых у человека всего 7-12, сверху имеют вид круглых образований 1-3 мм в диаметре; они находятся в ограниченной зоне поперек спинки языка у его корня. Третий тип, листовидные сосочки, образуют тесно расположенные складки вдоль задних краев языка. Они хорошо развиты у детей, но гораздо менее выражены и менее многочисленны у взрослых.
Нитевидные сосочки, занимающие остальную поверхность языка, не показаны на рис. 7-1, потому что в них нет вкусовых почек. Название «почка» говорит о форме этих органов (рис. 7-2). Положение их на сосочках варьирует; в случае желобоватых и листовидных сосочков много вкусовых почек заложено в боковых стенках, а на верхушке их нет. В грибовидных сосочках вкусовые почки ограничены поверхностью «шляпки гриба», которая может достигать 1 мм в диаметре.
Отдельная вкусовая почка имеет около 70 мкм в высоту и около 40 мкм в диаметре. Всего у человека около 2000 вкусовых почек, из них
238
X. Альтнер
Рис. 7-1. Схема распределения вкусовых сосочков, их иннервации и зон максимальной чувствительности к разным вкусовым качествам на языке человека.
Рис. 7-2. Положение вкусовых почек (красные) на вкусовых сосочках трех типов.
7. Физиология вкуса
Пора
239
Рис. 7-3. Строение и иннервация вкусовой почки. Элементы органа вкуса, сенсорные, опорные и базальные клетки расположены подобно лепесткам. Вся структура лежит под эпителиальной поверхностью так, что микровилли сенсорных клеток выдаются в наполненное жидкостью пространство над ними. Показано (красным) только два афферентных волокна; в действительности в одну вкусовую почку входят около 50 волокон и разветвляются в ней.
около половины-на желобоватых сосочках. Каждая вкусовая почка содержит 40-60 отдельных клеток.
В соединительную ткань под желобоватыми и листовидными сосочками погружены серозные железы, протоки которых открываются в углубления у основания сосочка (рис. 7-2); их секрет служит для смывания частиц пищи и микроорганизмов. Кроме того, он понижает концентрацию стимулирующего вещества вблизи вкусовых почек.
Внутри вкусовых почек различают три типа клеток: сенсорные, опорные и базальные (рис. 7-3). Растворимые в воде вещества, попадающие на поверхность языка, диффундируют через пору в наполненное жидкостью пространство над вкусовой почкой; здесь они соприкасаются с мембранами микровиллей, которые образуют наружные концы сенсорных клеток. Вкусовые рецепторы представляют собой вторичные сенсорные клетки без аксонов, которые проводят импульсы в центральном направлении. Их ответы передаются афферентными волокнами, которые образуют синапсы близ оснований сенсорных клеток. На рис. 7-3 показаны только два волокна, но в действительности в каждую вкусовую почку входят и разветвляются в ней около 50 волокон.
Продолжительность жизни сенсорных клеток во вкусовых почках невелика; происходит их непрерывная смена. В среднем одна сенсорная
240
X. Алыпнер
клетка замещается новой уже через 10 дней. За сменой клеток можно проследить, помечая их ядра 3Н-тимидином и определяя число меченых ядер, сохранившихся через некоторое время. Утраченные сенсорные клетки замещаются новыми, которые образуются из базальных клеток. При этой смене должны прерваться синапсы между афферентными волокнами и старыми клетками и возникнуть новые синапсы. В связи с такой перестройкой возникает много интересных вопросов, особенно если учесть тот факт, что сенсорные клетки различаются по своей чувствительности к разным стимулам. Так, смена сенсорных клеток может привести к изменению «вкусового профиля»-характерной формы ответов в афферентных волокнах, о чем пойдет речь в следующем разделе.
Центральные связи. Афферентные волокна, проводящие ответы от скоплений вкусовых луковиц, распределяются по двум черепномозговым нервам-лицевому (VII) и языкоглоточному (IX). Такое деление обычно соответствует областям языка, которые снабжаются этими волокнами (рис. 7-1). Так, волокна от желобоватых и листовидных сосочков идут преимущественно в составе языкоглоточного нерва, а волокна от грибовидных сосочков в передней части языка входят в барабанную струну (chorda tympani), ветвь лицевого нерва. У детей имеются добавочные вкусовые органы в эпителии мягкого нёба и задней стенки глотки до гортани; они иннервируются главным образом блуждающим нервом (X).
В головном мозгу вкусовые волокна на каждой стороне объединяются в солитарный тракт. Он оканчивается в продолговатом мозгу, в ядре солитарного тракта, где афферентные волокна образуют синапсы с нейронами второго порядка. Аксоны этих нейронов идут к вентральному таламусу в составе медиального лемниска. Третья совокупность нейронов связывает эту область с корой больших полушарий. Вкусовые зоны коры расположены в латеральной части постцентральной извилины.
Основные вкусовые ощущения. В обычных условиях, например при еде, слизистая ротовой полости подвергается действию сложных стимулов, включающих несколько модальностей. Благодаря тому что ротовая полость сообщается с носовой, пахучие вещества могут достичь обонятельных рецепторов в носу и вызвать другие ощущения. Кроме того, в слизистой оболочке рта и языка имеются терморецепторы, механорецепторы и болевые волокна, которые тоже подвергаются стимуляции. То, что обычно называют «вкусом», в действительности является мультимодальным ощущением, в котором на собственно вкусовые ощущения накладываются ощущения запаха, тепла или холода, давления и, возможно, даже боли.
Что касается запахов, то трудно распределить множество различных стимулов по группам входящих в них веществ, которые отличались бы определенными запаховыми качествами. В отличие от этого существуют четыре четко различимых основных вкусовых ощущения: сладкое, кислое, соленое и горкое.
7. Физиология вкуса
241
Таблица 7-1. Примеры веществ четырех основных вкусовых категорий
Сладкое Кислое Соленое Горькое
Глюкоза Соляная кислота Столовая соль Сернокислый хинин
Сахароза Уксусная кислота Хлористый Никотин
Сахарин Лимонная кислота аммоний Кофеин
D-лейцин Хлористый бериллий Винная кислота Хлористый магний Фтористый натрий L-лейцин Сернокислый магний
Кроме этих основных качеств различаются два дополнительных -щелочной и металлический. Щелочной (или мыльный) возникает при стимуляции поташем (углекислым калием). А некоторые металлы и металлические соли обладают специфическим «металлическим» вкусом.
В табл. 7-1 приведены некоторые вещества, относящиеся к каждой из этих основных категорий. Из таблицы видно, что фактором, определяющим кислый вкус, служат ионы Н+. Но что касается остальных трех качеств, то практически невозможно предсказать на основании физических или химических свойств данного вещества, каким качеством оно должно обладать. Вещества со сходным вкусом могут сильно различаться по химической структуре, и в то же время оптические изомеры могут быть совершенно разного вкуса; некоторые аминокислоты имеют в D-форме сладкий вкус, а в L-форме горький. Ясно, что стимулы, действующие в естественных условиях, сложны, включают несколько качеств, и, таким образом, большинство вкусовых ощущений смешанные.
Пороги обнаружения для разных качеств приходятся на разные концентрации. Пороговая концентрация сернокислого хинина (8 мкмоль/л, или 0,006 г/л) служит хорошим примером того, что вещества с горьким вкусом обнаруживаются при очень низких концентрациях. Порог обнаружения для сахарина составляет 23 мкмоль/л (0,0055 г/л), для виноградного сахара-0,08 моль/л, а для тростникового сахара-0,01 моль/л (соответственно 14,41 и 3,42 г/л). Эти данные характерны, и они показывают, что пороги для моно- и дисахаридов значительно выше, чем для синтетических сладостей. Пороги для уксусной кислоты (0,18 моль/л, или 0,108 г/л) и столовой соли (0,01 моль/л, или 0,585 г/л) служат иллюстрацией того общего правила, что пороги для кислого и соленого приблизительно того же порядка, что и для указанных выше сахаридов. Пороги для кислот приблизительно отражают степень их диссоциации. Сравнение порогов для виноградного и тростникового сахаров говорит о том, что раствор виноградного сахара должен быть более концентрированным, чем раствор тростникового сахара, для того чтобы они были одинаково сладкими. Экспериментальная проверка растворов разных надпороговых концентраций соответствует этому различию.
Но польза от таких точных пороговых данных ограничена, потому что для большинства веществ пороги подвержены значительной инди
242
X. Алътнер
видуальной вариабельности. Разумнее было бы говорить о диапазоне пороговых значений.
Интенсивность ощущений. Простое сравнение разных растворов показывает, что интенсивность вкусового ощущения зависит от концентрации вещества. При определении порогов обнаружено, что эффект от разбавления раствора стимулирующего вещества может быть компенсирован стимуляцией большей поверхности языка, т.е. большего числа рецепторов. Вероятно, это происходит благодаря пространственной суммации. В пороговой области существует входное соотношение между концентрацией и продолжительностью действия стимула. Эти данные можно суммировать в выражениях САП = к и t-Ca = k, где С - концентрация вещества в растворе; 4-площадь стимулируемой части языка; t-время действия стимула; /с-константа для каждого вещества; показатели степени п тоже специфичны для стимулирующего вещества. Эти выражения не включают всех параметров, влияющих на интенсивность вкусового ощущения; определенную роль играет температура стимулирующего раствора. Кроме того, следует помнить, что чувство вкуса подвержено определенной адаптации-при длительном действии стимула интенсивность ощущения снижается. Еще одним фактором является секреция серозных желез, которая разжижает действующее у вкусовых луковиц вещество и тем самым влияет на интенсивность ощущения.
Испытание ряда разведений солевых растворов в околопороговой области во многих случаях показывает, что ощущение может менять свое качество в зависимости от концентрации. Растворы столовой соли 0,02-0,03 моль/л имеют сладкий вкус, а в концентрации 0,04 моль/л или больше-соленый. Этот сдвиг качества, вероятно, можно понять, исходя из того, что вкусовые волокна обладают широким спектром чувствительности в пределах каждого качества (см. разд. 7.2).
Как видно из рис. 7-1, разные области языка у человека варьируют по чувствительности к четырем основным качествам. Кончик языка особенно чувствителен к сладким веществам, средние части краев отвечают лучше всего на кислые стимулы. Соленые стимулы всего эффективнее в области края языка, которая частично перекрывает первые две. Горькие вещества сильнее всего действуют на рецепторы близ корня языка, в области желобоватых сосочков. Поэтому повреждение языкоглоточного нерва понижает способность к обнаружению горечей, а после блокады проведения в лицевом нерве обнаруживаются только они одни.
В 7.1. Афферентные волокна от вкусовых луковиц являются
а) аксонами вкусовых сенсорных клеток;
б) неветвягцимися волокнами, которые образуют синаптические контакты на вкусовых сенсорных клетках;
в) ветвящимися волокнами, которые образуют синаптические контакты на вкусовых сенсорных клетках;
7. Физиология вкуса
243
г) сенсорными волокнами, оканчивающимися на поверхности эпителия между вкусовыми сенсорными клетками.
В 7.2. Перечислите на память названия нервов, идущих от вкусовых органов, и два высших центра переключения вкусового пути.
В 7.3. Свяжите следующие вещества с соответствующими вкусовыми качествами: 1) кофеин, 2) хлористый магний, 3) лимонная кислота, 4) глюкоза, 5) никотин, 6) винная кислота и 7) сернокислый хинин.
а) Сладкое.
б) Кислое.
в) Соленое.
г) Горькое.
В 7.4. Какая концентрация в относительных единицах должна быть у раствора тростникового сахара, чтобы он был таким же сладким, как 1) раствор виноградного сахара? 2) раствор сахарина? а) Соответственно более или же менее концентрированным, б) Более концентрированным в обоих случаях.
в) Соответственно менее и более концентрированным.
г) Менее концентрированным в обоих случаях.
В 7.5. На какие вкусовые качества реагируют сильнее всего 1) кончик языка и 2) область непосредственно впереди корня языка?
а) Сладкое.
б) Кислое.
в) Соленое.
г) Горькое.
7.2. Объективная физиология вкуса
В этом разделе речь пойдет о том, что способность к различению вкусовых качеств, весьма возможно, зависит от специфичности рецепторных молекул в мембранах сенсорных клеток. Может показаться удивительным, чтобы такой механизм отражался лишь диффузно или совсем не отражался в специфичности ответа отдельной сенсорной клетки и афферентного волокна. Как в таком случае может кодироваться качество и концентрация? На этот вопрос можно ответить, если принять во внимание совокупность ответов множества клеток или волокон. Поэтому сначала мы опишем их, а в заключение вкратце рассмотрим биологическое значение чувства вкуса.
Для регистрации активности как отдельных сенсорных клеток, так и афферентных волокон можно воспользоваться микроэлектродами. Такие записи показывают, что ни сами рецепторы, ни волокна, идущие к ЦНС, не дают качественно специфических ответов; как правило, эффективными оказываются стимулы более чем одной категории. На рис. 7-4 приведено изменение частоты импульсации в четырех афферентных волокнах (А-Г) при стимуляции веществами, которые представляют четыре основных качества. Очевидно, что каждое волокно реаги-
244
X. Алътнер
Рис. 7-4. Ответы четырех вкусовых волокон (Л-Г) на растворы стимулирующих веществ разного качества. Типичные конфигурации разных возбуждений называются вкусовыми профилями.
рует на стимулы нескольких категорий, но при рассмотрении разных градаций чувствительности выявляются различия. Иными словами, стимуляция раствором вещества в определенной концентрации активирует различные волокна в разной степени. Характер возбуждения, типичный для каждого отдельного волокна при ответах на ряд веществ, называется вкусовым профилем. Волокнами, наиболее близкими к качественной специфичности, являются те, которые реагируют на растворы сахаров увеличением частоты разрядов. Сравнительные исследования показали, что такие относительно специфичные волокна особенно характерны для обезьян.
Регистрация активности отдельных сенсорных клеток показала, что они обладают градуальной относительной специфичностью. Ответы волокон, идущих от этих клеток, в этом отношении отражают ответы клеток. Но афферентные волокна ветвятся во вкусовых луковицах (рис. 7-3), так что каждое волокно получает возбуждение от многих сенсорных клеток, которые, надо полагать, различаются по степени специфичности. Кроме того, обнаружено, что сенсорные клетки в разных сосочках образуют синапсы с коллатералями от одного афферентного волокна. Это значит, что вкусовые волокна получают входы от сенсорных клеток, распределенных по значительным участкам языка. Эти участки называются рецептивными полями (рис. 7-5). Ситуация с рецептивными полями усложняется тем, что отдельные сенсорные клетки могут получать иннервацию от нескольких различных волокон.
Суммируя, надо сказать, что градуальная относительная специфичность вкусовых волокон создается 1) градуальной относительной специфичностью сенсорных клеток и 2) ветвлением вкусовых волокон, создающим рецептивные поля. Частота импульсации в одиночном
7. Физиология вкуса
245
Рис. 7-5. Рецептивные поля языка крысы. Отдельные афферентные волокна отвечают на стимуляцию обширных участков, которые могут многократно перекрываться, причем каждый содержит несколько грибовидных сосочков.
афферентном волокне (рис. 7-4) поэтому меняется как от качества стимула, так и от его концентрации. Разумеется, важным фактором является также степень, в какой стимулируемая область покрывает рецептивное поле волокна. Очевидным выводом в отношении кодирования
стимула является то, что активность одного волокна не может дать однозначную информацию о качестве или концентрации. Только сравнение уровня возбуждения в нескольких волокнах может выявить характерные распределения (паттерны) активности, которые говорят что-то о качестве стимула. При условии что качество известно, частота им-пульсации в каждом отдельном волокне может служить мерой концентрации стимулирующего вещества. Отличительные черты вещества, следовательно, кодируются таким образом, что сложный, но характерный паттерн возбуждения создается одновременными, но разными ответами множества нейронов.
Первичный процесс. Условием возбуждения вкусового рецептора является взаимодействие между молекулами стимулирующего вещества и специально дифференцированными точками в мембране сенсорной клетки, где лежат рецепторные молекулы. Это взаимодействие называется первичным процессом; как полагают, он начинается с адсорбции молекулы вещества-стимула. Предполагают, что, когда это происходит, рецепторная-вероятно белковая-молекула изменяет свою структуру. Такое конформационное изменение рецепторной молекулы приводит в свою очередь к локальному изменению проницаемости мембраны клетки. Этот клеточный «усилительный механизм» мог бы послужить причиной генерации рецепторного потенциала.
К свидетельствам существования специфических рецепторных молекул относится наблюдение, что некоторые растительные вещества и препараты, например кокаин и гимневая кислота (получаемая из ин
246
X. Алътнер
дийского растения Gymnema sylvestre), избирательно блокируют некоторые вкусовые ощущения. Очевидно, эта кислота связывается с рецепторными молекулами для сладких веществ, поскольку ее нанесение делает такие вещества безвкусными. Первичный процесс в мембранах вкусовых сенсорных клеток еще по-настоящему не объяснен, но, согласно рабочей гипотезе, он сходен с процессом в холинергических синапсах, где особые молекулы меняют проницаемость в особых точках мембраны.
Роль вкусовой чувствительности. Вкусовые луковицы на языке реагируют на стимулы, локализованные во рту. Иными словами, вкусовая чувствительность у всех позвоночных участвует в ориентации на близком расстоянии. В то же время у рыб чувство вкуса может служить также ориентации на далеком расстоянии. В воде вкусовые вещества перемещаются благодаря диффузии и конвекции из очень далеких источников к вкусовым луковицам, которые могут быть рассеяны по всей поверхности тела рыбы.
Помимо своей роли в ориентации на близком расстоянии чувство вкуса у человека выполняет важную функцию, запуская ряд рефлексов. Например, отмывание языка секретом из серозных желез контролируется рефлексом, который находится под действием вкусовых луковиц. Секреция слюны тоже запускается рефлекторно соответствующей стимуляцией вкусовых рецепторов. Даже состав слюны варьирует в зависимости от характера стимулов, действующих на сенсорные клетки, и вкусовые стимулы влияют также на выделение желудочного сока. Наконец, доказано, что рвота вызывается при участии вкусовой чувствительности.
В 7.6. Как данное афферентное волокно, идущее от вкусовых луковиц, реагирует на вкусовые вещества?
а) Одинаково сильно на вещества одного и того же качества, б) В разной степени, хотя вещества обладают одним и тем же качеством.
в) С одинаковой силой на вещества разных качеств.
г) В разной степени на вещества разных качеств.
В 7.7. Что лежит в основе градуальной специфичности. отдельных вкусовых волокон?
а) Отсутствие абсолютной специфичности у сенсорных клеток, б) Ветвление волокон и наличие рецептивных полей.
в) Перекрывание рецептивных полей.
В 7.8. Которая из следующих реакций может быть вызвана рефлекторно возбуждением вкусовых рецепторов?
а) Кашель.
б) Секреторная активность серозных желез.
в) Секреция слюны.
г) Поток слез.
д) Удушье.
8. ФИЗИОЛОГИЯ ОБОНЯНИЯ
X. Альтнер
Нос человека способен различать тысячи разных пахучих веществ, но эта способность весьма скромна по сравнению с возможностями других организмов. Такая относительная слабость обонятельной функции наряду с чрезвычайно важной ролью других органов чувств, быть может, является одной из причин того малого интереса, какой проявлялся к исследованию обонятельной системы. Другая причина-это трудность экспериментирования с ней-несомненная в отношении людей, но также касающаяся и животных. В результате нет ни субъективных, ни объективных критериев, которые позволили бы четко определить качества запахов; поразительное разнообразие обонятельных стимулов мало упорядочено. Между тем, как будет видно из настоящей главы, обонятельные проводящие пути у млекопитающих изучены довольно хорошо. Структурный и физиологический анализ выявил ряд интересных свойств этой системы, например тот факт, что сигналы, посылаемые в мозг, находятся под эфферентным контролем. Много сведений получено также о значении чувства обоняния для автономных регуляторных процессов и эмоциональных состояний.
8.1. Обонятельная слизистая; периферические механизмы обонятельной рецепции
Первый предмет этого раздела составляет морфология органа обоняния и клеток, из которых он состоит. Затем речь пойдет о разнообразии стимулирующих веществ; они не поддаются классификации по качеству ни на основе вызываемых ими ощущений, ни на основе ответов одиночных сенсорных клеток.
Локализация и клеточная организация обонятельного эпителия. Носовая полость разделена на две части, левую и правую, стенкой, называемой носовой перегородкой. Поверхность обеих частей увеличена тем, что образует складки, раковины, которые вдаются внутрь из наружных стенок. Как видно на рис. 8-1, у взрослого человека с каждой стороны одна над другой лежат три раковины. Вся носовая полость выстлана слизистой оболочкой, но обонятельные сенсорные клетки расположены лишь на небольшом ее участке, в обонятельной области (на рис. 8-1 показана красным). Эта область занимает всю верхнюю раковину и образует островки на средней раковине. Обонятельный эпителий имеется также на прилежащих частях перегородки. Дыхательная область-та часть слизистой носа, в которой нет обонятельных клеток,-представляет собой ресничный эпителий, состоящий из двух слоев клеток, в том числе бокаловидных, которые продуцируют слизь.
248
X. Алыпнер
Рис. 8-1. Положение обонятельного эпителия и направление потока воздуха в носу человека; вид сбоку. Обонятельная область (Об, отмечена красным) ограничена верхней и средней раковинами. Аксоны сенсорных клеток образуют здесь обонятельные волокна fila olfactoria (черные), идущие к обонятельной луковице (ОЛ). Показаны также участки, откуда волокна проходят к тройничному (V), языкоглоточному (IX) и блуждающему (X) нервам.
У плода человека слизистая на перегородке содержит замкнутые трубочки, которые, по-видимому, никуда не ведут. Это рудименты якобсонова органа, обонятельного органа, который играет вспомогательную, а в некоторых случаях - важную роль у многих амфибий, рептилий и млекопитающих. У человека его нет.
На рис. 8-2 показано строение обонятельной слизистой, многослойного эпителия с двумя преобладающими типами клеток-обонятельными и опорными. Как и во вкусовых почках (гл. 7), здесь имеются также базальные клетки; они могут развиться в обонятельные и, таким образом, представляют собой незрелые сенсорные клетки. Обонятельные клетки (в отличие от вкусовых) являются первичными сенсорными клетками, посылающими аксоны от своего базального полюса. Эти волокна образуют толстые пучки под сенсорным эпителием (называемые также обонятельными волокнами, fila olfactoria), которые идут к обонятельной луковице.
На своем апикальном полюсе обонятельные клетки несут видоизмененные цилии, погруженные в слой слизи, которая покрывает обонятельный эпителий. Молекулы пахучего вещества диффундируют сквозь часть этого слизистого слоя, прежде чем достигнут самой периферической части обонятельных клеток, мембран цилий. Слизь происходит из
8. Физиология обоняния
249
Аксоны
Рис. 8-2. Элементы обонятельного эпителия. А. Слизистая оболочка обонятельной области. Б. Клетки обонятельной слизистой.
трех источников: 1) боуменовых желез, 2) бокаловидных клеток дыхательной области и 3) опорных клеток обонятельного эпителия, которые таким образом выполняют двойную функцию. Киноцилии клеток в дыхательной области регулируют ток слизи.
Обонятельные ощущения исходят не только от сенсорных клеток в обонятельной области. Дыхательная область содержит помимо уже упомянутых клеточных элементов свободные окончания сенсорных волокон тройничного нерва (V); они также реагируют на пахучие вещества. Поэтому даже при случайном полном перерыве обонятельных волокон ощущение запаха до некоторой степени сохраняется.
Запахи. Повседневный опыт показывает, что мы можем различать тысячи разных веществ по их запаху. Но в отличие от физиологии вкуса
250
X. Альтнер
в субъективных опытах по обонянию не удалось выявить четкого различия между запахами. В любой попытке создать такую классификацию есть нечто произвольное. Но для практических целей был определен ряд качеств и составлен список классов запахов, или первичных запахов. Ниже приводятся некоторые из них, и для каждого даны два примера химически чистых пахучих веществ данной категории.
Цветочный: ос-ионон, Р-фенилэтиловый спирт Эфирный: 1,2-дихлорэтан, бензилацетат
Мускусный: Циклические кетоны (С15_17), например цибетон и мускусный кетон
Камфорный: 1,8-цинеол, камфора
Запах пота: Изо валериановая кислота, масляная кислота Гнилостный: Сероводород, этилмеркаптан Едкий: Муравьиная кислота, уксусная кислота
Уже из этих нескольких примеров ясно, что химически сходные вещества могут оказаться в разных запаховых классах и что члены одного и того же класса могут значительно различаться по своей химической структуре. Запахи, которые встречаются в естественных условиях и по которым названы первичные классы (например, аромат цветов, запах пота или гниющего мяса), как правило, представляют собой смеси запахов, в которых преобладают определенные компоненты.
Показано, что 0,1-1% населения Европы отличается особой «обонятельной слепотой». Симптом такой частичной аносмии, которая в некоторых случаях может быть наследственной, состоит в том, что немногие вещества со сходными запахами (например, некоторые мускусные запахи) обнаруживаются только при очень высоких концентрациях. Иными словами, порог для таких веществ специфически повышен. Наличие частичных аносмий показывает, что разграничение запахов по их качеству до некоторой степени возможно.
Кодирование. Как и для вкусовой чувствительности (см. гл. 7), лежащий в основе обоняния первичный процесс не разгадан, хотя и в этом случае считается, что молекулы пахучего вещества взаимодействуют с особыми молекулами в мембране обонятельной клетки-с рецепторными молекулами. Наличие множества эффективных пахучих веществ не позволяет думать, что сенсорная мембрана содержит по отдельной рецепторной молекуле для каждого пахучего вещества. Вероятно, несколько близких пахучих веществ вступают в реакцию с одной и той же рецепторной молекулой. Свидетельством существования рецепторных молекул, которые могут взаимодействовать с несколькими пахучими веществами, служит явление упомянутой выше частичной аносмии. Однако отдельные стимулы должны как-то кодироваться, поскольку, как известно, существует различение отдельных запахов.
Каким образом кодируются параметры стимула, в частности природа и концентрация пахучего вещества, казалось бы, легче всего установить по регистрации ответов отдельных рецепторных клеток. Однако
8. Физиология обоняния
251
в действительности попытки выявить такие корреляции у позвоночных животных очень редко удавались. Пока что известно только, что обонятельные клетки обладают характерными спектрами ответов. Возбуждение каждой отдельной клетки возможно в разной степени под влиянием многих веществ, но относительная чувствительность к разным активным веществам (при данных концентрациях) варьирует от клетки к клетке. Очевидно, стимулы, действующие на обонятельные клетки, так же как на вкусовые луковицы (см. гл. 7), кодируются таким образом, что при данной концентрации каждое пахучее вещество вызывает свой особый паттерн возбуждения, охватывающего много сенсорных клеток. Иными словами, информация о природе вещества содержится в относительном уровне возбуждения. При повышении концентрации в большинстве случаев частота импульсов растет, хотя некоторые пахучие вещества тормозят спонтанную активность сенсорных клеток.
Обонятельные сенсорные клетки у насекомых, обычно расположенные на щетинковидных выступах кутикулярного экзоскелета, значительно более доступны для электрофизиологических исследований. Оказалось, что в отличие от позвоночных функции рецепторов насекомых чрезвычайно специфичны по отношению к определенным пахучим веществам. Другой интересный результат, полученный в опытах на тутовом шелкопряде, состоит в том, что распространяющийся импульс может быть вызван действием на мембрану сенсорной клетки самца одной-единственной молекулы аттрактанта, выделяемого самкой. Нет никаких оснований полагать, что в отношении обонятельных клеток человека и других позвоночных ситуация в корне отличается от описанной.
В 8.1. Сколько раковин в каждой половине носовой полости (А) и на каких из них расположена обонятельная область (Б)?
а) Три или четыре. Б. а) На всех.
б) Три. б) На верхней.
в) Больше трех. в) На нижней.
г) Две. г) На двух верхних.
д) Одна или две. д) На двух нижних.
В 8.2. Какие из следующих типов клеток имеются в обонятельной области слизистой носа?
а) Обонятельные клетки.
б) Бокаловидные клетки.
в) Опорные клетки.
г) Базальные клетки.
В 8.3. Назовите на память четыре класса запахов.
В 8.4. Что означает термин «частичная аносмия»?
а) Разрушение обонятельной области.
б) Повышение порога для небольшого числа близких запахов.
в) Общее повышение порога для пахучих стимулов.
г) Пониженная чувствительность к мускусным запахам.
252
X. Алътнер
В 8.5. Как кодируются пахучие вещества в ответах сенсорных клеток? а) Ответом клеток, специфических для данного качества.
б) Ответом клеток, специфических для данного вещества.
в) Образованием специфических паттернов возбуждения в большой популяции сенсорных клеток.
8.2. Субъективная физиология обоняния; центральные связи
Вначале мы обсудим вопрос о различии между порогом обнаружения запаха и порогом его опознания. Затем будет показано, что сила таких ощущений возрастает относительно медленно по мере повышения интенсивности стимулов. Анализ обонятельного проводящего пути показал, что эффективный контроль возбудительных входов осуществляется на периферии. Кроме того, обоняние вовсе не служит только для ориентации; оно действует еще и на течение вегетативных процессов, а также может сказываться на эмоциональных состояниях.
Порог обнаружения и порог опознания. При низких концентрациях пахучего вещества, едва достаточных, чтобы вызвать ощущение «какого-то запаха», человек, как правило, не может сказать, что это за запах. Он просто чувствует, что чем-то пахнет. Только при более высоких концентрациях запах вещества становится опознаваемым, и его можно определить. Поэтому полезно говорить о порогах обнаружения и порогах опознания.
Нос человека очень чувствителен ко многим веществам. Масляная кислота, которая относится к классу запахов пота, обнаруживается при концентрации всего лишь 2,4 1012 молекул на 1 л воздуха, что эквивалентно разведению чистого вещества в 1010 раз. Порог обнаружения бу-тилмеркаптана, вещества с неприятным гнилостным чесночным запахом, равен Ю10 молекул на 1 л воздуха (разведение 2,7 -1012).
Если известен порог обнаружения того или иного вещества и число рецепторов, то можно вычислить соответствующее число молекул пахучего вещества на обонятельную клетку за один вдох (или «принюхивание»). Такое вычисление для бутилмеркаптана дает на пороге максимум восемь молекул на клетку.
У животных установлены пороги гораздо ниже, чем у человека. Хорошо известно поразительно тонкое обоняние у собак. Угри обнаруживают бета-фенилэтиловый спирт в разведении 1:2,86-1018. Шелкопряд тоже замечателен в этом отношении. Если всего лишь 1% антеннальных рецепторов, чувствительных к аттрактанту самки, возбуждается молекулами этого вещества-причем в этом случае в каждой из двух антенн возникают около 200 импульсов-самец дает характерную поведенческую реакцию-трепетание крыльев.
Сила ощущения. При надпороговой стимуляции ощущение усиливается по мере повышения концентрации пахучего вещества. Эта зависимость описывается степенным законом Стивенса: ощущение пропорционально (С — Со)" (см. разд. 1.3). Если построить график в логарифми-
8. Физиология обоняния
253
Рис. 8-3. Зависимость силы ощущения запаха от концентрации вещества (в данном случае пентанола). Обе шкалы логарифмические. Коэффициент наклона меньше, чем для других модальностей.
Интенсивность стимула (концентрация, мг/л)
ческом масштабе, то получим прямую линию, коэффициент наклона которой показывает скорость усиления ощущения с повышением концентрации (рис. 8-3). Показатель степени п, который определяет этот коэффициент наклона, составляет для обоняния 0,5-0,6. Такой наклон невелик по сравнению с наклоном для некоторых других сенсорных модальностей; иными словами, обонятельные ощущения меняются по мере изменений стимула относительно медленно.
При длительном действии стимула ощущение слабеет-наступает адаптация. При продолжительной интенсивной стимуляции адаптация может быть даже полной, т.е. ощущение исчезает совсем. Адаптацию можно измерить по повышению порога после определенного периода стимуляции.
Стимуляция волокон тройничного нерва. Считается, что волокна тройничного нерва реагируют предпочтительно на едкие или горелые запахи. Но в опытах на животных эти волокна отвечают также на вещества, которые для человека вовсе не имеют едкого запаха. Наблюдения над больными с односторонним разрушением этого нерва тоже говорят о том, что тройничный компонент принимает более широкое участие в обонятельных ощущениях.
Центральные связи. В предыдущем разделе было указано, что объединенные в пучок аксоны обонятельных клеток, fila olfactoria, идут к обонятельной луковице. По пути они проходят сквозь lamina cribrosa решетчатой кости. На рис. 8-4 видно, что клеточные элементы в обонятельной луковице расположены слоями. Считая по направлению к центру, это следующие слои: 1) слой fila olfactoria, 2) слой гломерул, 3) наружный плексиформный слой, 4) слой тел митральных клеток и 5) слой клеток-зерен. Весьма крупные митральные клетки представляют собой нейроны второго порядка обонятельного пути. Они отдают один главный дендрит, дистальные веточки которого образуют синапсы с во-
254
X. Альтнер
Рис. 8-4. Схема слоев в обонятельной луковице; показаны связи клеток. Клетки между гломерулами, перигломерулярные, могут передавать латеральные влияния между митральными клетками; через перигломерулярные клетки и клетки-зерна осуществляется эфферентный контроль. Fila olfactoria и эфферентные волокна показаны красным. У дендро-дендритных синапсов красные стрелки означают торможение, черные -возбуждение.
локнами обонятельных клеток. Дендритные ветвления митральных клеток вместе с окончаниями приходящих сюда волокон образуют сферические массы, гломерулы. На каждой митральной клетке конвергирует около 1000 волокон. Аксоны обонятельных клеток синаптически контактируют также с перигломерулярными клетками, которые образуют латеральные связи между гломерулами.
Аксоны митральных клеток составляют обонятельный тракт. Этот тракт или непосредственно, или через свои связи с другими трактами передает обонятельные сигналы во многие области мозга, в том числе в обонятельную луковицу противоположной стороны, препириформную область и пириформную долю, гиппокамп и через миндалевидный комплекс к автономным ядрам гипоталамуса.
Выход возбудительных сигналов из обонятельной луковицы находится под эфферентным контролем, действующим на периферическом уровне. На рис. 8-4 центробежные волокна показаны красным. Эти аксоны оканчиваются на клетках-зернах и перигломерулярных клетках и таким образом в состоянии видоизменять возбудительную активность, приходящую от fila olfactoria на уровень митральных клеток. На
8. Физиология обоняния
255
рис. 8-4 дана очень упрощенная схема сложных связей в обонятельной луковице. Она показывает, что имеются дендро-дендритные контакты между митральными клетками и клетками-зернами, а также между митральными и перигломерулярными клетками. По таким контактам поток информации передается в обе стороны-от митральных к перигло-мерулярным клеткам и клеткам-зернам и в обратном направлении-от них к митральным клеткам. Рис. 8-4 указывает также на возможность латерального взаимодействия между митральными клетками через перигломерулярные клетки.
Как упомянуто выше, возбудительные сигналы от обонятельного органа передаются к автономным ядрам гипоталамуса. Функция этой связи еще недостаточно изучена. Несомненно, что у разных млекопитающих активность в этих проводящих путях влияет на процессы, связанные с размножением. Так, установлено, что активность элементов в обонятельном проводящем пути варьирует при изменении уровней половых гормонов.
Но полностью значение обоняния для млекопитающих выходит за пределы как его участия в регуляции размножения, так и его очевидной роли в поисках пищи. Например, пахучие вещества служат сигналами в общественных отношениях между группами и особями. Группа, к которой принадлежит животное, может быть обозначена обонятельным сигналом, точно так же как животное, занимающее некоторую территорию, метит ее границы определенным запахом. У людей обоняние утратило' свою важную роль по сравнению с другими ощущениями, но все же его значение недооценивается. Так, на эмоции оно может действовать в гораздо большей степени, чем мы себе представляем; знакомый аромат может сильно повлиять на настроение. Такое действие, возможно, определяется упомянутыми выше связями между органом обоняния и лимбической системой. Но эти отношения привлекали к себе мало внимания; лучше известно, что некоторые неприятные запахи вызывают у человека такие защитные рефлексы, как чиханье и задержка дыхания. Вещества с резким запахом, например аммиак, приводят к рефлекторной остановке дыхания.
В 8.6. Что связано с более высокими концентрациями-а) порог обнаружения или б) порог опознания?
В 8.7. Который из следующих показателей говорит о степени адаптации обоняния?
а) Повышенная чувствительность к пахучим веществам.
б) Повышение порога обнаружения.
в) Понижение порога опознания.
В 8.8. Который из следующих элементов осуществляет эфферентный контроль в обонятельной луковице?
a) Fila olfactoria.
б) Центробежные волокна в обонятельном тракте.
в) Клетки-зерна.
г) Перигломерулярные клетки.
9. ЖАЖДА И ГОЛОД
Р. Шмидт
Чувство жажды, которое мы испытываем, когда не выпили достаточно жидкости, и чувство голода-когда давно не ели, нельзя отнести к какому-то конкретному органу или части тела. Поэтому они называются «общими чувствами». Другими примерами общих чувств являются усталость, ощущение духоты и половое влечение. С точки зрения сенсорной физиологии все они характеризуются тем, что вызываются одним или несколькими адекватными стимулами, возникающими в самом организме, а не в окружающей среде. Эти стимулы обнаруживаются рецепторами (часть из которых еще неизвестна) и вызывают соответствующие общие ощущения (рис. 9-1, А). Например, ниже мы расскажем, как «сгущение» жидкости в организме из-за недостатка воды воспринимается осморецепторами, и в результате возникает чувство жажды (рис. 9-1, Б). Можно представить себе также, что в течение дня в крови накапливаются конечные продукты обмена и они вызывают чувство усталости, или же некоторые гормоны вызывают половое влечение или благоприятствуют его появлению, когда их концентрация в организме достаточно высока.
Стимулы, способные вызывать общие ощущения, вызывают также разные виды активности, направленные на снижение таких ощущений или их устранение. Иными словами, стимулы, вызывающие общие ощущения, приводят к драйвам (drive-побуждение)-мотивационным состояниям, которые побуждают организм добывать то, чего ему недостает. Хотя такие драйвы контролируются ощущениями, они могут возникать также и независимо (рис. 9-1, А). Недостаточное количество воды в организме ведет не только к ощущению жажды, но и к поискам воды, и если эти поиски успешны, то недостаток ее устраняется (рис. 9-1, Б). В самом общем виде это значит, что удовлетворение побуждения устраняет причину, вызвавшую общее чувство.
Драйвы, связанные с общими чувствами, способствуют выживанию индивидуума или вида. Поэтому, как правило, они должны удовлетворяться. Это врожденные состояния, не требующие обучения. Но в течение жизни многочисленные влияния видоизменяют их, в особенности на высших филогенетических уровнях. Эти влияния действуют в разные моменты всего процесса. Описание драйвов и их изменений не входит в задачу настоящей книги; здесь будут рассмотрены как примеры общих чувств только те аспекты жажды и голода, которые связаны с сенсорной физиологией.
9. Жажда и голод
257
Рис. 9-1. Отношения между общими чувствами и драйвами. А. Общий характер взаимоотношений между ощущениями и драйвами. Б. Возникновение чувства жажды и активация поиска воды при ее нехватке. В ощущении жажды помимо осморецепторов участвуют и другие рецепторы (см. рис. 9-2). Вертикальные красные полоски на кончиках верхних стрелок означают, что удовлетворение драйва ведет к устранению стимула.
9.1. Жажда
Условия возникновения чувства жажды. Организм взрослого человека на 70-85% (по весу) состоит из воды (не считая отложений жира). Это высокое содержание воды колеблется в очень узких пределах. В норме оно колеблется только на + 0,22% веса тела, так что у человека весом 70 кг колебания эти составляют + 150 мл. При потере воды больше
258
Р. Шмидт
Таблица 9-1. Суточный водный баланс у взрослого человека (Muntwyler, 1973)
Поступление воды МЛ Потери воды МЛ
Питье 1200 С мочой 1400
С твердой пищей 900 Через легкие и кожу 900
В процессе метаболизма 300 С калом 100
Всего 2400 2400
0,5% веса тела (около 350 мл у человека весом 70 кг) возникает жажда. Физиологически организм может терять воду четырьмя путями: с мочой, с потом, в виде водяных паров в выдыхаемом воздухе и (обычно в ограниченном количестве) с калом. Эта вода должна быть возмещена, чтобы не нарушилось жизненно важное тонко сбалансированное равновесие между множеством растворенных веществ. Как показывают данные по водному балансу (табл. 9-1), в умеренном климате человек, потребляющий обыкновенную пищу, получает воду главным образом в виде питья, а также как составную часть твердой пищи и в меньшей степени-как продукт окислительного распада этой пищи в организме. Суммарные цифры выражают суточный кругооборот воды у взрослого человека, составляющий около 3-4% веса тела.
Еще одним фактором, влияющим на чувство жажды, является распределение воды в организме на две части (компартмента) с разными свойствами. Около двух третей жидкости тела содержится в клетках, во внутриклеточном пространстве, а одна треть-во внеклеточном пространстве. Около трех четвертей внеклеточной воды находится в промежутках между клетками, в интерстициальном пространстве, а остальная-в сосудистой системе, где она образует водную фазу плазмы крови (крови без кровяных телец). Вне- и внутриклеточные жидкости разделены клеточными мембранами и имеют разный состав, в особенности в отношении катионов и анионов. В интересующем нас аспекте главное различие состоит в том, что внеклеточная жидкость содержит большие количества Na + , а внутриклеточная богата К+, но содержит мало Na+. Плазма крови и интерстициальная жидкость сообщаются через стенки капилляров; они почти не различаются по содержанию солей, но белка в плазме значительно больше, чем в интерстициальной жидкости.
Клеточные мембраны, разделяющие внутри- и внеклеточные пространства, легко проницаемы для воды, но для солей их проницаемость значительно ниже. Для простоты изложения мы примем, что мембраны непроницаемы для солей. В конечном счете диффузия воды через такие полупроницаемые мембраны (т. е. проницаемые только для воды) всегда направлена к пространству с более высокой концентрацией солей (т.е. с меньшей «концентрированностью воды»). При равной концентрации соли в обоих пространствах вода диффундирует сквозь клеточную мембрану одинаково в обе стороны, в итоге перемещения не происходит.
9. Жажда и голод
259
Но если из одного пространства удалить воду, так что концентрация соли в нем повысится, то вода будет перемещаться сюда из второго пространства, пока не выравняются концентрации. (Этот процесс называется осмосом, а разница в гидростатическом давлении, которая точно противодействовала бы стремлению воды диффундировать через полупроницаемую мембрану между пространствами, содержащими жидкости с разной концентрацией солей, называется осмотическим давлением. Солевой раствор с таким осмотическим давлением, как у нормальной жидкости тела, называется изотоническим. Отсюда гипертоническим и гипотоническим называют растворы солей соответственно более и менее концентрированные, чем нормальная тканевая жидкость.) Например, если вода удаляется из плазмы вследствие образования мочи в почке, то произойдет диффузия воды из интерстициального пространства в плазму, а из клеток в интерстициальное пространство. Если же клетка в потовой железе секретирует пот (т.е. теряет воду), то вода должна диффундировать из интерстициального пространства в клетку железы.
Физиологические потери воды организмом (с мочой, потом, влагой в выдыхаемом воздухе) все вместе приводят к потере воды как из внеклеточного, так и из внутриклеточного пространства. Одновременно повышается концентрация солей в этих растворах; они становятся гипертоническими, но обычно лишь в незначительной степени. Дальнейшим следствием является уменьшение секреции слюны, что приводит к ощущению сухости во рту и в глотке, столь характерному для жажды. Если бы существовали соответствующие рецепторы, можно было бы измерить потерю воды организмом а) по объему или осмотическому давлению клеток, б) по объему или осмотическому давлению внеклеточного пространства или в) косвенно по снижению секреции слюны и возникающей при этом сухости слизистых оболочек.
Адекватные стимулы для чувства жажды. Чтобы решить, которое из перечисленных изменений может быть причиной чувства жажды, каждый из этих факторов нужно изучить экспериментально по отдельности; иначе говоря, содержание воды и солей в одном пространстве надо менять, не затрагивая второго, или же надо менять одну только секрецию слюны. Такие опыты проведены на животных, причем количество выпиваемой воды служило показателем степени экспериментально вызванной жажды. Наиболее важные результаты этих опытов изложены ниже.
После внутривенного вливания гипертонического раствора собака выпивает воды вдвое больше, чем после внутривенного вливания осмотически эквивалентного раствора мочевины. В первом случае из-за непроницаемости клеточных мембран для ионов Na+ вода выходит из клеток. Но для мочевины клеточные мембраны легко проницаемы; поэтому, когда ее вводят, концентрации во внутри- и внеклеточных пространствах выравниваются при заметно меньшем изменении объема клеток и их тоничности (концентрации соли). Этот факт подтвердился в повторных опытах со многими модификациями на разнообразных
260
Р. Шмидт
млекопитающих. Отсюда следует вывод, что уменьшение объема клеток без изменения содержания в них солей (но не концентрации, которая возрастает) вызывает жажду.
Если количество Na+ во внеклеточном пространстве снизить в опыте (например, изменив состав пищи или посредством искусственной почки), то оно теряет воду; часть ее выходит из тела, а часть диффундирует в клетки. В этих условиях, даже несмотря на то что объем клеток возрастает, появляется жажда. (Возникает также потребность в соли, но этого мы здесь касаться не будем). Кроме того, можно экспериментально снизить объем внеклеточной жидкости, не меняя концентрацию NaCl в ней, так что объем клеток не меняется; в этом случае также возникает жажда. Отсюда можно сделать вывод, что уменьшение объема внеклеточной жидкости тоже вызывает жажду. Опыты показали, что эффекты этих двух факторов аддитивны; при одновременном понижении объема клеток и объема внеклеточной жидкости жажда становится особенно сильной.
Сухость во рту, сопровождающая все формы жажды, вызывается, как уже было указано, снижением секреции слюны. Это явление отражает недостаток воды. Но в отличие от прежних толкований оно, по-видимому, представляет собой не причину, а скорее симптом, связанный с чувством жажды. Об этом говорят следующие данные. Смачивание поверхности рта и глотки не устраняет жажды, хотя может немного облегчить ее. Жажду также не ослабляет и не предотвращает местная анестезия слизистой рта или даже полная денервация области рта и глотки. Врожденное отсутствие слюнных желез (у людей) или их оперативное удаление (у животных) не оказывает заметного действия на потребление воды; количество выпиваемой воды остается нормальным. Наконец, внутривенное внедрение воды немедленно приносит облегчение, хотя сухость во рту и глотке какое-то время может сохраняться.
Рецепторы и центральные механизмы. Нервные структуры, преимущественно регулирующие водно-солевой баланс, находятся в промежуточном мозгу (диэнцефалон), в основном в гипоталамусе и около него. Эта часть головного мозга имеет прямое отношение к регуляции вегетативных функций. Опытами здесь выявлено наличие осморецепторов, главным образом в областях впереди гипоталамуса. Это сенсорные элементы, реагирующие на повышение концентрации соли внутри клеток при потере ими воды. Например, после введения козе очень малых количеств (меньше 0,2 мл) гипертонического раствора NaCl в некоторые части этой области животное через 30-60 с начинало пить и в течение 2-5 мин поглощало 2-8 л воды. Электрическая стимуляция этих же нервных структур тоже заставляет животных много пить. Хирургическое удаление или коагуляция некоторых участков гипоталамуса может ослабить или совсем прекратить питье даже при нехватке воды в организме. Все эти данные показывают, что осморецепторы в промежуточном мозгу служат сенсорными аппаратами жажды, вызываемой дефицитом воды в клетках (рис. 9-2). Нейронные структуры в гипоталамусе несом-
9. Жажда и голод
261
Рис. 9-2. Возникновение чувства жажды. Участвующие в этом рецепторы показаны серым. Прямоугольники над ними означают их адекватные стимулы. Сухость во рту является косвенным следствием внутриклеточного и внеклеточного дефицита вода.
ненно играют решающую роль в переработке информации от этих осморецепторов.
Что касается сенсорных элементов, на которых основано чувство жажды, вызываемое дефицитом воды во внеклеточном пространстве, то здесь имеются только предположения и косвенные данные. В настоящее время представляется вероятным, что рецепторы растяжения в стенках крупных вен и в предсердиях не только влияют на кровообращение, но также участвуют в регуляции водного баланса и возникновения жажды (рис. 9-2). Гипоталамус служит важным центром переработки сигналов, приходящих от рецепторов растяжения (по афферентным волокнам блуждающего нерва). Внеклеточная дегидратация связана также с секрецией ренина и образованием ангиотензина II. Ангиотензин II-вещество, весьма действенно вызывающее жажду при введении его в кровоток или непосредственно в разные отделы гипоталамуса, в том числе в субфор-никальный орган. Ангиотензин несомненно способствует возникновению жажды, связанной с уменьшением объема жидкости (гиповолемия), но пока что относительное значение ангиотензина и других механизмов для такого рода жажды еще не определено.
Об уменьшении слюноотделения, связанном с дефицитом воды и приводящем к сухости во рту, сигнализуют рецепторы в слизистых оболочках рта и глотки. В опытах на животных обнаружены здесь раз
262
Р. Шмидт
ные виды рецепторов-механорецепторы, холодовые и тепловые рецепторы и, возможно, водяные рецепторы. Неизвестно, в какой степени каждый из них участвует в создании этого периферического компонента жажды. Если эти рецепторы стимулируются, когда в организме нет общего недостатка воды, что возможно, когда человек говорит, курит, дышит через рот или ест очень сухую пищу, то вызываемую ими «ложную жажду» можно утолить, смочив слизистую рта. Но сигналы, идущие из области рта и глотки, совсем не обязательны, если чувство жажды вызывается действительным дефицитом воды в организме. При истинной жажде, как указано выше, смачивание глотки может уменьшить, но не утолить жажду.
Таким образом, жажда-это общее ощущение, основанное на интегрированной реакции многих типов рецепторов, из которых одни находятся на периферии, а другие в ЦНС. Они приведены на рис. 9-2. Промежуточный мозг, и в частности гипоталамус, по-видимому, играет доминирующую роль в интеграции этого множества афферентных входов. Мы не знаем, насколько точно результаты опытов на животных приложимы к человеку, и также не знаем, какие центральные структуры вызывают чувство жажды. Но можно думать, что отношения, изображенные на рис. 9-2, распространяются и на человека.
Для чувства жажды не наступает адаптации. Этот субъективный опыт опять-таки подтверждается экспериментами на животных. Показано, что количество воды, потребляемое после внутривенного вливания гипертонического раствора соли, не зависит от быстроты вливания. Иными словами, жажда, вызываемая введением определенного количества раствора, оставалась неизменной независимо от того, возрастала концентрация NaCl очень медленно или очень быстро. Поскольку адаптация к жажде не наступает, единственный способ утолить ее-это выпить воды (рис. 9-1, Б).
Утоление жажды. Между моментом начала питья и устранением дефицита воды проходит значительное время; вода в желудке и кишечнике сначала должна всосаться (т.е. перейти в кровоток), что происходит главным образом в тонком кишечнике. Но как известно из обычных наблюдений и как неоднократно подтверждали опыты на животных, чувство жажды исчезает (т. е. питье прекращается) задолго до того, как дефицит воды во вне- и внутриклеточном пространствах мог быть возмещен. Это значит, что утолению жажды, наступающему после всасывания (absorption) воды,-так называемому постабсорбтивному-предшествует преабсорбтивное утоление; таким образом предотвращается потребление чрезмерного количества воды за период, пока усвоенная вода не окажет своего действия (рис. 9-3). Опыты на животных показывают, что этот преабсорбтивный механизм действует с большой точностью: количество выпитой воды очень близко соответствует тому, какое требуется в конечном счете.
Рецепторы и механизмы, лежащие в основе преабсорбтивного утоления жажды, неизвестны. Собака с фистулой в пищеводе пьет почти
9. Жажда и голод
263
Рис. 9-3. Утоление жажды (преабсорбтивное и постабсорбтивное). Рецепторы, показанные на рис. 9-2, представлены здесь одним прямоугольником (рецепторы жажды).
вдвое больше воды, чем нормальная собака с таким же дефицитом воды, а затем перестает пить на 20-60 мин. Следовательно, само по себе питье или связанные с ним моторные и сенсорные процессы вызывают некоторое временное утоление жажды. Растяжение желудка проглоченной жидкостью, по-видимому, тоже имеет значение. Вода, введенная крысам и другим животным непосредственно в желудок, соответственно снижает количество выпиваемой воды. Но связанные с этим нервные механизмы остаются неясными.
После того как дефицит воды действительно компенсирован (постабсорбтивное утоление жажды), проходит некоторое время, пока чувство жажды возобновится, даже если происходит непрерывная (хотя и медленная) физиологическая потеря воды. Это значит, что жажда имеет свой порог. Как уже было сказано выше, этот порог эквивалентен потере воды, приблизительно равной 0,5% веса тела. Порог ограничивает появление жажды определенными интервалами; без этого неэффективно уходило бы много времени на питье малых количеств воды. Итак, содержание воды в организме человека колеблется между максимумом после постабсорбтивного утоления жажды и минимумом, который в идеальном случае лежит немного ниже порога. Но нередко колебания в содержании воды в организме выходят за эти пределы. Одна из причин этого состоит в том, что часто мы потребляем воды больше, чем
264
Р. Шмидт
нужно для возмещения ее потерь; другая причина та, что не всегда есть возможность утолить жажду сразу же, как только она становится заметной.
Первичное и вторичное питье. Питье, вызываемое абсолютным или относительным дефицитом воды в одном из пространств тела, содержащих жидкость, называется первичным питьем, а питье без видимой нужды в восполнении потерянной воды-вторичным питьем. Собственно говоря, вторичное питье служит обычным путем поступления воды. Обычно мы (и другие млекопитающие) выпиваем физиологически потребную воду заранее. Например, жидкость пьют за едой и после еды-очевидно, мы научились приноравливать выпиваемое количество к характеру пищи. Когда еда соленая, мы пьем больше, даже не ощущая жажды. Известную роль, по-видимому, играет также привычка, но мы располагаем лишь весьма скудными сведениями о механизмах, посредством которых потребность в воде оценивается заранее. Во всяком случае, первичное питье -это реакция, осуществляемая в особых случаях, и к нему редко прибегают люди, ведущие размеренный образ жизни.
Патологическая жажда. Усиление жажды во время болезни может быть следствием ненормально большой потери воды без нарушения механизмов жажды. Но оно может также означать нарушение этих механизмов или регуляции водно-солевого баланса. Ярким примером первого случая служит потеря воды, связанная с длительной рвотой или сильным поносом, как это бывает, например, при холере. Еще одним хорошо известным примером того же порядка является несахарный диабет, при котором из-за недостатка антидиуретического гормона (АДГ) из организма выводится ежесуточно много литров гипотонической мочи. Такие больные страдают от неутолимой жажды, и все дневные дела связаны у них с постоянной необходимостью пить. Детали многих сторон патологической жажды можно найти в руководствах по патофизиологии и клинической медицине.
В 9.1. Вода в организме содержится в двух больших пространствах-внутриклеточном и внеклеточном. Отношение внутриклеточного объема к внеклеточному составляет а) 5:1;
б) 2:1;
в) 1:1;
г) 1:3;
д) 1 : Ю.
В 9.2. Тело человека состоит на 70-75% (по весу) из воды. Потеря какой части веса тела является порогом для появления чувства жажды? а) 0,05%.
б) 0,1%.
в) 0,5%.
г) 1%-
д) 3%.
9. Жажда и голод
265
В 9.3. Какие факторы участвуют в возникновении жажды?
а) Повышение тоничности внутриклеточной жидкости.
б) Понижение тоничности внутриклеточной жидкости.
в) Увеличение объема клетки.
г) Уменьшение объема внеклеточной жидкости.
д) Понижение тоничности внеклеточной жидкости.
В 9.4. Какие рецепторы сигнализируют о дефиците воды в организме? а) Хеморецепторы в каротидной железе.
б) Осморецепторы в промежуточном мозгу, в Рецепторы растяжения в желудке.
г) Рецепторы растяжения мыщц, участвующих в акте глотания, д) Рецепторы растяжения в крупных венах.
В 9.5. Которая из следующих форм питья чаще возмещает потери жидкости у человека?
а) Первичное питье.
б) Вторичное питье.
в) Первичное и вторичное питье используются одинаково часто.
9.2. Голод
Потребность в пище. Энергетический баланс сохраняется у любого животного, если содержание энергии в пище соответствует энергии, расходуемой при мышечной работе, в метаболических процессах (рост, перестройка) и при теплопотерях. Чрезмерное потребление пищи приводит к отложению жира и тем самым к увеличению веса, а недостаточное количество пищи ведет сначала к мобилизации и использованию отложений жира, а при длительном недоедании нарушает функции организма и иногда кончается смертью.
Человек и животные в норме приноравливают количество потребляемой пищи к меняющимся потребностям в зависимости от производимой работы, климата и питательной ценности пищи. Кратковременная регуляция принятия пищи накладывается на долговременную регуляцию, которая возмещает временные неправильности питания и обеспечивает возвращение к нормальному весу тела. Например, когда животные жиреют из-за насильственного кормления, а затем их возвращают в нормальные условия, они едят меньше, чем контрольные. Когда же они восстановят свой истинный контрольный вес, поедание пищи постепенно увеличивается. И наоборот, после периода голодания первоначальный вес тела приобретается вновь путем временного усиленного потребления пищи.
Недостаток пищи вызывает чувство голода, и связанный с ним пищевой драйв ведет к приему пищи и в конечном счете к сытости (рис. 9-1,4). Какие механизмы вызывают голод и чувство насыщения? Возникает также вопрос, лежат ли в основе кратковременной и долговременной регуляции потребления пищи одни и те же или разные механизмы? Несмотря на многочисленные исследования, на эти вопросы
266
Р. Шмидт
еще не получены исчерпывающие ответы. Пробелы в наших знаниях станут очевидными из дальнейшего изложения.
Одно установлено с очевидностью, а именно что в ощущениях голода и сытости участвует несколько факторов. Но совершенно неизвестно, каково их относительное значение, а также неясно, все ли относящиеся сюда факторы уже обнаружены.
Факторы, вызывающие голод. Субъективный опыт показывает, что голод представляет собой общее чувство, локализующееся в области желудка (или проецирующееся на эту область); оно возникает, когда желудок пустой, и исчезает или уступает место чувству насыщения, как только желудок наполняется пищей. Некоторые прежние исследователи этого вопроса считали, что чувство голода вызывается сокращениями пустого желудка. По мнению этих авторов, такое представление согласуется с тем фактом, что кроме обычных сокращений, посредством которых пища перерабатывается и перемещается, сокращается также и пустой желудок. Такие сокращения, по-видимому, тесно связаны с голодом и поэтому могли бы способствовать возникновению этого чувства. Возможно, что о них сигнализируют в ЦНС механорецепторы в стенке желудка (рис. 9-4, слева). Но действие сокращений пустого желудка на голод не стоит переоценивать; при денервации желудка или полном его удалении в опыте на животных их пищевое поведение практически не меняется. Следовательно, такие сокращения могут быть одним из факторов, приводящих к ощущению голода, но фактором не обязательным.
Решающую роль в возникновении ощущения голода, по-видимому, играет глюкоза (виноградный сахар). Этот сахар служит главным источником энергии для клеток организма. Уровень глюкозы в крови и доступность глюкозы для отдельных клеток контролируются гормонами. Экспериментально показано, что снижение доступности глюкозы (не сам по себе уровень сахара в крови) очень хорошо коррелирует с чувством голода и мощными сокращениями желудка, т.е. фактор «наличия глюкозы» является решающим параметром в развитии голода.
Эта «глюкостатическая» гипотеза подтверждается различными экспериментальными данными, показывающими, что в промежуточном мозгу, печени, желудке и тонком кишечнике существуют глюкорецепторы. Например, при инъекции мышам золото-тиоглюкозы (золото является ядом для клеток) многие клетки в промежуточном мозгу, которые, очевидно, поглощают особенно большие количества глюкозы, разрушаются; при этом резко нарушается пищевое поведение. Иными словами, глюкорецепторы обычно сигнализируют о снижении количества наличной глюкозы и таким образом вызывают голод (рис. 9-4).
Выдвинуто и другое представление о том, каким путем вызывается голод, но в его пользу имеется меньше экспериментальных данных, чем для глюкостатической гипотезы. Это термостатическая гипотеза, основанная на наблюдении, что теплокровные животные поедают пищу в количествах, обратно пропорциональных температуре среды. Чем ни-
9. Жажда и голод
267
Дефицит пищи
i^h ' i е н ?зЩ ни и ц ноли j
' 1ЭДЛОГО желадна ' | '('?b’tsd •W’V'hou
Снижение
W( ,Ы1ЧЪ
ДйЙзЖйеЯйёДД-] жйрошвшйОёйа]
Рис. 9-4. Возникновение чувства голода. Связанные с этим рецепторы показаны под их адекватными стимулами.
же температура окружающей среды, тем больше они едят, и наоборот. Согласно этой гипотезе, датчиками в процессе интеграции общего энергетического баланса служат внутренние (центральные) терморецепторы (см. разд. 3.3). В таком случае снижение общей теплопродукции влияет на внутренние терморецепторы, которые вызывают чувство голода (рис. 9-4). Можно показать экспериментально, что локальное охлаждение или нагревание в промежуточном мозгу, средоточии центральных терморецепторов, может изменить пищевое поведение, как это предсказывает гипотеза, но не исключены и другие толкования тех же самых данных.
Чрезмерное потребление пищи ведет к отложению жира в тканях, а когда пищи недостаточно, жировые отложения используются. Если предположить существование липорецепторов, то о таких отклонениях от идеального веса тела могли бы сигнализировать промежуточные продукты жирового обмена, которые появляются при отложении или использовании жира; так могли бы возникать сигналы чувства голода или сытости (рис. 9-4). В пользу липостатической гипотезы говорят не
268
Р. Шмидт
которые убедительные экспериментальные данные, в частности упомянутые выше данные о том, что после насильственного кормления животные едят меньше, чем контрольные, пока не израсходуются их жировые отложения.
Согласно этому толкованию, липостатический механизм голода действует главным образом при долговременной регуляции потребления пищи, а сокращения пустого желудка и глюкостатический механизм принимают участие прежде всего в кратковременной регуляции. Термостатический механизм, возможно, участвует и там, и там (см. рис. 9-4). При таком разнообразии физиологических механизмов, создающих ощущение голода, даже в самых сложных условиях это ощущение и пищевой драйв обеспечивают потребление пищи в должных количествах.
Насыщение. Как и при питье, человек и животные прекращают поедать пищу задолго до того, как ее всасывание из пищеварительного тракта устранит тот дефицит энергии, который первоначально вызвал голод и потребление пищи. Все процессы, в результате которых животное перестает есть, имеют общее название насыщение. Как всякий знает по собственному опыту, ощущение того, что пищи съедено достаточно,-это нечто большее, чем только исчезновение голода; одним из других его проявлений (часть из которых связана с удовольствием) является отчетливое чувство переполнения, если пищи съедено слишком много. По мере того как после принятия пищи проходит время, ощущение насыщения постепенно ослабляется и в конце концов после более или менее долгого нейтрального периода вновь уступает место голоду. По аналогии с процессами, приводящими к утолению жажды, можно принять как предпосылку, что ощущение в начале насыщения преаб-сорбтивно- оно наступает до усвоения пищи, т.е. в результате процессов, связанных с самим актом еды, а происходящее позднее усвоение питательных веществ вызывает постабсорбтивное насыщение и препятствует немедленному возобновлению голода. Обратимся теперь к процессам, лежащим в основе этих двух видов насыщения.
Вероятно, преабсорбтивное насыщение создается многими факторами. Животные с фистулой пищевода, через которую проглоченная пища выходит, не попав в желудок, едят значительно дольше, чем до операции. и через более короткие интервалы. По-видимому, преабсорбтивно-му насыщению способствует стимуляция во время еды обонятельных, вкусовых и механорецепторов в слизистой носа, рта, глотки и пищевода, а возможно, также сам акт жевания (рис. 9-5), хотя имеющиеся в настоящее время данные указывают, что эти влияния на возникновение и сохранение чувства насыщения невелики. Еще одним фактором, по-видимому, является растяжение желудка пищей (рис. 9-5). Если желудок экспериментального животного наполнить через фистулу еще до того, как его накормят, то потом оно поедает меньше пищи. Степень компенсации связана не с питательной ценностью пищи, а с объемом первоначального содержимого желудка и с временем, когда оно туда введено. В крайних случаях потребление пищи через рот может быть полностью
9. Жажда и голод
269
Рис. 9-5. Возникновение чувства насыщения при приеме пищи. Жевательные движения могут способствовать преабсорбтивному насыщению через прямую центральную эфферентационную копию моторных программ жевания, через рецепторы, активируемые при жевании (например, мышечные веретена и сухожильные органы), или через те и другие.
заторможено на недели, если в желудок непосредственно закладываются большие количества пищи незадолго до того, как животному полагается питаться. Следовательно, здесь безусловно играет роль растяжение желудка (и, возможно, прилегающей части кишечника). Наконец, хеморецепторы в желудке и верхних отделах тонкого кишечника (рис. 9-5), очевидно, чувствительны к содержанию глюкозы и аминокислот в пище. Наличие соответствующих «глюкозных» и «аминокислотных» рецепторов в кишечной стенке показано электрофизиологически.
Постабсорбтивное насыщение тоже может быть связано с указанными хеморецепторами, потому что они способны сигнализировать о концентрациях еще не использованных питательных веществ, оставшихся в пищеварительном тракте. К этому добавляются все энтероцептивные сенсорные процессы, рассмотренные при обсуждении кратковременной
270
Р. Шмидт
и долговременной регуляции голода. Увеличение количества глюкозы и усиление теплопродукции по мере переработки пищи, а также изменения жирового обмена действуют на соответствующие центральные рецепторы (рис. 9-5); возникающие эффекты противоположны тем, которые вызывают голод (обозначены красным на рис. 9-4). В этом смысле голод и насыщение составляют две стороны одной монеты. Чувство голода побуждает к еде, а чувство насыщения (преабсорбтивно-го) заставляет прекратить еду. Однако количество съеденной пищи и продолжительность пауз между приемами пищи определяются также процессами, которые мы назвали «долговременной регуляцией приема пищи» и «постабсорбционным насыщением»,-процессами, которые, как мы теперь понимаем, в большей или меньшей степени накладываются друг на друга (ср. рис. 9-4 и 9-5).
Психологические факторы, участвующие в регуляции потребления пищи. В дополнение к перечисленным физиологическим факторам в регуляции пищевого поведения участвует ряд психологических факторов. Например, время приема пищи и количество съеденной пищи зависят не только от чувства голода, но также от установившихся привычек, количества предлагаемой пищи, ее вкуса и т. п. Животные, как и человек, регулируют количество поедаемого корма в зависимости от того, когда ожидается следующее кормление, и от того, сколько энергии, вероятно, будет до этого затрачено. Этот элемент планирования пищевого поведения, благодаря которому энергия поступает заранее, аналогичен «вторичному питью» (см. разд. 9-1), т.е. обычному порядку потребления воды.
Наше желание поесть определенную пищу, т.е. стремление повторить полученное удовольствие, называется аппетитом. Он может возникать из ощущения голода (т. е. пищевого драйва) или же появляться независимо (когда человек видит или ему предлагают что-нибудь особенно вкусное). Аппетит часто имеет соматическую основу-например, стремление к соленой пище, когда организм потерял много соли; но он может также не зависеть от физических потребностей и отражать врожденные или приобретенные индивидуальные предпочтения. Такое приобретенное подедение, а также отказ от некоторых видов пищи объясняется наличием той или иной пищи и установившимися привычками, порой связанными с религиозными соображениями. С этой точки зрения «аппетитность» блюда-основные элементы которой составляют запах, вкус, консистенция, температура и то, как оно приготовлено и подано,-в большой мере зависит от нашей аффективной реакции на него. Примеров этому много, и их легко найти на местном, национальном и международном уровнях.
Почти любой человек, оказавшись перед соблазнительной пищей, иной раз съест ее больше, чем нужно. Механизмы кратковременной регуляции тут не справляются. После этого надо было бы сократить прием пищи, но в современном материально обеспеченном обществе не все ведут себя так. Причины несостоятельности долговременной регуляции,
9. Жажда и голод
271
к сожалению, мало понятны. Программы предупреждения тучности и борьбы с ней трудно разработать, и часто они не дают результатов; тучность и все связанные с ней опасности для здоровья во многих западных странах достигли, по-видимому, эпидемических размеров.
В заключение надо указать на связь между потреблением пищи и неврозами и психозами. Усиленная еда или отказ от пищи часто служат эквивалентом удовольствия или протеста у психически больных, когда на самом деле беспокойство связано с другими видами мотивации. Наиболее известным примером является anorexia nervosa, форма отказа от пищи, весьма распространенная у девочек в период созревания; это нарушение психического развития может быть настолько серьезным, что приводит к смерти от голода.
Центральные механизмы голода и насыщения. Гипоталамус-структура, тесно связанная с регуляцией вегетативных функций (см. разд. 9.1), по-видимому, является главной центральной перерабатывающей и интегрирующей структурой также для голода и насыщения. Двустороннее разрушение ткани в определенных вентромедиальных областях гипоталамуса вызывает у экспериментального животного чрезвычайную тучность в результате переедания. В то же время разрушение более латеральных областей может привести к отказу от пищи и в конце концов к смерти от голода. С этими данными сопоставимы результаты локальной стимуляции гипоталамуса через вживленные электроды и опыты с золототиоглюкозой. Таким образом, в течение некоторого времени внимание исследователей было направлено почти исключительно на гипоталамус. В результате роль других структур мозга в регуляции потребления пищи известна очень мало. Безусловно будет упрощением заключить на основании только что упомянутых опытов, что вся центральная переработка информации локализована в двух «центрах», один из которых действует как «центр насыщения», а другой как «центр голода». Согласно этой гипотезе, разрушение центра насыщения должно привести к растормаживанию центра голода и отсюда-к развитию волчьего аппетита; если же разрушен центр голода, то это должно вызывать постоянное чувство насыщения и отказ от всякой пищи. Однако положение явно намного сложнее. Например, упомянутые выше еда и питье «авансом» связаны с участием высших уровней головного мозга (лимбической системы, ассоциативной коры). Не следует также упускать из виду, что еда и питье-это сложные двигательные акты, которые соответственно требуют широкого участия двигательной системы.
В 9.6. Какой(ие) из следующих процессов, вероятно, участвует(ют) в возникновении чувства голода?
а) Увеличение количества промежуточных продуктов метаболического распада жировых отложений.
б) Понижение теплопродукции.
в) Мощные сокращения пустого желудка.
272
Р. Шмидт
г) Сухость во рту и в области глотки.
д) Увеличение количества глюкозы.
В 9.7. Какие из следующих рецепторов участвуют в кратковременной регуляции приема пищи и какие-в долговременной регуляции? а) Внутренние терморецепторы.
б) Механорецепторы в стенке желудка.
в) Липорецепторы.
г) Глюкорецепторы.
В 9.8. Какие из следующих рецепторов участвуют в преабсорбтивном насыщении и какие-в постабсорбтивном? (Возможно некоторое перекрывание.)
а) Глюкорецепторы в промежуточном мозгу.
б) Механорецепторы в желудке.
в) Аминокислотные рецепторы в желудочно-кишечном тракте, г) Обонятельные и вкусовые рецепторы.
д) Центральные термо- и липорецепторы.
е) Глюкорецепторы в желудочно-кишечном тракте.
Ответы на вопросы
Глава 1
В 1.1: Модальность, качества
В 1.2: а) М; б) I; в) Q; г) М; д) М; е) Q; ж) Q; з) I; и) М
В 1.3: в
В 1.4: Выработанный или приобретенный рефлекс, выражающийся в том, что данный стимул регулярно вызывает данную активность.
В 1.5: а, г
В 1.6: б, г
В 1.7: а, б
В 1.8: Время, пространство, качество, интенсивность (количество) (в любом порядке)
В 1.9: а, в, г
В 1.10: 5
В 1.11: в
В 1.12: д
Глава 2
В 2.1: Механо-, термо-, хемо- и фоторецепторы
В 2.2: б, в
В 2.3: а, в, г
В 2.4: в
В 2.5: б, г
В 2.6: б, д
В 2.7: г
В 2.8: Семь основных отделов следующие: спинной мозг, продолговатый мозг, мост, средний мозг, промежуточный мозг, конечный мозг и мозжечок. Продолговатый мозг, мост и средний мозг вместе называются стволом мозга
В 2.9: а, г
В 2.10: в, г
В 2.11: Дифференциальный порог нейрона-это минимальное изменение параметра стимула, которое вызывает измеримое изменение частоты импульсации
В 2.12: 1 г
В 2.13: Как на рис. 2-10
В 2.14: б, в, г
В 2.15: в
В 2.16: б, в
В 2.17: г
274
Ответы на вопросы
В 2.18: б
В 2.19: а, в
В 2.20: в
В 2.21: а, в
В 2.22: г
Глава 3
В 3.1: в, б, а, г, е, д
В 3.2: б
В 3.3: в
В 3.4: в
В 3.5: в, г
В 3.6: д
В 3.7: Ощущения положения, движения и силы
В 3.8: в
В 3.9: б, д, е, ж, и
В 3.10: в, д, е
В 3.11: г
В 3.12: а, б, г
В 3.13: б, д
В 3 14: а
В 3.15: в
В 3.16: а
В 3.17: г
В 3.18: в
В 3.19: г, д
В 3.20: б, в, д
В 3.21: д
В 3.22: в
В 3.23: б
В 3.24: а
В 3.25: см. с. 98-99
Глава 4
В 4.1: в
В 4.2: Читать, близорукими, вогнутые
В 4.3: б, в, г, е
В 4.4: 1е + ж, 2г, 36, 4д, 5в, 6з
В 4.5: б, г
В 4.6: г
В 4.7: а, б, в, г, д, е
В 4.8: 1,25: (дуговых минут)-1
В 4.9: а, г, д
В 4.10: б
В 4.11: в
Ответы на вопросы
275
В 4.12: а, б, в, д
В 4.13: б, е
В 4.14: а, г, е
В 4.15: Активацией, торможению, торможение, активацию, антагонистически, меньше
В 4.16: а, в, д
В 4.17: б, в
В 4.18: а
В 4.19: в, Д
В 4.20: в, г
В 4.21: б, в
Глава 5
В 5.1: б, а, г, в
В 5.2: г
В 5.3: в
В 5.4: а, в
В 5.5: а, в, е
В 5.6: б
В 5.7: б
В 5.8: в
В 5.9: в
В 5.10 В 5.11 В 5.12 В 5.13 В 5.14 В 5.15 В 5.16 : б : б, в : в : б : а : а, г, е, ж : а
Глава 6
В 6.1: а, б, г, д, е
В 6.2: б
В 6.3: а, б, в
В 6.4: в
В 6.5: а, б, в, г
Глава 7
В 7.1: в
В 7.2: Лицевой и языкоглоточный нервы; ядро одиночного тракта, вентральный таламус, постцентральная извилина
В 7.3: 1г, 2в, 36, 4а, 66, 7г
В 7.4: в
В 7.5: 1а, 2г
В 7.6: б, г
276
Ответы на вопросы
В 7.7: а, б
В 7.8: б, в, д
Глава 8
В 8.1: А: б, Б: г
В 8.2: а, в, г
В 8.3: Например, цветочный, эфирный, мускусный, камфорный, пота, гнилостный, едкий
В 8.4: б
В 8.5: в
В 8.6: б
В 8.8: б
В 8.8: б, в, г
Глава 9
В 9.1: б
В 9.2: в
В 9.3: а, г
В 9.4: б, д
В 9.5: б
В 9.6: а, б, в
В 9.7: Кратковременная регуляция: а, б, г; долговременная регуляция: а, в
В 9.8: Преабсорбтивное насыщение: б, в, г, е; постабсорбтивное насыщение: а, в, д, е
Рекомендуемая литература
Глава 1
Cartrette Е. С., Friedman М. Р. (eds.), 1974. Handbook of Perception, vol. 1 + 2, Academic Press, New York and London.
Grossman P„ 1967. A Textbook of Physiological Psychology, John Wiley and Sons, Inc., New York-London-Sidney, pp. 1-932. Handbook of Sensory Physiology, 1971, Vol. 1. Principles of Receptor Physiology (ed. W. R. Loewenstein); vol. IV. Olfaction (ed. L. M. Beidler); vol. VII/4. Visual Psychophysics (eds. D. Jameson, L. M. Hurvich), Berlin-Heidelberg-New York, Springer.
Marks LE„ 1974. Sensory Processes, New York-London, Academic Press.
Milner P.M., 1971. Physiological Psychology, Holt, Rinehart and Winston, London-New York-Sidney-Toronto, pp. 1-531. [Имеется перевод: Милнер П. Физиологическая психология.-М. Мир, 1973.]
Ruch F.L., Zimbardo P.G., 1975. Psychology and Life, 9th ed., Scott, Foresman and Company, Glenview-Illinois-London. Stevens S. S., 1975. Psychophysics, New York-London-Sidney-Toronto, John Wiley.
Глава 2
Brodal A., 1969. Neurological Anatomy in Relation to Clinical Medicine, 2nd ed., New York-London-Toronto, Oxford University Press.
Garner V.R., 1962. Uncertainty and Structure as Psychological Concepts, New York, John Wiley.
Iggo A. (ed.), 1973. Somatosensory System, Handbook of Sensory Physiology, Berlin-Heidelberg-New York, Springer, vol. II.
Kenshalo D. R. (ed.), 1968. The Skin Senses, Springfield/Ill., Charles C. Thomas.
Loevenstein W.R. (ed.), 1971. Principles of Receptor Physiology. Handbook of Sensory Physiology, Berlin-Heidelberg-New York, Springer, vol. I.
Milner P.M., 1970. Physiological Psychology, London-New York-Sidney-Toronto, Holt, Rinehart and Winston. [Имеется перевод: Милнер П. Физиологическая психология-М.: Мир, 1973.]
Mountcastle V.B. (ed.), 1974. Medical Physiology, 13th ed., St. Louis, Mosby, vol. I. Penfield W., Rasmussen T, 1950. The Cerebral Cortex of Man, New York, Macmillan. Ruch T. C., Patton H. D., 1966. Physiology and Biophysics, Philadelphia-London, Saunders.
Shannon С. E., Weaver W., 1949. The Mathematical Theory of Communication, Urbana, The University of Illinois Press.
Stein R.B. (1967). The information capacity of nerve cells, a frequency code, Biophys. J., 7, 797.
Wiener N., 1948. Cybernetics, Cambridge/Mass., MIT Press.
Zimmermann M., 1976. Neurophysiology of Nociception. In: International Review of Physiology, Neurophysiology II, Porter R. (ed.), University Park Press, Baltimore, vol. X, pp. 179-221.
Zotterman Y. (ed.), 1976. Sensory Function of the Skin in Primates, Oxford, Pergamon Press.
278
Рекомендуемая литература
Глава 3
De Reuck А. V. S., Knight J., 1969. Touch, Heat and Pain, London, J. and A. Churchill Ltd.
Fulton J. F., 1943. Physiology of the Nervous System, London-New York-Toronto, Oxford University Press.
Goodwin G.M., McCloskey D.I., Matthews P.B. C. (1972). The contribution of muscle afferents to kinesthesia shown by vibration induced illusions of movement and by the effects of paralysing joint afferents, Brain, 95, 705-748.
Iggo A. (ed.), 1973. Handbook of Sensory Physiology. Somatosensory System, Berlin-Heidelberg-New York, Springer, vol. II.
Janzen R., Keidel W. D., Herz A., Steichele C. (eds.), 1972. Pain, Stuttgart, Thieme.
Kornhuber H. H., Aschoff J. C. (eds.), 1976 Somato-Sensory System, Stuttgart, Thieme.
Matthews P. В. C., 1972. Mammalian Muscle Receptors and their Central Actions, London, Edward Arnold (Publishers) Ltd.
Mountcastle V.B., 1974. Medical Physiology, 13th ed. Saint Louis, The С. V. Mosby Company, vol. I.
Schmidt R.F., Thews G. (eds.), 1977. Physiologie des Menschen, 19th Ausg., Berlin-Heildelberg-New York, Springer.
Sherrington C.S., 1900. The muscular sense. In: Schafer’s Textbook of Physiology, London-Edinburgh, Pentland, vol. II, pp. 1002-1025.
Sweet W.H., 1959. Pain. In: Handbook of Physiology; Section I: Neurophysiology 1, Washington D. C., American Physiological Society, pp. 459-506.
Zotterman Y. (ed.), 1976. Sensory Functions of the Skin in Primates, Oxford, Pergamon Press.
Глава 4
Dartnall (ed.), 1972. Photochemistry of Vision. Handbook of Sensory Physiology, Berlin-Heidelberg-New York, Springer, vol. VII/1.
Helmholtz H. von, 1924. Physiological Optics (Translat. of 3rd ed.), New York, Optical Society of America.
Hering E., 1964. Outlines of a Theory of the Light Sense (Engl. Transl.), Cambridge Mass., Harvard Univ. Press.
Hurvich L. M., Jameson D., 1966. The Perception of Brightness and Darkness, Boston, Allyn and Bacon.
Jung R. (ed.), 1973. Handbook of Sensory Physiology, Berlin-Heidelberg-New York, Springer, vol. VII/3A, III/B.
Kuffler F. W, Nicholls J. G., 1976. From Neuron to Brain, Sunderland Mass., Sinauer Ass.
Lennerstrand G., Bach-Y-Rita P (eds.), 1975. The Oculomotor System, New York-London, Academic Press.
Lennerstrand G., Bach-Y-Rita P„ 1975. Basic Mechanisms of Ocular Motility and their Clinical Implications, Oxford-New York, Pergamon Press.
Mach E., 1959. The Analysis of Sensations, New York, Dover Publications Inc. Rodieck R.W., 1973. The Vertebrate Retina, San Francisco, W. H. Freeman and Co. Ruch F. L„ Patton H. D„ 1976. Physiology and Biophysics, 20th ed., Philadelphia, Saunders, vol. I.
Walls G. L., 1967. The Vertebrate Eye and its Adaptive Radiations, New York-London, Hafner.
Walsh F. B., Hoyt W.F., 1969. Clinical Neuroophthalmology, Baltimore, Williams and Wilkins Co., vol. III.
Yarbus A.L., 1967. Eye Movements and Vision, New York, Plenum Press.
Рекомендуемая литература
279
Глава 5
Dallas Р„ 1973. The Auditory Periphery, New York-London, Academic Press.
Evans E. F., Wilson J. P., 1977. Psychophysics and Physiology of Hearing, London, Academic Press.
Henderson D., Hamernik R.P., Darshan S.D., Mills J. H. (eds.), 1976. Effects of Noise on Hearing, New York, Raven Press.
Keidel W. D., Neef W. D. (eds.), 1974-1976. Handbook of Sensory Physiology, Berlin-Heidelberg-New York, Springer Verlag, vol. V/1-V/3.
Roederer J. G., 1975. Introduction to the Physics and Psychophysics of Music, 2nd ed., Berlin-Heidelberg-New York, Springer.
Tobias J. V. (ed.), 1970, 1972. Foundations of Modern Auditory Theory, New York, Academic Press, vol. 1, vol. 2.
Webster W.R., Atkin L.M., 1975. Central auditory processing. In: Handbook of Psychobiology, Gazzaniga M.S., Blakemore Colin (eds.), New York, Academic Press.
Глава 6
Brodal A., Pompeiamo O. (eds.), 1972. Basic Aspects of Central Vestibular Mechanisms, Amsterdam, Elsevier Publishing.
Kornhuber H. H. (ed.), 1974. Handbook of Sensory Physiology, Berlin, Springer-Verlag, vol. VI/1, VI/2.
Главы 7 и 8
Serie: Olfaction and Taste.
Zotterman Y. (ed.), 1963. Oxford-London-New York-Paris, Pergamon Press.
Hayashi T. (ed.), 1967. Oxford-London-New York-Paris, Pergamon Press.
Pfaffmann C. (ed.), 1969. New York City, Rockefeller University Press.
Schneider D. (ed.), 1972. Stuttgart, Wissenschaftliche Verlagsgesellschaft.
Denton D.A., Coghlan J.P. (eds.), 1975. New York-San Francisco-London, Academic Press.
Le Magnen J., Mac Lead P. (eds.), 1977. London-Washington D. C, Information Retrieval.
Beidler L.M. (ed.), 1971. Taste. In: Handbook of Sensory Physiology, Berlin-Heidelberg-New York, Springer, vol. IV/1.
Beidler L.M. (ed.), 1971. Olfaction. In: Handbook of Sensory Physiology, Berlin-Heidelberg-New Work, Springer, vol. IV/2.
Cain W.S. (ed.) (1974). Odors: Evaluation, Utilization and Control, Ann. N.Y. Acad. Sci, 237, 1-439.
Moulton D. G., Turk A., Johnston J. W. (eds.), 1975. Methods in Olfactory Research, London-New York-San Francisco, Academic Press.
Shepherd G.M., 1970. The Olfactory Bulb as a Simple Cortical System: Experimental Analysis and Functional Implications. In: Schmitt F.O. (ed.), The Neurosciences; Second Study Program, New York, Rockefeller University Press, pp. 539-552.
Shepherd G.M. (1972). Synaptic Organization of the Mammalian Olfactory Bubl, Physiol. Rev., 52, 864-917.
Shepherd G. M., 1974. The Synaptic Organization of the Brain, New
York-London-Toronto, Oxford University Press.
Wolstenholme G. E. W, Knight J., 1970. Taste and Smell in Vertebrates (Ciba Foundation Symposium), London, Churchill.
280 Рекомендуемая литература
Глава 9
Anand В. К. (1961). Nervous regulation of food intake, Physiol. Rev., 41, 677.
Andersson B., 1972. Receptors subserving hunger and thirst. In: Handbook of Sensory Physiology, Neil E. (ed.), Berlin-Heidelberg-New York, Springer, vol. III/l, p. 187.
Code C.F. (ed.), 1967. Handbook of Physiology. Section 6: Alimentary Canal, vol. I: Control of Food and Water Intake, Washington, American Physiological Society.
Fitzsimons J. T. (1972). Thirst. Physiol. Rev., 52, 468.
Novin D., Wyrwicka W., Bray G. A. (eds.), 1976. Hunger. Basic Mechanisms and Clinical Implications, New York, Raven Press.
Peters G., Fitzsimons J. T„ Peters-Haefeli L., 1975. Control Mechanisms of Drinking, Berlin-Heidelberg-New York, Springer.
Pilgri/n F.J., 1967. Human food attitudes and consumption. In: Handbook of Physiology, Sect. 6: Alimentary Canal, vol. I: Control of Food and Water Intake Code C. F., (ed.) Washington, American Physiological Society, p. 139.
Wolf A. V., 1958. Thirst: Physiology of the Urge to Drink and Problems of Water Lack, Springfield/Ill., Ch. C. Thomas.
Список авторов
Helmut Altner
Fachbereich Biologie der Universitat UniversitatsstraBe 31, D-8400 Regensburg
Josef Dudel
Physiologisches Institut der Technischen Universitiit
Biedersteiner StraBe 29, D-8000 Munchen 40
Otto-Joachim Grusser
Physiologisches Institut der Freien Universitat
Arnimallee 22, D-1000 Berlin 33
Ursula Grusser-Cornehls
Physiologisches Institut der Freien Universitat
Arnimallee 22, D-1000 Berlin 33
Rainer Klinke
Zentrum der Physiologic
Theodor-Stern-Kai 7, D-6000 Frankfurt 70
Manfred Zimmermann
II. Physiologisches Institut der Universitat Im Neuenheimer Feld 326, D-6900 Heidelberg
Предметный указатель
Агнозия (зрительная) 75, 186
Адаптация 41-43, 108, 129, 222, 242, 253, 262
- глаза 18-21, 156, 159-160, 178-180 - обонятельная 253
- терморецепторов 116, 117, 123
Аккомодация 147
«Активация» 75
Акупунктура см. Иглоукалывание
Амакриновые клетки 152, 153, 174, 179, 180
Анальгезия 139
Ангиотензин 261
Анестезия 133, 140-141
Аносмия 250
Антидиуретический гормон 262
Аппетит 271
Ассоциативная зона 74-75, 196, 271
Астигматизм 149
Аудиограмма 208
Аудиометрия 208
Афферентное торможение 62
Афферентные волокна 104, 112-114, 132-134
Базальные клетки 248
Базилярная (основная) мембрана 200, 212, 213
Барабанная перепонка 198
- струна 240
Бегущие волны 212-213
Бел 204
Белый шум 205
Бензилацетат 250
Бинокулярная интеграция 183
- конкуренция 168
- суммация 185
Бинокулярное взаимодействие 183
- зрение 166-168
- слияние 166, 168
Биполярные клетки 150-153
Близорукость см. Миопия
Блоковый нерв 50
Блуждающий нерв 50, 133, 240
Бодрствование 75, 76
Болевые рецепторы 129-134
Боль 126441
- проводящие пути 64
- фантомная 114
Боуменовы железы 249
Бугорки четверохолмия, верхние 191, 192, 196
— нижние 220
Вкус 91, 237-246
Вкусовые зоны 240
- почки 237-240, 248
- профили 244
- пути 240
- рецепторы 243, 261, 262
- сосочки 237-239
Водный баланс 257, 261
Волосковые клетки 200-202, 213-216, 218
Волосяных фолликулов рецепторы 99, 106
Восприятие 7, 12, 13, 73-80, 169-171, 173-174
Восходящая ретикулярная активирующая система 75
Вебера правило 29
Вебера проба 217
Вебера-Фехнера закон 29, 75
Вернике зона 196
Верхняя .олива 210
Вестибулоспинальный тракт 231
Вестибулярная мембрана 200, 212
Вестибулярное ядро 231, 232
Вестибулярный ганглий 231
Вестибулярный нистагм 232-235
Вибрация 93, 98, 103-104, 106
Витамин А 172
Внутреннее ухо, анатомия 199-201 — функция 212-217
Вызванные потенциалы 72, 73
Ганглиозные клетки 152, 175-183
Гипералгезия 139
Гиперметропия 150-151
Гиперпатия 136-137
Гипноз 128
Гипоталамус 77, 231, 235
- и потребление пищи 271
V- обонятельный путь 254, 255
- осморецепторы 260, 261
Гиппокамп 254
Гистамин 138, 139
Геликотрема 200, 213
Генераторный потенциал 42
Гештальт-психология 170
Глаз 144-153
- движения 188-196
Глазодвигательные рефлексы 192
Глазодвигательный нерв 50, 147
Глиальные клетки 150
Гломерулы 254
Глухота 216-217
Глюкоза 241, 267
Глюкорецепторы 267
Предметный указатель
283
Глюкостатическая гипотеза 267
Головная боль 127, 129, 138, 148
Головокружение 192, 235
Голод 265-271
Горизонтальная диспаратность 169
Горизонтальные клетки 150-152, 174, 178, 180
Гороптер 166, 167, 186
Громкость 205-207, 222
Давления ощущение 93, 94, 106
Датчики 101-105
Движения глаз 188-196
Двоичные знаки 81-83
Декодирование 79
Дерматомы 48, 136
Детекторы 106
Децибел 204, 206
Диоптрический аппарат 145, 146, 150
Дифференциальные пороги 26-28, 59-
60, 205, 222
Дихроматы 164
Добавочное ядро 210, 220
Добавочный нерв 50
Драйв (пробуждение) 257
Евстахиева труба 198
Жажда 257-264
Задние корешки 47-49
- рога (спинного мозга) 60-62
- столбы (спинного мозга) 61-64
Запахи 250, 252, 255
Затылочная кора 183, 186
Звук, физические свойства 202-205
Звуковое давление 203-204, 215, 222
Зональная теория 165
Зрачок 144
- реакции 148, 149
Зрение 142-196. См. также Глаз, Цветовое зрение
Зрительная агнозия 75, 186
- кора 182-186, 192, 195, 221
Зрительные нейроны 182-186
- пути 180-183, 186
Зрительный нерв 50, 146, 153, 158, 180
- тракт 181
Зуд 138
Иглоукалывание (акупунктура) 141, 213
Избыточность 88-89
Изофон 202. 206
Иллюзии (зрительные) 170
Импеданс акустический 212
Информация, кодирование 79. 82, 84
- количество 80
- теория 78-91
Инфразвук 207
Кинетоз см. Морская болезнь
Киноцилии 224
Клетки-зерна 254, 255
Кожа, рецепторы 90, 93. См. также отдельные виды рецепторов
Колбочки 150, 152, 159, 172, 174, 180
Коленчатые тела латеральные 181-186 — медиальные 220
Конвергенция 67, 152, 180
- глаз 189
Контраст 31-34, 36, 155-158, 177-179, 183
Конъюнктива 144, 145
Кора (большого мозга). См. также названия отдельных областей
- ассоциативная 74-75
- зрительная 183-186, 195
- нейтронная организация 73
- слуховая 210, 221
- соматосенсорная 70-73
- топография 182-183
Кориолиса ускорение 235
Кортиев орган 200, 201
Косоглазие 150, 190
Кохлеарное ядро 218, 220, 221
Кривая настройки 218, 219
Купула 228-230
Лабиринт см. Равновесия орган
Ландольта кольца 157
Латеральное коленчатое тело 181-186
- торможение 55, 183
Латеральный мениск 220
Легкое, рецепторы 133
Лемниск латеральный 220
- медиальный 240
Лемнисковая система 51, 63
Лестница барабанная 199, 212, 213
- средняя 199, 212
- преддверия 199
Лимбическая система 76, 77, 139, 192
— и голод 271
— роль в обонянии 255
Липорецепторы 267
Липостатическая гипотеза 267-268
Лицевой нерв 50, 240, 242
Лобная доля 139
Лоботомия 139
Локтевой нерв 135
Макулярный орган 224-225, 232
Маха полосы 155
Мезотопическое зрение 159
Мейснера тельца 99, 101, 103
284
Предметный указатель
Меркеля диски 99, 101-102, 106
Механорецепция 93-107
Механорецепторы, классификация 93 - структура 98-101
Микросаккады 190
Микрофонные потенциалы 212, 216
Микроэлектроды 98, 106
Миндалевидный комплекс 254
Миопия (близорукость) 149-151
Митральные клетки 254
Модальнось 8
- межмодальное сравнение 25-26
Мозжечок 192, 231
Мозолистое тело 182
Монохромат 164
Молоточек 198
Морская болезнь 231, 235
Мост (варолиев) 65
Мотивация 77
Мышечные веретена 109, 112, 232
Мышцы глаза 147, 149, 188-189
- слуховых косточек 212
Мюллеровские опорные клетки 150
Наковальня 198
Насыщение при голоде 265
Насыщенность цвета 163
Нейронные колонки 73
Нейроны зрительной коры 183-186 - рецептивные поля 56-59, 175-180, 183-186
- сенсорные 55-60
- слуховые 221-222
Неокортекс 75
Неоспинно-таламический тракт 64
Нервный импульс. См. также Потенциал действия
— содержание информации 83, 84
Нистагм 192-194, 232-235
Новокаин 133
Носовые раковины 247
Ноцицепторы 131-134
«Облегчение» 131
Обоняние 247-255
- у насекомых 251
Обонятельная луковица 248, 253-254
Обонятельные волокна 248
- клетки 248-251
- пути 253-255
- рецепторы 42
Обонятельный нерв 50
- тракт 254
- эпителий 247-248
Общее чувство 257
Овальное окно 199
Оливарный комплекс 218, 220
Оперантные условные рефлексы 16-21
Оппонентная теория цветов 165
Опсин 172
Ослепление 34
Осморецепторы 257, 261
Острота зрения 143, 146, 156-158
Отводящий нерв 50
Отолитовая мембрана 224
Ощущение 7, 13
- интенсивность 23-29
- шкала 23
- эмоциональный аспект 37-38
Палеоспинно-таламический тракт 64
Палочки 99, 104, 106, 112, 150, 152, 159, 172, 176, 179
Пачини тельца 99, 104, 106, 112
Передне-боковой пучок 61, 63-65 Перигломерулярные клетки 255 .
Периферические нервы 47
Печеночная колика 133
Пигментные клетки 150, 152
Пириформная доля 254
Пласидо диск 149, 150
Плечевое сплетение 48
Поведение 14-21, 140, 266, 267, 270
Подъязычный нерв 50
Покровная (текториальная) мембрана 200, 201, 203
Поле зрения 142-143, 196
Полисинаптические пути 64
Положение тела 114
Полукружные каналы 225, 226, 229
Порог(и), абсолютный 18, 19, 23, 59
- болевой 129, 207
- вкусовые 241, 242
- дифференциальные 26-28, 59-60, 205— 222
- зрительные 18-20
- обонятельные 252
- пространственные (кожные) 95-97, 120-122
- слуховые 20, 204-208, 222
- терморецепторные 121
Пороговый стимул 11
Последовательные образы 156
Постцентральная извилина 70, 240
Потенциал действия 13, 42, 98
— ганглиозных клеток сетчатки 175
— слухового нерва 216, 219
- покоя 40
Препириформная область 254
Пресбиакузис 208
Пресбиопия 147
Претектальная область 191
Прикосновение, рецепторы 93, 102-103, 106
Предметный указатель
285
Продолговатый мозг 61, 62, 231, 240
Промежуточный мозг 261, 267, 268
Проприоспинальные пути 61
Проприоцепторы 109, 111-112
Проприоцепция 108-115
Психозы 272
Психотерапия 138
Психохирургия 140
Психофизика 7, 24, 89-91
- зрения 155-171
Психофизическая проблема 7
Психофизический момент 34
Равновесие 224-235
Равновесия орган (лабиринт) 192, 199, 224-230
Радужная оболочка (радужка) 144, 145, 149
Рейснерова мембрана 200
Ренин 262
Ресничный поясок 145, 147
Ресничные мышцы 145, 147
Ретикулярная формация 65-67, 75, 231
Ретиналь 172
Рефлексы безусловные 15
- статические и статокинетические 232— 233
- условные 15-21
Рефракция глаза 147-150
Рецептивная единица 175
- мембрана 41
Рецептивные поля 56-59
- - вкусовые 244, 245
-- механорецепторных нейронов 105— 106
— нейронов сетчатки 58, 153, 175-180
— сложные 184, 186, 195
— терморецепторные 121
Рецепторные молекулы 245
Рецепторный потенциал 9, 40-42, 44 — фоторецепторов 173-174 Рецепторы 9
- передача информации 84-98
- определение и классификация 39-40, 99, 101-105
- структура 99-101
Речь 75, 90, 196, 221
Рикко закон 34
Роговица 144-146
Родопсин 174, 176, 172, 174
Руффини окончания 99, 101, 102, 112
Саккады 190, 193, 195
Саккулус 225, 229
Сахар (в крови) 267
Светлота 163
Сенситизация 129
Сенсомоторная интеграция 188-196
Сенсорная депривация 7
Сенсорное впечатление 9, 11, 12
Сенсорные нейроны (клетки) 41—42, 55-
60, 239, 248, 251
- стимулы 8, 9
- функция 47-53
Серая шкала 154-155
Сердце, рецепторы 133, 261
Сетчатка 146, 150-153, 181, 182
- нейроны 174-180
Синапсы 451, 55, 254
Скелетные мышцы (как источник боли) 133, 138
Склера 146
Скотопическое зрение 159
Следящие движения (глаз) 190, 192-194
Слезная жидкость 145
Слезные железы 145
Слепое пятно 158-159
Слияние мельканий 34, 160, 161, 195
Слух 198-222. См. также Ухо
Слуховая кора 210, 221
Слуховой нерв 50, 201-202, 216, 218— 222
- порог 20, 205, 222
- проход 198
Слуховые нейроны 221
- пути 210, 218-222
Слюна 246, 260, 261
Слюнные железы 261
Смешение цветов 163, 164
Снеллена отношение 157
Сознание 73-78, 91-92
Солитарный тракт 240
Соль 241, 260, 261
Соматовисцеральная система 50
- чувствительность 93-141 -
Соматосенсорная система 60-78
— информационные аспекты 79-91
- - рецептивные поля 56-60
— специфическая 51-53
- - торможение 65, 67
Соматосенсорные области (коры) 51, 70-72
Соматотопическая организация 51
Сосудистая оболочка 152
Сосудистый слой 146
Спинальные нервы 47
Спинной мозг 47-49, 60-65, 140
Спинно-мозжечковый тракт 64
Спинно-ретикулярный тракт 64
Спинно-таламический тракт 64, 135
Спинно-шейный тракт 64
Спиральный ганглий 201
Среднее ухо 198-201, 211-212, 217
Статолитовый орган 225
286
Предметный указатель
Ствол мозга 182, 192, 195
Стекловидное тело 146
Стенокардия 133
Стереоскопическое зрение 168-169
Стивенса закон 253
- функция 24, 29, 45
Стимул 9, 12, 15, 18, 35
- интенсивность 44-47
- информационный аспект 83
- определение 39
Стоячие потенциалы 215
Стремечко 198, 212, 213
Стрихнин 55
Субфорникальный орган 261
Субъективность 7, 20
Суставные рецепторы 109-112
Сухожильные органы Гольджи 109, 112-113, 232
Схема тела 114
Тактильные диски 102
Таламическая боль 143
Таламо-кортикальная система 73-74
Таламус 68-70
- и боль 139
- вестибулярный аппарат 231
- вкусовые пути 240
- латеральное торможение 55
- рецептивные поля 56-58, 70
Текториальная (покровная) мембрана 200, 201, 213
Темновая адаптация 18-21, 178-179 — кривая 159-160
Теория места 212-213, 222
Тепла и холода точки 120-122
Терморецепторы 39, 40, 116, 121-123
- внутренние 268
Терморецепция 115-125
Термостатическая гипотеза 267
Тоничность клеток 260
Тоны 204, 207-209
- порог различения 20
Тошнота 235
Трансдукция 41-42, 213, 215
Трансформация (стимула) 42, 44
Трихроматическая теория 164-165, 174
Трихроматы 164
Тройничный нерв 49, 50, 65, 249, 253
Улитка 199-201, 212-216
Ускорение 224, 227-229
Условные рефлексы 15-21
Утрикулус 225, 229
Ухо, анатомия 198-222. См. также Внутреннее ухо, Слух
Фантомные боли 114
Фехнера закон см. Вебера-Фехнера закон
Фиксация (зрительная) 166, 190, 192, 196
Фон 206, 207
Фоторецепторы 39, 41-42
- «квантовая эффективность» 172
Фрея волоски 94, 101
Хеморецепторы 39
Хиазма 180-181
Хордотомия 64
Хроматичность 163
Хрусталик 145-146
Цветовая слепота 164
Цветовое зрение 161-165,
- тело 164
- уравнение 163
Цветовой круг 161-164
- треугольник 164, 165
Центральная ямка 146, 150, 153, 156
179, 189, 190
Циклопический глаз 167
Цингулотомия 140
Частота (звука) 204
Черепные нервы 49-50
Шелкопряд 252
Шум (при передаче информации) 85-89
Щекотки ощущение 93, 105
Экстралемнисковая система 53
Электроокулография 192
Эндокохлеарный потенциал 215
Эндолимфа 199-200, 212, 225, 229, 235
Эфферентная копия 114
Язык 237
Языкоглоточный нерв 50
Яркость 33, 34
Оглавление
Предисловие редактора перевода............................................... 5
Предисловие ко второму изданию............................................... 6
Предисловие к первому изданию................................................ 6
1. Общая сенсорная физиология, психофизика. Дж. Дудел....................... 7
1.1. Основные понятия общей сенсорной физиологии.......................... 7
1.2. Связь между стимулом и поведением; условный рефлекс................. 15
1.3. Измерение интенсивности ощущений. Психофизика....................... 22
1.4. Пространственные, временные и эмоциональные аспекты ощущения ... 30
2. Нейрофизиология сенсорных систем. М. Циммерманы......................... 39
2.1. Преобразование стимулов в рецепторах............................... 39
2.2. Сенсорные функции ЦНС. Общее рассмотрение......................... 47
2.3. Свойства и функционирование сенсорных нейронов и нейронных ансамблей 55
2.4. Соматосенсорная система: спинной мозг, восходящие пути и ствол мозга 60
2.5. Соматосенсорная система в таламусе и коре........................... 68
2.6. Сенсорная система в свете теории информации......................... 78
3. Соматовисцеральная чувствительность. Р. Шмидт........................... 93
3.1. Механорецепция...................................................... 93
3.2. Проприоцепция...................................................... 108
3.3. Терморецепция...................................................... 115
3.4. Соматические и висцеральные боли................................... 126
3.5. Особые и аномальные виды болей; терапия при болях................. 135
4. Физиология зрения. О. Грюссер, У. Грюссер-Корнельс ....... 142
4.1. Глаз............................................................... 144
4.2. Психофизика зрительного восприятия................................. 154
4.3. Нейрофизиология зрения............................................. 172
4.4. Движение глаз и сенсомоторная интеграция при зрительном восприятии . . 188
5. Физиология слуха. Р. Клинке............................................ 198
5.1. Анатомия органа слуха.............................................. 198
5.2. Слух человека...................................................... 202
5.3. Функции среднего и внутреннего уха................................. 211
5.4. Слуховой нерв и высшие уровни слухового пути....................... 218
6. Физиология чувства равновесия. Р. Клинке............................... 224
6.1. Анатомия и физиология периферического органа....................... 224
6.2. Центральные механизмы чувства равновесия........................... 231
7. Физиология вкуса. X. Альтнер........................................... 237
7.1. Морфология органов вкуса; субъективная физиология вкуса............ 237
7.2. Объективная физиология вкуса....................................... 243
8. Физиология обоняния. X. Альтнер........................................ 247
8.1. Обонятельная слизистая; периферические механизмы обонятельной рецепции 247
8.2. Субъективная физиология обоняния; центральные связи................ 252
9. Жажда и голод. Р. Шмидт................................................ 256
9.1. Жажда............................................................. 257
9.2. Голод.............................................................. 265
Ответы на вопросы.......................................................... 273
Рекомендуемая литература................................................... 277
Список авторов............................................................. 281
Предметный указатель....................................................... 282
УВАЖАЕМЫЙ ЧИТАТЕЛЬ!
Ваши замечания о содержании книги, ее оформлении, качестве перевода и другие просим присылать по адресу:
129820, Москва, И-110, ГСП 1-й Рижский пер., д. 2, издательство «Мир».
основы СЕНСОРНОЙ ФИЗИОЛОГИИ
Под редакцией Р. Шмидта
Ст. научи, редактор Е. А. Яновская
Мл. научн. редактор О. А. Горгун
Художник В. Е. Карпов
Художественный редактор Л. М. Кузнецова
Технический редактор 3. И. Резник Корректор М. М. Колотнлнна
ИБ №3637
Сдано в набор 17.08.83.
Подписано к печати 21.02.84.
Формат 60 х 90/16.
Бумага офсетная № 1.
Гарнитура тайме. Печать офсетная.
Объем 9,0 бум. л.
Усл. печ. л. 18,00. Усл. кр.-отт. 41,00.
Уч.-нзд. л. 19,60. Изд. № 3637.
Тираж 5000 экз. Зак. 703. Цена 3 р. 40 к.
ИЗДАТЕЛЬСТВО «МИР»
Москва, 1-й Рижский пер., 2.
Можайский полиграфкомбннат Союзполиг-рафпрома при Государственном комитете СССР по делам издательств, полиграфии и книжной торговли.
г. Можайск, ул. Мира, 93.