/














































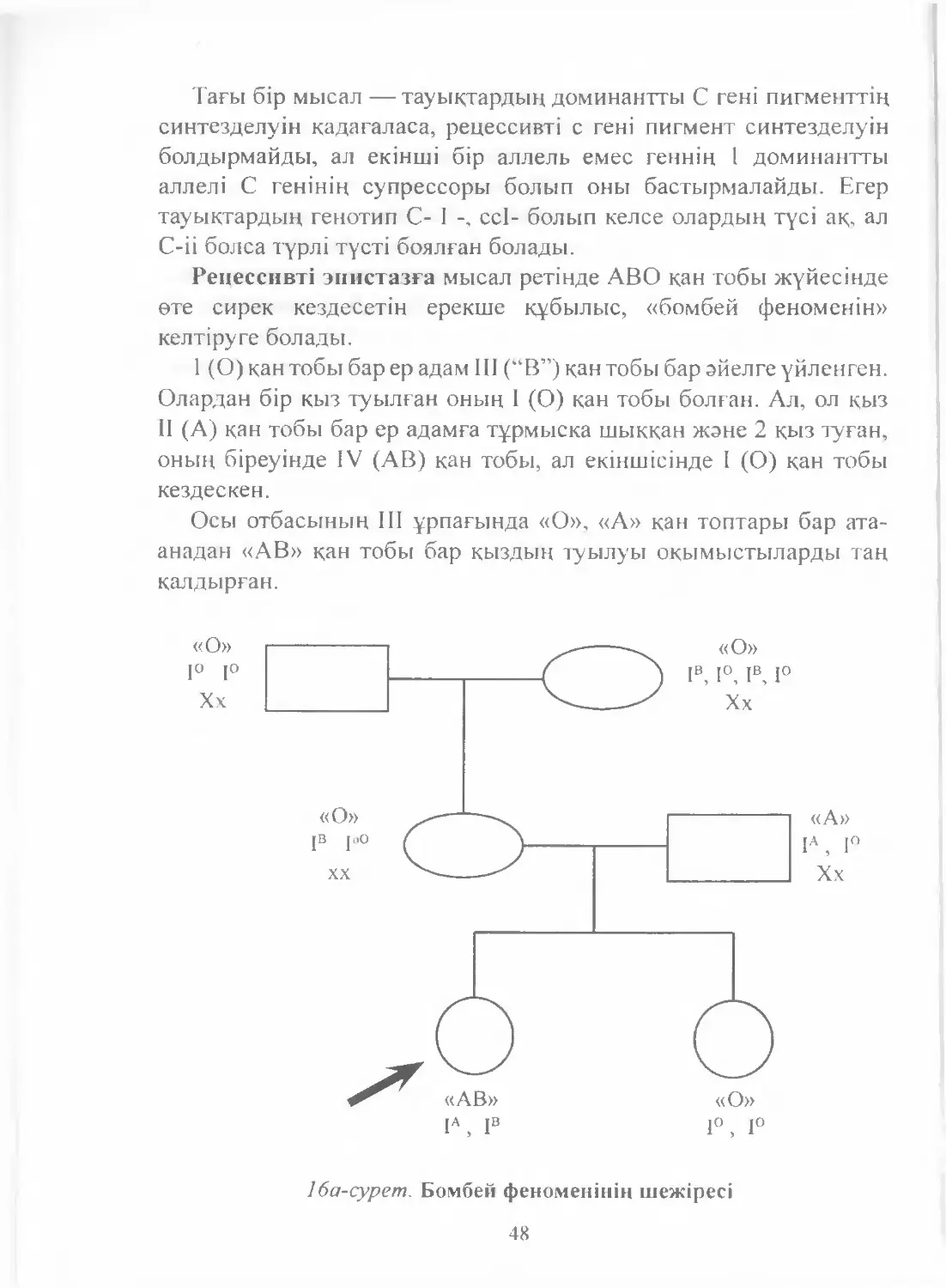










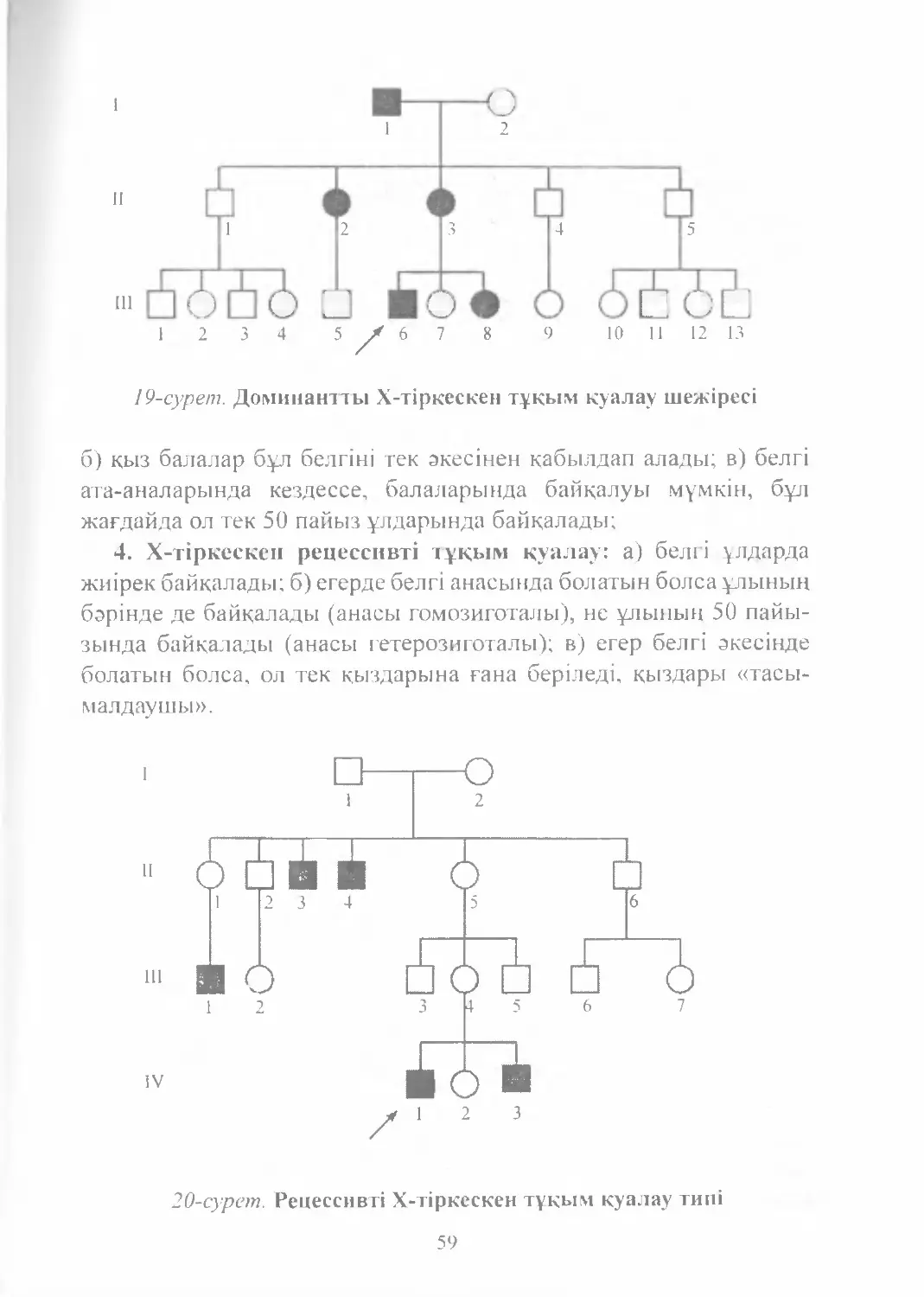

















































































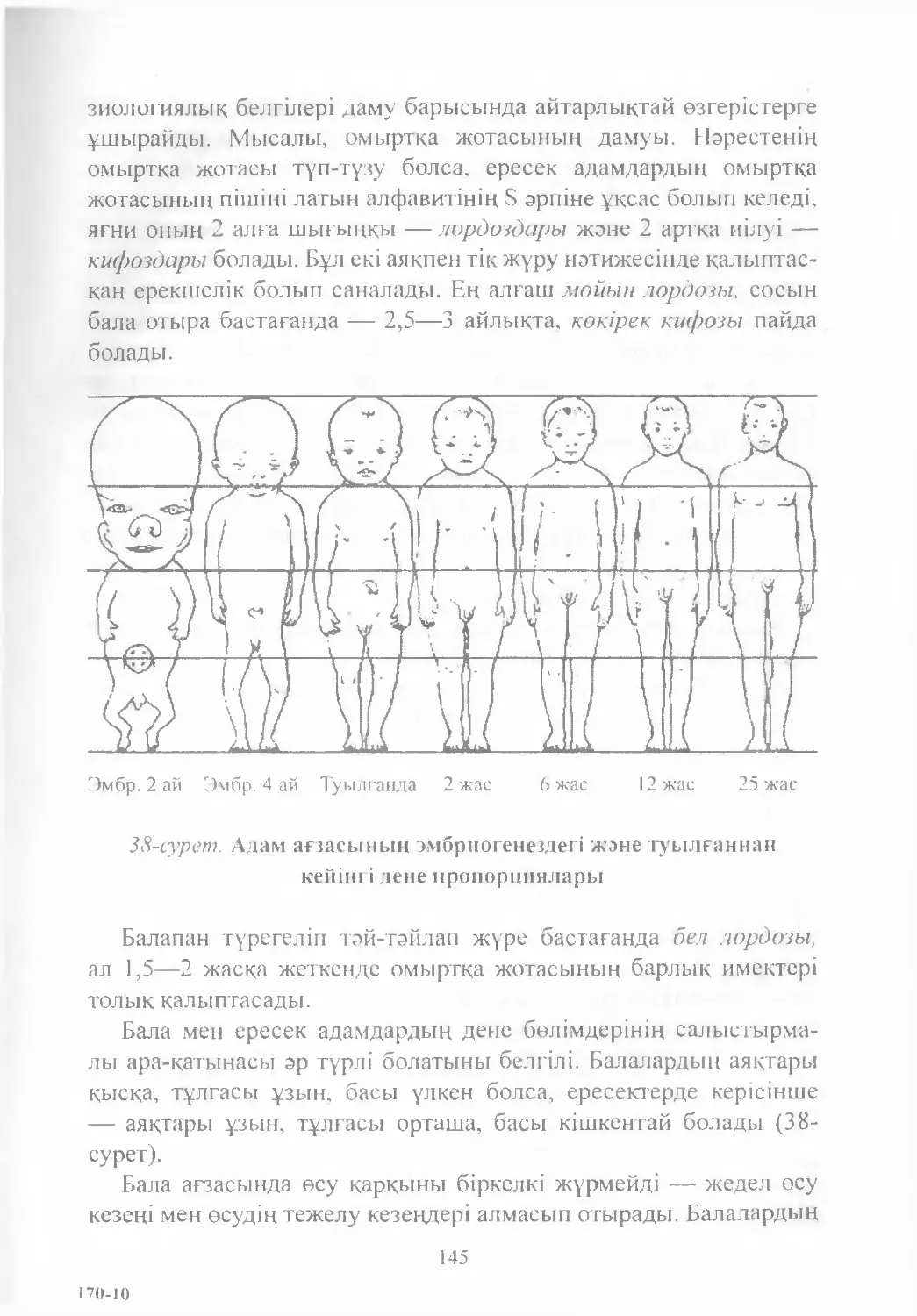
















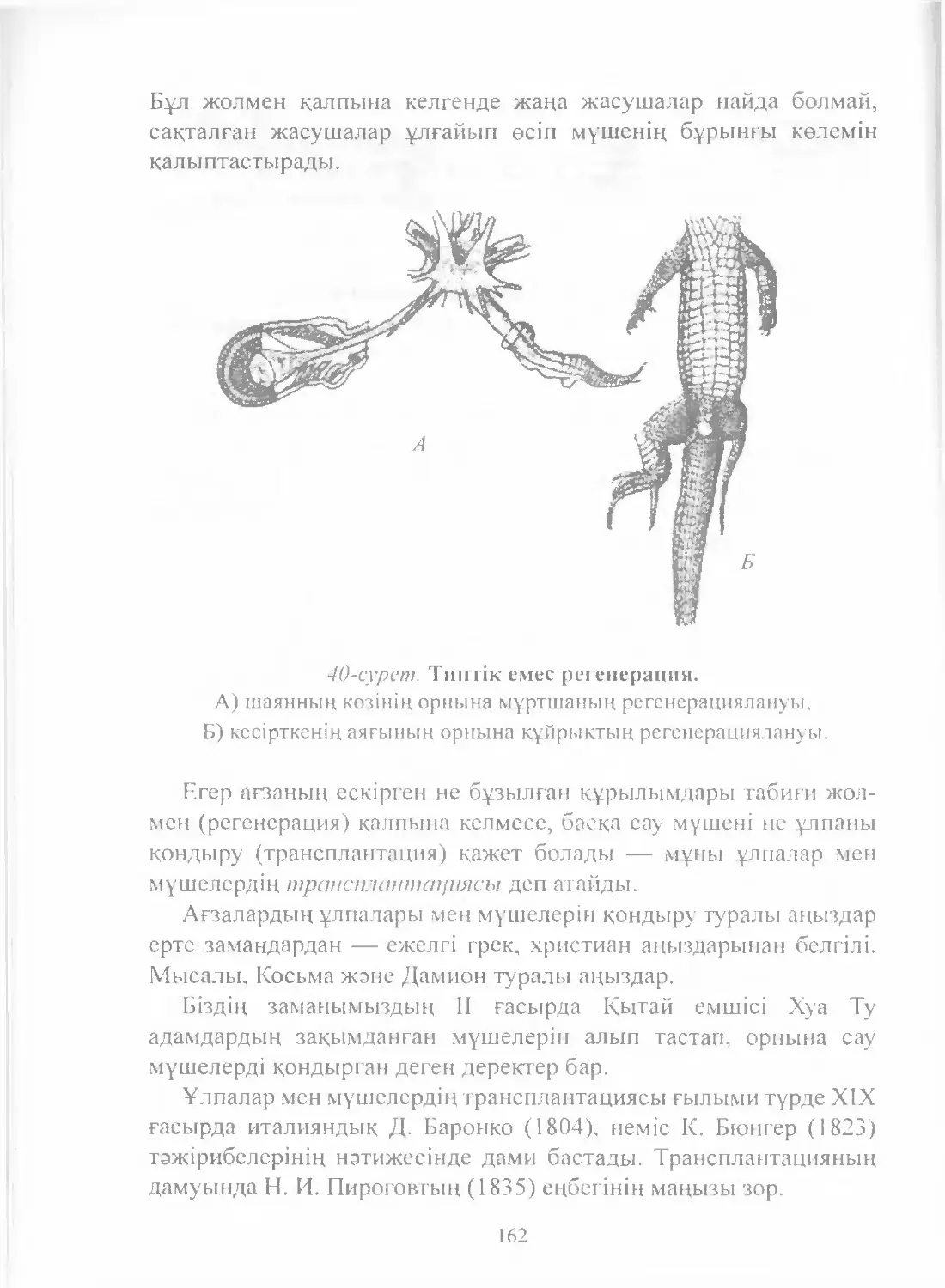

















































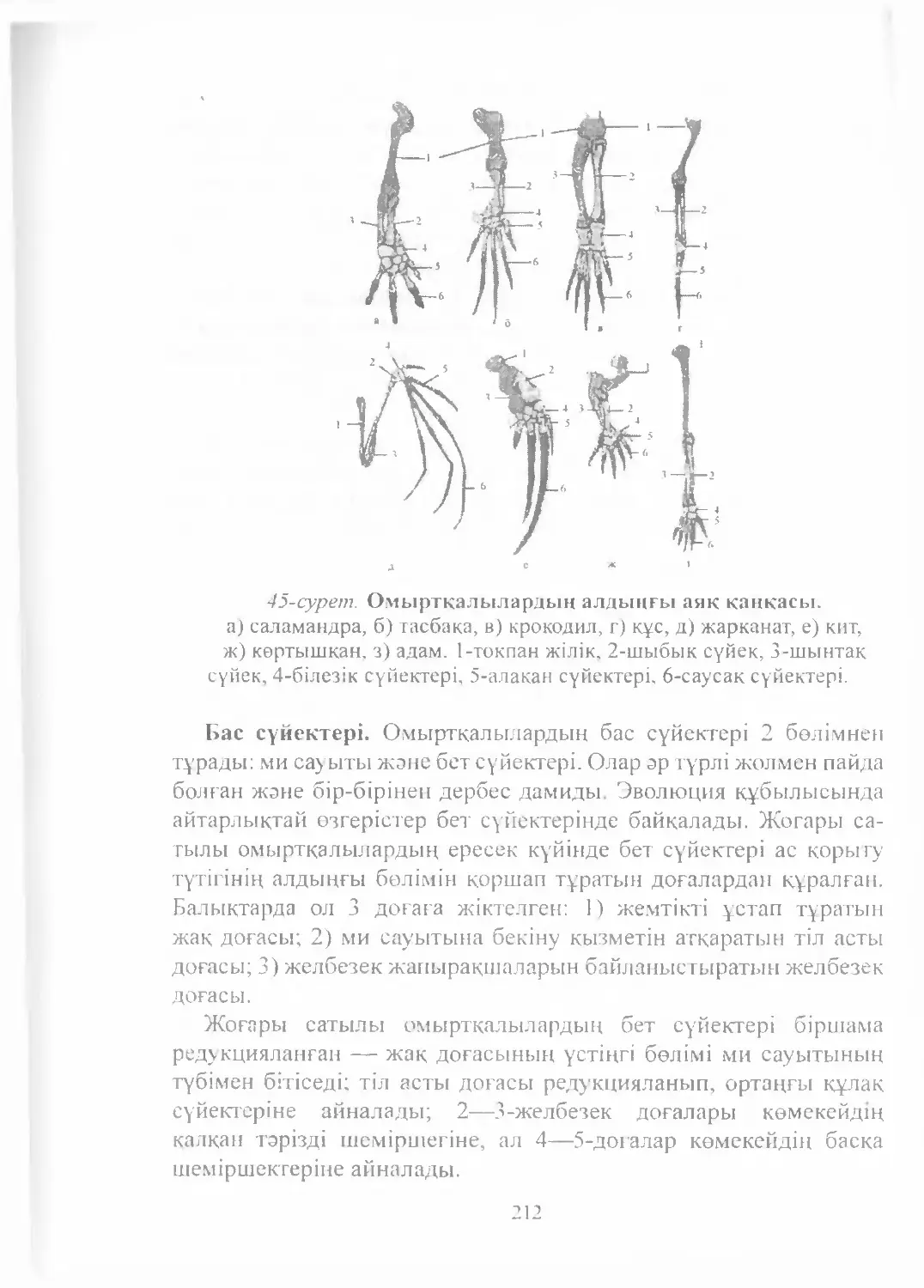




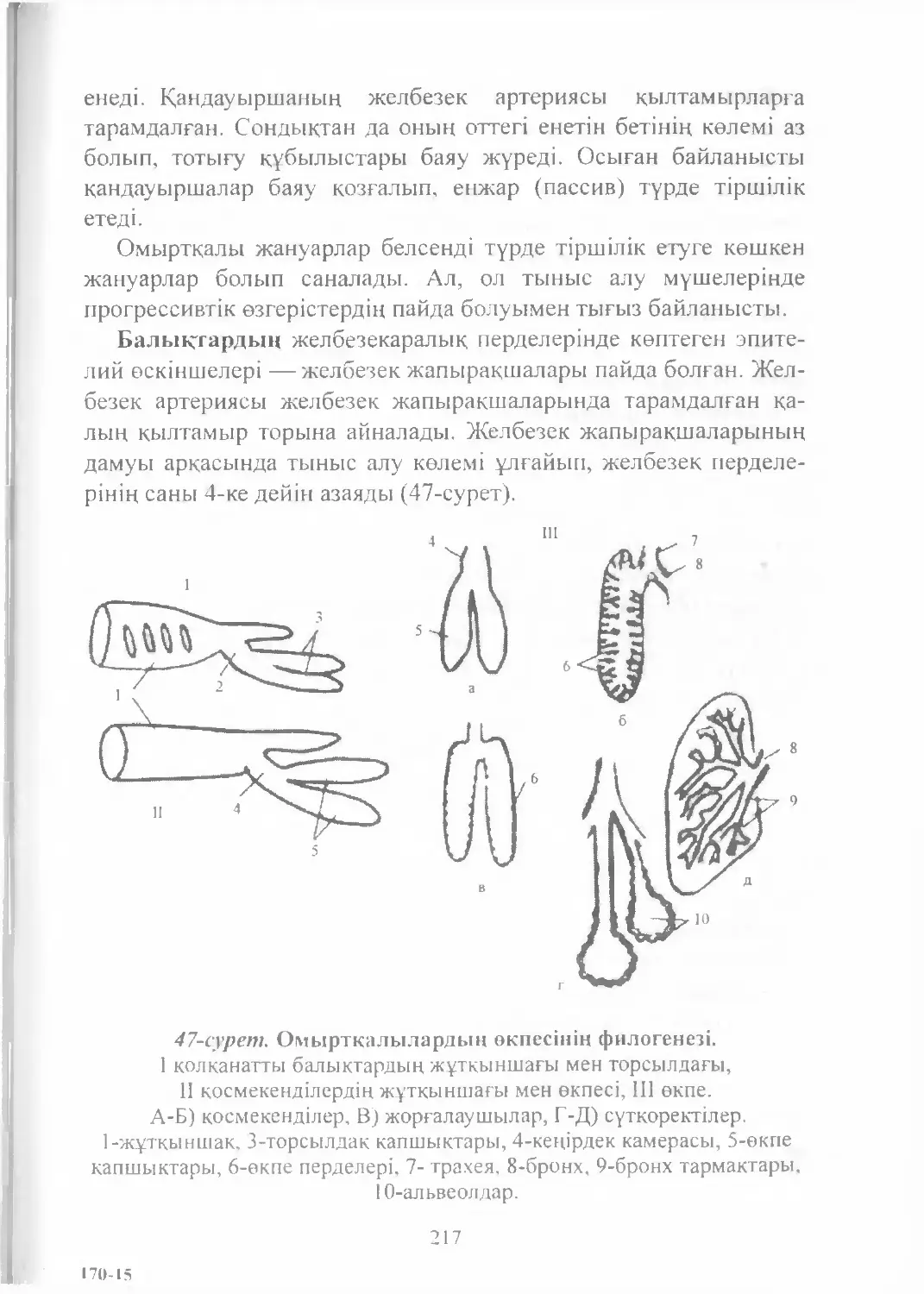



















































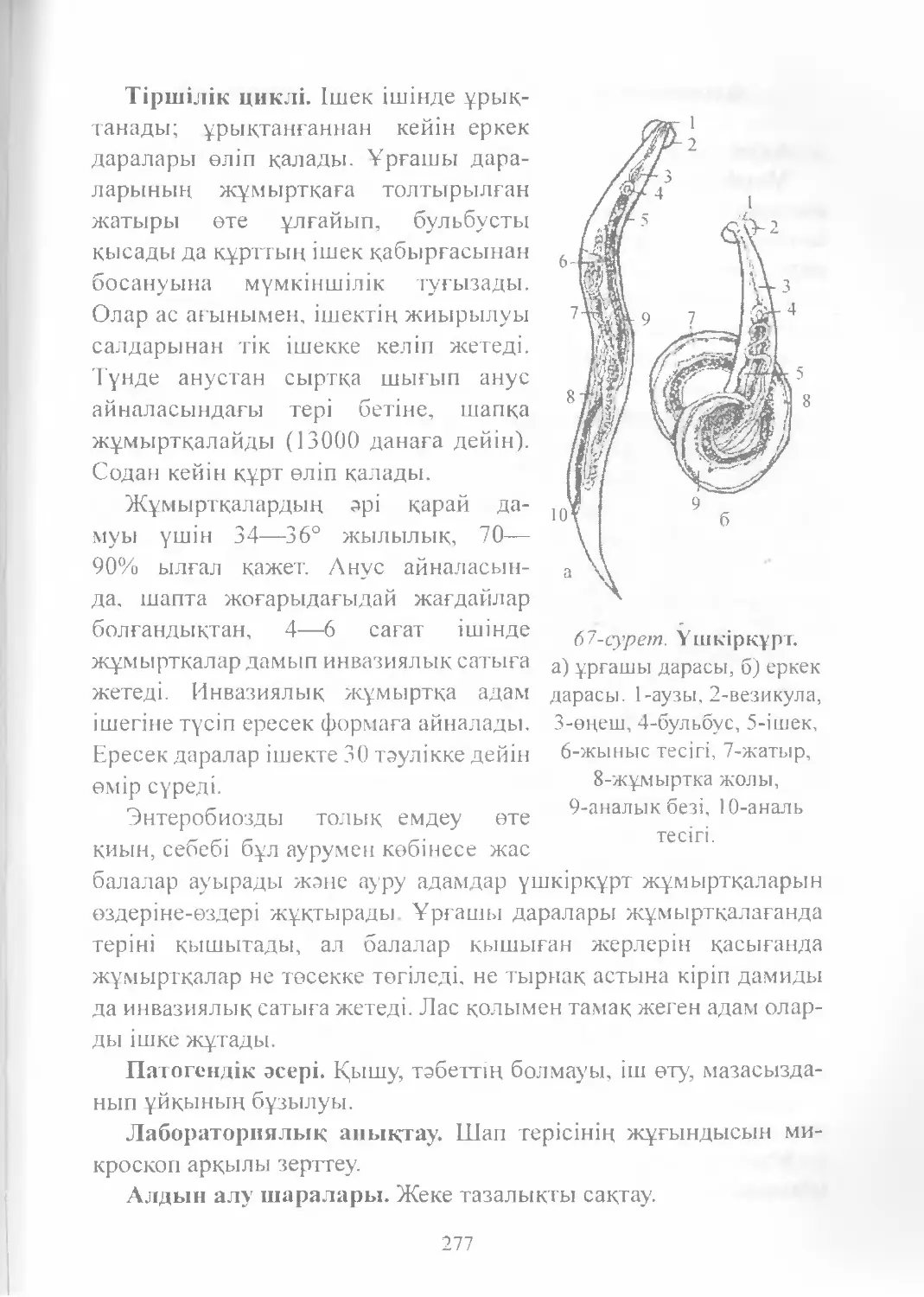







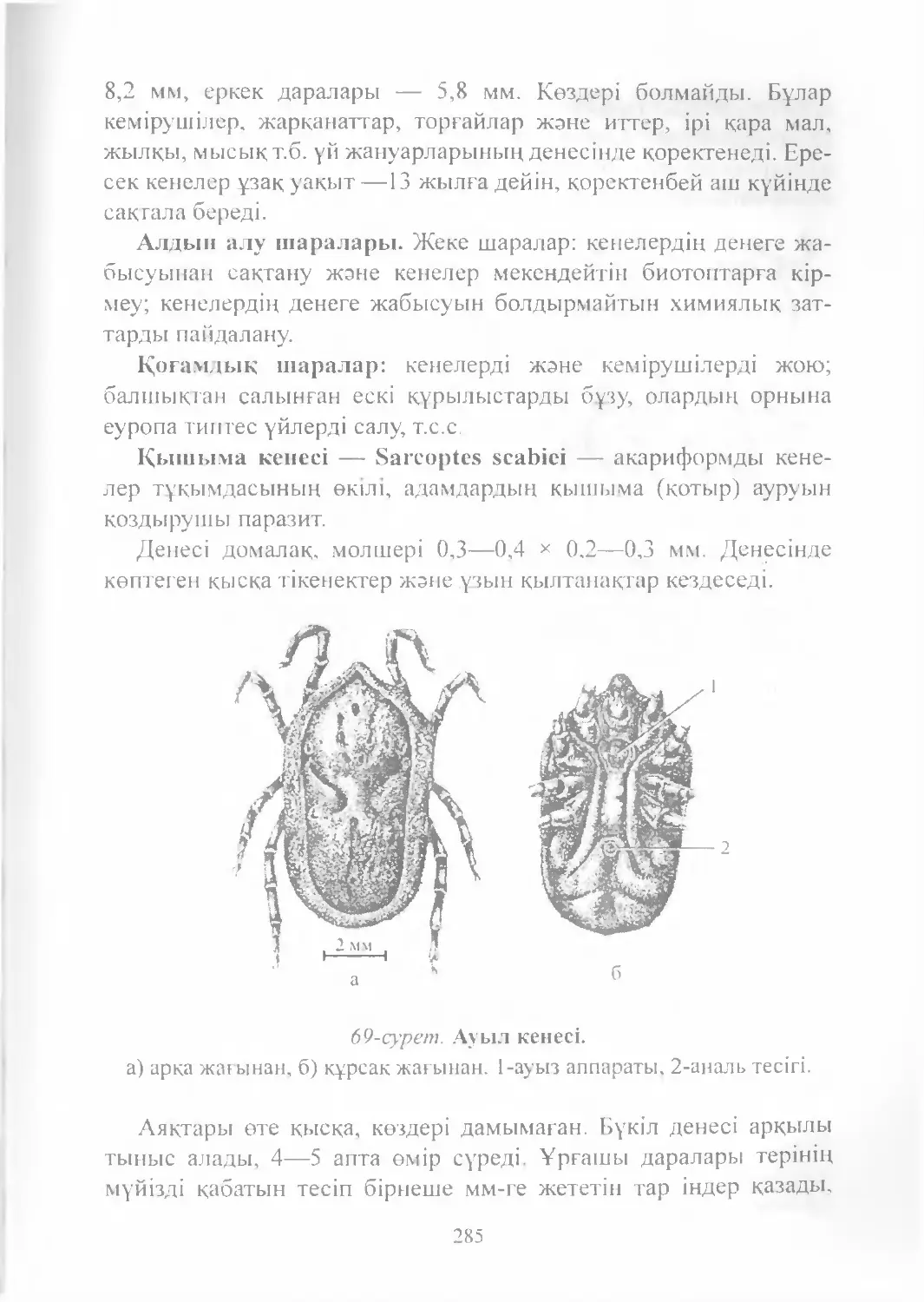
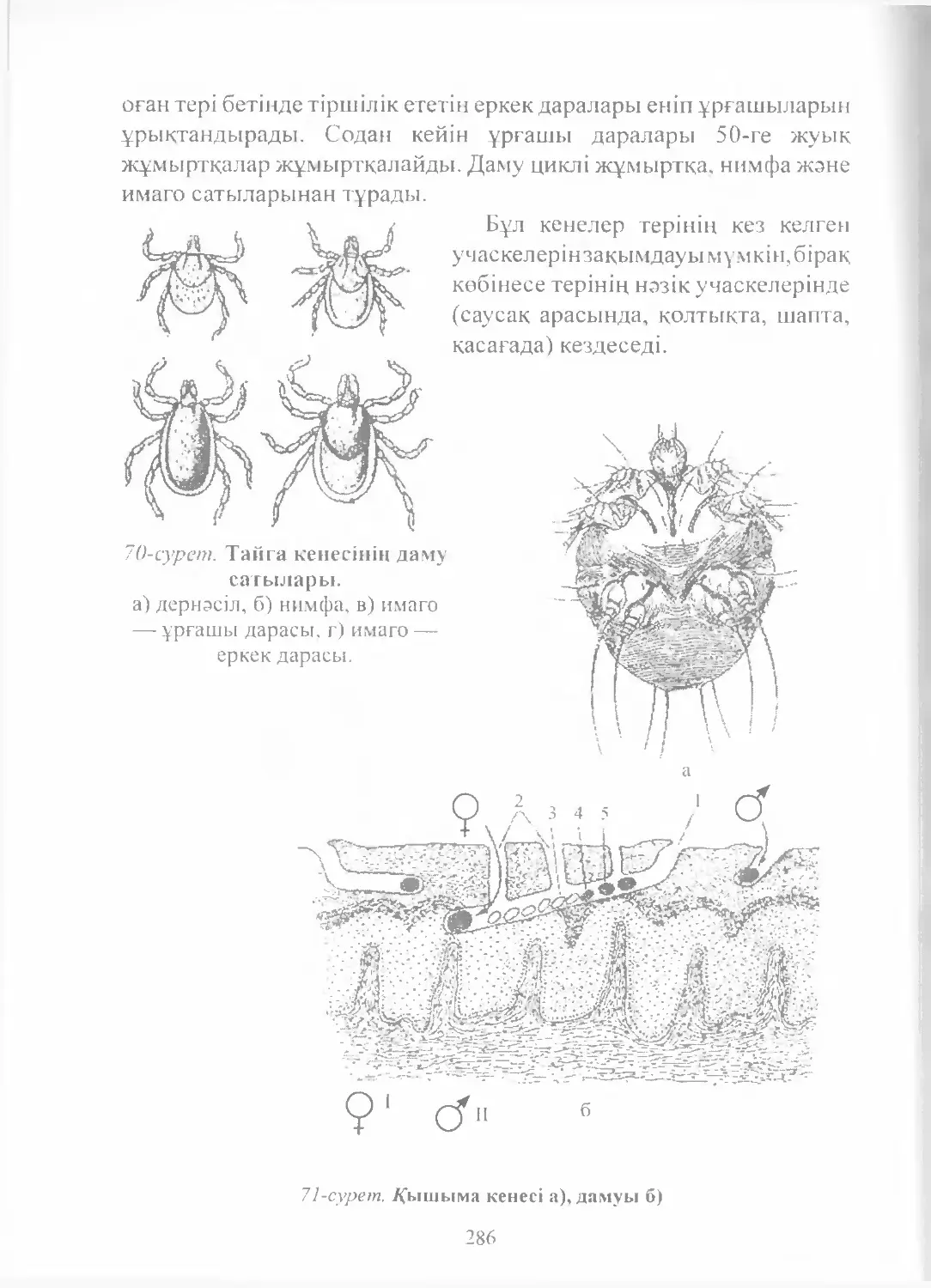




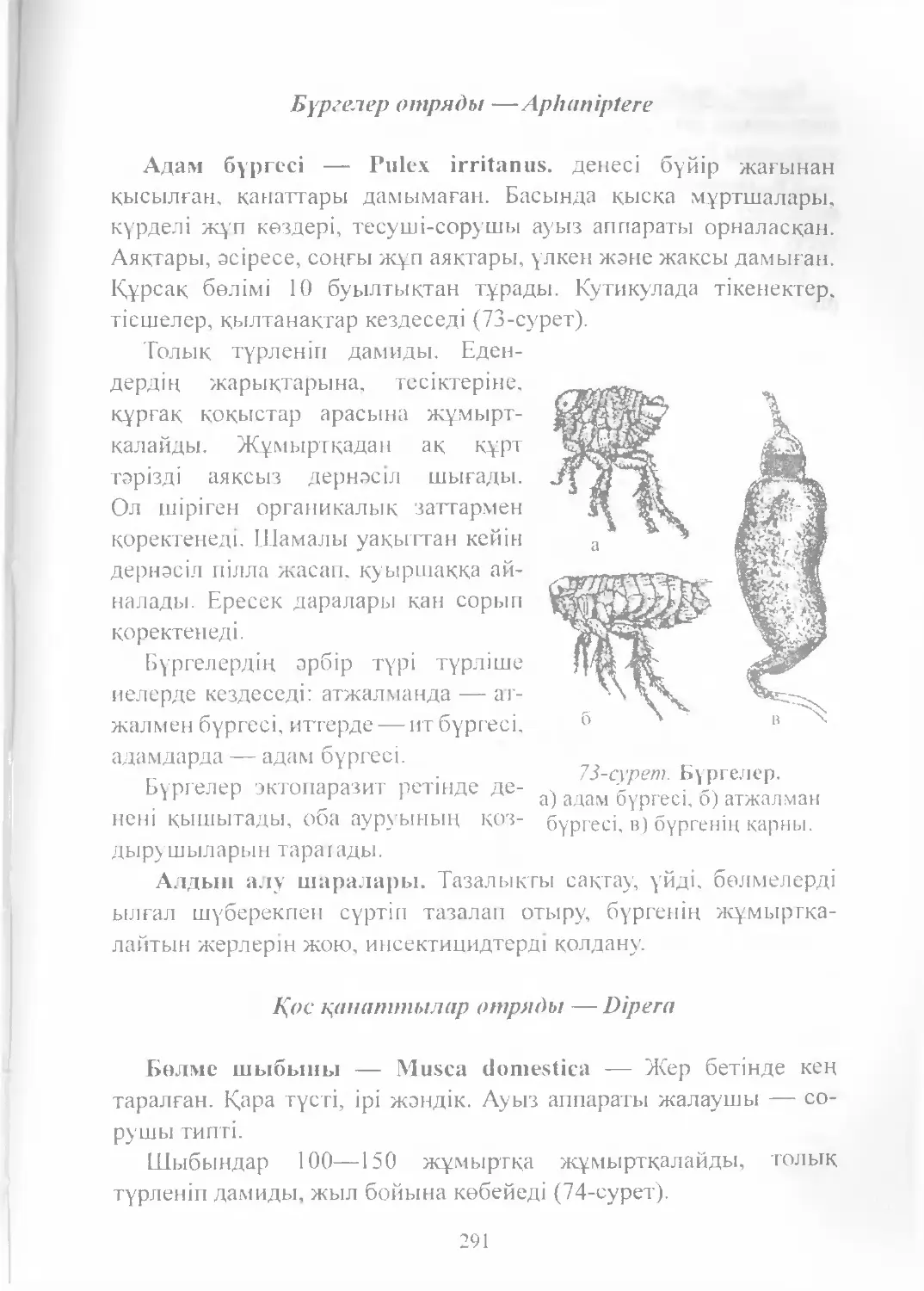






















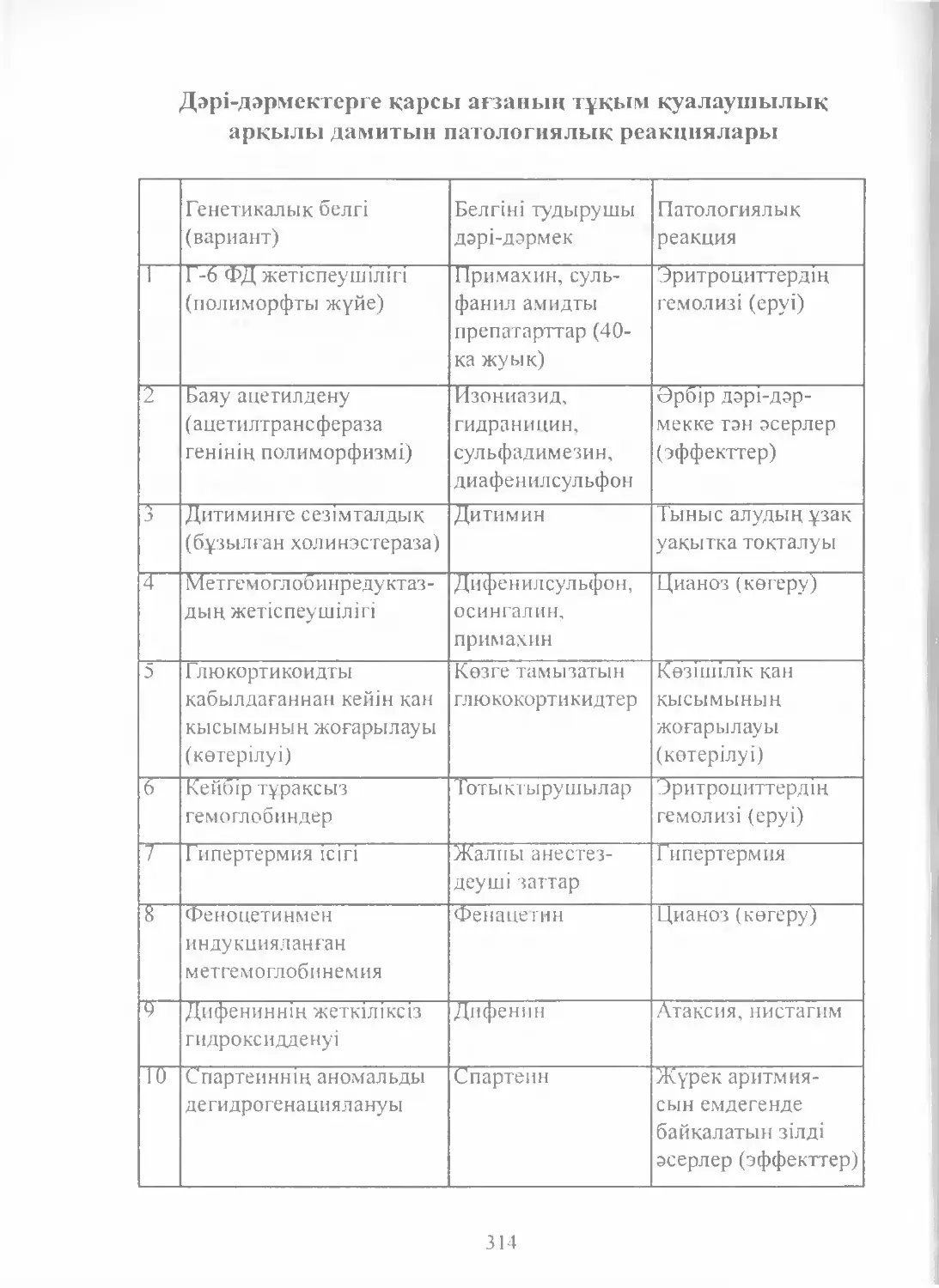


















































Text
Е. Ө. Қуандықов
С. А. Әбілаев
Медициналық
БИОЛОГИЯ
және
ГЕНЕТИКА
NURPRESS
Алматы
2011
УДК 575+612
ББК 52.5
К 71
Рецензенттер:
ОҚММА фармакология жэне фармакотерапия кафедрасының
меңгерушісі, ҚР МҒА мүше-корреспонденті, м.ғ.д.,
профессор ОРМАНОВ Н.Ж.
Қ.А.ЯССАУИ атындағы ХҚТУ Шымкент институтының биология
кафедрасының меңгерушісі, б.х.к., доцент МҮТӘЛИЕВ А.
Қ 71 Е. Ө. Қуандыков —медицина гылымдарыныц докторы,
профессор
С. А. Әбілаев — биология гылымдарыныц кандидаты, профессор
Медициналык биология жэне генетика. Окулык.—
Алматы: «NURPRESS» баспасы, 2011.— 356 бет.
ISBN 9965-830-11-8
Окулыкта Қазакстан Республикасының Денсаулык сактау министрлігі
бекіткен Мемлекеттік жалпыға бірдей міндетті білім берудің жаңа стан-
дартына (03.07-456-2003) сәйкес тіршіліктің негізгі касиеттері жэне
эволюциялык кұбылыстар бірізділікпен сипатталган. Адам онтогенезінде
және популяцияларында жалпы биологиялык заңдылыктардың байка-
лу ерекшеліктері камтылып, олардың медицина практикасындағы рөлі
керсетілген, генетика гылымының жаңа салалары — экогенетика жэне
фармакогенетика мәселелеріне де коңіл аударылған.
Оқулыктың соңғы жагында шэкірттердің өз бетінше окып ұйренуі
(білім алуы), бакылауы үшін сұрактар мен есептер және кыскаша тер-
миндер сөздігі келтірілген.
Окулык медициналык жоғары жэне орта кәсіби оку орындарының,
университеттердің биология факультеттерінің студенттеріне, аспирантта-
рына, дәрігерлер жэне биология мен генетика мәселелері кызыктыратын
мамандардың пайдалануына арналган.
УДК 575+612
ББК 52.5
ISBN 9965-830-11-8
© Қуандыков Е. Ө., Әбілаев С. А., 2011.
© «NURPRESS» баспасы, 2011.
С. Д. Асфендияров атындагы
Қазацтың Үлттық медицина
университетінің 75 жылдыгына
арналады.
Кіріспе
Биология — тіршілікті зерттейтін ғылым (биос — тіршілік,
логос — білім). Биология гылымы тіршіліктің барлық нысанда-
рын жан-жақты, кең ауқымды зерттейді, яғни ол тірі агзалардың
жэне олардың табиғи қауымдастықтарының құрылысын, функ-
циясын таралуын, шығу тегін, дамуын, олардың бір-бірімен жэне
тірі ағзалар мен өлі дүние арасындағы күрделі байланыстарды
зерттейді.
Биологиялықзандылықтарды білу табиғатты қорғау, оны үнемді
пайдалану, ластанудан сақтау және табиғатпен ғылыми негізде
қарым-қатынас жасау үшін қажет.
Биология терминін 1802 жылы Ж. Ламарк жэне Г. Тревиранус
бір мезгілде, бір-бірінен тәуелсіз енгізген болатын.
Биология ғылымының негізгі зерттеу әдістеріне:
1) сипаттап жазу эдісі;
2) салыстыру әдісі;
3) тарихи эдіс;
4) тэжірибе жасау әдістері жатады.
Сипаттап жазу әдісі — өте ерте кезде пайда болған ежелгі
әдіс. Кез келген кұбылысты зергтеу үшін оны толық сипаттау
қажет, ал ол үшін нақтылы деректер жинақтау керек. Бұл әдісті
қазіргі кездегі зерттеулерде де ғалымдар кеңінен қолданады.
XVIII ғ. салыстырмалы эдіс кең өріс алып дамыды — бұл әдістің
мәні ағзаларды не олардың мүшелерін бір-бірімен салыстырып
зерттеп, олардың ұқсастығын не аиырмг
інылыктарын анықтау бо-
систематикасы қүрастырылды.
лып саналады. Осы эдіс көмегімен eciisa^fe9ce>l!VНІН
Оқу-клиникалық бззасы
Кітапхака
Тарихи әдіс — ертеде өмір сүріп, жойылып кеткен агзаларды
зерттей отыра, қазіргі кезде тіршілік ететін агзалардың құрылысын,
шығу тегін жэне даму зандылықтарын анықтауда қолданылады.
Бұл эдіс Ч. Дарвиннің эволюциялық ілімі пайда болғаннан кейін
қалыптасқан.
Тәжірибе жасау әдісі — табиғат күбылыстарын тәжірибе жа¬
сау арқылы зерттеу болып табылады. Бұл эдіс химия, физика т.б.
табиғаттану гылымдарының жетістіктерін кеңінен қамтиды.
Биология — өте ертеде пайда болған ғылым. Ол адамдардың
практикалық қызметі нэтижесінде қалыптаскан. Ежелгі адамдар
өсімдіктермен, жануарлармен тамақтанған, оларды киім жэне ба-
спана жасауда пайдаланған. Сондықтан олар туралы деректерді
жинақтай бастаған. Одан эрі қарай адамдар жабайы өсімдіктерді
егіп, жануарларды қолға үйреткен. Ал, ол үшін осы ағзалардың
тіршілігін тереңірек білу қажет болған. Жинақталған тәжірибе,
білім үрпақтан-үрпакка үзақ жылдар бойына ауызша, аңыз күйінде
беріліп келген болса, жазу пайда болғаннан кейін, адамзаттың
табигат туралы білімі кеңінен жинакталып, сараланып, эрі қарай
жүйелі түрде беріліп отырған. Бұл осы ғылымның пайда болуына
тікелей жол ашкан.
Биологияның алғашқы қалыптасқан салалары ретінде ботаника
— өсімдіктер туралы ғылым жэне зоология — жануарлар туралы
гылымдарды атауға болады.
Ботаника гылымы б.ж.с. дейінгі IV—111 ғғ. калыптасқан, оның
атасы болып Теофраст (372—287 б.ж.с.д.) танылды. Ол өсімдіктер
туралы 11 кітап жазған. Олардың кейбіреулері біздің заманымызға
дейін жетіп сақталған.
Зоология гылымы да б.ж.с. дейінгі IV—III гғ. қалыптаскан
жэне оның атасы болып Аристотель (384—322 б.ж.с.д.) саналады.
Ол 500-ден астам жануарлар түрлерін сипаттап жазып, олардьщ
алғашқы жүйесін жасаған.
Биология ғылымының қалыптасуында ертедегі грек дәрігері
Гиппократтың рөлі ерекше (460—377 б.ғ.д.). Ол жануарлар
мен адамның қүрылысы туралы көптеген деректер қалдырған;
сүйектерді, бүлшықеттерді, сіңірлерді, миды жэне жүлынды си¬
паттап жазган. Гиппократ үнемі «...эрбір дәрігер табиғатты жете
білуі кажет» деп айтып отырган.
Адам денесінің құрылысын зерттеуде Рим дәрігері Галеннің
(130—200 б.ж.с.д.) еңбегі өте үлкен. Ол маймыл мен шошканы
сойып оның ішкі қүрылысын зерттеген.
Орта ғасырларда діни көзқарас басым болды. Ол, гылыми
көзкарасты қолдайгын адамдарды қатал жазалап, ғылымның да-
муына жол бермеді.
XV—XVI ғасырлардан бастап биология гылымы жаңадан өрлеп
дами бастады. 1583 жылы А. Чезальпино өсімдіктердің алғашқы
жасанды жүйесін (системасын) құрастырды. 1543 жылы А. Веза-
лий адам денесінің құрылысын зерттеп, «Адам денесінің күрылысы
туралы» атты кітап жазган; 1628 жылы У. Гарвей кіші қан айналым
шеңберін зерттеп, кан айналым туралы ілімді қалыптастырған;
1665 жылы Р. Гук жасушаны, (1673) Левенгук сперматозоидтар-
ды жэне микроағзалар дүниесін ашқан. XVIII ғасырда К. Линней
(1735) өсімдіктер дүниесінің жасанды жүйесін жасап, ботаниканың
дамуына зор үлес қосты.
1809 жылы Ж. Ламарк алгашқы эволюциялық теорияны
қүрастыруға талпыныс жасады; 1838—1839 жылдары Шванн жэне
Шлейден жасуша теориясын: Ч. Дарвин (1859) шынайы эволю-
циялық ілімді қалыптастырды, оныңкозғаушы күштерін анықтады;
1865 жылы Г.Мендель тұқым қуалаушылық зандылықтарын
ашты.
XX гасырда биология гылымы жедел дамыды, оган ат са-
лыскандар: эволюция теориясы саласында — С. С. Четве-
ряков, Р. А. Фишер, С. Райт, Г. Хаксли, Ф. Б. Добржанский,
А. Н. Северцев, Н. И. Вавилов, И. И. Шмальгаузен, Н. В. Тимофеев-
Ресовский т.б.
Молекулалық биология негізін қалагандар — Ф. Крик, Дж. Уот¬
сон (1953), ал, эрі карай дамытушылар — А. А. Баев, А. Н. Бе¬
лозерский, А. С. Спирин, В. А. Энгельгард, М. А. Айтхожин т.б.
болып саналады.
В. В. Вернадский (1928) — биосфера туралы ілімнің негізін
қалаган; В. И. Сукачев (1942) — биогеоценоз, Тенсли (1935) —
экосистема туралы ілімді қалыптастырган.
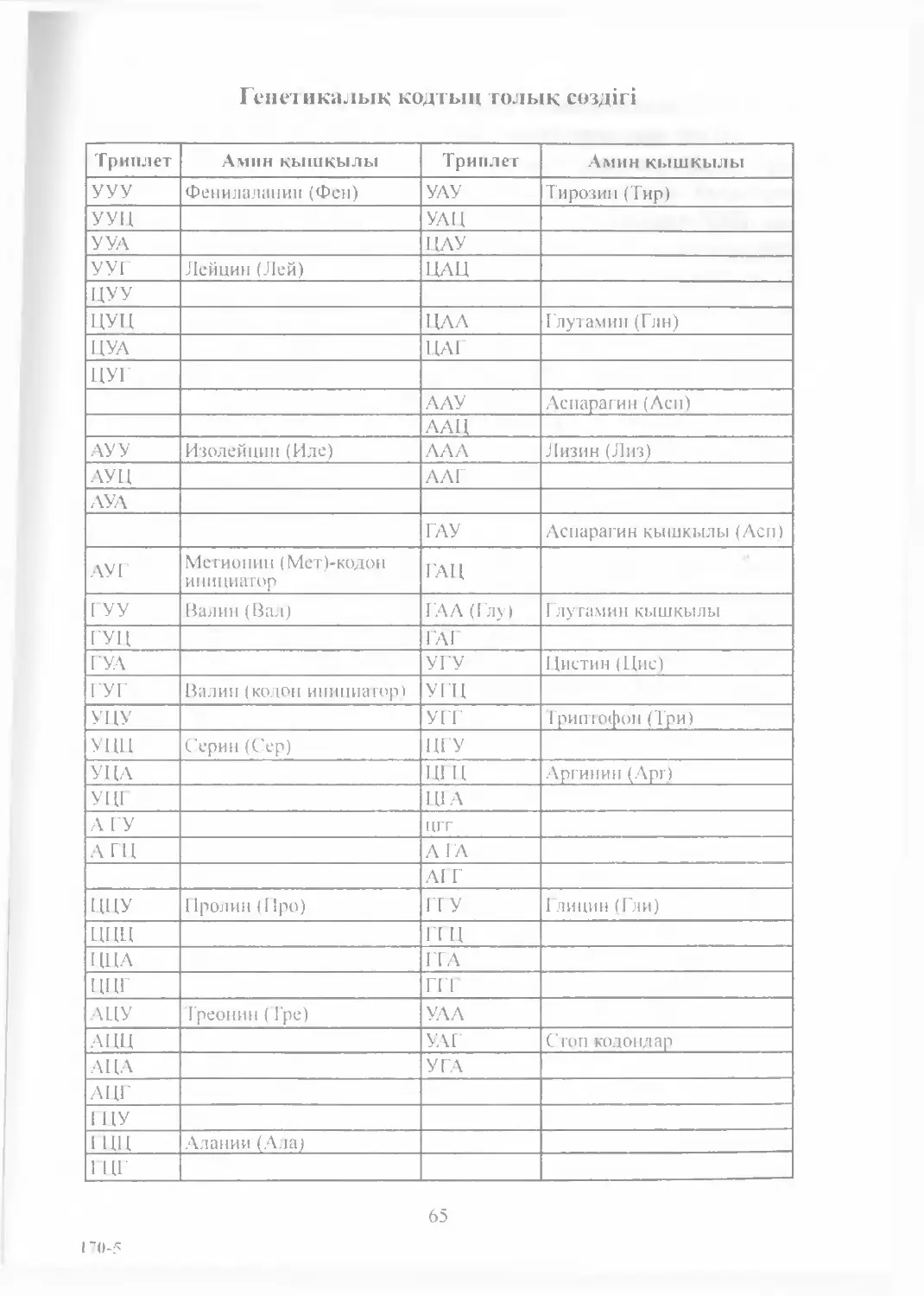
1953 жылы Ф. Крик жэне Дж. Уотсон нуклеин кышқылдарының
құрылысын ашты. 1961—1964 жылдары С. Очао, X. Корана,
М. Ниренберг т.б. оқымыстылардың еңбектерінің нәтижесінде
5
генетикалық кодтыц толық «сөздігі» құрастырылып, оның барлык
тірі ағзаларда бірдей болатындығы аныкталды. Осы жылдары
жасушада жүретін негізгі биохимиялық құбылыстардыц — ДНҚ
репликациясы, транскрипция жэне ақуыз синтезі — трансляция
механизмдері зерттелініп, оның прокариоттарда жэне эукариот-
тарда үқсас жоба күйінде жүретіндігі белгілі болды.
1970 жылы молекулалық биологияның жаңа саласы —
генетикалык инженерия, биотехнология қалыптасып, жасанды
генотипті ағзаларды (трансгенді агзаларды) жасауга, көптеген
гормондарды лабораториялық жағдайларда синтездеуге қол
жетті. Мысалы: осылайша 1977 жылы соматотропин, 1978-—
1979 жылдары инсулин, интерферон, простагландин гормондары
синтезделінді. XX ғасырдың аяғында ғалымдар жоғары сатылы
ағзалардың сомалық жасушаларын клондауға кол жеткізді. Мыса¬
лы: 1997 жылы Англияда алғаш рет клондау әдісі аркылы «Долли»
атты козы, 1998 жылы торай, 1999 жылы маймыл дүниеге келген.
Қазіргі кезде ғалымдар ДНҚ — тесттер арқылы адам геномын
анықтады. Осылайша молекулалық генетиканың жаңа саласы —
геномика қалыптасты. Мүның медицина үшін маңызы өте зор,
себебі, генодиагностика, генотерапия, жасушалык терапия т.б. ау-
руды емдеудің жэне анықтаудың жаңа әдістері медицина практи-
касына енгізілуде.
XXI гасыр — биология, оның ішінде генетика гасыры деп
аталуда. Оныц болашагы медицина үшін орасан зор, себебі жаңа
жетістіктер, жаңалықтар нэтижесінде осы уақытқа дейін емдеу-
ге болмайтын адамның түқым куалайтын ауруларын емдеуге қол
жетеді, адамдардыц өмір сүру үзактыгын едәуір ұзартуга мүмкіндік
туады.
Қазіргі кезде биология гылымы осы әдістердің бәрін де пайда-
ланып, тірі агзаларды кешенді түрде зерттейді.
Биология гылымы коп салалы, кешенді табигаттану гылымы бо¬
лып табылады. Оныц ішінде болашақ дэрігерлер үшін ең кажеттісі
— ол медициналық биология саласы болып саналады.
Медициналық биология — биологиялық зандылықтарды
адам кауымдастыгы түргысынан карастыратын жалпы биология
гылымының бір тармагы болып табылады.
6
Биологияның медицина үшін маңызы өте зор — ол медицина
үшін теориялык негіз болып саналады. Медицинаның көрнекті
теоретигі И. В. Давыдовскийдің айтуынша «... медицинаны теория
түрғысынан қарастыратын болсақ, ол, ең алдымен, жалпы биоло¬
гия деп білуіміз керек...».
Медицина жетістіктері биологиялық зерттеулермен тікелей
байланысты, сондықтан да дәрігерлер биология жаңалықтарын
үнемі біліп отырулары шарт. Бұған көптеген мысалдар келтіруге
болады: 1862 жылы J1. Пастердің тіршілік ездігінен, кенеттен пай¬
да бола алмайтындығын дәлелдеген зерттеулері, заттардың шіруі
мен ашуының негізгі себебі микроағзалар екенін дәлелдеуі, меди-
цинада үлкен өзгерістер енгізіп, хирургияныц дамуына жол ашты.
Медициналық практикаға алғаш антисептика (жараны химиялық
заттармен өңдеп, микробтарды жүқтырмау), ал, кейін асептика
(жараны таңуға арналган заттарды стерилдеу) әдістері енгізіледі.
Жасушаның ашылуы, тіршіліктің физиологиялық жэне
биохимиялық зандылықтарын зерттеулер аурудың пайда болу
себептерін тереңірек түсінуге, медицина практикасына кеселді
анықтаудың жэне емдеудің жаңа эдістерін енгізуге мүмкіншілік
берді.
И. Мечниковтың төменгі сатылы көпжасушалы жануарлардың
ас корыту ерекшеліктерін зерттеулері, фагоцитоз қүбылысының
ашылуына, иммунитет күбылысының мэнін айқындауға мүм-
кіншілік берді.
Адамдардыцденекұрылысы,қызметі,эсіресе,оныңқорғаныстык
механизмдері — адам тегініц миллиондаған жылдарға созылган
эволюциялық өзгерістерінің нэтижесі екенін естен шыгармауымыз
керек. Сондықтан да адамдардың патологиясының себептері де
жалпы биологиялық зандылықтарға байланысты екені айдан
анық.
Адамның ауруларының бәрі — белгілер болып саналады,
ал, ол белгілердің калыптасып дамуы адам генотипіне байланы¬
сты. Қазіргі кезде адамның 14000-нан астам ауруларының түқым
қуалайтындығы анықталып отыр. Кейбір аурулар тұкым қуаламаса
да, оларға бейімділіктің түқым қуалайтындығы анықталды (қант
ауруы, дизентерия т.б.). 'Гіпті, түқым қуаламайтын жүкпалы
аурулардың өзі адамдар генотипіне байланысты болатыны белгілі.
7
Мысалы: бір адам бір аурулармен қатал ауырса, екіншісі жеңіл
түрде, ал үшіншісі мүлдем ауырмауы мүмкін, сондықтан да адам-
дарды емдеу үшін эр түрлі әдістер қолданылуы қажет.
Дүниежүзілік денсаулық сактау үйымының (ВОЗ) деректеріне
Караганда адамның денінің сау болуы 50 пайыз жағдайда оның
өмір сүру ерекшелігіне, 20 пайыз жагдайда — экологиялық
факторларға, 20 пайыз жагдайда — генотипке, 10 пайыз жағдайда
медицина жетістіктеріне байланысты.
«Тәні саудың—дені сау». Ауырмай сау болу, ауырганға қарағанда
коғамға да, адамга да элдекайда арзанға түсетіні белгілі.
Сондықтан да тәуелсіз Қазақстанда адамдардың салауатты өмір
сүруін қалыптастыру мәселелері мемлекеттік мэселе дәрежесіне
көтерілген. Қазақстан Президенті Н. Назарбаевтың «Қазақстан —
2030» атты Жолдауында, оның жыл сайынғы Қазақстан халқына
арналган дәстүрлі жолдауларында осы мәселелер Қазақстанның
стратегиялық дамуының басты багыттарының бірі ретінде аталган.
Бүл мәселе 1999 жылдың басында болып әткен Республика
дәрігерлерінің бірінші қүрылтайында да жан-жақты талқыланды;
Үкімет қаулысымен «Салауатты омір сүруді қалыптастыру
мәселелері» атты ұлтгық орталық үйымдастырылып жүмыс
істеуде.
Қазіргі кезде биология гылымы мен медицина соншалықты
тыгыз байланысып, соңгысын көптеген жагдайларда, биомедицина
деп атауда.
Қазақстанда биология ғылымының жүйелі түрде дамуы
1932 жылы Алматыда КСРО Ғылым Академиясының филиа¬
лы ұйымдастырылуынан басталады. Бүган дейін, XIX гасырда,
Қазақстанның бай табигатын Ресей жэне баска да шетел галымдары
оқтын-оқтын келіп зерттеген.
Қазіргі кезде елімізде 10-га жуық биологиялық гылыми зерттеу
институттары бар. Олардың құрамында жүздеген гылым доктор-
лары, мындаган гылым кандидаттары еңбек етеді. Сонымен катар,
биологиялык зерттеулер ондаган университеттер мен институттар-
да да жүргізілуде.
Қазақстан биологтерінің көптеген еңбектері элемге эйгілі бо¬
лып, сыйлыктарга ие болган. Мысалы: С. Н. Боевтің «Легочные
нематоды копытных животных Казахстана», И. Г. Галузоның
«Токсоплазмоз животных», «Природная очаговость болезней и
вопросы паразитологии» атты монографиялары 1964—1965 жыл-
дары ағылшын тіліне аударылған. М. А. Айтқожин 1974 жылы бір
топ орыс молекулалық биохимиктерімен бірге жасушаның ерек-
ше органелласы — информосомаларды тауып, оның күрылысын,
кызметін аныктауға арналган еңбектері үшін Лениндік сыйлықтың
иегері атанды.
Қазақстанда биология ғылымының дамуына көптеген жергілікті
жэне орыс ғалымдары үлес коскан, олардың ішінен: ботаник-
тер — Н. В. Павлов, Б. В. Быков, Л. К. Қылышов, В. П. Кузьмин,
A. М. Габбасов, Е. X. Үзенбаев, Г. 3. Бияшов, Н. Л. Удольская,
B. С. Корнилова, Б. О. Байтулин; зоологтер — Б. А. Домбровский,
C. Н. Боев, И. Г. Галузо, В. В. Гвоздев, В. Шевченко; физиологтер
— А. П. Полосухин, Н. О. Базанова, Ф. Мүхаметкалиев; биохи-
миктер — П. А. Верболович, В. И. Якубовская, Т. Б. Дарқанбаев,
М.А.Айтқожин, Г. Т. Тәшенов, X. Жұматов, т.б. атауға болады.
Тіршіліктің мәнін анықтау — биологияның негізгі мақсат-
тарының бірі.
Тіршілік анықтамасы өте көп, бірак, солардың ішінде кең
таралғаны Ф. Энгельстің берген анықтамасы болып саналады.
Ф. Энгельс «Тіршілік — ақуыз денелердің өмір сүру формасы,
оның негізгі мәні сыртқы ортамен зат алмасуы» деп көрсеткен бо-
латын.
Қазіргі кездегі көзқарас бойынша тіршіліктің негізін 2 ірі биопо-
лимерлер — ақуыз жэне нуклеин қышқылдары қүрайды. Сондық-
тан да тіршіліктің барлық қүбылыстары осы екі биополимерлердің
кешенді касиеттеріне байланысты.
Tipi ағзалар өлі дүниеде кездесетін химиялық элементтерден
түрады, бірақ, сол химиялық заттардың тірі ағзалардағы күйі,
атқаратын қызметі, активтігі өлі дүниедегіден өзгеше болады.
Тірі ағзалар денесінде заттардың тек физико-химияық озге-
рістері мен күрделі физиологиялык күбылыстары жүріп қоймай,
сол сияқты онда сапалық биологиялық зандылыктар байқалады
(ыдырау-синтезделу; заттар, энергия, ақпараттар ағыны т.б.).
Тіршіліктің түпкілікті касиеттеріне мыналар жатады:
1) оздігінен жацару — ол заттар мен энергия агындарына бай¬
ланысты;
9
2) өздігінен көбею — бұл бірін-бірі алмастырып отыратын
ұрпақтар жалгасын қамтамасыз ететін процесс жэне ол акпараттар
ағынына байланысты болады;
3) өздігінен реттелу — ол заттардың, энергия жэне ақпараттар
ағындарына байланысты.
Тіршіліктің жоғарыда келтірілген түпкілікті қасиеттері
оның негізгі белгілерін қалыптастырады, олар: заттардың жэне
энергияның алмасуы, тітіркену, гомеостаз, көбею, тұқым қуалау
мен өзгергіштік, жеке және филогенетикалық даму, тіршіліктің
дискреттілігі жэне біртұтастыгы.
1) Заттардыц және энергияның алмасуы. Заттардың алмасуы
тіршіліктің ең негізгі белгілерінің бірі болып саналады. Ол кейде
қарапайым күйде өлі дүниеде де кездеседі. Бірақ, ол тірі ағзалардан
өзгеше жүреді, мысалы: көмір жанғанда О, пен СО, алмасады,
темірдің бетін тат басқанда да О, алмасуы байқалады. Өлі дүниеде
бүл процестер негізінде заттар ыдырап жойылып, жоқ болады, ал,
тірі ағзалардың тіршілік етуі үшін міндетті түрде зат алмасу болуы
қажет. Зат алмасу процесінде тірі агзаларда ыдыраған заттар жой-
ылмай қайтадан калпына келіп отырады, яғни өздігінен жаңару
жэне өздігінен көбею байқалады.
Тірі ағзалар ашық биологиялық жүйе болып табылады. Ол
арқылы үзіліссіз заттар, энергия жэне акпараттар агыны өтіп от¬
ырады.
2) Тітіркену — тіршіліктің негізгі белгілерінің бірі. Ол сыртқы
ортадан берілетін кез келген ақпаратқа тірі ағзалардың кері жауап
реакциясы болып саналады. Сыртқы ортадан қажетті ақпарат алу
арқылы биологиялық жүйелердің (жасуша, үлпа, мүше, агза т.б.)
өздігінен ретгелуі қамтамасыз етіледі.
Өздігінен реттелу негізінде ағзалардың қүрылым түрақтылығы
— гомеостаз бірқалыпты сақталынады.
3) Көбею — агзалардың негізгі белгілерінің бірі, онсыз тіршілік
болмайды, үрпақтар жалғаспайды.
4) Түқым қуалаушылық пен өзгергіштік — тірі ағзалардың
басты белгілері болып саналады.
5) Даму — тіршіліктің ең негізгі белгісі. Ол жеке даму (онтоге¬
нез) жэне тарихи даму (филогенез) болып болінеді.
6) Дискреттік жэне біртүтастық.
10
'Гіршілік бір жағынан біртұтас, ал, екінші жағынан дискретті
(бөлшектенген) болып келеді. Тірі дүние біртұтас -— бір агзаның
тіршілігі екінші ағза тіршілігімен тығыз байланысты, мысалы:
өсімдіктер, өсімдікқоректі жануарлар — жыртқыштар т.с.с.
Сондай-ақ, тіршілікжеке-жеке, бір-бірінен дербес бөлшектерден
(элементтерден) тұрады, мысалы: өсімдіктер, жануарлар дүниесі
— түрлерден; түрлер — популяциялардан, жеке ағзалардан; ағза
— мүшелерден, үлпалардан, жасушалардан түрады.
Тіршіліктіңдискреттілігі туралы үғым оны бірнеше қүрылымдық
деңгейлерге бөлуге мүмкіндік береді. Қазіргі кезде тіршіліктің 7
ірі қүрылым деңгейлері белгілі:
1) тіршіліктіц молекулалык, цүрылым деңгейі — ДНҚ, РНҚ,
ақуыздар (барлық тірі ағзаларда біркелкі болып келеді);
2) тіриііліктің жасушалъщ цұрылъш деңгейі (барлық тірі
ағзаларда біркелкі болып келеді);
3) тіриііліктіц үлпалыц ңурылым деңгейі жануарларда 5,
өсімдіктерде — 6 түрлі үлпалар кездеседі;
4) тіршіліктің агзальщ (онтогенездік) цүрылым децгейі — көп
түрлі болып келеді;
5) тіршіліктің популяг/ияльщ — түрлік қурылым деңгейі —
көп түрлі болып келеді; жануарлардың 1,5 миллионная астам,
өсімдіктердің 500 мыңнан астам түрлері белгілі;
6) тіршіліктің биогеоуеноздық ңурылым децгеііі — биоце¬
ноз дегеніміз бір-бірімен жэне сыргкы ортамен үнемі карым-
қатынаста болатын тірі ағзалардың түрақты, тарихи қалыптасқан
қауымдастығы;
7) тіршіліктің биосфералъщ цүрылым децгейі.
1. ЖАСУША БИОЛОГИЯСЫ
1.1. Жасушаның курылысы
Жасушаның ашылуы оптикалық құрал — микроскоптың
қүрастырылуымен тығыз байланысты. Оптикалық әйнектер тура¬
лы деректер өте ерте кезден-ақ калыптаса бастаған. XV ғасырдың
басында оптикалык эйнектерді адамдар көзәйнек ретінде
пайдаланған. Екі линзадан тұратын жэне ұсак денелерді үлкейтіп
көрсететін ең қарапайым оптикалық кұралды XVI ғасырдың
аяғында ағайынды Янсендер құрастырган, бірақ ол арқылы жасу-
шаны көруге мүмкіндік болмаган.
Жасушаны көруге мүмкіндік берген алғашқы микроскопты
1665 жылы физик Р. Гук күрастырып, сол микроскоп аркылы жа¬
сушаны ашқан. 1671 жылы М. Мальпиги, Н. Грю жасушаларды
зерттеп, олардың ен негізгі кұрамдық бөлігі — кабықшасы деп
болжамдаган.
XVIII—XIX гасырларда микроскоптың күрделіленуі,
микроскопиялық зерттеулер әдісінің жетілуі нәтижесінде бірте-
бірте жасушаның тірі заттары ашылған.
1831 жылы ағылшын ғалымы Р. Броун жасушаның түйіршік-
тенген қүрылымын ашып, оны ядро (nucleus) деп атаған; 1841
жылы чех галымы Ян Пуркинье жасушаның қоймалжың заты —
тірі загын ашып, оны протоплазма деп атаган. Осылайша XIX
ғасырда ғалымдардың жасуша туралы пікірі бірте-бірте өзгеріп,
оның негізгі заты қабықшасы емес, тірі загпы — деген үғым
қалыптасқан.
1838—1839 жылдары неміс ғалымдары Т. Шванн жэне М. Шлей-
ден еңбектерінде өсімдіктер мен жануарлар жасушаларының 200
жылға жуық созылган зерттеулері қорытындыланып, жасуша те-
ориясы қалыптасқан. Сол сияқты жасуша теориясының эрі қарай
дамуына неміс дәрігері Р. Вирхов та (1858) өз үлесін қосқан.
12
Қазіргі кезде жасуша теориясының мынадай негізгі кағидалары
белгілі:
1) жасуша тіршіліктің ең ұсақ құрылым бірлігі болып та-
былады, себебі барлық тірі ағзалар (өсімдіктер, жануарлар,
саңырауқұлақтар, бөлшектенушілер) жасушалардан түрады;
2) өсімдіктер мен жануарлардың жасушаларының қүрылысы,
жалпы алғанда бір-біріне ұқсас болады;
3) жасуша тек жасушадан, оның бөлінуі нәтижесінде пайда бо¬
лады (Р. Вирхов). «Omni cellula acellula»;
4) жасуша — ашық биологиялық жүйе, ол арқылы үнемі
заттардың, энергияның және ақпараттар ағыны өтіп отырады.
Қазіргі кездегі түжырым бойынша тіршіліктің 2 формасы
белгілі:
1) Тіріиіліктіц жасушасыз формасы — оған вирустар жа-
тады. Вирустар — өте кішкентай, тіпті жай микроскоп арқылы
көрінбейтін, денелер. Олар нуклеин кышқылдарынан және
ақуыздан түрады. Олардың тіршілігі тек жасушаға енгеннен кейін
гана байқалды, ал, өз беттерінше оларда тіршілік күбылыстары
байқалмайды. Вирустарды 1892 жылы орыс ғалымы Д. И. Иванов¬
ский ашкан.
2) Тіршіліктің жасуишлы формасы. Оның 2 түрі белгілі, а)
прокариотты жасушалар (бактериялар, көкжасыл балдырлар) ци-
топлазмасы қос қабатты липидтік мембранамен қоршалған, ядро-
сы болмайды, түқым қуалайтын материал сақина тэрізді ДНҚ мо-
лекуласынан тұрады; рибосомадан баска органоидтары болмай¬
ды, мөлшері жагынан өте үсақ болып келеді 0,1—0,5 мкм, митоз
кездеспейді.
3) Эукариотпіы жасуиіалар — цитоплазмасы қос қабатты
липидтік мембранамен қоршалған, ядросы болады; тұкым
куалайтын материалы хромосомаларда орналасады, барлық орга¬
ноидтары болады; жасуша мөлшері біршама ірі болып келеді —
15—65 мкм., митоз жолымен бөлінеді.
Эукариоттарды бір жасушалы агзалар жасушасы жэне көп
жасушалы агзалар жасушалары деп жіктейді. Ал, соңғыларын
өсімдіктер жэне жануарлар жасушалары деп бөледі (1 -сурет).
Бір жасушалы ағзалардың жасушасы (қарапайымдылар) —
өздері бір жасуша бола түрып, түтас ағзаға тэн қызметтер атқарады:
13
қозғалу, тітіркену, көбею, зэр шығару, ас қорыту жэне т.б. Ал,
көпжасушалы ағзалардың жасушалары белгілі бір қызмет атқаруға
маманданады, оларда эр түрлі ақуыздар синтезделінеді, мысалы:
эпителий ұлпасының жасушаларында -— меланин, бұлшық ет жа-
сушаларында — миозин т.б.
Өсімдіктер жасушасының жануарлар жасушасынан ерекшелігі
мынадай:
1-сурет. Жасуша түрлері
а) прокариоттар; б) эукариоттык өсімдік жасушасы;
в) эукариотгык жануар жасушасы.
1) жасуша сыртын қалың целлюлоза қабығы каптап түрады;
2) цитоплазмада пластидтер (хлоропласттар, хромопласттар,
лейкопласттар) кездеседі;
3)вакуолялары болады.
Қазіргі деректер бойынша жасушаның негізгі заты болып прото¬
пласт саналады. Протопласт цитоплазмаға жэне ядроға жіктеледі.
Ол сыртқы ортадан шеткі мембрана — плазмолемма арқылы шек-
телген. Цитоплазма өз кезегінде гиалоплазмаға (цитоплазманың
негізгі заты — матриксы) жэне органеллаларға жіктелген.
Гиалоплазма — 2 мембранамен (плазмолемма, тонопласт) шек-
телген қоймалжыңсүйықтық. Ол орган икал ы қ ж эн е бейорганикалъщ
заттардан түрады. Оның 80—90 пайызын су кұрайды. Органикалық
затгардың ішінен негізгілері — ақуыз, нуклеин кышқылдары, май-
лар, көмірсулар, АТФ т.б.
а
б
в
14
2-сурет. Жасушаның
мембраналык жүйесі
1 -ядро, 2-кедір-бұдыр
эндоплазмалык тор,
3-цитоплазмалык
көпіршік, 4-Гольджи
кешені, 5-гранула,
6-митохондрия,
7-лизосома
Гиалоплазмада органеллалар бы-
тыраңқы орналасады. Органелла¬
лар — жасушада белгілі бір қызмет
атқарып, цитоплазмада тұрақты түрде
кездесетін құрылымдар. Оларга —
митохондриялар, Гольджи комплек-
сі, эндоплазмалык тор, пластидтер,
рибосомалар, лизосомалар, жасуша
орталыгы, микроденешіктер, микро-
түтікшелер т.б. жатады (2-сурет).
Эндоплазмалықторды 1945 жылы
Портер ашқан. Ол өте ұсак, тек кана
электрондық мироскоп арқылы көруге
болатын, қос қабат мембранамен шек-
телген жэне тарамданып гиалоплаз-
маны өне бойына торлап тесіп өтіп
орналасқан микроарнашықтар мен
микроқуыстар жүйесі болып табыла-
ды. Оның 2 түрі белгілі: гранулалы не-
месе кедір-бүдырлы жэне агранулалы
немесе тегіс эндоплазмалык тор. Гра¬
нулалы (кедір-бұдыр) эндоплазмалык
тордың мембранасына рибосомалар
бекінген, ал, тегіс эндоплазмалык тор-
да рибосомалар болмайды.
Цитоплазмада, әдетте гранулалы тор
агранулалы торга Караганда элдеқайда жақсы жетілген. Агранула¬
лы тор кейбір ерекше қызмет атқаратын, ягни майлы заттарды көп
синтездейтін жасушаларда гана жақсы жетілген болады. Гранулалы
тордың қызметі — акуыз синтездеу, жасуша мембраналарын пайда
ету орталыгы болып саналады. Сол сияқты, ол вакуоля, лизосома,
микроденешіктерді де пайда ете алады. Эндоплазмалык арналар
аркылы макромолекулалар, иондар тасымалданады. Агранулалы
тор липидтерді жэне көмірсуларды синтездеуге қатынасады.
Гольджи кешенін 1878 жылы итальян галымы Гольджи жа¬
нуарлар жасушасынан ашкан жэне соңгы кездерге дейін ол тек
жануарлар жасушаларына гана тэн деп келінген. Бірак, кейінірек
15
ол өсімдіктер жасушаларынан да табылған. Сондықтан Гольджи
кешені барлық эукариотты жасушаларға тэн органелла болып
саналады. Гольджи кешені өте жүқа, жалпақ, бірінің үстіне бірі
орналасқан 5—20 қалташықтардан—диктиосомалардан қүралған.
Әрбір қалташықтың диаметрі 1 мкм, ал қалындығы не бары 20—
25 нм болып келеді. Қалташықтардың жиектері тесіліп біртебірте
торға айналған. Гольджи кешенінің кызметі — полисахаридтерді
синтездеу, жинактау жэне тасымалдау болып саналады.
Митохондриялар — жасушаның міндетті органеллаларыныц
бірі. Оның пішіні түрліше болып келеді: таяқша тәрізді, дөңгелек,
сопақша т.с.с, ал мөлшері 0,5—7 мкм тең. Митохондриялар кос
қабат мембранамен шектелген. Сыртқы мембранасы тегіс, түйык.
ал ішкі мембранасы митохондрияның ішіне қарай қатпарлар
пайда етеді. Оларды кристтер дегі атайды. Кристтер арасын-
да митохондрияның негізгі заты — матриксі орналаскан. Онда
ДНҚ, рибосомалар, ақуыздар т.б. кездеседі. Митохондриялар
органикалық заттарды ыдырату, АТФ синтездеу қызметтерін
атқарады.
Рибосомаларды 1955 жылы Палладе ашқан. Ол екі бөлшектен
(кіші бөлшегі, үлкен бөлшегі) түрады. Олар өте үсак, тек
электрондық микроскоп арқылы көруге болатын — органеллалар.
Оның мөлшері не бары 15—35 нм болады. Рибосомалар р-РНҚ-
дан жэне акуыздан тұрады, оның негізгі қызметі ақуызды синтез¬
деу болып саналады.
Лизосомалар — диаметрі 2 мкм, эр түрлі ферменттерден
түратын көпіршіктер. Олар органикалык заттарды гидролиздеу-
ыдырату процесіне қатынасады, ягни жасушаішілік ас корыту
қызметін атқарады.
Жасуша орталығы — жануарлар жасушаларына тэн орга¬
нелла. Ол 2 центриолядан тұрады. Әрбір центриоля диаметрі 150
нм, ұзындыгы 300—500 нм болып келетін куыс цилиндр. Оның
қабырғасы үш-үштен 9 топка топтасқан 27 микротүтікшелерден
қүрылған. Жасуша орталығының қызметі митоздың қалыпты
жүруін қамтамасыз ету, ягни, анафаза кезінде хроматидалардың
полюстерге ажырауын қамтамасыз ету болып табылады.
Микротүтікшелер — түрліше болып келетін үзын түтіктер,
оның диаметрі 24 нм-ге тең. Олар тірек кызметін атқарады.
16
Пластидтер тек өсімдік жаушаларына тэн органеллалар.
Олардың 3 түрі белгілі: жасыл пластидтер — хлоропласттар (Ком-
паретти. 1791) сары, кызыл пластидтер — хромопласттар (Берце¬
лиус 1837), түссіз пластидтер — лейкопласттар (Крюгер, 1854).
Цитологияның соңғы кездердегі ең маңызды жетістіктеріне мы-
наларды жатқызуга болады:
1. Цитоплазманың мембраналық құрылыс принциптерін тұжы-
рымдау;
2. Жасушаның ашық биологиялық жүйе екендігін түжырымдау,
ягни заттар, энергия жэне ақпарат ағындары туралы түжырымның
калыптасуы.
Цитоплазма және оның органеллалары биологиялық мембрана-
лардан түрады, оның қызметі, қасиеттері сол мембраналарға бай¬
ланысты болады. Шынында да, цитоплазманың құргақ затының 90
пайызын биологиялық мембраналар қүрайды; гиалоплазма 2 мем-
бранамен (плазмолемма, тонопласт) шектелген; органеллалардың
кобісі мембраналардан түрады.
Биологиялық мембрана өте жүқа, қалындығы не бары 5—10 нм
болып келетін қабықша. 1972 С. Сингер мен Г. Никольсон ұсынган
биомембрананың сүйықтық — мозаикалык моделіне сэйкес, ол 2
биополимерден түрады: ақуыз жэне липидтер (3-сурет).
2
3-сурет. Биологиялык мембрананын күрылысы
1-фосфолипид кабаттары, 2-акуыз молекулалары
Липидтер молекуласы биологиялык мембраналардың қаңкасын
күрайды, ол екі кабат сүйық фаза күйінде болады, ал, оның бетінде
17
170-2
немесе оған еніп, кейде оны түгел тесіп өтіп ақуыз молекулалары
орналасқан. Биологиялық мембраналар жартылай өткізгіштік (тан-
дамалы өткізгіштік) касиетке ие, оньң ұштары үнемі тұйықталган.
Биологиялық мембраналар арқасында жасушада көптеген дербес,
тұйық қуыстар (органеллалар) түзіледі. Мембраналар арқасында
осы қуыстардатекөздеріне ғанатэн химиялық қүрамы калыптасып,
бір мезгілде түрліше химиялық реакциялардың жүруіне мүмкіндік
туады.
Жасуша ашық биологиялық жүйе болып саналады. Ол арқылы
үнемі заттар, энергия жэне ақпараттар ағыны өтіп отырады.
Заттар ағыны — жасуша тіршілігі үшін өте қажет қүбылыс.
Жасушаға үнемі заттар еніп, олар зат алмасу процесінде өзгеріп
(ыдырап, синтезделіп), сыртқа шығарылып отырады. Бүл про¬
цесс үздіксіз жүреді, себебі заттар агыны тоқталса, жасуша да өз
тіршілігін тоқтатады. Жасушаға түйіршік тэрізді заттар — фаго¬
цитоз, ал сүйық заттар — пиноцитоз арқылы енеді. Пиноцитоз
осмос қүбылысы нәтижесінде жүреді. Оған мысал ретіңде жа¬
сушада байқалатын плазмолиз — деплазмолиз қүбылыстарын
келтіруге болады. Заттардың мембранадан белсенді тасымалдануы
ферменттердің қатысуы жэне энергия жүмсалуы арқылы жүреді.
Энергия агыны — жасуша тіршілігі үшін энергия кажет. Ол
энергия органикалық заттардың ыдырауы нәтижесінде бөлініп
шығадыдаАТФмолекуласындажинақталады. АТФ-тажинақталған
энергия оның эрі қарай АДФ, АМФ-қа ыдырауы арқылы болініп әр
түрлі тіршілік әрекеттеріне пайдаланылады. Энергия синтезі жа¬
сушада анаэробтық гликолиз, фотосинтез, хемосинтез, аэробтық
гликолиз реакциялары нәтижесінде іске асады. Энергияның
синтезделуі өсімдіктер жасушаларында пластидтерде, ал, жануар-
ларда — митохондрияларда жүреді.
Ақпараттар ағыны — жасуша тіршілігі туралы ақпарат
ядрода, хромосомаларда, ДНҚ молекуласында биологиялык
(генетикалық) код күйінде жазылган. Міне осы акпараг ДНҚ-
ДНҚ-ға, ДНҚ-РНҚ-га, РНҚ-дан ақуызга беріліп отырады. Арнайы
зертхана жағдайларында ревертаза ферментінің көмегімен РНҚ-
нан-ДНҚ-ны синтездеуге мүмкіндік пайда болды. Ал, ақуыздар
(ферменттер) жасушаның барлық тіршілік процестерін басқарып,
реттеп отырады. Ақпараттар ағынысыз жасуша тіршілігінің болуы
мүмкін емес.
18
1.2. Жасуша тіршілігіндегі мерзімдік озгерістер
(Жасушаның бөлінуі)
Ядро — эукариотты жасушалардың ең негізгі кұрамдык
бөлімдерініңбірі.Оны 1931 жылыағылшынғалымыГ.Броунашқан.
Ол цитоплазмаға Караганда тығыздау болып келеді. Оның пішіні
жасуша пішініне сәйкес болады, мөлшері 10—25 мкм шамасында:
эрбір жасушада 1, кейде 2 не одан да көп ядролар кездеседі. Ядро-
сыз жасуша болмайды, тек адам эритроциттері мен өсімдіктердің
сүзгілі түтіктерінің жасушаларында ядро кездеспейді.
Ядроның қүрылысы. Ядро екі мембранадан түратын қабығы
аркылы цитоплазмамен шектелген. Оның ішінде ядро шырыны,
ядрошыц жэне хроматин орналасады. Ядроның қүрылымы жасу¬
ша циклінің эр түрлі кезеқцерінде өзгеріп отырады. Осыған орай
оның интерфаза кезіндегі ядро (яғни ядроның кызметтік күйі) жоне
митоздық ядро күйі белгілі.
Ядроның интерфазалық күйінің негізгі бөлігі —хроматин. Ол
өте жіңішке, үзын, шумақталған хроматин жіпшесінен түрады. Хро¬
матин — хромосоманың белсенді — активті күйі. Хроматин ДНҚ
және гистондық белоктардан түрады. Ол жіңішке, диаметрі 10—13
нм болатын жіпше. Оның каңқасын 4 түрлі гистондык ақуыздардың
(Н2А, Н2В, НЗ, Н4)молекулаларынан қүрылған дөңгелек октомер
денешіктер — нуклеосомалар қүрайды. Нуклеосомаларға ДНҚ
молекуласы тыгыз ширатылып орналасқан. ДНҚ молекуласының
нуклеосомалық оралымдарын бір-біріне гистондық ақуыздардың
бесінші түрі — Н1 жалгайды. Сонымен, гистондық ақуыздардың
5 түрі белгілі. Олар қүрылымдық, ретгеуші қызметтер атқарады.
Хроматиннің ең негізгі бөлігі — ДНҚ молекуласы. Ол түқым
қуалаушылықтың материалы болып есептелінеді, себебі оның мо-
лекуласында түқым қуалаушылық туралы ақпарат «жазылған».
Эукариоттардың ДНҚ молекуласы 3 түрлі фракциядан түрады:
1) қайталаіібайгын нуклеотидтер тізбегі — ол ДНҚ бойын-
да бір дана күйінде кездеседі. Адамдардың гаплоидтық геномы
3—109 нуклеотидтер катарынан түратын болса, оның 70 пайызы,
яғни 2—109 нуклеотидтер катары қайталанбайтын нуклеотидтер
тізбегі болып саналады. Бүл жерде барлық қүрылымдық гендер
орналасқан;
19
2) орташа қайталанатын нуклеотидтер тізбектері — олар
ДНҚ молекуласының ұзына бойында 100-ден 100000 ретке дейін
қайталануы мүмкін. Олардың үлесіне геномның 22—23 пайыз
мөлшері жатады. Бүл жерде құрылымдық гендер кездеспейді,
бірақ рРНҚ, тРНҚ, гистондық белоктардың гендері кездеседі;
3) өте жиі кайталанатын нуклеотидтер тізбектері — олар
100000-нан 1000000 ретке дейін қайталанатын қысқа нуклео¬
тидтер тізбектері. Олардың үлесіне геномның 8 пайызы жатады.
Бүл бөлшектерде ешқандай гендер болмайды, олар гендердің
интрондық бөліктері болып табылады.
ДНҚ молекуласының қасиеттерінің бірі — оның тығыз шира-
тылуы. ДНҚ молекуласы нуклеосома денешіктеріне ширатылып
нуклеогистондық микрофибрилла соленоид түзеді, ал ол эрі қарай
ширатылып, жуандап митоздық хромосомаға айналады. ДНҚ
молекуласының осылайша ширатылуының нәтижесінде түқым
қуалаушылық материалы тыгыздалып, шагын денелерде (хромо-
сомаларда) сақталуына мүмкіншілік туады. Мысалы, адамның
дене жасушасының 1 ядросында 6 пг ДНҚ кездеседі десек, оның
жалпы үзындығы 2м тең. Адамның барлық жасушаларындағы
үзындығы не бары 150 мкм болатын хромосомаларыңдағы ДНҚ
молекулаларын бір-біріне тізіп жалғайтын болсақ, оның үзын-
дығы 2,5х 1010 км жетеді. Бүл Жер мен Күннің ара қашықтығынан
100 есе артық.
Митоздық (бөлінуші) ядроның негізгі бөлігі — митоздъщ хро-
мосомалар болып есептелінеді. Митоздық хромосомалар хрома¬
тин жіпшелерінің ширатылып, жуандап қысқаруы нәтижесінде
түзіледі. Хромосома профазада таяқша, ал метофазада X пішіндес
болып келеді. Хромосома бірінші реттік тартылыс (центромера)
аркылы жалғанған екі иіннен түрады. Хромосома иінінің үшьн
теломвра деп атайды. Хромосомалар үзына бойына 2 тепе-тең
бөлікке бөлінген, оларды хроматидачар деп атайды. Әрбір хро-
матида 2 тығыз ширатылған жіпшелерден — хромонемалардан
түрады.
Хромосомалардың мөлшеріне жэне центромерасының орна-
ласуына байланысты оларды — метоуентрлі, субметацентрлі,
акроцептрлі және телоцентрлі деп жіктейді.
20
4-сурет. Хромосомалар формасы
1-центромера,
2-серік,
I телоцентрлік,
II акроцентрлік,
3-кыска иін,
4-үлкен иін,
III субметацентрлік,
IV метацентрлік,
5-хроматидалар
Метацентрлік хромосоманың центромерасы дэл ортасында
орналасқан иіндері тең болады, субметацентрлік — центромерасы
бір шетіне сәл ғана ығысып орналасады, бір иіні ұзын, екінші иіні
кыскалау, акроцентрлік центромерасы бір ұшына жақын орналаса¬
ды, бір иіні өте ұзын, екіншісі өте қысқа, телоцентрлік центромера¬
сы ұшында орналасады, хромосоманыц бір ғана иіні болады. Кей¬
де хромосомада екінші реттік тартылыс пайда болады, иіндерінде
қосымша бөлік — серік пайда болады. Мұңдай хромосомаларды
серікті хромосомалар деп атайды.
Хромосома бойында ДНҚ молекуласының ширатылу дэрежесі
тұрліше болып келеді. Осыған орай хромосоманың эухрома-
тин, гетерохроматин учаскелерін ажыратады. Эухроматин —
хромосоманың митоз кезінде ширатылып тығыздалатын, спираль-
данатын, ал митоздан кейін ширатылуы ашылатын, деспиральда-
натын бөлігі. Бүл хромосоманыц ең белсенді — активті транскрип-
цияланатын, структуралық гендер кездесетін бөлігі; гетерохрома¬
тин — хромосоманыц митоз кезінде де, митоздан кейін де тыгыз
ширатылған — спиральданган, конденсацияланған, ашылмайтын
учаскесі. Онда структуралык гендер болмайды.
Ядро өте маңызды рөл атқарады. Ол жасуша тіршілігінің барлык
құбылыстарын басқарып, бағдарлап отырады.
ООО
в
Ш
Ж
°о°
ш
tl
щ
5-сурет. Митоздык цикл
М-митоз Gj-пресинтездік кезең, S-синтездік кезең,
02-постсинтездік кезең
6-сурет. Митоз жобасы
А-профаза, Б-метафаза, В-анафаза, Г-телофаза
Онда тұқым куалаушылық материалы — ДНҚ шоғырланған, ол
жасушаның өсуін, бөлінуін (көбеюін) басқарып отырады.
Жасушаның тіршілік циклі деп оның пайда болуы,
кұрылысының күрделіленуі, атқаратын қызметіне икемделуі,
бөлінуі немесе тіршілігін жою (өліп қалу) кезеңдері жиынтыгын
айтамыз. Яғни жасушаның циклі оның пайда болуынан бөлінуіне
немесе өлуіне дейінгі уақытты қамтиды, мысалы, ұдайы болінетін
жасушалардың тіршілік жиынтығын митоздьщ цикл (5-сурет) деп
атауға болады. Сонымен, митоздық цикл дегеніміз жасушаның
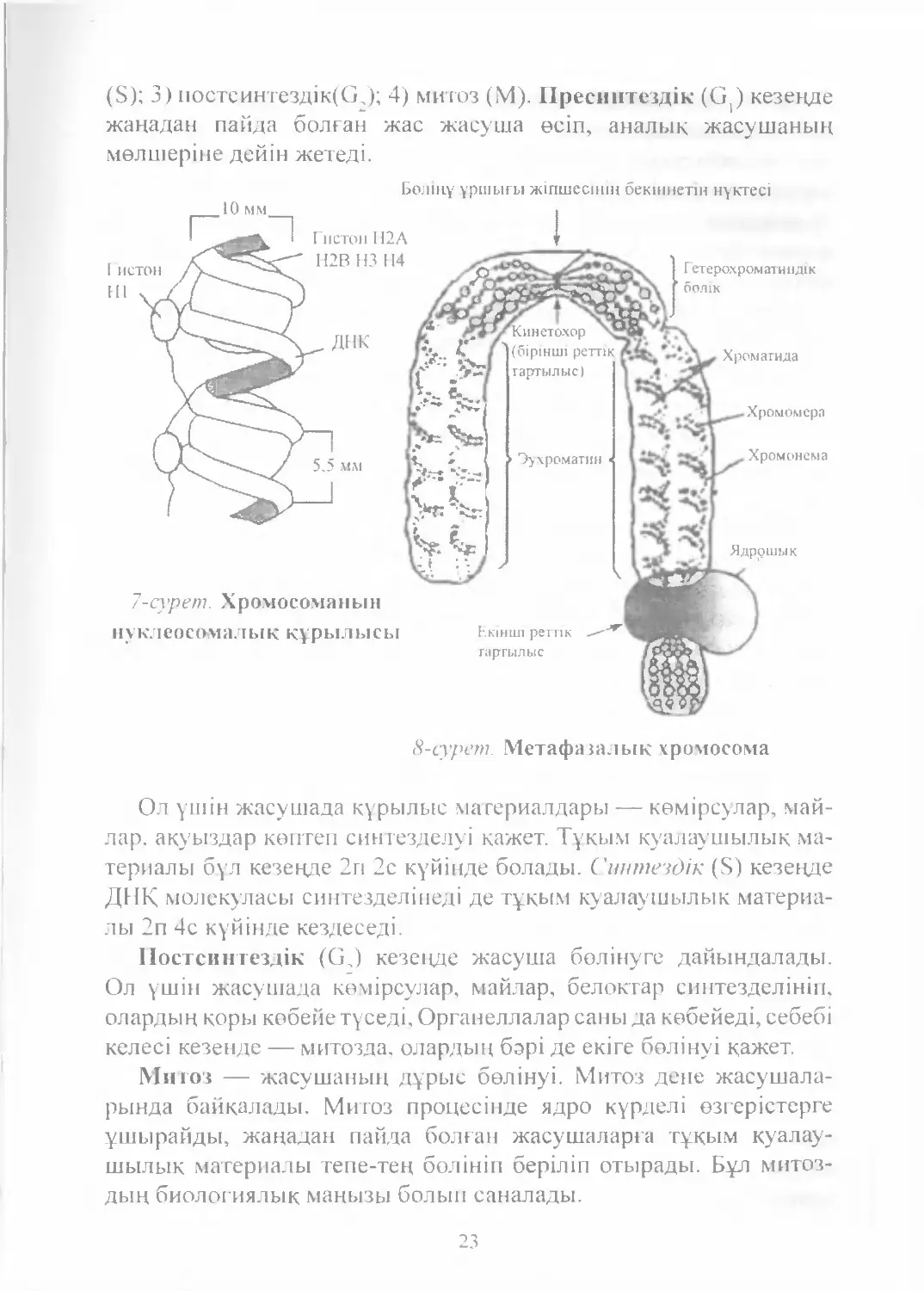
митоздық жолмен екі болінуі арасындагы байқалатын кұбылыстар
жиынтығы. Ол 4 кезеңге болінеді: 1) пресинтездік (G,); 2) синтездік
22
(S); 3) постсинтездік(С2); 4) митоз (М). Пресинтездік (G^ кезенде
жаңадан пайда болған жас жасуша өсіп, аналық жасушаның
мөлшеріне дейін жетеді.
Велінү ұршыгы жіпшесінің бекіннетін нүктесі
Гетерохроматиндік
болік
Хроматида
Хромомера
Хромонема
Ядрошык
7-сурет. Хромосоманың
нуклеосомалық күрылысы
8-сурет. Метафазалык хромосома
Ол үшін жасушада кұрылыс материалдары — көмірсулар, май-
лар, ақуыздар көптеп синтезделуі кажет. Тұқым куалаушылық ма¬
териалы бұл кезеңде 2п 2с күйінде болады. Синтездік (S) кезеңде
ДНҚ молекуласы синтезделінеді де түқым куалаушылык материа¬
лы 2п 4с күйінде кездеседі.
Постсинтездік (GJ кезеңде жасуша бөлінуге дайындалады.
Ол үшін жасушада көмірсулар, майлар, белоктар синтезделініп,
олардың қоры көбейе түседі, Органеллалар саны да көбейеді, себебі
келесі кезенде — митозда, олардың бэрі де екіге бөлінуі қажет.
Митоз — жасушаның дүрыс бөлінуі. Митоз дене жасушала-
рында байқалады. Митоз процесінде ядро күрделі өзгерістерге
үшырайды, жаңадан пайда болған жасушаларга түқым куалау¬
шылык материалы тепе-тең бөлініп беріліп отырады. Бүл митоз-
дың биологиялык маңызы болып саналады.
23
Митоздың негізгі себептері: 1) ядро — цитоплазмалық
ара қатынасының өзгеруі (V—Vg ден V69—‘/g9 ге дейін); 2)
«митогенетикалық сэулелер» — бөлінуші жасушалар көршілес
жатқан жасушалардың митоздық жолмен бөлінуіне эсер етеді;
3) «жарақат гормондарының» әсері — жарақаттанған жасушалар
жарақаттанбаған жасушалардың бөлінуіне ықпал ететін ерекше
заттар бөліп шығарады.
Біртүтас митоз кұбылысы 4 фазаға бөлінеді: профаза, .метафа¬
за, анафаза, телофаза. Профазада ядро көлемі үлғайып, хрома¬
тин жіпшелері тығыз ширатылып, жуандап, қысқарып митоздық
хромосомаларға айналады. Профазаның аяғында бөлінуші
жасушаның екі полюсіне центросоманың (жасуша орталығы) цен-
триолялары ажырап, арасында ахроматин жіпшелері пайда бола¬
ды.
Метафазада хромосомалар өздерінің центромрралары арқы-
лы ахроматин жіпшелеріне бекініп, бөлінуші жасушаның эк-
ваторына реттеліп орналасады. Бұл кезеңде әрбір хромосома
хроматидаларының теламерлік бөліктері ажырап, тек центроме¬
расы аркылы байланысып түрады. Бұл кезде хромосомалардың
пішіні анық көрінеді. Сондықтан, кариотипті анықтау және хро-
мосомаларды жіктеу осы метафазада (метафазалық пластинкада)
жүргізіледі.
Анафазада хромосомалардың хроматидалары бір-бірінен то-
лык ажырасады да эр түрлі полюстерге қарай тартыла бастайды.
Телофазада бөлінуші жасушаның полюстеріне топтасқан хро¬
мосомалар жіңішкеріп, үзарып хроматин жіпшелеріне айналады,
ядрошық, ядро кабықшасы пайда болып, 2 жаңа ядро түзіледі.
Ядро 2-ге бөлінгеннен кейін (кариокинез), цитоплазма да 2-ге
бөлінеді (цитокинез). Сөйгіп, митоз жэне цитокинез негізінде бір
жасушадан жаца 2 жасуша пайда болады.
Жасушалар митоздан басқа жолдармен де бөлінуі мүмкін, мы¬
салы: амитоз — жасушаныц тікелей (бүрыс) бөлінуі жэне мейоз
— жыныс жасушаларыныц бөлінуі.
Амитоз аркылы негізінен прокариоттар жэне регенерациялану-
шы эукариотты жасушалар бөлінеді. Бүл кезде көзге көрінетін хро¬
мосомалар жэне бөліну ұршығы түзілмейді. Амитоз ядроныц жэне
цитоплазманыц созылып екіге бөлінуі арқылы жүреді.
24
Митоздың ерекше түрлеріне эндомитоз жэне политенияларды
да жатқызамыз. Эндомитозда ядро бөлінбей, тек хромосомалар екі
еселеніп, полиплоидты жасушалар түзіледі.
Политенияда ДНҚ молекуласы бірнеше рет еселеніп, бірақ бір-
бірінен ажыраспайды, политенді (көп жіпшелі) хромосомалар пай¬
да болады (мысалы: жеміс шыбынында).
Хромосомалардың ережелері:
1. Хромосома санының түрақты болуы;
2. Хромосомалардың жүпты (гомологті) болуы;
3. Хромосомалардың жеке даралық ерекшелігі;
4. Хромосомалардың үздіксіздігі.
Хромосомалар жасушалардағы түқым қуалаушылық материа¬
лы болғандықтан, кез-келген биологиялык түрлерде олардың саны
түрақты болады. Хромосома жиынтығының диплойдтық саны (2п)
— дене жасушаларында, гаплоидтык саны (п) — жыныс жасуша-
ларында болады.
Кейбір ағзалардың хромосома жиынтығы (2п,п) төменде
келтірілген:
2п
п
1. Безгек плазмодиясы
2
1
2. Өзен шаяны
116
58
3. Гидра
32
16
4. Үй шыбыны
12
6
5. Бака
26
13
6. Кептер
80
40
7. Тышкан
40
20
8. Коян
44
22
9. Шимпанзе
48
24
10. Саналы адам
46
23
11. Жеміс шыбыны
8
4
Биологиялык түрлерде тек қана хромосома саны түрақты болып
қоймай, сол сияқты, олардың пішіні мен мөлшері де түрақты бола¬
ды. Биологиялык түрлердін хромосома жиынтыгының, пішінінің,
мөлшерінің кешенді сипаттамасын каршщцишд атаймыз. Адам
кариотипі 46 хромосомадан түрады, 'штелогаік >^П-нің
күрайды. Оның 22 жүбы екі жыныста да эірдёйводады, сондь).қт^ң;і
25 I КІтапхана
I ТіҒК£У N® JLSS2&,
оларды аутосомалар деп атайды, ал 23 жұп хромосомалары бір-
бірінен өзгеше болып келеді, оларды У жэне Xхромосома немесе
жыныс хромосомалары (гоносомалар) деп атайды. Кариотипті,
әдетте метафазалық препараттар дайындап зерттейді. Адам
кариотипін жіктеудің (классификациялаудың) 2 түрі белгілі: Ден¬
вер классификацпясы (1960) жэне Париж классификациясы (1971).
Денвер классификациисы бойынша хромосомаларды үлкенінен
кішкентайына қарай орналастырып, олардың идиограммасын
қүрастырады да әрқайсысын нөмірлейді, хромосома морфология-
сына қарай бірнеше топқа бөледі: мүнда негізгі көрсеткіш ретінде
центромералық индексті (ЦИ) пайдаланады. Центромералық ин¬
декс дегеніміз — хромосоманыц қысқа иіні ұзындығының бүкіл
хромосома ұзындыгына ара қатынасы (%) болып табылады:
А тобы: 1-2-3 хромосомалар; ең ірі метацентрлі хромосомалар,
(ЦИ=38-49);
Втобы:4-5;ірісубметацен-
трлі хромосомалар, (ЦИ=24-30);
С тобы; 6-12; орташа суб-
метацентрлі хромосомалар,
(ЦИ=27-35);
Д тобы:
акроцентрлі
(ЦИ=15);
Е тобы: 16-18; үсақ
субметацентрлі хромосома¬
лар, (ЦИ=26-40);
Ғ тобы: 19-20; ең ұсақ
метацентрлі хромосомалар,
(ЦИ=36-46);
G 21 -22;еңұсақакроцентр-
лі хромосомалар, (ЦИ=13-23);
X хромосома — орташа
субметацентрлі (С тобы) хро¬
мосома.
Ү хромосома — ең үсақ
акроцентрлі (G тобы) хромо¬
сомалар болып табылады.
13-15; орташа
хромосомалар.
9-сурет. Адам кариотипі (Париж
классификациясы бойынша)
26
Денвер жіктеуінің кемшілігіне бір топқа жататын хромосома-
ларды ажыратудың киындығы жатады.
Париж классификациясыО 971)хромосомалардыңтаңцамалы
боялуына байланысты жүргізіледі (9-сурет). Ол үшін хромосо-
маларды түрліше бояулармен бояйды (G, R, S). Сонда эр түрлі
(гомологтік емес) хромосомалар түрліше боялады, ал гомологтік
хромосомалар бірдей боялады, сондықтан хромосомалардың
гомологтік жүптарын табу жеңілдейді. Сонымен қатар, бүл класси¬
фикация бойынша хромосомалардағы нақтылы локустарды ажы-
ратып белгілеуге, хромосома картасын жасауға болады. Ол үшін
кейбір символдарды пайдаланады, мысалы: хромосоманыц қысқа
иінін р, үзын иінін q әрпімен белгілейді. Боялу интенсивтігіне
карай хромосоманыц эрбір иінін центромерадан теломераға қарай
аудандарға, ал аудандарды сегменттерге бөледі де, араб санда-
рымен белгілейді. Мысалы, Ір 22 хромосоманың қысқа иінініц 2
ауданындагы 2 сегмент дегенді білдіреді.
2. КӨБЕЮ БИОЛОГИЯСЫ
Tipi ағзалардың негізгі белгілерінің бірі — олардың көбеюге
кабілеттілігі. Осы қасиеттің арқасында миллиардтаған жылдар бой-
ына ұрпақтар жалғасып, Жер бетінде тіршілікбайқалыпкеледі, келе
де бермек. Ағзалардың көбею қасиеті тіршіліктің дискреттілігімен
тікелей байланысты. Мысалы, ағзалардың денелері көптеген жа-
сушалардан түрады, ал жеке жасушалардың тіршілік ұзақтығы
ағзалардың тіршілік ұзактыгымен салыстырғанда элдеқайда қысқа
болады, сондықтан да ағзалардың тіршілігі оның жасушаларының
үздіксіз көбеюі нәтижесінде ғана байқалуы мүмкін. Кез кел-
ген биологиялык түрлер жекелеген даралардан түрады, ал жеке
даралардың тіршілік үзақтыгы түрлердің тіршілік үзақтығына
Караганда әлдеқайда кысқа. Демек, түрлердің түрақты тіршілік етуі
оның дараларының үздіксіз көбеюінің нэтижесінде ғана байқалады
жэне т.с.с.
Ағзалардың көбею формалары түрліше болып келеді, ол жоба
күйінде төменде келтірілген.
Жыныссьп көбею
Жыныстык көбею
1. Бір жасушалы ағзалар:
а) жасушаның жай екіге бөлінуі
аркылы (прокариоттар, бір жасушалы
эукариоттар);
б) шизогония (бір жасушалы
эукариоттар — споралылар);
в) бүршіктену аркылы (ашыткы
саңыраукұлағы);
г) спора түзу аркылы (өсімдіктер);
2. Кеп жасушалы агзалар
(вегетативтік):
а) каламшалар, түйнектер аркылы;
б) бүршіктену аркылы;
в) сомалык эмбриогенез;
г) бөлшектену аркылы (жауынкұрты);
1. Агаметогамия: конъюгация
(инфузория).
2. Гаметогамия: копуляция
изогамия (хламидамонада);
анизогамия (карапайым-
дылар); оогамия.
28
Жыныссыз көбею жыныстык көбеюге қарағанда қарапайым
жэне ол эволюция кұбылысында алғаш пайда болған. Оның
негізгі ерекшелігі осы жолмен пайда болған үрпақтардыц аналық
ағзалардан ешбір айырмашылығы болмайды, себебі екеуінің
де тұқым қуалаушылық материалдары өзгеріссіз, бірдей. Ал
жыныстық көбею — күрделі процесс, себебі болашақ ұрпақтың
генетикалық материалы екі ата-ананың тұқым қуалаушылық
ақпаратгарының қосылуы нәтижесінде қалыптасады. Жыныстық
көбеюдің ең қарапайым түрі — конъюгация кейбір бір жасушалы
ағзаларға тэн. Конъюгация кезінде арнайы жыныс жасушаларын
(гаметаларды) пайда етпей, өздері өзара қосылады не генетикалық
ақпараттармен алмасады (қарапайымдылар, бір жасушалы бал-
дырлар), мысалы, кірпікшелі кебісше.
Копуляция. Tipi ағзалардыц басым копшілігініц жыныстық
көбеюіерекшежасушалар—жынысжасушалардыц(гаметалардыц)
косылып, зигота түзуі аркылы жүреді. Кейбір ағзалар мөлшері,
қозғалуы жағынан біркелкі болып келеді, мысалы хламидамонада,
оныц қүрылымы бірдей екі жасушасыныц қосылуын изогамия деп
атайды.
Енді бір агзалар қозғалуы жағынан эртүрлі гаметалар түзеді.
Олардыц бірі үсақ, жылдам қозгалады. Мысалы, аталық гамета,
ал екіншісі одан гөрі үлкендеу. баяу қозғалады, ол — аналық га¬
металар. Мүндай гаметалардыц өзара қосылуын анизоогамия деп
атайды, Оган мысал: безгек плазмодийлері.
Жыныстык процестіц ец күрделі түрі — оогамия. Бүнда ата-
аналар эр тұрлі гаметаларды пайда етеді. Бірі — үсақ, қозгалгыш
аталық гаметалар — сперматозоидтар пайда етсе, екіншісі өте
аз мөлшерде, тіпті тек бір дана күйінде үлкен, қозгалмайтын
аналық гамета — жүмыртқа жасушасын түзеді. Міне. осылардың
қосылуын оогамия (көбею) деп атайды.
Гаметалардыц түзілу процесін гаметогенез (сперматогенез, не
оогенез) деп атайды. Аталық гаметалар аталық бездерде, аналық
гаметалар — аналық бездерде пісіп жетіледі.
Сперматогенез 4 кезеңнен (көбею, өсу, пісіп жетілу, қалыптасу),
ал оогенез 3 кезецнен түрады (көбею, осу, пісіп жетілу).
Гаметаларды (сперматозоид жэне жүмыртқа жасушасы) пайда
ететін диплоидты жасушаларды сперматогониялар жэне ового-
пиялар деп атайды.
29
Гаметогенездің бірінші — көбею сатысътда жыныс бездерінде
сперматогониялар мен овогониялар митоз жолымен бөлініп.
ол ардың саны көбейеді. Ер адамдарда сперматогониялардың көбеюі
жыныстық жетілген кезден басталып қартайғанға дейін созыла-
ды, ал әйелдерде овогониялардың көбеюі олардың эмбриондық
дамуының алғашқы 2 айлығында басталып, жатырдағы дамудың
7-ші айларында овоциттердің көбісі миоздық профаза — І-де бо¬
лады.
10-сурет. Жыныс жасушалары
I сперматозоидтар:
1 -басы, 2-мойны, 3-ортаңгы бөлім, 4-кұйрық.
II аналык бездің көлденең кесіндісі:
1 -алғашкы фолликулалар, 2-есуші фолликулалар,
3-жетілген фолликула, 4-овуляция, 5-сары дене, 6-ак дене.
Ill жұмыртка жасушасы:
1-ядро, 2-цитоплазма, 3-кортикальдык кабат, 4-мембрана,
5-мөлдір аймак, 6-фолликула жасушалары.
30
Екіниіі өсу сатысында жаңадан түзілген жас жасушалар өсіп
бірінші реттік сперматоциттер мен бірінші реттік овоциттерге ай-
налады
Пісіп жетілу сатысында бірінші реттік сперматоциттер мен
бірінші реттік овоциттер мейоз жолымен екі рет бөлінеді. Бірінші
бөлінудің нәтижесінде екінші реттік сперматоциттер мен екінші
реттік овоцит жэне бір редукциялық денешік түзіледі, ал екінші
бөлінудің нәтижесінде әрбір сперматоциттен төрт сперматидтер
жэне эрбір екінші реттік овоциттен — бір жұмыртқа жасушасы
мен үш редукциялық (бағытгаушы) денешіктер пайда болады.
Сперматогенез — цалыптасу сатысымен аяқталады. Бүл
сатыда сперматидтер сперматозоидқа айналады. Бүл кезде
сперматидтің цитоплазмасы азаяды, хромосомалардың тығыз ши¬
ратылып инерттік күйге көшуіне байланысты ядро тыгыздалып
сперматозоидтың басы пайда болады. Эндоплазмалык тор, Гольд¬
жи комплексі сперматидтің бас бөлімінің алдыңғы полюсіне ығы-
сып акросомалық аппаратқа айналады, ал центросома қарама-қарсы
полюске орналасып сперматозоидтыц қүйрығын қалыптастырады,
ал оныц түбінде митохондриялар топтасып, сперматозоид мойнын
пайда етеді.
Гаметогенез процесініц ец негізгі мэнінің бірі — мейоз. Мейоз
— жасушаныц күрделі бөлінуі, оныц нәтижесінде бір диплоидтық
аналық жасушадан төрт гаплоидтық жыныс жасушалары пайда бо¬
лады. Мейоз — жасушаныц ДНҚ еселенуінсіз (қысқа интерфаза)
екі рет бөлінуі: оныц бірінші бөлінуін — редукциялъщ, екіншісін
— эквациялыц бөліну деп атайды. Мейоздыц эрбір бөлінуі митоз
сияқты терт фазадан (профаза, метафаза, анафаза, телофаза)түрады.
Мейоздыц негізгі корытындылары — гаплоидтық жасушалардыц
пайда болуы, түқым куалаушылық материалдарының рекомбина-
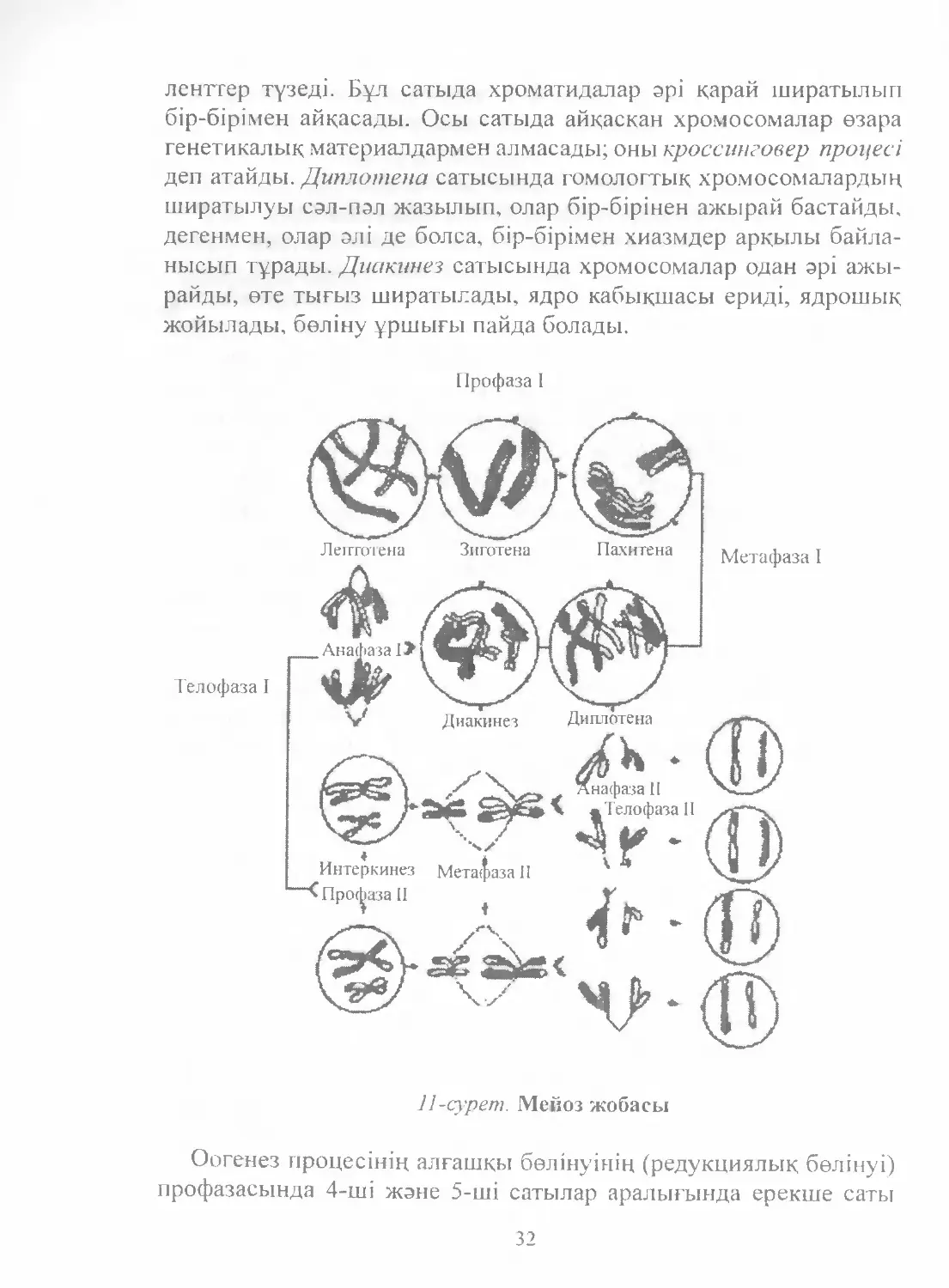
циялануы оның бірінші бөлінуінде жүзеге асады. Профаза-1 бес
сатыга жіктеледі: лептотена, зиготена, пахитена, диплотена жэне
диакинез. Лептотена сатысында диплоидтық хромосомалар ши-
ратыла бастайды, нашар тығыздалған жіпке үқсайды. Зиготе-
нада гомологтік хромосомалар эрі қарай ширатылып бір-біріне
жақындап қосылып, жүптар қүрайды (конъюгацияланады). Хро¬
мосома жүптарыныц саны гаплоидтық хромосома санына, яғни
адамдарда 23-ке тең болады. Пахитена сатысында гомологтік
хромосомалар толық қосылып төрт хроматидадан түратын бива-
31
лентгер түзеді. Бүл сатыда хроматидалар эрі қарай ширатылып
бір-бірімен айқасады. Осы сатыда айқасқан хромосомалар өзара
генетикалық материалдармен алмасады; оны кроссинговер процесі
деп атайды. Диплотена сатысында гомологтық хромосомалардың
ширатылуы сәл-пәл жазылып, олар бір-бірінен ажырай бастайды.
дегенмен, олар әлі де болса, бір-бірімен хиазмдер арқылы байла-
нысып тұрады. Диакинез сатысында хромосомалар одан әрі ажы-
райды, өте тығыз ширатылады, ядро кабықшасы ериді, ядрошық
жойылады, бөліну ұршығы пайда болады.
Профаза I
Тел оф аза I
^ «Телофаза II
ч/ 4 ¥ -
Интеркинез Мета|іаза II 4
Пролаза II
Метафаза I
11-сурет. Мейоз жобасы
Оогенез процесінің алғашқы бөлінуінің (редукциялық бөлінуі)
профазасында 4-ші жэне 5-ші сатылар аралығында ерекше саты
32
— диктиотена сатысы болады. Бұл саты эмбриогенезде басталып
қыздар 11—13 жасқа, яғни жыныстық жетілген жасқа, келгенге
дейін созылады. Ьүл кезде хромосомалар ерекше морфологиялық
формаға айналып эрі қарай бөлінуін уақытша тоқтатады. Ал, жы-
ныстықжетілу кезінде гипофиздіңлютеиндеушігормоныныңәсері-
нен ай сайын бір овоцит өзінің бөлінуін эрі қарай жалғастырады.
Метафаза 1-де биваленттер бөлінуші жасушаньщ экваторына
топтасады. Анафаза 1-де биваленттер екі хромосомаларға ажыра-
сып, олардың әрқайсысы карама-қарсы полюске қарай тартылады.
Телофаза І-де пайда болған ядролардағы хромосомалардың саны
екі есе азаяды (редукцияланады), жасуша цитокинез нәтижесінде
екіге бөлінеді. Қысқа интерфазадан (интеркинезден) кейін бірден
мейоздың екінші бөлінуі — эквациялық бөліну басталады. Ин-
теркинез кезінде екінші бөлінуге кажет ақуыздар жэне энергия
коры жиналады, бірақ ДНҚ еселенбейді. Екінші бөліну метафаза
ІІ-ден басталады. Бұл фазада митоздағыдай эрбір хромосома екі
хроматидаға ажырайды.
Анафаза П-де ажырасқан хроматидалар жасуша полюсіне қарай
тартылады. Телофаза І1-де хроматидалар деспиральданады. ядро
қабығы жэне ядрошык түзіледі, цитокинезден соц хроматидалар
жиынтығы гаплоидты 4 жасуша пайда болады.
Мейоздын биологиялык мацызы:
1) мейоздыц жэне ұрықтанудың нәтижесінде биологиялык
түрлердің хромосома санының тұрақтылығы сақталынады; 2)
мейоз нэтижесінде гаметаларда хромосомалардың еркін комби-
нациялану ыктималдығы кұшейе түседі, себебі әрбір гомологтік
хромосомалардың (әкесінен және шешесінен алынған) екі
гаметага ажырау ықтималдығы '/г-ге тең, ал барлық хромосома са-
нын алатын болсақ, олардың гаметаларда еркін комбинациялану
ықтималдығы 2 п дәрежесіне, ягни адамдарда 223 дәрежесіне тең.
Былайша айтқанда, бір гаметаға тек қана әкесінен алынган хро¬
мосомалар, ал екіншісіне шешесінен алынған хромосомалардың
топтасу мүмкіншілігі өте аз мөлшерде болады (1/2)22 =1/4 194304,
ягни 1/4000000-на тең; 3) мейоз процесінде кроссинговер салда-
рынан хромосомаларда генетикалык материалдар (гендер) ре-
комбинациаланады, түқым қуалаушылық материалдың ұрпақтар
жалгасында комбинативтік өзгергіштік байқалады.
33
170-3
Жыныстық көбеюдің негізгі мэні ұрықтану екені белгілі, бірақ
кейде жаңа ағза ұрықтанбаган жұмыртка жасушасынан да да-
муы мүмкін. Бұл құбылысты партеногенез деп атайды. Бұл жол-
мен пайда болған агзалар гаплоидты болады. Партеногенездің
екі түрі белгілі: андрогенез жэне гиногеиез. Андрогенезде жаңа
ағза сперматозоидтың ядросы негізінде дамиды, ягни спермато¬
зоид жұмыртка жасушасына енеді де оның ядросын ауыстырады,
ягни жұмыртқа жасушасының ядросы өліп жойылады; ал гиноге-
незде сперматозоидтар арқылы активтенген (бірақ қосылмаған)
жұмыртқа жасушасының ядросы жаңа агзаның дамуына алып
келеді.
3. ГЕНЕТИКА НЕГІЗДЕРІ
3.1. Туқым қуалаушылықтың негізгі зандылықтары
Генетика — тұқым қуалаушылық пен өзгергіштікті зерттейтін
гылым. Оның қалыптасуы 1900 жылдан басталады, ал негізін
қалаушысы, «атасы» — чех ғалымы Г. Мендель.
Тұкым қуалаушылық тірі ағзалардың негізгі қасиеттерінін
бірі — ол ұрпакган-үрпақка ата-аналарының белгілерінің,
қасиеттерінің үздіксіз беріліп отырылуын камтамасыз етеді. Гұқым
қуалаушылықтың екі мэні белгілі: 1) оның тұрақты, консервативті
болуы, яғни ұрпақтан-ұрпакқа ағзалардың негізгі белгілері мен
қасиеттерінің өзгеріссіз беріліп отырылуы. Оған мысал ретінде
қойдан қозының, түйеден ботаның, биеден құлынның, иттен
күшіктің туылуын атауға болады; бидай сепсек бидай жинаймыз,
жүгеріден жүгері өнеді; асқабақтан асқабақ жетіледі т.с.с. Тұкым
қуалаушылықтың консервативтілігінің нэтижесінде биологиялық
түрлердің, тіршіліктің тұрақтылығы, біртұтастыгы калыптасады;
2) түқым қуалаушылықтың өзгергіштігі, ягни әр түрлі
себептердің салдарынан агзалардың белгілері мен қасиеттері
азды-көпті өзгеріске ұшырайды. Оган мындаған мысал келтіруге
болады. Бір отбасының балалары бір-бірінен аз да болса ерекше,
өзгеше болады; қорадағы қой, ешкі, сиыр, жылқылар бір-бірінен
сзл де болса өзгеше болады; егістіктегі бидай, жүгері, арпа тағы
сол стяқты өсімдіктер биіктігі, өнімділігі жағынан түрліше болвп
келеді. Түқым қуалайтын өзгергіштіктің нэтижесінде тіршіліктің
сан алуан түрлері пайда болган.
Генетика ғылымының негізгі мақсаты — тұқым қуалаушылық
пен өзгергіштікті зерттеп, тіршіліктің негізгі зандылықтарының
сырын ашу, анықтау. Оның медицина, ауыл шаруашылығы, т.б.
салалар үшін маңызы өте зор. Адам ауруларының көбісі тұқым
қуалайды, ал инфекциялық, инвазиялық және т.б. аурулардың
зілділігі тұқымқуалашылықпен анықталады. Оларды анықтап ем-
35
деп, ауруды болдырмай алдын алу үшін генетиканы жақсы игеру
қажет.
Ағзалардың тұқым қуалаушылық қасиеті ертедегі грек
оқымыстыларына белгілі болған, бірақ олар оның мэнін дұрыс
түсіндіре алмаған.
Олардың болжамдары бойынша үрпақтардың белгілері, қасиет-
тері атасы мен анасының тұқым қуалаушылық материалдарының
өзара қосылуы нәтижесіңде қалыптасады. Осы сияқты болжамды
Ч.Дарвин де дамытқан. Ол өзінің эволюциялық ілімін генетикалық
тұргыдан түсіндіру үшін «пангенезис» теориясын үсынган. Бүл
болжам бойынша агзалардың әрбір мүшесі, үлпалары ерекше зат
—жыныстық жасушалар «пангендер» бөліп шығарады, ал олар қан
арқылы жыныс жасушаларына жеткізіліп, (гаметалар) үрықтанган
кезде өзара қосылады,— деген (12-сурет).
1865 жылы чех галымы Г. Мендель түқым қуалаушылықтың
дискретті (тәуелсіз түцым цуалау) теориясын қалыптастырды.
Бүл теория бойынша ата-аналарының түқым қуалаушылык фак-
торлары (гендер) жыныстық жасушалар қосылган кезде бір-
бірімен өзара араласпайды, керісінше олар дербес, дискретті
«таза күйінде» болады. Ата-аналарының белгілері ерте ме, кеш пе
үрпақтарда байқалып, қайталанып отырады (13-сурет).
ООО
ООО
„ Ғ
ООО
X
ООО
-► 1
ООО
ООО
<ё€)Ф
12-сурет. «Кіріккен» тұкым куалаушылык жобасы
ООО
ООО
F
ОООО
ООО
X
ООО
► Г1
ООО©
ООО
ООО
ОООО
ОООО
13-сурет. Тәуелсіз түкым куалаушылык жобасы
36
Г. Мендель ашқан тұқым қуалаушылық зандылықтары 1900
жылга дейін гылыми қауымға, көпшілікке белгісіз болып келді.
Ал 1900 жылы оқымыстылар — Г. Фриз, К. Корренс, Э. Чермак,
бір-бірінен тәуелсіз, Г. Мендель заңдарын қайта ашқаннан кейін
барып Г. Мендельдің еңбектері өз дәрежесінде багаланды. Бірақ
элі де болса тұқым қуалаушылықтың материалы белгісіз еді. 1902
жылы АҚШ галымы Вальтер Саттон жэне неміс — Теодор Бовери
тұқым қуалаушылық хромосомалармен байланысты болуы мүмкін
деген болжам жасаган. Оған негіз болған себеп, жасушаның ми¬
тоз жэне мейоз жолдарымен бөліну жэне ұрықтану процесінде
олар хромосомалардың қимыл-әрекеттерінің ұқсас болатындығын
байқаған.
Бұл болжамның растыгын, ягни тұқым қуалаушылықтың
хромосомалармен байланысты екенін, түрліше тәжірибелермен
дэлелдеген Т.Морган жэне оның шәкірттері болды. Олар 1910—
1911 жылдары жеміс шыбынын (Drosophila melanogaster) будан-
дастырып, түқьш қуалаушылыцтың хромосомалыц теориясын
қалыптастырды, тіркес түқым қуалау қүбылысын ашты.
XX ғасырдың басында К.Корренс (1908) цитоплаталыц тү-
қым цуалаушылыңты ашты, 1909 жылы В. Иогансен Г. Мендель-
дің «Түқым қуалаушылық факторларын» —ген деп атады.
XX ғасырдың 30—40 жылдары орыс галымдары Н. К. Кольцов,
А. С. Серебровский, Н. П. Дубинин, т.б. еңбектерінің нәтижесінде
ген теориясы қалыптасты.
1944 жылы О. Эйвери, К. Мак Леод, М. Мак Карта тұқым
қуалаушылықтың материалдық негізі — нуклеин қышқылдары
(ДНҚ) екендігін дәлелдеді, ал 1953 жылы Дж. Уотсон жэне
Ф. Крик ДНҚ молекуласы қүрылымының моделін анықтады.
XX ғасырдың екінші жартысынан бастап ғалымдардың
зерттеулері негізінен нуклеин қышқылдарының қасиетін
анықтауга, олардың молекуласында түқым қуалаушылық
акпараттың жазылу жэне жүзеге асу механизмдерін анықтауга,
генетикалық кодтың қүрылысын жэне қасиеттерін анықтауға,
ағзалардың нақтылы белгілері мен ересек (дефинитивтік)
фенотипінің қалыптасуында гендердің белсенділігінің ретгелу
механизмдерін зерттеуге бағытталады. XX ғасырдың 60-жылдары
М. Ниренберг, С. Очао, X. Корана жэне басқа да биохимиктердің
37
еңбектері нәтижесінде генетикалық код түгелдей анықталды, ал
70-жылдары генетикалық инженерия әдістері қалыптасып, тірі
ағзалардың түкым қуалаушылығын мақсатқа сай өзгертуге, жасан¬
ды тұқым қуалаушылық материалдары бар (трансгенді) ағзаларды
күрастыруға, жоғары сатылы омырткалы ағзаларды клондауға
мұмкіншілік туды.
Г. Мендель гендері эр түрлі хромосомаларда орналаскан
белгілердің тұқым қуалаушылыгын зерттеп дискреттік (тэуелсіз
түқым куалау) теориясын қалыптастырса, Т. Морган гендері бір
хромосомада орналасқан белгілердің тұқым қуалаушылығын зерт¬
теп, тіркес түқым қуалау кұбылысын ашқан.
Г. Мендель тэжірибелері
1865 жылы Чехословакияның Брно қаласының шагын ғылыми
ұйымының кезекті мэжілісінде Г. Мендель өзінің тәжірибелерінің
қорытындыларын баяндады. Ол моно-ди-потгибридтік будаида-
стыру әдістерін қолданған. Г. Мендельдің тэжірибелік эдістері сол
кездегі басқа оқымыстылардың эдістерінен мүлдем өзгеше бола¬
ды:
1) ол будандастыру үшін бір-бірінен айқын ажыратуға бола-
тын балама белгілерді колданған, мысалы: тұқымның түсінің
сары не жасыл болуы; жемістерінің түсінің сары не жасыл болуы;
гүлдерінің түсінің кызыл не ақ болуы; өсімдік сабақтарының ұзын
не қысқа болуы; гүлдерінің сабақ гөбесіне не жапырақ қойнына
орналасуы және т.с.с;
2) будандастыруға дейін өсімдіктерді бірнеше жыл бойы-
на өздігінен тозаңдатып, белгілердің тазалыгына (гомозиготалы
күйіне) жеткізген;
3) белгілердің үрпақтарда байкалуын ерінбей бір-бірлеп есеп-
теп санаған;
4) гибридология (будандастыру) нәтижесін талдау үшін
математикалық аппаратты кеңінен пайдаланған жэне үрпақтардыц
белгілерінің бәрін бірдей емес, тек нақтылы 1, 2 не 3 белгілердің
тұқым қуалауын ғана талдаған;
Моногибридті будандастыру. Г. Мендель бүршақтың тұқы-
мының түсі сары не жасыл болып келетін екі сортын өзара
38
будандастырған. Сонда бірінші үрпақтың тұкымдары бэрінде
де сары болып келген. Бірінші үрпақтың өкілдерін өздігінен
тозандандырғанда екінші ұрпакта сары түсті бүршақтармен қатар
жасыл түсті бүршақтар да байқалған. Г. Мендель бірінші ұрпақта
байқалған сары белгіні доминантты, ал бірінші ұрпақта байқалмай,
тек екінші ұрпақта ғана байкалатын жасыл түсті рецессивті белгі
деп атаған. Г. Мендель бүршақтың түсі (сары, жасыл) екі тәуелсіз,
дискретті түқым қуалаушылық факторлары арқылы беріледі
деп болжамдап, доминантты белгіні АА, рецессивті белгіні
аа әріптерімен жазып бейнелеген, сонда оның моногибридті
будандастыруының нэтижесі мынадай болады:
P. АА х аа
Г. А а
Ғ, Аа; Аа
Бірінші үрпақ өкілдерінің түқымдарының бэрінде бірдей (сары
түс) белгінің байқалуын Г. Мендельдің 1-заңы — гибридтер
белгілерінің біркелкілік зацы деп аталады.
Г. Мендельдің 2-заңы — белгілердің ажырау зацы деп атала¬
ды. Бірінші үрпақ өкілдерін (сары түсті бүршақтар) өздігінен
тозаңдастырсақ екінші үрпақта белгілердің ажырауын байқаймыз.
3 бөлігі сары (АА; Аа; Аа), ал бір бөлігі (аа) жасыл түсті болады.
Р. Аа хАа
Г. А а А а
Ғ, АА; Аа; Аа; аа
Г. Мендель дигибридтті будандастыруга да тэжірибе жасаган,
ягни ол сары түсті тегіс бүршақты (доминантты белгілер) жасыл
түсті кедір-бүдыр (рецессивті белгілер) бүршақпен будандастырган.
Сонда мынадай нәтиже алынган.
P. ААВВ х аавв
Г. АВ ав
?! АаВв; АаВв
Бірінші үрпақ дигетерозиготалы бүршақтарды өздігінен
тозандандырғанда төмендегідей ажырау байкалған.
Р. АаВв х АаВв
Г. АВ Ав аВ ав
Ғ2 Пеннет кестесіне қара:
39
Бірінші ұрпақтың өкілдерінің бэрінің тұқымдары сары және
тегіс болып Г. Мендельдің бірінші заңы — біркелкілік заңы
қайталанган. Ал екінші ұрпақта белгілер 9:3:3:1 арақатынасындай
ажырайды. Бұл жерде ата-аналарына тэн белгілер (сары тегіс. жа¬
сыл кедір-бұдыр) мен қатар жаңа, ата-аналарында кездеспейтін
белгілер де пайда боған (сары кедір-бүдыр, жасыл тегіс). Мұны
біз кроссинговер құбылысының нэтижесі деп білуіміз керек.
Сол сияқты, эрбір жұп белгілерді өз алдына дербес талдай-
тын болсақ, олар бір-бірінен тәуелсіз ажырасады жэне олар 3:1
арақатынасындай мөлшерде ажырасады. Мысалы, Г. Мендель
тәжірибесінде сары бұршақтар саны 12, жасыл бұршақтар 4 (12:4
=3:1); тегіс бұршақтар 12. кедір-бұдыр бұршақтар 4 (12:4 =3:1).
Міне осыған байланысты Г. Мендельдің 3-заңы — белгілердің
тәуелсіз тұқым цуалау зсщы деп аталады, ягни диполигибридті
будандасуларда эрбір жұп белгілер бір-бірінен тәуелсіз тұқым
қуалайды; тәуелсіз ажырасады.
Т. Морган тпәжірибелері
1908 жылы У. Сэттон жэне Р. Пеннет кей жағдайларда екінші
ұрпақ белгілерінің ажырауы Г. Мендельдің 3-заңы — белгілердің
еркін комбинациялануынан өзгеше болатындыгын байқаған.
1910—1911 жылдары Т. Морган жэне оның шәкірттері жеміс шы-
бындарына тэжірибе жасап, олардың белгілерінің ажырасуы
Г. Мендельдің дискреттік тұқым қуалау заңына қайшы келетіндігін
байқаган. Мысалы: қызыл көзді ұргашы шыбынды (қызыл көз до¬
минантты белгі), ақ көзді еркек шыбынмен (ақ көз рецессивті белгі)
40
будандастырғанда бірінші ұрпақтың бэрінің көздері қызыл бол¬
тан. Бұл Г. Мендельдің 1-заңына сэйкес келеді. Ал қызыл көзді Ғ
шыбындарын бір-бірімен будандастырғанда Ғ,-де шыбындардың
3-бөлігінің көзі қызыл, ал 1 -бөлігінің көзі ақ болып ажырасуы керек
(Мендель заңы бойынша). Бірак, еркек жэне ұрғашы шыбындарды
бөлек талдағанда ұрғашы шыбындардың бірінің көздері қызыл,
ал еркек шыбындардың жартысыньщ көзі қызыл, жартысының
көздері ақ болып келген.
Егер де ақ көзді (рецессивті белгі) ұргашы шыбынды қызыл
көзді (доминантты белгі) еркек шыбынмен будандастырғанда Ff
шыбындарының жартысының көздері қызыл, ал екінші жарты-
сы ақ көзді болған. Оның үстіне, қызыл көзді шыбындардың бэрі
ұрғашы, ал ақ көзділердің бэрі еркек шыбындар болған.
Бұларды Г. Мендель зандары арқылы түсіндіруге мүмкін емес.
Сондықтан Т. Морган мынадай болжам жасады: шыбындардың
көзінің түсін анықтайтын ген жыныс хромосомасында (X хромо-
сомада) орналасқан жэне онымен бірге түқым қуалайды, ал У хро-
мосомада ол кездеспейді.
25%
25%
25 %
25%
Р.
В.
V
V
14-сурет. Гендердін еркін комбинациялануы
Т. Морган гомозиготалы сұр түсті қалыпты қанатты (BBVV) шы¬
бынды қаратүсті кысқақанатты (bbvv) шыбынмен будандастырганда
бірінші үрпақтың бэрінде де доминантты белгі (сұр түсті қалыпты
қанатты шыбындар) байқалган, ягни. Мендельдің 1-заңы —
Біркелкілік заңы байқалган.
P. BBVV х bbvv
Г. BV bv
*Ү
BbVv
Бірінші ұрпақ шыбындарының генотипін анықтау үшін Т. Мор¬
ган талдаушы будандастыру жүргізген; яғни рецессивті гомозигота-
41
лы ұрғашы шыбынды (bbvv) дигетерозиготалы еркек шыбынымен
(BbVv) будандастырған. Сонда бірінші үрпақтың 50 %-ы сұр түсті
қалыпты қанатты —дигетерезиготалы (BbVv), 50 %-ы рецессивті
гомозиготалы (bbvv), яғни қара түсті қысқа капатты болған.
15-сурет. Толык тіркесу кезінде гаметалардың түзілу жобасы
Бұл жерде Г. Мендельдің 3-заңына сэйкес 4 түрлі фенотип
пайда болуы керек еді (14-сурет), бірақ тек 2 түрлі ғана фенотип
байқалған. Себебі дигетерозиготалы шыбынның 2 доминантты
гені (BV) бір хромосомада, ал 2 рецессивті гені екінші хромосо-
маларда орналасып, мейоз нәтижесінде олар әр түрлі гаметаларга
(BV), (bv), таралып тек қана 2 түрлі гаметалар түзілген (15-сурет).
Сондықтан да оларда тек ата-аналарына гана үқсас фенотиптер
байқалган. Бүл жағдайда гендер толық тіркескен.
Ал, дигетерозиготалы үргашы шыбынды (BbVv) рецессивті
гомозиготалы еркек шыбынмен (bvbv) будандастырғанда Г. Мен-
дельдің 3-заңына сэйкес 4 түрлі фенотип байқалған, бірақ Г. Мен-
дельдің еркін комбинациялану заңындагыдай эрбір фенотип 25%
емес, өзгеше мөлшерде, ягни 41,5 % + 8,5 % + 8,5 % + 41,5 %
мөлшерінде ажырасқан (16-сурет).
V
V
P. BbVv х bbvv
Г. BV Bv bv
vB vb
F2. BbVv; BbVv; bbVv; bbvv
41,5% 8,5% 8,5% 41,5%
42
”1 г
- G
У
7 I
У
—
J
j
■
■ в
* V і
b
3 v E
b
v
: В
: V
■ В ■
J
ь
A
b
и
: В
В VP
« в
V
■ b
: V
b
■ V
41,5% 8,5% 8,5% 41,5%
j 16-cypem. Кроссинговер жобасы |
Екінші тәжірибеде гендер толық тіркеспегендіктен, дигетеро¬
зиготалы дараларда 4 түрлі гаметалар түзіліп ата-аналарына ұқсас
41,5% сұр түсті қалыпты қанатты, 41,5 % қара түсті қысқа қанатты
жэне 8,5 %-тен гибридтік түрлер — сүр түсті қысқа қанатты жэне
қара түсті қалыпты қанатты шыбындар байқалган. Бұл тәжірибеде
бір хромосомадағы гендердің толық тіркесуі бүзылады, себебі
мейоз кезінде гомолог хромосомалар коньюгацияланады, аллельді
локустарымен алмасады (кроссинговер), хромосомалардагы
гендер жаңаша топтасады, тұқым қуалайтын материал реком-
бинацияланады. Сондықтан, екінші тәжірибеде гендер толық
тіркеспегендікген, дигетерозиготалы дараларда 4 түрлі гаметалар
түзіліп ата-аналарына ұксас 41,5 % сүр түсті қалыпты қанатты,
41,5% қара түсті қысқа қанатты жэне 17 %-ында кроссоверлік түр-
лер — сүр түсті қысқа қанатты жэне қара түсті қалыпты қанатты
шыбындар байқалган. Бұл шыбындардагы белгілердің топтасуы
ата-аналарына үксамайды, себебі кроссинговердің нэтижесінде
гендер жаңаша топтасады.
Осы тәжірибелердің нәтижесінде Т. Морган 1911—1912 жыл-
дары тұқым куалаушылықтың хромосомалық теориясын ашты.
Оның негізгі қагидалары:
1) Гендер хромосомаларда сызық бойымен тізіліп орналасады.
Әр түрлі хромосомаларда гендердің түрліше жиынтыгы болады, ал
гомологтық хромосомаларда гендер жиынтыгы бірдей болады;
2) Аллельді гендер гомологтік хромосомалардың бірдей локу-
сында (орында) орналасады;
43
3) Бір хромосомада орналасқан гендер тіркесіп тұқым куалай-
ды, тіркесу тобын құрайды. Тіркесу тобының саны гаплоидтық
хромосомалар санына тең;
4) Гендердің тіркесу күші олардың ара қашықтығына кері про-
порционал болады;
5) Кейде гендердің тіркесуі бұзылады, оның бірден-бір себебі
— кроссинговер;
6) Гендердің ара қашықтығының өлшем бірлігі ретінде морга-
нида қолданылады. 1 морганида тең 1% кроссинговерге;
7) Егер гендердің ара қашықтығы 50 морганидаға тең болса не¬
месе 50 морганидадан артық болса, онда Г. Мендель зандарына
сэйкес тәуелсіз тұқым қуалайды.
Тіркес тұқым қуалау аутосомаларда жэне жыныс хромосомала-
рында орналасқан гендерге тэн, соңгысын жыныспен (X, V хромо¬
сомалары) тіркес тұқым куалау деп атайды.
Адамдардың 150-ге жуық белгілері жыныспен тіркес тұқым
қуалайды (мысалы, гемофилия).
3.2. Ағзалардың генетикалық жүйесі.
Гендердің өзара әрекеттесуі
Агзалардың эрбір белгілерінің дискретті гендер аркылы
анықталуына қарамастан, олардың жеке даму процесінде (онто¬
генез) нақтылы биологиялык түрлердің морфофизиологиялық
типтеріне тэн өзара үйлескен белгілер мен қасиеттер кешені
түзіледі. Осылайша, заңды түрде безгек плазмодиі, адам ішекқүрты,
көдімгі қарағай, үнді пілі, саналы адам т.б. пайда болады. Ал бүл,
ағзалардың көптеген дискреттік гендерінің қызметтік түргыдан
алғанда біртүтас жүйеге — генотипке (геномга) топтасуының
нәтижесінде жүзеге асады.
Генотип дегеніміз агзалардың диплоидтық хромосома санын-
дағы гендер (аллельдер) жиынтығы, ал геном — гаплоидтық хро¬
мосома санындағы гендердің (аллельдердің) жиынтыгы.
Фенотип — ағзалардың белгілерінің жэне қасиеттерінің
жиынтығы. Генотип, фенотип деген терминдерді ғылымга 1909
жылы В. Иогансен енгізген.
44
Фенотип эр кезде генотипке байланысты болады жэне онын
қалыптасуы генотиптегі гендердің өзара эрекеттесуінің жэне
генотиптің орта факторларымен әрекеттесуінің нэтижесінде
жүзеге асады. Бірдей генотипке не ағзаларда эр түрлі фенотип
қалыптасуы мүмкін, ягни фенотип өзгермелі келеді, оны моди-
фикацііялар деп атайды. Ал ұқсас, бірдей фенотипке ие агзаларда
міндетті түрде бірдей генотиптің болуы шарт емес, мысалы сары
түсті бүршақтардың генотипі гомозиготалы (АА), не гетерозигота-
лы (Аа) болуы мүмкін.
Гомозиготалы деп геннің біркелкі аллельдерінен тұратын не¬
месе бір типті гаметаларды түзетін жасуша, не ағзаны айтамыз,
ал гетерозиготалы деп геннің эр түрлі аллельдерінен тұратын, не
эр түрлі типті гаметалар түзетін жасуша, не агзаны айтамыз. Со-
нымен қатар, аллельдер гемозиготалы күйінде де болуы мүмкін.
Гемозиготалы деп аллельі сыңар, тек бір хромосомада болып
екіншісінде кездеспейтін жасушаны не агзаны айтамыз. Қалыпты
жагдайда ол тек жыныс хромосомаларында ғана кездеседі.
Гендердің әрекеттесуі эр түрлі деңгейлерде болуы мүмкін:
1) генотип деңгейінде;
2) а — РНҚ мен ақуыз биосинтезінде түзілетін полипептидтер
арасында;
3) бір метаболизм цикліне катысатын әр түрлі ақуыз — фер-
менттер арасында.
Гендердің екінші деңгейдегі эрекеттесуінің, ягни олардың функ-
ционалдық актив өнімдері — а-РНҚ жэне полипептид арасындагы
эрекеттесуінің нэтижесінде кейбір күрделі белгілер қалыптасады,
мысалы, Моррис синдромы. Бүл синдром ХУкариотипіне ие, бірак
эйелдерге тэн фенотипте болатын, яғни эйелдерге сәйкес соңгы
жыныс белгілері қалыптасқан агзаларда байқалады. Оның себебі —
еркектерге сэйкес соңгы жыныстық белгілер кешені онтогенездің
белгілі бір сатысында 2 фактордың аталық жыныс гормонының
(тестостерон) жэне дене жасушаларын тестостеронга сезімтал
ететін ақуыз — рецептордың синтезделуіне байланысты. Моррис
синдромымен ауыратын адамдарда тестостерон қажетті мөлшерде
синтезделгенімен, дене жасушаларын оган сезімтал ететін ақуыз-
рецептор синтезделмейді. Сонымен, еркектерге сэйкес соңгы
жыныстық белгілер кешенінің калыпты дамуын 2 ген айқындайды
жэне олар өздерінін өнімдері деңгейінде әрекеттеседі.
45
Гендердің әрекеттесу деңгейіне қарамастан, олардың өзара
әрекеттесуінің 2 түрін ажыратады: аллелъді гендердің эрекеттесуі
жэне аллелъсіз гендердіц эрекеттесуі.
Аллельді гендер деп гомологтык хромосомалардың бірдей
локусында орналасқан жэне карама-қарсы (балама) белгілерді
анықтайтын гендерді айтамыз. Аллельді гендердің эрекеттесуіне
доминанттылык, рецессивтік, толық емес доминанттылық (тұқым
қуалаушылықтың аралық сипаты), аса жоғары доминанттылык,
кодоминанттылық (бірлесіп доминанттылык көрсету), аллель-
дер аралық комплементация (бірлесу), аллельдің біреуінің істен
шығарылуы жатады.
Доминанттылык, дегеніміз — аллельді бір геннің екінші
аллельді басып тастауы. Доминанттылыктың 3 түрі белгілі:
толық доминанттылык, толык емес доминанттылык, аса жоғары
доминанттылык.
Толыц доминаттылыц — дегеніміз доминантты аллельдің
рецессивті аллельді толык басып тастауы, мысалы: бүршақтың
(Аа) түсінің сары болуы.
Толыц емес доминанттылык, дегеніміз—доминантты аллельдің
рецессивті аллельді толык басып тастай алмауы салдарынан рецес-
сивті аллель аз да болса өзінің фенотипін көрсетеді, белгі аралык
сипатта дамиды. Мысалы, кызыл гүлді намазшамгүлін (АА) ак
гүлді намазшамгүлмен (аа) будандастырғанда бірінші үрпақтың
гүлінің түсі (Аа) не қызыл, не ақ емес, қызғылт болады.
Аса жогары доминанттылык, дегеніміз — Ғ,-дің гетерозиго-
талы (Аа) дараларында доминантты белгінің гомозиготалы (АА)
күйіне Караганда әлдеқайда күшті байқалуы, яғни Аа>АА.
Рецессивтік дегеніміз — бір аллельді геннің фенотиптік
байқалуының екінші бір аллель арқылы бастырмалануы.
Кодоминанттылық (бірлесіп доминанттылык,, корсету)
дегеніміз — екі гетерозиготалы доминантты аллельдің бірлесіп
бір белгіні дамытуы, ал жеке-жеке олар эр түрлі белгілерді дамы-
тады. Мысалы, адамдардың АВ (IV) қан тобыныңтүқым қуалауы.
Адамдардың АВО жүйесі 3 аллель арқылы — 1 °, 1А, 1 “анықталады.
Олардың екеуі 1А, 1в, 1°-ге қарағанда доминанттылык байқатады.
1° 1° — О (I) кан тобын, ІА ІА, 1А, 1° — II (А) қан тобын; Ів Ів, Ів 1° — III
(В) кан тобын анықтаса, ІА, Ів- АВ (IV) қан тобын анықтайды.
46
Аллелъаралыц сәйкестілік комплементация — аллельді ген-
дердің сирек кездесетін эрекеттесу түріне жатады. Бұл жағдайда
қалыпты доминантты (Д) белгінің дамуы осы геннің екі доминант¬
ты мутантгы гендерінің (Д1 Д11) өзара әрекеттесуі нәтижесінде
байкалады. Мысалы: бір қалыпты белгінің дамуы молекула
құрылымы күрделі, яғни 4 дәрежелі күрылысқа ие, бірдей екі по-
липептидтен түратын ақуыз молекуласының синтезделуіне бай¬
ланысты делік. Онда. мутантты Д1 гені (Д‘Д, Д‘д), мутантты Д1
полипептидінің. ал екінші мутантты Д" гені (ДПДП, Дпд) өзгерген
екінші Ди полипептидінің синтезделуіне алып келіп мутант¬
ты белок молекуласы түзіледі, яғни олар жеке-жеке осы белок
молекуласынын 4-ші дэрежелі құрылысын пайда етіп, қалыпты
белгіні дамытуы мүмкін.
Аллельдердіц біреуінің істен шыгарылуы дегеніміз гомогамета-
лы жыныстарда (мысалы әйелдерде) екі X хромосоманыц біреуі
активсізденіп жыныс хроматиніне (Барр денешігіне) айналуы
салдарынан онда орналасқан гендердін активсізденіп істен шыга¬
рылуы болып табылады. Әйелдердіц эмбриондық дамуының
16 тэулігінде екі X хромосоманыц біреуі активсізденіп, жыныс
хроматиніне айналады. Түрліше жасушаларда эр түрлі X хромо¬
сома (әкесінен алынган, не шешесінен алынган) активсізденетін-
діктен, ағза жасушаларында X хромосомасының мозаикасы пайда
болады.
Гендердің осы сияқты әрекеттесуі, яғни аллельдің істен шыга¬
рылуы кейбір ерекше антигендерге сэйкес келетін антиденелерді
синтездейтін В-лимфоциттерде де кездеседі.
Аллель емес гендердің әрекеттесуіне эпистаз, комплиментар-
лық, полимерия жатады. Эпистаз дегеніміз — бір аллель емес
геннің екінші бір аллель емес генді бастырмалауы. Ол доминант¬
ты жэне рецессивті эпистаз болып екіге бөлінеді. Доминант¬
ты эпистазға мысал ретінде асқабак жемісінің түқым куалауын
келтіруге болады. Асқабак жемісінің сары түсті болуы доминант¬
ты (В) аллельіне, жасыл түсті болуы рецессивті (в) аллеліне байла¬
нысты. Егер асқабақ генотипінде тагы бір доминантты (А) аллельі
кездесетін болса екінші аллельдің кайсысына болмасын эсер ететін
(ААВВ, ААВв, Аавв т.с.с.) жемістің түсі ақ болып келеді, себебі А
аллелі В аллельдеріне (сары не жасыл түс) эпистаздық эсер етеді.
47
Тағы бір мысал — тауықтардың доминантты С гені пигменттің
синтезделуін қадағаласа, рецессивті с гені пигмент синтезделуін
болдырмайды, ал екінші бір аллель емес геннің 1 доминантты
аллелі С генінің супрессоры болып оны бастырмалайды. Егер
тауыктардың генотип С- I ccl- болып келсе олардың түсі ақ, ал
С-іі болса түрлі тусті боялган болады.
Рецессивті эпистазға мысал ретінде ABO қан тобы жүйесінде
өте сирек кездесетін ерекше кұбылыс, «бомбей феноменін»
келтіруге болады.
1 (О) қан тобы бар ер адам III (“В”) қан тобы бар эйелге үйленген.
Олардан бір қыз туылган оның I (О) қан тобы болган. Ал, ол қыз
II (А) кан тобы бар ер адамга тұрмыска шыққан жэне 2 қыз туган,
оның біреуінде IV (АВ) қан тобы, ал екіншісінде I (О) қан тобы
кездескен.
Осы отбасының III ұрпагында «О», «А» қан топтары бар ата-
анадан «АВ» қан тобы бар қыздың туылуы оқымыстыларды таң
қалдырган.
«О»
1° 1°
Хх
«А»
1А, 1°
Хх
1А, 1в
1°, 1°
16а-сурет. Бомбей феноменінін шежіресі
48
Олай болуы мүмкін емес, себебі ол тұқым қуалаушылық
заңдылықтарына қайшы келеді. Оны тек эпистаздық эсер арқылы
түсіндіруге болады (1ба-сурет): Шынында да ABO жүйесінің
қан топтарын анықтайтын доминантты ІА Ів аллельдерінен басқа
тағы бір аллель емес рецессивті х гені гомозиготалы күйінде
(хх) I ұрпақтың геніне эпистаздық эсер етіп II үрпақтағы қыздың
генотипі I" 1°болуына қарамастан, доминантты В аллелі рецессивті
х гені арқылы бастырмаланып «О» қан тобының байқалуына
алып келген. Мұның себебі, А жэне В антигені синтезделуі үшін
олардың бастама заттары синтезделуі қажет. Ал антигендердің
бастама заттарының синтезделуі доминантты X геніне (XX, Хх)
байланысты болады. Осы геннің рецессивті гомозиготасы (хх) ор¬
ганизм генотипінде ІА, 1в гендерінің болуына қарамастан А жэне В
антигенінің синтезделуін болдырмайды. Сондықтан да бүл адам-
дарда «О» қан тобы байқалады.
Комплиментарлық дегеніміз екі не одан да көп аллель емес
доминантты гендердің бірлесіп жаңа белгіні дамытуы, ал тек бір
геннің доминатты аллелі болатын болса белгі дамымайды. Мы¬
салы, дала тышқандарының популяцияларында сүр түсті «агути»
даралары басым болады. Олардың жүндерінің сұр түсті болуы 2
доминантты аллель емес гендерге байланысты. Оның бірі А —
пигмент синтезін қадагаласа осы локустың рецессивті гомозиго¬
талы болуы (аа) альбинизмге алып келеді, ал екінші В гені сол
пигментгің түктің түбіне жэне ұшына шогырлануын кадагалайды
(осы локустың рецессивті гомозиготалы болуы вв — қара түсті бо¬
луына алып келеді).
Тагы бір мысал, адамдардың шаштарының реңі 2 компли-
ментарлық гендерге байланысты. М-гені кара пигмент —
меланиннің синтезін камтамасыз етсе (оның 3 аллелі бар: Мвк
меланиннің көп мөлшерде, MBW — орташа мөлшерде, MBd —
меланиннің аз мөлшерде синтезделуіне алып келеді). R-гені (R,
R,) қызыл пигменттің ситезделуін камтамасыз етеді. Осы геннің
өнімдері — қара пигмент меланинмен қызыл пигменттің өзара
араласуы нәтижесінде адамдардың шаштарының реңі алуан түрлі
болып келеді.
Күрделі белгі — есту 2 түрлі комплиментарлы геннің эрекет-
тесуіне байланысты калыптасады. Бір доминантты ген (А) ішкі
49
170-4
күлақтың иірімдерінің жетілуін бақыласа, екінші доминантты ген
(В) есту нервінің жетілуін қадағалайды — ААВВ, ААВв, АаВВ.
АаВв — > есітеді;
ааВВ, ааВв аавв ААвв, Аавв -> естімейді.
Тұқым қуалаушылықтың белгілер күйінде байкалуы генотиптің
сипатына жэне орта факторларына байланысты екенін гендердің
экспрессивтілігі мен пенетрстттылыгынан байқауға болады.
Гендердіц экспрессивтілігі дегеніміз — бірдей аллельге ие
болатын дараларда белгінің байқалу дәрежесі. Экспрессивтілік
тұрақты не құбылмалы болып келеді. Түракты экспрессивтілікке
мысал ретінде бұршақтың тұқымының түсінің сары не жасыл бо-
луын жатқызуга болады, ал қүбылмалы экспрессивтілікке Гентинг-
тон хореясын (доминантты ген), көздің түсін анықтайтын аллельді,
жеміс шыбынының көзінің дамуын қадагалайтын гендерді т.б.
жатқызуга болады.
Гендердің пенетранттылыгы дегеніміз — бірдей аллель¬
ге ие дараларда белгінің фенотиптік байқалу жиілігі. Ол пай-
ызбен өлшенеді. Ол толық жэне толық емес пенентранттылык
деп жіктелінеді. Толық пенетрантты аллель 100 пайыз жагдайда
байқалады, ал толык емес пенетрантты аллельдің фенотиптік
байқалуы 100 пайыздан кем болады. Мысалы, Гентингтен Хоре-
ясы (адамның басы, денесі, аяқ қолдары қалтырап, нерв жүйесі
бұзылып, ақыл-есінің кем болуы, агза қажып түбінде өлуге
алып келетін ауру). Осы ауру доминантты аллель арқылы тұкым
қуалайды жэне осы аллельге ие адамдардың кейбіреулері мүлдем
ауырмайды, ал ауырган адамдардың өзі эр түрлі жас шамасын-
да сыркаттануы мүмкін; бірі бала кезінде, екінші біреулері орта
жасқа келгенде, үшіншілері кэрілік кезенде сырқаттанады. Демек,
Гентингтон Хореясының аллелі (А) толык емес пенетранттылықка
ие болып оның экспрессивтігі күбылмалы болады.
Плейотропия дегеніміз — бір геннің бірнеше белгінің даму¬
ын қамтамасыз ететін ерекше қасиеті. Бұл құбылысты Г. Мендель
байқаган болатын. Ол бір геннің бұршак гүлінің түсін, тұқымының
түсін жэне жапырақ қынабының түсін аныктайтынына көніл
аударган. Жеміс шыбынының бір гені — оның канатының
дамуының (үзын не қысқа болуын), ызылдау аппаратының даму¬
ын, көбею қабілеттілігін, өмірініңұзақтыгын анықтайтыны белгілі.
50
Адамдарда фенилкетонурия ауруын анықтайтын рецессивті ген
осы аурумен қатар сол адамдардың шашының түсінің жирен түсті
болуын, микроцефалия (бастарының кіші болуын), акыл-естерінің
азды-копті кеміс болуын да аиыктайды.
Аллель емес гендердің әрекеттесуіне полимерияны да
жатқызуға болады. Полимерия дегеніміз — бірнеше аллель емес
гендердің бірлесіп бір белгіні дамытуы. Сол аллель емес гендердің
эрқайсысы белгінің дамуына шамалы ғана үлес қосады, ал бэрі
бірігігі бір белгіні дамытады. Мүндай жолмен сандық белгілер
түқым қуалайды. Мысалы, адамдардың бойының ұзындыгы,
салмағы, ақылдылығы, бидайдың дэнінің түсі, т.с.с. Мыса¬
лы: адамдардың терісінің түсінің боялу интенсивтілігі меланин
пигментінің синтезделуін қадағалайтын 4 аллельсіз ген арқылы
анықталады — Р,, Р„, Р,, Р4. Егерде генотипте осы полигендердің
8 доминантты аллельдері кездессе пигменттің өте көп мөлшерде
синтезделуінен адам терісінің түсі шымқай кара болады (Афри¬
ка негрлері — Р ?! Р2 Р2 Р, Р3 Р4 Р4), ал генотипте доминантты
аллельдердің болмауы, ягни тек рецессивті аллельдердің кездесуі
(р, р, р, р, р3рз Р4 Р4) пигменттің аз мөлшерде синтезделуі салда-
рынан адам терісі түсінің ашык ақшыл болуына алып келеді (еуро-
пеоидтар). Доминантты аллельдердің көп не аз болуы тері түсінің
түрліше боялу интенсивтілігіне алып келеді.
Сол сиякты, полимерия жолымен адамның кейбір белгілері жэне
аурулары — ақылдылык, кант диабеті, шизофрения, гипертония
(қан қысымының) көтерілуі жэне т.б. белгілері түкым қуалайды.
3.3. Адам генетикасы
Тұқым куалаушылық басқа тірі агзалар сияқты, адамдарга да
тэн касиет. Адамдардың көптеген белгілері Г.Мендель заңцарына
сэйкес тұқым қуалайды, оларды мендельденуші белгілер деп атай¬
ды. Қазіргі кезде адамдардың 4000-нан астам тұқым қуалайтын ау¬
рулары анықталган, олардың ішінде: тері аурулары — 250-ден; көз
аурулары — 200 астам т.с.с.
Адамдардың тұқым қуалаушылыгын зертгейтін генети¬
ка гылымының саласын адам генетикасы деп атайды. Ол ерте
51
кезден-ақ, XIX гасырдың аяғынан бастап Ф. Гальтон, А. Гэррод
еңбектерінің нәтижесінде дами бастаған.
Адам генетикасының адамдардың денсаулығын сақтаудағы рөлі
өте зор, себебі кез келген ауру — зат алмасудың, не зат алмасуга
катынасып оны реттейтін, жеделдететін ферменттердің кызметінің
бұзылуына байланысты. Ал фермент дегеніміз ақуыз, ол гендердің
экспрессиялануының өнімі болып саналады. Демек, нақтылы
бұзылған гендерді зерттеп, оны эр түрлі жолдармен жөндеп —
генетикалық инженерия әдістерімен, түрліше аурулармен күресуге
болады.
Қазіргі кезде емханалардағы ауру адамдардың 30—40 пайызға
жуығы тұқым қуалайтын аурулармен ауырады, жас нәрестелердің
жастайынан өлуінің 25—30 пайыз жағдайдағы себебі де сол тұқым
қуалайтын аурулар. Әйелдердің күні бүрын өздігінен түсік тастауы
50—70 пайыз жағдайында хромосомалар мутациясына байланы¬
сты.
Адамдардың бедеу болуының, балаларының болмауының
себебі де 20 пайыз жағдайында түқым қуалайтын аппараттың
бүзылуынан екендігі белгілі.
Осыншама көп аурудың болуы, оларды емдеу, коғам үшін
әлеуметтік жэне материалдық үлкен мағынасы бар проблема бо¬
лып табылады.
Ауруларды емдеуден гөрі сол аурудың алдын алып, оны бол-
дырмау әлдеқайда тиімді. Міне, осы проблеманы шешу адам
генетикасының негізгі міндеті.
Әдетте, генетика ғылымының ең негізгі зерттеу әдісі болып
гибридологиялық эдіс саналады, яғни зерттеушілер күні бұрын
будандастыру үшін ата-ана жүптарын тандап алып эксперимент
жасайды (мысалы, Г.Мендель тәжірибелері).
Бұл тұрғыдан алғанда адамдар генетикалық объекті ретінде
колайсыз болып келеді, себебі: 1) адамдарда күні бұрын мақсатқа
сай ата-ана жұптарын таңдау, эксперименттік некелер қүрастыру
мүмкін емес, себебі ол қоғамдык-әлеуметтік жағдайларға кайшы
келеді; 2) қазіргі кезде көптеген отбасында балалар саны аз, ягни
2—3 баладан гана болады, ал ол белгілердің түқым куалау шылыгын
объективті түрде талдауға жеткіліксіз; 3) адамдардың өмірі қысқа
болады, яғни бір үрпақтың тіршілік үзактығы шамамен 65—70
52
жылга тең. Ал олардың жыныстық жетілуі кешірек басталады,
яғни 13—15 жасқа келгенде гана. Сондықтан ғалым-генетик өз
өмірінің ішінде тікелей 1—2 ұрпактың ғана алмасуын бақылап
зерттей алады; 4) адамдардың хромосомаларында «тіркесу»
топтарының саны көп — 23, ал дрозофилада ол не бары 4; 5)
адамдардың генотипі көп түрлі болып келеді. Мысалы, егер эрбір
жұп гомологтық хромосомаларда тек бір жұп аллель болады де-
сек, онда 23 жұп хромосомалардың мейоз кезінде еркін комбина-
циялануы нәтижесінде 8.388.608 (223) әр түрлі гаметалар пайда бо-
лар еді. Ал шын мэнінде адам хромосомаларындағы аллельдердің
жалпы саны элдеқайда көп, мысалы, тек X хромосомада 92 локус
кездеседі. Адамның геномындағы гендердің жалпы саны 100.000
дейін жетеді. (Соңғы кездерде адам геномындағы гендердің саны
30 000 көлемінде деп айтылуда).
Адамдардың генетикалық көп түрлілігі жэне оның тіршілігіне
эсер ететін экологиялықжәне элеуметтік факторлардың әр түрлілігі
олардың көптеген фенотиптік өзгергіштігіне алып келеді. Осы
жэне басқа да кедергілер гибридологиялық әдісті адам генетика-
сын зерттеу үшін қолдануга мүмкіндік бермейді.
Дегенмен, казіргі кезде, адамның тұкым қуалаушылыгын зерт¬
теу жедел дамуда, оның фенотипі анатомиялық, физиологиялық,
биохимиялық, иммунологиялық жэне мінез-құлқы тұргысынан
алганда жан-жақты жэне терең зерттелген.
Адам генетикасын зерттеу үшін бірнеше ерекше эдістер
қолданылады:
1. Генеалогиячъщ әдіс — бүл адам шежіресін қүрастыру жэне
талдау арқылы жүргізіледі. Бұл эдісті XIX гасырдың аягында
Ф.Гальтон енгізген.
Шежіре күрастыруды пробандтан бастайды. Пробанд
дегеніміз шежіре кұрастыруга себепші адам. Адам генетикасын
зерттеуде генеалогиялық әдістің рөлі өте зор, себебі осы эдіс
арқылы: 1) зерттелетін белгілердің тұкым қуалайтынын, түқым
қуаламайтынын; 2) белгілердің тұқым қуалау типін (аутосомды
— доминантты, аутосомды — рецессивті, жыныспен тіркес түқым
қуалау); 3) аллельдің экспрессивтілігі мен пенетранттылығын
анықтауга болады; 4) белгілердің (аурулардың) ұрпақтарда байқа-
лу мүмкіндігін анықтауға болады.
53
□ 1 O' [W)
О 2 Q dn
7
s
p JO’
0 ®10
6
17-cypem. Шежіре күрастыру үшін
колданылатын символдар
■ т ц
В <1)12
1 -ер адам, 2-әйел адам, 3-неке, 4-сибстер,
5-монозиготалы егіздер, 6-дизиготалы
егіздер, 7-бедеу адамдар, 8-туыстык
неке, 9-пробанд, 10-тасымалдаушылар,
11-өлі туылу, 12-өлгендері.
5
2. Егіздерді зерттеу әдісі — белгілердің қалыптасуындағы
генотиптің жэне орта факторларының салысгырмалы рөлін анық-
тау үшін жүргізіледі. Ол үшін белгінің монозиготалы (М), дизиго-
талы (Д) егіздерде жэне популяцияның баска да өкілдерінде байқа-
луын салыстырады. Осының нәтижесінде: 1) бірдей генотиптердің
ұқсас экологиялық жағдайда; 2) эр түрлі генотиптердің үқсас
экологиялық жағдайларда немесе әр түрлі генотиптердің эр түрлі
экологиялық жағдайларда даму заңцылықтарын анықтайды.
Кейде монозиготалы егіздерді бір-бірінен ажыратып эр түрлі
экологиялық жэне әлеуметтік жағдайларда тәрбиелеп бірдей
генотиптің дамуындағы зандылықтарды зерттейді.
Егіздерді зерттеу нэтижесінде олардың сапалы белгілерінің
конкорданттылыгын, не дискорданттылыгын жэне сандық белгі-
лерінің дисперсиясын анықтайды.
Зерттелінетін белгі егіздің екеуінде де кездессе, оларды кон-
кордантты (үксас), ал егерде белгі егіздердің біреуінде гана болып
екіншісінде кездеспесе, онда оларды дискордантты деп атай-
мыз.
Белгінің конкорданттылық дэрежесі неғұрлым жоғары болса,
согүрлым белгінің дамуында генотиптің рөлінің жогары болган-
дығын көрсетеді.
Егерде конкорданттылық көрсеткіш монозиготалы жэне дизи-
готалы егіздерде бірдей болса, онда осы белгінің дамуында орта
факторлары әсерінің басым болгандығын көрсетеді.
54
Кейбір белгілер мен аурулардың монозиготалы жэне дизигота-
лы егіздердегі конкорданттылығы төмендегідей болып келеді:
Белгілер не аурулар
Конкорданттылык пайыэы
монозиготалы
егіздер (т)
диіиготалы
егіздер (d)
1. Көздің түсі
99,5
28,0
2. Шаштың реңі
97,0
23,0
3. Терінін реңі
100
45,0
4. Мүрынның пішіні
100
30,0-35,0
5. Қүлак калканшасының пішіні
28,0
40,0
6. Үстіңгі еріннің жырык болуы
33,0
5,0
7. Туберкулез
37,0
15,0
8. Демікпе ауруы (бронхиальная
астма)
47,0
24,0
9. Қант ауруы
65,0
18,0
10. Қызылша
98,0
94,0
Белгінің дамуында тұқым қуалаушылық пен орта факторлары-
ның рөлін аныктау ұшін Хольцингер формуласын қолданады:
Cvz - Cdz
Н 100;
100-Cdz
мұнда Н — тұқым қуалаушылық үлесі, mz % — монозиготалы-
лардың конкорданттылығы, dz % — дизиготалылардың конкор-
данттылығы.
Сандық белгілердің дисперсиясын статистикалық әдістер
арқылы аныктауга болады. Ол үшін мынадай формуланы қолда-
нады:
с £(х- пх)г
6= — :
Дисперсия дегеніміз — белгінің өзгергіштік шегін көрсететін
статистикалық көрсеткіш. Егер дисперсия мөлшері аз болса, онда
сол белгінің дамуында генотиптің рөлінің басым болғаны, ал,
керісінше, дисперсия мөлшері көп болса, онда ол белгінің даму¬
ында орта факторларының рөлінің басым болғандығын көрсетеді.
55
3. Цитогенетикалыц әдіс — микроскоп арқылы хромосомалар-
ды зерттеуден басталады.
Бұл эдіс арқылы: 1) агзалардың кариотипін анықтайды; 2) хро¬
мосома санын анықтайды; 3) хромосомалардың морфологиясын
зерттеп, олардың құрылысында өзгерістер бар-жоғын анықтайды,
яғни хромосомалық ауруларды анықтайды; 4) хромосомалардың
генетикалық карталарын жасау үшін пайдаланады — ол үшін
хромосомаларды тандамалы түстерге бояйды; 5) геномдық не
хромосомалық мутацияларды, олардың жиілігін анықтау үшін
пайдаланады.
4. Сомалъщ жасуиіалардыц генетикасы. Зерттеуге лейкоцпт-
терді, сүііектіц қызьл кемігі жасушсіларын немесе фибробласт-
тарды стады. Жасушаларды ағзадан тыс, жасанды қоректік
орталарда өсіреді. Оларды өсіру кезінде қажет болған немесе
зерттеушінің көзіне түскен ерекше жасушаны бөліп алып клондай-
ды, яғни көбейтеді жэне сол жасушалар тобын алып зерттейді. Ол
жасушаларды әрі карай сұрыптап будандастырады.
Бұл эдіс нәтижесінде: 1) гендердің тіркесуін жэне олардың хро¬
мосома бойында орналасу орнын анықтайды; 2) кейбір гендердің
активтілігінің нәтижесі ретінде синтезделінетін олардың нақтылы
өнімдерін зерттейді; 3) гендердің эрекеттесуін жэне олардың
активтілігінің реттелу механизмдерін анықтайды; 4) гендік мута¬
цияларды зерттеу үшін қолданады.
Осы әдісті пайдаланып медицина практикасында элі туылмаған
баланың жынысын, 60-қа жуық тұқым куалайтын ауруларын
анықтауға болады. Ол үшін жатырдан сүйықтық алып ондагы ұрық
жасушаларын өсіріп зерттейді (амниоцентез).
5. Популяцияіық-статистикалық әдіс. Бүл эдіс арқылы: 1)
популяциядағы белгілі бір генотиптің не аллельдің таралу жиілігін
анықтауға; 2) болашақ үрпақтарда нақтылы фенотиптің дамуын
болжауға; 3) рецессивті аллельдердің гетерозиготалы күйінде та-
сымалдану жиілігін есептеуге болады.
Ол үшін Харди-Вайнберг заңын қолданады.
1) р + q = 1
2) р2 + 2pq + q2 = 1
р — доминантты аллельдің жиілігі;
q — рецессивті аллельдің жиілігі;
56
р- — доминантты гомозиготалылар жиілігі;
2pq — гетерозиготалылар жиілігі;
q2 — рецессивті гомозиготалылар жиілігі.
Осылайша гемоглобин генінің кейбір елдердегі жиілігі анық-
талған.
Мысалы, Белорусияда HbS гені бойынша гомозиготалы адам¬
дар кездеспесе, Батые Африка елдерінде оның жиілігі: Камерун-
да 25 %-ға тең, Танзанияда 40 % дейін өзгеріп отырады. Әр түрлі
географиялық аймақтарда гендердің таралу ерекшеліктерін зерт-
теп түрліше этникалық топтардың шығу тегін жэне олардың ми-
грациялануын, жеке адамдардың тұқым куалайтын аурулармен
ауыру деңгейін анықтауға болады.
6. Биохимиялъщ әдістер—ферменттік жүйелердің белсенділігін
зерттеу арқылы жүргізіледі. Олар зат алмасу ауруларының себебі
— гендік мутацияларды анықтауға, сол сияқты биохимиялык
тесттер арқылы патологиялық гендердің гетерозиготалы тасымал-
даушыларын да анықтауга мүмкіндік береді. Ол үшін зерттелуші
адамның канына (венасына) белгілі бір аминқышқылының (мыса¬
лы фенилаланиннің) біршама мөлшерін енгізеді де, оның қандагы
концентрациясын мезгіл-мезгіл анықтайды. Егер зертгелуші адам
доминантты ген бойынша гомозиготалы (АА) болса, онда амин
қышқылының концентрациясы тез арада қалыпты (оны енгізгенге
дейінгі) деңгейге келеді, ал егер гетерозиготалы (Аа) болса, онда
амин кышқылының концентрациясының төмендеуі екі есе баяу
жүреді.
Осы сияқты тесттерді адамның мультифакторлы аурулармен,
мысалы, кант ауруы, гипертония ауруларымен ауыруға бейімділігін
анықтау үшін де жүргізеді.
Тұқым куалаушылық жэне тұкым қуалау терминдері бір-
біріне синоним емес. Туцым қуаіаушылық, дегеніміз — ағ-
залардың қүрылымдык-функционалдық біртұтастыгын ұрпақтан-
ұрпақ-ка жалгастырып отыратын қасиет, бүл — генетикалык
тетіктер жиынтығы. Ал түцым цуалау дегеніміз — нақтылы
биологиялық түрлердің үрпактарында өздеріне тэн структуралық
— функционалдық құрылымының, жекелеген белгілері мен
қасиеттерінің берілуі болып табылады. Тұқым қуалаудың 2 түрі
белгілі: моногендік жэне полигендік.
57
Моногендік механизмдер
аутосомды
жыныспен тіркескен
доминантты
рецессивті
Ү-тіркескен
доминантты
рецессивті
Моногендік түқым қуалау типтерінің өздеріне тэн ерекше
белгілері болады, олар мыналар:
1. Аутосомды-доминантты т^қым қуалау: а) белгі бірінші
үрпақган бастап кем дегенде 50 пайыз дараларда байқалады; б) ер¬
кек жэне үрғашы жыныстарда бірдей байқалады; в) ата-аналары
белгіні балаларына бірдей бере алады;
2. Аутосомды-рецессивті т^қым қуалау: а) белгі бірінші
ұрпақта байқалмай, келесі ұрпақтарда байқалуы мүмкін; б) белгі
ата-аналарында болмаса да балаларында байқалуы мүмкін, бұл
жағдайда белгінің байқалу мүмкіншілігі 25 пайызға тең; в) егер
белгі ата-аналардьң екеуінде де болатын болса, барлық балаларын¬
да да байқалады; г) егер белгі ата-аналарының біреуінде болатын
болса балаларында оның байқалу мүмкіншілігі 50 пайызға тең бо¬
лады; д) белгі еркек жэне ұрғашы жыныстарда бірдей дәрежеде
байқалады;
3. Х-тіркескен доминантты түқым куалау: а) белгі ұл бала-
ларда да, кыздарда да байқалады;
0
1
ъ
12 3 4
9 10 11 12 13 14 15 16
18-сурет. Аутосомды-доминантты түқым куалау шежіресі
58
-о
6 і
™йо£кі> □ во* о 666
12 3 4 5 У 6 7 8
9 10 11 12 13
19-сурет. Доминантты Х-тіркескен тұкым куалау шежіресі
б) қыз балалар бұл белгіні тек экесінен қабылдап алады; в) белгі
ата-аналарында кездессе, балаларында байқалуы мүмкін, бүл
жағдайда ол тек 50 пайыз ұлдарында байқалады;
4. Х-тіркескен рецессивті түкым куалау: а) белгі үлдарда
жиірек байқалады; б) егерде белгі анасында болатын болса үлының
бэрінде де байқалады (анасы гомозиготалы), не үлының 50 пайы-
зында байқалады (анасы гетерозиготалы); в) егер белгі экесінде
болатын болса, ол тек кыздарына ғана беріледі, қыздары «тасы-
малдаушы».
-о
б~йТГ ~ъ
2 3 4
III
IV
О
1 2
3
4 5
.
\ ^
1 2 3
20-сурет. Рецессивті Х-тіркескен түкым куалау типі
59
*rO
ОтІГ "IbO
III
IV
һоЬтО ІІҺО
3 f4 5 6
оЪ о
21-cypem. У-тіркескен тұкым куалау типі
5. У-тіркескен түкым қуалаудың ерекшелігі белгі тек еркек
жынысқа беріліп отырады, себебі У хромосома экесінен тек ұл
балаларына ғана беріледі. Оны голандриялыц гпұқым куалау деп
атайды, У хромосомамен тіркескен 3 ген белгілі: 1) гипертрихоз
гені — ұл балалардың құлақ қалқаншасын қалың түк басуы, ол
17 жастан кейін байкалады; 2) аталық безді дамытатын ген; 3) ер
адамдардың тіс жүйесінің дамуын қадагалайтын ген.
3.4. Гендердің кұрылысы мен қызмет етуі туралы
молекул алық-биологиялық түсінік
Молекулалық биология өз алдына дербес ғылым ретінде
XX ғасырдың 40—50 жылдары бөлініп шықты. Ол эр түрлі
гылымдардьщ (биология, химия, физика, т.с.с.) түйісуінің нәти-
жесінде пайда болды, сондықтан да молекулалык биология аталған
жэне басқа да ғылымдардың жетістіктерін, зерттеу эдістерін
кеңінен пайдаланып талдау жасайды.
Молекулалық биология — тіршілікті молекулалық деңгейде
зерттейтін биологияның ең маңызды салаларының бірі. Ол
биологиялык макромолекулалардың нуклеин қышқылдары мен
ақуыздардың құрылысын, қызметін зерттейді.
60
Қазіргі кезде молекулалық биология идеялары биологияның
барлық салаларында кеңінен таралган жэне ол теориялық,
эксперименталдық биологияның дамуына елеулі ықпал етеді.
Молекулалық биология идеялары генетика саласында XX
ғасырдың басында колданыла бастады. 1908 жылы ағылшын
емдеушісі А. Гэррод адамдардың алкаптонурия (адам несебінің
түсінің ауада қараюы) ауруын зерттеп, ол ауру адам агзасында го-
могентезин кышкылы алмасуының бұзылуы салдарынан болаты-
нын жэне бүл аурудың тұқым куалайтындыгын анықтаған.
1945 жылы Д. Бидл жэне Э. Татум зерттеулерінің нәтижесінде
бір ген — бір ақуыз (фермент) — бір белгі деген ұғым қалыптасып,
генетиканың биохимиямен тікелей байланысы бар екендігі
көрсетілді.
1950—1960 жылдары тұқым қуалаушылықтың материалы
— нуклеин қышқылдарының кұрылысы зерттелініп, 1953 жылы
Ф. Крик жэне Дж. Уотсон ДНҚ молекуласы қүрылысының моделін
анықтады; генетикалық ақпараттық ДНҚ молекуласындагы ну¬
клеотидтер тізбегінде генетикалық код кұйінде жазылатындыгы
анықталды; ДНҚ молекуласының өздігінен екі еселену (реплика-
циялану) механизмі белгілі болды; жасушада ақпараттың берілу
жолдары айқындалды.
1954 жылга дейін ген тұқым қуалаушылықтың ең кіші, әрі карай
бөлінбейтін қызметтік, мутациялық жэне рекомбинациялық өлшем
бірлігі деп келінген. Бірак, кейінгі зерттеулер ген құрылысының
күрделі екенін, оның мутацияланып, рекомбинацияланатындығын
көрсетті. Сондықтан да бүгінгі күні ген тұқым қуалаушылықтың
ең кіші қызметтік өлшем бірлігі деп қарастырылуда. Ген дегеніміз
— бір полипептид молекуласын анықтауга арналған аппаратқа ие
ДНҚ молекуласының бір бөлігі. Ген деген терминді 1909 жылы
Дания галымы В. Иогансен енгізген. Геннің бірнеше қасиетгері
белгілі: 1) ген әрекетінің дискреттілігі — эр түрлі белгілердің
қалыптасуын түрліше гендер айқындайды; 2) ген түрақты бола¬
ды — мутация болмаған жагдайда ол үр п а қтан -ұрп аққа өзгеріссіз
беріліп отырады; 3) гендердің эрекеті нақтылы болады, ягни
олардың эрқайсысы нақтылы бір белгінің дамуын қамтамасыз
етеді, бір ген — бір белгіні, немесе бір ген бірнеше белгілерді да-
мытуы мүмкін — плейотропия.
61
Гендердің бірнеше жіктелу жүйесі белгілі: аллельді, аллельді
емес гендер; летальды, жартылай летальды гендер, т.с.с. Сонымен
қатар гендерді 3 топқа бөледі: 1) структуралық гендер (цистрон-
дар); 2) модуляторлық гендер; 3) реттеуші гендер.
Қүрылымдъщ гендерге — құрылымдық жэне ферменттік акуыз-
дарды, рибосома ақуыздарын, гистонды ақуыздарды, р-РНҚ,
т-РНҚ молекулаларын анықтайтын гендер жатады. Құрылымдық
гендер нақтылы белгілерді қалыптастырады, оның қызметтік өнімі
ретінде аРНҚ, полипептид немесе р-РНҚ, т-РНҚ синтезделінеді.
Модуляторлың гендерге ингибиторлар не супрессорлар, интен-
сифакторлар жэне модификаторлар жатады. Модуляторлық гендер
белгінің даму процесіне эсер етеді, не структуралық гендердің му-
тациялану жиілігін күшейтеді.
Реттеугиі гендерге едфылымдык гендердің активтілігін рет-
тейтін гендер жатады. Мысалы регулятор, оператор гендері.
XX гасырдың 20-жылдары хромосомалардың ақуыз жэне ну¬
клеин қышқылдарынан түратындыгы белгілі болды. 1928 жылы
Н.К.Кольцов ген қызметін ақуыз молекуласы атқаруы мүмкін де¬
ген болжам жасады. Бірак, кейінірек, түқым куалаушылық ақпарат
ақуыз молекуласында емес, нуклеин қышқылдарының (ДНҚ) мо-
лекуласында болатындыгы дәлелденді. 1928 жылы Ф.Гриффитс
бактериялардың трансформациялану қабілетіне тэжірибе жасап
ДНҚ молекуласының тұқым қуалаушылықтың материалдық негізі
екенін алгаш рет айтқан.
Трансформация дегеніміз — бактерияның бір штаммының
екінші бір штаммның ДНҚ молекуласының бір бөлігін өзіне қосып
алып, оның қасиеттеріне ие болуы.
Ф. Гриффитс тышқандарга вирулентті және вирулентті емес
пневмококк штаммдарын енгізіп, пневмококк штаммдарының
вируленттік қасиеті ДНҚ молекуласының фрагменттері арқылы
беріледі деп болжамдаған. 1944 жылы О. Эйвери, К. Мак Леод
жэне М. Мак Карта осы тэжірибені жаңа әдістемелік децгейде
жүргізіп, Ф. Граффитс болжамын растады.
1952 жылы Н. Циндер жэне Д. Ледерберг трансдукция
кұбылысын ашып (трансдукі/ия — бактериофаггардың бакте-
рияның бір штаммының ДНҚ фрагментін екінші штаммына көші-
ре алу қасиеті) ДНҚ молекуласының түқым куалаушылықтағы
рөлі туралы тагы бір дэлелдемеге қол жеткізді.
62
1950 жылы X.Френкель — Конрат темекі өсімдігіне темекі
мозайкасы вирусының вирулентті жэне вирулентті емес штамм-
дарының белок жэне РНҚ молекулаларын жеке-жеке жэне бірге
енгізіп, тәжірибе жасап, тұқым қуалаушылық ақпарат ақуыз мо-
лекуласында емес нуклеин қышқылдарында болатындығын
үзілді-кесілді дэлелдеді.
X. Френкель — Конрат тәжірибелерінің жобасы:
Вирулентті вирус штаммының
V
Ауру дамымайды
акуыз молекуласы
f
Вирулентті вирус штаммының
а Ауру дам иды
РНҚ молекуласы
I
Вирулентті вирус штаммының
V
а Ауру дамымайды
вирулентті РНҚ
Вирулентті емес вирус
штаммының акуыз
а Ауру дам иды
молекуласы + вирулентті РНҚ
молекуласы
Сонымен түқым куалаушылык акпарат ДНҚ молекуласының
нуклеотидтер тізбегінде генетикалық код күйінде жазылған.
Генетикалык код — дегеніміз нуклеин қышқылдарының
молекуласындағы түкым қуалаушылық ақпараттың 4 нуклеотид
аркылы кысқаша жазылу, сақталу жэне эрі карай жүзеге асу жүйесі
болып саналады.
Нуклеин қышқылдарының (ДНҚ, РНҚ) молекуласында 5 ну¬
клеотид кездеседі пуриндік негіздер — аденин (А), гуанин (Г)
және гіиримидиндік негіздер — цитозин (Ц), тимин (Т) жэне ура-
цил (У). Ал, нақтылы нуклеин қышкылдарының молекуласында
тек 4 азоттық негіз болады: ДНҚ-да А-Т; Г-Ц; РНҚ молекуласында
тиминнің (Т) орнында урацил (У). Сонымен, 4 нуклеотид арқылы
20 амин қышқылдары аныкталады, демек генетикалық код 1 нукле-
отидтен коп болуы керек, себебі, код жалғыз нуклеотидтен түрады
десек, 4 нуклеотид не бары 4 амин кышқылын ғана аныктаған бо-
лар еді, амин қышқылдарының саны 20, демек ол жеткіліксіз. Ал,
егер генетикалык код 2 (жұп) нуклеотидтен түрады десек, 4 ну-
63
клеотидтен эр түрлі 16 жұп құрастыруга болар еді (42 =16), бұл да
жеткіліксіз, себебі амин қышкылдарының саны — 20.
1954 жылы американ ғалымы Г. Гамов теория күйінде генети-
калық код 3 нуклеотидтен түруы мүмкін деген болжам айтқан.
Шынында да 4 нуклеотидтен 64 эр түрлі үштік құрастыруға бола¬
ды (43—64), ал бұл амин қышқылдарының санынан (20) әлдеқай-
да көп, ягни жеткілікті.
1961 жылы Ф. Крик генетикалық код триплетті, яғни 3 ну¬
клеотидтен тұратындығын тәжірибе жасап дәлелдеді. Ал, 1964
жылы М. Ниренберг, С. Очао, X. Корана еңбектерінің нэтижесінде
барлық 64 кодонның мағынасы анықталды жэне оның негізгі
қасиеттері белгілі болады. Генетикалық кодтың қасиеттері мы¬
надай: 1) кодтың колинеарлылығы (сэйкестігі), яғни нукле¬
ин кышқылындағы нуклеотидтердің бірізділікпен орналасуы
полипептидтегі амин қышқылдарының орналасу ретіне сэйкес
келеді; 2) генетикалық кодтьщ транскрипциялануы белгілі бір
нүктеден басталып бір бағытга жүріп отырады; 3) генетикалық код
үзіліссіз, үтірсіз болады; 4) генетикалық код артық болады, ягни
1 амин қышқылы 2 не одан да көп кодон аркылы анықталады; 5)
код триплетті (үш өрімді) болады; 6) генетикалық код әмбебап —
(универсал) болады, яғни генетикалық код барлық тірі ағзаларда
бірдей амин қышқылын анықтайды.
61 кодон нақтылы амин қышқылдарды анықтайды, оларды
мағыналы кодондар, ал 3 кодон УАА, УАГ жэне УГА ешқандай
амин қышқылдарды анықтамайды, оларды мағынасыз кодон¬
дар деп атайды; олар транскрипцияның не трансляпияның (ақуыз
синтезінің) аяқталуын анықтайды (стоп кодондар). Сол сияқты,
ақуыз синтезінің басталуын инициациялайтын кодон — инициа-
торлар белгілі, олар: АУГ, ГУГ жэне УУГ.
Инициаторлық кодондар а-РНҚ-ның бас жағында орнала-
сып, синтезделінуші полипептидтің алғашқы амин қышқылы —
формилметионинді немесе валинді анықтап, белок биосинтезінің
басталуын анықтайды.
Генетикалық код тұжырымдамасына сэйкес жасушада ақпа-
раттың үзіліссіз берілу жүйесі болуы қажет. Жасушада генетика-
лык акпараттың берілуінің 3 жолы белгілі: 1) генетикалық ак-
параттың жалпы тасымалдану жолы немесе биологияның (Криктің)
64
Генетикалық кодтың толық сөздігі
Триплет
Амин қышқылы
Триплет
Амин кышкылы
УУУ
Фенилаланин (Фен)
УАУ
Тирозин (Тир)
УУЦ
УАЦ
УУА
ЦАУ
УУГ
Лейцин (Лей)
ЦАЦ
ЦУУ
ЦУЦ
ЦАА
Глутамин (Глн)
ЦУА
ЦАГ
ЦУГ
ААУ
Аспарагин (Асн)
ААЦ
АУУ
Изолейцин (Иле)
ААА
Лизин (Лиз)
АУЦ
ААГ
АУА
ГАУ
Аспарагин кышкылы (Асп)
АУГ
Метионин (Мет)-кодон
инициатор
ГАЦ
ІУУ
Валин(Вал)
ГАА (Глу)
Глутамин кышкылы
ГУЦ
ГАГ
Г'УА
УГУ
Цистин (Цис)
ГУГ
Валин (кодон инициатор)
УГЦ
УЦУ
УГГ
Триптофон (Три)
УЦЦ
Серин (Сер)
ЦГУ
УЦА
ЦГЦ
Аргинин (Apr)
УЦГ
ЦГА
А ГУ
цгг
А ГЦ
АГА
АГГ
ЦПУ
Пролин (Про)
ГГУ
Глицин (Гли)
ццц
ГГЦ
ЦЦА
ГГА
ццг
ГГГ
АЦУ
Треонин (Тре)
УАА
АЦЦ
УАГ
Стоп кодондар
АЦА
У ГА
АЦГ
ГЦУ
I цц
Алании (Ала)
гцг
65
17(1-5
негізгі постулаты. Оны Ф. Крик ашқан болатын. Оның негізгі мәні
— генетикалық ақпараттың ДНҚ-ДНҚ-РНҚ — акуыз бағытында
берілуі. ДНҚ молекуласындағы ақпарат ДНҚ молекуласына оның
жартылай консервативтік репликациялануы негізінде беріледі.
Ал, ДНҚ молекуласынан РНҚ молекуласына — транскрипция,
РНҚ-дан ақуызға — трансляция процесі арқылы беріліп отырады;
2) генетикалық ақпараттың ерекше тасымалдану жолы. Ол 1970
жылы ашылған. Кейбір вирустарда — ретровирустарда, тұқым
қуалаушылық ақпарат ДНҚ молекуласында емес, РНҚ молеку¬
ласында жазылған. Олардың кейбіреулері адамда рак (ісік) ауру-
ын тудырады. Сонда калайша, ненің көмегімен вирустың РНҚ-
сындағы ақпарат адам ДНҚ-сына беріледі. 1970 жылы американ
ғалымдары X. Темин жэне Д.Балтимор бір жіпшелі РНҚ моле¬
куласы негізінде қос жіпшелі ДНҚ молекуласының синтезделуін
катализдеуші фермент — кері транскриптаза (ревертаза) ферментін
ашқан. Осы ферменттің катысуымен генетикалық ақпарат РНҚ-дан
ДНҚ-ға одан ДНҚ-РНҚ — ақуыз бағытында беріледі. Ақпараттың
осындай ерекше тасымалдану жолы адамдар мен жануарлардың
кейбір калыпты жасушаларында да байқалған; 3) ақпарат тасы-
малдануының тыйым салынған жолы табигатга кездеспейтін
жолы — ақпараттың ақуыздан нуклеин қышқылдарына берілуі.
Ақпараттың тасымалдануының мұндай жолы Жер бетінде ешбір
тірі ағзаларда кездеспейді.
3.5. ДНҚ молекуласының репликациялану механизмдері
ДНҚ молекуласының ең маңызды қасиеттерінің бірі: оның
өздігінен екі еселенуі (репликациялануы) болып саналады. ДНҚ
репликациялануыныц негізінде тұқым куалаушылык туралы
ақпарат ұрпақтан-ұрпакка өзгеріссіз, тепе-тец мөлшерде беріліп
отырады жэне ұрпақтардыц жалғасуын қамтамасыз етеді.
ДНҚ молекуласыныц репликациялану қасиеті 1953 жылы
Дж.Уотсон жэне Ф. Криктіц ДНҚ молекуласы құрылысының қос
спиральді болатындығын ашқаннан кейін белгілі болды. Теория
күйінде ДНҚ молекуласыныц репликациялануыныц 3 түрлі әдісі
болжамданған: 1) консервативті; 2) жартылай консервативті;
3)дисперсті репликациялану.
66
Көптеген тәжірибелер нәтижесінде ДНҚ молекуласының ре-
пликациялануы жартылай консервативтік жолмен жүретіндігі
дәлелденді. Оны 1958 жылы М. Мезельсон жэне Ф. Сталь Е.соіі
жасушасында байқаган.
Қазіргі танда ДНҚ молекуласының сырт пішінінің 3 түрлі ны-
саны белгілі: түрақты сақиналы ДНҚ молекуласы (бактериофаг-
тер), қүбылмалы сақиналы ДНҚ молекуласы (бактериофагтер),
сызықты ДНҚ молекуласы (прокариоттар жэне эукариоттар).
Осыған сәйкес ДНҚ молекуласының жартылай консервативті
репликациялануының 3 түрлі әдісі белгілі: 1) тета репликация; 2)
сигма репликация; 3) У — тэрізді репликация.
Тета репликация түрақты сақиналы ДНҚ молекулаларына тэн.
Ол ДНҚ сақинасыныц бір нүктесінен басталып, карама-карсы
бағытына карай ісініп, репликацияланудыц белгілі бір кезецінде
грек алфавитінің тета — Ө әріпіне ұксас қүрылым пайда етеді.
Кейбір агзалардыц даму циклінде алгаш сақиналы ДНҚ мо¬
лекуласы жайлап сызықты пішінге өтеді. Мүндай өзгеріс ДНҚ
молекуласыныц сигма репликациялану әдісінде жүзеге асады.
Сигма репликация сақиналы молекуланыц бір спиралінде фос-
фодиэфир байланысыныц ыдырауынан үзілістіц екі жағында
«жалаңаш» 3‘-ОН жэне 5'-Р04 үштары пайда болуынан баста-
лады. Содан кейін ДНҚ сақинасыныц 3'-ОН үшы бар спиралі
комплиментарлық жаца спиральдіц синтезделуі үшін матрица бо¬
лып, 5‘-Р04 үшы сызыкшаланып сақинаныц қүйрығына айналады
да, оныц негізінде комплиментарлы жаца спираль синтезделінеді.
Репликацияныц бүл әдісінде оның аралык қүрылымы гректіц сиг¬
ма (б) эрпіне үқсас болып келеді.
Кейбір прокариоттардыц жэне барлык эукариоттардыц ДНҚ
молекулалары сызықша тэрізді болып келеді жэне олардыц репли-
кациялануы белгілі бір нүктеден, репликациялык ісінудің пайда бо¬
луынан басталып, карама-қарсы жагына қарай бағытталады. Кей¬
де эукариоттардыц ірі ДНҚ-да репликациялануда жүздеген ини¬
циация нүктелері пайда болуы мүмкін. Репликациялану негізінде
көршілес ісінділер бір-бірімен қосылып хромосома үшында У
эріпі тэрізді аралық кұрылымды тугызады. Мұны У тэрізді жарты¬
лай консервативті репликациялану дегі атайды.
67
Барлық эукариотгардың сызықша тэрізді ДНҚ молекулалары У
тэрізді жартылай консервативтік жолмен репликацияланады. Ре¬
пликация басталуы үшін аналық молекуланың қос спиралінің ши-
ратылуы жазылып, бір-бірінен ажырасуы қажет. Содан кейін бір-
бірінен ажырасқан спиральдар екінші спиральдің синтезделуі үшін
матрица (калып) қызметін атқарады. Екінші спиральдің синтезін,
яғни дезоксирибонуклеотидтердің бір-бірімен фосфодиэфирлік
байланыс арқылы байланысуын басқарушы фермент ДНҚ — по¬
лимераза ферменті болып саналады. Бірақ, ДНҚ —- полимераза
ферменті өз бетіндіе ДНҚ синтезін бастай алмайды, ол үшін бос
3'-0Н үшы бар праймер — «үйытқы» кажет. Ал ондай «праймер»
рөлін «РНҚ-праймер атқарады. «РНҚ-праймер» ДНҚ синтезін ба-
стайды да жойылады, себебі эрі карай оның кажеті жоқ. Сонымен
ол ДНҚ синтезін иницияциялаушы болып табылады.
22-сурет. Репликация жобасы.
ДНҚ молекуласының спиральдері Д. Уотсон — Ф. Криктің кос
спиральді моделіне сэйкес бір-біріне антипаралельді болып келеді,
яғни репликативтік айырда оныц З1 жэне 5' үштары бір-біріне
қарама-қарсы орналасады. Ал, ДНҚ синтезі белгілі бір бағытта
жүреді, ягни ДНҚ полимераза дезоксирибонуклеотидтерді «РНҚ,
праймерге» және бір-біріне 3‘-0Н үштары арқылы жалғайды.
Демек, ДНҚ синтезі спиральдің 51 — үшынан З1 _үшына қарай
жүреді. Сондықтан репликативтік ашаның бір спиралінде ДНҚ
үзіліссіз тізбек арқылы синтезделінеді, оны лидерлік тізбек, ал
екіншісінде үзіліп-үзіліп жүреді, оны ілесуиіі тізбек деп атайды.
Екінші ілесуші тізбекте синтезделінетін ДНҚ үзінділерін Оказа-
68
ки фрагменттері деп атайды, себебі бүл кұбылысты 1968 жылы
Р. Оказаки ашқан болатын. Оказаки фрагменпітері ДНҚ лигаза
ферментгің қатынасуымен бір-біріне жалганады.
Сонымен, бүгінгі танда ДНҚ синтезі, ягни ДНҚ репликациясы
өте күрделі, коп сатылы процесс екендігі белгілі болып отыр. Оған
көптеген ферменттер жэне баска да белоктар қатынасады. ДНҚ ре-
пликациясына мынадай белоктар қатынасады:
1) хеликаза — ДНҚ молекуласының кос спиралінің ширатылу-
ын ажыратады, репликативтік айыр пайда болады;
2) SSB — ақуыздар ажырасқан ДНҚ жіпшелеріне жабысып,
олардың бір-бірімен күні бүрын жанасуын болдырмайды;
3) топоизомераза — ДНҚ молекуласындағы тізбектердің ажы-
рауынан пайда болатын үлкенді-кішілі түйіндерді жояды;
4) ДНҚ — полимераза ДНҚ синтезін, ягни дезоксирибо-
нуклеотидтерді бір-бірімен фосфодиэфирлік байланыс арқылы
жалғаушы фермент;
5) праіімаза — «РНҚ праймерді» синтездейді;
6) 5'-3‘ — экзонуклеаза — «РНҚ праймерді» жояды, репара-
цияға қатынасады;
7) 3'-5' — эндонуклеаза репликация қателіктерін жондеуге
катынасады;
8) ДНҚ — лигаза — Оказаки фрагменттерін бір-біріне 3'-ОН
жэне 5'-Р04 үштарымен жалғайды;
9) эндонуклеаза — ДЫҚ репарациясына қатынасады;
10) Гликолизаза — ДНҚ репарациясына катынасады.
ДНҚ репликациясы нактылы, қателіксіз жүретін процесс. Де-
генмен, ІОМО10 нуклеотидтен біреуі қате синтезделуі мүмкін. ДНҚ
синтезі прокариотгарда да эукариоттарда да бірдей жоба бойын¬
ша жүреді, дегенмен прокариоттарда ол жылдам -1 секундта 1000
нуклеотид, ал эукариоттарда баяулау — не бары 100 нуклеотид
синтезделінеді.
3.6. Гендердің экспрессиялануының (активтенуініц)
реттелу механизмдсрі
Тұқым куалаушылықтың материалы — ДНҚ молекуласы,
негізінен ядрода шоғырланған, ал белок биосинтезі цитоплаз-
69
мада — рибосомада жүреді. Сонда, ДНҚ молекуласының бойы-
на «жазылған» ақпарат қалайша цитоплазмаға жеткізіледі? ДНҚ
молекуласындағы генетикалық акпараттың цитоплазмада белок-
фермент синтезі күйінде жүзеге асуы тіршіліктің түпкілікті
қасиеттерінің бірі болып саналады.
Цитоплазмада белгілі бір белоктың синтезделу активтігі
гендердің активтігіне (экспрессиялануына) байланысты екені айдан
анық. Жасушада белок биосинтезі екі саты арқылы жүзеге асады —
транскрипция жэне трансляция. Транскрипция дегеніміз — генніц
экспессиялануы (активтенуі) нэтижесінде а-РНҚ молекуласыныц
синтезделуі, яғни ДНҚ молекуласындағы ақпараттыц а-РНҚ мо-
лекуласына көшіріліп жазылуы (транскрипция — көшіріліп жа-
зылу деген үгымды береді), ал трансляция а-РНҚ молекуласыныц
негізінде цитоплазмада — рибосомада, полипептид молекула¬
сыныц синтезделуі болып табылады.
Транскрипция — өте күрделі процесс. ДНҚ бойына орналасқан
гендердің бэрі бірдей, бір мезгілде транскрипцияланбайды. Ол
біріншіден жасуша тіршілігінің белсенділігіне жэне даму кезеціне,
екіншіден, гендердіц экспрессиялануыньц реттелу механизмде-
ріне байланысты болады. Сондықтан да, бір мезгілде эр түрлі
жасушаларда түрліше гендер транскрипцияланады жэне ағзаныц
дамуыныц әр түрлі кезеңдерінде бір жасушаныц түрліше гендері
экспрессияланады.
Әдетте біз, бір ген — бір белок (фермент) деген үгым бой¬
ынша эрбір ген оз алдына жеке транскрипцияланады деп ойлай-
мыз. Ал, шын мәнінде, бір белгіні дамытатын бірнеше гендер
ДНҚ бойына қатар орналасып, бірге транскрипцияланады. Ондай
гендерді кластерлі гендер деп атайды. Кластерлі гендердіц бәрі
бірдей транскрипцияланып ортақ полицистронды а-РНҚ түзіледі.
Осыныц негізінде бір белгінің дамуына қажет барлық ферменттер
бір мезгілде синтезделінеді. Кластерлі гендердің экспрессиялану-
ын ерекше реттеуші гендер реттеп отырады.
Гендердіц экспрессиялануыныц реттелу механизмдерін зерт¬
теу үшін прокариоттар өте қолайлы объект болып саналады,
себебі олардыц геномдары не бары бірнеше гендерден құралган
жэне олар оте тез кобейе алады. Сонымен қатар, гендердіц
экспрессиялануыныц реттелу механизмдері прокариоттарда да,
эукариоттарда да ұқсас жүретіндігі анықталды.
70
Гендердің экспрессиялануының реттелу механизмін зерттеуде
ішек бактериясы — Escherichia соіі өте колайлы объект болды.
Е. соіі тіршілігі үшін қалыпты энергия көзі болып глюкоза сана¬
лады. Егер де тіршілік ортасында глюкоза болмаса ол лактозаны
пайдалануга көшеді. Осы кезде жасушада лактозаны ыдырата-
тын Р-галактозидаза ферменті синтезделуі қажет. р-галактозидаза
ферменті дисахарид-лактозаны галактозага жэне глюкозага ыды-
ратады.
акуыз-репрессор
молекуласы
23-сурет. Опероннын кұрылысы.
Е. coli жасушасында р-галактозидаза ферментінің синтезделуі
коректік ортада лактоза болган жагдайда индукцияланады, ал
оның молшері азайса, не мүлдем болмаса бүл ферменттің синтез-
делу қарқыны да азаяды, не тоқталады. Р-галактозидаза ферменті
синтезделу үшін Е. coli ДНҚ-сындагы Lac-Z деген ген транс-
крипцияланып, оның а-РНҚ,-сы түзілуі кажет, р-галактозидаза
ферментінің синтезделу қарқыны индукцияланғаннан кейін 1000
есеге дейін көбейеді жэне ол қоректік ортада индукторлакто-
за болса бір деңгейде үзақ уақыт сақталынады. р-галактозидаза
ферментінің лактозадан басқа индукторы ретінде оның ыды-
рауында пайда болатын аралық зат — аллолактоза да саналады.
Ортада индуктордың (лактоза не аллолактоза) азаюы не жойылуы
Р-галактозидаза а-РНҚ-сыньщ нуклеотидтерге ыдырап жойылуы-
на алып келеді. А-РНҚ-ньщ тіршілік үзақтығы бірнеше минутқа
ғана тең, сондықтан да оның бір деңгейде синтезделіп тұруы
71
үшін жэне жасушада үнемі [З-галактозидаза ферменті синтезделуі
үшін ол үнемі индукцияланып түруы қажет, яғни ортада лакто¬
за не аллолактоза болуы қажет. Сонда ғана а-РНҚ-ның ыдырауы
мен синтезделуі теңеседі, жасушада р-галактозидаза ферменті бір
қалыпты синтезделінеді.
Бактерия гендерінің экспрессиялануының реттелуін зерттеу-
де Ф.Жакоб жэне Ж. Моно еңбектерінің маңызы өте зор. Олар
өздерінің тәжірибелерінің нэтижесінде 1961 жылы гендердің
экспрессиялануының реттелуінің оперондъщ теориясын ] сынды.
Е. coliжасушасында лактозаны толық ыдырату үшін бір-бірімен
тығыз байланысқан екі геннің өнімі қажет. Олар — Р-галактозидаза
ферментін анықтайтын — Lac-Z+ гені жэне пермеаза ферментін
анықтайтын Lac-Y+ гені. в-галактозидаза ферментінің қызметі
белгілі, ал пермеаза ферменті Е. coli жасушасына лактозаны
белсенді тасымалдау қызметін атқарады. Осы екі генмен қатар
орналаскан үшінші ген де болады ол Lac-A+ гені. Lac-A+ гені тио-
галактозидтрансацетилаза ферментін анықтайды. Ол жогарыда
келтірілген 2 генмен — Lac-Z+, Ьас-У+, тығыз байланысып
кластерлік гендер кешенін күрайды, бірге реттелінеді, бірге транс¬
крипцияланады, бірақ лактозаның ыдырауына қатынаспайды. Бүл
үш ген мынадай тәртіппен орналасады Z-У-А. Осы үш геннің
бірге индукциялануының нэтижесінде бір полицистронды а-РНҚ
синтезделінеді.
ДНК
л
полтіистрондық а-РНҚ
A
активсвденген
репрессор
индуктор р-Галактозндаза
молекуласы
t
лЛд*
Пермеаза
о°о°оо
А-акуыз
24-сурет. Гендердін экспрессиялануынын реттелу механизмдері
Ф. Жакоб жэне Ж. Моно өздерінің тәжірибелері нэтижесінде мы¬
надай болжам жасады: Z+,y+,A+ кластерлі гендердің транскрип-
72
циялануы оператор (0+) деп аталатын геннің бақылауында бола¬
ды. ал оператордың кызметі, өз кезегінде. репрессор деп аталатын
(1+) ген арқылы реттелінеді. Репрессор екі түрлі қызмет атқарады:
1) егер ортада индуктор (лактоза, аллолактоза) болмаса оның өнімі
— репрессор-ақуыз молекулалары, операторды (0+) «тыгындап»,
әрі қарай Z+, У+, А+ гендеріне ақпаратты өткізбей, олардың транс-
крипциялануын болдырмайды; 2) ал егер ортада индуктор (лакто¬
за, аллолактоза) болатын боса, онда индуктор молекулалары (лак¬
тоза, не аллолактоза) репрессор-акуыз молекулаларына жабысып,
косылып оларды активсіздеңдіреді, сондықтан активсізденген ре¬
прессор молекулалары операторды (0+) «тыгындап» жаба алмай,
ақпарат әрі карай Z+,y+,A+ гендеріне өтіп олардың транскрип-
циялануына «рұқсат» беріледі. Осы гендердің транскрипциялануы
гіромотор учаскесінен басталады.
Ф.Жакоб, Ж.Моно- Z+,y+,A+ гендерін жэне оператор, промо¬
тор учаскелерін оперон деп атаған.
Сонымен. гендердің экспрессиялануының реттелуіне 3 түрлі
реттеуші элементтер қатынасады.
1)реттеуиіі ацуыздар — транскрипцияның инициациялануын-
да не тежелуінде РНҚ полимераза ферментінің активтілігі-не эсер
ететін акуыздар (репрессор);
2) эффекторлар — ұсак, белок емес заттар, олар реттеуші
акуыз молекулаларымен косылып олардың активтілігіне эсер етеді
(индукторлар — лактоза, аллолактоза);
3) реттеуші нуклеотид тізбектері — ДНҚ молекуласында
болатын кейбір реттеуші нуклеотид тізбектері (промотор, терми¬
натор). Реттеуші акуыздардың осы учаскелерге тигізетін эсерінің
нэтижесінде а-РНҚ синтезінің активтігі реттелінеді.
Ф. Жакоб жэне Ж. Моно өздерінің оперондық гипотезасын
теория күйінде болжамдаған болатын. Ал қазіргі кезде опе¬
рон теориясының шындығы коптеген тәжірибелер аркылы
дэлелденген. Мысалы, таза күйінде репрессорды бөліп алып зерт-
теп оның акуыз екендігі дэлелденді; репрессор молекуласындағы
амин қышқылдарының реті анықталып, оның 1+ геніндегі нуклео¬
тидтер тізбегінің кодына колинеарлы болатындыгы анықталды.
Сонымен катар, реттеуші ақуыздардың ДНҚ молекуласының
реттеуші аймактарымен нақтылы байланысатындығы анықталды.
73
Мысалы, 3,2 xl06 нуклеотид жұбынан тұратын Е. coli ДНҚ-сының
ішінен Lac — репрессор тандап, тек қана белгілі бір нуклеотидтер
тізбегімен (оператор) байланысады, ал оның ұзындығы не бары 24
нуклеотид жұбына тең.
Прокариоттар негізінде зерттелініп, анықталган гендердің
экспрессиялануының реттелуінің оперондьщ механизмі
эукариоттарға да тэн. Тек, эукариоттардың айырмашылығы, көп
жасушалы ағзаларда эр түрлі жасушалар бірдей жэне түрліше
ақуыздарды синтездейді. Яғни эукариоттардың гендері екі топқа
бөлінеді: 1) бір жасушаныңтүпкілікті, әмбебапты тіршілік кызметін
қамтамасыз ететін, яғни барлық жасушалар типінде кездесетін
гендер. Оларды «түрмыстың « гендер деп атайды, оларсыз кез
келген жасушаның тіршілігі мүмкін емес; 2) жасушаның ерекше
қызметтерін қамтамасыз ететін жэне тек кейбір жасушаларда ғана
актив болатын гендер. Оларды «молитлыц» гендері деп атайды.
Эукариотгар гендерінің экспрессиялануының реттелу жобасын
1972 жылы Г.П.Георгиев жасаған.
Коп жасушалы ағзалардың ДНҚ молекуласының ұзындығы
бір жасушалыларға қарағанда элдеқайда үзын, бірақ сол ДНҚ
молекуласының бэрі бірдей а — РНҚ-ға айналмайды, себебі
алгашқы про-а-РНҚ (Гя-РНҚ) түзіліп, оның тек азғана бөлігі -10%
ғана, а-РНҚ-ға айналады. Адам геномы 2,9 хІО6 нж түрады де¬
сек, одан пайда болган а-РНҚ мөлшері 2 хЮ8 нж-га тең. А-РНҚ
гендерінің орташа үзындығы 1800 нж-ға тең десек адам геномын-
да 110000-га жуық қүрылымдық гендер кездеседі (соңғы кездері
адам геномындағы гендердің саны 30000 деп айтылуда).
Эукариоттаргендерініңэкспрессиялануыныңретгелумеханизмі
прокариотгарга үқсас. Дегенмен, оның кейбір ерекшеліктері де
белгілі. Эукариоттар гендері экзон-интрон учаскелерінен түрады.
А-РНҚ-ның пісіп-жетілуінде (процессинг, сплайсинг) интрондар
үзіліп түсіп, экзондар бір-бірімен жалғанады. Сол сияқты, эукари¬
оттар жасушасында ДНҚ молекуласы гистонды белоктармен өте
тығыз байланысқан, ал бүл көп жагдайларда РНҚ — полимераза
ферментінің әсеріне беріле бермейді де, а-РНҚ синтезделмейді.
Эукариоттар гендерінің транскрипциялануының инициа-
циясы прокариоттар сияқты промотордан басталады. Промотор
учаскесіне жақын жерлерде А-Т нуклеотидтері жиі кездесетін
74
ген орналасқан, оның ұзындығы 20-30 нуклеотидтерге тең. Бұл
аймақты Гольдберг-Хогнесс боксы немесе «ТАТА...” тізбегі деп
атайды. «ТАТА...” тізбегі РНҚ-полимераза ферментімен байланы-
сып транскрипцияның басталу нүктесін анықтайды.
3.7. Транскрипциялану немесе РНҚ синтезінің
механизмдері
ДНҚ молекуласының транскрипциялану механизмі прокариот-
тар жасушасында жақсы зерттелінген.
Транскрипция жүру үшін ДНҚ молекуласының жіпшелері кем
дегенде 2 өрім бойына бір-бірінен ажырауы қажет. Транскрип¬
ция промоторлық учаскеден басталып терминаторлық учаскемен
аяқталады. РНҚ синтезін жүргізуші, яғни рибонуклеотидтерді бір-
біріне жалгастырушы ферментті — РНҚ — полимераза ферменті
деп атайды. РНҚ — полимераза молекуласыныц негізін 4 компо-
ненттен (б , б,, Яу, Я.,) түратын кофермент қүрайды. Коферментке
бір ақуыз — сигма (б) ақуыз, жалғанса, ол РНҚ — полимеразаныц
холоферментіне ягни актив күйіне айналады.
Сигма белок ДНҚ молекуласының промоторлык учаскесін дэл
табады және бір-бірінен ажырасқан ДНҚ жіпшелерініц қайсысы
матрица (қалып) қызметін атқаратынын анықтайды. РНҚ синтезі
басталганнан кейін сигма (б) ақуыздыц кажеті жоқ, сондықтан ол
холоферменттен ажырап тусіп қалады, ал оныц орнына басқа бір
ақуыз-nus-A генініц өнімі келіп қосылады. Nus-A гені өнімінің
қызметі — ДНҚ молекуласыныц терминациялық учаскесін та-
нып, транскрипция процесін тоқтату. Синтезделінген РНҚ
молекуласыныц ДНҚ спиральдерінен ажырап бөлініп шыгуы үшін
тагы бір ақуыз қажет, ол ро (р) фактор деп аталынады.
Эукариоттар ДНҚ-сының транскрипциясы, жалпы алғанда,
прокариоттарғаұқсас,дегенменбіршамаерекшеліктердекездеседі.
Мысалы, прокариоттарда РНҚ-ныц 3 түрі де (а-РНҚ, т-РНҚ,
р-РНҚ) бір РНҚ полимераза ферменті аркылы синтезделінеді, ал
эукариоттарда олардың эрқайсысы тек өздеріне ғана сэйкес РНҚ-
полимеразалар арқылы синтезделінеді: р-РНҚ-РНҚ-полимераза-І
арқылы; а-РНҚ-РНҚ-полимераза-ІІ арқылы; т-РНҚ-РНҚ-полиме-
раза-ІІІ арқылы синтезделінеді.
75
25-сурет. Транскрипция жобасы
Прокариоттар гендерінің құрылысы карапайым, ол тек магына-
лы нуклеотид тізбектерінен түрады жэне түгел транскрипцияла-
нып бірден цитоплазмаға өтіп трансляция үшін матрица (қалып)
бола алады. Ал, эукариоттар гендерінің құрылысы күрделі, ол
мағыналы және мағынасыз учаскелерден түрады. Мағыналы учас-
келерін экзондар, мағнасыз учаскелерін интрондар деп атайды.
Транскрипция процесінде гендердіц экзондық жэне интрондық
учаскелерініц бәрі де бірдей синтезделінеді, оны про-а-РНҚ (Гя-
РНҚ) деп атайды. Бірақ ол цитоплазмага өтіп, белок синтезі үшін
матрица (қалып) кызметін атқара алмайды, себебі ол пісіп-жетілуі
қажет, яғни нағыз а-РНҚ-ға айналуы қажет. Про-а-РНҚ немесе Гя-
РНҚ-дан нағыз а-РНҚ пайда болуы үшін оның интрондары қиылып
алынып тасталып, экзондары бір-бірімен жалғануы қажет. Бұл
процесті процессинг жэне сплайсинг деп атайды. Пісіп-жетілген
а-РНҚ ядродан цитоплазмага өтіп акуыз синтезі үшін матрица
(калып) қызметін атқарады.
3.8. Ақуыз биосинтезі
Ақуыз биосинтезі тіршіліктің ең мацызды қасиеттерініц
бірі, себебі тіршілік процесінде ақуыз молекулалары эр түрлі
биохимиялық реакцияларға түсіп, ыдырап жойылып отырады,
ал олардыц орнын толтыру тек жаца акуыз молекулаларыныц
синтезделуі нэтижесінде жүзеге асады.
76
А-РНҚ молекуласыныц нуклеотидтер тізбегінде жазылған
акпараттың колинеарлы полипептид молекуласыныц аминқыш-
қылдары ретіне берілуін, яғни акуыз синтезін трансляция деп
атаймыз.
Трансляция процесі полипептидтіц N ушынан басталып С
ұшына қарай жүреді жэне ол метионин амин қышқылынан ба-
сталады. Яғни, кез келген а-РНҚ молекуласыныц бас жағында
метионин амин қышкылына сэйкес келетін (колинеарлы) код-
инициаторлық код болады (АУТ). Полипептид синтезініц иниция-
лануы ерекше т-РНҚ-инициаторлық т-РНҚ-ның қатынасуымен
жүзеге асады. Бұл процеске 3 инициациялаушы факторлар — IF,
IF,, IF, катынасады. Ақуыз биосинтезініц жалғасуы немесе элон¬
гация сатысы 2 фактордың қатысуымен жүреді — EF-1, EF-2, ал
белок синтезінің терминациялануы кодон-терминаторларға жэне
терминациялық факторларга — RF-1, RF-2-ге байланысты. Кодон
терминаторлардың 3 түрі белгілі — УАА, УАГ, УГА.
Трансляция процесіне 3 РНҚ — үшеуі де қатынасады.
А-РНҚ-ақуыз синтезі үшін матрица (қалып) қызметін атқарса,
рибосомалық РНҚрибосомалардыц қүрамына кіреді. Рибосомалар
цитоплазманыц ец үсак субмикроскопиялық органеллалары бо¬
лып табылады. Акуыз синтезі жүруі үшін рибосомалар а-РНҚ-мен
байланысады. Рибосома 2 бөлшектен түрады: рибосоманың кіші
бөлшегі — 30S, рибосоманың үлкен бөлшегі — 50S. Әдетте, ақуыз
синтезі жүрмеген кезде, рибосоманың екі бөлшегі бір-бірінен
бөлек кездеседі, олардыц бір-бірімен қосылып біртүтас органо¬
ид — рибосома түзілуі ақуыз биосинтезі басталган кезде жүзеге
асады. А-РНҚ алғаш рибосоманыц кіші бөлшегімен қосылады, со-
дан кейін оларға иницияция факторлары IF, IF2, IF, жэне т-РНҚ —
аминоацилметионин қосылып инициаторлық кешен түзіледі.
Инициаторлық кешен түзілгеннен кейін оган рибосоманыц
үлкен бөлшегі келіп қосылады. Рибосоманыц үлкен бөлшегінде 2
учаске болады: пептидилдік бөлік — П бөлік, аминоацилдік бөлік
—А бөлік. Рибосоманыц үлкен бөлшегініц П бөлігіне формильден-
ген метионинмен инициаторлық т-РНҚ орналасады, ал А-учаскеге
а-РҒІҚ-ныц екінші кодонына сэйкес келетін антикодоны бар т-РНҚ
өзінің амин қышқылымен орналасады (26-сурет).
77
Трансляция процесінде а-РНҚ кодондары амин қышқылдары-
мен тікелей эрекеттесе алмайды. Олардың арасындағы байланыс
тасымалдаушы РНҚ (т-РНҚ) арқылы жүзеге асады. Әрбір амин
қышқылының тек өзіне ғана сэйкес т-РНҚ-сы болады, сонымен кем
дегенде т-РНҚ белгілі — 20-амин қышқылының 20 т-РНҚ-сы және
инициаторлық Т-РНҚ. Т-РНҚ-лар -т-РНҚФсн, т-РНҚАлан, т-РНҚСер,
т-РНҚИзол, т-РНҚАсп т.с.с деп аталынады. Барлық прокариоттардың
жэне эукариоттардың г-РНҚ молекулалары 80-ге жуық нуклео-
тидтерден құралған жэне олардың соңғы құрылысы бір-біріне
ұксас болып келеді. Олардың молекулаларының соңғы кұрылысы
беденің 3 құлақты жапырағына ұқсас. Т-РНҚ молекуласыныц 2
ерекше қасиеті белгілі: 1) т-РНҚ-ның 3'-ұшында ерекше рЦ рЦ
рА-ОН-тан туратын бос нуклеотид тізбегі болады, осы ұшымен
т-РНҚ-ға сэйкес келетін амин қышқылы ковалентті байланысады;
2) т-РНҚ-ның қарама-қарсы ұшында 3' нуклеотидтен тұратын ан¬
тикодон болады.
■ • в
26-сурет. Акуыз биосинтезінін жобасы
Т-РНҚ-ның негізгі қызметі — өзіне сэйкес амин қышқылын «та-
нып» 3'-ОН үшыиа қондыру және а-РНҚ-ның антикодонга компли-
78
ментарлы кодонын «танып» өзі алып келген амин қышқылын поли¬
пептид тізбегіне орналастыру болып саналады Амин қышқылының
өзіне сэйкес т-РНҚ молекуласының 3'-ОН ұшына ковалентті байла-
нысуы ерекше аминоацил т-РНҚ-синтетаза ферментінің қатысуы
арқылы жүреді.
20 амин қышқылына сэйкес 20 аминоацил т-РНҚ синтетаза
ферментгері болады. Аминоацил т-РНҚ, синтетаза ферменттері
өздеріне сэйкес келетін амин қышқылдарымен т-РНҚ-ның бір-
бірін «танып», амин қышқылыныц т-РНҚ молекуласыныц 3'-ОІ1
ұшына коваленттік байланысуын қамтамасыз етеді. Бұл процесс 2
сатыдан тұрады. Алғашқы сатыда амин қышқылдары активтенеді:
Мысалы, Серилацил т-РНҚ-синтетаза+серин+АТФа (серилацил-
АМФ) Серил-т-РНҚ-синтетаза+Ррі; Екінші сатыда активтенген
амин қышқылы-өзіне сәйкес т-РНҚ-ның 3'ОН ұшымен байланы-
сады. (Серилацил-АМФ) серил-т-РНҚ-синтетаза+т-РНҚ а серил-
т-РНҚ Сер+серил-т-РНҚ-синтетаза+АМФ түзіледі.
Аминоацил-т-РНҚ-синтетаза ферменттері трансляция про-
цесінің қателіксіз жүруін, яғни а-РНҚ кодына колинеарлы (сэйкес)
полипептид молекуласыныц синтезделуін қамтамасыз етеді. Де¬
мек, трансляцияның кателіксіз жүруі аминоацил-т-РНҚ-синтетаза
ферменттерінің өздеріне сэйкес амин кышқылдарын «танып» олар¬
ды тиісті т-РНҚ-ға қондыруына байланысты болады. Сонымен
қатар, аминоацил-т-РНҚ — синтетаза ферменттері бақылаушылық
қызметін де атқарады. Мысалы, изолейцил-т-РНҚ — синтетаза
ферменті өзіне сәйкес т-РНҚ орнына басқа—«жат» т-РНҚ, айталық
т-РНҚФен-ді, «таныды» делік. Осының нәтижесінде изолейцин-т-
рнқнзм орнына изолейцил-т-РНҚфен кешені пайда болар еді. Осы
қателіктіц нэтижесінде синтезделуші ақуыз молекуласында фени¬
лаланин амин қышқылының орнына изолейцин амин қышқылы ор-
наласар еді. Бірақ, фенилаланин-т-РНҚ-синтетаза ферменті өзініц
т-РНҚФЕН-сымен жат амин қышкылының қосылғанын сезіп калып
қате кешенді — изолейцин амин қышқылына жэне т-РНҚфен-ге
ыдыратады.
Аминоацил-т-РНҚ-синтетаза ферменттерініц екінші бақылау-
шылық қызметі амин кышқылдардыц активтену сатысында
байқалады. Айталық, изолейцил-т-РНҚ-синтетаза ферменті изо¬
лейцин амин қышқылыныц орнына валин амин қышкылын
активтендірді делік.
79
Изолейцил-т-РНҚ-синтетаза+валин+АТФ —»(валиладил-АМФ)
изол ейцил-т-РНҚ-синтетаза+Рр і.
Сонда, келесі сатыда «қате» түзілген кешен т-РНҚИзол-мен
әрекеттескенде валил-т-РНҚИзол пайда болуының орнына вали-
лацил-АМФ, еркін валин амин кышқылына жэне АМФ-ге гидро-
лизденеді.
(Валилацил-АМФ) изолейцил-т-РНҚ-синтетаза+т-РНҚИзол —>
валин +АМФ+т-РНҚИзсш+изолейцил-т-РНҚ-синтетаза.
Сонымен, аминоацил-т-РНҚ-синтетаза ферменттері нақтылы
амин қышқылдарын өздеріне тиісті антикодондармен байланы-
стырып генетикалық ақпараттың трансляциялану процесінде
маңызды рөл атқарады. Сонымен қатар, бұл ферменттердің қос
қабат бақылаушылық қызметі нэтижесінде трансляция процесінің
қателіксіз жүру деңгейі едэуір жоғарылайды. Мысалы, амин
қышкылдардың «қате» активтенуі шамамен 100 изолейцин моле-
куласынан 1 валинга тең болса, активтенген аминқышқылының
«жат» т-РНҚ-мен байланысуы 1/180-ге тең, сонда трансляцияның
жалпы қателігі 1/100 х 1/180= 1 /18000-нан аспайды.
Аминоацил-т-РНҚ кешені пайда болганнан кейін олар рет-
реттерімен тізіліп орналасып, амин қышқылдар арасында
пептидтік байланыс пайда болады. Оны рибосоманың үлкен
бөлшегінде кездесетін ерекше фермент — пептидил — трансфе-
раза ферменті қалыптастырады. Екі амин қышқылдары арасында
пептидтік байланыс пайда болганнан кейін рибосома бір кодонға
солға қарай жылжиды, осының нәтижесінде инициаторлық ке¬
шен рибосомадан сыртқа шығады, А учаскедегі кешен П учаскеге
өтеді, ал А учаскесіне а-РНҚ-ның үшінші кодонына сәйкес келетін
т-РНҚ, өзінің амин қышқылымен келіп орналасады, содан кейін
екінші жэне үшінші амин қышқылдары арасында пептидтік байла¬
ныс пайда болады. Бүл жерде элонгация факторлары — ЕҒ-1, ЕҒ-2
қатынасады. Содан кейін рибосома тағы бір кодонға солғажылжып
А учаске босайды. Осылайша ақуыз синтезі А учаскеге а-РНҚ-ның
терминаторлық кодондары келіп жеткенге дейін жалгаса береді.
Терминаторлық кодондарды (УАА, УАГ, УГА) арнайы терминация
факторлары — RF-1, RF-2 «таниды». Терминаторлык кодондарға
сэйкес келетін амин қышқылдары болмагандықтан А — учаске
бос қалып, пептидилтрансфераза ферменті пептидтік байланыс
80
жасай алмайды, сондықган рибосома 2 бөлшекке ыдырайды, син-
тезделген полипептид бөлініп шығады. Осылайша ақуыз синтезі
аяқталады.
3.9. Цитоплазмалық туқьш қуалаушылық.
Генетикалық инженерия. Жасушаның мутацияға қарсы
антимутациялық биологиялық механизмдері
Генетика ғылымынын негізін қалаушылар — Г. Мендель,
Т.Морган жэне олардың шәкірттерінің еңбектері нәтижесінде
калыптасқан тұкым қуалаушылық теориясын хромосомалық не¬
месе ядролық тұқым қуалаушылық деп атаймыз, себебі бұл тұкым
қуалаушылық ядромен байланысты.
Дегенмен, генетика ғылымының дамуында ядролық тұқым
қуалаушылық теориясы түрғысынан түсіндіруге болмайтын
көптеген деректер жинақталған. Осы деректерді түсіндіру үшін
тұқым қуалаушылық процесіне тек қана ядро емес, сол сияқты,
цитоплазма да катынаса ма деген пікір пайда болды.
Егер тұқым қуалаушылық материалы — ДНҚ молекуласы бо¬
лып саналатын болса, ол негізінен (90 пайызга жуыгы) ядрода
шогырланган, ал 10 пайыздай болігі цитоплазмада — митохондри-
яда, хлоропласттарда кездеседі. Митохондрияда, хлоропласттарда
ДНҚ-мен бірге рибосомалар да кездеседі, демек осы органелла-
ларда өздігінен ақуыз синтезделініп, олар оз бетінше көбейіп, жа-
сушадан жасушага беріле алады.
Мысалы, тек күн сэулесінің әсерімен протопластидтерден хло-
ропласттар түзілетіні белгілі, ал қараңғыда олар протопластид
күйінде қала береді. Егер радиация әсерімен протопластидтердің
ядросын жойсақ, онда олар хлоропласттарга айнала алады, ал
егер цитоплазмасын жойсақ, хлоропласттарга айнала алмайды.
Демек, протопластидтердіц хлоропластқа айналуы, ягни хлоро-
фил синтезделуі, ядро емес цитоплазма қасиетіне байланысты
болганы. Осылайша 1908 жылы К. Корренс цитоплазмалык,
(пластидтік) тұқым куалаушыльщты, ал 1949 жылы Б.Эфрусси
ашыткы саңыраукұлагын зерттеп, митохондриялық туқым
қуалауіиылыцты ашты. Цитоплазмалык тұқым қуалаушылық осы
органоидтарда кездесетін сақиналы ДНҚ-га.байланысты.
81
170-6
Адам митохондриясының геномы 16569 нж-дан тұратын
сақиналы ДНҚ куйінде болады, онда р-РНҚ, т-РНҚ, с,в-цитохром
т.б. белоктардың гендері кездеседі.
Цитоплазмалық тұқым қуалаушылық жасушада ядродан тыс
орналасқан гендер арқылы беріледі, оларды плазмогендер, ал
олардың жиынтығын плазмон деп атайды. Тұқым қуалаушылық-
тың мұндай түрі аналық жыныс жасушасы арқылы беріледі, себебі
сперматозоидтарда цитоплазма болмайды. Аналық жыныс жасу¬
шасы арқылы хламидомонаданың стрептомицин жэне т.б. анти-
биотиктерге төзімді болу қасиеті, өсімдік жапырақтарында ақ
дақтардың болуы, кейбір өсімдіктердің гүлдерінде аталықтардың
жетілмеуі, т.с.с. белгілер беріледі.
Кейбір отбасыларда үш үрпақ бойына 79 қыз туылып, бірде-бір
ұл баланыц туылмауын да осы цитоплазмалыктұқым қуалаушылық
арқылы түсіндіруге болады.
Плазмогендерді екі топка белуге болады: 1) ДНҚ-сы болатын
жасуша органеллаларының гендері (митохондриялар, хлоропласт-
тар); 2) жасуша симбионттарыныц (вирустар, плазмидалар) жэне
инфекция аркылы жасушага енген гендер.
Екі топтыц да плазмогендері ядролык гендерге үксас бо¬
лады, олар өздігінен екі еселенеді, үрпақтан-үрпаққа тепе-тең
молшерде беріліп отырады, мутацияланады. Олар кейбір өте
қажетті ферменттердіц синтезделуін реттеп, маңызды белгілерді
қалыптастыру үшін қажет.
Молекулалық биология, биохимия жэне генетика гылымдары-
ныц соцгы онжылдықтарда қарқынды дамуы нэтижесінде оның
жаца саласы — генетикалыц инженерия қалыптасты.
Генетикалық инженерия дегеніміз жасушада өздігінен
көбейе алатын, белгілі бір затты синтездеуге қабілетті, тұқым
қуалаушылық материалдарын қолдан жасайтын молекулалық
биологияныц жаца саласы. Генетикалық инженерия 1972 жылдан
бастап дамып келеді. Осы жылы Берг алгаш рет ішек бактериясы
мен бактериофаг ДНҚ-ларынан қүрылган жасанды гибридтік ДНҚ
молекуласын қүрастырган. Генетикалық инженерия ғылымыныц
өзіне тэн әдістері болады, олар: 1) кажетті гендерді ДНҚ молеку-
ласынан бөліп алу; 2) оларды қолдан көбейту; 3) ол генді басқа
жасушаға — иесіне енгізіп жүмыс істету. Осы операциялардың
бэрін плазмиданы пайдалану арқылы жүргізеді.
82
Плазмидалар дегеніміз жасушада тұрақты күйде кездесетін
жэне хромосомамен байланыссыз, дербес тұқым қуалау факторы.
Плазмида деген терминді 1952 жылы Ледерберг енгізген болатын.
Плазмидалар сақина тэрізді ДНҚ молекуласынан тұрады. Оларға
митохондрия, хлоропласттар, бактериялардың тұқым қуалаушы-
лық аппараты жатады. Плазмидалар хромосомамен қосылып
эписоманы туындатады да, етеді де активтенеді. Плазмидалар
генетикалық инженерия саласында жиі пайдаланылады, себебі
оған кез келген қажетті генді жалғауға, содан кейін оны бактерия
жасушасына ендіруге жэне көбейтуге болады.
Алгашқы плазмиданы ерекше ферменттер — рестриктазалар
арқылы «кеседі» жэне қажет генді бөліп алады. Содан кейін сол
генді плазмида — векторга «тігіп» енгізеді. Оны «тігу» үшін ДНҚ
— лигаза ферменттерін пайдаланады. Содан кейін сол гибридтік
плазмиданы «иесінің» жасушасына, эдетге ішек бактериясы жасу¬
шасына енгізеді.
Генетикалык инженерия эдісімен 1977 жылы соматотро-
пин гормонын, 1978 жылы инсулин, интерферон т.б. гормон-
дарын биотехнологиялық жолмен ала бастады. Генетикалық
инженерияның, эсіресе денсаулық сақтау саласында — тұқым
қуалаушылық аурулармен күресу үшін (гендік терапия әдісімен)
болашагы өте мол.
Гендік терапия дегеніміз — агзаның сомалык жасушаларын-
да жэне гаметаларында не зиготаның дамуының бастапқы саты-
ларында пайда болған генетикалық бұзылыстарды жөндеу болып
табылады.
Қазіргі кезде гендерді енгізу үшін тек сүйек кемігінің жасу-
шаларын немесе фибробласттарды пайдалануга болады. Оларды
ағзадан бөліп алып. өсіріп, оларға қажетті гендерді енгізіп паци-
енттерге көшіруге болады.
Гендік терапияны кең қолдану үшін оның адам ағзасына
қауіпсіз екендігіне көз жеткізіп барып жүргізу қажет, себебі адам
онкогендері ретровирустарға өте үқсас, ал оларды жасушаға
енгізгенде адам онкогені модификацияланып, рак онкогендеріне
айналуы эбден мүмкін.
Көпшілік жағдайда мутациялардың агза үшін зиянды бола-
тындыгы белгілі. Олар популяцияда мутациялық қысым салдары-
нан үнемі пайда болып отырады.
83
Жасушада мутациялардың пайда болу жиілігін реттеп отыра-
тын не олардың әсерлерін байқатпайтын мутацияға қарсы табиғи
механизмдер болады. Оларға мыналар жатады:
1) диплоидты эукариоттық ағзалардың хромосомаларының
жұп болуы. Екі гомологтық хромосомадағы аллельдің біреуі
мутацияланганымен, рецессивті күйінде оның әсері байқалмайды;
2) кейбір гендік мутацияларды болдырмайтын механизм ол
р-РНҚ, т-РНҚт.б. гендерінің бірнеше көшірме күйінде кездесуі;
3) мутацияны болдырмайтын тағы бір антимутациялық қасиет
ол ДНҚ молекуласының қателіксіз репликациялануы;
4) биологиялық кодтың триплетті жэне артық болуы; тіпті
триплеттің бір нуклеотиді өзгерген күнде де ол кодон-синонимге
айналып генетикалық ақпарат мағынасын бүзбайды, мысалы: адам
гемоглобинінің Я-полипептидінің гені 438 нуклеотидтен түрады.
Егер сол 438 нуклеотидтің 25 пайызына жуық нулеотидтерінің
біреуі өзгергенімен полипептид синтезі бүзылмайды, ал 2—3
нуклеотид өзгерсе қысқа полипептид синтезделінеді; ал 73 пай¬
ыз нуклеотидтерінің бір жүбы ауысса полипептидте бір амин
қышқылының алмасуы байқалады. Триплегтің үшінші нуклеотиді
ауысса, ол 64 пайыз жағдайда өз мағынасын бұзбайды.
5) полипептидтегі амин кышқылдарының қасиеттері де бірдей
бола бермейді. Егер де жаңадан қосылган амин қышқылы өз
қасиеті бойынша ауыстырылған амин қышқылына үқсас болса
белоктың кұрылысы жэне қасиеті болар-болмас қана өзгереді.
Мысалы, адамның мутантты S жэне С гемоглобиндерінің калыпты
гемоглобин молекуласымен салыстырғандагы айырмашылыгы
бір-ақ амин қышкылының — 6-амин қышқылы-глутамин амин
қышқылының алмасуында; S-гемоглобинде 6-амин қышқылы
-валин, С-гемоглобинінде — лизин болады. Глутамин амин
қышқылымен лизиннің қасиеттері үқсас — екеуі де гидрофильді,
сондықтан да С-гемоглобині адамдарда қан аздықтың жеңіл түрін
тудырады, ал глутамин амин қышқылы мен валиннің касиеттері
қарама-қарсы, біреуі гидрофильді екіншісі — гидрофобты.
Сондықтан S-гемоглобиннің қасиеті күрт өзгеріп адамдарда қан
аздықтың зілді, ауыр түрі байкалады.
6) ДНҚ молекуласынын қүрылысы бүзылган жағдайда ол репа¬
рация механизмдері арқылы жонделеді.
84
Репарация дегеніміз — ДНҚ молекуласында пайда болған
мутацияньщ ерекше ферменттердің қатынасуымен жөнделуі.
Репарацияның бірнеше түрлері белгілі: а) фотореактивация
немесе жарық репарация. Оны 1962 жылы К.Руперт ашқан.
Ультракүлгін сәулелері ДНҚ молекуласына эсер етіп онда тиминдік
димерлерді туғызады, ягни 2 көршілес тиминдер — адениндермен
байланысын үзіп, өзара байланысады. Бүл кезде фотореактивтеуші
ферменттер жарықтың (күн сәулесінің) эсерімен тиминдік
димерлерді ыдыратып ДНҚ молекуласының бүрынғы, қалыпты
күйіне келтіреді.
б) эксцизиалық немесе қараңғылық репарация — XX
ғасырдың 50-жылдары А. Геррен ашқан, ол жарықтың қаты-
насынсыз жүреді жэне 4 сатыдан түрады: 1. Эндонуклеаза
ферментгері пайда болған димерлерді «тауып», оларды «танып»,
қияды; 2. Кесілген ДНҚ молекулалары жіпшелерінің үштарын
экзонуклеаза ферменттері «танып» арасын алшақтатады да
ығыстырып шығарады; 3. ДНҚ — полимераза ферменті кесіліп
алынып тасталған нуклеотидтер орнына ДНҚ-ның үзілмеген
екінші жіпшесі негізінде комплиментарлық принциппен қалыпты
нуклеотидтерді синтездейді; 4. Лигаза ферменттері синтезделінген
нуклеотидтерді ДНҚ-ның кесілген жіпшесіне жалғайды.
в) Репликациядан кейінгі репарация. Егерде репарацияның
бірінші не екінші жолдары арқылы ДНҚ молекуласындагы
қателіктер репликация кезінде жөнделмесе, онда ол келесі репли-
кацияда матрица қызметін толық атқара алмайды. Сондықтан пай¬
да болған қателіктер репликациядан кейін жөнделуі тиіс. Ондай
жөнделуді репликациядан кейінгі репарация деп атайды.
3.10. Өзгергіштік
Өзгергіштік тұқым қуалаушылык сияқты тірі агзалардың ең
негізгі касиеттерінің бірі.
Өзгергіиітік дегеніміз — эр түрлі себептердің нәтижесінде
ағзалардың белгілерінің жэне қасиеттерінің азды-копті өзгеруі бо¬
лып табылады.
85
Егер тұқым қуалаушылық арқылы биологиялық түрлердің,
тіршіліктің біркелкілігі дамитын болса, өзгергіштік арқылы
тіршіліктің алуан түрлілігі қалыптасады. Қазіргі кезде Жер бетінде
500000-нан астам өсімдік түрлері, 1,5 миллионная астам жануар¬
лар түрлері кездеседі. Ал, ерте замандарда өмір сүрген түрлердің
саны қазіргі тірі ағзалардың санынан әлдеқайда көп екенін ескер-
сек, онда Жер бетінде тіршілік пайда болғалы бері 5 миллионная
астам тірі ағзалар түрлерінің өмір сүргені белгілі. Тірі ағзалардын
осындай алуан түрлілігі олардың өзгергіштігінің нәтижесі болып
саналады. Ал, әрбір биологиялық түрлер миллиондаган даралардан
түратынын және олардың әрқайсысы бір-біріне мүлдем ұксамай,
өзгеше болатынын ескерсек, табиғатта өзгергіштіктің рөлінің өте
зор екенін айқын байқаймыз.
Сонымен өзгергіштік тірі ағзалардың ең басты қасиеттерінің
бірі, ол органикалық дүниенің дамуына, сүрыптауға материал бо¬
лып табылады.
XIX ғасырға дейін табиғат, тірі дүние өзгермейді деген
метафизикалық үғым басым болған. Тек 1809 жылы француз
ғалымы Ж.Б.Ламарк алғашқы эволюциялық ілімді қалыптастырып,
тірі ағзалар үнемі өзгеріп отырады деп айтқан. Ж. Б. Ламарк
өзгергіштіктің тек 1 түрін ажыратқан жэне кез келген өзгерулер
түқым қуалайды деп ойлаған.
1859 жылы Ч. Дарвин өзінің эволюциялық ілімінде өзгергіштік-
тің 2 формасын — айқындалған жэне айқындалмаған өзгергіштікті
ажыратқан.
Қазіргі кезде өзгергіштікті 2 үлкен топка бөледі (сызбанүсқаны
қара).
Фенотиптік өзгергіштік орта факторларының эсерлері салда-
рынан туындайды жэне түқым қуаламайды. Ол модификациялық
жэне кездейсоқ өзгергіштік болып 2-ге бөлінеді. Егер фенотиптік
өзгергіштік «реакция нормасынан» шықпайтын жэне нақтылы бір
орта факторларының эсерінен болатын болса оны модификациялык
өзгергіштік деп атаймыз. Мысалы, ақ түсті коянның арқасының
жүнін қиып, салқын жерде біршама уақыт үстасақ оның арқасында
кара түсті жүндердің пайда болғанын байкаймыз, ал сол коянды
жылы жерге көшірсек, жылылық көтерілген сайын аркасындағы
қара түсті түктер бірте-бірте ақ түске айналатынын көреміз. Бүл
жерде жылылықтың, яғни температураның, әсерінен қоянның
86
түгінің түсінің ақтан — қараға -аққа өзгеретінін байқаймыз. «Ре¬
акция нормасы» дегеніміз — фенотиптік өзгергіштіктің шегі.
1. Фенотиптік өзгергіштік
модификациялық өзгергіштік кездейсоқ өзгергіштік
2. Генотиптік өзгергіштік
Комбинативтік өзгергіштік мутациялық өзгергіштік
сомалық
генеративтік
гендік Z' геномдық
хромосомалық
хромосомалық
Кездейсоқ фенотиптік өзгергіштік деп — ортаның көптеген
факторларының бірлесіп эсер етуі салдарынан пайда болатын
өзгергіштікті айтамыз.
Фенотиптік өзгергіштіктіңзандылықтарын зерттеп білу медици¬
на практикасы үшін өте маңызды, себебі оны дүрыс білген жағдайда
ғана ортаның эрбір факторларының, не олардың жиынтығының
фенотиптің қалыптасып дамуындагы рөлін нақтылы анықтауға
мүмкіндік бере тіпі анықталған. Оның үстіне, модификациялық
өзгергіштік кейде фенокопиялардың (фенокөшірмелердің) пайда
болуына да алып келеді. Фенокопиялар (фенокөшірмелер) дегеніміз
— генотиптері эр түрлі ағзаларда үксас, бірдей фенотиптің пайда
болуы, оның негізгі себебі орта факторларының әсері болып табы-
лады. Мысалы, адамдардың туа біткен ақаулықтарының бір түрі —
гипосподия (несеп шығаратын сыртқы тесіктің жыныс мүшесінің
үшынан оның түбіне карай ығысып орналасуы) — 2 жолмен: 1)
генетикалық материалдың өзгеруі (мутация) нэтижесінде; 2)
орта факторларының әсерінен калыпты эмбриогенездің бүзылуы
нэтижесінде пайда болады.
87
Соңғы жолмен пайда болғанда ол фенокошірме болып сана¬
лады. Бұл ақаулықты емдеу үшін оның қалайша пайда болганын
білудің кажеті жоқ, әдетте оны хирургиялық операция жасап
емдейді. Бірақ, болашақ үрпақтарда осы ақаулыктың қайталану
ықтималдығын анықтау үшін оның қандай себептермен (генотиптік
не фенотиптік) пайда болғанын білу өте қажет.
Генотиптік өзгергіштік — түқым куалаушылық материалда-
рының өзгеруі нэтижесінде пайда болады жэне ол міндетті түрде
фенотипті де өзгертеді. Генотиптік өзгергіштіктің негізгі себептері
мутациялар, не аллельдердің мейоз немесе үрықтану кезінде
жаңаша комбинациялануы болып саналады.
Генотиптік өзгергіштік комбинативтік жэне мутациялық
өзгергіштік болып екіге бөлінеді:
1) комбинативтік өзгергіиітік — тұқым қуалаушылық матери-
алдары өзгермей сол калпында қалып, олардың жаңаша комбина¬
циялануы (топтасуы) негізінде пайда болады. Оның негізгі көздері
кроссинговер, гомологтік хромосомалардың гаметаларға еркін,
тәуелсіз ажырасуы жэне кездейсоқ үрықтану қүбылыстары болып
саналады. Комбинативтік өзгергіштік үрпақтарда ата-аналарына
тэн емес белгілер комбинациясының пайда болуына алып келеді.
Мысалы, сары тегіс бүршақпен жасыл кедір-бүдыр бүршақты
будандастырғанда: сары, кедір-бүдыр, жасыл тегіс бұршактардың
пайда болуы, т.с.с.
2) мутациялық өгергіштік тұқым қуалаушылық материалда-
рының (ген, хромосома) қүрылысының не мөлшерінің озгеруі сал-
дарынан туындайды.
Мутация дегеніміз — ДНҚ молекуласыныц мөлшерінің, не
оныц қүрылысыныц озгеруі.
Мутациялар сомалық не генеративтік жасушаларда пайда бо¬
луы мүмкін. Сомалық мутациялар ағзалардыц дене жасушала-
рында пайда болады жэне оныц эволюция үшін мацызы шамалы.
Генеративтіқ мутациялар жыныс жасушаларда пайда болады, ол
түқым қуалайды жэне эволюцияныц материалы болып саналады.
Мутациялардыц 3 түрі белгілі: гендік, урамосомалъщ, ге-
номдыц.
Гендік мутациялар
Гендік мутациялар деп — жай көзге көрінбейтін, тіпті микро¬
скоп арқылы да байқауға болмайтын геннің кұрамында болатын
өзгерістер. Гендік мутациялар геннің химиялық құрамының өзге-
руі, яғни ДНҚ молекуласындағы нуклеотидтердің орналасу ретінің
өзгеруі, бір немесе бірнеше нуклеотид жұбының түсіп калуы, не
жаңадан бір нуклеотид жұбыныц қосылуы күйінде байқалады.
Осының салдарынан геннің мағынасы, яғни онда «жазылған»
генетикалык ақпарат, озгереді. Ал ол қалыпты белок-ферменттің
синтезделуінің орнына өзгерген, мутацияланган белок-ферменттің
синтезделуіне алып келеді. Молекулалық биологияда бір фермент
— бір белгі деген ұғымныц бар екенін ескерсек, гендік мутациялар
белгілердің озгеруіне (метаболизмнің бұзылуына) алып келетініне
көзіміз жетеді.
Мутациялануға қабілетті геннің ең кіші бөлігін.муию// деп атай¬
ды, ол 1 нуклеотид жүбына тең.
Гендік мутациялардың 2 тобы: 1) геннің нуклеотид жұптарының
алмасуы; 2) ген ақпаратының көшірілу рамкасының жылжуы
белгілі. Мутациялардың бірінші тобына гендік мутациялардың 20
пайызы, ал екінші тобына 80 пайызы жатады.
Нуклеотидтер жұптары алмасуының 2 түрі белгілі: транзиция-
лар жэне трансверсиялар. Транзициялар дегеніміз — бір пуриндік
негіздің (А) екінші бір пуринмен Г), немесе бір пиримидиндік
негіздің (Ц) екінші пиримидиндік (Т) негізбен, не керісінше, (А Г;
ГА) (ЦТ; ТЦ) алмасуы.
Трансверсиячар дегеніміз — пуриндік негіздің (А,Г) пири-
мидиндік негізбен (ЦТ), не керісінше алмасуы.
Ген ақпаратының көшірілу рамкасының жылжуы геннің
құрамында бір нуклеотид жұбыныц түсіп қалуының (делеция) не
бір немесе бірнеше нуклеотидтердің кыстырмаланып қосылуының
нэтижесінде пайда болады. Бүл екі себептердің қайсысы болмасын
геннің мағынасын бүзады.
Гендік мутациялар өздігінен, ешбір себепсіз, кенеттен пай¬
да болуы мүмкін, не олар белгілі бір физикалық не химиялық
факторлардыц эсерінен тууы мүмкін. Бірінші жолмен пайда болған
мутацияларды кенеттен, өздігінен пайда болған мутациялар, ал
екіншісін индукцияланган мутациялар деп атайды.
89
Гендік мутациялар зат алмасу құбылысының түрліше бұзылуы
күйінде байқалады. Қазіргі кезде адамдардың 3 мыңнан астам
гендік аурулары белгілі. Олардың популяциялардағы орташа
жиілігі 2—4 пайызға тең.
Гендік аурулардың бір тобы — амин қышқылдарының
метаболизмінің бұзылуына байланысты. Олардың бірі — фенил-
кетонурия жэне альбинизм.
Фенилаланин амин қышқылы ауыстыруға болмайтын амин
қышқылдары қатарына жатады. Жасушадағы фенилаланиннің
шамалы бөлігі белок синтезіне жұмсалады, ал көптеген бөлігі
эрі қарай өзгереді, ягни фенилаланингидроксилаза ферментінің
қатынасуымен ол алғаш тирозинге, содан кейін меланин пигментіне
айналады.
Фенилаланин ► Тирозин ► Меланин
фенилаланин тирозиназа
гидроксилаза
Фенилпирожүзім фенилкетонурия ► альбинизм
қышкылы ауруы
Фенилкетонурия ауруы аутосомды — рецессивті жолмен
тұқым қуалайды, оның гені 12 q22 24-де анықталган. Оның орташа
жиілігі 1:10000-ға тең.
Егер жасушада фенилаланингидроксилаза ферменті болмаса не
оның белсенділігі төмен болса фенилаланин амин қышқылы ти¬
розинге айнала алмайды. Ол фенилпирожүзім қышқылы күйінде
қанда көптеп жиналады жэне несеп арқылы сыртқа шығарылып
отырады. Сондықтанда бүл аурумен ауыратын адамдардан
«тышқандардың» иісіне үқсас иіс шығып түрады.
Фенилпирожүзім қышқылы миды зақымдайтын у болып табы-
лады, сондықтан ауру адамдар тез қозгыш болады, оларда гипер¬
рефлексия, бүлшық еттері тонусының жоғары болуы, талып қалу
т.б. жагымсыз қүбылыстар байқалады.
Альбинизм — аутосомды-рецессивті жолмен тұкым куалайды,
оның жиілігі 1-5000; 1-25000-ге тең. Бүл аурудың дамуы тирози¬
наза ферментінің белсенділігін бүзатын гендік мутацияга байла-
90
нысты, осының нәтижесінде тирозин амин қышқылы меланин
пигментіне айналмай, адамның тері жасушаларында меланин
пигменті болмағандықтан терінің, шаштардың, кірпіктің, сақал-
мүрттың түсі сүттей аппақ болады.
Алксттонурш — аутосомды-рецессивті жолмен тұқым
қуалайды. Жиілігі 3—5:100000-ға тең. Бүл ауру жасушада гомо-
гентезин қышкылының малеилацетосірке қышқылына айналуына
қатынасатын оксидаза ферментін синтездеуге жауап беретін геннің
мутациялануына байланысты. Ол ген 3 ц2-де орналасқан.
Галактоземия — аутосомды-рецессивті жолмен түқым
қуалайды, жиілігі — 1:100000-га тең. Бүл ауру галактозо-1-
фосфатты уридин-дифосфагалактозаға айналдыруға қатынасатын
галактозо-іфосфатуридилтрансфераза ферменті генінің мутация-
сына байланысты.
Муковисцидоз (үйкы безінің кистофиброзы), аутосомды-
рецессивті жолмен түқым қуалайды, жиілігі 1:2500-ге тең. Бүл ауру
7 хромосомадағы 7 q2131 генінің мутациялануына байланысты.
Ахондроплизия — аутосомды-доминантты жолмен түқым
қуалайды, жиілігі 1:100000-ға тең. Оның гені 4р 1416 орналасқан.
Бүл аурудың клиникалык сипаты — бойдың өте аласа болуы (Мы-
салы, ересек адамдарда— 120-130см).
Гемофилия А — қан үюының VIII факторының кемістігіне бай¬
ланысты дамитынөтезілді түқым қуалайтын ауру. Х-хромосомамен
тіркес рецессивті жолмен түқым қуалайды, жиілігі 1—6500 үл ба-
лаларда. Оның гені Х-хромосоманың үзын иінінде орналасқан —
Xq28.
Гемофилия В — қан ұюының IX факторының белсенділігінің
төмендеуіне байланысты дамитын өте зілді түқым қуалайтын ауру.
X — хромосомамен тіркес рецессивті жолмен тұкым қуалайды.
Оның гені X — хромосоманың үзын иінінде орналасқан — X q27.
Гемоглобипопатиялар—гемоглобин молекуласы құрылысының
бұзылуы салдарынан дамитын аурулар. Оның ең көп таралған
түрлері:
1) орац — жасуиіалы анемия (қаназдық) (HbS). Мұнда гемо¬
глобин молекуласының р-жіпшесінің 6-амин қышқылы-глутамин
қышқылы валинмен алмасқан. Бүл гемоглобиннің ерітілуін
төмендетеді. Гомозиготалыларда эритроциттер орақ пішінді бо-
лып келеді.
91
2) Талассемия — глобиндер мөлшерінің азаюына не мүлдем бол-
мауына алып келетін глобиндік гендердің мутациялануына байла¬
нысты дамитын гендік ауру. Оның а-талассемия, р-талассемия де¬
ген түрлері белгілі. Олардың гендері 16 хромосомада орналасқан
— 16р13,3.
Хромосомалыц мупіациялар
Хромосомалық мутацияларға олардың құрылысында пайда
болған өзгерістерді жатқызады.
Хромосомалық мутациялар хромосомаішілік жэне хромосома-
аралық болып екіге бөлінеді.
Хромосомаішілік мутацияларға делеция, дупликация, инвер¬
сия жатады, ал хромосомааралық мутацияларға транслокация,
робертсондық қайта құрылымдарды жатқызады.
Делеция дегеніміз — хромосоманың бір учаскесінің жойылып
түсіп қалуы.
Дупликация — хромосоманың бір учаскесінің екі еселенуі.
Инверсия хромосоманың бір учаскесінің қиылып 180°-қа айна-
лып орналасуы. Оның 2 түрі белгілі: перицентрлік инверсия жэне
парацентрлік инверсия.
Перицентрлік инверсия центромера арқылы жүреді.
Парацентрлік инверсия центромерасыз хромосоманың бір
иінінде жүреді.
Транслокация дегеніміз — гомологтік емес хромосомалардың
учаскелерімен алмасуы. Оның екі түрі белгілі: реципрокты және ре-
ципрокты емес. Реципрокты транслокация дегеніміз — гомологтік
емес хромосомалардыц өзара учаскелерімен алмасуы, ал реципрок¬
ты емес транслокация гомологтік емес хромосомалардыц біреуініц
учаскесініц үзіліп екіншісіне жалғануы.
Делеция жэне дупликациялар хромосомадағы гендер саныныц
өзгеруіне (кемуіне не көбеюіне) алып келсе, транслокациялар мен
инверсиялар хромосома гендерінің санын өзгертпей, олардың ор-
наласу ретін өзгертеді.
92
ABCDEFGH
П ГИГЖГП
11
ABCDEFGH
СЛИОХГО
t t
ABCDE FGH
azr*>4CD
t t
ABCDE FGH
ДГМХЕР
t t
ABCDE FGH
азЕгскш
A
i
MNOP Q R
t
ABCDE FGH
t "t t
GET
ABCEFGH
-^azaxm
ABCBCDEFGH
-*аиишхш
ABC GH
-* ССЕ»<ЖХЗ
FED
A E FGH
-* anb’
DCB
cm
MNOCDEFGH
шшшихт
ABPO R
aj»«
ADE FBCGH
'TP t~TT
27-cypem. Хромосомалык мутациялар
Робертсондық қайта қүрылымдарға центромералык косылу
және центромералық ажыраулар жатады. Центромералық қосылу
дегеніміз — екі акроцентрлік хромосомалардың центромера
арқылы қосылып, бір метацентрлік хромосоманы пайда етуі жата¬
ды. Осылайша адамдардың 2-хромосомасы пайда болған.
28-сурет. Робертсондык кайта
қүрылымдар
х i(xq) і(хр)
29-сурет.
X — хромосоманын
изохромосомалары
Центромералық косылу аркылы кейде 21 хромосоманың
транслокациялық трисомиясы да түзіледі (21/14).
93
Кейде хромосома центромера аркылы үзіліп, оның әрбір иіні
екі еселеніп (репликацияланып) бір-бірімен өзара жалғанып изо-
хромосомаларды пайда етеді (29-сурет). Олардың иіндері бірдей
гендер жиынтығынан тұрады.
Хромосомалық мутациялар кейбір гендердің мөлшерін, ор-
наласу ретін, тіркесу топтарын өзгертеді. Осының нәтижесінде
жасушада үйлесімді гендік тепе-тендік бұзылып ағза денесінің,
психикасыныц қалыпты дамуы бүзылады. Хромосомалық мута¬
циялар адамдар миының дамуының, тірек-қимыл, қан айналу, зэр
шығару жүйелерінің кемістігіне экеліп согады. Ондай адамдардың
ақыл-есі кеміс болады, элсіз болады, әйелдер түсік тастайды, ал
бала туған күнде де нәрестелер өте кішкентай, элсіз болып туы-
лып, жастайынан дүние салады.
Делециялар гомозиготалы күйінде жэне Х-хромосомада кездес-
се летальды болады. Адамдарда кездесетін «мысықша мияулау»
синдромы 5 хромосоманың кысқа иінінің делециясына байланы¬
сты. 4, 13, 18 хромосомалардьщ делециялары да гетерозиготалы
күйінде адамдардың ақыл-есінің кем болуына, әлсіз болуына алып
келеді.
Инверсиялар гетерозиготалы болса ағзалар үрпақ қалдыра ал-
майды, себебі пайда болган гаметалардың тең жартысы жүмыртка
жасушасын ұрыкгандыра алмайды.
Транслокациялар гетерозиготалы күйінде стерильді болады,
себебі мейоз нәтижесінде бүзылган (кеміс) гаметалар түзіледі.
Геномдық мутациялар
Геномдық мутациялар деп хромосома саныныц 1, 2, 3
хромосомага өзгеруін (кемуі не көбеюі), не хромосома санының
еселеп өсуін айтамыз. Мутацияның бірінші түрін аиеуплоидия, ал
екіншісін полиплоидия деп атаймыз.
Анеуплоидия — нуллисомик, моносомик, три, тетро жэне по-
лисомик күйлерінде кездеседі.
Нулчисомик дегеніміз — диплоидты хромосома жиынтыгында
бір жүп хромосоманың болмауы — 2п-2.
Моносомия дегеніміз — диплоидты хромосома жиынтыгында
бір хромосоманың сыцар күйінде болуы (2п-1; ХО).
94
Трисомия дегеніміз — диплоидты хромосома жиынтығында бір
хромосоманың 3 дана күйінде болуы (2п+1; 13+; 18+; 21+).
Анеуплоидия мейоз процесінде хромосомалардың дұрыс ажы-
распауы салдарынан пайда болады.
Моносомиктер, әсіресе нуллисомиктер, тіршілік ете алмайды.
Трисомиктер эр түрлі хромосомалық ауруларға алып келеді.
Полиплоидия хромосома санының еселеп өсуі нәтижесінде
пайда болады. Оның гаплоидия, триплоидия, тетра, пента, т.с.с.
плоидия деген түрлері белгілі. Полиплоидия өсімдіктер дүниесінде
жиі кездеседі жэне ол пайдалы қасиеттерге алып келеді (өнімді болу
т.с.с), ал жануарлар дүниесінде, эсіресе адамдарда, өте сирек бола¬
ды. Адамдарда полиплоидты ұрықтар күні бұрын түсіп қалады, ал
тірі туылған күнде де олар бірер минут кана өмір сүреді.
3.11. Адамдардың хромосомалық аурулары
Қазіргі кездегі ғылыми деректерге қарағанда дүниеге кел-
ген нәрестелердің 5 пайызы эр түрлі генетикалық өзгерістермен
туылады, ал олардың ішінен 0,5 пайызы шамасындағы бала-
ларда хромосомапық аурулар байқалады. Бүгінгі таңца 700-ге
жуық хромосомалық аберрациялар (бұзылыстар) сипатталып
жазылган, олардың ішінен 100-ге жуығы адамдардың ақыл-есінің
кеміс болуына, денелерінің дамуының бүзылуына, эр түрлі зілді
хромосомалық аурулардың дамуына алып келеді. Адамдардың
хромосомалық ауруларының негізгі клиникалық сипаты ретінде
туа біткен ақаулыктарды, ақыл-естерінің кем болуын, жыныстық
дамуының бүзылуы нэтижесінде бедеу болуын, яғни үрпақ қалдыра
алмауын, өздігінен түсік тастауын т.с.с атауға болады.
Хромосомальщ схурулар деп — клиникалық сипаттары жағынан
түрліше болып келетін адамдар патологиясының үлкен бір тобын
айтамыз. Олардың бэрінің себептері бір — ол хромосомалық не
геномдықмутацияларболыптабылады.Хромосомалықаурулардың
басқа түқым қуалайтын аурулардан ерекшелігі — олар Г. Мендель
зандарынан өзгеше жолмен түқым қуалайды.
Хромосомалық аурулар ата-аналарының гаметаларында пайда
болған мутациялар, не үрықтың дамуының алғашқы кезеңдерінде
95
пайда болған мутациялар салдарынан қалыптасуы мүмкін. Гаме-
таларда пайда болтан мутациялар бүл аурудыц толык нысаныныц,
ал үрық жасушаларында пайда болған мутациялар аралас (мо-
заикалық) формасының дамуына алып келеді. Аралас формалы
ағзалардың кейбір жасушаларында қалыпты кариотип болатын
болса, кейбіреулерінде бүзылған кариотип кездеседі.
Адамдар гаметаларында болатын хромосомалық ауытқушы-
лықтыңжалпы саны 750-ге жуық, ал оныц 700-і хромосомалардыц
қүрылымыныц бұзылуларыныц (аберрация) үлесіне тиеді.
Хромосомалық аурулардың пайда болу механизмдері
Көптеген хромосомалық аурулардыц пайда болуыныц басты
себебі — тарихи, эволюциялық қалыптасқан жүйеніц — кариотип-
тің езгеруі, яғни хромосома сандарыныц не хромосомалардыц
күрылымының бүзылуы болып табылады.
Ағзалардың хромосома сандарыныц ауытқуы жасушаныц
дүрыс бөлінбеуінің не эр түрлі мутагендік факторлардың әсерінен
жасуша бөлінуінде хромосомалардың бір-бірінен ажыраспауыныц
салдарынан болады. Бүл хромосома санының еселеп өсуіне (по-
лисомия — Зп, 4п, 5п, т.с.с.), не қалыпты кариотиптің бір немесе
бірнеше хромосомаға көбейіп не кемуіне алып келеді (анеуплои-
дия) — моносомия 2п-1; трисомия 2п+1.
Толық триплоидия (Зп) жэне тетраплоидия (4п) нысандары
адамдарда тек кенеттен, оздігінен өліп, түсіп қалған түсіктерде
ғана байқалған, ягни полиплоидты үрықтар тірі туылмай, дамудың
алгашқы кезеңдерінде-ақ өліп қалады. Ал, өсімдіктерде поли¬
плоидия (Зп, 4п, 5п) кұнды қасиеттерді -өміршең болуын, өнімді
болуын т.с.с. қалыптастырады. Сондықтан да селекционерлер
полиплоидтық нысандарды мэдени өсімдіктердіц жаца сорттарын
алу үшін кеңінен колданады.
Анеуппоидия — аутосомды не жыныс хромосомалар сандары¬
ныц ауытқуы салдарынан болуы мүмкін. Жыныс хромосомасы-
ның саны өзгергенде эрбір қосымша Х-хромосома өте тығыз
ширатылған гетерохроматин күйінде болып, оның гендері ак-
тивсіз болады. Дегенмен, гетерохроматин күйіндегі қосымша
Х-хромосомалар түгелдей дерлік инертті болмайды. Олар жасу-
96
шаларға, жасуша метаболизміне жэне ағзаның дамуына эсер
етеді. Гетерохроматинденген қосымша Х-хромосомаларда сандық
белгілерді анықтайтын полигендер болуы мүмкін.
1-12 жүп ірі хромосомалардыц ауытқулары бар үрықтар әдетте
өте ерте кезде-ак өліп қалады, яғни летальды болады. 13-18 жүп
хромосомалардыц трисомиялары (13+; 18+) жартылай летальды
(сублетальды) болады да, нэрестелер не өлі туылады, не балалық
шағында-ақ өліп қалады. Жыныс хромосомалары ауытқуларының
(ХО, XXV, XXXV, т.с.с.) жэне кейбір аутосомды трисомиялардың
(13+, 18+, 21+) тіршілік қабілеті айтарлықтай дәрежеде болуы
мүмкін.
Қазіргі кездегі сипатталған 100-ге жуық хромосомалык ауру-
лардың 95-і негізінен 5 хромосомалық ауытқу нысандарына тэн
болады: 13,18,21 хромосома трисомиялары, Шерешевский-Тернер
синдромы (45, ХО), Клайнфельтер синдромы (47, XXV).
1. Даун синдромы (21+)
Синдром Деп — белгілі бір ауруға жатпайтын бірнеше ауру
белгілерінің бір адамда қатар келуін айтамыз.
Бүл ауруды алғаш рет 1855 жылы J1. Даун сипатгап жазған,
бірақ оның себептері 100 жылдан кейін барып анықталған. Бүл
ауру екі жыныста да бірдей жиілікпен кездеседі, оның орташа
жиілігі 1/700-дей шамасындай.
Даун синдромының негізгі клиникалық сипаты — ақыл-есінің
туа біте кем болуында. Оларды оқытып үйретуге болады, бірақ
жазуға, санауға үйрету мүмкін емес. Орталық нерв жүйелерінде
айтарлықтай ауытқушылықтар болмаса да олар икемсіз, епсіз,
жайсыз болып келеді.
Бүл аурудың негізгі фенотиптік сипаттарына мыналарды
жатқызуга болады; бойлары аласа, шүйдесі тегіс, бас сүйектері
кішкентай трахицефальды, эпикант дамыған, көздері қысыңқы,
мүрындарының түбі жалпақ, кең кеңсірікті болып келеді.
Олардың жүрек-тамыр жүйесінің бүзылуы, сол сияқты, барлық
ішкі секреция бездері қызметтерінің бүзылыстары байқалады.
Дерматоглификасы — алақандарында терец көлденең сызық-
тарының және шынашагында 2-жұмылатын бүгілу сызығыныц ор-
нына тек 1 ғана сызықтың болуымен сипатталады.
97
170-7
2. Эдвардс синдромы (18+)
Бұл ауруды 1960 жылы Эдвардс айқындап тапқан, оның жиілігі
1/4500-ден 1/650-ге дейін болады. Бұл аурумен ерлерге қарағанда
эйелдер жиірек ауырады. Бұл — ұл балалардың эмбриональдық
даму кезінде не өмірінің алғашқы апталарында көптеп өлігі
қалатындығын көрсетеді.
Бұл аурудың негізгі сипатгамаларына мыналар жатады:
нәрестелердің салмағы өте жеңіл, бойлары кішкентай болады,
иектері тегіс, жақтары нашар дамыган, бас сүйегі кішкентай,
кұлақтары кішкентай жэне олар бас сүйегінен төмендеу орна-
ласқан, түмсықтары шығыңқы, күстүмсық болып келеді. Птоз,
экзофтальм, эпикант дамыған, көздерінің мөлдір кабығының
бүлдырлануы, көру нерв дискісінің семуі сияқты кору мүшелерінің
мүкістігі айқын байқалып түрады. Қол саусақтары оте үзын не-
месе өте кысқа болып, 2—5 саусақтары ерекше орналасқан бола¬
ды. Табандарының пішіні өзгереді: Жүрек-қантамыр жүйесінің,
бүйректерінің мүкістігі байқалады. Ересек жасқа дейін жеткен
балалардың ақыл-естерінің кем болатындығы байқалган. Эдвардс
синдромын нәресте туылған кезде бала жолдасының (плацента)
кішкентай болуы және жалғыз кіндік артериясының болуы арқылы
күні бүрын анықтауға болады.
3. Патау синдромы (13+)
1961 жылы Патау жэне онын әріптестері өте кемтар, сұрықсыз
баланың кариотипін зерттегенде оның Д тобында артык 1
хромосоманың болатынын анықтап, осы ауруды сипаттап жазган.
Бүл синдромның популяциялардагы жиілігін анықтау қиын,
себебі осы синдроммен ауырған балалар ерте өліп қалады; деген-
мен де оның орташа жиілігі 1:3500—4000 жуық.
Бүл синдромның клиникалық сипаттары — балалардың
салмағы өте жеңіл, бойлары кысқа жэне олар күні жетпей туы-
лады. Сол сияқты, осы синдромның ерекше белгілеріне жүмсақ
жэне қатты тандайларының жырық, көздерінің өте кішкентай
— эр түрлі дәрежеде микроофтальмиялы болып келуін де атауға
болады. Оларда туа біткен катаракта, беттерінің ангиомасы, поли¬
дактилия, синдактилия жэне табандарының өзгерулері байкалады.
Жүрегінің, бүйректерінің қызметтері бүзылады. Қыз балалар-
да жатырдың имек болуы, үлдардың үмаларының өзгерулері
байқалады. Гипотония жэне гипертония, ақыл-естері кем, тоқ
ішектің ауытқуы, қосымша көкбауыр кездеседі. Бүл синдроммен
ауырған нэрестелер өмірінің алғашқы күндерінде немесе алғашқы
аптада-ақ өліп қалады. Дегенмен, кейде олар 2-3 жыл өмір сүруі
де мүмкін.
4. Кдайнфельтер синдромы (ХХУ, ХХХУ, ХХХХУ, XXX У, ХУУ)
Клайнфельтер синдромы ер адамдарда кездеседі жэне ол
қосымшаХжыныс хромосомасының болуымен сипатталады (ХХУ,
ХХХУ, т.с.с). Оның орташа жиілігі 1:500-ге тең.
Бүл синдромның негізгі сипатына мыналарды жатқызуға бола¬
ды: бойлары өте үзын, иықтары тар, бөкселері кең, бүлшық еттері
нашар дамыған астеник немесе әтек (пішілген адам) типтес бо¬
лып келеді. Беттерінде және қолтықтарында мардымсыз, өте си-
рек түктері болады, ал қасағаның түктері әйелдерге үқсас болады;
олардың шәует жолдары семіп (атрофия) калган, сперматогенез
жоктыктан бедеу болып келеді. Акыл-естері кемістеу, өте сенгіш,
көңіл-күйі тез өзгергіш, қызбалау болады.
Клайнфельтер синдромымен ауырган адамдардың дермато-
глификасы өзгерген — қол саусақтарының өрнегінде догалар жиі
кездесіп. қырлар (гребень) саны азаяды.
5. Шереіиевский — Тернер синдромы (ХО)
Бүл синдромды 1925 жылы Н.А. Шерешевский жэне 1938 жылы
Тернер тауып сипатгап жазган. Оның орташа жиілігі 1: 3000-ге тең
жэне тек эйелдерде кездесіп, эсіресе аласа бойлы қыздар арасында
жиі байқалады.
Шерешевский-Тернер синдромын жаңа туылган қыз нәре-
стелерде айқын байқауга болады, себебі моносомия X (ХО)
кейбір мүшелер мен үлпалардың жатырда дамуын бүзатындықтан
99
нәрестелер бірнеше аномалиялармен туылады, яғни салмақтары
өте жеңіл, бойлары қысқа, табандарында жэне қолдарында
лимфоидтық ісіктер, тырнактарының гипоплазиясы (толық
жетілмеуі) байқалады. Жүректерінің туа біткен ақаулықтары,
қолқа (аорта), окпе артериясының тарылуы (стеноз, коаркта-
ция) байкалып, эпикант дамыған, шаштары қысқа, мойыны
қысқа жэне жуан болып келеді. Қаңқа дамуының аномалиялары,
көкірек қуысының өзгеруі, 4—5 саусақтарының қысқаруы да бүл
ауруға тэн белгілер болып табылады. Бойларының қысқа болуы¬
на байланысты аяқтары да қысқа, түлғалары ұзындау болып дене
кұрылысында диспропорция байқалады. Иықтары кең, бөкселері
тар болып өздерінің сыртқы құрылысы жағынан ер адамдарға
ұқсас келеді.
Ауруларда ішкі жэне сыртқы жыныс мүшелері дамымай, соңгы
жыныс белгілері — сүт бездері, қолтықтарында, қасага үстінде
түктер болмайды. Олар бедеу болады, себебі жыныс бездері
дамымаған.
Бүл аурумен ауырған әйелдерде жыныс хроматині кездеспейді,
олардың кариотипі 45 (ХО) тең болады. Сол сияқты X
хромосомасының басқа да аномалиялары үзын иінінің немесе
қысқа иінінің делециялары, екі X хромосомалардың транслокаци-
ясы, сақиналы X хромосома, т.с.с. байқалуы мүмкін.
6. «Мысық айқайы» синдромы (5р двлеииясы)
Бүл синдром 5р хромосоманың қысқа иінінің делециясымен
байланысты екенін 1965 жылы Герман дәлелдеген. Оның жиілігі
толық анықталмаған. Дегенмен соңғы кездері бүл синдром жиі
кездесетін болып жүр. Оның клиникалық сипаты — бүл аурумен
ауыратын балалардың дыбыс тембрі ерекше, мысықша «мияулап»,
жалынышты күйде болады. Сол сияқты олардың ақыл-есі кем,
денесінің дамуы нашар болады. Өсе келе бүл белгілер жойылуы
мүмкін.
Негізгі фенотиптік белгілері — беті доңгелек, эпикант дамыған,
микроцефалия және жүрегінің ақаулықтары айқын байқалады.
100
3.12. Мутагенез жэне канцерогенез
Мутациялардыц пайда болу процесін мутагенез, ал мута-
циялардыц пайда болуына алып келетін физикалық не химиялық
факторларды мутагендер деп атайды. Мутациялар ешбір себепсіз,
табиғи жолмен пайда болуы мүмкін — оларды өздігінен, кенеттен
пайда болған мутациялар дейді; ал кейбіреулері эр түрлі физика-
лык не химиялық мутагендік факторлардың эеері салдарынан пай¬
да болады — оларды индукцияланган мутациялар деп атайды.
Өздігінен пайда болатын гендік мутациялардыц жиілігі эр түрлі
агзаларда түрліше болып келеді, мысалы, адамдарда ол 10'4-10'6
дэрежесіне тең. Егер адамдардың бір үрпағыныц орташа тіршілік
үзақтығы 25—30 жыл, ал мутациянын орташа жиілігі 1x10'5
дәрежесіне тең, адамдардың генотипіндегі гендер саны 100000—
110000 деп алсақ, онда адамдардыц гаплоидтық хромосома санын-
да (гаметаларда) эрбір үрпақ сайын 1—10 жаца мутация пайДа бо¬
лып отырады. Бұл көп пе аз ба? Жеке бір адамды алатын болсақ
ол әрине аз, себебі адамдардыц бір үрпағында, яғни 25—30 жыл
ішінде түзілетін гаметаларының тек 1—10-ында ғана мутация пай¬
да болады. Ал егер бүкіл адамзатты, ягни тұтас адам популяция-
ларын алатын болсак жэне адамдар гаметаларында не бары 2-ақ
мутациядан пайда болды десек, онда мутациялардың жалпы саны
8x109 ягни 8 000 000 000 тец болады. Ал, бұл, эрине, өте көп.
Гендік мутациялар ағза белгілерініц түрліше озгерулеріне алып
келеді. Олар доминантгы, жартылай доминантты, рецессивті бо¬
лып келуі мүмкін. Мутанггы гендер, қалыпты гендерге қараганда,
оздерініц онімдерінің — ферменттерінің аз молшерде, немесе оте
коп молшерде синтезделуіне алып келуі мүмкін немесе фермент
мүлдем синтезделмеуі мүмкін. Ягни, мутация негізінде қалыпты
ген езініц магынасын біржолата жойып нонсенс — (мағынасыз)
кодонга, не оның магынасы өзгеріп миссенс — кодонга айна-
луы мүмкін. Гендік мутациялардыц агзалар үшін зияндылығы
— олар тіршілік процесініц бүзылуына (патология), тіршілігініц
төмендеуіне не кәбею кабілетінің кемуіне алып келеді, ягни
агзалардыц ортага бейімделуін төмендетеді. Кейде мутантты ген
агзалардың өліп қалуына да алып келеді (летальді мутациялар).
101
Дегенмен, мутацияларды өте сирек және тосыннан (кенегген)
пайда болатын, белгілі бір багытқа бағытталмаған құбылыс деп
қарастыруымыз керек:
1) мутациялар өте сирек пайда болады, себебі олар реплика-
циялану процесінде ДНҚ синтезінің қателігі күйінде пайда болуы
мүмкін. Ал, ДНҚ синтезі қателіксіз жүреді. Тек 100 000 нуклео-
тидтен біреуі ғана қате синтезделуі мүмкін. Тіпті ДНҚ синтезінде
қателік болған күннің өзінде, пайда болған қателік репарация
тетіктері арқылы жөнделіп отырады;
2) мутациялардыц өте сирек жэне кездейсоқ болатын тагы бір
себебі — олардыц нақтылы бір генде, нақтылы үрпақта пайда
болу-болмауын күні бүрын болжау мүмкін емес;
3) мутациялар ағзалардыц ортаға бейімделушілігін жогарылат-
пайды, сондыктан да оларды белгілі бір бағытқа багытталмаған
дейміз.
¥зақ жылдар бойына хромосомалық жэне гендік мутациялар¬
дыц пайда болу себептері белгісіз болып келеді. Сондықтан
коптеген ғалымдар мутациялар, әсіресе өздігінен пайда болатын
мутациялар, табиғатта сыртқы орта факторларыныц әсерінсіз
пайда болады деген жалған көзқараста болып келді. Тек, гендік
мутациялардыц санының есебін анықтау әдістері қалыптасқаннан
кейін ғана оларды эр түрлі мутагендік факторлар арқылы туғызу
мүмкін екендігі белгілі болды. Оларды физикалық, химиялықжәне
биологиялық мутагендік факторлар деп боледі.
Физикалық радиациялық мутагенез дегеніміз — әр түрлі
физикалық факторлардыц — иондаушы сәулелердің, ультракүлгін
сэулелердіц, т.б. эсерінен пайда болатын мутациялар. Иондау¬
шы сәулелердіц мутацияларды туғыза алатындығын алгаш рет
1925 жылы орыс ғалымдары Г.Надсон жэне С.Филипповтар
ашқан. 1927 жылы Г.Меллер рентген сәулелерінің әсерінен дро-
зофилаларда (жеміс шыбыны) коптеген мутациялардыц пайда
болатындығын тәжірибе жасап аныктаған. Қазіргі кезде радиа-
циялаушы сэулелердіц мутагендік эсері көптеген биологиялық
объектілерде дәлелденген.
Радиациялаушы сәулелердіц мутагендік механизмі түрліше бо¬
лып келеді. Олар гендік, хромосомалык, геномдық мутацияларды
туғыза алады.
102
Кейбір химиялық заттардың мутагендік әсерінің болатынды-
гын алғаш рет КСРО-да 1928 жылы Н. Мейсель, 1933 жылы В. Са¬
харов, 1934 жылы М. Лобашовтар ашкан болатын. Ең алғашқы
күшті мутагенді (жасушаға жат ДНҚ) 1939 жылы С. Гершензон
ашқан болатын. 1946 жылы генетик И. Рапопорт формалиннің
жэне этилениминнің күшті мутагендік эсері бар екенін аныктады.
Сол сияқты, осы жылдары ағылшын генетиктері UL1. Ауэрбах жэне
Д. Робсон иприттің мутагендік эсер ететінін анықтаған.
Қазіргі кезде жүздеген химиялық заттардың мутагендік эсерлері
айқындалды. Олардың ішінде ауыл шаруашылыгында жиі пай-
даланатын гербицидтер, дефолианттар және инсектицидтер
(ДДТ), кейбір дэрі-дәрмектер (әсіресе, нитрофурондар), эр түрлі
өнеркэсіпте пайдаланатын, не өнеркәсіпте бөлініп шыгатын зат-
тар, мысалы: токыма комбинаттарында пайдаланылатын — афок-
сид; жасанды шайырларды алуға пайдаланатын — формальде¬
гид; кағаз шығаруга пайдаланатын — гидроксиламин; азық-түлік
өнеркэсібінде шараптарды дайындауга пайдаланылатын — натрий
бисульфиті; азоттық иприт, 5-бромурацил жэне т.б.
Барлық химиялық мутагендер гендік, хромосомалық жэне
геномдық мутацияларды тудыра алады.
Химиялық заттардың мутагендік механизмі элі толық анық-
талған жоқ, дегенмен, олардың кейбіреулері жасушаның бөлінуі
кезеңінде ахроматин жіпшелерін үзіп, олардың кызметін жояды,
ал екінші біреулері, мысалы 5-бромурацил ДНҚ молекуласының
репликациялануында азоттык негіздердің орнын ауыстырып гран-
зициялар мен грансверсияларды пайда болгызады.
Химиялық мутагендер, радиация сэулелері сияқты, мейоздың
бүзылуына алып келіп, хромосомалардың дүрыс ажырасуын
болдырмайды не хромосомалардыц үзілуіне алып келеді. Кейбір
химиялық мутагендер тікелей ДНҚ молекуласына эсер етпей,
оныц репликациялану механизмін озгертеді, осыныц салдарынан
ДНҚ синтезінде қателіктер пайда болады. Ондай заттардыц бірі —
кофеин.
Канцерогенез дегеніміз — рак (ісік) ауруыныц пайда болу
процесі. Көптеген мутагендік факторлар канцерогендік те түрғы-
дан эсер етеді. Рак (қатерлі ісік) ауруы — XX ғасырдагы адамдар-
дың ең қауіпті ауруларыныц бірі.
103
Рак (ісік) ауруы жасушаларының кэдімгі дене жасушаларынан
2 ерекшеліктері болады: 1) кэдімгі дене жасушалары санаулы рет
қана бөлініп, содан кейін бөлінуін тоқтатып өліп қалатын болса,
рак (ісік) жасушалары шексіз бөлініп өсуге қабілетті. Сондықтан
да олар ағзага механикалық зиян келтіріп, өсіп көбейе береді де
ақырында оның өлуіне алып келеді; 2) рак жасушалары жайы-
лып метастаз береді, яғни оның кейбір жасушалары лимфа не
қантамырлар аркылы басқа мүшелерге, ұлпаларға таралып, сол
жерлерде жаңа ісіктер пайда болады.
Агзалардың бөлінуге қабілетті кез келген жасушалары ісікке
айнала алады. Бірақ, олар калайша, ненің әсерінен рак (қатерлі
ісік) жасушаларына айналады, элі толық шешілмеген мәселелер.
Дегенмен, қазіргі кезде белгілі болғаны — рак (қатерлі ісік) жасу¬
шаларында гендер қызметінің реттелу механизмінің бұзылуыныц
салдары екендігі. Екінші біртолықшешілмеген мэселе — рак(ісік)
ауруы тұқым қуалай ма, жоқ па? Бүл түрғыдан екі түрлі гипотеза-
лар айтылып жүр: бірі — рак (ісік) ауруы түқым қуалайды десе,
екіншісі — тұкым қуаламайды деп болжамдайды. Қалай болганда
да рак (ісік) ауруының пайда болуыныц негізгі себептері — сома-
лык мутациялар жэне вирустардың әсері екендігі анықталып
отыр.
Қазіргі кезде рак (ісік) ауруының кем дегенде 3—4 түрінің
түқым қуалайтындыгы дэлелденген, олар: ретинобластоманың бір
түрі, Фанкони анемиясы (кан аздылығы), пигменттік ксеродерма
жэне нейрофиброматоз.
Рак (ісік) ауруының көпшілігі сомалық мутациялардан баста-
лады.
Сомалык мутациялар адамдардыц дене жасушаларында эр түрлі
мутагендік факторлардыц — радиация сэулелерініц, ультра-күлгін
сәулелерініц, химиялық заттардыц әсерінен түзіледі. Сомалық
мутациялар алғаш жеке жасушада пайда болып, оның тағдыры
сол жасуша тіршілігіне байланысты болады. Егер осы мутация
эсерінен жасуша өліп қалса, пайда болған сомалық мутация да сол
жасушамен бірге жойылады, ал егер мутацияланган жасуша әрі
қарай бөлінігі көбейетін болса, онда сол жасушаныц үрпақтарыныц
бәрінде де мутация сақталынады.
104
30-сурет. Ісік (рак) ауруынын пайда болу жобасы
Сомалық мутация негізінде ағза денесінде жасуша мозаикасы
қалыптасады, яғни калыпты сау жасушалармен катар мутантты
жасушалар да кездеседі. Сомалык мутациялар негізінде мутант¬
ты жасушада хромосома тұрақтылығы бұзылады, ал ол эр түрлі
мутацияларға (делеция, транслокация, дупликация т.с.с.) алып
келеді. Осыныц нәтижесінде сол жасушалар жедел өсіп, көбейіп
ісік туғызады, мысалы, осылайша Фанкони анемиясы — (жас ба-
лаларда кездесетін ауру — оныц белгілері кацқа, эсіресе саусақ
сүйектері, кэрі жілік қүрылысы бүзылады, сүйектерде кемік жа¬
сушалары жетілмейді, сондықтан эритроциттер аз мөлшерде
түзіледі) дамиды.
Сомалық мутациялардыц рак (ісік) ауруын пайда қылуыныц
жалпы механизмініц жобасы келтірілген (30-сурет).
105
Түкым қуалайтын рак (ісік) ауруының тағы бірі — ретино-
бластома. Ол балалардың көздерінде болатын ісік ауруы. Егер
дер кезінде дұрыс емделмесе ісік миға беріліп балалардың өліп
қалуына алып келеді.
Ретинобластоманың екі түрі белгілі, бірі түқым куалайды,
екіншісі түқым қуаламайды.
Ретинобластоманың түқым қуалайтын түрі аутосомды — доми-
нантты жолмен беріледі. Оның пайда болуы екі сатыдан түрады:
1) мутация алғаш жыныс жасушаларында пайда болады, ал ол
сомалық мутациялардыц тууына алып келеді;
2) сомалық мутация пайда болған жасушалардыц біреуі өсіп,
көбейіп ретинобластомаға айналады. Ретинобластоманыц гені 13-
хромосоманыц үзын иінінде орналасқан.
Канцерогенездіц сомалық мутация болжамымен қатар вирустық
болжамы да белгілі.
Вирустардыц рак (ісік) ауруын туғыза алатындығы туралы бол-
жамдар ерте кезден-ақ белгілі. Бірақ, тек 1950 жылы американ
ғалымы Р. Делебекко кемірушілерді маймыл вирусымен зақымдап,
оларда рак (ісік) ауруыныц пайда болғанын зерттеп анықтаған.
Соцғы кездері көптеген рак (ісік) жэне лейкоз ауруларыныц ви-
рустар эсерінен болатындығы анықталды. Бірақ коптеген рак
туғызушы вирустарда түқым қуалаушылық материалдары ретінде
ДНҚ емес РНҚ молекуласы болатыны белгілі. Сонда, қалайша ви¬
рус РНҚ-сы жасуша ДНҚ-сымен қосыла алады?
1970 жылы Г. Темин жэне Д. Балтимор РНҚ молекуласы
негізінде ДНҚ молекуласыныц синтезделуін қамтамасыз ететін
ерекше ферменттіц кері транскриптаза (ревертаза) ферментін
ашты. Осындай РНҚ-лы вирустардыретпровирустар деп атайды.
Вирустар РНҚ күйінде жасушада үзақ уақыт зиянсыз өмір сүріп
жүре алады, ал сол РНҚ негізінде, кері транскриптаза ферментініц
қатысуымен, ДНҚ синтезделіп жасуша ДНҚ-сына жалғанса бол-
ды, жасуша «қүтырып», тез бөлініп, рак жасушасына айналады.
Вирус РНҚ-сында өздерініц тіршілігіне кажет гендермен қатар
жасушаны «қүтыртып» рак жасушасына айналдыратын гендер де
кездеседі. Оларды онкогендер V-onc деп атайды, олардыц жалпы
саны 20-га жуық. Бір қызығы, вирус онкогендеріне үқсас ген¬
дер адамныц қалыпты эукариотты жасушаларында да кездеседі,
106
бірак олар өз беттерінше қалыпты жасушаны рак жасушасына
айналдыра алмайды, керісінше, олар адам онтогенезінің алгашқы
кезеңцерінде өте маңызды рөл атқарады. Ол гендерді протоонко¬
гендер — с-опс деп атайды.
Вирус онкогендері мен жасуша онкогендері (протоонкоген¬
дер) ұксас белоктарды синтездейді. Дегенмен, с-опс интрон-
экзон учаскелерінен, ал V-onc тек экзондардан кұрылган. С-опс
активтенуі үшін оған вирус онкогенінің (V-onc) промоторы -вирус
LTR жалгануы кажет.
3.13. Медициналық генетика л ық кеңес беру негіздері
Тұқым қуалаушылық механизмдерінің тірі агзаның қалыпты
дамуы үшін маңызы өте зор, себебі түкым қуалаушылық мате-
риалдарында пайда болган озгерістер адамдардың денсаулығын
бүзады, түкым қуалайтын ауруларга альш келеді. Гендік ауру¬
лар структуралық гендердің мутациясы салдарынан дамиды, ал
хромосомалық ауруларға — хромосомалық не геномдық мутация¬
лар алып келеді. Адамдардыц тұкым қуалайтын патологиясына,
сол сиякты, түқым қуалауға бейім болып келетін ауруларды да
жатқызады. Олардың дамуына генетикалық факторлармен бірге
орта факторлары да эсер етеді.
Түқым қуалайтын ауруларды анықтау үшін клиникалық
лабораториялык зерттеулердің нэтижелері кеңінен пайдаланы-
лады. Кейде, осы ауруларды анықтау үшін арнайы генетикалық
әдістерді пайдалануға тура келеді. Көптеген гендік, хромосомалық
аурулардың бір ерекшелігі — олардың үрпақтарда қайталануы
жэне клиникалық дені сау адамдардыц гетерозиготалы күйінде
жагымсыз рецессивті аллельді тасымалдауы болып саналады.
Аталган екі жағдайларда да үрпақтыц денсаулығын болжау үшін
арнайы генетикалық зерттеулер қажет.
Түқым куалайтын ауруларды түбегейлі емдеу мүмкін емес,
олардыц тек алдын алуга болады. Ал олардыц алдын алудың ец
тиімді жолдарыныц бірі — дер кезінде медициналық генетикалық
кецес беру болып саналады.
107
Медициналық генетикалық кенес берудің негізгі міндеттері:
1. Тұқым қуалайтын аурулар байқалған отбасында оның гене-
тикалық қаупін анықтау.
2. Тұқым қуалайтын аурулардың диагнозын нақтылау жэне
қажет болған жағдайда оның генетикалық қаупін анықтау.
Ол үшін кеңес беруші дәрігер-генетик:
1. Ауру адамдарды не олардың туыстарын қосымша
лабораториялық әдістер (цитогенетикалык, биохимиялық, иммуно-
логиялық, рентгенологиялық т.с.с.) арқылы зерттеудің қажеттілігін
анықтайды.
2. Ауру адамдарға не олардьщ туысқандарына басқа маман
дәрігерлердің (эндокринолог, невропатолог, окулист т.с.с.) кеңес
беру қажеттілігін анықтайды.
Медициналық генетикалық кеңес алу үшін дамудың туа біткен
ақаулыктарының дараланған не көпшілікті түрлері байқалған,
ақыл-естері кем, жүйке-психикалық ауруларымен сырқаттанған, эр
түрлі мүшелері мен жүйелерінің үзак уақытқа созылған аурулары
байқалған адамдарға, жыныстық дамуы бұзылған балаларға, бедеу
жүбайларга, балаларын үнемі өлі туатын, түсікті жиі тастайтын не
нәрестелері ерте өліп қала беретін әйелдерге жолдама беріледі.
Медициналық генетикалық кеңеске жолдама бергенде аурудың
болжам күйінде болса да диагнозы көрсетілуі қажет. Тіпті, түқым
қуалайтын аурудың диагнозы нақтылы анықталган күннің өзінде,
кейде оны косымша басқа әдістер арқылы зерттеп нақтылау қажет
болады. Мысалы, көпшілік жагдайда Даун синдромын тәжірибелі
дәрігер-педиатр жеп-жеңіл аныктай алады. Дегенмен, оның
үрпақтарда генетикалық қаупін анықтау үшін сол аурудың пайда
болу себептерін дэл білу қажет, яғни ол жай трисомия (21+) жо-
лымен не транслокациялық трисомия жолымен пайда болады ма?
Себебі олардың генетикалык қаупі түрліше.
Медициналық генетикалык кеңес беру, көпшілік жагдайда ауру
адамдарды клиникалық-генеалогиялық талдау (шежірені талдау)
жэне сол аурудың генетикалык қаупін анықтаумен шектеледі.
Ал қайтадан жүкті болган эйелдерге пренатальдық (бала
туылғанға дейін) диагностика жасау тек баланың туа біткен
ақаулыктарын (нерв түтігінің кемістігі, еріннің не таңдайдың
жырыгы, гидроцефалия, дамудың туа біткен ақаулықтарының
108
көпшілікті түрі, т.с.с.) анықтау мүмкіншілігі күмәнсіз болған
жағдайда ғана қажет. Дегенмен, қайтадан жүкті болған әйелдердің
барлығы дерлік жүктіліктің 15-16 аптасынан бастап 2-3 рет ультра-
дыбыс арқылы зерттеулерден өту керектігін түсіндіру кажет, себебі
осы эдіс арқылы көлденең белгілерге сүйеніп (жатырда судың
кеп болуы, жатырда судың аз болуы, үрықтың гипотрофиясы,
үрық мөлшерінің не онын кейбір бөлімдері мөлшерінің жүктілік
мерзіміне сәйкес болмауы, үрықтың баяу козғалуы т.с.с.) үрықтың
кейбір ақаулықтарын анықтауға жэне үрықтың қүрсақ ішінде да-
муын кадағалап отыруға мүмкіндік туады.
Жыныстык дамуы бүзылған, бедеу, гермафродит адамдарға
медициналық-генетикалық зертгеулерді де жүргізу қажет. Ал
орталық нерв жүйесінің патологиялары бар адамдарга медици-
налық-генетикалық кеңес беру алдында олардың жан-жақты
клиникалық-биохимиялық зерттеулерден өтулері қажет екендігін
түсіндірген жөн.
Дамудың туа біткен ақаулықтарының көпшілікті түрлері
кездесетін адамдарга кеңес беру үшін олардың барлык ақаулық-
тарына жэне дамудың микроаномалияларына мүқият фенотиптік
сипаттама беру қажет. Сонымен қатар, пробандты — қажет болған
жағдайда оның гуыстарын, суретке түсіру шарт. Сол сияқты,
дамудың туа біткен ақаулықтарының көпшілікті түрлері кездесетін
балалардың бэрін дерлік цитогенетикалык эдіс аркылы зерттеу
кажет.
Лурулирдың генетикалық қатерін анықтау принциптері:
Медициналык генетикалык кеңес бергенде мендельденуші
(моногендік) жэне мендельденбейтін (полигендік, мультифак-
торлы) аурулар арасында біршама айырмашылық болатындыгын
ескерген жөн. Егер де мендельденуші аурулардың генетикалык
каупін аныктаудың теориялык негізі толық шешілген десек,
мендельденбейтін (мультифакторлы) аурулардың генетикалык
қаупін анықтау тек ойша, болжам кұйінде анықталуы мүмкін,
себебі оның генетикасы элі күнге дейін толық зерттелінбеген.
Мендельденуші аурулардың генетикалык қаупін анықтаганда
оның түкым куалау типі басшылыққа алынады, сондықтан кеңес
109
берушінің негізгі мақсаты — пробандтың, не оның ата-аналарының
нақтылы генотиптерін анықтаумен шектеледі. Ал мендельден-
бейтін (мультифакторлы) ауруларды алатын болсақ, қазіргі танда
оларға тэн дискретті генотиптерді атау мүмкін емес. Себебі ондай
аурудың пайда болуында генетикалық жэне орта факторларының
әсерлері байқалады. Сондықтан да мендельденбейтін белгілерді
генетикалык тұрғыдан талдаганда көптеген қиындықтар кездеседі
жэне күрделі математикалық эдістерді пайдалануды талап етеді.
Моногендік аурулардың қатеріп анықтау
Моногендік ауруларға кеңес беру барысындағы байқалатын
эр түрлі жағдайларды негізінен 3 топқа белуге болады: 1) ата-
аналардың генотипі белгілі; 2) ата-аналардың генотипін нақтылы
болжауға болады; 3) ата-аналардың генотипі белгісіз.
Ата-аналардың генотипі белгілі, не нақтылы болжауға болатын
болса генетикалық кауіпті оп-оңай, Мендельдің екінші заңына
сэйкес анықтайды. Ол үшін ата-аналарының жэне олардың мүмкін
болган үрпақтарының генотиптерін гамета типтеріне сэйкес тізіп
жазу қажет.
Кеңес беру процесіндегі негізгі проблема ретінде аурудың
түқым куалау типін анықтау болып табылады. Оны отбасы
шежіресін талдай отырып, кейбір белгілер негізінде, анықтауга
эбден болады. Мысалы, аутосомды-доминантты түқым қуалау: 1)
егер белгі үрпақтан-үрпақка беріліп отырса; 2) үрпақгардагы ауру
буындардың саны 50 пайыз шамасындай болатын болса; 3) ауру
әкеден үл балага берілетін болса; 4) үрпақтардың екі жынысы да
(үл балалар жэне қыздар) бірдей ауыратын болса. Бүл жагдайда
ата-аналары ауыратын, өзі де ауру индивид cay адаммен қосылса
олардың балаларының екеуінің біреуі ауру болып туылады. Ал
оның cay іні-карындастары не ага-апалары генотипі жэне фенотипі
жагынан алсақ та cay болып есептеледі жэне ауруды үрпақтарына
бере алмайды. Бүл, әрине, толық пенетрантты доминантты
ауруларга тән ережелер.
Ал толық емес пенетрантты аутосомды — доминантты ауру-
лардың генетикалык қатерін аныктаганда оның корсеткіші мүл-
дем басқаша болады. Бүл жагдайда отбасында жэне үрпақтарда
110
Кесте
Екі ата-ана генотиптері аныкталған жағдайдағы үрпактардың
мүмкін болған фенотиптері
Ay руд ын
түкы.м куалау
ТИПІ
Әкесінін
геногипі
Шешесі-
нін
генотині
¥лдары
Кыздары
я*
и
ауру
сау, бірак
тасылмал-
даушы
ауру
Я
W
сау, бірақ
тасымал-
даушы
/
2
3
4
5
б
7
8
9
Аутосомды-
АА
Аа
—
—
—
—
доминантты
АА
Аа
1
—
—
1
—
—
АА
Аа
1
—
—
1
—
—
Аа
АА
1
—
—
1
—
—
Аа
Аа
3/4
—
1/4
3/4
—
1/4
Аа
Аа
1/2
—
1/2
1/2
—
1/2
аа
АА
1
—
—
1
—
—
аа
Аа
1/2
—
1/2
1/2
—
1/2
аа
Аа
—
—
1
—
—
1
Аутосомды-
АА
АА
—
1
—
—
[
рецессивті
АА
Аа
—
1/2
1/2
—
1/2
1/2
аа
Аа
—
1
—
—
1
—
Аа
АА
—
1/2
1/2
—
1/2
1/2
аа
Аа
1/4
1/2
1/4
1/4
1/2
1/4
Аа
Аа
1/2
1/2
—
1/2
1/2
—
аа
АА
—
1
—
—
1
—
аа
Аа
1/2
1/2
—
1/2
1/2
—
аа
Аа
1
—
—
1
—
—
Доминантты
А
аА
і
—
—
;
—
Х-тіркескен
А
Аа
1/2
—
1/2
1
—
—
А
Аа
—
—
1
1
—
—
А
АА
1
—
—
1
—
—
а
Аа
1/2
—
1/2
1/2
—
1/2
а
Аа
—
—
1
—
—
1
Рецессивті
А
АА
—
Г
—
—
1
Х-тіркесксн
А
Аа
1/2
—
1/2
—
1/2
1/2
А
Аа
1
—
—
—
1
—
а
Аа
—
—
1
—
1
—
а
Аа
1/2
—
1/2
1/2
1/2
—
а
Аа
1
—
—
1
—
—
ЕСКЕРТУ: А — доминантты аллель;
а — рецессивті аллель.
аурулардың пайда болу жиілігі теориялық тұргыда күткендегі-
ден аз болады. Егер пенетранттылықты К-ге деп белгілесек, онда
бірінші буында ауру ұрпақтың туылуы 1/2 К тең болады. Егер
дендері сау жұбайлардан ауру бала туылса жэне жұбайлардың ата-
аналарының біреуі не жақын туыстары сол аурумен сырқаттанған
болса, оның генотипі гетерозиготалы болатындығын үлкен
сенімділікпен айтуға болады, бірақ бір отбасында өздігінен бірдей
екі мутация пайда болды деп болжау керек, ал бүл жағдай тіпті
мүмкін емес.
Аутосомды-доминанттыаурулардыңмедициналық-генетикалық
кеңес сүрауларыныц басты себептерінің бірі — ол жүбайлардың
біреуінің науқас болуы. Айталық, нейрофиброматоз ауруымен ау-
ыратын ер адам үрпақтарының денсаулыгын болжау үшін кеңес
беруді өтінді делік.
Нейрофиброматоз ауруынын шежіре схемасынын бір бөлігі
Нейрофиброматоз ауруы 80 % пенетранттылықпен ауто-
сомды — доминантты тұкым қуалайтыны белгілі. II үрпақтағы
2-нөмірлі ауру ер адамды гетерозиготалы (Аа), ал оның жүбайын
(ІІ-І) қалыпты ген бойынша (а) гомозиготалы (аа) деуге болады.
Сонда, олардың үрпақтарында кездесетін 3 типті генотиптер мен
фенотиптердің ара қатынасы мынадай болмақ:
1) ауру гетерозиготалылар (Аа) 1/2 К= 1 /2 К=3 8/10=0,4
2) сау гомозиготалылар (аа) — 0,5
3) сау гетерозиготалылар (Аа) — 1 (І-К)=0,1.
112
Сонымен, бірінші не кез келген келесі балаларының нейрофи¬
броматоз ауруымен ауыру мүмкіншілігі 40 %, ал сау балалардың
туылу мүмкіншілігі — 60 %-га тең болады.
Аутосомды-рецессивті түқым куалайтын аурулардың шежі-
релері төмендегідей белгілермен сипатталады: 1) ауру ата-
аналардың балалары сау, бірақтасымалдаушы болып келеді; 2) осы
некенің қалған балаларының ауру болып туылу мүмкіншілігі 25
%-ға тең жэне үл балалары да, қыз балалары да бірдей ауру болуы
мүмкін; Әдетте отбасында балалар саны санаулы-ақ болады, кейде
ауру бала сол отбасындағы жалғызы болуы мүмкін; Осы отбасы-
дағы баланың екеуінің де ауру болып туылуы (1/4)2=1/16=6,25
%-га тең; ал екеудің тек біреуінің ауру болып туылуы Ньютон
биномының жайылу заңына сэйкес 2x3/4x1/4=87,5 % тең болады;
3) ауру адамның балалары эдетте сау болып туылуы мүмкін;
4) кейде ауру балалардың ата-аналары бір-бірімен туыс болуы
мүмкін (туыстық неке), бүл жағдайда отбасының екі мүшесінің
де бір түрлі рецессивті ген бойынша тасымалдаушы болу
мүмкіншілігі, туыс емес некелерге қарағанда элдеқайда коп бо¬
лады. Оның үстіне отбасында ауру негүрлым сирек байқалатын
болса, оның түқым куалауында туыстық некелесудің (инбридинг)
эсерінің соншалықты жоғары болгандыгын көрсетеді. Егер гете¬
розиготалы индивид өзінің немере қарындасына үйленсе, онда
сол жігіттің мутантты генді тасымалдаушы гетерозиготалы әйелге
үйлену мүмкіншілігі 1/8-дей болады, ягни екеуі де тасымалдаушы,
ал мутантты ген бойынша гомозиготалы индивид өзінің немере
қарындасына үйленсе, онда эйелдің сол ген бойынша гетерозиго¬
талы болу мүмкіншілігі 1/8-ден 1/4-ке дейін оседі, ягни олардың
тек біреуі гана тасымалдаушы. Егер де отбасы шежірелерінде ауру
адамдар туралы деректер болмаса онда рецессивті ауру балалар-
дың туылу қаупінң мүмкіншілгін мынадай эмпирикалык формула
бойынша есептейді:
Р = -у Ғхп.
Ғ— инбридинг коэффициент;
п — популяцияныц кез келген адамдарының патологиялық
рецессивтік гендерінің орташа саны, ол 4-5 генге тец.
X — тіркескен рецессивті аурулардың шежірелеріне тэн
| белгілер төмендегідей: 1) ауру эке өзінің патологиялық генін
113
170-8
тек қыздарына береді, бірақ олар сау, тасымалдаушы болады; 2)
тасымалдаушы гетерозиготалы эйел өзінің патологиялық генін
балаларының 50%-на ягни тек жартысына ғана бере алады; 3) ауру
ер бала өзінің патологиялық генін тек анасынан ғана қабылдап ала¬
ды, себебі ол X— хромосомамен тіркескен; 4) тасымалдаушы әйел
өзінің патологиялық генін шешесінен де, әкесінен де қабылдап
алуы мүмкін, онда ол ауру әйелдің ұл балаларының бәрі ауру, ал
қыздары гетерозиготалы — тасымалдаушы болып келеді.
Сонымен, егер ер адам ауру болса, оньщ Х-хромосомасында
патологиялық геннің болганы. Сау әйелдің генотипі қалыпты ал¬
лель бойынша гомозиготалы не гетерозиготалы болуы мүмкін.
Егер эйелдің экесінде X — тіркескен ауру болса, онда ол эйел
100% жагдайда гетерозиготалы болады да, одан туылган үлдардьщ
ауру болуы 1/2 тең; 2) дендері сау жүбайлардан 2 ауру үл бала
туылса, оның анасының гетерозиготалы болгандыгын көрсетеді;
3) дендері сау жүбайлардан 1 ауру үл бала туылса жэне әйелдің
ағалары не інілері ауру болса, онда ол әйелді гетерозиготалы деп
санауга эбден болады.
Ата-ананың белгісіз генотипін анықтау
Бүл жағдайларда шежіре ерекшеліктеріне қарай бірнеше бол-
жам (гипотеза) түжырымдалып басшылықка алынады да, оның
қайсысы шындыққа жақын болатынын жорамалдап анықтайды.
Кейбір белгілердің бірнеше типті түқым қуалауы мүмкін — ауто-
сомды — доминантты, аутосомды — рецессивті, X — тіркескен
т.с.с. Міне. осы жагдайларда оның барлык түқым куалау типтерін
саралап талдап барып оның отбасындағы генетикалық қаупін
анықтайды. іМысалы, эдебиеттердегі деректерге қараганда,
пигментгік ритинит ауруы 88 % жагдайда аутосомды — рецес-
сивтік, 10 % жагдайда аутосомды — доминантты жэне 2 % X —
хромосомамен тіркес түқым қуалайды.
Пигменттік-ретинит ауруымен ауырагын баласы бар жүбайлар
(II-1, П-2) болашақ балаларында оның қайталану қаупін білмек бо¬
лып кеңес сүраған. Бүл жагдайда осы аурудың түқым қуалауының
3-типін де талдау кажет.
114
[ гипотеза — аутосомды-рецессивті тип; априрорлық мүм-
кіншілігі — 0,88 (88 %); шартты мүмкіншілігі — 0,25 (25 %)
бірлесе түқым қуалау мүмкіншілігі — 0,88 х 0,25 = 0,22 (22 %).
II гипотеза — аутосомды-доминантты тип: априорлық
мүмкіншілік — 0,1 (10 %); шартты мүмкіншілігі — 0, бірлесе
түқым қуалау мүмкіншілігі — 0.
III гипотеза — Х-тіркескен тип: априорлық мүмкіншілік — 0,02
(2 %).
Бүл жагдайда шартты мүмкіншілік ауру баланың анасының
(II—I) гомозиготалы не гетерозиготалы болу мүмкіншілігін
көрсетуі тиіс.
Оның генетикалык қатерін төмендегідей жолмен есептейді:
30 б-сурет
4. ДАМУ БИОЛОГИЯСЫ
4.1. Онтогенез. Ағзалардың жеке дамуы
Кез келген тірі ағза өсіп дамиды. Ағзалардын өсуі дегеніміз
оларда байқалатын сандық өзгерістер, яғни олардың денесінің не
жекелеген мүшелерінің мөлшерінің, салмағының өсуі. Ал, даму
дегеніміз — ағза денесінде, не оның мүшелерінде байқалатын кері
қайтпайтын сапалық өзгерістер болып табылады. Ағзалардың өсуі
мен дамуы бір-бірімен тығыз байланысқан, бірінсіз-бірі болмайтын
күрделі физиологиялық қүбылыстар болып саналады. Олардың
екеуі де генетикалык. гуморальдык және жүйкелік механизмдер
арқылы реттелініп отырады.
Даму құбылысын, эдетге, екі түрлі мағынада түсінеді: 1)
ағзалардың жеке дамуы — онтогенез; 2) агза топтарының, ягни
сиетематикалық топтардың немесе агзаның мүшелері мен мүшелер
жүйесінің тарихи дамуы — филогенез.
Онтогенез турлері. Онтогенез — (грекше ontos — тіршілік,
genesis — даму) тірі даралардың жеке дамуы, ягни зиготадан ба-
стап адам дүние салғанға (өлгенге) дейінгі уакыттар аралығында
байқалатын күрделі өзгерістер жиынтыгы болып табылады. Онто¬
генез терминін алгаш рет 1866 жылы Э. Геккель енгізген болатын.
Әр түрлі ағзалардың онтогенезі түрліше жолдармен жүреді.
Осыган орай, оның екі түрін ажыратады: 1) тура даму; 2) тура
емес (түрленіп) даму.
Тура даму дегеніміз — жүмыртқадан не ұрык қабығынан
барлық мүшелері қалыптасқан, ересек өкілдеріне өте үқсас, тек
олардан дене пропорциялары арқылы гана ерекшеленетін жэне өз
бетінше дербес тіршілік ете алатын ұрпактардың дүниеге келуін
айтамыз. Даралардың тура дамуы ана денесінен тыс жагдайларда
(мысалы, жүмырткаларда — қүстар, жорғалаушы: ?.р) не тікелей
ана агзасында (мысалы, жатырда — сүтқоректілер) жүруі мүмкін.
116
Турленіп даму (метаморфоза) деп жұмыртқадан ересек форма-
ларына мүлдем ұқсамайтын аралық формалардың (дернәсілдер,
куыршақ, т.с.с.) пайда болуы арқылы дамуды айтамыз. Оның екі
түрі белгілі: 1) толық турленіп даму — жүмыртқадан бір немесе
бірнеше формалы дернәсілдер, содан кейін қуыршак, ересек дара
— имаго дамып жетіледі (бунакденелілерде);
2) шала түрленіп даму — бүл жагдайда жүмыртқадан —
дернәсіл, ал одан ересек дара дамып жетіледі (бунақденелілер).
Жогарыда сипатталган онтогенез түрлері үрықтану нәтижесінде
жүзеге асады. Кейде даралардың дамуы үрықтанбай-ақ жұмыртқа
жасушасының ядросының, не сперматозоид ядросының бөлінуі
нәтижесінде жүзеге асуы мүмкін (жалпақ күрттар, бунакденелілер).
Дамудың бүндай жолын, ягни даралардың үрықтанбай дамуын —
партеногенез (гр. parthenos — қыз; genesis — даму) деп атайды.
Партеногенез жыныстық көбеюдің .ерекше түрі болып табыла-
ды. Мүндай жолмен көбейетін даралар үрпақтары гаплоидТы (п),
дара жынысты болып келеді. Партеногенездің екі түрі белгілі: 1)
гиногенез — жүмыртқа жасушасына сперматозоидтар еніп, бірақ
қосылмай оның эрі қарай дамуына түрткі болады; 2) андрогенез
— жүмыртқа жасушасына енген сперматозоид ядросы жүмыртқа
жасушасының оліп жойылган ядросын алмастырып жаңа агзаның
дамуын камтамасыз етеді.
4.1.1. Онтогенез болжамдары
Онтогенездің мэнін толык анықтайтын түпкілікті теория элі
күнге дейін жоқ. Бірақ XIX гасырдың басынан бастап оның мэнін
анықтау мақсатында бірнеше болжамдар (гипотезалар) айтылуда.
Олардың негізгілері — преформизм жэне эпигенез гипотезалары.
Преформизм болжамы бойынша болашак агза жыныс жасуша¬
ларында барлық мүшелері толық калыптасқан, бірақ өте кішкентай
күйде күні бүрын анықталган. Сол кішкентай агза ішінде олардың
болашак үрпақтары да айқындалган, ягни болашақ агзалар жыныс
жасушалары қосылмай түрып қалыптаскан. Ал, онтогенез (жеке
даму) дегеніміз — сол кішкентай, күні бүрын қалыптасқан агза
мүшелерінің, денесінің жай өсуі гана болып табылады, ягни онто-
117
J
генез барысында ешқандай жаңа құрылымдар — ұлпалар, мүшелер
түзілмейді деп уагыздаган.
Мүндай болжамды ең алғаш ежелгі грек дэрігері Гиппократ
айтқан болатын. Преформизмді қолдаушы оқымыстылардың
өздері дайын кішкентай ағза қандай жыныс жасушаларда кездеседі
(жұмыртқа жасушасында ма, әлде сперматозоидтарда ма) деген
сұракқа байланысты екі топқа бөлінген: бірі қалыптасқан кішкентай
ағза жүмыртқа жасушасында болады деп уагыздаса — овисттер;
екіншісі — сперматозоидтарда кездеседі деп болжамдаған — ани-
малкулисттер.
Қазіргі деректер бойынша жыныс жасушаларында барлық
мүшелері қалыптасқан кішкентай агза болады деп айту қате, де-
генмен, соңғы кездерге дейін преформизм идеясын жаңғыртуға
тырысушылар оқтын-оқтын табылып жатады, оларды неопрефор-
мисттер деп атайды (В. Ру, Г. Друш, Я. Свамердам т.б.).
Ерте замандардың өзінде-ақ преформизм гипотезасына
қарама-қарсы пікірлер айтылган. Бүл пікірлерді эпигенез болжа-
мы деп атайды. Эпигенез болжамы бойынша жыныс жасушала¬
рында (жүмыртқа жасушасы, сперматозоид) ешқандай дайын,
күні бүрын айқындалган кішкентай ағза болмайды, тіпті жыныс
жасушаларында ешқандай күрделі қүрылымдар да кездеспейді.
Олар қүрылыссыз, гомогенді —- біркелкі массадан түрады, ал
болашақ ағза үрықтанғаннан кейін күрделі жаңа кұрылу құбы-
лыстарының нәтижесінде дамып жетіледі деп айтқан. Ягни жыныс
жасушаларының агза онтогенезі үшін ешқандай рөлі болмайды
деп уағыздаған.
Эпигенез идеяларының өріс алып дамуы орыс ғалымы К. Во-
льфтың еңбегімен тығыз байланысты.
XVII-XVIII гасырларда осы екі болжам жақтастары арасында
үзаққа созылған ымырасыз пікірталас жүріп келген. Бірақ XIX
ғасырдың бас жагында К. Бэр өз тәжірибелері негізінде жогарыда
келтірілген гипотезалардың ешқандай шындыққа жанаспайтынын
дэлелдеп көрсеткен.
К. Бэрдің айтуынша, жыныс жасушаларында дайын күні бүрын
айқындалған кішкентай агза болады деп айту эбестік, сол сияқты
жыныс жасушалары болашақ ағзаның дамуына ешқандай үлес
қоспайды деу де дүрыс емес. Онтогенез өте күрделі қүбылыс,
118
оның эрбір кезеңдері мен сатыларында күрделі өзгерістер, жаңа
құрылымдар пайда болып отырады; оның келесі кезеңі мен
сатысының қалыпты жүруі оған дейінгі кезеңдер мен сатыларда
болып өткен кұбылыстар жиынтығына тығыз байланысты бола¬
ды. Сондықтан да онтогенезді — преформацияланган эпигенез деп
қарастыруымыз керек.
К. Бэрдің осы болжамы қазіргі кезде де көптеген оқымыстылар
тарапынан қолдау табуда.
4.1.2. Онтогенездің кезендерге бөлінуі
Онтогенез үзіліссіз жүріп отыратын біртүтас күбылыс болып та-
былады, бірақ оны зерттеуді, окып-үйренуді жеңілдету мақсатымен
бірнеше кезеңдерге, сатыларға, фазаларга жіктейді.
Әдетте, онтогенезді 2 кезеңге бөледі: эмбрионалдык кезең және
постэмбрионалдық кезең. Бірақ, жогары сатылы жануарлардың
жэне адам онтогенезін 3 кезеңге боледі: 1) үрык пайда болғанға
дейінгі кезең; 2) дүниеге келгенге дейінгі (антенатальдық,
эмбрионалдық) кезең; 3) туылғаннан кейінгі (постнатальдық,
постэмбрионалдық) кезең (1-кесте).
¥рық пайда болғанға дейінгі кезен. Бүл кезең шын мэнінде
тікелей онтогенезге жатпайды, себебі ол жыныс жасушаларының
пайда болу (гаметогенез — сперматогенез, овогенез) құбылыста-
рын гана қамтиды. Дегенмен, бүл кезеңнің онтогенездің
қалыпты жүруі ұшін маңызы өте зор, себебі: 1) жыныс жасу-
шаларынсыз үрықтану болмайды, демек онтогенез басталмай-
ды; 2) зиготаның жэне болашақ үрпақтың тектік түрі (генотипі)
жыныс жасушаларының геномдарының қосылуы нәтижесінде
қалыптасады; 3) жүмыртқа жасушасында зиготаның эрі қарай
дамуы үшін қажет қоректік заттар (сары уыз) жинақталады;
4) жүмыртқа жасушасының цитоплазмасында күрделі жіктелу
— ооплазмалық сегрегация, гендердіц амплификациясы, т.с. с.
қүбылыстар жүреді.
Әр түрлі агзалардың жүмыртқа жасушаларында сары уыз
түрліше жинақталуы мүмкін. Осыған орай, олардың 3 түрін ажы-
ратады:
119
1-кесте
Адам онтогенезінің кезендерге бөлінуі
Кезендер
Сатылар
Фазалар
1. Ұрык пайда
болғанга дейінгі кезең
1. Сперматогенез
2. Онтогенез
Сперматозоидтардың,
жұмыртка жасушасының
пайда болуы
2. Дүниеге келгенге
дейінгі /антенатал-
дык/ кезең
1. Ұрыктык саты /
ұрықтанғаннан -8
аптаға дейін/
а/ зигота
б/ бөлшектену
в/ бластула
г/ гаструланың пайда
болуы
д/ гисто-органогенез
2. Құрсактағы
бала сатысы /9
аптадан — дүниеге
келгенге дейін/
өсу
3. Дүниеге келген-
нен кейінгі /постна-
талдык/ кезең
1. Ёалалык шак —
13—15 жаска дейін
а/ нәрестенің дүниеге келуі
б/ емізулі бала -11 күннен
1 жас в/ сәби — 2-3 жаска
дейін
г/ бөбек 4-6 жас д/
бүлдіршін 7-11 жас е/
жеткіншек 11-13 /12-15/
жас
2. Жетілген шак
/жыныстык
жетілген кез/
а/ бозбала 14-20 /16-20/жас
б/ жігіт/бойжеткен/ 21—35
жас
в/ орта жас — 36-50 жас г/
жігіт ағасы 51 -60 жас
3. Кэрілік шак
а/кария 61-75 жас б/ егде
адам — 75-90 жас в/ ұзак
жасаушылар -90 жастан
артык г/ дүние салу
1) 1ізолецетапьды жүмырпщалар — сары уызы аз жэне цито-
плазмада біркелкі таралған. М.үндай жұмыртқалар төменгі сатылы
сүтқоректілерде, инетерілілерде кездеседі.
120
2) телолецеталъды жүмыріщалар — сары уызы өте көп жэне
ол вегетативтік полюсінде орналасқан, ал анимальдық полюсте
сарыуыз болмайды. Бұндай жұмыртқалар былқылдақденелілерде,
қосмекенділерде, жорғалаушыларда жэне құстарда кездеседі.
3) центрюлецетальды жүмырігщалар — сары уызы жұмырт-
қаның оргасына орналаскан, ал цитоплазма оны қоршап жатады.
Жұмыртқа жасушаларының түзілу (гаметогенез) құбылысын-
да қоректік заттар — сарыуыздың жинақталуымен қатар басқа
да күрделі процестер жүреді. Мысалы, ооплазмалық сегрегация,
гендердің амплификациясы, жүмыртқа жасушасының полярлығы,
т.с.с.
Жүмыртқа жасушасының ооплазмалық сегрегсп/иялануы де-
геніміз — сарыуыздың, гликоген, РНҚ молекулаларының т.б. зат-
тардың әркелкі таралуы салдарынан цитоплазманың эр түрлі сапа-
лы учаскелерге жіктелуі болып табылады.
а б в г
31-сурет. Ооплазмалык сегрегация.
а) төменгі сатылы хордалылар, б) балыктар, в) космекенділер,
г) жоргалаушылар жэне күстар. 1-эктодерма, 2-нерв тактасы,
3-хорда, 4-ішек энтодермасы, 5-мезодерма.
Осындай эркелкі сапалы учаскелерден болашақта түрліше
үлпалар мен мүшелер дамып жетіледі. Ягни, ересек даралардың
үлпалары мен мүшелерінің дамуы жүмыртқа жасушасында
үрықтануга дейін бастама күйінде айкындалган.
Гендердің стплификациясы дегеніміз — кейбір гендердің
көптеген көшірмелер күйінде көбейтіліп жинақталуы. Ол
зиготаның бөлшектенуі үшін өте қажет, себебі алгашқы кезендерде
ДНҚ молекуласының екі еселенуі (репликациялану) зиготаның
бөлшектену қаркынына ілесе алмайды. Осы кезде байқалатын
121
гендер «тапшылығы», жетіспеушілігі, күні бұрын көшірмеленіп
жинақталған гендер арқылы сезілмейді.
4.2. Дүниеге келгенге дейінгі
(эмбриондық, антенаталдық) кезең
Эмбриондық кезең зиготаның бөлшектенуінен басталады.
Зигота — ұрықтану нәтижесінде түзілетін үрықтың бір жасу¬
шалы, диплоидты фазасы болып табылады. Зигота жүмыртқа жа-
сушасы мен сперматозоидтың өзара қосылуы нэтижесінде пайда
болгандықтан, онда екі жүп хромосома саны (2п) кездеседі. Олар-
дын бір жартысы (п) аналық дарадан, ал екінші жартысы (п) аталық
дарадан келіп түседі.
Ұрықтанганнан кейін жүмыртқа жасушасының ооплазмалық
сегрегациялануы эрі қарай күшейе түседі.
Зиготаныц болшектенуі дегеніміз оның ядросынын 2, 4, 8, 16,
32, 64 т.б. жасушаларга (бластомерлерге) бөлінуі болып табыла¬
ды. Зигота бөлшектенуінің жай митоздық бөлінуден ерекшелігі
бөлшектену нэтижесінде түзілген бластомералар оспей эрі қарай
бөліне береді.
Бөлшектенудің биологиялыкк маңызы: 1) бөлшектену нэти-
жесінде бір жасушалы зиготадан көп жасушалы үрық түзіледі;
2) бөлшектену кезінде ДНҚ молекуласының қайта-қайта екі
еселенуі (репликациялануы) салдарынан зигота генотипі көбейеді;
3) зигота цитоплазмасының эр түрлі учаскелерінің кұрылысы
мен жіктелуі түрліше болып, ақырында бір-бірінен күрамы жэне
даму багыттары ерекше болып келетін бластомерлер жиынтыгы
түзіледі.
Жүмыртқа жасушасының түріне қарай зигота бөлшектенуінің
2 түрін ажыратады: 1) толық бөлшектену; 2) толык емес бел-
шектену.
Зиготаның толъщ бөлшектенуінде оның тірі заты түгелдей,
қалдықсыз бластомерлерге таралады, ал толык емес бөлшектенуде
зиготаның тек ақуызы гана бөлініп, сары уызы бөлінбейді.
Толық бөлшектенудің біркелкі толъщ бөлшектену, әркелкі
толық бөлшектену, ал толык емес бөлшектенудің дискі тәрізді
бөлшектену, үстіцгі бетінің бөлшектенуі деген түрлері белгілі.
122
Жұмыртканын типтері
қандауырша бака кұс
32-сурет. Жумыртқанын бөлшектенуі жэне бластула
Сол сияқты, бөлшектенудің спиральді, сэулелі (радиальді) син-
хронды және асинхронды бөлшектену деген түрлері де болады.
Зиготаның толық, біркелкі бөлшектенуінде (қандауырлар)
бірінші және екінші бөліну зиготаның анимальдық полюсінен
вегетативтік полюсі бағытында меридиан жазығында жұмыртқаны
2-ге, содан кейін 4-ке боледі. Үшінші боліну экватор жа-зығында,
яғни келденең бағытта жүріп жұмыртқаны 8 бластомерлерге
бөледі. Келесі болінулер меридиан жэне экватор бағыттарында
кезектесіп жүреді де, осылайша 6—7 рет болінуден кейін бласто-
мерлер саны 64-128-ге жетеді.
Болшектену — бір қабатты ұрык бластулапыц пайда болуымен
аяқталады. Бластула — іші сұйықтыққа толтырылған көпіршік.
Оның қабырғасын бластодерма, ал сұйықтыққа толтырылған
куысын бластоцелъ деп атайды.
Болшектену типіне карай бластуланың — кәдімгі бластула не-
месе целобласптула, сімфибласіпула, дискобластула және морула
деген түрлері белгілі.
123
Кэдімгі бластула немесе целобластула толық, біркелкі
бөлшектену нәтижесінде тұзіледі. Ол бластомерлердің бірдей бо¬
лу ымен сипатталады. Амфибластула толық, әркелкі бөлшектену
нэтижесінде түзіледі. Оның бластомерлері әркелкі болып
келеді: анимальді полюстің бластомерлері ұсақ микромерлер,
ал вегетативтік полюстің бластомерлері ірі макромерлер. Дис¬
кобластула — толық емес, диск тәрізді бөлшектену нәтижесінде
түзіледі. Морула — бластомерлері тығыз орналасқан, бластоцелі
болмайтын бластула. Оның сырт пішіні тұт агашының жемісіне
ұқсас болады.
Жогары сатылы сүтқоректілердің, соның ішінде адамдардың
зиготасында сарыуыз тапшы болып, толық, сәулелі не спиральді
жолмен бөлшектеніп бластоциста түзіледі. Оның қабырғасының
бір бөлігінде бластомерлер тығыз орналасып ұрық дискісі — эм-
бриобласт пайда болады, ал қалған бөлігінен қорғаныстық, қорек,
бекіну қызметтерін атқаратын трофобласт түзіледі.
Гаструляция. Гаструляция дегеніміз — 2 не 3 қабатты ұрық —
гаструланың пайда болу құбылысы. Гаструла пішіні кең, жалпақ
кұмыраға ұксас болып келеді. Оның қабыргасы алғаш 2 қабаттан
(эктодерма, энтодерма), кейінірек үшінші кабат — мезодермадан
кұрылган. Оның ішкі куысын — гастроцелъ немесе алгаиіцы
іиіек цуысы деп атайды. Гастроцель сырткы ортамен кішкене
тесік — бластопор немесе алгаиіқы ауыз арқылы байланысады.
Бластопордың жиектерін еріндер деп атайды. Эктодерма жэне эн¬
тодерма алғашқы, ал мезодерма соңгы ұрық жапырақшасы (қабаты)
деп аталады. Мезодерманы пайда ететін бастама қүрылымдар
алғаш не эктодерманың, не энтодерманың кұрамына кіреді.
Гаструланыц пайда болуының бірнеше жолдары белгілі: 1)
инвагинация немесе ішке ңараіі қайрылу — бір полюстің, әдетте
вегетативтік полюстіц бластомерлері ішке қарай тартылып ішкі
қабат — энтодерманы туындатады.
2) эпиболия немесе қапталу — вегетативтік полюстің ірі,
сарыуызға бай жасушалары қозғала алмағандыктан, анимальдык
полюстің микромерлері тез бөлініп, көбейіп оларды коршап
қаптайды да эктодермаға айналады.
3) иммиграция — бластодерманың бір полюсінің бластомерлер,
әдетте вегетативтік полюстің, бір-бірінен ажырасып үрық ішіне
шөгіп, үстіцгі қабаттың астына орналасып энтодермаға айналады.
124
а
э
б
в
33-сурет. Гаструла түрлері:
а) инвагинация, э) эпиболия, б) иммиграция, в) деляминация.
1-эктодерма, 2-энтодерма, 3-бластоцель
4) деляминация — морула типті бластулаларға тэн. Бұл
үрықтарда моруланың сыртқы бластомерлері эктодермаға, ішкі
бластомерлері ығысып энтодермаға айналады.
Деляминация жэне иммиграция негізінен ішекқуыстыларға тэн,
ал жануарлардыц басым көпшілігінде гаструляция аралас жолмен
жүреді, мысалы, инвагинация + эпиболия, кейде инвагинация +
эпиболия + иммиграция.
Екі қабатты гаструла тек ішекқуыстыларга гана тэн, ал
калган коп жасушалы жануарлардыц ұрыгында үшінші ұрық
жапырақшасы (кабаты) — мезодерма қалыптасады. Сондыктан да
оларды 3 қабатты жануарлар деп атайды.
Мезодерма негізінен 3 түрлі эдіс арқылы қалыптасады:
1) телобласттың әдіс — бұл көптеген омыртқасыздарға тэн.
Бұл эдісте мезодерма ұрықтыц артқы жагында эктодерма мен эн¬
тодерма арасында орналасқан ірі жасушалар — телобласттардан
пайда болады.
2) энтероцельдік әдіс — бұл инетерілілерге, қолаяқтыларға,
қылтанақаяқтыларға, бассүйексіздерге, дөцгелекауыздыларга,
балықтарга, космекенділерге тән. Бұл эдіс бойынша мезодер¬
ма бастамасы гаструляциясыныц алғашқы сатысында энтодер¬
ма қүрамына кіреді, сосын гаструляцияныц соцғы сатыларында,
ягни мезодерманың қалыптасу сатысында, энтодермадан бөлініп
шыгып мезодерманы қалыптастырады.
3) жогары сагпылы хордалыларда — жоргалаушылар, құстар
жэне сүтқоректілер, бластула сатысында мезодерма бастамала-
125
ры алғаш эктодерма құрамында болып, гаструляция ныц соңғы
сатысында олар эктодермадан бөлініп мезодерма кабатын калып-
тастырады.
Көптеген омырткалы жануарлардың мезодермасының дамуы
ұксас жобамен жүреді. Үрықтың арқа жағында хорда бастама-
сы қалыптасады. Оның екі жағында мезодерма буылтыкталған
(метамерлі) сомиттерге бөлінеді. Олар мезодерманың қүрсақ
боліміндегі буылтықталмаған бүйір тақталарымен (спланхнотом-
дармен) жіңішке аяқшалар немесе нефротомдар арқылы байланы-
сып тұрады. Әрі қарай эрбір сомит қабырғасы склеротом, дерма-
том жэне миотомдарға жіктеледі. Склеротомдар білік қаңкасын
(омыртқа жотасы) жэне дәнекер ұлпаны, дерматомдар — терінің
дәнекер үлпалы қабатын (дерма), миотомдар қанқа бұлшық еттерін
қалыптастырады.
Ұрық дамуында гаструляция сатысының болатындыгы жэне
үрық жапырақшалары туралы теорияны қалыптастырғандар орыс
ғалымдары: К.Ф. Вольф, Х.И. Пандер, К. Бэр, әсіресе А.О. Кова¬
левский жэне И.И. Мечниковтар болатын.
Гистогенез жэне органогенез. Гистогенез (гр. histos — ұлпа,
genesis — даму) — агза ұлпаларының пайда болуын, дамуын,
тіршілігін жэне қалпына келуін қамтамасыз ететін қүбылыстар
жиынтыгы болып табылады. Гистогенез негізі болып эмбриоге-
нездін ең алгашқы сатыларында басталатын жасушалар жіктеуі
(дифференцировка), ягни маманданушы жасушалар арасындагы
үдемелі морфо-физиологиялық ерекшеліктер саналады. Бұл
жасушалар эр түрлі гендердің активтенуін не активсізденуін
қамтамасыз ететін күрделі молекулалық-генетикалық қүбылыстар
болып табылады. Жасушалардың көбеюі, олардыц орын алмасты-
рып қозгалып жылжулары (миграция) жэне басқа да қүбылыстар
эмбриондық бастамалардыц қалыптасуына алып келеді. Эмб-
риондық бастамалардыц ұлгіалық жіктелуі нэтижесінде эр түрлі
мүшелердің ұлпалары дамиды.
Органогенез — жануарлар мүшелері мен мүшелер жүйесініц
пайда болу жэне даму сатысы болып табылады. Органогенез саты-
сы 2 кезецге болінеді: 1) нейруляция — бұл фазада нерв түтігі жэне
хорда қалыптасады: 2) екінші фазада ұрықтың қалган мүшелері
пайда болып, олардыц оздеріне тэн пішіндері мен құрылысыныц
өзара үйлесімділігі қалыптасады.
126
Сонымен гисто-органогенез сатысында ұрық жапырақшала-
рынаң (қабаттарынан) төмендегідей улпалар мен мүшелер түзі-
леді.
Эктодермадан — жүйке жүйесі, тері эпидермисі және оның
туындылары (тырнақ, шаш, түктер, май жэне тері бездері), есту,
көру, сезу мүшелері дамиды. Хордалыларда жүйке жүйесі өзінің
дамуында бірнеше кезендерден өтеді. Алғаш эктодермадан хорда
үстінде жүйке тақтасы пайда болады. Әрі қарай оның жиектері
кайрылып астауша түзіледі. Астауша жиектерінің қайрылуы эрі
қарай жалгасып ақырында олар бір-бірімен бітісіп жүйке түтігін
түзеді.
Энтодермадан — хорда, ас корыту жолдарының, бауыр, өкпе
эпителийлері, гипофиз, ұйқы безі, ішек-қарын бездері дамиды.
Мезодермадан — терінің дәнекер ұлпалы қабаты (дерма),
қаңқа бұлшық еттері, дәнекер ұлпалар, зэр шыгару, жыныс, жүрек-
кантамыр жүйелері т.б. дамиды.
4.3. Эмбриогенездің жасушалық
жэне генетикалық механизмдері
Эмбриогенез — өте күрделі процесс. Ол бір жагынан үнемі
жаңа жасушалар кешенінің түзілуі, екінші жағынан, алгаш тең
мүмкіндікті (тотипотентті) жасушалардың детерминацияла-
нып ақырында маманданған ұлпаларга және мүшелерге жіктелуі
арқылы жүреді. Ол жасушалық жэне генетикалык механизм-
дер аркылы басқарылып, реттеліп отырады. Эмбриогенездің
жасушалық механизмдеріне үрық жасушаларының көбеюін
(болінуін), жасушалардың миграциялануын, жасушалардың тан-
дамалы іріктелуін, жіктелуін жэне заңды түрде өліп жойылуын
жатқызуга болады. Жасушалардың көбеюі (бөлінуі) эмбриоге¬
нез процесінде маңызды рөл аткарады, себебі, тек жасушалардың
көбеюі нәтижесінде гана ұлпалар мен мүшелердің калыптасуы,
үрықтың өсуі мүмкін. Жасушаның көбеюі нэтижесінде ұлпалар
мен мүшелерге кажет күрылыс материалдары — жасушалар
түзіледі.
127
Жасушалардың көбеюі нәтижесінде түзілген жаңа жасуша¬
лар активті түрде жылжып, қозғалып, миграцияланады. Осының
арқасында жасушалар құрылыс материалдары ретінде ұрықтыц
қажет болған жерлеріне жеткізіледі. Жасушалар позициялық
ақпаратты пайдаланып амеба тэрізді қозғалыс нәтижесінде жыл¬
жып миграцияланады. Позт/гіяльщ ақпарат дегеніміз жасуша-
лардың жылжып қозғалып өздеріне тиісті орнын тауып орнала-
суына бағытталған ооплазмалық сигнал болып саналады.
Ал, позициялық ақпарат механизмдері ретінде жасушалардың
контактты (түйісу) әрекеттесуі немесе хемотаксис қүбылысы бо¬
луы мүмкін.
Эмбриогенез барысында көптеген жасушалар ішінен біркелкі
жасушалар іріктеліп алынып белгілі бір ұлпа не мүшелердің
қалыптасуына багытталынады.
Эмбриогенез барысында жасушалардың көбеюімен қатар
кейбір жасуша топтарының заңды түрде өліп жойылуы — апоптоз
да байқалады. Осылайша саусақтар бір-бірінен ажырасады, қуыс
мүшелердің (қантамырлар, ішектер) қуыстары тесіледі. Эмбрио¬
генез процесінде қол бастамасы тұтас тақта күйінде қалыптасады,
кейінірек саусақтар арасындагы паренхималық жасушалар өліп
жойылып, саусактар бір-бірінен ажырайды.
Жасушаныц жіктелуі дегеніміз — алгаш тотипотентті ұрық
жасушаларының детерминацияланып маманданган ұлпа не мүше
жасушаларына айналуы.
Тотгтотенттілік, ягни тең мүмкіншіліктілік дегеніміз —
кез келген ұрық жасушасының бір-бірінен ажыраса, өз алдына
жаңа ұрық дамыта алу қасиеті. Адам ұрығының бластомерлері
2-4 жасушалы күйінде тотипотентті болады. Ал зиготаның
келесі бөлшектенуінде, ягни 8 жасушалы күйінде бластомерлер
тотипотентті болмайды, себебі, олар детерминациялануға көшкен
болып саналады. Детерминациялану дегеніміз — жасушалардың
белгілі бір даму жолына түсуі. Ол күні бүрын генетикалык
бағдарланган жэне көрші жасушалардың өзара әрекеттесуінің,
гормондардың немесе сырткы факторлардың әсері нәтижесінде
қалыптасады. Детерминациялану түрақты жэне қүбылмалы детер¬
минациялану болып бөлінеді. Түрақты детерминацияланған жасу¬
шалар еш уақытга да қайтадан тотипотентгі күйге көше алмайды,
128
ал құбылмалы детерминацияланған жасушалар кейде қайтадан
тотипотенттілік қасиетке ие бола алады, оны трансдетерминация
деп атайды.
Эмбриогенездің бастапқы сатыларында жасушалардың детер-
минациялануын жасушаның ядродан тыс цитоплазмада орналас-
қан факторлары (ооплазмалық сегрегация, ұрық жасушаларының
детерминанттары, позициялық ақпарат) бағдарлап басқарып огы-
рады. Жасушалардың эрі қарай детерминациялануы жэне жіктелуі
ядро бағдарламасына тәуелді болады жэне ол гендердің тацдама-
лы экспрессиялану теориясы негізінде жүзеге асады.
Агзалардың жеке дамуы зигота ядросындагы генетикалық
багдарлама негізінде жүзеге асады. Бірақ, қалайша алгаш тең
қабілетті тотипотентті ұрық жасушаларынан түрліше құрылысты
жэне эр түрлі қызмет атқаруга маманданган ұлпалар мен
мүшелер, ересек агзалар қалыптасады? Мүны алғаш тотипотентті
эмбриондық жасушалардың эр түрлі даму жолдарына түсіп
жіктелуінің (дифференциялануыныц) салдары деп түсіну қажет.
Дифференцияланган (жіктелген) жасушалар эдетте өте күрделі
қүрылысты болып келеді. Мысалы: жүйке жасушалары, көздің
торлы кабыгының таяқшалары мен колбашықтары т.с.с. Сол
сияқты, әр түрлі ұлпа жасушаларында тек өздеріне гана тэн бе-
локтар синтезделінеді, мысалы: бұлшық ет жасушаларында -мио¬
зин; көздің көру рецепторларында — опсин; эритроциттерде —
гемоглобин т.с.с. Осы ақуыздар ұлпалардың ерекше белгілерін
анықтайды. Қазіргі кезде кез келген белок белгілі бір геннің
кызметтік активтігінің нәтижесінде синтезделінетіні баршага аян,
ал агзаның барлық жасушаларында гендер жиынтығы бірдей бола¬
ды, мысалы адам жасушаларында 30000-га жуық гендер кездеседі.
Сонда, белгілі бір даму жолына түскен жасушалардың жіктелуі
кейбір гендердің іріктеліп жойылуына алып келе ме, әлде олар үне-
мі активсізденіп отыра ма? Әр түрлі тэжірибелер нәтижесінде да¬
му процесінде міндетті түрде гендердің жойылмайтындыгы бел-
гілі болды. Сонымен қатар, осы тэжірибелер даму процесінде үрық
жасушаларында гендердің таңдамалы экспрессияланатындығын
айқындады. Гендердіц тацдамапы экспрессияпануы (белсенділе-
нуі) дегеніміз дамудың эрбір кезеңінде, эр түрлі жасушалар то-
129
170-9
бында, түрліше гендердің активтенуі не активсізденуі. Бұл болжам
бойынша әрбір жасушада гендердің 3 белсенді тобы болады:
1) барлық жасушаларда актив күйінде болатын жэне жасуша-
ның негізгі тіршілік процестерін (зат, энергия т.б. алмасуды)
қамтамасыз ететін гендер. Бұл гендерсіз кез келген жасуша
тіршілігі мүмкін емес, олар жасушага өте қажет гендер: оларды
«тұрмыстық» гендер деп атайды;
2) бір ұлпа жасушаларының морфофизиологиялық ұксасты-
гын қалыптастыратын гендер (мысалы, жүйке жасушаларының
тітіркенуін, олардың түрақты өскіншелерін дамытатын гендер);
3) нақтылы үлпалар жасушаларына гана тэн ерекше қызмет-
терін, қасиеттерін қалыптастыратын жэне басқа үлпаларда ак¬
тив болмайтын гендер (мысалы: эритроциттерде гемоглобин
синтезделуін, бүлшык ет жасушаларда миозин синтезделуін, көз-
дің көру рецепторларында опсин синтезделуін камтамасыз ететін
гендер). Бүл гендер тобын «молшылық» гендері деп атайды.
Гендердің активтену-активсіздену механизмдері элі күнге
дейін толык шешілмеген. Дегенмен, бүл процесте хромосоманың
физикалық-химиялық касиеттері өзгеретіндігі сөзсіз, ягни
ДНҚ молекуласы мен гистонды белоктардың ара қатынасы өз-
гереді. Активсіз хромосомаларда ДНҚ молекуласы нуклеосома
денешіктерімен тыгыз байланыскан, ал олар активтенгенде ДНҚ
молекуласымен нуклеосома денешіктері арасындагы байланыс
едәуір босап элсірейді.
Әр түрлі тәжірибелер нэтижесінде гендердің активтенуі зигота
цитоплазмасының эсеріне байланысты болатындыгы анықталды.
Шамасы, ооплазма хромосомамен эрекеттесетін эр түрлі зат-
тарды бөліп шыгарып, ондагы ДНҚ молекуласын нуклеосома
денешіктерінен босатып, гендерді активтендіретін болуы ке-
рек. Хромосоманың актив учаскесі микроскоппен қараганда
активсіз учаскелерінен біршама жуандау, ісіңкі болып келеді.
Хромосоманың ондай учаскелерін пуффачар деп атайды. Пуффа-
ларда ДНҚ молекуласы нуклеосомалардан босап ажырап, шираты-
луы жазылып транскрипцияланады (а-РНҚ синтезделінеді).
Зерттеулер нәтижесінде насекомдар дамуының эр түрлі
кезеңдерінде хромосоманың эр түрлі учаскелерінде түрліше пуф-
фалар пайда болатыны анықталды. Мысалы, жеміс шыбынының
130
дамуының дернәсілдік (личинкалық) сатысында бір хромосомада
3 пуффа пайда болса, дернәсілден қуыршақ сатысына өткенде сол
хромосомадағы алғашқы 3 пуффалар жойылып, оның басқа бір
жерінен жаңадан пуффа пайда болатыны байқалған.
Эмбриогенез процесінде гендердің тандамалы белсенділігі
болатындыгына дәлел ретінде ұрық дамуының эртұрлі кезенде-
рінде түрліше мутациялардың пайда болатындығын айтуға бола¬
ды. Бүл мутациялар гомозиготалы күйінде ұрықтың өліп қалуына
алып келеді, мысалы: t12 мутация бойынша гомозиготалы ұрықтар
морула сатысына дейін жетіп бластоцистаға өте алмай өліп
қалады; tw73 бойынша гомозиготалы ұрықтар бластоциста саты¬
сына өтіп жатырга толық бекіне алмай өліп қалады; t° бойынша
гомозиготалы ұрықтар гаструла сатысында өліп қалады; t-'vl — ор¬
ганогенез сатысында өліп қалады. Демек, мутантты t аллельдері
үрықтыц дамуыныц әр түрлі сатыларында пайда болып отыра-
тын жасушааралық әрекеттесуді тежейді десек, онда Т-локусы
қалыпты жагдайларда жасуша қабықшасының бетінің сезімталдық
қасиеттерін өзгерту арқылы дамуды қадагалап отырады деген бол-
жам жасауга әбден болады.
Гендердіц тандамалы экспрессиялануына тагы бір мысал —
эр түрлі жынысты агзалар жынысының анықталуы 2 кезеңнен
тұрады: 1) ядроның хромосомалык күрамы — гонадалардың
жыныстық жіктелуіне алып келеді, егер XX хромосомалары бол¬
са аналық безді, ХУ хромосомалары болса аталық безді қалып-
тастырады; 2) егер аталық без қалыптасса ол тестостерон гормонын
бөліп шыгарады. Ол ұрық денесіне таралып, сома жасушаларыныц
еркек ағзаларга тэн багытта дамуына мүмкіншілік тугызады.
Ал, егер аналык без қалыптасса тестостерон гормоныныц син-
тезделмеуі үрық жасушаларының эйелдерге тэн багытында даму¬
ына алып келеді.
Көптеген деректерге Караганда, аталык бездің дамуы Ү хромо¬
сомада орналасқан гендердің эрекетіне байланысты. Адамдардың
жыныс хромосомаларының мейоз кезінде дүрыс ажыраспауы мы-
надай зиготалардың пайда болуына алып келеді. ХО (2А) немесе
ХХУ (2А). ХО (2А) — ұргашы фенотипті дамытады (Тернер син¬
дромы), олардың аналық бездері өте нашар дамыган. ХХУ (2А)
— еркек фенотипті дамытады (Клайнфельтер синдромы), олардыц
131
аталық бездері дамиды, бірақ сперматогенез болмайды. Осының
бэрі адамдарда жыныстың қалыптасуы Ү хромомсомага байланы¬
сты болатындығын дәлелдейді.
Сонымен, ағзалардың соңғы жыныстық белгілерінің дамуы
гонадалардың жыныстық жіктелуіне, ал ол хромосома құрамына
(XX, XV) байланысты болады. Дамып келе жатқн аталық без-
дер тестостерон синтездейді, ал ол белгі (сигнал) ретінде еркек
фенотиптің дамуына алып келеді. Сол белгі болмаса үрық ұргашы
фенотипке сэйкес дамиды. Үрықтың еркек типіне сэйкес дамуы
X хромосомасымен тіркескен жалғыз геннің (Tfm+) бақылауында
болады. Ол ген тестостерон гормонымен байланысатын ақуыз
молекуласының синтезделуін қамтамасыз етеді жэне еркек,
ұргашы ағзалардың барлық жасушаларында кездеседі. Бұл ақуыз
реттеуші қызмет атқарады, ягни ол тестостеронмен байланысып
активтенеді, содан кейін активтенген белок-тестостерон кешені
ядроға еніп жасушаның еркек типке сэйкес багытта дамуы үшін
қажет гендерін активтендіреді. Адамдарда Tfm+ генінің мутациясы
белгілі (Tfhv). Ол тестикулярлы феминизация белгілерін тудыра-
ды (генотипі ХУ болғанымен, фенотип әйелдерге ұқсас болады).
Tfm- /У генотипі бар эмбрион жасушалары тестостерон гормо-
нын мүлдем сезбейді, ягни тестостерон гормонымен қосылып
активтенетін белок синтезделмейді, сондықтан тестостерон бол-
ганымен, ол ядрога еніп еркек типті фенотиптің дамуына кажет
гендерді активтендіре алмайды, осының нәтижесінде соңгы жы¬
ныс белгілері эйелдер фенотипіне сэйкес дамиды. Бірақ, мүндай
әйелдерде аналық бездің орнына аталық безі дамып, өздерінін
бөліп шыгарган гормондары — х-факторлары арқылы фалло-
пий түтікшелерінің жэне жатырдың дамуын тежеп түйық қынап
қалыптасады.
34-сурет. Алгашкы эмбриондык индукция:
а) донор материалы, э) донор материалын реципиент ұрыкка кондыру.
б) екінші жүйке (нерв) түтігінің пайда болуы, в) екінші урыктын пайда болуы.
а
э
б
в
132
Эмбриондьщ индукция. ¥рық дамуында эмбриогенездің жоға-
рыда айтылған жасушалық жэне генетикалық механизмдерімен
қатар эмбриональдық индукциялардың да рөлі зор. дегеніміз
дамып келе жатқан ұрықтың бір бөлімінің (индуктор) басқа
бөлімдеріне (эффектор) тигізетін әсері болып саналады. Эмб-
риондық индукцияны алғашқы рет 1924 жылы неміс биологы
Г. Шпеман қосмекенділер ұрыгында көз буршагының эктодер¬
ма қабатынан пайда болуын зерттеу кезінде ашқан. Егер ұрық
эктодермасынан көз бастамасын алып тастағанда көз бұршағы
қалыптаспаған.
Керісінше, ұрықтың бір бүйіріне көз бастамасын қондырса
эктодермадан көз бұршағы пайда болған, ал қалыпты жағдайда
одан тері эпидермисі түзілуі қажет болатын. Кейінірек Г. Шпеман
хордомезодерманың орталық жүйке жүйесі бастамасының (жүйке
тақтасының) эктодермадан пайда болуына индукциялық ықпал
ететіндігін байкаған. Бүл қүбылысты ол алгаиіңы эмбриональдьщ
индукция, ал индуктор — хордомезодерманы үйымдастыруиіы
деп атаған. Дамушы үрықтың кейбір бөлімдерін кесіп алып олар¬
ды бөлек-бөлек не бірге үрықтың басқа жерлеріне қондыру
негізінде эмбриондық индукцияның көптеген омыртқасыз жэне
барлық омыртқалы жануарлар арасында кең таралған қүбылыс
екендігі анықталды.
Эмбриондық индукция қүбылысы тек қүзырлы эффектор жа¬
сушаларында байқалады, ягни индукция ықпалын қабылдауға
қабілетті жэне тиесілі кері жауап бере алатын жағдайларда ғана
жүзеге асады.
Даму барысында көпсатылы эмбриондык индукциялар жүзеге
асады: жіктелуге (дифференцировка) сигнал (стимул) алган эффек¬
тор жасушалары дамып, эрі карай баска компонент жасушалар
үшін индуктор қызметін атқарады т.с.с, ягни алғашқы эмбриондық
индукция, екінші, үшінші, төртінші т.б. ретті индукцияларға
жалғаса береді.
4.4. ¥рыктың провшорлық мүшелері
Көптеген омыртқалылар эмбриогенезінде үрықтың тіршілігі
үшін өте қажет уакытша мүшелер — провизорлық мүшелер қа-
133
лыптасады, себебі ұрықтың элі толық жетіле қоймаған мүшелері
өздеріне тэн қызметтерді атқара алмайды. Ал, олар толық жетіліп
өздеріне тэн қызметтерді аткаруға кірісе бастағанда провизорлық
мүшелер семіп жойылады. Провизорльщ мүиіелер дегеніміз
омыртқалылардың ересек дараларында болмайтын, тек ұрықтың
дамуы үшін ғана қажет уақытша мүшелерді айтамыз. Оларға сары-
уызды қапшық және ұрық қабықтары — қағанақ (амнион), хорион,
аллантойстар жатады.
Провизорлық мүшелер үрықты механикалық зақымданудан
сақтайды, оның дамуы үшін қолайлы жағдайлар — сүйықтық
орта калыптастырады жэне тыныс алу, қоректену, бөліп шығару.
қозғалу, қантүзу т.б. қызметтерді атқарады.
Адамдардың провизорлық
мүшелерінің дамуы өте ерте
гаструляциямен бір мезгілде
басталады. Қағанақ (амнион)
пен хорионның дамуы ұрык
дамуының 7—8 тәулігінде ба¬
сталады. Хорион трофобласт-
тан, ал қағанақ (амнион) эпи-
бласт жасушаларының ығысуы
салдарынан пайда болады. Адам
үрығының қағанағы (амни¬
он) шизамнион деп аталады.
Қағанак қуысы шамалы уақыт
эпибласт жасушаларымен жэне
трофобласт учаскелерімен шек-
теледі. Кейінірек эпибластгың
бүйір қабырғасы қатпарлар
туғызып бір-бірімен кірігеді.
Қағанақ куысы түгелдей эпи¬
бласт жасушаларымен астар-
ланып түрады, ал оның сыртқы жағы үрықтан тыс мезодермалы
жасушалармен қапталған болады.
Қаганақ (амнион) үрық қабаттарының ең ішкі қабаты. Ол
үрықты тікелей қоршап түратын жэне сүйықтыққа толтырылған
эпидермалы капшық болып табылады. Қағанақ ұрықты кеуігі кету-
35-сурет. Омыртқалардын
провизорлык мүшелері:
а) балыцтар, ә) цүстар,
6) сүтцоректілер, 1-сарыуыздыц
қапиіық, 2-а.мнион, 3-аллантоис,
4-хорион
134
ден жэне механикалық зақымданудан сақтайды, оның дамуы үшін
ең қолайлы жэне табиғи ылғалды ортаны қалыптастырады.
Хорион — ұрык кабықтарының ең сыртқысы болып табылады
жэне ол қағанақ сияқты эктодермадан түзіледі. Хорион ұрықпен
оны қоршаған орта арасындағы зат алмасу қызметін атқарады.
Сүтқоректілерде ол тыныс алу, қоректену, бөліп шығару, сүзу
(фильтрация) жэне кейбір заттарды (мысалы, гормондарды) син-
тездеу қызметтерін де атқарады.
Сарыуызды цапшыц ішкі жасушалық массадан гипобласттың
жүқа қабаты мен ұрықтан тыс эпидермалы жасушалардың бір-
бірінен ажырасқан кезінде түзіледі. Алғашқы сарыуызды қапшық
үрық дамуының 12—13 тэулігінде гіайда болып, әрі қарай үрықпен
байланысқан екінші реттік сарыуызды капшыққа айналады.
Сарыуызды цапшыц эктодерма туындысы жэне ұрықтың ішек
түтігімен тікелей байланысқан. Сүтқоректілерде сарыуызды
қапшық энтодермасы — алғашқы жыныс жасушаларының, мезо-
дермасы — қан түйіршіктерінің түзілетін орны болып саналады.
Сол сияқты, сүтқоректілердің сарыуызды қапшығы сүйықтықпен
толтырылған, ал онда амин кышқылдарының, глюкозаның көптеген
қорлары жинақталады. Осыған байланысты ол акуыз алмасуына
қатынасуы мүмкін деген болжамдар айтылуда.
Аллантоис (сарыуыз қап) — провизорлық мүшелердің ең
соңынан түзілетін қабыгы болып саналады.
Адам аллантоисы артқы ішектің вентральды қабырғасының
қалтасы күйінде пайда болады. Оның қабырғасында үрықтың
негізгі қан тамырларымен байланысқан көптеген тор тэрізді қан
тамырлар дамиды. Аллантоис мезодермасы хорион мезодермасы-
мен қосылып өзінің қан тамырларын соңғысына береді. Осылайша
хорион — аллантоисты бала орны (плацента) дамиды.
Плацента (бала орны) дегеніміз ұрық қабықтарының ана
ағзасының үлпаларымен кірігуі болып табылады. Приматтар-
да жэне кейбір сүтқоректілерде аллантойстың энтодермасы
редукцияланған, ал мезодерма жасушалары тығыз созылмалы
жіпше пайда етеді. Ол клоака бөлімінен хорионға дейін созы-
лып жатады. Аллантойс мезодермасы арқылы хорионға сосудтар
өтеді, ал олар бөліп шығару, тыныс алу, қоректену қызметгерін
атқарады.
135
4.5. Дамудын қатерлі кезеңдері
Жануарлар дамуын зерттеу нәтижесінде дамудың қатерлі
кезеңдері болатындығы анықталды. Дамудыц катерлі кезеңдері
деп ұрықтың қоршаған ортаның эр түрлі зиянды факторларына
өте сезімтал кезендерін айтамыз. Бұл кезеңдерде зиянды фак-
торлар эсерінен онтогенездің қалыпты жүруі бүзылып, ұрык не
өліп қалады, не міндетті түрде оның бойында эр түрлі ақаулықтар
қалыптасады. Дамудың қатерлі кезеңдері терминін 1897 жылы
Браун енгізген болатын.
А. Н. Трифонов (1834) балықтардың дамуында 3 қатерлі кезең-
дерді ажыратқан: біріншісі — бөлшектенудің басы немесе орта-
сы; екіншісі — гаструляция басы; үшіншісі — білік мүшелерінің
(омыртқа жотасының) қалыптасу сатысы.
Дамудың эр түрлі қатерлі кезеңдерінде үрықтар түрліше
факторларға, мысалы дамушы уылдырықтар отгегі жетіспеу-
шілігіне, жылылыққа, механикалық эрекеттерге сезімтал болады.
П.Г. Светловтың айтуынша, үрық бір қатерлі кезеңде температура
эсеріне сезімтал болса, екінші кезеңде — химиялық заттар әсеріне
сезімтал болуы мүмкін, т.с.с.
Дамудын қатерлі кезеңдерінде үрықта метаболизм, ты¬
ныс алу кұбылыстары күшейеді, РНҚ қасиетгері, иммундық
мэртебесі өзгереді, өсу қарқыны томендейді. Оттектің жэне ну¬
клеин қышкылдарының жеткілікті болуы қиын кезеңдер салдарын
байқатпайтын факторлар болып табылады. Сол сияқты, дамудың
қатерлі кезеңдері эмбриогенездің бір кезеңінен екінші кезеңіне
өтуімен жэне морфологиялық жіктелу сатыларымен түспа-түс
келеді.
Адам үрығы дамуының киын кезеңдері бірінші аптаның аяғы
— екінші апта басы аралыгы жэне жүктіліктің 3—6 апта аралығы
болып саналады.
Дамудың қатерлі кезеңдерін ажыратуымен қатар эмбриогенезде
тератогендік терминациялық кезецдердің де маңызы зор екенін
ескерген жөн. Тератогендік терминациялық кезең дегеніміз
— зақымдаушы факторлардың эсер етуі нэтижесінде нақтылы
ақаулықтың пайда болу мүмкіншілігінің шегі.
136
4.6. Дамудың туа біткен ақаулықтары
Эмбриогенез құбылысының қалыпты жүруінің бұзылуы эр
түрлі акаулықтарға алып келеді, оларды дамудың туа біткен
ақаулықтары (ДТБА) деп атайды. Дамудың туа біткен ақаулықтары
немесе даму ақаулықтары деп агзаның қалыпты кұрылысының
морфологиялық бұзылуын айтамыз. ДТБА негізінен үрықтык
(антенатальдық) дамудың бұзылуы нэтижесінде немесе сирек,
туылғаннан кейін кейбір мүшелердің эрі қарай қалыптасуының
ауытқуы салдарынан (мысалы, тістің ақаулықтары, артериялык
боталл өзегінің ашық болуы, т.с.с.) пайда болады.
Адам ақаулықтарын зерттейтін ғылымды тератология, ал сол
ақаулықтарды тудыратын факторларды тератогендік факторлар
деп атайды.
Ғылыми деректерге қарағанда, қазіргі кездері өркениетті елдер
тұрғындарының, эсіресе жас балалардың сырқаттануы мен қаза
болуының басты себептерінің бірі ретінде адамдардың туа біткен
ақаулықтары саналады. ДТБА жаңадан туылған нәрестелердің
кем дегенде 2—3 пайызында кездеседі; нәрестелердің алғашқы
апталарда-ақ өліп қалуының себептерінің 25—30 пайызы да осы
ДТБА-ға байланысты. Кейбір елдерде (АҚШ, Жапония, Германия
т.б.) бөбектердің қаза болуының басты себебі де осы ДТБА болып
есептелінеді. Дамудың туа біткен ақаулыктарына төмендегідей
өзгерістерді жатқызады:
Агенезия — мүшенің мүлдем дамымауы, болмауы;
Аплазия — мүшенің өте нашар дамыған бастамасы гана болуы
мүмкін;
Туа біткен гипоплазия — мүшенің толық жетілмеуі (дамы¬
мауы);
Туа бігкен гипертрофия — мүше салмағының (не мөлшерінің)
салыстырмалы түрде үлғайып өсуі;
Туа біткен гипотрофия — нәрестенің не қүрсақтагы баланың
дене салмагының аз (кішкентай) болуы;
Макросомия —- нәресте денесінің үзын болуы;
Гетерогопия — кейбір мүшелерде өздеріне тэн емес жасу-
шалардың, ұлпалардың немесе басқа бір мүшенің түгелдей бір
бөлігінің кездесуі. Мысалы, мишык қабығында алмүрт пішіндес
137
невроциттердің кездесуі; кеңірдектен тыс, өкпеде кейбір шемір-
шектердің кездесуі т.с.с.
Гетероплазия — кейбір ұлпалардың жіктелуінің бұзылуы;
Эктопия — мүше орнының ыгысуы, яғни оның өзіне тән емес
жерлерде орналасуы. Мысалы, бүйректің жамбас куысында орна-
ласуы, жүректің көкірек қуысынан сыртта орналасуы;
Атрезия — қуыс мүше арнасының не табиғи тесікшесінің мүл-
дем болмауы;
Стеноз — куыс мүше арнасының не тесігінің тарылуы (ішек-
тердің стенозы, қан тамырлардың стенозы);
Бітісу (кірігу) — екі монозиготалы егіздердің бір-бірімен бі-
тісуі (кірігуі);
Дисхрония — ағза мүшелерінің даму қарқынының бүзылуы
(күшеюі не баяулауы).
36-сурет. Дамудын туа біткен акаулыктары.
а) үстіңгі еріннің жэне катты тандайдьщ бітіспеуі, б) аяк-кол
акаулыктары
ДТБА-ның жалпы саны өте көп, мындап саналады Олар¬
ды түқым куалайтын, экзогендік жэне мультифакторлы деп 3
топқа бөледі. Тұқым қуалайтын ақаулықтарға мутациялар
негізінде пайда болған ақаулықтар жатады. Оларды гендік және
хромосомалық түқым қуалайтын ақаулыктар дегі бөледі.
б
138
Экзогендік ақаулықтар
ұрықгыц не құрсақтағы ба-
ланың тератогендік фактор-
лармен тікелей зақымдануы
нэтижесінде калыптасады.
Мультифакторлы ақау-
лықтар генетикалык жэне
экзогендік факторлардың
бірлесе эрекет етуінің нэ-
тижесінде пайда болады.
ДТБА-ның пайда болу се-
бепгерінің 40 пайызы мута¬
циялар, 50 пайызы мульти¬
факторлы, 5 пайызы тера-
тогендік, оның ішінде 2 пай¬
ызы инфекция факторлары
болып есептелінеді.
ДТБА тератогендік фак-
торлардьщ эсер ету форма-
сында, ақаулықтардың агза
мүшелерінде таралуына ка¬
рай жіктеледі.
Тератогендік факторлардыц эсер ету уақытына қарай ақау-
лықтарды: 1) гаметопатия — гаметалардың зақымдануы. Бұл
кездері кіріккен егіздерде циклопия (жалгыз көзділік), сиреноме-
лия, т.с.с. акаулықтар пайда болады; 2) эмбриопатия — ұрықтың
16 күннен — 8 апта аралығында зақымдануы; 3) фетопатия —
ұрықтың 9 апталығынан нәресте дүниеге келгенге дейінгі ара-
лықта зақымдануы. Бұл кездері эмбриондық құрылымдардың әрі
қарай сақталып қалуы (пересистирование) (урахус, крипторхизм)
сияқты ақаулықтар пайда болады.
Агза денесінде таралуына карай ақаулықтарды былайша бөледі:
1) жеке акаулыцтар — бір мүшеде кездессе; 2) жуйелі ақаулықтар
— бір мүшелер жүйесінде кездессе; 3) көптеген ацаулыцтар — екі
не одан да коп мүшелер жүйесінде кездессе.
Ақаулықтардың жогарыда келтірілген жіктелуі этиологиялык
принципке негізделген жэне олар қазіргі тацда көп қолданыла
37-сурет. Біріккен егіздер.
А—Б—В — бастарымен біріккен,
Г—Д—Е — төстерімен біріккен,
Ж — 3—И — бөкселерімен біріккен
139
бермейді. Оның есесіне қазіргі кезде ДТБА бойынша адам денесін
анатомиялық — физиологиялық жүйеге бөлу принциптеріне сэй¬
кес жіктеу кеңінен қолдау табуда. Дүниежүзілік денсаулық сақтау
үйымының (ДДС¥) XXIX Ассамблеясында 1975 жылы қабылдан-
ған жіктелу де осы принципке негізделген. Ол төмендегідей:
А. Мүшелердің және мүшелер жүйелерінің туа біткен
ақаулықтары:
Орталық жүйке жүйесінің, сезу мүшелерінің ақаулықтары;
Мойын жэне бет ақаулықтары;
Жүрек-қантамыр жүйесінің ақаулықтары;
Тыныс алу жүйесінің ақаулықтары;
Ac қорыту жүйесінің ақаулықтары;
Тірек-қимыл жүйесінің ақаулықтары;
Зәр шығару жүйесінің ақаулықтары;
Жыныс мүшелерінің ақаулыктары;
Ішкі секреция бездерінің ақаулықтары;
Тері жэне оның туындыларының ақаулықтары;
Плацента (бала орны) ақаулықтары;
Басқа да акаулықтар;
Б. Көптеген ақаулықтар:
1. Хромосомалық ақаулықтар;
2. Гендік синдромдар;
3. Экзогендік факторларға байланысты синдромдар;
4. Пайда болуы анықталмаған синдромдар;
5. Айқындалмаган көпшілікті синдромдар;
Тератогендік факторларды 3 топқа бөледі:
1) Эндогендік факторлар:
а) мутациялар;
б) ішкі секреция бездері қызметініц бүзылуы;
в) жыныс жасушаларыныц пісіп кетуі;
г) ата-аналар жасы.
2) Экзогендік факторлар:
а) физикалық факторлар;
б) ионданушы радиация;
в) механикалық факторлар;
г) химиялық факторлар:
140
— дәрі-дәрмектер
— тұрмыста жэне өнеркәсіпте пайдаланылатын заттар
— дұрыс тамақтанбау (аштық)
— гипоксия.
3) Биологиялық факторлар:
а) виру стар;
б) микоплазмалар т.с.с.
Мутациялар — ақаулықтардың пайда болуыныц басты
себептерінің бірі болып табылады. Н. П. Дубининнің айтуын-
ша, адамдардыц барлық ақаулықтарын мутацияныың салдары
деп түсіну қажет. Мутациялардыц түрлері, пайда болуы, зиянды
әсерлері туралы алдыңғы тарауларда толык айтылган.
Ішкі секреция бездері қызметініц бұзылуы көптеген ақаулық-
тарга, мысалы, өздігінен тұсік тастауга не құрсақтағы баланыц
морфологиялық жэне физиологиялық ақаулықтары немесе нә-
рестенің көп өмір сүрмей өліп қалуына алып келеді. Әсіресе, кант
ауруларымен ауыратын әйелдерде бұл патология жиі байқалады.
Ішкі секреция бездері кызметініц бұзылу салдары ретінде
диабеттік эмбриопатия жэне диабеттік фетопатияларды айту-
ға болады. Аталған ақаулықтармен туылған нэрестелердіц дене
салмағы үлкен — 5 кг, миокардында (жүрек бұлшық етінде), бау-
ырда жэне каңка бұлшық еттерінде гликоген аз мөлшерде болады.
Олардыц эрі қарай дамуы баяулап кешігеді, ақыл-естері кеміс бо¬
лады. Диабеттік эмбриопатиямен ауырган балалардың 37 %-ының
тірек-қимыл жүйесініц, 24 %-ының жүрек-кан тамыр жүйесініц,
14 -ында ОНЖ ақаулықтары байқалған. Жыныс жасушаларыпыц
«пісіп» кетуі дегеніміз — жүмыртқа жасушасыныц не спермато-
зоидтардың жатыр түтікшесінде бір-бірімен дер кезінде кездесіп
қосылмай, үзақ уақыт түрып қалуы салдарынан әлсіреп, олар-
да өзгерістер болуы нәтижесінде үрықтану қабілетініц кемуі не
бүзылган зиготаларды түзуі болып табылады. Тәжірибелер аркылы
кейбір сүтқоректілердіц жыныс жасушаларыныц «пісіп» кетуініц
үрықтыц дамуына жэне ақаулықтардың пайда болуына тигізетін
септігі зерттелген жэне толық дэлелденген. Мысалы, жүмыртқа
жасушасы 12 сағаттан үзақ түрып қалса, оныц үрықтану қабілеті
76 %-га кеміген, ол үрықтанған күнде 43 % жагдайда эр түрлі
ақаулықтарга алып келген болып есептеледі.
141
Сол сияқты, сперматозоидтардың да «пісіп» кетуі, яғни аналық
жыныс жолдарында 1—2 тэулік бойына жұмыртқа жасушасы-
мен қосылмай тұрып қалуы, олардың әлсіреуіне алып келеді.
Егер элсіреген сперматозоидтар жүмыртқа жасушасымен қосыла
қойса, эр түрлі ақаулықтардың болатындығы байқалған. Жыныс
жасушаларының «пісіп» кетуінің негізгі тератогендік механизмі
хромосомалардыц бір-бірінен дұрыс ажырасуының бұзылуында
болса керек. Ал, ол өз кезегінде эр түрлі анеуплоидияға алып
келеді.
Ата-аналар жасы да балалар денсаулығына септігін тигізетін
фактор болып табылады, себебі адамдардыц кобею қабілеті жал-
пы биологиялық заңдылықтарға тэуелді. Сондықтан өте жас ата-
аналар орта жастағы жұбайларга қарағанда ауру балаларды жиірек
дүниеге келтіретіндігі түсінікті.
Өте жас аналар 22-35 жастағы аналарға қарағанда тірек-қимыл,
тыныс алу жүйелерінің ақаулықтары кездесетін нэрестелерді
жиірек туады. Ал, ананың жасы 35-тен жоғары болған сайын
нәрестелердің көпкездесетін ақаулықгары жэне ОНЖ ақаулық-
тары, эсіресе, әр түрлі анеуплоидиялар — трисомия 1318-21
жиі байқалады. Мысалы, олардың орташа жиілігі 30-34 жастагы
аналардың балаларында 1/150; 35-38 жаста 1/185; 40-44 жаста
1/63; 45 жастан үлкен аналарда 1/24 тең болады.
Жастары үлғайған ата-аналардың эр түрлі ақаулыктары бар
нәрестелерді дүниеге келтіруі эндогендік жэне экзогендік фак¬
торлар әсерінен болуы мүмкін, мысалы, сперматоциттердің жэне
овоциттердің «қартайып» қалуы. Ал, олардыц қажуы мутация
жиілігін жоғарылатады.
Экзогендік тератогендік факторлардыц ішінен ец маңыздысы
— ионданушы радиация болып саналады. Егер енді ғана екіқабат
болған (1-6 күн) әйел бір рет 0,1 гр мөлшерінде рентген сәулесімен
әрекеттессе зигота түсіп қалады, ал жүктіліктіц 2-6 аптасын-
да осыншама мөлшердегі рентген сэулесімен эсерленсе эр түрлі
акаулықтар пайда болады.
Кейбір дэрі-дэрмектердіц де тератогендік әсері белгілі, мы¬
салы, олардыц қатарында транквилизаторлар (қолдан жасалған
гормондық препараттар), талидамид жэне диазепамды атауга
болады. Талидамид ересек адамдарга зиянсыз, бірақ 4-10 ап¬
142
та аралыгындагы жүкті әйелдерге зиян келтіріп, үрықтың
талидамидті эмбриопатиясьша алып келеді. Бұл патология-
мен туылған балалардың аяқ-қолдары өте қысқа немесе қолдары
мүлдем дамымайды, ішкі мүшелерінің қүрылысы да бүзылады,
ал мүндай нәрестелер не өлі туылады, не 1—2 айдан кейін өліп
қалады. Диазепам — балалардың еріндерінің жырық болуына
алып келеді. Тератогендік дэрі-дэрмектерге тырысқаққа жэне рак
ауруына карсы қолданылатын дэрілерді де жатқызуга болады.
Вирустардың, эсіресе, қызылша вирусының, тератогендік
эсері 1941 жылы белгілі болды. Жүкті аналар қызылшамен ауыр-
са үрық не қүрсақтағы бала күні бұрын өздігінен түсіп қалады,
не ақаулықтармен туылады, 50% жагдайда олардың тістері да¬
мымайды. Тератогенездің жасушалык механизмдеріне жасуша
бөлінуінің, миграциялануынын жэне жіктелуінің ауытқуларын
жатқызуға болады.
4.7. Дамудың постэмбриондық кезені
Ағза туылғаннан соң немесе жүмыртқа не үрық қабығынан бо-
санып шыққаннан кейін постэмбриондық кезең басталады.
Әр түрлі жануарлар түрлерінде постэмбриондық кезең бірнеше
күннен ондаған жылдарға дейін созылуы мүмкін.
Постэмбриондық онтогенезді 3 кезеңге бөледі: 1) жастық
(ювенальдік) кезең; 2) жыныстық жетілген (репродуктивтік) кезең;
3) кэрілік кезең. Постнатальдық онтогенез агзаның дүние салуы-
мен аяқталады.
Онтогенез типіне байланысты жастық кезең екі түрлі жолмен
жүреді: тура даму жэне тура емес немесе түрленіп (метаморфоз —
metamorphosis — түрлену) даму.
Тура дамуда жүмыртқа қабыгынан босанып шыққан не¬
месе жанадан туылган жас агзаньщ ересек агзадан ешқандай
айырмашылыгы болмайды, олар тек дене бітімінің пропорциясы-
мен ғана ерекшеленеді. Мүндай даму көптеген жануарларга жэне
адамдарга тэн.
Түрленіп даму жұмыртқадан ересек агзаларга мүлдем
үқсамайтын бір немесе бірнеше дернэсілдік формалардың ал-
143
масуы арқылы жүреді. Метаморфоз арқылы даму әр түрлі жа-
нуарлар өкілдерінде (ішеккуыстылар, жалпақ, жүмыр құрттар,
былқылдақценелілер, бунақденелілер, қосмекенділер т.б.) кеңінен
таралған.
Постэмбриондық дамудың бірден-бір сипаты ағзаның өсуі бо¬
лып саналады. Өсу дегеніміз ағза мөлшерінің, салмағының сандық
өзгеруі болып табылады.
Ағзалардың өсуін екі топқа бөледі: шектеулі жэне шексіз
өсу. Шексіз өсу — ағзалардың өмірінің аягына дейін өсуі, бүл
былқылдақценелілерге, шаянтэрізділерге, балықтарға, жорғалау-
шыларға, қосмекенділерге және басқа да жануарларға тэн.
Шектеулі өсу — ағза өсуі белгілі бір жас шамасына жеткенге
дейін байқалады да тоқтайды. Ол шыбын-шіркейлерге, қүстарга,
сүтқоректілерге тэн.
Өсудің бірқатар жалпы заңдылықтары белгілі. Оларды зерт-
теуге ат салысқандар қатарынан американ зоологі К. Майнотты,
орыс ғалымы И. Шмальгаузенді ерекше атауга болады.
Онтогенездің бастапқы сатыларында даралардың өсуі қаркын-
ды түрде жүреді, ал кейінірек ол біртіндеп төмендейді және
дамудың эр түрлі кезеңдерінде өсу карқыны бірдей бола бермейді.
Онтогенезде өсу мен даму катар жүріп отыратын жэне бірінсіз-
бірі болмайтын күбылыстар болып саналады. Бірақ онтогенездің
эр түрлі сатыларында олардың ара-қатынасы тепе-тең болмайды,
яғни бір сатыда өсу қүбылысы дамудан басымдау болып келсе,
екіншісінде керісінше.
Адамдардың жастық кезеңі нәрестенің дүниеге келгенінен ба-
сталып жыныстық жетілген шаққа, яғни жыныс жасушаларының
түзіле басталуына (12-15 жас), дейін созылады. Оны бірнеше
сатыға: нәресте (емізулі бала), бәбек, бүлдіршін, жеткіншек т.б.
(кесте) бөледі.
Адамның постэмбриондық дамуының басқа сүтқоректілерден,
сонымен катар, приматтардан ерекшелігі — оның балалық шағы-
ның үзақ болуы, мысалы, кыздарда 11-13 жасқа, үл балаларда 13-
15 жасқа дейін. Мүның тек қана биологиялық емес, сол сияқты.
элеуметтік те маңызы бар, себебі осы кезде ағзаның физикалық
дамуымен қатар оның үжымдық өмір сүру жагдайларына
бейімделушіліктері қалыптасады. Адамның көптеген морфофи-
144
зиологиялық белгілері даму барысында айтарлықтай өзгерістерге
ұшырайды. Мысалы, омыртқа жотасының дамуы. Нәрестенің
омыртқа жотасы түп-түзу болса, ересек адамдардың омыртқа
жотасының пішіні латын алфавитінің S эрпіне ұқсас болып келеді,
яғни оның 2 алға шығыцқы — лордоздары жэне 2 артқа иілуі —
кифоздары болады. Бұл екі аяқпен тік жүру нәтижесінде қалыптас-
қан ерекшелік болып саналады. Ең алғаш мойын лордозы, сосын
бала отыра бастағанда — 2,5—3 айлықта, көкірек кифозы пайда
болады.
38-сурет. Адам ағзасынын эмбриогенездегі жэне туылғаннан
кейінгі дене пропорциялары
Балапан түрегеліп тәй-тәйлап жүре бастағанда бел лордозы,
ал 1,5—-2 жасқа жеткенде омыртқа жотасының барлық имектері
толық қалыптасады.
Бала мен ересек адамдардың дене бөлімдерінің салыстырма-
лы ара-қатынасы эр түрлі болатыны белгілі. Балалардың аяқтары
қысқа, түлғасы үзын, басы үлкен болса, ересектерде керісінше
— аяқтары ұзын, тұлгасы орташа, басы кішкентай болады (38-
сурет).
Бала ағзасында өсу қарқыны біркелкі жүрмейді — жедел өсу
кезеңі мен өсудің тежелу кезендері алмасып огырады. Балалардың
145
170-10
ең жедел өсуі бірінші жылы байқалады, бүл кезде оның денесі
23—25 см-ге ұзарады. Екінші жылы өсу қарқыны сэл төмендейді,
бірақ элі де болса ол жоғары деңгейде сақталады: 10—15 см;
үшінші жылы — 8 см, 4—7 жасқа дейін жылына 5—7 см өседі.
Төменгі мектеп жасында өсу біршама баяулайды, жылына 4—-5
см ғана өседі. Ал қыздар 11—12 жасқа, ұлдар 13—14 жасқа кел-
генде олардың өсу қарқыны қайтадан жоғарылайды, жылына 7—8
см дейін өседі. Оны пубертатты секіру деп атайды. Бұл құбылыс
балалардың жыныстық жетілуінің басталуымен тұспа-тұс келеді.
Балалар денесінің ұзарып өсуімен бірге оның салмағының ар-
туы да байқалады. Мысалы, алгашқы 5 айда нәресте салмағы екі
еселенеді, ал 1 жасқа жеткенде 3 есе көбейеді. Екі жасган кейін дене
салмагының өсуі баяулайды. 10 жасқа дейін қыздар мен ұлдардың
дене салмақтарында айтарлықтай айырмашылық байқалмайды, ал
11—12 жаста қыздарда ол біршама жылдамдайды, 15 жастан кейін
ұл балалар бұл көрсеткіш бойынша қыздарды басып озады жэне
бұл үрдіс эрі карай да сақталады.
Адамдар мен жануарлардың өсуі көптеген эндогендік және
экзогендік факторларға тәуелді болады.
Өсу—сандық белгі болып, полигендік жолмен тұқым қуалайды.
Сандық белгілердің қоршаған орта жагдайларына тәуелді бола-
тыны белгілі. Ағзаның өсуі генетикалық, эндокриндік, жүйкелік
т.б. жағдайлар арқылы бағдарланып отырады. Сонымен қатар,
қалыпты өсу жэне даму үшін толық қоректену қажет. Ac кұрамында
ағзаға қажет барлық амин қышқылдары жеткілікті мөлшерде бо¬
луы тиіс. Егер де олардың біреуі жетіспесе өсу тоқталады, даму
бүзылады жэне агза өліп қалады. Сол сияқты, өсу кұбылысы үшін
дэрумендердің, әсіресе А-дэрумені — ретинол. Д-дэрумені — каль¬
циферол, В дәрумендер тобының рөлі өте маңызды. Қалыпты өсу
үшін, сол сияқты минерал түздары мен микроэлементтер де қажет.
Өсу мен дамуға эсер ететін орта факторларынан — оттекті, темпе-
ратураны, жарықты айтуға болады. Ағзаның өсуі үшін жарықтың
бірден-бір рөлі — ол Д-дәруменінің синтезделуін қамтамасыз ету
болып табылады.
Ағзаның өсуі гипофиз безінің гормоны — соматотропин гормо¬
ны арқылы реттелініп отырады. Соматотропин гормоны үрықтық
даму кезінде де бөлінеді, бірақ оның әсері тек постэмбрионалдық
146
кезенде ғана байқалады. Гипофиздің соматотропин гормонын син-
тездеу карқыны ауытқыса (азайса не көбейсе) адамның қалыпты
өсуі бұзылады. Мысалы, балалық шақта гипофиз бездің гипо-
функциясы, яғни гормонды жеткіліксіз мөлшерде синтездеу,
гипофизарлыц ергежешікке — нанизмға алып келеді. Нанизмде
ағза өсуі баяулайды, бірақ дене бөлімдерінің дамуының салы-
стырмалы ара қатынасы өзгермейді. Гипофиздің гиперфункциясы,
ягни гормонды артық мөлшерде синтездеуі алыптыққа алып келеді
(бойы 2 метрден артық болады). Егер гипофиз гормонының гипер¬
функциясы агзаның өсуі тоқталганнан кейін болатын болса, онда
адамның кейбір мүшелерінің шектен тыс өсіп кетуі, үлғаюы, мы¬
салы қол саусақтарының, өкпенің, беттің өсуі байқалады. Мүндай
қүбылысты акромегалия деп атайды.
Соматотропин гормонының әсері калқанша бездің гормонда-
ры болган жагдайларда күшейе түседі. Бүл гормондар митохон-
дрияда тотыгу күбылысын күшейтіп энергетикапық алмасудың
артуына алып келеді. Қалқанша бездің гормоны — тироксин
әсерінен үлпалардың глюкозаны пайдалануы жогарылайды.
Жас жануарлардың калқанша безін алып тастаса агзада негізгі
алмасудың жэне жыныстық жетілудің тежелуі байқалады, орталық
жүйке жүйесінін тітіркенушілігі төмендейді, рефлекстің түзілуі
баяулайды. Адамдарда қалқанша бездің гипофункциясы балалық
шақта байкалатын болса кретинизм ауруы дамиды. Ол психикалық
кемтарлықпен, өсудің жэне жыныстық дамудың тежелуімен, дене
бөлімдерінің даму диспропорциясымен (түтікті суйектердің қысқа
жэне қалың болуы) сипатталады.
Акселерация (лат. acceleratio — жылдамдау) — балалар мен
жасөспірімдердің өсуі мен дамуының жылдамдауы. Соңгы 100—
150 жылда балалар мен жасөспірімдердің физиологиялық жетілуі
мен сомалық дамуының жылдамдауы байқалады.
Бүл қүбылыс XIX гасырдың 30-жылдарында жэне XX
гасырдың басында балалар мен жасөспірімдердің сомалық жэне
физиологиялық даму көрсеткіштерін салыстырып зерттеулер
негізінде байқалган. Акселерация кұбылысы тіпті қүрсақ ішіндегі
баланың дамуында да байқалган, мысалы XIX ғасырдың жылдары-
мен салыстырганда XX гасыр басында жаңатуылғын нәрестелердің
денесінің орташа ұзындыгы 0,5—1 см-ге, ал, салмагының 50—
147
100 грамнан артық болғаны анықталды. Емізулі балада да өсудің
біршама жеделдеуі байқалады. Ертеде нәрестенің дене салмағы-
ның екі еселенуі 6 айда байқалған болса, XX ғасырда 4—5 ай
аралығында байқалған. Сол сияқты, нәрестелерде сүт тістерінің
шыға бастауы да бұрынгы кезбен салыстырғанда ертелеу байқал-
ған. Бір жасқа жеткен балалардың салмағы осыдан 50 жыл бұрын
қазіргі кезбен салыстырғанда 1,5—2 кг кем болған. 1880 жыл-
дан 1950 жылға дейін Еуропа мен Солтүстік Америкада эрбір 10
жыл сайын 5—7 жасар балалардың денесінің үзындығы орташа
алганда 1,5 см-ге көбейіп, ал салмағы 0,5 кг-ға артқан. 13—15
жасар жасөспірімдерде тиісінше 2,5 см жэне 2 кг өсіп отырған.
XIX ғасырмен салыстырғанда XX ғасырда адам ағзасының өсуі
де ертерек тоқталған. Қазіргі уақытта қыздардың өсуі көпшілік
жағдайларда 16—18 жаста, ал үл балаларда 18—19 жаста
тоқталады. ¥зын сүйектердің өсуі эпифиз бен диафиз арасында
сүйек үлпасының «өсу жолағы» деп аталатын шеміршекті қабаты
сақталғанша жүреді. Қашан да оның орнына сүйек үлпасы да-
мыса, онда сүйектің ұзарып өсуі гоқталады. XX ғасырдың 70-
жылдарындағы әскер жасындағы үлдардың бойы 40-жылдармен
салыстырғанда орташа 8 см үзын болған. Акселерация себептері
туралы бірнеше пікірлер айтылуда. Бір болжам бойынша осу
мен дамудың жылдамдауын тамақтанудың жақсаруы, ағзага
дәрумендер мен ақуыздардың көптеп келіп түсуінен деп түсіндіреді
(Таннер, Ленц). Балалар ауруының азаюы, оларға медициналык
жәрдем берудің жақсаруы, гигиена жэне аурудың алдын алу ша-
ралары да дамудың жеделдеуіне өз ықпалын тигізбей қоймайды.
Кейбір пікірлер бойынша осу мен дамудың жылдамдауы жердің
магнит аймагының, иондаушы сәуле шашулардың жэне күн
радиацияларыньщ әрекетінің жылдан-жылга күшеюінен де бо¬
луы мүмкін. Сол сияқты, көптеген теле- радио қондыргылардан
шығарылатын электромагнитгі толқындардың әсерлері де дамуды
тездетуі мүмкін деген пікірлер айтылуда.
Урбанизация пікірі бойынша акселерацияныц негізгі себебі қа-
ладагы тіршілік жағдайларыныц балалардыц жүйке жүйесін жогарьі
дэрежеде тітіркендірулері болуы мүмкін деп болжамдайды.
Дегенмен, жогарыда келтірілген факторлардыц ешқайсысын
да акселерацияныц негізгі себебі деп қарастыруга болмайды. Ша-
148
масы акселерация көптеген факторлардың бірлескен эрекетінің
нэтижесі болуы эбден мүмкін. Сондықтан да оны эрі қарай эр
түрлі мамандар — дәрігерлер, педагогтер, биологтер, юристтер т.б.
терең зерттеулері қажет.
Жыныстық жетілу (репродуктивтік) кезең қыздарда 11—13 жа¬
ста, ұл балаларда 12—14 жаста басталады. Оның бірден-бір белгісі
қыздарда алғаш рет етеккірдің келуі, ал ұлдарда поллюцияның,
яғни түнде ұйықтап жатқанда шәуеттің еріксіз бөлінуі болып са-
налады. Репродуктивтік кезенді алғашқы репродуктивтік кезең —
35 жасқа дейін, соңғы репродуктивтік кезең — 36—55 (60) жасқа
дейін деп бөлінеді.
Алғашқы репродуктивтік кезенде де адам ағзасы эрі қарай —
18—20 жасқа дейін өсе береді. Осы кезде оның соңгы жыныстық
белгілері дамып жетіледі, дене түрпаты (конституциясы) қалып-
тасады.
Репродуктивтік кезеңнің аяқталуы әйелдерде менопаузаның
(климакс) келуі, ер адамдарда сперматогенездің тоқталуымен бай¬
ланысты. Көптеген адамдарда бұл кезде кәсіби еңбек ету қабілеті
толық сақталған, ақыл-естері кемелденген, белсенді тіршілік ету
мүмкіншілігі мол болып келеді.
4.8. Ағзаның қартаю кұбылыстары
жэне геронтология мәселелері
Қартаю барлық тірі ағзаларға тэн жалпы биологиялық күбылыс
болып саналады. Қартаю — онтогенез қорытындысы, агзада ерек¬
ше құрылымдық, қызметтік жэне биохимиялық өзгерістердің пай¬
да болуымен сипатталатын құбылыс.
Қартаю проблемаларын зерттейтін ғылымды геронтология (гр.
geron — шал) деп атайды. Геронтология қартаю қүбылысының
негізгі зандылықтарын молекулалық және жасушалық деңгейден
бастаптұтасагзаденгейінде анықтайды, оныңдаму ерекшеліктерін,
емдеу жэне аурудың алдын алу проблемаларын зерттейді.
Қартаю және ұзақ өмір сүру мәселелері барлык уақытта
ғалымдарды ойландырып келген мәселе. Ол, эсіресе, XX
ғасырда халық кұрамында терең демографиялық өзгерістердің
149
белең алуына байланысты ерекше мэнге ие болып отыр, себебі
көптеген экономикалык дамыган елдерде кэрі адамдар саны
өсуде. Геронтологияның міндеті — тек қана адам өмірін ұзарту
емес, сол сияқты, кэрі адамдардың қоғамдық өмірге жэне еңбек
қатынастарына белсенді араласуына көмектесу, яғни олардың
белсенді жэне толыққанды өмір сүру мүмкіншіліктерін үзарту бо¬
лып табылады.
4.8.1. Қартпюдың морфофизиологияпъщ сипапткшасы
Қартаю — жастық өзгерістердің занды қүбылысы болып та¬
былады. Қартаю құбылыстары ертеден басталады жэне ағзаның
қызмет ету мүмкіншілігін бірте-бірте қысқартады.
Ағза денесінде қартаю белгілері эр түрлі қүрылымдық дең-
гейлерде байқалады: молекулалық, жасушалық, үлпалық, жүйелік
жэне ағзалық. Ағзалық деңгейде картаю өзгерістері алдымен
сырткы белгілерден байқалады: дене пішіні, отыруы өзгереді, дене
көлемі азаяды, шаштары ағарады, түседі; терінің созылғыштық
қасиеті жойылып, әжімдер пайда болады. Ағзаның көру жэне есту
қабілеті нашарлайды, есте сақтауы төмендейді.
Қартаю кезеңінде адам ағзасының барлық мүшелері мен
мүшелер жүйесінде айтарлықтай өзгерістер байқалады.
50 жастан кейін адам терісінде түрақты өзгерістер—тыртықтар.
меңдер, сүйелдер, эжімдер пайда болады. Бүның негізгі себебі —
осы кезде тері асты май қабаты бірте-бірте жүкарып, жойылады.
Бүл терінің қүрғақтануына, оның иілімділігінің төмендеуіне алып
келеді.
Ac қорыту жүйесінде мынадай өзгерістер байқалады — тістер
босап, қаусап түсе бастайды, ас корыту солдерінің бөлінуі азаяды.
40 жастан өткеннен кейін артериялық қан қысымы жоғары-
лайды, қан тамырлар қабырғасына холестерин сіңіп жинақталып,
оның иілімділігі төмендейді.
Кэрілік кезенде бүйректің фильтрациялау қарқыны азаяды.
40 жастан өткеннен кейін өкпенің тіршілік сыйымдылығы азая¬
ды. Бүлшық еттер босап, оның тіршілік күші төмендейді, осының
нэтижесінде адам тез шаршайтын болады.
150
Ішкі секреция бездерінің қызметінде де айтарлықтай өзгерістер
байқалады — кейбір бездердің мысалы, жыныс бездері, калқанша
безі, бүйрек үсті безі, т.с.с. қызметі төмендеп, гормондарды аз
мөлшерде бөліп шығарады, ал кейбіреулерінің қызметі мысалы
гипофиз керісінше, жогарылайды.
Жас ұлгайган сайын агзаның иммундық реакциясы өзгереді,
гуморальдық және жасушалық иммунитет төмендейді.
Жүйке жасушаларының көбейе алмайтындыгын ескерсек, жас
үлғайған сайын олардыц саны бірте-бірте азаятындығы өздігінен-
ақ түсінікті. Мысалы, 100 жылда мысықтың нейрондар саны 25
пайызга кеміген.
Кэрілік сатыда адамдардың барлық сезім мүшелерінін
қызметтері төмендейді.
Ағзаның қартаю кезінде түқым қуалаушылық аппараттың қү-
рылысында да, кызметінде де өзгерістер болатыны анықталды.
Мысалы, РНҚ, ДНҚ мөлшері азаяды, хроматин ақуызының
физикалық-химиялық қасиеттері озгереді, гистонды ақуыздардың
ДНҚ молекуласымен байланысы қатая түседі, ал бүл көптеген
гендерді активсіз күйге көшіреді, себебі олар транскрипцияла-
на алмайды. Жасушалардың негізгі молекулалық генетикалық
механизмдері — транскрипция, трансляция, ДНҚ репликациясы
жэне репарациясы бұзылады.
Қартаю кезінде жасушалық децгейдегі озгерістер ішінен цито-
плазмада судыц азаюын, оныц иондарды тасымалдау белсенділі-
гініц өзгеруін атауға болады. Эндоплазмалык тор құрылысы
өзгереді.
Агзаның ескі жасушаларында бірқатар ферменттердіц бел-
сенділігі азаяды. цитоплазмада бос радикалдар жинақталады, осы-
ның салдарынан ассимиляция диссимиляцияның шығындарын
толық өтей алмайды.
¥зақ уақыт бойына геронтологияда мынадай пікір айтылып
келген: қартаю кезінде ағза қызметіңіц кері дамуы, яғни инволю-
циясы байқалады.
Жоғарыда келтірілген қартаю сипаттамалары шынында да осы
пікірді растайды. Дегенмен көптеген гомеостаздық қүбылыстар
қартаю кезецінде шұғыл түрде өзгермейді. Сол сияқты барлық
мүшелер бірдей озгермейді, айталық, бір мүшеніц қызметі кэрілік
151
сатысында төмендесе, екінші біреулерінің қызметі, керісінше,
жогарылайды, үшіншілерінің қызмет белсенділігі айтарлықтай
өзгермейді. Мысалы, бірінші типке жүректің жиырылуын,
гормондардың белсенділігін, ішкі секреция бездерінің (қалқанша
без, жыныс бездері) қызметін, көру, есту т.б.; екінші типке ги¬
пофиз қызметін, қандагы холестерин денгейін, жасушалардың
гуморальдық жэне химиялық факторларға сезімталдығын т.б.;
үшінші типке кандағы қант деңгейін, эритроциггер мен лейкоцит-
тер санын, гемоглобин мөлшерін т.б. жатқызуга болады. Агзаның
әр түрлі мүшелері мен мүшелер жүйесінде жастық өзгерістер
онтогенездің эр түрлі кезендерінде бір мезгілде байқалмайды —
оны гетерохрониялъщ қабілет деп атайды. Мысалы, тимустың
семуі 13—15 жаста басталса, аналық бездің қызметінің тоқталуы
48—52 жаста байқалады.
Жастық өзгерістер бір мүшенің эр түрлі қүрамдық бөлімдерін-
де бірдей бола бермейді, мысалы мидың эр түрлі бөлімдерінде
қартаю күбылыстары бірдей болмайды — оны гетеротопия
цүбылысы деп атайды.
Жастық өзгерістер бір жүйеде ерте пайда болып, жай дамыса
(мысалы, сүйек үлпасы), екінші біреулерінде кештеу байқалып тез
дамып, кейінірек алга шыгады (мысалы, орталық жүйке жүйесі).
Жастық озгерістердің пайда болуы тек күнтізбелік жаска гана бай¬
ланысты емес, ол бірқатар факторларга, оның ішінде әлеуметтік
факторларга да байланысты. Биологиялық жасты анықтау үшін
эр түрлі тесттік жүйелер пайдаланылады: артерия қысымы, кан
холестеринінің молшері, өкпенің тіршілік сыйымдылығы, бүлшык
ет күші т.б.
Ерте кездердегі геронтологтердің пікірінше (Пархон) қар-
таю — емдеуге болатын ауру. Бірақ қартаю — ауру емес, ол
онтогенездің занды нәтижесі екенін естен шыгармауымыз
кажет.
Ғылымның негізгі мақсаты биологиялық қүбылыстарды кері
қайтару емес, ол мүмкін де емес. Қартаю қүбылыстарын зерт-
теудің негізгі мақсаты — ерте қартаюдың алдын алу, болдырмау,
адамдарга физиологиялық қартаюга дейін толык әрі белсенді өмір
сүруге мүмкіндіктер жасау болып табылады.
152
4.8.2. Қартаюдың иегізгі теоршиары
Адам ағзасының қартаю себептері туралы 300-ге жуық бол-
жамдар айтылган. Олардың көбінің тек тарихи тұрғыдан ғана
маңызы бар. Қартаю теориялары ішінен М. Рубнердің (1908)
«цуаттык, қартаю теориясын» атауға болады. Бүл теория бойын¬
ша эрбір ағзаның қуат коры болады. Қуат қоры таусылса, сағаттың
серіппесі босагандай, тіршілік те баяулап, дүние салуға алып
келеді. Ағзаның қуат қорын жұмсауы дененің сыртқы мөлшеріне
тэуелді болады, яғни ағза дене сыртына жылуды қаншалықты көп
шығарса, соншалықты энергия алмасуы белсенді түрде болып,
оның қоры тез таусылады және ондай агзалар көп өмір сүрмейді.
¥сақ жануарлардың дене салмағына қарағанда оның үстіңгі
бетінің көлемі үлкен, сондықтан да олардың тіршілік үзақтығы
қыска болады (мысалы, егеуқүйрық 2—3 жыл, ит 20 жыл, піл 80
жыл өмір сүреді). Бірақ бүл түжырымды барлық жануарлар үшін
қолдана беруге болмайды, кейбіреулерінің тіршілік үзақтығы бүл
түжырымға қайшы келеді. Мысалы, егеуқұйрықтарга туыс бо¬
лып келетін тиіндер салмағы да, дене бетінің көлемі де, онымен
барабар, бірак тиіндер белсенді қозгалып 15—20 жыл тіршілік
етеді, яғни егеуқүйрықтардан 10 есе артык өмір сүреді. Белсенді
қозғалып тіршілік ететін сүр қояндар үй кроликтеріне қараганда
екі есе артық өмір сүреді.
И. И. Мечниковтың интоксикациялық (улану) теориясы
бойынша қартаю қүбылысы тек қана биологиялық факторга —
физиологиялық, патологиялық т.с.с. тэуелді болмай, сол сияқты
элеуметтік факторларға да тәуелді болады. И. И. Мечниковтың
пайымдауынша, агзаның өсуімен қатар эр турлі заттардың алма¬
суы, мысалы, азоттың алмасуы нәтижесінде жинақталган ыды-
рау енімдері — аммиак ағзаны улайды, тоқ ішекте шіру өнімдері
көптеп жинақталады. Бүл теория бойынша улы ыдырау өнімдері
кейбір мүшелер мен ұлпаларды, мысалы, бауыр, ми жасушаларын
көбірек улайды, ал дәнекер үлпа жасушалары керісінше көбейеді.
И. И. Мечников қартаю қүбылысын тек фагоцитоз ілімі негізінде
түсіндірмек болды. Ол ішекте шіру қүбылысын болдырмау үшін
шіріту' бактерияларының тіршілігіне қолайсыз орта жасау керек,
сондықтан сүт өнімдерімен көбірек қоректену қажет деп айтқан.
153
170-11
Академик А. А. Богомольц (1922) қартаю себептері ағзадагы
үлпааралық қатынастардың бүзылуы деп болжамдаған. Ол
жасушалар мен үлпалардың қоректенуін қамтамасыз ететін жэне
реттейтін дэнекер ұлпалардың картаю құбылысындағы маңызы
ерекше деп түсінген.
Қартаю құбылысының себептерін түсіну үшін И. П. Павлов
(1912) ілімінің де маңызы зор, себебі орталық жүйке жүйесі
ағзаның барлық мүшелері мен мүшелер жүйесінің қызметін
реттеуші, басқарушы орталық болып табылады. Әр түрлі
тәжірибелер нәтижесінде И. П. Павлов жүйке күйзелісі мен үзақ
уақыт жүйкенің шаршауы ерте қартаюды тудыратын факторлар
екенін анықтаған.
1940 жылы Нагорный «қартаю жасушада ақуыз молекула-
сының синтезделуінің бүзылуы салдарынан болады» деп бол-
жамдаган.
Қартаюдың қазіргі теориялары бойынша қартаю жасушаның
генетикалық аппаратының бүзылуы салдарынан болады. Онтоге¬
нез барысында нуклеопротеид кешенінің сандық және сапалық
өзгерістері байқалады, ал ол тіршіліктің түпкілікті қасиеттерінің
бірі — өзін-өзі жаңарту, ақуыз синтезі сияқты қүбылыстардың
өзгеруіне алып келеді.
Қартаю кезінде гистондар мөлшері көбейеді, олардың ДНҚ
молекуласымен байланысы қатая түседі, ал бүл көптеген гендер
мен гендер кешенінің актив күйінен активсіз күйіне кошуіне алып
келеді. Бүл қүбылысты Г. Д. Бердышев (1972), В. Н. Никитин (1972)
өз тәжірибелерінен байқаган.
XX гасырдың 90-жылдары Америка галымдары адам
агзасының «кэрілік» генін тауып оны «клото» гені деп атаган. Бүл
ген адамзаттың шамамен 25 пайызында кездеседі. Ол агзада екі
не одан да көп дана күйінде кездесуі мүмкін және геномда оның
саны негүрлым көп болса адамдар солғүрлым ерте қартайып дүние
салады.
«Клото» генінің екі данасы жаңадан туылган нәрестелердің 3
пайызында кездессе, 65 жастағы адамдардың тек 1,1 пайызында
гана кездескен. Демек, осыншама «клото» гені бар адамдар ерте
кезде, ягни 65 жасқа жетпей, дүние салган.
154
«Клото» генімен ағзаның ерте қартаюы арасында тікелей бай-
ланыс болатындығына ғалымдар әлі де күмандануда. Әйтсе де,
осы геннің адамдардың тіршілік ұзақтығын реттеуге қатынасатын-
дыгы сөзсіз.
Адамдарда ерте қартаю белгісі — прогерия дейтін ауру екені
белгілі. Бұл аурумен ауыратын адамдарда балалық шақтан бастап
(мүмкін жыныстық жағынан жетілер шақта — 13—15 жаста) жа-
сушаларда жедел картаю қүбылыстары байқалады.
Олардың терісінде қыртыстар пайда болып, шаштары ағарады.
көздері нашар көреді, тамырлардың атеросклерозасы т.б. дами¬
ды, яғни 20 жасқа жетпей-ак нагыз кэрі адамдарға тэн қартаю
қүбылыстары дамиды. Tan осындай қүбылыс 2001 жылы Аты-
рау облысының түрғыны 9—10 жастағы Нүржан атты балада
байқалган.
Бүгінгі танда медицина мүндай генетикалык ауру белгісімен ау¬
ыратын адамдарға түбегейпі көмек корсете алмайды, себебі оның
пайда болу жэне даму механизмдері элі толық анықталмаған. Де-
генмен, цитологиялық жэне молекулалық-биологиялық зерттеу-
лер нэтижесінде жасушаның не түтас ағзаның тіршілік үзақтығын
«таразылап» өлшеп отыратын кейбір механизмдер анықталган.
Олардың ішінен J1. Хейфликтің лимитін жэне оған негізделген А.
Оловниковтің теломерлік гипотезасын айтуға болады.
XX ғасырдың 70-жылдары Америка дәрігері JI. Хейфлик тірі
агзалар жасушаларының санаулы рет қана бөлінетіндігіне қоніл
аударған. Кейінірек эр түрлі түрлердің жасушаларының бөліну
максимумы түрліше болатындығы белгілі болды және ол агзаның
тіршілік үзақтығына тура пропорционал болатындығы анықтал-
ды. Мысалы, жүз жылға дейін өмір сүретін адамдарда (Homo sa¬
piens) Хейфлик лимиті — 50-re, 3 жыл өмір сүретін тышқандарда
— 15—20-ға, 175 жыл өмір сүретін галапагосс тасбақаларында —
110 -ға тең.
Ерте қартаю ауру белгілері (прогерия) байқалатын адамдардыц
жасушаларында Хейфлик лимиті 50-ден 10—15 ке дейін азайған.
Ғалымдардың пікірінше, жасуша бөлінген сайын онда кейбір
нәрселер біржолата жойылып не жинақталып отыра ма деген ой
пайда болды.
155
XX ғасырдың 80-жылдары А. М. Оловников жасушалардың
эрбір бөлінуінде олардың хромосомаларының ұштары — те-
ломералар азды-көпті үзіліп қысқарып отырады, ал хромосома
теломерлерінің үзындыгы минимальды мөлшерге жеткенде жасу¬
ша бөлінуін тоқтатады деген болжам айтқан. Кейінірек бұл бол-
жам тэжірибе күйінде дэлелденді.
Хромосома теломерлері маңызды механикалық (хромосома-
ларды ядро матриксіне жабыстырады; хроматидалар ұштарын бір-
біріне жалгайды;) хромосома қүрылымын түракггандырушы,
гендердің экспрессиялануына эсер етуші жэне жасуша бөлінуін
есептеуші қызметтерді атқарады.
Кейбір жасушаларда хромосома теломерлерінің үзілген
боліктерін қалпына келтіріп жалғап отыратын арнайы фермент —
теломераза ферменті синтезделінетіні аныкталды (1985).
Рак жэне жыныс жасушаларында теломераза ферменті үнемі
синтезделініп олардың шексіз бөлінуін қамтамасыз етеді.
Сонымен, қартаюдың нақтылы бір себептерін (механизмдерін)
боле-жара қарастыруға болмайды жэне ағзаның кәрілік өзгерістері
бір мүшеде жайлап басталып, эрі қарай тасқындап үдей дамып,
көптеген мүшелер мен мүшелер жүйесін қамтиды.
4.8.3. Агзалардың тіршілік ұзақтыгы
Біржасушалы өсімдіктер мен жануарлардың тіршілік үзақтығы
бірнеше сағатқа тең. Шөптесін өсімдіктердің тіршілік үзақтыгы
бір маусым ғана болады. Ағаш тэрізді өсімдіктер ондаған, жүзде-
ген жыл өмір сүреді. Мысалы, шие ағашы — 100 жыл, акация —
200 жыл, үйеңкі ағашы — 250 жыл, қарағай — 600 жыл, шырша
— 1000 жыл, ал секвоя — бірнеше мың жылға дейін өмір сүре
алады.
Жануарлар дүниесінде мүндай үзақ өмір сүрушілер кездеспей-
ді, дегенмен кейбір түрлердің өкілдері ондаған жылдан бірнеше
жүз жылға дейін өмір сүруі мүмкін. Мысалы, ішекқуыстылар —
50 жыл, жүмыр кұрттар (сүлік) — 25—30 жыл, жауын қүрттары
— 10 жылдай өмір сүретіні белгілі.
Жалпы алғанда, ірі жануарлар үсақ жануарларға қарағанда
үзақтау өмір сүреді, бірақ бүл абсолютті бола бермейді, мысалы;
156
кейбір былқылдақценелілер (моллюскалар) — 100 жыл өмір сүруі
мүмкін, көлбақалар — 16, қүрбақалар — 35—40 жыл, кептерлер
— 30, қаздар — 65, бүркіттер, тотықүстар — 70, тышқандар — 5
қояндар — 10, егеуқүйрықтар — 2—3 жыл, көртышқандар — 45—
50 жыл, жылқылар — 40, сиырлар — 20—-35 жыл, аюлар — 50,
арыстандар — 35, пілдер — 80 жылдан артық өмір сүреді.
Әр түрлі ағзалардың өмір сүру ұзақтығыныц ара қатынасы
туралы ортақ зандылық жоқ, дегенмен ағзалардың тіршілік
ұзактығының өмір сүруінің минималдық шегінің, жыныстық
пісіп-жетілуінің жэне үрпақ қалдыру белсенділіктерінің эволюция
барысында табиги сүрыптау арқылы қалыптасқандығына шүбэ
келтіруге болмайды.
XIX ғасырдың басында француз ғалымы Ж. Бюффон ағзалар-
дыц өмір сүру үзақтығы, олардыц өсу мерзімінен 5—7 есе артық
болатындығын есептеп шығарған. Ж. Бюффон коэффициентін
барлық жағдайларда қолдана беруге болмайды, әйтсе де көцтеген
жагдайларда оны пайдаланып түрлердіц өмір сүру үзақтығын
теория күйінде есептеуге әбден болады. Мысалы, иттердіц өсуі 2
жылға, ал өмір сүру үзақтығы 15 жылга тец; ірі кара мал -4 жыл
жэне 20 жыл; жылқы — 5 жэне 30—40 жыл; түйе — 8 жэне 40
жыл т.б. Осылайша А. Богомольц, И. Шмальгаузен, тағы басқалар
адамдардың өмір үзактығы — теория күйінде 120—150 жылға тең
болуы мүмкін деген болжам айтқан.
4.8.4. Адам өмірінің ұзақтыгы
Адам өмірініц орташа үзақтығы қүбылмалы болатындыгы
белгілі. Ерте замандарда адам өмірініц орташа үзақтығы қазіргі
кездегіден әлдеқайда қысқа болған. Мысалы, неандертальдықтар-
дың қазба күйінде табылған қацқаларын зерттегенде, олардыц 40
пайызы 14 жас шамасында дүние салғанын, 15 пайызы — 15—
20 жас арасында, тек 15 пайызы 40 жастан көбірек өмір сүргенін
көреміз. Тас дэуірі адамдарыныц 50 жасқа дейін өмір сүргендері
өте сирек. Мыцдаған жылдар бойына адамдар қартаю жасына жет-
пей өлген.
Адамдардыц орташа жасы Еуропада XVI ғасырда 21, XVII
ғасырда — 26, XVIII ғасырда — 34, XX ғасырдың басында — 50,
XX ғасырдыц аяғында — 60—65 жас шамасында болған.
157
Әйелдер ер адамдарға қарағанда біршама ұзагырақ өмір
сүретіні статистикалық түрғыдан анықталған. Бүл бұрыннан да
белгілі болатын, бірақ соңғы онжылдықтарда ол әсіресе айқын
байқалады. Мысалы, Ресейде 1896—1897 жылдары әйелдер мен
ер адамдардың орташа жасының айырмашылығы 2 жылды кира¬
са, 1926—1927 жылдары — 5 жылга, 1970-1971 жылдары — 9
жылға жеткен. Мүндай құбылыс көптеген елдерде де байқалады.
Бұл құбылысты кейбір ғалымдар әлеуметгік факторлардың әсері:
ер адамдардың еңбек ерекшелігі, көп жарақаттануы, жағымсыз
әдеттерге күштарлыгы (ішімдік, темекі тарту, нашақорлык т.б.) деп
болжамдайды. Дегенмен, бұл факторлар ер адамдар мен әйелдер
арасындағы орташа жас айырмашылыгын толық түсіндіре алмай¬
ды.
Бүл қүбылыс көптеген жануарлар түрлеріне де (насеком-
дар, балықтар, қүстар, сүтқоректілер) тән. Мынадай тәжірибе
жасалынган: адам аскаридасының еркек жэне үрғашы дараларын
бір-бірлерінен бөлек үстағанда, олардың екеулері де 20 күн шама-
сында омір сүрген. Ал, оларды араластырып үстағанда үргашы да-
ралары 20 күн, еркек даралары не бары 16 күн өмір сүрген. Осыған
қарап галымдар мынадай жорамал жасаған: еркек даралар сперма¬
тогенез қүбылысында көптеп (жүздеген, мыңдаган, миллиондаған)
шәуеттер бөліп шыгарады, ал сперматогенез болса, энергияны
коп жүмсауды қажет ететін кұбылыс екені белгілі. Демек, еркек
даралардың энергия шыгыны көп болгандықтан, үргашы дара-
лардан аздау өмір сүреді. Сонымен қатар бүл жагдай үргашы
даралардың нейрогуморальдық ретгелу ерекшеліктері мен жыныс
гормонының ерекше қорганыстық эрекетінің эсерінен де болуы
мүмкін деген пікірлер айтылуда. Мүмкін әйелдер каротипінде
екі X хромосоманың болуы олардың тіршілік қабілетін арттыра
түсетін фактор болар.
Адамдардың омір сүруінің биологиялық мүмкіншіліктері 120-
150 жыл, дегенмен осы жасқа дейін өмір сүрушілер — үзақ өмір
сүрушілер саны оте коп емес. Жалпы — 90 жастан көп өмір сүретін
адамдарды үзақ өмір сүрушілер деп атайды. Үзак омір сүрушілер
барлық елдерде де, барлық үлттарда да, сол сияқты Қазақстанда
да кездеседі. Мысалы, үлы Жамбылдың 100 жыл өмір сүргені
158
^эрімізге аян. Жүзді алқымдап қалған немесе 100-ден асып кет-
кен қариялар Қазакстанның түпкір-түпкірінен көптеп кездеседі.
Дегенмен, үзақ өмір сүрушілер, эсіресе Абхазияда, Аджарияда,
Грузияда, Әзірбайжанда, Солтүстік Кавказда, Таулы Алтайда, т.б.
жерлерде жиі кездеседі.
Дүниеде ең ұзак өмір сүрген, Гиннестің рекордтар кітабына ен-
ген адам — Шерэлі Мүслімов (эзірбайжан) болған. Ол 168 жыл, ал
әкесі 110, шешесі 90 жыл өмір сүрген.
4.8.5. Өлім — жеке дамудың (онтогенездіц) заңды
ңорытындысы
Әрбір ағзаның жеке тіршілігінің ақырғы фазасын — өлім деп
атайды. «Өмір барда, елім бар» деп халық бекер айтпаған, өлімнен
ешбір пенде қүтыла алмаган, қүтыла алмайды да.
Ағзалардың тіршілік құбылысында оның дене жасушалары бір
жағынан өліп жойылып отырса, екінші жағынан оның орны толты-
рылып (физиологиялық регенерация) қалпына келіп отырады.
Агзаның зат алмасу қүбылысы және сыртқы ортамен қарым-
қатынасы бұзылған кезде оның өлімі басталады. Ағза өлімінің
табиғи себептері болып қартаю қүбылыстарының тереңдеп үдей
түсуі, патологиялық қүбылыстар, сыртқы орта факторларының зи¬
янды әсерлері т.б. болуы мүмкін.
Жогары сатылы жануарларда жэне адамдарда өлімнің 2 түрін
ажыратады: физиологиялык, өлім жэне патологиялыц өлім.
Физиологиялъщ өлі.м — табиғи өлім, қартаю қүбылыстары
нәтижесінде пайда болады; патологиялык, өлім — мезгілсіз
өлім, адам ағзасының үзақ сырқаттану салдарынан мүшелерінің
қажып, қызмет өте алмау жағдайларында орын алады. Сол сияқты,
мезгілсіз өлім әр түрлі апат жағдайларының салдарынан да болуы
мүмкін.
Жогары сатылы көпжасушалы ағзаларда өлім клиникальщ өлім
жэне биологиялыц өлім болып бөлінеді.
Тіршіліктің негізгі кызметтерінің тоқталуы клиникальщ өлімге
жатады. Ол 5—6 минутқа созылады. Бүл кезде көптеген жасу¬
шалар мен мүшелер өздерінің тіршілік қабілетін элі толық жоя
159
коймағандықтан, олар өзін-өзі жаңарту құбылыстарын жалғастыра
береді, метаболизм жалгасады, бас миы өз қызметін элі толық
жоғалтпаған. Сондықтан да осы өлім жағдайындағы адамдарды
қайтадан тірілтуге болады. Оны реанимация деп атайды.
Тыныс алудың жэне қан айналымның ұзақ уақытқа (10—15
минут) тоқтатылуы үлкен ми сыңарларының жасушаларында кері
кайтпайтын өзгерістер пайда етеді, ол өз қызметгерін түгел жояды.
Оны қайта тірілту мүмкін емес. Өлімнің мүндай түрін биологиялъщ
өлім деп атайды.
4.9. Ағзаның қалпына келу қүбылыстары. Мүшелер мен
үлпалардыц регенерациясы және трансплантациясы
Ағза онтогенезінде көптеген дене жасушалары табиги не
патологиялық жағдайларға байланысты ескіреді, не бүзылады.
Олар жасушалардың бөлінуі нәтижесінде үнемі қалпына келіп от-
ырады, себебі онсыз кез келген ағза біртүтас жүйе ретінде тіршілік
ете алмайды. Оны регенерация деп атаймыз. Сонымен, регене¬
рация деп ағзаның жарақаттанған, не жойылган үлпалары мен
мүшелерінің қалпына келуін айтамыз. Оның физиологиялық реге¬
нерация жэне репаративтік регенерация деген 2 түрі белгілі.
Физиологиялъщ регенерация дегеніміз қалыпты тіршілік
жагдайларында агзаның ескірген не бүзылган қүрылымдарының
өліп қалып не түлеп түсіп отыруы жэне олардыц қалпына келуі.
Мысалы, шаянтәрізділер мен жэндіктердіц қуыршақтары өздерінің
қатты хитин қабатын тастап, түлеп өседі. Сол сияқты, жыландар да
жылына бір рет түлеп отырады, қүстардың қауырсындары түсіп,
оның орнына жаца қауырсындар пайда болады. Сүтқоректілер
мен адамдар терісініц эпителий жасушаларыныц түлеп түсуі,
ішектердіц сілемейлі қабығы жасушаларының түлеуі т.б. осыган
мысал бола алады. Қанның қызыл түйіршіктері — эритроциттер
күніне 1 пайыз мөлшерінде ауысып отырады. Шаштың, тырнақтыц
өсуі, үй жануарларыныц түктерініц түлеуі т.б. физиологиялық
регенерацияга мысал бола алады.
160
а э б
39-сурет. Аксолотль аяғынын регенерациясы
а) жаракат, э) бластеманьщ пайда болуы, б) аяктьщ регенерациялануы,
1-эпидермис, 2-дерма, 3-бұлшыкет, 4-сүйек.
Репаративтік регенераг/ш дегеніміз эр түрлі патологиялық
жағдайларда жойылған не жарақаттанған мүшелердің қалпына
келуі болып табылады. Репаративтік регенерация эр түрлі жану-
арларда түрліше деңгейде байқалады. Қарапайым ағзалардың
(ішек-қуыстылар, құрттар) кез келген жасушаларынан жаңа
дара өсіп дами алады. ИІаянтәрізділер жойылган қол-аяқтарын,
мұртшаларын, көздерін калпына келтіре алады. Қосмекенділер
жарақаттанған қол-аяқтарын, күйрығын жэне кейбір мүшелерін
қалпына келтіреді. Сүтқоректілер мен адамдардың күрделі
мүшелері толық қалпына келе алмайды, дегенмен бұл оларда
репаративтік регенерациялану болмайды деген сөз емес.
Сүтқоректілер мен адамдардыц әр түрлі ұлпалары түрліше
дәрежеде қалпына келеді, мысалы, тері, сілемейлі қабық эпителиі
жоғары дәрежеде регенерацияланса, дэнекер ұлпалар біршама
төменгі дэрежеде, ал шеміршек жэне нерв ұлпасы нашар регене-
рацияланады.
Регенерация процесі тіршіліктіц эр түрлі құрылым децгей-
лерінде байқалады — жүйелік, мүшелік, үлпалық, жасушалық
жэне жасушаішілік. Олардыц бэрінің негізі болып жасушаішілік
кұрылымдардыц жацаруы (регенерациялануы) саналады. Жасу-
шаішілік регенерацияныц мынадай түрлері белгілі: молекула-
лык регенерация; органоидішілік регенерация; органоидтық ре¬
генерация — органоидтардың санының көбеюі (цитоплазманың
гиперплазиясы).
Эндоморфоз — немесе регенерациялық гипертрофия деп
мүшелер ішінде жүретін толық емес регенерацияны айтамыз.
161
Бұл жолмен қалпына келгенде жаңа жасушалар пайда болмай,
сақталган жасушалар ұлғайып өсіп мүшенің бұрынгы көлемін
қал ы птастырад ы.
40-сурет. Типтік емес регенерация.
А) шаянның көзінің орнына мұртшаның регенерациялануы,
Б) кесірткенің аягының орнына кұйрыктың регенерациялануы.
Егер агзаның ескірген не бұзылган құрылымдары табиги жол¬
мен (регенерация) қалпына келмесе, басқа сау мүшені не үлпаны
қондыру (трансплантация) қажет болады — мұны үлпалар мен
мүшелердің трсінсплсттациясы деп атайды.
Агзалардың ұлпалары мен мүшелерін қондыру туралы аңыздар
ерте замандардан — ежелгі грек, христиан аңыздарынан белгілі.
Мысалы, Косьма жэне Дамион туралы аңыздар.
Біздіц заманымыздыц II гасырда Қытай емшісі Хуа Ту
адамдардыц зақымданган мүшелерін алып тастап, орнына сау
мүшелерді қондырган деген деректер бар.
Үлпалар мен мүшелердіц трансплантациясы гылыми түрде XIX
гасырда италияндық Д. Баронко (1804), неміс К. Бюнгер (1823)
тәжірибелерініц нәтижесінде дами бастады. Трансплантацияныц
дамуында Н. И. Пироговтыц (1835) ецбегініц мацызы зор.
162
Ресейде 1840 жылы Ж. Штраух, 1867 жылы Н. Фей көздің қасаң
қабыгын трансплантациялаған. 1865 жылы В. Антоневич тіс-
терді, 1892 жылы К. И. Сапожко сілемейлі қабықшаны қондыруға
тәжірибе жасаған.
1897 жылы В.Г. Григорьев ұлпаларды қондырудан (транс¬
плантация) мүшелерді қондыруға көшкен: ол аналық безді бір
адамнан екіншісіне қондыруга операция жасаған.
Қазіргі кезде ұлпалармен мүшелерді кондыру көптеген елдер¬
де жүргізілуде. 1980—1990 жылдары әлемде бүйректі қондыратын
301 орталық болып 25000 операция жасалса, жүректі қондыратын
64 орталықта 500-ге жуык операциялар жасалган.
Қазақстанда мүшелерді трансплантациялау операцияла-
ры академик М. Әлиев есімімен байланысты. М. Әлиевтіц
басшылыгымен XX ғасырдыц 80—90 жылдары бауырга, үйқы
безіне т.б. мүшелерге операциялар жасалынған. 2002—2003
жылдары Қазақстан дәрігерлері трансплантация саласында
жаца жетістіктерге қол жеткізді, себебі ұлпалармен мүшелерді
қондырудан жасушалық трансплантацияга көшті. Мысалы, 2002
жылы М. Алиевтің басшылығымен бауырға, 2003 жылы әлемде
бірінші болып М. Әлиев, Ж. Досқалиевтар қалқанша безге шприц-
пен эмбриондық жасушаларды енгізу арқылы сэтті операциялар
жасады. Қазіргі кезде Қазақстанда бауыры ауыратын адамдарға
эмбриондық гепотоциттерді енгізу аркылы жылына 200—300 сәтті
операциялар жасалуда. Бұл эдістіц болашағы мол.
Трансплантологияда мүшесі алынатын агзаны — донор, ал
сол мүшені қондыратын агзаны — рег/епиент, қондырушы мү-
шені трансплантант деп атайды.
Трансплантацияны — ау готрансплантация, аллотрансплан¬
тация жэне ксенотрансплантация деп үшке бөледі.
Аутотрансплантация деп бір агза мүшесін сол агзаның екінші
бір жеріне қондыруды айтамыз.
Аллотрансплантация дегеніміз бір түрге жататын агза мүшесін
екінші бір агзага қондыру.
Ксенотрансплантация деп донор жэне рецепиент эр түрлі
түрлерге жататын болса айтамыз.
Трансплантологияныц коптеген проблемалары белгілі:
1) трансплантация сэттілігі; 2) дер кезінде донор табу, т.с.с.
163
Трансплантация нәтижесі барлық уақытта сәтті бола бермейді,
себебі көпшілік жағдайда трасплантантпен рецепиент арасында
иммундық үйлесімділік болмайды. Яғни, егер донор мүше мен
рецепиент үлпалары арасында гистоүйлесімділік болмаса, реце¬
пиент ағзасында донор мүшесінің үлпаларына карсы бағытталған
антидене түзіледі де, соцғысын кері қайтарып трансплантацияны
болдырмайды. Ал егер, рецепиент пен трансплантант арасында
гистоүйлесімдік болса, онда трансплантация нәтижесі сэтті бо¬
лады. Бұл түрғыдан алғанда аутотрансплантация ец сэтті, ал ал¬
лотрансплантация, әсіресе ксенотрансплантация сэтсіздеу болып
келеді.
Аталған проблеманы шешу үшін көптеген әдістер — иммуно-
депрессанттарды қолдану, эмбриональдық қүрылымдарды пайда-
лану, ағза мүшелерін клондау т.б. қолданылады.
Қажетті донор мүшелерін дер кезінде табу екінші негізгі про¬
блема болып табылады. Оны мүшелер мен үлпалар банкін құру,
жасанды мүшелерді қүрастыру, ағза мүшелерін клондау арқылы
шешуге болады. Қазіргі кезде тері, қан банкілері қүрылған жэне
адамның барлык мүшелерініц жасанды баламалары қүрасты-
рылған. Дегенмен, трансплантологияныц болашағы ағза мүше-
лерін клондаумен байланысты.
5. ГОМЕОСТАЗ ТУРАЛЫ ІЛІМ НЕПЗДЕРІ
Tipi агзалар өздерін қоршаған сыртқы орта факторларымен
үнемі тығыз байланыста болады. Олар сыртқы ортадан эр түрлі
заттарды, энергия т.б. алып тіршілік етеді. Сыртқы орта фактор-
лары құбылмалы болып келеді, бірак орта қаншалықты өзгерсе
де тірі агзалар оган тез бейімделіп, өздерінің морфологиялық
біртүтастығын, негізгі қызметтерін, қасиеттерін, мінез-құлқын,
жасушалардың, ұлпаларының негізгі физико-химиялық көрсет-
кіштерін тұрақты күйде сақтап қала алады. Сыртқы ортаның
өзгеруіне қарай тірі агзалар сол өзгеріске сэйкес (адекватты) жауап
қайтара алады, яғни сырткы ортаның қолайсыз әсерлерін жойып,
не оның эсерін элсіретіп қалыпты тіршілік ете алады. Осының
нәтижесінде агзалардың тек қана ішкі ортасының тұрақтылыгы
сақталынып қоймай, сол сияқты олардың ағза ретінде біртұтастығы
сакталынады.
Сыртқы орта факторларының өзгеруіне қарамастан, тірі
ағзалардың өздеріне тэн ішкі ортасының тұрақтылығын, мор-
фологиялық, функционалдық біртұтастығын сақтап қалу қасиетін
гомеостаз деп атаймыз.
Гомеостаз — ағзалардың ең негізгі қасиеттерінің бірі жэне ол
ағзалардың қалыпты тіршілік етуі үшін өте маңызды, себебі тек
гомеостаз нәтижесінде агза қүбылмалы сыртқы ортага бейімделіп
тіршілік ете алады. Тірі ағзалардың мұндай қасиетінің маңызын
алгашқы рет 1878 жылы француз галымы К. Бернар байқаган.
Бірақ гоместаз деген терминді биологияга 1929 жылы америка
физиологі В. Кеннон енгізген болатын.
К. Бернардың айтуынша, сыртқы орта факторларының әсерлері
салдарынан эр түрлі агзалар және тіршіліктің түрліше форма-
лары қалыптасады. Ол тіршілікті 3 түрге бөлген: тіршіліктің жа-
сырын (латентті), тәуелді жэне тұрақты не еркін формалары.
Тіршіліктіц жасырын формасында тіршілік процестері
байқалмайды, зат алмасу толық тежеледі. Бұған мысал ретінде
165
өсімдіктердің тұқымдарын алуға болады. Түқымдар жүздеген,
мындаған жылдар бойына тіршіліктерін байкатпай сақталады,
қолайлы жагдай туса өніп, өсіп, жеке өсімдікке айнала алаты-
ны белгілі. Жануарлар дұниесінде тіршіліктің жасырын түрі
қарапайымдыларда циста күйінде байқалады. Қолайлы жағдай-
ларда циста қалыпты ағзаға айналады.
Тіршіліктің тәуелді формасы сыртқы орта факторларына
тәуелді болып келеді. Оларға салқын қанды —- пойкилотермді
жануарлардың бэрі, яғни омыртқасыздаржәне кейбір омыртқалылар
жатады. Олардыц тіршілігі сыртқы орта факторларына (су,
жылылық т.с.с.) тәуелді болады.
Тіршіліктіц теракты не еркін формасы жоғары сатылы
жануарларга тән. Бүл жануарлардыц тіршілігі сыртқы орта фак-
торлары күрт өзгерген күнде де тоқтамай жалғаса береді, себебі
оның мүшелері мен ұлпаларыныц қызмет активтілігі айтарлықтай
өзгермейді. Оның басты себебі мүшелерді, ұлпаларды қоршаған
ағзаныц ішкі ортасы өзгермейді. Нэтижесінде агза жылыжай-
да тіршілік еткен сияқты болып, түрақты, еркін өмір сүреді.
Сонымен, К. Бернардыц айіуынша, кез келген жоғары саты¬
лы жануарлардыц тіршілігі 2 ортада жүреді: ағзаны қоршаған
сырпщы орта жэне ұлпаларды, мүшелерді қоршаған ішкі орта.
К. Бернар ағзаныц ішкі ортасы деп ұлпаларды үнемі жуып-шай-
ып түратьш сұйықтықтарды — қан сұйықтығын, лимфа жэне үл-
па сұйықтықтарын айтқан. Қорыта келгенде, ол ағзаныц ішкі орта
тұрақгылығы тіршіліктіц тұрақгы, еркін формасыныц негізі бола¬
ды.
В. Кеннон көптеген физиологиялық зерттеулер нэтижесінде
жануарлардыц тіршілігі қоршаған ортаныц қолайсыз, зиянды
факторларының әсеріне қарамастан, ұзақ жылдар бойына қалыпты
күйде жүретінін байқаған.
Ағзалардың қалыпты тіршілігін ұзак уақыт өзгертпей сақтап
гүру қасиеті мыңдаған жылдар бойына күпия болып келді. Әйтсе
де, Гиппократтан бері қарай көптеген ғалымдар ағзалардың ауру-
лары олардың ішкі табиғи күштері негізінде емделетінін байқа-
ған. Бірақ, тек қазіргі кезде ғана ол күштерді ағзалардыц гомео-
стазды калыптастыру жэне зиянды факторларға бейімделуші-
лікті (адаптация пануды) дамыту мүмкіншіліктерімен байланысты
166
екендігі анықталды. Сонымен, гомеостаз дегеніміз ағзалардың
орта факторларына адаптациялануының (бейімделуінің) тарихи
қалыптасқан, тұкым қуалайтын қасиеті болып саналады. Орта
факторлары қысқа мерзімге, не ұзақ уакыт бойына өзгеруі мүмкін.
Бұл жағдайларда ағзалардың адаптациялануы (бейімделуі) ішкі
ортаның қасиеттерін қалпына келтіріп қана қоймай, қысқа мер-
зімге оның қызметтік активтігінің өзгеруіне де алып келеді (мыса¬
лы, жүрек қызметі ыргагыныц жиіленуі жэне тыныс алу жиілігінің
жылдамдануы). Қолайсыз жағдайлардың зиянды әсері үзақ
уақытқа созылатын, не қайталана беретін болса, ағзада түрақты
қүрылымдық өзгерістер пайда болуы мүмкін. Ал, егер бір мүше
жарақаттанатын болса, онда оны қалпына келтіру механизмдері
(регенерация) іске қосылады, не басқа мүшелердің қатынасуымен
жарақаттанған мүшенің қызметі қалыптастырылады. Гомеостаз-
ды калыптастыратын ағзаның реакциялары олардыц кұрылымдық
түрақтылыгын сактау, зиянды факторлардың әсерлерін жою не
шектеу процестерін үйлестіруге, құбылмалы орта факторла¬
ры мен агзалар арасында оңтайлы қарым-қатынас формаларын
сақтап калуга не жаңадан қалыптастыруға бағытталады. Осы
процестердің бәрін адаптация деп атаймыз. Медицинада адапта¬
ция деп ағзалардың қолайсыз жағдайларда тіршілік етуіне байла¬
нысты қалыптасқан бейімделушілігін айтады жэне адаптацияның
кез келген түрлері гомеостаз механизмдері негізінде пайда болады.
Ол механизмдер түрліше: шагын бір мүше не ұлпалар деңгейінде;
тұтас агза деңгейінде болып келуі мүмкін. Гомеостаздың шагын
не жүйелі формалары агзаның физиологиялық көрсеткіштерінің
сақталуында елеулі рөл атқарса, тұтас агза деңгейіндегі гомеостаз
механизмдері зияңды эрекеттерді жоюга бағытталады.
Гомеостаздың негізгі құрамдық бөліктеріне мыналар жатады:
A) Жасуша тіршілігін калыптастыратын материалдар:
Жасушаның өсуіне қажет заттар, энергия көздері — көмірсулар,
белоктар, майлар;
Су;
Натрий хлориді, кальцийда баска да бейорганикалык заттар;
Оттегі;
Ішкі секреция гормондары.
B) Жасуша активтігіне эсер ететін сыртқы факторлар:
167
1. Осмос қысымы;
2. Жылылық;
3. Сутек ионыныц концентрациясы (РН-көрсеткіш).
Г) Ағзаның қызметтік жэне қүрылымдық біртүтастығын
қалыптастыратын механизмдер:
1. Тұқым қуалаушылық;
2. Регенерация жэне репарация;
3. Иммунобиологиялық қасиеті.
Сыртқы ортаның қолайсыз жағдайларына агзаның нервтік
және эндокриндік реттелу механизмдерінің өзара әрекеттесуі сал¬
дарынан туындайтын ағзаның жалпы жауап реакциясына мысал
ретінде стресс-реакцияларды жатқызуға болады. Ол қолайсыз
тіршілік жағдайларының қалыптасқан ағза гомеостазын бұзу қаупі
туған кездерінде дамиды.
Стресс (күйзеліс-реакция) дегеніміз — ең алдымен күнделікті
өмірде ерекше, төтенше немесе экстремальды эсерлер нэтиже-
сінде пайда болатын ағзаның адаптивтікреакциясы болып санала-
ды. Сонымен қатар, күйзеліс кейде жүрек-қантамыр ауруларының,
жүйке бұзылуының, психикалық бұзылыстардың дамуыныц себебі
де болуы мүмкін.
Күйзеліс құбылысыныц мүндай бір-біріне қарама-қарсы
қасиеттерін табиғаттың белгісіз күштерініц барлық уақытта
мақсатқа сай эсер ете бермейтіндігімен түсіндіруге болады. Мы¬
салы, И. И. Мечниковтен бері қарай қабыну кұбылысы ағзаның
қорғаныстық реакциясы екендігі белгілі. Әйтсе де көптеген адам-
дар қабыну салдарынан дүние салады. Күйзеліс те сол сияқты.
Бүл реакция бір жағынан ағзаныц қорғаныстық реакциясы болса,
екінші жағынан ол гіатологиялық қүбылыстардыц пайда болуына
алып келуі мүмкін.
Стресс (stress) — құбылысын оқымыстылар түрліше мағынада
түсінеді: кейбіреулері ағзаныц кез келген түршігу күйін стресс деп
карастырады. Екінші біреулері, стресс Г. Сельеніц аурулар тура¬
лы іліміне сэйкес келеді деп есептейді. Олардыц пікірлерінше,
стресс — ағзада кез келген қолайсыз тіршілік жағдайларыныц
(стрессорлар) эсерлерінен пайда болатын кез келген аурулардың
өзіндік ерекшеліктеріне тэн (спецификалық) емес коріністері
деп есептейді. Ал аурудың өзіндік ерекшеліктеріне тән емес
168
көріністерінің тұрлері өте көп, мысалы, бас ауруы, қалтырау,
қабыну, невроз, шок, (есеңгіреу) парабиоз т.б.
Сондықтан да, қазіргі кезде стресс (күйзеліс) деп ағза го-
меостазының бұзылу қаупі төнген кезде пайда болатын ағзаның
спецификалык емес жалпы нейрогуморальдык реакциясын айтады.
Кесте
t t
Төтенше қоздырғыштар, экстремальды факторлар
Стресс реакциялардың ең негізгі сипаттамаларына мы-
налар жатады: АКТГ (адено-кортикотропты гормон) және
кортико-стероидтардың секрециясының күшеюі; бүйрекүсті без
қыртысының гипотрофиясы; тимус-лимфалық жүйенің бүзылуы
жэне кішіреюі; асқазан-ішек жолдарының қабынуы, жараның да¬
муы.
Стресс-реакциялардың дамуында 3 кезең байқалады:
1) ағзаның қорғаныстық күштерінің іске жүмылдырылуы (да-
был кезеңі);
2) агза резистенттілігінің (төзімділігінің) жогарылауы;
169
170-12
3) Қажу, стресс-реакцияның ақырғы кезеңі ағзаның өліміне
әкеледі.
Стресс-реакциялардыңақырғы нэтижесітүрлішеболуы мүмкін:
1) стресс-адаптивтік механизм ретінде ағзаны бүрынғы күйіне не
жаңа күйге көшіріп, гомеостазды қалпына келтіреді; 2) стресс-
реакция аурудың даму механизміне айналып, ауруды қоздырады.
Адам гомеостазының қалыптасуы мен реттелуінің бірнеше
механизмдері белгілі: нервтік, эндокриндік, биохимиялық,
физикахимиялық, генетикалык, күрылымдық, иммунологиялықт.б.
Олар туралы толық жэне нақтылы деректерді арнайы биологиялық,
клиникалық эдебиеттерден табуға болады (Гомеостаз. // Под ре¬
дакцией П. Д. Горизонтова, 1981. Москва).
6. ЭВОЛЮЦИЯЛЫҚ ІЛІМ НЕГІЗДЕРІ
6.1. Эволюциялық теорияның даму тарихы
Эволюциялық ілім биологияның ең ірі, түпкілікті тұжырымы
болып есептелінеді. Міне, 145 жылдан астам уақыт ішінде ол
биологияның барлық салаларының дамуына елеулі ықпал етіп
келеді.
Эволюция дегеиіміз — табиғаттың тарихи дамуы, тіршіліктің
қарапайым түрлерінің күрделі формаларының пайда болуы. Эво¬
люция процесі негізінде ағзалардың бейімделушілігі, биологиялық
түрлердің түзілуі жэне жойылуы, биогеоценоздың жэне био-
сфераның өзгеруі болады. Биологиялық эволюция нәтижесінде
Жер бетінде қоғамдық өмірдің негізі — адамның пайда болып,
өмір сүруіне қолайлы жағдайлар туды.
Ерте замандарда табигат өзгермейді, тіршілік құдайдың
әмірімен пайда болган деген діни, креационистік көзқарас басым
болды. Әйтсе де, кейбір ертедегі грек галымдары қалыптаскан
креационистік көзқарасқа қайшы көзқарас, тіршілік өлі дүниеден
оның дамуы негізінде пайда болды деген пікірлер айтқан. Мы¬
салы, Фалес (624—578 б.г.д.) дүние судан дамиды деп ойлаған.
Анаксимандр (610—546 б.г.д.) адам балықтардан келіп шыққан
деген пікір айтқан. Эмпедокл (483—423 б.г.д.) тірі дүние 4 баста-
мадан — от, су, ауа жэне топырақтан жаралган деп болжамдаган.
Гераклит (IV гасырдың аягы — III гасырдың басы б.г.д.) дүниенің
негізі қозгалыс деп ойлаган. Дүние унемі өзгеріп отырады...»суга
екі рет түсуге болмайды...» деген үғым айтқан.
Осындай даму туралы өте қарапайым пікірлерді эволюциялық
пікір деп қарастыруга болмайды, сондықтан да оларды транс¬
формизм деп атайды, себебі олар қарапайым формалардан күрделі
формалардың қалайша пайда болатынын түсіндіре алмаган.
Орта гасырларда ғылымның дамуына шіркеу тыйым салып,
креационистік көзқарас үстем болган. Дегенмен, XVI гасырдан
171
бастап ғылыми деректер жинақтала бастады. XVIII ғасырда
жасуша ашылады, К. Линней өсімдіктердіц алғашқы жүйесін
құрастырады.
Эволюциялық ілімнің қалыптасуына үлы француз материалис-
тері Ж. Ламетри, Д. Дидро жэне Гельвеций көп үлес қосты. Олар
табиғат дамудың нәтижесінде қалыптасты деген пікір айтқан. Ж.
Ламетри адамдар мен жануарлардыц қүрылысыныц жэне шығу
тегініц үқсастығына көңіл аударған. Даму туралы пікірлер Ж.
Бюффон ецбектерінде де кеңінен орын алған. Ол тірі дүние өлі
дүниеніц дамуыныц нэтижесінде пайда болды жэне олар орта
факторларыныц эсерімен өзгереді деген ғылыми пікір айтқан.
Жануарлар қүрылысыныц ұқсастығын олардыц шығу тегінің
ортақтығымен түсіндірген.
Алғашқы эволюциялық ілім 1809 жылы пайда болған. Оныц
авторы француз ғапымы — Ж. Б. Ламарк. Оның айтуынша, тірі
ағзалар үнемі өзгеріп отырады. Олардың өзгергіштігініц негізгі
себебі — орта факторларыныц әсері. Ол екі зац қалыптастырған:
1. Агзалар сыртқы орта факторлары эсерінен үнемі өзгерігі оты¬
рады, кез келген өзгерістер түқым қуалайды;
2. Жиі пайдаланылатын (жаттыгатын) мүшелер дамиды, ал пай-
даланылмайтын (жаттықпайтын) мүшелер жойылады (семеді).
Ж. Ламарк қарапайым формалардан күрделі формалардың пай¬
да болуын (ягни, нагыз эволюция процесін) градация принципі
арқылы түсіндірді. Ол барлық тірі ағзаларды бірнеше сатыларга
орналастырган. Оныц айтуынша, агзалардыц дамуы оларды үнемі
өздігінен күрделенуге жетелейтін белгісіз ішкі қозгаушы күшке
байланысты, соныц арқасыңда дамиды деген пікір айтқан.
Ж. Ламарк теориясы жалган теория, сондықтан да ол галымдар
арасында көп қолдау таппаган, дегенмен, бұл креационизмге қарсы
багытталган бірден-бір теория болатын.
Нагыз гылыми-эволюциялық теория 1859 жылы пайда болды,
оның авторы агылшын галымы — Ч. Дарвин. Оның эволюциялық
теориясы мынадай табиғи қүбылыстарга негізделген: өзгергіштік,
түқым қуалаушылық, тіршілік үшін күрес, табиги сүрыптау,
белгілердің ажырауы (дивергенция).
Ч. Дарвин агзалардың өзгергіштігініц 2 түрін ашты:
172
1. Айқындалган өзгергіштік көптеген дараларда бірдей пай¬
да болады жэне оның негізгі себебі сыртқы орта факторлары.
Айқындалған өзгергіштік тұқым қуаламайды.
2. Айқындалмаған өзгергіштік жекелеген, тіпті жеке бір дарада
ғана пайда болады. Айқындалмаған өзгергіштік тұкым қуалайды
жэне оның эволюция қүбылысы үшін маңызы өте зор.
Ағзалардың өзгергіштігі олардыц сыртқы ортаға түрліше
бейімделуіне алып келеді. Олардыц жақсы бейімделгендері өсіп,
дамып, ұрпақ қалдырып көбейе береді, ал бейімделмегендері, не
нашар бейімделгендері ұрпак қалдыра алмайды, өліп жойылады.
Міне, осы құбылысты, яғни тіршілік үшін күрес құбылысында
бейімделген даралардыц тірі қалып, бейімделмеген даралардыц
өліп қалуын Ч. Дарвин табиги сүрыпталу деп атаған. Тіріиілік
үіиін күрес деп Ч.Дарвин даралардыц бір-бірімен немесе орта
факторларымен қатынасу түрлерін айтқан. Неліктен табигатта да-
ралар арасында тіршілік үшін күрес болады? Оның негізгі себебі
тірі ағзалардың көбею интенсивтілігі (қарқындылығы) болып
табылады. Ағзалардың кобею қарқындылығы дегеніміз —
ағзалардың мейлінше көп мөлшерде ұрпақ қалдыруға тырысуы.
Бұл тірі ағзалардыц бэріне де тэн. Мысалы, бір бақбақ өсімдігініц
себетгүлінде мындаған тұқымшалар пісіп жетіледі, олардан не
бары 100 бақбақ өсіп шықты десек, бір жылда бір бақбақтан 100
бақбақ пайда болады, екі жылда 10 000, ал үш жылда 1000 000
т.с.с. Балықтар миллиондаған уылдырык шашады (мысалы, тре¬
ска 2 миллионға жуық), бірақ оныц бэрі ересек балыққа айнала
бермейді, себебі олардыц дамуыныц эр түрлі кезецдерінде тіршілік
үшін күрес болып отырады.
Ч. Дарвин тіршілік үшін күрестің 3 түрін ажыратқан: түрішілік
күрес, түраралъщ күрес, агзалардыц ортаның қолийсыз фактор¬
ларымен күресі.
Түрішілік күрес бір түрдіц даралары арасында жүреді, пгүра-
ральщ күрес — екі түрдіц даралары арасындағы күрес. Осылар-
дың ішінен ең қатапы — түрішілік күрес болып саналады, себебі
бір түрдің даралары ортақ жерді мекендеп, бір түрлі қорекпен
қоректенеді. Ортаныц цолайсыз жагдаііларымен күреске Ч. Дар¬
вин мысал ретінде, теңізден соққан желдің аралда тіршілік ететін
құстар жэне насекомдар арасында элсіз қанаттыларын жойып, тек
173
қанаттары жақсы жетілгендері мен қанатсыздарды тірі қалды-
руын келтірген.
Тіршілік үшін күрес, табиғи сұрыптау негізінде ортаға жақсы
бейімделген жаңа формалар пайда болады. Tipi дүниенің ортаға
бейімделуі алуан түрлі болып келеді, бірақ кез келген бейімделу-
шілік салыстырмалы түрде ғана болады, ягни бір бейімделушілік
барлық жағдайлардан сақтай алмайды, орта факторы өзгерсе
бүрынғы бейімделушілік дараларды қорғай алмайды.
Ч. Дарвин эволюцияның қозғаушы күштері — өзгергіштік.
тұқым қуалаушылық, тіршілік үшін күрес жэне табиги сүрыптау-
мен қатар белгілердің ажырауының (дивергенция) түртүзілудегі
рөлін айқындап көрсете білді. Баяу өзгеретін түрлер табигаттың
қүбылмалы жағдайларына бейімделе алмагандықтан күрып
жойылады, ал, керісінше, өзгергіш түрлер эр түрлі экологиялық
орталарга бейімделіп дамиды. Әдетте, қүбылмалы жагдайларда
белгілердің ең шеткі көрсеткіштеріне ие болган даралар тірі қалып,
орташа мэніне ие болгандары өліп отырады. Белгілердің диверген-
циясы нәтижесінде пайда болган болар-болмас айырмашылықтар
сақталынып, үрпақтан-үрпаққа жалгасып көбейіп, алгаш нәсілдер,
сосын түр тармагы, содан кейін барып жаңа түрлер түзіледі.
6.2. Биологиялық түрлер жэне олардың
популяциялық қүрамы
Түр деп морфологиялық жэне қызметтік белгілері, кариотипі
бірыңгай ұқсас, белгілі бір жерді үзақ уақыт мекендейтін, шыгу
тегі бірдей, өзара еркін будандасып, көбеюге қабілетті үрпақтар
қалдыра алатын даралар жиынтыгын айтамыз.
Түр деген терминді XVII гасырда Джон Рэй енгізген, бірак
түр туралы нақтылы деректер келтіріп, олардыц гылыми аталу
принциптерін (бинарлы номенклатура) қалыптастырган К. Линней
(XVII г.) болатын.
К. Линнейден бері қарай оқымыстылардың түрге деген көзқа-
растары өзгеріп отырган. К. Линней түрлер өзгермейді, түракты
болады деп болжамдаган.
174
Ж. Ламарк, керісінше, тұрлер үнемі өзгереді, табиғатта шынайы
түрлер кездеспейді, оларды адамдар ойлап тапкан, табиғатта тек
даралар ғана кездеседі деген.
Ч. Дарвин түрлер үнемі өзгеріп отырады, бірақ белгілі бір та-
рихи уақыт аралығында табиғатта олар кездеседі деп айтқан. Кез
келген түрлердің өздеріне тэн морфологиялық, физиологиялық,
цитогенетикалық, экологиялық, биохимиялық жэне т.б. крите-
рийлері болады.
Табиғи жагдайларда бір түрдің даралары өздерінін ареалда-
рында біркелкі таралмаған, бір жерлерде жиі кездесіп топтасып
орналасса, екінші бір жерлерде аз кездеседі, тіпті, кездеспеуі де
мүмкін. Түр дараларының үлкенді-кішілі топтарын попупяцияяар
деп атайды.
//о/гуляг/шг деп белгілібіржердіұзақуақытмекендеп,бір-бірімен
еркін будандасатын биологиялық түр дараларының жиынтыгын
айтамыз.
Популяциялар түрдің тіршілік ету, эволюциялану формасы
болып табылады. Олар ағзалардың сыртқы орта факторларымен
эрекеттесуінің нәтижесінде, тұкым қуалаушылык, өзгергіштік
және табиғи сүрыгітау салдарынан қалыптасады. Популяция
терминін биологияға 1903 жылы Иогансен енгізген болатын. Бір
түр бірнеше популяциялардан тұрады.
Популяциялардың экологиялық, генетикалык сигіаттамалары
белгілі.
Популяцияның экологиялық сипаттамасына мыналар жатады:
1. Ареал мөлшері;
2. Даралар саны;
3. Жастык күрамы;
4. Жыныстық қүрамы.
Популяцияның мекендейтін ареалының мөлшері түрліше бола¬
ды, ол ағзалардың жекелей белсенділігінің радиусына байланысты.
Мысалы, ұлу өте баяу қозғалады, ол бірнеше метрге ғана жылжи
алады, демек оның ареалының мөлшері кішкентай, ал су тышқаны
жүздеген метрге барып келе алады, түлкілер, қасқырлар—ондаған,
жүздеген километрге таралады, демек олардыц популяциялары-
ның ареалдары үлкен. Сол сияқты популяциядагы даралар саны да
эр түрлі болуы мүмкін. Мысалы, көлдерді мекендейтін инеліктің
175
популяциясында 30 000-ға жуық даралар болатын болса, ұлу по-
пуляциясында не бары 1000-ға жуық даралар кездеседі. Деген-
мен, популяциядағы даралар санының ең төменгі деңгейі болады.
Ол деңгейден азайса, популяциялар жойылады.
Адамдардың сан жағынан шағын, 1500—4000 адамнан аспай-
тын кішкентай популяцияларын демдер, ал одан да кішкентай
1500-ден аз адамдардан тұратын популяцияларды изоляттар деп
атайды. Демдер мен изолятгар өте баяу көбейеді, демдер — 20,
изоляттар — 25 пайыз мөлшерінде. Сол сияқты, демдер мен изо-
ляттарда туыстық некелесу жиілігі өте жоғары дәрежеде бола¬
ды, ал ол рецессивті аллельдердің гомозигота күйіне өтіп, кейбір
аурулардың дамуына алыгі келеді. Егер изоляттар 4 буыннан астам
уақыт бір жерде өмір сүретін болса, оның эрбір мүшелері бір-
бірімен кем дегенде шөберелес агалы-інілі немесе апалы-сіңілілі
болып келеді.
Әр түрлі түрлер популяцияларының жастық жэне жыныстық
құрамдары күбылмалы болып келеді, олар популяцияның тіршілік
ұзақтыгына, көбею белсенділігіне, жыныстык жетілу мерзімде-
ріне байланысты болады.
Популяцияны генетикалык түргыдан қарастыратын болсақ
оның өзіне тэн генофонды болады. Табиғи популяциялардың
генетикалык сипаттамалары:
1. Генофонд (ген қоры) дегеніміз популяция дараларының
генотиптерінің (аллельдерінің) жиынтыгы.
2. Генетикалык гетерогенділігі немесе генетикалык полимор¬
физм! (көптүрлілігі).
3. Генетикалык біртұтастыгы.
4. Әр түрлі генотипті даралардыц өзара динамикалык тепе-
теңдігі болып саналады.
Популяциялар генофондыныц генетикалык көптүрлілігі бір
генніц эр түрлі аллельдерінің бір мезгілде кездесуі нәтижесінде
байқалады. Ол мутациялық күбылыс негізінде қалыптасады.
Мутациялар, әдетте рецессивті болып гетерозиготалы агзалар
фенотипіне айтарлықтай ықпал етпей, популяция генофон-
дында табиги сұрыптауга ұшырамай, ұзақ уакыт сакталынады.
Олар жинақталынып «тұқым қуалаушы өзгергіштіктің қорын
қүрайды».
176
Комбинативтік өзгергіштіктің арқасында бұл «қор» эрбір буын-
да аллельдердің жаңа комбинацияларының пайда болуына алып
келеді. Ал, ол «қордың» мүмкіншілігі өте үлкен. Мысалы, егер
бір-бірімен 1 ООО локус бойынша ерекшеленетін, ал эрбір локуста
10 аллельдері бар ағзаларды өзара будандастырсақ, олардан пайда
болатын генотиптер варианттары 10 000-ға тең болар еді.
Популяцияның генетикалық біртұтастығы олардың кең мөл-
шердегі ттмиксиясымен байланысты, ягни популяция даралары
өзара еркін будандасып ұрпақтар генотиптерінің, аллельдерінің
көзі ретінде популяция генофондын «пайдаланады». Популяция
генофондында белгілі жагдайларда эр түрлі аллельдерден тұратын
генотиптер мөлшері ұрпақтан-үрпаққа тұрақты болады. Ол Д. Хар¬
ди — Г. Вайнберг заңы аркылы сипатталады.
Популяцияларда тұқым қуалаушылықты зерттеу олардыц
генотиптік құрамын, ягни эр түрлі генотиптің жэне аллельдердіц
жиілігін анықтаудан басталады.
Популяцияныц генотиптер не аллельдер жиілігі деп процент
мөлшерімен белгіленетін осы генотипке ие аллельдер, не даралар
жиынтығын айтамыз. Популяция дараларыныц жалпы саны 100
процент деп есептелінеді.
Жынысты, жолмен көбейетін агзаларды өздігінен ұрықтанатын
жэне айқас ұрықтанатын деп екі топқа белуге болады. Осы топ
агзалардыц популяцияларында түқым қуалау заңдылықтары
түрліше болады.
Өздігінен ұрықтанатын, түкымының түсі сары — гетерозиго¬
талы (Аа) болып келетін, бұршақтың (А доминантты — сары түсті;
а-рецессивті — ақ түсті); екінші үрпагыныц Ғ, тұқымының түсі
былайша ажырайды. ІАА: 2Аа: Іаа, ягни 25 пайыз — АА; 50 пайыз
— Аа; 25 пайыз — аа (Мендельдіц 2 зацы). Сонымен Ғ,-де — 50 %
гомозиготалы даралар (25 пайыз АА+25 пайыз аа) жэне 50 пайыз
гетерозиготалы даралар болады.
Келесі жылы, ягни Ғ,-те, егер көбею коэффиценті 4-ке тец
десек, гомозиготалы нысандардан ІАА-дан 4АА, 1аа-дан 4-аа;
ал гетерозиготалы даралардан -2АА+4Аа+2аа генотиптер пай¬
да болады. Осы генотиптерді өзара қоссақ, 2АА+4АА=6АА,
4Аа, 2аа+4аа=6аа, ягни 6АА+4Аа+6аа пайда болады, оларды
екіге қысқартсақ, ЗАА+2Аа+Заа генотиптердіц пайда болатынын
177
байқаймыз. Пайыздық жағына келетін болсақ Ғ3-те 37.5 пайыз
АА; 37.5 пайыз аа; 25 пайыз Аа кездеседі, яғни гомозиготалар (АА;
аа) — 75 пайыз, гетерозиготалар 25 пайызға тең болады. Ғ ұрпақта
гомозиготалар саны өсіп (50 пайыздан 75 пайызға дейін), ал гете¬
розиготалар керісінше кемиді (50 пайыздан 25 пайызға дейін). Ал
Ғ ұрпакта гомозиготалар 98.8 пайызга дейін өседі, гетерозигота¬
лар 0,2 пайызга дейін кемиді.
Панмиксиялық популяцляларда тұқым қуалау Харди-Вайнберг
заңына сэйкес болады жэне ол төмендегі шарттарга сэйкес келетін
популяцияларда байқалады:
1. Даралардың бір-бірімен еркін будандасуы қажет (панмик-
сия);
2. Популяцияда сүрыптаудың болмауы, ягни сүрыптаудың сал¬
дарынан гендердің жойылып кетпеуі қажет;
3. Миграция салдарынан жаңа гендердің келіп енбеуі қажет;
4. Гомозиготалы жэне гетерозиготалы даралар бірдей мөлшерде
көбеюі қажет;
5. Популяция көлемі ірі. ягни даралар саны өте көп болуы
қажет.
Харди-Вайнберг заңыныц 3 қагидасы белгілі: I. Нақтылы
популяциядагы бір геннің жиілігінің жиынтыгы тұрақты болады.
Егер популяциядагы доминантгы геннің (А) жиілігінің жиынты-
гын р деп, ал рецессивті аллелдіц «а» жиілігін — q деп белгілісек,
онда р + q=I; р — доминантты генніц жиілігі, q— рецессивті ген-
ніц жиілігі.
Егер популяцияда 100 000 дара болатын болса, бір локустыц
аллельдік гендерініц саны 200 000-га тең. Бірак доминантты жэне
рецессивті гендердіц саны міндетті түрде тепе-тец болмауы мүм-
кін. Доминантты ген 60%, рецессивті ген 40% немесе 90% жэне
10% т.с. болуы мүмкін, бірақ екеуініц қосындысы 1-ге (немесе
100%) тец болады (60%+40% =100%; 90%+10%= 100 т.с.с).
А=р
a=q
А=р
АА-р2
Aa-pq
a=q
Aa-pq
aa-q2
178
ғ
2. Нақтылы популяцияда бір аллельдіц генотиптер жиілігінің
жиынтығы тұрақты жэне ол Ньютон биномына сэйкес болады.
P2+2pq+q2=l (100%). Р2—АА генотипінің жиынтыгы. 2 pq — ге¬
терозиготалы генотиптер (Аа) жиынтыгы; q2 — рецессивті гомо¬
зиготалы (аа) генотиптердін жиынтыгы; 1 (100%) популяциядагы
даралар саны.
3. Тепе-тең популяцияларда гендердің жэне генотиптердің
жиілігі ұрпақтан-ұрпаққа тұрақты болады.
Егер, Ғр-де доминантты ген р=0.6 (60%) рецессивті ген=0.4
(40 %) деп алатын болсақ, олардың генотиптерінің жиілігі АА
(р2) =0.36 (36%). Аа (2pq)=0.48 (48%); аа=0.16 (16%) тең болады.
Келесі ұрпақта Ғ2 доминатты «А» гені бойынша гомозиготалылар-
да 36%, ал гетерозиготалыларда 24% осындай гаметалар түзіледі.
р=0.3+0.24=0.6 (60 %). Рецессивті гендерден тұратын гаметалар-
дың 24 пайызы гетерозиготалы дараларда, ал 16% рецессивті го-
мозиготаларда түзіледі q=0.24+0.16=0.4 (40%), ягни, екінші үр-
пақта да бірінші үрпаққа тэн генотиптер ара катынасы сақталады.
Бүл қүбылыс Ғ—Ғ10 т.с. сақталынып қайталанып отырады.
Қалыптасқан гендер мен генотиптер ара қатынасының өзгеруі
мүмкін бе? Мүмкін, егер де популяция тепе-тендігі бүзылса.
Популяция тепе-теңдігі әр түрлі себептермен, мысалы — орта
жагдайының өзгеруі салдарынан, сүрыптау нәтижесінде ген¬
дер саны азайса немесе жаңа мутациялар пайда болса бүзылуы
мүмкін.
6.3. Түртүзілу күбылысы. Табиғи иопуляциялардағы
карагіайым эволюциялык факторлардың әрекеті
Харди-Вайнберг заңы популяциялардыц генетикалық түрақ-
тылык жагдайларын сипаттайды. Бірнеше үргіақтар бойына ге-
нофондтары өзгермей, түрақты болып келетін популяцияларды
Мендель популяциялары деп атайды. Мендель популяциялары эво¬
люция қүбылысынан тыс қалып отырады, себебі табиги сүрып-
тау болмайды. Табигатга мүндай популяциялар кездеспейді. Хар¬
ди-Вайнберг зацында популяциялардыц генофондтарын озгерте-
тін жагдайлар келтірілген, олар:
179
1) панмиксияны шектеуші факторлар;
2) популяция дараларының санының күрт азаюы жэне ең
төменгі деңгейге жетуі;
3) оқшаулану кедергілері (барьерлері) т.с.с.
Эволюциялық құбылыс нэтижесінде нақтылы ортага бейім-
делген даралар жиынтығы (түрлер) пайда болатыны белгілі. Бүл
бейімделушілік биологиялық ақпарат күйінде үрпақтан-ұрпаққа
сақталынып беріліп отырады. Егер орта баяу өзгеретін бол¬
са түрдің үзақ уақыт сақталып өмір сүруі оның генофондының
түрақтылығына байланысты. Екінші жагынан түрақты генофонд
орта факторлары күрт өзгерген жагдайларда олардың сақталып,
тірі қалуын қамтамасыз ете алмайды. Мүндай генофонд түрлердің
ареалдарының кеңейіп, жаңа экологиялық орталарды игеруіне
де мүмкіншілік бермейді. Бір сөзбен айтқанда, түрлердің үзақ
уақыт біртүтас сақталуы оның генофондының түрақталуына
байланысты десек, екінші жагынан түрлердің генофондының
түрақтылыгы олардың эволюциялануын (дамуын) болдырмай-
ды. Себебі эволюция нәтижесінде түрлер озгеріп, жаңа белгілер
мен қасиеттерге ие болып, 2 не одан да коп жаңа түрлерге айна-
лады. Ал эволюцияның жалгыз бағыттаушы факторы — табиги
сүрыптау болып саналады.
Түрлердің эволюциясының мүндай қайшылықтарын олардың
популяциялық қүрылымы түргысынан түсіндіруге болады. Түрлер
көптеген үсақ популяциялардан түрады. Түр генофонды оның по-
пуляциялар генофондыныц жиынтыгы болып табылады. Әрбір по¬
пуляция генофонды эр түрлі багытта өзгеріп дамиды. Популяция-
лар түр шеңберінде ашық топтар болып саналады. Даралардыц
популяция аралык миграциясы сол популяцияларда пайда
болган азды-көпті ерекшеліктерін жойып. түрдіц біртүтастыгын
қамтамасыз етеді. Егер де кейбір популяциялар түрдіц калган по-
пуляцияларынан үзақ уақыт географиялық оқшауланатын болса,
онда бастапқы болар-болмас ерекшеліктер күшейе түсіп, ақырында
олардыц арасында генетикалык оқшаулану пайда болып, жаца түр
түзілуіне алып келеді. Эволюциялық қүбылысқа жекелеген попу¬
ляциялар қатынасады, ол жаңа түрдің пайда болуымен аяқталады.
Сонымен, популяция түрдіц қарапайым кұрылымы, эволюциялану-
шы бірлігі, ал түр эволцияныц сапалық кезеці болып есептелінеді.
180
'
Эволюцияның синтетикалык теориясына сәйкес түр түзілу
құбылысы қарапайым эволюциялық қүбылыстан басталады.
Қарапайым эволюциялык, құбылыс дегеніміз популяция гено-
фондының (генетикалық қүрылымыныц) өзгеруі. Қарапайым
эволюциялық құбылысқа, яғни популяция генофондының өзге-
руіне алып келетін қүбылыстар мен оқиғаларды қарапайым
эволюциялык, факторлар деп атаймыз.
Қарапайым эволюциялық факторларға мыналар жатады:
1) мутациялық процесс;
2) популяциялық толқындар;
3) оқшаулану;
4) миграция(гендер «ағыны»);
5) гендер дрейфі;
6) табиғи сұрыптау.
Мутациялық күбылыс. Ағзалардыц жыныс жасушаларында
үнемі гендік, хромосомалық не геномдық мутациялар пайда болып
түрады. Олар жыныс жасушаларының тұқым куалаушылық мате-
риалын өзгертеді. Осы мутациялардың ішінен ең жиі кездесетіні
жэне мацыздысы гендік мутациялар. Гендік мутациялар көптеген
аллельдердің пайда болуына алып келіп биологиялық ақпараттыц
көптүрл іл і гі н калы птастырады.
Түртүзілуде мутациялык қүбылыстыц екі түрлі әсері белгілі: 1)
бір аллельдін екінші аллельге қараганда жиілігін өзгертіп, популя¬
ция генофондының өзгеруіне тікелей эсер етеді;
2) мутантты аллельдердің пайда болуы нәтижесінде тұқым-
куалаушылық өзгергіштіктің қоры (резерв) түзіледі.
Тұқым қуалайтын өзгергіштіктің қоры келесі ұрпақтарда
комбинативтік өзгергіштік салдарынан агзалар генотиптерінің
аллельдік құрамының қүбылмалы болуына алып келеді. Мута¬
циялык құбылыс негізінде табиги популяциялардыц генетика¬
лык көптүрлілігінің жогары децгейде болуы қамтамасыз еті-
леді. Мутациялық құбылыс негізінде пайда болган аллельдер
жиынтыгы қарапайым эволюциялык құбылыстың материалы бо¬
лып, түртүзілу қүбылысында басқа да факторлардыц әрекет етуі
үшін негіз болып табылады.
Жекелеген мутацияның пайда болуы өте сирек қүбылыс
болганымен, популяцияда оныц жалпы саны өте коп болады.
181
Мысалы, егер 100 ООО гаметаның біреуінде мутация пайда бол¬
ды десек, геномында 10 ООО локусы бар 100 миллион даралардыц
әрқайсысы 1 ООО гаметадан түзетін болса, жалпы мутациялар саны
10'° тең болар еді. Ал түрдің орташа тіршілік уақытында (ондаған
мыц ұрпақтар жалгасында) мутациялар саны 10й тең болады.
Мутациялардыц көпшілігі даралар фенотипіне зиянды эсер етеді,
бірақ мутантты аллельдер рецессивті болып келіп гетерозиготалы
күйінде генофондта ұзақ уақыт байқалмай сақталады. Осыныц
нәтижесінде:
1) мутантты рецессивті аллельдердіц даралар фенотипіне зи¬
янды эсері байқалмайды;
2) қазіргі кезде бейімделушілік қүндылыгы болмаганымен
болашақта, орта факторлары күрт өзгерсе не жаца экологиялық
жагдайларды игерген жагдайларда олар пайдалы болуы мүмкін;
3) гетерозис қүбылысы арқасында көптеген мутациялар гете¬
розиготалы күйінде даралардыц тіршілік қабілетін жогарылатады
(өнімді көп береді). Пайдалы мутациялар саны аз болганымен,
олардың ұрпақтардагы не түрдің тіршілік уақытындағы жалпы
саны көп болуы мүмкін. Миллион мутациядан тек 1 гана пайдалы
болады десек, бір ұрпақта пайда болатын 10ю мутацияның 104 пай¬
далы болып келеді. Популяция генофондына үнемі мутациялық
кұбылыс қысымына эсер етеді, ал, ол урпактарда жеке дара
мутациялардыц жойылу ыктималдыгыныц орнын толтырады.
Адамдарда да мутациялық құбылыс басқа тірі агзалардагыдай
болады. Бірақ, қазіргі кездері адам генофондында мутациялық
құбылыстыц кысымы күшеюде. Оган бірден-бір себеп — гылыми
техникалық революция жагдайларында адамдардың қызметтік-
кәсіби іс-эрекеті нэтижесінде индукцияланган мутациялардыц
көптеп пайда болуы.
Популяциялыц толқындар — бұл популяция даралары саны-
ның өзгеруі (бірде азайып бірде көбейуі). Кейбір түрлердіц
(шыбын-шіркейлердіц, балықтардыцт.б.) популяция саны бірнеше
рет өзгеруі мүмкін. Популяциялар даралары санының кезецдік,
кезецге байланыссыз өзгеруін С. С. Четверяков популяциялық
толқындар немесе тіршілік толқындары деп атады.
Популяция даралары санының өзгеруі маусымдық құбылы-
старга, қоректену түріне байланысты болуы мүмкін. Кейде апат
182
■
құбылыстары да (су тасқыны, орман мен дала өрттері жэне т.б.)
популяция даралары санының өзгеруіне алып келеді. Өсімдіктер
мен жануарлар популяциялары санының өзгеруіне алып келетін
бірден-бір себеп адамныц белсенді іс-эрекеті болып саналады. По¬
пуляция даралары саныныц өзгеруі салдарынан гендер қоры (ге¬
нофонд) өзгереді.
Популяция даралары саныныц
ауытқуы негізінен экологиялық
жағдайларғабайланысты болады.
Мысалы, шөппен қоректенетін
жануарлар мен жыртқыштар
даралары саныныц ауытқуы.
Егер шөппен қоректенетін жа¬
нуарлар (қоян) популяциясы-
на жыртқыштар (қасқыр, түл-
кі. сілеусін т.б.) тарапынан аз
қысым көрсетілсе, олардың да¬
ралар саны тез өседі. Ал, бұл
аталган жыртқыштардың жемтік
(қорек) корын көбейтіп, олардың
санының өсуіне алып келеді.
Жыртқыштар өз кезегінде шөп-
пен қоректенетіндерді (қоян-
дар) көптеп өлтіріп жойып, соц-
гыларыныц санын азайтады (41-
сурет).
Популяция генофонды популя-
циялық толқындардыц шыцына
(пик) жеткен кезде де, құлдырап
төмендеген кезде де өзгереді. Дараларь.нын сандык ауыткулары.
п а) сілеусін, б) түлкі, в) касқыр
Популяция дараларының саны J 1
өскен кезде бұрын бір-бірінен оқшауланган ұсақ популяциялар
жэне олардың генофондтары өзара қосылады. Ал, эрбір популя¬
ция генофонды бірегей ерекше болатындықтан, олардын қосылуы
нәтижесінде өзгерген жаңа генофонд түзілетіні сөзсіз.
Популяция дараларының саны азайган кезде ірі популяциялар
бірнеше ұсак популяцияларга ыдырайды. Жацадан пайда болган
183
1932 1940 1948 1954
41-сурет. Жырткыштар мен
«жемтіктер» популяциялары
ұсақ популяцияларда өздеріне гана тән генофондтар қалыпта-
сады. Ағзалардың жаппай өліп қырылуы кезінде кейбір сирек
кездесетін мутантты аллельдер генофондтан біржолата жойылуы
мүмкін, не сақталып қалып олардың жиілігі өсуі мүмкін.
Популяциялық толқындар популяция генофондының өзгеруіне
алып келетін эволюциялық факторлардың бірі болып табылады,
бірақ олар өз беттерінше жаңа формаларды (бейімделушіліктерді)
туғыза алмайды.
Популяциялық толқындар популяция генофондын кездейсоқ
өзгертіп сүрыптау үшін колайлы материал болып табылады.
Оқшаулану. Ағзалардың бір-бірімен еркін будандасуының
(панмиксия) шектелуін оциіау.чану деп атаймыз. Ол панмиксия
деңгейін төмендетіп туыстық некелесуді жиілетеді. Ал, бүл го-
мозиготалылар жиілігін көбейтіп популяция генофондтарыныц
айырмашылықтарын тереңдете түседі. Оқшаулану популяция
аралық генотиптік айырмашылықтардыц тецесуіне кедергі кел-
тіріп, популяцияларда тіршілік мүмкіншілігі жоғары болып келе-
тін генотиптердің сақталуына, таралуына мүмкіншілік туғызады.
Оқшауланудыц екі түрі белгілі: географиялық жэне биоло-
гиялық оқшаулану.
Географиялъщ оқшаулану деп жер қыртысының ерекшеліктері
салдарынан кецістікте оқшауланып орналасқан популяциялар-
ды айтамыз. Мысалы, сулар, теціздер, тау қыраттары аркылы
бөлшектенген популяциялар. Гавай аралдарында қүрлық ұлулары
таулардың ойлы алқаптарын мекендейді жэне бір-бірімен
қыраттар арқылы бөлінген. Олардыц қыраттардан асып өтуіне
топырақтын күрғақ болуы жэне ормандар кедергі келтіреді.
Осының нэтижесінде бүл үлулар популяциялары толық болмаса
да бір-бірінен оқшауланып, фенотиптік түрғыдан ерекшеленген.
Биологиялық оқшаулану түрішілік ерекшеліктердіц негізінде
пайда болады жэне оның бірнеше түрлері белгілі: 1) экологиялық
оқшаулану; 2) генетикалық оқшаулану.
Экологиялық оқшаулануға даралардыц түсінің, қоректе-
нуінің, көбеюініц ерекшеліктері алып келеді. Мысалы, Молда-
вияда тышқанның бір-бірімен араласпайтын (будандаспайтын) 2
популяциясы кездеседі: сары тамақты орман тышқандары жэне
дала тышқандары. Олардың бір-бірінен бөлінуіне себеп — қорек
184
'
күрамының эр түрлі болуында. Популяциялардыц ажырасуы
фенотиптік ерекшеліктерге алып келген дала тышқандары ұсак,
ми сауытының пішіні өзгеше болып келеді.
¥зақ уақытқа созылган экологиялық оқшаулану популяциялар
дивергенциясын кұшейтіп, жаңа түрлердің түзілуіне алып келеді.
Бүған мысал ретінде адам аскаридасы мен шошқа аскаридасын
алуға болады. Олар бір-бірімен морфологиялық жагынан ұқсас,
біртекті болып келеді. Экологиялық оқшаулану популяция аралық
будандасу мүмкіншілігін төмендеткенімен, толык жоя алмайды.
Генетикалык, оциіаулану даралардыц өзара будандасуын бол-
дырмайтын тосқауыл, бөгеттер қояды. Ол бөгеттерге мыналар: 1)
гаметалардың өзара үйлеспеуі; 2) үрықтана сала зиготалардың өліп
қалуы; 3) ағзалардың стерильді болуы не гибридтердің тіршілік
мүмкіншілігінің томен дәрежеде болуы жатады.
Кейде популяциялардың жіктелуі генетикалык оқшауланудан
басталады. Ал, генетикалык оқшаулануга мутантты гаметалардың
хромосомдық жиынтыгын күрт өзгертетін полиплоидия нёмесе
хромосомалардыц кайта қүрылулары жэне т.с.с. жагдайлар алып
келеді.
Түртүзілу қүбылысында оқшаулану басқа да карапайым
эволюциялық факторлармен бірлесіп әрекет етеді. Ол мутациялық
қүбылыс жэне комбинативтік өзгергіштік салдарынан пайда
болган генотиптік ерекшеліктерді терендете түседі. Оқшаулану
негізінде калыптасқан топтардың генетикалык кұрамы эр түрлі
болып табиғи сүрыптаудыц қысымынан эсерленеді.
Адам қауымының дамуы оқшауланган шагын өндірістік
үжымдар жиынтыгы ретінде жүріп отырган. Олардыц некелесуі
негізінен сол шагын үжым шецберінде гана болган. Адамзат
тарихыныц бастапқы кезецдерінде географиялық оқшауланудыц
маңызы үлкен болган. Окшаулану факторлары адамдар популя-
цияларының генофондына айтарлықтай ықпал еткен, мысалы,
кейбір шағын үлыстардыц антропологиялык ерекшеліктері —
бушмендердіц қүлақ қалқаны пішінінін ерекшеліктері, крояктар
мен ителмендердің төменгі жагыныц диаметрініц үлкен болуы
т.с.с. соның салдары.
Гендер дрейфі (генетикалық-автоматты қүбылыстар).
Мутациялар, комбинативгік өзгергіштік популяция даралары
санының оқтын-оқтын ауытқуы популяция генофондын өзгертіп
185
170-13
отырады. Олар сұрыпталумен бірлесіп әрекет етіп, түртүзілу
құбылысында биологиялық өзгергіштікке бейімделушілік сипат
береді.
Кейбір шағын популяцияларда табиғи сұрыптауға ілінбей,
кездейсоқ себептер нэтижесінде кейбір сирек аллельдер жиілігі
өзгеруі мүмкін. Бүл қүбылысты, яғни мутация қүбылысына
не табиғи сүрыптауға байланыссыз, кейбір сирек аллельдер
жиілігінің кездейсоқ өзгеруін, гендердіц дреііфі не генетикалыц-
автоматтыц цүбылыс деп атайды. Оны 1931—1932 жылда¬
ры орыс генетиктері Н. П. Дубинин, Д. Д. Ромашов, ағылшын
ғалымы Р. Фишер, американ ғалымы С. Райт ашқан. Гендердің
дрейфі нәтижесінде кейбір аллельдер қүрып біржолата жойы-
луы, не сақталып гомозиготалануы мүмкін. Ол популяцияның
негізгі бөлігінен оқшауланған шағын топтар генофондының
қалыптасуында маңызды рол атқарады. Гендер дрейфінің пай¬
да болуына изоляция, инбридинг жэне популяцияның сандық
молшерінің аз болуы эсер етеді.
Адамдар қауымдастығы аса үлкен болмайды. Осыдан 1—2 жүз
жыл бүрын адамдар 25—30 үйден аспайтын шагын топтар күйінде
тіршілік еткен. Соңғы жылдарга дейін адамдар популяцияларыныц
саны 400—3500 адамнан аспаган. Адамдар популяциясы арасын-
дағы географиялық, экономикалық, нэсілдік, діни, мэдени
ерекшеліктер олардыц некелесуін аудан, тайпа, ауыл, діни топ¬
тар көлемінде шектеп отырган. Адамдардыц шагын популяцияла-
рында үзаққа созылған жоғары дэрежелі репродуктивтік о к in ау¬
ла ну олардыц үрпақтарында гендердіц дрейфіне қолайлы жағ-
дай жасаган. Гендердің дрейфі популяция ішіндегі өзгергіштікті
теңестіріп, изоляттар арасында кездейсоқ айырмашылықтардың
пайда болуына алып келеді. Мысалы, Памир түргындары арасын¬
да теріс резусты адамдар Еуропа түргындарымен салыстырғанда
2—3 есе аз. Көптеген қыстақтарда теріс резусты адамдар мөл-
шері 3—5% болса, кейбір изоляттарда ол 15%-га жетеді.
Адамдар популяциясында гендердіц дрейфінің әсеріне «ру
қалыптастырушы эффекті» («эффект родоначальника») мы-
сал бола алады. Ол, егер бірнеше отбасы өз популяциясынан қол
үзіп, оңашаланып, басқа бір жерлерге қоныс тепкен жагдайларда
жэне шагын популяция бастамасында туыстық некелесу жогары
186
деңгейде болып, оның генофондында кейбір аллельдер кездейсоқ
сақталған жағдайларда байқалады. Нәтижесінде өте сирек
аллельдердің жиілігі күрт көбеюі мүмкін. Мыеалы, АҚШ-тың
Пенсильвания штатын мекендейтін мениоттар діни ұйымының
мүшелер саны XX ғасырдьщ ортасында 8000-ға жеткен, олардың
бэрінің түп негізі 1770 жылы Америкаға қоныс аударған 3 отба-
сынан келіп шыққан. Осы изоляттың 55 адамында көпсаусақты
ергежейліліктің ерекше формалары байқалған. Бұл ауру аутосомды-
рецессивті жолмен түқым қуалайды жэне АҚШ-тың Индиана,
Огайо штаттарында тұратын мениоттар арасында байқалмаған.
Адамзат тарихында гендердің дрейфі олардың популяциялар
генофондының қалыптасуына елеулі ықпал етіп отырган. Мысалы,
Сібірдің, Арктика, Байкал, Орталық Азия, Орал тұргындарының
кейбір шағын топтарының тар жерлерге бейімделу ерекшелікте-
рінің типтері гендердің дрейфі әрекетінің салдары деп түсіну
кажет. Әйтсе де, бұл қүбылыс адамдардың жіктелуі мен эволюция-
лануына шешуші эсер ете алмайды.
Гендердің дрейфі әсерінің салдарынан кейбір тұқым қуалайтын
аурулар адам топтары арасында әркелкі таралған. Мысалы, Кве¬
бек жэне Ньюфаундленд түрғындарында цебромакулярлық
регенерацияның салыстырмалы жиілігі жоғары дэрежеде бо¬
луын, Францияда балалардың цистиноз ауруының, Чехослова-
кияда алкаптонурия, Оңтүстік Африкада ақ нэсілдер арасында
порфириялардың бір түрінің, эскимостар арасында аденогенти-
льдік синдромның болуын оқшаулану мен гендердің дрейфі
эсерлері арқылы түсіндіруге болады.
Табиғи сүрыпталу. Жыныстық жолмен көбейетін агзалар по-
пуляциясында генотиптердің жэне фенотиптердің алуан түрлілігі
байқалады. Жеке өзгергіштіктің арқасында тіршілік ортасына қарай
эр түрлі генотиптің (фенотиптің) бейімделушілігі түрліше болып
келеді. Бейімделушілікті эволюциялық тұргыдан қарастыратын
болсақ, ол даралардыц тіршілік қабілетімен көбею қабілетінің
туындысына тең болады. Агзалар арасындагы бейімделушілік
айырмашылыгы табиги сүрыгпалу арқылы айқындалады жэне ол
келесі ұрпақтарга аллельдерді беру дэрежесі арқылы багаланады.
Сүрыпталудың негізгі нэтижесі тек тіршілікке қабілетті даралар-
дың тірі қалуымен шектелмей, келесі ұрпақ генофондына салы¬
стырмалы үлес қосатын даралардыц тірі қалуы болып саналады.
187
42-сурет. Табиғи сүрыпталудын формалары.
а) тұрактандырушы, б) козғаушы, в) бөлшектеуші, Ғ1-3 — үрпактар.
Сұрыпталу тіршілік үшін күрес нәтижесінде жүзеге асады.
Табиғи сүрыгіталу ағзалар онтогенезінің барлык сатыларында
жүріп отырады жэне олардың тандамалы көбеюін қамтамасыз
етеді. 'Габиғи сүрыпталу арқасында даралардың тірі қалуын
жэне көбею қабілетін жогарылататын аллельдер үрпақтар
жалғасында жинақталып популяциялардың генетикалық күра-
мын биологиялық максатқа сай өзгертеді. Табиғи сүрыпталу
қарапайым экологиялық фактор ретінде популяцияларда әрекет
етеді. Популядиялар сүрыптаудың әрекет алаңы, жеке даралар —
эрекет объектісі, ал нақты белгілер эрекет ету нүктелері болып
саналады.
Табиги сүрыпталудың үш түрі белгілі: түрақтандырушы,
қозғаушы жэне бөлшектендіруші (дизруптивтік).
Табиғи сүрыпталудың түрацтандыруиіы формасын И. Шма-
льгаузен (1946) ашқан, ол сыртқы орта факторлары өзгермей
бірқалыпты, тұрақты болған жағдайларда байқалады. Бұл кез¬
де жаңадан пайда болган мутациялар зиянды болып бұрыннан
қалыптасқан бейімделушілікті бұзады, бірақ жаңа бейімделу-
шілік пайда болмайды; бұрынғы бейімделушіліктер сақталынады.
Мысалы, қалың қар жауып, қатты жел тұрған кезде 136 торғай
есеңгіреп жерге құлап түскен. Олардың 72-сі тірі қалған, ал 64-
сі өліп қалған. Тірі қалған торғайлардың қанатгарының ұзын-
дығы орта мөлшерде болган, ал өлген торғайлардың қанаттары өте
ұзын не өте қысқа болып келген. Тұрақтандырушы форма табиғи
сұрыпталудыц консервативтік рөліне сэйкес келеді.
Табиги сүрыпталудың цозгауиіы формасын Ч. Дарвин аш-
қан, ол сыртқы орта факторларының құбылмалы болып келу
жагдайларында байқалады жэне фенотиптің белгілі бір багытта
өзгеруіне әкеліп соғады. Табиғи сұрыпталудың қозғаушы форма-
сы жаңа бейімделушіліктің пайда болуына алып келеді. Қозғаушы
сұрыптау екі түрлі бағытга жүреді: реакция нормасының өзгеруі
немесе кеңеюі.
Мысал ретінде XX ғасырдың басында Англияның ірі өнеркә-
сіпті қалаларының айналасындагы қайынды ормандарды мекен-
деген ақ түсті көбелектердің бірте-бірте күңгірт түсті көбелектер-
мен алмасуын айтуға болады.
Табиғи сүрыгіталудың бөлшектеуші (дизруптивтік) формасы
бірдей бейімделген бірнеше фенотиптердің сақталуын камтамасыз
етіп, алғаш біртүтас популяцияны бірнеше үсак популяцияларга
бөлшектейді. (42-сурет)
Дизруптивтік сүрыптау бұрынғы түрлердің ыдырауына
(дивергенцияга), бір немесе бірнеше реакция нормасыныц пайда
болуына алып келеді. Оған мысал ретінде мүхит жанындагы жер-
лерде қанатсыз және үзын қанатты шыбын-шіркейлердіц пайда
болуын айтуга болады. Дизруптивтік сұрыптау саналы адам —
Homo sapiens-тің пайда болуында, әсіресе нәсілдердің түзілуінде
үлкен рөл атқарған.
Табиғи сүрыпталу басқа да карапайым эволюциялық факторлар
сияқты популяция генофондыныц аллельдерінің арақатынасын
өзгертеді. Бірақ ол өзгерістер белгілі бір багытқа бағытталған бо¬
лады. Табиги сүрыпталу басқа да қарапайым эволюциялық фак-
торлармен бірлесіп эрекет етеді, бірак, мутациялық қүбылыс,
189
оңашалану, гендердің дрейфі оған қажетті материал дайын-
даса, табиғи сұрыпталу популяция генофондының өзгеруіне
биологиялық мақсатқа сай багыт беріп отырады. Сондықтан ол
қарапайым формалардан күрделі формалардың пайда болуына
алып келеді.
Түртүзілу құбылысында табиғи сүрыпталу кездейсоқ, жекелей
өзгергіштікті биологиялық пайдалы, топтық-популяциялық не
түрлік өзгергіштікке айналдырады .
Оның тұрақтандырушы нысаны алдыңғы кезеңдерде пай¬
да болған аллельдердің тиімді комбинацияларын сақтап қалады.
Сүрыптау, сол сияқты популяцияның генетикалык полимор-
физмін қалыптастырады.
Қазіргі кезде адамдар популяциясында табиғи сүрыптау өзінің
түртүзілу функциясын жойып, тек генофондтардың түрақты-
лыгын жэне генетикалык көптүрлілігін гана қалыптаетырады.
Бүган мысал ретінде уақыты жетпей не уақытынан өтіп туылган
нәрестелердің көптеп өлуін айтуга болады.
Адамдарда табиғи сүрыпталу кейде гетерозиготаларға карсы,
кейде гомозиготаларга қарсы бағытталады. Гетерозиготаларга
қарсы сұрыптау ретінде Rh антигенінің синтезделуін келтіруге
болады. Еуропа түргындарының 85%-ның эритроциттерінде
Rh антигені оң резус тобын қүрайды, ал популяцияның қалган
мүшелерінің эритроциттерінде Rh антигені болмагандықтан,
теріс резусты болып келеді. Rh антигені доминантты (Д) ал¬
лель арқылы анықталынады. Rh (+) адамдардыц генотипі ДД
не Дсі, ал Rh (-) dd күйінде болады. Егер Rh (-) эйел Rh (+) ер
адамнан жүкті болып, қүрсақтагы бала Rh (+) болатын болса,
қүрсақтагы баланың Д (+) эритроциттері ана агзасына өтіп оны
иммундайды. Ана агзасында ол антигенге қарсы антидене ретінде
синтезделінеді. Бірінші Rh (+) бала қалыпты туылады, ал келесі
Rh (+) балаға жүкті болса, ананың антирезус-антиденесі бала
орнына (плацентага) еніп, қүрсақтагы баланың эритроциттерін
ыдыратады да, нэресте гемолитикалық аурумен ауырады. Оларга
медициналық жэрдем көрсетіп емдемесе, оліп қалады. Бүл
жагдайда сүрыпталу гетерозиготага карсы багытталган.
Гомозиготаларга қарсы багытталган сүрыптауга мысал ретін-
де орақ жасушалы анемияның тұқым қуалауын келтіруге болады.
190
Бұл ауру S аллельі арқылы доминантты тұқым қуалайды. Осы ал¬
лель бойынша гомозиготалы (SS) балалар эдетте өліп қалады, ал
гетерозиготалылар осы аурумен сырқаттанса да өлмейді.
Түртүзілу қүбылысы қарапайым эволюциялық факторлардың
өзара әрекеттесуі нэтижесінде жүреді. Түртүзілудің үш жолы
белгілі:
1) бір түрден екі не одан да көп түрлердің түзілуі — шынайы
түртүзілу;
2) филетикалық түртүзілу — бір түрдің баяу дамып, екінші бір
түрге айналуы;
3) гибридтік түртүзілу.
Сонымен қатар, түртүзілудің аллопатриялық жэне симпат-
риялық түртүзілу деген жолдары да белгілі. Аллопатриялық
не географиялык түртүзілу құбылысында алғаш географиялык
оқшаулану, содан кейін генетикалык оқшаулану гіайда болады.
Симнатриялық түртүзілуде жаңа түрлер бүрынгы түрдің попу-
ляциясы шеңберінде түзіледі. Бүл жагдайда о бастан генетикалык
оқшаулану пайда болады. Жануарлардың көптеген түрлері
аллогіатриялық, ал өсімдіктердің түрлері симпатриялық жолдар-
мен түзілген.
6.4. Табиғи популяциялардын генетикалық
полиморфизмі (көитүрлілігі)
Түртүзілу қүбылысында, бір жагынан, тірі агзалардың
көптүрлілігі түзілсе, екінші жагынан мекен ортасына бейімделген
формалар пайда болады. Тұқым куалайтын өзгергіштіктің
резервінің жэне генетикалык комбинация салдарынан әрбір буын-
да пайда болатын эр түрлі генотиптер арасынан тек кейбіреулері
гана мекен ортасына шынайы бейімделген, ал қалгандары нашар
бейімделген, кейбіреулері тіпті бейімделмеген болуы мүмкін.
Табиги сүрыптау нәтижесінде шынайы бейімделген генотиптер
таңдамалы түрде көбейіп, ал калгандары үрпак калдыра алмай
өліп калатын болса, онда популяциялар генофонды тек «сэтті»
аллельдерден жэне олардыц комбинацияларынан кұрылган болар
еді. Нәтижеде түкым куалайтын өзгергіштік тоқталып, гомозиго-
191
талы генотиптер мөлшері өсіп кетер еді. Бірақ, табиги популяция-
ларда бұған қарама-қарсы құбылыс байқалады — популяцияның
көптеген даралары жоғары дэрежеде гетерозиготалы болып
келеді. Кейбір даралар бірнеше локустар бойынша гетерозиго¬
талы болады, ал бұл популяциялардың гетерозиготалылыгын
жогарылатады. Мысалы, Антарктидада тіршілік ететін киттердің
қорегі ұсақ шаян тэрізділердің 126 дарасының 36 локусын электро¬
форез әдісі арқылы зерттегенде, олардың 15 локусында өзгергіштік
байқалмаган (бэрінде бірдей болган) 21 локусының 3—4 аллель-
дік формалары анықталган.
Осы популяциялар даралары 58% локустары бойынша ге¬
терозиготалы болып, олардың 2-ден көп аллельдері болатыны
анықталган. Орташа эрбір дара 5,8% гетерозиготалы локусқа
ие болган. Өсімдіктердің орташа гетерозиготалылыгы — 17,
омыртқасыздардың— 13,4, омыртқалылардың — 6,6, адамдардың
— 6,7%-га тең. Популяциялардың осыншама үлкен дэрежеде ге¬
терозиготалы болуын тек мутациялар аркылы түсіндіруге бол-
майды, себебі мутациялар оте сирек кұбылыс.
Погіуляцияларда ұзак уақыт тепе-теңдік күйінде болып,
мөлшері жагынан ең сирек кездесетін генотиптен 1%-дан артық
болатын бірнеше генотиптің кездесуін генетикалык, полиморфизм
(көптүрлілік) деп атаймыз. Генетикалык полиморфизм мутациялар
жэне комбинативтік өзгергіштік салдарынан түзіледі жэне табиги
сұрыптау аркылы бірқалыпты децгейде сакталынады. Генетикалык
полиморфизмнің 2 түрі белгілі:
1. Адаптациялык полиморфизм;
2. Балансты, не гетерозиготалы полиморфизм.
Зацды түрде өзгеріп отыратын ортада сурыптау эр түрлі ге-
нотиптерге қолайлы болатын болса адаптациялык, полиморфизм
түзіледі. Мысалы, екі нүктелі қанқызыныц популяциясында күз-
діц ақырында кара түсті қоцыздар (доминантты белгі), ал көктемде
қызыл түсті қоцыздар саны басым болады. Мүныц себебі қызыл
түсті қоцыздар суыққа шыдамды болса, қара түсті қоцыздар
жаздыц күні жедел көбейе алады.
Егер сүрыптау рецессивті жэне доминантты гомозиготалыларга
Караганда гетерозиготалыларга қолайлы болатын болса, онда
балансты (гетерозиготалы) полиморфизм қалыптасады. Мы-
192
салы, жеміс шыбынының жасанды популяциясында алғаш му¬
тантты қара денелі (рецессивті белгі) шыбындар басым болып,
шамалы уақыттан кейін қара денелілердің саны азайып, 10 пайыз
мөлшеріне жетіп токтаған. Зерттеулер нэтижесінде рецессивті
гомозиготалылар мен доминантты гомозиготалылардың гетеро¬
зиготалы дараларга Караганда тіршілік қабілетінің төмен болаты-
ны анықталган.
Гетерозиготалылардың гомозиготалыларга қараганда артық-
шылыгын аса жогары доминанттылық деп атайды. Сұрыптау-
дың гетерозиготалыларга қолайлы болу механизмдері эр түрлі
болып келеді. Жалпы алганда, сұрыптау белсенділігі нақтылы
генотип (фенотип) жиілігіне тәуелді болады. Мысалы, балықтар,
кұстар, сүтқоректілер жемтіктерін аулаганда сирек кездесетін
фенотиптерді аулайды. Гетерозиготалылардың артықшылыгының
тагы бір себебі, ол гетерозис кұбылысына байланысты.
Тіршілік ортасы факгорларының алуан түрлі болуының сал¬
дарынан сұрыптау бір мезгілде бірнеше багытта жүреді, ал
оның ақыргы нәтижесі сұрыптау багыттарының белсенділігінің
ара катынасына байланысты болады. Мысалы, адамдардың
орақ пішінді эритроциттерінің жартылай летальды аллельінің
концентрациясының Жер шарының кейбір аудандарында жогары
деңгейде болуы осы аллельге багытталган теріс сұрыптау үстіне
белсенді оң контрсүрыптаудың қабатгасуы салдарынан болып та¬
былады. Себебі, орак пішінді эритроциттер безгек індеті жиі бо¬
латын тропикалық аймақтарда агзалардың осы ауруга төзімділігін
жогарылатады. Популяциялардагы табиги сұрыптаудың ақыргы
нәтижесі сұрыптау жэне контрсұрыптау багыттарының бір-бірінің
үстіне қабаттасуына байланысты болады жэне осы арқылы бір
мезгілде генофондтың тұрақтылыгымен генетикалык көптүрлілігі
қалыптасады.
Балансты полиморфизм популяцияларга бірнеше құнды қасиет
береді:
1) генетикалык көптүрлі популяциялар сыртқы ортаның кең
аукымды жагдайларын игере алады;
2) оның генофондында кең көлемді түқым қуалаушылық
өзгергіштіктің қоры жинақталады, осының нэтижесінде ол эволю-
циялық икемділікке ие болып, кез келген багытта өзгере алады.
193
Адамдар популяцияларында генетикалык полиморфизм
жогары деңгейде болады, сондықтан олардың алуан түрлі
фенотиптері байқалады. Адамдар бір-бірінен терісінің, көзінің,
шашының рендерінің эр түрлі болуымен, мұрын және құлақ
қалқаншасының пішіндерінің, т.б. белгілерінің әр түрлі болуы¬
мен ерекшеленеді. Адамдарда бір не бірнеше амин қышқылдар
арқылы ерекшеленетін жэне эр түрлі қызмет атқаратын ақуыздар
түрлері белгілі. Акуыздар — белгілер, сондықтан да олар
адам агзасының гікелей генетикалык қүрылымын анықтайды.
Адамдарда ABO, резус жүйелері бойынша эритроцитарлық
антигендерінің көптеген түрлері белгілі. Гемоглобиннің 130-га
жуық түрлері, глюкозаның — 6, фосфатдегидрогеназа фермен-
тінің 70-ке жуық түрі белгілі. Жалпы, адамдардың ферментте-
рініц синтезделуін қадагалайтын 30 пайыз гендерінің жиілігі
түрліше болады, бірі жиі кездессе, екіншілері сирек кездеседі.
Мысалы, гемоглобиннің 130 түрінен жиі кездесетіні 4-HbS
(тропикалық Африкада, Жерорта теңізінде), НвС (Батыс Аф-
рикада), НвД (Үндістанда), НвЕ (Оңтүстік Шыгыс Азияда).
Ал гемоглобиннің қалган аллельдерінің концентрациясы 0,01-
0,0001 %-дан аспайды. Адамдар популяциясында аллельдердің
таралуының әр түрлі болуы карапайым эволюциялық фактор-
ларга, оның ішінде мутациялық күбылыска, табиги сүрыптауга,
гендер дрейфіне жэне миграцияга байланысты.
Адамдар генофондындагы аллельдердің көптүрлілігін 2 топқа
бөледі:
1. Сирек кездесетін варианттар — олар барлык жерде кездеседі
жэне олардың кездесу жиілігі 1 %-дан аз болады;
2. Кейбір таңдамалы популяцияларда жиі кездесетін вариант-
тар. Мысалы, HbS, НвС, НвД, НвЕ.
Популяциялар арасында кейбір аллельдер концентрациясыньщ
ұзақ уакыт түрліше болуы, бір популяцияда бірнеше аллельдіц
концентрациясыныңжоғарыдеңгейде болуы, табиги сүрыгіталудыц
не гендер дрейфінің әсерінен болады.
Кейбір аллельдер концентрациясыньщ популяция аралық
айырмашылыгына табиги сүрыпталудыц түрақтандырушы фор-
масы алып келеді. ABO қан тобы жүйесініц эритроцитарлық ан¬
тигендер аллельдерініц Жер шарында жиі таралуы өте қауіпті ин-
194
фекциялар жиі болып тұратын жерлерде эр түрлі қан топтары бар
адамдардың түрліше тірі қалуының нәтижесі болуы мүмкін.
Мысалы, Азия тұрғындарында I аллельінің томен болуы бұл
жерлерде оба індетінің жиі болуымен байланысты. Оба ауруын ту-
дырушы микроағзалардың Н — типтес антигендері болады, ал бүл
«О» қан тобы бар адамдарды оба ауруына сезімтал етеді, себебі
оларда да Н — типтес антигендері болғандықтан, сол микробтарға
қарсы антидене жеткілікті мөлшерде синтезделінбейді. Керісін-
ше, Австралия мен Полинезияның орнықты тұрғындарында, Аме¬
рика үндістерінде I аллельінің концентрациясы жогары болады,
себебі бүл жерлерде оба індеті бұрын-сонды болмаған.
Адамдар популяциясында бір мезгілде бір геннің бірнеше
аллельінің түракты сақталуының себебі сүрыптаудың гетеро-
зиготалыларға тиімді (қолайлы) болуының нәтижесі деп түсінген
дүрыс. Бұған мысал ретінде гемоглобиннің S, С, Е аллельдерінің
тропикалық безгек ауруының ошақтарында таралуын алуға бола¬
ды. Табиғи жағдайларда ағза фенотипіне көптеген факторлар эсер
еткендіктен, сұрыптау бірнеше бағытта жүріп, нақтылы жағдайда
популяциялардың тірі қалуын камтамасыз ететін аллельдер саны
мен жиілігінің балансты генофондын қалыптастырады, Бұл
жағдай адам популяцияларында жиі кездеседі. Мысалы, «О» қан
тобы бар адамдар «В» қан тобы бар адамдарға қарағанда оба ау¬
руына төзімсіз келеді. «О» қан тобы бар адамдар «А» қан тобы бар
адамдарға қарағанда туберкулез ауруынан қиындықпен емделеді.
«О» қан тобы бар адамдар — мерез ауруынан жылдам айығады,
олардың қарынның рак ауруымен, жатыр мойнының рак (ісік) ау-
руымен, ревматизм, жүректің ишемия ауруымен, холецистит, өтте
тас болу ауруымен ауру ықтималдығы «А» кан тобы бар адамдарға
қарағанда 20 гіайызға томен болады.
6.5. Генетикалық жүк жэне оның
медициналық-элеуметтік маңьпы
Генетикалык полиморфты популяцияларда эрбір ұрпақта орта
факторларына түрліше бейімделген генотиптер пайда болып от¬
ырады. Олардың ортаға бейімделушілігі бірдей болмайды, бірі
195
жақсы бейімделген болса, екіншілері нашар бейімделген. Мұн-
дай популяциялардың тіршілік қабілеттілігі эр кезде тек «сәтті»
генотиптер жиынтығынан томен болады.
Бірдей генофондқа ие нақтылы табиғи полиморфты по-
пуляциялардың бейімделушілігінің тек «сэтті» генотиптерден
қүралган үлгілі популяциялардың бейімделушілігінен айыр-
машылыгын генетикалык жүк деп атайды. Ягни, генетикалык
жүк дегеніміз популяциялардың орта факторларына нашар
бейімделген дараларының генотиптерінің жиынтыгы. Ол мута¬
ция негізінде қалыптасады. Генетикалык жүктің мутациялық,
сегрегациялық және субситуациялық жүк деп аталатын 3 түрі
белгілі.
Мутациялъщ жүк — популяцияларда қайта пайда болган
мутанттық аллельдер нәтижесінде түзіледі. Табиги сүрыпталу бүл
аллельдерге қарсы бағытталгандыктан, олардыц жиілігі аз бола¬
ды. Олар популяцияда мутациялық қысым арқасында қайтадан
пайда болып отырады.
Сегрегацішлыц жүк — гетерозиготалы ата-аналардан ортага
нашар бейімделген гомозиготалы генотиптердің бөлініп шыгуы-
ның нәтижесінде түзіледі. Бұл жүктің түрін Ф. Г. Добжанский
ашқан. Бұл популяцияларда гетерозиготалылардыц тіршілік
мүмкіншілігі гомозиготалыларга Караганда жогары болып келеді.
Субситуацияпъщ жүк — даралардыц адаптивтік құндылы-
гының өзгеруі нэтижесінде түзіледі жэне ол бір аллельдіц екінші
аллельмен алмасуына дейін сақталынады.
Сонымен, генетикалык жүк популяцияныц жалпы бейім-
делушілігін төмендетіп көптеген даралар үшін зиянды болып
келеді. Бірақ, генетикалық жүк барлық уақытта зиянды бола
бермейді, себебі:
1) эволюцияның генетикалык қоры болып есептелінеді;
2) оның арқасында популяцияда генетикалык полиморфизм
популяциясыныц эволюциялық икемділігі қалыптасады.
Адамдардыц генетикалык жүгінің мөлшері біршама жогары
болады. Оныц зардабы летальды гендердіц эсеріне тец, ягни эр
адамныц генотипіндегі жагымсыз, зиянды аллельдер жиынтыгы
3—8 рецессивті аллельдердіц гомозиготалы күйіндегі зиянды
әсеріне тец болады. Ондай агзалар көбею жасына жетпей оліп
қалады.
196
Адамдардың эрбір буынында пайда болатын үрпақтардыц
50%-ы жағымсыз аллельдердің болуы нәтижесінде биологиялық
тұргыдан алғанда мағынасыз, яғни келесі үрпаққа гендерін бере
алмайды, көбейе алмай өліп қалады. Оның ішінен, пайда болған
үрықтыц 15%-ы туылғанға дейін, эмбриондық даму кезінде
өліп қалады; 2%-ы туылғаннан кейін 1 жыл ішінде өліп қалады;
3%-ы репродуктивтік жас шамасына жетпей өледі; 20%-ы неке-
лесе алмайды; ал 10%-ы бедеу болып келеді. Осының бэрі адам
популяциясының генетикалык жүгінің зардабының салдары бо¬
лып табылады.
6.6. Микро- және макроэволюция.
Ағза топтарының эволюциясы
Қарапайым эволюциялық факторлардың әсерінен популяция-
лардың дивергенциясы (айырмашылығы) қалыптасады. Ол
әрі қарай тереңдеп, жаңадан 2 тұртүзілуге алып келеді. Түр
— түзілгенге дейін эр түрлі популяциялардың даралары бір-
бірімен будандасып гендерімен алмасады, бүл популяция аралық
айырмашылықтарды теңестіреді. Жаңадан түр-түзілгеннен кейін
олардың арасында репродуктивтік оқшаулану пайда болады да
олар будандаса алмайды.
Органикалық дүние өзінің дамуының эрбір кезеңдерінде
бір-бірінен репродуктивтік оқшауланган, бір-бірімен азды-
көпті туыстық байланыспен байланысқан агзалар топтарының
(таксондардың) күрделі иерархиялық жүйесі болып табылады.
Бүл табиғаттың біртұтас эволюциялық қүбылысының екі түрлі
қырларының нэтижесі болып саналады:
1) түртүзілу қүбылысы (микроэволюция);
2) түрден ірі жүйелілік таксондардың — туыс, түқымдас отряд,
т.с.с. пайда болу қүбылысы — макроэволюция.
Макроэволюция құбылысы мындаған, миллиондаған жыл-
дарға созылатындықтан, оны тікелей байқау не тэжірибе жасап
зерттеу мүмкін емес. Оныц зандылықтарын анықтауда палеон¬
тология, салыстырмалы эмбриология жэне салыстырмалы морфо¬
логия деректері мацызды рөл атқарады.
197
Макроэволюцияның қозғаушы факторлары туралы жалпыға
ортақ пікір, тұжырым элі күнге дейін жоқ. Көптеген болжамдардың
бірі—микро- жэне макроэволюция процестері ортақ эволюциялық
қозғаушы күштер арқылы жүреді деп болжамдайды, ол күштер —
қарапайым эволюциялық факторлар — мутация, оқшаулану, ген¬
дер дрейфі, сүрыптау.
Тірі ағзалар топтары эволюциясының бірнеше формалары
белгілі:
1) филетикалъщ эволюция — бір таксон өкілдері біртүтас бір
багытта өзгеріп дамып, жаңа таксонга айналуы. Мысалы, жылқы
тұқымдасының дамуы. Олардың дамуында ағаш бұталарымен
қоректенетін формалардың ақырындап шөптермен қоректенуге
көшкенін жэне түяқтары саныныц біреуге дейін азайғанын
байқауга болады;
2) дивергенттік эволюция — бір таксоннан бірнеше жаца
топтардыц (таксондардыц) пайда болуы. Эволюцияныц бүл
формасы ірі таксондардыц пайда болуына алып келеді, мыса¬
лы: класс ішінде бірнеше класс тармақтарының, отрядтардыц,
түқымдастардың пайда болуы.
Бірнеше таксондардыц салыстырмалы даму формаларына
эволюцияныц параллелизмін жэне конвергенциясын жаткызуга
болады.
Бірінші жагдайда бір ата-тектен пайда болган 2 таксон бірдей
багытта филетикалық жолмен дамиды. Топтар эволюциясын-
да катар (параллель) эволюциялану кезеңі конвергенциямен
ауысуы мүмкін. Конвергентті эволюция дегеніміз туыс емес 2
гаксонның бірдей филетикалық багытта қатар дамуы болып табы¬
лады. Конвергенция эр түрлі топтың өкілдері үқсас экологиялық
жагдайларда тіршілік етсе байқалады. Мысалы, акулалардыц және
киттэрізділердіц дене қалпының үқсас болуы конвергенттік эво¬
люция нэтижесі болып саналады.
Бейімделу ауқымына қарай эволюцияныц 2 типін ажыратады:
аллогенез (идиоадаптация) жэне арогенез (ароморфоз). Аллогенез
(идиоадаптация) дегеніміз — бір ірі таксон өкілдерініц (класс,
отряд, түқымдас) тіршілік кұрылымыныц деңгейі езгермей эр
түрлі экологиялық жагдайларды игеріп, жайылып дамуы. Бүган
мысал ретінде сүтқоректілер класыныц насекомқоректілер
отрядының эр түрлі экологиялық формаларын айтуга болады.
198
Арогенез (ароморфоз) деп — ірі таксондардыц кейбір ағза
топтарының қүрылым деңгейлері күрделіленіп жоғары деңгейге,
жаңа адаптивтік аймаққа көтерілуіне алып келетін эволюциялық
өзгерістерді айтамыз. Ароморфоз негізінде ағзалар сыртқы орта
қорларын игеру үшін жаңа мұмкіншіліктерге ие болды. Бүл
мүмкіншілік аллогенез (идиоадаптация) негізінде жүзеге асады,
ал ол жаңа ароморфозды пайда етеді. Сонымен, ағза топтарының
эволюциясының типтері- аллогенез жэне арогенез бір-бірімен
тыгыз байланысты болады. Аллогенез (идиоадаптация) жэне
арогенез (ароморфоз) биологиялық прогресс құбылысына алып
келеді. Кейде агзалар дамуында олардыц құрылысыныц, фи-
зиологиясының қарапайымдануы байқалады, мысалы, паразит-
тер. Эволюцияныц бүл түрін морфофизиологиялық регресс (де¬
генерация) деп атайды.
Агза топтары эволюциясыныц бірнеше ережелері белгілі:
1) эволюция бағыгының кері кайтпайтындығы — оны 1883
жылы Л. Долло түжырымдаган. Бүл ереже бойынша тарихи даму
барысында қандай да болсын бір топ өздері жүріп өткен бүрынгы
қалпына қайтып оралмайды. Бүл ереженіц генетикалық негізі бо¬
лып бір-біріне үқсас екі генофондтардың (генотиптердіц) пайда
болу ықтималдыгының өте аз мөлшерде болуы, тіпті болмауы са-
налады. Дегенмен, филогенез барысында кері мутациялардыц пай¬
да болуы арқасында ертеде кездескен кейбір қарапайым белгілердіц
қайталануы мүмкін.
2) агзалардың прогрессивтік мамандануы — оны 1876
жылы Ш. Депре түжырымдаган. Осы типпен эволюцияланушы
топ эрі қарай мамандануды терендетігі дамиды. Бүл ереженің
генетикалык негізі ретінде бір топ генофондыныц шектеулі болу¬
ын жэне оныц нактылы адаптивтік аймақ факторларыныц эсері-
нен калыптасатындыгын айтуга болады.
3) жаңа топтың (таксонның) қараиайым, маманданбаған
топтардан келіп шығуы — оны 1904 жылы Э. Коп тұжырым-
даган. Бүл ереже бойынша жаңа топтар тек қарапайым,
маманданбаган топтардан келіп шыгады. Оныц негізі ретінде
маманданбаган топтарда тіршілік үшін күрес эр алуан болатын-
дыгын жэне маманданбаган топтардан эр түрлі адаптациялар-
дыц пайда болатындыгын айтуга болады.
199
Қазіргі органикалық дүние тірі ағзалардың көптеген топта-
рынан тұрады. Олардың кұрылыстары, физиологиясы тұрліше
болғанымен, бэрі бірге тіршілік етеді, демек, олардың бәрінде
де тіршілік үшін күрес сәтті болғаны. Тарихи дамудың әр түрлі
кезеңдерінде органикалық дүниенің бір тобы басым болып, ал
екіншісі аз мөлшерде кездесіп отырған. Дегенмен де, шағын топ
ағзаларына да экспансия, яғни кең таралу, жаңа мекен орталарын
жаулап алып игеру қасиеті тэн. Ал үшінші біреулері қүрып жойы-
лу күйінде болады.
Кең таралған, басым топ ағзалары биологиялық прогресс
күйінде болады. Биологиялық прогрестің негізгі белгілері:
1) топ өкілдерінің жалпы санының шексіз көп болуы;
2) ірі таксондар ішінде ұсақ таксондардыц көп болуы (класс,
класс тармағы — отряд);
3) ареалдарының кеңеюі.
Жер тарихының эр түрлі кезеңдерінде эр түрлі жануарлар топта-
ры басым болган. Мысалы, космекенділер палеозой эрасының де¬
вон дәуірінде пайда болып, келесі 75 миллион жыл бойына басым
болган. Таскөмір дәуірінде жоргалаушылар пайда болып мезозой
эрасының триас дәуірінде басым болған; юра, бор дәуірлерінде
ірі диназаврлар басым болган; олар осыдан 65 млн. жыл бұрын
жойылып, олардан кейін сүтқоректілердің жаппай басымдылыгы
калыптасты.
Қазіргі агзалардың морфологиялық жэне физиологиялық
ерекшеліктері белгілі бір таксондардың тарихи дамуының
(филогенезінің) нәтижесі болып саналады. Олар кептеген үрпақтар
жалгасу барысында ата тектерінде пайда болган озгерістерді
бейнелейді. Әйтсе де, әрбір үрпақ дараларының қүрылымдық
— қызметтік белгілері олардың жеке дамуының (онтогенез)
нәтижесінде калыптасады.
Түрдің тарихи даму қүбылысы бір-бірін алмастырып отыратын
даралардың үрпақтары болып саналады. Биологиялық түргыдан
қарастырганда дара және оның онтогенезі үқсас, бірдей екені
белгілі, сондықтан тарихи даму бірізділікпен алмасып отыра¬
тын онтогенез қатарлары болып табылады. Демек, тарихи даму
мен онтогенез арасында тыгыз байланыс болады, ал оның мэнін
анықтау эволюциялық қүбылыстардың механизмдерін түсіну үшін
маңызды болып есептеледі.
200
Өткен ғасырдың аяғында биологтар ағзалардың жеке даму
құбылысында (онтогенез) оның кейбір мүшелері өздерінің
ататектерінің қарапайым өкілдерінің мүшелеріне ұқсас сатылар-
дан өтетіндігіне көңіл аударған. Мысалы, адам эмбриогенезінде
алғаш пронефрос, содан кейін мезонефрос және метанефрос
қалыптасады.
Салыстырмалы эмбриологиялық зерттеулер негізінде К. Бэр
үръщ үцсастыгы заңын тұжырымдады. Бұл заң бойынша
үрықтардыц дамуында туыс жануарлар топтарының ортақ
белгілері ерекше белгілеріне қарағанда ерте қалыптасады, ягни
жануарлардыц урықтык дамуында алгаш класқа тэн белгілер, со¬
дан кейін отрядқа, түқымдасқа, туысқа, ең ақырында тұрге тән
белгілер дамиды. Жеке дамудың алғашқы сатыларында эр түрлі
жануарлардыц үрықтары бір-біріне ұқсас болып келеді.
Филогенез жэне онтогенез арасындагы байланыс проблемала-
ры эрі қарай Ф.Мюллер, Э.Геккель, А.Н.Северцов еңбектерінде
дамытылған.
Ф. Мюллер шаянтәрізділер филогенезін зерттей отыра олардыц
кейбіреулерінің дернэсілдік формалары жойылып, құрып кеткен
ата-тектерініц түрлеріне ұқсас болатынына көңіл аударған. Осы
зерттеулер нәтижесінде ол мынадай қорытынды жасады: казіргі
шаянтәрізділер өздерінің эмбриогенезінде ата-тектерінің даму
тарихында өтіп кеткен жолдарын кайталайды.
Э.Геккель негізгі биогенетикалық занды тұжырымдады. Бүл
заңга сэйкес, онтогенез дегеніміз — филогенездіц қысқа жэне
жылдам қайталануы болып саналады.
Биогенетикалық заңныц растыгына дэлел ретінде рекапитуля¬
ция (қайталану) мысалдарын келтіруге болады. Айталық, күстар
мен сүтқоректілер эмбриогенезінде желбезек сацылаулары және
оган сэйкес қацқа күрылымдары, қантамырлары пайда болады.
Адам эмбриогенезінде тері эпидермисі алгашқыда бір қабатты,
содан кейін көп қабатты мүйізденбеген, көп қабатты болар болмас
мүйізденген, ақырында қалыпты мүйізденген эпителий күйінде
қалыптасады. Эпителийдіц аталган түрлері ересек қандауыршага,
сүйекті балықтарга, қүйрықты амфибияларга тэн. Э. Геккельдіц
айтуы бойынша, эволюциялық мацызга ие жаца белгілер тек ере¬
сек агзаларда пайда болады.
Онтогенез бен филогенез арасындагы байланысты ашуда
А.И. Северцовтың еңбектерінің маңызы зор. А. И. Северцовтың
пікірінше, эволюциялық өзгерістердің көзі болып ересек форма-
ларында емес, онтогенездің бастапқы сатыларында пайда бола¬
тын өзгерістер саналады. Егер олар ересек күйінде пайда болып
тұқым қуаласа, ұрпақтан-ұрпаққа беріліп сақталынады жэне сол
ағза топтарының филогенезіне кыстырылып қосылады. Ере¬
сек формалардың кұрылысында байқалатын жэне эволюциялық
маңызы бар эмбриондық өзгерістерді филэмбриогенездер деп
атайды. Оның 3 түрі белгілі: анаболия — эмбриогенездің бұзылуы
оның аяқ жағына қосымша сатының қосылуы салдарынан бола¬
ды; девиация — эмбриогенездің бұзылуы оның ортаңгы сатыла¬
рында болады; архаллаксис — эмбриогенездің бұзылуы оның
бастапқы сатыларында басталады.
Филэмбриогенез теориясына сэйкес жеке дамудын бастапқы
сатыларында пайда болган өзгерістер мүшелердің филогенети-
калык қайта құрылуларының негізі болып саналады.
г
7. ОМЫРТҚАЛЫЛАРДЫҢ МҮШЕЛЕР ЖҮЙЕСІНІҢ
ФИЛОГЕНЕЗІ
7.1. Филогенездің жалпы зандылықтары
Филогенетикалық өзгерістердің пайда болу зандылықтарын
зерттейтін ғылымды эволюциялыц морфология деп атайды. Ол
XIX ғасырдың аяғында салыстырмалы анатомия, салыстырма¬
лы эмбриология жэне палеонтология гылымдарының негізінде
қалыптасқан.
Эволюциялық морфология гылымының негізгі зерттеу әдісі бо¬
лып салыстырмалы анатомия, салыстырмалы эмбриология жэне
палеонтология әдістерін камтитын «уштік параллелизм» эдісі
саналады.
Үштік параллелизм эдісі аркылы эволюциялык морфология
гылымы мүшелердің гомологиялықжэне аналогиялық ұқсастығын
анықтайды жэне оларды жіктейді.
Гомологиялык үқсастык (грекше homologia — бірдей) деге-
німіз:
1) мүшелердің күрылысының, олардың бөлімдерінің арақаты-
насының, орналасуынын бірдей болуы;
2) мүшенің агза денесінде бірдей орналасуы;
3) мүшелердің бір ұрык бастамасынан пайда болуы.
Бұл мүшелердің атқаратын қызметгері бірдей не эр түрлі бо¬
луы мүмкін. Ягни гомологиялық мүшелер деп — шыгу тегі бірдей,
қүрылысы не атқаратын қызметтері бірдей не эр түрлі болып келе-
тін мүшелерді айтамыз. Бүган мысал ретінде омыртқалылардың
алдыңгы аяқтарын жатқызуга болады. Гомологиялық ұқсастық-
тың негізгі себебі — тарихи туыстық болып саналады, ягни олар
бір (ортақ) ата-тектен не оның мүшесінен пайда болган.
Апалогііялъщ щсастъщ (грекше analogia — бірдей) немесе
аналогиялық мүшелер деп шыгу тегі эр түрлі, бірак сырт пішіндері
бірдей, үқсас жэне бірдей қызмет атқаратын мүшелерді айтамыз.
203
Олар тек морфологиялық жагынан гана ұқсас болып, ортақ бір
қызмет атқару арқылы айқындалады, ал шығу тектері эр түрлі.
Аналогиялық мүшелердің үқсастыгы тіршілік ортасының ортақ
болуымен сипатталады жэне олардың туыстық байланыстары
болмайды. Мысал ретінде көбелектердің жэне құстардың қанатга-
рын келтіруге болады.
Филогенез заңдылықтарын дұрыс түсіну ұшін мүшелердің
құрылысының өзгерулерін зерттеумен қатар атқаратын қызметін де
зерттеген дүрыс. Бір мүшенің қүрылысының өзгеруі міндетті түрде
оның қызметінің де өзгеруіне алып келеді немесе керісінше.
Тарихи даму барысында мүше қызметінің алмасуы үнемі оның
формасының алмасуына үласады. Әдетте, мүшелердің эволюция-
сы оның қызметінің алмасуы арқылы жүзеге асады. Бүл жагдайда
мүшенің бүрынгы негізгі қызметі өз мэнін жойып, оның қосалқы
қызметі маңызды рөл атқаруы мүмкін. Қалай болганда да, барлық
уақытта, мүше қызметінің алмасуы оның күрылысын да өзгертеді.
Мысалы, омыртқалылардың ата-тектерінің тері қабыршақтары
жаққа қарай ыгысып өтіп тіске айналган, қол қанатты балықтардың
торсылдагы эрі қарай дамып ауамен тыныс алу мүшесіне айналган
жэне т.с.с.
Филогенетикалық өзгерістердің пайда болу жолдары ішінен
маңыздылары ретінде:
1) мүше кызметінің кеңеюін;
2) мүше қызметінің күшеюін жэне белсенділігінің артуын;
3) мүше мен оның қызметінің субснтуациясын (алмасуын)
атауга болады.
Мүше қызметінің кеңеюіне мысал ретінде балықтардың көкірек
канаттарының дамуын келтіруге болады. Алгаш олар балык
денесін бірқалыпты, түрақты үстап түру кызметін гана атқарган
болса, кейін терендікке сүңгу жэне жүзу багытының рулі қызметін
де қоса атқарган.
Мүше кызметінің күшеюі оның негізгі қызметтік бөлімдерінің
санының кобеюімен бірге жүреді. Ол мүше өлшемінің осуіне және
гистологиялық күрылымдарының қайта қүрылуына алып келеді.
Мысалы, омыртқалылардың алдынгы миының дамуы, сүт бездері
бөліктерінің көбеюі салдарынан аталган мүшелердің молшерінің
үлгаюын айтуга болады.
204
Мүше қызметі белсенділігінің артуы дегеніміз — бүрын
белсенді емес болып келген мүшелердің белсенді түрге айналуы
болып табылады.
Субситуация немесе мүше мен оның қызметінің алмасуы фило¬
генез қүбылысында бір мүшенің екінші бір мүшемен алмастыры-
луы болып табылады. Алмастырылатын мүше мүлдем жойылып
жоқ болады немесе рудимент күйінде сақталынады.
Мысалы, хорданың омыртқамен алмастырылуы, пронефрос-
тың цезонефроспен, ал оның метанефроспен алмастырылуы т.с.с.
Эволюция барысында бір мүше бастамасының пайда болу
орнының өзгеруі немесе мүшенің дене білеуінен (ось) салыстырма¬
лы түрде ығысып орналасуы да байқалады. Оны гетеротопия деп
атайды. Мысалы, құстар мен сүтқоректілердің жүрегінің көкірек
қуысына ығысуы, жогаргы сатылы омыртқалылардың алдыңгы
аяқтарының балықтардың көкірек қанаттарымен салыстырганда
біршама кейінірек орналасуы т.с.с.
Филогенетикалық өзгерістерге гетерохрониялар да жатады.
Гетерохрония дегеніміз — мүшелердің қалыптасу мерзімінің
өзгеруі.
Эволюция қүбылысында гетеробатмия деген қүбылыс та
байқалады. Гетеробатмия дегеніміз — ағза денесінің не бір
мүшесініц бөлімдерініц әр түрлі эволюциялық даму деңгейінде
болуы, яғни біреулері қарапайым, төменгі деңгейде болса, екінші
біреулері жогарғы децгейде, күрделі болуы мүмкін.
Эволюция қүбылысында бір мүшелер жаңадан пайда болып,
екінші біреулері қүрып жойылып отырады. Жаңа мүшелердің пай¬
да болуының себебі олардыц дифференциялануы (жіктелуі) бо¬
лып саналады, ал оның алғы шарты болып ескі мүшеніц атқара-
тын қызметінің сан алуан (мультифункционалды) болуында.
Мүшелердіц жойылуы (редукциялануы) эволюция қүбылы-
сында алгаш пайдалы, кажет болып келген мүшенің қажетсіз,
тіпті зиянды мүшеге айналуымен байланысты. Мүндай өзге-
рістерге, эдетте сыртқы ортаның өзгеруі алып келеді. Мысалы,
омыртқалылардың дамуында түлга бүйрегі (мезонефрос) пайда
болганнан кейін дененіц алдыцгы бөліміндегі бүйректіц (пронеф-
рос) редукциялануын айтуга болады. Мүшелердің редукцияла¬
нуы олардыц кішіреюі жэне күрылысыныц қарапайымдануымен
205
байланысты. Редукцияланған мүшелер өте өзгергіш келеді, мы¬
салы, соқыр ішектің ұзындыгы 2 см-ден 20 см-ге дейін болады.
Сол сияқты, мүшелердің редукциялану дэрежесі де түрліше болып
келеді. Кейде редукцияланған мүше ересек формаларда да, үрықта
да кездеспей мүлдем жойылып кетеді; кейде ол ересек формаларда
кездеспей тек үрықта ғана, эмбриондық бастама күйінде сақталуы
мүмкін. Кейде ол белгілі бір сатыга дейін қалыпты дамып, содан
кейін дамуын тоқтатып, ересек формаларда жетілмеген күйде
сақталуы мүмкін. Бүл жағдайда ол ешқандай қызмет атқармайды.
Ондай мүшелерді рудгшенттер деп атайды.
7.2. Омыртқалылардың герісінің филогенезі
Омыртқалылардың терісі негізінен қорғаныштык қызмет
атқарады. Сол сияқты, ол тітіркенулерді қабылдайтын, тыныс алу,
зэр шығару кызметтерін де атқаратын мүше болып табылады. Ол
үстіңгі қабат — эпидермистен жэне нагыз тері қабатынан (кориум)
түрады.
Қандауыршалардың терісі нашар дамыған. Эпителииі бір
қабаттан түрады, ал кориум кілегейленген дәнекер үлпалардан
қүрылған.
Омыртқалылардың терілері айқын екі қабатқа жіктелген. Эпи¬
дермис — көп қабатгы, ал кориум негізгі заттардан, талшықтардан
жэне жасушалардан кұрылған. Тері эр түрлі туындыларды пай¬
да болғызады, олардың ішінен негізгілері — қорғаныштық
кұрылымдар жэне бездер.
Балъщтардын терісі өте жүқа қабыршақтармен қапталған.
Эпидермис бір қабатты тірі жасушалардан түрады, онда көптеген
біржасушалы сілемей бездер кездеседі. Кориум қатты талшыкты
қабаттан құрылған.
Космекенділердіц де терісі жүқа, жалаңаш, ылгалды болып
келеді. Эпидермис бірнеше қабаттірі жасушалардан түрады. Онда
көптеген сілемей бездері кездеседі. Кориум талшықты, дәнекер
үлпадан жэне жасушалардан құрылған.
Жоргалауиіылар қүрлық жануарлары болғандықтан олардың
терісі калың, құрғақ және қабыршақтармен қапталған бола-
206
ды. Эпидермис айкын екі қабатка бөлінген, үстіңгі қабаты өлі
мүйізденген жасушалардан, ал астыңғы (мальпигиев қабаты) тірі,
бөлінгіш жасушалардан тұрады. Эпидермистің үстіңгі қабаты
үнемі түлеп түсіп отырады. Ол астыңғы қабат жасушаларының
бөлінуі нәтижесінде қалпына келеді. Жоргалаушылар терісінде
бездер болмайды.
Сүпщоректілердің терісі қалың және күрделі болып келеді.
Оның екі қабаты да (эпидермис жэне кориум) жақсы жетілген.
Эпидермис көп қабатты болып оның үстіңгі қабат жасушала¬
ры мүйізденген өлі жасушалардан, ал астыңғы қабаты тірі, тез
бөлінгіш жасушалардан тұрады. Эпидермистен көптеген туынды-
лар пайда болады. Олар — түктер, тырнақтар, түяқтар. мүйіздер,
қабыршақтар, әр түрлі бездер. Кориум қалың болып, негізінен
талшықты дәнекер ұлпадан тұрады. Кориумның астында тері асты
май қабаты жаксы дамыған.
Сүтқоректілердін ерекше-
ліктерінің бірі — денесінде
қалың түктердің болуы (43-
сурет, в). Оның негізгі қызметі
денені ыстықтан қорғайды,
суды аз буландыру болып
табылады. Ересек адамдар
денесінде алақаны мен табаны-
нан баска жерлерінің бәрінде
редукцияланған түктер кезде-
седі.
Сүткоректілер терісінде көп-
теген бездер — тер бездері, май
бездері, сүт бездері кездеседі.
Тер бездерінің секретгері ку¬
рам ы жағынан сілемейлі не
ақуызды болып келеді, кей¬
де онда май да болуы мүмкін.
Тері бездері зәр шығару жэне
дене температурасын реттеуде
маңызды рөл атқарады.
43-сурет. Құстар мен
сүткоректілердін тері
туындылары.
а) кауырсын, б) кауырсын бастама-
сы, в) түк бастамасы. 1-желпуіш,
2-каламша, 3-эпидермис, 5-бүршік.
6-дерма, 7-тер безі, 8-май безі,
9-өсу аймағы, 10—12-түктің орта¬
сы, 13-бұлшык етті капшык,
14—15-түк қалтасы.
207
Май бездерінің секреттері тері бетін жэне түктерді майлап
тұрады жэне оларды сыртқы орта факторларының әсерлерінен
қоргайды. Май бездері тек сүткоректілерге ғана тэн.
Сүт бездері тер бездеріне гомологиялық мүшелер болып сана¬
лады.
Сонымен, омыртқалылардың терісінің эволюциясы бір қа-
батты эпителийден көп қабатты эпителийге, тері бездерінің,
тері туындыларынын күрделіленуі, жіктелуі бағытында жүріп
отырған.
Адамдардың тері онтогенезінің алғашқы сатыларының бүзы-
луы салдарынан мынадай атавистік ақаулыктар пайда болуы
мүмкін:
гипертрихоз — қүлақ қалқаншасын жүн басуы; полимастия
— емшектің екеуден коп болуы; полителия — емшек емізігі саны-
ның көп болуы, денені түгел түк басуы жэне т.с.с.
7.3. Омыртқалылар қаңқасының филогенеіі
Омыртқалылар қанкасының маңызы өте зор. Біріншіден,
ол зақымданудан қорғайды, екіншіден тірек-қимыл кызметін
атқарады.
Қаңқа 3 бөлімнен түрады: білік қаңқасы (омыртқа жотасы), аяқ-
қол қаңқасы жэне бас сүйектері.
Білік қацқасы (омыртқа жотасы). Эволюция қүбылысында
омыргқалылардың білік қаңкасы біршама өзгеріске ұшыраған.
Алгаш білеу қаңқасы хорда күйінде пайда болып, кейінірек ол
жануарлардыц дамуы негізінде өзгеріп бірте-бірге омыртқа жота-
сымен алмасқан.
Қазіргі кездің өзінде кейбір төменгі сатылы омыртқалыларда
(мысалы, шеміршекті балықтарда) ересек күйінде хорда кездеседі.
Ал, қалған омыртқалыларда хорда тек дамудың бастапқы
кезендерінде болып, омыртқалармен алмастырылған. Омыртқа
жотасы көптеген омыртқалардан түрады. Омыртқа — денеден,
үстіцгі жэне астыңғы доғалардан кұрылган. Қандауырларда хор¬
да өмірінің ақырына дейін сақталынады, бірақ хорда үстінде
буылтықтанып жүптасып орналасқан шеміршекті омыртқа бас-
тамалары пайда болады, оларды устіңгі догалар деп атайды.
208
Төмепгі сатылы бсілъщтарда (шеміршекті балықтар) ұстіңгі
догамен қатар астыңгы догалар да қалыптасады, ал жогары са¬
тылы балыңтарда — сүйекті балықтарда омыртқалардың денесі
де пайда болады. Үстіңгі жэне астыңғы догалар бір үштарымен
омыртқа денесімен бітіседі, ал ұстіңгі доғаның екінші үшы бір-
бірімен қосылып жүлын орналасатын арна пайда етеді. Астыңғы
доғада өскіншелер пайда болып, оган қабырғалар бекінеді.
Сүйекті балықтарда хорда қалдығы омыртқалар араларын-
дағы шеміршек күйінде сақталған. Олардың омыртқа жотасы
2 бөлімге: кеуде жэне құйрық бөліміне бөлінеді. Кеуде бөлімі
омыртқаларының бэріне қабырғалар бекінген, ал қүйрық бөлі-
мінде қабыргалар болмайды.
Қосмекенділердің дамуының бастапқы кезінде-ақ хорда
омыртқалармен алмасады. Олардың омыртқа жотасы 4 бөлімге
бөлінген: мойын, кеуде, сегізкөз жэне қүйрық. Мойын бөлімінде
1 омыртқа, кеуде бөлімінде 5 омыртка кездеседі. Оның бәріне
қабырға бекінген. Қабырғалары кішкентай болып тоске жетпейді.
Сегізкоз бөлімінде 1 омыртқа болады, олжамбасжэне аяқсүйектері
үшін тірек болып табылады. Құйрықты қосмекенділердің қүйрық
бөлімінде көптеген омыртқалар кездессе, қүйрықсыздарында олар
бір сүйекке кіріккен.
Жоргалауиіылардыц омыртқа жотасы 5 бөлімнен түрады: мой¬
ын, кеуде, бел, сегізкөз жэне қүйрық. Мойын бөлімінде 8-ге дейін
омыртқалар кездеседі, оның біріншісі (атлант) сақина тәрізді, ал
екіншісінің оған кіріп тұратын өскіншесі болады. Сондықтан да
жорғалаушылардың басы эр түрлі багытта бүрыла алады. Кеуде
бөлімінде көптеген омыртқалар кездеседі, оларға жақсы жетілген
қабырғалар бекінеді. Қабырғалардың кейбіреулері төспен жал-
ғасады да кеуде қуысын қалыптастырады. Бел омыртқаларына
және сегізкоз бөлімінің алгашкы екі омыртқасына да қабырғалар
бекінеді. Қүйрык бөлімінің омыртқалар саны әр түрлі.
Сүтқоректілерде хорда калдығы омыртқалар арасындағы
шеміршек күйінде сақталынады. Қүйрык бөлімінің омырткалар
саны эр түрлі.
Омыртқа жотасы 5 бөлімге бөлінген: мойын, кеуде, бел, сегіз-
коз жэне қүйрық: мойын бөлімінде 7 омыртқа; кеуде бөлімінде
9—24 (12—13) омыртқалар кездеседі. Кеуде омыртқаларына
209
кабырғалар бекінеді, олардың көпшілігі төспен жалғасқан. Бел
омыртқаларының саны 3—9; сегізкөз бір-бірімен кіріккен 4—5
омыртқадан тұрады. Құйрық омыртқаларының саны тұрақсыз.
Омыртқалылардың филогенезіне сэйкес адамдардың эмб-
риональдық дамуында барлық омыртқаларда қабырга бастамала-
ры пайда болады, кейінірек олар тек кеуде омыртқаларында гана
сақталып дамиды, ал қалғандары редукцияланады. Кейде ең соң-
ғы мойын омыртқасында және бірінші бел омырткасында косым-
ша қабырғалар дамуы мүмкін — бұл атавизмге жатады. Адам-
ның 1,5—3 айлық ұрығында 8—11 бастама омыртқалардан тура-
тын күйрық болады, кейін олардың көпшлігі редукцияланады,
ал калган 4—5 нашар дамыған омыртқалар кірігіп құйымішак
пайда болады. Кейде кұйрық бөлімінің омыртқалары өсуін
тоқтатпайды, бұл үзындығы 20 см-ге жететін құйрықты
балалардың туылуына алып келеді (атавизм).
Сол сияқты білік каңқасының (омыртқа жотасының)
онтогенезінің бүзылуы басқа да атавистік ақаулықтарға алып келуі
мүмкін. Мысалы, омыртқалардың так өскіншелерініц кірікпеуі
салдарынан омырпща арнасыныц кемістігі (дефект) — Spina
bifida туындайды, мұндай жағдайда жырық арқылы жұлын қа-
бығы сыртқа шығып жүлыи жырыгы (грыжа) пайда болады.
Қол-сіяц цаңцасы. Омырткалылардың қол-аяқтарының 2
түрі белгілі: балықтардың жұп қанаттары (кеуде жэне қүрсақ
қанаттары) жэне қүрлық жануарларының 5 саусақты қол-аяқтары.
Балықтардын жүп қанаттарынан кұрлық жануарларының 5
саусақты қол-аяқтары пайда болған. Көптеген балықтардың жүп
қанаттарының қаңқасы ірі шеміршек тақтасынан тұратын ішкі
бөлімнен және радиалды орналасқан көптеген жіңішке сәулелер-
ден қүрылган шеткі бөлімдерден түрады. Жүзбе канат қаңқасы-
ның бөлімдері бір-бірімен қозғалыссыз байланысып біртұтас
жазықтық пайда болады. Ол иық белдеуімен де қозғалыссыз
байланысқан, сондықтан көптеген балықтардың қанаттары тірек
қызметін атқара алмайды.
Керісінше Девон дэуірінде өмір сүріп кұрып кеткен қолңа-
иатты балыцтардың жүзбе канаттарында 5 саусақты қол-аяққа
үқсас прогрессивтік өзгерістер пайда болған (44-сурет). Бүлардың
қанаттарының сүйектері бір-бірімен кіріккен жэне олардың саны
210
азайған. Қанаттың ішкі бөлімінде тек бір ұлкен сүйек (ол тоқпан
жілікке сэйкес келеді), ал шет бөлімінде 2 ірі сүйек сақталған (олар
кэрі жілікке сэйкес келеді), оларға сәулеленіп 7-12 жіңішке сүйек-
тер бекінген (бүлар саусақтарға сэйкес келеді). Сонымен қатар,
олардың қанаттары иық белдеуімен қозғалмалы байланысқан,
сондықтан олар денеге тірек болып, қайырылып қалған жағдай-
ларда жылжып козгалуы үшін де пайдаланылған.
Алғашқы күрлык жануарларының (стегоцефалдар) 5—7 сау-
сақты қол-аяқтары болган. Олардың қаңқасының кұрылысының
жобасы жэне сүйектердің өзара қатынасы қолқанаттылардың
қанаттарына өте ұқсас. Мысалы, қол-аяқтың ішкі бөлімі бір үлкен
сүйектен (тоқпан жілік), одан кейін 2 сүйектен (кэрі жілік), 3—4
катар орналасқан үсақ сүйектерден (білезік сүйектері) жэне сау-
сақ сүйектерінен күрылган. Барлық қүрлык жануарларының кол-
аяқ каңқалары осы жобага сэйкес болып келеді (45-сурет). Тек
омыртқалылардың қол-аяқ қанқасының эволюциясында мынадай
өзгерістерді атауга болады:
44-сурет. Балыктардың жүзбе канатгарынын бессаусакты
кол-аякка айналуы
1. Қаңқаның шеткі бөлімі қарапайымданады, яғни 7 саусақтың
2 шеткісі редукцияланып, 5 саусақ қалады;
2. Білезік сүйектерінің саны азаяды — қосмекенділерде 3 қатар,
жорғалаушылар мен сүтқоректілерде 2 қатар болып келеді;
3. Ішкі бөлімнің сүйектері (тоқпан жілік, кэрі жілік) бірте-бірте
үзарып, созылады.
211
45-сурет. Омырткалылардын алдынғы аяк канкасы.
а) саламандра, б) тасбака, в) крокодил, г) кұс, д) жарканат, е) кит,
ж) көртышкан, з) адам. 1-токпан жілік, 2-шыбык сүйек, 3-шынтак
сүйек, 4-білезік сүйектері, 5-алақан сүйектері, 6-саусак сүйектері.
Бас сүйектері. Омыртқалылардың бас сүйектері 2 бөлімнен
түрады: ми сауыты жэне бет сүйектері. Олар әр түрлі жолмен пайда
болған жэне бір-бірінен дербес дамиды. Эволюция кұбылысында
айтарлықтай өзгерістер бет сүйектерінде байқалады. Жоғары са-
тылы омыртқалылардың ересек күйінде бет сүйектері ас корыту
түтігінің алдыңғы бөлімін қоршап тұратын доғалардан қүралған.
Балықтарда ол 3 доғага жіктелген: 1) жемтікті үстап түратын
жақ доғасы; 2) ми сауытына бекіну қызметін атқаратын тіл асты
доғасы; 3) желбезек жапырақшаларын байланыстыратын желбезек
доғасы.
Жоғары сатылы омыртқалылардың бет сүйектері біршама
редукцияланған — жақ доғасының үстіңгі бөлімі ми сауытының
түбімен бітіседі; тіл асты доғасы редукцияланып, ортаңғы қүлақ
сүйектеріне айналады; 2—3-желбезек доғалары көмекейдің
қалқан тэрізді шеміршегіне, ал 4—5-доғалар көмекейдің баска
шеміршектеріне айналады.
212
Адам қаңқасының құрылысы сүтқоректілердің қаңқасына өте
ұқсас, онда сүтқоректілерде кездеспейтін бірде-бір сүйек бол-
майды. Дегенмен, антропогенез құбылысында адам қаңқасында
біршама өзгерістер қалыптасқан, олардың көпшілігі адамның тік
жүруімен байланысты, мысалы: 1) табанда күмбез дамып, оның
күрылысы өзгерген. Ол үстау қызметін жойып, тек тірек қызметін
ғана атқарады; 2) аяқтың бас саусағының қалған саусақтарға
Караганда өте мықты дамуы; 3) омыртқа жотасының түп-түзу
емес S әріпі — тәрізді иіліп келуі; 4) омыртка жотасы жамбасқа
60° бүрышпен орналасқан; 5) бас сүйегі омыртқа жотасына өзгеше
бекінген.
7.4. Омыртқалылардың ас қорыту жүйесінің филогенезі
Төменгі сатылы хордалылардың (жақсүйексіздер) жақтары бол-
майды, асқорыту түтігі бөлімдерге жіктелмеген; асқазаны болмай-
ды. Желбезек саңылауларымен тесілген жүтқыншақ бірден ішекке
жалгасады. Ішек түтігі түп-түзу, бөлімдерге бөлінбеген жэне қысқа
болып келеді. Бауыр ішектің өсіндісі ретінде дамыған, қарапайым
күрылысты болады; үйқы безі нашар дамыган.
Балықтардың ас корыту жүйесі ауыз қуысынан басталады.
Үстіңгі жэне астыңғы жақтарының жиектерінде көптеген біркелкі
үсак тістер орналасқан, ягни тістері гомодонтты болып келеді.
Бүлардың тістері шайнау қызметін атқармай, тек асты үстап түру
қызметін ғана атқарады. Ауыз қуысында қарапайым тілі болады,
бездер кездеспейді.
Ауыз қуысынан кейін желбезек саңылауларымен тесілген
жүтқыншақ қысқа өңеш, асқазан жэне ішекке жалғасады. Балық-
тардың ішегі аш ішекке жэне тоқ ішекке жіктеліп, ануспен
аяқталады. Ішектің жалпы ұзындығы біршама болады жэне ол
ирелеңдеп имектер туғызады. Ал ішектің ілмегінде үйқы безі
орналасқан, бауыр жақсы дамыған, өт қабы кездеседі.
Қосмекенділердің ауыз қуысы жүтқыншақтан бөлінген, тістері
гомодонтты болып келеді. Ауыз қуысында көптеген үсақ сілекей
бездері кездеседі, бірақ олардың секреттері асқа ешқандай хи-
миялық эсер етпей, тек оны ылғалдап түрады. Ауыз-жұткыншақ
213
қуысына хоаналар, евстахиев түтігі жэне көмекей саңылаулары
ашылады. Ол қуыс эрі қарай өңешке жэне асқазанға жалғасады.
Ішектері ұзын, аш ішекке жэне тоқ ішекке жіктеліп, клоакаға
жалғасады. Тарамдалған ұйқы безі аш ішектің ілмегінде орна-
ласқан, бауыры үлкен болып келеді.
Жоргалауиіылардыц ауыз қуысы жұтқыншақтан толық бө-
лінген. Тістері негізінен гомодонтты болады, бірақ кейбір өліп кет¬
кен формаларының тістері жіктеле бастаған. Тілі жақсы дамыған.
Көптеген үсақ жэне ірі сілекей бездері (тіл асты, тіс асты, ерін
асты) жақсы дамыған.
Жұтқыншақтың, өңештің жэне асқазанның қүрылысы
қосмекенділерге үқсас. Ішектері аш ішекке жэне тоқ ішекке
жіктелген. Аш ішектің тоқ ішекке өтетін жерінде кішкентай
түйық өскінше — бүйен пайда болады. Ішектің жалпы үзындығы
қосмекенділермен салыстырғанда біршама үзын. Ішек аш ішекке
жэне тоқ ішекке жіктеліп, клоакаға ашылады.
Сүтцоректілердің аскорыту жолының жіктелуі өте күрделі. Ол
еріндер, ұрт жэне жақтармен шектелген ауыз алды қуыстан бас-
талады. Бүлшық етті еріндер тек қана сүткоректілерге тэн жэне
олар жемтікті қармау қызметін атқарады.
Сүтқоректілердің тістері күрделі, гетеродонтты болады, яғни
олар күрек тістерге, ит тістеріне, кіші азу тістеріне (премоляр-
лар) жэне үлкен азу тістеріне (молярлар) жіктелген. Олардың
қүрылысы және аткаратын кызметтері түрліше болып келеді.
Сүтқоректілердің тістерінің жалпы саны эр түрлі — төменгі саты-
лы сүтқоректілерде 44, ал приматтардың көпшілігінде жэне адам¬
дарда 32-ге тең: 2, 1, 2, 3.
Сүтқоректілердің ауыз қуысында көптеген үсақ жэне ірі
сілекей бездері кездеседі: тіл асты, тіл түбіндегі, жақ асты, кұлақ
айналасындағы бездер. Жүтқыншаққа мүрын-жұткыншақ жол-
дары, евстахиев түтікшесі, көмекей саңылаулары ашылады.
Сүтқоректілердің асқазаны асқорыту жолының басқа бөлімдерінен
айқын бөлінген, оның кұрылысы әр түрлі жануарларда түрліше
болып келеді. Асқазанның сілемейлі қабатында көптеген асқазан
бездері кездеседі (46-сурет). Ішек — үлтабар, аш ішек, ток
ішек, бүйен және тік ішекке бөлінген. Ішектің жалпы үзындыгы
жорғалаушыларға қарағанда элдеқайда үзын. Адам онтогенезінің
214
бұзылуы салдарынан қалыптасатын асқорыту жүйесінің ата-
вистік ақаулықтары ретінде мыналарды атауға болады: 1) тістер
санының 32-ден аз не көп болуы; артық тістер жақ сүйегінде не¬
месе таңдайда пайда болуы мүмкін; кейде «ақыл тістер» мүлдем
шықпауы немесе кешігіп барып, яғни 25 жастан кейін, шығуы
мүмкін; немесе ол рудимент күйінде кішкентай, жіктелмеген бо¬
луы мүмкін; сиректеу тістер гомодонтты болып бэрі бірдей, үшкір
болып келуі мүмкін, көпшілік жагдайда азу тістердің бетінде 5
төмпешіктің орнына 3 төмпешік дамуы мүмкін.
46-сурет. Омырткалылардың ауыз куысындағы
секреторлык аппарат.
а) царапайым омыріщалыларда, 6) дөңгелек ауыздыларда,
в) балықтарда, г) космекенділерде, 1-сіле.меші жасушалар, 2-сілекей
бездер, д) жоргаіаушыларда, І-сілемей жасушалары, 2—3-тутік
тэрізді сілекей бездері, е) сүпщоректілерде, 2-ақуызды сілекей бездері,
3-сілемей, 4-аралас бездер, 5-ұсақ сілекей бездері.
2) Жұткыншак ақаулықтары ретінде мойынның шеткі саңы-
лаулары (латеральные свищи шеи) жэне мойынның шеткі
215
түйінді көпіршіктерін (кистасы) атауға болады. Біріншісі адам
эмбриогенезіндегі желбезек саңылауларының постэмбриондық
кезеқде сакталуы болып табылады.
Екіншісі поетэмбриондық кезеңде желбезек қапшықтарының
эмбриондық материалдарының сақталуымен байланысты.
3) ішек эмбриогенезінің бұзылуы салдарынан ас қорыту
жолының гипоплазиясы нашар дамуы байқалуы мүмкін, мысалы:
а) ішектердің кысқаруы; б) ішек бөлімдерінің нашар дамуы
не мүлдем дамымауы; в) ас қорыту бездерінің нашар дамуы не
мүлдем дамымауы; г) үйқы безі үлпаларының ішек не асқазан
қабырғасына ығысып орналасуы (гетеротопия); д) ересек адам¬
дарда клоаканың сақталуы. Қалыпты жағдайда 8 аптадан кейін
үрықтарда клоака тік ішекке, несеп жолына жэне үрпінің сыртқы
тесігіне жіктеліп жойылады.
7.5. Омыртқалылардың тыныс алу жүйесінің филогенезі
Төменгі сатылы омыртқасыз жануарлардың (ішеккуыстылар
типі, жалпақ қүрттар типі, жүмыр қүрттар типтерінің өкілдері) ар¬
найы тыныс алу мүшесі болмайды. Бүл агзалардың үлпалары мен
қоршаган орта арасында газ алмасуы дененің үстіңгі беті арқылы
жүзеге асады. Мүндай тыныс алуды диффузияпыц тыныс алу деп
атайды.
Жоғары сатылы омыртқасыз жәндіктерде, барлық омыртқалы
жануарларда газ алмасу қүбылысын арнайы тыныс алу мүшесі —
желбезек, кеңірдек жэне өкпе атқарады.
Желбезектер алғаш рет суды мекендейтін буылтық қүрттар
өкілдерінде, ал трахея мен өкпе насекомдармен өрмекшітәрізділерде
пайда болған.
Төменгі сатылы хордалыларда (кандауыршалар) тыныс алу
қызметін ішек түтігінің алдыңгы бөлімі —жүтқыншақ атқарады.
Оның қабыргасында 100—150 жүп тесіктер немесе желбезек
саңылаулары кездеседі. Тыныс алу мүшесі болып желбезекаралық
перделер саналады. Ол арқылы желбезек артериясы өтеді. Жел¬
безек саңылаулары аркылы су өткенде аталған перделерді
жуып-шаяды. Судағы оттегі артерия кабыргасы арқылы қанға
216
енеді. Қандауыршаның желбезек артериясы қылтамырларға
тарамдалған. Сондықтан да оның оттегі енетін бетінің көлемі аз
болып, тотыгу құбылыстары баяу жүреді. Осыған байланысты
қандауыршалар баяу қозғалып, енжар (пассив) түрде тіршілік
етеді.
Омырткалы жануарлар белсенді түрде тіршілік етуге көшкен
жануарлар болып саналады. Ал, ол тыныс алу мүшелерінде
прогрессивтік өзгерістердің пайда болуымен тыгыз байланысты.
Балықтардың желбезекаралық перделерінде көптеген эпите¬
лий өскіншелері — желбезек жапырақшалары пайда болған. Жел¬
безек артериясы желбезек жапырақшаларында тарамдалған қа-
лың қылтамыр торына айналады. Желбезек жапырақшаларының
дамуы арқасында тыныс алу көлемі үлғайып, желбезек перделе-
рінің саны 4-ке дейін азаяды (47-сурет).
47-сурет. Омырткалылардын өкпесінін филогенезі.
I колқанатты балыктардың жұтқыншағы мен торсылдағы,
II космекенділердің жұтқыншагы мен өкпесі, III өкпе.
А-Б) космекенділер, В) жорғалаушылар, Г-Д) сүтқоректілер.
1-жұгкыншақ, 3-торсылдак капшықтары, 4-кеңірдек камерасы, 5-өкпе
капшыктары, 6-өкпе перделері, 7- трахея, 8-бронх, 9-бронх тармактары,
10-альвеолдар.
217
170-15
Қолқанатгы балықтарда желбезектермен қатар атмосфералық
оттекпен тыныс алатын мүше пайда болады. Ол — торсылдақ.
Оның қабырғасында көптеген қылтамырлар кездеседі.
Қосмекенділер өкпесі қолқанатты балықтардың торсылдагына
гомологиялық мүше болып табылады. Қосмекенділер өкпесінің
қүрылысы өте қарапайым: ол 2 қапшықтан түрады; тыныс алу
көлемі өте кішентай. Өкпе арқылы денеге не бары 30—40%
мөлшерінде ғана оттегі енеді. Сондықтан да қосмекенділердің
негізгі тыныс алу мүшесі болып терісі саналады, онда көптеген
қылтамырлар кездеседі.
Қүрлықта тіршілік етуіне байланысты жоргалауиіылардыц
тыныс алу жүйесі эрі қарай күрделіленген. Олар тек өкпесі
арқылы тыныс алады. Өкпеде көптеген перделердің пайда болуы
нәтижесінде тыныс алу көлемі күрт ұлғайған. Сонымен қатар,
тыныс алу жолдарында да прогрессивтік өзгерістер қалыптасқан:
кеңірдекте шеміршекті сақиналар пайда болған; кеңірдек 2
бронхыға бөлінген; ал ол өкпеде әрі қарай тарамдалған.
Сүтқоректілер өкпесінің қүрылысы өте күрделі. Кеңірдек
2 бронхыға бөлініп өкпеде 3—4 дәрежелі ұсақ бронхыларға
тарамдалған. Бронхылар эрі қарай тарамдалып жүқа түтікшелі
бронхиоларға жіктелген, олардың ұштарында жүка эпителиймен
қапталған ұсақ копіршіктер немесе альвеолдар болады. Әрбір
альвеолдар қабырғалары калың қылтамыр торымен қапталған,
осы жерде газ алмасу кұбылысы жүреді. Өкпедегі альвеолдардың
жалпы саны өте көп, сондықтан тыныс алу көлемі күрт өсіп дене
бетінің көлемінен 50—100 есе артық болады.
Адамдардың тыныс алу жүйесінің онтогенезінің бұзылуы
салдарынан пайда болатын акаулыктарға томендегілерді жатқы-
зуға болады: 1) өңеш пен кеңірдектің (трахеяның) туа біткен
ақаулықтары — эзофаготрахеальды саңылаулар; 2) өкпе үлпасы-
ның альвеолярлық жіктелуінің бұзылуы — бронхының (ауатамыр)
дизонтогенетикалық түйінді көпіршіктері (кистасы); 3) өкпенің
түйінді көпіршіктері тектес гипоплазиясы (жалпы өкпенің нашар
дамуы).
218
7.6. Омыртқалылардың қанайналым жүйесінің
филогенезі
Жануарлар ұлпаларында зат алмасу қүбылысы үздіксіз жүріп
отыруы үшін оған үнемі оттегі пен қоректік заттар жеткізіліп, ал¬
масу құбылысында түзілген керексіз заттар мен көміркышқыл газы
шыгарылып түруы қажет.
Ағзада бұл заттардың алмасуы әр түрлі жануарларда түрліше
жүреді. Төменгі сатылы омыртңасыздарда (ішекқуыстылар,
жалпақ қүрттар) қоректік заттар мен оттегі ұлпа сұйықтыгының
диффузия құбылысы нәтижесінде жеткізіледі. Омыртқасыздар
түрлерінің көпшілігінде ұлпа сүйықтыгы эр түрлі багыттарда ағуы
мүмкін, ал кейбіреулерінің ұлгіаларында арнайы агын жолдары
пайда болып сұйықтық солар арқылы ағады. Осылайша қарапай-
ым агын жолдар пайда болған. Агын жолдарының эрі қарай эво-
люциялануы олардың қабырғасында бұлшық ет үлпасының даму¬
ына алып келген. Бұлшык еттердін қалыптасуы нэтижесінде ағын
жолдары жиырыла алады жэне оның ішіндегі сұйықтык ерекше
ұлпа — қанға айналады. ¥лпаларға оттегі қан жасушаларында
кездесетін ерекше заттар арқылы жеткізіледі. Қанайналым түйық
не ашыц болады. Туйыц ңанаііналым деп канның тек кантамырлар
аркылы агуын айтамыз; ал аіиъщ қанайналым деп кантамырлар
дене куысына ашылатын болса, ягни қанның қантамырлар арқылы
дене куысына құйылып, мүшелерді тікелей шайып тұратын қан ай-
налуды айтамыз.
Қанайналым алғаш буылтык кұрттарда пайда болған. Олар-
дың қан айналымы тұйык болып келеді. Қанайналым жүйесі
арқа жэне күрсақ қантамырлардан жэне оларды байланыстырып
тұратын сақиналы жолдардан тұрады.
Буынаяқтылардыц қанайналым жүйесі ашық. Ол жиырылғыш
көпбүрышты жүректен, артериялардан түрады. Жүрек камера-
ларының арасында қакпақшалар болады. Жүректің бірізділікпен
жиырылуы салдарынан қан қантамырларға келіп жетеді, ал олар
дене қуысына ашылып құйылады. Қан ұлпаларды шайып газ ал-
масады, содан кейін қалпақшаларда болатын жүп тесікшелер
арқылы жүрекке сорылады.
219
Төменгі сатылы хордалылар — қандауыршаның қан айналым
жүйесі тұйық. Жүрегі болмайды, оның қызметін ірі ағын жолдары
— құрсак аортасы (қолқасы) атқарады.
Балықтардың жүрегі 2 камералы (бір жүрекше, бір қарынша)
болып, бір қанайналым шеңбері кездеседі. Жүректе вена қаны бо¬
лады. Ол желбезектерге жеткізіліп, оттегімен қаныгып, бүкіл де-
неге таралады, ал жүрекке қан көктамырлар (вена) арқылы келіп
құяды.
Қосмекенділердің жүрегі 3 камералы — 2 жүрекше, бір
қарынша. Екі жүрекше қарыншага ортақ бір тесік аркылы ашы¬
лады, сондықтан да қарыншада артерия қанымен вена қаны ара-
ласады. Қосмекенділерде өкпесі арқылы тыныс алуына байланы¬
сты, кіші қанайналым шеңбері қалыптасқан. Сол жақ жүрекшеде
өкпеден келіп қүйылған (өкпе венасы) артерия каны, ал оң жак
жүрекшеде дене мүшелерінің үлпаларынан келіп кұйылатын
(қуыс көктамырлар) вена қаны болады. Қарыншада 3 қан бола¬
ды: оның сол жақ бөлімінде — артерия қаны, оң жақ бөлімінде
— вена қаны; ал орта бөлімінде — аралас қан. Қарыншадан бір
ірі тамыр — артерия конусы басталып, бірден 3 жүп тамырларга
тарамдалады: 1) тері-өкпе аргериясы; 2) қолқа (аорта) доғасы жэне
3) күретамыр. Тері-өкпе артериясы вена қанын теріге жэне өкпеге,
аорта догасы аралас қанды бүкіл дене мүшелеріне, күретамыр ар¬
терия канын мига жеткізеді.
Жорғалаушылардың қанайналым жүйесі бүдан да эрі қарай
күрделілене түседі. Олардың жүрегі 3 камералы, бірақ жүрекше-
лер бір-бірінен толық бөлінген жэне қарыншаға әрқайсысы
жеке тесікшелер арқылы ашылады. Қарынша жартылай перде
арқылы оң жэне сол жаққа жартылай бөлінген, ал крокодилдердің
жүрегінің қарыншасы толық екіге бөлінген. Сондықтан да
жорғалаушылардың қаны аз мөлшерде араласады.
Артерия баганасы 3 тамырга бөлінген жэне олар карыншадан
өз бетінше дербес басталады. Қарыншаның сол жагынан қолқа-
ның (аорта) оң жақ догасы басталады, ол арқылы артерия қаны
агады. Одан миына жэне алдыңгы аяқтарга баратын тамырлар
таралады. Қарыншаның орта бөлімінен, ягни жартылай перде
үстінен, аортаның сол жақ догасы басталады. Ол аралас қанды
бүкіл денеге таратады. Қарыншаның оң жагынан өкпе артериясы
басталып, вена қанын өкпеге жеткізеді.
220
Сүтқоректілердің жүрегі 4 камералы, бір-бірімен байланы-
спайтын 2 бөлікке бөлінген, сондықтан артерия қаны мен вена
қаны араласпайды. Жорғалаушыларда кездесетін қолқаның сол
жақ доғасы редукцияланады, қарыншадан 2 тамыр ғана баста¬
лады: оң жақ қарыншадан өкпе артериясы, сол жақ қарыншадан
қолқаның сол жақ догасы. Ол арқылы дене мүшелеріне,
үлпаларына оттегімен қаныққан артерия қаны жеткізіледі. Со-
нымен омыртқалылардың қан айналым жүйесінің эволюция-
сы жүрек камералары санының көбеюі (2 камералы, 3 камера¬
лы, 4 камералы), қан тамырларының жіктелуі жэне қанда оттегі
мөлшерінің өсуі (жогарылауы) бағытында жүреді.
I II III IV V
48-сурет. Омырткалылардын жүрегінін жэне артерия
доғаларынын күрылысы.
I балыктар, II космекенділер, III жорғалаушылар, IV кұстар,
V сүткоректілер.
Адамның көптеген артериялары желбезек артериясының ба-
стамаларынан дамиды. Омыртқалылар өкілдерінің эмбриондық
даму кезеңінде жүтқыншақ айналасындағы қантамырлардың
калыптасуы бірдей жобамен жүреді. Жүректен алға қарай тақ
тамыр-қүрсак қолқасы басталады, ал одан желбезек перделеріне
6 жүп (балықтарда 6—-7 жүп) ірі тамырлар тарамдалып, жұт-
қыншақты қоршап, арқа бөлімінде 2 тамыр бір-бірімен қосылып
арқа қолқасын қалыптастырады. Бүл тамырлар артерия немесе
желбезек догалары деп аталады.
221
Балықтардың ұрықтарында алдыңгы 2 жұп тамырлар тез арада
жойылады, себебі олар бас сүйегінің бет бөлімінің құрамына енеді,
қалған 4—5 ясүгт артерия тамырлары желбезек артериясы ретінде
қызмет етеді.
Қүрлықта тіршілік ететін омыртқалыларда да, балықтар
сияқты, алдыңғы 2 жүп артерия тез жойылады, ал қалған 4 жүбы
атмосферамен тыныс алуға байланысты ерекше өзгерістерге
үшырайды. Желбезек доғасының үшінші жүбы басқа жұп аргерия-
лармен байланысын үзіп, арқа аортасының тамырымен қосылып,
күретамырға (сонная артерия) айналады. Төртінші тамырлар
өте жақсы дамып арқа аортасының тамырларымен бірге ересек
күйінде қанайналымның үлкен шеңберінің негізгі тамырлары
болып саналады. Қосмекенділер мен жорғалаушыларда төртінші
жүп тамырларының екеуі де бірдей дамыған, ал сүтқоректілерде
сол жақ тамыр жойылады. Желбезек доғасының бесінші жүп та¬
мырлары редукцияланады, 6-жұбы өкпе артериясы на айналып
аорта доғаларымен жэне басқа да қантамырлармен байланысын
үзеді. Сонымен, қүрлықта тіршілік ететін омыртқалылардың
күретамырлары балықтардың желбезек доғасының үшінші
жүбына, аорта доғалары төртінші жүбына, өкпе артериясы алтын-
шы жүбына гомологиялык мүше болып саналады.
Жүректің дамуы. Адамдардың жүрегінің дамуында ататек-
теріне тән кейбір белгілер қайталанады: алғаш 2 камералы, содан
кейін 3 камералы (2 жүрекше, 1 қарынша) содан барып 4 камералы
жүрек дамиды.
Адам жүрегінің дамуында жиі кездесетін ауытқулар ретінде
төмендегі акаулықтарды атауға болады: 1) сиректеу жағдайда
нәрестенің туылу уақытында екі камералы жүректің сақталуы
мүмкін, мүндай нәрестелер өмір сүре алмайды; 2) жүрекше аралық
перденің кемістігі (1/1000 туылуға); 3) карынша аралық перденің
кемістіктері (2,5—5/1000 туылуға) жиірек кездеседі; 4) жүректің
мойын эктопиясы (мойынға ығысып орналасуы) — нәресте туыла
сала өліп қалады.
Қан тамырлардың дамуының атавистік ақаулықтарына мына-
лар жатады:
1) Қолқаның екі догасының да сақталуы (1/200 өлі нәрестелерде
кездеседі).
222
2) Салатамыр (артерия) не боталлов өзегінің сақталуы (0.5-
7.2/1000 туылуға) — арқа қолқасының түбімен сол жақтағы
4—6 салатамыр жүптарының кірігуі. Бұл кезде артерия каны
қанайналымының үлкен шеңберінен кіші қанайналым шеңберіне
құйылады.
3) Алғашқы эмбрионалдық діңгектің (ствол) сақталуы —
жүректен тек бір тамыр бастау алады — бүл өлімге алып келеді.
4) Тамырлар транспозициясы — қолка мен өкпе артериялары-
ның орын алмасуы, ягни қолқаның он жақ карыншадан, ал окпе
артериясының сол жақ қарыншадан басталуы — өлімге алып
келеді.
5) Үстіңгі екі куыс көктамырдың (вена) сақталуы. Егер екеуі
де оң жак жүрекшеге құйылса ауру байқалмайды, ал сол жақ қуыс
вена сол жақ журекшеге, оң жақ қуыс вена оң жақ жүрекшеге
құйылса үлкен қан айналым шеңберіне веналық кан келіп
құйылады: кейде екі қуыс вена да сол жақ жүрекшеге құйылуы
мүмкін — бүл жүрек қантамыр жүйесінің ақаулықтарының 1 %-га
жуығы. олар өлімге алып келеді.
7.7. Омыртқалылардың жүйке жүйесінің филогенезі
Жануарлардың жүйке жүйесі эктодермадан қалыптасады.
Оның негізгі қызметі тітіркенулерді қабылдау жэне өткізу болып
саналады.
Омыртқасыздардың жүйке жүйесі қарапайым — диффутя-
лық жуііке жүхіесі деп аталады; ол жіңішке өскіншелермен өзара
торланып байланысқан жүйке жасушаларының жиынтыгы болып
табылады.
Жүйке жүйесінің эрі қарай эволюциялануы жүйке жасуша-
ларының дененің бір жеріне шоғырланып жүйке түйіндерін пай¬
да етуіне алып келген. Мысалы, жалпақ жэне жүмыр күрттарда
жүйке түйіндері күрсак жэне арқа бөлімдерінде бір-бірімен бай-
ланысып, жүтқыншақ айналасында сақина пайда етеді; одан 3
жүп баганалар таралады; буылтық құрттарда жүйке тізбегі пайда
болады, ягни денесінің әрбір буылтыгында жүп жүйке түйіндері
кездеседі жэне олар бір-бірімен көлденең және үзына бойына
орналасқан тізбектер арқылы жалганган.
223
Хордалылардың жүйке жүйесінің дамуы бірнеше сатылардан
өтеді: алғаш жүйке тацтасы пайда болып, ол жүйке нсіуасына,
содан кейін жүйке түтігіне айналады.
Хордалылардың қарапайым өкілдерінде — (дөңгелекауыз-
дылар) жүйке жүйесі жіктелмеген, жүйке түтігінен ғана түрады,
ал барлық омыртқалылар онтогенезінде жүйке түтігінің бас
жағында алғаш 3 ми көпіршігі пайда болады: алдыңгы, ортаңгы
жэне аріщы көпіршіктер. Әрі қарай алдыңгы көпіршік екі бөлімге
бөлінеді де одан алдыңгы ми жэне аралық ми қалыптасады,
ортаңғы көпіршіктен ортаңгы ми түзіледі, артқы ми көпіршігі де
екіге бөлініп мишық жэне сопақша миды дамытады.
Омыртқалы жануарлардыц миы бес бөлімнен тұрады: алдынгы
ми, арстъщ ми, ортацгы ми, мишъщ және сопацила ми. Бірақ, әр
түрлі жануарларда бүл бөлімдер түрліше дәрежеде дамыған (49-
сурет).
Бальщтардыц миының қүрылысы өте қарапайым, көлемі
жағынан кішкентай және мидың алдыңгы бөлімдері нашар
дамыған. Алдыңғы ми басқа ми бөлімдерімен салыстырғанда өте
кішкентай, ми сыңарларына жіктелмеген, онда жүйке жасуша¬
лары болмайды, тек екі иіс сезгіш талшыктан түрады. Аралық
мидың үстіңгі бетінде эпифиз, ал астыңгы бетінде гипофиз безі
жэне гипоталамус орналасқан. Ортаңғы ми салыстырмалы түрде
жақсы дамыған, онда көру орталығы орналасқан. Сонымен қатар,
ортаңгы ми интегративтік (үйлестіруші) қызмет атқарады, ми
бөліктерінің қызметтерін реттейді. Сол сияқты балықтардың
мишығы да жақсы дамыған. Алдыңғы бөлімдеріне қараганда
артқы бөлімдері жақсы дамыған миды — ихтиопсидті ми типі
деп атайды.
Қосмекенділердің миының көлемі балыктарга қарағанда
біршама үлкен және жақсы дамыған, әсіресе, алдыңғы миы.
Алдыңгы ми екі ми сыңарларына бөлінген жэне оның үстіңгі
бетінде жүйке жасушалары кездеседі. Алдыңғы мидың түбінде
жолақты дене орналасқан. Алдыңгы ми иіс сезу қызметін
атқарады. Аралық ми біршама дамыған. Оның үстіңгі қабаты —
эпифиз, ал астыңгы қабатында — гипофиз орналаскан. Ортаңгы
ми салыстырмалы түрде үлкен, бірақ балықтардың ортаңгы ми-
ымен салыстырганда кішілеу болып келеді. Ортаңгы мида көру
224
орталығы орналасқан және ол мидың бөлімдерін үйлестіруші
бөлім болып саналады.
Қосмекенділердің мишығы нашар дамыған. Сонымен қосме-
кенділердің бас миы да ихтиопсидті ми типіне жатады.
Жоргалауіиыпардыц алдыңғы миы басқа бөлімдеріне кара-
ганда едәуір үлкен және жақсы дамыған болады. Оның көлемінің
үлкеюі негізінен жолақ денелер немесе астыңғы қабатының
дамуына байланысты. Алдыңгы мидың үстіңгі бетінде алғаш
рет ми қыртысы пайда болады, ол ми сыңарларын түгел жауып
тұрмайды, тек шетінде екі аралшық күйінде жауып тұрады. Ми
қыртысының қүрылысы карапайым, оны ежелгі ми қыртысы
(архикортекс) деп атайды. Алдыңғы мидың жолак денелері
жақсы дамыған жэне ол жүйке жүйесінің үйлестірушісі қызметін
атқарады. Мүндай миды заурапсидті ми типі деп атайды.
Ортаңгы ми көлемі кішірейіп, ол үйлестірушілік қызметін жояды.
Мишық жақсы дамыган.
49-сурет. Омырткалылардын миы.
а) сүйекті балыктар, б) скат, в) күрбака, г) жорғалаушылар, д) кұстар,
е) сүтқоректілер. 1-алдыңгы ми, 2-эпифиз, 3-гипофиз, 4-ортаңгы ми,
5-мишык, 6-сопакша ми, 7-аралык ми, 8-мантия, 9-жолак дене.
225
Сүтқоректілердің миының көлемі өте үлкен жэне құрылысы
қүрделі, эдетте алдыңғы ми өте жақсы дамыған. Алдыңгы мидың
дамуы астыңғы бетіне емес, үстіңгі бетінің мықты дамуына
байланысты. Ми сыңарларының бетін түтас ми қыртысы жауып,
қаптап тұрады. Оны жаңа ми қыртысы (неокортекс) деп атайды.
Жаңа ми қыртысы көп қабатты, эр түрлі жүйке жасушаларынан
қүрылған.
Төменгі сатылы сүтқоректілердің ми қыртысы тегіс, ал жоғары
сатылы сүтқоректілерде қатпарлар пайда болып, оның жалпы
көлемі күрт өседі. Ми қыртыстары ми бөлімдерінің қызметін
үйлестіруші болып табылады, онда сезу, көру, есту, қозғалу
орталықтары, сол сияқты жоғары жүйке қызметімен байланысты
орталықтар орналасқан. Мұндай ми типін маммальды ми типі
деп атайды. Аралық мида эпифиз, гипофиз бездері кездеседі.
Ортаңғы ми кішкентай, онда кору және есту болімдері болады.
Мишық жақсы дамыған.
Адамның оргалық нерв жүйесі жеке даму қүбылыстарын
үйлестіруші, басқарушы мүше болгандықтан, оның көптеген
ақаулықтары ағзаның оліп қалуына алып келетіні сөзсіз. Адамның
жүйке жүйесінің ақаулықтары:
— жүлынның ақаулығы ретінде рахисхиз (платиневрия) атауға
болады. Бүл нерв түтігі жиектерінің кірікпеуі.
— алдыңғы мидың ақаулығы — прозэнцефалия — ми сыңар-
ларының бөлінбеуі жэне қыртыстың дамымауы. Жогарыда
келтірілген екі ақаулық та агзаның өліп қалуына алып келеді.
— агирии — ми жүлгелерінің болмауы және олигогирия —
ірі жүлгелер санының аз болуы.
7.8. Омыртқалылардың зәр шығару жэне жыныс
жүйелерінің фнлогенезі
Ішекқуыстылардың арнайы зэр шығару мүшесі болмайды.
Олардын денесі екі қабатты болып диссимиляция өнімдері сыртқа
диффузиялық жолмен шығарылады.
Зэр шығару мүшесі алғаш рет жалпақ қүрттарда кездеседі, ол
протонефридиялар күйінде болады. Жүмыр қүрттарда өзгерген
226
протонефридиялармен катар зәр шығару қызметін кейбір алып
тері жасушачары да атқарады. Буылтық құрттардың зәр шығару
мүшесі бөлшектенген метанефридиялар күйінде болады. Әрбір
буылтықта жұп метанефридиялар орналасқан. Олар целомға ашы-
латын воронкадан жэне одан басталатын зэр шығару тесігінен (по-
расынан) тұрады.
Буынаяқтылардың зэр шығару жуйесі түрі өзгерген метанеф-
ридиялардан, ерекше түтікшелерден (Мальпиги түтіктері) жэне
майлы денеден тұрады.
Омыртқалылардың зәр шығару мүшесі болып бүйрек сана¬
лады. Бүйректер сыртқы пішіні жагынан омыртқасыздардың
жэне төменгі сатылы хордалылардың нефридияларынан ерек-
шеленген, бірақ кұрылысы жағынан оларга ұқсас болып келеді.
Буйректердің негізгі қүрылымдық бірлігі болып, нефридиялар
сияқты дене қуысына ашылатын воронка (шанақ) жэне одан ба¬
сталатын зэр шығару арнашықтары саналады. Бүйректің барлык
зәр шығару арнашықтары бір ортак арнаға — несеп ағар жолына
кұйылады.
Төменгі сатыдағы омыртқалылар (балықгар, космекенділер)
эмбриогенезінде дененің алдыңғы (бас) болімінде алғашқы буй-
рек, (пронефрос), одан кейін тұлға бөлімінде бірінші реттік буй-
рек (мезонефрос) калыптасады. Жоғарғы сатылы омыртқалыларда
буйректің аталған 2 түрінен баска дененің жамбас бөлімінде екін-
ші реттік, соңғы бүйрек, (метанефрос) қалыптасады. Бүйректер-
дің бірізділікпен қалыптасуында алғаш пайда болғандары бірте-
бірте редукцияланып жойылады (50-сурет).
Пронефростың немесе бас бүйрегінің қүрылысы өте
қарапайым. Ол барлық омыртқалылардың эмбриогенезінің
алғашқы сатыларында дененің бас бөлімінде қалыптасады. Ол
не бары 6-12 нефроннан қүрылған. Нефрон целомға ашылатын
кірпікшелі воронкадан (нефростом) басталып, қысқа жэне түзу
зэр шығару арнашыктары арқылы ортақ несеп жолына келіп
қүяды. Ал, несеп жолы омыртқа жотасын бойлай отыра клоакаға
ашылады. Воронка жанында, целом пердесініц сыртында бірнеше
қылтамырлы шумақ дамиды. Диссимиляция өнімдері шумақтан
дене қуысындағы сүйықтыкка келіп енеді, сосын онымен арала-
сып нефростомға, арнашықтарға, несеп жолына келіп күйылады.
227
Пронефростың қарапайымдылығы қан айналу жүйесі мен зэр
шығару жүйелері арасында тікелей байланыстың болмауында бо¬
лып саналады.
Қазіргі кездегі омыртқалыларда пронефрос тек эмбриональдық
даму кезінде ғана байқалады, ал ересек күйінде ол тек дөңге-
лекауыздыларда ғана кездеседі.
Мезонефрос омыртқалылар бүйрегінің эволюциясының келесі
кезеңі болып табылады. Ол дененің түлға бөлімінде қалыптасады.
Оның нефрондарының қүрылысы біршама күрделенген. Зэр
шығару арнашықтарының арқа (үстіңгі) бетінде екі қабатты
тостағанша тэрізді өскінше (капсула) пайда болады. Осы кап-
сулада қылтамырлы шумақ орналасып, онымен бірге бүйрек
денешігін түзеді. Осының арқасында қан айналу жүйесі мен зэр
шыгару жүйелері арасында тікелей байланыс орнайды. Енді дис¬
симиляция өнімдері дене қуысына құйылмай, тікелей бүйрекке
өтеді. Сондықтан да ыдырау онімдері ағзадан толық жэне тез
шығарылады. Алғашқы бүйректің воронкасы өз қызметін жоя-
ды және кейбір нефрондар бірте-бірте редукцияланып жойыла¬
ды. Нефронның прогрессивтік дамуының тағы бір көрінісі ол
зэр шығару арнашықтарының әжептеуір үзарып созылуы және
имектеліп бірнеше бөлімдерге жіктелуі. Осының нәтижесінде
алгашкы бүйректің зэр шығару арнашықтарында су, глюкоза, т.б.
кері сүзіліп. несеп концентрациясы күшейе түседі.
Алғашкы бүйрек ересек күйінде балықтарда, космекенділерде
қызмет етсе, жогары сатылы омырткалыларда (жоргалаушылар,
қүстар, сүтқоректілер) тек эмбриондық даму кезінде кездеседі.
Метанефрос жогары сатылы омыртқалыларга тэн. Оныц
нефронының қүрылысының ерекшелігі — воронкасының мүлдем
болмауы, сондықтан да дене куысымен байланыс біржолата
үзілген.
Нефрон тікелей бүйрек денешігінен басталады. Зэр шыгару
арнашыгы 3 бөлімге жіктеледі: проксималды арнашық, дис-
тальды арнашық жэне нефрон имегі (Генле имегі). Шумақты
аппарат қарапайымданады, бірақ арнашықтар қабыргалары
жасушаларының ыдырау өнімдерін арнашыққа сүзу қызметі
күшейеді. Осыныц нэтижесінде жамбас бүйрегінде диссимиля¬
ция өнімдері 2 жол арқылы: шумақтагы қан плазмасыныц сүзілуі
228
аркылы жэне арнашықтар жасушаларыныңзаттарды өткізуі кезінде
сүзіледі. Сонымен қатар арнашықтарда кері сүзілу қүбылыстары да
қатар жүріп отырады. Капсула қуысындағы несепте ағза үшін пай¬
далы көптеген заттар — қанттар, витаминдер, амин қышқылдары,
хлоридтер т.с.с. болады. Олардың көпшілігі арнашықтар арқылы
тасымалданғанда қанға кері сорылады. Соңғы буйректе 1 ООО 000-
га жуық нефрондар болады.
50-сурет. Нефрон эволкшиясы.
а) пронефрос, б) мезанефрос, в) метанефрос, г) адам бүйрегі.
1-нефростом, 2-шығарушы арнашык, 3-зәр шыгару арнасы.
4-кылтамырлар, 5-целом, 6-бүйрек шумағының капсуласы, 7-иілген
арнашык, 9-нефрон имегі.
Зэр шығару жүйесінің эволюциясының негізгі бағыты бүйрек-
тің зэр шыгару беттерінің өсуі, яғни алмасу өнімдерін тез жэне
толық шыгару болып табылады.
229
Омыртқалылардың зэр шығару жүйесі жыныс жүйесінің мүше-
лерімен байланысқан. Көптеген омыртқалыларда пронефростың
және алгашқы бүйректің бөлімдері жыныс бездерінің өнімдерін
шығару қызметін атқарады.
Омыртқалылардың жыныс бездері мезонефростың шеткі
жиектерінде жүп қатпарлар күйінде қалыптасады. Гонадалар ба-
стамалары қалың эпителийден жэне көптеген дэнекер үлпалардан
түрады. Алғаш аталық жэне аналык бездердің қүрылысы бірдей
болады. Кейінірек олар маманданып, әр түрлі жыныстарда зәр
шыгару жүйесінің түрліше бөлімдерімен байланысады.
Омыртқалыларда алғашқы бүйрек (мезанефрос) қалыптасқан
кезде пронефрос арнасы екі арнаға бөлінеді, оның бірін — Во.пьф
арыасы, екіншісін — Мюллер арнасы деп атайды. Вольф арна¬
сы алғашқы бүйректің нефрондарымен байланысып зэр шыгару
қызметін атқарса, Мюллер арнасы алдыңгы жагымен пронефро-
стын бір нефронымен байланысып жүмыртқа жолын (яйцевод)
пайда етеді, ал артқы жагымен клоакага ашылады.
Омыртқалылардың бэрінде жыныстарына қарамай, міндетті
түрде Вольф жэне Мюллер арналары түзіледі, бірақ олардыц эрі
қарайгы тагдырлары эр түрлі жыныстарда жэне эр түрлі класс
өкілдерінде түрліше болады.
Балыктардыц жэне қосмекенділердің үргашы дараларында
Вольф арнасы несепагар қызметін, Мюллер арнасы жұмыртқа
жолы (яйцевод) кызметтерін атқарады. Ал, еркек дараларында
Мюллер арнасы редукцияланып, Вольф арнасы несепагар жэне
жыныс қызметтерін коса атқарады.
Жоргалаушылар мен сүтқоректілерде Вольф арнасыныц
көптеген бөлімдері несеп шыгаруга катынаспайды, еркек дара¬
ларында ол шэует шыгару қызметін гана атқарады. Мюллер ар¬
насы толық редукцияланады. Ұргашы дараларда Вольф арнасы
редукцияланып, Мюллер арнасы жүмыртқа жолына, жатыр жэне
қынапқа жіктелген.
Жогары сатылы омыртқалылардыц жүмыртқа жолыныц
бөлімдері бір-бірімен эр түрлі децгейде бірігеді. Кейбіреулерініц
қынап бөлімі бірігіп, жатыр жүп күйінде болады (кемірушілер),
кейбіреулерінде жатырлар түп жагымен бірігіп қуыс пайда бола¬
ды да, үстіцгі жагы кірікпей дербес болады — мүны қос мүйізді
230
жатыр деп атайды (жыртқыштар, жүптүяқтылар), үшінші біреуле-
рінде жатырлар бір-бірімен түтас бірігіп, жай жатыр туғызады
(маймылдар, адамдар, жарқанаттар).
Адам эмбриогенезінің алғашқы кезендерінде жүп Вольф және
Мюллер арналары калыптасып, кейін олар эр түрлі жыныстар-
да түрліше редукцияланады. Ер адамдарда Мюллер арнасының
рудименті қуық безде еркек жатырша күйінде кездеседі.
Адамдардың бүйрегінің ақаулықтарына төмендегілерді жатқы-
замыз:
1) мезонефрос сакталып соңғы бүйректің (метанефрос) біреу-
інің болмауы — өте сирек кездеседі;
2) бір немесе бірнеше несепағары болатын бунақталған соңғы
бүйректің (метанефрос) дамуы;
3) бүйректің жамбас куысында орналасуы, бүл ұрық дамуы-
ның 2—4 айында бүйректің бел бөліміне ығысуының бүзылуы
салдарынан болатын ақаулық.
8. АНТРОПОГЕНЕЗ НЕГІЗДЕРІ
Антропогенез деп саналы адам түрінің пайда болуы мен оныц
қоғамдық өмірінің қалыптасу процестері жиынтығын атайды. Бұл
процестің қозғаушы факторларын, бағыттарын, заңдылыктарын
зерттейтін ғылымды антропология деп атайды. Антропология —
адамдар туралы гылым. Ол адамдардыц шыгу тегін, тарихи даму¬
ын жэне тұрпатын (конституциясын), өсуін жэне жеке, жастык,
жыныстық өзгергіштігін, нәсілдерге бөлінуін т.б. зерттейді.
Адамның өзінің табигаты жагынан биологиялық жэне әлеу-
меттік мэні бар. Адамдардыц кұрылысы, тіршілігі жануарларга
үқсас жэне биологиялық зандылыктарга багынады.
Адамдар — нагыз саналы адам (Homo sapiens sapiens) деп атала-
тын түрге, адам (Ношо) туысына, гоминидтер түқымдасына, при-
маттар отрядына, сүтқоректілер класына, хордалылар типіне жа-
тады. Сонымен қатар, адамдар тек қогамдық өмірдіц нәтижесінде,
қогамдық ортада гана адам болып калыптасқан. Адамды адам ет-
кен — ецбек пен қогамдық өмір.
Антропология гылымы бірнеше бөлімдерге бөлінеді: 1) антро¬
погенез — адамдардыц пайда болган жерін, уақытын, нақтылы
ата-тегін, адамның пайда болуыныц негізгі кезецдерін, қозгаушы
күштерін, адамдардыц алгашқы қогамыныц жэне сойлеуініц
дамуын т.б. зерттейді; 2) адам морфологиясы — адамдардыц
қалыпты сыртқы қүрылысын зерттейді; 3) адам физиология-
сы — адамдардыц қалыпты физиологиялық жэне биохимиялық
процестерін жэне олардыц көптүрлілігін зерттейді. Мысалы, по¬
пуляция қан тобыныц, гемоглобинніц көптүрлілігін, адам түрпаты-
мен сыртқы орта факторларыныц өзара байланысын, агзаныц әр
түрлі ауруларга бейімділігін т.б. зертгейді; 4) этникалық антрополо¬
гия — адам нәсілдерін, олардыц таралу ерекшеліктерін зерттейді.
Антропология гылымыныц негізгі зерттеу эдістері: 1) антропо¬
метрия — өлшеу; 2) деректерді вариациялық статистика эдісімен
өндеу (биометрия) болып табылады.
Антропология гылымынын медицина үшін теориялық жэне
практикалық мацызы зор: 1) адамдардыц шығу тегін, эволюция-
сын түсіндіреді. Атавистік жэне рудименттік мүшелерді зерттеп
кейбір ауруларды анықтауға көмектеседі; 2) кейбір этникалық
варианттардыц, адамныц физикалық дене құрылысыныц ерек-
шеліктерін түсіндіреді.
Адамныц дене тұрпаты немесе конституциясы туралы түсінік
ерте кезден-ақ қалыптасқан. Мысалы, Гиппократ адамдарды дене
құрылысына қарай бірнеше топқа бөлген: 1) күшті тип; 2) тыгыз
тип; 3) кұргақ тип; 4) ылгал тип; 5) майлы тип. Бұл типтердіц
әрқайсысы түрліше аурулармен ауруға бейімді болып келген.
Гален адамдар конституциясын 4 топқа бөлген жэне оларды
белгілі бір сұйықтықпен байланыстырган: 1) қан; 2) сұйық сілемей;
3) от; 4) қара от. Осы сұйықтықтардыц болуы эр түрлі тұрпаттагы
типтіц қалыптасуына алып келеді: 1) холерик — жеңілтек, қызба
адамдар; 2) сангвиник — көцілді, тез қимылдайтын адамдар; 3)
флегматик — тұрақты, сабырлы адамдар; 4) меланхолик — өте
сылбыр, баяу қимылдайтын адамдар.
Адам конституциясы тек морфологиялык-физиологиялық
ерекшеліктермен шектеліп қоймай, ол адамдардын мінез-
кұлқын, оныц ауруларга бейімділігін, зат алмасу процестерініц
ерекшеліктерін де калыптастырады.
Адамныц бойы, салмагы жэне көкірек куысы көлемініц
арақатынасына негізделінетін физикалық даму индексіне сүйеніп
М.В. Черноруцкий адамдарды 3 конституциялык типке бөлді: ас-
тениктер — жіцішке, ұзын бойлы; пормостениктер -— орта бой¬
лы, мыгым денелі; гиперстениктер -— аласа бойлы, өте мыгым,
кец иықты, қысқа мойынды.
Француз галымы Д. Сиго адамдарды 4 конституциялык
топка бөлді: 1) тынысалушы; 2) асқорытушы; 3) бұлшықетті;
4) церебральді типтер. Неміс галымы К. Кречмер адамдарды
астеникалъщ, пикникалъщ, атлеттер типі деп бөлді.
Адамдардыц пайда болуы туралы 2 теория белгілі: Ч. Дарвин-
ніц биологиялык теориясы жэне Ф. Энгельстіц ецбек теориясы.
Адамдардыц жануарлар дүниесінен шыққандыгын алгашқы-
лардыц бірі болып Ч. Дарвин дэлелдеген болатын. Ч. Дарвинніц
теориясын симиалъдык, теория деп атайды. (Симиа — маймыл).
233
170-16
Ол адамдар ертеде тіршілік еткен жогары сатылы маймылдардан
келіп шыққан деп ойлаган. Шынында да, адамдардың анатомиялық
құрылысы жэне дамуы, биохимиялық жэне физиологиялық
көрсеткіштері қазіргі кездегі адам тэрізді маймылдарға (шим¬
панзе, горилла) өте ұқсас. Ч. Дарвин өзінің теориясында
антропогенездің қозгаушы күші ретінде тек биологиялық фактор-
ларды — табиги сұрыптауды гана қарастырган.
XIX гасырдың 70-жылдары Ф. Энгельс антропогенездің ең-
бек теориясын қалыптастырды. Оның негізгі мэні бойынша ант¬
ропогенез процесінде адамның прогрессивтік эволюциясының
жэне тарихи дамуының негізгі факторы болып — еңбек, қогам-
дық өмір, сөйлеу, ягни әлеуметтік факторлар саналады. Еңбек
кұралдарын жасау және пайдалану, бұл қызметтіц мэнін түсініп
ұгу адамдардың табигатқа тигізетін эсерінің еселеп өсуіне алып
келіп, бірте-бірте эволюцияныц биологиялық факторларын
ыгыстырып шыгарган. Антропогенез құбылысының органика-
лык дүниенің дамуынан бірден-бір сапалық ерекшелігі де осында
болып саналады. К. Маркс, Ф. Энгельс адамды адам еткен, адам
когамын қалыптастырган ецбек деп санаган.
Адамдардыц алгашқы пайда болган жері ретінде эр түрлі мате-
риктер айтылып жүр, мысалы, Африка, Еуразия, Жерорта тецізініц
айналасы т.б. Дегенмен, бұл проблема элі күнге дейін өзініц
шешімін таппай отыр.
Антропогенез адамныц пайда болуы жэне адам когамыныц
тарихи қалыптасуын зерттейді. Ол негізінен палеонтологиялық
деректерге сүйенеді. Қазіргі тацда антропогенез мэселелері
толык шешілмеген, дегенмен, көптеген болжамдар баршылық.
Адам (Homo sapiens) сүтқоректілер класыныц приматтар
отрядына жатады. Приматтардыц алгашқы қарапайым түрлері
плиоцен дэуірініц басында (55—60 млн. жыл бүрын) пайда
болган. Ал, жогары сатылы маймылдар (Antropoidea) үшінші
дэуірдіц басында (эоцен) жартылай маймылдардан (Prosimii)
келіп шыққандыгы палеонтологиялық деректерден белгілі.
Антропоидтардыц отряд тармагы 3 түқымдасқа бөлінеді: 1)
кец танаулы маймылдар (Ceboiolea); 2) тар танаулы маймылдар
(Cercopitheeoiolea);
234
51-сурет. Ертедегі гоминидтердің филогенетикалык байланыстары
3) гоминидтер — ‘'адам тектес маймылдар”. Соңғы тұқымдасқа
қазіргі кездегі адам тәрізді маймылдар жэне адам туысы
жатқызылады.
Алғашқы екі тұқымдастың адамның пайда болуына ешқандай
қатынасы жоқ, ал үшінші тұқымдастың эволюциясы саналы адам
— Homo sapiens sapiens пайда болуына алып келген.
Палеонтологиялық деректер негізінде XX ғасырдың 70-
жылдарына дейін адамдардың ең ертедегі ата-тектері ретінде
плиоцен дэуірінде тіршілік еткен рамипитектер (Ramipithecus
pinjabicus) саналып келген. Оның қазба қалдыктары 1934 жылы
Үндістанда табылған. Олар бұдан 14 млн. жыл бұрын өмір сүрген.
Бойлары аласа — 110 см, екі аяғымен тік жүрген, ашық не бұталы
орманды жерлерді мекендеген.
1970—1980-ші жылдарда жүргізілген цитогенетикалык, гене¬
тикалык жэне биохимиялык зерттеулер нэтижесінде рамипитек¬
тер адамдарга қарағанда орангутангтарга жақын екендігі бел-
235
гілі болды, сондықтан да қазіргі кезде олар адамдардыц емес
орангутангтардың ата-тегі болуы мүмкін деген болжамдар айты-
луда.
Осы зерттеулер адам мен шимпанзенің ұқсас екендігін көрсет-
ті, мысалы адамдар мен шимпанзелердің 99 % белоктары бірдей.
1925 жылы Оңтүстік Африкада (Кения) адамдарга ұксас
жэндіктің бас сүйегі табылган. Оны оңтүстік маймылы —
австралопитек (Australopithecus africanus) деп атаған. Қазіргі
кезде австралопитектердің бірнеше түрлерінің қазба қалдықтары
табылган. Олардыц ішіндегі ең қарапайым түрі — афар австра-
лопитегі —Australopithecus afarensis (52-сурет).
Австролопитектер осыдан 2—3 млн.
жыл бүрын өмір сүрген. Олардыц бойла¬
ры — 120—150 см, ми көлемі 380—550
см’ екі аяқтарымен тік жүрген. Олар ашық
жерлерді, орманды далаларды мекендеген.
Олардыц адамдарга ец үқсас белгілерініц
бірі —жақтарыныц күрылысы: күрек тістері,
ит тістері салыстырмалы түрде үсақтау, ал
азу тістері ірі болган. Австралопитектердіц
жақтары адамдардыкінен элдеқайда үлкен
52-сурет. Афар және алга қарай шыгыццы болган, ми көлемі
австралопитегі адамдардан 3 есе кем, ал қазіргі адам тэрізді
маймылдардан (шимпанзе, горилла) екі есе үлкен.
Сондыктан да оларды адамдардыц ата-тегі ретінде қарас-
тырады. Австралопитектердіц ішінде афар австралопитегі
(Australopithecus afarensis) адамдардыц ата-тегі болуы мүмкін
деп болжамдануда.
1960 жылы Танзания елініц Олдувай шатқалынан австрало-
питектердіц сүйектерімен бірге адамга өте үқсас казба сүйектер
табылган. Олардыц бойлары шамамен 120 см, салмагы 40—50
килограмм, ми көлемі 650—750 см3, жақтарыныц құрылысы жэне
тістері австралопитектермен салыстырганда нэзіктеу болып кел¬
ген. Сонымен қатар, қазба сүйектермен бірге қолдан жасалынган
қарапайым ецбек қүралдары — пышақтар, сойылдар т.б. табылган.
Оларды Олдувай мәдениеті деп атайды. Осы қарапайым ецбек
кұралдарын жасаган қазба сүйектердіц иесі деп есептелініп, олар-
236
ды адам туысының алғашқы өкілі — іскер адам — Homo habilis
деп атаған (51-сурет). Епті адамдардың бас сүйегінің ішкі бетінде
бірнеше прогрессивтік нейроморфологиялық өзгерістердің пай¬
да болғаны анықталды: 1) ми сыңарларының асинхронды бо¬
луы; 2) екі сөйлеу орталығының бастамасының дамуы. Сонымен
қатар, аяқтарының бас бармағы табанға көлденең орналаспай
тура орналасқан. Бұл епті адамдарда тік жүруге байланысты
морфологиялык өзгерістердің толық аяқталғанын көрсетеді. Епті
адамдардың өмір сүрген уақыты 2—1,7 млн. жыл. Сонымен адам
туысы осыдан 2 млн. жыл бүрын пайда болған.
Адам пайда болғаннан кейінгі эволюцияны 3 кезеңге бөледі:
ежелгі адамдар — архантроптар; ертедегі адамдар — палеоан-
троптар; қазіргі типті адамдар — неоантроптар.
Архантроптар осыдан 2 млн жыл бүрын пайда болып 200—
100 мың жылға дейін тіршілік еткен. Оларға, тік жүретін адамдар
деп аталатын қүрама түр Homo erectus — питекантроп, синан¬
троп, Гейдельберг адамы т.б. жатқызылады. Питекантроптардың
сүйектері Шығыс және Солтүстік Африкадан, синантроптар Ази-
ядан табылган. Бүлардың тіршілік еткен уақыты 1 млн. жылдан
600-400 мың жыл аралықтарында. Олардың ми сауытының көлемі
үлкен болған — 850—1100 см\ Антропологтардың жорамалдау-
ынша, ең карапайым сөйлеу ми көлемі 750—800 см3 жеткен кез¬
де пайда болган, демек питекантроптар, синантроптар өте кара¬
пайым сөйлей білген. Олар отты пайдаланган, оны үзақ уақыт
сактай білген, бірақ өздері отты ала алмаган, Олардың шагын топ-
тары үңгірлерді мекендеген.
Тік жүретін адамдар қазіргі Қазақстан, Өзбекстан мемле-
кеттері аймагын мекендеген. Мысалы, Тешікташтан (Өзбекстан)
питекантроптың баласының қазба сүйегі, Қаратау, Ұлытау
өңірлерінен (Қазақстан) тік жүретін адамның мекен орыны, еңбек
қүралдары, тасқа салган суреттері көптеп табылган. Олар осы¬
дан 500 мың жыл бүрын өмір сүрген.
Палеоантроптар — ертедегі адамдар 100 мың жылдан 25-
35 мың жылга дейін омір сүрген. Оларга неандерталь ада¬
мы — Homo sapiens neanderthaliensis жатады. Бүлардың ми
көлемі 1500 см3 дейін болып, күрделі қауымдық тіршілік ет¬
кен. Олар өздері отты ала білген, сөздері күрделірек, былдыр-
237
лап сөйлейтін болған. Палеоантроптар өздерінің жақындарына,
жас балаларға, қарияларға қамқорлық жасаған, өлген адамдар¬
ды жерлеп отырған. Палеоантроптардың эволюциясында элі де
болса табиғи факторлар — сұрыптау елеулі рөл атқарған. Де¬
генмен, бірте-бірте палеоантроптардың әлеуметгік құрылымы,
еңбек қызметтері күрделеніп антропогенездің ең ақырғы —- 3-ші
кезеңіне — неоантроптарға көшкен. Бұл процесте адамдардың бір-
бірімен сөйлесу арқылы байланыстарының күшеюі, жинақталған
тәжірибені, білімді ұрпақтарга жалғастыруда сөздің күрделіленуі
шешуші рөл атқарған. Адамдардың сыртқы орта факторларына
тэуелсіздігінің күшеюі, жасанды тіршілік ортасының пайда бо¬
луы, қоғамның қалыптасуы табиғи сүрыптаудың творчестволық
(түртүзілу) мэнінің жойылуына алып келіп, адамдардың биоло-
гиялық эволюциясын біржолата токтатты.
Неоантроптар — қазіргі типті адамдардың — Homo sapiens
sapiens — қарапайым формалары осыдан 40—35 мың жыл бүрын
пайда болған. Оларды — кроманъонецтер деп атайды. Олардың
қазба сүйектері Жер шарының барлык жерлерінен — Америка,
Африка, Азия, Еуропа, Австралияда табылган. Олардың сырт
пішіні қазіргі адамдарга ұқсас.
Қазіргі адамдар бірнеше нэсілдерге бөлінеді. Нәсілдер деге-
німіз адамдардың үлкен популяциялары болып табылады. Нәсіл-
дердің физиологиялық, психологиялық үқсастығы олардың айыр-
машылықтарынан әлдеқайда жогары болады, олардыц көптеген
гендері де бірдей, ортақ болып келеді.
Адамдардыц 3 үлкен нәсілдері белгілі: негроидтыц, еуро-
пеоидтың жэне монголоидтьщ нәсілдер. Негроидтық (австра-
ло-негроидтық) нәсілдердіц шаштары бүйра, қара түсті, көздері
қара, мүрындары қысқа жэне кец, қалыц ерінді, терісі қоңыр-қара
болып келеді. Еуропеоидтардың шаштары жұмсак, толқынды,
эр түрлі рецді, терісі ашық түсті, сақал-мүрттары жақсы дамыған,
мұрындары қысыцқы, үлкен, жүқа ерінді болып келеді. Монголо-
идтар шаштары қатты, қара түсті, сакал-мүрты нашар дамыған,
терісі сары реңці, мүрындары жуан, орташа, қабақтары (эпикант)
жақсы дамыған.
Нәсілдер бір-бірінен негізінен терілерініц түстері аркылы ажы-
расады, ол адамдардыц эволюциясыныц бастапқы кезеңдерінде
238
табиги сүрыптау нәтижесінде ортаға бейімделушілік ретінде
қалыптасқан. Ежелгі адамдар — архантроптар 2 үлкен популя-
цияға — нәсілдерге бөлінген: монголоидтар және еуропеоидты-
негроидтар болып. Кейінірек еуропеоидты-негроидты популяция
тағы да — евразиялық (еуропеоидтық) жэне австрало-негроидтық
нәсілдер болып, 2-ге бөлінген.
XV ғасырга дейін, ягни жаңа құрлыктар. жаңа елдер ашылғанга
дейін эрбір нэсілдер бір-бірімен араласпай белгілі бір жерлерді;
мысалы, монголоидтар — Азияда, Солтүстік жэне Оңтүстік Аме-
рикада, австралонегроидтар — экваторлық тропика аудандарда;
еуропеоид — Еуропа, Солтүстік Африкада, Солтүстік Үндістанда,
т.б. мекендеген. Ал, кейінірек, әр түрлі көліктердің дамуына байла¬
нысты олар миграцияланып, бір-бірімен араласып кеткен.
9. МЕДИЦИНАЛЫҚ ПАРАЗИТОЛОГИЯ НЕГІЗДЕРІ
9.1. Жалпы мәселелер
Паразитология — паразитизм құбылысын, паразиттер био-
логиясын, экологиясын жэне олардьщ тудыратын ауруларын, па-
разиттермен күрес шараларын зерттейтін кешенді биологиялық
ғылым.
Паразитизм (паразиттік тіршілік ету) биотикалық байланы-
стардың бірі болып табылады, ол бір ағзаның (паразит) екінші бір
ағзаны (иесін) баспана, қорек ретінде пайдаланып зиян келтіруі.
Медициналық паразитология адам паразиттерін зерттейтін
жалпы паразитология гылымының бір саласы. Ол, сол сияқты,
паразиттерге қарсы күрес шараларын, паразиттер тудыратын ау¬
руларды анықтау, емдеу жэне алдын алу шараларын да зертгейді.
Медициналық паразитология 3 бөлімге бөлінеді: медициналық
протозоология — қарапайымдылар типінің адам паразиттерін;
медициналық гельминтология — жалпак жэне жұмыр құртгар
типінің адам паразиттерін; медициналық арахноэнтомоло-
гня — буынаяқтылар типінің адам ауруларын тудырушы, не табиги
резервуарлары болып келетін өкілдерін (кенелер) зерттейді.
Медициналык паразитологияның негізгі мақсаттарына мына-
лар жатады:
— паразиттердің түрлерін дэл анықтау үшін олардың даму
кезеңдеріндегі күрылыс ерекшеліктерін зерттеу. Олардың түрлерін
нақты анықтау дәрігерлер үшін өте маңызды, себебі тек осының
арқасында паразиттер қоздыратын ауруларды дұрыс анықтап ем-
деуге мүмкіншілік туады;
— паразиттердің және таратушылардың даму ерекшеліктерін
зертгеу; бұл паразиттердің табигатта таралуын жэне адам агзасына
ену эдістерін анықтауга мүмкіндік береді;
— паразит — ие арасындагы өзара байланыстарды зерттеу;
240
— паразиттердің зиянды әрекеттерін анықтау нәтижесінде
паразиттік ауруларды ғылыми негізде анықтау және емдеу эдістерін
қалыптастыру; паразиттер мен оларды таратушыларға қарсы күрес
жэне алдын алу шараларын жасау;
— аталған шаралардың бэрі медициналық паразитологияның
негізгі мақсаты — паразиттер қоздыратын ауруларды болдыр-
мау жәие алдын алу жүйесіи цүруга багытталады.
Медициналық паразитология ғылымы клиникалықжәне санита-
риялық-гигиена пәндерімен тығыз байланысқан.
Паразиттер адамнын барлык мүшелерін зақымдайды, сон-
дықтан да кез келген мамандықтағы дәрігерлер өздерінің күнде-
лікті жұмыс тэжірибелерінде паразиттік аурулармен кездесіп оты¬
рады, мысалы:
Терапевттерге ішек-қарын, өт жолдары зақымдалған (лям¬
блия, амеба, балантидий, ішеккүрт, үшкіркұрт т.б.), бауыры (эхи¬
нококк, трематодтар, альвекокк), өкпесі (өкпе сорғышы), висце-
ралды лейшманиозбен, безгек т.б. аурулармен ауыратын адамдар
көрінулері мүмкін;
Хирургтерге бауыр мен өкпе эхинококкозы, ішектері тыгын-
далып қалган (ішеккұрт) т.б. аурулар көрінуі мүмкін.
Паразиттік аурулармен, әсіресе, педиатрлар жиі кездеседі,
себебі иммундық жүйелері элі толык калыптаспаган балалар осы
аурулармен жиі ауырады.
Паразитология өз алдына дербес гылым ретінде XIX гасырдың
екінші жартысында бөлініп шыккан. Бұл кезде адамның кейбір
қауіпті жэне кең таралган ауруларын (безгек, лейшмания, амеби-
аз т.б.) тудыратын паразиттер және таратушылар ашылып, анық-
талып, олардың даму циклдары зерттелген болатын.
Паразитология гылымының калыптасуына жэне дамуына
орыс галымдары үлкен үлес қосқан. Мысалы, Г. Гросс адамның
паразиттік амебасын, Д. Ф. Лямбль лямблияны ашты, А. П. Фед-
ченко бірнеше паразиттік кұрттарды зерттеген. И. А. Порчинский
сонаның, масаның, шыбындардың паразиттік ауруларды тара-
тудагы рөлін анықтаған, Н. А. Холодовский биттерді зерттеген.
Паразитология гылымының, әсіресе гельминтология сала-
сының дамуында, К. И. Скрябиннің ролі өте зор. Ол дүние
жүзінде тұңгыш болып КСРО-да гельминтология институ-
241
тын ұйымдастырган. Сол сияқты, паразитологияның дамуын-
да Е.Н.Павловскийдің де еңбегі ерекше. Ол арахноэнтомоло-
гия саласында көп еңбек еткен, паразитоценоз, трансмиссивтік
аурулардың табиғи ошақтары туралы ұгымды қалыптастырған.
В. А. Догель экологиялық паразитологияны дамытқан.
Паразитология ғылымы Қазақстанда 1925 жылдан бастап
дамып келеді. Осы жылы Алматыда өлкелік мал дәрігерлік-
бактерологиялық институты құрылып, 1926 жылы оның кұра-
мында гельминтология, 1935 жылы протозоология бөлімі ұйым-
дастырылды. Қазақстанда паразитология гылымының дамуына ат
салысқан зоологтер — J1. С. Берг, И. А. Долгушин, А. А. Слудский,
М. И. Мариковский, М. Д. Зверев, Т. Н. Досжанов, И. Г. Галузо,
Е. Н. Боев, А. Бекенов т.б.
Табигатта паразитизм түрлері көптеп кездеседі. Паразиттер —
нагыз паразиттер, жалган паразиттер, уақытиіа жэне түрац-
ты паразиттер болып бөлінеОі. Нагыз паразиттер иесінің
денесімен едәуір уақыт бойына байланысқан. Бұл жагдайда
паразиттік тіршілік ету тарихи қалыптасқан жэне түрдің ерекше
белгісі болып саналады. Жалган паразит дегеніміз еркін тіршілік
ететін түрдің кейбір өкілдері кездейсоқ басқа бір түрдің дара-
сына еніп оның қалыпты тіршілігін бұзуы. Мысалы, шыбындар
дернәсілдерінің адам ішегіне енуі.
Уақытша паразиттер (негізінен қан соргыш буынаяқтылар)
өздерінің өмірінің шамалы гана уақытын иесінің денесінде өт-
кізеді, ал қалган уақытта еркін тіршілік етеді.
Түрацты паразиттер — салыстырмалы тұрақты жэне шарт¬
ты тұрақты паразиттер болып бөлінеді. Салыстырмалы тұрақты
паразиттер иесінің денесінде өзінің даму циклінің бір-ак кезеңін
өткізеді, ал қалган кезендерінде еркін тіршілік етеді.
Шартты тұрақты паразиттер — өздерінің барлық өмірін
иесінің денесінде өткізеді; олар еркін, ягни қожайын денесінен
тыс, тіршілік ете алмайды. Иесінің денесінде орналасуына
қарай паразиттерді эктопаразиттер жэне эндопаразиттер деп
бөледі.
Эктопаразиттер иесінің дене жамылгысының сыртында (тері,
түктер) тіршілік етеді. Оларга қансоргыш буынаяқтылар -— кене-
лер жатады.
242
Эндропаразиттер — иесінің ішкі мүшелерінде тіршілік етеді.
Паразиттердің иелерін аралық, негізгі деп бөледі. Иесі деге-
німіз — паразиттердің қорек жэне мекен ортасы ретінде пайда¬
ланатын тірі ағзалар. Кейбір паразиттер өздерінің даму циклінде
бір иесінен екіншісіне кошіп отырады, оны иесін алмастыру деп
атаймыз.
Аралъщ иесі деп паразиттің дернәсілдік сатысы кездесетін, не
жыныссыз жолмен көбейетін ағзаны айтамыз.
Негізгі иесі деп паразиттің ересек сатысы болатын не жыны-
стық жолмен көбейетін ағзаны айтамыз.
Осымен қатар резервуарлыц иесі деген ұғым да белгілі. Оларда
паразит тіршілігін жоймай ұзак уақыт сақталады, бірак әрі қарай
дамымайды.
Паразиттік тіршілік етудің пайда болу жолдары түрліше болып
келеді. Эктопаразиттердің көпшілігі алғаш жыртқыштық тіршілік
еткен, сосын олардың қожайын денесінде коректену, онымен бай-
ланысу мерзімі ұзарып, эктопаразиттіктіршілік етуге көшкен. Эво¬
люция процесінде кейбір эктопаразиттердің қоректену мерзімініц
ұзаруын байқауға болады, мысалы, шіркейдің ұргашы дарала¬
ры иесінің денесінде 1 минуттан аз уақыт қоректенсе, иксодалы
кенелердің дернэсілдері (личинкалары) 5—8 тәулік, ересек түрлері
5—14 тэулік қоректенеді, бүргелер иелерінің денесінде өздерінің
өмірінің көп бөлігін өткізеді, ал биттер өмір бойына иесінің
денесінде кездеседі.
Паразитизмге өтудің екінші жолы комменсализм (селбесу) бо¬
лып табылады. Мысалы, күстарда кездесетін кенелер. Олардыц
ата тектері қүстармен селбесіп тіршілік етіп кұстардыц корегініц
қалдыкгарымен коректенген. Бірте-бірте олар қүстардың денесіне
өтіп, мамықтарымен қоректенуге көшкен.
Эндопаразиттер эктопаразиттерден келіп шыкқап.
Ауру тудырушы паразиттердің таралу көзі болып адамдар не¬
месе жануарлар саналады, себебі, паразиттер адамдар не жануар¬
лар денесін мекендеп көбейеді жэне басқа иелерге беріледі.
Паразит ұзақ уақыт сақталатын ағзаларды резервуар деп
атайды. Кейбір паразиттер үшін резервуар болып адамдар (без-
гек, бөртпе сүзегі, іш сүзегі), кейбіреулері үшін жануарлар сана¬
лады. Жабайы жануарларды табиғи резервуарлар деп атайды.
243
Мысалы, кемірушілер лейшмания, оба, туляремия ауруларыньщ
қоздырғыштарына табиги резервуар болып саналады.
Көптеген паразиттердің таралуы үшін резервуарлардан
басқа таратушылар да қажет. Таратушылар қызметін қан
соргыш буын аяқтылар атқарады. Таратушылардың белсенділігі
нэтижесінде паразиттер табиғатта кең таралады. Оларды ар¬
найы таратушылар, механикалъщ таратушылар деп бөледі.
Арнайы таратушылар денесінде паразиттер өздерінің дамуының
бір кезеңін өткізеді (безгек масасы). Механикалық таратушылар
денесінде паразиттер дамымай тек олардың көмегімен кеңістікте
таралады. Мысалы, шыбын денесіне әр түрлі паразиттер жабы-
сып таралуы мүмкін.
Кейбір таратушылар паразиттердің табиғи резервуарлары
да болуы мүмкін. Мысалы, аргас кенелері қайталама сүзектің
қоздыргыштарын 20 жылға дейін денелерінде сақтай ала¬
ды, бүргелер 37° жылылықта оба паразиттерін 27 күн, ал 0-5°
жылылықта 358 күнге дейін сақтай алады.
Паразиттердің таратушылар денесінде сақталу мерзімінің со-
зылуына олардыц трансовариалдық жолмен берілуі маңызды
рөл атқарады. Паразиттердің трансовариалдъщ жолмен берілу
дегеніміз таратушылардың жүмыртка жасушалары (гамета)
аркылы паразиттердің үрпактан-үрпакка беріліп отырылуы болып
табылады. Паразиттер таратушылардың эр түрлі үлпаларымен
мүшелеріне, соның ішінде жыныс мүшелеріне — аналық безге,
жүмыртқа жасушаларына өтуі мүмкін. Осындай жүмыртқа жасу¬
шалары үрықтанғанда үрық қалыпты дамиды, бірақ оның барлық
сатыларында, ересек дараларында паразиттер болады жэне оны
үрпактан-ұрпаққа беріп отырады. Е.Н. Павловский лаборатория-
сында аргас кенелерінің осындай жолмен қайталама сүзегінің
паразиттерін 3 буын бойына беріп отырғаны байқалган, иксод
кенелері риккетсийлерді 12 буын бойына беріп отырған.
Пайда болу себептеріне қарай адамдар мен жануарлардыц ау¬
руларын инфекциялық (жүқпалы ауру, індет) жэне инвазиялық
(паразиттік) аурулар деп бөледі.
Инфекуиялық аурулар эр түрлі микроағзалардың — бактерия-
лар, спирохетгер, вирустар, саңыраукұлақтардың эсерінен пайда
болады.
244
Инвазиялъщ аурулар түрліше патогендік қарапайымдар (про-
тозооноздар), гельминтгер (гельминтоздар), насекомдар әсерінен
пайда болады.
Таратушылар арқылы берілетін ауруларды трансмиссивтік ау¬
рулар деп атайды. Оларға инфекциялық және инвазиялық аурулар
жатады.
Паразиттер мен иесінің арасындағы ерекше байланыстарға
қарай төмендегідей трансмиссивтік ауруларды ажыратады: зоо-
ноздар — тек қана жануарларға тэн аурулар (құстар безгегі); ан-
тропоноздар — тек қана адамдарға тэн аурулар (трихомонадоз,
амебиаз), антропозооноздар — адамдарда да, жануарларда да
кездесетін аурулар (тайга энцефалиті, лейшманиоз, оба, т.б.)
Е.Н. Павловский 1939 жылы экспедициялық, лабораториялық,
эксперименталдық зерттеулер нәтижесінде табиғи ошақтық ау¬
рулар деген ұгымды калыптастырды. Табиги ошақтық аурулар-
дың бірнеше ерекше белгілері болады:
1) олардың паразиттері адамның катынасынсыз табигатта өз
бетінше ұзақ уақыт айналып жүреді;
2) олардың таратушылары болып жабайы жануарлар саналады;
3) олар табигатта кең таралмай белгілі бір шагын жерлерде —
ошақтарда кездеседі.
Табиги ошақтық ауруларга мысал ретінде тайга энцефалитін
алуга болады. Тайганы мекендейтін жабайы жануарлар (тиін,
қоян, кірпі, құстар) — табиги резервуарлар; жануарлардың
паразиттері — иксод кенелері таратушылар болып саналады да,
табигатта адамнан тәуелсіз, өз беттерінше бірінен-біріне өтіп ұзак
жылдар бойына айналып жүреді. Олардың ареалдары қылқан
жапырақты ормандармен шектелген, одан тыс кездеспейді. Пара-
зиттің табиги резервуарларының, таратушыларының таралган
жерлерін (тайга) ошағы деп атайды.
Лейшманиоздың табиги ошагы — лейшманиямен зақымданган
тышқандар мен олардың індерін мекендейтін бэкене шыбындар
таралган шөлді жэне шөлейт жерлер саналады.
Табиги ошақтар адамдар үшін өте қауіпті. Егер кездейсоқ адам¬
дар сол ошақтарга бара қалса (экспедиция, аң аулау, орманшылар
т.б.) онда таратушылар сол паразиттерді адамдарга жұқтыруы
әбден мүмкін.
245
9.2. Қарапайымдылар типі — Protozoa
Бұл типке 20—25000-ға жуық түрлер жатады. Олар табиғатта
кең таралған жэне мұхиттарда, теңіздерде, тұщы суларда, топы-
рақтарда кездеседі. Олардың көптеген түрлері басқа ағзалар
денесінде тіршілік етуге (паразиттер) бейімделіп, жануарлардыц
жэне адамдардыц зілді ауруларын тудырады.
Қарапайымдар дене кұрылысы жағынан бір жасушадан (цито¬
плазма жэне бір немесе бірнеше ядродан) тұрады, ал қызметтік
жағынан ол біртүтас, дербес ағза болып саналады, яғни ол
қозғалады, жемтігін қармайды, көбейеді, жауларьшан қорғанады
т.с.с.
Қарапайымдардыц денесі өте кішкентай: 3 мкм-ден 160 мкм
аралығында болады. Ол сыртқы мембранадан, цитоплазмадан,
ядродан жэне органоидтардан түрады. Сыртқы мембрананыц
күрылысы жасушалардыц мембранасына ұқсас; цитоплазмасы
2 қабатқа бөлінген — сыртқы қабаты —- эктоплазма, тығыздау,
біркелкі жэне мөлдір, ал ішкі қабаты — эндоплазма, түйіршікті
жэне сүйықтау болып келеді. Эндоплазмада жалпы қызмет
атқаратын органоидтары болады: митохондриялар, эндоплазмалық
тор, Гольджи кешені т.с.с. Сонымен қатар, қарапайымдыларда ерек¬
ше қызмет атқаратын органоидтарда кездеседі. Оларға қозғалу, ас
қорыту, зэр шығару, қорғаныстық жэне т.с.с. органоидтары жата¬
ды.
Қарапайымдардыц қозғалу органоиды болып жалған аяқ-
тары, талшықтары, кірпікшелері саналынады.
Ac қорыту органоиды болып ас қорыту вакуольдері, зэр
шыгару органоиды — жиырылгыш вакуольдер саналады; ал
кейбіреулерінде ерекше тесікшелер — қылаулатқыштар кездеседі.
Қолайсыз жагдайларга орай коптеген карапайымдар ци¬
ста түзеді. Бүл кезде олар қозгалуын тоқтатып, қалыц қабықпен
қапталып, тыныштык күйіне көшеді. Олардыц зат алмасуы
мүлдем тоқталады. Циста күйінде олар қиын кезецдерді өткізеді,
таралады. Қолайлы жагдай туса олар қайтадан тіршілігін эрі қарай
жалгастыра береді.
Қарапайымдар жыныссыз жэне жынысты жолдармен көбейеді.
Жыныссыз көбеюі жасушаныц жай 2-ге бөлінуі арқылы, не
246
жасушаныд көптеп бөлінуі (шизогония) арқылы жүруі мүмкін.
Жыныстық процесс копуляция не конъюгация күйінде кездеседі.
Қарапайымдар типініц жіктелуі түрліше болып келеді. Оныц
кец таралган жіктелуі бойынша олар 4 класқа бөлінеді:
1) саркодалылар класы (Sarcodina);
2) талшықтылар класы (Flagellata);
3) споралылар класы (Sporozoa);
4) инфузория класы (Jnfusoria).
9.2.1. Саркодалылар класы (Sarcodina)
Бүларга 10000-га жуык гүрлер жатады; олар негізінен теціз-
дерде, тұщы суларда және топырақтарда кездеседі. Бұлар — ец
қарапайым жануарлар. Олардыц цитоплазмасы жіктелмеген, бір
немесе бірнеше ядролары болады. Дене пішіні түрақсыз, козгалу
органоидтары болып жалган аяқтары саналады. Жалган аяқтары
қозгалумен катар жемтікті қармау қызметін де атқарады. Ауыз
қуысы болмайды, жасушага заттардыц енуі жэне жасушадан
заттардыц сыртқа шыгарылуы денесініц кез келген жері арқылы
жүзеге асады. Жиырылгыш вакуольдері — біреу. Көбеюі негізінен
жыныссыз жолмен — жасушаныц 2-ге бөлінуі арқылы жүреді.
Бүл кластыц өкілдерініц ішінен бір отряд өкілдерініц меди-
циналық мацызы белгілі. Ол — Амеба отряды (Amoebina).
Амеба отряды —Amoebina
Адам денесінде амебаның бірнеше түрлері кездеседі, олардыц
ішінен мацыздысы — қантышқақ (дизентерия) амебасы (Entam¬
oeba histolytica) (53-сурет).
Дизентерия амебасы — Entamoeba histolytica адамдардыц
зілді ауруы — амеба дизентериясы немесе амебиаз ауруыныц
коздырушысы болып табылады. Ол адамныц тоқ ішегінде
паразиттік тіршілік етеді. Ол барлық жерлерде, әсіресе, климаты
ыстық болып келетін жерлерде, кездеседі.
Бүлар тек адам паразиті болып табылады. Оныц тіршілік
циклінде мынадай формалары кездеседі: циста, үсақ вегатативтік
формасы (forma minuta), ірі вегетативтік формасы (forma magna)
жэне ұлпалык формасы.
247
5
г
53-сурет. Дизентерия жэне ішек амебалары.
а) ірі вегетативтік формасы, б,в) ұсак вегетативтік формасы:
1-эктоплазма, 2-эндоплазма, 3-жалған аяктары, 4-ядро, 5-кариосома,
6-ас корыту вакуоляларындагы эритроциттер; г) циста.
Инвазиялык сатысы болып 4 ядролы циста саналады. Ол лас қол
аркылы, не жуылмаған жеміс-жидектерді жегенде адамға жұғады.
Адам ішегінде циста қабығы еріп 4 ядролы амебаға айналады. 4
ядролы амеба тез бөлініп, 4 бөлек бір ядролы ұсақ вегетативтік
форманы (7—15 мкм) (forma minuta) түзеді. Үсақ вегетативтік
амеба ток ішекте бактериялармен коректеніп, ұзақ уакыт зиянсыз
тіршілік етеді. Тоқ ішектің төменгі болімдеріне өтіп ол цистаға ай¬
налады да нэжіспен сыртқа шыгарылады.
Кейбір адамдарда, белгілі бір жағдайларда (мысалы, салқын-
дағанда, авитаминозда, диета бүзылғанда, гельминтоздарда) ұсак
вегетативтік форма ішек қабыргасына еніп ішектің сілемейлі
қабығында жара туғызады. Бұл оның ұлпалык формасы. Бұл кез¬
де кылтамырлар қабырғасы жараланып, ішек қуысына қан ағуы
байқалады.
248
54-сурет. Дизентерия амебасынын тіршілік циклі.
а) цистотасымалдаушы, б) амебиазбен ауыратын адам; 1-циста,
2-вегетативтік формасы, 3-ұсак вегетативтік формасы, 4-ірі вегетативтік
формасы, 5-ұлпалык формасы, 6—7-нэжіспен сыртка шығарылуы.
I шектің амеба аркылы зақымдануы салдарынан ұсақ вегетативтік
форма (forma minuta) ірі вегетативтік формаға (forma magna) айна-
лады. Оның өлшемі ірілеу 30—40 мкм жэне ядро құрылысы ерек¬
ше болып келеді. Олар эритроциттермен қоректенеді, сондықтан
оларды эритрофагтар деп атайды.
Ішек кабырғасының ұлпасында көбеюші амебалар (үлпалык
формасы) ішек қуысына өтуі мүмкін. Олардың қүрылысы жэне
көлемі ірі вегетативтік формаға ұқсас болады, бірак олар эритро¬
циттермен қоректене алмайды.
Емдеу немесе ағзаның қорғаныстьщ реакциясының күшеюі
нэтижесінде ірі вегетативтік форма қайтадан ұсақ вегетативтік
формаға өтіп цистаға айналады.
Кейбір адамдарда үсақ вегетативтік форма еш уақытта да ірі
вегетативтік формаға айналмайды. Ондай адамдарды уистота-
сымалдаушыпар деп атайды. Олар өздері ауырмайды, бірақ басқа
адамдар үшін өте қауіпті, себебі басқа адамдардыц зақымдану
көзінің бірі болып табылады. Циста тасымалдаушы 1 гәулікте 600
млн цисталарды бөліп шығарады. Оларды дер кезінде тауып емдеу
қажет.
Амебиаздың жұгуының жалғыз көзі болып адам саналады.
Нәжіспен бөлініп шыққан цисталар топырақты, суларды ластай-
ды, бау-бақшаларға жетіп жеміс-жидектерді ластайды. Ластанған
жеміс жидектерді жумай жеген кезде амеба цистасы адамға
жұғады. Олардың механикалық таратушылары болып шыбындар,
тарақандар саналады.
Клиникалық сипаты. Негізгі белгілері (симптомдары) ішектің
қансырап тұратын жарасы, қан мен сілемей араласқан сұйық үлкен
дәреттің жиі келуі болып табылатын зілді ауруды қоздырады.
Дұрыс емделмесе 40 % жагдайда дүние салуға апарып соғады.
Лабораториялык анықтау. Нәжісті микроскопиялық зерт¬
теу арқылы жүргізіледі. Аурудың зілді кезінде үлкен дәретте
эритроциттерді жұтқан ірі вегетативтік формасын, ал созылмалы
ауру кезінде не цистотасымалдаушыларда 4 ядролы цисталарды
іздейді.
Алдын алу шаралары ретінде жеміс-жидектерді жуып жеу,
қайнаған суды ғана ішу, тамақтану алдында, дэрет алғаннан кейін,
қолды сабындап жуу, яғни жеке тазалықты сақтау болып табы¬
лады, сондай-ақ механикалық тасымалдаушыларды (шыбындар,
тарақандар) жою жэне санитарлық ағартушылық жұмыс (үгіт-
насихат) жүргізу әдістерін атауға болады.
Адамның ас қорыту жолында дизентерия амебасымен қатар
басқа да патогенді емес амебалар кездеседі. Ол — ішек амебасы
(Entamoeba соіі), ауыз амебасы (Entamoeba dingivalis). Олар адам
денсаулығына зиянсыз болып табылады.
9.2.2. Талшьщтылар класы — Flagellata
Бұл класқа 6—8 мыңға жуық түрлер жатады, олардың көпшілігі
адам паразиттері болып табылады (55-сурет).
Денесі кішкентай, сопақша, домалақ не жіпше тэрізді, ол
сыртқы мембранамен, жүқа қабықшамен (пелликула) қапталып,
дене пішіні тұрақты болып келеді. Қозғалу органоиды бір неме¬
се бірнеше талшықтар. Талшықтар — цитоплазманың үзын жэне
жіңішке өскіншесі болып саналады. Ол, әдетте, дененің алдыңғы
жагынан басталады. Талшық еркін бөліктен жэне эктоплазмаға
сіңірілген бөлімнен, цилиндр пішінді базальдық денешіктен неме-
250
се кинетосомадан тұрады. Кейбір талшықтылардың (лейшмания,
трипоносома) талшыгының түбінде ерекше органоид-кинетопласт
кездеседі, ал кейбіреулерінің талшығы денесін ұзына бойына
бойлап өтіп, сыртқа шығып тұрады. Цитоплазма онымен жұқа
өскінше арқылы жалғанып желпуші жарғақша — (ундилденуші
жарғақша) тгғызады. Осы жарғақшаның толқынданып желпінуі
жэне талшықтың қозгалуы нәтижесінде олар қозғалады.
55-сурет. Паразит талшыктылар.
а) лейшмания (лептомонадалык формасы), б) лейшмания
(лейшманиялык формасы), в) трихомонада, г) лямблия.
1-талшык, 2-ядро, 3-кинетопласт, 4-аксостиль, 5-желпуші жарғакша.
Талшықтылардың көпшілігі гетеротрофты, кейбіреулері авто-
трофты немесе миксотрофты түрде қоректенеді. Паразит ныса-
ны қоректі денесінің бүкіл бетімен пиноцитоз арқылы сіңіреді;
гетеротрофтылардың ас қорыту вакуольдері болады. Беліп
шығару органоиды-жиырылғыш вакуольдері еркін өмір сүретін
формаларында ғана кездеседі, ал паразит формалары болмайды.
Талшықтыларда негізінен бір, кейде екі ядро кездесуі мүмкін.
Кобеюі жыныссыз жолмен, кейде жыныстық жолдармен де
жүреді.
251
Алғашқы монадалылар отряды
Лейшмания туысы —Leishmania
Лейшмания трипоносомалылар тұқымдасына жатады. Бұл
тұқымдастың ерекше белгісі ретінде даму циклінде, қоршаған орта
ерекшеліктеріне қарай, бірнеше морфологиялық формалардың
түзілуін айтуға болады: трипоносомалық, критидиалдық, лептомо-
надалық, лейшманиялық (жасуша ішілік) жэне метациклдік.
Трипоносамалыц формасының жалпақ, таспа тэрізді денесі
болады, оның ортасында сопақша ядро орналасқан. Талшық
денесінің артқы жағынан, ядродан кейіндеу, басталады, желпуші
перде жақсы жетілген.
Критидиалдыц формасының талшыгы денесінің ортасынан,
ядродан ілгерілеу, басталады: желпуші перде кысқа болып келеді.
Лептомонадалыц формасының талшығы денесінің алдыңғы
жағының жиегінен басталады, желпуші перде дамымаған.
Лейшманиялың (немесе жасуша ішілік) формасының денесі
домалақ, ядросы ірі, талшық мүлдем дамымаған не өте нашар
дамыған, оның тек базаль денесі ғана болады.
Метациклдік формасы критидиалдық формага ұқсас, бірақ
талшықтың еркін белігі дамымаған.
Лейшмания туысының өкілдерінде 2 морфологиялық фор¬
ма кездеседі: лептомонадалық жэне лейшманиялық (немесе
клеткаішілік).
Тіршілік циклі қожайын алмастыру арқылы жүреді. Лейш-
маниялық нысаны адамдарда және омыртқалыларда паразиттік
тіршілік етеді; лептомонадалық формасы — таратушы насеком-
дарда кездеседі. Лейшмания тудыратын ауруды лейшмаииоздар
деп атайды, ол табиғи-ошақтық, трансмиссивтік аурулар қатарына
жатады.
Лейшманиялар дерматотропты (теріні зақымдайды) жэне вис-
церотропты (ішкі мүшелерді зақымдайды) түрлер деп бөлінеді.
Висцералді пейшманиозды цоздырушы— Leishmania donovani. Ол
бауыр, көкбауыр, сүйек кемігі жасушаларында, лимфа түйінінде,
тері астындағы ретикулоэндотелиалды жасушаларда кездеседі.
Ол Жерорта теңізі елдерінде, Азия, тропикалық Африка,
Оңтүстік Америка, Орта Азия, Кавказ елдерінде таралған.
252
Даму цикпі. Бұлардың резервуарлары болып адамдар жэне
эр түрлі сүтқоректілер (иттер, шиебөрілер) саналады. Тарату-
шылары болып ұсақ қансорғыш насекомдар-бэкене шыбын-
дар есептелінеді. Лейшмания бэкене шыбынның ас қорыту
жолына еніп сілекей безіне өтеді. Ондай шіркей адамды шақса
лейшманияның лептомонадалы түрін жұқтырады. Олар қан,
лимфа арқылы таралып ішкі мүшелер жасушаларына өтіп
лейшманиялық формаға айналады да көбейеді. Бір жасушадағы
паразиттер саны 100—200-ге дейін жетуі мүмкін. Олар жасуша-
ны бүзып жаңа жасушаларға өтіп отырады.
Патогендік әсері. Көкбауыр жэне бауыр мөлшері үлкейіп
өседі. Ағзада қаназдылық, элсіздену, жүдеу процестері байка-
лады. Ауру зілді түрде не созылмалы түрде жүруі мүмкін. Дүрыс
емделмесе өлімге алып келеді. Бүл аурумен негізінен балалар
жиі ауырады.
Лабораторинлык анықтау — тос сүйегі кемігінің жағын-
дысын микроскоп арқылы зерттеп, паразиттің лейшманиялық
түрін табу болып саналады.
Алдын алу шарлары: жеке шаралар — бэкене шыбындар-
дан сақтану; қоғамдық шаралар — табиги резервуарларды жою;
санитариялық-ағартушьшық жұмыстар жүргізу, ауруларды емдеу
т.с.с.
Тері лейшманиозын қоздырушы — Leishmania tropica
Лейшманияның дерматотропты түрінің 3 түрлі тармагы бел-
гілі: L. tropica minor; L. tropica major (Жер шарының шығысында)
жэне L. tropica mexicana (Жер шарының батысында).
Олар тері жасушаларында кездеседі.
Еуропа, Азия, Америка, Африка, Орта Азия жэне Кавказ елде-
рінде кең таралған.
Лептомонадалық жэне лейшманиялық формаларының вис-
церотропты лейшманиядан айырмашылығы болмайды.
Даму циклі. Висцералды лейшманиозға ұқсас. Жүғу көзі болып
адам жэне жабайы жануарлар (үсақ кемірушілер) саналынады. Та-
ратушылары болып шіркейлер есептелінеді.
Патогендік эсері. Ағзаның ашык жерлерінде үзақ жазылмай-
тын жаралар пайда етеді. Жара жазылғаннан кейін оның орнында
сүйкімсіз тыртык қалады.
Лабораториялық анықтау — жара бетіндегі ылғалды микро-
скопиялық зерттеу арқылы жүргізіледі.
Алдын алу шаралары: жеке шаралар шіркейлердің шагуынан
с акта ну; қоғамдық шаралар шіркейлерді, табиғи резервуарларды
жою.
Көпталшыктылар отряды — Polymastagina
Урогениталды (қынап) трихомонада — Trichomonas vagina¬
lis. Урогениталды трихомонадоз ауруын қоздырушы — паразит, ол
әйелдер мен ер адамдардыц зэр шығару жэне жыныс жолдарында
кездеседі. Ол барлык жерлерде таралған. Бұлар ішек трихомона-
дасына ұқсас, бірақ ірілеу болып келеді, өлшемі 7—30 мкм. Дене
пішіні алмұрт тәрізді; 4 талшыгы болады, оның бірі денесінің ар-
тына карай бағытталган.
Патогендік (зардаптық) әсері. Оларжыныс-несеп жолдарының
қабыргасына бекініп механикалық эсер егеді.
Бұл ауруды жыныс-несеп жолдарының болінділерін микро-
скоппен зерттеу аркылы анықтайды.
Аурудың алдын алу шаралары. Жеке шаралар — гигиеналык
шарттарды сақтау болып табылады.
9.2.3. Споралылар кяасы — Sporozoci
Бұл класқа тек паразиттік формалар жатады. Бұлардың козғалу
органоидтары, ас корыту жэне жиырылғыш вакуольдері болмай-
ды. Жыныссыз жэне жыныстық жолдармен көбейеді. Жыныс¬
сыз көбеюі шизогония (көпшілікті түрде бөліну) күйінде болады.
Тіршілік циклі күрделі, кожайын алмастыру арқылы жүреді.
Адам паразиттері қан споралары жэне кокцидий отрядтарына
жатады.
Қан споралылар отряды — Heamasporidia
Қан споралылары өздерінің дамуының белгілі бір сатысын
адамның не эр түрлі омыртқалылардың эритроциттерінде өткізеді.
Олардың даму циклі иесін алмастыру арқылы жүреді. Жыныс¬
сыз көбеюі адам денесінде, жыныстық көбеюі омыртқасыз жану-
арларда (несекомдарда) жүреді. Бүлар спора түзбей бір иесінен
екіншісіне тікелей беріліп отырады. Адам паразиті болып безгек
платодиясы саналады — ол безгек ауруын қоздырады. Адамдарда
безгек плазмодийінің 4 түрі паразиттік тіршілік етеді:
254
Plasmodium vivax, Pl.malariae, Pl.falciparium, Pl.ovale. Соңғы
түрдің өкілдері бізде кездеспейді. Олардың бэрінің даму циклі
жэне кұрылысы ұқсас: аралық иесі -— адамдар, ақыргы иесі —
безгек масасы болып табылады.
а
б
в
56-сурет. Безгек плазмодийлерінің түрлері.
a) PI.vivax, б) Pl.malariae, в) PI.falciparum.
Тіршілік циклі үш кезеңге бөлінеді, оның екеуі адам денесінде,
үшіншісі маса денесінде өтеді.
I. Преэритроцитарлық шизогония. Плазмодий адам денесіне
закымданған маса сілекейінде болатын спорозоиттар күйінде
енеді, яғни бұлардың инвазиялық сатысы болып спорозоиттар са¬
налады. Спорозоиттар қан арқылы бүкіл денеге, бауыр жасушала-
рына келіп жетеді. Бұл жерде олар осіп, үлкейіп шизонтқа айнала-
ды. Содан кейін шизонт шизогония жолымен (көпшілікті бөліну)
бөлініп, көптеген (1000—5000) ұсақ бір ядролы мерозоиттар пайда
етеді. Бүл процесс преэритроцитарлыц немесе цлпалыц шизогония
деп аталады. Бауыр жасушалары бұзылғанда мерозоиттар шығып
қанға өтіп эритроциттерге енеді.
II. Эндоэритроцитарлық шизогония. Эритроциттерге енген
мерозоиттар гемоглобинмен қоректеніп өсіп, шизонтқа айналады.
Өздерінің дамуында мерозоиттар бірнеше сатыдан өтеді: алгаш
олардыц өлшемдері өте кішкентай, эритроцит көлемінің 1/3—1/6
бөлігін алып, цитоплазмасы сақина тэрізді болады — бүл сақиналы
255
шизонт сатысы; әрі қарай сақиналы шизонт ұлгайып, жалғанаяқ-
тар пайда болып, қозғалып, амеба тэрізді шизонтқа айналады.
Шизонт бірте-бірте өсіп, эритроцит көлемін түгел толтырады,
гемоглобинді толықжояды жэне шизогония жолымен бөлінеді. Әр
түрлі түрлерде түрліше мөлшерде мерозоиттар түзіледі. Pl.vivax-
22, Pl.malariae-6-12, PI.falciparum 12-18. Бүл кезде эритроциттер
қабықшасы жыртылып мерозоиттар қанга өтеді де зақымданбаған
жаңа эритроциттерге еніп жаңа цикл басталады. Мерозоиттармен
бірге қанға олардыц улы өнімдері де өтеді. Осыныц нэтижесінде
ағзада қалтырау үстамасы басталады. Эндоэритроцитарлық
кезец Pl.vivax, Pl.falciparum, PI.ovale-де 48 сагатка (3 тэуліктік
безгек), Pl.malariae — 72 сагатка (4 тәуліктік безгек) созылады.
Шизогонийдіц қайталануы нэтижесінде адам агзасындагы паразит¬
тер саны тез көбейеді. Бірнеше дүркін жыныссыз көбеюден кейін
жыныстык жолмен көбеюге дайындық басталады. Кейбір мерозо¬
иттар эритроциттерге еніп жетілмеген жыныстық формаларга —
гаметоциттерге айналады. Олардыц кейбіреулері аналык жыныс
формаларын — макрогаметоциттерді, енді біреулері — аталык
жыныс формаларын — микрогаметоциттерді тугызады. Гаме-
тоциттер иіизонттарга Караганда ірілеу, домалақ болып келеді,
олардыц қою кара ядросы болады. Адам агзасында гаметоциттер
эрі карай дамымайды. Олардыц әрі карай дамуы маса денесінде
өтеді.
III. Жыныстык кобеюі жэне спорогония. Маса агзасына гаме¬
тоциттер ауру адамныц қанымен бірге енеді. Маса асқазанында олар
жетілген жыныстық нысанга немесе гаметаларга айналады. Ми-
крогаметоциттер болініп 5—6 талшык тэрізді микрогаметаларга,
ал макрогаметоциттердіц колемі үлкейіп 1 макрогаметага айнала¬
ды. Ұрықтанганнан кейін зигота козғалгыш оокннетаға айналып.
асқазан кабырғасын тесіп өтіп оныц сыртқы қабатына бекінеді. Ол
кабықшамен қапталып ооцистаға айналады да тез осе бастайды.
Содан кейін спорогония процесі басталады, ягни ядро мен ци¬
топлазма бөлінігі ооциста ішінде көптеген (1000) спорозоиттар
түзіледі. Ооциста қабықшасы жарылып спорозоиттар масаныц
дене куысына, гемолимфага, сілекей безіне етеді.
Патогендік әсері. Ауруларда мезгіл-мезгіл калтырау үстамасы
(қызбасы) байкалады. Әрбір қызбада 6—12 сагатқа созылган
256
қалтырау жэне дене температурасының 40°С дейін көтерілуі
байқалады.
Бауыр мен талақ ұлгаяды, каназдық байқалады. Емделмесе,
әдетте, дүние салуга алып келеді.
Лабораторинлық анықтау үшін қан жүгындысын микроскоп
арқылы зерттеп паразитті табу кажет.
Аурудың алдын алу шаралары — безгек резервуары бо¬
лып саналатын ауру адамдарды дер кезінде тауып емдеу; ауру
паразиттерін тасымалдаушы — безгек масасын жою болып табы¬
лады.
Кокцидий отряды — Cocciilia
Бұл отряд өкілдерінің ішінен маңыздысы
— токсоплазма.
Токсоплазма — Toxoplasma gondii —
адамның жэне жануарлардың токсоплазмоз
ауруын тудырушы, кең таралган паразит.
Токсоплазма адам агзасының эр түрлі
мүшелерінде — бас миында, скелет (кап¬
ка) жэне жүрек бұлшық еттерінде, көз ұл-
пасында, өкпеде, жатыр қабыргасында жэне
ұрық кабықшаларында кездеседі.
Токсоплазма барлық жерлерде таралган.
Морфофизиологиялық ерекшелікгері.
Токсоплазма эндозоит сатысында апельсин
жемісінің бір бөліміне немесе жарты айға
ұқсас болады; оның өлшемі — ұзындыгы
4—7 мкм, ені — 2—4 мкм. Оның алдыңгы
жагы кысыңқы, ал артқы жагы кең жэне
домалактанып келген. Денесінің алдыңгы
жагында конус тэрізді құрылым — коноид
болады. Оның қабыргасында ширатылган
фибриллдар кездеседі. Коноид паразиттің
иесінің денесіне енуі кезінде тірек қызметін
атқарады деп есептелінеді. Коноидтан кей-
ін токсоплазма денесінін артына қарай бір-
те-бірте кеңейген калта торізді органоид —
57-сурет. Токсоплаз¬
ма эндозоидынын
кұрылысы.
1 -коноид,
2-микрожіпшелер,
3-микротүтікшелер,
4-микропоралар,
5-май тамшылары,
6-ядро, 7-эндоплазмалык
тор, 8-Гольджи кешені,
9-митохондрия,
10-роптрии.
257
ponmpuit бастама алады. Роптрийда паразиттің жасушаға енуін
жеңілдететін заттар болады. Роптрий айналасында, дененің
алдыңгы жағында, онымен байланысқан жэне ирелендеген жіп-
шелер — микронемалар кездеседі. Аталған органоидтармен катар
токсоплазмада басқа да жасуша органоидтары болады. Дене сыр-
тын 3 қабат мембранадан тұратын пелликула қаптап тұрады, оның
астында түтік тэрізді фибрилдер жүйесі орналасқан. Пелликула
мен фибрилдер жүйесі паразиттің сыртқы қаңқасы болып табы-
Тіршілік циклі.
Акырғы, негізг і иесі
болып мысық; ара-
лык иесі — қүстар,
сүтқоректілер жэне
адамдар саналады.
Аралық иесінің
ішегіне спорозоит-
тары бар ооциста
енеді. Спорозоиттар
ооцистадан босанып
шыгып ішектің эпи¬
телий жасушаларына
енеді және бөлініп
көбейе бастайды.
Оның қайта-қайта
көбеюі нэтижесінде иесінің денесінде паразит саны еселеп өседі.
Олар жалпы бір қабықшамен қапталып цистага айналады.
Циста ақыргы иесінің денесіне өтсе, ондағы спорозоиттар
домалақтанып шизонтқа айналады. Ол шизогония жолымен
көбейеді. Осының нэтижесінде көптеген талшық тэрізді мерозо¬
иттар түзіледі. Эндозоиттар ішек қуысына өтіп жаңа жасушаларға
енеді де қайтадан шизонтқа айналып көбейеді. Бірнеше дүркін
жыныссыз көбеюден кейін жыныстық көбею басталады. Ол
үшін кейбір мерозоиттар жасушаға еніп жетілмеген жыныстық
нысандарға — микро-макрогаметоциттерге айналады. Микро-
гаметоциттерден 2 жіпше тәрізді қос талшықты сперматозоидтар
лады.
58-сурет. Токсоплазманын тіршілік циклі.
1-түпкілікті иесі, 2-3-4-ооцистаның даму
сатылары, 5-6-аралык ие, 7-тышкандардың
трансовариальдык закымдануы.
258
түзіледі, ал макрогаметоциттер өсіп ұлғайып, макрогаметаларға
айналады. Үрықтанганнан кейін зигота 2 қабат қабықшамен
қапталып ооцистага үласады. Ооцистада спорогония басталып
әркайсысында 4 спорозоиттары бар 2 спора түзіледі. Ооцисталар
аралық иесі үшін де, түпкілікті иесі үшін де, инвазиялық сатысы
болып табылады.
Ооцисталар үлкен немесе кіші дәрет арқылы сыртқа
шыгарылады. Адамдар токсоплазманы үй жануарларынан, әсіресе,
мысықтардан жүктырады.
Патогендік әсері. Оның клиникалық белгілері түрліше бо¬
лып келеді. Әдетте нерв, жыныс, лимфа жүйелері, көру мүшелері
зақымданады.
Алдын алу шаралары: үй жануарларын күтіп-баққанда жеке
гигиенаны сақтау қажет.
9.2.4. Инфузория класы — Infusoria
Күрделі қарапайымдылар болып табылады. Дене пішіні түрліше
болып келеді. Өлшемі 30—40 мкм-ден 1000-2000 мкм дейін.
Қозгалу органоиды болып кірпікшелері саналады. Кейбіреулерінің
қорганыстық қызмет атқаратын ерекше қүрылымдары — трихо-
цисталары болады. Оларда қарапайым ауыз қуысы (цистостом),
жасушалық жүтқыншак (цитофаринкс) кездеседі. Қорытылмаган
ас қалдыгы ерекше тесікше — қылаулатқыш арқылы сыртқа
шыгарылады.
Инфузорияларда 2 ядро кездеседі: біреуі — үлкен макронук¬
леус, екіншісі — кіші микронуклеус. Макронуклеус зат алмасу,
өсу т.б. вегетативтік қызметтер аткарады, ал микронуклеус —
жыныстық көбеюге қагыпасады.
Бүлардың көбеюі жыныссыз жэне жыныстық жолдармен
жүреді. Жыныстық процесс конъюгация күйінде болады. Ол үшін
2 дара бір-бірімен жанасып аталык жыныс ядроларымен алмаса-
ды. Содан кейін аталық жэне аналық ядролар өзара қосылады, ал
инфузория даралары бір-бірінен ажырасады.
Бүл кластыц өкілдері ішінен адам паразиті болып тек баланти-
дий саналады.
259
Балантидий — Balantidium coli
Бұлар адамның балантидиаз ауруын тудырады.
Ол тоқ ішекте кездеседі жэне барлық жерлерде таралган. Ба¬
лантидий денесі жұмырлау немеее жұмыртқа пішінді болып, өл-
шемі — үзындығы 30—200 мкм, ені 20 мкм-ге тең. Денесінің
алдыңғы қысыңқы жағында перистом, содан кейін воронка
тэрізді цитофаринкс, ал денесінің артқы жағында аналь тесігі
— қылаулатңыш орналасқан. Балантидий қорытылмаған ас
қалдығымен, эритроциттермен қоректенеді.
Жиырылғыш вакуолялары — екеу. Цисталары сопақша не
домалақ (дм 50—60 мкм) пішінді болып келеді.
Тіршілік циклі. Инвазиялық сатысы болып циста санала¬
ды. Ac қорыту жолында цистадан вегетативтік формасы түзіледі.
Вегетативтік форма күйінде балантидий ішекте ұзақ уақыт зиян-
сыз тіршілік етеді. Бірақ кейде белгісіз жагдайлардың әсерінен
(аскаридоз), олар ішектің қабырғасына еніп, оны зақымдап, соның
салдарынан үлкен жара пайда болады. Ішектің артқы бөлімінде
вегетативтік форма цистаға айналады да үлкен дәретпен сыртқа
шығарылады.
Балантидийдің негізгі резервуарлары болып жабайы және
үй шошқалары саналады. Олардың нәжісімен цисталар сыртқа
шыгарылып қоршаған ортаны ластайды. Адамдар шошқаларды
күтіп-баққанда цисталарды жүқтыруы мүмкін, сонымен қатар
жеміс-жидектер, лас қол, қайнатылмаган су, т.с.с. арқылы да жүгуы
мүмкін.
Патогендік әсері. Ішек қабырғасында қансыраған жара
туғызады, қан аралас іш өткізеді. Емделмесе дүние салуға алып
келеді.
Лабораториялық анықтау әдісі— нәжісті микроскоп арқылы
зерттеп паразит цисталарын табу.
Аурудың алдын алу шаралары — жеке шаралар — жеке ги¬
гиена нормаларын сақтау; қоршаған ортаның шошқа қиымен ла-
стануын болдырмау; шошқа фермаларында тиісті еңбек тэртібін
үйымдастыру, т.б. болып табылады.
260
II
59-сурет. Балантидий.
а) вегетативтік формасы: I бактерияны жұткан, II эритроциттерді
жұткан. І-цитостом, 2-цитофарринкс, 3-ас корыту вакуолі, 4-бөліп
шығарушы вакуоль, 5-макронуклеус, 6-микронуклеус; б) циста.
9.3. Жалпақ қүрттар типі — Plathelminthes
Бұл типке 7300-ге жуық түрлер жатады. Олар теңіздерде, тұщы
суларда, топырақта кездеседі; көпшілігі паразиттік тіршілік етуге
көшкен.
Бұл типтің өкілдерінің ерекшеліктеріне мыналар жатады:
1) бүлар 3 қабатты жануарлар болып табылады;
2) денесі тері — бұлшық ет қапшығымен қапталған;
3) дене қуысы болмайды, яғни мүшелер арасы паренхимамен
толтырылған;
4) денесі екі жақты симметриялы (билатеральды) болып
келеді;
5) денесі арка-кұрсақ бағытында жалпақтанган;
261
1
6) бұларда ас қорыту, зэр шығару, жүйке жэне жыныс мүшелері
дамып жетілген, кан айналым, тыныс алу мүшелері болмайды.
Жалпақ күрттар типі 3 класқа бөлінеді: 1) кірпікшелі қүрттар
(Turbellaria); 2) сорғыштар класы (Trematoda); 3)таспа қүртгар кла¬
сы (Cestoda). Адам паразиттері соңгы екі класқа кіреді, сондықтан
да біз енді соларға тоқталамыз.
9.3.1. Соргыштар класы — Trenuitodes
Бүл класқа 3000-ға жуық түрлер жатады, олардың бэрі де пара¬
зиттер болып саналады.
Бүлардың денесі жалпақ, жапырақ тэрізді болып, өлшемі 2-ден
800 мм дейін жетеді. Денесі тері — бүлшық ет қапшыгымен
қапталған. Ол сыртқы қабаттан — тегумент жэне 3 қабаты
(сақиналы, диагональ, үзына бойына орналасқан) біркелкі салалы
бұлшық еттен қүралган.
Соргыштар класынын ерекше белгісінің бірі — иесінің
денесіне бекінуге жэне қоректік затты соруга арналган ерек¬
ше соргыштарының болуы. Әдетте, 2 соргышы болады — ауыз
соргышы және кұрсак соргышы. Ауыз соргышы дененің алдыңгы
жагында орналасады жэне ол ауыз тесігімен байланысқан, ал
қүрсақ соргышы дененің құрсак бетінде орналасып бекіну қыз-
метін атқарады.
Асцорытужүйесі—ауызтесігінен, бүлшық еттіжүтқыншактан,
өңештен жэне екіге тарамдалган ішектен түрады. Ішектері тұйық
аяқталады, аналь тесігі болмайды. Қорытылмаган ас қалдыгы ауыз
тесігі арқылы кері шыгарылады.
Соргыштар иесі ішегінің жасушаларымен, сілемей жэне қан
плазмасымен қоректенеді.
Зэр шыгару жүйесі — протонефридиялардан түрады. Олар
көптеген жүлдызша тэрізді (терминалды) жасушалардан бастала¬
ды. Жүлдызша тэрізді жасушалардың кірпікшелі арнашықтары бо¬
лады, ал олар көпжасушалы зэр шыгару арнасына келіп қүйылады.
Зэр шыгару арнасы зэр шыгару тесігі арқылы сыртқа ашылады.
Жүйке жүііесі — жүтқыншақ маңындагы нерв сақинасынан
жэне одан таралган 3 жүп жүйке баганаларынан — арқа, бүйір
жэне құрсақ жүйке баганаларынан түрады. Жүйке баганалары бір-
262
r
бірімен көлденең жіпшелер арқылы байланысқан, сондықтан да
олар тор тэрізді болып келеді.
Жыныс жүйесі. Сорғыштардың бэрі дерлік гермофродиттер
болып саналады. Аталық жыныс жүйесі — жұп аталық бездерден,
2 шэует жолынан, шәует қүйылатын арнадан жэне жыныс мүшесі
— циррустан тұрады.
Аналықжынысжүйесі—тақаналыкбезден,жүмыртқажолынан
жэне сарыуыз безінен тұрады. Олар кең оотипке келіп құйылады.
Оотипте жұмыртқа жасушасы ұрықтанады жэне сарыуыз безінен,
Мелис денешігінен келіп жететін коректік заттарды пайдаланып
әрі қарай дамиды. Жүмыртқалар оотиптен жатырға өтеді де жы¬
ныс тесігі арқылы сыртқа шығарылады. Жатыр, сол сияқты, қынап
қызметін де атқарады, яғни ол арқылы оотипке аталық жыныс жа¬
сушалары келіп жетеді. Соргыштар айқас ұрықтанады.
Тіршілік циклі — күрделі, иесін алмастыру жэне бірнеше
дернәсілдік (личинкалық) сатысының алмасуы арқылы жүреді.
Сорғыштардың ақырғы, негізгі иелері — адамдар жэне
омыртқалылар, аралық иесі болып міндетті түрде моллюскілер са¬
налады. Кейбір соғыштардың екінші аралық иелері де болады — ол
балықтар не омыртқасыз жануарлар. Тіршілік циклінің ерекшелігі
— дернәсілдік сатыларының партеногенез жолымен көбейеді.
Соргыштар тудыратын ауруларды трематоздар деп атайды.
Биуыр соргышы — Fasciola hepatica
Фасциолез ауруын қоздырушы — паразит. Ол адамдардың от
жолдарында, от қабында, кейде ұйқы безінде не басқа мүшелерінде
кездесуі мүмкін (60-сурет).
Ересек формасын марита деп атайды. Ол үлкен қүрт,
яғни үзындығы 3—5 см болып келеді. Өзге сорғыштардан
айырмашылығы мөлшерінің үлкен болып келуі жэне жыныс
мүшелерінің күрделілігі. Көптеп тарамдалған жатыр құрсак
сорғышынан кейін орналасады; сосын аналық без, ал денесінің
бүйірлерінде саруыз бездері орналасқан. Денесінің ортасында
аталық безі кездеседі.
Тіршілік циклі. Бүлардың ақырғы иесі — ірі, үсақ қарамал,
жылқылар, шошқалар, қояндар, кейде адамдар. Аралық иесі болып
үлу саналады.
263
60-сурет. Трематодалар.
а) бауыр сорғышы, б) зэр шығару жүйесі, 1-протонефридия
арнашыктары, 2-орталық зэр шығару арнасы.
Бауыр сорғышының жұмыртқасы дамуы үшін суға келіп түсуі
кажет (61-сурет). Одан дернэсіл шығады — оны мирацидиіі дел
атайды. Оның жүйке түйіндері, жарық сезгіш мүшесі, зэр шыгару
мүшелері жэне денесінің артында ұрық жасушалары болады.
Денесінің алдыңгы жагында аралық иесіне енгенде оның тірі
үлпаларын ерітетін фермент бөліп шыгаратын ерекше без болады.
Мирацидий денесі кірпікшелермен қапталган, сондықтан ол суда
еркінжүзіп белсенді түрде аралықиесініңденесіне өтеді. Әрі қарай
паразит келесі дернәсілдік сатысы — спороцистаға айналады.
Спороциста қалыпсыз қалта тәрізді болып келеді және оның ешбір
мүшесі (нерв, зэр шыгару) болмайды. Ол тек көбеюге қабілетті.
Спороцистада үрық жасушаларынан партеногенез (ягни
үрықтанбай) жолымен жаңа дернэсілдік сатысы — редийлер да¬
мып жетіледі. Спороцистадан редийлер шыгады да иесінің жасу¬
шаларында паразштік тіршілік егеді.
Редийдің кейбір мүшелері дамыган, атап айтқанда: ауызы,
жүтқыншақ, ас қорыту түтігі жэне жаңадан түзілетін үрпақ да-
раларын сыртқа шыгаратын тесікше. Редийдің үрық жасушала¬
рынан партеногенез жолымен келесі дернэсілдік үрпақ — цер-
264
карий дамиды. Церкарий денесінде маритаға тэн көптеген
мүшелер кездеседі: сорғыштары, ішектер, нерв жэне зэр шыгару
мүшелері. Оның маритадан айырмашылығы — ұзын бұлшық етті
кұйрығының болуы. Ол суда қозғалу қызметін атқарады. Цер¬
карий ұлу денесінен суга өтіп жүзіп жүреді. Шамалы уақыттан
кейін ол су өсімдіктеріне бекініп, жабысып, қалың қабықпен
қапталып адолескарийга айналады. Егер адолескарийді ақырғы
иесі жұтса ол ас қорыту жолында ересек формата — маритага
айналып бауырға өтеді.
61-сурет. Трематодтардын дернәсілдік сатылары.
а) мирацидий, б) спороциста, в) редия, г) церкарий, д) метацерка-
рий. 1-кірпікшелер, 2-кезі, 3-үрык жасушалары, 4-редий ұрыктары,
5-церкарий, 6-сорғышы, 7-күйрығы, 8-кабығы.
Сонымен, бауыр сорғышының аралық иесі үшін инвазиялық
сатысы болып мирацидий, ал түпкілікті иесі үшін адолескарий са-
265
170-18
налады. Жануарлар шөп жегенде, су ішкенде, ал адамдар көкөніс
арқылы адолескарийлерді жүқтыруы мұмкін.
Патогендік әсері. Фасциола иесіне механикалық эсер етеді.
Олардың тіршілігінде түзілетін улы заттар аллергиялық эсер етеді.
Олар — эритроциттерді, лейкоциттерді жэне өт жолдарының эпи¬
телий жасушаларын жұтып қоректенеді. Сондықтан да бауырда
цирроз байқалуы мүмкін. Мұны лабораториялық анықтау үшін
нәжісті микроскоп арқылы зерттейді.
Алдыналушаралары.Қайнатылмағансудыішпеу,көконістерді
жуып жеу жэне санитариялык ағарту жүмыстарын үйымдастыру
болып табылады.
Мысық немесе сібір соргышы — Opisthorchis felineus
Бұл — описторхоз ауруын қоздырушы паразит. Ол адамдар
мен үй жануарларының (мысық, ит) бауырында, от қабында, ұйқы
безінде кездеседі.
Описторхоз негізінен Сібір өзендерінің жағалауындағы аудан-
дарда кең таралған.
метацеркария Морф оф И 3 И ОЛ О ГИ Я Л Ы Қ
62-сурет. Мысык сорғышынын гінде жүмыртқадан мираци-
Ішінде мирацидийі бар
жүмыртқа суға түседі, оны
моллюска жұтса оның іше-
Тіршілік циклі. Бүлардың
ақырғы иелері болып адам¬
дар жэне балықтармен қорек-
тенетін жыртқыш сүтқорек-
тілер саналады (62-сурет).
ерекшеліктері. Сібір сорғы-
шының денесінің ұзындыгы
4—13 мм. Денесінің орта-
сында тарамдалған жатыр,
содан кейін дөңгелек аналық
безі жэне денесінің артқы
бөлімінде екі аталық безі
орналасқан.
тіршілік циклі.
діш шығады. Бауыр жа-
266
сушаларында мирацидий спороцистага айналады. Спороцистада
партеногенез жолымен редий, ал олардан церкарияяар дамиды.
Церкарий моллюскі денесінен суға өтеді де балық денесіне енеді.
Балық денесінде ол теріасты жасушаға және бұлшық еттерге
өтіп, кос қабықшамен капталып метацаркарийга айналады. Егер
ақыргы иелері піспеген балық етін жесе паразиттің метацаркарийін
жүтады, ал ол ас қорыту жолында қабықшадан босанып өт жолы¬
на, бауырға енеді де ересек формата — маритага айналады.
Сонымен, Сібір сорғышының дамуы 2 аралық иесін алмасты-
ру арқылы жүреді. Оның біріншісі үшін инвазиялық сатысы —
жұмыртқа, екіншісі (балык) үшін церкарий; ал ақырғы иесі үшін
метацеркарий саналады.
Патогендік әсері. Описторхоз зілді ауру, ол кейде өлімге алып
келеді. Паразиттердің көптеп жиналуы оттің ағуын болдырмайды,
бауыр циррозына алып келеді. Ауруды анықтау нәжісті жэне дуо-
денум сөлін микроскоп аркылы зерттеумен шектеледі.
Алдын алу шаралары — балықты жақсы пісіріп не қуырып
жеу жэне санитарлық-агарту жұмыстарын үйымдастыру.
9.3.2. Таспа қурттар класы — Cestoidea
Бұл кластың 1800-ге жуық түрлері белгілі. Олардың бэрі де
паразиттер болып саналады.
Бұлардың денесі арқа-қүрсақ багытында жалпактанып, таспа
тэрізді болып келеді. Денесінің алдыңғы ұшында басы — ско-
лекс, одан эрі мойыны жэне бунақталган денесі -— стробиласы
орналасқан. Стробила бунактарын проглоттидалар деп атайды.
Сколекс жүмыр немесе жалпақтау болып келеді. Онда ішек
кабырғасына бекінуге арналган түрліше соргыштар, кармақшалар
кездеседі. Жаңа жас бунақтар (проглоттидалар) үнемі мойынын-
да түзіліп отырады жэне оған дейін пайда болғандарын денесінің
артына карай ығыстырады. Сонымен, мойнынан денесінің артына
қарай бунақтар (проглоттидалар) бірте-бірте ескіре береді.
Таспа кұрттар денесінің сыртын тері-бүлшық ет қапшығы
каптап түрады. Ол тегументтен — сыртқы кабаты жэне үш кабат
бұлшық еттерден (сақиналы, ұзына бойына орналасқан жэне диа¬
гональ) құрылган.
267
Зәр шығару жүйесі протонефридиялардан, нерв жүйесі бас
бөлімінде орналасқан алдыңғы жүйке түйінінен (ганглия) және
денесін бойлай созылган 2 бүйірлік жүйке баганаларынан түрады.
Ac қорыту жүйесі дамымаган.
Әрбір буылтықта жыныс мүшелері болады. Таспа қүрттар гер-
мафродиттер. Мойынга жақын орналасқан, жас буылтықтарда
жыныс мүшелері элі қалыптаспаған; денесінің (стробиланың)
ортасындагы буылтықтарда алғаш аталык жыныс мүшелері, содан
кейін аналық жыныс мүшелері қалыптасады. Қос жыныс мүшелері
кездесетін проглоттидаларды гермафродит буылтъщтар деп атай¬
ды. Онда аналық безі, саруыз бездері, оотип, нашар дамыган жа¬
тыр, аталық бездер, шәует құйылатын арна жэне циррус кездеседі.
Бұл құрттар айқас үрықтанады. Кейбір өкілдерінің денесінің арткы
бөліміндегі буылтықтарындагы жатыр жүмыртқалармен толты-
рылып, өте жақсы дамиды да жыныс жүйесінің өзге мүшелерін
ығыстырып, буылтықты түгел алып түрады. Оларды жетілген
буылтъщтар деп атайды. Жетілген буылтықтар үзіліп нәжіспен
бірге сыртқа шыгарылады. Бүл қүрттар айқас үрықтанады.
¥рықтанған жүмыртқаның дамуы жатырда басталады.
Жүмыртқа ішінде 6 қармақшалы үрық — онкосфера дамиды.
Онкосфераның әрі қарай дамуы үшін жүмыртқа аралық иесінің
денесіне енуі қажет. Иесінің асқазанында жүмыртқадан онкосфе¬
ра босанып шыгып, қантамырлар арқылы бүлшык еттерге отеді де
келесі дернэсілдік сатысы — финнага айналады. Финна — құрт-
тың басы, мойны кері қайырылып орналасқан көпіршік, оның
бірнеше түрлері белгілі: 1) цистицерк — сұйықтықпен толты-
рылған копіршік; оның ішінде паразиттің теріс айналдырылған 1
сколексі (мойны) болады; 2) ценур — бірнеше сколексі болатын
копіршік; 3) цистицеркоид — көпіршік екі болікке бөлінген; а)
сколексі болатын кеңейген жері; б) қүйрық тәрізді болімі. 4) эхино¬
кокк — бірнеше үсақ көпіршіктерден күрылған аналық көпіршік.
Әрбір копіршіктерде жеке сколекстер болады; 5) процеркоид және
плероцеркоид қүрт тэрізді финна. Оның алдыңғы жагында екі
соргыш саңылауы болады.
Дернәсілдердің ересек формата дамуы ақырғы иелерінің
ішегінде жүреді. Бүл жерде асқорыту ферменттерінің әсерінен
268
көпіршік еріп жойылып, сколекс кері айналып ішек қабырғасына
бекінеді. Ал мойыннан буылтықтар бөлініп стробила жетіледі.
Ақырғы иелері зақымданған аралық иесін қорек ретінде
пайдаланғанда өздеріне паразитті жұқтырады.
Шоіиқа иемесе қаруланган цепень — Taenia solium
Бұл — тениоз ауруын қоздырушы паразит. Оның ересек форма¬
сы аш ішекте, ал финналары негізінен, бұлшық еттерде кездеседі.
Сол сияқты, ол коздеде, мида да кездеседі.
Морфофизиологиялық ерекшеліктері. Ересек формалары-
ның ұзындығы 2—3 м. Басы (сколекс) өте кішкентай (2—3 мм),
онда көптеген қармақшалар жэне 4 сорғыш орналасқан. Гермаф¬
родит буылтықтары квадрат тэрізді болады. Аналық жыныс безі
ортасына орналасып, 2 үлкен және бір кішкентай бөлімдерге
бөлінген. Көптеген аталық бездері проглаттиданың бүйір бөлімінде
орналасқан. Жетілген буылтықтарда жатыр 7—12 тармақтарға
тармақталған (63-сурет).
63-сурет. Шошка цепенінің тіршілік цнклі.
а) басы: 1-кармакшалар, 2-сорғыштар, 3-мойыны, б) гермафродит буын:
1-аналык безі, 2-сарыуыз, 3-оотип, 4- жатыр, 5-аталык без, 6-шэует
еткізуші, 7-кынап, 8-клоака, 9-зәр шыгару арнасы, в) жетілген буын:
1-жатырдың негізгі тармағы, 2-бүйір тармақтары,
г) шошка цепенінің стробиласы.
Тіршілік циклі. Ақырғы иесі — адам, ал аралық иелері шошка
жэне адам.
269
Ауру адам нәжісімен бірге жүмырткаға толы жетілген
буылтықтар сыртқа шығарылады. Жұмыргканын эрі қарай дамуы
үшін ол шошқа ішегіне өтуі қажет. Әдетте, шошқалар лас, қоқыс
жерлерде жайылып нәжіспен бірге паразит жұмыртқаларын да
жүтады. Қарында олардан 6 қармақшалы онкосфера түзіледі. Он¬
косфера қармақшалары арқылы ішек кабырғасын тесіп өтіп қан
тамырларға енеді де, канмен бүкіл денеге таралады, эсіресе қаңқа
бүлшық еттеріне. Бүл жерде олар цистицеркке (финнаға) айна¬
лады. Финналар бүлшық еттерде үзақ уақыт сақталады. Финналар
ақырғы иесінде — адам ішегінде ересек нысанга айналады. Шала
піскен шошқа етін жегенде адамдар финналарды жұктырады.
Патогендік әсері. Механикалық зақымдау (иесінің қоректік
затын пайдалану) және улы заттарымен улау. Тениоз кезінде
асқорытудың бүзылуы, қаназдык, жалпы әлсіздік байқалады.
Тениоздың тағы бір қауіптілігі ол адамның ақырғы жэне аралық
иесі ретінде болуы, ягни адамда финналық сатысының дамуы (ци-
стицеркоз).
Тениоз ауруымен ауырған адамдар лоқсып қүсқанда тоқ ішек-
тегі жетілген буылтықтар асқазанға, өңешке жэне жүтқыншаққа
өтуі мүмкін. Олардың кейбіреулерін адам кері жұтады. Асқазанда
жүмыртқалардан онкосфера пайда болып, кантамырлар арқылы
коз, ми, бүлшық ет үлпаларында цистицеркке (финнаға) айналуы
мүмкін. Бүл кезде тениоз цистицеркоз ауруына ұласады. Цисти-
церкоз өте қауіпті ауру, әсіресе, ми цистицеркозы, ол өлуге алып
келеді, коз цистицеркозы адамдарды соқыр етеді. Цистицеркозды
тек хирургиялық жолмен емдейді.
Алдын алу шаралары: жеке шаралар — піспеген, шала піскен
шошқаетінжемеу;қоғамдық шаралар—санитарлық-ветеринарлық
жүмыстарды дүрыс үйымдастыру.
Сиыр немесе қарулаибаган цепень — Taenicirhynchus saginatus
Бүл—тениоринхоз ауруын қоздырушы паразит. Ересек күйінде
адамдардыц аш ішегінде паразиттік тіршілік етеді.
Морфофизиологиялық ерекшеліктері. Адамның ец ірі
паразиттік қүрты, оныц үзындығы 10—18 м жетеді. Сколексінде 4
соргышы болады, ал қармақшалары болмайды, сондықтан оларды
царуланбаган цепень деп атайды (64-сурет).
270
Стробиланың орта бөлігіндегі гермафродит буылтықтарында
1 мыңга жуық көпіршіктенген аталык бездер кездеседі, аналық
безі екі бөлімге бөлінген. Жетілген буылтықтарында жатыр
жұмыртқалармен толтырылып 17—35 бүйір өскіншелерін
(тармақтарын) пайда қылада. Әрбір проглоттидадағы жүмыртқалар
саны 175 мыңға жетеді. Бір жылда паразит 2500 проглотгида бө-
ліп шығарады.
64-сурет. Сиыр цепені.
1-сколекс, 2-жетілмеген буын, 3-гермафродиттік буын,
4-пісіп жетілген буын.
Нәжіспен сыртқа шығарылған жұмыртқаларда қалыптасқан
онкосфералар болады жэне проглоттида ішінде олар ұзак уақыт
өзгеріссіз сақталады.
Тіршілік циклі. Сиыр цепенінің түпкілікті иесі — адам, ал
аралықиесі — ірі қарамал. Ірі қарамал шөпжегенде нэжістегі про-
глоттидаларды жұтады. Мал қарнындажүмыртқадан 6 қармақшалы
онкосфера шығады да кан аркылы бүкіл денеге, соның ішінде
қаңқа бүлшық етіне таралады. Бүл жерде олар цистиг/ерк типтес
финиша айналады. Адамдар шала пісірілген сиыр етін жегенде
оны өздеріне жұқтырады.
271
ІІатогендік әсері. Ауру тениозга ұқсас, бірақ адамда финналык
сатысы дамымайды. Бұл паразитті анықтау нэжісті микроскоппен
зерттеу арқылы жүргізіледі.
Алдын алу шаралары: Жеке шаралар — піспеген сиыр етін
жемеу, қогамдық шаралар — тениозға ұқсас.
Ергежейлі цепень —Hymenolepis папа
Гименолепидоз ауруын коздырушы паразит. Ол адамдардың аш
ішегінде кездеседі.
Морфофизиологиялық ерекшеліктері. Бұл цепеннің ұзын-
дығы 1—5 см, стробиласында 200-ге жуық буылтықтар бола¬
ды (65-сурет). Алмұрт пішінді
қармақшалары орналаскан.
65-сурет. Ергежейлі цепень,
а) жалпы түрі, б) тіршілік циклі.
Патогендік әсері. Бұл цепень
кездеседі, 7—14 жастағы балалар
ауырады.
Патогендік әсері ішектің бүрлі
өнімдері арқылы агзаны улау б
сколексінде 4 сорғышы жэне
Тіршілік циклі. Ергежей-
лі цепеннің ақырғы және
аралық иелері болып адам¬
дар саналады. Адам ішегінен
жетілген жүмыртқалар сырт-
қа шыгарылады. Егер оны
жүтатын болсақ, ас қорыту
сөлінің эсерімен одан онкос¬
фера шығып ішек бүрлеріне
бекінеді. Бүл жерде одан ци-
стицеркоид типтес финна-
пар дамиды. Бірнеше күннен
кейін зақымдалған ішек бүр-
лері жойылып цистицерко-
идтар ішек қуысына өтеді.
Ішекте асқорыту сөлінің эсе-
рінен цепеннің басы кері
қайырылып ішек қабырғасы-
на бекінеді, 14—15 күннен
кейін ол көбейе бастайды.
негізінен жас бөбектерде жиі
жэне ересек адамдар сиректеу
қабығын зақымдау жэне улы
элып табылады. Гименолепи-
дозда бас ауруы, асқазан ауруы, ішек, нерв қызметінің бұзылуы
байқалады, балалар тез шаршайды, жалпы әлсіздік байқалады. Ба¬
лалар мазасызданып бей-жай болады.
Алдын алу шаралары. Жеке гигиена нормаларын қатал сақтау,
балаларды жеке тазалықты сақтауга машықтандыру, санитарлық-
ағарту жүмыстарын жүргізу, балабақшаларда тазалық сақтау,
ойыншықтарды стерильдеу, т.с.с. Бұл паразит тек лас қол арқылы
жұгады.
Эхинококк — Echinococcus granulosus
Эхинококкоз ауруын қоздырушы паразит. Дернәсілдік са-
тысы адамның бауырында,
кездеседі.
Морфофизиологиялык
ерекшеліктері. Денесінің
ұзындығы 2—6 мм, 3—4
буылтықтан тұрады.
Оның ен соңгысы — ен
үлкен, жетілген, ал онын
алдындағысы гермафро¬
дит буылтык болып табы¬
лады. Жатырда 5000-га
дейін жүмыртқалар бола¬
ды. Сколексінде 4 соргышы
жэне екі катар орналасқан
қармақшалары кездеседі.
Тіршілік циклі. Иелері
ит, қасқыр, шиборілер, ал
аралык иелері адам, ірі жэне
үсақ кара мал, шошқа, түйе,
қояндар, т.б. сүтқоректілер.
Ақыргы иесінің нэжісімен
жұмыртқалар сыртқа шыга-
рылады (66-сурет).
Сонымен қатар, жетілген
буылтықтары иттердің
аналь тесігінен шыгып жүн-
өкпеде, миында, ұзын сүйектерінде
66-сурет. Эхинококтын
морфологиясы және тіршілік циклі.
1-басы, 2-мойыны, 3-гермафродит
буын, 4-жетілген буын, 5-жатыр.
273
деріне ж үмыртқаларды қалдырады, ал олар шабындықтарды
(өсімдіктерді) ластайды. Шабындықта жайылған, аунаған мал-
дар (сиыр, қой) кұрт жұмыртқаларын жүндеріне жабыстыра-
ды. Адамдар үй малдарын бағып күткенде қолдары арқылы
жүмыртқалар-ды жұтады, жұктырады. Аралық иесінің ас қорыту
жолында эхинококк жүмыртқасынан онкосфера шығады, ол қан
тамырлар арқылы бауырға, өкпеге, мига таралады да эхинококк
типтес финнаеа айналады. Ол оте үлкен болады, адамдарда жас
баланың басындай, тіпті кейбір жануарларда 64 кг дейін жетеді.
Оның құрылысы да күрделі. Көпіршік 2 қабатпен капталған:
сыртқы қабаты (хитинді), ішкі қабаты (үрықты), іші сүйықтықпен
толтырылған. Ұрықтық қабаттан көптеген ұсақ көпіршіктер
түзіледі, олардың іштерінде кері айналган сколекстер болады.
Аналық көпіршік ішінде 2 үрпақ көпіршіктері кездеседі.
Патогендік әсері. Эхинококк адам ағзасына айтарлықтай эсер
етеді, себебі оның финнасы үлпаларды закымдап, мүшелердің
кызмет етуін қиындатады жэне улайды.
Оны тек хирургиялык жолмен емдеуге болады. Қазақстанда
эхинококкоз ауруына байланысты жылына бірнеше жүздеген опе¬
рациялар жасалады.
Лаборагориялык анықтау. Эхинококкоз финнасы адамның
ішкі мүшелерінде болатындықтан, оны тікелей анықтау мүмкін
емес. сондыктан қосымша көлденең иммунологиялык эдістер рен-
геноскопия, томография, сканерлеу қолданылады.
Алдын алу шаралары: Жеке тазалықты сақтау, қогамдық ша¬
ралар — иесіз, каңгып жүрген иттерді жою; сойылган ірі және үсақ
қара малдардың еттерін сараптамадан өткізу т.с.с.
Альвеококк —Alveococcus multilocularis
Альвеококкоз ауруын коздырушы паразит. Адам денесінде та-
ралуы эхинококкозга үксас, бірақ алдымен бауырды зақымдайды.
Географиялык таралуы. Эхинококкоздан ерекшелігі —
ошақты болып келеді. Батыс Сібір, Якутия, Орта Азия, Башқұрт-
стан т.б. елдерде кездеседі.
Морфофизиологиялық ерекшеліктері. Ересек сатысы эхи¬
нококка ұқсас, айырмашылыгы басында қармақшалар саны аздау,
жатырының дөнгелек болуы, жыныс тесігінің жыныс мүшесінің
артқы жагында емес алдыңгы жағында орналасуы.
274
Финналық сатысы ерекшелеу — финнасы көптеген ұсак
көпіршіктерден құрылган жэне сұйықтығы болмайды, үнемі
бүршіктеніп өсіп отырады да метастаз пайда етеді.
Тіршілік циклі. Ақырғы иелері — түлкі, қасқыр, ит, аралық
иелері кемірушілер (тышқандар) жэне адам. Жалпы алғанда
тіршілік циклі эхинококқа ұқсас. Адамдар түлкі, ит, т.б. терілерінде
болатын жұмыртқаларды жұтып зақымданады.
Патогендік әсері. Альвеококкоз эхинококкозға қарағанда си-
ректеу кездеседі, бірақ ол рак ауруы сияқты жайылып (метастаз-
данып) мүшелерді, эсіресе, бауырды зақымдайды. Альвеококк
көпіршігі мөлшерінің диаметрі 15 см жетеді.
Алдын алу шаралары. Эхинококкозга үқсас.
9.4. Жүмыр қүрттар типі — Nemathelminthes
Денесі буылтықтанбаған, көлденең кесіндісі жүмыр болып
келетін қүрттар. Тері-бүлшық ет капшығының ішінде алғашкы
дене қуысы болады. Асқорыту түтігінде артқы бөлім дамып, аналь
тесігімен аяқталады. Зэр шығару жүйесі — протонефридий типтес
не тері бездері күйінде; көпшілігі — дара жынысты. Нерв жүйесі
нерв түйіндерінен жэне олардан таралган нерв тізбектерінен
түрады. Қан айналу, тыныс алу мүшелері болмайды.
Бүл тип 5 класка болінеді, бірак олардың ішінде медициналық
маңызы бар өкілдері — адам паразиттері, нағыз жүмыр қүрттар
класында кездеседі.
Нагыз жұмыр ңұрттар кіасы —Nemaloda
Көптеген еркін жэне паразиттік тіршілік ететін түрлерді
қамтиды.
Морфофизиологиялық ерекшеліктері. Денесі үзын, жүмыр
болып үштарына қарай бірте-бірте жіңішкеленген. Денесінің
алдыңғы жағында ауыз тесігі, ал арткы жағына жақын, қүрса-
ғында анус орналасқан.
Тері-бүлшық ет қапшығы кутикуладан, гиподермадан
жэне бүлшық ет қабаттарынан түрады. Кутикула тірек кызметін
275
атқарады, оның астында гиподерма жэне бір қабат бұлшық етгер
орналасады. Өте баяу қозгалады.
Тері-бұлшық ет қапшығының ішінде сүйықтыққа толтырылган
алғашқы дене қуысы болады. Қуыс сұйықтыгы жогары кы-
сымды болып қосымша тірек қызметін атқарады.
Ас корыту жүйесі ауыздан басталып ануспен аякталады. Ауыз
қуысы кутикулалы еріндермен қоршалган. Ac бір багытта жылжи-
ды, сондықтан да ол жақсы корытылып сіңіріледі.
Зэр шыгару жүйесі. 1 не 2 алып жасушалардан (тері бездері)
қүрылган. Оның денесі нематодтардың алдыңгы бөлімінде
орналасқан, одан артқы жэне алдыңгы жагына қарай арнашық-
тармен тесілген өсінділер таралган. Артқа қарай багытталган
арнашыктар түйық аяқталады, алдыңгы жагына багытталган
арнашықтар ортақ бір арнага топтасып сыртқа экскреторлық тесік
арқылы ашылады.
Нерв жүйесі. Жүтқыншак айналасындагы нерв сақинасынан
жэне одан тарамдалган нерв баганаларынан түрады.
Немагодтар дара жынысты жануарлар, оларда жыныстық ди¬
морфизм жақсы дамыган — еркек даралары үсақ, қүйрықтары
ширатылган, ал үргашы даралары үлкен, ірі болып келеді.
Тіршілік циклі. Нематодтардың көпшілігі иесін алмастырмай
дамиды. Ересек даралары паразиттік тіршілік етсе, жұмыртқа-
лары және дернэсілдері иесінен тыс — сыртқы ортада дамиды.
Оларды геогельминттер, ал кейбіреулерінің жүмыртқалары мен
дернәсілдері аралық иелерінің денесінде дамиды, оларды био-
гельминттер деп атайды.
Үшкірқұрт (аққұрт) — Enterobius vermicularis
Энтеробиоз ауруын тудыратын, кең таралган жэне жиі
кездесетіні — адам паразиті. Үшкірқүрт адамның аш ішегінің
(жіңішке ішек) төменгі, тоқ ішектің алдыңгы бөлімінде кездеседі.
Морфофизиологиялық ерекшеліктері. Үрғашылары 10—12
мм, еркек даралары 2—5 мм болып келетін ақ түсті қүрт. Ауыз
тесігін кутикула ісіндісі — везикула қоршап түрады, ол ішек
қабыргасына бекіну үшін қажет. Жүтқыншақтың артқы жагында
да шар тәрізді ісінді — бульбус болады, ол да бекіну қызметін
атқарады. Үшкіркұрт ішекте қорытылған аспен қоректенеді (67-
сурет).
276
Тіршілік циклі. Ішек ішінде ұрық-
танады; ұрықтанғаннан кейін еркек
даралары өліп қалады. Ұрғашы дара-
ларының жұмыртқаға толтырылған
жатыры өте ұлғайып, бульбусты
қысады да кұрттың ішек қабырғасынан
босануына мүмкіншілік туғызады.
Олар ас агынымен, ішектің жиырылуы
салдарынан тік ішекке келіп жетеді.
Түнде анустан сыртқа шығып анус
айналасындағы тері бетіне, шапқа
жұмыртқалайды (13000 данага дейін).
Содан кейін құрт өліп калады.
Жұмыртқалардың эрі қарай да¬
муы үшін 34—36° жылылық, 70—
90% ылғал қажет. Анус айналасын-
да, шапта жоғарыдағыдай жағдайлар
болғандықтан, 4—6 сагат ішінде
жұмыртқалар дамып инвазиялық сатыға
жетеді. Инвазиялық жұмыртқа адам
ішегіне түсіп ересек формаға айналады.
Ересек даралар ішекте 30 тэулікке дейін
өмір сүреді.
Энтеробиозды толық емдеу өте
қиын, себебі бұл аурумен көбінесе жас
балалар ауырады жэне ауру адамдар үшкірқүрт жұмыртқаларын
өздеріне-өздері жұқтырады. Үрғашы даралары жүмыртқалағанда
теріні қышытады, ал балалар қышыған жерлерін қасығанда
жүмыртқалар не төсекке төгіледі, не тырнақ астына кіріп дамиды
да инвазиялық сатыға жетеді. Лас қолымен тамақ жеген адам олар¬
ды ішке жүтады.
Патогендік әсері. Қышу, тәбеттің болмауы, іш оту, мазасызда-
нып ұйқының бұзылуы.
Лабораториялық анықтау. Шап терісінің жүғындысын ми¬
кроскоп аркылы зерттеу.
Алдын алу шаралары. Жеке тазалықты сақтау.
67-сурет. Үшкіркүрг.
а) үргашы дарасы, б) еркек
дарасы. 1-аузы, 2-везикула,
3-өңеш, 4-бульбус, 5-ішек,
6-жыныс тесігі, 7-жатыр,
8-жүмыртка жолы,
9-аналык безі, 10-аналь
тесігі.
277
Адам ііиекқұрты (аскарида) —Ascaris lumbricoides
Адамдардыц аскаридоз ауруының қоздырушысы.
Морфофизиологиялық ерекшелігі. ¥рғашы даралары 20—40
см, еркек даралары — 15—20 см жететін ірі қүрт. Ауыз тесігі 3
кутикалы еріндерімен қоршалған. Жұмыртқалары ірі, домалақ не
сопақшалау болып келеді.
Аш ішекте паразиттік тіршілік етеді. Энтеробиозға қарағанда
сиректеу кездеседі.
Тіршілік циклі. Тек қана адам ағзасында паразиттік тіршілік
ететіні —геогельминт. ¥рықтанған ұргашы даралары аш ішекте
тэулігіне 240000 жүмыртка жүмыртқалайды. Жүмыртқалардың эрі
карай дамуы үшін сыртқа шығуы қажет, себебі олар оттегі, ылғал
және 20—25° жылылық болған жагдайда дамиды. 21—24 тәулікте
жүмыртқада қозғалгыш дернәсіл пайда болып инвазиялық сатыға
жетеді. Көкөніс, жеміс-жидектерді жумай жегенде инвазиялық
жұмыртқалар ішекке өтеді. Онда жүмыртқа қабығы еріп
дернәсіл босанып шығады. Дернәсіл ішек кабырғасын тесіп, кан
гамырларға өтеді де қанмен бауырға, жүрекгің оң жақ бөліміне (оң
жақ жүрекше, оң жақ қарынша), өкпе артериясы арқылы өкпеге,
өкпе кылтамырларына өтіп ауысады. Осы кезде дернәсіл қозғала
бастайды. Қозғалғыш дернэсіл кылтамыр кабырғасын тесіп өкпе
көпіршіктеріне, бронхиолдарға, ауатамырға (бронх), кеңірдекке,
жүтқыншакка өтеді. Осы жерден дернэсіл сілекеймен бірге екінші
рет жұтылып ішекке келіп жетеді де, ересек формаға айналады.
Дернәсілдің көшіп қонуы 2 аптаға, ал ересек формага айналуы
70—75 тэулікке созылады.
Ересек даралардың тіршілігі 10—12 айға созылады.
Патогендік әсері. Дернәсілдік сатысы өкпе, бауыр ұлпаларын
бұзады жэне ағзаның аллергиялық реакцияларын туғызады.
Ересек формалары ағзаны улы заттармен улайды, ішекті
тығындап, механикалық эсер етеді.
Лабораториялық аныктау. Жүмыртқаларын табу үшін нәжісті
микроскоп арқылы зерттейді.
Алдын алу шаралары. Жеке тазалықты сақтау, жеміс-
жидектерді. кокөністерді жуып жеу.
Қоғамдық шаралар — санитарлық ағарту жүмыстарын
ұйымдастыру, ортаны ластанудан сақтау.
278
Қылбасқұрт — Trichocepltalus trichiurus
Адамдардың трихоцефалез ауруының қоздырушысы. Бүйенде,
соқыр ішек өсіндісінде, тоқ ішектің бас жағында кездеседі.
Морфофизиологиялық ерекшеліктері. Ұрғашы дараларының
ұзындығы 5,5 см, еркек даралары 1,5—3 см болады. Денесінің
алдыңғы жагы жіпше не қылға ұқсас өте жіңішке, ал артқы
жағы жуан болып келеді. Денесінің алдыңғы жіңішке бөлігінде
тек жұтқыншақ, ал калған асқорыту мүшелері артқы бөлімінде
орналасқан. Қылбасқұрт денесінің алдыңғы, жіңішке бөлімімен
иесінің ішек қабырғасына еніп, қанмен қоректенеді (68-сурет).
Тіршілік циклі. Ұрықтанғаі
ұргашы даралары ішек қуысын
жүмыртқаларын жүмыртқалайды
олар нәжіспен сыртқа шығарылады
Сыртқы ортада 26—28° жылылықта, -
апта ішінде жұмыртқалар инвазиялы
сатысына жетеді. Лас қол, жуылмағаі
жеміс-жидек, көкөністер арқылі
инвазиялық жүмыртқалар ішекке ке
ліп түседі де бүйенге жетіп, ересе:
формата айналады. Қылбасқұртта]
адам ағзасында 5 жылға дейін тіршілі:
ете алады.
Патогендік әсері. Ішектердіі
шаншып ауруы, тәбеттің бүзылуы
іш өту, нерв жүйесінің бұзылуы, ба
айналу, талып қалу, т.б. қүбылыстар"
байқалады.
Лабораториялык аныктау. Нәжі-
сті зерттеп, жұмыртқаларын табу.
Алдын алу шаралары. Жеке тазалықты сақтау.
68-сурет. Қылбаскүрт.
а) ұрғашы дарасы,
б) еркек дарасы.
Трихинелла — Trichinella spiralis
Табиги ошақ — трихинеллез ауруын қоздырушы паразит.
Ересек формасы иесінің аш ішегінде, дернәсілдері — бүлшык
еттерде тіршілік етеді.
279
Морфофизиологиялық ерекшеліктері. Өте кішкентай,
ұрғашы даралары 3-4-0,6 мм, еркек даралары 1,5x2-0,004 мм бо¬
лып келетін күрт. Бұлар дернэсілдерден тірі туады.
Тіршілік циклі. Адамдарда, үй жануарларында (шошқа, ит)
және жабайы жануарларда (жабайы шошқалар, атжалмандар,
тышқандар, аю, түлкілер т.б.) паразиттік тіршілік ететін биогель-
минттер. Бір ағза негізгі жэне аралык иесі бола алады.
Ерееек формалары аш ішекте 4 аптаға дейін тіршілік етеді.
Үрықтанғаннан кейін еркек даралары өліп қалады, ал ұрықтанган
ұрғашы даралары денесінің алдыңғы жағымен иесінің ішек
қабырғасына еніп, 2000-ға жуық тірі дернэсілдерді шығарады.
Олар қан не лимфа арқылы бүкіл денеге таралып, қаңка бұлшық
еттерінде, эсіресе, көкетте жинақталады. Қабырғааралық, шай-
нау, сан бұлшық еттерінде шамалы уақыттан кейін дернәсіл
шиыршықталып ширатылады. 2—2,5 айдан кейін дернәсіл
айналасындағы ұлпалар капсула қалыптастырады, оның мөлшері
0,25x0,66 мм жетеді. Бір жылдан кейін капсула қабырғасы
әктастанын, катаяды.
Дернэсілдің ересек формага айналуы үшін ол басқа иесінің
ағзасына өтуі кажет. Екінші иесінің ішегінде капсула еріп ол боса-
нып шығады да 2—3 күннен кейін ересек формаға айналады.
Патогендік әсері. Дернәсіл ағзаны улайды, сондықтан бет,
кабақ ісіп, дене температурсы 40° дейін көтеріледі, асқазан, ішек
қызметі бұзылады. Кейінірек бұлшык еттердің шаншып ауруы
байқалады.
Лабораториялық анықтау. Бұлшык еттерде дернәсілдерді
табу (биопсия) жэне иммунологиялык реакциялар арқылы.
Алдын алу шаралары. Қоғамдық шаралар — мал сойылатын
орындарда санитарлық-ветеринарлық нормаларды сақтау, мал-
дарды таза ұстау, атжалмандарды күрту.
9.5. Буынаяқтылар типі — Anthrpoda
Буынаяқтылар типінің көптеген өкілдері адам ауруларының
қоздырғыштары, таратушылары деп саналады, мекен ету орталары
аралык иелерінде және табиғи резервуарлар болып табылады.
280
Буынаяқтылар жоғары сатылы омыртқасыз жануарлар, оларға
1,0—1,5 миллиондай түрлер топтастырылады.
Буынаяқтылардьщ денесі буылтықтарға бөлінген. Буылтық-
тары бір-бірімен кірігіп дене бөлімдерін — бас, көкірек, құрсақты
кұрайды.
Буынаяқтылардың аяқтары да буылтықтардан түрады.
Аяқтарының кызметі түрліше — олар тек кана қозғалу қызметін
атқарып қоймай, сол сияқты сезу, ауыз аппараты, қорғаныс жэне
шабуылдау қызметтерін де атқарады.
Денесі хитин қабатымен қапталған. Хитин сыртқы қаңқа,
қорғаныс қызметтерін атқарады. Бұлшық еттері көлденең жолақты
болып, күшті жэне әр түрлі қимыл-қозғалыстарды камтамасыз
етеді.
Дене қуысы аралас немесе миксоцель күйінде. Эмбриогенез
барысында бірінші болып алғашқы дене қуысы қалыптасады, со-
сын соңғы дене қуысы пайда болады, бірак алғашқы дене қуысы
толық жойылмай, шамалы бөлігі болса да сақталынып калады.
Мүндай дене қуысын аралас дене цуысы деп атайды.
Ас корыту жүйесі — алдыңғы, ортаңғы, артқы бөлімдерден
түрады. Алдыңғы болімінде хитинді тістері (үнтақтаушы аппа¬
рат), ортангы бөлімде жүп өскінше — асқорыту безінің (бауыр)
бастамасы болады. Ауыз аппараты 3 жүп түрі өзгерген аяқтарынан
құрылган.
Қан айналуы ашық типті, 5 бүрышты қалта тэрізді жүректен
жэне қысқа қан тамырларынан түрады.
Тыныс алу мүшелері — желбезек, жапырақша тэрізді окпе
жэне трахеядан (демтүтік) түрады.
Зәр шығару жүйесі — метанефридиялардан, мальпигий
түтіктерінен немесе жасыл бездерден қүрылған.
Жүйке жүйесі жүтқыншақ манындағы сақинадан жэне кор¬
сак, жүйке тізбегінен түрады.
Буынаяқтылардың көпшілігі дара жынысты. Буынаяқтылар
типі — шаянтәрізділер, ормекшітэрізділер жэне бунақценелілер
(насекомдар) кластарына болінеді. Олардың ішінен өрмекші-
тэрізділердің жэне насекомдардың медициналык маңызы белгілі.
170-19
281
9.5.1. Өрмекіиітәрізділер класы —Arachnida
Өрмекшітәрізділердің кейбіреулерінің дене бөліктері бірігіп
2 бөлімге баскөкірек, кұрсаққа — (бүзаубастар) бөлінген, екін-
шілерінің барлық бөліктері өзара біріккен, бөлімдерге бөлінбеген.
мысалы, кенелер. Бұлардың 6 жұп аяқтары, оның ішінде 4 жұп
жұру аяқтары болады.
Ac қорыту жүйесініц құрылысы түрліше, бірақ барлық
өрмекшітәрізділердің қоректену ерекшелігі — сұйық, жартылай
қорытылған асты сорып қоректенеді. Ол үшін жүтқыншақ мықты
дамыған бұлшық еттерден құрылган. Алдыңғы ішекке жұп
сілекей бездері ашылады жэне ортаңғы ішекте жұп өскін — бауыр
дамыған.
Зәр шығару жүйесі түрі өзгерген метанефридиялардан не
мальпигий тамырларынан (сосудтарынан), тыныс алу жүйесі
жапырақ тэрізді өкпеден не трахеядан (демтүтік) түрады.
Жүйке жүйесі жүтқыншақ маңындагы сақинадан жэне кур-
сак, жүйке тізбектерінен түрады.
Өрмекшітэрізділер өкілдерінен тек кенелер отрядының меди-
циналық маңызы белгілі.
Кенелер отряды —Асигі
Кенелердің мөлшері өте кішкентай — 0,1 мм ден 10 мм-ге
дейін. Денесі бөлімдерге бөлінбеген, хитин қабаты тері тәрізді
созылған, ал кейбіреулерінде хитин қабатының учаскелері қатай-
ып қалқаншаға айналған.
Кенелердің 6 жұп аяктары, олардың екеуі (хелицерлер жэне
педипальпалар) ауыз аппаратына айналған, ал 4 жүбы жүру
аяқтары.
Тыныс алу мүшесі — трахея (демтүтіктер), зэр шығару мүшесі
— мальпигий түтіктері.
Кенелер дара жынысты насекомдар.
Тіршілік циклі. Өрмекшітәрізділер толық түрленіп (мета¬
морфоз) дамиды — жүмыртқа, дернәсіл, нимфа жэне имаго.
Дернэсілдерінің 3 жүп жүру аяқтары болады жэне бүкіл денесі-
мен тыныс алады. Олар түлеп нимфаға айналады.
Нимфалардың 4 жұп жүру аяқтары, тыныс алу мүшесі — тра-
хеялары болады, бірақ жыныс мүшелері элі қалыптаспаған. Ним¬
фа түлеп имагоға (ересек формага) айналады.
Кенелер таратушылар, паразиттер жэне табиги резервуарлар
болуы мүмкін.
Иксод кенелер тұқымдасы — Ixodidae
Ірі, мөлшері 4—5 мм болатын кенелер (70-сурет). Тойынған
ұргашы кенелердің мөлшері 10 мм ден артық болады. Еркек да-
раларда денесінің үстіңгі бетін толык жауып түратын қалқанша
кездеседі, ал ұргашы дараларда жэне дернэсілдерде қалқанша
тек денесінің алдыңғы бөлігінде ғана болады, ал қалған бөліктері
жұқа, созылмалы хитин қабатымен капталған. Мүның маңызы өте
зор, себебі ұрғашы даралар көп мөлшерде, массасынан 200—400
есе артық мөлшерде, қан сорып ісінеді.
Иксод кенелері тіршілігіне қолайсыз жерлерді — ормандарды
мекендейді. Ол жерлерге ұзақ уақыт қоректендіруші жануарлар
енбеуі мүмкін, сондыктан олар аштықка шыдауға бейімделген.
Сонымен қатар, мекен ортаеына коректендіруші жануарлардың
еніп келгенін сездіретін ерекше бейімделушіліктері болады: 1)
ірі жануарлардың (бұғы, аю, маралдар т.б.) жүруі салдарынан
топырақтың азды-көпті тербелісін (вибрациясын) сезетін өте
нәзік сезімтал құрылымдары дамыган; 2) мекен ортасына енген
жануарлардың тыныс алуы салдарынан орта температурасының
болар-болмас өзгеруін сезетін терморецепторлары дамыған.
Сонымен катар мекен ортасы олардың дамуы үшін қолайсыз
болғандыктан, көп мөлшерде жұмыртқалайды.
Тайга кенесі — Ixodes persulcatus — көктемгі-жазғы энцефа¬
лит ауруының тасымалдаушысы жэне табиғи резервуары. Сібір-
де, Шығыс Қазақстанда кең таралған.
Еркек дараларының мөлшері — 2,5 мм, ұрғашылары 4 мм.
Дернәсілдері кемірушілердің, кірпілердің, құстардың қанын сора-
ды және топыраққа өтіп түлейді. Нимфалары ірі кемірушілердің,
тиіндердің, қояндардың канын сорады. Ересек формалары — ірі
қара малдың, бұгылардың, маралдардың қанын сорады.
283
Ит кенесі — Ixodes ricinus — туляремия, шотландия энце-
фалитінің қоздырушысын тасымалдайды. Қылқан жапырақты,
бұталы аймақтарды мекендейді. Мөлшері, даму циклі тайга
кенесіне ұқсас.
Аргас кенелері тұқымдасы —Argasidae
Аргас кенелері жабық табиғи немесе жасанды мекендерді,
мысалы, үңгірлерді, жануарлардыц індерін, апандарды
жэне балшықтан салынған үйлерді, қора-қопсыларды т.с.с.
мекендейді. Бұлардың тіршілік жағдайлары иксод кенелерге
Караганда біршама қолайлы болып келеді, сондықтан да олар
көптеп өлмейді. Бұл кенелер мекен-жайына енген жануарлар-
мен ғана қанагаттанады, сондықтан олардың қожайындарының
саны өте көп (жоргалаушылардан бастап адамдарга дейін).
Қоректендірушілердің аз уақытқа болса да мекенге енгенін пай-
даланып тез қоректенеді де, 3—30 мин. арасында тойынады. Бұл
кенелер аз мөлшерде қоректенеді жэне мекен-жайлары дамуы
үшін қолайлырақ болгандыктан, санаулы гана (100) жүмыртқа
жұмыртқалайды, бірақ аз өмірінде бірнеше рет жүмыртқалайды
(69-сурет).
Бүлардың мекен-жайына ұзақ уакыт ешбір жануар енбеуі
мүмкін, сондыктан да кенелер көп жылдап ашығады, олардың да¬
муы 20—28 жылға созылады. Бұлардың дамуында жүмыртқа, ли¬
чинка сатыларынан кейін бірнеше нимфалық сатылары (нимфа-1,
нимфа-2, нимфа-3, т.б.) ауысып, содан кейін ересек формата (има¬
го) өтеді. Бүл тұкымдастың өкілінің бірі — ауыл (поселке) кенесі.
Ауыл кенесі — Ornithodorus papillipes
Қайталама кене сүзегінің таратушысы жэне табиги резервуары
болып табылады. Олар негізінен Орта Азия елдерінде таралган,
үңгірлерді, сүтқоректілер мен қүстардың індерін жэне ұяларын,
балшықтан салынган үйлердің, қоралардың едендегі тесіктерін
мекендейді.
Морфофизиологиялық ерешеліктері. Бұл кененің түсі —
күңгірт-сүр болып келеді. ¥ргашыларының денесінің үзындыгы
284
8,2 мм, еркек даралары — 5,8 мм. Көздері болмайды. Бұлар
кемірушілер, жарқанаттар, торғайлар жэне иттер, ірі қара мал,
жылқы, мысық т.б. үй жануарларының денесінде қоректенеді. Ере¬
сек кенелер ұзак уақыт —13 жылға дейін, коректенбей аш күйінде
сақтала береді.
Алдын алу шаралары. Жеке шаралар: кенелердің денеге жа-
бысуынан сақтану жэне кенелер мекендейтін биотоптарга кір-
меу; кенелердің денеге жабысуын болдырмайтын химиялық зат-
тарды пайдалану.
Қоғамдық шаралар: кенелерді жэне кемірушілерді жою;
балшықтан салынған ескі құрылыстарды бұзу, олардың орнына
еуропа типтес үйлерді салу, т.с.с.
Қышыма кенесі — Sarcoptes scabiei — акариформды кене¬
лер тұқымдасының өкілі, адамдардың қышыма (қотыр) ауруын
қоздырушы паразит.
Денесі домалак, молшері 0,3—0,4 х 0,2—0,3 мм. Денесінде
көптеген кысқа тікенектер жэне ұзын қылтанақтар кездеседі.
69-сурет. Ауыл кенесі.
а) арка жағынан, б) кұрсак жагынан. 1-ауыз аппараты, 2-аналь тесігі.
Аяқтары өте қысқа, көздері дамымаған. Бүкіл денесі арқылы
тыныс алады, 4—5 апта өмір сүреді. ¥рғашы даралары терінің
мүйізді қабатын тесіп бірнеше мм-ге жететін тар індер қазады,
285
оған тері бетінде тіршілік ететін еркек даралары еніп ұрғашыларын
ұрықтандырады. Содан кейін ұргашы даралары 50-ге жуық
жұмыртқалар жұмыртқалайды. Даму циклі жұмыртқа. нимфа жэне
имаго сатыларынан тұрады.
W\ / Бұл кенелер терінін кез келген
учаскелерінзақымдауымүмкін,бірақ
көбінесе терінің нэзік учаскелерінде
(саусак арасында, қолтықта, шапта,
касағада) кездеседі.
70-сурет. Тайга кенесінін даму
сатылары.
а) дернәсіл, б) нимфа, в) имаго
— ұргашы дарасы, г) имаго —
еркек дарасы.
71-сурет. А'ышымп кенесі а), дамуы б)
286
Кенелер теріні өте күшті қышытады, әсіресе, түнде. Адам¬
дар кышыган жерлерін қасығанда сол жерлерге микробтар түсіп
қабыну, іріндеу қүбылыстары байқалуы мүмкін.
Алдын алу шаралары — тазалықты сақтау, ауру адамдарды
емдеу.
9.5.2. Бунақдепелілер (насекомдар) класы — Insecta
Насекомдар ең жоғарғы сатылы омыртқасыз жануарлар болып
табылады. Сонымен қатар, насекомдар ең үлкен класс, яғни оларга
1 млн-нан астам түрлер кіреді.
Морфофизиологиялық ерекшеліктері. Насекомдардың дене-
сі 3 болімге болінген — бас, көкірек, қүрсақ. Көкірек бөлімі 3
буылтықтан түрады, эрбір буылтығында 1 жүп жүру аяқтары
болады. Демек, насекомдардың 3 жүп жүру аяқтары болады.
Қүрсак бөлімі 6—12 буылтыққа бөлінген. Басы көкірек бөлімімен
қозгалмалы түрде байланысқан. Онда жүп мүртшалары; ауыз ап¬
параты жэне 2 қарапайым не күрделі көздері орналасқан. Ауыз ап¬
параты 3 жүп аяқтарынан пайда болған. Ол үстіңгі және астыңғы
жақтардан, төменгі жэне үстіңгі еріндерден түрады. Ауыз ап-
паратына, сол сияқты, ауыз қуысының төменгі бетінің хитинді
өскіншесі — ті.п немесе гипофаринкста жатады.
Қоректену ерекшеліктеріне қарай — кеміруші, шағушы, жалау-
шы, сорушы ауыз аппараттары ажыратылады.
Насекомдардың копшілігінің көкірек болімінің екінші жэне
үшінші буылтықтарында жүп канаттары кездеседі. Кейбір насе-
комдардың қанаттары болмауы мүмкін. Қанатсыз насекомдар
алғашкы канатсыздар (қарапайым түрлері) жэне соңғы қанатсыз-
дар (биттер, бүргелер) болып бөлінеді. Соңгылары паразитгік
тіршілік етуге бейімделу салдарынан қанаттарын жойған.
Насекомдар денесінің сыртын хитин қабаты каптап түрады,
оның астында бір қабат гиподермалы эпителий орналасқан.
Терісінде түрліше бездер кездеседі. Бүл шық еттері көлденең
жолақты болып келеді.
Аскорыту жүйесі алдыңғы, ортаңгы және артқы ішектерге
жіктелген. Алдыңғы ішек ауыз қуысынан басталып, жүтқыншақ
жэне оңешке бөлінген. Өңештің артқы бөлігі кецейіп, жемсау
287
жэне шайнайтын карын қалыптасқан. Алдыңғы ішекке 3 жұп
сілекей бездері де кіреді. Олардың секреттері ас қорыту қызметіне
қатысады: асты ылғалдап тұрады жэне әр тұрлі ферменттер бө-
леді. Бал арасының сілекей бездерінің бір жұбының секреті жемса-
уда гүл шырындарымен араласып қосылып, бал қалыптастырады.
Ортаңғы ішекте ешқандай бездер болмайды, дегенмен, осы жерде
ас қорытыльш, сіңіріледі. Артқы ішек аналь тесігімен аяқталады.
Тыныс алу мүшесі болып трахеялар саналады. Ол дененің
барлық мүшесіне, үлпасына тарамдалып таралған түтіктер жүйе-
сінен түрады. Трахеялар сыртқа стигмалар арқылы ашылады.
Зәр шығару мүшесі болып Мальпигиев түтіктері саналады.
Ол ортаңгы жэне артқы ішектер аралыгында ішекке келіп қүятын
көптеген түтікшелерден түрады.
Қанайналым мүшелері — жүрек жэне аорта — денесінің
арқа бетінде орналасқан. Қанайналым жүйесі ашық жэне нашар
дамыған. Ол арқылы ағылатын сүйықтыкты гемолимфа деп атай¬
ды. Онда ақ қан түйіршіктері болады. Кейбір насекомдардың гемо-
лимфасы улы болып келеді.
Жүйке жүйесі жақсы дамыған. Ол жүтқыншақ маңындағы
түйіндерден жэне құрсақ-жүйке тізбегінен түрады. Ми 3 бөлімге
бөлінген: алдыңғы, ортаңгы және артқы. Жүтқыншак үстіндегі
түйіннің күрылысы өте күрделі, онда ерекше қызмет аткаратын
— ассоциативтік орталық болып табылатын саңырауқүлақ тәрізді
дене кездеседі.
Сезу мүшелері де жақсы дамыган — оның ішінде иіс сезу, кору,
дэм сезу және есту мүшелері.
Насекомдар — дара жынысты жануарлар. Олар шала тур-
лент, толық түрленіп, не түрленбей дамиды. Шала түрленіп
дамыганда жүмыртқадан ересек формаларына үксас дернэсіл
шыгады. Олар түлеп ересек формага — имагога айналады. Толық
түрленіп даму — жүмыртқа, дернэсіл, куыршақ жэне ересек фор¬
масы — имаго сатылары арқылы жүреді. Дернәсілдер ересек фор-
масынан морфологиялық қүрылысы, тіршілігі жагынан өзгеше
болып келеді, ал қуыршақ сатысында олар қоректенбейді. Дернэсіл
мүшелері ересек насекомдардың мүшелерімен алмастырылады.
Насекомдардың практикалық маңызы оте зор: олар гүлді
өсімдіктердің айқас тозандануын қамтамасыз етеді, кейбіреулері
288
топырақ түзу процесіне қатысады, кейбіреулері адамдарга пайда¬
лы өнімдер өндіреді, мысалы бал арасы, жібек кұрты, т.с.с. олар
колға үйретілген. Кейбір жыртқыш насекомдар ауыл шаруашы-
лык зиянкестерімен биологиялық тұргыда күресу үшін пайдала-
нылады.
Насекомдар арасында адам не жануарлар паразиттері немесе
ауру қоздырушы паразиттерді таратушылары да кептеп кезде-
седі. Сол сияқты, насекомдардың улы түрлері де белгілі.
Тарақандар отряды — Blattodea
Медициналық маңызы бар түрлеріне қара тарақан (Blatta огі-
entalis), шубар тарақан (Blatta germanica) жатады. Біріншісінің
олшемі 20—26 мм, екіншісінікі 8—11 мм. Олар үйлерде, әсіресе,
ас болмелерде, қоқыстар арасында кездеседі. Ас калдығымен,
нэжіспен коректенеді.
Тарақандар эр гүрлі аурулардың, әсіресе, асқазан — ішек
ауруларының іш сүзегі, туляремия, дифтерия, т.с.с. қоздырушы-
ларын таратады.
Алдын алу шаралары: Болмелерде тазалық сақтау, ас
калдықтарын дер кезінде жинап отыру, таракандарды жою.
Биттер отряды —Anoplure
Биттер — ұсак, қанатсыз насекомдар. Олардың көптеген
түрлері белгілі. Иесінің денесінде паразиттік тіршілік етеді. Адам¬
дарда бас биті, киім биті және қасага биттері кездеседі (72-сурет).
Бас биті — Pediculus humanus capitis, адамдардыц шашында
кездеседі. Молшері2—3 мм. Басы кішкентай, екі мұртшасы,екіжай
коздері, тесіп сорушы ауыз аппараты болады. Көкірек бөлімінде 3
жүп аяқтары кездеседі, аяқтарының ең соңғы буылтығының мық-
ты дамыған гырнагы болады, ол алдыңғы буылтықтын өсіндісі-
мен бірігіп, қысқышка ұксас қансоратын құрылым пайда етеді.
Осы құрылым аркылы биттер шаштарға да жабысып тұрады.
Қүрсағы көкірек бөліміне карағанда кеңдеу, 10 буылтықтан
тұрады.
289
72-сурет. Биттер.
а, б) бас биттері, в) киім биті, г) касаға биті, д) жұмырткалары.
Шала түрленіп дамиды. Жұмыртқалары шаштарга жабысып
тұрады (сіркелер). Жүмыртқадан ересек дараларға үксас дернэсіл
түзіледі, олар түлеп имагоға айналады.
Тіршілік ұзақтығы — 38 күн.
Бас биті — сүзек ауруының қоздырушысын таратады жэне эк¬
топаразит ретінде педикулез ауруын дамытады.
Алдын алу шаралары. Тазалықты сақтау -— мезгіл-мезгіл
моншаға түсу, киімдерді, бөлмелерді таза ұстау.
Киім биті — Pediculus humanus. Іш киімдердің тігістерінде
кездеседі, қан copy үшін теріге отеді. Сыртқы пішіні бас битіне
ұқсас, бірақ біршама үлкендеу — 4,7 мм, болып 48 күн өмір
сүреді.
Киім биті — эктопаразит жэне қайталама сүзегінің тарату-
шысы болып саналады.
Алдын алу шаралары. Бас битіне ұқсас.
Қасаға биті — Phthirus pubis, касағадағы жүндер арасында,
қаста, кірпік арасында кездеседі.
Мөлшері 1—1,5 мм, 26 күн өмір сүреді. Эктопаразит, ауру
қоздырғыштарын таратады.
290
Бургелер отряды —Aphaniptere
Адам бүргесі — Pulex irritanus. денесі бүйір жағынан
қысылган, қанаттары дамымаған. Басында қысқа мүргшалары,
күрделі жұп көздері, тесуші-сорушы ауыз аппараты орналасқан.
Аяқтары, әсіресе, соңғы жүп аяқтары, үлкен жэне жақсы дамыған.
Қүрсақ бөлімі 10 буылтықтан түрады. Кутикулада тікенекгер,
тісшелер, қылтанақтар кездеседі (73-сурет).
Толық түрленіп дамиды. Еден-
дердің жарықтарына, тесіктеріне,
қүрғақ қоқыстар арасына жұмырт-
қалайды. Жүмыртқадан ақ қүрт
тэрізді аяқсыз дернәсіл шығады.
Ол шіріген органикалық заттармен
қоректенеді. Шамалы уақыттан кейін
дернәсіл пілла жасап, қуыршаққа ай¬
налады. Ересек даралары кан сорып
қоректенеді.
Бүргелердің әрбір түрі түрліше
иелерде кездеседі: атжалманда — ат-
жалмен бүргесі, иттерде — ит бүргесі,
адамдарда — адам бүргесі.
Бүргелер эктопаразит ретінде де- а) адам бүргесі, б) ахжалман
нені қышытады, ооа ауруының қоз- бүргесі, в) бүргенің карны.
дырушыларын таратады.
Алдын алу шаралары. Тазалыкты сақтау, үйді, бөлмелерді
ылғал шүберекпен сүртіп тазалап отыру, бүргенің жұмыртқа-
лайтын жерлерін жою, инсектицидтерді колдану.
Қос қанаттылар отряды — Dipera
Бөлме шыбыны — Musca domestica — Жер бетінде кең
таралған. Қара түсті, ірі жэндік. Ауыз аппараты жалаушы — со-
рушы типті.
Шыбындар 100—150 жүмыртқа жүмыртқалайды, толық
түрленіп дамиды, жыл бойына көбейеді (74-сурет).
73-сурет. Бүргелер.
291
Бөлме шыбыны — ішек инфекциясының — тырысқақ,
қантышқақ, іш-сүзегі т.б. қоздырушыларының механикалық тара-
тушылары болып табылады.
Үй шыбыны — Muscina stabulans — барлық жерлерде
кездеседі. Денесінің түсі қоңыр, аяқтары, қылтанақтары сары
түсті. Адамның тамақтарымен, нәжістерімен қоректенеді.
Ішек ауруларының қоздырғыштарының механикалык тарату-
шысы.
74-сурет. Бөлме шыбыны.
а) жалпы түрі. б) жұмыртқалары, в) дернәсіл, г) қуыршақ.
Вольфарт шыбыны — Wohlfahrtia magnifica — миаз ауруын
қоздырушы паразит. Кавказда, Орта Азияда кең таралган.
Ақшыл сары түсті ірі шыбын. Жара бетіне өздерінің
дернэсілдерін тастайды, ал олар жүмсақ үлпаларға еніп паразиттік
тіршілік етеді. Қуыршақтану алдында иесін тастап, топыраққа
өтеді. Ересек формалары гүлдердің шырынымен қоректенеді.
292
10. ЭКОЛОГИЯ НЕГІЗДЕРІ
10.1. Жалпы экология мәселелері
Tipi агзалар белгілі бір ортада тіршілік етеді. Оларға бір
жагынан, тіршілік ортасының әр түрлі жагдайлары эсер етсе,
екінші жагынан өздері, ягни тірі агзалар қоршаган ортага кері эсер
етіп отырады.
Агзалар мен тіршілік ортасы қарым-катынасының заццы-
лықтарын, биогеоценоздың дамуы мен тіршілігінің заңдылық-
тарын зерттейтін гылымды экология деп атайды. Экология (OICOS
— үй, LOGOS — ілім) деген терминді 1866 жылы Э.Геккель
енгізген.
Экология заңдылыктары тіршіліктің эр түрлі кұрылым дең-
гейлерінде: даралар деңгейінде, популяциялар деңгейінде, биоце¬
ноз деңгейінде, биогеоценоз децгейінде байқалады.
Биоценоз (тірі агзалар қауымдастыгы) дегеніміз — бір жерді
ұзак уақыт мекендейтін тірі агзалар жиынтыгы.
Биогеоценоз дегеніміз — өсімдіктердіц, жануарлардыц,
микроагзалардыц өзара қарым-катынаста болып, атмосфера,
гидросфера, литосфера компоненттерімен тікелей байланы¬
сып, тұрақты не қүбылмалы қауымдастық күйінде тіршілік етуі.
Бұл терминді В. Н. Сукачев енгізген. Биогеоценоз-экосистема
терминіне сэйкес келеді (Тенсли, 1935), бірак экосистеманыц мәні
құбылмалы болады (кец не тар ауқымды).
Орта (тіршілік ортасы) дегеніміз — даралардын мекен-жайын-
да оларга эсер ететін элементгер жиынтыгы.
Даралардыц жеке дамуында оларга тікелей эсер ететін орта
элементтерін экологиялық факторлар деп атаймыз. Экологиялык
факторларды биотикалыц жэне абиотикалъщ деп 2-ге бөлеміз.
Абиотикалық факторлар — климаттық, эдафикалық (топырак)
жэне топографиялық (орографиялық) деп бөлінеді.
293
Биотикалық факторлардың — фитогендік (өсімдіктер әсері),
зоогендік (жануарлар эсері), антропогендік (адамдар әсері) деген
түрлері белгілі.
Ағзалар, популяциялар жэне биоценоздар тіршілігіне көпте-
ген факторлар жиынтыгы эсер етеді, дегенмен, эркайсысының
әсерлері түрліше болады. Агзаларға басты эсер ететін факторлар
тіршілік жагдайын қалыптастырады. Табигатта бір фактордың
әсерін екіншісінен бөліп, ажыратып қарастыру мүмкін емес, себебі
олар агзаларга бірлесіп эсер етеді.
Әр түрлі агзалар тіршілік ортасыныц факторларына түрліше
жауап қайтарады. Экологиялық факторлар әсерінен байқалатын
даралардың өзгергіштігінің шегі олардың генетикалык ерек-
шеліктеріне байланысты жэне оны реакция нормасы деп атайды.
Эр түрлі биологиялық түрлердің реакция нормасының мөлшері
түрліше болып келеді, бірі кең ауқымды, ал екіншісі тар аукымды
болуы мүмкін. Осыган байланысты олардыц ортага бейімделу
мүмкіншілігі, ягни экологиялық потенциялары түрліше болады.
Экологиялық потенциалы тар болып келетін түрлерді стено-
топты, ал кец ауқымды экологиялық потенциалды түрлерді — эв-
ритопты түрлер деп атайды.
Климаттық факторлардыц ішінен агзалар үшін ец мацыздылары
— жарык, жылылык, ылгалдылық т.с.с. болып есептелінеді.
Жарықтыц (күн сэулесінің) эсері. Жарық (күн сэулелері)
Жер бетіндегі энергияныц негізгі көзі болып табылады. Жарық
бір жагынан қарапайым физикалық бөлшектерден — корпу-
скулдардан тұрса, екінші жагынан ол толқындар қасиетіне ие.
Сәулелер толқындарыныц үзындыгы қаншалықты қысқа болса,
ол соншалықты энергияга бай болады жэне керісінше. Фотондар
энергиясы жасыл өсімдіктердіц фотосинтез процесі үшін қажет.
Ал фотосинтез негізінде синтезделінген органикалық заттар жэне
бөлініп шыққан оттегі жер бетінде тіршілік ететін барлық агзалар
үшін өте қажет.
Агзалардыц күнніц үзақтыгына реакциясын фотопериодизм
деп атайды. Ол агзалардыц тіршілігінде (агза түктерініц түлеп
түсуі, денеде май жинақталуы, көшіп-қонуы, көбеюі т.с.с) елеулі
рол атқарады.
294
Кұн сәулесінің кысқа толқынды сэулелері — ультракүлгін
сәулелері (0,29 мкм дейін) энергияга бай болғанымен жер-
ге жетпейді, оларды озон қабаты ұстап өткізбейді. Бұл сэулелер
тіршілік үшін өте қауіпті, ал жұмсак, энергияға тапшылау болып
келетін ультракүлгін сәулелері (0,3—0,4 мкм), озон қабатынан өтіп
жерге жетеді жэне ағзаларға екі түрлі эсер етеді:
1) пайдалы әсері — ағза денесінде провитаминнен Д дэруме-
нінің синтезделуін қамтамасыз етеді; 2) зиянды әсері — көптеген
мутацияларды туғызады.
Орташа толқынды, қызыл түсті сәулелерді (0,6—0,7 мкм)
өсімдіктер фотосинтез процесінде сіңіреді.
Адамдар мен жануарлардыц күн мен түннің алмасуына
бейімделу реакциясы ретінде зат алмасудыц тэуліктік ыргақ-
тылығыныц, тыныс алу жиілігініц, жүректіц соғу жиілігінің,
қанқысымы децгейінің, дене температурасының, жасушалардыц
бөліну жылдамдығыныц түрліше болуын т.с.с. атауға болады.
Жылулық. Ағзалардың қалыпты тіршілігі үшін жылулық-
тыц рөлі зор. Жасуша ферменттерінің активтігі 10—40°С
аралығында болады. Көптеген насекомдар салқын түссе топы-
раққа, ағаш діңіне, тау-тастар арасына тығылып паналайды,
бақалар үйыкқа кіріп кетеді, кейбір сүтқоректілер аю үзақ уақыт
ұйқыға кетеді.
Ағзалардыц ыстыкка бейімделу реакциясы ретінде өсімдік-
тердіц жапырақтары, жануарлардыц тыныс алу жолдары, терілері
аркылы суды буландыруларын айтуга болады.
Жасушаларында су мөлшері аз болып келетін — споралар,
тұқымдар, кейбір қарапайымдардыц цисталары ұзақ уақыт анаби¬
оз күйінде сақталады.
Ылғалдылық. Ағзалар тіршілігі үшін судыц рөлі өте жогары,
себебі цитоплазманыц 80—90% су күрайды. Ылгалдыкка
бейімделуіне карай өсімдіктерді гидрофиттер, гигрофиттер, мезо-
фиттер, ксерофиттер деп 4 топқа бөледі.
Шөлді жерлерді мекендейтін жануарлардыц су резервуар-
лары ретінде өркештеріндегі майлар (түйе), тері асты майлары
(кемірушілер), майлы денелері (насекомдар) саналады.
Абиотикалық факторларға, сол сияқты, атмосфера газдары, ми¬
нерал заттар, гидросфера, судыц не топыракгыц тұзды болуы т.б.
жатады.
295
Минералдық элементтер -— агзаларда тұз күйінде болып суда
диссоциаланады да иондарға ыдырайды:
Калий — бұлшық еттердің жиырылғыш ақуызы миозинмен
қосылып, оның босаңсуына эсер етеді. Кальций — бұлшық
еттердіц жиырылуын күшейтеді. Магний — хлорофилл, темір
— гемоглобин құрамына кіреді. Калий, натрий, кальций, маг¬
ний иондарының жетіспеушілігі не болмауы жасушаның қозу
қабілетінің тежелуіне жэно оның өліп қалуына алып келеді.
Табиги жагдайларда микроэлементтердіц жетімсіздігі адам¬
дардыц жергілікті ауруларына алып келеді.
Биотикалық факторларға өсімдіктер мен жануарлардыц
қауымдастыгыныц өзара әрекетгесуі жатады. Оларды түрішілік
жэне тураралық эрекеттесу деп бөледі.
Түрішілік әрекеттесуге бір түрдіц дараларыныц қорек, су,
энергия, кецістік т.б. үшін күресін жатқызуга болады. Түраралық
әрекеттесуге: мутуализм, комменсиализм, жыртқыштық, пара-
зиттік гіршілік етуді (паразитизм) жатқызады.
Мутуализм — екі түр агзаларының селбесіп тіршілік етіп,
өзара пайда келтіруі. Бүган мысал ретінде бүршақ тұқымдастар
өкілдерініц тамырындагы түйнек бактерияларын, қынаны,
адамныц тоқ ішегіндегі бактерияларды т.б. келтіруге болады.
Комменсиализм — екі түрге жататын агзалардьщ біреуінің
екіншісін зиян келтірмей пайдалануы, ал екіншісініц тіршілігі
үшін біріншісініц не пайдасы не зияны болмайды. Мысалы, кейбір
моллюскалардыц қабыршақтары үсақ жануарлар үшін пана бола¬
ды.
Жыртқыштық — бір агзаныц екінші агзаны өлтіріп, қорек
ретінде пайдалануы.
Паразитизм — бір агзаныц екінші агзаны зақымдап зиян
келтіруі, бірақ өлтірмейді.
Биогеоценоз — биосфераныц ең кіші өлшем бірлігі болып са¬
налады.
Tipi агзалардыц өзара әрекеттесуін жэне олардыц өлі дүние
элементтерімен көптүрлі байланысын биогеоценоз тұжырым-
дамасы айқын ашып көрсетеді.
Биогеоценоз дегеніміз — тірі агзалардыц — өсімдіктер, жа¬
нуарлар жэне микроагзалардыц озара жэне өлі дүние элементте-
296
рімен динамикалы не тұрақты байланыста болатын қауымдастығы
болып табылады.
Биогеоценоз биотикалық (биоценоз) жэне абиотикалық (эко¬
тип) бөлімдерден тұрады. Бұл екі бөлім арасында үзіліссіз зат,
энергия алмасуы болып тұрады. Ол ашық жүйе болып санала¬
ды. Оган үнемі күн сэулесінің энергиясы, минерал заттар, атмос¬
фера газдары келіп енеді, ал одан жылулық, көмірқышқыл газы,
биогендік заттар т.б. бөлініп шыгып отырады. Биогеоценоздың
негізгі қызметі табигатта энергия мен заттардыц алмасуын
қалыптастыру болып табылады. Биогеоценоз экономикасын-
да қорек тізбегі үлесініц мацызы өте зор. Қорек тізбегі бірнеше
трофикалық (қоректік) децгейлерден қүрылады. Энергияныц
алгашқы көзі болып — күн сэулесініц энергиясы саналады, ол 4,6
х 1026 Дж/с тец. Оныц тек 1-2000000 бөлігі, ягни 2,3—1020 Дж/с
Жерге келіп жетеді де, не бары 0,1-0.2% жасыл өсімдіктердің
фотосинтез процесінде пайдаланылады. Осімдіктер денесінде
жинақталган энергия биогеоценоздыц алгашкы таза енімі фи¬
тобиомасса деп аталынады. Б:ул фитобиомасса өсімдікқоректі
жануарлар үшін энергия көзі болып, олардың биомассасын түзу
үшін жүмсалынады. ал осімдікқоректі жануарларды жыртқыш-
тар (етқоректілер) қорек ретінде пайдаланады.
Трофикалық децгей жогарылаган сайын оның өнімділігі
бүрынгы (төменгі) деңгейдіц өнімділігімен салыстырганда азая
береді, ягни келесі деңгейге не бары 5-20% (10%) өтеді, себебі
әрбір децгейдіц дараларыныц тіршілігі үшін жұмсалатын энергия
мөлшері көбейеді де, өнімділігі кемиді. Мысалы, құрлықта тірші-
лік ететін агзалар биомассасы шамамен 3x1012 тец болса, оныц тек
1—3 пайызы гана жануарлар биомассасыныц (зоомассасыныц)
үлесіне тиеді.
Биогеоценоздын коректік тізбегініц трофикалық децгейі
жогарылаган сайын сіңірілетін энергия мөлшері прогрессивті
тұргыдан азаяды жэне ол экологиялық пирамида щрайды.
Биогеоценоздыц негізгі бөлігі болып биоценоз саналады. Ал
биоценоздардыц мацызды сипаттамасы агзалар популяцияла-
рыныц бір-бірімен тікелей не қосалқы эрекеттесуі болып табыла¬
ды.
297
170-2(1
10.2. Адам экологиясы
Адам экологиясы адамдардың сыртқы ортамен қарым-қаты-
насын зерттейтін жалпы экология ғылымының бір саласы.
Адам экологиясы кешенді гылым болып табылады, себебі онда
қогамтану, географиялык, табигаттану, биологиялық, медици-
налықт.б. проблемалар қарастырылады.
Сонымен, адам экологиясы антропоэкологиялық жүйелердің
пайда болу, даму заңдылықтарын зерттейтін гылым. Антро-
поэкологияльщ жуйелер деп ортамен құбылмалы байланыста
болып, өздеріне қажетті заттарды алып тіршілік ететін адам
кауымдастыгын айтамыз. Бұл жүйелердің көлемі түрліше бола¬
ды. Ол адам популяцияларының санына жэне құрылымына бай¬
ланысты. Адамдардың популяцияларын өндіріс ерекшеліктеріне,
тұрмыс-тіршіліктеріне карай изоляттар, демдер, ұлттар жэне
ұлыстар, ұлттан да үлкен құрылымдар, жалпы адамзат деп бөледі.
Антропоэкологиялык жүйелердің көлемін анықтауда табиги
жагдайлардың рөлі өте зор. Қазіргі кезде адамзаттың 80%-ын
қамтитын адамдардың ірі популяциялары құрлықтың ең қолайлы
44% жерлерін мекендейді. Оган тропикалық орманды аймак, са-
ванналар, коңыржай аймактың бұгалы не орманды өңірлері жата¬
ды.
Керісінше, шөлді жэне шөлейт жерлерде (құрлықтың 18%)
адамдардың не бары 4% мекендейді.
Антропоэкологиялык жүйелердің табиги экосистемалардан
бірден-бір айырмашылыгы — олардың құрамында барлык уақыт-
та басымдык, үстемдік көрсететін адам кауымдастығыньщ болу-
ында.
Антропоэкологиялык жүйелерде адамдар мен табиги орта
арасындагы байланыс екі багытта жүріп отырады: 1) орта
факторларының эсерінің нэтижесінде жеке индивидтердің жэне
когамның кейбір биологиялык, әлеуметгік көрсеткіштері өзгереді;
2) ортаны адамдар өз талаптарына сэйкес өзгертеді.
Адам қогамының даму тарихында осы екі багыттың соңгысы
күштірек өзгеріп отырган. Алгаш мэдени өсімдіктерді өсіріп
егіншілікпен айналысу, жануарларды қолга үйретіп, мал ша-
руашылыгымен айналысу нэтижесінде орта шамалы гана өзгер-
298
ген, ал кейінірек, өнеркәсіптің дамуы, қалалардың пайда бо¬
луы нәтижесінде адам қоғамы ортаны толық өзгертіп, өздерінің
тіршілік мүмкіншіліктерін едэуір кеңейтті.
Антропоэкологиялык жүйелердегі биологиялық жэне
әлеуметтік процестердің негізгі нэтижесі адам қоғамының эр түр-
лі ортаға жекелей жэне топтасып бейімделушілігін арттыру бо¬
лып саналады. Мүндай бейімделушіліктің қалган тірі ағзалардың
бейімделушіліктерінен негізгі ерекшелігі — адам ортага тек
физиологиялык жагынан бейімделіп қана қоймай, сол сияқты,
экономикалық, техникалық, эмоциялык жагынан да бейімделеді.
Адам өзінің мекен ортасында бір жагынан ортаның экологиялык
факторларының әсеріне ұшырайтын объект болып есептелінсе,
екінші жагынан, оның өзі ортага елеулі түрде эсер етеді.
Адамның экологиялық фактор ретіндегі ерекшелігі — оның
табигатқа тигізетін әсерінің саналы, нысаналы жэне қарқынды
түрде болуында. fX>. Энгельс «... жануарлар қоршаган ортадан тек
өздеріне қажет заттарды алып пайдаланып, оган өздерінің сол ор-
тада тіршілік етуі нәтижесінде гана эсер ететін болса, адам өзінің
іс-әрекеттері арқылы оны күрт өзгертіп, өзінің мүддесі үшін қыз-
мет етуге зорлайды, үстемдік етеді»,— деп жазган. ^jl
Кез келген биологиялық түрлердің энергия ресурстары шек-
теулі болады, сондықтан да оның ортага тигізетін әсері де шектеулі.
Мысалы, жасыл өсімдіктер күн сәулесінің энергиясын пайдала¬
нып органикалық зат түзеді. Жануарлар жасыл өсімдіктер син-
тездеген органикалық заттардың энергиясымен гана қоректенеді.
Ал, адамдар болса өзінің кол жэне ойлау еңбегінің нәтижесінде
энергиялық ресурстар көзін көбейтіп, оның жаңа көздерін ашып,
айталык, ядролық, термоядролық реакциялардың энергияларын
пайдаланады. Бұл адамдар санының өсуіне кедергі келтіретін
кейбір табиги жагдайларды жоюга мүмкіндік береді. Адамзаттың
санының өсуі, олардың энергиямен қамтамасыз етілуінің ар-
туы, техникадагы қол жеткен жетістіктер адамдардың кез кел¬
ген экологиялық жагдайларды игере алуына мол мүмкіншіліктер
тугызады. Жер бетінде шексіз кең таралган жалгыз түр — Homo
sapiens, саналы адам. Сондыктан адам элемдік маңызы бар
экологиялык факторга айналды. Адамдардың эсері биосфераның
барлық негізгі компоненттерін қамтитындыктан, ол планетаның
299
ең қиыр шегіндегі экологиялық аймақтарға дейін жетеді. Мыса¬
лы, Анграктидадағы пингвиндер мен тюленьдердің бауырлары-
нан инсектицид — ДДТ табылған, ал бұл жерлерде ешуақытта да
бірде-бір инсектицид қолданылған емес.
Адамныц экологиялық фактор ретіндегі тағы бір ерекшелігі
— олардыц іс-эрекетініц белсенді, творчестволық күйінде бо¬
луы. Өздері қолданатын энергияны адамдар ортаны өзгертуге
бағыттайды. Биологиялық механизмдерге негізделген адамдар¬
дыц тіршілік етуініц экологиялық оптимумы, барлық тірі ағзалар
сияқты, шектеулі болады. Олардыц кец таралу мүмкіншілігі
адамдардыц биологиясыныц өзгеруі нэтижесінде емес, ортаны
өзгертіп өздеріне қолайлы жагдай туғызып — жасанды орта жа-
сау аркылы жүзеге асады.
Табиғи жэне жасанды орта факторлары адамға үнемі эсер
етіп отырады. Адамзаттыц даму тарихында табиғи факторлар
әсерлерініц нәтижесі бүгінгі күні олардыц экологиялық жікте-
луіне, эр түрлі адаптивтік типтерге, нәсілдерге бөлінуіне алып
келген. Ал, әлеуметтік факторлардыц әсерлерініц нэтижесі ретінде
адамдар қауымдастыгыныц мэдени-шаруашылык типтерініц пай¬
да болуын жэне олардыц занды түрде алмасуын жатқызуга бола¬
ды. Мэдени-шаруашылык типі дегеніміз — нақтылы адамдар
кауымдастығына тэн шаруашылық жэне мәдениет кешені болып
саналады.
Қазіргі кезде планетада адамдардыц эр түрлі мэдени-
шаруашылык типтері кездеседі. Олар бір-бірінен пайда болу
мерзімініц түрліше болуымен, өндіріс децгейімен, халықтың эл-
ауқатыныц мөлшерімен жэне демографиялык ерекшеліктерімен
өзгешеленеді. Оларга мыналар жатады:
1) «иемденуші» тип — қазіргі кезде сирек кездесетін тип. Бүл
типтіц ерекшелігі — олардыц экономикасында ац аулау, балык
аулау, жеміс-жидек жинау сияқты қарапайым экономикалық
катынастар басым болады. Бүл тип Заирдіц ацшы-пигмейлеріне,
оцтүстік-шығыс Азия тауларын мекендейтін аэта жэне кубу тай-
паларына, Амазонка өзенініц бойын мекендейтін кейбір үндіс-
терге тэн;
2) экономикалық негізі болып қолмен жерді өндеп не соқамен
жерді жыртып егін егетін, мал шаруашылыгымен айналысатын
мәдени-шаруашылық типтері;
300
3) өркениетті елдерде ғылыми-техникалык революция жеті-
стіктеріне негізделген жоғары өнімді егін шаруашылығы мен мал
шаруашылығы басым болатын мәдени-шаруашылық типтері.
Мәдени-шаруашылық типтерінің қалыптасуы табиги орта фак-
торларына тәуелді болады. Ол эсіресе адам қоғамының дамуының
алғашқы кезеңдерінде өте жогары дэрежеде болған. Дегенмен, сол
кездерде де, эсіресе, кейінірек, мәдени-шаруашылық типтердің
қалыптасуына халықтардың элеуметтік-экономикалық дамуының
деңгейі елеулі эсер етіп отырған.
Жасанды орта табиғи экологиялық факторлардың адамдарга
тигізетін эсерлерін теңестіріп, әлсіретіп отырады. Бұл, эсіресе,
өркениетгі елдерде жогары дэрежеде байқалады. Адам когамының
дамуында ғылыми-техникалык эра үзақгыгы 1 % уақыт алады, яғни
100 жылдай. Ал, саналы адамныц — Homo sapiens дамуының 99%
уақытында оган табиғи ортаның эртүрлі климаттық, геохимиялық,
биологиялық, микробтық, паразиттік экстремалды факторлары
айтарлықтай эсер етіп келген жэне ол факторлардың багыты мен
қысымы Жер шарының эр түрлі жерлерінде түрліше болган. Сол
табиги қысымның салдарынан қазіргі адамдар бірнеше адаптивтік
типтерге бөлінеді.
Адаптивтік тип дегеніміз — мекен ортасының басым фактор-
ларына бейімделу ретінде қалыптасқан адамдардың биологиялық
реакция нормасы болып табылады. Ол адамдардың белгілі
бір физикалық ортага шынайы биологиялық бейімделушілігін
қалыптастырушы кешенді морфофункционалдық, биохимиялық,
иммунологиялық белгілердіц дамуы аркылы байқалады. Нақтылы
адаптивтік типке тән белгілер жиынтыгы адамдардың нәсілдік не
ұлгтық ерекшеліктеріне байланысты болмайды.
Адамдардың бірнеше адаптивтік типтері белгілі:
1) Арктикалық адантивтік тип — Арктика, Антрактида
тұргындарына тэн. Бұл типтің калыптасуына төменгі темпера¬
тура, орманныц болмауы, өсімдіктердіц тапшы болуы мен жа¬
нуарлар дүниесінің бай болуы (балықтар, сүткоректілер — аю,
түлкілер, қояндар, бүгылар т.б.) өз септігін тигізген. Сондықтан
да бүл жерде тамақ кұрамында ақуыз көп, көмірсу аз мөлшерде
болады. Адам организмі А, В, С, D дэрумендермен жеткілікті
мөлшерде камтамасыз етіледі, ауада микроорганизмдер аз бо-
301
лады. Бул типтің кешенді белгілері дененің қаңқа — бұлшық ет
компонентерінің мықты дамуы, көкірек клеткасы мөлшерінің
үлкен болуы, гемоглобиннің жоғарғы деңгейде, қанда ақуыздың,
холестериннің көп болуымен сипатталады.
2) Тропикалык адаптивтік тип — тропика, субтропика
түрғындарына тэн. Бұл типтің қалыптасуына ауаның өте ыстык
болуы, жауын-шашынның көп мөлшерде түсуі, маусымдық
өзгерістердің айтарлықтай байқалмауы сияқты экстремалды
факторлар эсер еткен. Ылгалды тропикалық ормандарда жану¬
арлар аз болса, субтропикада, керісінше, коп болады жэне бул
жерлерде ірі-ірі үйірлі андар кездеседі. Адамдардың тамағының
құрамында жануарлар ақуызы аз мөлшерде болады. Сол сияқты,
бул жерлерде экологиялык жағдай өте қубылмалы болып келеді.
Ол да өз эсерін тигізбей қоймайды. Тропикалық, субтропикалық
аймақтарда эр турлі нәсілдердің, улттардың жэне экономикалары
түрліше дамыған адамдардың топтары кездеседі. Бул жерлердегі
адамдардың сомалык белгілері өте қубылмалы болады, дәлірек
айтсақ, осы жерлерде ең аласа бойлы жэне ең ұзын бойлы тайпа-
лар кездеседі.
Дегенмен, тропиктер мен субтропика турғындарының ерекше
белгілері мынадай: денелері узын, булшық ет массасы аздау бо¬
лып келеді. Дене салмагы салыстырмалы түрде аздау болып, кол-
аяқтары ұзын, көкірек қуысының шеңбері тарлау болып келеді.
Терісінде тер бездері көп болғандықтан, тершең келеді, қанда хо¬
лестерин концентрациясы томен болады.
3) Қоцыржай аймақтың адапгивтік типі — арктикалык.
тропикалық типтермен салыстырганда, көптеген белгілері бойын¬
ша — сомалық. негізгі алмасу деңгейі, т.б. аралық көрсеткішке ие
болады. Бул осы жерлердегі биогеографиялык орта жағдайларына
сэйкес келеді.
4) Таулы жерлердің адаптивтік типі — бул жерлерде
адамдардың белгілерінің қалыптасуына эсер ететін ерекше фак¬
торлар -гипоксия, атмосфера қысымының томен болуы, суықтық,
тамақтың бір түрлі болуы. Бул типтің ерекшелігі зат алмасу
жогары деңгейде болады, қаңкасының ұзын сүйектері оте узын
болып келеді. Кокірек қуысы кең, қанында эритроцитгер молшері
коп болады.
302
Антропогендік экосистемаларда жасанды орта табиги ортага
Караганда басым болады. Қазіргі кездері негізгі антропогендік
экосистемаларга қалаларды жатқызуга болады. Адамзат тарихын-
да қалалар ерекше мекен ортасы болып есептелінеді. Олар бұдан
7000 жыл бұрын пайда болган. 1950 жылы калаларда адамдардьщ
28%, 1970 жылы — 40% мекендеген. 2000 жылы калаларда эр түрлі
деректерге қараганда 56-62-ден 70-90%-га түргындар мекендейді.
Қазіргі таңда қала тұргындарының 1/3 бөлігі ірі, тұргындары 1
млн-нан кем болмайтын қалаларда тұрады. Қалаларда өндіріс,
гылыми жэне мәдени ошақтар шоғырланган. Сондықтан да, қала-
да жүмысқа орналасу, білім алу, азық-түлікпен камтамасыз етілу,
медициналық жэрдем көрсету т.б. тұрмыстық мәселелер жақсы
жолга қойылган.
Сонымен катар, қалаларда табиги орта күшті өзгерген.
Қалада өндірістік жэне тұрмыстық мекемелер коп болгандықтан
топырақта, ауада, суларда және осімдіктерде көптеген микроэле-
менттер ерекше таралган. Қалаларда жұқпалы аурулардың таралу¬
ына қолайлы жагдайлар туады. Қала ауасының аэрозольдармен ла-
стануы салдарынан, өндіріс мекемелерінің, коліктердіңжылу бөлігі
шыгаруы салдарынан ауаның орташа жылдық, айлық, тәуліктік
температурасы қаланы қоршаган аумақтарга Караганда жогары
дәрежеде болады. Қалалардың ауасы түтінді болып, ультракүлгін
радиациясының деңгейін кыстың күндері 30%, жаздың күндері
5% төмендетеді, ал бүл өз кезегінде Д авитаминозына алып келіп,
адамдардьщ тез шаршауына, жұқпалы жэне тұмау ауруларына
төзімділігін төмендетеді. Бүлардан басқа қалада шу жэне вибра¬
ция жогары деңгейде болады.
Сыртқы ортага бейімделу ерекшеліктеріне карай адамдарды
стаііерлер жэне спринтерлер деп бөледі.
Стайерлер — сыртқы ортаныц ұзакқа созылган өзгерістеріне
бірте-бірте бейімделеді, ал қысқа мерзімді күрт өзгерістерге
бейімделе алмайды. Спринтерлер — керісінше, қысқа мерзімді
күрт өзгерістерге тез бейімделіп, ұзаққа созылатын озгерістерге
шыдай алмайды.
303
11. АДАМНЫҢ ЭКОЛОГИЯЛЫҚ ГЕНЕТИКАСЫ,
ФАРМАКОГЕНЕТИКА
11.1. Жалпы мәселелер
Адамның экологиялық генетикасы — мекен орта факторла-
рының тұқым қуалаушылыққа тигізетін эсерлерін зерттейді. Орта
факторлары генотиптің құрылымына жэне қызметіне екі тұрлі
эсер етуі мүмкін: 1) арнайы (спецификалық) факторлардың агзаға
эсер етуі салдарынан белгілі бір аллельдер әрекетінің байқалуы
өзгереді; 2) даралардын жэне популяциялардың генетикалық ма-
териалдары өзгереді.
Бірінші типті әсерлер жеке адамдар деңгейінде патологиялық
реакциялардың (аурулардың), ал популяциялық деңгейде — ортаға
жақсы не нашар бейімделуі (адаптациялануы, жерсінуі) күйінде
байқалуы мүмкін.
Орта факторлары эсерлерінен патологиялык аллельдердің
байқалуын экогенетикалық реакциялар немесе экогенетикалық ау¬
рулар деп атайды.
Екінші типті әсерлерге — қоршаған орта факторлары
индукциялаған мутациялық күбылыс пен сұрыптауды жатқыза-
мыз. Бүл екі құбылыс адамдардың тұқым қуалайтын өзгергіштігі
қарқынының жеке дара жэне популяциялық деңгейлерде
жоғарылауына алып келеді.
Адамның экологиялық генетикасының негізі болып эволю¬
ция құбылысының жалпы биологиялык зацдылықтары санала¬
ды. Генотиптердің өзгеруі ағзалардың (популяцияның) озгеруіне
алып келеді, ал популяциядағы сұрыптау популяция генофон-
дын қалыптастырады. Демек, бір биологиялық түрдіц, сол сияқты
адамдардыц эволюциясы — оныц генотиптерініц эволюциясы бо¬
лып табылады.
Эволюция қүбылысыныц негізі болып саналатын өзгергіш-
тіктіц бірден-бір көзі болып мутациялар саналады. Биологиялық
304
түрлердің маңызды жэне тұрақты сипаттамаларының бірі —
нақтылы орта жағдайларында олардың мутациялық қүбылы-
сының тұрақты жэне оптимальды деңгейде болуы.
Қоршаған орта популяциялардыц, не даралар тобының тұқым
қуалаушылық ерекшеліктеріне қарай олардың сұрыпталуына, тірі
қалуына, «гүлденіп» дамуына алып келеді. Биологиялық түрақгы
түрлерде мутациялық құбылыс пен сұрыптау арасында үнемі тепе-
тендік байқалып түрады.
Адам эволюциясы барысында оныц мекен ортасы (кли¬
мат, корек, от, киім, мекен-жай т.б.). үнемі өзгеріп отырған.
Бұл өзгерістерге адам агзасы бір жағынан кец колемді реакция
нормасы арқасында, екінші жагынан генотипгерінің өзгеруі
нэтижесінде бірте-бірте жайлап бейімделігі келген. Мұның бэрі
жүзмыңдаған жылдар бойына адамныц биологиялык болмысын
қалыптастырып, адамныц қоршаған ортага жеткілікті дәрежеде
бейімделуіне алып келді.
Қазіргі кезде адамньщ тіршілік ортасы тез қаркынмен жэне
кец көлемде өзгеруде, ал адамның тұқым куалаушылыгы,
популяциялық децгейде, осыншама тез өзгере алмайды.
Сондықтан да қазіргі кезде адам популяцияларында мутациялык
құбылыс пен сұрыптау деңгейі едэуір жоғарылады.
Эволюция барысында адам популяцияларында үнемі байқа-
лып отыратын мутациялык, генетикалық-автоматтық (гендердіц
дрейфі) күбылыс жэне сұрыптау салдарынан кец көлемді балан¬
сты полиморфизм қалыптаскан. Қазіргі адам популяцияларында
оның көлемі өте үлкен. Мысалы, адамныц антигендік, ферменттік,
рецепторлык жэне басқа да молекулалық-биохимиялық қасиетін
анықтайтын гендердіц кем дегенде 25%, яғни 25 000 гені, 2 не
одан да көп аллельдерден тұратын полиморфтық жүйе күйінде
кездеседі, демек жеке генотиптер вариациясы 225000 тец. Мұндай
көптүрліліктіц каншалықты үлкендігіне көз жеткізу үшін мына-
ны ескерген жон: гек 25 полиморфты жүйеніц вариациялары (225)
бүкіл жер шарын мекендейтін адамдар санына тең (6 миллиардтан
астам) эр түрлі жеке генотиптерді пайда етер еді.
Адамныц ферменттік жүйелерініц, тасымалдаушы ақу-
ызда-рыныц, антигендерініц жэне жасуша рецепторларыныц
коптеген вариациялары агзадагы химиялық заттардыц
305
метаболизмдерінің, биологиялық агенттерге не физикалық
факторларға кері жауап реакцияларының жеке ерекшеліктерін
туғызады. Осылардың бэрі адам экогенетикасының зерттеу
объектілері болып саналады.
Адам экогенетикасы XX гасырдың 50-жылдарынан бастап да¬
мып келеді. Бұл жылдары адам ағзасында кейбір ферменттердің
жетіспеушілігі салдарынан дэрі-дэрмектерге қарсы тұқым
куалайтын патологиялық реакциялар байқалған. Бұл құбылысты
сипаттау үшін неміс генетигі Ф. Фогель фармакогенетика деген
терминді колдануды ұсынды.
Адам экогенетикасын зерттеулер соңгы жылдары адамның
мекен ортасының «жаңа», бүрын кездеспеген факторлармен
(дәрі-дэрмектер, пестицидтер, тамақ қоспалары т.б.) ластануы
нәтижесінде жеделдеді. Бүрын адамдар бұл заттармен, тіпті,
жанаспаган, сондықтан да бұл заттарга қарсы сүрыптау болма-
ган. Кейбір аллельдер гендердің дрейфі не баска да себептер
нэтижесінде популяцияда жинакталуы мүмкін. бірақ олар ұзақ
уақыт «үнсіз» күйде болып фенотиптік байқалмаган. Ал, жаңа
жагдайларда олар активтеніп фенотиптік байқалуын көрсетуі
мүмкін.
Бұрын «үнсіз» күйде болып келген гендердің жаңа эко-
логиялық факторлар әсерінен активтенуін — факторлардың
экогенегикалық эсері деп атайды.
Экогенетикалық аурулардыц қалыптасуыныц популя-
циялық-генетикалық түсіндірмесі.
Қазіргі кезде агзалардың орта факторлары әсерлерінен түқым
куалайтын реакциялары тек қана дэрі-дэрмектерге емес, сол
сиякты физикалық факторларга, тамақтарга. әсіресе, тамақтарга
косылагын қоспаларга, атмосфера ластануына, кэсіби зиянды
факторларга да қатысты байқалган.
Сыртқы орта факторларының әрекеттеріне агзаның ге¬
нетикалык реакцияларының ерекшеліктерін клиникалык-
генеалогиялық, егіздерді зерттеу немесе популяциялық-ста-
тистикалық эдістер арқылы анықтауга болады.
306
11.2. Сыртқы орта әрекеттеріне ағзаның түқым
қуалайтын пагологиялық рсакциялары
Ағзаның орта факторлары әсерлеріне өте жогары сезімтал
болуының негізі болып кейбір арнайы (спецификалык) мутация¬
лар саналады. Ортаның зиянды факторлары барлық адамдарды
закымдамай, тек кейбір осы мутацияларга генетикалык бейімді
ағзаларды гана зақымдайды. Адам ағзасына енген химиялық
қосылыстар метаболизмінің (биотрансформациясының) генети¬
калык тұргыдан бақыланатыны (басқарылатыны) белгілі.
Агзага енген химиялык заттардың биотрансформациясына
қатынасып патологиялық экогенетикалық реакцияларды қалып-
тастыратын полиморфтық локустарга — цитохром Р-450,
N-ацетилтрансфераза, қан сарысуынын параксоназасы жэне
холинэстеразасы, глюкоза-6-фосфат-дегидрогеназа, лактоза,
протеиназ ингибиторы т.б. жатады.
Экогенетикалық реакциялар кейбір сирек кездесетін мутантты
аллельдер не полиморфты жүйелер аркылы, сол сияқты бір немесе
бірнеше гендер әрекеті нэтижесінде байқалуы мүмкін.
307
Атмосфераның ластануы
Автокөліктерден, көптеген зауыт, фабрикалардан бөлініп
шыққан газдармен және басқа да өнімдермен атмосфераның ла¬
стануы әлемдік көлемдегі гигиеналық проблемаға айналып отыр-
ғаны белгілі. Химиялық қосылыстар жэне шандар ағзаға екпе,
тері, кілегейлі қабықша т.с. арқылы еніп, онда эр түрлі реакция-
ларды тудырады. Олардың кейбіреулерімен адам үнемі жанасып
жүрсе, кейбіреулерімен сирек кездеседі. Агзаныц тұқым қуалай-
тын реакцияларының вариациялары (түрлері) кез келген фактор¬
лар әсерінен пайда болады.
Атмосфераның ластануы салдарынан агзаның реакция-
сын тудыратын мутациялардыц бірі — а! — антитрипсинніц
жетіспеушілігі болып табылады. Қан плазмасыныц бұл акуызын
протеиназ ингибиторы деп те атайды. Қалыпты жагдайда
оның концентрациясы екіқабат эйелдерде жэне қабыну
кезінде жогарылайды. Осы ақуыздың генетикалык вариантта-
ры көптеген популяцияларда байқалган. Оныц жетіспеушілігі
Z аллелі (рецессивті белгі) аркылы анықталады. ZZ — гомо¬
зигота жиілігі еуропеоидтарда 0,05%, гетерозиготалар (MZ;
SZ) жиілігі — 4,5%. Тұқым куалайтын протеиназ ингибито¬
ры жетіспеушілігі кездесетін адамдар гомозиготалы (ZZ) бол¬
са, созылмалы кабынуга жэне өкпе эмфиземасына бейім бо¬
лып келеді. Ғалымдардыц болжауынша, антитрипсин жүйесі
агзада кабыну құбылыстарыныц дамуын тежеуде маңызды рол
атқарады. Өкпе ұлпасыныц болар-болмас зақымдануы (қабыну,
микроциркуляцияныц бұзылуы) нэтижесінде ыдыратушы (про-
теолиздеуші) ферменттер зақымданган учаскені бұза бастай-
ды. Қалыпты жагдайда протеиназ ферменті синтезделіп, ыды¬
ратушы (протеолиздеуші) ферменттердіц эсерлерін жояды да,
өкпе ұлпасыныц бұзылыстары тоқталады. Ал, протеиназ ин¬
гибиторы өнімініц жетіспеушілігі (мутантты генотип) ыдыра¬
тушы (протеолиздеуші) ферменттердіц өкпеніц зақымданган
учаскелерін бұзып өкгіеде көптеген тесіктердіц дамуына алып
келеді. Темекі тарту, ауаныц ластануы эмфиземаныц дамуын
едэуір тездетеді.
308
г
Адамның мекен ортасында көмірсутектер, оның ішінде
полициклдік көмірсутектер, көптегі кездеседі. Олар ағзада арил-
гидрокарбонгидроксилазамен гидроксилденгеннен кейін бел-
сенді эпоксидтерге айналады, Ал, эпоксидтер канцерогендік
заттарға жатады. Адамда осы ферменттің (арилгидрокарбон-
гидроксилаза) синтезделуінің индукциялануының көптеген ва-
риациялары белгілі. Мысалы, фермент көп мөлшерде синтез-
делетін адамдарды — доминантты гомозиготалар, орташа жэне
аз мөлшерде синтезделетіндерді — гетерозиготалылар жэне
рецессивті гомозиготалылар деп қарастыру керек. Өкпе ісігімен
ауыратын адамдардьщ 30% ферменттің коп синтезделінетін то-
бына жатады, ал популяцияда бүл белгі сирек кездеседі. Он-
дай адамдарга темекіні тартудың жэне кэсіби қызметінде
көмірсутектермен жанасудың оте қауіпті екенін түсіндіру кажет.
Сол сияқты, қуықтын зілді ісігіне генетикалық бейімділік
те белгілі болды. Ол бауырдың N-ацетилтрансфераза генінің
мутациясымен байланысты. Осы ферменттің қатынасуымен
ксенобиотикгер (зиянды жат заттар) ацетилденігі (зиянсызда-
нып) ағзадан сыртқа шығарылады. Ксенобиотиктердің ацетил-
дену жылдамдыгына қарай 3 түрлі фенотипті ажыратады: тез
ацетилдеушілер (қалыпты аллель бойынша гомозиготалылар),
баяу ацетилдеушілер (мутантты аллель бойынша гомозиготалы¬
лар) жэне орташа ацетилдеушілер (аралық формасы) гетерозиго¬
талылар.
Тамақтар жэне тамаққа қосатын заттар
Генетикалык сезімтал адамдарда кейбір тамақтың түрлері
қолайсыз реакцияларды тудыруы мүмкін. Мысалы, лактоза-
ны көтере алмау. Осындай кемістігі бар адамдарда сүт жэне сут
өнімдерін пайдаланганнан кейін асқорыту жолының «дискомфор¬
та» жэне іштің отуі байқалады. Бүл кемістіктің себебі — ішекте
лактозаны (сүт қанты) ыдырататын ферментгің синтезделмеуі,
болмауы. Осының нәтижесінде шіріту микрофлорасының кө-
беюіне қолайлы жагдай туады. Бүл геннің мутантты формалары
309
шығыс халықтарында кең таралған (жиілігі 95—100%), амери-
ка негрлері мен үндістерінде (70—75%), ал европеоидтарда —
5—10%.
Сыр (ірімшік) кұрамында кездесетін катехоламиндер кейбір
ағзаларда бас сақинасы (мигрень) ауруын тудырады. Бұл тира-
миннің нашар конъюгациялануымен байланысты. Осы ауруды ту¬
дыратын тағы бір тамақ (азық) түрі — шоколад. Мұнда моноами-
ноксидаза ферментінің активтігі томен болады.
Адамдардьщ алкогольге деген арнайы (спецификалык) реак-
циялары да белгілі. Кейбір адамдарда алкогольді өте аз мөлшерде
қабылдаудың өзі беттің тез қызарып, жүрек согуын жиілетеді
(тахикардия), асқазаны бүріп ауырады, бұлшық еттің босаңсуы
т.б. уланудың белгілері байқалады. Ағзаның бұл реакциясы
спиртті ыдыратушы екі ферменттің вариацияларына байланы¬
сты: 1) бауырдын алкогольдегидрогеназа гендеріне — оның 3
аллелі белгілі (АДН-1, АДН-2, АДН-3); 2) альдегиддегидрогеназа
гендеріне — оның 2 аллелі белгілі (АЬДН-1, АЬДН-2). Жоғарыда
келтірілген алкогольдін болар-болмас дозасына деген агза реак¬
циясы АЬДН-1 аллелінің гетерозиготалы күйіне байланысты да¬
миды.
Физикалық факторлар жэне металдармеп улану
Суыққа, ыстыкка жэне күн сәулесі әсерлеріне адамдардьщ
сезімталдыгының түрліше болатындыгы белгілі болганымен,
бұл реакциялардың генетикалық механизмдері элі толық зерт-
телмеген. Дегенмен, суықтың әсеріне әр түрлі нэсілдердің
реакциясы түрліше болатындыгы анықталган, мысалы: нег-
рлер кавказ нәсілдеріне қараганда суыққа өте сезімтал келеді.
Мұны оларда жылуды өткізу жэне қан тамырларының кецею
деңгейлері түрліше болуымен түсіндіреді. Ультракүлгін сәуленің
эсерлеріне адамдардыц жекелей және нәсілдік реакцияларының
ерекшеліктері анықталган. Оган мысал ретінде — пигменттік ксе-
родерма ауруының дамуын келтіруге болады. Бұл ауру аутосом¬
ды — рецессивті жолмен тұкым қуалайды, мұнда күн сәулесінің
310
әсерінен тері күйіп, жаралар пайда болады да терінің зілді ісігіне
айналады. Пигменттік ксеродерма ауруында ДНҚ молекула-
сының бұзылыстарын жөндеуші репарациялық жүйе зақым-
данады, яғни ДНҚ репарациясына қатысатын бірнеше локус-
тардың (экзонуклеаза, эндонуклеаза, полимеразалар, лигазалар
т.б.) мутацияларына байланысты. Адамдардьщ ауыр металдар
(қорғасын, сынап, кадмий, т.б.) түздарына сезімталдығының
түрліше болатындығы анықталды. Мысалы, сынаптың орга-
никалық қосылыстарымен уланған адамдарда түрліше дәрежеде
нейро-психикалық бүзылыстар байқалған.
Қорғасын деңгейінің «улы» болмай-ақ, жай көтеріңкі деңгей-
інің өзі жас балаларда «беймаза» мінез-қүлықтың дамуының се-
бебі болуы мүмкін.
11.3. Фармакогенетика
Фармакогенетика — адам ағзасының дәрі-дәрмек әсерлеріне
қарсы түқым қуалайтын реакцияларын зерттейді. Адамдардьщ
кез келген фармакогенетикалық реакциялары адам популяция-
сында қазіргі кезде қолданылатын фармакологиялық заттарды
колданғанға дейін эволюция гіроцесінде қалыптасқан кең көлемді
генетикалык полиморфизмі негізінде дамиды.
Қолданылатын дәрі-дәрмектердің тиімділігі мен қосымша
әсерлері эр түрлі топтарда жэне ағзаларда түрліше болатындығы
дәрігерлік практикадан белгілі. Ағзаға дәрі-дәрмектің стандарт-
ты дозасын енгізгеннен кейін оның қандагы концентрациясы бір
адамдарда оптимумнан (тиімді доза) томен болса, яғни эсер етпе-
се, екіниіі біреулерде улау деңгейіне дейін жетеді.
Дэрі-дэрмекгің ағзадағы тағдыры оның биотрансформация-
лануына немесе оның сіңірілу, таралу (мүшелерге, үлпаларға,
жасушаларға, органеллаларға), рецепторлармен өзара эрекеттесу,
метаболизм жэне ағзадан шығарылу қүбылыстарына бай¬
ланысты. Осы фармакокинетикалық кұбылыстардың эрбір
кезендері генетикалық түрғыдан бақылауда болатыны, яғни,
311
тэн (спецификалық) жэне тэн емес (спецификалық емес) фер-
менттердің қатынасуымен жұретіні сөзсіз. Адам популяция¬
ларында кең көлемді балансты полиморфизм болатынын ескер-
сек, эрбір дэрі-дэрмектің фармакокинетикалық кезендердегі
тағдыры полиморфты ферменттер немесе ақуыз жүйесімен бай¬
ланысты екені өзінен-өзі түсінікті.
Агзаның тұқым қуалайтын реакцияларының ерекшеліктері
фармакокинетикалық кұбылыстың барлык құрамдық бөлімде-
рінде (сорылу, мүшелерде жэне үлпаларда таралу, рецепторлар-
мен әрекеттесу, метаболизм, шығарылу) сипатталған.
Қазіргі кезде дәрі-дәрмек қабылдағаннан кейін туындайтын
патологиялық реакциялардыц көптеген мутациялары табылған.
Олардыц тұқым қуалау типтері жэне алғашқы биохимиялық
бүзылыстары зерттелінген. Олардыц ішінде Г-б-ФД (глюкоза
-6-фосфатдегидрогеназа) жетіспеушілігі жақсы зерттелінген.
Безгекке қарсы препараттарды (примахин, дифенилсульфан,
сульфаниламидтер, көк толуидин т. б.) қабылдау эритроциттердіц
гемолизденуіне (еруіне) алып келеді.
Метгемоглобинредуктаза жетіспеушілігі моногендік жолмен
түқым қуалайды, гомозиготалы күйінде метгемоглобинемия ауруы
дамиды. Гетерозиготалы агзаларда примахин, хингамин, диафе-
нилсульфон дәрі-дәрмектерін кабылдаганнан кейін метгемоглоби¬
немия жэне цианоз (терініц когеруі) дамиды.
Суксаметонинга сезімталдык сарысу холинэстеразасы генініц
мутациясымен байланысты. Бүзылган холинэстераза дитилинді
активсіздендіре алмагандықтан, осы ақуыз иелерінде үзақ уақыт
(1 сагат дейін) тыныс алу тоқталады. Бүл генніц 2 мутантты
аллелі (Е5 жэне Е') түрліше эсер етеді, Еи —- қалыпты аллель. Е5, Е'
аллельдері бойынша гетерозиготаларда (Es Е1) патологиялық эсер
толық байқалады, ал Eu Es немесе Eu Ef, ягни қалыпты генмен ге-
терозиготалыларда холинэстераза активтігі қалыпты болып, ауру
дамымайды.
Фармакогенетикалық аурудыц тагы бір түрі — гипертер¬
мия ісігі. Ол доминантты тип арқылы түқым қуалайды деп бол-
жамдалады. Бүл ауруды қоздырушы факторлар болып кейбір
312
ингаляциялық анестетиктер (фторэтан, этил эфирі, метоксифлу-
ран) саналады. Ауру адамдардьщ дене температурасы 44С°-қа
дейін көтеріледі жэне тахикардия, дем алудыңжиіленуі, гипоксия,
ацидоз, гиперкалиемия, гипокальцемия т.б. байқалады. Осындай
180 ауру анықталып, олардьщ 60 пайызы жүректің тоқтауы сал-
дарынан қайтыс болган.
Фармакогенетикалық бұзылыстардың тағы бір түрі — дәрі-
дәрмектіц метаболизмінің бүзылуы салдарынан олардың ағзадан
шыгарылуындагы кедергілер. Бұған мысал ретінде бауырдың
N-ацетилтрансфераза генінің мутацияларын келтіруге болады.
Туберкулезге (өкпе кабынуы) қарсы қолданылатын изонио-
зидті стандартты дозада қабылдаған кейбір адамдарда 6 сагаттан
кейін оның сарысудағы концентрациясы күрт көтеріледі. Бұл осы
препараттың ацетилденбей агзадан баяу шығарылуына байланы¬
сты. Мұндай адамдарда изониазидке қосалқы реакция ретінде
перифериялық нервтердің зақымдануы байқалады. Патологиялық
аллель аутосомды-рецессивті тип аркылы тұқым қуалайды.
Кейбір отбасында агзалардың кейбір дэрі-дэрмектерге резис-
тенттілігі (төзімділігі), мысалы кумаринге резистенттілігі; ди-
фениннің гидроксилденуінің бұзылуы: амобарбиталдың гидро-
ксилденуінің жетіспеушілігі, ацетофенитидпен индукцияланган
метгемоглабинемия; эритроциттерде литий мен натрийдің тасы-
малдануының бұзылыстары т.б. анықталған.
Болашақта адамдардыц денсаулығын сақтауда адам экоге-
нетикасының маңызы еселеп өсетіні сөзсіз, себебі, профилакти-
калық медицина — адам экогенетикасы тұжырымына сэйкес,
әрбір даралардыц қалыпты тіршілік етуі биохимиялық полимор-
физмнің патологиялық экогенетикалық эсерлерініц алдын алу
үшін ец қолайлы (оптимальды) жағдайларды (тамак, дэрі-дэрмек,
жүмыс) қалыптастыруға багытталады.
Дәрі-дәрмектерге қарсы ағзаның түқым қуалаушылық
арқылы дамитын патологиялық реакциялары
Генетикалык белгі
(вариант)
Белгіні тудырушы
дэрі-дэрмек
Патологиялык
реакция
I
Г-6 ФД жетіспеушілігі
(полиморфты жүйе)
Примахин, суль-
фанил амидты
препатарттар (40-
ка жуык)
Эритроциттердің
гемолизі (еруі)
2
Баяу ацетилдену
(ацетилтрансфераза
генінің полиморфизмі)
Изониазид,
гидраницин,
сульфадимезин,
диафенилсульфон
Әрбір дэрі-дэр-
мекке тэн әсерлер
(эффекттер)
Дитиминге сезімталдык
(бұзылган холинэстераза)
Дитимин
Тыныс алудың ұзак
уакытка токталуы
4
Метгемоглобинредуктаз-
дың жетіспеушілігі
Дифенилсульфон,
осингалин,
примахин
Цианоз (көгеру)
э
Глюкортикоидты
кабылдағаннан кейін кан
кысымының жогарылауы
(көтерілуі)
Көзге тамызатын
глюкокортикидтер
Көзішілік кан
кысымының
жогарылауы
(көтерілуі)
6
Кейбір тұраксыз
гемоглобиндер
Тотыктырушылар
Эритроциттердің
гемолизі (еруі)
/
Гипертермия ісігі
Жалпы анестез-
деуші заттар
Гипертермия
8
Феноиетинмен
индукцияланған
метгемоглобинемия
Фенацетин
Цианоз (көгеру)
Дифениннің жеткіліксіз
гидроксидденуі
Дифенин
Атаксия, нистагим
іо
Спартеиннің аномальды
дегидрогенациялануы
Спартеин
Жүрек аритмия-
сын емдегенде
байкалатын зілді
әсерлер (эффекттер)
314
СТУДЕНТТЕРДІҢ ӨЗ БЕТІНШЕ ОҚЫП-ҮЙРЕНУШЕ,
БАҚЫЛАУЫНА АРНАЛҒАН СҰРАҚТАР МЕН ЕСЕПТЕР
1. Сүрақтар:
1. Дұрыс бір жауапты тап.
Жасуша қашан ашылды?
а) 1859;
б) 1809;
б) 1838-1839;
г) 1665;
д) 1865.
2. Жасушаны кім ашты?
а) Ч. Дарвин:
о) Ж. Ламарк;
в) Р. Гук;
г) Шванн-Шлейден;
д) Г. Мендель.
3. Жасуша теориясын кім қалыптастырды?
а) Ч. Дарвин;
б) Ж. Б. Ламарк;
в) Р. Гук;
г) Шванн-Шлейден;
д) Г. Мендель.
4. Жасуша теориясы қашан қалыптасты?
а) 1859;
б) 1809;
в) 1838-1839;
г) 1665;
д) 1865.
5. Дұрыс екі жауапты тап.
Жасуша қандай жолдармен бөлінеді?
а) эндамитоз;
315
б) политения;
в) митоз;
г) мейоз;
д) партеногенез.
6. Қате бір жауапты тап.
Жасушаның құрылымдық компоненттері:
а) ядро;
б) митохондрия;
в) генотип:
г) рибосома;
д) цитоплазма.
7. Дұрыс бір жауапты тап.
Дене жасушалары қандай жолмен бөлінеді?
а) бүршіктену;
б) мейоз;
в) митоз;
г) эндомитоз;
д) политения.
8. Жыныс жасушалары кандай жолмен бөлінеді?
а) бүриііктену;
б) мейоз;
в) митоз;
г) эндомитоз; д) политения.
9. Жасушаның қандай бөлінуінде хромосома саны тұрақты сол
күйінде қалады?
а) эндомитоз;
б) полителия;
в) мейоз;
г) митоз.
10. Жасушаның қандай бөлінуінде хромосома саны екі есеге
азаяды (кемиді)?
а) эндомитоз;
б) политения;
316
в) мейоз;
г) митоз.
11. Қате бір жауапты тап.
Митоздық хромосомалардыц қандай түрлерін білесің?
а) метацентрлі;
б) субметацентрлі;
в) акроцентрлі;
г) субцентрлі;
д) телоцентрлі.
12. Дұрыс бір жауапты тап.
Жасуша циклінің қандай кезеңінде ДНҚ молекуласы екі
еселенеді?
а) пресинтездік;
б) синтездік;
в) постсинтездік;
г) эмбриондыц;
д) мшпоз.
13. Қалыпты адам кариотипі қанша хромосомалардан тұрады?
а) 46;
б) 48;
в) 44;
г) 45;
д) 47.
14. Адам кариотипінің А тобына қандай хромосомалар топта-
стырылады?
а) орташа субметацентрлік;
б) ец ірі метацентрлік;
в) ірі субметацентрлік;
г) ұсақ акроцентрлік;
д) усаң метацентрлік.
15. Адам кариотипінің В тобына қандай хромосомалар топта-
стырылады?
а) орташа субметацентрлік;
317
б) ең ірі метацентрлік;
в) ірі субметацентрлік;
г) үсац акроцентрлік;
д) үсац метацентрлік.
16. Адам кариотипінің С тобына қандай хромосомалар топта-
стырылады?
а) орташа субмепіаг/ентрлік:
б) ең ірі метацентрлік;
в) ірі субметацентрлік;
г) үсаң акрогіептрлік:
д) үсақ метацентрлік.
17. Адам кариотипінің Д тобына қандай хромосомалар топта-
стырылады?
а) ең ірі метацентрлік;
б) ірі субметацептрлік;
в) орташа субметацентрлік;
г) орташа акроцеитрлік;
д) үсақ субметацентряік.
18. Адам кариотипінің Е тобына кандай хромосомалар топта-
стырылады?
а) ең ірі метацентрлік;
б) ірі субметацентрлік;
в) ортаіиа субметацентрлік;
г) орташа акрог/ентрлік;
д) үсак, субметацентрлік.
19. Адам кариотипінің Ғ тобына қандай хромосомалар топта-
стырылады?
а) ортаіиа субмепхацентрпік;
б) үсақ субметацентрлік;
в) үсақ метацентрлік;
г) ең үсақ акроцентрлік;
д) ец ірі метацентрлік.
20. Адам кариотипінің G тобына қандай хромосомалар топта-
стырылады?
а) орташа субметацентрлік; б) ұсақ субметацентрлік; в) усақ
метацентрлік; г) ец усақ акроцентрлік;
д) ең ірі метацентрлік.
21. Ер адамдар кариотипінің орташа субметацентрлік хромосо¬
малар саны қанша?
а) 14;
б) 16:
в) 15;
д) 10.
22. Әйелдер кариотипінің орташа субметацентрлік хромосома¬
лар саны қанша?
а) 14;
б) 16;
в) 15;
г) 12;
д) 10.
23. Нуклеосомалар дегеніміз:
а) микроскоп ирцылы корінетін хромотин туіпршіктері;
б) гистондыц ақуыздар жиынтыгы;
в) ДНҚ молекуласыныц бір бөлігі;
г) активсізденген Х-хромосома;
д) РНҚ молекуласыныц бір бөлігі.
24. Геном дегеніміз:
а) диплоидтық хромосомалар жиынтыгы;
б) диплоидтыц хромосомалардагы гендер жиынтыгы;
в) гаплоидтьщ хромосомалар жиынтыгы;
г) гаплоидтыц хромосомалардагы гендер жиынтыгы;
д) бір хромосомадагы гендер жиынтыгы.
25. Генотип дегеніміз:
а) диплоидтыц хромосомалар жиынтыгы;
319
б) диплоидтық хромосомалардагы гендер жиынтыгы;
в) гаплоидтыц хромосомалар жиынтыгы;
г) гаплоидтьщ хромосомалардагы гендер жиынтыгы;
д) бір хромосомадагы гендер жиынтыгы.
26. Эухроматин хромосоманьщ:
а) центромера айналасында;
б) теломера бөлігінде;
в) узын иінінде;
г) қысқа иінінде орналасқан.
27. Гетерохроматин хромосоманың:
а) центромера айналасында;
б) теломера бөлігінде;
в) узын иінінде;
г) қысқа иінінде орналасқан.
28. Прокариоттардьщ гендері:
а) тек интрондардан;
б) тек экзондардан;
в) интрон жэне экзондардан;
г) РНҚучаскесінен тұрады.
29. Эукариоттардың гендері:
а) тек интрондардан;
б) тек экзондардан;
в) интрон жэне экзондардан;
г) РНҚучаскесінен турады.
30. ДНҚ синтезі жасуша циклінің:
а) интерфаза;
б) пресинтездік;
в) синтездік;
г) постсинтездік;
О) митоз фазаларында жүреді.
31. Процессинг дегеніміз:
а) транскрипция;
320
б) трансляция;
в) интрондык, учаскелердің қиылуы;
г) экзондъщ учаскелердің жалгануы;
д) репликация.
32. Сплайсинг дегеніміз:
а) транскрипция;
б) трансляция;
в) интрондык, учаскелердіц циылуы;
г) экюндық учаскелердің жалгануы;
д) репликация.
33. Репрессор гені:
а) цүрылымдъщ ген;
б) кластерлі ген;
в) реттеуиіі ген;
г) ақпараттың көшірілуініц басталу нүктесі.
34. Промотор дегеніміз:
а) қурылымдьщ ген;
б) кластерлі ген;
в) реттеуиіі ген;
г) ақпараттың көиіірілуінің басталу нүктесі.
35. Оператор гені:
а) цурылымдьщ ген;
б) кластерлі ген;
в) реттеуші ген;
г) ақпараттың көшірілуінің басталу нүктесі.
36. Бір қате жауапты тап. Реттеуші гендерге жататыны:
а) репрессор гені;
б) оператор гені;
в) қүрылымОық ген;
г) промотор гені.
321
37. Құрылымдық гендер ДНҚ молекуласының:
и) цайталанбайтын;
б) орташа цайталанатын;
в) өте жиі цайталанатын учаскелерінде;
г) гетерохроматин учаскесінде орналасцан.
38. Нуклеогистонда ДНҚ молекуласы:
а) хроматинге;
б) нуклеосома денешіктеріне;
в) гистонды емес ацуыздарга;
г) цитоплазма ақуыздарына ишратылады.
39. Дұрыс екі жауапты ran. Хроматиннің құрамы:
а) РНҚ молекуласынан;
б) ДНҚ молекуласынан;
в) гистонды емес ацуыздардан;
г) гистонды ацуыздардан;
д) нуклеотидтерден түрады.
40. Дұрыс бір жауапты тап.
Қандай белгілердің гендері тәуелсіз гұкым қуалайды?
а) цитоплазма гендері;
б) ядро гендері;
в) әр турлі хромосомаларда орналасцан гендер;
г) бір хромосомаларда орналасцан гендер;
д) сандыц белгілер.
41. Қандай белгілердің гендері тіркес тукым қуалайды?
а) цитоплазма гендері;
б) ядро гендері;
в) әр түрлі хромосомаларда орналасцан гендер;
г) бір хромосомаларда орналасцан гендер;
д) сандыц белгілер.
42. Қате жауаптарды тап. Мутация түрлері:
а) хромосомалыц;
б) генотиптік;
в) геномдыц;
г) сомалыц.
322
43. Гендік мутациялар:
а) ген цүрамында;
б) нуклеотидтер алмасуында;
в) анеуплоидияда;
г) транзициялыц;
д) трансверсияларда болады.
44. Қате жауапты тап. Хромосомалық мутациялар:
а) геннің цурамында;
б) транслокацияда;
в) инверсияда;
г) анеуплоидняда;
д) транслокацияда болады.
45. Дұрыс бір жауапты тап. Геномдык мутациялар:
а) геннің цурамында;
б) транслокацияда;
в) инверсияда;
г) анеуплоидняда;
д) транслокацияда болады.
46. Адам генетикасын зерттеу әдістері:
а) гибридологиялыц;
б) цитогенетикалыц;
в) эксперименталдыц;
г) тарихи;
О) филогенетикалыц.
47. Дұрыс жауаптарды тап. Кариотип дегеніміз:
а) хромосома саны;
б) хромосомалар пііиіні;
в) гендер жиынтыгы;
г) генотип;
д) хромосомалар мөлшері.
48. Адам кариотипініц 45 болуы қандай өзгерістерге жатады?
а) мутациялыц;
323
б) гендік;
в) хромосомалыц;
г) геномдъщ;
д) моносомиялык,.
49. Адам кариотипінің 47 болуы қандай өзгерістерге жатады?
а) фенотиптік;
б) хромосомалыц;
в) геномдыц;
г) гендік;
д) трисомиялыц.
50. Дұрыс бір жауапты тап. Полиплоидия:
а) цачыпты кариотип;
б) хромосома саныныц 1 хромосомага кемуі;
в) хромосома саныныц 1 хромосомага кобеюі;
г) хромосома саныныц еселеп өсуі.
51. Шерешевский-Тернер синдромына тэн кариотипті тап.
а) 46, хх;
б) 46 ху;
в) 45 хо;
г) 47 хху;
д) 47-21+.
52. Даун синдромына тэн кариотипті тап:
а) 46, хх;
б) 46 ху;
в) 45 хо;
г) 47 хху;
д) 47-21+.
53. Патау синдромына тэн кариотипті тап:
а) 45 хо;
б) 47 ххх;
в) 47-13+;
г) 47-18+;
д) 47-21+.
324
54. Эдвардс синдромына тэн кариотипті тап:
а) 45 хо;
б) 47-13+;
в) 47-18+;
г) 47-21+;
д) 46-5рд.
55. «Мысықша мияулау» синдромына тэн кариотипті тап:
а) 45 хо;
б) 47-13+;
в) 47-18+;
г) 47-21+;
д) 4б-5рд.
56. Клайнфельтер синдромына тэн кариотипті тап:
а) 47 ххх;
б) 45 хо;
в) 47 хху;
?) 47-18+;
д) 46-5рд.
57. Қате бір жауапты тап. Бластула түрлері:
а) целобластула;
б) бластопора;
в) сшфибластула;
г) дискобластула;
д) морула.
58. Гаструланың құрылысы:
а) энтодерма;
б) эктодерма;
в) бластодерма;
г) гастроцелъ;
д) бластопора.
59. Дұрыс бір жауапты тап. Гаструляция жолдары:
а) эпиболия;
325
б) гомоморфоз;
в) эпиморфоз;
г) бластодерма;
д) гастроцель.
60. Қате бір жауапты тап. Эмбриогенездің жасушалық меха-
низмдері:
а) жасушачардыц бөлінуі;
б) миграциялану (көиііп-цону);
в) таңдамалы іріктелу;
г) нейруляция;
д) зацды турде өліп цалуы.
61. Қате бір жауапты тап.
Эмбриондык жасушалар өздерінің дамуында мынадай сатылар-
дан өтеді:
а) тотипотенттілік:
б) детерминациялану;
в) репликация;
г) дифференциация.
62. Эмбриогенез механизмдері:
а) жасушальщ;
б) генетикалык,;
в) эмбриондыц индукция;
г) транскрипция.
63. Дүрыс бір жауапты тап. Пубертаттық секіру қай кезде байка-
лады:
а) 3—5 жаста;
б) 8—10 жаста;
в) 11—13 жаста;
г) 18—20 жаста;
д) 40—50 жаста.
64. Пубертаттық секіру кезінде:
а) сүт тістері шыгады;
326
б) түрсщты тістері шыгады;
в) агзаның өсуі күрт жогарылайды;
г) агзаныц өсуі тоцтайды.
65. Дамудың туа біткен ақаулықтарына жатады:
а) гендік аурулар;
б) эмбриогенез барысында пайда болатын турсщты морфо-
логиялъщ бузылыстар;
в) туылганнан кейін пайда болатын өзгерістер;
г) моднфикациячар.
66. Дұрыс екі жауапты тап.
Бластула кұрылысы:
а) эктодерма;
б) бластодерма;
в) энтодерма;
г) бластоцель;
д) гастроцель.
67. Дуры с емес екі жауапты тап. Гаструла кұрылысы:
а) гастроцель;
б) бластопора:
в) бластоцель;
г) мезодерма;
д) морула.
68. Дұрыс бір жауапты тап.
Нерв түтігі қандай ұрық кабатынан түзіледі:
а) эктодерма;
б) энтодерма;
в) бластодерма;
г) мезодерма;
д) морула.
69. Хорда қандай ұрық қабатынан түзіледі:
а) эктодерма;
б) энтодерма;
327
в) бластодерма;
г) мезодерма; д)морула.
70. Тері эпителиі қандай ұрык кабатынан түзіледі:
а) эктодерма;
б) энтодерма;
в) бластодерма;
г) мезодерма;
д) морула.
71. Ас корыту жолдарының эпителийі қандай ұрық кабатынан
түзіледі:
а) эктодерма;
б) энтодерма;
в) бластодерма;
г) мезодерма;
д) морула.
72. Сезім мүшелері қандай ұрык қабатынан түзіледі:
а) эктодерма;
б) энтодерма;
в) бластодерма;
г) мезодерма;
д) морула.
73. Бүлшыкеттер қандай ұрык кабатынан түзіледі:
а) эктодерма;
б) энтодерма;
в) бластодерма;
г) мезодерма;
д) .морула.
74. Қаңқа қандай үрық кабатынан түзіледі:
а) эктодерма;
б) энтодерма;
в) бластодерма;
г) мезодерма;
д) морула.
328
75. Ішкі секреция бездері қандай ұрық қабатынан түзіледі:
а) эктодерма;
б) энтодерма;
в) бластодерма;
г) мезодерма;
д) морула.
76. Жүрек-қантамыр жүйесі қандай үрық кабатынан түзіледі
а) эктодерма;
б) энтодерма;
в) бластодерма;
г) мезодерма;
д) морула.
77. Жыныс мүшелері кандай ұрык кабатынан түзіледі:
а) эктодерма;
б) энтодерма;
в) бластодерма;
г) мезодерма;
д) морула.
78. Зэр шығару жүйесі кандай үрык кабатынан түзіледі:
а) эктодерма;
б) энтодерма;
в) бластодерма;
г) мезодерма;
д) морула.
79. Дұрыс екі жауапты тап.
Адам жынысыныц дамуы кандай факторларга байланысты:
а) экологиялыц;
б) аутосомдарга;
в) жыныс хромосомаіарына;
г) белок-рецепторларга;
д) физикалыц.
329
170-22
80. Дұрыс бір жауапты тап. Генофонд дегеніміз:
а) бір агзаныц гендерінің жиынтыгы;
б) популяция дараларыныц генотиптерініц жиынтыгы;
в) популяция дараларыныц генотиптері мен фенотиптерінің
жиынтыгы;
г) популяция дараларыныц фенотиптерініц жиынтыгы.
81. Популяция генофондына эсер ететін факторлар:
а) экологиялыц;
б) климаттыц;
в) гендердіц дрейфі;
г) физикалыц;
д) химиялыц.
82. Популяция генофондына эсер етпейтін факторлар:
а) мутация;
б) оцаиіалану;
в) климаттыц;
г) гендердіц дрейфі;
д) популяциялыц толцындар.
83. Популяция генофондыныц өзгеруі калай аталады?
а) царапайым экологиялыц цубылыс;
б) царапайым эволюциялыц факторлар:
в) царапайым эволюциялыц цубылыс;
г) мутация;
д) тіршілік толцындары.
84. Қате бір жауапты тап.
Популяцияныц экологиялық сипаттамасына жатады:
а) генофонд;
б) ареал мөлшері;
в) жастыц цурамы;
г) жыныстыц цүрамы;
д) даралар саны.
330
85. Дұрыс бір жауапты тап. Демдерде даралар саны:
а) 1400;
б) 1000;
в) 3500;
г) 10000;
д) 100000.
86. Изоляттарда даралар саны:
а) 1500;
б) 1000;
в) 4500;
г) 10000;
д) 100000
87. Адамдардьщ тұқым куалайтын ауруларыньщ пайда болу
жолы:
а) сырпщы орта факторларыньщ әсері;
б) генетикалык, факторлар;
в) сырпщы орта жэне генетикалык, фактор;
г) тотенше жагдатар.
88. Адамдардьщ мультифакторлы ауруларыньщ пайда болу
жолы:
а) сырпщы орта факторларыньщ әсері;
б) генетикалык, факторлар;
в) сырпщы орта жэне генетикалык, фактор;
г) төтениіе жагдатар.
89. Дұрыс бір жауапты тап. Генетикалык инженерия дегеніміз:
а)хро\юсомаларды зерттеу;
б) ДНҚ нуклеотидтерін аныцтау;
в) биологияльщ актив заттарды лабораториялык, жагдайда
өндіру;
г) жасанды тұқым қуалаушылық материалдарын цурастыру;
д) ауруларды емдеу.
331
90 Биотехнология дегеніміз:
а) хромосомаларды зерттеу;
б) ДНҚ иуклеотидтерін анъщтау;
в) биологиялъщ актив заттарды лабораториялъщ жагдайда
өндіру;
г) жасанды түцымцуалаушылъщ материалдарын цұрастыру;
д) ауруларды емдеу.
91. Трансгенді агзалар дегеніміз:
а) гибридтер;
б) мутацитанган өсімдіктер;
в) жасанды генотипке ие агзалар;
г) мутацііяланган бактергіялар;
д) мутациялапган жануарлар.
92. Клондау дегеніміз:
а) будандастыру;
б) сурыптау;
в) будандастыру жэне сурыптау;
г) бір жасуша не агза ұрпацтары;
О) цолдан өсіру.
93. Эмбриогенездің ұрықтық сатысы қай уақытқа дейін созы-
лады?
а) 8 аптага;
б) 10 аптага;
в) 15 аптага;
г) 30 аптага;
д) 40 аптага.
94. Эмбриогенездің кұрсақтағы бала сатысы қай уақытта баста¬
лады?
а) 1 апта; б) 7апта;
в) 9 апта;
г) 15 апта;
д) 30 аптадан.
332
95. Дұрыс үш жауапты тап. Тератогендік факторларға жатады
а) физикалыц;
б) топырац;
в) биологиялыц;
г) антропогендік;
д) химиятқ.
96. Қате бір жауапты тап. Канцерогендік факторларға жатады
а) химиялыц;
б) физикалыц;
в) антропогендік;
г) биологиялыц;
д) топырац.
97. Экологиялық апат аймағы:
а) адамдардыц тіриіілігіне цолайсыз;
б) адамдардыц тіриіілігіне цолайлы;
в) экологиялыц жагдайы томен;
г) әлеуметтік жагдаіілары нашар.
98. Дұрыс бір жауапты тап. Экогенетикалық эсер дегеніміз:
а) мутациялар әсері;
б) орта жагдайларыныц әсері;
в) орта жэне мутация әсерлері;
г) антропогендік эсерлер;
д) зоогендік эсерлер.
99. Гомологиялык мүшелер:
а) қүры.чыстары бірдей;
б) цызметтері бірдей;
в) цүрылысы. цызметтері әр түрлі;
г) иіыгу тегі бірдей;
д) иіыгу тегі эр түрлі.
100. Аналогиялық мүшелер:
а) цұрылыстары бірдей;
б) цызметтері бірдей;
333
в) қүрылысы, цышеттері эр түрлі;
г) шыгу тегі бірдей;
д) шыгу тегі әр түрлі.
101. Рудименттер:
а) қапыпты мүшелер;
б) кішіреііген мүшелер;
в) цалдыц мүиіелер;
г) ңаііталанган ата-тектерінің белгілері;
д) алып мүшелер.
102. Атавистік мүшелер:
а) цалыпты мүшелер;
б) кішіреііген мүиіелер;
в) цалдыц мүшелер;
г) цаіппаланган ата-тектерінің белгілері;
д) алып мүшелер.
103. Рудимент мүшелер:
а)) қол-аяқтар;
б) түктер;
в) цүііымшак;
г) ңүйрық;
д) тістер.
104. Дұрыс екі жауыпты тап.
Атавистік мүшелер:
а) цүйымиіац;
б) адамда қүйрықтың дамуы;
в) соңыр ішек өсіндісі;
г) денені түк басуы;
д) тістер.
105. Дүрыс бір жауапты тап.
Түр түзілудің қандай жолдарын білесің?
а) фтетнкалъщ;
б) эмбриондыц;
334
в) партеногенездік;
г) нейруляциялыц;
д) фетопатиялъщ.
Дүрыс жауаптар:
1г; 2в; Зг; 4в; 5в,г; 6в; 7в; 86; 9г; 10в; 11 г; 126; 13а; 146; 15в; 16а;
17г; 18д; 19в; 20г; 21в; 226; 236; 24г; 256; 266; 27а; 286; 29в; ЗОв;
31 в; 32г; ЗЗв; 34г; 35в; 36в; 37а; 386; 396, г; 40в; 41г; 42в; 43в; 44а;
45г; 466; 47а, б, д; 48д; 49д; 50г; 51 в; 52д; 53в; 54в; 55д; 56в; 576;
58в; 59а; 60г; 61 в; 62г; 63в; 64в; 656; 66 б, г; 67 в, д; 68а; 696; 70а;
716; 72г; 73г: 74г; 75а; 766; 77г; 78г; 79г; 80в, г; 816; 82в; 83д; 84а;
85в; 86а; 876; 88в; 89г; 90в; 91 в; 92г; 93а; 94в; 95а, в, д; 96д; 976;
98в; 99г; ЮОд; 101в; 102г: 1036; 104 6, г; 105а.
2. Есептер:
1. ДНҚ молекуласыныц бір жіпшесіндегі нуклеотидтер тізбегі
ААГГЦТЦТАГГТАЦЦАГТ куйінде орналасқан.
а) осы жіпшеге комплиментарлы жіпше нуклеотидтерін тап;
б) осы жіпше негізінде синтезделінетін а-РНҚ нукпеотидтерінің
тізбегін анықта;
в) осы жіпшедегі ақпарат негізінде синтезделінетін полипептид
молекуласын анықта.
2. Цистинурия ауруымен (несеп арқылықалыпты жағдайдағыдан
көп амин кышқылдарының бөлініп шығуы) ауыратын адам
несебімен бөлініп шыгатын амин қышқылдарына а-РНҚ-ның
мынадай кодондары сэйкес келеді: АЦУ; У ТУ; ГЦУ; ГГУ; ЦАА;
АБА; ААА. Сау адамдардыц несебінде аланин, серин, глутамин
қышқылы жэне глицин кездесетіні белгілі.
а) цистинурия ауруына тэн амин қышқылдарын тап;
б) сау адамдардыц несебінен бөлініп шыгатын амин
қышқылдарына сэйкес келетін а-РНҚ триплеттерін жаз.
3. Адамда қара көзд іц болуы көк көзге қараганда доминанттылық
байқатады. Көк көзді ер адам қара көзді эйелге үйленген. Әйелдіц
әкесінің козі көк, шешесініц көзі қара. Осы некеден кара көзді бала
туылган.
335
а) осы отбасының келесі балаларының көздері қандай болмақ;
б) көк көзді балалардьщ туылу ықтималдылығын тап;
в) жоғарыда келтірілген адамдардың генотиптері қандай?
4. Фенилкетонурия аутосомды-рецессивті белгі ретінде түқым
қуалайды.
а) егер ата-аналары гетерозиготалы болса балалары кандай
болмақ?
б) фенилкетонурия ауруымен ауыратын балалардьщ туылу
ықтималдылыгы кандай?
5. Шизофренияның (қояншык) кейбір формалары аутосомды-
доминантты жолмен тұқым қуалайды. Гомозиготаларда бүл геннің
пенетранттылығы 100%, ал гетерозиготалыларда — 20%. Ата-
аналарының екеуі де гетерозиготалы отбасында ауру балалардьщ
туылу ықтималдылыгын тап.
6. Адамдарда соқырлыктың екі түрі белгілі жэне олардың
әрқайсысы өзінің аутосомды-рецессивті гендері аркылы дамиды.
Екі белгінің гендері әр түрлі хромосомаларда орналасқан.
а) егер ата-аналары соқырлықтың бір түрімен ауырып екінші
түрі бойынша сау жэне гетерозиготалы болса, балаларының соқыр
болып туылу ықтималдылыгы кандай?
б) егер ата-аналары соқырлыктың эр түрлі түрлері бойынша
ауырган болса (екі ген бойынша да гомозиготалы) балалардьщ
сокыр болып туылуын аныкта.
7. Адамдарда дальтонизм — кызыл, жасыл түстерді ажырата ал-
мау, Х-хромосомамен тіркес рецессивті жолмен тұқым қуалайды.
а) қалыпты көретін қыз, әкесі дальтонизммен ауырады,
қалыпты көретін ер адамга күйеуге шыққан. Осы некеден туылган
балалардьщ түсті ажыратуы қандай болмак?
б) қалыпты көретін жұбайлардың: 1) дальтоник ұлы жэне
қалыпты көретін кызы бар; 2) калыпты көретін қызы, ал оның бір
дальтоник жэне 1 қалыпты көретін ұлдары бар; осы жанұяньщ
отбасыларының, балаларының, немерелерінің генотиптерін
анықга.
8. Адамда дальтонизм Х-хромосомамен тіркес рецессивті тұ-
қым қуалайды, ал талассемия аутосомды — доминантты белгі
ретінде тұқым қуалайды жэне оның екі формасы белгілі; гомози¬
готалы күйінде ол өте ауыр, зілді жүреді, тіпті өлімге экеледі, ал
гетерозиготалыларда жеңіл субклиникалық күйінде болады.
336
Қалыпты көретін жэне талассемияның жеңіл түрімен ауыра-
тын эйел, дальтоник, бірақ талассемиямен ауырмайтын ер адамға
күйеуге шыққан. Олардан талассемияның жеңіл түрімен ауыра-
тын жэне дальтоник үл бала туылған. Келесі үл баласының екі
белгі бойынша сау болып туылу ықтималдылығын анықта.
9. Ата-аналарында гетерозиготалы II жэне III қан тобы болса,
баласында қандай кан топтары байқалады?
10. Перзентханада екі нэрестені шатастырып алған. Бірінші
баланың ата-аналарында I жэне II қан тобы, екіншісінің ата-
аналарында I жэне IV кан топтары анықталған. Балалардьщ
біреуінде I, екіншісінде II кан тобы болган. Балалардьщ ата-
аналарын аныкта.
11. Альбинизм аутосомды-рецессивті белгі ретінде тұқым
куалайды. Бұл аурудың жиілігі 1:20000 тең. Популяциядагы гете-
розиготалылар санын тап.
12. Аниридия аутосомды-доминантты белгі ретінде тұқым
куалайды жэне оның орташа популяциялық жиілігі 1:10000 тең.
Популяциядагы гетерозиготалылар санын анықта.
13. Полипептид мынадай амин қышқылдарынан құрылган:
валин-аланин-глицин-лизин-триптофан-валин-серин-глутамин
кышқылы.
Осы полипептидті анықтайтын ДНҚ молекуласының учас-
кесін тап.
14. Полипептид молекуласын кодтаушы қалыпты ДНҚ
молекуласының учаскесі мынадай азоттык негіздерден тұрады:
ААААЦЦААААТАЦТТАТАЦАА. Репликациялану кезінде сол
жакгагы үшінші аденин түсіп қалган.
Қалыпты ДНҚ молекуласының учаскесін жэне репликация-
ланганнан кейінгі ДНҚ учаскесін аныктайтын полипептид моле-
кулаларын тап.
15. Ретинобластома (көздің тор кабатының ісігі) аутосомды
доминантты ген арқылы беріледі, оның пенетранттылыгы 70%.
Медициналық-генетикалық кеңес алу үшін дәрігерге жүкті эйел
келген. Анамнезден белгілі болганы — жұбайлардың екеуі де
сау, бірақ үлкен ұлы осы аурумен ауырады. Әйелдің шежіресінде
ауру адамдар кездеспеген, ал күйеуінің экесіне бала кезінде
ретинобластомага хирургиялык операция жасалган. Осы жанүяда
ауру баланың туылу ықтималдылыгы қандай?
337
16. Адамдардьщ қалыпты естуі 2-А жэне В комплиментар-
лы гендердің өзара эрекеттесуіне байланысты. Керең — мылқау
жұбайлардан (Смиттер) 4 керең — мылқау балалар, ал керең —
мылқау Вессондардан 5 керең — мылқау балалар туылған. Әйелі
дуние салғаннан кейін Смит жесір Вессонға үйленген. Бұл неке-
ден 6 қалыпты есітетін балалар туылған. Смиттердің (жұбайлар),
Вессондардың (жұбайлар) жэне олардың бірінші, екінші
некелерінен туылган балаларының генотиптерін анықта.
17. Үзын бойлы адамдардьщ генотипі А1А1А2А2АЗАЗ, ал
қысқа бойлылардікі — а1а1а2а2аЗаЗ. Орта бойлы адамдардьщ
генотиптерінің варианттарын жаз.
18. Адамдарда интерферон синтезі 2 тенге байланысты, оның
біреуі 2, ал екіншісі 5 хромосомада орналасқан.
а) осы гендердің әрекеттесуі қалай аталады?
б) егер жүбайлар осы гендер бойынша гетерозиготалы болса,
интерферон синтездей алмайтын баланьщ туылу ықтималдылығын
анықта.
19. Дрозофиланың дигетерозиготалы ұргашы дарасын
рецессивті еркек дарасымен будандастырғанда төмендегідей
ұрпақтар пайда болған:
а) АВ + Ав + Ав + ав = 25%+25%+25%+25%
б) АВ + Ав + аВ + ав = 45%+5%+5%+45%
в) АВ + Ав + аВ + ав = 5%+45%+45%+5%
1) кандай жагдайда тэуелсіз жэне тіркес тұқым куалау
байқалады?
2) а, б, в —- варианттарында гендер қалай орналасқан?
3) б, в — жагдайларында А жэне В гендерінің ара қашықтыгы
неге тең?
20. Ауру адамның кейбір жасушаларында қалыпты кариотип
кездессе, баскаларында 47 немесе 45 хромосомалар кездескен. Осы
өзгерістер қалай аталады жэне оньщ пайда болу механизмдерін
ата.
21. Дрозофилада, сүтқоректілерде жэне адамдарда үргашы жы¬
ныс — гомозиготалы, ал еркек жыныс — гетерозиготалы болып
келеді. Құстарда еркек жыныс — гомозиготалы, ал ұргашылары
— гетерозиготалы.
338
Төмендегі жануарлардыц сомалық жасушаларында қандай жы¬
ныс хромосомалар кездеседі?
1) ұргашы дрозофилада —
2) еркек дрозофилада -
3) тауықта —
4) әтеште —
5)әйелде —
6) ер адамда —
22. Америка банкирі Твистердің үлы 3 бірдей тұқым куалайтын
аурумен ауырган: гемофилия, дальтонизм жэне тістерінің болма¬
уы. Бұл аурулар Х-хромосомамен тіркескен. Твистердің ұлы ұзақ
уақыт жанұясынан бөлек, Парижде тұрған жэне 1944 жылы сол
жерде дүние салған. Ол өлгеннен кейін банкир Твистерге 15 жа¬
сар ұл баланы ертіп бір француз эйел келеді де, мына бала сіздіц
немереңіз дейді. Ол бала Твистердің ұлында аталган 3 аурумен
бірдей ауырады, бірак баланың туылуы туралы құжаттары бол¬
майды. Твистер жанұялык дәрігердің кеңесімен, туылуы туралы
кұжаты болмаса да, сол баланы немерем деп мойындаган, себебі
дэрігер осыншама сирек кездесетін тұқым куалайтын аурулардың
қатар келуі кездейсок емес деп оны сендіреді. Сіз дэрігердің ойы-
мен келісесіз бе?
23. Патау, Эдвардс, Даун, Клайнфельтер, Тернер жэне «Мы-
сықша мияулау» синдромдарының кариотиптерін жаз?
24. Құрамында күрделі, тіршілікті болдырмайтын, генети¬
калык бұзылыстары бар ұрыктар неліктен бөлшектену сатысынан
түгел өте алады?
25. Жасушаларында артык хромосомалары бар ұрықтар
бөлшектену сатысында тірі болып. эрі қарай өліп қалады. Мұны
немен түсіндіруге болады.
Дүрыс жауаптар
1. а) ТТЦЦГАГАТЦЦАТГГТЦА — екінші жіпше
б) ААГГЦУЦУАГГУАЦЦАГУ — а-РНҚ
в) лиз-ала-лей-гли-тре-сер-полипептид
2. а) цистеин, глютамин, аргинин, лизин
339
б) ГЦУ, ГЦА, ГЦА, ГЦГ, ГЦЦ — аланин; УЦУ, УЦА, УЦЦ,
УЦГ, АГУ, АГЦ — серин; ГАА, ГАГ — глютамин қышқылы; ГГУ,
ГГА — глицин.
3. а) көк көзді жэне қара көзді балалардыц туылу ықтималдығы
50% тец.
б) 50%;
в) экесі — аа, шешесі — Аа.
4. а) 75% — сау, 25% — ауру; б) 25%.
5. а) 35% — тец.
6. а) 75% — балалары 1 түрімен, 25% балалары 2 түрімен ауру
болады;
6)0.
7. а) қыздарыныц бэрініц көзі сау, ұл балалары 50% сау, 50%
дальтоник; б)Р:ХАУ; ХАХа Г,. ХАХА; ХАХа; ХАУ; ХАУ Ғ,. ХАУ;
ХаУ.
8. а) 25%.
9. a) I, II, III, IV — кан топтары.
10. а) I топ қаны бар бала бірінші жұп ата-аналардыц, 11-топ
қаны бар бала екінші жұп ата-аналардыц балалары.
11. а) 1,41%.
12. а) 0.012%.
13. а-РНҚГУУ, ГЦЦ, ГГУ, ААА, УГГ, ГУГ, УЦЦ, ГАА
ДНҚ-1 ЦААЦГГЦЦАТТТАЦЦЦАЦАГГЦТТ ДНҚ-2-
ГТТГЦЦГГТАААТГГЦТГТЦЦГАА
14. а) калыпты а-РНҚ молекуласы: УУУУГГУУУУАУГАА-
УАУГУУ полипептид молекуласы: фенилаланин-триптофон-
фенилаланин-триозин-глютамин кышқылы-триозин-валин.
б) мутацияланғаннан кейінгі а-РНҚ молекуласы: УУУГ-
ГУУУУАУГААУАУГУУ полипептид-фенилаланин-глицин-фени-
лаланин-метионин-аспарагин-метионин.
15. а) ауру баланыц туылу ыктималдыгы 35% тец.
16. а) ерлі-зайыпты Смиттердіц генотипі ААвв, ал Вессондар-
дікі — ааВВ; олардыц балаларыныц генотиптері — бірінші неке-
ден — ААвв жэне ааВВ, ал екінші некеден — АаВв.
17. а) орташа бойлылардыц генотипі — А, А, а, а2; а, а, А, А2;
А, а,А, а2.
18. а) комплиментарлық; б) 7/16 бөлігі.
340
19. 1-а — тәуелсіз; б, в — тіркес тұқым қуалау, 2-а — гендер эр
түрлі хромосомаларда орналасқан 3-10 морганида.
20. а) мозаикалық, эмбриогенездің бастапқы сатыларында бір
жұп хромосоманың ажыраспауы.
21. 1-ХХ 4-ХХ 2-ХУ 5-XX 3-ХУ 6-ХУ.
22. Жоқ, себебі Твистердің үлы өз баласына X хромосоманы
бере алмайды.
23. 1 —47, 13+; 4 — 47, ХХУ; 2-47, 18+; 5- 45, ХО; 3-47, 21+;
6- 46, 5 p.
24. Бөлшектену кезінде гендер фенотиптік белгі күйінде
байқалмайды.
25. Бөлшектену кезінде гендер гетеросинтетикалық қызмет
атқармайды, сондықтан артық хромосома бластомерлердің тір-
шілігіне зиян келтірмейді.
ТЕРМИН ДЕР СӨЗДІГІ
Аденозинтрифосфор қышқылы (АТФ) — құрамында аденин,
рибоза жэне фосфор қышқылының 3 қалдығы бар нуклеотид; тірі
жасушалардагы энергияның эмбебап аккумуляторы.
Акселерация — балалардьщ жэне жасөспірімдердің жедел
физикалык жэне физиологиялық дамуы.
Алкаптонурия — аутосомды-рецессивті жолмен тұқым куа-
лайтын гендік ауру; оньщ даму себебі — гемогентезин қышқылы-
ның малелацетатсірке кышқылына айналуын катализдеуші окси-
даза ферментінің синтезделуінің бұзылуы.
Аллель — бір геннің эр түрлі формалары.
Аллельді гендер — гомологтік хромосомалардыц бірдей локу-
сында орналаскан гендер.
Аллель емес гендер — гомологтік хромосомалардыц эр түрлі
локустарына орналаскан гендер.
Альбинизм — аутосомды-рецессивті жолмен тұқым куалай-
тын гендік ауру; оныц себебі триозиннің меланинге айналуын
катализдеуші триозиназа ферментінің синтезделуініц бүзылуы.
Амниоиентез — пренатальдық диагностика әдісі; мұнда жа-
тырдан сұйықтық алып, ондагы үрық жасушаларын генетикалық
зерттеулер үшін пайдаланады.
Анеуплоидия (гетероплоидия) — геномдық мутациялардыц
бір түрі; диплоидтық хромосома саныныц 1 немесе бірнеше
хромосомаларга өзгеруі.
Антибиоз — екі агзаныц (түрдіц) бірге тіршілік етуінін мүм-
кін болмауы, мысалы, бактериялар — саңырауқүлақтар.
Антикодон — а-РНҚ, молекуласыныц бос кодонына компли-
ментарлы т-РНҚ триплеті; олардыц өзара байланысуы полипеп¬
тид жіпшесіндегі амин қышкылыныц орнын аныктайды.
Антимутагенез — мутациялардыц агзада (жасушада) тұрақ-
талып сақталуын, не мутация әсерлерінен болдырмайтын
(байқатпайтын) биологиялық механизмдер жиынтыгы.
Антимутагендер — кенеттен пайда болатын жэне индук-
цияланган мутациялардыц жиілігін азайтатын факторлар.
Аралық ие — паразиттіц дернэсілдік (личинкалық) сатылары
кездесетін, ягни жыныссыз жолмен көбейетін агза.
342
Аутбридинг — туыс емес даралардыц будандасуы; гетерозиго-
талылар жиілігін жоғарылатады.
Аутосомалар — эйел жэне ер адамдарда бірдей болып келе-
тін 22 жұп хромосомалар.
Аутосомды-доминантты туқым қу>алау — аутосомаларда
орналасқан доминантты ген арқылы тұқым қуалау типі.
Аутосомды-рецессивті тұқым куалау — аутосомаларда
орналасқан рецессивті ген арқылы тұқым қуалау типі.
Ахондроплазия — аутосомды-доминантты жолмен тұқым
қуалайтын гендік ауру; 5-нуклеотидаза жэне глюкоза —
6-фосфотаза ферменттері белсенділігінің бүзылуы салдарынан
ұзын сүйектердің эпифизіндегі шеміршек ұлпасының өсуінің
бұзылуы.
Бивалент — екі конъюгацияланушы гомологтік хромосома¬
лар; эрбір биваленттер 4 хроматидадан тұрады.
Биологиялық түрлер — бір ареалды ұзак уақыт мекен-
деп, генетикалык, морфологиялық, физиологиялық жэне мінез-
күлықтары тұргысынан'ұқсас, еркін будандасып, өскелең ұрпак
беретін даралар жиынтыгы.
Будандастыру — генетиканың негізгі зерттеу әдісі.
Галактозем ия — аутосомды-рецессивті жолмен тұқым
қуалайтын гендік ауру.
Гаметогенез — жыныс жасушаларының түзілуі.
Гаметалар — жыныс жасушалары (жұмыртка жасушасы,
сперм атозоидтар).
Гаплочдчя — дене жасушаларында 1 п хромосома жиынтыгы
болатын геномдык мутадияның бір түрі.
Гемизиготалар — Х-не У-хромосомалардың гомологтік емес
учаскелерінде бір дана күйінде кездесетін аллель.
Гемофилия (А, В) — Х-хромосомамен тіркес рецессивті жол¬
мен тұқым қуалайтын зілді гендік ауру; қанның ұйымауы.
Ген — полипептид не нуклеин қышқылының синтезделуін
анықтаушы ДНҚ молекуласының бір учаскесі.
Гендердің амплификациясы — жұмыртқа жасушасының цито-
плазмасында кейбір гендердің көшірмеленіп көбеюі.
Гендердіц дрейфі — өте шагын популяцияларда сирек
аллельдердің жиілігінің кездейсоқ өзгеруі.
343
Генетикалык, инженерия — жасушадан (не ағзадан) белгілі
бір генді бөліп алып, рекомбинантты РНҚ жэне ДНҚ молекулала-
рын құрастыру жэне гендерді басқа жасуша не ағзаларға енгізуге
багытталған жана әдістер мен технологиялар жиынтыгы.
Гендік терапия — түқым қуалаушылық материалдарын (ДНҚ
не РНҚ) жасушаға не ағзага енгізу арқылы оның қасиеттерін
(қызметтерін) өзгерту.
Геном — гаплоидтық хромосома санындагы гендер жиын¬
тыгы.
Генотип — диплоидтық хромосома санындагы гендер жиын¬
тыгы.
Генофонд — популяция дараларыныц гендер не генотиптер
жиынтыгы.
Гетерозигота — бір геннің екі түрлі аллелі кездесетін жасуша
не агза.
Гетерохромотин —- интерфазада тыгыз ширатылган хромосо¬
ма учаскесі.
Гомологтіхромосомалар — жұп хромосомалар.
Генетикалык, жүк — популяция дараларының бейімделуші-
лігін төмендететін мутациялар жиынтыгы.
Генетикалык, полиморфизм — популяция даралары генотип-
терінің көптүрлілігі.
Дальтонизм — Х-хромосомамен тіркес рецессивті түкым
қуалайтын гендік ауру, түстерді ажырата алмау.
Даун синдромы — 21 хромосоманыц трисомиясына байланы¬
сты өте зілді ауру.
Делеция — хромосоманыц бір учаскесініц түсіп қалуына бай¬
ланысты мутация.
Демдер — 1500—4000 дарадан тұратын адамдардьщ шагын по-
пуляциясы.
Диакинез — I мейоздыц профазасыныц ец акыргы сатысы.
Диплоид — сомалық жасушалардагы хромосома саны (2п).
Диплотена — I мейоздыц профазасыныц 4 сатысы.
Диктиотена — оогенез процесінде мейоздыц I бөлінуініц
профазасыныц ерекше сатысы.
Дискорданттылық — егіздердіц бір белгі бойынша ерекшелік
дәрежесі.
344
ДНҚ-полимераза — ДНҚ синтезін қамтамасыз ететін, ягни
дезоксирибонуклеотидтерді фосфодиэфирлік байланыс арқылы
бір-біріне байланыстырушы фермент.
Доминанттылық — бір геннің басымшылык көрсетуі.
Дупликация — хромосомалардыц бір учаскесінің екі еселе-
нуіне байланысты мутация.
Жасуша циклі — жасушаның митоздық екі бөлінуі арасын-
дағы құбылыстар жиынтығы.
Зигота — бір жасушалы ұрық.
Зиготена — I мейоздыц профазасыныц 2 сатысы.
Идиограмма — хромосомаларды үлкенінен кішкентайына
қарай тізіп орналастыру.
Изоляттар — адам саны 1500-ден аспайтын адамдардыц өте
шагын топтары.
Ие — паразиттердіц бір сатысы мекендейтін тірі агзалар.
Импринпшнг (геномдық) — ата-ана жынысына байланысты
гомологтік гендердің (не хромосома учаскелерініц) активтігініц
айырмашылыгын қалыптастыратын механизм. Мысалы, бір
аурудыц ауру әкеден туылған балаларда ауру анадан туылганда-
рына қараганда жеңілдеу формада байқалуы.
Инбридинг — туыстык некелесу, ол рецессивтік белгілердіц
фенотиптік байқалу жиілігін көбейтеді.
Инвазия — адам агзасына сырттан енетін факторлар (паразит¬
тер).
Инвазиялық аурулар — паразит жәндікгердің тіршілігі нәти-
жесінде дамитын адам аурулары.
Инверсия — хромосоманыц бір учаскесінің 1800 айналып
қайта жалгануына байланысты мутация.
Индуктор — ақуыз-репрессормен байланысып транскрип¬
ция процесініц басталуына жол ашатын зат.
Инициация — трансляция (ақуыз синтезінің) басталу кезеці,
бұл кезде рибосома а-РНҚ-мен байланысып, рибосоманыц улкен
бөлшегіндегі А-учаскеге т-РНҚ-мен — метионинніц келіп орна-
ласуы.
Интрондар — эукариотгы гендердіц магынасыз учаскелері.
Интерфаза — жасушаныц 2 бөлінуі аралыгындагы кезец.
345
170-23
Кариотип — биологиялық түрлердің хромосома саны, пішіні,
мөлшері туралы кешенді сипаттамасы.
Клайпфельтер синдромы — ер адамдар кариотипінде қосым-
ша Х-хромосоманың болуына байланысты ауру.
Код (генетикалык) — ДНҚ (РНҚ) молекуласында
нуклеотидтердің бірізділікпен орналасуы арқылы тұқым қуалау-
шылық ақпараттың жазылу жүйесі.
Кодон (триплет) — геннің ең ұсақ, 3 нуклеотидтен тұратын
жэне бір амин қышқылын анықтайтын қызметтік бірлігі.
Кодоминанттылық — екі доминантты аллельдің бірлесіп
доминанттылық байқатуы.
Колинеарлық — ДНҚ молекуласында нуклеотидтердің орна-
ласуының полипептид молекуласыныц амин қышқылдарына
сэйкес болуы.
Комплиментарлық — 2 доминантты аллель емес геннің бір-
лесіп бір белгіні дамытуы.
Комменсализм — екі ағзаның (түрдің) біреуінің екіншісін пай-
даланып (баспана ретінде) тіршілік етуі, ал екіншісі үшін бірінші
агзаның (түрдің) не пайдасы не зияны жоқ.
Конкорданттыльщ — егіздердің бір белгі бойынша уқсас бо¬
луы.
Көпшілікті аллелизм — бір геннің екіден көп формада
кездесуі.
Кроссинговер — пахитенада хроматидалардың бір-бірімен
айқасып, учаскелерімен алмасуы.
Клондау — жасуша не генді көшірмелеп көбейту.
Қуыршақ — буынаяқтылардың түрленіп, дамуындағы бір
саты.
Лептотена — мейоздың бірінші бөлінуінің профазасының
1-сатысы.
Лигаза — нуклеин кышкылдарының молекула белшектерін
бір-бірімен «тігуші» фермент.
Локус — геннің хромосомада орналасқан орны.
Леталъ — жасушаның не ағзаның еліп қалуына алып келетін
мутация.
Мейоз — жыныс жасушаларыныц бөліну жолы, оныц салда¬
рынан бір аналық жасушадан 4 гаплоидтық жыныс жасушалар
түзіледі.
346
Миграция (көшіп-қону) — популяцияга сырттан жаңа гено
типтердің келіп қосылуы не шыгып кетуі.
Митоз — дене жасушаларының бөліну жолы.
Моиосомия — бір хромосоманың сьщар куйінде кездесуі;
анеуплоидияның бір түрі.
«Мысық айқайы» синдромы — 5 хромосоманың кысқа иінінің
делециясына байланысты ауру.
Мутагенез — мутациялардың пайда болу процесі.
Мутагендер — мутациялардың пайда болуына алып келетін
факторлар.
Мутация — генетикалык материалдың өзгеруі.
Мутон — геннің мутациялануга қабілетті ең кіші бөлігі.
Мутализм — екі ағзаның (түрдің) бір-біріне пайда келтіріп
тіршілік етуі, бір-бірінен бөлек олар тіршілік ете алмайды, мы¬
салы, жасыл балдырлар мен саңырауқүлақтар — қына денесінде,
адам ішегіндегі кейбір бактериялар.
Нуклеосома — гистонды ақуыздар кұрылған жэне ДНҚ моле¬
куласы ширатылатын денешік.
Нуклеоид — прокариоттардың генетикалык аппараты.
Нуклеотид — нуклеин қышқылдарының мономері.
Овогенез — жұмыртқа жасушасының пайда болу жэне жетілу
процессі.
Онкогендер — дене жасушасының рак жасушаларына айна-
луына алып келетін акуыздарды аныктайтын гендер.
Онтогенез — агзалардьщ жеке дамуы.
Ооплазмальщ сегрегация — жүмыртқа жасушасында хи-
миялык заттардың түрліше таралуына байланысты цитоплаз-
манын эр түрлі сапалы учаскелерге жіктелуі.
Оперон — генетикалык ақпараттың транскрипциялану бірлігі.
Өзгергіштік — агзалардьщ белгілері мен қасиеттерінің
өзгеруі.
Панмиксия — популяция дараларының бір-бірімен еркін бу-
дандасуы.
ГІатау синдромы — 13 хромосоманың трисомиясына байланы¬
сты ауру.
Похищена — мейоздың бірінші бөлінуінің профазасыныц 3-ші
сатысы; бұл кезде кроссинговер болады.
Пенетранттылъщ (гендердіц) — геннің белгі күйінде бай-
қалу мөлшері; ол % арқылы есептелінеді.
Плазмидалар — бактерия жасушаларында, эукариоттар жасу-
шасының кейбір органеллаларында (пластидтер, митохондриялар)
өз алдына дербес (автономды) кездесетін сақиналы ДНҚ молеку¬
ласы.
Плейотропия — бір геннің бірнеше белгіні дамыту қасиеті.
Полимерия — бірнеше аллельсіз гендердің бірлесіп бір белгіні
дамытуы (полигендік түқым қуалау).
Полиплоидия — хромосома санының еселеп өсуіне байланы¬
сты геномдық мутация (Зп, 4п, 5п, т.б.).
Политения — митоздың бір түрі; интерфазада хромати-
да көбейіп, бір-бірінен ажыраспай политендік (көп жіпшелі)
хромосомалардыц түзілуі.
Популяция — бір ареалды ұзақ уақыт мекендеп, ортақ
генофондқа ие, бір-бірімен еркін будандасатын, бір түрдің дара¬
лар жиынтыгы.
Популяциялық толқыидар — табиги популяциялардыц да¬
ралар саныныц мезгілді-мезгілсіз өзгеруі; қарапайым эволюция-
лык фактор.
Пробанд — шежіре құрастыруга себепші адам.
Прокариоттар — ядросы толық қалыптаспаған біржасу-
шалы агзалар.
Промотор — транскрипция басталатын ДНҚ молекуласыныц
бір учаскесі.
Процессинг — про-а-РНҚ интрондық учаскелерінің қиылып
түсіп калуы.
Реакция нормасы — модификациялық өзгергіштіктіц шегі.
Реанимация — ағзаны клиникалық өлім жагдайынан қайта
тірілту.
Резервуарлық иелер — паразиттердіц бір агзадан екіншісіне
өтіп, бірақ оларды зақымдамай, табигатта ұзақ сақталуын
қамтамасыз ететін тірі агзалар.
Резистенттілік — агзалардьщ белгілі бір факторга төзім-
ділігі.
Рекон — генніц рекомбинациялануга кабілетті ец кіші бөлігі,
ол 1 нж-га тец.
348
Репарация — ДНҚ молекуласының бұзылған құрылымда-
рының қалпына келуі.
Репликациялық аша — репликация басталатын учаске.
Репликация (ДНК) — ДНҚ молекуласыныц өздігінен екі
еселенуі (ДНҚ синтезі).
Репрессор — ген-операторды «тығындап», транскрипцияны
болдырмайтын ақуыз.
Рестриктазалар — нуклеин қышқылдарының белгілі бір ну¬
клеотид тізбектерін «танып», кесуші ферменттер («қайшылар»).
Рецессивтік — тек гомозиготалы күйінде байқалатын белгі не
аллель.
Сибстер — бір ата-анадан туылған балалар.
Симбиоз (селбесіп тіршілік ету) — екі агзаның (түрдің) бірге
(бірлесіп) тіршілік етуі.
Синдром — ағзаның көптеген мүшелері мен мүшелер жүйесін
қамтитын патологиялык белгілер кешені. Олар бір патогенез
негізінде калыптасады.
Спейсерлер — қүрылымдық гендерді бір-бірінен ажыратып
түратын ДНҚ молекуласыныц кысқа учаскелері.
Сперматогенез — спрематозоидтардыц пайда болуы жэне
пісіп-жетілу процесі.
Сплайсинг — про-а-РНҚ-ныц экзондарыныц бір-бірімен «жал-
гану» процесі.
Супрессорлар (ингибиторлар) — бір аллель емес генніц эре-
кетін бастырмалайтын ген.
Сүрыптау (табиги) — популяцияларда бейімделген дара¬
лардыц сақталуын камтамасыз ететін тіршілік үшін күрес
нәтижесі.
Табиги оіиақтық аурулар — табиги ошагы, таратушылары
жэне резервуарлық иелері болатын паразиттік аурулар, мысалы,
оба ауруы, лейшманиоздар т.б.
Табиги резервуарлар — паразиттердің жабайы резервуарлық
иелері.
Талассимия — қан аздықтың бір түрі; гендік ауру.
Теломер — хромосомалардыц үшы.
Теломераза — жасушаныц бөліну кезінде бұзылган хромо¬
сома үшыныц қүрылысын қалпына келтіруші фермент.
349
Таратушылар — паразиттердің бір сатысын (жұмыртқаларын
не дернэсілдерін) адамдарға жұқтыратын жәндіктер (кенелер, на¬
секомдар).
Тератология — туа біткен ақаулықтарды зерттейтін ғылым.
Терминация — транскрипция жэне трансляция құбылысыныц
аяқталуы.
Тіркесу тобы — бір жұп гомологтік хромосома гендері.
Тотипотенттілік — эмбриогенездің бастапқы сатыларын¬
да бластомералардың қызметтік тең мүмкінділігі; бұл кезде эрбір
бластомера жеке ағзаны дамыта алады.
Транзиция — гендік мугацияның бір түрі; бүл кезде бір
пуриндік негіз екінші пуринмен (А не Г) немесе бір пиримидиннің
екіншісімен алмасуы.
Трансверсия — гендік мутацияның бір түрі; бүл кезде пуриндік
негіз (А, Г) пиримидиндік негізбен (Ц, Т), не керісінше алмасады.
Трансдукиия — бактериофагтер арқылы бактериялардың бір
штаммының ДНҚ молекуласыныц бір учаскесінен екіншісіне
көшірілуі.
Транскрипция — ДНҚ молекуласындағы акпараттыц РНҚ мо-
лекуласына көшірілуі (РНҚ синтезі).
Транслокация — гомологтік емес хромосомалардыц учаске-
лер алмастыруы.
Трансмиссивтік аурулар — таратушылар арқылы берілетін
инвазиялык аурулар.
Тернер (Шерешевский-Терпер) синдромы — эйелдердіц бір
Х-хромосомасыныц жетіспеушілігіне байланысты ауру.
Трансляция (ақ\’ыз синтезі) — а-РНҚ негізінде полисомада
полипептид молекуласыныц синтезделуі.
Транспозондар ( «секіруші «гендер) — ДНҚ молекуласында
тұрақты орындары болмайтын, «секіріп» кешіп жүретін гендер —
геном «паразиттері».
Трансформация — бактерия штаммдарыныц өзара ДНҚ
учаскелерімен алмасып, қасиегтерін өзгертуі.
Трисомия — анеуплоидияныц бір түрі; кариотиптіц бір
хромосомага артық болуы, 2п+1.
Тұқым куалаушылық — агзалардьщ белгілері мен қасиетте-
рініц үрпақтарга берілуі.
350
Тұқым куалайтын аурулар (белгілер) — әр түрлі мутациялар
(гендік, хромосомалық, геномдық) салдарынан қалыптасатын
адамдардыц патологиялары.
Тұқым куалауга бейім аурулар (белгілер) — тұқым
қуалаушылығы нақтылы анықталмаса да, белгілі бір орта
факторларының әсерінен дамитын аурулар (белгілер).
Түпкілікті иесі — паразиттің ересек формасы кездесетін, яг¬
ни жыныстық жолмен көбейетін ағзасы.
Фенилкетонурия — фенилаланин амин қышқылын тиро-
зинге айналдыруға қатынасатын фенилаланин-гидрооксилаза
ферментінің синтезделінуі бұзылуыныц салдарынан байқалатын
гендік ауру.
Фенокөшірмелер — генотиптері эр түрлі ағзаларда ұқсас
фенотиптің байқалуы.
Фенотип — ағзаның белгілері мен қасиеттерінің жиынтыгы.
Хиазма — мейоздық бөлінудің профаза- 1-дегі хромотида-
лардың конъюгацияланып байланысуы.
Хроматин —• ДНҚ молекуласынан жэне гистонды ақуыз-
дардан құрылган интерфазалық хромосоманыц деспиральданган
күйі.
Хр омо сом ал i, i қ аурулар — хромосомалар саныныц жэне
құрылысыньщ өзгеруі салдарынан байқалатын аурулар.
Центромера — хромосома иіндерін жалгап тұратын орталық
керме.
Цистрон •— құрылымдық ген.
Экзондар — құрылымдық гендердіц магыналы учаскелері.
Экогенетика (экологиялық генетика) — қоршаган орта
факторларыньщ агзаныц түкым куалаушылыгына эсер етуі
негізінде қалыгітасатын түрліше реакцияларыныц ерекшеліктерін
зерттейтін генетика гылымыныц бір саласы.
Экспрессивтік — геннің белгі күйінде байқалу дэрежесі.
Элонгация — полипегітидтік тізбектіц үзаруы. акуыз синтезініц
эрі қарай жалгасуы.
Эмбриондық индукция — ұрықтыц дифференцияланган
бөлігінің көршілес жасушалар тобына әсері.
Эндомитоз — митоздыц бір түрі; хромосома екі еселеніп, ядро
бөлінбейді де полиплоидтық жасуша түзіледі.
351
Эпистаз — аллель емес гендердің әрекеггесуінің бір түрі; бір
аллель емес ген екінші аллель емес геннің әрекетін бастырмалай-
ды.
Эукариоттар — ядросы қалыптасқан жасушалы агзалар.
Ядро — эукариоттық жасушалардың негізгі компоненті.
Ядрошық — ядроның құрамдық бөлігі, онда рибосома
суббірліктері синтезделінеді.
ӘДЕБИЕТТЕР
1. Айала Ф., Кайгер Д. Ж. Современная генетика. В 3-томах.—
Москва, 1988.
2. Әбілаев С. Л., Қуандықов Е. Ө., Төлебаев Р. К. Медициналық
биология жэне генетика.—Шымкент, 1996.
3. Биология. / Под редакцией В. Н. Ярыгина.— Москва, 1984.
4. Биология. В 2-х книг., // Под редакцией В. Н. Ярыгина.— Мо¬
сква, 2001.
5. Биотехнология. Отв. ред. Баев А. А.— Москва, 1984.
6. Бочков Н. П. Клиническая генетика.— Москва, 1997.
7. Гофман-Кадашников П. Б. «Задачник по общей и медицин¬
ской генетике».— М., 1969.
8. Заяі/ Р. Г., Бутвииловский В. Э., Рачковская К. В., Давы¬
дов В. В. Общая и медицинская генетика. Лекции и задачи.— Ро¬
стов на Дону, 2002.
9. Қазымбет П. К, Аманжолова П. Л., Нүртаева Қ. Меди-
циналық биология.— Алматы, 2000.
10. Қуандъщов Е. Ө., Төлебаев Р. К., Әбілаев С. Л., Медицина-
лык биология жэне генетика терминдерініц орысша-қазакаіа
түсіндірме сөздігі.— Алматы, 1996.
11. Мушкамбаров Н. Н, Кузнецов С. Л. Молекулярная биоло¬
гия.— Москва, 2003.
12. Слюсарев А. А., Жукова С. В. Биология.— Киев, 1987.
13. Тератология человека. // Под редакцией Лазюка Г. И.— Мо¬
сква, 1991.
14. Фогель Ф., Мотулъский А. Генетика человека. В 3-томах.—
Москва, 1989.
МАЗМ¥НЫ
Кіріспе 3
1. Жасуша биологиясы 12
1.1. Жасушаның кұрылысы 12
1.2. Жасуша тіршілігіндегі мерзімдік өзгерістер
(жасушаның бөлінуі) 19
2. Көбею биологиясы 28
3. Генетика негіздері 35
3.1. Тұкым куалаушылыктың негізгі заңдылыктары 35
3.2. Ағзалардың генетикалык жүйесі. Гендердің өзара әрекеттесуі...44
3.3. Адам генетикасы 51
3.4. Гендердің кұрылысы мен кызмет етуі туралы
молекулалык-биологиялык түсінік 60
3.5. ДНҚ молекуласының репликациялану механизмдері 66
3.6. Гендердің экспрессиялануының (активтенуінің)
реттелу механизмдері 69
3.7. Транскрипция немесе РНҚ синтезінің механизмдері 75
3.8. Акуыз биосинтезі 76
3.9. Цитоплазмалык түкым куалаушылык. Генетикалык инженерия.
Жасушаның мутацияға карсы антимутациялык механизмдері 81
3.10. Өзгергіштік 85
3.11. Адамдардың хромосомалык аурулары 95
3.12. Мутагенез жэне канцерогенез 101
3.13. Медициналык-генетикалык кенес беру негіздері 107
4. Даму биологиясы 116
4.1. Онтогенез. Агзалардьщ жеке дамуы.. 116
4.2. Дүниеге келгенге дейінгі (эмбриондык) кезең 122
4.3. Эмбриогенездің жасушалык жэне генетикалык механизмдері.,127
4.4 Ұрыктың провизорлык мүшелері 133
4.5 Дамудың катерлі кезеңдері 136
4.6. Дамудың туа біткен акаулыктары 137
4.7. Дамудың постэмбриондык кезеңі 143
4.8. Агзаның картаю күбылыстары жэне геронтология
мәселелері 149
4.9. Агзаның калпына келу кұбылыстары. Мүшелер мен
үлпалардың регенерациясы және трансплантациясы 160
5. Гомеостаз туралы ілім негіздері 165
6. Эволюциялык ілім негіздері 171
354
6.1. Эволюциялык теорияның даму тарихы 171
6.2. Биологиялык түрлер жэне оның популяциялык кұрылысы 174
6.3. Түртүзілу күбылысы. Табиғи популяциялардағы
карапайым эволюциялык факторлардың әрекеті 179
6.4. Табиғи популяциялардың генетикалык полиморфизмі 191
6.5. Генетикалык жүк және оның медициналык-әлеуметтік
маңызы 195
6.6. Микро- жэне макроэволюция. Ағза топтарының
эволюциясы 197
7. Омыргкалылардың мүшелері мен мүшелер жүйесінің
филогенезі 203
7.1 .Филогенездің жалпы заңдылыктары 203
7.2. Омырткалылардың терісінің филогенезі 206
7.3. Омырткалылар каңкасының филогенезі 208
7.4. Омыртқалылардың ас корыту жүйесінін филогенезі 213
7.5. Омырткалылардың тыныс алу жүйесінің филогенезі 216
7.6. Омырткалылардың канайналым жүйесінід филогенезі 219
7.7. Омырткалылардың жүйке жүйесінің филогенезі 223
7.8. Омырткалылардың зэр шыгару, жыныс жүйелерінің
филогенезі 226
8. Антропогенез негіздері .....232
9. Медициналык паразитология негіздері 240
9.1. Жалпы мэселелер 240
9.2. Қарапайымдар типі 246
9.3. Жалпак күрттар типі 261
9.4. Жұмыр күрттар типі 275
9.5. Буынаяктылар тигіі 280
10. Экология негіздері 293
10.1. Жалпы экология мәселелері 293
10.2. Адам экологиясы 298
11. Адамның экологиялык генетикасы. Фармакогенетика 304
11.1. Жалпы мэселелер 304
11.2. Сырткы орта эрекеттеріне агзаның тұкым куалайтын
патологиялык реакиялары 307
11.3. Фармакогенетика 311
Студенттердін өз бетінше окып-үйренуіне, бакылауына
арналған сұрактар мен есептер 315
Терминдер сөздігі 342
Әдебиеттер 353
355
Есенгелді Өсербайұлы Қуандықов
Сэтбай Ахатжыұлы Әбілаев
Медициналық биология жэне генетика
Оқулық
Корректор Б. Т. Мекеев
Беттеуші А. А. Сляднева
Дизайн А. В. Милованов
Басуға 06.12.2010 кол койылды. Офсеттік басылыс.
Пішімі 60х80'/16. Қағазы офсеттік. Қаріп түрі «Таймс».
Шартты баспа табағы 22,5.
Таралымы 500. Тапсырыс № 80.
ISBN 9965-830-11-8
789965 830112
«NUR.PR.ESS» баспасы
050042 Алматы к.,
Таугуль ы/а, 51 үй, 8 оф.
Тел/факс: (727) 226-03-29.
E-mail: nurpress@mail.ru
«Курсив» ЖШС баспаханасында басылған
Издательство «NURPRESS»
НОВИНКИ!!!
С. Т. Культелеев
Практикум по экологическому праву РК:
Учебное пособие.— Алматы, 2011.— 17ё с.
В учебном пособии освещается содержание курса
лекций и семинарских занятий. По основным темам
курса приведены задачи, которые помогают студен¬
там глубже усвоить учебную дисциплину. В пособии
приводится методический материал необходимый при
изучении курса, в том числе тестовые занятия, схемы
по отдельным темам, а также рекомендуемые для изу¬
чения нормативно-правовые акты и литература.
К. А. Алишева
Экология: Учебник-
- Алматы, 2011.— 344 с.
ЭКОЛОГИЯ
В учебнике изложены, помимо основ экологии, по¬
следние данные по экологической действительности
на сегодняшний день, учитывающей реалии настоя¬
щего времени. Экологическое состояние окружающей
природной среды является одним из самых актуаль¬
ных и приоритетных направлений устойчивого разви¬
тия Республики Казахстан в XXI веке.
Рекомендуется в качестве учебника для студентов
вузов, где введен курс «Экология», являющийся обя¬
зательным на всех факультетах разных направлений
специализации.
С. К. Жүрсімбаев
Қазақстан Республикасының қүқық корғау
органдары: Окулық.— Алматы, 2010.— 360 бет.
«Қазакстан Республикасының кұкык коргау орган¬
дары» курсы — заң оку орындарында студенттерге
кұқық қоргау және құқык қолдану органдарының
қызметі туралы негізгі мәліметтерді беретін пән.
Курсты оқудың мақсаты — болашак зацгерлерді
кұкык қоргау қызметін жүзеге асыратын соттың,
прокуратураның жэне баска да органдардың құкықтык
негіздері, міндеттері, күрылымы мен кұзырегіері
жөнінде қажетті ақпаратпен қамтамасыз ету.
Толықтырылып ұсынылып отырған осы екінші
басылым — зац оку орындарынын студенттеріне,
окытушыларға жэне күкық корғау органдарыньщ
кызметкерлеріне арналған.
• ifA* *Ці-
... //Лцософц»
H. Ч. Джолдасбаева
Квалификация убийств: теоретические и прак¬
тические аспекты.— Алматы, 2010.— 292 стр.
В данной работе на основе анализа действующего
законодательства Республики Казахстан, законода¬
тельства ряда стран СНГ, основанном на едином Мо¬
дельном Уголовном кодексе и объединенных общей
историей, теоретической литературы, посвященной
проблемам квалификации убийств, выборочного из¬
учения судебной практики и материалов обобщений
Верховного Суда Республики Казахстан, включая соб¬
ственный многолетний правоприменительный опыт,
сделана попытка дать правильное, по мнению автора,
решение ряда спорных в практике вопросов по квали¬
фикации отдельных видов убийств.
Т. X. Габитов
Философия права. Учебное пособие для юридиче¬
ских вузов.— Алматы, 2010.— 228 стр.
В данном учебном пособии в соответствии с
требованиями государственного образовательного
стандарта даны основы философии права. В нем
использованы отдельные материалы из следующих
опубликованных книг: Нуржанов Б. Г., Габитов Т. X.,
Ибраева А. С., Айдарбаев С. Ж. и др. На пути к культуре
мира. Алматы: КазГУ, 2000; Профессиональная этика
сотрудников правоохранительных органов — Москва,
Щит и меч, 1998; Букреев В. И., Римская И. Н. Этика
права — М., 1998; Габитов Т. X. и др. Основы этики
— А., 1997.
С. К. Журсимбаев
Правоохранительные органы Республики Казах¬
стан. Учебник.— Алматы, 2010.— 400 стр.
Вводная дисциплина в юридическом учебном заве¬
дении, в рамках которого студенты получают основ¬
ные сведения о деятельности правоохранительного и
правоприменительных органов. Цель изучения курса
— это обеспечение необходимой информацией буду¬
щих юристов о правовой основе, задачах, структуре и
компетенции суда, прокуратуры и других органов, осу¬
ществляющих правоохранительную деятельность.
Издательство
«NURPRESS»
050042 г. Алматы, мкр. Таугуль, д. 51, оф. 8, т./ф.: (727) 226-03-29
E-mail: nurpress@mail.ru
Издательство «NURPRESS» впервые в Казахстане
выпустило в свет аудиокнигу для детей
«Қазақ ертегілері»
со звуковым и музыкальным сопровождением
В нее включены такие сказки, как:
1. Батыр қойшы (Отважный пастух) — 10:34;
2. Қасиетті құдық (Волшебный колодец) — 7:44;
3. Уәзір мен шал (Визирь и старик) — 3:55;
4. Бақыт іздеушілер (Искатели счастья) — 7:22;
5. Түлкі мен қырғауыл (Лиса и фазан) — 3:58;
Общее время — 34:27
Текст читает актер Казахского
государственного академического театра драмы
им. М. Ауэзова — Бахтияр Кожа