



































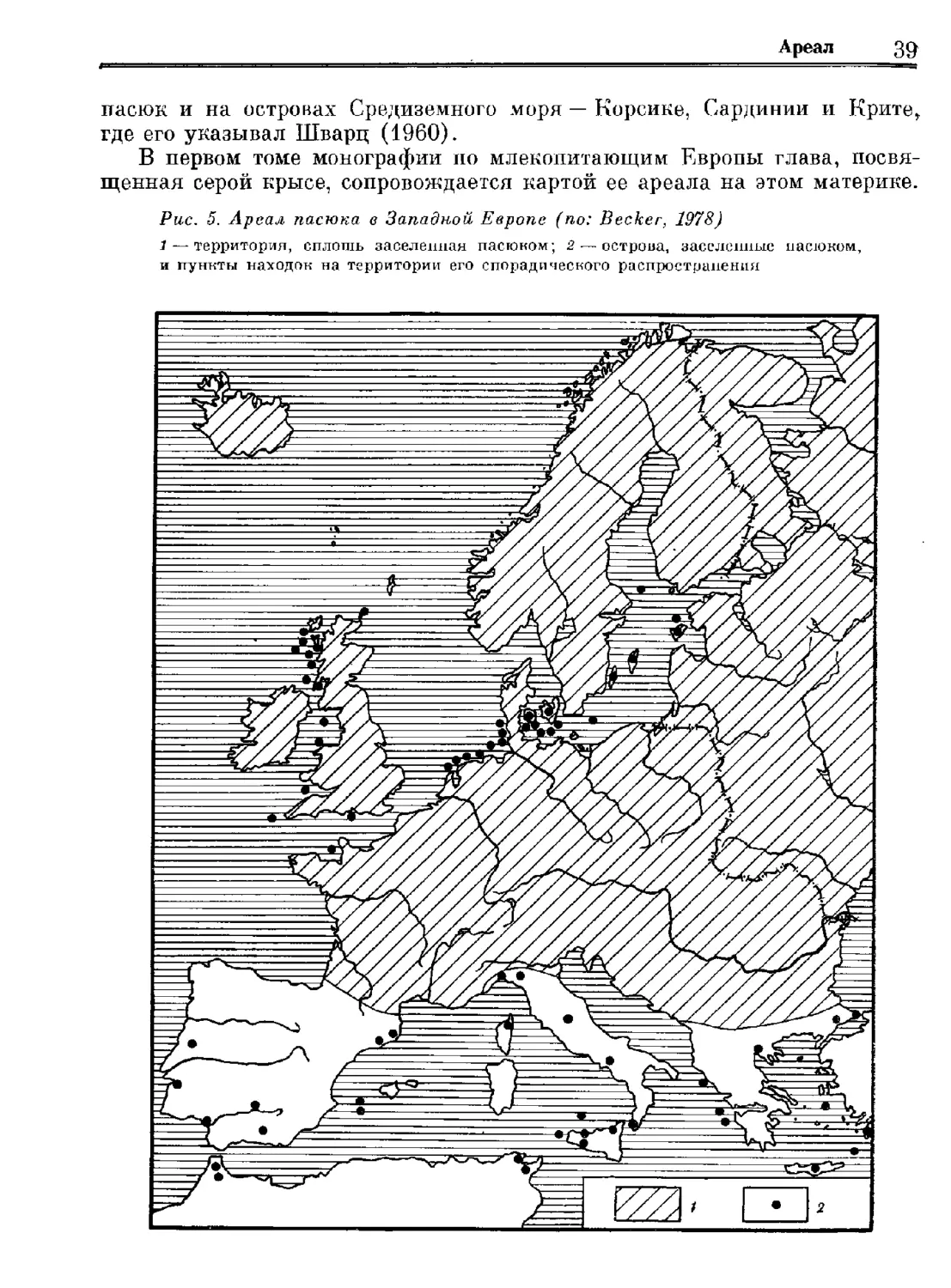







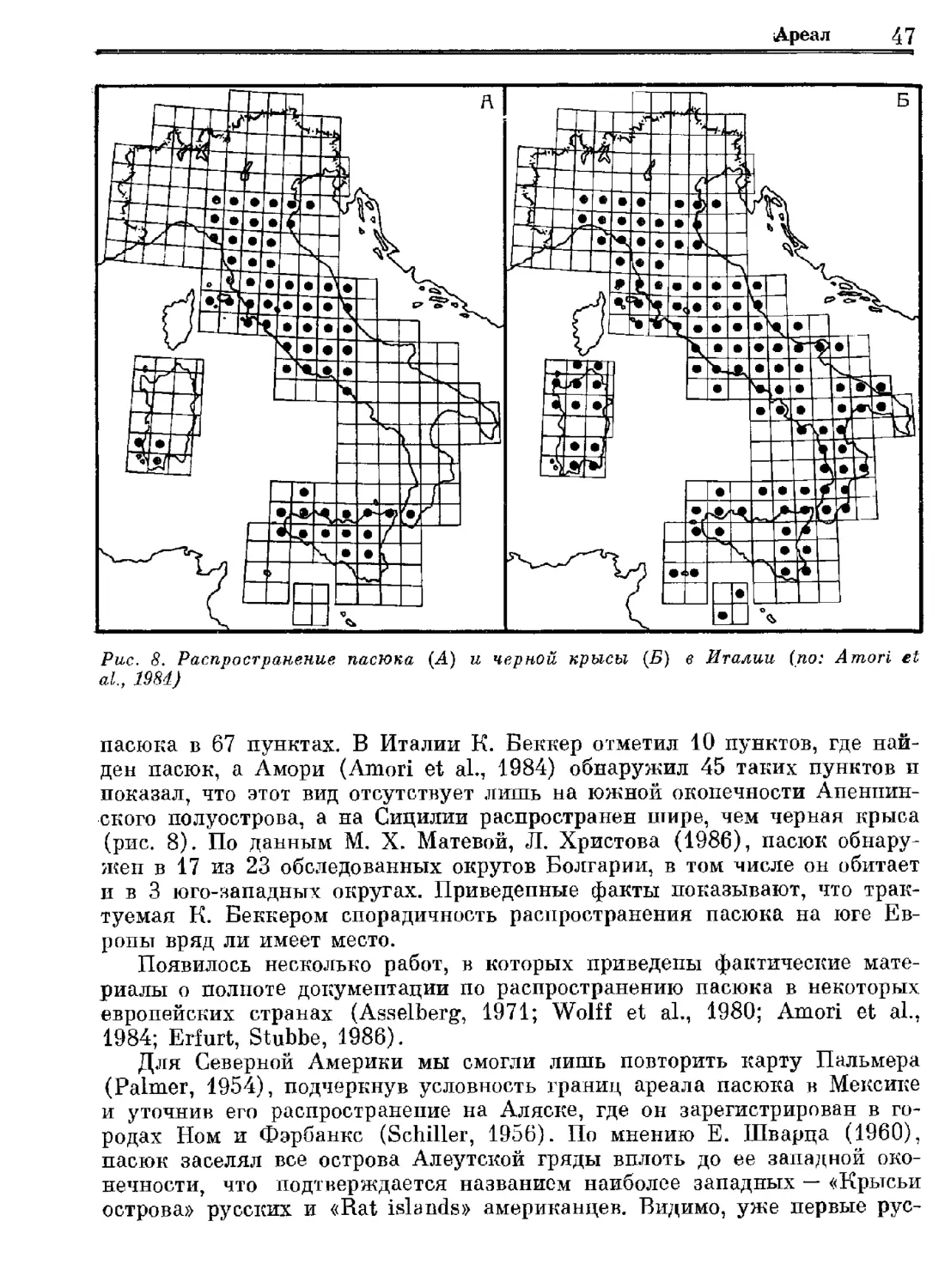

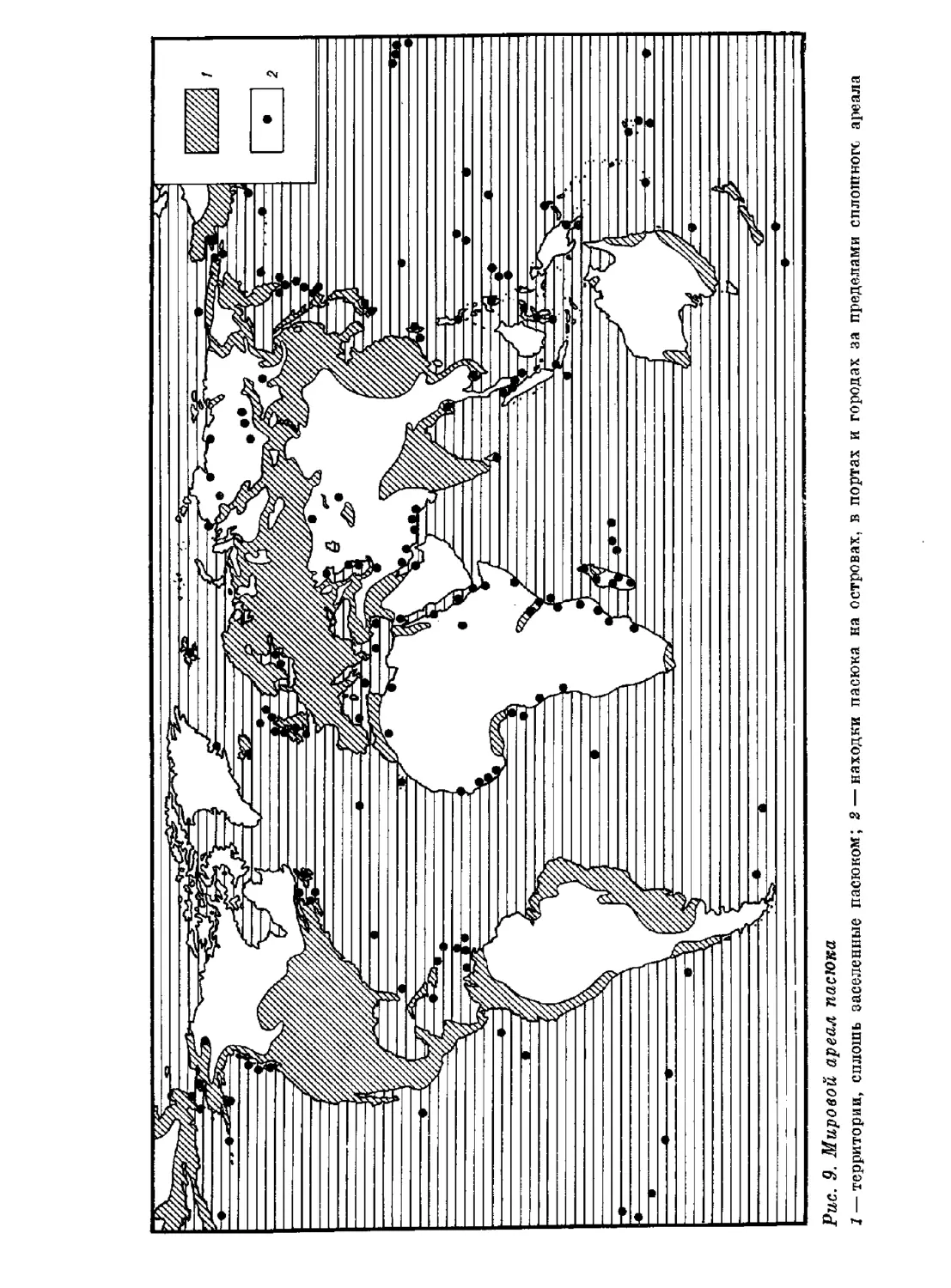


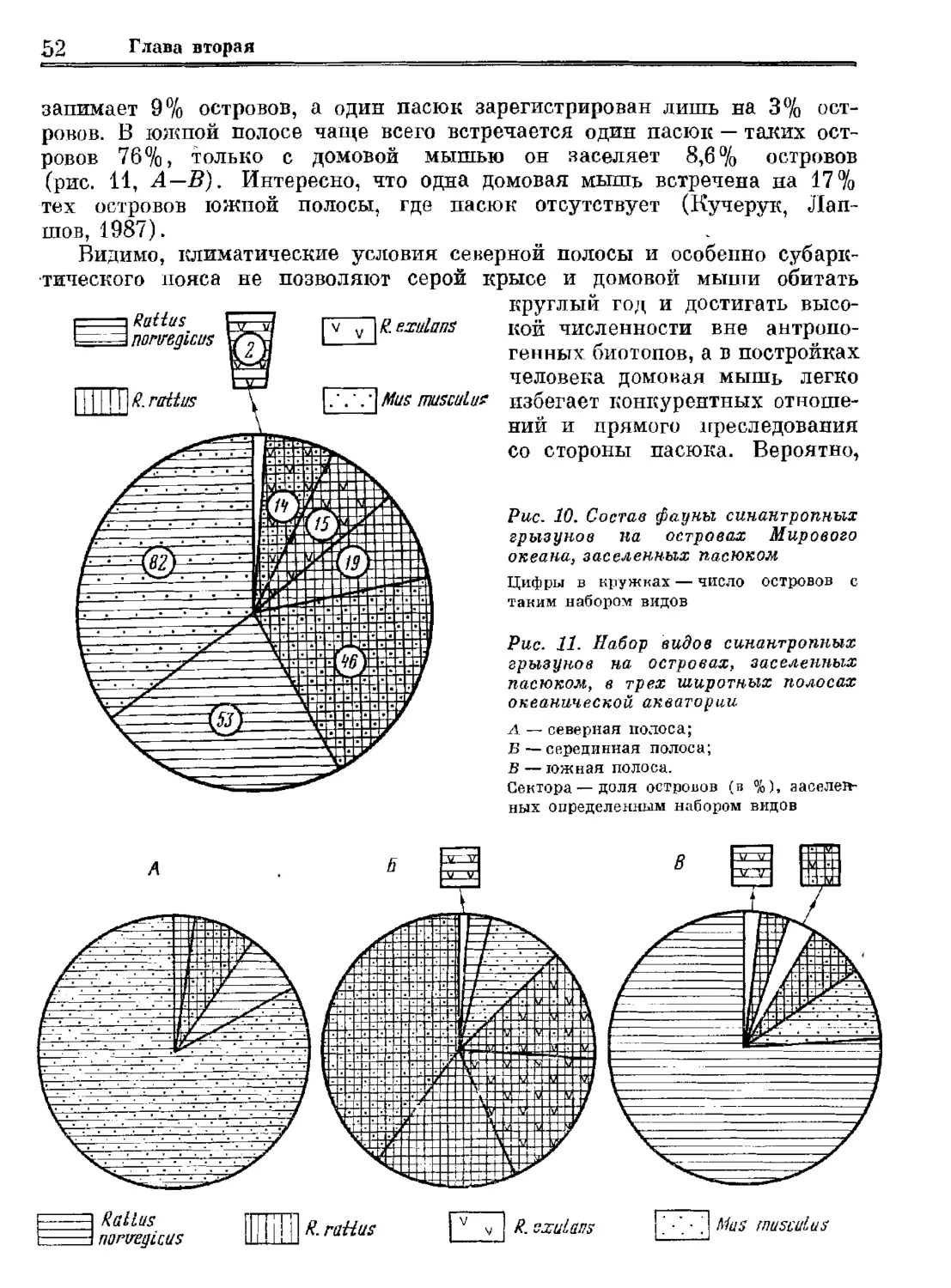















































































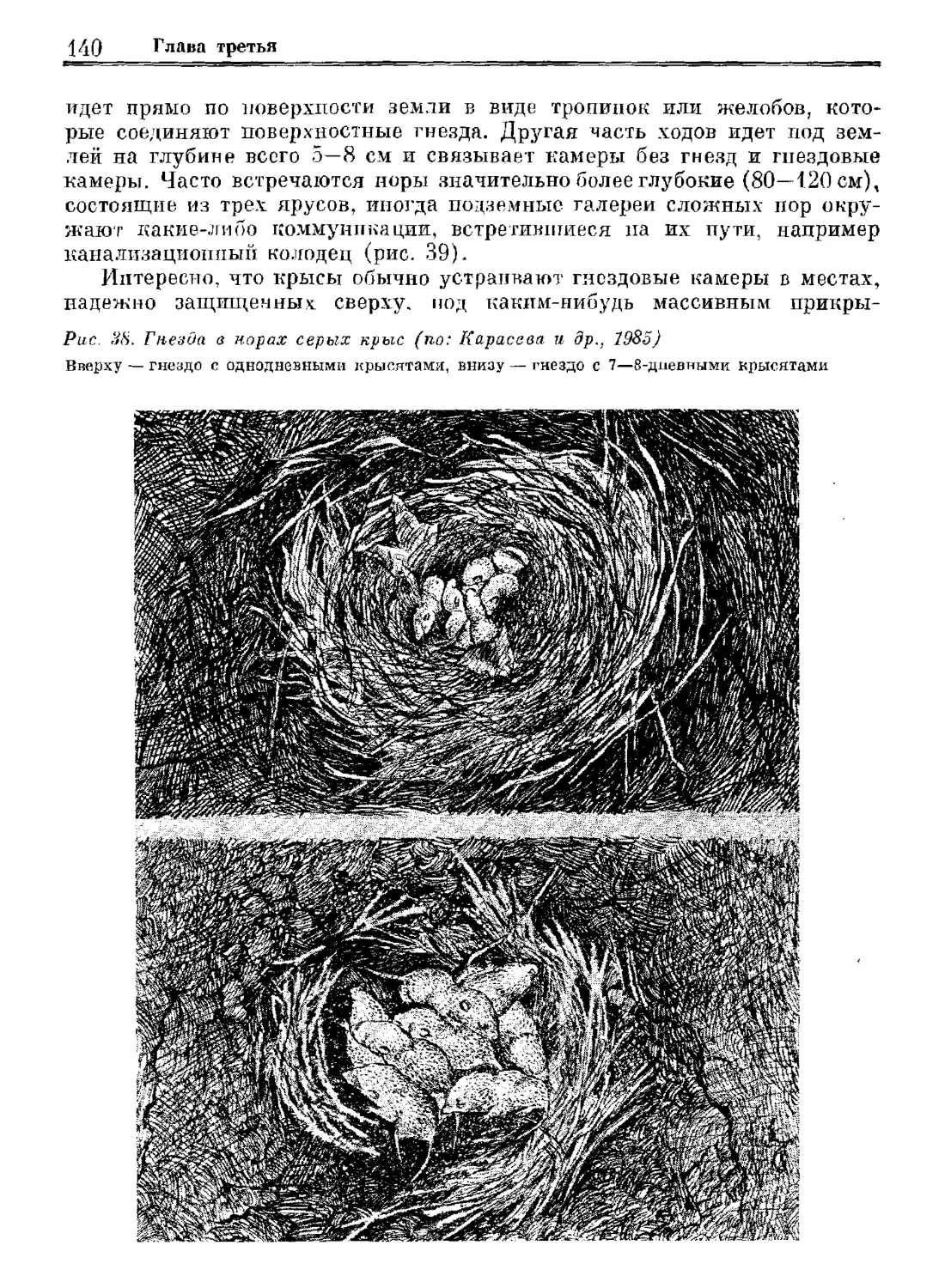
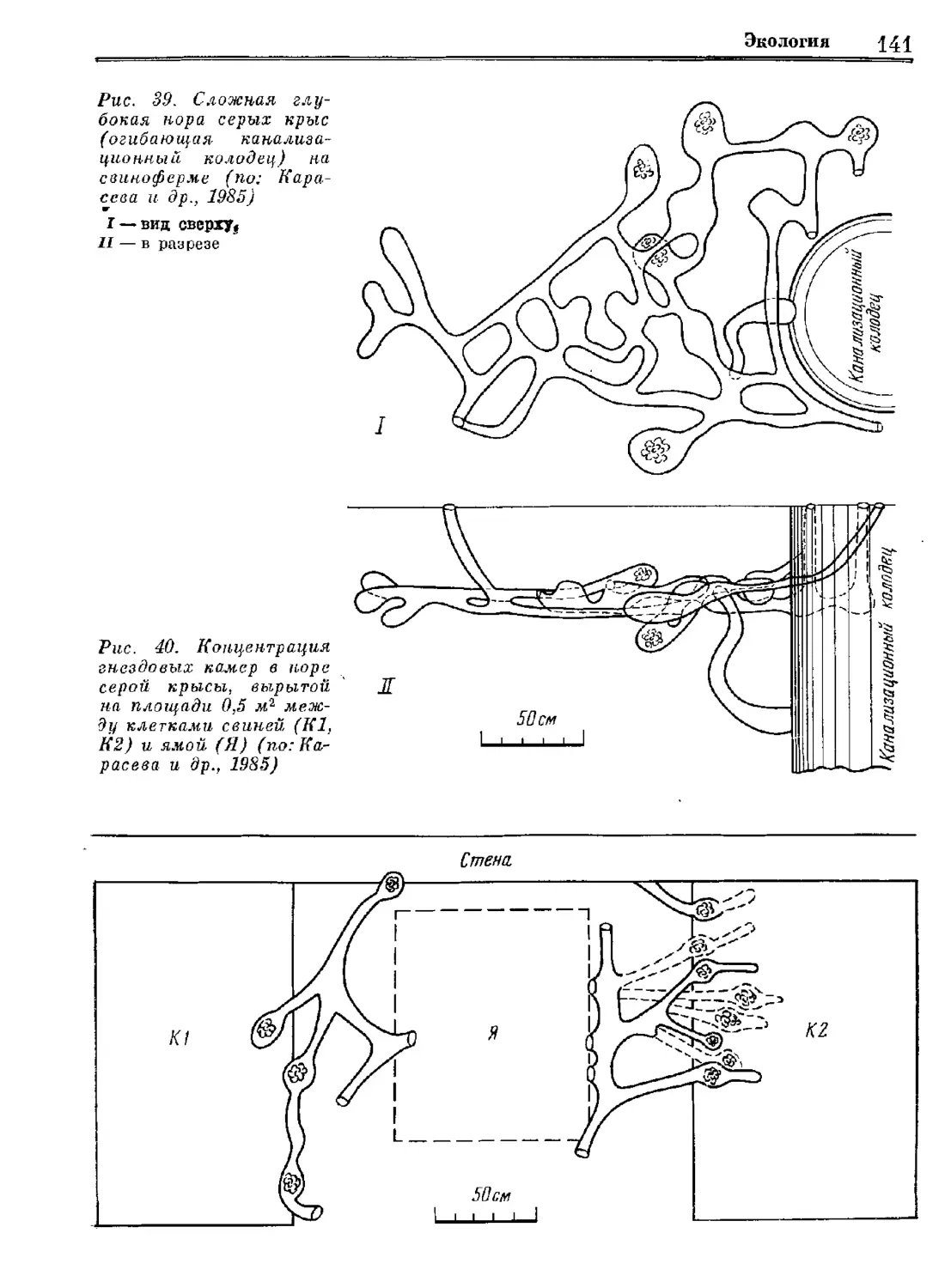











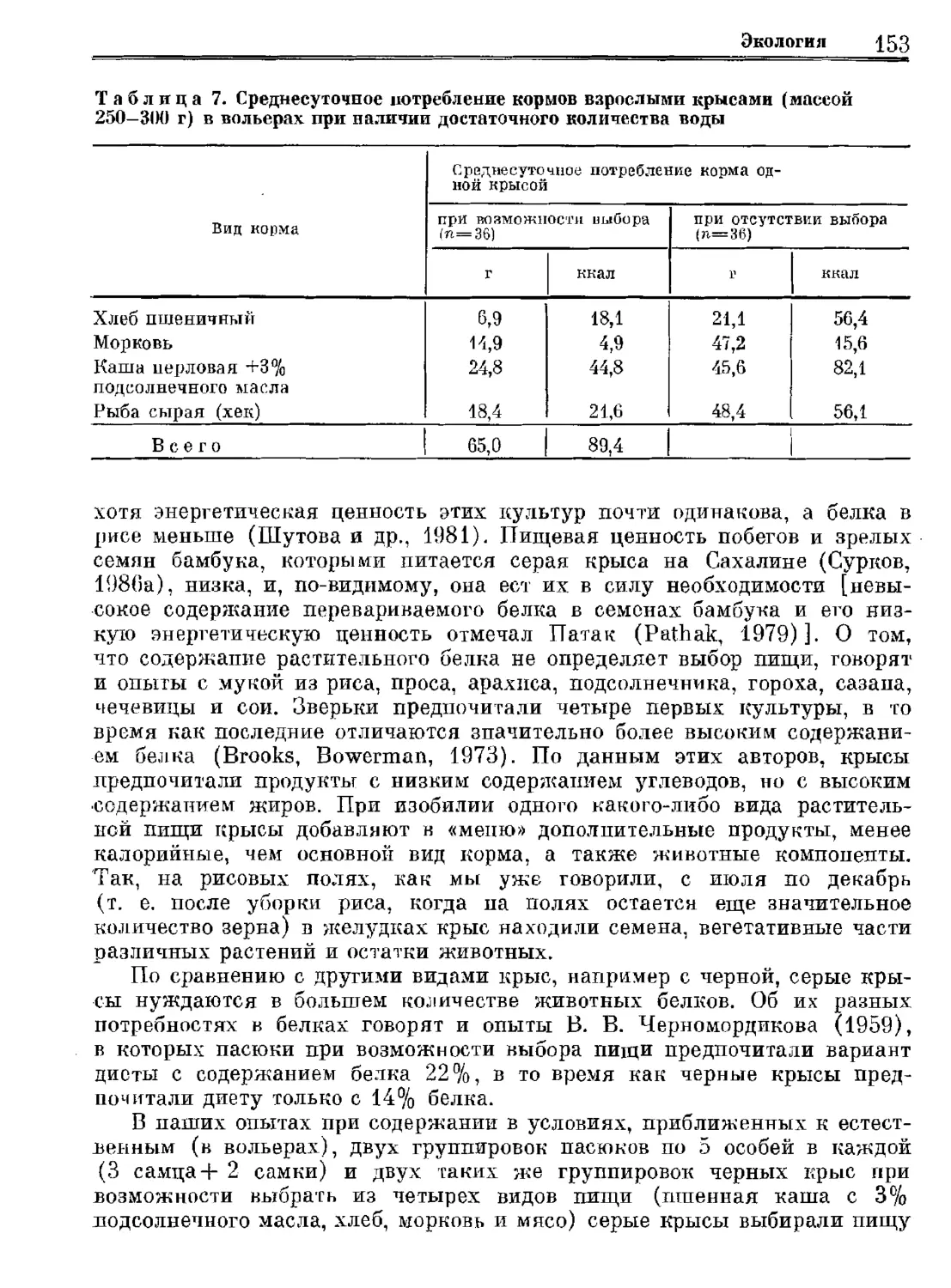











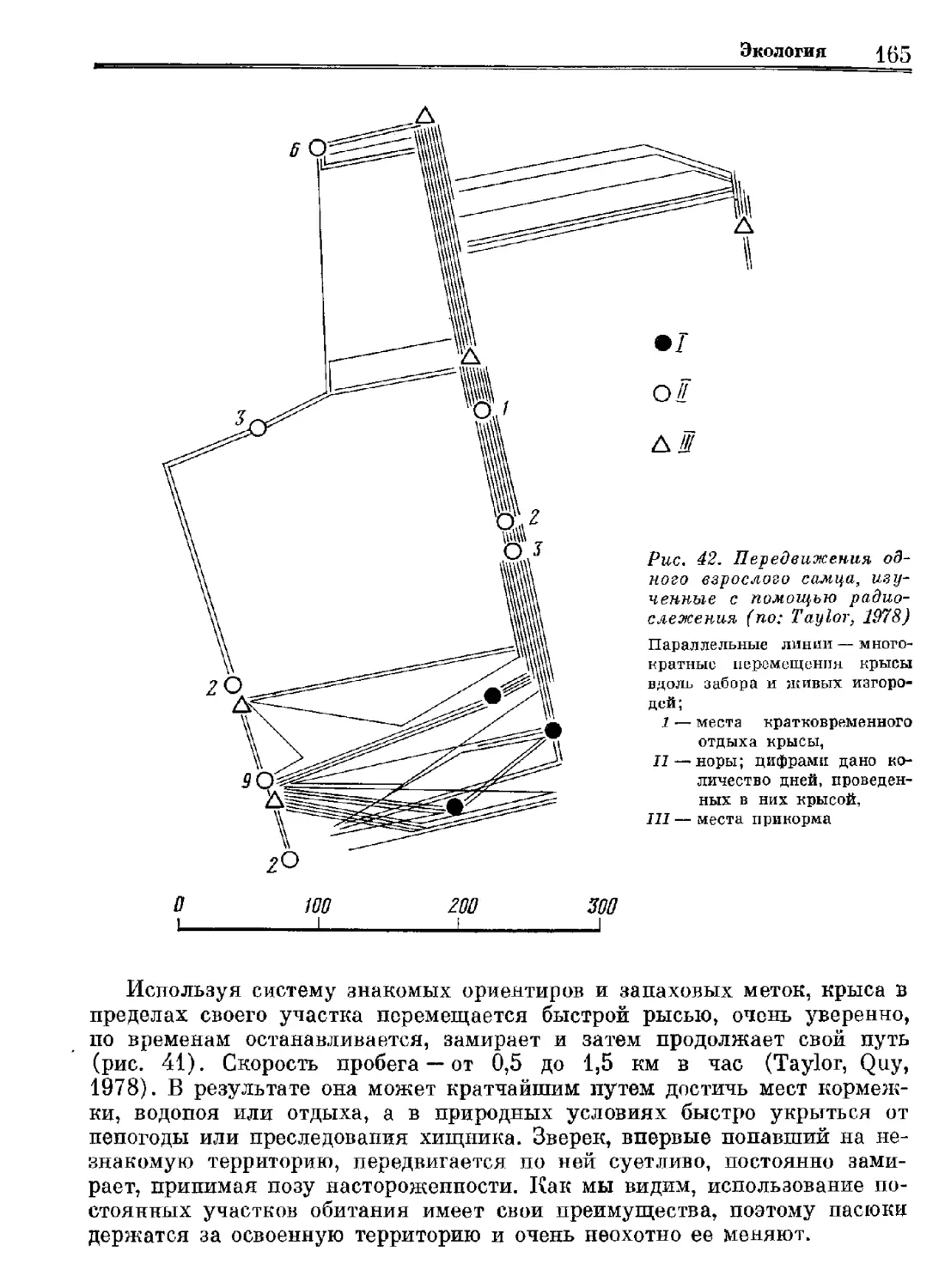













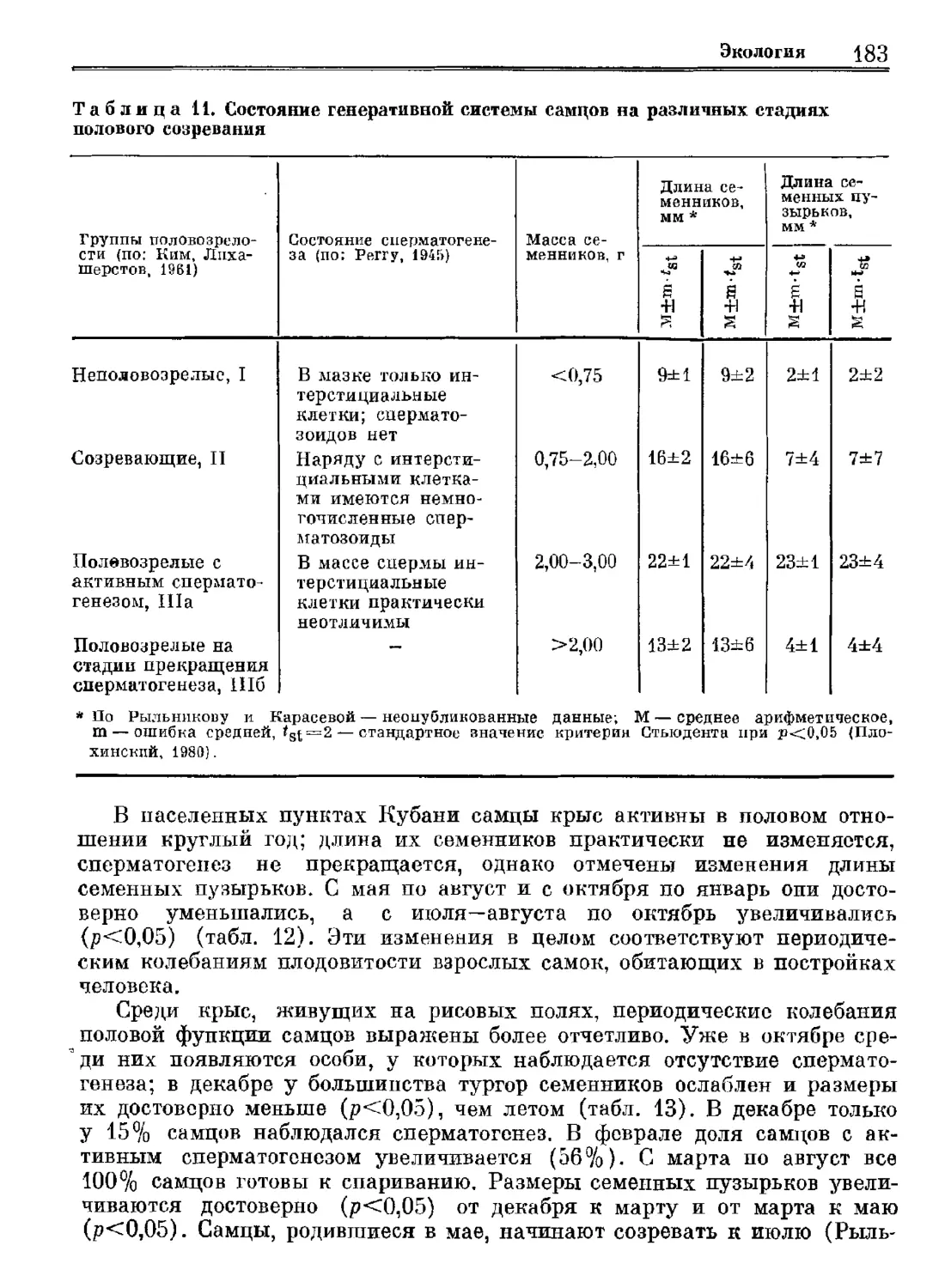
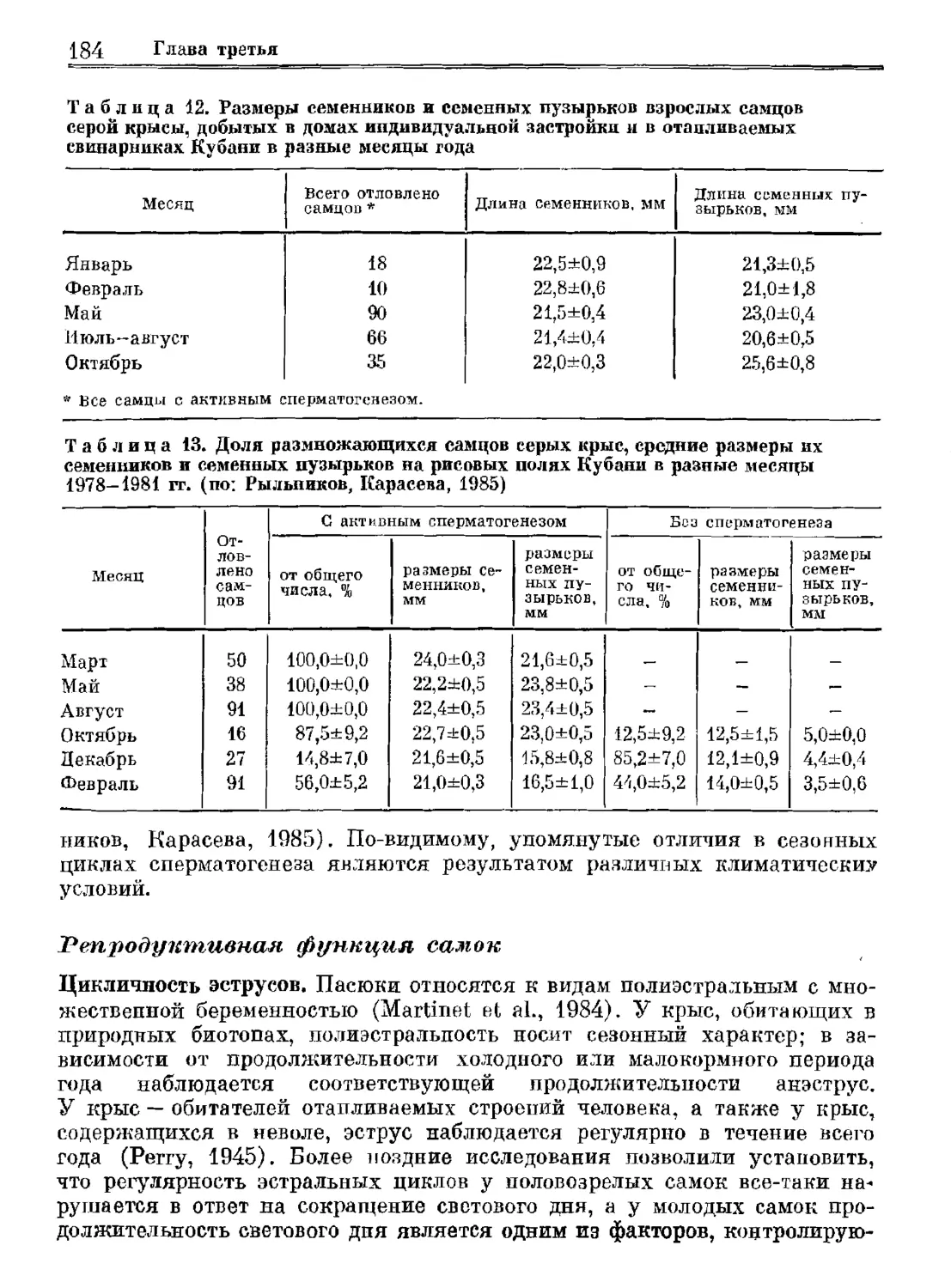
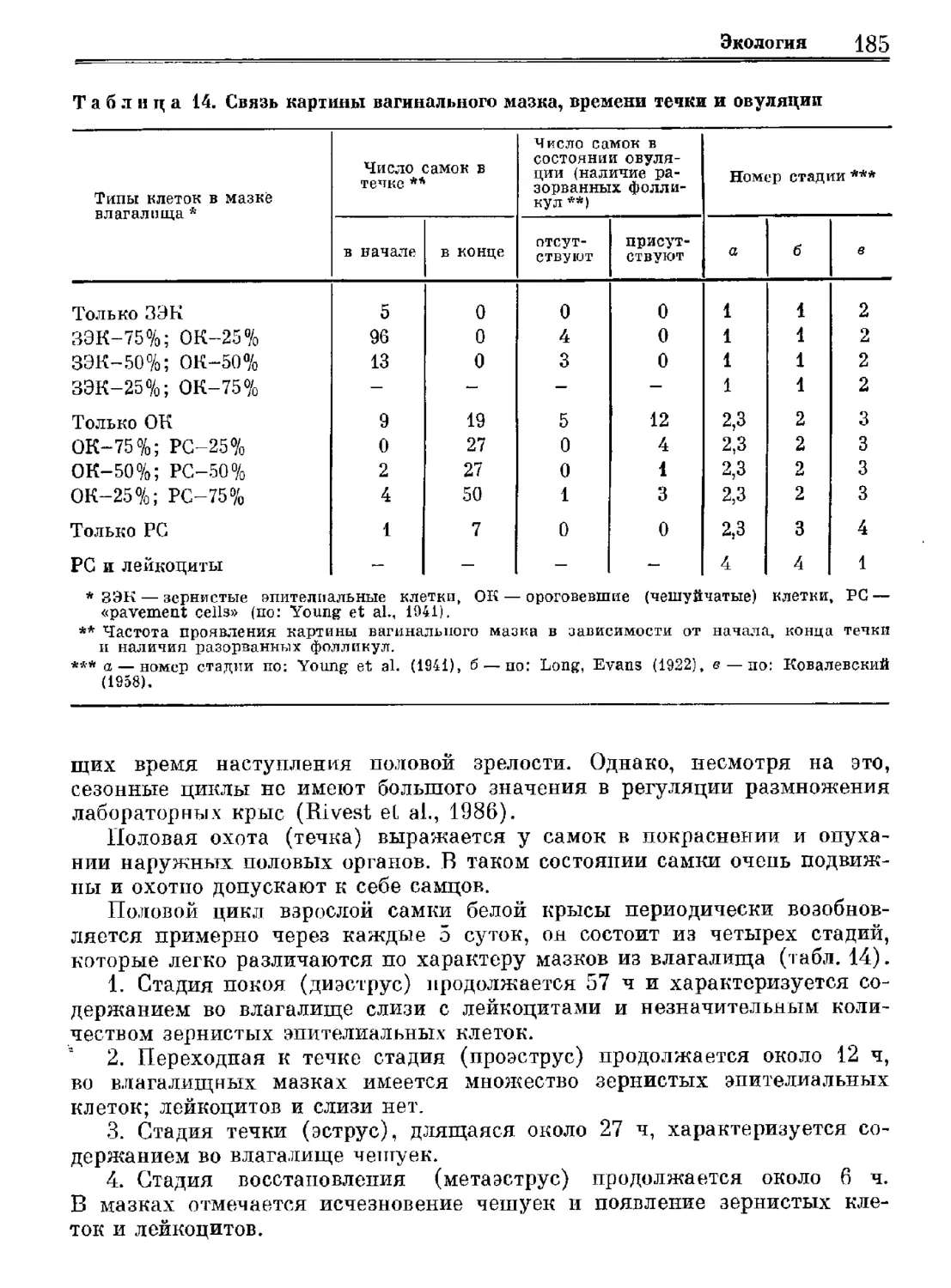




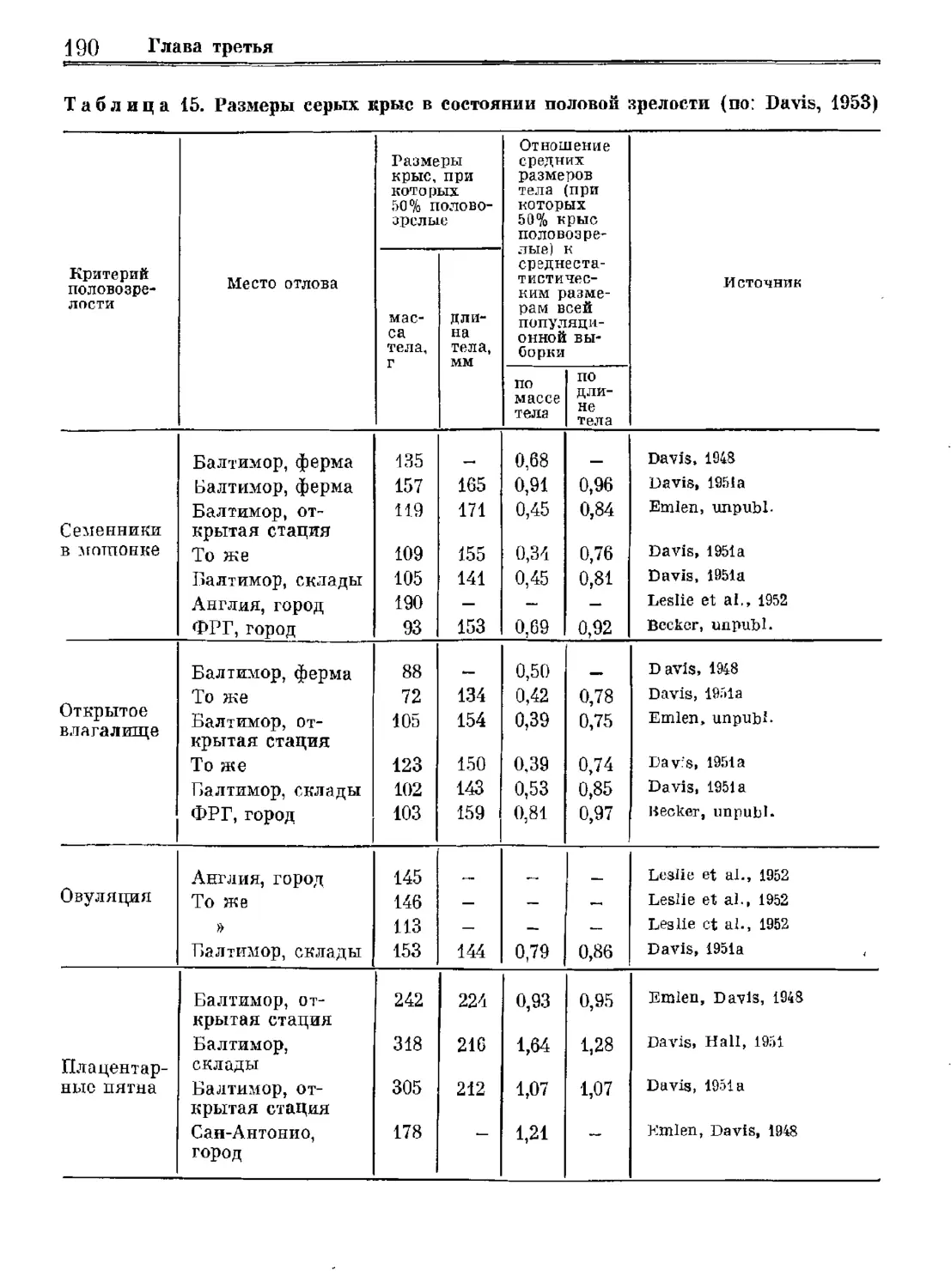





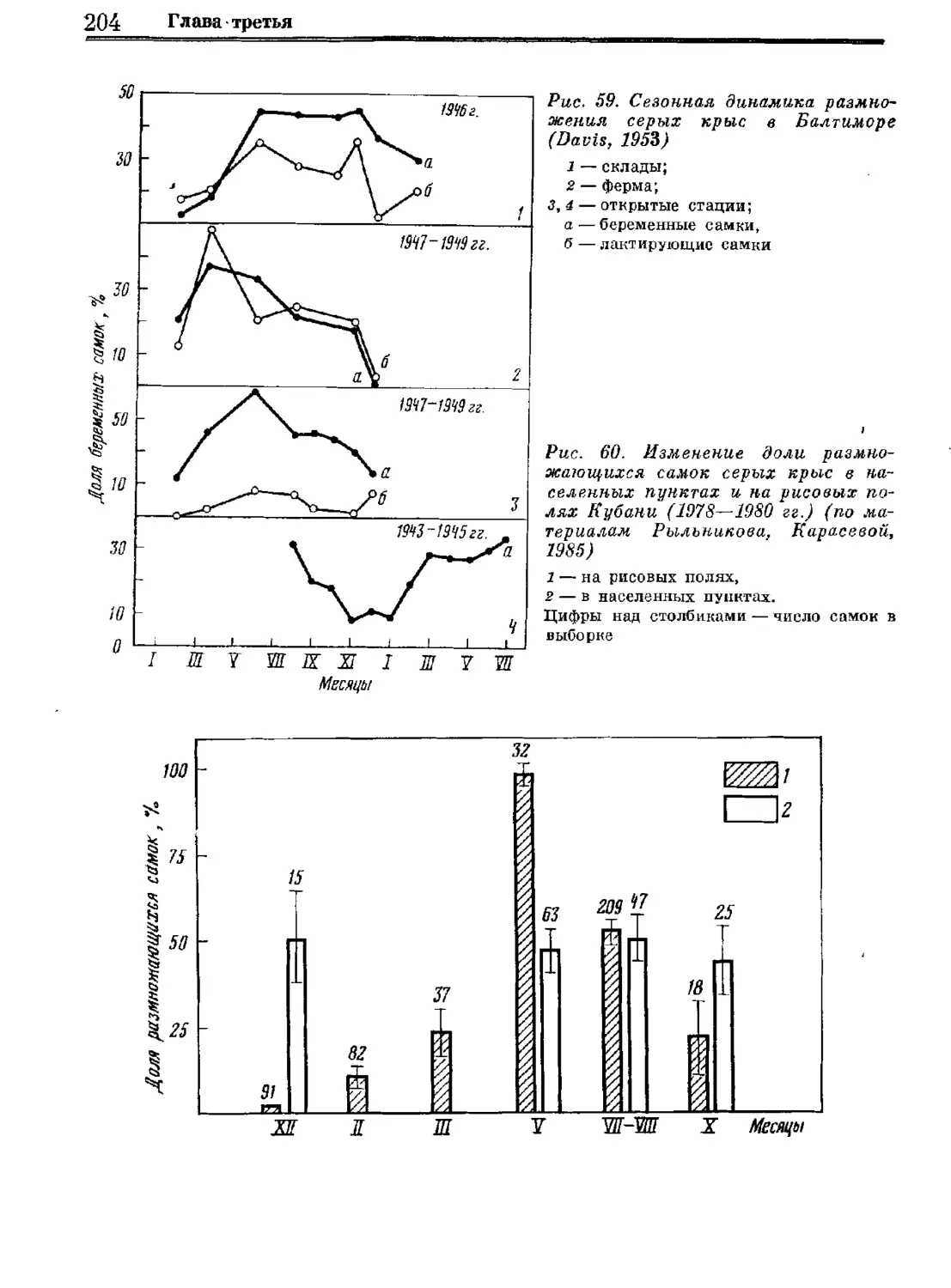
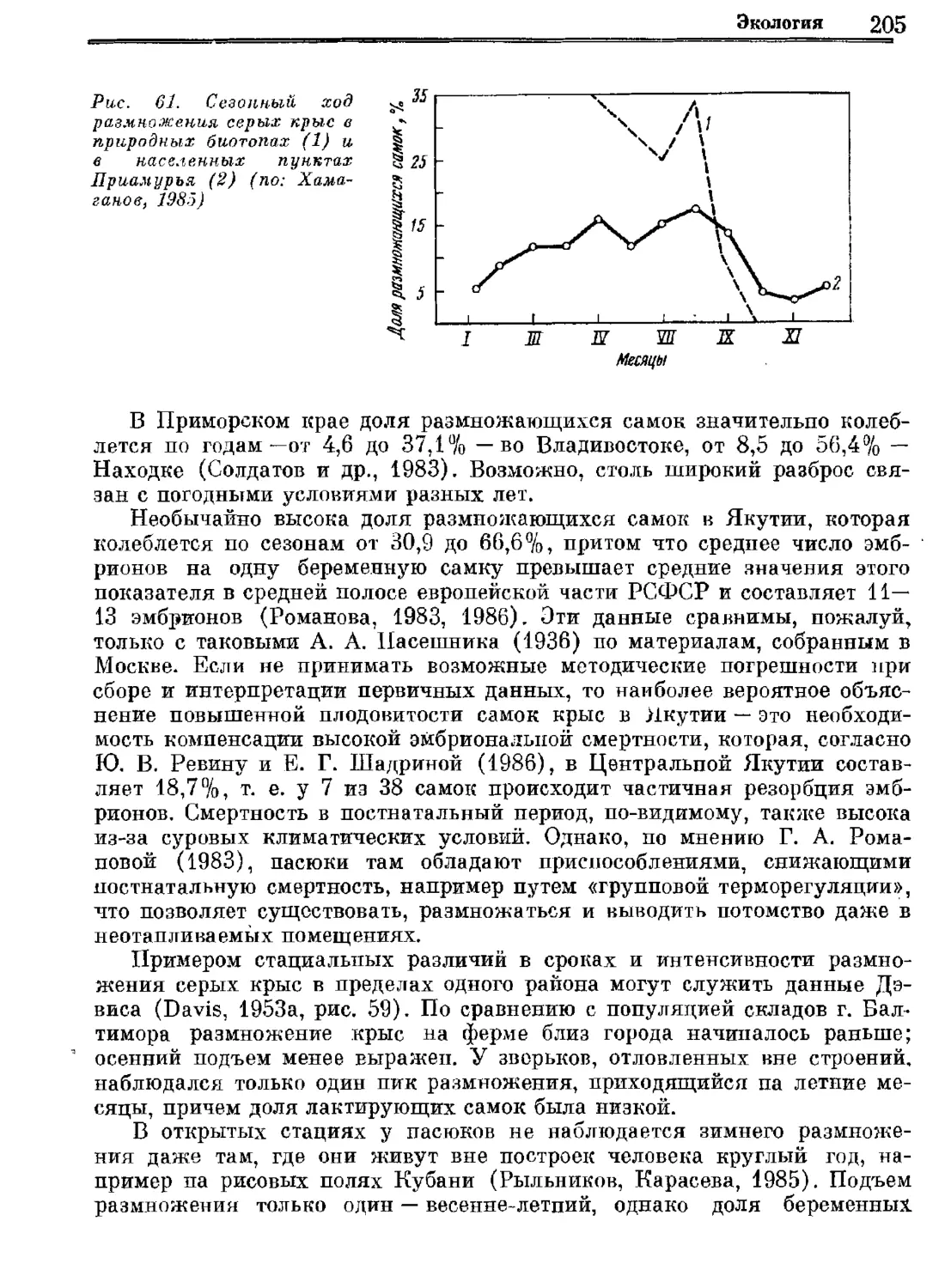


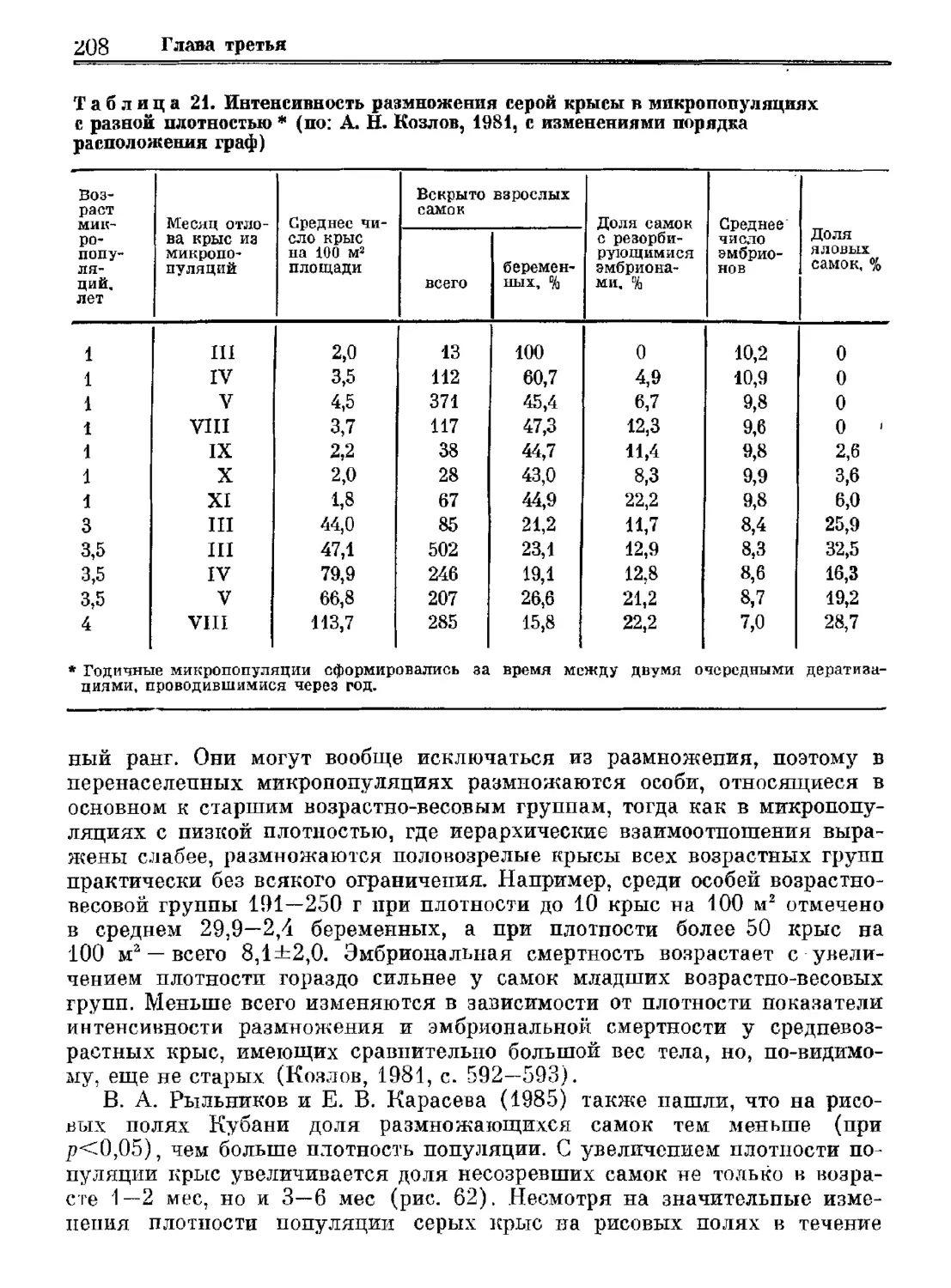
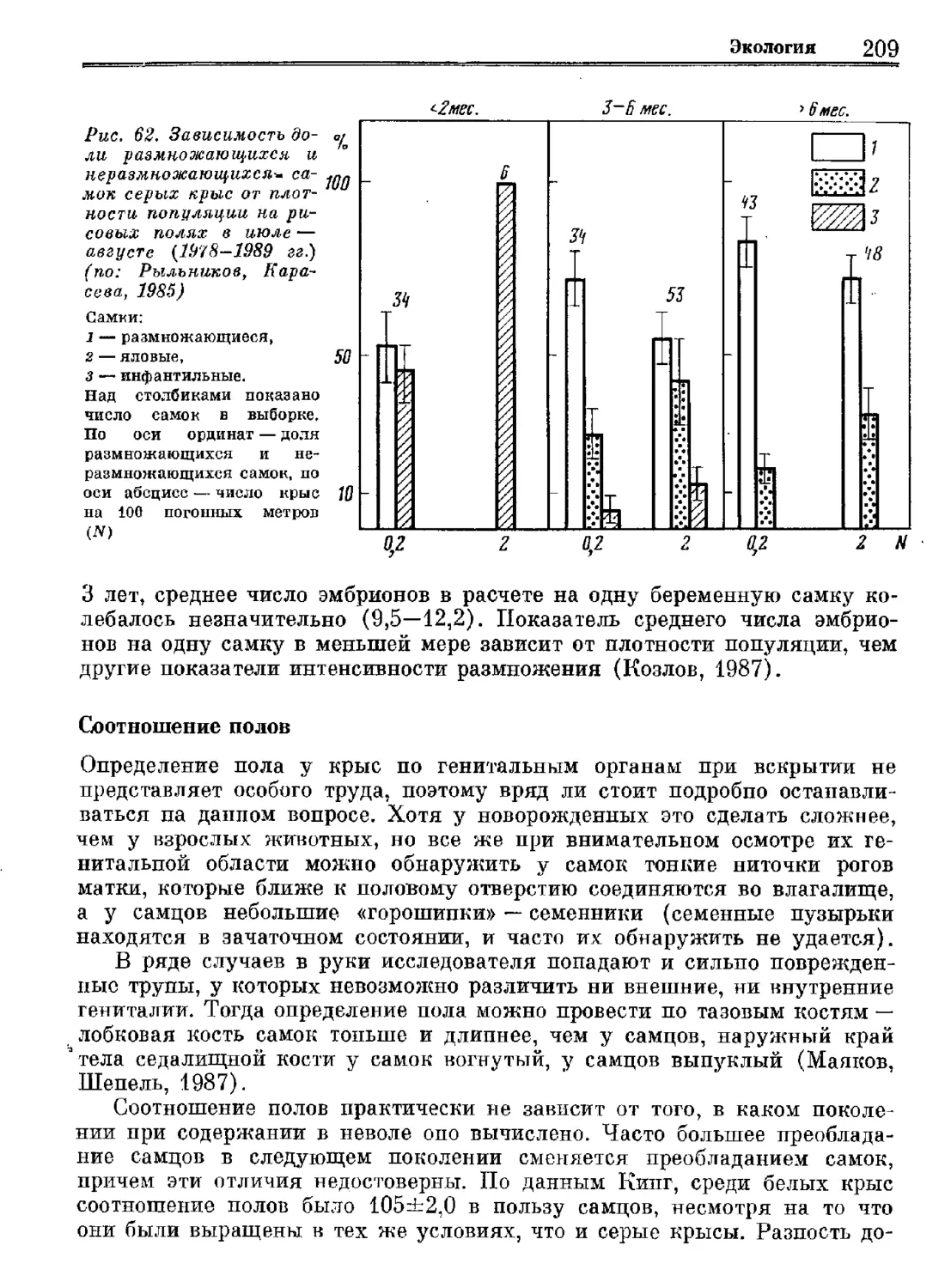


































































































































































































































Author: Орлова Г.М.
Tags: rodentia lagomorpha грызуны образ жизни животных поведение животных экология этология животные и окружающая среда (биономия) биология животный мир мир животных зоология фауна
ISBN: 5-02-005230-2
Year: 1990




Академия наук СССР
Институг
эволюционной морфологии
и экологии животных
им. А. Н. Северцова
Научный совет по проблеме
«Биологические основы
освоения, реконструкции
и охраны животного мира»
Виды
фауны СССР
и сопредельных
стран
Советский комитет
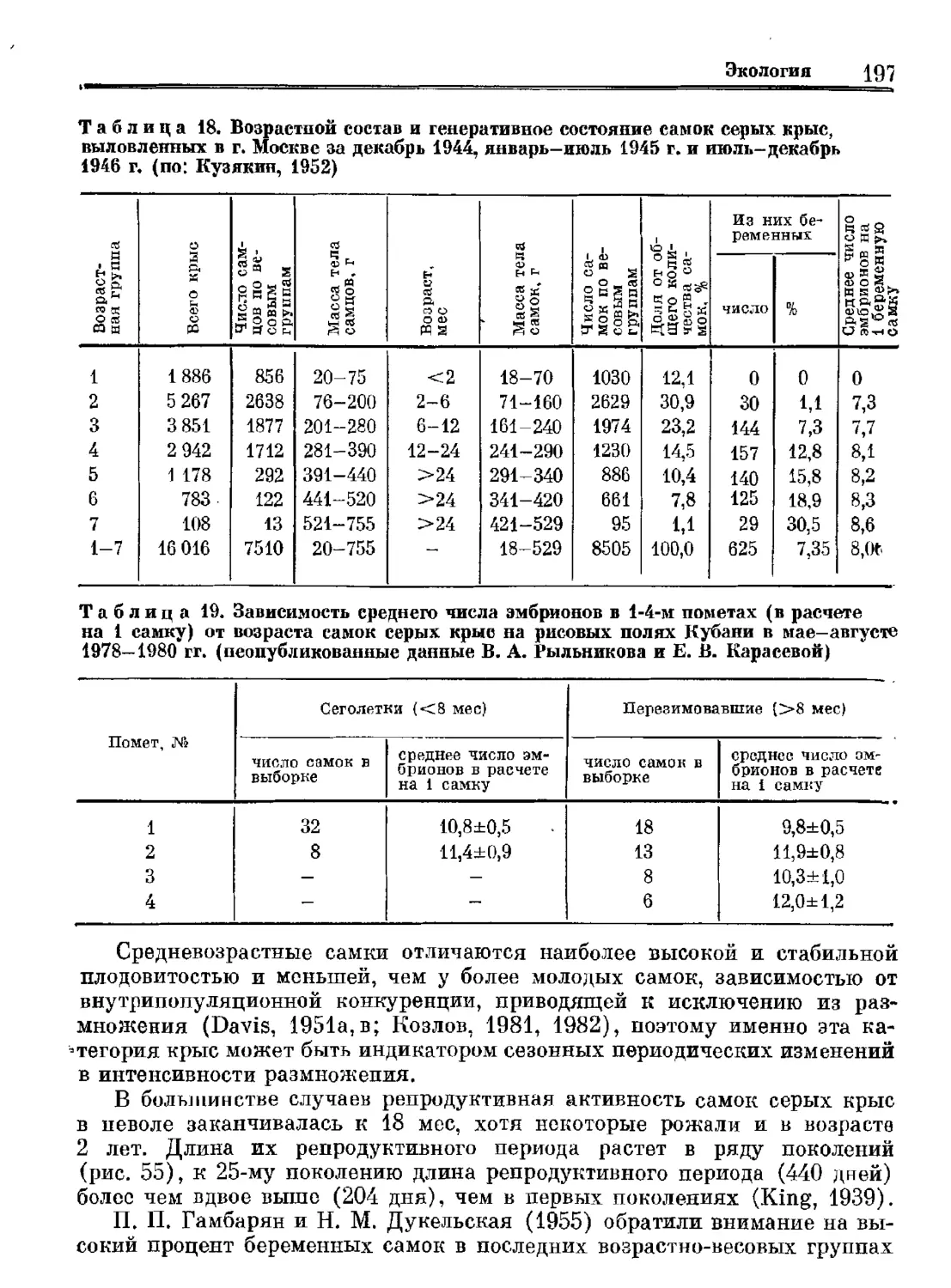
по программе ЮНЕСКО
„Человек и биосфера“
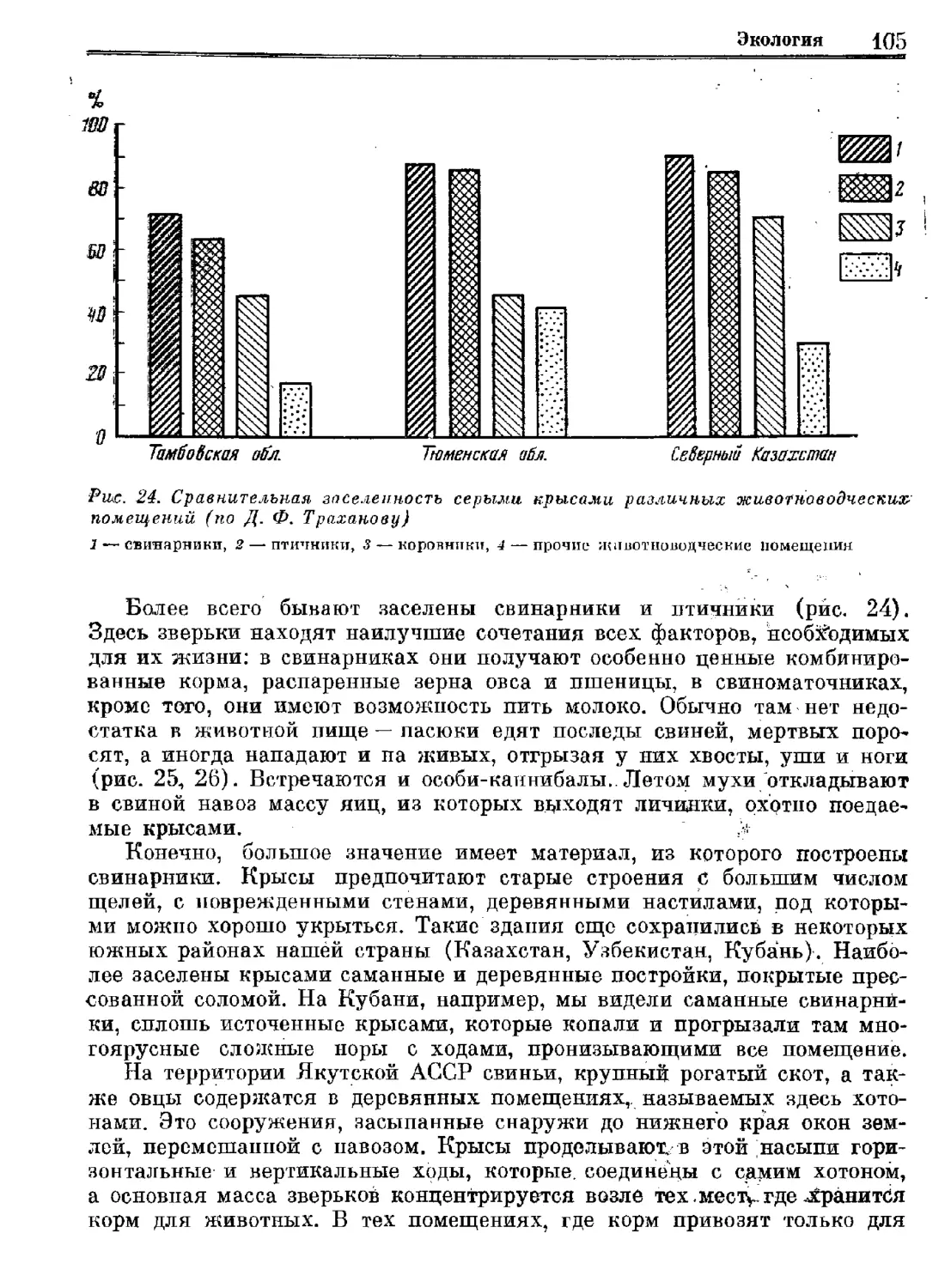
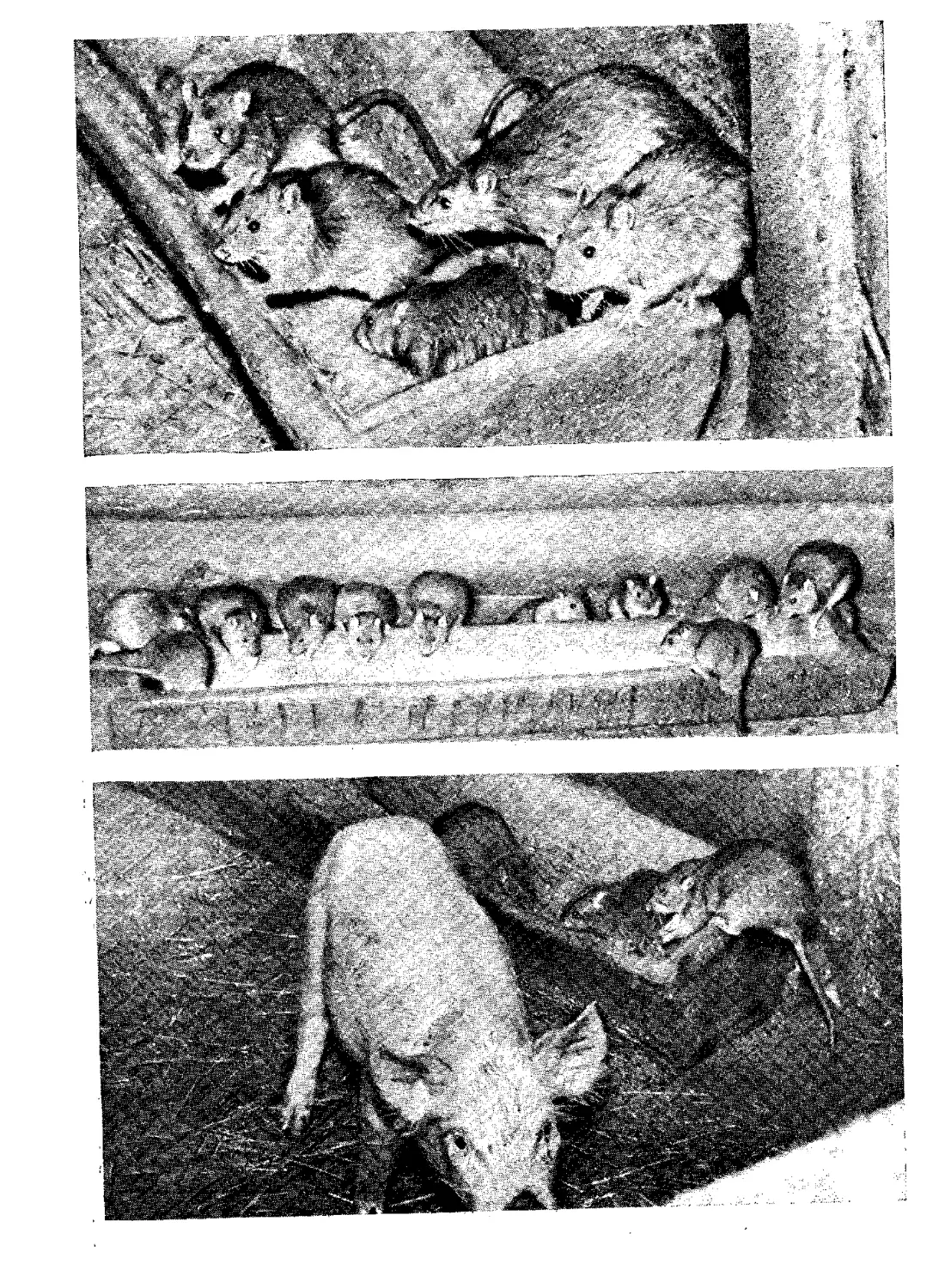
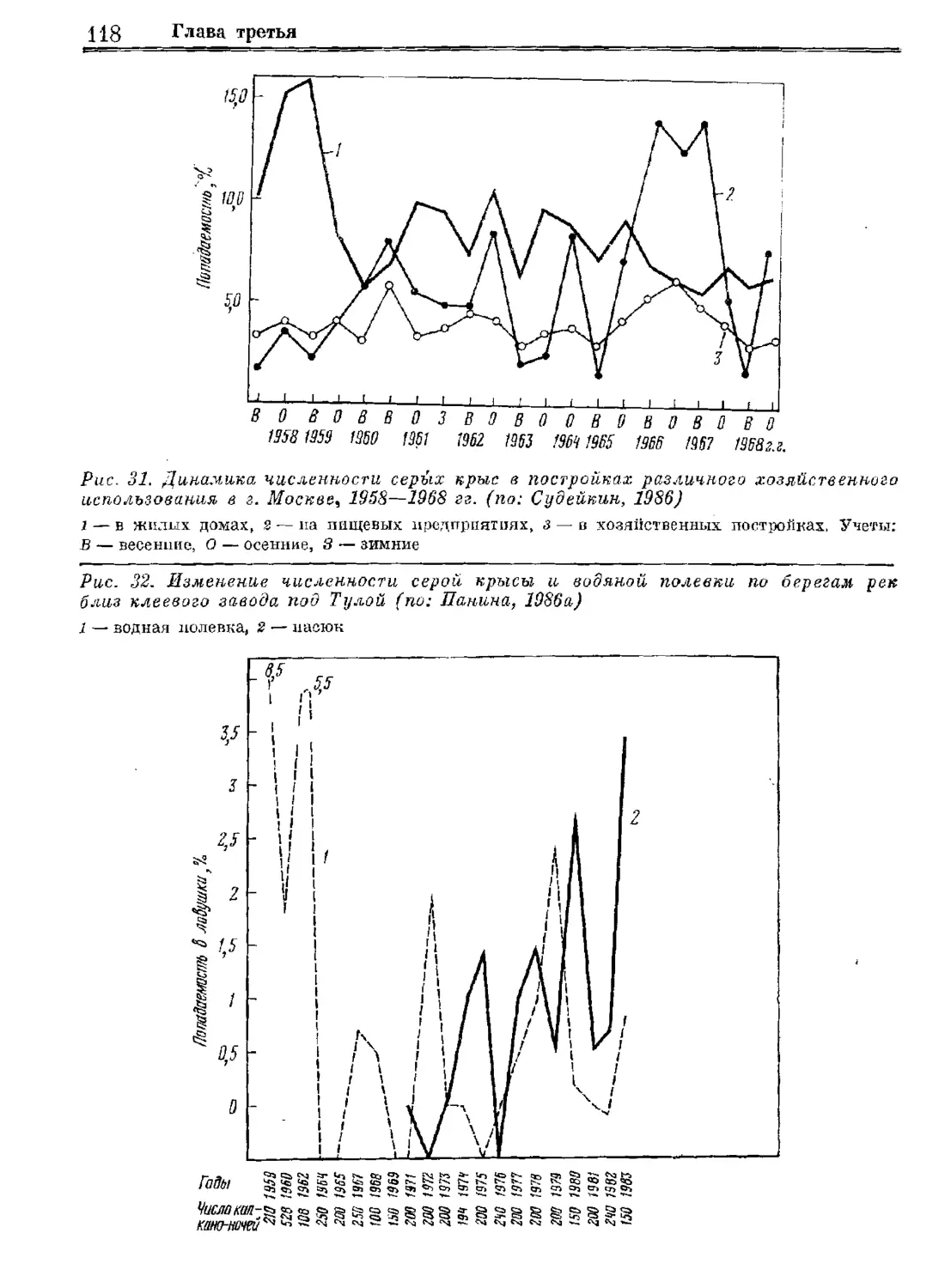
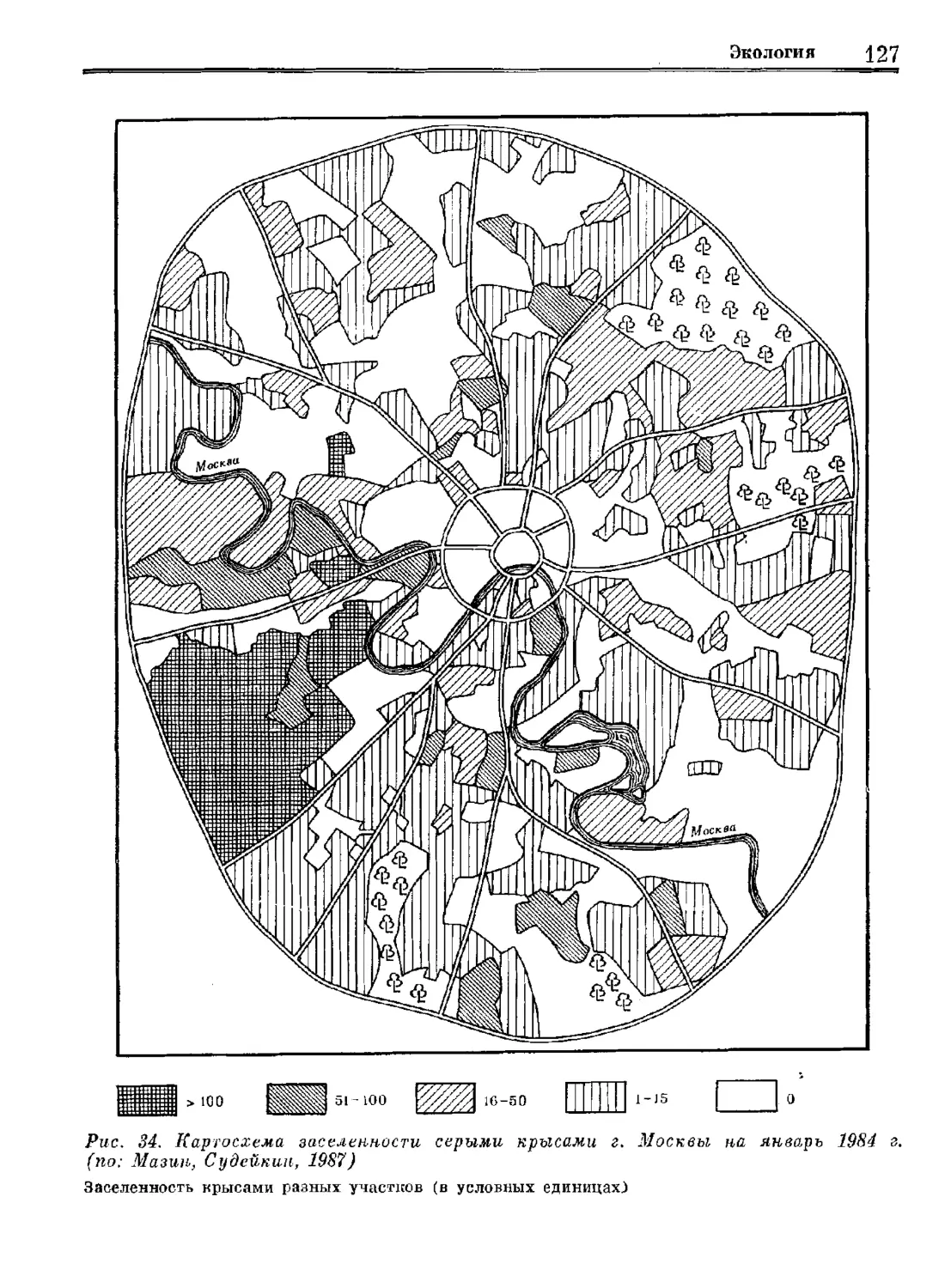
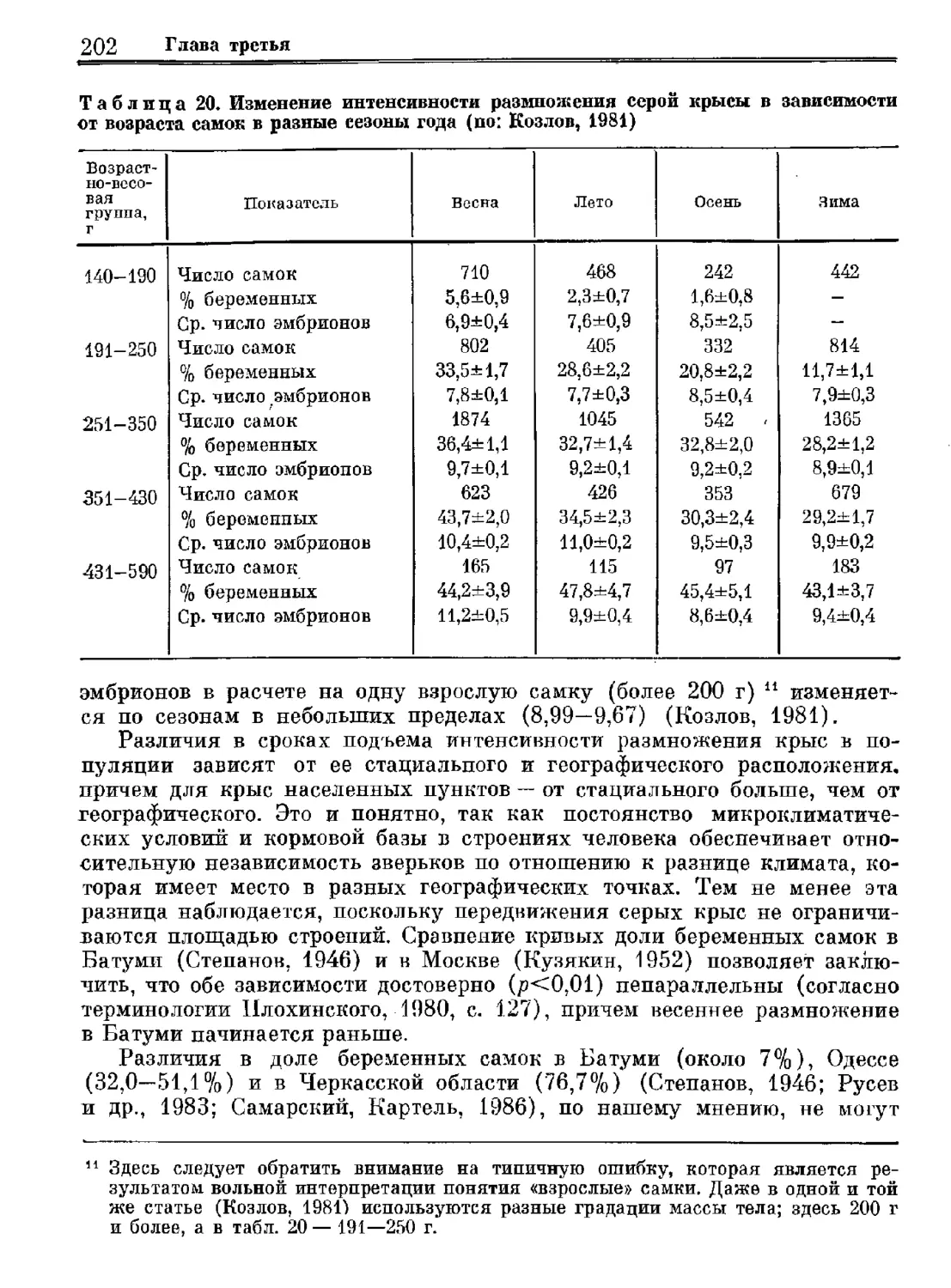
Серая крыса
Систематика
Экология
„Регуляция численности
Издательство «Наука»
Серая крыса
Систематика Эвология Регуляция численности
Могуау Вай
Зузетайс$ Есооку Роршаноп Соптя
Серая крыса
Систематика Эвология Регуляция численности
Могуау Вай
Зузетайс$ Есооку Роршаноп Соптя
03$ Асаехлу оЁ Зе1епсе
А. М. Зехегыох Гази ище
оЁ Ехо опагу
Апиа! Могрво]ову
ап@ Есоюсу
Заепыс Сопие
фог В1оюр1са! Стопа@з
0Ё Мапасешеп®,
Весопзигасйоп
ап СопзехуаНоп
о? Апира]з
Бресез
0# Ще Каппа оЁ Ще 9588
апд Ве Соп1о0$
Сопиие8
Те Зег!ез
Зоу1её СошиыИее {ог Ше отв 11 1979
ОМЕЗСО Ртооташише
«Мап ап@ В1озрКеге»
No Noшгуау Ка
фбуяетайс5
ЕсОову
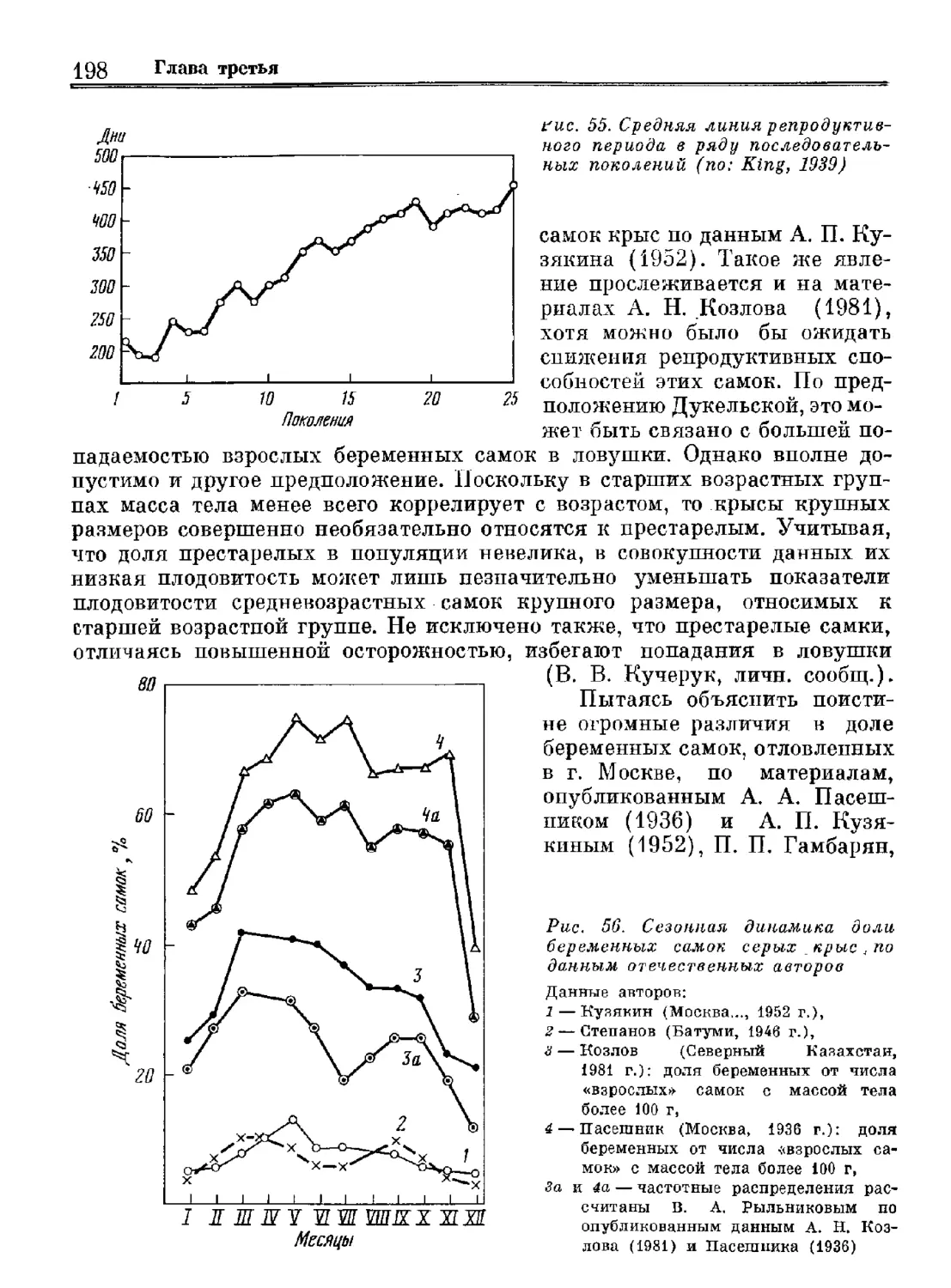
Роршайоп сот]
Ед Иот8-ш-СШей
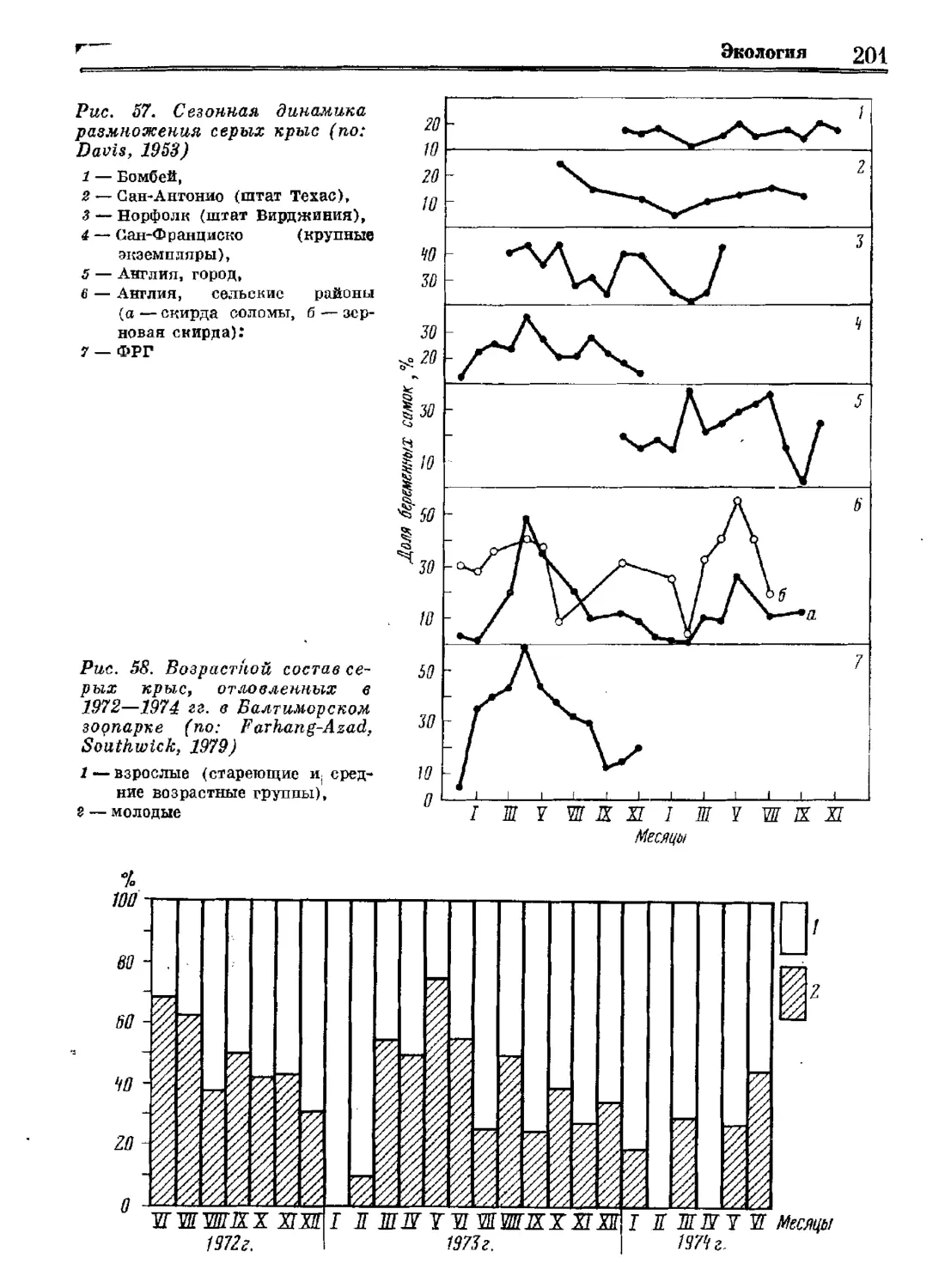
Асад. У. Е. ЗОКОГОХ,
Пг. зс. Е. У. КАВАЗОУА
Мапцка Ра1 Вегз
Мозео\ 1990
Академия паук СССР
Институт
эволюционной морфологии
и экологии животных
им. А. Н. Северцова
Научный совет
по проблемам
«Биологические осповы
освоения, реконструкции
и охраны
животного мира»
Виды фауны СССР
°
и сопредельных стран
.
Серия основана
в 1979 г.
Советский комитет
по программе ЮНЕСКО
«Человек и биосфера»
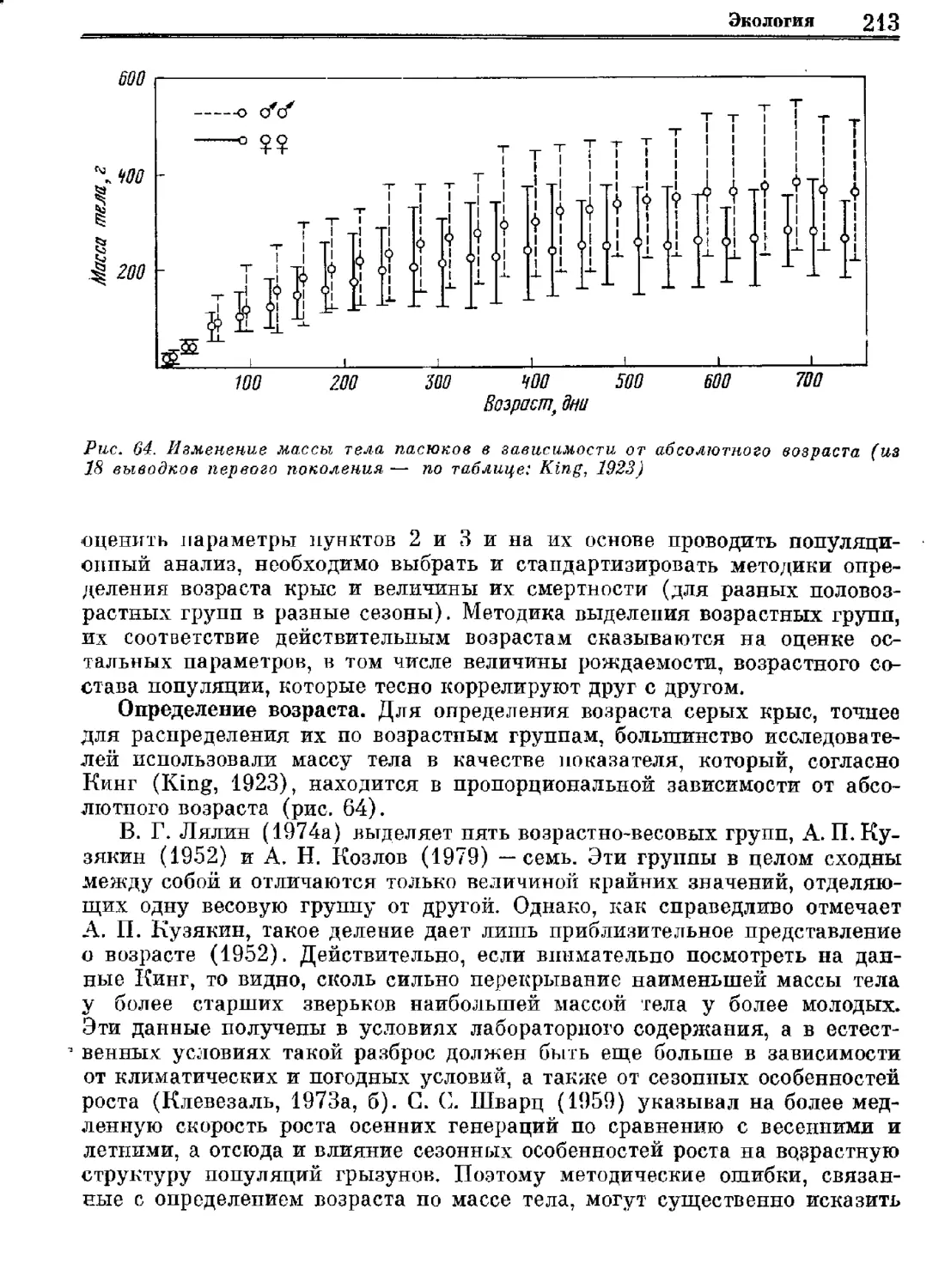
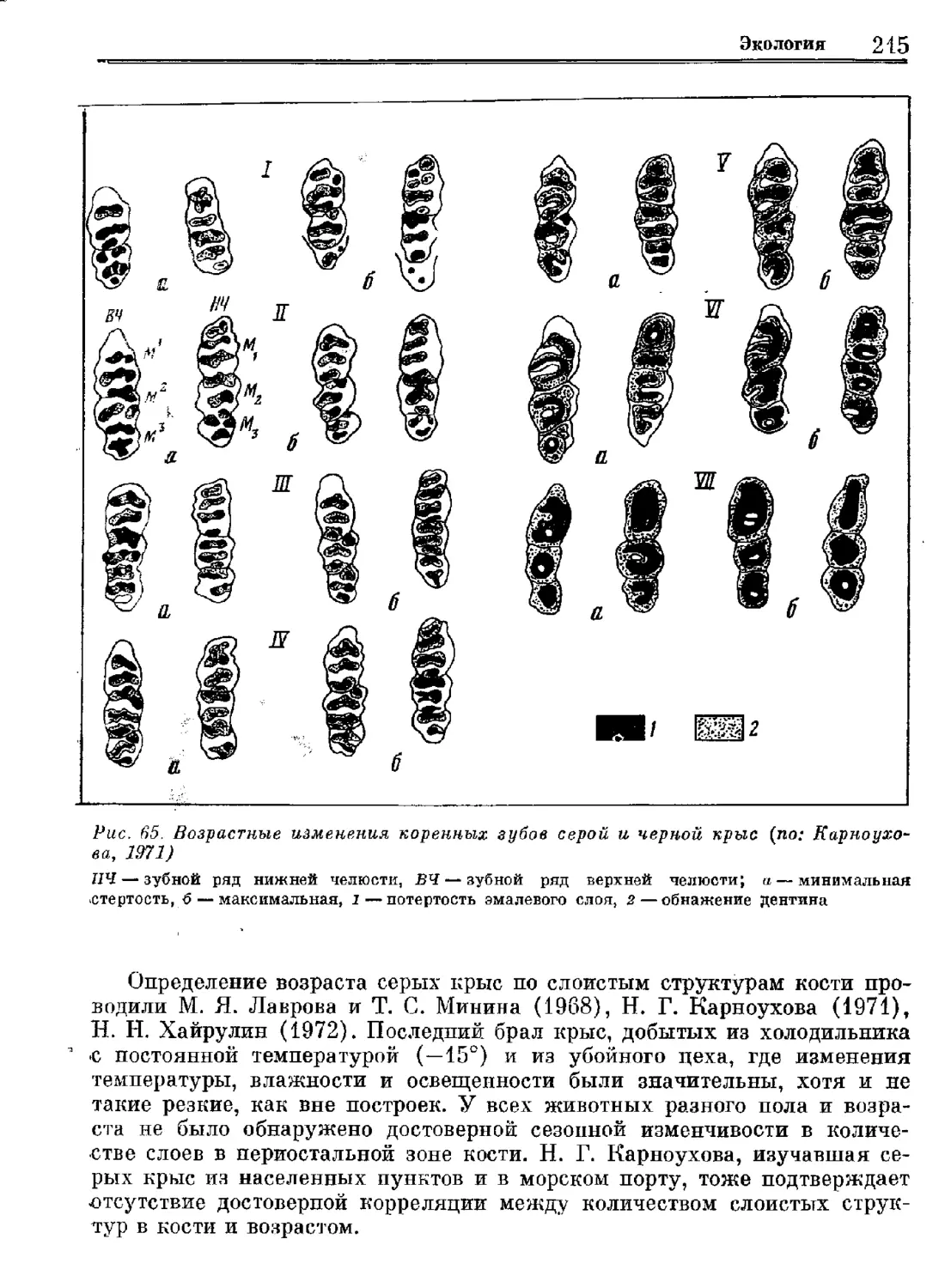
Серая крыса.
Систематика
Экология'
Регуляция численности
Ответственные редакторы тома
академик В. Е. СОКОЛОВ,
доктор биологических наук Е. В. КАРАСЕВА
ВНаукаМосква 1990
03$ Асаехлу оЁ Зе1епсе
А. М. Зехегыох Гази ище
оЁ Ехо опагу
Апиа! Могрво]ову
ап@ Есоюсу
Заепыс Сопие
фог В1оюр1са! Стопа@з
0Ё Мапасешеп®,
Весопзигасйоп
ап СопзехуаНоп
о? Апира]з
Бресез
0# Ще Каппа оЁ Ще 9588
апд Ве Соп1о0$
Сопиие8
Те Зег!ез
Зоу1её СошиыИее {ог Ше отв 11 1979
ОМЕЗСО Ртооташише
«Мап ап@ В1озрКеге»
No Noшгуау Ка
фбуяетайс5
ЕсОову
Роршайоп сот]
Ед Иот8-ш-СШей
Асад. У. Е. ЗОКОГОХ,
Пг. зс. Е. У. КАВАЗОУА
Мапцка Ра1 Вегз
Мозео\ 1990
Академия паук СССР
Институт
эволюционной морфологии
и экологии животных
им. А. Н. Северцова
Научный совет
по проблемам
«Биологические осповы
освоения, реконструкции
и охраны
животного мира»
Виды фауны СССР
°
и сопредельных стран
.
Серия основана
в 1979 г.
Советский комитет
по программе ЮНЕСКО
«Человек и биосфера»
Серая крыса.
Систематика
Экология'
Регуляция численности
Ответственные редакторы тома
академик В. Е. СОКОЛОВ,
доктор биологических наук Е. В. КАРАСЕВА
ВНаукаМосква 1990
УДК 599.323.4:591.52
Серая крыса: Систематика, экология, регуляция численности. М.: Наука, 1990. 456 с-
Т3ВМ 5-02-005230-2
.
Книга представляет собой обзор современных знаний по систематике, экологии, мировом
распространению вида и методам ограничения его численности. Этот обзор должен соста-
вить научную основу для разработки более рациональных и эффективных мер по контролю.
численности серой крысы— вреднейшего из грызунов.
Могуау Вае бузешайсз, ЕсоЮву, Вегщацов 0# Нз Роршайоп. М.; Маика, 1990.
456 р.
ТЗВМ 5-02-005230-2
—
в
ТВе Боок 15 а тезем оп зуфешайсь, ес0105у ап@ тота Фетаоп 0 Ве зресфез апа те о6ь
оР {с темаоп 01 Из роршаНопз. ТЫ 8 течет 15 $0 рто\!Че а Ъавз Тот е1афогаМоп о{ тоге таНо-
па! апб еесёте глеазитез ой сопгоШо5 ве Мог\уау гай, (пе тпо5Т Патио! рез.
Редакционная коллегия серии
СОКОЛОВ В. Е.
ИЛЬИЧЕВ В. Д.
СТАРОБОГАТОВ Я. И.
{отв. редактор)
ЛЕБЕДКИНА Н. С.
СУХАНОВ В. Б.
БОЛЬШАКОВ В. Н.
ЛУКИН Е. Ф.
СТЕПАНЯН Л. С.
ВОЛЬСКИС Р. С.
НЕРОНОВ В. М.
ХАБАРМАН Х. Х.
ГРОМОВ И. М.
РАСС Т. С.
ШАТУНОВСКИЙ М. И.
ДАРЕВСКИЙ И. С.
СВЕШНИНОВ В. А.
ЩЕБАК Н. Н,
ЗАЯНЧКАУСКАС ЦП. л.
СМИРНОВ Н. Н.
ЯБЛОНОВ А. В.
ИВАНОВ А. В.
Рецензенты:
доктор биологических наук Д. И. БИБИКОВ,
донтор биологических наук С. А. ШИЛОВА
Едиот1а! Воаг@ оЁ 41е Эег1ез
ЗОКОГ.ОУ У. Е.
ТРУЕНЕУ У. О.
ЗТАВОВОбАТОУ Та. Т-
(ЕаМот-ш-СЫеЕ)
ТЕВЕО КМА М. 8.
ЗОКНАНМОХ У. В. |
ВОРЗНАКОУ У, М.
ОКМ Е. #.
ЗТЕРАМУАМ Г.. 8.
УО1.8К1З В. 8.
МЕВОМОУ У. М.
КНАВАВМАМ КВ. КВ.
акомоу 1. м.
ВА8$ Т. 8.
ЗНАТОМОУЗКУ М. 1.
РАКЕУЗКУ 1. 8.
ЗУЕЗНМКОУ У. А.
ЗСНЕВВАК М. М.
2АХАМСНКАОВКА$ Р.А.
ЗМТВМОХ М. М.
УАВРОКОХ А. У.
ТУАМОУ А. У.
КБез1езуега:
Г. 1. ВТВЕКОХ, РгоГ., Ог. 8с. (В101057у),
8. А. ЗНИОУА, Рго{., Ог. Зс. (8101089)
1907 000 000-555
055(02)-90
Т5ВМ 5-02-005280-2
589-89, кн. 2
© Институт эволюционной морфологии.
и экологии животных им. А. Н. Северцова, 1990
Предисловие
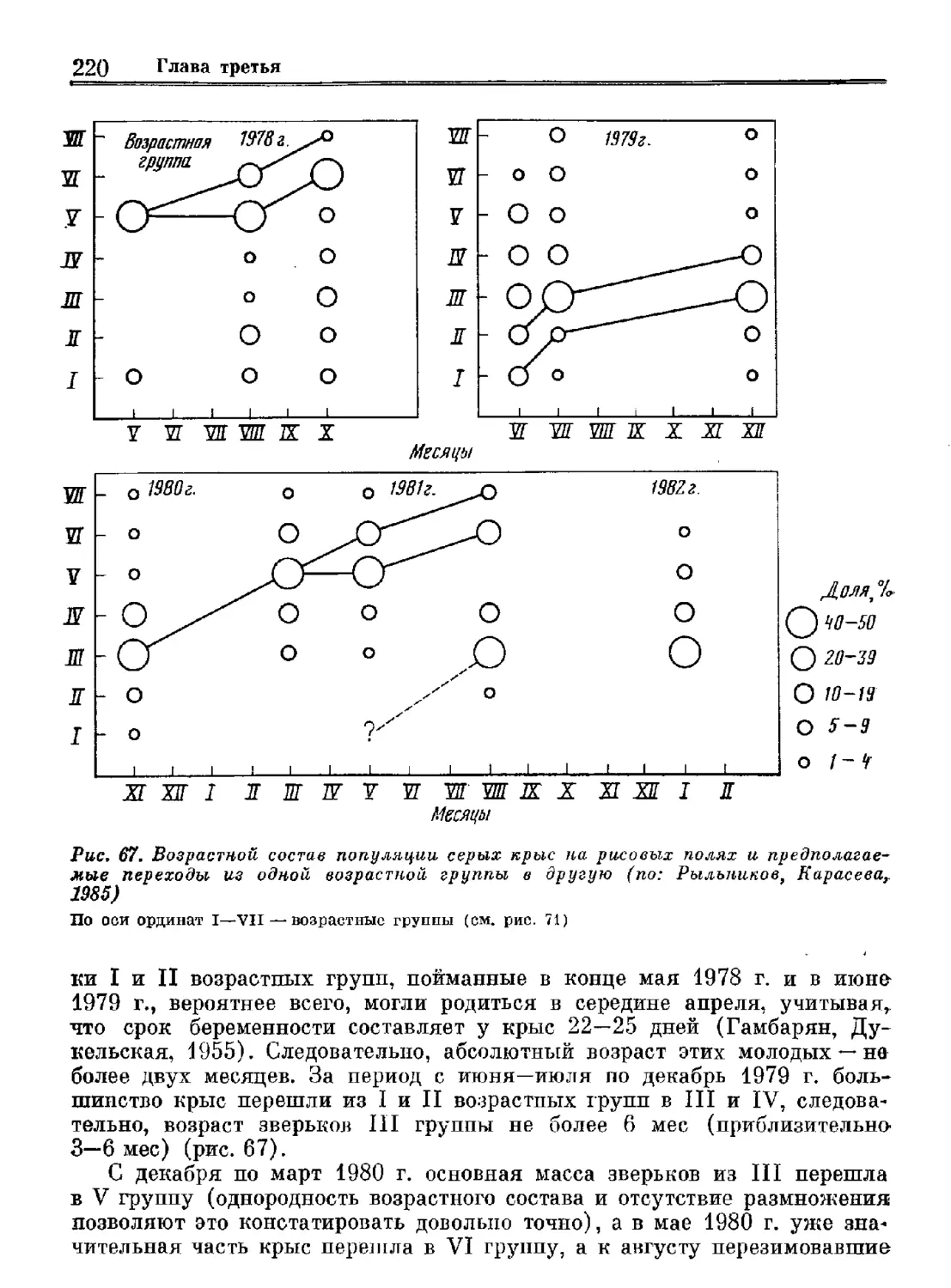
В серии «Виды фауны СССР и сопредельных стран» вышло уже три мо-
нографии, посвященных млекопитающим,— «Зубр», «Рыжая полевка» и
«Волк». Задача этой серии— дать на современном уровне наиболее пол-
ное и всестороннее описание вида: особенностей его систематики, рас-
пространения в мире, морфологии, экологии, поведения и практической
значимости. В издании этих монографий мы видим большой смысл, так
как они представляют собой не только обобщение и анализ имеющейся
информации, но и служат стимулом дальнейшего изучения вида. В из-
дании монографии по серой крысе назрела особая необходимость, пото-
му что этот вид является одним из самых злейших врагов человечества
среди животных.
В последние десятилетия резко увеличилась антропогенная трансфор-
мация среды: сведение лесов, устройство водохранилищ, появление но-
вых населенных пунктов, а также железных дорог, активизация живот-
новодства и пр. Все это привело к значительному расширению ареала
крысы, уплотнению его «кружева» и увеличению численности вида.
Специальной службой с пасюком ведется постоянная борьба. У нас в
стране это отделы профилактической дератизации Министерства здраво-
охранения. Но, несмотря на большие усилия и затрату немалых средств,
крыс остается еще очень много. Для разработки более эффективных мер
отраничения численности этого грызуна нсобходимо детальное знание его
особенностей, образа жизни и повадок. На необходимость изучения нпа-
сюка указывали специалисты многих стран мира еще в 1966 г.
в Женеве на семинаре Весмирной организации здравоохранения. В 1973 г.
проблеме изучения биологии крыс и методов борьбы с ними был поевя-
щен специальный Международный конгресс (Будапешт).
В 1983 г. при Всесоюзном териологическом обществе (ВТО) была со-
здана координационная группа по изучению пасюка. В нее вошли со-
трудники Илститута эволюционной морфологии и экологии животных
им. А. Н. Северцова АН СССР (ИЭМЭЖ), Всесоюзного научно-исследо-
вательского института ветеринарпой гигиены (ВНИИВГ), Веесоюзного
научно-исследовательского института дезинфекции и стерилизации
(ВНИИДиС) Министерства здравоохранения СССР.
В задачи группы входит не только привлечение зоологов разных вс-
домств (особенно молодежи) к изучению серой крысы, но и координация
их работы. Группа провела ряд рабочих совещаний (всесоюзных), в ре-
зультате которых вытпли следующие публикации под редакцией В. Е. Со-
колова и Е. В. Карасевой:
1. Экология и медицинское значение серой крысы (Ваз потуев1сиз).
М.: АН СССР, 1983.
2. Распространение и экология серой крысы и методы ограничения ее
численности. М.: Наука, 1985.
УДК 599.323.4:591.52
Серая крыса: Систематика, экология, регуляция численности. М.: Наука, 1990. 456 с-
Т3ВМ 5-02-005230-2
.
Книга представляет собой обзор современных знаний по систематике, экологии, мировом
распространению вида и методам ограничения его численности. Этот обзор должен соста-
вить научную основу для разработки более рациональных и эффективных мер по контролю.
численности серой крысы— вреднейшего из грызунов.
Могуау Вае бузешайсз, ЕсоЮву, Вегщацов 0# Нз Роршайоп. М.; Маика, 1990.
456 р.
ТЗВМ 5-02-005230-2
—
в
ТВе Боок 15 а тезем оп зуфешайсь, ес0105у ап@ тота Фетаоп 0 Ве зресфез апа те о6ь
оР {с темаоп 01 Из роршаНопз. ТЫ 8 течет 15 $0 рто\!Че а Ъавз Тот е1афогаМоп о{ тоге таНо-
па! апб еесёте глеазитез ой сопгоШо5 ве Мог\уау гай, (пе тпо5Т Патио! рез.
Редакционная коллегия серии
СОКОЛОВ В. Е.
ИЛЬИЧЕВ В. Д.
СТАРОБОГАТОВ Я. И.
{отв. редактор)
ЛЕБЕДКИНА Н. С.
СУХАНОВ В. Б.
БОЛЬШАКОВ В. Н.
ЛУКИН Е. Ф.
СТЕПАНЯН Л. С.
ВОЛЬСКИС Р. С.
НЕРОНОВ В. М.
ХАБАРМАН Х. Х.
ГРОМОВ И. М.
РАСС Т. С.
ШАТУНОВСКИЙ М. И.
ДАРЕВСКИЙ И. С.
СВЕШНИНОВ В. А.
ЩЕБАК Н. Н,
ЗАЯНЧКАУСКАС ЦП. л.
СМИРНОВ Н. Н.
ЯБЛОНОВ А. В.
ИВАНОВ А. В.
Рецензенты:
доктор биологических наук Д. И. БИБИКОВ,
донтор биологических наук С. А. ШИЛОВА
Едиот1а! Воаг@ оЁ 41е Эег1ез
ЗОКОГ.ОУ У. Е.
ТРУЕНЕУ У. О.
ЗТАВОВОбАТОУ Та. Т-
(ЕаМот-ш-СЫеЕ)
ТЕВЕО КМА М. 8.
ЗОКНАНМОХ У. В. |
ВОРЗНАКОУ У, М.
ОКМ Е. #.
ЗТЕРАМУАМ Г.. 8.
УО1.8К1З В. 8.
МЕВОМОУ У. М.
КНАВАВМАМ КВ. КВ.
акомоу 1. м.
ВА8$ Т. 8.
ЗНАТОМОУЗКУ М. 1.
РАКЕУЗКУ 1. 8.
ЗУЕЗНМКОУ У. А.
ЗСНЕВВАК М. М.
2АХАМСНКАОВКА$ Р.А.
ЗМТВМОХ М. М.
УАВРОКОХ А. У.
ТУАМОУ А. У.
КБез1езуега:
Г. 1. ВТВЕКОХ, РгоГ., Ог. 8с. (В101057у),
8. А. ЗНИОУА, Рго{., Ог. Зс. (8101089)
1907 000 000-555
055(02)-90
Т5ВМ 5-02-005280-2
589-89, кн. 2
© Институт эволюционной морфологии.
и экологии животных им. А. Н. Северцова, 1990
Предисловие
В серии «Виды фауны СССР и сопредельных стран» вышло уже три мо-
нографии, посвященных млекопитающим,— «Зубр», «Рыжая полевка» и
«Волк». Задача этой серии— дать на современном уровне наиболее пол-
ное и всестороннее описание вида: особенностей его систематики, рас-
пространения в мире, морфологии, экологии, поведения и практической
значимости. В издании этих монографий мы видим большой смысл, так
как они представляют собой не только обобщение и анализ имеющейся
информации, но и служат стимулом дальнейшего изучения вида. В из-
дании монографии по серой крысе назрела особая необходимость, пото-
му что этот вид является одним из самых злейших врагов человечества
среди животных.
В последние десятилетия резко увеличилась антропогенная трансфор-
мация среды: сведение лесов, устройство водохранилищ, появление но-
вых населенных пунктов, а также железных дорог, активизация живот-
новодства и пр. Все это привело к значительному расширению ареала
крысы, уплотнению его «кружева» и увеличению численности вида.
Специальной службой с пасюком ведется постоянная борьба. У нас в
стране это отделы профилактической дератизации Министерства здраво-
охранения. Но, несмотря на большие усилия и затрату немалых средств,
крыс остается еще очень много. Для разработки более эффективных мер
отраничения численности этого грызуна нсобходимо детальное знание его
особенностей, образа жизни и повадок. На необходимость изучения нпа-
сюка указывали специалисты многих стран мира еще в 1966 г.
в Женеве на семинаре Весмирной организации здравоохранения. В 1973 г.
проблеме изучения биологии крыс и методов борьбы с ними был поевя-
щен специальный Международный конгресс (Будапешт).
В 1983 г. при Всесоюзном териологическом обществе (ВТО) была со-
здана координационная группа по изучению пасюка. В нее вошли со-
трудники Илститута эволюционной морфологии и экологии животных
им. А. Н. Северцова АН СССР (ИЭМЭЖ), Всесоюзного научно-исследо-
вательского института ветеринарпой гигиены (ВНИИВГ), Веесоюзного
научно-исследовательского института дезинфекции и стерилизации
(ВНИИДиС) Министерства здравоохранения СССР.
В задачи группы входит не только привлечение зоологов разных вс-
домств (особенно молодежи) к изучению серой крысы, но и координация
их работы. Группа провела ряд рабочих совещаний (всесоюзных), в ре-
зультате которых вытпли следующие публикации под редакцией В. Е. Со-
колова и Е. В. Карасевой:
1. Экология и медицинское значение серой крысы (Ваз потуев1сиз).
М.: АН СССР, 1983.
2. Распространение и экология серой крысы и методы ограничения ее
численности. М.: Наука, 1985.
6
Предисловие
3. Экология серой крысы: 4-й съезд ВТО. М., 1986. Т. 3.
А. Серая крыса: в 2 т. М.: АН СССР, 1956.
.
.
5. Материалы по экологии и ограничению численности серой крысы:
вэт. М.: АН СССР, 1987.
Немалое внимание серой крысе уделялось на международных конфе-
ренциях по борьбе с вредными трызунами, проведенных в Кувейто
(Ргосеебзаз ой Ве Е зутрозиии оп Весеш Адуапсез ш Воде соо,
Кизай, 1982); Зесопа Зутрозиииа оЁ Вссей АЧуапсез п Водет Сошо,
Кимай, 1985).
.
Настоящий том подготовлен коллективом авторов: сотрудниками
ИЭМЭЖ им. А. Н. Северцова АН СССР, ВНИИДиС Мииздрава СССР,
Зоологического музея Тартуского университета и МГУ, ВНИИВГ, Ин-
ститута эпидемиологии и микробиологии (ИЭМ) им. Н. Ф. Гамалеи
АМН СССР, Института полиомиелита Минздрава СОСР, противочумной
станции (Одесса) и Института систематической и экологической биоло-
тии (Брно, Чехословакия).
й
.
Морфологии пасюка посвящен следующий том монографии, который
предполагается выпустить в недалеком будущем.
.
.
Эта книга о серой крысе является фактически первой капитальной
сводкой на русском языке по данному виду. У нас в стране по пасюку
была издана лишь одна небольшая книга (Гамбарян, Дукельская, 1955),
но по экологии крысы в ней имеются только отрывочные данные.
У зарубежных авторов нам известны сводки лишь по поведению се-
рой крысы, в основном в лабораторных условиях (Ватое, 1975) или па
отороженной территории (СаШоип, 1968). Имеются книги по ограниче-
нию численности синантронных трызунов, в том числе серых крыс
(Сыму, 1954; Меевал, 1984), а также сводка по эпидемиолотическому
значепию крыс и мышей (\УУаЦет, 1982).
Мы надеемся, что эта книга даст более полную и разностороннюю
характеристику иптереснейшего и высокооргапизованного трызуна. Мо-
нографический обзор современных знаний о серой крысе составит науч-
ную основу для проведения более рациональных и эффективных мер по
ограничению численности пасюка.
При составлении монографии мы пользовались ценными советами
проф. В. В. Кучерука, проф. Ю. А. Мясникова, д. б.н. С. Н. Варшавско-
тоикб. п. Н. Н. Мешковой. Нами были использованы неопубликован-
ные данные, любезно предоставленные нам зоологами Н. В. Ольковой,
В Я Антонюком и И. Р. Мерзликиным, которым мы выражаем искрен-
нюю благодарность.
Фотографии для обложки выполнены П. Роделем {Чехословакия).
Академик В. Е. Соколов
Доктор биологических паук Е. В. Карасева
Глава первая.
Систематика
Систематика рола Ва@лаз
Невозможно получить правильное представление о систематическом по-
ложении серой крысы ВаНиз погуеяеиз (Веткепроц®, 1769), структурной
и функциональной специфике этого вида, ето эволюционной судьбе без
обращения к систематике рода Вабиз в целом. В связи с этим мы сочли
необходимым обобщить имеющуюся к моменту написания монографии
литературу по этому вопросу. Такая компиляция оправдана еще и тем,
что зоологи-несистематики, а также специалисты по систематике других
трупп часто обращаются к устаревшим сводкам, будучи не в состоянии
следить за многочисленными узкоспециальными публикациями послед-
них лет, коренным образом изменившими таксономию сем. МанЧае и
особенно рода ВаМиз. Предлагаемое обобщение позволит избежать пута-
ницы в номенклатуре и основанных на ней неверных филогенетических
и зоотеотрафических выводов.
Описание рода ВаМлз осложнено тем, что пока не существует ого
таксономической ревизии в объеме мировой фаупы. Предложенные в
ходе ревизий таксонов более высокого ранга варианты (ЕПеттап, 1941,
1949; М1зопре, 1969) оказались неудовлетворительными. Обитание в
труднодоступных районах земного шара, рассеянность типовых экземп-
ляров по музсям четырех континентов, обилие форм при их слабой мор-
фологической дифференциации, номенклатурный хаос
— все это услож-
няет работу специалистов по рассматриваемой группе. До сих пор не
существует единого диагноза рода ВаНиз, и только начинают прояснять-
ся ото истинные таксономические границы. Тем не менее благодаря ин-
тенсивным исследованиям систоматиков ссылки на работы которых бу-
дут дапы ниже, можно надеяться, что в скором времени таксономичоская
ревизия рода Вайлз в объеме мировой фауны появится. Пока же для
того, чтобы сделать возможным описание, нам приходится использовать
предварительный список видов, обоснование которому дано в первом
разделе главы, посвященном истории изучения рода.
История изучения и востав рода
Европейцы были знакомы с крысами, по крайней мере с черной кры-
сой В. тайиз (Т,., 1758), задолго до выхода в свет в 1758 г. 10-го изда-
ния «Системы природы» Карла Линнея (Тлппаеиз, 1758), положившего
начало систематике живых организмов. Утверждение о том, что древние
греки и римляне пе знали крыс (см., например, Кесслер, 1866), ошибоч-
но. Данные археологии свидетельствуют об обратном (Антипина, Мас-
лов, 1984: Зсвлиу9, 1984). Отсутствие упоминаний о крысах в обширной
6
Предисловие
3. Экология серой крысы: 4-й съезд ВТО. М., 1986. Т. 3.
А. Серая крыса: в 2 т. М.: АН СССР, 1956.
.
.
5. Материалы по экологии и ограничению численности серой крысы:
вэт. М.: АН СССР, 1987.
Немалое внимание серой крысе уделялось на международных конфе-
ренциях по борьбе с вредными трызунами, проведенных в Кувейто
(Ргосеебзаз ой Ве Е зутрозиии оп Весеш Адуапсез ш Воде соо,
Кизай, 1982); Зесопа Зутрозиииа оЁ Вссей АЧуапсез п Водет Сошо,
Кимай, 1985).
.
Настоящий том подготовлен коллективом авторов: сотрудниками
ИЭМЭЖ им. А. Н. Северцова АН СССР, ВНИИДиС Мииздрава СССР,
Зоологического музея Тартуского университета и МГУ, ВНИИВГ, Ин-
ститута эпидемиологии и микробиологии (ИЭМ) им. Н. Ф. Гамалеи
АМН СССР, Института полиомиелита Минздрава СОСР, противочумной
станции (Одесса) и Института систематической и экологической биоло-
тии (Брно, Чехословакия).
й
.
Морфологии пасюка посвящен следующий том монографии, который
предполагается выпустить в недалеком будущем.
.
.
Эта книга о серой крысе является фактически первой капитальной
сводкой на русском языке по данному виду. У нас в стране по пасюку
была издана лишь одна небольшая книга (Гамбарян, Дукельская, 1955),
но по экологии крысы в ней имеются только отрывочные данные.
У зарубежных авторов нам известны сводки лишь по поведению се-
рой крысы, в основном в лабораторных условиях (Ватое, 1975) или па
отороженной территории (СаШоип, 1968). Имеются книги по ограниче-
нию численности синантронных трызунов, в том числе серых крыс
(Сыму, 1954; Меевал, 1984), а также сводка по эпидемиолотическому
значепию крыс и мышей (\УУаЦет, 1982).
Мы надеемся, что эта книга даст более полную и разностороннюю
характеристику иптереснейшего и высокооргапизованного трызуна. Мо-
нографический обзор современных знаний о серой крысе составит науч-
ную основу для проведения более рациональных и эффективных мер по
ограничению численности пасюка.
При составлении монографии мы пользовались ценными советами
проф. В. В. Кучерука, проф. Ю. А. Мясникова, д. б.н. С. Н. Варшавско-
тоикб. п. Н. Н. Мешковой. Нами были использованы неопубликован-
ные данные, любезно предоставленные нам зоологами Н. В. Ольковой,
В Я Антонюком и И. Р. Мерзликиным, которым мы выражаем искрен-
нюю благодарность.
Фотографии для обложки выполнены П. Роделем {Чехословакия).
Академик В. Е. Соколов
Доктор биологических паук Е. В. Карасева
Глава первая.
Систематика
Систематика рола Ва@лаз
Невозможно получить правильное представление о систематическом по-
ложении серой крысы ВаНиз погуеяеиз (Веткепроц®, 1769), структурной
и функциональной специфике этого вида, ето эволюционной судьбе без
обращения к систематике рода Вабиз в целом. В связи с этим мы сочли
необходимым обобщить имеющуюся к моменту написания монографии
литературу по этому вопросу. Такая компиляция оправдана еще и тем,
что зоологи-несистематики, а также специалисты по систематике других
трупп часто обращаются к устаревшим сводкам, будучи не в состоянии
следить за многочисленными узкоспециальными публикациями послед-
них лет, коренным образом изменившими таксономию сем. МанЧае и
особенно рода ВаМиз. Предлагаемое обобщение позволит избежать пута-
ницы в номенклатуре и основанных на ней неверных филогенетических
и зоотеотрафических выводов.
Описание рода ВаМлз осложнено тем, что пока не существует ого
таксономической ревизии в объеме мировой фаупы. Предложенные в
ходе ревизий таксонов более высокого ранга варианты (ЕПеттап, 1941,
1949; М1зопре, 1969) оказались неудовлетворительными. Обитание в
труднодоступных районах земного шара, рассеянность типовых экземп-
ляров по музсям четырех континентов, обилие форм при их слабой мор-
фологической дифференциации, номенклатурный хаос
— все это услож-
няет работу специалистов по рассматриваемой группе. До сих пор не
существует единого диагноза рода ВаНиз, и только начинают прояснять-
ся ото истинные таксономические границы. Тем не менее благодаря ин-
тенсивным исследованиям систоматиков ссылки на работы которых бу-
дут дапы ниже, можно надеяться, что в скором времени таксономичоская
ревизия рода Вайлз в объеме мировой фауны появится. Пока же для
того, чтобы сделать возможным описание, нам приходится использовать
предварительный список видов, обоснование которому дано в первом
разделе главы, посвященном истории изучения рода.
История изучения и востав рода
Европейцы были знакомы с крысами, по крайней мере с черной кры-
сой В. тайиз (Т,., 1758), задолго до выхода в свет в 1758 г. 10-го изда-
ния «Системы природы» Карла Линнея (Тлппаеиз, 1758), положившего
начало систематике живых организмов. Утверждение о том, что древние
греки и римляне пе знали крыс (см., например, Кесслер, 1866), ошибоч-
но. Данные археологии свидетельствуют об обратном (Антипина, Мас-
лов, 1984: Зсвлиу9, 1984). Отсутствие упоминаний о крысах в обширной
8
Глава первая
античной литературе объясняется тем, что в древнегреческом и латин-
ском языках не существовало специального названия для этих животных
{Нагь 1982). Такой донаучный номенклатурный нюанс привел впослед-
ствии к неверным зоотеотрафическим выводам. Считалось, что черные
крысы обитали в Европе до расцвета древнегреческой цивилизации, за-.
тем по непонятным причинам вымерли, а во время крестовых походов
(Х!-ХИТ вв.) были завезены вновь из Палестины (Вела,
;
16
вание крыс в свропейской литературе принадлежит, ви-
димо, Альберту Великому (ХПТ в.). Старший синоним черной крысы из
списка, составленного Эркслебеном (Егхефеп, 1777), олубликован в
1693 г., а серой крысы— в 1736 г. Черная крыса была единственпым
видом современного рода ВаМиз, известным Линнею, который поместил
ее в один из зшести родов грызунов (СНгез). Помимо Миз гаМаз, род
включал сще 16 видов, среди которых значились морская свинка, аль-
пийский сурок, египетский тушкапчик (Гипаече, 1758).
6
Линнеевскому роду Миз суждено было выполнять своеобразную.
функцию. В него включали все более или менее мышеподобные формы,
не подходящие под диагнозы других родов. Несмотря на то что род не-
однократно дробился, обработка материалов многочисленных заморских
экспедиций пополняла его все новыми и новыми формами, и к началу
ХХ в. он превратился в искусственное образование, насчитывающее сотни
разных по происхождению видов и объединепных лишь поверхностным
сходством. Возникла настоятельная необходимость ревизии рода.
Сложная структура рода Миз была отражена номенклатурно в конце
ХГХ в. Трусаром. В своем «Каталоге млекопитающих» он, в частности,
выделяет подрод Ерйпуз, к которому были отнесены 58 видов грызунов,
в том числе встречающиеся в Европе крысы (Тгоцеззат, 1881). ак был
выделен таксон, который сейчас мы знаем как род Кация. Новый под-
род стал быстро разрастаться. В 1897 г. он насчитывал 142 вида, а в
1904 г. 225 (Тгочезват 1897, 1904—1905).
В 1910 г. Миллер дал четкий диагноз роду Миз. В результате в его
составе осталась небольшая часть видов, соответствующих диагнозу,
а основная их масса вошла в новый род Ерппуз (МШег, 4910; Твотпав,
1910), который унаследовал функцию «таксономического мусорного ящи-,
ка, в который выбрасываются все виды, не подходящие ни в один из дру-
гих родов» (ЕПегтаи, 1949, с. 189). Так как в начале нашего 2ек
описание повых видов грызунов шло особенно интенсивно, то объем
р
быстро увеличивался.
то
свое современное название— ВаИлз, Холлис-
тер (НоШзэет, 1916а) обнаружил, что еще в 1803 г. петербургский уче-
ный Г. Фишер фон Вальдхайм вместо традиционного Миз применил по
отпошению к серой крысе родовое название ВаИиз (в книге допущена
опечатка
— ВаНиз; ЕзсБег уоп \УаВейи, 1803, с. 128). Сотласно пра-
випу приоритета, в дальнейшем для обозначения рода стал использовать-
ся старший синоним— ВаИлз. В качестве типового вида Холлистер ука-
зал В. десатапиз РаПаз (=В. погуемеиз ВегКепВой{) — единственный
Систематика
9
вид, упоминаемый Фишером (Но ет, 1946Ъ). Следует отметить, что на-
звание ВаМиз употреблялось еще в 1775 г. Фритем (Рызср), однако его
работа не была строго биноминальна.
Род ВаЙмз продолжал разрастаться и в 30-е тоды насчитывал сотни
видов. Возникла необходимость ревизии рода или хотя бы каталогиза-
ции описанных форм. Отсутствие сводки затрудняло выполнение регио-
нальных исследований и способствовало вагромождению и без того запу-
танной номенклатуры повыми синонимами. С другой стороны, проведе-
ние региональных исследований было необходимо, так как они облегча-
ли ревизию таксонов в объеме мировой фауны. Именно в этот наиболее
трудпый период была составлена сводка по Мимдае СССР А. И. Арги-
ропуло (4940).
Чрезвычайно сложную работу по ревизии отряда Водепыа выиолнил
Дж. Эллерман. Его фундаментальная сводка «Семейства п роды рецент-
ных грызунов» выходит в двух томах в 1940 п 1944 тг. Второй том це-
ликом посвящен Мимдае зепзи 1а40 (ЕПегтам, 1940, 1941). Позже Эл.
лерман, обработав новые материалы, несколько пересмотрел свой взгля-
ды и изменения и дополнения были изданы отдельным ломом (ЕПег-
тап, 1949).
Род Цашиз Эллермана замечателен своим объемом. Он включает
554 формы (275 из них получили видовой статус). Это был самый боль-
шой род в отряде трызупов. Дж. Симисоп назвал его «самым многооб-
разпым родом млекопитающих...» (Зпирзоп, 1945, с. 89). Род имел слож-
ную структуру. Он был разделен на 38 групп и подродов, охватывающих
не только индо-австралийские, но и африканские виды. В третьем томе
Эллерман объединяет группы индо-австралийских НаНлз в подроды.
В результате в составе рода было выделено 15 подродов, из них 7 ин-
до-австралийских (Нафиз, Э\епотуз, Махошуз, Аротуз, ГеороЧатуз,
Вегуплуз, Сгеттошуз) и 8 африканских (З4оНотуз, Ргаотуз, Нуотуз-
сиз, Рерпошуз, Муотуз, Мазютуз, Меаеатуз, ОсЬтотуз).
Эллерман проделал колоссальную работу по каталогизации и клас-
сификации огромного количества форм трызунов и еще большего коли-
чества назвапий. Но в его концепции род ВаМиз представлял собой
контломерат форм разного происхождения, скрывающий истинные фило-
генетические связи между видами, не дающий ничего для попимания
географии и эволюции входящих в него таксонов. Сведение сотен видов
В один не поддающийся диагностике род крайне затрудняло определение
видов, Такое положение не могло удовлетворить последователей Эллер-
мана, и с 1969 г. пачался процесс дробления рода.
Первым, кто пересмотрел систему Эллермапа, был бельгийский зоолог
Мизон. Используя в качестве таксономических признаков особенности
строения коренпых зубов и опираясь на теорию эволюции их структуры,
он предложил свою классификацию сом. МимЧае (М1воппе, 1969).
В составе рода ВаНаз Мизон оставил только 4 из 45 подродов Эллерма-
на: ВаЙлз, Бепотуз, Георо]4атуз, Вали.
Состав их лишь частично совпадаст с составом одноименных подро-
дов Эллермана. Африканские формы исключены полностью. В составе
8
Глава первая
античной литературе объясняется тем, что в древнегреческом и латин-
ском языках не существовало специального названия для этих животных
{Нагь 1982). Такой донаучный номенклатурный нюанс привел впослед-
ствии к неверным зоотеотрафическим выводам. Считалось, что черные
крысы обитали в Европе до расцвета древнегреческой цивилизации, за-.
тем по непонятным причинам вымерли, а во время крестовых походов
(Х!-ХИТ вв.) были завезены вновь из Палестины (Вела,
;
16
вание крыс в свропейской литературе принадлежит, ви-
димо, Альберту Великому (ХПТ в.). Старший синоним черной крысы из
списка, составленного Эркслебеном (Егхефеп, 1777), олубликован в
1693 г., а серой крысы— в 1736 г. Черная крыса была единственпым
видом современного рода ВаМиз, известным Линнею, который поместил
ее в один из зшести родов грызунов (СНгез). Помимо Миз гаМаз, род
включал сще 16 видов, среди которых значились морская свинка, аль-
пийский сурок, египетский тушкапчик (Гипаече, 1758).
6
Линнеевскому роду Миз суждено было выполнять своеобразную.
функцию. В него включали все более или менее мышеподобные формы,
не подходящие под диагнозы других родов. Несмотря на то что род не-
однократно дробился, обработка материалов многочисленных заморских
экспедиций пополняла его все новыми и новыми формами, и к началу
ХХ в. он превратился в искусственное образование, насчитывающее сотни
разных по происхождению видов и объединепных лишь поверхностным
сходством. Возникла настоятельная необходимость ревизии рода.
Сложная структура рода Миз была отражена номенклатурно в конце
ХГХ в. Трусаром. В своем «Каталоге млекопитающих» он, в частности,
выделяет подрод Ерйпуз, к которому были отнесены 58 видов грызунов,
в том числе встречающиеся в Европе крысы (Тгоцеззат, 1881). ак был
выделен таксон, который сейчас мы знаем как род Кация. Новый под-
род стал быстро разрастаться. В 1897 г. он насчитывал 142 вида, а в
1904 г. 225 (Тгочезват 1897, 1904—1905).
В 1910 г. Миллер дал четкий диагноз роду Миз. В результате в его
составе осталась небольшая часть видов, соответствующих диагнозу,
а основная их масса вошла в новый род Ерппуз (МШег, 4910; Твотпав,
1910), который унаследовал функцию «таксономического мусорного ящи-,
ка, в который выбрасываются все виды, не подходящие ни в один из дру-
гих родов» (ЕПегтаи, 1949, с. 189). Так как в начале нашего 2ек
описание повых видов грызунов шло особенно интенсивно, то объем
р
быстро увеличивался.
то
свое современное название— ВаИлз, Холлис-
тер (НоШзэет, 1916а) обнаружил, что еще в 1803 г. петербургский уче-
ный Г. Фишер фон Вальдхайм вместо традиционного Миз применил по
отпошению к серой крысе родовое название ВаИиз (в книге допущена
опечатка
— ВаНиз; ЕзсБег уоп \УаВейи, 1803, с. 128). Сотласно пра-
випу приоритета, в дальнейшем для обозначения рода стал использовать-
ся старший синоним— ВаИлз. В качестве типового вида Холлистер ука-
зал В. десатапиз РаПаз (=В. погуемеиз ВегКепВой{) — единственный
Систематика
9
вид, упоминаемый Фишером (Но ет, 1946Ъ). Следует отметить, что на-
звание ВаМиз употреблялось еще в 1775 г. Фритем (Рызср), однако его
работа не была строго биноминальна.
Род ВаЙмз продолжал разрастаться и в 30-е тоды насчитывал сотни
видов. Возникла необходимость ревизии рода или хотя бы каталогиза-
ции описанных форм. Отсутствие сводки затрудняло выполнение регио-
нальных исследований и способствовало вагромождению и без того запу-
танной номенклатуры повыми синонимами. С другой стороны, проведе-
ние региональных исследований было необходимо, так как они облегча-
ли ревизию таксонов в объеме мировой фауны. Именно в этот наиболее
трудпый период была составлена сводка по Мимдае СССР А. И. Арги-
ропуло (4940).
Чрезвычайно сложную работу по ревизии отряда Водепыа выиолнил
Дж. Эллерман. Его фундаментальная сводка «Семейства п роды рецент-
ных грызунов» выходит в двух томах в 1940 п 1944 тг. Второй том це-
ликом посвящен Мимдае зепзи 1а40 (ЕПегтам, 1940, 1941). Позже Эл.
лерман, обработав новые материалы, несколько пересмотрел свой взгля-
ды и изменения и дополнения были изданы отдельным ломом (ЕПег-
тап, 1949).
Род Цашиз Эллермана замечателен своим объемом. Он включает
554 формы (275 из них получили видовой статус). Это был самый боль-
шой род в отряде трызупов. Дж. Симисоп назвал его «самым многооб-
разпым родом млекопитающих...» (Зпирзоп, 1945, с. 89). Род имел слож-
ную структуру. Он был разделен на 38 групп и подродов, охватывающих
не только индо-австралийские, но и африканские виды. В третьем томе
Эллерман объединяет группы индо-австралийских НаНлз в подроды.
В результате в составе рода было выделено 15 подродов, из них 7 ин-
до-австралийских (Нафиз, Э\епотуз, Махошуз, Аротуз, ГеороЧатуз,
Вегуплуз, Сгеттошуз) и 8 африканских (З4оНотуз, Ргаотуз, Нуотуз-
сиз, Рерпошуз, Муотуз, Мазютуз, Меаеатуз, ОсЬтотуз).
Эллерман проделал колоссальную работу по каталогизации и клас-
сификации огромного количества форм трызунов и еще большего коли-
чества назвапий. Но в его концепции род ВаМиз представлял собой
контломерат форм разного происхождения, скрывающий истинные фило-
генетические связи между видами, не дающий ничего для попимания
географии и эволюции входящих в него таксонов. Сведение сотен видов
В один не поддающийся диагностике род крайне затрудняло определение
видов, Такое положение не могло удовлетворить последователей Эллер-
мана, и с 1969 г. пачался процесс дробления рода.
Первым, кто пересмотрел систему Эллермапа, был бельгийский зоолог
Мизон. Используя в качестве таксономических признаков особенности
строения коренпых зубов и опираясь на теорию эволюции их структуры,
он предложил свою классификацию сом. МимЧае (М1воппе, 1969).
В составе рода ВаНаз Мизон оставил только 4 из 45 подродов Эллерма-
на: ВаЙлз, Бепотуз, Георо]4атуз, Вали.
Состав их лишь частично совпадаст с составом одноименных подро-
дов Эллермана. Африканские формы исключены полностью. В составе
10
Глава первая
рода Мизон упоминает 41 вид, но этот список неполон, так как автор
‘пе ставил своей целью ревизию видового состава.
В 70-е п 80-е годы выходит целый цикл публикаций Массера и Мас-
сера с соавторами по систематике мурид, изменивших трактовку рода
ВаНниз еще более. Знакомство с музейными коллекциями Америки и Ев-
ропы, личные сборы в Индонезии позволили Массеру, с одной стороны,
описать ряд новых видов мурид, в том числе два вида Вакиз (В. оз5009
и В цауажетз15), с другой стороны, еще более сузить таксономиче-
ские границы рода Вайиз, выделив из его состава ряд новых родов
(ЗгИавкатуз, Горо!датуз, Мгиуещег, Вегуплуз, Кадагвапотиуз, Зпи-
Чэшуз, Котодотуз) (Миззет, Вова@!, 1980; Миззег, 1981а, 19815; Миззег,
МсусошЪ, 1983).
Исследования Масссра приблизили нас к пониманию истинного раз-
пообразия мурид, позволили выявить скрытые ранее филогенетические
связи, привели к важным зоогеографическим обобщениям. В тот же пе-
риод появились две региональные ревизии, посвященные австралийским
(Тау1от, Ногпег, 1973) и новогвипейским (Тауог её а1., 1982) ВаИлз.
Таковы основные вехи истории систематики рода Вайлз. С момента
своего возникновения в качестве подрода Ерйпуз в 1881 г. до 19441 г.
число видов в нем увеличилось примерно в пять раз (с 58 до 275). По
современным представлениям в роде около 50 видов (Мизвег, МечсотЬ,
1983.) Конечно, менялось не только число видов, но и качественный во-
став рода. Можно с уверенностью прогнозировать, что оба эти параметра
будут меняться и впредь, особенно интенсивно в ближайшие годы. Будет
продолжаться описание новых видов, пересматриваться таксономический
статус уже известных форм, не исключено дальнойттее дробление рода.
Так, например, Массер высказал предположение, что индо-малайские и
австралийские Вайиз имеют разное происхождение, а их сходство в03-
никло копвергентно (Маззег, 1984с, с. 169).
.
Систематическое описание рода НВаЙиз дапное в настоящей главе
относится к видам, перечисленным в приведенном ниже списке. Он взят
из публикации Массера и Ньюкомба (Миззег, МезсошЬ, 1988) и допол-
пен двумя новыми видами.
В
ВаМав
43. В. о Полет! НШ, 1960
(о Мпеог МелусошЬ, 1983, 1985; Миз-
14. В. БоЁпат! (МаёзсШе, 1901)
|
15. В. Рообог\уегН Свазеп, 4939
зег, Неапеу, 1985)
Боорег\ ее,
4. В. адизбаз Зоду, 1940
16. в. Домепя би ее 1985
2. В. аппайда1е! (Вопвоее, 4903)
уВ.У,
1871)
3. ео
(ВоБлзоп её К1033,
19; В. шавиз (МШег, 1909)
4. В. Башепя!з (ТБотав, 1894)
5.н.ео в ат)
5. В. Бгопаеиз (Нодёзот, 1845)
22. В. шшаогепз:з (ТБоваз, 1898)
6. В. сегашисяз ТЬоштаз. 1920
23. В. шогаах (ТВошав, 190/)
1. П. оарь оз чоду, т
24. В. тлогоба{спз18 КеПоя, 1945
8. Н. спвашия 8
25, В. паНуЦаЫз (Твотаз, 1889)
.В.ем 1
Ут,
:м
10. В. ехйапз (Рез[е, 1848)
26. В. поро (Твошаз, 1906)
11. В. ГеНесаз (ТВотаз, 1920)
27. В. 01195, {Нодезоп, 1845)
12. В. Газегрез (\У/афегВоцзе, 1839)
28. В. погуер1сиз (Вегкепфой, 1769)
Систематика
44
29. В. поуаетеае Тау]ог её Са1аБу,
37. В. “еп Витек, 1935
1982
38. В. зюсиз (МШег, 1902)
30. В. озвоодт Миззег её МезусотЬ, 1985
39. В. 1а\На\!еоз1з Моззег её Неапеу,
34. В. рабиагит (йеерог, 1869)
1985
32. В. ргаеюг (ТНотваз, 1888)
40. В. Нотап1саз (МШег, 1900)
33. В. гаймз (Глипаеиз, 1758)
41, В. таппеу! (Топаз, 1904)
ЗА. В. ЧеБагдзот Тафе, 1949
42. В. пхке\жатеиз (Зафрала, 1903)
35. В. “ККииелз!з Ницов, 1949
43. В. усгесипдиз (ТБошаз, 1904)
36. В. зог диз (Сош@, 1858)
44, В. хашВитгиз (Стау, 1867)
В приведенный выше список не включены формы нсяеного таксоно-
мического статуса, отнесенные Массером к группе райпагиал (В. Батгав,
Биатгезсейз, зила|игелз!з, 1а$ае, фа, шещам!) и комплексу хащпогаз
{В. шагтозигиз, Гасефиз, омешаНз, реГагаз, Бовфапиз, Гогатизпеиз).
Некоторые исследователи склоппы рассматривать азиатскую форму
черной крысы (2п=42) как самостоятельный вид В. Фагди (Тешк,
1880) (Разепг её а|., 1982; ВаустэбосК её а1., 1983), другие предпочитают
не делать вывод до таксономической ревизии многочисленных форм
В. гамиз (Миззег, СаЦНа, 1982). Нами принята позиция последних.
Спорен статус трех австралийских форм: зот@Чиз, соПем и уШоз3-
зпичз. На основании морфологического сходства все они вместе с двумя
новогнинейскими формами (сезы1 и атапца) были объединены в один
политипический вид В. зог@1!Чиз (Тау]ог, Ногпег, 4973; Тау1ог её а1.,
1982, 1983).
Между тем оказалось, что австралийские формы отличаются друг
от друга хромосомными наборами: зог@9з — 2п=32, соПеф — 2и=42,
у от — 21=50 и дают гибридов с пониженной плодовитостьо.
Это послужило основанием для признания трех самостоятельных видов:
В. зог19а, В. соПея, В. уШоязапииз (ВауегзюсКк её а1., 1975, 1977).
Биохимические исследования показали, что все три формы очень
близки генетически, однако полученные дистанции пс позволяют опре-
деленно высказаться в пользу какого-либо одного номенклатурного ре-
шения. Здесь принята точка зрепия «объедипителей».
Номенклатура
Род Вамиз Е1зсВег уоп У/’аЮВет, 1803 — настоящие крысы! Е1зеБег
уоп \УаЧВеш С. Газ МайбопаПаазеии 9ег МайгсеземМеме ти Раг!8.
Ва. 2. Етапки а,/М., 8. 128 (опечатка — Виз).
1758. Маз [лппасиз, Зузета Мабагае, е4. Х, %. 1, р. 61.
1814. Мизсшаз Вайбпезаие, Ргесз Песопуегез, Тгауапх Бот1ю]081-
Чисз, р. 13.
1867. Асатротуз Огау (но не Те5зоп, 1842 или Токода, 1944), Ргос.
7001. Зос. Тюп4., р. 597.
+ Принятое в русском языке название рода «крысы» (Соколов, 1984; Громов, 1981)
недостаточно, так как оно применяется ко всем крупным муридам других родов,
грызунам других семейств (например, бамбуковые крысы, мешотчатые крысы) и
Даже представителям других отрядов млекопитающих (сумчатые крысы).
10
Глава первая
рода Мизон упоминает 41 вид, но этот список неполон, так как автор
‘пе ставил своей целью ревизию видового состава.
В 70-е п 80-е годы выходит целый цикл публикаций Массера и Мас-
сера с соавторами по систематике мурид, изменивших трактовку рода
ВаНниз еще более. Знакомство с музейными коллекциями Америки и Ев-
ропы, личные сборы в Индонезии позволили Массеру, с одной стороны,
описать ряд новых видов мурид, в том числе два вида Вакиз (В. оз5009
и В цауажетз15), с другой стороны, еще более сузить таксономиче-
ские границы рода Вайиз, выделив из его состава ряд новых родов
(ЗгИавкатуз, Горо!датуз, Мгиуещег, Вегуплуз, Кадагвапотиуз, Зпи-
Чэшуз, Котодотуз) (Миззет, Вова@!, 1980; Миззег, 1981а, 19815; Миззег,
МсусошЪ, 1983).
Исследования Масссра приблизили нас к пониманию истинного раз-
пообразия мурид, позволили выявить скрытые ранее филогенетические
связи, привели к важным зоогеографическим обобщениям. В тот же пе-
риод появились две региональные ревизии, посвященные австралийским
(Тау1от, Ногпег, 1973) и новогвипейским (Тауог её а1., 1982) ВаИлз.
Таковы основные вехи истории систематики рода Вайлз. С момента
своего возникновения в качестве подрода Ерйпуз в 1881 г. до 19441 г.
число видов в нем увеличилось примерно в пять раз (с 58 до 275). По
современным представлениям в роде около 50 видов (Мизвег, МечсотЬ,
1983.) Конечно, менялось не только число видов, но и качественный во-
став рода. Можно с уверенностью прогнозировать, что оба эти параметра
будут меняться и впредь, особенно интенсивно в ближайшие годы. Будет
продолжаться описание новых видов, пересматриваться таксономический
статус уже известных форм, не исключено дальнойттее дробление рода.
Так, например, Массер высказал предположение, что индо-малайские и
австралийские Вайиз имеют разное происхождение, а их сходство в03-
никло копвергентно (Маззег, 1984с, с. 169).
.
Систематическое описание рода НВаЙиз дапное в настоящей главе
относится к видам, перечисленным в приведенном ниже списке. Он взят
из публикации Массера и Ньюкомба (Миззег, МезсошЬ, 1988) и допол-
пен двумя новыми видами.
В
ВаМав
43. В. о Полет! НШ, 1960
(о Мпеог МелусошЬ, 1983, 1985; Миз-
14. В. БоЁпат! (МаёзсШе, 1901)
|
15. В. Рообог\уегН Свазеп, 4939
зег, Неапеу, 1985)
Боорег\ ее,
4. В. адизбаз Зоду, 1940
16. в. Домепя би ее 1985
2. В. аппайда1е! (Вопвоее, 4903)
уВ.У,
1871)
3. ео
(ВоБлзоп её К1033,
19; В. шавиз (МШег, 1909)
4. В. Башепя!з (ТБотав, 1894)
5.н.ео в ат)
5. В. Бгопаеиз (Нодёзот, 1845)
22. В. шшаогепз:з (ТБоваз, 1898)
6. В. сегашисяз ТЬоштаз. 1920
23. В. шогаах (ТВошав, 190/)
1. П. оарь оз чоду, т
24. В. тлогоба{спз18 КеПоя, 1945
8. Н. спвашия 8
25, В. паНуЦаЫз (Твотаз, 1889)
.В.ем 1
Ут,
:м
10. В. ехйапз (Рез[е, 1848)
26. В. поро (Твошаз, 1906)
11. В. ГеНесаз (ТВотаз, 1920)
27. В. 01195, {Нодезоп, 1845)
12. В. Газегрез (\У/афегВоцзе, 1839)
28. В. погуер1сиз (Вегкепфой, 1769)
Систематика
44
29. В. поуаетеае Тау]ог её Са1аБу,
37. В. “еп Витек, 1935
1982
38. В. зюсиз (МШег, 1902)
30. В. озвоодт Миззег её МезусотЬ, 1985
39. В. 1а\На\!еоз1з Моззег её Неапеу,
34. В. рабиагит (йеерог, 1869)
1985
32. В. ргаеюг (ТНотваз, 1888)
40. В. Нотап1саз (МШег, 1900)
33. В. гаймз (Глипаеиз, 1758)
41, В. таппеу! (Топаз, 1904)
ЗА. В. ЧеБагдзот Тафе, 1949
42. В. пхке\жатеиз (Зафрала, 1903)
35. В. “ККииелз!з Ницов, 1949
43. В. усгесипдиз (ТБошаз, 1904)
36. В. зог диз (Сош@, 1858)
44, В. хашВитгиз (Стау, 1867)
В приведенный выше список не включены формы нсяеного таксоно-
мического статуса, отнесенные Массером к группе райпагиал (В. Батгав,
Биатгезсейз, зила|игелз!з, 1а$ае, фа, шещам!) и комплексу хащпогаз
{В. шагтозигиз, Гасефиз, омешаНз, реГагаз, Бовфапиз, Гогатизпеиз).
Некоторые исследователи склоппы рассматривать азиатскую форму
черной крысы (2п=42) как самостоятельный вид В. Фагди (Тешк,
1880) (Разепг её а|., 1982; ВаустэбосК её а1., 1983), другие предпочитают
не делать вывод до таксономической ревизии многочисленных форм
В. гамиз (Миззег, СаЦНа, 1982). Нами принята позиция последних.
Спорен статус трех австралийских форм: зот@Чиз, соПем и уШоз3-
зпичз. На основании морфологического сходства все они вместе с двумя
новогнинейскими формами (сезы1 и атапца) были объединены в один
политипический вид В. зог@1!Чиз (Тау]ог, Ногпег, 4973; Тау1ог её а1.,
1982, 1983).
Между тем оказалось, что австралийские формы отличаются друг
от друга хромосомными наборами: зог@9з — 2п=32, соПеф — 2и=42,
у от — 21=50 и дают гибридов с пониженной плодовитостьо.
Это послужило основанием для признания трех самостоятельных видов:
В. зог19а, В. соПея, В. уШоязапииз (ВауегзюсКк её а1., 1975, 1977).
Биохимические исследования показали, что все три формы очень
близки генетически, однако полученные дистанции пс позволяют опре-
деленно высказаться в пользу какого-либо одного номенклатурного ре-
шения. Здесь принята точка зрепия «объедипителей».
Номенклатура
Род Вамиз Е1зсВег уоп У/’аЮВет, 1803 — настоящие крысы! Е1зеБег
уоп \УаЧВеш С. Газ МайбопаПаазеии 9ег МайгсеземМеме ти Раг!8.
Ва. 2. Етапки а,/М., 8. 128 (опечатка — Виз).
1758. Маз [лппасиз, Зузета Мабагае, е4. Х, %. 1, р. 61.
1814. Мизсшаз Вайбпезаие, Ргесз Песопуегез, Тгауапх Бот1ю]081-
Чисз, р. 13.
1867. Асатротуз Огау (но не Те5зоп, 1842 или Токода, 1944), Ргос.
7001. Зос. Тюп4., р. 597.
+ Принятое в русском языке название рода «крысы» (Соколов, 1984; Громов, 1981)
недостаточно, так как оно применяется ко всем крупным муридам других родов,
грызунам других семейств (например, бамбуковые крысы, мешотчатые крысы) и
Даже представителям других отрядов млекопитающих (сумчатые крысы).
42
Глава нервая
1881. Ерппуз Ттопеззатё, ВаЦ. бос. Ефа4ез Зс1. Апяетз,1. 10, р. 140 (подрод
рода Миз).
1910. Ззепотуз ТВотаз, Апп. Маз. Маё. Н134., зег. 8, уо|. 6, р. 506.
1947. Ертотуз ЕШоЬ А свеск-В5 оЁ таштиа]з 0? е Мог Атетсав
сопбпете..., бирр!., р. 41 (поправка названия Ерилуз Тгомеззаг).
1941. Сьчзющуз Зоду, ТтечЫа, у01. 18, No 2, р. 260.
4941. Стопотуз Зоду, ТтечЩа, у01. 18. No 2, р. 260.
4941. Сеготуз боду, Тгеаа, у01. 18, No 2, р. 260.
4941. МоШсотуз Зоду, 'Тгешыа, 01. 18, No 2, р. 260.
1944. Осютуз боду (но не Твотаз, 1920), ТгепБа, у01. 18, No 2, р. 261.
1941. РаШоштуз боду, ТгецЫа, уо|. 18, No 2, р. 260.
1941. Таегошуз боу, ТгепЫа, уо|. 18, No 2, р. 260.
Тин рода— Миз десотапоз РаПав=М. погуереиз Вегкепнои.
В некоторых публикациях в качестве типа рода выступает черная
крыса (Миз табов) (Та, 1936; Громов, 1984). В 1912 г. Дж. Миллер
указал па черную крысу как типовой вид рода Ерииуз (тип по после-
дующему обозпачению}. Однако после номенклатурных преобразований
(НоШзег, 1946а, 19165) Холлистер пересмотрел свои взгляды в пользу
серой крысы (МШЬг, 1923, с. 428). Тейт считает В. гаМлз типом по абоо-
лютной тавтономии. Между тем согласно пункту «@» статьи 68-го
Международного кодекса зоологической номенклатуры (4966) виды
тавтонимы становятся типами рода в том случае, если они «первоначаль-
но включены во вновь установленный номинальный род». Черная крыса
не упоминается в оригинальном описании рода, следовательно, она не
может быть типом по абсолютной тавтопимии. Единственным видом,
указанным Фишером фон Вальдхаймом, была серая крыса (Миз Чеса-
тапиз РаПаз, 1778=М. погусе1саз Вегкепвоп, 1769), что позволяет
считать ее типом рода по монотинии.
ОЭписание
Диагноз. Удовлетворительното диагноза рода, позволяющего иден-
тифицировать животных в любой части родового ареала, пока не сущеет-
вует.
Морфология. Мы не претендуем на исчерпывающую полноту‘
данного ниже морфологического описания рода. Но оно позволяет соста-
вить общее представление о роде, так как охватывает большую часть
указанных в списке видов. Иногда нам придется прибегать к описанию
одной типичной в смысле строения форме, т. е. той, признаки которой
характерны для большинства других видов. В качестве такой «типичной
Вайиз» (но пе тинового вида!) мы вслед за Массером и Ньюкомбом
(Миззег, Мезусошь, 1983) используем В. гайаз Фатай (Теомок, 1980).
Внешний вид. Грызуны мелких и средних размеров. Средняя
длина тела самого маленького вида, В. ехщапз 4145 мм (Тайланд,
МатзваП, 1977), масса тела самцов и самок того же вида в возрасте
20 нед в среднем соответственно 72,0 и 54,2 г (Гавайские острова, Нтаг-
Систематика
13
(а, Мазз, 1974). Максимальная длина тела представителей рода, видимо,
не превышает 270 мм, а масса тела 500 г. У трети видов средняя длина
тела превыптает 200 мм (виды группы раПпагит, В. ]оЫепз1з, В. погуе-
21спз, В. хаюигиз). Лишь у нескольких видов она меныне 150 мм
(В. ехщапз, В. 1озеа, В. шоБе). Большая же часть видов имест среднюю
длину тела от 150 до 200 мм. В зависимости от образа жизни туловище
крыс может быть либо компактным, вальковатым, либо стройным, вы-
тянутым. Так, на { см длины тела черных крыс из Западного Берлина
приходится 0,9 г массы тела, тогда как на 1 см длины серой крысы из
Бонна — 6,4 г (высчитано по таблицам 92 и 96: ВесКег, 1978).
Хвост длинный, чешуйчатый, с редкими короткими волосками, не
образующими на конце кисточки, его относительная длипа составляет
от 60 до 130%. У большей части видов этот показатель превышает 90%.
Меньший средний индекс хвоста имеют: В. Ра3с1рез, В. Иамепя5,
В. Татеов, В. шогдах, В. пИй9из, В. погуеюеиз, В. 0300091, В. ргаефог,
ЗВ. зог@1 из, В. мешь В. звоеиз, В. фауНа\метз1з, В. бллтеу!. Относитель-
ная длина хвоста весьма изменчива даже в пределах одной популяции.
Так, по нашим данным, у взрослых серых крыс Таллиннекого зоопарка
она колеблется от 70 до 90%. С возрастом у молодых животных этот.
показатель возрастает. Тем не менее относительная длина хвоста от“
ражает образ жизни вида
— древесные и лазающие формы имеют осо-
бенпо длинные хвосты.
Ушные раковины овальные, снаружи покрыты редкими, короткими
волосками, через которые просвечивает кожа. Длина их составляет
примерно 50% от длины черепа, незначительно варьируя от вида к
виду в ту или иную сторону в зависимости от их образа кизпи, среды
обитания и особенностей слуха.
‚
Стопа имеет вытянутую форму и превышает по размерам кисть.
Длипа стопы составляет 18—25% от длины тела. Конечности пятина-
лые, но на кисти первый (внутрепний) палец в разной степени реду-
цирован. На вентральной поверхности кисти и стопы имеются пальце-
вые, межпальцевые, пястные и плюсневые подошвенные бугорки. Сте-
пень их развития связана с образом жизни крыс. Самые большие и
высокие бугорки имеют древесные виды. Характерно наличие на стопе
четырех межпальцевых и двух плюеневых бугорков. Внутренний плюс-
новой бугорок вытянут и имеет форму запятой. Пальцы снабжены
когтями, форма которых также зависит от образа жизни крыс. У лазаю-
щих форм опи короткие, сильно изогнутые, острые, у роющих
— длин-
ные, слабо изогнутые. Дорсальпые поверхности кисти и стопы частично
покрыты короткими редкими волосками, подошвы голые.
Меховой покров широко варьирует по густоте, длине, наличию и
соотношению различных типов волос. Он бывает очень густым, особен-
но у горных видов (В. и сВагазоп, В. шоре, В. 0360041), или менее
густым, с длинными направляющими остями (В. тасеаг, В. епбапиз,
В. хапВигиав, В. зКПиепзз, В. гаМлз) или без пих (В. паНуцаи$),
тладким (В. уетеситдаз, В. агрепбуещет, В. {а\Ца\1е1$13) или пушис-
тым (В. збююме, В. аппарпдае!, В. 103еа). Многие виды имеют иглы
42
Глава нервая
1881. Ерппуз Ттопеззатё, ВаЦ. бос. Ефа4ез Зс1. Апяетз,1. 10, р. 140 (подрод
рода Миз).
1910. Ззепотуз ТВотаз, Апп. Маз. Маё. Н134., зег. 8, уо|. 6, р. 506.
1947. Ертотуз ЕШоЬ А свеск-В5 оЁ таштиа]з 0? е Мог Атетсав
сопбпете..., бирр!., р. 41 (поправка названия Ерилуз Тгомеззаг).
1941. Сьчзющуз Зоду, ТтечЫа, у01. 18, No 2, р. 260.
4941. Стопотуз Зоду, ТтечЩа, у01. 18. No 2, р. 260.
4941. Сеготуз боду, Тгеаа, у01. 18, No 2, р. 260.
4941. МоШсотуз Зоду, 'Тгешыа, 01. 18, No 2, р. 260.
1944. Осютуз боду (но не Твотаз, 1920), ТгепБа, у01. 18, No 2, р. 261.
1941. РаШоштуз боду, ТгецЫа, уо|. 18, No 2, р. 260.
1941. Таегошуз боу, ТгепЫа, уо|. 18, No 2, р. 260.
Тин рода— Миз десотапоз РаПав=М. погуереиз Вегкепнои.
В некоторых публикациях в качестве типа рода выступает черная
крыса (Миз табов) (Та, 1936; Громов, 1984). В 1912 г. Дж. Миллер
указал па черную крысу как типовой вид рода Ерииуз (тип по после-
дующему обозпачению}. Однако после номенклатурных преобразований
(НоШзег, 1946а, 19165) Холлистер пересмотрел свои взгляды в пользу
серой крысы (МШЬг, 1923, с. 428). Тейт считает В. гаМлз типом по абоо-
лютной тавтономии. Между тем согласно пункту «@» статьи 68-го
Международного кодекса зоологической номенклатуры (4966) виды
тавтонимы становятся типами рода в том случае, если они «первоначаль-
но включены во вновь установленный номинальный род». Черная крыса
не упоминается в оригинальном описании рода, следовательно, она не
может быть типом по абсолютной тавтопимии. Единственным видом,
указанным Фишером фон Вальдхаймом, была серая крыса (Миз Чеса-
тапиз РаПаз, 1778=М. погусе1саз Вегкепвоп, 1769), что позволяет
считать ее типом рода по монотинии.
ОЭписание
Диагноз. Удовлетворительното диагноза рода, позволяющего иден-
тифицировать животных в любой части родового ареала, пока не сущеет-
вует.
Морфология. Мы не претендуем на исчерпывающую полноту‘
данного ниже морфологического описания рода. Но оно позволяет соста-
вить общее представление о роде, так как охватывает большую часть
указанных в списке видов. Иногда нам придется прибегать к описанию
одной типичной в смысле строения форме, т. е. той, признаки которой
характерны для большинства других видов. В качестве такой «типичной
Вайиз» (но пе тинового вида!) мы вслед за Массером и Ньюкомбом
(Миззег, Мезусошь, 1983) используем В. гайаз Фатай (Теомок, 1980).
Внешний вид. Грызуны мелких и средних размеров. Средняя
длина тела самого маленького вида, В. ехщапз 4145 мм (Тайланд,
МатзваП, 1977), масса тела самцов и самок того же вида в возрасте
20 нед в среднем соответственно 72,0 и 54,2 г (Гавайские острова, Нтаг-
Систематика
13
(а, Мазз, 1974). Максимальная длина тела представителей рода, видимо,
не превышает 270 мм, а масса тела 500 г. У трети видов средняя длина
тела превыптает 200 мм (виды группы раПпагит, В. ]оЫепз1з, В. погуе-
21спз, В. хаюигиз). Лишь у нескольких видов она меныне 150 мм
(В. ехщапз, В. 1озеа, В. шоБе). Большая же часть видов имест среднюю
длину тела от 150 до 200 мм. В зависимости от образа жизни туловище
крыс может быть либо компактным, вальковатым, либо стройным, вы-
тянутым. Так, на { см длины тела черных крыс из Западного Берлина
приходится 0,9 г массы тела, тогда как на 1 см длины серой крысы из
Бонна — 6,4 г (высчитано по таблицам 92 и 96: ВесКег, 1978).
Хвост длинный, чешуйчатый, с редкими короткими волосками, не
образующими на конце кисточки, его относительная длипа составляет
от 60 до 130%. У большей части видов этот показатель превышает 90%.
Меньший средний индекс хвоста имеют: В. Ра3с1рез, В. Иамепя5,
В. Татеов, В. шогдах, В. пИй9из, В. погуеюеиз, В. 0300091, В. ргаефог,
ЗВ. зог@1 из, В. мешь В. звоеиз, В. фауНа\метз1з, В. бллтеу!. Относитель-
ная длина хвоста весьма изменчива даже в пределах одной популяции.
Так, по нашим данным, у взрослых серых крыс Таллиннекого зоопарка
она колеблется от 70 до 90%. С возрастом у молодых животных этот.
показатель возрастает. Тем не менее относительная длина хвоста от“
ражает образ жизни вида
— древесные и лазающие формы имеют осо-
бенпо длинные хвосты.
Ушные раковины овальные, снаружи покрыты редкими, короткими
волосками, через которые просвечивает кожа. Длина их составляет
примерно 50% от длины черепа, незначительно варьируя от вида к
виду в ту или иную сторону в зависимости от их образа кизпи, среды
обитания и особенностей слуха.
‚
Стопа имеет вытянутую форму и превышает по размерам кисть.
Длипа стопы составляет 18—25% от длины тела. Конечности пятина-
лые, но на кисти первый (внутрепний) палец в разной степени реду-
цирован. На вентральной поверхности кисти и стопы имеются пальце-
вые, межпальцевые, пястные и плюсневые подошвенные бугорки. Сте-
пень их развития связана с образом жизни крыс. Самые большие и
высокие бугорки имеют древесные виды. Характерно наличие на стопе
четырех межпальцевых и двух плюеневых бугорков. Внутренний плюс-
новой бугорок вытянут и имеет форму запятой. Пальцы снабжены
когтями, форма которых также зависит от образа жизни крыс. У лазаю-
щих форм опи короткие, сильно изогнутые, острые, у роющих
— длин-
ные, слабо изогнутые. Дорсальпые поверхности кисти и стопы частично
покрыты короткими редкими волосками, подошвы голые.
Меховой покров широко варьирует по густоте, длине, наличию и
соотношению различных типов волос. Он бывает очень густым, особен-
но у горных видов (В. и сВагазоп, В. шоре, В. 0360041), или менее
густым, с длинными направляющими остями (В. тасеаг, В. епбапиз,
В. хапВигиав, В. зКПиепзз, В. гаМлз) или без пих (В. паНуцаи$),
тладким (В. уетеситдаз, В. агрепбуещет, В. {а\Ца\1е1$13) или пушис-
тым (В. збююме, В. аппарпдае!, В. 103еа). Многие виды имеют иглы
44
Глава первая
(В. ргаеог, В. шотдах, В. 1виоориз, В. поуаеритеае, В. ]01е1515, Ц. зог-
919ив, В. ехШалз) или иглоподобные гнущиеся щетинки (В. 10зва,
В. агрепуетег, В. эеш, В. 01615). Часть видов имеет иглы только
на спине, другие на спине и брюшке. Количество игл с возрастом уве-
личивается. У новотвинейских Н. ехи]апз иглы имеются только
у животпых из равнинных и предгорных популяций, тогда как обитаю-
щие выше 1500 м крысы лишены их (Тау]ог с% а1., 1982).
Окраска изменчива, причем часто значительно варьирует внутри вида.
Окраска верха может быть черпой (табиз, раПпатит), серо-коричневой
различных оттенков от темного до светло-коричневого, рыжей. Брюшко
может быть чисто-белым, белым с кремовым оттенком, грязно-белым,
пепельно-серым, темно-серым с охриетым налетом или без него, темным
ссро-коричневым. Брюшко, как правило, окрашено светлее спины и бо-
ков. Переход от цвета спины к цвету брюшка может быть постепенным
или резким. Ушные раковины обычно окрашены под цвет спины или
телесного цвета. Покрытые волосом части кистей и стоп могут быть под
цвет спины или белыми, оголенные части телесного цвета. Хвост бывает
а) равномерно пигментирован по всей длине, сверху и снизу (В. габиз),
б) двухцветпым: темным сверху и светлым снизу (В. погуееаз),
в} темным в проксимальной части и белым или телесным дистально.
(В. Боовсг\уегй). Относительно часто встречаются цветовые аберрации.
Черные особи отмечались в природе у В. еха]апз, В. 1озеа, И. погуер1стз,
В. гаи, белые у В. агдепйуетег, В. погуе&1ств, желтые у В. агсепй-
уетиет, В. погусвиз. У ряда видов на груди (ипогда на брюшке) встре-
чаются белые либо охристые пятна. Белые грудные пятна зарсгистриро-
ваны У В. погуео1сия, В. габиз, В. ргаеют, В. шотдах, В. зе, В. уеге-
сипдиз, охристые у В. шогдах, В. ]оепз1з.
Количество сосков у самок варьирует от 4 (В. уегесчидиз чапдечзет!)
до 12 (табл. 1). Крысы равных видов, имеющие одинаковое количество
сосков, нередко отличаются их расположением.
Таблица 1. Количество сосков у самок некоторых видов рода ВаМлз
Чиело сосков
4—6
6
6—8
8.
10
40—12
42
уетесиидиз |плоБе эены Боодегуеот
фаГаеив! погуей{- |зот9198
1ецсориз |0 1еп31$ | 5401615
раПпагит |018
01495
аппапда[е?
мотает
агрепйуещег
исБагазот
таз
эт еп818
ргаеюг
Тозеа
Бтоппеиз
910913
0570001
{меК
с ат100$
поуаеритеае
ехщапз
{а На\1еп13
Систематика
15
Краниологические признаки. Пределы изменчивости наи-
большей длины черепа можно представить, исходя из средних значений
для самого маленького вида и одного из наиболее крупных. Так, для
В. ехшатз эта величина составляет 34,9 мм [о-в Новая Гвинея (Тау]от
её а1., 1982)], а для В. еуегей
— 53,8 мм [о-в Динагат, Филиппины
(Неапеу, ВаБог, 1982) |.
Череп имеет вытянутую Форму. Скуловая ширипа примерно вдвое
меньше его наибольшей длины (46—52%). Длина носовых костей со-
ставляет 35—44 от наибольшей длины черепа. Пределы изменчивости
закных в таксономическом отношении костных слуховых барабанов
(БаПа ‘ушрап!) и резцового отверстия (Готатеп зпезуит) совпадают
по величине и составляют 13—20% от наибольтей длины черена. Шири-
на резцового отверстия составляет 27—45 от его длины. Относитель-
пая альвеолярная длина верхних коренных зубов 16-20%.
Массер и Ньюкомб считают характерными для Вамяз следующие
особенности строения черепа и зубов В. газ Фаг@н (полный перечень
см.: Миззег, МезсотаЬ, 1983, с. 345—349).
1. Резцы покрыты оранжевой эмалью и не имеют бороздок. Верхние
резцы либо ортодонтные, либо опистодоптные, но не проодонтные.
2. На вссх верхних молярах отсутствует бугорок 47°.
3. Буторки МЫз и &2Ы5 на первых верхних молярах обычно отсутет-
вуют.
4. Позади бугорка 46 на нсех порвых и вторых верхних молярах от-
сутствует дополнительный бугорок. Позади бугорков %4 и (3 на всех мо-
лярах нет гребней.
5. Второй и третий нижние моляры имеют переднегубной бугорок.
6. На каждом первом нижнем моляре переднеязычный бугорок боль-
ше переднегубного. Образуемая ими петля слегка уже, чем следующая за
ней.
7. Интерорбитальная и посторбитальная области, а также дорсо-лате-
ральные части черепной коробки очерчены выпуклыми гребнями.
8. Межтеменная кость большая. Она формирует большую часть кры-
ши черепа в затылочной области.
9. Поверхности черенной коробки между скуловыми отростками ви
сочной кости и височными гребнями вертикальны или почти вертикальны.
40. Резцовое отверстие длинное. Его задний край достигает уровня
переднего края первых верхних моляров.
14. Задний край костного пёба выдается за линию, соединяющую зад-
ний край последних верхних моляров.
19. Костные слуховые барабаны круглые и по отношению к разме-
рам черепа большие. Евстахисвы трубы игирокие и короткие.
13. Верхние и нижние моляры брахиодонтные. В верхнем зубном
ряду первый моляр налегает на второй, а второй на третий. В нижнем
ряду третий моляр налегает на второй, а второй на первый. Моляры в
сравнении с площадью костного нёба и мандибулы маленькие.
= Номенклатура зубов мурид дана на рис. 1.
44
Глава первая
(В. ргаеог, В. шотдах, В. 1виоориз, В. поуаеритеае, В. ]01е1515, Ц. зог-
919ив, В. ехШалз) или иглоподобные гнущиеся щетинки (В. 10зва,
В. агрепуетег, В. эеш, В. 01615). Часть видов имеет иглы только
на спине, другие на спине и брюшке. Количество игл с возрастом уве-
личивается. У новотвинейских Н. ехи]апз иглы имеются только
у животпых из равнинных и предгорных популяций, тогда как обитаю-
щие выше 1500 м крысы лишены их (Тау]ог с% а1., 1982).
Окраска изменчива, причем часто значительно варьирует внутри вида.
Окраска верха может быть черпой (табиз, раПпатит), серо-коричневой
различных оттенков от темного до светло-коричневого, рыжей. Брюшко
может быть чисто-белым, белым с кремовым оттенком, грязно-белым,
пепельно-серым, темно-серым с охриетым налетом или без него, темным
ссро-коричневым. Брюшко, как правило, окрашено светлее спины и бо-
ков. Переход от цвета спины к цвету брюшка может быть постепенным
или резким. Ушные раковины обычно окрашены под цвет спины или
телесного цвета. Покрытые волосом части кистей и стоп могут быть под
цвет спины или белыми, оголенные части телесного цвета. Хвост бывает
а) равномерно пигментирован по всей длине, сверху и снизу (В. габиз),
б) двухцветпым: темным сверху и светлым снизу (В. погуееаз),
в} темным в проксимальной части и белым или телесным дистально.
(В. Боовсг\уегй). Относительно часто встречаются цветовые аберрации.
Черные особи отмечались в природе у В. еха]апз, В. 1озеа, И. погуер1стз,
В. гаи, белые у В. агдепйуетег, В. погуе&1ств, желтые у В. агсепй-
уетиет, В. погусвиз. У ряда видов на груди (ипогда на брюшке) встре-
чаются белые либо охристые пятна. Белые грудные пятна зарсгистриро-
ваны У В. погуео1сия, В. габиз, В. ргаеют, В. шотдах, В. зе, В. уеге-
сипдиз, охристые у В. шогдах, В. ]оепз1з.
Количество сосков у самок варьирует от 4 (В. уегесчидиз чапдечзет!)
до 12 (табл. 1). Крысы равных видов, имеющие одинаковое количество
сосков, нередко отличаются их расположением.
Таблица 1. Количество сосков у самок некоторых видов рода ВаМлз
Чиело сосков
4—6
6
6—8
8.
10
40—12
42
уетесиидиз |плоБе эены Боодегуеот
фаГаеив! погуей{- |зот9198
1ецсориз |0 1еп31$ | 5401615
раПпагит |018
01495
аппапда[е?
мотает
агрепйуещег
исБагазот
таз
эт еп818
ргаеюг
Тозеа
Бтоппеиз
910913
0570001
{меК
с ат100$
поуаеритеае
ехщапз
{а На\1еп13
Систематика
15
Краниологические признаки. Пределы изменчивости наи-
большей длины черепа можно представить, исходя из средних значений
для самого маленького вида и одного из наиболее крупных. Так, для
В. ехшатз эта величина составляет 34,9 мм [о-в Новая Гвинея (Тау]от
её а1., 1982)], а для В. еуегей
— 53,8 мм [о-в Динагат, Филиппины
(Неапеу, ВаБог, 1982) |.
Череп имеет вытянутую Форму. Скуловая ширипа примерно вдвое
меньше его наибольшей длины (46—52%). Длина носовых костей со-
ставляет 35—44 от наибольшей длины черепа. Пределы изменчивости
закных в таксономическом отношении костных слуховых барабанов
(БаПа ‘ушрап!) и резцового отверстия (Готатеп зпезуит) совпадают
по величине и составляют 13—20% от наибольтей длины черена. Шири-
на резцового отверстия составляет 27—45 от его длины. Относитель-
пая альвеолярная длина верхних коренных зубов 16-20%.
Массер и Ньюкомб считают характерными для Вамяз следующие
особенности строения черепа и зубов В. газ Фаг@н (полный перечень
см.: Миззег, МезсотаЬ, 1983, с. 345—349).
1. Резцы покрыты оранжевой эмалью и не имеют бороздок. Верхние
резцы либо ортодонтные, либо опистодоптные, но не проодонтные.
2. На вссх верхних молярах отсутствует бугорок 47°.
3. Буторки МЫз и &2Ы5 на первых верхних молярах обычно отсутет-
вуют.
4. Позади бугорка 46 на нсех порвых и вторых верхних молярах от-
сутствует дополнительный бугорок. Позади бугорков %4 и (3 на всех мо-
лярах нет гребней.
5. Второй и третий нижние моляры имеют переднегубной бугорок.
6. На каждом первом нижнем моляре переднеязычный бугорок боль-
ше переднегубного. Образуемая ими петля слегка уже, чем следующая за
ней.
7. Интерорбитальная и посторбитальная области, а также дорсо-лате-
ральные части черепной коробки очерчены выпуклыми гребнями.
8. Межтеменная кость большая. Она формирует большую часть кры-
ши черепа в затылочной области.
9. Поверхности черенной коробки между скуловыми отростками ви
сочной кости и височными гребнями вертикальны или почти вертикальны.
40. Резцовое отверстие длинное. Его задний край достигает уровня
переднего края первых верхних моляров.
14. Задний край костного пёба выдается за линию, соединяющую зад-
ний край последних верхних моляров.
19. Костные слуховые барабаны круглые и по отношению к разме-
рам черепа большие. Евстахисвы трубы игирокие и короткие.
13. Верхние и нижние моляры брахиодонтные. В верхнем зубном
ряду первый моляр налегает на второй, а второй на третий. В нижнем
ряду третий моляр налегает на второй, а второй на первый. Моляры в
сравнении с площадью костного нёба и мандибулы маленькие.
= Номенклатура зубов мурид дана на рис. 1.
16
Глава первая
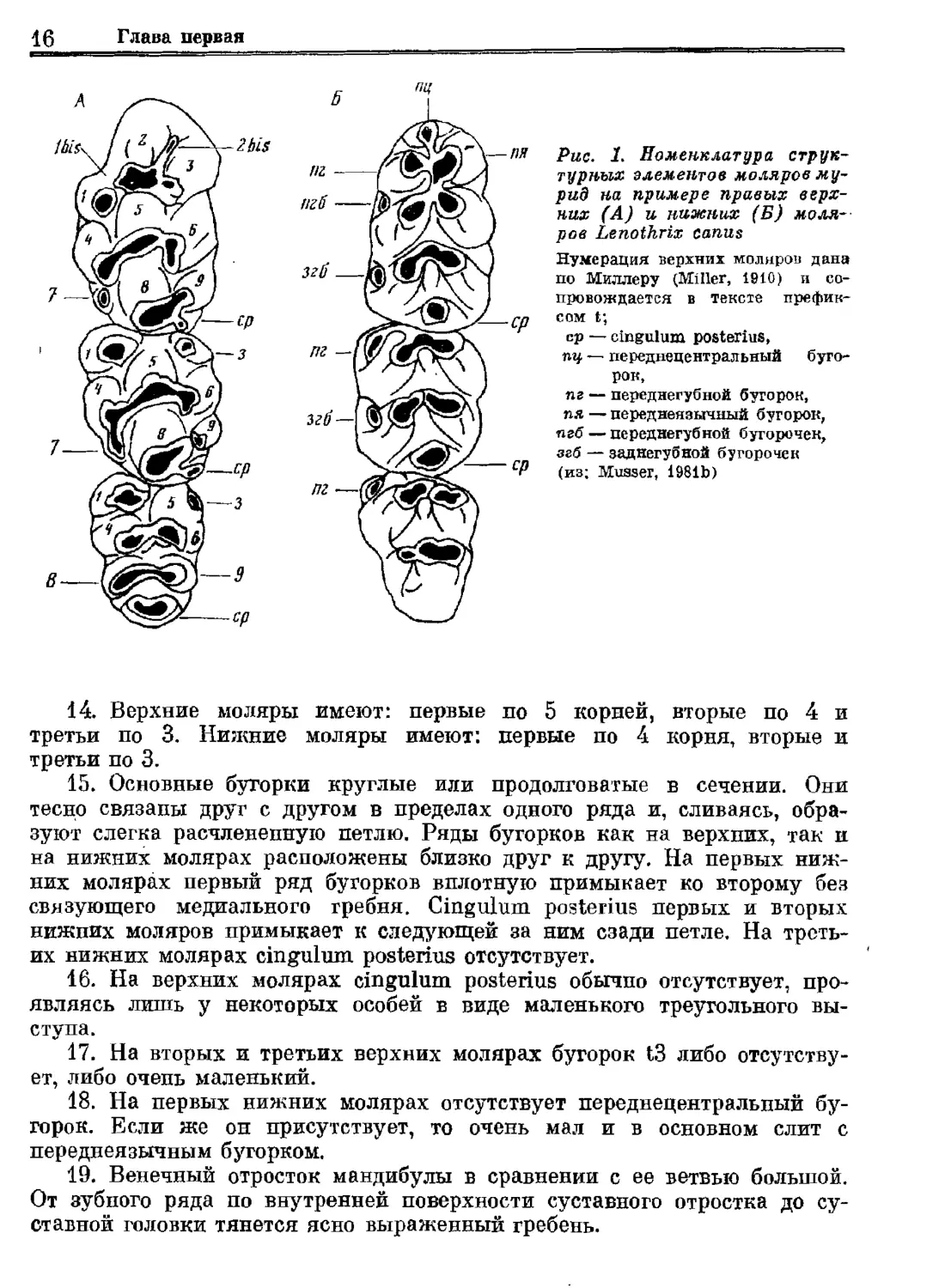
Рис. 1. Номенклатура струк-
турных элементов моляров му-
рид на примере правых верх-
ров Гепо фт сапиз
Нумерация верхних моляров дана
по Миллеру (МШег, 1910) и со-
провождается в тексте префик-
сом в;
ср — спеиция розбегйаз,
пу — переднецентральный буго-
рок,
пг — переднегубной бугорок,
пя — переднеязычный бугорок,
пгб — переднегубной бугорочек,
згб — задиегубной бугорочек
(из; Миязег, 19815)
а 14. Верхние моляры имеют: первые по 5 корней, вторые по 4 и
етьи по 3. Нижние моляры имеют: п: первые по 4
Я СЗ.
р
о 4 корня, вторые и
ве 15. Основные бугорки круглые или продолговатые в сечении. Они
но связаны друг с другом в пределах одного ряда и, сливаясь, обра-
зуют слегка расчлененную петлю. Ряды бугорков как на верхних, так и
на нижних молярах расположены близко друг к другу. На первых ниж-
пих молярах первый ряд бугорков вплотную примыкает ко второму без
ь язующего медиального гребня. Степи розбеггаз первых и вторых
ижних моляров примыкает к следующей за ним сзади петле. На треть-
их Ниних молярах сотеиПии розбегиз отсутствует.
я а верхних молярах от розбеиз$ обычно отсутствует, про-
ишь у некоторых особей в виде малень
ля
д
кого треугольного вы-
17. На вторых и тр
.
етьих верхних молярах бугорок {3 л
г
з
иб
-
ет, либо очень маленький.
Ут
о отоутотву
м 18. На первых нижних молярах отсутствует переднецентральный бу-
рок. Юсли же он присутствует, то очень мал и в основном слит с
переднеязычным бугорком.
о 9. Венечный отросток мандибулы в сравнении с ее ветвью большой.
т зубного ряда по внутренней поверхности суставного отростка до су-
ставной головки тянется ясно выраженный гребень.
них (А) ц нижних (Б) ноля--
Систематика
17
20. На первых нижних молярах переднегубной бугорочек обычно от-
сутствует. Заднегубные бугорочки присутствуют на первом и втором
нижних молярах.
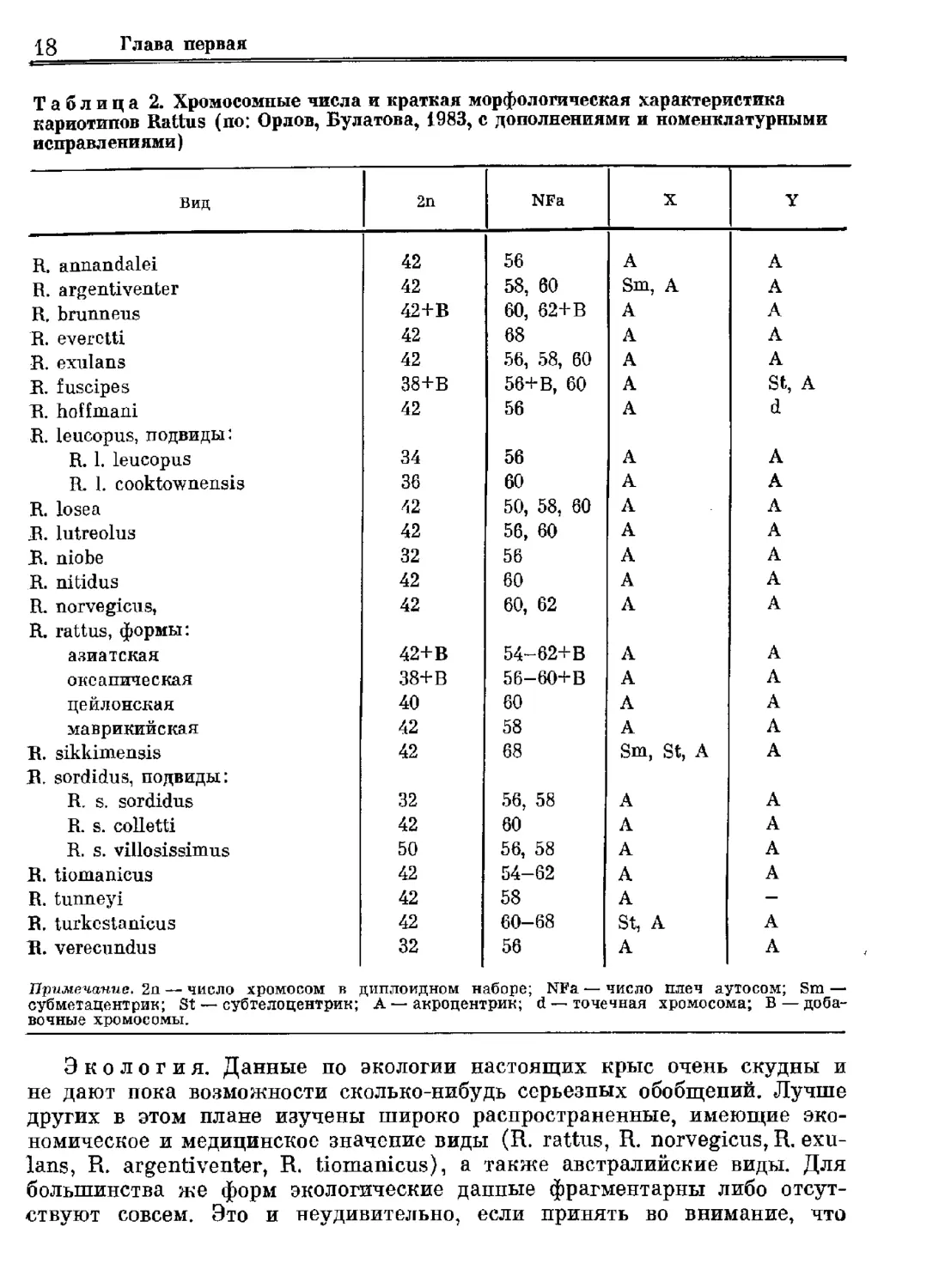
Кариология. Крысы рода ВаМиз довольно интенсивно исследова-
лись цитогенетиками. Результаты многочисленных работ неоднократно
обобщались (Вашал, ЗВатта, 1977; Уоз9а, 1980; Орлов, Булатова, 1983;
Бекасова, 1984).
В настоящее время известны кариотипы 20 видов. Большинство форм
имеет в диплоидном наборе 42 хромосомы. У некоторых Форм число
хромосом меньше: 32, 34, 36, 38, 40. Только у В. зог@ Чи У 08158 Пиз
число хромосом больше 42 (2и=50). Число хромосомных плеч колеб-
лется от 50 до 68. У трех видов обнаружены дополнительные хромосомы
(В. Бгиппеиз, В. Илзерез, В. таМиз). Половые хромосомы чаще всего
акроцентрические (табл. 2).
Предполагается, что исходный диплоидный набор Ваз состоял из
А? хромосом (18 пар акроцентриков, 7 пар метацентриков и акроцентри-
ческие Х и У хромосомы) (Ватал, бВагша, 1977; Уоз9а, 1980, 1985).
В дальнейшем хромосомная эволюция шла за счет инверсий, робертсо-
новских слияний и робертсоновских разделений. Различия в морфологии
хромосом, возникшие вследствие ипверсий, отражены в варьировании
плеч хромосом. Путем робертсононских слияний возникли кариотипы
цейлонских и океанических черных крыс, В. #No1зо:рез, В. Тепсориз,
В. зог@Чиз зот@ из. Робертсоновскими разделениями объясняется воз-
никновение хромосомных наборов маврикийской черной крысы и В. зог-
Чаи У Позитив. Иосида указывает на широкое распространение па-
раллелизмов в хромосомной эволюции настоящих крыс. Черная крыса,
названная Баверстоком «цитогенетическим чудом», имеет много карио-
типических форм, как правило, имеющих географическую приуроченность.
Часто хромосомные наборы других видов оказываются морфологически
идентичными какой-либо из этих форм (Уоз а, 1985).
В каждом конкретном случае трудно установить, объясняется ли
сходство кариотипов общностью происхождения или параллельной эво-
люцией, что осложняет филогенетическую интерпретацию данных по
морфологии хромосом. Выход из положения дает сочетание этих данных
с данными о внутреннем строении хромосом, классической морфологией
и биохимией.
В цитогенетическом и эволюционном плане особенно интересны таксо-
пы В. таназ и В. з0т@@ из. Биохимические, тибридологические и морфо-
логические исследования показали, что все хромосомные формы в преде-
лах каждого из этих таксонов, несмотря на кариотипические различия,
состоят в более тесном родстве друг с другом, чем с каким-либо другим
видом. Вместе с тем некоторые из хромосомных форм представляют со-
бой зарождающиеся виды (Тауюог, Ногпег, 1973; Тауог ей а1., 1983;
Вауегзоск еЁ а|., 1981, 1983). В результате хромосомных мутаций часто
возникает полная или частичная репродуктивная изоляция, способст-
вующая дивергепции вида.
16
Глава первая
Рис. 1. Номенклатура струк-
турных элементов моляров му-
рид на примере правых верх-
ров Гепо фт сапиз
Нумерация верхних моляров дана
по Миллеру (МШег, 1910) и со-
провождается в тексте префик-
сом в;
ср — спеиция розбегйаз,
пу — переднецентральный буго-
рок,
пг — переднегубной бугорок,
пя — переднеязычный бугорок,
пгб — переднегубной бугорочек,
згб — задиегубной бугорочек
(из; Миязег, 19815)
а 14. Верхние моляры имеют: первые по 5 корней, вторые по 4 и
етьи по 3. Нижние моляры имеют: п: первые по 4
Я СЗ.
р
о 4 корня, вторые и
ве 15. Основные бугорки круглые или продолговатые в сечении. Они
но связаны друг с другом в пределах одного ряда и, сливаясь, обра-
зуют слегка расчлененную петлю. Ряды бугорков как на верхних, так и
на нижних молярах расположены близко друг к другу. На первых ниж-
пих молярах первый ряд бугорков вплотную примыкает ко второму без
ь язующего медиального гребня. Степи розбеггаз первых и вторых
ижних моляров примыкает к следующей за ним сзади петле. На треть-
их Ниних молярах сотеиПии розбегиз отсутствует.
я а верхних молярах от розбеиз$ обычно отсутствует, про-
ишь у некоторых особей в виде малень
ля
д
кого треугольного вы-
17. На вторых и тр
.
етьих верхних молярах бугорок {3 л
г
з
иб
-
ет, либо очень маленький.
Ут
о отоутотву
м 18. На первых нижних молярах отсутствует переднецентральный бу-
рок. Юсли же он присутствует, то очень мал и в основном слит с
переднеязычным бугорком.
о 9. Венечный отросток мандибулы в сравнении с ее ветвью большой.
т зубного ряда по внутренней поверхности суставного отростка до су-
ставной головки тянется ясно выраженный гребень.
них (А) ц нижних (Б) ноля--
Систематика
17
20. На первых нижних молярах переднегубной бугорочек обычно от-
сутствует. Заднегубные бугорочки присутствуют на первом и втором
нижних молярах.
Кариология. Крысы рода ВаМиз довольно интенсивно исследова-
лись цитогенетиками. Результаты многочисленных работ неоднократно
обобщались (Вашал, ЗВатта, 1977; Уоз9а, 1980; Орлов, Булатова, 1983;
Бекасова, 1984).
В настоящее время известны кариотипы 20 видов. Большинство форм
имеет в диплоидном наборе 42 хромосомы. У некоторых Форм число
хромосом меньше: 32, 34, 36, 38, 40. Только у В. зог@ Чи У 08158 Пиз
число хромосом больше 42 (2и=50). Число хромосомных плеч колеб-
лется от 50 до 68. У трех видов обнаружены дополнительные хромосомы
(В. Бгиппеиз, В. Илзерез, В. таМиз). Половые хромосомы чаще всего
акроцентрические (табл. 2).
Предполагается, что исходный диплоидный набор Ваз состоял из
А? хромосом (18 пар акроцентриков, 7 пар метацентриков и акроцентри-
ческие Х и У хромосомы) (Ватал, бВагша, 1977; Уоз9а, 1980, 1985).
В дальнейшем хромосомная эволюция шла за счет инверсий, робертсо-
новских слияний и робертсоновских разделений. Различия в морфологии
хромосом, возникшие вследствие ипверсий, отражены в варьировании
плеч хромосом. Путем робертсононских слияний возникли кариотипы
цейлонских и океанических черных крыс, В. #No1зо:рез, В. Тепсориз,
В. зог@Чиз зот@ из. Робертсоновскими разделениями объясняется воз-
никновение хромосомных наборов маврикийской черной крысы и В. зог-
Чаи У Позитив. Иосида указывает на широкое распространение па-
раллелизмов в хромосомной эволюции настоящих крыс. Черная крыса,
названная Баверстоком «цитогенетическим чудом», имеет много карио-
типических форм, как правило, имеющих географическую приуроченность.
Часто хромосомные наборы других видов оказываются морфологически
идентичными какой-либо из этих форм (Уоз а, 1985).
В каждом конкретном случае трудно установить, объясняется ли
сходство кариотипов общностью происхождения или параллельной эво-
люцией, что осложняет филогенетическую интерпретацию данных по
морфологии хромосом. Выход из положения дает сочетание этих данных
с данными о внутреннем строении хромосом, классической морфологией
и биохимией.
В цитогенетическом и эволюционном плане особенно интересны таксо-
пы В. таназ и В. з0т@@ из. Биохимические, тибридологические и морфо-
логические исследования показали, что все хромосомные формы в преде-
лах каждого из этих таксонов, несмотря на кариотипические различия,
состоят в более тесном родстве друг с другом, чем с каким-либо другим
видом. Вместе с тем некоторые из хромосомных форм представляют со-
бой зарождающиеся виды (Тауюог, Ногпег, 1973; Тауог ей а1., 1983;
Вауегзоск еЁ а|., 1981, 1983). В результате хромосомных мутаций часто
возникает полная или частичная репродуктивная изоляция, способст-
вующая дивергепции вида.
18
Глава первая
Систематика
19
Таблица 2. Хромосомные числа и краткая морфологическая характеристика
кариотипов КаЙиз (по: Орлов, Булатова, 1983, с дополнениями и номенклатурными
исправлениями)
Вид
20
МЕа
х
У
В. аппапда!е
42
56
А
А
В. аглепиуещег
42
58, 60
Зш, А
А
В. Бгиппейз
42+В
60,62+В |А
А
В. еуегой1
42
68
А
А
В. ехшаоз
42
56,58,60 |А
А
В. озорез
38+В
56-В,60|А
ЗА
В. воЙшаш
42
56
А
9
В. 1еисориз, подвиды:
В. 1. 1еиеориз
34
56
А
А
В. 1. сооКю\пепзз
36
60
А
А
В. 10зеа
12
50,58,60 |А
А
В. Пигео[из
42
56, 60
А
А
ЗВ. шоре
32
56
А
А
В. в а9диз
42
60
А
А
В. погуе1елв,
42
60, 62
А
А
В. гаNoлз, формы:
азиатская
А2+В
54-62+В | А
А
оксапическая
38+ В
56-60+В | А
А
цейлонская
40
60
А
А
маврикийская
42
58
А
А
В. эККивепз1$
42
68
Зт,8%А
А
В. зог919$, подвиды:
В. в. зогда4из
32
56, 58
А
А
В. в. соНе
42
60
А
А
В. 3. Шо 8818
50
56, 58
А
А
В. Иоташетз
42
54—62
А
А
В. борлеу!
42
58
А
_
В. иткезаюеиз
42
60—68
З,А
А
ЗВ. уегеспидиз
32
56
А
А
Примечание. 2п — число хромосом в диплоидном наборе; Ма
— число плеч аутосом; эт —
субметацентрик, 5% — субтелоцентрик; А — акроцентрик; @ — точечная хромосома; В — доба-
вочные хромосомы.
Экология. Данные по экологии настоящих крыс очень скудны и
не дают пока возможности сколько-нибудь ссрьезпых обобщений. Лучше
других в этом плане изучены широко распространенные, имеющие эко-
номическое и медицинское значение виды (В. га биз, В. погуез1еле, В. ехи-
]апз, В. агеепуетщег, В. Нотатсиз), а также австралийские виды. Для
большинства же форм эколотические даппые фрагментарны либо отсут-
ствуют совсем. Это и неудивительно, если принять во внимание, что
часть видов известна лишь по одному или немногим коллекционным эк-
земплярам.
Обычно настоящих крыс рассматривают как группу, у членов которой
отсутствуют крайние формы специализации. Это действительно так, но
даже внутри такой деспециализированной группы отчетливо видна адап-
тивная радиация, идущая по традиционным для всего животного мира
паправлепиям. В качестве примера укажем несколько характерных для
рода экологических типов.
1. Древесные формы. Ирысы, принадлежащие к этому типу,
имеют вытянутое тонкое туловище. Хвост длиннее тела и у некоторых
видов цепкий. Подотвенные бугорки болышие и выпуклые. Когти не-
большие, сильно изогпутые, острые. К этому типу принадлежат В. ап-
папда]е1, В. гайлз и некоторые другие виды.
:
2. Наземные формы. Имеют более плотное телосложение, чем
древесные. Хвост нецепкий. Его длина короче длины тела или равна
ей. Подошвенные бугорки небольшие, плоские. К этому экологическому
типу относится значительное число видов. В качестве примера можно
привести равнинных В. 10зеа и В. астепЯхегиег и обитателей высокото-
рий Новой Гвинеи В. вПамепз!5, В. шофе, В асвагазот.
.
3. Роющие формы. Большинство видов настоящих крыс в той
или иной степени связаны с норами и их рытьем. Специализирован в
этом направлении вид © о-ва Рождества В. памуйа6з. Эти крупные
крысы имеют вальковатое тело, болыние кисти и ступни, вооруженные
широкими и длинными, особенло па передних лапах, когтями. Хвост
короткий и толстый.
4. Околоводные формы. Тесно связаны с водой В. погуерлсаз
и В. Гатео]аз. Однако их морфологическая специализация в этом направ-
лении но зашла далеко. У серой крысы кожа между пальцами задних
лап образует небольшую перепонку.
Известно, что есть виды, специализирующиеся на поедании богатых
клетчаткой вегетативных частей растений (В. ]итео]лз). Другие поеда-
ют преимущественно их плоды и семена (В. таМлз). Многие виды упо-
требляют в пищу животные корма. По данным Уаттса, в питании В. 1е1-
сориз и В. мзефез удельный вес насекомых составляет 20—90 (Уайз,
1977). ивотные корма охотно используют в пищу В. агрепбуещет,
В. ехШапз, В. потуер1сиз, В. тайиз. Все они в разной степени питаются
как беспозвоночными, так и позвоночными.
Удивительный пример хищничества описан Кеплером (Керег, 1967).
Самые маленькие из крыс, В. ехшапз, на Гавайских островах не только
поедали яйца и птенцов морских птиц, но и нападали ночью на спя-
щих взрослых альбатросов (О1отейеа пииииа3), выгрызая мясо у них
на спине. Согласно Неплеру, из-за крыс ежегодно гибло 1—2,5% пого-
ловья этих огромных птиц. На основании его наблюдений В. схи]апв
была обвинена даже в исчезповении целого ряда островных видов, в том
числе гигантского моа (Е1епупр, 1969).
Наличие и соотношение различных видов кормов в рационе конкрет-
ного вида меняются в зависимости от их наличия в природе, сезона года,
18
Глава первая
Систематика
19
Таблица 2. Хромосомные числа и краткая морфологическая характеристика
кариотипов КаЙиз (по: Орлов, Булатова, 1983, с дополнениями и номенклатурными
исправлениями)
Вид
20
МЕа
х
У
В. аппапда!е
42
56
А
А
В. аглепиуещег
42
58, 60
Зш, А
А
В. Бгиппейз
42+В
60,62+В |А
А
В. еуегой1
42
68
А
А
В. ехшаоз
42
56,58,60 |А
А
В. озорез
38+В
56-В,60|А
ЗА
В. воЙшаш
42
56
А
9
В. 1еисориз, подвиды:
В. 1. 1еиеориз
34
56
А
А
В. 1. сооКю\пепзз
36
60
А
А
В. 10зеа
12
50,58,60 |А
А
В. Пигео[из
42
56, 60
А
А
ЗВ. шоре
32
56
А
А
В. в а9диз
42
60
А
А
В. погуе1елв,
42
60, 62
А
А
В. гаNoлз, формы:
азиатская
А2+В
54-62+В | А
А
оксапическая
38+ В
56-60+В | А
А
цейлонская
40
60
А
А
маврикийская
42
58
А
А
В. эККивепз1$
42
68
Зт,8%А
А
В. зог919$, подвиды:
В. в. зогда4из
32
56, 58
А
А
В. в. соНе
42
60
А
А
В. 3. Шо 8818
50
56, 58
А
А
В. Иоташетз
42
54—62
А
А
В. борлеу!
42
58
А
_
В. иткезаюеиз
42
60—68
З,А
А
ЗВ. уегеспидиз
32
56
А
А
Примечание. 2п — число хромосом в диплоидном наборе; Ма
— число плеч аутосом; эт —
субметацентрик, 5% — субтелоцентрик; А — акроцентрик; @ — точечная хромосома; В — доба-
вочные хромосомы.
Экология. Данные по экологии настоящих крыс очень скудны и
не дают пока возможности сколько-нибудь ссрьезпых обобщений. Лучше
других в этом плане изучены широко распространенные, имеющие эко-
номическое и медицинское значение виды (В. га биз, В. погуез1еле, В. ехи-
]апз, В. агеепуетщег, В. Нотатсиз), а также австралийские виды. Для
большинства же форм эколотические даппые фрагментарны либо отсут-
ствуют совсем. Это и неудивительно, если принять во внимание, что
часть видов известна лишь по одному или немногим коллекционным эк-
земплярам.
Обычно настоящих крыс рассматривают как группу, у членов которой
отсутствуют крайние формы специализации. Это действительно так, но
даже внутри такой деспециализированной группы отчетливо видна адап-
тивная радиация, идущая по традиционным для всего животного мира
паправлепиям. В качестве примера укажем несколько характерных для
рода экологических типов.
1. Древесные формы. Ирысы, принадлежащие к этому типу,
имеют вытянутое тонкое туловище. Хвост длиннее тела и у некоторых
видов цепкий. Подотвенные бугорки болышие и выпуклые. Когти не-
большие, сильно изогпутые, острые. К этому типу принадлежат В. ап-
папда]е1, В. гайлз и некоторые другие виды.
:
2. Наземные формы. Имеют более плотное телосложение, чем
древесные. Хвост нецепкий. Его длина короче длины тела или равна
ей. Подошвенные бугорки небольшие, плоские. К этому экологическому
типу относится значительное число видов. В качестве примера можно
привести равнинных В. 10зеа и В. астепЯхегиег и обитателей высокото-
рий Новой Гвинеи В. вПамепз!5, В. шофе, В асвагазот.
.
3. Роющие формы. Большинство видов настоящих крыс в той
или иной степени связаны с норами и их рытьем. Специализирован в
этом направлении вид © о-ва Рождества В. памуйа6з. Эти крупные
крысы имеют вальковатое тело, болыние кисти и ступни, вооруженные
широкими и длинными, особенло па передних лапах, когтями. Хвост
короткий и толстый.
4. Околоводные формы. Тесно связаны с водой В. погуерлсаз
и В. Гатео]аз. Однако их морфологическая специализация в этом направ-
лении но зашла далеко. У серой крысы кожа между пальцами задних
лап образует небольшую перепонку.
Известно, что есть виды, специализирующиеся на поедании богатых
клетчаткой вегетативных частей растений (В. ]итео]лз). Другие поеда-
ют преимущественно их плоды и семена (В. таМлз). Многие виды упо-
требляют в пищу животные корма. По данным Уаттса, в питании В. 1е1-
сориз и В. мзефез удельный вес насекомых составляет 20—90 (Уайз,
1977). ивотные корма охотно используют в пищу В. агрепбуещет,
В. ехШапз, В. потуер1сиз, В. тайиз. Все они в разной степени питаются
как беспозвоночными, так и позвоночными.
Удивительный пример хищничества описан Кеплером (Керег, 1967).
Самые маленькие из крыс, В. ехшапз, на Гавайских островах не только
поедали яйца и птенцов морских птиц, но и нападали ночью на спя-
щих взрослых альбатросов (О1отейеа пииииа3), выгрызая мясо у них
на спине. Согласно Неплеру, из-за крыс ежегодно гибло 1—2,5% пого-
ловья этих огромных птиц. На основании его наблюдений В. схи]апв
была обвинена даже в исчезповении целого ряда островных видов, в том
числе гигантского моа (Е1епупр, 1969).
Наличие и соотношение различных видов кормов в рационе конкрет-
ного вида меняются в зависимости от их наличия в природе, сезона года,
20
Глава первая
возраста жинотных, генетической специфики популяций и даже пищевых
традиций. Так, в лесах Израиля, где произрастают совместно сосна
Рапиаз Веерепз!15 и кипарис Сиргезз зетрег\гепз, было обпаружено, что
одна популяция черных крыс поедала шишки сосны и не трогала шишки
кипариса, тогда как члены друтой популяции поступали наоборот
(Аззпег, 1984).
Активность крыс, изучавшихся в этом плане, падает в основном на
темное время суток. Для строго ночных видов характерны большие вы-
пуклые глаза, больпие ушные раковины и длинные вибриссы.
Стратегия размножения разпых видов крыс одинакова. Для них ха-
рактерны короткая беременность, большие выводки, быстрое созревание
детенышей. Репродуктивные параметры известны лишь для немногих ви-
дов. Большая их часть очень лабильна и характеризует не вид, а конк-
ретпую экологическую ситуацию, поэтому в описании рода мы их не
приводим. Средние размеры выводков, возможно, в какой-то мере кор-
релируют с количеством сосков у самок (см. табл. 1).
Замечательная особенность настоящих крыс— их склонность к си-
нантропии, которая проявляется у многих видов, особенно в подроде
Вабиз (В. габоз, В. потуез1сиз, В. ехшапз, В. агрепйуещег, В. миаз
и др.).
Один вид-— В. погуес1сяв
— одомашнен и широко используется как
лабораторное животное.
Распространение
Современное распространение рода Ва Маз близко к космополитическо-
му. Огромную роль в рассенении настоящих крыс сыграла их склонность
к синантропии. Люди расселились по всей планете благодаря созданию
оптимального микроклимата в своих жилищах и искусственной кормовой
базы за счет сельскохозяйственного производства. Этим самым были
еломлены климатические и трофические препятствия па пути расселе-
ния сипаптропных крыс. Географические барьеры, опять же, преодоле-
ваются крысами с помощью людей. Для видов, заселяющих суда, являю-
щиеся по существу плавающими домами, не существует водных преград.
С помощью транспортных средств, главным образом морских и речных
судов, черная и серая крысы за последние столетия необыкновенно рас-
ширили свои ареалы, достигнув семикосмополитизма. Современный аре-
ал рода в основном укладывается в рамки границ ареалов этих двух ви-
дов.
Несмотря на исключительную приспособленпость к обитанию в но-
стройках человека, на распространение черных и серых крыс оказывают
влияние и климатические факторы. Так, черные крысы широко распрост-
ранены в тропической и субтропической зонах всех континентов, на
север же не проникают дальше 60° с. ит. Серые крысы широко распрост-
ранены в умеренных зонах Евразии и Америки, проникая за Северный
полярный круг, а на юге до 54° ю. ш. (0-в Южная Георгия}, но прак-
тически отсутствуют в тропиках, встречаясь там спорадически в порто-
Сисхематика
941
вых городах. Оба вида с трудом заселяют области с аридным климатом.
Иное распространение у других синантропов-антропохоров. Третий по
величине ареал имеет малая крыса (В. ехцапз). Она распространена на
Азиатском материке от Бангладеш до Вьетнама, на островах Зондского
архипелага, Филиппинах, Новой Гвинее, в Новой Зеландии и на многих
тихоокеанских островах вплоть до Гавайских островов и о-ва Пасхи.
Любопытно, что ©с нет па Австралийском материке. Такой ареал, несом-
ненпо, возник в результате завоза крыс людьми: считается, что полине-
зийские мореплаватели завозили их случайно, с сельскохозяйственными
продуктами. Однако допустима и версия преднамеренной интродукции,
которой придерживается, в частности, Хейердал (Неуетда1, 1974). Из-
вестно, что крысы используются полинезийцами в пищу и служат объ-
ектом спортивной охоты (Элтон, 1960, с. 122—123).
Кроме упомянутых трех видов, по мнению Массера (Миззег, 1977),
еще два расширили свой ареал благодаря человеку. Это обитатели рисо-
вых полей В. аглепнусщег и В. 194з, заселяющие дома и сады. Они
распространены в материковой ТОго-Восточной Азии и на многочислен-
ных островах между Азией и Австралией до Новой Гвинеи включи-
тельно.
Для целей филогении и систематики, являющихся предметом настоя-
щей главы, важно установить естественный ареал рода, который распо-
ложен в основпом в пределах Индо-Малайской и Австралийской зоогео-
графических областей. В Палеарктику проникает лишь туркестанская
крыса (В. бигкеашсиз).
Распределение видов внутри родового ареала отражено в табл. 3,
причем широко распространенные виды указаны по месту их предпола-
гаемого происхождения.
П роисхождение и история расселения
Атетиз сЫшНпз5, обнаруженный в среднем миоцене Пакистана,
наиболее древний и примитивный из известных мурид. Согласно гино-
тезе Джекобса, он был исходной формой для многих фоссильных родов
мурид, найденных в позднемиоценовых
— ранненлейстоценовых слоях в
Пакистане и Афганистане. Один из этих родов, Кагиппайа, дал эволю-
ционную ветвь, включающую близкие друг другу роды Со|ал4а, Муюотуз
и Рыошуз, а также Атуеатиз и Кайлз (УасоЬз, 1978). Существуют и
другие точки зрения па раннюю эволюцию мурид. Они упоминаются
Карлетоном и Массером (Саеюп, Миззег, 1984, с. 356).
Наиболее древние находки Ваз относятся к средпему плейстоцену.
По остеологическому материалу, относимому к этому времени, собранно-
му в Триниле на о-ве Ява знаменитым Дюбуа, Массером был описан но-
вый фоссильный вид В. ШПейз!з (Мет, 1982). Известны мноточис-
ленные находки «Ерпиуз габиз» из плейстоцена Китая (Чоукоутин),
однако, согласно Мизону, костный матервал был неправильно идентифи-
цирован и принадлежит вилам, ныне относимым к другим родам (Ме
20
Глава первая
возраста жинотных, генетической специфики популяций и даже пищевых
традиций. Так, в лесах Израиля, где произрастают совместно сосна
Рапиаз Веерепз!15 и кипарис Сиргезз зетрег\гепз, было обпаружено, что
одна популяция черных крыс поедала шишки сосны и не трогала шишки
кипариса, тогда как члены друтой популяции поступали наоборот
(Аззпег, 1984).
Активность крыс, изучавшихся в этом плане, падает в основном на
темное время суток. Для строго ночных видов характерны большие вы-
пуклые глаза, больпие ушные раковины и длинные вибриссы.
Стратегия размножения разпых видов крыс одинакова. Для них ха-
рактерны короткая беременность, большие выводки, быстрое созревание
детенышей. Репродуктивные параметры известны лишь для немногих ви-
дов. Большая их часть очень лабильна и характеризует не вид, а конк-
ретпую экологическую ситуацию, поэтому в описании рода мы их не
приводим. Средние размеры выводков, возможно, в какой-то мере кор-
релируют с количеством сосков у самок (см. табл. 1).
Замечательная особенность настоящих крыс— их склонность к си-
нантропии, которая проявляется у многих видов, особенно в подроде
Вабиз (В. габоз, В. потуез1сиз, В. ехшапз, В. агрепйуещег, В. миаз
и др.).
Один вид-— В. погуес1сяв
— одомашнен и широко используется как
лабораторное животное.
Распространение
Современное распространение рода Ва Маз близко к космополитическо-
му. Огромную роль в рассенении настоящих крыс сыграла их склонность
к синантропии. Люди расселились по всей планете благодаря созданию
оптимального микроклимата в своих жилищах и искусственной кормовой
базы за счет сельскохозяйственного производства. Этим самым были
еломлены климатические и трофические препятствия па пути расселе-
ния сипаптропных крыс. Географические барьеры, опять же, преодоле-
ваются крысами с помощью людей. Для видов, заселяющих суда, являю-
щиеся по существу плавающими домами, не существует водных преград.
С помощью транспортных средств, главным образом морских и речных
судов, черная и серая крысы за последние столетия необыкновенно рас-
ширили свои ареалы, достигнув семикосмополитизма. Современный аре-
ал рода в основном укладывается в рамки границ ареалов этих двух ви-
дов.
Несмотря на исключительную приспособленпость к обитанию в но-
стройках человека, на распространение черных и серых крыс оказывают
влияние и климатические факторы. Так, черные крысы широко распрост-
ранены в тропической и субтропической зонах всех континентов, на
север же не проникают дальше 60° с. ит. Серые крысы широко распрост-
ранены в умеренных зонах Евразии и Америки, проникая за Северный
полярный круг, а на юге до 54° ю. ш. (0-в Южная Георгия}, но прак-
тически отсутствуют в тропиках, встречаясь там спорадически в порто-
Сисхематика
941
вых городах. Оба вида с трудом заселяют области с аридным климатом.
Иное распространение у других синантропов-антропохоров. Третий по
величине ареал имеет малая крыса (В. ехцапз). Она распространена на
Азиатском материке от Бангладеш до Вьетнама, на островах Зондского
архипелага, Филиппинах, Новой Гвинее, в Новой Зеландии и на многих
тихоокеанских островах вплоть до Гавайских островов и о-ва Пасхи.
Любопытно, что ©с нет па Австралийском материке. Такой ареал, несом-
ненпо, возник в результате завоза крыс людьми: считается, что полине-
зийские мореплаватели завозили их случайно, с сельскохозяйственными
продуктами. Однако допустима и версия преднамеренной интродукции,
которой придерживается, в частности, Хейердал (Неуетда1, 1974). Из-
вестно, что крысы используются полинезийцами в пищу и служат объ-
ектом спортивной охоты (Элтон, 1960, с. 122—123).
Кроме упомянутых трех видов, по мнению Массера (Миззег, 1977),
еще два расширили свой ареал благодаря человеку. Это обитатели рисо-
вых полей В. аглепнусщег и В. 194з, заселяющие дома и сады. Они
распространены в материковой ТОго-Восточной Азии и на многочислен-
ных островах между Азией и Австралией до Новой Гвинеи включи-
тельно.
Для целей филогении и систематики, являющихся предметом настоя-
щей главы, важно установить естественный ареал рода, который распо-
ложен в основпом в пределах Индо-Малайской и Австралийской зоогео-
графических областей. В Палеарктику проникает лишь туркестанская
крыса (В. бигкеашсиз).
Распределение видов внутри родового ареала отражено в табл. 3,
причем широко распространенные виды указаны по месту их предпола-
гаемого происхождения.
П роисхождение и история расселения
Атетиз сЫшНпз5, обнаруженный в среднем миоцене Пакистана,
наиболее древний и примитивный из известных мурид. Согласно гино-
тезе Джекобса, он был исходной формой для многих фоссильных родов
мурид, найденных в позднемиоценовых
— ранненлейстоценовых слоях в
Пакистане и Афганистане. Один из этих родов, Кагиппайа, дал эволю-
ционную ветвь, включающую близкие друг другу роды Со|ал4а, Муюотуз
и Рыошуз, а также Атуеатиз и Кайлз (УасоЬз, 1978). Существуют и
другие точки зрения па раннюю эволюцию мурид. Они упоминаются
Карлетоном и Массером (Саеюп, Миззег, 1984, с. 356).
Наиболее древние находки Ваз относятся к средпему плейстоцену.
По остеологическому материалу, относимому к этому времени, собранно-
му в Триниле на о-ве Ява знаменитым Дюбуа, Массером был описан но-
вый фоссильный вид В. ШПейз!з (Мет, 1982). Известны мноточис-
ленные находки «Ерпиуз габиз» из плейстоцена Китая (Чоукоутин),
однако, согласно Мизону, костный матервал был неправильно идентифи-
цирован и принадлежит вилам, ныне относимым к другим родам (Ме
22
Глава первая
Таблица 3. Аборигенные виды Ва физ различных географичееких облаетей (по:
Мизвег, Меуусошь, 1983, 1985; Мизвег, Неапеу, 1985)
Австралия
Филиппинские
Австралия
О-в Новая Гвинея пои Нохая Молукнские острова | острова
Газс1рез
поре
}епсориз
сегати1стя
ечетеЯ
Нитеовз
и1сВагазоп!
зога19з
1еНссив
119 0гепз13
{иппеу
уегесипд аз
того а1етз15
фауЦа\/1е015
ргаеют
с] арЫтиаз
шотгдах
Уепи
оЦиа\уе1813
поузегашеае
до еп
* Виды. распространенные сейчас и за пределами указанной области.
уетцег, Г.еоро!датуз). Плейстоценовые В. гаМлз из Китая не известны
(М1зоппе, 1969, с. 27, 128, 140).
Возраст серой крысы по «молекулярным часам» определен в
8 мли лет или в 2,5 млн лет в зависимости от метода исследования (со-
ответственно электрофорез и иммунология альбуминов) (Вауст®юосЕ сё а|.,
1983). Учитывая неравномерность темпов эволюции, зависимость ре-
зультатов исследования от соотпошепия «быстро» и «медленно» эволюи-
рующих локусов, а также отмеченное выше несовпадение в определении
возраста, мы пе припимаем всерьез этих цифр. К сожалению, из-за чрез-
вычайно слабой палеонтологической изученности Юго-Восточной Азии и
на палеонтологические данные в этом вопросе пока нельзя опереться.
Центром происхождения и расселения рода Ва Млз, видимо, является
материковая Юго-Восточная Азия (УозЧа, 1980; Миззег, Ме\усошЪ,
1983). История расселепия рода сейчас видится следующим образом.
1. Северное направление. Распространению на север пре-
пятствовали горы и пустыни Центральной Азии, обрамленпые с юга вы-
сочайшей грядой Гималаев, поэтому проникнуть на север крысы смогли,
только обойдя этот неприступный барьер с запада и востока.
2. Восточное направление. Распространение по тропикам и
субтропикам материковой Восточной Азии, должно быть, не встречало
сорьезных препятствий. У берегов Восточно-Китайского моря сейчас
обитают: Ш. п19из, В. 10зеа, В. ККииензз, В. табазв. Такой
своеобразпый вид, как серая крыса, возник, видимо, в Восточпом Китае.
Благодаря приспособленности к климату умеренной полосы пасюк обес-
печил продвижение рода па север, в обход Центральной Азии с востока.
3. Юго-Восточное направление. Расселение крыс по
островам Индо-Австралии и проникновение на Австралийский материк
происходило в периоды низкого уровня Мирового океана, когда возника-
Систематика
23
Мелкие острова к
й
востоку, югу и вапа-
Острова Зондеко
Датериковая Азия
О-в Сулавеси
. ду от Зондекого
го шельфа
А.
шельфа
.
:
7205
и
Вотан
3601613
Чоташеа$
погус21си8 ,
хап ити
рабвагат
Балет
атвепиусщег
аретз
Ъоорегуег И
ехи!ап$
.
..: +
аЧаза$
аппапда]е1
п9$
озеа
епрапиз
и
|
шас] саг
фагкезват1 18
туцац
топпеи$
пануйац$
ы|,
ЕКО с
тала *
озеоо4
ли перешейки между островами, между островами и маториками, пу
лись проливы. В такие периоды происходили петро м и одно.
кратно возникавшие в плиоцене н плоботощене,
ая
мался над поверхностью воды, образу
окр
‚ Новая
лась персшейком © Австралией (см. карты в ра :
ов 19876). В преднем плейстоцене Вайлз ре
ва озе Ява (Миззег, 1982), в позднем плейстоцене — в
., 1983).
|
Тема в проникновения настоящих крыс в Австралийскую
область. Естественио было бы предположить, то, ото и 29 де ке
Малых Зондеких островов, однако на них
В ромы
сильных, ни рецентных аборигенных ВаНиз (Миззег, 1 . с
у
: 1) Вай» на Малых Зондеких
три объяснепия этому обстоятельству
д
ых Зотдеких
есть. но пока не найдены; 2) заселение Авотралийск
т через о-в Сулавеси и Молуккские острова, где сейчас воть этдомич-
ные виды Ва\бо5; 3) австралийскио виды возникли нове ко от аааат
ских и имеют с пими лишь конвергентное ‚оходетво иззег, 1981с,
с. 169). Какая из трех версий правильна, покажет удущее о поминезий-
В голоцене шло расселение на восток В. ехч|ап5. Вмест ней
скими мореплавателями они проникли на острова тожжной ие
Пацифики от Новой Зеландии до Гавайских островов
Пас, бот.
А. Западное направление. Расселение па запад ло соот.
венно через Индийский субконтинент и по склонам им ал вов. та
п-ове Индостан произошли основные события в мое ее
черной крысы
— образование цеилонской и океаническо ом вая
двух последовательных робертоононеких слияний, о в, ем вы
съ только на о-ве Шри
,
й
но м рисселиться по всему свету. Азиатская форма черной крысы
22
Глава первая
Таблица 3. Аборигенные виды Ва физ различных географичееких облаетей (по:
Мизвег, Меуусошь, 1983, 1985; Мизвег, Неапеу, 1985)
Австралия
Филиппинские
Австралия
О-в Новая Гвинея пои Нохая Молукнские острова | острова
Газс1рез
поре
}епсориз
сегати1стя
ечетеЯ
Нитеовз
и1сВагазоп!
зога19з
1еНссив
119 0гепз13
{иппеу
уегесипд аз
того а1етз15
фауЦа\/1е015
ргаеют
с] арЫтиаз
шотгдах
Уепи
оЦиа\уе1813
поузегашеае
до еп
* Виды. распространенные сейчас и за пределами указанной области.
уетцег, Г.еоро!датуз). Плейстоценовые В. гаМлз из Китая не известны
(М1зоппе, 1969, с. 27, 128, 140).
Возраст серой крысы по «молекулярным часам» определен в
8 мли лет или в 2,5 млн лет в зависимости от метода исследования (со-
ответственно электрофорез и иммунология альбуминов) (Вауст®юосЕ сё а|.,
1983). Учитывая неравномерность темпов эволюции, зависимость ре-
зультатов исследования от соотпошепия «быстро» и «медленно» эволюи-
рующих локусов, а также отмеченное выше несовпадение в определении
возраста, мы пе припимаем всерьез этих цифр. К сожалению, из-за чрез-
вычайно слабой палеонтологической изученности Юго-Восточной Азии и
на палеонтологические данные в этом вопросе пока нельзя опереться.
Центром происхождения и расселения рода Ва Млз, видимо, является
материковая Юго-Восточная Азия (УозЧа, 1980; Миззег, Ме\усошЪ,
1983). История расселепия рода сейчас видится следующим образом.
1. Северное направление. Распространению на север пре-
пятствовали горы и пустыни Центральной Азии, обрамленпые с юга вы-
сочайшей грядой Гималаев, поэтому проникнуть на север крысы смогли,
только обойдя этот неприступный барьер с запада и востока.
2. Восточное направление. Распространение по тропикам и
субтропикам материковой Восточной Азии, должно быть, не встречало
сорьезных препятствий. У берегов Восточно-Китайского моря сейчас
обитают: Ш. п19из, В. 10зеа, В. ККииензз, В. табазв. Такой
своеобразпый вид, как серая крыса, возник, видимо, в Восточпом Китае.
Благодаря приспособленности к климату умеренной полосы пасюк обес-
печил продвижение рода па север, в обход Центральной Азии с востока.
3. Юго-Восточное направление. Расселение крыс по
островам Индо-Австралии и проникновение на Австралийский материк
происходило в периоды низкого уровня Мирового океана, когда возника-
Систематика
23
Мелкие острова к
й
востоку, югу и вапа-
Острова Зондеко
Датериковая Азия
О-в Сулавеси
. ду от Зондекого
го шельфа
А.
шельфа
.
:
7205
и
Вотан
3601613
Чоташеа$
погус21си8 ,
хап ити
рабвагат
Балет
атвепиусщег
аретз
Ъоорегуег И
ехи!ап$
.
..: +
аЧаза$
аппапда]е1
п9$
озеа
епрапиз
и
|
шас] саг
фагкезват1 18
туцац
топпеи$
пануйац$
ы|,
ЕКО с
тала *
озеоо4
ли перешейки между островами, между островами и маториками, пу
лись проливы. В такие периоды происходили петро м и одно.
кратно возникавшие в плиоцене н плоботощене,
ая
мался над поверхностью воды, образу
окр
‚ Новая
лась персшейком © Австралией (см. карты в ра :
ов 19876). В преднем плейстоцене Вайлз ре
ва озе Ява (Миззег, 1982), в позднем плейстоцене — в
., 1983).
|
Тема в проникновения настоящих крыс в Австралийскую
область. Естественио было бы предположить, то, ото и 29 де ке
Малых Зондеких островов, однако на них
В ромы
сильных, ни рецентных аборигенных ВаНиз (Миззег, 1 . с
у
: 1) Вай» на Малых Зондеких
три объяснепия этому обстоятельству
д
ых Зотдеких
есть. но пока не найдены; 2) заселение Авотралийск
т через о-в Сулавеси и Молуккские острова, где сейчас воть этдомич-
ные виды Ва\бо5; 3) австралийскио виды возникли нове ко от аааат
ских и имеют с пими лишь конвергентное ‚оходетво иззег, 1981с,
с. 169). Какая из трех версий правильна, покажет удущее о поминезий-
В голоцене шло расселение на восток В. ехч|ап5. Вмест ней
скими мореплавателями они проникли на острова тожжной ие
Пацифики от Новой Зеландии до Гавайских островов
Пас, бот.
А. Западное направление. Расселение па запад ло соот.
венно через Индийский субконтинент и по склонам им ал вов. та
п-ове Индостан произошли основные события в мое ее
черной крысы
— образование цеилонской и океаническо ом вая
двух последовательных робертоононеких слияний, о в, ем вы
съ только на о-ве Шри
,
й
но м рисселиться по всему свету. Азиатская форма черной крысы
24 Глава первая
проникла на запад до Джелалабадекой долины в Афганистане [если не
считать недавней находки ее в Мексике (!) (А]0пзо её а1., 1982)]. Через
Индию уже в историческое время проходил основной путь расселения на
запад серой крысы.
Гималаи с их высотной зональностью, представляющей широкий:
спектр экологических условий, послужили «коридором», по которому про-
никли на запад горные виды. Помимо азиатской формы черной крысы
сейчас здесь обитают В. 913, В. зШиаепзз, В. Бтапиеиз и В. ишКе-
361115, Первые два вида широко распространены в Юго-Восточной
и Восточной Азии. В. Бгапаси$ нигде, кроме Гималаев, пе встречается.
Туркестанская крыса обитает на склонах Гималаев и в 'торных системах
к западу от них. Она освоила Гиндукуш, горы Таджикистана, Западный
Тянь-Шань вплоть до южной части хребта Кара-Тау (Казахстан) на се-
вере. На востоке зарегистрирована в китайских провинциях ТОньнань
и Гуандун. Интересна паходка черепа туркестанской крысы в средне-
вековых погадках филина на юго-восточной окраине Сарыкамышской:
низменности, в 900 км западное современной границы ареала (Стальма-
НН юге Узбекистана она известна из среднего плейстоцена.
Филогенетическое положение
Филогенетические взаимоотношения внутри семейства Мин4ас далеко
еще не ясны. Их выявлению препятствует, помимо прочего, неопределен-
ность таксономических границ рода ВаИлз.
Согласно Массеру и Ныокомбу, наиболее близки к Ваз (из 13 изу-
ченных в этом плане родов) зондекие Раа\апотуз (0-в Палаван)
и Каатзапотуз (о-в Ява). Другие виды мурид образуют самостоятель-.
ные группы, не состоящие в близком родстве с Ваз. Это NoМ\уенег
Махотуз, ГеороЧашуз, Вегупуз, Гопойих, РИНесвей,, Нараотуз СЬг-.
тородотуз, Наегошуз, Зипдатуз, Миз. Возможно, к Вайаз близки афри-
канские АтусапИйз, филиппинские роды, например Татпотуз и Тагзо-
туз, сулавесские 'Тасготуз и Рагиготуз, Рарасотуз и Кошойдотуз Малых.
Зопдских островов (Миззет, Меусотшь, 1983, с. 577—578).
Таксономичесная структура
Эволюция представлений о структуре рода ВаНи$ отражена в первом:
разделе главы, посвященном истории его изучения. Попытки выделепия
подродов внутри рода в современном его понимании пока не предприни-
малось. Здесь при обсуждении внутриродовой структуры мы опирались.
на следующие работы: Тауог, Ногпег, 1973; Тау]ог её 21., 1982, 4983;
Миззег, МехусотаЪ, 1983, 1985; Миззег, Неалеу, 1985.
|у|
Определенно признается объединение крыс материковой Лзии
(см. табл. 3) в подрод Вайяв. К этому же подроду относят зондских
В. БаГаепз!з, В. Вооветуегй, В. Нотап!соз и филиппинскую В. пушдогеп-
Систематика
25
34в. Половина видов этого подрода раньше считалась подвидами черной
крысы.
Крупные крысы, населяющие мелкие острова возле Зондокого шель-
фа, за ето пределами, объединяются Массером в группу райтагииа
(В. ранпагит, В. 1велз, В. адизиз). К этой группе близки В. фа\уца-
\1е1515 и В. БоЁтал.
К сулавесским крысам из комплекса хатигив примыкают В. шас-
Зеам и В. епхапиз.
Новогвинейские крысы близки друг другу и менее близки, но обна-
руживают сходство с молуккскими В. сегапцств и В. {еЙсепз с одной
«тороны, и австралийскими видами с другой (см. табл. 3).
Систематика Вайиз погуе21е18
Номенклатура
Ваз потуее1сив (Вегкепрои, 1769} — серая крыса, пасюк. ВегКепноц 7. ОпИшез
Е Ве Мага! Езогу оЁ Сгеак Вгйаа ап@ Гге]апд, уо1. 1, р. 5.
Типовая местность: Великобритания.
Голотип: неизвестен.
Принятое ныне научное название серой крысы сложилось не сразу.
В связи с выделением настоящих крыс в самостоятельный род и ноиском
старших синонимов оно неоднократно менялось. Наибольшее распростра-
нение получили следующие сочетания (в скобках указаны даты номен-
‘клатурных преобразований, но по инерции недействительные названия
употреблялись и позже): Миз Чеситапиз РаЦаз (1778—1900 тг.), Маз
‘погуерлсив ЕгхеБеп (1900—1940 гг.), Ерипуз погуезств Етхефев (1910—
1946 гт.), Вами погуерсиз Етхефеп (с 1916 г.), В. потуесиз Вегкеп-
Фой* (видимо, с 30-х тг.).
Беркенхаут и Эркслебен пазвали серую крысу независимо друг от
друга «норвежской». Очевидно, это название укоренилось в ХУП! в.
во многих западноевропейских языках. В списке синонимов пасюка
у Эркслебена (Егфереп, 1977) оно встречается неоднократно начиная
с 50-х годов ХУШ в. (Маз ех Могуеза, Оз Могуез1сив, 1е Ваё 4е М т-
убое). В английском языке название «норвежская крыса» широко упо-
требляется до сих пор, сохранилось оно и во французском языке.
Ниже приводятся списки русских. английских, немецких и француз-
ских синонимов В. погуес1стз. Списки начинаются с наиболее употреби-
тельных названий и заканчиваются наименее употребительными и уста-
ревшими. Латияская синонимика дана при описании подвидов.
Русские названия: серая крыса, пасюк, карако (подвид), рыжая кры-
са, амбарная крыса, домовая крыса, обыкновенная крыса, кочевая крыса.
Английские названия: Мотуау га. Бгомжп гаь сотой таб, зе\ег таб,
ТзВ гаф \уаег-гаф, отау гаф, ууатЁ га, Баги гаь Напоуемаи га4.
Немецкие названия: \\апдеггаНе, З1ед1иезуапдеггаИе, Бгаипе Вайе,
ЭсвИЁгаНе, огаие ВаЧе.
24
Глава первая
проникла на запад до Джелалабадекой долины в Афганистане [если не
считать недавней находки ее в Мексике (!) (А]0пзо её а1., 1982)]. Через
Индию уже в историческое время проходил основной путь расселения на
запад серой крысы.
Гималаи с их высотной зональностью, представляющей широкий:
спектр экологических условий, послужили «коридором», по которому про-
никли на запад горные виды. Помимо азиатской формы черной крысы
сейчас здесь обитают В. 913, В. зШиаепзз, В. Бтапиеиз и В. ишКе-
361115, Первые два вида широко распространены в Юго-Восточной
и Восточной Азии. В. Бгапаси$ нигде, кроме Гималаев, пе встречается.
Туркестанская крыса обитает на склонах Гималаев и в 'торных системах
к западу от них. Она освоила Гиндукуш, горы Таджикистана, Западный
Тянь-Шань вплоть до южной части хребта Кара-Тау (Казахстан) на се-
вере. На востоке зарегистрирована в китайских провинциях ТОньнань
и Гуандун. Интересна паходка черепа туркестанской крысы в средне-
вековых погадках филина на юго-восточной окраине Сарыкамышской:
низменности, в 900 км западное современной границы ареала (Стальма-
НН
юге Узбекистана она известна из среднего плейстоцена.
Филогенетическое положение
Филогенетические взаимоотношения внутри семейства Мин4ас далеко
еще не ясны. Их выявлению препятствует, помимо прочего, неопределен-
ность таксономических границ рода ВаИлз.
Согласно Массеру и Ныокомбу, наиболее близки к Ваз (из 13 изу-
ченных в этом плане родов) зондекие Раа\апотуз (0-в Палаван)
и Каатзапотуз (о-в Ява). Другие виды мурид образуют самостоятель-.
ные группы, не состоящие в близком родстве с Ваз. Это NoМ\уенег
Махотуз, ГеороЧашуз, Вегупуз, Гопойих, РИНесвей,, Нараотуз СЬг-.
тородотуз, Наегошуз, Зипдатуз, Миз. Возможно, к Вайаз близки афри-
канские АтусапИйз, филиппинские роды, например Татпотуз и Тагзо-
туз, сулавесские 'Тасготуз и Рагиготуз, Рарасотуз и Кошойдотуз Малых.
Зопдских островов (Миззет, Меусотшь, 1983, с. 577—578).
Таксономичесная структура
Эволюция представлений о структуре рода ВаНи$ отражена в первом:
разделе главы, посвященном истории его изучения. Попытки выделепия
подродов внутри рода в современном его понимании пока не предприни-
малось. Здесь при обсуждении внутриродовой структуры мы опирались.
на следующие работы: Тауог, Ногпег, 1973; Тау]ог её 21., 1982, 4983;
Миззег, МехусотаЪ, 1983, 1985; Миззег, Неалеу, 1985.
|
у
|
Определенно признается объединение крыс материковой Лзии
(см. табл. 3) в подрод Вайяв. К этому же подроду относят зондских
В. БаГаепз!з, В. Вооветуегй, В. Нотап!соз и филиппинскую В. пушдогеп-
Систематика
25
34в. Половина видов этого подрода раньше считалась подвидами черной
крысы.
Крупные крысы, населяющие мелкие острова возле Зондокого шель-
фа, за ето пределами, объединяются Массером в группу райтагииа
(В. ранпагит, В. 1велз, В. адизиз). К этой группе близки В. фа\уца-
\1е1515 и В. БоЁтал.
К сулавесским крысам из комплекса хатигив примыкают В. шас-
Зеам и В. епхапиз.
Новогвинейские крысы близки друг другу и менее близки, но обна-
руживают сходство с молуккскими В. сегапцств и В. {еЙсепз с одной
«тороны, и австралийскими видами с другой (см. табл. 3).
Систематика Вайиз погуе21е18
Номенклатура
Ваз потуее1сив (Вегкепрои, 1769} — серая крыса, пасюк. ВегКепноц 7. ОпИшез
Е Ве Мага! Езогу оЁ Сгеак Вгйаа ап@ Гге]апд, уо1. 1, р. 5.
Типовая местность: Великобритания.
Голотип: неизвестен.
Принятое ныне научное название серой крысы сложилось не сразу.
В связи с выделением настоящих крыс в самостоятельный род и ноиском
старших синонимов оно неоднократно менялось. Наибольшее распростра-
нение получили следующие сочетания (в скобках указаны даты номен-
‘клатурных преобразований, но по инерции недействительные названия
употреблялись и позже): Миз Чеситапиз РаЦаз (1778—1900 тг.), Маз
‘погуерлсив ЕгхеБеп (1900—1940 гг.), Ерипуз погуезств Етхефев (1910—
1946 гт.), Вами погуерсиз Етхефеп (с 1916 г.), В. потуесиз Вегкеп-
Фой* (видимо, с 30-х тг.).
Беркенхаут и Эркслебен пазвали серую крысу независимо друг от
друга «норвежской». Очевидно, это название укоренилось в ХУП! в.
во многих западноевропейских языках. В списке синонимов пасюка
у Эркслебена (Егфереп, 1977) оно встречается неоднократно начиная
с 50-х годов ХУШ в. (Маз ех Могуеза, Оз Могуез1сив, 1е Ваё 4е М т-
убое). В английском языке название «норвежская крыса» широко упо-
требляется до сих пор, сохранилось оно и во французском языке.
Ниже приводятся списки русских. английских, немецких и француз-
ских синонимов В. погуес1стз. Списки начинаются с наиболее употреби-
тельных названий и заканчиваются наименее употребительными и уста-
ревшими. Латияская синонимика дана при описании подвидов.
Русские названия: серая крыса, пасюк, карако (подвид), рыжая кры-
са, амбарная крыса, домовая крыса, обыкновенная крыса, кочевая крыса.
Английские названия: Мотуау га. Бгомжп гаь сотой таб, зе\ег таб,
ТзВ гаф \уаег-гаф, отау гаф, ууатЁ га, Баги гаь Напоуемаи га4.
Немецкие названия: \\апдеггаНе, З1ед1иезуапдеггаИе, Бгаипе Вайе,
ЭсвИЁгаНе, огаие ВаЧе.
26
Глава первая
Французские названия: баги 10, Ва ас Могуёяе, Ва Бгип, Ва дт1,
Вак (’6с0и, Ваф сазреп, Ваё папда!5.
у
Описание
Поскольку детальное описание морфологии, кариотипа и экологии серой
крысы дано в соответствующих разделах пастоящей монографии, здесь
приводится, по существу, лишь раслниренный диагноз вида.
Морфология. Крупная крыса, телосложение плотное. Толстый
основания хвост короче длины тела (70—95%) и имеет в среднем
182 (163—201) кольца чешуек (ВесКег, 1978). Ушные раковины малень-
кие, овальной формы, снаружи покрыты короткими волосками. Кисти
и стопы с относительно маленькими и плоскими подошвенными бугор-
ками. На кисти первый (впутренний) палец сильно редуцирован. Хорошо:
развитые складки кожи между пальцами стоны образуют подобие плава-
тельной перепонки. Когти слабо изогнуты, относительно длинные.
Характер мехового покрова изменчив. Он зависит от образа жизни
крыс, их пола и возраста, сезона года. У экзоантропных форм меховой
покров гуще и мягче, у синантропных реже и грубее. Меховой покров:
сампов грубее, чем у самок. Зимний мех в 1,3—1,4 раза гуще летнего
(Козлов, 1983). Направляющие волосы на несколько миллиметров пре-
вышают длину остевых волос. Особенно хорото они выражены на хреб-
те у взрослых самцов, но пикогда пс достигают такого чрезвычайного
развития, как у черной крысы. Иглы и иглоподобные щетины отсут-
ствуют.
Волосяной покров верхней части головы, спины и боков окрашен
зопально. Волосы имеют серое основание, черную вершину и желтое
кольцо в промежутке, что в совокуппости даст коричнево-серуто окраску
верха тела животного. Тон окраски сильно варьирует в зависимости
от ширины отдельных цветовых зоп и их интенсивности, обусловленпой
характером расположения гранул меланинов в волосе и их количеством.
Эти призпаки, с одной стороны, находятся под генетическим контролем,
с другой стороны, подвержены внешним влияниям, например, зависят
от состава кормов (Во зоп, 1965). Оттенок окраски крыс, как и дру-
гих млекопитающих с зональной окраской волоса, зависит также от ста-
дии роста волос и, следовательно, неоднократно меняется в течение жиз-
ни индивидуума в связи с возрастпыми и сезопными линьками. Молодые.
пасюки, как правило, серее взрослых и по мере роста рыжеют.
Брюшко имеет грязно-белую окраску, обусловленную серым основа-
нием и белым окончанием волос. Граница между цветом боков и брюшка
не резкая, но, как правило, хорошо заметна.
Кожа серых крысе лишена пигмента, поэтому тс участки, где она.
оголена (кончик носа, внутренние поверхности ушных раковин, пальцы)
или покрыта редкими депитмептированными волосками (дорсальные по-
верхности плюсны и пясти, вентральная поверхность хвоста), имеют
телеспый цвет. Покрытые редкими и короткими пигментированными
волосками наружная поверхность узиных раковин и дорсальная поверх-
ность хвоста имеют серую окраску.
Систематика
27
В сстественных популяциях серых крыс нередко попадаются особи
с аберрантиой окраской (ВоБтзоп, 1965; Зеае, 1968; Милютин, 1981,
1982: Романова, 1985). Чаще всего встречается «ирландская погость» —
белое пятно на грули, обусловленное одним рецессивным аллелем (в).
Второе место по частоте встречаемости припадлежит мутации пе агути».
У гомозиготных животных волосы лишены желтой зоны, в силу чего
они имеют черный цвет. Эта окраска, обычно навываемая в зоологиче-
ской литературе «меланизмом», детерминирустся рецессивным аллелем
другого локуса (а). Относительно часто выщенляются пасюки с «капю-
пюнной» окраской (No6). У них окрашены только передняя часть туло-
ваща («капюшон») и верх спипы и хвоста («ремень»), все остальное
тело белое, т. е. лишено пигмента. Ширипа «ремня» изменчива, он мо-
жет прерываться или отсутствовать совсем.
Очень редко встречаются альбиносы
— полностью белые животные
< красными глазами (сс) (известны, правда, я‹изнеспособные дикие коло-
нии убежавших лабораторных альбиносов, например в Душанбинском
зоопарке). Чрезвычайно редко попадаются пасюки других окрасок, на-
пример желтые (Ееаа, 1963), с белым брюшком (Уазештз, 1972;
В. Е. Лошаков, личн. сообщ.).
.
Самки обычно имеют 12 сосков: 1 грудную пару, 2 постаксиллярных,
1 брюшную и 2 паховых. Количество сосков может варьировать от 10
до 14 (Аргиропуло, 4940; ВесКег, 1978).
Череп угловатый с хорошо выраженными гребнями. Массетерная
пластинка нерхнечелюстной кости крупная, © сильно выступающим впе-
фед верхпим углом и наклонениым в направлении назад передним краем.
Скуловые дуги паиболее птироко расставлены в их вадней трети или
в средпей части. Наибольшее сужение лобных костей совпадает прибли-
зительно с их серединой. Теменные кости у взрослых особей невыпуклы
и расположены приблизительно в одной плоскости с лобными и межте-
менной; они ограничены по бокам почти прямыми или слабо изогнуты-
ми гребнями, идущими параллельно или слабо расходящимися в направ-
пении пазад [описание черепа дано по Й. М. Громову с соавт. (1965,
<. 443)].
Кариотип. Цитогенетика серой крысы относительно хорошо изуче-
на. Стандартный диплоидный хромосомный набор состоит из 42 хромо-
<ом: 7 пар метацентриков, 4 пары субтелоцентриков, 9 пар акроцентри-
ков и акроцентрические Х и У хромосомы ({МЕа
= 62). Для диких
‘и лабораторных популяций зарегистрирован полиморфизм по положению
центромеры в Х хромосоме и многих зутосомах. В большинстве случаев
зн рассматривался как результат перицентрических инверсий.
Экология. Экзоантропные серые крысы — околоводные грызуны.
`Они хорошо плавают и ныряют. Для живущих у воды пасюков водоем
служит источником корма, убежищем от врагов и средой для передви-
жения. Серые крысы хорошо приспособлены к рытью и обычно живут
в норах, в которых проводят значительную часть суток (Карасева и др.,
1985; Милютин и др., 4986). При необходимости пасюки могут забирать-
ся на деревья и экить на чердаках, однако значительно уступают в спо-
26
Глава первая
Французские названия: баги 10, Ва ас Могуёяе, Ва Бгип, Ва дт1,
Вак (’6с0и, Ваф сазреп, Ваё папда!5.
у
Описание
Поскольку детальное описание морфологии, кариотипа и экологии серой
крысы дано в соответствующих разделах пастоящей монографии, здесь
приводится, по существу, лишь раслниренный диагноз вида.
Морфология. Крупная крыса, телосложение плотное. Толстый
основания хвост короче длины тела (70—95%) и имеет в среднем
182 (163—201) кольца чешуек (ВесКег, 1978). Ушные раковины малень-
кие, овальной формы, снаружи покрыты короткими волосками. Кисти
и стопы с относительно маленькими и плоскими подошвенными бугор-
ками. На кисти первый (впутренний) палец сильно редуцирован. Хорошо:
развитые складки кожи между пальцами стоны образуют подобие плава-
тельной перепонки. Когти слабо изогнуты, относительно длинные.
Характер мехового покрова изменчив. Он зависит от образа жизни
крыс, их пола и возраста, сезона года. У экзоантропных форм меховой
покров гуще и мягче, у синантропных реже и грубее. Меховой покров:
сампов грубее, чем у самок. Зимний мех в 1,3—1,4 раза гуще летнего
(Козлов, 1983). Направляющие волосы на несколько миллиметров пре-
вышают длину остевых волос. Особенно хорото они выражены на хреб-
те у взрослых самцов, но пикогда пс достигают такого чрезвычайного
развития, как у черной крысы. Иглы и иглоподобные щетины отсут-
ствуют.
Волосяной покров верхней части головы, спины и боков окрашен
зопально. Волосы имеют серое основание, черную вершину и желтое
кольцо в промежутке, что в совокуппости даст коричнево-серуто окраску
верха тела животного. Тон окраски сильно варьирует в зависимости
от ширины отдельных цветовых зоп и их интенсивности, обусловленпой
характером расположения гранул меланинов в волосе и их количеством.
Эти призпаки, с одной стороны, находятся под генетическим контролем,
с другой стороны, подвержены внешним влияниям, например, зависят
от состава кормов (Во зоп, 1965). Оттенок окраски крыс, как и дру-
гих млекопитающих с зональной окраской волоса, зависит также от ста-
дии роста волос и, следовательно, неоднократно меняется в течение жиз-
ни индивидуума в связи с возрастпыми и сезопными линьками. Молодые.
пасюки, как правило, серее взрослых и по мере роста рыжеют.
Брюшко имеет грязно-белую окраску, обусловленную серым основа-
нием и белым окончанием волос. Граница между цветом боков и брюшка
не резкая, но, как правило, хорошо заметна.
Кожа серых крысе лишена пигмента, поэтому тс участки, где она.
оголена (кончик носа, внутренние поверхности ушных раковин, пальцы)
или покрыта редкими депитмептированными волосками (дорсальные по-
верхности плюсны и пясти, вентральная поверхность хвоста), имеют
телеспый цвет. Покрытые редкими и короткими пигментированными
волосками наружная поверхность узиных раковин и дорсальная поверх-
ность хвоста имеют серую окраску.
Систематика
27
В сстественных популяциях серых крыс нередко попадаются особи
с аберрантиой окраской (ВоБтзоп, 1965; Зеае, 1968; Милютин, 1981,
1982: Романова, 1985). Чаще всего встречается «ирландская погость» —
белое пятно на грули, обусловленное одним рецессивным аллелем (в).
Второе место по частоте встречаемости припадлежит мутации пе агути».
У гомозиготных животных волосы лишены желтой зоны, в силу чего
они имеют черный цвет. Эта окраска, обычно навываемая в зоологиче-
ской литературе «меланизмом», детерминирустся рецессивным аллелем
другого локуса (а). Относительно часто выщенляются пасюки с «капю-
пюнной» окраской (No6). У них окрашены только передняя часть туло-
ваща («капюшон») и верх спипы и хвоста («ремень»), все остальное
тело белое, т. е. лишено пигмента. Ширипа «ремня» изменчива, он мо-
жет прерываться или отсутствовать совсем.
Очень редко встречаются альбиносы
— полностью белые животные
< красными глазами (сс) (известны, правда, я‹изнеспособные дикие коло-
нии убежавших лабораторных альбиносов, например в Душанбинском
зоопарке). Чрезвычайно редко попадаются пасюки других окрасок, на-
пример желтые (Ееаа, 1963), с белым брюшком (Уазештз, 1972;
В. Е. Лошаков, личн. сообщ.).
.
Самки обычно имеют 12 сосков: 1 грудную пару, 2 постаксиллярных,
1 брюшную и 2 паховых. Количество сосков может варьировать от 10
до 14 (Аргиропуло, 4940; ВесКег, 1978).
Череп угловатый с хорошо выраженными гребнями. Массетерная
пластинка нерхнечелюстной кости крупная, © сильно выступающим впе-
фед верхпим углом и наклонениым в направлении назад передним краем.
Скуловые дуги паиболее птироко расставлены в их вадней трети или
в средпей части. Наибольшее сужение лобных костей совпадает прибли-
зительно с их серединой. Теменные кости у взрослых особей невыпуклы
и расположены приблизительно в одной плоскости с лобными и межте-
менной; они ограничены по бокам почти прямыми или слабо изогнуты-
ми гребнями, идущими параллельно или слабо расходящимися в направ-
пении пазад [описание черепа дано по Й. М. Громову с соавт. (1965,
<. 443)].
Кариотип. Цитогенетика серой крысы относительно хорошо изуче-
на. Стандартный диплоидный хромосомный набор состоит из 42 хромо-
<ом: 7 пар метацентриков, 4 пары субтелоцентриков, 9 пар акроцентри-
ков и акроцентрические Х и У хромосомы ({МЕа
= 62). Для диких
‘и лабораторных популяций зарегистрирован полиморфизм по положению
центромеры в Х хромосоме и многих зутосомах. В большинстве случаев
зн рассматривался как результат перицентрических инверсий.
Экология. Экзоантропные серые крысы — околоводные грызуны.
`Они хорошо плавают и ныряют. Для живущих у воды пасюков водоем
служит источником корма, убежищем от врагов и средой для передви-
жения. Серые крысы хорошо приспособлены к рытью и обычно живут
в норах, в которых проводят значительную часть суток (Карасева и др.,
1985; Милютин и др., 4986). При необходимости пасюки могут забирать-
ся на деревья и экить на чердаках, однако значительно уступают в спо-
28
Глава первая
собности к лазанью ведущим древесный образ жизни п
,
Е:
редставителям
рода (НШ её а., 1983; Аристов и др., 1984; Милютин и др., 1986).
Пасюки исключительно хорошо приспособлены к синантропному образу:
жизни. Синантропные серые крысы предпочитают заселять нижние:
ярусы помещений, подземные коммупикации, канализации. Они обычные
обитатели шахт.
По образу жизни и адаптивным особенностям строения тела серая
крыса сходна с В. 105са и В. агрепИуещег.
Пасюки всеядны. От других представителей рода они отличаются по-
вышенной склонностью к употреблению животной пищи, в том числе
мяса позвоночных животных, которых способны добывать сами.
Характер распределения активности в точение суток лабилен. Серые
крысы активны преимущественно в темное время суток, хотя укесткой
связи с темным периодом нет. Для них, видимо, характерно наличие.
Вор предутренних пиков активности (Рраа, 1965; Козлов, Коз-
Происхождение и история расселения
#2
в
Е
Реконструкция путей расселения серой крысы, возможная на оспове зна-
ния центра и времени ее происхождения, палеогеотрафической и исто-
рической ситуации в период и в районе расселения, а также степепи ва-
гильности вида, дает ключ к пониманию его внутривидовой дифферен-
циации и в какой-то мере также филогенетических связей.
Современный мировой ареал серой крысы показан в настоящей книге.
на карте, составленной В. В. Кучеруком. Пасюк расселился по всем кон-
типоптам, кроме Антарктиды, но далеко не достиг космополитизма.
арактер его распространения и документальные свидетельства говорят"
о том, что большая часть современного ареала сформировалась путем
пассивного расселения с помощью человека
вне]
в исторически н
прошлом.
у
нем
Серая к
И
ИСКОЙ
р р рыса возникла, вероятно, на Великой Китайской равнине.
уда известны самые древние ее паходки, датируемые поздним плей-
стоценом (Громов, 1981), там она обитает в природе и сейчас. Резуль-
ты ета возраста серой крысы с помошью «молекулярных часов»
иворечивы. Возникновение на периферии а
д
еала рода указыв
то, что это молодой вид.
р РОЛА Унааиваот а
Исходя из природных условий Восточной и Юго-Восточной Азии
в плейстоцене-голоцене, истории этих региопов, физических возмож-
стей и экологии серой крысы, мы предполагаем, что ее расселение из.
вичного ареала началось в голоцене и шло в вух о
‹
-
первичи
двух осповных направ
Расестение в северном направлении шло в основном активным
путем. рысы заселили Маньчжурию, Корею, Приморье, Приамурье,
осточную Монголию, Забайкалье. В формировании населения этих тер-
риторий участвовали наиболее северные популяции вида. Нача-
лось оно еще в плейстоцене.
Систематика
29;
Расссление в западном направлении за пределы Китая шло поч-
ти исключительно или исключительно пассивпым способом — на морских
судах, с помощью которых серые крысы проникли на п-ов Индостан,
а оттуда распространились по всему свету вплоть до того же Забайкалья,
достигнутого в отом случае с запада. В формирования населения серых
крыс Индии участвовали восточные приморские популя-
ции. Проникпуть в Индию пасюки могли не ранее Тв. до нашей эры.
Оттуда в УП-ХУ вв. они, вероятно, завозились арабскими мореплава-
телями в порты Персидского залива, Красного моря, Восточной Африки.
Природные условия этих мест, за исключением Месопотамии, крайне
неблагоприятны для обитапия пасюка.
На рубеже ХУ-ХУТ вв. зарождается морская торговля между свро-
пейскими странами и Индией. Это событие повлекло за собой бурное рас-
селение по Европе серой крысы, нашедтей здесь благоприятные клима-
тические и хозяйствеппые условия. Европейские пасюки были затем.
завезены на северное и западиое побережье Африки, в Северную и Юж-
ную Америку, в Австралию и Новую Зеландию.
Детали истории формирования ареала серой крысы продолжают оста-
ваться неясными и спорными (Кузиков, 1983; Милютин, 1983а, б).
Здесь существенно подчеркнуть два момепта.
1. Филогенстически наиболее близкие к серой крысе формы следует
искать в Восточной Азии.
2. Внутри рода В. потуезюи$ можно выделить, по нашему мпению,
наиболее крупные филетические линии: восточноазиатскую и индийскую.
Представители первой обитают в Восточном Китае многие тысячи, воз-
можно, несколько миллионов лет и расширили свой ареал на прилегаю-
щие районы, особенно далеко продвинувшись в северном направлении
Индийская ветвь сформировалась из приморских популяций восточно-
азиатской, возможно, около 2 тыс. лет назад. Исходя из объема морской
торговли между странами и се географических и нолитических обобен-
ностей, можно предполагать, что все остальные регионы Земли, где сей-
час встречается серая крыса, были заселены преимущественно представи-
телями индийской ветви.
Филогенетическое положение
Систематики, опирающиеся па морфологические признаки, едиподушно
относят серых крыс к подроду ВаНиз (ЕПегтап, 1949; М!зоппе, 1969;
Миззег, МечусошЬ, 1985), правда, по-разному понимая его состав. Всеми
признана близость В. погуев1стз и Н. гайаз при ясно выраженном морфо-
логическом своеобразии первой. Эллерман (ЕПегтап, 1949) подчеркнул
оба момента, поместив серую крысу в труппу погуер1сиз подрода.
Вайлз.
Кариотип серой крысы, как и других видов рода с 2п = 42, имеет
сходство с кариотипом азиатской формы черной крысы и морфологическя
идентичен одной из вариаций последней. Предполагается, что хромосом-
ный набор серой крысы является производным этой вариации либо воз-
28
Глава первая
собности к лазанью ведущим древесный образ жизни п
,
Е:
редставителям
рода (НШ её а., 1983; Аристов и др., 1984; Милютин и др., 1986).
Пасюки исключительно хорошо приспособлены к синантропному образу:
жизни. Синантропные серые крысы предпочитают заселять нижние:
ярусы помещений, подземные коммупикации, канализации. Они обычные
обитатели шахт.
По образу жизни и адаптивным особенностям строения тела серая
крыса сходна с В. 105са и В. агрепИуещег.
Пасюки всеядны. От других представителей рода они отличаются по-
вышенной склонностью к употреблению животной пищи, в том числе
мяса позвоночных животных, которых способны добывать сами.
Характер распределения активности в точение суток лабилен. Серые
крысы активны преимущественно в темное время суток, хотя укесткой
связи с темным периодом нет. Для них, видимо, характерно наличие.
Вор предутренних пиков активности (Рраа, 1965; Козлов, Коз-
Происхождение и история расселения
#2
в
Е
Реконструкция путей расселения серой крысы, возможная на оспове зна-
ния центра и времени ее происхождения, палеогеотрафической и исто-
рической ситуации в период и в районе расселения, а также степепи ва-
гильности вида, дает ключ к пониманию его внутривидовой дифферен-
циации и в какой-то мере также филогенетических связей.
Современный мировой ареал серой крысы показан в настоящей книге.
на карте, составленной В. В. Кучеруком. Пасюк расселился по всем кон-
типоптам, кроме Антарктиды, но далеко не достиг космополитизма.
арактер его распространения и документальные свидетельства говорят"
о том, что большая часть современного ареала сформировалась путем
пассивного расселения с помощью человека
вне]
в исторически н
прошлом.
у
нем
Серая к
И
ИСКОЙ
р р рыса возникла, вероятно, на Великой Китайской равнине.
уда известны самые древние ее паходки, датируемые поздним плей-
стоценом (Громов, 1981), там она обитает в природе и сейчас. Резуль-
ты ета возраста серой крысы с помошью «молекулярных часов»
иворечивы. Возникновение на периферии а
д
еала рода указыв
то, что это молодой вид.
р РОЛА Унааиваот а
Исходя из природных условий Восточной и Юго-Восточной Азии
в плейстоцене-голоцене, истории этих региопов, физических возмож-
стей и экологии серой крысы, мы предполагаем, что ее расселение из.
вичного ареала началось в голоцене и шло в вух о
‹
-
первичи
двух осповных направ
Расестение в северном направлении шло в основном активным
путем. рысы заселили Маньчжурию, Корею, Приморье, Приамурье,
осточную Монголию, Забайкалье. В формировании населения этих тер-
риторий участвовали наиболее северные популяции вида. Нача-
лось оно еще в плейстоцене.
Систематика
29;
Расссление в западном направлении за пределы Китая шло поч-
ти исключительно или исключительно пассивпым способом — на морских
судах, с помощью которых серые крысы проникли на п-ов Индостан,
а оттуда распространились по всему свету вплоть до того же Забайкалья,
достигнутого в отом случае с запада. В формирования населения серых
крыс Индии участвовали восточные приморские популя-
ции. Проникпуть в Индию пасюки могли не ранее Тв. до нашей эры.
Оттуда в УП-ХУ вв. они, вероятно, завозились арабскими мореплава-
телями в порты Персидского залива, Красного моря, Восточной Африки.
Природные условия этих мест, за исключением Месопотамии, крайне
неблагоприятны для обитапия пасюка.
На рубеже ХУ-ХУТ вв. зарождается морская торговля между свро-
пейскими странами и Индией. Это событие повлекло за собой бурное рас-
селение по Европе серой крысы, нашедтей здесь благоприятные клима-
тические и хозяйствеппые условия. Европейские пасюки были затем.
завезены на северное и западиое побережье Африки, в Северную и Юж-
ную Америку, в Австралию и Новую Зеландию.
Детали истории формирования ареала серой крысы продолжают оста-
ваться неясными и спорными (Кузиков, 1983; Милютин, 1983а, б).
Здесь существенно подчеркнуть два момепта.
1. Филогенстически наиболее близкие к серой крысе формы следует
искать в Восточной Азии.
2. Внутри рода В. потуезюи$ можно выделить, по нашему мпению,
наиболее крупные филетические линии: восточноазиатскую и индийскую.
Представители первой обитают в Восточном Китае многие тысячи, воз-
можно, несколько миллионов лет и расширили свой ареал на прилегаю-
щие районы, особенно далеко продвинувшись в северном направлении
Индийская ветвь сформировалась из приморских популяций восточно-
азиатской, возможно, около 2 тыс. лет назад. Исходя из объема морской
торговли между странами и се географических и нолитических обобен-
ностей, можно предполагать, что все остальные регионы Земли, где сей-
час встречается серая крыса, были заселены преимущественно представи-
телями индийской ветви.
Филогенетическое положение
Систематики, опирающиеся па морфологические признаки, едиподушно
относят серых крыс к подроду ВаНиз (ЕПегтап, 1949; М!зоппе, 1969;
Миззег, МечусошЬ, 1985), правда, по-разному понимая его состав. Всеми
признана близость В. погуев1стз и Н. гайаз при ясно выраженном морфо-
логическом своеобразии первой. Эллерман (ЕПегтап, 1949) подчеркнул
оба момента, поместив серую крысу в труппу погуер1сиз подрода.
Вайлз.
Кариотип серой крысы, как и других видов рода с 2п = 42, имеет
сходство с кариотипом азиатской формы черной крысы и морфологическя
идентичен одной из вариаций последней. Предполагается, что хромосом-
ный набор серой крысы является производным этой вариации либо воз-
30
Глава нервая
Рис. 8. Гипотетицеские дис-
танции между четырьмя ви-
дами Паниз, основанные на
данных по развитию гиб-
ридных эмбрионов (из: Уоз-
да, 1985)
К. ПОРГЕЙГИЯ
Ю.аппатие
ник независимо от нее в результате перицентрических инверсий акро-
центрических хромосом осповного кариотипа черной крысы (Уозаа, 1980,
1985; ВауетзфосК её 21., 1983). Изучение внутренней структуры хромосом
продемонстрировало поверхностность сходства кариотипов азиатской чер-
ной и серой крыс. Оказалось, что они различаются по С-полосатости
(у серой крысы участки гетерохроматина маленькие и неясные) и по ло-
кализации зоны ядрышкового организатора (МОВ). Последняя находится
у черных крыс и хромосомных парах 3, 8 и 13, а у серых— 3, 12 и 13
{Уоза, 1985).
Мпогочисленные биохимические исследования белков и ДНК крыс
< последующим вычислением генетических дистанций между таксонами
пока не пролили свет на их филогенетические связи. Результаты этих
работ настолько не согласуются друг с другом, что количество предло-
женных филогепетических древ практически равно количеству публи-
каций.
Так, японские генетики (МоггуаЁК её а]., 1974; НауазН} её а[., 1979)
утверждали, что серая крыса ближе к азиатской черной крысе, чем
последняя к океанической форме свосго же вида, однако последующие
исследования других авторов этих выводов не подтвердили (Разеог
её а1., 1982; ВауегеюсК еф а]., 1983).
По Геммеке, Нитхаммеру и Баверстоку с соавторами, генетическая
дистанция между серой и черной крысами такая же, как между послед-
ней и В. ехп]апз, тогда как, по Частеру, с соавторами, серая крыса гораз-
до ближе к черной, чем В. ехи]ааз (Разфетг её э1., 1982; ВауетаюсК её
21., 1983; Сетштеке, Фе атшатег, 1984).
Согласно одному источнику (Разйеиг ей а]., 1982), генетическая дис-
танция между серой и черной крысами такая же, как между черной
к
з
Систематика
З1
крысой и Н. Ношашсиз, согласно другому (Сеттеке, Фе фатшииег, 1984),
В. Ноташсйз ближе к черной крысе, чем пасток.
По Баверстоку с соавторами (ВауегчюсК её г[., 1983), В. 1озса ближе
к черной крысе, чем серая.
Совершенно иначе подошли к решению проблемы Иосида и Тайя
(Тауа) (пит по: Уоз а, 1985). Они осеменили искусственпо самок серых
крыс спермой самцов Ц. га аз, В. аппапда!е! и В. 1озеа. Во всех трех
вариантах происходило оплодотворение, но эмбрионы погибали соответ-
ственно через две недели, одну неделю и на стадии морулы. На основе
этих данных построена схема гипотетических дистанций между этими
четырьмя видами (рис. 2). Мы видим, что чем короче линия, соединяю-
щая на рисунке два вида, тем ближе их родетво.
Внутривидовая структура. Опираясь па данпые по исто-
рии формирования ареала серой крысы, можно предположить наличие.
двух наиболее крупных внутривидовых структурных едипиц, упомянутых
выше как восточноазиатская и индийская филетические ветви. Эти
структурные единицы соответствуют двум номинальным подвидам:
В. и. погуерсиз и В. п. сагасо. Деление В. погуееиз на эти два под-
вида согласуется со взглядами Корбета (Сотфеь 1978) и Эллермана’
и Моррисона-Скотта (ЕПегтав, Могйзоп-Зе04, 1951). Последние авторы
упоминают также третий подвид: Н. п. 1опеаидиз Мом, но это назва-
нис пыне принадлежит синонимике черной крысы (\Уоп, \Noоо, 1960).
Джоне и Джонсон (Топез, Торизов, 1965), помимо погуезсиз и сагасо,
считают действительными названия зосег и 1150|аз, вое и тзо|айлз,
а Ву (\\п, 1982) — ВитШайаз и вое.
Принятая здесь трактовка отражает лишь самый верхний уровепь
сложной таксономической структуры В. погуезеиз, которая, можно
надеяться, будет вскрыта последующими исследованиями.
Помимо двух подвидов диких серых крыс, в этом разделе под назва-
нием В. п. {. дотезйсиз кратко рассматривается их одомашненная форма,
обычно называемая «лабораторной крысой».
ДаНиз погоесив потоесиз (ВегКкеппош, 1769)
1769. Маз погуе1стз Всткепроць Оп тез о {Ве Мака! Н15®юту о! Огеав
ВгНаш ап9 Гге]ал9, у01. 1, р, 5. Великобритания.
1778. Миз 4есмшапиз РаПаз, Моуае бресез Опадгаредит с СН
Ог@1ле: р. 94. Европа.
1779. М (из) загтоТобав Зеусгиоизв, Телйатеп 7001. Напрансае: р. 73.
Центральная Европа.
1800. М (из) а(есишавиз) ВуБмаиз Весвэ ют, Реппализ аПоеш. 'ОЪог-
58% 4. УзегРавз ТЫеге, ВЧ. 2. 8. 497, 713. Вальтерсхаузен, Восточ-
ная Германия (описан экземпляр черной окраски).
1816. Миз сазргз ОКеп, ГевЬ. 9. Мабагя., Ва. 3, 1аЁ. 2, $. 895 (предло-
жено вместо десатапиз).
1837. Миз БФегизсиз ТВотарзоп, Ргос. 7001. Зое. Топдоп. Ва. 52. (Раз-
фриленд, Даун, Ирландия).
1839. Миз шапгаз У’э4егВотсе, 7001. Веазе, р. 31. Америка.
30
Глава нервая
Рис. 8. Гипотетицеские дис-
танции между четырьмя ви-
дами Паниз, основанные на
данных по развитию гиб-
ридных эмбрионов (из: Уоз-
да, 1985)
К. ПОРГЕЙГИЯ
Ю.аппатие
ник независимо от нее в результате перицентрических инверсий акро-
центрических хромосом осповного кариотипа черной крысы (Уозаа, 1980,
1985; ВауетзфосК её 21., 1983). Изучение внутренней структуры хромосом
продемонстрировало поверхностность сходства кариотипов азиатской чер-
ной и серой крыс. Оказалось, что они различаются по С-полосатости
(у серой крысы участки гетерохроматина маленькие и неясные) и по ло-
кализации зоны ядрышкового организатора (МОВ). Последняя находится
у черных крыс и хромосомных парах 3, 8 и 13, а у серых— 3, 12 и 13
{Уоза, 1985).
Мпогочисленные биохимические исследования белков и ДНК крыс
< последующим вычислением генетических дистанций между таксонами
пока не пролили свет на их филогенетические связи. Результаты этих
работ настолько не согласуются друг с другом, что количество предло-
женных филогепетических древ практически равно количеству публи-
каций.
Так, японские генетики (МоггуаЁК её а]., 1974; НауазН} её а[., 1979)
утверждали, что серая крыса ближе к азиатской черной крысе, чем
последняя к океанической форме свосго же вида, однако последующие
исследования других авторов этих выводов не подтвердили (Разеог
её а1., 1982; ВауегеюсК еф а]., 1983).
По Геммеке, Нитхаммеру и Баверстоку с соавторами, генетическая
дистанция между серой и черной крысами такая же, как между послед-
ней и В. ехп]апз, тогда как, по Частеру, с соавторами, серая крыса гораз-
до ближе к черной, чем В. ехи]ааз (Разфетг её э1., 1982; ВауетаюсК её
21., 1983; Сетштеке, Фе атшатег, 1984).
Согласно одному источнику (Разйеиг ей а]., 1982), генетическая дис-
танция между серой и черной крысами такая же, как между черной
к
з
Систематика
З1
крысой и Н. Ношашсиз, согласно другому (Сеттеке, Фе фатшииег, 1984),
В. Ноташсйз ближе к черной крысе, чем пасток.
По Баверстоку с соавторами (ВауегчюсК её г[., 1983), В. 1озса ближе
к черной крысе, чем серая.
Совершенно иначе подошли к решению проблемы Иосида и Тайя
(Тауа) (пит по: Уоз а, 1985). Они осеменили искусственпо самок серых
крыс спермой самцов Ц. га аз, В. аппапда!е! и В. 1озеа. Во всех трех
вариантах происходило оплодотворение, но эмбрионы погибали соответ-
ственно через две недели, одну неделю и на стадии морулы. На основе
этих данных построена схема гипотетических дистанций между этими
четырьмя видами (рис. 2). Мы видим, что чем короче линия, соединяю-
щая на рисунке два вида, тем ближе их родетво.
Внутривидовая структура. Опираясь па данпые по исто-
рии формирования ареала серой крысы, можно предположить наличие.
двух наиболее крупных внутривидовых структурных едипиц, упомянутых
выше как восточноазиатская и индийская филетические ветви. Эти
структурные единицы соответствуют двум номинальным подвидам:
В. и. погуерсиз и В. п. сагасо. Деление В. погуееиз на эти два под-
вида согласуется со взглядами Корбета (Сотфеь 1978) и Эллермана’
и Моррисона-Скотта (ЕПегтав, Могйзоп-Зе04, 1951). Последние авторы
упоминают также третий подвид: Н. п. 1опеаидиз Мом, но это назва-
нис пыне принадлежит синонимике черной крысы (\Уоп, \Noоо, 1960).
Джоне и Джонсон (Топез, Торизов, 1965), помимо погуезсиз и сагасо,
считают действительными названия зосег и 1150|аз, вое и тзо|айлз,
а Ву (\\п, 1982) — ВитШайаз и вое.
Принятая здесь трактовка отражает лишь самый верхний уровепь
сложной таксономической структуры В. погуезеиз, которая, можно
надеяться, будет вскрыта последующими исследованиями.
Помимо двух подвидов диких серых крыс, в этом разделе под назва-
нием В. п. {. дотезйсиз кратко рассматривается их одомашненная форма,
обычно называемая «лабораторной крысой».
ДаНиз погоесив потоесиз (ВегКкеппош, 1769)
1769. Маз погуе1стз Всткепроць Оп тез о {Ве Мака! Н15®юту о! Огеав
ВгНаш ап9 Гге]ал9, у01. 1, р, 5. Великобритания.
1778. Миз 4есмшапиз РаПаз, Моуае бресез Опадгаредит с СН
Ог@1ле: р. 94. Европа.
1779. М (из) загтоТобав Зеусгиоизв, Телйатеп 7001. Напрансае: р. 73.
Центральная Европа.
1800. М (из) а(есишавиз) ВуБмаиз Весвэ ют, Реппализ аПоеш. 'ОЪог-
58% 4. УзегРавз ТЫеге, ВЧ. 2. 8. 497, 713. Вальтерсхаузен, Восточ-
ная Германия (описан экземпляр черной окраски).
1816. Миз сазргз ОКеп, ГевЬ. 9. Мабагя., Ва. 3, 1аЁ. 2, $. 895 (предло-
жено вместо десатапиз).
1837. Миз БФегизсиз ТВотарзоп, Ргос. 7001. Зое. Топдоп. Ва. 52. (Раз-
фриленд, Даун, Ирландия).
1839. Миз шапгаз У’э4егВотсе, 7001. Веазе, р. 31. Америка.
32
Глава первая
1841. Маз аеситато!ез
(пошеп пидит).
1842. Маз 1епеозюги
й
рм
тпит Вйрре!, Миз. ЗепсКепЬ., ВЧ. 3, $. 108, 146. Эри-
8. Миз шапетааз Ма
|
8
спег, АтсВ. Мабагр., Ва. 14, 8. 186. Е
1904. Миз {погус1сиз) БоЁтат! Тгоцеззаге (но не МайзеШе 1904) Сат
1904 шаттва]. Зпрр/., р. 365. Сулавеси,
’
|
- 48 ПОогУуе1сиз ргаезбатз 'Тргопеззать, 114. р.
4918 дя прооккупировапного названия оНшае 1) 540 (потеп поушшо
. уз зу|уаНсиз 41зс0]ог Моась, 7. Рог п Та
1
.
. ТазЧмезсп, Вет].
.!
194 5. 466. Эберсвальде близ Берлина, Восточная Германии то, 4. 50,
а: (?} Чесатут Стапа1 ет, ВиЦ. Миз. Раш, у01. 2, 6, р. 478. Мадагаска
. Вайиз погуся1сиз агоопаща Глоу, Уч. зап. Горьк. ун-та, т. 14, с 39.
д Полхидокая низменность, Грузия.
и
иагнов. среднем крупнее В. п. сагасо, ступня
длипнее, хвост менее густо покрыт волосками, меховой покров ры
аспрострапение. Семикосмополитическое.
ПаЙиз потоесиз сатасо (РаЦаз, 1778)
1778. Миз сагасо РаПаз, Мочае Зресу
‚
речез Опайгиред
У
тае:
Р. 91, р. 335. Китай, Восточная ие претит о ОННиа Оттаве:
Нойезов, 7. Азаь Зос. Вепя., у01. 10, р. 915
Рис. 3. Миз сатасо (=В. п. сагасо)
— копия
описания Палласа (РаИаз, 1778, ва. ХхХША)}
уветной таблицы из оригинального
Системахика
33
1865. Маз ВаШамаз МИле-Едужагаз, ПоспегсВез Н136. Ма. 4ез Мапии!-
{егез: р. 187. Окрестности Пекина, Китай.
1871. Миз вызе!рескав МИл-Едхуатв, Моцу. Атсв. Миз. ВоЦ., у01. 1, р. 93.
Сычуань, Китай.
1874. Маз рипафеиз МИпе-Ейуагсз, ВеспегсВез Н 15, Маё. 4ез МапапыЁе-
тез, р. 138. Хэбей, Китай.
41912. Миз погуесиз ргитатгз КазбзеВепко, Ежегодник Зоол. муз.
Импер. акад. наук. т. 17, с. 376. Забайкальс.
1914. Ерыпуз потуссиз зосег МШег, Ргое. Вю. бое. УУазьпидоп,
\01. 27, р. 90. Ганьсу, Витай.
1927. Вайав Вайпыайлз Шзо]афаз Но\меП, No9. уо]. 40, р. 44. Шаньси,
Китай.
1928. Вайав ВишЩатаз зожегру: Но\уеП, 119., ухо]. 44, р. 42. Гирин,
Китай.
Диагноз. В среднем мельче номинативного подвида, ступня относи-
тельно короче, хвост гуще покрыт волосками, меховой покров мягкий
(рис. 3).
Распространение. Восточный Китай, Корея, Восточная Монго-
лия, Сахалин и Хоккайдо.
Примечание. Интересно замечание А. И. Аргиропуло '(1940), что
карако лучше номинативного подвига приспособлена к рытью и в связи
© этим «отличается от типичной формы более коротким хвостом, массив-
ным черепом и итирокими скулами, признаками определенными, но столь
мелкими, что они иногда улавливаются лишь на сериях».
ВаНиз потоетсиз }. аотезИсиз
Робинсон (НоБМтзоп, 1965) описывает процесс доместикации серых
крыс следующим образом. По крайней мере с 1800 г.в Англии и Фран-
ции была популярна травля крыс собаками. Обученных терьеров сажа-
ли в яму, где находилось 100—200 крыс. Побеждала собака, быстрее
других убившая всех крыс. Для нужд этого «спорта» отлавливали огром-
ное количество крыс, а понадавшихся изредка альбипосов оставляли
для разведения и демонстрации.
Первая работа, в которой отмечено применение подопытных крыс,
была выполнена Филипо (РиШрезих) и опубликована в 1856 г. В 1877—
1885 гг. Крамп (Сгатшре) проводит эксперименты по разведению пасюков-
альбиносов и скрещиванию их с дикими. На рубеже ХХ и ХХ вв. белые
крысы европейского происхождения завозятся в Америку и дают нача-
ло ныне широко распространенной линии Вистар (КоМтзоп, 1965).
Доместицированные пасюки размножаются под контролем человека
и практически полностью изолированы от дикой формы. Жесткий отбор
и инбридирование в сочетании с быстрой сменой поколений оказали
сильнейшее воздействие на генетическую конституцию животных. Сов-
ременная «лабораторная крыса» такое же домашисе животное, как
собака или корова.
2 Серая крыса
32
Глава первая
1841. Маз аеситато!ез
(пошеп пидит).
1842. Маз 1епеозюги
й
рм
тпит Вйрре!, Миз. ЗепсКепЬ., ВЧ. 3, $. 108, 146. Эри-
8. Миз шапетааз Ма
|
8
спег, АтсВ. Мабагр., Ва. 14, 8. 186. Е
1904. Миз {погус1сиз) БоЁтат! Тгоцеззаге (но не МайзеШе 1904) Сат
1904 шаттва]. Зпрр/., р. 365. Сулавеси,
’
|
- 48 ПОогУуе1сиз ргаезбатз 'Тргопеззать, 114. р.
4918 дя прооккупировапного названия оНшае 1) 540 (потеп поушшо
. уз зу|уаНсиз 41зс0]ог Моась, 7. Рог п Та
1
.
. ТазЧмезсп, Вет].
.!
194 5. 466. Эберсвальде близ Берлина, Восточная Германии то, 4. 50,
а: (?} Чесатут Стапа1 ет, ВиЦ. Миз. Раш, у01. 2, 6, р. 478. Мадагаска
. Вайиз погуся1сиз агоопаща Глоу, Уч. зап. Горьк. ун-та, т. 14, с 39.
д Полхидокая низменность, Грузия.
и
иагнов. среднем крупнее В. п. сагасо, ступня
длипнее, хвост менее густо покрыт волосками, меховой покров ры
аспрострапение. Семикосмополитическое.
ПаЙиз потоесиз сатасо (РаЦаз, 1778)
1778. Миз сагасо РаПаз, Мочае Зресу
‚
речез Опайгиред
У
тае:
Р. 91, р. 335. Китай, Восточная ие претит о ОННиа Оттаве:
Нойезов, 7. Азаь Зос. Вепя., у01. 10, р. 915
Рис. 3. Миз сатасо (=В. п. сагасо)
— копия
описания Палласа (РаИаз, 1778, ва. ХхХША)}
уветной таблицы из оригинального
Системахика
33
1865. Маз ВаШамаз МИле-Едужагаз, ПоспегсВез Н136. Ма. 4ез Мапии!-
{егез: р. 187. Окрестности Пекина, Китай.
1871. Миз вызе!рескав МИл-Едхуатв, Моцу. Атсв. Миз. ВоЦ., у01. 1, р. 93.
Сычуань, Китай.
1874. Маз рипафеиз МИпе-Ейуагсз, ВеспегсВез Н 15, Маё. 4ез МапапыЁе-
тез, р. 138. Хэбей, Китай.
41912. Миз погуесиз ргитатгз КазбзеВепко, Ежегодник Зоол. муз.
Импер. акад. наук. т. 17, с. 376. Забайкальс.
1914. Ерыпуз потуссиз зосег МШег, Ргое. Вю. бое. УУазьпидоп,
\01. 27, р. 90. Ганьсу, Витай.
1927. Вайав Вайпыайлз Шзо]афаз Но\меП, No9. уо]. 40, р. 44. Шаньси,
Китай.
1928. Вайав ВишЩатаз зожегру: Но\уеП, 119., ухо]. 44, р. 42. Гирин,
Китай.
Диагноз. В среднем мельче номинативного подвида, ступня относи-
тельно короче, хвост гуще покрыт волосками, меховой покров мягкий
(рис. 3).
Распространение. Восточный Китай, Корея, Восточная Монго-
лия, Сахалин и Хоккайдо.
Примечание. Интересно замечание А. И. Аргиропуло '(1940), что
карако лучше номинативного подвига приспособлена к рытью и в связи
© этим «отличается от типичной формы более коротким хвостом, массив-
ным черепом и итирокими скулами, признаками определенными, но столь
мелкими, что они иногда улавливаются лишь на сериях».
ВаНиз потоетсиз }. аотезИсиз
Робинсон (НоБМтзоп, 1965) описывает процесс доместикации серых
крыс следующим образом. По крайней мере с 1800 г.в Англии и Фран-
ции была популярна травля крыс собаками. Обученных терьеров сажа-
ли в яму, где находилось 100—200 крыс. Побеждала собака, быстрее
других убившая всех крыс. Для нужд этого «спорта» отлавливали огром-
ное количество крыс, а понадавшихся изредка альбипосов оставляли
для разведения и демонстрации.
Первая работа, в которой отмечено применение подопытных крыс,
была выполнена Филипо (РиШрезих) и опубликована в 1856 г. В 1877—
1885 гг. Крамп (Сгатшре) проводит эксперименты по разведению пасюков-
альбиносов и скрещиванию их с дикими. На рубеже ХХ и ХХ вв. белые
крысы европейского происхождения завозятся в Америку и дают нача-
ло ныне широко распространенной линии Вистар (КоМтзоп, 1965).
Доместицированные пасюки размножаются под контролем человека
и практически полностью изолированы от дикой формы. Жесткий отбор
и инбридирование в сочетании с быстрой сменой поколений оказали
сильнейшее воздействие на генетическую конституцию животных. Сов-
ременная «лабораторная крыса» такое же домашисе животное, как
собака или корова.
2 Серая крыса
Глава вторая.
Ареал
Принято считать, что пасюк распространился по воему этру и аселит
` ‹и сплошь все материки, за ис
.
ав © тим авторы большей части сводок по млекопитающим, или.
грызупам избегают подробпого о
сколько а
пли ином регионе и пе приводят
‚Вото рим.
ров. В подробнейших монографиях по менотита И
де пОЕ-
Келсона и Холла (НаП, Кезоп, 1959; Най,
,
00 картосхем ареалов ее
„отеутотвуют кары
ка и пе даны описания
Америке. В сводке по грызунам Индии и прилежащих стран дерма
(ЕЦегтап, 1961) нет описания распространения пасюка ие ре
пы точки его находок на рассматриваемой территор ан он
(Зашё Сутопз, 1973) в монографии по млекопитающим рати рт
Кенилюкса посвятила описанию распространения пасюка вс ‹
„
<
к находок.
|
ое В тех случаях, ога авторы приводят карты ареала паса, ‚они
обычно сплошь заштриховывают всю. отисывасмую территорию кота
пространения пасюка в Ёвро
Ве о Я, 1956). Аналогична карта ия
в США (без Аляски), приводимая Брауном (Вгочтт, 1: о. д тет,
в 14 томе «Энциклопедии жизни животных» Гржимека ни терлем
(Г1еещет) привел карту ареала рода Ваниз (с. 356), на Которой оплот
ной штриховкой от северных до южных пределов материк вл а
Африки, Евразии, Северной и Южной Америки показано распр
авы совстских сводках также бытует представление о прак
тически всесветном распространении серой крысы. А. _Н. \ргиропуло
(1940, с. 52) пишет: «Тичичная форма пасюка, родина которой и ‚уста
повлена, сейчас распространена по всему земному шару, кро ани
северных пунктов, не заселенпых оседлым человеком». Б. С. отраоя
иА. Н. Аргиропуло (1944, с. 152—154} писали о пасюке: | „р спроста
нен в настоящее время по всему земному шару, кроме краин их ‚север
ных районов и области пустынь». Почти дословно эта формулировя о
вторена в следующем издании определителя трызупов Б. ‚Вито граде.
вым, а также И. М. Громовым (1952) в опредехителе ме о таощих
{Громов и др., 19658) ив «Систематике млекопитающих .. около.
вым (1977). Н. А. Бобринский и др. (1965, с. ) утвер да т более
решительно: «..распространен почти по всему земному шару». т
формулировка использована А. П. Кузякиным © ‚соавтор
И
в 6м томе «Жизнь животных». В «Каталоге млекопитающих, ССР
(1981) о серой крысе написано кратко и определенно: распростр
всесветно.
Ареал
35
На самом деле подобные формулировки весьма неточно отражают
истинное положение. От пасюка свободны большая часть Африки, Юж-
ной Америки, Австралии. В Азии им занято не более половины конти-
непта. Лишь в умеренном климате больтая часть Северной Америки и
подавляющая территория Европы заселены серой крысой.
История изучения ареала пасюка
Первое полноценное исследование ареала пасюка принадлежит А. П. Ку-
зякину (1954). Это большая статья, посвященная истории расселения,
современному распространению и местообитаниям пасюка в СССР. В ней
перечислены все имеющиеся в литературе данные о местах достоверных
находок пасюка па территории СССР и МНР. По сути дела, данная
работа явилась первым опытом составления кадастра ареала конкретного
вида млекопитающих на территории СССР. На приведенной картосхеме
‘очерчены предполагаемые границы ареала. Отсутствие пасюка впутри
ареала обозначено для Калмыкии, Волжско-Уральского междуречья,
к югу от Ладожского озера, в междуречьях Северной Двины и Печоры,
Печоры и Оби. Видимо, северная граница ареала пасюка проведена‘
А. П. Вузякиным пе совсем точно. Она показана по морскому побережью
от западной границы СССР до устья Печоры и по Северному полярному
кругу от Печоры до Оби. В то же время он сам пишет, что от пасюка
‹вободна вся зона тундр и лесотундр Евразии (за исключением Мур-
манска, нижней Печоры и Салехарда) и почти вся таежная полоса
северо-востока европейской части СССР, Урала и Западной Сибири.
Несмотря на указанные выше недостатки, исследование Л. П. Кузякина
явилось крупным шагом в изучении ареала пасюка на Рьропейском кон-
тиненте. Карта ареала пасюка, составленная А. П. Кузякиным, была
многократно повторно опубликована в слегка упрощенном виде (Гамба-
рян, Дукельская, 1955; Айзенттадт, 1959; Кузякин и др., 1971; Кучерук,
Кузиков, 1985).
В {950 г. Шварцем (Зем аги, 1960) впервые была опубликована кар-
та мирового ареала серой крысы. Он считал, что сс арсал занимает почти
всю Европу, включая средиземноморские острова. Южная траница ареала
здесь идет полосой вдоль северо-восточного побережья Средиземпого
моря и обрамляет южный берег Черного моря. От восточного окончания
последнего она идет к южному берегу Каспия. Внутренние части Тур-
ции и Восточная Сирия свободны от крысы. Западный и северный берега
Каспийского моря служат восточной границей ареала пасюка. Далее
южная граница подходит к райопу Уральска, от которого ареал узкой
полоской тянется к востоку.
Северо-восточная граница европейской части ареала пасюка целиком
лежит в пределах СССР. Она идет вдоль западного побережья Белого
моря, сворачивает на запад к Финскому заливу Балтийского моря. От
последнего резко поворачивает па юго-восток, пересекает Волгу на уров-
ие Саратова и р. Урал в районе Уральска. Восточнее Уральска ареал
идет узкой полосой, которая в представлении автора, видимо, тянется
2*
Глава вторая.
Ареал
Принято считать, что пасюк распространился по воему этру и аселит
`
‹и сплошь все материки, за ис
.
ав © тим авторы большей части сводок по млекопитающим, или.
грызупам избегают подробпого о
сколько а
пли ином регионе и пе приводят
‚Вото рим.
ров. В подробнейших монографиях по менотита И
де пОЕ-
Келсона и Холла (НаП, Кезоп, 1959; Най,
,
00 картосхем ареалов ее
„отеутотвуют кары
ка и пе даны описания
Америке. В сводке по грызунам Индии и прилежащих стран дерма
(ЕЦегтап, 1961) нет описания распространения пасюка ие ре
пы точки его находок на рассматриваемой территор ан он
(Зашё Сутопз, 1973) в монографии по млекопитающим рати рт
Кенилюкса посвятила описанию распространения пасюка вс ‹
„
<
к находок.
|
ое В тех случаях, ога авторы приводят карты ареала паса, ‚они
обычно сплошь заштриховывают всю. отисывасмую территорию кота
пространения пасюка в Ёвро
Ве о Я, 1956). Аналогична карта ия
в США (без Аляски), приводимая Брауном (Вгочтт, 1: о. д тет,
в 14 томе «Энциклопедии жизни животных» Гржимека ни терлем
(Г1еещет) привел карту ареала рода Ваниз (с. 356), на Которой оплот
ной штриховкой от северных до южных пределов материк вл а
Африки, Евразии, Северной и Южной Америки показано распр
авы совстских сводках также бытует представление о прак
тически всесветном распространении серой крысы. А. _Н. \ргиропуло
(1940, с. 52) пишет: «Тичичная форма пасюка, родина которой и ‚уста
повлена, сейчас распространена по всему земному шару, кро ани
северных пунктов, не заселенпых оседлым человеком». Б. С. отраоя
иА. Н. Аргиропуло (1944, с. 152—154} писали о пасюке: | „р спроста
нен в настоящее время по всему земному шару, кроме краин их ‚север
ных районов и области пустынь». Почти дословно эта формулировя о
вторена в следующем издании определителя трызупов Б. ‚Вито граде.
вым, а также И. М. Громовым (1952) в опредехителе ме о таощих
{Громов и др., 19658) ив «Систематике млекопитающих .. около.
вым (1977). Н. А. Бобринский и др. (1965, с. ) утвер да т более
решительно: «..распространен почти по всему земному шару». т
формулировка использована А. П. Кузякиным © ‚соавтор
И
в 6м томе «Жизнь животных». В «Каталоге млекопитающих, ССР
(1981) о серой крысе написано кратко и определенно: распростр
всесветно.
Ареал
35
На самом деле подобные формулировки весьма неточно отражают
истинное положение. От пасюка свободны большая часть Африки, Юж-
ной Америки, Австралии. В Азии им занято не более половины конти-
непта. Лишь в умеренном климате больтая часть Северной Америки и
подавляющая территория Европы заселены серой крысой.
История изучения ареала пасюка
Первое полноценное исследование ареала пасюка принадлежит А. П. Ку-
зякину (1954). Это большая статья, посвященная истории расселения,
современному распространению и местообитаниям пасюка в СССР. В ней
перечислены все имеющиеся в литературе данные о местах достоверных
находок пасюка па территории СССР и МНР. По сути дела, данная
работа явилась первым опытом составления кадастра ареала конкретного
вида млекопитающих на территории СССР. На приведенной картосхеме
‘очерчены предполагаемые границы ареала. Отсутствие пасюка впутри
ареала обозначено для Калмыкии, Волжско-Уральского междуречья,
к югу от Ладожского озера, в междуречьях Северной Двины и Печоры,
Печоры и Оби. Видимо, северная граница ареала пасюка проведена‘
А. П. Вузякиным пе совсем точно. Она показана по морскому побережью
от западной границы СССР до устья Печоры и по Северному полярному
кругу от Печоры до Оби. В то же время он сам пишет, что от пасюка
‹вободна вся зона тундр и лесотундр Евразии (за исключением Мур-
манска, нижней Печоры и Салехарда) и почти вся таежная полоса
северо-востока европейской части СССР, Урала и Западной Сибири.
Несмотря на указанные выше недостатки, исследование Л. П. Кузякина
явилось крупным шагом в изучении ареала пасюка на Рьропейском кон-
тиненте. Карта ареала пасюка, составленная А. П. Кузякиным, была
многократно повторно опубликована в слегка упрощенном виде (Гамба-
рян, Дукельская, 1955; Айзенттадт, 1959; Кузякин и др., 1971; Кучерук,
Кузиков, 1985).
В {950 г. Шварцем (Зем аги, 1960) впервые была опубликована кар-
та мирового ареала серой крысы. Он считал, что сс арсал занимает почти
всю Европу, включая средиземноморские острова. Южная траница ареала
здесь идет полосой вдоль северо-восточного побережья Средиземпого
моря и обрамляет южный берег Черного моря. От восточного окончания
последнего она идет к южному берегу Каспия. Внутренние части Тур-
ции и Восточная Сирия свободны от крысы. Западный и северный берега
Каспийского моря служат восточной границей ареала пасюка. Далее
южная граница подходит к райопу Уральска, от которого ареал узкой
полоской тянется к востоку.
Северо-восточная граница европейской части ареала пасюка целиком
лежит в пределах СССР. Она идет вдоль западного побережья Белого
моря, сворачивает на запад к Финскому заливу Балтийского моря. От
последнего резко поворачивает па юго-восток, пересекает Волгу на уров-
ие Саратова и р. Урал в районе Уральска. Восточнее Уральска ареал
идет узкой полосой, которая в представлении автора, видимо, тянется
2*
36
Глава вторая
Ареал
37
вдоль транссибирской железнодорожной магистрали до Байкала, нижнего
течения Аргуни и выходит к устью Амура. К югу здесь лежит обширное
пятно ареала, занимающее южную часть советского Дальнего Востока
и восточную часть Китая на юг до границ Бирмы, Лаоса и Вьетнама.
Я
Кроме того, указано сплошное распространение серой крысы на Камчат-
ке, Сахалине, Японских островах и Корейском полуострове. Свободен от
крысы Сихоте-Алинь (рис. 4).
Видимо, Е. Шзарцу не были известны вышеупомянутые публикации
советских специалистов, поэтому приведенные им очертания ареала пасю-
ка на территории СССР весьма далеки от истинных.
В Африке обитание серой крысы отмечено Е. Шзарцем только по
восточному и южному побережьям континента. Ее наличие показано
лишь в райопах крупных портов: Момбасы, Мозамбика, Мапуту, Дурба-
на, Порт-Елизабет, Кейптауна. Автор специально отмечает, что крыса
есть в Момбасе, но не заселила Найроби; есть в Кейптауне, но не про-
пикла в Йоханнесбург и Преторию. На Мадагаскаре обитание пасюка
5)
показано в одном месте— на восточном берегу острова в районе порта
Таматаве. Не показано наличия пасюка в Северо-Западной Африке по
побережью Атлантического океана и западной части Средиземного моря.
А
Распространение пасюка в Австралии Шварц изобразил шестью огра-
-1
Ь
ниченными, не связанными между собою пятнами, приуроченными к рас-
положению крупных портовых городов: Маунт-Морган, Брисбен, Сидней,
|
1
Мельбурн, Порт-Пири и Порт-Огаста, Перт. Кроме того, показано обита-
3 ||]
ние пасюка на южном и северном островах Новой Зеландии, на Новых
у
Гебридах и на Гавайских островах.
В Северной Америке этот автор следующим образом очерчивает ареал
мы
серой крысы. На Аляске пасюк заселил узкую полосу вдоль побережья
у
3
Чукотского моря на запад от мыса Барроу до границы с Канадой. Далее
такой же узкой полосой вдоль тихоокеанского побережья Канады ареал
спускается до Ванкувера. От последнего северная граница ареала пасюка.
круто поворачивает па северо-восток и идет вдоль Саскачевана до север-
ного окончания залива Джеймса. П-ов Лабрадор она пересекает по линии
от Форт-Джоржа на западе до устья р. Черчилл на востоке. К югу от
этой границы пасюк сплошь распространси по Канаде и США. В Мек-
сике занимает большую половипу страны, доходя на юг до 20° с. ш. Кро-
ме того, крыса заселила весь п-ов Юкатан, Багамские, Большие и Малые
Антильские, а также все Алеутские острова.
Следует отметить, что на карте в полевом определителе млекопитаю-
щих Северной Америки Р. Палмер (Рарлег, 1954) несколько иначе про-
водит северную границу ареала пасюка в Канаде. Этот вид лишь незна-
чительно заходит в юто-восточную часть провинции Британская Колум-
бия, занимает самые южные части провинций Саскачеван и Манитоба.
Далее граница ареала идет по северному побережью оз. Верхнее, рекам
Оттаве и Св. Лаврентия и выходит на побережьс Атлантического океана
южнее залива св. Лаврентия. Северпее пасюк обитает здесь лишь на
островах Антикости и Ньюфаундленд.
п
о
и
,
и
к
”
А
И
и
7
”
р
а
Р
и
с
.
4
.
М
и
р
о
в
о
й
а
р
е
а
л
п
а
с
ю
к
а
(
п
о
:
б
с
в
р
а
г
л
,
1
9
6
0
)
36
Глава вторая
Ареал
37
вдоль транссибирской железнодорожной магистрали до Байкала, нижнего
течения Аргуни и выходит к устью Амура. К югу здесь лежит обширное
пятно ареала, занимающее южную часть советского Дальнего Востока
и восточную часть Китая на юг до границ Бирмы, Лаоса и Вьетнама.
Я
Кроме того, указано сплошное распространение серой крысы на Камчат-
ке, Сахалине, Японских островах и Корейском полуострове. Свободен от
крысы Сихоте-Алинь (рис. 4).
Видимо, Е. Шзарцу не были известны вышеупомянутые публикации
советских специалистов, поэтому приведенные им очертания ареала пасю-
ка на территории СССР весьма далеки от истинных.
В Африке обитание серой крысы отмечено Е. Шзарцем только по
восточному и южному побережьям континента. Ее наличие показано
лишь в райопах крупных портов: Момбасы, Мозамбика, Мапуту, Дурба-
на, Порт-Елизабет, Кейптауна. Автор специально отмечает, что крыса
есть в Момбасе, но не заселила Найроби; есть в Кейптауне, но не про-
пикла в Йоханнесбург и Преторию. На Мадагаскаре обитание пасюка
5)
показано в одном месте— на восточном берегу острова в районе порта
Таматаве. Не показано наличия пасюка в Северо-Западной Африке по
побережью Атлантического океана и западной части Средиземного моря.
А
Распространение пасюка в Австралии Шварц изобразил шестью огра-
-1
Ь
ниченными, не связанными между собою пятнами, приуроченными к рас-
положению крупных портовых городов: Маунт-Морган, Брисбен, Сидней,
|
1
Мельбурн, Порт-Пири и Порт-Огаста, Перт. Кроме того, показано обита-
3 ||]
ние пасюка на южном и северном островах Новой Зеландии, на Новых
у
Гебридах и на Гавайских островах.
В Северной Америке этот автор следующим образом очерчивает ареал
мы
серой крысы. На Аляске пасюк заселил узкую полосу вдоль побережья
у
3
Чукотского моря на запад от мыса Барроу до границы с Канадой. Далее
такой же узкой полосой вдоль тихоокеанского побережья Канады ареал
спускается до Ванкувера. От последнего северная граница ареала пасюка.
круто поворачивает па северо-восток и идет вдоль Саскачевана до север-
ного окончания залива Джеймса. П-ов Лабрадор она пересекает по линии
от Форт-Джоржа на западе до устья р. Черчилл на востоке. К югу от
этой границы пасюк сплошь распространси по Канаде и США. В Мек-
сике занимает большую половипу страны, доходя на юг до 20° с. ш. Кро-
ме того, крыса заселила весь п-ов Юкатан, Багамские, Большие и Малые
Антильские, а также все Алеутские острова.
Следует отметить, что на карте в полевом определителе млекопитаю-
щих Северной Америки Р. Палмер (Рарлег, 1954) несколько иначе про-
водит северную границу ареала пасюка в Канаде. Этот вид лишь незна-
чительно заходит в юто-восточную часть провинции Британская Колум-
бия, занимает самые южные части провинций Саскачеван и Манитоба.
Далее граница ареала идет по северному побережью оз. Верхнее, рекам
Оттаве и Св. Лаврентия и выходит на побережьс Атлантического океана
южнее залива св. Лаврентия. Северпее пасюк обитает здесь лишь на
островах Антикости и Ньюфаундленд.
п
о
и
,
и
к
”
А
И
и
7
”
р
а
Р
и
с
.
4
.
М
и
р
о
в
о
й
а
р
е
а
л
п
а
с
ю
к
а
(
п
о
:
б
с
в
р
а
г
л
,
1
9
6
0
)
38
Глава вторая
В Южной Америке, по Е. Шварцу, насюк занимает узкую полосу
тихоокеанского побережья от Панамы на севере до 40° ю. иг. на юге.
По атлантическому побережью такой же узкой полосой распространен
от Венесузльского залива на севере до Ла-Платы на юге (см. рис. 4).
Статья Е. Шварца (1960) впервые показала превратность общеприня-
тых представлений об ареале пасюка
ских сведений по этому вопросу.
и явную недостаточность фактиче-
В 1965 г появились две публикации, в которых рассматривался
ареал пасюка во внетропической Евразии. В определителе млекопитаю-
щих СССР Н. А. Бобринский, Б. А.
привели карту ареала пасюка, котора
ликованной А. П. Кузякиным (1951),
Кузнецов, А. П. Кузякин (1965)
я существенно отличалась от олуб-
А. П. Кузякиным, Ц. П. Второвым,
Н. Н. Дроздовым (4971). В пей южная граница ареала на пространстве
от восточного побережья Каспийского моря до Алтайского края была
значительно сдвинута к югу, что в общих чертах подтвердилось и даль
нейшими исследованиями. К востоку
казан выходящим за пределы СССР.
на Южном Ллтае и к югу от Саян.
ареала проходит много севернее, чем
идет по Лене до впадения р. Оленек,
от Зайсанской котловины ареал по-
На самом деле крыса отсутствует
На карте в определителе граница
у А. П. Кузякина. От Братска она
через истоки Зеи и Буреи и далее
севернее Амтгуни выходит на побережье Охотского моря. Насколько
такие очертания ареала соответствуют действительности, мы рассмотрим
в разделе, посвященном современному ареалу пасюка в СССР. Новым
является показ на карте определителя обитапия серой крысы в Форте-
Шевченко, Ташкенте, на крайнем северо-востоке Сибири (Певех, бухта
Провидения, устье Анадыря), Магадане, Охотске, на Командорских и
Курильских островах.
В статье по синантропным грызунам (Киснегак, 1965) дана карта
ареала пасюка во внетронической Евразии, где объединены материалы
А. П. Кузякина (1951), Ван ден Бринка (1956), No. Шварца (1960)
и добавлены некоторые новые материалы об обитании крысы в Дудин-
ке — Норильске, Турухапске, по южному побережью Каспийского моря,
Месопотамской низменности и Турции.
Корбет (СотБеф, 1978) в сводке по млекопитающим Палеарктики дал
схематические карты ареалов всех видов, свойственных этому региопу,
в том числе насюка. Он объединил сведения по Западной Европе & мате-
риапами Н. А. Бобринского и др. (1965) по СССР, несколько схематизи-
ровав последние и повторив все недостатки указанной работы. Ироме
того, был уточнен ареал крысы в Иране и АРЕ по Мизону (М1зоппе,
1960) и Гаррисону (Нагзоп, 1972). На юте, в Нитае, граница ареала
показана не доходящей до 20° с. ш. Почему-то не отмечено обитание пасю-
ка на Камчатке, Чукотке и Курильских островах, неоднократно регист-
рируемое его предшественниками (Кузякин, 1951; Шварц, 1960; Бобрин-
ский и др., 1965; Кучерук, 1965). Не показана территория, заселения
пасюком в средиземноморской Северо-Западной Африке, о которой упо-
минают Петтер и Сан-Жирон (Рейег, Зай\ Суговз, 1965). Не отмечен
Ареал
39
пасюк и на островах Средиземного моря— Корсике, Сардинии и Крите
где его указывал Шварц (1960).
у
В первом томе монографии но млеконитающим Европы глава, посвя-
щенная серой крысе, сопровождается картой ее ареала на этом материке.
Рис. 5. Ареал пасюна в Западной Европе (по: ВесКег, 1978}
1 — территория, сплошь заселенная пасюком; 2 — острова, заселенные пасюком
и пункты находок на территории его спорадического распространения
р
о
У
ср
—
ь
О
о
т
ы
_
х
38
Глава вторая
В Южной Америке, по Е. Шварцу, насюк занимает узкую полосу
тихоокеанского побережья от Панамы на севере до 40° ю. иг. на юге.
По атлантическому побережью такой же узкой полосой распространен
от Венесузльского залива на севере до Ла-Платы на юге (см. рис. 4).
Статья Е. Шварца (1960) впервые показала превратность общеприня-
тых представлений об ареале пасюка
ских сведений по этому вопросу.
и явную недостаточность фактиче-
В 1965 г появились две публикации, в которых рассматривался
ареал пасюка во внетропической Евразии. В определителе млекопитаю-
щих СССР Н. А. Бобринский, Б. А.
привели карту ареала пасюка, котора
ликованной А. П. Кузякиным (1951),
Кузнецов, А. П. Кузякин (1965)
я существенно отличалась от олуб-
А. П. Кузякиным, Ц. П. Второвым,
Н. Н. Дроздовым (4971). В пей южная граница ареала на пространстве
от восточного побережья Каспийского моря до Алтайского края была
значительно сдвинута к югу, что в общих чертах подтвердилось и даль
нейшими исследованиями. К востоку
казан выходящим за пределы СССР.
на Южном Ллтае и к югу от Саян.
ареала проходит много севернее, чем
идет по Лене до впадения р. Оленек,
от Зайсанской котловины ареал по-
На самом деле крыса отсутствует
На карте в определителе граница
у А. П. Кузякина. От Братска она
через истоки Зеи и Буреи и далее
севернее Амтгуни выходит на побережье Охотского моря. Насколько
такие очертания ареала соответствуют действительности, мы рассмотрим
в разделе, посвященном современному ареалу пасюка в СССР. Новым
является показ на карте определителя обитапия серой крысы в Форте-
Шевченко, Ташкенте, на крайнем северо-востоке Сибири (Певех, бухта
Провидения, устье Анадыря), Магадане, Охотске, на Командорских и
Курильских островах.
В статье по синантропным грызунам (Киснегак, 1965) дана карта
ареала пасюка во внетронической Евразии, где объединены материалы
А. П. Кузякина (1951), Ван ден Бринка (1956), No. Шварца (1960)
и добавлены некоторые новые материалы об обитании крысы в Дудин-
ке — Норильске, Турухапске, по южному побережью Каспийского моря,
Месопотамской низменности и Турции.
Корбет (СотБеф, 1978) в сводке по млекопитающим Палеарктики дал
схематические карты ареалов всех видов, свойственных этому региопу,
в том числе насюка. Он объединил сведения по Западной Европе & мате-
риапами Н. А. Бобринского и др. (1965) по СССР, несколько схематизи-
ровав последние и повторив все недостатки указанной работы. Ироме
того, был уточнен ареал крысы в Иране и АРЕ по Мизону (М1зоппе,
1960) и Гаррисону (Нагзоп, 1972). На юте, в Нитае, граница ареала
показана не доходящей до 20° с. ш. Почему-то не отмечено обитание пасю-
ка на Камчатке, Чукотке и Курильских островах, неоднократно регист-
рируемое его предшественниками (Кузякин, 1951; Шварц, 1960; Бобрин-
ский и др., 1965; Кучерук, 1965). Не показана территория, заселения
пасюком в средиземноморской Северо-Западной Африке, о которой упо-
минают Петтер и Сан-Жирон (Рейег, Зай\ Суговз, 1965). Не отмечен
Ареал
39
пасюк и на островах Средиземного моря — Корсике, Сардинии и Крите
где его указывал Шварц (1960).
у
В первом томе монографии но млеконитающим Европы глава, посвя-
щенная серой крысе, сопровождается картой ее ареала на этом материке.
Рис. 5. Ареал пасюна в Западной Европе (по: ВесКег, 1978}
1 — территория, сплошь заселенная пасюком; 2 — острова, заселенные пасюком
и пункты находок на территории его спорадического распространения
р
о
У
ср
—
ь
О
о
т
ы
_
х
40
Глава вторая
Автор главы К. Беккер (Вескег, 1978) считает, что пасюк сплошь за
селяет Исландию, Великобританию, Ирландию и прилежащие к ним мел-
кие острова. На севере континентальной Европы, свободной от пасюка,
показана лить самая северо-восточная часть Порвегии к северу от
плоскогорья Финмаркен. Больптинство островов Балтийского, Северного
морей и Ла-Манша заселено пасюком.
Подавляющая часть Европейского континента сплошь заселена пасю-
ком. По мнению К. Беккера, только в самой южной ее части: па Пире-
нейском, Апеннинском и Балканском полуостровах— пасюх распростра-
нен спорадично. В Португалии показано 2 пункта регистрации пасюка,
в Испании —6, па Лпеннинах— 65, па Балканах—1{1 пункт в Греции
и 2— в европейской части Турции. Среди островов Средиземного моря
обнаружен па Фермептере, Ивисе, Сицилии, Устике, Эгадских островах,
Крите; в Ионическом море— на Закинфе, Кефалинии, Керкире; в Эгей-
ском море-— на Родосе, Нарпатосе, Икарии. По мнению К. Беккера,
пасюк отсутствует на островах: Мальорка, Менорка, Сардиния и Эвбея
{рис. 5).
Сравнительно недавно была опубликована специальная работа, посвя-
щенная современному ареалу серой крысы (Кучерук, Кузиков, 1985).
В ней были приведены карты ареала пасюка в СССР по А, П. Вузякину
(19541), по Н. А. Бобринскому, Б. А. Кузнецову, А. П. Кузякину (1965),
оригинальная карта современного распространения этого вида в СССР,
картосхемы изменений его ареала в СССР за 30 лет (4951—1984), про-
гноза вероятного расширения ареала в СССР на ближайшие годы и сте-
пепи вредоносности в СССР в 1958—1961 гг. по В. В. Груздеву (1963).
Кроме того; повторены карты мирового ареала пасюка из работы
Е. Шварца (1960) и распространения его в Палеарктике из книги
Г. Б. Корбета. Завериталась статья оригинальной картой мирового ареала
этого вида по данным на конец семидесятых годов. Все карты сопровож-
дались подробным описанием и сопоставлением.
Современные знания об ареале пасюка
Мы продолжали сбор и апализ фаунистической литературы © целью
уточнения ареала серой крысы, в том числе и распространения ее на
океанических островах. В последние 8—10 лет появился ряд специальных
статей, уточняющих ареал этого вида в нескольких странах Европы,
а также и в отдельных регионах СССР. В Советском Союзе было про-
ведепо несколько совещаний (1983, 1986, 1987 гг.) и издано четыре сбор-
ника, посвященных серой крысе. Все это позволило существенно уточ-
нить наши представления о распространении этого вида. Начнем © описа-
ния распространения пасюка за пределами СССР.
Зарубежная территория
В Северной Африке пасюк распространен в дельте Нила от Порт-Саида
и Исмалии на востоке до Даманхура, Абу-Хуммуса, Александрии на за-
паде. Вверх по Нилу он прослежен до Исны и Асуана (ВЁааё её а|.,
Ареал
41
1969; Озроги, Нешиу, 1980). Выше по Нилу изолированное поселепие
пасюка существует в Хартуме с начала нынешнего века (ВаКНоит, 1911 3;
Еюоууег, 1932). В 1945 г. мероприятия по ограничению численпости этого
вида в Хартуме проводили УУаов, Ретту (1954). Ф. Диетерлен в феврале
1973 г. наблюдал пасюков на набережной Нила. Он упоминает о паличии
Н Бриталоком музее естественной истории двух экземпляров пасюка, до-
а
РТУ в 1904 и одного в 1929 гг. (ГЛеецев, Мо! ап,
Восточнее по побережью Средиземного моря пасюк обитает в Триполи
{Вапк, 1968), широко распространен в области жестколистных, вечно-
зеленых средиземноморских лесов от Туниса до Марокко (Рейег баш
Сто, 1965). Он обнаружен в городах: Тунис, Алжир Бени-Упиф Фес
Гетуан, Мекнес, Рабат, Эль-Джадида, Атадир — ив устье Ширрата меж
ду „Касабланкой и Рабатом (Реапое, 1925; Толиттет, 1960; МаШоох
1965; МаШочх, Вет тане, 1970; Г.аитеп(, 1937: Бернар, 1974). ’
”
По атлантическому побережью Африки пасюк обитает в крупных
портах: Дакаре, Конакри, Фритауне, Монровии (Рекеузег, 1955; Уда
В1е] её а1., 1969). На участке, прилежащем к Гвинейскому заливу пасюк.
сплошь заселяет побережье от Абиджана до дельты Нигера обитает
в г. Дуала (Двала) и на островах Фернандо-По и Сан-Томе { АЦеп
1939; Усвеке, 1958; Уое]ске!, Уамегаз, 1960). Далее к югу паслок есть
в Пуэнт-Нуар и, по-видимому, в Луанде.
у
"
В ТОАР пасюк силошь заселяет побережье от Кейптауна до Порт-
Еливабет, восточнее он известен из Ист-Лондона, Дурбана. Далее, по
всей вероятности, обитает в Мануту, Иньямбане, Бейре Мозамбике
(Парк, 1960). Достоверно известен в Дар-эс-Саламе, Занзибаре, Садани
томбвсе (Кавдоп, 1974). Из последней по железной дороге проник
в Найроби (Нигз, 1926—1927) и достиг территории Заира на западном
берегу оз. Мобуту-Сесе-Сено.
^
Г. Д. Авери (Ауегу, 1985) считает, что пасюк с равной вероятностью
мог быть завезен в Восточную Африку как из Европы, так и из Азии
(из Йемена или стран Персидского залива в [--ГХ вв, нашей эры) По
мнению этого автора, пасюх отсутствует в Мозамбике, но заселяет по то-
вые города ЮАР, Памибии, Анголы и страп Западной Африки аныо
рапние сведения о пасюке в Нейптауне относятся к 1832 г. ав катал ге
Южно-Африканского музея, изданном в 1862 г., уже указано несколько
экземпляров этого вида. Авери считает, что пасюк плохо приспособлен ‘к
Условиям тропического климата.
В Сомали серая крыса, по всей вероятности, встречается в Могадишо
и Джибути. В Эфиопии достоверно обитает на о-ве Фатма и вг. Массава
(Уаеп её а1., 1976), на суданском побережье в Порт-Судане и Аб -Ра-
миде (Нооззтаа!, 1963; ОзЪоги, Нету, 1980).
у
Как видно из вышеизложенного, пасюк не смог освоить Африканский
материк. Он заселил преимущественно портовые города. В тлубь мате-
рика этот вид сумел распространиться на отдельных неболыших участ-
3 Цит. по; Сгаш, 1973.
40
Глава вторая
Автор главы К. Беккер (Вескег, 1978) считает, что пасюк сплошь за
селяет Исландию, Великобританию, Ирландию и прилежащие к ним мел-
кие острова. На севере континентальной Европы, свободной от пасюка,
показана лить самая северо-восточная часть Порвегии к северу от
плоскогорья Финмаркен. Больптинство островов Балтийского, Северного
морей и Ла-Манша заселено пасюком.
Подавляющая часть Европейского континента сплошь заселена пасю-
ком. По мнению К. Беккера, только в самой южной ее части: па Пире-
нейском, Апеннинском и Балканском полуостровах— пасюх распростра-
нен спорадично. В Португалии показано 2 пункта регистрации пасюка,
в Испании —6, па Лпеннинах— 65, па Балканах—1{1 пункт в Греции
и 2— в европейской части Турции. Среди островов Средиземного моря
обнаружен па Фермептере, Ивисе, Сицилии, Устике, Эгадских островах,
Крите; в Ионическом море — на Закинфе, Кефалинии, Керкире; в Эгей-
ском море-— на Родосе, Нарпатосе, Икарии. По мнению К. Беккера,
пасюк отсутствует на островах: Мальорка, Менорка, Сардиния и Эвбея
{рис. 5).
Сравнительно недавно была опубликована специальная работа, посвя-
щенная современному ареалу серой крысы (Кучерук, Кузиков, 1985).
В ней были приведены карты ареала пасюка в СССР по А, П. Вузякину
(19541), по Н. А. Бобринскому, Б. А. Кузнецову, А. П. Кузякину (1965),
оригинальная карта современного распространения этого вида в СССР,
картосхемы изменений его ареала в СССР за 30 лет (4951—1984), про-
гноза вероятного расширения ареала в СССР на ближайшие годы и сте-
пепи вредоносности в СССР в 1958—1961 гг. по В. В. Груздеву (1963).
Кроме того; повторены карты мирового ареала пасюка из работы
Е. Шварца (1960) и распространения его в Палеарктике из книги
Г. Б. Корбета. Завериталась статья оригинальной картой мирового ареала
этого вида по данным на конец семидесятых годов. Все карты сопровож-
дались подробным описанием и сопоставлением.
Современные знания об ареале пасюка
Мы продолжали сбор и апализ фаунистической литературы © целью
уточнения ареала серой крысы, в том числе и распространения ее на
океанических островах. В последние 8—10 лет появился ряд специальных
статей, уточняющих ареал этого вида в нескольких странах Европы,
а также и в отдельных регионах СССР. В Советском Союзе было про-
ведепо несколько совещаний (1983, 1986, 1987 гг.) и издано четыре сбор-
ника, посвященных серой крысе. Все это позволило существенно уточ-
нить наши представления о распространении этого вида. Начнем © описа-
ния распространения пасюка за пределами СССР.
Зарубежная территория
В Северной Африке пасюк распространен в дельте Нила от Порт-Саида
и Исмалии на востоке до Даманхура, Абу-Хуммуса, Александрии на за-
паде. Вверх по Нилу он прослежен до Исны и Асуана (ВЁааё её а|.,
Ареал
41
1969; Озроги, Нешиу, 1980). Выше по Нилу изолированное поселепие
пасюка существует в Хартуме с начала нынешнего века (ВаКНоит, 1911 3;
Еюоууег, 1932). В 1945 г. мероприятия по ограничению численпости этого
вида в Хартуме проводили УУаов, Ретту (1954). Ф. Диетерлен в феврале
1973 г. наблюдал пасюков на набережной Нила. Он упоминает о паличии
Н Бриталоком музее естественной истории двух экземпляров пасюка, до-
а
РТУ в 1904 и одного в 1929 гг. (ГЛеецев, Мо! ап,
Восточнее по побережью Средиземного моря пасюк обитает в Триполи
{Вапк, 1968), широко распространен в области жестколистных, вечно-
зеленых средиземноморских лесов от Туниса до Марокко (Рейег баш
Сто, 1965). Он обнаружен в городах: Тунис, Алжир Бени-Упиф Фес
Гетуан, Мекнес, Рабат, Эль-Джадида, Атадир — ив устье Ширрата меж
ду „Касабланкой и Рабатом (Реапое, 1925; Толиттет, 1960; МаШоох
1965; МаШочх, Вет тане, 1970; Г.аитеп(, 1937: Бернар, 1974). ’
”
По атлантическому побережью Африки пасюк обитает в крупных
портах: Дакаре, Конакри, Фритауне, Монровии (Рекеузег, 1955; Уда
В1е] её а1., 1969). На участке, прилежащем к Гвинейскому заливу пасюк.
сплошь заселяет побережье от Абиджана до дельты Нигера обитает
в г. Дуала (Двала) и на островах Фернандо-По и Сан-Томе { АЦеп
1939; Усвеке, 1958; Уое]ске!, Уамегаз, 1960). Далее к югу паслок есть
в Пуэнт-Нуар и, по-видимому, в Луанде.
у
"
В ТОАР пасюк силошь заселяет побережье от Кейптауна до Порт-
Еливабет, восточнее он известен из Ист-Лондона, Дурбана. Далее, по
всей вероятности, обитает в Мануту, Иньямбане, Бейре Мозамбике
(Парк, 1960). Достоверно известен в Дар-эс-Саламе, Занзибаре, Садани
томбвсе (Кавдоп, 1974). Из последней по железной дороге проник
в Найроби (Нигз, 1926—1927) и достиг территории Заира на западном
берегу оз. Мобуту-Сесе-Сено.
^
Г. Д. Авери (Ауегу, 1985) считает, что пасюк с равной вероятностью
мог быть завезен в Восточную Африку как из Европы, так и из Азии
(из Йемена или стран Персидского залива в [--ГХ вв, нашей эры) По
мнению этого автора, пасюх отсутствует в Мозамбике, но заселяет по то-
вые города ЮАР, Памибии, Анголы и страп Западной Африки аныо
рапние сведения о пасюке в Нейптауне относятся к 1832 г. ав катал ге
Южно-Африканского музея, изданном в 1862 г., уже указано несколько
экземпляров этого вида. Авери считает, что пасюк плохо приспособлен ‘к
Условиям тропического климата.
В Сомали серая крыса, по всей вероятности, встречается в Могадишо
и Джибути. В Эфиопии достоверно обитает на о-ве Фатма и вг. Массава
(Уаеп её а1., 1976), на суданском побережье в Порт-Судане и Аб -Ра-
миде (Нооззтаа!, 1963; ОзЪоги, Нету, 1980).
у
Как видно из вышеизложенного, пасюк не смог освоить Африканский
материк. Он заселил преимущественно портовые города. В тлубь мате-
рика этот вид сумел распространиться на отдельных неболыших участ-
3 Цит. по; Сгаш, 1973.
42
Глава вторая
ках. Он проник по долине Нила вверх до Ассуана и закрепился в Харту-
ме, куда попал, вероятно, по железной дороге из Порт-Судана. Однако
нет никаких сведений об обитании пасюка в городах вдоль этой дороги.
Широко распространен в области жестколистных вечнозеленых лесов и
кустарников африканского Средиземноморья (Петтер, Сан-Жирон, 1965).
Видимо, пасюк также обычен на территории, запятой жестколистными
вечнозелепыми кустарпиками Каиской области. Расселилоя вдоль желез-
пой дороги Момбаса, Найроби, Кампала, Касесе. Однако нет сведений о
том, насколько здесь многочисленны и постоянны его поселения. Также
нет сведений, насколько далеко проник пасюк от побережья Гвинейско-
то залива. Почти все попытки освоения пасюком внутриматериковых тер-
риторий приурочены к районам с субтропическим климатом или к сред-
негорьям, где мепее жарко.
Несомненно, тропический климат неблагоприятен для пасюка. Одна-
ко ето экспансия в глубь континента Южной Америки проходила значи-
тельно успеинее. Видимо, многочисленные африканские представители
семейстна мышиных (Миаш@ае) оказались более конкурентоспособными
по сравнению с южноамериканскими хомячьйми ({СнсейЧае).
Для Малой Азии, Аравии, Ирана есть несколько работ, в которых
приведены карты с точками фактической добычи пасюка на той или
иной территории. Такая карта, охватывающая территорию от Аденского
залива, Красного моря, Суэцкого канала, восточного побережья Среди-
земного моря, Восточной Турции, 1Ожного Кавказа и большей части
Ирана приведена в работе Гаррисона (Нагт1зоп, 1972), перечисление мест
находок и карты с очертаниями ареала даны по Ирану В. Мизоном
(М1зопие, 1956, 1960), а для Израиля— П. Г. Гратцом (1973). Кадхим
и Мустафа (Кали, Мозага, 1983) опубликовали карту распростране-
ния паслока, на которой значками отмечено 18 пунктов регистрации этого
вида в Ираке.
В общих чертах распространение пасюка в Юго-Западной Азии вы-
глядит следующим образом. На Аравийском полуострове по берегу Крас-
ного моря пасюк обнаружен в северной оконечности залива Акаба и
вг Эйлат. По-видимому, он есть и в г. Акаба. На Средиземноморском
побережье обитание серой крысы установлено повсюду от Аскалона на
юге до Хайфы и Бейрута на севере. В Сирии она найдена в Дамаске и
в Эт-Телльэль-Абьяде на грапице с Турцией. На востоке средиземно-
морского побережья Турции зарегистрирована в порту Мерсиан. Вероятно
заселяет все морское побережье Израиля, Лизана, Сирии и Турции.
На непрерывные поселения пасюка по морскому побережью Турции
указывают Е. Шварц (1960) и Корбет (Согьеф, 1978). Вдоль восточной
части южного берега Черного моря эти поселения идут узкой полосой,
уходя в пределы СССР, где между Большим и Малым Кавказом тяпутся
до Каспийского побережья. Все южное побережье Каспия от Ленкоран-
ской низменности до Гассан-Кули и южлые склоны Эльбурса заселены
пасюком. Оп найден в 40 населенных пунктах от Астары, Пехлеви, Реш-
та на западе до Горгана, Алиабада, Шахнесенда, Тилабада на востоке.
Изолированное поселение существует в Гегерапе (Мизон, 1956).
‘
Ареал
43
ы Оженая траница ареала этого вида в Юго-Западной Азии идет в Си-
ри и г раке ине Евфрата и спускается вдоль побережья Персид-
ана
ой границы Кувейта, где пасюк обнаружен и на
и у - Южнее его поселения известны в Бахрейне и Хухуфе
р осоверному берегу Персидского залива его регистрировали з Бен-
дер-А все и Бушире. На карте в работе К. Мизона (1956) показано, что
пасюк тает вдоль всего Иранского побережья Персидского залива.
оНАКО и личной беседе он отметил, что такая трактовка отибочна и па
ею
р
я здесь только в крупных портовых горо-
ад ие9 Персидского залива граница ареала пасюка идет на северо-
осточной границы провинции Хузестан в Иране, пересекает
Рис. 6. Распространение пасюка в Юго-Западной Азии
1—т ерритории, сплошь заселенные иасюном; 2 — пункты регистрации пасюка
<?
не,
==:
ъ
>.
=
$
и
@
®
.
-®
КИ,
С-}.
=
-
—_
42
Глава вторая
ках. Он проник по долине Нила вверх до Ассуана и закрепился в Харту-
ме, куда попал, вероятно, по железной дороге из Порт-Судана. Однако
нет никаких сведений об обитании пасюка в городах вдоль этой дороги.
Широко распространен в области жестколистных вечнозеленых лесов и
кустарников африканского Средиземноморья (Петтер, Сан-Жирон, 1965).
Видимо, пасюк также обычен на территории, запятой жестколистными
вечнозелепыми кустарпиками Каиской области. Расселилоя вдоль желез-
пой дороги Момбаса, Найроби, Кампала, Касесе. Однако нет сведений о
том, насколько здесь многочисленны и постоянны его поселения. Также
нет сведений, насколько далеко проник пасюк от побережья Гвинейско-
то залива. Почти все попытки освоения пасюком внутриматериковых тер-
риторий приурочены к районам с субтропическим климатом или к сред-
негорьям, где мепее жарко.
Несомненно, тропический климат неблагоприятен для пасюка. Одна-
ко ето экспансия в глубь континента Южной Америки проходила значи-
тельно успеинее. Видимо, многочисленные африканские представители
семейстна мышиных (Миаш@ае) оказались более конкурентоспособными
по сравнению с южноамериканскими хомячьйми ({СнсейЧае).
Для Малой Азии, Аравии, Ирана есть несколько работ, в которых
приведены карты с точками фактической добычи пасюка на той или
иной территории. Такая карта, охватывающая территорию от Аденского
залива, Красного моря, Суэцкого канала, восточного побережья Среди-
земного моря, Восточной Турции, 1Ожного Кавказа и большей части
Ирана приведена в работе Гаррисона (Нагт1зоп, 1972), перечисление мест
находок и карты с очертаниями ареала даны по Ирану В. Мизоном
(М1зопие, 1956, 1960), а для Израиля— П. Г. Гратцом (1973). Кадхим
и Мустафа (Кали, Мозага, 1983) опубликовали карту распростране-
ния паслока, на которой значками отмечено 18 пунктов регистрации этого
вида в Ираке.
В общих чертах распространение пасюка в Юго-Западной Азии вы-
глядит следующим образом. На Аравийском полуострове по берегу Крас-
ного моря пасюк обнаружен в северной оконечности залива Акаба и
вг Эйлат. По-видимому, он есть и в г. Акаба. На Средиземноморском
побережье обитание серой крысы установлено повсюду от Аскалона на
юге до Хайфы и Бейрута на севере. В Сирии она найдена в Дамаске и
в Эт-Телльэль-Абьяде на грапице с Турцией. На востоке средиземно-
морского побережья Турции зарегистрирована в порту Мерсиан. Вероятно
заселяет все морское побережье Израиля, Лизана, Сирии и Турции.
На непрерывные поселения пасюка по морскому побережью Турции
указывают Е. Шварц (1960) и Корбет (Согьеф, 1978). Вдоль восточной
части южного берега Черного моря эти поселения идут узкой полосой,
уходя в пределы СССР, где между Большим и Малым Кавказом тяпутся
до Каспийского побережья. Все южное побережье Каспия от Ленкоран-
ской низменности до Гассан-Кули и южлые склоны Эльбурса заселены
пасюком. Оп найден в 40 населенных пунктах от Астары, Пехлеви, Реш-
та на западе до Горгана, Алиабада, Шахнесенда, Тилабада на востоке.
Изолированное поселение существует в Гегерапе (Мизон, 1956).
‘
Ареал
43
ы Оженая траница ареала этого вида в Юго-Западной Азии идет в Си-
ри и г раке ине Евфрата и спускается вдоль побережья Персид-
ана
ой границы Кувейта, где пасюк обнаружен и на
и у - Южнее его поселения известны в Бахрейне и Хухуфе
р осоверному берегу Персидского залива его регистрировали з Бен-
дер-А все и Бушире. На карте в работе К. Мизона (1956) показано, что
пасюк тает вдоль всего Иранского побережья Персидского залива.
оНАКО и личной беседе он отметил, что такая трактовка отибочна и па
ею
р
я здесь только в крупных портовых горо-
ад ие9 Персидского залива граница ареала пасюка идет на северо-
осточной границы провинции Хузестан в Иране, пересекает
Рис. 6. Распространение пасюка в Юго-Западной Азии
1—т ерритории, сплошь заселенные иасюном; 2 — пункты регистрации пасюка
<?
не,
==:
ъ
>.
=
$
и
@
®
.
-®
КИ,
С-}.
=
-
—_
АА
Глава вторая
границу Ирака и Турции по левобережью р. Тигра и, круто повернув на
запад, выходит к турецкому побережью Средиземного моря. Очертания
этой границы в западном Иране и восточном Ираке не ясны, возможно,
что этот участок ареала соединяется с Прикаснийским. В Хузестане па-
сюк зарегистрирован в городах Абадан, Хорремшехр, Ахваз (Мизон,
1956), а в Ираке найден в Мухафазах: Басра. в 3 пунктах, Ди-Кар —
в 2. Майсан—в 3, Муттана— в 2, Кадисия—в 2, Васит —в 1, Ба-
биль —в 1, Багдад — в 3, Наинава —в2 (Кайши, Миаза{а, 1983).
Согласно рис. 6 ареал пасюка в Юго-Западной Азии имеет весьма
сложное очертание, территории, заселенные этим грызуном, приурочены
к морским побережьям, низменностям и долинам крупных рек. Горы и
равнинные пустыни свободны от него.
В Афганистане пасюка нет. В Пакистане он обнаружен в трех горо-
дах на берегу Индийского океана: Гвадаре, Пасни, Карачи и в Лахоре
у северо-восточной границы страны (ВЛашога, 1888—1891; ВоЪемз, 1977,
Мизон, 1956).
В Индии пасюк встречается во всех больших городах и деревнях,
расположенных по берегам судоходных рек (В]аз{от@, 1888). Однако
Мизоп считает, что в Ипдии пасюка можно обнаружить только в городах
и морских портах, но не в деревиях. Деорас (Оеогаз, 1964) отмечает на-
личие пасюка в 49 штатах Индии. Специальных работ по распростране-
нию пасюка в Индии мы пе нашли. По-видимому, он отсутствует в Се-
веро-Западной Индии, а северная граница его распространения в этом
регионе проходит по предгорьям Гималаев. Есть пасюк в Калькутте,
Дакке и Рангуне. В работе по мышиному тифу в Таиланде (ЗапКази\ап
сё а1.. 1969) есть указапия па обитание пасюка в этой стране. В Малай-
зии пасюк приурочен к портам (Нагмзот, 1957; Согдон ЗшИР её а|.,
1961а, Ь), достоверно зарегистрирован в Сипгапуре, Келанге, Джорж-
тауне на о-ве Пенанг и на о-ве Лапкави в г. Куах. В Кампучии он об-
паружен в Пномпене.
Пасюк широко распространен в городах и портах Въетнама (Као Ван
Шунг и др., 1982), его обитание зарегистрировано во всех зоогеотрафиче-
ских районах этой страны за исключением северо-западного (Дао Ван
Тьен, 1978). Пасюка добывали в Хошимине, Нячанге, Дананге, Винье,
Ханое, Тхайнгуене, Лапгшоне (Дао Ван Тьен, 1962; Петров, Нгуен,
Ван Нье, 1983; Попов, Тесленко, 1984).
Распространение пасюка в Индонезии, Папуа-Новой Гвинее и на Фи-
липпинах документировано чрезвычайно плохо. Мизон (М15оппе, 1959)
считает, что этот вид в Индонезии строго приурочен к морским портам.
Пасюк достоверно известен из Манилы, Минданао, Горонтало и Макас-
сара на Сулавеси, Джакарты на Яве, Кавиепга на о-ве Новая Ирландия,
Лаэ и Порта Мореби в Папуа-Новой Гвинее (Гаме, НЛЬ 1954; Мепяез,
Дери4з, 1979). Е. Шварц (1960) указывает на обитание пасюка па Соло-
моповых островах.
В Китае пасюк обитает практически во всех портовых городах Южно-
Китайского, Восточно-Китайского и Желтого морей. Он обычен на 'Тай-
зане и Пескадорских островах, многочислен в Сянгане (Гопконг) и Гуанч-
Ареал
45
жоу. Западная граница его распространения идет примерно от Куньмина
на Чжаотун, Баоцзи, Тайюань, Датун (АПеп, 1940) и далее на Чифын,
Кайлу, Чжаньюй, Байчэн, Улан-Хото и выходит на верховья р. Нумэр-
тин-Гол па восточной границе МНР. Мы добывали его на западе провин-
ции Гирин в городах Таонань и Байчоп, в автономном районе Внутрен-
няя Монголия в городах Улан-Хото и Солунь. К северу от оз. Буир-Нур
вдоль р. Орчун-Гол пасюк проник на восточное побережье оз. Далайнор
и заселил всю железную дороту на восток от ст. Маньчжурия. Восточнее
и севернее этой границы пасюк повсеместно распространен в автономном
районе Внутренняя Монголия и целиком заселяет провинцию Хэйлун-
цзян (Ма У-СШих её а1., 1986).
На крайнем востоке МНР пасюк встречается в естественных местооби-
тапиях всего бассейна р. Нумэргин-Гол и ло долине Халхин-Гол от устья
Дэгэ-Гол до Буир-Нура. Также он обнаружен в постройках по побережью
оз. Буир-Нур и в пос. Тамсаг-Булак (Кучерук, 1946). Указание А. Г. Бап-
никова (1954) об обитании пасюка па оз. Сангин-Далай-Нур ошибочно.
Второй участок ареала серой крысы в МНР приурочен к южным скло-
нам Хентся. Мы добывали пасюка в 1942—1944 тг. в пойме Онона про-
тив устья Кыры и у пересечения р. Хара-Гол автомобильным трактом от.
Кяхты в Улан-Батор. В. Н. Скалон (1949) добыл экземпляр серой крысы
в пойме Онона у сомона Биндэр, В. М. Линаев (1963) в 1955/56 г. нашел
паслока в среднем течении р. Бальджа, на берегах Опопа у сомона Бат-
Ширэт и в развалинах монастыря Хутагтэ-Хурэ в верховьях р. Мурон.
При обследовании ж. д. Сухэ-Батор — Улан-Батор Ж. Демберил (1962)
обнаружил пасюка во всех поселках от Алтан-Булак и Сухэ-Батор на се-
вере до Дзун-Хара и Мандал на юге (рис. 7).
Серая крыса занимает весь Корейский полуостров и Янонские остро-
ва: Хоккайдо, Хонсю, Сикоку, Кюсю (Корбет, 1978). Предполагается, что
в Япопию пасюк был случайно завезен во время периода 'Тайкаа (нача-
ло ХГУ в.) из Кореи. Впачале он появился на территории префектур Фу-
куй и Тояма (0-в Хонсю), а затем расселился по всему государству
(Каваути, 1930, цит. по Реймерсу и др., 1968). По нашему мнению, па-
сюк либо автохтон японских островов, либо был завезен, при заселении
островов японцами в первых веках нашей эры.
Распространение пасюка на территории Европы за пределами СССР
изображено в работах Ван ден Бринка (4958), Е. Шзарца (1960) и
Г. Б. Корбета (1978) практически тождественно. Показано сплошное рас-
пространение вида по всем европейским странам, включая Британские
острова и Исландию. Не указан пасюк на Ишицбертене и в Гренландии.
В то же время Ван ден Бринк (1956) считает, что пасюк не поднимается
в горы выше 1600 м. Следовательно, в Альпах, Апеннинах, Карпатах и
других горных системах Европы значительные площади не заселены па-
стоком, и это можно отразить на картах ареала. Видимо, пасюк должен
отсутетвовать на большей части тундр Скандинавского полуострова, что
не нашло отражения ни на одной из трех вышеупомянутых карт его
ареала. На карте, составленной К. Беккером (ВесКег, 1978), в пределах
Европы выделены области сплошного и впорадичного распространения
АА
Глава вторая
границу Ирака и Турции по левобережью р. Тигра и, круто повернув на
запад, выходит к турецкому побережью Средиземного моря. Очертания
этой границы в западном Иране и восточном Ираке не ясны, возможно,
что этот участок ареала соединяется с Прикаснийским. В Хузестане па-
сюк зарегистрирован в городах Абадан, Хорремшехр, Ахваз (Мизон,
1956), а в Ираке найден в Мухафазах: Басра. в 3 пунктах, Ди-Кар —
в 2. Майсан—в 3, Муттана— в 2, Кадисия—в 2, Васит —в 1, Ба-
биль —в 1, Багдад — в 3, Наинава —в2 (Кайши, Миаза{а, 1983).
Согласно рис. 6 ареал пасюка в Юго-Западной Азии имеет весьма
сложное очертание, территории, заселенные этим грызуном, приурочены
к морским побережьям, низменностям и долинам крупных рек. Горы и
равнинные пустыни свободны от него.
В Афганистане пасюка нет. В Пакистане он обнаружен в трех горо-
дах на берегу Индийского океана: Гвадаре, Пасни, Карачи и в Лахоре
у северо-восточной границы страны (ВЛашога, 1888—1891; ВоЪемз, 1977,
Мизон, 1956).
В Индии пасюк встречается во всех больших городах и деревнях,
расположенных по берегам судоходных рек (В]аз{от@, 1888). Однако
Мизоп считает, что в Ипдии пасюка можно обнаружить только в городах
и морских портах, но не в деревиях. Деорас (Оеогаз, 1964) отмечает на-
личие пасюка в 49 штатах Индии. Специальных работ по распростране-
нию пасюка в Индии мы пе нашли. По-видимому, он отсутствует в Се-
веро-Западной Индии, а северная граница его распространения в этом
регионе проходит по предгорьям Гималаев. Есть пасюк в Калькутте,
Дакке и Рангуне. В работе по мышиному тифу в Таиланде (ЗапКази\ап
сё а1.. 1969) есть указапия па обитание пасюка в этой стране. В Малай-
зии пасюк приурочен к портам (Нагмзот, 1957; Согдон ЗшИР её а|.,
1961а, Ь), достоверно зарегистрирован в Сипгапуре, Келанге, Джорж-
тауне на о-ве Пенанг и на о-ве Лапкави в г. Куах. В Кампучии он об-
паружен в Пномпене.
Пасюк широко распространен в городах и портах Въетнама (Као Ван
Шунг и др., 1982), его обитание зарегистрировано во всех зоогеотрафиче-
ских районах этой страны за исключением северо-западного (Дао Ван
Тьен, 1978). Пасюка добывали в Хошимине, Нячанге, Дананге, Винье,
Ханое, Тхайнгуене, Лапгшоне (Дао Ван Тьен, 1962; Петров, Нгуен,
Ван Нье, 1983; Попов, Тесленко, 1984).
Распространение пасюка в Индонезии, Папуа-Новой Гвинее и на Фи-
липпинах документировано чрезвычайно плохо. Мизон (М15оппе, 1959)
считает, что этот вид в Индонезии строго приурочен к морским портам.
Пасюк достоверно известен из Манилы, Минданао, Горонтало и Макас-
сара на Сулавеси, Джакарты на Яве, Кавиепга на о-ве Новая Ирландия,
Лаэ и Порта Мореби в Папуа-Новой Гвинее (Гаме, НЛЬ 1954; Мепяез,
Дери4з, 1979). Е. Шварц (1960) указывает на обитание пасюка па Соло-
моповых островах.
В Китае пасюк обитает практически во всех портовых городах Южно-
Китайского, Восточно-Китайского и Желтого морей. Он обычен на 'Тай-
зане и Пескадорских островах, многочислен в Сянгане (Гопконг) и Гуанч-
Ареал
45
жоу. Западная граница его распространения идет примерно от Куньмина
на Чжаотун, Баоцзи, Тайюань, Датун (АПеп, 1940) и далее на Чифын,
Кайлу, Чжаньюй, Байчэн, Улан-Хото и выходит на верховья р. Нумэр-
тин-Гол па восточной границе МНР. Мы добывали его на западе провин-
ции Гирин в городах Таонань и Байчоп, в автономном районе Внутрен-
няя Монголия в городах Улан-Хото и Солунь. К северу от оз. Буир-Нур
вдоль р. Орчун-Гол пасюк проник на восточное побережье оз. Далайнор
и заселил всю железную дороту на восток от ст. Маньчжурия. Восточнее
и севернее этой границы пасюк повсеместно распространен в автономном
районе Внутренняя Монголия и целиком заселяет провинцию Хэйлун-
цзян (Ма У-СШих её а1., 1986).
На крайнем востоке МНР пасюк встречается в естественных местооби-
тапиях всего бассейна р. Нумэргин-Гол и ло долине Халхин-Гол от устья
Дэгэ-Гол до Буир-Нура. Также он обнаружен в постройках по побережью
оз. Буир-Нур и в пос. Тамсаг-Булак (Кучерук, 1946). Указание А. Г. Бап-
никова (1954) об обитании пасюка па оз. Сангин-Далай-Нур ошибочно.
Второй участок ареала серой крысы в МНР приурочен к южным скло-
нам Хентся. Мы добывали пасюка в 1942—1944 тг. в пойме Онона про-
тив устья Кыры и у пересечения р. Хара-Гол автомобильным трактом от.
Кяхты в Улан-Батор. В. Н. Скалон (1949) добыл экземпляр серой крысы
в пойме Онона у сомона Биндэр, В. М. Линаев (1963) в 1955/56 г. нашел
паслока в среднем течении р. Бальджа, на берегах Опопа у сомона Бат-
Ширэт и в развалинах монастыря Хутагтэ-Хурэ в верховьях р. Мурон.
При обследовании ж. д. Сухэ-Батор — Улан-Батор Ж. Демберил (1962)
обнаружил пасюка во всех поселках от Алтан-Булак и Сухэ-Батор на се-
вере до Дзун-Хара и Мандал на юге (рис. 7).
Серая крыса занимает весь Корейский полуостров и Янонские остро-
ва: Хоккайдо, Хонсю, Сикоку, Кюсю (Корбет, 1978). Предполагается, что
в Япопию пасюк был случайно завезен во время периода 'Тайкаа (нача-
ло ХГУ в.) из Кореи. Впачале он появился на территории префектур Фу-
куй и Тояма (0-в Хонсю), а затем расселился по всему государству
(Каваути, 1930, цит. по Реймерсу и др., 1968). По нашему мнению, па-
сюк либо автохтон японских островов, либо был завезен, при заселении
островов японцами в первых веках нашей эры.
Распространение пасюка на территории Европы за пределами СССР
изображено в работах Ван ден Бринка (4958), Е. Шзарца (1960) и
Г. Б. Корбета (1978) практически тождественно. Показано сплошное рас-
пространение вида по всем европейским странам, включая Британские
острова и Исландию. Не указан пасюк на Ишицбертене и в Гренландии.
В то же время Ван ден Бринк (1956) считает, что пасюк не поднимается
в горы выше 1600 м. Следовательно, в Альпах, Апеннинах, Карпатах и
других горных системах Европы значительные площади не заселены па-
стоком, и это можно отразить на картах ареала. Видимо, пасюк должен
отсутетвовать на большей части тундр Скандинавского полуострова, что
не нашло отражения ни на одной из трех вышеупомянутых карт его
ареала. На карте, составленной К. Беккером (ВесКег, 1978), в пределах
Европы выделены области сплошного и впорадичного распространения
46
Глава вторая
пасюка (см. рис. 5). К территориям спорадичного распространения он от-
нес Пиренейский, Апеннинский и Балканский полуострова. В Португа-
лии им показано всего два пункта обитания пасюка: Коимбра и Порто-
Ково, а в статье о вредных грызунах Португалии Баумлер с соавт.
(ВёииЦег её а1., 1984) пишут, что пасюк обитает вне поселений чело-
века в течение всего года, многочислен на рисовых полях, посевах куку-
рузы, садах, животноводческих хозяйствах. Потери урожая кукурузы и
риса, связанные с этим грызуном, достигают 30%. Пасюк служит источ-
ником заражения людей лептоспирозом Вейля — болезнью, которой часто
заражаются рабочие рисовых полей.
В Испании К. Беккер отметил лишь семь мест регистрации пасюка,
а Гозальбез © соавт. (СозаШея ©ё а|., 1985) только в Каталонии нашли
Рис. 7. Распространение пасюка в Южной и Восточной Азии
1 — территории, сплошь заселенные пасюком; 2 -—— пункты достоверной регистрации пасюка
‘Ареал
47
,[-
&|
Б
АН.
грЖ
НЫ
РЕГ г
р
я
к 4®|®) |\
®|$
®[® $
|
|
ЗАСОЧАО
® во|©
49:
а."
<.
*
С5:С
{®]® | ер
®|6)| |® 2%
\ НОС
мь
и
ы
[.
.
д
Ф||®|
®
С]
|аре
ее
®
®©
—А
®2® ©
Н
Е
во
Рис. 8. Распространение пасюка (А) и черной крысы (Б) в Италии (по: Атон е
а1., 1984)
пасюка в 67 пунктах. В Италии К. Беккер отметил 10 пунктов, где най-
ден пасюк, а Амори (Атот! © а1., 1984) обнаружил 45 таких пунктов и
показал, что этот вид отсутствует лишь на южной оконечности Аненпин-
ского полуострова, а на Сицилии распространен шире, чем черная крыса
(рис. 8). По данным М. Х. Матевой, Л. Христова (1986), пасюк обнару-
жен в 47 из 23 обследованных округов Болгарии, в том числе он обитает
ив 3 юго-западных округах. Приведепные факты показывают, что трак-
туемая К. Беккером спорадичность распространения пасюка на юге Ев-
роны вряд ли имеет место.
Появилось несколько работ, в которых приведены фактические мате-
риалы о полноте докумептации по распространению пасюка в некоторых
европейских странах (АззеШета, 1971; \УаШ её а1., 1980; Ашот её а1.,
1984; Ета, Зее, 1986).
Для Северной Америки мы смогли лишь повторить карту Пальмера
(Рариег, 1954), подчеркнув условность границ ареала пасюка в Мексике
и уточнив его распространение на Аляске, где он зарегистрирован в го-
родах Ном и Фэрбанкс (ЗсВШег, 1956). По мнению Е. Шварца (1960),
пасюк заселял все острова Алеутской гряды вплоть до ее западной око-
нечности, что подтверждается названием наиболее западных — «Крысьи
острова» русских и «Ваф 131з145» американцев. Видимо, уже первые рус-
46
Глава вторая
пасюка (см. рис. 5). К территориям спорадичного распространения он от-
нес Пиренейский, Апеннинский и Балканский полуострова. В Португа-
лии им показано всего два пункта обитания пасюка: Коимбра и Порто-
Ково, а в статье о вредных грызунах Португалии Баумлер с соавт.
(ВёииЦег её а1., 1984) пишут, что пасюк обитает вне поселений чело-
века в течение всего года, многочислен на рисовых полях, посевах куку-
рузы, садах, животноводческих хозяйствах. Потери урожая кукурузы и
риса, связанные с этим грызуном, достигают 30%. Пасюк служит источ-
ником заражения людей лептоспирозом Вейля — болезнью, которой часто
заражаются рабочие рисовых полей.
В Испании К. Беккер отметил лишь семь мест регистрации пасюка,
а Гозальбез © соавт. (СозаШея ©ё а|., 1985) только в Каталонии нашли
Рис. 7. Распространение пасюка в Южной и Восточной Азии
1 — территории, сплошь заселенные пасюком; 2 -—— пункты достоверной регистрации пасюка
‘Ареал
47
,[-
&|
Б
АН.
грЖ
НЫ
РЕГ г
р
я
к 4®|®) |\
®|$
®[® $
|
|
ЗАСОЧАО
® во|©
49:
а."
<.
*
С5:С
{®]® | ер
®|6)| |® 2%
\ НОС
мь
и
ы
[.
.
д
Ф||®|
®
С]
|аре
ее
®
®©
—А
®2® ©
Н
Е
во
Рис. 8. Распространение пасюка (А) и черной крысы (Б) в Италии (по: Атон е
а1., 1984)
пасюка в 67 пунктах. В Италии К. Беккер отметил 10 пунктов, где най-
ден пасюк, а Амори (Атот! © а1., 1984) обнаружил 45 таких пунктов и
показал, что этот вид отсутствует лишь на южной оконечности Аненпин-
ского полуострова, а на Сицилии распространен шире, чем черная крыса
(рис. 8). По данным М. Х. Матевой, Л. Христова (1986), пасюк обнару-
жен в 47 из 23 обследованных округов Болгарии, в том числе он обитает
ив 3 юго-западных округах. Приведепные факты показывают, что трак-
туемая К. Беккером спорадичность распространения пасюка на юге Ев-
роны вряд ли имеет место.
Появилось несколько работ, в которых приведены фактические мате-
риалы о полноте докумептации по распространению пасюка в некоторых
европейских странах (АззеШета, 1971; \УаШ её а1., 1980; Ашот её а1.,
1984; Ета, Зее, 1986).
Для Северной Америки мы смогли лишь повторить карту Пальмера
(Рариег, 1954), подчеркнув условность границ ареала пасюка в Мексике
и уточнив его распространение на Аляске, где он зарегистрирован в го-
родах Ном и Фэрбанкс (ЗсВШег, 1956). По мнению Е. Шварца (1960),
пасюк заселял все острова Алеутской гряды вплоть до ее западной око-
нечности, что подтверждается названием наиболее западных — «Крысьи
острова» русских и «Ваф 131з145» американцев. Видимо, уже первые рус-
48
Глава вторая
—
й
ские землепроходцы в ХУШ в. столкнулись с изобилием крыс на Алеут-
ских островах‘. Раннее проникновение пасюка на северное побережье
Аляски подтверждается коллекционным экземпляром, добытым па о-ве
Кальяк в 1844 г. (ЗИН АН СССР), ва 23 года до продажи Аляски и
Алеутских островов США. Достоверное обитание пасюка у побережья
Аляски в пастоящее время установлено для следующих островов: Кадьяк,
||©
Нунивак, Св. Лаврентия, Уналашка, Адак (Зе Шег, 1956; Нааз её а1.,
о
1986; Кучерук, Лапшов, 1987).
ТВ.
В некоторых работах упомянуто, что в пределах США есть террито-
рии (пустыни, северные области Скалистых гор), не заселенные пасюком
(Еске, 1954; Нагиозюой, УУгььь 1960). Видимо, при более тщательном
изучении ареала пасюка в Северной Америке будет выявлепа достаточно
сложпая картина кружева ареала этого вида, которая уже установлена
в СССР.
Карта ареала пасюка в Южной Америке была составлена следующим
образом. Сначала по книге «Р]авие ш Ше Алемсаз, 1945» выбрали горо-
да, в которых документально установлено обитание пасюка. Их пунсона-
ми обозпачили на карте. Затем эти данные были экстраполированы на
прилежащие территории, основываясь на картах растительноети, плотно-
сти людского населения, густоты железнодорожной и автодорожной сети.
Понимая вею относительность подобного приема, мы уверены, что состав-
ленная таким образом карта ареала пасюка в Южной Америке ближе
соответствует действительному положению, чем карта для этого конти-
нента, опубликованная в работе Е. Шварца (1960).
Видимо, в связи с малой конкурентной способностью местных видов
насюк завоевывает территорию Южной Америки значительно успешнее,
чем в Африке.
В
В Австралии пасюк широко распространен в сельской местности про-
винций Квинсленд и Новый ТОжный Уэльс. Здесь он в массе встречает-
ся на плантациях сахарного тростника и обычен во всех крупных и пор-
товых городах (Торп8оп, 1950; Оегыск, Роре, 1960; Ващеу её а1., 1964;
Апаньип, Карасева, 1971). В провинции Южная Австралия пасюк есть
р
вг. Аделаида и на п-ове Эйр, в Западной Австралии па территории с от-
носительно плотным людеким населением от Джералдтона на севере до
Олбани па юге (Шварц, 1960). На Северной Территории этот вид отме-
чен в районе г. Дарвина. Пасюк достаточно обычен в Новой Зеландии
как на Северном, так и на Южном островах (рис. 9).
На обоих опубликованных картах мирового ареала пасюка (Шварц,
1960; Кучерук, Кузиков, 1985) отсутствовали сведения о размещении
этого вида на океанических островах. Мы попытались исправить это упу-
щение, подобрав фактические данные и подготовив статьи о синантроп-
ных грызунах океанических островов (Кучерук, Лапшов, 1987). Материа-
лы Для описания океанического ареала пасюка заимствованы из этих
публикаций.
Р
о
4
7
7
2
2
0
4
4
* Во время экспедиции В. Беринга (1725—1730 и 1733—4744 гг.) быпи открыты Але-
утские и Командорские острова.
1
—
т
е
р
р
и
т
о
р
и
и
,
с
п
л
о
ш
ь
з
а
с
е
л
е
н
н
ы
е
п
а
с
ю
к
о
м
;
8
—
н
а
х
о
д
к
и
п
а
с
ю
к
а
н
а
о
с
т
р
о
в
а
х
,
в
п
о
р
т
а
х
и
г
о
р
о
д
а
х
з
а
п
р
е
д
е
л
а
м
и
с
п
л
о
п
т
н
о
г
о
а
р
е
а
л
а
Р
и
с
.
9
.
М
и
р
о
в
о
й
а
р
е
а
л
п
а
с
ю
к
а
48
Глава вторая
—
й
ские землепроходцы в ХУШ в. столкнулись с изобилием крыс на Алеут-
ских островах‘. Раннее проникновение пасюка на северное побережье
Аляски подтверждается коллекционным экземпляром, добытым па о-ве
Кальяк в 1844 г. (ЗИН АН СССР), ва 23 года до продажи Аляски и
Алеутских островов США. Достоверное обитание пасюка у побережья
Аляски в пастоящее время установлено для следующих островов: Кадьяк,
||©
Нунивак, Св. Лаврентия, Уналашка, Адак (Зе Шег, 1956; Нааз её а1.,
о
1986; Кучерук, Лапшов, 1987).
ТВ.
В некоторых работах упомянуто, что в пределах США есть террито-
рии (пустыни, северные области Скалистых гор), не заселенные пасюком
(Еске, 1954; Нагиозюой, УУгььь 1960). Видимо, при более тщательном
изучении ареала пасюка в Северной Америке будет выявлепа достаточно
сложпая картина кружева ареала этого вида, которая уже установлена
в СССР.
Карта ареала пасюка в Южной Америке была составлена следующим
образом. Сначала по книге «Р]авие ш Ше Алемсаз, 1945» выбрали горо-
да, в которых документально установлено обитание пасюка. Их пунсона-
ми обозпачили на карте. Затем эти данные были экстраполированы на
прилежащие территории, основываясь на картах растительноети, плотно-
сти людского населения, густоты железнодорожной и автодорожной сети.
Понимая вею относительность подобного приема, мы уверены, что состав-
ленная таким образом карта ареала пасюка в Южной Америке ближе
соответствует действительному положению, чем карта для этого конти-
нента, опубликованная в работе Е. Шварца (1960).
Видимо, в связи с малой конкурентной способностью местных видов
насюк завоевывает территорию Южной Америки значительно успешнее,
чем в Африке.
В
В Австралии пасюк широко распространен в сельской местности про-
винций Квинсленд и Новый ТОжный Уэльс. Здесь он в массе встречает-
ся на плантациях сахарного тростника и обычен во всех крупных и пор-
товых городах (Торп8оп, 1950; Оегыск, Роре, 1960; Ващеу её а1., 1964;
Апаньип, Карасева, 1971). В провинции Южная Австралия пасюк есть
р
вг. Аделаида и на п-ове Эйр, в Западной Австралии па территории с от-
носительно плотным людеким населением от Джералдтона на севере до
Олбани па юге (Шварц, 1960). На Северной Территории этот вид отме-
чен в районе г. Дарвина. Пасюк достаточно обычен в Новой Зеландии
как на Северном, так и на Южном островах (рис. 9).
На обоих опубликованных картах мирового ареала пасюка (Шварц,
1960; Кучерук, Кузиков, 1985) отсутствовали сведения о размещении
этого вида на океанических островах. Мы попытались исправить это упу-
щение, подобрав фактические данные и подготовив статьи о синантроп-
ных грызунах океанических островов (Кучерук, Лапшов, 1987). Материа-
лы Для описания океанического ареала пасюка заимствованы из этих
публикаций.
Р
о
4
7
7
2
2
0
4
4
* Во время экспедиции В. Беринга (1725—1730 и 1733—4744 гг.) быпи открыты Але-
утские и Командорские острова.
1
—
т
е
р
р
и
т
о
р
и
и
,
с
п
л
о
ш
ь
з
а
с
е
л
е
н
н
ы
е
п
а
с
ю
к
о
м
;
8
—
н
а
х
о
д
к
и
п
а
с
ю
к
а
н
а
о
с
т
р
о
в
а
х
,
в
п
о
р
т
а
х
и
г
о
р
о
д
а
х
з
а
п
р
е
д
е
л
а
м
и
с
п
л
о
п
т
н
о
г
о
а
р
е
а
л
а
Р
и
с
.
9
.
М
и
р
о
в
о
й
а
р
е
а
л
п
а
с
ю
к
а
50
Глава вторая
В Северном Ледовитом океане мпого островов, однако в большинстве
они лишены постоянных поселений людей и не имеют условий для су-
яцествования синантропных грызунов. На северном побережье Канады и
на севере Сибири в СССР пасюк практически отсутствует. Поэтому засе-
лепие островов Ледовитого океана могло идти только через Норвежское,
Баренцево или Берингово моря. К настоящему времени поселения пасюка
в западной части океана извостны в Гренландии и на Шпиинбергене,
а в восточной — на островах Врангеля, Нунавак и Св. Лаврентия (Нуче-
рук, Лапшов, 1987). Вероятно проникновение пасюка на острова Колгуев
и Вайгач, но фактов, подтверждающих это предположение, нет. Судя по
последним исследованиям, на архипелаге Ссверпая Земля пасюк отсут-
ствует.
На севере Атлантического океана пасюк встречается на всех островах
Ирландии и Великобритании, включая Гебридские, Фарерские, Оркней-
скис (Таушт, 1977), и в Исландии. У побережья Канады оп найден на
островах Ньюфаундленд, Антикости, Принца Эдуарда, Кейп-Бретон.
Южнее пасюк населяет Бермудские, Багамские острова, Кубу, Ямайку,
Пуэрто-Рико, Анегаду, Гваделупу, Мартинику, Барбадос. У Сенеро-Афри-
канского побережья
— Сан-Мигуэль из Азорских островов (ГИтвь 1926—
1927} и Канарские острова. В тропической части океана пасюк обитает
па островах Биоко и Сан-Томе в Гвинейском заливе и на о-ве Св. Елены.
Нет его на Фернанду-ди-Норонья и о-ве Вознесения. Мы не нашли све-
дений о наличии или отсутствии синантропных грызунов на Островах
Зеленого Мысв (Республика Кабо-Верде), Сан-Паулу и Тринидад. На
Южно-Сандвичевых, Южно-Оркпейских и на о-ве Буве синантроппых
грызунов не обнаружено.
В Индийском океане пасюк проник на небольшое число острозов. Он
обнаружен на Занзибаре, Мадагаскаре, на Маскарепских островах (Ре-
юньон, Маврикий, Родригес), лежащих на важнейшем морском пути,
связывающем страны Персидского залива с Западной Европой и Север-
ной Америкой. В восточной части океана он обычен в городах Шри
Ланки, найдеп на островах Лапкави и Пинанг в Малаккском проливе,
в Джакарте на Яве и на о-ве Рождества. Вдоль Африканского побережья
его нет на Сокотре, Пембе, Мафии, Коморских и островах, лежащих к
северу от Мадагаскара, Амирантских и Сейшельских островах. Он отсут-
ствует на Лаккадивских, Мальдивских островах и на архипелаге Чагос,
вытянувшихся цепочкой от юго-западного побережья Индии в цептр
Индийского океана. Нет его на Андаманеких, Никобарских и островах,
прилежащих к западному побережью Суматры, а также на островах во-
слочнее Явы и па Кокосовых островах. Большинство перечисленных
выше островов заселены черной крысой (Кучерук, Лаишов, 1987). От-
сутствует пасюк на островах Амстердам, Сен-Поль, Кергелен, Крозе,
Принс Эдуард, лежащих в центральной и субантарктической частях океа-
на (см. рис. 9).
Слабое заселение пасюком островов Ипдийского океана объясняется
его малочисленпостью на восточном побережье Африки и юго-западных
берегах Азии, а также изобилием и копкурентоспособностью многочис-
Ареал
51
ленных опдемичных представителей рода Вайаз в Юго-Восточной Азии
и на о-ве Новая Гвинея.
В северной части Тихого океана гасюк обычен на островах Алеутской
гряды и на Нурильских островах. Многочислен на Сахалине, Хоккайдо,
Хонсю, Сикоку, Кюсю. Есть на о-ве Тайвань, Пескадорских островах и на
о-ве Кадзан (Волкано). Заселяет некоторые города на Лусоне, Минданао,
Сулавеси. Обнаружен на Шалау и Понаие
— Каролинские острова, на
Гуаме и Сайпане
— Марианские острова и на атолле Эниветок
— Мар-
шалловы острова. Южнее обитает на Новой Ирландии и в двух пунктах
Папуа-Новой Гвинеи. Из Гавайских островов заселил лишь четыре самых.
крупных: Гавайи, Мауи, Оаху и Кауаи. Южнее, в тропической части
океана, найден на Новой Каледонии, Фиджи, островах Тонга, Северных
Кука, Общества, Туамоту, Маркизских. У берегов Центральной Америки
встречен на Ревилья-Хихедо, Кокосе и Галапагосах. Южнее Южного
тропика обитает на о-ве Лорд-Хау, Антиподов, Кэмибел, Питкэрн, Пасхи,
Хуан-Фернандес.
Из 174 острова Тихого океана, на которых обнаружены синантроппые
грызуны, серая крыса заселила 65, полинезийская— 97 и черная —
140 островов (Кучерук, Лапшшов, 1987).
,
Пасюк заселил 231 остров Мирового океана. Мы проанализировали
состав их фауны настоящих или полных синантропов (пасюк, черная и
полинезийская крысы, домовая мышь). Из восьми возможных наборов
видов отсутствует только один— сочетание пасюка, полинезийской кры-
сы и домовой мыши (рис. 10). Видимо, это явление не случайно. Мелкая
полинезийская крыса малоконкурентоспособна и вытеснлется из жилых
помещений домовой мышью, а из естественных местообитаний и хозяй-
ственных построек
— агрессивным пасюком (Кучерук, Лапшюоз, 1987).
Один пасюк встречен на 23% заселенных им островов, зпачительно чаще
он обитает вместе с домовой мышью— 35,5% островов. Совместное оби-
тание с домовой мышью и черной крысой отмечено на 20% островов, а на
все остальтые паборы сочетаний видов приходится около 22$ заселен-
ных пасюком островов. Реже всего встречаются остроза, где обитают па-
сюк и полинезийская крыса — всего 0,9%.
Для оценки влияния природных условий, и в частности климата, мы
сопоставили состав синантропных грызунов островов, заселенных пасю-
ком, в трех тиротных полосах океанической акватории: северной, сере-
динной и южной (рис. 11). В северную полосу включены субарктический
и северный умеренный климатические пояса. В серединную полосу—
субтропические, тропические, субэкваториальный и экваториальный.
В южную полосу
— южный умеренный и субантарктический климатиче-
ские пояса.
На островах северной полосы обитают преимущественно пасюк с до-
мовой мышью
— таких островов 83%. Одип пасюк обнаружен на 7,2$
островов. Сочетанием пасюка с черной крысой или с черной крысой и
домовой мышью занято 9,6% островов северной полосы. Острова сере-
динной полосы заняты преимущественно наборами видов с участием чер-
ной крысы— таких островов 87%. Пасюк совместно с домовой мышью
50
Глава вторая
В Северном Ледовитом океане мпого островов, однако в большинстве
они лишены постоянных поселений людей и не имеют условий для су-
яцествования синантропных грызунов. На северном побережье Канады и
на севере Сибири в СССР пасюк практически отсутствует. Поэтому засе-
лепие островов Ледовитого океана могло идти только через Норвежское,
Баренцево или Берингово моря. К настоящему времени поселения пасюка
в западной части океана извостны в Гренландии и на Шпиинбергене,
а в восточной — на островах Врангеля, Нунавак и Св. Лаврентия (Нуче-
рук, Лапшов, 1987). Вероятно проникновение пасюка на острова Колгуев
и Вайгач, но фактов, подтверждающих это предположение, нет. Судя по
последним исследованиям, на архипелаге Ссверпая Земля пасюк отсут-
ствует.
На севере Атлантического океана пасюк встречается на всех островах
Ирландии и Великобритании, включая Гебридские, Фарерские, Оркней-
скис (Таушт, 1977), и в Исландии. У побережья Канады оп найден на
островах Ньюфаундленд, Антикости, Принца Эдуарда, Кейп-Бретон.
Южнее пасюк населяет Бермудские, Багамские острова, Кубу, Ямайку,
Пуэрто-Рико, Анегаду, Гваделупу, Мартинику, Барбадос. У Сенеро-Афри-
канского побережья
— Сан-Мигуэль из Азорских островов (ГИтвь 1926—
1927} и Канарские острова. В тропической части океана пасюк обитает
па островах Биоко и Сан-Томе в Гвинейском заливе и на о-ве Св. Елены.
Нет его на Фернанду-ди-Норонья и о-ве Вознесения. Мы не нашли све-
дений о наличии или отсутствии синантропных грызунов на Островах
Зеленого Мысв (Республика Кабо-Верде), Сан-Паулу и Тринидад. На
Южно-Сандвичевых, Южно-Оркпейских и на о-ве Буве синантроппых
грызунов не обнаружено.
В Индийском океане пасюк проник на небольшое число острозов. Он
обнаружен на Занзибаре, Мадагаскаре, на Маскарепских островах (Ре-
юньон, Маврикий, Родригес), лежащих на важнейшем морском пути,
связывающем страны Персидского залива с Западной Европой и Север-
ной Америкой. В восточной части океана он обычен в городах Шри
Ланки, найдеп на островах Лапкави и Пинанг в Малаккском проливе,
в Джакарте на Яве и на о-ве Рождества. Вдоль Африканского побережья
его нет на Сокотре, Пембе, Мафии, Коморских и островах, лежащих к
северу от Мадагаскара, Амирантских и Сейшельских островах. Он отсут-
ствует на Лаккадивских, Мальдивских островах и на архипелаге Чагос,
вытянувшихся цепочкой от юго-западного побережья Индии в цептр
Индийского океана. Нет его на Андаманеких, Никобарских и островах,
прилежащих к западному побережью Суматры, а также на островах во-
слочнее Явы и па Кокосовых островах. Большинство перечисленных
выше островов заселены черной крысой (Кучерук, Лаишов, 1987). От-
сутствует пасюк на островах Амстердам, Сен-Поль, Кергелен, Крозе,
Принс Эдуард, лежащих в центральной и субантарктической частях океа-
на (см. рис. 9).
Слабое заселение пасюком островов Ипдийского океана объясняется
его малочисленпостью на восточном побережье Африки и юго-западных
берегах Азии, а также изобилием и копкурентоспособностью многочис-
Ареал
51
ленных опдемичных представителей рода Вайаз в Юго-Восточной Азии
и на о-ве Новая Гвинея.
В северной части Тихого океана гасюк обычен на островах Алеутской
гряды и на Нурильских островах. Многочислен на Сахалине, Хоккайдо,
Хонсю, Сикоку, Кюсю. Есть на о-ве Тайвань, Пескадорских островах и на
о-ве Кадзан (Волкано). Заселяет некоторые города на Лусоне, Минданао,
Сулавеси. Обнаружен на Шалау и Понаие
— Каролинские острова, на
Гуаме и Сайпане
— Марианские острова и на атолле Эниветок
— Мар-
шалловы острова. Южнее обитает на Новой Ирландии и в двух пунктах
Папуа-Новой Гвинеи. Из Гавайских островов заселил лишь четыре самых.
крупных: Гавайи, Мауи, Оаху и Кауаи. Южнее, в тропической части
океана, найден на Новой Каледонии, Фиджи, островах Тонга, Северных
Кука, Общества, Туамоту, Маркизских. У берегов Центральной Америки
встречен на Ревилья-Хихедо, Кокосе и Галапагосах. Южнее Южного
тропика обитает на о-ве Лорд-Хау, Антиподов, Кэмибел, Питкэрн, Пасхи,
Хуан-Фернандес.
Из 174 острова Тихого океана, на которых обнаружены синантроппые
грызуны, серая крыса заселила 65, полинезийская— 97 и черная —
140 островов (Кучерук, Лапшшов, 1987).
,
Пасюк заселил 231 остров Мирового океана. Мы проанализировали
состав их фауны настоящих или полных синантропов (пасюк, черная и
полинезийская крысы, домовая мышь). Из восьми возможных наборов
видов отсутствует только один— сочетание пасюка, полинезийской кры-
сы и домовой мыши (рис. 10). Видимо, это явление не случайно. Мелкая
полинезийская крыса малоконкурентоспособна и вытеснлется из жилых
помещений домовой мышью, а из естественных местообитаний и хозяй-
ственных построек
— агрессивным пасюком (Кучерук, Лапшюоз, 1987).
Один пасюк встречен на 23% заселенных им островов, зпачительно чаще
он обитает вместе с домовой мышью— 35,5% островов. Совместное оби-
тание с домовой мышью и черной крысой отмечено на 20% островов, а на
все остальтые паборы сочетаний видов приходится около 22$ заселен-
ных пасюком островов. Реже всего встречаются остроза, где обитают па-
сюк и полинезийская крыса — всего 0,9%.
Для оценки влияния природных условий, и в частности климата, мы
сопоставили состав синантропных грызунов островов, заселенных пасю-
ком, в трех тиротных полосах океанической акватории: северной, сере-
динной и южной (рис. 11). В северную полосу включены субарктический
и северный умеренный климатические пояса. В серединную полосу—
субтропические, тропические, субэкваториальный и экваториальный.
В южную полосу
— южный умеренный и субантарктический климатиче-
ские пояса.
На островах северной полосы обитают преимущественно пасюк с до-
мовой мышью
— таких островов 83%. Одип пасюк обнаружен на 7,2$
островов. Сочетанием пасюка с черной крысой или с черной крысой и
домовой мышью занято 9,6% островов северной полосы. Острова сере-
динной полосы заняты преимущественно наборами видов с участием чер-
ной крысы— таких островов 87%. Пасюк совместно с домовой мышью
52
Глава вторая
запимает 9% островов, а один пасюк зарегистрирован лишь на 3% ост-
ровов. В южной полосе чаще всего ветречается один пасюк — таких ост-
ровов 76%, только с домовой мышью он заселяет 8,6% островов
(рис. 14, А-В). Интересно, что одна домовая мышь встречена на 17%
тех островов южной полосы, где пасюк отсутствует (Кучерук, Лап-
шов, 1987).
.
Видимо, климатические условия северной полосы и особенно субарк-
тического пояса не позволяют серой крысе и домовой мыши обитать
круглый год и достигать высо-
Кии,
[У уе 22ииу кой численности вне антропо-
ОНИ
генных биотопов, а в постройках
’
человека домовая мышь легко
Пе лав
[- 77] ив пивени" избегает конкурентных отноше-
ний и прямого преследования
со стороны пасюка. Вероятно,
иениежиня |
Ы
Рис. 10. Состав фауны синантропных
Е
р
грызунов на островах Мирового
океана, заселенных пасюком
Цифры в кружках
— число островов с
таким набором видов
Рис. 1. Набор видов синантропных
|
грызунов на островах, заселенных
7
пасюком, в трез широтных полосах
7
океанической акватории
А — северная полоса;
чи
.
<
Б — серединная полоса;
`
;
В — южная полоса.
Сектора— доля островов (в %$), ааселен-
ных определенным набором видов
Ю ети
1.15 ИИЗЕИУ
Кан
поРееЕСи5 Ш К. гаНи5
74
/ГаЁ
т
]
и
-
г.
\-
ту
Г
\
7
К—
у
ар
No
р
17:
м
и
/
х
Ё—юЮ
`6
я
Ареал
53
пасюк и домовая мышь, имея сходную инвазивность, легко избегают кон-
куренции, осваивая (и на судах) разные местообитания.
Серая крыса и домовая мышь обрели на субантарктических островах
свою вторую родину. Оба эти вида живут здесь как в естественных био-
топах, так и в постройках человека. Пасюк активно вытесняет другие
виды, в том числе и домовую мыть. Она в отсутствие других синантро-
пов обнаружена преимущественио на небольших островах. Наверное, си-
стема каботажной транспортировки малотоннажными судами препятству-
ет завозу крупных крыс, но обеспечивает запос © малогабаритным грузом
небольшой домовой мыши.
Современный ареал пасюка в СССР
Мы продолжили кадастр мест находок пасюка в СССР, начатый А. П. Ку-
зякиным (1951), приведя его в соответствие с современными требования-
ми построения кадастрово-справочных карт (Тупикова, Тесленко, 1967;
Тупикова, 1969). На первом этапе были проверены по первоисточникам
все пункты находок пасюка, описанные А. П. Нузякиным, им были при-`
своены порядковые номера и все места обнаружения вида отмечены на
бланковой карте масштаба 1:10 000 000 точками с номером из журнала
кадастра мест находок. Порядок ведения такого журнала подробно опи-
сан в вынеупомянутых работах. В дальнейшем эта работа была продол-
жена с использованием всех доступных нам публикаций, в которых уно-
мянуты конкретные места находок пасюка на территории СССР. Уста-
новлено около 1800 пунктов, для которых в результате специальных
исследований было установлено наличие или отсутствие пасюка. Состав-
ленные нами кадастрово-справочная карта и журнал кадастра хранятся
в Зоологическом музее Московского университета.
Путем сопоставления современного ареала с его очертаниями к нача-
лу пятидесятых годов (Кузякин, 1951) была сделана картосхема изме-
нения распространения пасюка в СССР за последние 40 лет (рие. 12).
Хорошо видно, что наиболее существенные сдвиги в распространении
этого вида объясняются освоением целинных земель в Казахстане, рас-
пространением свиноводства в районах с мусульманским населением,
развитием промышленности и строительством современных городов и по-
селков в северных районах страны. Большое значение для раситирения
грапиц и уплотнения кружева ареала имело увеличение грузопотоков в
бурно развивающихся районах и стремительный рост поливного земледе-
лия. Этими же причинами объясняется проникновение иасюка в южные,
засушливые части страны и возникновение там его устойчивых изолиро-
ванных поселений.
Вероятно, установленные нами изменения границ распространения
пасюка отражают не только фактическое расширение его ареала, но и
увеличение информации о недостаточно изученных в недалеком прош-
лом территориях СССР. Это в первую очередь касается Дальнего Востока
и Восточной Сибири.
52
Глава вторая
запимает 9% островов, а один пасюк зарегистрирован лишь на 3% ост-
ровов. В южной полосе чаще всего ветречается один пасюк — таких ост-
ровов 76%, только с домовой мышью он заселяет 8,6% островов
(рис. 14, А-В). Интересно, что одна домовая мышь встречена на 17%
тех островов южной полосы, где пасюк отсутствует (Кучерук, Лап-
шов, 1987).
.
Видимо, климатические условия северной полосы и особенно субарк-
тического пояса не позволяют серой крысе и домовой мыши обитать
круглый год и достигать высо-
Кии,
[У уе 22ииу кой численности вне антропо-
ОНИ
генных биотопов, а в постройках
’
человека домовая мышь легко
Пе лав
[- 77] ив пивени" избегает конкурентных отноше-
ний и прямого преследования
со стороны пасюка. Вероятно,
иениежиня |
Ы
Рис. 10. Состав фауны синантропных
Е
р
грызунов на островах Мирового
океана, заселенных пасюком
Цифры в кружках
— число островов с
таким набором видов
Рис. 1. Набор видов синантропных
|
грызунов на островах, заселенных
7
пасюком, в трез широтных полосах
7
океанической акватории
А — северная полоса;
чи
.
<
Б — серединная полоса;
`
;
В — южная полоса.
Сектора— доля островов (в %$), ааселен-
ных определенным набором видов
Ю ети
1.15 ИИЗЕИУ
Кан
поРееЕСи5
Ш К. гаНи5
74
/
Га
Ё
т
]
и
-
г.
\-
ту
Г
\
7
К—
у
ар
No
р
17:
м
и
/
х
Ё—юЮ
`6
я
Ареал
53
пасюк и домовая мышь, имея сходную инвазивность, легко избегают кон-
куренции, осваивая (и на судах) разные местообитания.
Серая крыса и домовая мышь обрели на субантарктических островах
свою вторую родину. Оба эти вида живут здесь как в естественных био-
топах, так и в постройках человека. Пасюк активно вытесняет другие
виды, в том числе и домовую мыть. Она в отсутствие других синантро-
пов обнаружена преимущественио на небольших островах. Наверное, си-
стема каботажной транспортировки малотоннажными судами препятству-
ет завозу крупных крыс, но обеспечивает запос © малогабаритным грузом
небольшой домовой мыши.
Современный ареал пасюка в СССР
Мы продолжили кадастр мест находок пасюка в СССР, начатый А. П. Ку-
зякиным (1951), приведя его в соответствие с современными требования-
ми построения кадастрово-справочных карт (Тупикова, Тесленко, 1967;
Тупикова, 1969). На первом этапе были проверены по первоисточникам
все пункты находок пасюка, описанные А. П. Нузякиным, им были при-`
своены порядковые номера и все места обнаружения вида отмечены на
бланковой карте масштаба 1:10 000 000 точками с номером из журнала
кадастра мест находок. Порядок ведения такого журнала подробно опи-
сан в вынеупомянутых работах. В дальнейшем эта работа была продол-
жена с использованием всех доступных нам публикаций, в которых уно-
мянуты конкретные места находок пасюка на территории СССР. Уста-
новлено около 1800 пунктов, для которых в результате специальных
исследований было установлено наличие или отсутствие пасюка. Состав-
ленные нами кадастрово-справочная карта и журнал кадастра хранятся
в Зоологическом музее Московского университета.
Путем сопоставления современного ареала с его очертаниями к нача-
лу пятидесятых годов (Кузякин, 1951) была сделана картосхема изме-
нения распространения пасюка в СССР за последние 40 лет (рие. 12).
Хорошо видно, что наиболее существенные сдвиги в распространении
этого вида объясняются освоением целинных земель в Казахстане, рас-
пространением свиноводства в районах с мусульманским населением,
развитием промышленности и строительством современных городов и по-
селков в северных районах страны. Большое значение для раситирения
грапиц и уплотнения кружева ареала имело увеличение грузопотоков в
бурно развивающихся районах и стремительный рост поливного земледе-
лия. Этими же причинами объясняется проникновение иасюка в южные,
засушливые части страны и возникновение там его устойчивых изолиро-
ванных поселений.
Вероятно, установленные нами изменения границ распространения
пасюка отражают не только фактическое расширение его ареала, но и
увеличение информации о недостаточно изученных в недалеком прош-
лом территориях СССР. Это в первую очередь касается Дальнего Востока
и Восточной Сибири.
З
а
м
“
м
—
А
т
г
=
.
Ё
г
е
р
р
а
б
Ареал
55
Подробнее изменения в распространении пасюка будут рассмотрены
при более детальном описании ареала этого вида по отдельным районам
страны.
При содействии созданной при всесоюзном териологическом обществе
координационной труппы по изучению серой крысы (председатель
Е. В. Карасева) мы получили от ряда смециалистов сведения, уточняю-
щие распространение этого вида в некоторых областях СССР. Такие
сведения были присланы: А. М. Головановым (Смоленская обл.),
Н. Н. Дапиловым (Карельская АССР), Р. И. Мерзликиным (Сумская
обл.), А. И. Милютиным (Эстонская ССР), М. Г. Михайленко ( Молдав-
ская ССР), Ю. К. Поповым (Удмуртская АССР), Т. И. Савиновой (Ноз-
городская обл.), В. Б. Семеновым (Архангельская обл.), В. И. Соловье-
вым (Правобережная Украина), 3. С. Сорокиной (Дагестанская АССР),
М. И. Тарасовым (Кавказ), А. И. Феоктистовым (Калужская обл.),
Е. С. Харитоновой (Калининская обл.), Н. В. Щепотьевым (Приволжьс).
Выражаю глубокую благодарность всем перечисленным выше лицам
(использование присланпых материалов оговаривается).
При онисании ареала мы выделяем следующие основные регионы
|
\
9
-
СССР:
-
4. Европейская часть.
М
2. Кавказ.
м
3. Западная Сибирь и Северный Вазахстан.
К
4. Средняя Сибирь.
5. Северо-восток Сибири и Дальний Восток.
6. Средняя Азия и Южный Казахстан.
В зависимости от размеров региона и степени изученности он может
быть подразделен на участки.
Прежде чем перейти к описанию ареала пасюка и сго изменений в
выделенных регионах, сделаем следующие оговорки. За редким исключе-
нием мы не будет ссылаться на литературу, опубликованную До 1950 т.,
так как статья А. П. Кузякипа (1951) содержит почти исчерпывающую
библиографию на это время. Также не будем упоминать многочисленные
“,
фаунистические работы с ‘указанием находок пасюка в том или ином
рый
месте. Будут приведены ссылки на специальные статьи по пасюку, си-
Ы
нантропным грызунам и региональные монографии по млекопитающим,
'
в которых рассмотрено распрострапепие пасюка в пределах конкретных
территорий.
Европейская часть СССР
Прибалтика. А. П. Кузякин (1951) практически не имел сведений о рас-
пространении пасюка в республиках Прибалтики. Он отметил лишь на-
личие этого грызуна в г. Риге и в Литве. В настоящее время в резуль-
тате исследований А. И. Милютина, 9. П. Езбрекене (1984, 1986) Эсто-
ния и Латвия стали территориями, где распространение крыс докумепти-
ровапо наиболее полно (рис. 18). Как хорошо видно на прилагаемой
Р
и
с
.
1
2
.
К
а
р
т
о
с
т
е
м
а
и
з
м
е
н
е
н
и
й
а
р
е
а
л
а
п
а
с
ю
к
а
в
С
С
С
Р
з
а
п
о
с
л
е
д
н
и
е
5
0
л
е
т
(
п
о
:
К
у
ч
е
р
у
к
,
К
у
з
и
к
о
в
,
1
9
8
5
,
с
д
о
п
о
и
н
е
-
н
и
я
м
и
)
{
-
—
а
р
е
а
л
п
а
с
ю
к
а
п
о
А
,
Н
.
К
у
з
я
к
и
н
у
(
1
9
5
1
)
;
2
—
у
ч
а
с
т
к
и
а
р
е
а
л
а
,
в
ы
я
в
л
е
н
н
ы
е
к
н
а
с
т
о
я
щ
е
м
у
в
р
е
м
е
н
и
карте, пасюк сплошь занимает северо-западную часть Эстонской ССР.
З
а
м
“
м
—
А
т
г
=
.
Ё
г
е
р
р
а
б
Ареал
55
Подробнее изменения в распространении пасюка будут рассмотрены
при более детальном описании ареала этого вида по отдельным районам
страны.
При содействии созданной при всесоюзном териологическом обществе
координационной труппы по изучению серой крысы (председатель
Е. В. Карасева) мы получили от ряда смециалистов сведения, уточняю-
щие распространение этого вида в некоторых областях СССР. Такие
сведения были присланы: А. М. Головановым (Смоленская обл.),
Н. Н. Дапиловым (Карельская АССР), Р. И. Мерзликиным (Сумская
обл.), А. И. Милютиным (Эстонская ССР), М. Г. Михайленко ( Молдав-
ская ССР), Ю. К. Поповым (Удмуртская АССР), Т. И. Савиновой (Ноз-
городская обл.), В. Б. Семеновым (Архангельская обл.), В. И. Соловье-
вым (Правобережная Украина), 3. С. Сорокиной (Дагестанская АССР),
М. И. Тарасовым (Кавказ), А. И. Феоктистовым (Калужская обл.),
Е. С. Харитоновой (Калининская обл.), Н. В. Щепотьевым (Приволжьс).
Выражаю глубокую благодарность всем перечисленным выше лицам
(использование присланпых материалов оговаривается).
При онисании ареала мы выделяем следующие основные регионы
|
\
9
-
СССР:
-
4. Европейская часть.
М
2. Кавказ.
м
3. Западная Сибирь и Северный Вазахстан.
К
4. Средняя Сибирь.
5. Северо-восток Сибири и Дальний Восток.
6. Средняя Азия и Южный Казахстан.
В зависимости от размеров региона и степени изученности он может
быть подразделен на участки.
Прежде чем перейти к описанию ареала пасюка и сго изменений в
выделенных регионах, сделаем следующие оговорки. За редким исключе-
нием мы не будет ссылаться на литературу, опубликованную До 1950 т.,
так как статья А. П. Кузякипа (1951) содержит почти исчерпывающую
библиографию на это время. Также не будем упоминать многочисленные
“,
фаунистические работы с ‘указанием находок пасюка в том или ином
рый
месте. Будут приведены ссылки на специальные статьи по пасюку, си-
Ы
нантропным грызунам и региональные монографии по млекопитающим,
'
в которых рассмотрено распрострапепие пасюка в пределах конкретных
территорий.
Европейская часть СССР
Прибалтика. А. П. Кузякин (1951) практически не имел сведений о рас-
пространении пасюка в республиках Прибалтики. Он отметил лишь на-
личие этого грызуна в г. Риге и в Литве. В настоящее время в резуль-
тате исследований А. И. Милютина, 9. П. Езбрекене (1984, 1986) Эсто-
ния и Латвия стали территориями, где распространение крыс докумепти-
ровапо наиболее полно (рис. 18). Как хорошо видно на прилагаемой
Р
и
с
.
1
2
.
К
а
р
т
о
с
т
е
м
а
и
з
м
е
н
е
н
и
й
а
р
е
а
л
а
п
а
с
ю
к
а
в
С
С
С
Р
з
а
п
о
с
л
е
д
н
и
е
5
0
л
е
т
(
п
о
:
К
у
ч
е
р
у
к
,
К
у
з
и
к
о
в
,
1
9
8
5
,
с
д
о
п
о
и
н
е
-
н
и
я
м
и
)
{
-
—
а
р
е
а
л
п
а
с
ю
к
а
п
о
А
,
Н
.
К
у
з
я
к
и
н
у
(
1
9
5
1
)
;
2
—
у
ч
а
с
т
к
и
а
р
е
а
л
а
,
в
ы
я
в
л
е
н
н
ы
е
к
н
а
с
т
о
я
щ
е
м
у
в
р
е
м
е
н
и
карте, пасюк сплошь занимает северо-западную часть Эстонской ССР.
56
Глава вторая
Здесь он обнаружен в 151 обследованном пункте. Граница сглоптного.
распространения пасюка в республике проходит с юго-запада на северо-
восток по следующим населенным пупцтам: Таагепера — Хольстре —
Вильянди— Пуурмани, Каарепере — Сааре — Вепевере — Йыуга — Куре-
мяе — Алайые. К востоку от этой линии пасюк распространен спорадич-
по, он обнаружен в 19 из 84 обследованных поселков, хуторов, ферм. Тут
он повсюду обитал совместно с черной крысой, которая встречалась
сплошь.
В Латвии пасюк имеет сплошное распространение в западной полови-
не республики, где он обнаружен во всех обследованных населенных
пунктах (151 пункт). В восточной половине республики пасюк встреча-
ется спорадично, он найден здесь в [5 из 154 обследованных населенных
пунктов, большинство которых заселено черной крысой.
В Литве до 60-х годов пасюк отсутствовал на крайнем северо-востоке
республики. В настоящее время он обитает повсюду и обнаружен в сот-
нях населенных пунктов всех 44 районах Литвы (Ликявичене, 1960;
Милютин, Изёрскене, 1986).
В Калининградской области пасюк был отловлен в 7 (Багратионов-
ский, Гурьевский, Краснознаменский, Пестеровский, Озерский, Полес-
ский, Славский) из 13 административных районов.
ранение пасюка и
черной крысы в
Эстонии (вверху)
и Латвии (вни-
зу) (по: Милютин,
Езёрскене, 1984,
1986)
Населенные пункты,
в которых найдены:
1 — только пасюк;
2 — только черная
крыса;
3—обавида
Рис. 18. Распрост-‘
Ареал
57
С запада к Прибалтике примыкает Северо-Западный район РСФСР,
х которому относятся Пековская, Ленинградская и Новгородская обла-
сти, А. П. Кузякин (1951) специально оговаривал отсутствие пасюка в
некоторых районах современных Псковской, Новгородской, Калининской
и Смоленской областей. В Поковекой области до 1964 г. регистрировали
только черную крысу, которая была найдена в 45 населенных пунктах
22 из 24 районов области. Пасюка в области пе было (Федосеева и др.,
1967; Милютин, 1987). В настоящее время насюк проник в Плюсский и
'Струго-Краспенокий райопы о стороны Ленинграда, в Дновский со
стороны Старой Руссы и в Питаловский— со стороны Латвийской ССР
(рис. 14).
Г. А. Новиков и др. (1970) предполагали отсутствие пасюка в отдель-
ных пунктах юго-запада Ленинградской области. В последней до настоя-
ре: Распространение крыс в областях Северо-Западного экономического района
1— населенные пункты, в которых обнаружен только пасюк; 2 — обнаружены пасюк и чер-
ная крыса; 3 — обнаружена только черная крыса; & — областные центры
т
-
77
56
Глава вторая
Здесь он обнаружен в 151 обследованном пункте. Граница сглоптного.
распространения пасюка в республике проходит с юго-запада на северо-
восток по следующим населенным пупцтам: Таагепера — Хольстре —
Вильянди— Пуурмани, Каарепере — Сааре — Вепевере — Йыуга — Куре-
мяе — Алайые. К востоку от этой линии пасюк распространен спорадич-
по, он обнаружен в 19 из 84 обследованных поселков, хуторов, ферм. Тут
он повсюду обитал совместно с черной крысой, которая встречалась
сплошь.
В Латвии пасюк имеет сплошное распространение в западной полови-
не республики, где он обнаружен во всех обследованных населенных
пунктах (151 пункт). В восточной половине республики пасюк встреча-
ется спорадично, он найден здесь в [5 из 154 обследованных населенных
пунктов, большинство которых заселено черной крысой.
В Литве до 60-х годов пасюк отсутствовал на крайнем северо-востоке
республики. В настоящее время он обитает повсюду и обнаружен в сот-
нях населенных пунктов всех 44 районах Литвы (Ликявичене, 1960;
Милютин, Изёрскене, 1986).
В Калининградской области пасюк был отловлен в 7 (Багратионов-
ский, Гурьевский, Краснознаменский, Пестеровский, Озерский, Полес-
ский, Славский) из 13 административных районов.
ранение пасюка и
черной крысы в
Эстонии (вверху)
и Латвии (вни-
зу) (по: Милютин,
Езёрскене, 1984,
1986)
Населенные пункты,
в которых найдены:
1 — только пасюк;
2 — только черная
крыса;
3—обавида
Рис. 18. Распрост-‘
Ареал
57
С запада к Прибалтике примыкает Северо-Западный район РСФСР,
х которому относятся Пековская, Ленинградская и Новгородская обла-
сти, А. П. Кузякин (1951) специально оговаривал отсутствие пасюка в
некоторых районах современных Псковской, Новгородской, Калининской
и Смоленской областей. В Поковекой области до 1964 г. регистрировали
только черную крысу, которая была найдена в 45 населенных пунктах
22 из 24 районов области. Пасюка в области пе было (Федосеева и др.,
1967; Милютин, 1987). В настоящее время насюк проник в Плюсский и
'Струго-Краспенокий райопы о стороны Ленинграда, в Дновский со
стороны Старой Руссы и в Питаловский— со стороны Латвийской ССР
(рис. 14).
Г. А. Новиков и др. (1970) предполагали отсутствие пасюка в отдель-
ных пунктах юго-запада Ленинградской области. В последней до настоя-
ре: Распространение крыс в областях Северо-Западного экономического района
1— населенные пункты, в которых обнаружен только пасюк; 2 — обнаружены пасюк и чер-
ная крыса; 3 — обнаружена только черная крыса; & — областные центры
т
-
77
58
Глава вторая
щего времени распрострапепие пасюка документировано совершенно не-
достаточно.
.
В пограничных © Ленинградской районах Новгородской области (Нов-
городском, Чудовском, Маловищерском, Хвойнинском, Боровичевском,
Мошенском, Пестовском) обнаружен только пасюк, также только насюк
найден в южном Маревском районе на границе с Калининской областью.
На севере в Любытинском, на западе в Солецком, Шимском, Волотовском,
Старорусском и на востоке в Валдайском районах распространены и па-
сюк, и черная крыса. Наконец, в Окуловском, Батецком, Поддорском,
Холмском и Демянском районах обнаружена только черная крыса {дан-
ные Т. А. Савиной).
.
В Белорусской ССР насюк достоверно зарегистрирован в Минской
области, в г. Любань, Пос. Смолевичи, в Березинском заповеднике; в Мо-
тилевской обл. широко распространен в Осиповичивском районе; в Го-
мельской обл.— в г. Турове; в Петриковском, Копаткевичеком, Чичерском
районах, в Брестком районе и Беловежской пуще. Так как черная крыса
распространена по всей территории Белоруссии и встречается значитель-
но чаще пасюка (Сержанин, 1961), а в некоторых пограничных с Бело-
руссией областях (Псковской, Смоленской, на западе Латвии) обитает
иреимущественно черная крыса (Милютин, Езёрскене, 1986), охаракте-
ризовать совремепное распространепие пасюка в пределах БССР без
проведения специальных исследований не представляется возможным.
А. П. Кузякин (1951), описывая распространение пасюка на Украи-
не. указал, что в правобережной части республики этот вид был найден
в Одессе и в пизовьях Южного Буга (с. Ивапово-Рыбальче). Кроме того,
пасюк указан для городов Киева, Днепропетронска, Харькова, Золочева,
Сум, Новгорода-Северского и дикоживущая популяция была обнаружена
на р. Обиточной, впадающей в Азовское море. В Крыму пасюк обитах
в г. Севастополе, но А. П. Кузякин специально отметил, что далеко не
весь полуостров заселен им. В то же время не было указано, что еще до
1889 г пасюка пашли в Евпатории (Тихомиров, Корчагин, 1889),
а Б. Воляпский (1929) описал его птирокое распространение в населен
пых пунктах и по морскому побережью Керченского полуострова. В кол-
лекциях Зоомузея МГУ есть экземпляр пасюка из Армянска сборов.
1929 г. И. И. Пузанов (4981) добыл его в 1927 г. на перевале к северу,
от Алушты. Для Феодосии этот вид указап Ф. А. Пушницей, П. И. Ши-
рановичем (1960).
Как это ни удивительно, но за время, прошедшее с выхода в снет
обзора А. П. Кузякина, отпосительно Украины не появилось ни одной
публикации, посвященной распространению пасюка на копкретной терри-
тории, а авторы общих работ по млекопитающим обычно ограничивались
указанием на повсеместное обитание этого вида.
В книге о зверях Украины (Мигулин, 1988) говорится, что пасюк
обитает на всей территории республики, но конкретных мест сбора кол-
лекционных материалов упомянуто всего лишь семь. По мнепию И. Т. Со-
кура (1960), пасюк водится во всех природных районах Украины, по
особенно многочислен в степной и лесостепной части ее. Обычен в при-
Ареал
59
родных биотопах пойм Дуная, Днестра, Днепра, Десны. В книге приведе-
но 46 карт распространения млекопитающих Украины, в том числе 20 ви-
дов грызунов, но карта мест находок насюка отсутствует.
В западных областях Украины К. А. Татаринов (1956) обнаружил
пасюка во всех’ обследованных им пунктах, отметив, что Карпаты этот
вид заселяет до высот 1350—1400 м. Распространению и экологии пасюка
в естественных биотопах верховьев Днестра посвящена специальная
статья (Татаринов, 1954).
Носительство пасюками лептоспир обпаружено в следующих городах
Украины: Киеве, Одессе, Черкассах, Днепропетровске, Херсоне, Воро-
зпиловграде, Донецке, Черновцах (Бернасовская и др., 1983).
В Сумской области обитание пасюка подтверждено для всех райопных
цептров за исключением Бурыни и Тростянца. Кроме того, пасюк обна-
ружен еще в 62 населенных пунктах районов, расположенных к западу
ик югу от Сум, в том числе и в трех населенных пунктах Тростянецко-
го района (данные И. Р. Мерзликина).
В Одесской области все районы заселены пасюком (данные В. И. Со-
ловьева).
По нашему мнению, уверенность украинских зоологов в повсеместном
распространении пасюка недостаточно обоснована. К восточным областям
Украины прилегают территории, на которых пасюк появился недавно
(Дмитриева, 1975) или отсутствует (ШЩепотьев, Ткачева, 1966; Козлов,
1986). На севере Украины, в Полесье, весьма обычна черная крыса (Ги-
ренко, 1950; Сокур, 1960), еще более многочисленна она на прилежащих
областях Белоруссии (Сержанин, 1961), возможно, что на часть террато-
рии, занятой черной крысой, пасюк еще не проник.
Уже давно назрела необходимость в хорошей обзорной работе по па-
сюку на Украине с хорошей фактической аргументацией, использовани-
ем коллекционных материалов и подробными картами распростра-
пения.
В Молдавии пасюки распространены повсюду, зверьки обпаружены
более чем в ста обследованных населенных пунктах. Дикоживущие по-
луляции обычны в низовьях Днестра, Дуная и Прута (Лозан, 1971).
В Молдавской ССР на основании отлова более тысячи пасюков со-
ставлена кадастровая карта мест его находок. Насюк был обнаружен в
тородах: Кишинев, Бельцы, Бендеры, Кагул, Оргеев, Сороки, Тирасполь,
Унгены и в 49 населенных пунктах в 22 из 38 районов республики.
В 19 точках 12 административных районов 138 экз. отловили в естествен-
ных местообитаниях (данные А. Г. Михайленко).
Северный район РСФСР (Карельская АССР, Мурманская, Архангель-
ская, Вологодская области и Коми АССР).
В работе А. П. Кузякина (1951) для рассматриваемого региона от-
мечены следующие достоверные пункты регистрации пасюка: Мурманск,
Архангельск, Медвежегорск, Троицко-Печорск. Кроме того, оконтурены
поймы Печоры и нижней Усы от Троицко-Печорска до Печорской губы,
где, по указаниям В. П. Теплова и Е. Н. Тепловой (1947), пасюк встре-
чается на пристанях. Кузякин считает, что пасюка, видимо, не было на
58
Глава вторая
щего времени распрострапепие пасюка документировано совершенно не-
достаточно.
.
В пограничных © Ленинградской районах Новгородской области (Нов-
городском, Чудовском, Маловищерском, Хвойнинском, Боровичевском,
Мошенском, Пестовском) обнаружен только пасюк, также только насюк
найден в южном Маревском районе на границе с Калининской областью.
На севере в Любытинском, на западе в Солецком, Шимском, Волотовском,
Старорусском и на востоке в Валдайском районах распространены и па-
сюк, и черная крыса. Наконец, в Окуловском, Батецком, Поддорском,
Холмском и Демянском районах обнаружена только черная крыса {дан-
ные Т. А. Савиной).
.
В Белорусской ССР насюк достоверно зарегистрирован в Минской
области, в г. Любань, Пос. Смолевичи, в Березинском заповеднике; в Мо-
тилевской обл. широко распространен в Осиповичивском районе; в Го-
мельской обл.— в г. Турове; в Петриковском, Копаткевичеком, Чичерском
районах, в Брестком районе и Беловежской пуще. Так как черная крыса
распространена по всей территории Белоруссии и встречается значитель-
но чаще пасюка (Сержанин, 1961), а в некоторых пограничных с Бело-
руссией областях (Псковской, Смоленской, на западе Латвии) обитает
иреимущественно черная крыса (Милютин, Езёрскене, 1986), охаракте-
ризовать совремепное распространепие пасюка в пределах БССР без
проведения специальных исследований не представляется возможным.
А. П. Кузякин (1951), описывая распространение пасюка на Украи-
не. указал, что в правобережной части республики этот вид был найден
в Одессе и в пизовьях Южного Буга (с. Ивапово-Рыбальче). Кроме того,
пасюк указан для городов Киева, Днепропетронска, Харькова, Золочева,
Сум, Новгорода-Северского и дикоживущая популяция была обнаружена
на р. Обиточной, впадающей в Азовское море. В Крыму пасюк обитах
в г. Севастополе, но А. П. Кузякин специально отметил, что далеко не
весь полуостров заселен им. В то же время не было указано, что еще до
1889 г пасюка пашли в Евпатории (Тихомиров, Корчагин, 1889),
а Б. Воляпский (1929) описал его птирокое распространение в населен
пых пунктах и по морскому побережью Керченского полуострова. В кол-
лекциях Зоомузея МГУ есть экземпляр пасюка из Армянска сборов.
1929 г. И. И. Пузанов (4981) добыл его в 1927 г. на перевале к северу,
от Алушты. Для Феодосии этот вид указап Ф. А. Пушницей, П. И. Ши-
рановичем (1960).
Как это ни удивительно, но за время, прошедшее с выхода в снет
обзора А. П. Кузякина, отпосительно Украины не появилось ни одной
публикации, посвященной распространению пасюка на копкретной терри-
тории, а авторы общих работ по млекопитающим обычно ограничивались
указанием на повсеместное обитание этого вида.
В книге о зверях Украины (Мигулин, 1988) говорится, что пасюк
обитает на всей территории республики, но конкретных мест сбора кол-
лекционных материалов упомянуто всего лишь семь. По мнепию И. Т. Со-
кура (1960), пасюк водится во всех природных районах Украины, по
особенно многочислен в степной и лесостепной части ее. Обычен в при-
Ареал
59
родных биотопах пойм Дуная, Днестра, Днепра, Десны. В книге приведе-
но 46 карт распространения млекопитающих Украины, в том числе 20 ви-
дов грызунов, но карта мест находок насюка отсутствует.
В западных областях Украины К. А. Татаринов (1956) обнаружил
пасюка во всех’ обследованных им пунктах, отметив, что Карпаты этот
вид заселяет до высот 1350—1400 м. Распространению и экологии пасюка
в естественных биотопах верховьев Днестра посвящена специальная
статья (Татаринов, 1954).
Носительство пасюками лептоспир обпаружено в следующих городах
Украины: Киеве, Одессе, Черкассах, Днепропетровске, Херсоне, Воро-
зпиловграде, Донецке, Черновцах (Бернасовская и др., 1983).
В Сумской области обитание пасюка подтверждено для всех райопных
цептров за исключением Бурыни и Тростянца. Кроме того, пасюк обна-
ружен еще в 62 населенных пунктах районов, расположенных к западу
ик югу от Сум, в том числе и в трех населенных пунктах Тростянецко-
го района (данные И. Р. Мерзликина).
В Одесской области все районы заселены пасюком (данные В. И. Со-
ловьева).
По нашему мнению, уверенность украинских зоологов в повсеместном
распространении пасюка недостаточно обоснована. К восточным областям
Украины прилегают территории, на которых пасюк появился недавно
(Дмитриева, 1975) или отсутствует (ШЩепотьев, Ткачева, 1966; Козлов,
1986). На севере Украины, в Полесье, весьма обычна черная крыса (Ги-
ренко, 1950; Сокур, 1960), еще более многочисленна она на прилежащих
областях Белоруссии (Сержанин, 1961), возможно, что на часть террато-
рии, занятой черной крысой, пасюк еще не проник.
Уже давно назрела необходимость в хорошей обзорной работе по па-
сюку на Украине с хорошей фактической аргументацией, использовани-
ем коллекционных материалов и подробными картами распростра-
пения.
В Молдавии пасюки распространены повсюду, зверьки обпаружены
более чем в ста обследованных населенных пунктах. Дикоживущие по-
луляции обычны в низовьях Днестра, Дуная и Прута (Лозан, 1971).
В Молдавской ССР на основании отлова более тысячи пасюков со-
ставлена кадастровая карта мест его находок. Насюк был обнаружен в
тородах: Кишинев, Бельцы, Бендеры, Кагул, Оргеев, Сороки, Тирасполь,
Унгены и в 49 населенных пунктах в 22 из 38 районов республики.
В 19 точках 12 административных районов 138 экз. отловили в естествен-
ных местообитаниях (данные А. Г. Михайленко).
Северный район РСФСР (Карельская АССР, Мурманская, Архангель-
ская, Вологодская области и Коми АССР).
В работе А. П. Кузякина (1951) для рассматриваемого региона от-
мечены следующие достоверные пункты регистрации пасюка: Мурманск,
Архангельск, Медвежегорск, Троицко-Печорск. Кроме того, оконтурены
поймы Печоры и нижней Усы от Троицко-Печорска до Печорской губы,
где, по указаниям В. П. Теплова и Е. Н. Тепловой (1947), пасюк встре-
чается на пристанях. Кузякин считает, что пасюка, видимо, не было на
60
Глава вторая
всем Кольском полуострове к востоку от железной дороги. Он не имел
сведений о пасюке в долине р. Мезепи, но на водораздельных простран-
ствах между Северной Двиной, Мезенью и Печорой пасюка, по всей ве-
роятпости, не было.
К настоящему времени, кроме перечисленных выше, поселения пасю-
ка в естественных биотопах зарегистрированы па северном берегу Ла-
дожекого озера. Обитание его в Карельской ССР установлено в Пудоже,
Кондопоге, Юкоозере. В Мурманской обл.—в Кандалакше
и Коле. В Ар-
хангельской обл.—в Каргополе, Онеге, Северодвинске, Верхней Тойме,
Красноборске. В бассейне р. Мезень он обнаружен в г. Мезени, в с. Ле-
шуканском, Вожгаре, в ряде пунктов дельты Печоры, в том числе и в
Нарьян-Маре. В Коми АССР вверх по Нечоре отмечен в Хабарихе, нА
ст. Печора, в Кожве и Воркуте, южнее в Айкине (Сосницкий, Некрасова,
1966; Сосницкий, 1973). По В. В. Груздеву (1963), северная граница мас-
совой вредоносной деятельности пасюка проходит между южными побе-
режьями Ладожского и Онежекого озер и языками вдается на север по
Онеге и Северной Двине. Видимо, эти очертания отражают сплошное
распространение пасюка в описываемом регионс.
9. В. Ивантер (1975) считает, что пасюк в Карелии теперь широко
расселился и встречается почти повсеместно даже в самых глухих лес-
ных поселках, причем обитает не только в человеческих постройках, но
и вне населенных пунктов. Обитание пасюка показано во всех населен-
ных пунктах, обозначенных на карте Карельской АССР масштаба
1:750 000 (данные П. И. Данилова).
В Архангельской области пасюк распространен повсюду, за исключе-
нием самых северных и глухих таежных поселков. Встречается во всех
городах, на речных пристанях, железподорожных станциях, а таже в
сельских крупных населенных пунктах. Появлепие пасюков в сельской
местности относится к тридцатым годам текущего столетия, с момента
создания в селах колхозных зерновых фуражных складов, с возникнове-
нием животноводческих ферм. В последнее десятилетие в Архангельской
области пасюка добывали в городах Опеге, Северо-Двинске, Каргоноле,
Шенкурске, Котласе, Мезепи, в районных центрах Холмогоры, Березник,
Октябрьский (Шангалы), Карпогоры, Лешуконское, Верхняя Тойма,
Красноборск, Ильинское-Подомское. В естественных биотопах его ловили:
в дельте Северной Двины, на заливных лугах в 5 км от Архангельска,
в 30 км от Архангельска по шоссе на Вологду, в восточной части Хол-
могорского района, в Верхнетоймском, Красноборском, Вилегодском, Лен-
ском районах. В поселке Усть-Кара Ненецкого автономного округа появ-
ление серых крыс впервые было отмечено в 1934 г. (данные В. Б. Се-
менова).
А. Г. Фаршатов (1986) пишет, что пасюк в массовом количестве стал
появляться на территории Номи АССР в 30-е годы нашего столетия и к
настоящему времепи заселил большую часть ее территории. Отсутствует
он па западе республики в населенных пунктах по р. Вашка, по некото-
рым рекам Приуралья, в тундровых поселках, удаленных от основных
водных путей и железных дорог. На юге республики в Прилуцком райо-
дреал
61
не пасюка нет в 40% населенных пунктов, в Приуралье в Троицко-Пе-
чорском районе в 94% населенных пунктов.
До настоящего времени отсутствуют полноценные публикации о рас-
пространении пасюка как в целом по региону, так и по отдельным об-
ластям и АССР.
Центральный экономический район. К нему относятся: Калининская,
Ярославская, Костромская, Смоленская, Московская, Ивановская, Влади-
мирская, Брянская, Калужская, Орловская, Тульская, Рязанская области.
А. П. Кузякин (1951) указывал на обитание пасюка в Калинине, Ря:е-
ве, Рыбинске, Москве, Серпухове, Брянске, Туле, Рязани, Сасове, Ми-
хайлове, на многочисленность этого вида во всех уездах юго-востока Ор-
ловской губернии. Отмечал отсутствие пасюка в Луковпиковском, Сели-
жаровеком, Осташковском районах Калининской области, в средних и
северных районах Смоленской области, вероятно, во многих районах Ве-
ликолукской и юга Ленинградской области.
В настоящее время пасюк заселил болыпую часть Калининской об-
ласти. Только этот вид отмечен в Фировском, Бологовском, Вышневолоц-
ком, Кувшиновском, Ржевском, Старицком, Торжокском, Лихославль-
ском, Калининском, Максатихинском, Молоковеком, Бежецком, Конаков-.
ском, Весъегонском, Нраснохолмском, Санковском, Несовогорском, Ка-
шинском и Кимрском районах. В Осташковском и Западнодвинском встре-
чается вместе с черной крысой. Только черная крыса зарегистрирована
в Нелидовском районе (данные Г. С. Харитоновой).
Д. В. Поярков (1961) обнаружил пасюков в Ростовском районе Яро-
славской области, Галичеком и Сусанинском районах Костромской об-
ласти.
Пасюк найден только в шести северо-восточных районах Смоленской
области: Гагаринском (Гжатском), Темкинском, Тумановском, Сычевском,
Холм-Жирковском и Вяземском. В последпем оп встречается вместе с
черной крысой. Все остальные районы области заселены только черной
крысой (дапные А. М. Голованова).
В Московской области пасюк обитает в населенных пунктах по всей
территории. В 20 обследованных городах на его долю пришлось 34,5%
всех отловленных мелких млекопитающих. В помещепиях сельских на-
селенных пунктов пасюк составлял всего 3,5% добытых зверьков. По
берегам водоемов паслока добывали в Москве, Серпуховском, Ступинском
и Одинцовском районах (Домбровский, 1983). По нашим наблюдениям,
пасюк заселяет берега больштинства мелких речушек, текущих в пределах
городской черты Москвы.
-
В. С. Усова (1986) обнаружила поселения пасюков в природных био-
топах 11 из 17 административных райопов Владимирской области (Ков-
ровском, Маленковском, Собинском, Гусь-Хрустальном, Вязниковском,
Юрьев-Польском, Селивановском, Судогодском, Кольчугинском, Горохо-
венком, Суздальском). Кроме того, она упоминает о выделении от пасю-
ка возбудителя исевдотуберкулеза в Четушинском районе.
В Калужекой области с 1952 по 1986 г. многократно обследовали 22
из 24 административных районов. В 104 населенном пункте была обна-
60
Глава вторая
всем Кольском полуострове к востоку от железной дороги. Он не имел
сведений о пасюке в долине р. Мезепи, но на водораздельных простран-
ствах между Северной Двиной, Мезенью и Печорой пасюка, по всей ве-
роятпости, не было.
К настоящему времени, кроме перечисленных выше, поселения пасю-
ка в естественных биотопах зарегистрированы па северном берегу Ла-
дожекого озера. Обитание его в Карельской ССР установлено в Пудоже,
Кондопоге, Юкоозере. В Мурманской обл.—в Кандалакше
и Коле. В Ар-
хангельской обл.—в Каргополе, Онеге, Северодвинске, Верхней Тойме,
Красноборске. В бассейне р. Мезень он обнаружен в г. Мезени, в с. Ле-
шуканском, Вожгаре, в ряде пунктов дельты Печоры, в том числе и в
Нарьян-Маре. В Коми АССР вверх по Нечоре отмечен в Хабарихе, нА
ст. Печора, в Кожве и Воркуте, южнее в Айкине (Сосницкий, Некрасова,
1966; Сосницкий, 1973). По В. В. Груздеву (1963), северная граница мас-
совой вредоносной деятельности пасюка проходит между южными побе-
режьями Ладожского и Онежекого озер и языками вдается на север по
Онеге и Северной Двине. Видимо, эти очертания отражают сплошное
распространение пасюка в описываемом регионс.
9. В. Ивантер (1975) считает, что пасюк в Карелии теперь широко
расселился и встречается почти повсеместно даже в самых глухих лес-
ных поселках, причем обитает не только в человеческих постройках, но
и вне населенных пунктов. Обитание пасюка показано во всех населен-
ных пунктах, обозначенных на карте Карельской АССР масштаба
1:750 000 (данные П. И. Данилова).
В Архангельской области пасюк распространен повсюду, за исключе-
нием самых северных и глухих таежных поселков. Встречается во всех
городах, на речных пристанях, железподорожных станциях, а таже в
сельских крупных населенных пунктах. Появлепие пасюков в сельской
местности относится к тридцатым годам текущего столетия, с момента
создания в селах колхозных зерновых фуражных складов, с возникнове-
нием животноводческих ферм. В последнее десятилетие в Архангельской
области пасюка добывали в городах Опеге, Северо-Двинске, Каргоноле,
Шенкурске, Котласе, Мезепи, в районных центрах Холмогоры, Березник,
Октябрьский (Шангалы), Карпогоры, Лешуконское, Верхняя Тойма,
Красноборск, Ильинское-Подомское. В естественных биотопах его ловили:
в дельте Северной Двины, на заливных лугах в 5 км от Архангельска,
в 30 км от Архангельска по шоссе на Вологду, в восточной части Хол-
могорского района, в Верхнетоймском, Красноборском, Вилегодском, Лен-
ском районах. В поселке Усть-Кара Ненецкого автономного округа появ-
ление серых крыс впервые было отмечено в 1934 г. (данные В. Б. Се-
менова).
А. Г. Фаршатов (1986) пишет, что пасюк в массовом количестве стал
появляться на территории Номи АССР в 30-е годы нашего столетия и к
настоящему времепи заселил большую часть ее территории. Отсутствует
он па западе республики в населенных пунктах по р. Вашка, по некото-
рым рекам Приуралья, в тундровых поселках, удаленных от основных
водных путей и железных дорог. На юге республики в Прилуцком райо-
дреал 61
не пасюка нет в 40% населенных пунктов, в Приуралье в Троицко-Пе-
чорском районе в 94% населенных пунктов.
До настоящего времени отсутствуют полноценные публикации о рас-
пространении пасюка как в целом по региону, так и по отдельным об-
ластям и АССР.
Центральный экономический район. К нему относятся: Калининская,
Ярославская, Костромская, Смоленская, Московская, Ивановская, Влади-
мирская, Брянская, Калужская, Орловская, Тульская, Рязанская области.
А. П. Кузякин (1951) указывал на обитание пасюка в Калинине, Ря:е-
ве, Рыбинске, Москве, Серпухове, Брянске, Туле, Рязани, Сасове, Ми-
хайлове, на многочисленность этого вида во всех уездах юго-востока Ор-
ловской губернии. Отмечал отсутствие пасюка в Луковпиковском, Сели-
жаровеком, Осташковском районах Калининской области, в средних и
северных районах Смоленской области, вероятно, во многих районах Ве-
ликолукской и юга Ленинградской области.
В настоящее время пасюк заселил болыпую часть Калининской об-
ласти. Только этот вид отмечен в Фировском, Бологовском, Вышневолоц-
ком, Кувшиновском, Ржевском, Старицком, Торжокском, Лихославль-
ском, Калининском, Максатихинском, Молоковеком, Бежецком, Конаков-.
ском, Весъегонском, Нраснохолмском, Санковском, Несовогорском, Ка-
шинском и Кимрском районах. В Осташковском и Западнодвинском встре-
чается вместе с черной крысой. Только черная крыса зарегистрирована
в Нелидовском районе (данные Г. С. Харитоновой).
Д. В. Поярков (1961) обнаружил пасюков в Ростовском районе Яро-
славской области, Галичеком и Сусанинском районах Костромской об-
ласти.
Пасюк найден только в шести северо-восточных районах Смоленской
области: Гагаринском (Гжатском), Темкинском, Тумановском, Сычевском,
Холм-Жирковском и Вяземском. В последпем оп встречается вместе с
черной крысой. Все остальные районы области заселены только черной
крысой (дапные А. М. Голованова).
В Московской области пасюк обитает в населенных пунктах по всей
территории. В 20 обследованных городах на его долю пришлось 34,5%
всех отловленных мелких млекопитающих. В помещепиях сельских на-
селенных пунктов пасюк составлял всего 3,5% добытых зверьков. По
берегам водоемов паслока добывали в Москве, Серпуховском, Ступинском
и Одинцовском районах (Домбровский, 1983). По нашим наблюдениям,
пасюк заселяет берега больштинства мелких речушек, текущих в пределах
городской черты Москвы.
-
В. С. Усова (1986) обнаружила поселения пасюков в природных био-
топах 11 из 17 административных райопов Владимирской области (Ков-
ровском, Маленковском, Собинском, Гусь-Хрустальном, Вязниковском,
Юрьев-Польском, Селивановском, Судогодском, Кольчугинском, Горохо-
венком, Суздальском). Кроме того, она упоминает о выделении от пасю-
ка возбудителя исевдотуберкулеза в Четушинском районе.
В Калужекой области с 1952 по 1986 г. многократно обследовали 22
из 24 административных районов. В 104 населенном пункте была обна-
62
Глава вторая
ружена черная крыса. По пасюку данных нет (материалы А. И. Феок-
тистова).
Ю. Л. Мясциков (4976) с 1956 п 1968 г. обнаружил пасюков в
11 населенных пунктах Арсеньевского, в 4 — Одоевского, в 3 — Чернь-
ского районов и в г. Узловой Тульской области. В 1972—1983 гг. пасюк
расширил свое распространение. Он был найден в 24 из .68 обеледован-
ных населенных лунктов в 8 из 10 административных районов лесостен-
ной части Тульской области. Не поймали пасюка в 6 населенных пунк-
тах Заокского и в 3 Одоенското районов. Среди 215 крыс было 86 пасю-
ков. В стенной части области обследовали 84 населенных пункта »
13 районах. Отсутствовал пасюк в 28 населенных пунктах 6 администра-
тивных районов (Воловского, Кфремовского, Каменского, Куркинского,
Тепло-Огаревского, Черньского). Из 289 поймапных крыс было 212 па-
сюков (Панина, 1986а). В Туле первый пасюк был пойман в апреле
1972 г., в 1975 г. среди отловленных в городе крыс на долю пасюка
приходилось 45$, в 1978—81%, в 1979— более 90%. а после 1982 г.—
96—98% (Панина, 1986). Таким образом, удалось наблюдать активный
процесс замены черной крысы пасюком.
.
Мн не нашли после 4954 г. никаких новых литературных сведений о
пасюке в Иваповской, Брянской, Орловской и Рязанской областях.
Центрально-Черноземный район. В него входят Липецкая, Тамбовская,
Курская, Белгородская и Воронежская области.
.
А. П. Кузякии (1951) считал, что сведений о пасюке в бассейпе верх-
него и среднего течения Дона почти нет, Это же подтверждает Т. В. Дми-
триева (1970), отмечая, что данные о размещении пасюка на территории
Воропежекой области в довоенное время отсутствуют. По ее мнению, в
первые годы после войны пасюк обитал преимущественно в городах, где
на него приходилось 18,3% добытых грызунов, в сельских населенных
пунктах его доля была равна 1,8%. Пасюк отсутствовал или был крайте
малочислен в южных и западных районах области. Много пасюка в
сельской местности было только в северо-восточных районах — в бассей-
не Хопра. В первые послевоенные годы ареал пасюка еще более сокра-
тился и принял пятнистый характер. Одновремепно заселенные этим ви-
дом пятна появились и па юго-востоке области, дойдя до левобережья До-
на у пересечения им границы области. Все правобережье Дона было прак-
тически свободно от пасюка.
.
С конца 50-х годов началось быстрое расселение этого вида.
В 1962—1963 тг. резко вырос удельный вес крысиного населепия в
сельской местности, на долю пасюка падало 29% выловленных грызунов.
Расселение и подъем численности пасюка были связаны с увеличением
сельскохозяйственной продукциии особенно © ростом животноводства.
К 1968 г. подавляющая часть области, хотя и с разной плотностью, была
заселена пасюком, отсутствовали зверьки только в части западных райо-
нов. Статья (Дмитриева, 1970) проиллюстрирована тремя весьма труднв
читаемыми картосхемами размещения пасюка в Воронежской области в
1946, 1948 и 1968 гг.
Ареал
63
В работе, посвященной областям Центральночерноземного экономиче-
ского райопа (Дмитриева, 1971), в общей форме сообщено о паличии
насюка в пределах Тамбовской, Курской и Линецкой областей. Отмечены
регулярные встречи этого вида во влажных биотопах Курской области.
В Линецкой области пасюк распространен более широко и особенно мно-
гочислен в юго-восточных, пограничных с Воронежской областью Усман-
ском и Добринском районах.
На основании обследования 8 тыс. построек и птирокого использова-
ния анкетных сведений территория Воронежской области была разделена
на три участка, различающихся размещением, обилием и вредоносной
делтельностью пасюка. Первый участок наименьшего вреда занимает пра-
вобережье Дона от Семилукского и Рамонского районов на севере до
Кантемировского и Богучаровского на юге. На севере участка пасюком
населено 13% сельских населенных пунктов. К югу пасюка стаповится
больше, многочисленны зверьки в поселках, расположенных по берегам
Дона, Богучара, Черной Калитвы. Процент населенных поселков здесь
увеличивается до 41. Появление пасюка и заселение им юга участка про-
ходили в конце 50-х — начале 60-х годов.
Участок средней вредопосности пасюока занимает полосу вдоль левого.
берега Дона и Калачевскую возвышенность на северо-восток до Бобров-
ского, Таловского и Новохоперского районов. Показатель заселенности
пасюком населенных пунктов лежит в пределах 0,50—0,85 баллов. В се-
верной половине участка им населено 75% поселков, а в южной — 69%.
Участок высокой вредоносности лежит на северо-востоке области, в
пределах южной части Окско-Донской низменности, занимая Эртильский,
Аннинский, Терновский, Грибаповский, Борисоглебский и Поворинский
районы. Здесь иасюк обитал издавна, численность его высока и постоян-
па.
По мнению А. Н. Козлова (1986), пасюк отсутствует в ряде населен-
ных пунктов Воронежской области, граничащих с Алексеевским районом
Белгородской области. Он же обнаружил только черную крысу в сельских
населенных пунктах (Верейка, Скляево, Айдарово) северной части Ра-
мопьского райопа Воропежской области. В райопном пентре Рамопь оби-
тают как черная крыса, так и пасюк.
Обитание черной крысы в Лебедянском и Елецком уездах (современ-
ная территория Липецкой области) было известно в середине прошлого
века. Первая специальная работа по изучению размещения крыс в пре-
делах Липецкой области ‘была осуществлена Т. В. Дмитриевой, Л. Д. Фи-
линым (1972). Они использовали как непосредственное обследование на-
селеппых пунктов, так и широкую рассылку и апализ анкет. Во многих
обследованных ими районах в начале текущего столетия пасюк был не-
известен жителям. В 1944 г. г. Елец был сплошь заселен черной кры-
сой. Пасюк практически отсутствовал.
Эти авторы дают следующую картину заселения пасюком Липецкой
области. Оп пропик в пее тремя путями. Первый шел с востока из Там-
бовской и Воронежской областей. В Добринском районе пасюк появился
в 1947 г. или несколько раньше. Далее расселение его шло но малым
62
Глава вторая
ружена черная крыса. По пасюку данных нет (материалы А. И. Феок-
тистова).
Ю. Л. Мясциков (4976) с 1956 п 1968 г. обнаружил пасюков в
11 населенных пунктах Арсеньевского, в 4 — Одоевского, в 3 — Чернь-
ского районов и в г. Узловой Тульской области. В 1972—1983 гг. пасюк
расширил свое распространение. Он был найден в 24 из .68 обеледован-
ных населенных лунктов в 8 из 10 административных районов лесостен-
ной части Тульской области. Не поймали пасюка в 6 населенных пунк-
тах Заокского и в 3 Одоенското районов. Среди 215 крыс было 86 пасю-
ков. В стенной части области обследовали 84 населенных пункта »
13 районах. Отсутствовал пасюк в 28 населенных пунктах 6 администра-
тивных районов (Воловского, Кфремовского, Каменского, Куркинского,
Тепло-Огаревского, Черньского). Из 289 поймапных крыс было 212 па-
сюков (Панина, 1986а). В Туле первый пасюк был пойман в апреле
1972 г., в 1975 г. среди отловленных в городе крыс на долю пасюка
приходилось 45$, в 1978—81%, в 1979— более 90%. а после 1982 г.—
96—98% (Панина, 1986). Таким образом, удалось наблюдать активный
процесс замены черной крысы пасюком.
.
Мн не нашли после 4954 г. никаких новых литературных сведений о
пасюке в Иваповской, Брянской, Орловской и Рязанской областях.
Центрально-Черноземный район. В него входят Липецкая, Тамбовская,
Курская, Белгородская и Воронежская области.
.
А. П. Кузякии (1951) считал, что сведений о пасюке в бассейпе верх-
него и среднего течения Дона почти нет, Это же подтверждает Т. В. Дми-
триева (1970), отмечая, что данные о размещении пасюка на территории
Воропежекой области в довоенное время отсутствуют. По ее мнению, в
первые годы после войны пасюк обитал преимущественно в городах, где
на него приходилось 18,3% добытых грызунов, в сельских населенных
пунктах его доля была равна 1,8%. Пасюк отсутствовал или был крайте
малочислен в южных и западных районах области. Много пасюка в
сельской местности было только в северо-восточных районах — в бассей-
не Хопра. В первые послевоенные годы ареал пасюка еще более сокра-
тился и принял пятнистый характер. Одновремепно заселенные этим ви-
дом пятна появились и па юго-востоке области, дойдя до левобережья До-
на у пересечения им границы области. Все правобережье Дона было прак-
тически свободно от пасюка.
.
С конца 50-х годов началось быстрое расселение этого вида.
В 1962—1963 тг. резко вырос удельный вес крысиного населепия в
сельской местности, на долю пасюка падало 29% выловленных грызунов.
Расселение и подъем численности пасюка были связаны с увеличением
сельскохозяйственной продукциии особенно © ростом животноводства.
К 1968 г. подавляющая часть области, хотя и с разной плотностью, была
заселена пасюком, отсутствовали зверьки только в части западных райо-
нов. Статья (Дмитриева, 1970) проиллюстрирована тремя весьма труднв
читаемыми картосхемами размещения пасюка в Воронежской области в
1946, 1948 и 1968 гг.
Ареал
63
В работе, посвященной областям Центральночерноземного экономиче-
ского райопа (Дмитриева, 1971), в общей форме сообщено о паличии
насюка в пределах Тамбовской, Курской и Линецкой областей. Отмечены
регулярные встречи этого вида во влажных биотопах Курской области.
В Линецкой области пасюк распространен более широко и особенно мно-
гочислен в юго-восточных, пограничных с Воронежской областью Усман-
ском и Добринском районах.
На основании обследования 8 тыс. построек и птирокого использова-
ния анкетных сведений территория Воронежской области была разделена
на три участка, различающихся размещением, обилием и вредоносной
делтельностью пасюка. Первый участок наименьшего вреда занимает пра-
вобережье Дона от Семилукского и Рамонского районов на севере до
Кантемировского и Богучаровского на юге. На севере участка пасюком
населено 13% сельских населенных пунктов. К югу пасюка стаповится
больше, многочисленны зверьки в поселках, расположенных по берегам
Дона, Богучара, Черной Калитвы. Процент населенных поселков здесь
увеличивается до 41. Появление пасюка и заселение им юга участка про-
ходили в конце 50-х — начале 60-х годов.
Участок средней вредопосности пасюока занимает полосу вдоль левого.
берега Дона и Калачевскую возвышенность на северо-восток до Бобров-
ского, Таловского и Новохоперского районов. Показатель заселенности
пасюком населенных пунктов лежит в пределах 0,50—0,85 баллов. В се-
верной половине участка им населено 75% поселков, а в южной — 69%.
Участок высокой вредоносности лежит на северо-востоке области, в
пределах южной части Окско-Донской низменности, занимая Эртильский,
Аннинский, Терновский, Грибаповский, Борисоглебский и Поворинский
районы. Здесь иасюк обитал издавна, численность его высока и постоян-
па.
По мнению А. Н. Козлова (1986), пасюк отсутствует в ряде населен-
ных пунктов Воронежской области, граничащих с Алексеевским районом
Белгородской области. Он же обнаружил только черную крысу в сельских
населенных пунктах (Верейка, Скляево, Айдарово) северной части Ра-
мопьского райопа Воропежской области. В райопном пентре Рамопь оби-
тают как черная крыса, так и пасюк.
Обитание черной крысы в Лебедянском и Елецком уездах (современ-
ная территория Липецкой области) было известно в середине прошлого
века. Первая специальная работа по изучению размещения крыс в пре-
делах Липецкой области ‘была осуществлена Т. В. Дмитриевой, Л. Д. Фи-
линым (1972). Они использовали как непосредственное обследование на-
селеппых пунктов, так и широкую рассылку и апализ анкет. Во многих
обследованных ими районах в начале текущего столетия пасюк был не-
известен жителям. В 1944 г. г. Елец был сплошь заселен черной кры-
сой. Пасюк практически отсутствовал.
Эти авторы дают следующую картину заселения пасюком Липецкой
области. Оп пропик в пее тремя путями. Первый шел с востока из Там-
бовской и Воронежской областей. В Добринском районе пасюк появился
в 1947 г. или несколько раньше. Далее расселение его шло но малым
84
Глава вторая
рекам на север до р. Матыра и на запад до р. Воронеж. На этом участ-
‹е обитает только пасюк.
С Второй путь пропикновения пасюка в Липецкую область шел из Туль-
ской области по р. Красивая Меча. Здесь в большом количестве этот вид
стал регистрироваться с 1950 г. Он расселялея вниз по реке, проник в
Краснинский район, дошел до Дона и вверх по последнему стал рассе-
ляться к северу.
.
"" Претий м пропикновения пасюка шел из Орловской области мо ре-
кам Сосна и Большая Чернава в Измалковский район. Здесь он появился
в начале 50-х годов, а к 1954 г. стал массовым видом. _
Н 70-м годам вея Липецкая область к западу от Дона была населена
пасюком и черной крысой. Численность крыс в средпей части этой терря-
тории (Измалковский, Становлянский, Красненский, Клецкий, Долгору-
ковский районы) очень велика. Северный — Данковский —и южные —
Воловский и Тербунский районы — относительно слабо заселены крысами.
Между Доном и Воронежем крысы малочисленны и распространены спо-
радично. Пасюк обнаружен в Лев-Толетовском, Липецком и Задонском
районах (Дмитриева, Филин, 1972).
Нам кажется сомнительным утверждение авторов о проникповепии
пасюка в Липецкую область по долине Красивой Мечи. В граничащих ©
Липоцкой районах Тульской области — Ефремовском, Куркинском — па-
сюка нет. Отсутствует этот вид и в верхнем течении Красивой Мечи—
Каменском, Воловском, Тепло-Огаревском районах (Панина, 1986а). Бо-
лее вероятен завоз пасюка в г. Елец, являющийся крупным железнодо-
рожным узлом, тем более что ст. Узловая в Тульской области уже в
50-х годах была заселена пасюком (Мясников, 1976). .
В 1981—1982 гг. распространение крыс в Липецкой области изучал
А. Н. Козлов (1986), уточнивший данные Т. В. Дмитриевой, А, Д. Фи-
лина (1972). По его данным, на юго-востоке в Добринском и Усманском
районах обитает только пасюк, время его появления здесь не установлено.
В расположенном севернее Грязинском районе пасюк появился в середи-
не 60-х годов. В сельских населенных пунктах района обитает только па-
сюк, а в районном центре г. Грязи отмечены ласюк и черная крыса.
В перечисленных районах сейчас пасюком заселены все населенные пунк-
ты, в половине их отмечена высокая и очень высокая численность крыс.
В Липецком районе в шести сельских населенных пунктах обнаружен
пасюк, а в двух — черная крыса, в г. Липецке обитают оба вида. В Доб-
ровском районе до 1974 г. крыс не было. Затем его заселили оба вида,
причем в районном центре обитает только пасюк. Кроме перечисленных
выше, пасюк обитает в сельских населенных пунктах еще пяти районов,
В четырех районах он встречается вместе с черной крысой только в рай-
онных центрах, а остальные населенные пункты заняты черпой крысой.
Наконец, в четырех западпых районах (Измалковеком, Долгоруковском,
ЗВоловском, Тербунском) отмечена только черная крыса.
Различия в описапиях (Дмитриева, Филин, 1972; Козлов, 1986),
вероятно, объясняются не изменениями ареалов крыс Липецкой области,
а уточнением сведений об их распространении.
Ареал
65
Пасюк издавна распространен в юго-западной части Белгородской
области, пограничной с Сумской и Харьковской областями УССР.
В эту часть целиком входят Ракитянский, Борисовский, Яковлевский,
Белгородский и Шебекинский районы. Здесь пасюк равпомерпо заселяет
большинство населенных пупктов — 94% обследованных. Отсюда он,
расселяясь на север, в 1960—1970 гг. проник в южные части Ивнянско-
го и Корочанского районов. Одновременно пасюк был зарегистрирован в
центральной и западной частях Новооскольского района. В Прохоров-
ском, Губкинском и Старооскольском районах оп появился в 1967 г.,
расселяясь со стороны Курской области. Во второй половине 60-х годов
пасюк проник из Ворошиловградской области в юго-восточные райопы
Белгородской области, заселив южпую половину Ровеньского и юг Вей-
делевского районов (Дмитриева, Ковалев, 1974). По мнению А. Н. Воз-
лова (1986), за период с 1969 по 1982 г. этот вид заселил Ивнянский,
Губкипский, Черпянский и Валуйский районы. До сих пор пасюк отсут-
ствует в восточных Алексеевском и Ирасногвардейском районах.
Авторы (Дмитриева, 1971; Козлов, 1989) , занимавшиеся раслттростране-
нием пасюка в Центральном Черноземье, конкретпых сведепий по Там-
бовской и Курской областям пе привели.
Поволжекий экономический район. В него входят Калмыцкая АССР,
Астраханская, Волгоградская, Саратовская, Пензенская, Куйбышевская,
Ульяновская области и Татарская АССР. По А. П. Кузякину (1951),
пасюка не было в восточных районах Ростовской области, западных и
юго-западных районах Астраханской области. В последней он заселял
почти всю дельту Волги на запад до п. Яндыки, селепия по правобережью
Волги на север до Черного Яра. В Волгоградекой области его отмечали в
Волгограде, Николаевске, Камышине. В Саратовской области он найден в
Саратове, Усть-Вурдюме, Аткареке, Пугачеве. Севернее зарегистрирован
в Куйбышеве и Пензе.
Распространению пасюка в Поволжье в настоящее время посвящена
серия обзорных статей (Варшавский Б. С. и др., 1983; Варшавский С. Н.
и др., 1986, 1987; Шилов и др., 1986), в которых есть карты размеще-
ния этого вида в Волго-Ахтубинской пойме к югу от Ахтубинска, в Ден-
гизском районе Гурьевской области и в нижнем течении р. Урала.
Современная восточная граница ареала пасюка в Калмыдкой АССР
идет от Волгограда на Сарпинский район, где он найден в Уманцове,
Канукове, Артан-Зельмене, но отсутствует в районном центре Садовое.
Нет пасюка и в расположенном северо-восточнее Малодербетовском райо-
не. Не обнаружен этот вид и в Приозерном районе в поселках Ергинин-
ский, Кегульта, нет его в поселке Овата и с. Троицком — районном цент-
ре Целинного района, однако на самом юге района пасюк встречен в
п. Вознесеновка. Обитает он в г. Элисте и поселках Лола, Бага-Бурул,
Ики-Бурул, Чулуп-Хамур Ики-Бурульского района (Варшавский и др.
1986, 1987). Восточнее перечисленных пунктов, за исключением Каспий-
ского побережья, пасюка в Калмыкии пока не обнаружили (рис. 15).
Примыкающие к Калмыцкой АССР юго-западные районы Волгоградской,
восточные Ростовской и северо-восточные Ставропольского края к на-
3 Серая крыса
84
Глава вторая
рекам на север до р. Матыра и на запад до р. Воронеж. На этом участ-
‹е обитает только пасюк.
С Второй путь пропикновения пасюка в Липецкую область шел из Туль-
ской области по р. Красивая Меча. Здесь в большом количестве этот вид
стал регистрироваться с 1950 г. Он расселялея вниз по реке, проник в
Краснинский район, дошел до Дона и вверх по последнему стал рассе-
ляться к северу.
.
"" Претий м пропикновения пасюка шел из Орловской области мо ре-
кам Сосна и Большая Чернава в Измалковский район. Здесь он появился
в начале 50-х годов, а к 1954 г. стал массовым видом.
_
Н 70-м годам вея Липецкая область к западу от Дона была населена
пасюком и черной крысой. Численность крыс в средпей части этой терря-
тории (Измалковский, Становлянский, Красненский, Клецкий, Долгору-
ковский районы) очень велика. Северный — Данковский —и южные —
Воловский и Тербунский районы — относительно слабо заселены крысами.
Между Доном и Воронежем крысы малочисленны и распространены спо-
радично. Пасюк обнаружен в Лев-Толетовском, Липецком и Задонском
районах (Дмитриева, Филин, 1972).
Нам кажется сомнительным утверждение авторов о проникповепии
пасюка в Липецкую область по долине Красивой Мечи. В граничащих ©
Липоцкой районах Тульской области — Ефремовском, Куркинском — па-
сюка нет. Отсутствует этот вид и в верхнем течении Красивой Мечи—
Каменском, Воловском, Тепло-Огаревском районах (Панина, 1986а). Бо-
лее вероятен завоз пасюка в г. Елец, являющийся крупным железнодо-
рожным узлом, тем более что ст. Узловая в Тульской области уже в
50-х годах была заселена пасюком (Мясников, 1976). .
В 1981—1982 гг. распространение крыс в Липецкой области изучал
А. Н. Козлов (1986), уточнивший данные Т. В. Дмитриевой, А, Д. Фи-
лина (1972). По его данным, на юго-востоке в Добринском и Усманском
районах обитает только пасюк, время его появления здесь не установлено.
В расположенном севернее Грязинском районе пасюк появился в середи-
не 60-х годов. В сельских населенных пунктах района обитает только па-
сюк, а в районном центре г. Грязи отмечены ласюк и черная крыса.
В перечисленных районах сейчас пасюком заселены все населенные пунк-
ты, в половине их отмечена высокая и очень высокая численность крыс.
В Липецком районе в шести сельских населенных пунктах обнаружен
пасюк, а в двух — черная крыса, в г. Липецке обитают оба вида. В Доб-
ровском районе до 1974 г. крыс не было. Затем его заселили оба вида,
причем в районном центре обитает только пасюк. Кроме перечисленных
выше, пасюк обитает в сельских населенных пунктах еще пяти районов,
В четырех районах он встречается вместе с черной крысой только в рай-
онных центрах, а остальные населенные пункты заняты черпой крысой.
Наконец, в четырех западпых районах (Измалковеком, Долгоруковском,
ЗВоловском, Тербунском) отмечена только черная крыса.
Различия в описапиях (Дмитриева, Филин, 1972; Козлов, 1986),
вероятно, объясняются не изменениями ареалов крыс Липецкой области,
а уточнением сведений об их распространении.
Ареал
65
Пасюк издавна распространен в юго-западной части Белгородской
области, пограничной с Сумской и Харьковской областями УССР.
В эту часть целиком входят Ракитянский, Борисовский, Яковлевский,
Белгородский и Шебекинский районы. Здесь пасюк равпомерпо заселяет
большинство населенных пупктов — 94% обследованных. Отсюда он,
расселяясь на север, в 1960—1970 гг. проник в южные части Ивнянско-
го и Корочанского районов. Одновременно пасюк был зарегистрирован в
центральной и западной частях Новооскольского района. В Прохоров-
ском, Губкинском и Старооскольском районах оп появился в 1967 г.,
расселяясь со стороны Курской области. Во второй половине 60-х годов
пасюк проник из Ворошиловградской области в юго-восточные райопы
Белгородской области, заселив южпую половину Ровеньского и юг Вей-
делевского районов (Дмитриева, Ковалев, 1974). По мнению А. Н. Воз-
лова (1986), за период с 1969 по 1982 г. этот вид заселил Ивнянский,
Губкипский, Черпянский и Валуйский районы. До сих пор пасюк отсут-
ствует в восточных Алексеевском и Ирасногвардейском районах.
Авторы (Дмитриева, 1971; Козлов, 1989) , занимавшиеся раслттростране-
нием пасюка в Центральном Черноземье, конкретпых сведепий по Там-
бовской и Курской областям пе привели.
Поволжекий экономический район. В него входят Калмыцкая АССР,
Астраханская, Волгоградская, Саратовская, Пензенская, Куйбышевская,
Ульяновская области и Татарская АССР. По А. П. Кузякину (1951),
пасюка не было в восточных районах Ростовской области, западных и
юго-западных районах Астраханской области. В последней он заселял
почти всю дельту Волги на запад до п. Яндыки, селепия по правобережью
Волги на север до Черного Яра. В Волгоградекой области его отмечали в
Волгограде, Николаевске, Камышине. В Саратовской области он найден в
Саратове, Усть-Вурдюме, Аткареке, Пугачеве. Севернее зарегистрирован
в Куйбышеве и Пензе.
Распространению пасюка в Поволжье в настоящее время посвящена
серия обзорных статей (Варшавский Б. С. и др., 1983; Варшавский С. Н.
и др., 1986, 1987; Шилов и др., 1986), в которых есть карты размеще-
ния этого вида в Волго-Ахтубинской пойме к югу от Ахтубинска, в Ден-
гизском районе Гурьевской области и в нижнем течении р. Урала.
Современная восточная граница ареала пасюка в Калмыдкой АССР
идет от Волгограда на Сарпинский район, где он найден в Уманцове,
Канукове, Артан-Зельмене, но отсутствует в районном центре Садовое.
Нет пасюка и в расположенном северо-восточнее Малодербетовском райо-
не. Не обнаружен этот вид и в Приозерном районе в поселках Ергинин-
ский, Кегульта, нет его в поселке Овата и с. Троицком — районном цент-
ре Целинного района, однако на самом юге района пасюк встречен в
п. Вознесеновка. Обитает он в г. Элисте и поселках Лола, Бага-Бурул,
Ики-Бурул, Чулуп-Хамур Ики-Бурульского района (Варшавский и др.
1986, 1987). Восточнее перечисленных пунктов, за исключением Каспий-
ского побережья, пасюка в Калмыкии пока не обнаружили (рис. 15).
Примыкающие к Калмыцкой АССР юго-западные районы Волгоградской,
восточные Ростовской и северо-восточные Ставропольского края к на-
3 Серая крыса
66
Глава вторая
Тк
1966;
стоящему времени сплопть заселены пасюком (Щепотьев, Ткачева,
;
шавский и др., 1987).
|
.
ва
вы Потраханокой области поселения пасюка приурочены & дольте Вот
ти и Волго-Ахтубинекой пойме. Между железной дорогой р
сюка в областях Поволжского экономического района
Рис. 15. Распространение па
1—т итори
тот аселенные пасюком; & — ПУН чз
зарегистрировано обитание па-
ерритории, сп ъ зас
асюком,
пункты, где
иср
‚
сюка; 3 — пункты, где пасюк не обнаружен
‚
Ареал
67
Кизляр, дельтой Волги и побережьем Каспийского моря в пределах Аст-
раханской области он заселяет всю территорию. По самому западному
протоку Волги— Бахтемиру— пасюк распространен па юг до Бирючьей
косы, Бударина, Рынка, Вышки. Еще южнее он обнаружен на террито-
рии Калмыкии в Джалыкове, Каспийске ив 27—30 км юго-восточнее и
восточнее стапции Артезиан. Вероятно, по побережью Каспийского моря
поселения пасюка смыкаются с Дагестанскими. В Денгизском районе
Гурьевской области по побережью Каспия пасюк распространен восточ-
нее протока Кигач до о-ва Новинского (Шилов и др., 1986).
Пасюк встречается практически во всех селах правобережья Нижней
Волги па север вплоть до Светлого Яра Волгоградской обл. Он обычен во
всех селах по левому берегу Ахтубы от Волжского на севере до Акса-
райского и Красного Яра на юге. Внутри Волго-Ахтубипекой поймы па-
хюк обычен как в населенных пунктах, так и в естественных биотопах
от Волгограда до Астрахани. За пределами поймы он встречается на
разъездах и станциях железной дороги от Аксарайской до Верхнего Бас-
кунчака (Варшавский и др., 1987; Опарина, 1987).
В настоящее время пасюк сплошь заселяет юго-западные, примыкаю-
щие к Цимлянскому водохранилищу районы Волгоградской области, куда`
он проник из Ростовской области. Этот вид встречается здесь как в при-
брежных, так и в удаленных от водохранилища сельских населенных
пунктах. Северная граница распространения пасюка на этом участке про-
ходила вблизи железной дороги Волгоград—Морозовск (Щепотьев, Тка-
чева, 1966). Его находили в Ильевке, Калаче-на-Дону, Суровкине, Облив-
ной. Севернее перечисленных пунктов пасюк не обпаружен.
Вверх по Волге обитание серой крысы строго приурочено к берегам
Волгоградского водохранилища, по правому берегу до Камышина, а по
левому — до устья Еруслана. От Камышина граница ареала поворачива-
ла на юго-запад и шла через населенные пункты Саломатино, Ольховка,
Зензеватка, Фролово, Отрадное, Михайловка, Карагичевский, Алексеев-
ская, Захоперский, Верхнереченский, за которыми пересекала границу
Воропежской области. К северу от этой линии начинается область сплон-
ного распространения пасюка (см. рис. 15).
По мнению Н. В. Щепотьева, Н. В. Ткачевой (1966), в 60-е годы па-
сюк отсутствовал между описанной границей и железпой дорогой Волго-
град— Морозовск (Подтелковский, Серафимовичский, Клетекий, Иловлин-
ский райопы Волгоградекой области). В юго-восточных районах Воро-
нежской области этот вид крайне редок (Дмитриева, 1975), а сведений
о распространении его в северных районах Ростовской области нет. Не
ясно, насколько изменилась в настоящее время ситуация с распростране-
нием пасюка на западе Волгоградской области.
По восточному берегу Волгоградского водохранилища пасюк был при-
урочен к берегу вплоть до устья Еруслана. По линии железной дороги
Верхний Баскупчак—Красный Кут его периодически регистрировали
только на ст. Джаныбек и Палласовка. Сплошь заселяет келезную доро-
гу серая крыса лишь к северу от Красного Кута.
3*
66
Глава вторая
Тк
1966;
стоящему времени сплопть заселены пасюком (Щепотьев, Ткачева,
;
шавский и др., 1987).
|
.
ва
вы Потраханокой области поселения пасюка приурочены & дольте Вот
ти и Волго-Ахтубинекой пойме. Между железной дорогой р
сюка в областях Поволжского экономического района
Рис. 15. Распространение па
1—т итори
тот аселенные пасюком; & — ПУН чз
зарегистрировано обитание па-
ерритории, сп
ъ зас
асюком,
пункты, где
иср
‚
сюка; 3 — пункты, где пасюк не обнаружен
‚
Ареал
67
Кизляр, дельтой Волги и побережьем Каспийского моря в пределах Аст-
раханской области он заселяет всю территорию. По самому западному
протоку Волги— Бахтемиру— пасюк распространен па юг до Бирючьей
косы, Бударина, Рынка, Вышки. Еще южнее он обнаружен на террито-
рии Калмыкии в Джалыкове, Каспийске ив 27—30 км юго-восточнее и
восточнее стапции Артезиан. Вероятно, по побережью Каспийского моря
поселения пасюка смыкаются с Дагестанскими. В Денгизском районе
Гурьевской области по побережью Каспия пасюк распространен восточ-
нее протока Кигач до о-ва Новинского (Шилов и др., 1986).
Пасюк встречается практически во всех селах правобережья Нижней
Волги па север вплоть до Светлого Яра Волгоградской обл. Он обычен во
всех селах по левому берегу Ахтубы от Волжского на севере до Акса-
райского и Красного Яра на юге. Внутри Волго-Ахтубипекой поймы па-
хюк обычен как в населенных пунктах, так и в естественных биотопах
от Волгограда до Астрахани. За пределами поймы он встречается на
разъездах и станциях железной дороги от Аксарайской до Верхнего Бас-
кунчака (Варшавский и др., 1987; Опарина, 1987).
В настоящее время пасюк сплошь заселяет юго-западные, примыкаю-
щие к Цимлянскому водохранилищу районы Волгоградской области, куда`
он проник из Ростовской области. Этот вид встречается здесь как в при-
брежных, так и в удаленных от водохранилища сельских населенных
пунктах. Северная граница распространения пасюка на этом участке про-
ходила вблизи железной дороги Волгоград—Морозовск (Щепотьев, Тка-
чева, 1966). Его находили в Ильевке, Калаче-на-Дону, Суровкине, Облив-
ной. Севернее перечисленных пунктов пасюк не обпаружен.
Вверх по Волге обитание серой крысы строго приурочено к берегам
Волгоградского водохранилища, по правому берегу до Камышина, а по
левому — до устья Еруслана. От Камышина граница ареала поворачива-
ла на юго-запад и шла через населенные пункты Саломатино, Ольховка,
Зензеватка, Фролово, Отрадное, Михайловка, Карагичевский, Алексеев-
ская, Захоперский, Верхнереченский, за которыми пересекала границу
Воропежской области. К северу от этой линии начинается область сплон-
ного распространения пасюка (см. рис. 15).
По мнению Н. В. Щепотьева, Н. В. Ткачевой (1966), в 60-е годы па-
сюк отсутствовал между описанной границей и железпой дорогой Волго-
град— Морозовск (Подтелковский, Серафимовичский, Клетекий, Иловлин-
ский райопы Волгоградекой области). В юго-восточных районах Воро-
нежской области этот вид крайне редок (Дмитриева, 1975), а сведений
о распространении его в северных районах Ростовской области нет. Не
ясно, насколько изменилась в настоящее время ситуация с распростране-
нием пасюка на западе Волгоградской области.
По восточному берегу Волгоградского водохранилища пасюк был при-
урочен к берегу вплоть до устья Еруслана. По линии железной дороги
Верхний Баскупчак—Красный Кут его периодически регистрировали
только на ст. Джаныбек и Палласовка. Сплошь заселяет келезную доро-
гу серая крыса лишь к северу от Красного Кута.
3*
68
Глава вторая
В восточных районах Волгоградской области южная граница расиро-
странения пасюка птла но правобережью Еруслана, уходя в Саратовскую
область. В последией он зарегистрирован в Луговском, Дьяков, сатово,
Журавлевке, Красном Куте. По железпой дороге Саратов— пральск вс те-
чался на всех крупных станциях на восток до Демьяса. осточнее о
следнего по железной дороге отсутствовал вплоть до Уральска. ор. М -
лый Увень зарегистрирован в Ершоне, рой Падине, Мортанке,
р
х
н епотьев, Ткачева,1 .
.
о, Малом Узее ПИ пасюк обнаружен на севере Дженыбекского ралона
Уральской области в поселках Таловка, Жаксыбай, Шерембетсай { ар-
шавский и др., 1987). Таким образом, соединились поселения Тото их
на ст. Палласовка с поселениями но Мадому Узеню (ем. рис. ). ока
нет сведений об обитании пасюка в Александрово-Гайском, овоузене ры
и Озинском районах Саратовской области. Сенеро-восточнее ст. Демьяо
он найден в населенных пунктах бассейна р. Камелик — левого прито
Иргиза.
й
—
т отрашотие пасюка в правобережных районах Саратовской обла”
сти документировано недостаточно. Ето обитапие’ зарегистрирован
Красноармейском, Саратовском, Аткарском и Само ловоком рай их я.
Конкретных сведений о размещении пасюта в уйбышевской,
кой °
зенской областях практ
.
поой ет АССР (см. рис. 45) пасюка добывали в г. Казани й
Вытегорском районе в постройках человека и в Агрызском, утопает.
ском, Клабужском и Мамадыжском районах в естественных био
у
<
4).
“Пиринсная, 1986). экономический район входят Кировская и Горько
ская области, Марийская, Чувашская и Мордовская АССР. А. п. Кузя.
кип (1951) отметия для отого региона обитание пасюка в г. Горько и
Мордовском заповедпике. В последующие годы пасюк был заре ис Тир .
ван в г. Саранске. Больше никаких конкретных оводоний о раз ежщении
этого вида в Мордовской, Чувашской, Марийской АС и Горь ко ской
области в литературе не обнаружено. Кировская область, видимо, сти
заселена пасюком, он зарегистрирован В 15 из 39 администрат вных
районов области, в том числе и и северпых: Лузском и Вер’
татев, 1967; Чарушина,
.
О
,
о
окономическото района в пределах европейской саста
РСФСР лежат Пермская область, Удмуртская и Башкирская УСС ’
Для этого региона А. П. Кузякин (1954) указывает пасюка только д тя
Пермской области— в городах Соликамске, Чердыне и в са г северо
восточном Ныробском районе, где зверьки местами лостаточно, того
численны. В дальнейшем насюк был найден В Г. Алекоапдровс <, Бар:
дымском, Гайнинском, Кунгурском и Лысьвенском районах, д в
области (Кучерук, Кузиков, 1985). В. И. Чабовский, В. Г. Дар
м
5 В конце прошлого века пасюк был многочислен в Ардатовском и Курмышско
здах Симбирской губернии (Житков, 1898).
8 беткот ртеную область мы будем рассматривать вместе с Казахстаном.
Ареал
69
(1964) сообщили, что в заброшенных поселках Александровского и
Лысьвенского райопов Пермекой области отсутствие людей в течение
одной зимы привело к полному исчезновению пасюков. На основании
анализа заготовок шкурок серой крысы и отлова зверьков было установ-
лено обитание этого вида во всех 25 административных районах Удмурт-
ской АССР (данные Ю. К. Попова).
Кавказ и Нредкавказье
А. П. Вузякин (1951) так описывает расиространение пасюка в этом
регионе. Южная граница его из Турции заходит в Южное Закавказье,
тде огибает пустынные районы долины Аракса, Зангезурский и Карабах-
ский хребты, и по верхнему поясу Талышских гор спова уходит за гра-
ницу вдоль каспийското побережья Ирана. Не было пасюка в пустын-
ных и полупустынных районах северо-западного Прикаспия, в северном
Дагестане, в восточных районах Ростовской области, в западных и юго-
западных районах Астраханской области. Отсутетвовал он между Волгой
и Уралом к северу от линии Камышин или даже Саратов Уральск.
На карте кружева ареала пасюка на Кавказе М. В. Шидловский’
(1976) следующим образом очерчивал распространение этого вида.
Ареал относительно узкой полосой идет вдоль Черноморского побережья
до Геленджика и Новороссийска. Затем вдоль железных дорог соединя-
ется с Северо-Кавказской частью ареала, которая простирается от север-
ных склонов Главного Кавказского хребта до побережья Азовского моря,
нижнего Дона и устья Терека. От последнего, спускаясь вдоль побережья
Каспийского моря, арсал пасюка занимает северную и приморскую части
Дагестана (Лавровский, Колесников, 1956), заходя по долинам рек глу-
боко в горы, уходит в Азербайджанскую ССР, смыкаясь в ней с закав-
казской частью ареала. Примерно так же показано распространение
пасюка на Северном Кавказе на карте, приведенной А. К. Темботовым
(1972).
Закавказская часть ареала пасюка на карте М. В. Шидловского
(1976) вытлядит следующим образом. По Колхиде она широким языком
вдается на восток, поднимаясь вверх мо долинам рек Цхенисцноли и
Риони. Далее на восток идет полосой вдоль железной дороги Кутаиси—
Тбилиси на север, заходя в пределы Юго-Осетинской АО (г. Цхинвали)
и к югу спускаясь по верховьям рек Куры и Цоблеани. Восточнее Тби-
лиси ареал идет неширокой полосой вдоль железной дороги, идущей на
Баку, не удаляясь от нее на расстояние, превышающее 30 км (Рябцева,
1980), и языком вдается па восток по долипам Алазана и Агричая.
В Армении ареал пасюка небольшим выступом заходит из Турции в
район Ленинакана, занимает большую часть Араратской долины и из
Грузинской и Азербайджанской ССР проникает в северо-восточные
районы республики.
Как уже упоминалось выше, пасюк отсутствует в полупустынных
райопах северо-западного Прикаспия. За последние 40 лет ареал его
здесь заметно расширился. Этому процессу посвящен ряд публикаций
68
Глава вторая
В восточных районах Волгоградской области южная граница расиро-
странения пасюка птла но правобережью Еруслана, уходя в Саратовскую
область. В последией он зарегистрирован в Луговском, Дьяков, сатово,
Журавлевке, Красном Куте. По железпой дороге Саратов— пральск вс те-
чался на всех крупных станциях на восток до Демьяса. осточнее о
следнего по железной дороге отсутствовал вплоть до Уральска. ор. М -
лый Увень зарегистрирован в Ершоне, рой Падине, Мортанке,
р
х
н епотьев, Ткачева,1 .
.
о, Малом Узее ПИ пасюк обнаружен на севере Дженыбекского ралона
Уральской области в поселках Таловка, Жаксыбай, Шерембетсай { ар-
шавский и др., 1987). Таким образом, соединились поселения Тото их
на ст. Палласовка с поселениями но Мадому Узеню (ем. рис. ). ока
нет сведений об обитании пасюка в Александрово-Гайском, овоузене ры
и Озинском районах Саратовской области. Сенеро-восточнее ст. Демьяо
он найден в населенных пунктах бассейна р. Камелик — левого прито
Иргиза.
й
—
т
отрашотие пасюка в правобережных районах Саратовской обла”
сти документировано недостаточно. Ето обитапие’ зарегистрирован
Красноармейском, Саратовском, Аткарском и Само ловоком рай их я.
Конкретных сведений о размещении пасюта в уйбышевской,
кой °
зенской областях практ
.
поой ет АССР (см. рис. 45) пасюка добывали в г. Казани й
Вытегорском районе в постройках человека и в Агрызском, утопает.
ском, Клабужском и Мамадыжском районах в естественных био
у
<
4).
“Пиринсная, 1986). экономический район входят Кировская и Горько
ская области, Марийская, Чувашская и Мордовская АССР. А. п. Кузя.
кип (1951) отметия для отого региона обитание пасюка в г. Горько и
Мордовском заповедпике. В последующие годы пасюк был заре ис Тир .
ван в г. Саранске. Больше никаких конкретных оводоний о раз ежщении
этого вида в Мордовской, Чувашской, Марийской АС и Горь ко ской
области в литературе не обнаружено. Кировская область, видимо, сти
заселена пасюком, он зарегистрирован В 15 из 39 администрат вных
районов области, в том числе и и северпых: Лузском и Вер’
татев, 1967; Чарушина,
.
О
,
о
окономическото района в пределах европейской саста
РСФСР лежат Пермская область, Удмуртская и Башкирская УСС ’
Для этого региона А. П. Кузякин (1954) указывает пасюка только д тя
Пермской области— в городах Соликамске, Чердыне и в са г северо
восточном Ныробском районе, где зверьки местами лостаточно, того
численны. В дальнейшем насюк был найден В Г. Алекоапдровс <, Бар:
дымском, Гайнинском, Кунгурском и Лысьвенском районах, д в
области (Кучерук, Кузиков, 1985). В. И. Чабовский, В. Г. Дар
м
5 В конце прошлого века пасюк был многочислен в Ардатовском и Курмышско
здах Симбирской губернии (Житков, 1898).
8 беткот ртеную область мы будем рассматривать вместе с Казахстаном.
Ареал
69
(1964) сообщили, что в заброшенных поселках Александровского и
Лысьвенского райопов Пермекой области отсутствие людей в течение
одной зимы привело к полному исчезновению пасюков. На основании
анализа заготовок шкурок серой крысы и отлова зверьков было установ-
лено обитание этого вида во всех 25 административных районах Удмурт-
ской АССР (данные Ю. К. Попова).
Кавказ и Нредкавказье
А. П. Вузякин (1951) так описывает расиространение пасюка в этом
регионе. Южная граница его из Турции заходит в Южное Закавказье,
тде огибает пустынные районы долины Аракса, Зангезурский и Карабах-
ский хребты, и по верхнему поясу Талышских гор спова уходит за гра-
ницу вдоль каспийското побережья Ирана. Не было пасюка в пустын-
ных и полупустынных районах северо-западного Прикаспия, в северном
Дагестане, в восточных районах Ростовской области, в западных и юго-
западных районах Астраханской области. Отсутетвовал он между Волгой
и Уралом к северу от линии Камышин или даже Саратов Уральск.
На карте кружева ареала пасюка на Кавказе М. В. Шидловский’
(1976) следующим образом очерчивал распространение этого вида.
Ареал относительно узкой полосой идет вдоль Черноморского побережья
до Геленджика и Новороссийска. Затем вдоль железных дорог соединя-
ется с Северо-Кавказской частью ареала, которая простирается от север-
ных склонов Главного Кавказского хребта до побережья Азовского моря,
нижнего Дона и устья Терека. От последнего, спускаясь вдоль побережья
Каспийского моря, арсал пасюка занимает северную и приморскую части
Дагестана (Лавровский, Колесников, 1956), заходя по долинам рек глу-
боко в горы, уходит в Азербайджанскую ССР, смыкаясь в ней с закав-
казской частью ареала. Примерно так же показано распространение
пасюка на Северном Кавказе на карте, приведенной А. К. Темботовым
(1972).
Закавказская часть ареала пасюка на карте М. В. Шидловского
(1976) вытлядит следующим образом. По Колхиде она широким языком
вдается на восток, поднимаясь вверх мо долинам рек Цхенисцноли и
Риони. Далее на восток идет полосой вдоль железной дороги Кутаиси—
Тбилиси на север, заходя в пределы Юго-Осетинской АО (г. Цхинвали)
и к югу спускаясь по верховьям рек Куры и Цоблеани. Восточнее Тби-
лиси ареал идет неширокой полосой вдоль железной дороги, идущей на
Баку, не удаляясь от нее на расстояние, превышающее 30 км (Рябцева,
1980), и языком вдается па восток по долипам Алазана и Агричая.
В Армении ареал пасюка небольшим выступом заходит из Турции в
район Ленинакана, занимает большую часть Араратской долины и из
Грузинской и Азербайджанской ССР проникает в северо-восточные
районы республики.
Как уже упоминалось выше, пасюк отсутствует в полупустынных
райопах северо-западного Прикаспия. За последние 40 лет ареал его
здесь заметно расширился. Этому процессу посвящен ряд публикаций
70
Глава вторая
{Миропов, 1964; Миронов и др. 1965; Шенотьев, Ткачева, 1966; Про-
кофьева, Изапов, 1974; Варшавский и др., 1983; Шилов и др. 1983).
Р настоящее время граница ареала пасюка на этой территории идет от
побережья Каспийского моря в устье Терека вверх по долине реки до
станции Галюгасвской. От нее круто поворачивает на север и через по-
селок 'Тукуй-Мектеб, Махмуд-Мектеб выходит в долину Кумы в районе
Нефтекумска и Озек-Сулата, вверх по Куме доходит до виадения в пее
Мокрой Буйвалы. Отсюда вповь поворачивает на север к впадению
р. Калауса в Маныч, далее идет в 40 км восточнее и 5—6 км севернее
Элисты, через Федосеевку, Заветное, Киселевку на Кануково, Уманцево
Садовского района Цалмыцкой АССР выходит через Абганерово и
п. Трудолюбие к правому берегу Волги (Варшавский и Др., 1987).
В настоящее время появилась возможность охарактеризовать тоиогра-
фию ареала пасюка на Кавказе и в Предкавказье. С этой целью все
опубликованные сведения о пунктах, в которых нашли пасюка (более
360) или в которых при специальном обследовании его пе удалось обна-
ружить (более 80), были напесены на карту. Одновременно по гипсомет-
рическим, геоботаническим и картам плотности людского населения окон-
турепы территории, на которых обитание пасюка маловероятно (рис. 16).
В низовьях Дона, на Черноморском побережье, Ставропольской в03-
вышенности, в Кабардино-Балкарии, Северной Осетии широкое распрост-
ранение пасюка было известно еще в прошлом веке (Россиков, 1887;
Сатунин, 1904; Губарев, 1941; Кузякин, 1951; Верещагин, 1959; Вурят-
пиков. 1986). Одпако, как видно из рис. 16, для приазовской части Рос-
товской области, северо-восточной половипы Красиодарского и западной
части Ставропольского краев явно недостает документации находок этого
вида. Но черноморскому побережью он зарегистрирован в Тамани и да-
лее в Анапе, Новороссийске, Геленджике, Дивноморском. Миогочислен
в плавнях и лиманах низовьев Кубани с ее левыми притоками. На север
плотные поселения пасюка достигают Приморско-А хтарского района
(Лаврова, 1962).
Нет сведений о наличии пасюка па Черпоморском побережье от
Дивпоморска до Туанее. К юго-востоку от последнего серая крыса за-
селяет практически все прибрежные и железподорожные поселки вплоть
до Батуми. До Сухуми ареал этого вида тянется вдоль побережья отно-
сительно узкой полосой (ПТидловский, 1947). По долинам наиболее зна-
читольных рек оп проникает относительно высоко в горы. Зарегистриро-
вано его обитание в верховьях Р. Шахэ-Бабук-Луле, в среднем и верх-
нем течении р. 'Мзымты
— Красная Поляна, кордоны Ачиспе, Пелух
(Кормилицина, 1980). Иет сведений о наличии этого вида на оз. Рица
п в верховьях р. Бзыбь.
Южнее Сухуми ареал пасюка расширяется, захватывая всю Колхид-
скую низменность и поднимаясь вверх по р. Ингури до п. Джавари,
по р. Цхенисцщкали до г. Цагери, по р. Риони и ее верхним притокам
до г. Опи и побелка Кваиси (Цихистави, Читиаитвили, 1986). Выше
пасюхк отсутствует в Сванетии в высокогорных Местийском и Лентех-
ском районах. При специальном обследовании не натили его в райцентре
Ареал
ТА
ия
селах. Местные жители не знали этого грызуна
р схистави, Дхмухадее, 1984). М. В. Шидловский (1964), обследовав
ных пунктов в криволесье и па альпийских лугах (1800—
2400 м над Ур. моря) Большого Кавказа вп
ой
не обпаружил в них пасюка.
редоелах Грузинской ССР,
Рис. 16. Топограф
. 40.
ия ареала пасюка на Ка
церук, Кузиков)
вказе и в Предкавказье (составители Ку-
1 — населенные п ункты, в которых пасюк найден; 8 — населенные пункты, в которых пасюн
отсутствует; 3 — места регист
2
е рации пасюка в естественных
;
циально пригодная для обитания нпасюка
Оиототах; 4— территория, потен-
©
Ростов-на-Дону
®
®.
+
©,
Элиста.
Астрахань.
,
®,
Краснодар.
Майне:
Ставрополь
&
[=
Я
а7
о
‘Черкёсск
——<
—*
—
.
—Е;
ри”
——
из
>
.
ж
о
Е
®, 'НАлЬЗин,
ф
——
о
Грозный
Е
ыы
[)
г
5
4
——
#
Орджлийи
Махачкала:
=—
<
—$
о
в,
[®)
о
=
©
997 оо
=:
2
©°>
4
хинвли : о
-
о8°
Батуми
©.
<,
Тбилиси,
[© ре
—
[®)
=
о«О
7 ®,\Ереван!
в
Баку
ры
ь уам
,
о НАхич! о
Ям
ь
70
Глава вторая
{Миропов, 1964; Миронов и др. 1965; Шенотьев, Ткачева, 1966; Про-
кофьева, Изапов, 1974; Варшавский и др., 1983; Шилов и др. 1983).
Р настоящее время граница ареала пасюка на этой территории идет от
побережья Каспийского моря в устье Терека вверх по долине реки до
станции Галюгасвской. От нее круто поворачивает на север и через по-
селок 'Тукуй-Мектеб, Махмуд-Мектеб выходит в долину Кумы в районе
Нефтекумска и Озек-Сулата, вверх по Куме доходит до виадения в пее
Мокрой Буйвалы. Отсюда вповь поворачивает на север к впадению
р. Калауса в Маныч, далее идет в 40 км восточнее и 5—6 км севернее
Элисты, через Федосеевку, Заветное, Киселевку на Кануково, Уманцево
Садовского района Цалмыцкой АССР выходит через Абганерово и
п. Трудолюбие к правому берегу Волги (Варшавский и Др., 1987).
В настоящее время появилась возможность охарактеризовать тоиогра-
фию ареала пасюка на Кавказе и в Предкавказье. С этой целью все
опубликованные сведения о пунктах, в которых нашли пасюка (более
360) или в которых при специальном обследовании его пе удалось обна-
ружить (более 80), были напесены на карту. Одновременно по гипсомет-
рическим, геоботаническим и картам плотности людского населения окон-
турепы территории, на которых обитание пасюка маловероятно (рис. 16).
В низовьях Дона, на Черноморском побережье, Ставропольской в03-
вышенности, в Кабардино-Балкарии, Северной Осетии широкое распрост-
ранение пасюка было известно еще в прошлом веке (Россиков, 1887;
Сатунин, 1904; Губарев, 1941; Кузякин, 1951; Верещагин, 1959; Вурят-
пиков. 1986). Одпако, как видно из рис. 16, для приазовской части Рос-
товской области, северо-восточной половипы Красиодарского и западной
части Ставропольского краев явно недостает документации находок этого
вида. Но черноморскому побережью он зарегистрирован в Тамани и да-
лее в Анапе, Новороссийске, Геленджике, Дивноморском. Миогочислен
в плавнях и лиманах низовьев Кубани с ее левыми притоками. На север
плотные поселения пасюка достигают Приморско-А хтарского района
(Лаврова, 1962).
Нет сведений о наличии пасюка па Черпоморском побережье от
Дивпоморска до Туанее. К юго-востоку от последнего серая крыса за-
селяет практически все прибрежные и железподорожные поселки вплоть
до Батуми. До Сухуми ареал этого вида тянется вдоль побережья отно-
сительно узкой полосой (ПТидловский, 1947). По долинам наиболее зна-
читольных рек оп проникает относительно высоко в горы. Зарегистриро-
вано его обитание в верховьях Р. Шахэ-Бабук-Луле, в среднем и верх-
нем течении р. 'Мзымты
— Красная Поляна, кордоны Ачиспе, Пелух
(Кормилицина, 1980). Иет сведений о наличии этого вида на оз. Рица
п в верховьях р. Бзыбь.
Южнее Сухуми ареал пасюка расширяется, захватывая всю Колхид-
скую низменность и поднимаясь вверх по р. Ингури до п. Джавари,
по р. Цхенисцщкали до г. Цагери, по р. Риони и ее верхним притокам
до г. Опи и побелка Кваиси (Цихистави, Читиаитвили, 1986). Выше
пасюхк отсутствует в Сванетии в высокогорных Местийском и Лентех-
ском районах. При специальном обследовании не натили его в райцентре
Ареал
ТА
ия
селах. Местные жители не знали этого грызуна
р схистави, Дхмухадее, 1984). М. В. Шидловский (1964), обследовав
ных пунктов в криволесье и па альпийских лугах (1800—
2400 м над Ур. моря) Большого Кавказа вп
ой
не обпаружил в них пасюка.
редоелах Грузинской ССР,
Рис. 16. Топограф
. 40.
ия ареала пасюка на Ка
церук, Кузиков)
вказе и в Предкавказье (составители Ку-
1 — населенные п ункты, в которых пасюк найден; 8 — населенные пункты, в которых пасюн
отсутствует; 3 — места регист
2
е рации пасюка в естественных
;
циально пригодная для обитания нпасюка
Оиототах; 4— территория, потен-
©
Ростов-на-Дону
®
®.
+
©,
Элиста.
Астрахань.
,
®,
Краснодар.
Майне:
Ставрополь
&
[=
Я
а7
о
‘Черкёсск
——<
—*
—
.
—Е;
ри”
——
из
>
.
ж
о
Е
®, 'НАлЬЗин,
ф
——
о
Грозный
Е
ыы
[)
г
5
4
——
#
Орджлийи
Махачкала:
=—
<
—$
о
в,
[®)
о
=
©
997 оо
=:
2
©°>
4
хинвли : о
-
о8°
Батуми
©.
<,
Тбилиси,
[© ре
—
[®)
=
о«О
7 ®,\Ереван!
в
Баку
ры
ь уам
,
о НАхич! о
Ям
ь
72
Глава вторая
Юзкнее Батуми по долине р. Чороха его ареал выходит за пределы
СССР, а по р. Аджарие-Цкали поднимается до райцентра еда и бл -
зкайших сел (Флеров, 1927; Цихистави, Читиашвили, 1986). одтанс и
„убальпийском поясе Аджарии, ие 2000 м та пеохетоком и
|<
{
:
лапия и др.,
‚
ском хробтах пасюка не нашли
1
и.
По всей версятности, пасюк заселяет все крупные Паобленные пу
же: Й
и шоссе Самтредиа-—'Тбилиси,
ты вдоль железной дороги
и, Но докумен
й
тот вид в Сачхере, коне
тировано это плохо. Найден э
оч
кте
попезнодорожной ветки па север от г. Зестафони. па ветке Хашури
Боржоми—Вале зарегистрирован в горолах Боржоми, Ахалцихе, по ос
св проник на восток в районный центр Аспиндзу, но отсутс вует Е
Ахалкалакском районе. Еще далее на восток в 1970 о ны ся в
‹
го времени не обнаружен
г. Богдановка, по до пастояще
у
Цихистави, 'Читиантвили, 1986). В Богдановку пасюк мог быть завезен
т
к из Тбилиси, так и со стороп
автотранспортом по шоссе ка:
то
и.
па етом 1984 г. был зарегистрироваи в Дманисском районе, считавтие
а.
ся до того свободным от пасюк
|,
3 восточной части Грузии распространение пасюка уе
льно ш
х
т
емя ареал его тут зпачите
,
плохо. Однако в настоящее вр
у
это я пнисашо М. В. Шидловским (1976). Но всей вероятности, он заселяот
подавляющее большинство населенных пунктов вдоль О
"
\ ак и веток от Тбилиси до Телави
от Тбилиси до Кировакана
ы
|
-
Цкаро. На них он зарегистрирован в Гурджаани и Сигнахи (Кузякии,
1951). В Цителцкаройском районе пасюк обнаружен В ра ор всей
у
ле Кура—
эт
отсутствует на водоразде
вероятности, этот грызун
„Кура ори и на
б
Й
<
плоскогорья, гие овцеводе
большей части ИШорского
,
т
-
правление хозяйства, мала плотность паселения людей и редко располо
ты.
яепы небольшие населенные иун
_
.
" На южные склоны Большого Кавказа пасюк подпимается до Г. и
-
кой д
ктов вдоль Военно-Грузине
ли. известен из многих пун
Ко
Коби. Сиони, Казбеги), многократно появлялся на Крестовом
а
вые: 2379 м (Курятников 1986). Отсутствует этот грызун
перевале па высоте
к
,
Е
Тушетии и Хевсуретии. 1. 2.
‚
вернее с. Квемо-Алвани в
|
В К Читиашвили (1986) пишут об обитании пасюта ниже о ая
пре
береж
лазани, не
:
пунктах левобережья А: и,
у
в предгорных населеплых
апи,
г
я
окротных мест находок. В пограничных частях, Авербайджеокой
;
:
хи
”
т
Белоканы и Закаталы. На терри
этот вид известен из сел
Н
у _Н.
итории Гру
й
Большого Кавказа на высота»
‹ой ССР в горно-несном поясе
1500
о 1800 м а, моря было обследовано девять населенных пунктов
и в одном из них пасюка не обпаружили, не было сго здесь и
‹ий, 1964).
осным сведениям (Шидловский, Е
.
а.
р Пасюк давно известен из западной части Армянской ССР. В ориввн
‹
всех
-
й губ
б более или менее обыкповенен во к
‹ой губернии оп был
в
ть
ых оселенных пунктах еще в начале пынешнего века (Пявотер, 1912).
й
@.
а
Для Ленинакана упоминается © 1919 г. (Кузякин, 1951). о
м
утверждение Н. В. Верещагина {1959, с. 301) о том, что в Ереново и
Джульфе пасюки появились лишь | 30-х годах нашего ст
,
Ареал
73
иостройки железной дороги, связавшей названные города с Тбилиси и
Баку», не выдерживает критики, тем более что железная дорога, связав-
шая Тбилиси с Ленинаканом и Карсом, Ереваном и Джульфой, действо-
вала уже в 1913 г. Видимо, насюк проник в Приараксинскую котловину
с юго-востока. В Нахичевани пасюк был добыт еще в 1917 г. (коллекции
ЗИН АН СССР).
Т. М. Соспихина (1946, 1950), изучавшая население грызунов городов
Армении в первой половине 40-х годов, отметила отсутствие пасюка в
Кировакане и Алаверди. В обзоре млекопитающих Армянской ССР
С. В. Даль (1954) пишет, что пасюк обычен в городах Ереване, Эчмиад-
зине, Ленинакане, а также в селениях Аштаракского, Эчмиадзинского,
Абовянского (Котайкского), Арташатского и Араратското (Вединского)
районов. Этот вид обитает в естественных биотопах по речкам, впадаю-
щим в Аракс к югу от Еревана (Шидловский, 1976).
В настоящее время пасюк заселяет всю западную часть Армении за
исключением северо-западных, пограничных с Грузией Амасийского,
Гукасянекого, Калининского райопов и высокогорий Арагацкого массива.
Он занимает целиком Араратскую низменность. Встречается во всех
круппых населенных пунктах на шоссе Ереван-Талин—Ленинакан и
вдоль железной дороги Ереван—Тбилиси, включая города Кировакан,
Алаверди. На северо-запад от железной дороги проник в г. Степанаван.
На север от Еревана вдоль железной дороги расселился до г. Раздан.
На восток доходит до Гарни, Веди, Арарата. В Ехегнадзор проник, ви-
димо, по широкой долине р. Арпа. На северс-востоке Армении найден в
четырех селах Носмберянского района. Существуют неопределенные
указания на возможность обитания этого вида по берегам правых при-
токов Куры— рек Агстев в Иджеванском, Ахун и Хндзрот в Шамшадин-
ском райопах (Варшавский и др., 1987).
Нет пасюка вокруг Севапа и в высокогорьях хребтов Армянского
нагорья и Малого Кавказа. Зарегистрировано отсутствие этого вида в
Кафане, Дастакерте, Сисиане, Базаурчае (Соспихина, 1946; Петров,
Адамян, 1966). В Нахичеванской АССР пасюк найден в Ильичевском
{ст. Ильичевск, сел. Ханлыклар, Верхний и Нижний Аралыг, Дашарх) и
Нахичеванском районах (Лобанова, Петросян, 1959; Рябцева, 1980;
Исмаилов и др., 1986).
В Азербайджане кости пасюка найдены в слоях позднего средневе-
ковья (ХУ-ХУГ вв. п. 2.) при раскопках дворца ширваншахов в Баку
(Верещагин, 1959). В настоящее время пасюк распространен на большей
части Азербайджана. Оп встречается в бассейнах Алазани и Агричая от
Белокан и Закатал на западе до Вартаптена и Куткашена на востоке. По
железной дороге Махачкала—Баку отмечен на большинетве круиных стан-
ций (Худат, Хачмае, Дивичи, Сиазань). Проникает далеко вверх по
долине Самура, по р. Кудиалчаю доходит до Кубы. Нет пасюка в высо-
когорьях Большого Кавказа на востоке ло массива Бабадага включи-
тельно (см. рис. 46).
Он найден на всех крупных станциях железных дорог Тбилиси—
Баку, Баку—Астара, начиная от г. Казах на западе и на ветке
72
Глава вторая
Юзкнее Батуми по долине р. Чороха его ареал выходит за пределы
СССР, а по р. Аджарие-Цкали поднимается до райцентра еда и бл -
зкайших сел (Флеров, 1927; Цихистави, Читиашвили, 1986). одтанс и
„убальпийском поясе Аджарии, ие 2000 м та пеохетоком и
|<
{
:
лапия и др.,
‚
ском хробтах пасюка не нашли
1
и.
По всей версятности, пасюк заселяет все крупные Паобленные пу
же: Й
и шоссе Самтредиа-—'Тбилиси,
ты вдоль железной дороги
и, Но докумен
й
тот вид в Сачхере, коне
тировано это плохо. Найден э
оч
кте
попезнодорожной ветки па север от г. Зестафони. па ветке Хашури
Боржоми—Вале зарегистрирован в горолах Боржоми, Ахалцихе, по ос
св проник на восток в районный центр Аспиндзу, но отсутс вует Е
Ахалкалакском районе. Еще далее на восток в 1970 о ны ся в
‹
го времени не обнаружен
г. Богдановка, по до пастояще
у
Цихистави, 'Читиантвили, 1986). В Богдановку пасюк мог быть завезен
т
к из Тбилиси, так и со стороп
автотранспортом по шоссе ка:
то
и.
па етом 1984 г. был зарегистрироваи в Дманисском районе, считавтие
а.
ся до того свободным от пасюк
|,
3 восточной части Грузии распространение пасюка уе
льно ш
х
т
емя ареал его тут зпачите
,
плохо. Однако в настоящее вр
у
это я пнисашо М. В. Шидловским (1976). Но всей вероятности, он заселяот
подавляющее большинство населенных пунктов вдоль О
"
\ ак и веток от Тбилиси до Телави
от Тбилиси до Кировакана
ы
|
-
Цкаро. На них он зарегистрирован в Гурджаани и Сигнахи (Кузякии,
1951). В Цителцкаройском районе пасюк обнаружен В ра ор всей
у
ле Кура—
эт
отсутствует на водоразде
вероятности, этот грызун
„Кура ори и на
б
Й
<
плоскогорья, гие овцеводе
большей части ИШорского
,
т
-
правление хозяйства, мала плотность паселения людей и редко располо
ты.
яепы небольшие населенные иун
_
.
" На южные склоны Большого Кавказа пасюк подпимается до Г. и
-
кой д
ктов вдоль Военно-Грузине
ли. известен из многих пун
Ко
Коби. Сиони, Казбеги), многократно появлялся на Крестовом
а
вые: 2379 м (Курятников 1986). Отсутствует этот грызун
перевале па высоте
к
,
Е
Тушетии и Хевсуретии. 1. 2.
‚
вернее с. Квемо-Алвани в
|
В К Читиашвили (1986) пишут об обитании пасюта ниже о ая
пре
береж
лазани, не
:
пунктах левобережья А: и,
у
в предгорных населеплых
апи,
г
я
окротных мест находок. В пограничных частях, Авербайджеокой
;
:
хи
”
т
Белоканы и Закаталы. На терри
этот вид известен из сел
Н
у _Н.
итории Гру
й
Большого Кавказа на высота»
‹ой ССР в горно-несном поясе
1500
о 1800 м а, моря было обследовано девять населенных пунктов
и в одном из них пасюка не обпаружили, не было сго здесь и
‹ий, 1964).
осным сведениям (Шидловский, Е
.
а.
р Пасюк давно известен из западной части Армянской ССР. В ориввн
‹
всех
-
й губ
б более или менее обыкповенен во к
‹ой губернии оп был
в
ть
ых оселенных пунктах еще в начале пынешнего века (Пявотер, 1912).
й
@.
а
Для Ленинакана упоминается © 1919 г. (Кузякин, 1951). о
м
утверждение Н. В. Верещагина {1959, с. 301) о том, что в Ереново и
Джульфе пасюки появились лишь | 30-х годах нашего ст
,
Ареал
73
иостройки железной дороги, связавшей названные города с Тбилиси и
Баку», не выдерживает критики, тем более что железная дорога, связав-
шая Тбилиси с Ленинаканом и Карсом, Ереваном и Джульфой, действо-
вала уже в 1913 г. Видимо, насюк проник в Приараксинскую котловину
с юго-востока. В Нахичевани пасюк был добыт еще в 1917 г. (коллекции
ЗИН АН СССР).
Т. М. Соспихина (1946, 1950), изучавшая население грызунов городов
Армении в первой половине 40-х годов, отметила отсутствие пасюка в
Кировакане и Алаверди. В обзоре млекопитающих Армянской ССР
С. В. Даль (1954) пишет, что пасюк обычен в городах Ереване, Эчмиад-
зине, Ленинакане, а также в селениях Аштаракского, Эчмиадзинского,
Абовянского (Котайкского), Арташатского и Араратското (Вединского)
районов. Этот вид обитает в естественных биотопах по речкам, впадаю-
щим в Аракс к югу от Еревана (Шидловский, 1976).
В настоящее время пасюк заселяет всю западную часть Армении за
исключением северо-западных, пограничных с Грузией Амасийского,
Гукасянекого, Калининского райопов и высокогорий Арагацкого массива.
Он занимает целиком Араратскую низменность. Встречается во всех
круппых населенных пунктах на шоссе Ереван-Талин—Ленинакан и
вдоль железной дороги Ереван—Тбилиси, включая города Кировакан,
Алаверди. На северо-запад от железной дороги проник в г. Степанаван.
На север от Еревана вдоль железной дороги расселился до г. Раздан.
На восток доходит до Гарни, Веди, Арарата. В Ехегнадзор проник, ви-
димо, по широкой долине р. Арпа. На северс-востоке Армении найден в
четырех селах Носмберянского района. Существуют неопределенные
указания на возможность обитания этого вида по берегам правых при-
токов Куры— рек Агстев в Иджеванском, Ахун и Хндзрот в Шамшадин-
ском райопах (Варшавский и др., 1987).
Нет пасюка вокруг Севапа и в высокогорьях хребтов Армянского
нагорья и Малого Кавказа. Зарегистрировано отсутствие этого вида в
Кафане, Дастакерте, Сисиане, Базаурчае (Соспихина, 1946; Петров,
Адамян, 1966). В Нахичеванской АССР пасюк найден в Ильичевском
{ст. Ильичевск, сел. Ханлыклар, Верхний и Нижний Аралыг, Дашарх) и
Нахичеванском районах (Лобанова, Петросян, 1959; Рябцева, 1980;
Исмаилов и др., 1986).
В Азербайджане кости пасюка найдены в слоях позднего средневе-
ковья (ХУ-ХУГ вв. п. 2.) при раскопках дворца ширваншахов в Баку
(Верещагин, 1959). В настоящее время пасюк распространен на большей
части Азербайджана. Оп встречается в бассейнах Алазани и Агричая от
Белокан и Закатал на западе до Вартаптена и Куткашена на востоке. По
железной дороге Махачкала—Баку отмечен на большинетве круиных стан-
ций (Худат, Хачмае, Дивичи, Сиазань). Проникает далеко вверх по
долине Самура, по р. Кудиалчаю доходит до Кубы. Нет пасюка в высо-
когорьях Большого Кавказа на востоке ло массива Бабадага включи-
тельно (см. рис. 46).
Он найден на всех крупных станциях железных дорог Тбилиси—
Баку, Баку—Астара, начиная от г. Казах на западе и на ветке
ТА
Глава вторая
—
лезных дорог удаляется в сторону не далее чем на
ая а 1980). обычен в Нагорном Карабахе (Шуша, Гадрут,
Мартуни, Мардакерт). Встречается на большинотве пристапей Ур ы ло
ее устья и юга Куринской Носы. Обнаружен на Оль зе островов
Апитеронского полуострова и Бакипского архипелага. Отсу р у ры
ок в верхней части Тальтиских гор, на наиболее высоких хре тах вр
мянского нагорья и Малого Кавказа, а также на Нооброе: ть в тоста
полынных полупустынь (Верещагин, 1959; Кузя, 195
тоа
Петросян, 1959; Петросян и др. 1962; Кададкий, вт учерук,
ков, 1985; Исмаилов и др., 1986: Варшавский и др.,
.
Поселения пасюка па севере Дагестана вдоль морского побережья
смыкаются с калмыдцкими. Им заселены все протоки и болота, дель гы
Терека, включая Прорву и Сулла-Чубушла. к югу от железной дор
сплошь заняты этим видом Казбековский, Буйнакский и бол
сть
Левашинского района. В Гергебильском районе паек найден в д ух
|
.
°
‹утинском районе он обнаружен
3
населенных пунктах, в Акуш
‚обнаружен в пятива
в. Отсюда пропик‘'на` запад
Ч
тести обследованных поселко
<
`
п
а
занал
т
кото ` (с. Укарах) р
{\
ерную часть Дахадаевс
Е
(с. Кумух, Кунды) и в северну
т
{Я
итагокий
кий. Хивский, Сулейман-Ст
,
на. Кайтагский, Табасаранский,
ле
к
ы
йоны, сплошь заселены ‘пасюком.
й
как и все приморские районы,
м. Ото
‹
проник в о Курах, а вверх по Самуру заселил Ахтьшекий район д пай
ден в поселках Шиназ и Аракул Рутульского района. р тульокий, лин.
й
бский
‹ульский, Гумбетовский и все
ский, Гунибский, Упцуку
,
падные
райопы горного Дагестана свободны от этого руна а о чад
‹
Ботлихский район (ем. рис: 10).
‘из Ведено насюк проник в 5:
гм
6. В сто.
`
‹
ах Дагестана ‘в обслед
т
бодных от пасюка горных раиоп
ух ве
жили. Все заселенные ‘ пасюком
менных пунктах его не обнару
рые паюком в т
:
в пределах 1000—
м над ур:
.
населенные пункты расположены
у
ад УР. моря.
‹
ка, в десяти поселках их бы
Численность зверьков здесь невелика,
то малои
асселение пасюков в горах осуще
в семи— среднее количество. р
уществиястоя
бильным транспортом. Как правило,
преимущественно автомо
к правило, засоля тоя
<
енные по наиболее оживле
населениые пункты, располож
й
и-
лям (Сорокина, Бацин, Коржов, 1986; Сорокина, Потапова, 1987; Вар-
ий
987)
-
,
шавский и др., 19 .
,
И
.ко
В равнинной части Чечено-Иигушеко, АССР`по: рекам Тороку, Су
‹
пасюк обитает. новвеместно. Ю}
и железнодорожным станциям
о
г тута
:
едепо, Шали, Уру
ил в республике идет через
,
ница его распроетранен
депо, Шали, Урус
ой
Н
никидзе. В горной ча
Мартан, Ачхой-Мартан на г. Орджо
‘асти етеко:
т
пийскКии»
{
й
аселенных пунктах (совхоз «Аль
,
п Веченского районов в пяти н
г
о
ины
Эр-1
‹<
бнаружили орок
Юр-Дар, Итум- Кале, рать Макажой) крыс не обнару
ки
на, Бации, Коржов
.
ко
ля по находкам костей в погадках филина древностью в носкоть ко
столетий. пасюк уже давно проник па территорию современной се ро
Осетинской АССР. В начале пынешиего столетия он был о ычтым оби
й
по Военно-Грузинекой дороге з
тателем равпинных районов, а
мился
высоко в горы. Активное расселение этого вида пачалось с момента ‘про
Ареал
75
ведепия колеспых дорог в горные районы Северной Осетии, Первые све-
дения о пасюке в горных ущельях относятся к концу 40-х годов.
Сплошной ареал пасюка, где он встречается практически во всех
сельских населенных пунктах и городах, а также живет круглый год по
берегам прудов, каналов и в заболоченных поймах рек, занимает всю
предгорную часть республики. Южная граница сплошного распростране-
ния этого вида с запада на восток идет примерно по линии Чикола, Дур-
Дур, Алагир, Балта, Тарское. По ущельям и автомобильным дорогам
насюк проникает далеко
к тогу от этой линии. В верховьях бассейна р. Урух
он достиг селений Ахсау и Дунта, вверх по Ардону он обычен в селе-
ниях Мизур, Архон, Бурон. Из последнего завезен в Нижний Цей, пе-
риодически появлялся и исчезал в Нижнем Зармаге. У Мизура на высо-
те 1300 м обнаружены поселения в естественных биотопах.
Вверх по р. Фиагдону устойчивая популяция пасюка сформировалась
в п. Верхний Фиагдон. Здесь на высоте 1400 м крысы встречаются на
огородах и в естественных биотопах. Из Верхнего Фиагдопа пасюк про-
ник в последние населенные пункты верховьев ущелья
— Хидикус и
Хариджин, Вверх по р. Гизельдон пасюк зарегистрирован в селеньях
Кобан, Даргавс и на курорте Кармадон (Курятников, 1986; Сорокина
и др., 1986; Варшавский и др., 1987).
|
В горной части республики ареал пасюка имеет пятнистый характер,
поселения его неустойчивы и могут многократно возникать и исчезать.
Н. Н. Курятников (1986), обеледовавигий в 1972—1984 гг. горные селе-
ния республики, нашел пасюка в 5 из них, а в 16 этот вид отсутствовал.
Из 21 горного поселка, обследованного в республике за последнее трид-
патилетие, пасюк был обнаружен в 5 (Сорокина и др., 1986). Одновре-
менно отмечено в 1986 г. появление пасюка в поселках, где его ранее
не наблюдали, курорт Кармадон и др. (Варшавский и др., 1987).
В Кабардино-Балкарской АССР юго-западная граница сплошного
распространепия пасюка идет от поселков Таши-Тала и Верхний Лескен
на Бабуген, Карасу, Нижний Чегем, Жанхотеко, Каменпомостское, Кич-
малку и выходит в пределы Карачаево-Черкесской АО, где в ауле Ха-
суат пасюк обитал в конце прошлого века (Россиков, 1887). Вверх по
Баксану пасюк найден в пос. Белым и г. Тырнауз, где с конца 60-х го-
дов сформировалась многочисленная и устойчивая популяция. Особенно
многочислен пасюк в зоне лесостепи в Советском и Зольном районах
(Темботов, 1972).
В Карачаево-Черкесской АО от Хасуата южная граница распростра-
нения пасюка идет через п. Эльбрусский, г. Карачаевск, станицу Зелен-
‚чукскую, Преградную, Курджиново на пос. Псебай в нижнем течении
р. Малой Лабы.
Распространение этого вида здесь цокументировано нлохо. Зарегист-
рированы его находки в Теберде и Домбае. Возможно, оп проник от
Эльбрусского вверх по р. Кубани до пос. Хурзук, а по Большому Зелен-
чуку вверх до Архыза.
В Краснодарском крае пасюка добывали в четырех паселенных пупк-
тах южной части Отрадненского района и в ст. Чернореченской на
ТА
Глава вторая
—
лезных дорог удаляется в сторону не далее чем на
ая а 1980). обычен в Нагорном Карабахе (Шуша, Гадрут,
Мартуни, Мардакерт). Встречается на большинотве пристапей Ур ы ло
ее устья и юга Куринской Носы. Обнаружен на Оль зе островов
Апитеронского полуострова и Бакипского архипелага. Отсу р у ры
ок в верхней части Тальтиских гор, на наиболее высоких хре тах вр
мянского нагорья и Малого Кавказа, а также на Нооброе: ть в тоста
полынных полупустынь (Верещагин, 1959; Кузя, 195
тоа
Петросян, 1959; Петросян и др. 1962; Кададкий, вт учерук,
ков, 1985; Исмаилов и др., 1986: Варшавский и др.,
.
Поселения пасюка па севере Дагестана вдоль морского побережья
смыкаются с калмыдцкими. Им заселены все протоки и болота, дель гы
Терека, включая Прорву и Сулла-Чубушла. к югу от железной дор
сплошь заняты этим видом Казбековский, Буйнакский и бол
сть
Левашинского района. В Гергебильском районе паек найден в д ух
|
.
°
‹утинском районе он обнаружен
3
населенных пунктах, в Акуш
‚обнаружен в пятива
в. Отсюда пропик‘'на` запад
Ч
тести обследованных поселко
<
`
п
а
занал
т
кото ` (с. Укарах) р
{\
ерную часть Дахадаевс
Е
(с. Кумух, Кунды) и в северну
т
{Я
итагокий
кий. Хивский, Сулейман-Ст
,
на. Кайтагский, Табасаранский,
ле
к
ы
йоны, сплошь заселены ‘пасюком.
й
как и все приморские районы,
м. Ото
‹
проник в о Курах, а вверх по Самуру заселил Ахтьшекий район д пай
ден в поселках Шиназ и Аракул Рутульского района. р тульокий, лин.
й
бский
‹ульский, Гумбетовский и все
ский, Гунибский, Упцуку
,
падные
райопы горного Дагестана свободны от этого руна а о чад
‹
Ботлихский район (ем. рис: 10).
‘из Ведено насюк проник в 5:
гм
6. В сто.
`
‹
ах Дагестана ‘в обслед
т
бодных от пасюка горных раиоп
ух ве
жили. Все заселенные ‘ пасюком
менных пунктах его не обнару
рые паюком в т
:
в пределах 1000—
м над ур:
.
населенные пункты расположены
у
ад УР. моря.
‹
ка, в десяти поселках их бы
Численность зверьков здесь невелика,
то малои
асселение пасюков в горах осуще
в семи— среднее количество. р
уществиястоя
бильным транспортом. Как правило,
преимущественно автомо
к правило, засоля тоя
<
енные по наиболее оживле
населениые пункты, располож
й
и-
лям (Сорокина, Бацин, Коржов, 1986; Сорокина, Потапова, 1987; Вар-
ий
987)
-
,
шавский и др., 19 .
,
И
.ко
В равнинной части Чечено-Иигушеко, АССР`по: рекам Тороку, Су
‹
пасюк обитает. новвеместно. Ю}
и железнодорожным станциям
о
г тута
:
едепо, Шали, Уру
ил в республике идет через
,
ница его распроетранен
депо, Шали, Урус
ой
Н
никидзе. В горной ча
Мартан, Ачхой-Мартан на г. Орджо
‘асти етеко:
т
пийскКии»
{
й
аселенных пунктах (совхоз «Аль
,
п Веченского районов в пяти н
г
о
ины
Эр-1
‹<
бнаружили орок
Юр-Дар, Итум- Кале, рать Макажой) крыс не обнару
ки
на, Бации, Коржов
.
ко
ля по находкам костей в погадках филина древностью в носкоть ко
столетий. пасюк уже давно проник па территорию современной се ро
Осетинской АССР. В начале пынешиего столетия он был о ычтым оби
й
по Военно-Грузинекой дороге з
тателем равпинных районов, а
мился
высоко в горы. Активное расселение этого вида пачалось с момента ‘про
Ареал
75
ведепия колеспых дорог в горные районы Северной Осетии, Первые све-
дения о пасюке в горных ущельях относятся к концу 40-х годов.
Сплошной ареал пасюка, где он встречается практически во всех
сельских населенных пунктах и городах, а также живет круглый год по
берегам прудов, каналов и в заболоченных поймах рек, занимает всю
предгорную часть республики. Южная граница сплошного распростране-
ния этого вида с запада на восток идет примерно по линии Чикола, Дур-
Дур, Алагир, Балта, Тарское. По ущельям и автомобильным дорогам
насюк проникает далеко
к тогу от этой линии. В верховьях бассейна р. Урух
он достиг селений Ахсау и Дунта, вверх по Ардону он обычен в селе-
ниях Мизур, Архон, Бурон. Из последнего завезен в Нижний Цей, пе-
риодически появлялся и исчезал в Нижнем Зармаге. У Мизура на высо-
те 1300 м обнаружены поселения в естественных биотопах.
Вверх по р. Фиагдону устойчивая популяция пасюка сформировалась
в п. Верхний Фиагдон. Здесь на высоте 1400 м крысы встречаются на
огородах и в естественных биотопах. Из Верхнего Фиагдопа пасюк про-
ник в последние населенные пункты верховьев ущелья
— Хидикус и
Хариджин, Вверх по р. Гизельдон пасюк зарегистрирован в селеньях
Кобан, Даргавс и на курорте Кармадон (Курятников, 1986; Сорокина
и др., 1986; Варшавский и др., 1987).
|
В горной части республики ареал пасюка имеет пятнистый характер,
поселения его неустойчивы и могут многократно возникать и исчезать.
Н. Н. Курятников (1986), обеледовавигий в 1972—1984 гг. горные селе-
ния республики, нашел пасюка в 5 из них, а в 16 этот вид отсутствовал.
Из 21 горного поселка, обследованного в республике за последнее трид-
патилетие, пасюк был обнаружен в 5 (Сорокина и др., 1986). Одновре-
менно отмечено в 1986 г. появление пасюка в поселках, где его ранее
не наблюдали, курорт Кармадон и др. (Варшавский и др., 1987).
В Кабардино-Балкарской АССР юго-западная граница сплошного
распространепия пасюка идет от поселков Таши-Тала и Верхний Лескен
на Бабуген, Карасу, Нижний Чегем, Жанхотеко, Каменпомостское, Кич-
малку и выходит в пределы Карачаево-Черкесской АО, где в ауле Ха-
суат пасюк обитал в конце прошлого века (Россиков, 1887). Вверх по
Баксану пасюк найден в пос. Белым и г. Тырнауз, где с конца 60-х го-
дов сформировалась многочисленная и устойчивая популяция. Особенно
многочислен пасюк в зоне лесостепи в Советском и Зольном районах
(Темботов, 1972).
В Карачаево-Черкесской АО от Хасуата южная граница распростра-
нения пасюка идет через п. Эльбрусский, г. Карачаевск, станицу Зелен-
‚чукскую, Преградную, Курджиново на пос. Псебай в нижнем течении
р. Малой Лабы.
Распространение этого вида здесь цокументировано нлохо. Зарегист-
рированы его находки в Теберде и Домбае. Возможно, оп проник от
Эльбрусского вверх по р. Кубани до пос. Хурзук, а по Большому Зелен-
чуку вверх до Архыза.
В Краснодарском крае пасюка добывали в четырех паселенных пупк-
тах южной части Отрадненского района и в ст. Чернореченской на
76
Глава вторая
й
й
у
мина
Гольшой Лабе в Псебайском раионе данные ооо г и и
Н. Г. Калашникова, И. Я. Классовского, Л. И. толя И. В. Марко
(1949) указал на заселение пасюком в 1925 г. и. мы к хо х
р. Белой, давиее обитание его в 1. Новопрохладное на р. тие т тсут
ствие крыс на корлонах Кавказского заповедника Киша и у
т.
елах заповедника, в верховьях р.
.
Видимо, где-то здесь, в пред
‚
Белой Киши. и закапчивается незаселенная пасюком территория Боль
7
ы
,
ттого Кавказа (см. рис. 16).
Западная Сибирь и Казахстан
Й
хин
Северную границу ареала пасюка в Западной Сибири А. П. Кузяцин
{19541) проводил от средпего течения Усы по Полярному руту ло Са.
Оби и ее правому притоку зулыш
.
лехарда, затем вверх по
у
у Пульшимащу, от ею
<
` ль транссибирской железной дор
верховьев на восток вдо
д
‚оо
-
мо отсутствует в тае;
‹
аривался, что пасюк, види
р ри ом панаду
т
аежных и степных
й
<
ижнего течения Оби и на т
ных районах к западу от н
атас
степи
Т:
Ишима и Иртьша, Иртыша
.
одоразделах Тобола и Ишима,
й "И, | Лаптев (1958) существенно уточнил северную часть ареала
пасока в Западной Сибири. На приведенной в его: работе картосхеме
северная граница силоптного распространения пасюка, идет вдоль, рек
а,Г
места впадения в него. Тобола д
Пелыма, Тавды, Иртыша от
па до устья
этой
з водораздел к месту впадеп
р
Тары. вверх ио этой реке и через в
я
в 06; Далее вдоль бассейна Чульшмана выходит ‘па Енисей. ее
<
к: ты
Й
{ грого приурочен к долинам рек:
этой границы ареал пасюка стр
„рек: Иртыту
<
- ийска, вверх по Конде до усть
от Тобольска до Ханты-Мане
‚
уетья полые
Е верх по ее притокам Кети,
,
Тана, по Оби до Салехарда и в й
и
ману,
: Цазыму и Сосьве. По Оби север
Тыму. в нижнем течении Югана, по
устья Ваха пасюк был достоверно известен из г. Сургута, ео
Салехарда. Его добывали в Салехарле (Оборот) их о
ов
—1939 гг. В. В..
5
лекции ЗММУ). Отметим, чт
т.|
поцветающую популяцию пасюка в домах района пристани г. Салехарда.
Две статьи о современном рапрострашении а ы Я
йоб
{
Г. Лялин
Аа,1.
-
‹ ласти опубликовал В. Г.
м
ед
$
енные на основании
} хемы ареалов, составл
их статьях приведены картос
К
нные па основании
5
лков Томской и Тюмепсекой обла
-
обследования более 150 посе
на-
лиза [0 ответов на специальные анкеты, разосланные в 5 областе
(табл. 4).
.
ри
<
пасюки заселили почти в
Автор отмечает, что к семидосятьм годам ое ое
Стре
:
ыи
поселки в нижнем течении Ирт ,
ровекое, Стре.
й
:
й
юганск и г. Сургут) и нижне
свой, Нижневартовсий, Нефте ь
ни
Оби (Урманный, Красноленинский, Карымкары, Октябрьское, Шерка =.
Березово). Приведенная в работе картосхема ареала иринцитиально
отличается от вышеонисапной схемы И. П. Лаптева (1958). иным отп.
Несомненно, строительство современных домов © централь тым отот-
пением позволит пасюку продвинуться еще дальие на совер. ет
сообщения об укоренении пасюка в пос. Краспоселькупе на Хх
Ареал
77
Таблица 4. Наличие паеюков в паселенных п
Е
Ч
унктах админиетративпых
территорий Западной Сибири (по: Лялин, 1974)
Число насе-
ленных
Доля населен-
пунктов, по | ных пунктов с
Область, округ
которым ио- | наличием па-
лучены све- | стока, %
дения
Гомекая
226
96,6
Кемеровская
342
97,6
Новосибирская
862
98,2
Омская
696
97,0
Тюменская (без автономных округов)
307
97,4
Ханты-Мансийский автономный округ
187
43,4
Ямало-Ненецкий автономный округ
63
6,3
Сале па Обской губе (Чистяков, 1965), в пос. Аксарка и Салемал на
нижней Оби, а также о проникновении этого вида в пос. Новый Порт.
и Мыс Каменный (Ельшин, 1983).
На севере Свердловской области пасюк заселил поселки вдоль желез-
ной дороги Ивдель-—Обь (Серегинский) и появился в населенных пунк-
тах по верховьям Лозьвы до 100 км севернее г. Ивдель. В природных
биотопах в теплое время года зверьков регулярно отлавливали в поймах
рек Исети, Пышмы, Туры. На севере области, в верховьях Лозьвы, бро-
шенные поселки лесозаготовителей уже через год освобождаются от
пасюков (Стадухин, 1975).
Южную границу ареала пасюка в Казахстапе и Западной Сибири
А. П. Кузякин (1951) проводил от устья Кумы на Актюбинск, оговари-
вая отсутствие крыс в междуречье нижней Волги и Урала. От Аклю-
бинска она пла на Аще-Булак около Орска, через Кустанай, Атбасар,
Целиноград (Акмолипск). От последнего граница круто поднималась к
северу, пересекая Иртыш южнее Омска, и, обойдя Кулупдинскую низ-
менность, вновь выходила па левобережье Иртыша у Семипалатинска.
В Семипалатинске пасюк был добыт В. 9. Мартино в 1915 г. (коллекция
ЗИН). В районе Усть-Каменогорска и Лепиногорска опять круто свора-
чивала п северо-востоку, пересекала Обь чуть ниже Бийска.
Следует отметить, что пасюк появился в Северпом Казахстане еще в
конце прошлого века. Он был добыт в г. Петропавловске в 1890 г. (кол-
лекция Зоомузея МГУ).
В западной части Цазахстапа к настоящему времени ареал пасюка
существенно сократился. Он отмечен в низовьях Урала на север до
106. Алмалы. Далее вверх по Уралу его посслепия отсутствуют на про-
тяжении 300 км до районного центра Чанаево (см. рис. 15). Подробные
сведения о распространении пасюка в низовьях р. Урал приведены
О. С. Опариной, М. Л. Опариным, Е. Г. Самариным (1985). На севере
Уральской области он отмечен в поймах р. Деркул Каменского района,
76
Глава вторая
й
й
у
мина
Гольшой Лабе в Псебайском раионе данные ооо г и и
Н. Г. Калашникова, И. Я. Классовского, Л. И. толя И. В. Марко
(1949) указал на заселение пасюком в 1925 г. и. мы к хо х
р. Белой, давиее обитание его в 1. Новопрохладное на р. тие т тсут
ствие крыс на корлонах Кавказского заповедника Киша и у
т.
елах заповедника, в верховьях р.
.
Видимо, где-то здесь, в пред
‚
Белой Киши. и закапчивается незаселенная пасюком территория Боль
7
ы
,
ттого Кавказа (см. рис. 16).
Западная Сибирь и Казахстан
Й
хин
Северную границу ареала пасюка в Западной Сибири А. П. Кузяцин
{19541) проводил от средпего течения Усы по Полярному руту ло Са.
Оби и ее правому притоку зулыш
.
лехарда, затем вверх по
у
у Пульшимащу, от ею
<
` ль транссибирской железной дор
верховьев на восток вдо
д
‚оо
-
мо отсутствует в тае;
‹
аривался, что пасюк, види
р ри ом панаду
т
аежных и степных
й
<
ижнего течения Оби и на т
ных районах к западу от н
атас
степи
Т:
Ишима и Иртьша, Иртыша
.
одоразделах Тобола и Ишима,
й "И, | Лаптев (1958) существенно уточнил северную часть ареала
пасока в Западной Сибири. На приведенной в его: работе картосхеме
северная граница силоптного распространения пасюка, идет вдоль, рек
а,Г
места впадения в него. Тобола д
Пелыма, Тавды, Иртыша от
па до устья
этой
з водораздел к месту впадеп
р
Тары. вверх ио этой реке и через в
я
в 06; Далее вдоль бассейна Чульшмана выходит ‘па Енисей. ее
<
к: ты
Й
{ грого приурочен к долинам рек:
этой границы ареал пасюка стр
„рек: Иртыту
<
- ийска, вверх по Конде до усть
от Тобольска до Ханты-Мане
‚
уетья полые
Е верх по ее притокам Кети,
,
Тана, по Оби до Салехарда и в й
и
ману,
: Цазыму и Сосьве. По Оби север
Тыму. в нижнем течении Югана, по
устья Ваха пасюк был достоверно известен из г. Сургута, ео
Салехарда. Его добывали в Салехарле (Оборот) их о
ов
—1939 гг. В. В..
5
лекции ЗММУ). Отметим, чт
т.|
поцветающую популяцию пасюка в домах района пристани г. Салехарда.
Две статьи о современном рапрострашении а ы Я
йоб
{
Г. Лялин
Аа,1.
-
‹
ласти опубликовал В. Г.
м
ед
$
енные на основании
} хемы ареалов, составл
их статьях приведены картос
К
нные па основании
5
лков Томской и Тюмепсекой обла
-
обследования более 150 посе
на-
лиза [0 ответов на специальные анкеты, разосланные в 5 областе
(табл. 4).
.
ри
<
пасюки заселили почти в
Автор отмечает, что к семидосятьм годам ое ое
Стре
:
ыи
поселки в нижнем течении Ирт ,
ровекое, Стре.
й
:
й
юганск и г. Сургут) и нижне
свой, Нижневартовсий, Нефте ь
ни
Оби (Урманный, Красноленинский, Карымкары, Октябрьское, Шерка =.
Березово). Приведенная в работе картосхема ареала иринцитиально
отличается от вышеонисапной схемы И. П. Лаптева (1958). иным отп.
Несомненно, строительство современных домов © централь тым отот-
пением позволит пасюку продвинуться еще дальие на совер. ет
сообщения об укоренении пасюка в пос. Краспоселькупе на Хх
Ареал
77
Таблица 4. Наличие паеюков в паселенных п
Е
Ч
унктах админиетративпых
территорий Западной Сибири (по: Лялин, 1974)
Число насе-
ленных
Доля населен-
пунктов, по | ных пунктов с
Область, округ
которым ио- | наличием па-
лучены све- | стока, %
дения
Гомекая
226
96,6
Кемеровская
342
97,6
Новосибирская
862
98,2
Омская
696
97,0
Тюменская (без автономных округов)
307
97,4
Ханты-Мансийский автономный округ
187
43,4
Ямало-Ненецкий автономный округ
63
6,3
Сале па Обской губе (Чистяков, 1965), в пос. Аксарка и Салемал на
нижней Оби, а также о проникновении этого вида в пос. Новый Порт.
и Мыс Каменный (Ельшин, 1983).
На севере Свердловской области пасюк заселил поселки вдоль желез-
ной дороги Ивдель-—Обь (Серегинский) и появился в населенных пунк-
тах по верховьям Лозьвы до 100 км севернее г. Ивдель. В природных
биотопах в теплое время года зверьков регулярно отлавливали в поймах
рек Исети, Пышмы, Туры. На севере области, в верховьях Лозьвы, бро-
шенные поселки лесозаготовителей уже через год освобождаются от
пасюков (Стадухин, 1975).
Южную границу ареала пасюка в Казахстапе и Западной Сибири
А. П. Кузякин (1951) проводил от устья Кумы на Актюбинск, оговари-
вая отсутствие крыс в междуречье нижней Волги и Урала. От Аклю-
бинска она пла на Аще-Булак около Орска, через Кустанай, Атбасар,
Целиноград (Акмолипск). От последнего граница круто поднималась к
северу, пересекая Иртыш южнее Омска, и, обойдя Кулупдинскую низ-
менность, вновь выходила па левобережье Иртыша у Семипалатинска.
В Семипалатинске пасюк был добыт В. 9. Мартино в 1915 г. (коллекция
ЗИН). В районе Усть-Каменогорска и Лепиногорска опять круто свора-
чивала п северо-востоку, пересекала Обь чуть ниже Бийска.
Следует отметить, что пасюк появился в Северпом Казахстане еще в
конце прошлого века. Он был добыт в г. Петропавловске в 1890 г. (кол-
лекция Зоомузея МГУ).
В западной части Цазахстапа к настоящему времени ареал пасюка
существенно сократился. Он отмечен в низовьях Урала на север до
106. Алмалы. Далее вверх по Уралу его посслепия отсутствуют на про-
тяжении 300 км до районного центра Чанаево (см. рис. 15). Подробные
сведения о распространении пасюка в низовьях р. Урал приведены
О. С. Опариной, М. Л. Опариным, Е. Г. Самариным (1985). На севере
Уральской области он отмечен в поймах р. Деркул Каменского района,
78
Тлава вторая
й
е
Барбастау Теректинского и р. Илек Бурминского районов. тоне
ы Уралу южная граница ареала пасюка уходит в Ореп ур у
область Южнее по Уялу, Темиру и Иргизу, где его в потом зо ы
мечал Н. А. Зарудный (1897), сейчас этого вида нет (Варшаве др.,
©
.
и
18) результате осноепия целины и организации многочиеленных, зерно.
вых совхозов, имеющих свиноформы, Пао аНаЧитОлЬо аи во
р
‹стане (Груздев, 1959; Слуд , .. ;
в Северном Казахстан
„ 19:
1
оалов 1979). Из 76 поселков Нустанайской области, в которых пасюк
был обнаружен в 1967 г. 43 населенных пункта были заселены он,
начала освоения целинных земель, с 1954 по 1967 г. (Борисенко, 1969).
В вастоящее время южная граница сплошного распространения асю
ходит до Нов
-
;
Е
так. По р. Илеку она доход
ка в Казахстане выглядит
ЕВА
1
у
т к верховьям Тобола, д
к’, Карабутак иде
}
евки и через Актюбинск’,
вьям Тобола, иле
:
кат, Докучаевку, Держа
,
,
востоку через Пушкино, Ор ,
\т-
У Делинограх. От последпего граница сплошного распростраления
сворачивает к северу и идет через Алексеевку, Стошняк, епинград ое
:
елаулы и ч
‹ья озер Селентытениз и
Кзылту на северные побережья
С
аулыи чер
‹одит в пойму Иртыша южнее “р у .
Андриановку, Разумовку выходи
Краснокутока.
й
ы изолированные поселения
ТОжнее и восточнее этой траниц,
Спаоока из
ховьях Тургая, по нижнему р
,
вестны в Карсакпае, в вер
о
ороаккану
и
оз. Тенгиз, южнее Нуры, в р
‚
северо-восточного побережья 03.
,
о
,
Я покаревке Осакаровке, к востоку от Алексесвки, районе Ерментау
*
Шидерты.
,
.
.
Вверх по Иртышу пасток распространен вплоть до оз Зайсан, где мно
гочислен на Тополевом мысу. В Алтайском крае распространех в ду,
кроме Кош-Агачского и Улагановского аймакон (Кирьянов, | Очв-
До конца 70-х годов пасюк в Тувинской АССР не был найден очи.
ров, Башанов, 1975; Бояркин, 1984). О
вали
1
гы
зал
хай
у
черук, Нузиков,
в
пасюка в ближайлие годы (Ку
.
,
ы продокават
торию Тувы. Действительно,
проникновение его па терря
тель
сентябре
1982 г. 3 экз. были впервые обнаружены з Уюкской кои и
сле 15 г. добыли п
.
,
ках Аржан и Хадыг. В апр
ь
я
у
линовка, Пий-Хемского района.
пос. Аржан, Хадын, Тарлас, Ма:
‚
ооо
у
‹е 195 зверьков, они вновь появ
квартале 1985 г. отловили уже
Е
в ос Уюк и Шевелик. Пасюк стал обычным обитателем домон м на
цворных построек перечисленных выше населенных пунктов ( ки о о,
Галацевич, 1986). Видимо, следует ожидать дальнейшего р р
ареала пасюка в 'Тувинской АССР.
Средняя Сибирь (бассейны Енисея и Лены)
\
ареала пасюка в Средней
к уже упоминалось зыше, северную границу
ее А) П. Кузякин (1951) проводил по правому берегу р. Чушьыа
и далее к востоку от Ачинска до Красноярска вдоль железной дороги.
ке (Набе-
7 Исть указание на отсутствие пасюка в настоящее время в г. Актюбинске (
нов, 1981).
Арезл
79
долине Енисея граница поднималась к’ северу до Енисейска или даже до
с. Подкаменная Тунгуска, но не дальше. Если в Игарке пасюк уже укре-
пился, то его обитание здесь правильнее трактовать как вновь возникшее
изолированное поселение. От Енисейска граница шла по Ангаре, Илиму,
захватывала неболыпой отрезок верхней Лены и поресекала Байкал
южнее о-ва Ольхон. Восточнее она шла от устья Селенги к Улан-Удэ
и далее вдоль железной дороги к Чите. При этом А. П. Кузякин ука-
зывал, что оставалось неясным, жили ли в это время пасюки между
транссибирской железпой дорогой и долиной средней Ангары и вотречал-
‹я ли этот вид в долине Подкаменной Тунгуски °.
С этого времени наши представления об ареале пасюка в описынаемом
регионе существенно изменились. Это объяспяется как расплирением ареа-
ла вида, так и обснедованием прежде недостаточно изученных территорий.
В настоящее время приходится считать, что северная граница ареала
пасюка от верховьев р. Кети выходит в долину Енисея у г. Лесосибир-
ска. Вниз по Еписею этот вид, несомненно, обитает во всех крупных на-
селенных пунктах по берегам реки (известен из Ёписейска, Ярцева, Бора,
Сумарокова, Верхнеимбатска, Верещагина, Туруханска, Игарки, Дудинки,
Усть-Порта, Караула, Диксона). По-видимому, из Туруханска он проник
в Ираспоселькуп. Стал обычным обитателем г. Норильска, Утольного и
окрестных шахт, встречается здесь и в естественных биотопах.
ПТГироко распространен в естественных биотопах бассейпа верхней
Ангары (Кузякип, 1951), известен из Братска, Усть-Илимска, Богучан.
Регулярно занозится в населенные пупкты на Нижней Тунгуске: Непа,
Ербогачен, Преображенка, Подволомино (Тарасов, 1965). Найден в
Ногинске (Хамаганов и др., 1986).
Подробные сведения о распространения пасока в южной половине
Иркутской области и западной части Бурятекой АССР привел А.`А. Мак-
симов (1937). Опубликована (Хамаганов и др., 1986) схематическая знач-
ковая карта распространения насюка на территория Восточной Сибири и
Дальнего Востока. В Иркутской области пасюк обнаружен в 36 населен-
ных пунктах 20 административных районов. Недостает сведений о его
распространении в западных и северных районах области. В западную
часть Бурятской АССР пасюк проникаст вверх по долипам рек Иркута
и Джиды. Видимо, нет его в восточных отрогах Большого Саяна и на
Хапгарульском хребте (рис. 17).
На средней Лене пасюк обитает издавна. А. П. Окладников (1953)
обнаружил его кости в верхних слоях неолитической стоянки человека в
100 км ниже устья Витима. В 50 кы ниже Пеледуя, па обрывах берега
Лены, субфоссильные остатки пасюка з погадках филина нашол
О. В. Егоров (1960).
В настоящее время по Лене паесюк встречается в Качуге, Жигалове,
Усть-Куте, Киренске, ПВитиме, Олекмипске (Кузякин, 1951; Тарасов,
1965). В 1970 г. поселение пасюка обнаружено в пос. Амга, на реке
того же назнания, левом притокс Алдана (Романова, 1974). К 1973 г. ов
8 А. П. Кузякин называет эту реку Верхней Тунгуской.
78
Тлава вторая
й
е
Барбастау Теректинского и р. Илек Бурминского районов. тоне
ы
Уралу южная граница ареала пасюка уходит в Ореп ур у
область Южнее по Уялу, Темиру и Иргизу, где его в потом зо ы
мечал Н. А. Зарудный (1897), сейчас этого вида нет (Варшаве др.,
©
.
и
18) результате осноепия целины и организации многочиеленных, зерно.
вых совхозов, имеющих свиноформы, Пао аНаЧитОлЬо аи во
р
‹стане (Груздев, 1959; Слуд ,
..;
в Северном Казахстан
„ 19:
1
оалов 1979). Из 76 поселков Нустанайской области, в которых пасюк
был обнаружен в 1967 г. 43 населенных пункта были заселены он,
начала освоения целинных земель, с 1954 по 1967 г. (Борисенко, 1969).
В вастоящее время южная граница сплошного распространения асю
ходит до Нов
-
;
Е
так. По р. Илеку она доход
ка в Казахстане выглядит
ЕВА
1
у
т к верховьям Тобола, д
к’, Карабутак иде
}
евки и через Актюбинск’,
вьям Тобола, иле
:
кат, Докучаевку, Держа
,
,
востоку через Пушкино, Ор ,
\т-
У Делинограх. От последпего граница сплошного распростраления
сворачивает к северу и идет через Алексеевку, Стошняк, епинград ое
:
елаулы и ч
‹ья озер Селентытениз и
Кзылту на северные побережья
С
аулыи чер
‹одит в пойму Иртыша южнее “р у .
Андриановку, Разумовку выходи
Краснокутока.
й
ы изолированные поселения
ТОжнее и восточнее этой траниц,
Спаоока из
ховьях Тургая, по нижнему р
,
вестны в Карсакпае, в вер
о
ороаккану
и
оз. Тенгиз, южнее Нуры, в р
‚
северо-восточного побережья 03.
,
о
,
Я покаревке Осакаровке, к востоку от Алексесвки, районе Ерментау
*
Шидерты.
,
.
.
Вверх по Иртышу пасток распространен вплоть до оз Зайсан, где мно
гочислен на Тополевом мысу. В Алтайском крае распространех в ду,
кроме Кош-Агачского и Улагановского аймакон (Кирьянов, | Очв-
До конца 70-х годов пасюк в Тувинской АССР не был найден очи.
ров, Башанов, 1975; Бояркин, 1984). О
вали
1
гы
зал
хай
у
черук, Нузиков,
в
пасюка в ближайлие годы (Ку
.
,
ы продокават
торию Тувы. Действительно,
проникновение его па терря
тель
сентябре
1982 г. 3 экз. были впервые обнаружены з Уюкской кои и
сле 15 г. добыли п
.
,
ках Аржан и Хадыг. В апр
ь
я
у
линовка, Пий-Хемского района.
пос. Аржан, Хадын, Тарлас, Ма:
‚
ооо
у
‹е 195 зверьков, они вновь появ
квартале 1985 г. отловили уже
Е
в ос Уюк и Шевелик. Пасюк стал обычным обитателем домон м на
цворных построек перечисленных выше населенных пунктов ( ки о о,
Галацевич, 1986). Видимо, следует ожидать дальнейшего р р
ареала пасюка в 'Тувинской АССР.
Средняя Сибирь (бассейны Енисея и Лены)
\
ареала пасюка в Средней
к уже упоминалось зыше, северную границу
ее А) П. Кузякин (1951) проводил по правому берегу р. Чушьыа
и далее к востоку от Ачинска до Красноярска вдоль железной дороги.
ке (Набе-
7 Исть указание на отсутствие пасюка в настоящее время в г. Актюбинске (
нов, 1981).
Арезл
79
долине Енисея граница поднималась к’ северу до Енисейска или даже до
с. Подкаменная Тунгуска, но не дальше. Если в Игарке пасюк уже укре-
пился, то его обитание здесь правильнее трактовать как вновь возникшее
изолированное поселение. От Енисейска граница шла по Ангаре, Илиму,
захватывала неболыпой отрезок верхней Лены и поресекала Байкал
южнее о-ва Ольхон. Восточнее она шла от устья Селенги к Улан-Удэ
и далее вдоль железной дороги к Чите. При этом А. П. Кузякин ука-
зывал, что оставалось неясным, жили ли в это время пасюки между
транссибирской железпой дорогой и долиной средней Ангары и вотречал-
‹я ли этот вид в долине Подкаменной Тунгуски °.
С этого времени наши представления об ареале пасюка в описынаемом
регионе существенно изменились. Это объяспяется как расплирением ареа-
ла вида, так и обснедованием прежде недостаточно изученных территорий.
В настоящее время приходится считать, что северная граница ареала
пасюка от верховьев р. Кети выходит в долину Енисея у г. Лесосибир-
ска. Вниз по Еписею этот вид, несомненно, обитает во всех крупных на-
селенных пунктах по берегам реки (известен из Ёписейска, Ярцева, Бора,
Сумарокова, Верхнеимбатска, Верещагина, Туруханска, Игарки, Дудинки,
Усть-Порта, Караула, Диксона). По-видимому, из Туруханска он проник
в Ираспоселькуп. Стал обычным обитателем г. Норильска, Утольного и
окрестных шахт, встречается здесь и в естественных биотопах.
ПТГироко распространен в естественных биотопах бассейпа верхней
Ангары (Кузякип, 1951), известен из Братска, Усть-Илимска, Богучан.
Регулярно занозится в населенные пупкты на Нижней Тунгуске: Непа,
Ербогачен, Преображенка, Подволомино (Тарасов, 1965). Найден в
Ногинске (Хамаганов и др., 1986).
Подробные сведения о распространения пасока в южной половине
Иркутской области и западной части Бурятекой АССР привел А.`А. Мак-
симов (1937). Опубликована (Хамаганов и др., 1986) схематическая знач-
ковая карта распространения насюка на территория Восточной Сибири и
Дальнего Востока. В Иркутской области пасюк обнаружен в 36 населен-
ных пунктах 20 административных районов. Недостает сведений о его
распространении в западных и северных районах области. В западную
часть Бурятской АССР пасюк проникаст вверх по долипам рек Иркута
и Джиды. Видимо, нет его в восточных отрогах Большого Саяна и на
Хапгарульском хребте (рис. 17).
На средней Лене пасюк обитает издавна. А. П. Окладников (1953)
обнаружил его кости в верхних слоях неолитической стоянки человека в
100 км ниже устья Витима. В 50 кы ниже Пеледуя, па обрывах берега
Лены, субфоссильные остатки пасюка з погадках филина нашол
О. В. Егоров (1960).
В настоящее время по Лене паесюк встречается в Качуге, Жигалове,
Усть-Куте, Киренске, ПВитиме, Олекмипске (Кузякин, 1951; Тарасов,
1965). В 1970 г. поселение пасюка обнаружено в пос. Амга, на реке
того же назнания, левом притокс Алдана (Романова, 1974). К 1973 г. ов
8 А. П. Кузякин называет эту реку Верхней Тунгуской.
80
Глава вторая
расселился вниз по течепию и заселил поселки Чимикулан, Абага,
Михайловское. В 1976—1978 гг. пасюк закрепился в Якутоко и началось
стремительное раслиирение ареала вида. На правобережье ены эр я
появились напротив Якутска в пос. Цавловск и Нижний Бестях, в ы .
их обнаружили в селе Огородтах севернее Якутска,в селах Кыеыл Сыр
и Антоновка па р. Вилюй (Вилюйский и Ленинский районы), а ПИ г.
в нос. Харабалах на нижней Амге и селе булун на р. Алдан.
ры.
были заселепы все животповодческие комплексы и перевалочные базы,
расположенные по р. Лене от Якутска до Синска. В это же время пасюк
Рис. И. Распространение пасюка в Восточной Сибири и на Дальнем Востоке
‹
и8—
юк не
1— территории, сплошь заселенные пасюком; 2 — точки находок; 8 пункты, где пас
найден
—#9иг.
-——>
".
==
2
<
7
45
—No
_.
й
|
м
ь
|
й
‚
Ц
>
©
©
.
О
о
м
т
`
|
«5 ОЕ
®Ей
й
и[© Аини.
г
С
©)
м
м
|
П
И
[
й
сх
Ареал
81
проник в населенные пункты по р. Яне (Нижнеянск, Кулар, Уеть-Куй-
га, Батагай). Устойчивое поселение возникло в низовьях р. Оленек в
ос. Таймылар. Началось заселение пасюком автотрассы Ленск—Мирный,
тде в пос. Арылах впервые он появился в 1984 г. К настоящему време-
ни пасюк населяет порты Тикси, Быковский, Черский на Колыме и пос.
Зырянку в среднем течении этой реки (Романовна, 1986; Романовна и др.,
1986; Егоров, Хмелева, 1987).
В пределах Олекмо-Витимской горной страны пасюк обитает в пойме
правого притока Витима р. Каренги. Найден в верховьях р. Амалат в
с. Монгон и в пойме верховьев р. Нерчи. По Витиму найден в естеетвеп-
ных биотопах и постройках с. Неляты. Широко распространен в природ-
ных биотопах Муйско-Куандииской котловипы (Дагопчан, среднее тече-
ние р. Парам). Обычен в Бодайбо. Ветречается в пойме р. Тунгир, пра-
вого притока Верхней Олекмы (Тарасов, 1965; Очиров, 1968 а, б, 1980;
Немченко, Быков, 1983). В северной части Байкала пасюк есть зв
пос. Байкальское, Нижнеангарск и в населенных пунктах долины Верх-
ней Ангары (Швецов, 1977). Видимо, бассейпы р. Селенги, Шилки,
верховьев Витима и Олекмы относятся к естественному (исходному)
ареалу ласлока.
Мы можем констатировать быстрое распространение пасюка в райо-
ны промышленного. освоения Средней Сибири. Если вся южная часть
региона входила в естественный, исконный ареал пасюка, то сейчас
наблюдается стремительное расселение этого вида на сезер в районы
промышленных новостроек. Видимо, недалеко то время, когда пасюк
васелит большую часть населенных пунктоз долин Виалюя и Алдана,
проникнет в населенные пункты вдоль железной и автомобильной дорог
от Тынды до Беркакита и Якутска. Заселит поселки вдоль автотрассы
Сусуман—Хандыга— Якутск.
Южное Забайкалье, Дальний Восток,
Северо-Восточная Сибирь
Северную границу ареала пасюка в этом регионе А. П. Кузякин (1951)
проводил от устья Селенги па Улан-Удо и далее по железной дороге
через Читу, Скозородино, Тыгду к пристани Зея, Нижнюю Бурею на
Амур у Хабаровска. По долине Амура пасюк распространен до устья.
Изолированные поселения пасюка были указаны для Камчатки и отмс-
чены два пункта завоза крысы на крайний северо-восток Сибири.
Как мы уже отмочали выше, в пастолщее время установлено птяро-
кое распространение пасюка в естественных местообитаниях Олекмо-Ви-
тимской горной страпы. К его исконному ареалу относится территория
бассейнов Верхней Ангары, Селенги, Онона, Ингоды, Шилки и Аргупя.
Здесь он повсюду обычен в природных биотопах (Фетисов, 1945; Ха-
маганов, Греков, 1983).
Подробные сведения о распространении пасюка в южных районах
Читинской области приведены А. С. Фетисовым (1945, 1946), Н. В. Не-
хииеловым (1958, 1960), Н. В. Ольковой и др. (1987). Пасюк обычен по
80
Глава вторая
расселился вниз по течепию и заселил поселки Чимикулан, Абага,
Михайловское. В 1976—1978 гг. пасюк закрепился в Якутоко и началось
стремительное раслиирение ареала вида. На правобережье ены эр я
появились напротив Якутска в пос. Цавловск и Нижний Бестях, в ы .
их обнаружили в селе Огородтах севернее Якутска,в селах Кыеыл Сыр
и Антоновка па р. Вилюй (Вилюйский и Ленинский районы), а ПИ г.
в нос. Харабалах на нижней Амге и селе булун на р. Алдан.
ры.
были заселепы все животповодческие комплексы и перевалочные базы,
расположенные по р. Лене от Якутска до Синска. В это же время пасюк
Рис. И. Распространение пасюка в Восточной Сибири и на Дальнем Востоке
‹
и8—
юк не
1— территории, сплошь заселенные пасюком; 2 — точки находок; 8 пункты, где пас
найден
—#9иг.
-——>
".
==
2
<
7
45
—No
_.
й
|
м
ь
|
й
‚
Ц
>
©
©
.
О
о
м
т
`
|
«5
ОЕ
®Ей
й
и[© Аини.
г
С
©)
м
м
|
П
И
[
й
сх
Ареал
81
проник в населенные пункты по р. Яне (Нижнеянск, Кулар, Уеть-Куй-
га, Батагай). Устойчивое поселение возникло в низовьях р. Оленек в
ос. Таймылар. Началось заселение пасюком автотрассы Ленск—Мирный,
тде в пос. Арылах впервые он появился в 1984 г. К настоящему време-
ни пасюк населяет порты Тикси, Быковский, Черский на Колыме и пос.
Зырянку в среднем течении этой реки (Романовна, 1986; Романовна и др.,
1986; Егоров, Хмелева, 1987).
В пределах Олекмо-Витимской горной страны пасюк обитает в пойме
правого притока Витима р. Каренги. Найден в верховьях р. Амалат в
с. Монгон и в пойме верховьев р. Нерчи. По Витиму найден в естеетвеп-
ных биотопах и постройках с. Неляты. Широко распространен в природ-
ных биотопах Муйско-Куандииской котловипы (Дагопчан, среднее тече-
ние р. Парам). Обычен в Бодайбо. Ветречается в пойме р. Тунгир, пра-
вого притока Верхней Олекмы (Тарасов, 1965; Очиров, 1968 а, б, 1980;
Немченко, Быков, 1983). В северной части Байкала пасюк есть зв
пос. Байкальское, Нижнеангарск и в населенных пунктах долины Верх-
ней Ангары (Швецов, 1977). Видимо, бассейпы р. Селенги, Шилки,
верховьев Витима и Олекмы относятся к естественному (исходному)
ареалу ласлока.
Мы можем констатировать быстрое распространение пасюка в райо-
ны промышленного. освоения Средней Сибири. Если вся южная часть
региона входила в естественный, исконный ареал пасюка, то сейчас
наблюдается стремительное расселение этого вида на сезер в районы
промышленных новостроек. Видимо, недалеко то время, когда пасюк
васелит большую часть населенных пунктоз долин Виалюя и Алдана,
проникнет в населенные пункты вдоль железной и автомобильной дорог
от Тынды до Беркакита и Якутска. Заселит поселки вдоль автотрассы
Сусуман—Хандыга— Якутск.
Южное Забайкалье, Дальний Восток,
Северо-Восточная Сибирь
Северную границу ареала пасюка в этом регионе А. П. Кузякин (1951)
проводил от устья Селенги па Улан-Удо и далее по железной дороге
через Читу, Скозородино, Тыгду к пристани Зея, Нижнюю Бурею на
Амур у Хабаровска. По долине Амура пасюк распространен до устья.
Изолированные поселения пасюка были указаны для Камчатки и отмс-
чены два пункта завоза крысы на крайний северо-восток Сибири.
Как мы уже отмочали выше, в пастолщее время установлено птяро-
кое распространение пасюка в естественных местообитаниях Олекмо-Ви-
тимской горной страпы. К его исконному ареалу относится территория
бассейнов Верхней Ангары, Селенги, Онона, Ингоды, Шилки и Аргупя.
Здесь он повсюду обычен в природных биотопах (Фетисов, 1945; Ха-
маганов, Греков, 1983).
Подробные сведения о распространении пасюка в южных районах
Читинской области приведены А. С. Фетисовым (1945, 1946), Н. В. Не-
хииеловым (1958, 1960), Н. В. Ольковой и др. (1987). Пасюк обычен по
82
Глава вторая
всему бассейну рек Онона и Борзи. К югу от них известен из Красной
Ималки, Кулусутая, Соловъевска. Зарегистрирован на всех крупных
станциях Забайкальской железной дороги. Заселяет все приаргунские
села до Нерчепского завода. Встречен в остественных биотопах поймы
р. Уров. В стенной части Юго-Восточного Забайкалья численность его не-
велика, в некоторых паселенных пунктах он появлялся лишь периоди-
чески (Пекинелов, 1958; Олькова и др., 1987).
Вверх ло 3ез пасюк прослежен до Зейского заповедника, по Бурее
доходил до Ургала в Чегдомына, ость в Кондоне, в пизовьях Амгунй и
устье Уды (Кучерук, Кузиков, 1985). На восточном отрезке БАМа пасюк
зарегистрирован в Февральском, Звонком, Ургале, Дуссе-Алине, Сулуке,
Герби, Баджале, Березовом (Ковалевский, Корепберг, 1983).
Вдоль побережья Охотского моря пасок обитает во многих населеп-
ных пунктах от Ульи и г. Охотска на юге до Магадана и пос. Цушки,
Таяти и нижиуего течепия р. Пенжины на севере. На север от Магадана
но Колымской трассе проник в пос, Палатка, Атка, Ягодное и ряд дру-
тих поселков верховьев Колымы (Хамаганов, 1972; Юдин и др., 1976;
Липаев и др., 1983). Зарегистрирован в ряде населенных пунктов запал-
ного побережья Камчатки, в г. Петропавловек-Камчатский, обычен по
долине р. Камчатки от устья до верховьев. Пасюк Зарегистрирован на
многих из Курильских островов и распространен практически по всему
Сахалину, достигая здесь весьма высокой численности (Тимофеева, Сур-
ков, 1968; Воронов, 1974; Костенко, 1976). Встречается почти во всех
населенных пунктах, расположенных В бассойне Анадыря (села Краене-
но, Усть-Белая, Снежное, Ерпол, Ламутское, Танорер и др.). К северу от
Анадыря обычен в пос. Эгвекинот, па Чукотском полуострове в пос. Про-
видения, Лаврентия, Уэлен. Далее к западу известен в пос. Иультин,
мые Шмидта, г. Певек. Завозвился в порты Зеленый Мыс и Черский в
низовьях Колымы (Хамаганов, 1972; Юдин и др., 1976; Костенко, 1976).
Т(ирокое распрострапение пастока’ на побережьях Восточно-Сибиреко-
то и Бериогова морей как в Азии, так и з Америке заставляет считать
его достаточно давним обитателем этих мест. Возможно, как это считает
С А. Хамаганов (1972), Ападырь и Чукотка входят в первичный ареал
«дикоживущих» крыс.
о региону опубликовано несколько карт ареала пасюка. А. А. Тимо-
фесва, В. С. Сурков (1968) привели карту размещения этого видав на-
селенных пунктах и природных мостообитаниях на 0-ве Сахалип, где от-
мсчено более 200 точек регистрации пасюка. С. А Хамаганов (1972)
оттубликовал карту распространения пасюка на Дальпем Востоке (Амур-
ская, Камчатская, Магаданская, Сахалниская области, Приморский и Ха-
баровский края). На ней разными пунсопами обозначены: 1} паселенные
пункты, в которых пасюк постоянно обитает; 2) места обитания пасока
в природных биотопах; 3) пункты, где крысы не найдены. В тексте ска-
зано, что обитание пасюка зарегистриронано в 450 населенных пунктах
Дальнего Востока, в том числе в Амурской области— в 51, Камчат-
ской —24, Магаданской
— 40, Сахалинской— 54, Приморском крае—
91, Хабаровском —490. На карте значками таких пунктов показано:
Ареал
82
в Амурской области — 8, Камчатской — 13, Магаданской — 17, Сахали
ской — 24, Приморском крас — 18, Хабаровском
—37.ВАмускойоб.
ласти в 15% населенных пунктон пасюк отсутствовал, но на а те пока-
ван лишь один пункт © отсутствием крысы. Как отбирались я Нвчки ля
показа на карте, з работе не упомянуто. На карте размещения масок
на Дальнем Востоке (Костенко, 1976) тоновым ареалом показано бита.
ние вида в естественных биотопах. Даны две градации: а) вид едок —
менее 1 зверька па 100 ловушко-ночей и б) вид обычен — более во ь-
ка на 100 ловушко-ночей. Пунсоном показаны места, где вид вст Чен
только в насслениом пункте. Пунктиром обозначены основные т но т-
ные трассы, как сухопутные, так и морские. Карта вата тот е
регион и по степени подробности практически не отличается
6..
кованной С. А. Хамагановым.
от опт
Недавно в Южном Приморье обнаружено позднеплейстоценовое и го-
лоценовое скопление остатков грызунов— пещера Близнец близ г. Па
тизапска. Из 38,5 тыс. костных остатков 2,1% (не менее 250 обобей),
приходится на пасюка. Единичные кости этого вида встречены на глуби-
то 88 м, затем на 6,2 мис 6 м до поверхности встречаются ностояпно.
шее число костей пасюка приходится на горизонт от 6 до 3,2 м
затем относительная численность их резко сокращается, и в таком коли-
честве „они доходят до поверхности. На глубине 3 м возраст отложений
п церы по содержанию С“ составлял 14 965-65 лет (Алексеева, 1986)
Следовательно, проникновение пасюка на эту территорию не б о
.
но с деятельностью человека.
р
ло ов
Последние публикации (Хамаганов и др., 1986; Олькова и др., 1987)
не внесли существенных изменений в наити представления о рас] остра-
нении пасюка па Дальнем Востоке и Северо-Восточной Сибири ры
Изолированные участки ареала пасюка
в Средней Азии и Южном Казахстане
Впервые на появление пасюка в Ташкенте указал И. И, Колеснико
рен пасюка в этом райопе посвящены работы В. П. ром.
в в 32), В. п. Промитова и др. (41981, 1983), Г. С. Давыдова (1964,
м
оящ время на севере пасюк достиг пос. Янгибазар на
р. лесе в Чимкентской области и заселил юго-восточное побережье
ардаринского водохрапилища. На юго-запад он распространился
границы Самаркандской области, к югу— до поселков Ура-Тюбе и ау
на севере Таджеякистана. На восток вдоль южного побережья Кайрак ы
ского водохранилища заселил г. Канибадам и, двигаясь по олезной до.
роге, достиг пос. Киров — районного центра Ферганской области. Можно
предполагать быстрое растпирение ареала пасюка в этом регионе практи-
по всем паправлениям. Карта ареала пасюка в Узбекистане опуб-
ликована В. П. Промптовым и др. (1986).
у
р
ще об обнаружении пасюка ва юго-западе Таджикистапа
й ‚ 1986), видимо, есть следствие ошибки в определении вида
первые па территории г. Алма-Аты трех пасюков добыли в октябре
82
Глава вторая
всему бассейну рек Онона и Борзи. К югу от них известен из Красной
Ималки, Кулусутая, Соловъевска. Зарегистрирован на всех крупных
станциях Забайкальской железной дороги. Заселяет все приаргунские
села до Нерчепского завода. Встречен в остественных биотопах поймы
р. Уров. В стенной части Юго-Восточного Забайкалья численность его не-
велика, в некоторых паселенных пунктах он появлялся лишь периоди-
чески (Пекинелов, 1958; Олькова и др., 1987).
Вверх ло 3ез пасюк прослежен до Зейского заповедника, по Бурее
доходил до Ургала в Чегдомына, ость в Кондоне, в пизовьях Амгунй и
устье Уды (Кучерук, Кузиков, 1985). На восточном отрезке БАМа пасюк
зарегистрирован в Февральском, Звонком, Ургале, Дуссе-Алине, Сулуке,
Герби, Баджале, Березовом (Ковалевский, Корепберг, 1983).
Вдоль побережья Охотского моря пасок обитает во многих населеп-
ных пунктах от Ульи и г. Охотска на юге до Магадана и пос. Цушки,
Таяти и нижиуего течепия р. Пенжины на севере. На север от Магадана
но Колымской трассе проник в пос, Палатка, Атка, Ягодное и ряд дру-
тих поселков верховьев Колымы (Хамаганов, 1972; Юдин и др., 1976;
Липаев и др., 1983). Зарегистрирован в ряде населенных пунктов запал-
ного побережья Камчатки, в г. Петропавловек-Камчатский, обычен по
долине р. Камчатки от устья до верховьев. Пасюк Зарегистрирован на
многих из Курильских островов и распространен практически по всему
Сахалину, достигая здесь весьма высокой численности (Тимофеева, Сур-
ков, 1968; Воронов, 1974; Костенко, 1976). Встречается почти во всех
населенных пунктах, расположенных В бассойне Анадыря (села Краене-
но, Усть-Белая, Снежное, Ерпол, Ламутское, Танорер и др.). К северу от
Анадыря обычен в пос. Эгвекинот, па Чукотском полуострове в пос. Про-
видения, Лаврентия, Уэлен. Далее к западу известен в пос. Иультин,
мые Шмидта, г. Певек. Завозвился в порты Зеленый Мыс и Черский в
низовьях Колымы (Хамаганов, 1972; Юдин и др., 1976; Костенко, 1976).
Т(ирокое распрострапение пастока’ на побережьях Восточно-Сибиреко-
то и Бериогова морей как в Азии, так и з Америке заставляет считать
его достаточно давним обитателем этих мест. Возможно, как это считает
С А. Хамаганов (1972), Ападырь и Чукотка входят в первичный ареал
«дикоживущих» крыс.
о региону опубликовано несколько карт ареала пасюка. А. А. Тимо-
фесва, В. С. Сурков (1968) привели карту размещения этого видав на-
селенных пунктах и природных мостообитаниях на 0-ве Сахалип, где от-
мсчено более 200 точек регистрации пасюка. С. А Хамаганов (1972)
оттубликовал карту распространения пасюка на Дальпем Востоке (Амур-
ская, Камчатская, Магаданская, Сахалниская области, Приморский и Ха-
баровский края). На ней разными пунсопами обозначены: 1} паселенные
пункты, в которых пасюк постоянно обитает; 2) места обитания пасока
в природных биотопах; 3) пункты, где крысы не найдены. В тексте ска-
зано, что обитание пасюка зарегистриронано в 450 населенных пунктах
Дальнего Востока, в том числе в Амурской области— в 51, Камчат-
ской —24, Магаданской
— 40, Сахалинской— 54, Приморском крае—
91, Хабаровском —490. На карте значками таких пунктов показано:
Ареал
82
в Амурской области — 8, Камчатской — 13, Магаданской — 17, Сахали
ской — 24, Приморском крас — 18, Хабаровском
—37.ВАмускойоб.
ласти в 15% населенных пунктон пасюк отсутствовал, но на а те пока-
ван лишь один пункт © отсутствием крысы. Как отбирались я Нвчки ля
показа на карте, з работе не упомянуто. На карте размещения масок
на Дальнем Востоке (Костенко, 1976) тоновым ареалом показано бита.
ние вида в естественных биотопах. Даны две градации: а) вид едок —
менее 1 зверька па 100 ловушко-ночей и б) вид обычен — более во ь-
ка на 100 ловушко-ночей. Пунсоном показаны места, где вид вст Чен
только в насслениом пункте. Пунктиром обозначены основные т но т-
ные трассы, как сухопутные, так и морские. Карта вата тот е
регион и по степени подробности практически не отличается
6..
кованной С. А. Хамагановым.
от опт
Недавно в Южном Приморье обнаружено позднеплейстоценовое и го-
лоценовое скопление остатков грызунов— пещера Близнец близ г. Па
тизапска. Из 38,5 тыс. костных остатков 2,1% (не менее 250 обобей),
приходится на пасюка. Единичные кости этого вида встречены на глуби-
то 88 м, затем на 6,2 мис 6 м до поверхности встречаются ностояпно.
шее число костей пасюка приходится на горизонт от 6 до 3,2 м
затем относительная численность их резко сокращается, и в таком коли-
честве „они доходят до поверхности. На глубине 3 м возраст отложений
п церы по содержанию С“ составлял 14 965-65 лет (Алексеева, 1986)
Следовательно, проникновение пасюка на эту территорию не б о
.
но с деятельностью человека.
р
ло ов
Последние публикации (Хамаганов и др., 1986; Олькова и др., 1987)
не внесли существенных изменений в наити представления о рас] остра-
нении пасюка па Дальнем Востоке и Северо-Восточной Сибири ры
Изолированные участки ареала пасюка
в Средней Азии и Южном Казахстане
Впервые на появление пасюка в Ташкенте указал И. И, Колеснико
рен пасюка в этом райопе посвящены работы В. П. ром.
в в 32), В. п. Промитова и др. (41981, 1983), Г. С. Давыдова (1964,
м
оящ время на севере пасюк достиг пос. Янгибазар на
р. лесе в Чимкентской области и заселил юго-восточное побережье
ардаринского водохрапилища. На юго-запад он распространился
границы Самаркандской области, к югу— до поселков Ура-Тюбе и ау
на севере Таджеякистана. На восток вдоль южного побережья Кайрак ы
ского водохранилища заселил г. Канибадам и, двигаясь по олезной до.
роге, достиг пос. Киров — районного центра Ферганской области. Можно
предполагать быстрое растпирение ареала пасюка в этом регионе практи-
по всем паправлениям. Карта ареала пасюка в Узбекистане опуб-
ликована В. П. Промптовым и др. (1986).
у
р
ще об обнаружении пасюка ва юго-западе Таджикистапа
й ‚ 1986), видимо, есть следствие ошибки в определении вида
первые па территории г. Алма-Аты трех пасюков добыли в октябре
84
Глава вторая
1982 г. Их появление связывали с масеовым заводом картофеля из Севоро-
Казахстанской области. За 1983—1984 тг. в том же районе город был
добыт 61 зверек. При выборочном обследовании тысячи часты к домо-
владений более 100 из них оказались заселеппыми насюхом. а ьшая
плотность населения зверьков наблюдалась н ностроиках, где содер ли
свиней, нутрий и домашнюю птицу. Поселения этого вида встреча тоя то
берегам ручьев и речек Октябрьского и Московского ра
р
“Стогов, ВВ) `пасюка в г. Красповодске известны © то основания.
В дальнейшем его отмечали там неоднократно (Бондарь, ь 16: а р,
1958). Современная харавторнотика популяции пасюка в Вр
м
().
вы. м "полы, кроме Красноводека, пасюка добывали в портах
Уфра и Бекдапт. В последиие годы пасюк, кроме Красповолека, пы ви
наружен в Уфре, па п-ове Челскен (г. Челекен, и аратель, о
Аладжа) и в пос. Аджи-Яб Гасан-Кулийского района ар
й
р сопоставлении современных данных о распроораноти пасюка м
картой его ареала, опубликованной нами ранее р
а
1985), видно, что во многих местах удалось уточнить топограф ва
этого вида, а также зафиксировать дальнейшее расселение
тории СССР.
Глава третья.
Экология
Места обитания
Неравномерность распределения животных на местности определяется ее
пеодпородностью: одни участки территории полностью отвечают запросам
того или иного вида, другие в меньшей степени, а третьи могут быть сов-
сем для него непригодпы. Как известно, серая крыса, являясь экологиче-
ски пластичным видом, может жить, приспосабливаясь к самым различ-
ным условиям, но тем не менее она предъявляет совершенпо определен-
ныс требования к среде— се распределение всегда зависит от наличия
достаточного количества воды. В связи с этим пасюк в пределах ареала
распространеп далеко пе повосместпо, как пишут многие авторы (Арги-
ропуло, 1940; Громов и др, 1963), а распределяется мозаично, составляя
так называемое «кружево ареала».
Мозаичные поселения вида могут быть двух типов: линейными или
островными (Наумов, 1954). Для пасюков в одпих случаях характерен.
линейный тип, в других островной.
Крысы обитают в населенных пунктах и в природных биотопах, при
этом в постройках человека опи живут в пределах всего ареала, а в при-
роде только в отдельных его частях. В процессе эволюции эти грызуны
так приспособились к жизии за счет человска, что обитание в постройках
стало для них наиболее типично.
Мы, как и Б. Росицкий и И. Кратохвил (1953), будем подразделять
крыс на две экологические формы: на экзоантропных, обитающих круг-
лый год в открытых стациях, и синантроппых, паселяющих жилища че-
ловска и постройки, в той или иной степени связанные с его деятель-
ностью. Такое деление хотя и вполне правомочно, по несколько условно,
так как иногда крысы, обитающие з прироче, охотно переходят к пита-
нию за счет человека и становятся сипаптронными, а крысы-сипантропы
могут переселяться в естествеппые стации,
При описании биотопического распределения всегда необходимо иметь сведения
об обилии особей на территории и, следовательно, надо располагать данными учета
численности.
По количественному учету млекопитающих существуют два способа, дающие
относительные и абсолютные показатели. При относительном учете получаются дан-
ные, показывающие не только во сколько раз численность вида в одном местообита-
нии отличается от таковой в другом, но также и в каком направлении меняется
обилие особей по сезонам и годам.
Абсолютный учет в отличие от относительного более объективен и дает воз-
можность определить численность вида на единицу площади, но проводить его нам-
ного сложнее.
В, В. Кучерук и 9. И. Коренберг (1964) делят учеты численности на три катего-
рии: 1) относительный косвенный учет, 2) относительный прямой и 3) абсолютный
учет.
Эта классификация разработана для животных, обитающих в природе, но все
применяемые в настоящее время методы учета синантропных крыс подходят к ее
традациям.
84
Глава вторая
1982 г. Их появление связывали с масеовым заводом картофеля из Севоро-
Казахстанской области. За 1983—1984 тг. в том же районе город был
добыт 61 зверек. При выборочном обследовании тысячи часты к домо-
владений более 100 из них оказались заселеппыми насюхом. а ьшая
плотность населения зверьков наблюдалась н ностроиках, где содер ли
свиней, нутрий и домашнюю птицу. Поселения этого вида встреча тоя то
берегам ручьев и речек Октябрьского и Московского ра
р
“Стогов, ВВ) `пасюка в г. Красповодске известны © то основания.
В дальнейшем его отмечали там неоднократно (Бондарь, ь 16: а р,
1958). Современная харавторнотика популяции пасюка в Вр
м
().
вы. м "полы, кроме Красноводека, пасюка добывали в портах
Уфра и Бекдапт. В последиие годы пасюк, кроме Красповолека, пы ви
наружен в Уфре, па п-ове Челскен (г. Челекен, и аратель, о
Аладжа) и в пос. Аджи-Яб Гасан-Кулийского района ар
й
р сопоставлении современных данных о распроораноти пасюка м
картой его ареала, опубликованной нами ранее р
а
1985), видно, что во многих местах удалось уточнить топограф ва
этого вида, а также зафиксировать дальнейшее расселение
тории СССР.
Глава третья.
Экология
Места обитания
Неравномерность распределения животных на местности определяется ее
пеодпородностью: одни участки территории полностью отвечают запросам
того или иного вида, другие в меньшей степени, а третьи могут быть сов-
сем для него непригодпы. Как известно, серая крыса, являясь экологиче-
ски пластичным видом, может жить, приспосабливаясь к самым различ-
ным условиям, но тем не менее она предъявляет совершенпо определен-
ныс требования к среде— се распределение всегда зависит от наличия
достаточного количества воды. В связи с этим пасюк в пределах ареала
распространеп далеко пе повосместпо, как пишут многие авторы (Арги-
ропуло, 1940; Громов и др, 1963), а распределяется мозаично, составляя
так называемое «кружево ареала».
Мозаичные поселения вида могут быть двух типов: линейными или
островными (Наумов, 1954). Для пасюков в одпих случаях характерен.
линейный тип, в других островной.
Крысы обитают в населенных пунктах и в природных биотопах, при
этом в постройках человека опи живут в пределах всего ареала, а в при-
роде только в отдельных его частях. В процессе эволюции эти грызуны
так приспособились к жизии за счет человска, что обитание в постройках
стало для них наиболее типично.
Мы, как и Б. Росицкий и И. Кратохвил (1953), будем подразделять
крыс на две экологические формы: на экзоантропных, обитающих круг-
лый год в открытых стациях, и синантроппых, паселяющих жилища че-
ловска и постройки, в той или иной степени связанные с его деятель-
ностью. Такое деление хотя и вполне правомочно, по несколько условно,
так как иногда крысы, обитающие з прироче, охотно переходят к пита-
нию за счет человека и становятся сипаптронными, а крысы-сипантропы
могут переселяться в естествеппые стации,
При описании биотопического распределения всегда необходимо иметь сведения
об обилии особей на территории и, следовательно, надо располагать данными учета
численности.
По количественному учету млекопитающих существуют два способа, дающие
относительные и абсолютные показатели. При относительном учете получаются дан-
ные, показывающие не только во сколько раз численность вида в одном местообита-
нии отличается от таковой в другом, но также и в каком направлении меняется
обилие особей по сезонам и годам.
Абсолютный учет в отличие от относительного более объективен и дает воз-
можность определить численность вида на единицу площади, но проводить его нам-
ного сложнее.
В, В. Кучерук и 9. И. Коренберг (1964) делят учеты численности на три катего-
рии: 1) относительный косвенный учет, 2) относительный прямой и 3) абсолютный
учет.
Эта классификация разработана для животных, обитающих в природе, но все
применяемые в настоящее время методы учета синантропных крыс подходят к ее
традациям.
86
Глава третья
4. Отноеительный косвенный учет численности паеюков.
а. Глазомерная оценка численности по следам жизнедеятельности крыс
—#0
трызам, входным отверотиям в норы, калу ит. п. Численность в этих случая опреде-
ляют как много, средне и Мало. Этот метод очень субъективен и дает весьма от-
восительные данные.
б. Учет численности по количеству съеденной приманки за сутки. Обычно для
этого используют зерповую приманку (овсянку) с растительным маслом, расклады-
вая ее ежедневно в течение 10 дней, каждый раз взвептивая порцию и се остатки.
Расчет обитающих в постройке крыс проводится по максимальному суточному ко-
личеству съеденной приманки, поделенному на норму потребления одним зверьком за
сутки (СыИУ, 1942; Дукельская, 1948).
К сожалению, о суточном потреблении корма крысами в литературе приводятся
различные данные, составляющие от 20 до 60 г (Дукельская, 1948; Турок, 1970;
"Граханов, 1983; Соколов и др., 1986). В. А. Судейкин указывает на большую разницу
в поедании корма молодыми крысами (30 г) и взрослыми {60 г}. Величина суточной
нормы крыс зависит еще и от состава кормов в помещении, тде они обитают (Пасеш-
ник, 1947; Мирзоев, 1984). Таким образом, различные показатели суточной ноедаемо-
сти крысами приманки приводят к трудно сравнимым между собой результатам
учетов.
в. Определение численности © помощью пылевых площадок. Ю. В. Тощигин
(1983) предложил использовать процент площадок со следами крыс по отношению
к числу выставленных площадок. При этом методе площадки (лотки из фанеры или
картона) засынают слоем (2 мм) какого-либо сыпучего материала {тальк, мука,
зола, мелкий песок) и располагают их в местах, наиболее посещаемых крысами.
тот метод в последнее время широко применяется как средство обнаружения
крыс в помещении, но он недостаточно разработан — нет данных о размере площадок,
сколько их должно быть выложено на определенной площади и т. Д.
г. Определение свободных и заселенных грызунами построек. При этом методе
рассчитывают, какой процент зданий от числа обследованных заселен крысами.
Несомненно, чем больше строений свободно от трызунов, тем ниже численность в
данном регионе и наоборот. Полученный показатель может быть использован для
изучения пространственного распределения грызунов на больших территориях на-
селенного пункта. Его недостаток заключается в том, что при низкой численности
наличие крыс трудно установить и здания часто оптибочно попадают в число сво-
бодных от пасюков, что снижает показатели общей численности на всей территории.
2 Относительный прямой учет численности паеюков.
|
а. Учет с помощью ловушек Геро (наживленных стандартной приманкой— короч
кой хлеба с подсолнечным маслом). Способ учета на ловупко-линиях, широко при-
меняемый зоологами в природе, в большинстве случаев непригоден В помещениях,
так как для размещения линии ловуптек не хватает места. Ловушки ставят, как пра.
вило, в определенном количестве на единицу площади. Показателем относительной
численности служит число пойманных зверьков на 1000 м? (Вишняков и Др. 1955)
или на 100 ловушко-суток (Леви, Судейкин, 1977). Метод имеет явные недостатки,
так как осторожные крысы часто избегают ловушек, часть их обладает неофобией; а
в ряде случаев, только задев приманку, зверек спускает ловугику, пугается и потом
начинает ее избегать.
В настоящее время отсутствует единый подход к учету крыс ловушкамя Геро.
В различных инструкциях рекомендуются не только разное количество ловушек на
единицу площади, но и различное количество дней проведения учета, а также и
различные принципы расчета средних показателей. Тем не менее этот метод широко
применяется и поэтому заслуживает внимания.
.
б. Число собранных трупов после проведения истребительных мероприятий.
Труты подечитывают и относят на единицу площади (объчно на 4000 м?). Недоетаток
метода заключается в том, что часть крыс, беря приманку, погибает в норах, под
полом, в перекрытиях и т. п., в результате чего их трупы нельзя учесть. Другая же
часть не берет приманку вообще. Этот метод позволяет сказать, какая была чиос-
ленность, но не какая она есть в настоящее время.
Экология
87
т
я
- 1900 +.
3. я)
21208 5;
о,
21000 =
ъх
Е
33 30.
]8
$3
<
ЗВ
7 600
53
ь
ЕВ зд
Я 900ыы
р
- 203
0
Рис. 18. Сопоставление резуль-
татов учета численности крыс,
полученных разными метода- 5% 00
‚ми, в подвале жилого много-
‚
|
м
ы
„
”
х
и
д
а
ы
З
„
>
х
.
.
>
Р
о
й
=
|
—
р
ы
х
этажного дома (по материалам З
мх`
и‘
3
В.К.м
ы
т8|
`меи\
<
А — сравнение заслежелности Ч
ры
площадок и поедаемости зер- 9 8 г
05
новых приманок;
з|-
7
Е5
Б — сравнение заслеженности пло- $ 40 Е
еб
щадок и попадаемости в ло- х
8
вушки Геро.
а
Г
2к
1-— попадаемость в ловушки 20|
з
Геро,
.
.
<
|
З
2 — количество съеденной при- З
8
ле
<
ООО
О
ОО
О
О
5
ГН|Л|
3 — процент заслеженны: пло-
ГРЯЗИ
Е
щадок
.
„Дни наблюдений
в. Глазомерна
:
с
а
рая опека численности крыс. Пастюки в основном активны в сумер-
ках и ве но при очен ь высокой численности все зверьки не могут насытиться
ато Рромят ооо ит ают выходить из укрытий днем. Считается, что если
а
теля в светлое время суток находятся единичные особи
о
ое чел ы группы, К зверьков очень много (Вегпаг4, СоНл,
использован только как орпентировочный., 67). Копетно, этот мотод может быть
ероятно, наибол
:
ного орояти р а того фотка при учете, можно достигнуть, вводя коррекцию од-
В Абеотютный учет численности пасюков.
ен 902 т ые четы отличаются большой сложностью и трудоемкостью. По отно-
ени к серым Зрыба ; практически их не применяют. Но мы все же остановимся
двух ИЗ пих ко ры ‚могут ыть использованы при научных исследованиях
ен
орк ‚ Суть метода сводится к следующему: в течение 8-7
вы
стороженным ловушкам * с приманкой, после чего они
о
аивтоя и а 2—3 ночи удается выловить в помещениях если не
а
отных и составить себе представление об их численности
б. Оценка исленвоети, о учету 119
О
т зверьков с помощью выпуска меченых особей. Этот ме-
ав
И рее бухова и В. В. Раевского (1933), применявитих его для
учета полевокв омотах.
учета в помещениях серых крыс их отлавливают жи-
,
выпускают на Месте поимки. После этого все помещение *5б-
При спл Н!
ыл
эффек ивно применение
овых кацканов No 0 или
э
оном в ове более
у
м
"т:
риме
д
86
Глава третья
4. Отноеительный косвенный учет численности паеюков.
а. Глазомерная оценка численности по следам жизнедеятельности крыс
—#0
трызам, входным отверотиям в норы, калу ит. п. Численность в этих случая опреде-
ляют как много, средне и Мало. Этот метод очень субъективен и дает весьма от-
восительные данные.
б. Учет численности по количеству съеденной приманки за сутки. Обычно для
этого используют зерповую приманку (овсянку) с растительным маслом, расклады-
вая ее ежедневно в течение 10 дней, каждый раз взвептивая порцию и се остатки.
Расчет обитающих в постройке крыс проводится по максимальному суточному ко-
личеству съеденной приманки, поделенному на норму потребления одним зверьком за
сутки (СыИУ, 1942; Дукельская, 1948).
К сожалению, о суточном потреблении корма крысами в литературе приводятся
различные данные, составляющие от 20 до 60 г (Дукельская, 1948; Турок, 1970;
"Граханов, 1983; Соколов и др., 1986). В. А. Судейкин указывает на большую разницу
в поедании корма молодыми крысами (30 г) и взрослыми {60 г}. Величина суточной
нормы крыс зависит еще и от состава кормов в помещении, тде они обитают (Пасеш-
ник, 1947; Мирзоев, 1984). Таким образом, различные показатели суточной ноедаемо-
сти крысами приманки приводят к трудно сравнимым между собой результатам
учетов.
в. Определение численности © помощью пылевых площадок. Ю. В. Тощигин
(1983) предложил использовать процент площадок со следами крыс по отношению
к числу выставленных площадок. При этом методе площадки (лотки из фанеры или
картона) засынают слоем (2 мм) какого-либо сыпучего материала {тальк, мука,
зола, мелкий песок) и располагают их в местах, наиболее посещаемых крысами.
тот метод в последнее время широко применяется как средство обнаружения
крыс в помещении, но он недостаточно разработан — нет данных о размере площадок,
сколько их должно быть выложено на определенной площади и т. Д.
г. Определение свободных и заселенных грызунами построек. При этом методе
рассчитывают, какой процент зданий от числа обследованных заселен крысами.
Несомненно, чем больше строений свободно от трызунов, тем ниже численность в
данном регионе и наоборот. Полученный показатель может быть использован для
изучения пространственного распределения грызунов на больших территориях на-
селенного пункта. Его недостаток заключается в том, что при низкой численности
наличие крыс трудно установить и здания часто оптибочно попадают в число сво-
бодных от пасюков, что снижает показатели общей численности на всей территории.
2 Относительный прямой учет численности паеюков.
|
а. Учет с помощью ловушек Геро (наживленных стандартной приманкой— короч
кой хлеба с подсолнечным маслом). Способ учета на ловупко-линиях, широко при-
меняемый зоологами в природе, в большинстве случаев непригоден В помещениях,
так как для размещения линии ловуптек не хватает места. Ловушки ставят, как пра.
вило, в определенном количестве на единицу площади. Показателем относительной
численности служит число пойманных зверьков на 1000 м? (Вишняков и Др. 1955)
или на 100 ловушко-суток (Леви, Судейкин, 1977). Метод имеет явные недостатки,
так как осторожные крысы часто избегают ловушек, часть их обладает неофобией; а
в ряде случаев, только задев приманку, зверек спускает ловугику, пугается и потом
начинает ее избегать.
В настоящее время отсутствует единый подход к учету крыс ловушкамя Геро.
В различных инструкциях рекомендуются не только разное количество ловушек на
единицу площади, но и различное количество дней проведения учета, а также и
различные принципы расчета средних показателей. Тем не менее этот метод широко
применяется и поэтому заслуживает внимания.
.
б. Число собранных трупов после проведения истребительных мероприятий.
Труты подечитывают и относят на единицу площади (объчно на 4000 м?). Недоетаток
метода заключается в том, что часть крыс, беря приманку, погибает в норах, под
полом, в перекрытиях и т. п., в результате чего их трупы нельзя учесть. Другая же
часть не берет приманку вообще. Этот метод позволяет сказать, какая была чиос-
ленность, но не какая она есть в настоящее время.
Экология
87
т
я
- 1900 +.
3. я)
21208 5;
о,
21000 =
ъх
Е
33 30.
]8
$3
<
ЗВ
7 600
53
ь
ЕВ зд
Я900ыы
р
- 203
0
Рис. 18. Сопоставление резуль-
татов учета численности крыс,
полученных разными метода- 5% 00
‚ми, в подвале жилого много-
‚
|
м
ы
„
”
х
и
д
а
ы
З
„
>
х
.
.
>
Р
о
й
=
|
—
р
ы
х
этажного дома (по материалам З
мх`
и‘
3
В.К.м
ы
т8|
`меи\
<
А — сравнение заслежелности Ч
ры
площадок и поедаемости зер- 9 8 г
05
новых приманок;
з|-
7
Е5
Б — сравнение заслеженности пло- $ 40 Е
еб
щадок и попадаемости в ло- х
8
вушки Геро.
а
Г
2к
1-— попадаемость в ловушки 20|
з
Геро,
.
.
<
|
З
2 — количество съеденной при- З
8
ле
<
ООО
О
ОО
О
О
5
ГН|Л|
3 — процент заслеженны: пло-
ГРЯЗИ
Е
щадок
.
„Дни наблюдений
в. Глазомерна
:
с
а
рая опека численности крыс. Пастюки в основном активны в сумер-
ках и ве но при очен ь высокой численности все зверьки не могут насытиться
ато Рромят ооо ит ают выходить из укрытий днем. Считается, что если
а
теля в светлое время суток находятся единичные особи
о
ое чел ы группы, К зверьков очень много (Вегпаг4, СоНл,
использован только как орпентировочный., 67). Копетно, этот мотод может быть
ероятно, наибол
:
ного орояти р а того фотка при учете, можно достигнуть, вводя коррекцию од-
В Абеотютный учет численности пасюков.
ен 902 т ые четы отличаются большой сложностью и трудоемкостью. По отно-
ени к серым Зрыба ; практически их не применяют. Но мы все же остановимся
двух ИЗ пих ко ры ‚могут ыть использованы при научных исследованиях
ен
орк ‚ Суть метода сводится к следующему: в течение 8-7
вы
стороженным ловушкам * с приманкой, после чего они
о
аивтоя и а 2—3 ночи удается выловить в помещениях если не
а
отных и составить себе представление об их численности
б. Оценка исленвоети, о учету 119
О
т зверьков с помощью выпуска меченых особей. Этот ме-
ав
И рее бухова и В. В. Раевского (1933), применявитих его для
учета полевокв омотах.
учета в помещениях серых крыс их отлавливают жи-
,
выпускают на Месте поимки. После этого все помещение *5б-
При спл Н!
ыл
эффек ивно применение
овых кацканов No 0 или
э
оном в ове более
у
м
"т:
риме
д
88
Глава третья
лавливают дуговыми канканами, а также уничтожают зверьков © помощью отравлен-
ной приманки,
Численность популяции (Х) определяют по формуле ХЫ—ас[®, где а— число по-
меченных крыс, с число трупов, 8 — число меченных крыс среди трунов.
В природных условиях, там, где позволяет растительный покров, учитывают число
открывптихся входных отверстий в норы, предварительно прикапывая их. Это дает
возможность при подсчете на 4 га площади получить внолне объективные данные по.
численности жилых нор. Если при этом часть нор обловить капканами или раскопать.
(выловив при этом вее население крыс), то можно, сделав соответствующий пере-
счет :°, получить данные об абсолютной численности крыс (Рыльников, Карасева,
1985; Метод. указания... 1984).
К сожалению, в большинстве работ, характеризующих обитание крыс в природ-
ных биотопах, приводится только процент их попадания в ловушки.
Несмотря на все несовершенство перечисленных вые методов учета численности.
крыс, мы, рассматривая распространение их 8 различных местах обитания, вынуж-
дены пользоваться теми данными, которые нриводят в литературе, так как некоторые
представления о численности пасюков они все же дают.
Экзоантропные крысы
Крысы, обитающие в природе круглый год («дикие»), по своей экология
несколько отличаются от синантропных форм. Они не размножаются в
холодное время года, ведут себя как мелкие хищники, несмотря на то,
что в их рацион входит значительная доия растительных кормов. Велед-
ствие более жестокой борьбы за существование «дикие» крысы не дожи-
вают до такого возраста, как синантроиные, и смена населепия у них
происходит скорее. Зверьки никогда пе бывают такими жирными, как
разъевитиесл и малоподвижные пасюки населепных пунктов (особенно на
животноводческих фермах).
В естественных биотопах крысы чаще всего встречаются в поймах
равнинных рек, стариц, речуитек и ручьев с медленным течением, а так-
же по берегам озер и прудов. Наиболее типичные стации этих грызу-
нов — заросли гидрофитов, луговой травянистой растительности, кустар-
ников, ив и др., а также берега с отдельно стоящими деревьями е мощ-
ной корпевой системой. Иногда крысы населяют берега водоемов, зарос-
ших тростником и осокой с включениями камыша, рогоза, хноща и пр.
(рис. 19). Предпочитают они места, заваленные сухими сучьями, сухой
травой и т. п., большое значение для пих имеют не только состав расти-
тельности и захламленность места, но и характер берега. Охотнее всего
зверьки поселяются на борегах с уклоном в 10--30° с крытыми уступа-
ми. избегая сильно заболоченных мест, водораздельных лесов (как хвой-
ных, так и лиственных) и гор.
Там, где пасюки обитают в природе, обычно большая их часть не свя-
зана с крысами населенных пунктов. Они могут жить на расстоянии 20—
25 км и более от ближайших построек, нс соверлая в них сезонных миг-
раций и не перемешиваясь, таким образом, с синантронными формами.
Но если на территории, где обитают дикие популяции крыс, хотя бы
временпо поселяется человек, то зверьки очень быстро концентрируются
15 Зная коэффициент заселения (К), т. е. число входных отверстий, приходящихся
на одну крысу, и среднее число входных отверстии на единицу площади (а),„
можно получить данные по численности на ней крыс (Х): Х-=а/К.
Экология
89
около его жилищ или стоянок (рыболовецких станов, сенокосных стой-
бищ, туристических палаток и т. п.). Поэтому в ирироде наибольшая
плотность населения диких крыс (их экологический оптимум) почти всег-
да приурочена к местам фупкционирования людей— зверьков привлека-
ют остатки еды и фекалии человека, но опи охотпо селятся ив заб 0-
шенных постройках (Хамаганов, 1985), часто устраивают норы и убожи.
ща под деревянными мостиками и переправами через речки и ЗИ
под настилами, брошенными около дороги предметами домашнего обихо
‚да (колесами, ведрами, тряпками и т. д.). Такое тяготение к человеку и
ко всему, что связано с его деятельностью, возможно, объясняется ем
что пасюк на значительной части ареала ранее был синантропным и по-
том уже вторично выселился в природу.
Снециальных исследований поселений экзоантроиных крыс проводи-
‚лось немного — часто авторы просто приводят даппые о распространении
и численпости пасюков, случайно попавших в капканы, выставленные для
уе воднной полевки (Яковлев, 1949; Кондрашкии, 1949; Тихвинская,
А). Целенаправлениое изучение серых крыс в естественных биотопах
воли А. С. Фетисов (1946), Г. И. Львов (1949), К. А. Татаринов (1954)
. Д. Бернштейн (1959), С. А. Хамаганов (1985), М. Я. Лаврова, Т. С. Ми-.
Рис. 19. Нижнее Приднестровье
— места обитания серой крысы. Фото И. Т. Русева
88
Глава третья
лавливают дуговыми канканами, а также уничтожают зверьков © помощью отравлен-
ной приманки,
Численность популяции (Х) определяют по формуле ХЫ—ас[®, где а— число по-
меченных крыс, с число трупов, 8 — число меченных крыс среди трунов.
В природных условиях, там, где позволяет растительный покров, учитывают число
открывптихся входных отверстий в норы, предварительно прикапывая их. Это дает
возможность при подсчете на 4 га площади получить внолне объективные данные по.
численности жилых нор. Если при этом часть нор обловить капканами или раскопать.
(выловив при этом вее население крыс), то можно, сделав соответствующий пере-
счет :°, получить данные об абсолютной численности крыс (Рыльников, Карасева,
1985; Метод. указания... 1984).
К сожалению, в большинстве работ, характеризующих обитание крыс в природ-
ных биотопах, приводится только процент их попадания в ловушки.
Несмотря на все несовершенство перечисленных вые методов учета численности.
крыс, мы, рассматривая распространение их 8 различных местах обитания, вынуж-
дены пользоваться теми данными, которые нриводят в литературе, так как некоторые
представления о численности пасюков они все же дают.
Экзоантропные крысы
Крысы, обитающие в природе круглый год («дикие»), по своей экология
несколько отличаются от синантропных форм. Они не размножаются в
холодное время года, ведут себя как мелкие хищники, несмотря на то,
что в их рацион входит значительная доия растительных кормов. Велед-
ствие более жестокой борьбы за существование «дикие» крысы не дожи-
вают до такого возраста, как синантроиные, и смена населепия у них
происходит скорее. Зверьки никогда пе бывают такими жирными, как
разъевитиесл и малоподвижные пасюки населепных пунктов (особенно на
животноводческих фермах).
В естественных биотопах крысы чаще всего встречаются в поймах
равнинных рек, стариц, речуитек и ручьев с медленным течением, а так-
же по берегам озер и прудов. Наиболее типичные стации этих грызу-
нов — заросли гидрофитов, луговой травянистой растительности, кустар-
ников, ив и др., а также берега с отдельно стоящими деревьями е мощ-
ной корпевой системой. Иногда крысы населяют берега водоемов, зарос-
ших тростником и осокой с включениями камыша, рогоза, хноща и пр.
(рис. 19). Предпочитают они места, заваленные сухими сучьями, сухой
травой и т. п., большое значение для пих имеют не только состав расти-
тельности и захламленность места, но и характер берега. Охотнее всего
зверьки поселяются на борегах с уклоном в 10--30° с крытыми уступа-
ми. избегая сильно заболоченных мест, водораздельных лесов (как хвой-
ных, так и лиственных) и гор.
Там, где пасюки обитают в природе, обычно большая их часть не свя-
зана с крысами населенных пунктов. Они могут жить на расстоянии 20—
25 км и более от ближайших построек, нс соверлая в них сезонных миг-
раций и не перемешиваясь, таким образом, с синантронными формами.
Но если на территории, где обитают дикие популяции крыс, хотя бы
временпо поселяется человек, то зверьки очень быстро концентрируются
15 Зная коэффициент заселения (К), т. е. число входных отверстий, приходящихся
на одну крысу, и среднее число входных отверстии на единицу площади (а),„
можно получить данные по численности на ней крыс (Х): Х-=а/К.
Экология
89
около его жилищ или стоянок (рыболовецких станов, сенокосных стой-
бищ, туристических палаток и т. п.). Поэтому в ирироде наибольшая
плотность населения диких крыс (их экологический оптимум) почти всег-
да приурочена к местам фупкционирования людей— зверьков привлека-
ют остатки еды и фекалии человека, но опи охотпо селятся ив заб 0-
шенных постройках (Хамаганов, 1985), часто устраивают норы и убожи.
ща под деревянными мостиками и переправами через речки и ЗИ
под настилами, брошенными около дороги предметами домашнего обихо
‚да (колесами, ведрами, тряпками и т. д.). Такое тяготение к человеку и
ко всему, что связано с его деятельностью, возможно, объясняется ем
что пасюк на значительной части ареала ранее был синантропным и по-
том уже вторично выселился в природу.
Снециальных исследований поселений экзоантроиных крыс проводи-
‚лось немного — часто авторы просто приводят даппые о распространении
и численпости пасюков, случайно попавших в капканы, выставленные для
уе воднной полевки (Яковлев, 1949; Кондрашкии, 1949; Тихвинская,
А). Целенаправлениое изучение серых крыс в естественных биотопах
воли А. С. Фетисов (1946), Г. И. Львов (1949), К. А. Татаринов (1954)
. Д. Бернштейн (1959), С. А. Хамаганов (1985), М. Я. Лаврова, Т. С. Ми-.
Рис. 19. Нижнее Приднестровье
— места обитания серой крысы. Фото И. Т. Русева
90
Глава третья
нина (1968), М. Н. Лозан (1971). В последние годы появились о
пия В. С. Суркова в, Т. Русева и В. И. Соловьева
,
. В. Читиаливили
идр. \! .
ь рилье следователи тоже немного ухеляли, олимания изучен
крыс в природе, (Зиафол её а[., 1977; Ното[Ка,
; атаг,
;
30).
.
ов } о очень важный раздел экологии серой крысы, потому
что пасюки, обитающие в природе, являются постоянным резорнуаром,
из которого могут быть заселены вновь построенные паселенные пу ты,
а также вояможен приток зверьков в старые поселки и города. Несм тря
на то что в природе серые крысы редко достигают высокой плотно ти
нассления, они широко распределены по территории и поэтому в цел В
многочисленны. Эти грызуны могут быть носителями многих инфекции,
опасных для человека, и формируют природные очаги болезней. Бороть я
же с ними в естественных условиях, где они рассредоточены, значител
но труднее, чем в населенных пунктах.
Природные биотопы
Рсем известные на сегодияшний день районы обитания оквоантронных
крыс в Европе характеризуются мягким климатом, относительно тотл Е ии
зимами и богатой кормовой базой. Заведомо круглогодичное обитание
крыс в природе пока зарегистрировано только в южной части Европы.
Рассмотрим последовательно с запада на носток все места обитан
РЫТ "сообщению Вольфа с соавт. (МОШЕ ей а1., 1980), серые крысы ши-
роко распространены в вотественных биотопах Придун ай ской ни .
менности, обитают по берегам рек Дуная и Марха. Встречаю гся в
дельте Дуная, где устраивают норы чате зсего под корнями т
(Бутабоп её а[., 1977), заселяют некоторые острова на Дунае
фаг,
198.
6
и плавней
Низовья р. Прут. До осушения заболоченных берегов
ея
реки основными местами обитания крыс были Качульские плавни пло
щадь 2500 га). Силавины под действием ветра свободно перемещались.
На пих местами были густые тростниковые крепи или заросли рогов,
рдестов, кубышки, стрелолиста и друтих болотных растений, а темпера
тура воздуха здесь достигала +35° С. Зимой на плавнях скап палось
много снега, под действием ветров ом уплотнялся, и крысы в п ках
пищи легко передвигались по его понерхности, Температура воздуха +
заросллх болотных растений и зимой была нытие, чем в открытых тах.
Численность крыс в этих условиях было трудно учесть. Но, по-вики-
мому, она была немалой, так как на 100 капканов в среднем то лось
12 крыс. На одной из сплавин при сплошном вылове на поел
было выловлено 32 пасюка (Лозан, Кучук, 1969; Лозан,
ии.
После осущения планней (1978 г.) численность крыс резко пои
лась, и в настоящее время она очень невелика (Лозан. устн. сообщ.)
Пойма р. Днестр. Здесь крысы обитали и до строител
Экология
94
Дубоссарской ГЭС и Мотилево-Подольской плотины (Татаринов, 1954; Со-
кур, 1960). В последние годы в связи со строительством этих соору-
жений большой размах получили работы, связанные с хозяйственным ос-
воснием Дпестровской поймы. Велед за завершением строительства ГЭС
последовало обвалование берегов и создание пойменных агроценозон об-
цей площадью 7000 га.
В настоящее время наиболее благоприятной зоной обитания пасюков
является центральная часть поймы, где гидрологический режим позволя-
ет им строить постоянные норы, не затопляемые водой.
В Нижнем Приднестровье серые крысы обитают по берегам
реки, в пойменном лесу и особенно многочисленны в местах, где летом
концентрируются отдыхающие люди (Русев, Соловьев, 1987).
Корченский полуостров. Б. Ролянский (1929) указывал,
что серые крысы очень многочислепиы в естественных биотопах полу-
острова, обитая между камнями в скалах. Особенно их много по берегу
Черного моря, где они в значительной степени питаются его выбросами.
В настоящее время в Крыму крысы селятся около пляжей.
Пойма р. Дон. О круглогодичном обитании здесь серых крыс со-
общал М. Г, Яковлев (1949): «Паесюкизжили в одних биотопах © водяной
крысой, но в отличие от нее норы серых крыс всегда располагались на
возвышенных местах, редко заливаемых водой». Больше всего отлавлива-
ли пасюков па рыболовных тонях (39% попадания в капканы), на в0оз-
вышенных берегах Дона попадаемость была в среднем равна 14,3%, в за-
рослях тростника— 7,3% и по берегам озер— 118%. По берегам дель-
ты р. Дона, ериков и озер в пастолщее время серые крысы многочис-
ленны и обитают там круглогодично (Кондратенко и др., 1986).
Приазовские плавни. В 50-х годах нашего столетия поселе-
ния крыс были зарегистрированы на протяжепии от г. Темрюка до
г. Приморско-Ахтарска. Здесь в некоторых местах зверек был многочис-
лен, особенно в глухих обширных тростниковых зарослях, куда выпусти-
ли путрию, которую систематически подкармлизали, что, конечно, обес-
нечивало полноценное питание и крысам.
М. П. Павлов и И. Б. Кирис (1956) сообщают, что серая крыса была
наиболее распространенным видом из всех млекопитающих обитателей
приазовских плавней— она буквально наводняла лиманы и ерики. Посе-
ления ее встречались круглогодично, особенно густо были населены узкие
пространства земли (иерешейки) между водоемами, где удобно устраивать
норы.
В настоящее время нутриевая ферма там не работает и крыс немного.
Зоолог Г. И. Мажникова сообщила нам, что попадаемость крыс в каика-
ны не превышает 1—2%.
Наши наблюдения в августе 1975 г. в приазовских плавнях между го-
родами Темрюком и Приморско-Ахтарском показали, что численпость се-
рых крыс в это зремя была невелика — 0,1—0.5 крысы на 100 погонных
метров. Почва здесь сильпо заболочена, и зверьки могут селиться только
вдоль сухих гринок (лиманных гряд), зозвышающихся над болотами.
К таким гривам обычно приурочены проселочные дороги, и, следователь-
90
Глава третья
нина (1968), М. Н. Лозан (1971). В последние годы появились о
пия В. С. Суркова в, Т. Русева и В. И. Соловьева
,
. В. Читиаливили
идр. \! .
ь рилье следователи тоже немного ухеляли, олимания изучен
крыс в природе, (Зиафол её а[., 1977; Ното[Ка,
; атаг,
;
30).
.
ов } о очень важный раздел экологии серой крысы, потому
что пасюки, обитающие в природе, являются постоянным резорнуаром,
из которого могут быть заселены вновь построенные паселенные пу ты,
а также вояможен приток зверьков в старые поселки и города. Несм тря
на то что в природе серые крысы редко достигают высокой плотно ти
нассления, они широко распределены по территории и поэтому в цел В
многочисленны. Эти грызуны могут быть носителями многих инфекции,
опасных для человека, и формируют природные очаги болезней. Бороть я
же с ними в естественных условиях, где они рассредоточены, значител
но труднее, чем в населенных пунктах.
Природные биотопы
Рсем известные на сегодияшний день районы обитания оквоантронных
крыс в Европе характеризуются мягким климатом, относительно тотл Е ии
зимами и богатой кормовой базой. Заведомо круглогодичное обитание
крыс в природе пока зарегистрировано только в южной части Европы.
Рассмотрим последовательно с запада на носток все места обитан
РЫТ "сообщению Вольфа с соавт. (МОШЕ ей а1., 1980), серые крысы ши-
роко распространены в вотественных биотопах Придун ай ской ни .
менности, обитают по берегам рек Дуная и Марха. Встречаю гся в
дельте Дуная, где устраивают норы чате зсего под корнями т
(Бутабоп её а[., 1977), заселяют некоторые острова на Дунае
фаг,
198.
6
и плавней
Низовья р. Прут. До осушения заболоченных берегов
ея
реки основными местами обитания крыс были Качульские плавни пло
щадь 2500 га). Силавины под действием ветра свободно перемещались.
На пих местами были густые тростниковые крепи или заросли рогов,
рдестов, кубышки, стрелолиста и друтих болотных растений, а темпера
тура воздуха здесь достигала +35° С. Зимой на плавнях скап палось
много снега, под действием ветров ом уплотнялся, и крысы в п ках
пищи легко передвигались по его понерхности, Температура воздуха +
заросллх болотных растений и зимой была нытие, чем в открытых тах.
Численность крыс в этих условиях было трудно учесть. Но, по-вики-
мому, она была немалой, так как на 100 капканов в среднем то лось
12 крыс. На одной из сплавин при сплошном вылове на поел
было выловлено 32 пасюка (Лозан, Кучук, 1969; Лозан,
ии.
После осущения планней (1978 г.) численность крыс резко пои
лась, и в настоящее время она очень невелика (Лозан. устн. сообщ.)
Пойма р. Днестр. Здесь крысы обитали и до строител
Экология
94
Дубоссарской ГЭС и Мотилево-Подольской плотины (Татаринов, 1954; Со-
кур, 1960). В последние годы в связи со строительством этих соору-
жений большой размах получили работы, связанные с хозяйственным ос-
воснием Дпестровской поймы. Велед за завершением строительства ГЭС
последовало обвалование берегов и создание пойменных агроценозон об-
цей площадью 7000 га.
В настоящее время наиболее благоприятной зоной обитания пасюков
является центральная часть поймы, где гидрологический режим позволя-
ет им строить постоянные норы, не затопляемые водой.
В Нижнем Приднестровье серые крысы обитают по берегам
реки, в пойменном лесу и особенно многочисленны в местах, где летом
концентрируются отдыхающие люди (Русев, Соловьев, 1987).
Корченский полуостров. Б. Ролянский (1929) указывал,
что серые крысы очень многочислепиы в естественных биотопах полу-
острова, обитая между камнями в скалах. Особенно их много по берегу
Черного моря, где они в значительной степени питаются его выбросами.
В настоящее время в Крыму крысы селятся около пляжей.
Пойма р. Дон. О круглогодичном обитании здесь серых крыс со-
общал М. Г, Яковлев (1949): «Паесюкизжили в одних биотопах © водяной
крысой, но в отличие от нее норы серых крыс всегда располагались на
возвышенных местах, редко заливаемых водой». Больше всего отлавлива-
ли пасюков па рыболовных тонях (39% попадания в капканы), на в0оз-
вышенных берегах Дона попадаемость была в среднем равна 14,3%, в за-
рослях тростника— 7,3% и по берегам озер— 118%. По берегам дель-
ты р. Дона, ериков и озер в пастолщее время серые крысы многочис-
ленны и обитают там круглогодично (Кондратенко и др., 1986).
Приазовские плавни. В 50-х годах нашего столетия поселе-
ния крыс были зарегистрированы на протяжепии от г. Темрюка до
г. Приморско-Ахтарска. Здесь в некоторых местах зверек был многочис-
лен, особенно в глухих обширных тростниковых зарослях, куда выпусти-
ли путрию, которую систематически подкармлизали, что, конечно, обес-
нечивало полноценное питание и крысам.
М. П. Павлов и И. Б. Кирис (1956) сообщают, что серая крыса была
наиболее распространенным видом из всех млекопитающих обитателей
приазовских плавней— она буквально наводняла лиманы и ерики. Посе-
ления ее встречались круглогодично, особенно густо были населены узкие
пространства земли (иерешейки) между водоемами, где удобно устраивать
норы.
В настоящее время нутриевая ферма там не работает и крыс немного.
Зоолог Г. И. Мажникова сообщила нам, что попадаемость крыс в каика-
ны не превышает 1—2%.
Наши наблюдения в августе 1975 г. в приазовских плавнях между го-
родами Темрюком и Приморско-Ахтарском показали, что численпость се-
рых крыс в это зремя была невелика — 0,1—0.5 крысы на 100 погонных
метров. Почва здесь сильпо заболочена, и зверьки могут селиться только
вдоль сухих гринок (лиманных гряд), зозвышающихся над болотами.
К таким гривам обычно приурочены проселочные дороги, и, следователь-
92
Глава третья
но, зверьки имеют возможность ие только строить норы в сухих пезатои-
ляемых мостах, но и пользоваться рассыпанным при перевозке по дороге
верном.
В таких условиях крысы обычно обитают пе кучно, Чаще живут самка
с дотенылнами и еще две-три крысы, а следующее подобное поселение мо-
жет быть расположено вдоль гривы на расстоянии 500—600 м и более.
М. Я. Лаврова и Т. С. Минина (1968) сообщают о высокой численности
крыс в низовьях Кубани, отмечая наибольшую концептрацию их пор на
незатопляемых грядах. Относительно более высокая числелность крыс в
районе исследований М. Ч. Лавровой и Т. С. Мининой, вероятно, объяс-
няется близостью ностроек животноводческих ферм станицы Гризинекой.
В приазовских плавиях поселения крыс распределены мозаично н
плотность их значительно выше близ населенных пунктов, особенно жи-
вотноводческих ферм.
В Колхидской низменности типичными местами обитания
крыс в естественных биотопах являются территории, которые з прежние
времена были сплошь заболочены и покрыты широколистненными густы-
ми лесами, состоящими из дуба, граба, каштапа, ольхи и других деревь-
ев, густо перевитых лиапами (Сосновский, 1930). В настоящее время
большая часть заболоченной территории осушена, леса сведены и на их
местах вовделывают винограл, кукурузу и другие культуры. Но там и
сейчас сохранилась система озер (Бебисири, Палеостоми, Партоцоми)
с заболоченными берегами, на которых есть участки типичных Для
Колхиды очень густых, сырых, труднопроходимых широколиственных ле-
сов, где обитают крысы. Есть они и в полинах рек Риони и Печоры.
Вероятно, серая крыса в природных биотопах обитает на всем протя-
жений черноморского побережья от Гагр до Батуми, но конкретные опи-
сания се биотопического распределения мы имеем только в прибрежных
лесах вышеназванных озер и рек (УИитков, 1944; Львов, 1949; Шидлов-
ский, 1947; Верещагин, 1959; Читиашвили и др., 1987).
Помимо серой крысы, здесь весьма многочисленны черные крысы, л6с-
ные, а местами полевые и домовые мыши. С 1955 но 1956 г. в районе
оз. Бебисири при отлове плаликами (добыто 629 зверьков перечисленных
видов) серая крыса составляла 45,1% и преобладала (80,3%) ино самым
берегам озера (Бернштейн, 1959). В прибрежных зарослях она состав-
ляла 29,3%, а в лесу на ее долю приходилось всего 4—8, где она усту-
пала место черной крысе.
В настоящее время близ 0з. Бебисири, несмотря на увелизивающуюся
мелиорацию, численность зверьков мало изменилась: по нашиям наблюде-
ниям, в 4984 г. их на его берегах и сейчас немало— в средпем на
100 капкано-ночей, выставленных здесь, приходится 10—12, особей.
Па побережье оз. Палеостоми (близ города Поти) результаты осутие-
ния на жизни зверьков сказались больше. Там работают машины, рою-
щие осушительные каналы, лес во многих местах сильно вырублен,
и численность крыс здесь колеблется от 5 до 15% попадания. Поселения
пасюков приурочены в основном к мостам, гдс ловят рыбу, и к одиноч-
ным домам (например. около домика егеря заповедника).
Экология
93
Талышско-Ленкоранскал низменность. Пасюки с дав
них пор здесь населяют заболоченные леса, берега оз. Морцо и рек ы -
о
(Житков, 1944; Кузякии, 1951; Верещагин, 1959; ке
О они ы Соле мы не располагаем более подробными и
ое
.
этом интересном месте обитания серых
в Кура А ра ксин ок ая провинция. В основном пасюк отмечен
орых островах Каспия (Эйгелие, 1980). По мере ожегодног
уменьшения акватории, связанного с отступлением Каспийского мо я,
образуются обсохшие бугры, которые заселяются грызунами разных ны
дов, в том числе серой крысой (Шилов и др., 1986).
р
в
пе
"крыс р я пом ров ин ЦИЯ — упоминаются поселения экзо-
Не).
менных лесах рек Самур и Суле (Лавровский,
Терско-Кумская пизменность. Серая крыса встречается в
зарослях тростника в устье рек Кумы и Подкумки, в дельте р. Терека
а также по берегам рек Сулак, Талец, Злавка, Кизляр (Прокофьева,
Иванов, 1974; Сорокина и др., 1986).
Предкавказье и северные склоны Главного Кав-
каза. Весть отдельные указания на обитание серых крыс в приро
а именно по берегам р. Западного Маныча (Кайзер, 1941) речек Пур
Дур и Чикола (Нрокофьева, Иванов, 1974; Сорокина и др `1986) ок.
рестностях г. Нальчика (Темботов, 1972). Интересен случай обитания
тих грызунов по берегам прудов и каналов, где они часто заселяют норы
водяной полевки у пос. Редант (в 9 км от г. Орджоникидзе). Это ве
ние было описано Д. В. Красовскям еще в 1929 г. затем Л Б емо
в 1936 г. и существует по настоящее время (Варшавский и пр 1987)
ПВолго-Ахтубинская пойма. В природных биотопах крыса
широко распространена, но, по-видимому, прежде нигде пе была много-
численна. Из работы Г. А. Кондрашкина (1949) мы знаем, что в дель
Волги обитало небольшое количество зверьков (1—10 особей па и м
береговой линии), которые были равпомерно расселены по всей те ито.
рии дельты. Автор пишет, что отдельные носеления их иногда тоя
друг от друга на 5-10 км, а распространение связано главным образом
ми берсгами, в которых они роют норы, часто размещая их меж-
ду и под корнями деревьев, особенно под старыми одиночными ветлами
Близость груиптовых вод исключает возможность выкапывания очень гл и
боких нор. В период паводка прибывающая вода вытеспяет я аоков 6
нор и опи выселяются на деревья. В настоящее время в результате ант-
ропогенного преобразования пойма дельты существенно изменилась. За-
регулирование речного стока привело к уменьшению поступления талых
ИВ на территорию поймы. Площадь затопления сократилась на 25—
55%, длительность паводка уменьшилась в среднем на 20 дней. Все эти
преобразования не могли не сказаться на распределении и образе жизни
серых крыс. Но сообщению О. С. Опаряной (1987), для них (в районе
Краспого Яра, дер. Сетаевки) оптимальными условиями являются высо-
кие берега ериков, проток, небольших речек, как правило, заросших
92
Глава третья
но, зверьки имеют возможность ие только строить норы в сухих пезатои-
ляемых мостах, но и пользоваться рассыпанным при перевозке по дороге
верном.
В таких условиях крысы обычно обитают пе кучно, Чаще живут самка
с дотенылнами и еще две-три крысы, а следующее подобное поселение мо-
жет быть расположено вдоль гривы на расстоянии 500—600 м и более.
М. Я. Лаврова и Т. С. Минина (1968) сообщают о высокой численности
крыс в низовьях Кубани, отмечая наибольшую концептрацию их пор на
незатопляемых грядах. Относительно более высокая числелность крыс в
районе исследований М. Ч. Лавровой и Т. С. Мининой, вероятно, объяс-
няется близостью ностроек животноводческих ферм станицы Гризинекой.
В приазовских плавиях поселения крыс распределены мозаично н
плотность их значительно выше близ населенных пунктов, особенно жи-
вотноводческих ферм.
В Колхидской низменности типичными местами обитания
крыс в естественных биотопах являются территории, которые з прежние
времена были сплошь заболочены и покрыты широколистненными густы-
ми лесами, состоящими из дуба, граба, каштапа, ольхи и других деревь-
ев, густо перевитых лиапами (Сосновский, 1930). В настоящее время
большая часть заболоченной территории осушена, леса сведены и на их
местах вовделывают винограл, кукурузу и другие культуры. Но там и
сейчас сохранилась система озер (Бебисири, Палеостоми, Партоцоми)
с заболоченными берегами, на которых есть участки типичных Для
Колхиды очень густых, сырых, труднопроходимых широколиственных ле-
сов, где обитают крысы. Есть они и в полинах рек Риони и Печоры.
Вероятно, серая крыса в природных биотопах обитает на всем протя-
жений черноморского побережья от Гагр до Батуми, но конкретные опи-
сания се биотопического распределения мы имеем только в прибрежных
лесах вышеназванных озер и рек (УИитков, 1944; Львов, 1949; Шидлов-
ский, 1947; Верещагин, 1959; Читиашвили и др., 1987).
Помимо серой крысы, здесь весьма многочисленны черные крысы, л6с-
ные, а местами полевые и домовые мыши. С 1955 но 1956 г. в районе
оз. Бебисири при отлове плаликами (добыто 629 зверьков перечисленных
видов) серая крыса составляла 45,1% и преобладала (80,3%) ино самым
берегам озера (Бернштейн, 1959). В прибрежных зарослях она состав-
ляла 29,3%, а в лесу на ее долю приходилось всего 4—8, где она усту-
пала место черной крысе.
В настоящее время близ 0з. Бебисири, несмотря на увелизивающуюся
мелиорацию, численность зверьков мало изменилась: по нашиям наблюде-
ниям, в 4984 г. их на его берегах и сейчас немало— в средпем на
100 капкано-ночей, выставленных здесь, приходится 10—12, особей.
Па побережье оз. Палеостоми (близ города Поти) результаты осутие-
ния на жизни зверьков сказались больше. Там работают машины, рою-
щие осушительные каналы, лес во многих местах сильно вырублен,
и численность крыс здесь колеблется от 5 до 15% попадания. Поселения
пасюков приурочены в основном к мостам, гдс ловят рыбу, и к одиноч-
ным домам (например. около домика егеря заповедника).
Экология
93
Талышско-Ленкоранскал низменность. Пасюки с дав
них пор здесь населяют заболоченные леса, берега оз. Морцо и рек ы -
о
(Житков, 1944; Кузякии, 1951; Верещагин, 1959; ке
О они ы Соле мы не располагаем более подробными и
ое
.
этом интересном месте обитания серых
в Кура А ра ксин ок ая провинция. В основном пасюк отмечен
орых островах Каспия (Эйгелие, 1980). По мере ожегодног
уменьшения акватории, связанного с отступлением Каспийского мо я,
образуются обсохшие бугры, которые заселяются грызунами разных ны
дов, в том числе серой крысой (Шилов и др., 1986).
р
в
пе
"крыс р я пом ров ин ЦИЯ — упоминаются поселения экзо-
Не).
менных лесах рек Самур и Суле (Лавровский,
Терско-Кумская пизменность. Серая крыса встречается в
зарослях тростника в устье рек Кумы и Подкумки, в дельте р. Терека
а также по берегам рек Сулак, Талец, Злавка, Кизляр (Прокофьева,
Иванов, 1974; Сорокина и др., 1986).
Предкавказье и северные склоны Главного Кав-
каза. Весть отдельные указания на обитание серых крыс в приро
а именно по берегам р. Западного Маныча (Кайзер, 1941) речек Пур
Дур и Чикола (Нрокофьева, Иванов, 1974; Сорокина и др `1986) ок.
рестностях г. Нальчика (Темботов, 1972). Интересен случай обитания
тих грызунов по берегам прудов и каналов, где они часто заселяют норы
водяной полевки у пос. Редант (в 9 км от г. Орджоникидзе). Это ве
ние было описано Д. В. Красовскям еще в 1929 г. затем Л Б емо
в 1936 г. и существует по настоящее время (Варшавский и пр 1987)
ПВолго-Ахтубинская пойма. В природных биотопах крыса
широко распространена, но, по-видимому, прежде нигде пе была много-
численна. Из работы Г. А. Кондрашкина (1949) мы знаем, что в дель
Волги обитало небольшое количество зверьков (1—10 особей па и м
береговой линии), которые были равпомерно расселены по всей те ито.
рии дельты. Автор пишет, что отдельные носеления их иногда тоя
друг от друга на 5-10 км, а распространение связано главным образом
ми берсгами, в которых они роют норы, часто размещая их меж-
ду и под корнями деревьев, особенно под старыми одиночными ветлами
Близость груиптовых вод исключает возможность выкапывания очень гл и
боких нор. В период паводка прибывающая вода вытеспяет я аоков 6
нор и опи выселяются на деревья. В настоящее время в результате ант-
ропогенного преобразования пойма дельты существенно изменилась. За-
регулирование речного стока привело к уменьшению поступления талых
ИВ на территорию поймы. Площадь затопления сократилась на 25—
55%, длительность паводка уменьшилась в среднем на 20 дней. Все эти
преобразования не могли не сказаться на распределении и образе жизни
серых крыс. Но сообщению О. С. Опаряной (1987), для них (в районе
Краспого Яра, дер. Сетаевки) оптимальными условиями являются высо-
кие берега ериков, проток, небольших речек, как правило, заросших
94
Глава третья
о-
бурьяном, кустарником или отдельными деревьями. По кромке воды про-
из астают тростник, рогов и осока. В этих местах крысы ны пот бога
ую кормовую базу и места для устройства убежищ. Т ПОЩВДЬ отах т
у езультате антр
й,К
ло сказано, сократилась в р
ий, кок уже о
Й
начительно увеличилась за
Й
ь орошаемой земли 3
г
во
зверьков в пойме. Валы
особствовало расселению звер
последние годы, что сп
ойме. Валы
'ростником и другими р
алов быстро зарасталот трос
Витя,
тих грызунов в природе
у
ми для обитания э
р
ий и становятся пригодны
у
роде
круглый тод. Численность их подвержена сезонным колебаниям и дос
1987).
максимума осенью (Опарина,
р
о более Северной части Ахтубы (р-н Лепинска) ыы, по видимому,
ото
-
50-х годах они обитали по берегам пр
менее многочисленны. В о -х
т
о
седству © водяной полевкой, но после того как оттуда, и ото
и исчезли, и в
г.В.В.
.
зодческие фермы, зверьк
,
сообщ ) их поселения в этих местах не мог обнаружить.
ванных
европейской части СССР пасюки в естсет
ве
отита
й
за исключением тех
биотопах нигде не достигают высокой численности, за
мест, где явно сказывается влияние деятельности человека. реал паибо-
ИЕ)
ст
ы в азиатской ча
Экзоантропные поселения крыс
‚зат
паибо-
лее типичны для Амуро-Байкальского участка. А. С. Ксении В я
Й ибир
-
о для Западнои
лин (1986) указывают, чт
3
чи
оселений этого грызуна в природе.
ствие круглогодичных п
о
ащии крыс
пия (с мая по октябрь) Ч
только сезонные перемеще
крыс
из населенных пунктов в природные и п обратно. То же вамо
азахстану (Козлов, Козлов,
.
ри б К кал ье и Забайкалье. Рыло показано обитание серых
зверь-
крыс по рекам Иро, Кыра, Онопу, Борзеи Ангаре. Сообщалось, что зверь.
ки живут здесь не только летом, но # хорошо переносят з у аибонее
типичны для них ивово-тополево-черемуховые и осоково трости ов
заросли во берегам рек старик и озер (Скалон, 1933; отисов, 15,
1946; Кузякин, 1951; Хамаханов, 1985 и др.) В пастоящее эре п по ро
общению зоологов противочумного отделения Б. о е мал т
това, поселения крыс обнаружены только по р. Онону, гд
Ы
в таскной зоне Восточной Сибири «диких, поселения серой
крысы отсутствуют (Романова, 1986; Олькова и др.., д : 7. я ем Восто-
ых биотопах на
Об обитании крыс в естественн
,
м
т. по: Хамаганов,
1861) и Радде (1862) (цит.
ке сообщали еще Маак (
мы
сы речных долин края
1985}. Богатые кормовые ресур
зы.
:
астительным кор
— их привлекает нарнду Ср
живанию этих грызунов
тм
оллюсков, рыбы, лягушек
Ы
обилие двустворчатых м
‚
а
ки могут находить кру
Рыбу в этих местах пасют
МОЙ она
у
ах, часть ее гибн
от русла реки водоемах,
е
остается в отшиурованных
С
‚ре.
зультате промерзания проток и стаповится легкой добычей для нас
осов, 1945
не Ь пет подо льдом и глубокие снежные заносы, поаволяют
вверьку спасаться от морозов и поддерживать активный обра
Экология
95
в зимний период (Хамагапов, 1983, 1985; Хамагапов и др., 1986). Наи-
большая численность крыс отмечена но берегам протоков и стариц.
Вероятно, в азиатской части ареала в отличие от европейской крысы
исходно жили в открытых биотопах и, оттуда вселяясь в постройки,
становились сннантропами (Кузякин, 1951; Хамаганов и др., 1986).
В. В. Кучерук (1946) указывает, что в Монголия в условиях сурового
континентального климата в районе Халхин-Гола, когда ранее необитае-
мал территория стала заселяться людьми, крысы быстро появились в но-
вых населенных пунктах (куда они не могли понаеть с транспортом),
т. е., по-видимому, они переселились туда из природных биотопов.
Особый интерес представляют места обитания крыс на различных
островах, например на о-ве Сахалине и па южных Курильских островах
(Кунашир, Шикотан), где они имеют великолепные условия существо-
вания.
На территории островного края насчитывается свыше 6500 рек и
ручьев (протяженностью около 105000 км) и 17120 озер с общей пло-
щадью водной поверхности 1100 км” (Бродский, Ножкина, 1967). Берс-
га почти всех водоемов, как правило, заселены крысами, причем не толь-
ко в низинных участках, но и в горах до 500—700 м над ур. моря. Не-
редко зверьки обитают вдоль мелких ручьев.
Весьма благоприятные условия создаются в густых травяных зарос-
лях: дудника, сахалинской гречихи и др., которые под воздействием теп-
лаи обильной влаги могут подниматься в два и более человеческих роста
и образовывать труднопроходимые заросли. Крысы находят здесь надеж-
ные укрытия и охотно питаются семенами и сочными корнями этих ра-
стений.
Болыное значение в жизни пасюков имеет литораль. Во время штор-
мов на берег выбрасываются крабы, моллюски, морские звезды и рыба.
Этим питаются многие животные, и в том числе серые крысы.
На Кунашире, Шикотане и Итуруне численность крыс выше, чем на
Сахалине. На о-ве Итуруп из грызупов обитает только серая крыса, ко-
торую японцы специально разводили здесь для подкармливания ЛИСИЦ.
По сообщению В. С. Суркова (1986а), в августе 1962 г. на этом острове
средняя попадаемость крыс в ловушки составляла 83%. Такая высокая
численность зверька объясняется чрезвычайно благоприятными для него
условиями обитания, а также тем, что был допущен перепромысел лисиц.
Л. М. Беньковский (1986) пишет, что в последние годы на о-ве
Итуруп числеппость серых крыс уже пе так велика. В районе населенных
пунктов Буревестник — Касатка — Куйбышевск и Курильск на 100 ловуш-
ко-суток в среднем ловится 5—7 крыс, и только около Лесозаводека (по
берегам рек и ручьев) процент попадания зверьков часто превышает 10.
С помощью человека, который транспортирозал крыс на судах различ-
ного типа, крыса заселила мнотие другие океанические острова.
На южных границах полярного бассейна пастоки расселяются в есте-
ственных биотопах островов (Кучерук, Лапшов, 1987).
На остров Гельголанд пасюк проник в 1730 г., в 1890 г. он был
уже мпогочислеп в постройках, а поздпее заселил открытые биотопы ост-
94
Глава третья
о-
бурьяном, кустарником или отдельными деревьями. По кромке воды про-
из астают тростник, рогов и осока. В этих местах крысы ны пот бога
ую кормовую базу и места для устройства убежищ. Т ПОЩВДЬ отах т
у езультате антр
й,К
ло сказано, сократилась в р
ий, кок уже о
Й
начительно увеличилась за
Й
ь орошаемой земли 3
г
во
зверьков в пойме. Валы
особствовало расселению звер
последние годы, что сп
ойме. Валы
'ростником и другими р
алов быстро зарасталот трос
Витя,
тих грызунов в природе
у
ми для обитания э
р
ий и становятся пригодны
у
роде
круглый тод. Численность их подвержена сезонным колебаниям и дос
1987).
максимума осенью (Опарина,
р
о более Северной части Ахтубы (р-н Лепинска) ыы, по видимому,
ото
-
50-х годах они обитали по берегам пр
менее многочисленны. В о -х
т
о
седству © водяной полевкой, но после того как оттуда, и ото
и исчезли, и в
г.В.В.
.
зодческие фермы, зверьк
,
сообщ ) их поселения в этих местах не мог обнаружить.
ванных
европейской части СССР пасюки в естсет
ве
отита
й
за исключением тех
биотопах нигде не достигают высокой численности, за
мест, где явно сказывается влияние деятельности человека. реал паибо-
ИЕ)
ст
ы в азиатской ча
Экзоантропные поселения крыс
‚зат
паибо-
лее типичны для Амуро-Байкальского участка. А. С. Ксении В я
Й ибир
-
о для Западнои
лин (1986) указывают, чт
3
чи
оселений этого грызуна в природе.
ствие круглогодичных п
о
ащии крыс
пия (с мая по октябрь) Ч
только сезонные перемеще
крыс
из населенных пунктов в природные и п обратно. То же вамо
азахстану (Козлов, Козлов,
.
ри б К кал ье и Забайкалье. Рыло показано обитание серых
зверь-
крыс по рекам Иро, Кыра, Онопу, Борзеи Ангаре. Сообщалось, что зверь.
ки живут здесь не только летом, но # хорошо переносят з у аибонее
типичны для них ивово-тополево-черемуховые и осоково трости ов
заросли во берегам рек старик и озер (Скалон, 1933; отисов, 15,
1946; Кузякин, 1951; Хамаханов, 1985 и др.) В пастоящее эре п по ро
общению зоологов противочумного отделения Б. о е мал т
това, поселения крыс обнаружены только по р. Онону, гд
Ы
в таскной зоне Восточной Сибири «диких, поселения серой
крысы отсутствуют (Романова, 1986; Олькова и др.., д : 7. я ем Восто-
ых биотопах на
Об обитании крыс в естественн
,
м
т. по: Хамаганов,
1861) и Радде (1862) (цит.
ке сообщали еще Маак (
мы
сы речных долин края
1985}. Богатые кормовые ресур
зы.
:
астительным кор
— их привлекает нарнду Ср
живанию этих грызунов
тм
оллюсков, рыбы, лягушек
Ы
обилие двустворчатых м
‚
а
ки могут находить кру
Рыбу в этих местах пасют
МОЙ она
у
ах, часть ее гибн
от русла реки водоемах,
е
остается в отшиурованных
С
‚ре.
зультате промерзания проток и стаповится легкой добычей для нас
осов, 1945
не Ь пет подо льдом и глубокие снежные заносы, поаволяют
вверьку спасаться от морозов и поддерживать активный обра
Экология
95
в зимний период (Хамагапов, 1983, 1985; Хамагапов и др., 1986). Наи-
большая численность крыс отмечена но берегам протоков и стариц.
Вероятно, в азиатской части ареала в отличие от европейской крысы
исходно жили в открытых биотопах и, оттуда вселяясь в постройки,
становились сннантропами (Кузякин, 1951; Хамаганов и др., 1986).
В. В. Кучерук (1946) указывает, что в Монголия в условиях сурового
континентального климата в районе Халхин-Гола, когда ранее необитае-
мал территория стала заселяться людьми, крысы быстро появились в но-
вых населенных пунктах (куда они не могли понаеть с транспортом),
т. е., по-видимому, они переселились туда из природных биотопов.
Особый интерес представляют места обитания крыс на различных
островах, например на о-ве Сахалине и па южных Курильских островах
(Кунашир, Шикотан), где они имеют великолепные условия существо-
вания.
На территории островного края насчитывается свыше 6500 рек и
ручьев (протяженностью около 105000 км) и 17120 озер с общей пло-
щадью водной поверхности 1100 км” (Бродский, Ножкина, 1967). Берс-
га почти всех водоемов, как правило, заселены крысами, причем не толь-
ко в низинных участках, но и в горах до 500—700 м над ур. моря. Не-
редко зверьки обитают вдоль мелких ручьев.
Весьма благоприятные условия создаются в густых травяных зарос-
лях: дудника, сахалинской гречихи и др., которые под воздействием теп-
лаи обильной влаги могут подниматься в два и более человеческих роста
и образовывать труднопроходимые заросли. Крысы находят здесь надеж-
ные укрытия и охотно питаются семенами и сочными корнями этих ра-
стений.
Болыное значение в жизни пасюков имеет литораль. Во время штор-
мов на берег выбрасываются крабы, моллюски, морские звезды и рыба.
Этим питаются многие животные, и в том числе серые крысы.
На Кунашире, Шикотане и Итуруне численность крыс выше, чем на
Сахалине. На о-ве Итуруп из грызупов обитает только серая крыса, ко-
торую японцы специально разводили здесь для подкармливания ЛИСИЦ.
По сообщению В. С. Суркова (1986а), в августе 1962 г. на этом острове
средняя попадаемость крыс в ловушки составляла 83%. Такая высокая
численность зверька объясняется чрезвычайно благоприятными для него
условиями обитания, а также тем, что был допущен перепромысел лисиц.
Л. М. Беньковский (1986) пишет, что в последние годы на о-ве
Итуруп числеппость серых крыс уже пе так велика. В районе населенных
пунктов Буревестник — Касатка — Куйбышевск и Курильск на 100 ловуш-
ко-суток в среднем ловится 5—7 крыс, и только около Лесозаводека (по
берегам рек и ручьев) процент попадания зверьков часто превышает 10.
С помощью человека, который транспортирозал крыс на судах различ-
ного типа, крыса заселила мнотие другие океанические острова.
На южных границах полярного бассейна пастоки расселяются в есте-
ственных биотопах островов (Кучерук, Лапшов, 1987).
На остров Гельголанд пасюк проник в 1730 г., в 1890 г. он был
уже мпогочислеп в постройках, а поздпее заселил открытые биотопы ост-
96
Глава третья
рова, в основном концентрируясь на побережье. Эта часть популяции
крыс на зяму в помещения не переселялась, основной корм ес составляли
выбросы моря, но охотно поедались также птицы и их яйца. К 1944 г.
тнездивитиеся з дюнах популяции желтой трясогузки были полностью
истреблены крысами (Уапк, 1958).
Серые крысы сумели адаптироваться к суровым климатическим усло-
зиям многих антарктических островов (Кучерук, Лапшон, 1987), в оенов-
ном они закрепились и выжили там, где обитают колониальные птицы
и, следовательно, пет недостатка
в питце (яйца, птенцы, взрослые особи).
На один из антарктических остронов — Ю жная Георгия —кры-
сы были случайно завезены в 1800 г. и расселились там в тустых прибреж-
пых зарослях, на кочковатых лугах, где могли строить норы. В начале
ХХ в. здесь промышляли китов и яверьки питались в основном их мясом.
Впоследствии они перешли на питание главным образом за счет птиц в
колопиях буревостников. Мелкие воробъиные птицы, в частности антарк-
тические коньки, не гнездятся там, тде жхивут крысы. Буревестники хотя
и страдают от крыс, но могут © ними совместно существовать (Рус,
Воппет, 1980).
Биотопы, созданные человеком, или места обитания,
на которые человек оказал большое влияние
Места постоянного сбрасывания пищевых отходов,
создающих кормовую базу. Самый простой пример биотопа, создан-
ного руками челозека,—это свалки, куда обычно выбрасывают не только
остатки пищи, но также различный хлам, который создает надежные
укрытия для пасюков. Часто такие свалки находятся в нескольких кило-
метрах от паселенного пункта, и зверьки обитают там круглый год (Ба-
кутис, 1956)
А. Н. Формозов (1945) описал поселение крыс в 7 км от г. Горького
на свалке, расположенной на речке Старко среди полей и небольших 60-
лот. Интересно, что и сейчас здесь имеется популяция серых крыс, кото-
рая обитает вдоль этой речки и впадающих в нее ручьев и существует за
счет отходов пищевой промышленности, сплавляемых из города по воде
(Петров, Леонтьев, 1977).
В нескольких километрах от г. Тулы большая свалка окопапа тлубо-
кой каназой. Крысы имеют здесь великолепные защитные и кормовые ус-
ловия, некоторые устраивают короткие норы или гнезда прямо на свалке
под различным хламом, другяе же роют норы по склонам каназы, которые
сплошь ископаны.
Аналогичное поселение крыс описано в Северной Моравии, где боль-
тпое количество пасюков обитает в яме-свалке, в которую свозят испор-
ченные продукты. Норы располагаются преимущественно в степах ямы.
а обитающие здесь зверьки часто посещают также и расположенные
вблизи поля свеклы и других культур (‘Лар1ейа1, Саег, 1983).
В небольшом узбекском городе Паркенте протекает река Паркент-сай,
которая весной сильно разливается. Для укрепления ве берегов из круп-
Экология
97
ных камней
у
с
й
ных к иней, закрепленных сеткой, сложена дамба, которую посещают
рители орда, а в некоторых местах выбрасывают золу и остатки пищи
На хровалках постоянно обитают крысы, находя укрытия между кам-
.П.
я здесь зверьков
5
:
авнялась —4
б
на 100 м? (рис. 20; Квашнин, Карасева, 1985) р
39-16 овобям
а свалках, гле много пи
:
евых отх
;
свалках, тле мно
щ
отходов, крысы способны переносять
ие
ратуры воздуха. Например, на Аляске близ г. Нома
И
ной
свалке, песмотря на сильные морозы
.
1 и можно было судит
(от
присуте
|
удить по пару, поднимающе-
у ад свалкой. Часть пасюков гибли от мороза, у т бкоторых и
Рис
рые
ис. 20. Серые крысы на свалке-дамбе в Узбекистане
Ввейху- - крыса обнюхивает
}, внизу —
са пьзт из рзки. Фото
В. Карасовой
са обню
хлеб,
крыса г
фо
ар:
А Серая крыса
96
Глава третья
рова, в основном концентрируясь на побережье. Эта часть популяции
крыс на зяму в помещения не переселялась, основной корм ес составляли
выбросы моря, но охотно поедались также птицы и их яйца. К 1944 г.
тнездивитиеся з дюнах популяции желтой трясогузки были полностью
истреблены крысами (Уапк, 1958).
Серые крысы сумели адаптироваться к суровым климатическим усло-
зиям многих антарктических островов (Кучерук, Лапшон, 1987), в оенов-
ном они закрепились и выжили там, где обитают колониальные птицы
и, следовательно, пет недостатка
в питце (яйца, птенцы, взрослые особи).
На один из антарктических остронов — Ю жная Георгия —кры-
сы были случайно завезены в 1800 г. и расселились там в тустых прибреж-
пых зарослях, на кочковатых лугах, где могли строить норы. В начале
ХХ в. здесь промышляли китов и яверьки питались в основном их мясом.
Впоследствии они перешли на питание главным образом за счет птиц в
колопиях буревостников. Мелкие воробъиные птицы, в частности антарк-
тические коньки, не гнездятся там, тде жхивут крысы. Буревестники хотя
и страдают от крыс, но могут © ними совместно существовать (Рус,
Воппет, 1980).
Биотопы, созданные человеком, или места обитания,
на которые человек оказал большое влияние
Места постоянного сбрасывания пищевых отходов,
создающих кормовую базу. Самый простой пример биотопа, создан-
ного руками челозека,—это свалки, куда обычно выбрасывают не только
остатки пищи, но также различный хлам, который создает надежные
укрытия для пасюков. Часто такие свалки находятся в нескольких кило-
метрах от паселенного пункта, и зверьки обитают там круглый год (Ба-
кутис, 1956)
А. Н. Формозов (1945) описал поселение крыс в 7 км от г. Горького
на свалке, расположенной на речке Старко среди полей и небольших 60-
лот. Интересно, что и сейчас здесь имеется популяция серых крыс, кото-
рая обитает вдоль этой речки и впадающих в нее ручьев и существует за
счет отходов пищевой промышленности, сплавляемых из города по воде
(Петров, Леонтьев, 1977).
В нескольких километрах от г. Тулы большая свалка окопапа тлубо-
кой каназой. Крысы имеют здесь великолепные защитные и кормовые ус-
ловия, некоторые устраивают короткие норы или гнезда прямо на свалке
под различным хламом, другяе же роют норы по склонам каназы, которые
сплошь ископаны.
Аналогичное поселение крыс описано в Северной Моравии, где боль-
тпое количество пасюков обитает в яме-свалке, в которую свозят испор-
ченные продукты. Норы располагаются преимущественно в степах ямы.
а обитающие здесь зверьки часто посещают также и расположенные
вблизи поля свеклы и других культур (‘Лар1ейа1, Саег, 1983).
В небольшом узбекском городе Паркенте протекает река Паркент-сай,
которая весной сильно разливается. Для укрепления ве берегов из круп-
Экология
97
ных камней
у
с
й
ных к иней, закрепленных сеткой, сложена дамба, которую посещают
рители орда, а в некоторых местах выбрасывают золу и остатки пищи
На хровалках постоянно обитают крысы, находя укрытия между кам-
.П.
я здесь зверьков
5
:
авнялась —4
б
на 100 м? (рис. 20; Квашнин, Карасева, 1985) р
39-16 овобям
а свалках, гле много пи
:
евых отх
;
свалках, тле мно
щ
отходов, крысы способны переносять
ие
ратуры воздуха. Например, на Аляске близ г. Нома
И
ной
свалке, песмотря на сильные морозы
.
1 и можно было судит
(от
присуте
|
удить по пару, поднимающе-
у ад свалкой. Часть пасюков гибли от мороза, у т бкоторых и
Рис
рые
ис. 20. Серые крысы на свалке-дамбе в Узбекистане
Ввейху- - крыса обнюхивает
}, внизу —
са пьзт из рзки. Фото
В. Карасовой
са обню
хлеб,
крыса г
фо
ар:
А Серая крыса
98
Глава третья
ты или лапы, но тем не менее осповная масса не толь-
жены увое
овЫ
множалась, и численность их,
ко выживала, по и весьма интенсивно раз
особенно летом, была высока (Зев\Щет, 1956).
ть.
Особенно благоприятные услония для крыс создаются са окотомотиль-
никах при недостаточном соблюдении ветеринарными ра отниками сани-
й скот свозят в определенное место, отстоя
тарных норм. Обычно павший
щее от животповодческих ферм на несколько километров. Обилие корма
принлекаст сюда пасюков, и они во множестве там. размножаются. и
Например, в Краснодарском крае близ совхоза « лавянох ь ‹ уг
скотомотильника вся почва была изрыта крысамина м: в ср й ом
приходилось 5—6 входных отверстии в их норы. Аналогичные ситу пи
известны в Кировской, Ростовской, Воронежской и других област
биков и др., 1987).
Отвалы земли по берегам иску
п арыков играют больную роль в жизни крыс.
сственных каналов
Обычно с одной сто-
Рис. 91. Распределение крыс в разные месяцы по дамбам рыборазводных прудов
{по материалам И. Т. Русева)
1— кормовая будка, 2 — дамба, 3 — пруды, 4 — территория, зассленная крысами;
1—Х17 — месяцы
1-П
попиш
.,
ОООО
х
зЯ
и
с
$7д
и
р
<ЕИ
П
8
|]
080: 05] Е} Е!
Экология
99
роны канала от бульдозера вырастают отвалы из выброшенной земли и
отдельные пласты неплотно прилегают друг к другу, образуя пустоты,
используемые крысами, перестраивающими их под свои норы. Насыпь
зарастает растительностью, создающей хорошие защитные условия,
а среди растений зверьки протаптывают поверхностные дорожки. Такие
каналы являются экологическими руслами, по которым крысы широко
перемещаются.
В Волго-Уральском междуречье после выкапывапия каналов крысы
быстро заселили отвалы земли и, хотя численность их тут была ненели-
ка (1—1,54 попадания в ловушки), обитали вне связи с населенными
пунктами круглый год (Шилов и др., 1986).
В Северном Казахстане зверьки заселили земляные отвалы части ка-
нала Иртыш—Карагапда, где опи встречаются главным образом в трост-
никово-рогозовых зарослях (1—2% поцадания в капканы) (Стогов, Бай-
танаев, 1983).
В Узбекистане и северо-западном Таджикистане крысы встречаются
вдоль оросительных каналов (которых там очень много), иногда распо-
ложенных в 10—15 км от населенных пунктов, но численность их там
невелика. Они очень многочисленны вдоль каналов и арыков, которые °
проходят в пределах паселеппых пупктов или недалеко от них.
Рыборазводные пруды. Это одно из излюбленных мест оби-
тания крыс, созданных человеком. Зверьков привлекают здесь корм для
рыб, а также снулая рыба, периодические заморы которой на прудах при-
водят к ее болыним скоплениям в прибрежной части. Работники рыбных
хозяйств часто отстреливают рыбоядных птиц, трупы которых крысы
тоже очень охотно поедают. Кроме того, на этих прудах немало и есте-
ствеппых кормов— личинок водяпых насокомых, моллюсков, головасти-
ков и пр.
Норы крысы устраивают по берегам прудов и каналов в искусствен-
ных насыпях. Поселения их чаще всего встречаются по берегам сброс-
ных каналов, вдоль которых насыпи особенно высоки. В Ташкентской
области на рыборазводных прудах пос. Бапыкчи, вдоль ебросных кана-
лов, по нашим учетам, в среднем было 28 входных отверстий в норы на
100 погонных метров, тогда как по дамбам вдоль берегов пруда их насчи-
тали только 10. Пространственное распределение серых крыс на рыбораз-
водных прудах сильно меняется по сезонам. По наблюдениям И. Т. Русе-
ва, в нижнем Приднестровье зимой крысы держатся локальпо только па
дамбе, около будки, в которой хранится корм для рыбы. Здесь они нахо-
лят пищу, а убежищем для них служат постоянные норы’на дамбе. Ле-
том обилие легкодоступного корма приводит к тому, что эти грызуны
расселяются по всему периметру берегов пруда. Наибольшей численности
насюки достигают в октябре — в это время пижияя часть склонов дамб
бывает ими почти полностью заселена (рис. 21). После вылова рыбы,
в конце ноября — в декабре, крысы уходят с прежних мест обитания и
концентрируются только на дамбе у кормовой будки. Они заселяют не
все рыборазводные пруды, а, как правило, только дамбы тех прудов, где
бывают часто заморы рыбы (Русев, Соловьев, 1987).
4*
98
Глава третья
ты или лапы, но тем не менее осповная масса не толь-
жены увое
овЫ
множалась, и численность их,
ко выживала, по и весьма интенсивно раз
особенно летом, была высока (Зев\Щет, 1956).
ть.
Особенно благоприятные услония для крыс создаются са окотомотиль-
никах при недостаточном соблюдении ветеринарными ра отниками сани-
й скот свозят в определенное место, отстоя
тарных норм. Обычно павший
щее от животповодческих ферм на несколько километров. Обилие корма
принлекаст сюда пасюков, и они во множестве там. размножаются. и
Например, в Краснодарском крае близ совхоза « лавянох ь ‹ уг
скотомотильника вся почва была изрыта крысамина м: в ср й ом
приходилось 5—6 входных отверстии в их норы. Аналогичные ситу пи
известны в Кировской, Ростовской, Воронежской и других област
биков и др., 1987).
Отвалы земли по берегам иску
п арыков играют больную роль в жизни крыс.
сственных каналов
Обычно с одной сто-
Рис. 91. Распределение крыс в разные месяцы по дамбам рыборазводных прудов
{по материалам И. Т. Русева)
1— кормовая будка, 2 — дамба, 3 — пруды, 4 — территория, зассленная крысами;
1—Х17 — месяцы
1-П
попиш
.,
ОООО
х
зЯ
и
с
$7д
и
р
<ЕИ
П
8
|]
080: 05] Е} Е!
Экология
99
роны канала от бульдозера вырастают отвалы из выброшенной земли и
отдельные пласты неплотно прилегают друг к другу, образуя пустоты,
используемые крысами, перестраивающими их под свои норы. Насыпь
зарастает растительностью, создающей хорошие защитные условия,
а среди растений зверьки протаптывают поверхностные дорожки. Такие
каналы являются экологическими руслами, по которым крысы широко
перемещаются.
В Волго-Уральском междуречье после выкапывапия каналов крысы
быстро заселили отвалы земли и, хотя численность их тут была ненели-
ка (1—1,54 попадания в ловушки), обитали вне связи с населенными
пунктами круглый год (Шилов и др., 1986).
В Северном Казахстане зверьки заселили земляные отвалы части ка-
нала Иртыш—Карагапда, где опи встречаются главным образом в трост-
никово-рогозовых зарослях (1—2% поцадания в капканы) (Стогов, Бай-
танаев, 1983).
В Узбекистане и северо-западном Таджикистане крысы встречаются
вдоль оросительных каналов (которых там очень много), иногда распо-
ложенных в 10—15 км от населенных пунктов, но численность их там
невелика. Они очень многочисленны вдоль каналов и арыков, которые °
проходят в пределах паселеппых пупктов или недалеко от них.
Рыборазводные пруды. Это одно из излюбленных мест оби-
тания крыс, созданных человеком. Зверьков привлекают здесь корм для
рыб, а также снулая рыба, периодические заморы которой на прудах при-
водят к ее болыним скоплениям в прибрежной части. Работники рыбных
хозяйств часто отстреливают рыбоядных птиц, трупы которых крысы
тоже очень охотно поедают. Кроме того, на этих прудах немало и есте-
ствеппых кормов— личинок водяпых насокомых, моллюсков, головасти-
ков и пр.
Норы крысы устраивают по берегам прудов и каналов в искусствен-
ных насыпях. Поселения их чаще всего встречаются по берегам сброс-
ных каналов, вдоль которых насыпи особенно высоки. В Ташкентской
области на рыборазводных прудах пос. Бапыкчи, вдоль ебросных кана-
лов, по нашим учетам, в среднем было 28 входных отверстий в норы на
100 погонных метров, тогда как по дамбам вдоль берегов пруда их насчи-
тали только 10. Пространственное распределение серых крыс на рыбораз-
водных прудах сильно меняется по сезонам. По наблюдениям И. Т. Русе-
ва, в нижнем Приднестровье зимой крысы держатся локальпо только па
дамбе, около будки, в которой хранится корм для рыбы. Здесь они нахо-
лят пищу, а убежищем для них служат постоянные норы’на дамбе. Ле-
том обилие легкодоступного корма приводит к тому, что эти грызуны
расселяются по всему периметру берегов пруда. Наибольшей численности
насюки достигают в октябре — в это время пижияя часть склонов дамб
бывает ими почти полностью заселена (рис. 21). После вылова рыбы,
в конце ноября — в декабре, крысы уходят с прежних мест обитания и
концентрируются только на дамбе у кормовой будки. Они заселяют не
все рыборазводные пруды, а, как правило, только дамбы тех прудов, где
бывают часто заморы рыбы (Русев, Соловьев, 1987).
4*
100
Глава третья
О поселении крыс на рыборазволных прулах в Доно-Аксайской поймо
сообщают В. Е. Кондратенко с соавторами (1986). Отдельные поссления
пасюков вотречаются в Авотри та рыборазводных прудах в окрест
х
линга (УТоЦЕ её а1., | .
стих Пафтинте С9С Особый интерес представляют поселепия крыс па
различных агроценозах. В одних случаях опи живут там постояипо я зе
дут себя как представители экзоантропной формы, в других ст ко
временно выселяются на поля из населенных пунктов в теплое вр
года.
Наиболее оптимальные условия существования для крыс создаются
на полях, заливаемых водой, это обычио рисовые поля и плантации
н го сахара.
о аномерное Чорбдовавие земляных валиков и рисовых деко ‚ пред-
ставлиет для крыс исключительно благоприятное сочетание богатой кор-
мовой базы и мест, удобных для норелия (рис. 22). Мелкие, хорошо про
треваемые солнцем водоемы изобилуют различными бесповвопочными,
здесь обычио мпого рыбы, земноводных — нее это составляет остов аки
вотных кормов крыс. На наликах, окаймляющих чеки, держатся тр `
ченные животным кормом птицы — чайки, цапли, кулики. Они удо ряют
испражнениями почву и благодаря этому и обилию воды по валика
буйно разрастаются различные растения— в одних местах преоблад
—_
,
й
Рис. 99. Валики оросительной системы на рисовыт полят места обитания серо
крысы. Фото В. Ф. Семенова
Слева —- канал, справа — нора
Экология
101
разнотравье, в других— бурьян, в наиболее увлажненпых местах—
тростник. Поры крыс бывают расположены по валикам, а растительность
является для них хорошим укрытием.
Распределение зверьков на рисовых полях лимитируется возмож
ностью рыть норы, т. е. дрепажем почвы валиков.
Осенью после вспашки крысы обитают уже не только па валиках, но’
делают поры и под болышими пластами перевернутой земли, передвига-
ясь под ними. На некоторых участках рисовых полей пе проводят осен-
нюю распашку, и там остается много обмолоченной соломы. Часть пасю-
ков остается зимовать на валиках в норах, где они с осени углубляют
и чистят их. Но большая их часть концентрируется около куч соломы
риса, где оспову их питапия в это время составляют оставшиеся зерна
(рис. 23).
Численпость крыс па рисовых полях значительно вые, чем в при-
родных етациях. По нашим данным, па Кубани по берегам лиманов
число крыс на 100 погонных метров составляло всего 0,1—0,2 особи, тог-
да как по валикам рисовых чеков оно доходило до 10 и более особей (Ка-
расева и др., 1986).
Численность пасюков па рисовых полях меняется {о сезонам, а также
в разные годы. С ноября по февраль количество их убывает за счет вы-
сокой смертности и отсутствия размножения, весной наблюдается неболь-
шой подъем, а наибольшая скорость
Рис. 93. Россыпи риса после уборки. Прироста численности имеет место с
Фото В. Ф. Семенова
июня ло август. В годы нашей рабо-
ты на Цубани ее наименышая величи-
на отмечена в 1978 г., несколько
большая—в 1979 г., а в 1980 и
1981 тг. численность крыс была еще
выше (Рыльников, Карасева, 1985).
К. А. Илятер-Плохицкий (1931,
1936), работавший на рисовой систе-
мев Приханкайской пизменности, от-
мечает, что в 1920-х годах числен-
пость крыс там была высокой и с0-
ставляла в среднем 50 нор на 1 га..
Зверьки приносили немалый . вред,
плавая в разных направлениях по че-
кам и поедая молодые ростки риса,
что при их высокой численности ока-.
зывало заметное влияние на пониже-
ние урожая.
В последние годы на высокую чие-.
ленность серых крыс на рисовых по-
лях Приханкайской пизменности ука-
зывает В. А. Нестеренко (1985).
Наиболее существенный зред
зверьки припосят, роя поры в вали-
100
Глава третья
О поселении крыс на рыборазволных прулах в Доно-Аксайской поймо
сообщают В. Е. Кондратенко с соавторами (1986). Отдельные поссления
пасюков вотречаются в Авотри та рыборазводных прудах в окрест
х
линга (УТоЦЕ её а1., | .
стих Пафтинте С9С Особый интерес представляют поселепия крыс па
различных агроценозах. В одних случаях опи живут там постояипо я зе
дут себя как представители экзоантропной формы, в других ст ко
временно выселяются на поля из населенных пунктов в теплое вр
года.
Наиболее оптимальные условия существования для крыс создаются
на полях, заливаемых водой, это обычио рисовые поля и плантации
н го сахара.
о аномерное Чорбдовавие земляных валиков и рисовых деко ‚ пред-
ставлиет для крыс исключительно благоприятное сочетание богатой кор-
мовой базы и мест, удобных для норелия (рис. 22). Мелкие, хорошо про
треваемые солнцем водоемы изобилуют различными бесповвопочными,
здесь обычио мпого рыбы, земноводных — нее это составляет остов аки
вотных кормов крыс. На наликах, окаймляющих чеки, держатся тр `
ченные животным кормом птицы — чайки, цапли, кулики. Они удо ряют
испражнениями почву и благодаря этому и обилию воды по валика
буйно разрастаются различные растения— в одних местах преоблад
—_
,
й
Рис. 99. Валики оросительной системы на рисовыт полят места обитания серо
крысы. Фото В. Ф. Семенова
Слева —- канал, справа — нора
Экология
101
разнотравье, в других— бурьян, в наиболее увлажненпых местах—
тростник. Поры крыс бывают расположены по валикам, а растительность
является для них хорошим укрытием.
Распределение зверьков на рисовых полях лимитируется возмож
ностью рыть норы, т. е. дрепажем почвы валиков.
Осенью после вспашки крысы обитают уже не только па валиках, но’
делают поры и под болышими пластами перевернутой земли, передвига-
ясь под ними. На некоторых участках рисовых полей пе проводят осен-
нюю распашку, и там остается много обмолоченной соломы. Часть пасю-
ков остается зимовать на валиках в норах, где они с осени углубляют
и чистят их. Но большая их часть концентрируется около куч соломы
риса, где оспову их питапия в это время составляют оставшиеся зерна
(рис. 23).
Численпость крыс па рисовых полях значительно вые, чем в при-
родных етациях. По нашим данным, па Кубани по берегам лиманов
число крыс на 100 погонных метров составляло всего 0,1—0,2 особи, тог-
да как по валикам рисовых чеков оно доходило до 10 и более особей (Ка-
расева и др., 1986).
Численность пасюков па рисовых полях меняется {о сезонам, а также
в разные годы. С ноября по февраль количество их убывает за счет вы-
сокой смертности и отсутствия размножения, весной наблюдается неболь-
шой подъем, а наибольшая скорость
Рис. 93. Россыпи риса после уборки. Прироста численности имеет место с
Фото В. Ф. Семенова
июня ло август. В годы нашей рабо-
ты на Цубани ее наименышая величи-
на отмечена в 1978 г., несколько
большая—в 1979 г., а в 1980 и
1981 тг. численность крыс была еще
выше (Рыльников, Карасева, 1985).
К. А. Илятер-Плохицкий (1931,
1936), работавший на рисовой систе-
мев Приханкайской пизменности, от-
мечает, что в 1920-х годах числен-
пость крыс там была высокой и с0-
ставляла в среднем 50 нор на 1 га..
Зверьки приносили немалый . вред,
плавая в разных направлениях по че-
кам и поедая молодые ростки риса,
что при их высокой численности ока-.
зывало заметное влияние на пониже-
ние урожая.
В последние годы на высокую чие-.
ленность серых крыс на рисовых по-
лях Приханкайской пизменности ука-
зывает В. А. Нестеренко (1985).
Наиболее существенный зред
зверьки припосят, роя поры в вали-
102 Глава третья
ках, что способствует размыванию последних и требует затраты значитель-
ных средств для ремонта системы.
В нокоторых районах СССР на рисовых чеках крыс очень немното.
Так, ио нашим наблюдениям, в Узбекистапе осенью 1982 и 1988 тг.
численность насюков в среднем была равна всего 0,5 зверька на 1 га.
Это, вероятно, объясняется тем, что они сравнительно недавно веселились
па территорию Ташкентского оазиса (см. гл. об, арезлах). За последние
30—40 лет зверьки достигли здесь очень высокой численности, но только
в помещениях, главным образом па животноводческих фермах. В откры-
тых биотопах, и в том числе на рисовой системе, их немного.
На рисовых плантациях Италии и Испании показана высокая числен-
ность серых крыс (8—10% попадашия в ловутики; ВаБафег, 1954). Бар-
нес Брукс (Вагпез Вгоокз) пишет об их массовом появлении на рисовых
полях в Северной Америке, но пе приводит конкретных данных (цит. по:
Сноив, 1982). Серые крысы многочисленны на нлаптациях сахарного
тростника в Квинсленде, Новом Южном Уэльсе, а также па Гавайях
(Ловпзоп, 1950; Рорр, 1950; Топуев, 1969).
.
На зерновых культурах крысы обычно не живут круглый
тод, а выселяются туда только в теплое время года в период поспевания
зерна. У нас в стране (во всяком случае в больших количествах) на та-
ких полях эти грызуны не встречаются.
Есть интересные сведения о заселении крысами хлебного жнивья в
Британии. Авторы (Во\е, 1962: Втоб1е её а1., 1983) указывают, что пасю-
ки во время летних месяцев буквально наводняют посевы хлеба, где они
питаются созревающими колосьями. После уборки урожая часть зверьков
выселяются на окраины полей в бурьяны, покрывающие земляпые вали-
ки. По сообщению Винойбле и Лесли (Уепаез, ГезНе, 1942), пасюки
скапливаются под скирдами пеобмолоченпого хлеба в ометах соломы
(цит. по: Кузякин, 1951).
В Шотландии крысы мпогочисленны на полях ячменя, овса и на ого-
родах, откуда они после уборки урожая выселяются в фермерские пост-
ройки (Вгобс ей а1., 1983).
.
О заселенности крысами зерновых полей и огородов в Бельгии с00б-
щают Бернар и Колен (Вегпага, СоШа, 1966). Во Франции крысы посе-
ляются на полях кукурузы, где причиняют немалый вред. Особенно дни
многочисленны на полях, близко расположенных к берегам рек и прудов
(ЗЖибан, 1977).
` Места отдыха людей, пляжи, Как уже говорилось, крыс,
обитающих в природе, всегда привлекают временные стоянки людей, где
остается много неиспользовапных продуктов. По сообщению И. Т. Русе-
ва и В И. Соловьева (1987), характер распределения зверьков в При-
диестровье определяется главным образом степенью освоения леса отды-
хающими. В теплое время года па выходные дни, а также и па длитель-
ный срок собирается очень много народу. Приезжающие (особенно только
на выходные дни), как правило, оставляют на своих отояпках обильные
остатки еды. Отдыхающие более долгий срок расходуют продукты эко-
номней, но и от пих остаются объедки. Авторы облавливали капканами в
Экология
103
Таблица 5. Соотношение числа еерых крые
и водяных полевок, выловленных в котловине 03. Неро
в разные годы (но: Карасева и др., 1986а)
Годы
всего Х
Доли серых крыс,
ных полевок
ыы
1950—1955
329
8,2-1,5
1960-1964
173
9,5=2,2
1970—1974
450
32.2438
1981
65
90,7+3,6
разные сезоны года 26 участков стоянок отдыхающих. В апреле, когда
людей практически не было, удалось отловить всего трех крыс. В мае,
когда отдыхающие уже пачипали появляться, пойманы уже 13 крыс,
и, наконец, в июле и августе, в сезон наибольшего скопления людей,
крыс было поймано соответственно 33 и 36.
Подобных примеров можно приводить много. В Ярославской области
в приозерной котловине оз. Неро в течение 30 лет велись наблюдения за‘
численностью мелких млекопитающих. В пачале 50-х годов по берегам
ручьев и реки, впадающих в озеро, серых крыс в летнее время было срав-
нительно немного
— преобладали водяные полевки. Но в последующие
годы в прнозерной котловине значительно увеличилось число отдыхаю-
щих, которые приходили сюда собирать ягоды, купаться в реке, ловить
рыбу ит. п. В результате этих походов ностояпио оставались объедки от
завтраков, в некоторых местах появлялись свалки мусора. В результате
резко изменилось соотношение численпости двух видов — водяной полев-
ки стало мало (отдыхающие вытаптывали траву), а лоля серых крыс з
стловах значительно возросла (табл. 5) (Карасева и др., 1986а).
Большое значение в использовании крысами мест отдыха людей име-
ют пляжи и особенно места вблизи них, гие скапливается много отды-
хающих. Крыс мы часто наблюдали на пляжах в Сочи (курорт Лазарев-
ское па р. Псезуапсе), Батуми и Сухуми, встречаются они и на пляжах
Крыма (Коковин и др., 1978; Арутюнян, устн. сообщ.).
Синантропные ирысы
Как мы уже говорили, благодаря высокой эколого-физмологической пла-
стичности серая крыса смогла адаптироваться к жизни в самых различ-
пых паселенных пунктах. Она не только выдерживает низкие темпера-
туры воздуха (—10°...—12°), по и успешно размножается при наличии
корма (в холодильниках, где хранится мясо). Переносят и достаточно
высокие температуры воздуха (-28...--30°), поселяясь в обшивках нпа-
ровых котлов. Однако температурный оптимум вида +18...+20°, т. е.
тот же, что паиболее приемлем для человека.
Кроме того, вследствие многовековой адаптации пасюка к жизни око-
ло человека, стремящегося уничтожить крысу, зид приобрел ряд этоло-
102 Глава третья
ках, что способствует размыванию последних и требует затраты значитель-
ных средств для ремонта системы.
В нокоторых районах СССР на рисовых чеках крыс очень немното.
Так, ио нашим наблюдениям, в Узбекистапе осенью 1982 и 1988 тг.
численность насюков в среднем была равна всего 0,5 зверька на 1 га.
Это, вероятно, объясняется тем, что они сравнительно недавно веселились
па территорию Ташкентского оазиса (см. гл. об, арезлах). За последние
30—40 лет зверьки достигли здесь очень высокой численности, но только
в помещениях, главным образом па животноводческих фермах. В откры-
тых биотопах, и в том числе на рисовой системе, их немного.
На рисовых плантациях Италии и Испании показана высокая числен-
ность серых крыс (8—10% попадашия в ловутики; ВаБафег, 1954). Бар-
нес Брукс (Вагпез Вгоокз) пишет об их массовом появлении на рисовых
полях в Северной Америке, но пе приводит конкретных данных (цит. по:
Сноив, 1982). Серые крысы многочисленны на нлаптациях сахарного
тростника в Квинсленде, Новом Южном Уэльсе, а также па Гавайях
(Ловпзоп, 1950; Рорр, 1950; Топуев, 1969).
.
На зерновых культурах крысы обычно не живут круглый
тод, а выселяются туда только в теплое время года в период поспевания
зерна. У нас в стране (во всяком случае в больших количествах) на та-
ких полях эти грызуны не встречаются.
Есть интересные сведения о заселении крысами хлебного жнивья в
Британии. Авторы (Во\е, 1962: Втоб1е её а1., 1983) указывают, что пасю-
ки во время летних месяцев буквально наводняют посевы хлеба, где они
питаются созревающими колосьями. После уборки урожая часть зверьков
выселяются на окраины полей в бурьяны, покрывающие земляпые вали-
ки. По сообщению Винойбле и Лесли (Уепаез, ГезНе, 1942), пасюки
скапливаются под скирдами пеобмолоченпого хлеба в ометах соломы
(цит. по: Кузякин, 1951).
В Шотландии крысы мпогочисленны на полях ячменя, овса и на ого-
родах, откуда они после уборки урожая выселяются в фермерские пост-
ройки (Вгобс ей а1., 1983).
.
О заселенности крысами зерновых полей и огородов в Бельгии с00б-
щают Бернар и Колен (Вегпага, СоШа, 1966). Во Франции крысы посе-
ляются на полях кукурузы, где причиняют немалый вред. Особенно дни
многочисленны на полях, близко расположенных к берегам рек и прудов
(ЗЖибан, 1977).
` Места отдыха людей, пляжи, Как уже говорилось, крыс,
обитающих в природе, всегда привлекают временные стоянки людей, где
остается много неиспользовапных продуктов. По сообщению И. Т. Русе-
ва и В И. Соловьева (1987), характер распределения зверьков в При-
диестровье определяется главным образом степенью освоения леса отды-
хающими. В теплое время года па выходные дни, а также и па длитель-
ный срок собирается очень много народу. Приезжающие (особенно только
на выходные дни), как правило, оставляют на своих отояпках обильные
остатки еды. Отдыхающие более долгий срок расходуют продукты эко-
номней, но и от пих остаются объедки. Авторы облавливали капканами в
Экология
103
Таблица 5. Соотношение числа еерых крые
и водяных полевок, выловленных в котловине 03. Неро
в разные годы (но: Карасева и др., 1986а)
Годы
всего Х
Доли серых крыс,
ных полевок
ыы
1950—1955
329
8,2-1,5
1960-1964
173
9,5=2,2
1970—1974
450
32.2438
1981
65
90,7+3,6
разные сезоны года 26 участков стоянок отдыхающих. В апреле, когда
людей практически не было, удалось отловить всего трех крыс. В мае,
когда отдыхающие уже пачипали появляться, пойманы уже 13 крыс,
и, наконец, в июле и августе, в сезон наибольшего скопления людей,
крыс было поймано соответственно 33 и 36.
Подобных примеров можно приводить много. В Ярославской области
в приозерной котловине оз. Неро в течение 30 лет велись наблюдения за‘
численностью мелких млекопитающих. В пачале 50-х годов по берегам
ручьев и реки, впадающих в озеро, серых крыс в летнее время было срав-
нительно немного
— преобладали водяные полевки. Но в последующие
годы в прнозерной котловине значительно увеличилось число отдыхаю-
щих, которые приходили сюда собирать ягоды, купаться в реке, ловить
рыбу ит. п. В результате этих походов ностояпио оставались объедки от
завтраков, в некоторых местах появлялись свалки мусора. В результате
резко изменилось соотношение численпости двух видов — водяной полев-
ки стало мало (отдыхающие вытаптывали траву), а лоля серых крыс з
стловах значительно возросла (табл. 5) (Карасева и др., 1986а).
Большое значение в использовании крысами мест отдыха людей име-
ют пляжи и особенно места вблизи них, гие скапливается много отды-
хающих. Крыс мы часто наблюдали на пляжах в Сочи (курорт Лазарев-
ское па р. Псезуапсе), Батуми и Сухуми, встречаются они и на пляжах
Крыма (Коковин и др., 1978; Арутюнян, устн. сообщ.).
Синантропные ирысы
Как мы уже говорили, благодаря высокой эколого-физмологической пла-
стичности серая крыса смогла адаптироваться к жизни в самых различ-
пых паселенных пунктах. Она не только выдерживает низкие темпера-
туры воздуха (—10°...—12°), по и успешно размножается при наличии
корма (в холодильниках, где хранится мясо). Переносят и достаточно
высокие температуры воздуха (-28...--30°), поселяясь в обшивках нпа-
ровых котлов. Однако температурный оптимум вида +18...+20°, т. е.
тот же, что паиболее приемлем для человека.
Кроме того, вследствие многовековой адаптации пасюка к жизни око-
ло человека, стремящегося уничтожить крысу, зид приобрел ряд этоло-
4104
Глава третья
тических и психологических свойств, помогающих зверькам скрываться
от людей, находить даже в непосредственной близости от них особые
скрытые убежища, а также избегать различных ловушек и отравленных
приманок.
Таким образом, серая крыса стала классическим синантропом, с одпой
стороны, в связи © исключительной экологической тибкостью, а с лру-
той — вследствие воздействия антропогенных факторов, которые, в свою
очередь, способствовали ее синантропизму (Соколов, Карасева, 1985).
Сельская местность
В сельской местпости серые крысы заселяют поселки городского и дачно-
го типа, села, деревни, населенные пункты нового типа (садово-огородные
участки), аулы, кишлаки и, наконец, одиночно стоящие строения.
В некоторых местах все паселенные пункты бывают сплошь заселены
этими грызунами. Например, в Словакии.из 126 обследованных деревень
пе было ни одной не заселенной крысами. А при обследовании 56 293 зда-
ний крысы оказались в 29,6%, при этом в среднем на одно здание при-
ходилось 37 особей (Оеуаг, 1986). Зверьки заселяют жилые дома и 0со-
беняо падворные постройки (помещения для. скота и домашней птицы),
уборные, риги, сараи, сеновалы, мельницы, различные склады, бойни, по
очень многочисленны на животноводческих фермах.
Килые дома и надворпые постройки. В сельских
усадьбах серые крысы заселяют главным образом падворные постройки,
т. е. сарай, кладовые, пристройки, где обычно держат свиней, скот, кур,
‚кроликов и т. п. В сараях лежат дрова, свалены разные предметы домаш-
него обихода (ведра, корыта, ящики, тряпки, старые доски, сено и т. п.),
в кладовых хранятся продукты иитания, зерпо, фураж. Все эти. помеще-
ния обячно сообщаются межу собой густой сетью крысиных ходов,
и зверьков здесь бывает много. Антисанитарное состояние надверных по-
строск создает. для них отличные условия обитания. А. Н.. Козлов
(1983а) сообщает, что посие истребительных работ в одном сарайчике
ему пришлось подобрать 70 трупов крыс. В самих жилых домах зверь-
ков обычно бывает значительно мопьше, чем в надворных постройках, но
если они там есть, то чаще всего живут в подполье, иногда устраивая
поры в земляном полу нод половицами.
|
Наиболее благоприятны для заселения серыми крысами тлинобитные
и деревянные постройки, в кирпичных и каменных домах этих животных
бывает меньше (ОТе]аг, 1980, 1986). Как правило, в домах, содержанцих-
ся чисто, где нет свалок, крыс не бывает. Так, в некоторых населенных
пунктах ФРГ благодаря хорошему санитарному состоянию жилых домов
в них крысы не вотречаются, хотя в окрестностях их много (ТеЦе, 1962).
Животноводческие фермы. На многих животноводческих
фермах крысы имеют оптимальные условия существования, обилие еды,
постоянный источник воды и удобные места для устройства убежищ (По-
лежаев и др., 1959; Миронов и др.. 1961; Козлов, 1964; Хамаганов, 1968;
Траханов, 1983; Ое]&г, 1980; и др.).
|
Экология
405
|
| 2222 122
|
ты
8
[<Я
Я:
169591
”
оС
|
р<
4
]
к
`No;
а
[53%
ох
ро
ы
5:
Я
ее
и
|
ех
о
мен
ро
х
о
ро
к.
РЯ
|
хх
Косо<о
во
.
Во
ее
мы
к
ко
Вох
|
рос
4.
р
|
я
ря
10
ро
и
р
вое,
252
ро
ро
хх
и
|
я
на
вы
99°
45
К
|-
РСЯ
ео
о
о
ха
о
ра
Ко
ро
м
нее
р
р
>>.
Хх
ро
те
[хо
Е
рых
хх
о
29
хх
ся
ХХ
роса
44
СС
ыы
С
не
|
ыы
РОЯ
РС
20°
то
ое
=
а
р
ыы
х
}
ео
р
ры
я
о
ро
[4
амбовская вл
Поменская дбл.
СЕбЕДНЫЙ КЕЗОЕСТаН
Рис. 24. Сравнительная заселеиность серыми крысами различных животноводческих:
помещений (по Д. Ф. Траханову}
1-— свинарники, 2 — птичники, 3 — коровники, 4 — прочие животноводческие помещения
Более всего бынают заселены свинарники и птичники (рис. 24).
Здесь зверьки находят наилучшие сочетания всех факторов, необ%одимых
для их жизни: в свинарниках они получают особенно ценные комбиниро-
ванные корма, распаренные зерна овса и пшеницы, в свиноматочниках,
кроме того, они имеют возможность пить молоко. Обычно там. нет недо-
остатка в животной пище— пасюки едят последы свиней, мертвых поро-
сят, а иногда нападают и па живых, отгрызая у них хвосты, уши и ноги
(рис. 25, 26). Встречаются и особи-каннибалы.
Летом мухи откладывают
в свиной навоз массу яиц, из которых выходят личинки, охотно поедае-
мые крысами.
-к
Конечно, большое значение имеет материал, из которого построены
свинарники. Крысы предпочитают старые строения © большим числом
щелей, с новрежденными стенами, деревянными настилами, под которы-
ми можно хорошо укрыться. Такие здапия още сохранились в некоторых
южных районах нашёй страны (Казахстан, Узбекистан, Кубань). Наибо-
лее заселены крысами саманные и деревянные постройки, покрытые прес-
сованной соломой. На Кубани, например, мы видели саманные свинарнй-
ки, сплошь источенные крысами, которые копали и прогрызали там мно-
тоярусные сложные норы с ходами, пронизывающими все помещение.
На территории Якутской АССР свинья, крупный рогатый скот, а так-
же овцы содержатся в деревянных помещениях, называемых здесь хото-
нами. Это сооружения, засыпанные снаружи до нижнего края окон зем-
лей, перемешанной с павозом. Крысы проделывают.в этой ‘насыпи гори-
зонтальные. и вертикальные ходы, которые. соединены с самим хотоном,
а основная масса зверьков концентрируется возле тех ‚месть где Хранитбя
корм для животных. В тех помещениях, где корм привозят только для
4104
Глава третья
тических и психологических свойств, помогающих зверькам скрываться
от людей, находить даже в непосредственной близости от них особые
скрытые убежища, а также избегать различных ловушек и отравленных
приманок.
Таким образом, серая крыса стала классическим синантропом, с одпой
стороны, в связи © исключительной экологической тибкостью, а с лру-
той — вследствие воздействия антропогенных факторов, которые, в свою
очередь, способствовали ее синантропизму (Соколов, Карасева, 1985).
Сельская местность
В сельской местпости серые крысы заселяют поселки городского и дачно-
го типа, села, деревни, населенные пункты нового типа (садово-огородные
участки), аулы, кишлаки и, наконец, одиночно стоящие строения.
В некоторых местах все паселенные пункты бывают сплошь заселены
этими грызунами. Например, в Словакии.из 126 обследованных деревень
пе было ни одной не заселенной крысами. А при обследовании 56 293 зда-
ний крысы оказались в 29,6%, при этом в среднем на одно здание при-
ходилось 37 особей (Оеуаг, 1986). Зверьки заселяют жилые дома и 0со-
беняо падворные постройки (помещения для. скота и домашней птицы),
уборные, риги, сараи, сеновалы, мельницы, различные склады, бойни, по
очень многочисленны на животноводческих фермах.
Килые дома и надворпые постройки. В сельских
усадьбах серые крысы заселяют главным образом падворные постройки,
т. е. сарай, кладовые, пристройки, где обычно держат свиней, скот, кур,
‚кроликов и т. п. В сараях лежат дрова, свалены разные предметы домаш-
него обихода (ведра, корыта, ящики, тряпки, старые доски, сено и т. п.),
в кладовых хранятся продукты иитания, зерпо, фураж. Все эти. помеще-
ния обячно сообщаются межу собой густой сетью крысиных ходов,
и зверьков здесь бывает много. Антисанитарное состояние надверных по-
строск создает. для них отличные условия обитания. А. Н.. Козлов
(1983а) сообщает, что посие истребительных работ в одном сарайчике
ему пришлось подобрать 70 трупов крыс. В самих жилых домах зверь-
ков обычно бывает значительно мопьше, чем в надворных постройках, но
если они там есть, то чаще всего живут в подполье, иногда устраивая
поры в земляном полу нод половицами.
|
Наиболее благоприятны для заселения серыми крысами тлинобитные
и деревянные постройки, в кирпичных и каменных домах этих животных
бывает меньше (ОТе]аг, 1980, 1986). Как правило, в домах, содержанцих-
ся чисто, где нет свалок, крыс не бывает. Так, в некоторых населенных
пунктах ФРГ благодаря хорошему санитарному состоянию жилых домов
в них крысы не вотречаются, хотя в окрестностях их много (ТеЦе, 1962).
Животноводческие фермы. На многих животноводческих
фермах крысы имеют оптимальные условия существования, обилие еды,
постоянный источник воды и удобные места для устройства убежищ (По-
лежаев и др., 1959; Миронов и др.. 1961; Козлов, 1964; Хамаганов, 1968;
Траханов, 1983; Ое]&г, 1980; и др.).
|
Экология
405
|
| 2222 122
|
ты
8
[<Я
Я:
169591
”
оС
|
р<
4
]
к
`No;
а
[53%
ох
ро
ы
5:
Я
ее
и
|
ех
о
мен
ро
х
о
ро
к.
РЯ
|
хх
Косо<о
во
.
Во
ее
мы
к
ко
Вох
|
рос
4.
р
|
я
ря
10
ро
и
р
вое,
252
ро
ро
хх
и
|
я
на
вы
99°
45
К
|-
РСЯ
ео
о
о
ха
о
ра
Ко
ро
м
нее
р
р
>>.
Хх
ро
те
[хо
Е
рых
хх
о
29
хх
ся
ХХ
роса
44
СС
ыы
С
не
|
ыы
РОЯ
РС
20°
то
ое
=
а
р
ыы
х
}
ео
р
ры
я
о
ро
[4
амбовская вл
Поменская дбл.
СЕбЕДНЫЙ КЕЗОЕСТаН
Рис. 24. Сравнительная заселеиность серыми крысами различных животноводческих:
помещений (по Д. Ф. Траханову}
1-— свинарники, 2 — птичники, 3 — коровники, 4 — прочие животноводческие помещения
Более всего бынают заселены свинарники и птичники (рис. 24).
Здесь зверьки находят наилучшие сочетания всех факторов, необ%одимых
для их жизни: в свинарниках они получают особенно ценные комбиниро-
ванные корма, распаренные зерна овса и пшеницы, в свиноматочниках,
кроме того, они имеют возможность пить молоко. Обычно там. нет недо-
остатка в животной пище— пасюки едят последы свиней, мертвых поро-
сят, а иногда нападают и па живых, отгрызая у них хвосты, уши и ноги
(рис. 25, 26). Встречаются и особи-каннибалы.
Летом мухи откладывают
в свиной навоз массу яиц, из которых выходят личинки, охотно поедае-
мые крысами.
-к
Конечно, большое значение имеет материал, из которого построены
свинарники. Крысы предпочитают старые строения © большим числом
щелей, с новрежденными стенами, деревянными настилами, под которы-
ми можно хорошо укрыться. Такие здапия още сохранились в некоторых
южных районах нашёй страны (Казахстан, Узбекистан, Кубань). Наибо-
лее заселены крысами саманные и деревянные постройки, покрытые прес-
сованной соломой. На Кубани, например, мы видели саманные свинарнй-
ки, сплошь источенные крысами, которые копали и прогрызали там мно-
тоярусные сложные норы с ходами, пронизывающими все помещение.
На территории Якутской АССР свинья, крупный рогатый скот, а так-
же овцы содержатся в деревянных помещениях, называемых здесь хото-
нами. Это сооружения, засыпанные снаружи до нижнего края окон зем-
лей, перемешанной с павозом. Крысы проделывают.в этой ‘насыпи гори-
зонтальные. и вертикальные ходы, которые. соединены с самим хотоном,
а основная масса зверьков концентрируется возле тех ‚месть где Хранитбя
корм для животных. В тех помещениях, где корм привозят только для
Экология
107
одпо-двухдневного кормления, этих грызунов или пет, или они встреча-
ются редко. Видимо, для ностояпного обитания крыс в животноводческих
помещениях Восточной Сибири только болыние запасы кормов могут со-
ставить благоприятные условия для образования устойчивых популяций
(Антонюк и др., 1987).
|
Пемалое значение для крыс имеет п температурный фактор — в тече-
ние года в свипарниках температура воздуха меняется менее резко, чем
снаружи. Под деревянными настилами, на которых лежат свипьи, зимой
температура воздуха (благодаря теплу от свиней) достигает --24...--26° С.
Именно здесь крысы чаще всего держатся сами и устраивают гнезда
(рис. 27).
Интересно, что в некоторых свинарниках носеления крыс на северной
и южной сторопах расноложены неравномерно. С южной стороны нор’
в 3—4 раза больше, чем с северной. То же наблюдается в соломенных и
трэстниковых крышах, где с солнечной стороны располагается до 80%,
всех наружных нор.
В холодное время года крысы концентрируются и в других номеще-
инях свипофермы — они любят сидеть под паровыми котлами, на кухне,
<_
Рис. 25. Серые крысы в свинарнике. Фото А. Н. Козлова
Рис. 26. Крысы, грызущие собрата. Фото П. Роделя
”
Экология
107
одпо-двухдневного кормления, этих грызунов или пет, или они встреча-
ются редко. Видимо, для ностояпного обитания крыс в животноводческих
помещениях Восточной Сибири только болыние запасы кормов могут со-
ставить благоприятные условия для образования устойчивых популяций
(Антонюк и др., 1987).
|
Пемалое значение для крыс имеет п температурный фактор — в тече-
ние года в свипарниках температура воздуха меняется менее резко, чем
снаружи. Под деревянными настилами, на которых лежат свипьи, зимой
температура воздуха (благодаря теплу от свиней) достигает --24...--26° С.
Именно здесь крысы чаще всего держатся сами и устраивают гнезда
(рис. 27).
Интересно, что в некоторых свинарниках носеления крыс на северной
и южной сторопах расноложены неравномерно. С южной стороны нор’
в 3—4 раза больше, чем с северной. То же наблюдается в соломенных и
трэстниковых крышах, где с солнечной стороны располагается до 80%,
всех наружных нор.
В холодное время года крысы концентрируются и в других номеще-
инях свипофермы — они любят сидеть под паровыми котлами, на кухне,
<_
Рис. 25. Серые крысы в свинарнике. Фото А. Н. Козлова
Рис. 26. Крысы, грызущие собрата. Фото П. Роделя
”
108 Глава третья
.
Рис. 97. Изменение температуры 803-
.
духа по месяцам в помещении для саи-
ней (1) и в подполье (2) (по А. Н. Коз-
лову, 1980а)
где готовится корм, возле обогрева-
тельных труб, калориферов, на окнах
с солнечной стороны и т. д. Летом
часть зверьков выселяется из свинар-
ника и обитает в норах © наружной
их стороны. Некоторые пасюки ухо-
дят из помещений на зпачительные
расстояния.
Численность крыс зв некоторых
свинарниках достигает колоссальных
от-
размеров — на Кубани в марте 1982 г. в р
свинариико, те о
Е
ы
.Д.),
пия зверьков (кал, погрызы,
мечали много следов пребыва
в
торм и т. Жо,
о объективные ноказатели.
пами были получены достаточн
Моя
5
тили их отрезанием пальцев
живоловками 58 крыс, поме
‚
и
выустнии
ке на месте поимки. Далее нами
обратно в том же свипарии
г Дале т были обло.
ы иииздобытых
крыс
лепы все помещения капкапам
Крыс моче ими
свинариике обитали 955 взр
зались 13. Следовательно, в
|
К этому количеству следует прибавить и число молодых крысят, которыс
еще пе покипули материнских гнезд: из 124 отловленпых капканачи
взрослых крыс самок было 54, из пих 41 заведомо патио
дедов
р
`
из которых
-
ии было 227 взрослых самок,
тельно, всего в помещен
к, Иа которых 5 35
й
-
ыводки (формулу расчет , . ВТ,
бей лактировали, т. е. имели в
.
г
ВЫ.
й
ый по вскрытию беремент
}
Средний выводок (устаповленн
1
й
Е мки имелось
у каждой лактирующей са
был 10,4. Если считать, что У
Не самки по
х составляло 490 особей. Та
разом,
10 хрысят, то поголовье молоды
ВИ
бе
зом,
ли 553 взрослые кр
лощадью 900 м’ обитали
1
в помещении свинофермы и
ь
плюс 490 молодых, т. е. все поголовье составляло 1043 особи и на 1
холилось более одной крыс.
|
" мене ирише исследования мы провели в июле 1984 г. в Узбевиота то
Е
й
б
‹олхоза. Там числеп-
‹
‹озяйства рыборазводного к ва.
в свинарнике подсобного хо
<
:от
ность рые была еще выше— па 1 м’ приходилось 4 особи. Такие. ие
данные 0 численности крыс па свинофермах Украины привод
—
би на1м..
И. Абеленцев (1956) — 4 осо
ь О весьма высокой численности этих грызунов на свинофермах товорят
и визуальные наблюдения в Узбекиотане, где пам дпем ино Приходи
590 особей, а на Кубани— 2—5.
еть одновременно 15
,
.
м
СОС боташева (1967: табл. 8), из всех животноводческих помещений
.
;
у
о
,
о больший процент зданий,
встречается значительн
среди свинарников
ой
:
идеть одипочных крыс и д
в которых днем можно в
тЫ Ихет
й
крыс в свинарниках д
Об очень большой численности
„ствует
число их трупов, которые подбирают после применения ядовитых | ри =.
нок. В. С. Никул (1978) указывает, что в свинарниках Молд
Экология 109
Таблица 6. Заселенноеть пасюками животноводческих
помещений (Кировекая область) но визуальным
наблюдениям (по: Остатев, 1967)
Доля зцаний, в которых
днем, обнаружены кры-
Ра
сы, %
Объекты
Все оваНо
зданий
отдельные
труппы
особи
особей
Свинарники
784
37,2
6,8
Птичники
321
14.8
34
Коровники
2127
9,6
81
Телятники
1663
10,0
1,0
Конютни
2480
3,8
0,8
Овчарни
439
4,4
0
дератизации собирали до полутора тысяч трупоз. Н. П. Козлов (4961)
в Казахстане лишь в одном здании собрал более тысячи трупов (рие. 28).
Конечно, такое обилие крыс наблюдается не повсеместно: там, где.
дератизация проводится регулярно, опи не достигают большой численно-
сти, хотя дератизационные мероприятия в большинстве случаев приводят
лишь к временному спижению плотности поселения зверьков, которая в
течение ряда лет колеблется в пебольших пределах. Полностью ликвиди-
ровать пасюков чрезвычайно трудно.
Современные животноводческие комплексы, построенные из железобе-
тона, где бетонирован пол и нет подполья и деревянных настилов, каза-
лось бы, менее благоприятны для обитания крыс, чем деревянные и са-
манные постройки, однако этих грызунов в них бывает также очень
много: они хорошо приспосабливаются к жизни и в таких условиях, ис-
пользуя все трещины и места, где разрушастся цемепт. Их поры часто
находятся под кормушками и там, где имеются отверстия между основа-
пием стен и полом. Поэтому в давно построенных зданиях, где появились
уже трещины и дефекты стен, численность крыс может быть особенно
велика. Пасюки обитают в канализационных каналах свинарников и в
вентиляционной системе, особенно если она бездействует. Здесь зверьки
часто спасаются от дератизационных мероприятий. В современных комп-
лексах эти грызуны держатся в подсобных помещениях, где готовят корм
свиньям, несмотря на то что там во время нагревания котлов стоит шум
от электромоторов и сильный запах от дизельного топлива. Много их бы-
вает в постройках, под приемным бункером для корма, под кормовым
транспортером и т. п. В тамбурах серые крысы селятся под весами для
свиней.
В одном из хозяйств были проведены истребительные работы. На про-
тяжении полутора месяцев собрано на поверхности 6827 крыс, при подия-
тий кровли их подобрано еще 42 758 штук. Часть трупов не была обна-
ружена, так как они остались в подземных коммупикациях, в траве,
108 Глава третья
.
Рис. 97. Изменение температуры 803-
.
духа по месяцам в помещении для саи-
ней (1) и в подполье (2) (по А. Н. Коз-
лову, 1980а)
где готовится корм, возле обогрева-
тельных труб, калориферов, на окнах
с солнечной стороны и т. д. Летом
часть зверьков выселяется из свинар-
ника и обитает в норах © наружной
их стороны. Некоторые пасюки ухо-
дят из помещений на зпачительные
расстояния.
Численность крыс зв некоторых
свинарниках достигает колоссальных
от-
размеров — на Кубани в марте 1982 г. в р
свинариико, те о
Е
ы
.Д.),
пия зверьков (кал, погрызы,
мечали много следов пребыва
в
торм и т. Жо,
о объективные ноказатели.
пами были получены достаточн
Моя
5
тили их отрезанием пальцев
живоловками 58 крыс, поме
‚
и
выустнии
ке на месте поимки. Далее нами
обратно в том же свипарии
г Дале т были обло.
ы иииздобытых
крыс
лепы все помещения капкапам
Крыс моче ими
свинариике обитали 955 взр
зались 13. Следовательно, в
|
К этому количеству следует прибавить и число молодых крысят, которыс
еще пе покипули материнских гнезд: из 124 отловленпых капканачи
взрослых крыс самок было 54, из пих 41 заведомо патио
дедов
р
`
из которых
-
ии было 227 взрослых самок,
тельно, всего в помещен
к, Иа которых 5 35
й
-
ыводки (формулу расчет , . ВТ,
бей лактировали, т. е. имели в
.
г
ВЫ.
й
ый по вскрытию беремент
}
Средний выводок (устаповленн
1
й
Е мки имелось
у
каждой лактирующей са
был 10,4. Если считать, что У
Не самки по
х составляло 490 особей. Та
разом,
10 хрысят, то поголовье молоды
ВИ
бе
зом,
ли 553 взрослые кр
лощадью 900 м’ обитали
1
в помещении свинофермы и
ь
плюс 490 молодых, т. е. все поголовье составляло 1043 особи и на 1
холилось более одной крыс.
|
" мене ирише исследования мы провели в июле 1984 г. в Узбевиота то
Е
й
б
‹олхоза. Там числеп-
‹
‹озяйства рыборазводного к ва.
в свинарнике подсобного хо
<
:от
ность рые была еще выше— па 1 м’ приходилось 4 особи. Такие. ие
данные 0 численности крыс па свинофермах Украины привод
—
би на1м..
И. Абеленцев (1956) — 4 осо
ь О весьма высокой численности этих грызунов на свинофермах товорят
и визуальные наблюдения в Узбекиотане, где пам дпем ино Приходи
590 особей, а на Кубани— 2—5.
еть одновременно 15
,
.
м
СОС боташева (1967: табл. 8), из всех животноводческих помещений
.
;
у
о
,
о больший процент зданий,
встречается значительн
среди свинарников
ой
:
идеть одипочных крыс и д
в которых днем можно в
тЫ Ихет
й
крыс в свинарниках д
Об очень большой численности
„ствует
число их трупов, которые подбирают после применения ядовитых | ри =.
нок. В. С. Никул (1978) указывает, что в свинарниках Молд
Экология 109
Таблица 6. Заселенноеть пасюками животноводческих
помещений (Кировекая область) но визуальным
наблюдениям (по: Остатев, 1967)
Доля зцаний, в которых
днем, обнаружены кры-
Ра
сы, %
Объекты
Все оваНо
зданий
отдельные
труппы
особи
особей
Свинарники
784
37,2
6,8
Птичники
321
14.8
34
Коровники
2127
9,6
81
Телятники
1663
10,0
1,0
Конютни
2480
3,8
0,8
Овчарни
439
4,4
0
дератизации собирали до полутора тысяч трупоз. Н. П. Козлов (4961)
в Казахстане лишь в одном здании собрал более тысячи трупов (рие. 28).
Конечно, такое обилие крыс наблюдается не повсеместно: там, где.
дератизация проводится регулярно, опи не достигают большой численно-
сти, хотя дератизационные мероприятия в большинстве случаев приводят
лишь к временному спижению плотности поселения зверьков, которая в
течение ряда лет колеблется в пебольших пределах. Полностью ликвиди-
ровать пасюков чрезвычайно трудно.
Современные животноводческие комплексы, построенные из железобе-
тона, где бетонирован пол и нет подполья и деревянных настилов, каза-
лось бы, менее благоприятны для обитания крыс, чем деревянные и са-
манные постройки, однако этих грызунов в них бывает также очень
много: они хорошо приспосабливаются к жизни и в таких условиях, ис-
пользуя все трещины и места, где разрушастся цемепт. Их поры часто
находятся под кормушками и там, где имеются отверстия между основа-
пием стен и полом. Поэтому в давно построенных зданиях, где появились
уже трещины и дефекты стен, численность крыс может быть особенно
велика. Пасюки обитают в канализационных каналах свинарников и в
вентиляционной системе, особенно если она бездействует. Здесь зверьки
часто спасаются от дератизационных мероприятий. В современных комп-
лексах эти грызуны держатся в подсобных помещениях, где готовят корм
свиньям, несмотря на то что там во время нагревания котлов стоит шум
от электромоторов и сильный запах от дизельного топлива. Много их бы-
вает в постройках, под приемным бункером для корма, под кормовым
транспортером и т. п. В тамбурах серые крысы селятся под весами для
свиней.
В одном из хозяйств были проведены истребительные работы. На про-
тяжении полутора месяцев собрано на поверхности 6827 крыс, при подия-
тий кровли их подобрано еще 42 758 штук. Часть трупов не была обна-
ружена, так как они остались в подземных коммупикациях, в траве,
140
Глава третья
Рис. 28. Кучи мертвых крыс, собранных после истребительных работ. Фото А. Н.Коз-
лова
Рис. 29. Крысы, поедающие яйца в курятнике. Фото Д. Ф. Траханова
Экология 114
около зданий и т. д. Оставшимся живым крысам продолжали выклады-
вать приманку, и они съедали ее около 1 кг в сутки в каждом здании,
следовательно, живых зверьков осталось еще достаточное количество.
Почти так же сильно, как свинарпики, бывают заселены крысами и
птичники. В них относительно постоянный микроклимат, изобилие кор-
ма (в том числе и куриных яиц; рис. 29), воды и много удобных мест
для устройства гнезд. Однако в отличие от свинарников корм в птични-
кс сухой, что менее привлекательно для влаголюбивых крыс. На птице-
фермах зверьков всогда больше в помещении с цыплятами, где темпера-
тура поддерживается в пределах 25°. В Кустанайской области в одном
курятнике после истробительных работ на площади 1800 м” было собра-
но 900 трупов крыс (Борисенко, 1969).
В птичниках крысы, по наблюдениям А. Н. Козлова (1983а), в ос-
новном селятся в трещинах бетонного пола, за металлическими баками и
бочками, в местах ввода в помещение водонроводных, обогревательных и
канализационных труб, по углам, в калориферных отсеках, вокруг кор-
мосмесителей. Особенно много крысиных нор внутри пометного короба в
слое высохшего помета, тде он по бокам короба скребками транспортера
плохо удаляется. В помещениях с клоточным содержанием кур крысы
живут вдоль лотка, по которому проходит лента пометного транспортера;
в вентиляционных коробах, устаповлепных вдоль стен и открывающихся
впутрь помещения отдушинами с металлической сеткой, в неглубоких
выемках вдоль степ, где проложены обогревательные трубы.
В телятниках крыс несколько меньше, чем в свинарпиках и птични-
ках, численность их в этих помещениях лимитируется количеством кор-
ма. Телятам в течение дня, кроме зеленого корма, дают сравнительно
немпого зерновой подкормки, которую они быстро съедают, и крысам
достается незначительное количество зерен, просыпавшихея сквозь щели
кормушек.
В коровпиках пригодного для крыс корма еще меньше. Короз кормят
зеленым кормом, зерна не дают совсем, а комбикорма дают в очень не-
большом количестве, и скот быстро его съедает. Поэтому пасюки в телят-
никах и коровниках концентрируются вокруг кладовых и ларей с ком-
бикормом. В тех помещениях, где комбикорм не доступен для крыс, их
очень мало или нет совсем.
Мест для устройства убежищ крыс в коровниках также пемного. В од-
ном из подмосковных колхозов, где в кормовом отсеке пол был зацемен-
тиронан, крысы вынуждены были устраивать свои убежища между по-
толком и крышей. Они лазили туда по углам вертикальных кирпичных
стен. Интересно. что в таких помещениях жили только молодые, некруп-
ные крысы, т. е., вероятно, те особи, которым легче было лазать.
В конюшнях и овчарнях крыс обычно мало. Здесь, как правило, для
них нет хороших условий существования. Таким образом, по заселенно-
сти пасюками, несомненно, на первом месте стоят свянарники и птич-
ники. По численпости же крыс в помещениях с другими сельскохозяй-
ственными животными конкретных данных нет, так как в копюпшях,
овчарнях, да и в коровниках зверьков обычно не истребляют и соответст»
140
Глава третья
Рис. 28. Кучи мертвых крыс, собранных после истребительных работ. Фото А. Н.Коз-
лова
Рис. 29. Крысы, поедающие яйца в курятнике. Фото Д. Ф. Траханова
Экология 114
около зданий и т. д. Оставшимся живым крысам продолжали выклады-
вать приманку, и они съедали ее около 1 кг в сутки в каждом здании,
следовательно, живых зверьков осталось еще достаточное количество.
Почти так же сильно, как свинарпики, бывают заселены крысами и
птичники. В них относительно постоянный микроклимат, изобилие кор-
ма (в том числе и куриных яиц; рис. 29), воды и много удобных мест
для устройства гнезд. Однако в отличие от свинарников корм в птични-
кс сухой, что менее привлекательно для влаголюбивых крыс. На птице-
фермах зверьков всогда больше в помещении с цыплятами, где темпера-
тура поддерживается в пределах 25°. В Кустанайской области в одном
курятнике после истробительных работ на площади 1800 м” было собра-
но 900 трупов крыс (Борисенко, 1969).
В птичниках крысы, по наблюдениям А. Н. Козлова (1983а), в ос-
новном селятся в трещинах бетонного пола, за металлическими баками и
бочками, в местах ввода в помещение водонроводных, обогревательных и
канализационных труб, по углам, в калориферных отсеках, вокруг кор-
мосмесителей. Особенно много крысиных нор внутри пометного короба в
слое высохшего помета, тде он по бокам короба скребками транспортера
плохо удаляется. В помещениях с клоточным содержанием кур крысы
живут вдоль лотка, по которому проходит лента пометного транспортера;
в вентиляционных коробах, устаповлепных вдоль стен и открывающихся
впутрь помещения отдушинами с металлической сеткой, в неглубоких
выемках вдоль степ, где проложены обогревательные трубы.
В телятниках крыс несколько меньше, чем в свинарпиках и птични-
ках, численность их в этих помещениях лимитируется количеством кор-
ма. Телятам в течение дня, кроме зеленого корма, дают сравнительно
немпого зерновой подкормки, которую они быстро съедают, и крысам
достается незначительное количество зерен, просыпавшихея сквозь щели
кормушек.
В коровпиках пригодного для крыс корма еще меньше. Короз кормят
зеленым кормом, зерна не дают совсем, а комбикорма дают в очень не-
большом количестве, и скот быстро его съедает. Поэтому пасюки в телят-
никах и коровниках концентрируются вокруг кладовых и ларей с ком-
бикормом. В тех помещениях, где комбикорм не доступен для крыс, их
очень мало или нет совсем.
Мест для устройства убежищ крыс в коровниках также пемного. В од-
ном из подмосковных колхозов, где в кормовом отсеке пол был зацемен-
тиронан, крысы вынуждены были устраивать свои убежища между по-
толком и крышей. Они лазили туда по углам вертикальных кирпичных
стен. Интересно. что в таких помещениях жили только молодые, некруп-
ные крысы, т. е., вероятно, те особи, которым легче было лазать.
В конюшнях и овчарнях крыс обычно мало. Здесь, как правило, для
них нет хороших условий существования. Таким образом, по заселенно-
сти пасюками, несомненно, на первом месте стоят свянарники и птич-
ники. По численпости же крыс в помещениях с другими сельскохозяй-
ственными животными конкретных данных нет, так как в копюпшях,
овчарнях, да и в коровниках зверьков обычно не истребляют и соответст»
412
Глава третья
{
чис-
веппо не полечитывают трупы. Но если бы были такио данные по си
ленности крыс, то разница была бы еще более значительна, чем
з
еленности зданий.
.
° ” Летом крысы могут жить вне животноводческих строений, р
а пометного т
-
колодцев, в местах выход
вокруг канализационных
,
ы
‚пор.
те Ве т. п. Чаеть их выселяются из помещений и обитают рядом с ним :
|: адах, огородах, зарослях бурьяна, на свалках, возле а
1х
<
ако питаться большинство ;
ах дров и мусора. Оди
о
о
НЫ
свиней и птиц. В это вре
х
икам — к кормушкам
ходит к прежним источи
-
я они часто поселяются в яме под весами и в будке весовщика, тд
ения холодов.
тут жить вплоть до наступл
..
мо Общие биологические особенности крыс, населяющих животноводче
:
хоро
-
к следующему: благодаря
скис помещения, сводятся
оротой обеовельа-
ы размножаются здесь кру
ности кормами эти грызуп
.
-
И жительность жизни у них выше, чем у эквовытропной формы. В род
оживают до 1,5—2 лет. Нром
,
нем в помещениях крысы д
Оотор
й итанностью и более круппыми размерами.
отличаются большой уп
)
и
сравнении одновозрастных крыс экзоантропной и синантропной фор
ь
пнее.
инантропные обычно бывают кру
.
,
С Наличие животноводческих помещений имеет КООальНОО очен
й
н
с. Попав случайпо вместе с тр
при расширении ареала крыс.
в
вые районы, зверьки быстро заселяют наиболее НИЙ ‚стан
х
тде начинают интенсивно
ии — животноводческие фермы,
-
ся В ряде территорий нашей страны— Голодная степь, Ростомекая 0б-
пасть Калмыцкая АССР, Узбекистаи
— расселение пою было ‹ т
с
ми
р
р
водческими строения
но исключительно с животно
иями
1
|
1961). Таким же путем им удалось освоить районы о
Сенера,
Гыдалский полуостров, Таймыр и др. (Кучерук, „Кузиков, ва р ерыь
сельс
}
.
Прочие помещения в
ыо
рыбы нередко заселяют складские помещения для хранения пище ‚х
продуктов, где устраивают гпезда прямо н мешках с сыпучими пр ук
у
й
н
3
Бывают они многочисленны и
тами, например с мукой.
(Очиров Бондарчук, 1963; Степин, 1952; Хамаганов, 1968) и Крыс ош
р
_-.
.
зерна, не являютея д
-
Склады, где содержатся запасы
,
ают влажный
межки, так как опи предпочит
мальными местами для корь
,
корм а не сухой. Обычно от таких складов тянутся длинные, хорото
се склад.
-
Г
;
ою. Однако в Словакии в
протоптанные дорожки к водой
>
мещения сольскоховяйственных кооперативов были заражены крысам
ОТе1Аг, 1980).
я
Крысы бывают многочисленны в овощехранилищах и на звероформ
Очиров, Бондарчук, 1963).
.
В, Сродной Азии пасюки заселяют пункты, в которых хранятся хло
пок или сто семена. Заселяются также старые стога сена (Тонышева,
ПШаяндяпан, 1968).
Экология
113
Город
Малые города (до 100 тыс. населения) по условиям обитания для крыс
ненамного отличаются от населенных пунктов в сельской местности.
Крупные же города (до 1 млн жителей) и крупнейшие (свыше 1 млн)
представляют собой очень сложную экологическую систему, где наряду
с человеком обитает множество других живых организмов, в большей
или менышей степени связанных друг с другом и зависящих от среды
обитания.
Среда обитания города
— это комплекс разпообразных как по архи-
тектуре, так и по хозяйственному использованию строепий, инженерных
сооружений и коммуникаций. Применение нозых строительных материа-
лов, сооружение крупных многоэтажных зданий, мнотоцелевое назначе-
ние строепий, густая сеть подземных коммуникаций создают совершенно
особые условия существования синантроппых грызунов в современных
городах.
Подземные системы (канализация, сеть тепло- и газопроводов, элект-
рокабель) создают для обитания крыс особенно благоприятные условия.
В городах всегда есть незастроенные участки: парки, бульвары, сады,
пустыри, а также неудобные для строительства места
— овраги, косого-
ры, поймы речушек и пр. Здания, заасфальтированные улицы и площа-
ди излучают накопленное теплю, строения задерживают ветер. Дождевые
воды отводятся непосредственио в канализационпые каналы, вследствие
чего значительно снижастся поверхность испарения. В результате в го-
родах более теплый и менее влажный микроклимат, характеризующийся
меньшей скоростью ветра.
Еще Т. В. Дмитриева (1971) отмечала большую приуроченность се-
рых крыс к крупным городам. По ее данным, крысы в них встречаются
чаще, чем в небольших поселках городского типа. Например, в г. Воро-
неже они составляют 21,7% от общего числа выловленных грызунов, а в
поселках городского типа — всего 18,3%.
Многие территории, не занятые ранее крысой, стали заселяться ес
именно с городов. Например, в Западной Сибири пасюк в первую оче-
редь заселил города Омск и Томек (Кащенко, 1912; Зверев, 1930; Ля-
лин, 1973), в Средней Азии — г. Ташкент (Колесников, 1952).
илая
— самая большая зона в современных городах— занимает по
площади две трети ноего города. Ее основные компоненты — жилые дома,
которые крысы заселяют по-разному, что зависит от многих причин:
1) от материала, из которого построены дома; 2) от архитектурных осо-
бенностей (в частности, от числа этажей); 3) от типа хозяйственного
использования; 4) санитарного состояния; 5) от взаимного расположе-
ния зданий.
Более всего бызают заселены крысами, как и в сельской местности,
деревянные строения, стены хоторых зверьки легко прогрызают.
В начале века ббльшая часть городов состояла из деревянных или
смешанных (дерево и «дикий» камень) зданий и пасюков в них было
очень много. Например, в Москве в это время деревянные и смешанные
412
Глава третья
{
чис-
веппо не полечитывают трупы. Но если бы были такио данные по си
ленности крыс, то разница была бы еще более значительна, чем
з
еленности зданий.
.
° ” Летом крысы могут жить вне животноводческих строений, р
а пометного т
-
колодцев, в местах выход
вокруг канализационных
,
ы
‚пор.
те Ве т. п. Чаеть их выселяются из помещений и обитают рядом с ним :
|: адах, огородах, зарослях бурьяна, на свалках, возле а
1х
<
ако питаться большинство ;
ах дров и мусора. Оди
о
о
НЫ
свиней и птиц. В это вре
х
икам — к кормушкам
ходит к прежним источи
-
я они часто поселяются в яме под весами и в будке весовщика, тд
ения холодов.
тут жить вплоть до наступл
..
мо Общие биологические особенности крыс, населяющих животноводче
:
хоро
-
к следующему: благодаря
скис помещения, сводятся
оротой обеовельа-
ы размножаются здесь кру
ности кормами эти грызуп
.
-
И жительность жизни у них выше, чем у эквовытропной формы. В род
оживают до 1,5—2 лет. Нром
,
нем в помещениях крысы д
Оотор
й итанностью и более круппыми размерами.
отличаются большой уп
)
и
сравнении одновозрастных крыс экзоантропной и синантропной фор
ь
пнее.
инантропные обычно бывают кру
.
,
С Наличие животноводческих помещений имеет КООальНОО очен
й
н
с. Попав случайпо вместе с тр
при расширении ареала крыс.
в
вые районы, зверьки быстро заселяют наиболее НИЙ ‚стан
х
тде начинают интенсивно
ии — животноводческие фермы,
-
ся В ряде территорий нашей страны— Голодная степь, Ростомекая 0б-
пасть Калмыцкая АССР, Узбекистаи
— расселение пою было ‹ т
с
ми
р
р
водческими строения
но исключительно с животно
иями
1
|
1961). Таким же путем им удалось освоить районы о
Сенера,
Гыдалский полуостров, Таймыр и др. (Кучерук, „Кузиков, ва р ерыь
сельс
}
.
Прочие помещения в
ыо
рыбы нередко заселяют складские помещения для хранения пище ‚х
продуктов, где устраивают гпезда прямо н мешках с сыпучими пр ук
у
й
н
3
Бывают они многочисленны и
тами, например с мукой.
(Очиров Бондарчук, 1963; Степин, 1952; Хамаганов, 1968) и Крыс ош
р
_-.
.
зерна, не являютея д
-
Склады, где содержатся запасы
,
ают влажный
межки, так как опи предпочит
мальными местами для корь
,
корм а не сухой. Обычно от таких складов тянутся длинные, хорото
се склад.
-
Г
;
ою. Однако в Словакии в
протоптанные дорожки к водой
>
мещения сольскоховяйственных кооперативов были заражены крысам
ОТе1Аг, 1980).
я
Крысы бывают многочисленны в овощехранилищах и на звероформ
Очиров, Бондарчук, 1963).
.
В, Сродной Азии пасюки заселяют пункты, в которых хранятся хло
пок или сто семена. Заселяются также старые стога сена (Тонышева,
ПШаяндяпан, 1968).
Экология
113
Город
Малые города (до 100 тыс. населения) по условиям обитания для крыс
ненамного отличаются от населенных пунктов в сельской местности.
Крупные же города (до 1 млн жителей) и крупнейшие (свыше 1 млн)
представляют собой очень сложную экологическую систему, где наряду
с человеком обитает множество других живых организмов, в большей
или менышей степени связанных друг с другом и зависящих от среды
обитания.
Среда обитания города
— это комплекс разпообразных как по архи-
тектуре, так и по хозяйственному использованию строепий, инженерных
сооружений и коммуникаций. Применение нозых строительных материа-
лов, сооружение крупных многоэтажных зданий, мнотоцелевое назначе-
ние строепий, густая сеть подземных коммуникаций создают совершенно
особые условия существования синантроппых грызунов в современных
городах.
Подземные системы (канализация, сеть тепло- и газопроводов, элект-
рокабель) создают для обитания крыс особенно благоприятные условия.
В городах всегда есть незастроенные участки: парки, бульвары, сады,
пустыри, а также неудобные для строительства места
— овраги, косого-
ры, поймы речушек и пр. Здания, заасфальтированные улицы и площа-
ди излучают накопленное теплю, строения задерживают ветер. Дождевые
воды отводятся непосредственио в канализационпые каналы, вследствие
чего значительно снижастся поверхность испарения. В результате в го-
родах более теплый и менее влажный микроклимат, характеризующийся
меньшей скоростью ветра.
Еще Т. В. Дмитриева (1971) отмечала большую приуроченность се-
рых крыс к крупным городам. По ее данным, крысы в них встречаются
чаще, чем в небольших поселках городского типа. Например, в г. Воро-
неже они составляют 21,7% от общего числа выловленных грызунов, а в
поселках городского типа — всего 18,3%.
Многие территории, не занятые ранее крысой, стали заселяться ес
именно с городов. Например, в Западной Сибири пасюк в первую оче-
редь заселил города Омск и Томек (Кащенко, 1912; Зверев, 1930; Ля-
лин, 1973), в Средней Азии — г. Ташкент (Колесников, 1952).
илая
— самая большая зона в современных городах— занимает по
площади две трети ноего города. Ее основные компоненты — жилые дома,
которые крысы заселяют по-разному, что зависит от многих причин:
1) от материала, из которого построены дома; 2) от архитектурных осо-
бенностей (в частности, от числа этажей); 3) от типа хозяйственного
использования; 4) санитарного состояния; 5) от взаимного расположе-
ния зданий.
Более всего бызают заселены крысами, как и в сельской местности,
деревянные строения, стены хоторых зверьки легко прогрызают.
В начале века ббльшая часть городов состояла из деревянных или
смешанных (дерево и «дикий» камень) зданий и пасюков в них было
очень много. Например, в Москве в это время деревянные и смешанные
114
Глава третья
ы
*
щ
у
ены
ания занимали 70
пл ади и грыз нами были заселен: на 90
Л
у
[о
здани
ые мел-
)
вянными являются олько сам
Теперь в больших городах дере
КИ. остро
ар
алатки бараки. Они часто бывают и кар
—
е
р йки: ларьки, п.
.
аркасно
засыпными, почти никогда не имеют углубленного фундамента, и кры-
сы. легко прогрызая деревянные степки, устраивают гнезда в грунте под
полом, в проетрапстве между чистым и черным полом или в пустотах
стен.
Олеяр
городах Словакии деревянных Потро
978%, а постройки из камня —н
ф
’
коз — всего на 4%.
Кирпичные здания
ленность крысами в
15
: ет на большую засе
О,
По его данным, они заселены на
и. наконец, из бетонных бло-
з
П-
обычно заселены крысами меньгие,, чем дерев
азуют
-
Олнако з старых зданиях, где кирпич раврушается, образу
шины=т
м тва. дапий Уцотребляют главным
ля строитель
авнмм
настоящее время д
потребляют т
б бы железобетонные конструкции. я
пВы
о т минием а также синтетические и другие ед,
На.
в
|
ы легко
ерые крые
-
ыгие материалам с
тя,
я
устройства снойх гнезд они с успехом ИополнУ от поно.
Рю вуКо и топлоизоляционные прослойки, а вк ра
Г
р
апт
.
товь ой прокладывают ходы (Мелкова, Кун: ши 198). боспочива-
Вт Полежаев (4947) выделяет пять условий, рые обои
‹
.
. В. е оступность («крысонепропицаемость») помещен Я алые
ют Нн т ие бетонированных или цемоптироваппых не о тя ыы
1) нали и на окнах или вентиляционных отверстиях, толь.
том хоста сопряжений трубопроводов со стенами; , удовл и
ое ение дверей (т. е. отсутствие возможности пр
ное соедин
.в.
можно
НЫ
бра м Москве свидетельствует о том, что в
в
,
ы
з 498 анкет, собранных
ото том,
что
и
‹отда все приведенные выше показатели отсу уют, по
о
лены
ами почти на 100%. При наличии всех по я
Й
селены крыс
Ио
те
ИИ оопопроницаемости» зассленными оказались т
,
леи «<
|
'
ании зданий
и
б
олучены при обследов
таты были полу
зданий
налогичные резуль т
а
Л р ада (Судейкип, 1986). Автор полагает, т о пабе оу
ое стнисм, влияющим на заселение крыс
троек, являет
пят
2
Ио бетонирозапного или цомоптированного по а
одних нии,
ше емкость местообитания и
й, тем бол
Й
тих нарушении, т
ен
ем, ие ре можст в нем поселиться. Показано, что Во одах С -
ак более заселенными пасюками оказались дома, построонтые в
м
уда В военные и послевоенные годьт, когда качество матер зла и ра.
вы6
изкими, поэтому здания имели зн
ичест-
боты были более н
рые крысы могли использовать. В дома т
й
стот, кото
оРМ
зо щелей Е 41920 гг., были заселены знерькамт ма в)
О
на 29%, а после 1965 г. всего ма 17,8% (О1е]Аг,
тг.
‚
Экология
115
Как правило, вие зависимости от того, из какого материала здапис
построено, серые крысы заселяют в большей степени подвальные поме-
щения (78,7%) и первый этаж (8,5%) (Кузякин, 1950).
Приуроченность пасюков к подвалам и первым этажам отмечается и
в настоящее нремя в домах современной конструкции. Доля зверьков от
числа всех выловлепных особей в подвалах и на первых этажах состав-
ляла 93,5%, а на вторых этажах — только 6,5% (Судейкин, 1986).
Серая крыса находит оптимальные условия обитания в подвалах.
Люди относительно редко их посещают, там полумрак, повышениая влаж-
ность, а щели, через которые можно проникнуть в пустоты, обеспечива-
ют пасюкам удобные места для устройства гнозд. Правда, есть сообще-
пия о том, что крысы иногда не только проникают в верхние этажи,
но и размножаются там. В Ленинграде гостиница, сданная в экеплуата-
цию в 1970 г., оказалась заселенной серыми крысами от подвалов до
чердака (Лысов, 1984).
В. А. Судейкин (1986) описывает случай, когда в Москве в 1947 г.
семья серых крыс была обпаружена на 9-м этаже гостиницы «Москва».
Иногда поселение крысе на чердаках определяется наличием там то.
лубей, птенцы и яйца которых привлекают их (Романов, 1963). Но, ве-.
роятно, эти случаи падо рассматривать как исключения. Заселенность
зданий крысами зависит не только от материала, из которого они по-
строены, но в значительной степени от их этажности.
Современная жилая зона характеризуется смешанной застройкой зда-
пиями различной этажности. В крупнейших и крупных городах предус-
матривается смешанная застройка зданиями в девять и более этажей с
застичным возведением пятиэтажных зданий. В побольших городах и
поселках строятся, как правило, пятиэтажные жилые дома.
Важно подчеркнуть, что характерным и неотъемлемым - элементом
многоэтажного жилого дома (свыше 5 этажей) является система мусо-
ропроводов, что оказалось весьма важным фактором, влияющим на за-
осленность зданий крысами.
В зданиях с малым и средним количеством этажей иместся бачковая
или контейнерная система сбора бытовых отходов. Бачки располагаются
на площадке не далос 100 м от жилых домов, причем одна площадка
обычно обеспечивает несколько домов. Следовательно, крысы, населяю-
щие подвалы, должны пробегать до места копцентрации корма пемалое
расстояние по открытой местности, подвергая себя определенному риску.
В многоэтажных жилых домах (от 6 этажей и зыше) бытовые отхо-
ды собираются в мусорокамеры, расположенные внутри здания. Таким
образом, в этом случае крысы имеют возможность кормиться, не выходя
из подвала, где они яхивут.
Вывоз бытовых отходов осуществляется далеко не регулярно, но
даже в случае ожедневной очистки мусорокамер отходы скапливаются в
них и крысы обеспечены кормом. Весьма важно и т0, что в многоэтаж-
ных домах сбор бытовых отходов осуществляется в каждой секции дома,
следозательно, количество точек, где могут кормиться крысы, во много
раз больше, чем на той же площади, где располагаются малоэтажные
114
Глава третья
ы
*
щ
у
ены
ания занимали 70
пл
ади и грыз нами были заселен:
на 90
Л
у
[о
здани
ые мел-
)
вянными являются олько сам
Теперь в больших городах дере
КИ. остро
ар
алатки бараки. Они часто бывают и кар
—
е
р йки: ларьки, п.
.
аркасно
засыпными, почти никогда не имеют углубленного фундамента, и кры-
сы. легко прогрызая деревянные степки, устраивают гнезда в грунте под
полом, в проетрапстве между чистым и черным полом или в пустотах
стен.
Олеяр
городах Словакии деревянных Потро
978%, а постройки из камня —н
ф
’
коз — всего на 4%.
Кирпичные здания
ленность крысами в
15
: ет на большую засе
О,
По его данным, они заселены на
и. наконец, из бетонных бло-
з
П-
обычно заселены крысами меньгие,, чем дерев
азуют
-
Олнако з старых зданиях, где кирпич раврушается, образу
шины=т
м тва. дапий Уцотребляют главным
ля строитель
авнмм
настоящее время д
потребляют т
б бы железобетонные конструкции. я
пВы
о т минием а также синтетические и другие ед,
На.
в
|
ы легко
ерые крые
-
ыгие материалам с
тя,
я
устройства снойх гнезд они с успехом ИополнУ от поно.
Рю вуКо и топлоизоляционные прослойки, а вк ра
Г
р
апт
.
товь ой прокладывают ходы (Мелкова, Кун: ши 198). боспочива-
Вт Полежаев (4947) выделяет пять условий, рые обои
‹
.
. В. е оступность («крысонепропицаемость») помещен Я алые
ют Нн т ие бетонированных или цемоптироваппых не о тя ыы
1) нали и на окнах или вентиляционных отверстиях, толь.
том хоста сопряжений трубопроводов со стенами; , удовл и
ое ение дверей (т. е. отсутствие возможности пр
ное соедин
.в.
можно
НЫ
бра м Москве свидетельствует о том, что в
в
,
ы
з 498 анкет, собранных
ото том,
что
и
‹отда все приведенные выше показатели отсу уют, по
о
лены
ами почти на 100%. При наличии всех по я
Й
селены крыс
Ио
те
ИИ оопопроницаемости» зассленными оказались т
,
леи «<
|
'
ании зданий
и
б
олучены при обследов
таты были полу
зданий
налогичные резуль т
а
Л р ада (Судейкип, 1986). Автор полагает, т о пабе оу
ое стнисм, влияющим на заселение крыс
троек, являет
пят
2
Ио бетонирозапного или цомоптированного по а
одних нии,
ше емкость местообитания и
й, тем бол
Й
тих нарушении, т
ен
ем, ие ре можст в нем поселиться. Показано, что Во одах С -
ак более заселенными пасюками оказались дома, построонтые в
м
уда В военные и послевоенные годьт, когда качество матер зла и ра.
вы6
изкими, поэтому здания имели зн
ичест-
боты были более н
рые крысы могли использовать. В дома т
й
стот, кото
оРМ
зо щелей Е 41920 гг., были заселены знерькамт ма в)
О
на 29%, а после 1965 г. всего ма 17,8% (О1е]Аг,
тг.
‚
Экология
115
Как правило, вие зависимости от того, из какого материала здапис
построено, серые крысы заселяют в большей степени подвальные поме-
щения (78,7%) и первый этаж (8,5%) (Кузякин, 1950).
Приуроченность пасюков к подвалам и первым этажам отмечается и
в настоящее нремя в домах современной конструкции. Доля зверьков от
числа всех выловлепных особей в подвалах и на первых этажах состав-
ляла 93,5%, а на вторых этажах — только 6,5% (Судейкин, 1986).
Серая крыса находит оптимальные условия обитания в подвалах.
Люди относительно редко их посещают, там полумрак, повышениая влаж-
ность, а щели, через которые можно проникнуть в пустоты, обеспечива-
ют пасюкам удобные места для устройства гнозд. Правда, есть сообще-
пия о том, что крысы иногда не только проникают в верхние этажи,
но и размножаются там. В Ленинграде гостиница, сданная в экеплуата-
цию в 1970 г., оказалась заселенной серыми крысами от подвалов до
чердака (Лысов, 1984).
В. А. Судейкин (1986) описывает случай, когда в Москве в 1947 г.
семья серых крыс была обпаружена на 9-м этаже гостиницы «Москва».
Иногда поселение крысе на чердаках определяется наличием там то.
лубей, птенцы и яйца которых привлекают их (Романов, 1963). Но, ве-.
роятно, эти случаи падо рассматривать как исключения. Заселенность
зданий крысами зависит не только от материала, из которого они по-
строены, но в значительной степени от их этажности.
Современная жилая зона характеризуется смешанной застройкой зда-
пиями различной этажности. В крупнейших и крупных городах предус-
матривается смешанная застройка зданиями в девять и более этажей с
застичным возведением пятиэтажных зданий. В побольших городах и
поселках строятся, как правило, пятиэтажные жилые дома.
Важно подчеркнуть, что характерным и неотъемлемым - элементом
многоэтажного жилого дома (свыше 5 этажей) является система мусо-
ропроводов, что оказалось весьма важным фактором, влияющим на за-
осленность зданий крысами.
В зданиях с малым и средним количеством этажей иместся бачковая
или контейнерная система сбора бытовых отходов. Бачки располагаются
на площадке не далос 100 м от жилых домов, причем одна площадка
обычно обеспечивает несколько домов. Следовательно, крысы, населяю-
щие подвалы, должны пробегать до места копцентрации корма пемалое
расстояние по открытой местности, подвергая себя определенному риску.
В многоэтажных жилых домах (от 6 этажей и зыше) бытовые отхо-
ды собираются в мусорокамеры, расположенные внутри здания. Таким
образом, в этом случае крысы имеют возможность кормиться, не выходя
из подвала, где они яхивут.
Вывоз бытовых отходов осуществляется далеко не регулярно, но
даже в случае ожедневной очистки мусорокамер отходы скапливаются в
них и крысы обеспечены кормом. Весьма важно и т0, что в многоэтаж-
ных домах сбор бытовых отходов осуществляется в каждой секции дома,
следозательно, количество точек, где могут кормиться крысы, во много
раз больше, чем на той же площади, где располагаются малоэтажные
116
Глава третья
дома, где бытовые отходы от нескольких зданий концентрируются в од-
з
ном месте.
.
ичество бытовых отхо
О
ых ах "В значительной степени
мусороприемных
.
дов скаиливается в
р
ПОЙ отерени
чисток (85,71%), немалую д
это отходы из картофельных о
г
аВяЮт остатки весьма питательных иродуктов (мясо 3,4, рыба
й 3,1%)
230 остатки других овощей 8,1%).
.
в К Мелкова провела расчет на количество кормовых единиц в пи-
тевых отходах. Около одного мусоропровода в 12-этажном доме могут
прокормиться от 44 по 62 крыс. Результаты добычи крысе и г
льтаты
0со-
‹
одов показали близкие резу.
около одпого их мусоропров
-
би). Таким образом, собираемые в мусорокамерах бытовые отходы могу
нность крые.
поддерживать высокую числе
|
“р подвалах многоэтажных домов крысы всегда концентрируются око
ло мусороприемных камер. Это хорошо прослежено при отлов пасюков
и при сборе их трупов после проведения истребительных работ. ноже
По поедаемости приманки число обитающих в здания крыс может
достигать 106 особей на 1000 м?. Постоянно проводимые дератизац о
ныс мероприятия позволяют держать численность крыс на гораздо
лее низком уровне.
—
В жилых домах зверьки почти ноегда находят место для питья —
в подвалах нередко стоят лужи воды, но животным достаточно и сов
небольшого количества влаги, которая просачивается из тру . ти их
Заселенность домов крысами зависит не только от о еспеченн их
кормом и водой, но также и от условий, благоприятствующих устр ь
.
Й
я
и
‚-
ну убежищ. Нятиэтажные постройки обычно возводятея не на затлуб-
ленных фундаментах, а на сваях. В подвалах этих домон полы в от о
от многоэтажных строений не бетонирозаны. Крысы легко в ы пр т
кают и имеют возможность рыть поры з земляном трунте (рис.
Судейкин, 1986).
,.
В многоэтажных домах, несмотря на то что там условия для, устро -
ства убежищ крысами хуже, обычно имеется много технических ру-
шений (нарушений целостности порокрытий, мест сопряжений ве г
й
яту-
й
аций и пр.) и зверьки наход
нотрукций, вывода коммуник
-
но мес Я своих убожищ. В. К. Мелкова провела в Москве оболодома
ние 64 мусорокамер многоэтажных Домов и установила, о о ь
илось
н
.
еру в среднем приход
,
случаев на одну мусорокам
Во мпогих случаях были установлены повреждения, обеспечивающие
то знач
с
камер в полости перекрытии, ч
крысам проход от мусоро
е
В
но
йства нор и убежищ.
мест, пригодных для устро
увеличивает число
,
и
рощи
13 жилых домов (площадь
,
следования подвалов
780 м) пот
одвале приходилось 14, р
что на поверхность пола в ГП
ы
100 м”. Это обследование показало также и очень большую захламлон
ность подвалов, в которых часто держат макулатуру, ненужные р тики.
Большую роль в жизни крыс в жилых зданиях играют трубы с р
мы горячего водоснабжения. Трубы большого диаметра яеляютол ил -
пленными местами, где пасюки сидят как поодиночке, так и труп .
Экология 117
Таким образом, в жилых домах городов имеются весьма благоприят-
ные условия для обитания крыс. По сообщению В. С. Судейкина (1986),
в Москве с 1959 по 1968 г. более всего были заселены ими именно жи-
лые дома— на 5644, пищевые предириятия крысы заселяли на 4,9%,
лечебные учреждения— на 0,2%, детские (без школ) — па 0,5% и
прочие 38,3%.
Если в столовых прозодится интенсивная дератизация, то числен-
ность серых крыс в них может быть ниже, чем в жилых помещениях,
но она быстро восстановится из-за обильной и постоянпой кормовой
базы (рис. 31). В прежние годы, когда дератизация не была интенсиз-
на, пищевые предприятия отличались особенно большой численностью
крыс (Дмитриева, 1971).
Промышленная зона. В этой зоне крыс бывает особенно мно-
го в помещениях мясокомбинатов, боен, рыбокомбинатов, на складах и
территориях заводов и фабрик. Великолепная кормовая база, которой
здесь обеспечены пасюки, подчас сочетается с недостаточным соблюдени-
ем санитарных условий. Если, например, территория завода не оборудо-
вана, т. е. не заасфальтирована, загружена свалкой, отдельные участки
заросли бурьяном, то крысы заселяют ес с высокой плотностью. Напри-
мер, в Ярославской области в пос. Поречье-Рыбное на территории коп-`
сервпого завода в 1950-х годах была высокая численность крыс. Специ-
альная комиссия, расследовавшая причины, вызвавшие столь высокую
плотность паселения этих грызунов (10—15% попадания в ловушки),
установила следующее: с территории завода подолгу не вывозили отходы
Рис. 80. Крыса у норы в подвале жилого дома. Фото В. К. Мелковой
116
Глава третья
дома, где бытовые отходы от нескольких зданий концентрируются в од-
з
ном месте.
.
ичество бытовых отхо
О
ых ах "В значительной степени
мусороприемных
.
дов скаиливается в
р
ПОЙ отерени
чисток (85,71%), немалую д
это отходы из картофельных о
г
аВяЮт остатки весьма питательных иродуктов (мясо 3,4, рыба
й 3,1%)
230 остатки других овощей 8,1%).
.
в К Мелкова провела расчет на количество кормовых единиц в пи-
тевых отходах. Около одного мусоропровода в 12-этажном доме могут
прокормиться от 44 по 62 крыс. Результаты добычи крысе и г
льтаты
0со-
‹
одов показали близкие резу.
около одпого их мусоропров
-
би). Таким образом, собираемые в мусорокамерах бытовые отходы могу
нность крые.
поддерживать высокую числе
|
“р подвалах многоэтажных домов крысы всегда концентрируются око
ло мусороприемных камер. Это хорошо прослежено при отлов пасюков
и при сборе их трупов после проведения истребительных работ.
ноже
По поедаемости приманки число обитающих в здания крыс может
достигать 106 особей на 1000 м?. Постоянно проводимые дератизац о
ныс мероприятия позволяют держать численность крыс на гораздо
лее низком уровне.
—
В жилых домах зверьки почти ноегда находят место для питья —
в подвалах нередко стоят лужи воды, но животным достаточно и сов
небольшого количества влаги, которая просачивается из тру . ти их
Заселенность домов крысами зависит не только от о еспеченн их
кормом и водой, но также и от условий, благоприятствующих устр ь
.
Й
я
и
‚-
ну убежищ. Нятиэтажные постройки обычно возводятея не на затлуб-
ленных фундаментах, а на сваях. В подвалах этих домон полы в от о
от многоэтажных строений не бетонирозаны. Крысы легко в ы пр т
кают и имеют возможность рыть поры з земляном трунте (рис.
Судейкин, 1986).
,.
В многоэтажных домах, несмотря на то что там условия для, устро -
ства убежищ крысами хуже, обычно имеется много технических ру-
шений (нарушений целостности порокрытий, мест сопряжений ве г
й
яту-
й
аций и пр.) и зверьки наход
нотрукций, вывода коммуник
-
но мес Я своих убожищ. В. К. Мелкова провела в Москве оболодома
ние 64 мусорокамер многоэтажных Домов и установила, о о ь
илось
н
.
еру в среднем приход
,
случаев на одну мусорокам
Во мпогих случаях были установлены повреждения, обеспечивающие
то знач
с
камер в полости перекрытии, ч
крысам проход от мусоро
е
В
но
йства нор и убежищ.
мест, пригодных для устро
увеличивает число
,
и
рощи
13 жилых домов (площадь
,
следования подвалов
780 м) пот
одвале приходилось 14, р
что на поверхность пола в ГП
ы
100 м”. Это обследование показало также и очень большую захламлон
ность подвалов, в которых часто держат макулатуру, ненужные р тики.
Большую роль в жизни крыс в жилых зданиях играют трубы с р
мы горячего водоснабжения. Трубы большого диаметра яеляютол ил -
пленными местами, где пасюки сидят как поодиночке, так и труп .
Экология 117
Таким образом, в жилых домах городов имеются весьма благоприят-
ные условия для обитания крыс. По сообщению В. С. Судейкина (1986),
в Москве с 1959 по 1968 г. более всего были заселены ими именно жи-
лые дома— на 5644, пищевые предириятия крысы заселяли на 4,9%,
лечебные учреждения— на 0,2%, детские (без школ) — па 0,5% и
прочие 38,3%.
Если в столовых прозодится интенсивная дератизация, то числен-
ность серых крыс в них может быть ниже, чем в жилых помещениях,
но она быстро восстановится из-за обильной и постоянпой кормовой
базы (рис. 31). В прежние годы, когда дератизация не была интенсиз-
на, пищевые предприятия отличались особенно большой численностью
крыс (Дмитриева, 1971).
Промышленная зона. В этой зоне крыс бывает особенно мно-
го в помещениях мясокомбинатов, боен, рыбокомбинатов, на складах и
территориях заводов и фабрик. Великолепная кормовая база, которой
здесь обеспечены пасюки, подчас сочетается с недостаточным соблюдени-
ем санитарных условий. Если, например, территория завода не оборудо-
вана, т. е. не заасфальтирована, загружена свалкой, отдельные участки
заросли бурьяном, то крысы заселяют ес с высокой плотностью. Напри-
мер, в Ярославской области в пос. Поречье-Рыбное на территории коп-`
сервпого завода в 1950-х годах была высокая численность крыс. Специ-
альная комиссия, расследовавшая причины, вызвавшие столь высокую
плотность паселения этих грызунов (10—15% попадания в ловушки),
установила следующее: с территории завода подолгу не вывозили отходы
Рис. 80. Крыса у норы в подвале жилого дома. Фото В. К. Мелковой
148
Глава третья
5
=
>
П
о
п
а
й
т
е
м
о
с
т
ь
9
,
я
=
и|11|
80808
1958 1959 1960 1961 1962 1963 1964 1965 1966 1967 19682.г.
©
|
-
=
|
-
&
в
о
[
-
<
=
©
[
-
>
Е
5
5
[
-
>
|
5
Е
=
в
|
-
=
>
в
-
-
м
=
7
ё
йственного
Рис. 31. Динамика численности серих врыс в о
иное дозя
Ё
пользования в г Москве, 1958—1968 а. (по: Субейкин,
—
илых до:
— на пищевых иредприятиях —вх
стройках. четы:
1 вжил
мах, 2
и х ен чятпях, 8 в хозяйственных постро
У
В — весениие, О — осенние, 8 — зимние
й
Г.
грегам рек
Рис. 32. Изменение численности серой крысы водяной полевки по бер
р
близ клеевого завода под Тулой (по: Панина, 1986а)
1— водная полевка, 8 — насюк
85
Г? 9:
|
9
5-
2
25Г
=ы
$2
25Г
,
ы
.—Г
|
-°
ЗИ!
.
ви!
Г
|о!
0|-
м
У
хаюкноос
о зав
пы ЗВ ЕВЕ ЕЗЕЕБЕЕЕЕЕЕНаНЫ
ЕЕ
ЕВ
Число кии-= 53 3 В В ЗВ ВЗЕ НЕЕ НЕНЕВ
Экологил
119
зеленого горошка, воторые представляли собой витамипизированный
корм, способствующий интенсивному размножению крыс. Здесь же от-
крыто хранился большой запас бумажных этикеток и, таким образом, не
было недостатка в материале для подстилок гнезд. Крысы мелко пере-
трызали бумагу и в этой массе устраивали гнезда. Кроме того, неподале-
ку, здесь же, на территории завода, были высокие заросли полыни и
крапивы. Земля там, где рос этот бурьян, была сплошь перекопана зверь-
ками, а в гнездовых камерах нор часто встречались изгрызенные эти-
кетки,
Близ г. Тулы в 1967 г. начал функционировать клеевой завод. Кости
для переработки в клей привозили на автомашинах и по железной доро-
ге из разпых областей страны, сваливая их в штабеля на земле под от-
крытым небом. Вместе с костями случайно были завезены серые крысы.
Неремещаясь между костями, устраивая там в хороших защитных усло-
виях гисзда и питаясь мясом, которое сохранилось на костях, животные
имели благоприятные условия существования, быстро размножились и
стали расселяться.
.
Учеты водяных полевок дуговыми капканами на ближайшей от
свалки речке Шат. показали, как начиная с 1974 г. здесь стали попа-
даться крысы и в последующие годы численность их росла (рис. 32; Па-
нина, 19686).
Ком мунально-складская зопа. В коммунально-складской
зоне города крысы населяют различные базы и склады; особенно много
их дерлхится на складах животноводческого сырья, различных продуктов
питания.
В Иркутске в 1985—1986 гг. паибольшая концентрация крыс обнару-
жена в пригороде Жилкипо, где находятся крупные продовольственные
базы: мясокомбинаты, маслосырбаза, комбикормовый завод и т. д. По
учету плашками численность зверьков здесь достигает больших разме-
ров — 15% попадапия, а максимально
— 27% (Олькова и др., 1987).
В городах, особенно раньше, когда дератизация была поставлепа очень
слабо, большие скопления крыс отмечали на территории рынков. В Одес-
се до Октябрьской революции зверьки были чрезвычайно многочислен-
ны. особенно много их было на рынке, где они держались в куче отбро-
сов, пенужной тары и т. п. Столь высокая численность привела к рас-
пространению в городе эпизоотии чумы.
На высокую численность крыс на рынках Балтимора и Вашингтона
указывали Дэвис (Пау!з, 1953а) и Ларсон (Гатзоп, 1944).
Крыс на рынках отмечают и сейчас в некоторых южных тородах на-
нтей страпы — в Ереване, Краснодаре, Красноводске.
Крысы бывают мноточисленны в наземных уборпых с выгребными
ямами (Моргиловская и др., 1979).
Впутренний транспорт. В крупных городах иногда немалую
роль в существовании крыс играет метрополитен. Зверьки, как известно,
оклонны к обитанию в подземпых сооружениях, но степень заселенности
ими метрополитена всецело зависит от того, как он строится.
148
Глава третья
5
=
>
П
о
п
а
й
т
е
м
о
с
т
ь
9
,
я
=
и|11|
80808
1958 1959 1960 1961 1962 1963 1964 1965 1966 1967 19682.г.
©
|
-
=
|
-
&
в
о
[
-
<
=
©
[
-
>
Е
5
5
[
-
>
|
5
Е
=
в
|
-
=
>
в
-
-
м
=
7
ё
йственного
Рис. 31. Динамика численности серих врыс в о
иное дозя
Ё
пользования в г Москве, 1958—1968 а. (по: Субейкин,
—
илых до:
— на пищевых иредприятиях —вх
стройках. четы:
1 вжил
мах, 2
и х ен чятпях, 8 в хозяйственных постро
У
В — весениие, О — осенние, 8 — зимние
й
Г.
грегам рек
Рис. 32. Изменение численности серой крысы водяной полевки по бер
р
близ клеевого завода под Тулой (по: Панина, 1986а)
1— водная полевка, 8 — насюк
85
Г? 9:
|
9
5-
2
25Г
=ы
$2
25Г
,
ы
.—Г
|
-°
ЗИ!
.
ви!
Г
|о!
0|-
м
У
хаюкноос
о зав
пы ЗВ ЕВЕ ЕЗЕЕБЕЕЕЕЕЕНаНЫ
ЕЕ
ЕВ
Число кии-= 53 3 В В ЗВ ВЗЕ НЕЕ НЕНЕВ
Экологил
119
зеленого горошка, воторые представляли собой витамипизированный
корм, способствующий интенсивному размножению крыс. Здесь же от-
крыто хранился большой запас бумажных этикеток и, таким образом, не
было недостатка в материале для подстилок гнезд. Крысы мелко пере-
трызали бумагу и в этой массе устраивали гнезда. Кроме того, неподале-
ку, здесь же, на территории завода, были высокие заросли полыни и
крапивы. Земля там, где рос этот бурьян, была сплошь перекопана зверь-
ками, а в гнездовых камерах нор часто встречались изгрызенные эти-
кетки,
Близ г. Тулы в 1967 г. начал функционировать клеевой завод. Кости
для переработки в клей привозили на автомашинах и по железной доро-
ге из разпых областей страны, сваливая их в штабеля на земле под от-
крытым небом. Вместе с костями случайно были завезены серые крысы.
Неремещаясь между костями, устраивая там в хороших защитных усло-
виях гисзда и питаясь мясом, которое сохранилось на костях, животные
имели благоприятные условия существования, быстро размножились и
стали расселяться.
.
Учеты водяных полевок дуговыми капканами на ближайшей от
свалки речке Шат. показали, как начиная с 1974 г. здесь стали попа-
даться крысы и в последующие годы численность их росла (рис. 32; Па-
нина, 19686).
Ком мунально-складская зопа. В коммунально-складской
зоне города крысы населяют различные базы и склады; особенно много
их дерлхится на складах животноводческого сырья, различных продуктов
питания.
В Иркутске в 1985—1986 гг. паибольшая концентрация крыс обнару-
жена в пригороде Жилкипо, где находятся крупные продовольственные
базы: мясокомбинаты, маслосырбаза, комбикормовый завод и т. д. По
учету плашками численность зверьков здесь достигает больших разме-
ров — 15% попадапия, а максимально
— 27% (Олькова и др., 1987).
В городах, особенно раньше, когда дератизация была поставлепа очень
слабо, большие скопления крыс отмечали на территории рынков. В Одес-
се до Октябрьской революции зверьки были чрезвычайно многочислен-
ны. особенно много их было на рынке, где они держались в куче отбро-
сов, пенужной тары и т. п. Столь высокая численность привела к рас-
пространению в городе эпизоотии чумы.
На высокую численность крыс на рынках Балтимора и Вашингтона
указывали Дэвис (Пау!з, 1953а) и Ларсон (Гатзоп, 1944).
Крыс на рынках отмечают и сейчас в некоторых южных тородах на-
нтей страпы — в Ереване, Краснодаре, Красноводске.
Крысы бывают мноточисленны в наземных уборпых с выгребными
ямами (Моргиловская и др., 1979).
Впутренний транспорт. В крупных городах иногда немалую
роль в существовании крыс играет метрополитен. Зверьки, как известно,
оклонны к обитанию в подземпых сооружениях, но степень заселенности
ими метрополитена всецело зависит от того, как он строится.
120 „Глава третья
В послевоенные годы в Москве крысы были постоянными обитателями
метрополитена (Дукельская, 1947, 1948). В 1950-х годах строительство
его шло главным образом закрытым методом. В это время основным
путем провикповения этих грызунов В тупнели были вентиляциопные
трубы, которые часто выводились в виде так называемых «киосков» 30
дворы, вблизи сараев и других надворных построек, где почти всегда
были крысы, и из 87 обследованных «киосков» они были обнаружены в
64. В то время в Москве наиболее засоленными зверьками были траесы
от Крымской площади до Сокольликов, а также привокзальные стапции
метро: Курская, Павелецкая, Киевская и Белорусская.
В последующие годы с ликвидацией надворных построек, различных
сараев и других строений, из которых крысы проникали во вновь строя-
щиеся липии метрополитена, этих грызунов в метро стало очепь мало
(Судейкин, 1986).
Внешний транспорт. Железнодорожные вокзалы и морские
порты— это ворота, через которые пасюки могут проникнуть в город.
РВ Западной Сибири они в первую очередь заселили большие железно-
дорожные станции городов Омска, Томска и станцию Тайга, а затем.
расселились уже по самому городу (Кащенко, 1942; Зверев, 1930). Су-
ществует болышая опасность завоза ипфицированных крыс из других
стран, особенно на морских судах.
На вокзалах и в портах постоянно сосредоточето много людей, 060-
бенно в периоды народных бедствий и энергичного освоения новых эко-
номических районов, что при высокой численности крыс уже само по
себе представляет больттую опасность. В связи с этим па вокзалах, в пор-
тах, а также па прибывающих судах постоянио проводится весьма ип-
тенсивная дератизация, и тем ие менее крысы там периодически бывают.
Железнодорожные вокзалы. В пределах ареала серой кры-
сы она обитает не на всех вокзалах, и в заселении сю этих зданий хоро-
по выражена сезонность
— она появляется там в холодное время года,
причем максимум зверьков приходится на осень. Резкое уменьшение
количества заселенных воквалов зимой связано, по-видимому, с интенсив-
но проводящимися после осеннего заселения дератизационными меро-
приятиями. Весной число заселенных вокзалов увеличивается за счет
ослабления дератизации, а летом они полностью освобождаются от крыс.
вследствие истребительных мероприятий и из-за выселения зверьков В-
открытые стации. Вокзалов, которые были бы постоянно заселены крыса-
ми, нет — все они заселяются периодически по сезонам и даже по годам.
Наиболее часто крысы появляются на вокзалах плохого санитарно-техни--
ческого состояния. Это, как правило, старые здапия довоенпой постройки,
Например, в городе Ростове Ярославском старый вокзал периодически
засслялся крысами, а новый, построенный в 1979 г. свободен от них
(Дубровский, 1987).
На вокзалах серые крысы предпочитают селиться в пищевых объектах
и в подвалах. Реже их отмечают в административных и подсобных поме--
щениях. В подавляющем большинстве случаев (88%) источником поступ-
ления пасюка на вокзалы являются близлежащие строения: элеваторы. ,
Экология’ 124
склады, жилые дома ‚ свинарники, городская к
ск
и
, дская канализация и пр.
стаций заселение крысами вокзалов отмечено я иш
а вен
завоз с поездами — в 2%
эишь р 10%
Ирысы селя‚елятся также непосредс твенпо на желе:
используя обычно насыпи (Айзенштадт, 1950а)
о
рские и речные
`|
р
порты, с а. ЦП
и
:
‚ суда. орты горо
о варажеты серыми крысами, которые оржатоя в ера
мо етиях, д складах, где хранятся грузы. В пекоторых регионах р ды.
сы обитают тол ко в портовых городах, где они сосредоточиваются пре.
их портах. Шо Атлантическому побережью Африки з
Нр.
р
сюк 00
А—
,
р
у
.
к
ртах
д
априме
насю ооитае
олько в порта
акаре Конакри Ф
Й
итауне
и др. (РеКеузет, 1965; ВШ у
1985).
у
; В уап © а[., 1969, цит. по: Кучерук, Кузиков,
случаев,
дорожных путях,
На крайнем северо-востоке се
м
рые крысы заселяют в
то торода, ор на Камчатке — Петропавловск Камчалекий порто
Сухуми, Поти (Пушница и пр. оо
49. а Батуми,
1983), так и Белого и Барен ева рей — и Дро98О: Гуцель и др.
В
Ц морей — Рига, Архангельск, Ленинград
Пауштица т р р звестпы они и на Дальнем Востоке в портах.
Численность, их оная
В ВМО, 1985)
ииелениое
.
Ч
<
из-за периодически п
-
ри а,
ии
Большинство авторов отмечают за после.
тие
нижние, однако в Советской Гавани по учетам за
поели : р трируют высокую численность крыс — в с
, ы ооадания в ловушки (Олькова и др., 1987).
редем
ея ций порт пасюки попадают не только с судами, но и вместе
о пе то об: тм транснортом ‘и по железной дороге (Гуцель
и др. 1988) та 0 х типах судов заграничного плавания, прибывающих
па Сахалин, иногда воз речаются крысы, они есть и на каботажных судах
веет их. ма ры оловецких траулерах (Тимофеева, 1962). —
р но ие Зорокик судах тораздо чаще встречаются черные
нию, более ловки и ат
щыт
аа.
более многочисленны, чем серые крысы (Солов ито у ца
О
ьев и др., 1986; Пушница
Для Сибири типичел завоз большого
пой
чеи
количества продоволь
50-
тор «лолтое время хранится на пристанях (до навигации }. то прино
м
оплениям и интенсивному размножению здесь крыс
которые о
дах широко распространяются по территории,
«пакетирование» грузов которое било веде, ры 978 в, И аварию
«пакоти
в,к
нос
г. По 45—70 -
т руаку Н трюмы © ово в огромные пакеты, что облегчает их по.
У Втр ты У ов, так как при этом методе с помощью подъемпого
крапа можно одно ременно загрузить сразу много мешков. Пакеты скла-
т
Бо табеля (ста оры» — местное название) высотой в 78 мн
О
Е рые в течение всой зимы ‚часто остаются лежать
небом, прикрытые брезентом. Содержимое мешков (ком-
120 „Глава третья
В послевоенные годы в Москве крысы были постоянными обитателями
метрополитена (Дукельская, 1947, 1948). В 1950-х годах строительство
его шло главным образом закрытым методом. В это время основным
путем провикповения этих грызунов В тупнели были вентиляциопные
трубы, которые часто выводились в виде так называемых «киосков» 30
дворы, вблизи сараев и других надворных построек, где почти всегда
были крысы, и из 87 обследованных «киосков» они были обнаружены в
64. В то время в Москве наиболее засоленными зверьками были траесы
от Крымской площади до Сокольликов, а также привокзальные стапции
метро: Курская, Павелецкая, Киевская и Белорусская.
В последующие годы с ликвидацией надворных построек, различных
сараев и других строений, из которых крысы проникали во вновь строя-
щиеся липии метрополитена, этих грызунов в метро стало очепь мало
(Судейкин, 1986).
Внешний транспорт. Железнодорожные вокзалы и морские
порты— это ворота, через которые пасюки могут проникнуть в город.
РВ Западной Сибири они в первую очередь заселили большие железно-
дорожные станции городов Омска, Томска и станцию Тайга, а затем.
расселились уже по самому городу (Кащенко, 1942; Зверев, 1930). Су-
ществует болышая опасность завоза ипфицированных крыс из других
стран, особенно на морских судах.
На вокзалах и в портах постоянно сосредоточето много людей, 060-
бенно в периоды народных бедствий и энергичного освоения новых эко-
номических районов, что при высокой численности крыс уже само по
себе представляет больттую опасность. В связи с этим па вокзалах, в пор-
тах, а также па прибывающих судах постоянио проводится весьма ип-
тенсивная дератизация, и тем ие менее крысы там периодически бывают.
Железнодорожные вокзалы. В пределах ареала серой кры-
сы она обитает не на всех вокзалах, и в заселении сю этих зданий хоро-
по выражена сезонность
— она появляется там в холодное время года,
причем максимум зверьков приходится на осень. Резкое уменьшение
количества заселенных воквалов зимой связано, по-видимому, с интенсив-
но проводящимися после осеннего заселения дератизационными меро-
приятиями. Весной число заселенных вокзалов увеличивается за счет
ослабления дератизации, а летом они полностью освобождаются от крыс.
вследствие истребительных мероприятий и из-за выселения зверьков В-
открытые стации. Вокзалов, которые были бы постоянно заселены крыса-
ми, нет — все они заселяются периодически по сезонам и даже по годам.
Наиболее часто крысы появляются на вокзалах плохого санитарно-техни--
ческого состояния. Это, как правило, старые здапия довоенпой постройки,
Например, в городе Ростове Ярославском старый вокзал периодически
засслялся крысами, а новый, построенный в 1979 г. свободен от них
(Дубровский, 1987).
На вокзалах серые крысы предпочитают селиться в пищевых объектах
и в подвалах. Реже их отмечают в административных и подсобных поме--
щениях. В подавляющем большинстве случаев (88%) источником поступ-
ления пасюка на вокзалы являются близлежащие строения: элеваторы. ,
Экология’ 124
склады, жилые дома ‚ свинарники, городская к
ск
и
, дская канализация и пр.
стаций заселение крысами вокзалов отмечено я иш а вен
завоз с поездами — в 2%
эишь р 10%
Ирысы селя‚елятся также непосредс твенпо на желе:
используя обычно насыпи (Айзенштадт, 1950а)
о
рские и речные
`|
р
порты, с а. ЦП
и
:
‚ суда. орты горо
о варажеты серыми крысами, которые оржатоя в ера
мо етиях, д складах, где хранятся грузы. В пекоторых регионах р ды.
сы обитают тол ко в портовых городах, где они сосредоточиваются пре.
их портах. Шо Атлантическому побережью Африки з
Нр.
р
сюк 00
А—
,
р
у
.
к
ртах
д
априме
насю ооитае
олько в порта
акаре Конакри Ф
Й
итауне
и др. (РеКеузет, 1965; ВШ у
1985).
у
; В уап © а[., 1969, цит. по: Кучерук, Кузиков,
случаев,
дорожных путях,
На крайнем северо-востоке се
м
рые крысы заселяют в
то торода, ор на Камчатке — Петропавловск Камчалекий порто
Сухуми, Поти (Пушница и пр. оо
49. а Батуми,
1983), так и Белого и Барен ева рей — и Дро98О: Гуцель и др.
В
Ц морей — Рига, Архангельск, Ленинград
Пауштица т р р звестпы они и на Дальнем Востоке в портах.
Численность, их оная
В ВМО, 1985)
ииелениое
.
Ч
<
из-за периодически п
-
ри а,
ии
Большинство авторов отмечают за после.
тие
нижние, однако в Советской Гавани по учетам за
поели : р трируют высокую численность крыс — в с
, ы ооадания в ловушки (Олькова и др., 1987).
редем
ея ций порт пасюки попадают не только с судами, но и вместе
о пе то об: тм транснортом ‘и по железной дороге (Гуцель
и др. 1988) та 0 х типах судов заграничного плавания, прибывающих
па Сахалин, иногда воз речаются крысы, они есть и на каботажных судах
веет их. ма ры оловецких траулерах (Тимофеева, 1962). —
р но ие Зорокик судах тораздо чаще встречаются черные
нию, более ловки и ат
щыт
аа.
более многочисленны, чем серые крысы (Солов ито у ца
О
ьев и др., 1986; Пушница
Для Сибири типичел завоз большого
пой
чеи
количества продоволь
50-
тор «лолтое время хранится на пристанях (до навигации }. то прино
м
оплениям и интенсивному размножению здесь крыс
которые о
дах широко распространяются по территории,
«пакетирование» грузов которое било веде, ры 978 в, И аварию
«пакоти
в,к
нос
г. По 45—70 -
т руаку Н трюмы © ово в огромные пакеты, что облегчает их по.
У Втр ты У ов, так как при этом методе с помощью подъемпого
крапа можно одно ременно загрузить сразу много мешков. Пакеты скла-
т
Бо табеля (ста оры» — местное название) высотой в 78 мн
О
Е рые в течение всой зимы ‚часто остаются лежать
небом, прикрытые брезентом. Содержимое мешков (ком-
422
Глава третья
©)
й температуре — 30
вет, и при внешней т
ка, крупа и т. п.) преет,
98.
м ых бывает тепло и, следовательно, создаются очень хорошие
условия для обитания и размножения крыс. Из-за того что ме Е к плот.
и связаны в пакеты, зверьки легко перемещаются из и,
используют простр
ают в них гнезда и
котками.
Полавленные на места грузы в виде неразобранных т
к,
ло
перевозят на склады. В. Я. Аптонюк © соавт.
общак,
крысами я аборке пакетов опи видели потрясающее количество ее
щи о крыс Содержимое мешков, которые обычно снаружи ие поврежле.
вы я нотда полностью изгрызено, а следо крыс не видно,
у
з штабелей.
и не выходят на мороз и
я ан поиах
т очень большая численность крыс была аще ох
й
. Несомненно, ч
а
Олекыинск, Мындогей, Нюрба и др о оино, ой
серой крысы и заселепие этим видом Яку
водным путем.
ет
Ка О пизация Особое место в жизни крыс города рые
Н
.
п
наливационная система, которая связывает разные зоны. , ля ры
то магистрали, по которым они мотут перемещаться иногд
Я
?
.
_
их
а лок
асполагаются на тлубине 1 7 м, ноэто
илекторы капализации р
тому
м а воздуха в них отличается стабильностью и бывает р и
О
1986), что соответствует оптимальной
О
С
шчкина 4957). В зимнее время температура в
серой крысы и
‚
.
И
р. лекторах поддерживается на относительно высоком уров о таке
а счет гниения ортанических остатков с выделением те г. Каша ия
со бучается с поверхностью сотнями И тысячами коммуникац и
атается непосредственно под постройками города, многие и
ь
лаг
нон
й степени
и оотранение пасюков в канализации в вначительной стене
ано у ес устройством. Наиболее благоприятна для хи зализацих
в песплавного типа, предназначенная для отвода как ат Е ферных, так
ие тарных вод и слагающаяся из единой системы СУ ‚ беру м тва
ив до
вые водостоки состоят из труб не ольшого д
6—2тр
х в более крупные водостоки (45—60 см), несу-
Оомеюо
160 см), внадающие в подземные
‹оллекторы (90— ем),
ные
ы в уличные коллект
даем
альт, оторые иногда имеют вид туннеля с подукругтым свод о
ДЕ
ходит желоб для
,.\
ны
т апимают пе более 10—12% объема жело-
<
мя года сточные воды
ма жел.
ба, роль Которого проходят банкеты, по которым легко перед
,
частся
“РЕ апализация общесплавного типа сравнительно редко потречае т ы
нас в стране и широко распространена в США и Вах
но,
ами,—ч
‚
‚
сивно заселена крысами,
оны, где она интен
я
)
тузтарт (Тощигии, 1968, 1986; ВеюЦеу, 1960). ой предотаваяет со-
Раздельная канализация в отличие от общеспла.
редотаваяет ве.
бой не параллельные системы труб небольшого диам тр НИ
для санитарпых и атмосферных вод. Эта система о
Экология 123
нена жидкостью и гораздо менее благоприятна для заселения крысами.
Ю. В. 'Гощигин (1986) проводил сравнительные обследования канали-
заций обоих типов. По его дапным, в общесплавной канализации в Одес-
се, Черповцах, Львове заселено крысами от 32 до 72% общесплавной
территории, тогда как канализация раздельцого типа в Саратове, Калу-
ге, Ярославле, Смоленске, Перми оказалась свободной от пих. Только в
Волгограде (раздельный тип канализации) крыс было обнаружено 6,4%.
Благодаря периодическим изменениям уровня воды в канализацион-
ной системе распределение зверьков там неравномерно. Следы их пребы-
зания часто обнаруживаются на банкетах, а также в сухих бездействую-
щих притоках (Айзенштадт, Арсеньев, 1954; ВепИеу, 1960). Крысы часто
интенсивно роют норы в осповпом там, где канализационные трубы пов-
реждепы (Тощигин, 1986). По данным этого автора, зверьки засоляют ка-
нализацию неравномерно, но максимально используют удобные для но-
рения места. По сго наблюдениям, в Одессе в среднем на 100 м кана-
лизационной системы приходилось 2,86 пормы.
Для распространения пасюков по канализации большое значение име-
ют разрушенные участки системы. Этому способствуют газы, выделяю-
щиеся при гниении отбросов, особенно в тех участках, которые отли-
чаются очень медленным течением. Кирпичные и цемептпые стенки кол-
лекторов разрыхляются и стаповятся доступными для повреждения
крысами.
Крысиные норы в канализациях обычно располагаются невысоко,
в нескольких сантиметрах над водой, а иногда и прямо над уровием
воды. В случае затопления пор животные паходят себе дополнительные
убежища па высоте 30—80 см от дна коллектора на выступах стен. Пи-
таются крысы в канализациях ‘пищевыми отбросами и экскрементами
(ВепШеу, 1960), которые, по мнению Ю. В. Тощигина (1986), являются
основпой пищей.
При дератизации коллекторов большое значение имеет вопрос о го-
ризонтальном и вертикальном перемещении крыс, так как от величины
этих перемещений должны зависеть размеры обрабатываемого участка.
Горизонтальные перемещения обычно невелики — от 30 до 200 м и лишь
иногда доходят до 600 м. Малая величина горизоптальпых перемещений
в первую очередь связана © колебаниями уровня воды, не позволяю-
щими зверькам удаляться на большие расстояпия (Тощигин, Харченко,
1967).
Проведенные опыты позволяют считать, что в условиях Одессы назем-
ные и канализационные популяции крыс мало сообщаются между собой.
Но в других городах, например во Львове, ири обильных дождях и, сле-
довательно, ири повышении уровня воды в канализации резко возраста-
ет численность крыс в домах (Тощитин, 1986).
Вероятно, не только повышение уровня воды в канализации, но и
увеличение плотности паселения крыс способствует их выселению на по-
верхность земли в наземные строевия. На эту сторону связи крыс, оби-
тающих в канализационной системе и в паземпых постройках, указывают
многие авторы (Айзенштадт, Арсеньев, 1954; Зенишег, 1960; и др.).
422
Глава третья
©)
й температуре — 30
вет, и при внешней т
ка, крупа и т. п.) преет,
98.
м ых бывает тепло и, следовательно, создаются очень хорошие
условия для обитания и размножения крыс. Из-за того что ме Е к плот.
и связаны в пакеты, зверьки легко перемещаются из и,
используют простр
ают в них гнезда и
котками.
Полавленные на места грузы в виде неразобранных т
к,
ло
перевозят на склады. В. Я. Аптонюк © соавт.
общак,
крысами я аборке пакетов опи видели потрясающее количество ее
щи о крыс Содержимое мешков, которые обычно снаружи ие поврежле.
вы я нотда полностью изгрызено, а следо крыс не видно,
у
з штабелей.
и не выходят на мороз и
я ан поиах
т очень большая численность крыс была аще
ох
й
. Несомненно, ч
а
Олекыинск, Мындогей, Нюрба и др о оино, ой
серой крысы и заселепие этим видом Яку
водным путем.
ет
Ка О пизация Особое место в жизни крыс города рые
Н
.
п
наливационная система, которая связывает разные зоны. , ля ры
то магистрали, по которым они мотут перемещаться иногд
Я
?
.
_
их
а лок
асполагаются на тлубине 1 7 м, ноэто
илекторы капализации р
тому
м а воздуха в них отличается стабильностью и бывает р и
О
1986), что соответствует оптимальной
О
С
шчкина 4957). В зимнее время температура в
серой крысы и
‚
.
И
р. лекторах поддерживается на относительно высоком уров о таке
а счет гниения ортанических остатков с выделением те г. Каша ия
со бучается с поверхностью сотнями И тысячами коммуникац и
атается непосредственно под постройками города, многие и
ь
лаг
нон
й степени
и оотранение пасюков в канализации в вначительной стене
ано у ес устройством. Наиболее благоприятна для хи зализацих
в песплавного типа, предназначенная для отвода как ат Е ферных, так
ие тарных вод и слагающаяся из единой системы СУ ‚ беру м тва
ив до
вые водостоки состоят из труб не ольшого д
6—2тр
х в более крупные водостоки (45—60 см), несу-
Оомеюо
160 см), внадающие в подземные
‹оллекторы (90— ем),
ные
ы в уличные коллект
даем
альт, оторые иногда имеют вид туннеля с подукругтым свод о
ДЕ
ходит желоб для
,.\
ны
т апимают пе более 10—12% объема жело-
<
мя года сточные воды
ма жел.
ба, роль Которого проходят банкеты, по которым легко перед
,
частся
“РЕ апализация общесплавного типа сравнительно редко потречае т ы
нас в стране и широко распространена в США и Вах
но,
ами,—ч
‚
‚
сивно заселена крысами,
оны, где она интен
я
)
тузтарт (Тощигии, 1968, 1986; ВеюЦеу, 1960). ой предотаваяет со-
Раздельная канализация в отличие от общеспла.
редотаваяет ве.
бой не параллельные системы труб небольшого диам тр НИ
для санитарпых и атмосферных вод. Эта система о
Экология 123
нена жидкостью и гораздо менее благоприятна для заселения крысами.
Ю. В. 'Гощигин (1986) проводил сравнительные обследования канали-
заций обоих типов. По его дапным, в общесплавной канализации в Одес-
се, Черповцах, Львове заселено крысами от 32 до 72% общесплавной
территории, тогда как канализация раздельцого типа в Саратове, Калу-
ге, Ярославле, Смоленске, Перми оказалась свободной от пих. Только в
Волгограде (раздельный тип канализации) крыс было обнаружено 6,4%.
Благодаря периодическим изменениям уровня воды в канализацион-
ной системе распределение зверьков там неравномерно. Следы их пребы-
зания часто обнаруживаются на банкетах, а также в сухих бездействую-
щих притоках (Айзенштадт, Арсеньев, 1954; ВепИеу, 1960). Крысы часто
интенсивно роют норы в осповпом там, где канализационные трубы пов-
реждепы (Тощигин, 1986). По данным этого автора, зверьки засоляют ка-
нализацию неравномерно, но максимально используют удобные для но-
рения места. По сго наблюдениям, в Одессе в среднем на 100 м кана-
лизационной системы приходилось 2,86 пормы.
Для распространения пасюков по канализации большое значение име-
ют разрушенные участки системы. Этому способствуют газы, выделяю-
щиеся при гниении отбросов, особенно в тех участках, которые отли-
чаются очень медленным течением. Кирпичные и цемептпые стенки кол-
лекторов разрыхляются и стаповятся доступными для повреждения
крысами.
Крысиные норы в канализациях обычно располагаются невысоко,
в нескольких сантиметрах над водой, а иногда и прямо над уровием
воды. В случае затопления пор животные паходят себе дополнительные
убежища па высоте 30—80 см от дна коллектора на выступах стен. Пи-
таются крысы в канализациях ‘пищевыми отбросами и экскрементами
(ВепШеу, 1960), которые, по мнению Ю. В. Тощигина (1986), являются
основпой пищей.
При дератизации коллекторов большое значение имеет вопрос о го-
ризонтальном и вертикальном перемещении крыс, так как от величины
этих перемещений должны зависеть размеры обрабатываемого участка.
Горизонтальные перемещения обычно невелики — от 30 до 200 м и лишь
иногда доходят до 600 м. Малая величина горизоптальпых перемещений
в первую очередь связана © колебаниями уровня воды, не позволяю-
щими зверькам удаляться на большие расстояпия (Тощигин, Харченко,
1967).
Проведенные опыты позволяют считать, что в условиях Одессы назем-
ные и канализационные популяции крыс мало сообщаются между собой.
Но в других городах, например во Львове, ири обильных дождях и, сле-
довательно, ири повышении уровня воды в канализации резко возраста-
ет численность крыс в домах (Тощитин, 1986).
Вероятно, не только повышение уровня воды в канализации, но и
увеличение плотности паселения крыс способствует их выселению на по-
верхность земли в наземные строевия. На эту сторону связи крыс, оби-
тающих в канализационной системе и в паземпых постройках, указывают
многие авторы (Айзенштадт, Арсеньев, 1954; Зенишег, 1960; и др.).
124
Глава третья
астки города. Изучение распространо-
жно, так как ое
я зверьков в пезастроенной части города особешно нак, та ак о
в ются резерватами, из которых грызуны мотут т ты тв адания.
явля
венпых биотопах, в пределах тород
атотею к
а
ажиени
:
местам с постоянным нали зием
сырым увлажпенным у часткам и к
р!
}
р
гав.
оой зар
Не1нЕечастиород
В
Я
3.
ода обычно представ тяю со р аро
частки с неровным рельефом («иеудоби»), а при подоотатон
о
ртом надзоре население выбрасывает туда му ор. Все
вместе ввятее сода пра
мае восредоточивастся а
Поля фи и в е `отстанваются : очищаются печистоты, выведен
полях а" лизацией. На окраинах Москвы в Люблииских д
тЫ ВЫХ ПОЛЯХ фильтрации процент попадания в плашки насю
у
ое
КИ»
х
20
дит до 20.
_
По своей структуре эти пол
ные карты, в которые по палив
Незастроенные уч
сие
рисовые плантации. Квадрат-
я похожи На
орода печис-
ным каналам поступают из г
нежная траншея крыс зимои.
Рис. 88. Тропа и разрытая подс
Фото Н. В. Степановой
Экология
125
тоты, окружепы земляными валиками. Во многих местах валики зараста-
ют кустарниками, преимущественно бузиной, черсмухой и ивой. Местами
буйно разрастается бурьяп и травянистая растительность. Норы крыс
распределены по валикам неравномерно, большее их число встречастся в
менее увлажненных частях. Часто входные отверстия в норы открыва-
ются между корпями кустов. Крысы живут здесь круглый год, и основная
их масса не переселяется в помещения. Зимой пасюки передвигаются по
поверхности снега, но чаще делают подземные транитеи, около входов в
которые обычно бывает много крысиного кала (рис. 33).
Места отдыха. Парки, бульвары, скверы привлекают отдыхающих
во все времена года. В Москве круглогодичное обитание крыс пока за-
регистрировано только в Измайловском парке па р. Серебрянке, где живут
дикие утки, которых постоянно кормят посетители парка, а поэтому кры-
сы не имеют недостатка в нище. Там же, у мостика через речку, сосре-
доточены их норы (Сильвестров и др. 1986).
На пебольших речках в Царицынском и других лесопарках с водяны-
ми полевками обитают вместе и пасюки, причем в последние годы
чиеленпость последних возрастает, а водяных полевок резко уменьшается.
В 1968—1970 гг. процент попадания в ловушки водяных полевок был ра-
вен 6—12%, а серых крысе— всего 0,5—1,7%. В 1983—1984 тг. водяпых
полевок не стало совсем, а процент попадания серых крыс возрос до 6—
12% (Степанова и др., 1986). Это объясняется, с одной стороны, тем,
что отдыхающие вытаптывают траву, а в некоторых местах ее специаль-
но выкашивают или выжигают, ухудшая тем самым условия обитания
водяных нолевок, а с другой — больное количество брошенпых пищевых
остатков способствует привлечению пасюков.
Угольные знахты
Как и канализационная система, угольные шахты очень бизгонриятны
для обитания крыс, по микроклимат шахт, располагающихся на большей
глубине, значительно стабильнее. Относительная влажность воздуха до-
стигает 90% и более, па мпогих участках скаиливается н стоит вода,
Температура воздуха колеблется от +15 до 25". Реже встречаются шахты
с пониженной (+10°) или повышиенной (-30°) температурой (Кузьми-
чев п др., 1958). Таким образом, микроклимат в них примерно тот же,
что и в норах насюков и, следовательно, восьма благоприятен для послед-
них (Тощигин, 1986).
В шахтах поры крыс встречаются редко — паличие здесь больлтого ко-
личества нии и щелей исключает в них необходимость (Тумое, 1961,
1980).
Ю.В, Тощигин, (1968) сообщает, что, но анкетным данным, в СССР
из 673 тахт серой крысой оказались заселенными 277, т. с. 42%. В Анг-
лии из 562 шахт крысы были обнаружены в 253, т. е. в 454%.
Есть сведения о наличии пасюков в шахтах Дальнего Востока (Змеев,
1931), Донбасса (Гажиев и др., 1986а), Караганды (Кузьмичев и др.,
1958). Есть они и в шахтах Кировоградской области (Соловьев и др..
124
Глава третья
астки города. Изучение распространо-
жно, так как ое
я зверьков в пезастроенной части города особешно нак, та ак о
в ются резерватами, из которых грызуны мотут т ты тв адания.
явля
венпых биотопах, в пределах тород
атотею к
а
ажиени
:
местам с постоянным нали зием
сырым увлажпенным у часткам и к
р!
}
р
гав.
оой зар
Не1нЕечастиород
В
Я
3.
ода обычно представ тяю со р аро
частки с неровным рельефом («иеудоби»), а при подоотатон
о
ртом надзоре население выбрасывает туда му ор. Все
вместе ввятее сода пра
мае
восредоточивастся а
Поля фи и в е `отстанваются : очищаются печистоты, выведен
полях а" лизацией. На окраинах Москвы в Люблииских д
тЫ ВЫХ ПОЛЯХ фильтрации процент попадания в плашки насю
у
ое
КИ»
х
20
дит до 20.
_
По своей структуре эти пол
ные карты, в которые по палив
Незастроенные уч
сие
рисовые плантации. Квадрат-
я похожи На
орода печис-
ным каналам поступают из г
нежная траншея крыс зимои.
Рис. 88. Тропа и разрытая подс
Фото Н. В. Степановой
Экология
125
тоты, окружепы земляными валиками. Во многих местах валики зараста-
ют кустарниками, преимущественно бузиной, черсмухой и ивой. Местами
буйно разрастается бурьяп и травянистая растительность. Норы крыс
распределены по валикам неравномерно, большее их число встречастся в
менее увлажненных частях. Часто входные отверстия в норы открыва-
ются между корпями кустов. Крысы живут здесь круглый год, и основная
их масса не переселяется в помещения. Зимой пасюки передвигаются по
поверхности снега, но чаще делают подземные транитеи, около входов в
которые обычно бывает много крысиного кала (рис. 33).
Места отдыха. Парки, бульвары, скверы привлекают отдыхающих
во все времена года. В Москве круглогодичное обитание крыс пока за-
регистрировано только в Измайловском парке па р. Серебрянке, где живут
дикие утки, которых постоянно кормят посетители парка, а поэтому кры-
сы не имеют недостатка в нище. Там же, у мостика через речку, сосре-
доточены их норы (Сильвестров и др. 1986).
На пебольших речках в Царицынском и других лесопарках с водяны-
ми полевками обитают вместе и пасюки, причем в последние годы
чиеленпость последних возрастает, а водяных полевок резко уменьшается.
В 1968—1970 гг. процент попадания в ловушки водяных полевок был ра-
вен 6—12%, а серых крысе— всего 0,5—1,7%. В 1983—1984 тг. водяпых
полевок не стало совсем, а процент попадания серых крыс возрос до 6—
12% (Степанова и др., 1986). Это объясняется, с одной стороны, тем,
что отдыхающие вытаптывают траву, а в некоторых местах ее специаль-
но выкашивают или выжигают, ухудшая тем самым условия обитания
водяных нолевок, а с другой — больное количество брошенпых пищевых
остатков способствует привлечению пасюков.
Угольные знахты
Как и канализационная система, угольные шахты очень бизгонриятны
для обитания крыс, по микроклимат шахт, располагающихся на большей
глубине, значительно стабильнее. Относительная влажность воздуха до-
стигает 90% и более, па мпогих участках скаиливается н стоит вода,
Температура воздуха колеблется от +15 до 25". Реже встречаются шахты
с пониженной (+10°) или повышиенной (-30°) температурой (Кузьми-
чев п др., 1958). Таким образом, микроклимат в них примерно тот же,
что и в норах насюков и, следовательно, восьма благоприятен для послед-
них (Тощигин, 1986).
В шахтах поры крыс встречаются редко — паличие здесь больлтого ко-
личества нии и щелей исключает в них необходимость (Тумое, 1961,
1980).
Ю.В, Тощигин, (1968) сообщает, что, но анкетным данным, в СССР
из 673 тахт серой крысой оказались заселенными 277, т. с. 42%. В Анг-
лии из 562 шахт крысы были обнаружены в 253, т. е. в 454%.
Есть сведения о наличии пасюков в шахтах Дальнего Востока (Змеев,
1931), Донбасса (Гажиев и др., 1986а), Караганды (Кузьмичев и др.,
1958). Есть они и в шахтах Кировоградской области (Соловьев и др..
126
Глава третья
Экология
127
1986). Кроме серых крысе, шахты часто бывают заселены домовыми мы-
шами, есть случая обнаружения в них черной в туркестанской крыс.
Ю. В. Тощигин (1986) указывает, что питапие всех этих грызунов состо-
ит из двух компонентов
— пищевых отбросов и человеческого кала.
Очевидно, что количество корма определяет возможность обитания
крыс в шахтах. Там, где много подземных рабочих (более 200), крысы
з 95,5% случаев встречаются в шахтах. Шахты с мепьтим количеством
рабочих заселены зверьками только в 33.1% случаев (Тощигин, 1986).
|
Зверьки в условии шахт мало перемещаютсн и обитают только на тех
участках, где работают люди, но удаляясь от них более чем на 300—
350 м. Вода всегда имеется в избытке. По наблюдениям Твитга ('Т\188,
1964), крысы перемещаются только вслед за переходом людей с одного
участка на другой. На поверхность земли из шахт крысы переселяются
сравнительно редко. Опыты Ю. В. Тоигина (1968) показали, что они
подпимались 10 наклонным стволам шахт только до их середины. Этот
же автор сообщает, что около входов в шахты крысиных нор он ие об-
наруживал, а заселенпость зверьками поселков, расположенных рядом,
оказалась весьма умеренной. Таким образом, нет оснований считать, что
крысы (во всяком случае систематически) выходят из шахт наружу.
Гиологические особенпости пасюков, населяющих шахты, изучены
недостаточно. Но все-таки можио сказать, что это в основном обособлен-
ные, не смешивающиеся © наземпыми группировками крыс поселения.
Благодаря стабильному микроклимату и наличито корма крысы в шахтах
размножаются круглый год— среди самок, пойманных в шахтах в Мар-
те—апреле, было семь молодых зверьков, что свидетельствует о том, что
в шахте шло зимнее размножение (Тощигин, 1968).
Таким образом, рассмотрев места обитания серых крыс, мы видим,
что этот вид всегда распространен мозаично и распределение его, как
правило, определяется структурой территории и, конечно, вниянием дея-
тельности человека и особенио дератизации. Но для экзоантропных крыс
в основном характерны ленточные или линейные поселепия, а для синан-
тропных — островные.
«Дикие поселения» серых крыс бывают главным образом вытянуты
вдоль берегов рек и ручьев или встречаются на плавнях вдоль сухих грив
«бровок», где они могут рыть норы.
,
На риеовых полях, например, липейные поселения пасюков расположе-
ны вдоль оросительных и сбросных каналов. Встречаются оши и вдоль
осушительных каналов и берегов рыборазводпых прудов.
Паселенные пункты и особенно болыпие города представляют собой
крайне сложную и пеструю мозанку из неоднородных экологических ус-
ловий, что и определяет особенности распространения здесь пасюка—
х
—
,
в большинстве тородов всегда имеются какие-то территории — острова,
5|ы-юИЛА-в0|
1-15 Г|о
наиболее интенсивно заселенные крысами, и участки, где их вообще нет
Рис. 84. Картосхема засел
(рис. 34) (Мелкова, Туров, 1986).
(по; Мазин, Судейкии, у
серыми крысами г. Москвы на январь 1984 г.
З аселенность крысами разных участиов (в условных единицах)
126
Глава третья
Экология
127
1986). Кроме серых крысе, шахты часто бывают заселены домовыми мы-
шами, есть случая обнаружения в них черной в туркестанской крыс.
Ю. В. Тощигин (1986) указывает, что питапие всех этих грызунов состо-
ит из двух компонентов
— пищевых отбросов и человеческого кала.
Очевидно, что количество корма определяет возможность обитания
крыс в шахтах. Там, где много подземных рабочих (более 200), крысы
з 95,5% случаев встречаются в шахтах. Шахты с мепьтим количеством
рабочих заселены зверьками только в 33.1% случаев (Тощигин, 1986).
|
Зверьки в условии шахт мало перемещаютсн и обитают только на тех
участках, где работают люди, но удаляясь от них более чем на 300—
350 м. Вода всегда имеется в избытке. По наблюдениям Твитга ('Т\188,
1964), крысы перемещаются только вслед за переходом людей с одного
участка на другой. На поверхность земли из шахт крысы переселяются
сравнительно редко. Опыты Ю. В. Тоигина (1968) показали, что они
подпимались 10 наклонным стволам шахт только до их середины. Этот
же автор сообщает, что около входов в шахты крысиных нор он ие об-
наруживал, а заселенпость зверьками поселков, расположенных рядом,
оказалась весьма умеренной. Таким образом, нет оснований считать, что
крысы (во всяком случае систематически) выходят из шахт наружу.
Гиологические особенпости пасюков, населяющих шахты, изучены
недостаточно. Но все-таки можио сказать, что это в основном обособлен-
ные, не смешивающиеся © наземпыми группировками крыс поселения.
Благодаря стабильному микроклимату и наличито корма крысы в шахтах
размножаются круглый год— среди самок, пойманных в шахтах в Мар-
те—апреле, было семь молодых зверьков, что свидетельствует о том, что
в шахте шло зимнее размножение (Тощигин, 1968).
Таким образом, рассмотрев места обитания серых крыс, мы видим,
что этот вид всегда распространен мозаично и распределение его, как
правило, определяется структурой территории и, конечно, вниянием дея-
тельности человека и особенио дератизации. Но для экзоантропных крыс
в основном характерны ленточные или линейные поселепия, а для синан-
тропных — островные.
«Дикие поселения» серых крыс бывают главным образом вытянуты
вдоль берегов рек и ручьев или встречаются на плавнях вдоль сухих грив
«бровок», где они могут рыть норы.
,
На риеовых полях, например, липейные поселения пасюков расположе-
ны вдоль оросительных и сбросных каналов. Встречаются оши и вдоль
осушительных каналов и берегов рыборазводпых прудов.
Паселенные пункты и особенно болыпие города представляют собой
крайне сложную и пеструю мозанку из неоднородных экологических ус-
ловий, что и определяет особенности распространения здесь пасюка—
х
—
,
в большинстве тородов всегда имеются какие-то территории — острова,
5 |ы-юИЛА-в0| 1-15Г|о
наиболее интенсивно заселенные крысами, и участки, где их вообще нет
Рис. 84. Картосхема засел
(рис. 34) (Мелкова, Туров, 1986).
(по; Мазин, Судейкии, у
серыми крысами г. Москвы на январь 1984 г.
З аселенность крысами разных участиов (в условных единицах)
128
Глава третья
Убежища
Жизнь серой крысы, так же как и большинства видов грызупов, связана
‹ убежищами. В них она находит не только защиту от неблатоприятпых
воздействий внешпой среды и нападения хищников, но опи ей служат для
родов и выкармливания потомства,
и
0б убежинщах серых крыс как у отечественных авторов (Козлов, Коз-
—
7
#
х
лов, 1974; Хамаганов, 1975; Карасева и др., 1985), так и зарубежных
(Восс! её а1., 1948; "Р/запо, Зфогог, 1948) данных пемного. В сводках по
млекопитающим Дависа и Джолли (Бау1з, ТоПу, 1963) о норах. серой кры-
сы вообще пет упоминания. Калхоун (СаШоцп, 1962} исследовал особен-
ности поведения этого грызуна в связи с устройством нор ‘на огоро-
эженной торритории. Роющую деятельность серых крысе в искусственных
условиях изучали Броунелл и Чан ил Чун (Втомупей, СБап Н-Сва,
1977).
`
Е
|
<1.
Норы и убежища животных обычно. дифференцируют как ид обобен-
ностям их строения, так и по целевому назначению (Тихвинский,
1932; Кучерук, 1983}. Применительно к серой крысе ее жилниа по свое-
му строению могут быть подразделены на. не норовые убежища,
кула входят 1} поверхностные гнезда, 2) убежища различных емкостях
(пустотах) под навесами и в’ укрытиях, и на норы, подразделяемые на
1) бескамерные, 2) с камерами (без гнезд) 'и.3) гнездовые норы (про-
стые и сложные). По целевому. назначению мы зыделяем, норы 1) `за-
цитные и кормовые, 2) выводковые, 3) зимовочные, 4}. полуфункциональ-
ные.
.
|
и
Для изучения строения норы перед ее раскопкой нанослт на план все
входные отверстия, выбросы земли, поверхпостные ‘дорожки и другие
следы жизнедеятельности крыс. После этого для выявления хознев в
большинстве случаев нору облавливают дуговыми капканами-или, когда
это удается, вылавливают зверьков в процессе раскопки -и.зна том: же
плане обозначают обнаживишуюся систему галерей, тупики, пустые гнездо-
вые камеры и пр. (Карасева и др., 1985}. В помещёниях животноводче-
ских ферм норы чаще всего раскапывают во время ремонта здания (Коз-
лов, Козлов, 1971). © _
о
По своему строению норы ‘крысе, обитающих в природе и в домах,
очень разнообразны, но все они не имеют снециальных камер для запа-
сов и уборных. Роющая способность крыс велика, роют они ‘очень быст-
ро. По наблюдениям, в неволе нора длиной в 2,25 м с двумя камерами
может быть выкопана за два дня (Вго\пей, Свав -Сроз, 1977).
В. Г. Лялин (1972) указывает, что в свинарнике за одну почь два зверь-
ка выбросили на поверхность пола дна ведра земли.
8
Пизано и Сторер (Р4запо, Эюгег, 1948), изучая норовую деятельность
крыс, помещали их в клетку без дна, которую устанавливали вад ящи-
ком с землей. Зверьки очень быстро рыли передними лапами, ударами
одпой или двух передних же лап выбрасывали землю под озживот и уже
потом выкидывали ее задними копечностями. После того как крыса про-
капывала туннель, равный длине своего тела, она поворачивалась и вы-
Экология
129
Рис. 35. Схема строения
нор серых крыс
@а—г—норы
синантрон-
ных крыс (по: Карасева и
др., 1985):
а — бескамерная,
б —с камерой без гнезда,
в — простая гнездовая,
2 — сложная многоярусная
гнездовая;
9—3: норы экзоантропных
крыс:
98 — в бугре,
е — в береге реки,
ж,а—вполе
1 — камера без гнезда, .
2 — гнездовая камера,
3 — земляная пробка
талкивала землю передними лапами, помогая себе головой. В. Г. Лялин
(1972) пишет, что ипогда эти грызуны перетаскивают землю во рту.
Самки роют больше, чем самцы, с возрастом их роющая деятельность
понижается (РгоупеН, Свап П-Свиа, 1977). Калхоун (Сао, 1962)
сообщаст, что высокоранговые самки копают больше, чем низкорапговые
Крысы роют норы не только в почве, по и в разнообразных субстра-
тах: в глинобитпых стенах, окружающих кошары и другие животновод-
ческие фермы, в шлаколитных помещениях (И. 3. Климченко, устн со.
общ.) в слежавшихся опилках (В. А. Судейкин, устн. сообщ.), |: кучах
перетиоя, соломы. В твердом грунте рытье сочетается с грызущей деятель-
Норы крыс по своему строению, протяженности ходов, глубипе зале-
гания, а также по взаимнсму расположению очень различны (рис. 85).
Бескамерные норы имеют самое примитивное устройство
диаметр их ходов зависит от грунта. В твердом грунте он равен 5—7 см,
5 Серал крыса
128
Глава третья
Убежища
Жизнь серой крысы, так же как и большинства видов грызупов, связана
‹ убежищами. В них она находит не только защиту от неблатоприятпых
воздействий внешпой среды и нападения хищников, но опи ей служат для
родов и выкармливания потомства,
и
0б убежинщах серых крыс как у отечественных авторов (Козлов, Коз-
—
7
#
х
лов, 1974; Хамаганов, 1975; Карасева и др., 1985), так и зарубежных
(Восс! её а1., 1948; "Р/запо, Зфогог, 1948) данных пемного. В сводках по
млекопитающим Дависа и Джолли (Бау1з, ТоПу, 1963) о норах. серой кры-
сы вообще пет упоминания. Калхоун (СаШоцп, 1962} исследовал особен-
ности поведения этого грызуна в связи с устройством нор ‘на огоро-
эженной торритории. Роющую деятельность серых крысе в искусственных
условиях изучали Броунелл и Чан ил Чун (Втомупей, СБап Н-Сва,
1977).
`
Е
|
<1.
Норы и убежища животных обычно. дифференцируют как ид обобен-
ностям их строения, так и по целевому назначению (Тихвинский,
1932; Кучерук, 1983}. Применительно к серой крысе ее жилниа по свое-
му строению могут быть подразделены на. не норовые убежища,
кула входят 1} поверхностные гнезда, 2) убежища различных емкостях
(пустотах) под навесами и в’ укрытиях, и на норы, подразделяемые на
1) бескамерные, 2) с камерами (без гнезд) 'и.3) гнездовые норы (про-
стые и сложные). По целевому. назначению мы зыделяем, норы 1) `за-
цитные и кормовые, 2) выводковые, 3) зимовочные, 4}. полуфункциональ-
ные.
.
|
и
Для изучения строения норы перед ее раскопкой нанослт на план все
входные отверстия, выбросы земли, поверхпостные ‘дорожки и другие
следы жизнедеятельности крыс. После этого для выявления хознев в
большинстве случаев нору облавливают дуговыми капканами-или, когда
это удается, вылавливают зверьков в процессе раскопки -и.зна том: же
плане обозначают обнаживишуюся систему галерей, тупики, пустые гнездо-
вые камеры и пр. (Карасева и др., 1985}. В помещёниях животноводче-
ских ферм норы чаще всего раскапывают во время ремонта здания (Коз-
лов, Козлов, 1971). © _
о
По своему строению норы ‘крысе, обитающих в природе и в домах,
очень разнообразны, но все они не имеют снециальных камер для запа-
сов и уборных. Роющая способность крыс велика, роют они ‘очень быст-
ро. По наблюдениям, в неволе нора длиной в 2,25 м с двумя камерами
может быть выкопана за два дня (Вго\пей, Свав -Сроз, 1977).
В. Г. Лялин (1972) указывает, что в свинарнике за одну почь два зверь-
ка выбросили на поверхность пола дна ведра земли.
8
Пизано и Сторер (Р4запо, Эюгег, 1948), изучая норовую деятельность
крыс, помещали их в клетку без дна, которую устанавливали вад ящи-
ком с землей. Зверьки очень быстро рыли передними лапами, ударами
одпой или двух передних же лап выбрасывали землю под озживот и уже
потом выкидывали ее задними копечностями. После того как крыса про-
капывала туннель, равный длине своего тела, она поворачивалась и вы-
Экология
129
Рис. 35. Схема строения
нор серых крыс
@а—г—норы
синантрон-
ных крыс (по: Карасева и
др., 1985):
а — бескамерная,
б —с камерой без гнезда,
в — простая гнездовая,
2 — сложная многоярусная
гнездовая;
9—3: норы экзоантропных
крыс:
98 — в бугре,
е — в береге реки,
ж,а—вполе
1 — камера без гнезда, .
2 — гнездовая камера,
3 — земляная пробка
талкивала землю передними лапами, помогая себе головой. В. Г. Лялин
(1972) пишет, что ипогда эти грызуны перетаскивают землю во рту.
Самки роют больше, чем самцы, с возрастом их роющая деятельность
понижается (РгоупеН, Свап П-Свиа, 1977). Калхоун (Сао, 1962)
сообщаст, что высокоранговые самки копают больше, чем низкорапговые
Крысы роют норы не только в почве, по и в разнообразных субстра-
тах: в глинобитпых стенах, окружающих кошары и другие животновод-
ческие фермы, в шлаколитных помещениях (И. 3. Климченко, устн со.
общ.) в слежавшихся опилках (В. А. Судейкин, устн. сообщ.), |: кучах
перетиоя, соломы. В твердом грунте рытье сочетается с грызущей деятель-
Норы крыс по своему строению, протяженности ходов, глубипе зале-
гания, а также по взаимнсму расположению очень различны (рис. 85).
Бескамерные норы имеют самое примитивное устройство
диаметр их ходов зависит от грунта. В твердом грунте он равен 5—7 см,
5 Серал крыса
430
Глава третья
в сыпучем 12—15 см. Длина галерей может быть самой различной (до
10 м и более), в основном она определяется расстоянием от укрытия до
корма. Обычно такие поры выполняют кормовые и защитные функции —
пасюки в них прячутся при любой опасности (например, при резких не-
знакомых звуках), а если поедапие корма там, где он находится, почему-
либо опаспо, то они песут его в свои подземные галереи, где спокоино
съедают. При высокой плотности населения бескамерпые норы сливаются
нередко вместе с гиездовыми, создавая сплошные сложные лабиринты.
Норы © камерой без гнезда имеют разнообразное строепие.
Наиболее простой случай, когда наклонный ход уходит в землю на глу-
бину 30—40 см и ведет к округлой или овальной, несколько приплюсну-
той сверху пустой камере. Такая нора может иметь песколько входных
отверстий, но чаще бывает одно. Норы с камерой, но без гнезда могут
быть и более сложного строения— иногда они имеют несколько камер.
Назначение таких нор неясно, возможно, они служат для длительного
отдыха самцов, но мы на свинофермах многократно наблюдали самцов,
спящих прямо на полу около кормушек или внутри них.
Грездовые норы по своему строению бывают простыми и сложтыми,
хотя деление это условно. Простая гнездовая нора имеет
1—2 входных отверстия, общая длина ходов обычно не превышает 2,5—
3 м, тиездовых камер одна или две. Такие гнездовые норы обычно при-
надлежат одной самке, которая выкармливает в ней свой выводок.
Сложные гнездовые норы —это сооружения, предотавляю-
щие собой лабиринты, в которых длина ходов варьирует от 3 до 35 м.
Входных отверстий в них бывает самое различное количество— от 3 до
нескольких десятков. В норе всегда есть 3—8 тнездовых камер, но мо-
жет быть и значительно больше (максимум 25). Камер без гнезда в них
меньшее количество (1—15), и они, как и в простых норах, расположены
не так глубоко.
Деление нор серых крыс на типы несколько условно. Между ними мо-
гут быть переходы, и не всегда каждую нору можно четко отнести к
тому или иному типу.
Экзоантропные зрысы
Природные биотопы
В природе поселения крыс, как влаголюбивых животпых, часто приуро-
чены к берегам различных водоемов: рек, стариц, ручьев, озер, морских
лиманов, а также к плавиям. В этих условиях зверьки ведут себя как
мелкие хищиики, рассредоточиваясь на местности.
Неноровые убежища
Поверхностные гнезда. В сырых, заболоченных местах, где
затруднено рытье нор, серые крысы устраивают поверхностные гнезда,
располагая их у основания кочек, в скоплениях сухих болотпых растений,
или же строят висячие гнезда, закрепляя их между стеблями тростника
Экология 4134
па высоте 10—20 см от поверхности земли. Все эти гнезда закрытые,
округлой формы, диаметром в 22—26 см и состоят из сухой растительно-
сти (Лозан, 1971). П. М Павлов и И. Б. Кирис (1956) находили гнезда
крыс в завалах тростника, на лежках кабанов. Иногда зверьки устраивают
поверхностные гнезда и на отпосительно сухой почве. 'Гак, В. С. Петров
п М. П. Леонтьев (1977) пишут, что летом расселяющиеея по пойме
р. Старка (Горьковская обл.) пасюки наряду © осповпым типом убе-
жищ — подземными порами — устраивают и надземные гнезда, размещая
их у осоковых кочек, перевитых стеблями вьющихся растений (под-
маренники и др.). Такое гнездо обычпо силетено из сухих стеблей
осоки.
Описано, что в низовьях р. Дуная, Прута, Днестра, в дельте Волги
весной крысы спасаются от половодья на деревьях, устраивая гнезда в
дуплах, а там, где нет дуплистых деревьев, подвешивают их в развилках
ветвей на высоте 0,25—1,5 м от поверхпости воды. Такие гнезда бывают
также шарообразной формы или несколько сплюснутые, диаметром 35—
80 см. В этом случае крысы делают их из свежесрезанных веточек дли-
пой 15—20 см. В основании гнезда имеются 1—3 входа, ведущие во
внутреннюю камеру, выложенную листьями (Кондрашкин, 1949; Айзен-
титадт, 19506). Описанные выше гнезда, по словам этих авторов, могут
быть использованы крысами в основном как защитные
— они сидят в
них, по словам Д. С. Айзеиштадта, дпем и покидают в сумерках. Вероят-
но, в случаях преследования зверька хищниками такие гнезда не могут
служить надежными убежищами, так как при нападении крысы выпуж-
депы покидать их, часто даже прыгая в воду. Н. Р. Кондрашкин опиеы-
вает случаи, когда в гнездах на деревьях эти грызуны размно-
жаются.
Кроме закрытых шарообразных поверхностных тпезд, по сообщению
И.Р. Мерзликина (19876), крысы устраивают своеобразные лежки (под-
стилки), которые состоят из травянистых растений и располагаются на
земле обычно в небольших углублениях. Эти подстилки зверьки делают
пе с целью предохранить себя от чрезмерной увлажненности почвы, а,
возможно, используют их для отдыха в наиболее комфортных условиях.
Убежища в различных емкостях (пустотах), под
навесами, в укрытиях. Пасюки, живущие в открытых 'биотопах,
охотно используют для своих убежищ различные пустоты. В пойме р. Пар-
кент (Узбекистан) они жили в дамбе, состоящей из крупного булыжни-
ка, укрепленного металлической сеткой. Длительные наблюдения за ме-
чепыми крысами показали, что они ностоянно использовали одни и те
же промежутки между камнями в качестве входных отверстий. Отсутст-
вие выбросов земли говорит о том, что, по всей вероятности, внутри дам-
бы они не рыли нор. В период наблюдения (июль-август) удавалось ви-
деть беременных самок и периодически появлялись подроспшие крысята,
следовательно, пасюки использовали убежища внутри дамбы для родов и
воспитания детенышей (Квашнин, Карасева, 1985). Есть указания на
то, что они живут в каменистых россыпях недалеко от поселков (Фети-
сов, 1945; Курятников, 1986).
5+
430
Глава третья
в сыпучем 12—15 см. Длина галерей может быть самой различной (до
10 м и более), в основном она определяется расстоянием от укрытия до
корма. Обычно такие поры выполняют кормовые и защитные функции —
пасюки в них прячутся при любой опасности (например, при резких не-
знакомых звуках), а если поедапие корма там, где он находится, почему-
либо опаспо, то они песут его в свои подземные галереи, где спокоино
съедают. При высокой плотности населения бескамерпые норы сливаются
нередко вместе с гиездовыми, создавая сплошные сложные лабиринты.
Норы © камерой без гнезда имеют разнообразное строепие.
Наиболее простой случай, когда наклонный ход уходит в землю на глу-
бину 30—40 см и ведет к округлой или овальной, несколько приплюсну-
той сверху пустой камере. Такая нора может иметь песколько входных
отверстий, но чаще бывает одно. Норы с камерой, но без гнезда могут
быть и более сложного строения— иногда они имеют несколько камер.
Назначение таких нор неясно, возможно, они служат для длительного
отдыха самцов, но мы на свинофермах многократно наблюдали самцов,
спящих прямо на полу около кормушек или внутри них.
Грездовые норы по своему строению бывают простыми и сложтыми,
хотя деление это условно. Простая гнездовая нора имеет
1—2 входных отверстия, общая длина ходов обычно не превышает 2,5—
3 м, тиездовых камер одна или две. Такие гнездовые норы обычно при-
надлежат одной самке, которая выкармливает в ней свой выводок.
Сложные гнездовые норы —это сооружения, предотавляю-
щие собой лабиринты, в которых длина ходов варьирует от 3 до 35 м.
Входных отверстий в них бывает самое различное количество— от 3 до
нескольких десятков. В норе всегда есть 3—8 тнездовых камер, но мо-
жет быть и значительно больше (максимум 25). Камер без гнезда в них
меньшее количество (1—15), и они, как и в простых норах, расположены
не так глубоко.
Деление нор серых крыс на типы несколько условно. Между ними мо-
гут быть переходы, и не всегда каждую нору можно четко отнести к
тому или иному типу.
Экзоантропные зрысы
Природные биотопы
В природе поселения крыс, как влаголюбивых животпых, часто приуро-
чены к берегам различных водоемов: рек, стариц, ручьев, озер, морских
лиманов, а также к плавиям. В этих условиях зверьки ведут себя как
мелкие хищиики, рассредоточиваясь на местности.
Неноровые убежища
Поверхностные гнезда. В сырых, заболоченных местах, где
затруднено рытье нор, серые крысы устраивают поверхностные гнезда,
располагая их у основания кочек, в скоплениях сухих болотпых растений,
или же строят висячие гнезда, закрепляя их между стеблями тростника
Экология 4134
па высоте 10—20 см от поверхности земли. Все эти гнезда закрытые,
округлой формы, диаметром в 22—26 см и состоят из сухой растительно-
сти (Лозан, 1971). П. М Павлов и И. Б. Кирис (1956) находили гнезда
крыс в завалах тростника, на лежках кабанов. Иногда зверьки устраивают
поверхностные гнезда и на отпосительно сухой почве. 'Гак, В. С. Петров
п М. П. Леонтьев (1977) пишут, что летом расселяющиеея по пойме
р. Старка (Горьковская обл.) пасюки наряду © осповпым типом убе-
жищ — подземными порами — устраивают и надземные гнезда, размещая
их у осоковых кочек, перевитых стеблями вьющихся растений (под-
маренники и др.). Такое гнездо обычпо силетено из сухих стеблей
осоки.
Описано, что в низовьях р. Дуная, Прута, Днестра, в дельте Волги
весной крысы спасаются от половодья на деревьях, устраивая гнезда в
дуплах, а там, где нет дуплистых деревьев, подвешивают их в развилках
ветвей на высоте 0,25—1,5 м от поверхпости воды. Такие гнезда бывают
также шарообразной формы или несколько сплюснутые, диаметром 35—
80 см. В этом случае крысы делают их из свежесрезанных веточек дли-
пой 15—20 см. В основании гнезда имеются 1—3 входа, ведущие во
внутреннюю камеру, выложенную листьями (Кондрашкин, 1949; Айзен-
титадт, 19506). Описанные выше гнезда, по словам этих авторов, могут
быть использованы крысами в основном как защитные
— они сидят в
них, по словам Д. С. Айзеиштадта, дпем и покидают в сумерках. Вероят-
но, в случаях преследования зверька хищниками такие гнезда не могут
служить надежными убежищами, так как при нападении крысы выпуж-
депы покидать их, часто даже прыгая в воду. Н. Р. Кондрашкин опиеы-
вает случаи, когда в гнездах на деревьях эти грызуны размно-
жаются.
Кроме закрытых шарообразных поверхностных тпезд, по сообщению
И.Р. Мерзликина (19876), крысы устраивают своеобразные лежки (под-
стилки), которые состоят из травянистых растений и располагаются на
земле обычно в небольших углублениях. Эти подстилки зверьки делают
пе с целью предохранить себя от чрезмерной увлажненности почвы, а,
возможно, используют их для отдыха в наиболее комфортных условиях.
Убежища в различных емкостях (пустотах), под
навесами, в укрытиях. Пасюки, живущие в открытых 'биотопах,
охотно используют для своих убежищ различные пустоты. В пойме р. Пар-
кент (Узбекистан) они жили в дамбе, состоящей из крупного булыжни-
ка, укрепленного металлической сеткой. Длительные наблюдения за ме-
чепыми крысами показали, что они ностоянно использовали одни и те
же промежутки между камнями в качестве входных отверстий. Отсутст-
вие выбросов земли говорит о том, что, по всей вероятности, внутри дам-
бы они не рыли нор. В период наблюдения (июль-август) удавалось ви-
деть беременных самок и периодически появлялись подроспшие крысята,
следовательно, пасюки использовали убежища внутри дамбы для родов и
воспитания детенышей (Квашнин, Карасева, 1985). Есть указания на
то, что они живут в каменистых россыпях недалеко от поселков (Фети-
сов, 1945; Курятников, 1986).
5+
132
Глава третья
=
т
Л_
Как известно, крысы охотно заселяют свалки и помойки. В ото и у.
чае они используют различные предметы (старую обувь, о Уду, ищи
в качестве защитных убе
и другие брошенные вещи)
ди Для утро
чаях они часто перемещаю
я
ства под ними гнезд. В этих слу
т
яотое-
и том же проходам, образовавшимся между старой ат
те
нты, когда эти тры
чаются комбинированные варианты,
ут дия
устройства убежиц какие-либо укрытия, вокруг или около которых р
систему нор.
Норы
.
срых крыс в природных биотонах, мы
не можем согласиться с мпением д. С. Айвениитадта 955 от то
эти грызуны ведут бродячий обр
-
я иа посят зременный характер и не имеют определенного плана строе”
пия. Ничем не нотревоженные крысы достаточно долго ‚ точение 2
3 месяцев (срок наблюдения) — живут в одних и тех же нора;
вынужденного выселения вновь возвразцаются в них. я от ло, де они
Устройство и взаимное расположение нор завися | ТОГО, ТД ови
вырыты, а именно от микрорельефа, то
и
та, защитных условий, в
льНо
-
У обоберто м еятольности человека. Поэтому норы крыс, обитаощих
в природе, по своему строепию более вариабельны, чем у синантр
пасюков, живущих в менее разнообразных условиях. ах в па других
В сырых местах зворьки роют норы на бутрах, гри д на других
сухих участках почвы, не заливаемых во время паводна, часто рыкаы
вают поры по склонам крутых берегов рок, каналов. При
2
гда выходят только над в .
первую оасредь верьки занимают искусствепно созданные челове-
ком нарушепия рельефа. Например, среди приазовских пла этой в о
хих грив тянутся проселочные дороги, которые во мног гостах укроп
лены насыпями. Именно вдоль этих дорог, на склоне насыпе й прочат
ся норы крыс. Иногда но ходу дорог через ручьи п канавы, пр жены
мостики и простые переправы — норы крыс часто можно н ти под оти-
ми укрытиями. В перечисленных выше биотопах встречаются 1 ры вова
мерные и с камерами. Интересно, что входные отверстия их чото
бывают совершенно открыты, т. е. не защищены какой-лиоо ра тительно-
стью. Бескамерные норы иногда проложены на десятки метров пл ль 66
рега (папример, на р. Пеёл на Украине). Гнездовые воры пр я то стро.
ния обычно имеют одно-два входных отверстия, а общая про лисе ть
ходов доходит до 2—3 м, но иногда в мягком, сыпучем грунте они мотут
быть очень короткими и фактически гнездовая камера прям рвет
ся входным отверстием. Диаметр тиездовых камер колеоле си в продела»
15—20, высота
— 12—15 см. Крысы, обитающие в открыт > биотоп ох,
делают гнезда преимущественно из сухой травы и листьев, ато тре-
чаются перья. Если зверек находит тряпку, кусок веревки, зум т
что-либо подобное, то обычпо он все подбирает и тащит в с
.
Переходя к характеристике нор ©
Экология
133
В поре имеются одна, реже две камеры с гнездами и, как правило, ость
пустые камеры. Последние чаще находятся рядом с гнездовыми, но могут
располагаться и в противоложном конце поры, В большинстве случаев
такие норы неглубоки, а гнездовые камеры располагаются па глубине
35—40 см.
Весьма типично для гнездовых пор паличие земляных пробок, которы:
ми забита часть ходов. Мы встречали норы, где отнорок не лоходил до
поверхности земли на 7—10 см. При онаспости крыса быстро прокапы-
вает это расстояние и спасается бегством. Обычно вокруг гисздовой норы
находятся песколько бескамерных нор 25—50 см длиной, играющих роль
защитных.
Для природпых биотопов характерно преобладание гнездовых нор
простого строения, и липть по берегам рек иногда встречаются сложные
тнездовые норы, вырытые под защитой больших деревьев.
В приазовских плавнях гиездовые норы располагались на разных рас-
стояниях друг от друга. Иногда можно было вотретить их рядом в 1—
2 м одну от другой, но бывало и так, что, найдя одлу нору, при тщатель-
пом обследовании территории другую удавалось обнаружить только в.
1—2 км от первой. Паиболее типично расположение групп нор (тнездо-
вых, с пустыми камерами и бескамерных) на расстоянии 50-100 м друг
от друга.
Для крысе, живущих в открытых биотопах, особенно типично исполь-
зование поверхностных дорожек, хорошо заметных на залитых водой ри-
совых полях, где зверьки могут рыть норы только на земляных валиках
и то не везде, а там, где это позволяет дренаж почвы. Поэтому простые
норы часто соединяются пс подземными галереями, а поверхностными до-
рожками. Длительные наблюдения и картирование входных отверстий, вы.
бросов земли, поверхностных дорожек на опытном участке на Кубани
ноказали, что последние идут вдоль уреза воды и в основном играют роль
охотничьих троп. На них часто можно найти остатки лягушек, кости рыб,
раковины моллюсков и пр. Обычно они беспрерывно тянутся на 150—
200 м.
Так как в природе поселения крыс в основном рассредоточены и не
бывает больших скоплений зверьков, длительно из поколения в поколение
живущих на одном месте, то среди гнездовых нор преобладают норы про-
стого типа. Однако иногда встречаются поры, вырытые, по-видимому, пе
одпим поколением крыс и имеющие сложную структуру. Часть ходов и ка-
мер бывает забита землей.
Крысы часто используют пустоты под корнями деревьев, расширяя и
усложняя их. В этом случае благодаря очень хороптим защитным услови-
ям здесь могут быть сложные гнездовые норы ({Кондраикин, 1949; Фе-
тисов, 1945; Хаматанов, 1985).
При наличии укрытий, например фундамепта из больших кусков дер-
на разваливитегося рыбацкого домика, крысы устраивали поры гораздо
более сложного строения, причем ходы проходили между пластами дерна.
При рытье оросительных каналов бульдозерами (Узбекистан) большие
пласты земли складывают вдоль каналов и между пеплотно слежавшими-
132
Глава третья
=
т
Л_
Как известно, крысы охотно заселяют свалки и помойки. В ото и у.
чае они используют различные предметы (старую обувь, о Уду, ищи
в качестве защитных убе
и другие брошенные вещи)
ди Для утро
чаях они часто перемещаю
я
ства под ними гнезд. В этих слу
т
яотое-
и том же проходам, образовавшимся между старой ат
те
нты, когда эти тры
чаются комбинированные варианты,
ут дия
устройства убежиц какие-либо укрытия, вокруг или около которых р
систему нор.
Норы
.
срых крыс в природных биотонах, мы
не можем согласиться с мпением д. С. Айвениитадта 955 от то
эти грызуны ведут бродячий обр
-
я иа посят зременный характер и не имеют определенного плана строе”
пия. Ничем не нотревоженные крысы достаточно долго ‚ точение 2
3 месяцев (срок наблюдения) — живут в одних и тех же нора;
вынужденного выселения вновь возвразцаются в них. я от ло, де они
Устройство и взаимное расположение нор завися | ТОГО, ТД ови
вырыты, а именно от микрорельефа, то
и
та, защитных условий, в
льНо
-
У обоберто м еятольности человека. Поэтому норы крыс, обитаощих
в природе, по своему строепию более вариабельны, чем у синантр
пасюков, живущих в менее разнообразных условиях. ах в па других
В сырых местах зворьки роют норы на бутрах, гри д на других
сухих участках почвы, не заливаемых во время паводна, часто рыкаы
вают поры по склонам крутых берегов рок, каналов. При
2
гда выходят только над в .
первую оасредь верьки занимают искусствепно созданные челове-
ком нарушепия рельефа. Например, среди приазовских пла этой в о
хих грив тянутся проселочные дороги, которые во мног гостах укроп
лены насыпями. Именно вдоль этих дорог, на склоне насыпе й прочат
ся норы крыс. Иногда но ходу дорог через ручьи п канавы, пр жены
мостики и простые переправы — норы крыс часто можно н ти под оти-
ми укрытиями. В перечисленных выше биотопах встречаются 1 ры вова
мерные и с камерами. Интересно, что входные отверстия их чото
бывают совершенно открыты, т. е. не защищены какой-лиоо ра тительно-
стью. Бескамерные норы иногда проложены на десятки метров пл ль 66
рега (папример, на р. Пеёл на Украине). Гнездовые воры пр я то стро.
ния обычно имеют одно-два входных отверстия, а общая про лисе ть
ходов доходит до 2—3 м, но иногда в мягком, сыпучем грунте они мотут
быть очень короткими и фактически гнездовая камера прям рвет
ся входным отверстием. Диаметр тиездовых камер колеоле си в продела»
15—20, высота
— 12—15 см. Крысы, обитающие в открыт > биотоп ох,
делают гнезда преимущественно из сухой травы и листьев, ато тре-
чаются перья. Если зверек находит тряпку, кусок веревки, зум т
что-либо подобное, то обычпо он все подбирает и тащит в с
.
Переходя к характеристике нор ©
Экология
133
В поре имеются одна, реже две камеры с гнездами и, как правило, ость
пустые камеры. Последние чаще находятся рядом с гнездовыми, но могут
располагаться и в противоложном конце поры, В большинстве случаев
такие норы неглубоки, а гнездовые камеры располагаются па глубине
35—40 см.
Весьма типично для гнездовых пор паличие земляных пробок, которы:
ми забита часть ходов. Мы встречали норы, где отнорок не лоходил до
поверхности земли на 7—10 см. При онаспости крыса быстро прокапы-
вает это расстояние и спасается бегством. Обычно вокруг гисздовой норы
находятся песколько бескамерных нор 25—50 см длиной, играющих роль
защитных.
Для природпых биотопов характерно преобладание гнездовых нор
простого строения, и липть по берегам рек иногда встречаются сложные
тнездовые норы, вырытые под защитой больших деревьев.
В приазовских плавнях гиездовые норы располагались на разных рас-
стояниях друг от друга. Иногда можно было вотретить их рядом в 1—
2 м одну от другой, но бывало и так, что, найдя одлу нору, при тщатель-
пом обследовании территории другую удавалось обнаружить только в.
1—2 км от первой. Паиболее типично расположение групп нор (тнездо-
вых, с пустыми камерами и бескамерных) на расстоянии 50-100 м друг
от друга.
Для крысе, живущих в открытых биотопах, особенно типично исполь-
зование поверхностных дорожек, хорошо заметных на залитых водой ри-
совых полях, где зверьки могут рыть норы только на земляных валиках
и то не везде, а там, где это позволяет дренаж почвы. Поэтому простые
норы часто соединяются пс подземными галереями, а поверхностными до-
рожками. Длительные наблюдения и картирование входных отверстий, вы.
бросов земли, поверхностных дорожек на опытном участке на Кубани
ноказали, что последние идут вдоль уреза воды и в основном играют роль
охотничьих троп. На них часто можно найти остатки лягушек, кости рыб,
раковины моллюсков и пр. Обычно они беспрерывно тянутся на 150—
200 м.
Так как в природе поселения крыс в основном рассредоточены и не
бывает больших скоплений зверьков, длительно из поколения в поколение
живущих на одном месте, то среди гнездовых нор преобладают норы про-
стого типа. Однако иногда встречаются поры, вырытые, по-видимому, пе
одпим поколением крыс и имеющие сложную структуру. Часть ходов и ка-
мер бывает забита землей.
Крысы часто используют пустоты под корнями деревьев, расширяя и
усложняя их. В этом случае благодаря очень хороптим защитным услови-
ям здесь могут быть сложные гнездовые норы ({Кондраикин, 1949; Фе-
тисов, 1945; Хаматанов, 1985).
При наличии укрытий, например фундамепта из больших кусков дер-
на разваливитегося рыбацкого домика, крысы устраивали поры гораздо
более сложного строения, причем ходы проходили между пластами дерна.
При рытье оросительных каналов бульдозерами (Узбекистан) большие
пласты земли складывают вдоль каналов и между пеплотно слежавшими-
134 Глава третья
ся пластами образуются полости, в которых крысы устраивают норы.
Очень типично использование ими ниш, часто образующихся между пла-
стами земли. Зверьки прокапывают в задией стенке нипти ходы, которые
ведут в норы, и в таких случаях галереи часто тяпутся межлу пластами
земли на протяжении 30—40 м, соединяя пустые и гнездовые камеры.
На рисовых полях после уборки урожая тракторами перепахиваются
чеки. Огромпые неплотно слежавшиеся пласты земли создают хорошие
укрытия для крыс, которые свободно перемещаются в образовавшихся пу-
стотах, кое-где разрывая пласты и проделывая входные отверстия.
По целевому назначению сложные гнездовые норы могут быть отнесе-
ны к выводковым и зимовочным. В выводковых норах обитает чаще всего
одна самка с выводком, реже две самки. По нашим наблюдениям, крысы
не роют специально зимовочных нор, а остаются зимовать в тех, же но-
рах, где оши жили и размножались летом, сильно расчищая и углублия
их осенью перед зимовкой. В октябре-ноябре па опытном участке (на ри-
совом поле) число свежих выбросов по сравнению © июлем августом
увеличивается более чем в 4—5 раз.
|
|
Раскопка зимовочных нор в приазовских плавиях показала, что гнез-
довые камеры находятся па значительно большей глубине (70—80 см),
чем летом, диаметр их увеличивается до 30 см, а высота до 22 см (Ка-
расева и др.. 1985).
.
Особый интерес представляет способность крысе поселяться в норах
других животных, перестраивая их па свой лад.
Серая крыса
— сильтый, агрессивный грызун. Нередко опа загрызает
хозлина норы или вынуждает покинуть свое жилище, часто засоляет и
старые, брошенные норы. Известно, что серые крысы поселяются в порах
даурской пишухи (Оспобопа дата РаН.)‚. даурского суслика ( я
Зашстз Вгап@!), бурундука (Ешат1аз зутеиз Гахт.) (Фетисов, 9,
водяной полевки (Аг еоа фегтезиМ8 р.) (Павлов, Кирис, т 956;
Лялин, 1972), обыкновенного хомяка (Седсейлз омсебиз Г.) ( ат
её а1., 1968а, No), земляной белки (Хегаз шат!з) (Р1запо, Звогет, 1948),
Помимо этого, крысы часто, расширяя ходы и гнездовые камеры полевок,
селятся в их норах, а именно в норах обыкновенных нолевок (Мусгойлз
агуаНз РаН.), полевок-экопомок (Мтегомя оесопошиз) , больших полевок
(М. Тотыз ВисБлег) и унгурских (М. ппеигеоз!з Эойг.) (Фетисов, 1945;
Хамаганов, 1975). Пасюки нередко устраивают гнезда прямо па хатках он-
датры (Опдайга фе са Г..).
_.
На Украине, на крутых отвесных берегах реки Пеёл, крысы осваивают
норы береговых ласточек (рама грата Ра|.), поедая яйца и птенцов,
а подчас и взрослых птиц. Сами норы они несколько расширяют и удли-
няют, а иногда пристраивают к ним свои сложные норы (рис. 36). Ин-
тересно, что крысы поселяются только в нижнем ярусе колонии ласточек
и, живя среди птиц, продолжают совершать набеги на их гнезда, опусто-
итая их (Мерзликин, 1986а).
|
Мы ни разу не встречали у крыс, живущих в открытых биотопах,
запасов и, следовательно, пе видели в их порах кладовых (Карасева и
др., 1985). Об отсутствии запасов у крыс писали многие исследователи
Экология
135
(Плятер-Плохоцкий, 1931; Формозов, 1945; Кондразикин, 1949; Хамага-
нов, 1975; Лозан, 1971; Русев, Соловьев, 1987; Боге, 1980; и др.). Про-
тивоположные сведения мы паходим в двух работах: А. С. Фетисов (1945)
сообщает о том, что пасюки на Дальнем Востоке делают запасы из кор-
ней диких и из корнеплодов культурных растений, а М. Хамар с соав-
торами (Нашаг еб а1., 19685) пишут о запасе зерна (кукурузы) в 5,4 кг,
обнаруженном в норе крысы. Но можно предположить, что в том и в
другом случае эти запасы припадлежали не крысам, а другим грызупам.
В той же работе А. С. Фетисов пишет, что крысы часто поселяются в
порах полевок (М. 1огиз и М. ппбагеп513) и используют их запасы.
Для крые, живущих в открытых биотопах, типична роющая деятель-
ность. В сырых местах они используют любой небольшой участок сухой
Рис. 86. Норы береговых ласточек, переоборудованные серыми крысами в сложные
норы
1— норы ласточен, 2 — ходы серой крысы, $ — гнезца серой крысы, 4 — земляная пробка (по
И. Р. Мерзликину)
земли для того, чтобы устроить в ном нору. И, по-видимому, только в
самых экстремальных условиях (наводнение, сплоитная заболоченность
и т. п.) зверьки переходят к устройству поверхиостных гиезд или соору-
жают их, как мы уже говорили, на деревьях.
Крысы, обитающие в природе, быстро реагируют на появление людей
па заселеппой ими территории и фактически становятся синантропными.
В этом случае расположение и устройство их нор в значительной степени
зависят от тех изменепий, которые вносит в природу хозяйственная дея-
тельность человека (наличие мостов, переправ, насыпей и тр.).
Синантропные крысы
Населенные пункты
Серые крысы могут обитать буквально во всех типах населенных пунктов,
начиная от одиночно стоящих домиков, деревень, аулов и кончая круп-
нейшими современными городами. Этот вид обладает удивительной спо-
собностью селиться и устраивать свои убежища иногда даже в самых,
казалось бы, неподходящих местах: в обшивке паровых котлов, в холо-
134 Глава третья
ся пластами образуются полости, в которых крысы устраивают норы.
Очень типично использование ими ниш, часто образующихся между пла-
стами земли. Зверьки прокапывают в задией стенке нипти ходы, которые
ведут в норы, и в таких случаях галереи часто тяпутся межлу пластами
земли на протяжении 30—40 м, соединяя пустые и гнездовые камеры.
На рисовых полях после уборки урожая тракторами перепахиваются
чеки. Огромпые неплотно слежавшиеся пласты земли создают хорошие
укрытия для крыс, которые свободно перемещаются в образовавшихся пу-
стотах, кое-где разрывая пласты и проделывая входные отверстия.
По целевому назначению сложные гнездовые норы могут быть отнесе-
ны к выводковым и зимовочным. В выводковых норах обитает чаще всего
одна самка с выводком, реже две самки. По нашим наблюдениям, крысы
не роют специально зимовочных нор, а остаются зимовать в тех, же но-
рах, где оши жили и размножались летом, сильно расчищая и углублия
их осенью перед зимовкой. В октябре-ноябре па опытном участке (на ри-
совом поле) число свежих выбросов по сравнению © июлем августом
увеличивается более чем в 4—5 раз.
|
|
Раскопка зимовочных нор в приазовских плавиях показала, что гнез-
довые камеры находятся па значительно большей глубине (70—80 см),
чем летом, диаметр их увеличивается до 30 см, а высота до 22 см (Ка-
расева и др.. 1985).
.
Особый интерес представляет способность крысе поселяться в норах
других животных, перестраивая их па свой лад.
Серая крыса
— сильтый, агрессивный грызун. Нередко опа загрызает
хозлина норы или вынуждает покинуть свое жилище, часто засоляет и
старые, брошенные норы. Известно, что серые крысы поселяются в порах
даурской пишухи (Оспобопа дата РаН.)‚. даурского суслика ( я
Зашстз Вгап@!), бурундука (Ешат1аз зутеиз Гахт.) (Фетисов, 9,
водяной полевки (Аг еоа фегтезиМ8 р.) (Павлов, Кирис, т 956;
Лялин, 1972), обыкновенного хомяка (Седсейлз омсебиз Г.) ( ат
её а1., 1968а, No), земляной белки (Хегаз шат!з) (Р1запо, Звогет, 1948),
Помимо этого, крысы часто, расширяя ходы и гнездовые камеры полевок,
селятся в их норах, а именно в норах обыкновенных нолевок (Мусгойлз
агуаНз РаН.), полевок-экопомок (Мтегомя оесопошиз) , больших полевок
(М. Тотыз ВисБлег) и унгурских (М. ппеигеоз!з Эойг.) (Фетисов, 1945;
Хамаганов, 1975). Пасюки нередко устраивают гнезда прямо па хатках он-
датры (Опдайга фе са Г..).
_.
На Украине, на крутых отвесных берегах реки Пеёл, крысы осваивают
норы береговых ласточек (рама грата Ра|.), поедая яйца и птенцов,
а подчас и взрослых птиц. Сами норы они несколько расширяют и удли-
няют, а иногда пристраивают к ним свои сложные норы (рис. 36). Ин-
тересно, что крысы поселяются только в нижнем ярусе колонии ласточек
и, живя среди птиц, продолжают совершать набеги на их гнезда, опусто-
итая их (Мерзликин, 1986а).
|
Мы ни разу не встречали у крыс, живущих в открытых биотопах,
запасов и, следовательно, пе видели в их порах кладовых (Карасева и
др., 1985). Об отсутствии запасов у крыс писали многие исследователи
Экология
135
(Плятер-Плохоцкий, 1931; Формозов, 1945; Кондразикин, 1949; Хамага-
нов, 1975; Лозан, 1971; Русев, Соловьев, 1987; Боге, 1980; и др.). Про-
тивоположные сведения мы паходим в двух работах: А. С. Фетисов (1945)
сообщает о том, что пасюки на Дальнем Востоке делают запасы из кор-
ней диких и из корнеплодов культурных растений, а М. Хамар с соав-
торами (Нашаг еб а1., 19685) пишут о запасе зерна (кукурузы) в 5,4 кг,
обнаруженном в норе крысы. Но можно предположить, что в том и в
другом случае эти запасы припадлежали не крысам, а другим грызупам.
В той же работе А. С. Фетисов пишет, что крысы часто поселяются в
порах полевок (М. 1огиз и М. ппбагеп513) и используют их запасы.
Для крые, живущих в открытых биотопах, типична роющая деятель-
ность. В сырых местах они используют любой небольшой участок сухой
Рис. 86. Норы береговых ласточек, переоборудованные серыми крысами в сложные
норы
1— норы ласточен, 2 — ходы серой крысы, $ — гнезца серой крысы, 4 — земляная пробка (по
И. Р. Мерзликину)
земли для того, чтобы устроить в ном нору. И, по-видимому, только в
самых экстремальных условиях (наводнение, сплоитная заболоченность
и т. п.) зверьки переходят к устройству поверхиостных гиезд или соору-
жают их, как мы уже говорили, на деревьях.
Крысы, обитающие в природе, быстро реагируют на появление людей
па заселеппой ими территории и фактически становятся синантропными.
В этом случае расположение и устройство их нор в значительной степени
зависят от тех изменепий, которые вносит в природу хозяйственная дея-
тельность человека (наличие мостов, переправ, насыпей и тр.).
Синантропные крысы
Населенные пункты
Серые крысы могут обитать буквально во всех типах населенных пунктов,
начиная от одиночно стоящих домиков, деревень, аулов и кончая круп-
нейшими современными городами. Этот вид обладает удивительной спо-
собностью селиться и устраивать свои убежища иногда даже в самых,
казалось бы, неподходящих местах: в обшивке паровых котлов, в холо-
136
Глава третья
дильциках. Но и для синантропных, так же как и для пасюков, живущих
в природе, справедливо подразделение всех убежищ на категории, ко
рые приведепы выше.
Неноровые убежища
Поверхностные гнезда. Серым крысам свойственно быстро ока
ливаться в мостах концентрации корма. В этих случаях они часто сонеем
не роют нор, а устраивают гнезда, например, нопосредотвонио в кулях
с комбикормом, мукой, сахаром и другими продуктами. незда ‹ стоят
из нагрызенных материалов, находящихся поблизости о эти ‚трят
ки, бумага, пакля, вата, мочало, шерсть, перья и пр.). Бну ти тнезда
бывает более мягкая подстилка из мелко нагрызенных матер лов. | й
ипогда слепые крысята лежат совсем без подстилки прямо на ко икор-
ме, муке и т. д. (Романова, 1986). В холодильниках, папример, гнезда
крыс находятся в замороженных тушах и, за отсутствием прут
риалов, бывают сделаны из сухожилий этих туш ( икитин,
о
В новых многоэтажных городоких домах пасюки обычно держатся в
подвалах, около камер мусоропроводов. В. К. Мелкова обнаружииа гиоздо
со слепыми крысятами, расположенное открыто примерно в 4 смо ь усо-
роприемной камеры. Защитные гиезда, в которых крысы отдыхают, ие
сколько меньших размеров и на складах часто, так жо как и выводко
вые камеры, располатаются между мешками или ящиками с продуктами.
В одном из складов на Украине по следам крыс, оставленным на ра :ы:
панной муке, удалось установить наличие постоянно используе:
рп,
связывающих пять тнезд, два из которых былис дотеньштами, а три -
пустые (защитные). Общая площадь, па которой находились оти гнезд
‹ностные дорожки, была равна
.
” "В оольской зови ‘например на свинофермах, крысы живут огром-
ными сообществами, по иногда поселяются небольшими труппами, пара-
ми и одипочками. Осенью одна-две крысы обычно селятся во време, о
пежилых домах (дачах) и устраивают гиезда в постелях, диванах и дру-
гой мягкой мебели, измельчая вату и материю для гпезда. Б данном слу-
чае тнезда несут функцию зимовочных убежищ, так как эворьки, как
правило, в неотапливаемых помещениях, где пет больного колич ‘ва
твия, не размножаются.
о аким образом, "самое примитивное убежище синантропных крыс —
это гнездо, которое находится прямо внутри «массы корма» или тепло
изолирующих материалов. Оно может пост функцию как выводк ,
‹
го и зимовочного убежища.
убежища В Зазличных емкостях (пустотах). Несколь-
ко более сложный случай мы видим, когда крысы переоборудуют под свое
жилище какие-либо емкости: различные ящики, бочки, коробки и р.
Г. А. Романова (1986) сообщает, например, о гнездах крыс в пустых бу
оброс крыс грызть очень велика. Принято считать, что опи гры-
зут в основном для того, чтобы стачивать отрастающие резцы, или в тех
Экология
137
случаях, когда между пими и кормом находится препятствие (Емельяно-
ва, 1983). Однако часто зверьки прибегают к использованию своих гры-
зущих способностей и для того, чтобы переоборудовать какой-либо пред-
мет под свое убежище. Начав что-либо грызть, крыса делает это неуклон-
но и систематически. По нашим наблюдениям, она способна грызть, если
ей не мешают, буквально в течение всех суток с побольшими перерыва-
ми для еды и отдыха.
Нам многократно приходилось находить в пустых ящиках гнезда па-
сюков, расположенпые, как правило, в углах. Зверек чаще начинает про-
трызать снизу одну из боковых степок ящика.
В Калининской области в оставленном на зиму доме крысы прогрыз-
ли в дверце деревянного шкафа входное отверстие диаметром 5,5 см ив
углу на нижней полке устроили гнездо из мелко нагрызенной бумаги и
мочала, вытащепного из матраца, На второй полке шкафа было спизу про-
грызено другое отверстие, через которое зверьки постоянно посещали
«второй ярус» своего убежища.
Очень часто крысы устраивают убежища в емкостях стен современных
строений. Например, прогрызают многослойпые панели, из которых
построепы животноводческие комплексы, и делают тнезда в среднем слое
пенопласта и других материалов. Зверьки устраивают убежища также и
в теплоизоляционном слое крыш. Особенно интересно использование ими
в свинарниках электрических щитов, за которыми и даже внутри кото-
рых они часто устраивают гнезда (Козлов, 1980а).
Убежища под навесами и в укрытиях. Крысы весьма
охотно селятся в укрытиях, созданных человеком. Нри перекладывании
поленницы дров, куч хвороста, различной тары (ящиков, бочек и пр.),
находящихся поблизости от источников корма (например, от курятника
или крольчатпика), под ними часто паходят гнезда крыс, расположенные
обычно на земле.
В. Я. Антонюк (устн. сообщ.) сообщает о случае, когда он обнаружил
гнездо серой крысы в мешке
с крупной солью. Гнездо находилось в нише,
образовавшейся в слежавшейся и затвердевшей соли, и состояло из
обрывков бумаги, этикеток, мешковины и пр.
В постройках свиноводческих комплексов, где полы бетонированы, кры-
сам негде выкапывать норы. В большинстве случаен они устраивают гнез-
да в нишах, сделанных для мусоропроводных камер, а также в системе
подземной вентиляции (Козлов, 1980а). Строения, в которых только на»
мечаются повреждения бетопированного пола и образуются трещины в
стенах, крысы быстро осваивают и приступают к рытью нор. Преобла-
дающее число нор входных отверстий выходит в углах помещения или в
местах повреждения полов. Зверьки проникают под бетонные покрытия
также в местах ввода отопительной и водопроводной систем. В розульта-
те их интенсивной роющей деятельности за ночь может быть выброшено
до трех-четырех водер земли, шлака и щебня. Под бетоном образуются
большие пустоты, над которыми покрытия легко проваливаются, и, та-
ким образом, для зверьков складываются благоприятные условия для
дальнейшего устройства подземных убежищ.
136
Глава третья
дильциках. Но и для синантропных, так же как и для пасюков, живущих
в природе, справедливо подразделение всех убежищ на категории, ко
рые приведепы выше.
Неноровые убежища
Поверхностные гнезда. Серым крысам свойственно быстро ока
ливаться в мостах концентрации корма. В этих случаях они часто сонеем
не роют нор, а устраивают гнезда, например, нопосредотвонио в кулях
с комбикормом, мукой, сахаром и другими продуктами. незда ‹ стоят
из нагрызенных материалов, находящихся поблизости о эти ‚трят
ки, бумага, пакля, вата, мочало, шерсть, перья и пр.). Бну ти тнезда
бывает более мягкая подстилка из мелко нагрызенных матер лов. | й
ипогда слепые крысята лежат совсем без подстилки прямо на ко икор-
ме, муке и т. д. (Романова, 1986). В холодильниках, папример, гнезда
крыс находятся в замороженных тушах и, за отсутствием прут
риалов, бывают сделаны из сухожилий этих туш ( икитин,
о
В новых многоэтажных городоких домах пасюки обычно держатся в
подвалах, около камер мусоропроводов. В. К. Мелкова обнаружииа гиоздо
со слепыми крысятами, расположенное открыто примерно в 4 смо ь усо-
роприемной камеры. Защитные гиезда, в которых крысы отдыхают, ие
сколько меньших размеров и на складах часто, так жо как и выводко
вые камеры, располатаются между мешками или ящиками с продуктами.
В одном из складов на Украине по следам крыс, оставленным на ра :ы:
панной муке, удалось установить наличие постоянно используе:
рп,
связывающих пять тнезд, два из которых былис дотеньштами, а три -
пустые (защитные). Общая площадь, па которой находились оти гнезд
‹ностные дорожки, была равна
.
” "В оольской зови ‘например на свинофермах, крысы живут огром-
ными сообществами, по иногда поселяются небольшими труппами, пара-
ми и одипочками. Осенью одна-две крысы обычно селятся во време, о
пежилых домах (дачах) и устраивают гиезда в постелях, диванах и дру-
гой мягкой мебели, измельчая вату и материю для гпезда. Б данном слу-
чае тнезда несут функцию зимовочных убежищ, так как эворьки, как
правило, в неотапливаемых помещениях, где пет больного колич ‘ва
твия, не размножаются.
о аким образом, "самое примитивное убежище синантропных крыс —
это гнездо, которое находится прямо внутри «массы корма» или тепло
изолирующих материалов. Оно может пост функцию как выводк
,
‹
го и зимовочного убежища.
убежища В Зазличных емкостях (пустотах). Несколь-
ко более сложный случай мы видим, когда крысы переоборудуют под свое
жилище какие-либо емкости: различные ящики, бочки, коробки и р.
Г. А. Романова (1986) сообщает, например, о гнездах крыс в пустых бу
оброс крыс грызть очень велика. Принято считать, что опи гры-
зут в основном для того, чтобы стачивать отрастающие резцы, или в тех
Экология
137
случаях, когда между пими и кормом находится препятствие (Емельяно-
ва, 1983). Однако часто зверьки прибегают к использованию своих гры-
зущих способностей и для того, чтобы переоборудовать какой-либо пред-
мет под свое убежище. Начав что-либо грызть, крыса делает это неуклон-
но и систематически. По нашим наблюдениям, она способна грызть, если
ей не мешают, буквально в течение всех суток с побольшими перерыва-
ми для еды и отдыха.
Нам многократно приходилось находить в пустых ящиках гнезда па-
сюков, расположенпые, как правило, в углах. Зверек чаще начинает про-
трызать снизу одну из боковых степок ящика.
В Калининской области в оставленном на зиму доме крысы прогрыз-
ли в дверце деревянного шкафа входное отверстие диаметром 5,5 см ив
углу на нижней полке устроили гнездо из мелко нагрызенной бумаги и
мочала, вытащепного из матраца, На второй полке шкафа было спизу про-
грызено другое отверстие, через которое зверьки постоянно посещали
«второй ярус» своего убежища.
Очень часто крысы устраивают убежища в емкостях стен современных
строений. Например, прогрызают многослойпые панели, из которых
построепы животноводческие комплексы, и делают тнезда в среднем слое
пенопласта и других материалов. Зверьки устраивают убежища также и
в теплоизоляционном слое крыш. Особенно интересно использование ими
в свинарниках электрических щитов, за которыми и даже внутри кото-
рых они часто устраивают гнезда (Козлов, 1980а).
Убежища под навесами и в укрытиях. Крысы весьма
охотно селятся в укрытиях, созданных человеком. Нри перекладывании
поленницы дров, куч хвороста, различной тары (ящиков, бочек и пр.),
находящихся поблизости от источников корма (например, от курятника
или крольчатпика), под ними часто паходят гнезда крыс, расположенные
обычно на земле.
В. Я. Антонюк (устн. сообщ.) сообщает о случае, когда он обнаружил
гнездо серой крысы в мешке
с крупной солью. Гнездо находилось в нише,
образовавшейся в слежавшейся и затвердевшей соли, и состояло из
обрывков бумаги, этикеток, мешковины и пр.
В постройках свиноводческих комплексов, где полы бетонированы, кры-
сам негде выкапывать норы. В большинстве случаен они устраивают гнез-
да в нишах, сделанных для мусоропроводных камер, а также в системе
подземной вентиляции (Козлов, 1980а). Строения, в которых только на»
мечаются повреждения бетопированного пола и образуются трещины в
стенах, крысы быстро осваивают и приступают к рытью нор. Преобла-
дающее число нор входных отверстий выходит в углах помещения или в
местах повреждения полов. Зверьки проникают под бетонные покрытия
также в местах ввода отопительной и водопроводной систем. В розульта-
те их интенсивной роющей деятельности за ночь может быть выброшено
до трех-четырех водер земли, шлака и щебня. Под бетоном образуются
большие пустоты, над которыми покрытия легко проваливаются, и, та-
ким образом, для зверьков складываются благоприятные условия для
дальнейшего устройства подземных убежищ.
138
Глава третья
В подвалах жилых многоэтажных домов, где пол заботонирован, кры-
сы также пе могут рыть норы и устраивают расположенные отчрь я
верхпостные гнезда. По чаще они их располагают под как либо р
тием, а захламлонность подвалов увеличивает количество уе’ и лая
них. А. В. Судейкип, например, сообщил о гнезде с крысятами, 1 дет
ном в подвале мпотоэтажного жилого дома под укрыти теплопровода,
заваленного битым кирпичом и кусками бетона. Крысы устра то жили.
ца также во внутренней полости перекрытий, иопольвуя режд
поверхности в качестве входных отверстий (Мелкова,
.
Норы
Килые норы отличаются чистотой, только ипотда в ходах можно обиа-
ружить небольшое количество экскрементов (Карасева и До. $85;
Тоте, 1980). Беременные самки, найдя подходящий корм, „методически
у
:
однак
недалеко от входа, где, ол ,
переносят его в свою нору н
ы
‹
т
я орциальных камер для запасов. Иногда самки окладываюе корм в м
||
ает неболь
200—
Е
т норы. Занас корма быв
либо укрытии поблизости о
12.
(200--
300 г). Нередко зверьки для того, чтобы спокойно питаться, затаск ват
в нору какой-нибудь относительно крупный кусок пищи. мер, т
ик
но потом остатки удаляют. /1ор ое,
павшей курицы, рыбу и т. п.),
р
1980) ет, что в Г. Пью-Джерси около мусорооборщика он обедов
ч
}
{
ыхкамер : -
5
ных
оных пор. В 16 из 18 раскопанн
пар сложных гнезд
пани:
ен т ездовой материал (трава, бумага, остатки тряпок, полиэтиленовых
мешков и пр.), по ни в одной из пих не был обнаружен запас корм .
)
.),
Гнезда крыс в норах, так же как и поверхностные, состоят из ото
имеющегося поблизости пригодного материала. В одном из о (Се
И
&
;
шили березовые веники и
верный Казахстан) в сарае су
я
ор в этом помещении обнаружили, что все И почти целиком состо
к
Ковлов, 1978).
т из сухих березовых листьев (Козлов,
” Простые гнездовые поры (кроме выводковых) по целевому назначению
бывают и зимовочными, но также выполняют функции защитных и кор
мовых. Встречаются опи там, где нет больыпото скопления крыс,—в жи
ых домах сельской местности, в новых строениях ИВОТНОВОЧеСКИ?
форм В зданиях пасюки часто роют норы в земляном полу под полов .
цами, в этом случае часто ходы идут вблизи от поверхности и иногд
имеют вид желоба, но подземные галереи могут ить в ину,
‹
С
ис. 37). -
ся в 30—50 ем от поверхности (р
и тогда камеры располагают
рис. 37)
:
х нор могут быть вытянуты р
е
тда ходы простых гнездовых
:
з
10—12 м Крысы часто устраивают их в парниках в слое унароженно
земли, где сами ходы идут вдоль степок парника (Антонюк, устн. с00 щ.).
В гнездах, где ость крысята, материал всегда свежий, никогда не бы
заст заплесневелым и затхлым. Само ето при наличии ие ДО ты-
И
ис. . По мере роста кры
‚
игей круглое, закрытое сверху (р
увеличивается, стенки утончаются и гнездо, приобретает форму лотка,
открытого сверху. Такие норы бывают неглубоки. гнездовая камера р
положена па глубине 20—30 см от поверхпости. В большинстве случаев
Экология
139
Рис. 37. Сложная по-
верхностная нора серых
крыс на свиноферме под
деревянным настилом
(по: Карасева и др.
1985}
1— поверхностные дорож-
ки-желоба,
2 — поверхностные гнезда,
$ — подземные галереи,
4 — подземные пустые ка-
меры,
5 — камеры с гнездами
{вид сверху)
[ЕЕ Е [Е]з [Ем [535
в норе есть и пустая камера, которая находится ближе к поверхности
земли. Обычно есть несколько сленых отнорков. Часть ходов и входных
отверстий иногда, вероятно с целью утепления норы, бывает забита зем-
лей или соломой, Но, может быть, крысы делают это и для защиты от
вторжения чужих особей (Лялин, 1972). Калхоун (СаШоив, 1962) ука-
зывает, что затыкать изнутри ходы более свойственно подчиненным о0со-
бям.
Сложные гнездовые норы встречаются в местах большого скопления
крыс, в частпости в старых зданиях животноводческих ферм, особенно в
свинарниках, где постоянно имеется избыток корма и воды. Хамар с со-
авторами (Натаг её а|., 1968а, Ъ) пишут, что в некоторых уездах Румы-
нии при очень высокой численности крыс они прогрызали стены глино-
битных зданий, которые быстро после этого разрушались. Земляной пол
под деревянными настилами весь был изрыт крысами, изрешечены даже
деревянные балки и доски.
В Казахстане в степах самапных стросний ферм крысы проделывают
многочисленные вертикальные и горизонтальные ходы, образующие
сплошной лабиринт, который тянется вдоль всей стены па песколько
десятков метров и открывается множеством входных отверстий, особенно
в местах кормления и водопоя домашних животных (Козлов, 19805).
В Якутия свиньи и другой скот содержатся в деревянных помеще-
ниях, засыпанных снаружи до нижнего края окон (до высоты 150—
160 см) землей, поремешанной с навозом. Нрысы проделывают в этой
насыни вдоль и поперек ходы, которые соединены с самим помещением.
Внутри они прогрызают ходы в деревянном полу возле стен по всему
нериметру помещения. Сложные гнездовые норы в свинарниках часто
располагаются под деревянными настилами. В этих случаях часть ходов
138
Глава третья
В подвалах жилых многоэтажных домов, где пол заботонирован, кры-
сы также пе могут рыть норы и устраивают расположенные отчрь я
верхпостные гнезда. По чаще они их располагают под как либо р
тием, а захламлонность подвалов увеличивает количество уе’ и лая
них. А. В. Судейкип, например, сообщил о гнезде с крысятами, 1 дет
ном в подвале мпотоэтажного жилого дома под укрыти теплопровода,
заваленного битым кирпичом и кусками бетона. Крысы устра то жили.
ца также во внутренней полости перекрытий, иопольвуя режд
поверхности в качестве входных отверстий (Мелкова,
.
Норы
Килые норы отличаются чистотой, только ипотда в ходах можно обиа-
ружить небольшое количество экскрементов (Карасева и До. $85;
Тоте, 1980). Беременные самки, найдя подходящий корм, „методически
у
:
однак
недалеко от входа, где, ол ,
переносят его в свою нору н
ы
‹
т
я орциальных камер для запасов. Иногда самки окладываюе корм в м
||
ает неболь
200—
Е
т норы. Занас корма быв
либо укрытии поблизости о
12.
(200--
300 г). Нередко зверьки для того, чтобы спокойно питаться, затаск ват
в нору какой-нибудь относительно крупный кусок пищи. мер, т
ик
но потом остатки удаляют. /1ор ое,
павшей курицы, рыбу и т. п.),
р
1980) ет, что в Г. Пью-Джерси около мусорооборщика он обедов
ч
}
{
ыхкамер : -
5
ных
оных пор. В 16 из 18 раскопанн
пар сложных гнезд
пани:
ен т ездовой материал (трава, бумага, остатки тряпок, полиэтиленовых
мешков и пр.), по ни в одной из пих не был обнаружен запас корм .
)
.),
Гнезда крыс в норах, так же как и поверхностные, состоят из ото
имеющегося поблизости пригодного материала. В одном из о (Се
И
&
;
шили березовые веники и
верный Казахстан) в сарае су
я
ор в этом помещении обнаружили, что все И почти целиком состо
к
Ковлов, 1978).
т из сухих березовых листьев (Козлов,
” Простые гнездовые поры (кроме выводковых) по целевому назначению
бывают и зимовочными, но также выполняют функции защитных и кор
мовых. Встречаются опи там, где нет больыпото скопления крыс,—в жи
ых домах сельской местности, в новых строениях ИВОТНОВОЧеСКИ?
форм В зданиях пасюки часто роют норы в земляном полу под полов .
цами, в этом случае часто ходы идут вблизи от поверхности и иногд
имеют вид желоба, но подземные галереи могут ить в ину,
‹
С
ис. 37). -
ся в 30—50 ем от поверхности (р
и тогда камеры располагают
рис. 37)
:
х нор могут быть вытянуты р
е
тда ходы простых гнездовых
:
з
10—12 м Крысы часто устраивают их в парниках в слое унароженно
земли, где сами ходы идут вдоль степок парника (Антонюк, устн. с00 щ.).
В гнездах, где ость крысята, материал всегда свежий, никогда не бы
заст заплесневелым и затхлым. Само ето при наличии ие ДО ты-
И
ис. . По мере роста кры
‚
игей круглое, закрытое сверху (р
увеличивается, стенки утончаются и гнездо, приобретает форму лотка,
открытого сверху. Такие норы бывают неглубоки. гнездовая камера р
положена па глубине 20—30 см от поверхпости. В большинстве случаев
Экология
139
Рис. 37. Сложная по-
верхностная нора серых
крыс на свиноферме под
деревянным настилом
(по: Карасева и др.
1985}
1— поверхностные дорож-
ки-желоба,
2 — поверхностные гнезда,
$ — подземные галереи,
4 — подземные пустые ка-
меры,
5 — камеры с гнездами
{вид сверху)
[ЕЕ Е [Е]з [Ем [535
в норе есть и пустая камера, которая находится ближе к поверхности
земли. Обычно есть несколько сленых отнорков. Часть ходов и входных
отверстий иногда, вероятно с целью утепления норы, бывает забита зем-
лей или соломой, Но, может быть, крысы делают это и для защиты от
вторжения чужих особей (Лялин, 1972). Калхоун (СаШоив, 1962) ука-
зывает, что затыкать изнутри ходы более свойственно подчиненным о0со-
бям.
Сложные гнездовые норы встречаются в местах большого скопления
крыс, в частпости в старых зданиях животноводческих ферм, особенно в
свинарниках, где постоянно имеется избыток корма и воды. Хамар с со-
авторами (Натаг её а|., 1968а, Ъ) пишут, что в некоторых уездах Румы-
нии при очень высокой численности крыс они прогрызали стены глино-
битных зданий, которые быстро после этого разрушались. Земляной пол
под деревянными настилами весь был изрыт крысами, изрешечены даже
деревянные балки и доски.
В Казахстане в степах самапных стросний ферм крысы проделывают
многочисленные вертикальные и горизонтальные ходы, образующие
сплошной лабиринт, который тянется вдоль всей стены па песколько
десятков метров и открывается множеством входных отверстий, особенно
в местах кормления и водопоя домашних животных (Козлов, 19805).
В Якутия свиньи и другой скот содержатся в деревянных помеще-
ниях, засыпанных снаружи до нижнего края окон (до высоты 150—
160 см) землей, поремешанной с навозом. Нрысы проделывают в этой
насыни вдоль и поперек ходы, которые соединены с самим помещением.
Внутри они прогрызают ходы в деревянном полу возле стен по всему
нериметру помещения. Сложные гнездовые норы в свинарниках часто
располагаются под деревянными настилами. В этих случаях часть ходов
140
Глава третья
идет прямо по новерхпости земли в виде тролипок ИЛИ желобов, кото-
рые соединяют поверхностные гнезда. Другая часть ходов идет под зем-
лей на глубине всего 5—8 см и связывает камеры без гнезд и гиездовые
камеры. Часто встречаются норы значительно более глубокие (80—120 см),
состоящие из трех ярусов, ипогда подземные галереи сложных пор окру-
жаютг какие-либо коммуникации, встретивитиеся па их пути, например
‹анализационпый колодец (рис. 39).
Ииптересно, что крысы обычно устраивают гнездовые камеры в местах,
надежно защищенных сверху. нод каким-нибудь массивным прикры-
Рис. 88. Гнезда в норах серых врыс (по: Карасева и др., 1985)
р
р
—8-диевными крысятами
Вверху — гнездо с однодневными крысятами, внизу — гнездо с 7
Экология
141
Рис. 39. Сложная глу-
бокая нора серых крыс
{огибающая канализа-
ционный колодец) на
свиноферме (по: Кара-
сева и др., 1985)
Т— вид сверху,
И — в разрезе
А
п
н
а
л
и
з
а
н
и
в
н
н
ы
й
А
С
л
о
д
е
А
Яя
ПОЙа ЙЕ
НЕТ|Е
422 18
Рис. 40. Концентрация
Е
гнездовых камер в норе
Е
серой крысы, вырытой `
>
на площади 0,5 м? меж-
Е
Эу клетками свиней (КТ,
Е
КЗ) и ямой (Я) (по: Ка-
Е
расева и др., 1985)
&
1
К?
К!
140
Глава третья
идет прямо по новерхпости земли в виде тролипок ИЛИ желобов, кото-
рые соединяют поверхностные гнезда. Другая часть ходов идет под зем-
лей на глубине всего 5—8 см и связывает камеры без гнезд и гиездовые
камеры. Часто встречаются норы значительно более глубокие (80—120 см),
состоящие из трех ярусов, ипогда подземные галереи сложных пор окру-
жаютг какие-либо коммуникации, встретивитиеся па их пути, например
‹анализационпый колодец (рис. 39).
Ииптересно, что крысы обычно устраивают гнездовые камеры в местах,
надежно защищенных сверху. нод каким-нибудь массивным прикры-
Рис. 88. Гнезда в норах серых врыс (по: Карасева и др., 1985)
р
р
—8-диевными крысятами
Вверху — гнездо с однодневными крысятами, внизу — гнездо с 7
Экология
141
Рис. 39. Сложная глу-
бокая нора серых крыс
{огибающая канализа-
ционный колодец) на
свиноферме (по: Кара-
сева и др., 1985)
Т— вид сверху,
И — в разрезе
А
п
н
а
л
и
з
а
н
и
в
н
н
ы
й
А
С
л
о
д
е
А
Яя
ПОЙа ЙЕ
НЕТ|Е
422 18
Рис. 40. Концентрация
Е
гнездовых камер в норе
Е
серой крысы, вырытой `
>
на площади 0,5 м? меж-
Е
Эу клетками свиней (КТ,
Е
КЗ) и ямой (Я) (по: Ка-
Е
расева и др., 1985)
&
1
К?
К!
142
Глава третья
тием — бочкой, стеной сарая или бетонированным настилом и т. п. В тех
случаях, когда почему-нибо рытье нор особенно затруднено и животные
вынуждены копать их па ограниченной территории, гнездовые камеры
могут быть расположены в непосредственной близости друг от друга.
Например, в одном из совхозов Северного Казахстапа па площади
1,2 м? было 11 тнездовых камер, причем семь из них располагались
в два яруса на площади в 0,5 м’, первый на глубине А—9 см, второй —
30—45 см (рис. 40) (Карасева и др., 1985).
В Западной и Восточной Сибири, а также в Северном Казахстане
на свинофермах поры крыс помещаются главным образом в стенах зда-
пия и внутри них и только часть входных отверстий выходит наружу
(Лялин, 1972; Романова, 1986). Но в районах с более теплым климатом
(Северный Кавказ) они часто располагаются вне помещений и даже
в 2—5 мот них, а от входных отверстий к стене свинарника или к источ-
нику воды ведут хорошо протоптанные поверхностные дорожки.
Размеры камер и характер устройства гнезд в норах как простых, так
и сложных зависят от температурных условий. В теплых помещениях
(свинарники, телятники) камеры невелики: диаметр их 14—19 сы,
а высота 11—15 см. В неотапливаемых зданиях (кладовые, склады
ит. 1.) размеры гиездовых камер гораздо больше: диаметр 17—29 см,
а высота 14—19 см.
Масса гнезда также очень различна. В Северном Казахстане в теплых
помещениях она не превышает 30—60 г, в холодных— 100—160 г
(Козлов, 1980). В Якутии масса гнезда доходит до 300 г (Романова,
1986). Глубина залегапия камер зависит не только от температурных
условий, но и от плотности грунта. В твердом грунте они находятся на
меньшей глубине, чем в рыхлом. В холодных помещениях с рыхлым.
грунтом норы бывают относительно глубоки (140—400 см). В теплых
помещениях они более поверхностны (8—10 см).
.
Сложные норы выкапывают, несомненно, несколько поколений крыс.
Такие поры существуют только в старых здапиях свиноферм, в новых
помещениях зверьки не успевают построить столь сложные, требующие
больших усилий сооружения,
Если крысы вновь носелнются в помещениях или около них (папри-
мер, после дератизации или ремонта), то они в большинстве случаев не
роют новых нор, а заселяют старые, персделывая их по-своему — в этом
случае выбросы земли свидетельствуют о том, что животные роют новые
ходы, отнорки и камеры. По-видимому, приверженность пасюка к старым
норам объясняется тем, что сохраняются запаховые метки (Соколов
и ур., 19796) и различные следы их жизнедеятельности, которые служат
гарантией пригодности этого места для обитания.
По целевому назначению такие сложные норы должны быть отнесены
к нолуфункциональным — крысы используют их и как выводковые,
и как защитные. В них часто одновременно встречаются разновозраст-
ные выводки. Изпример, в Северной Осетии (Моздокский район) на
свиноферме в трехъярусной норе (общая протяженность ходов 18 м,
11 гнездовых камер, 5 пустых) было добыто 8 взрослых крыс (2 самца
Экология
143
и 6 кормящих самок), 3 выводка (25 слепых крысят) и 15 подросших
неноловозрелых особей — всего 48 крыс.
В другой подобной норе обитали 32 крысы, в том числе 3 взрослых
самца, 4 кормящие самки, 4 выводка крысят и 3 молодых неполовозрелых
самца (Царасева и др., 1985).
Вероятно, каждую нору населяет один клан (группировка) крыс, сре-
ди которых есть доминирующий самец, охраняющий данную нору и при-
легающую к ней территорию. Трудно себе представить, чтобы в одной
норе могло быть больше одпого клана, т. е. чтобы крысы могли внутри
норы делить территорию. Если в нашем случае в одной норе жили
48 крыс, то очень интересно было бы проследить, как они стали рас-
селяться, когда крысята выросли. К сожалению, у пас таких наблюде-
ний нет, но Калхоун (Капоии, 1962) пишет, что более высокораптовые
особи чаще покидают обжитые норы и роют или осваивают другие,
а молодые остаются в старых норах.
Сложные гнездовые поры, помимо животноводческих ферм, иногда
встречаются в надворных постройках частных домов. Большинство
их сконцентрировано в сараях, в которых содержат скот и птицу, 060-
бенно в помещениях для свиней и кладовых.
Несомненно, что нора — это нестабильное сооружение. Она постоянно
«живет», видоизменяется и может переходить из одного типа. нор в другой.
Простые гнездовые норы могут усложпяться по мере размножения
крыс и роста их численности. Калхоун (1962) пишет, что старые самки
на периферии норы роют новые камеры, где родят следующие выводки.
А. Н. Козлов (1980а) наблюдал, что молодые беременные самочки, от-
деляясь от материнского гнезда, в разных местах окраин поры делают
углубления и копают камеры. Вероятно, возможны тот и другой вариан-
ты. Иногда сложная пора образуется за счет того, что несколько простых
тисздовых нор соединяются подземными галереями.
Подводя итог всему изложенному выше, мы можем сказать, что
пасюк проявляет необычайную пластичность в характере устройства
и использования убежищ. Трудно согласиться с А. Н. Формозовым
(1945) и М. Хамар с соавторами (Нашаг еЁ а!., 1968а, Ь), которые
нишут, что серая крыса малороющий зверь. Как мы видели, этому виду
свойственна интенсивная розощая деятельность, но благодаря своей высо-
кой экологической гибкости он выживает там, где рытье пор невозможно,
устраивая поверхностные гнезда или используя для убежища укрытия
и пустоты
Прокапывая и прогрызая длинные ходы в деревянных, глинобитных
и шлаковых степах животноводческих ферм, а также выкапывая глубо-
кие и сложные норы под стенами зданий, крысы приводят их тем самым
в негодное состояпие, в результате чего они разрушаются значительно
рансе положенного срока (Козлов, 1980а,6) Выкапывая поры в берегах
оросительных каналов, пасюки способствуют их размыванию и разруше-
нию (Карасева и др., 1985). Гораздо более экономично тратить больше
ценег на дератизационные работы, чем па ликвидацию последствий рою-
щей деятельности этого опаснейшего из всех видов грызунов.
142
Глава третья
тием — бочкой, стеной сарая или бетонированным настилом и т. п. В тех
случаях, когда почему-нибо рытье нор особенно затруднено и животные
вынуждены копать их па ограниченной территории, гнездовые камеры
могут быть расположены в непосредственной близости друг от друга.
Например, в одном из совхозов Северного Казахстапа па площади
1,2 м? было 11 тнездовых камер, причем семь из них располагались
в два яруса на площади в 0,5 м’, первый на глубине А—9 см, второй —
30—45 см (рис. 40) (Карасева и др., 1985).
В Западной и Восточной Сибири, а также в Северном Казахстане
на свинофермах поры крыс помещаются главным образом в стенах зда-
пия и внутри них и только часть входных отверстий выходит наружу
(Лялин, 1972; Романова, 1986). Но в районах с более теплым климатом
(Северный Кавказ) они часто располагаются вне помещений и даже
в 2—5 мот них, а от входных отверстий к стене свинарника или к источ-
нику воды ведут хорошо протоптанные поверхностные дорожки.
Размеры камер и характер устройства гнезд в норах как простых, так
и сложных зависят от температурных условий. В теплых помещениях
(свинарники, телятники) камеры невелики: диаметр их 14—19 сы,
а высота 11—15 см. В неотапливаемых зданиях (кладовые, склады
ит. 1.) размеры гиездовых камер гораздо больше: диаметр 17—29 см,
а высота 14—19 см.
Масса гнезда также очень различна. В Северном Казахстане в теплых
помещениях она не превышает 30—60 г, в холодных— 100—160 г
(Козлов, 1980). В Якутии масса гнезда доходит до 300 г (Романова,
1986). Глубина залегапия камер зависит не только от температурных
условий, но и от плотности грунта. В твердом грунте они находятся на
меньшей глубине, чем в рыхлом. В холодных помещениях с рыхлым.
грунтом норы бывают относительно глубоки (140—400 см). В теплых
помещениях они более поверхностны (8—10 см).
.
Сложные норы выкапывают, несомненно, несколько поколений крыс.
Такие поры существуют только в старых здапиях свиноферм, в новых
помещениях зверьки не успевают построить столь сложные, требующие
больших усилий сооружения,
Если крысы вновь носелнются в помещениях или около них (папри-
мер, после дератизации или ремонта), то они в большинстве случаев не
роют новых нор, а заселяют старые, персделывая их по-своему — в этом
случае выбросы земли свидетельствуют о том, что животные роют новые
ходы, отнорки и камеры. По-видимому, приверженность пасюка к старым
норам объясняется тем, что сохраняются запаховые метки (Соколов
и ур., 19796) и различные следы их жизнедеятельности, которые служат
гарантией пригодности этого места для обитания.
По целевому назначению такие сложные норы должны быть отнесены
к нолуфункциональным — крысы используют их и как выводковые,
и как защитные. В них часто одновременно встречаются разновозраст-
ные выводки. Изпример, в Северной Осетии (Моздокский район) на
свиноферме в трехъярусной норе (общая протяженность ходов 18 м,
11 гнездовых камер, 5 пустых) было добыто 8 взрослых крыс (2 самца
Экология
143
и 6 кормящих самок), 3 выводка (25 слепых крысят) и 15 подросших
неноловозрелых особей — всего 48 крыс.
В другой подобной норе обитали 32 крысы, в том числе 3 взрослых
самца, 4 кормящие самки, 4 выводка крысят и 3 молодых неполовозрелых
самца (Царасева и др., 1985).
Вероятно, каждую нору населяет один клан (группировка) крыс, сре-
ди которых есть доминирующий самец, охраняющий данную нору и при-
легающую к ней территорию. Трудно себе представить, чтобы в одной
норе могло быть больше одпого клана, т. е. чтобы крысы могли внутри
норы делить территорию. Если в нашем случае в одной норе жили
48 крыс, то очень интересно было бы проследить, как они стали рас-
селяться, когда крысята выросли. К сожалению, у пас таких наблюде-
ний нет, но Калхоун (Капоии, 1962) пишет, что более высокораптовые
особи чаще покидают обжитые норы и роют или осваивают другие,
а молодые остаются в старых норах.
Сложные гнездовые поры, помимо животноводческих ферм, иногда
встречаются в надворных постройках частных домов. Большинство
их сконцентрировано в сараях, в которых содержат скот и птицу, 060-
бенно в помещениях для свиней и кладовых.
Несомненно, что нора — это нестабильное сооружение. Она постоянно
«живет», видоизменяется и может переходить из одного типа. нор в другой.
Простые гнездовые норы могут усложпяться по мере размножения
крыс и роста их численности. Калхоун (1962) пишет, что старые самки
на периферии норы роют новые камеры, где родят следующие выводки.
А. Н. Козлов (1980а) наблюдал, что молодые беременные самочки, от-
деляясь от материнского гнезда, в разных местах окраин поры делают
углубления и копают камеры. Вероятно, возможны тот и другой вариан-
ты. Иногда сложная пора образуется за счет того, что несколько простых
тисздовых нор соединяются подземными галереями.
Подводя итог всему изложенному выше, мы можем сказать, что
пасюк проявляет необычайную пластичность в характере устройства
и использования убежищ. Трудно согласиться с А. Н. Формозовым
(1945) и М. Хамар с соавторами (Нашаг еЁ а!., 1968а, Ь), которые
нишут, что серая крыса малороющий зверь. Как мы видели, этому виду
свойственна интенсивная розощая деятельность, но благодаря своей высо-
кой экологической гибкости он выживает там, где рытье пор невозможно,
устраивая поверхностные гнезда или используя для убежища укрытия
и пустоты
Прокапывая и прогрызая длинные ходы в деревянных, глинобитных
и шлаковых степах животноводческих ферм, а также выкапывая глубо-
кие и сложные норы под стенами зданий, крысы приводят их тем самым
в негодное состояпие, в результате чего они разрушаются значительно
рансе положенного срока (Козлов, 1980а,6) Выкапывая поры в берегах
оросительных каналов, пасюки способствуют их размыванию и разруше-
нию (Карасева и др., 1985). Гораздо более экономично тратить больше
ценег на дератизационные работы, чем па ликвидацию последствий рою-
щей деятельности этого опаснейшего из всех видов грызунов.
АА Глава третья
Поселяясь в норах многочисленных животных, серые крысы спобоб-
ствуют циркуляции различных инфекций с природной очаговостью
(псевдотуберкулез, иерсениоз, лептоспироз и др.), опасных для человека.
Питание
Обитание серых крыс в самых разнообразных экологических нишах за-
ставляет задумываться о том, какие факторы способствуют адаптации
этих грызунов к новым условиям. Питание крыс, изучением которого
занимались в последние десятилетия многие специалисты, безусловно,
может быть одним из этих факторов.
Изучение особенностей питания крыс в различных условиях (геогра-
фических, лабораторных, естественных и вольерных) показало, что ииро-
кий пабор кормов присущ пасюкам вне зависимости от привязанности
к географическим зонам. Однако данные, полученные многими исследо-
вателями, часто трудпосопоставимы как из-за биотопической, стациаль-
ной и сезоппой разницы во время сбора материала, так и из-за различия
в методиках определения отдельных видов корма.
Наиболее распрострапенными методами исследования питапия диких
видов грызунов являются анализы содержимого желудков, экскрементов,
кормовых запасов и остатков пищи на местах кормежки. Эти методы
реально отражают качественный состав пищи, хотя каждый из них не
лишен недостатков. Сильная измельченность корма в желудке затрудняет
его определение, а разница в скорости переваривания растительных и
животных кормов нередко снижает точность ‘информации за определен-
ные промежутки времени. Более совершенный метод определения содер-
жимого желудков описан В. А. Рыльниковым и Е. В. Карасевой (1985).
Ими былн получены эталонные образцы нереваренных кормов путем
скармливания лабораторным белым крысам различной растительной и
животной пищи. Через 2, 4, 6 ч после поедания этих кормов забивали
по две крысы для изучения отличительных особенностей нищи на раз-
ных стадиях переваривания. Содержимое желудков отловлеппых пасюков
сопоставляли © такими эталонными образцами, но предварительно его
обрабатывали смесью спиртового раствора йода с йодистым калием, бла-
тоцаря чему растительные остатки за счет включенного в них крахмала
окрашивались в синий цвет, а животные— в фиолетово-красный.
Попытки определения питания пасюка по содержимому экскрементов
не дают достаточно эффективного результата, так как в кале в основном
содержатся пепереваренные остатки растений в виде клетчатки и 0еезо-
звоночных в виде хитипа. Данпые по питанию этого грызуна, собранные
мпогими авторами в разных местообитаниях, говорят как о возможности
его существования в условиях с самой различной кормовой базой, так
и о его способности переходить с одного вида корма па другой.
В разных географически удаленных друг от друга районах у экзо-
антропных популяций серых крыс прослеживается сезопиость в смепе
кормов в зависимости от динамики и обилия фоновых видов мелких
позвоночных, беспозвоночных и растений в биоценозе. У пасюков, насе-
Экология
145
ляющих агроценозы, эта связь находится в более тесной зависимости от
произрастаюних там сельскоховяйственных культур, времени их посева,
созревания и уборки.
Питапие синаптроппых пасюков во всех географических зопах цели-
ком зависит от того, какие продукты они находят на хозяйственных
объектах человека, и здесь спектр их кормов практически не ограничен.
Экзоантропные крысы
Природные биотопы
Имеющийся в нашем распоряжении материал по питапию окзоантропных
серых крыс является в достаточной степени противоречивым. Нет удов-
летворительных сведений о различии в питапии по сезонам, почти пол-
ностью отсутствуют данные о составе пищи молодых особей. Нередко
авторы па основании своих исследований, проведенных только за один
какой-либо сезон года, делают вывод о том, что пасюк в основном яв-
ляется хищником или растительноядным.
С нашей точки зрения, паслок являстся воеядным, но процентный со-
став его пищи может меняться в зависимости от мест обитания и от
времени года.
В природе круглогодичное обитание серых крысе определяется в знз-
чительной степени наличием кормовой базы и воды.
На юге нашей страны по берегам оз. Бебисири (Абхазия), по данным
А. Д. Бериштейна (1959), в желудках крыс в 80% случаев преобладала
растительная пища, а в 30% — животная, т. е. остатки рыб, лягушек,
раков и грызунов. Там же находили разгрызенных моллюсков, а на нут-
риевой ферме, расположенной невдалеке, отмечены случаи пападения на
нутрий.
О преимущественном питании южных пасюков растительными корма-
ми писали Н. К. Верецатин (1959) и Г. Н. Львов (1949), описывая кол-
хидского пасюка. При изучении состава «кормовых столиков» ими было
установлено, что в летние месяцы вблизи водоемов зверьки в 88% слу-
чаев поедали листья осоки, молодые побеги тростника, плоды водяного
ореха. Животпые корма встречались реже: моллюски в 36% случаев,
а насекомые и раки —в 6%. В сентябре, октябре и ноябре основу нита-
ния составляли водяпые орехи. УЖитков (1944) писал, что в феврале
1939 г., по словам Н. В. Верещагина, крысы за ночь съели попавшую
в капкан нутрию, весящую 1,5 кг. Там же попадались остатки шкурок
и кости водяных полевок. Автор предполагает, что в зимнее время пика
насоков содержит пресноводных моллюсков и лягушек. Не исключено,
что нахождение авторами в разпое время года погрызов, а в нище—
растительных остатков носит сезонный характер, тем более что боль-
шинство исследователей считают пасюка преимуществепио плотоядным.
По данным Д. С. Айзенштадта (1955) и Г. В. Романова (1963),
южный пасюк— хищник. М. Т. Яковлев (1949) проанализировал желуд-
ки диких популяций пасюков дельты Дона и в 78—100% желудков нашел
АА
Глава третья
Поселяясь в норах многочисленных животных, серые крысы спобоб-
ствуют циркуляции различных инфекций с природной очаговостью
(псевдотуберкулез, иерсениоз, лептоспироз и др.), опасных для человека.
Питание
Обитание серых крыс в самых разнообразных экологических нишах за-
ставляет задумываться о том, какие факторы способствуют адаптации
этих грызунов к новым условиям. Питание крыс, изучением которого
занимались в последние десятилетия многие специалисты, безусловно,
может быть одним из этих факторов.
Изучение особенностей питания крыс в различных условиях (геогра-
фических, лабораторных, естественных и вольерных) показало, что ииро-
кий пабор кормов присущ пасюкам вне зависимости от привязанности
к географическим зонам. Однако данные, полученные многими исследо-
вателями, часто трудпосопоставимы как из-за биотопической, стациаль-
ной и сезоппой разницы во время сбора материала, так и из-за различия
в методиках определения отдельных видов корма.
Наиболее распрострапенными методами исследования питапия диких
видов грызунов являются анализы содержимого желудков, экскрементов,
кормовых запасов и остатков пищи на местах кормежки. Эти методы
реально отражают качественный состав пищи, хотя каждый из них не
лишен недостатков. Сильная измельченность корма в желудке затрудняет
его определение, а разница в скорости переваривания растительных и
животных кормов нередко снижает точность ‘информации за определен-
ные промежутки времени. Более совершенный метод определения содер-
жимого желудков описан В. А. Рыльниковым и Е. В. Карасевой (1985).
Ими былн получены эталонные образцы нереваренных кормов путем
скармливания лабораторным белым крысам различной растительной и
животной пищи. Через 2, 4, 6 ч после поедания этих кормов забивали
по две крысы для изучения отличительных особенностей нищи на раз-
ных стадиях переваривания. Содержимое желудков отловлеппых пасюков
сопоставляли © такими эталонными образцами, но предварительно его
обрабатывали смесью спиртового раствора йода с йодистым калием, бла-
тоцаря чему растительные остатки за счет включенного в них крахмала
окрашивались в синий цвет, а животные— в фиолетово-красный.
Попытки определения питания пасюка по содержимому экскрементов
не дают достаточно эффективного результата, так как в кале в основном
содержатся пепереваренные остатки растений в виде клетчатки и 0еезо-
звоночных в виде хитипа. Данпые по питанию этого грызуна, собранные
мпогими авторами в разных местообитаниях, говорят как о возможности
его существования в условиях с самой различной кормовой базой, так
и о его способности переходить с одного вида корма па другой.
В разных географически удаленных друг от друга районах у экзо-
антропных популяций серых крыс прослеживается сезопиость в смепе
кормов в зависимости от динамики и обилия фоновых видов мелких
позвоночных, беспозвоночных и растений в биоценозе. У пасюков, насе-
Экология
145
ляющих агроценозы, эта связь находится в более тесной зависимости от
произрастаюних там сельскоховяйственных культур, времени их посева,
созревания и уборки.
Питапие синаптроппых пасюков во всех географических зопах цели-
ком зависит от того, какие продукты они находят на хозяйственных
объектах человека, и здесь спектр их кормов практически не ограничен.
Экзоантропные крысы
Природные биотопы
Имеющийся в нашем распоряжении материал по питапию окзоантропных
серых крыс является в достаточной степени противоречивым. Нет удов-
летворительных сведений о различии в питапии по сезонам, почти пол-
ностью отсутствуют данные о составе пищи молодых особей. Нередко
авторы па основании своих исследований, проведенных только за один
какой-либо сезон года, делают вывод о том, что пасюк в основном яв-
ляется хищником или растительноядным.
С нашей точки зрения, паслок являстся воеядным, но процентный со-
став его пищи может меняться в зависимости от мест обитания и от
времени года.
В природе круглогодичное обитание серых крысе определяется в знз-
чительной степени наличием кормовой базы и воды.
На юге нашей страны по берегам оз. Бебисири (Абхазия), по данным
А. Д. Бериштейна (1959), в желудках крыс в 80% случаев преобладала
растительная пища, а в 30% — животная, т. е. остатки рыб, лягушек,
раков и грызунов. Там же находили разгрызенных моллюсков, а на нут-
риевой ферме, расположенной невдалеке, отмечены случаи пападения на
нутрий.
О преимущественном питании южных пасюков растительными корма-
ми писали Н. К. Верецатин (1959) и Г. Н. Львов (1949), описывая кол-
хидского пасюка. При изучении состава «кормовых столиков» ими было
установлено, что в летние месяцы вблизи водоемов зверьки в 88% слу-
чаев поедали листья осоки, молодые побеги тростника, плоды водяного
ореха. Животпые корма встречались реже: моллюски в 36% случаев,
а насекомые и раки —в 6%. В сентябре, октябре и ноябре основу нита-
ния составляли водяпые орехи. УЖитков (1944) писал, что в феврале
1939 г., по словам Н. В. Верещагина, крысы за ночь съели попавшую
в капкан нутрию, весящую 1,5 кг. Там же попадались остатки шкурок
и кости водяных полевок. Автор предполагает, что в зимнее время пика
насоков содержит пресноводных моллюсков и лягушек. Не исключено,
что нахождение авторами в разпое время года погрызов, а в нище—
растительных остатков носит сезонный характер, тем более что боль-
шинство исследователей считают пасюка преимуществепио плотоядным.
По данным Д. С. Айзенштадта (1955) и Г. В. Романова (1963),
южный пасюк— хищник. М. Т. Яковлев (1949) проанализировал желуд-
ки диких популяций пасюков дельты Дона и в 78—100% желудков нашел
146
Глава третья
шерсть, остатки насекомых и рыб. Автор сделал логичный вывод, что
характер питания зависит от наличия того или иного вида корма, и
вона.
то реимущестненное питание животными кормами ры ущих
в дельте Волги, описывает Г. А. Копдрашкин (1949, 1 р ; Там
в 93% желудков, содержащих пищу, обнаружили шерсть, Кост имя
мелких грызунон. Лишь в 5—26 % желудков были остатки ор и,
зелепые части и семена растений. Зимой здесь крысы питались молод-
няком водяных полевок, другими видами трызунов, итицами, снулой
рыбой. Весной и осенью они охотились за лятушками, рыбой и двуствор-
чатыми моллюсками, летом — за жуками-плавунцами, водолюбами и дру-
тими насекомыми. Особенно много они уничтожали лягушек. О преоб-
ладапии в рациопе крыс животпого корма свидетельствуют и остатки,
найденные в норах. В январе в одной из раскопанных нор нашли
35 шкурок и скелетов полевых мышеи, 14 черепов лягушек, остатки
сазанчиков, жуков, ящериц, а в другой норе
— остатки выеденных водн-
` ` илима).
.
ых орехов ий) (1945) обнаружил, что желудок крысы, добытой
в дельте Волти в ноябре, был туго набит остатками озерных ‘лягушек.
Интересно сообщение Н. Н. Бакеева (1967) о том, как зимой паки
могут добывать беспозвоночных: в приазовских плавнях дельты У ани
в феврале 1964 г. в зарослях сухой лебеды мнотие серые крысы взбира
лись по стеблям на высоту 1—1,5 м и выгрызали из них личинок огнев-
ки. В желудке пойманной там крысы их обнаружено 49 штук.
О преимуществепном питании крыс, обитающих в окоповодных ота-
циях Моравии, пищей животного происхождения сообщает . аплета
(ТарееЪ 1964). Апализ 21 желудка показал, что содержимое на
15% составляли части пасекомых, моллюсков, млекопитающих и другие
пеидентифицированные остатки мяса, и костей, а в 25—45% случаев
остатки семян злаков и непереваренная зелень.
,
.
На западном берегу Каспийского моря в плавнях р. Таловки весной
и летом пищей крысам служили яйца птиц, рыба, попавшая в вентери,
и мелкие грызуны (Афанасьев, 1956).
.
На севере Апшеронокого полуострова песок у воды был ен
крысами, подбиравиими выбросы моря, рыбу, ракообразных и др. ит
ков, 1944). М. В. Тихвинская (1964) в Татарии и Башкирии в желудках
серых крыс в основном обнаружила животные корма, хотя вегетативные
части растепий, семена ежеголовника и кубышки там тоже имелись.
Д. С. Айзенштадт, работая эпидемиологом на северо-западной части
фронта, на основании вскрытия желудков крыс установил, что в мае -
июне во время их интенсивного размножения п расселения молодняк
часть питания составляли зеленые части растении и семена хвоиных
(до 30%) даже у особей, живших в блиндажах. Расселившиеся молодые
зверьки целиком переходили на природный животный или растительный
корм (АЙйзенщтадт, 1945).
.в
Пищу животвото происхождения молодые нпасюки начинают употре
лять очепь рано. Даже в желудках крысят © массон тела 4 —57Т г
Экология
147
(что соответствует приблизительно 4—5 неделям), выкопанных из норы
вместе с родителями, обнаружена шерсть мышей с небольшой примесью
зелени.
Возрастные различия в питании пасюков, обитающих в котловине
оз. Неро (Ярославская область), отметили Е. В. Карасева с соавторами
(1986а). Апализ 50 желудков показал, что летом взрослые особи (стар-
ше 9 месяцев) здесь, как и в других ириродных биотопах, всеядпы, по
все же питаются опи преимущественно позвоночными. У более молодых
особей (до 2 месяцев) в содержимом желудков больше остатков беспо-
звоночных и растений, что, по-видимому, связано с тем, что они пока
еще не могут добывать позвоночных.
В. А. Рыльников и Е. В. Карасева (1985) паблюдали вариации
в питацпии крыс разных возрастных групп летом на рисовых полях
Краснодарского края. В пище молодняка в возрасте до 2 месяцев отчет-
ливо преобладали вегетативные наземные части растений и беспозвоноч-
ные, а доля других видов корма, особенно семян и позвоночных, была
незначительна. У крыс в возрасте от 2 месяцев и старше доля семян
и животных кормов возрастает, а вегетативных наземных частей расте-
ний убывает. Взрослые крысы питаются преимущественно животной
пищей и семенами растений. Таким образом, на разницу в составе кор-
мов у крыс, обитающих в разных регионах, кроме сезопа, может влиять
и возраст зверьков.
Пасюки, добытые на берегу р. Урал (Казахстан) и оз. Имантау
(Кокчетавская обл.), питались рыбой и моллюсками, а один из векры-
тых желудков содержал мясо и шерсть мелкого грызуна (Борисенко,
1977).
В Западной Сибири (Томский и Нарабелиский районы) вскрытые
желудки 14 крыс, отловленных в зарослях прибрежной растительности,
содержали как растительные, так и животные корма (Лялин, 1978), при-
чем последние были встречены во всех желудках (100%), а раститель-
ные— только в 6 (42.84) и их количественное содержание было неве-
лико— в пределах 10—17%. В состав содержимого желудков входили
рогоз, осоки, в также пырей ползучий. Из животных кормов больше все-
то попадались дождевые черви и моллюски—в 78,6% исследованпых
желудков; в 50% желудков встречались насекомые, чаще всего жуки-
листоеды. По-видимому, эти виды являются здесь наиболее доступными
для крыс. Из мышевидных грызунов отмечены два вида полевок (в двух
случаях — красная, и в четырех— экономка).
На востоке нашей страны, в Хабаровском крае, у обитающих в при-
роде серых крыс (карако) семена растений встречались в 68% желудков,
зелепых частей растений не находили, а животные корма найдены
в 89% желудков (Хаматанов, 1967, 1968). Эти крысы охотились за мел-
кими грызупами, лягушками, снулыми лососевыми рыбами, птицами,
двустворчатыми моллюсками и насекомыми. Автор на основании вскры-
тия 120 желудков крыс из поймы р. Амур показывает сезонные измене-
ния в их питании. Зимой основу питания составляли семепа растений,
грызуны и рыба. Летом больше половины исследованных желудков было
146
Глава третья
шерсть, остатки насекомых и рыб. Автор сделал логичный вывод, что
характер питания зависит от наличия того или иного вида корма, и
вона.
то реимущестненное питание животными кормами ры ущих
в дельте Волги, описывает Г. А. Копдрашкин (1949, 1 р ; Там
в 93% желудков, содержащих пищу, обнаружили шерсть, Кост имя
мелких грызунон. Лишь в 5—26 % желудков были остатки ор и,
зелепые части и семена растений. Зимой здесь крысы питались молод-
няком водяных полевок, другими видами трызунов, итицами, снулой
рыбой. Весной и осенью они охотились за лятушками, рыбой и двуствор-
чатыми моллюсками, летом — за жуками-плавунцами, водолюбами и дру-
тими насекомыми. Особенно много они уничтожали лягушек. О преоб-
ладапии в рациопе крыс животпого корма свидетельствуют и остатки,
найденные в норах. В январе в одной из раскопанных нор нашли
35 шкурок и скелетов полевых мышеи, 14 черепов лягушек, остатки
сазанчиков, жуков, ящериц, а в другой норе
— остатки выеденных водн-
` ` илима).
.
ых орехов ий) (1945) обнаружил, что желудок крысы, добытой
в дельте Волти в ноябре, был туго набит остатками озерных ‘лягушек.
Интересно сообщение Н. Н. Бакеева (1967) о том, как зимой паки
могут добывать беспозвоночных: в приазовских плавнях дельты У ани
в феврале 1964 г. в зарослях сухой лебеды мнотие серые крысы взбира
лись по стеблям на высоту 1—1,5 м и выгрызали из них личинок огнев-
ки. В желудке пойманной там крысы их обнаружено 49 штук.
О преимуществепном питании крыс, обитающих в окоповодных ота-
циях Моравии, пищей животного происхождения сообщает . аплета
(ТарееЪ 1964). Апализ 21 желудка показал, что содержимое на
15% составляли части пасекомых, моллюсков, млекопитающих и другие
пеидентифицированные остатки мяса, и костей, а в 25—45% случаев
остатки семян злаков и непереваренная зелень.
,
.
На западном берегу Каспийского моря в плавнях р. Таловки весной
и летом пищей крысам служили яйца птиц, рыба, попавшая в вентери,
и мелкие грызуны (Афанасьев, 1956).
.
На севере Апшеронокого полуострова песок у воды был ен
крысами, подбиравиими выбросы моря, рыбу, ракообразных и др. ит
ков, 1944). М. В. Тихвинская (1964) в Татарии и Башкирии в желудках
серых крыс в основном обнаружила животные корма, хотя вегетативные
части растепий, семена ежеголовника и кубышки там тоже имелись.
Д. С. Айзенштадт, работая эпидемиологом на северо-западной части
фронта, на основании вскрытия желудков крыс установил, что в мае -
июне во время их интенсивного размножения п расселения молодняк
часть питания составляли зеленые части растении и семена хвоиных
(до 30%) даже у особей, живших в блиндажах. Расселившиеся молодые
зверьки целиком переходили на природный животный или растительный
корм (АЙйзенщтадт, 1945).
.в
Пищу животвото происхождения молодые нпасюки начинают употре
лять очепь рано. Даже в желудках крысят © массон тела 4 —57Т г
Экология
147
(что соответствует приблизительно 4—5 неделям), выкопанных из норы
вместе с родителями, обнаружена шерсть мышей с небольшой примесью
зелени.
Возрастные различия в питании пасюков, обитающих в котловине
оз. Неро (Ярославская область), отметили Е. В. Карасева с соавторами
(1986а). Апализ 50 желудков показал, что летом взрослые особи (стар-
ше 9 месяцев) здесь, как и в других ириродных биотопах, всеядпы, по
все же питаются опи преимущественно позвоночными. У более молодых
особей (до 2 месяцев) в содержимом желудков больше остатков беспо-
звоночных и растений, что, по-видимому, связано с тем, что они пока
еще не могут добывать позвоночных.
В. А. Рыльников и Е. В. Карасева (1985) паблюдали вариации
в питацпии крыс разных возрастных групп летом на рисовых полях
Краснодарского края. В пище молодняка в возрасте до 2 месяцев отчет-
ливо преобладали вегетативные наземные части растений и беспозвоноч-
ные, а доля других видов корма, особенно семян и позвоночных, была
незначительна. У крыс в возрасте от 2 месяцев и старше доля семян
и животных кормов возрастает, а вегетативных наземных частей расте-
ний убывает. Взрослые крысы питаются преимущественно животной
пищей и семенами растений. Таким образом, на разницу в составе кор-
мов у крыс, обитающих в разных регионах, кроме сезопа, может влиять
и возраст зверьков.
Пасюки, добытые на берегу р. Урал (Казахстан) и оз. Имантау
(Кокчетавская обл.), питались рыбой и моллюсками, а один из векры-
тых желудков содержал мясо и шерсть мелкого грызуна (Борисенко,
1977).
В Западной Сибири (Томский и Нарабелиский районы) вскрытые
желудки 14 крыс, отловленных в зарослях прибрежной растительности,
содержали как растительные, так и животные корма (Лялин, 1978), при-
чем последние были встречены во всех желудках (100%), а раститель-
ные— только в 6 (42.84) и их количественное содержание было неве-
лико— в пределах 10—17%. В состав содержимого желудков входили
рогоз, осоки, в также пырей ползучий. Из животных кормов больше все-
то попадались дождевые черви и моллюски—в 78,6% исследованпых
желудков; в 50% желудков встречались насекомые, чаще всего жуки-
листоеды. По-видимому, эти виды являются здесь наиболее доступными
для крыс. Из мышевидных грызунов отмечены два вида полевок (в двух
случаях — красная, и в четырех— экономка).
На востоке нашей страны, в Хабаровском крае, у обитающих в при-
роде серых крыс (карако) семена растений встречались в 68% желудков,
зелепых частей растений не находили, а животные корма найдены
в 89% желудков (Хаматанов, 1967, 1968). Эти крысы охотились за мел-
кими грызупами, лягушками, снулыми лососевыми рыбами, птицами,
двустворчатыми моллюсками и насекомыми. Автор на основании вскры-
тия 120 желудков крыс из поймы р. Амур показывает сезонные измене-
ния в их питании. Зимой основу питания составляли семепа растений,
грызуны и рыба. Летом больше половины исследованных желудков было
148
Глава третья
наполнено остатками лягущек, но попадались и семена культурных зла-
ков. Весной преимущественное значение имели грызуны и рыба. Осенью
преобладали семена и мышевидные грызупы (Хамаганов, 1985).
В отличие от других авторов А. С. Фетисов (1945), описывая питание
восточной формы серых крыс— карако, считает, что растительная пища
занимает в их рационе более зпачительное место, чем животная. Этот
вывод сделан им на основании изучения остатков и запасов зверьков, но
при анализе содержимого их желудков этим автором обнаружены остат-
ки жуков и муравьев, а также шерсть изицухи, двух видов мышей и
полевок, даурского хомячка и монгольского тушканчика. Имелась также
и примесь стеблей рогоза и ивы. При раскопке крысиных пор А. С. Фе-
тисов обнаружил шкурки перечисленных выше видов грызунов и остатки
более 25 видов растений. Ему удалось установить, что крысы, обитающие
в тех же стациях, что и полевки-экономки и унгурские полевки, затас-
кивают в свои норы запасы корней смолевки и горнеца, сделанные
полевками. При этом они разрывают их поры и полностью забирают со-
держимое кормовых камер. К сожалению, автор не указывает сезон,
котла проводился анализ содержимого желудков. запасов и остатков
пищи из нор и кормовых камер. По-видимому, на основании таких мето-
дически разпоречивых данных певозможно однозначно решить вопрос
о преимущественном питании одним или другим видом пищи.
Дапные о составе животных и растительных кормов островных попу-
ляций серых крыс также весьма отличпы. Возможно, что в некоторой
степени
— это результат различий в методиках обора и анализа мате-
риала.
Так, на Сахалине и Курильских озтровах «дикие» крысы зимой
питаются в бамбуковых зарослях семенами и побегами курильского бам-
бука, а на побережье островов — выбросами моря (крабами, моллюсками,
морскими звездами, рыбой), в нерестовых же реках
— спулыми лососе-
зыми рыбами (Сурков, 1972, 1983, 1986а}. Автор, однако, при анализе
218 желудков зверьков обнаружил, что больше половины их были на-
полнены мясом и шерстью грызунов и землероек, 6 желудков содержали
мясо мелких итиц, а 7— яичный желток и частицы скорлупы. Н. Ф. Рей-
мерс с соавторами (1968) тоже отмечают преимуществениое питание
крыс на Южном Сахалине животными кормами
— желудки почти всех
отловленпых в природе зверьков были наполнены мясом и шерстью:
Рид иностранных авторов приводят данные о преимущественном пи-
тании серых крыс на островах растительной пищей. Так, в Южной
Атлантике, па о-ве Южная Джорджия, у крыс, обитакацих на кочкова-
тых лугах, анализ содержимого желудков показал преобладание расти-
тельных кормов, но регулярпо попадались и насекомые; реже встреча-
лись морские беспозвоночные, рыбы, мясо трупов ластонотих и яйца
птиц (Рус, Воппег, 1980). В Иплийском океане, на о-ве Европа (близ
Мадагаскара), крысы уничтожали гнездивиихся на земле птиц, репти-
лий, амфибий, а также завезенных человеком кур (Терепате, 1966).
В Северпом море крысы, живущие с 1930 г. на дюнах о-ва Гельго-
ланд, в основном питались выбросами моря и насекомыми. Но охотились
Экология
149
также за трясогузками, коньками, поедани яйца серебристых чаек
(Узи, 1958).
Драммонд (Огитиоп4, 1960а), анализируя экскременты крысе, обитав-
ших на о-ве Бриджмарш, пришел к выводу, что основу их питапия с0-
оставляют одподольпые растепия, а в осенний период
— семена двудоль-
ных, которыми они питаются и в течение всей зимы. Остатки насекомых
и ракообразных встречаются реже. Однако мы уже отмечали, что экекре-
менты дают весьма приблизительную картину питания, так как обычно
содержат наименее ценные для организма непереварепиые остатки
клетчатку, хитин и т. д.
С нашей точки зрения, серые крысы, обитающие в природе, повсе-
местпо всеядпы, но корма животного происхождения в их рационе пре-
обладают и состоят из фоновых видов мелких позвоночных, беспозвопоч-
ных (в ‚том числе насекомых), а также сомян и вегетативных частей
растении.
И. Р. Мерзликин предоставил пам свои материалы по детальному изучению пи-
тания серых крыс, выселившихся в теплое время года из помещений в естественные
биотопы. Обследование произведено им в 1983—1986 гг. на Украине, в Сумской об-
ласти. В результате анализа 167 желудков таких пасюков остатки позвоночных
(грызуны, птицы, лягушки, рыбы) были обнаружены в 7,3% желудков, а в 92,7%
желудков находились остатки беспозвоночных, среди которых осповное место занима-
ли членистоногие (92%), главным образом личиночные формы насекомых, а имен-
но личинки мух. В единичных случаях встречались остатки брюхоногих. моллюсков
малощетинковых червей.
‘
Из растений по частоте встреч в желудках преобладали семена дикорастущих
трав (46,7%), из которых наибольшее число относилось к семейству злаковых (24,1% }
далее шли гречишные (11,1%), розоцветные (44,1%), сложнодветные и маковые (по
7,44 каждое семейство). В остальные семейства входили от 1,9 до 5,6% наименований
растений. Зеленые части растений в желудках встречались в 25,79
$
ыеч
: случаев (кусоч-
ки листьев и стеблей).
т
"7% случ (вувоч
Семена встречались в желудках в различных количествах и соотношениях ©
другими пищевыми компонентами. В одних желудках они были представлены в од-
НОМ экземпляре, а в других составляли все содержимое их. Весной семена расте-
ний были встречены в двух желудках (1,2%), летом —в 42 (25,1%), а осенью— в
34 желудках (20,4%). Семена представителей семейства маковых ветречались толь-
ко летом (в июле—антусте), сомена злаковых и розоцветных — в августе и сентябре,
а семена, например, различных горцев и череды трехраздельной — только в сентябре-
ктябре.
Агроценозы
Питание крыс в агроценозах зависит не только от фоповых сельскохозяй-
ственных культур, сроков их посева и созревания, но также от заселен-
ности их другими видами животных (как позвоночных, чак и беспозво-
ночных), входящих в рациопы питалия зверьков. Прявязанность к агро-
цепозам с обильной кормовой базой приводит к сезонному переходу
пасюков на преимущественно растительный корм. Однако животная
пища всегда остается составной частью рациона, и зверьки ве упускают
возможности разнообразить его насекомыми или мелкими позвоночными.
Так. на юге (в Краснодарском крае) по содержимому желудков крыс.
живущих на рисовых полях, было установлено, что осснью и зимой они
148
Глава третья
наполнено остатками лягущек, но попадались и семена культурных зла-
ков. Весной преимущественное значение имели грызуны и рыба. Осенью
преобладали семена и мышевидные грызупы (Хамаганов, 1985).
В отличие от других авторов А. С. Фетисов (1945), описывая питание
восточной формы серых крыс— карако, считает, что растительная пища
занимает в их рационе более зпачительное место, чем животная. Этот
вывод сделан им на основании изучения остатков и запасов зверьков, но
при анализе содержимого их желудков этим автором обнаружены остат-
ки жуков и муравьев, а также шерсть изицухи, двух видов мышей и
полевок, даурского хомячка и монгольского тушканчика. Имелась также
и примесь стеблей рогоза и ивы. При раскопке крысиных пор А. С. Фе-
тисов обнаружил шкурки перечисленных выше видов грызунов и остатки
более 25 видов растений. Ему удалось установить, что крысы, обитающие
в тех же стациях, что и полевки-экономки и унгурские полевки, затас-
кивают в свои норы запасы корней смолевки и горнеца, сделанные
полевками. При этом они разрывают их поры и полностью забирают со-
держимое кормовых камер. К сожалению, автор не указывает сезон,
котла проводился анализ содержимого желудков. запасов и остатков
пищи из нор и кормовых камер. По-видимому, на основании таких мето-
дически разпоречивых данных певозможно однозначно решить вопрос
о преимущественном питании одним или другим видом пищи.
Дапные о составе животных и растительных кормов островных попу-
ляций серых крыс также весьма отличпы. Возможно, что в некоторой
степени
— это результат различий в методиках обора и анализа мате-
риала.
Так, на Сахалине и Курильских озтровах «дикие» крысы зимой
питаются в бамбуковых зарослях семенами и побегами курильского бам-
бука, а на побережье островов — выбросами моря (крабами, моллюсками,
морскими звездами, рыбой), в нерестовых же реках
— спулыми лососе-
зыми рыбами (Сурков, 1972, 1983, 1986а}. Автор, однако, при анализе
218 желудков зверьков обнаружил, что больше половины их были на-
полнены мясом и шерстью грызунов и землероек, 6 желудков содержали
мясо мелких итиц, а 7— яичный желток и частицы скорлупы. Н. Ф. Рей-
мерс с соавторами (1968) тоже отмечают преимуществениое питание
крыс на Южном Сахалине животными кормами
— желудки почти всех
отловленпых в природе зверьков были наполнены мясом и шерстью:
Рид иностранных авторов приводят данные о преимущественном пи-
тании серых крыс на островах растительной пищей. Так, в Южной
Атлантике, па о-ве Южная Джорджия, у крыс, обитакацих на кочкова-
тых лугах, анализ содержимого желудков показал преобладание расти-
тельных кормов, но регулярпо попадались и насекомые; реже встреча-
лись морские беспозвоночные, рыбы, мясо трупов ластонотих и яйца
птиц (Рус, Воппег, 1980). В Иплийском океане, на о-ве Европа (близ
Мадагаскара), крысы уничтожали гнездивиихся на земле птиц, репти-
лий, амфибий, а также завезенных человеком кур (Терепате, 1966).
В Северпом море крысы, живущие с 1930 г. на дюнах о-ва Гельго-
ланд, в основном питались выбросами моря и насекомыми. Но охотились
Экология
149
также за трясогузками, коньками, поедани яйца серебристых чаек
(Узи, 1958).
Драммонд (Огитиоп4, 1960а), анализируя экскременты крысе, обитав-
ших на о-ве Бриджмарш, пришел к выводу, что основу их питапия с0-
оставляют одподольпые растепия, а в осенний период
— семена двудоль-
ных, которыми они питаются и в течение всей зимы. Остатки насекомых
и ракообразных встречаются реже. Однако мы уже отмечали, что экекре-
менты дают весьма приблизительную картину питания, так как обычно
содержат наименее ценные для организма непереварепиые остатки
клетчатку, хитин и т. д.
С нашей точки зрения, серые крысы, обитающие в природе, повсе-
местпо всеядпы, но корма животного происхождения в их рационе пре-
обладают и состоят из фоновых видов мелких позвоночных, беспозвопоч-
ных (в ‚том числе насекомых), а также сомян и вегетативных частей
растении.
И. Р. Мерзликин предоставил пам свои материалы по детальному изучению пи-
тания серых крыс, выселившихся в теплое время года из помещений в естественные
биотопы. Обследование произведено им в 1983—1986 гг. на Украине, в Сумской об-
ласти. В результате анализа 167 желудков таких пасюков остатки позвоночных
(грызуны, птицы, лягушки, рыбы) были обнаружены в 7,3% желудков, а в 92,7%
желудков находились остатки беспозвоночных, среди которых осповное место занима-
ли членистоногие (92%), главным образом личиночные формы насекомых, а имен-
но личинки мух. В единичных случаях встречались остатки брюхоногих. моллюсков
малощетинковых червей.
‘
Из растений по частоте встреч в желудках преобладали семена дикорастущих
трав (46,7%), из которых наибольшее число относилось к семейству злаковых (24,1% }
далее шли гречишные (11,1%), розоцветные (44,1%), сложнодветные и маковые (по
7,44 каждое семейство). В остальные семейства входили от 1,9 до 5,6% наименований
растений. Зеленые части растений в желудках встречались в 25,79
$
ыеч
: случаев (кусоч-
ки листьев и стеблей).
т
"7% случ (вувоч
Семена встречались в желудках в различных количествах и соотношениях ©
другими пищевыми компонентами. В одних желудках они были представлены в од-
НОМ экземпляре, а в других составляли все содержимое их. Весной семена расте-
ний были встречены в двух желудках (1,2%), летом —в 42 (25,1%), а осенью— в
34 желудках (20,4%). Семена представителей семейства маковых ветречались толь-
ко летом (в июле—антусте), сомена злаковых и розоцветных — в августе и сентябре,
а семена, например, различных горцев и череды трехраздельной — только в сентябре-
ктябре.
Агроценозы
Питание крыс в агроценозах зависит не только от фоповых сельскохозяй-
ственных культур, сроков их посева и созревания, но также от заселен-
ности их другими видами животных (как позвоночных, чак и беспозво-
ночных), входящих в рациопы питалия зверьков. Прявязанность к агро-
цепозам с обильной кормовой базой приводит к сезонному переходу
пасюков на преимущественно растительный корм. Однако животная
пища всегда остается составной частью рациона, и зверьки ве упускают
возможности разнообразить его насекомыми или мелкими позвоночными.
Так. на юге (в Краснодарском крае) по содержимому желудков крыс.
живущих на рисовых полях, было установлено, что осснью и зимой они
150 Глава третья
питаются преимущественно семенами риса, куриного проса и др., & так
же вегетативными частями злаков и осок. УКивотные корма (насекомые,
мыши, земноводные) встречаются в меньшем количестве. Весной в пита-
нии также преобладают вегетативные части растепий. Летом, в июне—
июле, в рационе достоверно преобладает животная нища (мыши, земно-
водные, моллюски, насекомые). Особенно велика доля земноводных
(жерлянки, прудовые лятупки). В это время рисовые поля залиты водой
и доля земноводных в рационе втрое больше, чем в безводный период.
На суше около воды находили «кормовые столики» © остатками прудо-
виков, изредка беззубок и мелкой рыбы. Преобладание животного корма
над растительным сохранялось до сентября включительно, в этот же
период несколько увеличивалась доля подземных частей растений (Шу-
това и др., 1981; Соколов, Карасева, 1983; 1985; Рыльников, Карасе-
ва, 1985).
.
|
На Дальнем Востоке у ирые, обитающих на сельскохозяйствеппых
угодьях, по даппым А. М. Колосова (1978), в рациопе преобладает в
осповном растительная пища. Зверьки повреждают овощи, подоолнечник,
и нередко их желудки бывают до отказа наполнены соем. Но даже при
обилии корма на рисовых полях хорошей приманкой крысам могут быть
живая полевка или мышь, посаженные в вершу (Некипелов, 1957).
В Западной Сибири, по описанию В. Г. Лялина (1978), пасюки, по-
падая па поля и приусадебные участки, поедали корнеплоды моркови
п клубни картофеля, всходы пшеницы и овса, огурцы и молодые побеги
клевера, но во всех исследованных желудках встречались и животные
корма.
.
В Восточной Сибири в период уборки сельскохозяйственпых культур
крысы концентрируются на полях (Фетисов, 1945). Автор отмечает, что.
здесь в отношепии питания они не отличаются от зверьков, обитакяцих
в западной части ареала,—хорошю едят зерна ржи, пшеницы, семена
конопли, гречихи и гороха. Поселяясь на огородах, едят морковь, брюкву,
сахарную свеклу и сырой картофель.
.
М. Заплетал (7арека 1962), анализируя фекалии экзоантропных
насюков из агроцепозов Южной Моравии (Чехословакия), показал, что:
весь год их пища на 90% по объему состояла из семян ишеницы, куку-
рузы и сорняков. Судя но частоте встречаемости остатков пищи, она
на 81% была растительного происхождения. Это были зерна, вегетатив-
ные части трав. Лишь небольшую часть составляли остатки червеи,
насекомых, костей мелких млекопитающих.
Пасоки в агроценозах Добруджи (Румыния), судя по запасам в норе,
питались как растительной, так и животной пищей. При ее расконке
были найдены 5,4 кг кукурузы, 45 черепов и шкурки хомяка Мезостюсе-
15 пемюш. Весной 1975 г. на кукурузных полях Большого острова
Браилы (на Дунае) пасюки, составляющие 90% всех добытых грызу-
нов, кормились проросшими семенами кукурузы и мелкими грызунами
(Натахг, 1977).
Экология
1541
Синантропные крысы
Характер питания синантропных пасюков обычно определяется паличием
того или иного продукта в помещении. Чаще бывают заселены ими
животноводческие помещения. Проведенный В. Г. Лялиным (1978) ана-
лиз 490 желудков крыс, обитающих на животноводческих фермах Запад-
ной Сибири, показал, что основой пищевого рациона зверьков были
зерно, мука и комбикорм. В свипарпиках к перечисленным кормам добав-
иялись еще рыба, отходы из столовых и корнеплоды. В качестве второ-
степенных кормов пасюки летом поедали рубленый клевер, а снаружи —
дождевых червей и насекомых. Силос зимой они поедали только с низ-
ким содержанием кислот.
Сезонные колебания в питании пасюков в животноводческих помеще-
ниях незначительны и зависят от поступающих кормов. Только колеба-
ния в потреблепии второстепенных кормов связаны с сезоном, когда
зверьки имеют возможность выходить из помещения. Н. П. Козлов
(1961) отмечает у синантропных форм случаи каннибализма, которые
приходилось наблюдать и нам в разных местах.
С. А. Хамаганов (1985) детально изучил питание ссрых крыс в Хаба-
ровске и примыкающих к нему селах. В содержимом 235 исследованных
им желудков крыс животные и растительные корма встречались пример-
но в равной степени. Из растительных кормов преобладали зерновые
культуры и хлебные продукты (47,6%), из животных — мяеспые продук-
ты (31,2%) и грызуны (19%).
По сообщению А. Н. Козлова (1981), в свинарниках Северного Казах-
стана пищей крысам служат не только свиной корм, но и мясо павших
свиней, последы. Они загрызают новорождепиых поросят, ягнят, кроль-
чат и цыплят. У живых малоподвижиых сальных свиней эти грызуны
выгрызают в слое жира глубокие лунки (Натат её а|., 1968а).
Не пренебрегают крысы и трупами людей (Цорытип, 1976), экскре-
ментами в капализациопной системе, шахтах и на полях фильтрации
({ЗнепКег 1980; Тощитин, 1986).
Кроме основных кормов, которыми располатают синантропные крысы
в помещениях разного целевого назначепия, в их рацион входит много
дополнительных компонентов, которые опи паходят вблизи строений.
По материалам, предоставленным нам И. Р. Мерзликиным, в результате деталь-
ного определения содержимого 300 желудков крыс, отловленных на животноводчес-
ких фермах и в надворных ностройках частных домов (Украина, Сумская обл.),
складывается следующая картина: 1 - основу питания здесь составляет корм сель-
скохозяйственных животных и 2— один он встречен в среднем в 52% желудков,
в остальных в различных соотношениях присутствовали и другие виды пищи.
Наиболее часто попадались насскомые (28,74), зеленые части растений (9%);
реже присутствовали остатки мышевидных грызупов, дождевых червей, моллюсков,
‹емена культурных растений и трав.
Соотношение различных пищевых компонентов значительно изменяется по
сезонам. Доля желудков, в которых присутствует только основной корм с января
по апрель, в среднем составляет 63,7%. Начиная с апреля их доля падает и доети-
гает своего минимума в июне (19%), что связано с обилием других кормов в этот
а
Затем значение основного корма начинает возрастать и в ноябре достигает
86,2%.
150 Глава третья
питаются преимущественно семенами риса, куриного проса и др., & так
же вегетативными частями злаков и осок. УКивотные корма (насекомые,
мыши, земноводные) встречаются в меньшем количестве. Весной в пита-
нии также преобладают вегетативные части растепий. Летом, в июне—
июле, в рационе достоверно преобладает животная нища (мыши, земно-
водные, моллюски, насекомые). Особенно велика доля земноводных
(жерлянки, прудовые лятупки). В это время рисовые поля залиты водой
и доля земноводных в рационе втрое больше, чем в безводный период.
На суше около воды находили «кормовые столики» © остатками прудо-
виков, изредка беззубок и мелкой рыбы. Преобладание животного корма
над растительным сохранялось до сентября включительно, в этот же
период несколько увеличивалась доля подземных частей растений (Шу-
това и др., 1981; Соколов, Карасева, 1983; 1985; Рыльников, Карасе-
ва, 1985).
.
|
На Дальнем Востоке у ирые, обитающих на сельскохозяйствеппых
угодьях, по даппым А. М. Колосова (1978), в рациопе преобладает в
осповном растительная пища. Зверьки повреждают овощи, подоолнечник,
и нередко их желудки бывают до отказа наполнены соем. Но даже при
обилии корма на рисовых полях хорошей приманкой крысам могут быть
живая полевка или мышь, посаженные в вершу (Некипелов, 1957).
В Западной Сибири, по описанию В. Г. Лялина (1978), пасюки, по-
падая па поля и приусадебные участки, поедали корнеплоды моркови
п клубни картофеля, всходы пшеницы и овса, огурцы и молодые побеги
клевера, но во всех исследованных желудках встречались и животные
корма.
.
В Восточной Сибири в период уборки сельскохозяйственпых культур
крысы концентрируются на полях (Фетисов, 1945). Автор отмечает, что.
здесь в отношепии питания они не отличаются от зверьков, обитакяцих
в западной части ареала,—хорошю едят зерна ржи, пшеницы, семена
конопли, гречихи и гороха. Поселяясь на огородах, едят морковь, брюкву,
сахарную свеклу и сырой картофель.
.
М. Заплетал (7арека 1962), анализируя фекалии экзоантропных
насюков из агроцепозов Южной Моравии (Чехословакия), показал, что:
весь год их пища на 90% по объему состояла из семян ишеницы, куку-
рузы и сорняков. Судя но частоте встречаемости остатков пищи, она
на 81% была растительного происхождения. Это были зерна, вегетатив-
ные части трав. Лишь небольшую часть составляли остатки червеи,
насекомых, костей мелких млекопитающих.
Пасоки в агроценозах Добруджи (Румыния), судя по запасам в норе,
питались как растительной, так и животной пищей. При ее расконке
были найдены 5,4 кг кукурузы, 45 черепов и шкурки хомяка Мезостюсе-
15 пемюш. Весной 1975 г. на кукурузных полях Большого острова
Браилы (на Дунае) пасюки, составляющие 90% всех добытых грызу-
нов, кормились проросшими семенами кукурузы и мелкими грызунами
(Натахг, 1977).
Экология
1541
Синантропные крысы
Характер питания синантропных пасюков обычно определяется паличием
того или иного продукта в помещении. Чаще бывают заселены ими
животноводческие помещения. Проведенный В. Г. Лялиным (1978) ана-
лиз 490 желудков крыс, обитающих на животноводческих фермах Запад-
ной Сибири, показал, что основой пищевого рациона зверьков были
зерно, мука и комбикорм. В свипарпиках к перечисленным кормам добав-
иялись еще рыба, отходы из столовых и корнеплоды. В качестве второ-
степенных кормов пасюки летом поедали рубленый клевер, а снаружи —
дождевых червей и насекомых. Силос зимой они поедали только с низ-
ким содержанием кислот.
Сезонные колебания в питании пасюков в животноводческих помеще-
ниях незначительны и зависят от поступающих кормов. Только колеба-
ния в потреблепии второстепенных кормов связаны с сезоном, когда
зверьки имеют возможность выходить из помещения. Н. П. Козлов
(1961) отмечает у синантропных форм случаи каннибализма, которые
приходилось наблюдать и нам в разных местах.
С. А. Хамаганов (1985) детально изучил питание ссрых крыс в Хаба-
ровске и примыкающих к нему селах. В содержимом 235 исследованных
им желудков крыс животные и растительные корма встречались пример-
но в равной степени. Из растительных кормов преобладали зерновые
культуры и хлебные продукты (47,6%), из животных — мяеспые продук-
ты (31,2%) и грызуны (19%).
По сообщению А. Н. Козлова (1981), в свинарниках Северного Казах-
стана пищей крысам служат не только свиной корм, но и мясо павших
свиней, последы. Они загрызают новорождепиых поросят, ягнят, кроль-
чат и цыплят. У живых малоподвижиых сальных свиней эти грызуны
выгрызают в слое жира глубокие лунки (Натат её а|., 1968а).
Не пренебрегают крысы и трупами людей (Цорытип, 1976), экскре-
ментами в капализациопной системе, шахтах и на полях фильтрации
({ЗнепКег 1980; Тощитин, 1986).
Кроме основных кормов, которыми располатают синантропные крысы
в помещениях разного целевого назначепия, в их рацион входит много
дополнительных компонентов, которые опи паходят вблизи строений.
По материалам, предоставленным нам И. Р. Мерзликиным, в результате деталь-
ного определения содержимого 300 желудков крыс, отловленных на животноводчес-
ких фермах и в надворных ностройках частных домов (Украина, Сумская обл.),
складывается следующая картина: 1 - основу питания здесь составляет корм сель-
скохозяйственных животных и 2— один он встречен в среднем в 52% желудков,
в остальных в различных соотношениях присутствовали и другие виды пищи.
Наиболее часто попадались насскомые (28,74), зеленые части растений (9%);
реже присутствовали остатки мышевидных грызупов, дождевых червей, моллюсков,
‹емена культурных растений и трав.
Соотношение различных пищевых компонентов значительно изменяется по
сезонам. Доля желудков, в которых присутствует только основной корм с января
по апрель, в среднем составляет 63,7%. Начиная с апреля их доля падает и доети-
гает своего минимума в июне (19%), что связано с обилием других кормов в этот
а
Затем значение основного корма начинает возрастать и в ноябре достигает
86,2%.
452
Глава третья
Начиная с апреля отмечается резкое увеличение числа встреч насекомых в ра-
ционе крыс и достигает своего максимума в мае (71,4%), далее начинается ее снад.
Употребление зеленых частей растений достигает максимума в июне (33,3%), посте-
пенно снижается к сентябрю (91%) и далее нрекращается до начала весеннего не-
риода. Мышевидные грызуны встречаются
в желудках насюков почти на всем протя-
жении года. В январе-феврале они составляют 2,84, а в июне достигают 12,5%
и к октябрю снижаются До 2,3%. Птицы в желудках крыс встречаются в единичных
случаях.
Отмечены и популяции крыс, например в Балтиморском зоопарке
питающиеся в основном растительной нищей,— из 100 отловленпых па-
сюков в 90% случаев желудки содержали зерно ячменя, пшеницы, под-
солнечника, овощей и фруктов. Лишь у 2% крыс в желудках пашли
остатки членистоногих. Крысы, живущие рядом с медведями и пекари,
кормились разнообразнее, чем те, что обитали вблизи буйволов, а неко-
торая часть их, особенно зимой, были каннибалами (ЕатВапо-АзаЯ, Зош-
иск, 1979).
Изучение питания крыс в лабораторных условиях
О роли отдельных компонентов нищи крыс и степени их предпочтения
накоплен обншрный, по также весьма противоречивый материал. В ос-
новном исследования проводились при клеточном содержании зверьков.
Так, А. П. Кузякин (4950) пишет, что лучше всего крысы поедают пше-
ничный хлеб и столовую свеклу, Н. И. Фирстов (1958) ечитает, что они
лучите едят мясной фарш, А. К. Голещихин (1955) — капту из кукуруз-
пой муки, В. Т. Абеленцев (1959) — зерно пшеницы, ячменя, кукурузы
(хуже поедают мясо, творог, яйца); Н. И. Никифоров и др. (1964) пишут
о предпочтепии мяспого и рыбного фарша, В. К. Шепелева (1950) также
пишет о предпочтении животной нищи. Н. М. Дукельская (1948) счи-
тает, что лучше всего едят ржапой хлеб, колбасу, творог, сметану, мяс-
ной фарш, Н. И. Нечаева (1954} — хлебные продукты, С. А. Хамаганов,
Д. В. Шахуров (1959) — муку, поджаренную на жире, картофельное
пюре на масле, овсяную крупу, мясо. Таких примеров можно еще приве-
сти немало.
Поставленные пами в вольерах па группировках крысе опыты по по-
едаемости основных элементов кормов, упомянутых вышенеречислениыми
авторами, показали, что количество съеденпой пищи в большой степени
зависит как от наличия других видов кормов, так и от условий содер-
жания зверьков (табл. 7).
Однако крысы на диете, состоящей только из сырых овощей, долго
существовать не могут
— начинается отставание в весе, приводящее к
гибели (Шепелева, 1950). Об этом же говорит и минимальное количество
овощей в рационе предпочитаемых зверьками кормов в опытах Козлова
(1980а).
Эксперименты, проведенные в лабораторных, вольерных и естествен-
пых условиях, показали, что только энергетической ценностью нельзя
объяснить выбор продуктов, поедаемых этими трызунами. На рисовых
полях они предпочитали рис (более знакомый им) пшенице и ячменю,
Экология 153
Таблица 7. Среднесуточное потребление кормов взрослыми крысами (массой
250—300 г) в вольерах при наличии достаточного количества воды
Среднесуточное потребление корма од-
нои крысой
при возможности выбора
при отсутствии выбора
Вид корма
(п=36)
(и==35)
г
нкал
т
ккал
Хлеб пшеничный
6,9
18,1
21,1
56,4
Морковь
171,9
4,9
47,2
15,6
Каша перловая +3%
24,8
448
45,6
82,1
подсолнечного масла
Рыба сырая (хек)
18,4
21,6
48,4
56,1
Всего
|60|304|
хотя энергетическая ценность этих культур почти одинакова, а белка в
рисе меньше (Шутова и др., 1981). Пищевая ценность побегов и зрелых.
семяи бамбука, которыми питается серая крыса на Сахалине (Сурков,
1986а), низка, и, по-видимому, она ест их в силу необходимости [невы-
сокое содержание переваринаемого белка в семенах бамбука и ето низ-
кую энертетическую ценность отмечал Патак (Рафак, 1979)]. О том,
что содержапие растительного белка не определяет выбор пищи, говорят
и опыты с мукой из риса, проса, арахиса, подсолнечника, гороха, сазана,
чечевицы и сои. Эверьки предпочитали четыре первых культуры, в то
время как последние отличаются зпачительно более высоким содержани-
ем белка (Вгоок$, Вожегтаю, 1973). По данным этих авторов, крысы
предпочитали продукты © низким содержанием углеводов, но © высоким
содержанием жиров. При изобилии одного какого-либо вида раститель-
ной пищи крысы добавляют в «мепю» дополнительные продукты, менее
калорийные, чем основной вид корма, а также животные компопенты,
Так, на рисовых полях, как мы уже говорили, с июля по декабрь
(т. ©. после уборки риса, когда па полях остаетсн еще значительное
количество зерна) в желудках крые находили семена, вегетативные части
различных растений и остатки животных.
По сравнению с другими видами крыс, например с черной, серые кры-
сы нуждаются в большем количестве животных белков. 0б их разных
потребностях в белках говорят и опыты В. В. Черноморднкова (1959),
в которых пасюки при возможности выбора пищи предпочитали вариант
цисты с содержанием белка 22%, в то время как черные крысы пред-
почитали диету только с 14% белка.
В паших опытах при содержании в условиях, приближенных к естест-
венным (в вольерах), двух группировок пасюков по 5 особей в каждой
{3 самца-+ 2 самки) и двух таких же группировок черных крыс при
возможности выбрать из четырех видов пищи (ишенная каша с 3%
подсолнечного масла, хлеб, морковь и мясо) серые крысы выбирали пищу
452
Глава третья
Начиная с апреля отмечается резкое увеличение числа встреч насекомых в ра-
ционе крыс и достигает своего максимума в мае (71,4%), далее начинается ее снад.
Употребление зеленых частей растений достигает максимума в июне (33,3%), посте-
пенно снижается к сентябрю (91%) и далее нрекращается до начала весеннего не-
риода. Мышевидные грызуны встречаются
в желудках насюков почти на всем протя-
жении года. В январе-феврале они составляют 2,84, а в июне достигают 12,5%
и к октябрю снижаются До 2,3%. Птицы в желудках крыс встречаются в единичных
случаях.
Отмечены и популяции крыс, например в Балтиморском зоопарке
питающиеся в основном растительной нищей,— из 100 отловленпых па-
сюков в 90% случаев желудки содержали зерно ячменя, пшеницы, под-
солнечника, овощей и фруктов. Лишь у 2% крыс в желудках пашли
остатки членистоногих. Крысы, живущие рядом с медведями и пекари,
кормились разнообразнее, чем те, что обитали вблизи буйволов, а неко-
торая часть их, особенно зимой, были каннибалами (ЕатВапо-АзаЯ, Зош-
иск, 1979).
Изучение питания крыс в лабораторных условиях
О роли отдельных компонентов нищи крыс и степени их предпочтения
накоплен обншрный, по также весьма противоречивый материал. В ос-
новном исследования проводились при клеточном содержании зверьков.
Так, А. П. Кузякин (4950) пишет, что лучше всего крысы поедают пше-
ничный хлеб и столовую свеклу, Н. И. Фирстов (1958) ечитает, что они
лучите едят мясной фарш, А. К. Голещихин (1955) — капту из кукуруз-
пой муки, В. Т. Абеленцев (1959) — зерно пшеницы, ячменя, кукурузы
(хуже поедают мясо, творог, яйца); Н. И. Никифоров и др. (1964) пишут
о предпочтепии мяспого и рыбного фарша, В. К. Шепелева (1950) также
пишет о предпочтении животной нищи. Н. М. Дукельская (1948) счи-
тает, что лучше всего едят ржапой хлеб, колбасу, творог, сметану, мяс-
ной фарш, Н. И. Нечаева (1954} — хлебные продукты, С. А. Хамаганов,
Д. В. Шахуров (1959) — муку, поджаренную на жире, картофельное
пюре на масле, овсяную крупу, мясо. Таких примеров можно еще приве-
сти немало.
Поставленные пами в вольерах па группировках крысе опыты по по-
едаемости основных элементов кормов, упомянутых вышенеречислениыми
авторами, показали, что количество съеденпой пищи в большой степени
зависит как от наличия других видов кормов, так и от условий содер-
жания зверьков (табл. 7).
Однако крысы на диете, состоящей только из сырых овощей, долго
существовать не могут
— начинается отставание в весе, приводящее к
гибели (Шепелева, 1950). Об этом же говорит и минимальное количество
овощей в рационе предпочитаемых зверьками кормов в опытах Козлова
(1980а).
Эксперименты, проведенные в лабораторных, вольерных и естествен-
пых условиях, показали, что только энергетической ценностью нельзя
объяснить выбор продуктов, поедаемых этими трызунами. На рисовых
полях они предпочитали рис (более знакомый им) пшенице и ячменю,
Экология 153
Таблица 7. Среднесуточное потребление кормов взрослыми крысами (массой
250—300 г) в вольерах при наличии достаточного количества воды
Среднесуточное потребление корма од-
нои крысой
при возможности выбора
при отсутствии выбора
Вид корма
(п=36)
(и==35)
г
нкал
т
ккал
Хлеб пшеничный
6,9
18,1
21,1
56,4
Морковь
171,9
4,9
47,2
15,6
Каша перловая +3%
24,8
448
45,6
82,1
подсолнечного масла
Рыба сырая (хек)
18,4
21,6
48,4
56,1
Всего
|60|304|
хотя энергетическая ценность этих культур почти одинакова, а белка в
рисе меньше (Шутова и др., 1981). Пищевая ценность побегов и зрелых.
семяи бамбука, которыми питается серая крыса на Сахалине (Сурков,
1986а), низка, и, по-видимому, она ест их в силу необходимости [невы-
сокое содержание переваринаемого белка в семенах бамбука и ето низ-
кую энертетическую ценность отмечал Патак (Рафак, 1979)]. О том,
что содержапие растительного белка не определяет выбор пищи, говорят
и опыты с мукой из риса, проса, арахиса, подсолнечника, гороха, сазана,
чечевицы и сои. Эверьки предпочитали четыре первых культуры, в то
время как последние отличаются зпачительно более высоким содержани-
ем белка (Вгоок$, Вожегтаю, 1973). По данным этих авторов, крысы
предпочитали продукты © низким содержанием углеводов, но © высоким
содержанием жиров. При изобилии одного какого-либо вида раститель-
ной пищи крысы добавляют в «мепю» дополнительные продукты, менее
калорийные, чем основной вид корма, а также животные компопенты,
Так, на рисовых полях, как мы уже говорили, с июля по декабрь
(т. ©. после уборки риса, когда па полях остаетсн еще значительное
количество зерна) в желудках крые находили семена, вегетативные части
различных растений и остатки животных.
По сравнению с другими видами крыс, например с черной, серые кры-
сы нуждаются в большем количестве животных белков. 0б их разных
потребностях в белках говорят и опыты В. В. Черноморднкова (1959),
в которых пасюки при возможности выбора пищи предпочитали вариант
цисты с содержанием белка 22%, в то время как черные крысы пред-
почитали диету только с 14% белка.
В паших опытах при содержании в условиях, приближенных к естест-
венным (в вольерах), двух группировок пасюков по 5 особей в каждой
{3 самца-+ 2 самки) и двух таких же группировок черных крыс при
возможности выбрать из четырех видов пищи (ишенная каша с 3%
подсолнечного масла, хлеб, морковь и мясо) серые крысы выбирали пищу
154
Глава третьх
со значительно большим содержанием животных белков (61,7% от обще-
го количества белков), чем черные крысы (до 42,5).
Наиболее стабильно серые крысы в отличие от черпых поедали имен-
по мясо. По всем остальным кормам индивидуальная изменчивость
очень велика. Лишенные на длительное время мяспой и рыбной пищи
пасюки проявляют хищнические наклонности. Если в это время их с0-
единить с черными крысами в одной вольерс, то опи воспринимают по-
следних как добычу, подстерегают их, убивают и съедают. Но нам при-
ходилось наблюдать и другую картипу: когда у пасюков было достаточно
мясной пищи, то при соединении их в вольере с черными крысами опи:
лишь оттесняли их от кормушек и частично занимали их территорию.
Никаких отношений хищник— жертва в этом случае не наблюдалось.
Н. В. Башенина (1977) считает, что животный корм у грызунов слу-
жит источником белка и особенно ценен тем, что в организме каждого.
животного имеется полпый набор незаменимых аминокислот. Кроме того,
оп пополняет недостаток некоторых минеральных веществ, микроэлемен-
тов, солей. Физиологическую адаптивность крые к переходу на животную.
пищу отмечал А. М. Уголев (1959) —у крыс, питающихся мясной и мо-
лочной нищей преобладает зоолитическая активность в слюне (активная
по гликогену), как у плотоядных.
Наши опыты по содержанию крыс в клетках дали возможность уста-
новить большие индивидуальные различия в выборе еды зверьками.
Усредненные данные показывают только самые общие склонности этих
грызунов к определенпым видам корма. Если же рассчитать для каждого:
из дней опыта коэффициент вариации выбранной и съеденной {12 зверь-
ками пищи, то хорошо заметна индивидуальная изменчивость их
«вкусов».
Коэффициент вариации у пасюков при поедании каши и хлеба коле-
бался от 12 до 27%, а по количеству съеденной рыбы — от 22 до 67$.
К таким же результатам. пришел П. А. Свириденко (1940), изучая в
ольфактометре поедаемость крысами большого ассортимента пищевых
продуктов (ржаной хлеб, овес, шшеница, морковь, вареный картофель).
Перечисленпые корма поедались подопытными животными в разных ко-
личествах, и были отмечены резкие индивидуальные различия в выборе
пищи.
Различная реакция на пищевые продукты и разнообразие их в есте-
ственных местах обитания крыс пе позволяют составить список кормов
в порядке их предпочитаемости, тем более что крысы их выбирают в за-
висимости от условий и могут при пеобходимости переходить © одного:
вида корма па другой. Они способны выбирать пищевые продукты, пеоб-
ходимые для поддержапия энергетического равновесия (Вагпей, 1963).
Механизм этого явления еще недостаточно изучен, по именно он дает
дополнительную возможность обитать пасюку в среде с самой различной
кормовой базой.
Н. Н. Воронцов (1967а) и Н. В. Башенина (1977) считают всеядность.
приспособительной реакцией на неустойчивость кормовой базы. В этом
отпошении прислособительные способности серых крыс проявляются зпа-
Экология
45
с
л
чительно шире, чем у других видов грызунов (от растительноядности до
активного хищничества). Проявление такой пластичности может быть
одним из объяснений распирения ареала пасюка и в последнее время
‘очень быстрого его распрострапения по земпому шару. Именно пластич-
ность в выборе пищи и пищевого поведения создает наибольшие трудно-
сти в снижении численности этого вида, имеющего большое экопомиче-
ское и эпидемиологическое значезие.
Ритм суточной пищевой активности и пищевое
поведение
Серые крысы активны в сумеречно-ночные часы, и это особенно заметно
у зверьков, живущих в природных биотопах (Хамаганов, 1968; Лозан,
1971; Ворцайе, Вочей 1976).
Изучая влияние светового режима па пищевую активность серых
крые, В. Г. Полежаев (1950) приходит к выводу, что крыса ночное жи-
ротное, по суточный ритм ее не стабилен и может измениться при изме-
нении внешних экологических причин. Суточный ритм может меняться
даже при изменении одного только света при прочих равных условиях.
температуры и влажности. Автор определял активность по круглосуточ-
но автоматически регистрируемым нодходам крысе к кормушкам. Живот-
пых в опыте содержали одипочно в садках. Действительно, в затемненных
садках (интенсивность освещения падала в 30 раз по сравнению с нор-
мальным дневным светом) ритм пищевой активности менялся: повыша-
лась дневная активность за счет некоторого снижения ночной и актив-
ность в целом распределялась более равномерно в течение суток. Воз-
можно, в этой обстановке лишепные всякой информации стрессирован-
вые крысы следовали своему природному ритму питания, вызываемому
чувством голода. В этой же работе, описывая влияние светового режима
на пищевую активность свободноживущих на различных объектах крыс,
которая регистрировалась по количеству съеденной ими пищи, Нолежаев
показал, что изменение светового режима вызывало только некоторое из-
менение пищевой активности. Как круглосуточное освещение, так и круг-
лосуточпое затемнение несколько увеличивали нотреблепие пищи в днев-
ные часы. Основная же часть активности продолжала оставаться в вечер-
ние и ночные часы.
Анализируя эти данные по отдельным категориям помещений, мож-
но заметить, что искусственное изменение освещения оказывало наиболь-
шес влияпие на пищевую активность на нежилых объектах, где изначаль-
но дневная активность была выше, чем на других объектах. Еще более
значительное воздействие на этот процесс оказывает характер суточной
активности человека. Но даже и в этих условиях основным временем пи-
тания оставались вечерне-ночные часы.
Преимущественио ночпую пищевую активность синантропных крыс
отмечают и другие исследователи (Черномордиков, Горячева, 1961;
Н. Козлов, А. Козлов, 1977). А. Н. Козлов (1980а), изучая суточную
‚активность серых крыс в свинарниках, описывает наибольшее число пи-
154
Глава третьх
со значительно большим содержанием животных белков (61,7% от обще-
го количества белков), чем черные крысы (до 42,5).
Наиболее стабильно серые крысы в отличие от черпых поедали имен-
по мясо. По всем остальным кормам индивидуальная изменчивость
очень велика. Лишенные на длительное время мяспой и рыбной пищи
пасюки проявляют хищнические наклонности. Если в это время их с0-
единить с черными крысами в одной вольерс, то опи воспринимают по-
следних как добычу, подстерегают их, убивают и съедают. Но нам при-
ходилось наблюдать и другую картипу: когда у пасюков было достаточно
мясной пищи, то при соединении их в вольере с черными крысами опи:
лишь оттесняли их от кормушек и частично занимали их территорию.
Никаких отношений хищник— жертва в этом случае не наблюдалось.
Н. В. Башенина (1977) считает, что животный корм у грызунов слу-
жит источником белка и особенно ценен тем, что в организме каждого.
животного имеется полпый набор незаменимых аминокислот. Кроме того,
оп пополняет недостаток некоторых минеральных веществ, микроэлемен-
тов, солей. Физиологическую адаптивность крые к переходу на животную.
пищу отмечал А. М. Уголев (1959) —у крыс, питающихся мясной и мо-
лочной нищей преобладает зоолитическая активность в слюне (активная
по гликогену), как у плотоядных.
Наши опыты по содержанию крыс в клетках дали возможность уста-
новить большие индивидуальные различия в выборе еды зверьками.
Усредненные данные показывают только самые общие склонности этих
грызунов к определенпым видам корма. Если же рассчитать для каждого:
из дней опыта коэффициент вариации выбранной и съеденной {12 зверь-
ками пищи, то хорошо заметна индивидуальная изменчивость их
«вкусов».
Коэффициент вариации у пасюков при поедании каши и хлеба коле-
бался от 12 до 27%, а по количеству съеденной рыбы — от 22 до 67$.
К таким же результатам. пришел П. А. Свириденко (1940), изучая в
ольфактометре поедаемость крысами большого ассортимента пищевых
продуктов (ржаной хлеб, овес, шшеница, морковь, вареный картофель).
Перечисленпые корма поедались подопытными животными в разных ко-
личествах, и были отмечены резкие индивидуальные различия в выборе
пищи.
Различная реакция на пищевые продукты и разнообразие их в есте-
ственных местах обитания крыс пе позволяют составить список кормов
в порядке их предпочитаемости, тем более что крысы их выбирают в за-
висимости от условий и могут при пеобходимости переходить © одного:
вида корма па другой. Они способны выбирать пищевые продукты, пеоб-
ходимые для поддержапия энергетического равновесия (Вагпей, 1963).
Механизм этого явления еще недостаточно изучен, по именно он дает
дополнительную возможность обитать пасюку в среде с самой различной
кормовой базой.
Н. Н. Воронцов (1967а) и Н. В. Башенина (1977) считают всеядность.
приспособительной реакцией на неустойчивость кормовой базы. В этом
отпошении прислособительные способности серых крыс проявляются зпа-
Экология
45
с
л
чительно шире, чем у других видов грызунов (от растительноядности до
активного хищничества). Проявление такой пластичности может быть
одним из объяснений распирения ареала пасюка и в последнее время
‘очень быстрого его распрострапения по земпому шару. Именно пластич-
ность в выборе пищи и пищевого поведения создает наибольшие трудно-
сти в снижении численности этого вида, имеющего большое экопомиче-
ское и эпидемиологическое значезие.
Ритм суточной пищевой активности и пищевое
поведение
Серые крысы активны в сумеречно-ночные часы, и это особенно заметно
у зверьков, живущих в природных биотопах (Хамаганов, 1968; Лозан,
1971; Ворцайе, Вочей 1976).
Изучая влияние светового режима па пищевую активность серых
крые, В. Г. Полежаев (1950) приходит к выводу, что крыса ночное жи-
ротное, по суточный ритм ее не стабилен и может измениться при изме-
нении внешних экологических причин. Суточный ритм может меняться
даже при изменении одного только света при прочих равных условиях.
температуры и влажности. Автор определял активность по круглосуточ-
но автоматически регистрируемым нодходам крысе к кормушкам. Живот-
пых в опыте содержали одипочно в садках. Действительно, в затемненных
садках (интенсивность освещения падала в 30 раз по сравнению с нор-
мальным дневным светом) ритм пищевой активности менялся: повыша-
лась дневная активность за счет некоторого снижения ночной и актив-
ность в целом распределялась более равномерно в течение суток. Воз-
можно, в этой обстановке лишепные всякой информации стрессирован-
вые крысы следовали своему природному ритму питания, вызываемому
чувством голода. В этой же работе, описывая влияние светового режима
на пищевую активность свободноживущих на различных объектах крыс,
которая регистрировалась по количеству съеденной ими пищи, Нолежаев
показал, что изменение светового режима вызывало только некоторое из-
менение пищевой активности. Как круглосуточное освещение, так и круг-
лосуточпое затемнение несколько увеличивали нотреблепие пищи в днев-
ные часы. Основная же часть активности продолжала оставаться в вечер-
ние и ночные часы.
Анализируя эти данные по отдельным категориям помещений, мож-
но заметить, что искусственное изменение освещения оказывало наиболь-
шес влияпие на пищевую активность на нежилых объектах, где изначаль-
но дневная активность была выше, чем на других объектах. Еще более
значительное воздействие на этот процесс оказывает характер суточной
активности человека. Но даже и в этих условиях основным временем пи-
тания оставались вечерне-ночные часы.
Преимущественио ночпую пищевую активность синантропных крыс
отмечают и другие исследователи (Черномордиков, Горячева, 1961;
Н. Козлов, А. Козлов, 1977). А. Н. Козлов (1980а), изучая суточную
‚активность серых крыс в свинарниках, описывает наибольшее число пи-
156
Глава третья
тающихся зверьков в вечерние часы, после раздачи корма свиньям, ког-
ла уходит обслуживающий персонал. Эта активность продолжается почти
всю ночь, хотя вечерний пик значительно выше ночного. В помещениях,
тде работники ие преследуют крыс, их высокая активность отмечается
уже во время раздачи корма. Автор считает, что периодичность кормле-
ния животпых на фермах также оказывает влияцие на суточную актив-
ность крыс. Там, тде проводилось дневное кормление животных, отмечен
и третий пик активности пасюков— в середиие дня. Длилсн он около
часа.
_
М. Н. Лозан с соавторами (1980) провел интересную работу, наблю-
дая меченых зверьков в животноводческих помещениях Молдавии. Там
пищевая активность крыс характеризовалась четким бимодальным рит-
мом. Утренний пик пищевой активности был иптеисивнее вечернего и
продолжался с 5 до 9 ч. Вечерний длился © 19 до 23 ч, достигая макси-
мума к 20 ч. Минимальная пищевая активность наблюдалась в светлые
часы суток.
Фигала (Рода, 1965), изучая активность крыс в животноводческих
помещениях, применял прибор, регистрирующий число посещений ими
мест, в которых раскладывали приманку. Оп отмечал два суточных пика
пищевой активности: первый пик — носле захода солица и второй—
в 9 ч ночи. Ритм пищевой активности в значительном степени зависел
от деятельности человека.
р
У сипантропной серой крысы — вида, живущего постоянно вблизи че-
ловека, сложился стереотип поведения, проявляющийся в ночной актив-
ности, т. е. тогда, когда деятельность человека сведопа к минимуму.
В то же время он довольно легко мепялел с изменением ритма этой.
деятельности.
Однако ритм пищевой активности может меняться и под влиянием
других факторов. Ими могут быть резксе изменение кормовой базы,
а также высокая плотность популяции крыс. Н. Ц. Козлов, А. П. Козлов
(1977) отмечают, что в свинарниках с высокой численностью крыс неза-
висимо от других факторов в дневные часы ветречается всегда больше
активных зверьков, чем в помещениях с их небольшой численностью.
В этих случаях крысы могут реагировать на присутствие человека как
на сигнал подачи корма. Мы наблюдали, как в свинарнике днем (около
46 ч) крысы значительно активизировались, высовывались наружу из-
под досок пола, пищали, а во время раздачи корма свиньям не отбегали.
от свинарки, стараясь схватить кусочки из-под носа свиней. В одиночно
расположенном в степи свипарнике в Добрулже (Румыния),по пашим на-
блюдениям, после эвакуации оттуда свиней крысы, лишенные корма,
днем реатировали на открывание дверей человеком, собираясь у свиных
кормушек и перегородок. То же самое отмечает и А. Н. Козлов для жи-
вотноводческих помещений Казахстана. Там у крыс после вывода живот-
ных в летние лагеря наблюдался подъем дневной активности, по опа ни-
когда не достигала вечернего пика.
С. А. Квашнин и Е. В. Карасева (1985) в Узбекистане отмечали пи-
щевую активпость крыс на дамбе в светлые часы суток как летом, так
Экология 457
и зимой. Причем вечерний пик был выражен несколько слабее утреппего.
Авторы связыватт это явление со временем поступления корма с мусором
на свалку. Наши опыты, проведенные на трех сложившихся группиров-
ках серых крыс
в вольерах (3 самца и 2 самки в каждой вольере), пока-
зали, что при круглосуточном «сумеречном» освещении ири обилии пищи
и воды крысы сохраняли стереотип вечерне-ночной пищевой активности,
несмотря на то, что наблюдатели находились вблизи вольер и ночью.
Регистрацию пищевой активности в этом случае проводили как автома-
тически, по пересечепию ипфракраспого луча у входа в камеру с кор-
мушкой, так и визуально, отмечая время нахождения каждого зверька
у кормушки. В этих условиях свет пе влиял на ритм пищевой активпости.
Судя по данным, полученным в природе, в различных местах обита-
ния синантропных крыс, а также в вольерах и в лабораторных условиях,
трудпо устаповить, каким факторам наиболее подвержено изменение су-
точного ритма пищевой активности. Весьма вероятно, что на изначаль-
ную сумеречно-почпую активпость крыс в зпачитольной степени оказы-
вает влияние изменение светового режима, однако, по нашему мнению,
основным фактором, воздействующим на характер пищевой суточной
активности, служит ритм деятельности человека. Быстрая адаптация к
его хозяйственной деятельности позволяет крысам жить вблизи пего, в са-
мые короткие сроки осваивать новые места обитания и добывать пищу
в самых разнообразных условиях.
Помимо одного, двух или трех пиков суточной пищевой активности
крыс (Полежаев, 1950; Ешайа, 1965; Н. Козлов, А. Козлов, 1977), неко-
торые авторы отмечают и более тонкую двухчасовую ритмику (ГосКагд,
Ноаеге, 1969). В наших опытах в вольерах при наблюдении за группи-
ровками крыс, состоящими каждая из пяти зверьков, в течение двух
месяцев была рассчитана частота актов еды и питья в среднем на одно
животное в сутки, а также средняя длительность интервалов между ними
{табл. 8).
Эти данные, полученные в контролируемых условиях вольер, соответ-
ствуют результатам, полученным Фигала (Еша]а, 1965) в естественных
условиях, где автор пользовался методикой раскладки приманки и о6б-
наружил среднюю двухчасовую ритмику пищевой активности крыс. Пили
воду крысы, как и у нас в опытах, в два раза реже, чем ели.
Однако наши наблюдения за отдельными мечеными зверьками в
вольерах показали наряду со средпими по группировке результатами
более частый ритм кормежки некоторых зверьков. Частота подходов к
кормушке и длительность нахождения около нее зависели от иерархиче-
ского положения самца в группировке. Высокоранговые самцы могли
подходить к кормушкам в любое (в основном в «удобное» для них время,
т. е. в вечерие-почные часы). Они подолгу оставались у кормушки,
питаясь на месте (до 27 мин). Вместе с ними у кормушек могли нахо-
диться неполовозрелые молодые зверьки и самки. Низкорапговые самцы
кормилясь в основном в отсутствие самца-доминанта, часто забегая з
кормовую камеру, чтобы вытащить оттуда кусочки и съесть их в укром-
ном месте. Часто они бросали недоеденный кусок и вновь бежали к кор-
156
Глава третья
тающихся зверьков в вечерние часы, после раздачи корма свиньям, ког-
ла уходит обслуживающий персонал. Эта активность продолжается почти
всю ночь, хотя вечерний пик значительно выше ночного. В помещениях,
тде работники ие преследуют крыс, их высокая активность отмечается
уже во время раздачи корма. Автор считает, что периодичность кормле-
ния животпых на фермах также оказывает влияцие на суточную актив-
ность крыс. Там, тде проводилось дневное кормление животных, отмечен
и третий пик активности пасюков— в середиие дня. Длилсн он около
часа.
_
М. Н. Лозан с соавторами (1980) провел интересную работу, наблю-
дая меченых зверьков в животноводческих помещениях Молдавии. Там
пищевая активность крыс характеризовалась четким бимодальным рит-
мом. Утренний пик пищевой активности был иптеисивнее вечернего и
продолжался с 5 до 9 ч. Вечерний длился © 19 до 23 ч, достигая макси-
мума к 20 ч. Минимальная пищевая активность наблюдалась в светлые
часы суток.
Фигала (Рода, 1965), изучая активность крыс в животноводческих
помещениях, применял прибор, регистрирующий число посещений ими
мест, в которых раскладывали приманку. Оп отмечал два суточных пика
пищевой активности: первый пик — носле захода солица и второй—
в 9 ч ночи. Ритм пищевой активности в значительном степени зависел
от деятельности человека.
р
У сипантропной серой крысы — вида, живущего постоянно вблизи че-
ловека, сложился стереотип поведения, проявляющийся в ночной актив-
ности, т. е. тогда, когда деятельность человека сведопа к минимуму.
В то же время он довольно легко мепялел с изменением ритма этой.
деятельности.
Однако ритм пищевой активности может меняться и под влиянием
других факторов. Ими могут быть резксе изменение кормовой базы,
а также высокая плотность популяции крыс. Н. Ц. Козлов, А. П. Козлов
(1977) отмечают, что в свинарниках с высокой численностью крыс неза-
висимо от других факторов в дневные часы ветречается всегда больше
активных зверьков, чем в помещениях с их небольшой численностью.
В этих случаях крысы могут реагировать на присутствие человека как
на сигнал подачи корма. Мы наблюдали, как в свинарнике днем (около
46 ч) крысы значительно активизировались, высовывались наружу из-
под досок пола, пищали, а во время раздачи корма свиньям не отбегали.
от свинарки, стараясь схватить кусочки из-под носа свиней. В одиночно
расположенном в степи свипарнике в Добрулже (Румыния),по пашим на-
блюдениям, после эвакуации оттуда свиней крысы, лишенные корма,
днем реатировали на открывание дверей человеком, собираясь у свиных
кормушек и перегородок. То же самое отмечает и А. Н. Козлов для жи-
вотноводческих помещений Казахстана. Там у крыс после вывода живот-
ных в летние лагеря наблюдался подъем дневной активности, по опа ни-
когда не достигала вечернего пика.
С. А. Квашнин и Е. В. Карасева (1985) в Узбекистане отмечали пи-
щевую активпость крыс на дамбе в светлые часы суток как летом, так
Экология 457
и зимой. Причем вечерний пик был выражен несколько слабее утреппего.
Авторы связыватт это явление со временем поступления корма с мусором
на свалку. Наши опыты, проведенные на трех сложившихся группиров-
ках серых крыс
в вольерах (3 самца и 2 самки в каждой вольере), пока-
зали, что при круглосуточном «сумеречном» освещении ири обилии пищи
и воды крысы сохраняли стереотип вечерне-ночной пищевой активности,
несмотря на то, что наблюдатели находились вблизи вольер и ночью.
Регистрацию пищевой активности в этом случае проводили как автома-
тически, по пересечепию ипфракраспого луча у входа в камеру с кор-
мушкой, так и визуально, отмечая время нахождения каждого зверька
у кормушки. В этих условиях свет пе влиял на ритм пищевой активпости.
Судя по данным, полученным в природе, в различных местах обита-
ния синантропных крыс, а также в вольерах и в лабораторных условиях,
трудпо устаповить, каким факторам наиболее подвержено изменение су-
точного ритма пищевой активности. Весьма вероятно, что на изначаль-
ную сумеречно-почпую активпость крыс в зпачитольной степени оказы-
вает влияние изменение светового режима, однако, по нашему мнению,
основным фактором, воздействующим на характер пищевой суточной
активности, служит ритм деятельности человека. Быстрая адаптация к
его хозяйственной деятельности позволяет крысам жить вблизи пего, в са-
мые короткие сроки осваивать новые места обитания и добывать пищу
в самых разнообразных условиях.
Помимо одного, двух или трех пиков суточной пищевой активности
крыс (Полежаев, 1950; Ешайа, 1965; Н. Козлов, А. Козлов, 1977), неко-
торые авторы отмечают и более тонкую двухчасовую ритмику (ГосКагд,
Ноаеге, 1969). В наших опытах в вольерах при наблюдении за группи-
ровками крыс, состоящими каждая из пяти зверьков, в течение двух
месяцев была рассчитана частота актов еды и питья в среднем на одно
животное в сутки, а также средняя длительность интервалов между ними
{табл. 8).
Эти данные, полученные в контролируемых условиях вольер, соответ-
ствуют результатам, полученным Фигала (Еша]а, 1965) в естественных
условиях, где автор пользовался методикой раскладки приманки и о6б-
наружил среднюю двухчасовую ритмику пищевой активности крыс. Пили
воду крысы, как и у нас в опытах, в два раза реже, чем ели.
Однако наши наблюдения за отдельными мечеными зверьками в
вольерах показали наряду со средпими по группировке результатами
более частый ритм кормежки некоторых зверьков. Частота подходов к
кормушке и длительность нахождения около нее зависели от иерархиче-
ского положения самца в группировке. Высокоранговые самцы могли
подходить к кормушкам в любое (в основном в «удобное» для них время,
т. е. в вечерие-почные часы). Они подолгу оставались у кормушки,
питаясь на месте (до 27 мин). Вместе с ними у кормушек могли нахо-
диться неполовозрелые молодые зверьки и самки. Низкорапговые самцы
кормилясь в основном в отсутствие самца-доминанта, часто забегая з
кормовую камеру, чтобы вытащить оттуда кусочки и съесть их в укром-
ном месте. Часто они бросали недоеденный кусок и вновь бежали к кор-
158
Глава третья
Таблица 8. Структура пищевого поведения крые разных видов во времени
Длительность интервалов между акта- Средняя частота дейст-
ми (мин)
вий одной крысы в 1 ч
Вид
еды
питья
занпаса-
еды ПИТЬЯ ния коп-
ма
среднее |модальное| среднее |модальное
Ласюк
21,1
15
27,1
10
0,47 0,22
0,17
Черная крыса
43,2,
5
19,3
10
0,90 0,66
0,05
мушке, словно опасаясь, что туда болыше ие будет доступа. Посещение
камеры с кормушкой низкоранговыми самцами было в 2—3 раза чаще,
чем доминантом, а затрата времени на каждое посещение кормушки—
значительно меньшей. Подсаженным самцам-чужакам доступ к корму
был практически закрыт. Мы наблюдали в течение двух суток, как в
одной из вольерных группировок самец-доминант не подпускал чужака
® корму, загнал его на сетку вольеры и сторожил. При малейшей но-
пытке приблизиться к кормушке он набрасывался на чужака, кусал его
за лапы, хвост и морду. Зверька пришлось снимать с сетки совершенно
обессиленным.
Разпицу в пищевой активности самцов и самок в естественных ме-
стообитаниях наблюдали М. Н. Лозан © сотрудниками (1980). Вечерний
пик активности у самок но продолжительности питания и но числу кор-
мящихся особей преобладал над таковым у самцов, а в период низкой
активности питались преимущественно самки. Продолжительноеть одно-
разового пищевого поведения отдельных особей достигала максимального
значения ‘у самок н период с 9 до 12 ч, составляла 26 мин 31 сау
самцов —с 21 до 24 ч и равнялась 20 мин 42 с. Минимальное значение
одноразового пищевого поведения зарегистрировано с 12 до 15 ч и 0©0-
ставляло 18 с. Максимальный одноразовый присом пищи был отмечен у
самки — 13 мин 24 с. По пашим данным, в вольерах самец-доминант мог
питаться у кормушки до 27 мин.
.
Наблюдая в вольерах за питанием молодых крысят, нам не удалось
установить фактов научения подражанием, что соответствует данным
Барнетта (Вагпей, 1963). Не наблюдали мы и проявления предупреждаю-
щего поведения отдельных животных по отношению к остальным членам
труппировки в случае контакта с отравленной пищей, как это отмечал
В. Г. Лялин (1974а), песмотря на то, что проводили несколько месяцев
круглосуточные наблюдения, давая крысам два тина ратицидов: моно-
фторин и дифенацин. Может быть, нодобная информация передается
другими формами коммуникации между животными. Возможно, прояз-
ления болевых ощущений подтравившихся крыс могли быть приняты за
предупреждающее поведение. Барнетт (Ваглей, 1963) считает, что нзаи-
Экология
159
моотношения как диких, так и лабораторных серых крыс влияют на их
пищевое новедение лишь эпизодически. На наш взгляд, это проявляется
лишь в стабильных группировках при невысокой численпости крыс, ког-
да разнорантовые особи могут рассредоточиться во времени ири доста-
точном количестве и доступности корма. Мы скорее склониы согласиться
с В. Е. Соколовым с соавторами (1977), что низкоранговые самцы на ос-
нове проптлого опыта избегают подходить к кормушкам в присутствии
домипаита, поэтому их питание пе совпадает по времени. О разновре-
менных подходах к кормушкам доминанта и подчиненных особей в ис-
кусственных условиях говорит и С. А, Квашпшии (1983а).
Как мы уже отмечали, отличительной чертой рациона серых крыс
является его разнообразие. В связи с отсутствием специализации к ка-
кому-нибудь виду корма вся система поведения зверьков позволяет им
найти еду в определенном районе вокруг гнезда и выбрать согласно сво-
им потребностям и «вкусам». У пасюков пет каких-либо специфических
приемов в манипулировании пищей, но очень велики индивидуальные
вариации в процессах питания и кормодобывания. В зависимости от
структуры и размера продукта они едят его на месте (сыпучие продукты,
мелкие зерна, жидкие и вязкие корма) или перетаскивают в укрытия
или в нору, где могут спокойно, не подвергаясь опасности, его есть.
Зверьки могут слизывать сыпучую и жидкую пищу, откусывать кусочки,
а мелкие предметы и зерна могут брать одной или двумя лапками и
есть, сидя на задних лалках.
Способы поедания растений весьма разнообразны. В. Г. Лялин (1978)
описывает питапие крыс картофелем на полях. В первую очередь поеда-
лись крупные и здоровые клубни, у которых пасюки выгрызали внутрен-
нюю часть, в результате чего образовывалась лунка диаметром в 2—3 ем
и глубиной в 8 см. Обычно поедались нетронутые клубни, но при недо-
статке их подбирались уже подгрызенные другими особями. В несколь-
ких случаях автор наблюдал, как зверьки очищали клубень от кожуры,
снимая ее небольиими кусочками, и только потом поедали. Вететатив-
ные части растений крысы ели в местах их произрастания и лишь при
испуге, убегая, захватывали с собой сорванные растения. Откусив сте-
бель или вырвав его с корнем, пасюки съедали более сочную нижнюю:
часть, совершенно не трогая верхнюю.
По-другому они едят различные семена — они быстро заталкивают их
в рот и, убегая в укрытие, их там высыпают и едят. Куски до 20 г
хватают зубами и бегом уносят в укрытия, более тяжелую добычу (100—
200 г) держат в зубах и, поддерживая передними лапами, передвигают:
ея прыжками на задних. Если маеса предмета превосходит втрое массу
тела зверька, то крысы волокут его за собой, пятясь.
Яйца птиц пасюки держат в лапках вертикально и прогрызают в них
сверху дырку, затем засовывают туда морду и выпивают содержимое.
При транспортировке яйца они передвигаются прыйкками, зажимая его
передними лапами и зубами.
Так, Ф. Штейнигер (Зепишет, 1952) описывал, как трохсотграммовая
крыса перетаскивала убитую крякву. Если на пути ее передвижения с
158
Глава третья
Таблица 8. Структура пищевого поведения крые разных видов во времени
Длительность интервалов между акта- Средняя частота дейст-
ми (мин)
вий одной крысы в 1 ч
Вид
еды
питья
занпаса-
еды ПИТЬЯ ния коп-
ма
среднее |модальное| среднее |модальное
Ласюк
21,1
15
27,1
10
0,47 0,22
0,17
Черная крыса
43,2,
5
19,3
10
0,90 0,66
0,05
мушке, словно опасаясь, что туда болыше ие будет доступа. Посещение
камеры с кормушкой низкоранговыми самцами было в 2—3 раза чаще,
чем доминантом, а затрата времени на каждое посещение кормушки—
значительно меньшей. Подсаженным самцам-чужакам доступ к корму
был практически закрыт. Мы наблюдали в течение двух суток, как в
одной из вольерных группировок самец-доминант не подпускал чужака
® корму, загнал его на сетку вольеры и сторожил. При малейшей но-
пытке приблизиться к кормушке он набрасывался на чужака, кусал его
за лапы, хвост и морду. Зверька пришлось снимать с сетки совершенно
обессиленным.
Разпицу в пищевой активности самцов и самок в естественных ме-
стообитаниях наблюдали М. Н. Лозан © сотрудниками (1980). Вечерний
пик активности у самок но продолжительности питания и но числу кор-
мящихся особей преобладал над таковым у самцов, а в период низкой
активности питались преимущественно самки. Продолжительноеть одно-
разового пищевого поведения отдельных особей достигала максимального
значения ‘у самок н период с 9 до 12 ч, составляла 26 мин 31 сау
самцов —с 21 до 24 ч и равнялась 20 мин 42 с. Минимальное значение
одноразового пищевого поведения зарегистрировано с 12 до 15 ч и 0©0-
ставляло 18 с. Максимальный одноразовый присом пищи был отмечен у
самки — 13 мин 24 с. По пашим данным, в вольерах самец-доминант мог
питаться у кормушки до 27 мин.
.
Наблюдая в вольерах за питанием молодых крысят, нам не удалось
установить фактов научения подражанием, что соответствует данным
Барнетта (Вагпей, 1963). Не наблюдали мы и проявления предупреждаю-
щего поведения отдельных животных по отношению к остальным членам
труппировки в случае контакта с отравленной пищей, как это отмечал
В. Г. Лялин (1974а), песмотря на то, что проводили несколько месяцев
круглосуточные наблюдения, давая крысам два тина ратицидов: моно-
фторин и дифенацин. Может быть, нодобная информация передается
другими формами коммуникации между животными. Возможно, прояз-
ления болевых ощущений подтравившихся крыс могли быть приняты за
предупреждающее поведение. Барнетт (Ваглей, 1963) считает, что нзаи-
Экология
159
моотношения как диких, так и лабораторных серых крыс влияют на их
пищевое новедение лишь эпизодически. На наш взгляд, это проявляется
лишь в стабильных группировках при невысокой численпости крыс, ког-
да разнорантовые особи могут рассредоточиться во времени ири доста-
точном количестве и доступности корма. Мы скорее склониы согласиться
с В. Е. Соколовым с соавторами (1977), что низкоранговые самцы на ос-
нове проптлого опыта избегают подходить к кормушкам в присутствии
домипаита, поэтому их питание пе совпадает по времени. О разновре-
менных подходах к кормушкам доминанта и подчиненных особей в ис-
кусственных условиях говорит и С. А, Квашпшии (1983а).
Как мы уже отмечали, отличительной чертой рациона серых крыс
является его разнообразие. В связи с отсутствием специализации к ка-
кому-нибудь виду корма вся система поведения зверьков позволяет им
найти еду в определенном районе вокруг гнезда и выбрать согласно сво-
им потребностям и «вкусам». У пасюков пет каких-либо специфических
приемов в манипулировании пищей, но очень велики индивидуальные
вариации в процессах питания и кормодобывания. В зависимости от
структуры и размера продукта они едят его на месте (сыпучие продукты,
мелкие зерна, жидкие и вязкие корма) или перетаскивают в укрытия
или в нору, где могут спокойно, не подвергаясь опасности, его есть.
Зверьки могут слизывать сыпучую и жидкую пищу, откусывать кусочки,
а мелкие предметы и зерна могут брать одной или двумя лапками и
есть, сидя на задних лалках.
Способы поедания растений весьма разнообразны. В. Г. Лялин (1978)
описывает питапие крыс картофелем на полях. В первую очередь поеда-
лись крупные и здоровые клубни, у которых пасюки выгрызали внутрен-
нюю часть, в результате чего образовывалась лунка диаметром в 2—3 ем
и глубиной в 8 см. Обычно поедались нетронутые клубни, но при недо-
статке их подбирались уже подгрызенные другими особями. В несколь-
ких случаях автор наблюдал, как зверьки очищали клубень от кожуры,
снимая ее небольиими кусочками, и только потом поедали. Вететатив-
ные части растений крысы ели в местах их произрастания и лишь при
испуге, убегая, захватывали с собой сорванные растения. Откусив сте-
бель или вырвав его с корнем, пасюки съедали более сочную нижнюю:
часть, совершенно не трогая верхнюю.
По-другому они едят различные семена — они быстро заталкивают их
в рот и, убегая в укрытие, их там высыпают и едят. Куски до 20 г
хватают зубами и бегом уносят в укрытия, более тяжелую добычу (100—
200 г) держат в зубах и, поддерживая передними лапами, передвигают:
ея прыжками на задних. Если маеса предмета превосходит втрое массу
тела зверька, то крысы волокут его за собой, пятясь.
Яйца птиц пасюки держат в лапках вертикально и прогрызают в них
сверху дырку, затем засовывают туда морду и выпивают содержимое.
При транспортировке яйца они передвигаются прыйкками, зажимая его
передними лапами и зубами.
Так, Ф. Штейнигер (Зепишет, 1952) описывал, как трохсотграммовая
крыса перетаскивала убитую крякву. Если на пути ее передвижения с
150
Глава третья
добычей встречалось препятствие, пишет он, то она затаскивала добычу
на препятствие, а затем стаскивала с него. У норы зверек сначала пы-
тался затолкать туда принесенную добычу, а если это не удавалось,
то проникал в нору через другой вход, пытаясь затащить добычу из-
путри.
Описывая характер питания серых крыс, мы отмечали в их рационе
значительное содержание животных кормов, которые они добывают, не-
релко нанадая на позвоночных животных, Наиболее частой их жертвой
в естественных биотопах бывают. лягупки, остатки которых на кормовых
столиках насюков находили многие авторы, однако способ добывания
этих земноводных интереспо описан И. Р. Мерзликиным (1986). По его
словам, характер охоты на лягушек, преимущественио прудовых, зависел
от плотности их поселепия и от сезона. Па берегу реки при пизкой плот-
ности лягушек крысы специально на них не охотились, а при оболедова-
нии берега схватывали тех, что попадались ина пути в укрытия. Нри
изобилии лягушек (после сброса воды в пруду) зверьки заходили в
воду но брюхо и, двигаясь по мелководью, ощупывали дно передними
лапами. Обнаружив лягушку, крыса быстро погружала голову под воду
и хватала ее. После заморозков, когда земноводные зарылись в ил, кры-
сы начали за ними пырять. Автор отмечает, что зверьки отлавливали их
значительно больше, чем необходимо было для питания. После неудач-
ных охот насюки отыскивали на берегу давние остатки и съедали их.
Многолетние наблюдения В. С. Суркова (19866) на Сахалине показали,
что в природных условиях пасюки активно преследуют лесных нолевок
и в период генеративиого цикла раскапывают их выводковые гнезда и
поедают детенышей. Этому же автору неоднократно приходилось сталки-
ваться с фактом разорения насюком гнезд рябчиков, болотной курочки,
вальдшнепов, а также некоторых мелких птиц, гиездивигихся на земле и
в траве. Отмечено также, что в годы массового размножения крыс зча-
чительная часть зайчат-беляков уничтожается ими, зверьки нередко по-
едают в норах ондатрят, когда их родители выходят на кормежку из
норы. В природных условиях лесостенной зоны крысы нападали на птен-
цов ласточек-береговушек в гнездах, а также на полевок и землероек
(Мерзликин, 1986).
В агроценозах Добруджи (Румыния) в норе насюка были обнаружены
более 40 черенов и шкурок хомяка (Мезостеейз ре\{071), который по
размерам мало уступает серой крыее (Пашаг её а\., 1968а, Б). |
Но сообщению Г. В. Романова (1963), крысы, обитая на чердаке жи-
лого дома, систематически нападали на голубей, поедая не только яйца
и птенцов, но также и взрослых птиц,
Пасюки, обитающие з жилище человека, нередко делают болыние
запасы пищи, состоящие из десятков килограммов картофеля и овощей,
куриных яиц (до 38 1ит.), а в одном случае нашли запас около 3 кг
мяспых пельменей (Сурков, 1986,6). Но устному сообщению В. И. Со-
ловъева, крысы за одну ночь унесли и сложили несколько сот грецких
орехов. Мы находили занасы, состоящие из 3—4 кг моркови, яблок и дру-
тих продуктов. Стремление к запасанию у синантропных крыс можно
Экология 161
объяснить невозможностью подходить к источнику пищи в любое время
суток вследствие деятельности человека. Однако у этих зверьков хорошо
развиты челночные передвижения, так характерные для животных, за-
насающих корм.
Наши опыты показали, что лабораторные белые, равно как и серые,
крысы в двухкамерных прозрачных клетках (размером 1 мХ4 мх80 см),
горизонтально разделенных на «тнездовую» и «пищевую» камеры, из
трех видов корма (зерна кукурузы, кусочки хлеба и кукурузная мука)
два первых полностью переносили в гнездовую камеру в течение 2 чи
только потом приступали к еде. Наблюдения в вольерах показали, что
запасы делают в осповпом доминанты и субдомпнанты. Подчиненные
зверьки лишь оттаскивают в сторону пищу, поедая или бросая ее там.
Эти данные соответствуют тому, что описывал Калхоун (СаШФопп, 1962).
Пищу запасают ие только голодные крысы, но даже при обилии корма
часть зверьков неретаскивают его в тнездовую камеру.
Большая экспериментальная работа, проведенная многими исследова-
телями, показала, что запасапие зависит от целого ряда факторов, среди
которых основную роль играют предварительное голодание крыс и размер
убежища. Однако запасенный корм далеко не всегда полностью исполь-
зуется, так как зверьки постоянно кормятся вне убежища. Сезонных по-
стояпных запасов у «диких» крыс нет, но в зависимости от сезона в ка-
мерах может преобладать пища растительного или животного происхож-
дения. У синантропных пасюков камеры могут быть наполнены любыми
кормами, достаточно удобными для переноски.
Барнетт считает (Вагое, 1963), что биологическая значимость запа-
сання корма в том, что оно потенциально важно для выживания особи
или группы особей. Но природа такого поведения и непосредственные
стимулы к запасанию изучены недостаточно и пуждаются в дополнитель-
ном исследовании.
Подвижность
Одни из ословных элементов экологии животных— их подвижность и
отношение к территории, что, в свою очередь, преимущественно опреде-
ляется характером питания и особенностями поведения вида.
Об отношении серой крысы к территории в литературе есть очень
много противоречивых сведений. Одни авторы считают, что она все время
перемещается, не имея постоянного местообитания, т. е. ведет, как писал
ДД. С. Айзенштадт (41945), бродячий образ жизни. Такото же мнения при-
лерживались ГК. Л. Ковдрашкин (1949), В. Г. Полежаев и др. (1962),
Б. Л. Шура-Бура и В. Ц. Харламов (1966), В. С. Судейкин (1976).
Другие авторы полагают, что миграционные способности серой крысы
невелики и что в осповиом она ведет оседлый образ жизни (Оа\15 её а|.,
1948; Дукельская, 1959; Нонов, 1970; и др.). На самом деле этот грызун
в отношенпи пспользования территории проявляет пеобычайпую гиб-
кость.
6 (Серая крыса
150
Глава третья
добычей встречалось препятствие, пишет он, то она затаскивала добычу
на препятствие, а затем стаскивала с него. У норы зверек сначала пы-
тался затолкать туда принесенную добычу, а если это не удавалось,
то проникал в нору через другой вход, пытаясь затащить добычу из-
путри.
Описывая характер питания серых крыс, мы отмечали в их рационе
значительное содержание животных кормов, которые они добывают, не-
релко нанадая на позвоночных животных, Наиболее частой их жертвой
в естественных биотопах бывают. лягупки, остатки которых на кормовых
столиках насюков находили многие авторы, однако способ добывания
этих земноводных интереспо описан И. Р. Мерзликиным (1986). По его
словам, характер охоты на лягушек, преимущественио прудовых, зависел
от плотности их поселепия и от сезона. Па берегу реки при пизкой плот-
ности лягушек крысы специально на них не охотились, а при оболедова-
нии берега схватывали тех, что попадались ина пути в укрытия. Нри
изобилии лягушек (после сброса воды в пруду) зверьки заходили в
воду но брюхо и, двигаясь по мелководью, ощупывали дно передними
лапами. Обнаружив лягушку, крыса быстро погружала голову под воду
и хватала ее. После заморозков, когда земноводные зарылись в ил, кры-
сы начали за ними пырять. Автор отмечает, что зверьки отлавливали их
значительно больше, чем необходимо было для питания. После неудач-
ных охот насюки отыскивали на берегу давние остатки и съедали их.
Многолетние наблюдения В. С. Суркова (19866) на Сахалине показали,
что в природных условиях пасюки активно преследуют лесных нолевок
и в период генеративиого цикла раскапывают их выводковые гнезда и
поедают детенышей. Этому же автору неоднократно приходилось сталки-
ваться с фактом разорения насюком гнезд рябчиков, болотной курочки,
вальдшнепов, а также некоторых мелких птиц, гиездивигихся на земле и
в траве. Отмечено также, что в годы массового размножения крыс зча-
чительная часть зайчат-беляков уничтожается ими, зверьки нередко по-
едают в норах ондатрят, когда их родители выходят на кормежку из
норы. В природных условиях лесостенной зоны крысы нападали на птен-
цов ласточек-береговушек в гнездах, а также на полевок и землероек
(Мерзликин, 1986).
В агроценозах Добруджи (Румыния) в норе насюка были обнаружены
более 40 черенов и шкурок хомяка (Мезостеейз ре\{071), который по
размерам мало уступает серой крыее (Пашаг её а\., 1968а, Б). |
Но сообщению Г. В. Романова (1963), крысы, обитая на чердаке жи-
лого дома, систематически нападали на голубей, поедая не только яйца
и птенцов, но также и взрослых птиц,
Пасюки, обитающие з жилище человека, нередко делают болыние
запасы пищи, состоящие из десятков килограммов картофеля и овощей,
куриных яиц (до 38 1ит.), а в одном случае нашли запас около 3 кг
мяспых пельменей (Сурков, 1986,6). Но устному сообщению В. И. Со-
ловъева, крысы за одну ночь унесли и сложили несколько сот грецких
орехов. Мы находили занасы, состоящие из 3—4 кг моркови, яблок и дру-
тих продуктов. Стремление к запасанию у синантропных крыс можно
Экология 161
объяснить невозможностью подходить к источнику пищи в любое время
суток вследствие деятельности человека. Однако у этих зверьков хорошо
развиты челночные передвижения, так характерные для животных, за-
насающих корм.
Наши опыты показали, что лабораторные белые, равно как и серые,
крысы в двухкамерных прозрачных клетках (размером 1 мХ4 мх80 см),
горизонтально разделенных на «тнездовую» и «пищевую» камеры, из
трех видов корма (зерна кукурузы, кусочки хлеба и кукурузная мука)
два первых полностью переносили в гнездовую камеру в течение 2 чи
только потом приступали к еде. Наблюдения в вольерах показали, что
запасы делают в осповпом доминанты и субдомпнанты. Подчиненные
зверьки лишь оттаскивают в сторону пищу, поедая или бросая ее там.
Эти данные соответствуют тому, что описывал Калхоун (СаШФопп, 1962).
Пищу запасают ие только голодные крысы, но даже при обилии корма
часть зверьков неретаскивают его в тнездовую камеру.
Большая экспериментальная работа, проведенная многими исследова-
телями, показала, что запасапие зависит от целого ряда факторов, среди
которых основную роль играют предварительное голодание крыс и размер
убежища. Однако запасенный корм далеко не всегда полностью исполь-
зуется, так как зверьки постоянно кормятся вне убежища. Сезонных по-
стояпных запасов у «диких» крыс нет, но в зависимости от сезона в ка-
мерах может преобладать пища растительного или животного происхож-
дения. У синантропных пасюков камеры могут быть наполнены любыми
кормами, достаточно удобными для переноски.
Барнетт считает (Вагое, 1963), что биологическая значимость запа-
сання корма в том, что оно потенциально важно для выживания особи
или группы особей. Но природа такого поведения и непосредственные
стимулы к запасанию изучены недостаточно и пуждаются в дополнитель-
ном исследовании.
Подвижность
Одни из ословных элементов экологии животных— их подвижность и
отношение к территории, что, в свою очередь, преимущественно опреде-
ляется характером питания и особенностями поведения вида.
Об отношении серой крысы к территории в литературе есть очень
много противоречивых сведений. Одни авторы считают, что она все время
перемещается, не имея постоянного местообитания, т. е. ведет, как писал
ДД. С. Айзенштадт (41945), бродячий образ жизни. Такото же мнения при-
лерживались ГК. Л. Ковдрашкин (1949), В. Г. Полежаев и др. (1962),
Б. Л. Шура-Бура и В. Ц. Харламов (1966), В. С. Судейкин (1976).
Другие авторы полагают, что миграционные способности серой крысы
невелики и что в осповиом она ведет оседлый образ жизни (Оа\15 её а|.,
1948; Дукельская, 1959; Нонов, 1970; и др.). На самом деле этот грызун
в отношенпи пспользования территории проявляет пеобычайпую гиб-
кость.
6 (Серая крыса
162
Глава третья
Таблица 9. Частота повторных ловов у меченых грызунов разных видов
Среднее
Число
исло по
Вид
особей И
Автор
особь
Серая крыса
459
1,75
Пау!5 (1948)
Полевая мыть
458
4,1
Никитина (1980)
Лесная мышь
107
5,5
Ильсико, Зубчанинова (1963)
Обыкновенный хомяк
486
5,5
Карасева (1962)
Водяная полевка
1000
До 15 раз Пантелеев (1968)
_
Полевка-экономка
786
5,4
Карасева, Ильснко (1957)
Рыжая полевка
118
Л
Меркова (1955)
Подвижность как серых крыс, так и других животных изучают глав-
ным образом при помощи индивидуального и массового мечения (Пу-
черук, Никитина, 1980). В первом случае каждое животное имеет свою
метку и многократные отловы одното и того же зверька или визуальное
паблюдение за ним дают возмояатюость установить характер использова-
ния территории. Во втором случае производится массовое мечепие зверь-
ков в каком-либо месте и их вылов в другом. Это гораздо менее трудоем-
кий сп060б, но результаты его применения обычно сводятся только к
определению пройденного расстояния одпим зверьков или группой их.
При мечении серых крыс возникают большие осложнения, так Как
эти животные в отличие от других грызунов имеют хорошую память,
а нопав один раз в ловушку-живоловку и будучи номеченными, они вто-
рой раз в нее идут очень неохотно, ноэтому повторных ловов бывает мало
(табл. 9).
й
Даже в первый раз в живоловку серых крыс поймать нелегко, они
очень осторожны, а некоторые из них более или менее выраженные пео-
фобы (Тауюг, Очу, 1973, 1978). Еели к пенастороженным ловупкам
предварительно приучить крыс прикормом, то они, привыкнув, будут
беспрепятственно входить и выходить из них. После этого в насторожен-
ные ловушки поймать зверьков уже значительно легче (Сыбу, 1974; Ву-
зякин, 1950; Квалинин, Карасева, 1985).
Индивидуально крыс метят чаще всего, отрезая им пальцы в разных
комбипациях или нанося на шкурку пятна красящего вещества (обычно
урзола).
.
В последнее лесятилетие стали применять наиболее эффективный
метод индивидуального мечения серых крыс — радиопрослеживание.
Пойманное животное апестезируют и надевают ему ошейник с радиодат-
чиком весом 8—9 г. Антенна состоит из петли с сердечником из желез-
ных опилок. Неремещения пасюка улавливаются с помощью приемника
с антенной, установленного на треножнике, а для уточнения его место-
нахождения применяют дна переносных нриемника с антеннами. Для
Экология 163
визуального наблюдения за крысами на ошейник одновременно наносят
флюоресцирующий состав (Тау1ог, Опу, 4978; Тауог, 1978).
Гораздо легче пасюков метить массовым способом без предваритель-
ного отлова. Таких приемов есть несколько. Обычно в одном месте для
зверьков выставляют приманку с каким-либо метчиком, а затем их от-
лавливают на разных расстояниях от места мечения капканами или тра-
вят ядом и уже у мертвого животного устанавливают наличие метки.
В отдельных случаях удовлетворяются сбором испражнений, которые мо-
гут пметь следы метки.
В качестве метчика употребляют изотолы, а именно радиоактивный
фосфор (Р®) или радиоактивный кальций (Са“) (0,6—0,8 мг на 1 кг
приманки). Нойманную крысу умерщьляют, ампутируют у нее часть
хвоста, которую сжигают, и определяют наличие в ней изотопа (Шура-
Бура и др., 1962; Шура-Бура, Харламов, 1966; Полежаев и др., 1982;
Судейкин, 1976; Судейкин и др., 1962).
Более простой и эффективный снособ массового мечения заключается
в применении антибиотиков. Добавление, например, 1% тетрациклина к
приманке приводит к тому, что кальций соединяется © ним в растущих
частях костей и в ультрафиолетовых лучах дает специфическое свечение
(Тауог, Опу, 1978). Применяя различвые антибиотики грунпы тетра-
циклина (окситетрациклин, хлортетрациклин, метациклин и др.), можно
получить метки, при которых свечение будет разных цветов. Добавляя в
приманку гистологические красители, не вредные для крысе (эозин, ме-
тиленовая синяя, трипанблау и др.), удается по окрашенному калу уста-
повить, как далеко переместилось животное (Ра\13 еф а[., 1948; Рыльни-
ков и др., 1981). А. Н. Козлов (1978) не только собирал окрашенные
фекални, но и извлекал их у убитой крысы из прямой кишки. В. А. Рыль-
ников © соавторами (1983) усовершенствовали этот метод, введя хрома-
тографию, что позволило выявлять, не ела ли одна особь приманку ©
разпыми красителями.
Наконец, проводили опыты, при которых пропыливали входы в норы
крыс цветным порошком (обычно родамином), Вылезая из нор, крысы
пачкали шкурку и затем оставляли небольшие частицы порошка на раз-
пых предметах, что позволяло определять направление их передвижений
{Дукельская, 1957; Тау]ог, Оцу, 1978).
Помимо мечения, исдвижность крысе может быть изучена по заслежен-
ным пылевым площадкам (Мелкова, 1987), а также при сравнении во
времени результатов учетов их численности. В ряде случаев, сопоставляя
числеппость в различных участках, удается установить, куда именно перс-
‘местились крысы. Иногда приходят на помощь и прямые наблюдения за
персселяющимися зверьками (Козлов, 1978; Эюд4агь, 1981).
Все перечисленные методы позволяют судить о характере использова-
ния территории оседлыми крысами и о передвижении мигрантов.
6+
162
Глава третья
Таблица 9. Частота повторных ловов у меченых грызунов разных видов
Среднее
Число
исло по
Вид
особей И
Автор
особь
Серая крыса
459
1,75
Пау!5 (1948)
Полевая мыть
458
4,1
Никитина (1980)
Лесная мышь
107
5,5
Ильсико, Зубчанинова (1963)
Обыкновенный хомяк
486
5,5
Карасева (1962)
Водяная полевка
1000
До 15 раз Пантелеев (1968)
_
Полевка-экономка
786
5,4
Карасева, Ильснко (1957)
Рыжая полевка
118
Л
Меркова (1955)
Подвижность как серых крыс, так и других животных изучают глав-
ным образом при помощи индивидуального и массового мечения (Пу-
черук, Никитина, 1980). В первом случае каждое животное имеет свою
метку и многократные отловы одното и того же зверька или визуальное
паблюдение за ним дают возмояатюость установить характер использова-
ния территории. Во втором случае производится массовое мечепие зверь-
ков в каком-либо месте и их вылов в другом. Это гораздо менее трудоем-
кий сп060б, но результаты его применения обычно сводятся только к
определению пройденного расстояния одпим зверьков или группой их.
При мечении серых крыс возникают большие осложнения, так Как
эти животные в отличие от других грызунов имеют хорошую память,
а нопав один раз в ловушку-живоловку и будучи номеченными, они вто-
рой раз в нее идут очень неохотно, ноэтому повторных ловов бывает мало
(табл. 9).
й
Даже в первый раз в живоловку серых крыс поймать нелегко, они
очень осторожны, а некоторые из них более или менее выраженные пео-
фобы (Тауюг, Очу, 1973, 1978). Еели к пенастороженным ловупкам
предварительно приучить крыс прикормом, то они, привыкнув, будут
беспрепятственно входить и выходить из них. После этого в насторожен-
ные ловушки поймать зверьков уже значительно легче (Сыбу, 1974; Ву-
зякин, 1950; Квалинин, Карасева, 1985).
Индивидуально крыс метят чаще всего, отрезая им пальцы в разных
комбипациях или нанося на шкурку пятна красящего вещества (обычно
урзола).
.
В последнее лесятилетие стали применять наиболее эффективный
метод индивидуального мечения серых крыс — радиопрослеживание.
Пойманное животное апестезируют и надевают ему ошейник с радиодат-
чиком весом 8—9 г. Антенна состоит из петли с сердечником из желез-
ных опилок. Неремещения пасюка улавливаются с помощью приемника
с антенной, установленного на треножнике, а для уточнения его место-
нахождения применяют дна переносных нриемника с антеннами. Для
Экология 163
визуального наблюдения за крысами на ошейник одновременно наносят
флюоресцирующий состав (Тау1ог, Опу, 4978; Тауог, 1978).
Гораздо легче пасюков метить массовым способом без предваритель-
ного отлова. Таких приемов есть несколько. Обычно в одном месте для
зверьков выставляют приманку с каким-либо метчиком, а затем их от-
лавливают на разных расстояниях от места мечения капканами или тра-
вят ядом и уже у мертвого животного устанавливают наличие метки.
В отдельных случаях удовлетворяются сбором испражнений, которые мо-
гут пметь следы метки.
В качестве метчика употребляют изотолы, а именно радиоактивный
фосфор (Р®) или радиоактивный кальций (Са“) (0,6—0,8 мг на 1 кг
приманки). Нойманную крысу умерщьляют, ампутируют у нее часть
хвоста, которую сжигают, и определяют наличие в ней изотопа (Шура-
Бура и др., 1962; Шура-Бура, Харламов, 1966; Полежаев и др., 1982;
Судейкин, 1976; Судейкин и др., 1962).
Более простой и эффективный снособ массового мечения заключается
в применении антибиотиков. Добавление, например, 1% тетрациклина к
приманке приводит к тому, что кальций соединяется © ним в растущих
частях костей и в ультрафиолетовых лучах дает специфическое свечение
(Тауог, Опу, 1978). Применяя различвые антибиотики грунпы тетра-
циклина (окситетрациклин, хлортетрациклин, метациклин и др.), можно
получить метки, при которых свечение будет разных цветов. Добавляя в
приманку гистологические красители, не вредные для крысе (эозин, ме-
тиленовая синяя, трипанблау и др.), удается по окрашенному калу уста-
повить, как далеко переместилось животное (Ра\13 еф а[., 1948; Рыльни-
ков и др., 1981). А. Н. Козлов (1978) не только собирал окрашенные
фекални, но и извлекал их у убитой крысы из прямой кишки. В. А. Рыль-
ников © соавторами (1983) усовершенствовали этот метод, введя хрома-
тографию, что позволило выявлять, не ела ли одна особь приманку ©
разпыми красителями.
Наконец, проводили опыты, при которых пропыливали входы в норы
крыс цветным порошком (обычно родамином), Вылезая из нор, крысы
пачкали шкурку и затем оставляли небольшие частицы порошка на раз-
пых предметах, что позволяло определять направление их передвижений
{Дукельская, 1957; Тау]ог, Оцу, 1978).
Помимо мечения, исдвижность крысе может быть изучена по заслежен-
ным пылевым площадкам (Мелкова, 1987), а также при сравнении во
времени результатов учетов их численности. В ряде случаев, сопоставляя
числеппость в различных участках, удается установить, куда именно перс-
‘местились крысы. Иногда приходят на помощь и прямые наблюдения за
персселяющимися зверьками (Козлов, 1978; Эюд4агь, 1981).
Все перечисленные методы позволяют судить о характере использова-
ния территории оседлыми крысами и о передвижении мигрантов.
6+
464
Глава третья
Характер использования территории
оседло живущими прысами
Экзоантропные и синантропные формы крыс, имеющие достаточное ко-
личество корма, не будучи ничем потревоженными, могут нодолгу
(во всяком случае но нескольку месяцев) жить на одпом месте.
Как мы увидим дальше, такой консерватизм в использовании терри-
тории может быть временным и период оседлого существования этих
грызунов весьма вариабелен. Тем не менее, нока пасюки живут оседло,
им свойственна, как и другим видам грызунов, пространственная струк
тура населения, которая выражается в закономерном размещении особей
п их группировок по отпошению к определенным элементам внешней
среды и друг к друту. Эта структура необходима, с сдной стороны, для
снижения конкурентных отношений между собой, а с другой— для
обеспечения необходимого уровня контактов между особями (Шилов,
1985).
,
Серым крысам в болыпинстве случаев снойствеппо жить груннами или
кланами (СаШоми, 1962). Каждая группировка занимает участок обита-
ния, т. е. территорию, в пределах которой зверьки находят все неооходи-
мые условия для существования: пищу, воду, места для устройства убе-
жи и естественные укрытия. Участки отдельных группировок в значи“
тельной степени обособлены друг от друга и охраняются от вторжения
особей из других кланов. Для серых крыс типично очень активное 000-
рудование своего участка, который они метят главиым образом вдоль
границ, запаховыми метками (в основном мочой} для охраны территории.
от вторжения особей из других группировок. Но такие же метки зверькн
оставляют и внутри участка — на поверхностных дорожках, около вхолов
внорыит.п.
Рис. 41. Крыеа на своем участке. Фото Е. В. Карасевой
Экология
165
Р
е
—
=
=
р
о. {©
5
О./
]
АХ
ул
Рис. 42. Передвижения 0д-
ного взрослого самца, изу-
ченные с помощью радио-
слежения (по: Гашог, 1978)
Параллельные линии — много-
кратные перемещения крывы
вцоль забора и живых изгоро-
дей;
1 — места кратковременного
отдыха крысы,
Н — норы; цифрами дано ко-
личество дней, проведен-
ных в них крысой,
ПП — места прикорма
0
108
200
309]
Используя систему знакомых ориентиров и запаховых меток, крыса в
пределах своего участка перемещается быстрой рысью, очень уверенно,
по временам останавливается, замирает и затем продолжает свой путь
(рис. 44). Скорость пробега — от 0,5 до 1,5 км в час (Тауот, Оцу,
1978). В результате она может кратчайшим путем достичь мест кормеж-
ки, водопоя или отдыха, а в природных условиях быстро укрыться от
пелогоды или преследования хищника. Зверек, впервые понавший на не-
знакомую территорию, передвигается по ней суетливо, постоянно зами-
рает, припимая позу настороженности. Как мы видим, использование по-
стоянных участков обитания имеет свои преимущества, поэтому пасюки
держатся за освоенную территорию и очень неохотно ее меняют.
464
Глава третья
Характер использования территории
оседло живущими прысами
Экзоантропные и синантропные формы крыс, имеющие достаточное ко-
личество корма, не будучи ничем потревоженными, могут нодолгу
(во всяком случае но нескольку месяцев) жить на одпом месте.
Как мы увидим дальше, такой консерватизм в использовании терри-
тории может быть временным и период оседлого существования этих
грызунов весьма вариабелен. Тем не менее, нока пасюки живут оседло,
им свойственна, как и другим видам грызунов, пространственная струк
тура населения, которая выражается в закономерном размещении особей
п их группировок по отпошению к определенным элементам внешней
среды и друг к друту. Эта структура необходима, с сдной стороны, для
снижения конкурентных отношений между собой, а с другой— для
обеспечения необходимого уровня контактов между особями (Шилов,
1985).
,
Серым крысам в болыпинстве случаев снойствеппо жить груннами или
кланами (СаШоми, 1962). Каждая группировка занимает участок обита-
ния, т. е. территорию, в пределах которой зверьки находят все неооходи-
мые условия для существования: пищу, воду, места для устройства убе-
жи и естественные укрытия. Участки отдельных группировок в значи“
тельной степени обособлены друг от друга и охраняются от вторжения
особей из других кланов. Для серых крыс типично очень активное 000-
рудование своего участка, который они метят главиым образом вдоль
границ, запаховыми метками (в основном мочой} для охраны территории.
от вторжения особей из других группировок. Но такие же метки зверькн
оставляют и внутри участка — на поверхностных дорожках, около вхолов
внорыит.п.
Рис. 41. Крыеа на своем участке. Фото Е. В. Карасевой
Экология
165
Р
е
—
=
=
р
о. {©
5
О./
]
АХ
ул
Рис. 42. Передвижения 0д-
ного взрослого самца, изу-
ченные с помощью радио-
слежения (по: Гашог, 1978)
Параллельные линии — много-
кратные перемещения крывы
вцоль забора и живых изгоро-
дей;
1 — места кратковременного
отдыха крысы,
Н — норы; цифрами дано ко-
личество дней, проведен-
ных в них крысой,
ПП — места прикорма
0
108
200
309]
Используя систему знакомых ориентиров и запаховых меток, крыса в
пределах своего участка перемещается быстрой рысью, очень уверенно,
по временам останавливается, замирает и затем продолжает свой путь
(рис. 44). Скорость пробега — от 0,5 до 1,5 км в час (Тауот, Оцу,
1978). В результате она может кратчайшим путем достичь мест кормеж-
ки, водопоя или отдыха, а в природных условиях быстро укрыться от
пелогоды или преследования хищника. Зверек, впервые понавший на не-
знакомую территорию, передвигается по ней суетливо, постоянно зами-
рает, припимая позу настороженности. Как мы видим, использование по-
стоянных участков обитания имеет свои преимущества, поэтому пасюки
держатся за освоенную территорию и очень неохотно ее меняют.
466
Глава третья
Границы участков могут быть очень различны и меняются в зависи-
мости от изменивлтихся условий. Часто участки состоят из двух частей -
в одной крыса живет (т. е. там находится ее основная жилая нора), ав
другую ходит кормиться. Иногда зверьки обеспечены кормом и хор ми
условиями для убежищ, ио не имеют воды. Тогда они регул; р солят
на водопой, и, следовательно, в этом случае их участок также с
т
к
.
В
ыы умы ходят в пределах участка, а также переходя из одной его
части в другую, обычно по одним и тем же хорошо протоптантым Яс-
рожкам, часто вдоль какого-либо укрытия. По на людепию Тейлора
(Тау!ог, 1978) с помощью радиопрослеживания, крыса поредвитв: ус
вдоль заборов или ых иоторолой по многу раз, иногда пер
эграпства (рис. .
р аы участков битаний крыс зависят от ряда причин и могуг
быть очень изменчивы. Их величина определяется запасом корма иго
распределением по территории, количеством мест, удобных для устройст_
ва убежищ, и численностью зверьков. Большое значение имеет степень
неодпородиости территории
— чем сложнее участок, тем у крысь яв
ляется больше возможностей его переоборудования в своих интересах,
и в результате на относительно небольшом пространстве может
бо;
чество особей.
у рые. обитающих в природных условиях, в большинстве олучзев
пища рассредоточена, зверьки в поисках ее должны отовительшо ти.
роко передвигаться, в связи с чем и участки обитания бывают ть ии.
ми. У синантропных крыс, хорошо обеспеченных кормом, участки ме
В литературе данных о величине участков крыс и их взаимном
{
много, и они противоречивы.
и родны е б п ото ыы Консерватизм пасюков в испольаовании
территории при экизни в природных биотопах хорошо прослеживается ‚ а
рисовых полях Нубапи, где они обитают круглый год, пе перемещаяс
насоленные пункты и, следовательно, не смешиваясь с оинаптропными
формами. В пределах самой рисовой системы Значительная часть отих
грызунов живет как оседлые животные. В ноябре-декабре опи конце
трируются на участках рисовых полей, там, где кормовые условия наиб
лее благоприятны (остается много педомолоченной соломы).
По нашим многолетним наблюдениям удалось показать, что размеры
участков различны при разной численности крыс. Поселения их вытяну-
ты влоль земляных валиков, окаймляющих чеки, заполнениые зодой.
Протяженность групповых участков составляет в среднем около
м
При снижении плотности населения. крыс в 15 раз (отлов па
длина участков увеличивалась до 552 м (Рыльников, арасева, Но .
Нами в ноябре 1984 г. на территории рисовых полей площадью га
в четырех участках были расставлены кормушки с приманкой. в которую
добавили разные антибиотики грунты тетрациклипа. Заким ооразом,
крысы в каждом участке приобретали свою метку. Зимой, в феврале
1982 г., при сплошном облове территории калканами было отловлено
Экология 167
Июль 79672.
кабрь
— февраль 1982г.
——_—.
се
®
+
т
э
о®
[<]®›©
|)ы|
Гюо| 1
зозо®||
от
оо1
[2[]зВ
1 <].
||
—
ие
Е—-=:
В
+
г
|
||
|
1|
®
Е--
|
И
+
е-
Е
-
—
НФобос[|1ево
о
|
Гюое|1
|©9106©|||сь
оо
|оО
©
е||
|
[65]
|6 [6]
о
о
г [бо
6||||
ооо
|
|05||
с
—
———
=—
Е Е2 ЕЗзЕ=3*Е-35[26[е]7[8[$]3
9"
Рис. 43. Передвижение серых крыс на рисовых полях Кубани (по: Рыльников,
Карасева, 1985)
1 — земляные валики, 2—5 — границы участка меченил (2 — тетрациклин, 3 -— тетрациклин +
+ окситетрациклин, 4 — хлортетрациклин + метациклин, 5-- хлортетрациклин); 6—9 — зве-
рек, получивший метку (6 — тетрациклин, 7 — тетрациклин--окситетрациклин, & — хлор-.
тетрациклин + метациклин, 9 — хлортетрациклин)
Рис. 44. Расположение групповых участков крыс на рисовых полях Кубани по
результатам мечения кала крыс гистологическими красителями (по; Рыльников и
др., 1981)
Места прикормки +: 1— эозином, 2 — трипан-блау, 3 — брильянт-грюн.
Точки обнаружения кала крыс +: 1а — эозин, 32а — трипан-блау, 3а — брильянт-грюн.
Предполагаемые границы кланов: 16 — первого, 26 — второго, 36 — третьего
(
)
О
л
я
©!см——м@24. Фуа ед
140 крыс, из которых 120 (86%) оказались мечеными, т. е. на протязже-
нии 3,5 мес зверьки не ушли с подопытной территории. Часть их пере-
местились лишь в пределах этой территории © одного участка на другой
(рис. 43).
Среднее расстояние перемещения равнялось 72=2 м.
Использование территории крысами летом было там же изучено на
других четырех участках с применением той мс методики. Через лва
месяца после раскладки примапки поймали всего 68 насюков, из кото-
рых 30 (44%) оказались номеченными тетрациклином. При этом только
466
Глава третья
Границы участков могут быть очень различны и меняются в зависи-
мости от изменивлтихся условий. Часто участки состоят из двух частей -
в одной крыса живет (т. е. там находится ее основная жилая нора), ав
другую ходит кормиться. Иногда зверьки обеспечены кормом и хор ми
условиями для убежищ, ио не имеют воды. Тогда они регул; р солят
на водопой, и, следовательно, в этом случае их участок также с
т
к
.
В
ыы умы ходят в пределах участка, а также переходя из одной его
части в другую, обычно по одним и тем же хорошо протоптантым Яс-
рожкам, часто вдоль какого-либо укрытия. По на людепию Тейлора
(Тау!ог, 1978) с помощью радиопрослеживания, крыса поредвитв: ус
вдоль заборов или ых иоторолой по многу раз, иногда пер
эграпства (рис. .
р аы участков битаний крыс зависят от ряда причин и могуг
быть очень изменчивы. Их величина определяется запасом корма иго
распределением по территории, количеством мест, удобных для устройст_
ва убежищ, и численностью зверьков. Большое значение имеет степень
неодпородиости территории
— чем сложнее участок, тем у крысь яв
ляется больше возможностей его переоборудования в своих интересах,
и в результате на относительно небольшом пространстве может
бо;
чество особей.
у рые. обитающих в природных условиях, в большинстве олучзев
пища рассредоточена, зверьки в поисках ее должны отовительшо ти.
роко передвигаться, в связи с чем и участки обитания бывают ть ии.
ми. У синантропных крыс, хорошо обеспеченных кормом, участки ме
В литературе данных о величине участков крыс и их взаимном
{
много, и они противоречивы.
и родны е б п ото ыы Консерватизм пасюков в испольаовании
территории при экизни в природных биотопах хорошо прослеживается ‚ а
рисовых полях Нубапи, где они обитают круглый год, пе перемещаяс
насоленные пункты и, следовательно, не смешиваясь с оинаптропными
формами. В пределах самой рисовой системы Значительная часть отих
грызунов живет как оседлые животные. В ноябре-декабре опи конце
трируются на участках рисовых полей, там, где кормовые условия наиб
лее благоприятны (остается много педомолоченной соломы).
По нашим многолетним наблюдениям удалось показать, что размеры
участков различны при разной численности крыс. Поселения их вытяну-
ты влоль земляных валиков, окаймляющих чеки, заполнениые зодой.
Протяженность групповых участков составляет в среднем около
м
При снижении плотности населения. крыс в 15 раз (отлов па
длина участков увеличивалась до 552 м (Рыльников, арасева, Но .
Нами в ноябре 1984 г. на территории рисовых полей площадью га
в четырех участках были расставлены кормушки с приманкой. в которую
добавили разные антибиотики грунты тетрациклипа. Заким ооразом,
крысы в каждом участке приобретали свою метку. Зимой, в феврале
1982 г., при сплошном облове территории калканами было отловлено
Экология 167
Июль 79672.
кабрь
— февраль 1982г.
——_—.
се
®
+
т
э
о®
[<]®›©
|)ы|
Гюо| 1
зозо®||
от
оо1
[2[]зВ
1 <].
||
—
ие
Е—-=:
В
+
г
|
||
|
1|
®
Е--
|
И
+
е-
Е
-
—
НФобос[|1ево
о
|
Гюое|1
|©9106©|||сь
оо
|оО
©
е||
|
[65]
|6 [6]
о
о
г [бо
6||||
ооо
|
|05||
с
—
———
=—
Е Е2 ЕЗзЕ=3*Е-35[26[е]7[8[$]3
9"
Рис. 43. Передвижение серых крыс на рисовых полях Кубани (по: Рыльников,
Карасева, 1985)
1 — земляные валики, 2—5 — границы участка меченил (2 — тетрациклин, 3 -— тетрациклин +
+ окситетрациклин, 4 — хлортетрациклин + метациклин, 5-- хлортетрациклин); 6—9 — зве-
рек, получивший метку (6 — тетрациклин, 7 — тетрациклин--окситетрациклин, & — хлор-.
тетрациклин + метациклин, 9 — хлортетрациклин)
Рис. 44. Расположение групповых участков крыс на рисовых полях Кубани по
результатам мечения кала крыс гистологическими красителями (по; Рыльников и
др., 1981)
Места прикормки +: 1— эозином, 2 — трипан-блау, 3 — брильянт-грюн.
Точки обнаружения кала крыс +: 1а — эозин, 32а — трипан-блау, 3а — брильянт-грюн.
Предполагаемые границы кланов: 16 — первого, 26 — второго, 36 — третьего
(
)
О
л
я
©!см——м@24. Фуа ед
140 крыс, из которых 120 (86%) оказались мечеными, т. е. на протязже-
нии 3,5 мес зверьки не ушли с подопытной территории. Часть их пере-
местились лишь в пределах этой территории © одного участка на другой
(рис. 43).
Среднее расстояние перемещения равнялось 72=2 м.
Использование территории крысами летом было там же изучено на
других четырех участках с применением той мс методики. Через лва
месяца после раскладки примапки поймали всего 68 насюков, из кото-
рых 30 (44%) оказались номеченными тетрациклином. При этом только
168 Глава третья
два меченых самца переместились из своего участка на два других на
расстояние 100 и 300 м.
В Узбекистане с помощью наблюдений за мечепыми зверьками было
показано, что на дамбе крысы жили круглый год и даже в зимний пе-
риод не переселялись в дома (Квашнин, Карасева, 1985).
В природных биотонах Великобритании крысы обитали по берегу ‘од-
ного из островов. Их отлавливали живоловками, метипи и там же вы-
пускали. При повторных отловах большая часть зверьков была поймана
не далее 12 м от места мечения. Когда поспела итиеница, часть живот
ных стали ходить на поля кормиться, но жить они продолжали на берегу.
Таким образом, в этом случае крысы проявили определенный терри-
ториальный консерватизм
— прибавив к своему участку новую террито-
рию, они все же не переселились на нее (Пау!з еЁ а]., 1948).
В Среднем Уэльсе, но данным мечения, длина индизидуального участ“
ка самцов в среднем равнялась 66,1 м, а самок — 54,8 м. Наиболее даль-
нне переходы самцов составляли 954 м, а самок — 850 м (Нат@еу, В1зЪор,
1979).
Для выяснения взаимного расположения участков отдельных кланов
было предпринято изучение распределения по земляным валикам рисо-
вых полей Кубани кала крыс, окрашенного гистологическими красителя-
ми (эозин, брильянт-грюн, трипанблау). Оказалось, что участки кланов
ме перекрываются (рис. 44) (Рыльников и др. 1981). В Приазовеких
плавнях протяженность группового участка крысе примерно такая же и
зверьки имеют возможность устраивать норы только вдоль сухих грив.
Тщательное обследование таких грив показало, что жилые норы расно-
лагадись на них по прямой линии на протяжении 250 м, а на ответв-
лепии грив— на 100 м. Можно предположить, что все обитатели нор
принадлежали к одному клану. Среди добытых здесь зверьков был один
взрослый самец, обитавший в типичной для самцов норе, а три другие
норы были выводковые. На других гривах норы не были обнаружены
(рис. 45).
В средней полосе России с помощью гистологических красителей было
установлено, что клан крыс (8 особей) обитал на участке, вытянутом
вдоль ручья, длиной также 250—300 м (Карасева и др. 1986а).
Крысы не всегда живут сообществами (кланами). В природных усло-
виях, выселившись несной из помещения, опи иногда живут одиночно и
имеют ирдивидуальный участок территории. На ием можно выделить
особо охрапяемую часть
— ядро участка и посещаемую, но пе охраняе“
мую часть. Но наблюдениям И. Р. Мерзликина (19876), на р. Сумке (Ук-
раина) одиночно живущий самец использовал территорию участка нерав-
номерно. Несмотря на то что он постоянно перемещалея по всей терри-
тории (500 м длиной), он активно охранял только ее часть длиной В
70 м, патрулируя, метя ее границы и преследуя появляющихся здесь
других крыс (рис. 46).
Населенные нункты. Образ жизни синантропных крыс, насе-
„яющих постройки человека, значительно отличается от такового в при-
родных биотонах. Крысы экзоантропиой экологической формы ведут себя
Экология
169
и
И
/
й
и
/
/
/
1
|
|
|
1
м
1
К]
=
|
—2
\
[4]:
ии
=
‚ы
с
у
58
Л
293
=
|
5
|
МИ
]
ОООООТООО
Рис. 45. Расположен
р
‚30.
#
@
ние жилых нор в их обитателей н
йе
плавней
р
а сутой гриве Приазовских
три 2 — проселочная дорога, $ — норы крыс (на врезке рядом схемы их строения и
исло пойманных в них крыс}, 4 — камера без гнезда, 5 — камера с гнездом
168 Глава третья
два меченых самца переместились из своего участка на два других на
расстояние 100 и 300 м.
В Узбекистане с помощью наблюдений за мечепыми зверьками было
показано, что на дамбе крысы жили круглый год и даже в зимний пе-
риод не переселялись в дома (Квашнин, Карасева, 1985).
В природных биотонах Великобритании крысы обитали по берегу ‘од-
ного из островов. Их отлавливали живоловками, метипи и там же вы-
пускали. При повторных отловах большая часть зверьков была поймана
не далее 12 м от места мечения. Когда поспела итиеница, часть живот
ных стали ходить на поля кормиться, но жить они продолжали на берегу.
Таким образом, в этом случае крысы проявили определенный терри-
ториальный консерватизм
— прибавив к своему участку новую террито-
рию, они все же не переселились на нее (Пау!з еЁ а]., 1948).
В Среднем Уэльсе, но данным мечения, длина индизидуального участ“
ка самцов в среднем равнялась 66,1 м, а самок — 54,8 м. Наиболее даль-
нне переходы самцов составляли 954 м, а самок — 850 м (Нат@еу, В1зЪор,
1979).
Для выяснения взаимного расположения участков отдельных кланов
было предпринято изучение распределения по земляным валикам рисо-
вых полей Кубани кала крыс, окрашенного гистологическими красителя-
ми (эозин, брильянт-грюн, трипанблау). Оказалось, что участки кланов
ме перекрываются (рис. 44) (Рыльников и др. 1981). В Приазовеких
плавнях протяженность группового участка крысе примерно такая же и
зверьки имеют возможность устраивать норы только вдоль сухих грив.
Тщательное обследование таких грив показало, что жилые норы расно-
лагадись на них по прямой линии на протяжении 250 м, а на ответв-
лепии грив— на 100 м. Можно предположить, что все обитатели нор
принадлежали к одному клану. Среди добытых здесь зверьков был один
взрослый самец, обитавший в типичной для самцов норе, а три другие
норы были выводковые. На других гривах норы не были обнаружены
(рис. 45).
В средней полосе России с помощью гистологических красителей было
установлено, что клан крыс (8 особей) обитал на участке, вытянутом
вдоль ручья, длиной также 250—300 м (Карасева и др. 1986а).
Крысы не всегда живут сообществами (кланами). В природных усло-
виях, выселившись несной из помещения, опи иногда живут одиночно и
имеют ирдивидуальный участок территории. На ием можно выделить
особо охрапяемую часть
— ядро участка и посещаемую, но пе охраняе“
мую часть. Но наблюдениям И. Р. Мерзликина (19876), на р. Сумке (Ук-
раина) одиночно живущий самец использовал территорию участка нерав-
номерно. Несмотря на то что он постоянно перемещалея по всей терри-
тории (500 м длиной), он активно охранял только ее часть длиной В
70 м, патрулируя, метя ее границы и преследуя появляющихся здесь
других крыс (рис. 46).
Населенные нункты. Образ жизни синантропных крыс, насе-
„яющих постройки человека, значительно отличается от такового в при-
родных биотонах. Крысы экзоантропиой экологической формы ведут себя
Экология
169
и
И
/
й
и
/
/
/
1
|
|
|
1
м
1
К]
=
|
—2
\
[4]:
ии
=
‚ы
с
у
58
Л
293
=
|
5
|
МИ
]
ОООООТООО
Рис. 45. Расположен
р
‚30.
#
@
ние жилых нор в их обитателей н
йе
плавней
р
а сутой гриве Приазовских
три 2 — проселочная дорога, $ — норы крыс (на врезке рядом схемы их строения и
исло пойманных в них крыс}, 4 — камера без гнезда, 5 — камера с гнездом
170
Глава третья
как мелкие хищники. Они, как уже было сказано, должны широко не-
ремещалься в ноисках корма, скрываться от преследования врагов, пере-
носить неблагоприятную погоду и т. н. Синантропные крысы в большия-
стве случаев обеспечены кормом, им не надо далеко ходить в поисках
его. Живя под крышей, опи не подвергаются неблагоприятным воздейст-
виям погоды, врагов среди животных у них мало. Все эти экологические
отличия накладывают отпечаток на пространственную структуру их на-
селения. В основном это касается размеров, конфигурации и взаимного
расположения участков обитания. В целом же характер использования
территории крысами обеих экологических форм нидослецифичен.
На свинофермах крысы всегда с избытком обеспечены кормом, повэто-
му участки их обитания бывают очень небольшими (30—50 м“), чаще
располагаются по клетям, в которых держат свиней. Численность этих
грызупов здесь достигает иногда колоссальных размеров, и для увеличе-
ния емкости пространства своего обитания пасюки начинают селиться и
перемещаться в пустотах стен, лазают по трубам, стропилам, бегают под
настилами, используют подземные помещения, передвигаются под кры-
шей. Таким образом, их местообитание становится как бы трехмерным,
способствуя тем самым как уменьшению контактов между особями, так и,
следовательно, степени стресса.
Отлов индивидуально меченных крыс показал, что их перемещения в
пределах участка невелики — 15—25 м (Соколов, Карасева, 1985}
(рис. 47). Групповые участки относительно изолированы друг от друга
благодаря тому, что доминанты защищают их грапицы и между ними
располагается нейтральная территория. На свинофермах это тамбуры,
проходы между загонами и коридоры (Лозан, Никул, 1983).
Территория участка группировки крыс но ее использованию, как и у
экзоантронных пасюков, неодпородна. На ней выделяется центральная
часть, наиболее защищаемая (30—50% площади всего участка), границы
которой доминант метит мочевыми точками и активно защищает от втор-
жения чужих самцов. Периферия участка обычно самцом-доминантом не
посещается — там чаще держатся подчиненные особи. Участки соседних
группировок перекрываются только на их нериферии (Михайленко, 1982),
Крысы редко без причины переселяются из одпого корпуса животно-
водческой фермы в другие места (Нозлов, 1978). Они настолько бывают
привязаны к своей территории, что часть их иногда длительное время не
покидают даже пустые помещения, из которых выведен скот и, следова-
тельно, отсутетвует корм. По почам зверьки ходят питаться в соседние
корпуса, расположенные на расстоянии 25—50 и даже 200 м.
Иногда сила этой привязанности поразительна. Например, в одном из
рядом стоящих свинарников во время ремонта были вскрыты полы и сня-
та часть крыши. Пищи для крыс в этом номещении не было, убежища
в значительной степени были разрушены. Несмотря на это, животные
жили там, а кормились за 45 м в соседнем свинарнике.
В условиях большого города крысы также часто консервативны в ис-
пользовании территории (Дукельская, 1957; Рау!$ её а1., 1948; Мелкова,
1987). И. И. Траут (1950), например, нишет, что в отдельных кварта-
Экология
174
5баЕ=8[97]?
Рис. 46. Использование одиночным самцом территории вдоль р. Сумки (Украина)
(по: Мерзликин, 1987)
А — план всего участка, Б — охраняемая часть участка; 1— жилые норы, 2 — естественные
убежища, 8 — направление переправ крысы через реку, 4 — граница всего участка, 5 — грани-
ца охраняемой его части, 6 — деревья, 7 — нереправа
Рис. 47. Перемещения индивидуально меченных крыс в свинарнике
1 — место мечения крысы, 2? — место поимки меченой крысы
\
1.\
=.
—__
[
о.
.-
Г|„20,|й
<] ®2
‚ мах крысы придерживаются определенных строений, годами не давая о
себе знать в других постройках.
Нри сносе старых, ветхих домов с надворными постройками зверьки
никуда далеко не уходят и стремятся заселить новые многоэтажные
дома, расположенные вблизи сломаппых, часто даже раньше, чем их сда-
дут в эксплуатацию (Дмитриева, 1958).
О постоянстве обитания крыс в одних и тех же зданиях свидетельст-
вуют многочисленные исследования Дэвиса с соавторами (Рау:5 её а|.,
170
Глава третья
как мелкие хищники. Они, как уже было сказано, должны широко не-
ремещалься в ноисках корма, скрываться от преследования врагов, пере-
носить неблагоприятную погоду и т. н. Синантропные крысы в большия-
стве случаев обеспечены кормом, им не надо далеко ходить в поисках
его. Живя под крышей, опи не подвергаются неблагоприятным воздейст-
виям погоды, врагов среди животных у них мало. Все эти экологические
отличия накладывают отпечаток на пространственную структуру их на-
селения. В основном это касается размеров, конфигурации и взаимного
расположения участков обитания. В целом же характер использования
территории крысами обеих экологических форм нидослецифичен.
На свинофермах крысы всегда с избытком обеспечены кормом, повэто-
му участки их обитания бывают очень небольшими (30—50 м“), чаще
располагаются по клетям, в которых держат свиней. Численность этих
грызупов здесь достигает иногда колоссальных размеров, и для увеличе-
ния емкости пространства своего обитания пасюки начинают селиться и
перемещаться в пустотах стен, лазают по трубам, стропилам, бегают под
настилами, используют подземные помещения, передвигаются под кры-
шей. Таким образом, их местообитание становится как бы трехмерным,
способствуя тем самым как уменьшению контактов между особями, так и,
следовательно, степени стресса.
Отлов индивидуально меченных крыс показал, что их перемещения в
пределах участка невелики — 15—25 м (Соколов, Карасева, 1985}
(рис. 47). Групповые участки относительно изолированы друг от друга
благодаря тому, что доминанты защищают их грапицы и между ними
располагается нейтральная территория. На свинофермах это тамбуры,
проходы между загонами и коридоры (Лозан, Никул, 1983).
Территория участка группировки крыс но ее использованию, как и у
экзоантронных пасюков, неодпородна. На ней выделяется центральная
часть, наиболее защищаемая (30—50% площади всего участка), границы
которой доминант метит мочевыми точками и активно защищает от втор-
жения чужих самцов. Периферия участка обычно самцом-доминантом не
посещается — там чаще держатся подчиненные особи. Участки соседних
группировок перекрываются только на их нериферии (Михайленко, 1982),
Крысы редко без причины переселяются из одпого корпуса животно-
водческой фермы в другие места (Нозлов, 1978). Они настолько бывают
привязаны к своей территории, что часть их иногда длительное время не
покидают даже пустые помещения, из которых выведен скот и, следова-
тельно, отсутетвует корм. По почам зверьки ходят питаться в соседние
корпуса, расположенные на расстоянии 25—50 и даже 200 м.
Иногда сила этой привязанности поразительна. Например, в одном из
рядом стоящих свинарников во время ремонта были вскрыты полы и сня-
та часть крыши. Пищи для крыс в этом номещении не было, убежища
в значительной степени были разрушены. Несмотря на это, животные
жили там, а кормились за 45 м в соседнем свинарнике.
В условиях большого города крысы также часто консервативны в ис-
пользовании территории (Дукельская, 1957; Рау!$ её а1., 1948; Мелкова,
1987). И. И. Траут (1950), например, нишет, что в отдельных кварта-
Экология
174
5баЕ=8[97]?
Рис. 46. Использование одиночным самцом территории вдоль р. Сумки (Украина)
(по: Мерзликин, 1987)
А — план всего участка, Б — охраняемая часть участка; 1— жилые норы, 2 — естественные
убежища, 8 — направление переправ крысы через реку, 4 — граница всего участка, 5 — грани-
ца охраняемой его части, 6 — деревья, 7 — нереправа
Рис. 47. Перемещения индивидуально меченных крыс в свинарнике
1 — место мечения крысы, 2? — место поимки меченой крысы
\
1.\
=.
—__
[о.
.-
Г|„20,|й
<] ®2
‚ мах крысы придерживаются определенных строений, годами не давая о
себе знать в других постройках.
Нри сносе старых, ветхих домов с надворными постройками зверьки
никуда далеко не уходят и стремятся заселить новые многоэтажные
дома, расположенные вблизи сломаппых, часто даже раньше, чем их сда-
дут в эксплуатацию (Дмитриева, 1958).
О постоянстве обитания крыс в одних и тех же зданиях свидетельст-
вуют многочисленные исследования Дэвиса с соавторами (Рау:5 её а|.,
472
Глава третья
1948) в г. Балтиморе, где пасюко» отлавливали, метили и отпускали па
месте поимки. Через 3 месяца более 80% меченых зверьков ловили в
тех же зланиях, где они были выпущены. Интересны и другие опыты
этих же авторов, доказывающие высокую привязанность крые к своим
местам обитапия.— животных метили в северной и южной частях г. Бал-
лимора, а выпускали в центре города. Последующие отловы показали,
что из 181 помеченной крысы 53 особи, т. е. 29%, вернулись обратно,
на прежние места, т. е. проявили «инстинкт дома».
О размерах участков крыс в г. Балтиморе можно судить липть по рас-
стояниям, на которые перемещались индивидуально меченные зверьки
(табл. 10). Судя по данным таблицы, половина крыс уходила не более
чем па 6 м и только едипичные особи удалялись от места мечения на
36 м. Авторы считают, что эти расстояния есть радиусы участков крыс.
Данлые В. К. Мелковой (1987), полученные другой методикой (грунпо-
вое мечение крыс тетрациклином в подвале многоэтажного дома), также
говорят о невысокой подвижности оседло живущих пасюков. Из 31 ме-
ченой особи 27 были обнаружены на территории мечепия и только 4 за
ее пределами на расстоянии 4—5 м. Самое дальнее обнаружение одной
крысы составляло 37,5 м.
Таким образом, в подвалах жилых многоэтажных домов крысы лере-
мещаются на небольшие расстояния, причем в основном они держатся
около источника пищи — мусороприемных камер. Вероятно, около каж-
дой из камер обитает группировка крыс, которая сдерживает натиск осо-
бей соседних кланов. Регистрация следов крые на равпомерно разложен-
ных пылевых площадках ноказывает, что после проведения истребитель-
ных мероприятий, когда часть пасюков около одной камеры уничтожена,
несколько дней здесь следы крыс отсутствуют, затем появляются снова,
т. е. освободившаяся територия занолняетея новыми зверьками (Мел-
кова, 1987).
Таблица 190. Дальность перемещения меченых крыс в условиях города по данным
повторных отловов (по: Рау!$ ей а|., 1948)
Расстояние, м
Возраст крыс
0—6
1,5—12
13,5 —19 19,5—24 25,5—30 31,5—36
Всего
Самцы
Взрослые
21 ИИ
9
1
_
2
АА
Молодые
10
1
—
1
1
-
13
Самки
Взрослые
81
14
7
3
-
1
53
Молодые
3
3
—
_
_
-
6
Всего
65
26
16
5
1
3
116
ф
55
224
43,6
4,2
1,6
2,6
100
‹
Экология
4173
В подвальных помещениях многоэтажных домов емкость пространст-
ва, населенного крысами, как в свинарниках, увеличинается за счет
«усложненности» территории. Крысы используют трубы горячего и хо-
плодного водоснабжения, расположенные как в горизонтальном, так и в
вертикальном направлении. В. К. Мелкова указывает, что пасюки бегают
по трубам группами или поодиночке, на торизонтальных трубах больтю-
го диаметра иногда спят. Захламленность подвалов, посторонние предме-
ты еще больше увеличивают емкость прострапства.
В подвалах домов при высокой численности крысы используют в 0с-
повном всю поверхность пола. Они перемещаются не только вдоль степ,
но и на расстоянии от них. При низкой численности пасюки чаще бегают
только вдоль стен, редко совершают перебежки по всему подвалу. Эверь-
ки, обитающие в нодвалах летом, выходят из зданий на улицу, расттиряя
территорию своих участков. Визуальные наблюдения Мелковой показа-
ли, что крысы регулярно используют полосы земли, заросшие травой, ко-
торые обычно бывают расположены между наружной стопой и подъезда-
ми в домах.
Подвижность крыс
Несмотря на высокий уровень территориального консерватизма, серые
крысы в природе, сельской местности и в условиях города бывают очень
подвижны и совершают значительные поремещения, способствующие их
боле? широкому расселепию. Если в сельской местности насюки пере-
двигаются но поверхности земли вдоль какого-либо укрытия {повален-
ного дерева, стены, вдоль канавы, насыпи, живой изгороди), иногда пе-
ресская и открытое пространство (200—300 м; Лялин, 1973; Тау|от,
1978), то в городе они обычно ходят по подземным сооружениям, в кото-
рых проложены водопроводные и канализационные трубы, кабельная
сеть, газопроводы и т. д. (Оа\1з её а1., 1948). Есть указания и на то,
что в случае ликвидации источников нитания или при ремонте зданий
крысы пересекают даже шумные, многолюдные улицы с большим коли-
чеством транспорта (Дукельская, 1959).
Причины, вызывающие перемещение крыс, многообразны и далеко
не достаточио изучены. В ряде случаев подвижность может быть вызва-
на не одной. а несколькими причинами. Большое значение имеет сильно
развитая исследовательская активпость зверьков (СаШоии, 1962; Ваглей,
1963).
В природных условиях крысы зимой не размножаются. Весной, с на-
ступлением теплой погоды, у самцов вновь возникает сперматогенез,
самки начинают овулировать и в связи с этим возрастает подвижность
зверьков. В это время при отловах капканами иногда оши попадаются на
значительном расстояпии от своих нор. Особенно увеличивается весенняя
подвижность крыс при их низкой численности, когда опи вынуждены ис-
кать половых партнеров.
Все, кто ловил крыс канканами в открытых биотонах, знают, что вес-
ной в норах, в которых полностью выловлено население этих грызунов,
472
Глава третья
1948) в г. Балтиморе, где пасюко» отлавливали, метили и отпускали па
месте поимки. Через 3 месяца более 80% меченых зверьков ловили в
тех же зланиях, где они были выпущены. Интересны и другие опыты
этих же авторов, доказывающие высокую привязанность крые к своим
местам обитапия.— животных метили в северной и южной частях г. Бал-
лимора, а выпускали в центре города. Последующие отловы показали,
что из 181 помеченной крысы 53 особи, т. е. 29%, вернулись обратно,
на прежние места, т. е. проявили «инстинкт дома».
О размерах участков крыс в г. Балтиморе можно судить липть по рас-
стояниям, на которые перемещались индивидуально меченные зверьки
(табл. 10). Судя по данным таблицы, половина крыс уходила не более
чем па 6 м и только едипичные особи удалялись от места мечения на
36 м. Авторы считают, что эти расстояния есть радиусы участков крыс.
Данлые В. К. Мелковой (1987), полученные другой методикой (грунпо-
вое мечение крыс тетрациклином в подвале многоэтажного дома), также
говорят о невысокой подвижности оседло живущих пасюков. Из 31 ме-
ченой особи 27 были обнаружены на территории мечепия и только 4 за
ее пределами на расстоянии 4—5 м. Самое дальнее обнаружение одной
крысы составляло 37,5 м.
Таким образом, в подвалах жилых многоэтажных домов крысы лере-
мещаются на небольшие расстояния, причем в основном они держатся
около источника пищи — мусороприемных камер. Вероятно, около каж-
дой из камер обитает группировка крыс, которая сдерживает натиск осо-
бей соседних кланов. Регистрация следов крые на равпомерно разложен-
ных пылевых площадках ноказывает, что после проведения истребитель-
ных мероприятий, когда часть пасюков около одной камеры уничтожена,
несколько дней здесь следы крыс отсутствуют, затем появляются снова,
т. е. освободившаяся територия занолняетея новыми зверьками (Мел-
кова, 1987).
Таблица 190. Дальность перемещения меченых крыс в условиях города по данным
повторных отловов (по: Рау!$ ей а|., 1948)
Расстояние, м
Возраст крыс
0—6 1,5—12
13,5 —19 19,5—24 25,5—30 31,5—36
Всего
Самцы
Взрослые
21 ИИ
9
1
_
2
АА
Молодые
10
1
—
1
1
-
13
Самки
Взрослые
81
14
7
3
-
1
53
Молодые
3
3
—
_
_
-
6
Всего
65
26
16
5
1
3
116
ф
55
224
43,6
4,2
1,6
2,6
100
‹
Экология
4173
В подвальных помещениях многоэтажных домов емкость пространст-
ва, населенного крысами, как в свинарниках, увеличинается за счет
«усложненности» территории. Крысы используют трубы горячего и хо-
плодного водоснабжения, расположенные как в горизонтальном, так и в
вертикальном направлении. В. К. Мелкова указывает, что пасюки бегают
по трубам группами или поодиночке, на торизонтальных трубах больтю-
го диаметра иногда спят. Захламленность подвалов, посторонние предме-
ты еще больше увеличивают емкость прострапства.
В подвалах домов при высокой численности крысы используют в 0с-
повном всю поверхность пола. Они перемещаются не только вдоль степ,
но и на расстоянии от них. При низкой численности пасюки чаще бегают
только вдоль стен, редко совершают перебежки по всему подвалу. Эверь-
ки, обитающие в нодвалах летом, выходят из зданий на улицу, расттиряя
территорию своих участков. Визуальные наблюдения Мелковой показа-
ли, что крысы регулярно используют полосы земли, заросшие травой, ко-
торые обычно бывают расположены между наружной стопой и подъезда-
ми в домах.
Подвижность крыс
Несмотря на высокий уровень территориального консерватизма, серые
крысы в природе, сельской местности и в условиях города бывают очень
подвижны и совершают значительные поремещения, способствующие их
боле? широкому расселепию. Если в сельской местности насюки пере-
двигаются но поверхности земли вдоль какого-либо укрытия {повален-
ного дерева, стены, вдоль канавы, насыпи, живой изгороди), иногда пе-
ресская и открытое пространство (200—300 м; Лялин, 1973; Тау|от,
1978), то в городе они обычно ходят по подземным сооружениям, в кото-
рых проложены водопроводные и канализационные трубы, кабельная
сеть, газопроводы и т. д. (Оа\1з её а1., 1948). Есть указания и на то,
что в случае ликвидации источников нитания или при ремонте зданий
крысы пересекают даже шумные, многолюдные улицы с большим коли-
чеством транспорта (Дукельская, 1959).
Причины, вызывающие перемещение крыс, многообразны и далеко
не достаточио изучены. В ряде случаев подвижность может быть вызва-
на не одной. а несколькими причинами. Большое значение имеет сильно
развитая исследовательская активпость зверьков (СаШоии, 1962; Ваглей,
1963).
В природных условиях крысы зимой не размножаются. Весной, с на-
ступлением теплой погоды, у самцов вновь возникает сперматогенез,
самки начинают овулировать и в связи с этим возрастает подвижность
зверьков. В это время при отловах капканами иногда оши попадаются на
значительном расстояпии от своих нор. Особенно увеличивается весенняя
подвижность крыс при их низкой численности, когда опи вынуждены ис-
кать половых партнеров.
Все, кто ловил крыс канканами в открытых биотонах, знают, что вес-
ной в норах, в которых полностью выловлено население этих грызунов,
174
Глава третья
через несколько дней они часто снова попадаются. „Тетом обловленпые
норы подолгу остаются необитаемыми, так как в этот период подвижность
крыс снижается, самки привязаны к норам с выводками. По мере под-
растания детенышей семьи начинают распадаться. В нервую очередь вы-
водок покидает самка, которая обычно выселяется на небольшое расетоя-
ние (20—30 м) (СаШопа, 1962), далее следует расселение молодых с0-
зревающих особей. В природных условиях там, где нет скоплений запасов
корма, поселения крыс редко достигают высокой плотности, следователь-
но, причиной выселения молодых не является агрессивность доминантных
особей (на теле молодых почти никогда нет следов от их покусов). Но-
о
*
С
:
о
о
б
о
5
с
о
Ф
о
5
5
0
С
С
Т
е
]
р
Ф
9
0
О
Рис. 43. Расположение нор серых крыс на рисовом поле Кубани (подопытная пло-
щадка)} в мае (черные кружки) и в июле (светлые кружки) 1980 г.
видимому, расселение происходит на оспове врожденной программы по-
ведения (Шилов, 1985).
Конечным результатом расселения может оказаться расширение пло-
щади обитания популяции с возможным перемещением зверьков на зна-
чительные расстояпия (800—1500 м) (Лялин, 1973). Однако если рядом
есть незанятая территория, то молодые крысы выселяются на небольшие
расстояния.
Результаты систематического обследования на Кубани (1979—1980)
земляных валиков на подопытной карте рисовой системы и картирование
расположения жилых нор показали, что весной, до начала размножения
взрослых крыс, их норы рассредоточены и находятся на значительном
расстоянии друг от друга. Летом при интенсивном размпожепии зверь-
ков и увеличении их численности вокруг этих нор появляются группы
нор (рис. 48). Это позволяет предположить, что молодые созревающие
зверьки в большинстве случаев при расселении не перемещаются далеко
от родительских нор.
У синантропных крыс, обитающих в отапливаемых помещепиях и
обеспеченных кормом, нет зимнего нерерыва в размножении. Тем не ме-
нее весной и у них увеличивается интенсивность половой активности и
в связи с этим резко возрастает подвижность. Значительная часть зверь-
ков как в сельской, так и в городской местности весной выселяются из
помещений в природные биотопы (Поярков, 1961; Тихвинская, 1964;
Судейкин, 1976). Некоторые авторы считают, что весеннее выселение
крысе объясняется не только проявлением их половой активности, но и
потребностью в витаминизированном корме (Мелкова, 1987).
Экология
175
В Ярославской области в 1981 г. в начале мая в деревне на живот-
поводческой ферме, где была средияя численность крыс, разложили при-
манку с тетрациклином. В июле этого же года произвели их отлов каи-
канами на ферме и в ее окрестностях, в тех местах, где были обнаружены
следы зверьков (их кал или норы). Следы пребывания крыс были ветре-
чены в обновном по берегам ручья, заросшего ивой, ольхой, орешником
и другим кустарником. На ферме было отловлено 16 крыс, из которых
9 было меченых (564-12), на ручье 52 крысы, из них 8 меченых
(15,3% ==8). Меченые крысы на ручье были пойманы в 2 км от деревни
(т. с. места, где они были помечены), рядом с палаточным лагерем.
Можно было бы думать, что крысы продолжали постоянно жить на фер-
ме и берег ручья входил в их участок, который они носешали, возвра-
щаясь обратно в свои норы в населенном пупкте. Но анализ возвраетного
состава и участия в размножении меченых некрытых крыс, пойманных
на ручье, говорит о другом. Из 6 взрослых самок 3 были кормящими.
Цак известно, кормящие самки у грызунов не отходят далеко от своих
нор с дотенышами и размеры их участков во время лактации уменьша-
ются (Карасева, Ильенко, 1957). Следовательно, крысь, переселивииеся
на берег ручья, остались там жить (Карасева и др. 198ба). В этой же
области обнаружили весеннее переселение крыс и на большие расстоя-
ния. Весной зверьки регулярно выселялись из поселка на расстояние до
7 км вдоль р. Сары вплоть до ее устья. В норах крыс часто встречались
выводки. Осенью с паступлением холодов этих грызунов на берегу реки
обнаружить не удавалось.
Повышение нодвижности крыс весной, связанное с активизацией по-
ловых инстинктов, отмечено и в городе. Например, в Москве весной пе-
реселенцы составляют 82%, а осенью только 54% (Судойкин, 1976).
Мелкова (1987) также указывает на то, что весной крысы часто высе-
ляются из подвалов многоэтажных жилых домов в открытые биотопы
города.
Половая активность крыс стимулирует повышенную подвижность
зверьков при их очень низкой численности, которая возникает в нометце-
ниях после дератизации, и животные интсепсивно перемещаются в по-
иске половых партнеров (Дмитрисва, 1958). Очень высокая плотность
паселения крыс также способствует увеличению их подвижности.
Синантропные пасюки, как мы видели, на складах, мясокомбинатах и
особенно на животноводческих фермах могут достигать колоссальной
численности (4 особи на 1 м"). В таких случаях мы имеем дело с явным
„перенаселением крыс, которое приводит к стрессовому состоянию, вынуж-
дающему животных покидать свои места обитания и переселяться. Вы-
сокая плотность населения синаптронных крысе повьшшает агрессивность
взрослых самцов, при этом особенно страдают подчиненные особи. Обыч-
по молодой самец в панике убогает от преследующего его самца, находя-
щегося на более высокой ступени иерархии. Последний цепляется моло-
дому самцу в основание хвоста, нанося ему глубокие кровоточащие раны.
В населенных пунктах, особенно в животноводческих фермах, среди
взрослых самцов покусаппые составляли 50%. Чем больше плотность на-
174
Глава третья
через несколько дней они часто снова попадаются. „Тетом обловленпые
норы подолгу остаются необитаемыми, так как в этот период подвижность
крыс снижается, самки привязаны к норам с выводками. По мере под-
растания детенышей семьи начинают распадаться. В нервую очередь вы-
водок покидает самка, которая обычно выселяется на небольшое расетоя-
ние (20—30 м) (СаШопа, 1962), далее следует расселение молодых с0-
зревающих особей. В природных условиях там, где нет скоплений запасов
корма, поселения крыс редко достигают высокой плотности, следователь-
но, причиной выселения молодых не является агрессивность доминантных
особей (на теле молодых почти никогда нет следов от их покусов). Но-
о
*
С
:
о
о
б
о
5
с
о
Ф
о
5
5
0
С
С
Т
е
]
р
Ф
9
0
О
Рис. 43. Расположение нор серых крыс на рисовом поле Кубани (подопытная пло-
щадка)} в мае (черные кружки) и в июле (светлые кружки) 1980 г.
видимому, расселение происходит на оспове врожденной программы по-
ведения (Шилов, 1985).
Конечным результатом расселения может оказаться расширение пло-
щади обитания популяции с возможным перемещением зверьков на зна-
чительные расстояпия (800—1500 м) (Лялин, 1973). Однако если рядом
есть незанятая территория, то молодые крысы выселяются на небольшие
расстояния.
Результаты систематического обследования на Кубани (1979—1980)
земляных валиков на подопытной карте рисовой системы и картирование
расположения жилых нор показали, что весной, до начала размножения
взрослых крыс, их норы рассредоточены и находятся на значительном
расстоянии друг от друга. Летом при интенсивном размпожепии зверь-
ков и увеличении их численности вокруг этих нор появляются группы
нор (рис. 48). Это позволяет предположить, что молодые созревающие
зверьки в большинстве случаев при расселении не перемещаются далеко
от родительских нор.
У синантропных крыс, обитающих в отапливаемых помещепиях и
обеспеченных кормом, нет зимнего нерерыва в размножении. Тем не ме-
нее весной и у них увеличивается интенсивность половой активности и
в связи с этим резко возрастает подвижность. Значительная часть зверь-
ков как в сельской, так и в городской местности весной выселяются из
помещений в природные биотопы (Поярков, 1961; Тихвинская, 1964;
Судейкин, 1976). Некоторые авторы считают, что весеннее выселение
крысе объясняется не только проявлением их половой активности, но и
потребностью в витаминизированном корме (Мелкова, 1987).
Экология
175
В Ярославской области в 1981 г. в начале мая в деревне на живот-
поводческой ферме, где была средияя численность крыс, разложили при-
манку с тетрациклином. В июле этого же года произвели их отлов каи-
канами на ферме и в ее окрестностях, в тех местах, где были обнаружены
следы зверьков (их кал или норы). Следы пребывания крыс были ветре-
чены в обновном по берегам ручья, заросшего ивой, ольхой, орешником
и другим кустарником. На ферме было отловлено 16 крыс, из которых
9 было меченых (564-12), на ручье 52 крысы, из них 8 меченых
(15,3% ==8). Меченые крысы на ручье были пойманы в 2 км от деревни
(т. с. места, где они были помечены), рядом с палаточным лагерем.
Можно было бы думать, что крысы продолжали постоянно жить на фер-
ме и берег ручья входил в их участок, который они носешали, возвра-
щаясь обратно в свои норы в населенном пупкте. Но анализ возвраетного
состава и участия в размножении меченых некрытых крыс, пойманных
на ручье, говорит о другом. Из 6 взрослых самок 3 были кормящими.
Цак известно, кормящие самки у грызунов не отходят далеко от своих
нор с дотенышами и размеры их участков во время лактации уменьша-
ются (Карасева, Ильенко, 1957). Следовательно, крысь, переселивииеся
на берег ручья, остались там жить (Карасева и др. 198ба). В этой же
области обнаружили весеннее переселение крыс и на большие расстоя-
ния. Весной зверьки регулярно выселялись из поселка на расстояние до
7 км вдоль р. Сары вплоть до ее устья. В норах крыс часто встречались
выводки. Осенью с паступлением холодов этих грызунов на берегу реки
обнаружить не удавалось.
Повышение нодвижности крыс весной, связанное с активизацией по-
ловых инстинктов, отмечено и в городе. Например, в Москве весной пе-
реселенцы составляют 82%, а осенью только 54% (Судойкин, 1976).
Мелкова (1987) также указывает на то, что весной крысы часто высе-
ляются из подвалов многоэтажных жилых домов в открытые биотопы
города.
Половая активность крыс стимулирует повышенную подвижность
зверьков при их очень низкой численности, которая возникает в нометце-
ниях после дератизации, и животные интсепсивно перемещаются в по-
иске половых партнеров (Дмитрисва, 1958). Очень высокая плотность
паселения крыс также способствует увеличению их подвижности.
Синантропные пасюки, как мы видели, на складах, мясокомбинатах и
особенно на животноводческих фермах могут достигать колоссальной
численности (4 особи на 1 м"). В таких случаях мы имеем дело с явным
„перенаселением крыс, которое приводит к стрессовому состоянию, вынуж-
дающему животных покидать свои места обитания и переселяться. Вы-
сокая плотность населения синаптронных крысе повьшшает агрессивность
взрослых самцов, при этом особенно страдают подчиненные особи. Обыч-
по молодой самец в панике убогает от преследующего его самца, находя-
щегося на более высокой ступени иерархии. Последний цепляется моло-
дому самцу в основание хвоста, нанося ему глубокие кровоточащие раны.
В населенных пунктах, особенно в животноводческих фермах, среди
взрослых самцов покусаппые составляли 50%. Чем больше плотность на-
176
Глава третья
селения крыс, тем выше процент пострадавших самцов (Козлов, 1980а;
Рыльников, Карасева, 1985).
А. ПН. Козлов (1978) сообщает, что в одном из современных совхозов
Павлодарской области в двух свинарпиках, расположенных друг от друга
на расстоянии 15 м, после истребительных работ были собраны трупы
крыс — в одпом 123, в другом 19 трунов. 49 трупов были с многочислен-
ными следами покусов, т. е. можно думать, что пострадавшие крысы пе-
реселились в другое здапие.
Часто подчиненные особи, не дожидаясь нападения, уклоняются от
преследования домипантов, эмитрируя из помещений с высокой плот-
ностью. Об этом свидетельствуют наблюдения многих этологов (СЬШу,
1954: СТоцвВ, 1965; ЕайЪай, 1978; и др.).
.
При высокой скученности крыс им часто не хватает места для устрой-
ства гнезд, и они начинают переселяться в другие помещения.
Часто насюки выпуждены переселяться в связи с измепившимися ус-
ловиями обитания, особенно при разрушении их убежищ. Это мы можем
наблюдать во время весенних паводков, когда вода заливает норы, и на
агроцепозах. Так, на рисовых полях весной во время гидромедиоратив-
ных и агротехпических работ разрушаются убежища крыс, и они вынуж-
депы покидать места своего зимнего обитания, тереселяясь ипогда на рас-
стояния 1,5-2 км. Летом, если в каналах рисовой системы поднимается
вода, заливаются норы крыс. Выравнивание поверхпости земли трактора-
ми приводит к разрушению нор и к выселению их обитателей (Рыльни-
ков, Карасева, 1985).
Большую роль в подвижности крыс играют недостаток корма или его
перемещение.
Экзоантропные крысы не в такой степени обеспечены полноценным
кормом, как пасюки синантропной формы, поэтому им более свойствепна
ирокая подвижность в поисках корма.
Во время войны зоолог Д. С. Айзенштадт (1945) наблюдал, как кры-
сы, привлоченные остатками нищи военпых частей, расположенных в
лесу в блинлажах, переместились туда на 20 км от ближайшего паселен-
ного пункта.
|
На Кубани пасюки, обитая в плавнях, перемещались к полевой
кухне охотничьего хозяйства, где выбрасывали отходы стрянни (в основ-
ном остатки рыбы) прямо в заросли тростпика.
.
На Украине мы наблюдали, как к трупу лошади, который долго не
убирали, быстро стали собираться крысы, привлеченные обилием пищи,
и рыли поблизости поры. В первые дни в сумерках наблюдали липть еди-
ничных особей, а через десять дней их были уже целые скопления. Ве-
роятно, такая концентрация крыс в местах, изобилующих кормом, может
быть объяснена тем, что отдельные особи, обладая повышенной исследо-
вательской активностью, оставляют мочевые метки на путях своего пере-
мещения и вслед за ними, ориентируясь на эти метки, приходят другие
зверьки.
Что касается синантропных крыс, то здесь мы имеем большее количе-
ство данных, чем по экзоантропным формам, свидетельствующих об их
Экология 177
поведении при измепепии условий обитания. Так, А. Н. Козлов (1978)
сообщает, что в Северном Казахстане в зданиях свинарников, находя-
щихея около р. Каракал (Есильский райоп), была высокая численность
крыс. Когда па реке прорвало дамбу и затонило свинарпик, то на другой
же день зверьки появились в коровнике в 500 м от места затопления,
тде их ранее не было. Их обнаружили также на расстоянии 1 км в на-
селенном пункте, который прежде был свободен от них.
В Южной Моравии после паполнения водохранилища была затоплена
деревия, из которой серые крысы перешли на поля кукурузы, находя-
щиеся от нее в 5 км, заселили их с высокой плотностью и долго там дер-
жались (Ното!ка, 1983).
В городах в связи с нарушением водопроводной системы передко бы-
вают затоплены подвалы жилых многоэтажных домов, что приводит к
выселению оттуда крысе (Мелкова, 1987). Затонление нор в канализаци-
онпой системе городов снособствует выселению пасюков на поверхность,
ипогда опи пропикают в квартиры через унитазы (Тощигин, 1986).
Но причиной нереселения крыс часто может быть и разрушение по-
строек, в которых они обитают. При ремонте помещений, как уже было
сказано, часть крыс могут не покидать своих мест обитания, по в боль-
шинстве случаев пасюки уходят из зданий, где разрушены их убежища,
при этом они переселяются обычно в другие помещения.
В Северном Казахстане в мае был разрушен старый свипарник с вы-
сокой плотностью паселения крыс (20—30 особей на 100 м). В 600 м
от свинарника находился населенный пункт, где через 2 дия после раз-
рушения свинарника из 26 домов в 19 заметили резкое увеличение чис-
ленности крыс (Козлов, 1978).
Помимо вынужденного выселения, связанного с разрушением убежищ,
крысы часто мигрируют и в связи с другими причинами, например з
связи с недостатком корма или его перемещением.
Как бы ни была высока численпость крыс в свинарниках, пока в но-
мещениях есть свиньи, крысы обеспечены кормом. Но, когда весной жи-
вотных переводят из ферм в летние помещения, зверьки остаются без
пищи. Многие из них и уходят велед за свиньями на расстояние 400—
600 ми даже 1,5—2 км. А. Н. Козлов (1978) считает, что крысы опре-
деляют направление пути по следам свиней и их помету.
Есть наблюдения, показывающие, что крысы покидают и помещения
коровников после перевода скота на пастбища. В Северном Казахетане
в апреле, накануне выгона коров на летние настбища, крысами было
заселено 34,9% коровников, а в автусте
— всего 6,1% (Козлов, 1978).
В некоторых зарубежных странах синантропные крысы регулярно
выселяются на пахотные поля, когда начинают поспевать возделываемые
па пих зерновые культуры. При высокой числениости передвижение
идет вдоль экологических русел — каналов к местам сброса воды, откуда
зверьки переходят уже на поля (Вто@е, Еаз®, Ста, 1983). Под Брюс-
селем пасюки покидали склад и, передвигаясь по берегу реки, собирались
сперва у озерка и уже оттуда переходили на ноля пшеницы, которые за-
селяли с большой плотностью (Вегпаг@, СоНп, 1966).
176
Глава третья
селения крыс, тем выше процент пострадавших самцов (Козлов, 1980а;
Рыльников, Карасева, 1985).
А. ПН. Козлов (1978) сообщает, что в одном из современных совхозов
Павлодарской области в двух свинарпиках, расположенных друг от друга
на расстоянии 15 м, после истребительных работ были собраны трупы
крыс — в одпом 123, в другом 19 трунов. 49 трупов были с многочислен-
ными следами покусов, т. е. можно думать, что пострадавшие крысы пе-
реселились в другое здапие.
Часто подчиненные особи, не дожидаясь нападения, уклоняются от
преследования домипантов, эмитрируя из помещений с высокой плот-
ностью. Об этом свидетельствуют наблюдения многих этологов (СЬШу,
1954: СТоцвВ, 1965; ЕайЪай, 1978; и др.).
.
При высокой скученности крыс им часто не хватает места для устрой-
ства гнезд, и они начинают переселяться в другие помещения.
Часто насюки выпуждены переселяться в связи с измепившимися ус-
ловиями обитания, особенно при разрушении их убежищ. Это мы можем
наблюдать во время весенних паводков, когда вода заливает норы, и на
агроцепозах. Так, на рисовых полях весной во время гидромедиоратив-
ных и агротехпических работ разрушаются убежища крыс, и они вынуж-
депы покидать места своего зимнего обитания, тереселяясь ипогда на рас-
стояния 1,5-2 км. Летом, если в каналах рисовой системы поднимается
вода, заливаются норы крыс. Выравнивание поверхпости земли трактора-
ми приводит к разрушению нор и к выселению их обитателей (Рыльни-
ков, Карасева, 1985).
Большую роль в подвижности крыс играют недостаток корма или его
перемещение.
Экзоантропные крысы не в такой степени обеспечены полноценным
кормом, как пасюки синантропной формы, поэтому им более свойствепна
ирокая подвижность в поисках корма.
Во время войны зоолог Д. С. Айзенштадт (1945) наблюдал, как кры-
сы, привлоченные остатками нищи военпых частей, расположенных в
лесу в блинлажах, переместились туда на 20 км от ближайшего паселен-
ного пункта.
|
На Кубани пасюки, обитая в плавнях, перемещались к полевой
кухне охотничьего хозяйства, где выбрасывали отходы стрянни (в основ-
ном остатки рыбы) прямо в заросли тростпика.
.
На Украине мы наблюдали, как к трупу лошади, который долго не
убирали, быстро стали собираться крысы, привлеченные обилием пищи,
и рыли поблизости поры. В первые дни в сумерках наблюдали липть еди-
ничных особей, а через десять дней их были уже целые скопления. Ве-
роятно, такая концентрация крыс в местах, изобилующих кормом, может
быть объяснена тем, что отдельные особи, обладая повышенной исследо-
вательской активностью, оставляют мочевые метки на путях своего пере-
мещения и вслед за ними, ориентируясь на эти метки, приходят другие
зверьки.
Что касается синантропных крыс, то здесь мы имеем большее количе-
ство данных, чем по экзоантропным формам, свидетельствующих об их
Экология 177
поведении при измепепии условий обитания. Так, А. Н. Козлов (1978)
сообщает, что в Северном Казахстане в зданиях свинарников, находя-
щихея около р. Каракал (Есильский райоп), была высокая численность
крыс. Когда па реке прорвало дамбу и затонило свинарпик, то на другой
же день зверьки появились в коровнике в 500 м от места затопления,
тде их ранее не было. Их обнаружили также на расстоянии 1 км в на-
селенном пункте, который прежде был свободен от них.
В Южной Моравии после паполнения водохранилища была затоплена
деревия, из которой серые крысы перешли на поля кукурузы, находя-
щиеся от нее в 5 км, заселили их с высокой плотностью и долго там дер-
жались (Ното!ка, 1983).
В городах в связи с нарушением водопроводной системы передко бы-
вают затоплены подвалы жилых многоэтажных домов, что приводит к
выселению оттуда крысе (Мелкова, 1987). Затонление нор в канализаци-
онпой системе городов снособствует выселению пасюков на поверхность,
ипогда опи пропикают в квартиры через унитазы (Тощигин, 1986).
Но причиной нереселения крыс часто может быть и разрушение по-
строек, в которых они обитают. При ремонте помещений, как уже было
сказано, часть крыс могут не покидать своих мест обитания, по в боль-
шинстве случаев пасюки уходят из зданий, где разрушены их убежища,
при этом они переселяются обычно в другие помещения.
В Северном Казахстане в мае был разрушен старый свипарник с вы-
сокой плотностью паселения крыс (20—30 особей на 100 м). В 600 м
от свинарника находился населенный пункт, где через 2 дия после раз-
рушения свинарника из 26 домов в 19 заметили резкое увеличение чис-
ленности крыс (Козлов, 1978).
Помимо вынужденного выселения, связанного с разрушением убежищ,
крысы часто мигрируют и в связи с другими причинами, например з
связи с недостатком корма или его перемещением.
Как бы ни была высока численпость крыс в свинарниках, пока в но-
мещениях есть свиньи, крысы обеспечены кормом. Но, когда весной жи-
вотных переводят из ферм в летние помещения, зверьки остаются без
пищи. Многие из них и уходят велед за свиньями на расстояние 400—
600 ми даже 1,5—2 км. А. Н. Козлов (1978) считает, что крысы опре-
деляют направление пути по следам свиней и их помету.
Есть наблюдения, показывающие, что крысы покидают и помещения
коровников после перевода скота на пастбища. В Северном Казахетане
в апреле, накануне выгона коров на летние настбища, крысами было
заселено 34,9% коровников, а в автусте
— всего 6,1% (Козлов, 1978).
В некоторых зарубежных странах синантропные крысы регулярно
выселяются на пахотные поля, когда начинают поспевать возделываемые
па пих зерновые культуры. При высокой числениости передвижение
идет вдоль экологических русел — каналов к местам сброса воды, откуда
зверьки переходят уже на поля (Вто@е, Еаз®, Ста, 1983). Под Брюс-
селем пасюки покидали склад и, передвигаясь по берегу реки, собирались
сперва у озерка и уже оттуда переходили на ноля пшеницы, которые за-
селяли с большой плотностью (Вегпаг@, СоНп, 1966).
178
Глава третья
%60
Рис. 49. Распределение
.
нрыс в течение года в
`
разных складских поме-
88|
щениях (по; Эюддам,
1981)
10
1— хранилища сена и со-
ломы,
307
2 — склады продовольст-
вия,
29е
8 — зернохранилища,
4 — склады фуража
а
МЕСяцы
Интересные данные о сезонных переселениях крыс в постройки ферм
и из них приводит Штоддарт (З%юЧ9аг, 1981), собиравший материал в
1966 п 1987 гг. За это время ежемесячно обследовали различные склады
(продовольственные, зернохранилища, фураж) на 1584 фермах. В каждом
помещении учитывали свежие следы жизнедеятельности крыс и затем
рассчитывали процент заселенных зданий от всех обследованных. Во всех
зданиях пик численпости приходился на апрель, затем начинался спад,
достигавший очень низкого уровня в июле (рис. 49). Апрельский лик
может быть объяспен как результат роста популяции во время репродук-
тивного периода, так как подъем иптенсивности размножения регистри-
ровали в марте. В апреле-июне не отмечали повьшшенной смертности,
и низкий уровень численности в июле можно отнести только за счет вы-
солепия пасюков, совпадающего с поспеванием зерновых культур. После
уборки урожая на полях наблюдается обратное переселение этих грызу-
нов в помещения (Низоп, Вептизов, 1981). То же явление в открытых
биотопах мы наблюдали с наступлением осенью похолодания (Козлов,
1978; Карасева и др., 198ба).
Таким образом, мы видим, что в сельской местности подвижность крыс
может быть вызвана разными причинами, в том числе недостатком ос-
повного корма. Это очень наглядно показано в описаниом нами ниже
эксперименте Тейлора и Киу (Тау]ог, Опу, 1978), из которого видно, что
крыса в поисках переместившегося источника корма может за ночь прой-
ти больше трех километров.
Эксперимент был прозеден в Британии в сельской местности: вдоль
зеленой изгороди из кустарников расставили 12 кормушек с зерном. Че-
рез несколько дней, когда крысы нрикормились, выставили живоловки.
Был пойман один самец © массой тела 300 г. На него падели радиодат-
чик в виде ошейника и после этого отпустили на том же месте, в поеле-
дующие дни проводилось радиослежение. В нервые дни меченая крыса,
выходя из своей рядом расположенной норы, никуда не уходила от кор-
мушек, но через 6 дней их убрали. Тогда она стала ходить гораздо даль-
шс, передвигаясь до конца изгороди на 300—400 м в одну и в другую
Экология 179
сторону, но всегда возвращалась в свою нору. Через 7 дней осле изъятия
кормушек крыса посетила необитаемую пору в 1,5 км от своей норы,
но там она провела 3,5 ч, а затем вернулась в свою собственпую нору.
Следующие 7 дней крыса провела в другой поре (рис. 50). В течение
одной ночи она ‘двигалась по кругу (3,5 км) и верпулась в свою нору.
Этот эксперимент показывает, с одной стороны, каких значительных раз-
Рис. 50. Перемещение взрослого сам-
ца в сельской местности (по: Тау1ог,
Оиу, 1978)
1 — дороги,
2 — живая изгороль,
3 — участки леса,
4 — направление перемещения крысы,
5 — жилая нора подопытной крысы,
6 — норы, которые крыса посещала вре-
менно,
7 — места прикормки
100 200 300
ГОН ОН
Рис. 51. Места периодического по-
явления крыс (по: Мелкова, 1957)
1— стойкие длительно существующие
поселения крыс,
2 — места периодического появления
крыс,
3— знания, постоянно свободные от
крыс,
4 — железная дорога
ПИ2 Э500и|
178
Глава третья
%60
Рис. 49. Распределение
.
нрыс в течение года в
`
разных складских поме-
88|
щениях (по; Эюддам,
1981)
10
1— хранилища сена и со-
ломы,
307
2 — склады продовольст-
вия,
29е
8 — зернохранилища,
4 — склады фуража
а
МЕСяцы
Интересные данные о сезонных переселениях крыс в постройки ферм
и из них приводит Штоддарт (З%юЧ9аг, 1981), собиравший материал в
1966 п 1987 гг. За это время ежемесячно обследовали различные склады
(продовольственные, зернохранилища, фураж) на 1584 фермах. В каждом
помещении учитывали свежие следы жизнедеятельности крыс и затем
рассчитывали процент заселенных зданий от всех обследованных. Во всех
зданиях пик численпости приходился на апрель, затем начинался спад,
достигавший очень низкого уровня в июле (рис. 49). Апрельский лик
может быть объяспен как результат роста популяции во время репродук-
тивного периода, так как подъем иптенсивности размножения регистри-
ровали в марте. В апреле-июне не отмечали повьшшенной смертности,
и низкий уровень численности в июле можно отнести только за счет вы-
солепия пасюков, совпадающего с поспеванием зерновых культур. После
уборки урожая на полях наблюдается обратное переселение этих грызу-
нов в помещения (Низоп, Вептизов, 1981). То же явление в открытых
биотопах мы наблюдали с наступлением осенью похолодания (Козлов,
1978; Карасева и др., 198ба).
Таким образом, мы видим, что в сельской местности подвижность крыс
может быть вызвана разными причинами, в том числе недостатком ос-
повного корма. Это очень наглядно показано в описаниом нами ниже
эксперименте Тейлора и Киу (Тау]ог, Опу, 1978), из которого видно, что
крыса в поисках переместившегося источника корма может за ночь прой-
ти больше трех километров.
Эксперимент был прозеден в Британии в сельской местности: вдоль
зеленой изгороди из кустарников расставили 12 кормушек с зерном. Че-
рез несколько дней, когда крысы нрикормились, выставили живоловки.
Был пойман один самец © массой тела 300 г. На него падели радиодат-
чик в виде ошейника и после этого отпустили на том же месте, в поеле-
дующие дни проводилось радиослежение. В нервые дни меченая крыса,
выходя из своей рядом расположенной норы, никуда не уходила от кор-
мушек, но через 6 дней их убрали. Тогда она стала ходить гораздо даль-
шс, передвигаясь до конца изгороди на 300—400 м в одну и в другую
Экология 179
сторону, но всегда возвращалась в свою нору. Через 7 дней осле изъятия
кормушек крыса посетила необитаемую пору в 1,5 км от своей норы,
но там она провела 3,5 ч, а затем вернулась в свою собственпую нору.
Следующие 7 дней крыса провела в другой поре (рис. 50). В течение
одной ночи она ‘двигалась по кругу (3,5 км) и верпулась в свою нору.
Этот эксперимент показывает, с одной стороны, каких значительных раз-
Рис. 50. Перемещение взрослого сам-
ца в сельской местности (по: Тау1ог,
Оиу, 1978)
1 — дороги,
2 — живая изгороль,
3 — участки леса,
4 — направление перемещения крысы,
5 — жилая нора подопытной крысы,
6 — норы, которые крыса посещала вре-
менно,
7 — места прикормки
100 200 300
ГОН ОН
Рис. 51. Места периодического по-
явления крыс (по: Мелкова, 1957)
1— стойкие длительно существующие
поселения крыс,
2 — места периодического появления
крыс,
3— знания, постоянно свободные от
крыс,
4 — железная дорога
ПИ2 Э500и|
180 Глава третья
меров достигает подвижность крыс при неремещении корма, а с дру-
й—
к ее консерватизм.
ви" родах недостаток корма или его перемещение тоже приводят к
а
ыС.
В осажденном Ленинграде было очень мпого серых крыс. Длительная
осада города, гибель запасов продовольствия в результате действий про
тивника и в связи с этим значительное сокращение пайков в зиму
1944/42 тг., конечно, отразились и на этих грызунах, питающихся за счет
рациона людей. Весной 1942 г. крысы совершали массовые миграции,
совпадающие по времени с переселением жителей из района в район,
и покидали те части города, из которых уходило население и где, сле-
довательно, пе оставалось пищи. Переселение крыс в новые районы ста-
новилось очень заметно
— они нанадали на пищевые объекты, огороды,
кладбища, участилось число случаев их непосредственного нападения и
людей (по регистрации числа укушенных граждан, обращавшихся в Па-
стеровский институт). Часть пасюков выселялись за городскую черту,
на передний край обороны (Токаревич, ‚4947).
|
Но поиск крысами лучших условий жизни, в том числе и корма,
в значительной степени может определяться их усиленной исследователь-
ской активностью. Мы, например, знаем случаи (Молкова, 1987), когда
зверьки доходили до девятого этажа и подгрызали там двери в квартиры.
В. А. Судейкии (1976) описывает передвижение насюков в городе по
поверхности на расстояние в 4 км, а в одном случае даже на | км,
но преобладающее большинство (82,5%) особей, но словам автора, пере
ходили на расстояния меньше 4 км.
р
В. К. Мелкова (1987) провела очень интересное наблюдение на экс-
периментальном участке города, где, несмотря на интенсивную и система-
тическую дератизацию, сохранялись в течение ряда лет два очень стой-
ких поселения крыс на расстоянии примерно 2 км друг от друга (рис. 51).
На территории между этими поселениями еженедельно проверяли нали-
чие (или отсутствие) крыс с помощью площадок, засыпанных тальком,
и по мере надобности проводили дератизацию, благодаря. которой эти
грызуны там отсутствовали по нескольку месяцев. Но оказалось, что на
этой освобождасмой от крыс территории они периодически опять появ-
лялись группами по 5—8 особей на расстоянии в 300—500 ми болое от
мест их стойкого обитания. Мы можем только предположить, что такие
дальние выходы совершаются зверьками в связи с желанием получить
информацию о возможности заселения нового места обитания.
Отношение серых крыс к территории в настоящее время изучено не-
достаточно, но уже можно сказать, что, несмотря на высокую подр.
ность, этих грызунов нельзя назвать кочующим видом, ведущим бродя-
чий образ жизни (Айзенштадт, 1957). В большинстве случасв они ведут
как территориальные животные.
са + особны перемещаться на большие расстояния, и это позво-
ляет им обнаруживать скопления пищи, скучиваться около нее и при
благоприятных условиях подолгу пребывать на одном месте, сокращая,
таким образом, риск гибели от врагов и других причин.
Экология
181
Размноженис, возрастной состав и смертность
Рожавемость и смертность среди пасюков, относящихся к разным поло“
вым и возрастным группам, характеризуются различными величинами в
зависимости от географических и стациальных условий, уровня числен-
ности и времени тода, что в конечном счете определяет динамику воз-
растного состава популяции — одной из важнейших характеристик попу-
ляционной динамики.
Соотношение половозрастных групи в популяции может определять
не только репродуктивный потенциал полуляции, но и направление раз-
вития адаптаций, изменение ее генетической структуры.
Репродуктивная функция самцов
Состояние генеративной системы самцов обычно устанавливают путем
их векрытия, визуального осмотра, измерения семенников, семенных пу-
зырьков и эпидидимиса. На основании такого обследования выделяют
четыре груплы половозрелости в соответствии с делением, предложенным
Т. А. Ким и В. М. Лахашерстовым (1964) (табл. 11). О наличии спер-
матогенсза судят по присутствию зрелых сперматозоидов (Ретгу, 1945),
однако для определения половозрелости иногда используют только массу
и размеры половых органов, которые довольно жестко коррелируют с со-
стоянием сперматогенеза (рис. 52). Наибольшую трудность при опреде-
лении половой зрелости и половой активности составляет различение П
и 1116 груни. У самцов П группы лишь небольшое количество спермато-
зоидов, причем масса семенников быстро увеличивается с возрастом.
Крысы Гб группы, как правило, имеют массу тела более 200 г, однако
Тургор семенников ослаблен, семенные пузырьки опавшие, длиной око-
ло 4—5 мм. Появление в нопуляции крыс П16 грунны обычно связано с
депрессией размножения зимой,
С возрастом ропродуктивная функция самцов практически не ослабе-
вает. По всей видимости, лишь немногие из них доживают до такого воз-
раста, когда они становятся сексуально неактивными (Регту, 1945; Коз-
лов, 1984). Старшие самцы выделяют максимальное количество спермы
и оставляют наибольшее число эмбрионов при третьей эякуляции. У мо-
лодых самцов количество выбрасываемой спермы последовательно снижа-
ется от первой до седьмой эякуляции, но количество зачатий до шестой
эякуляции остается постоянным. Таким образом, возраст самца и поряд-
ковый номер эякуляции не влияют на фертильность снариваний (Топег,
‚ АЧюг, 1985). Исследовапие изменений гаметогенеза и продукции стероид-
ных гормонов в семенниках самцов лабораторных крыс также свидетель-
ствует о том, что снижения репродуктивной фупкции с возрастом не наб-
людается (Э1ерег, Ниапо, 1983).
Половая активность самцов наблюдается большую часть года, снижа-
ясь только зимой. Среди особей, отловленных в феврале в строениях
г. Красноярска, активный сперматогенез наблюдали в 56% случаев (Ким,
Лихашерстов, 1964).
180 Глава третья
меров достигает подвижность крыс при неремещении корма, а с дру-
й—
к ее консерватизм.
ви" родах недостаток корма или его перемещение тоже приводят к
а
ыС.
В осажденном Ленинграде было очень мпого серых крыс. Длительная
осада города, гибель запасов продовольствия в результате действий про
тивника и в связи с этим значительное сокращение пайков в зиму
1944/42 тг., конечно, отразились и на этих грызунах, питающихся за счет
рациона людей. Весной 1942 г. крысы совершали массовые миграции,
совпадающие по времени с переселением жителей из района в район,
и покидали те части города, из которых уходило население и где, сле-
довательно, пе оставалось пищи. Переселение крыс в новые районы ста-
новилось очень заметно
— они нанадали на пищевые объекты, огороды,
кладбища, участилось число случаев их непосредственного нападения и
людей (по регистрации числа укушенных граждан, обращавшихся в Па-
стеровский институт). Часть пасюков выселялись за городскую черту,
на передний край обороны (Токаревич, ‚4947).
|
Но поиск крысами лучших условий жизни, в том числе и корма,
в значительной степени может определяться их усиленной исследователь-
ской активностью. Мы, например, знаем случаи (Молкова, 1987), когда
зверьки доходили до девятого этажа и подгрызали там двери в квартиры.
В. А. Судейкии (1976) описывает передвижение насюков в городе по
поверхности на расстояние в 4 км, а в одном случае даже на | км,
но преобладающее большинство (82,5%) особей, но словам автора, пере
ходили на расстояния меньше 4 км.
р
В. К. Мелкова (1987) провела очень интересное наблюдение на экс-
периментальном участке города, где, несмотря на интенсивную и система-
тическую дератизацию, сохранялись в течение ряда лет два очень стой-
ких поселения крыс на расстоянии примерно 2 км друг от друга (рис. 51).
На территории между этими поселениями еженедельно проверяли нали-
чие (или отсутствие) крыс с помощью площадок, засыпанных тальком,
и по мере надобности проводили дератизацию, благодаря. которой эти
грызуны там отсутствовали по нескольку месяцев. Но оказалось, что на
этой освобождасмой от крыс территории они периодически опять появ-
лялись группами по 5—8 особей на расстоянии в 300—500 ми болое от
мест их стойкого обитания. Мы можем только предположить, что такие
дальние выходы совершаются зверьками в связи с желанием получить
информацию о возможности заселения нового места обитания.
Отношение серых крыс к территории в настоящее время изучено не-
достаточно, но уже можно сказать, что, несмотря на высокую подр.
ность, этих грызунов нельзя назвать кочующим видом, ведущим бродя-
чий образ жизни (Айзенштадт, 1957). В большинстве случасв они ведут
как территориальные животные.
са + особны перемещаться на большие расстояния, и это позво-
ляет им обнаруживать скопления пищи, скучиваться около нее и при
благоприятных условиях подолгу пребывать на одном месте, сокращая,
таким образом, риск гибели от врагов и других причин.
Экология
181
Размноженис, возрастной состав и смертность
Рожавемость и смертность среди пасюков, относящихся к разным поло“
вым и возрастным группам, характеризуются различными величинами в
зависимости от географических и стациальных условий, уровня числен-
ности и времени тода, что в конечном счете определяет динамику воз-
растного состава популяции — одной из важнейших характеристик попу-
ляционной динамики.
Соотношение половозрастных групи в популяции может определять
не только репродуктивный потенциал полуляции, но и направление раз-
вития адаптаций, изменение ее генетической структуры.
Репродуктивная функция самцов
Состояние генеративной системы самцов обычно устанавливают путем
их векрытия, визуального осмотра, измерения семенников, семенных пу-
зырьков и эпидидимиса. На основании такого обследования выделяют
четыре груплы половозрелости в соответствии с делением, предложенным
Т. А. Ким и В. М. Лахашерстовым (1964) (табл. 11). О наличии спер-
матогенсза судят по присутствию зрелых сперматозоидов (Ретгу, 1945),
однако для определения половозрелости иногда используют только массу
и размеры половых органов, которые довольно жестко коррелируют с со-
стоянием сперматогенеза (рис. 52). Наибольшую трудность при опреде-
лении половой зрелости и половой активности составляет различение П
и 1116 груни. У самцов П группы лишь небольшое количество спермато-
зоидов, причем масса семенников быстро увеличивается с возрастом.
Крысы Гб группы, как правило, имеют массу тела более 200 г, однако
Тургор семенников ослаблен, семенные пузырьки опавшие, длиной око-
ло 4—5 мм. Появление в нопуляции крыс П16 грунны обычно связано с
депрессией размножения зимой,
С возрастом ропродуктивная функция самцов практически не ослабе-
вает. По всей видимости, лишь немногие из них доживают до такого воз-
раста, когда они становятся сексуально неактивными (Регту, 1945; Коз-
лов, 1984). Старшие самцы выделяют максимальное количество спермы
и оставляют наибольшее число эмбрионов при третьей эякуляции. У мо-
лодых самцов количество выбрасываемой спермы последовательно снижа-
ется от первой до седьмой эякуляции, но количество зачатий до шестой
эякуляции остается постоянным. Таким образом, возраст самца и поряд-
ковый номер эякуляции не влияют на фертильность снариваний (Топег,
‚ АЧюг, 1985). Исследовапие изменений гаметогенеза и продукции стероид-
ных гормонов в семенниках самцов лабораторных крыс также свидетель-
ствует о том, что снижения репродуктивной фупкции с возрастом не наб-
людается (Э1ерег, Ниапо, 1983).
Половая активность самцов наблюдается большую часть года, снижа-
ясь только зимой. Среди особей, отловленных в феврале в строениях
г. Красноярска, активный сперматогенез наблюдали в 56% случаев (Ким,
Лихашерстов, 1964).
132
Глава третья
В животноводческих помещениях Северного Казахстана зимой, судя
по уменьшению массы семенников, также наступает депрессия спермато-
гопеза. У многих самцов зимой семенники дряблые. Однако даже в сере-
дине зимы встречаются крысы старших возрастно-весовых групп © хоро-
шо развитыми семенпиками размером 19—22
х 11—14 мм (Козлов, 1981).
Рис. 62. Гистологические срезы семенников (по: Тауог её в., 1981)
Вверху— семяироводятцие канальцы неполовоарелых самцов в возрасте одного месяца; видны
сперматоциты и многочисленные картины мейоза. Х 315; внизу — семяпроводящие канальцы
половозрелых животных; видны стенки двух канальцев со зрелыми сперматозоидами по бокам
Экология
183
Таблида 11. Состояние генеративной системы самцов на различных стадиях
полового созревания
Длина се- Длина се-
менников, менных пу-
мм
зырьков,
мм*
Группы половозрело- Состояние сиерматогене- | Масса се-
сти (по: Ким, Лиха-
за (по; Реггу, 1945}
менников, г
.>
-
+
-
шерстов, 1961)
.в
.?
Ра
„р
Е
|:
Е
Я
-Н
Н
Н
Я
>
я
=
Е:
Неноловозрелыс, 1
В мазке только ин-
< 0,75
9+1|9=2|21| 2+2
терстициальные
клетки; спермато-
зоидов нет
Наряду с интерсти-
циальными клетка-
ми имеются немно-
точисленные спер-
Созревающие, П
0,75—2,00 |146=2 | 166 | 7+4 | 7=7
матозоиды
Полевозрелые с
В массе спермы ин- 2,00—3,00 | 22-1 | 22-54 | 23-51 [23-4
активным спермато- | терстициальные
генезом, ИТа
клетки практически
неотличимы
Половозрелые на
-
>2.00 13-2 | 43=6 | 4+1 | 4-4
стадии прекращения
сперматогенеза, 1116
* По Рыльникову и Карасевой— неонубликованные данные; М — среднее арифметическое,
ш — ошибка средней, 1;4=2
— стандартное значение критерия Стьюдента ири 20,05 (Пло-
хинский, 1980).
В населенных пунктах Кубани самцы крыс активны в ноловом отно-
шений круглый год; длина их семенников практически не изменястся,
сперматогепез не прекращается, однако отмечены изменения длины
семенных пузырьков. С мая по автуст и с октября по январь опи досто-
верно уменьшались, а с июля—августа по октябрь увеличивались
{р=0,05) (табл. 12). Эти изменения в целом соответствуют периодиче-
ским колебаниям плодовитости взрослых самок, обитающих в постройках
человека.
Среди крыс, живущих на рисовых полях, периодические колебания
половой функции самцов выражены более отчетливо. Уже в октябре сре-
`ди них появляются особи, у которых наблюдается отсутетвие спермато-
генеза; в декабре у большинства тургор семенников ослаблен и размеры
их достоверно меньше (р<0,05), чем летом (табл. 13). В декабре только
у 15% самцов наблюдался сперматогенез. В феврале доля самцов с ак-
тивным сперматогенезом увеличивается (56%). С марта но август все
100% самцов готовы к спариванию. Размеры семенных пузырьков увели-
чиваются достоверно (р<0,05) от декабря к марту и от марта к маю
(р<0,05). Самцы, родивлтиеся в мае, начинают созревать к июлю (Рыль-
132
Глава третья
В животноводческих помещениях Северного Казахстана зимой, судя
по уменьшению массы семенников, также наступает депрессия спермато-
гопеза. У многих самцов зимой семенники дряблые. Однако даже в сере-
дине зимы встречаются крысы старших возрастно-весовых групп © хоро-
шо развитыми семенпиками размером 19—22
х 11—14 мм (Козлов, 1981).
Рис. 62. Гистологические срезы семенников (по: Тауог её в., 1981)
Вверху— семяироводятцие канальцы неполовоарелых самцов в возрасте одного месяца; видны
сперматоциты и многочисленные картины мейоза. Х 315; внизу — семяпроводящие канальцы
половозрелых животных; видны стенки двух канальцев со зрелыми сперматозоидами по бокам
Экология
183
Таблида 11. Состояние генеративной системы самцов на различных стадиях
полового созревания
Длина се- Длина се-
менников, менных пу-
мм
зырьков,
мм*
Группы половозрело- Состояние сиерматогене- | Масса се-
сти (по: Ким, Лиха-
за (по; Реггу, 1945}
менников, г
.>
-
+
-
шерстов, 1961)
.в
.?
Ра
„р
Е
|:
Е
Я
-Н
Н
Н
Я
>
я
=
Е:
Неноловозрелыс, 1
В мазке только ин-
< 0,75
9+1|9=2|21| 2+2
терстициальные
клетки; спермато-
зоидов нет
Наряду с интерсти-
циальными клетка-
ми имеются немно-
точисленные спер-
Созревающие, П
0,75—2,00 |146=2 | 166 | 7+4 | 7=7
матозоиды
Полевозрелые с
В массе спермы ин- 2,00—3,00 | 22-1 | 22-54 | 23-51 [23-4
активным спермато- | терстициальные
генезом, ИТа
клетки практически
неотличимы
Половозрелые на
-
>2.00 13-2 | 43=6 | 4+1 | 4-4
стадии прекращения
сперматогенеза, 1116
* По Рыльникову и Карасевой— неонубликованные данные; М — среднее арифметическое,
ш — ошибка средней, 1;4=2
— стандартное значение критерия Стьюдента ири 20,05 (Пло-
хинский, 1980).
В населенных пунктах Кубани самцы крыс активны в ноловом отно-
шений круглый год; длина их семенников практически не изменястся,
сперматогепез не прекращается, однако отмечены изменения длины
семенных пузырьков. С мая по автуст и с октября по январь опи досто-
верно уменьшались, а с июля—августа по октябрь увеличивались
{р=0,05) (табл. 12). Эти изменения в целом соответствуют периодиче-
ским колебаниям плодовитости взрослых самок, обитающих в постройках
человека.
Среди крыс, живущих на рисовых полях, периодические колебания
половой функции самцов выражены более отчетливо. Уже в октябре сре-
`ди них появляются особи, у которых наблюдается отсутетвие спермато-
генеза; в декабре у большинства тургор семенников ослаблен и размеры
их достоверно меньше (р<0,05), чем летом (табл. 13). В декабре только
у 15% самцов наблюдался сперматогенез. В феврале доля самцов с ак-
тивным сперматогенезом увеличивается (56%). С марта но август все
100% самцов готовы к спариванию. Размеры семенных пузырьков увели-
чиваются достоверно (р<0,05) от декабря к марту и от марта к маю
(р<0,05). Самцы, родивлтиеся в мае, начинают созревать к июлю (Рыль-
184
Глава третья
Таблица 12. Размеры семенников и семенных пузырьков взроелых самцов
серой крысы, добытых в домах индивидуальной застройки и в отанливаемых
свинарниках Кубани в разные месяцы года
Месяц
всего отловлено
Длина семенников, мм оных пу
Январь
18
22,5+0,9
24,3=0,5
Февраль
10
22,8-=0,6
21,0+1,8
Май
90
21,5=0,А
23,0-0,4
Июль--август
66
21,4=0,4
20,6=0,5
Октябрь
35
22,0+0,3
25,6=0,8
* Все самды с активным сперматогенезом.
Таблица 13. Доля размножающихся самцов серых крыс, средние размеры их
семенников и семенных пузырьков на рисовых полях Кубани в разные месяцы
1978—1984 гг. (по: Рыльников, Карасева, 1985)
С активным сперматогенезом
Без сперматогенеза
ов
размеры
овамеры
Месни
само|Отобщего ое |ныхшу-|5очи
|
семениы. | ных пу-
дов
.
мм
зырьков, | сла, % ков, мм зырьков,
мм
мм
Март
50 100,0=0,0
24,0-0,3 21,6=0,5
_
_
—_
Май
38 400,0-0,0 22,2-=0,5 23,8+0,5
-
-
-
Август
94 400,0=0,0
22,4—0,5 23,4=0,5
—
—
—
Октябрь
16
87,5=9,2
22,7-=0,5 23,0=0,5 | 12,5=9,2 | 12,515 5,0-0,0
Цекабрь
27
44,8=7,0 21.6—0,5 15,8-=0,8 | 85,2=7,0 | 12,1=0,9 | 4,4-0,4
Февраль
91
56,0=5,2 21,0=0,3 416,5—=1,0 | 44,0-55,2 | 14,0--0,5 | 3,5=0,6
ников, Карасева, 1985). По-видимому, упомянутые отличия в сезонных
циклах сперматогенеза являются результатом различных климатическиу
условий.
Репродуктивная функция самов
,
Цикличноеть эструсов. Пасюки относятся к видам полиэстральным с мно-
жествепной беременностью (Магпе её а1., 1984). У крыс, обитающих в
природных биотопах, полизстральпость носит сезонный характер; в з8-
висимости от продолжительности холодного или малокормного периода
тода наблюдается соответствующей продолжительности анэструс.
У крыс— обитателей отапливаемых строепий человека, а также у крыс,
содержащихся в неволе, эструс наблюдается регулярно в течение всего
года (Реггу, 1945). Более поздние исследования позволили устаповить,
что регулярность эстральных циклов у ноловозрелых самок все-таки на-
рушается в ответ на сокращение светового дня, а у молодых самок про-
должительность светового дпя является одним из факторов, контролирую-
а
к
,
Экология 185
Таблица 14. Связь картины вагинального мазка, времени течки и овуляции
Число самок в
состоянии овуля-
И
Типы клеток в мазке
кул *#)
влагалища *
в вачале | в конце ри
р
а
[2
в
Только 395
5
0
0
0
1
1
2
ЗЭК-75%; ОК-25%
96
0
А
0
1
1
2
ЗЭК-50$; ОК-50%
13
0
3
01
12
39-25%; ОК-75%
_
-
_
_
1
1
2
Только ОК
э
19
5
122323
ОК-75%; РС-25$
0
27
0
4
2,3
2
3
ОК-50%; РС-50%
2
27
0
4
2,3
2
3
ОК—25%; РС-75%
4
50
1
3
23
2
3
Только РС
1
7
0
02,334
РС и лейкоциты
—
—
_
-
4
А
А
с—
* ЗЭК — зернистые эпителпиальные клетки, ОК — ороговевшие (чешуйчатые) клетки, Р
«рауетей$ сеПз» (по: Уоция её а]., 1941).
** Частота проявления картины вагинального мазка в зависимости от начала, конца течки
и наличия разорванных фолликул.
ыыы пы номер стадии по: Уоипе еф а1. (1941), 6 — по: Топа, Еуаиз (1922), в — по: Ковалевский
58).
щих время наступления половой зрелости. Однако, несмотря на это,
сезонные циклы не имеют большого значения в регуляции размножения
лабораторных крысе (В!уезф е[ а|., 1986).
Ноловая охота (течка) выражается у самок в покраенении и опуха-
нии наружных ноловых органов. В таком состояпии самки очепь подвиж-
ны и охотно допускают к себе самцов.
Половой цикл взрослой самки белой крысы периодически возобнов-
ляется примерно через каждые 5 суток, он состоит из четырех стадий,
которые легко различаются по характеру мазков из влагалища (табл. 14).
1. Стадия покоя (диэструс) продолжается 57 ч и характеризуется со-
держанием во влагалище слизи с лейкоцитами и незначительным коли-
чеством зернистых эпителиальных клеток.
`° 2. Переходная к течке стадия (проэструе) продолжается около 12 ч,
во влагалищных мазках имеется множество зернистых эпителиальных
клеток; лейкоцитов и слизи нет.
3. Стадия течки (эструс), длящаяся около 27 ч, характеризуется со-
держанием во влагалище чептуек.
А. Стадия восстаповлепия (метаэструс) продолжается около 6 ч.
В мазках отмечается исчезновение чешуек и появление зернистых кле-
ток и лейкоцитов.
184
Глава третья
Таблица 12. Размеры семенников и семенных пузырьков взроелых самцов
серой крысы, добытых в домах индивидуальной застройки и в отанливаемых
свинарниках Кубани в разные месяцы года
Месяц
всего отловлено
Длина семенников, мм оных пу
Январь
18
22,5+0,9
24,3=0,5
Февраль
10
22,8-=0,6
21,0+1,8
Май
90
21,5=0,А
23,0-0,4
Июль--август
66
21,4=0,4
20,6=0,5
Октябрь
35
22,0+0,3
25,6=0,8
* Все самды с активным сперматогенезом.
Таблица 13. Доля размножающихся самцов серых крыс, средние размеры их
семенников и семенных пузырьков на рисовых полях Кубани в разные месяцы
1978—1984 гг. (по: Рыльников, Карасева, 1985)
С активным сперматогенезом
Без сперматогенеза
ов
размеры
овамеры
Месни
само |Отобщего ое |ныхшу-|5очи
|
семениы. | ных пу-
дов
.
мм
зырьков, | сла, % ков, мм зырьков,
мм
мм
Март
50 100,0=0,0
24,0-0,3 21,6=0,5
_
_
—_
Май
38 400,0-0,0 22,2-=0,5 23,8+0,5
-
-
-
Август
94 400,0=0,0
22,4—0,5 23,4=0,5
—
—
—
Октябрь
16
87,5=9,2
22,7-=0,5 23,0=0,5 | 12,5=9,2 | 12,515 5,0-0,0
Цекабрь
27
44,8=7,0 21.6—0,5 15,8-=0,8 | 85,2=7,0 | 12,1=0,9 | 4,4-0,4
Февраль
91
56,0=5,2 21,0=0,3 416,5—=1,0 | 44,0-55,2 | 14,0--0,5 | 3,5=0,6
ников, Карасева, 1985). По-видимому, упомянутые отличия в сезонных
циклах сперматогенеза являются результатом различных климатическиу
условий.
Репродуктивная функция самов
,
Цикличноеть эструсов. Пасюки относятся к видам полиэстральным с мно-
жествепной беременностью (Магпе её а1., 1984). У крыс, обитающих в
природных биотопах, полизстральпость носит сезонный характер; в з8-
висимости от продолжительности холодного или малокормного периода
тода наблюдается соответствующей продолжительности анэструс.
У крыс— обитателей отапливаемых строепий человека, а также у крыс,
содержащихся в неволе, эструс наблюдается регулярно в течение всего
года (Реггу, 1945). Более поздние исследования позволили устаповить,
что регулярность эстральных циклов у ноловозрелых самок все-таки на-
рушается в ответ на сокращение светового дня, а у молодых самок про-
должительность светового дпя является одним из факторов, контролирую-
а
к
,
Экология 185
Таблица 14. Связь картины вагинального мазка, времени течки и овуляции
Число самок в
состоянии овуля-
И
Типы клеток в мазке
кул *#)
влагалища *
в вачале | в конце ри
р
а
[2
в
Только 395
5
0
0
0
1
1
2
ЗЭК-75%; ОК-25%
96
0
А
0
1
1
2
ЗЭК-50$; ОК-50%
13
0
3
01
12
39-25%; ОК-75%
_
-
_
_
1
1
2
Только ОК
э
19
5
122323
ОК-75%; РС-25$
0
27
0
4
2,3
2
3
ОК-50%; РС-50%
2
27
0
4
2,3
2
3
ОК—25%; РС-75%
4
50
1
3
23
2
3
Только РС
1
7
0
02,334
РС и лейкоциты
—
—
_
-
4
А
А
с—
* ЗЭК — зернистые эпителпиальные клетки, ОК — ороговевшие (чешуйчатые) клетки, Р
«рауетей$ сеПз» (по: Уоция её а]., 1941).
** Частота проявления картины вагинального мазка в зависимости от начала, конца течки
и наличия разорванных фолликул.
ыыы пы номер стадии по: Уоипе еф а1. (1941), 6 — по: Топа, Еуаиз (1922), в — по: Ковалевский
58).
щих время наступления половой зрелости. Однако, несмотря на это,
сезонные циклы не имеют большого значения в регуляции размножения
лабораторных крысе (В!уезф е[ а|., 1986).
Ноловая охота (течка) выражается у самок в покраенении и опуха-
нии наружных ноловых органов. В таком состояпии самки очепь подвиж-
ны и охотно допускают к себе самцов.
Половой цикл взрослой самки белой крысы периодически возобнов-
ляется примерно через каждые 5 суток, он состоит из четырех стадий,
которые легко различаются по характеру мазков из влагалища (табл. 14).
1. Стадия покоя (диэструс) продолжается 57 ч и характеризуется со-
держанием во влагалище слизи с лейкоцитами и незначительным коли-
чеством зернистых эпителиальных клеток.
`° 2. Переходная к течке стадия (проэструе) продолжается около 12 ч,
во влагалищных мазках имеется множество зернистых эпителиальных
клеток; лейкоцитов и слизи нет.
3. Стадия течки (эструс), длящаяся около 27 ч, характеризуется со-
держанием во влагалище чептуек.
А. Стадия восстаповлепия (метаэструс) продолжается около 6 ч.
В мазках отмечается исчезновение чешуек и появление зернистых кле-
ток и лейкоцитов.
186
Глава третья
Покрытие самцом самки возможно только в нервой половине эструса,
когда самка находится в возбуждениом состояпии. Во время других ста-
дий полового цикла она не допускает к себе самца. После акта покрытия
самки во влагалище образуется пробка, состоящая из секрета семенных
пузырьков. При изучении содержимого мазка из влагалища под микро-
сколом можно обнаружить присутствие сперматозоидов (Ковалевский,
1958, с. 98).
По мнению некоторых других авторов (бобома, цит. по НаЪег, 1945),
у белых крыс овуляция имеет место споктанно и независимо от копуля-
ции, действие которой может предшествовать или следовать за овуляци-
ей. Одновремепно с этим Суботта приводит очевидные свидетельства при-
вязки копуляции к овуляции, а именно: спаривания были более много-
численны через 29 дней после родов, т. е. совпадали со вторым периодом
овуляции.
Осеменение самок крыс чаще всего происходит в первые часы ночи
(МШег, 1911).
Оплодотворение осуществляется в фаллопиевых трубах, куда попада-
ет отпосительно пебольшое количество сперматозоидов. Их жизнь в гени-
тальных органах самок белых крыс составляет около 10 ч (НоЪет, 4915).
Беременность у диких крыс часто возникает лосле первой послеродо-
вой овуляции, что является причиной случаев совмещения беременности
и лактации. У лабораторных крыс эта овуляция лишь изредка кончалась
беременностью (Реггу, 1945).
Протекание и продолжительность беременноети. Продолжительность
беременности от копуляции до родов составляет в 90% случаев 24-—
22 дня (НиЪег, 1945). Но Миллеру (МШег, 1944), она длится 23,5—
25,5 дня, по Эмлену и Давису (Епеп, Оау1з, 1948) — 24 дня и разделена
на 2 стадии: 1) оплодотворение-имнлантация (5—6 дней, причем до
А-го дня оплодотворенные яйца находятся в фаллониевых трубах, 1—
2 дня в виде свободных бластоциетов в матке); 2) визуально различимая
беременность— 18 дней (НиЪег, 1945; КиВаш, 1916; цит. ло: ГезИе её
а1., 1952; Еп еп, Бау!в, 1948; ТезЦе её а1., 1952). В литературе есть ука-
вания и на более короткие сроки видимой беременности: 47 дней (Реггу.
1945), 16 дней (Еафюп, Эвитей, 1928) и 14 дней (Кузякин, 1952; Вишня-
ков, 1970), что, по мнению Лесли с соавторами (ТлезЦе её 21., 1952), свя-
зано с возможностями конкретных исследований по идентификации бере-
меплости после имплантации. Имплантировавшихся эмбрионов различают
макроскопически, свободные маточные бластоцисты обнаруживают с по-
мощью выдавливания из матки; идентификацию проводят на гистологи-
ческих срезах. Оплодотворенные яйца в фаллонпиевых трубах обнаружи-
вали в их серийных срезах (Роггу, 1945).
Желтые тела формируются в том самом месте, где высвобождается
яйцо, они хорозпо визуально различимы. Поэтому чиело зитот может
быть приблизительно определено путем подсчета желтых тел. Яичники
крые содержат обычно желтые тела разных возрастов. относящиеся к не-
скольким последовательным овуляциям. Несмотря на то что желтые тела
разных возрастов практически неотличимы друг от друга, желтые тела
Экология 187
текущей беременности характеризуются крупными размерами (рис. 53)
внлоть до родов (Реггу, 1945; Рыльников, Карасева, 1985).
Старые плацентарные пятна четко различимы как темные пятна на
матке около мезометриума в течение длительного времени после родов у
крыс — обитателей сельских населенных пунктов (Реггу, 1945).
У многих видов грызунов обнаружено явление совмещения лактации
и беременности, изучение которого оказалось интересным по двум при-
чинам. Во-первых, с точки зрения оценки энергетических возможностей
организма матери по совместимости одновременпого кормлепия новорож-
денпых и вынаптивания следующего потомства, во-вторых, с точки зре-
ния влияния лактации на продолжительность беременности, что необхо-
димо учитывать при оценке доли беременных самок в популяции.
Для целого ряда видов грызунов было показано, что вероятность оп-
лодотворения беременных самок мала (За@ет, 1984), по если они все же
оплодотворепы, то продолжительность беременности увеличивается за
счет задержки имплантации (Туникова, 1964). Сначала материалы Крех-
биль (Клевые, 1941), а затем детальные исследования Перри (Регту,
1945) на серых крысах показали, что увеличение нродолжительности бе-
ремепности оплодотворенной, лактирующей самки до 34 дней происходит
за счет замедления роста зародышей на стадиях до имплантации. Число
животных со свободными бластоцистами среди кормящих самок выше
ожндаемого (по сравнению с некормящими). Это связано с тем, что фак-
тически среднее для каждого бластоциста время пребывания в свободном
состоянии в полости матки больше (приблизительно на 4 дня} у кормя-
ших, чем у некормящих (1—2 дпя), из чего Перри заключает, что увели-
Рис. 53. Яичники НаНиз азятИ5 (по: Ташог еф а1., 1981)
Слева
— яичники неполовозрелых самок в возрасте {1 меслца; видны растущие п зачаточные
фолликулы. справа
— янчники половозрелых самок с тремя полностью сформированными
желтыми телами. Х 22
186
Глава третья
Покрытие самцом самки возможно только в нервой половине эструса,
когда самка находится в возбуждениом состояпии. Во время других ста-
дий полового цикла она не допускает к себе самца. После акта покрытия
самки во влагалище образуется пробка, состоящая из секрета семенных
пузырьков. При изучении содержимого мазка из влагалища под микро-
сколом можно обнаружить присутствие сперматозоидов (Ковалевский,
1958, с. 98).
По мнению некоторых других авторов (бобома, цит. по НаЪег, 1945),
у белых крыс овуляция имеет место споктанно и независимо от копуля-
ции, действие которой может предшествовать или следовать за овуляци-
ей. Одновремепно с этим Суботта приводит очевидные свидетельства при-
вязки копуляции к овуляции, а именно: спаривания были более много-
численны через 29 дней после родов, т. е. совпадали со вторым периодом
овуляции.
Осеменение самок крыс чаще всего происходит в первые часы ночи
(МШег, 1911).
Оплодотворение осуществляется в фаллопиевых трубах, куда попада-
ет отпосительно пебольшое количество сперматозоидов. Их жизнь в гени-
тальных органах самок белых крыс составляет около 10 ч (НоЪет, 4915).
Беременность у диких крыс часто возникает лосле первой послеродо-
вой овуляции, что является причиной случаев совмещения беременности
и лактации. У лабораторных крыс эта овуляция лишь изредка кончалась
беременностью (Реггу, 1945).
Протекание и продолжительность беременноети. Продолжительность
беременности от копуляции до родов составляет в 90% случаев 24-—
22 дня (НиЪег, 1945). Но Миллеру (МШег, 1944), она длится 23,5—
25,5 дня, по Эмлену и Давису (Епеп, Оау1з, 1948) — 24 дня и разделена
на 2 стадии: 1) оплодотворение-имнлантация (5—6 дней, причем до
А-го дня оплодотворенные яйца находятся в фаллониевых трубах, 1—
2 дня в виде свободных бластоциетов в матке); 2) визуально различимая
беременность— 18 дней (НиЪег, 1945; КиВаш, 1916; цит. ло: ГезИе её
а1., 1952; Еп еп, Бау!в, 1948; ТезЦе её а1., 1952). В литературе есть ука-
вания и на более короткие сроки видимой беременности: 47 дней (Реггу.
1945), 16 дней (Еафюп, Эвитей, 1928) и 14 дней (Кузякин, 1952; Вишня-
ков, 1970), что, по мнению Лесли с соавторами (ТлезЦе её 21., 1952), свя-
зано с возможностями конкретных исследований по идентификации бере-
меплости после имплантации. Имплантировавшихся эмбрионов различают
макроскопически, свободные маточные бластоцисты обнаруживают с по-
мощью выдавливания из матки; идентификацию проводят на гистологи-
ческих срезах. Оплодотворенные яйца в фаллонпиевых трубах обнаружи-
вали в их серийных срезах (Роггу, 1945).
Желтые тела формируются в том самом месте, где высвобождается
яйцо, они хорозпо визуально различимы. Поэтому чиело зитот может
быть приблизительно определено путем подсчета желтых тел. Яичники
крые содержат обычно желтые тела разных возрастов. относящиеся к не-
скольким последовательным овуляциям. Несмотря на то что желтые тела
разных возрастов практически неотличимы друг от друга, желтые тела
Экология 187
текущей беременности характеризуются крупными размерами (рис. 53)
внлоть до родов (Реггу, 1945; Рыльников, Карасева, 1985).
Старые плацентарные пятна четко различимы как темные пятна на
матке около мезометриума в течение длительного времени после родов у
крыс — обитателей сельских населенных пунктов (Реггу, 1945).
У многих видов грызунов обнаружено явление совмещения лактации
и беременности, изучение которого оказалось интересным по двум при-
чинам. Во-первых, с точки зрения оценки энергетических возможностей
организма матери по совместимости одновременпого кормлепия новорож-
денпых и вынаптивания следующего потомства, во-вторых, с точки зре-
ния влияния лактации на продолжительность беременности, что необхо-
димо учитывать при оценке доли беременных самок в популяции.
Для целого ряда видов грызунов было показано, что вероятность оп-
лодотворения беременных самок мала (За@ет, 1984), по если они все же
оплодотворепы, то продолжительность беременности увеличивается за
счет задержки имплантации (Туникова, 1964). Сначала материалы Крех-
биль (Клевые, 1941), а затем детальные исследования Перри (Регту,
1945) на серых крысах показали, что увеличение нродолжительности бе-
ремепности оплодотворенной, лактирующей самки до 34 дней происходит
за счет замедления роста зародышей на стадиях до имплантации. Число
животных со свободными бластоцистами среди кормящих самок выше
ожндаемого (по сравнению с некормящими). Это связано с тем, что фак-
тически среднее для каждого бластоциста время пребывания в свободном
состоянии в полости матки больше (приблизительно на 4 дня} у кормя-
ших, чем у некормящих (1—2 дпя), из чего Перри заключает, что увели-
Рис. 53. Яичники НаНиз азятИ5 (по: Ташог еф а1., 1981)
Слева
— яичники неполовозрелых самок в возрасте {1 меслца; видны растущие п зачаточные
фолликулы. справа
— янчники половозрелых самок с тремя полностью сформированными
желтыми телами. Х 22
188
Глава третья
чение продолжительности беременности может быть связано с параллель-
но протекающей лактацией, которая продлевает первую стадию беремен-
ности до 10 дней. Развитие зародышей до имплантации и во время на-
хождения в фаллопиевых трубах задерживалось в болышей мере, чем
свободных бластоцистов в матке. Лактация наблюдалась у каждой треть-
ей самки со свободпыми бластоцистами в матке и у каждой пятой—
с яйцеклетками в фаллопиевых трубах. Сроки видимой беременности не
увеличиваются. Основанием для такого заключения послужило сравнение
фактической и ожидаемой частоты встречаемости случаев совмещения
лактации с видимой беременностью.
Рост и развитие врые
Масса тела, развитие и возраст эмбрионов. Специальных работ, посзя-
щенных этому вопросу, оказалось немного, однако среди них есть одна
достаточно полная сводка Геннеберга (Непперего, 1937 — цит. по: Тупи-
кова, Медведева, 1956).
Масса тела и развитие новорожденных. Окот у крысе в большинстве
случаев происходит ночью, реже утром или днем. Нормально роды про-
должаются 20—35 мин. В первые дни после рождения крысы кормят де-
тенышей через каждые 40—55 мин (Ковалевский, 1958).
Масса эмбрионов самцов в выводках диких пасюков, состоящих из
5—13 зародышей, перед рождением составляла 3,9—5,6 г при длине тела
87—42 мм, а самок
— 3,7—5,9 г при длине тела 36—42 мм. Масса тела
самцов в средпем при рождепии превышала таковую самок па 5,8=1,0%.
Чем меныше число эмбрионов, тем больше масса их тела при рождении.
У новорожденных крысят эта зависимость отчетливо проязляется. Напри-
мер, в выводке из 4 детенышей масса тела одного крысенка в среднем
равна 8,1 г, а в выводке из 14 детенышей того же возраста — 6,3 г. При
асимметричном расположении эмбрионов в матке средняя масса их в
роге с меныним числом зародьцией, как правило, больше на 2—17%;
при этом чем сильнее асимметрия, тем существеннее разница в массе
эмбрионов
— она зависит от их расположения в матке. Наиболее круп-
ные эмбрионы находятся, как правило, в середине, рядом с влагалищем
(Козлов, 19866).
,
Крысята насюков покрываются шерстью через 9—10 дней после рож-
дения при массе тела 17—19 г (Ковалевский, 1958, с. 102). Глаза откры-
вались на 16—17-й день по Миллеру (МШег, 1914 — цит, по: Попа1450п,
1924), между 14-м и 17-м днем ло Ковалевскому (4958), на 15-й день
при массе тела самцов 26,3 г, самок 25,0 г_по А. Н. Козлову (1984).
Половое созревание
Половозрелыми можно считать самок, потенциально способных принести
потомство. Гдинственным надежным критерием половозрелости самок
крыс можно считать наличие у них зрелых фолликулов и острального
цикла (овуляции). Однако лишь немногие авторы (Реггу, 1945; КагВапо-
Экология
189
Аза, ЗошучсЕ, 1979) пользовались этим критерием. Зрелые фолликулы
хорошо различимы только у нерожазних самок; у рожавших самок ока-
зывается трудным визуально отличить фолликулы от желтых тел, отно-
сящихся к разным пометам. Приготовление же гистологических срезов
ничников оказывается очень трудоемким. Определение стадий эстрально-
то цикла по мазкам практиковалось в осповном па лабораторных живот-
вых. По-видимому, именно поэтому больпинство исследователей ислоль-
зовали косвенный показатель половозрелости, такой, как наличие откры-
того влагалища (Оау!з, 1953Ъ).
Поскольку созревание крыс происходит в определенном возрастном
интервале, это послужило основанием для того, чтобы использовать при-
надлежность к определенной возрастной трунне в качестве среднестати-
стического показателя половозрелости данной особи. Однако явно идя по
пути наименытего сопротивленйй, большинство авторов опирались на воз-
растио-весовые группы, а срок жизни рассчитывали по липии регрессии
между массой тела и возрастом лабораторных крыс.
В большинстве случаев половозрелость наступает при массе тела
100—200 г, что соответствует возрасту от 45 до 147 дней, в среднем 77—
80 дням (по: Гоп, Еуапё 1922
— цит. по: ГезНе её а1., 4952; Ретту,
1945; ГезНе её а|., 1952; Еаграпб-АзаЯ, Зои мсек, 1979), 90 дням (по:
Еаюп, Эбитей, 1928 — цит. по: Кше, Попа19з0п, 1929). В редких случаях
половая зрелость паступаст при массе тела 95 г (Регту, 1945) и даже
72 г (Хамаганов, 1985) и 51 г (Випняков, 1970),
Исходя из этого половозрелыми считали самок с массой тела более
145 г (50% среди них были половозрелыми
— ГезИе еф а|., 1952);
А. А. Пасешник (1936) в г. Москве и В. С. Сурков (1986) на о-ве Са-
халин процент беременных самок рассчитывали среди самок с массой
тела более 100 г, П. П. Гамбарян, Н. М. Дукельская (1955) — среди крыс
старше 6 месяцев (или с массой тела более 170—190 г), В. Г. Лялин
(19746) в Западной Сибири и А, Н. Козлов (1984) в Северном Казах-
стане — среди крыс массой более 200 г.
Такой разнобой в показателях половозрелости вносит большие трудно-
сти при сравпепии материалов перечисленпых авторов. Дэвис (Раз,
1953) также обратил внимание на то, что доля беременных самок варьн-
рует в зависимости от того, какую категорию самок каждый из авторов
принимал за «взрослых».
Методически такой подход позволяет судить о половом созревании са-
мок по их фактическому вступлепию в размножение, что нс одно и то
же. Сроки пастуиления половой зрелости и сроки вступления в размно-
*жепие могут отличаться на несколько месяцев. В разреженных популяци-
ях половозрелость крыс настунает раньше, в переуплотненных
— позже
(Козлов, 1981; Рыльников, Карасева, 1985), а весной крысы становятся
половозрелыми при меньшем возрасте и массе тела, чем осенью (Козлов,
1981). В популяции, состоящей из крыс с болышими среднестатистиче-
скими размерами тела, самки созревают при более крупных размерах,
чем в популяции, где крысы мельче (Па\!з, 1953). Недавно проведенные
исследования нодтверждают эту закопомерпость. Пасюки в райоце БАМа,
188
Глава третья
чение продолжительности беременности может быть связано с параллель-
но протекающей лактацией, которая продлевает первую стадию беремен-
ности до 10 дней. Развитие зародышей до имплантации и во время на-
хождения в фаллопиевых трубах задерживалось в болышей мере, чем
свободных бластоцистов в матке. Лактация наблюдалась у каждой треть-
ей самки со свободпыми бластоцистами в матке и у каждой пятой—
с яйцеклетками в фаллопиевых трубах. Сроки видимой беременности не
увеличиваются. Основанием для такого заключения послужило сравнение
фактической и ожидаемой частоты встречаемости случаев совмещения
лактации с видимой беременностью.
Рост и развитие врые
Масса тела, развитие и возраст эмбрионов. Специальных работ, посзя-
щенных этому вопросу, оказалось немного, однако среди них есть одна
достаточно полная сводка Геннеберга (Непперего, 1937 — цит. по: Тупи-
кова, Медведева, 1956).
Масса тела и развитие новорожденных. Окот у крысе в большинстве
случаев происходит ночью, реже утром или днем. Нормально роды про-
должаются 20—35 мин. В первые дни после рождения крысы кормят де-
тенышей через каждые 40—55 мин (Ковалевский, 1958).
Масса эмбрионов самцов в выводках диких пасюков, состоящих из
5—13 зародышей, перед рождением составляла 3,9—5,6 г при длине тела
87—42 мм, а самок
— 3,7—5,9 г при длине тела 36—42 мм. Масса тела
самцов в средпем при рождепии превышала таковую самок па 5,8=1,0%.
Чем меныше число эмбрионов, тем больше масса их тела при рождении.
У новорожденных крысят эта зависимость отчетливо проязляется. Напри-
мер, в выводке из 4 детенышей масса тела одного крысенка в среднем
равна 8,1 г, а в выводке из 14 детенышей того же возраста — 6,3 г. При
асимметричном расположении эмбрионов в матке средняя масса их в
роге с меныним числом зародьцией, как правило, больше на 2—17%;
при этом чем сильнее асимметрия, тем существеннее разница в массе
эмбрионов
— она зависит от их расположения в матке. Наиболее круп-
ные эмбрионы находятся, как правило, в середине, рядом с влагалищем
(Козлов, 19866).
,
Крысята насюков покрываются шерстью через 9—10 дней после рож-
дения при массе тела 17—19 г (Ковалевский, 1958, с. 102). Глаза откры-
вались на 16—17-й день по Миллеру (МШег, 1914 — цит, по: Попа1450п,
1924), между 14-м и 17-м днем ло Ковалевскому (4958), на 15-й день
при массе тела самцов 26,3 г, самок 25,0 г_по А. Н. Козлову (1984).
Половое созревание
Половозрелыми можно считать самок, потенциально способных принести
потомство. Гдинственным надежным критерием половозрелости самок
крыс можно считать наличие у них зрелых фолликулов и острального
цикла (овуляции). Однако лишь немногие авторы (Реггу, 1945; КагВапо-
Экология
189
Аза, ЗошучсЕ, 1979) пользовались этим критерием. Зрелые фолликулы
хорошо различимы только у нерожазних самок; у рожавших самок ока-
зывается трудным визуально отличить фолликулы от желтых тел, отно-
сящихся к разным пометам. Приготовление же гистологических срезов
ничников оказывается очень трудоемким. Определение стадий эстрально-
то цикла по мазкам практиковалось в осповном па лабораторных живот-
вых. По-видимому, именно поэтому больпинство исследователей ислоль-
зовали косвенный показатель половозрелости, такой, как наличие откры-
того влагалища (Оау!з, 1953Ъ).
Поскольку созревание крыс происходит в определенном возрастном
интервале, это послужило основанием для того, чтобы использовать при-
надлежность к определенной возрастной трунне в качестве среднестати-
стического показателя половозрелости данной особи. Однако явно идя по
пути наименытего сопротивленйй, большинство авторов опирались на воз-
растио-весовые группы, а срок жизни рассчитывали по липии регрессии
между массой тела и возрастом лабораторных крыс.
В большинстве случаев половозрелость наступает при массе тела
100—200 г, что соответствует возрасту от 45 до 147 дней, в среднем 77—
80 дням (по: Гоп, Еуапё 1922
— цит. по: ГезНе её а1., 4952; Ретту,
1945; ГезНе её а|., 1952; Еаграпб-АзаЯ, Зои мсек, 1979), 90 дням (по:
Еаюп, Эбитей, 1928 — цит. по: Кше, Попа19з0п, 1929). В редких случаях
половая зрелость паступаст при массе тела 95 г (Регту, 1945) и даже
72 г (Хамаганов, 1985) и 51 г (Випняков, 1970),
Исходя из этого половозрелыми считали самок с массой тела более
145 г (50% среди них были половозрелыми
— ГезИе еф а|., 1952);
А. А. Пасешник (1936) в г. Москве и В. С. Сурков (1986) на о-ве Са-
халин процент беременных самок рассчитывали среди самок с массой
тела более 100 г, П. П. Гамбарян, Н. М. Дукельская (1955) — среди крыс
старше 6 месяцев (или с массой тела более 170—190 г), В. Г. Лялин
(19746) в Западной Сибири и А, Н. Козлов (1984) в Северном Казах-
стане — среди крыс массой более 200 г.
Такой разнобой в показателях половозрелости вносит большие трудно-
сти при сравпепии материалов перечисленпых авторов. Дэвис (Раз,
1953) также обратил внимание на то, что доля беременных самок варьн-
рует в зависимости от того, какую категорию самок каждый из авторов
принимал за «взрослых».
Методически такой подход позволяет судить о половом созревании са-
мок по их фактическому вступлепию в размножение, что нс одно и то
же. Сроки пастуиления половой зрелости и сроки вступления в размно-
*жепие могут отличаться на несколько месяцев. В разреженных популяци-
ях половозрелость крыс настунает раньше, в переуплотненных
— позже
(Козлов, 1981; Рыльников, Карасева, 1985), а весной крысы становятся
половозрелыми при меньшем возрасте и массе тела, чем осенью (Козлов,
1981). В популяции, состоящей из крыс с болышими среднестатистиче-
скими размерами тела, самки созревают при более крупных размерах,
чем в популяции, где крысы мельче (Па\!з, 1953). Недавно проведенные
исследования нодтверждают эту закопомерпость. Пасюки в райоце БАМа,
190
Глава третья
Таблица 15. Размеры серых крыс в воетоянии половой зрелости (по: Оам!з, 195$)
Отношение
Размеры
средних
крыс, при
размеров
которых
тела (при
50% полово- | которых
зрелые
50% крыс
половозре-
зые) к
среднеста-
ритерий.
Место отлова
я рае
Источник
лости
рам всей
мас- | цли- популяци-
са на онной вы-
тела, | тела, | борки
г
мм
по
по
массе ди-
Тела | тела
Балтимор, ферма 135 - 0,68 _ рау!в, 1948
Балтимор, ферма 457 | 165 | 0,954 | 0,96 | разв, 19а
Балтимор, от-
119 474 | 0,45 | 0,84 Епцеп, эпруЪ1.
Семенники | крытая стация
.
в мотаонке | То же
109 155 | 0,34 | 0,76 ау, 1951а
Палтимор, склады | 105 | 141 | 0,45 | 0,81 Рав. 1951а
Англия, город
190 _
-
- ТевНе её а, 1952
ФРГ, город
93 153 | 0,69 | 0,92 веског, зараЪ1.
Балтимор, ферма 88 _ 0,50 - 2, 148
То же
72 134 | 0,42 | 0,78 Рау!з, 19а
Открытое
5
Ешеп, ипрар!.
Балтимор, от-
405 154 | 0,39 | 0,75
‚
влагалище крытая стация
То же
123|150|0,39|0,74|Рау’в,19а
Палтимор, склады | 102 | 143 | 0,53 | 0,85 | Рау, 19а
| ФРГ, город
403 | 159 10,84 | 0,97 | Нескег, иарцы!.
Англия, город
145 —
—
- ТезПе еф а1., 1952
Овуляция То же
146 _
_
- ТезИе е% а1., 1952
р
113 _
-_
—
ТезИе сф а1., 1952
Палтимор, склады | 153 4АА | 079 | 0,86 | Рау, 19а
р
Балтимор, от-
242 | 224 | 0,93 | 0,95 | Еимеш, Ба\з, 1948
крытая стация
|
р
Балтимор,
348|246|1,64|128|ра\ь,Нац,1953
Плацентар- | Склады
И
ные пятна | Балтимор, от-
305|212|4,07|1,07|Раз,19в
крытая стация
Сан-Антонио,
178
— | 4,21 _ хииеп, Рау, 1948
город
Экология
191
Рис. 54. Различие в массе тела крысе 3
—
разного возраста 1-го (1) и 25-го (2) 300 |-
ея”
поколений
„-=7
2
"3250 —
„77“
з
и
5
|
8 200
и
З150|-,/
*100и
20 Г,
1
1
1
1
|
501920394%550
Возраст , ни
которых принято относить к подвиду ЦаМав погуе21ем$ сагасо РаП.,
мельче размерами тела, чем европейские серые крысы В. п. погуер1сия
ВегкК. При этом они начинают размножаться в среднем при размерах
тела, соответствующих 1,5—2-месячным насюкам номинативного подвида
(Москва) (Вигоров, 1985; Шутова и др., 1985).
Дэвие (Ра\з, 1953) предлагал количественно оценивать отношение
между размерами, при которых 50% особей достигают половозрелости,
и средними размерами всей полуляции крыс. Чем больше это отношение,
тем крысы относительно круннее при достижении половозрелости
(табл. 15).
.
В лабораторных условиях благодаря извостному абсолютному возра-
сту крыс все рассмотрепные методические трудности при определепии
времени вступления в размножение остаются в стороне.
Самки серых крыс первого поколения от скрещивания диких родите-
лей в лабораторных условиях размножаются, как правило, в возрасте не
моложе 4 мес. В порядке появлепия последующих генераций наблюда-
лось заметное увеличепие репродуктивного периода (с 227 до 369 дней).
В последующих генерациях наблюдались значительные вариации време-
ни начала размножения самок.
Средций возраст самок, в котором они начинают размножаться, носто-
янно уменынается в последовательных поколениях и к 25-му поколению
становится равпым 119 дням, что на 147 дней мепьше возраста начала
размножения самок первых выводков, которые размножались при сред-
нем возрасте 266 дней. По мнению Кинг (Кр, 1939), песмотря на то
что половые органы самок серых крыс в возрасте до 3 мес. смособны
функционировать, они еще псдостаточно развились и еще не в полной
мере способны питать эмбрионы, чем в болес половозрелом состоянии.
Некоторые дикие формы крыс никогда не размножаются после пере-
несения их из естественных условий в неволю; другие приносят помет,
однако отказываются его кормить. Помет дикой самки приходится пере-
носить к самке-кормилице, обычно альбиноске, поскольку самка-мать
чаще всего ие обращает внимания на своих детенышей, что приводит их
к гибели. Стерильность у диких, недавно отловленных крыс, так же как
иу индивидов ранпих генераций в неволе, вызвана нарушением функции
190
Глава третья
Таблица 15. Размеры серых крыс в воетоянии половой зрелости (по: Оам!з, 195$)
Отношение
Размеры
средних
крыс, при
размеров
которых
тела (при
50% полово- | которых
зрелые
50% крыс
половозре-
зые) к
среднеста-
ритерий.
Место отлова
я рае
Источник
лости
рам всей
мас- | цли- популяци-
са на онной вы-
тела, | тела, | борки
г
мм
по
по
массе ди-
Тела | тела
Балтимор, ферма 135 - 0,68 _ рау!в, 1948
Балтимор, ферма 457 | 165 | 0,954 | 0,96 | разв, 19а
Балтимор, от-
119 474 | 0,45 | 0,84 Епцеп, эпруЪ1.
Семенники | крытая стация
.
в мотаонке | То же
109 155 | 0,34 | 0,76 ау, 1951а
Палтимор, склады | 105 | 141 | 0,45 | 0,81 Рав. 1951а
Англия, город
190 _
-
- ТевНе её а, 1952
ФРГ, город
93 153 | 0,69 | 0,92 веског, зараЪ1.
Балтимор, ферма 88 _ 0,50 - 2, 148
То же
72 134 | 0,42 | 0,78 Рау!з, 19а
Открытое
5
Ешеп, ипрар!.
Балтимор, от-
405 154 | 0,39 | 0,75
‚
влагалище крытая стация
То же
123|150|0,39|0,74|Рау’в,19а
Палтимор, склады | 102 | 143 | 0,53 | 0,85 | Рау, 19а
| ФРГ, город
403 | 159 10,84 | 0,97 | Нескег, иарцы!.
Англия, город
145 —
—
- ТезПе еф а1., 1952
Овуляция То же
146 _
_
- ТезИе е% а1., 1952
р
113 _
-_
—
ТезИе сф а1., 1952
Палтимор, склады | 153 4АА | 079 | 0,86 | Рау, 19а
р
Балтимор, от-
242 | 224 | 0,93 | 0,95 | Еимеш, Ба\з, 1948
крытая стация
|
р
Балтимор,
348|246|1,64|128|ра\ь,Нац,1953
Плацентар- | Склады
И
ные пятна | Балтимор, от-
305|212|4,07|1,07|Раз,19в
крытая стация
Сан-Антонио,
178
— | 4,21 _ хииеп, Рау, 1948
город
Экология
191
Рис. 54. Различие в массе тела крысе 3
—
разного возраста 1-го (1) и 25-го (2) 300 |-
ея”
поколений
„-=7
2
"3250 —
„77“
з
и
5
|
8 200
и
З150|-,/
*100и
20 Г,
1
1
1
1
|
501920394%550
Возраст , ни
которых принято относить к подвиду ЦаМав погуе21ем$ сагасо РаП.,
мельче размерами тела, чем европейские серые крысы В. п. погуер1сия
ВегкК. При этом они начинают размножаться в среднем при размерах
тела, соответствующих 1,5—2-месячным насюкам номинативного подвида
(Москва) (Вигоров, 1985; Шутова и др., 1985).
Дэвие (Ра\з, 1953) предлагал количественно оценивать отношение
между размерами, при которых 50% особей достигают половозрелости,
и средними размерами всей полуляции крыс. Чем больше это отношение,
тем крысы относительно круннее при достижении половозрелости
(табл. 15).
.
В лабораторных условиях благодаря извостному абсолютному возра-
сту крыс все рассмотрепные методические трудности при определепии
времени вступления в размножение остаются в стороне.
Самки серых крыс первого поколения от скрещивания диких родите-
лей в лабораторных условиях размножаются, как правило, в возрасте не
моложе 4 мес. В порядке появлепия последующих генераций наблюда-
лось заметное увеличепие репродуктивного периода (с 227 до 369 дней).
В последующих генерациях наблюдались значительные вариации време-
ни начала размножения самок.
Средций возраст самок, в котором они начинают размножаться, носто-
янно уменынается в последовательных поколениях и к 25-му поколению
становится равпым 119 дням, что на 147 дней мепьше возраста начала
размножения самок первых выводков, которые размножались при сред-
нем возрасте 266 дней. По мнению Кинг (Кр, 1939), песмотря на то
что половые органы самок серых крыс в возрасте до 3 мес. смособны
функционировать, они еще псдостаточно развились и еще не в полной
мере способны питать эмбрионы, чем в болес половозрелом состоянии.
Некоторые дикие формы крыс никогда не размножаются после пере-
несения их из естественных условий в неволю; другие приносят помет,
однако отказываются его кормить. Помет дикой самки приходится пере-
носить к самке-кормилице, обычно альбиноске, поскольку самка-мать
чаще всего ие обращает внимания на своих детенышей, что приводит их
к гибели. Стерильность у диких, недавно отловленных крыс, так же как
иу индивидов ранпих генераций в неволе, вызвана нарушением функции
192
Глава третья
нервной системы под действием страха, испытываемого животными от
нахождения внутри ограниченного пространства. Страх вызывает секре-
цию желез, нодавляющих активность репродуктивных органов. После от-
носительно короткого срока неволи крысы начинают нормально размно-
жаться. В течение первых восьми генераций стерильность среди самок
уменьшается от 37,29 до 5,88%. В последних генерациях все самки пло-
довиты.
Более раннее размножение самок поздних гепераций может быть
также связано с их более быстрым ростом (рис. 54). Можно допустить
также, что некоторые индивиды данной липии несут тены, которые имеют
отношение к более раннему половому созреванию. В результате инбри-
диига такие гены мотут со временем аккумулироваться (Кв, 1939).
Тогда закономерным оказывается факт, что крысы-альбипосы становятся
половозрелыми при средней массе тела 100 г, т. е. в возрасте 50 дней
(Оопа140п, 1924), хотя есть данные, что в лабораторных условиях неко-
торые самки могут быть покрыты в возрасте 30—35 дней (Сахаров, 1937).
Анализ плодовитости самок
Плодовитость и рождаемость. Показатели плодовитости. Прежде чем пере-
ходить к определению плодовитости и рождаемости, пеобходимо наном-
нить, какое место нроцессы размножения занимают в популяционной ди-
намике.
Измепение плотности 2,;({Г) происходит в результате взаимодействия
четырех процессов: скорости размножения (В;), скорости гибели (0;
иммиграции (1;) и эмиграции (Ё;), которые имеют размерность особъ/
нлощадь/время. Конечно, разностное уравнение изменения плотности
(р за единицу времени: А; =(&(Н-Т)-ж()/в(р =В-ОТ-Е,.
Как и в монографии В. Д. Федорова и Т. Г. Гильманова (1980), под «рож-
даемостью» и «смертностью» мы подразумеваем не В; и Д,, а соответет-
венно их удельные скорости 6; и 4, которые мы будем называть «удель-
ной рождаемостью» и «удельной смертностью». 6; и 4;, согласно упомя-
нутым выше авторам, имеют размерность 1/время и от площади не
зависят.
Абсолютные скорости размножения и гибели можно представить в
виде произведения плотности (2;) на соответствующие удельные скорости
(6,@;):В, Буй
-у
Представляется необходимым выделить в этих функциях пе завися-
щие от плотности х; аддитивные компоненты, т. е. представить их в виде
сумм вида 65,-=6»-Н6» и а=а»Н4и. Осуществив такос разбиение, можно
выяснить, насколько динамика данной популяции зависит от исключи-
тельно внешних по отношению к ней факторов.
`
Удельная скорость изменения численности популяции (г;) равпа
г.=0;-4; (при условии, что удельные значения иммиграции и эмиграции
равны нулю), тогда значение изменения плотности Алу(К =т().
Удельную скорость изменения популяции необходимо отличать от ко-
нечной скорости ). которая выражается отношением А;=х;(#1)/2(В)
и связана с г; соотношением г; ^..
Экология
193
Таблица 16. Показатели плодовитости еамок и удельная рождаемость в популяции
серых крые рисовых полей Кубани за 12 мес (1978—1980 гг.)
Число самок
Число эмбрионов
+
в расчете
ав
ь
Е
еРИЕ|=ыВ
Е
Месяцы отлова, 1 Е 3
8
=
аЕВВЕ
РЗ
вы5р
|
о
$
юао
5;
мк|2ЕЕ
ЕЕ
ЕЕоЕЕ
ы&
о бобр |2 ЕЕК
ЗЕ|Бо|Чо
ЕЕ
Е ЕЕ-Е=|98&ВЕ
2|в |345|58 |8688|«ВЕ.
ХЕ
442 91
0
0,00
_
142} -—
-
-
п
4154 82
2
0,02
15 169|7,5|0,18|0,089
ш
55 Ри
3
0,08
26 81 87 | 0,70 0,324
У|
803247
0,53
199 219 | 44,7 | 6,22 0,743
УТ-УП
385 196 70
0,41
751|1136|10,9|3,84|0,664
1х-х
51 45| 4 0,27
АЗ 78| 86|2,87|0,550
Сумма
992 453 96
-
1034 | 1718 —
_
—
Невзвешенное
-
_
- Ре-=0,1871
_
9,5 | 2,3 0,524
среднее
Примечание.
продолжительность видимой беременности (дни)
18
.б
средняя продолжительность сезони размножения (ини) = 270 — 0061; 0,067 =
0,187
= 00 = 2,8 (число пометов в расчете на 1 беременную самку за 270 дней);
Е:
Мо-0,07
— 34,5 (число эмбрионов в расчете на 1 самку из числа отловленных за 270 дней).
Число поворожденных рассчитано по Числу эмбрионов (за вычетом постимплантационных
потерь), которые появляются на свет в течение срока, в среднем равного
дня
периода истинной беременности).
ты
р
ы
м -бтоловине
Величипа принлода в популяции крыс складывается из трех показа-
телей плодовитости: 1) средней доли беременных самок Ру (пределы
варьирования от 7 до 70%), 2) среднего числа эмбрионов в помете (8,0—
9,0 в естественных условиях и 6,1 в неволе, пределы варьирования 1—7),
3) числа пометов в году, которое, в свою очередь, определяетел средней
продолжительностью сезона размножения (от 200 до 365 дней в году).
Первые два показателя могут быть заменены одним показателем — сред-
ним числом поворожденных крысят (или эмбрионов за вычетом постим-
плантационных потерь) в расчете на одну самку из числа активных чле-
нов популяции или самку данной возрастной группы за определенный
промежуток времени (ЛУ. Этот же показатель, отнесенный к общему чи-
слу животных в популяции (включая новорожденных), позволяет оце-
нить удельную рождаемость (5,). Рассчитывая число новорожденных на
число животных данной возрастной групны, можно оценить удельную
рождаемость в соответствующей возрастной группе (табл. 16, 17).
7 Серая крыса
192
Глава третья
нервной системы под действием страха, испытываемого животными от
нахождения внутри ограниченного пространства. Страх вызывает секре-
цию желез, нодавляющих активность репродуктивных органов. После от-
носительно короткого срока неволи крысы начинают нормально размно-
жаться. В течение первых восьми генераций стерильность среди самок
уменьшается от 37,29 до 5,88%. В последних генерациях все самки пло-
довиты.
Более раннее размножение самок поздних гепераций может быть
также связано с их более быстрым ростом (рис. 54). Можно допустить
также, что некоторые индивиды данной липии несут тены, которые имеют
отношение к более раннему половому созреванию. В результате инбри-
диига такие гены мотут со временем аккумулироваться (Кв, 1939).
Тогда закономерным оказывается факт, что крысы-альбипосы становятся
половозрелыми при средней массе тела 100 г, т. е. в возрасте 50 дней
(Оопа140п, 1924), хотя есть данные, что в лабораторных условиях неко-
торые самки могут быть покрыты в возрасте 30—35 дней (Сахаров, 1937).
Анализ плодовитости самок
Плодовитость и рождаемость. Показатели плодовитости. Прежде чем пере-
ходить к определению плодовитости и рождаемости, пеобходимо наном-
нить, какое место нроцессы размножения занимают в популяционной ди-
намике.
Измепение плотности 2,;({Г) происходит в результате взаимодействия
четырех процессов: скорости размножения (В;), скорости гибели (0;
иммиграции (1;) и эмиграции (Ё;), которые имеют размерность особъ/
нлощадь/время. Конечно, разностное уравнение изменения плотности
(р за единицу времени: А; =(&(Н-Т)-ж()/в(р =В-ОТ-Е,.
Как и в монографии В. Д. Федорова и Т. Г. Гильманова (1980), под «рож-
даемостью» и «смертностью» мы подразумеваем не В; и Д,, а соответет-
венно их удельные скорости 6; и 4, которые мы будем называть «удель-
ной рождаемостью» и «удельной смертностью». 6; и 4;, согласно упомя-
нутым выше авторам, имеют размерность 1/время и от площади не
зависят.
Абсолютные скорости размножения и гибели можно представить в
виде произведения плотности (2;) на соответствующие удельные скорости
(6,@;):В, Буй
-у
Представляется необходимым выделить в этих функциях пе завися-
щие от плотности х; аддитивные компоненты, т. е. представить их в виде
сумм вида 65,-=6»-Н6» и а=а»Н4и. Осуществив такос разбиение, можно
выяснить, насколько динамика данной популяции зависит от исключи-
тельно внешних по отношению к ней факторов.
`
Удельная скорость изменения численности популяции (г;) равпа
г.=0;-4; (при условии, что удельные значения иммиграции и эмиграции
равны нулю), тогда значение изменения плотности Алу(К =т().
Удельную скорость изменения популяции необходимо отличать от ко-
нечной скорости ). которая выражается отношением А;=х;(#1)/2(В)
и связана с г; соотношением г; ^..
Экология
193
Таблица 16. Показатели плодовитости еамок и удельная рождаемость в популяции
серых крые рисовых полей Кубани за 12 мес (1978—1980 гг.)
Число самок
Число эмбрионов
+
в расчете
ав
ь
Е
еРИЕ|=ыВ
Е
Месяцы отлова, 1 Е 3
8
=
аЕВВЕ
РЗ
вы5р
|
о
$
юао
5;
мк|2ЕЕ
ЕЕ
ЕЕоЕЕ
ы&
о бобр |2 ЕЕК
ЗЕ|Бо|Чо
ЕЕ
Е ЕЕ-Е=|98&ВЕ
2|в |345|58 |8688|«ВЕ.
ХЕ
442 91
0
0,00
_
142} -—
-
-
п
4154 82
2
0,02
15 169|7,5|0,18|0,089
ш
55 Ри
3
0,08
26
81 87 | 0,70 0,324
У|
803247
0,53
199 219 | 44,7 | 6,22 0,743
УТ-УП
385 196 70
0,41
751|1136|10,9|3,84|0,664
1х-х
51 45| 4 0,27
АЗ 78| 86|2,87|0,550
Сумма
992 453 96
-
1034 | 1718 —
_
—
Невзвешенное
-
_
- Ре-=0,1871
_
9,5 | 2,3 0,524
среднее
Примечание.
продолжительность видимой беременности (дни)
18
.б
средняя продолжительность сезони размножения (ини) = 270 — 0061; 0,067 =
0,187
= 00 = 2,8 (число пометов в расчете на 1 беременную самку за 270 дней);
Е:
Мо-0,07
— 34,5 (число эмбрионов в расчете на 1 самку из числа отловленных за 270 дней).
Число поворожденных рассчитано по Числу эмбрионов (за вычетом постимплантационных
потерь), которые появляются на свет в течение срока, в среднем равного
дня
периода истинной беременности).
ты
р
ы
м -бтоловине
Величипа принлода в популяции крыс складывается из трех показа-
телей плодовитости: 1) средней доли беременных самок Ру (пределы
варьирования от 7 до 70%), 2) среднего числа эмбрионов в помете (8,0—
9,0 в естественных условиях и 6,1 в неволе, пределы варьирования 1—7),
3) числа пометов в году, которое, в свою очередь, определяетел средней
продолжительностью сезона размножения (от 200 до 365 дней в году).
Первые два показателя могут быть заменены одним показателем — сред-
ним числом поворожденных крысят (или эмбрионов за вычетом постим-
плантационных потерь) в расчете на одну самку из числа активных чле-
нов популяции или самку данной возрастной группы за определенный
промежуток времени (ЛУ. Этот же показатель, отнесенный к общему чи-
слу животных в популяции (включая новорожденных), позволяет оце-
нить удельную рождаемость (5,). Рассчитывая число новорожденных на
число животных данной возрастной групны, можно оценить удельную
рождаемость в соответствующей возрастной группе (табл. 16, 17).
7 Серая крыса
194 _ Глава третья
Таблина 17. Показатели плодевитости самок и удельная рождаемость в популяции
серых крые рисовых полей. Кубани за 90 дней (июль-август 1978—1980 гг.)
Экология
195
0
Число самок
ыы
,
,
Е
Е
ы
8
Е
Е
ЕР
Зозраст (мес)
ЕЯ
8
Е
БЕ
&
[>=
Ф
я
ЕРЕ2
5Е
эм
ви
о+
=
я
НЕакЕеЕ
ВЕ
=<1
3
0,008
1
0
0,00
1—2
40
0,404
18
0
0,00
3—6
180
0,468
86
23
0,27
7—8
53
0,140
27
1А
0,52
9—11
25
0,065
13
9
0,69
12—46
82
0,213
42
19
0,52
Больше 47
Ат
0,044
9
5
0,17
Сумма
385
1,000
196
70
_
Невзвешенное сред-
-
_
_
_
Ре-=0,43
нее
Взвешенное среднее
-
_
-
-
Ре=0,44 ,
продолжительность видимой беременности (дни) —_ 48 по Рб __ 0,43 _
Примечание.
90 дней
=0=0,2 =
=2,15
нов в расчете на 1 самку из числа отловленных за 90 дней.
* Число новорожденных рассчитано по числу эмбрионов (за вычетом постимплантационных
периода истинной беременности).
При расчете итогового показателя — числа эмбрионов, приходящих-
ся на 1 самку за определенный промежуток времени, необходимо учиты-
вать следующее замечание Лесли с соавторами (ТезЦе её а1., 1952). Если
Е, — общее число эмбрионов, а No — число самок в выборке
— беремен-
ных и небеременных, то Г,/No — чиело эмбрионов на 1 самку. Если No —
з
исто ВЫ
Общее коли-
Число эмбрионов в расчете
чество крыс
—
густа (мес
58
Е
>
м
мы
м
тесново- В
Рав
ЕЕ
98 Е й ы рожденные |62©|8310| ВЕ
ЕЕЕ
М|ды, |НЕ
|
Зы
ая
#
И
=
Век|СЯ
858.°
-
_
_
0,000
3
0,00 0,00
0,000
-
-_
-
0,00
40
0,00 0,00
0,000
8
1
236 0,314
446
10,30
2,14
0,567
4
10
6
м
т
12
5
10
И
159
0,212
212
14,40
5,89
0,750
113
42
5
13
1
45
1
7
4
92 0,123
117 10,30 7,10
0,786
40
4
11
2
12
2
8
:
241
0,280
293
41,10
5,00
0,720
10
1
и
1
12
5
13
3
1А
3
15
2
10
1
53 0,071
70
41,20
6,20
0,757
11
3
13
1
-
-
751
_
1136
_
-_
-
-
-
-
_
_
10,90 - 3,84
0,661
Е
—чиИсло пометов в расчете на {1 беременную самку за 90 дней; и
В 19,5 — число эмбрио-
No 0,2
потерь), которые появляются на свет в течение срока, в среднем равного 12 дням (половина
число беременных самок, то Е/No=МNo/МРИNo или М/МNoМ==рх,
где р=МNo'/МNo и х==Р./ Ме. Тогда при условии, что р и х были независимы,
дисперсия (7) оценки Г/М, могла быть равна У (Р/МNo) ==?Т (р)-+р?У(1).
Однако фактически р и х связаны друг с другом, поэтому любые диспер-
‹ии оценки Р‚/No. будут занижепы и при их сравнении с помощью стан-
7
194 _ Глава третья
Таблина 17. Показатели плодевитости самок и удельная рождаемость в популяции
серых крые рисовых полей. Кубани за 90 дней (июль-август 1978—1980 гг.)
Экология
195
0
Число самок
ыы
,
,
Е
Е
ы
8
Е
Е
ЕР
Зозраст (мес)
ЕЯ
8
Е
БЕ
&
[>=
Ф
я
ЕРЕ2
5Е
эм
ви
о+
=
я
НЕакЕе
Е
ВЕ
=<1
3
0,008
1
0
0,00
1—2
40
0,404
18
0
0,00
3—6
180
0,468
86
23
0,27
7—8
53
0,140
27
1А
0,52
9—11
25
0,065
13
9
0,69
12—46
82
0,213
42
19
0,52
Больше 47
Ат
0,044
9
5
0,17
Сумма
385
1,000
196
70
_
Невзвешенное сред-
-
_
_
_
Ре-=0,43
нее
Взвешенное среднее
-
_
-
-
Ре=0,44 ,
продолжительность видимой беременности (дни) —_ 48 по Рб __ 0,43 _
Примечание.
90 дней
=0=0,2 =
=2,15
нов в расчете на 1 самку из числа отловленных за 90 дней.
* Число новорожденных рассчитано по числу эмбрионов (за вычетом постимплантационных
периода истинной беременности).
При расчете итогового показателя — числа эмбрионов, приходящих-
ся на 1 самку за определенный промежуток времени, необходимо учиты-
вать следующее замечание Лесли с соавторами (ТезЦе её а1., 1952). Если
Е, — общее число эмбрионов, а No — число самок в выборке
— беремен-
ных и небеременных, то Г,/No — чиело эмбрионов на 1 самку. Если No —
з
исто ВЫ
Общее коли-
Число эмбрионов в расчете
чество крыс
—
густа (мес
58
Е
>
м
мы
м
тесново- В
Рав
ЕЕ
98 Е й ы рожденные|62©|8310| ВЕ
ЕЕЕ
М|ды, |НЕ
|
Зы
ая
#
И
=
Век|СЯ
858.°
-
_
_
0,000
3
0,00 0,00
0,000
-
-_
-
0,00
40
0,00 0,00
0,000
8
1
236 0,314
446
10,30 2,14
0,567
4
10
6
м
т
12
5
10
И
159
0,212
212
14,40
5,89
0,750
113
42
5
13
1
45
1
7
4
92 0,123
117 10,30 7,10 0,786
40
4
11
2
12
2
8
:
241 0,280
293
41,10
5,00
0,720
10
1
и
1
12
5
13
3
1А
3
15
2
10
1
53 0,071
70
41,20 6,20
0,757
11
3
13
1
-
-
751
_
1136
_
-_
-
-
-
-
_
_
10,90 - 3,84
0,661
Е
—чиИсло пометов в расчете на {1 беременную самку за 90 дней; и
В 19,5 — число эмбрио-
No 0,2
потерь), которые появляются на свет в течение срока, в среднем равного 12 дням (половина
число беременных самок, то Е/No=МNo/МРИNo или М/МNoМ==рх,
где р=МNo'/МNo и х==Р./ Ме. Тогда при условии, что р и х были независимы,
дисперсия (7) оценки Г/М, могла быть равна У (Р/МNo) ==?Т (р)-+р?У(1).
Однако фактически р и х связаны друг с другом, поэтому любые диспер-
‹ии оценки Р‚/No. будут занижепы и при их сравнении с помощью стан-
7
198
Глава третья
дартного отклонения возможны значительные ошибки. Подытоживая,
Лесли с соавторами приходят к выводу о том, что легче работать с пока-
зателем доли беременных самок, поскольку статистическим методом для
сравнения различий в долях является критерий г.
Остается непонятным, зачем нужно рассчитывать Ё,/No через р их,
если эту оценку можно нолучить непосредственно из полевого материала
(табл. 16, 17). Тем не менее необходимость замечания, сделанного Лес-
ли, очевидна, поскольку некоторые исследователи (ЕпЦеп, Пау!з, 1948;
Вишняков, 1970; Самарский, Картель, 1986) допускают эту ошибку, рас-
считывая через долю беременных самок число новорожденных на 1 самку
в популяции.
Расчет плодовитости на одну «взрослую» самку, как это предлагает
Лесли с соавторами (ГезШе её а|., 1952),— величина столь же субъектив-
ная, как и само нонятие «взрослая». Этот расчет должен быть донолнен
расчетом плодовитости но отдельным возрастным группам (желательно,
чтобы эти возрастные группы были выделены не на основе возрастно-ве-
совых или основанных на измерении длины тела показатолей). Для анз-
лиза размерных отношений Дэвис (Рал!з, 1953») предлагает использо-
вать группирование по массе тела или его длине. Например, беременны:
5,6% из 231 самок с длиной тела, равной 130—179 мм; 13,9% из 180 са-
мок — длиной тела 180—190 мм; 21,1% из 199 самок
— более 199 мм дли-
ной. Потенциальный репродуктивный вклад самок разных возрастных
групп может быть онределен так, как предлагают Н. В. Тупикова и
Ю. Г. Швецов (1956) для водяных крыс и Е. В. Иванкина (1987) для
рыжих полевок— ло отношению количества выводков, приходящихся на
данную группу, ко всем выводкам, припесенным за сезон размножения.
Хотя более полным является показатель числа новорожденных, прихо-
дящихся на самок данной возрастной группы но отношению к общему
числу новорожденных в популяции, где учитывается не только число вы-
водков, но и их размер.
Связь между размером тела (масса тела, длина), возраетом и плодови-
тостью. Как следует из данных большинства авторов, плодовитость уже
созревших зв половом отношении самок крыс увеличивается с увеличени-
ем массы их тела. Причем возрастает не только доля беременных самок,
но и число эмбрионов
в расчете на одну самку данных веса или длины тела
(табл. 18) (Регу, 1945; Фетисов, 1945; Ра\!з, 1953а, в; Кузякин, 1952;
Вишняков, 1970; Козлов, 1981; Хамаганов, 1985). Однако, как указывали
Лесли с соавторами (ТезНе еф а|., 1952), масса тела не единственный
фактор, от которого зависит плодовитость самок, не менее важное значе-
ние может иметь их возраст, даже, более вероятно, предшествующий
опыт их участия в размножении. Это подтверждается данными Кинг
(Кто, 1924; Кая, Оопа1азоп, 1929), которая показала, что в лабораторных
условиях среднее число молодых в помете уволичивается (до 68,55) с воз-
растом самок, что, внрочем, сопровождается увеличением массы их тела,
При анализе размножения серых крыс Кубани установлено, что во
втором помете число эмбрионов было больше, чем в первом (р<0,05)
(табл. 19).
Экология
197
Таблица 18. Возрастной состав и генеративное еостояние самок серых крыс,
выловленных в г. Москве за декабрь 1944, январь-июль 1945 г. и июль-декабрь
1946 г, (по: Кузякин, 1952)
и
бе-
..|.
р
,
.. роменных 8 28
ВЕ
я |288 Зы
Е 5% |ЗН4
бе
|2
С:
н.
{
ан
ы
Е
25
од
я
-з
р
ре
но
Зо_& 58
ЗЕЯ
ЕР888:38ЕЗЕ|328врЕ
ВЕР
ЕЕ 5 |Эв88 38 ЕЕ ВЕ | ЗЕЕЫ З85Ы чиело| % |5684
ЗЕ
я [993=| #58
ЕЕ
5 |588 НЯ
5-5
1 1886|856|20—75 <2|1810|1030| 124 о|о |0
2 5267|2638|76-200|2-6|71-460|2629|309|30| 44|73
3 3851|1877|201-280|6-42 |164—240|4974|23,2|лад|73|17
4 2942|1742|281-390|42-24 |244-290|4230|ЧАБ|457|428|81
5
1178|292|391-440|224 |294-340|886|404|140|458|82
6
783. | 122 | 444-520 | 224 |344-420| 6в61| 78| 125 | 189 | 83
7
108 43 | 524-755 >24 | 424-5299] | 14| 29| 305 | 86
17 | 46016 | 7540 | 20-755 - 418—529 | 8505 | 100,0 | 625 | 7,35| 8,0%
Таблица 19. Зависимость среднего числа эмбрионов в 1-4-м пометах (в расчете
на 1 самку} от возраста самок серых крыс на рисовых полях Кубани в мае-авгуехе
1978—1980 гг. (пеошубликованные данные В. А. Рыльникова и Е. В. Карасевой)
Сеголетки (<8 мес}
Перезимовавитие {>>8 мес)
Помет, No
среднее число эм-
среднее число ом-
число самок в
число самок в
брионов в расчете
брионов в расчете
выборке
на {1 самку
выборке
на+ами
И
32
10,805 .
18
9,8=0,5
2
8
А1,А-=0,9
13
11,9=0,8
3
_
_
8
40,3= 1,0
4
-
-
6
12,0=1,2
Средневозрастные самки отличаются наиболее высокой и стабильной
плодовитостью и меньшей, чем у более молодых самок, зависимостью от
внутрипопуляционной конкуренции, приводящей к исключению из раз-
множения (ШОа\1з, 1951а,в; Козлов, 1981, 1982), поэтому именно эта ка-
.тегория крыс может быть индикатором сезонных периодических изменений
в интенсивности размножепия.
В большинстве случаев репродуктивная активность самок серых крыс
в неволе заканчивалась к 18 мес, хотя нокоторые рожали и в возрасте
2 лет. Длина их репродуктивного периода растет в ряду поколений
(рис. 55), к 25-му поколению длина ренродуктивного периода (440 дней)
болес чем вдвое выше (204 дня), чем в первых поколениях (Клшо, 1939).
П. П. Гамбарян и Н. М. Дукельская (1955) обратили внимание на вы-
сокий процент беременных самок в последних возрастно-весовых группах
198
Глава третья
дартного отклонения возможны значительные ошибки. Подытоживая,
Лесли с соавторами приходят к выводу о том, что легче работать с пока-
зателем доли беременных самок, поскольку статистическим методом для
сравнения различий в долях является критерий г.
Остается непонятным, зачем нужно рассчитывать Ё,/No через р их,
если эту оценку можно нолучить непосредственно из полевого материала
(табл. 16, 17). Тем не менее необходимость замечания, сделанного Лес-
ли, очевидна, поскольку некоторые исследователи (ЕпЦеп, Пау!з, 1948;
Вишняков, 1970; Самарский, Картель, 1986) допускают эту ошибку, рас-
считывая через долю беременных самок число новорожденных на 1 самку
в популяции.
Расчет плодовитости на одну «взрослую» самку, как это предлагает
Лесли с соавторами (ГезШе её а|., 1952),— величина столь же субъектив-
ная, как и само нонятие «взрослая». Этот расчет должен быть донолнен
расчетом плодовитости но отдельным возрастным группам (желательно,
чтобы эти возрастные группы были выделены не на основе возрастно-ве-
совых или основанных на измерении длины тела показатолей). Для анз-
лиза размерных отношений Дэвис (Рал!з, 1953») предлагает использо-
вать группирование по массе тела или его длине. Например, беременны:
5,6% из 231 самок с длиной тела, равной 130—179 мм; 13,9% из 180 са-
мок — длиной тела 180—190 мм; 21,1% из 199 самок
— более 199 мм дли-
ной. Потенциальный репродуктивный вклад самок разных возрастных
групп может быть онределен так, как предлагают Н. В. Тупикова и
Ю. Г. Швецов (1956) для водяных крыс и Е. В. Иванкина (1987) для
рыжих полевок— ло отношению количества выводков, приходящихся на
данную группу, ко всем выводкам, припесенным за сезон размножения.
Хотя более полным является показатель числа новорожденных, прихо-
дящихся на самок данной возрастной группы но отношению к общему
числу новорожденных в популяции, где учитывается не только число вы-
водков, но и их размер.
Связь между размером тела (масса тела, длина), возраетом и плодови-
тостью. Как следует из данных большинства авторов, плодовитость уже
созревших зв половом отношении самок крыс увеличивается с увеличени-
ем массы их тела. Причем возрастает не только доля беременных самок,
но и число эмбрионов
в расчете на одну самку данных веса или длины тела
(табл. 18) (Регу, 1945; Фетисов, 1945; Ра\!з, 1953а, в; Кузякин, 1952;
Вишняков, 1970; Козлов, 1981; Хамаганов, 1985). Однако, как указывали
Лесли с соавторами (ТезНе еф а|., 1952), масса тела не единственный
фактор, от которого зависит плодовитость самок, не менее важное значе-
ние может иметь их возраст, даже, более вероятно, предшествующий
опыт их участия в размножении. Это подтверждается данными Кинг
(Кто, 1924; Кая, Оопа1азоп, 1929), которая показала, что в лабораторных
условиях среднее число молодых в помете уволичивается (до 68,55) с воз-
растом самок, что, внрочем, сопровождается увеличением массы их тела,
При анализе размножения серых крыс Кубани установлено, что во
втором помете число эмбрионов было больше, чем в первом (р<0,05)
(табл. 19).
Экология
197
Таблица 18. Возрастной состав и генеративное еостояние самок серых крыс,
выловленных в г. Москве за декабрь 1944, январь-июль 1945 г. и июль-декабрь
1946 г, (по: Кузякин, 1952)
и
бе-
..|.
р
,
.. роменных 8 28
ВЕ
я |288 Зы
Е 5% |ЗН4
бе
|2
С:
н.
{
ан
ы
Е
25
од
я
-з
р
ре
но
Зо_& 58
ЗЕЯ
ЕР888:38ЕЗЕ|328врЕ
ВЕР
ЕЕ 5 |Эв88 38 ЕЕ ВЕ | ЗЕЕЫ З85Ы чиело| % |5684
ЗЕ
я [993=| #58
ЕЕ
5 |588 НЯ
5-5
1 1886|856|20—75 <2|1810|1030| 124 о|о |0
2 5267|2638|76-200|2-6|71-460|2629|309|30| 44|73
3 3851|1877|201-280|6-42 |164—240|4974|23,2|лад|73|17
4 2942|1742|281-390|42-24 |244-290|4230|ЧАБ|457|428|81
5 1178|292|391-440|224 |294-340|886|404|140|458|82
6
783. | 122 | 444-520 | 224 |344-420| 6в61| 78| 125 | 189 | 83
7
108 43 | 524-755 >24 | 424-5299] | 14| 29| 305 | 86
17 | 46016 | 7540 | 20-755 - 418—529 | 8505 | 100,0 | 625 | 7,35| 8,0%
Таблица 19. Зависимость среднего числа эмбрионов в 1-4-м пометах (в расчете
на 1 самку} от возраста самок серых крыс на рисовых полях Кубани в мае-авгуехе
1978—1980 гг. (пеошубликованные данные В. А. Рыльникова и Е. В. Карасевой)
Сеголетки (< 8 мес}
Перезимовавитие {>>8 мес)
Помет, No
среднее число эм-
среднее число ом-
число самок в
число самок в
брионов в расчете
брионов в расчете
выборке
на {1 самку
выборке
на+ами
И
32
10,805 .
18
9,8=0,5
2
8
А1,А-=0,9
13
11,9=0,8
3
_
_
8
40,3= 1,0
4
-
-
6
12,0=1,2
Средневозрастные самки отличаются наиболее высокой и стабильной
плодовитостью и меньшей, чем у более молодых самок, зависимостью от
внутрипопуляционной конкуренции, приводящей к исключению из раз-
множения (ШОа\1з, 1951а,в; Козлов, 1981, 1982), поэтому именно эта ка-
.тегория крыс может быть индикатором сезонных периодических изменений
в интенсивности размножепия.
В большинстве случаев репродуктивная активность самок серых крыс
в неволе заканчивалась к 18 мес, хотя нокоторые рожали и в возрасте
2 лет. Длина их репродуктивного периода растет в ряду поколений
(рис. 55), к 25-му поколению длина ренродуктивного периода (440 дней)
болес чем вдвое выше (204 дня), чем в первых поколениях (Клшо, 1939).
П. П. Гамбарян и Н. М. Дукельская (1955) обратили внимание на вы-
сокий процент беременных самок в последних возрастно-весовых группах
198
Глава третья
„Дни
500
-150
400
350
300
250
200
ЛДжоления
ис. 55. Средняя линия репродуктив-
ного периода в ряду последователь-
ных поколений (по; Юта, 1989)
самок крыс по данным А. П. Ву-
зякина (1952). Такое же явле-
ние прослеживается и на мате-
риалах А. Н. Козлова (1981),
хотя можно было бы ожидать
спижения репродуктивных спо-
собностей этих самок. По пред-
положению Дукельской, это мо-
жет быть связано с большей по-
падаемостью взрослых беременных самок в ловушки. Однако вполне до-
пустимо и другое предположение. Поскольку в старших возрастных груп-
пах масса тела менее всего коррелирует с возрастом, то крысы крупных
размеров совершенно необязательно относятся к престарелым. Учитывая,
что доля престарелых в популяции невелика, в совокупности данных их
низкая плодовитость может лишь пезпачительно уменьшать показатели
плодовитости средневозрастных самок крупного размера, относимых к
старшей возрастной группе. Не исключено также, что престарелые самки,
отличаясь повышенной осторожностью, избегают попадания в ловушки
80
З
т
„
Д
о
л
я
б
е
р
е
м
е
н
н
ы
х
с
а
м
о
к
,
%
=
>
5
С
=
)
(В. В. Кучерук, личн. сообщ.).
Пытаясь объяснить поисти-
не отромные различия в доле
беременных самок, отловленных
в г. Москве, по материалам,
опубликованным А. А. Пасеш-
ником (1936) и А. П. Вузя-
киным (1952), П. П. Гамбарян,
Рис. 56. Сезонная дицамика доли
беременных самок серых крыс, по
данным отечественных авторов
Данные авторов:
1— Кувзякин (Москва..., 1952 т.),
8 — Степанов (Батуми, 1946 г.),
3 — Козлов
{Северный
Казахстан,
1981 г.): доля беременных от числа
«взрослых» самок с массой тела
более 100 г,
4 — Пасешник (Москва, 1936 г.): доля
беременных от числа взрослых са-
мок» с массой тела более 100 г,
За и 4а — частотные распределения рас-
считаны В. А. Рыльниковым по
опубликованным данным А. Н. Коз-
лова (1981) и Насешника (1936)
з
Экология 199
Н. М. Дукельская (1955) и С. В. Вишняков (4970) считают, что непра-
вильно было рассчитывать долю беремеппых на всех самок выборки, так
как это и послужило причиной занижения данных в расчетах А. П. Цу-
зякина (1952) и И. В. Степанова (41946). Мы провели пересчет числа бе-
ременных самок по отпошению к общему числу пойманных, используя
материалы А. А. Пасешника (1936) и А. Н. Козлова (1984) (рис. 56}.
Сравнение полученных кривых динамики плодовитости самок в течение
тода свидетельствует о том, что причина упомянутых различий пе только
в методике обработки материалов. Неудивительно, что доля беременных
самок по материалам И. В. Степанова и А. П. Кузякина совпала. Оба
автора использовали не только единую методику обработки материала,
но и отлова серых крыс ловушками Геро. П. Ц. Гамбарян и Н. М. Ду-
кельская (1955) замечают, что молодые зверьки легче пападаются в раз-
личные ловушки и по сравиепию с ними взрослые отличаются гораздо
большей осторожностью, поэтому выловить их труднее. Этим и объясня-
ется завышенный процент молодых крыс в уловах. Так, по материалам,
использованпым А. П. Кузякиным (4952), 30,9% (т. е. доля, которая
вносит существенный вклад в среднестатистические показатели процента
беременных самок,— замечание В. А. Рыльникова) от общего числа са-
мок составляли крысы в возрасте от 2 до 6 мес.
Это замечание справедливо, одпако, только по отношению к ловушкам
Геро, а не ко всем ловушкам. Серые крысы, отловлеппые А. А. Пасеш-
ником (4936), были добыты вершами и дуговыми капканами. По нашему
собственному опыту известно, что доля взрослых крыс очень велика в
отловах дуговыми канканами при их иепродолжительной экспозиции в
одном и том же месте (не более 2—3 дней). Молодые крысы пачипали
ловиться липть спустя 1—2 недели после начала отлова вершами (Пау1в,
Епщеп, 1956).
А. Н. Козлов (1981) использовал сборы трупов крыс после их истреб-
ления фосфидом цинка на приманке. Учитывая, что эффективность дера-
тизации составляла 95—98% (Козлов, 19806}, репрезептативность вы-
борки, полученной путем сбора трупов крыс на поверхности (даже если
они составляют мепьптую долю от общего числа погибших), оказывается
напбольшей по сравнению с другими способами сбора материала. Линть
с этими отоворками можно приступать к анализу материалов по сезонной
динамике плодовитости самок,
Сезонный ход размножения
Дипамика плодовитости складывается из показателей плодовитости самок
из выборок, взятых в отдельные месяцы одного года или за ряд лет.
Вместе с тем кривая изменений плодовитости самок является результа-
том тех изменений, которые происходят в возрастном составе самок из
выборок, взятых из популяции в последовательные интервалы звре-
мени.
Средняя в течение года доля беременных самок формируется в основ-
198
Глава третья
„Дни
500
-150
400
350
300
250
200
ЛДжоления
ис. 55. Средняя линия репродуктив-
ного периода в ряду последователь-
ных поколений (по; Юта, 1989)
самок крыс по данным А. П. Ву-
зякина (1952). Такое же явле-
ние прослеживается и на мате-
риалах А. Н. Козлова (1981),
хотя можно было бы ожидать
спижения репродуктивных спо-
собностей этих самок. По пред-
положению Дукельской, это мо-
жет быть связано с большей по-
падаемостью взрослых беременных самок в ловушки. Однако вполне до-
пустимо и другое предположение. Поскольку в старших возрастных груп-
пах масса тела менее всего коррелирует с возрастом, то крысы крупных
размеров совершенно необязательно относятся к престарелым. Учитывая,
что доля престарелых в популяции невелика, в совокупности данных их
низкая плодовитость может лишь пезпачительно уменьшать показатели
плодовитости средневозрастных самок крупного размера, относимых к
старшей возрастной группе. Не исключено также, что престарелые самки,
отличаясь повышенной осторожностью, избегают попадания в ловушки
80
З
т
„
Д
о
л
я
б
е
р
е
м
е
н
н
ы
х
с
а
м
о
к
,
%
=
>
5
С
=
)
(В. В. Кучерук, личн. сообщ.).
Пытаясь объяснить поисти-
не отромные различия в доле
беременных самок, отловленных
в г. Москве, по материалам,
опубликованным А. А. Пасеш-
ником (1936) и А. П. Вузя-
киным (1952), П. П. Гамбарян,
Рис. 56. Сезонная дицамика доли
беременных самок серых крыс, по
данным отечественных авторов
Данные авторов:
1— Кувзякин (Москва..., 1952 т.),
8 — Степанов (Батуми, 1946 г.),
3 — Козлов
{Северный
Казахстан,
1981 г.): доля беременных от числа
«взрослых» самок с массой тела
более 100 г,
4 — Пасешник (Москва, 1936 г.): доля
беременных от числа взрослых са-
мок» с массой тела более 100 г,
За и 4а — частотные распределения рас-
считаны В. А. Рыльниковым по
опубликованным данным А. Н. Коз-
лова (1981) и Насешника (1936)
з
Экология 199
Н. М. Дукельская (1955) и С. В. Вишняков (4970) считают, что непра-
вильно было рассчитывать долю беремеппых на всех самок выборки, так
как это и послужило причиной занижения данных в расчетах А. П. Цу-
зякина (1952) и И. В. Степанова (41946). Мы провели пересчет числа бе-
ременных самок по отпошению к общему числу пойманных, используя
материалы А. А. Пасешника (1936) и А. Н. Козлова (1984) (рис. 56}.
Сравнение полученных кривых динамики плодовитости самок в течение
тода свидетельствует о том, что причина упомянутых различий пе только
в методике обработки материалов. Неудивительно, что доля беременных
самок по материалам И. В. Степанова и А. П. Кузякина совпала. Оба
автора использовали не только единую методику обработки материала,
но и отлова серых крыс ловушками Геро. П. Ц. Гамбарян и Н. М. Ду-
кельская (1955) замечают, что молодые зверьки легче пападаются в раз-
личные ловушки и по сравиепию с ними взрослые отличаются гораздо
большей осторожностью, поэтому выловить их труднее. Этим и объясня-
ется завышенный процент молодых крыс в уловах. Так, по материалам,
использованпым А. П. Кузякиным (4952), 30,9% (т. е. доля, которая
вносит существенный вклад в среднестатистические показатели процента
беременных самок,— замечание В. А. Рыльникова) от общего числа са-
мок составляли крысы в возрасте от 2 до 6 мес.
Это замечание справедливо, одпако, только по отношению к ловушкам
Геро, а не ко всем ловушкам. Серые крысы, отловлеппые А. А. Пасеш-
ником (4936), были добыты вершами и дуговыми капканами. По нашему
собственному опыту известно, что доля взрослых крыс очень велика в
отловах дуговыми канканами при их иепродолжительной экспозиции в
одном и том же месте (не более 2—3 дней). Молодые крысы пачипали
ловиться липть спустя 1—2 недели после начала отлова вершами (Пау1в,
Епщеп, 1956).
А. Н. Козлов (1981) использовал сборы трупов крыс после их истреб-
ления фосфидом цинка на приманке. Учитывая, что эффективность дера-
тизации составляла 95—98% (Козлов, 19806}, репрезептативность вы-
борки, полученной путем сбора трупов крыс на поверхности (даже если
они составляют мепьптую долю от общего числа погибших), оказывается
напбольшей по сравнению с другими способами сбора материала. Линть
с этими отоворками можно приступать к анализу материалов по сезонной
динамике плодовитости самок,
Сезонный ход размножения
Дипамика плодовитости складывается из показателей плодовитости самок
из выборок, взятых в отдельные месяцы одного года или за ряд лет.
Вместе с тем кривая изменений плодовитости самок является результа-
том тех изменений, которые происходят в возрастном составе самок из
выборок, взятых из популяции в последовательные интервалы звре-
мени.
Средняя в течение года доля беременных самок формируется в основ-
200 Глава третья
ном в зависимости от климатических условий, обеспеченности кормами и
уровня численности популяции, а также от таких субъективных причин,
как способ отлова крые и квалификация ловцов.
Наибольшая доля беременных самок наблюдается весной, иногда ле-
том и осенью (рис. 57).
Ответ на вопрос, почему весеннее размножение начинается так
дружно и с такой высокой интенсивностью, нужно искать не только в
периодических явлениях в физиологии самок, но и в возрастном составе
популяции (рис. 58). Вследствие пониженной интенсивности размноже-
ния в зимние месяцы и повышенной смертности в популяции в это вре-
мя преобладают зверьки средних и старших возрастных групп, в подавля-
ющем большинстве своем половозрелые. Доля молодых наиболее низка по
сравнению с другими сезонами года, поэтому весной в размножепии
участвуют практически все самки.
В результате массового выхода молодняка в мае-июне доля неполово-
зрелых самок резко возрастает, и в зависимости от того, какую долю мо-
лодых каждый из авторов включает в расчеты доли беременных самок,
значение этого показателя имеет более или менее выраженный осеппий
пик, который связан со вступлением в размножение сеголеток весенних
пометов. Известны и отклонепия от этого правила, к которым можно при-
числить ход сезонной динамики размножения крыс в лесостепной зоне
УССР (Черкасская обл.). Осепью наблюдается повышение плодовитости
самок серой крысы даже по сравнению с весенним подъемом, что, по-
видимому, является региональной особенностью, поскольку не наблюда-
ется в степной зоне УССР (Одесская обл.). В условиях лесостепи выше
влажность, что сказывается на размещении грызунов в природе ина
длительности их обитания там. Отчетливо выраженный осеппий подъем
доли размножающихся самок крыс наблюдался в Сумской обл. УССР.
Однако величина осеппего пика меныше весеннего, причем доля размно-
жающихся самок крыс закономерно увеличивается от осени к зиме, до-
стигая максимума в марте—мае. Летом показатели плодовитости самок
спижаются. По мнению автора этих материалов — И. Р. Мерзликина
(1987Та), самки серых крыс, забеременевшие еще в природных биотопах,
рождают детенышей уже в постройках человека, куда мигрируют при по-
холодапии. Как он считает, именно за счет этих самок происходит вто-
рой нодъем размножения крыс, который отмечался рапее рядом авторов.
В настоящее время, по-видимому, трудно дать однозначное толкование
этим результатам, так же как и необъяспимому увеличению числа бере-
менных самок серых крыс в Англии зимой 1941/42 г. (Оау!з, 1953а).
Наблюдаемое явление заслуживает серьезного анализа, поскольку отлича-
ется от наиболее распространенной схемы сезонной динамики размно-
жения.
Среднее число эмбрионов в расчете па беременную самку также
подвержено сезонным измепениям, связанным с периодическими явле-
ниями в физиологии самок (табл. 20). Наибольшее число эмбрионов у
взрослых самок (с массой тела более 251 г) наблюдается веслой и летом.
У молодых половозрелых самок (140—190 г) — осенью. Среднее число
Экология 204
Рис. 57. Сезонная динамика
р
размножения серых крыс (по:
д
—.———>
Ваз, 1953)
И
1 — Бомбей,
20-
2
2 — Сан-Антонио (штат Техас),
Е
3 — Норфолк (штат Вирджиния),
0
4 — Сан-Франциско
(крупные
экземпляры),
0Е
3
5 — Англия, город,
З0Е
6 — Англия, сельские районы
{а — скирда соломы, б — зер-
новая скирда):
ЗЕ
}
7— ФРГ
5Е
х
яя
ы
з
|
”
ЁИ
$
Зя |-
5
$51
з
Ча
=
5
ПЕ
а
Рис. 58. Возрастйой состав се- 0
7
рых крыс, отловленных в
|
1972—1974 гг. в Балтиморском
зоопарке (по: Ратвапи-Агаа,
30 |-
бошфилсЕ, 1979)
Е
1— взрослые (стареющие и сред-
Па
ние возрастные группы),
|)
1
|
111ЙдЙ11|й
2 — молодые
ТПУИЖЯТГПШЕИШЕЯ
МЕсяци
%100 |
[]
80-
-Г7
т
77
РВ
50 РА
Я
-й
А
0-
27
й
АРТА
й
#7
20-
77/777 РА
йр
й
ИИШЕХ ИГ Л ИПУИШИХХ ХИТ 1 ИМТ И Меж
1972г.
1975г.
797%г.
200 Глава третья
ном в зависимости от климатических условий, обеспеченности кормами и
уровня численности популяции, а также от таких субъективных причин,
как способ отлова крые и квалификация ловцов.
Наибольшая доля беременных самок наблюдается весной, иногда ле-
том и осенью (рис. 57).
Ответ на вопрос, почему весеннее размножение начинается так
дружно и с такой высокой интенсивностью, нужно искать не только в
периодических явлениях в физиологии самок, но и в возрастном составе
популяции (рис. 58). Вследствие пониженной интенсивности размноже-
ния в зимние месяцы и повышенной смертности в популяции в это вре-
мя преобладают зверьки средних и старших возрастных групп, в подавля-
ющем большинстве своем половозрелые. Доля молодых наиболее низка по
сравнению с другими сезонами года, поэтому весной в размножепии
участвуют практически все самки.
В результате массового выхода молодняка в мае-июне доля неполово-
зрелых самок резко возрастает, и в зависимости от того, какую долю мо-
лодых каждый из авторов включает в расчеты доли беременных самок,
значение этого показателя имеет более или менее выраженный осеппий
пик, который связан со вступлением в размножение сеголеток весенних
пометов. Известны и отклонепия от этого правила, к которым можно при-
числить ход сезонной динамики размножения крыс в лесостепной зоне
УССР (Черкасская обл.). Осепью наблюдается повышение плодовитости
самок серой крысы даже по сравнению с весенним подъемом, что, по-
видимому, является региональной особенностью, поскольку не наблюда-
ется в степной зоне УССР (Одесская обл.). В условиях лесостепи выше
влажность, что сказывается на размещении грызунов в природе ина
длительности их обитания там. Отчетливо выраженный осеппий подъем
доли размножающихся самок крыс наблюдался в Сумской обл. УССР.
Однако величина осеппего пика меныше весеннего, причем доля размно-
жающихся самок крыс закономерно увеличивается от осени к зиме, до-
стигая максимума в марте—мае. Летом показатели плодовитости самок
спижаются. По мнению автора этих материалов — И. Р. Мерзликина
(1987Та), самки серых крыс, забеременевшие еще в природных биотопах,
рождают детенышей уже в постройках человека, куда мигрируют при по-
холодапии. Как он считает, именно за счет этих самок происходит вто-
рой нодъем размножения крыс, который отмечался рапее рядом авторов.
В настоящее время, по-видимому, трудно дать однозначное толкование
этим результатам, так же как и необъяспимому увеличению числа бере-
менных самок серых крыс в Англии зимой 1941/42 г. (Оау!з, 1953а).
Наблюдаемое явление заслуживает серьезного анализа, поскольку отлича-
ется от наиболее распространенной схемы сезонной динамики размно-
жения.
Среднее число эмбрионов в расчете па беременную самку также
подвержено сезонным измепениям, связанным с периодическими явле-
ниями в физиологии самок (табл. 20). Наибольшее число эмбрионов у
взрослых самок (с массой тела более 251 г) наблюдается веслой и летом.
У молодых половозрелых самок (140—190 г) — осенью. Среднее число
Экология 204
Рис. 57. Сезонная динамика
р
размножения серых крыс (по:
д
—.———>
Ваз, 1953)
И
1 — Бомбей,
20-
2
2 — Сан-Антонио (штат Техас),
Е
3 — Норфолк (штат Вирджиния),
0
4 — Сан-Франциско
(крупные
экземпляры),
0Е
3
5 — Англия, город,
З0Е
6 — Англия, сельские районы
{а — скирда соломы, б — зер-
новая скирда):
ЗЕ
}
7— ФРГ
5Е
х
яя
ы
з
|
”
ЁИ
$
Зя |-
5
$51
з
Ча
=
5
ПЕ
а
Рис. 58. Возрастйой состав се- 0
7
рых крыс, отловленных в
|
1972—1974 гг. в Балтиморском
зоопарке (по: Ратвапи-Агаа,
30 |-
бошфилсЕ, 1979)
Е
1— взрослые (стареющие и сред-
Па
ние возрастные группы),
|)
1
|
111ЙдЙ11|й
2 — молодые
ТПУИЖЯТГПШЕИШЕЯ
МЕсяци
%100 |
[]
80-
-Г7
т
77
РВ
50 РА
Я
-й
А
0-
27
й
АРТА
й
#7
20-
77/777 РА
йр
й
ИИШЕХ ИГ Л ИПУИШИХХ ХИТ 1 ИМТ И Меж
1972г.
1975г.
797%г.
202
Глава третья
Таблица 20. Изменение интенсивности размножения серой крысы в зависимости
от возраета самок в разные сезоны года (по: Козлов, 1984)
Возрает-
но-весо-
вая
Показатель
Весна
Лето
Осень
Зима
грунна,
т
140—190 | Число самок
710
468
242
442
4 беременных
5,6+0,9
2,3=07
1,6-0,8
-
Ср. зисло эмбрионов
6,9-0,4
7,6-0,9
8,5=2,5
—
191—250 | Число самок
802
405
332
814
$ беременных
33,517 28,6—2,2 20,8+2,2 11,7-=1,4
Ср. число эмбрионов
7,8=0,1
7,1=0,3
8,5=0,А
7,9=0,3
251-350 | Число самок
1874
1045
542, 1365
% беременных
36,4=1,1 32,114 32,8=2,0 28,2+1,2
Ср. число эмбрионов
9,7—=0,1
9,2=0,1
9,2-0,2
8,9-=0,1
351—430 | Число самок
623
426
358
679
Ф беременных
43,7=2,0
34552,83
30,3=2,4
29,217
Ср. число эмбрионов
10,4—0,2
11,0=0,2
9,5-0,3
9,9-0,2
431-590 | Число самок.
165
115
97
183
4 беременных
44,2+3,9 47,841 45,4-5,1 43,1+3,7
Ср. число эмбрионов
11,2-=0,5
9,9-0,4
8,6=0,4
9,4—0,4
эмбрионов в расчете на одну взрослую самку (более 200 г) * изменяет-
ся по сезонам в небольших пределах (8,99—9,67) (Козлов, 19814).
Различия в сроках подъема интенсивности размножения крыс в по-
пуляции зависят от ее стациальпого и географического расположения.
причем для крыс населенных пунктов — от стациального больше, чем от
теографического. Это и понятно, так как постоянство микроклиматиче-
ских условий и кормовой базы з строениях человека обеспечивает отно-
сительную независимость зверьков по отношению к разнице климата, ко-
торая имеет место в разных географических точках. Тем не менее эта
разница наблюдается, поскольку передвижения серых крыс не ограничи-
заются площадью строепий. Сравпение крязых доли беременных самок в
Батуми (Степанов, 1946) и в Москве (Кузякин, 1952) позволяет заклю-
чить, что обе зависимости достоверно (р<0,01) непараллельны (согласно
терминологии Нлохинского, 1980, с. 127), причем весеннее размножение
в Батуми пачинается раньше.
Различия в доле беременных самок в Батуми (около 7$), Одессе
(32,0—51,1%) и в Черкасской области (76,7%) (Степанов, 1946; Русев
и др., 1983; Самарский, Картель, 1986), по нашему мнению, не могут
11 Здесь следует обратить внимание на тапичную ошибку, которая является ре-
зультатом вольной интерпретации понятия «взрослые» самки. Даже в одной и той
же статье (Козлов, 19841) используются разные градации массы тела; здесь 200 г
и более, а в табл. 20— 191—250 г.
Экология 203
являться результатом различий, связанных с географическим расположе-
нием. Вероятнее всего, это результат применения разных способов сбора
и обработки первичных данных.
В аридных райопах Северного Таджикистана главным лимитирую-
щим фактором для крыс является вода. При наличии ее и калорийной
пищи зверьки размножаются с конца февраля по начало поября. Доля
размнпожающихся самок особенно высока в периоды с умеренными тем-
пературами, т. е. весной и осенью, и составляет 50—73, однако число
эмбрионов па 1 беременную самку относительно невелико, в среднем
6,0—7,3 (Давыдов, 1986), что сопоставимо с таковым Узбекистана —
7,0 эмбрионов (Митропольский и др., 1986), но меньше, чем в Северном
Казахстане, 9,0—9,7 (Козлов, 1981), в средней полосе— 8 эмбрионов
{в среднем; Кузякин, 1952), на Украине— 9,7-0,3 (в среднем; Самар-
ский, Картель, 1986) и на Сахалине — 9,5 эмбриона (в среднем; Сурков,
1986 а). В Узбекистане по сравпению с Северным Таджикистаном ниже
доля беременных самок (в среднем их около 20%; Митропольский и др.,
1986) в те же периоды года. Учитывая географическую близость средне-
азиатских республик, наблюдаемые различия с региональных позиций
малообъяснимы. Интересно, что современные показатели плодовитости
самок серых крыс (1978—1983 тг.) в Узбекистане отличаются от таковых
в 1951—1953 тт., когда был начальный период освоения Узбекистапа
этим видом; произошло повышение доли размножающихся самок, в то
время как среднее число эмбрионов на 1 беременную самку снизилось
© 7,9 (начальный период) до 7,0 (последующий). Предполагают, что за
тридцатилетний период крысы адаптировались к новым условиям, что
повлекло за собой увеличение числа размножающихся самок при спиже-
нии плодовитости отдельных особей. Не исключено, что эти изменения —
результат преобразования возрастного состава в сторону преобладапия
молодых животных (Митропольский и др., 1986).
В. С. Сурков, С. В. Рябов (1983) в ©. А. Хаматанов (1985) сравни-
вают плодовитость половозрелых самок крыс в Приамурье и па о-ве Са-
халин. Па Сахалине оказались выше доля беременных самок и число
эмбрионов в расчете на одну половозрелую самку. По сравнению с евро-
нейскими сроками подъема размножения (апрель—сентябрь) на Дальнем
Востоке сроки активного размножения сокращены
— май—август (Ха-
маганов, 1985) и июнь—сентябрь (Сурков, Рябов, 1983). Эти отличия
авторы объясняют главным образом климатическими особенностями Даль-
него Востока. Близость моря и холодного течения обусловливает медлен-
ное наступление весны. Она запаздывает но сравнению с европейскими
сроками почти на месяц.
Обращают на себя внимание расхождения между В. С. Сурковым,
С. В. Рябовым (1983), с одной сторопы, и С. А. Хамагановым (1985) —
с другой, в оценке доли размножающихся самок (22—24 и 53,8—55,3%
соответственно) на о-ве Сахалин с мая по автуст, притом что значитель-
ных колебапий в размножении по голам эти авторы не отмечают, Можел
быть, опять сказались отличия в методикс отлова крыс и обработки
данных?
202
Глава третья
Таблица 20. Изменение интенсивности размножения серой крысы в зависимости
от возраета самок в разные сезоны года (по: Козлов, 1984)
Возрает-
но-весо-
вая
Показатель
Весна
Лето
Осень
Зима
грунна,
т
140—190 | Число самок
710
468
242
442
4 беременных
5,6+0,9
2,3=07
1,6-0,8
-
Ср. зисло эмбрионов
6,9-0,4
7,6-0,9
8,5=2,5
—
191—250 | Число самок
802
405
332
814
$ беременных
33,517 28,6—2,2 20,8+2,2 11,7-=1,4
Ср. число эмбрионов
7,8=0,1
7,1=0,3
8,5=0,А
7,9=0,3
251-350 | Число самок
1874
1045
542, 1365
% беременных
36,4=1,1 32,114 32,8=2,0 28,2+1,2
Ср. число эмбрионов
9,7—=0,1
9,2=0,1
9,2-0,2
8,9-=0,1
351—430 | Число самок
623
426
358
679
Ф беременных
43,7=2,0
34552,83
30,3=2,4
29,217
Ср. число эмбрионов
10,4—0,2
11,0=0,2
9,5-0,3
9,9-0,2
431-590 | Число самок.
165
115
97
183
4 беременных
44,2+3,9 47,841 45,4-5,1 43,1+3,7
Ср. число эмбрионов
11,2-=0,5
9,9-0,4
8,6=0,4
9,4—0,4
эмбрионов в расчете на одну взрослую самку (более 200 г) * изменяет-
ся по сезонам в небольших пределах (8,99—9,67) (Козлов, 19814).
Различия в сроках подъема интенсивности размножения крыс в по-
пуляции зависят от ее стациальпого и географического расположения.
причем для крыс населенных пунктов — от стациального больше, чем от
теографического. Это и понятно, так как постоянство микроклиматиче-
ских условий и кормовой базы з строениях человека обеспечивает отно-
сительную независимость зверьков по отношению к разнице климата, ко-
торая имеет место в разных географических точках. Тем не менее эта
разница наблюдается, поскольку передвижения серых крыс не ограничи-
заются площадью строепий. Сравпение крязых доли беременных самок в
Батуми (Степанов, 1946) и в Москве (Кузякин, 1952) позволяет заклю-
чить, что обе зависимости достоверно (р<0,01) непараллельны (согласно
терминологии Нлохинского, 1980, с. 127), причем весеннее размножение
в Батуми пачинается раньше.
Различия в доле беременных самок в Батуми (около 7$), Одессе
(32,0—51,1%) и в Черкасской области (76,7%) (Степанов, 1946; Русев
и др., 1983; Самарский, Картель, 1986), по нашему мнению, не могут
11 Здесь следует обратить внимание на тапичную ошибку, которая является ре-
зультатом вольной интерпретации понятия «взрослые» самки. Даже в одной и той
же статье (Козлов, 19841) используются разные градации массы тела; здесь 200 г
и более, а в табл. 20— 191—250 г.
Экология 203
являться результатом различий, связанных с географическим расположе-
нием. Вероятнее всего, это результат применения разных способов сбора
и обработки первичных данных.
В аридных райопах Северного Таджикистана главным лимитирую-
щим фактором для крыс является вода. При наличии ее и калорийной
пищи зверьки размножаются с конца февраля по начало поября. Доля
размнпожающихся самок особенно высока в периоды с умеренными тем-
пературами, т. е. весной и осенью, и составляет 50—73, однако число
эмбрионов па 1 беременную самку относительно невелико, в среднем
6,0—7,3 (Давыдов, 1986), что сопоставимо с таковым Узбекистана —
7,0 эмбрионов (Митропольский и др., 1986), но меньше, чем в Северном
Казахстане, 9,0—9,7 (Козлов, 1981), в средней полосе— 8 эмбрионов
{в среднем; Кузякин, 1952), на Украине— 9,7-0,3 (в среднем; Самар-
ский, Картель, 1986) и на Сахалине — 9,5 эмбриона (в среднем; Сурков,
1986 а). В Узбекистане по сравпению с Северным Таджикистаном ниже
доля беременных самок (в среднем их около 20%; Митропольский и др.,
1986) в те же периоды года. Учитывая географическую близость средне-
азиатских республик, наблюдаемые различия с региональных позиций
малообъяснимы. Интересно, что современные показатели плодовитости
самок серых крыс (1978—1983 тг.) в Узбекистане отличаются от таковых
в 1951—1953 тт., когда был начальный период освоения Узбекистапа
этим видом; произошло повышение доли размножающихся самок, в то
время как среднее число эмбрионов на 1 беременную самку снизилось
© 7,9 (начальный период) до 7,0 (последующий). Предполагают, что за
тридцатилетний период крысы адаптировались к новым условиям, что
повлекло за собой увеличение числа размножающихся самок при спиже-
нии плодовитости отдельных особей. Не исключено, что эти изменения —
результат преобразования возрастного состава в сторону преобладапия
молодых животных (Митропольский и др., 1986).
В. С. Сурков, С. В. Рябов (1983) в ©. А. Хаматанов (1985) сравни-
вают плодовитость половозрелых самок крыс в Приамурье и па о-ве Са-
халин. Па Сахалине оказались выше доля беременных самок и число
эмбрионов в расчете на одну половозрелую самку. По сравнению с евро-
нейскими сроками подъема размножения (апрель—сентябрь) на Дальнем
Востоке сроки активного размножения сокращены
— май—август (Ха-
маганов, 1985) и июнь—сентябрь (Сурков, Рябов, 1983). Эти отличия
авторы объясняют главным образом климатическими особенностями Даль-
него Востока. Близость моря и холодного течения обусловливает медлен-
ное наступление весны. Она запаздывает но сравнению с европейскими
сроками почти на месяц.
Обращают на себя внимание расхождения между В. С. Сурковым,
С. В. Рябовым (1983), с одной сторопы, и С. А. Хамагановым (1985) —
с другой, в оценке доли размножающихся самок (22—24 и 53,8—55,3%
соответственно) на о-ве Сахалин с мая по автуст, притом что значитель-
ных колебапий в размножении по голам эти авторы не отмечают, Можел
быть, опять сказались отличия в методикс отлова крыс и обработки
данных?
р
а
,
204 Глава третья
Экология 205
Я
у
Рис. 59. Сезонная динамика размно-
Рис. 61. Сезонный ход
#
<.
г]
|
1946г.
жения серых крыс в Балтиморе
размножения серых крые в
|
хи!
{Валлз, 1953)
природных биотопах (1) и
хи\
Г
а
1— склады;
8 населенных пунктах 25
у\
6
2— ферма;
Приамурья (2) (по: Хама-
\
з*
ы
д
-
! 3,4 — открытые стации;
ганов, 1985}
\
а — беременные самки,
1947-1949 ге.
б — лактирующие самки
„
Д
а
л
я
р
а
з
м
н
о
н
а
н
щ
и
с
т
с
я
с
а
м
о
к
,
%
я
т
„
Д
о
л
я
б
е
р
е
м
е
н
н
ы
х
с
а
м
о
к
,
%
з
Т
7
ИВ га
,
В Приморском крае доля размножающихся самок значительно колеб-
и
Рис. 60. Изменение доли разино-
лется по годам —от 4,6 до 37,1% — во Владивостоке, от 8,5 до 56,4% —
|.
жающился самок серых крыс в на-
Находке (Солдатов и др., 1983). Возможно, столь широкий разброс свя-
ЮГ
в
35| селенных пунктах и на рисовых по-
зан с погодными условиями разных лет.
ляз Кубани (1978—1980 °е.) й по ма-
Необычайно высока долл размножающихся самок в Якутии, которая
т
1943-1945 гг. ” 8 Рыльникова, Карасевой,
колеблется по сезонам от 80,9 до 66,6%, притом что средпее число эмб-`
1— на рисовых полях,
рионов на одну беременную самку превышает средние значения этого
г
2—в населенных пунктах.
показателя в средней полосе европейской части РСФСР и составляет 11—
ве
.| Цифры изд столбиками
— число самок в
13 эмбрионов (Романова, 1983, 1986). Эти данные сравнимы, пожалуй,
еек
| 660
только с таковыми А. А. Насешника (1936) по материалам, собранным в
Г ШУШКЯГТГШТИ
Москве. Если не принимать возможные методические погрешности при
Месяцы
сборе и интерпретации первичных данных, то наиболее вероятное объяс-
нение повышенной плодовитости самок крыс в Якутии — это необходи-
мость компенсации высокой эмбриональной смертности, которая, согласно
РА1
Ю. В. Ревину и Е. Г. Шадриной (1986), в Центральной Якутии состав-
ляет 18,7%, т.е. у 7 из 38 самок происходит частичная резорбция эмб-
Г12
рионов. Смертность в постнатальный период, по-видимому, также высока
,
из-за суровых климатических условий. Однако, по мнению Г. А. Рома-
новой (1983), пасюки там обладают приспособлениями, снижающими
в
|
постнатальную смертность, например путем «групповой терморегуляции»,
в 297
25
что позволяет существовать, размножаться и выводить потомство даже в
.
неотапливаемых помещениях.
|
у
|
Примером стациальных различий в сроках и интенсивности размно-
УТ
100 |-
е
я
5
т
18
жения серых крыс в пределах одного района могут служить данные Дэ-
виса (Пау!з, 1953а, рис. 59). По сравнению с популяцией складов г. Бал-
тимора размножение крыс на ферме близ города начипалось раньше;
осенний подъем менее выражен. У зверьков, отловленных вне строений.
наблюдался только один пик размножения, приходящийся па летние ме-
|
сяцы, причем доля лактирующих самок была низкой.
Хх МNoжщи
В открытых стациях у пасюков не наблюдается зимнего размноже-
ния даже там, где они живут вне построек человека круглый год, на-
пример па рисовых полях Кубани (Рыльников, Карасева, 1985). Подъем
размножения только олин — весенне-летний, однако доля беременных
„
Д
о
л
я
р
а
з
м
н
о
м
а
ю
щ
и
т
с
я
с
а
м
о
к
,
5
=
<
>
>
т
т
ы
с
в
н
я
Е
Я
м
ы
р
а
,
204 Глава третья
Экология 205
Я
у
Рис. 59. Сезонная динамика размно-
Рис. 61. Сезонный ход
#
<.
г]
|
1946г.
жения серых крыс в Балтиморе
размножения серых крые в
|
хи!
{Валлз, 1953)
природных биотопах (1) и
хи\
Г
а
1— склады;
8 населенных пунктах 25
у\
6
2— ферма;
Приамурья (2) (по: Хама-
\
з*
ы
д
-
! 3,4 — открытые стации;
ганов, 1985}
\
а — беременные самки,
1947-1949 ге.
б — лактирующие самки
„
Д
а
л
я
р
а
з
м
н
о
н
а
н
щ
и
с
т
с
я
с
а
м
о
к
,
%
я
т
„
Д
о
л
я
б
е
р
е
м
е
н
н
ы
х
с
а
м
о
к
,
%
з
Т
7
ИВ га
,
В Приморском крае доля размножающихся самок значительно колеб-
и
Рис. 60. Изменение доли разино-
лется по годам —от 4,6 до 37,1% — во Владивостоке, от 8,5 до 56,4% —
|.
жающился самок серых крыс в на-
Находке (Солдатов и др., 1983). Возможно, столь широкий разброс свя-
ЮГ
в
35| селенных пунктах и на рисовых по-
зан с погодными условиями разных лет.
ляз Кубани (1978—1980 °е.) й по ма-
Необычайно высока долл размножающихся самок в Якутии, которая
т
1943-1945 гг. ” 8 Рыльникова, Карасевой,
колеблется по сезонам от 80,9 до 66,6%, притом что средпее число эмб-`
1— на рисовых полях,
рионов на одну беременную самку превышает средние значения этого
г
2—в населенных пунктах.
показателя в средней полосе европейской части РСФСР и составляет 11—
ве
.| Цифры изд столбиками
— число самок в
13 эмбрионов (Романова, 1983, 1986). Эти данные сравнимы, пожалуй,
еек
| 660
только с таковыми А. А. Насешника (1936) по материалам, собранным в
Г ШУШКЯГТГШТИ
Москве. Если не принимать возможные методические погрешности при
Месяцы
сборе и интерпретации первичных данных, то наиболее вероятное объяс-
нение повышенной плодовитости самок крыс в Якутии — это необходи-
мость компенсации высокой эмбриональной смертности, которая, согласно
РА1
Ю. В. Ревину и Е. Г. Шадриной (1986), в Центральной Якутии состав-
ляет 18,7%, т.е. у 7 из 38 самок происходит частичная резорбция эмб-
Г12
рионов. Смертность в постнатальный период, по-видимому, также высока
,
из-за суровых климатических условий. Однако, по мнению Г. А. Рома-
новой (1983), пасюки там обладают приспособлениями, снижающими
в
|
постнатальную смертность, например путем «групповой терморегуляции»,
в 297
25
что позволяет существовать, размножаться и выводить потомство даже в
.
неотапливаемых помещениях.
|
у
|
Примером стациальных различий в сроках и интенсивности размно-
УТ
100 |-
е
я
5
т
18
жения серых крыс в пределах одного района могут служить данные Дэ-
виса (Пау!з, 1953а, рис. 59). По сравнению с популяцией складов г. Бал-
тимора размножение крыс на ферме близ города начипалось раньше;
осенний подъем менее выражен. У зверьков, отловленных вне строений.
наблюдался только один пик размножения, приходящийся па летние ме-
|
сяцы, причем доля лактирующих самок была низкой.
Хх МNoжщи
В открытых стациях у пасюков не наблюдается зимнего размноже-
ния даже там, где они живут вне построек человека круглый год, на-
пример па рисовых полях Кубани (Рыльников, Карасева, 1985). Подъем
размножения только олин — весенне-летний, однако доля беременных
„
Д
о
л
я
р
а
з
м
н
о
м
а
ю
щ
и
т
с
я
с
а
м
о
к
,
5
=
<
>
>
т
т
ы
с
в
н
я
Е
Я
м
ы
206
Глава третья
самок в течение этого периода даже выше, чем в населенных пунктах
(рис. 60). В природных биотопах Приамурья (Хамаганов, 1985) паблю-
дается сходная картина (рис. 61). Цо мнению авторов обоих сообщений,
более высокая интенсивность размножения крыс в природных биотопах
в весенне-летний период, которая определяется особо блатоприятными
условиями, компенсирует его ограниченность в сроках.
В открытых стациях лесостепной зопы УССР (Сумская обл.) наблю-
даются два подъема интенсивности размножения самок — весной и
осенью. Это оказалось отчетливо заметным как по числу размножающих-
ся самок, так и по среднему количеству эмбрионов в расчете на одну
беремеппую самку; летом —81, а осенью (сентябрь-октябрь) —
410,1 эмбриона (Мерзликин, 1987а). Сопоставляя эти материалы с полу-
ченными нами на рисовых полях Кубани (Рыльников, Карасева, 1985),
можно предположить следующее: в Сумской области влажность ноздуха
в летние месяцы и количество выпадающих осадков значительно меньше,
чем на Кубани. Высыхают мелкие, с сочной растительностью водоемы,
служащие местом концентрации водных насекомых и других беспозво-
ночных, являющихся основой питания серых крыс. Это, видимо, приво-
дит к резкому сужению площади, заселенной зверьками, ухудшению ус-
ловий их обитания, следствием чего может быть снижение плодовитости
самок. Средние показатели числа эмбрионов могут быть снижены, как
это уже обсуждалось выше, в результате участия в размножении моло-
дых самок, размер помета которых меньше, чем у взрослых. Осенью при
еще достаточно высоких положительных температурах влажность и ко-
личество осадков увеличиваются, что, вероятно, приводит к рассредото-
чению крыс по природным биотопам, снижению среднего уровня плотно-
сти и, как следствие этого, к подъему размножения. Хотелось бы, чтобы
эти наши предположения были подтверждены или опровергнуты полевы-
ми наблюдениями. Миграция взрослых крыс и их потомства в отапливае-
мые помещения и другие постройки человека обеснечивает защиту от
неблагоприятных условий в период похолодания (Карасева и др., 198 ба;
Мерзликин, 19876).
В средних и более северных широтах сроки размпожепия крыс в при-
родных биотопах сжаты — обычно с первой половины мая до конца сен-
тября. За это время самки успевают принести один-два помета. Не ис-
ключена возможность участия в размпожении и прибылых самок, по-
скольку в возрасте 2,5—3,0 мес они уже становятся половозрелыми (Тих-
винская, 1964; Осташтев, 1967), однако в литературе мы не нашли дан-
ных, подтверждающих это предположение. Наптими собственными иссле-
дованиями в котловине оз. Неро Ярославской области также пе удалось
выявить случаев размножения сеголеток.
Зависимость плодовитости от плотности популя-
ции. Этому вопросу посвящены классические работы Дэвиса конца
40-х
— начала 50-х годов, который показал, что зависимость плодовито-
сти от плотности населения крысе нельзя рассматривать без учета того,
на какой фазе численности находится популяция и каков средний уро-
вень ее насыщения.
`
Экология 207
Влияние плотности популяции па плодовитость самок крысе определя-
ется тем, к какому из трех типов относится исследуемая популяция:
растущая, постоянная или убывающая (Пау13, 4954а—с). Без этого по-
нятие высокой или пизкой плотности популяции не несет биологического
смысла, а является лишь относительной величиной, выражающей обилие.
Принадлежность к одному из трех упомянутых тинов популяции мо-
жет быть установлена по фазе логистического роста популяции, который,
согласно Эмлену с соавторами (Еп ел её а1., 1948), описывается так:
у=А/(41-е), гле А —уровень численности, соответствующий насыще-
нию, Ё-- параметр, &— время, измеренное от точки с координатами,
в которой у достигает среднего значения своего роста. Перлом (Реа,
1925} установлено, что скорость роста численности популяции резко спи-
жается при ее насыщепии. Согласно Эмлену с соавторами, это происхо-
дит в результате снижения плодовитости самок. При искусствеппом ра=-
режении плотности плодовитость самок увеличивается, причем тем боль-
ше, чем выше уровень ее насыщения, т. е. чем больше емкость среды
обитания.
у
В работе Дэвиса (ау!з, 1951а—с) приведены материалы эксперимен-
та по удалению известной доли крыс из понуляции с помощью орудий’
лова; это сказывается прежде всего на увеличении (на 60$) доли бере-
менных самок среди оставитихся в живых. При этом число эмбрионов в
расчете па одпу беремепную самку практически не изменяется в зависи-
мости от фазы численности: при постоянной численности — 10,3 эмбрио-
на, при растущей— также 10,3, при убывающей — 9,9. Мало изменяется
доля лактирующих самок: 43,0, 42,4, 39,9 соответственно. Дэвис предпо-
лагает, что это обусловлено повышенной смертностью крысят в растущей
популяции в гнездовой период. Больший прирост численности крыс в ра-
<тущей популяции объясняется большей выживаемостью молодняка пос-
ле их выхода из пор. Автор не находит объяснения тому факту, что в
убывающей популяции доля беременных самок выше, чем в стацио-
нарной.
В отечественной литературе наиболее полные данпые по вопросу о
плодовитости серых крыс в условиях различной плотности поселения
представлены в работах А. Н. Козлова (4981, 1982), где он пишет, что
при высокой численности крыс в помещении в условиях перенаселения
резко умепьшается доля самок, участвующих в размножении, снижается
плодовитость, повышается эмбриональная смертпость, задерживается по-
ловое созревапие молодняка (табл. 21) и что сильнее всего зависит от
плотности процент беременных (г=—0,97-0,03). Коэффициепт корреля-
ции между плотностью и процептом яловых г==-[0,84--0,17. Существует
связь между плотностью и долей самок с резорбирующимися эмбриона-
ми (г=--0,84=0,05), при которой наряду со значительным возрастани-
ем эмбриональной смертности в условиях высокой плотности наблюдает-
ся некоторое увеличение частоты резорбции в микропопуляциях с очень
низкой плотностью.
Особенно сильно подавляется репродуктивная функция в условиях
высокой плотности у молодых крыс, видимо, имеющих низкий социаль-
206
Глава третья
самок в течение этого периода даже выше, чем в населенных пунктах
(рис. 60). В природных биотопах Приамурья (Хамаганов, 1985) паблю-
дается сходная картина (рис. 61). Цо мнению авторов обоих сообщений,
более высокая интенсивность размножения крыс в природных биотопах
в весенне-летний период, которая определяется особо блатоприятными
условиями, компенсирует его ограниченность в сроках.
В открытых стациях лесостепной зопы УССР (Сумская обл.) наблю-
даются два подъема интенсивности размножения самок — весной и
осенью. Это оказалось отчетливо заметным как по числу размножающих-
ся самок, так и по среднему количеству эмбрионов в расчете на одну
беремеппую самку; летом —81, а осенью (сентябрь-октябрь) —
410,1 эмбриона (Мерзликин, 1987а). Сопоставляя эти материалы с полу-
ченными нами на рисовых полях Кубани (Рыльников, Карасева, 1985),
можно предположить следующее: в Сумской области влажность ноздуха
в летние месяцы и количество выпадающих осадков значительно меньше,
чем на Кубани. Высыхают мелкие, с сочной растительностью водоемы,
служащие местом концентрации водных насекомых и других беспозво-
ночных, являющихся основой питания серых крыс. Это, видимо, приво-
дит к резкому сужению площади, заселенной зверьками, ухудшению ус-
ловий их обитания, следствием чего может быть снижение плодовитости
самок. Средние показатели числа эмбрионов могут быть снижены, как
это уже обсуждалось выше, в результате участия в размножении моло-
дых самок, размер помета которых меньше, чем у взрослых. Осенью при
еще достаточно высоких положительных температурах влажность и ко-
личество осадков увеличиваются, что, вероятно, приводит к рассредото-
чению крыс по природным биотопам, снижению среднего уровня плотно-
сти и, как следствие этого, к подъему размножения. Хотелось бы, чтобы
эти наши предположения были подтверждены или опровергнуты полевы-
ми наблюдениями. Миграция взрослых крыс и их потомства в отапливае-
мые помещения и другие постройки человека обеснечивает защиту от
неблагоприятных условий в период похолодания (Карасева и др., 198 ба;
Мерзликин, 19876).
В средних и более северных широтах сроки размпожепия крыс в при-
родных биотопах сжаты — обычно с первой половины мая до конца сен-
тября. За это время самки успевают принести один-два помета. Не ис-
ключена возможность участия в размпожении и прибылых самок, по-
скольку в возрасте 2,5—3,0 мес они уже становятся половозрелыми (Тих-
винская, 1964; Осташтев, 1967), однако в литературе мы не нашли дан-
ных, подтверждающих это предположение. Наптими собственными иссле-
дованиями в котловине оз. Неро Ярославской области также пе удалось
выявить случаев размножения сеголеток.
Зависимость плодовитости от плотности популя-
ции. Этому вопросу посвящены классические работы Дэвиса конца
40-х
— начала 50-х годов, который показал, что зависимость плодовито-
сти от плотности населения крысе нельзя рассматривать без учета того,
на какой фазе численности находится популяция и каков средний уро-
вень ее насыщения.
`
Экология 207
Влияние плотности популяции па плодовитость самок крысе определя-
ется тем, к какому из трех типов относится исследуемая популяция:
растущая, постоянная или убывающая (Пау13, 4954а—с). Без этого по-
нятие высокой или пизкой плотности популяции не несет биологического
смысла, а является лишь относительной величиной, выражающей обилие.
Принадлежность к одному из трех упомянутых тинов популяции мо-
жет быть установлена по фазе логистического роста популяции, который,
согласно Эмлену с соавторами (Еп ел её а1., 1948), описывается так:
у=А/(41-е), гле А —уровень численности, соответствующий насыще-
нию, Ё-- параметр, &— время, измеренное от точки с координатами,
в которой у достигает среднего значения своего роста. Перлом (Реа,
1925} установлено, что скорость роста численности популяции резко спи-
жается при ее насыщепии. Согласно Эмлену с соавторами, это происхо-
дит в результате снижения плодовитости самок. При искусствеппом ра=-
режении плотности плодовитость самок увеличивается, причем тем боль-
ше, чем выше уровень ее насыщения, т. е. чем больше емкость среды
обитания.
у
В работе Дэвиса (ау!з, 1951а—с) приведены материалы эксперимен-
та по удалению известной доли крыс из понуляции с помощью орудий’
лова; это сказывается прежде всего на увеличении (на 60$) доли бере-
менных самок среди оставитихся в живых. При этом число эмбрионов в
расчете па одпу беремепную самку практически не изменяется в зависи-
мости от фазы численности: при постоянной численности — 10,3 эмбрио-
на, при растущей— также 10,3, при убывающей — 9,9. Мало изменяется
доля лактирующих самок: 43,0, 42,4, 39,9 соответственно. Дэвис предпо-
лагает, что это обусловлено повышенной смертностью крысят в растущей
популяции в гнездовой период. Больший прирост численности крыс в ра-
<тущей популяции объясняется большей выживаемостью молодняка пос-
ле их выхода из пор. Автор не находит объяснения тому факту, что в
убывающей популяции доля беременных самок выше, чем в стацио-
нарной.
В отечественной литературе наиболее полные данпые по вопросу о
плодовитости серых крыс в условиях различной плотности поселения
представлены в работах А. Н. Козлова (4981, 1982), где он пишет, что
при высокой численности крыс в помещении в условиях перенаселения
резко умепьшается доля самок, участвующих в размножении, снижается
плодовитость, повышается эмбриональная смертпость, задерживается по-
ловое созревапие молодняка (табл. 21) и что сильнее всего зависит от
плотности процент беременных (г=—0,97-0,03). Коэффициепт корреля-
ции между плотностью и процептом яловых г==-[0,84--0,17. Существует
связь между плотностью и долей самок с резорбирующимися эмбриона-
ми (г=--0,84=0,05), при которой наряду со значительным возрастани-
ем эмбриональной смертности в условиях высокой плотности наблюдает-
ся некоторое увеличение частоты резорбции в микропопуляциях с очень
низкой плотностью.
Особенно сильно подавляется репродуктивная функция в условиях
высокой плотности у молодых крыс, видимо, имеющих низкий социаль-
208 Глава третья
Таблица 21. Ивнтенеивноеть размножения серой крысы в микропопуляциях
с разной плотноетью * (по: А. Н. Козлов, 1984, е изменениями порядка
расположения граф)
Воз-
Векрыто взрослых
раст
самок
мик-
Месяц отно- | Среднее чи-
Доля самок | Среднее”
ро-
ва крыс из | сло крыс
с резорби- число Доля
попу-
микропо-
на 100 м?
рующимися | эмбрио- | Яяловых
ня-
пуляций
площади
беремен- | эмбриона- нов
самок, %
ций,
всего
ных, 9
ми. й
лет
Я
Ш
2,0
13
100
0
10,2
0
1
ТУ
3,5
112
60,7
4,9
10,9
0
1
У
45
371
45,4
6,7
9,8
0
1
УШ
3,7
117
473
12,3
9,6
0‚
1
Хх
2,2
38
44,7
41,4
9,8
2,6
1
х
20
28
43,0
8,3
9,9
3,6
1
Хх
18
67
44,9
22,2
9,8
6,0
3
И
44,0
85
24,2
АТ
8,4
25,9
3,5
ш
411
502
23,1
12,9
8,3
32,5
3,5
ТУ
79,9
246
19,1
12,8
8,6
16,3
3,5
У
66,8
207
26,6
21,2
8,7
49,2
4
УНР
113,7
285
15,8
22,2
7,0
287
* Годичные микропопуляции сформировались за время между двумя очередными дератиза-
циями, проводившимися через год.
ный ранг. Они могут вообще исключаться из размножения, поэтому в
перенаселепных микропопуляциях размножаются особи, относящиеся в
основном к старшим возрастно-весовым группам, тогда как в микропопу-
ляциях с пизкой плотностью, где иерархические взаимоотпошения выра-
жены слабее, размножаются половозрелые крысы всех возрастных групп
практически без всякого ограничения. Например, среди особей возрастно-
весовой группы 191—250 г при плотности до 10 крыс на 100 м? отмечено
в среднем 29,9—2, беременных, а при плотпости более 50 крыс на
100 м?-— всего 8,1-2,0. Эмбриональная смертность возрастает с -увели-
чением плотности гораздо сильнее у самок младших возрастпо-весовых
групи. Меньше всего изменяются в зазисимости от плотности показатели
интенсивности размножения и эмбриональной смертности у средпевоз-
растных крыс, имеющих сравнительно большой вес тела, но, по-видимо-
му, еще не старых (Козлов, 1984, с. 592—593).
В. А. Рыльников и Е. В. Карасева (1985) также пашли, что на рисо-
вых полях Кубани доля размножающихся самок тем меньше (при
р<0,05), чем больше плотность популяции. С увеличением плотности по-
пуляции крыс увеличивается доля несозревших самок не только в возра-
сте 1—2 мес, но и 3—6 мес (рис. 62). Несмотря на значительные изме-
пения плотности популяции серых крыс ва рисовых полях в течение
Экология
209
«мег.
8—6 мес.
>бмес.
Г}
|-
2
7773
|18
Рис. 62. Зависимость д0- ч’
ли размножающихея и
неравиножающится са- по |
мов серых крыс от плот-
|
ности популяции на ри-
совых полях в июле—
Я
,
августе (1978—1989 гг.)
{по: Рыльников, Кара-
й
сева, 1985)
я
Самки:
1 — размножающиеся,
2 — яловые,
3 — инфантильные.
Над столбиками показано
число самок в выборке.
Но оси ординат
— доля
размножающихея и не-
размножающихся самок, по
оси абсцисс
— число крыс 10
|
на 100 погонных метров
мыы
02
2
2
3 лет, среднее число эмбрионов в расчете на одну беременную самку ко-
лебалось незначительно (9,5—12,2). Показатель среднего числа эмбрио-
нов на одну самку в меньшей мере зависит от плотности популяции, чем
другие показатели интенсивности размножения (Козлов, 1987).
Соотношение полов
Определение пола у крыс по генитальным органам при вскрытии не
представляет особого труда, поэтому вряд ли стоит подробно остапазли-
ваться па данпом вопросе. Хотя у новорожденных это сделать сложнее,
чем у взрослых животных, но все же при внимательном осмотре их ге-
нитальной области можно обнаружить у самок тонкие ниточки рогов
матки, которые ближе к половому отверстию соединяются во влагалище,
а у самцов неболыние «горошинки»
— семенники (семенные пузырьки
находятся в зачаточном состоянии, и часто их обнаружить не удается).
В ряде случаев в руки исследователя попадают и сильно поврежден-
ныс трупы, у которых невозможно различить ни внешние, ни внутренние
гениталии. Тогда определение пола можно провести по тазовым костям —
‚лобковая кость самок тоньше и длипнее, чем у самцов, наружный край
“тела седалищной кости у самок вогнутый, у самцов выпуклый (Маяков,
Шепель, 1987).
Соотношение полов практически не зависит от того, в каком поколе-
нии при содержании в неволе опо вычислено. Часто болышее преоблада-
ние самцов в следующем поколении сменяется преобладанием самок,
причем эти отличия недостоверны. По данным Кинг, среди белых крыс
соотношение полов было 105--=2,0 в пользу самцов, несмотря на то что
они были выращены в тех же условиях, что и серые крысы. Разность до-
208 Глава третья
Таблица 21. Ивнтенеивноеть размножения серой крысы в микропопуляциях
с разной плотноетью * (по: А. Н. Козлов, 1984, е изменениями порядка
расположения граф)
Воз-
Векрыто взрослых
раст
самок
мик-
Месяц отно- | Среднее чи-
Доля самок | Среднее”
ро-
ва крыс из | сло крыс
с резорби- число Доля
попу-
микропо-
на 100 м?
рующимися | эмбрио- | Яяловых
ня-
пуляций
площади
беремен- | эмбриона- нов
самок, %
ций,
всего
ных, 9
ми. й
лет
Я
Ш
2,0
13
100
0
10,2
0
1
ТУ
3,5
112
60,7
4,9
10,9
0
1
У
45
371
45,4
6,7
9,8
0
1
УШ
3,7
117
473
12,3
9,6
0‚
1
Хх
2,2
38
44,7
41,4
9,8
2,6
1
х
20
28
43,0
8,3
9,9
3,6
1
Хх
18
67
44,9
22,2
9,8
6,0
3
И
44,0
85
24,2
АТ
8,4
25,9
3,5
ш
411
502
23,1
12,9
8,3
32,5
3,5
ТУ
79,9
246
19,1
12,8
8,6
16,3
3,5
У
66,8
207
26,6
21,2
8,7
49,2
4
УНР
113,7
285
15,8
22,2
7,0
287
* Годичные микропопуляции сформировались за время между двумя очередными дератиза-
циями, проводившимися через год.
ный ранг. Они могут вообще исключаться из размножения, поэтому в
перенаселепных микропопуляциях размножаются особи, относящиеся в
основном к старшим возрастно-весовым группам, тогда как в микропопу-
ляциях с пизкой плотностью, где иерархические взаимоотпошения выра-
жены слабее, размножаются половозрелые крысы всех возрастных групп
практически без всякого ограничения. Например, среди особей возрастно-
весовой группы 191—250 г при плотности до 10 крыс на 100 м? отмечено
в среднем 29,9—2, беременных, а при плотпости более 50 крыс на
100 м?-— всего 8,1-2,0. Эмбриональная смертность возрастает с -увели-
чением плотности гораздо сильнее у самок младших возрастпо-весовых
групи. Меньше всего изменяются в зазисимости от плотности показатели
интенсивности размножения и эмбриональной смертности у средпевоз-
растных крыс, имеющих сравнительно большой вес тела, но, по-видимо-
му, еще не старых (Козлов, 1984, с. 592—593).
В. А. Рыльников и Е. В. Карасева (1985) также пашли, что на рисо-
вых полях Кубани доля размножающихся самок тем меньше (при
р<0,05), чем больше плотность популяции. С увеличением плотности по-
пуляции крыс увеличивается доля несозревших самок не только в возра-
сте 1—2 мес, но и 3—6 мес (рис. 62). Несмотря на значительные изме-
пения плотности популяции серых крыс ва рисовых полях в течение
Экология
209
«мег.
8—6 мес.
>бмес.
Г}
|-
2
7773
|18
Рис. 62. Зависимость д0- ч’
ли размножающихея и
неравиножающится са- по |
мов серых крыс от плот-
|
ности популяции на ри-
совых полях в июле—
Я
,
августе (1978—1989 гг.)
{по: Рыльников, Кара-
й
сева, 1985)
я
Самки:
1 — размножающиеся,
2 — яловые,
3 — инфантильные.
Над столбиками показано
число самок в выборке.
Но оси ординат
— доля
размножающихея и не-
размножающихся самок, по
оси абсцисс
— число крыс 10
|
на 100 погонных метров
мыы
02
2
2
3 лет, среднее число эмбрионов в расчете на одну беременную самку ко-
лебалось незначительно (9,5—12,2). Показатель среднего числа эмбрио-
нов на одну самку в меньшей мере зависит от плотности популяции, чем
другие показатели интенсивности размножения (Козлов, 1987).
Соотношение полов
Определение пола у крыс по генитальным органам при вскрытии не
представляет особого труда, поэтому вряд ли стоит подробно остапазли-
ваться па данпом вопросе. Хотя у новорожденных это сделать сложнее,
чем у взрослых животных, но все же при внимательном осмотре их ге-
нитальной области можно обнаружить у самок тонкие ниточки рогов
матки, которые ближе к половому отверстию соединяются во влагалище,
а у самцов неболыние «горошинки»
— семенники (семенные пузырьки
находятся в зачаточном состоянии, и часто их обнаружить не удается).
В ряде случаев в руки исследователя попадают и сильно поврежден-
ныс трупы, у которых невозможно различить ни внешние, ни внутренние
гениталии. Тогда определение пола можно провести по тазовым костям —
‚лобковая кость самок тоньше и длипнее, чем у самцов, наружный край
“тела седалищной кости у самок вогнутый, у самцов выпуклый (Маяков,
Шепель, 1987).
Соотношение полов практически не зависит от того, в каком поколе-
нии при содержании в неволе опо вычислено. Часто болышее преоблада-
ние самцов в следующем поколении сменяется преобладанием самок,
причем эти отличия недостоверны. По данным Кинг, среди белых крыс
соотношение полов было 105--=2,0 в пользу самцов, несмотря на то что
они были выращены в тех же условиях, что и серые крысы. Разность до-
210
Глава третья
лей составила 7,9-2,1, что может определяться либо отибкой исходных
данных, либо эти различия паследственпо закреплепы (Киша, 1924, 1939).
Таким образом, в лабораторных условиях у серых крыс и крыс-альбино-
ов соотпошепие полов пеностоянно.
Если допустить, что в среднем соотношение, самцов и самок в популя-
ции остается приблизительно равным в течение года (1:1), что имеет
место в ряде случаев (Жирнов, 1983; Давыдов, 1986; Митропольский
и др., 1986), то это не должно влиять на сезонную и годовую продук-
тивность, прирост численности популяции и полозой отбор. Однако фак-
тически соотношение меняется в разных возрастных грунпах крысе по го-
дам, по сезонам и в разных местообитатиях.
Доля самцов среди 434 эмбрионов, согласно данным Перри (Гегту,
1945), оказалась равной 49,0-9,6%ф. Среди новорожденных крысят, оби-
тающих в скирдах соломы, самцы составляли 51,87%, причем их преоб-
ладапие наблюдали во все месяцы, кроме мая. При массе тела особей
45-194 г самцы также преобладали (51,87%), а при массе тела более
195 г опи составляли 41,34% (ГезЦе её а|., 1952). Есть данпые и об об-
ратном соотношении полов среди новорожденных. Миллер (МЩег, 1911)
нашел, что среди 54 новорожденного было самцов 82,1%, притом что
число самок принято за 100%. По 9. В. Митропольскому с соавторами
{1986), среди неполовозрелых самки составляли 52,3%, а среди взрос-
лых — 50,2%.
Установлено, что соотношение полов не имеет определенной связи с
размером пометов серых крысе (Ктр, БюмепЬиго, 1916 —цит. по: Кох,
1939). В то же время возраст матерей в некоторой степени влияет на со-
отвошение полов у новорожденных, а именно увеличивается доля самок
по отношению к самцам (Кто, 1924, 1939). Кронинг (Кгбоше, 1934,
цит. по: Кше 1939) также показал, что соотношение полов у новорож-
денных среди молодых имеет с возрастом тепдепцию к уменьшению доли
хамцов по отношению к доле самок. Это явление объясняют увеличением
пренатальной смертности самцов по мере старения их матерей, однако с
чем это связано, неизвестно. Возрастные изменения метаболизма организ-
ма, структуры матки или гормональной секреции, воздействующей на ре-
продуктивные процессы самок из группы «зепех», вероятно, снижают
жизнеспособность эмбрионов, причем в большей мере мужского пола,
чем женского (Ко, 1939).
По данным трехлетних наблюдений в Балтиморском зоопарке, доля
самцов в средпем ниже, чем доля самок. Паибольшая их доля наблюда-
лась в феврале—марте 1973 г., а наименьшая— в сентябре-октябре и
январе—феврале 1974 г. (рис. 63) (Еахпапя-Азай, Зои еК, 1979).
Изменение соотношения полов от года к году было отмечено еще
Ч. Дарвином для ряда видов, а именно: «Отношение между полами слег-
ка колеблется в последовательном ряде годов; так, у чистокровных лошеа-
дей на 100 родивитихся кобыл число жеребцов колебалось от 107,1 в од-
ном году до 92,6 в другом, а у борзых собак от 146,3 до 95,3. Но если
бы наблюдения записывались на большем пространстве, чем Англия, то
Экология 211
3
Рис. 68. Соотношение полов
по результатам ежемесячных
отловов серых крыс в Балти-
морском зоопарке (по: Рат-
рапа-Азай, бопилеь, 1979)
з
а
т
т
дАл ЛА
АУТ
„
Д
а
л
я
с
а
м
ц
о
в
,
3
,
з
0
КВ
10
ее’
ЕЕ
ОЕ
ЕЕ
ишхтпишхи и
1972г.
19732
1979г.
при большем числе эти колебания сгладились бы» (Ч. Дарвин. Происхож-
дение человека и половой отбор, с. 175).
Кэндэл (КепааП, 1984) установил, что в отловах крыс на ферме доля
самцов несколько увеличивалась зимой и уменыпалась летом. По его
мнению, эти изменения связаны с социальными взаимоотношениями
зверьков и половыми различиями в подвижности. Согласно Дэвису (Оа-
у13, 1953а), в период высокой репродуктивной активности доля самцов
составляла 47,0%, а в период низкой
— 51,3%, что соответствует мате-
риалам Кэндэла, так как зима — это период пизкой репродуктивной ак-
тивности.
Согласно наблюдепиям И. Р. Мерзликина (19876), в период летнего
выселения крыс в природные стации самцы там превосходят самок по
численности, поэтому доля самцов, обитающих в постройках, летом
уменьшается. В октябре, в период осенних миграций крыс в постройки
из открытых стаций, в средней полосе европейской части СССР наблю-
дается резкое преобладание самок в природе над самцами, которые пер-
выми уходят в отапливаемые помещения (Мерзликин, 19876}. Возможно,
труднообъяснимые колебапия в соотношении полов, отмечаемые многими
авторами, являются результатом дифференцированной гибели, а также
временного оттока или притока определенных категорий крысе из тех
мест, где производился их вылов.
Полуторакратное преобладание самцов над самками в природных
блотопах побережья Янонского моря наблюдала М. С. Девяткина (14966).
В природных биотопах Приамурья среди половозрелых особей самцы со-
ставляли 57,5% (Хамаганов, 1985).
Дэвис (ау, 1953в) не нашел отличий в соотношении полов в зави-
симости от уровня численности популяции. Напротив, А. Н. Козлов
(1981) приводит материалы о том, что в условиях высокой плотности
рождаемость самцов несколько выше. Так, в микропопуляциях с высокой
плотностью среди 805 эмбрионов самцов было 54,3%, а в микропопуля-
циях с низкой плотностью— 51,9%. По нашему мнению, в основе этих
отличий скорее всего лежат чисто случайные явления.
Необходимо иметь в виду, что способ отлова крыс может существенно
изменять действительное соотношение полов в популяции. Попадаемость
самцов и самок наверняка отличается по отношению к разным орудиям
210
Глава третья
лей составила 7,9-2,1, что может определяться либо отибкой исходных
данных, либо эти различия паследственпо закреплепы (Киша, 1924, 1939).
Таким образом, в лабораторных условиях у серых крыс и крыс-альбино-
ов соотпошепие полов пеностоянно.
Если допустить, что в среднем соотношение, самцов и самок в популя-
ции остается приблизительно равным в течение года (1:1), что имеет
место в ряде случаев (Жирнов, 1983; Давыдов, 1986; Митропольский
и др., 1986), то это не должно влиять на сезонную и годовую продук-
тивность, прирост численности популяции и полозой отбор. Однако фак-
тически соотношение меняется в разных возрастных грунпах крысе по го-
дам, по сезонам и в разных местообитатиях.
Доля самцов среди 434 эмбрионов, согласно данным Перри (Гегту,
1945), оказалась равной 49,0-9,6%ф. Среди новорожденных крысят, оби-
тающих в скирдах соломы, самцы составляли 51,87%, причем их преоб-
ладапие наблюдали во все месяцы, кроме мая. При массе тела особей
45-194 г самцы также преобладали (51,87%), а при массе тела более
195 г опи составляли 41,34% (ГезЦе её а|., 1952). Есть данпые и об об-
ратном соотношении полов среди новорожденных. Миллер (МЩег, 1911)
нашел, что среди 54 новорожденного было самцов 82,1%, притом что
число самок принято за 100%. По 9. В. Митропольскому с соавторами
{1986), среди неполовозрелых самки составляли 52,3%, а среди взрос-
лых — 50,2%.
Установлено, что соотношение полов не имеет определенной связи с
размером пометов серых крысе (Ктр, БюмепЬиго, 1916 —цит. по: Кох,
1939). В то же время возраст матерей в некоторой степени влияет на со-
отвошение полов у новорожденных, а именно увеличивается доля самок
по отношению к самцам (Кто, 1924, 1939). Кронинг (Кгбоше, 1934,
цит. по: Кше 1939) также показал, что соотношение полов у новорож-
денных среди молодых имеет с возрастом тепдепцию к уменьшению доли
хамцов по отношению к доле самок. Это явление объясняют увеличением
пренатальной смертности самцов по мере старения их матерей, однако с
чем это связано, неизвестно. Возрастные изменения метаболизма организ-
ма, структуры матки или гормональной секреции, воздействующей на ре-
продуктивные процессы самок из группы «зепех», вероятно, снижают
жизнеспособность эмбрионов, причем в большей мере мужского пола,
чем женского (Ко, 1939).
По данным трехлетних наблюдений в Балтиморском зоопарке, доля
самцов в средпем ниже, чем доля самок. Паибольшая их доля наблюда-
лась в феврале—марте 1973 г., а наименьшая— в сентябре-октябре и
январе—феврале 1974 г. (рис. 63) (Еахпапя-Азай, Зои еК, 1979).
Изменение соотношения полов от года к году было отмечено еще
Ч. Дарвином для ряда видов, а именно: «Отношение между полами слег-
ка колеблется в последовательном ряде годов; так, у чистокровных лошеа-
дей на 100 родивитихся кобыл число жеребцов колебалось от 107,1 в од-
ном году до 92,6 в другом, а у борзых собак от 146,3 до 95,3. Но если
бы наблюдения записывались на большем пространстве, чем Англия, то
Экология 211
3
Рис. 68. Соотношение полов
по результатам ежемесячных
отловов серых крыс в Балти-
морском зоопарке (по: Рат-
рапа-Азай, бопилеь, 1979)
з
а
т
т
дАл ЛА
АУТ
„
Д
а
л
я
с
а
м
ц
о
в
,
3
,
з
0
КВ
10
ее’
ЕЕ
ОЕ
ЕЕ
ишхтпишхи и
1972г.
19732
1979г.
при большем числе эти колебания сгладились бы» (Ч. Дарвин. Происхож-
дение человека и половой отбор, с. 175).
Кэндэл (КепааП, 1984) установил, что в отловах крыс на ферме доля
самцов несколько увеличивалась зимой и уменыпалась летом. По его
мнению, эти изменения связаны с социальными взаимоотношениями
зверьков и половыми различиями в подвижности. Согласно Дэвису (Оа-
у13, 1953а), в период высокой репродуктивной активности доля самцов
составляла 47,0%, а в период низкой
— 51,3%, что соответствует мате-
риалам Кэндэла, так как зима — это период пизкой репродуктивной ак-
тивности.
Согласно наблюдепиям И. Р. Мерзликина (19876), в период летнего
выселения крыс в природные стации самцы там превосходят самок по
численности, поэтому доля самцов, обитающих в постройках, летом
уменьшается. В октябре, в период осенних миграций крыс в постройки
из открытых стаций, в средней полосе европейской части СССР наблю-
дается резкое преобладание самок в природе над самцами, которые пер-
выми уходят в отапливаемые помещения (Мерзликин, 19876}. Возможно,
труднообъяснимые колебапия в соотношении полов, отмечаемые многими
авторами, являются результатом дифференцированной гибели, а также
временного оттока или притока определенных категорий крысе из тех
мест, где производился их вылов.
Полуторакратное преобладание самцов над самками в природных
блотопах побережья Янонского моря наблюдала М. С. Девяткина (14966).
В природных биотопах Приамурья среди половозрелых особей самцы со-
ставляли 57,5% (Хамаганов, 1985).
Дэвис (ау, 1953в) не нашел отличий в соотношении полов в зави-
симости от уровня численности популяции. Напротив, А. Н. Козлов
(1981) приводит материалы о том, что в условиях высокой плотности
рождаемость самцов несколько выше. Так, в микропопуляциях с высокой
плотностью среди 805 эмбрионов самцов было 54,3%, а в микропопуля-
циях с низкой плотностью— 51,9%. По нашему мнению, в основе этих
отличий скорее всего лежат чисто случайные явления.
Необходимо иметь в виду, что способ отлова крыс может существенно
изменять действительное соотношение полов в популяции. Попадаемость
самцов и самок наверняка отличается по отношению к разным орудиям
212
Глава третья
лова, в разные сезоны и в разных местообитаниях. Как было показано
ранее, с возрастом реакция на орудия лова также изменяется. Учесть
все эти методические погрешности и оценить уровень их значимости не
представляется возможным. Поэтому в болыпинстве случаев ошибкой с
‘разбросом примерпо в =10% можно препебречь, за исключением тех
случаев, когда отклонения оказываются значительно больше и сохраня-
ются в течепие длительного срока. Такие изменепия происходят в попу-
ляциях крыс, которые подвергаются истребительным мероприятиям.
По мнению Ч. Дарвина, «у некоторых, немногих животных в диком
востояний числеппые отпошения, как показано в прибавлении, колеблют-
ся в разные годы или в разных местностях, ио-видимому, достаточпо
сильно, чтобы вызвать действие полового отбора» (Ч. Дарвин. Происхож-
дение человека и половой отбор, с. 175). Здесь уместно напомнить, что
серые крысы — полигамы, а «многоженство ведет к тем же результатам,
как и действительное численное неравенство полов, потому, что, если
каждый самец обладает двумя или тремя самками, многие самцы не бу-
дут иметь возможности спариваться, и в этих последних условиях будут,
конечно, находиться более слабые и менее привлекательные особи» (Там
же). Таким образом, у серых крыс действие полового отбора может про-
исходить даже в условиях численного равенства полов. С увеличением
доли самцов по сравнению с самками значимость нолового отбора долж-
на возрастать. Это замечапие особенно важно в аспекте того, что самцы
более изменчивы, чем самки, как свидетельствуют данные как самого
Ч. Дарвина (Происхождение человека и половой отбор, с. 178), так и бо-
лее поздних исследователей (Кир, 1939). Иначе говоря, самцы являются
носителями более богатого материала для отбора.
Возрастной состав и смертность
Соотношение рождаемости, смертности и их влияние на возрастной со-
став популяции, Когда мы говорим о возрастном составе популяции се-
рых крыс, то должны иметь в виду, что это некоторое, фиксированное в
данный момент времени частотное соотношение числеппости (выражен-
ное в процентах от общей числеппости выборки) возрастных групп, ко-
торое складывается в результате взаимодействия трех векторов за опре-
деленпый промежуток времени (#), а именно: 1} удельной рождаемости
(6 — число новорожденных на одну особь из числа активных членов #10-
пуляции), 2) удельной смертности (4 — число погибающих в расчете на
одпу особь из числа активных членов популяции) и 3) удельной скоро-
сти прироста численности (г). Иначе говоря, возрастное распределение
популяции (5.) является результатом взаимодействия векторов въыживае-
мости ([.) и скорости прироста численности (67”“), т. е. 5.=[-е—"=
(Коли, 1979, с. 144—146), и отражает измепепия в выживаемости отдель-
ных возрастных групи.
Величина г зависит в естественных условиях от; 1) величины рождае-
мости, ее связи с возрастом; 2) величины смертности молодняка; 3) ожи-
даемого срока жизни взрослых самок (ГезЦе её а]., 1952). Для того чтобы
ы
Экология
213
600
т
=“
ТЕТ
ыы
ТТ:
“
|
ГОО ЗО
|
Е1
5
НН
т"
|
|
|
ь
ИРЕ
ы
лье
моя
ЯЕ|119
з
|[е
а
их
9
91ТЕ91| Е|+ 1
Зм1 тои
|
в[1
о
ааа
5
|
1
1
1
|
1
|
108
200
800
400
200
600
70
возраст,ни
Рис. 64. Изменение массы тела пасюков в зависимости от абсолютного возраста (из
8 выводков первого поколения — по таблице: Ктд, 1983}
оценить параметры пунктов 2 и Зи на их основе проводить популяци-
оппый анализ, необходимо выбрать и стапдартизировать методики опре-
деления возраста крыс и величины их смертности (для разных половоз-
растных групп в разные сезоны). Методика выделепия возрастных групи,
их соответствие действительным возрастам сказываются на оценке ос-
тальных параметров, в том числе величины рождаемости, возрастного со-
става популяции, которые тесно коррелируют друг с другом.
Определение возраста. Для определения возраста серых крыс, точнее
для распределения их по возрастным группам, больттинетво исследовате-
лей использовали массу тела в качестве показателя, который, согласно
Кинг (Кше, 1923), находится в пропорциональной зависимости от абсо-
лютного возраста (рис. 64).
В. Г. Лялин (1974а) выделяет пять возрастно-весовых групп, А. П. Ву-
зякин (1952) и А. Н. Козлов (1979) — семь. Эти группы в целом сходны
между собой и отличаются только величиной крайних значений, отделяю-
щих одну весовую группу от другой. Однако, как спразедливо отмечает
А. П. Бузякин, такое деление дает лишь приблизительное представление
о возрасте (1952). Действительно, если внимательно посмотреть на дан-
ные Кинг, то видно, сколь сильно перекрывание наименьшей массы тела
у более старших зверьков наибольшей массой тела у более молодых.
Эти данные получены в условиях лабораторного содержания, а в естест-
венных условиях такой разброс должен быть еще больше в зависимости
от климатических и погодных условий, а также от сезонпых особенностей
роста (Илевезаль, 197За, 6). Ц. С. Шварц (1959) указывал на более мед-
ленную скорость роста осенних генераций по сравнению © весенними и
летними, а отсюда и влияние сезонных особенностей роста на возрастную
структуру популяций грызунов. Поэтому методические оптибки, связан-
ные с определением возраста по массе тела, могут существенно исказить
212
Глава третья
лова, в разные сезоны и в разных местообитаниях. Как было показано
ранее, с возрастом реакция на орудия лова также изменяется. Учесть
все эти методические погрешности и оценить уровень их значимости не
представляется возможным. Поэтому в болыпинстве случаев ошибкой с
‘разбросом примерпо в =10% можно препебречь, за исключением тех
случаев, когда отклонения оказываются значительно больше и сохраня-
ются в течепие длительного срока. Такие изменепия происходят в попу-
ляциях крыс, которые подвергаются истребительным мероприятиям.
По мнению Ч. Дарвина, «у некоторых, немногих животных в диком
востояний числеппые отпошения, как показано в прибавлении, колеблют-
ся в разные годы или в разных местностях, ио-видимому, достаточпо
сильно, чтобы вызвать действие полового отбора» (Ч. Дарвин. Происхож-
дение человека и половой отбор, с. 175). Здесь уместно напомнить, что
серые крысы — полигамы, а «многоженство ведет к тем же результатам,
как и действительное численное неравенство полов, потому, что, если
каждый самец обладает двумя или тремя самками, многие самцы не бу-
дут иметь возможности спариваться, и в этих последних условиях будут,
конечно, находиться более слабые и менее привлекательные особи» (Там
же). Таким образом, у серых крыс действие полового отбора может про-
исходить даже в условиях численного равенства полов. С увеличением
доли самцов по сравнению с самками значимость нолового отбора долж-
на возрастать. Это замечапие особенно важно в аспекте того, что самцы
более изменчивы, чем самки, как свидетельствуют данные как самого
Ч. Дарвина (Происхождение человека и половой отбор, с. 178), так и бо-
лее поздних исследователей (Кир, 1939). Иначе говоря, самцы являются
носителями более богатого материала для отбора.
Возрастной состав и смертность
Соотношение рождаемости, смертности и их влияние на возрастной со-
став популяции, Когда мы говорим о возрастном составе популяции се-
рых крыс, то должны иметь в виду, что это некоторое, фиксированное в
данный момент времени частотное соотношение числеппости (выражен-
ное в процентах от общей числеппости выборки) возрастных групп, ко-
торое складывается в результате взаимодействия трех векторов за опре-
деленпый промежуток времени (#), а именно: 1} удельной рождаемости
(6 — число новорожденных на одну особь из числа активных членов #10-
пуляции), 2) удельной смертности (4 — число погибающих в расчете на
одпу особь из числа активных членов популяции) и 3) удельной скоро-
сти прироста численности (г). Иначе говоря, возрастное распределение
популяции (5.) является результатом взаимодействия векторов въыживае-
мости ([.) и скорости прироста численности (67”“), т. е. 5.=[-е—"=
(Коли, 1979, с. 144—146), и отражает измепепия в выживаемости отдель-
ных возрастных групи.
Величина г зависит в естественных условиях от; 1) величины рождае-
мости, ее связи с возрастом; 2) величины смертности молодняка; 3) ожи-
даемого срока жизни взрослых самок (ГезЦе её а]., 1952). Для того чтобы
ы
Экология
213
600
т
=“
ТЕТ
ыы
ТТ:
“
|
ГОО ЗО
|
Е1
5
НН
т"
|
|
|
ь
ИРЕ
ы
лье
моя
ЯЕ|119
з
|[е
а
их
9
91ТЕ91| Е|+ 1
Зм1 тои
|
в[1
о
ааа
5
|
1
1
1
|
1
|
108
200
800
400
200
600
70
возраст,ни
Рис. 64. Изменение массы тела пасюков в зависимости от абсолютного возраста (из
8 выводков первого поколения — по таблице: Ктд, 1983}
оценить параметры пунктов 2 и Зи на их основе проводить популяци-
оппый анализ, необходимо выбрать и стапдартизировать методики опре-
деления возраста крыс и величины их смертности (для разных половоз-
растных групп в разные сезоны). Методика выделепия возрастных групи,
их соответствие действительным возрастам сказываются на оценке ос-
тальных параметров, в том числе величины рождаемости, возрастного со-
става популяции, которые тесно коррелируют друг с другом.
Определение возраста. Для определения возраста серых крыс, точнее
для распределения их по возрастным группам, больттинетво исследовате-
лей использовали массу тела в качестве показателя, который, согласно
Кинг (Кше, 1923), находится в пропорциональной зависимости от абсо-
лютного возраста (рис. 64).
В. Г. Лялин (1974а) выделяет пять возрастно-весовых групп, А. П. Ву-
зякин (1952) и А. Н. Козлов (1979) — семь. Эти группы в целом сходны
между собой и отличаются только величиной крайних значений, отделяю-
щих одну весовую группу от другой. Однако, как спразедливо отмечает
А. П. Бузякин, такое деление дает лишь приблизительное представление
о возрасте (1952). Действительно, если внимательно посмотреть на дан-
ные Кинг, то видно, сколь сильно перекрывание наименьшей массы тела
у более старших зверьков наибольшей массой тела у более молодых.
Эти данные получены в условиях лабораторного содержания, а в естест-
венных условиях такой разброс должен быть еще больше в зависимости
от климатических и погодных условий, а также от сезонпых особенностей
роста (Илевезаль, 197За, 6). Ц. С. Шварц (1959) указывал на более мед-
ленную скорость роста осенних генераций по сравнению © весенними и
летними, а отсюда и влияние сезонных особенностей роста на возрастную
структуру популяций грызунов. Поэтому методические оптибки, связан-
ные с определением возраста по массе тела, могут существенно исказить
214
Глава третья
Е
тата.
действительное соотношение возрастных групи при анализе возрастной
структуры популяции.
Е. Н. Смирпов (1968) для определения возраста серых крыс, отлов-
ленных им в свинарниках Северпой Осетин, пользовался тремя методика-
ми: 1) по стертости жевательной поверхности зубов (Наумов, 1934,
1935; Кирис, 1937), 2} по слоистым структурам периостальной зоны ман-
дибулы (Клевезаль, Клейненберг, 1967) и 3) по размерам (высоте, копди-
лобазальной длине) и другим морфологическим особенностям черепа.
Е. Н. Смирновым была обнаружена тесная связь между перечисленными
показателями, которые он и положил в основу деления па пять возраст-
ных групи.
Определение возраста зверьков по стертости жевательной поверхно-
сти зубов верхнего ряда проводят обычно по методикам Е. Н. Смирнова
(1968) или Н. Г. Карноуховой (1974, рис. 65), отличающимся лишь тем,
что в первом случае выделено пять возрастных трупп (1—\), а во вто-
ром семь (1-УП). Е. Н. Смирнов указывал, что ГУ грунпа качественаяо
отличается от ПТ тем, что у животных ТУ группы па жевательной но:
верхности зубов дентин замыкается в петли, а У группа отличается от
ТУ тем, что у нее дентин занимает всю жевательную поверхность
(рис. 66). На основашии стого В. А. Рыльников и Е. В. Карасева (1985)
делят серых крыс по стертости жевательной поверхности зубов на пять
возрастных групи с подгруппами в [Ш и ТУ труппах, так как в них на-
ходятся зверьки, сильно отличающиеся по возрасту. Морфологически эти
отличия выражаются в виде разной степени обнажения дентина, которые
и дают право на выделение подтруни внутри этих групи. С точки зрения
этих авторов, такое деление более удобно для практического определения,
так как сначала позволяет обратить впимание на наиболее существенные
критерии стадии стертости зубов и на этой основе вести дальнейшее, бо-
лее детальное определение. По сути же предложенное этими авторами
деление на пять возрастных групп © подгрупиами в ПТ и ТУ трупиах
ничем не отличается от деления на семь возрастных групп, предложенно-
го Н. Г. Карноуховой (1971), которыми мы и будем пользоваться в даль-
нейшем.
В основу определения абсолютного возраста серых крыс по стертости
жевательной поверхности зубов Н. Г. Карноухова положила наблюдение
за стирапием зубов у крыс известиого возраста в лабораторных условиях.
Однако очевидно, что стирание зубов здесь будет проходить медленнее,
чем у диких зверьков, питающихся более грубой пищей. Могущие влиять
на скорость стирания зубов обменные процессы также протекают несрав-
нимо медленнее. Кроме того, срок жизни лабораторных крыс намного
больше, чем у диких. Все это может быть причиной завышения Карно-
уховой абсолютного возраста крыс в группах У— УП.
Изучение слоистых структур в периостальной зоне мандибулы обычна
проводят на ее поперечных срезах, начиная от участка с середины по-
следнего коренного зуба (Мз) и копчая сочлеповым отростком (Клеве-
заль, Илейненберг, 1967). Количество слоев в периостальной зоне должно
отражать число перезимовок.
з
Экология
245
Рис. 65. Возрастные изменения коренных зубов серой и черной крыс (по: Карноухо-
ва, 1971)
ИЧ — зубной ряд нижней челюсти, БЧ — зубной ряд верхней челюсти; аи — минимальная
стертость, о — максимальная, 1 — потертость эмалевого слоя, 2 — обнажение дентина
Определение возраста серых крыс по слоистым структурам кости про-
водили М. Я. Лаврова и Т. С. Минина (1968}, Н. Г. Карноухова (1971),
Н. Н. Хайрулин (1972). Последний брал крыс, добытых из холодильника
© постоянной температурой (—15°) и из убойного цеха, где изменения
температуры, влажности и освещенности были значительны, хотя и не
такие резкие, как вне построек. У всех животных разного пола и возра-
ста не было обнаружено достоверной сезопной изменчивости в количе-
стве слоев в периостальной зоне кости. Н. Г. Карноухова, изучавшая се-
рых крыс из населенных пунктов и в морском порту, тоже подтверждает
отсутствие достоверной корреляции между количеством слоистых струк-
тур в кости и возрастом.
214
Глава третья
Е
тата.
действительное соотношение возрастных групи при анализе возрастной
структуры популяции.
Е. Н. Смирпов (1968) для определения возраста серых крыс, отлов-
ленных им в свинарниках Северпой Осетин, пользовался тремя методика-
ми: 1) по стертости жевательной поверхности зубов (Наумов, 1934,
1935; Кирис, 1937), 2} по слоистым структурам периостальной зоны ман-
дибулы (Клевезаль, Клейненберг, 1967) и 3) по размерам (высоте, копди-
лобазальной длине) и другим морфологическим особенностям черепа.
Е. Н. Смирновым была обнаружена тесная связь между перечисленными
показателями, которые он и положил в основу деления па пять возраст-
ных групи.
Определение возраста зверьков по стертости жевательной поверхно-
сти зубов верхнего ряда проводят обычно по методикам Е. Н. Смирнова
(1968) или Н. Г. Карноуховой (1974, рис. 65), отличающимся лишь тем,
что в первом случае выделено пять возрастных трупп (1—\), а во вто-
ром семь (1-УП). Е. Н. Смирнов указывал, что ГУ грунпа качественаяо
отличается от ПТ тем, что у животных ТУ группы па жевательной но:
верхности зубов дентин замыкается в петли, а У группа отличается от
ТУ тем, что у нее дентин занимает всю жевательную поверхность
(рис. 66). На основашии стого В. А. Рыльников и Е. В. Карасева (1985)
делят серых крыс по стертости жевательной поверхности зубов на пять
возрастных групи с подгруппами в [Ш и ТУ труппах, так как в них на-
ходятся зверьки, сильно отличающиеся по возрасту. Морфологически эти
отличия выражаются в виде разной степени обнажения дентина, которые
и дают право на выделение подтруни внутри этих групи. С точки зрения
этих авторов, такое деление более удобно для практического определения,
так как сначала позволяет обратить впимание на наиболее существенные
критерии стадии стертости зубов и на этой основе вести дальнейшее, бо-
лее детальное определение. По сути же предложенное этими авторами
деление на пять возрастных групп © подгрупиами в ПТ и ТУ трупиах
ничем не отличается от деления на семь возрастных групп, предложенно-
го Н. Г. Карноуховой (1971), которыми мы и будем пользоваться в даль-
нейшем.
В основу определения абсолютного возраста серых крыс по стертости
жевательной поверхности зубов Н. Г. Карноухова положила наблюдение
за стирапием зубов у крыс известиого возраста в лабораторных условиях.
Однако очевидно, что стирание зубов здесь будет проходить медленнее,
чем у диких зверьков, питающихся более грубой пищей. Могущие влиять
на скорость стирания зубов обменные процессы также протекают несрав-
нимо медленнее. Кроме того, срок жизни лабораторных крыс намного
больше, чем у диких. Все это может быть причиной завышения Карно-
уховой абсолютного возраста крыс в группах У— УП.
Изучение слоистых структур в периостальной зоне мандибулы обычна
проводят на ее поперечных срезах, начиная от участка с середины по-
следнего коренного зуба (Мз) и копчая сочлеповым отростком (Клеве-
заль, Илейненберг, 1967). Количество слоев в периостальной зоне должно
отражать число перезимовок.
з
Экология
245
Рис. 65. Возрастные изменения коренных зубов серой и черной крыс (по: Карноухо-
ва, 1971)
ИЧ — зубной ряд нижней челюсти, БЧ — зубной ряд верхней челюсти; аи — минимальная
стертость, о — максимальная, 1 — потертость эмалевого слоя, 2 — обнажение дентина
Определение возраста серых крыс по слоистым структурам кости про-
водили М. Я. Лаврова и Т. С. Минина (1968}, Н. Г. Карноухова (1971),
Н. Н. Хайрулин (1972). Последний брал крыс, добытых из холодильника
© постоянной температурой (—15°) и из убойного цеха, где изменения
температуры, влажности и освещенности были значительны, хотя и не
такие резкие, как вне построек. У всех животных разного пола и возра-
ста не было обнаружено достоверной сезопной изменчивости в количе-
стве слоев в периостальной зоне кости. Н. Г. Карноухова, изучавшая се-
рых крыс из населенных пунктов и в морском порту, тоже подтверждает
отсутствие достоверной корреляции между количеством слоистых струк-
тур в кости и возрастом.
216
Глава третья
Экология
2417
Рис. 66. Возрастные группы по стадиям стертости жевательной поверхности зубов
верхней челюсти серой крысы. Фото В. А. Лапшова по материалам В. А. Рыльни-
вова
216
Глава третья
Экология
2417
Рис. 66. Возрастные группы по стадиям стертости жевательной поверхности зубов
верхней челюсти серой крысы. Фото В. А. Лапшова по материалам В. А. Рыльни-
вова
218
Глава третья
Г. А. Елевезаль (1973а, б; 1974) неоднократно указывала, что в но-
пуляцин животных, обитающих в районах со слабоконтинентальным кли-
матом, годовые линии склеивания выражены менее четко, поэтому падо
осторожно подходить к определению возраста у этой категорин.. Очевид-
но, что ее замечание в такой же мере должно относиться и к серым кры-
сам, обитающим в постройках человека, где круглый год колебания тем-
пературы, влажности и других неблагоприятных факторов в значительной
мере сглажены.
Наши собственные исследования, проведенные на серых крысах рисо-
вых полей и в населенных пунктах Кубани (Рыльников, Карасева, 1985),
лишь иллюстрируют замечание, сделаппое Цлевезаль.
Нами обнаружена невысокая корреляция между количеством слоев в
периостальной зоне мапдибулы, которая должна отражать число перези-
мовок, и показателем возраста по стертости коронок зубов у пасюков ©
рисовых полей (г=0,6, при р<0,01). Полное отсутствие такой корреля-
ции обнаружено у зверьков из построек человека ("=—0,06). Это можно
объяснить не только мягкими условиями перезимовывания, но также и
зимпим размножением крыс в постройках. У молодых животных Т, Пи
Ш возрастных групп, кости которых имеют губчатое строение, полосы
склеивания не закладываются или резорбируются (Клевезаль, Клейнен-
берг, 1967). Этим можно объяснить, почему мвогие зверьки УП возраст“
ной группы имеют лишь одну линию склеивания, а мпогие зверьки
У иУ!| групп не имеют линий вообще. Будучи отловленными обенью или
зимой, до того как успевает заложиться очередная линия склеизания,
они вполне могли быть в возрасте не менее двух лет при условии, что.
родились зимой или ранней весной. Исходя из этого можно объяснить,
почему в постройках среди зверьков УГ и УП возрастных групп (пере.
зимовавших) отношение числа крые с одной линией склеивания к
числу крыс с двумя линиями больше (0,75), чем в открытых стациях
(0,23). Аналогичные данные по числу линий склеивания у серых крыс
на Кубани получены М. Я. Лавровой и Т. С. Мининой (1968). Среди пе-
резимовавших крыс, отловленпых па лиманах Азовского моря, преоблада-
ли двухлетки, в то время как среди крыс, отловленных на открытых ста-
циях, прилегающих к станице, годовалые. Одпако авторы объясняют
это наличием миграционных путей, проходящих по околоводным биото-
пам (примыкающим к станице), по которым расселяются молодые кры-
сы. С нашей точки зрения, эти отличия определяются условиями перези-
мовывания, при которых у пасюков закладываются линии склеивания.
Таким образом, анализ слоистых структур кости у серых крыс необ-
ходимо проводить с болыной осторожностью. Однако согласиться с Кар-
воуховой (1971) в том, что эта методика совершенно непригодна для
пзучения возраста серых крыс, нельзя.
В качестве критерия относительного возраста используют также мор-
фологические особенности черепа: длину и высоту черепа, выраженность
и развитие, форму и размеры отдельных костей черепа (Смирнов, 1968).
Н. П, Наумов (1935) подчеркивал, что по этим признакам, а также по
скуловой ширине, широкоскулости (отношению скуловой ширины к кон-
г
Экология 219
дилобазальной длине), курносости (отношению длины мозговой коробки
к длине носовых костей) и т. д. можно достаточно падежно отделить
прибылых зверьков (рождения текущего года) от взрослых, но нельзя
на основе перечисленных критериев осуществлять разбивку по возраст-
вым группам из-за высокой степени трапегрессии рядов.
По нашим данным (неопубл. материалы Рыльникова и Карасевой),
кондилобазальная длина черепа обнаруживает зпачительный разброс в
зависимости от показателя возраста по степени стертости жевательной
поверхности зубов как среди самцов, так и самок, отловленных в одпом
и том же месте в одно и то же время. Но кондилобазальная длина черепа
серых крыс одного возраста, отловленных в 1978 г. в свинарниках (отап-
ливаемые помещения) и в 1980 г. в частном секторе (в неотапливаемых
сараях), достоверно (р<0,05) отличается у зверьков из отапливаемых и
холодных помещений. Эти различия скорее всего связаны с условиями
перезимовывания под влиянием разных микроклиматических факторов.
Внешние условия обитания и внутрипопуляционные факторы оказывают
существенное влияние на морфометрические показатели (Клевезаль
19736).
Существуют четыре метода оценки соответствия между относитель-
ным и абсолютным возрастом в полевых условиях: 1} мечение индивиду-
альными метками (отрезание пальцев, папесепие насечек на ушах, вы-
стригание или окрашивание шерсти и др.) с повторным отловом; 2) мас-
<овое мечение тетрациклином в приманках с повторпым или однократным
отловом; 3) определение числа линий склеивания в периостальной зоне
манлибулы; 4) многолетний анализ возрастной структуры сезонно раз-
множалщейся популяции серых крыс путем установления скорости пере-
хода модальной группы из менынего относительного возраста в больший.
Первый метод хотя и трудоемок, по позволяет наиболее точно опреде-
лить абсолютный возраст зверьков. Второй метод менее трудоемок. Мече-
ние тетрациклином производится путем однократной раскладки приман-
ки с тетрациклином. Если отлов произведен на протяжении не более
30 дней для крыс с массой тела приблизительно до 150 г, а для более
взрослых в течение срока не более 40 дней (соответственно сроки обнов-
ления резцов), время, прошедшее с момента мечения до момента отлова,
может быть с точностью до 2 дней установлено па поперечном срезе рез-
ца по числу суточных слоев, находящихсн между меткой и пульпой
(Клевезаль, 1984; Рыльников, Звонарев, 1986).
Третий метод подробпо обсуждался выше и, по всей вероятности, все
зке малонригоден.
Четвертый метод имеет свой безусловные ограничения и практически
пригоден только для изученин популяций серых крыс природных биото-
пов при условии их круглогодичного обитапия там. Тем не менее мы
подробно остановимся на нем, так как в ряде случаев мы им руководст-
вовались.
В качестве примера мы приводили произведенный нами анализ воя-
растного состава и размножения серых крыс на рисовых полях Кубани
{Рыльпиков, Карасева, 1985), позволивший установить следующее: зверь-
’
218
Глава третья
Г. А. Елевезаль (1973а, б; 1974) неоднократно указывала, что в но-
пуляцин животных, обитающих в районах со слабоконтинентальным кли-
матом, годовые линии склеивания выражены менее четко, поэтому падо
осторожно подходить к определению возраста у этой категорин.. Очевид-
но, что ее замечание в такой же мере должно относиться и к серым кры-
сам, обитающим в постройках человека, где круглый год колебания тем-
пературы, влажности и других неблагоприятных факторов в значительной
мере сглажены.
Наши собственные исследования, проведенные на серых крысах рисо-
вых полей и в населенных пунктах Кубани (Рыльников, Карасева, 1985),
лишь иллюстрируют замечание, сделаппое Цлевезаль.
Нами обнаружена невысокая корреляция между количеством слоев в
периостальной зоне мапдибулы, которая должна отражать число перези-
мовок, и показателем возраста по стертости коронок зубов у пасюков ©
рисовых полей (г=0,6, при р<0,01). Полное отсутствие такой корреля-
ции обнаружено у зверьков из построек человека ("=—0,06). Это можно
объяснить не только мягкими условиями перезимовывания, но также и
зимпим размножением крыс в постройках. У молодых животных Т, Пи
Ш возрастных групп, кости которых имеют губчатое строение, полосы
склеивания не закладываются или резорбируются (Клевезаль, Клейнен-
берг, 1967). Этим можно объяснить, почему мвогие зверьки УП возраст“
ной группы имеют лишь одну линию склеивания, а мпогие зверьки
У иУ!| групп не имеют линий вообще. Будучи отловленными обенью или
зимой, до того как успевает заложиться очередная линия склеизания,
они вполне могли быть в возрасте не менее двух лет при условии, что.
родились зимой или ранней весной. Исходя из этого можно объяснить,
почему в постройках среди зверьков УГ и УП возрастных групп (пере.
зимовавших) отношение числа крые с одной линией склеивания к
числу крыс с двумя линиями больше (0,75), чем в открытых стациях
(0,23). Аналогичные данные по числу линий склеивания у серых крыс
на Кубани получены М. Я. Лавровой и Т. С. Мининой (1968). Среди пе-
резимовавших крыс, отловленпых па лиманах Азовского моря, преоблада-
ли двухлетки, в то время как среди крыс, отловленных на открытых ста-
циях, прилегающих к станице, годовалые. Одпако авторы объясняют
это наличием миграционных путей, проходящих по околоводным биото-
пам (примыкающим к станице), по которым расселяются молодые кры-
сы. С нашей точки зрения, эти отличия определяются условиями перези-
мовывания, при которых у пасюков закладываются линии склеивания.
Таким образом, анализ слоистых структур кости у серых крыс необ-
ходимо проводить с болыной осторожностью. Однако согласиться с Кар-
воуховой (1971) в том, что эта методика совершенно непригодна для
пзучения возраста серых крыс, нельзя.
В качестве критерия относительного возраста используют также мор-
фологические особенности черепа: длину и высоту черепа, выраженность
и развитие, форму и размеры отдельных костей черепа (Смирнов, 1968).
Н. П, Наумов (1935) подчеркивал, что по этим признакам, а также по
скуловой ширине, широкоскулости (отношению скуловой ширины к кон-
г
Экология 219
дилобазальной длине), курносости (отношению длины мозговой коробки
к длине носовых костей) и т. д. можно достаточно падежно отделить
прибылых зверьков (рождения текущего года) от взрослых, но нельзя
на основе перечисленных критериев осуществлять разбивку по возраст-
вым группам из-за высокой степени трапегрессии рядов.
По нашим данным (неопубл. материалы Рыльникова и Карасевой),
кондилобазальная длина черепа обнаруживает зпачительный разброс в
зависимости от показателя возраста по степени стертости жевательной
поверхности зубов как среди самцов, так и самок, отловленных в одпом
и том же месте в одно и то же время. Но кондилобазальная длина черепа
серых крыс одного возраста, отловленных в 1978 г. в свинарниках (отап-
ливаемые помещения) и в 1980 г. в частном секторе (в неотапливаемых
сараях), достоверно (р<0,05) отличается у зверьков из отапливаемых и
холодных помещений. Эти различия скорее всего связаны с условиями
перезимовывания под влиянием разных микроклиматических факторов.
Внешние условия обитания и внутрипопуляционные факторы оказывают
существенное влияние на морфометрические показатели (Клевезаль
19736).
Существуют четыре метода оценки соответствия между относитель-
ным и абсолютным возрастом в полевых условиях: 1} мечение индивиду-
альными метками (отрезание пальцев, папесепие насечек на ушах, вы-
стригание или окрашивание шерсти и др.) с повторным отловом; 2) мас-
<овое мечение тетрациклином в приманках с повторпым или однократным
отловом; 3) определение числа линий склеивания в периостальной зоне
манлибулы; 4) многолетний анализ возрастной структуры сезонно раз-
множалщейся популяции серых крыс путем установления скорости пере-
хода модальной группы из менынего относительного возраста в больший.
Первый метод хотя и трудоемок, по позволяет наиболее точно опреде-
лить абсолютный возраст зверьков. Второй метод менее трудоемок. Мече-
ние тетрациклином производится путем однократной раскладки приман-
ки с тетрациклином. Если отлов произведен на протяжении не более
30 дней для крыс с массой тела приблизительно до 150 г, а для более
взрослых в течение срока не более 40 дней (соответственно сроки обнов-
ления резцов), время, прошедшее с момента мечения до момента отлова,
может быть с точностью до 2 дней установлено па поперечном срезе рез-
ца по числу суточных слоев, находящихсн между меткой и пульпой
(Клевезаль, 1984; Рыльников, Звонарев, 1986).
Третий метод подробпо обсуждался выше и, по всей вероятности, все
зке малонригоден.
Четвертый метод имеет свой безусловные ограничения и практически
пригоден только для изученин популяций серых крыс природных биото-
пов при условии их круглогодичного обитапия там. Тем не менее мы
подробно остановимся на нем, так как в ряде случаев мы им руководст-
вовались.
В качестве примера мы приводили произведенный нами анализ воя-
растного состава и размножения серых крыс на рисовых полях Кубани
{Рыльпиков, Карасева, 1985), позволивший установить следующее: зверь-
’
22)
Глава третья
УТГ Ярастная — 7976.
Е
О 1974.
и|арии
пЕоо
о
УГ
о
оо
о
и
о
[®)
оо
ш|
о©)
по
|
О {®)
ШЕ
о
то
[®) (©)
ТЕ
о
о
|
1
1
1
|
|
|
1
|
1
|
1
1
у\иШш
х
яияиикхяи
Е
19672.
УЕ
о
Е
(©)
т
„доля, >
М
О
Ои-я
ИЕ
@)
О-з
и.
О 0-13
ТЕ
О 5-7
о[7
14
|
1
|
1
|
1
|
]
|и
[[
|
|
диптих
ЯЯтГТТЛ
МЕСЯЦЫ
Рис, 67. Возрастной состав популяции серых крыс на рисовых полях и предполагае-
мые переходы из одной возрастной группы в другую (по: Рыльников, Карасева,
1985)
По оси ординат 1Т-УН — возраетные группы (см. рис. 71)
хи Ги П возрастцпых групп, пойманные в конце мая 1978 г. и в июне
1979 г., вероятнее всего, могли родиться в середине апреля, учитывая,
что срок беременности составляет у крыс 22—25 дней (Гамбарян, Ду-
кельская, 1955). Следовательно, абсолютный возраст этих молодых— нв
более двух месяцев. За период с июня—июля по декабрь 1979 г. боль-
шинство крыс перешли из Ги П возрастпых трупп в ПТ и ГУ, следова-
тельно, возраст зверьков [Ш группы не более 6 мес (приблизительно:
3—6 мес) (рис. 67).
С декабря по март 1980 г. основная масса зверьков из ПТ перешла
в У группу (однородность возрастного состава и отсутствие размножения
позволяют это констатировать довольно точно), а в мае 1980 г. уже зна
чительная часть крыс перешла в УТ группу, а к августу перезимовавшие
Экология
291
Таблица 22. Соответствие относительного возраста по
стертости коронок зубов абеолютному возрасту серых крые
Возрастная
Преднолагаемый абсолютный возраст, мес
группа по стер-
ов коронок
по: Карноухова, по: Смир- | 10: Ррльни-
1971
нов, 1968
сева, 41985
Т
—<1
<3
4
ИП
1—4
4—6
1-2
пьан)
3—4—6-7
3—6
п (У)
6—7 12
17—12
7—8
ТУ, (У)
12—24
9—11
ТУ У)
24—42
13—24
12—16
У (УП)
>42
>25
т
крысы составляли УТ—УП группы, отчетливо обособивииеся от сеголе-
ток (Т-ТУ). В феврале 1981 г. зверьки старших возрастных групи почти
исчезли из популяции. Гаким образом, можно считать, что возраст зверь-
ков У группы в среднем не более 11 мес (9—11), а возраст зверьков.
УГ группы в средпем пе более 16 мес. Согласно данным 1978 г., зверьки
У группы перешли в УТ грунну за период с июня по октябрь (т. е. за
4—5 мес), что подтверждают изложенные выше выводы.
Учитывая, что дважды (в 1979 и 1980 тт.) на рисовых полях Кубани
наблюдали практически полную смепу состава популяции, можно с уве-
репностью сказать, что средний срок жизни серых крыс там не превыша-
ет 2 лет с одной перезимовкой. Это, конечно, не исключает наличия в по-
пуляции пебольшого числа крысе УП группы, нерезимовавших 2 и 3 раза,
о чем свидетельствуют линии склеивания в периостальной зоне мандибу-
лы. Доля таких крыс колеблется в зависимости от скорости обновления
возрастного состава популяции.
|
На основании полученных данных составлена таблица соответствия’
возрастных групп приблизительному абсолютному возрасту (табл. 22).
Как видно, данные Рыльникова и Карасевой (1985) соответствуют
данным Карноуховой (1971) только для возрастных групп 1-—1У и рас-
ходятся с данными Е. Н. Смирнова (1968) за счет пропорционального
смещения абсолютного возраста по всем возрастным группам.
*
Величина смертности
Смертность в общем балансе сил, определяющих динамику численности,
является отрицательной составляющей, которая имеет место на всех
этапах от момента рождения молодняка. Пренатальная смертность
квалифицируется как потери до момента рождения, а поэтому должна
учитываться при расчете продуктивности самок путем вычитания
потерь до имплаптации и после нее.
22)
Глава третья
УТГ Ярастная — 7976.
Е
О 1974.
и|арии
пЕоо
о
УГ
о
оо
о
и
о
[®)
оо
ш|
о©)
по
|
О {®)
ШЕ
о
то
[®)
(©)
ТЕ
о
о
|
1
1
1
|
|
|
1
|
1
|
1
1
у\иШш
х
яияиикхяи
Е
19672.
УЕ
о
Е
(©)
т
„доля, >
М
О
Ои-я
ИЕ
@)
О-з
и.
О 0-13
ТЕ
О 5-7
о[7
1
4
|
1
|
1
|
1
|
]
|
и
[
[
|
|
диптих
ЯЯтГТТЛ
МЕСЯЦЫ
Рис, 67. Возрастной состав популяции серых крыс на рисовых полях и предполагае-
мые переходы из одной возрастной группы в другую (по: Рыльников, Карасева,
1985)
По оси ординат 1Т-УН — возраетные группы (см. рис. 71)
хи Ги П возрастцпых групп, пойманные в конце мая 1978 г. и в июне
1979 г., вероятнее всего, могли родиться в середине апреля, учитывая,
что срок беременности составляет у крыс 22—25 дней (Гамбарян, Ду-
кельская, 1955). Следовательно, абсолютный возраст этих молодых— нв
более двух месяцев. За период с июня—июля по декабрь 1979 г. боль-
шинство крыс перешли из Ги П возрастпых трупп в ПТ и ГУ, следова-
тельно, возраст зверьков [Ш группы не более 6 мес (приблизительно:
3—6 мес) (рис. 67).
С декабря по март 1980 г. основная масса зверьков из ПТ перешла
в У группу (однородность возрастного состава и отсутствие размножения
позволяют это констатировать довольно точно), а в мае 1980 г. уже зна
чительная часть крыс перешла в УТ группу, а к августу перезимовавшие
Экология
291
Таблица 22. Соответствие относительного возраста по
стертости коронок зубов абеолютному возрасту серых крые
Возрастная
Преднолагаемый абсолютный возраст, мес
группа по стер-
ов коронок по: Карноухова, по: Смир- | 10: Ррльни-
1971
нов, 1968
сева, 41985
Т
—<1
<3
4
ИП
1—4
4—6
1-2
пьан)
3—4—6-7
3—6
п (У)
6—7 12
17—12
7—8
ТУ, (У)
12—24
9—11
ТУ У)
24—42
13—24
12—16
У (УП)
>42
>25
т
крысы составляли УТ—УП группы, отчетливо обособивииеся от сеголе-
ток (Т-ТУ). В феврале 1981 г. зверьки старших возрастных групи почти
исчезли из популяции. Гаким образом, можно считать, что возраст зверь-
ков У группы в среднем не более 11 мес (9—11), а возраст зверьков.
УГ группы в средпем пе более 16 мес. Согласно данным 1978 г., зверьки
У группы перешли в УТ грунну за период с июня по октябрь (т. е. за
4—5 мес), что подтверждают изложенные выше выводы.
Учитывая, что дважды (в 1979 и 1980 тт.) на рисовых полях Кубани
наблюдали практически полную смепу состава популяции, можно с уве-
репностью сказать, что средний срок жизни серых крыс там не превыша-
ет 2 лет с одной перезимовкой. Это, конечно, не исключает наличия в по-
пуляции пебольшого числа крысе УП группы, нерезимовавших 2 и 3 раза,
о чем свидетельствуют линии склеивания в периостальной зоне мандибу-
лы. Доля таких крыс колеблется в зависимости от скорости обновления
возрастного состава популяции.
|
На основании полученных данных составлена таблица соответствия’
возрастных групп приблизительному абсолютному возрасту (табл. 22).
Как видно, данные Рыльникова и Карасевой (1985) соответствуют
данным Карноуховой (1971) только для возрастных групп 1-—1У и рас-
ходятся с данными Е. Н. Смирнова (1968) за счет пропорционального
смещения абсолютного возраста по всем возрастным группам.
*
Величина смертности
Смертность в общем балансе сил, определяющих динамику численности,
является отрицательной составляющей, которая имеет место на всех
этапах от момента рождения молодняка. Пренатальная смертность
квалифицируется как потери до момента рождения, а поэтому должна
учитываться при расчете продуктивности самок путем вычитания
потерь до имплаптации и после нее.
222
Глава третья
На основе теоретических выкладок, приведенных В. Д. Федоровым,
Т. Г. Гильмановым (1980) и Коли (1979), можно выделить основные
попятия, характеризующие смертность в популяции. Величина удель-
ной смертности (4;} в популяции равна
а;— (пупа) Ап п=Ш—5,
где 4; — доля животных погибающих, а 5, — доля животных, выживаю-
щих с возраста 7 до 71, п-.- численность возрастного класса /-1;
п, 5;— численность и удельная выживаемость возрастного класса.
Выживаемость &;—=п/и и смертность 1—1,„ которые характеризуют
вероятность выжить или соответственно умереть в возрастном интервале
от пулевого класса до возрастного класса ], где п. — численность нуле-
вого возрастного класса.
Необходимо ввести еще одно понятие
— средняя удельная смертность,
которая характеризует среднюю долю крыс всех возрастов, погибающих
т
в популяции в возрастном интервале от 0 до т, а именно @’== р ат —
т
а=У
}—0
(п—п...)/М — взвешенное среднее, которое наиболее часто используется
при практических расчетах, где М— численность популяции,
Величина смертности (средняя удельная смертность
— замечание
Рыльникова) может быть определена как число случаев гибели в расчете
на одну крысу в год и вычисляется отношением числа случаев гибели в
год к среднему числу особей в популяции.
Потери до имплантации. Преимплантационные потери подсчитывают
по отношению числа имплантировавшихся эмбриопов (включая резор-
бирующиеся) к числу желтых тел текущей беременности в расчете на
одну самку. По данным Перри (Реггу, 1945), у обследованных самок сред-
нее число овулировавших яиц было 10,84, а среднее число здоровых эм-
брионов — 8,66, т.е. потери составляли 20,5%. У 56,7% оплодотворенных
самок было потеряно по одному или по два яйца.
Иногда (7% случаев) паблюдается полиовуляция, которую рекомен-
дуется исключать из преимплантационных потерь. Число желтых ‚тел
текущей беременности в расчете на одну беременную самку в среднем
составляет 10,25. Наблюдаются изменения доли потеряпных зигот от
года к году. Зимой их доля немного увеличивается но сравнению с
летом или осенью (табл. 23). За 1,5 г. 61% самок крысе в Балтиморском
зоопарке нотеряли 11% зигот.
Одной из причин преимплантационпых потерь является пезакончив-
зпаяся лактация (ТезНе её а]., 1952).
Потери после имплантации. Постимплантационные потери опреде-
ляют обычно: 1} путем подсчета числа резорбирующихся эмбрионов по
отношению к общему числу найденных у всех беременных самок (взве-
шенное среднее); 2) по доле беременных самок, у которых обнаружены
случаи резорбции эмбрионов.
невзвешенное среднее, где 72 -— число возрастных классов, или
Экология
223
Таблица 23. Преимилантационные потери у серых крые в Балтиморском
зоопарке (19721974 гг,) (по: Еаграпя-Аха@, Зошулек, 1979)
1972 г.
1973 г.
1974 г.
Показатель
лето
осень зима весна лето осень зима
всего
Число самок
6
5
4
6
8
6
4
39
Число желтых тел
60
514260806137
394
Число эмбрионов
58
463550715432
346
Число самок с нотеря-
2
435433
24
ми зигот
Число потерянных зигот
2
5
710
9
7
5
45
Доля самок, потеряв-
33,3 80,0 | 75,0 | 83,3 | 50,0 | 50,0 | 75,0 61,0
ших зиготы, $
Доля потерянных зи-
3,3
9,8|16,6|46,7|11.2|14,55|13,5
11,0
тот, $
В Балтиморском зоопарке случаи резорбции эмбрионов наблюдались
у 20,0% самок, а доля резорбирующихся эмбрионов составила 4,6%.
Процент резорбирующихся эмбрионов был ниже весной и летом и выше
осенью и зимой (до 16,2% — табл. 24). Это пе случайно, поскольку пост-
имплантационные потери в большинстве случаев связаны со стрессом,
вызванным неблагоприятными условиями внешней среды, а также с
переуплотненностью поселений серых крыс. Осенью наблюдается зна-
чительный приток мигрантов в постройки, где численность популяции и
без того высока в результате весенне-летнего размножения. Процент
резорбирующихся эмбрионов среди молодых (200—299 г) и самых ста-
рых (500—599 г) самок (хотя и педостоверно) несколько ниже, чем
среди средневозрастных (300—499 г) (Еагвапя-Ахад, Зои иск, 1979).
Увеличенная доля самок старших возрастных групп, имеющих резер-
бирующиеся эмбрионы, по сравнению с младшими наблюдалась и среди
крыс открытых стаций Кубани и Украины (Рыльников, Карасева, 1985;
Мерзликин, 1987а), что как явление противоречит основной гипотезе,
согласно которой в процессе регуляции численности первыми из размно-
жения исключаются молодые животные, запимающие обычно низкие
иерархические ступени (Шилов, 1977, с. 174). Подтверждение этой
закономерности у серых крыс наблюдал А. Н. Козлов (1984) в животно-
,водческих помещениях Северного Казахстана, где эмбриональная смерт-
ность в условиях увеличения плотности чаще наблюдалась У самок
младиих возрастпо-весовых групп. Отмеченные противоречия, скорее
всего, являются свидетельством неоднозначности в способах регулирова-
ния численности серых крыс разных популяций.
Величина эмбриональной смертности может быть связана с условия-
ми пренатального развития. Например, вероятность резорбции зароды-
шей оказалась наименьшей на средних стадиях беременности и наиболь-
222
Глава третья
На основе теоретических выкладок, приведенных В. Д. Федоровым,
Т. Г. Гильмановым (1980) и Коли (1979), можно выделить основные
попятия, характеризующие смертность в популяции. Величина удель-
ной смертности (4;} в популяции равна
а;— (пупа) Ап п=Ш—5,
где 4; — доля животных погибающих, а 5, — доля животных, выживаю-
щих с возраста 7 до 71, п-.- численность возрастного класса /-1;
п, 5;— численность и удельная выживаемость возрастного класса.
Выживаемость &;—=п/и и смертность 1—1,„ которые характеризуют
вероятность выжить или соответственно умереть в возрастном интервале
от пулевого класса до возрастного класса ], где п. — численность нуле-
вого возрастного класса.
Необходимо ввести еще одно понятие
— средняя удельная смертность,
которая характеризует среднюю долю крыс всех возрастов, погибающих
т
в популяции в возрастном интервале от 0 до т, а именно @’== р ат —
т
а=У
}—0
(п—п...)/М — взвешенное среднее, которое наиболее часто используется
при практических расчетах, где М— численность популяции,
Величина смертности (средняя удельная смертность
— замечание
Рыльникова) может быть определена как число случаев гибели в расчете
на одну крысу в год и вычисляется отношением числа случаев гибели в
год к среднему числу особей в популяции.
Потери до имплантации. Преимплантационные потери подсчитывают
по отношению числа имплантировавшихся эмбриопов (включая резор-
бирующиеся) к числу желтых тел текущей беременности в расчете на
одну самку. По данным Перри (Реггу, 1945), у обследованных самок сред-
нее число овулировавших яиц было 10,84, а среднее число здоровых эм-
брионов — 8,66, т.е. потери составляли 20,5%. У 56,7% оплодотворенных
самок было потеряно по одному или по два яйца.
Иногда (7% случаев) паблюдается полиовуляция, которую рекомен-
дуется исключать из преимплантационных потерь. Число желтых ‚тел
текущей беременности в расчете на одну беременную самку в среднем
составляет 10,25. Наблюдаются изменения доли потеряпных зигот от
года к году. Зимой их доля немного увеличивается но сравнению с
летом или осенью (табл. 23). За 1,5 г. 61% самок крысе в Балтиморском
зоопарке нотеряли 11% зигот.
Одной из причин преимплантационпых потерь является пезакончив-
зпаяся лактация (ТезНе её а]., 1952).
Потери после имплантации. Постимплантационные потери опреде-
ляют обычно: 1} путем подсчета числа резорбирующихся эмбрионов по
отношению к общему числу найденных у всех беременных самок (взве-
шенное среднее); 2) по доле беременных самок, у которых обнаружены
случаи резорбции эмбрионов.
невзвешенное среднее, где 72 -— число возрастных классов, или
Экология
223
Таблица 23. Преимилантационные потери у серых крые в Балтиморском
зоопарке (19721974 гг,) (по: Еаграпя-Аха@, Зошулек, 1979)
1972 г.
1973 г.
1974 г.
Показатель
лето
осень зима весна лето осень зима
всего
Число самок
6
5
4
6
8
6
4
39
Число желтых тел
60
514260806137
394
Число эмбрионов
58
463550715432
346
Число самок с нотеря-
2
435433
24
ми зигот
Число потерянных зигот
2
5
710
9
7
5
45
Доля самок, потеряв-
33,3 80,0 | 75,0 | 83,3 | 50,0 | 50,0 | 75,0 61,0
ших зиготы, $
Доля потерянных зи-
3,3
9,8|16,6|46,7|11.2|14,55|13,5
11,0
тот, $
В Балтиморском зоопарке случаи резорбции эмбрионов наблюдались
у 20,0% самок, а доля резорбирующихся эмбрионов составила 4,6%.
Процент резорбирующихся эмбрионов был ниже весной и летом и выше
осенью и зимой (до 16,2% — табл. 24). Это пе случайно, поскольку пост-
имплантационные потери в большинстве случаев связаны со стрессом,
вызванным неблагоприятными условиями внешней среды, а также с
переуплотненностью поселений серых крыс. Осенью наблюдается зна-
чительный приток мигрантов в постройки, где численность популяции и
без того высока в результате весенне-летнего размножения. Процент
резорбирующихся эмбрионов среди молодых (200—299 г) и самых ста-
рых (500—599 г) самок (хотя и педостоверно) несколько ниже, чем
среди средневозрастных (300—499 г) (Еагвапя-Ахад, Зои иск, 1979).
Увеличенная доля самок старших возрастных групп, имеющих резер-
бирующиеся эмбрионы, по сравнению с младшими наблюдалась и среди
крыс открытых стаций Кубани и Украины (Рыльников, Карасева, 1985;
Мерзликин, 1987а), что как явление противоречит основной гипотезе,
согласно которой в процессе регуляции численности первыми из размно-
жения исключаются молодые животные, запимающие обычно низкие
иерархические ступени (Шилов, 1977, с. 174). Подтверждение этой
закономерности у серых крыс наблюдал А. Н. Козлов (1984) в животно-
,водческих помещениях Северного Казахстана, где эмбриональная смерт-
ность в условиях увеличения плотности чаще наблюдалась У самок
младиих возрастпо-весовых групп. Отмеченные противоречия, скорее
всего, являются свидетельством неоднозначности в способах регулирова-
ния численности серых крыс разных популяций.
Величина эмбриональной смертности может быть связана с условия-
ми пренатального развития. Например, вероятность резорбции зароды-
шей оказалась наименьшей на средних стадиях беременности и наиболь-
294
Глава третья
Таблица 24. Постимплантационные потери у серых крые (1972—1974 гг.)
(по: РагВапб-Азай, ЗоиК, 1979)
1972 г,
1973 г,
1974 г.
Показатель
лето осень зима весна лето
озень
зима
всего
Число самок
8
9
5
14
9
11
А
60
Число жизнеспособных
6677Я12879
100
32
519
О поорбирующих-
3
5
6
3
1
4
2
24
я тмотоьсре-
222
21
2
1
42
до робит
4,9 6,5 | 16,2 24] 13
4,0 6,3
4,62
Доля пометов с
25,0|22,2|40|143|14/1 18,2 |250 20,0
резорбциями, %
шей в ее начале и конце. Величина эмбриональной смертности связана
< первоначальным размером выводка геометрической прогрессией
х=0,7950", где х— вероятность того, что любой помет из п зародышей
останется без потерь (Реггу, 1945).
Смертность крысят в период пребывания в гнезде
К настоящему времени в литературе упоминается только один способ
оценки Этого показателя— по отношению числа новорожденных крысят,
обнаруженных в гнездах, к числу эмбрионов в расчете на одну самку
в год.
В неволе среди 31 670 белых крыс смертность крысят в гнезде состаз-
ляла 13% (Ко, 1921, цит. по: Кор, 1939), а среди 6295 лабораторных
серых крыс
— 1,2% (Кс, 1935). За 26 генераций серых крыс при их
содержании в неволе среди 53077 новорожденных мертвыми оказались
только 268. Смертность среди молодых крысят в период их выкармли-
вания значительно снизилась после 10 генераций. Ц этому времени
самки так хорошо приспособились к жизни в неволе, что редко умерщв-
ляли свое потомство после манипуляций (взвешивание, промеры и т. п.),
проводимых с пими исследователями. Изредка кормящие самки уничто-
жали свое потомство после того, как часть сосков была поранена молод-
няком во время кормления (К!ав, 1939), или в том случае, если условия
содержания были крайне плохие. Гнезда редко разрушались другими
животными, обитающими в той же клетке. Согласно дапным Лесли с
соавторами (ГезЦе её а1., 1952), среднее число крысят в одном гнезде 7,0,
а число эмбрионов на одну самку
— 9,03. Таким образом, вскоре после
родов по неизвестным причинам оказываются потерянными два новорож-
денных.
Дэвис (Оау!з, 1953а, Б) предлагал следующую схему расчета вероят-
Экология 225
ности выживания молодняка во время выкармливания в популяции
крыс, обитающей в складах г. Балтимора. На одну беременную самку
приходится 9,4 эмбриона. Число раз в году, когда одна взрослая самка
беременеет, составляет 4,8. Тогда число эмбрионов, которое будет про-
извелено в популяции за 16 дней. (ерок беременности по Довису), 9,4Х
Х4,8=45. В среднем 3,5 раза в году самка бывает кормящей. Если допу-
стить, что к середине срока лактации два новорожденных погибают (или
0,5 от ежегодного числа выводков), то до середины лактации выживает
детепымтей 7,4Х3,0=22. Тогда вероятность. выживания крысят от 16-го
дня беременности до середины срока лактации составит 22:45^220,49 в
течение 25 дней.
При использовании этого способа расчета самое трудное — опреде-
лить число новорожденных в помете, поскольку обнаружить гнездо крыс
сы в большинстве случаев невозможно из-за его недоступности. В таких
случаях для характеристики смертности крысят во время выкармлива-
ния постаточно воспользоваться отношением числа лактирующих самок
к числу беременных, рассчитанных за один и тот же срок наблюдения.
Так и поступает сам Дэвие (Рау, 1954а—с) при анализе характери-
стик популяций крыс. Им показано, что наибольшая смертпость крысят
во время выкармливания паблюдается в растущей популяции при срав-
нении с убывающей и стационарной, а увеличение численпости крыс
в растущей популяции происходит преимущественно за счет лучшего
выживания крысят после их выхода па поверхность ив нор (Пау1в, 1954а).
Дэвис (Рау!з, 1953а, Ь) рассматривает четыре возможных способа
определения величины смертности у жизотных в естественных условиях:
1) по возрастному составу среди погибитих животных; 2) по скорости
исчезновения из популяции маркированных животных путем мечеция
< повторным отловом; 3) по пропорции взрослых крыс в постоянных по
численности популяциях, поскольку к моменту, когда каждая молодая
крыса достигает половозрелости, погибает равное число взрослых живот-
ных; 4) по разности между известными значениями величины прироста
численности и величины рождаемости в популяции.
Вее случаи применимости этих и других методов обсуждаются в
монографии Коли (1979), однако возможности их использования по от-
ношению к серой крысе весьма небольшие.
Первые три метода Дэвис (Гау1з, 1953а) применил для определения
величины смертности серых крыс на ферме близ Балтимора. Им, в част-
ности, показано, что около 5% крыс доживали до 1 года при следующих
оказателях выживания с ранней весны (пачало отлова), где первопа-
чальное число зверьков принималось за 100%. Доля выживших поздней
весной составляла 74%, летом —60%, поздним летом
— 54%, ранпей
осенью
— 27%, поздней осенью— 15%, зимой—54. Эти дапные полу-
чепы на основе маркирования 1036 крыс, из которых 366 были отловле-
ны повторно в следующие сезоны. Величина гибели крыс на осповании
апализа возраста 60 подобранных трупов существенно пе отличалась от
таковой, установленной мечением с повторным отловом. Самки жили в
среднем на 20% дольше самцов. Сравнение продолжительности их жизни
8 Серая крыса
294
Глава третья
Таблица 24. Постимплантационные потери у серых крые (1972—1974 гг.)
(по: РагВапб-Азай, ЗоиК, 1979)
1972 г,
1973 г,
1974 г.
Показатель
лето осень зима весна лето
озень
зима
всего
Число самок
8
9
5
14
9
11
А
60
Число жизнеспособных
6677Я12879
100
32
519
О поорбирующих-
3
5
6
3
1
4
2
24
я тмотоьсре-
222
21
2
1
42
до робит
4,9 6,5 | 16,2 24] 13
4,0
6,3
4,62
Доля пометов с
25,0|22,2|40|143|14/1 18,2 |250 20,0
резорбциями, %
шей в ее начале и конце. Величина эмбриональной смертности связана
< первоначальным размером выводка геометрической прогрессией
х=0,7950", где х— вероятность того, что любой помет из п зародышей
останется без потерь (Реггу, 1945).
Смертность крысят в период пребывания в гнезде
К настоящему времени в литературе упоминается только один способ
оценки Этого показателя— по отношению числа новорожденных крысят,
обнаруженных в гнездах, к числу эмбрионов в расчете на одну самку
в год.
В неволе среди 31 670 белых крыс смертность крысят в гнезде состаз-
ляла 13% (Ко, 1921, цит. по: Кор, 1939), а среди 6295 лабораторных
серых крыс
— 1,2% (Кс, 1935). За 26 генераций серых крыс при их
содержании в неволе среди 53077 новорожденных мертвыми оказались
только 268. Смертность среди молодых крысят в период их выкармли-
вания значительно снизилась после 10 генераций. Ц этому времени
самки так хорошо приспособились к жизни в неволе, что редко умерщв-
ляли свое потомство после манипуляций (взвешивание, промеры и т. п.),
проводимых с пими исследователями. Изредка кормящие самки уничто-
жали свое потомство после того, как часть сосков была поранена молод-
няком во время кормления (К!ав, 1939), или в том случае, если условия
содержания были крайне плохие. Гнезда редко разрушались другими
животными, обитающими в той же клетке. Согласно дапным Лесли с
соавторами (ГезЦе её а1., 1952), среднее число крысят в одном гнезде 7,0,
а число эмбрионов на одну самку
— 9,03. Таким образом, вскоре после
родов по неизвестным причинам оказываются потерянными два новорож-
денных.
Дэвис (Оау!з, 1953а, Б) предлагал следующую схему расчета вероят-
Экология 225
ности выживания молодняка во время выкармливания в популяции
крыс, обитающей в складах г. Балтимора. На одну беременную самку
приходится 9,4 эмбриона. Число раз в году, когда одна взрослая самка
беременеет, составляет 4,8. Тогда число эмбрионов, которое будет про-
извелено в популяции за 16 дней. (ерок беременности по Довису), 9,4Х
Х4,8=45. В среднем 3,5 раза в году самка бывает кормящей. Если допу-
стить, что к середине срока лактации два новорожденных погибают (или
0,5 от ежегодного числа выводков), то до середины лактации выживает
детепымтей 7,4Х3,0=22. Тогда вероятность. выживания крысят от 16-го
дня беременности до середины срока лактации составит 22:45^220,49 в
течение 25 дней.
При использовании этого способа расчета самое трудное — опреде-
лить число новорожденных в помете, поскольку обнаружить гнездо крыс
сы в большинстве случаев невозможно из-за его недоступности. В таких
случаях для характеристики смертности крысят во время выкармлива-
ния постаточно воспользоваться отношением числа лактирующих самок
к числу беременных, рассчитанных за один и тот же срок наблюдения.
Так и поступает сам Дэвие (Рау, 1954а—с) при анализе характери-
стик популяций крыс. Им показано, что наибольшая смертпость крысят
во время выкармливания паблюдается в растущей популяции при срав-
нении с убывающей и стационарной, а увеличение численпости крыс
в растущей популяции происходит преимущественно за счет лучшего
выживания крысят после их выхода па поверхность ив нор (Пау1в, 1954а).
Дэвис (Рау!з, 1953а, Ь) рассматривает четыре возможных способа
определения величины смертности у жизотных в естественных условиях:
1) по возрастному составу среди погибитих животных; 2) по скорости
исчезновения из популяции маркированных животных путем мечеция
< повторным отловом; 3) по пропорции взрослых крыс в постоянных по
численности популяциях, поскольку к моменту, когда каждая молодая
крыса достигает половозрелости, погибает равное число взрослых живот-
ных; 4) по разности между известными значениями величины прироста
численности и величины рождаемости в популяции.
Вее случаи применимости этих и других методов обсуждаются в
монографии Коли (1979), однако возможности их использования по от-
ношению к серой крысе весьма небольшие.
Первые три метода Дэвис (Гау1з, 1953а) применил для определения
величины смертности серых крыс на ферме близ Балтимора. Им, в част-
ности, показано, что около 5% крыс доживали до 1 года при следующих
оказателях выживания с ранней весны (пачало отлова), где первопа-
чальное число зверьков принималось за 100%. Доля выживших поздней
весной составляла 74%, летом —60%, поздним летом
— 54%, ранпей
осенью
— 27%, поздней осенью— 15%, зимой—54. Эти дапные полу-
чепы на основе маркирования 1036 крыс, из которых 366 были отловле-
ны повторно в следующие сезоны. Величина гибели крыс на осповании
апализа возраста 60 подобранных трупов существенно пе отличалась от
таковой, установленной мечением с повторным отловом. Самки жили в
среднем на 20% дольше самцов. Сравнение продолжительности их жизни
8 Серая крыса
296
Глава третья
может быть сделапо также на основании изменений в соотношении нолов
(Пау!з, 1948).
Сходные дапные получены Хартли
и Бинюпом (Наг@еу, В1зПор, 1979).
Согласпо их предварительной оценке, продолжительность жизни серых
крыс на фермах Уэльса, полученная с помощью мечения с повторным
отловом,— от 1,5 до 9,5 мес. Некоторые крысы, жившие на полях около
фермы, достигли возраста 24—35 мес.
Однако, если принять во внимание данные как самого Дэвиса (Пау13,.
1953Ь), так и других авторов (Тапака, 1956; Тау|ог её а|., 1974, 1984;
Рыльников, 1983) о том, что вероятность повторного отлова серой крысы
резко уменьшается, можно предполагать, что величина смертности, по-
лученная этим способом, оказывается сильно завышенной.
В популяциях серых крыс с выраженным сезонным размножением
оказывается возможным при анализе динамики возрастпого состава вы-
делить группу перезимовавиих животных и проследить за изменением.
их доли в популяции в течение года.
На рисовых полях Кубани возрастной состав популяции обновляет-
ся з течение срока, не превышающего 2, а то и 1,5 лет (см. рис. 67).
Об этом же свидетельствуют и линии склеивания н периостальной зоне
мандибулы. Зверьки, пережившие 2—3 зимы, составляли ничтожную.
долю в нопуляции.
В. А. Рыльниковым и Е. В. Карасевой (1985) сделан вывод о ежегод-
ной смене значительной части состава популяции крыс за счет гибели
зверьков старших возрастных груни. Чем более суровыми оказываются
условия перезимовки, тем большая доля старых зверьков гибнет. В на-
селенных пунктах Кубани доля старших возрастных групп практически
больше во все месяцы года, что, по мнению авторов, свидетельствует о’
более медленной сменяемости поколений и может быть связано с мень-
шей смертностью от неблагоприятных факторов среды (Лаврова, Мини-
на, 1968).
В населенных пунктах температурные условия предшествовавшего
тода могут оказывать существенное влияние на долю молодняка в попу-
ляции крыс; в Ленинграде она в весенне-летний период была тем боль-
те, чем холоднее прошедшая зима (Жирнов, 1980, 1983).
Рождаемость и выживаемость за 1 год. Споеобы раечета. Поскольку‘
период видимой беременности составляет около 18 дней, то это и есть та
единица времени, по отпомению к которой выражают среднее число пол-
ных беременностей (или принесенных пометов) и среднее число крысят
(эмбриопов) в год.
Согласно Г. Коли (1979), величина, обратная средней продолжитель-
ности истинной беременности в течение сезона размножения, составит
18/365=0,05 (для серых крыс — обитателей населенных пунктов и дру-
гих местообитаний с круглогодичным размножением), а, например, для
серых крыс — обитателей рисовых полей и других природных биотонов
Кубани — 18/270=0,03, тогда число пометов в течение этих сроков соста-
вит Ро/0,05 или Рь/0,03, где Ро — доля самок с видимой беременностью, т.е.
3,1 и 2,8 соответственно (см. табл. 16, 17).
Экология 297
Дадим необходимые пояснения по технике вычисления параметра Ре.
Согласно рекомендациям Г. Коли (1979, с. 137), для усреднения доли
беременных самок и числа эмбрионов па одну самку, разбитых по воз-
растным групнам, может применяться невзвешенное среднее (Р%), при-
пт пи птиц|1
м + ии)<›ГДть,пи,ти—числоэмбрионовв
помете самок Т, П, ПТ возрастных групы, М, Ма, Мн — число самок
Т, П, ПТ возрастных групп, для которых производятся расчеты, С — число
возрастных групп.
Когда усреднение производится по месяцам года, в формуле цифры
1, П, 11... будут означать месяцы года, а С — число месяцев, в которые
зарегистрировано размножение.
В тех случаях, когда разбивка по возрастам не производится, удобнев
фассчитывать взвешенное среднее:
3—1Ри,
М+М п-т
Число случаев выкармливания (лактации) рассчитывается так же,
только при расчете в числителе используют значение срока лактации, .
который равен 36 дням (Ра\у!з, 1953), т. е. в пересчете на 1 год 36/365=
0,0986. Тогда число случаев лактации у самок за 365 дней составит:
Р/0,0986, где Р„ — доля лактирующих самок.
Пределы варьирования значений числа случаев беременности и числа
случаев лактации при 5% -ном уровне значимости могут быть оценены пу-
тем добавления к ним 20.
Среднее число эмбрионов, имеющихся у одной самки (Р,), за 1 Тод
рассчитывают следующим образом: Р/No/0,05 или No/МNo/0,08— у самок,
живущих в строениях, и у самок, живущих в природных биотопах соот-
ветственпо.
В населенных пунктах и па рисовых полях Кубани самки имеют в
среднем по 29,8 и 34,5 эмбриона в год при расчете на кафкдую независи-
мо от возраста (из числа активных членов популяции).
Поскольку многие авторы проводили расчет этих показателей па одну
взрослую самку, то можно ожидать, что цифры будут сильно завышены.
По данным Эмлена и Дэвиса (Епеп, Оау!з, 1948), в г. Балтиморе
взрослые самки (с массой тела более 250 г) приносят в среднем 3,34 поме-
та за 218 дней (с учетом 3 внутриутробных потерь), среднее число эм-
брионов составит 9,0, число эмбрионов на одну взрослую самку за
218 дней составляет 3,34Ж9,0+30,0. На одну половозрелую самку (с раз-
витыми фолликулами) в Балтиморском зоопарке приходилось в среднем
41,9 эмбриона, а с учетом ностимилантационных потерь (5,3%) — 39 но-
ворожденных в год (ЕагВапе-Аза4, ЗоВмск, 1979).
Для подсчета числа пометов и эмбрионов в них могут быть испольяо-
ваны плацентарные пятна, которые сохраняются на стенках матки в те-
чение 8—10 мес (Реггу, 1945; Смирнов, 19868).
Согласно неонубликованным данным В. А. Рыльникова и Е. В. Ка-
расевой, на рисовых полях Кубани самки-сеголетки (в возрасте до 8 мес)
чемРо=
8+
296
Глава третья
может быть сделапо также на основании изменений в соотношении нолов
(Пау!з, 1948).
Сходные дапные получены Хартли
и Бинюпом (Наг@еу, В1зПор, 1979).
Согласпо их предварительной оценке, продолжительность жизни серых
крыс на фермах Уэльса, полученная с помощью мечения с повторным
отловом,— от 1,5 до 9,5 мес. Некоторые крысы, жившие на полях около
фермы, достигли возраста 24—35 мес.
Однако, если принять во внимание данные как самого Дэвиса (Пау13,.
1953Ь), так и других авторов (Тапака, 1956; Тау|ог её а|., 1974, 1984;
Рыльников, 1983) о том, что вероятность повторного отлова серой крысы
резко уменьшается, можно предполагать, что величина смертности, по-
лученная этим способом, оказывается сильно завышенной.
В популяциях серых крыс с выраженным сезонным размножением
оказывается возможным при анализе динамики возрастпого состава вы-
делить группу перезимовавиих животных и проследить за изменением.
их доли в популяции в течение года.
На рисовых полях Кубани возрастной состав популяции обновляет-
ся з течение срока, не превышающего 2, а то и 1,5 лет (см. рис. 67).
Об этом же свидетельствуют и линии склеивания н периостальной зоне
мандибулы. Зверьки, пережившие 2—3 зимы, составляли ничтожную.
долю в нопуляции.
В. А. Рыльниковым и Е. В. Карасевой (1985) сделан вывод о ежегод-
ной смене значительной части состава популяции крыс за счет гибели
зверьков старших возрастных груни. Чем более суровыми оказываются
условия перезимовки, тем большая доля старых зверьков гибнет. В на-
селенных пунктах Кубани доля старших возрастных групп практически
больше во все месяцы года, что, по мнению авторов, свидетельствует о’
более медленной сменяемости поколений и может быть связано с мень-
шей смертностью от неблагоприятных факторов среды (Лаврова, Мини-
на, 1968).
В населенных пунктах температурные условия предшествовавшего
тода могут оказывать существенное влияние на долю молодняка в попу-
ляции крыс; в Ленинграде она в весенне-летний период была тем боль-
те, чем холоднее прошедшая зима (Жирнов, 1980, 1983).
Рождаемость и выживаемость за 1 год. Споеобы раечета. Поскольку‘
период видимой беременности составляет около 18 дней, то это и есть та
единица времени, по отпомению к которой выражают среднее число пол-
ных беременностей (или принесенных пометов) и среднее число крысят
(эмбриопов) в год.
Согласно Г. Коли (1979), величина, обратная средней продолжитель-
ности истинной беременности в течение сезона размножения, составит
18/365=0,05 (для серых крыс — обитателей населенных пунктов и дру-
гих местообитаний с круглогодичным размножением), а, например, для
серых крыс — обитателей рисовых полей и других природных биотонов
Кубани — 18/270=0,03, тогда число пометов в течение этих сроков соста-
вит Ро/0,05 или Рь/0,03, где Ро — доля самок с видимой беременностью, т.е.
3,1 и 2,8 соответственно (см. табл. 16, 17).
Экология 297
Дадим необходимые пояснения по технике вычисления параметра Ре.
Согласно рекомендациям Г. Коли (1979, с. 137), для усреднения доли
беременных самок и числа эмбрионов па одну самку, разбитых по воз-
растным групнам, может применяться невзвешенное среднее (Р%), при-
пт пи птиц|1
м + ии)<›ГДть,пи,ти—числоэмбрионовв
помете самок Т, П, ПТ возрастных групы, М, Ма, Мн — число самок
Т, П, ПТ возрастных групп, для которых производятся расчеты, С — число
возрастных групп.
Когда усреднение производится по месяцам года, в формуле цифры
1, П, 11... будут означать месяцы года, а С — число месяцев, в которые
зарегистрировано размножение.
В тех случаях, когда разбивка по возрастам не производится, удобнев
фассчитывать взвешенное среднее:
3—1Ри,
М+М п-т
Число случаев выкармливания (лактации) рассчитывается так же,
только при расчете в числителе используют значение срока лактации, .
который равен 36 дням (Ра\у!з, 1953), т. е. в пересчете на 1 год 36/365=
0,0986. Тогда число случаев лактации у самок за 365 дней составит:
Р/0,0986, где Р„ — доля лактирующих самок.
Пределы варьирования значений числа случаев беременности и числа
случаев лактации при 5% -ном уровне значимости могут быть оценены пу-
тем добавления к ним 20.
Среднее число эмбрионов, имеющихся у одной самки (Р,), за 1 Тод
рассчитывают следующим образом: Р/No/0,05 или No/МNo/0,08— у самок,
живущих в строениях, и у самок, живущих в природных биотопах соот-
ветственпо.
В населенных пунктах и па рисовых полях Кубани самки имеют в
среднем по 29,8 и 34,5 эмбриона в год при расчете на кафкдую независи-
мо от возраста (из числа активных членов популяции).
Поскольку многие авторы проводили расчет этих показателей па одну
взрослую самку, то можно ожидать, что цифры будут сильно завышены.
По данным Эмлена и Дэвиса (Епеп, Оау!з, 1948), в г. Балтиморе
взрослые самки (с массой тела более 250 г) приносят в среднем 3,34 поме-
та за 218 дней (с учетом 3 внутриутробных потерь), среднее число эм-
брионов составит 9,0, число эмбрионов на одну взрослую самку за
218 дней составляет 3,34Ж9,0+30,0. На одну половозрелую самку (с раз-
витыми фолликулами) в Балтиморском зоопарке приходилось в среднем
41,9 эмбриона, а с учетом ностимилантационных потерь (5,3%) — 39 но-
ворожденных в год (ЕагВапе-Аза4, ЗоВмск, 1979).
Для подсчета числа пометов и эмбрионов в них могут быть испольяо-
ваны плацентарные пятна, которые сохраняются на стенках матки в те-
чение 8—10 мес (Реггу, 1945; Смирнов, 19868).
Согласно неонубликованным данным В. А. Рыльникова и Е. В. Ка-
расевой, на рисовых полях Кубани самки-сеголетки (в возрасте до 8 мес)
чемРо=
8+
223
Глава третья
приносят 1—2, реже 3 помета, а перезимовавшие самки в большинстве
случаев припосят 2—3, реже 4 помета. Среди крыс населенных пунктов.
Кубани самок, принесших 3 и 4 помета, в возрасте старше 9 мес было
больше, чем на рисовых полях (рис. 68). Вак видно, эти данные находят-
ся в соответствии с произведенными выше расчетами и выводами
В. А. Рыльникова и Е. В. Карасевой (1985) о том, что, несмотря па
большую продолжительность периода размножения серых крыс в насе-
ленных пунктах, интенсивность их размножения на рисовых полях выше.
Таким образом, у серых крыс оба показателя плодовитости (число по-
метов и число детенышей в расчёте на одну самку за 1 год) популяции
зпачительпо варьируют в различных стациях и географических зонах.
Черные крысы приносят большее число выводков — в среднем 5 в год.
Число прибылых на одну самку у них 27 за один год (Оа\!з, 1953а, Ъ).
По данным К. Бейшебаева (1962), одна самка туркестанской крысы
в природных биотопах за один год жизни может дать не более 2—3 по-
метов. Это подтверждается числом плацептарных пятен па стенках мат-
ки — их не более 17—18 при средней величине выводка 410,8.
Показатель числа новорожденных в расчете на одну самку за опреде-
ленный промежуток времени (например, за один год), характеризуя по-
тенциал воспроизводства, не может быть использован для оценки попу-
ляционной динамики, в частности годовой продуктивности. Для этих.
целей используют показатели: 1) «удельной рождаемости» (5,;), т. е. от-
ношение числа новорожденных (или эмбрионов, исключая постимплап-
тационные потери) к числу животных в популяционной выборке или к
числу животных данной возрастной группы из этой популяционной вы-
борки, 2) «удельной смертности» (4,:+1) или «удельной выживаемости»
(5;,;-1). Причем последний показатель рассчитывают отношением числен-
ности последующего класса (7+1) к числепности предыдущего (7).
К сожалению, данные о выживаемости
— вероятности выжить в воз-
растном интервале от пулевого класса (0) до возрастного класса (7), обо-
значаемой 1, серых крыс в естественных условиях весьма скудны.
С учетом оговорок, упомянутых в разделе «Величина смертности»,
выживаемость серых крыс на сельскохозяйственных фермах Англии и
США можно считать примерно равной 0,05 за 1 год (5% пасюков, дожи-
вающих до возраста 1 год) (Пау!з, 1953а, 5; НагИеу, В13Ъор, 1979).
С учетом того, что удельная рождаемость за 1 год среди пасюков на фер-
мах Кубани (паши данные} составляет примерно 0,5, удельная скорость
изменения численности (г) будет равна г=0,50-— (1—0,05) =—0,40, т. е. она
отрицательна, в результате почти двукратного преобладания смертности
над рождаемостью. При таком значении удельной скорости прироста чис-
ленности серые крысы на животноводческих фермах вымерли бы в корот-
кие сроки, однако числепность там весьма высока и регулируется
преимущественно путем ограничения размножения посредством внутри-
популяционпых отпошений. Эти расчеты лишний раз убеждают в том,
что показатели выживаемости, полученные Дэвисом, а также Хартли и
Бишопом, сильно занижены. Исходя из данных самого же Дэвиса, сви*
детельслвующих, что в условиях отсутствия истребительных мероприя“
А
Числе ПОМЕТОВ
‚
ши. г
7+
[2
М
8—15|--
9—1|-
_16 8
7-8 |--
No
ца
Сг
|
|
]
|
|
д
У
ИИ
Х-М
Л
Й
8 17+
МЕСЯЦИ
<“ Г8
Челд потетов
|и
8-в|-
Е
М
9—2
в
СС`_
«
'
|
|
|
|
У
ИИ
Хх
М-Л
ВЕСА
Рис. 68. Доля самок серых крыс (от числа отловленных, 4$), принесших от 1 д0
4 пометов в зависимости от возраста и времени года
А — на рисовых полях Кубани, Б— в населенных пунктах. Цифры над кружками обозна-
чают число исследованных самок
тий численность крыс более или менее стационарпа (т. е. г=0), а, сле-
довательно, показатели удельной рождаемости и смерности примерно
равпы, вероятпость новорожденных дожить до 1 года должна быть равна
Ь, 1—4, —=1—0,50=0,50, т. е. через год остается в живых 50% пасюков.
223
Глава третья
приносят 1—2, реже 3 помета, а перезимовавшие самки в большинстве
случаев припосят 2—3, реже 4 помета. Среди крыс населенных пунктов.
Кубани самок, принесших 3 и 4 помета, в возрасте старше 9 мес было
больше, чем на рисовых полях (рис. 68). Вак видно, эти данные находят-
ся в соответствии с произведенными выше расчетами и выводами
В. А. Рыльникова и Е. В. Карасевой (1985) о том, что, несмотря па
большую продолжительность периода размножения серых крыс в насе-
ленных пунктах, интенсивность их размножения на рисовых полях выше.
Таким образом, у серых крыс оба показателя плодовитости (число по-
метов и число детенышей в расчёте на одну самку за 1 год) популяции
зпачительпо варьируют в различных стациях и географических зонах.
Черные крысы приносят большее число выводков — в среднем 5 в год.
Число прибылых на одну самку у них 27 за один год (Оа\!з, 1953а, Ъ).
По данным К. Бейшебаева (1962), одна самка туркестанской крысы
в природных биотопах за один год жизни может дать не более 2—3 по-
метов. Это подтверждается числом плацептарных пятен па стенках мат-
ки — их не более 17—18 при средней величине выводка 410,8.
Показатель числа новорожденных в расчете на одну самку за опреде-
ленный промежуток времени (например, за один год), характеризуя по-
тенциал воспроизводства, не может быть использован для оценки попу-
ляционной динамики, в частности годовой продуктивности. Для этих.
целей используют показатели: 1) «удельной рождаемости» (5,;), т. е. от-
ношение числа новорожденных (или эмбрионов, исключая постимплап-
тационные потери) к числу животных в популяционной выборке или к
числу животных данной возрастной группы из этой популяционной вы-
борки, 2) «удельной смертности» (4,:+1) или «удельной выживаемости»
(5;,;-1). Причем последний показатель рассчитывают отношением числен-
ности последующего класса (7+1) к числепности предыдущего (7).
К сожалению, данные о выживаемости
— вероятности выжить в воз-
растном интервале от пулевого класса (0) до возрастного класса (7), обо-
значаемой 1, серых крыс в естественных условиях весьма скудны.
С учетом оговорок, упомянутых в разделе «Величина смертности»,
выживаемость серых крыс на сельскохозяйственных фермах Англии и
США можно считать примерно равной 0,05 за 1 год (5% пасюков, дожи-
вающих до возраста 1 год) (Пау!з, 1953а, 5; НагИеу, В13Ъор, 1979).
С учетом того, что удельная рождаемость за 1 год среди пасюков на фер-
мах Кубани (паши данные} составляет примерно 0,5, удельная скорость
изменения численности (г) будет равна г=0,50-— (1—0,05) =—0,40, т. е. она
отрицательна, в результате почти двукратного преобладания смертности
над рождаемостью. При таком значении удельной скорости прироста чис-
ленности серые крысы на животноводческих фермах вымерли бы в корот-
кие сроки, однако числепность там весьма высока и регулируется
преимущественно путем ограничения размножения посредством внутри-
популяционпых отпошений. Эти расчеты лишний раз убеждают в том,
что показатели выживаемости, полученные Дэвисом, а также Хартли и
Бишопом, сильно занижены. Исходя из данных самого же Дэвиса, сви*
детельслвующих, что в условиях отсутствия истребительных мероприя“
А
Числе ПОМЕТОВ
‚
ши. г
7+
[2
М
8—15|--
9—1|-
_16 8
7-8 |--
No
ца
Сг
|
|
]
|
|
д
У
ИИ
Х-М
Л
Й
8 17+
МЕСЯЦИ
<“ Г8
Челд потетов
|и
8-в|-
Е
М
9—2
в
СС`_
«
'
|
|
|
|
У
ИИ
Хх
М-Л
ВЕСА
Рис. 68. Доля самок серых крыс (от числа отловленных, 4$), принесших от 1 д0
4 пометов в зависимости от возраста и времени года
А — на рисовых полях Кубани, Б— в населенных пунктах. Цифры над кружками обозна-
чают число исследованных самок
тий численность крыс более или менее стационарпа (т. е. г=0), а, сле-
довательно, показатели удельной рождаемости и смерности примерно
равпы, вероятпость новорожденных дожить до 1 года должна быть равна
Ь, 1—4, —=1—0,50=0,50, т. е. через год остается в живых 50% пасюков.
Глава четвертая.
Поведение
Онтогенез поведения
Необходимый этап при изучении процессов, связанных с развитием нове-
дения,— количественное и качественное их описание и выделение таких
периодов, в течение которых воздействие различных факторов среды мо-
жжот привести к существенпым изменениям в поведении животных. Всего
можно выделить три категории изменений, которые происходят в течение
жизни животных, особенно в ранний период: 1) некоторые формы пове-
дения исчезают по мере созревания организма, 2) с возрастом ноявляют-
ся новые поведенческие акты, 3) в ходе развития изменяется относи-
тельная частота выполнения поведенческих, в том числе и социальных,
действий. Как правило, на основе таких изменепий и с учетом характера
чувствительности животных к влиянию внешних факторов выделяются
критические (или чувствительные) перноды в их онтогенезе (Эсом, 1958;
Еох, 1966).
В этом плане наиболее обычные объекты исследований
— лаборатор-
ные крысы (Стоззеф, РоНи?, 1982). Относительно их диких родоначаль-
ников сведений значительно меньше, к тому же почти все они получены
за счет наблюдений и экспериментов в искусствеппых условиях. В дан-
пом разделе мы ограничимся качественным анализом общих закономерно-
стей в развитии поведения, акцентируя внимание на социальных взаимо-
действиях животных.
Онтогенез поведения в ранний постнатальный
период
Первые сведения о развитии детенышей лабораторных крыс мы паходим
уже в ранних работах (ЗтаП, 1899; ТИюеу, 1933). Значительно нозже по-
явились исследования, в которых становление поведенческих реакций
описывается подробно. Болле и Вуде (ВоМез, \Noооз, 1964), наблюдая
ежелневно за крысятами линии Бргавае-Оаз]еу с момепта их рождения
и до 36-дневного возраста (до 22-дневного детально), проследили динами-
ку различных форм активности детенышей (рис. 69). Анализ особенно-
стей поведения зверьков в ото время дает нам основание выделить ряд
периодов в раннем онтогенеяе лабораторных крыс подобно тому, что сде-
лано для лабораторных мышей (\У’ИНашз, 5е0й, 1953), больших несча-
пок (Бухникашвили, Крученкова, 1976) и собак (Бсой, 1958; Зеой,
Еи|ег, 1965).
1. Неонатальный период-— от рождения до 5—6-дневного возраста.
Жрысята сопершенно беспомощны и полностью зависят от матерей, ко-
торые их кормят, чистят и согревают. Движения их просты и ограничи-
ваются ползанием, сосанием и изгибапием тела. Основные формы контак-
Поведение 231
А
к
т
и
в
н
о
с
т
ь
‚
9
,
&
л
|
1
Й
1
02Й68102о6182022
Возраст, дни
Рис. 69. Динамика некоторых форм активности крысят в раннем возрасте (по: ВоПез,
Й’оо4$, 1964)
1— игра, 2 — ходьба, бег, прыжки, 3 — ползание, 4 — чистки (п том числе социальные)
тов — тактильшгые контакты друг с другом и с матерями и конкуренция
во время кормежки. В этот период в зачаточной форме появляются
чистки и первые признаки ориентации в гнезде. Спят беспокойно.
2. Переходный период
— возраст от 6 до 14 дней. Движения стано-
вятся более координированными и совершенными. Детеныши перестают
ползать и пачипают ходить. Развиваются перзые олемепты иселедова-
тельского поведения. На 13-й день открываются наружные слуховые про-
ходы. Хорошо развиты ипдивидуальные чистки. На 14-й день у пекото-
рых крысят открываются глаза. Появляется интерес к предметам и дру-
гим детенышам, отмечаются взаимные чистки. Сон более спокойный.
3. Социальный период
— возраст от 15 де 21 дня. У всех крысят от-
крываются глаза. Постепенно они начинают переходить на твердую
пишу. Общая активность резко возрастает. Наблюдается активное иссле-
довательекое поведение и манинулирование предметами. Возрастает чис-
ло игровых контактов. Характерна постоянная смена деятельпости.
С 18-дневного возраста начинают лазить по стенкам клетки. На 19-й день
крысята начинают реагировать на наблюдателя, и первая их реакция—
это приближение к дверцам клетки, даже во время их открывания. Нели
на 21-й день матерей отсадить от детенышей, то их активность несколь-
ко падает.
4. Ювенильный период— возраст от 22. до 36 дней (в 36 дней на-
блюдения были прекращены). Общий уровень активности (после отсажи-
вания матерей) быстро восстанавливается. Основной вид деятельности —
игра. Детенытти реагируют на малейние звуки. По-видимому, в это вре-
мя формируются отдельные элементы оборонительного поведения, начи-
нается перетаскивание пищи в гнезда, появляются первые несовернтенные
элементы полового новедения в виде апогетитальной инспекции и поны-
ток садок.
Глава четвертая.
Поведение
Онтогенез поведения
Необходимый этап при изучении процессов, связанных с развитием нове-
дения,— количественное и качественное их описание и выделение таких
периодов, в течение которых воздействие различных факторов среды мо-
жжот привести к существенпым изменениям в поведении животных. Всего
можно выделить три категории изменений, которые происходят в течение
жизни животных, особенно в ранний период: 1) некоторые формы пове-
дения исчезают по мере созревания организма, 2) с возрастом ноявляют-
ся новые поведенческие акты, 3) в ходе развития изменяется относи-
тельная частота выполнения поведенческих, в том числе и социальных,
действий. Как правило, на основе таких изменепий и с учетом характера
чувствительности животных к влиянию внешних факторов выделяются
критические (или чувствительные) перноды в их онтогенезе (Эсом, 1958;
Еох, 1966).
В этом плане наиболее обычные объекты исследований
— лаборатор-
ные крысы (Стоззеф, РоНи?, 1982). Относительно их диких родоначаль-
ников сведений значительно меньше, к тому же почти все они получены
за счет наблюдений и экспериментов в искусствеппых условиях. В дан-
пом разделе мы ограничимся качественным анализом общих закономерно-
стей в развитии поведения, акцентируя внимание на социальных взаимо-
действиях животных.
Онтогенез поведения в ранний постнатальный
период
Первые сведения о развитии детенышей лабораторных крыс мы паходим
уже в ранних работах (ЗтаП, 1899; ТИюеу, 1933). Значительно нозже по-
явились исследования, в которых становление поведенческих реакций
описывается подробно. Болле и Вуде (ВоМез, \Noооз, 1964), наблюдая
ежелневно за крысятами линии Бргавае-Оаз]еу с момепта их рождения
и до 36-дневного возраста (до 22-дневного детально), проследили динами-
ку различных форм активности детенышей (рис. 69). Анализ особенно-
стей поведения зверьков в ото время дает нам основание выделить ряд
периодов в раннем онтогенеяе лабораторных крыс подобно тому, что сде-
лано для лабораторных мышей (\У’ИНашз, 5е0й, 1953), больших несча-
пок (Бухникашвили, Крученкова, 1976) и собак (Бсой, 1958; Зеой,
Еи|ег, 1965).
1. Неонатальный период-— от рождения до 5—6-дневного возраста.
Жрысята сопершенно беспомощны и полностью зависят от матерей, ко-
торые их кормят, чистят и согревают. Движения их просты и ограничи-
ваются ползанием, сосанием и изгибапием тела. Основные формы контак-
Поведение 231
А
к
т
и
в
н
о
с
т
ь
‚
9
,
&
л
|
1
Й
1
02Й68102о6182022
Возраст, дни
Рис. 69. Динамика некоторых форм активности крысят в раннем возрасте (по: ВоПез,
Й’оо4$, 1964)
1— игра, 2 — ходьба, бег, прыжки, 3 — ползание, 4 — чистки (п том числе социальные)
тов — тактильшгые контакты друг с другом и с матерями и конкуренция
во время кормежки. В этот период в зачаточной форме появляются
чистки и первые признаки ориентации в гнезде. Спят беспокойно.
2. Переходный период
— возраст от 6 до 14 дней. Движения стано-
вятся более координированными и совершенными. Детеныши перестают
ползать и пачипают ходить. Развиваются перзые олемепты иселедова-
тельского поведения. На 13-й день открываются наружные слуховые про-
ходы. Хорошо развиты ипдивидуальные чистки. На 14-й день у пекото-
рых крысят открываются глаза. Появляется интерес к предметам и дру-
гим детенышам, отмечаются взаимные чистки. Сон более спокойный.
3. Социальный период
— возраст от 15 де 21 дня. У всех крысят от-
крываются глаза. Постепенно они начинают переходить на твердую
пишу. Общая активность резко возрастает. Наблюдается активное иссле-
довательекое поведение и манинулирование предметами. Возрастает чис-
ло игровых контактов. Характерна постоянная смена деятельпости.
С 18-дневного возраста начинают лазить по стенкам клетки. На 19-й день
крысята начинают реагировать на наблюдателя, и первая их реакция—
это приближение к дверцам клетки, даже во время их открывания. Нели
на 21-й день матерей отсадить от детенышей, то их активность несколь-
ко падает.
4. Ювенильный период— возраст от 22. до 36 дней (в 36 дней на-
блюдения были прекращены). Общий уровень активности (после отсажи-
вания матерей) быстро восстанавливается. Основной вид деятельности —
игра. Детенытти реагируют на малейние звуки. По-видимому, в это вре-
мя формируются отдельные элементы оборонительного поведения, начи-
нается перетаскивание пищи в гнезда, появляются первые несовернтенные
элементы полового новедения в виде апогетитальной инспекции и поны-
ток садок.
232
Глава четвертая
Таким образом, даппая периодизация раннего постнатального онтоге-
неза лабораторных крыс осповывается преимущественно па поведенче-
ских признаках. Несмотря на определенную условность (особенно в на-
званиях периодов), она тем не менее отражает стенень зависимости де-
тенышей от окружающих факторов и позволяет более тщательно иссле-
довать влияние раннего опыта на последующее поведение взрослых
особей.
,
Как происходит рост и развитие детенышей серых крыс в период
гнездовой жизни, до сих пор мало известно. Отдельные наблюдепия ряда
авторов (СаШопп, 1963; Ляпунова, 1980; Квашнин, 1983а) можно сум-
мировать следующим образом. В первые 5 дней жизни крысята издают
слышимые звуки, если их беспокоят. В 6—11 дней у них появляется во-
лосяной покров (если матери оставляют гнездо с крысятами в возрасте
до 12—14 дней в прохладную погоду, то перед уходом они их «утспляют»,
заваливая бумагой, сеном и т. п. (Квашнин, неопубл. данные)), что, оче-
видно, связано с особеппостями механизмов терморегуляции крысят. На
14-й — 21-й день у детенытей открываются глаза. Первые их выходы из
тнезд и убежищ па поверхность наблюдаются з возрасте 20—29 дней.
Тогда же отмечается их самостоятельное питание твердыми кормами
(возможно, переход на немолочное питание происходит и раньше, по-
скольку матери активно перетаскивают пишу в гпезда задолго до выхода
крысят из укрытий). В возрасте 27—30 дней крысята начинают перетас-
кивать в гнезда кусочки пищи и строительпый материал. И пакопец, по
данным Колхауна, дикие крысы перестают питаться молоком матери в
возрасте 30—38 дней (у лабораторных крыс лактация прекращается
раньше — к 28 дням (ВаБсКу её а]., 1970).
Сопоставляя изложенные выше данные, можно отметить, что в инди-
видуальном развитии диких и доместицированных крыс имеется ряд 0б-
щих черт — примерно в одно и то же время происходит открывание глаз,
начинается переносе в убежища предметов, в том числе и пищи. Лабора-
торные животные замечают наблюдателя (т. е. постороннии раздражи-
тель) в тот период, когда дикие зверьки впервые выходят за пределы
гнезд. Однако, по-видимому, ранний период жизни у серых крыс более
продолжительтый, чем у лабораторных, поскольку они дольше питаются
молоком матери. Возможно, и формирование нозеденческих реакций у
доместицированных крыс происходит раньше, о чем, в частности, свиде-
тельствует полное отсутствие у них реакции страха на новые стимулы,
тогда как у серых крыс наблюдается ярко выраженная пугливость, 0со-
бенно заметная при их первых выходах из укрытий.
Онтогенез поведения в послегнездовой период
Начиная с ювенильного нериода и вплоть до взрослого возраста в поведе-
нии молодых крыс происходят существенные изменения, аналогичные
тем, что характерны для раннего постнатального периода: с одной сторо-
ны, появляются или исчезают отдельные элементы поведения, а с дру-
гой — изменяются характер и частота социальных коптактов между всеми
Поведение
733
животными. Последние изменения являются пе менее важными для оп-
ределения так называемых «критических» периодов в позднем постна-
тальном онтогенезе молодняка (Ногулсй, 1972).
Динамика поведенческих реакций в это время достаточно полно про-
слежена у диких серых крыс, поэтому им и будет уделено основное вни-
мание. Здесь следует оговорить, что основной формой активности доме-
стицироваипых и серых крыс в «послегнездовой» период является игра,
которую мы рассмотрим пиже.
Долговремеппые наблюдения Колхауна {СаФопл, 1963) за колонией
серых крыс в условиях большой выгородки позволили ему выявить неко-
торые особенности поведения молодых зверьков и их взаимоотношений
со взрослыми животными вплоть до формирования у них полового пове-
дения (рис. 70). Однако эти данные носят отрывочный характер. Вос-
полнить этот пробел нам удалось за счет длительных и тщательных на-
блюдепий за динамикой поведенческих реакций у молодых серых крыс
в условиях небольших вольер (Соколов, Квашнин, 1983а, 1985; Квашнин,
1983а). На осноне этих результатов в послегнездовом онтогенезе пасюков
нами были выделены следующие возрастпые периоды, каждый из кото-
рых характеризовался своими особенностями поведения молодняка и спе-
цифическими формами социальных отношений между всеми членами экс-
периментальных групп (рис. 74):
1. Ювенильный период; по-видимому, начипается с момента выхода
крысят из гнезд в возрасте 20—25 дней“, когда они пачинают осваи-
вать повую для них среду, и заканчивается, когда крысята начинают фи-
зиологически созревать (в 65—70 дней, по Колхауну немпого позже).
Родители и другие взрослые особи мало внимания обращают на детепы-
шей вне гнезд, как бы пе замечают их присутствия. Для крысят, напро-
тив, характерен усиленный интерес к взрослым особям, особенно к мате-
рям— они постоянно следуют за ними, повторяют их движения и их
реакцию на предметы внептней среды. В этот период для детеньшней се-
рых крыс характерны, с одной стороны, повышенная пугливость и осто-
рожность, а с другой
— слепое подражапие действиям матери. Молодые
звервки много и охотно играют друг с другом и с матерью. В их поведен-
ческом репертуаре отсутствуют элементы полового, агонистического и
маркировочного поведения, характерные для взрослых особей.
Поведение лабораторных крыс этого возраста уже заметно отличается
от таконого у диких животных. Начало репродуктивного поведения ха-
рактерно для самцов в возрасте 36—40 дней, когда они впервые проявля-
ют влечение к самкам. В возрасте 41—45 дней у самцов отмечаются сад-
‘ки, а у самок впервые появляются позы лордоза (ВоПез, У’оойв, 1964;
Меоапеу, Эезагь 1981Ъ). Морфологические признаки полового созревания
наступают несколько раньше, чем проявляется половое влечение (Мегс!-
ет © а|., 1982). Б этом возрасте у лабораторных животных резко увели-
12 Это следует из периодизации раннего постнатального онтогенеза лабораторных
крыс. Однако накопление Данных об этом периоде развития У серых крысе, воз-
можно, внесет поправки и уточнения в возрастную классификацию пасюков.
232
Глава четвертая
Таким образом, даппая периодизация раннего постнатального онтоге-
неза лабораторных крыс осповывается преимущественно па поведенче-
ских признаках. Несмотря на определенную условность (особенно в на-
званиях периодов), она тем не менее отражает стенень зависимости де-
тенышей от окружающих факторов и позволяет более тщательно иссле-
довать влияние раннего опыта на последующее поведение взрослых
особей.
,
Как происходит рост и развитие детенышей серых крыс в период
гнездовой жизни, до сих пор мало известно. Отдельные наблюдепия ряда
авторов (СаШопп, 1963; Ляпунова, 1980; Квашнин, 1983а) можно сум-
мировать следующим образом. В первые 5 дней жизни крысята издают
слышимые звуки, если их беспокоят. В 6—11 дней у них появляется во-
лосяной покров (если матери оставляют гнездо с крысятами в возрасте
до 12—14 дней в прохладную погоду, то перед уходом они их «утспляют»,
заваливая бумагой, сеном и т. п. (Квашнин, неопубл. данные)), что, оче-
видно, связано с особеппостями механизмов терморегуляции крысят. На
14-й — 21-й день у детенытей открываются глаза. Первые их выходы из
тнезд и убежищ па поверхность наблюдаются з возрасте 20—29 дней.
Тогда же отмечается их самостоятельное питание твердыми кормами
(возможно, переход на немолочное питание происходит и раньше, по-
скольку матери активно перетаскивают пишу в гпезда задолго до выхода
крысят из укрытий). В возрасте 27—30 дней крысята начинают перетас-
кивать в гнезда кусочки пищи и строительпый материал. И пакопец, по
данным Колхауна, дикие крысы перестают питаться молоком матери в
возрасте 30—38 дней (у лабораторных крыс лактация прекращается
раньше — к 28 дням (ВаБсКу её а]., 1970).
Сопоставляя изложенные выше данные, можно отметить, что в инди-
видуальном развитии диких и доместицированных крыс имеется ряд 0б-
щих черт — примерно в одно и то же время происходит открывание глаз,
начинается переносе в убежища предметов, в том числе и пищи. Лабора-
торные животные замечают наблюдателя (т. е. постороннии раздражи-
тель) в тот период, когда дикие зверьки впервые выходят за пределы
гнезд. Однако, по-видимому, ранний период жизни у серых крыс более
продолжительтый, чем у лабораторных, поскольку они дольше питаются
молоком матери. Возможно, и формирование нозеденческих реакций у
доместицированных крыс происходит раньше, о чем, в частности, свиде-
тельствует полное отсутствие у них реакции страха на новые стимулы,
тогда как у серых крыс наблюдается ярко выраженная пугливость, 0со-
бенно заметная при их первых выходах из укрытий.
Онтогенез поведения в послегнездовой период
Начиная с ювенильного нериода и вплоть до взрослого возраста в поведе-
нии молодых крыс происходят существенные изменения, аналогичные
тем, что характерны для раннего постнатального периода: с одной сторо-
ны, появляются или исчезают отдельные элементы поведения, а с дру-
гой — изменяются характер и частота социальных коптактов между всеми
Поведение
733
животными. Последние изменения являются пе менее важными для оп-
ределения так называемых «критических» периодов в позднем постна-
тальном онтогенезе молодняка (Ногулсй, 1972).
Динамика поведенческих реакций в это время достаточно полно про-
слежена у диких серых крыс, поэтому им и будет уделено основное вни-
мание. Здесь следует оговорить, что основной формой активности доме-
стицироваипых и серых крыс в «послегнездовой» период является игра,
которую мы рассмотрим пиже.
Долговремеппые наблюдения Колхауна {СаФопл, 1963) за колонией
серых крыс в условиях большой выгородки позволили ему выявить неко-
торые особенности поведения молодых зверьков и их взаимоотношений
со взрослыми животными вплоть до формирования у них полового пове-
дения (рис. 70). Однако эти данные носят отрывочный характер. Вос-
полнить этот пробел нам удалось за счет длительных и тщательных на-
блюдепий за динамикой поведенческих реакций у молодых серых крыс
в условиях небольших вольер (Соколов, Квашнин, 1983а, 1985; Квашнин,
1983а). На осноне этих результатов в послегнездовом онтогенезе пасюков
нами были выделены следующие возрастпые периоды, каждый из кото-
рых характеризовался своими особенностями поведения молодняка и спе-
цифическими формами социальных отношений между всеми членами экс-
периментальных групп (рис. 74):
1. Ювенильный период; по-видимому, начипается с момента выхода
крысят из гнезд в возрасте 20—25 дней“, когда они пачинают осваи-
вать повую для них среду, и заканчивается, когда крысята начинают фи-
зиологически созревать (в 65—70 дней, по Колхауну немпого позже).
Родители и другие взрослые особи мало внимания обращают на детепы-
шей вне гнезд, как бы пе замечают их присутствия. Для крысят, напро-
тив, характерен усиленный интерес к взрослым особям, особенно к мате-
рям— они постоянно следуют за ними, повторяют их движения и их
реакцию на предметы внептней среды. В этот период для детеньшней се-
рых крыс характерны, с одной стороны, повышенная пугливость и осто-
рожность, а с другой
— слепое подражапие действиям матери. Молодые
звервки много и охотно играют друг с другом и с матерью. В их поведен-
ческом репертуаре отсутствуют элементы полового, агонистического и
маркировочного поведения, характерные для взрослых особей.
Поведение лабораторных крыс этого возраста уже заметно отличается
от таконого у диких животных. Начало репродуктивного поведения ха-
рактерно для самцов в возрасте 36—40 дней, когда они впервые проявля-
ют влечение к самкам. В возрасте 41—45 дней у самцов отмечаются сад-
‘ки, а у самок впервые появляются позы лордоза (ВоПез, У’оойв, 1964;
Меоапеу, Эезагь 1981Ъ). Морфологические признаки полового созревания
наступают несколько раньше, чем проявляется половое влечение (Мегс!-
ет © а|., 1982). Б этом возрасте у лабораторных животных резко увели-
12 Это следует из периодизации раннего постнатального онтогенеза лабораторных
крыс. Однако накопление Данных об этом периоде развития У серых крысе, воз-
можно, внесет поправки и уточнения в возрастную классификацию пасюков.
234
Глава четвертая
Возраст, дни
78090100101
1И20340Е5й
:!
0
—!
7
57
ня
у
5
ёк—
м
5
7
&
8
3
=
© — матери перетаскивают детенышей,
пределах норы,
связанные с запасанием корма,
тельное норение отдельно от матери и сибсов,
раютел вместе, но в основном не в родной о, лы
взрослыми особями, которые терпимо относятся к ›
,
поведения, смена юзенильного шерстного п
взрослыми особями
чивается уровень гормонов (Пезя-Е
наблюдается эрекция пениса (Засй$, М
вительность центральной нервн
т
число клеток Лейдига, секретирующих го
2— первые выходы на поверхность около норы,
4— прекращение питания молоком матери,
еамостоя
6 — следование за матерями, 7 —
еи0сы
8 — согласование взаимодействий со
Рис. 70. Временная пос-
ледовательность проявле-
ния отдельных форм по
ведения у молодых се-
рых крыс до формиро-
вания у них полового
поведения (по: СаТонп,
1968)
1 — исследовательское поведение вне гнезда, но В
3 — первые передвижения,
5 — самостоя-
9 — начало взрослого полового
окрова па варослый, заметный антагонизм с более
еЪег, Газигало[, 1979), у самцов
е1зе1, 1979), увеличивается чувст-
ой системы к андрогенам и нарастает
рмоны в семенниках (Клотг
1 а1., 4970). Начиная с 54—55-дневного возраста, молодые крысы уже де-
9%а[.,41.
монстрируют половое поведение, ха.
пеу, тать 1981а, 5; Адашз, Во1се, 1983).
ы
т
32
2.
ВЕ
72
®
|
|
1
!
|
1
|
}
—10 91-59
й-80 91-100 111-120
ы 91—50 51-П 81-9
0-10
Возраст, дну
Формы контактов:
рактерное для взрослых особей (Меа-
Noв
Рис. 71. Динамика со-
циальных
контактов
межбу молодыми серы-
ми крысами в семейных
группах (по: Соколов,
Квашнин, 1985)
1-— пазо-назальные и обнюхивания участков тела, 2 — перелезания через
—
ь
—
—
6—
партнеров, 3 — назо-анальные, 4 — попытки садок и преследованил, 5 — агрессивные,
‚
игровые
— кри-
По оси ординат
— число контактов за 1 ч: слева (No) — на кривой 1, справа {No2) на кр!
вых 2—6
Поведение
235
Развитие атонистических форм поведения у доместицированных крыс
также имеет свои особенности. Например, первые агрессивные реакции
по отношению к чужим особям у них проявляются в возрасте 45—
50 дней (ЗсоН, Ргедочевот, 1951; Зе, 1966; Тоге, ЗИро-Еапегфу, 1984).
С наступлением половой зрелости у молодых крыс в их играх, которые
состоят в основном из элементов агонистического поведения, заметно уве-
лнчивается количество этих элементов. Постепенно игровые драки приоб-
ретают отношения доминирования—подчинения между партнерами одпо-
то пола и сохраняются неизменными длительное время (Ваепишоег,
1966). Однако в другой работе показано, что иерархические отношения,
возникающие в играх лабораторных крыс, нестабильны во времени и не
коррелируют с системой доминировапия у взрослых особей (А4атз,
Вогсе, 1983). Эти авторы считают, что полная социальная зрелость у ла-
боратортых крыс наступает к пятимесячному возрасту.
Для лабораторных крые характерной формой взаимодействия являет-
ся и социальный труминг, который расценивается как проявление друже-
любия (Стапь МаскимюозВ, 1963; Роше, Е1зЬ, 1975, 4976; Меапеу, 5е-
зать 19845; Огелуз, 1982). В ювенильный период но сравнению с пред-
шествующими у детенышей заметно возрастает частота взаимных чисток’
как друг с другом, так и с матерями, причем молодые самки чистят ма-
терей почти в лва раза чаще, чем самцы-сверстники (Меапеу, Зезуагь,
19815). Груминг свойствен и взрослым лабораторным крысам, что дало
основание использовать эту форму поведения, например, для изучения до-
минантных отношений между животными в различных конкурентных те-
стах (Зруре|, Егазег, 1974). Здесь можно отметить, что социальные чист-
ки у серых крыс (молодых и взрослых) встречались крайне редко и были
характерны в основпом для самцов-доминантов по отношению к избран-
ным самкам.
Конечно, различия в поведении между дикими и доместицированными
крысами не ограпичиваются только указанными вьипе. Но и этих данных,
касающихся основных форм социального поведения, достаточно, чтобы
сделать вывод 0 том, что детенынти пасюков развиваются значительно
медленнее (по крайней мере в послегиездовое время), а время формиро-
вания отдельных поведенчоских реакций у них длится мпого дольше, чем
у лабораторных крыс. Причины таких явлений пока остаются неясными,
хотя, несомненно, они связапы с условиями жизни животных.
В естественных условиях крысята в возрасте до 30—35 дней редко
появляются на поверхности, предпочитая проводить время в порах. По-
стененно их передвижения становятся более активными, хотя и ограни-
чиватотся близлежащей к родной поре территорией (З{епикег, 1950; Са1-
Воит, 1963). В это время детеньнии одного выводка могут попасть в вер-
шу вслед за матерью или отвергнуть какую-либо пищу, также повторяя
действия матери. Сходные реакции были иродемонстрированы и в экепе-
риментальных условиях у лабораторных крысе (СаеГ, СЛагк, 197Ча,Ь;
СаеЁ, 1976). Неудивительно поэтому, что эффективность дератизацион-
ных мероприятий по отношению к диким крысам ювенильного возраста
может значительно варьировать.
234
Глава четвертая
Возраст, дни
78090100101
1И20340Е5й
:!
0
—!
7
57
ня
у
5
ёк—
м
5
7
&
8
3
=
© — матери перетаскивают детенышей,
пределах норы,
связанные с запасанием корма,
тельное норение отдельно от матери и сибсов,
раютел вместе, но в основном не в родной о, лы
взрослыми особями, которые терпимо относятся к ›
,
поведения, смена юзенильного шерстного п
взрослыми особями
чивается уровень гормонов (Пезя-Е
наблюдается эрекция пениса (Засй$, М
вительность центральной нервн
т
число клеток Лейдига, секретирующих го
2— первые выходы на поверхность около норы,
4— прекращение питания молоком матери,
еамостоя
6 — следование за матерями, 7 —
еи0сы
8 — согласование взаимодействий со
Рис. 70. Временная пос-
ледовательность проявле-
ния отдельных форм по
ведения у молодых се-
рых крыс до формиро-
вания у них полового
поведения (по: СаТонп,
1968)
1 — исследовательское поведение вне гнезда, но В
3 — первые передвижения,
5 — самостоя-
9 — начало взрослого полового
окрова па варослый, заметный антагонизм с более
еЪег, Газигало[, 1979), у самцов
е1зе1, 1979), увеличивается чувст-
ой системы к андрогенам и нарастает
рмоны в семенниках (Клотг
1 а1., 4970). Начиная с 54—55-дневного возраста, молодые крысы уже де-
9%а[.,41.
монстрируют половое поведение, ха.
пеу, тать 1981а, 5; Адашз, Во1се, 1983).
ы
т
32
2.
ВЕ
72
®
|
|
1
!
|
1
|
}
—10
91-59
й-80
91-100 111-120
ы 91—50 51-П
81-9
0-10
Возраст, дну
Формы контактов:
рактерное для взрослых особей (Меа-
Noв
Рис. 71. Динамика со-
циальных
контактов
межбу молодыми серы-
ми крысами в семейных
группах (по: Соколов,
Квашнин, 1985)
1-— пазо-назальные и обнюхивания участков тела, 2 — перелезания через
—
ь
—
—
6—
партнеров, 3 — назо-анальные, 4 — попытки садок и преследованил, 5 — агрессивные,
‚
игровые
— кри-
По оси ординат
— число контактов за 1 ч: слева (No) — на кривой 1, справа {No2) на кр!
вых 2—6
Поведение
235
Развитие атонистических форм поведения у доместицированных крыс
также имеет свои особенности. Например, первые агрессивные реакции
по отношению к чужим особям у них проявляются в возрасте 45—
50 дней (ЗсоН, Ргедочевот, 1951; Зе, 1966; Тоге, ЗИро-Еапегфу, 1984).
С наступлением половой зрелости у молодых крыс в их играх, которые
состоят в основном из элементов агонистического поведения, заметно уве-
лнчивается количество этих элементов. Постепенно игровые драки приоб-
ретают отношения доминирования—подчинения между партнерами одпо-
то пола и сохраняются неизменными длительное время (Ваепишоег,
1966). Однако в другой работе показано, что иерархические отношения,
возникающие в играх лабораторных крыс, нестабильны во времени и не
коррелируют с системой доминировапия у взрослых особей (А4атз,
Вогсе, 1983). Эти авторы считают, что полная социальная зрелость у ла-
боратортых крыс наступает к пятимесячному возрасту.
Для лабораторных крые характерной формой взаимодействия являет-
ся и социальный труминг, который расценивается как проявление друже-
любия (Стапь МаскимюозВ, 1963; Роше, Е1зЬ, 1975, 4976; Меапеу, 5е-
зать 19845; Огелуз, 1982). В ювенильный период но сравнению с пред-
шествующими у детенышей заметно возрастает частота взаимных чисток’
как друг с другом, так и с матерями, причем молодые самки чистят ма-
терей почти в лва раза чаще, чем самцы-сверстники (Меапеу, Зезуагь,
19815). Груминг свойствен и взрослым лабораторным крысам, что дало
основание использовать эту форму поведения, например, для изучения до-
минантных отношений между животными в различных конкурентных те-
стах (Зруре|, Егазег, 1974). Здесь можно отметить, что социальные чист-
ки у серых крыс (молодых и взрослых) встречались крайне редко и были
характерны в основпом для самцов-доминантов по отношению к избран-
ным самкам.
Конечно, различия в поведении между дикими и доместицированными
крысами не ограпичиваются только указанными вьипе. Но и этих данных,
касающихся основных форм социального поведения, достаточно, чтобы
сделать вывод 0 том, что детенынти пасюков развиваются значительно
медленнее (по крайней мере в послегиездовое время), а время формиро-
вания отдельных поведенчоских реакций у них длится мпого дольше, чем
у лабораторных крыс. Причины таких явлений пока остаются неясными,
хотя, несомненно, они связапы с условиями жизни животных.
В естественных условиях крысята в возрасте до 30—35 дней редко
появляются на поверхности, предпочитая проводить время в порах. По-
стененно их передвижения становятся более активными, хотя и ограни-
чиватотся близлежащей к родной поре территорией (З{епикег, 1950; Са1-
Воит, 1963). В это время детеньнии одного выводка могут попасть в вер-
шу вслед за матерью или отвергнуть какую-либо пищу, также повторяя
действия матери. Сходные реакции были иродемонстрированы и в экепе-
риментальных условиях у лабораторных крысе (СаеГ, СЛагк, 197Ча,Ь;
СаеЁ, 1976). Неудивительно поэтому, что эффективность дератизацион-
ных мероприятий по отношению к диким крысам ювенильного возраста
может значительно варьировать.
236 Глава четвертая
2. Период полового созревания— возраст молодых серых крыс от
65—70 до 90—95 дней. В этот период происходит физиологическое созре-
вапие крысят: у самцов появляется сексуальный интерес к самкам,
ау самок в копце периода начинаются эстральные циклы. У молодых
особей формируются снецифические элементы поведения, связанные с
маркировкой территории и партнеров по группе мочой. Проявляется аг-
рессивность по отношению к чужим особям. Сохраняется высокий уро-
вень игровых контактов между молодыми и между ними и матерями.
В конце периода взрослые особи начинают проявлять антагонизм к мо-
й
ку.
т данным Колхауна, первые раны на теле молодых крыс в осповном
появлялись. когла они достигали 85—90-дневного возраста, что совпада-
ет с вашими наблюдениями. Однако в этот период агрессивность взрос-
лых животных по отношению к молодым проявлялась слабо, поскольку
с «поведенческой точки зрения» членов группировки они не достигли по-
ловой и социальной зрелости — молодые особи еще только накапливали
сексуальный опыт через общение с сородичами. В этот период отчетливо
проявлялись различия в активности у разнополых зверьков: самцы ста-
повились более самостоятельными и удалялись от выводковой норы зна-
чительно дальше и на более длительное время, чем самки. |
3. Период поведенческого созревания
— наиболее важный и продол-
жительпый в развитии молодняка. Он начинается с 95—100-дневного
возраста и характеризуется следующими особенностями: в это время про-
исходят значительные изменения в индивидуальной активности молодых
особей и в характере социальных взаимодействий между всеми членами
групп. У молодых самцов резко возрастает частота всех форм контактов
с самками-сверстницами. У последних появляются первые выводки, в свя-
зи с чем увеличивается их агрессивность. В этом периоде у молодняка
резко сокращается количество игр, и они практически исчезают к четы-
фехмесячному возрасту. У взрослых животных усиливается антагонизм к
молодняку. Наблюдается «выравнивание» социальной активности взрос-
лых и молодых особей и «включение» последних во внутригрупповые от-
ношения, в результате чего формируются повые иерархические связи
между членами группы. В условиях вольер такое включение выражалось
в том, что у одного из молодых самцов стали регулярно проявляться эле-
менты агопистического поведения (позы напряжения, затаивапие, вздыб-
ливание шерсти, хлестание хвостом и т. д.) ® по отношению к взрослым
самцам, которые вначале не замечали измепений в поведении молодняка,
но затем стали проявлять агрессивность по отношению к ним. Одновре-
13 Для таких самцов были характерны и другие особенности поведения: они зна-
чительно чаще начинали игры, Достоверно больше маркировали территорию и
партнеров по группе, по были менее сексуально активны, чем однополые сибсы.
Им также были свойственны уверенность движений и смелость, т. е. черты высо-
коранговых особей. Остальные молодые самцы также отличались друг от друга
по характеру поведения во внутригрупповых контактах и по отношению к чу-
жим особям (Соколов, Квашнин, 1985).
Поведение 237
менпо у таких самцов проявлялась атрессивность по отношению к дру-
тим самцам-сверстникам, у которых соответственно развивалась реакция
избегания при столкновении с ними. Во всех экспериментальных груп-
пах молодые самцы занимали подчиненное положение отпосительно сам-
ца-доминанта. Интересно отметить, что иерархические отношения в таких
труппах по форме были аналогичными тому, что описано для естествен-
ных группировок серых крыс.
Барнетт (Ваглей, 1975) считает, что этот период развития серых крыс,
‚по крайней мере для самцов, соответствует их «зрелости». Исследования
на лабораторных животных показали, что этот период у них наступает
раньше и характеризуется тем, что у них увеличивается содержание анд-
рогенов в крови, семенники полностью опускаттся в мошонку и появля-
ются такие элементы поведения, как, папример, нетерпимость, свойствен-
ные взрослым особям (Втопзоп, Оездага!тз, 1971).
Впервые о необходимости выделения в возрастной классификации се-
рых крыс категории особей, проходящих стадию поведенческого созрева-
ния, высказался Колхаун (СаФопл, 1948а, Ь; 1963). По его мнению, этот
период заканчивается у самцов к шестимесячному возрасту, а у самок к
семимесячному, после чего животные становятся «полностью взрослыми».
Значение периода поведенческого созревания для молодых крыс заклю-
чается в приобретении ими социального опыта — они обучаются избегать
угроз и пападений взрослых особей и сдерживать такое поведение, кото-
рое могло бы вызвать агрессию со стороны взрослых крыс. В эта время
поведение молодых зверьков определяется уже не столько возрастом,
<колько принадлежностью к полу и индивидуальными характеристиками.
По наблюдениям Колхауна, в его колоний молодые самки предпочита-
ли оставаться в пределах участков обитания матерей вплоть до начала
своего размножения, тогда как взаимоотноптения между самцами в этот
период резко обострялись, и это, по-видимому, определяло дальнейшую
судьбу молодняка. С одной стороны, в результате агреесии со стороны
нзрослых животных происходило расселение молодых крыс за пределы
участков обитания, что характерио и для других видов грызунов (Свтз-
Цал, 1970; Зешег, 1972; Векой, 1977; Ктебз, 1978), ас другой — остав-
шиеся в составе группировок особи становились полноправпыми ее чле-
нами, т. е. происходил процесс «социализации» (МеВнае, 1964; Можем,
Атшйаре, 1979).
Влияние раннего опъита на формирование поведения
взрослых животных
Многочисленные эксперименты показали, что влияние факторов внешней
среды в различные периоды роста и развития крысят приводит к разно-
образным изменениям в морфологии, физиологии и поведении взрослых
животных.
Детеныши лабораторных крыс оказались чувствительными к различ-
ным воздействиям уже на ранней стадии онтотенеза, т. е. в пренатальный
период. Экспериментальным путем показано, что если самку во время
236 Глава четвертая
2. Период полового созревания— возраст молодых серых крыс от
65—70 до 90—95 дней. В этот период происходит физиологическое созре-
вапие крысят: у самцов появляется сексуальный интерес к самкам,
ау самок в копце периода начинаются эстральные циклы. У молодых
особей формируются снецифические элементы поведения, связанные с
маркировкой территории и партнеров по группе мочой. Проявляется аг-
рессивность по отношению к чужим особям. Сохраняется высокий уро-
вень игровых контактов между молодыми и между ними и матерями.
В конце периода взрослые особи начинают проявлять антагонизм к мо-
й
ку.
т данным Колхауна, первые раны на теле молодых крыс в осповном
появлялись. когла они достигали 85—90-дневного возраста, что совпада-
ет с вашими наблюдениями. Однако в этот период агрессивность взрос-
лых животных по отношению к молодым проявлялась слабо, поскольку
с «поведенческой точки зрения» членов группировки они не достигли по-
ловой и социальной зрелости — молодые особи еще только накапливали
сексуальный опыт через общение с сородичами. В этот период отчетливо
проявлялись различия в активности у разнополых зверьков: самцы ста-
повились более самостоятельными и удалялись от выводковой норы зна-
чительно дальше и на более длительное время, чем самки. |
3. Период поведенческого созревания
— наиболее важный и продол-
жительпый в развитии молодняка. Он начинается с 95—100-дневного
возраста и характеризуется следующими особенностями: в это время про-
исходят значительные изменения в индивидуальной активности молодых
особей и в характере социальных взаимодействий между всеми членами
групп. У молодых самцов резко возрастает частота всех форм контактов
с самками-сверстницами. У последних появляются первые выводки, в свя-
зи с чем увеличивается их агрессивность. В этом периоде у молодняка
резко сокращается количество игр, и они практически исчезают к четы-
фехмесячному возрасту. У взрослых животных усиливается антагонизм к
молодняку. Наблюдается «выравнивание» социальной активности взрос-
лых и молодых особей и «включение» последних во внутригрупповые от-
ношения, в результате чего формируются повые иерархические связи
между членами группы. В условиях вольер такое включение выражалось
в том, что у одного из молодых самцов стали регулярно проявляться эле-
менты агопистического поведения (позы напряжения, затаивапие, вздыб-
ливание шерсти, хлестание хвостом и т. д.) ® по отношению к взрослым
самцам, которые вначале не замечали измепений в поведении молодняка,
но затем стали проявлять агрессивность по отношению к ним. Одновре-
13 Для таких самцов были характерны и другие особенности поведения: они зна-
чительно чаще начинали игры, Достоверно больше маркировали территорию и
партнеров по группе, по были менее сексуально активны, чем однополые сибсы.
Им также были свойственны уверенность движений и смелость, т. е. черты высо-
коранговых особей. Остальные молодые самцы также отличались друг от друга
по характеру поведения во внутригрупповых контактах и по отношению к чу-
жим особям (Соколов, Квашнин, 1985).
Поведение 237
менпо у таких самцов проявлялась атрессивность по отношению к дру-
тим самцам-сверстникам, у которых соответственно развивалась реакция
избегания при столкновении с ними. Во всех экспериментальных груп-
пах молодые самцы занимали подчиненное положение отпосительно сам-
ца-доминанта. Интересно отметить, что иерархические отношения в таких
труппах по форме были аналогичными тому, что описано для естествен-
ных группировок серых крыс.
Барнетт (Ваглей, 1975) считает, что этот период развития серых крыс,
‚по крайней мере для самцов, соответствует их «зрелости». Исследования
на лабораторных животных показали, что этот период у них наступает
раньше и характеризуется тем, что у них увеличивается содержание анд-
рогенов в крови, семенники полностью опускаттся в мошонку и появля-
ются такие элементы поведения, как, папример, нетерпимость, свойствен-
ные взрослым особям (Втопзоп, Оездага!тз, 1971).
Впервые о необходимости выделения в возрастной классификации се-
рых крыс категории особей, проходящих стадию поведенческого созрева-
ния, высказался Колхаун (СаФопл, 1948а, Ь; 1963). По его мнению, этот
период заканчивается у самцов к шестимесячному возрасту, а у самок к
семимесячному, после чего животные становятся «полностью взрослыми».
Значение периода поведенческого созревания для молодых крыс заклю-
чается в приобретении ими социального опыта — они обучаются избегать
угроз и пападений взрослых особей и сдерживать такое поведение, кото-
рое могло бы вызвать агрессию со стороны взрослых крыс. В эта время
поведение молодых зверьков определяется уже не столько возрастом,
<колько принадлежностью к полу и индивидуальными характеристиками.
По наблюдениям Колхауна, в его колоний молодые самки предпочита-
ли оставаться в пределах участков обитания матерей вплоть до начала
своего размножения, тогда как взаимоотноптения между самцами в этот
период резко обострялись, и это, по-видимому, определяло дальнейшую
судьбу молодняка. С одной стороны, в результате агреесии со стороны
нзрослых животных происходило расселение молодых крыс за пределы
участков обитания, что характерио и для других видов грызунов (Свтз-
Цал, 1970; Зешег, 1972; Векой, 1977; Ктебз, 1978), ас другой — остав-
шиеся в составе группировок особи становились полноправпыми ее чле-
нами, т. е. происходил процесс «социализации» (МеВнае, 1964; Можем,
Атшйаре, 1979).
Влияние раннего опъита на формирование поведения
взрослых животных
Многочисленные эксперименты показали, что влияние факторов внешней
среды в различные периоды роста и развития крысят приводит к разно-
образным изменениям в морфологии, физиологии и поведении взрослых
животных.
Детеныши лабораторных крыс оказались чувствительными к различ-
ным воздействиям уже на ранней стадии онтотенеза, т. е. в пренатальный
период. Экспериментальным путем показано, что если самку во время
238 Глава четвертая
беременности беспокоить или подвергать действию стресс-факторов, то
это приводит к существенным сдвигам в поведении ее потомства. Крыся-
та, родившиеся от перенеситих стресс самок, отличались от контрольных
повышенной «эмоциональностью» (характеристика поведения в тесте
«открытого поля», оспованная на измерении латентного периода и уров-
ня активности, а также частоты дефекации) и пугливостью (НосКтап,
1964; Гьотарзоп сё а1., 1962; ТоНе, 1965; Мотга, 1965; РАжег, Гу в,
1983). Подобный эффект наблюдалея даже у внуков стрессированных са-
мок (Виззе[, 1971). Также было установлено, что у взрослых самцов, ко-
торые родились от перенесших стресс во время беременпости самок, была.
понижена полозая активность и их новедение было более феминизирован-
ным (\ата, 1972).
Экспериментальные воздействия на детенышей крыс в постнаталь-
ный период развития непосредственно или через их матерей существеп-
ным образом сказывались на их физиологии и поведении в более стар-
тем возрасте. Для изучения таких воздействий Нинг (Кшо, 1958; цит.
по: Рау, 1973) было предложено выделить три периода в онтогенезе
крыс — инфантильный (от рождения до 21-дневного возраста“), юве-
нильный (от 22-дневного возраста и до половой зрелости) и взрослый.
Наиболес чувствительным периодом развития является инфантильный,
тогда как стимуляция в остальные периоды уже менее эффективна.
(см., например: Вагоп е4 а|., 4957; Бау, Ообу, 1967).
В исследовании Зейтца (Зейя, 1954) было показано, что количество
крысят в помете (от 3 до 12) влияло на материнское поведение самки
и затем на последующее поведение детенышей: крысы из больших поме-
тов были более эмоциональными и болыпе запасали корма, а самцы из
таких выводков меньше весили и были более сексуально активными, чем
выроситие в малых пометах.
В том случае, если детенышей вынуждали питаться молоком матери
дольше обычного, во взрослом состоянии опи хуже обучались избегапию,
чем контрольные (ТГеуше, 4958). Раннее прекращение кормления моло-
ком матери приводило, в частности, к пеобратимым измепениям фулкций
почек (КтесеЕ, 1970) и пониженной сопротивляемости к различным за-
болевапиям (АсКегтап её а]., 1975).
Стимуляция детенышей в раннем возрасте, связанная с усложнением
обычных лабораторных условий или введением новых факторов, приводит
к различным эффектам. Наиболее часто наблюдаемые из них следующие
(Рау, 1973): понижение эмоциональности, улучшение обучения, более
адекватное реагирование па стрессовые воздействия, ускорепие роста и
развития, изменения в физиологических функциях.
11 У лабораторных крыс в это время обычно отделяют матерей от детенышей, так
как последние уже способны вести самостоятельную жизнь — они могут питаться
твердыми кормами (а у самок почти прекращается выделение молока). У диких
крысе, как считает Барнетт (Вагпей, 1958а), отделение матерей от детенышей
можно проводить не ральше, чем крысята достигнут 28-дневного возраста.
Поведение
239
Взятие крысят в руки (хэндлинг) в течение первых трех недель их
жизни приводило к тому, что они во взрослом возрастс оказывались ме-
исе эмоциопальными (Пеппепето, Зимйь, 1968; МоБапбу, М1зИга, 1984)
демонстрировали повышенную ориентирозочно-исследовательскую рсак-
цию (ПОеппепЪего, Стофа, 1964) и более адекватную реакцию на стрессо-
вые воздействия (более умеренным выбросом кортикостероидов в новой
ситуации и более интенсивной реакцией надпочечников в ситуациях
угрозы) (Геуше, МиШиз, 1966), чем контрольные животные. Интересно
отметить, что крысята, которых брали в руки в течение первых 10 дней
жизни, лучше обучались избеганию, чем детепьши, которых подвергли
хэндлингу в возрасте 11—20 дней (РеппепЪеге, Кагаз, 1960). Хэндлинг
приводил также к ускоренному росту и развитию крысят (хотя и не
всегда) и к повышенной выживаемости (Геуше, ОШз, 1958; ПеппепЪего,
Кагаз, 1961). Попытки выделения критических периодов у крысят в пер-
вые 10 дней их жизни путем хэпдлинга привели Дененберга (Оепнеп-
Ъегр, 1962) к заключению, что и возраст детенынтей, и число дней хэнд-
линга являются важными параметрами, воздействующими па последую-
щес поведение крысят.
Воздействие на крысят в инфантильном возрасте различными стресс-
факторами также приводило, как правило, к заметным изменениям в по-
следующем их поведении. Гак, Дененберг и Смит (ОеппепЪеге, ЗтИЪ,
1963) показали, что крысы, на которых воздействовали в детстве элек-
трическим током, отличались от контрольных пониженной эмоциональ-
ностью. Аналогичные результаты получены и другими исследователями
(Вагоп сё а1., 1957; Со]Чтап, 1964). Однако не во всех работах были по-
лучены определенные данные. Так, Гриффитс и Стрингер (СтИЁз, Зиш-
сег, 1952) не обнаружили по сравпению с контрольными различий в по-
ведении в лабиринте и в тесте открытого поля у взрослых крыс, которых
в течение первых девяти дней жизни подвергали воздействию электрото-
ка, звуковой стимуляции, быстрому вращению и т. д.
Таким образом, период наибольшей чувствительности к внешним воз-
действиям у крысят приходится на первые 5—10 дней жизни, хотя неко-
торые эффекты были обнаружены и при стимуляции в более поздние сро-
ки. Как считает Депеиберг (Оеппепрего, 1968), ранций постнатальный
(инфантильный) период развития детенышей крые является периодом
‘максимальной чувствительности к качеству и количеству воздействий,
хотя истинного критического периода в раннем детстве не существует.
Для объяснения описанных выше эффектов «сверхстимуляции» было
выдвинуто несколько гипотез. Согласно «гипотермической» гипотезе, эти
эффекты вызываются непосредственным воздействием на детенышей за
счет измепепия температуры тела, когда их вынимают из гнезда
и берут в руки. Основанием для этого предположения послужили,
в частности, эксперименты, в которых животных подвергали действию
только холода и в которых были получены соответствующие результаты
(ЗеВеаЁег, 1978, цит. по: Дьюсбери, 4981). Вторая гипотеза объясняет
наблюдаемые эффокты через изменение гормональных связей у детены-
шей и в связи с этим более адекватное реагирование на стрессовые
238 Глава четвертая
беременности беспокоить или подвергать действию стресс-факторов, то
это приводит к существенным сдвигам в поведении ее потомства. Крыся-
та, родившиеся от перенеситих стресс самок, отличались от контрольных
повышенной «эмоциональностью» (характеристика поведения в тесте
«открытого поля», оспованная на измерении латентного периода и уров-
ня активности, а также частоты дефекации) и пугливостью (НосКтап,
1964; Гьотарзоп сё а1., 1962; ТоНе, 1965; Мотга, 1965; РАжег, Гу в,
1983). Подобный эффект наблюдалея даже у внуков стрессированных са-
мок (Виззе[, 1971). Также было установлено, что у взрослых самцов, ко-
торые родились от перенесших стресс во время беременпости самок, была.
понижена полозая активность и их новедение было более феминизирован-
ным (\ата, 1972).
Экспериментальные воздействия на детенышей крыс в постнаталь-
ный период развития непосредственно или через их матерей существеп-
ным образом сказывались на их физиологии и поведении в более стар-
тем возрасте. Для изучения таких воздействий Нинг (Кшо, 1958; цит.
по: Рау, 1973) было предложено выделить три периода в онтогенезе
крыс — инфантильный (от рождения до 21-дневного возраста“), юве-
нильный (от 22-дневного возраста и до половой зрелости) и взрослый.
Наиболес чувствительным периодом развития является инфантильный,
тогда как стимуляция в остальные периоды уже менее эффективна.
(см., например: Вагоп е4 а|., 4957; Бау, Ообу, 1967).
В исследовании Зейтца (Зейя, 1954) было показано, что количество
крысят в помете (от 3 до 12) влияло на материнское поведение самки
и затем на последующее поведение детенышей: крысы из больших поме-
тов были более эмоциональными и болыпе запасали корма, а самцы из
таких выводков меньше весили и были более сексуально активными, чем
выроситие в малых пометах.
В том случае, если детенышей вынуждали питаться молоком матери
дольше обычного, во взрослом состоянии опи хуже обучались избегапию,
чем контрольные (ТГеуше, 4958). Раннее прекращение кормления моло-
ком матери приводило, в частности, к пеобратимым измепениям фулкций
почек (КтесеЕ, 1970) и пониженной сопротивляемости к различным за-
болевапиям (АсКегтап её а]., 1975).
Стимуляция детенышей в раннем возрасте, связанная с усложнением
обычных лабораторных условий или введением новых факторов, приводит
к различным эффектам. Наиболее часто наблюдаемые из них следующие
(Рау, 1973): понижение эмоциональности, улучшение обучения, более
адекватное реагирование па стрессовые воздействия, ускорепие роста и
развития, изменения в физиологических функциях.
11 У лабораторных крыс в это время обычно отделяют матерей от детенышей, так
как последние уже способны вести самостоятельную жизнь — они могут питаться
твердыми кормами (а у самок почти прекращается выделение молока). У диких
крысе, как считает Барнетт (Вагпей, 1958а), отделение матерей от детенышей
можно проводить не ральше, чем крысята достигнут 28-дневного возраста.
Поведение
239
Взятие крысят в руки (хэндлинг) в течение первых трех недель их
жизни приводило к тому, что они во взрослом возрастс оказывались ме-
исе эмоциопальными (Пеппепето, Зимйь, 1968; МоБапбу, М1зИга, 1984)
демонстрировали повышенную ориентирозочно-исследовательскую рсак-
цию (ПОеппепЪего, Стофа, 1964) и более адекватную реакцию на стрессо-
вые воздействия (более умеренным выбросом кортикостероидов в новой
ситуации и более интенсивной реакцией надпочечников в ситуациях
угрозы) (Геуше, МиШиз, 1966), чем контрольные животные. Интересно
отметить, что крысята, которых брали в руки в течение первых 10 дней
жизни, лучше обучались избеганию, чем детепьши, которых подвергли
хэндлингу в возрасте 11—20 дней (РеппепЪеге, Кагаз, 1960). Хэндлинг
приводил также к ускоренному росту и развитию крысят (хотя и не
всегда) и к повышенной выживаемости (Геуше, ОШз, 1958; ПеппепЪего,
Кагаз, 1961). Попытки выделения критических периодов у крысят в пер-
вые 10 дней их жизни путем хэпдлинга привели Дененберга (Оепнеп-
Ъегр, 1962) к заключению, что и возраст детенынтей, и число дней хэнд-
линга являются важными параметрами, воздействующими па последую-
щес поведение крысят.
Воздействие на крысят в инфантильном возрасте различными стресс-
факторами также приводило, как правило, к заметным изменениям в по-
следующем их поведении. Гак, Дененберг и Смит (ОеппепЪеге, ЗтИЪ,
1963) показали, что крысы, на которых воздействовали в детстве элек-
трическим током, отличались от контрольных пониженной эмоциональ-
ностью. Аналогичные результаты получены и другими исследователями
(Вагоп сё а1., 1957; Со]Чтап, 1964). Однако не во всех работах были по-
лучены определенные данные. Так, Гриффитс и Стрингер (СтИЁз, Зиш-
сег, 1952) не обнаружили по сравпению с контрольными различий в по-
ведении в лабиринте и в тесте открытого поля у взрослых крыс, которых
в течение первых девяти дней жизни подвергали воздействию электрото-
ка, звуковой стимуляции, быстрому вращению и т. д.
Таким образом, период наибольшей чувствительности к внешним воз-
действиям у крысят приходится на первые 5—10 дней жизни, хотя неко-
торые эффекты были обнаружены и при стимуляции в более поздние сро-
ки. Как считает Депеиберг (Оеппепрего, 1968), ранций постнатальный
(инфантильный) период развития детенышей крые является периодом
‘максимальной чувствительности к качеству и количеству воздействий,
хотя истинного критического периода в раннем детстве не существует.
Для объяснения описанных выше эффектов «сверхстимуляции» было
выдвинуто несколько гипотез. Согласно «гипотермической» гипотезе, эти
эффекты вызываются непосредственным воздействием на детенышей за
счет измепепия температуры тела, когда их вынимают из гнезда
и берут в руки. Основанием для этого предположения послужили,
в частности, эксперименты, в которых животных подвергали действию
только холода и в которых были получены соответствующие результаты
(ЗеВеаЁег, 1978, цит. по: Дьюсбери, 4981). Вторая гипотеза объясняет
наблюдаемые эффокты через изменение гормональных связей у детены-
шей и в связи с этим более адекватное реагирование на стрессовые
240
Глаза четвертая
воздействия у взрослых животпых (Геуше, МаШиз, 1966). Наиболее
убедительной зыглядит третья точка зрения, согласно которой «еверх-
стимуляция» действует не столько пепосредственно на молодых зверьков,
сколько через их матерей. Действительно, после возвращения стрессиро-
ванных детенышей в гнезда матери обращают на них повышенное вни-
мание— тщательно их обпюхивают и облизывают (Вагпей, Виги, 1967;
Мепдоза еф а|., 1980), что может быть вызвано интенсивными ультразву-
ковыми сигналами, которые издают такие детепыши. 'Гакже показано,
что если подвергнуть электрораздражению самок во время лактации, то их
детеныши впоследствии будут более эмоциональными, чем контрольные
(ПеппепБегя её а]., 1962). .
Другой экснериментальный подход к изучению влияния рапнего опы-
та связан с обеднением среды. Изоляция детенышей в период, предше-
ствующий отнятию от матери, приводила к различным эффектам. Такие
крысята в более позднем возрасте были более возбудимы и активнее в
исследовательском поведении, чем росшие в нормальных условиях
(Нас её а]., 1963; Ваепшшеег, 1967). Изоляция в раннем возрасте при-
водила к заметному увеличению агрессивности, а также к повышенной
чувствительности к электрораздражению и тактильной стимуляции у
взрослых животных (\!а!]егап@ её а]., 1983; Мовау, М!зВга, 1984).
Отдельное содержание (вне контактов с матерью) оказывало существен-
ное влияние на рост детенышей (НабН сё а]., 1963) и на некоторые фи-
зиологические фупкции (Нофег, 1973; Аскегтап ей а1., 1975).
Особое значение в жизни крысят, по-видимому, имеет возраст с 45-го
по 24-й день (социальный период). Именно в это время у них формиру-
ются устойчивые поведенческие связи с матерями и друг с другом. В есте-
ственных условиях развитие детепышей крыс с момента рождения и до
копца этого периода происходит в норах или укрытиях, т. е. в относи-
тельной (или даже полной) темноте. Поэтому в становлении взаимоотно-
шений между матерью и ее потомством важную роль должны играть
ольфакторные сигналы. На доместицированных крысах показано, что у
самок-матерей в кале происходит выделение феромона, когда детеныши
достигают 14-дневного возраста и у них, в свою очередь, проявляется к
нему аттрактивность (Теоп, МоМ=, 1974, 1972, 1973; Теоп, 1974, 1975;
НоНпка, Савоп, 1976). Мы предполагаем, что у детенышей диких серых
крыс именно в этот период происходит запечатление на обонятельные
сигналы, исходящие от матери. Результатом импринтинта может быть
хорошо выраженная реакция следования в основном за матерями у мо-
лодых зверьков, которые в конце этого периода начинают осваивать про-
странство вне убежищ (СаШNoопи, 1963; Соколов, Квашнин, 1983а, 6; 1985).
Во всяком случае, реакция следования обеспечивает сплоченность семьи,
синхронизацию активности ее членов, освоение крысятами новой для них
среды и выработку у них адаптивных реакций, а также способствует
преодолению детеньшами чувства страха (Мох, 1960; Куае, 4966; Вгоп-
воп, 1968а, Ь), которое особенно сильно развито у пих в ювенильном
возрасте.
Новедение 241
На формирование поведения детенышей крыс в ювенильный период
их онтогенеза значительное влияние оказывают уже не только матери,
но и сверстники. Для оценки этого влияния крысят отнимали от мате.
рей в возрасте 21—23 дня, подвергали действию различных факторов
и оценивали их последотвия у взрослых животных. Детеныши, роспие
с трехнедельного возраста в усложненных условиях (в больших клетках
© различными предметами, лесенками и т. п.), во взрослом возрасте по-
казывали лучшую обучаемость в лабиринте и были менее эмоциональны-
ми, чем росшие в обычных условиях (Вше\йат, Ст Из, 1952). Крысята,
подвергавшиеся джентлингу (взятие в руки и поглаживание по спине)
с момента отпятия их от матери, обучались во взрослом возрасте решать
задачи на различение быстрее контрольных (Вегимешт, 1957), а также
значительно чаще доминировали пад животными из контрольной группы
в ситуациях пищевой конкуренции (Мезе!, Возеп, 1960).
На формирование поведения зверьков в ювенильный период сущест-
венное влияние оказывают условия, связанные с социальной деприваци-
ей. Изолированное содержание крысят с 23- до 51-дневного возраста при-
водило к тому, что во взрослом возрасте они хуже обучались и были
более активны в лабириите и в открытом поле (Ватутр]е-АШогд, Вегот,
1984). Эти авторы предположили, что такие измепения обусловлены раз-
витием у крысят синдрома «раскрепощения» (независимости от сибсов).
Изоляция оказывает значительное влияние на полозое и агрессивное
поведение. Отдельное содержание приводило к новышенной сексуальной
активности и обусловливало неадекватное половое поведение у самцов
(Веасв, 1942; СогаШ её а1., 1967; Стиепае!, Атпоа, 1969). У взрослых
самок, которые были изолированы с 30-дневного возраста, отмечали по-
вышенную активность. Однако начало полового созревания и особенно.
сти эстральных циклов у них были сходными с самками, содержавшимися
труппами (Напзеп, 1977). Изоляция в ювенильном возрасте приводила
также к повышенной агрессивности у взрослых животных и к повыццен-
ной чувствительности к стрессу (Тоге, ЕаплеПу, 1984; У а экана е1 а1,
1983; Тюге, Зиро-анену, 1984; Тапа её а., 1984). Изолированное со-
держапие крысят в 41—68-дневном возрасте уже не сказывалось на их
агонистическом поведении во взрослом возрасте (Ма ахана о а]., 1983).
Изоляция у лабораторных крыс отражалась также на многих физио-
логических реакциях, на чувствительности к различным химическим
веществам и на ряде других форм поведения (Возепя\уе её а1., 1972;
Слопим, 1976; Уждавини, 1980; и др.).
Подобных эксперимептов с дикими серыми крысами проводилось
храйпе мало. Только в нескольких работах приводятся данные о послед-
ствиях частичной социальной депривации па развитие крысят и на фор-
мировацие их поведения (Соколов, Квашнин, 1983а,6: 1985; Квапшин,
1983а, 6). Суть опыта сводилась к тому, что из вольер были изъяты
взрослые жкивотные (родители) в ювепиальный период развития детепы-
шей, когда им было 36—40 дней от роду. Дальнейшие наблюдеция в те-
чение 2,5 мес ноказали, что характер и частота социальных контактов
между крысятами, росшими без родителей, были сходными с таковыми
240
Глаза четвертая
воздействия у взрослых животпых (Геуше, МаШиз, 1966). Наиболее
убедительной зыглядит третья точка зрения, согласно которой «еверх-
стимуляция» действует не столько пепосредственно на молодых зверьков,
сколько через их матерей. Действительно, после возвращения стрессиро-
ванных детенышей в гнезда матери обращают на них повышенное вни-
мание — тщательно их обпюхивают и облизывают (Вагпей, Виги, 1967;
Мепдоза еф а|., 1980), что может быть вызвано интенсивными ультразву-
ковыми сигналами, которые издают такие детепыши. 'Гакже показано,
что если подвергнуть электрораздражению самок во время лактации, то их
детеныши впоследствии будут более эмоциональными, чем контрольные
(ПеппепБегя её а]., 1962). .
Другой экснериментальный подход к изучению влияния рапнего опы-
та связан с обеднением среды. Изоляция детенышей в период, предше-
ствующий отнятию от матери, приводила к различным эффектам. Такие
крысята в более позднем возрасте были более возбудимы и активнее в
исследовательском поведении, чем росшие в нормальных условиях
(Нас её а]., 1963; Ваепшшеег, 1967). Изоляция в раннем возрасте при-
водила к заметному увеличению агрессивности, а также к повышенной
чувствительности к электрораздражению и тактильной стимуляции у
взрослых животных (\!а!]егап@ её а]., 1983; Мовау, М!зВга, 1984).
Отдельное содержание (вне контактов с матерью) оказывало существен-
ное влияние на рост детенышей (НабН сё а]., 1963) и на некоторые фи-
зиологические фупкции (Нофег, 1973; Аскегтап ей а1., 1975).
Особое значение в жизни крысят, по-видимому, имеет возраст с 45-го
по 24-й день (социальный период). Именно в это время у них формиру-
ются устойчивые поведенческие связи с матерями и друг с другом. В есте-
ственных условиях развитие детепышей крыс с момента рождения и до
копца этого периода происходит в норах или укрытиях, т. е. в относи-
тельной (или даже полной) темноте. Поэтому в становлении взаимоотно-
шений между матерью и ее потомством важную роль должны играть
ольфакторные сигналы. На доместицированных крысах показано, что у
самок-матерей в кале происходит выделение феромона, когда детеныши
достигают 14-дневного возраста и у них, в свою очередь, проявляется к
нему аттрактивность (Теоп, МоМ=, 1974, 1972, 1973; Теоп, 1974, 1975;
НоНпка, Савоп, 1976). Мы предполагаем, что у детенышей диких серых
крыс именно в этот период происходит запечатление на обонятельные
сигналы, исходящие от матери. Результатом импринтинта может быть
хорошо выраженная реакция следования в основном за матерями у мо-
лодых зверьков, которые в конце этого периода начинают осваивать про-
странство вне убежищ (СаШNoопи, 1963; Соколов, Квашнин, 1983а, 6; 1985).
Во всяком случае, реакция следования обеспечивает сплоченность семьи,
синхронизацию активности ее членов, освоение крысятами новой для них
среды и выработку у них адаптивных реакций, а также способствует
преодолению детеньшами чувства страха (Мох, 1960; Куае, 4966; Вгоп-
воп, 1968а, Ь), которое особенно сильно развито у пих в ювенильном
возрасте.
Новедение 241
На формирование поведения детенышей крыс в ювенильный период
их онтогенеза значительное влияние оказывают уже не только матери,
но и сверстники. Для оценки этого влияния крысят отнимали от мате.
рей в возрасте 21—23 дня, подвергали действию различных факторов
и оценивали их последотвия у взрослых животных. Детеныши, роспие
с трехнедельного возраста в усложненных условиях (в больших клетках
© различными предметами, лесенками и т. п.), во взрослом возрасте по-
казывали лучшую обучаемость в лабиринте и были менее эмоциональны-
ми, чем росшие в обычных условиях (Вше\йат, Ст Из, 1952). Крысята,
подвергавшиеся джентлингу (взятие в руки и поглаживание по спине)
с момента отпятия их от матери, обучались во взрослом возрасте решать
задачи на различение быстрее контрольных (Вегимешт, 1957), а также
значительно чаще доминировали пад животными из контрольной группы
в ситуациях пищевой конкуренции (Мезе!, Возеп, 1960).
На формирование поведения зверьков в ювенильный период сущест-
венное влияние оказывают условия, связанные с социальной деприваци-
ей. Изолированное содержание крысят с 23- до 51-дневного возраста при-
водило к тому, что во взрослом возрасте они хуже обучались и были
более активны в лабириите и в открытом поле (Ватутр]е-АШогд, Вегот,
1984). Эти авторы предположили, что такие измепения обусловлены раз-
витием у крысят синдрома «раскрепощения» (независимости от сибсов).
Изоляция оказывает значительное влияние на полозое и агрессивное
поведение. Отдельное содержание приводило к новышенной сексуальной
активности и обусловливало неадекватное половое поведение у самцов
(Веасв, 1942; СогаШ её а1., 1967; Стиепае!, Атпоа, 1969). У взрослых
самок, которые были изолированы с 30-дневного возраста, отмечали по-
вышенную активность. Однако начало полового созревания и особенно.
сти эстральных циклов у них были сходными с самками, содержавшимися
труппами (Напзеп, 1977). Изоляция в ювенильном возрасте приводила
также к повышенной агрессивности у взрослых животных и к повыццен-
ной чувствительности к стрессу (Тоге, ЕаплеПу, 1984; У а экана е1 а1,
1983; Тюге, Зиро-анену, 1984; Тапа её а., 1984). Изолированное со-
держапие крысят в 41—68-дневном возрасте уже не сказывалось на их
агонистическом поведении во взрослом возрасте (Ма ахана о а]., 1983).
Изоляция у лабораторных крыс отражалась также на многих физио-
логических реакциях, на чувствительности к различным химическим
веществам и на ряде других форм поведения (Возепя\уе её а1., 1972;
Слопим, 1976; Уждавини, 1980; и др.).
Подобных эксперимептов с дикими серыми крысами проводилось
храйпе мало. Только в нескольких работах приводятся данные о послед-
ствиях частичной социальной депривации па развитие крысят и на фор-
мировацие их поведения (Соколов, Квашнин, 1983а,6: 1985; Квапшин,
1983а, 6). Суть опыта сводилась к тому, что из вольер были изъяты
взрослые жкивотные (родители) в ювепиальный период развития детепы-
шей, когда им было 36—40 дней от роду. Дальнейшие наблюдеция в те-
чение 2,5 мес ноказали, что характер и частота социальных контактов
между крысятами, росшими без родителей, были сходными с таковыми
242
Глава четвертая
между семейными животными. Исключение составляло только маркиро-
вочное поведение: молодые самцы в период поведепческого созревания
метили партперов по группе и предметы среды значительно чаще, чем их
сверстники, росшие со взрослыми животными. Не выявлены различия в
сроках физиологического созревания молодых самок в разных типах
трунт. Существенные различия были установлены в скорости роста—
крысята из семейных групи росли значительно быстрее. Зверьки, кото-
рые росли без родителей, значительно долыпе сохраняли пугливость,
и у них впоследствии ярче проявлялась пассивпо-оборонительная реакция
и агрессивность но отношению к чужим особям. По-видимому, эти изме-
нения в новедении крысят были обусловлены отсутствием матери, хотя и
не ясны их механизмы.
Роль и значение взрослых самцов (отцов) в формировании новедепия
молодых особей не определены. Для лабораторных животных показано,
что молодые крысы в возрасте от 25 до 86 дней часто взаимодействовали
© отцами в семейных группах. Предполагастсл, что присутствие отцов
влияет на процессе социализации молодняка (Отелуз, 1982). Результаты
паблюдений за дикими крысами отличны от описанных выше — крысята
в этом возрасте относительно редко вступали в коптакты с отцами (Со-
колов, Квашнин, 1985). Однако преднолатается, что присутетвие взрослых
самцов оказывает сдерживающее влияние на формирование агрессивного
и маркировочного поведения у молодых самцов.
В заключение хотелось бы отметить, что различные эффекты, проде-
монстрированные в экоперимептах, трудно переносить па диких живот-
пых в естественных условиях обитания. С одной стороны, потому, что
мы нс можем полностью сопоставить условия, в которых проводятся ла-
бораторные опыты, с факторами, существенными для развития, выжива-
ния и размножения животных в природе, а с другой стороны, потому,
что обтсктами исследовапий служат в основном лабораторные крысы,
которые значительно отличаются от своих родоначальников.
Игровое поведение
Игры животных привлекали и продолжают привлекать внимание ученых
разных специальностей, что нашло свос отражение в значительном коли-
честве публикаций по этому вопросу. Однако при попытках дать опреде-
ление игре, выявить ве критерии и характеристики, а также установить
ее природу и функциопальное значение исследователи столкнулись со
многими трудпостями (Веась, 1945; Готов, 1966: Каоеп, 1974, 1981;
Фабри, 1976, 1982; Эльконин, 1978; Векой, 1984; и др.). Все это спра-
ведливо и по отношению к играм лабораторных и серых крыс.
Детеныши доместицированных крыс пачинают играть вскоре после
того, как у них открываются глаза. (Тауог, 1980; Меапеу, Зета, 19ВЛа;
РапКзерр, 1981). То же самое свойствепно, по-видимому, и диким крысам:
при паблюдениях в вольерах за группами пасюков мы неоднократно от-
мечали шум и сотрясение домиков, в которых находились детенынти тако-
го возраста. Постепенио частота игр увеличивается: у лабораторных крыс
Поведение
243
она достигает максимума ко времени их нолового созревания, а у диких
вскоре после выхода из убежищ. Игровые взаимодействия между серыми
крысами были характерны для них и в период их полового созревания,
поэтому здесь уместно отметить, что неправомочно игровой период онто-
тепеза поведепия и психики животных считать синонимом лишь ювениль-
ного периода, как это делают некоторые психологи (Фабри, 1976, 1982;
Крымов, 1984, 1983). У лабораторных крыс частота игр заметно снижа-
ется к 2,5—3-месячному возрасту (РапКзерр, 1984; Тпог, НоПожау,
1984а, Ъ), ау серых —к 3—3,5-месячному возрасту, т. е. к периоду пове-
денческого созревапия (Соколов, Иваптниы, 1985). У взрослых серых и
лабораторных крыс действия, которые можно охарактеризовать как игро-
вые, встречаются уже крайне редко.
Игровая активпость молодых животных проявляется вн трех формах.
Во-первых, это локомоторная или индивидуальная игра, которая выража-
ется в эпергичных прыжках и стремительных пробежках. Во-вторых, жи-
вотные могут играть с различными предметами. И наконец, широко рас-
пространены социальные игры, которыс привлекают наибольтиее внимание
исследователей. Для молодых крыс характерны в основном социальные
игры, хотя нередки и индивидуальные, которым свойственны импульеив-
пые скачкообразные движения, происходящие ние контактов с другими
зверьками, т. е. детеныш играет как бы сам по себе. Если играюний
таким образом крысенок сталкивается с другим зверьком, то индивидуаль-
пая игра чаще всего переходит в социальную (Рое, Е1зЪ, 1975; Соколов,
Квашнин, 1985; Квашнии, 1983а). Очевидно, что в данном случае прыж-
ки и наскоки одного детеныша на другого являются сигпалами, вызываю-
щими игру. Наличие таких «метакоммуникационных» сигналов — сигна-
лов, приглатающих к игре и указывающих на «несерьезность»
намерений, является необходимым условием возникновения межиндиви-
дуальных игр (Вафезоп, 1955; АШешапи, 1967; Еазеп, 1981). У крыс к
игровой сигнализации относятся и такие элементы поведения, как хвата-
ние за птерсть или за хвост, перелезания или подлезания под партпера,
а также взаимные чистки (Роое, ЕазН, 1975; Меапеу, Зце\мгагь 1981; ТВот,
НоПожау, 1983а, Ь).
Социальные игры у детенышей крыс происходят в основиом между
двумя зверьками, но иногда случаются и групповые игры, которые на-
поминают «кучу малу». Изучение половых и индивидуальных различий
между детенышами крысе в социальных играх дало противоречивые ре-
зультаты. В ряде работ, в которых использовали метод парных встреч,
не выявлены половые различия в играх лабораторных животных (Рапк-
зерр, Всайу, 1980; Рапкзерр, 1981; ТакафазЫ, Гоге, 1983). Однако в дру-
гих работах была показапа существенная разница в частоте игр, начатых
по инициативе самцов и самок — наиболее «игривыми» были самцы (Роо-
1е, ЗВ, 1986; ОПОН, Збеууат4, 1978; Меапеу, Эфемаге, 1981а; Вике, Мезапеу,
1983; Рапкзерр её 21., 1984; ТБог, НоПозмау, 1984а). Как считают последние
авторы, эти различия обусловлены различными условиями эксперимептов.
Наблюдения за поведением детенышей серых крыс показали, что ко-
личество ипдивидуальных игр пе различалось у молодых самцов и самок
242
Глава четвертая
между семейными животными. Исключение составляло только маркиро-
вочное поведение: молодые самцы в период поведепческого созревания
метили партперов по группе и предметы среды значительно чаще, чем их
сверстники, росшие со взрослыми животными. Не выявлены различия в
сроках физиологического созревания молодых самок в разных типах
трунт. Существенные различия были установлены в скорости роста—
крысята из семейных групи росли значительно быстрее. Зверьки, кото-
рые росли без родителей, значительно долыпе сохраняли пугливость,
и у них впоследствии ярче проявлялась пассивпо-оборонительная реакция
и агрессивность но отношению к чужим особям. По-видимому, эти изме-
нения в новедении крысят были обусловлены отсутствием матери, хотя и
не ясны их механизмы.
Роль и значение взрослых самцов (отцов) в формировании новедепия
молодых особей не определены. Для лабораторных животных показано,
что молодые крысы в возрасте от 25 до 86 дней часто взаимодействовали
© отцами в семейных группах. Предполагастсл, что присутствие отцов
влияет на процессе социализации молодняка (Отелуз, 1982). Результаты
паблюдений за дикими крысами отличны от описанных выше — крысята
в этом возрасте относительно редко вступали в коптакты с отцами (Со-
колов, Квашнин, 1985). Однако преднолатается, что присутетвие взрослых
самцов оказывает сдерживающее влияние на формирование агрессивного
и маркировочного поведения у молодых самцов.
В заключение хотелось бы отметить, что различные эффекты, проде-
монстрированные в экоперимептах, трудно переносить па диких живот-
пых в естественных условиях обитания. С одной стороны, потому, что
мы нс можем полностью сопоставить условия, в которых проводятся ла-
бораторные опыты, с факторами, существенными для развития, выжива-
ния и размножения животных в природе, а с другой стороны, потому,
что обтсктами исследовапий служат в основном лабораторные крысы,
которые значительно отличаются от своих родоначальников.
Игровое поведение
Игры животных привлекали и продолжают привлекать внимание ученых
разных специальностей, что нашло свос отражение в значительном коли-
честве публикаций по этому вопросу. Однако при попытках дать опреде-
ление игре, выявить ве критерии и характеристики, а также установить
ее природу и функциопальное значение исследователи столкнулись со
многими трудпостями (Веась, 1945; Готов, 1966: Каоеп, 1974, 1981;
Фабри, 1976, 1982; Эльконин, 1978; Векой, 1984; и др.). Все это спра-
ведливо и по отношению к играм лабораторных и серых крыс.
Детеныши доместицированных крыс пачинают играть вскоре после
того, как у них открываются глаза. (Тауог, 1980; Меапеу, Зета, 19ВЛа;
РапКзерр, 1981). То же самое свойствепно, по-видимому, и диким крысам:
при паблюдениях в вольерах за группами пасюков мы неоднократно от-
мечали шум и сотрясение домиков, в которых находились детенынти тако-
го возраста. Постепенио частота игр увеличивается: у лабораторных крыс
Поведение
243
она достигает максимума ко времени их нолового созревания, а у диких
вскоре после выхода из убежищ. Игровые взаимодействия между серыми
крысами были характерны для них и в период их полового созревания,
поэтому здесь уместно отметить, что неправомочно игровой период онто-
тепеза поведепия и психики животных считать синонимом лишь ювениль-
ного периода, как это делают некоторые психологи (Фабри, 1976, 1982;
Крымов, 1984, 1983). У лабораторных крыс частота игр заметно снижа-
ется к 2,5—3-месячному возрасту (РапКзерр, 1984; Тпог, НоПожау,
1984а, Ъ), ау серых —к 3—3,5-месячному возрасту, т. е. к периоду пове-
денческого созревапия (Соколов, Иваптниы, 1985). У взрослых серых и
лабораторных крыс действия, которые можно охарактеризовать как игро-
вые, встречаются уже крайне редко.
Игровая активпость молодых животных проявляется вн трех формах.
Во-первых, это локомоторная или индивидуальная игра, которая выража-
ется в эпергичных прыжках и стремительных пробежках. Во-вторых, жи-
вотные могут играть с различными предметами. И наконец, широко рас-
пространены социальные игры, которыс привлекают наибольтиее внимание
исследователей. Для молодых крыс характерны в основном социальные
игры, хотя нередки и индивидуальные, которым свойственны импульеив-
пые скачкообразные движения, происходящие ние контактов с другими
зверьками, т. е. детеныш играет как бы сам по себе. Если играюний
таким образом крысенок сталкивается с другим зверьком, то индивидуаль-
пая игра чаще всего переходит в социальную (Рое, Е1зЪ, 1975; Соколов,
Квашнин, 1985; Квашнии, 1983а). Очевидно, что в данном случае прыж-
ки и наскоки одного детеныша на другого являются сигпалами, вызываю-
щими игру. Наличие таких «метакоммуникационных» сигналов — сигна-
лов, приглатающих к игре и указывающих на «несерьезность»
намерений, является необходимым условием возникновения межиндиви-
дуальных игр (Вафезоп, 1955; АШешапи, 1967; Еазеп, 1981). У крыс к
игровой сигнализации относятся и такие элементы поведения, как хвата-
ние за птерсть или за хвост, перелезания или подлезания под партпера,
а также взаимные чистки (Роое, ЕазН, 1975; Меапеу, Зце\мгагь 1981; ТВот,
НоПожау, 1983а, Ь).
Социальные игры у детенышей крыс происходят в основиом между
двумя зверьками, но иногда случаются и групповые игры, которые на-
поминают «кучу малу». Изучение половых и индивидуальных различий
между детенышами крысе в социальных играх дало противоречивые ре-
зультаты. В ряде работ, в которых использовали метод парных встреч,
не выявлены половые различия в играх лабораторных животных (Рапк-
зерр, Всайу, 1980; Рапкзерр, 1981; ТакафазЫ, Гоге, 1983). Однако в дру-
гих работах была показапа существенная разница в частоте игр, начатых
по инициативе самцов и самок — наиболее «игривыми» были самцы (Роо-
1е, ЗВ, 1986; ОПОН, Збеууат4, 1978; Меапеу, Эфемаге, 1981а; Вике, Мезапеу,
1983; Рапкзерр её 21., 1984; ТБог, НоПозмау, 1984а). Как считают последние
авторы, эти различия обусловлены различными условиями эксперимептов.
Наблюдения за поведением детенышей серых крыс показали, что ко-
личество ипдивидуальных игр пе различалось у молодых самцов и самок
244
Глава четвертая
Рис. 72. Игровая активность молодых серых
крыс в семейных группаз (по: Соколов,
Евашнин, 1985)
р
А — индивидуальные игры,
Б — социальные
д:
игры —с0 всеми членами группы, взрослыми и
молодыми, инициаторами ноторых были молодые
особи, В — социальные игры только между моло-
дыми крысами (количество игр, инициаторами
которых были самцы и самки}. По оси ординат —
число игр за 1 ч наблюдений, по оси абециес—
возраст крыеят Р:Ж> 0,05, ЖЗИ ЖЖЖ <
ь
—
—
>
*
ж
|
*
+
к
В
!
Е
й
во все периоды их развития, кроме пе-
риода поведепческого созревания (Со-
7.
колов, Нвашнин, 1985; — Квашнин,
1983а, 6) (рис. 72). Аналогичные ре-
зультаты получены и по социальным
играм, если учитывать общую игровую
активность зверьков, т. е. количество
Е
.
игр со всеми членами (молодыми и
В
взрослыми) семейных групп, инициато-
рами которых были крысята обоего
т
пола. Однако при анализе социальных
игр, встречавшихся только между мо-
лодняком, оказалось, что самцы значи-
т
р
тельто чаще пачипали играть со свер-
р”
стниками, чем самки. Более того, среди
й
сибеов самцы предпочитали играть с
самками, а нес другими самцами. В свою
9-й Н-П 391—120 очередь, молодые самки значительно
_
р бо и чаще «заигрывали» с матерями, чем это
Г о” ЕА-6$
делали самцы. Для всех крысят было
характерпо почти полное отсутствие «заигрываний» со взрослыми сам-
цами.
В пределах одного выводка все молодые крысы, как серые, так и ла-
бораторные, проявляли высокую игровую активность. Однако отдельные
зверьки чаще и интенсивнее других участвовали в играх. Именно такие
особи (в осповном самцы) во взрослом возрасте демонстрировали повы-
шенную агрессивность по отношению к знакомым и чужим зверькам и
стаповились доминантными по отношению к сверстникам (Тауог, 1980;
Квашнин, 1983а; Соколов, Квашнин, 1985). Индивидуальные различия
могли выражаться и в том, что у отдельных крысят во время игр про-
являлись в разной степени элементы агрессивного и подчиненного пове-
дения (Роое, #1зВ, 1976).
У млекопитающих на игровую активность молодых животных могут
оказывать влияние различные факторы среды. В естественных условиях
обитания на частоту и продолжительность игр оказывают влияние преж-
де всего экологические факторы. Наши наблюдения за серыми крысами
Р
а
Поведение
245
в естественных условиях обитания показали, что игры между детепьыша-
ми встречались крайне редко по сравнению с вольерными условиями.
Таким образом, очевидно, что необходимым условием возникновения игры
является комфортное состояпие
— отсутствие голода, жажды, беспоноя-
щих или стрессирующих факторов среды.
В последпие годы начались интенсивные исследования эксперимен-
тальных воздействий на игровую активность животных. На лабораторных
крысах показано, что частота игр значительно увеличивается после изо-
ляции детеныша от сверстников на песколько дней и, напротив, обога-
щенная социальная среда снижает уровень игровой активности (Тауог,
1980; Рапкзерр, Веаму, 1980; РапКкзерр её а1., 1984}. Существенное влия-
ние на игру оказывают фармакологические средства (ВеаМу её а|., 1982,
19822), аносмия (Твог, НоПо\жау, 1982), нарушение мозговой нерегородки
(ВеаЦу её а|., 1982) и введение нейролептических веществ (скополами-
на) (ТЬог, НоЙо\жау, 1983а, Б). Также было установлено, что игровая
активность крые связана с уровнем гормонов в крови, в частности поло-
вых стероидов (Меапеу, З:е\ать 1981а; Меапеу её а|., 1982, 1983). На-
пример, введение тестостерона или его неароматизированного метаболита
За-дигидротестостерона увеличивало частоту социальных игр у зверьков
перед их половым созреванием. Эстрадиол не влиял на частоту итр,
а прогестерон и медроксипрогестеронацетат снижали уровень игр у сам-
пов и самок (ВиКе, Меапеу, 1983). Подобные исследования еще только
приносят первые результаты, поэтому преждевременно делать какие-либо
заключения.
Социальные игры являются видоспецифическими и состоят из эле-
ментов позедения, характерных для разных форм поведения взрослых
животных. Структурный подход к апализу игр делает упор на форму и
последовательность действий в игре (Еасеп, 1974, 1981). При этом пред-
полагается, что эта последовательность отлична от лоследовательности
неигровых движений (или случайна). Такой точки зрения придерживает-
ся, папример, Лойзос (1.01205, 1966). Однако исследования, проведенные
на разных видах млекопитающих, в том числе и на лабораторных крысах,
показали, что игровые последовательности имеют определепиую структу-
ру (Роше, Е1зЪ, 1975; Момлек: АтшИасе, 1979). Характерными особенно-
стями игр являются «преувеличенность» некоторых движений и целых
последовательностей и их многократная повторяемость. Существенным
признаком игры является также отсутствие очевидной цели или харак-
терного завершающего состояния. Она совершается как бы «вхолостую»,
хотя и отличается от «вакуумпой»» активности (Гогепт, 4956; ТБогре,
1966).
Элементы поведения, составляющие социальные игры, являются, как
правило, эквивалентами действий взрослых животных. Более того, у боль-
шииства изученных видов млекопитающих основная часть игровых эле-
мептов сходна по форме с агонистическим поведением взрослых живот-
ных (Рофе, 1966; Наточу, 1969; ВеКо#1, 1974а, 5; Мозле, АтшИаве, 1979).
У лабораторных крыс доля таких действий в социальных играх составляет
две трети (ВоПез, У!’ о0д3, 1964; Роше, Е1зВ, 1975). Это дало основание
244
Глава четвертая
Рис. 72. Игровая активность молодых серых
крыс в семейных группаз (по: Соколов,
Евашнин, 1985)
р
А — индивидуальные игры,
Б — социальные
д:
игры —с0 всеми членами группы, взрослыми и
молодыми, инициаторами ноторых были молодые
особи, В — социальные игры только между моло-
дыми крысами (количество игр, инициаторами
которых были самцы и самки}. По оси ординат —
число игр за 1 ч наблюдений, по оси абециес—
возраст крыеят Р:Ж> 0,05, ЖЗИ ЖЖЖ <
ь
—
—
>
*
ж
|
*
+
к
В
!
Е
й
во все периоды их развития, кроме пе-
риода поведепческого созревания (Со-
7.
колов, Нвашнин, 1985; — Квашнин,
1983а, 6) (рис. 72). Аналогичные ре-
зультаты получены и по социальным
играм, если учитывать общую игровую
активность зверьков, т. е. количество
Е
.
игр со всеми членами (молодыми и
В
взрослыми) семейных групп, инициато-
рами которых были крысята обоего
т
пола. Однако при анализе социальных
игр, встречавшихся только между мо-
лодняком, оказалось, что самцы значи-
т
р
тельто чаще пачипали играть со свер-
р”
стниками, чем самки. Более того, среди
й
сибеов самцы предпочитали играть с
самками, а нес другими самцами. В свою
9-й Н-П
391—120 очередь, молодые самки значительно
_
р бо и чаще «заигрывали» с матерями, чем это
Г о” ЕА-6$
делали самцы. Для всех крысят было
характерпо почти полное отсутствие «заигрываний» со взрослыми сам-
цами.
В пределах одного выводка все молодые крысы, как серые, так и ла-
бораторные, проявляли высокую игровую активность. Однако отдельные
зверьки чаще и интенсивнее других участвовали в играх. Именно такие
особи (в осповном самцы) во взрослом возрасте демонстрировали повы-
шенную агрессивность по отношению к знакомым и чужим зверькам и
стаповились доминантными по отношению к сверстникам (Тауог, 1980;
Квашнин, 1983а; Соколов, Квашнин, 1985). Индивидуальные различия
могли выражаться и в том, что у отдельных крысят во время игр про-
являлись в разной степени элементы агрессивного и подчиненного пове-
дения (Роое, #1зВ, 1976).
У млекопитающих на игровую активность молодых животных могут
оказывать влияние различные факторы среды. В естественных условиях
обитания на частоту и продолжительность игр оказывают влияние преж-
де всего экологические факторы. Наши наблюдения за серыми крысами
Р
а
Поведение
245
в естественных условиях обитания показали, что игры между детепьыша-
ми встречались крайне редко по сравнению с вольерными условиями.
Таким образом, очевидно, что необходимым условием возникновения игры
является комфортное состояпие
— отсутствие голода, жажды, беспоноя-
щих или стрессирующих факторов среды.
В последпие годы начались интенсивные исследования эксперимен-
тальных воздействий на игровую активность животных. На лабораторных
крысах показано, что частота игр значительно увеличивается после изо-
ляции детеныша от сверстников на песколько дней и, напротив, обога-
щенная социальная среда снижает уровень игровой активности (Тауог,
1980; Рапкзерр, Веаму, 1980; РапКкзерр её а1., 1984}. Существенное влия-
ние на игру оказывают фармакологические средства (ВеаМу её а|., 1982,
19822), аносмия (Твог, НоПо\жау, 1982), нарушение мозговой нерегородки
(ВеаЦу её а|., 1982) и введение нейролептических веществ (скополами-
на) (ТЬог, НоЙо\жау, 1983а, Б). Также было установлено, что игровая
активность крые связана с уровнем гормонов в крови, в частности поло-
вых стероидов (Меапеу, З:е\ать 1981а; Меапеу её а|., 1982, 1983). На-
пример, введение тестостерона или его неароматизированного метаболита
За-дигидротестостерона увеличивало частоту социальных игр у зверьков
перед их половым созреванием. Эстрадиол не влиял на частоту итр,
а прогестерон и медроксипрогестеронацетат снижали уровень игр у сам-
пов и самок (ВиКе, Меапеу, 1983). Подобные исследования еще только
приносят первые результаты, поэтому преждевременно делать какие-либо
заключения.
Социальные игры являются видоспецифическими и состоят из эле-
ментов позедения, характерных для разных форм поведения взрослых
животных. Структурный подход к апализу игр делает упор на форму и
последовательность действий в игре (Еасеп, 1974, 1981). При этом пред-
полагается, что эта последовательность отлична от лоследовательности
неигровых движений (или случайна). Такой точки зрения придерживает-
ся, папример, Лойзос (1.01205, 1966). Однако исследования, проведенные
на разных видах млекопитающих, в том числе и на лабораторных крысах,
показали, что игровые последовательности имеют определепиую структу-
ру (Роше, Е1зЪ, 1975; Момлек: АтшИасе, 1979). Характерными особенно-
стями игр являются «преувеличенность» некоторых движений и целых
последовательностей и их многократная повторяемость. Существенным
признаком игры является также отсутствие очевидной цели или харак-
терного завершающего состояния. Она совершается как бы «вхолостую»,
хотя и отличается от «вакуумпой»» активности (Гогепт, 4956; ТБогре,
1966).
Элементы поведения, составляющие социальные игры, являются, как
правило, эквивалентами действий взрослых животных. Более того, у боль-
шииства изученных видов млекопитающих основная часть игровых эле-
мептов сходна по форме с агонистическим поведением взрослых живот-
ных (Рофе, 1966; Наточу, 1969; ВеКо#1, 1974а, 5; Мозле, АтшИаве, 1979).
У лабораторных крыс доля таких действий в социальных играх составляет
две трети (ВоПез, У!’ о0д3, 1964; Роше, Е1зВ, 1975). Это дало основание
246 Глава четвертая
называть данную форму активности молодых животных игровой борьбой.
Большинство исследователей считают, что игровые драки молодых не то
же самое, что драки между взрослыми особями. Однако пекоторые уче-
ные полагают, что слово «игровая» только вводит в заблуждение. На са-
мом деле взаимоотношения между детенышами по сути являются агопи-
стическими, но характерными для сексуально пезрелых животных
(Ваепишеег, 1974; Тауог, 1980; Меапеу, З1емаг, 1984а). Существует и
такая точка зрения, что социальные игры вообще нельзя называть игровой:
борьбой, поскольку составляющие их элементы поведения относятся к
различиым (ис только агонистическим) формам взаимодействий взрослых
животных (Соколов, Квашнин, 1985). Эту точку зрения подтверждает и
то, что по мере роста крысят отдельные элементы игры постепенно «пре-
образовывались» в элементы собственно полового, дружелюбного и аго-
нистического поведепия. В копце концов это приводило к тому, что игры
молодых крыс все чаще оканчивались серьезными столкновениями. Ана-
логичное «преобразование» элементов игры в элементы взрослого пове-
депия отмечали у крыс и другие исследователи (СаШопи, 19683: Тьог
НоПо\жау, 1984а). Все это дает основание считать, что игра есть не обо
бая категория поведения, а составная часть онтотенеза, в которой обыч-
ные элементы поведения проявляются в специфических ювенильных
(предвзрослых) формах и в течение которой происходит формирование
социальных и других реакций, характерных для взрослых животных
(Веасв, 1945; Фабри, 1976, 1982; Крымов, 1981, 1983; Соколов, Квашнин
1985).
|
О природе игровой активности животных высказывались различные
точки зрения. Лоренц (Гогеп», 1950, 1956) ввел предетавление о «влече-
нии» к игре. Темброк (Тетьгоск, 1960, цит. по: Фабри, 1976) также по-
лагал, что игры являтотся своеобразными инстинктивными действиями с
самостоятельным мотивационным механизмом (центром игры). Однако
специальные исследования это не подтвердили (Незз, цит. по: Фабри
1976). Исходя из концепции Н. 9. Фабри (1976, 1982), которую под
тверждают эксперименты, проведенные А. А. Крымовым (1984, 1983),
игра есть развивающаяся доятельность и ее особенности связаны в пер-
вую очередь с развитием мотиваций, характерных для взрослых живот-
ных. Вначале отдельные элементы игры имеют нефункциональный ха-
рактер и лишь впоследствии происходит «наполнение» активности адек-
ватным ей предметным содержанием, т. е. игровая активность преобра-
зуется во взрослое поведепис.
До настоящего времени остается спорным и неяспым биологическое
значение игр. Одни исследователи отрицают их функциональное значе-
пие, считая, что «избыток энергии» молодых животных реализуется в по-
лучении удовольствия— им просто приятио играть. Однако большинство
зоологов и пеихологов полагают, что игры выполняют важную роль в
социализации животных (социализация
— результирующая всего с0-
циального опыта, который изменяет развитие особи
— \УПзоп, 1982).
С точки зрения «функционалистов» (Равеп, 4974, 1981), в игре развива-
ются и совершенствуются последующие адантивные реакции на физиче-
Поведение
247
ские и социальные факторы среды. Игра способствует развитию физиче-
ской силы, выносливости и ловкости; через игру происходит регуляция
скорости роста, приобретение чувствепиого опыта, развитие поведенческой
гибкости и изобретательности, необходимые для внутривидовой конкурен-
ции; игра усиливает социальные связи в группах и способствует форми-
рованию нормальных половых, дружелюбных и агонистических взаимо-
действий (Нато\ж, 1969; Рорчег, Зав, 1974; Равеп, 1984; У’Цвоп, 1982;
Веко!!, 1984; и др.). Также возможно, что через игру происходит «па-
стройка» молодых животцых к конкретным социальным И популяцион-
ным условиям и таким образом осуществляются последующие измепения
в социальной оранизации вида (Вагазв, 1974; ВекоН, 1974а, Ъ, 1975;
АгтИаое, 1975, 1977).
Мы описали общие закономерности онтотспеза поведения серых и ла-
бораторных крыс и на их основе выделили четко различающиеся ноз-
растные категории животных. Несмотря на относительную неполноту иа-
тих знаний об особенностях развития серых. крыс и па ряд существен-
ных различий между ними и дДомостицированными зверьками, мы
надеемся, что изложенные данные могут быть использованы как при эк-
спериментальных работах е крысами, так и при разработке проблем по-
пуляционной экологии этого вида. Возрастная классификация постна-’
тальпого периода жизни крыс может быть учтена при изучецпии влияния
раннего опыта на формирование поведения взрослых животных. Сопо-
ставление данной классификации с классификацией крыс, основанной на
‘морфологических признаках (Рыльников, Царасева, 1985), показывает,
что эти классификации не всегда и не в полной мере согласуются. По-
этому нам представляется весьма перспективным нахождение корреляции
между ними с тем, чтобы более адекватно изучать возрастную структуру
природпых популяций крые и ес динамику.
- Взаимоотношения между матерью и детенышами
Экспериментальное изучение взаимоотношений между матерью и дете-
нышами у лабораторной формы серой крысы началось еще в прошлом
веке (Зша|, 1899). В 1933 г. была опубликована сводка, где обобщались
данные полевых наблюдеций за дикой серой крысой с результатами экс-
перимептальных иселедований ее лабораторных форм (У езпег, ЗЪеаг@,
1933). Лабораторная крыса стала модельным объектом для изучения со-
циальных связей и особенно отпошений между матерью и детенышем.
Крыса — единственный вид, у которого изучены столь различные аспек-
ты такой связи, как поведенческий, физиологический (эндокринологиче-
ский и невральный) и энергетический, и заложены основы современных
представлений не только об общих нринципах отпошений между матерью
и детеньшием у млеконитающих, по и о механизмах, лежащих в их осно-
ве, Мы не ставим перед собой задачу с исчерпывающей полнотой изло-
жить этот разнородный материал, но рассмотрим лишь паиболее интен-
сивно разрабатываемые аспекты взаимоотношений между матерью и де-
тенышами у лабораторных крыс.
246 Глава четвертая
называть данную форму активности молодых животных игровой борьбой.
Большинство исследователей считают, что игровые драки молодых не то
же самое, что драки между взрослыми особями. Однако пекоторые уче-
ные полагают, что слово «игровая» только вводит в заблуждение. На са-
мом деле взаимоотношения между детенышами по сути являются агопи-
стическими, но характерными для сексуально пезрелых животных
(Ваепишеег, 1974; Тауог, 1980; Меапеу, З1емаг, 1984а). Существует и
такая точка зрения, что социальные игры вообще нельзя называть игровой:
борьбой, поскольку составляющие их элементы поведения относятся к
различиым (ис только агонистическим) формам взаимодействий взрослых
животных (Соколов, Квашнин, 1985). Эту точку зрения подтверждает и
то, что по мере роста крысят отдельные элементы игры постепенно «пре-
образовывались» в элементы собственно полового, дружелюбного и аго-
нистического поведепия. В копце концов это приводило к тому, что игры
молодых крыс все чаще оканчивались серьезными столкновениями. Ана-
логичное «преобразование» элементов игры в элементы взрослого пове-
депия отмечали у крыс и другие исследователи (СаШопи, 19683: Тьог
НоПо\жау, 1984а). Все это дает основание считать, что игра есть не обо
бая категория поведения, а составная часть онтотенеза, в которой обыч-
ные элементы поведения проявляются в специфических ювенильных
(предвзрослых) формах и в течение которой происходит формирование
социальных и других реакций, характерных для взрослых животных
(Веасв, 1945; Фабри, 1976, 1982; Крымов, 1981, 1983; Соколов, Квашнин
1985).
|
О природе игровой активности животных высказывались различные
точки зрения. Лоренц (Гогеп», 1950, 1956) ввел предетавление о «влече-
нии» к игре. Темброк (Тетьгоск, 1960, цит. по: Фабри, 1976) также по-
лагал, что игры являтотся своеобразными инстинктивными действиями с
самостоятельным мотивационным механизмом (центром игры). Однако
специальные исследования это не подтвердили (Незз, цит. по: Фабри
1976). Исходя из концепции Н. 9. Фабри (1976, 1982), которую под
тверждают эксперименты, проведенные А. А. Крымовым (1984, 1983),
игра есть развивающаяся доятельность и ее особенности связаны в пер-
вую очередь с развитием мотиваций, характерных для взрослых живот-
ных. Вначале отдельные элементы игры имеют нефункциональный ха-
рактер и лишь впоследствии происходит «наполнение» активности адек-
ватным ей предметным содержанием, т. е. игровая активность преобра-
зуется во взрослое поведепис.
До настоящего времени остается спорным и неяспым биологическое
значение игр. Одни исследователи отрицают их функциональное значе-
пие, считая, что «избыток энергии» молодых животных реализуется в по-
лучении удовольствия— им просто приятио играть. Однако большинство
зоологов и пеихологов полагают, что игры выполняют важную роль в
социализации животных (социализация
— результирующая всего с0-
циального опыта, который изменяет развитие особи
— \УПзоп, 1982).
С точки зрения «функционалистов» (Равеп, 4974, 1981), в игре развива-
ются и совершенствуются последующие адантивные реакции на физиче-
Поведение
247
ские и социальные факторы среды. Игра способствует развитию физиче-
ской силы, выносливости и ловкости; через игру происходит регуляция
скорости роста, приобретение чувствепиого опыта, развитие поведенческой
гибкости и изобретательности, необходимые для внутривидовой конкурен-
ции; игра усиливает социальные связи в группах и способствует форми-
рованию нормальных половых, дружелюбных и агонистических взаимо-
действий (Нато\ж, 1969; Рорчег, Зав, 1974; Равеп, 1984; У’Цвоп, 1982;
Веко!!, 1984; и др.). Также возможно, что через игру происходит «па-
стройка» молодых животцых к конкретным социальным И популяцион-
ным условиям и таким образом осуществляются последующие измепения
в социальной оранизации вида (Вагазв, 1974; ВекоН, 1974а, Ъ, 1975;
АгтИаое, 1975, 1977).
Мы описали общие закономерности онтотспеза поведения серых и ла-
бораторных крыс и на их основе выделили четко различающиеся ноз-
растные категории животных. Несмотря на относительную неполноту иа-
тих знаний об особенностях развития серых. крыс и па ряд существен-
ных различий между ними и дДомостицированными зверьками, мы
надеемся, что изложенные данные могут быть использованы как при эк-
спериментальных работах е крысами, так и при разработке проблем по-
пуляционной экологии этого вида. Возрастная классификация постна-’
тальпого периода жизни крыс может быть учтена при изучецпии влияния
раннего опыта на формирование поведения взрослых животных. Сопо-
ставление данной классификации с классификацией крыс, основанной на
‘морфологических признаках (Рыльников, Царасева, 1985), показывает,
что эти классификации не всегда и не в полной мере согласуются. По-
этому нам представляется весьма перспективным нахождение корреляции
между ними с тем, чтобы более адекватно изучать возрастную структуру
природпых популяций крые и ес динамику.
- Взаимоотношения между матерью и детенышами
Экспериментальное изучение взаимоотношений между матерью и дете-
нышами у лабораторной формы серой крысы началось еще в прошлом
веке (Зша|, 1899). В 1933 г. была опубликована сводка, где обобщались
данные полевых наблюдеций за дикой серой крысой с результатами экс-
перимептальных иселедований ее лабораторных форм (У езпег, ЗЪеаг@,
1933). Лабораторная крыса стала модельным объектом для изучения со-
циальных связей и особенно отпошений между матерью и детенышем.
Крыса — единственный вид, у которого изучены столь различные аспек-
ты такой связи, как поведенческий, физиологический (эндокринологиче-
ский и невральный) и энергетический, и заложены основы современных
представлений не только об общих нринципах отпошений между матерью
и детеньшием у млеконитающих, по и о механизмах, лежащих в их осно-
ве, Мы не ставим перед собой задачу с исчерпывающей полнотой изло-
жить этот разнородный материал, но рассмотрим лишь паиболее интен-
сивно разрабатываемые аспекты взаимоотношений между матерью и де-
тенышами у лабораторных крыс.
248 `Глава четвертая
Общая характеристика связи между матерью
% детенышами
Беременность крысы длится 22—23 дня. В это время (особенно в пер-
вой половипе беременности) самка в экспериментальных условиях ипдиф-
ферентно относится к подкладываемым в ее клетку крысятам. Носле
рождения детенышей у нее появляется весь комплекс адекватных мате-
ринских реакций, обеспечивающих сохранение и затиту потометва !5.
В пернод рождения и выкармливания детенышей глубоко меняется фи-
зиолотия и поведение самки: для этого времени характерен особый гор-
мональный профиль (ем. например: Возеп а, З1ее1, 1980), меняются
скорость метаболических процессов, уровни нотреблепия пищи и воды
(Сгозуепог, Мепа, 1974; Мешше, 1978: Емейтап ©! аГ., 1984), баланс
теплопродукции и теплоотдачи (Теоп © а|., 1978), режим активности
(Стоа, Адет, 1969), повышается агрессивность по отношению к членам
собственной группы и к чужакам (НезКш её а1., 1978) 9.
У незрелорождающегося крысенка связи с окружающей средой и ре-
сурсы, которые он использует, принципиально иные, чем у взрослой кры-
сы. Основной источник его энергетических ресуреов и основной компо-
нент окружающей среды— мать, которая в первую неделю после родов
около 80% своего времени проводит в гнезде, в непосредственном коп-
такте с выводком (Стгофа, АЪег, 1969). Важнейшие черты поведепия де-
тенышей обусловливаются этой принципиально важной для его выжива-
ния связью © самкой. Новеденческие возможности поворожденного неве-
лики: ориентация и движепие к материпокому соску, акты сосания,
восстановление физического контакта с самкой или другими детенышами.
Соответственио ограничены и перцептивные способности. Осповные из
них — восприятие запахов и температурного градиента — контролируют
движение детепыша по направлепию к самке и достижение сосков (см.,
например: Афемз, 1976; АЪегз, Мау, 1984).
Первое кормление молоком происходит сразу же после родов — самка
закрывает детенышей сверху, как бы нависая над ними, так что область
15 В то же время у крыс (по крайней мере лабораторных) самки иногда поедают
новорожденных детенышей. Если мать уничтожает часть выводка, то за остав-
шимися детенышами она ухаживает не хуже, чем самка, у которой каннибализм’
не наблюдался (Пе Зап5, ЗеВта]и, 1984).
Атрессивное поведение самки в период выкармливания выводка отличаетея от
агрессивного поведения самцов по структуре, функциям и по гормональным ме-
ханизмам. В частности, для «материнской агрессии» характерны очень малые
латентные периоды до и между атаками, большое количество коротких, малоин-
тенсивных укусов, изправленных на голову противника; локализация ран на те-
ле противника более разнообразна, чем ран, наносимых во время схваток самцов
(ОПуег её а1., 1985). Для изучения механизмов «материнской агрессии» модельным
видом являются мыши. Мы существенно меньше знаем о факторах, определяю-
щих это поведение у крыс. Однако вполне резонно предположить, что резкое воз-
растание агрессивности у самки крысы во вторую неделю выкармливания вывод-
ка также зависит от повышения уровня пролактина в крови и стимуляции дете-
нышами сосков матери при сосании.
®
Поведение
249
сосков оказывается в непосредственном контакте с детенышами. Коли-
чество молока, продуцируемого самкой, зависит от стимуляции сосков
детенышами во время сосания (Стозуепог её а|., 1970). Температура мо-
лока и режим его поступления в значительной степени определяют нор-
мальпое функционирование пищеварительной и других физиологических
‹истем крысенка. Это сказывается, например, на скорости сердечных
<окращений (НоЁег, 1981), на ритме «сон- бодротвование» крысят (Но-
Чег, ЗВа!г, 1982).
В первые дии жизни детеньши не способен самостоятельно испраж-
няться. Самка, вылизывая его аногенитальную область, стимулирует
уринацию н дефекацию и очищает тело от экскрементов. С помощью
введепия в кровь детенышам изотопов, переходящих затем вследствие
слизывания мочи крысят в оргацпизм самки, было показано, что сущест-
венная часть ее потребности в воде (которая в период лактации значи-
тельно повышена) удовлетворяется именно за счет слизывания мочи дете-
нышей (Емефтаи еб а], 1984; СоЪегиек, А1Ъег, 1985). Стимул, при-
влекающий самку к обнюхиванию и вылизыванию аногенитальной обла-
сти крысят, представляет собой секрет их препуциальных желез, после
оперативного удаления которых 75% новорожденных погибает из-за то-
то, что самка их почти не вылизывает (Уегпеф-Маигу, Вгопеме, 1985).
`У самок, экспериментально лишенных возможности слизывать мочу де-
тенышей, резко повышается степень риска обезвоживания организма
({СаБегшск, А]Ъегз, 1983).
Детеныпи-самцы существенно больню вылизываются матерью, чем
детеныши-самки (Мооге, 1981). Это различение крысят по полу опреде-
ляется, по-видимому, особенностями запаха апогенитальной области сам-
цов и самок, который, в свою очередь, зависит от гормонального профи-
ля детенышей. Тактильная стимуляция апогенитальной области имеет
существенное значение для развития комплекса копуляторного поведения
самцов. Детепыши-самцы, чьи матери вследствие экспериментального на-
рушения восприятия запахов мало их вылизывали, во взрослом соетоянии
по некоторым характеристикам полового поведения (латептный период
эйякуляций и интромиесий, интервалы между интромиссиями и др.) от-
личаются от выращенных интактными самками (Мооге, 1984).
Кроме молока, самка предоставляет детенышу и другой столь же вазк-
ный для выживания энергетический ресурс
— тепло, необходимое ему
для нормальпого роста. Пойкилотермные детеныши получают его при
физическом контакте с самкой в гнезде, когда она кормит выводок или
‚ просто скучивается с ним. Самка, определяя свой режим пребывания в
гнезде, в контакте с детенышами регулирует этим и количество отдавае-
мого им тепла (Реоп её а]., 1985). Первый период своей жизни крысята
проводят в материнском гнезде, самка строит его незадолго перед рода-
ми, в нем детеныши появляются. па свет. Гиездо с его особым микрокли-
матом также представляет защиту детеньшам от неблагоприятных воз-
действий внешней среды (Во{се, 1977).
Ирысята на первых стадиях постнатального развития не способны
самостоятельно передвигаться на сколько-нибудь значительные расстоя-
248 `Глава четвертая
Общая характеристика связи между матерью
% детенышами
Беременность крысы длится 22—23 дня. В это время (особенно в пер-
вой половипе беременности) самка в экспериментальных условиях ипдиф-
ферентно относится к подкладываемым в ее клетку крысятам. Носле
рождения детенышей у нее появляется весь комплекс адекватных мате-
ринских реакций, обеспечивающих сохранение и затиту потометва !5.
В пернод рождения и выкармливания детенышей глубоко меняется фи-
зиолотия и поведение самки: для этого времени характерен особый гор-
мональный профиль (ем. например: Возеп а, З1ее1, 1980), меняются
скорость метаболических процессов, уровни нотреблепия пищи и воды
(Сгозуепог, Мепа, 1974; Мешше, 1978: Емейтап ©! аГ., 1984), баланс
теплопродукции и теплоотдачи (Теоп © а|., 1978), режим активности
(Стоа, Адет, 1969), повышается агрессивность по отношению к членам
собственной группы и к чужакам (НезКш её а1., 1978) 9.
У незрелорождающегося крысенка связи с окружающей средой и ре-
сурсы, которые он использует, принципиально иные, чем у взрослой кры-
сы. Основной источник его энергетических ресуреов и основной компо-
нент окружающей среды— мать, которая в первую неделю после родов
около 80% своего времени проводит в гнезде, в непосредственном коп-
такте с выводком (Стгофа, АЪег, 1969). Важнейшие черты поведепия де-
тенышей обусловливаются этой принципиально важной для его выжива-
ния связью © самкой. Новеденческие возможности поворожденного неве-
лики: ориентация и движепие к материпокому соску, акты сосания,
восстановление физического контакта с самкой или другими детенышами.
Соответственио ограничены и перцептивные способности. Осповные из
них — восприятие запахов и температурного градиента — контролируют
движение детепыша по направлепию к самке и достижение сосков (см.,
например: Афемз, 1976; АЪегз, Мау, 1984).
Первое кормление молоком происходит сразу же после родов — самка
закрывает детенышей сверху, как бы нависая над ними, так что область
15 В то же время у крыс (по крайней мере лабораторных) самки иногда поедают
новорожденных детенышей. Если мать уничтожает часть выводка, то за остав-
шимися детенышами она ухаживает не хуже, чем самка, у которой каннибализм’
не наблюдался (Пе Зап5, ЗеВта]и, 1984).
Атрессивное поведение самки в период выкармливания выводка отличаетея от
агрессивного поведения самцов по структуре, функциям и по гормональным ме-
ханизмам. В частности, для «материнской агрессии» характерны очень малые
латентные периоды до и между атаками, большое количество коротких, малоин-
тенсивных укусов, изправленных на голову противника; локализация ран на те-
ле противника более разнообразна, чем ран, наносимых во время схваток самцов
(ОПуег её а1., 1985). Для изучения механизмов «материнской агрессии» модельным
видом являются мыши. Мы существенно меньше знаем о факторах, определяю-
щих это поведение у крыс. Однако вполне резонно предположить, что резкое воз-
растание агрессивности у самки крысы во вторую неделю выкармливания вывод-
ка также зависит от повышения уровня пролактина в крови и стимуляции дете-
нышами сосков матери при сосании.
®
Поведение
249
сосков оказывается в непосредственном контакте с детенышами. Коли-
чество молока, продуцируемого самкой, зависит от стимуляции сосков
детенышами во время сосания (Стозуепог её а|., 1970). Температура мо-
лока и режим его поступления в значительной степени определяют нор-
мальпое функционирование пищеварительной и других физиологических
‹истем крысенка. Это сказывается, например, на скорости сердечных
<окращений (НоЁег, 1981), на ритме «сон- бодротвование» крысят (Но-
Чег, ЗВа!г, 1982).
В первые дии жизни детеньши не способен самостоятельно испраж-
няться. Самка, вылизывая его аногенитальную область, стимулирует
уринацию н дефекацию и очищает тело от экскрементов. С помощью
введепия в кровь детенышам изотопов, переходящих затем вследствие
слизывания мочи крысят в оргацпизм самки, было показано, что сущест-
венная часть ее потребности в воде (которая в период лактации значи-
тельно повышена) удовлетворяется именно за счет слизывания мочи дете-
нышей (Емефтаи еб а], 1984; СоЪегиек, А1Ъег, 1985). Стимул, при-
влекающий самку к обнюхиванию и вылизыванию аногенитальной обла-
сти крысят, представляет собой секрет их препуциальных желез, после
оперативного удаления которых 75% новорожденных погибает из-за то-
то, что самка их почти не вылизывает (Уегпеф-Маигу, Вгопеме, 1985).
`У самок, экспериментально лишенных возможности слизывать мочу де-
тенышей, резко повышается степень риска обезвоживания организма
({СаБегшск, А]Ъегз, 1983).
Детеныпи-самцы существенно больню вылизываются матерью, чем
детеныши-самки (Мооге, 1981). Это различение крысят по полу опреде-
ляется, по-видимому, особенностями запаха апогенитальной области сам-
цов и самок, который, в свою очередь, зависит от гормонального профи-
ля детенышей. Тактильная стимуляция апогенитальной области имеет
существенное значение для развития комплекса копуляторного поведения
самцов. Детепыши-самцы, чьи матери вследствие экспериментального на-
рушения восприятия запахов мало их вылизывали, во взрослом соетоянии
по некоторым характеристикам полового поведения (латептный период
эйякуляций и интромиесий, интервалы между интромиссиями и др.) от-
личаются от выращенных интактными самками (Мооге, 1984).
Кроме молока, самка предоставляет детенышу и другой столь же вазк-
ный для выживания энергетический ресурс
— тепло, необходимое ему
для нормальпого роста. Пойкилотермные детеныши получают его при
физическом контакте с самкой в гнезде, когда она кормит выводок или
‚ просто скучивается с ним. Самка, определяя свой режим пребывания в
гнезде, в контакте с детенышами регулирует этим и количество отдавае-
мого им тепла (Реоп её а]., 1985). Первый период своей жизни крысята
проводят в материнском гнезде, самка строит его незадолго перед рода-
ми, в нем детеныши появляются. па свет. Гиездо с его особым микрокли-
матом также представляет защиту детеньшам от неблагоприятных воз-
действий внешней среды (Во{се, 1977).
Ирысята на первых стадиях постнатального развития не способны
самостоятельно передвигаться на сколько-нибудь значительные расстоя-
250
Тлава четвертая
р
%
р
Е
А
90-
86 |-
30—
ве
Г
20—
50-
40Е
ОЕ
30|
-
26
0
д
1
О ООО
О
ОТТЗООООВ ОО
0
0
20
36
40 50
ГУИН
ИВЛ
[ олова,
г
4
\
р
А
Е
\
т
А
урл
обама е
Рис. 73. Характеристика периода лактации
А — количество молока, выделяемого самкой в период кормления детенышей, измеренное ме--
тодом меченых изотопов. По оси абсцисс
— дни после родов (А—Г), по оси ординат
— отно-
Сительное количество высасываемого детенышами молока.
В — время, проводимое кормящей самкой в гнезде в контакте с выводком. Но оси ординат—
доля времени, проводимого с выводком.
В— выраженность у самки реакций транспортировки детенышей
1— реакция возвращения детенышей в гнездо после их удаления экспериментатором, 2 —
реакция переноса в новое гнездо носле разрушения старого. По оси ординат
— процент самок,
у которых выражена реакция транспортировки всего выводка полностью.
Г — гнездостроительная активность кормящих самок. По оси ординат
— доля самок, занимаю-
щихся гнездостроением (%). Все из (быеЁ, 1981)
ния. Если в этом возникает необходимость (когда детеныши в силу ка-
ких-нибудь обстоятельств оказываются вне гнезда), то их переносит мать.
Новорожденных самки осторожно берут зубами поперек тела, старитих
держат за кожу загривка или спины. В распоряжении детенышей есть
средетво активно воздействовать на поведение самки: оказавшись вие
г
а
Поведение 251
гнезда, крысенок издает ультразвуковые крики, основная частота кото-
рых (около 32 кГц) лежит в области максимальной чувствительности к
ультразвукам взрослых крыс (СонгеуйеЬ, Наск, 1966). Эти крики пе
только привлекают внимание самки, но и стимулируют проявление реак-
ций трапспортировки. В экспериментальных условиях в отсутствие де-
тенышей самки под воздействием таких звуков могут перепосить и нс-
живые объекты (АШт, Вапк$, 1972; Втеуузог, Геоп, 1980).
По мере увеличения самостоятельности крысенка мепяются обтем и
специфика материпской заботы. Ниже мы приводим графики, отражаю-
щис изменспис уровня проявления различных видов материнского пове-
дения крысы по мере роста детенышей. Так, уменьшение количества мо-
лока у самки происходит в то время, когда у крысенка формируются
способности, делающие возможным питание твердыми кормами (16—
40 дней); время, проводимое самкой в коптакте с детенышами, сокраща-
ется, когда они становятся гомойтермными; число переносов идет на
убыль, когда у крысят формируются локомоторпые способности, харак-
териые для взрослых животных; гнездостроительная активность матери
(рис. 73) падает ио мере роста детенышей. Поиск механизмов, обеспечи-
вающих такую сипхронность отношений,— одна из важнейпгих задач со-
временных исследований.
Изменение взаимоотношений между матерью и растущим детенышем
не сводится к простому ослаблению и разрушению комплекса связей,
характерных для отношений между самкой и новорожденпым. Онтогенез
детепылта— не только постепенное созревание всех систем и функций,
но и смена среды обитания. Детеныят в процессе своего развития прохо-
дит через ряд экологических пиш (Са]сТ, 1981), каждая из которых
предъявляет растущему организму свои требования, На 3—4-й неделе
детеныит переходит от питания материнским молоком к самостоятельно-
му питапию твердыми кормами. С этим связаны серъезные перестройки
физиологии пищеварения и пищевого поведения. Меняется также исполь-
зование пространства: крысепок пачипаст самостоятельно осваивать тер-
риторию. С этим процессом связана глубокая перестройка перцептивных
и локомоторных способностей животного, ведущая к изменению отноше-
пия к пространству и к сородичам. Материнская забота помогает детены-
иту «справиться» со всеми этими задачами. Возникают новые связи меж-
ду пим и матерью. Грысят пачицает привлекать запах экскрементов
матери. Это усиливает дистантную связь между самкой и подвижными
детенышами, облегчает их ориентацию в прострапстве. Поедацие крыся-
тами фекалий матери (которые в это время имеют особый состав)
способствует переходу на питание твердыми кормами, ускоряет развитие
детенышей (Теоп, 1974; Гес, Мо, 1984а). В этот период онтогенеза де-
тенышей мать определяет выбор кормов, обучает их сиецифическим при-
емам добывания пищи, коптролирует режим использовапия территории
(Сао, 1977). Рассмотрим подробнее наиболее изученные аспекты этих
отношений.
250
Тлава четвертая
р
%
р
Е
А
90-
86 |-
30—
ве
Г
20—
50-
40Е
ОЕ
30|
-
26
0
д
1
О ООО
О
ОТТЗООООВ
ОО
0
0
20
36
40 50
ГУИН
ИВЛ
[ олова,
г
4
\
р
А
Е
\
т
А
урл
обама е
Рис. 73. Характеристика периода лактации
А — количество молока, выделяемого самкой в период кормления детенышей, измеренное ме--
тодом меченых изотопов. По оси абсцисс
— дни после родов (А—Г), по оси ординат
— отно-
Сительное количество высасываемого детенышами молока.
В — время, проводимое кормящей самкой в гнезде в контакте с выводком. Но оси ординат—
доля времени, проводимого с выводком.
В— выраженность у самки реакций транспортировки детенышей
1— реакция возвращения детенышей в гнездо после их удаления экспериментатором, 2 —
реакция переноса в новое гнездо носле разрушения старого. По оси ординат
— процент самок,
у которых выражена реакция транспортировки всего выводка полностью.
Г — гнездостроительная активность кормящих самок. По оси ординат
— доля самок, занимаю-
щихся гнездостроением (%). Все из (быеЁ, 1981)
ния. Если в этом возникает необходимость (когда детеныши в силу ка-
ких-нибудь обстоятельств оказываются вне гнезда), то их переносит мать.
Новорожденных самки осторожно берут зубами поперек тела, старитих
держат за кожу загривка или спины. В распоряжении детенышей есть
средетво активно воздействовать на поведение самки: оказавшись вие
г
а
Поведение 251
гнезда, крысенок издает ультразвуковые крики, основная частота кото-
рых (около 32 кГц) лежит в области максимальной чувствительности к
ультразвукам взрослых крыс (СонгеуйеЬ, Наск, 1966). Эти крики пе
только привлекают внимание самки, но и стимулируют проявление реак-
ций трапспортировки. В экспериментальных условиях в отсутствие де-
тенышей самки под воздействием таких звуков могут перепосить и нс-
живые объекты (АШт, Вапк$, 1972; Втеуузог, Геоп, 1980).
По мере увеличения самостоятельности крысенка мепяются обтем и
специфика материпской заботы. Ниже мы приводим графики, отражаю-
щис изменспис уровня проявления различных видов материнского пове-
дения крысы по мере роста детенышей. Так, уменьшение количества мо-
лока у самки происходит в то время, когда у крысенка формируются
способности, делающие возможным питание твердыми кормами (16—
40 дней); время, проводимое самкой в коптакте с детенышами, сокраща-
ется, когда они становятся гомойтермными; число переносов идет на
убыль, когда у крысят формируются локомоторпые способности, харак-
териые для взрослых животных; гнездостроительная активность матери
(рис. 73) падает ио мере роста детенышей. Поиск механизмов, обеспечи-
вающих такую сипхронность отношений,— одна из важнейпгих задач со-
временных исследований.
Изменение взаимоотношений между матерью и растущим детенышем
не сводится к простому ослаблению и разрушению комплекса связей,
характерных для отношений между самкой и новорожденпым. Онтогенез
детепылта— не только постепенное созревание всех систем и функций,
но и смена среды обитания. Детеныят в процессе своего развития прохо-
дит через ряд экологических пиш (Са]сТ, 1981), каждая из которых
предъявляет растущему организму свои требования, На 3—4-й неделе
детеныит переходит от питания материнским молоком к самостоятельно-
му питапию твердыми кормами. С этим связаны серъезные перестройки
физиологии пищеварения и пищевого поведения. Меняется также исполь-
зование пространства: крысепок пачипаст самостоятельно осваивать тер-
риторию. С этим процессом связана глубокая перестройка перцептивных
и локомоторных способностей животного, ведущая к изменению отноше-
пия к пространству и к сородичам. Материнская забота помогает детены-
иту «справиться» со всеми этими задачами. Возникают новые связи меж-
ду пим и матерью. Грысят пачицает привлекать запах экскрементов
матери. Это усиливает дистантную связь между самкой и подвижными
детенышами, облегчает их ориентацию в прострапстве. Поедацие крыся-
тами фекалий матери (которые в это время имеют особый состав)
способствует переходу на питание твердыми кормами, ускоряет развитие
детенышей (Теоп, 1974; Гес, Мо, 1984а). В этот период онтогенеза де-
тенышей мать определяет выбор кормов, обучает их сиецифическим при-
емам добывания пищи, коптролирует режим использовапия территории
(Сао, 1977). Рассмотрим подробнее наиболее изученные аспекты этих
отношений.
252
Глава четвертая
Материнекное поведение
При изучении материнского поведепия впимацие исследователей концен-
трировалось на следующих основных проблемах: какие гормональные и
певральные механизмы обеспечивают материпское поведение? Какую
роль в организации материнского поведения играет стимуляция, исходя-
щая от детенышей? Как эти внутренние и внешние по отношению к
самке факторы взаимодействуют друг с другом? Каково значение родов.
в появлении материнского поведения?
Для выработки многих приемов регистрации материнского ловеде-
ния крысы важное значение имеют две предпосылки. Первая из них 0с-
нована на том, что по крайней мере три основные характеристики мате-
рииского поведения (гнездостроение, время поддержания физического:
контакта с выводком и реакция возвращения детенышей в гпездо) кор-
релируют между собой по времени и частоте проявления. Хотя эти кор-
реляции относительны, они дают основание условно ввести одну пере-
менную— комплекс материнского поведения— и оценивать его по одно-
му из трех наиболее удобных для измерения параметров (чаще всего’
используется характеристика «возвращение крысят в гпездо»).
Вторая предпосылка осповапа на том, что на материнское позедение:
крысы (по крайней мере лабораторной) не влияет индивидуальное зна-
кометво с детенышами: она ведет себя практически одинаково, нодкла-
дывает ли ей экспериментатор ее собственных или чужих детенышей *".
Действенными являются лишь возрастные особеппости детенышей: дете-
ныши одного возраста вызывают у самки одни и те же реакции. Матери:
не реагируют на замену своих детенышей чужими, даже если эта замена
осуществляется ежедневно па протяжении всего периода вскармливалия-
(см. например: Геоп, 1974). Именно благодаря этому обстоятельству
был выработан простой и универсальный метод для измерения чувстви-
тельности крысы к стимулам, вызывающим материнское поведение. Он
заключается в следующем: в клетку к крысе подкладывают детенышей
определенного возраста и регистрируют латентный период до момента
переноса первого детеныша в гнездо, количество перепесенных детенышей
за определенный промежуток времени, период времени между последую-
щими переносами и пеноторые другие количественные характеристики.
Этот тест оказался пригоден не только для оценки изменения мотивацион-
ного состояния данного индивидуума, но и для анализа межиндивидуаль-
пых и межлинейных различий и стал использоваться для широкого круга
других задач.
Сразу же после появления на свет детенышей самка, даже если она
рожала впервые, проявляет по отпошению к ним все необходимые фор-
11 Это вовсе не означает, что крысы не способны отличать своих детенышей от
чужих. Напротив, было показано (ВеасН, Уаупез, 1956), что крысы способны по
запаху отличать свой выводок от чужого. Однако вполне оправдано предположе-
ние, что на исследуемый сейчас уровень организации материнекого поведения
эта способность крысы практически не влияет.
Новедение
253
мы материнской заботы. Какие факторы управляют материнским поведе-
нием крысы? Уже вскоре после открытия гормонов было высказано пред-
положепие, что стероидные гормоны
— прогестерон, эстрогены, а также
тормон передней доли гипофиза — пролактин, управляющие беремен-
ностью, родами и лактацией, определяют и «инстинкты заботы о потом-
ствс» (см., например: Уезпег, ЗВеага, 1933). Исследования гормональ-
ного субстрата материнского поведения крысы велись начиная с 40-х го-
дов, параллельно с работами по гормональной регуляции беременности,
родов и лактации. Их результаты с описанием условий гормональной
стимуляции материнского поведения можно найти в сводке Розепблатта
и Сейджела (ВозепЫан, З1есе1, 1980).
Согласно современным представлениям, в развитии материнского по-
ведеция можно выделить две фазы, различающиеся по принципам фи-
зиологического контроля. Первая
— «включение» материнского поведе-
ния — соответствует по времени периоду родов, когда в поведении сам-
ки внезапно и комплексно проявляются все зиды заботы о детеньишах.
Вторая фаза — «поддержапие» материнского поведения
— протекает
в период выкармливания выводка (см., например: Нозеп ай еб а1.,
1985).
у
Гормональное включение материнского поведения. Период, когда про-
исходит включение материцского поведения, характеризуется резким из-
мепением гормонального баланса организма самки (рис. 74). За относи-
тельно короткий период (20—23-й депь) падает уровень прогестерона,
быстро возрастает уровень эстрогенов (19—23-й день) и пролактипа.
(21—23-й день). Особое «предродовое» состояцие организма выражается
и во многих других измепепиях в секреции гормонов и чувствительности
в пим. Возникает предположение, что включение материнского поведе-
ния определяется имепно этим особым гормональным профилем.
Действительно, было показано, что в крови самки в период включения
материнского поведения — за сутки до родов и сутки после них — содер-
держится пекий активный компонент, который способен стимулировать
материнское поведение. Инъекция плазмы крови, взятой у самок-мате-
рей, нерожавшим и небеременпым самкам вызывает у них комплекеное
включение материнского поведения, Особенно успешпым в этом отноше-
нии оказалось перекрестное переливание крови: самкам вживляли в серд-
це катетер с длинной трубкой, которая выходила на затылке и давала
возможность объединить кровеносные системы двух животных. Носле
нескольких часов такой транефузии крови между самкой, которая толь-
ко что родила, и небеременной самкой кровь этих животных полностью
*перемептивалась и у небеременной самки «включалось» материнское по-
ведение (Тегке], Нозеп Ман, 41968, 1972). Однако сам активный компо-
нент крови остался неидентифицированным.
Для его определепия были сделаны попытки искусственио создать
предродовой гормональный профиль различными экспериментальными
вмешательствами. Так, было показано, что, если вводить небеременным
самкам по специальной схеме только три гормона: эстроген, прогестерон
и пролактин, можно вызвать комплексное включение материнского пове-
252
Глава четвертая
Материнекное поведение
При изучении материнского поведепия впимацие исследователей концен-
трировалось на следующих основных проблемах: какие гормональные и
певральные механизмы обеспечивают материпское поведение? Какую
роль в организации материнского поведения играет стимуляция, исходя-
щая от детенышей? Как эти внутренние и внешние по отношению к
самке факторы взаимодействуют друг с другом? Каково значение родов.
в появлении материнского поведения?
Для выработки многих приемов регистрации материнского ловеде-
ния крысы важное значение имеют две предпосылки. Первая из них 0с-
нована на том, что по крайней мере три основные характеристики мате-
рииского поведения (гнездостроение, время поддержания физического:
контакта с выводком и реакция возвращения детенышей в гпездо) кор-
релируют между собой по времени и частоте проявления. Хотя эти кор-
реляции относительны, они дают основание условно ввести одну пере-
менную— комплекс материнского поведения— и оценивать его по одно-
му из трех наиболее удобных для измерения параметров (чаще всего’
используется характеристика «возвращение крысят в гпездо»).
Вторая предпосылка осповапа на том, что на материнское позедение:
крысы (по крайней мере лабораторной) не влияет индивидуальное зна-
кометво с детенышами: она ведет себя практически одинаково, нодкла-
дывает ли ей экспериментатор ее собственных или чужих детенышей *".
Действенными являются лишь возрастные особеппости детенышей: дете-
ныши одного возраста вызывают у самки одни и те же реакции. Матери:
не реагируют на замену своих детенышей чужими, даже если эта замена
осуществляется ежедневно па протяжении всего периода вскармливалия-
(см. например: Геоп, 1974). Именно благодаря этому обстоятельству
был выработан простой и универсальный метод для измерения чувстви-
тельности крысы к стимулам, вызывающим материнское поведение. Он
заключается в следующем: в клетку к крысе подкладывают детенышей
определенного возраста и регистрируют латентный период до момента
переноса первого детеныша в гнездо, количество перепесенных детенышей
за определенный промежуток времени, период времени между последую-
щими переносами и пеноторые другие количественные характеристики.
Этот тест оказался пригоден не только для оценки изменения мотивацион-
ного состояния данного индивидуума, но и для анализа межиндивидуаль-
пых и межлинейных различий и стал использоваться для широкого круга
других задач.
Сразу же после появления на свет детенышей самка, даже если она
рожала впервые, проявляет по отпошению к ним все необходимые фор-
11 Это вовсе не означает, что крысы не способны отличать своих детенышей от
чужих. Напротив, было показано (ВеасН, Уаупез, 1956), что крысы способны по
запаху отличать свой выводок от чужого. Однако вполне оправдано предположе-
ние, что на исследуемый сейчас уровень организации материнекого поведения
эта способность крысы практически не влияет.
Новедение
253
мы материнской заботы. Какие факторы управляют материнским поведе-
нием крысы? Уже вскоре после открытия гормонов было высказано пред-
положепие, что стероидные гормоны
— прогестерон, эстрогены, а также
тормон передней доли гипофиза — пролактин, управляющие беремен-
ностью, родами и лактацией, определяют и «инстинкты заботы о потом-
ствс» (см., например: Уезпег, ЗВеага, 1933). Исследования гормональ-
ного субстрата материнского поведения крысы велись начиная с 40-х го-
дов, параллельно с работами по гормональной регуляции беременности,
родов и лактации. Их результаты с описанием условий гормональной
стимуляции материнского поведения можно найти в сводке Розепблатта
и Сейджела (ВозепЫан, З1есе1, 1980).
Согласно современным представлениям, в развитии материнского по-
ведеция можно выделить две фазы, различающиеся по принципам фи-
зиологического контроля. Первая
— «включение» материнского поведе-
ния — соответствует по времени периоду родов, когда в поведении сам-
ки внезапно и комплексно проявляются все зиды заботы о детеньишах.
Вторая фаза — «поддержапие» материнского поведения
— протекает
в период выкармливания выводка (см., например: Нозеп ай еб а1.,
1985).
у
Гормональное включение материнского поведения. Период, когда про-
исходит включение материцского поведения, характеризуется резким из-
мепением гормонального баланса организма самки (рис. 74). За относи-
тельно короткий период (20—23-й депь) падает уровень прогестерона,
быстро возрастает уровень эстрогенов (19—23-й день) и пролактипа.
(21—23-й день). Особое «предродовое» состояцие организма выражается
и во многих других измепепиях в секреции гормонов и чувствительности
в пим. Возникает предположение, что включение материнского поведе-
ния определяется имепно этим особым гормональным профилем.
Действительно, было показано, что в крови самки в период включения
материнского поведения — за сутки до родов и сутки после них — содер-
держится пекий активный компонент, который способен стимулировать
материнское поведение. Инъекция плазмы крови, взятой у самок-мате-
рей, нерожавшим и небеременпым самкам вызывает у них комплекеное
включение материнского поведения, Особенно успешпым в этом отноше-
нии оказалось перекрестное переливание крови: самкам вживляли в серд-
це катетер с длинной трубкой, которая выходила на затылке и давала
возможность объединить кровеносные системы двух животных. Носле
нескольких часов такой транефузии крови между самкой, которая толь-
ко что родила, и небеременной самкой кровь этих животных полностью
*перемептивалась и у небеременной самки «включалось» материнское по-
ведение (Тегке], Нозеп Ман, 41968, 1972). Однако сам активный компо-
нент крови остался неидентифицированным.
Для его определепия были сделаны попытки искусственио создать
предродовой гормональный профиль различными экспериментальными
вмешательствами. Так, было показано, что, если вводить небеременным
самкам по специальной схеме только три гормона: эстроген, прогестерон
и пролактин, можно вызвать комплексное включение материнского пове-
254
Глава четвертая
Ае/МЯ
О
Рис. 74. Концент-
и
рация гормонов в
плазме крови вры-
ве мн
Е
сы во время бе-
ременности
=&100 Е
Стрелка слева
—оп
яя [8
5
лодотворение, спра-
аЗЕЙ
1 ва—роды.
з
аз
1— пролактин,
з0Гз805.50Г
2 — эстрадиол,
8 — прогестерон. Из
9
(Возепай ©
2
20 15|
а1., 1979)
20Е
1
0|8пе0ПООЕ
О
О
ЧО
О
О
о
1
ть
197916
ПВЯЯ
|
беременность , дни
дения (Мо её а., 1970; Галтом её а1., 1971). Однако для успешной
стимуляции необходимо придерживаться сложного режима введения гор-
мопов, сильно зависящего от экспериментальных условий, линии крыс
и других факторов. Часто попытки повторить эти опыты оказывались
неудачными (Козеп ай, Зее], 1980).
Предродовой гормопальный баланс можно воссоздать и другими сно-
собами. Так, удаление матки или кесарево сечение стимулируют вклю-
ченис материнского поведения. Эти операции вызывают гормональные
события, сходные с теми, что наблюдаются при родах: уменьшение со-
держания прогестерона, рост уровня эстрогенов. Увеличение продукции
эстрогенов после удаления матки приводит к тому, что через 48 ч
после операции наступают эструс и овуляция (как и после родов).
Чем позже во время беремепиости проведены удалепие матки или кеса-
рево сечение, тем быстрее и комплекснее проявляется материнское пове-
денис. Это показывает, что важны не только взаимный баланс эстрогенов
и прогестерона, но и основные их уровни (Возен ай, 1есе], 1975).
Гели кроме матки удалить яичники, то материнское поведение не вклю-
чается, так как роет эстрогенов, продуцирусмых яичпниками, отсутствует.
Введение эстрогенов в такой ситуации стимулирует появление материн-
ского поведения, а введепие значительных доз прогестерона подавляет
его (З1есе|. ВозепМай, 1975; Вм@зез её а1., 1978а, Ъ). Все это свидетель-
ствует о том, что понижение уровня прогестеронов при повышении уров-
ня эстрогенов и вызывает включение материнского поведения. В даль-
цейшем было показано, что некий базовый уровень пролактина является
также необходимым фактором для включения (Матап её а1., 1972). Так,
у самок с удаленным гипофизом вызвать включение материнского пове-
дения одними манипуляциями 0 стероидпыми гормопами не удается.
Однако введение пролактина таким самкам позволяет включить материн-
Поведение
255
ское поведение (Вт1Ч вез еЁ аё., 1985). Уровепь окситоцина (гормона зад-
ней доли гипофиза, играющего важную роль в процессе родов и стиму-
лирующего молокоотдачу) также существенно важен для включения
материпского поведения (Повфегот её а1., 1977; Редегзен, Ргапое, 1979).
Влияние процесса родов на включение материнекого поведения. Итак,
включение материнского поведения самки, происходящее приблизитель-
но в период родов (пачипая за 1—2 суток до родов), осуществляется
под влиянием гормональных событий. Важен ли при этом сам процесс
родов? Оказываетсл, что его влияние несущественно. Так, после кесарева
сечения даже те самки, которые были беременны впервые, проявляют
нормальпое материнское поведение (Том, Возеп ай, 1969). Тщательные
наблюдения за поведением самок позволили установить, что основные
изменения их реактивности но отношению к детепышам происходят еще
до родов. Начипаются они за 1—2 сут и становятся особенно резкими
в последние часы перед родами.
За 24—48 ч перед родами в ответ на проигрывание магнитофонной
записи звуков, издаваемых двухдневными детеныптами, у самки возрас-
тает двигательная активность и она ориентируст внимапие в направле-
нии источника звуков (Когапу! её а|., 1976). За несколько часов перед,
родами во много раз усиливается гнездостроительная активность (510{-
п1сК её а|., 1973), прежнее безразличие к подкладываемым в клетку
детенышам исчезает, самка перетаскивает их в гнездо и, располагаясь
с ними, принимает позы, характерные для кормления молоком (Козеп-
аи, З1еае|, 1975; Вт вез, 1978). Выраженность этих реакций неуклонно
растет но мере приближепия момента родов, и за несколько часов перед
ними крысы некоторых липий могут комплексно проявлять к подклады-
ваемым детеныитам все компоненты материиского поведепия так же ин-
тенсивно, как и по отношению к собственным после родов (Мауег,
ВозепЫ а, 1984). В последние сутки перед родами изменяются и другие
аспекты поведения самки: резко увеличивается ее агрессивность по отно-
тнению к незнакомым животным (ГезВш в а1., 4978), учащаются ультра-
звуковые крики (Геууз, Зей"Иег, 1985, цит. но: Возев ай её а1., 1985),
а в экспериментальпых тестах на различение самки теперь предпочи-
тают запах гнезда самок, находящихся на поздпей стадии беременности,
запаху гнезд пебеременных самок (Вацег, 1983).
Таким образом, сам процесс родов оказывается малосущественным
для подготовки крысы к проявлению материнского поведепия и самка
еще до родов оказывается готовой проявлять его. Интересно, что эта.
готовность «стать матерью» появляется не постепенпо, а как бы «вклю-
чается» внезапно.
Влияние опыта на процесе включения материнского поведения. Опре-
деленнос влияние на характеристики включения материнского поведсиия
перед родами оказывает предварительный опыт самки в воспитании
детенышей. Так, у самок, которые предварительно рожали и выкармли-
зали детенышей, овариоэктомия па 20-м дне беременноети не является
препятствием для включения материнского поведения, а у самок, бере-
менных впервые. материнское поведение после овариоэктомии не вклю-
254
Глава четвертая
Ае/МЯ
О
Рис. 74. Концент-
и
рация гормонов в
плазме крови вры-
ве мн
Е
сы во время бе-
ременности
=&100 Е
Стрелка слева
—оп
яя [8
5
лодотворение, спра-
аЗЕЙ
1 ва—роды.
з
аз
1— пролактин,
з0Гз805.50Г
2 — эстрадиол,
8 — прогестерон. Из
9
(Возепай ©
2
20 15|
а1., 1979)
20Е
1
0|8пе
0ПООЕ
О
О
ЧО
О
О
о
1
ть
197916
ПВЯЯ
|
беременность , дни
дения (Мо её а., 1970; Галтом её а1., 1971). Однако для успешной
стимуляции необходимо придерживаться сложного режима введения гор-
мопов, сильно зависящего от экспериментальных условий, линии крыс
и других факторов. Часто попытки повторить эти опыты оказывались
неудачными (Козеп ай, Зее], 1980).
Предродовой гормопальный баланс можно воссоздать и другими сно-
собами. Так, удаление матки или кесарево сечение стимулируют вклю-
ченис материнского поведения. Эти операции вызывают гормональные
события, сходные с теми, что наблюдаются при родах: уменьшение со-
держания прогестерона, рост уровня эстрогенов. Увеличение продукции
эстрогенов после удаления матки приводит к тому, что через 48 ч
после операции наступают эструс и овуляция (как и после родов).
Чем позже во время беремепиости проведены удалепие матки или кеса-
рево сечение, тем быстрее и комплекснее проявляется материнское пове-
денис. Это показывает, что важны не только взаимный баланс эстрогенов
и прогестерона, но и основные их уровни (Возен ай, 1есе], 1975).
Гели кроме матки удалить яичники, то материнское поведение не вклю-
чается, так как роет эстрогенов, продуцирусмых яичпниками, отсутствует.
Введение эстрогенов в такой ситуации стимулирует появление материн-
ского поведения, а введепие значительных доз прогестерона подавляет
его (З1есе|. ВозепМай, 1975; Вм@зез её а1., 1978а, Ъ). Все это свидетель-
ствует о том, что понижение уровня прогестеронов при повышении уров-
ня эстрогенов и вызывает включение материнского поведения. В даль-
цейшем было показано, что некий базовый уровень пролактина является
также необходимым фактором для включения (Матап её а1., 1972). Так,
у самок с удаленным гипофизом вызвать включение материнского пове-
дения одними манипуляциями 0 стероидпыми гормопами не удается.
Однако введение пролактина таким самкам позволяет включить материн-
Поведение
255
ское поведение (Вт1Ч вез еЁ аё., 1985). Уровепь окситоцина (гормона зад-
ней доли гипофиза, играющего важную роль в процессе родов и стиму-
лирующего молокоотдачу) также существенно важен для включения
материпского поведения (Повфегот её а1., 1977; Редегзен, Ргапое, 1979).
Влияние процесса родов на включение материнекого поведения. Итак,
включение материнского поведения самки, происходящее приблизитель-
но в период родов (пачипая за 1—2 суток до родов), осуществляется
под влиянием гормональных событий. Важен ли при этом сам процесс
родов? Оказываетсл, что его влияние несущественно. Так, после кесарева
сечения даже те самки, которые были беременны впервые, проявляют
нормальпое материнское поведение (Том, Возеп ай, 1969). Тщательные
наблюдения за поведением самок позволили установить, что основные
изменения их реактивности но отношению к детепышам происходят еще
до родов. Начипаются они за 1—2 сут и становятся особенно резкими
в последние часы перед родами.
За 24—48 ч перед родами в ответ на проигрывание магнитофонной
записи звуков, издаваемых двухдневными детеныптами, у самки возрас-
тает двигательная активность и она ориентируст внимапие в направле-
нии источника звуков (Когапу! её а|., 1976). За несколько часов перед,
родами во много раз усиливается гнездостроительная активность (510{-
п1сК её а|., 1973), прежнее безразличие к подкладываемым в клетку
детенышам исчезает, самка перетаскивает их в гнездо и, располагаясь
с ними, принимает позы, характерные для кормления молоком (Козеп-
аи, З1еае|, 1975; Вт вез, 1978). Выраженность этих реакций неуклонно
растет но мере приближепия момента родов, и за несколько часов перед
ними крысы некоторых липий могут комплексно проявлять к подклады-
ваемым детеныитам все компоненты материиского поведепия так же ин-
тенсивно, как и по отношению к собственным после родов (Мауег,
ВозепЫ а, 1984). В последние сутки перед родами изменяются и другие
аспекты поведения самки: резко увеличивается ее агрессивность по отно-
тнению к незнакомым животным (ГезВш в а1., 4978), учащаются ультра-
звуковые крики (Геууз, Зей"Иег, 1985, цит. но: Возев ай её а1., 1985),
а в экспериментальпых тестах на различение самки теперь предпочи-
тают запах гнезда самок, находящихся на поздпей стадии беременности,
запаху гнезд пебеременных самок (Вацег, 1983).
Таким образом, сам процесс родов оказывается малосущественным
для подготовки крысы к проявлению материнского поведепия и самка
еще до родов оказывается готовой проявлять его. Интересно, что эта.
готовность «стать матерью» появляется не постепенпо, а как бы «вклю-
чается» внезапно.
Влияние опыта на процесе включения материнского поведения. Опре-
деленнос влияние на характеристики включения материнского поведсиия
перед родами оказывает предварительный опыт самки в воспитании
детенышей. Так, у самок, которые предварительно рожали и выкармли-
зали детенышей, овариоэктомия па 20-м дне беременноети не является
препятствием для включения материнского поведения, а у самок, бере-
менных впервые. материнское поведение после овариоэктомии не вклю-
256 Глава четвертая
чается (Возепа еф а|., 1985). Таким образом, материнский опыт пони-
жает порог включения и делает его возможным при гормональных уров-
нях более низких, чем те, в которых нуждается неопытпая самка. При
первом экспериментальном предъявлении детенышей беременным самкам
многократно рожавшие особи пачинают проявлять материнское поведе-
ние значительно быстрее, чем беременные впервые. Реактивность пеопыт-
ных самок по отношенио к детенышам изменяется за несколько часов
перед родами, много раз рожавшие самки стаповятся чувствительтыми
к детепышам за 1—2 суток перед родами. За 3,5—6 ч перед родами
большая часть из них уже готова проявлять полный комплекс материн-
<кого поведения. Однако эти различия между опытными и неопытными
самками исчезают по мере приближения срока родов, т. е. по мере уси-
ления выраженности специфического гормонального профиля. Таким об-
разом, к момепту рождения собственных детенышей опытные и неопыт-
ные самки в равной степени готовы к проявлепию материиского поведе-
ния (Мауег, Возеп ай, 1980).
Включение материнского поведения сенситизацией самки. В нормаль-
ных условиях готовность самки к материнскому поведению, как мы уже
товорили, обусловлепа эпдогепными изменениями ее гормонального про-
филя. Однако интересно, что существует и другой путь «включения»
материпеского поведения, при котором ведущую роль играют не эндоген-
ные изменения гормонального статуса, а стимулы внешней среды. Если
з клетке У самки, не готовой к проявлению материнского поведения
(небеременной или девственной), постоянно присутствуют детеныши
«младенческого» (до 10-дневного) возраста, то ее первоначальное равно-
душие к пим через песколько дней исчезает. Появляются все учащаю-
пиеся обнюхивания и вылизывания детенышей, а затем внезапно вклю-
чается весь комплекс материнского поведения (за исключением лакта-
ции) (ЕНетшо, Козеп ам, 1974а). Этот фепомен, известпый под
названием «сенситизации», или «конкавеации» (У езпег, ЗНеага, 1933),
можно наблюдать и у самцов. Материнскому поведению, вызванному
сенситизацией, свойственны все особенности поведения, вызванного есте-
ственным эпдокринным путем: оно сходно с таковым по всем главным
характеристикам и аналогично меняется в зависимости от возраста и
количества детенышей (Мото, 1972а). Латентный период сенситизации
зависит от гепетических факторов (крысы определенных линий более
зтувствительны к такой стимуляции, чем другие), возраста и физиологи-
ческого состояния особей (Мауег, Возеп ай, 1979; Возеп ай её а].,
1985). Большое значение имеет режим стимуляции. Гак, детеныши в воз-
расте 3 дней являются наиболее, а старше 12 дней
— наименее эффек-
тивными для стимуляции материнского поведения (З4еги, МеЮштоп,
1978). Латентный период сенситизации зависит от количества стимуля-
ции — непрерывная стимуляция (когда детеныши находятся в клетке
постоянно) является более эффективной, чем стимуляция перемежаю-
щаяся (когда детенышей периодически убирают); в маленьких клетках
(площадь 200 см?) крысы быстрее «становятся матерями», чем в боль-
ших (площадь 3000 см?) (Тегке|, Возеп ай, 1971). Сенситизация совер-
Поведение
257
тя еее, если в соседней клетке находится самка с выводком
Физиологические механизмы сепситизации неизвестпы; хотя есть
предположения, что при сенситизации несколько меняется гормональный
профиль самки, но данные, подтверждающие их, противоречивы. Так
Маринари и Мольти (Магизат, Мой, 1978) сообщают, что постоянное
присутствие детенышей вызывает как у рожавших, так и у девственных
самок нарушение эстральной циклики и включение материнского поведе-
ния чаще происходит в экстральной и проэстральной фазах цикла. Одна-
ко другие исследователи таких связей не обнаружили (Э1етп, З1ее,
1978). Леон и Мольтц (Теоп, Мой=, 1975) сообщили, что введение
эстрогена и прогестерона может влиять на латентный период сенситиза-
ции, однако повторные исследования не подтвердили этих эффектов
(Возеп ай, её а|., 1979). У самок с удалеппыми гипофизом и яичниками
сенситизация проходит так же или несколько быстрее, чем у интактных
(Возеп ай, 1967). Кровь сенситизированной самки не содержит «актив-
ного элемента», который влияет на продолжительность латентного перио-
да сепситизации материнского поведения другой самки (Тегке!, Возеп-
Маз, 1968, 1972). Таким образом, при сенситизации материнского поведе-
ния влияпие гормональных процессов не очевидно.
Наблюдаются закономерные изменения в процессе сенситизации в за-
висимости от возраста крысы. Уже в 18-дневном возрасче у крысят
обоих полов проявляется тенденция контактировать с детенышами
3—5-дневного возраста. У крысят 22-дневного возраста сенситизацией
можно вызвать комплексное проявление материпского поведения, в том
числе и позу кормления. Как и взрослые животные, 22-дневные крысята
предпочитают двухдневных детенышей восьмидпевным, живых детены-
шей мертвым (Мауег, Возеп ам, 1979) и реагируют на двухдневных
детенышей не так, как на пластиковые муляжи такой же формы и веса.
Детенышей они переносят в гнездо, обогревают и т. п., а с муляжами
играют и грызут их (ТакоБо\зВ, Тегке], 1984; Стау, СВефеу, 1984).
Илтересно, что у самцов этого возраста «материнское» поведение вызвать
легче, чем у самок.
Большое значение для сенситизации имеет опыт предварительного
общения с детенышами. Так, взрослые самцы, которые не имели пред-
варительного опыта общения с 1—5-дневными детепышами, восприпи-
мают их при первом предъявлении как корм и поедают, применяя те
же специфические движения, что и при принятии пищи (Раш, КЙрег-
зови, 1975, цит. но: АозепЫаЕЬ её а|., 1979). Однако, постоянно под-
кладывая дотепьшюй, можно добиться сенситизации материнского пове-
цения и у таких самцов. Если самцы были предварительно сенситизиро-
ваны в возрасте 22 дней, то во взрослом состояний, при новой
сенситизации, «материнское» поведение включается у них быстрее, чем
У неопытных. Аналогичное влияние предыдущей сенситизации известно
и для взрослых самок (Мепите, Возеп а, 1974).
Механизм ‚включения материнского поведения. Связь между эндо-
кринной базой и сенеорным входом. Какой может быть связь между
9 Серая крыса
256 Глава четвертая
чается (Возепа еф а|., 1985). Таким образом, материнский опыт пони-
жает порог включения и делает его возможным при гормональных уров-
нях более низких, чем те, в которых нуждается неопытпая самка. При
первом экспериментальном предъявлении детенышей беременным самкам
многократно рожавшие особи пачинают проявлять материнское поведе-
ние значительно быстрее, чем беременные впервые. Реактивность пеопыт-
ных самок по отношенио к детенышам изменяется за несколько часов
перед родами, много раз рожавшие самки стаповятся чувствительтыми
к детепышам за 1—2 суток перед родами. За 3,5—6 ч перед родами
большая часть из них уже готова проявлять полный комплекс материн-
<кого поведения. Однако эти различия между опытными и неопытными
самками исчезают по мере приближения срока родов, т. е. по мере уси-
ления выраженности специфического гормонального профиля. Таким об-
разом, к момепту рождения собственных детенышей опытные и неопыт-
ные самки в равной степени готовы к проявлепию материиского поведе-
ния (Мауег, Возеп ай, 1980).
Включение материнского поведения сенситизацией самки. В нормаль-
ных условиях готовность самки к материнскому поведению, как мы уже
товорили, обусловлепа эпдогепными изменениями ее гормонального про-
филя. Однако интересно, что существует и другой путь «включения»
материпеского поведения, при котором ведущую роль играют не эндоген-
ные изменения гормонального статуса, а стимулы внешней среды. Если
з клетке У самки, не готовой к проявлению материнского поведения
(небеременной или девственной), постоянно присутствуют детеныши
«младенческого» (до 10-дневного) возраста, то ее первоначальное равно-
душие к пим через песколько дней исчезает. Появляются все учащаю-
пиеся обнюхивания и вылизывания детенышей, а затем внезапно вклю-
чается весь комплекс материнского поведения (за исключением лакта-
ции) (ЕНетшо, Козеп ам, 1974а). Этот фепомен, известпый под
названием «сенситизации», или «конкавеации» (У езпег, ЗНеага, 1933),
можно наблюдать и у самцов. Материнскому поведению, вызванному
сенситизацией, свойственны все особенности поведения, вызванного есте-
ственным эпдокринным путем: оно сходно с таковым по всем главным
характеристикам и аналогично меняется в зависимости от возраста и
количества детенышей (Мото, 1972а). Латентный период сенситизации
зависит от гепетических факторов (крысы определенных линий более
зтувствительны к такой стимуляции, чем другие), возраста и физиологи-
ческого состояния особей (Мауег, Возеп ай, 1979; Возеп ай её а].,
1985). Большое значение имеет режим стимуляции. Гак, детеныши в воз-
расте 3 дней являются наиболее, а старше 12 дней
— наименее эффек-
тивными для стимуляции материнского поведения (З4еги, МеЮштоп,
1978). Латентный период сенситизации зависит от количества стимуля-
ции — непрерывная стимуляция (когда детеныши находятся в клетке
постоянно) является более эффективной, чем стимуляция перемежаю-
щаяся (когда детенышей периодически убирают); в маленьких клетках
(площадь 200 см?) крысы быстрее «становятся матерями», чем в боль-
ших (площадь 3000 см?) (Тегке|, Возеп ай, 1971). Сенситизация совер-
Поведение
257
тя еее, если в соседней клетке находится самка с выводком
Физиологические механизмы сепситизации неизвестпы; хотя есть
предположения, что при сенситизации несколько меняется гормональный
профиль самки, но данные, подтверждающие их, противоречивы. Так
Маринари и Мольти (Магизат, Мой, 1978) сообщают, что постоянное
присутствие детенышей вызывает как у рожавших, так и у девственных
самок нарушение эстральной циклики и включение материнского поведе-
ния чаще происходит в экстральной и проэстральной фазах цикла. Одна-
ко другие исследователи таких связей не обнаружили (Э1етп, З1ее,
1978). Леон и Мольтц (Теоп, Мой=, 1975) сообщили, что введение
эстрогена и прогестерона может влиять на латентный период сенситиза-
ции, однако повторные исследования не подтвердили этих эффектов
(Возеп ай, её а|., 1979). У самок с удалеппыми гипофизом и яичниками
сенситизация проходит так же или несколько быстрее, чем у интактных
(Возеп ай, 1967). Кровь сенситизированной самки не содержит «актив-
ного элемента», который влияет на продолжительность латентного перио-
да сепситизации материнского поведения другой самки (Тегке!, Возеп-
Маз, 1968, 1972). Таким образом, при сенситизации материнского поведе-
ния влияпие гормональных процессов не очевидно.
Наблюдаются закономерные изменения в процессе сенситизации в за-
висимости от возраста крысы. Уже в 18-дневном возрасче у крысят
обоих полов проявляется тенденция контактировать с детенышами
3—5-дневного возраста. У крысят 22-дневного возраста сенситизацией
можно вызвать комплексное проявление материпского поведения, в том
числе и позу кормления. Как и взрослые животные, 22-дневные крысята
предпочитают двухдневных детенышей восьмидпевным, живых детены-
шей мертвым (Мауег, Возеп ам, 1979) и реагируют на двухдневных
детенышей не так, как на пластиковые муляжи такой же формы и веса.
Детенышей они переносят в гнездо, обогревают и т. п., а с муляжами
играют и грызут их (ТакоБо\зВ, Тегке], 1984; Стау, СВефеу, 1984).
Илтересно, что у самцов этого возраста «материнское» поведение вызвать
легче, чем у самок.
Большое значение для сенситизации имеет опыт предварительного
общения с детенышами. Так, взрослые самцы, которые не имели пред-
варительного опыта общения с 1—5-дневными детепышами, восприпи-
мают их при первом предъявлении как корм и поедают, применяя те
же специфические движения, что и при принятии пищи (Раш, КЙрег-
зови, 1975, цит. но: АозепЫаЕЬ её а|., 1979). Однако, постоянно под-
кладывая дотепьшюй, можно добиться сенситизации материнского пове-
цения и у таких самцов. Если самцы были предварительно сенситизиро-
ваны в возрасте 22 дней, то во взрослом состояний, при новой
сенситизации, «материнское» поведение включается у них быстрее, чем
У неопытных. Аналогичное влияние предыдущей сенситизации известно
и для взрослых самок (Мепите, Возеп а, 1974).
Механизм ‚включения материнского поведения. Связь между эндо-
кринной базой и сенеорным входом. Какой может быть связь между
9 Серая крыса
258 Глава четвертая
включением материнского поведения эндокринным путем и сенситиза-
цией? Чтобы обсудить этот вопрос, следует внимательно рассмотреть.
природу сенсорной стимуляции, воздействующей на включение материн-
ского поведения. Согласно наиболее обоспованной к настоящему времени
гипотезе (Репа, Возеп ай, 1974. с), основная сенсорная модаль-
ность, управляющая материнским поведением крысы,— хемореценция.
Предполагается, что самка крысы в любом физиологическом состоянии
готова проявить материнское поведение, но эта готовность блокируется
впеншим фактором
— запахом детенышей. Гормональные события, про-
исходящие на последних стадилх беременности, меняют реакции крысы
на ольфакторные стимулы, и это снимает иптибирующий эффект запаха
новорожденных. Можно предположить, что сенситизация другим путем
спимает ту же ингибицию. Происходит постепенное привыкание к за-
паху детенышей, ослабляющее и в итоге снимающее ее действие.
Основные факты в пользу этой гипотезы следующие. Аноемия, вы-
званная выжиганием обонятельного эпителия 70$0., или перерезка
латерального ольфакториого тракта, как и вомероназальная деафферен-
тация, резко сокращают латентный период сенситизации, и материнское:
поведение проявляется необычайно быстро (Решше, Возеп аи, 19745, с;
Ееш!то её а]., 1979). Гормональная перестройка неред родами меняет
реакцию на запахи. Например, на поздних стадиях беременпости
(за 1—2 дня перед родами) самки начипают предпочитать запах гнезда
беременных самок запаху гнезд небеременных (Вацег, 1983). Этот факт,
кроме свидетельства об измепепии реактивности на запах, позволяет
предположить и прямую блокировку каким-то привлекательным субетра-
том первичной аверсивной реакции на запах новорожденных детенышей.
Невральная база материнского позедепия, на которую могут воздейст-
вовать как хемосенсорпый вход, так и соответствующие гормональные
перестройки, структурно связана с медиальной преоптической областью.
типоталамуса, а также с вентральной тегментальной областью и черной
субстанцией среднего мозга (Миша, 1983).
|
Фаза поддержания материнского поведения. Какими факторами
управляется материнское поведение после того, как оно уже «включено»
(точнее, устранена его ингибиция)? Роль гормональных факторов стано-
вится гораздо менее существенной: если материнское поведепие уже
«включено», повлиять на пего какими-либо гормональными манипуля-
циями пе удается. Удаление яичников и матки (\У7Лезпег, Бпеага,
41933), надночечников и гипофиза (Возеп ай, её а1., 1979; Нгзкш, 1978)
не влияет на материнское поведение, если самка уже пачала его прояв-
лять. Введение прогестерона в дозах, достаточных для блокировапия
«включения» материнского поведения (НеттепКо#, Глзк, 1973), блокада.
секреции окситоцина (УоКкоуата, Оа, 1959) также не влияют на него.
Если блокировать секрецию пролактина эргокринтином у самки, которая
несколько дней назад родила, то количество молока у пее резко умень-
шится, но все виды материнского поредения, включая пребывание в позе
кормления, при контакте с крысятами останутся прежними (54егп, 1977).
Самки с удаленными перед спариванием молочными железами или соска-
Поведение
259
ми проявляют все виды материнского поведения (Мох еф а1., 1967).
"Таким образом, поведение кормления детепьшей непосредственно не за-
висит от влияния гормонов, хотя секреция молока и контролируется про-
лактином.
Вместе с тем в период выкармливация детенышей самке присущ
высокоспецифичный гормональный статус, характеризующийся прежде
всего высоким уровнем пролактица, обусловливающего не только секре-
цию молока, но и подавление эетральных ЦИКЛОВ, В этом отношении
интересно, что в отличие от других стадий репродуктивного цикла гор-
мональный статус самки во время лактации непосредственно зависит от
стимулов внешней среды, прежде всего от детеньштей. Стимуляция с0с-
ков во время кормления (ВоеЬП4, 1960; Решите, 1976), ультразвуки
4—7-дневных детенышей (ТегКе] ей а1., 1979), звуки детенышей во время
сосания (Г1ез, 1968) и само присутствие детенышей в клетке матери
(Стозуепот, Мепа, 1974) повышают секрецию пролактина и других гор-
монов лактогеппого комплекса. Одновременно интибируется секреция
фолликулостимулирующих гормонов и развитие эстральных циклов
у большинства лактирующих самок (Во ева, 1960). Диэструс, вызван-
ный лактацией, длится 2—4 недели и зависит от внешней стимуляции:
у самок, имеющих небольшие выводки (1—4 детеныша) эстральные цик-
лы восстанавливаются быстрее, чем у самок с 5 и более детепышами
{Тискег, ТвабсВег, 1968). Если детенышей удалить сразу же после родов,
эстральная циклика самки восстанавливается через 4—5 или 9—10 дней
{Вобпевйа, 1960). У оплодотворенной во время послеродового эструса
самки во время лактации имплантация яиц может задерживаться на
12—13 дней и новый выводок появляется лишь через 35 дней после спа-
ривания (см., например, ОПЬетв её а|., 1983).
Внешние факторы не просто повышают, по регулируют уровень про-
лактина. Если уровень стимуляции понижается (например, сосание со-
вершается нерегулярно), секреция пролактипа уменьшается (Мепа,
1971). Однако даже если лактация прервана отделением детенышей на
9 дней, то она через несколько дней после их возвращения восстанавли-
вается (Ош, УоКоуаша, 1965). Внешний фактор
— размер выводка—
определяет как количество функционирующих молочных желез (Мепа,
Сгозуепог, 1968), так и гормональную реакцию самки па стрессирующие
воздействия (ЗтоШегтаи еф а]., 1976).
Таким образом, гормопальный статус самки после рождения детены-
шей и лактация в значительной степени определяются внешней относи-
тельно самки, исходящей от выводка стимуляцией. Эта же внешняя
стимуляция контролирует развитие всех форм материнского поведения
{Зейя, 1954а; Стова, 1973), причем главные факторы, определяющие воз-
действие,— возраст детенышей и их количество.
Интенсивность и специфика материнского поведения зависят в пер-
вую очередь от возрастных особенпостей крысят, с которыми взаимодей-
ствует самка, и в значительно меньшей степени от других факторов,
например от того, насколько давно произошли роды. То есть материнское
поведение определяется спецификой взаимодействия с крысятами даппо-
9*
258 Глава четвертая
включением материнского поведения эндокринным путем и сенситиза-
цией? Чтобы обсудить этот вопрос, следует внимательно рассмотреть.
природу сенсорной стимуляции, воздействующей на включение материн-
ского поведения. Согласно наиболее обоспованной к настоящему времени
гипотезе (Репа, Возеп ай, 1974. с), основная сенсорная модаль-
ность, управляющая материнским поведением крысы,— хемореценция.
Предполагается, что самка крысы в любом физиологическом состоянии
готова проявить материнское поведение, но эта готовность блокируется
впеншим фактором
— запахом детенышей. Гормональные события, про-
исходящие на последних стадилх беременности, меняют реакции крысы
на ольфакторные стимулы, и это снимает иптибирующий эффект запаха
новорожденных. Можно предположить, что сенситизация другим путем
спимает ту же ингибицию. Происходит постепенное привыкание к за-
паху детенышей, ослабляющее и в итоге снимающее ее действие.
Основные факты в пользу этой гипотезы следующие. Аноемия, вы-
званная выжиганием обонятельного эпителия 70$0., или перерезка
латерального ольфакториого тракта, как и вомероназальная деафферен-
тация, резко сокращают латентный период сенситизации, и материнское:
поведение проявляется необычайно быстро (Решше, Возеп аи, 19745, с;
Ееш!то её а]., 1979). Гормональная перестройка неред родами меняет
реакцию на запахи. Например, на поздних стадиях беременпости
(за 1—2 дня перед родами) самки начипают предпочитать запах гнезда
беременных самок запаху гнезд небеременных (Вацег, 1983). Этот факт,
кроме свидетельства об измепепии реактивности на запах, позволяет
предположить и прямую блокировку каким-то привлекательным субетра-
том первичной аверсивной реакции на запах новорожденных детенышей.
Невральная база материнского позедепия, на которую могут воздейст-
вовать как хемосенсорпый вход, так и соответствующие гормональные
перестройки, структурно связана с медиальной преоптической областью.
типоталамуса, а также с вентральной тегментальной областью и черной
субстанцией среднего мозга (Миша, 1983).
|
Фаза поддержания материнского поведения. Какими факторами
управляется материнское поведение после того, как оно уже «включено»
(точнее, устранена его ингибиция)? Роль гормональных факторов стано-
вится гораздо менее существенной: если материнское поведепие уже
«включено», повлиять на пего какими-либо гормональными манипуля-
циями пе удается. Удаление яичников и матки (\У7Лезпег, Бпеага,
41933), надночечников и гипофиза (Возеп ай, её а1., 1979; Нгзкш, 1978)
не влияет на материнское поведение, если самка уже пачала его прояв-
лять. Введение прогестерона в дозах, достаточных для блокировапия
«включения» материнского поведения (НеттепКо#, Глзк, 1973), блокада.
секреции окситоцина (УоКкоуата, Оа, 1959) также не влияют на него.
Если блокировать секрецию пролактина эргокринтином у самки, которая
несколько дней назад родила, то количество молока у пее резко умень-
шится, но все виды материнского поредения, включая пребывание в позе
кормления, при контакте с крысятами останутся прежними (54егп, 1977).
Самки с удаленными перед спариванием молочными железами или соска-
Поведение
259
ми проявляют все виды материнского поведения (Мох еф а1., 1967).
"Таким образом, поведение кормления детепьшей непосредственно не за-
висит от влияния гормонов, хотя секреция молока и контролируется про-
лактином.
Вместе с тем в период выкармливация детенышей самке присущ
высокоспецифичный гормональный статус, характеризующийся прежде
всего высоким уровнем пролактица, обусловливающего не только секре-
цию молока, но и подавление эетральных ЦИКЛОВ, В этом отношении
интересно, что в отличие от других стадий репродуктивного цикла гор-
мональный статус самки во время лактации непосредственно зависит от
стимулов внешней среды, прежде всего от детеньштей. Стимуляция с0с-
ков во время кормления (ВоеЬП4, 1960; Решите, 1976), ультразвуки
4—7-дневных детенышей (ТегКе] ей а1., 1979), звуки детенышей во время
сосания (Г1ез, 1968) и само присутствие детенышей в клетке матери
(Стозуепот, Мепа, 1974) повышают секрецию пролактина и других гор-
монов лактогеппого комплекса. Одновременно интибируется секреция
фолликулостимулирующих гормонов и развитие эстральных циклов
у большинства лактирующих самок (Во ева, 1960). Диэструс, вызван-
ный лактацией, длится 2—4 недели и зависит от внешней стимуляции:
у самок, имеющих небольшие выводки (1—4 детеныша) эстральные цик-
лы восстанавливаются быстрее, чем у самок с 5 и более детепышами
{Тискег, ТвабсВег, 1968). Если детенышей удалить сразу же после родов,
эстральная циклика самки восстанавливается через 4—5 или 9—10 дней
{Вобпевйа, 1960). У оплодотворенной во время послеродового эструса
самки во время лактации имплантация яиц может задерживаться на
12—13 дней и новый выводок появляется лишь через 35 дней после спа-
ривания (см., например, ОПЬетв её а|., 1983).
Внешние факторы не просто повышают, по регулируют уровень про-
лактина. Если уровень стимуляции понижается (например, сосание со-
вершается нерегулярно), секреция пролактипа уменьшается (Мепа,
1971). Однако даже если лактация прервана отделением детенышей на
9 дней, то она через несколько дней после их возвращения восстанавли-
вается (Ош, УоКоуаша, 1965). Внешний фактор
— размер выводка—
определяет как количество функционирующих молочных желез (Мепа,
Сгозуепог, 1968), так и гормональную реакцию самки па стрессирующие
воздействия (ЗтоШегтаи еф а]., 1976).
Таким образом, гормопальный статус самки после рождения детены-
шей и лактация в значительной степени определяются внешней относи-
тельно самки, исходящей от выводка стимуляцией. Эта же внешняя
стимуляция контролирует развитие всех форм материнского поведения
{Зейя, 1954а; Стова, 1973), причем главные факторы, определяющие воз-
действие,— возраст детенышей и их количество.
Интенсивность и специфика материнского поведения зависят в пер-
вую очередь от возрастных особенпостей крысят, с которыми взаимодей-
ствует самка, и в значительно меньшей степени от других факторов,
например от того, насколько давно произошли роды. То есть материнское
поведение определяется спецификой взаимодействия с крысятами даппо-
9*
260
Глаза четвертая
го возраста, а не длительностью периода времени, прошедшего с момента
включения материнского поведения. Естественный процесс угашения ма-
теринского поведения, который в пормальных условиях паблюдается
тогда, когда детеныши достигают 3—4-педельного возраста, можпо
предотвратить с помощью соответствующим образом подобранного режи-
ма подмены детенышей. Так, заменяя каждые 5 дней детенышей на
поворожденных, можно добиться высокого уровня материнского поведе-
ния, характерного для обращения с детеньшами этого нозраста, т ущи-
непия срока лактации на несколько недель (МУПезпег, ЗВеага, 1933) или
даже до года (Втисе, 1961). С другой стороны, процессе угашепия мате-
рипского поведения можно аналогичным способом экспериментально.
ускорить: при замене 5-дневных детепышей на 12-дневных у самок про-
исходит резкое сокращение материнской заботы до уровня, соответотвую-
щего взаимодействию с детенышами этого возраста (Сгофа, 1973).
Установление связи между натерью и детеньишем
в первые дни его постнатального развития
Первые дни после родов — переходный период в регуляции материнского
поведения. К началу этого периода гормональный контроль достигает
максимальной силы, после чего он начинает ослабляться, а действие
факторов внешней среды усиливается (ВозепМаи, З1езе, 1980).
Поведение самки крысы во время и непосредственно после родов
хорошо структурировано и состоит из определенной последовательности
стереотипных действий. В организации этой последонательности можно
выделить чередующиеся фазы
— фазы, в которых действия матери на-
правлены па детенышей, сменяются фазами, в которые мать не обращает
па них никакого внимания, занимаясь, например, чисткой или едои.
Продолжительность различных фаз (всего их выделено пять) довольно
стереотипна. Вполне возможно, что правильный режим изменения кон
центрации внимания пеобходим для пормальной организации материн-
ского поведения (НоНочхау е а1., 1980).
Новорожденный детеныш способен влиять на поведение матери во все
фазы ее активности, но это влияние особеппо эффективно, когда ее вни-
мание ориентировано на него. Ультразвуковые крики новорожденных
стимулируют мать к освобождению их от плодных оболочек р продупреж-
дают случайные ранения детенышей при плацентофагии. Особое зиа °-
ние в развивающейся связи между матерью и детенышем имеет хем -
рецепция. Бульбоэктомия беременной самки парушает ее поведотие во
время родов, патология поведения проявляется именно в те фазы акти
ности, когда ее внимание должно быть направлено на детенышей. ы кое
парушение режима внимания в период родов приводит к весьма важ ым
последствиям: детеныши, которые находились в первые часы своей жи:
ни с бульбоэктомированными самками, выживают хуже и к моменту
отлучения их поведение в «открытом поле» отличается от поведения
контрольных (Нооугау е% а1., 1980).
„.
Первые дни контакта самки с новорожденными можно назвать «кр
тическими» для дальнейшего формирования связи между матерью и
Поведение
261
детеныптем. Если детенышей убрать от самки сразу же после родов,
то склонность ее к материпскому поведению резко падает: только 30%
самок проявляют материнское поведение по отношению к детенытам,
возвращенным спустя 2 дня после рождепия, материнское поведение во-
все исчезает, если эту разлуку продлить до 4 дней. Если же детенышей
убрать после трехдневного общения с матерью, то разлучение на те же
А дня понижает материнскую реакцию только у 30—40% самок (Возеп-
Ыаи, Гебттап, 1963; Возеп Май, 1967).
Если самка на протяжении 4—6 ч после родов сстается © детеныша-
ми, то спустя 25 дней (при восстановлении эстральной циклики) латент-
ный период сенситизации у нее будет существенно короче, чем у тех
самок, детеныши которых были удалены сразу же после родов. Даже
кратковременные контакты — обнюхивания и облизывания самкой дете-
нышейво время родов
— имеют существенные «следовые» влияния на ла-
тентный период сенситизации материнского поведения через 25 дней
после родов. У самок, которым было сделано кесарево сечение, этот
фепомен также проявляется: достаточно небольшого периода коптактов
с детенышами сразу после операции, чтобы существенно облегчить
«восстановлепие» материнского повецения через 25 дней (Вм@еез, 1977,
1978).
На детеныша коптакт с матерью оказывает еще большее влияние,
чем на мать. Условия, в которых крысята проводят первые 12 ч своей
жизни, определяют и темп их роста, и выживаемость. Если спустя час
после рождения детенышей отделить от матери и перенести к друтой
самке, которая лактирует 10 дней, то это ухудшит их выживаемость и
к месячному возрасту их масса будет ниже, чем у контрольных. Этот
эффект пропадает, если перенос отсрочить на 10 ч (Пеппепфегр ей а[.,
1976).
Одна из важнейпих и наиболее изученных связей, возникающих не-
посредственно после родов,— взаимные действия матери и детеныша,
обеспечивающие детеныша молоком. Сразу же после родов, обнюхав
и вылизав крысят, мать приступает к их кормлению. Движения поиска
соска сформированы у детеныша уже к моменту рождения. Первое корм-
ление совершается по инициативе самки. [Ше поведение оранизовано
таким образом, что предельно облегчает детенышу поиск соска
— ему
нужно лишь взять его в рот и начать сосать. Чтобы четче определить
возможности крысенка, была разработана следующая методика: самка
временно обездвиживается и укладывается на спину таким образом, что-
бы область сосков была доступной для детеныша. Новорожденный пны-
тается достичь сосков «своими силами» без помощи матери. Оказалось,
что в этой ситуации важнейшую роль в ориентации крысенка играют
ольфакторные стимулы, исходящие от соска. Если сосок помыть, то де-
теныш не прикрепляется к нему, даже если находится совсем рядом *;
1 Если соски мыть постоянно, то детеныши в первые дни жизни не способны отыс-
кивать их без помощи самки. К аналогичным результатам приводит и аносмил
дехенышей (ТесВег её а]., 1978).
260
Глаза четвертая
го возраста, а не длительностью периода времени, прошедшего с момента
включения материнского поведения. Естественный процесс угашения ма-
теринского поведения, который в пормальных условиях паблюдается
тогда, когда детеныши достигают 3—4-педельного возраста, можпо
предотвратить с помощью соответствующим образом подобранного режи-
ма подмены детенышей. Так, заменяя каждые 5 дней детенышей на
поворожденных, можно добиться высокого уровня материнского поведе-
ния, характерного для обращения с детеньшами этого нозраста, т ущи-
непия срока лактации на несколько недель (МУПезпег, ЗВеага, 1933) или
даже до года (Втисе, 1961). С другой стороны, процессе угашепия мате-
рипского поведения можно аналогичным способом экспериментально.
ускорить: при замене 5-дневных детепышей на 12-дневных у самок про-
исходит резкое сокращение материнской заботы до уровня, соответотвую-
щего взаимодействию с детенышами этого возраста (Сгофа, 1973).
Установление связи между натерью и детеньишем
в первые дни его постнатального развития
Первые дни после родов — переходный период в регуляции материнского
поведения. К началу этого периода гормональный контроль достигает
максимальной силы, после чего он начинает ослабляться, а действие
факторов внешней среды усиливается (ВозепМаи, З1езе, 1980).
Поведение самки крысы во время и непосредственно после родов
хорошо структурировано и состоит из определенной последовательности
стереотипных действий. В организации этой последонательности можно
выделить чередующиеся фазы
— фазы, в которых действия матери на-
правлены па детенышей, сменяются фазами, в которые мать не обращает
па них никакого внимания, занимаясь, например, чисткой или едои.
Продолжительность различных фаз (всего их выделено пять) довольно
стереотипна. Вполне возможно, что правильный режим изменения кон
центрации внимания пеобходим для пормальной организации материн-
ского поведения (НоНочхау е а1., 1980).
Новорожденный детеныш способен влиять на поведение матери во все
фазы ее активности, но это влияние особеппо эффективно, когда ее вни-
мание ориентировано на него. Ультразвуковые крики новорожденных
стимулируют мать к освобождению их от плодных оболочек р продупреж-
дают случайные ранения детенышей при плацентофагии. Особое зиа °-
ние в развивающейся связи между матерью и детенышем имеет хем -
рецепция. Бульбоэктомия беременной самки парушает ее поведотие во
время родов, патология поведения проявляется именно в те фазы акти
ности, когда ее внимание должно быть направлено на детенышей. ы кое
парушение режима внимания в период родов приводит к весьма важ ым
последствиям: детеныши, которые находились в первые часы своей жи:
ни с бульбоэктомированными самками, выживают хуже и к моменту
отлучения их поведение в «открытом поле» отличается от поведения
контрольных (Нооугау е% а1., 1980).
„.
Первые дни контакта самки с новорожденными можно назвать «кр
тическими» для дальнейшего формирования связи между матерью и
Поведение
261
детеныптем. Если детенышей убрать от самки сразу же после родов,
то склонность ее к материпскому поведению резко падает: только 30%
самок проявляют материнское поведение по отношению к детенытам,
возвращенным спустя 2 дня после рождепия, материнское поведение во-
все исчезает, если эту разлуку продлить до 4 дней. Если же детенышей
убрать после трехдневного общения с матерью, то разлучение на те же
А дня понижает материнскую реакцию только у 30—40% самок (Возеп-
Ыаи, Гебттап, 1963; Возеп Май, 1967).
Если самка на протяжении 4—6 ч после родов сстается © детеныша-
ми, то спустя 25 дней (при восстановлении эстральной циклики) латент-
ный период сенситизации у нее будет существенно короче, чем у тех
самок, детеныши которых были удалены сразу же после родов. Даже
кратковременные контакты — обнюхивания и облизывания самкой дете-
нышейво время родов
— имеют существенные «следовые» влияния на ла-
тентный период сенситизации материнского поведения через 25 дней
после родов. У самок, которым было сделано кесарево сечение, этот
фепомен также проявляется: достаточно небольшого периода коптактов
с детенышами сразу после операции, чтобы существенно облегчить
«восстановлепие» материнского повецения через 25 дней (Вм@еез, 1977,
1978).
На детеныша коптакт с матерью оказывает еще большее влияние,
чем на мать. Условия, в которых крысята проводят первые 12 ч своей
жизни, определяют и темп их роста, и выживаемость. Если спустя час
после рождения детенышей отделить от матери и перенести к друтой
самке, которая лактирует 10 дней, то это ухудшит их выживаемость и
к месячному возрасту их масса будет ниже, чем у контрольных. Этот
эффект пропадает, если перенос отсрочить на 10 ч (Пеппепфегр ей а[.,
1976).
Одна из важнейпих и наиболее изученных связей, возникающих не-
посредственно после родов,— взаимные действия матери и детеныша,
обеспечивающие детеныша молоком. Сразу же после родов, обнюхав
и вылизав крысят, мать приступает к их кормлению. Движения поиска
соска сформированы у детеныша уже к моменту рождения. Первое корм-
ление совершается по инициативе самки. [Ше поведение оранизовано
таким образом, что предельно облегчает детенышу поиск соска
— ему
нужно лишь взять его в рот и начать сосать. Чтобы четче определить
возможности крысенка, была разработана следующая методика: самка
временно обездвиживается и укладывается на спину таким образом, что-
бы область сосков была доступной для детеныша. Новорожденный пны-
тается достичь сосков «своими силами» без помощи матери. Оказалось,
что в этой ситуации важнейшую роль в ориентации крысенка играют
ольфакторные стимулы, исходящие от соска. Если сосок помыть, то де-
теныш не прикрепляется к нему, даже если находится совсем рядом *;
1 Если соски мыть постоянно, то детеныши в первые дни жизни не способны отыс-
кивать их без помощи самки. К аналогичным результатам приводит и аносмил
дехенышей (ТесВег её а]., 1978).
262
Глава четвертая
Однако если на вымытый сосок нанести мазок с невымытого соска, то
детеныш быстро находит его. Оказалось, что на сосках содержится сме-
шанная со слюной амниотическая жидкость, наносимая на них самкой
при вылизывании себя после родов (Теёслег, В\азз, 1977а, Ъ). Занах
эмниотической жидкости хоротпо воспринимается крысенком и привле-
кает ого, вероятно, благодаря обучению, которое происходит еще в эм-
бриональном состоянии. Так, естественную реакцию на запах амниоти-
ческой жидкости можно изменить, если в амниотическую полость
эмбрионов вводить раствор с запахом лимона и напосить этот раствор
на крысят сразу же после рождения. 'Гаких детенышей привлекают лишь
ле соски, на которые нанесен запах лимона, и опи не сосут нормально
пахнущие соски (В1а55, Редегзеп, 1982). Начиная с 17—19-го дия после
зачатия у эмбрионов крыс можно выработать вкусовые и запаховые
аверсии на оощества, которые вводятся в амниотическую полость
совместно с хлоридом лития. Новорожденные крысята избегают сосать
те соски, на которые нанесено вещество, аверсия к которому вырабаты-
залась в эмбриональном состоянии (см. папример: ЗтоНегтаи, 1985).
Если мы будем оценивать «успех достижения соска» по тому, лостит
ли крысенок соска обездвиженной самки за. какой-либо определенный
интервал времени или не достиг, то окажется, что па протяжении пер-
зых 12 ч жизни детеныша это поведение сильно изменяется. Взятые от
матери через 1 ч после рождения крысята в опытах с неподвижной
самкой самостоятельно достигают соска только в небольшой части слу-
чаев. Гели их лишить доступа к соску в первые 12 ч жизни (например,
отделив их от матери или удалив у нее грудные железы), то их способ-
иссти к самостоятельному поиску соска развиваются существенно хуже.
Само подкрепление молоком здесь не играет роли— детеньшии, сосавшие
самок, молочные железы которых были перевязаны, достигали сосков
с таким же успехом, как и контрольные, сосавттие молоко. Таким обра-
зом, подкреплением в данном случае служит сам акт сосапия и в первые
12 ч жизни крысята обучаются активно вести поиск соска. Уже с первого
дня жизни проявляется суточная ритмика: детеныши успешней и быст-
рее прикрепляются к соску ночью, а пе днем. Использование самок
с реверсированными циклами активности показало, что дела здесь не
в изменении поведеция самки— трехдиевные детеныши успешней при-
крепляются к соску «своей почью», а не «ночью матери». Оказывается
также, что детепыши уснешней достигают соеков самки, которая имеет
детенышей, сходных по возрасту с тестируемыми, чем той, что имеет
детенышей на 7 дней старше (РоШлаег © а]., 1980).
Таким образом, в первые часы и дни после родов мать и детеныш
крысы оказывают друг на друга особенно сильные взаимные воздейст-
вия, имеющие не только непосредственные, но и весьма отдаленные па
времени следствия в поведении и физиологии как матери, так и дете-
ныша.
Прямые следствия влияния детеньшиа
— окончательная подготовка
организма матери к проявлению материнских реакций. Среди связей, воз-
никающих в этот период между матерью и детенышем, одна из важней-
Поведение
263
ших, оспована на обучепии детеныша поиску соска и прикреплению.
к нему при ориентации на химические стимулы матери. Важную роль.
в этом обучении играет поведение матери.
Взаимоотношения между матерью и детенышежн
в процеесе становления гомойотермии детеньиия
До определенного возраста детепыши крысы пе способны самостоятель-
но поддерживать постоянную температуру тела. Эта временная пойкило-
термия обусловливает многие важные черты поведения новорожденных
как, например, положительный термотаксис — постоянное стремление к
скучиванию с матерью и собратьями по выводку. Детеныш должен поль-
зоваться термальными ресурсами окружающей среды, и главным их
источником является мать. В этом отношении крысенок столь же сильно
зависит от матери, как и в отношении питания. Однако неверно было бы
рассматривать пойкилотермию как некое несовершенство организма, не-
доразвитие физиологических функций. Среда, определяющая биологию
детеныша, совсем иная, чем у взрослых того же вида, и пойкило-
термия— это приспособление к особой экологической нише (см., папри-`
мер: СаеЁ, 1981).
”
После рождения детеныши постоянно находятся в гнезде, куда перио-
дически приходит самка. Там она их кормит и поддерживает с ними
физический контакт, после чего на определенный промежуток времени
покидает выводок. Таким образом, в отношении теплоспабжения можно
выделить две резко различающиеся между собой фазы в состоянии сре-
ды, окружающей детеныша. Фаза, когда самка отсутствует, характери-
зуется низкой температурой среды, а фаза, когда самка находится в кон-
такте с детенышами, характеризуется стабильной температурой тела
‚самки. В этих условиях пойкилотермия оказывается энергетически выгод-
ной. В периоды, когда самки нет в гнезде, температура тела детенышей
падает, уменьшаются энергетические траты на теплопродукцию, пони-
жается уровень метаболизма. В это время детеньши неактивен и "обычно
спит. Когда самка приходит в гнездо, у детенышей появляется возмож-
ность извне получать энергию от тепла ее тела и от теплого молока.
Температура его тела, согреваемого матерью, повышается, одповремепио.
резко усиливаются метаболические процессы
— вся энергия, получаемая
от матери, интенсивно используется для роста. Расчеты показывают, что
если бы в этот период онтогенеза детеныши были бы томойотермными то.
для поддержания температуры тела (-33°) и сохранения обычного тем-
па роста при комнатной температуре окружающей среды крысятам по-
требовалось бы по крайней мере вдвое больше молока, чем пойкилотерм-
ным детенышам (Нави её а|., 1964).
На второй неделе жизни детенышей время, проводимое самкой
в гнезде в контакте с пими, резко уменьшается (Стоба, Адег, 1969).
Это уменьшение продолжительности контактов и соответственное умепь-
шение теплоснабжения выводка происходит за счет сокращения продол-
жительности «визитов» самки в гнездо, хотя общее число посещений
262
Глава четвертая
Однако если на вымытый сосок нанести мазок с невымытого соска, то
детеныш быстро находит его. Оказалось, что на сосках содержится сме-
шанная со слюной амниотическая жидкость, наносимая на них самкой
при вылизывании себя после родов (Теёслег, В\азз, 1977а, Ъ). Занах
эмниотической жидкости хоротпо воспринимается крысенком и привле-
кает ого, вероятно, благодаря обучению, которое происходит еще в эм-
бриональном состоянии. Так, естественную реакцию на запах амниоти-
ческой жидкости можно изменить, если в амниотическую полость
эмбрионов вводить раствор с запахом лимона и напосить этот раствор
на крысят сразу же после рождения. 'Гаких детенышей привлекают лишь
ле соски, на которые нанесен запах лимона, и опи не сосут нормально
пахнущие соски (В1а55, Редегзеп, 1982). Начиная с 17—19-го дия после
зачатия у эмбрионов крыс можно выработать вкусовые и запаховые
аверсии на оощества, которые вводятся в амниотическую полость
совместно с хлоридом лития. Новорожденные крысята избегают сосать
те соски, на которые нанесено вещество, аверсия к которому вырабаты-
залась в эмбриональном состоянии (см. папример: ЗтоНегтаи, 1985).
Если мы будем оценивать «успех достижения соска» по тому, лостит
ли крысенок соска обездвиженной самки за. какой-либо определенный
интервал времени или не достиг, то окажется, что па протяжении пер-
зых 12 ч жизни детеныша это поведение сильно изменяется. Взятые от
матери через 1 ч после рождения крысята в опытах с неподвижной
самкой самостоятельно достигают соска только в небольшой части слу-
чаев. Гели их лишить доступа к соску в первые 12 ч жизни (например,
отделив их от матери или удалив у нее грудные железы), то их способ-
иссти к самостоятельному поиску соска развиваются существенно хуже.
Само подкрепление молоком здесь не играет роли— детеньшии, сосавшие
самок, молочные железы которых были перевязаны, достигали сосков
с таким же успехом, как и контрольные, сосавттие молоко. Таким обра-
зом, подкреплением в данном случае служит сам акт сосапия и в первые
12 ч жизни крысята обучаются активно вести поиск соска. Уже с первого
дня жизни проявляется суточная ритмика: детеныши успешней и быст-
рее прикрепляются к соску ночью, а пе днем. Использование самок
с реверсированными циклами активности показало, что дела здесь не
в изменении поведеция самки— трехдиевные детеныши успешней при-
крепляются к соску «своей почью», а не «ночью матери». Оказывается
также, что детепыши уснешней достигают соеков самки, которая имеет
детенышей, сходных по возрасту с тестируемыми, чем той, что имеет
детенышей на 7 дней старше (РоШлаег © а]., 1980).
Таким образом, в первые часы и дни после родов мать и детеныш
крысы оказывают друг на друга особенно сильные взаимные воздейст-
вия, имеющие не только непосредственные, но и весьма отдаленные па
времени следствия в поведении и физиологии как матери, так и дете-
ныша.
Прямые следствия влияния детеньшиа
— окончательная подготовка
организма матери к проявлению материнских реакций. Среди связей, воз-
никающих в этот период между матерью и детенышем, одна из важней-
Поведение
263
ших, оспована на обучепии детеныша поиску соска и прикреплению.
к нему при ориентации на химические стимулы матери. Важную роль.
в этом обучении играет поведение матери.
Взаимоотношения между матерью и детенышежн
в процеесе становления гомойотермии детеньиия
До определенного возраста детепыши крысы пе способны самостоятель-
но поддерживать постоянную температуру тела. Эта временная пойкило-
термия обусловливает многие важные черты поведения новорожденных
как, например, положительный термотаксис — постоянное стремление к
скучиванию с матерью и собратьями по выводку. Детеныш должен поль-
зоваться термальными ресурсами окружающей среды, и главным их
источником является мать. В этом отношении крысенок столь же сильно
зависит от матери, как и в отношении питания. Однако неверно было бы
рассматривать пойкилотермию как некое несовершенство организма, не-
доразвитие физиологических функций. Среда, определяющая биологию
детеныша, совсем иная, чем у взрослых того же вида, и пойкило-
термия— это приспособление к особой экологической нише (см., папри-`
мер: СаеЁ, 1981).
”
После рождения детеныши постоянно находятся в гнезде, куда перио-
дически приходит самка. Там она их кормит и поддерживает с ними
физический контакт, после чего на определенный промежуток времени
покидает выводок. Таким образом, в отношении теплоспабжения можно
выделить две резко различающиеся между собой фазы в состоянии сре-
ды, окружающей детеныша. Фаза, когда самка отсутствует, характери-
зуется низкой температурой среды, а фаза, когда самка находится в кон-
такте с детенышами, характеризуется стабильной температурой тела
‚самки. В этих условиях пойкилотермия оказывается энергетически выгод-
ной. В периоды, когда самки нет в гнезде, температура тела детенышей
падает, уменьшаются энергетические траты на теплопродукцию, пони-
жается уровень метаболизма. В это время детеньши неактивен и "обычно
спит. Когда самка приходит в гнездо, у детенышей появляется возмож-
ность извне получать энергию от тепла ее тела и от теплого молока.
Температура его тела, согреваемого матерью, повышается, одповремепио.
резко усиливаются метаболические процессы
— вся энергия, получаемая
от матери, интенсивно используется для роста. Расчеты показывают, что
если бы в этот период онтогенеза детеныши были бы томойотермными то.
для поддержания температуры тела (-33°) и сохранения обычного тем-
па роста при комнатной температуре окружающей среды крысятам по-
требовалось бы по крайней мере вдвое больше молока, чем пойкилотерм-
ным детенышам (Нави её а|., 1964).
На второй неделе жизни детенышей время, проводимое самкой
в гнезде в контакте с пими, резко уменьшается (Стоба, Адег, 1969).
Это уменьшение продолжительности контактов и соответственное умепь-
шение теплоснабжения выводка происходит за счет сокращения продол-
жительности «визитов» самки в гнездо, хотя общее число посещений
2ВА
Глава четвертая
остается прежним (Ёеоп ей а|., 1978). ЕК этому времени у детенышей
резко усиливается способность к гомойотермии. Возникает вопрос: чем
регулируется эта согласованность в изменении потребпостей детенышей
и реакций матери? Было показано, что время контактов самки с детены-
шами хорошо скоррелировано © температурой окружающей среды:
при +18° самка проводит с детеньишами большие времени, чем при +22°,
а при +26°
— меньше, чем при 22° (Стозкеггу © а1., 1978). Таким
образом, термальные характеристики являются важными определяющими
контактов. Возникло предположение, что самка кончает период контак-
тов (и тем самым регулирует время, проводимое © детенышами) в тот
момент, когда температура тела детенышей достигает определенной
величины, например стаповится равной температуре тела самки. Этим
можно объяспить постепенное сокращение периодов коптактов по мере
роста гомойотермии крысят. Действительно, если экспериментально ох-
лаждать детенышей, то продолжительность периодов коитактов у матери
с ними увеличивается. Однако, если припять зту гипотезу, нагревание
детенышей до той температуры, которая бывает в момент окончания
контактов, должно было бы резко уменьшить время иребывапия самки
с детопышами. Этого не происходит (Геоп её а|., 1978). Таким образом,
температура детенышей является не единственным фактором, определяю-
щим режим контактов. Возникло предположение, что окончание периода
контактов определяется некими особенностями работы системы теплопро-
дукции-—теплоотдачи самки. Действительно, изменение режима фупкцио-
нирсвания этой системы оказывает влияние на время, проводимое сам-
кой с детенышами. Так, если эксперимептально увеличить теплоотдачу
самки, обрив ей шерсть, продолжительность периодов контактов с крыся-
тами резко возрастает. И наоборот, экспериментальное умепьшецие спо-
собпости к теплоотдаче (отрезание хноста) вызывает сокращение перио-
дов контактов самки © детенышами. Более того, время, проводимое
с крысятами самкой, у которой отрезана половина хвоста, промежуточ-
ное между тем, что проводят бесхвостые и интактные самки (Геоп её а|.,
1978).
С помощью вживления температурных датчиков под кожу (в области
сосков) и в преоптическую зону переднего мозга самки было установле-
но, что момент окончапия контакта характеризуется резким повышецием
температуры тела и мозга (УУоодя4е, Геоп, 1980). Возможно, это и яв-
ляется стимулом, вызывающим прерывание контакта. Кроме того, было
показано, что для самки во вторую неделю лактации характерно резкое
увеличение теплопродукции (на 17,3% по сравнению с нелактирующими
самками) и хроническое повышение температуры тела (Тапез, Геоп,
1983; Геоп её а1., 1985). Это делает самку весьма чувотвительной к до-
млолнительному повьштению температуры во время контактов © детены-
шами, заставляя ее раньше прерывать контакт для того, чтобы сохранить
термальный гомеостаз. Гормональные механизмы этого феномепа пока
изучены далеко не достаточно. Однако ясно, что хроническое повышение
температуры обусловлено, по крайцей мере частично, повышением уров-
ия циркулирующего пролактина. Предполагается, во-первых, что пролак-
=
Поведение
265
тин стимулирует секрецию прогестеропа, который поднимает на повый
уровень термальные требования лактирующей самки. Во-вторых, в это
время повышается секреция адренальных глюкокортикоидов которые
усиливают тканевый метаболизм в такой степени, чтобы были удовле-
творены эти новые требования (см., папример: Геоп её а|., 1985).
Если на следующий день после родов экспериментально подавить
уровень циркулирующего пролактина, то температура тела кормящей
самки хронически попизится, а контакт © детенытами удлипяется
(Геоп её а|., 1985). Оцнако такие эффекты проявляются лишь на вто-
рой неделе лактации. К этому времсни оказывается подавленным уро-
вень прогестеропа. Герапия прогестероном у самок с подавлепным уров-
нем пролактина восстанавливает и темпоратуру, типичную для лакти-
рующих самок и нормальную продолжительность коптактов с детенышами
(\\содз14е еф а|1., 1981). В то же время адрепалэктомия лактирующих
самок также приводит к хроническому снижению температуры тсола
и удлинению продолжительности контактов с детенышами (Созуе, Ео]-
Теу, 1961). Восстанозительная терапия с помощью глюкокортикоидов под-
нимает температуру тела и нормализует продолжительность контактов
с детенышами (УУоодз14е, [еоп, 1980).
Таким образом, основная характеристика термальной связи между
матерью и детенытем крысы
— интенсивный обогрев детеньипей в пер-
вую неделю постнатального развития и постепенное уменьшение тепло-
спабжепия по мере того, как у детеныша уменьнтается потребность
в нем. Существенную роль в этом процессе играет внешне простой меха-
низм термоконтроля, определяющий прерывание коптакта самки с крыся-
тами. Однако этот механизм коптролируется сложным взаимодействием
двух процессов: физиологическими изменениями у детенышей в связи со
становлением гомойотермии и гормопальными изменениями в организме
самки, активно регулирующими механням термоконтроля. Восьма важно
что термальные связи между матерью и детепышами непосредственно
определяют и другие типы их взаимоотношений, например продолжи-
тельность кормлений детенышей молоком.
Хотя в рассмотренный период оптогенеза детенышей их термальная
зависимость от матери проявляется гораздо наглядней, чем в другое вре-
мя, это не означает, что детеныити стараиего возраста становятся пол-
ностью независимыми в этом отношении. Даже у 18—21-дневпых крысят
способных поддерживать постоянную температуру тела, эти Возможности
не столь соверптенны, как у взрослых крыс (Сопки, Несрепезз, 1974)
Поэтому зверьки данного возраста большую часть времени проводят
ы ое дано стабильным и благоприятным температурным режи-
Если двухнедельных крысят содержать в течение нескольких дией
без матери при комнатной температуре, то у них существенно замед-
ляется созревапие мозга, скелета, внутренних органов, повышается общая
смертность (Э\юпе ей 21., 1976). Однако если содержать крысят без мате-
ри при температуре жилого гнезда (+35°), то эти отклонения исчезают
Таким образом, хотя зверьки этого возраста в общем способны самостоя-
2ВА
Глава четвертая
остается прежним (Ёеоп ей а|., 1978). ЕК этому времени у детенышей
резко усиливается способность к гомойотермии. Возникает вопрос: чем
регулируется эта согласованность в изменении потребпостей детенышей
и реакций матери? Было показано, что время контактов самки с детены-
шами хорошо скоррелировано © температурой окружающей среды:
при +18° самка проводит с детеньишами большие времени, чем при +22°,
а при +26°
— меньше, чем при 22° (Стозкеггу © а1., 1978). Таким
образом, термальные характеристики являются важными определяющими
контактов. Возникло предположение, что самка кончает период контак-
тов (и тем самым регулирует время, проводимое © детенышами) в тот
момент, когда температура тела детенышей достигает определенной
величины, например стаповится равной температуре тела самки. Этим
можно объяспить постепенное сокращение периодов коптактов по мере
роста гомойотермии крысят. Действительно, если экспериментально ох-
лаждать детенышей, то продолжительность периодов коитактов у матери
с ними увеличивается. Однако, если припять зту гипотезу, нагревание
детенышей до той температуры, которая бывает в момент окончания
контактов, должно было бы резко уменьшить время иребывапия самки
с детопышами. Этого не происходит (Геоп её а|., 1978). Таким образом,
температура детенышей является не единственным фактором, определяю-
щим режим контактов. Возникло предположение, что окончание периода
контактов определяется некими особенностями работы системы теплопро-
дукции-—теплоотдачи самки. Действительно, изменение режима фупкцио-
нирсвания этой системы оказывает влияние на время, проводимое сам-
кой с детенышами. Так, если эксперимептально увеличить теплоотдачу
самки, обрив ей шерсть, продолжительность периодов контактов с крыся-
тами резко возрастает. И наоборот, экспериментальное умепьшецие спо-
собпости к теплоотдаче (отрезание хноста) вызывает сокращение перио-
дов контактов самки © детенышами. Более того, время, проводимое
с крысятами самкой, у которой отрезана половина хвоста, промежуточ-
ное между тем, что проводят бесхвостые и интактные самки (Геоп её а|.,
1978).
С помощью вживления температурных датчиков под кожу (в области
сосков) и в преоптическую зону переднего мозга самки было установле-
но, что момент окончапия контакта характеризуется резким повышецием
температуры тела и мозга (УУоодя4е, Геоп, 1980). Возможно, это и яв-
ляется стимулом, вызывающим прерывание контакта. Кроме того, было
показано, что для самки во вторую неделю лактации характерно резкое
увеличение теплопродукции (на 17,3% по сравнению с нелактирующими
самками) и хроническое повышение температуры тела (Тапез, Геоп,
1983; Геоп её а1., 1985). Это делает самку весьма чувотвительной к до-
млолнительному повьштению температуры во время контактов © детены-
шами, заставляя ее раньше прерывать контакт для того, чтобы сохранить
термальный гомеостаз. Гормональные механизмы этого феномепа пока
изучены далеко не достаточно. Однако ясно, что хроническое повышение
температуры обусловлено, по крайцей мере частично, повышением уров-
ия циркулирующего пролактина. Предполагается, во-первых, что пролак-
=
Поведение
265
тин стимулирует секрецию прогестеропа, который поднимает на повый
уровень термальные требования лактирующей самки. Во-вторых, в это
время повышается секреция адренальных глюкокортикоидов которые
усиливают тканевый метаболизм в такой степени, чтобы были удовле-
творены эти новые требования (см., папример: Геоп её а|., 1985).
Если на следующий день после родов экспериментально подавить
уровень циркулирующего пролактина, то температура тела кормящей
самки хронически попизится, а контакт © детенытами удлипяется
(Геоп её а|., 1985). Оцнако такие эффекты проявляются лишь на вто-
рой неделе лактации. К этому времсни оказывается подавленным уро-
вень прогестеропа. Герапия прогестероном у самок с подавлепным уров-
нем пролактина восстанавливает и темпоратуру, типичную для лакти-
рующих самок и нормальную продолжительность коптактов с детенышами
(\\содз14е еф а|1., 1981). В то же время адрепалэктомия лактирующих
самок также приводит к хроническому снижению температуры тсола
и удлинению продолжительности контактов с детенышами (Созуе, Ео]-
Теу, 1961). Восстанозительная терапия с помощью глюкокортикоидов под-
нимает температуру тела и нормализует продолжительность контактов
с детенышами (УУоодз14е, [еоп, 1980).
Таким образом, основная характеристика термальной связи между
матерью и детенытем крысы
— интенсивный обогрев детеньипей в пер-
вую неделю постнатального развития и постепенное уменьшение тепло-
спабжепия по мере того, как у детеныша уменьнтается потребность
в нем. Существенную роль в этом процессе играет внешне простой меха-
низм термоконтроля, определяющий прерывание коптакта самки с крыся-
тами. Однако этот механизм коптролируется сложным взаимодействием
двух процессов: физиологическими изменениями у детенышей в связи со
становлением гомойотермии и гормопальными изменениями в организме
самки, активно регулирующими механням термоконтроля. Восьма важно
что термальные связи между матерью и детепышами непосредственно
определяют и другие типы их взаимоотношений, например продолжи-
тельность кормлений детенышей молоком.
Хотя в рассмотренный период оптогенеза детенышей их термальная
зависимость от матери проявляется гораздо наглядней, чем в другое вре-
мя, это не означает, что детеныити стараиего возраста становятся пол-
ностью независимыми в этом отношении. Даже у 18—21-дневпых крысят
способных поддерживать постоянную температуру тела, эти Возможности
не столь соверптенны, как у взрослых крыс (Сопки, Несрепезз, 1974)
Поэтому зверьки данного возраста большую часть времени проводят
ы ое дано стабильным и благоприятным температурным режи-
Если двухнедельных крысят содержать в течение нескольких дией
без матери при комнатной температуре, то у них существенно замед-
ляется созревапие мозга, скелета, внутренних органов, повышается общая
смертность (Э\юпе ей 21., 1976). Однако если содержать крысят без мате-
ри при температуре жилого гнезда (+35°), то эти отклонения исчезают
Таким образом, хотя зверьки этого возраста в общем способны самостоя-
266
Глава четвертая
=
и.
тельно поддерживать постоянную температуру тела, неоольнтое количест
во тепла, которое периодически поставляется им матерью при. ее визи-
тах в гнездо (примерно 15 мин каждый час), оказывается необходимым
для их нормального развития (Нофег, 1975).
Взаимоотношения между матерью и детеньиами,
в период прекращения питания молоком
и их перехода в самостоятельному питанию
Для незрелорождающегося детеныша крысы материнское молоко —
единственный источник питательных веществ. Многие важ с к рты ето
физиологии и поведения определяются особенностями молочи
кары-
ливапия. Поэтому переход к питанию твердои пищеи сопровозкда ‚ся
существенной перестройкой не только физиологии пинцоварительной сп
стемы детеныша и его пищевого поведепия, но и отношен й ‚матерью.
На первых стадиях постнатального развития у детеныша, оянно
выражено стремление при любой возможности тянуться к соску, удерки-
рать ето во рту и сосать молоко до тех пор, лока оно ие ачинает
выливаться из глотки (ПаП, Вовеп ай, 1977). Если лищить самку 1 -
можности прервать контакт с выводком, то детеныши самостояте ьно пе
отрываются от сосков и могут оставаться прикрепленными к ним чт ами
(На| её а1., 1975). До 15-дневного возраста у крысят практ чески пол
ностью отсутетвуют эпзимы, необходимые для усвоения твердых к рмов,
зато лактаза, требующаяся для адсорбции и перера отки лак совы то.
ка, производится в больших количествах. Это можно ‚рассма в т
«физиологическую адаптацию» к молочному питанию (А]уаг г 385, т.
Стимуляция сосков самки при сосании приводит к повышен по се р о
пролактина, что, в свою очередь, ведетк увеличению пе тол т пр цук-
ции молока, но и потребления самкой кормов, нео ходимы ия
полнения энергетических трат на лактацию (Стозуепог е ы 970). в.
От рождепия и до 15-дневного возраста детенышей на па ря пе-
прерывное прибывапие количества т
ОЙ мото те
этого продукция его постепенно 001 ает
-
м
аету
Й
олностью исчезает (ВаБсКу её 21., 1970). Таким обр
а
раоте 45-27 дней осуществляется отлучение крысят от молоч-
ного вскармливания и переход к иитанию твердыми кормами. то д а от.
носительно самостоятельных процесса, но оба они развиваются то иль.
ным влиянием матери. Самка воздействует на процесс отлучения, меняя
свое отношение к попыткам детеньштеи сосать ее. Если па первых ‹ а
диях онтогенеза кормление происходит по инициативе матери, к торая
приходит в гнездо и принимает особую позу, то в период "луч тия
ипициативу перехватывают детеныши, активно преследу тие ие
пылках начать сосание. У самки же появляется и усиливае тепден-
ция избегать их. В лабораторных условиях кормление трехнедел! ых
зверьков происходит уже не в гнезде, а в любой НЫ м
они пастигнут мать и вынудят ее начать кормление (11056 ь 69).
Эти изменения в поводении самки обусловлены изменениям
Поведение
267
характеристик самих детенышей (возможно, их активности), так как,
подкладывая ей крысят младшего возраста, можно добиться, как уже
говорилось, продления лактации и стремления самки кормить выводок.
В лабораторных условиях до 19-дневного возраста детенышей ритм
питания определяется матерью. Детепьхии, чьи матери имеют естествен-
пый ритм ночного питания, также питаются ночью. Нри реверсии ритма
матери ритм реверсируется и у детенышей, а детспыши ослепленных ма-
терей, не имеющих четко выраженного суточного ритма питания, также
утрачивают этот ритм. После 19-дневного возраста пормальный ритм
ночного питания появляется у детенышей даже тогда, когда взрослые
отсутетвуют или когда у взрослых нет четкого ритма. Одпако при нару-
мении зрения детенышей их ритм соответствует материнскому ив более
поздлем возрасте (Геуш, Э4егп, 1975).
У 2—3-недельных крысят происходят существенные физиологические
перестройки. Резко сокращается синтез лактазы — настолько, что удли-
нение молочной диеты приводит к расстройствам пищеварения (ТлБег-
тап М, Г4Ъегтап О., 1978). Изменяются и способноети к обучению при
пищевом подкреплении. Например, у детенышей этого возраста можно
выработать аверсию (т. е. отрицательную реакцию) к соску, тогда как
крысят младше 15-дневного возраста этому обучить практически невоз-
можно (МагЫп АЮегв, 1979). Вполие возможно, что желудочно-кишеч-
ный дистресс, вызванный недостатком лактазы, ассоциируется у детепы:
шей с сосанием молока и это побуждаст их искать другую пищу
(СзеЁ, 1981). Такого рода обучение явно имеет очень большое значение
в биологии крые и по многим характеристикам отличается от других
форм обучения, например по быстроте выработки реакций, их устойчи-
вости и больному латентному периоду от момента предъявления стиму-
ла (т.е. поедания корма) до получепия подкрепления (т. е. оценки при-
годности пищи) (см., например: Сатза, Кое, 1966).
В естественных условиях в период, когда крысята переходят на твер-
дую пищу, взрослые растиряют норы и запасают в них корм. В оспоз-
пом эти запасы съедают детеныши (Ватлей, 1975). Примерно до 21-
дневного возраста детеныши пе покидают нору и едят только то, что при-
посят им родители (СаШоип, 1962). В 22-дневном возрасте крысята уже
выходят из норы, но их пищевые привычки но-прежнему складываются
под влилнием взрослых, в первую очередь матери. Иследования, прове-
денные на диких серых крысах, содержащихся в полувольных условиях,
показали, что крысята едят только те виды пищи, которыми питаются
взрослые члены группы и практически не потребляют те, к которым было
экспериментально выработано отвращение у взрослых (СяеЁ Сатк,
1971а). Каким образом это достигается? Во-первых, наблюдение за едой
взрослых животных является для крысят мощным стимулом для начала
их собственной пищевой активности, причем детеньшии стремятся есть
именно в тех местах и из тех кормушек, где едят взрослые (СаеЁ,
Сагк, 1971Ъ). Во-вторых, даже в отсутствие взрослых крысята предно-
читают есть в тех местах, где раньше ели взрослые, ориентируясь по за-
наховым меткам (Са]её, Нефег, 1976). Кроме того, есть указания (Са]еЁ,
266
Глава четвертая
=
и.
тельно поддерживать постоянную температуру тела, неоольнтое количест
во тепла, которое периодически поставляется им матерью при. ее визи-
тах в гнездо (примерно 15 мин каждый час), оказывается необходимым
для их нормального развития (Нофег, 1975).
Взаимоотношения между матерью и детеньиами,
в период прекращения питания молоком
и их перехода в самостоятельному питанию
Для незрелорождающегося детеныша крысы материнское молоко —
единственный источник питательных веществ. Многие важ с к рты ето
физиологии и поведения определяются особенностями молочи
кары-
ливапия. Поэтому переход к питанию твердои пищеи сопровозкда ‚ся
существенной перестройкой не только физиологии пинцоварительной сп
стемы детеныша и его пищевого поведепия, но и отношен й ‚матерью.
На первых стадиях постнатального развития у детеныша, оянно
выражено стремление при любой возможности тянуться к соску, удерки-
рать ето во рту и сосать молоко до тех пор, лока оно ие ачинает
выливаться из глотки (ПаП, Вовеп ай, 1977). Если лищить самку 1 -
можности прервать контакт с выводком, то детеныши самостояте ьно пе
отрываются от сосков и могут оставаться прикрепленными к ним чт ами
(На| её а1., 1975). До 15-дневного возраста у крысят практ чески пол
ностью отсутетвуют эпзимы, необходимые для усвоения твердых к рмов,
зато лактаза, требующаяся для адсорбции и перера отки лак совы то.
ка, производится в больших количествах. Это можно ‚рассма в т
«физиологическую адаптацию» к молочному питанию (А]уаг г 385, т.
Стимуляция сосков самки при сосании приводит к повышен по се р о
пролактина, что, в свою очередь, ведетк увеличению пе тол т пр цук-
ции молока, но и потребления самкой кормов, нео ходимы ия
полнения энергетических трат на лактацию (Стозуепог е ы 970). в.
От рождепия и до 15-дневного возраста детенышей на па ря пе-
прерывное прибывапие количества т
ОЙ мото те
этого продукция его постепенно 001 ает
-
м
аету
Й
олностью исчезает (ВаБсКу её 21., 1970). Таким обр
а
раоте 45-27 дней осуществляется отлучение крысят от молоч-
ного вскармливания и переход к иитанию твердыми кормами. то д а от.
носительно самостоятельных процесса, но оба они развиваются то иль.
ным влиянием матери. Самка воздействует на процесс отлучения, меняя
свое отношение к попыткам детеньштеи сосать ее. Если па первых ‹ а
диях онтогенеза кормление происходит по инициативе матери, к торая
приходит в гнездо и принимает особую позу, то в период "луч тия
ипициативу перехватывают детеныши, активно преследу тие ие
пылках начать сосание. У самки же появляется и усиливае тепден-
ция избегать их. В лабораторных условиях кормление трехнедел! ых
зверьков происходит уже не в гнезде, а в любой НЫ
м
они пастигнут мать и вынудят ее начать кормление (11056 ь 69).
Эти изменения в поводении самки обусловлены изменениям
Поведение
267
характеристик самих детенышей (возможно, их активности), так как,
подкладывая ей крысят младшего возраста, можно добиться, как уже
говорилось, продления лактации и стремления самки кормить выводок.
В лабораторных условиях до 19-дневного возраста детенышей ритм
питания определяется матерью. Детепьхии, чьи матери имеют естествен-
пый ритм ночного питания, также питаются ночью. Нри реверсии ритма
матери ритм реверсируется и у детенышей, а детспыши ослепленных ма-
терей, не имеющих четко выраженного суточного ритма питания, также
утрачивают этот ритм. После 19-дневного возраста пормальный ритм
ночного питания появляется у детенышей даже тогда, когда взрослые
отсутетвуют или когда у взрослых нет четкого ритма. Одпако при нару-
мении зрения детенышей их ритм соответствует материнскому ив более
поздлем возрасте (Геуш, Э4егп, 1975).
У 2—3-недельных крысят происходят существенные физиологические
перестройки. Резко сокращается синтез лактазы — настолько, что удли-
нение молочной диеты приводит к расстройствам пищеварения (ТлБег-
тап М, Г4Ъегтап О., 1978). Изменяются и способноети к обучению при
пищевом подкреплении. Например, у детенышей этого возраста можно
выработать аверсию (т. е. отрицательную реакцию) к соску, тогда как
крысят младше 15-дневного возраста этому обучить практически невоз-
можно (МагЫп АЮегв, 1979). Вполие возможно, что желудочно-кишеч-
ный дистресс, вызванный недостатком лактазы, ассоциируется у детепы:
шей с сосанием молока и это побуждаст их искать другую пищу
(СзеЁ, 1981). Такого рода обучение явно имеет очень большое значение
в биологии крые и по многим характеристикам отличается от других
форм обучения, например по быстроте выработки реакций, их устойчи-
вости и больному латентному периоду от момента предъявления стиму-
ла (т.е. поедания корма) до получепия подкрепления (т. е. оценки при-
годности пищи) (см., например: Сатза, Кое, 1966).
В естественных условиях в период, когда крысята переходят на твер-
дую пищу, взрослые растиряют норы и запасают в них корм. В оспоз-
пом эти запасы съедают детеныши (Ватлей, 1975). Примерно до 21-
дневного возраста детеныши пе покидают нору и едят только то, что при-
посят им родители (СаШоип, 1962). В 22-дневном возрасте крысята уже
выходят из норы, но их пищевые привычки но-прежнему складываются
под влилнием взрослых, в первую очередь матери. Иследования, прове-
денные на диких серых крысах, содержащихся в полувольных условиях,
показали, что крысята едят только те виды пищи, которыми питаются
взрослые члены группы и практически не потребляют те, к которым было
экспериментально выработано отвращение у взрослых (СяеЁ Сатк,
1971а). Каким образом это достигается? Во-первых, наблюдение за едой
взрослых животных является для крысят мощным стимулом для начала
их собственной пищевой активности, причем детеньшии стремятся есть
именно в тех местах и из тех кормушек, где едят взрослые (СаеЁ,
Сагк, 1971Ъ). Во-вторых, даже в отсутствие взрослых крысята предно-
читают есть в тех местах, где раньше ели взрослые, ориентируясь по за-
наховым меткам (Са]её, Нефег, 1976). Кроме того, есть указания (Са]еЁ,
968
Глава четвертая
ЗВегту, 1973), что крысята могут распознавать диету матери по, су
молока и при переходе па твердые корма даже в отсутствие матер
почитают ту пищу, которую ела она, когда кормила их ом
примап.
Штайнигер (Зленуеег, 1950) первым описал, как отравление, ‚примат
ка, вначале успешно применяющаяся для борьбы е крысами, о тепенио
стаповится малоэффективной. Он считал, что это происходит ва ‹ ет
того, что крысята обучаются избегать той пищи, которую, избе т
родители и родственники (получившие вкусовое отвращени ‚к ной вслед
ствие сублетальных доз яда). Современные исследования, | Е зо, п ка-
зывают, что этот эффект может достигаться только за счет осу отия пи.
щевым предпочитаниям, существование же специфического м
изиеганию пищи пока не обнаружено (СаеЁ, ВескК, 5). |
Запии в
Известно, что между колониями диких крыс наблюдаются вари к и
потреблении различных видов пищи; так, есть «кланы», специализ тру о-
щиеся на питапии рыбой, придонными моллюсками, молкими воробьи
ными, утятами, яйцами, птенцами из гнезд и т. д. (см., петро ор, свод.
ку: СаеЁ, 1980). Добывание всех этих видов пищи тре ует ‚обуч шя
крысят специфическим моторным навыкам. Чем обуслов твветея ото
обучение, пока не ясно. Возможно, что В условиях И
етоятельно
зых ресурсов на территории клана каждый крысенок сам тоятельто
обучается добыванию наиболее доступных видов пищи и в су мо
приводит к пищевой специализации клана. льтерпативн " ут я
обучение крысят у взрослых животных в результате на людопи» а
действиями, т. е. поддержание своеобразных «культурных радиций».
В первом случае пищевая специализация полностью определяет
или.
ом тех или ипых пищевых ресурсов, т. е. внешней по отношению к ла
пу эколотической средой. Во втором — она определяется не только этим,
но и социальными воздействиями сородичей. Хотя наблюдения р урали-
стов создавали впечатление о высоком развитии способностей ко учению
наблюдением у серой крысы, эксперимептальное выявление тих особ-
ностей оказалось делом непростым. Галеф (СаеЁ, 1980) о учат кр т
плавать и доставать пищу со дна водоема. Число крысят, о учиниихея
этому, было примерно равпым в тех случаях, когда их мать умел пла
вать и плавала при них, и в тех, когда мать плавать не умела, тя
крысята с «плавающей» матерью обучались в более раннем возрас ыы
успех обучения детенышей сильно влияла их пищевая мотивац
нехватке пищи число обучивитихся резко возрастало.
.
Интересные данные в этом отношении были получены при т
печерные крысы, пит
ы
.
оченио оомяни
зпишек включает сложную последовательность поведенческих т :повсте ие
не врожденное, а передается обучением от родителей детеннипам. В ея ое о в
крысы, не живущие в лесах, умирали от и
а
нойое©
отно шитаклцейся семенами. о нако если в клетку к такой ‹умеющей».
крысе помещали детеныша, то он быстро обучался специфическим моторным навыг
кам и полностью обеспечивал себя едой (А1зпег, Тегке], 1985).
=
Поведение
269
Значение капрофагии за время перехода "рысенка
на твердые корма
В формировании способности крысенка к перевариванию твердых кормов
большое значение имеет поедапие экскрементов матери, привленатель-
ность которых для иего резко повышается с двухнедельного возраста,
Изучение этой специфической капрофагии всирыло целую цепь синхроп-
ных изменений поведенческих и физиологических реакций матери и де-
тепытна.
На первых этапах исследования было обнаружено, что помещенные в
лабиринт крысята 14—27-дневного возраста чаще выбирают рукав, веду-
щий к матери, чем рукав, ведущий к нерожавшей самке. Ориептация их
совершается по запаху, так как «целевые камеры» лабиринта можно
сделать звуко- и светонепроницаемыми, но ток воздуха обязательно дол-
щен быть направлен от самок
к дотеньииу, а не наоборот. Леон и Мольтц
(Тео, Моя, 1971) предиоложили наличие в экскрементах химического
вещества— «материнского феромона», привлекающего детепышей. Наб-
людаемое предпочтение запаха матери оказалось не связанным © ее ип-
дивидуальными особенностями. Любая самка, имеющая детенышей сход-
ного возраста, может быть почти столь же привлекательной дия крысен-
ка, как и его собственная мать. Особенно привлекают крысят те самки,
рацион которых совпадает с рационом их матери (Теоп, 1975).
Возрастное повыптение чувствительности крысенка к действию мате-
ринекого феромона точпо соответствует периоду времени, когда самка
продуцирует этот феромон. Это было обнаружено при сопоставлепии ре-
акций 16-дыевного крысенка на самок, отличающихся по возрасту их де-
тенышей, с реакциями крысят различных возрастоз на самку, имеющую
16-дневных детенышей. Как реакция на феромон, так и его секреция в
нормальных условиях появляется через 14 дней после родов. Максималь-
пую реакцию проявляют детеныши 21-дневного возраста, и к 21-му дню
после родов самки достигают для них наибольшей привлекательности.
У детепышей 27-дневного возраста реакция резко ослабевает, а у самок,
имеющих детенышей этого возраста, прекращается секреция феромона
(Геоп, Мо, 1972).
Никакими экспериментальными воздействиями на крысенка этот пе-
риод реагировапия на феромон изменить не удается (например, сместить
‘его в сторону более раннего или более позднего включения). У самки же
срок секреции феромона можно менять, продлив его до 100 дней, если
ей, после того как она вырастит детенышей до месячного возраста, перио-
дически заменять их на 16-дневных детенытией (Мо]4и © а1., 1974). Если
же крысе, имеющей двухнедельных детенышей, заменить их на детены-
шей 1--5-дневного возраста, то секреция феромона исчезает. Если самку
содержать 16 дией после родов с однодневными Детенышами, ежедневно
заменяя их, то она не становится привлекательной для 16-дневных дете-
нышей, т, в. секреция феромона у нее не происходит. Если же, ежеднев-
но мепяя детеньшшей, увеличивать их возраст так, чтобы спустя 16 дней
после родов с самкой паходились 16-дневные детепьиги, она будет столь
968
Глава четвертая
ЗВегту, 1973), что крысята могут распознавать диету матери по, су
молока и при переходе па твердые корма даже в отсутствие матер
почитают ту пищу, которую ела она, когда кормила их ом
примап.
Штайнигер (Зленуеег, 1950) первым описал, как отравление, ‚примат
ка, вначале успешно применяющаяся для борьбы е крысами, о тепенио
стаповится малоэффективной. Он считал, что это происходит ва ‹ ет
того, что крысята обучаются избегать той пищи, которую, избе т
родители и родственники (получившие вкусовое отвращени ‚к ной вслед
ствие сублетальных доз яда). Современные исследования, | Е зо, п ка-
зывают, что этот эффект может достигаться только за счет осу отия пи.
щевым предпочитаниям, существование же специфического м
изиеганию пищи пока не обнаружено (СаеЁ, ВескК, 5). |
Запии в
Известно, что между колониями диких крыс наблюдаются вари к и
потреблении различных видов пищи; так, есть «кланы», специализ тру о-
щиеся на питапии рыбой, придонными моллюсками, молкими воробьи
ными, утятами, яйцами, птенцами из гнезд и т. д. (см., петро ор, свод.
ку: СаеЁ, 1980). Добывание всех этих видов пищи тре ует ‚обуч шя
крысят специфическим моторным навыкам. Чем обуслов твветея ото
обучение, пока не ясно. Возможно, что В условиях И
етоятельно
зых ресурсов на территории клана каждый крысенок сам тоятельто
обучается добыванию наиболее доступных видов пищи и в су мо
приводит к пищевой специализации клана. льтерпативн " ут я
обучение крысят у взрослых животных в результате на людопи» а
действиями, т. е. поддержание своеобразных «культурных радиций».
В первом случае пищевая специализация полностью определяет
или.
ом тех или ипых пищевых ресурсов, т. е. внешней по отношению к ла
пу эколотической средой. Во втором — она определяется не только этим,
но и социальными воздействиями сородичей. Хотя наблюдения р урали-
стов создавали впечатление о высоком развитии способностей ко учению
наблюдением у серой крысы, эксперимептальное выявление тих особ-
ностей оказалось делом непростым. Галеф (СаеЁ, 1980) о учат кр т
плавать и доставать пищу со дна водоема. Число крысят, о учиниихея
этому, было примерно равпым в тех случаях, когда их мать умел пла
вать и плавала при них, и в тех, когда мать плавать не умела, тя
крысята с «плавающей» матерью обучались в более раннем возрас ыы
успех обучения детенышей сильно влияла их пищевая мотивац
нехватке пищи число обучивитихся резко возрастало.
.
Интересные данные в этом отношении были получены при т
печерные крысы, пит ы
.
оченио оомяни
зпишек включает сложную последовательность поведенческих т :повсте ие
не врожденное, а передается обучением от родителей детеннипам. В ея ое о в
крысы, не живущие в лесах, умирали от и
а
нойое©
отно шитаклцейся семенами. о нако если в клетку к такой ‹умеющей».
крысе помещали детеныша, то он быстро обучался специфическим моторным навыг
кам и полностью обеспечивал себя едой (А1зпег, Тегке], 1985).
=
Поведение
269
Значение капрофагии за время перехода "рысенка
на твердые корма
В формировании способности крысенка к перевариванию твердых кормов
большое значение имеет поедапие экскрементов матери, привленатель-
ность которых для иего резко повышается с двухнедельного возраста,
Изучение этой специфической капрофагии всирыло целую цепь синхроп-
ных изменений поведенческих и физиологических реакций матери и де-
тепытна.
На первых этапах исследования было обнаружено, что помещенные в
лабиринт крысята 14—27-дневного возраста чаще выбирают рукав, веду-
щий к матери, чем рукав, ведущий к нерожавшей самке. Ориептация их
совершается по запаху, так как «целевые камеры» лабиринта можно
сделать звуко- и светонепроницаемыми, но ток воздуха обязательно дол-
щен быть направлен от самок
к дотеньииу, а не наоборот. Леон и Мольтц
(Тео, Моя, 1971) предиоложили наличие в экскрементах химического
вещества— «материнского феромона», привлекающего детепышей. Наб-
людаемое предпочтение запаха матери оказалось не связанным © ее ип-
дивидуальными особенностями. Любая самка, имеющая детенышей сход-
ного возраста, может быть почти столь же привлекательной дия крысен-
ка, как и его собственная мать. Особенно привлекают крысят те самки,
рацион которых совпадает с рационом их матери (Теоп, 1975).
Возрастное повыптение чувствительности крысенка к действию мате-
ринекого феромона точпо соответствует периоду времени, когда самка
продуцирует этот феромон. Это было обнаружено при сопоставлепии ре-
акций 16-дыевного крысенка на самок, отличающихся по возрасту их де-
тенышей, с реакциями крысят различных возрастоз на самку, имеющую
16-дневных детенышей. Как реакция на феромон, так и его секреция в
нормальных условиях появляется через 14 дней после родов. Максималь-
пую реакцию проявляют детеныши 21-дневного возраста, и к 21-му дню
после родов самки достигают для них наибольшей привлекательности.
У детепышей 27-дневного возраста реакция резко ослабевает, а у самок,
имеющих детенышей этого возраста, прекращается секреция феромона
(Геоп, Мо, 1972).
Никакими экспериментальными воздействиями на крысенка этот пе-
риод реагировапия на феромон изменить не удается (например, сместить
‘его в сторону более раннего или более позднего включения). У самки же
срок секреции феромона можно менять, продлив его до 100 дней, если
ей, после того как она вырастит детенышей до месячного возраста, перио-
дически заменять их на 16-дневных детенытией (Мо]4и © а1., 1974). Если
же крысе, имеющей двухнедельных детенышей, заменить их на детены-
шей 1--5-дневного возраста, то секреция феромона исчезает. Если самку
содержать 16 дией после родов с однодневными Детенышами, ежедневно
заменяя их, то она не становится привлекательной для 16-дневных дете-
нышей, т, в. секреция феромона у нее не происходит. Если же, ежеднев-
но мепяя детеньшшей, увеличивать их возраст так, чтобы спустя 16 дней
после родов с самкой паходились 16-дневные детепьиги, она будет столь
270 Глава четвертая
же привлекательна в тестах, как и самка, зыращивающая свой собствен-
ный выводок. Однако если сразу же после родов к самке поместить
10-дневных детеньиией, то привлекать коитрольных через 6 дней она не
будет. Таким образом, секреция феромона находится под контролем сти-
мулов, исходящих от крысят, однако важную роль играет и специфиче-
ское состояние самки, которая 2 недели назад родила и выращивает
выводок.
Было высказано предположение, что секрецией феромона управляют
гормональные механизмы (Теоп, Мо|, 1973). Для проверки этого пред-
положения проверяли реакции детенышей па самок с удаленными эндо,
крипными железами и измененным вследствие этого гормопальным про
филем. Самки с удаленными яичниками и падпочечниками вырабатывают
феромон. Однако при эртокорниновой блокаде секреции пролактина при-
влекательность самок исчезает. Если после блокады вводить пролактин,
привлекательпость восстанавлинается. Был сделап вывод, что выработка
феромона обусловлена высоким уровнем секреции пролактина.
Дальнейшие исследования показали, что для реакции детеныпта соб-
ственное присутствие самки в целевой камере несущественно, достаточно
оставлять ее фекалии. В естествениых ситуациях фекалии матери очень
привлекательны для детенышей. Как мы уже говорили, крысят пе только
привлекает их запах, но они чаето поедают их (КИрайтеК её а1., 1983).
Таким образом, был сделан вывод, что «материнский феромон» — хими-
ческое вещество, содержащееся в испражнениях самки.
Поиек этого вещества привлек знимапие исследователей к факту,
что как объем, так и состав фекальных массе самки, когда крысята дости-
гают двухнедельного возраста, сильно изменяются. Вес фекальных масс
в этот период в 2—3 раза превытиает таковой девственпых самок. Соот-
ветственно увеличивается и потребление ими кормов. В фекалиях резко
повышается содержание летучих компонентов (оцениваемое по соотно-
шению весов свежих и высушенных экскрементов), они светлее обычных,
сильнее пахнут и менее структурированы (Геоп, 1974; Цеоп, Мо|Нз, 1971).
На этом этапе пути двух исследователей, открывших «материнский феро-
моп», разошлись. Леон предположил, что основное отличие фекалий ма-
тери состоит в присутствии особой ботатой микрофлорой субстанции, на-
званной «цекотрофом» и вырабатываемой слепой кишкой у всех особей
вврослых крыс. Автор предполагает, что, помимо обычных темноократен-
ных и сформированных в бульбочки фекалий, взрослые животные экс-
кретируют и цекотроф, но сразу же после выделения из анального от-
верстия поедают его. У самки, имеющей двухнедельных детспышей, ко-
личество испражнений такого тина пастолько увеличивается, что значи-
тельная часть их остается несъеденной. Таким образом, именно цекотроф
матери, а не ее обычные фекалии привлекательны для детенышей. Кроме
того, Леоном было показано, что детенышей привлекают фекальные мас-
сы, взятые из слепой кишки любой взрослой крысы. Слепая кишка — ме-
сто усиленного размножения бактерий. Если ингибировать кинечную
флору введением антибиотика, то содержимое слепой кишки и фекалии
самки теряют свою привлекательпость для детеньшией (Теоп, 4994).
Поведение
274
Автор сделал вывод, что цекотрофный компонент фекалий определяет
их запаховую привлекательность для крысят. Можно предположить, что,
поедая экскременты матери, крысята получают бактериальную флору ее
кишечника, специфическую для предпочитасмых ею кормов. Поскольку
у крысят вырабатывается предпочтение тех же кормов, что и матери,
то специализация микрофлоры может существенно блатоприятствовать
оптимальному использованию кормов.
Однако Мольтц
— второй автор открытия «материнского феромонах —
не согласился с предположением о наличии цекотрофа, так как прямые
доказательства существования двух форм экскрементов у крыс (обычных
и цекотрофиых) отсутствуют. После детального исследования механизмов
продукции экскрементов лактирующей самкой он предложил другое объ-
яспение их привлекательности для крысят. Увеличение уровня пролакти-
на в крови лактирующей самки приводит к постепенному увеличению
концентрации этого гормона в печени (Ёее еф а1., 1982), а это, в свою
очередь, приводит к увеличению синтеза печенью основной желчной—
холиевой кислоты. В слепой кишке крыс под воздействием определенных
бактерий из холиевой кислоты синтезируется вторичная желчная
— де-
оксихолиеная кислота. Потребность В ней и заставляет крысят поедать
экскременты матери, а привлекает их феромон-содержащий компонент °
фекалий — производное деоксихолиевой кислоты.
В болышой серии экспериментов было показано, что вплоть до 28-
дневного возраста крысят уровень секреции деоксихолиевой кислоты у
них значительно ниже, чем у взрослых крыс. С другой стороны, в фека-
лиях самки-матери уровель деоксихолневой кислоты значительно выше,
чем у нерожавптих самок (КПрабтсК её а1., 1983). Причины недостаточ-
ной продукции деоксихолиевой кислоты крысятами неяспы.
Потребность в деоксихолиевой кислоте может объясняться по крайней
мере двумя основными причинами. Первая— повышение иммупозащиты
у крысят, вторая
— мислинизация мозга. Показано, что среди крысят,
лишенных доступа к экекрементам матери и подвергшихся холодовому
стрессу, увеличивается гибель от острого энтерита. Этот эффект пропада-
ет, если к их корму добавить деоксихоливвую кислоту. Иредполагается,
что опа инактивирует эпдотоксины, участвующие в патотепезе острых
энтеритов, т. е. обусловливает «физико-химическую» защиту, расщепляя
токсичные полимеры, производимые такими микрооргапизмами, как
Езовет1сШа соц, на более мелкие нетоксичные структурные элементы (Гее,
Мо, 19845).
При сравнении крысят, имеющих доступ к экскрементам матери и
лиленных его, обнаружилось, что у последних уровни триглицеридов и
свободных жирных кислот в плазме крови значительно ниже. У этих же
крысят. меньше масса миелина относительно массы мозга и ниже кон-
` центрация фосфолипидов в миелине, чем у коптрольтой групны. При до-
бавлении в корм деоксихолиевой кислоты все перечисленные эффекты
пропадают. Предполатается, что деоксихолиевая кислота способствует
абсорбции липидов с длинными цепями, которые необходимы для разви-
тия миелиновых структур мозга (Тее, Мол, 1984а, Ъ).
270 Глава четвертая
же привлекательна в тестах, как и самка, зыращивающая свой собствен-
ный выводок. Однако если сразу же после родов к самке поместить
10-дневных детеньиией, то привлекать коитрольных через 6 дней она не
будет. Таким образом, секреция феромона находится под контролем сти-
мулов, исходящих от крысят, однако важную роль играет и специфиче-
ское состояние самки, которая 2 недели назад родила и выращивает
выводок.
Было высказано предположение, что секрецией феромона управляют
гормональные механизмы (Теоп, Мо|, 1973). Для проверки этого пред-
положения проверяли реакции детенышей па самок с удаленными эндо,
крипными железами и измененным вследствие этого гормопальным про
филем. Самки с удаленными яичниками и падпочечниками вырабатывают
феромон. Однако при эртокорниновой блокаде секреции пролактина при-
влекательность самок исчезает. Если после блокады вводить пролактин,
привлекательпость восстанавлинается. Был сделап вывод, что выработка
феромона обусловлена высоким уровнем секреции пролактина.
Дальнейшие исследования показали, что для реакции детеныпта соб-
ственное присутствие самки в целевой камере несущественно, достаточно
оставлять ее фекалии. В естествениых ситуациях фекалии матери очень
привлекательны для детенышей. Как мы уже говорили, крысят пе только
привлекает их запах, но они чаето поедают их (КИрайтеК её а1., 1983).
Таким образом, был сделан вывод, что «материнский феромон» — хими-
ческое вещество, содержащееся в испражнениях самки.
Поиек этого вещества привлек знимапие исследователей к факту,
что как объем, так и состав фекальных массе самки, когда крысята дости-
гают двухнедельного возраста, сильно изменяются. Вес фекальных масс
в этот период в 2—3 раза превытиает таковой девственпых самок. Соот-
ветственно увеличивается и потребление ими кормов. В фекалиях резко
повышается содержание летучих компонентов (оцениваемое по соотно-
шению весов свежих и высушенных экскрементов), они светлее обычных,
сильнее пахнут и менее структурированы (Геоп, 1974; Цеоп, Мо|Нз, 1971).
На этом этапе пути двух исследователей, открывших «материнский феро-
моп», разошлись. Леон предположил, что основное отличие фекалий ма-
тери состоит в присутствии особой ботатой микрофлорой субстанции, на-
званной «цекотрофом» и вырабатываемой слепой кишкой у всех особей
вврослых крыс. Автор предполагает, что, помимо обычных темноократен-
ных и сформированных в бульбочки фекалий, взрослые животные экс-
кретируют и цекотроф, но сразу же после выделения из анального от-
верстия поедают его. У самки, имеющей двухнедельных детспышей, ко-
личество испражнений такого тина пастолько увеличивается, что значи-
тельная часть их остается несъеденной. Таким образом, именно цекотроф
матери, а не ее обычные фекалии привлекательны для детенышей. Кроме
того, Леоном было показано, что детенышей привлекают фекальные мас-
сы, взятые из слепой кишки любой взрослой крысы. Слепая кишка — ме-
сто усиленного размножения бактерий. Если ингибировать кинечную
флору введением антибиотика, то содержимое слепой кишки и фекалии
самки теряют свою привлекательпость для детеньшией (Теоп, 4994).
Поведение
274
Автор сделал вывод, что цекотрофный компонент фекалий определяет
их запаховую привлекательность для крысят. Можно предположить, что,
поедая экскременты матери, крысята получают бактериальную флору ее
кишечника, специфическую для предпочитасмых ею кормов. Поскольку
у крысят вырабатывается предпочтение тех же кормов, что и матери,
то специализация микрофлоры может существенно блатоприятствовать
оптимальному использованию кормов.
Однако Мольтц
— второй автор открытия «материнского феромонах —
не согласился с предположением о наличии цекотрофа, так как прямые
доказательства существования двух форм экскрементов у крыс (обычных
и цекотрофиых) отсутствуют. После детального исследования механизмов
продукции экскрементов лактирующей самкой он предложил другое объ-
яспение их привлекательности для крысят. Увеличение уровня пролакти-
на в крови лактирующей самки приводит к постепенному увеличению
концентрации этого гормона в печени (Ёее еф а1., 1982), а это, в свою
очередь, приводит к увеличению синтеза печенью основной желчной—
холиевой кислоты. В слепой кишке крыс под воздействием определенных
бактерий из холиевой кислоты синтезируется вторичная желчная— де-
оксихолиеная кислота. Потребность В ней и заставляет крысят поедать
экскременты матери, а привлекает их феромон-содержащий компонент °
фекалий — производное деоксихолиевой кислоты.
В болышой серии экспериментов было показано, что вплоть до 28-
дневного возраста крысят уровень секреции деоксихолиевой кислоты у
них значительно ниже, чем у взрослых крыс. С другой стороны, в фека-
лиях самки-матери уровель деоксихолневой кислоты значительно выше,
чем у нерожавптих самок (КПрабтсК её а1., 1983). Причины недостаточ-
ной продукции деоксихолиевой кислоты крысятами неяспы.
Потребность в деоксихолиевой кислоте может объясняться по крайней
мере двумя основными причинами. Первая— повышение иммупозащиты
у крысят, вторая
— мислинизация мозга. Показано, что среди крысят,
лишенных доступа к экекрементам матери и подвергшихся холодовому
стрессу, увеличивается гибель от острого энтерита. Этот эффект пропада-
ет, если к их корму добавить деоксихоливвую кислоту. Иредполагается,
что опа инактивирует эпдотоксины, участвующие в патотепезе острых
энтеритов, т. е. обусловливает «физико-химическую» защиту, расщепляя
токсичные полимеры, производимые такими микрооргапизмами, как
Езовет1сШа соц, на более мелкие нетоксичные структурные элементы (Гее,
Мо, 19845).
При сравнении крысят, имеющих доступ к экскрементам матери и
лиленных его, обнаружилось, что у последних уровни триглицеридов и
свободных жирных кислот в плазме крови значительно ниже. У этих же
крысят. меньше масса миелина относительно массы мозга и ниже кон-
` центрация фосфолипидов в миелине, чем у коптрольтой групны. При до-
бавлении в корм деоксихолиевой кислоты все перечисленные эффекты
пропадают. Предполатается, что деоксихолиевая кислота способствует
абсорбции липидов с длинными цепями, которые необходимы для разви-
тия миелиновых структур мозга (Тее, Мол, 1984а, Ъ).
272 Глава четвертая
Следовательно, в период перехода крысят на твердые корма происхо-
дит согласованная перестройка физиологии и поведепия крысят и мате-
ри. В фекалиях матери появляется запаховый агент, привлекающий дете-
нышей 2—4-недельного возраста.
|
Гормональная перестройка организма матери приводит к резкому уве-
личению в ее экскрементах деоксихолиевой кислоты. Капрофатия ликви-
дирует нехватку в организме крысенка этого вещества, повышающего им-
мунпозащиту и способствующего процессам миелинизации мозга.
Кроме того, можно предположить, что фекалии матери обогащены и
кишечной микрофлорой, специфичной для предпочитаемых ею кормов.
Таким образом, капрофагия способствует физиологической пищевой спе-
циализации крысят.
Общие неспецифические влижния натери
на физиологию и поведение детенышей
Существует целый ряд четко выявляемых изменений в поведении и фи-
зиологии крысят, обусловливающие факторы которых пока пеизвестны,
хотя точно установлено, что они определяются матерью.
Факты такого рода были собраны в основном с помощью двух экспе-
риментальных подходов. Первый состоит в экспериментальных изменени-
ях поведения и физиологии матери и носледующей оценке следствий
этого воздействия на какие-либо характеристики крысят. В этом случае из-
вестно экспериментальное воздействие на мать и результирующие изме-
нения у крысят, но не известно, какие именно факторы имели решающее
значение. Второй подход оспован на лишении детенышей возможности
общения с матерью и на оценке последствий такой депривации. И здесь
мы сталкиваемся с трудностями в идептификации факторов, вызываю-
щих изменения в поведении крысенка.
Первый подход был наиболее распространен в 60-е годы и использо-
вался в осповцом для оцепки передачи эффектов от стресс-факторов. Так,
было показано, что если самку подвергать стрессу в период беременности
или даже перед копуляцией, то это оказывает долговременное влияние па
ее потомство. Это влияние оценивали в известном тесте «открытого
поля», измеряя длину пробежек крыс в пустой камере, число оставлен-
ных ими фекалий и еще ряд параметров. Характеристика, выводимая из
оценки, получила название «эмоциональности». Оказалось, что эмоцио-
нальность крыс, матери которых были стрессированы, статистически до-
стоверно отличается от таковой у интактных и тот же эффект можно об-
наружить через поколение, т. е. у внуков стрессированных самок (Пеп-
пепфега, ВозепЪега, 1967; Виззе], 1971; и др.). Аналогичные феномены
можно получить и при использовании других экспериментальных звоздей-
ствий. Например, если детеныша в период молочного вскармливания под-
вергать различным «стимулирующим процедурам» (взятие в руки, легкие
удары электрическим током, поглаживание) (см. например: Геуше,
1969), то это воздействие оказывает существенное влияние на ето пове-
дение (оцененное в «открытом поле») и физиологические (стресс) реак-
=
Поведение
273
ции во взрослом состоянии (ПеппепЬего вф а1., 1969). Оказалось, что в
этом случае важны не только (а возможно, и не столько) стимуляция де-
тенышта эксперимептатором, но и воздействие на пего самки после воз-
вращения в гнездо. Этот «второй период стимулнции», в котором самка
интенсивно вылизывает, обнюхивает и таскает детеныша, может оказы-
вать долговременное воздействие на его поведение (Е еБаг4з, 1966; Ваг-
пей, Виги, 1967). Поведение матери после возвращения детеньшпа в гпез-
до зависит от степени предшествующей стимуляции детеныша, например
от стрессирующей силы воздействия (отсаживание в чистую клетку, взя-
тие в руки, легкий ток) (Меп4оза её а1., 1980). Возможно, интенсиь-
ность реакций матери определяется ультразвуковыми криками воззра-
щенных детенышей. Во всяком случае, интенсивность их криков зависит
от силы предшествующего стресса, а эта иптенеивность положительно
коррелирует с усилением материнских реакций к стрессировапиому дете-
пышу (Ве её а]., 1974).
Второй экспериментальный подход основан на использовании мето-
дов депривации
— разлучения самки с детенышами и оценки влияния
этой процедуры на различные физиологические и поведенческие харак-
теристики детенышей. Известно довольно большое количество феноменов
такого рода, но мы остановимся лишь па некоторых.
Если 15-дневных крысят отделить от матери, то у них резко повыша-
ется риск заболевания язвой желудка. На первый взгляд кажется, что
«критическим» фактором в данном случае является преждевременное
(па 1—2 недели раньше положенного срока) окончание молочного вскар-
мливания. Однако если крысят пачиная с 15-дневного возраста содержать
с нелактирующей самкой, проявляющей материнское поведение, риск за-
болевания существенно уменьшается. Был сделан вывод, что в данном
случае важны сами взаимодействия между матерью и детенышем, а не
снабжение детенышей молоком (Аскегтап её а|., 1975).
Сразу ке после отделения от самки у двухнедельных крысят изменя-
ются гормональный профиль и котщентрация энзимов мозга, что важно
для нормального синтеза протеинов (ВиШег её а1., 1978; Коби её а1.,
1978). Если поеле отделения от матери детенышей содержать с самкой,
У которой перевязаны молочные протоки, то этих отклонений не будет,
по если вместо этого детенышам предоставить возможность кормиться от
обездвиженной самки, нарушения развития происходят, Таким образом,
как и в предыдущем случае, оказываются важными какие-то аспекты
взаимодействия с активной матерью.
У отделепных от матери 15-дневных крысят происходит резкое замед-
ление ритма сердечных сокращений (Но{ег, 1973). Такое измепение со-
храняется, если детенышей содержать с нелантирующей самкой, и ока-
‘залось что оно происходит из-за парушения ритма кормления молоком.
В этих экспериментах детенышей кормили каждые 4 ч, внодя молоко
непосредотвенно в желудок в количестве, достаточном для нормального
увеличения массы тела, по частота сердечных сокращений все равно
уменьшалась. Если же вводить молоко в том же временнбмы режиме,
272 Глава четвертая
Следовательно, в период перехода крысят на твердые корма происхо-
дит согласованная перестройка физиологии и поведепия крысят и мате-
ри. В фекалиях матери появляется запаховый агент, привлекающий дете-
нышей 2—4-недельного возраста.
|
Гормональная перестройка организма матери приводит к резкому уве-
личению в ее экскрементах деоксихолиевой кислоты. Капрофатия ликви-
дирует нехватку в организме крысенка этого вещества, повышающего им-
мунпозащиту и способствующего процессам миелинизации мозга.
Кроме того, можно предположить, что фекалии матери обогащены и
кишечной микрофлорой, специфичной для предпочитаемых ею кормов.
Таким образом, капрофагия способствует физиологической пищевой спе-
циализации крысят.
Общие неспецифические влижния натери
на физиологию и поведение детенышей
Существует целый ряд четко выявляемых изменений в поведении и фи-
зиологии крысят, обусловливающие факторы которых пока пеизвестны,
хотя точно установлено, что они определяются матерью.
Факты такого рода были собраны в основном с помощью двух экспе-
риментальных подходов. Первый состоит в экспериментальных изменени-
ях поведения и физиологии матери и носледующей оценке следствий
этого воздействия на какие-либо характеристики крысят. В этом случае из-
вестно экспериментальное воздействие на мать и результирующие изме-
нения у крысят, но не известно, какие именно факторы имели решающее
значение. Второй подход оспован на лишении детенышей возможности
общения с матерью и на оценке последствий такой депривации. И здесь
мы сталкиваемся с трудностями в идептификации факторов, вызываю-
щих изменения в поведении крысенка.
Первый подход был наиболее распространен в 60-е годы и использо-
вался в осповцом для оцепки передачи эффектов от стресс-факторов. Так,
было показано, что если самку подвергать стрессу в период беременности
или даже перед копуляцией, то это оказывает долговременное влияние па
ее потомство. Это влияние оценивали в известном тесте «открытого
поля», измеряя длину пробежек крыс в пустой камере, число оставлен-
ных ими фекалий и еще ряд параметров. Характеристика, выводимая из
оценки, получила название «эмоциональности». Оказалось, что эмоцио-
нальность крыс, матери которых были стрессированы, статистически до-
стоверно отличается от таковой у интактных и тот же эффект можно об-
наружить через поколение, т. е. у внуков стрессированных самок (Пеп-
пепфега, ВозепЪега, 1967; Виззе], 1971; и др.). Аналогичные феномены
можно получить и при использовании других экспериментальных звоздей-
ствий. Например, если детеныша в период молочного вскармливания под-
вергать различным «стимулирующим процедурам» (взятие в руки, легкие
удары электрическим током, поглаживание) (см. например: Геуше,
1969), то это воздействие оказывает существенное влияние на ето пове-
дение (оцененное в «открытом поле») и физиологические (стресс) реак-
=
Поведение
273
ции во взрослом состоянии (ПеппепЬего вф а1., 1969). Оказалось, что в
этом случае важны не только (а возможно, и не столько) стимуляция де-
тенышта эксперимептатором, но и воздействие на пего самки после воз-
вращения в гнездо. Этот «второй период стимулнции», в котором самка
интенсивно вылизывает, обнюхивает и таскает детеныша, может оказы-
вать долговременное воздействие на его поведение (Е еБаг4з, 1966; Ваг-
пей, Виги, 1967). Поведение матери после возвращения детеньшпа в гпез-
до зависит от степени предшествующей стимуляции детеныша, например
от стрессирующей силы воздействия (отсаживание в чистую клетку, взя-
тие в руки, легкий ток) (Меп4оза её а1., 1980). Возможно, интенсиь-
ность реакций матери определяется ультразвуковыми криками воззра-
щенных детенышей. Во всяком случае, интенсивность их криков зависит
от силы предшествующего стресса, а эта иптенеивность положительно
коррелирует с усилением материнских реакций к стрессировапиому дете-
пышу (Ве её а]., 1974).
Второй экспериментальный подход основан на использовании мето-
дов депривации
— разлучения самки с детенышами и оценки влияния
этой процедуры на различные физиологические и поведенческие харак-
теристики детенышей. Известно довольно большое количество феноменов
такого рода, но мы остановимся лишь па некоторых.
Если 15-дневных крысят отделить от матери, то у них резко повыша-
ется риск заболевания язвой желудка. На первый взгляд кажется, что
«критическим» фактором в данном случае является преждевременное
(па 1—2 недели раньше положенного срока) окончание молочного вскар-
мливания. Однако если крысят пачиная с 15-дневного возраста содержать
с нелактирующей самкой, проявляющей материнское поведение, риск за-
болевания существенно уменьшается. Был сделан вывод, что в данном
случае важны сами взаимодействия между матерью и детенышем, а не
снабжение детенышей молоком (Аскегтап её а|., 1975).
Сразу ке после отделения от самки у двухнедельных крысят изменя-
ются гормональный профиль и котщентрация энзимов мозга, что важно
для нормального синтеза протеинов (ВиШег её а1., 1978; Коби её а1.,
1978). Если поеле отделения от матери детенышей содержать с самкой,
У которой перевязаны молочные протоки, то этих отклонений не будет,
по если вместо этого детенышам предоставить возможность кормиться от
обездвиженной самки, нарушения развития происходят, Таким образом,
как и в предыдущем случае, оказываются важными какие-то аспекты
взаимодействия с активной матерью.
У отделепных от матери 15-дневных крысят происходит резкое замед-
ление ритма сердечных сокращений (Но{ег, 1973). Такое измепение со-
храняется, если детенышей содержать с нелантирующей самкой, и ока-
‘залось что оно происходит из-за парушения ритма кормления молоком.
В этих экспериментах детенышей кормили каждые 4 ч, внодя молоко
непосредотвенно в желудок в количестве, достаточном для нормального
увеличения массы тела, по частота сердечных сокращений все равно
уменьшалась. Если же вводить молоко в том же временнбмы режиме,
274
Глава четвертая
как это делает самка, то изменений в работе сердца не наблюдается
(Нотег, 4973; Ноег, \Уешег, 1974; Нофег, Зват, 1982).
Через 18 ч после отделения 15-дневных крысят тестами «открытого
поля» обнаруживаются существенные изменения поведения, парушается
режим сна—бодротвования. Оказалось, что эти нарутения можно предо-
твратить, подвергая детенышей каждый час на 15 мин (что примерно
соответствует режиму приходов самки в гнездо) легкой тактильной сти-
муляции или стимуляции слабым электрическим током. Таким образом,
в дапиом случае для поддержания нормального функционирования фи-
зиологических и поведенческих систем детеныша важны сам режим, а не
форма стимуляции (НоЁег, 1984). Мать выступает внешним регулятором
работы этих систем.
Общее влияние матери па активацию физиологических и поведенче-
ских систем детеныша выявляют и опыты с «перекрестным воспитанием».
В этом случае разлученных с матерью крысят дают па воспитание дру-
гой кормящей самке, которая отличается от матери по некоторому набору
характеристик. Так, в линии УЯщаг есть две подлинии. Представители
одпой из них неизменно атакуют и убивают мышей, у представителей
другой этой реакции нет. Реакция на мынтей передается по наследетву.
Одпако когда детеныши одпой сублинии с рождения воспитываются мате-
рями другой, то они приобретают черты «воспитателей». Так, детеныши,
принадлежащие к подлинии «неубийц», но воспитанные приемными ма-
терями-«убийцами», в 30-дневном возрасте в тестовых ситуациях убивают
мышей. Интересно, что их реакция определяется пе прямым обучением:
крысята убивают мышей, даже если опи до этого не имели возможности
наблюдать, как это делают взрослые крысы и сами пикогда не контак-
тировали с мышами. Возможность наблюдения за реакциями взрослых
крыс, убивающих мышей, еще более усилизает эти реакции у детенышей.
Авторы показали, что в данном случае происходит не просто выработка
специфической реакции убийства, а измепение широкого диапазона пове-
денческих и физиологических характеристик, связь между которыми пока
недостаточно ясна. Реакция убийства мышей сочетается с более высоким
уровнем неспецифичоской возбудимости, более интенсивными действия-
ми при драках с сородичами, измспением в предкопуляторном поведепии,
ббльшим потреблением воды и солевых растворов (Еапаега, Моуасоча,
1974).
,
Влияние матери оказывается существенным и при межвидовом пере-
крестном воспитании. Усыновить новорожденного мышонка может и не-
лактирующая самка крысы. Даже если такая приемная мать будет про-
сто находиться в одной клетке с мышью-матерью, ее присутствие окажет
существенный и долговремеппый эффект на поведение мышах. Став
взрослыми, такие мыши менее активны в «открытом поле», менее агрес-
сивны, у них менее выражен адренокортикальный ответ на новизпу по
сравнению с мышами, выращенными приемпыми матерями своего вида
(еппенЪегр её а1., 1964; 1969; Возепфеге еЁ а1., 1970). Этот эффект пере-
дастся пе через молоко, так как сохрапяется в опытах с нелактирующи-
ми крысами, и, видимо, определяется не ольфакторными сигиалами, так
Поведение
275
как если поведенческие взаимодействия крысы с мышатами предотвра-
тить проволочной сеткой, то присутствие крысы перестает оказывать
влияние на детенышей. Нредполатается, что влияние крысы на мышопка
осуществляется благодаря их активным взаимодействиям (чисткам, иг-
рам и пр.), хотя конкретный механизм воздействия неизвестен.
Таким образом, существуют четко выявляемые измепепия в поведо-
пии и физиологии крысят, вызываемые матерью. 'Гочные факторы, обус-
ловливающие эти взаимодействия, неизвестны.
#& жж
Мы дали обзор феноменов связи между матерью и детеныттем, наиболее
интенсивно изучаемых у лабораторных крыс. Даже если судить по цити-
рованным работам, использованным в разделах этой главы, становится
ясно, что описанные направления исследований разрабатывались отиоси-
тельно независимо друг от друга разными научными группами. Интегра-
ция этих данных есть дело ближайшего будущего, так как очевидно, что
большинетво исследованпых феномепов имеет общие нейроэндокринные
механизмы. Прогресс, достигнутый в последние годы в эндокринологии и
нейроэтологии,— залог успехов и в рассмотренпой нами области исследо-
вапий. Дапные экспериментаторов— физиологов, эндокринологов, психо-
логов— несомненно создают фундамент для будущих зоологических ис-
следовапий поведенческой и экологической специфики связи между ма-
терью и детенышами у серой крысы.
Социальная организация
Серые крысы несомпеиио относятся к высокосоциальным видам, для ко-
торых характерен групповой образ жизни как на самых ранних этапах
жизни, так и во взрослом возрасте пезависимо от сезона тода и разно-
образия мест обитания. Известно, что взаимодействие отдельных живот-
‚ных и их группировок является одним из эффективных механизмов.
поддерживающих устойчивость популяционной структуры и ее интегри-
рованную адаптивную реакцию в ответ на влияние факторов среды
{\УУуппе-Е@\тагаз, 1965; Стоок, 1970; Шилов, 1977; КгеЬз, 1978; У Пвоп,
1982; Панов, 1983). Поэтому изучение социального поведения как средст-
за осуществления таких взаимодействий на основе определенной терри-
ториальной структуры представляется важным не только в теоретическом
плане, но и для соверптенствования контроля за популяциями экономиче-
ски важных видов. К сожалению, в этом плане серая крыса не может
быть отнесена к хорошо исследованному объекту, что отчасти обусловле-
но ее преимущественно почным и сумеречным образом жизни, скрыт-
ностью и осторожностью, обитанием в малодоступных для наблюдений
местах. Исходно поведение серых крыс изучали в ограниченных, контро-
` лируемых условиях. Мы постараемся представить социальную организа-
цию естественных поселений ® серых крыс, осповываясь на эксперимен-
тальных и пемпогочисленных данных полевых исследований.
1 Под социальной организацией мы понимаем систему информационных связей меж-
ду особями, входящими в состав конкретной популяции и отдельных внутрипо-
274
Глава четвертая
как это делает самка, то изменений в работе сердца не наблюдается
(Нотег, 4973; Ноег, \Уешег, 1974; Нофег, Зват, 1982).
Через 18 ч после отделения 15-дневных крысят тестами «открытого
поля» обнаруживаются существенные изменения поведения, парушается
режим сна—бодротвования. Оказалось, что эти нарутения можно предо-
твратить, подвергая детенышей каждый час на 15 мин (что примерно
соответствует режиму приходов самки в гнездо) легкой тактильной сти-
муляции или стимуляции слабым электрическим током. Таким образом,
в дапиом случае для поддержания нормального функционирования фи-
зиологических и поведенческих систем детеныша важны сам режим, а не
форма стимуляции (НоЁег, 1984). Мать выступает внешним регулятором
работы этих систем.
Общее влияние матери па активацию физиологических и поведенче-
ских систем детеныша выявляют и опыты с «перекрестным воспитанием».
В этом случае разлученных с матерью крысят дают па воспитание дру-
гой кормящей самке, которая отличается от матери по некоторому набору
характеристик. Так, в линии УЯщаг есть две подлинии. Представители
одпой из них неизменно атакуют и убивают мышей, у представителей
другой этой реакции нет. Реакция на мынтей передается по наследетву.
Одпако когда детеныши одпой сублинии с рождения воспитываются мате-
рями другой, то они приобретают черты «воспитателей». Так, детеныши,
принадлежащие к подлинии «неубийц», но воспитанные приемными ма-
терями-«убийцами», в 30-дневном возрасте в тестовых ситуациях убивают
мышей. Интересно, что их реакция определяется пе прямым обучением:
крысята убивают мышей, даже если опи до этого не имели возможности
наблюдать, как это делают взрослые крысы и сами пикогда не контак-
тировали с мышами. Возможность наблюдения за реакциями взрослых
крыс, убивающих мышей, еще более усилизает эти реакции у детенышей.
Авторы показали, что в данном случае происходит не просто выработка
специфической реакции убийства, а измепение широкого диапазона пове-
денческих и физиологических характеристик, связь между которыми пока
недостаточно ясна. Реакция убийства мышей сочетается с более высоким
уровнем неспецифичоской возбудимости, более интенсивными действия-
ми при драках с сородичами, измспением в предкопуляторном поведепии,
ббльшим потреблением воды и солевых растворов (Еапаега, Моуасоча,
1974).
,
Влияние матери оказывается существенным и при межвидовом пере-
крестном воспитании. Усыновить новорожденного мышонка может и не-
лактирующая самка крысы. Даже если такая приемная мать будет про-
сто находиться в одной клетке с мышью-матерью, ее присутствие окажет
существенный и долговремеппый эффект на поведение мышах. Став
взрослыми, такие мыши менее активны в «открытом поле», менее агрес-
сивны, у них менее выражен адренокортикальный ответ на новизпу по
сравнению с мышами, выращенными приемпыми матерями своего вида
(еппенЪегр её а1., 1964; 1969; Возепфеге еЁ а1., 1970). Этот эффект пере-
дастся пе через молоко, так как сохрапяется в опытах с нелактирующи-
ми крысами, и, видимо, определяется не ольфакторными сигиалами, так
Поведение
275
как если поведенческие взаимодействия крысы с мышатами предотвра-
тить проволочной сеткой, то присутствие крысы перестает оказывать
влияние на детенышей. Нредполатается, что влияние крысы на мышопка
осуществляется благодаря их активным взаимодействиям (чисткам, иг-
рам и пр.), хотя конкретный механизм воздействия неизвестен.
Таким образом, существуют четко выявляемые измепепия в поведо-
пии и физиологии крысят, вызываемые матерью. 'Гочные факторы, обус-
ловливающие эти взаимодействия, неизвестны.
#& жж
Мы дали обзор феноменов связи между матерью и детеныттем, наиболее
интенсивно изучаемых у лабораторных крыс. Даже если судить по цити-
рованным работам, использованным в разделах этой главы, становится
ясно, что описанные направления исследований разрабатывались отиоси-
тельно независимо друг от друга разными научными группами. Интегра-
ция этих данных есть дело ближайшего будущего, так как очевидно, что
большинетво исследованпых феномепов имеет общие нейроэндокринные
механизмы. Прогресс, достигнутый в последние годы в эндокринологии и
нейроэтологии,— залог успехов и в рассмотренпой нами области исследо-
вапий. Дапные экспериментаторов— физиологов, эндокринологов, психо-
логов— несомненно создают фундамент для будущих зоологических ис-
следовапий поведенческой и экологической специфики связи между ма-
терью и детенышами у серой крысы.
Социальная организация
Серые крысы несомпеиио относятся к высокосоциальным видам, для ко-
торых характерен групповой образ жизни как на самых ранних этапах
жизни, так и во взрослом возрасте пезависимо от сезона тода и разно-
образия мест обитания. Известно, что взаимодействие отдельных живот-
‚ных и их группировок является одним из эффективных механизмов.
поддерживающих устойчивость популяционной структуры и ее интегри-
рованную адаптивную реакцию в ответ на влияние факторов среды
{\УУуппе-Е@\тагаз, 1965; Стоок, 1970; Шилов, 1977; КгеЬз, 1978; У Пвоп,
1982; Панов, 1983). Поэтому изучение социального поведения как средст-
за осуществления таких взаимодействий на основе определенной терри-
ториальной структуры представляется важным не только в теоретическом
плане, но и для соверптенствования контроля за популяциями экономиче-
ски важных видов. К сожалению, в этом плане серая крыса не может
быть отнесена к хорошо исследованному объекту, что отчасти обусловле-
но ее преимущественно почным и сумеречным образом жизни, скрыт-
ностью и осторожностью, обитанием в малодоступных для наблюдений
местах. Исходно поведение серых крыс изучали в ограниченных, контро-
` лируемых условиях. Мы постараемся представить социальную организа-
цию естественных поселений ® серых крыс, осповываясь на эксперимен-
тальных и пемпогочисленных данных полевых исследований.
1 Под социальной организацией мы понимаем систему информационных связей меж-
ду особями, входящими в состав конкретной популяции и отдельных внутрипо-
276
Глава четвертая
Вопрос о социальной организации у серых крыс стал предметом спе-
циального изучения вначале благодаря блестящей работе Колхауна.
В конце 40-х годов оп соорудил большую выгородку площадью около
1000 м? со всеми подходящими для животных условиями, имитируя хо-
зяйственное подворье в г. Балтиморе, и выпустил в нее 10 разнополых
пезпакомых между собой взрослых крыс (рис. 75). В таких приближеп-
ных к естоственным условиях этот исследователь наблюдал развитие
«колонии» в течение 27 месяцев. Результаты этих наблюдений изложепы
в ряде публикаций (СаШФони. 1948а, Ь, 1949) и подробно в классической
монографии (СаШФоти, 1963).
|
Между исходными животными, выпущенными в вытородку, в резуль-
тате агрессивных столкновений возникли разобщепные во времени и
пространстве соподчиненные группировки, которые существовали в тече-
ние всего периода наблюдений. Наиболее отчетливо территориальная
структура колонии проявилась з конце эксперимента. Несмотря на ис-
кусственность условий, которые наверияка наложили отпечаток на полу-
ченные данные, Колхаун полагал, что аналогичным образом организова-
ны и естественные поселения серых крыс. Однако подтверждения этому
были получены лишь в конце 70-х— начале 80-х годов. Так, на восста-
повленных лугах в Японии посолепия крыс с мая по декабрь состояли из
отдельных группировок (Ттопй, 1979). Аналогичные результаты получе-
пы при наблюдениях за животными, обитавшими на мусорных свалках
(Гл, 1975) и на речной дамбе в пебольшом городе в Узбекистане
(Квашнин, Карасева, 1983, 1985). Такая организация поселений оказа-
лась характерной не только для зверьков, обитающих в открытых биото-
пах, но и для крыс, которые живут в постройках человека, в частности
па свинофермах (Лозан, Никул, 1981; Михайленко, 1982, 1987).
Таким образом, естественные поселения серых крыс в стабильных ус-
ловиях ** представлены рядом отдельных группировок, которые в лите-
ратуре имеют различные названия [парцеллы— по Н. Н. Нзумову (1967,
41971), знутрипопуляционные труппировки— ло И. А. Шилову (1977,
1985), локальные («тотоирз») группы — по СаШоии (1963)]. В дальней-
тем для описания таких естественных объединений животных мы будем
использовать термин группировка, а для искусственных объединений—
труппа.
4
пуляционных группировок. Таким образом, территориальное размещение особей
отражает форму организации таких связей, а социальное поведепис — средства и
способ их обуществления. Понятие популяция у серых крыс, чрезвычайно рас-
плывчато, что отразилось, в частности, в названиях естественных скоплений этих
животных (сообщества, поселения, колонии и т. п.). Мы будем использовать тер-
мин «поселение»,
Нод стабильными условиями мы понимаем отпосительную неизменность обнов-
ных параметров среды, необходимых и достаточных для нормального существо-
вания животных. Нарушение какого-либо параметра приводит к дезорганизации
поселений и миграциям особей. Вопрос о социальной организации временных по-
селений крыс, которые возникают, например, при зимовке на рисовых полях или
возле нового, но не постоянного источника корма, остается открытым, и здесь
мы не будем его касаться.
2
>
Поведение
277
Рис. 75. Локализация аруппиро-
вок серых крыс в искусственной
колонии (по: Саоип, 1968)
Размер выгородки:
100х100 футов.
С, Ю, В, 3 — стороны горизонта,
Социальные ранги группировок:
1— высокий,
2 — средний,
3 — низкий
(в пределах границ группировок ука-
зано количество варослых самцов и
самок),
4 — ограждение из сетки © прохо-
дами,
5 — кормушка
7222
Я;
6}
ГВ
Размеры участков, которые запимают грунпировки, и характер ис.
пользования общей территории отдельными особями изучены Нока недо-
статочно полно. По имеющимся литературным данным, складывается
следующая картина. В открытых биотонах площадь группового участка
составляет 78—157 м? на восстановленных лутах (Т2аии, 1979) и 120-—
240 м° на речпой дамбе (Квашнин, Карасева, 1983, 1985), причем разме-
ры территорий, по-видимому, не зависят от плотности паселепия. Важно,
что групповые участки пе перекрывались (расстояние между ними на
речной дамбе составляло 40—70 м) как в этих условиях, так и в услови-
ях большой выгородки. Весьма пеоднозначные результаты получены при
наблюдениях за синантропными грызунами. Так, по данным А. Н. Коз-
лова (19836), площадь групповых участков в животноводческих помеще-
ниях составляет 50—80 м’, причем она также не зависит от плотности
зверьков, а сами участки изолированы друг от друга. Иные сведепия мы
находим у А. Г. Михайленко (1982, 1987). Размеры участков, занятых
групнировками крыс на свинофермах, варьировали от 430 ло 830 м’.
Участки соседпих группировок перекрывались так, что граница участка
обитания одной группировки обычно определялась границей защищае-
мой территории, составляющей 30—50% площади участка друтой.
Таким образом, имеются существенные расхождения в данных по раз-
мерам и взаимному расположению групповых участков серых крыс, 0со-
*бепно в синантропных условиях обитания.
Численный и половой состав отдельных группировок серых крыс от-
личается большим разпообразием в различных условиях обитания живот-
ных, что отражает адаптивные возможности вида (Большаков, Нубан-
цев, 1984). По мнению Цолхауна, оптимальная численность группиронок
не должна превышать 10—15 взрослых особей. В этом случае возможие
поддержание и упорядочивание информационлых связей как между чле-
нами одной группировки, так и между членами разных группировок,
276
Глава четвертая
Вопрос о социальной организации у серых крыс стал предметом спе-
циального изучения вначале благодаря блестящей работе Колхауна.
В конце 40-х годов оп соорудил большую выгородку площадью около
1000 м? со всеми подходящими для животных условиями, имитируя хо-
зяйственное подворье в г. Балтиморе, и выпустил в нее 10 разнополых
пезпакомых между собой взрослых крыс (рис. 75). В таких приближеп-
ных к естоственным условиях этот исследователь наблюдал развитие
«колонии» в течение 27 месяцев. Результаты этих наблюдений изложепы
в ряде публикаций (СаШФони. 1948а, Ь, 1949) и подробно в классической
монографии (СаШФоти, 1963).
|
Между исходными животными, выпущенными в вытородку, в резуль-
тате агрессивных столкновений возникли разобщепные во времени и
пространстве соподчиненные группировки, которые существовали в тече-
ние всего периода наблюдений. Наиболее отчетливо территориальная
структура колонии проявилась з конце эксперимента. Несмотря на ис-
кусственность условий, которые наверияка наложили отпечаток на полу-
ченные данные, Колхаун полагал, что аналогичным образом организова-
ны и естественные поселения серых крыс. Однако подтверждения этому
были получены лишь в конце 70-х— начале 80-х годов. Так, на восста-
повленных лугах в Японии посолепия крыс с мая по декабрь состояли из
отдельных группировок (Ттопй, 1979). Аналогичные результаты получе-
пы при наблюдениях за животными, обитавшими на мусорных свалках
(Гл, 1975) и на речной дамбе в пебольшом городе в Узбекистане
(Квашнин, Карасева, 1983, 1985). Такая организация поселений оказа-
лась характерной не только для зверьков, обитающих в открытых биото-
пах, но и для крыс, которые живут в постройках человека, в частности
па свинофермах (Лозан, Никул, 1981; Михайленко, 1982, 1987).
Таким образом, естественные поселения серых крыс в стабильных ус-
ловиях ** представлены рядом отдельных группировок, которые в лите-
ратуре имеют различные названия [парцеллы— по Н. Н. Нзумову (1967,
41971), знутрипопуляционные труппировки— ло И. А. Шилову (1977,
1985), локальные («тотоирз») группы — по СаШоии (1963)]. В дальней-
тем для описания таких естественных объединений животных мы будем
использовать термин группировка, а для искусственных объединений—
труппа.
4
пуляционных группировок. Таким образом, территориальное размещение особей
отражает форму организации таких связей, а социальное поведепис — средства и
способ их обуществления. Понятие популяция у серых крыс, чрезвычайно рас-
плывчато, что отразилось, в частности, в названиях естественных скоплений этих
животных (сообщества, поселения, колонии и т. п.). Мы будем использовать тер-
мин «поселение»,
Нод стабильными условиями мы понимаем отпосительную неизменность обнов-
ных параметров среды, необходимых и достаточных для нормального существо-
вания животных. Нарушение какого-либо параметра приводит к дезорганизации
поселений и миграциям особей. Вопрос о социальной организации временных по-
селений крыс, которые возникают, например, при зимовке на рисовых полях или
возле нового, но не постоянного источника корма, остается открытым, и здесь
мы не будем его касаться.
2
>
Поведение
277
Рис. 75. Локализация аруппиро-
вок серых крыс в искусственной
колонии (по: Саоип, 1968)
Размер выгородки:
100х100 футов.
С, Ю, В, 3 — стороны горизонта,
Социальные ранги группировок:
1— высокий,
2 — средний,
3 — низкий
(в пределах границ группировок ука-
зано количество варослых самцов и
самок),
4 — ограждение из сетки © прохо-
дами,
5 — кормушка
7222
Я;
6}
ГВ
Размеры участков, которые запимают грунпировки, и характер ис.
пользования общей территории отдельными особями изучены Нока недо-
статочно полно. По имеющимся литературным данным, складывается
следующая картина. В открытых биотонах площадь группового участка
составляет 78—157 м? на восстановленных лутах (Т2аии, 1979) и 120-—
240 м° на речпой дамбе (Квашнин, Карасева, 1983, 1985), причем разме-
ры территорий, по-видимому, не зависят от плотности паселепия. Важно,
что групповые участки пе перекрывались (расстояние между ними на
речной дамбе составляло 40—70 м) как в этих условиях, так и в услови-
ях большой выгородки. Весьма пеоднозначные результаты получены при
наблюдениях за синантропными грызунами. Так, по данным А. Н. Коз-
лова (19836), площадь групповых участков в животноводческих помеще-
ниях составляет 50—80 м’, причем она также не зависит от плотности
зверьков, а сами участки изолированы друг от друга. Иные сведепия мы
находим у А. Г. Михайленко (1982, 1987). Размеры участков, занятых
групнировками крыс на свинофермах, варьировали от 430 ло 830 м’.
Участки соседпих группировок перекрывались так, что граница участка
обитания одной группировки обычно определялась границей защищае-
мой территории, составляющей 30—50% площади участка друтой.
Таким образом, имеются существенные расхождения в данных по раз-
мерам и взаимному расположению групповых участков серых крыс, 0со-
*бепно в синантропных условиях обитания.
Численный и половой состав отдельных группировок серых крыс от-
личается большим разпообразием в различных условиях обитания живот-
ных, что отражает адаптивные возможности вида (Большаков, Нубан-
цев, 1984). По мнению Цолхауна, оптимальная численность группиронок
не должна превышать 10—15 взрослых особей. В этом случае возможие
поддержание и упорядочивание информационлых связей как между чле-
нами одной группировки, так и между членами разных группировок,
278 Глава четвертая
Таблица 25. Характериетика группировок серых крые, обитавших
на речной дамбе, в летние и зимние периоды года (по; Квашнин,
Карасева, 1985)
Общее чис- Количество
ло взров-
индивиду-
лых особей ально ме-
Число
на участ-
ченных
Учас-
Чис
206
Сезон работ
том. ке*
особей
выВОДКов ро ей м?
Г
©
99 [уе 9$
Лето 1982 г.
1
8
7
8
т
1
100
2
6
2
А
1
1
4,0
3
5
2
3
0
1
3,5
4
7
А
6
3
3
4,6
5
1
А
4
2
3
4,1
Всего
27192213
9
ср. 5,2
Зима 1983 г.
4
0
0
0
0
—
_
2
2
3
2
2
—
_
3
2
1
0
1
—
-
4
1
1
0
0
—
_
5
1
1
0
0
—
—
Всего
6623
-
-
* Количество немеченых особей опрецеляли по числу животных, одновременно
наблюдавшихся на поверхности дамбы, а также по внешнему виду и особен-
ностим поведения,
что обеспечивает наиболее эффективную регуляцию в пределах локаль-
ных объединений и поселения в целом (СаШоип, 1963а). Действительно,
исказапо, что количество взрослых особей, входящих в состав групииро-
вок в открытых биотопах, пе превытиает 15, причем независимо от сезо-
па года (Таша, 1975; Тао, 1979; Квашнин, Карасева, 1985; а также
ем. рис. 75 и табл. 25).
В животноводческих помещениях, однако, численность отдельных
группировок могла достигать и значительно больной величины — при вы-
сокой плотности до 50 взрослых особей, хотя наиболее часто в состав
группировок входило 10—20 животных (Лозан, Никул, 1981; Михайлен-
ко, 1987; Козлов, 19886).
Половая структура локальных группировок животных варьирует в
широких пределах. Так, для поселений крыс на свинофермах, по-види-
мому, свойственно примерное равепство самцов и самок в состане раз-
личных групцировок (Лозан, Никул, 1981). Однако, по дапным Колхауна,
наряду с группировками, в которые входило равное число разнополых
особей, были и группировки, в которых многократно преобладали либо
самки, либо самцы (в последпем случае — вплоть до полного отсутствия
Поведение
279
Таблица 26. Характеристика группировок, принадлежащих к разны
социальным рамкам, у верых крые (по: СаБоип, 1963)
.
‚
м
-
ч
Число
Масса тела (орех Среднее число ран ео х
Ранг
группиров-
ыы
ся|°° ос’ 99 ое
99 общее|на!9
Высокий
724
510
Аба
10,4
5,2
95
3,96
Средний
12 47
479
431
11,4
5,5
37
2,18
Низкий
42 18
441
419
15,4
6,4
0
0,0
самок). Аналогичпые результаты приводят и другие авторы (Квашнин,
Карасева, 1983, 1985; см. табл. 25).
Локальные группировки серых крыс неравноцеппы между собой не
только по описанным выше признакам. По наблюдениям Колхауна, от-
дельные группировки имели даже свой социальный ранг. Члены высоко-
ранговых труппировок имели преимущество в удовлетворении своих
потребностей над членами низкорантовых группировок. Члены нысоко-
ранговых группировок характеризовались и другими особенпостями —
у них была больше масса тела и выше скорость роста, на их телах было
меньше рац; в состав высокоранговых группировок входило большее чис-
ло самок, которые успешнее размпожались и выращивали детепышей
(табл. 26). К этому можно добавить, что высокоранговые группировки з9-
нимали более блатоприятные местообитания и состояли в основном из
особей, родившихся на территории данной группировки (Саотт, 1963).
Функциональная перавпоценность (в частности, по эффективпости
размножения) отдельных группировок такие была отмечена в поселении
крыс на речной дамбе (Квашнин, Карасева, 1985) и даже в условиях не-
болыних вольер среди искусственпо созданных групп (Квашнин, 1983а).
Отличительными чертами естественных труппировок и искусственных
труни, в которых самки размножались слабо или не имели выводков сов-
сем, были также отсутствие иерархической соподчипенности среди сам-
цов, отсутствие хорошо выраженной агрессивной реакции на чужих 0060-
бей, изменения в суточной активности и ряд других особенностей (см.
например: СаВотл, 1963; Квашнин и др., 1986).
Таким образом, группировки серых крысе характеризуются значитель-
ным разнообразием по многим параметрам и в первую очередь по своему
* «вкладу» во внутрипопуляционные процессы.
Взаимоотношения между членами отдельных группировок в естест-
венных условиях практически мало изучены. По наблюдениям Колхаупа,
столкновения особей, принадлежащих к группировкам разных рангов,
в любых участках выгородки, как правило, заканчивались победой живот-
ных из высокоранговых группировок. Это дало основание автору предпо-
лагать, что между разпыми группировками существуют соподчипепнные
отпошения. Такого же мнения придерживаются М. Н. Лован и В. С. Ни-
278 Глава четвертая
Таблица 25. Характериетика группировок серых крые, обитавших
на речной дамбе, в летние и зимние периоды года (по; Квашнин,
Карасева, 1985)
Общее чис- Количество
ло взров-
индивиду-
лых особей ально ме-
Число
на участ-
ченных
Учас-
Чис
206
Сезон работ
том. ке*
особей
выВОДКов ро ей м?
Г
©
99 [уе 9$
Лето 1982 г.
1
8
7
8
т
1
100
2
6
2
А
1
1
4,0
3
5
2
3
0
1
3,5
4
7
А
6
3
3
4,6
5
1
А
4
2
3
4,1
Всего
27192213
9
ср. 5,2
Зима 1983 г.
4
0
0
0
0
—
_
2
2
3
2
2
—
_
3
2
1
0
1
—
-
4
1
1
0
0
—
_
5
1
1
0
0
—
—
Всего
6623
-
-
* Количество немеченых особей опрецеляли по числу животных, одновременно
наблюдавшихся на поверхности дамбы, а также по внешнему виду и особен-
ностим поведения,
что обеспечивает наиболее эффективную регуляцию в пределах локаль-
ных объединений и поселения в целом (СаШоип, 1963а). Действительно,
исказапо, что количество взрослых особей, входящих в состав групииро-
вок в открытых биотопах, пе превытиает 15, причем независимо от сезо-
па года (Таша, 1975; Тао, 1979; Квашнин, Карасева, 1985; а также
ем. рис. 75 и табл. 25).
В животноводческих помещениях, однако, численность отдельных
группировок могла достигать и значительно больной величины — при вы-
сокой плотности до 50 взрослых особей, хотя наиболее часто в состав
группировок входило 10—20 животных (Лозан, Никул, 1981; Михайлен-
ко, 1987; Козлов, 19886).
Половая структура локальных группировок животных варьирует в
широких пределах. Так, для поселений крыс на свинофермах, по-види-
мому, свойственно примерное равепство самцов и самок в состане раз-
личных групцировок (Лозан, Никул, 1981). Однако, по дапным Колхауна,
наряду с группировками, в которые входило равное число разнополых
особей, были и группировки, в которых многократно преобладали либо
самки, либо самцы (в последпем случае — вплоть до полного отсутствия
Поведение
279
Таблица 26. Характеристика группировок, принадлежащих к разны
социальным рамкам, у верых крые (по: СаБоип, 1963)
.
‚
м
-
ч
Число
Масса тела (орех Среднее число ран ео х
Ранг
группиров-
ыы
ся|°° ос’ 99 ое
99 общее|на!9
Высокий
724
510
Аба
10,4
5,2
95
3,96
Средний
12 47
479
431
11,4
5,5
37
2,18
Низкий
42 18
441
419
15,4
6,4
0
0,0
самок). Аналогичпые результаты приводят и другие авторы (Квашнин,
Карасева, 1983, 1985; см. табл. 25).
Локальные группировки серых крыс неравноцеппы между собой не
только по описанным выше признакам. По наблюдениям Колхауна, от-
дельные группировки имели даже свой социальный ранг. Члены высоко-
ранговых труппировок имели преимущество в удовлетворении своих
потребностей над членами низкорантовых группировок. Члены нысоко-
ранговых группировок характеризовались и другими особенпостями —
у них была больше масса тела и выше скорость роста, на их телах было
меньше рац; в состав высокоранговых группировок входило большее чис-
ло самок, которые успешнее размпожались и выращивали детепышей
(табл. 26). К этому можно добавить, что высокоранговые группировки з9-
нимали более блатоприятные местообитания и состояли в основном из
особей, родившихся на территории данной группировки (Саотт, 1963).
Функциональная перавпоценность (в частности, по эффективпости
размножения) отдельных группировок такие была отмечена в поселении
крыс на речной дамбе (Квашнин, Карасева, 1985) и даже в условиях не-
болыних вольер среди искусственпо созданных групп (Квашнин, 1983а).
Отличительными чертами естественных труппировок и искусственных
труни, в которых самки размножались слабо или не имели выводков сов-
сем, были также отсутствие иерархической соподчипенности среди сам-
цов, отсутствие хорошо выраженной агрессивной реакции на чужих 0060-
бей, изменения в суточной активности и ряд других особенностей (см.
например: СаВотл, 1963; Квашнин и др., 1986).
Таким образом, группировки серых крысе характеризуются значитель-
ным разнообразием по многим параметрам и в первую очередь по своему
* «вкладу» во внутрипопуляционные процессы.
Взаимоотношения между членами отдельных группировок в естест-
венных условиях практически мало изучены. По наблюдениям Колхаупа,
столкновения особей, принадлежащих к группировкам разных рангов,
в любых участках выгородки, как правило, заканчивались победой живот-
ных из высокоранговых группировок. Это дало основание автору предпо-
лагать, что между разпыми группировками существуют соподчипепнные
отпошения. Такого же мнения придерживаются М. Н. Лован и В. С. Ни-
280 Глава четвертая
кул (1981) в отношении поселений крыс на свинофермах. Однако в
тех же условиях другой исследователь не наблюдал подобной иерархик
между разными группировками (Михайленко, 1982, 1987). Но, с его точ-
ки зрения, отдельные группировки объединены в труппировки более вы-
сокого порядка — мерусы, в пределах которых особи из разпых группи-
ровок контактируют значительно чаще, чем с представителями других
мерусов. По пашим представлениям, вопрос о взаимоотношениях между
крысами, принадлежащими к локальным группировкам, и о существова-
нии мерусов нуждается в дополнительных исследованиях, тем более что.
в открытых биотопах не отмечено межгрупповой соподчиненности (Шхи-
пи, 1979; Кваптнин, Карасева, 1983, 1985).
В этой же связи представляет интерес вопрос о характере передвиже-
пий отдельных членов группировок как в пределах групповой территории,
так и вне се. Изучению активности животных и размеров индизидуаль-
пых участков обитания было посвящено немало работ. Важность таких
данных несомненна. Одпако в отих исследованиях, как правило, пе учи-
тывался групповой образ жизни крыс, который оказывает влияние на все
стороны их жизнедеятельности и в частпости на характер использования
территории отдельными особями. Только в одной работе приводятся све-
дения о связи индивидуальной активности крыс, занимающих различное
социальное положение в системе внутригрупповых связей, с интенсив-
ностью передвижений в пределах группового участка обитания (Квали-
нин, Карасева, 1985). Судя по рис. 76, высокорантовые самцы (доминант
и субдоминант) свободно передвигались по всему участку, причем цент-
ры их активности не совпадали. Для подчиненных самцов характерно
использование лишь какой-либо части общего участка, причем их инди-
видуальные участки значительно перекрывались. То же было свойствен-
но и самкам.
Совершенно очевидно, что группировки серых крыс не являются замк-
нутыми сообществами, как считали некоторые ученые (Тогоп2, 1966).
Даже в стабильных условиях обитания у пекоторых особей время от вре-
мени наблюдаются исследовательские выходы за пределы групполой тер-
ритории, хотя для большинства зверьков, по-видимому, характерны огра-
ниченные передвижения, нс превышающие 100--150 м (Гамз св а|.,
1948; Таут, Оцу, 1978). Эту категорию животных Колхаун назвал «но-
сителями информации» (теззепеетв). Возможно, такие рейды могут про-
исходить у многих членов группировок независимо от их социальной при-
надлежности. В условиях речной дамбы подобные выходы достигали
400 м и продолжались до 15 дней (Квашнин, Карасева, 1985). Дополни-
тельная информация по этому вопросу получена и в опытах с насильст-
венным перенесением взрослых особей из одних участков обитания в дру-
гие. Установлено, что часть таких зверьков возвращалась обратно
(расстояние переноса не превышало 400 м), тогда как другие даже «при-
живались» в составе новых группировок и занимали в них различное
социальное положение. (от домипирующего до подчиненного). Наконец,
стоит отметить, что характер передвижений отдельных крыс, отличаю-
щихся индивидуальными особенностями, полом, возрастом и социальным
Поведение
281
80|
„@
с’о’
/\
й\
30Е
/
\
<< 20
&
Еп
5з
зЯ
е|
Е5$
0
|
)уИЙум
``
|
7234.в
78эп
ип
Сектор
Рис. 76. Использование участка обитания отдельными членами группировки (по:
Квашнин, Карасева, 1985)
Самцы: 1 — доминант, 2 — субдоминант, 3—6 — подченениые. Для самок (7—410) эти обозна-
чения условны. В кружках номера крысе
положением, за пределы «родных» мест и причины, их обусловливающие,
чрезвычайно важный, но слабо изученный вопрос, который тесно связан
с. процессами внутрицопуляционной регуляции, и в частности с процесса-
ми формирования новых поселений крыс.
Возникновение новых группировок в естественных условиях, по-види-
мому, происходит различными путями в зависимости от конкретной эко-
логической ситуации. По мнению Штайнигера (З4ениег, 1950) и Бар-
нетта (Вагпей, 1967, 1975), в новом месте маловероятно объединение
незнакомых друг с друтом взрослых животных, а новые поселения обра-
зуются из пескольких зпакомых и(или) родственных особей, каким-либо
образом попавших в подходящие условия. Однако наблюдения Колхауна
свидетельствуют об обратпом — мпогие группировки в колонии, особенно
«низкоранговые, сформировались из взрослых незнакомых между собой
крыс, которые по разным причипам покинули моста своего рождения
(СаШочцп, 1963). В природе наиболее вероятным представляется образова-
ние новых группировок на незанятых территориях из молодых, нолово-
зрелых особей, вынужденных покинуть места своего рождения в резуль-
тате конкуренции с более взрослыми животными (Вагпей, 1955, 1956;
СаШоип, 1963; Козлюв, 1983). Таким образом, во вновь образовавитихся
поселениях их члены мотут быть и не родственными между собой, С уче-
том того, что и в постоянные группировки периодически «внедряются»
280 Глава четвертая
кул (1981) в отношении поселений крыс на свинофермах. Однако в
тех же условиях другой исследователь не наблюдал подобной иерархик
между разными группировками (Михайленко, 1982, 1987). Но, с его точ-
ки зрения, отдельные группировки объединены в труппировки более вы-
сокого порядка — мерусы, в пределах которых особи из разпых группи-
ровок контактируют значительно чаще, чем с представителями других
мерусов. По пашим представлениям, вопрос о взаимоотношениях между
крысами, принадлежащими к локальным группировкам, и о существова-
нии мерусов нуждается в дополнительных исследованиях, тем более что.
в открытых биотопах не отмечено межгрупповой соподчиненности (Шхи-
пи, 1979; Кваптнин, Карасева, 1983, 1985).
В этой же связи представляет интерес вопрос о характере передвиже-
пий отдельных членов группировок как в пределах групповой территории,
так и вне се. Изучению активности животных и размеров индизидуаль-
пых участков обитания было посвящено немало работ. Важность таких
данных несомненна. Одпако в отих исследованиях, как правило, пе учи-
тывался групповой образ жизни крыс, который оказывает влияние на все
стороны их жизнедеятельности и в частпости на характер использования
территории отдельными особями. Только в одной работе приводятся све-
дения о связи индивидуальной активности крыс, занимающих различное
социальное положение в системе внутригрупповых связей, с интенсив-
ностью передвижений в пределах группового участка обитания (Квали-
нин, Карасева, 1985). Судя по рис. 76, высокорантовые самцы (доминант
и субдоминант) свободно передвигались по всему участку, причем цент-
ры их активности не совпадали. Для подчиненных самцов характерно
использование лишь какой-либо части общего участка, причем их инди-
видуальные участки значительно перекрывались. То же было свойствен-
но и самкам.
Совершенно очевидно, что группировки серых крыс не являются замк-
нутыми сообществами, как считали некоторые ученые (Тогоп2, 1966).
Даже в стабильных условиях обитания у пекоторых особей время от вре-
мени наблюдаются исследовательские выходы за пределы групполой тер-
ритории, хотя для большинства зверьков, по-видимому, характерны огра-
ниченные передвижения, нс превышающие 100--150 м (Гамз св а|.,
1948; Таут, Оцу, 1978). Эту категорию животных Колхаун назвал «но-
сителями информации» (теззепеетв). Возможно, такие рейды могут про-
исходить у многих членов группировок независимо от их социальной при-
надлежности. В условиях речной дамбы подобные выходы достигали
400 м и продолжались до 15 дней (Квашнин, Карасева, 1985). Дополни-
тельная информация по этому вопросу получена и в опытах с насильст-
венным перенесением взрослых особей из одних участков обитания в дру-
гие. Установлено, что часть таких зверьков возвращалась обратно
(расстояние переноса не превышало 400 м), тогда как другие даже «при-
живались» в составе новых группировок и занимали в них различное
социальное положение. (от домипирующего до подчиненного). Наконец,
стоит отметить, что характер передвижений отдельных крыс, отличаю-
щихся индивидуальными особенностями, полом, возрастом и социальным
Поведение
281
80|
„@
с’о’
/\
й\
30Е
/
\
<< 20
&
Еп
5з
зЯ
е|
Е5$
0
|
)уИЙум
``
|
7234.в
78эп
ип
Сектор
Рис. 76. Использование участка обитания отдельными членами группировки (по:
Квашнин, Карасева, 1985)
Самцы: 1 — доминант, 2 — субдоминант, 3—6 — подченениые. Для самок (7—410) эти обозна-
чения условны. В кружках номера крысе
положением, за пределы «родных» мест и причины, их обусловливающие,
чрезвычайно важный, но слабо изученный вопрос, который тесно связан
с. процессами внутрицопуляционной регуляции, и в частности с процесса-
ми формирования новых поселений крыс.
Возникновение новых группировок в естественных условиях, по-види-
мому, происходит различными путями в зависимости от конкретной эко-
логической ситуации. По мнению Штайнигера (З4ениег, 1950) и Бар-
нетта (Вагпей, 1967, 1975), в новом месте маловероятно объединение
незнакомых друг с друтом взрослых животных, а новые поселения обра-
зуются из пескольких зпакомых и(или) родственных особей, каким-либо
образом попавших в подходящие условия. Однако наблюдения Колхауна
свидетельствуют об обратпом — мпогие группировки в колонии, особенно
«низкоранговые, сформировались из взрослых незнакомых между собой
крыс, которые по разным причипам покинули моста своего рождения
(СаШочцп, 1963). В природе наиболее вероятным представляется образова-
ние новых группировок на незанятых территориях из молодых, нолово-
зрелых особей, вынужденных покинуть места своего рождения в резуль-
тате конкуренции с более взрослыми животными (Вагпей, 1955, 1956;
СаШоип, 1963; Козлюв, 1983). Таким образом, во вновь образовавитихся
поселениях их члены мотут быть и не родственными между собой, С уче-
том того, что и в постоянные группировки периодически «внедряются»
282 Глава четвертая
чужие особи (СаШойл, 1963; Квашнин, Карасева, 1985), эти процессы
представляются эффективным механизмом поддержапия генетического
полиморфизма и, следовательно, повышения адантивных возможностей
вида,
р
Но как конкретно происходит формирование новых груплировок в ес-
тественных условиях, остается не внолне ясным. Можно только отметить,
что восстановление структуры группировок после дератизации или при
изменении мест обитания, что является частным случаем формирования
труппировок, сопровождается повышением частоты агрессивных контак-
тов. Так, при определении количества ран на теле крые, которых отлови-
ли на поле со зрелым сахарным тростником (стабильная группировка)
и на поле, где недавно убрали тростник (нестабильная группировка),
оказалось, что у самцов и самок из нестабильной групиировки их было.
значительно больше (ВТапевагЯ её а1., 1985). Аналогичные наблюдения
по восстановлению поселения серых крыс в евинарнике после эффектив-
ной дератизации приводит и С. А. Квашнин (1986). Однако внолно воз-
можен и «мирный» процесс образования группировок, поскольку он за-
висит от конкретных условий среды и от того, какие именно животные:
вошли в их состав (СаШоти, 1968; Ватгпе&, 1975).
Ввиду трудностей наблюдений за серыми крысами в природе процессы
формирования группировок паиболее подробно изучены в ограниченных
условиях. Штайнигер (З4епиюег, 1950) выпускал несколько пар взрослых
пезнакомых между собой животпых в вольеру площадью 64 м? которая,
по его мнению, соответствовала минимуму «естественного жизненного:
пространства» у крыс. Вначале образовывалась пара (самец и самка),
которая начинала преследовать других зверьков вплоть до их гибели.
Только после этого у них появлялись выводки и формировалось сооблще-
ство, члены которого относились друг к другу миролюбиво. Этот иссле-
дователь считает, что крысы, населяющие одно местообитание, принадле-
жат к одной семье.
-
В других работах было показано, что при ссаживании незнакомых
друг с другом взрослых диких крыс обоего пола в небольших вольсрах
(площадью 1—6 м”) основными формами контактов между ними были в
большинстве опытов агонистические взаимодействия (Вагпей, 1956,
1958а,5; 1964, 1975; Квашнин, Суров, 1976; Квашнин, 1975, 4978, 1983).
В результате таких взаимодействий между самцами устанавливались от-
пошения домипирования-подчинения, а весь процессе формирования
иерархической структуры группы не превышал 5—6 дней и характери-
зовался резким снижением частоты агрессивпых контактов к концу этого-
периода. В таких группах агрессия доминантов была столь сильно выра-
жена, что приводила, как правило, к гибели наиболее слабых самцов.
В первые дни жизни таких групи взаимоотношения между самками носи-
ли в целом нейтральный характер. Лишь у некоторых из них в этот
период проявлялась агрессивпость к другим членам группы, но она воет-
да была менее выражена, чем у самцов, и вскоре пропадала.
В тех случаях, когда в вольеры помещали знакомых друг с другом
взрослых особей (выловленных в одном естественном местообитании или.
Поведение
283
выросших вместе в лаборатории), агрессивные контакты между вамцами
‘были выражены в целом мпого слабее, а между самками отсутствовали
полностью (их взаимоотношения были миролюбивыми).
В нодобных опытах инотда возникали группы (причем независимо от
того, знакомые или незнакомые между собой зверьки входили в их оо-
став), в которых взаимоотноптения между самцами не носили характер
соподчинения и характеризовались отсутствием агонистических форм но-
ведения (Вагпей, 1958а, Ь; 1975; Квашнин, 1983). В таких группах не
‘формировалась устойчивая структура отношений между животными.
Таким образом, у серых крыс как в искусственных, так и в естест-
веппых условиях наблюдаются ‘и нестабильные объединения особей. Ме-
хапизмы их образования наиболее подробно изучены в лабораторных
условиях на другом синантропе — домовой мьгтти (Каменов, 1973; Маслов
и др., 1974; Шилов, 1977). В этих работах было показано, что формиро-
вапие устойчивой структуры внутригрупповых отношений возможно
только при достаточной степени физиологической и поведенческой разно-
качественности взрослых животных (основанной в первую очередь на
уровне агрессивности и типе нервной системы отдельных особей), объ-
единенных на новой территории. Очевидно, что нестабильные объедине-
ПИЯ серых крыс в вольерных условиях возникли за счет случайного под-
‘бора зверьков с низким уровпем ипдивидуальной агрессивности и слабым
типом нервной системы, тотда как в природе их происхождение носит
«естественный» характер.
Наиболее подробно процессы формировапия социальной структуры ис-
кусственных трупп описаны для крыс, которые являлись потомками
пасюков в течение нескольких поколений разводимых в лаборатории и
отбиравшихся по признаку пониженной агрессивности к человеку (Со-
колов и др., 1977; Ляпунова, 1980; Соколов, Ляпунова, 1982). На основв
значительного материала в процессе формирования трупп, образованных
из взрослых пезпакомых между собой животных, было выделено два по-
«ледовательных этапа — период становления иерархической структуры и
период ее стабилизации.
В период становления социальной структуры между зверьками на-
блюдали разной степени выраженности агрессивные столкновения, в ре-
зультате которых каждый самец занимал определенное положение в ие-
рархии. Для второго периода
— периода стабилизации —была характер-
на выработка определенного стереотипа поведения всеми самцами.
Если время до появления первой агрессии пе превышало 3 ч, то в боль-
лиинстве групп (40 из 50) продолжительность периода становления была
не больше 1—2 сут, а периода стабилизации — 4 суток (Соколов, Ляпу-
пова, 1982). В таких группах социальная структура оставалась стабиль-
ной на протяжении длительного времени. В группах, где период с момен-
та запуска на территорию до появления агрессии составлял от 3 до 24 ч,
‘продолжительнссть становления социальной структуры растягивалась до
1—2 недель, а иногда иерархическая структура пе формировалась совсем.
Социальная структура в стабильных группах поддерживалась за’ счет оп-
‘'ределенпых, характеризующихся слабой степенью агрессивности (часто
282 Глава четвертая
чужие особи (СаШойл, 1963; Квашнин, Карасева, 1985), эти процессы
представляются эффективным механизмом поддержапия генетического
полиморфизма и, следовательно, повышения адантивных возможностей
вида,
р
Но как конкретно происходит формирование новых груплировок в ес-
тественных условиях, остается не внолне ясным. Можно только отметить,
что восстановление структуры группировок после дератизации или при
изменении мест обитания, что является частным случаем формирования
труппировок, сопровождается повышением частоты агрессивных контак-
тов. Так, при определении количества ран на теле крые, которых отлови-
ли на поле со зрелым сахарным тростником (стабильная группировка)
и на поле, где недавно убрали тростник (нестабильная группировка),
оказалось, что у самцов и самок из нестабильной групиировки их было.
значительно больше (ВТапевагЯ её а1., 1985). Аналогичные наблюдения
по восстановлению поселения серых крыс в евинарнике после эффектив-
ной дератизации приводит и С. А. Квашнин (1986). Однако внолно воз-
можен и «мирный» процесс образования группировок, поскольку он за-
висит от конкретных условий среды и от того, какие именно животные:
вошли в их состав (СаШоти, 1968; Ватгпе&, 1975).
Ввиду трудностей наблюдений за серыми крысами в природе процессы
формирования группировок паиболее подробно изучены в ограниченных
условиях. Штайнигер (З4епиюег, 1950) выпускал несколько пар взрослых
пезнакомых между собой животпых в вольеру площадью 64 м? которая,
по его мнению, соответствовала минимуму «естественного жизненного:
пространства» у крыс. Вначале образовывалась пара (самец и самка),
которая начинала преследовать других зверьков вплоть до их гибели.
Только после этого у них появлялись выводки и формировалось сооблще-
ство, члены которого относились друг к другу миролюбиво. Этот иссле-
дователь считает, что крысы, населяющие одно местообитание, принадле-
жат к одной семье.
-
В других работах было показано, что при ссаживании незнакомых
друг с другом взрослых диких крыс обоего пола в небольших вольсрах
(площадью 1—6 м”) основными формами контактов между ними были в
большинстве опытов агонистические взаимодействия (Вагпей, 1956,
1958а,5; 1964, 1975; Квашнин, Суров, 1976; Квашнин, 1975, 4978, 1983).
В результате таких взаимодействий между самцами устанавливались от-
пошения домипирования-подчинения, а весь процессе формирования
иерархической структуры группы не превышал 5—6 дней и характери-
зовался резким снижением частоты агрессивпых контактов к концу этого-
периода. В таких группах агрессия доминантов была столь сильно выра-
жена, что приводила, как правило, к гибели наиболее слабых самцов.
В первые дни жизни таких групи взаимоотношения между самками носи-
ли в целом нейтральный характер. Лишь у некоторых из них в этот
период проявлялась агрессивпость к другим членам группы, но она воет-
да была менее выражена, чем у самцов, и вскоре пропадала.
В тех случаях, когда в вольеры помещали знакомых друг с другом
взрослых особей (выловленных в одном естественном местообитании или.
Поведение
283
выросших вместе в лаборатории), агрессивные контакты между вамцами
‘были выражены в целом мпого слабее, а между самками отсутствовали
полностью (их взаимоотношения были миролюбивыми).
В нодобных опытах инотда возникали группы (причем независимо от
того, знакомые или незнакомые между собой зверьки входили в их оо-
став), в которых взаимоотноптения между самцами не носили характер
соподчинения и характеризовались отсутствием агонистических форм но-
ведения (Вагпей, 1958а, Ь; 1975; Квашнин, 1983). В таких группах не
‘формировалась устойчивая структура отношений между животными.
Таким образом, у серых крыс как в искусственных, так и в естест-
веппых условиях наблюдаются ‘и нестабильные объединения особей. Ме-
хапизмы их образования наиболее подробно изучены в лабораторных
условиях на другом синантропе — домовой мьгтти (Каменов, 1973; Маслов
и др., 1974; Шилов, 1977). В этих работах было показано, что формиро-
вапие устойчивой структуры внутригрупповых отношений возможно
только при достаточной степени физиологической и поведенческой разно-
качественности взрослых животных (основанной в первую очередь на
уровне агрессивности и типе нервной системы отдельных особей), объ-
единенных на новой территории. Очевидно, что нестабильные объедине-
ПИЯ серых крыс в вольерных условиях возникли за счет случайного под-
‘бора зверьков с низким уровпем ипдивидуальной агрессивности и слабым
типом нервной системы, тотда как в природе их происхождение носит
«естественный» характер.
Наиболее подробно процессы формировапия социальной структуры ис-
кусственных трупп описаны для крыс, которые являлись потомками
пасюков в течение нескольких поколений разводимых в лаборатории и
отбиравшихся по признаку пониженной агрессивности к человеку (Со-
колов и др., 1977; Ляпунова, 1980; Соколов, Ляпунова, 1982). На основв
значительного материала в процессе формирования трупп, образованных
из взрослых пезпакомых между собой животных, было выделено два по-
«ледовательных этапа — период становления иерархической структуры и
период ее стабилизации.
В период становления социальной структуры между зверьками на-
блюдали разной степени выраженности агрессивные столкновения, в ре-
зультате которых каждый самец занимал определенное положение в ие-
рархии. Для второго периода
— периода стабилизации —была характер-
на выработка определенного стереотипа поведения всеми самцами.
Если время до появления первой агрессии пе превышало 3 ч, то в боль-
лиинстве групп (40 из 50) продолжительность периода становления была
не больше 1—2 сут, а периода стабилизации — 4 суток (Соколов, Ляпу-
пова, 1982). В таких группах социальная структура оставалась стабиль-
ной на протяжении длительного времени. В группах, где период с момен-
та запуска на территорию до появления агрессии составлял от 3 до 24 ч,
‘продолжительнссть становления социальной структуры растягивалась до
1—2 недель, а иногда иерархическая структура пе формировалась совсем.
Социальная структура в стабильных группах поддерживалась за’ счет оп-
‘'ределенпых, характеризующихся слабой степенью агрессивности (часто
284 Глава четвертая
ритуализированных) взаимоотношений между самцами. Для групи с не-
стабильной структурой был характерен высокий уровень агрессивных
взаимоотношений между доминирующим и подчиненными самцами. Цак
правило, в таких группах подчиненные самцы погибали, а в ряде случаев
происходила смена доминанта.
Таким образом, из приведенных данных следует, что процессы фор-
мировапия как искусственных, так и естественных группировок серых
крыс отличаются значительным разнообразием и в конечном итоге при-
водят к возникновению разных типов взаимоотношений между животны-
ми в их пределах.
Территориальная структура поселений серых крыс и состав отдель-
ных группировок определяют особенности поведения их членов и, в свою
очередь, являются результатом всей совокупности взаимоотношений о6о-
бей как в пределах группировок, так и между животными разных груп-
пировок.
В пределах группировки взаимоотношения между ое члепами строят-
ся, как правило, по иерархическому принципу. Отношения доминирова-
пия—подчинения, основанные на агонистических контактах, характерны
для взрослых самцов, что наблюдали как в синантропных (Лозан, Ни-
кул, 19841; Михайленко, 1987), так и в экзоантронных (Квашнин, 1982:
Квашнин, Карасева, 1983, 1985) поселениях крыс. Апалогичные данные
получены и для искусственных групл животных. При этом форма иерар-
хических отношений (если они наблюдались) нарьировала в значитель-
ных пределах — от деспотического доминирования одной или пары о6о-
бей (З\енишег, 1950; Вагпей, 1964; Квашнин, 1988) при относительно
равноценном подчинении остальных до линейно-групповой соподчиненно-
сти, когда выделяются отдельные особи (или их группы), занимающие
определенное социальное положение (Вагпей, 1958а, Ъ, 1975; Соколов
и др., 1977; Соколов, Ляпунова, 1982; Квашнин, 1983; Катраи@, 1984).
Взаимоотношения между самками в пределах естественных группировок
носят миролюбивый характер (вне периода размножений). Между ними,
как правило, не возникают устойчивые отношения соподчинения (Са]-
Воцп, 1963; Квашнин, Карасева, 1983; Михайленко, 1987). Аналогичные
выводы можно сделаль и из лабораторных исследований. Однако при елу-
чайном подборе животных для формирования искусственных групп между
самками иногда возпикали иерархические отношения, и они могли как
сохрапяться длительное время (Эфе!сет, 1950; Ляпунова, 1980; Ватра,
1984), так и ограничиваться непродолжительными периодами (Вагпей
1958а, Ь; Саотл, 1963; Квашнин, 41983).
у
Общая система иерархии с участием самцов и самок, по-видимому,
не своиственна серым крысам в отличие от черных крыс (ТеПо, 1966:
Еузет, 1971). Только в одной работе сообщается о существовании единого
ряда соподчипенпости для обоих полов в группировках синантропных
серых крыс (Лозан, Никул, 1981). Однако отсутствие фактических лан-
ных в этой работе делает ее трудносопоставимой с наблюдениями других
исследователсй.
Поведение
285
Взаимоотношения взрослых особей в пределах группировок, описап-
ные выше, носят обобщенный (статичный) характер. Но в природе по-
стоянно происходят измепения различных абиотических и биотических
факторов, что влияет, в частности, и на поведение животных. Конкрет-
ные пути и механизмы влияния таких изменений на структуру взаимоот-
ношений изучены еще крайне недостаточно, поэтому мы коенемся только
некоторых факторов. Один из них связан с размножением самок. Как в
естественных, так и в искусственпых условиях у беременных и особенно.
лактирующих самок заметно повышается агрессивность. В экспериментах
было показано, что на взаимоотношения между самцами значительное
влияние оказывает само по себе присутствие самок —в однополых груп-
пах, образованных из одних самцов, уровни агрессивности и смертности
были существенно ниже, чем в обосполых группах (Вагпей, 1955,
1958а, Ъ; Е!аппеПу, Тюте, 1977). На отношения между членами групи зна-
чительное влияние оказывают самки в течке и беременные самки (Са]-
Бои, 1963; Вагпей, 1975; Соколов, Ляпунова, 1982). В эксперименталь-
ных условиях было также показано, что агрессия доминирующих сам-
цов на подчиненных особей вскоре после родов самок возрастает столь
резко, что приводит к гибели оставшихся в живых (после периода фор-
мирования) подчиненных самцов (Цвашиин. 1983). Интересно, что в ус-
лориях вольер не у всех самок отмечалось повышение агрессивности в
репродуктивный период, а также не все размножавшиеся самки стиму-
лировали агрессивное поведение самцов. Можно предположить, что в
естественных условиях такие изменения в поведении животных имеют
несомненное биологическое значение — агрессия беременпых и лактирую-
щих самок обеспечивает защиту потомства, а усиление агрессивности у
самцов в связи с размножением самок способствует лучшей охране участ-
ка обитания от чужих особей и служит дополнительным стимулом к их
расселению.
° Другой фактор, который существенно влияет па впутригрупповые
отношения, связан с появлением молодых зверьков. Взрослые крысы, как
правило, игнорируют детенышей до достижепия ими 3—3,5 мес. Исключе-
пия наблюдаются лишь в нескольких случаях. Например, лактирующие
самки мотут проявить аптагонизм к крысятам в возрасте более 1,5 мес,
но не раньтте, что, очевидно, имеет значение для выживания молодпяка
(СаШоцп, 1963). Так же нетерпимо взрослые особи могут относиться к
детенышам, которые к ним пристают или паходятся около кормушки.
Агрессия взрослых особей по отношению к крысятам может происходить
и при дезорганизации поселения (Квашпин, 1986). В период полового и
особенно поведенческого созревания молодые особи «включаются» в си-
стему взаимоотношений между взрослыми крысами, причем часть из них
‚ действительно входит в эту систему, а другая покидает пределы груп-
повой территории под влиянием не столько взрослых, сколько более стар-
ших молодых животных (СаФопп, 1963). Цак происходят эти процессы
у серых крыс в естественных условиях, остается пока неясным.
Взаимоотношения, которые складываются между членами естествен-
ных «типичных» группировок и искусственных групи серых крыс, опре-
284 Глава четвертая
ритуализированных) взаимоотношений между самцами. Для групи с не-
стабильной структурой был характерен высокий уровень агрессивных
взаимоотношений между доминирующим и подчиненными самцами. Цак
правило, в таких группах подчиненные самцы погибали, а в ряде случаев
происходила смена доминанта.
Таким образом, из приведенных данных следует, что процессы фор-
мировапия как искусственных, так и естественных группировок серых
крыс отличаются значительным разнообразием и в конечном итоге при-
водят к возникновению разных типов взаимоотношений между животны-
ми в их пределах.
Территориальная структура поселений серых крыс и состав отдель-
ных группировок определяют особенности поведения их членов и, в свою
очередь, являются результатом всей совокупности взаимоотношений о6о-
бей как в пределах группировок, так и между животными разных груп-
пировок.
В пределах группировки взаимоотношения между ое члепами строят-
ся, как правило, по иерархическому принципу. Отношения доминирова-
пия—подчинения, основанные на агонистических контактах, характерны
для взрослых самцов, что наблюдали как в синантропных (Лозан, Ни-
кул, 19841; Михайленко, 1987), так и в экзоантронных (Квашнин, 1982:
Квашнин, Карасева, 1983, 1985) поселениях крыс. Апалогичные данные
получены и для искусственных групл животных. При этом форма иерар-
хических отношений (если они наблюдались) нарьировала в значитель-
ных пределах — от деспотического доминирования одной или пары о6о-
бей (З\енишег, 1950; Вагпей, 1964; Квашнин, 1988) при относительно
равноценном подчинении остальных до линейно-групповой соподчиненно-
сти, когда выделяются отдельные особи (или их группы), занимающие
определенное социальное положение (Вагпей, 1958а, Ъ, 1975; Соколов
и др., 1977; Соколов, Ляпунова, 1982; Квашнин, 1983; Катраи@, 1984).
Взаимоотношения между самками в пределах естественных группировок
носят миролюбивый характер (вне периода размножений). Между ними,
как правило, не возникают устойчивые отношения соподчинения (Са]-
Воцп, 1963; Квашнин, Карасева, 1983; Михайленко, 1987). Аналогичные
выводы можно сделаль и из лабораторных исследований. Однако при елу-
чайном подборе животных для формирования искусственных групп между
самками иногда возпикали иерархические отношения, и они могли как
сохрапяться длительное время (Эфе!сет, 1950; Ляпунова, 1980; Ватра,
1984), так и ограничиваться непродолжительными периодами (Вагпей
1958а, Ь; Саотл, 1963; Квашнин, 41983).
у
Общая система иерархии с участием самцов и самок, по-видимому,
не своиственна серым крысам в отличие от черных крыс (ТеПо, 1966:
Еузет, 1971). Только в одной работе сообщается о существовании единого
ряда соподчипенпости для обоих полов в группировках синантропных
серых крыс (Лозан, Никул, 1981). Однако отсутствие фактических лан-
ных в этой работе делает ее трудносопоставимой с наблюдениями других
исследователсй.
Поведение
285
Взаимоотношения взрослых особей в пределах группировок, описап-
ные выше, носят обобщенный (статичный) характер. Но в природе по-
стоянно происходят измепения различных абиотических и биотических
факторов, что влияет, в частности, и на поведение животных. Конкрет-
ные пути и механизмы влияния таких изменений на структуру взаимоот-
ношений изучены еще крайне недостаточно, поэтому мы коенемся только
некоторых факторов. Один из них связан с размножением самок. Как в
естественных, так и в искусственпых условиях у беременных и особенно.
лактирующих самок заметно повышается агрессивность. В экспериментах
было показано, что на взаимоотношения между самцами значительное
влияние оказывает само по себе присутствие самок —в однополых груп-
пах, образованных из одних самцов, уровни агрессивности и смертности
были существенно ниже, чем в обосполых группах (Вагпей, 1955,
1958а, Ъ; Е!аппеПу, Тюте, 1977). На отношения между членами групи зна-
чительное влияние оказывают самки в течке и беременные самки (Са]-
Бои, 1963; Вагпей, 1975; Соколов, Ляпунова, 1982). В эксперименталь-
ных условиях было также показано, что агрессия доминирующих сам-
цов на подчиненных особей вскоре после родов самок возрастает столь
резко, что приводит к гибели оставшихся в живых (после периода фор-
мирования) подчиненных самцов (Цвашиин. 1983). Интересно, что в ус-
лориях вольер не у всех самок отмечалось повышение агрессивности в
репродуктивный период, а также не все размножавшиеся самки стиму-
лировали агрессивное поведение самцов. Можно предположить, что в
естественных условиях такие изменения в поведении животных имеют
несомненное биологическое значение — агрессия беременпых и лактирую-
щих самок обеспечивает защиту потомства, а усиление агрессивности у
самцов в связи с размножением самок способствует лучшей охране участ-
ка обитания от чужих особей и служит дополнительным стимулом к их
расселению.
° Другой фактор, который существенно влияет па впутригрупповые
отношения, связан с появлением молодых зверьков. Взрослые крысы, как
правило, игнорируют детенышей до достижепия ими 3—3,5 мес. Исключе-
пия наблюдаются лишь в нескольких случаях. Например, лактирующие
самки мотут проявить аптагонизм к крысятам в возрасте более 1,5 мес,
но не раньтте, что, очевидно, имеет значение для выживания молодпяка
(СаШоцп, 1963). Так же нетерпимо взрослые особи могут относиться к
детенышам, которые к ним пристают или паходятся около кормушки.
Агрессия взрослых особей по отношению к крысятам может происходить
и при дезорганизации поселения (Квашпин, 1986). В период полового и
особенно поведенческого созревания молодые особи «включаются» в си-
стему взаимоотношений между взрослыми крысами, причем часть из них
‚ действительно входит в эту систему, а другая покидает пределы груп-
повой территории под влиянием не столько взрослых, сколько более стар-
ших молодых животных (СаФопп, 1963). Цак происходят эти процессы
у серых крыс в естественных условиях, остается пока неясным.
Взаимоотношения, которые складываются между членами естествен-
ных «типичных» группировок и искусственных групи серых крыс, опре-
286 Глава четвертая
деляют особеппости поведения особей, занимающих различное социаль-
ное положение. Особенно отчетливо это проявляется у самцов.
Для доминирующих самцов, или альфа-самцов (по: Вагоей, 1975),
характерна определенная манера поведения, которая отличается неза-
висимостью и «расковапностью». Опи патрулируют групповую террито-
рию и свободно по ней нередвигаются. Для них характерно территориаль-
ное поведение
— мечение участка обитания (или территории вольер)
мочой и ярко выраженная агрессивная реакция на чужих особей—
в основном самцов (Вагпей, 1958а, Ъ, 1975; Квашнин, Суров, 1976; Соко-
лов и др., 1977, 1978; Соколов, Ляпунова, 1982; Квашнин, Карасева,
1985). В естественных условиях доминанты, по-видимому, одними из
первых обнаруживают повые источники пищи и незнакомые предметы,
и их реакция может определять отношение к ним других членов груп-
пировки (СаФопо, 1963; Мисве], 1976; Лозан, Пикул, 1984). Домини-
рующие самцы, по-видимому, могуг являться «регуляторами» состава
группировок [в условиях вольер они наиболее агрессивны во внутригрун-
повых конфликтах (Ляпунова, 1980)], вынуждая покинуть групповую
территорию определенных самцов (но не самок; почему некоторые из
них мигрируют за пределы участков обитания, неизвестно) (СаШопп,
1963). По одним наблюдениям, их агрессия направлена в основном на
подчиненных и полувзрослых особей (Лозан, Никул, 1981; Козлов,
1983), согласно другим —на субдоминантов (Квашнин, Карасева, 1983,
1985). Вероятно, стратегия их поведения может меняться в зависимо-
сти от конкретных условий
— плотности населения, достатка корма и
убежищ, индивидуальных особенностей и т. п. В экспериментальтых груп-
нах для доминирующих самцов характерна маркировка мочой членов
своей группы в результате такой формы поведения, как перелезапие.
Самцы демонстрируют перелезания в основпом по отношению к избран-
ным самкам, что можно рассматривать как создание группового запаха
(Квашнин, 1983). Однако в естественных условиях такая форма поведе-
ния наблюдалась крайне редко. В условиях вольер доминанты имеют пре-
имущество в спаривании с самками; собственно, от них и появлялись
детеныши в стабильных группах (Ляпунова, 1980; Квашнин, 1983). Сте-
пень участия таких самцов в размножении в естественных группировках
пока неизвестна.
Остальные самцы в группировках относятся к подчиненным особям.
Однако они неравноценны между собой по особенностям поведения, по-
этому их можно подразделить на различные категории. Наибольшие раз-
ногласия между исследователями проявились в отношении так пазывае-
мых субдоминантов. Не вдаваясь в дискуссию, мы будем исходить из
того, что сами наблюдали в природе и в лаборатории.
В некоторых естественных группировках, помимо домипантов, выде-
лялись крупные сильные самцы, поводенис которых было сходным с по-
ведепием высокоранговых особей (Квашнин, 1982; Квашнин, Карасева,
1985). В таких группировках их количество не превышало 1—2 особи.
В других группировках они могли вообще отсутствовать. В синантрол-
ных условиях также наблюдали таких зверьков, и опи были отнесены к
=
Поведение
287
категории «угнетенных» (Михайленко, 1987). Действительно, мы отмеча-
ли, что в условиях речной дамбы агрессивная реакция доминантов была
адресована в основном субдоминирующим самцам. Несмотря на это, для
них было характерно свободное передвижение по всей групповой терри-
тории (см. рис. 76). Они также отличались уверенным поведением,
а «скованность» в движениях у них проявлялась только при встречах с
доминантами, которых они избегали. У субдомипаптов наблюдали агрес-
сивную реакцию по отношению к чужим знерькам, а в пределах груп-
пировок их агрессия была направлепа в основном на подчиненных сам-
цов. Исходя из этого, можно предположить, что самцы-субдомипапты
являются основными конкурентами для доминантов.
Такая категория особей не была обнаружена в искусствеппых груп-
пах, образованных из взрослых самцов и самок диких крысе (Вагпе\й,
1975; Квашнин, 1983). В этих группах четко выделялись только две
категории самцов —доминанты и подчиненные. В связи с обсуждаемым
вопросом интересны лаблюдения ряда авторов. Так, Барнетт (Вагпей,
1958Ъ, 1975), формируя группы только из одних самцов пасюков, выде-
лил в их составе категорию так называемых бета-особей. Аналогичная
категория самцов была отмечена и в группах, образованных из потомков
серых крыс, разводимых в ряду поколений в лаборатории, названная аз-
торами «субдоминантами» (Ляпунова, 1980; Соколов, Ляпунова, 1982).
Такие животпые хотя и избегали доминантов, но сумели приспособиться
к их агрессии. Они не только не теряли в весе, а иногда и прибавляли;
могли одновременно с доминантом питаться на кормушке; принимали
участие в спаривании с самками в эструсе. Для таких самцов не было
характерно территориальное поведение. Эти факты говорят о том, что
крысы, разводимые в течение нескольких поколений в лаборатории, от-
личаются от своих родоначальциков попиженной агрессивностью не
только к человеку, но и к особям своего вида. К таким же выводам при-
шли и другие исследователи, которые специально изучали влияние доме
стикации диких крыс на их поведение, в частпости агрессивное (Юта.
1929, 1939; Вагиеи, 1975; Васе, 1972; 1лоте, ЕТаппеПу, 1981). Во всяком
случае, особи бета-категории по своим поведенческим характеристикам
не сходны © субдоминантами, которых мы наблюдали в остественных
группировках.
Для второй категории собственно подчинениых самцов (омега— по
Ватпей, 1975, субординантные — по Михайленко, 1987) характерны пол-
пая (или почти полная) утрата агрессивных реакций и хорошо выра-
женные путгливость, скованность в движениях, постоянная готовпость
убежать. В естественных труппировках они избегают контактов © други-
ми особями, а их перемещения ограничены какой-либо частью групповой
‘территории. В искусственных группах активность подчиненных самцов
приурочена к часам, когда доминанты не активны (Ляпунова, 1980). Для
них не свойственно тсрриториальное поведение. С самками в течке они
спаризались только в отсутствие домипирующего самца или вообще не
обращали на них внимания. Как правило, они теряли в весе и многие
из них в конце концов погибали. Интересно отметить, что в некоторых
286 Глава четвертая
деляют особеппости поведения особей, занимающих различное социаль-
ное положение. Особенно отчетливо это проявляется у самцов.
Для доминирующих самцов, или альфа-самцов (по: Вагоей, 1975),
характерна определенная манера поведения, которая отличается неза-
висимостью и «расковапностью». Опи патрулируют групповую террито-
рию и свободно по ней нередвигаются. Для них характерно территориаль-
ное поведение
— мечение участка обитания (или территории вольер)
мочой и ярко выраженная агрессивная реакция на чужих особей—
в основном самцов (Вагпей, 1958а, Ъ, 1975; Квашнин, Суров, 1976; Соко-
лов и др., 1977, 1978; Соколов, Ляпунова, 1982; Квашнин, Карасева,
1985). В естественных условиях доминанты, по-видимому, одними из
первых обнаруживают повые источники пищи и незнакомые предметы,
и их реакция может определять отношение к ним других членов груп-
пировки (СаФопо, 1963; Мисве], 1976; Лозан, Пикул, 1984). Домини-
рующие самцы, по-видимому, могуг являться «регуляторами» состава
группировок [в условиях вольер они наиболее агрессивны во внутригрун-
повых конфликтах (Ляпунова, 1980)], вынуждая покинуть групповую
территорию определенных самцов (но не самок; почему некоторые из
них мигрируют за пределы участков обитания, неизвестно) (СаШопп,
1963). По одним наблюдениям, их агрессия направлена в основном на
подчиненных и полувзрослых особей (Лозан, Никул, 1981; Козлов,
1983), согласно другим —на субдоминантов (Квашнин, Карасева, 1983,
1985). Вероятно, стратегия их поведения может меняться в зависимо-
сти от конкретных условий
— плотности населения, достатка корма и
убежищ, индивидуальных особенностей и т. п. В экспериментальтых груп-
нах для доминирующих самцов характерна маркировка мочой членов
своей группы в результате такой формы поведения, как перелезапие.
Самцы демонстрируют перелезания в основпом по отношению к избран-
ным самкам, что можно рассматривать как создание группового запаха
(Квашнин, 1983). Однако в естественных условиях такая форма поведе-
ния наблюдалась крайне редко. В условиях вольер доминанты имеют пре-
имущество в спаривании с самками; собственно, от них и появлялись
детеныши в стабильных группах (Ляпунова, 1980; Квашнин, 1983). Сте-
пень участия таких самцов в размножении в естественных группировках
пока неизвестна.
Остальные самцы в группировках относятся к подчиненным особям.
Однако они неравноценны между собой по особенностям поведения, по-
этому их можно подразделить на различные категории. Наибольшие раз-
ногласия между исследователями проявились в отношении так пазывае-
мых субдоминантов. Не вдаваясь в дискуссию, мы будем исходить из
того, что сами наблюдали в природе и в лаборатории.
В некоторых естественных группировках, помимо домипантов, выде-
лялись крупные сильные самцы, поводенис которых было сходным с по-
ведепием высокоранговых особей (Квашнин, 1982; Квашнин, Карасева,
1985). В таких группировках их количество не превышало 1—2 особи.
В других группировках они могли вообще отсутствовать. В синантрол-
ных условиях также наблюдали таких зверьков, и опи были отнесены к
=
Поведение
287
категории «угнетенных» (Михайленко, 1987). Действительно, мы отмеча-
ли, что в условиях речной дамбы агрессивная реакция доминантов была
адресована в основном субдоминирующим самцам. Несмотря на это, для
них было характерно свободное передвижение по всей групповой терри-
тории (см. рис. 76). Они также отличались уверенным поведением,
а «скованность» в движениях у них проявлялась только при встречах с
доминантами, которых они избегали. У субдомипаптов наблюдали агрес-
сивную реакцию по отношению к чужим знерькам, а в пределах груп-
пировок их агрессия была направлепа в основном на подчиненных сам-
цов. Исходя из этого, можно предположить, что самцы-субдомипапты
являются основными конкурентами для доминантов.
Такая категория особей не была обнаружена в искусствеппых груп-
пах, образованных из взрослых самцов и самок диких крысе (Вагпе\й,
1975; Квашнин, 1983). В этих группах четко выделялись только две
категории самцов —доминанты и подчиненные. В связи с обсуждаемым
вопросом интересны лаблюдения ряда авторов. Так, Барнетт (Вагпей,
1958Ъ, 1975), формируя группы только из одних самцов пасюков, выде-
лил в их составе категорию так называемых бета-особей. Аналогичная
категория самцов была отмечена и в группах, образованных из потомков
серых крыс, разводимых в ряду поколений в лаборатории, названная аз-
торами «субдоминантами» (Ляпунова, 1980; Соколов, Ляпунова, 1982).
Такие животпые хотя и избегали доминантов, но сумели приспособиться
к их агрессии. Они не только не теряли в весе, а иногда и прибавляли;
могли одновременно с доминантом питаться на кормушке; принимали
участие в спаривании с самками в эструсе. Для таких самцов не было
характерно территориальное поведение. Эти факты говорят о том, что
крысы, разводимые в течение нескольких поколений в лаборатории, от-
личаются от своих родоначальциков попиженной агрессивностью не
только к человеку, но и к особям своего вида. К таким же выводам при-
шли и другие исследователи, которые специально изучали влияние доме
стикации диких крыс на их поведение, в частпости агрессивное (Юта.
1929, 1939; Вагиеи, 1975; Васе, 1972; 1лоте, ЕТаппеПу, 1981). Во всяком
случае, особи бета-категории по своим поведенческим характеристикам
не сходны © субдоминантами, которых мы наблюдали в остественных
группировках.
Для второй категории собственно подчинениых самцов (омега— по
Ватпей, 1975, субординантные — по Михайленко, 1987) характерны пол-
пая (или почти полная) утрата агрессивных реакций и хорошо выра-
женные путгливость, скованность в движениях, постоянная готовпость
убежать. В естественных труппировках они избегают контактов © други-
ми особями, а их перемещения ограничены какой-либо частью групповой
‘территории. В искусственных группах активность подчиненных самцов
приурочена к часам, когда доминанты не активны (Ляпунова, 1980). Для
них не свойственно тсрриториальное поведение. С самками в течке они
спаризались только в отсутствие домипирующего самца или вообще не
обращали на них внимания. Как правило, они теряли в весе и многие
из них в конце концов погибали. Интересно отметить, что в некоторых
288 Глава четвертая
группах, образованных из диких серых крыс, иногда наблюдали такую
ситуацию, когда у доминанта по каким-то причинам агрессивность по
отношению к подчиненным самцам значительно уменьшалась (или даже
пропадала) (Квашнин, 1983). Именпо в такие периоды «затишья» под-
чиненные самцы «оживали» и начинали принимать активное участие в
жизни групиы
— спаривались с самками, маркировали территорию, за-
пасали корм и даже «выясняли отношения» между собой, т. е. проявля-
лись различия между ними. Обычно эти периоды не превьипали двух не-
дель и заканчивались из-за того, что у доминаптов вновь проявлялась
агрессивность цо отношению к ним в связи с размножением самок. Та-
ким образом, очевидно, что в ограниченных групцах агрессивное поведе-
ние доминирующих самцов подавляет социальную активность подчинен-
ных зверьков, тогда как в естественных условиях «подчиненное»
поведение определяется, по-видимому, индивидуальными особенностями
животных.
Взаимоотношения между самками в пределах сстественных группи-
ровок носят, как уже упоминалось, в целом миролюбивый характер.
Между ними не возникают отношения соподчинения в отличие от того,
что наблюдали в некоторых искусственных группах. Атгрессивное пове-
дение характерно для них только в репродуктивный период— во время
эструса и лактации (охрана гнезда). В условиях вольер все самки наби-
рают в весе, одновременно с высокоранговыми самцами активны вне
убежищ, запасают корм и строят гнезда, демонстрируют половое и тер-
риториальное поведение (СаШопп, 1963; Ватпем, 1975; Ляпунова, 1980;
Квашнин, 19883). Однако они не равноценны между собой по социальной
активпости — в разпой степени у них проявляются все формы деятельно-
сти (вплоть до отсутотвия некоторых из них, например маркировочного
поведения или агрессивной реакции на чужих особей}. К тому же самцы-
доминанты избирательно реагируют на самок, предпочитая контактиро-
вать с определенными из них даже во время течки (Квашнин, 1983).
В естественных условиях самки также отличаются друг от друга, в част-
ности по характеру перемещений в пределах групповой территории и по
реакции на ловушки (Квашнин, Карасева, 1985). Для тех еамок, кото-
рые свободно передвигались но всему участку подобно высокораяговым
‹самцам, было характерпо также территориальное поведение (агрессияна
чужих особей). У них же зарегистрировано наибольшее число детенышей
(Саопл, 1963; Квашнин, Карасева, 1985). Таким образом, самки в пре-
делах группировок неравнозначны между собой как по особенностям
поведения, так и по своему «вкладу» в общегруиповые функции,
Таким образом, социальная организация у серых крыс представлена
локальными группировками, которые относительно независимы друг от
друга и чрезвычайно разнообразны по своему происхождению, составу и
характеру взаимоотношений между отдельными особями. Именно это
разнообразие группировок, оспованное на индивидуальных особенностях
их членов, определяет снецифичность организации поселений пасюка и,
следовательно, пластичность и быструю приспособляемость животных к
постоянно меняющимся условиям обитания. Конкретные механизмы адап-
Поведение
289
тации серых крыс к реальным условиям среды мотут быть различными,
они требуют дополнительных и всесторонних исследований. Несомненно
одно: именно на уровне отдельных грулиировок разворачиваются авторе-
туляторные процессы, которые обеспечивают существование и процвета-
пие поселений этих грызунов, несмотря на интенсивную борьбу, веду-
щуюся с ними,
Обонятельная коммуникация
Химичоский способ передачи информации универсален и осуществляется
у животных самых различных систематических групп. Наиболес сложным
образом хемочуветвительные органы построены у позвоночных; они вклю-
чают основную, донолнительную обонительные системы, вкусовую и си-
стему общего химического чувства. Слрогие доказательства участия этих
систем в химической коммуникации млекопитаютцих существуют только
для органов обоняпия.
Осповная обонятельная система (обопятольные реценторы и оконча-
ния тройничного нерва) реагирует только на летучие компопенты выде-
лений животных, а дополнительная (вомероназальный орган) восприни-
мает малолетучие и нелетучие компоненты. Пахучие вощества могут
стимулировать несколько интраназальных структур: обонятельные рецец-
торы, свободные нервные окончания тройничного нерва и специализиро-
ванные эпителиальные рецепторы вомероназального органа (орган
Якобсона) и септального органа Мазера.
.
Поведение крыс находится под контролем всех сенсорных систем,
при этом одна и та же информация, как правило, дублируется сигналами
различных модальностей. Вопрос об относительном вкладе каждого из
анализаторов в процесс обработки информации об окружающей среде изу-
чен недостаточно. Тем не менее есть основания говорить об особой зна-
чимости обоилтельных сигналов в поведении крыс. главным образом в
тех случаях, когда восприятие сигналов других модальностей затруднено
или вообще невозможно (Робу, 1986). Примером затрудненного восприя-
тия слуховых сигналов могут быть звуковые помехи различного рода,
а зрительных
— темные норы. В таких ситуациях химический способ
коммупикации приобретает серьезные преимущества по сравнению со
всеми другими. Во-первых, обонятельные сигналы могут восприниматься
и анализироваться крысятами в раннем онтогенезе, когда их глаза еще
не функционируют, а слуховой апализатор физиологически незрел. Во-
вторых, химические сигналы могут быть легко распределены в определен-
‘ных точках пространства, в значительной степени упрощая организацию
территорнального поведения у крыс. В-третьих, химические сигналы об-
падают более длинными периодами жизни по сравнению © сигналами
другой природы. В-четвертых, химический епособ коммуникаций не тре-
бует теспого контакта между животными и обеспечивает передачу инфор-
мации на большие расстояния.
Рассмотрим роль обонятельных сигналов в организации поведенческих
актов у крыс.
10 Серая крыса
288 Глава четвертая
группах, образованных из диких серых крыс, иногда наблюдали такую
ситуацию, когда у доминанта по каким-то причинам агрессивность по
отношению к подчиненным самцам значительно уменьшалась (или даже
пропадала) (Квашнин, 1983). Именпо в такие периоды «затишья» под-
чиненные самцы «оживали» и начинали принимать активное участие в
жизни групиы
— спаривались с самками, маркировали территорию, за-
пасали корм и даже «выясняли отношения» между собой, т. е. проявля-
лись различия между ними. Обычно эти периоды не превьипали двух не-
дель и заканчивались из-за того, что у доминаптов вновь проявлялась
агрессивность цо отношению к ним в связи с размножением самок. Та-
ким образом, очевидно, что в ограниченных групцах агрессивное поведе-
ние доминирующих самцов подавляет социальную активность подчинен-
ных зверьков, тогда как в естественных условиях «подчиненное»
поведение определяется, по-видимому, индивидуальными особенностями
животных.
Взаимоотношения между самками в пределах сстественных группи-
ровок носят, как уже упоминалось, в целом миролюбивый характер.
Между ними не возникают отношения соподчинения в отличие от того,
что наблюдали в некоторых искусственных группах. Атгрессивное пове-
дение характерно для них только в репродуктивный период— во время
эструса и лактации (охрана гнезда). В условиях вольер все самки наби-
рают в весе, одновременно с высокоранговыми самцами активны вне
убежищ, запасают корм и строят гнезда, демонстрируют половое и тер-
риториальное поведение (СаШопп, 1963; Ватпем, 1975; Ляпунова, 1980;
Квашнин, 19883). Однако они не равноценны между собой по социальной
активпости — в разпой степени у них проявляются все формы деятельно-
сти (вплоть до отсутотвия некоторых из них, например маркировочного
поведения или агрессивной реакции на чужих особей}. К тому же самцы-
доминанты избирательно реагируют на самок, предпочитая контактиро-
вать с определенными из них даже во время течки (Квашнин, 1983).
В естественных условиях самки также отличаются друг от друга, в част-
ности по характеру перемещений в пределах групповой территории и по
реакции на ловушки (Квашнин, Карасева, 1985). Для тех еамок, кото-
рые свободно передвигались но всему участку подобно высокораяговым
‹самцам, было характерпо также территориальное поведение (агрессияна
чужих особей). У них же зарегистрировано наибольшее число детенышей
(Саопл, 1963; Квашнин, Карасева, 1985). Таким образом, самки в пре-
делах группировок неравнозначны между собой как по особенностям
поведения, так и по своему «вкладу» в общегруиповые функции,
Таким образом, социальная организация у серых крыс представлена
локальными группировками, которые относительно независимы друг от
друга и чрезвычайно разнообразны по своему происхождению, составу и
характеру взаимоотношений между отдельными особями. Именно это
разнообразие группировок, оспованное на индивидуальных особенностях
их членов, определяет снецифичность организации поселений пасюка и,
следовательно, пластичность и быструю приспособляемость животных к
постоянно меняющимся условиям обитания. Конкретные механизмы адап-
Поведение
289
тации серых крыс к реальным условиям среды мотут быть различными,
они требуют дополнительных и всесторонних исследований. Несомненно
одно: именно на уровне отдельных грулиировок разворачиваются авторе-
туляторные процессы, которые обеспечивают существование и процвета-
пие поселений этих грызунов, несмотря на интенсивную борьбу, веду-
щуюся с ними,
Обонятельная коммуникация
Химичоский способ передачи информации универсален и осуществляется
у животных самых различных систематических групп. Наиболес сложным
образом хемочуветвительные органы построены у позвоночных; они вклю-
чают основную, донолнительную обонительные системы, вкусовую и си-
стему общего химического чувства. Слрогие доказательства участия этих
систем в химической коммуникации млекопитаютцих существуют только
для органов обоняпия.
Осповная обонятельная система (обопятольные реценторы и оконча-
ния тройничного нерва) реагирует только на летучие компопенты выде-
лений животных, а дополнительная (вомероназальный орган) восприни-
мает малолетучие и нелетучие компоненты. Пахучие вощества могут
стимулировать несколько интраназальных структур: обонятельные рецец-
торы, свободные нервные окончания тройничного нерва и специализиро-
ванные эпителиальные рецепторы вомероназального органа (орган
Якобсона) и септального органа Мазера.
.
Поведение крыс находится под контролем всех сенсорных систем,
при этом одна и та же информация, как правило, дублируется сигналами
различных модальностей. Вопрос об относительном вкладе каждого из
анализаторов в процесс обработки информации об окружающей среде изу-
чен недостаточно. Тем не менее есть основания говорить об особой зна-
чимости обоилтельных сигналов в поведении крыс. главным образом в
тех случаях, когда восприятие сигналов других модальностей затруднено
или вообще невозможно (Робу, 1986). Примером затрудненного восприя-
тия слуховых сигналов могут быть звуковые помехи различного рода,
а зрительных
— темные норы. В таких ситуациях химический способ
коммупикации приобретает серьезные преимущества по сравнению со
всеми другими. Во-первых, обонятельные сигналы могут восприниматься
и анализироваться крысятами в раннем онтогенезе, когда их глаза еще
не функционируют, а слуховой апализатор физиологически незрел. Во-
вторых, химические сигналы могут быть легко распределены в определен-
‘ных точках пространства, в значительной степени упрощая организацию
территорнального поведения у крыс. В-третьих, химические сигналы об-
падают более длинными периодами жизни по сравнению © сигналами
другой природы. В-четвертых, химический епособ коммуникаций не тре-
бует теспого контакта между животными и обеспечивает передачу инфор-
мации на большие расстояния.
Рассмотрим роль обонятельных сигналов в организации поведенческих
актов у крыс.
10 Серая крыса
290 Глава четвертая
Обоняние и половое поведение крыс
Существенная роль в организации полового поведения серой крысы при-
надлежит обонятельным сигналам. Ц настоящему времепи накоплен зна-
чительный экспериментальный материал, подтверждающий зпачимость.
химических сигналов выделений крыс для успешного поиска брачного
партнера, синхропизации физиологических и поведенческих реакции ири
спаривации (Вегталь, Тау|ог, 1969; Р/аН, РаЙтап, 1969), для полового
созревания (Зафю её а}, 1984) и нормальной организации эстральных цик-
лов (МеСИюоск, 1981, 1983а—‹). Значительная часть работ посвящена.
выяснению роли основной И дополнительной обонятельной систем в в06-
приятии обонятельных сигналов и реализации их воздействия на различ-
ные элементы полового поведения (СаПезо её а|., 1982; ВеапсВатр её а1.,
1983: КееВе, Агоп, 1984).
.
Начало исследованию аттрактивности выделений самок крыс для
особей противоположного пола было положено работами Ле-Маньяна
(Ге Мартеп, 1952), который в Т-образном лабиринте самцам крысе линии
Мэг предъявлял два источника запаха: от рецептивпой и нерецептив-
пой самок. Самцы достоверно чаще посещали рукав лабиринта с запахом
рецептивной самки. Если же в один из рукавов продували струю чистого
воздуха, а в другом находился источник запаха рецептивной или нере-
цептивной самки, то в 90% случаев самцы предпочитали рукав лабиринта
с запахом самки. На основании этих экспериментов автором был сделан.
вывод о том, что самцов крыс привлекает запах самок независимо от их
физиологического состояния, однако запах рецептивной самки более:
предпочтителен.
Исследовательская активность является необходимым элементом по-
ведения, направлепного на поиск возможного брачного партнера, Именно.
поэтому проблема аттрактилности тех или иных запахов для крыс при-
влекла к ссбе такой широкий круг исследователеи (Соколов и др.,
1978, 1979а, 6, 1983а, 6, 1985, 1986; Вто, 1985; Сагт, 1974; Самлепо\зВ..
1977; и др.)..
.
Колхаун (СаТопш, 1963), наблюдая поведение самки перед наступле-
нием рецептивности, отмечал, что в это кремя двигательная активность ее
возрастает. Она отходит на значительное расстояние от норы, часто трет-
ся апогенитальной областью о субстрат и окружающие предметы, остав-
ляя на них свои метки, которые исследуются самцами, что стимулирует”
у них поисковую активность, приводящую к обпаружению самки. О по-
вышенной маркировочной активности самок также и в период рецептив-
ности свидетельствует целый ряд работ (Ве, 1978; Ви\е, За ег, 1984).
Па пачальных этапах взаимодействия брачных партнеров поведение:
самки в присутствии самца амбивалептио: она то приближается к пему,
то убегает, когда он пачинает преследование (Руегсе, Миа, 1964).
В этот период стремление рецептивной самки к запаху самца может быть.
компонентом реакции приближения, способствующим объединению парт-
неров.
:
\
Поведение 291
Разработка проблемы аттрактивпости выделений крыс для особей сво-
<го вида шла по пути выяспения возможной роли различных факторов в
проявлении предпочтения запахов. Первым шагом в этом направлении
стало изучение роли сексуального опыта самцов при различении физио-
логического состояния самок. В клетку к одиночным самцам линии [.оп5-
Еуапз помещали два картоппых цилиндра, в одном из которых ранее
хидели самки в состоянии эструса, а в другом
— нерецептивные. В те-
чение получаса каждые 5 мин фиксировали время исследования самцом
одпого и другого цилиндра. Сексуально неопытные, а также кастрирован-
ные в раннем возрасте самцы не обнаруживали достоверной разницы во
времепи обследовапия цилипдров. Запах рецентивной самки предпочитали
лишь особи, имеющие половой опыт (Сагг её а1., 1965). Этот факт был
подтвержден эксперимептально рядом авторов (Р!аМ, Р!аНзиао, 1969;
Эцеги, 1970), хотя в экспериментах Хитта с сотрудниками (НИ её а1.,
1970) самцы крыс, не имеющие полового опыта, обнаруживали слабое
предпочтение запаху самки в состоянии эструса по сравнепию с нере-
цептивной самкой. Однако предпочтение угасало, если оно не сопро-
вождалоеь последующим приобретением полового опыта. Это послужило
предиосылкой для дальнейшего, более детальпого исследования вопроса
предпочтения запахов (Сагг, 1974; Сапфапег её а1., 1977; Тудей, Бобу,
1972). Оказалось, что уже достаточным условием для возникновения
предпочтения запаха самки в состоянии эструса является частичный по-
ловой опыт самцов (садки без интромиссий; б4егпи, 1970). Более того,
существуют экспериментальные подтверждения тому, что способность
самцов различать физиологическое состояние самки носит времеппой ха-
рактер и зависит от возраста самца. Карр (Сагг, 1974) пришел к анало-
тичпому выводу: предпочтение запаха эстральной самки проявляют мо-
лодые половозрелые самцы (в данном случае 7—15 мес). Самцы в воз-
расте 4—7 мес, как правило, не различают физиологического состояния
самки. Отсутетвие у них полового опыта способствует в дальнейшем уга-
санию такого предпочтения.
Таким образом, некоторые разногласия в результатах исследований
можно объяснить разновозрастностью использовапных животных.
На проявление предпочтения запаха рецептивной самки может также
‘оказывать влияние не только половой опыт как таковой, по и его харак-
тер: спаривание с одной или песколькими самками (Сагг её а|., 1970).
Так, самцы линии Гопя-Еуапз, спаривавшиеся только с одной самкой,
проявляют достоверное предпочтение запаху пезнакомой эстральной
самки по сравпепию со знакомой ренентивной самкой, тогда как для
самцов, имеющих опыт спаривания с несколькими самками, такого явле-
ния не обпаружено (Сагг 0ё а1., 1980).
Брауном (Вго\п, 1977а) была предпринята попытка составить ряд
запахов по их предпочитаемости для самцов капютонных крыс линии
Топе-Еуапз. Были использованы Интактные и кастрированные самцы,
имеющие и не имеющие полового опыта, содержащиеся только с самцами
или в смешанных лабораторных колониях. В парных сочетаниях предъ-
10*
290 Глава четвертая
Обоняние и половое поведение крыс
Существенная роль в организации полового поведения серой крысы при-
надлежит обонятельным сигналам. Ц настоящему времепи накоплен зна-
чительный экспериментальный материал, подтверждающий зпачимость.
химических сигналов выделений крыс для успешного поиска брачного
партнера, синхропизации физиологических и поведенческих реакции ири
спаривации (Вегталь, Тау|ог, 1969; Р/аН, РаЙтап, 1969), для полового
созревания (Зафю её а}, 1984) и нормальной организации эстральных цик-
лов (МеСИюоск, 1981, 1983а—‹). Значительная часть работ посвящена.
выяснению роли основной И дополнительной обонятельной систем в в06-
приятии обонятельных сигналов и реализации их воздействия на различ-
ные элементы полового поведения (СаПезо её а|., 1982; ВеапсВатр её а1.,
1983: КееВе, Агоп, 1984).
.
Начало исследованию аттрактивности выделений самок крыс для
особей противоположного пола было положено работами Ле-Маньяна
(Ге Мартеп, 1952), который в Т-образном лабиринте самцам крысе линии
Мэг предъявлял два источника запаха: от рецептивпой и нерецептив-
пой самок. Самцы достоверно чаще посещали рукав лабиринта с запахом
рецептивной самки. Если же в один из рукавов продували струю чистого
воздуха, а в другом находился источник запаха рецептивной или нере-
цептивной самки, то в 90% случаев самцы предпочитали рукав лабиринта
с запахом самки. На основании этих экспериментов автором был сделан.
вывод о том, что самцов крыс привлекает запах самок независимо от их
физиологического состояния, однако запах рецептивной самки более:
предпочтителен.
Исследовательская активность является необходимым элементом по-
ведения, направлепного на поиск возможного брачного партнера, Именно.
поэтому проблема аттрактилности тех или иных запахов для крыс при-
влекла к ссбе такой широкий круг исследователеи (Соколов и др.,
1978, 1979а, 6, 1983а, 6, 1985, 1986; Вто, 1985; Сагт, 1974; Самлепо\зВ..
1977; и др.)..
.
Колхаун (СаТопш, 1963), наблюдая поведение самки перед наступле-
нием рецептивности, отмечал, что в это кремя двигательная активность ее
возрастает. Она отходит на значительное расстояние от норы, часто трет-
ся апогенитальной областью о субстрат и окружающие предметы, остав-
ляя на них свои метки, которые исследуются самцами, что стимулирует”
у них поисковую активность, приводящую к обпаружению самки. О по-
вышенной маркировочной активности самок также и в период рецептив-
ности свидетельствует целый ряд работ (Ве, 1978; Ви\е, За ег, 1984).
Па пачальных этапах взаимодействия брачных партнеров поведение:
самки в присутствии самца амбивалептио: она то приближается к пему,
то убегает, когда он пачинает преследование (Руегсе, Миа, 1964).
В этот период стремление рецептивной самки к запаху самца может быть.
компонентом реакции приближения, способствующим объединению парт-
неров.
:
\
Поведение 291
Разработка проблемы аттрактивпости выделений крыс для особей сво-
<го вида шла по пути выяспения возможной роли различных факторов в
проявлении предпочтения запахов. Первым шагом в этом направлении
стало изучение роли сексуального опыта самцов при различении физио-
логического состояния самок. В клетку к одиночным самцам линии [.оп5-
Еуапз помещали два картоппых цилиндра, в одном из которых ранее
хидели самки в состоянии эструса, а в другом
— нерецептивные. В те-
чение получаса каждые 5 мин фиксировали время исследования самцом
одпого и другого цилиндра. Сексуально неопытные, а также кастрирован-
ные в раннем возрасте самцы не обнаруживали достоверной разницы во
времепи обследовапия цилипдров. Запах рецентивной самки предпочитали
лишь особи, имеющие половой опыт (Сагг её а1., 1965). Этот факт был
подтвержден эксперимептально рядом авторов (Р!аМ, Р!аНзиао, 1969;
Эцеги, 1970), хотя в экспериментах Хитта с сотрудниками (НИ её а1.,
1970) самцы крыс, не имеющие полового опыта, обнаруживали слабое
предпочтение запаху самки в состоянии эструса по сравнепию с нере-
цептивной самкой. Однако предпочтение угасало, если оно не сопро-
вождалоеь последующим приобретением полового опыта. Это послужило
предиосылкой для дальнейшего, более детальпого исследования вопроса
предпочтения запахов (Сагг, 1974; Сапфапег её а1., 1977; Тудей, Бобу,
1972). Оказалось, что уже достаточным условием для возникновения
предпочтения запаха самки в состоянии эструса является частичный по-
ловой опыт самцов (садки без интромиссий; б4егпи, 1970). Более того,
существуют экспериментальные подтверждения тому, что способность
самцов различать физиологическое состояние самки носит времеппой ха-
рактер и зависит от возраста самца. Карр (Сагг, 1974) пришел к анало-
тичпому выводу: предпочтение запаха эстральной самки проявляют мо-
лодые половозрелые самцы (в данном случае 7—15 мес). Самцы в воз-
расте 4—7 мес, как правило, не различают физиологического состояния
самки. Отсутетвие у них полового опыта способствует в дальнейшем уга-
санию такого предпочтения.
Таким образом, некоторые разногласия в результатах исследований
можно объяснить разновозрастностью использовапных животных.
На проявление предпочтения запаха рецептивной самки может также
‘оказывать влияние не только половой опыт как таковой, по и его харак-
тер: спаривание с одной или песколькими самками (Сагг её а|., 1970).
Так, самцы линии Гопя-Еуапз, спаривавшиеся только с одной самкой,
проявляют достоверное предпочтение запаху пезнакомой эстральной
самки по сравпепию со знакомой ренентивной самкой, тогда как для
самцов, имеющих опыт спаривания с несколькими самками, такого явле-
ния не обпаружено (Сагг 0ё а1., 1980).
Брауном (Вго\п, 1977а) была предпринята попытка составить ряд
запахов по их предпочитаемости для самцов капютонных крыс линии
Топе-Еуапз. Были использованы Интактные и кастрированные самцы,
имеющие и не имеющие полового опыта, содержащиеся только с самцами
или в смешанных лабораторных колониях. В парных сочетаниях предъ-
10*
292 Глава четвертая
являлись следующие запахи: 1 — мочи эстральной самун (а); 2— вари
оэктомированной самки (6); 3 — мочи членов собственной группы в
4 — кастрированного самца (2); 5 — интактного самца (9); 6 — воды °).
Для сексуально опытных самцов, живших в колониях е сородичам
обоего пола, запахи по их предпочитаемости расположились в следую-
щий ряд: а>б=в=е=д>е. Сексуально неопытные и кастрированные сам-
цы не оказывали предпочтения запаху мочи эстральной самки.
Сравнение реакций самок па запах кастрированных и иптактных сам-
цов показало, что независимо от их физиологического состояния (вструс
или диэструс) самки, имеющие половой опыт, достоворпо дольше иосле-
довали запах интактных самцов по сравнению с запахом кастратов. ам-
ки, пе имеющие полового опыта, дольше исследовали запах иитактных
самцов только в состоянии эструса, а у самок в состоянии диэструса та
кого предночтения не обнаружено (Сати её а1., 1965).
Изучение исследовательского поведения крыс в условиях вольерного
содержания позволлет получать более полные данные, отличающиеся от
таковых при тестировании животных, содержащихся в стапдартных кю
ветах в условиях вивария, и выделять дополнительный фактор (сопиал
ный ранг), влияющий на предпочтение различных запахов у крыс. этих
экспериментах использовали сексуальпо опытных самцов и самок в воз
расте 7—10 мес —потомков диких серых крыс, рожденных в виварии
(Соколов, Ляпунова, 1979, 1982). В вольору площадью 4 м, пасоленную
группой крыс из 6—10 особей с устойчивой иерархической структурой,
в качестве источника запаха помещали поодиночке в специально окон
струированных камерах-коробках самок, усыпленных нембуталом. дна
из них находилась в состолнии эструса. Контролем служила такая же,
но пустая коробка. Самцы достоверно дольше (р<0,01) исследовали ко-
робку с эстральной самкой, однако уровень их исследовательской актив-
ности был разным. Всегда максимальное время исследования домопстри-
ровал доминант. Более того, он начинал усилеппо патрулировать терри-
торню, припимал угрожающие позы и атаковал подчиненных самцов,
интенсивно метил мочой коробку © эстральшой самкой и близлежащее
пространство. Субдомивант проявлял аналогичные реакции, однако к
источнику запаха подходил реже и только в отсутствие доминанта.
од.
чипенцые самцы в такой ситуации редко покидали свои убежища и прРд-
почитали принюхиваться к коробке с самкой с большого расстояния.
Если же доминантного самца изымали из группы за 4 дня до экопери-
мента, то субдоминант занимал сго место и демонстрировавал реакции на
запах самки, аналогичные для доминанта. При парном предъявлении за-
пахов самок (как знакомых, так и незнакомых) в состоянии эструса
сексуально опытным самцам разиого социального ранга доминирующий
самец всегда предпочитал запах эстральной самки (оценка производилась
по времени исследования). Нодчиненные самцы избегали запаха самки
как в состоянии эструса, так и диэструса, если оп предъявлялся в паре
с запахом самца-доминанта, и при этом демонстрировали реакцию испу-
га. В 10 же время тестирование подчиненпых самцов в камере-ольфак-
тометре показало, что они предпочитают запах эстральной самки при
=
Поведение
293
предъявлении сго в паре с запахом одиночно содержавиегося самца.
Использование запаха социально доминантного самца в паре с запахом
эстральной самки измепяет реакцию подчинепных самцов (Соколов, Ля-
пунова, 1979).
Наблюдение в вольере за самками показало, что все они проявляют
слабую исследовательскую реакцию при внесепии на их территорию ко-
робки с запахом домипантвого самца. При этом они предпочитают скры-
ваться в убежищах. Предъявление запаха подчиненного самца или его
подстилки вызывает у всех самок возбуждение, интенсивную исследова-
тельскую реакцию, повышенную двигательную активность, перетаскива-
ние корма. При тестировании этих же самок в камере-ольфактометре в
течение 5 мин все самки достоверно дольше исследуют запах «чужого»
одиночного самца по сравнению с самцами различных социальных ран-
гов из своей группы. Наибольшая разница во времени исследования на-
блюдалась при предъявлении запаха «чулюго» одиночного самца и зна-
комого подчиненного (Соколов, Ляпунова, 1979).
Таким образом, реакция на запах у члепов группировки серых крыс
зависит не только от сексуального опыта особей, физиологического состоя-
ния самок, предварительного знакомства с донором запаха, но и от со-
ниального ранга самца.
Способность различать химические сигпалы партнера важна для ус-
нешпого спаривания крыс. Аногенитальное обследование, как правило,
предшествует садкам, поэтому его можно рассматривать как неотъемле-
мую часть поведения самцов при спаривапии. Если кастрированным в
раннем возрасте самцам ввести тестостерон, то у них время аногениталь-
ного обследовапия рецептивных самок возрастаст по сравнению с кастри-
рованными самцами (Этрег, 1972).
Экспонирование запаха рецептивной самки самцам, живущим в одно-
полых груплах, увеличивает процент животных, делающих садки на с0-
жителей по клетке, по срависнию с экспонировапием запаха нерецентив-
ной самки. Это свидетельствует о более высоком уровне возбуждения
реципиентов (РГа\, Ра тап, 1969). Вместе с тем известно, что аноеми-
рованные самцы, будучи несколько менее активными, тоже спариваются,
если их создить с рецентивной самкой (Вегтап, Тау]от, 1969). Поведе-
ние крыс при спаривапии практически не изменяется после перерезки
эфферентпых ольфаклорных нутей (Бич, 1960). Аносмированные сам-
ки продолжают циклировать и проявлять лордозис-рефлексе в парах с
с самцом (Атоп её а1., 1970). У циклирующих самок или у овариоэктоми-
рованных на фоне введения эстрогена и прогестерона проявление лорло-
вис-рефлекса даже облегчается при удалении обонятельных луковиц
(Ко]све: Агоп, 1984). В группах как полностью, так и частично бульбо-
эктомированных крыс частота лордоза выше, чем у ложнооперированных
и интактных крыс (БНлеНег её а1., 1982). Причем больше всего рецеп-
тивных самок (определяемых в тесте на спариванис с самцом) обнаружи-
вается в группе полностью бульбоэктомировапных крыс. Это указывает
на тот факт, что функциопирование обонятельной системы в норме ока-
зывает тормозные воздействия на рецептивность самок. Для реализации
292 Глава четвертая
являлись следующие запахи: 1 — мочи эстральной самун (а); 2— вари
оэктомированной самки (6); 3 — мочи членов собственной группы в
4 — кастрированного самца (2); 5 — интактного самца (9); 6 — воды °).
Для сексуально опытных самцов, живших в колониях е сородичам
обоего пола, запахи по их предпочитаемости расположились в следую-
щий ряд: а>б=в=е=д>е. Сексуально неопытные и кастрированные сам-
цы не оказывали предпочтения запаху мочи эстральной самки.
Сравнение реакций самок па запах кастрированных и иптактных сам-
цов показало, что независимо от их физиологического состояния (вструс
или диэструс) самки, имеющие половой опыт, достоворпо дольше иосле-
довали запах интактных самцов по сравнению с запахом кастратов. ам-
ки, пе имеющие полового опыта, дольше исследовали запах иитактных
самцов только в состоянии эструса, а у самок в состоянии диэструса та
кого предночтения не обнаружено (Сати её а1., 1965).
Изучение исследовательского поведения крыс в условиях вольерного
содержания позволлет получать более полные данные, отличающиеся от
таковых при тестировании животных, содержащихся в стапдартных кю
ветах в условиях вивария, и выделять дополнительный фактор (сопиал
ный ранг), влияющий на предпочтение различных запахов у крыс. этих
экспериментах использовали сексуальпо опытных самцов и самок в воз
расте 7—10 мес —потомков диких серых крыс, рожденных в виварии
(Соколов, Ляпунова, 1979, 1982). В вольору площадью 4 м, пасоленную
группой крыс из 6—10 особей с устойчивой иерархической структурой,
в качестве источника запаха помещали поодиночке в специально окон
струированных камерах-коробках самок, усыпленных нембуталом. дна
из них находилась в состолнии эструса. Контролем служила такая же,
но пустая коробка. Самцы достоверно дольше (р<0,01) исследовали ко-
робку с эстральной самкой, однако уровень их исследовательской актив-
ности был разным. Всегда максимальное время исследования домопстри-
ровал доминант. Более того, он начинал усилеппо патрулировать терри-
торню, припимал угрожающие позы и атаковал подчиненных самцов,
интенсивно метил мочой коробку © эстральшой самкой и близлежащее
пространство. Субдомивант проявлял аналогичные реакции, однако к
источнику запаха подходил реже и только в отсутствие доминанта.
од.
чипенцые самцы в такой ситуации редко покидали свои убежища и прРд-
почитали принюхиваться к коробке с самкой с большого расстояния.
Если же доминантного самца изымали из группы за 4 дня до экопери-
мента, то субдоминант занимал сго место и демонстрировавал реакции на
запах самки, аналогичные для доминанта. При парном предъявлении за-
пахов самок (как знакомых, так и незнакомых) в состоянии эструса
сексуально опытным самцам разиого социального ранга доминирующий
самец всегда предпочитал запах эстральной самки (оценка производилась
по времени исследования). Нодчиненные самцы избегали запаха самки
как в состоянии эструса, так и диэструса, если оп предъявлялся в паре
с запахом самца-доминанта, и при этом демонстрировали реакцию испу-
га. В 10 же время тестирование подчиненпых самцов в камере-ольфак-
тометре показало, что они предпочитают запах эстральной самки при
=
Поведение
293
предъявлении сго в паре с запахом одиночно содержавиегося самца.
Использование запаха социально доминантного самца в паре с запахом
эстральной самки измепяет реакцию подчинепных самцов (Соколов, Ля-
пунова, 1979).
Наблюдение в вольере за самками показало, что все они проявляют
слабую исследовательскую реакцию при внесепии на их территорию ко-
робки с запахом домипантвого самца. При этом они предпочитают скры-
ваться в убежищах. Предъявление запаха подчиненного самца или его
подстилки вызывает у всех самок возбуждение, интенсивную исследова-
тельскую реакцию, повышенную двигательную активность, перетаскива-
ние корма. При тестировании этих же самок в камере-ольфактометре в
течение 5 мин все самки достоверно дольше исследуют запах «чужого»
одиночного самца по сравнению с самцами различных социальных ран-
гов из своей группы. Наибольшая разница во времени исследования на-
блюдалась при предъявлении запаха «чулюго» одиночного самца и зна-
комого подчиненного (Соколов, Ляпунова, 1979).
Таким образом, реакция на запах у члепов группировки серых крыс
зависит не только от сексуального опыта особей, физиологического состоя-
ния самок, предварительного знакомства с донором запаха, но и от со-
ниального ранга самца.
Способность различать химические сигпалы партнера важна для ус-
нешпого спаривания крыс. Аногенитальное обследование, как правило,
предшествует садкам, поэтому его можно рассматривать как неотъемле-
мую часть поведения самцов при спаривапии. Если кастрированным в
раннем возрасте самцам ввести тестостерон, то у них время аногениталь-
ного обследовапия рецептивных самок возрастаст по сравнению с кастри-
рованными самцами (Этрег, 1972).
Экспонирование запаха рецептивной самки самцам, живущим в одно-
полых груплах, увеличивает процент животных, делающих садки на с0-
жителей по клетке, по срависнию с экспонировапием запаха нерецентив-
ной самки. Это свидетельствует о более высоком уровне возбуждения
реципиентов (РГа\, Ра тап, 1969). Вместе с тем известно, что аноеми-
рованные самцы, будучи несколько менее активными, тоже спариваются,
если их создить с рецентивной самкой (Вегтап, Тау]от, 1969). Поведе-
ние крыс при спаривапии практически не изменяется после перерезки
эфферентпых ольфаклорных нутей (Бич, 1960). Аносмированные сам-
ки продолжают циклировать и проявлять лордозис-рефлексе в парах с
с самцом (Атоп её а1., 1970). У циклирующих самок или у овариоэктоми-
рованных на фоне введения эстрогена и прогестерона проявление лорло-
вис-рефлекса даже облегчается при удалении обонятельных луковиц
(Ко]све: Агоп, 1984). В группах как полностью, так и частично бульбо-
эктомированных крыс частота лордоза выше, чем у ложнооперированных
и интактных крыс (БНлеНег её а1., 1982). Причем больше всего рецеп-
тивных самок (определяемых в тесте на спариванис с самцом) обнаружи-
вается в группе полностью бульбоэктомировапных крыс. Это указывает
на тот факт, что функциопирование обонятельной системы в норме ока-
зывает тормозные воздействия на рецептивность самок. Для реализации
294
Глава четвертая
этого эффекта пеобходима интактность как осповной обонятельной систе-
мы, так и вомероназального органа. Экспонирование самкам мочи самца
учащает у них проявление лордозис-рефлекса. Но всей вероятности,
в моче взрослых самцов содержатся вещества, способные снять тормоз-
ные эффекты на рецептивиость самок от химических стимулов иного
рода, воспринимаемых обонятельной системой.
Попытка проверить это предположение была предприпята Кельше и
Ароном (Кесве, Агоп, 1984). Они в экспериментах брали трех-четырох-
месячных сексуально опытных самок крыс пинии Маг и для того, что-
бы выяснить роль основной и дополнительной обонятельных систем
(Е1етушр её а1., 1979; баПево еб а1., 1982; Веаисватр еф а1., 1988), ис-
пользовали два способа предъявления запахов. Одной группе самок предо-
ставлялась возможность контакта с источником запаха, а другую поме-
щали на площадку, приподнятую на 5 см от обрабатываемой тестируе-
мыми веществами поверхности. В первом случае пол клетки обрызтивал-
ся мочой взрослых самцов. Тестирование самок па рецептивность
производилось через 8 ч. Все самки были в состоянии позднего проэстру-
са. Независимо от способа предъявления запаха в группе самок, подвер-
гавшихоя воздействию запаха мочи самцов, частота спариваний в опыт-
ной группе была достоверно выше, чем в контрольной (р<0,05), а частота
проявления лордозис-рефлекса в этих группах не различалась. Во втором
случае были тестированы на рецептивность самки с разрушенным воме-
роназальным органом. Частота спариваний и проявления лордоза ме от-
личалась в групие оперированных п ложнооперированных животных.
Экспонирование самкам с разрушенным вомероназальным органом мочи
самцов пе сказалось на исследуемых параметрах полового поведения
независимо от способа предъявления запаха. Результаты этой работы
свидетельствуют о том, что основная и дополнительная обопятельные
системы несут различную функциональную нагрузку. Вомероназальным
органом воспринимаются химические сигналы, стимулирующие проявле-
ние лордозис-рефлекса, а основная обонятельная система, по всей вероят- `
ности, оказывает тормозное воздействие на половое поведение самок в
отсутствие адекватного стимула. Таким образом, основная и дополнитель-
ная обонятельные системы связаны между собой реципрокными взаимо-
действиями (Кесве, Агоп, 1984).
.,
Гормональный контроль восприятия запаха мочи самцов серой крысы
был изучен по проявлению лордозис-позы на модели кастрированных в
3—4 мес самцов, которым вводили эстрадиол (75 мкг) и ссаживали через
48 ч с активными интактпыми самцами, регистрируя число лордозис-поз
в ответ на садки. Если кастрированным самцам через 40 ч после введе-
ния эстрадиола вводили 1 мг протестеропа, то на фоне его действия
увеличивалось число особей, демонстрирующих лордозис-позу. Экспози-
ция мочи сексуально активных самцов кастрированпым самцам за 9ч
до тестирования не увеличивала число особей, демопотрирующих лорло-
зис-позу. Более низкие дозы прогестерона (менее 1 мг) повышали чувот-
вительность кастрированных самцов к запаху мочи. Эти результаты сви-
детельствуют, что облегчение проявления лорлозис-реакции у кастриро-
Поведение
295
ванных самцов связано с взаимодействием гормональных и сенсорпых
механизмов (Сар её а|., 1985).
Обонятельные ситналы, исходлщио от самок, по-видимому, не влияют
на проявление лордозис-рефлекса у других самок. В норме ороговение
вагипального эпителия и лордозис-рефлекс возникают одновременно на
стадий позднего проэструса, на чем и основано тестирование самок на
рецептивность. Однако при длительной социальной изоляции происходит
диссоциация этих явлений. У значительной части одиночно живущих
самок лордоз перестает быть циклическим и может быть вызван в любой
день просто взятием в руки. Тем не мепое ороговение вагинального эпи-
телия будет по-прежнему наблюдаться одип раз в 4—5 дней. Продувание
в боксы с такими самками воздуха от других самок на различных ста-
диях эстрального цикла пе нормализует состояния реципиентов ( МеСПа-
фосКк, 1981, 1983а). Подача только акустических сигналов или предостав-
ление возможности видеть другую крысу (самку) в отсутствие сигпалов
других модальтостей также не приводит к синхронизации лордозис-реф-
лекса с началом эструса у социально изолированных самок. Макклинток
на основании этих экспериментов делает вывод о необходимости тактиль-
ных раздражений, исходящих от сородичей, для нормального циклическо-
го проявления лордозис-рефлекса.
Ряд работ посвящен изучению роли обонятельных сигналов в половом
созревании крысы. Вапденбергом (Уап4епЪегов, 1976) было. описано яв-
лепие ускорения полового созревания самок крыс на 6—7 дней под влия-
нием запаха мочи взрослых самцов. У самок линии \/15 ат, лишенных
контактов с самцами (контактирующих лишь
с запахом взрослой самки),
влагалище открывалось на 34,9-2,9 день, а первый эструс наступал на
35,2=3,4 день. У самок, выращенных в атмосфер запахов взрослых сам-
цов, эти показатели составили 28,1=1,9 и 29,2=2,3 дня соответственно
(Запепез-Смадо, СаПего, 1979). Периферическая диафферентация допол-
нительной обонятельной системы в 2[-дневном возрасте путем электроли-
тического разрушепия обоих вомероназальных нервов блокирует ускоре-
ние полового созревания под влиянием запаха мочи самца. Предполага-
ется, что функционирование дополнительной обонятельной системы
связапо с эндокринными реакциями на праймер-феромоны (Запсвез-
Стадо, 1982).
Задержка открытия влагалища у крыс была ранес обнаружена после
периферической аносмии. Бульбоэктомия или разрушение обонятельных
трактов в шестидновном возрасте приводят к задержке открытия влага-
лища на 10 дней по сравнению с контрольной группой (Заю её а1., 1984).
“Следует отметить, что сроки полового созревания лабораторных крыс
несколько отличаются от сроков, известных для их диких сородичей
(Мегетег её а1., 1982). Цак правило, в природе крысы достигают полово-
зрелости в возрасте околю 3 мес (Соколов, Карасева, 1985). Эти данные
подтверждаются наблюдениями за самками диких крыс, содержащихся в
условиях вольеры (Квашнин, 1983). В синантропиых популяциях серых
крыс, в населенных пунктах, половое созревание молодняка может
затятиваться до шестимесячного возраста (Рыльников, 1983). В регуля-
294
Глава четвертая
этого эффекта пеобходима интактность как осповной обонятельной систе-
мы, так и вомероназального органа. Экспонирование самкам мочи самца
учащает у них проявление лордозис-рефлекса. Но всей вероятности,
в моче взрослых самцов содержатся вещества, способные снять тормоз-
ные эффекты на рецептивиость самок от химических стимулов иного
рода, воспринимаемых обонятельной системой.
Попытка проверить это предположение была предприпята Кельше и
Ароном (Кесве, Агоп, 1984). Они в экспериментах брали трех-четырох-
месячных сексуально опытных самок крыс пинии Маг и для того, что-
бы выяснить роль основной и дополнительной обонятельных систем
(Е1етушр её а1., 1979; баПево еб а1., 1982; Веаисватр еф а1., 1988), ис-
пользовали два способа предъявления запахов. Одной группе самок предо-
ставлялась возможность контакта с источником запаха, а другую поме-
щали на площадку, приподнятую на 5 см от обрабатываемой тестируе-
мыми веществами поверхности. В первом случае пол клетки обрызтивал-
ся мочой взрослых самцов. Тестирование самок па рецептивность
производилось через 8 ч. Все самки были в состоянии позднего проэстру-
са. Независимо от способа предъявления запаха в группе самок, подвер-
гавшихоя воздействию запаха мочи самцов, частота спариваний в опыт-
ной группе была достоверно выше, чем в контрольной (р<0,05), а частота
проявления лордозис-рефлекса в этих группах не различалась. Во втором
случае были тестированы на рецептивность самки с разрушенным воме-
роназальным органом. Частота спариваний и проявления лордоза ме от-
личалась в групие оперированных п ложнооперированных животных.
Экспонирование самкам с разрушенным вомероназальным органом мочи
самцов пе сказалось на исследуемых параметрах полового поведения
независимо от способа предъявления запаха. Результаты этой работы
свидетельствуют о том, что основная и дополнительная обопятельные
системы несут различную функциональную нагрузку. Вомероназальным
органом воспринимаются химические сигналы, стимулирующие проявле-
ние лордозис-рефлекса, а основная обонятельная система, по всей вероят- `
ности, оказывает тормозное воздействие на половое поведение самок в
отсутствие адекватного стимула. Таким образом, основная и дополнитель-
ная обонятельные системы связаны между собой реципрокными взаимо-
действиями (Кесве, Агоп, 1984).
.,
Гормональный контроль восприятия запаха мочи самцов серой крысы
был изучен по проявлению лордозис-позы на модели кастрированных в
3—4 мес самцов, которым вводили эстрадиол (75 мкг) и ссаживали через
48 ч с активными интактпыми самцами, регистрируя число лордозис-поз
в ответ на садки. Если кастрированным самцам через 40 ч после введе-
ния эстрадиола вводили 1 мг протестеропа, то на фоне его действия
увеличивалось число особей, демонстрирующих лордозис-позу. Экспози-
ция мочи сексуально активных самцов кастрированпым самцам за 9ч
до тестирования не увеличивала число особей, демопотрирующих лорло-
зис-позу. Более низкие дозы прогестерона (менее 1 мг) повышали чувот-
вительность кастрированных самцов к запаху мочи. Эти результаты сви-
детельствуют, что облегчение проявления лорлозис-реакции у кастриро-
Поведение
295
ванных самцов связано с взаимодействием гормональных и сенсорпых
механизмов (Сар её а|., 1985).
Обонятельные ситналы, исходлщио от самок, по-видимому, не влияют
на проявление лордозис-рефлекса у других самок. В норме ороговение
вагипального эпителия и лордозис-рефлекс возникают одновременно на
стадий позднего проэструса, на чем и основано тестирование самок на
рецептивность. Однако при длительной социальной изоляции происходит
диссоциация этих явлений. У значительной части одиночно живущих
самок лордоз перестает быть циклическим и может быть вызван в любой
день просто взятием в руки. Тем не мепое ороговение вагинального эпи-
телия будет по-прежнему наблюдаться одип раз в 4—5 дней. Продувание
в боксы с такими самками воздуха от других самок на различных ста-
диях эстрального цикла пе нормализует состояния реципиентов ( МеСПа-
фосКк, 1981, 1983а). Подача только акустических сигналов или предостав-
ление возможности видеть другую крысу (самку) в отсутствие сигпалов
других модальтостей также не приводит к синхронизации лордозис-реф-
лекса с началом эструса у социально изолированных самок. Макклинток
на основании этих экспериментов делает вывод о необходимости тактиль-
ных раздражений, исходящих от сородичей, для нормального циклическо-
го проявления лордозис-рефлекса.
Ряд работ посвящен изучению роли обонятельных сигналов в половом
созревании крысы. Вапденбергом (Уап4епЪегов, 1976) было. описано яв-
лепие ускорения полового созревания самок крыс на 6—7 дней под влия-
нием запаха мочи взрослых самцов. У самок линии \/15 ат, лишенных
контактов с самцами (контактирующих лишь
с запахом взрослой самки),
влагалище открывалось на 34,9-2,9 день, а первый эструс наступал на
35,2=3,4 день. У самок, выращенных в атмосфер запахов взрослых сам-
цов, эти показатели составили 28,1=1,9 и 29,2=2,3 дня соответственно
(Запепез-Смадо, СаПего, 1979). Периферическая диафферентация допол-
нительной обонятельной системы в 2[-дневном возрасте путем электроли-
тического разрушепия обоих вомероназальных нервов блокирует ускоре-
ние полового созревания под влиянием запаха мочи самца. Предполага-
ется, что функционирование дополнительной обонятельной системы
связапо с эндокринными реакциями на праймер-феромоны (Запсвез-
Стадо, 1982).
Задержка открытия влагалища у крыс была ранес обнаружена после
периферической аносмии. Бульбоэктомия или разрушение обонятельных
трактов в шестидновном возрасте приводят к задержке открытия влага-
лища на 10 дней по сравнению с контрольной группой (Заю её а1., 1984).
“Следует отметить, что сроки полового созревания лабораторных крыс
несколько отличаются от сроков, известных для их диких сородичей
(Мегетег её а1., 1982). Цак правило, в природе крысы достигают полово-
зрелости в возрасте околю 3 мес (Соколов, Карасева, 1985). Эти данные
подтверждаются наблюдениями за самками диких крыс, содержащихся в
условиях вольеры (Квашнин, 1983). В синантропиых популяциях серых
крыс, в населенных пунктах, половое созревание молодняка может
затятиваться до шестимесячного возраста (Рыльников, 1983). В регуля-
296 Глава четвертая
ции эстральных циклов самок крыс обонятельным сигналам принадлеячит
весьма ответственная роль. Работами Шато © сотрудниками (Свафеам е
а1., 1972) было показано, что моча ках от интактных, так и от кастри-
рованных самцов и овариозктомированных самок крыс линии \У/15ват
обладает сходным действием на эстральные циклы, укорачивая их © 5
до 4 дней. Авторы достигали указанного эффекта обрызгиванием мочо
пола клеток одипочно содержащихся животных. Измерение уровня про-
тестерона в крови и гистологическое исследование яичников у опытных
и контрольных животных показали, что укорочение острального цикла
является результатом спижения уровня прогестерона в крови нод воздей-
ствием запаха мочи сородичей (СЬабеам ек а1., 1976). В группе питантных
самок спонтанное укорочение цикла с 5 до 4 дней отмечается в 5 No слу-
чаев (Агол, СЬзеаи, 1971). Существуют экспериментальные подтвержде-
пия того, что в регуляции длительности эстрального цикла участвуют
сигналы, воспринимаемые дополнительной обопятельной системой.
У 60-лневных самок крыс с разрушенным вомероназальным органом не
обнаруживается укороченис длительности зотральпых циклов мод влия-
нием запаха мочи самцов. В группе ТО-дневных самок, подвергнутых
электролитическому разрушению вомероназальных нервов, воврастаот
процент самок, приходящих в состояние эструса при экспозиции мот
самцов (СаПесо её а|., 1982). При раздельном содержании самцов, и я
мок линии УЧиаг ежедневно в состоянии эструса находится 29,1+0,8%
самок. Экспонирование общего запаха самцов увеличивает этот процент
ко 48,9+4,4 (СаПево, Запснез-Ста4о, 1979}.
6
В
Изучение воздействия запаха самок на самих самок было пачат
такого простого эксперимента. Молодых самок липии Эргазие- ом еу к
регулярными четырехдневными циклами содержали в группах по осо
бей. В контрольной группе животных содержали изолированно в клетках,
отстоящих одпа от друтой на 70 см. Оказалось, что эотральные циклы
самок при групповом способе содержания достоверно более оинхронты по
сравнению с таковыми у одиночных животных. Макклипток ( ИеСаво ,
1983а—с) была предпринята попытка проанализировать воздействие за
пахов самок с различным физиологическим состоянием а регуляцию
эстральных циклов. Эстральный цикл самки крысы условно был разлелен
на три фазы, отличающиеся по концентрации соответствующих омонор
в крови, а также по гистологической картине влагалищного мазка: ) во.
ликулярная стадия (диэструс-проэструс), 2) овуляторная стадия (про
эструс-эструс), 3) лютеальная стадия (метаэструс-дизотрус). казалось,
что запах самок, находящихся на фолликулярной стадии, нормализует. у
пругих самок эстральные циклы, сводя их у большинства к трех-чоты-
рехдневным. Запах самок на овуляторной стадии удтиняот оотр тыс
циклы, увеличивая долю самок с циклами от 5 до 11 дней. Запах сал <
в лютеальной фазе не оказывает достоверного воздействия на прололжи
тельность эстральных циклов. В последней групие животы дите етооть
эстральных циклов составляла от 4 до Т лней (МеСШмоскК,
›). No
Поскольку в нормальных условиях содержания большинство самог
либо беременные, либо кормящие, представляла интерес проверка воз
Поведение
297
действия запахов беременных и кормящих самок на эстральные циклы
других самок. Для этого Макклипток (1983а) молодых самок крыс ли-
нии бргарие-Оаз]еу содержала в боксах, через которые продувала воз-
дух от беременных или лактирующих самок. Запах от беременных самок
укорачивал эстральные циклы животных как по сравнению с контролем,
так и по сравнению с собственными циклами самок опытной группы до
начала эксперимента. Эффект запаха беременных самок аналогичен
таковому от самок в фолликулярной стадий, т. е. эти запахи увеличива-
ют долю самок с четырехдневными эстральтыми циклами, что связано с
подъемом эстрогена в крови. Запахи от лактирующих самок с детеныша-
ми удлипяют эстральные циклы у подопытпых самок таким образом, что
наряду с нормально циклирующими самками выделяется группа с 9—
16-дневными периодами. Эффект запаха лактирующих самок сопоставим
с таковым от самок на овуляторной стадии. Однако автор воздерживается
от вывода об однотицности химических сигналов, исходящих от беремен-
пых самок и самок на фолликулярной стадии, лактирующих и на ову-
ляторной стадии.
Обочлтельные сигналы в материневом поведении крысе
Только что рожденные крысята не видят и не слышат. Однако спустя
всего лишь несколько дней зверьки начинают анализировать некоторые
сигпалы, поступающие из внешнего мира. Дыхание через ное обеспечи-
вает наступление непрерывпого потока информации об окружающей их
среде. Критериями, позволяющими отличать обнюхивание от простого
вдыхания, служат изменение дыхательных ритмов, а также зависимость
этих ритмов от обонятельной стимуляции. У крысят в возрасте до 7 дней
обонятельная стимуляция вызывает беспорядочное изменение дыхатель-
ных ритмов. Но к 11—20-му дню их жизни формируются учащенные рит-
мы дыхапия, характерные как для направленного обследования, так и
для необходимого его компонента — обиюхивания (АШегз. Мау, 1980а, Ь).
С двухдиевпого возраста крысята способны к ассоциативному обуче-
нию на новый запах. При экспонированипи запаха лимона и последую-
щей инъекции раствора хлорида лития, вызывающего болевые ощущения,
онспериментальные крысята на 8-й день в отличие от контрольных из-
бегали предъявляемого им запаха (Виду, Спеайе, 1977). Также было ус-
таповлепо, что чувствительность к моче особей свосго вида и к типичио-
му обонятельному раздражителю
— амилацетату повышается у крысят
С возрастом. До пятидневного возраста они способны к анализу этих за-
пахов, однако различают лишь довольно высокие концентрации. Тем не
менее существует ряд экспериментальных доказательств, что у крысят в
раннем возрасте именно обонятельные сигналы являются наиравляющи-
ми в поиске сосков лактирующей самки. Так, при удалении шерсти у
матери на брюхе и изменении ее температуры тела зверьки продолжают
присасываться к соскам (Вгапо с а1., 1980).
Крысята могут реагировать на запах матери, ипгибирующий их чрез-
мерную двигательную активпость, со 2-го до 12-го дня (Зварго, За[ав,
296 Глава четвертая
ции эстральных циклов самок крыс обонятельным сигналам принадлеячит
весьма ответственная роль. Работами Шато © сотрудниками (Свафеам е
а1., 1972) было показано, что моча ках от интактных, так и от кастри-
рованных самцов и овариозктомированных самок крыс линии \У/15ват
обладает сходным действием на эстральные циклы, укорачивая их © 5
до 4 дней. Авторы достигали указанного эффекта обрызгиванием мочо
пола клеток одипочно содержащихся животных. Измерение уровня про-
тестерона в крови и гистологическое исследование яичников у опытных
и контрольных животных показали, что укорочение острального цикла
является результатом спижения уровня прогестерона в крови нод воздей-
ствием запаха мочи сородичей (СЬабеам ек а1., 1976). В группе питантных
самок спонтанное укорочение цикла с 5 до 4 дней отмечается в 5 No слу-
чаев (Агол, СЬзеаи, 1971). Существуют экспериментальные подтвержде-
пия того, что в регуляции длительности эстрального цикла участвуют
сигналы, воспринимаемые дополнительной обопятельной системой.
У 60-лневных самок крыс с разрушенным вомероназальным органом не
обнаруживается укороченис длительности зотральпых циклов мод влия-
нием запаха мочи самцов. В группе ТО-дневных самок, подвергнутых
электролитическому разрушению вомероназальных нервов, воврастаот
процент самок, приходящих в состояние эструса при экспозиции мот
самцов (СаПесо её а|., 1982). При раздельном содержании самцов, и я
мок линии УЧиаг ежедневно в состоянии эструса находится 29,1+0,8%
самок. Экспонирование общего запаха самцов увеличивает этот процент
ко 48,9+4,4 (СаПево, Запснез-Ста4о, 1979}.
6
В
Изучение воздействия запаха самок на самих самок было пачат
такого простого эксперимента. Молодых самок липии Эргазие- ом еу к
регулярными четырехдневными циклами содержали в группах по осо
бей. В контрольной группе животных содержали изолированно в клетках,
отстоящих одпа от друтой на 70 см. Оказалось, что эотральные циклы
самок при групповом способе содержания достоверно более оинхронты по
сравнению с таковыми у одиночных животных. Макклипток ( ИеСаво ,
1983а—с) была предпринята попытка проанализировать воздействие за
пахов самок с различным физиологическим состоянием а регуляцию
эстральных циклов. Эстральный цикл самки крысы условно был разлелен
на три фазы, отличающиеся по концентрации соответствующих омонор
в крови, а также по гистологической картине влагалищного мазка: ) во.
ликулярная стадия (диэструс-проэструс), 2) овуляторная стадия (про
эструс-эструс), 3) лютеальная стадия (метаэструс-дизотрус). казалось,
что запах самок, находящихся на фолликулярной стадии, нормализует. у
пругих самок эстральные циклы, сводя их у большинства к трех-чоты-
рехдневным. Запах самок на овуляторной стадии удтиняот оотр тыс
циклы, увеличивая долю самок с циклами от 5 до 11 дней. Запах сал <
в лютеальной фазе не оказывает достоверного воздействия на прололжи
тельность эстральных циклов. В последней групие животы дите етооть
эстральных циклов составляла от 4 до Т лней (МеСШмоскК,
›). No
Поскольку в нормальных условиях содержания большинство самог
либо беременные, либо кормящие, представляла интерес проверка воз
Поведение
297
действия запахов беременных и кормящих самок на эстральные циклы
других самок. Для этого Макклипток (1983а) молодых самок крыс ли-
нии бргарие-Оаз]еу содержала в боксах, через которые продувала воз-
дух от беременных или лактирующих самок. Запах от беременных самок
укорачивал эстральные циклы животных как по сравнению с контролем,
так и по сравнению с собственными циклами самок опытной группы до
начала эксперимента. Эффект запаха беременных самок аналогичен
таковому от самок в фолликулярной стадий, т. е. эти запахи увеличива-
ют долю самок с четырехдневными эстральтыми циклами, что связано с
подъемом эстрогена в крови. Запахи от лактирующих самок с детеныша-
ми удлипяют эстральные циклы у подопытпых самок таким образом, что
наряду с нормально циклирующими самками выделяется группа с 9—
16-дневными периодами. Эффект запаха лактирующих самок сопоставим
с таковым от самок на овуляторной стадии. Однако автор воздерживается
от вывода об однотицности химических сигналов, исходящих от беремен-
пых самок и самок на фолликулярной стадии, лактирующих и на ову-
ляторной стадии.
Обочлтельные сигналы в материневом поведении крысе
Только что рожденные крысята не видят и не слышат. Однако спустя
всего лишь несколько дней зверьки начинают анализировать некоторые
сигпалы, поступающие из внешнего мира. Дыхание через ное обеспечи-
вает наступление непрерывпого потока информации об окружающей их
среде. Критериями, позволяющими отличать обнюхивание от простого
вдыхания, служат изменение дыхательных ритмов, а также зависимость
этих ритмов от обонятельной стимуляции. У крысят в возрасте до 7 дней
обонятельная стимуляция вызывает беспорядочное изменение дыхатель-
ных ритмов. Но к 11—20-му дню их жизни формируются учащенные рит-
мы дыхапия, характерные как для направленного обследования, так и
для необходимого его компонента — обиюхивания (АШегз. Мау, 1980а, Ь).
С двухдиевпого возраста крысята способны к ассоциативному обуче-
нию на новый запах. При экспонированипи запаха лимона и последую-
щей инъекции раствора хлорида лития, вызывающего болевые ощущения,
онспериментальные крысята на 8-й день в отличие от контрольных из-
бегали предъявляемого им запаха (Виду, Спеайе, 1977). Также было ус-
таповлепо, что чувствительность к моче особей свосго вида и к типичио-
му обонятельному раздражителю
— амилацетату повышается у крысят
С возрастом. До пятидневного возраста они способны к анализу этих за-
пахов, однако различают лишь довольно высокие концентрации. Тем не
менее существует ряд экспериментальных доказательств, что у крысят в
раннем возрасте именно обонятельные сигналы являются наиравляющи-
ми в поиске сосков лактирующей самки. Так, при удалении шерсти у
матери на брюхе и изменении ее температуры тела зверьки продолжают
присасываться к соскам (Вгапо с а1., 1980).
Крысята могут реагировать на запах матери, ипгибирующий их чрез-
мерную двигательную активпость, со 2-го до 12-го дня (Зварго, За[ав,
298 Глава четвертая
1970). Их способность отвечать в достаточно раннем возрасте на мате-
ринские запахи интенсивными переворачивапиями по направлению к
гнезду и более тонкими, ориептирующими движениями при коптакте с
матерью в поиске соска была подтверждена как с применением‘ перифе-
рической аносмии
— обработкой обонятельной поверхиости сульфатом
цинка— так и центральной
— удалением обонятельных луковиц (РоНаск,
Сасрз, 1975; Зов, ТоЪасв, 1975; АШехв, 1976; Это в а[., 1976).
Тщательные наблюдения за поведением аносмированных крысят пока-
зывают, что они не только не ориентируются по отношению к гнезду и
к матери, но и при контакте с ней нелегко находят соски. По-видимому,
обнаружение сосков и присасывание к пим зависят от обонятельных си-
тналов. Крысята, аносмированные периферически, оказываются неспособ-
пыми присасываться к соскам анастезированной лактирующей матери,
теряют в весе и даже погибают (Зе её а1., 1976). Сосание у крысят
также может быть подавлено промыванием материнских сосков ацето-
ном, спиртом и водой. Возможно, что имеются какие-то компонечты,
секретируемые специализированными кожными железами, расположен-
ными в районе сосков, выделяющие химические сигналы, с помощью ко-
торых крысята ориептируются и присасываются. В материнском молоке
присутствие этих сигналов пе обпаружено: крысята не проявляют спо-
собности ориентироваться на экстракты материанского молока, нанесенные
па се шкурку (Ног её а1., 1976).
Влияние запаха матери на физиологическое состояние крысят было
показано по изменению у них частоты сердечных сокращений при отня-
тии у матери. Экспонирование запаха матери способотвует восстаповле-
нию исходной частоты сокращений. Запах пелактирующей самки таким
свойством не обладает (Сошрюп её а1., 1977). Самки серых крыс способ-
ны различать пол своих детенышей: опи дольше вылизывают аногени-
тальную область у детеныитей-самцов (Вс оопб, ЗасКз, 1984). Если де-
теньциамсамкам вводить мужокой половой гормон (тестостерон), то
мать вылизывает их более интенсивно первые девять суток со времени _
инъекции. Введение эстрадиола также в равной стенени увсличивает ин-
тенсивность вылизывания (Мооге, 1981). Эффект эстрадиола можно
объяснить наличием у самок крысят биохимической систомы деаромати-
зации кольца А у эстрадиола. Взрослые самки крыс также обнаруживают
предночтение запаха мочи крысят-самцов по сравнению с таковым кры-
сят-самок (Мооге, 1985).
По всей видимости, химические сигпалы выполняют определяющую
роль в процессе взаимодействия лактирующей самки с ое выводком, по-
скольку бульбооктомированные самки пе проявляют материнского пове-
дения, в результате чего выводок либо игнорируется ето, либо поедается.
Для выяспения происхождения и природы обонятельных сигналов лак-
тирующей самки крысы были предприняты мпогочисленные исследова-
ния. Один из таких сигналов получил название материпского феромона
(аттрактант для крысят), продуцируется самкой на 14-й день после ро-
дов, и именно с этого момента крысята начинают им привлекаться. Сро-
ки выделения этого аттрактанта не могут быть ускорены заменой собот-
Поведение
299
венного выводка матери крысятами более старшего возраста (Мот
оп, 1973).
Возможно, крысята могут ассоциировать запах каловых масс, специ-
фичных для материнской диеты, с соответствующим поведенческим отве-
том. В таком случае материнский феромон не является врожденным видо-
сисцифическим релизером. Таким образом, связь мать-—-детеныш может
быть построена па «обонятельном имипринтипге».
Ряду авторов удалось воспроизвести явление «обонятельного имприн-
тинга» в лабораторных условиях. Крысятам экспонировали искусствен-
ные пахучие вещества в точепие первых дней жизни. При тестировании
в более позднем возрасте они оказывали предпочтение особям свосго
вида, несущим эти запахи (Магг, ТАШэюп, 1969). Однако существует и
противоположное мнение: аттрактивность материнского феромона посит
врожденный характер и является обонятельным сигпалом, выработанным
в процессе эволюции (ЗсНашасВог, Мое, 1982). Крысятам в возрасте
14—27 дней предоставляли выбор между двумя сигналами, одним из
которых всегда были либо самка, продунирующая феромон, либо сам фе-
ромон, а другим 1) знакомый крысятам запах; 2) самка крысы, не нро-
дуцирующая феромон (в этой серии экспериментов крысята не имели
предварительного зпакомства с материнским феромоном, так как кормя-
щая их самка содержалась на специальной дисте, исключающей его по-
явление); 3) самка крысы, пе продуцирующая феромон, но несущая зна-
комый для крысят запах их гнезда (лимонное масло).
Во всех сериях экспериментов зверьки предпочитали запах либо ма-
теринского феромопа, либо продуцирующей его самки. Эти экоперимен-
ты подтверждают точку зрения о врожденном характере воздействия ма-
теринского феромопа. Вероятно, в первые дни жизни крысята запечатле-
вают различные запахи гнезда, не связанные © запахом материнского
феромона, что в дальнейшем способствует правильному выбору ими места
и продуктов питания, а также места постройки гнезда (ЕсВап@а сё а1.
1982).Таким образом, поведение крысят в ответ на материнский феромон,
характеризующееся ориентацией и движением к источнику запаха,
наиболее вероятно, является врожденным и уровень предварительного
знапия этого запаха (т. е. наличие его предварительной экспозиции) не
оказывает существенного влиянил на его предпочтение.
Химическая природа материнского феромона серых крыс
— состав и
строение аттрактивных для крысят компонентов — до настоящего времени
пе установлена.
Функции описанного выше материнского феромона были исследованы
на лабораторных крысах линии У$аг и Гопо-Еуапз, причем действие
его аналогично у крыс обеих генетических линий (Вгоуп, 41982). Одпако
в литературе содержатся сведения, что 21-дневпые крысята линии Гопр-
Еуап8 не обнаруживают предпочтения запаха анальных выделений лак-
тирующей самки по сравнению с черожавшей (Се Мозсиз, 1979).
В настоящее время имеется мало данных о роли обонятельных сигна-
лов в Коммуникации крысят между собой. Установлена способность уз-
“
298 Глава четвертая
1970). Их способность отвечать в достаточно раннем возрасте на мате-
ринские запахи интенсивными переворачивапиями по направлению к
гнезду и более тонкими, ориептирующими движениями при коптакте с
матерью в поиске соска была подтверждена как с применением‘ перифе-
рической аносмии
— обработкой обонятельной поверхиости сульфатом
цинка
— так и центральной
— удалением обонятельных луковиц (РоНаск,
Сасрз, 1975; Зов, ТоЪасв, 1975; АШехв, 1976; Это в а[., 1976).
Тщательные наблюдения за поведением аносмированных крысят пока-
зывают, что они не только не ориентируются по отношению к гнезду и
к матери, но и при контакте с ней нелегко находят соски. По-видимому,
обнаружение сосков и присасывание к пим зависят от обонятельных си-
тналов. Крысята, аносмированные периферически, оказываются неспособ-
пыми присасываться к соскам анастезированной лактирующей матери,
теряют в весе и даже погибают (Зе её а1., 1976). Сосание у крысят
также может быть подавлено промыванием материнских сосков ацето-
ном, спиртом и водой. Возможно, что имеются какие-то компонечты,
секретируемые специализированными кожными железами, расположен-
ными в районе сосков, выделяющие химические сигналы, с помощью ко-
торых крысята ориептируются и присасываются. В материнском молоке
присутствие этих сигналов пе обпаружено: крысята не проявляют спо-
собности ориентироваться на экстракты материанского молока, нанесенные
па се шкурку (Ног её а1., 1976).
Влияние запаха матери на физиологическое состояние крысят было
показано по изменению у них частоты сердечных сокращений при отня-
тии у матери. Экспонирование запаха матери способотвует восстаповле-
нию исходной частоты сокращений. Запах пелактирующей самки таким
свойством не обладает (Сошрюп её а1., 1977). Самки серых крыс способ-
ны различать пол своих детенышей: опи дольше вылизывают аногени-
тальную область у детеныитей-самцов (Вс оопб, ЗасКз, 1984). Если де-
теньциамсамкам вводить мужокой половой гормон (тестостерон), то
мать вылизывает их более интенсивно первые девять суток со времени _
инъекции. Введение эстрадиола также в равной стенени увсличивает ин-
тенсивность вылизывания (Мооге, 1981). Эффект эстрадиола можно
объяснить наличием у самок крысят биохимической систомы деаромати-
зации кольца А у эстрадиола. Взрослые самки крыс также обнаруживают
предночтение запаха мочи крысят-самцов по сравнению с таковым кры-
сят-самок (Мооге, 1985).
По всей видимости, химические сигпалы выполняют определяющую
роль в процессе взаимодействия лактирующей самки с ое выводком, по-
скольку бульбооктомированные самки пе проявляют материнского пове-
дения, в результате чего выводок либо игнорируется ето, либо поедается.
Для выяспения происхождения и природы обонятельных сигналов лак-
тирующей самки крысы были предприняты мпогочисленные исследова-
ния. Один из таких сигналов получил название материпского феромона
(аттрактант для крысят), продуцируется самкой на 14-й день после ро-
дов, и именно с этого момента крысята начинают им привлекаться. Сро-
ки выделения этого аттрактанта не могут быть ускорены заменой собот-
Поведение
299
венного выводка матери крысятами более старшего возраста (Мот
оп, 1973).
Возможно, крысята могут ассоциировать запах каловых масс, специ-
фичных для материнской диеты, с соответствующим поведенческим отве-
том. В таком случае материнский феромон не является врожденным видо-
сисцифическим релизером. Таким образом, связь мать-—-детеныш может
быть построена па «обонятельном имипринтипге».
Ряду авторов удалось воспроизвести явление «обонятельного имприн-
тинга» в лабораторных условиях. Крысятам экспонировали искусствен-
ные пахучие вещества в точепие первых дней жизни. При тестировании
в более позднем возрасте они оказывали предпочтение особям свосго
вида, несущим эти запахи (Магг, ТАШэюп, 1969). Однако существует и
противоположное мнение: аттрактивность материнского феромона посит
врожденный характер и является обонятельным сигпалом, выработанным
в процессе эволюции (ЗсНашасВог, Мое, 1982). Крысятам в возрасте
14—27 дней предоставляли выбор между двумя сигналами, одним из
которых всегда были либо самка, продунирующая феромон, либо сам фе-
ромон, а другим 1) знакомый крысятам запах; 2) самка крысы, не нро-
дуцирующая феромон (в этой серии экспериментов крысята не имели
предварительного зпакомства с материнским феромоном, так как кормя-
щая их самка содержалась на специальной дисте, исключающей его по-
явление); 3) самка крысы, пе продуцирующая феромон, но несущая зна-
комый для крысят запах их гнезда (лимонное масло).
Во всех сериях экспериментов зверьки предпочитали запах либо ма-
теринского феромопа, либо продуцирующей его самки. Эти экоперимен-
ты подтверждают точку зрения о врожденном характере воздействия ма-
теринского феромопа. Вероятно, в первые дни жизни крысята запечатле-
вают различные запахи гнезда, не связанные © запахом материнского
феромона, что в дальнейшем способствует правильному выбору ими места
и продуктов питания, а также места постройки гнезда (ЕсВап@а сё а1.
1982).Таким образом, поведение крысят в ответ на материнский феромон,
характеризующееся ориентацией и движением к источнику запаха,
наиболее вероятно, является врожденным и уровень предварительного
знапия этого запаха (т. е. наличие его предварительной экспозиции) не
оказывает существенного влиянил на его предпочтение.
Химическая природа материнского феромона серых крыс
— состав и
строение аттрактивных для крысят компонентов — до настоящего времени
пе установлена.
Функции описанного выше материнского феромона были исследованы
на лабораторных крысах линии У$аг и Гопо-Еуапз, причем действие
его аналогично у крыс обеих генетических линий (Вгоуп, 41982). Одпако
в литературе содержатся сведения, что 21-дневпые крысята линии Гопр-
Еуап8 не обнаруживают предпочтения запаха анальных выделений лак-
тирующей самки по сравнению с черожавшей (Се Мозсиз, 1979).
В настоящее время имеется мало данных о роли обонятельных сигна-
лов в Коммуникации крысят между собой. Установлена способность уз-
“
300
Глава четвертая
навания ими родни того же возраста в тестах с предпочтением запаха.
Ирысята в возрасте от 10 до 15 дней предпочитают запах зпакомых
крысят и незнакомых родственников по сравнению с незнакомыми не-
родственниками. С возрастом доля животных, предпочитающих незнако-
мых ровеспиков, возрастает (Неррег, 1983).
Марки ровочное поведение и поддержание
черархической структуры группы
Возможность быстрого опознания индивидуально каждой особи и ее фи-
зиологического состояния— необходимое условие существования соци-
альпой структуры.
Пахучие вещества, содержащиеся в моче крыс, несут информацию о
видовой, половой принадлежности особи, ее социальном рапге и физиоло-
гическом состояпии (Сагг сё а1., 4970; Афемз, Са1еЁ, 1973; Соколов и др.,
1982, 1985; п др.). Серые крысы способны различать своих сородичей по
обонятельным сигналам даже в отсутствие сигналов других модальностей.
Потеря обоняния снижает или полностью устраняет агрессивность у ди-
ких серых (Аелз, СаеЁ 1973) и лабораторных белых крые (Гаезпо,
Тюте, 1975). Способность различать индивидуальные и «групповые» запа-
хи рядом авторов рассматривается как основа для впутри- и межгруп-
повых взаимодействий (Наумов, Гольцман, 1972).
Как живуние синантропно, так и в экзоаптроиных популяциях кры-
сы метят мочой новые предметы на своей территории, особенно если они
несут информацию о сородичах (Адашз, 1976; Вто\уп, 1975; Рисе, 1975;
В:срагдз. З{еуепз, 1977; и др.), а также партнеров (Цвалинии, 1983;
Тауюг её а1., 1984). Самцы предпочитают метить предметы с меткой са-
мок, тогда как самки не обнаруживают такого выраженного предцочтения
к меткам самцов (Вго\уп, 1977а, Ь). В то же время самки чаще метят
предметы, несущие обонятельную ипформанию об интактных самцах по
сравнению с кастрированными (Тауог ей а1., 1982). Изолированное с0-
держание крыс снижает маркировочную активность как у самцов, так и
у самок. Самцы, содержавишеся в изоляции, избегают запахов сородичей
и метят в основном предметы с собственным запахом (Вто\ми, 1985).
Кастрированные самцы крыс не метят территории. Достоверно чаще ме-
тят территорию самки крыс в состоянии эструса по сравнению © самка-
ми в состоянии диэструса (Вго\и, 1977а, Б).
Пронесе взаимодействия двух самцов при вторжении одпого на тер-
риторию другого характеризуется активным маркировочным поведением.
Частота актов мечения предметов не коррелирует с уровнем агрессив-
ности животного. Одпако интенсивность маркировки со стороны «чужа-
ка» в значительной степени снижается после первой же атаки «хозяи-
на» (Лаашз, 1976). Для возникновения маркировочной активности крыс
половое возбуждение является более существенным фактором по сравне-
нию с ростом Уровня агрессивности. Вторжение самок па территорию
«хозяина» сопровождается большей частотой мечения иредметов при со-
поставлепии с таковой при вторжении самцов (Вто\ми, 1982). Самки
„
Поведение
301
практически никогда не подвергаются агрессивным атакам со стороны
самцов, даже если на их шкуру нанесена моча «чужака» (АЪегз, Са[е!,
1973). Вызвать агроссивпые реакции у самцов серых крыс изменением
обонятельных сигналов у знакомых самок удается лишь в том случае,
если самец находится в состоянии ожидания схватки с «чужаком» (Со-
колов п др., 4986).
«Запажи стресса»
Хорошо известен факт, что крысы способны различать по запаху стрес-
сированных и нестрессированных сородичей (Узетца, Noеъу, 1968; е-
уе0з, Кбзег, 1972; и лр.). Более того, у них можно выработать двига-
тельные (нажатие на педаль) дифференцировки па запах стрессирован-
ных (удар электрическим током) и нестрессированных крыс (Кшр её &1.,
1975). Существуют эксперимептальные подтверждения того факта, что
крысы различают запах сородичей в состоянии фрустрации (пропуск под-
крепления в опытах с положительным обусловливанием) от такового ин-
тактных особей (Могмзоп, Бадмезопт, 1970; ЕзНпосг, Гаду1озот, 1980;
и др.). Запах крысы в состоянии «фрустрации» может использоваться ее
сородичами как дискриминационный (Вутиз её а1., 1981).
Различие в ответах митральных клеток обонятельных луковиц на сти-
муляцию «запахом фрустрации», пищи и мочи интактных особей рас-
сматривается как доказательство химической специфичности сигпала
фрустрации (Сабаге! еб а1., 1977; УоотВеев, ВешЪеу, 19841).
‚Характер реакций, проявляемых особью-рецициептом запаха, зависит
как от способа стрессирования животного-донора (удар электрическим то-
ком, интенсивный бег, звуковой раздражитель и др.), так и от самих
Условий проведения эксперимента (жилая клетка, экспериментальная
камера, «открытое поле» и др.). Это может быть, например, исследова-
тельская реакция [известно, что крысы более активны в атмосфере запа-
ха стрессированных сородичей (МасКкау-Зил, Гат, 1980) ] или реакция
эмоционального возбуждения и связанцого с пей повышенного уровня
дефекации, замирания, испуга и попыток избежать запаха (Маскау-
Зла, Баше, 1981; Эфеусиз, @ет20с-Твотаз, 1977). По всей видимости,
конкретное проявление реакции на «запах стресса» зависит от уровпя
стрессировапиости животного-донора запаха. А сам «запах стресса» по
своей природе является неспецифическим и служит лишь своего рода
пусковым сигналом к перестройкам в поведении животного (Вго\уп, 1935).
Вопрос об источнике «запаха стресса» до сих пор остается нерешен-
пым. Так, существуют данные, что сигнал «стресса» присутствует в кро-
ви животных. Крысы избирательно реагируют на кровь, гомогенаты
‚мышц и гомогенаты мозга. При этом реакция на кровь и мышцы
етрессироваппых сородичей отличается от таковой на гомогенаты тканей
морских свинок (З1еуепз, Сег2ос-ТВошаз, 1977). Есть данные, указываю-
щис, что источником «феромона стресса» могут быль экокременты (Кир,
1989) и моча (Маскау-Зии, Гаше, 1981). Запах мочи стрессированных
белых крыс вызывает у их сородичей снижение потребления кислорода
300
Глава четвертая
навания ими родни того же возраста в тестах с предпочтением запаха.
Ирысята в возрасте от 10 до 15 дней предпочитают запах зпакомых
крысят и незнакомых родственников по сравнению с незнакомыми не-
родственниками. С возрастом доля животных, предпочитающих незнако-
мых ровеспиков, возрастает (Неррег, 1983).
Марки ровочное поведение и поддержание
черархической структуры группы
Возможность быстрого опознания индивидуально каждой особи и ее фи-
зиологического состояния— необходимое условие существования соци-
альпой структуры.
Пахучие вещества, содержащиеся в моче крыс, несут информацию о
видовой, половой принадлежности особи, ее социальном рапге и физиоло-
гическом состояпии (Сагг сё а1., 4970; Афемз, Са1еЁ, 1973; Соколов и др.,
1982, 1985; п др.). Серые крысы способны различать своих сородичей по
обонятельным сигналам даже в отсутствие сигналов других модальностей.
Потеря обоняния снижает или полностью устраняет агрессивность у ди-
ких серых (Аелз, СаеЁ 1973) и лабораторных белых крые (Гаезпо,
Тюте, 1975). Способность различать индивидуальные и «групповые» запа-
хи рядом авторов рассматривается как основа для впутри- и межгруп-
повых взаимодействий (Наумов, Гольцман, 1972).
Как живуние синантропно, так и в экзоаптроиных популяциях кры-
сы метят мочой новые предметы на своей территории, особенно если они
несут информацию о сородичах (Адашз, 1976; Вто\уп, 1975; Рисе, 1975;
В:срагдз. З{еуепз, 1977; и др.), а также партнеров (Цвалинии, 1983;
Тауюг её а1., 1984). Самцы предпочитают метить предметы с меткой са-
мок, тогда как самки не обнаруживают такого выраженного предцочтения
к меткам самцов (Вго\уп, 1977а, Ь). В то же время самки чаще метят
предметы, несущие обонятельную ипформанию об интактных самцах по
сравнению с кастрированными (Тауог ей а1., 1982). Изолированное с0-
держание крыс снижает маркировочную активность как у самцов, так и
у самок. Самцы, содержавишеся в изоляции, избегают запахов сородичей
и метят в основном предметы с собственным запахом (Вто\ми, 1985).
Кастрированные самцы крыс не метят территории. Достоверно чаще ме-
тят территорию самки крыс в состоянии эструса по сравнению © самка-
ми в состоянии диэструса (Вго\и, 1977а, Б).
Пронесе взаимодействия двух самцов при вторжении одпого на тер-
риторию другого характеризуется активным маркировочным поведением.
Частота актов мечения предметов не коррелирует с уровнем агрессив-
ности животного. Одпако интенсивность маркировки со стороны «чужа-
ка» в значительной степени снижается после первой же атаки «хозяи-
на» (Лаашз, 1976). Для возникновения маркировочной активности крыс
половое возбуждение является более существенным фактором по сравне-
нию с ростом Уровня агрессивности. Вторжение самок па территорию
«хозяина» сопровождается большей частотой мечения иредметов при со-
поставлепии с таковой при вторжении самцов (Вто\ми, 1982). Самки
„
Поведение
301
практически никогда не подвергаются агрессивным атакам со стороны
самцов, даже если на их шкуру нанесена моча «чужака» (АЪегз, Са[е!,
1973). Вызвать агроссивпые реакции у самцов серых крыс изменением
обонятельных сигналов у знакомых самок удается лишь в том случае,
если самец находится в состоянии ожидания схватки с «чужаком» (Со-
колов п др., 4986).
«Запажи стресса»
Хорошо известен факт, что крысы способны различать по запаху стрес-
сированных и нестрессированных сородичей (Узетца, Noеъу, 1968; е-
уе0з, Кбзег, 1972; и лр.). Более того, у них можно выработать двига-
тельные (нажатие на педаль) дифференцировки па запах стрессирован-
ных (удар электрическим током) и нестрессированных крыс (Кшр её &1.,
1975). Существуют эксперимептальные подтверждения того факта, что
крысы различают запах сородичей в состоянии фрустрации (пропуск под-
крепления в опытах с положительным обусловливанием) от такового ин-
тактных особей (Могмзоп, Бадмезопт, 1970; ЕзНпосг, Гаду1озот, 1980;
и др.). Запах крысы в состоянии «фрустрации» может использоваться ее
сородичами как дискриминационный (Вутиз её а1., 1981).
Различие в ответах митральных клеток обонятельных луковиц на сти-
муляцию «запахом фрустрации», пищи и мочи интактных особей рас-
сматривается как доказательство химической специфичности сигпала
фрустрации (Сабаге! еб а1., 1977; УоотВеев, ВешЪеу, 19841).
‚Характер реакций, проявляемых особью-рецициептом запаха, зависит
как от способа стрессирования животного-донора (удар электрическим то-
ком, интенсивный бег, звуковой раздражитель и др.), так и от самих
Условий проведения эксперимента (жилая клетка, экспериментальная
камера, «открытое поле» и др.). Это может быть, например, исследова-
тельская реакция [известно, что крысы более активны в атмосфере запа-
ха стрессированных сородичей (МасКкау-Зил, Гат, 1980) ] или реакция
эмоционального возбуждения и связанцого с пей повышенного уровня
дефекации, замирания, испуга и попыток избежать запаха (Маскау-
Зла, Баше, 1981; Эфеусиз, @ет20с-Твотаз, 1977). По всей видимости,
конкретное проявление реакции на «запах стресса» зависит от уровпя
стрессировапиости животного-донора запаха. А сам «запах стресса» по
своей природе является неспецифическим и служит лишь своего рода
пусковым сигналом к перестройкам в поведении животного (Вго\уп, 1935).
Вопрос об источнике «запаха стресса» до сих пор остается нерешен-
пым. Так, существуют данные, что сигнал «стресса» присутствует в кро-
ви животных. Крысы избирательно реагируют на кровь, гомогенаты
‚мышц и гомогенаты мозга. При этом реакция на кровь и мышцы
етрессироваппых сородичей отличается от таковой на гомогенаты тканей
морских свинок (З1еуепз, Сег2ос-ТВошаз, 1977). Есть данные, указываю-
щис, что источником «феромона стресса» могут быль экокременты (Кир,
1989) и моча (Маскау-Зии, Гаше, 1981). Запах мочи стрессированных
белых крыс вызывает у их сородичей снижение потребления кислорода
302
Глава четвертая
в большей степени, чем запах мочи пестрессированных животных. Дли-
тельность сохранения пониженного потребления кислорода достигает 1 ч
при прямом воздействии и не превышает 25 мин в случае применения
мочи нестрессированных крыс (Кунцевич, 1986). В то же время крысы
не избегают участков и источников пищи, меченных мочой и экскременп-
тами стрессироваиных особей (Вго\ти, 1985).
Вполне вероятно, что предполагаемый «сигнал стресса» может при-
сутствовать сразу в нескольких продуктах метаболизма в разных кон-
центрациях и что период жизни этих веществ в различных выделениях
может варьировать.
ПНахучие вещества ка дополнительный источник
информации о бистантно удаленных источниках пищи
Традиционно наибольшее впимание исследователей привлекают высоко-
специализированные формы внутривидовой коммуникации у животных,
отобранные в процессе длительной эволюции вида (ши, 1977). Такие
специализированные сигналы не удается выделить у позвоночных. Во-
нрос о возможности передачи дополнительной информации о дистантно
удаленных источниках пищи при помощи пахучих компонентов продуктов
жизненеятельноети крыс до сих пор остается малоизученным, хотя су-
ществуют достаточно убедительные доказательства передачи информации
именно таким способом с последующим облегчением поиска пищи у них
(Са]еГ, 1986). Нора крыс служит своего рода «информационпым цептром»
(\Noага, Ханау, 1973), где происходит непрерывный обмен информацией
между обитателями о пригодности пищи в окружающем пространстве
(СаеЁ 1983). Нассивпо подаваемые особъю обонятельные сигналы ©
поедаемой пище в дальнейшем ориентируют сородичей на выбор тех или
ипых источников питания.
Используемые в лабораториях модели для изучения таких способов
передачи информации, конечно, не в состоянии отразить всей сложности
наблюдаемого явления в природпых колониях серых крыс, но тем не ме-
нее они позволяют выделить некоторые закопомерности, весьма существен-
ные для понимания этого вопроса.
Неофобия — хоропю известная особопноеть поведения серых крыс
(Вагпей, 1975). Избегание новой пищи у них также можно рассматривать
как частный случай неофобии. Поедапие новой пищи со специфическим
запахом, например какао или корицы, в значительной степени облегчает-
ся, если крысе предоставляется возможность контакта в течение несколь-
ких минут со своим сородичем, уже имевшим опыт поедания такой пищи
(«демонстратором»). Явлепие предпочтения особъю-наблюдателем пищи,
поедасмой «демонстратором», при наличии возможности выбора подтверж-
дено рядом исследователей с использованием самых различных методик
(Са1еЁ УПототе, 1983; Розадаз-Апдхеув, Рорег, 1983; Зи'ирр, ГеуИзКу,
1984). Необходимым условием для воспроизведения результатов этой се-
рии экспериментов является интактпость обонятельной системы реципиен-
тов, а также возможность восприятия «наблюдателем» обонятельной
=
Поведение 303
информации. При выключении периферического звена обонятельной си-
стемы сульфатом цинка или при замепе сотки, разделяющей животных
в экспериментальной камере, на плексигласовую перегородку явление
предпочтения пищи, поедаемой «демонстратором», полностью исчезает
(А\Ъегз, СаеГ, 1973; бяе!, УЯстоге, 1983). Выбор пищи «наблюдате-
лем» не зависит от таких параметров, как пол животного, возраст, сте-
пень знакомства с «демонстратором», состояние сытости или голода, со-
став используемой пищи, а также от линии использованных крысе, степе-
ни доместикации диких серых крыс (СаеЁ, 1986). Более того, даже пред-
варительное знакомство с используемой в эксперименте пищей или ее
запахом в течение 1—2 суток не изменяет выбора крысы-«наблюдателя»
(Са1еЁ еЁ а[., 1985). Только длительное потребление опродоленного рода
пищи, например с запахом миндаля, способствует в дальнейшем ее пред-
почтению всем остальным (Розадаз-ЛАпаге\уз, Ророг, 41983), так же как
экспозиция ее в раннем периоде постнатального развития (Теоп еф а|.,
1977).
Таким образом, выбор той или иной пищи крысой-«наблюдателем»
нельзя объяснить только редукцией пеофобии под влиянием экспозиции
запаха пищи, исходящего от «демонстратора». Сама крыса-«демонстра-
тор» являстся пеобходимым коптекстом, на фоне которого воспринимает-
ся обонятельная информация о пище. По всей видимости, «демопотрато-
ру» в этой ситуации принадлежит пассивная роль, поскольку информация
о пише воспринимается ренициентами и от анастезированных животных,
как употреблявших пищу, так и просто с ее следами на кончиках лап или
морды. Однако неэффективно в дапной ситуации использование чучела
крысы (Са]еГ, 1986). Природа дополнительных сигналов, исходящих от
эживотпого-«демонстратора», не ясна, но результаты предыдущих экспе-
риментов позволяют предположить, что этими сигналами являются
пахучие компоненты выделений крыс, возможно, сигнализирующих о ви-
довой принаклежности животпого.
В естественной среде обитания крысы нопрерывно обмепиваются
обонятельной информацией сразу с несколькими сородичами, причем про-
межуток времени между поеданием пищи и коптактом с сородичами мо-
жет варьировать. Такая ситуация в ее упрощенном варианте может быть
воспроизведена в лабораторных условиях. Крысе-‹наблюдателю» предо-
ставлялась возможность кратковременного контакта с четырьмя крысами-
«демонстраторами», каждая из которых содержалась на определенной
диете (с добавлением кофе, како, корицы или уксуса). В достоверном ко-
личестве случаев «наблюдатель» предпочитал пищу только одного из
«демонстраторов», тогда как в условиях выбора одной диеты из двух кры-
сы всегда предпочитают пищу «демонстратора». Таким образом, они в
состоянии суммировать информацию о пище, поедаемой сородичами, ана-
лизировать эту информацию и на ее основе производить выбор. Проме-
жуток времени, прошедитего со времени поедания пищи до контакта с
крысой-«наблюдателем», пе оказывал существенного влияния на выбор
диеты «наблюдателем». Крысы-«наблюдатели» оказывали достоверпое
предпочтение пище «демонстратора» даже по прошествии 24 ч с момента
302
Глава четвертая
в большей степени, чем запах мочи пестрессированных животных. Дли-
тельность сохранения пониженного потребления кислорода достигает 1 ч
при прямом воздействии и не превышает 25 мин в случае применения
мочи нестрессированных крыс (Кунцевич, 1986). В то же время крысы
не избегают участков и источников пищи, меченных мочой и экскременп-
тами стрессироваиных особей (Вго\ти, 1985).
Вполне вероятно, что предполагаемый «сигнал стресса» может при-
сутствовать сразу в нескольких продуктах метаболизма в разных кон-
центрациях и что период жизни этих веществ в различных выделениях
может варьировать.
ПНахучие вещества ка дополнительный источник
информации о бистантно удаленных источниках пищи
Традиционно наибольшее впимание исследователей привлекают высоко-
специализированные формы внутривидовой коммуникации у животных,
отобранные в процессе длительной эволюции вида (ши, 1977). Такие
специализированные сигналы не удается выделить у позвоночных. Во-
нрос о возможности передачи дополнительной информации о дистантно
удаленных источниках пищи при помощи пахучих компонентов продуктов
жизненеятельноети крыс до сих пор остается малоизученным, хотя су-
ществуют достаточно убедительные доказательства передачи информации
именно таким способом с последующим облегчением поиска пищи у них
(Са]еГ, 1986). Нора крыс служит своего рода «информационпым цептром»
(\Noага, Ханау, 1973), где происходит непрерывный обмен информацией
между обитателями о пригодности пищи в окружающем пространстве
(СаеЁ 1983). Нассивпо подаваемые особъю обонятельные сигналы ©
поедаемой пище в дальнейшем ориентируют сородичей на выбор тех или
ипых источников питания.
Используемые в лабораториях модели для изучения таких способов
передачи информации, конечно, не в состоянии отразить всей сложности
наблюдаемого явления в природпых колониях серых крыс, но тем не ме-
нее они позволяют выделить некоторые закопомерности, весьма существен-
ные для понимания этого вопроса.
Неофобия — хоропю известная особопноеть поведения серых крыс
(Вагпей, 1975). Избегание новой пищи у них также можно рассматривать
как частный случай неофобии. Поедапие новой пищи со специфическим
запахом, например какао или корицы, в значительной степени облегчает-
ся, если крысе предоставляется возможность контакта в течение несколь-
ких минут со своим сородичем, уже имевшим опыт поедания такой пищи
(«демонстратором»). Явлепие предпочтения особъю-наблюдателем пищи,
поедасмой «демонстратором», при наличии возможности выбора подтверж-
дено рядом исследователей с использованием самых различных методик
(Са1еЁ УПототе, 1983; Розадаз-Апдхеув, Рорег, 1983; Зи'ирр, ГеуИзКу,
1984). Необходимым условием для воспроизведения результатов этой се-
рии экспериментов является интактпость обонятельной системы реципиен-
тов, а также возможность восприятия «наблюдателем» обонятельной
=
Поведение 303
информации. При выключении периферического звена обонятельной си-
стемы сульфатом цинка или при замепе сотки, разделяющей животных
в экспериментальной камере, на плексигласовую перегородку явление
предпочтения пищи, поедаемой «демонстратором», полностью исчезает
(А\Ъегз, СаеГ, 1973; бяе!, УЯстоге, 1983). Выбор пищи «наблюдате-
лем» не зависит от таких параметров, как пол животного, возраст, сте-
пень знакомства с «демонстратором», состояние сытости или голода, со-
став используемой пищи, а также от линии использованных крысе, степе-
ни доместикации диких серых крыс (СаеЁ, 1986). Более того, даже пред-
варительное знакомство с используемой в эксперименте пищей или ее
запахом в течение 1—2 суток не изменяет выбора крысы-«наблюдателя»
(Са1еЁ еЁ а[., 1985). Только длительное потребление опродоленного рода
пищи, например с запахом миндаля, способствует в дальнейшем ее пред-
почтению всем остальным (Розадаз-ЛАпаге\уз, Ророг, 41983), так же как
экспозиция ее в раннем периоде постнатального развития (Теоп еф а|.,
1977).
Таким образом, выбор той или иной пищи крысой-«наблюдателем»
нельзя объяснить только редукцией пеофобии под влиянием экспозиции
запаха пищи, исходящего от «демонстратора». Сама крыса-«демонстра-
тор» являстся пеобходимым коптекстом, на фоне которого воспринимает-
ся обонятельная информация о пище. По всей видимости, «демопотрато-
ру» в этой ситуации принадлежит пассивная роль, поскольку информация
о пише воспринимается ренициентами и от анастезированных животных,
как употреблявших пищу, так и просто с ее следами на кончиках лап или
морды. Однако неэффективно в дапной ситуации использование чучела
крысы (Са]еГ, 1986). Природа дополнительных сигналов, исходящих от
эживотпого-«демонстратора», не ясна, но результаты предыдущих экспе-
риментов позволяют предположить, что этими сигналами являются
пахучие компоненты выделений крыс, возможно, сигнализирующих о ви-
довой принаклежности животпого.
В естественной среде обитания крысы нопрерывно обмепиваются
обонятельной информацией сразу с несколькими сородичами, причем про-
межуток времени между поеданием пищи и коптактом с сородичами мо-
жет варьировать. Такая ситуация в ее упрощенном варианте может быть
воспроизведена в лабораторных условиях. Крысе-‹наблюдателю» предо-
ставлялась возможность кратковременного контакта с четырьмя крысами-
«демонстраторами», каждая из которых содержалась на определенной
диете (с добавлением кофе, како, корицы или уксуса). В достоверном ко-
личестве случаев «наблюдатель» предпочитал пищу только одного из
«демонстраторов», тогда как в условиях выбора одной диеты из двух кры-
сы всегда предпочитают пищу «демонстратора». Таким образом, они в
состоянии суммировать информацию о пище, поедаемой сородичами, ана-
лизировать эту информацию и на ее основе производить выбор. Проме-
жуток времени, прошедитего со времени поедания пищи до контакта с
крысой-«наблюдателем», пе оказывал существенного влияния на выбор
диеты «наблюдателем». Крысы-«наблюдатели» оказывали достоверпое
предпочтение пище «демонстратора» даже по прошествии 24 ч с момента
304
Глава четвертая
ее поедания «демонстратором» (Са1еТ, 1983). Однако восприятие ипфор-
мации о поедаемой сородичами пище, анализ и выбор на ее основе со-
ответствующей диеты еще не означают сами по себе облегчение ориента-
пни в поисках пищи в условиях сложноструктурировапной окружающей
среды. Некоторым приближением такой ситуации является модель с ис-
пользованием лабирипта с тремя отсеками, в которые помещалась различ-
ного рода пища. Животное должно было отыскать пищу © добавлением
сыра, корицы или какао в одпом из отсеков лабиринта. Эксперименталь-
ной групие животных предоставлялась возможность коптакта с «демон-
стратором», предварительно поедавшим соответствующий вид пищи перед
процедурой обучения. Контролем служили животные, не контактировав-
тние с «опытными» сородичами. Обонятельная информация о съеденной
ранее «демонстраторами» пище облегчала последующее отыскание этой
пищи у 4 животных из 7 (Са]еЁ, УЙетаоте, 41983).
Таким образом, имеющиеся к настоящему времени сведения о возмож
ных путях передачи обонятельной ипформации о дистаптно удаленных
источниках пищи у крыс свидетельствуют о том, что эта форма комму-
никации может служить дополнением или контекстом, уточняющим бо-
лее споциализированные сигналы. Представляется весьма перспективной
постановка такото рода экспериментов в полевых условиях.
Х и.ническое изучение
обонятельных сигналов и их источников
У серой крысы, как и у других видов млекопитающих, источниками обо-
нятельных сирпалов или их биохимических предшественников являются
секреты специфических экзокринных иолез и смеси компонентов из раз-
ных источников (кожный жир, слюна, вагинальные выделения, моча,
экскременты). Исследования липидов поверхности кожного покрова крыс
показали, что их основу составляют сложные эфиры стеринов — 27%,
воска (сложные моноэфиры— 17$ и диэфиры
— 21%, триглицериды—
7%, свободные спирты — 7%, свободные кислоты— 1%) (\ККам, 4974).
В состав отих линидов у серой крысы в отличие от других исследован-
ных видов млекопитающих в необычно высоком количестве входит лато-
стерин (до 40% от суммы стеринов), отнотение которого к другому глав-
ному стерину
— холестерину — меняется в зависимости от пололой при-
надлежности особи и се возраста (\/ВваЦеу, Тапсз, 1957). Найдены также
и некоторые другие стерины— агпостерин, ланостерин, производные де-
мостерина и др. (Noсо]а1Чез, 1965). В липидах самого кожного покрова
крыс было идентифицировано 15 различных стеринов, основными из ко-
торых являются холестерин — 32%, платостерин — 21%, 4,4-диметилхо-
лест-8-сн-3-ол — 10%, 4а-метилхолест-7-ен-3-ол — 71%, 4А-метилхолест
8-ен-3-ол —7% (С]ауюн еЁа|., 1973).
Секреты препуциальтых желез крыс содержат в основном триглице-
рилы — 60%, воска -- 14% и около 2% свободных алифатических кислот
и спиртов наряду с другими классами липидов, среди которых, однако,
не обнаружены сложные диэфиры (NoФоо1а ев, 1965). Препуциальные же-
лезы, как и другие дополнительные половые железы, чувствительны к
Поведение
305.
содержанию гормонов. Эти гормонально зависимые органы удачно расно-
ложепы для введения биологически значимых сигналов в мочу и таким
образом вместе с этим субстратом могут участвовать во многих коммуни-
кативных процессах. Липиды препуциальных желез показывают аттрак-
тивпые свойства, зависимые от пола особей, в отличие от липидов дру-
гих тканей. Кроме того, крысы различают липиды препуциальных желез
не только особей разного пола, но также животных одного и того же
пола, но различного гормонального состояния (Самтево\мзЕ, 1977; Сба\и-
епо\узКЕ еЁ а[., 1975, 4976). Самки крыс привлекаются экстрактами пре-
пуциальных желез половозрелых интактных самцов и пс привлекаются
экстрактами препуциальных желез кастрированных самцов, препуциаль-
ных желез самок, коагуляционных и подчелюстпой желез самцов, подош-
венной ткапи самцов. Самцы крыс привлекаются экстрактами препуциаль-
ных желез эстральных рецоптивпых самок (но не анэстральных самок),
препуциальных желез самцов, подчелюстной железы самок, подошвенной
тканью самок, Нейтральные липиды, ответственные за это привлечение,
могут быть отогнаны в вакууме из липидного окотракта и разделены с
помощью газовой хроматографии на фракции, отличающиеся биологиче-
ской активностью.
Диметилдисульфид и диметилсульфит, обнаруженные в экстрактах
препуциальной ткапи самок, были аттрактивны только для самцов крыс
(Саччепо\узК 1977).
Химическое изучение состава гардеровых желез, расположенных по-
зади глазного яблока, секрет которых вместе с секретом слезпых желез
смачиваст глаз, показало наличие трех порфиринов
— протопорфирина
ТХ — 64%, гардеропорфирипа — 29% и копропорфирина — 9% (Кеипеду,
1970). Из других изученных компонентов ткани гардеровых желез
нейтральные липиды составляют 66%, среди которых обнаружены слож-
ные эфиры с необычным положением двойной связи (А=7) как в спир-
тах, так и кислотах. Хотя длана цепи кислот и спиртов варьирует в ши-
роких пределах, в наибольших количествах присутствует кислота с
20 углеродными атомами, а спирт с 24 (Мигахзкт, Тозь, 1974). На воз-
можную функцию гардеровых желез указывают эксперименты с меченым
андрост-4,16-диен-3-оном — биосинтетическим предшественвиком мужеко-
то полового феромопа у свиней. При введении крысам этот стероид в 06-
новном аккумулируется в гардеровой и препуциальной железах (ВгооКз-
БапК её а1., 1973), в которых, следовательно, можно ожидать образование
летучих производных стероидов, апалогичных мужскому половому феро-
мону свиней.
Изучение связанных в виде солей или свободных органических осно-
ваний и кислот мочи крыс показало паличис мпогих моноаминов и моно-
карбоновых кислот, содержание которых меняется в широких пределах:
диметил-и (или) диэтиламин от 3 до 22% от суммы оснований, трет-бу-
тиламин от 0 до 50%, триэтиламин от 4 до 68%; алифатические кислоты
с числом углеродпых атомов С»—С., а также ароматические— бензойная
и фенилуксусная кислоты могут составлять до 85 от суммы кислот
(Зинкевич, Курятов, 1979). Установлено, что кислоты значимы при разли-
304
Глава четвертая
ее поедания «демонстратором» (Са1еТ, 1983). Однако восприятие ипфор-
мации о поедаемой сородичами пище, анализ и выбор на ее основе со-
ответствующей диеты еще не означают сами по себе облегчение ориента-
пни в поисках пищи в условиях сложноструктурировапной окружающей
среды. Некоторым приближением такой ситуации является модель с ис-
пользованием лабирипта с тремя отсеками, в которые помещалась различ-
ного рода пища. Животное должно было отыскать пищу © добавлением
сыра, корицы или какао в одпом из отсеков лабиринта. Эксперименталь-
ной групие животных предоставлялась возможность коптакта с «демон-
стратором», предварительно поедавшим соответствующий вид пищи перед
процедурой обучения. Контролем служили животные, не контактировав-
тние с «опытными» сородичами. Обонятельная информация о съеденной
ранее «демонстраторами» пище облегчала последующее отыскание этой
пищи у 4 животных из 7 (Са]еЁ, УЙетаоте, 41983).
Таким образом, имеющиеся к настоящему времени сведения о возмож
ных путях передачи обонятельной ипформации о дистаптно удаленных
источниках пищи у крыс свидетельствуют о том, что эта форма комму-
никации может служить дополнением или контекстом, уточняющим бо-
лее споциализированные сигналы. Представляется весьма перспективной
постановка такото рода экспериментов в полевых условиях.
Х и.ническое изучение
обонятельных сигналов и их источников
У серой крысы, как и у других видов млекопитающих, источниками обо-
нятельных сирпалов или их биохимических предшественников являются
секреты специфических экзокринных иолез и смеси компонентов из раз-
ных источников (кожный жир, слюна, вагинальные выделения, моча,
экскременты). Исследования липидов поверхности кожного покрова крыс
показали, что их основу составляют сложные эфиры стеринов — 27%,
воска (сложные моноэфиры— 17$ и диэфиры
— 21%, триглицериды—
7%, свободные спирты — 7%, свободные кислоты— 1%) (\ККам, 4974).
В состав отих линидов у серой крысы в отличие от других исследован-
ных видов млекопитающих в необычно высоком количестве входит лато-
стерин (до 40% от суммы стеринов), отнотение которого к другому глав-
ному стерину
— холестерину — меняется в зависимости от пололой при-
надлежности особи и се возраста (\/ВваЦеу, Тапсз, 1957). Найдены также
и некоторые другие стерины— агпостерин, ланостерин, производные де-
мостерина и др. (Noсо]а1Чез, 1965). В липидах самого кожного покрова
крыс было идентифицировано 15 различных стеринов, основными из ко-
торых являются холестерин — 32%, платостерин — 21%, 4,4-диметилхо-
лест-8-сн-3-ол — 10%, 4а-метилхолест-7-ен-3-ол — 71%, 4А-метилхолест
8-ен-3-ол —7% (С]ауюн еЁа|., 1973).
Секреты препуциальтых желез крыс содержат в основном триглице-
рилы — 60%, воска -- 14% и около 2% свободных алифатических кислот
и спиртов наряду с другими классами липидов, среди которых, однако,
не обнаружены сложные диэфиры (NoФоо1а ев, 1965). Препуциальные же-
лезы, как и другие дополнительные половые железы, чувствительны к
Поведение
305.
содержанию гормонов. Эти гормонально зависимые органы удачно расно-
ложепы для введения биологически значимых сигналов в мочу и таким
образом вместе с этим субстратом могут участвовать во многих коммуни-
кативных процессах. Липиды препуциальных желез показывают аттрак-
тивпые свойства, зависимые от пола особей, в отличие от липидов дру-
гих тканей. Кроме того, крысы различают липиды препуциальных желез
не только особей разного пола, но также животных одного и того же
пола, но различного гормонального состояния (Самтево\мзЕ, 1977; Сба\и-
епо\узКЕ еЁ а[., 1975, 4976). Самки крыс привлекаются экстрактами пре-
пуциальных желез половозрелых интактных самцов и пс привлекаются
экстрактами препуциальных желез кастрированных самцов, препуциаль-
ных желез самок, коагуляционных и подчелюстпой желез самцов, подош-
венной ткапи самцов. Самцы крыс привлекаются экстрактами препуциаль-
ных желез эстральных рецоптивпых самок (но не анэстральных самок),
препуциальных желез самцов, подчелюстной железы самок, подошвенной
тканью самок, Нейтральные липиды, ответственные за это привлечение,
могут быть отогнаны в вакууме из липидного окотракта и разделены с
помощью газовой хроматографии на фракции, отличающиеся биологиче-
ской активностью.
Диметилдисульфид и диметилсульфит, обнаруженные в экстрактах
препуциальной ткапи самок, были аттрактивны только для самцов крыс
(Саччепо\узК 1977).
Химическое изучение состава гардеровых желез, расположенных по-
зади глазного яблока, секрет которых вместе с секретом слезпых желез
смачиваст глаз, показало наличие трех порфиринов
— протопорфирина
ТХ — 64%, гардеропорфирипа — 29% и копропорфирина — 9% (Кеипеду,
1970). Из других изученных компонентов ткани гардеровых желез
нейтральные липиды составляют 66%, среди которых обнаружены слож-
ные эфиры с необычным положением двойной связи (А=7) как в спир-
тах, так и кислотах. Хотя длана цепи кислот и спиртов варьирует в ши-
роких пределах, в наибольших количествах присутствует кислота с
20 углеродными атомами, а спирт с 24 (Мигахзкт, Тозь, 1974). На воз-
можную функцию гардеровых желез указывают эксперименты с меченым
андрост-4,16-диен-3-оном — биосинтетическим предшественвиком мужеко-
то полового феромопа у свиней. При введении крысам этот стероид в 06-
новном аккумулируется в гардеровой и препуциальной железах (ВгооКз-
БапК её а1., 1973), в которых, следовательно, можно ожидать образование
летучих производных стероидов, апалогичных мужскому половому феро-
мону свиней.
Изучение связанных в виде солей или свободных органических осно-
ваний и кислот мочи крыс показало паличис мпогих моноаминов и моно-
карбоновых кислот, содержание которых меняется в широких пределах:
диметил-и (или) диэтиламин от 3 до 22% от суммы оснований, трет-бу-
тиламин от 0 до 50%, триэтиламин от 4 до 68%; алифатические кислоты
с числом углеродпых атомов С»—С., а также ароматические— бензойная
и фенилуксусная кислоты могут составлять до 85 от суммы кислот
(Зинкевич, Курятов, 1979). Установлено, что кислоты значимы при разли-
305
Глава четвертая
чении половой принадлежности как самками, так и самцами серой крысы
(Соколов и др., 1983а, 6, 1986). Сочетание определенных оснований и кис-
лот мочи серой крысы (н-бутиламин
— уксусная кислота и н-бутила-
мин — пропионовая кислота) несут для крыс информацию о принадлеж-
ности к своему виду (Соколов и др., 1979а, 6) и, таким образом, являются
феромоном своего вида. Впервые на таких примерах сложного состава
феромонов были сформулированы представления о том, что эти вещества
могут являться «обонятельными образами», т. е. содержат группы комно-
нентов, ключевых для распознавания (Соколов и др., 1978).
Обопятельпые сигналы экскрементов млекопитающих, как и мочи,
участвуют в организации их поведенческих реакций, однако опи практи-
чески пе изучены. Для серой крысы было установлено, что информация
о половой принадлежности содержится в экскрементах, вносится она сек-
ретом анальной железы (Скурат и др., 1979).
Была исследовапа реакция испуга серой крысы на запах экскремен-
тов лисицы и установлено, что повьшшение уровия кортикостерона в плаз-
ме крови крыс вызывалось двумя пахучими веществами, содержащими
серу и азот, которые присутотвуют в выделениях лисицы в следовых ко-
личествах. Эти компоненты были идентифицированы хроматомасс-спек-
трометрией как цис- и трапс-изомеры триметил-А*-тиазолины (Уегпе[-
Маигу, 1980). В дальнейшем было показано, что некоторые серосодержа-
щие соединения, такие, как 3,3-диметил-1,2-дитиолан (компонент секрета
анальной железы американской норки), цис- и транс-меркаптоментон,
4-мерканто-4-метил-пептан-2-он, А-меркаитопентан-2-он, 3-меркапто-3-
метилпентан-2-он и 2-метилбутан-2-тиол, вызывают стрессовую реакцию
в поведении самцов серой крысы линии \\13{аг в стапдартных условиях
«открытого ноля», а так же по данным изменения уровня кортикостерона
в плазме крови (Устве-Маигу ей а1., 1984).
Таким образом, исследовапия химической коммуникации серой крысы
выявили определенные закономерности передачи и приема информации
с помощью обонятельных сигналов и раскрыли некоторые механизмы .
изменения состояний и поведения животных. Использование этих. знаний
позволяет начать создание новых методов регуляции численности этого
животного, чрезвычайно адаптивного к деятельности человека (Соколов
и др., 1980, 1981; Судейкин и др., 1983).
‹
Акустическая коммуникация
Акустическая коммуникация серой крысы изучена значительно лучше
акустической коммуникации болыпинства других видов млекопитающих.
Однако практически все известпые наблюдения и эксперименты выпол-
нены в лабораторных условиях на лабораторных крысах. Это обетоя-
тельство обычно затрудпяет или делает невозможной экологическую
иптерпретацию получаемых результатов. Моделирование поведепчоских
ситуаций в лабораторных условиях создает, конечно, исключительно
благоприятные возможности для изучения ключевых моментов коммуни-
=ативного процосса и для выяснения сигнальной значимости акустиче-
Новедение
307
ских реакций, но вместе с тем экологически обедненная среда сильпо
маскирует экологическую специфику коммуникативного поведения. На-
копленпый опыт показывает, что окспериментальпые модели иногда
исключают очень важные поведенческие ситуации, обычные в естествен-
ных популяциях или характерные для них.
Кроме того, до настоящего времени основное внимание уделяется
ультразвуковым сигналам, в то время как слыпшимые звуки обсуждаются
крайне редко, хотя нот осповапий предполагать, что опи играют мень-
шую роль в коммуникативном процессе данного вида, чем ультразвуки.
В связи с этим пеобходимо заметить, что граница между звуками и ульт-
развуками, строго говоря, условна, а ее выделение в большинстве слу-
чаев неоправданно, поскольку создается ложное виечатление о двух
резко различных системах акустической коммупикации — звуковой и
ультразвуковой. Изучение ультразвуков у серой крысы имеет давнюю
историю и связапо прежде всего с именами Андерсона (Апдетзоп, 1954),
Сеуилл (Зе\е!, 1967) и Нойрот (Мото, 1968).
у
Учитывая изложенное выше и то, что собственным материалом мы не
располагаем, настоящая глава представляет собой обзор литературы пре-
имущественно по ультразвуковой коммуникации лабораторной формы се-
рой крысы.
Габота Уаттса (УУаИз, 1980) позволяет предполагать, что репертуар
акустических сигпалов лабораторных крыс не сильно отличается от ре-
пертуара звуков и ультразвуков, издаваемых дикими серыми крысами.
Уаттс описал осповпые акустические сигналы черной (ВаМиз гаи) и
серой (лабораторной) крыс и семи австралийских видов крыс. Несмотря
па разпую степень родства, у всех девяти видов рода Ваз в сходных
экспериментальных условиях оказался один и тот же репертуар акусти-
ческих сигналов, в основном не сильно различающихся между видами по
частотным и вромеппым характеристикам. Мало вероятно, чтобы раз-
личия в акустическом репертуаре между дикими и лабораторными кры-
сами оказались больше, чем между разными видами.
Акустическая активность детенышей и поведение
кормящих самок серых крысе
На ранних стадиях развития (в возрасте до трех недель) акустическая
активность детенышей серых крыс может быть вызвана различными
«стрессовыми» ситуациями, такими, как понижение температуры, изоля-
ция от гнезда, болевое раздражение. Издаваемые крысятами ультразву-
\‹и привлекают внимание кормящих самок и, по мнепию ряда авторов,
стимулируют у них различные поведенческие и физиологические реак-
ции, связанные с заботой о потомстве. Ультразвуковая активность кры-
сят сильно зависит не только от характера стимуляции, но и от возраста
детенышей. Ультразвуки, издаваемые ими в возрасте до трех недель, за-
нимают диапазон частот от 30 до 142 кГн (За]ез, БшиВ, 1978; Баез,
Руе, 1974). Обычно опи модулированы по частоте, глубина и особенно-
сти частотной модуляции различны. Частота мепяется медленно и посте-
305
Глава четвертая
чении половой принадлежности как самками, так и самцами серой крысы
(Соколов и др., 1983а, 6, 1986). Сочетание определенных оснований и кис-
лот мочи серой крысы (н-бутиламин
— уксусная кислота и н-бутила-
мин — пропионовая кислота) несут для крыс информацию о принадлеж-
ности к своему виду (Соколов и др., 1979а, 6) и, таким образом, являются
феромоном своего вида. Впервые на таких примерах сложного состава
феромонов были сформулированы представления о том, что эти вещества
могут являться «обонятельными образами», т. е. содержат группы комно-
нентов, ключевых для распознавания (Соколов и др., 1978).
Обопятельпые сигналы экскрементов млекопитающих, как и мочи,
участвуют в организации их поведенческих реакций, однако опи практи-
чески пе изучены. Для серой крысы было установлено, что информация
о половой принадлежности содержится в экскрементах, вносится она сек-
ретом анальной железы (Скурат и др., 1979).
Была исследовапа реакция испуга серой крысы на запах экскремен-
тов лисицы и установлено, что повьшшение уровия кортикостерона в плаз-
ме крови крыс вызывалось двумя пахучими веществами, содержащими
серу и азот, которые присутотвуют в выделениях лисицы в следовых ко-
личествах. Эти компоненты были идентифицированы хроматомасс-спек-
трометрией как цис- и трапс-изомеры триметил-А*-тиазолины (Уегпе[-
Маигу, 1980). В дальнейшем было показано, что некоторые серосодержа-
щие соединения, такие, как 3,3-диметил-1,2-дитиолан (компонент секрета
анальной железы американской норки), цис- и транс-меркаптоментон,
4-мерканто-4-метил-пептан-2-он, А-меркаитопентан-2-он, 3-меркапто-3-
метилпентан-2-он и 2-метилбутан-2-тиол, вызывают стрессовую реакцию
в поведении самцов серой крысы линии \\13{аг в стапдартных условиях
«открытого ноля», а так же по данным изменения уровня кортикостерона
в плазме крови (Устве-Маигу ей а1., 1984).
Таким образом, исследовапия химической коммуникации серой крысы
выявили определенные закономерности передачи и приема информации
с помощью обонятельных сигналов и раскрыли некоторые механизмы .
изменения состояний и поведения животных. Использование этих. знаний
позволяет начать создание новых методов регуляции численности этого
животного, чрезвычайно адаптивного к деятельности человека (Соколов
и др., 1980, 1981; Судейкин и др., 1983).
‹
Акустическая коммуникация
Акустическая коммуникация серой крысы изучена значительно лучше
акустической коммуникации болыпинства других видов млекопитающих.
Однако практически все известпые наблюдения и эксперименты выпол-
нены в лабораторных условиях на лабораторных крысах. Это обетоя-
тельство обычно затрудпяет или делает невозможной экологическую
иптерпретацию получаемых результатов. Моделирование поведепчоских
ситуаций в лабораторных условиях создает, конечно, исключительно
благоприятные возможности для изучения ключевых моментов коммуни-
=ативного процосса и для выяснения сигнальной значимости акустиче-
Новедение
307
ских реакций, но вместе с тем экологически обедненная среда сильпо
маскирует экологическую специфику коммуникативного поведения. На-
копленпый опыт показывает, что окспериментальпые модели иногда
исключают очень важные поведенческие ситуации, обычные в естествен-
ных популяциях или характерные для них.
Кроме того, до настоящего времени основное внимание уделяется
ультразвуковым сигналам, в то время как слыпшимые звуки обсуждаются
крайне редко, хотя нот осповапий предполагать, что опи играют мень-
шую роль в коммуникативном процессе данного вида, чем ультразвуки.
В связи с этим пеобходимо заметить, что граница между звуками и ульт-
развуками, строго говоря, условна, а ее выделение в большинстве слу-
чаев неоправданно, поскольку создается ложное виечатление о двух
резко различных системах акустической коммупикации — звуковой и
ультразвуковой. Изучение ультразвуков у серой крысы имеет давнюю
историю и связапо прежде всего с именами Андерсона (Апдетзоп, 1954),
Сеуилл (Зе\е!, 1967) и Нойрот (Мото, 1968).
у
Учитывая изложенное выше и то, что собственным материалом мы не
располагаем, настоящая глава представляет собой обзор литературы пре-
имущественно по ультразвуковой коммуникации лабораторной формы се-
рой крысы.
Габота Уаттса (УУаИз, 1980) позволяет предполагать, что репертуар
акустических сигпалов лабораторных крыс не сильно отличается от ре-
пертуара звуков и ультразвуков, издаваемых дикими серыми крысами.
Уаттс описал осповпые акустические сигналы черной (ВаМиз гаи) и
серой (лабораторной) крыс и семи австралийских видов крыс. Несмотря
па разпую степень родства, у всех девяти видов рода Ваз в сходных
экспериментальных условиях оказался один и тот же репертуар акусти-
ческих сигналов, в основном не сильно различающихся между видами по
частотным и вромеппым характеристикам. Мало вероятно, чтобы раз-
личия в акустическом репертуаре между дикими и лабораторными кры-
сами оказались больше, чем между разными видами.
Акустическая активность детенышей и поведение
кормящих самок серых крысе
На ранних стадиях развития (в возрасте до трех недель) акустическая
активность детенышей серых крыс может быть вызвана различными
«стрессовыми» ситуациями, такими, как понижение температуры, изоля-
ция от гнезда, болевое раздражение. Издаваемые крысятами ультразву-
\‹и привлекают внимание кормящих самок и, по мнепию ряда авторов,
стимулируют у них различные поведенческие и физиологические реак-
ции, связанные с заботой о потомстве. Ультразвуковая активность кры-
сят сильно зависит не только от характера стимуляции, но и от возраста
детенышей. Ультразвуки, издаваемые ими в возрасте до трех недель, за-
нимают диапазон частот от 30 до 142 кГн (За]ез, БшиВ, 1978; Баез,
Руе, 1974). Обычно опи модулированы по частоте, глубина и особенно-
сти частотной модуляции различны. Частота мепяется медленно и посте-
308
Глава четвертая
700
=
|
г——
к
ди
``г
5Еслим
|
й
Г
1
1
|
|
|
РА
99
60
й
0
90
Время, м
Рис. 77. Сонограмма ультразвунов детенышей серых крыс
а — ультразвук с медленным и илавным изменением частоты, издавасмый однодневным кры-
сенком (по: Зашз, ЗшИВ, 1978), 6. - ультразвук с быстрым и резким изменением частоты,
издаваемый двзнадцатидиневным крысенком (но; За!ез, Руе, 1974)
пенно или значительно и резко (рис. 77). Ситнал может занимать поло-
су частот от 1 до 20 кГи. В болынинстве известпых случаев на соно-
грамме удается выделить лишь одну спектральную составляющую, что,
по мнению Саль и Пий, (Заез, Руе, 1974, с. 155) отличает ультра-
звуки мышиных (Митдае) от ультразвуков хомяковых (СтоеНдае).
Ивдаваемые крысятами ультразвуки не организованы в серии, следу-
ют одиночно в ноопределепном ритме или почти с одипаковыми иптерва-
лами, их длительность колеблется от 1 до 500 мс, т. е. в очень итироких
пределах (Мальцов, 1974; За1ез, Рус, 1974; без, ЗшИ, 1978).
Интенсивность ультразвуков невелика и составляет обычно 60—
80 дБ (Окоп, 1974; За]ез, ЗшиВ, 1978).
Изменение частоты и длительности ультразвуков в раппем онтогенезе
серых крыс изучено недостаточно. В. П. Мальцев (1974) обнаружил с0-
пращение доли частотпо-модулированных ультразвуков у крысят более
старшего возраста, что особенно заметно по сокращению числа ультра-
звуков с резкой, ступенчатой модуляцией. Возрастные изменения дли-
тельности сигнала Мальцевым не обнаружены. Саль и Смит (1978) счи-
тают, что у новорожденных крысят в возрасте до трех дней наиболее
обычны относительно длительные ультразвуки с неглубокой, медленной
модуляцией частоты. Когда крысята становятся старше, в их репертуаре
начипают преобладать короткие крики длительностью менее 65 мс с резкой
и глубокой частотной модуляцией, напоминая ультразвуки взрослых жи-
вотных, издаваемые ими во время агрессивных взаимодействий.
Ультразвуки удается зарегистрировать уже в первые часы после
рождения крысят, но акустическая активность детенышей на протяжении
первых недель их жизни не остается постоянной. Меняется как число
ультразвуков в единицу времени (собственно акустическая активность),
так и интенсивность подаваемых сигналов. Первое исследование, посвя-
щенное данному вопросу, принадлежит Нойрот (Мото 1968), которая,
изолировав детенышей от гнезда, обнаружила у них низкую ультразву-
ковую активность в первые дни жизпи, которая резко возрастала в пе-
Поведение
309
третьей недели жизни детеныттей.
Рис. 7%. Иамепение с возристом интен- в;
_
сивности ультразвуков, издаваемых 9де-
тенышами серых крыс в ответ на из- 83|
менение температуры гнезда (по: Окоп,
1971, с изменениями)
=
=
т
риод открывания наружного слу-
хового прохода (на 2-й—5-й день).
„Достигнув на отой стадии макси-
мума, акустическая активность
падала почти до пуля к концу
>
<
з
е
т
з
а
з
а
т
т
т
Не обнаружено увеличения аку-
стичоской активности и к моменту
открывания (на 12-й, 17-й день)
тлаз у крысят. По наблюдениям
И
н
т
е
н
с
и
в
ы
в
с
р
л
в
у
л
ь
т
р
а
з
в
у
к
а
,
0
5
‹
>
Т
Нойрот, ультразвуковая актив- ИГ
`
ность была выше до этого момеп- Ель
Е рьь
та, чем во время открывания глаз’ 02800.86
Возраст,дни
или в последующие дни.
В дальнейшем более детальные
наблюдения показали, что динамика акустической активности крысе в пер-
вые педели жизни сильпо зависит от температуры окружающей среды и
связана с этапами развития у них терморегуляции. Наиболее показатель-
ны в этом отношении эксперименты Окоп (ОКоп, 1974, 1972), который
исходил из того, что температура гнезда крыс равна примерно 33°, а тем-
пература тела животных при достижении ими гомойтермии к 20-му дню —
.38°. С 1-го и до 20-го дня крысят ежедневно в течение часа выдерживали
при температурах 2—3, 412, 22 и 33°. Зверьки, выдерживаемые при темпе-
ратуре -+-2...-3°, только в возрасте 20 дней достигали через час томпе-
ратуры тела, равной 36°. До двухнедельного возраста температура их
‘тела вообще мало отличалась от температуры окружающей среды (вклю-
чая все 4 варнанта температур). Так, в возрасте 6 дней у крысят, выдер-
‘жанных прин 2°... + 3°, температура тела через час понижалась до 6,2°,
при12°—до18,6°ипри22°—до27,8°.
На рис. 78 показано, как меняется интенсивность излучаемых кры-
сятами ультразвуков по диям в зависимости от температуры внешней
среды. Как видио из рисунка, наиболее интенсивные „уъьтразнуки кры-
сята издавали при температурах виешией среды
„+3 и 12.
„ Ирнчем в первый день животные были молчаливы, мы 68 -го дня уро-
“вень интенсивности был невелик, а пик акустической активности наблю-
дался между 8-м и 12—15-м днями. Крысята, выдержанпые при темпе-
ратуре -2... +3°, продолжали издавать ультразвуки до 20-го дня, в то
время как при температуре -12°— до 16-го дпя, а при температуре
+22? — до 12-го или 13-го дня. Температура среды +33° вызывала слу-
чайные акустические реакции. Аналогичная динамика акустической ак-
тивности крысят наблюдалась и по числу ультразвуков, изцаваемых жи-
-ВвОоТнНыЫМиИ,
308
Глава четвертая
700
=
|
г——
к
ди
``г
5Еслим
|
й
Г
1
1
|
|
|
РА
99
60
й
0
90
Время, м
Рис. 77. Сонограмма ультразвунов детенышей серых крыс
а — ультразвук с медленным и илавным изменением частоты, издавасмый однодневным кры-
сенком (по: Зашз, ЗшИВ, 1978), 6. - ультразвук с быстрым и резким изменением частоты,
издаваемый двзнадцатидиневным крысенком (но; За!ез, Руе, 1974)
пенно или значительно и резко (рис. 77). Ситнал может занимать поло-
су частот от 1 до 20 кГи. В болынинстве известпых случаев на соно-
грамме удается выделить лишь одну спектральную составляющую, что,
по мнению Саль и Пий, (Заез, Руе, 1974, с. 155) отличает ультра-
звуки мышиных (Митдае) от ультразвуков хомяковых (СтоеНдае).
Ивдаваемые крысятами ультразвуки не организованы в серии, следу-
ют одиночно в ноопределепном ритме или почти с одипаковыми иптерва-
лами, их длительность колеблется от 1 до 500 мс, т. е. в очень итироких
пределах (Мальцов, 1974; За1ез, Рус, 1974; без, ЗшИ, 1978).
Интенсивность ультразвуков невелика и составляет обычно 60—
80 дБ (Окоп, 1974; За]ез, ЗшиВ, 1978).
Изменение частоты и длительности ультразвуков в раппем онтогенезе
серых крыс изучено недостаточно. В. П. Мальцев (1974) обнаружил с0-
пращение доли частотпо-модулированных ультразвуков у крысят более
старшего возраста, что особенно заметно по сокращению числа ультра-
звуков с резкой, ступенчатой модуляцией. Возрастные изменения дли-
тельности сигнала Мальцевым не обнаружены. Саль и Смит (1978) счи-
тают, что у новорожденных крысят в возрасте до трех дней наиболее
обычны относительно длительные ультразвуки с неглубокой, медленной
модуляцией частоты. Когда крысята становятся старше, в их репертуаре
начипают преобладать короткие крики длительностью менее 65 мс с резкой
и глубокой частотной модуляцией, напоминая ультразвуки взрослых жи-
вотных, издаваемые ими во время агрессивных взаимодействий.
Ультразвуки удается зарегистрировать уже в первые часы после
рождения крысят, но акустическая активность детенышей на протяжении
первых недель их жизни не остается постоянной. Меняется как число
ультразвуков в единицу времени (собственно акустическая активность),
так и интенсивность подаваемых сигналов. Первое исследование, посвя-
щенное данному вопросу, принадлежит Нойрот (Мото 1968), которая,
изолировав детенышей от гнезда, обнаружила у них низкую ультразву-
ковую активность в первые дни жизпи, которая резко возрастала в пе-
Поведение
309
третьей недели жизни детеныттей.
Рис. 7%. Иамепение с возристом интен- в;
_
сивности ультразвуков, издаваемых 9де-
тенышами серых крыс в ответ на из- 83|
менение температуры гнезда (по: Окоп,
1971, с изменениями)
=
=
т
риод открывания наружного слу-
хового прохода (на 2-й—5-й день).
„Достигнув на отой стадии макси-
мума, акустическая активность
падала почти до пуля к концу
>
<
з
е
т
з
а
з
а
т
т
т
Не обнаружено увеличения аку-
стичоской активности и к моменту
открывания (на 12-й, 17-й день)
тлаз у крысят. По наблюдениям
И
н
т
е
н
с
и
в
ы
в
с
р
л
в
у
л
ь
т
р
а
з
в
у
к
а
,
0
5
‹
>
Т
Нойрот, ультразвуковая актив- ИГ
`
ность была выше до этого момеп- Ель
Е рьь
та, чем во время открывания глаз’ 02800.86
Возраст,дни
или в последующие дни.
В дальнейшем более детальные
наблюдения показали, что динамика акустической активности крысе в пер-
вые педели жизни сильпо зависит от температуры окружающей среды и
связана с этапами развития у них терморегуляции. Наиболее показатель-
ны в этом отношении эксперименты Окоп (ОКоп, 1974, 1972), который
исходил из того, что температура гнезда крыс равна примерно 33°, а тем-
пература тела животных при достижении ими гомойтермии к 20-му дню —
.38°. С 1-го и до 20-го дня крысят ежедневно в течение часа выдерживали
при температурах 2—3, 412, 22 и 33°. Зверьки, выдерживаемые при темпе-
ратуре -+-2...-3°, только в возрасте 20 дней достигали через час томпе-
ратуры тела, равной 36°. До двухнедельного возраста температура их
‘тела вообще мало отличалась от температуры окружающей среды (вклю-
чая все 4 варнанта температур). Так, в возрасте 6 дней у крысят, выдер-
‘жанных прин 2°... + 3°, температура тела через час понижалась до 6,2°,
при12°—до18,6°ипри22°—до27,8°.
На рис. 78 показано, как меняется интенсивность излучаемых кры-
сятами ультразвуков по диям в зависимости от температуры внешней
среды. Как видио из рисунка, наиболее интенсивные „уъьтразнуки кры-
сята издавали при температурах виешией среды
„+3 и 12.
„ Ирнчем в первый день животные были молчаливы, мы 68 -го дня уро-
“вень интенсивности был невелик, а пик акустической активности наблю-
дался между 8-м и 12—15-м днями. Крысята, выдержанпые при темпе-
ратуре -2... +3°, продолжали издавать ультразвуки до 20-го дня, в то
время как при температуре -12°— до 16-го дпя, а при температуре
+22? — до 12-го или 13-го дня. Температура среды +33° вызывала слу-
чайные акустические реакции. Аналогичная динамика акустической ак-
тивности крысят наблюдалась и по числу ультразвуков, изцаваемых жи-
-ВвОоТнНыЫМиИ,
310
Глава четвертая
Иптереспые результаты получены Окон (ОКоп, 1971) при длительной
экспозинии крысят в условиях низкой температуры. При темпсратурах
+12 и +22° число ультразвуков за 5 мин к концу часа падало пример-
но © 120 до 20—40, экспозиция при +2... 23° приводила к тому, что
уже нк 25-й мин число ультразвуков падало с 150 до 0. Но в этом слу-
чае прекращалась пе только акустическая активность, но и резко сокра-
щалась частота дыхательных ритмов.
На основании полученных результатов Окон выделяет три стадии
акустической активности серых крыс в возрасте до 3 недель: 1) стадия
низкого акустического ответа в первые дни после рождения; 2) стадия
максимальной акустической активности в более старшем возрасте и
3) стадия понижения акустической активности даже при очень низких
температурах окружающей среды, совпадающая с периодом становления
гомойтермии. Выделенные Оконом стадии хоропто совпадают со стадиями
терморегуляции у крыс и с динамикой толерантности данного вида но
отношению к низким температурам. Ссылаясь на работы Викштайна и
Хана (\УеЕзюш, 1964; ПаБи, 1958), Окон подчеркивает, что резкий
скачок акустической активности у крыс совцадает с такими важными 060-
бенностями развития эндокринной и нервной системы, как включение в
процесс терморегуляции щитовидной железы к 14А-му дню и симпати-
ческих нервных ганглиев к 10—16-му дням после рождения.
Гейср (Сеуег, 1979) обратил впимапие на то, что у серых крыс и
других мышевидных грызунов резкое снижение акустической активности
детеныитей хорошо совпадает не только с развитием у пих гомойтермии,
но также и с развитием способности ориентироваться и активно переме-
щаться на запах. Относительная замена одного сенсорного капала дру-
гим происходит в тот момепт, когда спижается забота матери о нотом-
стве, а детеныши переходят к более самостоятельному образу жизни.
Тактильная стимуляция при комнатной температуре также вызывает
У крысят издавание ультразвуков. Динамика акустической активности
животных в этом случае аналогична той, которая связана с развитием
терморегуляции (Окоп, 1972; Заез, ЗшИЛ, 1978). Но уровень активности,
измеренный как интенсивность ультразвуков, зависит от характера сти-
муляции. Так, сжимание корня хвоста вызывает у крысят ультразвуко-
вую реакцию с интенсивностью до 77 дБ, вахват руками— до 72дБ,
а прихватывание детенышей самкой во вромя нперетаскивания-— всего:
около 60 дБ, причем в последнем случае не удалось обнаружить возраст-
ных изменений в акустической активности детеныттей. Уровепь звуко-
вого давления был одним и тем же во все дни наблюдений, от 1-го до
18-го (ОКоп, 1972).
Развивая предыдущие эксперименты, Освальт и Мейер (Озууа, Ме!-
ег, 1975) показали, что причиной акустической активности крысят яв-
ляется в значительной стецени изменение привычной для гиезда обета-
повки, создаваемой комплексом стимулов, куда входят температура, запа-
хи, особенности субстрата. Совершенно очевидно, что перечислепный
комплекс стимулов создаст характерпое для гнездового периода живот-
ных биологическое сигнальное поле (Наумов, 1971). По мнению Осваль-
Поведение
ЗИ
\
та и Мейер (Озмуай, Маег, 1975), в первые дни пребывания детеныптей
в гнезде у них развиваются «импринтинг-подобные» явления, способет-
зующие запечатлению обстановки гнезда. Было, в частности, показано,
что крысята в возрасте от 3 до 13 дней дольше издают ультразвуки, если
их поместить на пустой поддон, по сравнению с крысятами, помещенны-
ми на поддон с чистой подетилкой или же с подстилкой из гнезда. В дан-
ном случае стимулом, запускающим реакцию, было непривычное тактиль-
ное раздражение( твердый субстрат). С друзой сторопы, ультразвуко-
пая активность крысят была выше, когда их помещали на поддон с чис-
той подстилкой по сравнению с тем, когда они находились на поддоне с
подстилкой из гпезда, содержащей привычный запах, отсутствие которого
и было запускающим ультразвуковую реакцию стимулом. Еели крысят
помещали па поддон, покрытый ткапью, то их ультразвуковая актив-
ность становилась выше, когда под тканью отсутствовала подстилка, чем
в том случае, когда ткань прикрывала подстилку гнезда. В этой же
серии экспериментов была подтверждена способность крысят адекватно
выражать восприятие температуры гнезда через акустическую актив-
ность. При компатной температуре акустическая активность детенышей
вытие, чем в тех случаях, когда эксцериментальная установка подогрева-
лась до +32°, т, е. до температуры гиезда. Эти различия оказались ста-
тистически достоверны для крысят 3, 5 и 7-Дневного возраста, но не бо-
лее старших (10, 43 и 15-дневных).
Таким образом, в настоящее время известно, что стимулами, вызы-
зающими акустическую активность крысят, могут быть 1) понижение
температуры пиже температуры гнезда, 2) тактильное раздражение,
причиняющее боль, или непривычно твердый субстрат, 3) незнакомые
запахи или отсутствие привычного запаха гнезда. Но остается пеяспым,
влияет ли на акустическую активность крысят потеря социального, ком-
муникативного контакта с матерью. В большинстве экспериментов невоз-
можно отделить физические или химические стимулы от социальных.
Аллин и Бенкс (АШш, Ваокз, 1971), исследуя влияние температуры
на ультразвуковую активпость крысят, во время эксперимента изолиро-
вали детенытней от гнезда на 4,5 мин. Низкая температура, равная -+2°
и 20°, вызывала значительное число ультразвуков: температура, близкая
к нормальной (35°) и несколько более высокая (40°), вызывала их го-
раздо менышцее число. По мнению авторов, в последних двух случаях
акустическая активность крысят была связана не с температурным сти-
мулом, ас изоляцией. Авторы считают, что изоляция сама по себе отно-
сительно пезффективиа, вызывая у крысят пизкую акустическую актив-
ность.
Очень мало известпо о сигналах крысят в звуковом диапазоне частот
ниже 16 кГц. Первые 3—4 дня они реагируют на тактильное раздраже-
ние в основном слышимыми писками или визгами и очень редко издают
ультразвуки. В дальнейшем ультразвуки преобладают, по после
16-го дня их число резко сокращается, ас 20-го дня они становятся край-
не редкими и замещаются слышимыми звуками. В отличие от реакции на
тактильное раздражение на понижение температуры крысята реагируют
310
Глава четвертая
Иптереспые результаты получены Окон (ОКоп, 1971) при длительной
экспозинии крысят в условиях низкой температуры. При темпсратурах
+12 и +22° число ультразвуков за 5 мин к концу часа падало пример-
но © 120 до 20—40, экспозиция при +2... 23° приводила к тому, что
уже нк 25-й мин число ультразвуков падало с 150 до 0. Но в этом слу-
чае прекращалась пе только акустическая активность, но и резко сокра-
щалась частота дыхательных ритмов.
На основании полученных результатов Окон выделяет три стадии
акустической активности серых крыс в возрасте до 3 недель: 1) стадия
низкого акустического ответа в первые дни после рождения; 2) стадия
максимальной акустической активности в более старшем возрасте и
3) стадия понижения акустической активности даже при очень низких
температурах окружающей среды, совпадающая с периодом становления
гомойтермии. Выделенные Оконом стадии хоропто совпадают со стадиями
терморегуляции у крыс и с динамикой толерантности данного вида но
отношению к низким температурам. Ссылаясь на работы Викштайна и
Хана (\УеЕзюш, 1964; ПаБи, 1958), Окон подчеркивает, что резкий
скачок акустической активности у крыс совцадает с такими важными 060-
бенностями развития эндокринной и нервной системы, как включение в
процесс терморегуляции щитовидной железы к 14А-му дню и симпати-
ческих нервных ганглиев к 10—16-му дням после рождения.
Гейср (Сеуег, 1979) обратил впимапие на то, что у серых крыс и
других мышевидных грызунов резкое снижение акустической активности
детеныитей хорошо совпадает не только с развитием у пих гомойтермии,
но также и с развитием способности ориентироваться и активно переме-
щаться на запах. Относительная замена одного сенсорного капала дру-
гим происходит в тот момепт, когда спижается забота матери о нотом-
стве, а детеныши переходят к более самостоятельному образу жизни.
Тактильная стимуляция при комнатной температуре также вызывает
У крысят издавание ультразвуков. Динамика акустической активности
животных в этом случае аналогична той, которая связана с развитием
терморегуляции (Окоп, 1972; Заез, ЗшИЛ, 1978). Но уровень активности,
измеренный как интенсивность ультразвуков, зависит от характера сти-
муляции. Так, сжимание корня хвоста вызывает у крысят ультразвуко-
вую реакцию с интенсивностью до 77 дБ, вахват руками— до 72дБ,
а прихватывание детенышей самкой во вромя нперетаскивания-— всего:
около 60 дБ, причем в последнем случае не удалось обнаружить возраст-
ных изменений в акустической активности детеныттей. Уровепь звуко-
вого давления был одним и тем же во все дни наблюдений, от 1-го до
18-го (ОКоп, 1972).
Развивая предыдущие эксперименты, Освальт и Мейер (Озууа, Ме!-
ег, 1975) показали, что причиной акустической активности крысят яв-
ляется в значительной стецени изменение привычной для гиезда обета-
повки, создаваемой комплексом стимулов, куда входят температура, запа-
хи, особенности субстрата. Совершенно очевидно, что перечислепный
комплекс стимулов создаст характерпое для гнездового периода живот-
ных биологическое сигнальное поле (Наумов, 1971). По мнению Осваль-
Поведение
ЗИ
\
та и Мейер (Озмуай, Маег, 1975), в первые дни пребывания детеныптей
в гнезде у них развиваются «импринтинг-подобные» явления, способет-
зующие запечатлению обстановки гнезда. Было, в частности, показано,
что крысята в возрасте от 3 до 13 дней дольше издают ультразвуки, если
их поместить на пустой поддон, по сравнению с крысятами, помещенны-
ми на поддон с чистой подетилкой или же с подстилкой из гнезда. В дан-
ном случае стимулом, запускающим реакцию, было непривычное тактиль-
ное раздражение( твердый субстрат). С друзой сторопы, ультразвуко-
пая активность крысят была выше, когда их помещали на поддон с чис-
той подстилкой по сравнению с тем, когда они находились на поддоне с
подстилкой из гпезда, содержащей привычный запах, отсутствие которого
и было запускающим ультразвуковую реакцию стимулом. Еели крысят
помещали па поддон, покрытый ткапью, то их ультразвуковая актив-
ность становилась выше, когда под тканью отсутствовала подстилка, чем
в том случае, когда ткань прикрывала подстилку гнезда. В этой же
серии экспериментов была подтверждена способность крысят адекватно
выражать восприятие температуры гнезда через акустическую актив-
ность. При компатной температуре акустическая активность детенышей
вытие, чем в тех случаях, когда эксцериментальная установка подогрева-
лась до +32°, т, е. до температуры гиезда. Эти различия оказались ста-
тистически достоверны для крысят 3, 5 и 7-Дневного возраста, но не бо-
лее старших (10, 43 и 15-дневных).
Таким образом, в настоящее время известно, что стимулами, вызы-
зающими акустическую активность крысят, могут быть 1) понижение
температуры пиже температуры гнезда, 2) тактильное раздражение,
причиняющее боль, или непривычно твердый субстрат, 3) незнакомые
запахи или отсутствие привычного запаха гнезда. Но остается пеяспым,
влияет ли на акустическую активность крысят потеря социального, ком-
муникативного контакта с матерью. В большинстве экспериментов невоз-
можно отделить физические или химические стимулы от социальных.
Аллин и Бенкс (АШш, Ваокз, 1971), исследуя влияние температуры
на ультразвуковую активпость крысят, во время эксперимента изолиро-
вали детенытней от гнезда на 4,5 мин. Низкая температура, равная -+2°
и 20°, вызывала значительное число ультразвуков: температура, близкая
к нормальной (35°) и несколько более высокая (40°), вызывала их го-
раздо менышцее число. По мнению авторов, в последних двух случаях
акустическая активность крысят была связана не с температурным сти-
мулом, ас изоляцией. Авторы считают, что изоляция сама по себе отно-
сительно пезффективиа, вызывая у крысят пизкую акустическую актив-
ность.
Очень мало известпо о сигналах крысят в звуковом диапазоне частот
ниже 16 кГц. Первые 3—4 дня они реагируют на тактильное раздраже-
ние в основном слышимыми писками или визгами и очень редко издают
ультразвуки. В дальнейшем ультразвуки преобладают, по после
16-го дня их число резко сокращается, ас 20-го дня они становятся край-
не редкими и замещаются слышимыми звуками. В отличие от реакции на
тактильное раздражение на понижение температуры крысята реагируют
312
Глава четвертая
15
_
т
=
ыйГг
х
э
—
—
ь
—-
Рис. 79. Сонограммы виз-
ВНВ
М
га семидневного детены-
Е
ша серой крысы (а) и
8
—__
взрослого самца (6),
5| Реакция на тактильное
&
—__
“| раздражение (по: И’аИя,
0
|
й
И
|
1980)
Время
100 мс
__
только ультразвуками (ОКоп, 1971, 1972), Длительность визгов меняется
от 30 до 60 мс. Они имеют гармоническую структуру спектра с оенов-
пой частотой в диапазоне от 0,5 до 5 кГц. Доминирует обычно одна из
тармоник, а не основная частота. У взрослых животных по сравнению с
молодыми основная частота визгов ниже и практически отсутствует
частотная модуляция (УУаИз, 1980, рис. 79).
Считается, что ультразвуки крысят привлекают внимание самок, од-
нако сигнальная значимость их исследована далеко не достаточно и
нуждается в дальнейшей эксцериментальной проверке. В этом отношении
показательна критическая статья Белла (ВеП, 1974), которая появилась
непосредетвенно за первыми публикациями, посвящеппыми ультразву-
ковой активности мышевидпых грызунов, включая серую крысу (Зее!
1970; Мошоь 19725; Заез, Руе, 1974). Белл совершенно справедливо об-
ращает внимание на то, что ультразвуки мелких млекопитающих имеют
небольшую интенсивность и очень узкий спектр направленности из-за
коротких длин волн подаваемого сигнала. Опи быстро поглощаются и
рассеиваются средой, в результате чего не могут быть переданы на
большое расстояние. Общение с их помощью вряд ли возможно далее
нескольких десятков сантиметров, т. е. ограничено пределами гнезда или
его ближайших окрестностей. Далее Белл отмечает, что ультразвуки де-
тенышей отражают прежде всего уровень возбуждения источника сигна-
ла, могут передавать информацию об этом уровне и о местоположении
донора. По его мнению, возбуждение, выраженное ультразвуками, может
быть передапо реципиептам, привлекая их внимание и стимулируя их к
тем или иным действиям. Но конкретные формы активности рецициептов
зависят от различных факторов и не закодированы в параметрах сигна-
ла, «запускающего» лишь уровень возбуждения. Рассматривая ультра-
звуковую коммуникацию в широком аспекте, Белл подчеркивает, что
специфические формы активности реципиентов зависят от предшествую-
щего уровня возбуждения адресатов и их предшествующей конкретной
активпости, от того, в каком физиологическом состоянии они находятся
(эструс, лактация, уровень ‘андрогена, кортикостероидная активность),
Поведение
313
других стимульных свойств ситуации (контакт с партнерами по группе,
запах партнеров).
Первая экспериментальная проверка влияния ультразвуков, издавае-
мых крысятами, на поведепие взрослых животных принадлежит Аллину
и Бенксу (АП, ВашЕз, 1972; цит. по: ба1ез, Руе, 1974). Эксперимент
проводили па лактирующих самках, нерожавитих самках и взрослых сам-
цах белых крыс. В качестве стимула использовали 90-минутную магнит-
ную запись семидневных крысят. Контролем служила магнитная запись
шума. Гнездовая камера была устаповлена в шестиугольном полигоне.
Сигнал транслировали в случайном порядке с одной из шести стороп.
Аллин и Бепкс сообщают, что 20 из 40 лактирующих самок покидали
гнездо в ответ на ультразвуки, а большинство из пих ориептировались
на источник сигнала, Самцы и нерожавитие самки не покидали гнездо во
время воспроизведения ультразвуков, но пекоторыс самцы ориентирова-
ли головы в направлении источника звука.
Стимулирующее действие ультразвуковых сигналов детенышей повы-
шается в присутствии запаха крысят. Это было показано в эксперимеп-
тах Смотермапа с соавторами (Зтоегтап её а1., 4974). Лактирующим
самкам крыс предлагали выбрать один из рукавов лабиринта и верпуть
в гнездо находящегося в лабиринте детеныша. Привлекающими стиму-
лами были ультразвуки, издаваемые крысятами, которые в этом случае
были одновременно источником как акустического, так и химического
стимулов; магпитная запись ультразвуковых сигналов крысят (химиче-
ский стимул в этом случае исключался); запах крысят (детеныши, ох-
лажденные до коматозного состояния, не издавали ультразвуков). В ка-
честве контроля использовали пустой рукав лабиринта, свободный от
экспериментальных стимулов. Во всех случаях самки покидали гнездо-
вую коробку и направлялись в лабиринт, если присутствовал запах кры-
сят, а латентный период сокращалея, когда действовали одновременпо
Два стимула — акустический и химический. Это могли быть крысята, не-
посредственно издающие ультразвуки, или трансляция магпитпой запи-
си ультразвуков с одновременным предъявлением молчащего детеныша.
Ультразвуки крысят влияют пе только па поведение самок, но и на
их физиологическое состояние. Теркель с соавторами (Тегке| еф а1.,
1979) показали, что ультразвуковая активность крысят стимулирует сек-
рецию пролактина и таким образом регулирует нормальпую лактацию у
кормящих самок. Более того, ультразвуковая стимуляция незначитель-
но, но достоверно повышает количество пролактина в крови не только
кормящих, но и нерожавтних самок. Исследуя действие ультразвуковых
“сигналов детепьшней па секрецию пролактина, крысят удаляли из гнезда
на бчив течение 15 мин воспроизводили магнитную запись их голо-
сов. Коптролем служило отсутствие воспроизведения или воспроизведе-
ние в течение 15 мин магнитной записи шума. Таким образом, повыше-
ние пролактина вызывали только ультразвуки крысят. Существенно, что,
когда крысят возвращали в гпездо и они начинали сосать, т. е. когда
зключались несе сенсорные модальности (слуховая, ольфакторпая, визу-
альная и тактильпая), количество пролактина увеличивалось еще вдвое
312
Глава четвертая
15
_
т
=
ыйГг
х
э
—
—
ь
—-
Рис. 79. Сонограммы виз-
ВНВ
М
га семидневного детены-
Е
ша серой крысы (а) и
8
—__
взрослого самца (6),
5| Реакция на тактильное
&
—__
“| раздражение (по: И’аИя,
0
|
й
И
|
1980)
Время
100 мс
__
только ультразвуками (ОКоп, 1971, 1972), Длительность визгов меняется
от 30 до 60 мс. Они имеют гармоническую структуру спектра с оенов-
пой частотой в диапазоне от 0,5 до 5 кГц. Доминирует обычно одна из
тармоник, а не основная частота. У взрослых животных по сравнению с
молодыми основная частота визгов ниже и практически отсутствует
частотная модуляция (УУаИз, 1980, рис. 79).
Считается, что ультразвуки крысят привлекают внимание самок, од-
нако сигнальная значимость их исследована далеко не достаточно и
нуждается в дальнейшей эксцериментальной проверке. В этом отношении
показательна критическая статья Белла (ВеП, 1974), которая появилась
непосредетвенно за первыми публикациями, посвящеппыми ультразву-
ковой активности мышевидпых грызунов, включая серую крысу (Зее!
1970; Мошоь 19725; Заез, Руе, 1974). Белл совершенно справедливо об-
ращает внимание на то, что ультразвуки мелких млекопитающих имеют
небольшую интенсивность и очень узкий спектр направленности из-за
коротких длин волн подаваемого сигнала. Опи быстро поглощаются и
рассеиваются средой, в результате чего не могут быть переданы на
большое расстояние. Общение с их помощью вряд ли возможно далее
нескольких десятков сантиметров, т. е. ограничено пределами гнезда или
его ближайших окрестностей. Далее Белл отмечает, что ультразвуки де-
тенышей отражают прежде всего уровень возбуждения источника сигна-
ла, могут передавать информацию об этом уровне и о местоположении
донора. По его мнению, возбуждение, выраженное ультразвуками, может
быть передапо реципиептам, привлекая их внимание и стимулируя их к
тем или иным действиям. Но конкретные формы активности рецициептов
зависят от различных факторов и не закодированы в параметрах сигна-
ла, «запускающего» лишь уровень возбуждения. Рассматривая ультра-
звуковую коммуникацию в широком аспекте, Белл подчеркивает, что
специфические формы активности реципиентов зависят от предшествую-
щего уровня возбуждения адресатов и их предшествующей конкретной
активпости, от того, в каком физиологическом состоянии они находятся
(эструс, лактация, уровень ‘андрогена, кортикостероидная активность),
Поведение
313
других стимульных свойств ситуации (контакт с партнерами по группе,
запах партнеров).
Первая экспериментальная проверка влияния ультразвуков, издавае-
мых крысятами, на поведепие взрослых животных принадлежит Аллину
и Бенксу (АП, ВашЕз, 1972; цит. по: ба1ез, Руе, 1974). Эксперимент
проводили па лактирующих самках, нерожавитих самках и взрослых сам-
цах белых крыс. В качестве стимула использовали 90-минутную магнит-
ную запись семидневных крысят. Контролем служила магнитная запись
шума. Гнездовая камера была устаповлена в шестиугольном полигоне.
Сигнал транслировали в случайном порядке с одной из шести стороп.
Аллин и Бепкс сообщают, что 20 из 40 лактирующих самок покидали
гнездо в ответ на ультразвуки, а большинство из пих ориептировались
на источник сигнала, Самцы и нерожавитие самки не покидали гнездо во
время воспроизведения ультразвуков, но пекоторыс самцы ориентирова-
ли головы в направлении источника звука.
Стимулирующее действие ультразвуковых сигналов детенышей повы-
шается в присутствии запаха крысят. Это было показано в эксперимеп-
тах Смотермапа с соавторами (Зтоегтап её а1., 4974). Лактирующим
самкам крыс предлагали выбрать один из рукавов лабиринта и верпуть
в гнездо находящегося в лабиринте детеныша. Привлекающими стиму-
лами были ультразвуки, издаваемые крысятами, которые в этом случае
были одновременно источником как акустического, так и химического
стимулов; магпитная запись ультразвуковых сигналов крысят (химиче-
ский стимул в этом случае исключался); запах крысят (детеныши, ох-
лажденные до коматозного состояния, не издавали ультразвуков). В ка-
честве контроля использовали пустой рукав лабиринта, свободный от
экспериментальных стимулов. Во всех случаях самки покидали гнездо-
вую коробку и направлялись в лабиринт, если присутствовал запах кры-
сят, а латентный период сокращалея, когда действовали одновременпо
Два стимула — акустический и химический. Это могли быть крысята, не-
посредственно издающие ультразвуки, или трансляция магпитпой запи-
си ультразвуков с одновременным предъявлением молчащего детеныша.
Ультразвуки крысят влияют пе только па поведение самок, но и на
их физиологическое состояние. Теркель с соавторами (Тегке| еф а1.,
1979) показали, что ультразвуковая активность крысят стимулирует сек-
рецию пролактина и таким образом регулирует нормальпую лактацию у
кормящих самок. Более того, ультразвуковая стимуляция незначитель-
но, но достоверно повышает количество пролактина в крови не только
кормящих, но и нерожавтних самок. Исследуя действие ультразвуковых
“сигналов детепьшней па секрецию пролактина, крысят удаляли из гнезда
на бчив течение 15 мин воспроизводили магнитную запись их голо-
сов. Коптролем служило отсутствие воспроизведения или воспроизведе-
ние в течение 15 мин магнитной записи шума. Таким образом, повыше-
ние пролактина вызывали только ультразвуки крысят. Существенно, что,
когда крысят возвращали в гпездо и они начинали сосать, т. е. когда
зключались несе сенсорные модальности (слуховая, ольфакторпая, визу-
альная и тактильпая), количество пролактина увеличивалось еще вдвое
314 Глава четвертая
и достигало у лактирующих самок в среднем 895 ит/мл. Авторы считают,
что поскольку пролактин способствует росту детенышей, поступая к ним
с молоком матери, то ультразвуковая коммуникация опосредствованно-
выполняет роль стимулятора роста.
Но спустя 5 лет в том же журнале была опубликована работа ЛТтер-
на с соавторами (Э1еги её 21., 1984) под интригующим названием «Дей-
ствительпо ли ультразвуки детенышей вызызают секрецию пролактина
у лактирующих самок?», в которой результаты Теркеля с соавторами
(Тегке| её а1., 1979) не были подтверждены. В частности, самкам спустя
неделю после рождения детенышей транслировали магнитную запись.
ультразвуков крысят, прерывистый тон с частотой 45 кГц и шум. Пред-
варительно, как и в эксиерименте предшествеппиков, крысят удаляли
из гнезда на 6 ч. Ультразвуки детенышей и 45 кГц тон вызывали у всех
исследованных самок вполне адекватную поведенчоескую реакцию (ори-
онтацию на источник ультразвука, исследования клетки, чистку), но как
в опыте, так и в контроле не было обнаружено повышение пролактина.
И только после того, как крысята были подсажены к самкам и в тече-
ние 15 мин кормились их молоком, содержание пролактина в крови рез-
ко повысилось. Особенно сильно возросло содержание пролактина у тех
самок, которым предварительно в качестве стимула предлагали ультра-
звуки детенышей, в меньшей степени
— прерывистый тон и еще мень-
шей
— шум. По мнению авторов, вопрос о том, влияют ли ультразвуки
детенышей па содержание пролактина в крови, требует дальнейшей экс-
периментальной проверки, а результаты исследовация могут сильно за-
висеть от постановки эксперимента и особенностей самих животных.
Например, в опытах Теркеля с соавторами (Тегке] её а1., 1979) макси-
мальное содержание пролактина в крови самок, когда детенышей возвра-
щали в гнездо, достигало в среднем 895 нг/мл, в то время как в экспе-
риментах Штерна с соавторами (З4етп сё а1., 1984) оно составляло всего
213 нг/мл, хотя в обоих случаях крысы принадлежали одной гопетиче-
ской линии. Несмотря на трудности в интерпретации указапных вытие
работ, нельзя не отметить, что результаты исследования Шлерна с со-
авторами не абсолютно противоречат результатам, полученным Теркелем
с соавторами, так как возвращение крысят вызывало большее повышение
содержания пролактина у тех самок, которым предварительно предлагали
ультразвуки крысят.
'
Остается неясным зпачепие ультразвуков, издаваемых крысятами во
время их перетаскивания самками. Эксперименты по данному вопросу
отсутствуют, но некоторые авторы считают, что в подобных ситуациях
акустическая активность детенышей снижает агрессивность самок (За-
1ез, ши, 1978).
Леви и Шрифер (Го\1з, боевые ег, 1982) обнаружили повышенную
ультразвуковую активность у беременных самок серых крыс. С первого
и до последнего дня беременности опи издавали достоверно большес чис-
ло ультразвуков, чем небеременные самки. Основная частотпая компо-
нента этих ультразвуков лежит в области 40 кГи. Накануне родов резко
увеличивается размах индивидуальной изменчивости частоты повторений
Поведение
315
Ультразвуков в час и расширяется диапазон излучаемых частот от 15 до
60 кГц, что авторы связывают с предродовым стрессом. Не установлено,
имеют ли ультразвуки беременных самок сигнальное зпачепие или же
только отражают уровень и специфику возбуждения животных.
Акустическая коммуникация в брачном поведении
серых крыс
Первые исследования, посвященные акустической коммуникации в брач-
ном поведении серых крыс, припадлежат Саль (Заез, 1972а) и Барфил-
ду и Гейер (ВагНе!3, Сеуег, 1972). Самцы, помещенные в клетку к сам-
кам, издают ультразвуки, обнюхивая генитальную область самок, пре-
следуя их или совершая садки. Ультразвуки сопровождают также иптро-
миссии и эякуляции, Впервые попадая в клетку к самке, самцы издают
обычно ритмически не оргапизоваппые короткие ультразвуки. Длитель-
ность ультразвуков от 3 до 50 ме, диацазон частот— от 50 до 75 кГц.
Обнюхивая гепиталии самки, преследуя ее перед садкой и во время
садки, самцы издают серии более длительных, частотно-модулироваппых
ультразвуков. Обычно удается выделить два их типа — длительные, оди-
ночные, относительно низкочастотные ультразвуки и серии коротких,
частотно-модулированных сигналов. Первые имеют длительность от 100
до 800 мс и частоту от 30 до 55 кГц. Частота постененно понижается к
концу сигнала. Нередко он содержит отдельные короткие элементы.
В смектре выделяются первая, иногда вторая гармоники (рис. 80, а).
Длительность серий достигает 200 мс, длительность отдельных элемен-
тов 2—40 ме, интервалы между ними 3—15 мс. Каждый из отдельных
элементов содержит один, ипогда два цикла частотной модуляции, глуби-
на которой достигает нескольких десятков кГц (рис. 80,6; Ба1оз, 1972).
Ультразвуковой сигпал, описанный Барфилдом и Гейером (ВагНаЧ,
Сеуег, 1972), итироко вошел в литературу под названием «22-килогерце-
вый постоякуляторный сигнал (иногда
— песня) самца». Впервые опи-
сывая его, авторы отметили, что чаще всего сигнал удается зарегистри-
ровать вскоре после эякуляции. Он состоит из импульсов большой дли-
тельности (1-8 с) и отпосительно низкой частоты (22—23 кГц). Ин-
тервалы между отдельными ультразвуками около 0,25 с. Уровень звуко-
вого давлепия, замеренный на расстоянии 5—10 см от животного,
равен 80 дБ. Авторы подчеркивают, что этот сигнал им не удавалось за-
регистрировать ни в одной фазе брачного поведения, за исключением ре-
фракторного периода эякуляции. Подавая сигнал, самцы в большинстве
случаев спокойно лежат на субстрате, реже чистятся или медленно пере-
мещаются. Самки в это время стараются отделиться от них. Сходную
ультразвуковую реакцию Барфилд и Гейер обнаружили у подчиненных
самцов серых крыс. Авторы обращают внимание на то, что рефрактер-
ный период эякуляции, для которого характерен данный сигнал, соот-
зетствует стадии торможения. Это подтверждается, в частности, электро-
энцефалограммой. В рефрактерную фазу эякуляции она соответствует
состоянию, напоминающему состояние сна. В обычпом лабораторном
314 Глава четвертая
и достигало у лактирующих самок в среднем 895 ит/мл. Авторы считают,
что поскольку пролактин способствует росту детенышей, поступая к ним
с молоком матери, то ультразвуковая коммуникация опосредствованно-
выполняет роль стимулятора роста.
Но спустя 5 лет в том же журнале была опубликована работа ЛТтер-
на с соавторами (Э1еги её 21., 1984) под интригующим названием «Дей-
ствительпо ли ультразвуки детенышей вызызают секрецию пролактина
у лактирующих самок?», в которой результаты Теркеля с соавторами
(Тегке| её а1., 1979) не были подтверждены. В частности, самкам спустя
неделю после рождения детенышей транслировали магнитную запись.
ультразвуков крысят, прерывистый тон с частотой 45 кГц и шум. Пред-
варительно, как и в эксиерименте предшествеппиков, крысят удаляли
из гнезда на 6 ч. Ультразвуки детенышей и 45 кГц тон вызывали у всех
исследованных самок вполне адекватную поведенчоескую реакцию (ори-
онтацию на источник ультразвука, исследования клетки, чистку), но как
в опыте, так и в контроле не было обнаружено повышение пролактина.
И только после того, как крысята были подсажены к самкам и в тече-
ние 15 мин кормились их молоком, содержание пролактина в крови рез-
ко повысилось. Особенно сильно возросло содержание пролактина у тех
самок, которым предварительно в качестве стимула предлагали ультра-
звуки детенышей, в меньшей степени
— прерывистый тон и еще мень-
шей
— шум. По мнению авторов, вопрос о том, влияют ли ультразвуки
детенышей па содержание пролактина в крови, требует дальнейшей экс-
периментальной проверки, а результаты исследовация могут сильно за-
висеть от постановки эксперимента и особенностей самих животных.
Например, в опытах Теркеля с соавторами (Тегке] её а1., 1979) макси-
мальное содержание пролактина в крови самок, когда детенышей возвра-
щали в гнездо, достигало в среднем 895 нг/мл, в то время как в экспе-
риментах Штерна с соавторами (З4етп сё а1., 1984) оно составляло всего
213 нг/мл, хотя в обоих случаях крысы принадлежали одной гопетиче-
ской линии. Несмотря на трудности в интерпретации указапных вытие
работ, нельзя не отметить, что результаты исследования Шлерна с со-
авторами не абсолютно противоречат результатам, полученным Теркелем
с соавторами, так как возвращение крысят вызывало большее повышение
содержания пролактина у тех самок, которым предварительно предлагали
ультразвуки крысят.
'
Остается неясным зпачепие ультразвуков, издаваемых крысятами во
время их перетаскивания самками. Эксперименты по данному вопросу
отсутствуют, но некоторые авторы считают, что в подобных ситуациях
акустическая активность детенышей снижает агрессивность самок (За-
1ез, ши, 1978).
Леви и Шрифер (Го\1з, боевые ег, 1982) обнаружили повышенную
ультразвуковую активность у беременных самок серых крыс. С первого
и до последнего дня беременности опи издавали достоверно большес чис-
ло ультразвуков, чем небеременные самки. Основная частотпая компо-
нента этих ультразвуков лежит в области 40 кГи. Накануне родов резко
увеличивается размах индивидуальной изменчивости частоты повторений
Поведение
315
Ультразвуков в час и расширяется диапазон излучаемых частот от 15 до
60 кГц, что авторы связывают с предродовым стрессом. Не установлено,
имеют ли ультразвуки беременных самок сигнальное зпачепие или же
только отражают уровень и специфику возбуждения животных.
Акустическая коммуникация в брачном поведении
серых крыс
Первые исследования, посвященные акустической коммуникации в брач-
ном поведении серых крыс, припадлежат Саль (Заез, 1972а) и Барфил-
ду и Гейер (ВагНе!3, Сеуег, 1972). Самцы, помещенные в клетку к сам-
кам, издают ультразвуки, обнюхивая генитальную область самок, пре-
следуя их или совершая садки. Ультразвуки сопровождают также иптро-
миссии и эякуляции, Впервые попадая в клетку к самке, самцы издают
обычно ритмически не оргапизоваппые короткие ультразвуки. Длитель-
ность ультразвуков от 3 до 50 ме, диацазон частот— от 50 до 75 кГц.
Обнюхивая гепиталии самки, преследуя ее перед садкой и во время
садки, самцы издают серии более длительных, частотно-модулироваппых
ультразвуков. Обычно удается выделить два их типа — длительные, оди-
ночные, относительно низкочастотные ультразвуки и серии коротких,
частотно-модулированных сигналов. Первые имеют длительность от 100
до 800 мс и частоту от 30 до 55 кГц. Частота постененно понижается к
концу сигнала. Нередко он содержит отдельные короткие элементы.
В смектре выделяются первая, иногда вторая гармоники (рис. 80, а).
Длительность серий достигает 200 мс, длительность отдельных элемен-
тов 2—40 ме, интервалы между ними 3—15 мс. Каждый из отдельных
элементов содержит один, ипогда два цикла частотной модуляции, глуби-
на которой достигает нескольких десятков кГц (рис. 80,6; Ба1оз, 1972).
Ультразвуковой сигпал, описанный Барфилдом и Гейером (ВагНаЧ,
Сеуег, 1972), итироко вошел в литературу под названием «22-килогерце-
вый постоякуляторный сигнал (иногда
— песня) самца». Впервые опи-
сывая его, авторы отметили, что чаще всего сигнал удается зарегистри-
ровать вскоре после эякуляции. Он состоит из импульсов большой дли-
тельности (1-8 с) и отпосительно низкой частоты (22—23 кГц). Ин-
тервалы между отдельными ультразвуками около 0,25 с. Уровень звуко-
вого давлепия, замеренный на расстоянии 5—10 см от животного,
равен 80 дБ. Авторы подчеркивают, что этот сигнал им не удавалось за-
регистрировать ни в одной фазе брачного поведения, за исключением ре-
фракторного периода эякуляции. Подавая сигнал, самцы в большинстве
случаев спокойно лежат на субстрате, реже чистятся или медленно пере-
мещаются. Самки в это время стараются отделиться от них. Сходную
ультразвуковую реакцию Барфилд и Гейер обнаружили у подчиненных
самцов серых крыс. Авторы обращают внимание на то, что рефрактер-
ный период эякуляции, для которого характерен данный сигнал, соот-
зетствует стадии торможения. Это подтверждается, в частности, электро-
энцефалограммой. В рефрактерную фазу эякуляции она соответствует
состоянию, напоминающему состояние сна. В обычпом лабораторном
316 Глава четвертая
109Е
Рис. 80. Сонограммы
\
.
учьтразвуков, сопровож-
“ дающих брачное пове-
50Е
-
дение самцов серых
=
крыс (по: Эщез, Вуе,
<
1974)
<
Г/А
а0
1
:
1
—
а — последние 50 ме уль-
Е
‹
;
вое
„.
.
=
тразвука, в котором
5
Ро%=\#%7%/\—
медленное — измененпе
#
&. Ы7:
:#
г
г
-
частоты оканчивается
Е“
-=
_
2
ы
резкой частотной моду-
ляцией;
$ б — серия частотно-моду-
0р
т
р
з
р,
1
1
лированных ультразву-
$
50
ИЙ
й
ков
Время, ме
эксперименте наблюдается от 6 до 8 рефрактерпых фаз эякуляции, когда
самка и самец расходятся на 5—40 мин. В это время самцы проявляют
резко пониженный интерес к самкам и имепно в этот период чаще всего
издают 22-кГц сигнал (ЛАег, Ап!зКо, 1979). Для удобства изложения в
дальнейшем описанную выше акустическую реакцию мы будем пазызать
22-кГи сигпалом.
В 1975 г. Барфилд и Гейер (Вахе, Сеуег, 1975) выделили абсо-
лютную и относительную рефрактерпые фазы. Абсолютная фаза, во.
время которой самцы наиболее пассивны, а ЭЭГ показывает, что они па-
ходятся в состоянии, близком к состоянию спа, совпадает по времени
с 22-кГц сигналом. Замолкая, животные становятся активнее, начинают
чиститься и вновь переходят к брачному поведению. ЭЭГ подтверждает
переход самцов от заторможепного состояния к бодретвованию. Ультра-
звуковая активность (и абсолютная рефрактерная фаза) занимают около.
75% времени всего межэякуляторного периода, 25% занимает относи-
тельная рефрактерная фаза. Аниско с соавторами (АтзКо её а[., 1978}
подтвердили, что три четверти всего постэякуляторного периода запи-
мает ультразвуковая активность, но в ряде случаев 22-кГц сигнал уда-
валось зарегистрировать и в его последней четверти. Шо мнению авторов,
полученные различия могут быть вызваны индивидуальными особетно-
стями животных или спецификой экспериментов.
Рефрактерный период эякуляции находится цод гормональным ковт-
ролем. Аидрогены оказывают большее влияние на относительную реф-
рактерную фазу, чем на абсолютную. Это было показано Нэрротом (Раг-
то, 1976) в экспериментах па кастрированпных самцах. Значительное и
прогрессивное от первой недели после кастрации к третьей удлинение
рефрактерного периода эякуляции происходило преимущественно за
счет относительной фазы. Длительность ультразвуковой активности
уменьшалась гораздо медленнее, хотя у части кастрированных самцов
22-кГц сигнал вообще отсутствовал.
По мнению Френсис (Егапсз, 1977), 22-кГц сигнал встречается у се-
рых крыс значительно шире. Он обнаружен у самцов и самок, на месяц
изолированных от группы и таким образом лишенных социальных кон-
Поведение
317
тактов. Животные находились в темном помещении, где искусственно че-
редовались светлое и темпое время суток. Корм и вода давались в свет-
лое время. Наименьшее число 22-кГц сигналов было зарегистрировано в
первые часы темного времени, а наибольшее — в середине этого периода.
К концу его число их опять падало. У самок акустическая активпость
никак пе была связана в этом эксцеримелте с эстральными циклами.
Причиной появления 22-кГц сигналов были также стрессовые стимулы,
такие, как электрический шок или грубое манипулирование животными
в руках экспериментатора. Френсис отмечает, что социальная изоляция
тоже, видимо, может быть квалифицирована как состояние длительного
стресса. Те же ультразвуки были обнаружены и у лактирующих самок,
содержащихся вместе с выводком, у животных, соприкасающихся друг
с другом в индивидуальных клетках, а также в колониях как лаборатор-
ных, так и диких крыс. Френсис приходит к выводу, что данный сигнал
отражает прежде всего чередование периодов активности и покоя. Одиа-
ко нефункциопальность сигнала может быть, по мпению автора, только
кажущейся. Френсис не исключает, что 22-кГц сигнал способствует син-
хронизации ритмов активности в колониях крыс, а подаваемый лакти-
рующими самками способствует формированию у детеньшшей ассоциатив-
ных связей между акустическим сигналом и периодами активности и от-
дыха.
Пезависимо от Френсис отсутствие абсолютно жесткой связи между
22-кГц сигналом и постэякуляторной паузой обнаружили Аниско с с0-
авторами (АпизКо её а[., 1978). Прежде всего они отметили, что впечат-
пение о жесткости такой связи может быть вызвано условиями экспери-
мента с подсаживанием, когда сильно обедняется разнообразие внутри-
групповых контактов. Авторы проводили эксперимент в условиях,
приближенных к остественным, в большой вольере на многочисленной
группе животных. Они обнаружили, что в 45% случаев 22-кГц сигнал
подавался перед серией эякуляций, а пс только в постэякуляционные
рефрактерные фазы. Причем 26% преэякуляторных омиссий сигнала
были реакцией на агрессивные действия самок по отношению к самцам.
Более того, только 69% рефрактерных периодов эякуляции сопровожца-
лись ультразвуками на частоте 22 кГц. Обобщая полученные результа-
ты, Аниско с соавторами выделили 4 осповных типа чсоциосексуальных
состояний», в которых удается зарегистрировать даппый сигнал: 1) аг-
рессия с0 стороны особей своего вида; 2) презякуляторное состояние с
одновременным избегапием самки; 3) отдых во время постэякуляторного
интервала и 4) последующий за этим болезненный шок, вызываемый на-
падением особей стоего вида. Не исключая связи между сигналом и пост-
эякуляторным торможением самцов, Аниско с соавторами считают, что
тю характеру мотивации 22-кГи сигиал означает прежде всего, что его
поситель «выходит» из социальных контактов, демонстрируя подавлен-
ное состояпис. По мнению авторов, в нормальной групце крыс, обычно
состоящей из 4—5 самок и 2—3 самцов, 22-кГц сигнал способствует ко-
ординации поведения животных. В группе, состоящей из нескольких вам-
цов и нескольких эстральных самок, только один самец копулирует в
316 Глава четвертая
109Е
Рис. 80. Сонограммы
\
.
учьтразвуков, сопровож-
“ дающих брачное пове-
50Е
-
дение самцов серых
=
крыс (по: Эщез, Вуе,
<
1974)
<
Г/А
а0
1
:
1
—
а — последние 50 ме уль-
Е
‹
;
вое
„.
.
=
тразвука, в котором
5
Ро%=\#%7%/\—
медленное — измененпе
#
&. Ы7:
:#
г
г
-
частоты оканчивается
Е“
-=
_
2
ы
резкой частотной моду-
ляцией;
$ б — серия частотно-моду-
0р
т
р
з
р,
1
1
лированных ультразву-
$
50
ИЙ
й
ков
Время, ме
эксперименте наблюдается от 6 до 8 рефрактерпых фаз эякуляции, когда
самка и самец расходятся на 5—40 мин. В это время самцы проявляют
резко пониженный интерес к самкам и имепно в этот период чаще всего
издают 22-кГц сигнал (ЛАег, Ап!зКо, 1979). Для удобства изложения в
дальнейшем описанную выше акустическую реакцию мы будем пазызать
22-кГи сигпалом.
В 1975 г. Барфилд и Гейер (Вахе, Сеуег, 1975) выделили абсо-
лютную и относительную рефрактерпые фазы. Абсолютная фаза, во.
время которой самцы наиболее пассивны, а ЭЭГ показывает, что они па-
ходятся в состоянии, близком к состоянию спа, совпадает по времени
с 22-кГц сигналом. Замолкая, животные становятся активнее, начинают
чиститься и вновь переходят к брачному поведению. ЭЭГ подтверждает
переход самцов от заторможепного состояния к бодретвованию. Ультра-
звуковая активность (и абсолютная рефрактерная фаза) занимают около.
75% времени всего межэякуляторного периода, 25% занимает относи-
тельная рефрактерная фаза. Аниско с соавторами (АтзКо её а[., 1978}
подтвердили, что три четверти всего постэякуляторного периода запи-
мает ультразвуковая активность, но в ряде случаев 22-кГц сигнал уда-
валось зарегистрировать и в его последней четверти. Шо мнению авторов,
полученные различия могут быть вызваны индивидуальными особетно-
стями животных или спецификой экспериментов.
Рефрактерный период эякуляции находится цод гормональным ковт-
ролем. Аидрогены оказывают большее влияние на относительную реф-
рактерную фазу, чем на абсолютную. Это было показано Нэрротом (Раг-
то, 1976) в экспериментах па кастрированпных самцах. Значительное и
прогрессивное от первой недели после кастрации к третьей удлинение
рефрактерного периода эякуляции происходило преимущественно за
счет относительной фазы. Длительность ультразвуковой активности
уменьшалась гораздо медленнее, хотя у части кастрированных самцов
22-кГц сигнал вообще отсутствовал.
По мнению Френсис (Егапсз, 1977), 22-кГц сигнал встречается у се-
рых крыс значительно шире. Он обнаружен у самцов и самок, на месяц
изолированных от группы и таким образом лишенных социальных кон-
Поведение
317
тактов. Животные находились в темном помещении, где искусственно че-
редовались светлое и темпое время суток. Корм и вода давались в свет-
лое время. Наименьшее число 22-кГц сигналов было зарегистрировано в
первые часы темного времени, а наибольшее — в середине этого периода.
К концу его число их опять падало. У самок акустическая активпость
никак пе была связана в этом эксцеримелте с эстральными циклами.
Причиной появления 22-кГц сигналов были также стрессовые стимулы,
такие, как электрический шок или грубое манипулирование животными
в руках экспериментатора. Френсис отмечает, что социальная изоляция
тоже, видимо, может быть квалифицирована как состояние длительного
стресса. Те же ультразвуки были обнаружены и у лактирующих самок,
содержащихся вместе с выводком, у животных, соприкасающихся друг
с другом в индивидуальных клетках, а также в колониях как лаборатор-
ных, так и диких крыс. Френсис приходит к выводу, что данный сигнал
отражает прежде всего чередование периодов активности и покоя. Одиа-
ко нефункциопальность сигнала может быть, по мпению автора, только
кажущейся. Френсис не исключает, что 22-кГц сигнал способствует син-
хронизации ритмов активности в колониях крыс, а подаваемый лакти-
рующими самками способствует формированию у детеньшшей ассоциатив-
ных связей между акустическим сигналом и периодами активности и от-
дыха.
Пезависимо от Френсис отсутствие абсолютно жесткой связи между
22-кГц сигналом и постэякуляторной паузой обнаружили Аниско с с0-
авторами (АпизКо её а[., 1978). Прежде всего они отметили, что впечат-
пение о жесткости такой связи может быть вызвано условиями экспери-
мента с подсаживанием, когда сильно обедняется разнообразие внутри-
групповых контактов. Авторы проводили эксперимент в условиях,
приближенных к остественным, в большой вольере на многочисленной
группе животных. Они обнаружили, что в 45% случаев 22-кГц сигнал
подавался перед серией эякуляций, а пс только в постэякуляционные
рефрактерные фазы. Причем 26% преэякуляторных омиссий сигнала
были реакцией на агрессивные действия самок по отношению к самцам.
Более того, только 69% рефрактерных периодов эякуляции сопровожца-
лись ультразвуками на частоте 22 кГц. Обобщая полученные результа-
ты, Аниско с соавторами выделили 4 осповных типа чсоциосексуальных
состояний», в которых удается зарегистрировать даппый сигнал: 1) аг-
рессия с0 стороны особей своего вида; 2) презякуляторное состояние с
одновременным избегапием самки; 3) отдых во время постэякуляторного
интервала и 4) последующий за этим болезненный шок, вызываемый на-
падением особей стоего вида. Не исключая связи между сигналом и пост-
эякуляторным торможением самцов, Аниско с соавторами считают, что
тю характеру мотивации 22-кГи сигиал означает прежде всего, что его
поситель «выходит» из социальных контактов, демонстрируя подавлен-
ное состояпис. По мнению авторов, в нормальной групце крыс, обычно
состоящей из 4—5 самок и 2—3 самцов, 22-кГц сигнал способствует ко-
ординации поведения животных. В группе, состоящей из нескольких вам-
цов и нескольких эстральных самок, только один самец копулирует в
318 Глава четвертая
даппый момент, а его временный выход из социальных коптактов в ре-
зультате иодавленного состояния в рефрактерный нериод зякуляции
приводит к перестройке иерархической структуры группы.
Браун (Вгомт, 1979) обнаружил, что 22-кГц сигпал самцы довольно
часто подают и до серии интромиссий и копуляций, особенно в тех слу-
чаях, когда самки уклоняются от спаривания и проявляют новышенную
агрессивность. Более того, было замечено, что, издавая данный ультра-
звук, самцы могут преследовать самок. Браун считает, что его наблюде-
ние не противоречит гипотезе Аниско с соавторами о выходе самцов из
социальных контактов. По его мнепию, с помощью 22-кГц сигнала сам-
цы в различных ситуациях информируют членов группы о том, что они
не в состоянии нормально функционировать в данном взаимодействии,
По наблюдениям Гейера и Барфилда (Сеуег, ВагНе!4, 1978а), дли-
тельность презякуляторных сигналов меньше (менее 2 с), чем постаяку-
ляторных. Авторы подчеркивают роль фрустрации в случаях, когда сам-
цы издают 22-кГц ультразвуки. По их наблюдениям, самцы подают этот
сигнал в 45% случаев до эякуляторпых серий, при этом подвергаясь
атакам со стороны самок или же сами преследуя их. В подобных ситуа-
циях самцы сильно возбуждались, но пе могли реализовать половую мо-
тивацию, что и приводило к развитию фрустрации, сопровождающейся
22-кГц сигналом. Адлер и Аниско (АЧ]ег, АзтзКо, 1979) подтвердили,
что 22-кГц сигнал появляется у самцов крыс в различных ситуациях,
в которых животные испытывают сослояние утомления, торможения, ио-
давленности. Этот сигнал характеризует пе ситуацию, а состояние источ-
ника сигпала, по в половом поведении может выполнять и узко специ-
фическую функцию. Обычный в интервале между эякуляциями, 22-кГц
сигнал способствует поддержапию дистанции между брачными партие-
рами, препятствуя частым повторениям зякуляций. Рефрактерная фаза
эякуляций необходима для нормального прохождения спермы по матке.
В противпом случае вагино-цервикальная стимуляция препятствует это-
му процессу.
Как уже говорилось, помимо 22-кГц сигнала, самцы и самки серых
крыс сопровождают брачное поведение и более высокими ультразвука-
ми. Обычно их частота находится в диапазоне от 40 до 60 кГц, но мо-
жет быть и выше. В литературу эта ультразвуковая реакция вошла как
«50-кГц сигнал». В дальнейшем изложении мы также будем пользовать-
ся этим термином. 50 кГц сигнал чаще всего регистрируют в пачальных
<тадиях брачного поведения. В отличие от 22-кГц сигнала он соответст-
вует более активному состоянию животных. Хотя о 50 кГц сигнале впер-
вые сообщается в 1972 г. (Зайез, 1972 а, Ь), но до настоящего времени из-
вестно о нем очень мало. Барфилд с соавторами в обзорной статье, опуб-
ликоваппой в 1979 г. (ВагНе4 её а|., 1979), писали в связи с этим, что
неизвестно, представляют ли ультразвуки па частоте, близкой к 50-кГц,
отдельный, но сильно измепчивый сигнал, или же это несколько различ-
ных, но связанных между собой сигналов.
Влияние 50-кГц сигнала на поведение самок виервые было показапо
Макинтошем с соавторами (МеПиозН её а|., 1978). Ссаживая эстральных
Поведение
319
самок с кастрированными самцами, авторы обнаружили, что активность
полового поведения самок резко возрастает, если через громкоговоритель
транслировать ультразвуковой сигнал. Возрастает стремление самок к
самцам, сокращается латентный период между отдельными волнами по-
лового возбуждепия, чаще появляется лордоз. Причем моча интактных
самцов не стимулировала половую активность самок, а одновременное
предъявление мочи и ультразвуков вызывало не больший эффект, чем
только ультразвуковая стимуляция.
Несколько противоречивые результаты получены Барфилдом с соавто-
рами (ВагНе4 еф а|., 1979). Они сравнивали ультразвуковую активность,
крыс в случаях удачной и неудачной копуляции, т. е. закончившейся или
незакопчившейся эякуляцией. В обоих случаях оказалось, что число
50-кГц сигналов выше в активный период брачного поведения, когда сам-
ки не проявляют агопизма по отпошению к самцам, чем в подготовитель-
ный. Но в случаях удачных копуляций, закончившихся эякуляциями,
число ультразвуков оказалось в 2,5 раза мепьше, чем в случае неудачных.
Авторы, считая результаты эксперимента парадоксальными, отмечают,
что они заслуживают дальнейшего исследования. В этой же работе пока-
запо, что число 50-кГц сигналов неред интромиссиями было большим, чем
перед садками.
В эксперимептах Томаса с соавторами (Трошаз её а1, 1981) самцы,
лишенные голоса оперативным путем, не изменяли своего полового пове-
дения, но самки отдавали предпочтепие пормальпым самцам перед не-
мыми, хотя общий рисунок их поведения как с теми, так и с другими
оставался одним и тем же. Авторы считают, что 50-кГц сигпал способет-
вует половой восприимчивости самок по отношению к самцам. В другой
серии экспериментов Томас с соавторами (ТЬотаз еф а|., 1982) показали,
что самки проводят одно и то же количество времени как с немыми, так
и с нормальными самцами, но активнее ориентируются на нормальных
самцов или на пемых, позади которых размещены ультразвуковые источ-
ники, транслирующие 50-кГц сигнал. 1о мнению авторов, эти эксиеримен-
ты продемонстрировали, что данная акустическая реакция пе дает преи-
муществ в непосредственном физическом сближении брачных партнеров,
способствуя лишь ориентации эстральных самок на сексуально активных
самцов. Гейер с соавторами (Сеуег её а|., 1978а; Сеуег, ВагНе!а, 1978)
считают, что после сближения брачных партнеров 50-кГн ультразвуки
возбуждают самца и самку, повышая взаимпое влеченис.
Координации коммуникативных процессов в брачном поведении гры-
зунов, в том числе и серых крыс, в зпачительной степепи способствует
\гормопальпая регуляция акустической активности. Гейер и Барфилд
(Сеуег, Вагбе!4 1978), ссаживая кастрированных самок с интактпыми
самцами, обпаружили очень низкий уровень ультразвуковой активности
как тех, так и других. После введения самкам половых гормонов (эстро-
гена или прогестерона) ультразвуковая активность животных резко по-
высилась, как и другие элементы полового поведения. При этом макси-
мальный эффект был достигнут после введепил обоих гормопов. Меньшее
влияние па акустическую активность оказало введение только эстрогенов.
318 Глава четвертая
даппый момент, а его временный выход из социальных коптактов в ре-
зультате иодавленного состояния в рефрактерный нериод зякуляции
приводит к перестройке иерархической структуры группы.
Браун (Вгомт, 1979) обнаружил, что 22-кГц сигпал самцы довольно
часто подают и до серии интромиссий и копуляций, особенно в тех слу-
чаях, когда самки уклоняются от спаривания и проявляют новышенную
агрессивность. Более того, было замечено, что, издавая данный ультра-
звук, самцы могут преследовать самок. Браун считает, что его наблюде-
ние не противоречит гипотезе Аниско с соавторами о выходе самцов из
социальных контактов. По его мнепию, с помощью 22-кГц сигнала сам-
цы в различных ситуациях информируют членов группы о том, что они
не в состоянии нормально функционировать в данном взаимодействии,
По наблюдениям Гейера и Барфилда (Сеуег, ВагНе!4, 1978а), дли-
тельность презякуляторных сигналов меньше (менее 2 с), чем постаяку-
ляторных. Авторы подчеркивают роль фрустрации в случаях, когда сам-
цы издают 22-кГц ультразвуки. По их наблюдениям, самцы подают этот
сигнал в 45% случаев до эякуляторпых серий, при этом подвергаясь
атакам со стороны самок или же сами преследуя их. В подобных ситуа-
циях самцы сильно возбуждались, но пе могли реализовать половую мо-
тивацию, что и приводило к развитию фрустрации, сопровождающейся
22-кГц сигналом. Адлер и Аниско (АЧ]ег, АзтзКо, 1979) подтвердили,
что 22-кГц сигнал появляется у самцов крыс в различных ситуациях,
в которых животные испытывают сослояние утомления, торможения, ио-
давленности. Этот сигнал характеризует пе ситуацию, а состояние источ-
ника сигпала, по в половом поведении может выполнять и узко специ-
фическую функцию. Обычный в интервале между эякуляциями, 22-кГц
сигнал способствует поддержапию дистанции между брачными партие-
рами, препятствуя частым повторениям зякуляций. Рефрактерная фаза
эякуляций необходима для нормального прохождения спермы по матке.
В противпом случае вагино-цервикальная стимуляция препятствует это-
му процессу.
Как уже говорилось, помимо 22-кГц сигнала, самцы и самки серых
крыс сопровождают брачное поведение и более высокими ультразвука-
ми. Обычно их частота находится в диапазоне от 40 до 60 кГц, но мо-
жет быть и выше. В литературу эта ультразвуковая реакция вошла как
«50-кГц сигнал». В дальнейшем изложении мы также будем пользовать-
ся этим термином. 50 кГц сигнал чаще всего регистрируют в пачальных
<тадиях брачного поведения. В отличие от 22-кГц сигнала он соответст-
вует более активному состоянию животных. Хотя о 50 кГц сигнале впер-
вые сообщается в 1972 г. (Зайез, 1972 а, Ь), но до настоящего времени из-
вестно о нем очень мало. Барфилд с соавторами в обзорной статье, опуб-
ликоваппой в 1979 г. (ВагНе4 её а|., 1979), писали в связи с этим, что
неизвестно, представляют ли ультразвуки па частоте, близкой к 50-кГц,
отдельный, но сильно измепчивый сигнал, или же это несколько различ-
ных, но связанных между собой сигналов.
Влияние 50-кГц сигнала на поведение самок виервые было показапо
Макинтошем с соавторами (МеПиозН её а|., 1978). Ссаживая эстральных
Поведение
319
самок с кастрированными самцами, авторы обнаружили, что активность
полового поведения самок резко возрастает, если через громкоговоритель
транслировать ультразвуковой сигнал. Возрастает стремление самок к
самцам, сокращается латентный период между отдельными волнами по-
лового возбуждепия, чаще появляется лордоз. Причем моча интактных
самцов не стимулировала половую активность самок, а одновременное
предъявление мочи и ультразвуков вызывало не больший эффект, чем
только ультразвуковая стимуляция.
Несколько противоречивые результаты получены Барфилдом с соавто-
рами (ВагНе4 еф а|., 1979). Они сравнивали ультразвуковую активность,
крыс в случаях удачной и неудачной копуляции, т. е. закончившейся или
незакопчившейся эякуляцией. В обоих случаях оказалось, что число
50-кГц сигналов выше в активный период брачного поведения, когда сам-
ки не проявляют агопизма по отпошению к самцам, чем в подготовитель-
ный. Но в случаях удачных копуляций, закончившихся эякуляциями,
число ультразвуков оказалось в 2,5 раза мепьше, чем в случае неудачных.
Авторы, считая результаты эксперимента парадоксальными, отмечают,
что они заслуживают дальнейшего исследования. В этой же работе пока-
запо, что число 50-кГц сигналов неред интромиссиями было большим, чем
перед садками.
В эксперимептах Томаса с соавторами (Трошаз её а1, 1981) самцы,
лишенные голоса оперативным путем, не изменяли своего полового пове-
дения, но самки отдавали предпочтепие пормальпым самцам перед не-
мыми, хотя общий рисунок их поведения как с теми, так и с другими
оставался одним и тем же. Авторы считают, что 50-кГц сигпал способет-
вует половой восприимчивости самок по отношению к самцам. В другой
серии экспериментов Томас с соавторами (ТЬотаз еф а|., 1982) показали,
что самки проводят одно и то же количество времени как с немыми, так
и с нормальными самцами, но активнее ориентируются на нормальных
самцов или на пемых, позади которых размещены ультразвуковые источ-
ники, транслирующие 50-кГц сигнал. 1о мнению авторов, эти эксиеримен-
ты продемонстрировали, что данная акустическая реакция пе дает преи-
муществ в непосредственном физическом сближении брачных партнеров,
способствуя лишь ориентации эстральных самок на сексуально активных
самцов. Гейер с соавторами (Сеуег её а|., 1978а; Сеуег, ВагНе!а, 1978)
считают, что после сближения брачных партнеров 50-кГн ультразвуки
возбуждают самца и самку, повышая взаимпое влеченис.
Координации коммуникативных процессов в брачном поведении гры-
зунов, в том числе и серых крыс, в зпачительной степепи способствует
\гормопальпая регуляция акустической активности. Гейер и Барфилд
(Сеуег, Вагбе!4 1978), ссаживая кастрированных самок с интактпыми
самцами, обпаружили очень низкий уровень ультразвуковой активности
как тех, так и других. После введения самкам половых гормонов (эстро-
гена или прогестерона) ультразвуковая активность животных резко по-
высилась, как и другие элементы полового поведения. При этом макси-
мальный эффект был достигнут после введепил обоих гормопов. Меньшее
влияние па акустическую активность оказало введение только эстрогенов.
320 Глава четвертая
Самцы издавали не только 50-кГц, но и 22-кГц ультразвуки, что также
зависело от гормонального состояния партнери. Переход от 50-кГц к
22-кГц сигналам наблюдался в случае максимально восприимчивых самок,
обработанных обоими гормонами. Партнеры были разделены сеткой.
Церевозбуждение самцов вследствие нереализованной мотивации приводи-
ло затем к резкому утомлению, которое сопровождалось подачей 22-кГц
сигналов. В экспериментах Гейера и Барфилда нельзя было установить,
кто издает ультразвуки, самец или самка, но одипочные самцы обычно
подавали данный сигнал, находясь в клетке, испачканной эстральной
самкой, а одиночные самки подавали данный сигнал, реагируя на запах
мочи самца. Влияние запаховой стимуляции на акустическую активность
крыс вообще, вероятно, очень велико. В той же серии эксперимептов как
самцы, так и самки в чистых клетках не издавали ультразвуков, или же
они были редки и случайны. Правда, запах инициировал, но не поддер-
живал акустическую активность животпых. Для ее продолжения было
необходимо присутствие партнеров, сопровождаемое демонстрацией поло-
вого поведения. Самки, возбужденные половыми гормонами, но обездви-
женные анестезией, вызывали гораздо меньшую акустическую и вообще
половую активность у самцов, чем самки, находящиеся в бодротвующем
состоянии.
Самый общий вывод, к которому приитли Гейер с соавторами (Сеует,
ВагНе@, 1978; Сеуег её а1., 1978а, Ъ), состоит в том, что наиболее
быструю и сильную акустическую реакцию в брачном поведении крыс
вызывает комплекс стимулов
— акустических, химических, оптических и
тактильных.
Акустическая коммуникация в агонистическом поведении
серых крыс
Акустическая коммуникация в агонистическом поведении серых крыс
остается, пожалуй, наименее исследованной формой звуковой активпости
У данного вида. В литературе эта форма акустической коммуникации об-
суждается в основном в связи с брачным и территориальным поведепием,
поскольку в том и другом случае агрессивность у крыс проявляется на-
иболее часто. Первое исследование, специально посвященное роли акус-
тической активности в агрессивпом поведении крыс, принадлежит Саль
(Заез, 1972Ъ, цит. но: байез, Руе, 1974). Саль моделировала территори-
альпые колфликты между самцами, подсаживая чужаков в клетку хозяи-
на. После короткого взаимного обнюхивания и исследовательской реак-
ции чужака одна из крыс, чаще хозяин, атаковала другую. Стычка за-
канчивалась тем, что побежденная особь принимала подчиненную лозу,
ложась на спину или на бок, иногда со слегка раздвинутыми задними ко-
нечностями, а победитель стаповился пад ней. В течение эксперимента,
который длился один час, стычки могли повторяться, прерываясь нейт-
ральной активпостью обоих партнеров. 39 из 40 взаимодействий сопро-
вождались двумя типами ультразвуков, длительными и короткими. Дли-
тельность первых обычно равнялась 800—1600 мс и простиралась до
- Поведение
321
Рис. 81. Сопограммы
улотразвуков, сопровож-
дающих агонистическое
поведение серых крыс
в|
(по: ев, Эти, 1974)
а — сонограмма начальных
60 ме ультразвука,
издаваемого подчинен-
ной особью;
б — сонограмма ультразву-
7
Ю
Ч
е
е
т
о
в
т
а
е
,
к
у
З
о
)
ка, издаваемого сам-
|
Г
цами во время агрес-
р
1
к
,
Е
г
сивных контактов
Г!
РГ)
20
Е
40
ЗЕ
50
Дремя,ис
3400 ме. Единственная спектральная компонента занимала частотный
диапазон между 22 и 30 кГц. Частотная модуляция или отсутствовала
(рис. 81), или частота медленцо понижалась к концу сигнала. Длитель-
ные ультразвуки принадлежали подчиненным особям, обычно сопровождая
длительные выдохи животных, паходящихся в паиболее выраженной позе
подчинепил, Если домипирующий во встрече самец активно поддерживал
подчиненное состояние оппонента, то длительный сигнал мог продолжать-
ся до 15—20 мин, а иногда и после того, как животных рассаживали
Длительность коротких ультразвуков равнялась 3—65 Мс. частотный
диапазон — 40—70 кГц. Короткие ультразвуки, как правило "собраны в
серии. Ряд из них имеют глубокую частотную модуллцию. В отличие от
длительных короткие ультразвуки не были синхронизированы с дыханием
крыс, поэтому трудно было определить, кому из партнеров они принадле-
жат. Чаще всего короткие ультразвуки регистрировали в наиболее актив-
ную фазу стычек — атака, боксирование, борьба. Саль приводит рисунки
соответствующих поз (Заез, Руе, 1974). Очень мало коротких ультразву-
ков издавали крысы, демонстрируя явное подчинение и домипирование.
Чтобы уточнить, кому из партнеров принадлежат ультразвуки, особен-
по короткие, Томас с соавторами (ТБотаз её а|., 1983) ‘подсаживали к ин-
тактным самцам хозяевам иптактных чужаков и чужаков, лишенных го-
лоса оперативным путем. При подсаживании немых самцов резко сокра-
щалось число коротких 50-кГц ультразвуков и полностью исчезали дли-
тельные 22-кГц сигналы. Было, таким образом, показано, что как
короткие 50-кГц ультразвуки, так и длительные 22-кГк издают в основном
(а последпие исключительно) менее агрессивные подчиненные чужаки.
Если же голоса лишали хозяина клегки, то ультразвуковая активпость
продолжала оставаться высокой. что также подтвержцало, что ультразву-
ки издают подчиненные самцы (ТаКавазь! её а1., 1983).
|
небольшой серии экспериментов с дикими серыми крысами Саль и
Пи (За1ез, Руе, 1974) установили в осповном те же закономерности аку-
стической активности самцов. Дикие крысы оказались не более агрессив-
ны, чем лабораторные. Длительные ультразвуки (300—600 мс) были ха-
рактерны для самцов, находившихся в подчиненной позе. Короткие уль-
тразвуки самцы издавали при взаимном обнюхивании, еближая носы, или
11 Серая врыса
320 Глава четвертая
Самцы издавали не только 50-кГц, но и 22-кГц ультразвуки, что также
зависело от гормонального состояния партнери. Переход от 50-кГц к
22-кГц сигналам наблюдался в случае максимально восприимчивых самок,
обработанных обоими гормонами. Партнеры были разделены сеткой.
Церевозбуждение самцов вследствие нереализованной мотивации приводи-
ло затем к резкому утомлению, которое сопровождалось подачей 22-кГц
сигналов. В экспериментах Гейера и Барфилда нельзя было установить,
кто издает ультразвуки, самец или самка, но одипочные самцы обычно
подавали данный сигнал, находясь в клетке, испачканной эстральной
самкой, а одиночные самки подавали данный сигнал, реагируя на запах
мочи самца. Влияние запаховой стимуляции на акустическую активность
крыс вообще, вероятно, очень велико. В той же серии эксперимептов как
самцы, так и самки в чистых клетках не издавали ультразвуков, или же
они были редки и случайны. Правда, запах инициировал, но не поддер-
живал акустическую активность животпых. Для ее продолжения было
необходимо присутствие партнеров, сопровождаемое демонстрацией поло-
вого поведения. Самки, возбужденные половыми гормонами, но обездви-
женные анестезией, вызывали гораздо меньшую акустическую и вообще
половую активность у самцов, чем самки, находящиеся в бодротвующем
состоянии.
Самый общий вывод, к которому приитли Гейер с соавторами (Сеует,
ВагНе@, 1978; Сеуег её а1., 1978а, Ъ), состоит в том, что наиболее
быструю и сильную акустическую реакцию в брачном поведении крыс
вызывает комплекс стимулов
— акустических, химических, оптических и
тактильных.
Акустическая коммуникация в агонистическом поведении
серых крыс
Акустическая коммуникация в агонистическом поведении серых крыс
остается, пожалуй, наименее исследованной формой звуковой активпости
У данного вида. В литературе эта форма акустической коммуникации об-
суждается в основном в связи с брачным и территориальным поведепием,
поскольку в том и другом случае агрессивность у крыс проявляется на-
иболее часто. Первое исследование, специально посвященное роли акус-
тической активности в агрессивпом поведении крыс, принадлежит Саль
(Заез, 1972Ъ, цит. но: байез, Руе, 1974). Саль моделировала территори-
альпые колфликты между самцами, подсаживая чужаков в клетку хозяи-
на. После короткого взаимного обнюхивания и исследовательской реак-
ции чужака одна из крыс, чаще хозяин, атаковала другую. Стычка за-
канчивалась тем, что побежденная особь принимала подчиненную лозу,
ложась на спину или на бок, иногда со слегка раздвинутыми задними ко-
нечностями, а победитель стаповился пад ней. В течение эксперимента,
который длился один час, стычки могли повторяться, прерываясь нейт-
ральной активпостью обоих партнеров. 39 из 40 взаимодействий сопро-
вождались двумя типами ультразвуков, длительными и короткими. Дли-
тельность первых обычно равнялась 800—1600 мс и простиралась до
- Поведение
321
Рис. 81. Сопограммы
улотразвуков, сопровож-
дающих агонистическое
поведение серых крыс
в|
(по: ев, Эти, 1974)
а — сонограмма начальных
60 ме ультразвука,
издаваемого подчинен-
ной особью;
б — сонограмма ультразву-
7
Ю
Ч
е
е
т
о
в
т
а
е
,
к
у
З
о
)
ка, издаваемого сам-
|
Г
цами во время агрес-
р
1
к
,
Е
г
сивных контактов
Г!
РГ)
20
Е
40
ЗЕ
50
Дремя,ис
3400 ме. Единственная спектральная компонента занимала частотный
диапазон между 22 и 30 кГц. Частотная модуляция или отсутствовала
(рис. 81), или частота медленцо понижалась к концу сигнала. Длитель-
ные ультразвуки принадлежали подчиненным особям, обычно сопровождая
длительные выдохи животных, паходящихся в паиболее выраженной позе
подчинепил, Если домипирующий во встрече самец активно поддерживал
подчиненное состояние оппонента, то длительный сигнал мог продолжать-
ся до 15—20 мин, а иногда и после того, как животных рассаживали
Длительность коротких ультразвуков равнялась 3—65 Мс. частотный
диапазон — 40—70 кГц. Короткие ультразвуки, как правило "собраны в
серии. Ряд из них имеют глубокую частотную модуллцию. В отличие от
длительных короткие ультразвуки не были синхронизированы с дыханием
крыс, поэтому трудно было определить, кому из партнеров они принадле-
жат. Чаще всего короткие ультразвуки регистрировали в наиболее актив-
ную фазу стычек — атака, боксирование, борьба. Саль приводит рисунки
соответствующих поз (Заез, Руе, 1974). Очень мало коротких ультразву-
ков издавали крысы, демонстрируя явное подчинение и домипирование.
Чтобы уточнить, кому из партнеров принадлежат ультразвуки, особен-
по короткие, Томас с соавторами (ТБотаз её а|., 1983) ‘подсаживали к ин-
тактным самцам хозяевам иптактных чужаков и чужаков, лишенных го-
лоса оперативным путем. При подсаживании немых самцов резко сокра-
щалось число коротких 50-кГц ультразвуков и полностью исчезали дли-
тельные 22-кГц сигналы. Было, таким образом, показано, что как
короткие 50-кГц ультразвуки, так и длительные 22-кГк издают в основном
(а последпие исключительно) менее агрессивные подчиненные чужаки.
Если же голоса лишали хозяина клегки, то ультразвуковая активпость
продолжала оставаться высокой. что также подтвержцало, что ультразву-
ки издают подчиненные самцы (ТаКавазь! её а1., 1983).
|
небольшой серии экспериментов с дикими серыми крысами Саль и
Пи (За1ез, Руе, 1974) установили в осповном те же закономерности аку-
стической активности самцов. Дикие крысы оказались не более агрессив-
ны, чем лабораторные. Длительные ультразвуки (300—600 мс) были ха-
рактерны для самцов, находившихся в подчиненной позе. Короткие уль-
тразвуки самцы издавали при взаимном обнюхивании, еближая носы, или
11 Серая врыса
322 Глава четвертая
в случаях, когда одно животное било ногами другое. По мнению этих
авторов, короткие ультразвуки связаны скорее с агрессивной мотивацией,
а длительные с мотивацией подчинения. Имеющиеся наблюдения позво-
лили им высказать предположение, что длительные ультразвуки оказыва-
ют тормозящий эффект на агрессивное поведение партнеров по встрече.
Сотласно Грапту (Стапшь 1963), подчиненная поза сокращает агрессиз-
ность оплонента, но ее влияние должно быть времепным, пока длится
‘взаимодействие. Подавая длительные ультразвуковые сигналы, животные
‘постоянно или периодически подтверждают свое подчиненное положение,
одновременно имея возможность совершать текущую, жизненпо важную
активность (питание, потребление воды, исследование территории).
Акустическое сходство длительных ультразвуков с 22-кГц постэяку-
ляторным сигналом побудило Адлера и Аписко (АФег, Ат1зКо, 1979) про-
верить влияние последнего на агрессивное поведение самцов, используя
для этой цели животных, имевших перед ссаживанием контакт с самкой.
Чужака подсаживали к хозяину клетки после и до эякуляции. После
зякуляции самцы, подсажештые в клетку к агрессивному хозяину, изда-
вали длительные ультразвуки гораздо раньше (в среднем через 83,6 с
после подсаживания), чем в отсутствие полового коптакта с самкой
(227,8 с). С другой стороны, в первом случае активные агрессивные дей-
ствия хозяина начинались существенно позже, чем во втором. Если чу-
жак после эякуляции попадал в клетку к хозяину, вскоре начиная изда-
вать длительные ультрезвухи, 10 хозлии активпо атаковывал его через
334,8 с, если же чужак предварительно не контактировал © самкой и в
результате с большим латентным периодом начинал издавать ультразву-
ки, то был активпо атакован уже через 92,5 с. Адлер и Аписко, считают,
что результаты эксперимента подтверждают тормозящее действие длитель-
ных ультразвуков на агрессинное поведение самцов. Обсуждая структур-
ное сходство 22-кГц ультразвуков, сопровождающих брачное поведепие
и агрессивные контакты, авторы обращают впимание на связь возможной
сигнальной значимости ультразвуков с конкретным новедепческим коп-
текстом: один и тот же сигнал в зависимости от конкретной поведенче-
ской ситуации может восприниматься животными по-разному. Не при
этом Адлер и Аниско не исключают, что сходетво между сигналами,по-
даваемыми в различных ситуациях, может оказаться весьма поверхност-
ным. Чтобы выяснить этот вопрос, требуётся более детальное изучепие
акустических особенностей издаваемых крысами ультразвуков.
‘
Подавление агрессивности с помощью длительных ультразвуков было
показано Лором с соавторами (Т.оте её а1., 1976). Чужаков подсаживали
на 24 ч в клетку к хозяевам, регистрируя поведение животных в тече-
ние первых 30 мин. Спустя неделю эксперимент повторили с другими
хозяевами клетки. В первом эксперименте в шести из девяти случаев
чужаки были атакованы хозяевами и издавали длительные ультразвуки
только после этих энергичных атак. Во втором эксперименте ни один “из
девяти подсаженных самцов не был атакован в начале встречи с хозяи-
ном, но все девять самцов издавали ультразвуки, причем с меньшим ла-
тентпым периодом и более активно. Средияя длительность латентного
Поведение 323
периода в первом эксперименте равнялась 1459 с, а во втором— 687 с.
Средняя суммарная длительность в первом эксперименте составляла всего
46,1 с, а во втором — 431,0 с.
Нрямые эксперименты, посвященные выяснению сигнальной значимо-
сти ультразвуков, издаваемых крысами при агрессивных коптактах, по-
казали неопределенные результаты. В цитированной выше работе Томаса
© соавторами (ТВотаз её а|., 1983) была создана ситуация, когда из по-
веденческого репертуара партперов ло встрече исключались как длитель-
ный, так и короткий ультразвуки (чужаков лишали голоса оперативным
путем). Выключение акустического канала связи никак не повлияло ни
на активность поведения, ни на специфику двигательных реакций обоих
партнеров, агрессивного хозяина и ведущего себя подчипенно чужака.
Аналогичные результаты получены и в тех случаях, когда хозяина клет-
ки лишали слуха, заполняя наружный слуховой проход вазелином. По
<равнепию с контролем агрессивная реакция хозяев не менялась, хотя
они и не могли слышать ультразвуки, издаваемые чужаками. Таким об-
разом, акустическая активпость чужаков пе снимала агрессивности хозя-
ев клетки и контакты между партнерами ироходили без заметных изме-
нений двигательных реакций (ТакавазВ её аЁ., 1983). Такахаши с соав-
торами обращают внимание на значительные ограничения, вызываемые
условиями экспериментов. В число ограничений они включают короткий
период времени (пе более 1 ч) контактов между животными в зкспери-
ментальных условиях. Авторы не исключают, что в естественных группах
акустическая активность партнеров имеет осповное значение не в началь-
ный период установления иерархических отношений, а в будущем, когда
иерархия установлена. Возможный отсроченный эффект ультразвуков с0-
стоит в том, что опи, маркируя членов группы, способствуют индивилду-
альному опознаванию. Опыт же индивидуального опознавапия пакапли:
вается в первых контактах. В цитированной выше работе Лор с соавто-
рами (Тоте её аЁ., 1976) показали, что предварительный опыт влияет в
том числе и на ультразвуковую активность крыс. Как уже говорилоеь,
чужаки, подсажеппые в клетку к самцам-хозяевам, в первом эксперимен-
те издавали ультразвуки только после того, как были активно атакованы
агрессивным хозяином. Но педелю спустя уже с другим партнером они
подавали ультразвуковой сигнал независимо от того, атаковал их хозяин
клетки или нет. Таким образом, по крайней мере в течение недели у
крыс не только сохраняется условная связь на тревожную обстановку, но
и посредством акустической активности «маркируется» треволкное состоя-
ние.
‘ Вопрос о стимулах, вызывающих акустическую активность у крыс в
социально неблатоприятных условиях, в настоящее время почти не иссле-
дован. Наиболее детальные эксперименты, посвященные этой проблеме,
принадлежат Корригану и Фланнелли (Согнрап, РаппеПу, 1979). В част-
ности, показано, что ольфакторная стимуляция, хотя и влияет на акусти-
ческую активность самцов, в качестве единственной сенсорной модально-
сти недостаточна для стимуляции. Акустическая активность чужаков была
низкой в присутствии кастрированных или обездвиженных хозяев клетки
11*
322 Глава четвертая
в случаях, когда одно животное било ногами другое. По мнению этих
авторов, короткие ультразвуки связаны скорее с агрессивной мотивацией,
а длительные с мотивацией подчинения. Имеющиеся наблюдения позво-
лили им высказать предположение, что длительные ультразвуки оказыва-
ют тормозящий эффект на агрессивное поведение партнеров по встрече.
Сотласно Грапту (Стапшь 1963), подчиненная поза сокращает агрессиз-
ность оплонента, но ее влияние должно быть времепным, пока длится
‘взаимодействие. Подавая длительные ультразвуковые сигналы, животные
‘постоянно или периодически подтверждают свое подчиненное положение,
одновременно имея возможность совершать текущую, жизненпо важную
активность (питание, потребление воды, исследование территории).
Акустическое сходство длительных ультразвуков с 22-кГц постэяку-
ляторным сигналом побудило Адлера и Аписко (АФег, Ат1зКо, 1979) про-
верить влияние последнего на агрессивное поведение самцов, используя
для этой цели животных, имевших перед ссаживанием контакт с самкой.
Чужака подсаживали к хозяину клетки после и до эякуляции. После
зякуляции самцы, подсажештые в клетку к агрессивному хозяину, изда-
вали длительные ультразвуки гораздо раньше (в среднем через 83,6 с
после подсаживания), чем в отсутствие полового коптакта с самкой
(227,8 с). С другой стороны, в первом случае активные агрессивные дей-
ствия хозяина начинались существенно позже, чем во втором. Если чу-
жак после эякуляции попадал в клетку к хозяину, вскоре начиная изда-
вать длительные ультрезвухи, 10 хозлии активпо атаковывал его через
334,8 с, если же чужак предварительно не контактировал © самкой и в
результате с большим латентным периодом начинал издавать ультразву-
ки, то был активпо атакован уже через 92,5 с. Адлер и Аписко, считают,
что результаты эксперимента подтверждают тормозящее действие длитель-
ных ультразвуков на агрессинное поведение самцов. Обсуждая структур-
ное сходство 22-кГц ультразвуков, сопровождающих брачное поведепие
и агрессивные контакты, авторы обращают впимание на связь возможной
сигнальной значимости ультразвуков с конкретным новедепческим коп-
текстом: один и тот же сигнал в зависимости от конкретной поведенче-
ской ситуации может восприниматься животными по-разному. Не при
этом Адлер и Аниско не исключают, что сходетво между сигналами,по-
даваемыми в различных ситуациях, может оказаться весьма поверхност-
ным. Чтобы выяснить этот вопрос, требуётся более детальное изучепие
акустических особенностей издаваемых крысами ультразвуков.
‘
Подавление агрессивности с помощью длительных ультразвуков было
показано Лором с соавторами (Т.оте её а1., 1976). Чужаков подсаживали
на 24 ч в клетку к хозяевам, регистрируя поведение животных в тече-
ние первых 30 мин. Спустя неделю эксперимент повторили с другими
хозяевами клетки. В первом эксперименте в шести из девяти случаев
чужаки были атакованы хозяевами и издавали длительные ультразвуки
только после этих энергичных атак. Во втором эксперименте ни один “из
девяти подсаженных самцов не был атакован в начале встречи с хозяи-
ном, но все девять самцов издавали ультразвуки, причем с меньшим ла-
тентпым периодом и более активно. Средияя длительность латентного
Поведение 323
периода в первом эксперименте равнялась 1459 с, а во втором— 687 с.
Средняя суммарная длительность в первом эксперименте составляла всего
46,1 с, а во втором — 431,0 с.
Нрямые эксперименты, посвященные выяснению сигнальной значимо-
сти ультразвуков, издаваемых крысами при агрессивных коптактах, по-
казали неопределенные результаты. В цитированной выше работе Томаса
© соавторами (ТВотаз её а|., 1983) была создана ситуация, когда из по-
веденческого репертуара партперов ло встрече исключались как длитель-
ный, так и короткий ультразвуки (чужаков лишали голоса оперативным
путем). Выключение акустического канала связи никак не повлияло ни
на активность поведения, ни на специфику двигательных реакций обоих
партнеров, агрессивного хозяина и ведущего себя подчипенно чужака.
Аналогичные результаты получены и в тех случаях, когда хозяина клет-
ки лишали слуха, заполняя наружный слуховой проход вазелином. По
<равнепию с контролем агрессивная реакция хозяев не менялась, хотя
они и не могли слышать ультразвуки, издаваемые чужаками. Таким об-
разом, акустическая активпость чужаков пе снимала агрессивности хозя-
ев клетки и контакты между партнерами ироходили без заметных изме-
нений двигательных реакций (ТакавазВ её аЁ., 1983). Такахаши с соав-
торами обращают внимание на значительные ограничения, вызываемые
условиями экспериментов. В число ограничений они включают короткий
период времени (пе более 1 ч) контактов между животными в зкспери-
ментальных условиях. Авторы не исключают, что в естественных группах
акустическая активность партнеров имеет осповное значение не в началь-
ный период установления иерархических отношений, а в будущем, когда
иерархия установлена. Возможный отсроченный эффект ультразвуков с0-
стоит в том, что опи, маркируя членов группы, способствуют индивилду-
альному опознаванию. Опыт же индивидуального опознавапия пакапли:
вается в первых контактах. В цитированной выше работе Лор с соавто-
рами (Тоте её аЁ., 1976) показали, что предварительный опыт влияет в
том числе и на ультразвуковую активность крыс. Как уже говорилоеь,
чужаки, подсажеппые в клетку к самцам-хозяевам, в первом эксперимен-
те издавали ультразвуки только после того, как были активно атакованы
агрессивным хозяином. Но педелю спустя уже с другим партнером они
подавали ультразвуковой сигнал независимо от того, атаковал их хозяин
клетки или нет. Таким образом, по крайней мере в течение недели у
крыс не только сохраняется условная связь на тревожную обстановку, но
и посредством акустической активности «маркируется» треволкное состоя-
ние.
‘ Вопрос о стимулах, вызывающих акустическую активность у крыс в
социально неблатоприятных условиях, в настоящее время почти не иссле-
дован. Наиболее детальные эксперименты, посвященные этой проблеме,
принадлежат Корригану и Фланнелли (Согнрап, РаппеПу, 1979). В част-
ности, показано, что ольфакторная стимуляция, хотя и влияет на акусти-
ческую активность самцов, в качестве единственной сенсорной модально-
сти недостаточна для стимуляции. Акустическая активность чужаков была
низкой в присутствии кастрированных или обездвиженных хозяев клетки
11*
324
Глава четвертая
или же в их отсутствии. В последпем случае стимулом присутствия аг-
рессивного хозяина служила испачканная им подстилка. Напротив, наи-
большее число ультразвуков регистрировали в тех случаях, когда агрес-
сивпый хозяин клетки активно атаковал чужака. Обсуждая полученные
результаты, Карригап и Фланнелли также обращают внимание на отра-
ниченность экспериментальной обстаповки. По их мнению, в естественных
условиях лучшей стратегией было бы не издавапие ультразвуков, а из-
бегание агрессивного партпера, и лишь при высокой плотности популя-
ций акустическая активность имеет адаптивный смысл. При условии, что
она действительно тормозит агрессивность, число столкновений может
быть понижено.
Агопистические контакты между серыми крысами сопровождаются не
только ультразвуковыми сигналами, но и звуками в слышимом диапазо-
пе. Барнетт (Вагпей, 1975), ссылаясь на неопубликованные материалы
Еванса (Еуапз), упоминает о дуэте самцов серых крыс во время их не-
ожиданных встреч. Одно из животных издавало звуки с основной часто-
той 2400 Гц, с хорошо выраженными гармониками; у другого самца ос-
новная частота сигнала составляла 2600 Гц. Эти звуки издают скорее
подвержеппые пападению самцы, чем атакующие,— их слышали, напри-
мер, от самца, защищающего гпездо. Вероятно, они производят отпуги-
вающее действие независимо от двигательных реакций оборопяющегося
животного. Но это предположение требует дальнейней проверки. Анало-
тичные звуки используют также и самцы, защищая гнездо с детеныша-
ми, предупреждая ими приближение незнакомого самца.
Берг и Беннингер (Веге, Ваепотеег, 1973) описали нипепие крыс, из-
давасмое при близких контактах как самцами, так и самками. Шипение
отличается очень широким сцектром частот, примерно от 8 до 30 кГц.
Основной энергетический максимум на 12 кГц и два менее выраженных —
на 20 и 26 кГц. Длительность звуков около 1 с. Крысы шипели в ответ
на болезненное прищинывание хвоста, но только в том случае, если в
непосредственной близости от них находилась другая крыса. Одиночно
сидящие в экспериментальной клетке животные не издавали этих звуков.
Их также не удавалось вызвать и в тех случаях, когда крыс помещали в
большие клетки. В случае активных стычек между крысами воспроизве-
дение магнитной записи шипения никак не влияло на поведение партне-
ров, не уменьшая и не увеличивая число атак. Авторы считают, что ши-
пение появляется в результате конфликта между тенденциями бегства и
нападепия.
Уатс (Уаиз, 1980) в репертуаре семи видов крыс, в том числе и се-
рой, выделил визг. Визг сопровождает агрессивные контакты между сам-
цами. В нем обычно доминирует частотная область между Зи 4 кГц.
Основная частота визга у серой крысы равна 1,6—2,2 кГц. Длительность
отдельных звуков колеблется в диапазоне от 150 ‚до 850 мс. Частота не
модулирована. По мнению Уатса, визг более свойствен защищающимся
особям, чем атакующим. Для агрессивных контактов крыс характерно
также постукивание зубами. Наиболее часто этот звук издают животные,
Поведение
325
находящиеся в боксерской стойке. Оп состоит из серии коротких иелчков
с гармонической структурой спектра.
Единственное исследование, посвященное экспериментальной провер-
ке сигпальной значимости слытшимых звуков серых крыс, принадлежит
Цайнеру и Пику (7ештег, Реек, 1969; Рееке, Сешег, 1970), воспроизводив-
шим магпитпые записи криков, которые крысы издают при сильном бо-
левом раздражении. Реакция крыс на эти крики заметно отличается от
реакции на индифферентный раздражитель такой же интенсивности,
Крысы, приученные в определенное время нить, отказывались от воды го-
раздо чаще (на 30%), когда через громкоговоритель транслировали «дист-
ресс-крик», а не чистый тон. Причем привыкание к воспроизведению
«дистресс-крика» наступало значительно медленнее, чем к воспроизведс-
нию тона.
Таким образом, известные в настоящее время паблюдения и экспери-
менты говорят в пользу высокой акустической активности серых крыс.
Наиболее заметно она проявляется на ранних стадиях постэмбрионально-
го развития, а также в брачном и агопистическом поведении взрослых
животных, т. е. в ситуациях, обычпых для большинства видов млекопи-
тающих. Но экологическая специализация акустической активпости у
данного вида пока не обнаружена. Последнее в значительной степени
связано с тем, что подавляющее большинство работ выполнено в экспо-
риментальпых условиях на лабораторных животных,
Остастся открытым вопрос и о числе акустических сигналов у серой
крысы. По мнению Уатса (\!айз, 1980), основной акустический репер-
туар вида, как и некоторых других представителей рода Вайиз, состоит
из шести структурных типов: визг, писк, свист, кашель, нипение и по-
стукивание зубами. Но, как известно, выделение «сигналов» в значитель-
ной степени зависит от интерпретации соотношения дискретности и кон-
тинуальности акустических реакций. Весьма вероятно, что при детальном
исследовании ситуативных изменений издаваемых крысами звуков и
ультразвуков число выделенных Уатсом сигналов будет увеличено за
счет более дробного деления пекоторых из шести ‘перечисленных струк-
турных типов.
``
Позы и выразительные движения
Описание и классификация поз и выразительных движений животных
могут строиться на различных принципах, один из которых— подразде-
пепие на функциональные группы, т. е. группы элементов и их сочетаний,
объединенных общими причинными связями. Следует иметь в зиду, что
такая классификация относительна, поскольку отдельные элементы мо-
гут встречаться в различных ситуациях и в различных тинах взаимодей-
ствий и вызываются разными причипами. К тому же копкротные позы
весьма различаются в зависимости от социального, полового и возрастпо-
то статуса ‘партнеров и от индивидуальных особенностей зверьков.
Орнентировочно-исследовательекое поведение. Такое поведение вклю-
чает специфические позы и движения, направленные на получение ин-
324
Глава четвертая
или же в их отсутствии. В последпем случае стимулом присутствия аг-
рессивного хозяина служила испачканная им подстилка. Напротив, наи-
большее число ультразвуков регистрировали в тех случаях, когда агрес-
сивпый хозяин клетки активно атаковал чужака. Обсуждая полученные
результаты, Карригап и Фланнелли также обращают внимание на отра-
ниченность экспериментальной обстаповки. По их мнению, в естественных
условиях лучшей стратегией было бы не издавапие ультразвуков, а из-
бегание агрессивного партпера, и лишь при высокой плотности популя-
ций акустическая активность имеет адаптивный смысл. При условии, что
она действительно тормозит агрессивность, число столкновений может
быть понижено.
Агопистические контакты между серыми крысами сопровождаются не
только ультразвуковыми сигналами, но и звуками в слышимом диапазо-
пе. Барнетт (Вагпей, 1975), ссылаясь на неопубликованные материалы
Еванса (Еуапз), упоминает о дуэте самцов серых крыс во время их не-
ожиданных встреч. Одно из животных издавало звуки с основной часто-
той 2400 Гц, с хорошо выраженными гармониками; у другого самца ос-
новная частота сигнала составляла 2600 Гц. Эти звуки издают скорее
подвержеппые пападению самцы, чем атакующие,— их слышали, напри-
мер, от самца, защищающего гпездо. Вероятно, они производят отпуги-
вающее действие независимо от двигательных реакций оборопяющегося
животного. Но это предположение требует дальнейней проверки. Анало-
тичные звуки используют также и самцы, защищая гнездо с детеныша-
ми, предупреждая ими приближение незнакомого самца.
Берг и Беннингер (Веге, Ваепотеег, 1973) описали нипепие крыс, из-
давасмое при близких контактах как самцами, так и самками. Шипение
отличается очень широким сцектром частот, примерно от 8 до 30 кГц.
Основной энергетический максимум на 12 кГц и два менее выраженных —
на 20 и 26 кГц. Длительность звуков около 1 с. Крысы шипели в ответ
на болезненное прищинывание хвоста, но только в том случае, если в
непосредственной близости от них находилась другая крыса. Одиночно
сидящие в экспериментальной клетке животные не издавали этих звуков.
Их также не удавалось вызвать и в тех случаях, когда крыс помещали в
большие клетки. В случае активных стычек между крысами воспроизве-
дение магнитной записи шипения никак не влияло на поведение партне-
ров, не уменьшая и не увеличивая число атак. Авторы считают, что ши-
пение появляется в результате конфликта между тенденциями бегства и
нападепия.
Уатс (Уаиз, 1980) в репертуаре семи видов крыс, в том числе и се-
рой, выделил визг. Визг сопровождает агрессивные контакты между сам-
цами. В нем обычно доминирует частотная область между Зи 4 кГц.
Основная частота визга у серой крысы равна 1,6—2,2 кГц. Длительность
отдельных звуков колеблется в диапазоне от 150 ‚до 850 мс. Частота не
модулирована. По мнению Уатса, визг более свойствен защищающимся
особям, чем атакующим. Для агрессивных контактов крыс характерно
также постукивание зубами. Наиболее часто этот звук издают животные,
Поведение
325
находящиеся в боксерской стойке. Оп состоит из серии коротких иелчков
с гармонической структурой спектра.
Единственное исследование, посвященное экспериментальной провер-
ке сигпальной значимости слытшимых звуков серых крыс, принадлежит
Цайнеру и Пику (7ештег, Реек, 1969; Рееке, Сешег, 1970), воспроизводив-
шим магпитпые записи криков, которые крысы издают при сильном бо-
левом раздражении. Реакция крыс на эти крики заметно отличается от
реакции на индифферентный раздражитель такой же интенсивности,
Крысы, приученные в определенное время нить, отказывались от воды го-
раздо чаще (на 30%), когда через громкоговоритель транслировали «дист-
ресс-крик», а не чистый тон. Причем привыкание к воспроизведению
«дистресс-крика» наступало значительно медленнее, чем к воспроизведс-
нию тона.
Таким образом, известные в настоящее время паблюдения и экспери-
менты говорят в пользу высокой акустической активности серых крыс.
Наиболее заметно она проявляется на ранних стадиях постэмбрионально-
го развития, а также в брачном и агопистическом поведении взрослых
животных, т. е. в ситуациях, обычпых для большинства видов млекопи-
тающих. Но экологическая специализация акустической активпости у
данного вида пока не обнаружена. Последнее в значительной степени
связано с тем, что подавляющее большинство работ выполнено в экспо-
риментальпых условиях на лабораторных животных,
Остастся открытым вопрос и о числе акустических сигналов у серой
крысы. По мнению Уатса (\!айз, 1980), основной акустический репер-
туар вида, как и некоторых других представителей рода Вайиз, состоит
из шести структурных типов: визг, писк, свист, кашель, нипение и по-
стукивание зубами. Но, как известно, выделение «сигналов» в значитель-
ной степени зависит от интерпретации соотношения дискретности и кон-
тинуальности акустических реакций. Весьма вероятно, что при детальном
исследовании ситуативных изменений издаваемых крысами звуков и
ультразвуков число выделенных Уатсом сигналов будет увеличено за
счет более дробного деления пекоторых из шести ‘перечисленных струк-
турных типов.
``
Позы и выразительные движения
Описание и классификация поз и выразительных движений животных
могут строиться на различных принципах, один из которых— подразде-
пепие на функциональные группы, т. е. группы элементов и их сочетаний,
объединенных общими причинными связями. Следует иметь в зиду, что
такая классификация относительна, поскольку отдельные элементы мо-
гут встречаться в различных ситуациях и в различных тинах взаимодей-
ствий и вызываются разными причипами. К тому же копкротные позы
весьма различаются в зависимости от социального, полового и возрастпо-
то статуса ‘партнеров и от индивидуальных особенностей зверьков.
Орнентировочно-исследовательекое поведение. Такое поведение вклю-
чает специфические позы и движения, направленные на получение ин-
326 Глава четвертая
В:
Рис. 82. Ориентировочно-исследовательское поведение (рис. В. М. Смирина)
а — поза настороженности
— зверек напрязкен, замер на месте, его тело вытянуто и как бы
приземлено, хвост нараллелен земле, голова, уши и вибриссы направлены вперед; б — вер-
тикальная поза
— зверек прислунтивается, принюхивается и присматриваетвя к происходя-
щему на расстояний; в, г — манипуляционная и исследовательская активность по отношению
к новым предметам
Рис. 83. Поведение при обследовании партнера (рис. В. М. Смирина)
а — назо-назальный контакт между незнакомыми зверьнками; б — взаимное обследование ано-
генитальной области; в, г — обнюхивание участков тела
Поведение
327
формации об окружающей среде. Наиболее характерные из них представ-
лепы па рис. 82. Поза настороженности (рис. 82, а), которая отражает
конфликт между тенденцией приблизиться к чему-то или кому-то и тен-
денцией избежать возможной опасности, наиболее часто встречается в на-
чале активности, когда зверьки только выходят из укрытий, а также у
низкоранговых и молодых особей. Вертикальная поза (рис. 82, 6) харак-
терпа для крыс при освоении новой территории. При обнаружении на
Рис. 84. Дружелюбное поведение
а — скучивание зверьков низкоранговой грунпировки в гнезде (по Колхауну, 1963); б — пере-
лезание через лежащего партнера (чаще встречается у доминирующих самцов); в — подле-
зание под партнера (чаще встречается у молодых особей и взрослых самок); г— чистка
шерсти партнера (рис. 6—г по В. М. Смирину)
326 Глава четвертая
В:
Рис. 82. Ориентировочно-исследовательское поведение (рис. В. М. Смирина)
а — поза настороженности
— зверек напрязкен, замер на месте, его тело вытянуто и как бы
приземлено, хвост нараллелен земле, голова, уши и вибриссы направлены вперед; б — вер-
тикальная поза
— зверек прислунтивается, принюхивается и присматриваетвя к происходя-
щему на расстояний; в, г — манипуляционная и исследовательская активность по отношению
к новым предметам
Рис. 83. Поведение при обследовании партнера (рис. В. М. Смирина)
а — назо-назальный контакт между незнакомыми зверьнками; б — взаимное обследование ано-
генитальной области; в, г — обнюхивание участков тела
Поведение
327
формации об окружающей среде. Наиболее характерные из них представ-
лепы па рис. 82. Поза настороженности (рис. 82, а), которая отражает
конфликт между тенденцией приблизиться к чему-то или кому-то и тен-
денцией избежать возможной опасности, наиболее часто встречается в на-
чале активности, когда зверьки только выходят из укрытий, а также у
низкоранговых и молодых особей. Вертикальная поза (рис. 82, 6) харак-
терпа для крыс при освоении новой территории. При обнаружении на
Рис. 84. Дружелюбное поведение
а — скучивание зверьков низкоранговой грунпировки в гнезде (по Колхауну, 1963); б — пере-
лезание через лежащего партнера (чаще встречается у доминирующих самцов); в — подле-
зание под партнера (чаще встречается у молодых особей и взрослых самок); г— чистка
шерсти партнера (рис. 6—г по В. М. Смирину)
328
Глава: четвертая
знакомом участке незнакомых предметов у зверьков мотут проявляться
различные формы мапипуляционной и исследовательской активности
(рис. 82, в, г).
Поведение при обследовании партнера. Г этой группе выразительных
движений можно отнести назо-назальные и назо-апальпые контакты, а
также обнюхивания различных участков тела (рис. 83). Эти формы коп-
тактов обычно предшествуют и сопровождают все взаимодействия зверь-
ков, указывая на важность обопятельной информации в их жизни. Через
назо-назальный контакт и обнюхивания участков тела передаются сведе-
ния о принадлежности к зпакомым или незнакомым, своим или чужим
зверькам и, возможно, об индивидуальных особенностях особей. Через
пазо-назальный контакт возможна передача и пищевой информации,
Через назо-анальный контакт зверьки получают представление о фи-
зиологическом состоянии партнера, поскольку такая форма общения ха-
рактерна в основном для самцов по отношению к самкам.
Дружелюбное поведение. В эту функциональную группу объединены
выразительные движения, паправленные на поддержание социальных
связей и характеризующиеся отсутствием агрессивных намерении.
Серым крысам свойственно собираться группами, особенно в укрыти-
ях во время отдыха. Такое «скучивание» наблюдается даже в тех слу-
ь
Рис. 85. Поведение при спаривании и при мечении территории
а — назо-анальный контакт с вылизыванием самцом гениталий самки в течке; 6 — самка де-
монстрирует иозу лордоза, а самец совершает садну с эякуляцией (по: Барнетт, 1975); в —
посткопуляторная чистка; г-— мечение территории
— самец ироводит с небольшой задерж-
кой аногенитальной областью по полу, оставляя при этом кавельни мочи (рис. В. М. Сми-
рина)
:
Поведение
329
чаях, когда нет недостатка в убежищах. Наиболее характерно оно для
крыс, которые входят в состав низкорантовых группировок (рис. 84, а),
и для молодых зверьков.
Функциональное значение скучивания у взрослых крыс не вполне яс-
но. В естественных условиях в холодный период года подобные взаимо-
действия могут выполнять роль терморегулятора, однако они характерны
для крыс также и в теплое время года и в лабораторных условиях. Воз-
можно, что тактильные коптакты, которые зверьки имеют во время ску-
чивания, создают чувство уверенности и безолабности, а также служат
для формирования единого запаха, облегчающего распозпавание друг дру-
га в условиях вненоровой активности.
К этой же функциональной группе относятся перелезания через парт-
пера (рис. 84, 6), во время которых зверек проводит аногенитальной об-
ластью по шерсти партнера с одновременным выделнием капелек мочи.
Эти контакты характерны как ‘для взрослых особей, так и для крысят и
могут быть использованы для оценки характера межиндивидуальных свя-
зей.
Рис. 86. Агонистическое поведение (рис. В. М. Смирина)
а — самец (справа) угрожает самке, которая приняла защитно-усмиряющую позу; б — самка
(справа) пристает к самцу, который проявляет свое недовольство в виде боковой позы; в —
самец (слева) н позе угрозы перед нападением, правый зверек, прижавшись к стенке, демон-
стрирует подчинение и крайний испуг; г — самец (справа) пристает к самке, отталкивает
ее ногой
328
Глава: четвертая
знакомом участке незнакомых предметов у зверьков мотут проявляться
различные формы мапипуляционной и исследовательской активности
(рис. 82, в, г).
Поведение при обследовании партнера. Г этой группе выразительных
движений можно отнести назо-назальные и назо-апальпые контакты, а
также обнюхивания различных участков тела (рис. 83). Эти формы коп-
тактов обычно предшествуют и сопровождают все взаимодействия зверь-
ков, указывая на важность обопятельной информации в их жизни. Через
назо-назальный контакт и обнюхивания участков тела передаются сведе-
ния о принадлежности к зпакомым или незнакомым, своим или чужим
зверькам и, возможно, об индивидуальных особенностях особей. Через
пазо-назальный контакт возможна передача и пищевой информации,
Через назо-анальный контакт зверьки получают представление о фи-
зиологическом состоянии партнера, поскольку такая форма общения ха-
рактерна в основном для самцов по отношению к самкам.
Дружелюбное поведение. В эту функциональную группу объединены
выразительные движения, паправленные на поддержание социальных
связей и характеризующиеся отсутствием агрессивных намерении.
Серым крысам свойственно собираться группами, особенно в укрыти-
ях во время отдыха. Такое «скучивание» наблюдается даже в тех слу-
ь
Рис. 85. Поведение при спаривании и при мечении территории
а — назо-анальный контакт с вылизыванием самцом гениталий самки в течке; 6 — самка де-
монстрирует иозу лордоза, а самец совершает садну с эякуляцией (по: Барнетт, 1975); в —
посткопуляторная чистка; г-— мечение территории
— самец ироводит с небольшой задерж-
кой аногенитальной областью по полу, оставляя при этом кавельни мочи (рис. В. М. Сми-
рина)
:
Поведение
329
чаях, когда нет недостатка в убежищах. Наиболее характерно оно для
крыс, которые входят в состав низкорантовых группировок (рис. 84, а),
и для молодых зверьков.
Функциональное значение скучивания у взрослых крыс не вполне яс-
но. В естественных условиях в холодный период года подобные взаимо-
действия могут выполнять роль терморегулятора, однако они характерны
для крыс также и в теплое время года и в лабораторных условиях. Воз-
можно, что тактильные коптакты, которые зверьки имеют во время ску-
чивания, создают чувство уверенности и безолабности, а также служат
для формирования единого запаха, облегчающего распозпавание друг дру-
га в условиях вненоровой активности.
К этой же функциональной группе относятся перелезания через парт-
пера (рис. 84, 6), во время которых зверек проводит аногенитальной об-
ластью по шерсти партнера с одновременным выделнием капелек мочи.
Эти контакты характерны как ‘для взрослых особей, так и для крысят и
могут быть использованы для оценки характера межиндивидуальных свя-
зей.
Рис. 86. Агонистическое поведение (рис. В. М. Смирина)
а — самец (справа) угрожает самке, которая приняла защитно-усмиряющую позу; б — самка
(справа) пристает к самцу, который проявляет свое недовольство в виде боковой позы; в —
самец (слева) н позе угрозы перед нападением, правый зверек, прижавшись к стенке, демон-
стрирует подчинение и крайний испуг; г — самец (справа) пристает к самке, отталкивает
ее ногой
330
Глава четвертая
Сходное с описанным вые выразительное движение — подлезапие под
партнера (рис. 84, в) — наиболее характерно для детенышей крыс по от-
пошению к матерям, У взрослых особей подлезание встречается редко и
связапо, как правило, с провоцированием перелезания у партнера.
Перелезапия через партнера и подлезания под пего, по-видимому,
функционально связаны как между собой, так и с индивидуальным пове-
цением по мечению территории (рис. 85, г). Поэтому все эти элементы
поведения могут быть отнесены не только к дружелюбному, но и к мар-
кировочному поведению. Взаимные чистки (груминг) характерные для
лабораторных крыс, относительно редки во взаимодействиях между их
дикими родоначальниками (рис. 84, г).
Все элементы дружелюбного поведения служат для установления и
укрепления контактов между отдельными животными. Однако передки
случаи, когда у зверька, который пытается осуществить дружелюбное
действие и который наталкивается на сопротивление партнера, поведение
меняется на агрессивное.
Рис. 57. Агонистическое и неспецифическое поведение (рис. В. М. Смирина)
и — левый самец, нападая, опрокинул соперника на спину; б — правый зверек из боковой
стойки на задних лапах готовится к нападению, левый принял позу подчинения и падает
пронзительные звуки; в — «боксирование» в вертикальном полежении, левый, ее
дает, правый защищается; г —- завершившееся «боксирование» между миролюбино нг
ными партнерами
Поведение
331
Новедение при спаривании. Поведение ирн спаривании у крыс вклю-
чает пазо-апальные контакты с вылизыванием самцом тепиталий самки
(рис. 85, @), преследование самки самцом, совершение последним садок,
во время которых самка демонстрирует позу подставления (лордоза),
а также садок с серией интромиссий и садок с эякуляцией— интромис-
сий завершаются сильпым и затяжным последним толчком (рис. 85, 6),
после которого следует небольшая пауза, а затем чистка и вылизывание
гениталий (рис. 85, в) и посткопуляторный отдых.
Агонистическое поведение. В эту функциональную группу объедине-
ны все выразительные позы и движения, связанные с конкуренцией или
конфликтом между животцыми, т. е. состоящие из агрессивных и защит-
ных действий и других проявлений отношепий доминирования—нодчи-
нения.
У серых крыс любому конфликту, как правило, предшествуют позы
угрозы или боковые позы (рис. 86, а—2; 87, 6). Форма и интенсивпость
позы угрозы чрезвычайно лабильны. Это поведение демопстрируется в
основном по отношению к неподвижному партперу, для того чтобы выну-
дить его к бегству, и к тем зверькам, которые находятся в укрытиях.
Такая поза демонстрируется и тогда, когда зверек педоволен действиями
партнера (рис. 86, 6, г). _
Атрессивному поведению крыс свойственны также нападения, пресле-
дование убегаютцего зверька, схватки и взаимные укусы. В поведении
высокоагрессивных животных в случаях сильного возбуждения нередко
наблюдаются такие элементы, как «стрекотание» зубами и хлестание
хвостом. Для поведепия низкоранговых особей характерны пугливость и
напряженность, а при агонистических контактах
— бегство от партнера,
избегание контактов с ним и разнообразные формы подчинения (рис. 86,
а—в; 87, а—в). В ряде случаев подчиненные особи издают резкие звуки.
Однако принятие защитной позы и пронзительные крики лишь приоста-
навливают пападающего, но не меняют его агрессивных намерений.
Неспецифическое поведение. К неспецифическому поведению мы от-
носим такие олемепты поведения, как боксирование (рис. 87, в, г), по-
валивание на спину (рис. 87, а) и приставание (см. рис. 86, 0, г), т. е.
такие элементы, которые могут встречаться в различных типах взаимо-
действий. Примечательно, что игровые действия крысят состоят в основ-
ном из этих же выразительных движений.
Описанные элементы поведения серых крыс не претендуют на исчер-
пывающую полноту в силу значительной лабильности самих поведенче-
ских реакций животных.
330
Глава четвертая
Сходное с описанным вые выразительное движение — подлезапие под
партнера (рис. 84, в) — наиболее характерно для детенышей крыс по от-
пошению к матерям, У взрослых особей подлезание встречается редко и
связапо, как правило, с провоцированием перелезания у партнера.
Перелезапия через партнера и подлезания под пего, по-видимому,
функционально связаны как между собой, так и с индивидуальным пове-
цением по мечению территории (рис. 85, г). Поэтому все эти элементы
поведения могут быть отнесены не только к дружелюбному, но и к мар-
кировочному поведению. Взаимные чистки (груминг) характерные для
лабораторных крыс, относительно редки во взаимодействиях между их
дикими родоначальниками (рис. 84, г).
Все элементы дружелюбного поведения служат для установления и
укрепления контактов между отдельными животными. Однако передки
случаи, когда у зверька, который пытается осуществить дружелюбное
действие и который наталкивается на сопротивление партнера, поведение
меняется на агрессивное.
Рис. 57. Агонистическое и неспецифическое поведение (рис. В. М. Смирина)
и — левый самец, нападая, опрокинул соперника на спину; б — правый зверек из боковой
стойки на задних лапах готовится к нападению, левый принял позу подчинения и падает
пронзительные звуки; в — «боксирование» в вертикальном полежении, левый, ее
дает, правый защищается; г —- завершившееся «боксирование» между миролюбино нг
ными партнерами
Поведение
331
Новедение при спаривании. Поведение ирн спаривании у крыс вклю-
чает пазо-апальные контакты с вылизыванием самцом тепиталий самки
(рис. 85, @), преследование самки самцом, совершение последним садок,
во время которых самка демонстрирует позу подставления (лордоза),
а также садок с серией интромиссий и садок с эякуляцией— интромис-
сий завершаются сильпым и затяжным последним толчком (рис. 85, 6),
после которого следует небольшая пауза, а затем чистка и вылизывание
гениталий (рис. 85, в) и посткопуляторный отдых.
Агонистическое поведение. В эту функциональную группу объедине-
ны все выразительные позы и движения, связанные с конкуренцией или
конфликтом между животцыми, т. е. состоящие из агрессивных и защит-
ных действий и других проявлений отношепий доминирования—нодчи-
нения.
У серых крыс любому конфликту, как правило, предшествуют позы
угрозы или боковые позы (рис. 86, а—2; 87, 6). Форма и интенсивпость
позы угрозы чрезвычайно лабильны. Это поведение демопстрируется в
основном по отношению к неподвижному партперу, для того чтобы выну-
дить его к бегству, и к тем зверькам, которые находятся в укрытиях.
Такая поза демонстрируется и тогда, когда зверек педоволен действиями
партнера (рис. 86, 6, г). _
Атрессивному поведению крыс свойственны также нападения, пресле-
дование убегаютцего зверька, схватки и взаимные укусы. В поведении
высокоагрессивных животных в случаях сильного возбуждения нередко
наблюдаются такие элементы, как «стрекотание» зубами и хлестание
хвостом. Для поведепия низкоранговых особей характерны пугливость и
напряженность, а при агонистических контактах
— бегство от партнера,
избегание контактов с ним и разнообразные формы подчинения (рис. 86,
а—в; 87, а—в). В ряде случаев подчиненные особи издают резкие звуки.
Однако принятие защитной позы и пронзительные крики лишь приоста-
навливают пападающего, но не меняют его агрессивных намерений.
Неспецифическое поведение. К неспецифическому поведению мы от-
носим такие олемепты поведения, как боксирование (рис. 87, в, г), по-
валивание на спину (рис. 87, а) и приставание (см. рис. 86, 0, г), т. е.
такие элементы, которые могут встречаться в различных типах взаимо-
действий. Примечательно, что игровые действия крысят состоят в основ-
ном из этих же выразительных движений.
Описанные элементы поведения серых крыс не претендуют на исчер-
пывающую полноту в силу значительной лабильности самих поведенче-
ских реакций животных.
Глава пятая.
Практическое значение
Экономический ущерб
Несмотря на огромный иптерес, который проявляют исследователи к раз-
личным аспектам жизнедеятельности грызунов, реальный экономический
ущерб, приносимый этими животными, и в частпости серой крысой, изу-
чен крайпе педостаточно.
Пасюк — один из самых массовых но численпости синантропных видов
грызунов, занимающий среди пих ведущее место по приносимому эконо-
мическому ущербу. Вредоносная деятельность этого грызуна с каждым
годом расширяется.
И данным Деораса (Оеогаз, 1975), ущерб от серой крысы в США
составляет 300 млн долларов в год, Англия ежегодио теряет 15 млн фун-
тов стерлингов, Франция— 15 мли франков, Индия— 700 мли рупий.
На Кубе, шю подсчетам Эрнандеса и Драммопда (Нетпап4ея, Отититопа,
1984), только в провинции Вилла-Клара крысы и домовая мышь причи-
няют ежегодный ущерб в 80 тыс. песо. В СССР опубликовано лишь не-
сколько работ по этому вопросу (Лялин, 1983; Судейкип, 1983; Соколов
и др., 198ба).
Для оценки ущерба от пасюка берутся сведепил по следующим пара-
метрам: 1) ежедневный пищевой рацион зверька в весовых категориях,
2) средняя стоимость продуктов, 8) плотность крыс па единицу площади,
4) годовая и сезонная динамика численности, Для уточнения пищевого
рациона и среднего. количества корма, употребляемого крысой в сутки,
В. Г. Соколов с соавторами (4986а) провели серию экспериментов, при
которых зверьков содержали в вольерах группами, а в клетках парами.
В качестве корма предлагали наиболее стандартные продукты: хлеб, ка-
ши, мясо, овощи, а регистрацию их поедаемости проводили взвешиванием
остатков каждые 24 ч. Для обоих случаев средпее количество корма,
съеденпого одной крысой за сутки, составляло 56,7 г. А. А. Пасешник
(1947) и Ш. М. Марзоев (1974, 1978) по количеству съедаемых продух-
тов получили аналогичные результаты— 59—63 г. корма (в зависимости
от веса крысы). Следует особо отметить, что зверьки всегда предночита-
ют доброкачественные продукты (Дукельская, 1948), и это падо учиты-
вать при оргапизации дератизационных работ. Среднюю стоимость продук-
тов, съеденных пасюком, мы условно принимаем равной 20 коп. за 1 кг.
В. Е. Соколов с соавторами (1986) подсчитали, что если в среднем в
населенных нунктах на объектах обитает 1 крыса па 1000 м", то ма
площади в 1000000 м” живет 1000 крыс. За 1 день зверек съедает 60 г
продуктов, за 1 год— 21 кг 900 г. За год 1000 крыс съедают 219000 кг
{условно 22 т). Стоимость этих продуктов — 20 коп.Х22 000 кг--4400 руб.
Оценить экономический ущерб от насюков, обитающих по всему Совет-
скому Союзу, можно только с большими допущениями, поскольку реаль-
*)
Практическое значение
335
ная площадь всех строепий очепь изменчива и не поддается точному уче-
ту даже при анализе всей доступной справочной литературы. Жилые
помещения и другие постройки занимают не менее '10 млрд м”. Следо-
вательно, если па 1 млн м’ площади крысы уничтожают в год 22 т
продуктов, то на 10 млрд м* они способны уничтожить 220 000 т продук-
тов.
В местах повышенной концентрации пасюков экономический ущерб,
припосимый ими, может быть во много раз выше. По подсчетам тех же
авторов, на одном из мясокомбинатов площадью 970000 м’ жило
9000 крыс. По расчетным данным, при исключительном питании мясом
дневиая норма для крые может снижаться до 30 г. Соответственно за
год один зверек съедает 10 кг 900 г мясных продуктов (для удобства ав-
торы снизили цифру до 40 кг). 9000 крыс, живущих на мясокомбипате,
за год съедают 90 т мясных продуктов, и при минимальной стоимости
их в (р. 50 коп. за 1 кг убыток в денежном выражении составляет
135 000 руб. ежегодно.
,
В. Е. Соколовым с соавторами (1986) был оценен также ущерб,
причиняемый серой крысой в портах, площадь которых составляет около
49 млн м’. Ориентировочная численность крыс в них —2 особи на
1000 м?. Соответственно на этой площади может обитать 98 тыс. крыс,
съедающих там в год 2446 т 200 кг продуктов на сумму около 500 тыс. руб.
В. Г. Лялиным (1983) экономический ущерб, причиняемый серой кры-
сой за год в Западной Сибири, оценивается в миллионы рублей. Им
были учтены стоимость съеденной и испорченной сельскохозяйственной
продукции, испорченпых промышленных товаров, а также затраты па
проведение работ по борьбе с пасюком. В результате наблюдений, экспе-
римептов, сбора и анализа дапных автор установил, что ежегодно в Том-
ской и Тюменской областих крысами приводилось в негодность 0,1%
площади полов, числа кормушек и другого оборудования животноводче-
ских ферм: в среднем зверьки наносили ущерб в 1,8 коп. на одно живот-
ное. Ежегодные потери от порчи крысами промышленных товаров, по ис-
следованию в Томске, составляют в средпем 0,27 коп. на одного жителя.
За год один зверек поедает и приводит в негодность в среднем 9,8 кг
продуктов и кормов на сумму 78,4 коп.
В. Я. Антонюк и Н. В. Олькова предоставили нам свои материалы по
экопомическому ущербу, наносимому серой крысой в Сибири и на Даль-
нем Востоке. Так, в Иркутске на базе молочных продуктов крысами был
поврежден теплоизоляционный слой холодильной камеры стоимостью
150 тыс. руб. Они прогрызали отверстия в стенах холодильных камер,
тем самым вызывали большой расход электроэнергии для поддержания
нужной температуры. Здесь же опи грызли блоки сливочного масла и
сыра. Работники базы отмечают, что эти зверьки выбирают продукты
высшего качества. В бочках с полуфабрикатами сыра они выгрызают со-
держимое, оставляя сверху тонкую корочку, и при внешнем осмотре за-
мечаетсл только отверстие в сыроподобной массе.
Авторы при характеристике денежного ущерба в отдельных населен-
ных пупктах Якутской АССР приводят следующую таблицу (табл. 27).
Глава пятая.
Практическое значение
Экономический ущерб
Несмотря на огромный иптерес, который проявляют исследователи к раз-
личным аспектам жизнедеятельности грызунов, реальный экономический
ущерб, приносимый этими животными, и в частпости серой крысой, изу-
чен крайпе педостаточно.
Пасюк — один из самых массовых но численпости синантропных видов
грызунов, занимающий среди пих ведущее место по приносимому эконо-
мическому ущербу. Вредоносная деятельность этого грызуна с каждым
годом расширяется.
И данным Деораса (Оеогаз, 1975), ущерб от серой крысы в США
составляет 300 млн долларов в год, Англия ежегодио теряет 15 млн фун-
тов стерлингов, Франция— 15 мли франков, Индия
— 700 мли рупий.
На Кубе, шю подсчетам Эрнандеса и Драммопда (Нетпап4ея, Отититопа,
1984), только в провинции Вилла-Клара крысы и домовая мышь причи-
няют ежегодный ущерб в 80 тыс. песо. В СССР опубликовано лишь не-
сколько работ по этому вопросу (Лялин, 1983; Судейкип, 1983; Соколов
и др., 198ба).
Для оценки ущерба от пасюка берутся сведепил по следующим пара-
метрам: 1) ежедневный пищевой рацион зверька в весовых категориях,
2) средняя стоимость продуктов, 8) плотность крыс па единицу площади,
4) годовая и сезонная динамика численности, Для уточнения пищевого
рациона и среднего. количества корма, употребляемого крысой в сутки,
В. Г. Соколов с соавторами (4986а) провели серию экспериментов, при
которых зверьков содержали в вольерах группами, а в клетках парами.
В качестве корма предлагали наиболее стандартные продукты: хлеб, ка-
ши, мясо, овощи, а регистрацию их поедаемости проводили взвешиванием
остатков каждые 24 ч. Для обоих случаев средпее количество корма,
съеденпого одной крысой за сутки, составляло 56,7 г. А. А. Пасешник
(1947) и Ш. М. Марзоев (1974, 1978) по количеству съедаемых продух-
тов получили аналогичные результаты— 59—63 г. корма (в зависимости
от веса крысы). Следует особо отметить, что зверьки всегда предночита-
ют доброкачественные продукты (Дукельская, 1948), и это падо учиты-
вать при оргапизации дератизационных работ. Среднюю стоимость продук-
тов, съеденных пасюком, мы условно принимаем равной 20 коп. за 1 кг.
В. Е. Соколов с соавторами (1986) подсчитали, что если в среднем в
населенных нунктах на объектах обитает 1 крыса па 1000 м", то ма
площади в 1000000 м” живет 1000 крыс. За 1 день зверек съедает 60 г
продуктов, за 1 год— 21 кг 900 г. За год 1000 крыс съедают 219000 кг
{условно 22 т). Стоимость этих продуктов — 20 коп.Х22 000 кг--4400 руб.
Оценить экономический ущерб от насюков, обитающих по всему Совет-
скому Союзу, можно только с большими допущениями, поскольку реаль-
*)
Практическое значение
335
ная площадь всех строепий очепь изменчива и не поддается точному уче-
ту даже при анализе всей доступной справочной литературы. Жилые
помещения и другие постройки занимают не менее '10 млрд м”. Следо-
вательно, если па 1 млн м’ площади крысы уничтожают в год 22 т
продуктов, то на 10 млрд м* они способны уничтожить 220 000 т продук-
тов.
В местах повышенной концентрации пасюков экономический ущерб,
припосимый ими, может быть во много раз выше. По подсчетам тех же
авторов, на одном из мясокомбинатов площадью 970000 м’ жило
9000 крыс. По расчетным данным, при исключительном питании мясом
дневиая норма для крые может снижаться до 30 г. Соответственно за
год один зверек съедает 10 кг 900 г мясных продуктов (для удобства ав-
торы снизили цифру до 40 кг). 9000 крыс, живущих на мясокомбипате,
за год съедают 90 т мясных продуктов, и при минимальной стоимости
их в (р. 50 коп. за 1 кг убыток в денежном выражении составляет
135 000 руб. ежегодно.
,
В. Е. Соколовым с соавторами (1986) был оценен также ущерб,
причиняемый серой крысой в портах, площадь которых составляет около
49 млн м’. Ориентировочная численность крыс в них —2 особи на
1000 м?. Соответственно на этой площади может обитать 98 тыс. крыс,
съедающих там в год 2446 т 200 кг продуктов на сумму около 500 тыс. руб.
В. Г. Лялиным (1983) экономический ущерб, причиняемый серой кры-
сой за год в Западной Сибири, оценивается в миллионы рублей. Им
были учтены стоимость съеденной и испорченной сельскохозяйственной
продукции, испорченпых промышленных товаров, а также затраты па
проведение работ по борьбе с пасюком. В результате наблюдений, экспе-
римептов, сбора и анализа дапных автор установил, что ежегодно в Том-
ской и Тюменской областих крысами приводилось в негодность 0,1%
площади полов, числа кормушек и другого оборудования животноводче-
ских ферм: в среднем зверьки наносили ущерб в 1,8 коп. на одно живот-
ное. Ежегодные потери от порчи крысами промышленных товаров, по ис-
следованию в Томске, составляют в средпем 0,27 коп. на одного жителя.
За год один зверек поедает и приводит в негодность в среднем 9,8 кг
продуктов и кормов на сумму 78,4 коп.
В. Я. Антонюк и Н. В. Олькова предоставили нам свои материалы по
экопомическому ущербу, наносимому серой крысой в Сибири и на Даль-
нем Востоке. Так, в Иркутске на базе молочных продуктов крысами был
поврежден теплоизоляционный слой холодильной камеры стоимостью
150 тыс. руб. Они прогрызали отверстия в стенах холодильных камер,
тем самым вызывали большой расход электроэнергии для поддержания
нужной температуры. Здесь же опи грызли блоки сливочного масла и
сыра. Работники базы отмечают, что эти зверьки выбирают продукты
высшего качества. В бочках с полуфабрикатами сыра они выгрызают со-
держимое, оставляя сверху тонкую корочку, и при внешнем осмотре за-
мечаетсл только отверстие в сыроподобной массе.
Авторы при характеристике денежного ущерба в отдельных населен-
ных пупктах Якутской АССР приводят следующую таблицу (табл. 27).
ЗЗА
Глава пятая
Таблица 27. Количество еписанных товаров, испорченных и съеденных крысами
по Ленинскому райпотребсоюзу в ное. Нюрба Якутской АССР (1980-1987 гг.)
К-
к.-
то [|Приемные |чел, |Оум тод | поме |ети, |риа
4980 | Макаронные
353
197.80 |1984 Комбикорм
2576
854.66
изделия
Крупа
925 813.70
1981 | Мука
330 | 10244
Мата
353 | 241.82
Крупа
54 47.87
"Итого
за
Иу
54 4 910.18
Макаронные
335 28713
т
изделия
1985 Макаронные
4231 1573.91
Итого
719
388.04
изделия
Обувь
-
657.01
1982 | Мука
356 | 117.08
коаная
Крупа
116
13.33
Мешки (тара) | 3224 | 1386.32
Макаронные
395
305.97
(ттук)
изделия
Итого
4 231 3617.24
И
6
436.38
того 867
1386-| Сахар
355 | 33310
1983 | Мука
310 95.98
Крупа
2385 | 1079.08
Крупа
. 48 305.39 |апре-| Мука
4502 1 680.08
Макаронные
369
228.96 |ля) Мешки (тара) 4 126 2266.95
изделия
(штук)
Рыба
—
849.35
Обувь
_
81.00
Итого
727 | 1479.68
(уцененная)
Итого
14 368 | 5495.81
За все годы
25 344 | 26 1971.46
В связи © экономическим развитием севера Сибири идет быстрое уве-
личение производства пищевой, и в первую очередь мясной, продукции,
За последние 20 лет в Якутской АССР произошел рост поголовья свиней
в 3, а птиц в 9 раз. Резко увеличился поток грузов по Ленскому речно-
му бассейну из порта Осетрово в г. Усть-Кут. В частности, в 2,5 раза
возросли перевозки зернофуража и комбикорма для скота. Этот груз на-
чали хранить и перевозить пакетами по 50—60 мешков и в контейнерах.
Из-за недостатка складов комбикорм хранится на открытых площадках в
штабелях, старых крысопроницаемых складах и контейнерах, где созда-
ются благоприятные условия для жизни и размножения этих грызунов.
Ленский речпой бассейн стал основпым путем продвижения пасюка па
северо-восток Сибири. Здесь происходит большая порча им и приведение
в негодпость хранимого и транспортируемого комбикорма и зернофуража.
Во многих случаях повреждено от 50 до 100% мешков.
В 1984 т. на торговой базе пос. Осетрово находилось 125880 мешков
комбикорма. При высокой численности крыс (1000 экз. в одном штабеле)
почти все мешки были повреждены, т. е. 100 тыс. мептков стоимостью
в 150 тыс. руб. (стоимость одного мешка 4 руб. 50 коп.). Комбикорм
высыпался на землю или на пол склада, сментивался с мусором, пылью,
>
Практическое значение
335
снегом и к дальнейшему использованию был пепригоден. При количестве
100 тыс. поврежденных мешков эти просыни составили примерно 25 т,
а в деньгах — 4475 руб. (расчет сделан из наблюдений, что из 80 мешков
просыпается в среднем 20 кг). Следовательно, только 1 тыс. крыс на од-
ной крупной базе приносит убыток в 160 тыс. руб.
Оценка экономического ущерба, причиняемого серыми крысами в
Москве, сделана В. А. Судейкиным (41983). С 1951 по 1980 г. дезинфек-
ционной станцией было выловлено и подобрано 614,8 тыс. трупов отрав-
ленных крыс, которые, по расчетам автора, принесли бы ущерб за эти
годы в 12 544 руб.
В животповодческих помещениях, по данным Д. Ф. Траханова и
А. Ф. Кадирова (1981) и Ш. М. Мирзоева (1978), только в свинарниках
крысы ежегодно уничтожают 500 тыс. т кормов па сумму в 75 млн руб.
Общие ежегодные убытки от них в животноводстве эти авторы оценивают
в 157,6 млн руб.
Кроме того, что крысы поедают пищевые продукты, большое коли-
чество их они загрязняют своей мочой и калом. Оценить удельный вес
таких продуктов очень трудно. Имеются сведения, что один зверек выде-
ляет в год 25 тыс. катышков кала и 20—30 мл мочи за сутки (Пасеш-
ник, 1947). Но данным исследователей, одна крыса загрязняет мочой до
1000 зерен крупы в сутки.
Сведения об ущербе, причиняемом серой крысой, не ограничиваются
только порчей и уничтожением пищевых продуктов. Так, 20% пожаров
в США вызваны замыкапиями, произошедшими из-за повреждения этим
трызупом кабеля. Из-за повреждения кабеля и узлов автоматической
сигнализации крысами в Японии на длительное время парупалось дви-
жепие электропоездов (Наумов, Емельянова, 1979). Те же авторы с00б-
цают, что в г. Ахваз (Иран) крысы повредили линии связи в подзем-
пых коммуникациях, в результате чего в течение нескольких дпей во
многих учреждениях и жилых домах не работала телефонная снязь. При-
водятся снедения о новреждении зверьками проводов в самолетах, что
может повлечь за собой воздушнуто катастрофу (Емельянова, Данилки-
на, 1983). Значительный ущерб крысы иричиняют в подземных соору-
жепиях. В канализации сотни нор, сделанных ими в стенках коллекто-
ров, приводят к быстрому разрушению коллекторов, утечке сточных вод
и к обралам (Тощигин, 1986). Присутствие зверьков в шахтах представ-
ляет еще большую опасность, так как они повреждают изоляцию прово-
дов, упаковку и оболочку взрывчатых устройств (Иоффе, 1959). Во вре-
мя обследования шахт Донбасса неоднократно отмечали поврежденные
крысами оболочки детонаторов и потрызы в ящиках па складах взрыв-
чатых веществ (Тощигин, 1986). Ущерб от повреждений серой крысой
носит и косвенный характер: например, повреждения ею герметической
‘упаковки на приборах, храпящихся на складе, приводят к порче послед-
них. Поселяясь в животноводческих помещениях, крысы прогрызают
многочисленные отверстия в полу, и находящийся здесь скот ломает
ноги, проваливаясь в дыры; аварии на подстанциях приводят к остапэв-
ке работы на предприятиях (Емельянова, 1986) ит. д.
ЗЗА
Глава пятая
Таблица 27. Количество еписанных товаров, испорченных и съеденных крысами
по Ленинскому райпотребсоюзу в ное. Нюрба Якутской АССР (1980-1987 гг.)
К-
к.-
то [|Приемные |чел, |Оум тод | поме |ети, |риа
4980 | Макаронные
353
197.80 |1984 Комбикорм
2576
854.66
изделия
Крупа
925
813.70
1981 | Мука
330 | 10244
Мата
353 | 241.82
Крупа
54
47.87
"Итого
за
Иу
54 4 910.18
Макаронные
335
28713
т
изделия
1985 Макаронные
4231 1573.91
Итого
719
388.04
изделия
Обувь
-
657.01
1982 | Мука
356 | 117.08
коаная
Крупа
116
13.33
Мешки (тара) | 3224 | 1386.32
Макаронные
395
305.97
(ттук)
изделия
Итого
4 231 3617.24
И
6
436.38
того 867
1386-| Сахар
355 | 33310
1983 | Мука
310 95.98
Крупа
2385 | 1079.08
Крупа
. 48 305.39 |апре-| Мука
4502 1 680.08
Макаронные
369
228.96 |ля)
Мешки (тара) 4 126 2266.95
изделия
(штук)
Рыба
—
849.35
Обувь
_
81.00
Итого
727 | 1479.68
(уцененная)
Итого
14 368 | 5495.81
За все годы
25 344 | 26 1971.46
В связи © экономическим развитием севера Сибири идет быстрое уве-
личение производства пищевой, и в первую очередь мясной, продукции,
За последние 20 лет в Якутской АССР произошел рост поголовья свиней
в 3, а птиц в 9 раз. Резко увеличился поток грузов по Ленскому речно-
му бассейну из порта Осетрово в г. Усть-Кут. В частности, в 2,5 раза
возросли перевозки зернофуража и комбикорма для скота. Этот груз на-
чали хранить и перевозить пакетами по 50—60 мешков и в контейнерах.
Из-за недостатка складов комбикорм хранится на открытых площадках в
штабелях, старых крысопроницаемых складах и контейнерах, где созда-
ются благоприятные условия для жизни и размножения этих грызунов.
Ленский речпой бассейн стал основпым путем продвижения пасюка па
северо-восток Сибири. Здесь происходит большая порча им и приведение
в негодпость хранимого и транспортируемого комбикорма и зернофуража.
Во многих случаях повреждено от 50 до 100% мешков.
В 1984 т. на торговой базе пос. Осетрово находилось 125880 мешков
комбикорма. При высокой численности крыс (1000 экз. в одном штабеле)
почти все мешки были повреждены, т. е. 100 тыс. мептков стоимостью
в 150 тыс. руб. (стоимость одного мешка 4 руб. 50 коп.). Комбикорм
высыпался на землю или на пол склада, сментивался с мусором, пылью,
>
Практическое значение
335
снегом и к дальнейшему использованию был пепригоден. При количестве
100 тыс. поврежденных мешков эти просыни составили примерно 25 т,
а в деньгах — 4475 руб. (расчет сделан из наблюдений, что из 80 мешков
просыпается в среднем 20 кг). Следовательно, только 1 тыс. крыс на од-
ной крупной базе приносит убыток в 160 тыс. руб.
Оценка экономического ущерба, причиняемого серыми крысами в
Москве, сделана В. А. Судейкиным (41983). С 1951 по 1980 г. дезинфек-
ционной станцией было выловлено и подобрано 614,8 тыс. трупов отрав-
ленных крыс, которые, по расчетам автора, принесли бы ущерб за эти
годы в 12 544 руб.
В животповодческих помещениях, по данным Д. Ф. Траханова и
А. Ф. Кадирова (1981) и Ш. М. Мирзоева (1978), только в свинарниках
крысы ежегодно уничтожают 500 тыс. т кормов па сумму в 75 млн руб.
Общие ежегодные убытки от них в животноводстве эти авторы оценивают
в 157,6 млн руб.
Кроме того, что крысы поедают пищевые продукты, большое коли-
чество их они загрязняют своей мочой и калом. Оценить удельный вес
таких продуктов очень трудно. Имеются сведения, что один зверек выде-
ляет в год 25 тыс. катышков кала и 20—30 мл мочи за сутки (Пасеш-
ник, 1947). Но данным исследователей, одна крыса загрязняет мочой до
1000 зерен крупы в сутки.
Сведения об ущербе, причиняемом серой крысой, не ограничиваются
только порчей и уничтожением пищевых продуктов. Так, 20% пожаров
в США вызваны замыкапиями, произошедшими из-за повреждения этим
трызупом кабеля. Из-за повреждения кабеля и узлов автоматической
сигнализации крысами в Японии на длительное время парупалось дви-
жепие электропоездов (Наумов, Емельянова, 1979). Те же авторы с00б-
цают, что в г. Ахваз (Иран) крысы повредили линии связи в подзем-
пых коммуникациях, в результате чего в течение нескольких дпей во
многих учреждениях и жилых домах не работала телефонная снязь. При-
водятся снедения о новреждении зверьками проводов в самолетах, что
может повлечь за собой воздушнуто катастрофу (Емельянова, Данилки-
на, 1983). Значительный ущерб крысы иричиняют в подземных соору-
жепиях. В канализации сотни нор, сделанных ими в стенках коллекто-
ров, приводят к быстрому разрушению коллекторов, утечке сточных вод
и к обралам (Тощигин, 1986). Присутствие зверьков в шахтах представ-
ляет еще большую опасность, так как они повреждают изоляцию прово-
дов, упаковку и оболочку взрывчатых устройств (Иоффе, 1959). Во вре-
мя обследования шахт Донбасса неоднократно отмечали поврежденные
крысами оболочки детонаторов и потрызы в ящиках па складах взрыв-
чатых веществ (Тощигин, 1986). Ущерб от повреждений серой крысой
носит и косвенный характер: например, повреждения ею герметической
‘упаковки на приборах, храпящихся на складе, приводят к порче послед-
них. Поселяясь в животноводческих помещениях, крысы прогрызают
многочисленные отверстия в полу, и находящийся здесь скот ломает
ноги, проваливаясь в дыры; аварии на подстанциях приводят к остапэв-
ке работы на предприятиях (Емельянова, 1986) ит. д.
336
Глава пятая
Материалы по вредоноспой деятельности крыс пам передали
В. Я. Антонюк и Н. В. Олькова. Так, на Сахалине, по данным КНорса-
ковской пожарной части, в период с 1950 по 1960 г. причиной возник-
новения пожаров в 15% случаев были серые крысы. Авторы наблюдали
повреждение четырехжильного кабеля в резиновой упаковке на зерповом
токе в пос. Владимир-Петровка в Приморском крае. В Иркутске крысы
повреждают стены помещений, электропроводку, обувь, одежду и т. д.
В. С. Сурков (1971) описывает происшедшее ночью короткое замыка-
ние в результате того, что крысы обгрызли изоляцию проводов.
Причины повреждения серой врысой материалов
а техничесвих изделий
В. Е. Соколов с соавторами (1986) указывают следующие причипы по-
вреждения материалов и технических изделий пасоком:
1. Прогрызание ходов и отверстий на пути к пище, воде и убежищам.
Одна из самых распространенных причип повреждений, большая часть
которых возникает именно в результате желания зверька преодолеть
препятствия на пути к жизненно важным для него объектам. Известно,
что, добираясь до пищи, крысы повреждают тару и упаковку, прогры-
зают отверстия в полу и стенах складов, зернохранилищ, животповодчс-
ских построек.
|
2. Гнездостроительная деятельность. Серая крыса использует в каче-
стве гнездового материала все, что имеется поблизости: бумагу, ткани,
войлок, пенопласт, пленку и т. д. Д. С. Айзенштадт (19506) находил
в гнездах зверьков, населявигих насыпи вдоль железнодорожного полот-
на, обрывки бумаги, папиросные и спичечные коробки, трянки, которые
зверьки извлекали из мусора, выброшенного из вагонов. В эксперименте
серые крысы в качестве гпездового материала использовали изоляцию
проводов, резину, пенопласты, тонкую проволоку (Емельянова, Малыгип,
1973). При отсутствни или недостатке естественпых: материалов новреж-
дения, связаппые с гпездостроением усиливаются, особенно в период раз-
множения, так как у беременпых самок наблюдается повышение грызу-
щей активности (Данилкипа, 1984).
3. Необходимость стачивания резцов. По данным Ветцеля (Меца,
1927), у серой крысы недельный прирост верхних резцов составляет
2,4 мм, нижних
— 3,45 мм. Твердость эмали на резцах
— 5,0-5,5 еди-
ницы по шкале Мооса (Него, 1950). Для сравнепия укажем, что твер-
дость свинца по шкале — 1,5 сдипицы, цинка — 2,5, железа — 4,5, квар-
ца — 7 единиц. Естественно, что такой интенсивный прирост резцов
вызывает у зверьков потребность грызть окружающие предметы, чтобы
стачивать зубы.
А. Повреждение материалов в результате повышенной активности,
При возникновении стрессовых ситуаций в период резкого возрастапия
численпости, когда нарушается структура популяций и усиливается кон-
куренция за корм и убежища, возрастает и грызущая: активность зверь“
Практическое значение
337
ков и увеличивается вероятность панесения повреждений (Емельяно-
ва, 1986).
5. Повреждение материалов в результате интенсивного исследователь-
ского поведения. Повреждения серые крысы наносят ири исследовании
новых объектов. Грызение одной особи может также возбуждать грызу-
щую активность у других зверьков (Емельяпова, 1986). В эксперименте
показано, что при знакомстве с новой обстановкой крысы трызли пол и
стены, особое внимание обращая на стыки и углы, т. е. места, где удоб-
но начинать грызть (Меликова, 1981).
Все перечисленные выше причины, обусловливающие грызущую дея-
тельность крыс, вызывают необходимость точного знания не только стой-
кости материалов и изделий, используемых в промышленности, строи-
тельстве и т. д., к повреждению пасюком, по также и их привлекатель-
ности для этого грызупа.
Стойкость материалов к повреждениям крысой
Понятие стойкости (или устойчивости) материалов к повреждениям
теми или ипыми организмами имеет два значения. Под биологической
стойкостью понимается механическая прочность материала по сравнению
с прочностью резцов крысы при максимальных условиях, которые она
может развить в стрессовой ситуации. Нри отсутствии стрессов крыса
не будет повреждать материал даже менее механически прочный, если
он пе пмест для пее какой-либо привлекательности. В этом случае мож-
по говорить об экологической стойкости.
‚Для изучения стойкости пользуются тремя методами: 1) принудитель-
ный метод, 2) метод «свободного выбора» и 3) метод стимулирования.
Но принудительному методу (ГОСТ 9.057—75) взрослых крыс помещают
в клетку размером 70% 35Ж35 см, разделенную перегородкой па две рав-
ныё части. В перегородке имеется отверстие размером 10Х 10 см. В одной
половине клетки. находится гнездо, в другой корм. Пластинкой исследуе-
мого материала закрывается отверстие в перегородке, а снизу оставляет-
ся. щель в 15 мм. Корм крысы получают за 24 ч до опыта, воду опи
имеют без ограничения, сам опыт длится тоже 24 ч. Степень поврежие-
ния образцов оцепивают визуально по пятибалльной 1икале: 0 — образец
пе поврежден, 1 — на поверхности образца имеются следы зубов, 2 — по-
вреждены покрытие или поверхпость образца, 3 — образец значительно
поврежден, но не прогрызен, 4 — образец прогрызен.
‚ Материал считается неустойчивым, если степень повреждения соот-
вётствует баллам 2, 3 и4. Этим методом было испытано около 600 раз-
личных образцов мат ериалов.
„Сущность метода. «свободного выбора» (Дадонов и др., 1988) заклю-
чается в том, что в боксе 2Ж1,5Ж2 м поселяют группировку крыс
и имитируют природные условия; испытываемый материал на длитель-
пое время (15: суток) номещается в Тот же бокс и по истечепии срока
оценивается его устойчивость по ГОСТ 9.057—75. Дополнительно прово-
длтел. испытания методом «стимулирования». В тот же бокс заклады-
336
Глава пятая
Материалы по вредоноспой деятельности крыс пам передали
В. Я. Антонюк и Н. В. Олькова. Так, на Сахалине, по данным КНорса-
ковской пожарной части, в период с 1950 по 1960 г. причиной возник-
новения пожаров в 15% случаев были серые крысы. Авторы наблюдали
повреждение четырехжильного кабеля в резиновой упаковке на зерповом
токе в пос. Владимир-Петровка в Приморском крае. В Иркутске крысы
повреждают стены помещений, электропроводку, обувь, одежду и т. д.
В. С. Сурков (1971) описывает происшедшее ночью короткое замыка-
ние в результате того, что крысы обгрызли изоляцию проводов.
Причины повреждения серой врысой материалов
а техничесвих изделий
В. Е. Соколов с соавторами (1986) указывают следующие причипы по-
вреждения материалов и технических изделий пасоком:
1. Прогрызание ходов и отверстий на пути к пище, воде и убежищам.
Одна из самых распространенных причип повреждений, большая часть
которых возникает именно в результате желания зверька преодолеть
препятствия на пути к жизненно важным для него объектам. Известно,
что, добираясь до пищи, крысы повреждают тару и упаковку, прогры-
зают отверстия в полу и стенах складов, зернохранилищ, животповодчс-
ских построек.
|
2. Гнездостроительная деятельность. Серая крыса использует в каче-
стве гнездового материала все, что имеется поблизости: бумагу, ткани,
войлок, пенопласт, пленку и т. д. Д. С. Айзенштадт (19506) находил
в гнездах зверьков, населявигих насыпи вдоль железнодорожного полот-
на, обрывки бумаги, папиросные и спичечные коробки, трянки, которые
зверьки извлекали из мусора, выброшенного из вагонов. В эксперименте
серые крысы в качестве гпездового материала использовали изоляцию
проводов, резину, пенопласты, тонкую проволоку (Емельянова, Малыгип,
1973). При отсутствни или недостатке естественпых: материалов новреж-
дения, связаппые с гпездостроением усиливаются, особенно в период раз-
множения, так как у беременпых самок наблюдается повышение грызу-
щей активности (Данилкипа, 1984).
3. Необходимость стачивания резцов. По данным Ветцеля (Меца,
1927), у серой крысы недельный прирост верхних резцов составляет
2,4 мм, нижних
— 3,45 мм. Твердость эмали на резцах
— 5,0-5,5 еди-
ницы по шкале Мооса (Него, 1950). Для сравнепия укажем, что твер-
дость свинца по шкале — 1,5 сдипицы, цинка — 2,5, железа — 4,5, квар-
ца — 7 единиц. Естественно, что такой интенсивный прирост резцов
вызывает у зверьков потребность грызть окружающие предметы, чтобы
стачивать зубы.
А. Повреждение материалов в результате повышенной активности,
При возникновении стрессовых ситуаций в период резкого возрастапия
численпости, когда нарушается структура популяций и усиливается кон-
куренция за корм и убежища, возрастает и грызущая: активность зверь“
Практическое значение
337
ков и увеличивается вероятность панесения повреждений (Емельяно-
ва, 1986).
5. Повреждение материалов в результате интенсивного исследователь-
ского поведения. Повреждения серые крысы наносят ири исследовании
новых объектов. Грызение одной особи может также возбуждать грызу-
щую активность у других зверьков (Емельяпова, 1986). В эксперименте
показано, что при знакомстве с новой обстановкой крысы трызли пол и
стены, особое внимание обращая на стыки и углы, т. е. места, где удоб-
но начинать грызть (Меликова, 1981).
Все перечисленные выше причины, обусловливающие грызущую дея-
тельность крыс, вызывают необходимость точного знания не только стой-
кости материалов и изделий, используемых в промышленности, строи-
тельстве и т. д., к повреждению пасюком, по также и их привлекатель-
ности для этого грызупа.
Стойкость материалов к повреждениям крысой
Понятие стойкости (или устойчивости) материалов к повреждениям
теми или ипыми организмами имеет два значения. Под биологической
стойкостью понимается механическая прочность материала по сравнению
с прочностью резцов крысы при максимальных условиях, которые она
может развить в стрессовой ситуации. Нри отсутствии стрессов крыса
не будет повреждать материал даже менее механически прочный, если
он пе пмест для пее какой-либо привлекательности. В этом случае мож-
по говорить об экологической стойкости.
‚Для изучения стойкости пользуются тремя методами: 1) принудитель-
ный метод, 2) метод «свободного выбора» и 3) метод стимулирования.
Но принудительному методу (ГОСТ 9.057—75) взрослых крыс помещают
в клетку размером 70% 35Ж35 см, разделенную перегородкой па две рав-
ныё части. В перегородке имеется отверстие размером 10Х 10 см. В одной
половине клетки. находится гнездо, в другой корм. Пластинкой исследуе-
мого материала закрывается отверстие в перегородке, а снизу оставляет-
ся. щель в 15 мм. Корм крысы получают за 24 ч до опыта, воду опи
имеют без ограничения, сам опыт длится тоже 24 ч. Степень поврежие-
ния образцов оцепивают визуально по пятибалльной 1икале: 0 — образец
пе поврежден, 1 — на поверхности образца имеются следы зубов, 2 — по-
вреждены покрытие или поверхпость образца, 3 — образец значительно
поврежден, но не прогрызен, 4 — образец прогрызен.
‚ Материал считается неустойчивым, если степень повреждения соот-
вётствует баллам 2, 3 и4. Этим методом было испытано около 600 раз-
личных образцов мат ериалов.
„Сущность метода. «свободного выбора» (Дадонов и др., 1988) заклю-
чается в том, что в боксе 2Ж1,5Ж2 м поселяют группировку крыс
и имитируют природные условия; испытываемый материал на длитель-
пое время (15: суток) номещается в Тот же бокс и по истечепии срока
оценивается его устойчивость по ГОСТ 9.057—75. Дополнительно прово-
длтел. испытания методом «стимулирования». В тот же бокс заклады-
338
Глава пятал
вается образец материала таким образом, чтобы преградить путь к более
вкусной пище (при достаточном количестве обычной пищи и воды).
Возможность повреждения крысой материалов зависит от характера
их поверхности (гладкая или птершавая), твердости и структуры
(пористая, плотная, вязкая и др.). Крысы легко повреждали пенопласты,
когда пластинкой материала закрывали полностью окно в перегородке,
разделяющей кормовое и тнездовое отделения клетки, так как пористая
поверхность при грызении дает упор для резцов. Тот же пенопласт,
окралтенный эпоксидпой шпатлевкой, грызуны не могли прогрызть, так
как розцы скользили по гладкой поверхности. Пластинка была понреж-
дена лишь после того, как в ней были сделаны отверстия диаметром 5 мм
(Емельянова, 1986). Хук и Ла Брейн (Ниск, Га Вгцп, 1959) отмечали
то же явление при испытании пленки из поливинилхлорида. Наличие
вов, выступов, отверстий также способствует панесению повреждений.
Нейгауз (МепВалз, 1957) показал, что крысы не повреждали соедини-
тельные муфты из пластмассы до тех пор, пока им не стал доступеп
их край.
В опытах И. А. Емельяновой (1986) крысы перегрызали кабель диа-
метром до 17 мм. Значительные повреждения были ею отмечены на кабе-
не диаметром 24—29 мм и серьезные повреждения на кабеле диамет-
ром 60 мм в резиновой оболочке. Столь широкий слектр повреждаемых
сврой крысой материалов вызвал к жизни проблему защиты материалов
и технических изделий от этого грызуна.
Медицинское значение
Серая крыса
— классический носитель целого ряда инфекций, опаспых
для человека. Трудно найти другой вид животного, который нес бы
в себе такой огромный потенциал опаспости для здоровья людей.
В настоящее время подтверждено выделением культур носительство
крысами следующих инфекций и инвазий: бешенства. геморрагической
лихорадки с почечпым синдромом, крысиного сыпного тифа, клещевого
сыппого тифа, незикулезного, оспоподобного риккетсиозов, лихорадки
цуцугамущи, О-лихорадки, чумы, псевдотуберкулеза, кишечного иерси-
ниоза, пастереллезов, туляремии, бруцеллеза, эризипелоида, листериоза,
сальмонеллезов, лептоспирозов, содоку, токсоплазмоза и балантидиоза.
Из гельминтозов с крысой наиболее связан трихипеллез.
Описапие чумных эпизоотий, возникающих в больших городах,
на сегодняшний депь представляет лишь исторический интерес. Однако
быстрое увеличение ареала серой крысы и продвижение ее в Среднюю
Азию не может не беспокоить органы здравоохранения, так как проник-
повение этого грызуна в аридные регионы, где обитает большая песчан-
ка — один из основных носителей чумы, конечно, очень опасно.
В настоящее время больное эпидемиологическое значение имеют эпи-
зоотии среди крыс иктерогеморрагического леитоспироза (болезнь
Васильева—Вейля), псевдотуберкулеза, кишечпого иерсиниоза, эризине-
Практическое значение
339
лоида, а из вирусных заболеваний — геморрагическая лихорадка с почеч-
ным синдромом. Из инвазий распространен трихинеллез.
Пасюк является основным носителем болезпи содоку и крысиного
сыппого тифа.
При остальных инфекциях возбудитель попадает в популяции крысе
от других животных— основных носителей, но и среди пасюков возбу-
дитель может циркулировать достаточно долгое время.
Несмотря на очевидный территориальный консерватизм, который
способствует поддержанию стойких локальных очагов инфекций, серым
крысам свойственна большая подвижпость. Опи совершают сезонные
перемещения из населенных пунктов в открытые биотопы и обратно и
способствуют таким образом связи сипантронных грызунов с природны-
ми очагами инфекций.
Благодаря исключительной экологической пластичности, всеядности,
высокой интенсивности размножения пасюки часто достигают очепь
большой численпости, приводящей к тесному контакту между особями.
Зверьки ведут общественный образ жизни, им свойственно скучивание,
постояппое перемещение по одним и тем же путям, обитание многих
особей в одной норе и т. п. Все оти экологические особенности вида со-
здают высокую вероятность передачи инфекции от особи к особи, что
часто приводит к весьма интенсивным эпизоотиям. Серые крысы как
источники инфекции. представляют собой колоссальную опаспость для
людей из-за своего близкого контакта с ними. Обитая в населенных
пунктах, эти грызуны могут заражать своими выделениями продукты
питания, что приводит к инфицированию людей главным образом не-
трансмиссивными инфекциями (псевдотуберкулез, кишечный иерсиниоз,
листериоз, лептоспирозы и др.). Наличие на крысах большого .количе-
ства эктопаразитов (несколько видов блох, гамазовых клещей и др.)
делает возможным широкое расиространение в их популяции трапсмис-
сивных инфекций (чума, риккетсиозы).
Пасюки нередко кусают спящих людей или малепьких детей, что
опасно не только в силу напесения травм, но также может способство-
вать заражению, например бешенством и содоку (Судейкин, 1983а). Так,
в Москве лица, покусапные крысами, составили 7,0% всех обратившихся
за антирабической медицинской помощью. В Греции они составили 9,8%,
в Алжире — 11.87, а в Ливии — 47,6% (Селимов, 1978).
В городах в годы войны и в послевоеппые годы инфицирование людей
от крыс наблюдалось особенно часто. При частичном разрушении зда-
ний появляются многочисленные щели, через которые легко проникают
крысы. Нарушается водоснабжение, что может приводить к употребле-
вию людьми воды, загрязненной их выделениями. Элидемии ряда ипфек-
ций, вспыхивающие в траншеях среди солдат, часто бывают связаны со
скоплениями там крыс (Токаревич, 1947).
Помимо антропургических очагов, среди экзоантропных поселений
крыс возникают зпизоотии (например, лептоспироз), формируя природ-
ные очаги инфекций, с которыми особеппо трудно бороться. Эти очаги
имеют специфическую пространственную структуру, Которой определяют-
338
Глава пятал
вается образец материала таким образом, чтобы преградить путь к более
вкусной пище (при достаточном количестве обычной пищи и воды).
Возможность повреждения крысой материалов зависит от характера
их поверхности (гладкая или птершавая), твердости и структуры
(пористая, плотная, вязкая и др.). Крысы легко повреждали пенопласты,
когда пластинкой материала закрывали полностью окно в перегородке,
разделяющей кормовое и тнездовое отделения клетки, так как пористая
поверхность при грызении дает упор для резцов. Тот же пенопласт,
окралтенный эпоксидпой шпатлевкой, грызуны не могли прогрызть, так
как розцы скользили по гладкой поверхности. Пластинка была понреж-
дена лишь после того, как в ней были сделаны отверстия диаметром 5 мм
(Емельянова, 1986). Хук и Ла Брейн (Ниск, Га Вгцп, 1959) отмечали
то же явление при испытании пленки из поливинилхлорида. Наличие
вов, выступов, отверстий также способствует панесению повреждений.
Нейгауз (МепВалз, 1957) показал, что крысы не повреждали соедини-
тельные муфты из пластмассы до тех пор, пока им не стал доступеп
их край.
В опытах И. А. Емельяновой (1986) крысы перегрызали кабель диа-
метром до 17 мм. Значительные повреждения были ею отмечены на кабе-
не диаметром 24—29 мм и серьезные повреждения на кабеле диамет-
ром 60 мм в резиновой оболочке. Столь широкий слектр повреждаемых
сврой крысой материалов вызвал к жизни проблему защиты материалов
и технических изделий от этого грызуна.
Медицинское значение
Серая крыса
— классический носитель целого ряда инфекций, опаспых
для человека. Трудно найти другой вид животного, который нес бы
в себе такой огромный потенциал опаспости для здоровья людей.
В настоящее время подтверждено выделением культур носительство
крысами следующих инфекций и инвазий: бешенства. геморрагической
лихорадки с почечпым синдромом, крысиного сыпного тифа, клещевого
сыппого тифа, незикулезного, оспоподобного риккетсиозов, лихорадки
цуцугамущи, О-лихорадки, чумы, псевдотуберкулеза, кишечного иерси-
ниоза, пастереллезов, туляремии, бруцеллеза, эризипелоида, листериоза,
сальмонеллезов, лептоспирозов, содоку, токсоплазмоза и балантидиоза.
Из гельминтозов с крысой наиболее связан трихипеллез.
Описапие чумных эпизоотий, возникающих в больших городах,
на сегодняшний депь представляет лишь исторический интерес. Однако
быстрое увеличение ареала серой крысы и продвижение ее в Среднюю
Азию не может не беспокоить органы здравоохранения, так как проник-
повение этого грызуна в аридные регионы, где обитает большая песчан-
ка — один из основных носителей чумы, конечно, очень опасно.
В настоящее время больное эпидемиологическое значение имеют эпи-
зоотии среди крыс иктерогеморрагического леитоспироза (болезнь
Васильева—Вейля), псевдотуберкулеза, кишечпого иерсиниоза, эризине-
Практическое значение
339
лоида, а из вирусных заболеваний — геморрагическая лихорадка с почеч-
ным синдромом. Из инвазий распространен трихинеллез.
Пасюк является основным носителем болезпи содоку и крысиного
сыппого тифа.
При остальных инфекциях возбудитель попадает в популяции крысе
от других животных— основных носителей, но и среди пасюков возбу-
дитель может циркулировать достаточно долгое время.
Несмотря на очевидный территориальный консерватизм, который
способствует поддержанию стойких локальных очагов инфекций, серым
крысам свойственна большая подвижпость. Опи совершают сезонные
перемещения из населенных пунктов в открытые биотопы и обратно и
способствуют таким образом связи сипантронных грызунов с природны-
ми очагами инфекций.
Благодаря исключительной экологической пластичности, всеядности,
высокой интенсивности размножения пасюки часто достигают очепь
большой численпости, приводящей к тесному контакту между особями.
Зверьки ведут общественный образ жизни, им свойственно скучивание,
постояппое перемещение по одним и тем же путям, обитание многих
особей в одной норе и т. п. Все оти экологические особенности вида со-
здают высокую вероятность передачи инфекции от особи к особи, что
часто приводит к весьма интенсивным эпизоотиям. Серые крысы как
источники инфекции. представляют собой колоссальную опаспость для
людей из-за своего близкого контакта с ними. Обитая в населенных
пунктах, эти грызуны могут заражать своими выделениями продукты
питания, что приводит к инфицированию людей главным образом не-
трансмиссивными инфекциями (псевдотуберкулез, кишечный иерсиниоз,
листериоз, лептоспирозы и др.). Наличие на крысах большого .количе-
ства эктопаразитов (несколько видов блох, гамазовых клещей и др.)
делает возможным широкое расиространение в их популяции трапсмис-
сивных инфекций (чума, риккетсиозы).
Пасюки нередко кусают спящих людей или малепьких детей, что
опасно не только в силу напесения травм, но также может способство-
вать заражению, например бешенством и содоку (Судейкин, 1983а). Так,
в Москве лица, покусапные крысами, составили 7,0% всех обратившихся
за антирабической медицинской помощью. В Греции они составили 9,8%,
в Алжире — 11.87, а в Ливии — 47,6% (Селимов, 1978).
В городах в годы войны и в послевоеппые годы инфицирование людей
от крыс наблюдалось особенно часто. При частичном разрушении зда-
ний появляются многочисленные щели, через которые легко проникают
крысы. Нарушается водоснабжение, что может приводить к употребле-
вию людьми воды, загрязненной их выделениями. Элидемии ряда ипфек-
ций, вспыхивающие в траншеях среди солдат, часто бывают связаны со
скоплениями там крыс (Токаревич, 1947).
Помимо антропургических очагов, среди экзоантропных поселений
крыс возникают зпизоотии (например, лептоспироз), формируя природ-
ные очаги инфекций, с которыми особеппо трудно бороться. Эти очаги
имеют специфическую пространственную структуру, Которой определяют-
340
Глава пятая
ся особенности распределения крыс. Из-за способности зверьков скучи-
ваться образуются ядра очага, а вследствие подвижности пасюков эпи-
зоотия может широко распространиться (Карасева и др., 1986).
Вирусные заболевания
Роль серых крыс в энизоотологии и эпидемиологии вирусных инфенций
певелика и описана только при бешенстве и геморрагической лихорадке.
Бешенство — возбудитель ВаМез у1тиз. Все эпизоотии бешенства
принято делить на эпизоотии природного тина, поддерживаемые различ-
ными хищными животными, и эпизоотии городскогб
— поддерживаемые
домашними животпыми. Наряду с собаками (основным хранителем и
источником бешенства в очагах городского типа), рогатым скотом,
лошадьми, кошками было выявлено и поражение вирусом бешенства
серых крыс. В отчетах одной из русских пастеровских станций в 1909 г.
впервые сообщается о гибели укушенного крысой человека от паралити-
ческой формы бешепства (Али-Заде, 1958). Позже стал известен еще
ряд заболеваний гидрофобией людей, отмечавших в своем анамнезе
укусы крыс. Позднсе от крыс был выделен вирус бешенства, а в 1960—
1970 гг. в ряде стран Европы и Америки появились новые сообщения
о лабораторном подтверждении бешенства среди пих. В 1982 г. зареги-
стрирован случай бешенства у серой крысы в г. Донецке (Гажиев и др.,
1986), где она искусала школьников-подростков при разборе металлоло-
ма. Крыса была убита, и при лабораторном исследовании у пее под-
тверждеи диагноз бешенства.
Воирос о том, могут ли пасюки и другие грызуны поддерживать само-
стоятельно эпизоотии бешенства или они случайно включаются в элпи-
зоотическую цепь, до сих пор не ретзен.
Несмотря на высокую чувствительность серых крыс к эксперимен-
тальному заражению вирусом бешенства, эпизоотии среди них не отме-
чается. Очевидно, в естественпых условиях заражение их, как и других
грызунов, вирусом бешенства приводит к пемедленной гибели и на этом
эпизоотический процесс обрывается (Седимов, 1978).
Геморрагическая лихорадка с почечным синдр.о-
мом (ГЛИС) — возбудитель Наша \!тгиз. Особенностью инфекции
ГЛИС является отсутствие у животпых клинических проявлений болез-
ни. Признаками наличия инфекции у них служат обпаружение антигена
вируса ГЛИС в органах (преимущественно в легких) и антител в крови.
В организме серой крысы возможна длительная хроническая инфек-
ция ГЛПС, сопровождающаяся постоянным выделением вируса в окру-
жующую среду респираторным путем, со слюной, мочой и калом. Инди-
катором существования таких инфицированных популяций серой крысы
служат вспышки или спорадические случаи заболевания ГЛИС у чело-
века. Легкие формы ГЛИС в Китае, отсутствие клинически выраженных
случаев ГЛИС в Америке и на Гавайях при контактах людей с инфици-
рованными крысами позволили предположить, что в серых крысах цир-
Практическое значение
ЗАЛ
Таблица 28. Изоляция штаммов вируса ГЛИС из спонтанно инфицированных
серых крые
Число
Место и время от- выделен-| Наименова-
Эпидемическая
А.
1
ных
ам
.
втор штамма, лите-
лоза животных
ТАМним
ситуация в месте ратурный источник
мов
Сеул (жилые по-
11
80/39
Спорадические
Тее сб а1., 1986
стройки), Южная
случаи заболева-
Корея, 1982
ния людей
Шанхай иль
1
-
Всенышки ГЛИС Бопё бай её а!., 1982
постройки),
(легкое течение)
КНР, 1982
Саппоро (жилые
2
-
Спорадизческие
КИатига, цит. по:
постройки),
случаи заболева- | Гаврилонская, Бойно,
Япония, 1983
ния
1985
Токио (портовая
1
-
Случаев ГЛИС
Зитата, цит, по:
зона), Япопия,
нет
Гавриловская, Бойко,
1984
1985
Филадельфия,
2 Джирард
‘Го же
Те Рис еЁа|., 1985
Хьюстон (порто-
Поинт
вые зоны), США,
1984
Новый Орлеан
1 Чоупитулас | То же
Те рис сё &., 1985
(портовая зона),
.
США, 1985
Белем (портовая
1 Белем
р
Тза{, цит. по: 1е рае её
зона), Бразилия,
аГ., 198%
1985
кулирует аптигепто отличный, хотя и близкородственный прототипному,
“вирус ГЛИС (Могйа её а|., 1985).
В настоящее время все питаммы, изолироваппые от серых крыс
в Южной Корее, Японии, Китае, СПТА и Бразилии, отнесены к серотину
Вациз (ЭсвшаЦовл еф а1., 1985).
Антигенные свойства этих штаммов, изолированных в различных ре-
гионах мира, идентичны.
Возможность заражения человека при контакте с инфицированными
крысами была впервые установлена при обследовании служителей вива-
риев в Токийском университете. Ли с соавторами (Гее её а1., 1986)
сообщили, что с 1976 г. среди персонала, обслуживающего виварии
‚в Корее и Японии, зарегистрирована 24 вспышка ГЛИС (заболели
130 человек). Серологическое обследование лабораторных крыс в этих
‚ вивариях показало, что все линии крыс заражены вирусом ГЛИС.
Апалогичпые вспышки ГЛИС были описаны также и в Европе
в 1978 г. (Бельгия, Англия, Франция).
:
У серых крыс, отловлеппых в Китае и Южной Корее, был обнаружен,
кроме антител в крови, также антиген вируса ГЛПС в легких. Опреде-
ление антигена в органах этих грызупов свидетельствует о зпачительной
340
Глава пятая
ся особенности распределения крыс. Из-за способности зверьков скучи-
ваться образуются ядра очага, а вследствие подвижности пасюков эпи-
зоотия может широко распространиться (Карасева и др., 1986).
Вирусные заболевания
Роль серых крыс в энизоотологии и эпидемиологии вирусных инфенций
певелика и описана только при бешенстве и геморрагической лихорадке.
Бешенство — возбудитель ВаМез у1тиз. Все эпизоотии бешенства
принято делить на эпизоотии природного тина, поддерживаемые различ-
ными хищными животными, и эпизоотии городскогб
— поддерживаемые
домашними животпыми. Наряду с собаками (основным хранителем и
источником бешенства в очагах городского типа), рогатым скотом,
лошадьми, кошками было выявлено и поражение вирусом бешенства
серых крыс. В отчетах одной из русских пастеровских станций в 1909 г.
впервые сообщается о гибели укушенного крысой человека от паралити-
ческой формы бешепства (Али-Заде, 1958). Позже стал известен еще
ряд заболеваний гидрофобией людей, отмечавших в своем анамнезе
укусы крыс. Позднсе от крыс был выделен вирус бешенства, а в 1960—
1970 гг. в ряде стран Европы и Америки появились новые сообщения
о лабораторном подтверждении бешенства среди пих. В 1982 г. зареги-
стрирован случай бешенства у серой крысы в г. Донецке (Гажиев и др.,
1986), где она искусала школьников-подростков при разборе металлоло-
ма. Крыса была убита, и при лабораторном исследовании у пее под-
тверждеи диагноз бешенства.
Воирос о том, могут ли пасюки и другие грызуны поддерживать само-
стоятельно эпизоотии бешенства или они случайно включаются в элпи-
зоотическую цепь, до сих пор не ретзен.
Несмотря на высокую чувствительность серых крыс к эксперимен-
тальному заражению вирусом бешенства, эпизоотии среди них не отме-
чается. Очевидно, в естественпых условиях заражение их, как и других
грызунов, вирусом бешенства приводит к пемедленной гибели и на этом
эпизоотический процесс обрывается (Седимов, 1978).
Геморрагическая лихорадка с почечным синдр.о-
мом (ГЛИС) — возбудитель Наша \!тгиз. Особенностью инфекции
ГЛИС является отсутствие у животпых клинических проявлений болез-
ни. Признаками наличия инфекции у них служат обпаружение антигена
вируса ГЛИС в органах (преимущественно в легких) и антител в крови.
В организме серой крысы возможна длительная хроническая инфек-
ция ГЛПС, сопровождающаяся постоянным выделением вируса в окру-
жующую среду респираторным путем, со слюной, мочой и калом. Инди-
катором существования таких инфицированных популяций серой крысы
служат вспышки или спорадические случаи заболевания ГЛИС у чело-
века. Легкие формы ГЛИС в Китае, отсутствие клинически выраженных
случаев ГЛИС в Америке и на Гавайях при контактах людей с инфици-
рованными крысами позволили предположить, что в серых крысах цир-
Практическое значение
ЗАЛ
Таблица 28. Изоляция штаммов вируса ГЛИС из спонтанно инфицированных
серых крые
Число
Место и время от- выделен-| Наименова-
Эпидемическая
А.
1
ных
ам
.
втор штамма, лите-
лоза животных
ТАМним
ситуация в месте ратурный источник
мов
Сеул (жилые по-
11
80/39
Спорадические
Тее сб а1., 1986
стройки), Южная
случаи заболева-
Корея, 1982
ния людей
Шанхай иль
1
-
Всенышки ГЛИС Бопё бай её а!., 1982
постройки),
(легкое течение)
КНР, 1982
Саппоро (жилые
2
-
Спорадизческие
КИатига, цит. по:
постройки),
случаи заболева- | Гаврилонская, Бойно,
Япония, 1983
ния
1985
Токио (портовая
1
-
Случаев ГЛИС
Зитата, цит, по:
зона), Япопия,
нет
Гавриловская, Бойко,
1984
1985
Филадельфия,
2 Джирард
‘Го же
Те Рис еЁа|., 1985
Хьюстон (порто-
Поинт
вые зоны), США,
1984
Новый Орлеан
1 Чоупитулас | То же
Те рис сё &., 1985
(портовая зона),
.
США, 1985
Белем (портовая
1 Белем
р
Тза{, цит. по: 1е рае её
зона), Бразилия,
аГ., 198%
1985
кулирует аптигепто отличный, хотя и близкородственный прототипному,
“вирус ГЛИС (Могйа её а|., 1985).
В настоящее время все питаммы, изолироваппые от серых крыс
в Южной Корее, Японии, Китае, СПТА и Бразилии, отнесены к серотину
Вациз (ЭсвшаЦовл еф а1., 1985).
Антигенные свойства этих штаммов, изолированных в различных ре-
гионах мира, идентичны.
Возможность заражения человека при контакте с инфицированными
крысами была впервые установлена при обследовании служителей вива-
риев в Токийском университете. Ли с соавторами (Гее её а1., 1986)
сообщили, что с 1976 г. среди персонала, обслуживающего виварии
‚в Корее и Японии, зарегистрирована 24 вспышка ГЛИС (заболели
130 человек). Серологическое обследование лабораторных крыс в этих
‚ вивариях показало, что все линии крыс заражены вирусом ГЛИС.
Апалогичпые вспышки ГЛИС были описаны также и в Европе
в 1978 г. (Бельгия, Англия, Франция).
:
У серых крыс, отловлеппых в Китае и Южной Корее, был обнаружен,
кроме антител в крови, также антиген вируса ГЛПС в легких. Опреде-
ление антигена в органах этих грызупов свидетельствует о зпачительной
Глава пятая
342
интенсивности ипфекцнопного процесса в организме хозяина и об актив-
пом выделении инфекционного вируса во внешнюю среду (Гавриловская,
Бойко, 1985). Наличие только антител, как показали дальнейшие иссле-
дования, также свидетельствует об ипфицироваппости вирусом ГЛИС,
по с меньшей интенсивностью процесса. Чаще антитела обнаруживали
у крыс старше чести месяцев, что. по-видимому, является результатом
постепенного накопления хронически инфицированных особей в попу-
ляции.
Вирус ГЛИС был изолирован из органов серых крыс, отловленных
в различных регионах мира (табл. 28). Участие серой крысы в эпизоотии
ГЛИС привело, по-видимому, к распространению ипфекции в глобальном
масштабе.
В СССР первые находки антигена вируса с пизкими титрами анти-
гена (1:2) у единичных серых крыс, отловленных в природе, были
в Истринском районе Московской области (Повалишина, 1985). В Хаба-
ровском крае антиген найден у 1,1% исследованных серых крыс (Гапо-
пова и др., 1985). По данным Г. Д. Трофимчука (1985), в Приморском
крае этот процент достигал 10,3.
Педавно опубликовано сообщение о находках антигена ГЛНС у серых
крыс в Грузии (Иванидзе, 1987).
Ривкетвиозы
Роль серой крысы в эпидемиологии и эпизоотологии риккетсиозов уста-
новлена при крысином сыпном тифе, везикулезпом риккстсиозе, клеще-
зом сыппом тифе Северной Азии, лихорадке цуцугамути, Ку-лихорадке.
Наиболее подробно исследовано зпачепие серой крысы как источпика
ипфекции в очагах крысиного сыпного тифа. Нри других риккетсиозах
ев роль менее изучена и сводитсл к прокормлению переносчиков и источ-
ников инфекции— иксодовых, гамазовых или краснотелковых клещей.
Крысиный сыпной тиф (эндемический сыпной тиф, тород-
ской или магазинный тиф Малайи, блошиный сыпной тиф) — возбуди-
тель Пиускейча буры (по0зет!). Крысиный сыпной тиф распространен
в Западном и Восточном полушариях, случаи заболевапия у людей,
ипфицированность крыс и их эктопаразитов отмечены преимущественно
в бассейнах Средиземного, Черного, Каспийского и Северного морей,
Атлантического и Тихого, а также Индийского океанов.
Естественные побители возбудителя сыпного тифа — серые и черные
крысы, но большее значение в поддержании очагов, как установлено,
имеет серая крыса.
Естественная зараженность пасюка подтверждена выделением штам-
мов возбудителя в Батуми, Сухуми и Поти (Солитерман, 1948), а также
в Баку (Кулагин, Имамалиев, 1952).
Передача возбудителя в популяции крыс происходит как респиратор-
ным, так и алиментарпым (в результате капнибализма) путем. Зверьки
выделяют вирус с мочой до 16,5 месяца (Солитерман, 1948; Шарашид-
зе, 1958).
‘
®
‹
в Ирасноярском
343
Практическое значение
Большую роль в распространении инфекции среди крыс играет транс-
миссивный путь передачи, а именпо через блох. Главным передатчиком
являются космополитический вид ХепорзШа сВеорз1з, а также виды
С4епорВанииз зевтиз, СегаюриШиз ГЁазсафав, С. шизсай и некото-
рые другие.
Блохи заражаются при кровососании на больных крысах, выделяют
возбудителя с фекалиями, которые являются главным фактором пере-
дачи.
Сходпую, а возможно и ббльшуто роль, в поддержании очага крыси-
ного сыпного тифа играет крысиный клещ ОтоЙВоп155из фасой, у кото-
рого установлена трансфазовая и трансоварнальная передача возбуди-
теля (Поуе, ЗВе]пите, 1934). Штаммы возбудителя от Васой (в смешан-
ной пробе с Гаарз есШ@тшпиаз и Шегиаи1558 ва тае) выделены
в нашей стране в Новороссийске (Сомова и др., 1960).
Заболевания людей всегда возникают на фоне энизоотии среди сипан-
тропных грызунов
— крыс и мышей. Факторами передачи, как мы уже
говорили, являются моча крыс, фекалии блох, заражение возможно через
укусы гамазовых клацей. Основной путь передачи человеку— контакт-
ный при расчесывапии ранок, вызываемых укусами блох, через слизис-
тые оболочки, а также алиментарный
— через пищу, инфицированную
грызунами; возможно респираторное заражение.
Инфекция крысиного сыппого тифа стала космополитом в результате
ее заноса крысами в разные порты мира и имеет очаги как городского,
так и сельского типа. Так, в Закавказье очаги устаповлены не только
на Черноморском побережье, но и в районах, удаленных от моря
на 26—300 км (Бабалова, 1958). В США, где крысиный сыпной тиф при-
влекал внимание оргапов здравоохранения, в течение ряда лет просле-
жепо «перемещение» очагов крысиного сыпного тифа из городов Восточ-
ного побережья в сельскую местность.
Сходный процессе происходил, по-видимому, в Китае, где в настоящее
время крысипый ‘сынной тиф известен как в приморских районах, так
и в сельской местности в поскольких провинциях. В 1981—1982 гг. здесь
для предотвращения вспьшшек заболевания паселение было привлечено
к уничтожению крыс, что дало положительные результаты.
Клещевой сынной тиф Северной Азии
— возбудитель
Вискенз!а зфйЧса. Осповной источник и переносчик ипфекции — иксодо-
вые клещи родов Пегтасешюот, Настарпузаз и др., у которых установ-
лена трансфазовая и трансовариальная передача риккетсий.
х Зараженность серой крысы клещевым сыпным тифом устаповлепа
крае (Яцимирская-Кронтовская, 1940), антитела
к В. Ынеа у нее найдены в Азербайджане, в Ленкоранском райопе,
на ‘о-ве Глиняный, на Ашперонском полуострове (Стрехова, Ахундова,
1971). Можно предположить, что при хозяйственном освоении террито-
рий, потенциально опасных по клещевому сыпному тифу, при замене
диких животных — прокормителей иксодовых клещей — синантропными
трызунами возникнет ситуация, которая будет способствовать усилению
роли серой крысы в циркуляции В. зФр“са.
Глава пятая
342
интенсивности ипфекцнопного процесса в организме хозяина и об актив-
пом выделении инфекционного вируса во внешнюю среду (Гавриловская,
Бойко, 1985). Наличие только антител, как показали дальнейшие иссле-
дования, также свидетельствует об ипфицироваппости вирусом ГЛИС,
по с меньшей интенсивностью процесса. Чаще антитела обнаруживали
у крыс старше чести месяцев, что. по-видимому, является результатом
постепенного накопления хронически инфицированных особей в попу-
ляции.
Вирус ГЛИС был изолирован из органов серых крыс, отловленных
в различных регионах мира (табл. 28). Участие серой крысы в эпизоотии
ГЛИС привело, по-видимому, к распространению ипфекции в глобальном
масштабе.
В СССР первые находки антигена вируса с пизкими титрами анти-
гена (1:2) у единичных серых крыс, отловленных в природе, были
в Истринском районе Московской области (Повалишина, 1985). В Хаба-
ровском крае антиген найден у 1,1% исследованных серых крыс (Гапо-
пова и др., 1985). По данным Г. Д. Трофимчука (1985), в Приморском
крае этот процент достигал 10,3.
Педавно опубликовано сообщение о находках антигена ГЛНС у серых
крыс в Грузии (Иванидзе, 1987).
Ривкетвиозы
Роль серой крысы в эпидемиологии и эпизоотологии риккетсиозов уста-
новлена при крысином сыпном тифе, везикулезпом риккстсиозе, клеще-
зом сыппом тифе Северной Азии, лихорадке цуцугамути, Ку-лихорадке.
Наиболее подробно исследовано зпачепие серой крысы как источпика
ипфекции в очагах крысиного сыпного тифа. Нри других риккетсиозах
ев роль менее изучена и сводитсл к прокормлению переносчиков и источ-
ников инфекции— иксодовых, гамазовых или краснотелковых клещей.
Крысиный сыпной тиф (эндемический сыпной тиф, тород-
ской или магазинный тиф Малайи, блошиный сыпной тиф) — возбуди-
тель Пиускейча буры (по0зет!). Крысиный сыпной тиф распространен
в Западном и Восточном полушариях, случаи заболевапия у людей,
ипфицированность крыс и их эктопаразитов отмечены преимущественно
в бассейнах Средиземного, Черного, Каспийского и Северного морей,
Атлантического и Тихого, а также Индийского океанов.
Естественные побители возбудителя сыпного тифа — серые и черные
крысы, но большее значение в поддержании очагов, как установлено,
имеет серая крыса.
Естественная зараженность пасюка подтверждена выделением штам-
мов возбудителя в Батуми, Сухуми и Поти (Солитерман, 1948), а также
в Баку (Кулагин, Имамалиев, 1952).
Передача возбудителя в популяции крыс происходит как респиратор-
ным, так и алиментарпым (в результате капнибализма) путем. Зверьки
выделяют вирус с мочой до 16,5 месяца (Солитерман, 1948; Шарашид-
зе, 1958).
‘
®
‹
в Ирасноярском
343
Практическое значение
Большую роль в распространении инфекции среди крыс играет транс-
миссивный путь передачи, а именпо через блох. Главным передатчиком
являются космополитический вид ХепорзШа сВеорз1з, а также виды
С4епорВанииз зевтиз, СегаюриШиз ГЁазсафав, С. шизсай и некото-
рые другие.
Блохи заражаются при кровососании на больных крысах, выделяют
возбудителя с фекалиями, которые являются главным фактором пере-
дачи.
Сходпую, а возможно и ббльшуто роль, в поддержании очага крыси-
ного сыпного тифа играет крысиный клещ ОтоЙВоп155из фасой, у кото-
рого установлена трансфазовая и трансоварнальная передача возбуди-
теля (Поуе, ЗВе]пите, 1934). Штаммы возбудителя от Васой (в смешан-
ной пробе с Гаарз есШ@тшпиаз и Шегиаи1558 ва тае) выделены
в нашей стране в Новороссийске (Сомова и др., 1960).
Заболевания людей всегда возникают на фоне энизоотии среди сипан-
тропных грызунов
— крыс и мышей. Факторами передачи, как мы уже
говорили, являются моча крыс, фекалии блох, заражение возможно через
укусы гамазовых клацей. Основной путь передачи человеку— контакт-
ный при расчесывапии ранок, вызываемых укусами блох, через слизис-
тые оболочки, а также алиментарный
— через пищу, инфицированную
грызунами; возможно респираторное заражение.
Инфекция крысиного сыппого тифа стала космополитом в результате
ее заноса крысами в разные порты мира и имеет очаги как городского,
так и сельского типа. Так, в Закавказье очаги устаповлены не только
на Черноморском побережье, но и в районах, удаленных от моря
на 26—300 км (Бабалова, 1958). В США, где крысиный сыпной тиф при-
влекал внимание оргапов здравоохранения, в течение ряда лет просле-
жепо «перемещение» очагов крысиного сыпного тифа из городов Восточ-
ного побережья в сельскую местность.
Сходный процессе происходил, по-видимому, в Китае, где в настоящее
время крысипый ‘сынной тиф известен как в приморских районах, так
и в сельской местности в поскольких провинциях. В 1981—1982 гг. здесь
для предотвращения вспьшшек заболевания паселение было привлечено
к уничтожению крыс, что дало положительные результаты.
Клещевой сынной тиф Северной Азии
— возбудитель
Вискенз!а зфйЧса. Осповной источник и переносчик ипфекции — иксодо-
вые клещи родов Пегтасешюот, Настарпузаз и др., у которых установ-
лена трансфазовая и трансовариальная передача риккетсий.
х Зараженность серой крысы клещевым сыпным тифом устаповлепа
крае (Яцимирская-Кронтовская, 1940), антитела
к В. Ынеа у нее найдены в Азербайджане, в Ленкоранском райопе,
на ‘о-ве Глиняный, на Ашперонском полуострове (Стрехова, Ахундова,
1971). Можно предположить, что при хозяйственном освоении террито-
рий, потенциально опасных по клещевому сыпному тифу, при замене
диких животных — прокормителей иксодовых клещей — синантропными
трызунами возникнет ситуация, которая будет способствовать усилению
роли серой крысы в циркуляции В. зФр“са.
ЗАА
Глава пятая
Везикулезный (оспоподобный) риккетсиоз-— возбу-
дитель В. ака (татииаз). Очаги везикулезного риккетсиоза городского
типа описапы в цептральных районах Укранны, в США, в городах
на Атлантическом побережье (Риверс, 1955).
|
Источник п переносчик инфекции в очагах городского типа — гамазо-
вые клещи АПодегтапуззи$ запеитеиз и крысиный клещ О. Басой
(Кулагин, Земская, 1953). Потепциальная полифагия этих клещей при-
водит к их паразитированию на синантронных грызунах. в том числе
на серой крысе. Зараженность крыс (с большей долей вероятности мож-
но предположить, что серых крыс) в очаге в г. Донецке установлена
С. М. Кулагиным и С. А. Имамалиевым (1952).
Лихорадка цуцугамуши — возбудитель В. 1зшзиватазЕ.
Распространена на Дальнем Востоке, в Приморском крае и на Камчатке,
в страпах Юго-Восточной Азии, западной части Тихого океана, на севере
Австралии (Кулагин, Тарасевич, 1972. Тапцуа, 1962). Источник и пере-
носчик инфекции— личинки краспотелковых клещей родов Г.ербоготЬ-
Чит, МеотошЫ Чит и др. Мелкие млекопитающие, в том числе и серые
крысы, являются прокормителями краспотелок. Луди (Ап! 1968), изу-
чавший эволюцию очагов цуцугамуши, полагал, что Юго-Восточная
Азия была центром формировапия очагов этой инфекции, в которых
крысы играли главную роль. Серая крыса на севере ареала цуцугамуши
доминирует среди многих видов прокормителей краснотелок. По крайней
мере 10—50 или более личинок краснотелок ежедневно прокармливаются
на одной особи крысы. Естественная зараженность серой крысы В. 131-
{3исатисЬ1 установлена в Японии (Тапууа, 1962).
О-лихорадка (коксиеллез) — возбудитель СожеНа Ъитпей.
Гаспространена повсеместно. Основной источник инфекции — крупный и
мелкий рогатый скот, пути передачи возбудителя — респираторный, али-
ментарный, контактный, трансмиссивпый. Возбудитель выделяется во
внешнюю среду с околоплодными водами, мочой, фекалиями, молоком
животных, чрезвычайно устойчив к воздействию физических и химиче-
ских агентов, месяцами сохраняется на загрязненных выделениями жи-
вотных предметах.
Серая крыса может ипфицироваться вблизи помещений, где содер-
жится домашний скот. Естественная зараженность ее установлена в на-
птей стране в Крыму, в Азербайджане (Кулагин, Тарасевич, 1972). Анти-
тела к С. Багпей! найдены у серой крысы в Омской области. Естествен-
ная зараженность и антитела к С. Бигпей! обнаружены у пасюка в Туни-
се (Лишог её а|., 1980, цит. по: Федорова, 1968) н в ЧССР (ЗугасЕ
её а{., 1956; цит. по: Федорова, 1968).
Бактериозы
Чума (возбудитель чумпой микроб. Уегзица резиз) передается: от боль-
ного грызуна к здоровому главным образом трансмиссивпым путем, через
кровососущих насекомых— блох. Иногда возможно заражение грызунов
Практическое значение
345
респираторным путем при контакте зверьков друг с другом или при
копании ими почвы, зараженной микробами.
Природпые очаги чумы существуют на всех коптинентах мира, кроме
Австралии, но основная их масса нахоцится в Африке и Азии.
Для разных видов грызупов, носителей чумного микроба, характерны
специфические виды переносчиков — блохи. На серых крысах паразити-
руют преимущественно ХепорзШа сБеорз и -СегафюорВуЦиз {азс1абав.
Чумной микроб размножается в организме насекомых, при этом
у них образуется так называемый чумной блок преджелудка. Блокиро-
ванные блохи не могут нормально питаться и остаются голодными.
Поэтому они многократно кусают хозяина, пытаясь пасытиться, и зара-
жжают его чумой. Именно этот феномеп имеет колоссальное эпизоотоло-
гическое значение (Флегонтова, Малафеева, 1960; Бибикова, Класовский,
1975; и др.). У серых крыс часто бывает высокий паразитарный индекс,
и. в этом случае эпизоотия среди них быстро распространяется. Сами
зверьки не обладают высоцой чувствительностью к чуме, вернее сказать,
их степень чувствительности имеет большие индивидуальные вариа-
ции — одни 0с0би очень высоко чувствительны, другие резистентны
(Анисимова, 1957, 1959, 1962; Мамед-Заде и др., 1962). При эксперимен-
тальном заражении чумой у серых. крыс в большинстве случаев болезнь
протекает остро с бактериемией, которая может обеспечить заражение
большого количества блох. Возбудитель чумы начинает выделяться
У крыс после заражепия со второго по десятый день (Хвещенко, 1962,
1963). Показано, что менее чувствительны к чуме молодые крысы. Но
мпению А. П. Аписимовой (1962), существует изменение чувствитель-
ности популяции крыс в разные сезоны года, а именно: повышается опа
летом и зимой и становится меньше весной и осенью. У крыс возможно
хроническое носительство чумного микроба (Некипелов, 1959). Индий-
ские ученые показали возможность течения инфекции среди частично
иммунных грызупов в ипанараптной, или субклинической, форме (Зеа1,
1961).
.
В оргапизме крыс продуцируются антитела к чумному микробу в раз-
ных титрах, что свидетельствует о приобретенном иммунитете (Мамед-
Заде и др., 1962).
Роль серых крыс в эпидемиологии чумы была очень велика. Пассивно
распространяясь по земному нару, главным образом с морскими судами,
они способствовали завозу этой инфекции в портовые города.
В конце ХХ и начале ХХ в. с развитием международных морских
сцошений резко увеличилась вероятность завоза крыс в портовые города
мира. Зараженные чумой зверьки попали в порты Европы, Африки,
Америки, Азии и на Океанические острова. В Австралии очаги чумы
не’ укоренились.
.
Очаги чумы крысиного типа сформировались на Мадагаскаре, . Яве,
Гавайских островах, а также па юго-востоке Китая, во Вьетнаме, Бирме
и Тайланде. В подавляющем больитинстве случаев чума не. проникала
в глубину материков, а сосредоточивалась только в портовых городах.
С 1900 по 19% г. в порту Гамбурга с 6466 прибывающих из разных
ЗАА
Глава пятая
Везикулезный (оспоподобный) риккетсиоз-— возбу-
дитель В. ака (татииаз). Очаги везикулезного риккетсиоза городского
типа описапы в цептральных районах Укранны, в США, в городах
на Атлантическом побережье (Риверс, 1955).
|
Источник п переносчик инфекции в очагах городского типа — гамазо-
вые клещи АПодегтапуззи$ запеитеиз и крысиный клещ О. Басой
(Кулагин, Земская, 1953). Потепциальная полифагия этих клещей при-
водит к их паразитированию на синантронных грызунах. в том числе
на серой крысе. Зараженность крыс (с большей долей вероятности мож-
но предположить, что серых крыс) в очаге в г. Донецке установлена
С. М. Кулагиным и С. А. Имамалиевым (1952).
Лихорадка цуцугамуши — возбудитель В. 1зшзиватазЕ.
Распространена на Дальнем Востоке, в Приморском крае и на Камчатке,
в страпах Юго-Восточной Азии, западной части Тихого океана, на севере
Австралии (Кулагин, Тарасевич, 1972. Тапцуа, 1962). Источник и пере-
носчик инфекции— личинки краспотелковых клещей родов Г.ербоготЬ-
Чит, МеотошЫ Чит и др. Мелкие млекопитающие, в том числе и серые
крысы, являются прокормителями краспотелок. Луди (Ап! 1968), изу-
чавший эволюцию очагов цуцугамуши, полагал, что Юго-Восточная
Азия была центром формировапия очагов этой инфекции, в которых
крысы играли главную роль. Серая крыса на севере ареала цуцугамуши
доминирует среди многих видов прокормителей краснотелок. По крайней
мере 10—50 или более личинок краснотелок ежедневно прокармливаются
на одной особи крысы. Естественная зараженность серой крысы В. 131-
{3исатисЬ1 установлена в Японии (Тапууа, 1962).
О-лихорадка (коксиеллез) — возбудитель СожеНа Ъитпей.
Гаспространена повсеместно. Основной источник инфекции — крупный и
мелкий рогатый скот, пути передачи возбудителя — респираторный, али-
ментарный, контактный, трансмиссивпый. Возбудитель выделяется во
внешнюю среду с околоплодными водами, мочой, фекалиями, молоком
животных, чрезвычайно устойчив к воздействию физических и химиче-
ских агентов, месяцами сохраняется на загрязненных выделениями жи-
вотных предметах.
Серая крыса может ипфицироваться вблизи помещений, где содер-
жится домашний скот. Естественная зараженность ее установлена в на-
птей стране в Крыму, в Азербайджане (Кулагин, Тарасевич, 1972). Анти-
тела к С. Багпей! найдены у серой крысы в Омской области. Естествен-
ная зараженность и антитела к С. Бигпей! обнаружены у пасюка в Туни-
се (Лишог её а|., 1980, цит. по: Федорова, 1968) н в ЧССР (ЗугасЕ
её а{., 1956; цит. по: Федорова, 1968).
Бактериозы
Чума (возбудитель чумпой микроб. Уегзица резиз) передается: от боль-
ного грызуна к здоровому главным образом трансмиссивпым путем, через
кровососущих насекомых— блох. Иногда возможно заражение грызунов
Практическое значение
345
респираторным путем при контакте зверьков друг с другом или при
копании ими почвы, зараженной микробами.
Природпые очаги чумы существуют на всех коптинентах мира, кроме
Австралии, но основная их масса нахоцится в Африке и Азии.
Для разных видов грызупов, носителей чумного микроба, характерны
специфические виды переносчиков — блохи. На серых крысах паразити-
руют преимущественно ХепорзШа сБеорз и -СегафюорВуЦиз {азс1абав.
Чумной микроб размножается в организме насекомых, при этом
у них образуется так называемый чумной блок преджелудка. Блокиро-
ванные блохи не могут нормально питаться и остаются голодными.
Поэтому они многократно кусают хозяина, пытаясь пасытиться, и зара-
жжают его чумой. Именно этот феномеп имеет колоссальное эпизоотоло-
гическое значение (Флегонтова, Малафеева, 1960; Бибикова, Класовский,
1975; и др.). У серых крыс часто бывает высокий паразитарный индекс,
и. в этом случае эпизоотия среди них быстро распространяется. Сами
зверьки не обладают высоцой чувствительностью к чуме, вернее сказать,
их степень чувствительности имеет большие индивидуальные вариа-
ции — одни 0с0би очень высоко чувствительны, другие резистентны
(Анисимова, 1957, 1959, 1962; Мамед-Заде и др., 1962). При эксперимен-
тальном заражении чумой у серых. крыс в большинстве случаев болезнь
протекает остро с бактериемией, которая может обеспечить заражение
большого количества блох. Возбудитель чумы начинает выделяться
У крыс после заражепия со второго по десятый день (Хвещенко, 1962,
1963). Показано, что менее чувствительны к чуме молодые крысы. Но
мпению А. П. Аписимовой (1962), существует изменение чувствитель-
ности популяции крыс в разные сезоны года, а именно: повышается опа
летом и зимой и становится меньше весной и осенью. У крыс возможно
хроническое носительство чумного микроба (Некипелов, 1959). Индий-
ские ученые показали возможность течения инфекции среди частично
иммунных грызупов в ипанараптной, или субклинической, форме (Зеа1,
1961).
.
В оргапизме крыс продуцируются антитела к чумному микробу в раз-
ных титрах, что свидетельствует о приобретенном иммунитете (Мамед-
Заде и др., 1962).
Роль серых крыс в эпидемиологии чумы была очень велика. Пассивно
распространяясь по земному нару, главным образом с морскими судами,
они способствовали завозу этой инфекции в портовые города.
В конце ХХ и начале ХХ в. с развитием международных морских
сцошений резко увеличилась вероятность завоза крыс в портовые города
мира. Зараженные чумой зверьки попали в порты Европы, Африки,
Америки, Азии и на Океанические острова. В Австралии очаги чумы
не’ укоренились.
.
Очаги чумы крысиного типа сформировались на Мадагаскаре, . Яве,
Гавайских островах, а также па юго-востоке Китая, во Вьетнаме, Бирме
и Тайланде. В подавляющем больитинстве случаев чума не. проникала
в глубину материков, а сосредоточивалась только в портовых городах.
С 1900 по 19% г. в порту Гамбурга с 6466 прибывающих из разных
346
Глава пятая
концов мира судов были исследованы крысы на носительство чумы. Ока-
залось, что на 64 судах (т. е. в 1% случаев) были обпаружены зверьки,
заражеппые чумой.
В. Н. Федоров и В. П. Козакевич (1959) пишут: «Особенно широкий
обмен портовых городов мира зараженными чумой крысами происходил
в течение всей первой четверти текущего столетия. Об этом можно
судить по тому, что, например, в течение 1925 г. чума среди людей
была зарегистрирована в 59 портах. к пим относятся в Европе 14 портов,
в Азии —21, Африке— 14, Южной Америке —7, в Северной Амери-
ке —2ив Австралии — 1».
Такое интенсивное заселение крысами судов и портов объяснялось
в первую очередь исключительно скверным их санитарным состоянием
(Гамалея, 1903).
В нашей стране в 1901—1902 гг. вспыхнула эпидемия чумы в Одессе.
Источником заражения людей были крысы. Во время этой эпидемии
па судах вылавливали почти исключительно черных крыс, а в порту
и в городе преобладали серые крысы (Гамалея, 1903).
На созванной в Париже международной санитарлой конференции
(1903 г.) по ликвидации эпидемии чумы были разработаны меры про-
филактики портовой чумы, что в первую очередь сводилось к изысканию
эффективных методов уничтожения крыс, защите портовых сооружепий
и судов от заселения этим грызуном и к палаживанию удовлетворитель-
ного санитарного состояния всех территорий портов и портовых соору-
жений.
В связи с проведением в жизнь этих мероприятий число поражаемых
чумой портов резко сократилось.
Но в 1910 г. эпидемия в г. Одессе повторилась. Возбудитель чумы
был выделен от серых крыс, отловленных в жилых домах, на рынке
и в хлебных складах. Там же были зарегистрированы больные чумой
люди (Заболотный, 1912: Щастпый, 1912; Малиновский и др., 1912).
К 1945 г. заболевание людей чумой было зарегистрировано всего
в 22 портах, ав 1955 с.— только в 6.
Очаги чумы, в которых основную роль играют серые крысы, регист-
рируют и в настоящее время, главным образом в Юго-Восточной Азии.
В Ипдии в портовых городах многократно выявляли зараженных чумой
крыс (Малолетков, 1963). В Бирме одним из храпителей источников
инфекции для людей ‘являются также серые крысы (Вгоокз её а|.,
1977).
Заболевание чумой людей, связанное с заражением-от крыс, регистри-
ровали в 70-х годах в портах Ирака, Израиля и других стран Ближнего
Востока (Лавровский и др., 1973).
:
Очень велико эпидемиологическое зпачепие серой крысы в южной
части Вьетнама, особенно в приморских селениях. Несомненно, что это-
му способствовали военные действия, которые привели к ослаблению
профилактических мероприятий и к распространению ипфицированных
крыс по путям военных коммупикаций (Алиев и др., 1983).
Практическое значение
347
Эпизоотии чумы среди синантропных крыс представляют собой колос-
сальную эпидемиологическую опасность потому, что протекают непосред-
ственно в жилищах человека, что резко увеличивает вероятность зараже-
ния людей чумой. |
Кроме того, в ряде случаев серые крысы могут контактировать с гры-
зупами, населяющими природные очаги чумы, что еще в большей степе-
ни обусловливает эпидемиологическую опасность пасюка (Шилов и др.,
1986).
Комитет экспертов ВОЗ по чуме (1974) подчеркивал, что чума, сохра-
нявиаяся среди грызунов в различпых частях мира, и сейчас требует
пристального внимания со стороны медицинской службы и ВОЗ.
Псевдотуберкулез — возбудитель Тегзица рзепдоарегси10318.
У серых крыс имеет место непродолжительная латентная инфекция
с вовлечением в процесс только лимфатического аппарата кишечника.
В населенных пунктах зараженных серых крыс обпаруживают главпым
образом на плодоовощных базах, в холодильниках, животноводческих
фермах и в жилом секторе. Возбудитель псевдотуберкулеза накапливает-
ся па пищевых продуктах, загрязняемых выделениями инфицированных
пасюков, и размножается там. Заражение людей происходит при поеда-
нии пемытых продуктов, и заболеваемость носит характер вспышек
(Ющенко, 1964; Федоров и др., 1970; Сомов, 1979).
В СССР псевдотуберкулез у серых крыс впервые обнаружили
К. Н. Токаревич © соавторами (1940). В 1965 г. возбудитель инфекции
выделен из органов серой крысы, отловленной на Тбилисском мясоком-
бинате (Сакварелидзе и др., 1983). Псевдотуберкулез пасюков обнаружен
также в Риге, Ташкенте, Ереване, Мипгечауре и других местах
(табл. 29).
Эта инфекция среди крыс отмечена во многих странах Европы,
в США, Индии, Алжире (Ющенко, 1964). Так, в Польше при бактерио-
”`погическом исследовании 1184 серых крыс, отловленных на судах, в пор-
ту и на побережье Гданьского воеводства, изолирован 31 штамм.
Г. Н. Ющенко предполагает эпидемиологическую связь поражеп-
пых возбудителем исевдотуберкулеза серых крыс с заболеваниями этой
ипфенцией детей.
Возбудитель псевдотуберкулеза был обпаружеп у серых крыс и в при-
родных условиях, а именно по берегам рек и озер о-ва Сахалии.
В г. Владивостоке больниипство очагов псевдотуберкулеза находится на
окраинах города, что связывают с вероятным контактом серых крыс
с грызунами, живущими в природе, которые тоже играют определенную
„роль в формировании очагов этой болезни (Нацкий, Игнатович, 1970).
В. связи с тем что основной путь заражения людей алиментарный,
‚ антропургические очаги имеют большую эпидемиологическую валент-
ность, чем природные.
Кишечный иерсиниоз-— возбудитель Фегзина етцегосоН@са.
Возбудитель у крыс является условно-патогенным обитателем кишечни-
ка— заболевание и гибель животных наблюдаются лишь при резком
ослаблении организма. В антропургических очагах происходит циркуля-
346
Глава пятая
концов мира судов были исследованы крысы на носительство чумы. Ока-
залось, что на 64 судах (т. е. в 1% случаев) были обпаружены зверьки,
заражеппые чумой.
В. Н. Федоров и В. П. Козакевич (1959) пишут: «Особенно широкий
обмен портовых городов мира зараженными чумой крысами происходил
в течение всей первой четверти текущего столетия. Об этом можно
судить по тому, что, например, в течение 1925 г. чума среди людей
была зарегистрирована в 59 портах. к пим относятся в Европе 14 портов,
в Азии —21, Африке— 14, Южной Америке —7, в Северной Амери-
ке —2ив Австралии — 1».
Такое интенсивное заселение крысами судов и портов объяснялось
в первую очередь исключительно скверным их санитарным состоянием
(Гамалея, 1903).
В нашей стране в 1901—1902 гг. вспыхнула эпидемия чумы в Одессе.
Источником заражения людей были крысы. Во время этой эпидемии
па судах вылавливали почти исключительно черных крыс, а в порту
и в городе преобладали серые крысы (Гамалея, 1903).
На созванной в Париже международной санитарлой конференции
(1903 г.) по ликвидации эпидемии чумы были разработаны меры про-
филактики портовой чумы, что в первую очередь сводилось к изысканию
эффективных методов уничтожения крыс, защите портовых сооружепий
и судов от заселения этим грызуном и к палаживанию удовлетворитель-
ного санитарного состояния всех территорий портов и портовых соору-
жений.
В связи с проведением в жизнь этих мероприятий число поражаемых
чумой портов резко сократилось.
Но в 1910 г. эпидемия в г. Одессе повторилась. Возбудитель чумы
был выделен от серых крыс, отловленных в жилых домах, на рынке
и в хлебных складах. Там же были зарегистрированы больные чумой
люди (Заболотный, 1912: Щастпый, 1912; Малиновский и др., 1912).
К 1945 г. заболевание людей чумой было зарегистрировано всего
в 22 портах, ав 1955 с.— только в 6.
Очаги чумы, в которых основную роль играют серые крысы, регист-
рируют и в настоящее время, главным образом в Юго-Восточной Азии.
В Ипдии в портовых городах многократно выявляли зараженных чумой
крыс (Малолетков, 1963). В Бирме одним из храпителей источников
инфекции для людей ‘являются также серые крысы (Вгоокз её а|.,
1977).
Заболевание чумой людей, связанное с заражением-от крыс, регистри-
ровали в 70-х годах в портах Ирака, Израиля и других стран Ближнего
Востока (Лавровский и др., 1973).
:
Очень велико эпидемиологическое зпачепие серой крысы в южной
части Вьетнама, особенно в приморских селениях. Несомненно, что это-
му способствовали военные действия, которые привели к ослаблению
профилактических мероприятий и к распространению ипфицированных
крыс по путям военных коммупикаций (Алиев и др., 1983).
Практическое значение
347
Эпизоотии чумы среди синантропных крыс представляют собой колос-
сальную эпидемиологическую опасность потому, что протекают непосред-
ственно в жилищах человека, что резко увеличивает вероятность зараже-
ния людей чумой. |
Кроме того, в ряде случаев серые крысы могут контактировать с гры-
зупами, населяющими природные очаги чумы, что еще в большей степе-
ни обусловливает эпидемиологическую опасность пасюка (Шилов и др.,
1986).
Комитет экспертов ВОЗ по чуме (1974) подчеркивал, что чума, сохра-
нявиаяся среди грызунов в различпых частях мира, и сейчас требует
пристального внимания со стороны медицинской службы и ВОЗ.
Псевдотуберкулез — возбудитель Тегзица рзепдоарегси10318.
У серых крыс имеет место непродолжительная латентная инфекция
с вовлечением в процесс только лимфатического аппарата кишечника.
В населенных пунктах зараженных серых крыс обпаруживают главпым
образом на плодоовощных базах, в холодильниках, животноводческих
фермах и в жилом секторе. Возбудитель псевдотуберкулеза накапливает-
ся па пищевых продуктах, загрязняемых выделениями инфицированных
пасюков, и размножается там. Заражение людей происходит при поеда-
нии пемытых продуктов, и заболеваемость носит характер вспышек
(Ющенко, 1964; Федоров и др., 1970; Сомов, 1979).
В СССР псевдотуберкулез у серых крыс впервые обнаружили
К. Н. Токаревич © соавторами (1940). В 1965 г. возбудитель инфекции
выделен из органов серой крысы, отловленной на Тбилисском мясоком-
бинате (Сакварелидзе и др., 1983). Псевдотуберкулез пасюков обнаружен
также в Риге, Ташкенте, Ереване, Мипгечауре и других местах
(табл. 29).
Эта инфекция среди крыс отмечена во многих странах Европы,
в США, Индии, Алжире (Ющенко, 1964). Так, в Польше при бактерио-
”`погическом исследовании 1184 серых крыс, отловленных на судах, в пор-
ту и на побережье Гданьского воеводства, изолирован 31 штамм.
Г. Н. Ющенко предполагает эпидемиологическую связь поражеп-
пых возбудителем исевдотуберкулеза серых крыс с заболеваниями этой
ипфенцией детей.
Возбудитель псевдотуберкулеза был обпаружеп у серых крыс и в при-
родных условиях, а именно по берегам рек и озер о-ва Сахалии.
В г. Владивостоке больниипство очагов псевдотуберкулеза находится на
окраинах города, что связывают с вероятным контактом серых крыс
с грызунами, живущими в природе, которые тоже играют определенную
„роль в формировании очагов этой болезни (Нацкий, Игнатович, 1970).
В. связи с тем что основной путь заражения людей алиментарный,
‚ антропургические очаги имеют большую эпидемиологическую валент-
ность, чем природные.
Кишечный иерсиниоз-— возбудитель Фегзина етцегосоН@са.
Возбудитель у крыс является условно-патогенным обитателем кишечни-
ка— заболевание и гибель животных наблюдаются лишь при резком
ослаблении организма. В антропургических очагах происходит циркуля-
348 Глава нятая
Таблица 29. Выделение от серых крыс возбудителя песвдотуберкулеза
вСССР*
Число
Число ис- выделен-
Место
следован- ных
Автор. год
ных крые
культур
Москва
38 865
38
Ющенко, 1957
Сахалин
15 190
93
Сурков, Гайдукова, 1972
Владивосток
12 820
ТА
Якунина и др., 197%
Москва
7 930
Ющенко, 1976
Закарпатье
3816
30
Сакаль, 1976
Новгородекая область
22 145
28 . Покровская, 1980
Находка
2 801
4
Жиров и др., 1983
Нижнее Приамурье-
3882
47
Липаев, Бусоедова, 1983
* Выделение культур расположено в хронологическом поряяке.
ция возбудителя между домашпими животными, которые болетот перси-
пнозным энтероколитом.
Ведущая роль в передаче инфекций людям, очевидпо, припадлежит
инфицированным выделениям домашних животных и серых крыс. Зара-
жение людей происходит при поедании немытых продуктов. Наибольшая
интенсивность эпизоотии среди пасюков, например в Ленинграде, имеет
место в овощехранилищах, причем в холодное времл года (Лютов н др.,
1980; Ващенок и др., 1983).
В Японии при эпизоотологическом обследовании скотобойни установ-
лена высокая пораженность иерсиниозом туш свиней, при этом показа-
но, что выловленные здесь же серые крысы являются носителями возбу-
пителя. Вероятно, пасюки повинны во вторичном заражепии туп: и мяса
свиней, что способствует возпикповению среди людей заболеваний (Теп
Уой её а., 1974).
.
.
.
Роль серых крыс как хранителя возбудителя кишечного иерсиниоза
установлена в Нольше в порту в Гданьском воеводстве.
В СССР носительство возбудителя кишечного иерсиниоза среди серых
крыс установлено в Ленинграде (Лютов и др., 1980), Москве (Степано-
ва и др., 1986), Одессе (Андреева и др., 1983), на Чукотке (Сомов
и др., 1977; Шубин и др., 1978) и на Сахалине (Подболотов, Сурков,
1984).
.
Пастереллез — возбудитель РазегеПа тиЦос!:Ч9а. Многимн ис-
следователями установлено носительство возбудителя пастереллеза у се-
рых крыс. Т. Н. Попомарева и Л. В. Родкевич. (1964) при исследовании
55 342 серых крыс, отловленных в 1953—1961 гг. в Москве и ближнем
Подмосковье, выделили 107 культур возбудителя. Не их даниым, инфи-
цированность зверьков, поймапных на окраине города, в 4—5 раз выше,
чем в центральных районах. Она наиболее значительна среди грызунов
в пригородных животноводческих хозлйствах (4,9%), мясокомбинатах
Практическое значение
349
(3,1), открытых стациях (2.0%), в жилом секторе (1,7%). Пастерел-
лоносительство у серых крыс авторы наблюдали круглогодично с незпа-
чительным увеличением в зимний период. Г. В. Ющенко с соавторами
(1965), при анализе приведенных выше дапных, обратили внимание на
носительство пастерелл у серых крыс, отловленных также на железно-
дорожных и речпых вокзалах, в аэропортах.
Инфицированность возбудителем настереллеза серых крыс г. Лепин-
града, но данным исследований, проведенныхв 1961—1977 гг. (80 429 экз.),
составила 2,41 на 1000 особей и была выше, чем у других грызунов
(Лютов, 1980). На Сахалине Р. шаЦомЧа у серых крыс впервые изоли-
рована в начале 60-х годах (Климепко, Клименко, 1962). В. С. Сурков
(1971) подтвердил роль серых крыс как носителей возбудителя пасте-
реллеза на Сахалине и обосповал их большое эпидемиологическое зна-
чтение при этой инфекции, связанное с высокой численностью и заселен-
ностью различных объектов, а также с обитанием в поймах рек, на
побережье моря, в зарослях бамбука, на картофельных полях, откуда
происходит миграция грызунов в жилища человека.
Туляремия — возбудитель ЕгапезеМа иатепз1з (М. Соу её Сва-
рш). Серые крысы в очагах туляремии не играют существенной роли —
они относятся к группе высоковосприимчивых, но малочувствительных
к туляремии видов (Г труппа), для которых характерно доброкачест-
веннос тсечепие ипфекцин и формирование иммунитета, препятствующего
размножению возбудителя в их органах и ограничивающего этим воз-
можность передачи возбудителя через выделения кровососущим парази-
там или алиментарным путем (Айсели, 1951; Купресова, Левцов, 1951;
Олсуфьев, Дунаева, 1960).
.
Вследствие особенностей патогенеза туляремни у серых крыс оти
трызуны не могут служить источником заражения для других особей
своего и других видов. Обычно они заражаются туляремией от зверьков
других видов, высокочувствительных к ней.
Впервые туляремия у серых крыс обнаружена в 1924 г. в Калифор-
нии во время их обследования на зараженность чумой (Олефег, ВБодез,
1926).
.
В СССР пемногочисленные случаи вовлечения серых крыс в эпизо-
отии. туляремии в природных очагах отмечались преимущественно при
чрезвычайно высокой численности осповных хозяев инфекции (водяные
и обыкновенные полевки, домовые мыши). Контакт серых крыс с основ-
ными носителями происходит и в насслепных пунктах, куда в зимнее
время вселяются зверьки из природных очагов лугополевого и степного
тАпов.
В очагах пойменно-болотного типа серые крысы. могут постоянно кон-
тактировать с основными носителями туляремни — водяными полевками,
но заражение их регистрируется редко. Так, в дельте р. Допа, где серая
крыса постоянно встречается в открытых стациях, зараженность ее ту-
ляремией установлена лишь один раз в 1949 г., в то. время как у. водя-
ных половон эпизоотин обнаруживали ежегодно с 1949—1955 гг. (Вожен-
ко, 1957).
348 Глава нятая
Таблица 29. Выделение от серых крыс возбудителя песвдотуберкулеза
вСССР*
Число
Число ис- выделен-
Место
следован- ных
Автор. год
ных крые
культур
Москва
38 865
38
Ющенко, 1957
Сахалин
15 190
93
Сурков, Гайдукова, 1972
Владивосток
12 820
ТА
Якунина и др., 197%
Москва
7 930
Ющенко, 1976
Закарпатье
3816
30
Сакаль, 1976
Новгородекая область
22 145
28 . Покровская, 1980
Находка
2 801
4
Жиров и др., 1983
Нижнее Приамурье-
3882
47
Липаев, Бусоедова, 1983
* Выделение культур расположено в хронологическом поряяке.
ция возбудителя между домашпими животными, которые болетот перси-
пнозным энтероколитом.
Ведущая роль в передаче инфекций людям, очевидпо, припадлежит
инфицированным выделениям домашних животных и серых крыс. Зара-
жение людей происходит при поедании немытых продуктов. Наибольшая
интенсивность эпизоотии среди пасюков, например в Ленинграде, имеет
место в овощехранилищах, причем в холодное времл года (Лютов н др.,
1980; Ващенок и др., 1983).
В Японии при эпизоотологическом обследовании скотобойни установ-
лена высокая пораженность иерсиниозом туш свиней, при этом показа-
но, что выловленные здесь же серые крысы являются носителями возбу-
пителя. Вероятно, пасюки повинны во вторичном заражепии туп: и мяса
свиней, что способствует возпикповению среди людей заболеваний (Теп
Уой её а., 1974).
.
.
.
Роль серых крыс как хранителя возбудителя кишечного иерсиниоза
установлена в Нольше в порту в Гданьском воеводстве.
В СССР носительство возбудителя кишечного иерсиниоза среди серых
крыс установлено в Ленинграде (Лютов и др., 1980), Москве (Степано-
ва и др., 1986), Одессе (Андреева и др., 1983), на Чукотке (Сомов
и др., 1977; Шубин и др., 1978) и на Сахалине (Подболотов, Сурков,
1984).
.
Пастереллез — возбудитель РазегеПа тиЦос!:Ч9а. Многимн ис-
следователями установлено носительство возбудителя пастереллеза у се-
рых крыс. Т. Н. Попомарева и Л. В. Родкевич. (1964) при исследовании
55 342 серых крыс, отловленных в 1953—1961 гг. в Москве и ближнем
Подмосковье, выделили 107 культур возбудителя. Не их даниым, инфи-
цированность зверьков, поймапных на окраине города, в 4—5 раз выше,
чем в центральных районах. Она наиболее значительна среди грызунов
в пригородных животноводческих хозлйствах (4,9%), мясокомбинатах
Практическое значение
349
(3,1), открытых стациях (2.0%), в жилом секторе (1,7%). Пастерел-
лоносительство у серых крыс авторы наблюдали круглогодично с незпа-
чительным увеличением в зимний период. Г. В. Ющенко с соавторами
(1965), при анализе приведенных выше дапных, обратили внимание на
носительство пастерелл у серых крыс, отловленных также на железно-
дорожных и речпых вокзалах, в аэропортах.
Инфицированность возбудителем настереллеза серых крыс г. Лепин-
града, но данным исследований, проведенныхв 1961—1977 гг. (80 429 экз.),
составила 2,41 на 1000 особей и была выше, чем у других грызунов
(Лютов, 1980). На Сахалине Р. шаЦомЧа у серых крыс впервые изоли-
рована в начале 60-х годах (Климепко, Клименко, 1962). В. С. Сурков
(1971) подтвердил роль серых крыс как носителей возбудителя пасте-
реллеза на Сахалине и обосповал их большое эпидемиологическое зна-
чтение при этой инфекции, связанное с высокой численностью и заселен-
ностью различных объектов, а также с обитанием в поймах рек, на
побережье моря, в зарослях бамбука, на картофельных полях, откуда
происходит миграция грызунов в жилища человека.
Туляремия — возбудитель ЕгапезеМа иатепз1з (М. Соу её Сва-
рш). Серые крысы в очагах туляремии не играют существенной роли —
они относятся к группе высоковосприимчивых, но малочувствительных
к туляремии видов (Г труппа), для которых характерно доброкачест-
веннос тсечепие ипфекцин и формирование иммунитета, препятствующего
размножению возбудителя в их органах и ограничивающего этим воз-
можность передачи возбудителя через выделения кровососущим парази-
там или алиментарным путем (Айсели, 1951; Купресова, Левцов, 1951;
Олсуфьев, Дунаева, 1960).
.
Вследствие особенностей патогенеза туляремни у серых крыс оти
трызуны не могут служить источником заражения для других особей
своего и других видов. Обычно они заражаются туляремией от зверьков
других видов, высокочувствительных к ней.
Впервые туляремия у серых крыс обнаружена в 1924 г. в Калифор-
нии во время их обследования на зараженность чумой (Олефег, ВБодез,
1926).
.
В СССР пемногочисленные случаи вовлечения серых крыс в эпизо-
отии. туляремии в природных очагах отмечались преимущественно при
чрезвычайно высокой численности осповных хозяев инфекции (водяные
и обыкновенные полевки, домовые мыши). Контакт серых крыс с основ-
ными носителями происходит и в насслепных пунктах, куда в зимнее
время вселяются зверьки из природных очагов лугополевого и степного
тАпов.
В очагах пойменно-болотного типа серые крысы. могут постоянно кон-
тактировать с основными носителями туляремни — водяными полевками,
но заражение их регистрируется редко. Так, в дельте р. Допа, где серая
крыса постоянно встречается в открытых стациях, зараженность ее ту-
ляремией установлена лишь один раз в 1949 г., в то. время как у. водя-
ных половон эпизоотин обнаруживали ежегодно с 1949—1955 гг. (Вожен-
ко, 1957).
350
Глава пятая
Известны случаи обнаружения туляремии у серых крыс на мясоком-
бипате, на котором регистрировали заболевания людей при убое и раздел-
ке туш овец (Терещенко и др., 1956; Овасапян, 1957).
Таким образом, заражепие насюков туляремией носит случайный ха-
рактер и связано с развитием эпизоотий у грызунов других видов, высо-
кочувствительных к этой ипфекции.
Бруцеллез
— возбудители РгисеПа ше]Иепз!з, В. афомаз. Роль се-
рых крыс в очагах бруцеллеза до сих пор окопчательно не выяснена.
Восприимчивость к возбудителям этой болезни установлена более чем
у 60 видов позвоночных, в том числе у серых крыс, однако значение по-
следних в эпизоотологии бруцеллеза изучено мало.
.
И. А. Каркадиновская (1936), А. В. Панкратов (1939) и другие авто-
ры полагают, что крысы заражаются преимущественно при поедании кор-
ма, загрязненного послеродовыми выделениями сельскохозяйственных
животных, и, следовательно, они могут служить одним из факторов рас-
пространения бруцеллеза среди людей и скота. Подтверждением этого
служит выделение от пасюка культур В. афогиз.
Есть данные о выделении В. шеШепз15 от серых крыс в овцеводче-
ском хозяйстве в период абортов и массовых окотов овец и об отсутствии
ипфицированных зверьков в помещениях, где содержали здоровый скот
{Кучерова, 1957, 1959).
Несмотря на восприимчивость серых крыс к бруцеллезу, инфекцион-
ный процесс среди них длится недолго, следовательно. они только корот-
кое время выделяют возбудитель во внешнюю среду. Поэтому бруцеллез-
пая инфекция в популяциях крыс пе может протекать длительно
(быстро паступает выздоровление), следонательно, эпизоотологическая и
эпидемическая роль их очень певелика (Баландин, 1958; Миронов, 1964).
Однако есть и другая точка зрения. Н. И. Тихенко с соавторами (1982)
считают, что эпизоотия среди крыс способствует длительному сохране-
нию возбудителя во внешней среде, что приводит к заражению сельско-
хозяйственных животных. Культуры бруцелл у крыс удается чаще выде-
лять из крови и песколько реже из мочи.
Патолого-анатомические изменения У пасюков, зараженных В. тет}
1еп313, сводятся к увеличепию лимфатических желез, селезенки и появ-
лению в печени некротических узлов (Алтарева, 1952: Вершилова и др.,
1959).
Очевидно, что для выяснения истинной роли серых крыс в эпизоото-
лотии этой болезни среди сельскохозяйственпых животных и в эпидемио-
логии необходимы дальнейшие исследования.
.
Листериоз — возбудитель Мена топосуюет15. Серые крысы
часто явллются носителями возбудителя листериоза, но сами к нему
устойчивы. Полную смертельную дозу для пих устаповить не удалось,
а единичные экземпляры погибали лишь при заражении 10 млрд микроб-
ных клеток.
Зараженные зверьки выделяют листерии с мочой и калом, но больше
всего с носовым секретом. После заражения ипфицированппым калом они
выделяют листерии не только в течение заболевания, но и на протяжении
Практическое значение 351
Таблица 30. Выделение от серых крыс культуры возбудителя
листериоза в СССР
Число
Число ис- получен-
Место обнаружения
следован- ных
Автор, год
ных крыс
культур
Уссурийск
379
2
Хвещенко и др., 1956
Киев
—
33
Мороз, Хижинская, 1959
Москва
55 342
83
Огнева, 1961, 1964
Владивоеток
2 086
Михайловна, Якунина, 1962
7
Ярославская обл.
_
4
Малахов, 1962, 1964
еще 12 дней после клинического выздоровления (Триполитова и др.,
1965).
При вскрытии у больных крыс отмечают геморрагические очаги в лег-
ких, воспалительные изменепия в печени и некротические фокусы в селе-
зепке (Соломкин, 1959).
Среди крыс инфекция передается трансмиссивным, но чаще алимен-
тарным путем. В этом случае у них развивается генерализованная или
кишечная форма инфекционного процесса (Огнева, 1964).
Зараженных листериозом крыс отлавливают на окраинах города, на
млсокомбинатах, в пригородных совхозах (Терещенко, 1955; Терещенко
и др., 1957). Предполагают, что, возможно. они заражаются от домовых
мышей (Малахов, 1962, 1964) и обыкновениых полевок, формирующих
природные очаги этой инфекции (Огнева, 1961, 1964).
„Листериозньые эпизоотии среди крыс установлены во мпогих райопах
` и, городах пашей страны (табл. 30).
В последние годы проводят интенсивные. исследования крыс на листе-
риоз в Москве. Показано, что с 197) по 1984 г. выделение возбудителя
от них составило: 0,05% (Степанова и др., 1986). В Ленинграде заражен-
ность пасюков. примерно такая же — 0,24 на 1000 исследованных (Лютов
и др., 1980).
Сальмонеллезы — возбудители сальмопеллы, которые подразде-
ляются па ряд серологических вариантов. Серые крысы чаще всего носят
За топеЦа пат13 и 5. ещегИ 413.
носительстве сальмонелл пасюками в советской литературе впервые
сообщили Н. Н.. Клодницкий и Л. Я. Кац (1936). Инфицированность
зверьков может быть очень высока и на территории СССР в отдельные
тоды" составляет более’ 50% (Никитюк, 1970) (табл. 31). Зараженных
‘сальмонеллезами серых крыс чаще всего отлавливали в совхозах на ок-
раинах города и на пищевых объектах (Троп, Чуловский, 1960)..
Д. В. Поярков и Н. Н. Пояркова (1967) приводят данные о высокой
зараженности серых крыс в Москве (до. 34%) в парниках и на полях
фильтрации.
350
Глава пятая
Известны случаи обнаружения туляремии у серых крыс на мясоком-
бипате, на котором регистрировали заболевания людей при убое и раздел-
ке туш овец (Терещенко и др., 1956; Овасапян, 1957).
Таким образом, заражепие насюков туляремией носит случайный ха-
рактер и связано с развитием эпизоотий у грызунов других видов, высо-
кочувствительных к этой ипфекции.
Бруцеллез
— возбудители РгисеПа ше]Иепз!з, В. афомаз. Роль се-
рых крыс в очагах бруцеллеза до сих пор окопчательно не выяснена.
Восприимчивость к возбудителям этой болезни установлена более чем
у 60 видов позвоночных, в том числе у серых крыс, однако значение по-
следних в эпизоотологии бруцеллеза изучено мало.
.
И. А. Каркадиновская (1936), А. В. Панкратов (1939) и другие авто-
ры полагают, что крысы заражаются преимущественно при поедании кор-
ма, загрязненного послеродовыми выделениями сельскохозяйственных
животных, и, следовательно, они могут служить одним из факторов рас-
пространения бруцеллеза среди людей и скота. Подтверждением этого
служит выделение от пасюка культур В. афогиз.
Есть данные о выделении В. шеШепз15 от серых крыс в овцеводче-
ском хозяйстве в период абортов и массовых окотов овец и об отсутствии
ипфицированных зверьков в помещениях, где содержали здоровый скот
{Кучерова, 1957, 1959).
Несмотря на восприимчивость серых крыс к бруцеллезу, инфекцион-
ный процесс среди них длится недолго, следовательно. они только корот-
кое время выделяют возбудитель во внешнюю среду. Поэтому бруцеллез-
пая инфекция в популяциях крыс пе может протекать длительно
(быстро паступает выздоровление), следонательно, эпизоотологическая и
эпидемическая роль их очень певелика (Баландин, 1958; Миронов, 1964).
Однако есть и другая точка зрения. Н. И. Тихенко с соавторами (1982)
считают, что эпизоотия среди крыс способствует длительному сохране-
нию возбудителя во внешней среде, что приводит к заражению сельско-
хозяйственных животных. Культуры бруцелл у крыс удается чаще выде-
лять из крови и песколько реже из мочи.
Патолого-анатомические изменения У пасюков, зараженных В. тет}
1еп313, сводятся к увеличепию лимфатических желез, селезенки и появ-
лению в печени некротических узлов (Алтарева, 1952: Вершилова и др.,
1959).
Очевидно, что для выяснения истинной роли серых крыс в эпизоото-
лотии этой болезни среди сельскохозяйственпых животных и в эпидемио-
логии необходимы дальнейшие исследования.
.
Листериоз — возбудитель Мена топосуюет15. Серые крысы
часто явллются носителями возбудителя листериоза, но сами к нему
устойчивы. Полную смертельную дозу для пих устаповить не удалось,
а единичные экземпляры погибали лишь при заражении 10 млрд микроб-
ных клеток.
Зараженные зверьки выделяют листерии с мочой и калом, но больше
всего с носовым секретом. После заражения ипфицированппым калом они
выделяют листерии не только в течение заболевания, но и на протяжении
Практическое значение 351
Таблица 30. Выделение от серых крыс культуры возбудителя
листериоза в СССР
Число
Число ис- получен-
Место обнаружения
следован- ных
Автор, год
ных крыс
культур
Уссурийск
379
2
Хвещенко и др., 1956
Киев
—
33
Мороз, Хижинская, 1959
Москва
55 342
83
Огнева, 1961, 1964
Владивоеток
2 086
Михайловна, Якунина, 1962
7
Ярославская обл.
_
4
Малахов, 1962, 1964
еще 12 дней после клинического выздоровления (Триполитова и др.,
1965).
При вскрытии у больных крыс отмечают геморрагические очаги в лег-
ких, воспалительные изменепия в печени и некротические фокусы в селе-
зепке (Соломкин, 1959).
Среди крыс инфекция передается трансмиссивным, но чаще алимен-
тарным путем. В этом случае у них развивается генерализованная или
кишечная форма инфекционного процесса (Огнева, 1964).
Зараженных листериозом крыс отлавливают на окраинах города, на
млсокомбинатах, в пригородных совхозах (Терещенко, 1955; Терещенко
и др., 1957). Предполагают, что, возможно. они заражаются от домовых
мышей (Малахов, 1962, 1964) и обыкновениых полевок, формирующих
природные очаги этой инфекции (Огнева, 1961, 1964).
„Листериозньые эпизоотии среди крыс установлены во мпогих райопах
` и, городах пашей страны (табл. 30).
В последние годы проводят интенсивные. исследования крыс на листе-
риоз в Москве. Показано, что с 197) по 1984 г. выделение возбудителя
от них составило: 0,05% (Степанова и др., 1986). В Ленинграде заражен-
ность пасюков. примерно такая же — 0,24 на 1000 исследованных (Лютов
и др., 1980).
Сальмонеллезы — возбудители сальмопеллы, которые подразде-
ляются па ряд серологических вариантов. Серые крысы чаще всего носят
За топеЦа пат13 и 5. ещегИ 413.
носительстве сальмонелл пасюками в советской литературе впервые
сообщили Н. Н.. Клодницкий и Л. Я. Кац (1936). Инфицированность
зверьков может быть очень высока и на территории СССР в отдельные
тоды" составляет более’ 50% (Никитюк, 1970) (табл. 31). Зараженных
‘сальмонеллезами серых крыс чаще всего отлавливали в совхозах на ок-
раинах города и на пищевых объектах (Троп, Чуловский, 1960)..
Д. В. Поярков и Н. Н. Пояркова (1967) приводят данные о высокой
зараженности серых крыс в Москве (до. 34%) в парниках и на полях
фильтрации.
352 Глава пятая
Таблица 31. Выделенные от серых крые сальмонеллы в СССР
Число
Чи-
иесследо- | сло
Место обнаружения
ванных штам- Ссровар
Автор, год
крыс
мов
Пенинград
36 222 36
_
Гребенчук и др., 1965
Москва
7930 27
24.000
19
Поярков, Пояркова,
1967
Сахалин
25 093
8
Дзюбан и др., 1983
Нижнее Приамурье
2782
т
_
Пипаев, Бусоедова,
1983
Владивосток
257 | 6 |Различные
Якунина и др., 197%
|
.
`
`
серовары
Тбилиси
1547
16 |То же
Алания и др., 1986
Эризицелоинд — широко распространенпиое инфекционное заболе-
вание, вызываемое палочкой Егузре!о Вых тразорайсае и нередаваемое
человеку как домашними, так и дикими животными. Но биологическим и
серологическим свойствам заболевание может быть разделено на два ва-
рианта — эризипелоид свиной (уаг. 113) и мышиный (уаг. шие
зер ыса).
Серая крыса— это основной- носитель инфекции пе только в антро-
пургических очагах городского (мясокомбинаты, холодильники, зоопарки)
и сельского (животноводческие фермы) типов, но и в природпых очагах
(Мясников, 1973; Мясников, Белова, 1978). Сам зверек малочувствите-
лен к этой инфекции и не погибает даже при подкожном введении
1 млрд микробных клеток и более.
:
,
Крысы, больные эризипелоидом, выделяют возбудитель с мочой и ка-
лом. Не исключена возможность их заражения и при каппибализме (Мар-
тиновский, 19641, цит. по: Мясников, 1973), а также через кровососущих
членистоногих, у которых известны многочисленные находки возбудите-
ля. Широко распространено носительство эризипелотриксов в зеве (Ова-
сапян и др., 1964; Новиков и др., 1964, цит. по: Мясников, 1973).
Т. Н. Нономарева с соавторами (1962, цит. но: Мясников, 1973) выделя-
ла возбудитель из зева серых крыс в 7 раз чаще, чем из внутренних
органов.
.
Г. Д. Вилявин (1955, цит. по: Мясников, 1973) первый обратил вни-
мание на то, что на мясокомбинатах люди заражаются не столько от
мяса больных свиней, сколько от млса, обсемененного возбудителями
эризипелойда, занесенными через выделения грызунов, в основпом серых
крыс.
В зоопарках эта ипфекция становится настоящим бедствием, поражая
многие виды ценных млекопитающих и птиц. Разносчиками здесь .явля-
ются серые крысы, свободпо проникающие в клетки и вольеры. В Мос-
козском зоопарке зараженных крыс было 2% (Корчевская, 1946, цит. по:
Практическое значение
353
Мясников, 1973). В центре города пораженпость зверьков была меньше
(0,4%), чем на окраинах, 1,8% (Родкевич и др., 1955, цит. по: Мяени-
ков, 1978). О. В. Овасапян с соавторами (1964) пашли в Армении среди
крыс сельской местности 3,4% зараженпых, а среди зверьков, отловлен-
ных на мясокомбинате,— 346%.
Крысы, помимо проиэводственных {мясокомбинаты, холодильники)
и сельскохозяйственных (животноводческие фермы), вызывают также и
бытовые заражения (уборка помещений, приготовление пищи из инфи-
цированных крысами продуктов). Так, по данным М. А. Беловой (1972),
из 1723 обследованных ею случаев заболевания эризипелоидом рабочие
мясокомбинатов составили всего 14%, тогда как на рабочих предприятий,
не связанных с переработкой животного сырья, приходилось 34%, а на
домашних хозяев — 28% заболеваний.
у
В СССР от серых крые выделяли эризинелотрикс в Москве. Ленин-
граде, „енинтрадской, Ивановской, Вологодской и других областях
РСФСР, а также в Армении, Азербайдекане, на Украине (Мясников, Бе-
лова, 1978).
‚ Лептоспирозы, возбудители — патогенные лептоспиры. цирку-
лирующие в популяциях диких и домашних животных. Лептоспиры объ-
единены в ряд серогрупп, полразделяющихся па серовары. Серая крыса
связана с носительством лептоспир серогруппы Гееговаетогаае п
обычно в’очагах, обусловленных этим возбудителем, бывает основным
носителем. Полученные от нее культуры лептоспир неоднородны. Иден-
тификация штаммов до уровня серовара, проведенная с помощью много-
факторных сывороток, к сероварам СорепВакет и Теегопаетоттв аз1ае
показала, что у нас в стране наиболее часто встречаются У крыс лепто-
спиры Сореппазеп: (ервинская, 1968; Ананьина и др., 1987).
- Возбудитель вызывает у людей лептоспироз под названием болезнь
Васильева—Вейля, которая характеризуется особенно тяжелой КЛИНИКОЙ
‚ .И нередкими летальными исходами (Гольдентитейн, 1976; Антопова и др.,
1979; Бернасовская и др., 1986).
За последние годы во многих странах: в СССР, в Великобритании,
Италии, Франции, Румынии и других— сильно возросла доля этой 6бо-
лезни в общей структуре заболеваемости людей лентоспирозами (Берна-
совская, Копдратенко, #987; Ананьипа и др., 1987). В связи с этим мы
уделяем описанию роли серых крыс в этой болезни паибольшее вниманис.
Среди серых крыс эпизоотии иктерогеморрагического лептоспироза
часто протекают очень интенсивпо. Процент заражепиых зверьков дохо-
дит до 50—60: (Карасева, Свешникова, 1987). Такая высокая папряжен-
нобчь инфекционного процесса определяется больной восприимчивостью
пасюков к заражению лептоспирами Тсбегонаетогтваяае (ЗсВайтог,
Киепеп, 1923; Тыегтап, 1981). Лентосниры размножаются в корковом
. слое почек зверьков и эвакуируются наружу вместе с мочой. Эти микро-
бы — типичные гидробионты— попав на сухой субстрат, быстро гибнут,
но во влажной среде могут пекоторое время существовать, не теряя
свосй подвижности и патогенных свойств. Осповной путь передачи инфек-
ций от 060би к особи
— водный или контактный (Ананьин, Карасева,
1/212 Серая крыса
352 Глава пятая
Таблица 31. Выделенные от серых крые сальмонеллы в СССР
Число
Чи-
иесследо- | сло
Место обнаружения
ванных штам- Ссровар
Автор, год
крыс
мов
Пенинград
36 222 36
_
Гребенчук и др., 1965
Москва
7930 27
24.000
19
Поярков, Пояркова,
1967
Сахалин
25 093
8
Дзюбан и др., 1983
Нижнее Приамурье
2782
т
_
Пипаев, Бусоедова,
1983
Владивосток
257 | 6 |Различные
Якунина и др., 197%
|
.
`
`
серовары
Тбилиси
1547
16 |То же
Алания и др., 1986
Эризицелоинд — широко распространенпиое инфекционное заболе-
вание, вызываемое палочкой Егузре!о Вых тразорайсае и нередаваемое
человеку как домашними, так и дикими животными. Но биологическим и
серологическим свойствам заболевание может быть разделено на два ва-
рианта — эризипелоид свиной (уаг. 113) и мышиный (уаг. шие
зер ыса).
Серая крыса— это основной- носитель инфекции пе только в антро-
пургических очагах городского (мясокомбинаты, холодильники, зоопарки)
и сельского (животноводческие фермы) типов, но и в природпых очагах
(Мясников, 1973; Мясников, Белова, 1978). Сам зверек малочувствите-
лен к этой инфекции и не погибает даже при подкожном введении
1 млрд микробных клеток и более.
:
,
Крысы, больные эризипелоидом, выделяют возбудитель с мочой и ка-
лом. Не исключена возможность их заражения и при каппибализме (Мар-
тиновский, 19641, цит. по: Мясников, 1973), а также через кровососущих
членистоногих, у которых известны многочисленные находки возбудите-
ля. Широко распространено носительство эризипелотриксов в зеве (Ова-
сапян и др., 1964; Новиков и др., 1964, цит. по: Мясников, 1973).
Т. Н. Нономарева с соавторами (1962, цит. но: Мясников, 1973) выделя-
ла возбудитель из зева серых крыс в 7 раз чаще, чем из внутренних
органов.
.
Г. Д. Вилявин (1955, цит. по: Мясников, 1973) первый обратил вни-
мание на то, что на мясокомбинатах люди заражаются не столько от
мяса больных свиней, сколько от млса, обсемененного возбудителями
эризипелойда, занесенными через выделения грызунов, в основпом серых
крыс.
В зоопарках эта ипфекция становится настоящим бедствием, поражая
многие виды ценных млекопитающих и птиц. Разносчиками здесь .явля-
ются серые крысы, свободпо проникающие в клетки и вольеры. В Мос-
козском зоопарке зараженных крыс было 2% (Корчевская, 1946, цит. по:
Практическое значение
353
Мясников, 1973). В центре города пораженпость зверьков была меньше
(0,4%), чем на окраинах, 1,8% (Родкевич и др., 1955, цит. по: Мяени-
ков, 1978). О. В. Овасапян с соавторами (1964) пашли в Армении среди
крыс сельской местности 3,4% зараженпых, а среди зверьков, отловлен-
ных на мясокомбинате,— 346%.
Крысы, помимо проиэводственных {мясокомбинаты, холодильники)
и сельскохозяйственных (животноводческие фермы), вызывают также и
бытовые заражения (уборка помещений, приготовление пищи из инфи-
цированных крысами продуктов). Так, по данным М. А. Беловой (1972),
из 1723 обследованных ею случаев заболевания эризипелоидом рабочие
мясокомбинатов составили всего 14%, тогда как на рабочих предприятий,
не связанных с переработкой животного сырья, приходилось 34%, а на
домашних хозяев — 28% заболеваний.
у
В СССР от серых крые выделяли эризинелотрикс в Москве. Ленин-
граде, „енинтрадской, Ивановской, Вологодской и других областях
РСФСР, а также в Армении, Азербайдекане, на Украине (Мясников, Бе-
лова, 1978).
‚ Лептоспирозы, возбудители — патогенные лептоспиры. цирку-
лирующие в популяциях диких и домашних животных. Лептоспиры объ-
единены в ряд серогрупп, полразделяющихся па серовары. Серая крыса
связана с носительством лептоспир серогруппы Гееговаетогаае п
обычно в’очагах, обусловленных этим возбудителем, бывает основным
носителем. Полученные от нее культуры лептоспир неоднородны. Иден-
тификация штаммов до уровня серовара, проведенная с помощью много-
факторных сывороток, к сероварам СорепВакет и Теегопаетоттв аз1ае
показала, что у нас в стране наиболее часто встречаются У крыс лепто-
спиры Сореппазеп: (ервинская, 1968; Ананьина и др., 1987).
- Возбудитель вызывает у людей лептоспироз под названием болезнь
Васильева—Вейля, которая характеризуется особенно тяжелой КЛИНИКОЙ
‚ .И нередкими летальными исходами (Гольдентитейн, 1976; Антопова и др.,
1979; Бернасовская и др., 1986).
За последние годы во многих странах: в СССР, в Великобритании,
Италии, Франции, Румынии и других— сильно возросла доля этой 6бо-
лезни в общей структуре заболеваемости людей лентоспирозами (Берна-
совская, Копдратенко, #987; Ананьипа и др., 1987). В связи с этим мы
уделяем описанию роли серых крыс в этой болезни паибольшее вниманис.
Среди серых крыс эпизоотии иктерогеморрагического лептоспироза
часто протекают очень интенсивпо. Процент заражепиых зверьков дохо-
дит до 50—60: (Карасева, Свешникова, 1987). Такая высокая папряжен-
нобчь инфекционного процесса определяется больной восприимчивостью
пасюков к заражению лептоспирами Тсбегонаетогтваяае (ЗсВайтог,
Киепеп, 1923; Тыегтап, 1981). Лентосниры размножаются в корковом
. слое почек зверьков и эвакуируются наружу вместе с мочой. Эти микро-
бы — типичные гидробионты— попав на сухой субстрат, быстро гибнут,
но во влажной среде могут пекоторое время существовать, не теряя
свосй подвижности и патогенных свойств. Осповной путь передачи инфек-
ций от 060би к особи
— водный или контактный (Ананьин, Карасева,
1/212 Серая крыса
354
Глава пятая
1961). Заражению лептоспирозом здоровых крыс способствует их привер-
жениость к сырым местам обитания.
..
|
Характер поведения крыс также благоприятствует течепию инфекци-
онного процесса в популяции. Обычио крысы эвакуируют мочу 2—3 раза
в сутки, но доминант, стремясь пометить большую часть территории,
мочится гораздо чаще — до 4—5 раз в час; следовательно, если он лепто-
спироноситель, то широко рассеивает инфекцию (Карасева, 1983).
После заражения лентоспиры появляются в моче грызунов к концу вто-
рой или началу третьей недели (Карасева, Свешпикова, 1971). Чем стар-
ше крысы, тем больше среди них носителеи лентоспир (Амосенкова, Но-
пова, 1954; Карасева и др., 1986).
аи
.
Лептоспироз у пасюков протекает хронически, и опи.остаются носите-
лями лентоспир в течение всей своей жизни ,(Ва\!з, 1951а; Карасева
и др., 1986).
о
,:
Зараженность их лептоспирозом, вызываемым лентоспирами Тсеговае-
таоттВаз1ае, зарегистрирована почти во всем мире (на всех коптинентах,
кроме Антарктиды) и документировапа выделением культур. Однако,
кроме этих лептоспир, наиболее часто выделяемых от серых крыс и име-
ющих наибольшее эпидемиологическое значение, пасюки. бывают носите-
лями лептоспир и других серотрупи. В Европе. они иногда посят лепто-
спир серогруппы Ротопа, Вано, Небботад13, Вафамае..и; др. (Азот,
Вгоот, 1958). В Юго-Восточной Азии выделяются регионы (острова у-
матра, Язва, Сулавеси, Филиппины, а также Тайланд, Вьетнам), где-кры-
сы носят лептоспир серогруппы Вабам!ае и являются их осиовными носи-
телями (Торзт её а|., 1970; Петров, 1983). В Новой Зеландии показана
высокая инфицированность серых крыс лептоснирами серогруппы ВаПам
и устаповлена взаимосвязь зараженности пасюхов
с; плотностью их насе-
ления. Однако здесь основными носителями лептоснир серогруппы ВаПам
являются не серые, а черные крысы (Нафамалу,'В1аскитоте, 1981).
Эпизоотии лептоспироза встречаются как в. нашей стране, так и за
рубежом среди синантропных и экзоантропных крыс, образуя соответст-
венно антропургические и природные очаги... 2‘. °
—.
Города. Особенно много лептоспироносителей ‘обнаруживали в порто-
вых городах Европы — Лиссабоне, Копенгагене и. др (Ао её а1.,
1958). Много их было и в Израиле (Ноедеп уап'4ег; 4964, 1967). |
Вместе с судами крыс завезли в Африку. Они транспортировали' туда
п лентоспир, и в ряде портовых городов среди них обнаружены эпизоотии
лептоспироза (Тунис, Дакар и др.) (Ве. 1958). Зараженпых зверьков
там отлавливали главным образом в местах- их скопления: на рынках,
бойнях, в канализациях, на свалках.
ие.
.
В настоящее время среди крыс, обитающих во многих городах мира,
копстатируют, главным образом в жилых кварталах; этизоотии лептоспи-
роза. В восточных районах г. Детройта, например, ‘было _отловлено
358 серых крыс и культуры серогруины Теуеговастоггна1ае выделены
в 59,2% случаев (ТЫегтаил, 1977).
бои реа
Е
У нас в стране зпачительный процент ‚ лептоспироносителей. среди
крыс был обнаружен в разных городах (табл. 32).
ит
не
|
Практическое значение 855
Шахты. Благоприятные условия имеются для обитания серых крыс п
для течения среди них лептоспироза в шахтах. Эпизоотии лептоспироза в
популяциях пасюков в угольных шахтах вызывали заболеваемость людей
и описаны в ряде тран Европы (No, 1958).
У нас зарегистриррвано заболевапие лептоспирозом в шахтах Донбас-
са (Гажиев и др., 1986).
Сельская местность. В сельской местпости пасюки бывают особенно
многочисленны на животноводческих фермах. Высокая плотность паселе-
ния зверьков, а следовательно, большан напряженпость контакта между
особями и. вероятность их встречи с инфицированпыми мочевыми точка-
ми часто приводят.к интенсивным эпизоотиям лептоспироза среди зверь-
ков. На фермах они протекатт в течение всех периодов года (Карасева,
Свешникова, 1986, 1987). Зараженность лептоспирозом крыс в сельской
местности зарегистрирована во многих странах. В США, например,
в штате Иллинойс процент зараженных зверьков в среднем был равеп
60 (ЗсЬпитгепЪегесг. сё а1., 1970).
У нас в стране. эпизоотии иктерогеморрагического лептоспироза среди
пасюков, населяющих свинофермы, выявлены в основном в южных. райо-
нах страны, где.в среднем процент заражепиости зверьков составляет
20—22 (Коковин и.др., 1978; и др.). Надо отметить, что среди крыс, на-
селяющих свинарники, встречаются посители и летоспир серогруппы Ро-
топа, которыми они, вероятно, заражаются от свиней (табл. 33).
Природные биотопы. Среди крыс, живущих вне населенных пунктов,
как правило, интенсивность эпизоотии лептоспироза бывает выше, чем у
зверьков, обитающих в помещениях (по-видимому, в связи с лучшими
условиями переживания лентоспир во внешней среде).
Зараженные крысы в природе встречаются в естественных биотопах,
а именно в поймах и старицах южных рек с плавнями, берегами, зарос-
шими густой растительностью, на лиманах (Лаврова, Минина, 1968; Ка-
расева и др., 1986; Берпасовскал и др., 1986) (табл. 34).
Агроценозы. Особый интерес представляют очаги лентоспироза в аг-
роценозах. Наиболее оптимальные условия для течения эпизоотий созда-
ются на рисовых полях и на плантациях сахарного тростпика.
З Испании в окрестностях Кастелоне в связи с высокой заболевае-
мостью лептоспирозом рабочих, занимающихся уборкой риса вручную,
проводили исследования крыс, отловленных на полях. Зверьки там в пе-
которых местах были очепь. мпогочисленны (до 50 особей на 1 га) и про-
цент зараженных особей в разные годы колебался от 10 до 58.
З Восточной Испании— в Валенсии— были выделены от крыс куль-
туры Гоегопаетоттваятае. В Италии на рисовых полях лептоспироз среди
пасюков был установлен в районе Павия (Вари@ем, 1954), на о-ве Сар-
диния и в других местах (Вафа ет, Мозсо\1е, 1955).
Зараженность лентоспирозом крыс на рисовых полях констатировали
и па о-ве 'Гайвапь (КтезЬ е* а]., 1968).
В СССР на Кубани крысы-лептоснироносители встречаются во все вре-
мена года. Наибольшей интенсивности эпизоотический процессе среди них
достигает в конце лета (30—50% зараженных особей). К осени этот по-
12*
354
Глава пятая
1961). Заражению лептоспирозом здоровых крыс способствует их привер-
жениость к сырым местам обитания.
..
|
Характер поведения крыс также благоприятствует течепию инфекци-
онного процесса в популяции. Обычио крысы эвакуируют мочу 2—3 раза
в сутки, но доминант, стремясь пометить большую часть территории,
мочится гораздо чаще — до 4—5 раз в час; следовательно, если он лепто-
спироноситель, то широко рассеивает инфекцию (Карасева, 1983).
После заражения лентоспиры появляются в моче грызунов к концу вто-
рой или началу третьей недели (Карасева, Свешпикова, 1971). Чем стар-
ше крысы, тем больше среди них носителеи лентоспир (Амосенкова, Но-
пова, 1954; Карасева и др., 1986).
аи
.
Лептоспироз у пасюков протекает хронически, и опи.остаются носите-
лями лентоспир в течение всей своей жизни ,(Ва\!з, 1951а; Карасева
и др., 1986).
о
,:
Зараженность их лептоспирозом, вызываемым лентоспирами Тсеговае-
таоттВаз1ае, зарегистрирована почти во всем мире (на всех коптинентах,
кроме Антарктиды) и документировапа выделением культур. Однако,
кроме этих лептоспир, наиболее часто выделяемых от серых крыс и име-
ющих наибольшее эпидемиологическое значение, пасюки. бывают носите-
лями лептоспир и других серотрупи. В Европе. они иногда посят лепто-
спир серогруппы Ротопа, Вано, Небботад13, Вафамае..и; др. (Азот,
Вгоот, 1958). В Юго-Восточной Азии выделяются регионы (острова у-
матра, Язва, Сулавеси, Филиппины, а также Тайланд, Вьетнам), где-кры-
сы носят лептоспир серогруппы Вабам!ае и являются их осиовными носи-
телями (Торзт её а|., 1970; Петров, 1983). В Новой Зеландии показана
высокая инфицированность серых крыс лептоснирами серогруппы ВаПам
и устаповлена взаимосвязь зараженности пасюхов
с; плотностью их насе-
ления. Однако здесь основными носителями лептоснир серогруппы ВаПам
являются не серые, а черные крысы (Нафамалу,'В1аскитоте, 1981).
Эпизоотии лептоспироза встречаются как в. нашей стране, так и за
рубежом среди синантропных и экзоантропных крыс, образуя соответст-
венно антропургические и природные очаги... 2‘. °
—.
Города. Особенно много лептоспироносителей ‘обнаруживали в порто-
вых городах Европы — Лиссабоне, Копенгагене и. др (Ао её а1.,
1958). Много их было и в Израиле (Ноедеп уап'4ег; 4964, 1967). |
Вместе с судами крыс завезли в Африку. Они транспортировали' туда
п лентоспир, и в ряде портовых городов среди них обнаружены эпизоотии
лептоспироза (Тунис, Дакар и др.) (Ве. 1958). Зараженпых зверьков
там отлавливали главным образом в местах- их скопления: на рынках,
бойнях, в канализациях, на свалках.
ие.
.
В настоящее время среди крыс, обитающих во многих городах мира,
копстатируют, главным образом в жилых кварталах; этизоотии лептоспи-
роза. В восточных районах г. Детройта, например, ‘было _отловлено
358 серых крыс и культуры серогруины Теуеговастоггна1ае выделены
в 59,2% случаев (ТЫегтаил, 1977).
бои реа
Е
У нас в стране зпачительный процент ‚ лептоспироносителей. среди
крыс был обнаружен в разных городах (табл. 32).
ит
не
|
Практическое значение 855
Шахты. Благоприятные условия имеются для обитания серых крыс п
для течения среди них лептоспироза в шахтах. Эпизоотии лептоспироза в
популяциях пасюков в угольных шахтах вызывали заболеваемость людей
и описаны в ряде тран Европы (No, 1958).
У нас зарегистриррвано заболевапие лептоспирозом в шахтах Донбас-
са (Гажиев и др., 1986).
Сельская местность. В сельской местпости пасюки бывают особенно
многочисленны на животноводческих фермах. Высокая плотность паселе-
ния зверьков, а следовательно, большан напряженпость контакта между
особями и. вероятность их встречи с инфицированпыми мочевыми точка-
ми часто приводят.к интенсивным эпизоотиям лептоспироза среди зверь-
ков. На фермах они протекатт в течение всех периодов года (Карасева,
Свешникова, 1986, 1987). Зараженность лептоспирозом крыс в сельской
местности зарегистрирована во многих странах. В США, например,
в штате Иллинойс процент зараженных зверьков в среднем был равеп
60 (ЗсЬпитгепЪегесг. сё а1., 1970).
У нас в стране. эпизоотии иктерогеморрагического лептоспироза среди
пасюков, населяющих свинофермы, выявлены в основном в южных. райо-
нах страны, где.в среднем процент заражепиости зверьков составляет
20—22 (Коковин и.др., 1978; и др.). Надо отметить, что среди крыс, на-
селяющих свинарники, встречаются посители и летоспир серогруппы Ро-
топа, которыми они, вероятно, заражаются от свиней (табл. 33).
Природные биотопы. Среди крыс, живущих вне населенных пунктов,
как правило, интенсивность эпизоотии лептоспироза бывает выше, чем у
зверьков, обитающих в помещениях (по-видимому, в связи с лучшими
условиями переживания лентоспир во внешней среде).
Зараженные крысы в природе встречаются в естественных биотопах,
а именно в поймах и старицах южных рек с плавнями, берегами, зарос-
шими густой растительностью, на лиманах (Лаврова, Минина, 1968; Ка-
расева и др., 1986; Берпасовскал и др., 1986) (табл. 34).
Агроценозы. Особый интерес представляют очаги лентоспироза в аг-
роценозах. Наиболее оптимальные условия для течения эпизоотий созда-
ются на рисовых полях и на плантациях сахарного тростпика.
З Испании в окрестностях Кастелоне в связи с высокой заболевае-
мостью лептоспирозом рабочих, занимающихся уборкой риса вручную,
проводили исследования крыс, отловленных на полях. Зверьки там в пе-
которых местах были очепь. мпогочисленны (до 50 особей на 1 га) и про-
цент зараженных особей в разные годы колебался от 10 до 58.
З Восточной Испании— в Валенсии— были выделены от крыс куль-
туры Гоегопаетоттваятае. В Италии на рисовых полях лептоспироз среди
пасюков был установлен в районе Павия (Вари@ем, 1954), на о-ве Сар-
диния и в других местах (Вафа ет, Мозсо\1е, 1955).
Зараженность лентоспирозом крыс на рисовых полях констатировали
и па о-ве 'Гайвапь (КтезЬ е* а]., 1968).
В СССР на Кубани крысы-лептоснироносители встречаются во все вре-
мена года. Наибольшей интенсивности эпизоотический процессе среди них
достигает в конце лета (30—50% зараженных особей). К осени этот по-
12*
356 Глава нятая
Таблица 32. Города в СССР, в которых выделены лептоспиры серогрупл
ТебсговаетогтБазтае и Рошопа от серых крыс
Выделено культур
Число ис-
Город
следован-
Тобего-
Автор и год
ных крыс | раешог- | рошова
тваглае
Москва
300
1
Ананьин, 1941
1260
32
4
Николаев, 1945
Ленинград
398
25
Токаревич, Гавеман, 1936
1172
109
Амосенкова, Попова, 1954
1302
90
Лютов и др., 1983
Архангельск
375
6
Сосницкий, 1979
Казань
900
15
Каримова, 1954
Таллипн
143
—
19 Славин, Мединекой, 1956
Омск
—
3
Троп, Чуловский, 1959
Калининград
1619
4
1 Токалова и др., 1963
Львов
82
—
5 Ежова, 1955
Киев
20
8
- Балилевский, 1929
469
25
— Бернасовсная и др., 1986
Одесса
243
15
_ Шаферштейн, Айзен-
штадт, 1959
3611
90
_ Буров, Крицкая, 1964
469
12
- Зубко и др., 1983
Ялта
82
41
Подкорытов и др., 1986
Краснодар
410
21
З Мажникова, 1979
Сочи (Лазаревская}
9
2
- Коковин и др., 1978
Сухуми
9
3
—
Ананьин, Карасева, 1955
45
»
_
Кавтарадзе п др., 1957
Тбилиси
-
20
- Кулов, 1987
казатель несколько падает, по и зимой в средпем составляет 15—20%.
С ранней весны напряженность эпизоотии увеличивается.
|
Во все сезоны среди’ крыс-лентоспироносителей встречались особи,
не имеющие в крови антител, т. е. заразившиеся 2—3 недели назад.
Следовательно, заражение особей 1ило во все времена года, в том числе
и зимой.
|
О высокой степени зараженности почвы рисовых полей Кубани лепто-
спирами можно судить по результатам микроскопии смывов проб почвы:
из 146 проб лептосниры были обнаружены в 17 случаях, т.е. в 14,6%
(Карасева и др., 1986). Эпизоотии лептоснироза на плантациях сахарно-
го тростника известны па Газайских островах, где серые крысы были
заражены в среднем на 55,9% (Мтеце, 1964).
Несмотря па то что описаны весьма интенсивные и широкоразлитые
эпизоотии иктерогеморрагического лептоспяроза среди серых крыс, есть
Практическое значение
357
Таблица 33. Выделение лептоспир серогрупп Те(егоВаетоггВаяае и Ротопа от
серых крыс в сельекой местноети СССР (преимущественно на свинофермах) *
Число выделенных
В
культур
Место
сследо-
олего
Азтор, год
вано
Ваешог- | Рошопа
тВаз1ае
Московская обл,
_
—
1
Никифорова, 1957
Калипипградская обл.
_
9
_
Евдокимова и др., 1988
Славянский район
375
32
Карасева и др., 1986
Краснодарского края
Лабинек
45
42
Кокавин и др., 197$
Моздокский район (Северо-
533
-
6 Карасева и др., 1963
Осетинская АССР)
Чечено-Ингушская АССР
69
-
6 Коковин и др, 1962
Азербайджан
395
—
3
Алекнсров, 1983, 1986
Краснодарский край
379
6
1
дабовокий, Плотников,
Крымская обл., Бахчисарай-
20
9
- Подкорытов и др., 1986
ский райоп
Ташкентская обл.
71
4
_
Карасева и др., 1986
* На о-ве Сахалин от серых крыс была выделена культура лентаспир Сап1со]а (Безденежных,
Шаферштейн, 195%). а в Дагестане
— Тагазоу1 (Горшанова, 1962).
территории, где опи не являются носителями этих лептоспир. Несмотря
на тщательные исследования серых крыс, отловленных в свинарпиках в
Чечено-Ингушетии, Северной Осетии, в Азербайджане, у них лептоспиры
ТсегоВаетоттВас1ае выделены не были (см. табл. 33). Возможно, разные
популяции этих грызунов имеют неодинаковую восприимчивость к леп-
тоспирам ТеёеговаетоттВаяас.
За последние десятилетия серые крысы пшироко распрострапялись
и, увеличивая свой ареал, транспортировали и лептосиир [сйеговаезтюогтВа-
ятае. Вместе с грузами зверьки случайно были завезены на п-ов Таймыр,
где на животноводческих фермах среди них возпикли эпизоотии иктеро-
теморрагического лептоспироза (Райхлин, 1976).
Ранее серые крысы не обитали в Средней Азии, по, проникнув в
г. Ташкент и расселившись в пределах Ташкентской и Сыр-дарьинской
дбластей, они сформировали очаги лептоспироза типа ТсбетоваетогтВа-
отав.
Так, среди пасюков, отловленных в свинарниках близ Ташкента, леп-
тоспироносители составляли 42,2%, а антитела с лептоспирами [с4еговае-
тогграятас обпаружены в 54%. Ностоянно перемелаясь из помещений в
открытые биотопы, пасюки широко обсеменяют лентоспирами окружаю-
щую среду (Карасева и др., 1986).
Серая крыса быстро распространнется в регионы промытиленного ос-
воения Средней Сибири, заселяет повостройки. В связи с расширением
12* Серая крыса
356 Глава нятая
Таблица 32. Города в СССР, в которых выделены лептоспиры серогрупл
ТебсговаетогтБазтае и Рошопа от серых крыс
Выделено культур
Число ис-
Город
следован-
Тобего-
Автор и год
ных крыс | раешог- | рошова
тваглае
Москва
300
1
Ананьин, 1941
1260
32
4
Николаев, 1945
Ленинград
398
25
Токаревич, Гавеман, 1936
1172
109
Амосенкова, Попова, 1954
1302
90
Лютов и др., 1983
Архангельск
375
6
Сосницкий, 1979
Казань
900
15
Каримова, 1954
Таллипн
143
—
19 Славин, Мединекой, 1956
Омск
—
3
Троп, Чуловский, 1959
Калининград
1619
4
1 Токалова и др., 1963
Львов
82
—
5 Ежова, 1955
Киев
20
8
- Балилевский, 1929
469
25
— Бернасовсная и др., 1986
Одесса
243
15
_ Шаферштейн, Айзен-
штадт, 1959
3611
90
_ Буров, Крицкая, 1964
469
12
- Зубко и др., 1983
Ялта
82
41
Подкорытов и др., 1986
Краснодар
410
21
З Мажникова, 1979
Сочи (Лазаревская}
9
2
- Коковин и др., 1978
Сухуми
9
3
—
Ананьин, Карасева, 1955
45
»
_
Кавтарадзе п др., 1957
Тбилиси
-
20
- Кулов, 1987
казатель несколько падает, по и зимой в средпем составляет 15—20%.
С ранней весны напряженность эпизоотии увеличивается.
|
Во все сезоны среди’ крыс-лентоспироносителей встречались особи,
не имеющие в крови антител, т. е. заразившиеся 2—3 недели назад.
Следовательно, заражение особей 1ило во все времена года, в том числе
и зимой.
|
О высокой степени зараженности почвы рисовых полей Кубани лепто-
спирами можно судить по результатам микроскопии смывов проб почвы:
из 146 проб лептосниры были обнаружены в 17 случаях, т.е. в 14,6%
(Карасева и др., 1986). Эпизоотии лептоснироза на плантациях сахарно-
го тростника известны па Газайских островах, где серые крысы были
заражены в среднем на 55,9% (Мтеце, 1964).
Несмотря па то что описаны весьма интенсивные и широкоразлитые
эпизоотии иктерогеморрагического лептоспяроза среди серых крыс, есть
Практическое значение
357
Таблица 33. Выделение лептоспир серогрупп Те(егоВаетоггВаяае и Ротопа от
серых крыс в сельекой местноети СССР (преимущественно на свинофермах) *
Число выделенных
В
культур
Место
сследо-
олего
Азтор, год
вано
Ваешог- | Рошопа
тВаз1ае
Московская обл,
_
—
1
Никифорова, 1957
Калипипградская обл.
_
9
_
Евдокимова и др., 1988
Славянский район
375
32
Карасева и др., 1986
Краснодарского края
Лабинек
45
42
Кокавин и др., 197$
Моздокский район (Северо-
533
-
6 Карасева и др., 1963
Осетинская АССР)
Чечено-Ингушская АССР
69
-
6 Коковин и др, 1962
Азербайджан
395
—
3
Алекнсров, 1983, 1986
Краснодарский край
379
6
1
дабовокий, Плотников,
Крымская обл., Бахчисарай-
20
9
- Подкорытов и др., 1986
ский райоп
Ташкентская обл.
71
4
_ Карасева и др., 1986
* На о-ве Сахалин от серых крыс была выделена культура лентаспир Сап1со]а (Безденежных,
Шаферштейн, 195%). а в Дагестане
— Тагазоу1 (Горшанова, 1962).
территории, где опи не являются носителями этих лептоспир. Несмотря
на тщательные исследования серых крыс, отловленных в свинарпиках в
Чечено-Ингушетии, Северной Осетии, в Азербайджане, у них лептоспиры
ТсегоВаетоттВас1ае выделены не были (см. табл. 33). Возможно, разные
популяции этих грызунов имеют неодинаковую восприимчивость к леп-
тоспирам ТеёеговаетоттВаяас.
За последние десятилетия серые крысы пшироко распрострапялись
и, увеличивая свой ареал, транспортировали и лептосиир [сйеговаезтюогтВа-
ятае. Вместе с грузами зверьки случайно были завезены на п-ов Таймыр,
где на животноводческих фермах среди них возпикли эпизоотии иктеро-
теморрагического лептоспироза (Райхлин, 1976).
Ранее серые крысы не обитали в Средней Азии, по, проникнув в
г. Ташкент и расселившись в пределах Ташкентской и Сыр-дарьинской
дбластей, они сформировали очаги лептоспироза типа ТсбетоваетогтВа-
отав.
Так, среди пасюков, отловленных в свинарниках близ Ташкента, леп-
тоспироносители составляли 42,2%, а антитела с лептоспирами [с4еговае-
тогграятас обпаружены в 54%. Ностоянно перемелаясь из помещений в
открытые биотопы, пасюки широко обсеменяют лентоспирами окружаю-
щую среду (Карасева и др., 1986).
Серая крыса быстро распространнется в регионы промытиленного ос-
воения Средней Сибири, заселяет повостройки. В связи с расширением
12* Серая крыса
358
Глава пятая
Таблица 34. Выделение лептоспир серогруппы ТебегоВаетогеВахае от серых
крые в природных биотонах СССР
Всего ис-
Число выде-
Место
следовано ленных
Автор, год
крыс
культур
Низовья р. Кубани и плавни,
281
12
Лаврова, Минина, 1968
Краснодарский край,
ст. Гравинская
Плавни Азовского моря близ
58
6
Карасева и пр., 1986
Славянска на Кубани
Рисовые поля Краснодарского
647
47
То же
края
Берега р. Дупая, Измайловский
121
6
Бернасовская и др., 1986
район Одесской области
Дельта р. Дона, Октябрьский
50
10
Коциратенко и др., 1986
район Ростовской области
Абхазия оз. Бебисири
98
З
Кавтарадзе и др., 1957
Там же
_
1
Читиашвили и др., 1987
посевов риса и птироким развитием комплексного животноводства в педа-
леком будущем новые территории будут заселены серой крысой, и вслед
за этим могут вспыхнуть очаги лептоснироза. Необходимо предвидеть это
заранее и принимать соответствующие меры.
Болезнь содоку -- возбудитель риа пмтаз. Резервуар инфек-
ции — главным образом серые крысы, домовые мыши, кошки и собаки.
Встречается в разных странах мира, в том числе в СССР в виде еди-
ничных случаев.
П ротозойпые инфекции
Токсоплазмоз
— возбудитель Тосзор1азта 2оп41. В ряде работ советеких
и зарубежных авторов, относящихся к 50—60-м годам, иметотся упомипа-
ния о заражеппости токсоплазмами серых крыс. Так, Д. Н. Засухин
(1961) указывает на паразитирование токсоплазм у серых крыс и домо-
вых мылюй и полагает, что заражение свиней происходит при поедании
этих грызунов, а заражение человока— от инвазированных сельскохо-
зяйственпых животных. Впервые Д. Н. Засухин (4965) высказал также
предположение о возможности пепосредственного инвазирования токсо-
плазмами людей от серых крые и других синантропных грызунов.
В настоящее время имеется много сведений о зараженности серых
крыс токеоплазмозом в разных местах нашей страпы (Грузия, Туркме-
ния, Западная Сибирь, Украина, Хабаровский край и др.).
За рубежом эпизоотии токсоплазмоза в популяциях серых крыс уста-
новлены в США, Норвегии, ФРГ (ог 3,2 до 12% зараженных особей, по
данным Д. Н. Засухина и С. П. Васаной, 1955). Циркуляция токсоплазм
обнаружена у серых крыс в Хельсинки (Сгоотооз, Зашиутен, 1955). В Че-
Практическое значение
359
хословакии пораженность серых крыс токсоплазмами колебалась от 1,2
до 20%; возбудитель обнаружен у пих в мозге, сосудистой и лимфатиче-
ской системах. На Тайване токсоплазмы выделены от крыс методом био-
пробы па белых мышах (Уей Уао-СЬше, 1966). Зараженность токсоплаз-
мами серых крыс установлена на маленьких островах в западной части
Тихого океана (2,29% — по: УаПасе еб а[., 4972).
Болеют токсоплазмозом люди, контактирующие с домашними животны-
ми и серыми крысами. Часто случаи заболевания встречаются среди де-
ратизаторов, лаборантов и ветеринарных работников.
Балантидиоз — возбудитель ВаапиФа (паразитические инфу-
зории).
Почти во всех странах мира, преимущественио в районах развитого
свиповодства, отмечаются спорадические случаи балантидиоза среди лю-
дей. Основной резервуар
— свипьи, которые почти поголовпо ипфициро-
ваны балантидиями и постоянно выделяют с фекалиями цистные формы
этих паразитов.
На ряде территорий СССР выявлена зараженность балантидиями серых
крыс. Так, А. А. Авакян (1936) сообщил, что из 27 крыс, отловленных в
Московском зоопарке, 8 оказались ипзвазироваипыми. И. Д. Гвелисиани
(1949) исследовал 10 серых крыс, отловленных в пораженном свинарни-
ке, у одной из них обнаружены бэлантидии. Он не выявил существеппых
различий при изучение балаптидий, выделенных от людей, свиней и
крыс. При исследовании 79 серых крыс, отловленных на территории сви-
нарника в одном из сельских паселенных пунктов Днепропетровской об-
ласти, диагностировали балантидиоз у 4 экз. П. Л. Ианченко и А. Д. Тим-
ченко (1967) установили ипвазироваипость балантидиями 2,6% серых
крыс, отловленных в пригородах Оцессы. В ряде случаев при изучении
спонтанного балаптидиоза у крыс обнаруживали глубокие патологические
изменения стенки кишечника, сходные с поражениями, наблюдаемыми
при балантидиозе у человека. Однако прямых доказательств эпидемиоло-
гической роли ‘крыс при спорадияеском балаптидиозе еще пет.
Гельъминтозы
Энидемиологическая роль серых крысе в распространении тельминтозов
установлена при таких заболеваниях, как трихипеллез, гимеполепидоз,
шистозоматоз, описторхоз и др.
Трихинеллез. Развитие возбудителя трихинеллеза ТисьтеПа
5ригаН$ происходит с участием окончательного и промежуточного хозяев.
Круг хозяев трихиполл очень велик и включает множество домашних и
диких животных, в том числе серых крыс. По данным А. С. Бессонова
(1970). при исследовании 1522 серых крые на различных территориях
СССР было выявлено 323 зараженных особи (2,1%).
Распространение трихинеллеза среди основного носителя
— свиней
связано © распрострапением серой крысы, которая является не только
основным источником заражения свиней, но и основным переносчиком
трихинеллезной инвазии из дикой природы в антроиургические очаги.
12**
358
Глава пятая
Таблица 34. Выделение лептоспир серогруппы ТебегоВаетогеВахае от серых
крые в природных биотонах СССР
Всего ис-
Число выде-
Место
следовано ленных
Автор, год
крыс
культур
Низовья р. Кубани и плавни,
281
12
Лаврова, Минина, 1968
Краснодарский край,
ст. Гравинская
Плавни Азовского моря близ
58
6
Карасева и пр., 1986
Славянска на Кубани
Рисовые поля Краснодарского
647
47
То же
края
Берега р. Дупая, Измайловский
121
6
Бернасовская и др., 1986
район Одесской области
Дельта р. Дона, Октябрьский
50
10
Коциратенко и др., 1986
район Ростовской области
Абхазия оз. Бебисири
98
З
Кавтарадзе и др., 1957
Там же
_
1
Читиашвили и др., 1987
посевов риса и птироким развитием комплексного животноводства в педа-
леком будущем новые территории будут заселены серой крысой, и вслед
за этим могут вспыхнуть очаги лептоснироза. Необходимо предвидеть это
заранее и принимать соответствующие меры.
Болезнь содоку -- возбудитель риа пмтаз. Резервуар инфек-
ции — главным образом серые крысы, домовые мыши, кошки и собаки.
Встречается в разных странах мира, в том числе в СССР в виде еди-
ничных случаев.
П ротозойпые инфекции
Токсоплазмоз
— возбудитель Тосзор1азта 2оп41. В ряде работ советеких
и зарубежных авторов, относящихся к 50—60-м годам, иметотся упомипа-
ния о заражеппости токсоплазмами серых крыс. Так, Д. Н. Засухин
(1961) указывает на паразитирование токсоплазм у серых крыс и домо-
вых мылюй и полагает, что заражение свиней происходит при поедании
этих грызунов, а заражение человока— от инвазированных сельскохо-
зяйственпых животных. Впервые Д. Н. Засухин (4965) высказал также
предположение о возможности пепосредственного инвазирования токсо-
плазмами людей от серых крые и других синантропных грызунов.
В настоящее время имеется много сведений о зараженности серых
крыс токеоплазмозом в разных местах нашей страпы (Грузия, Туркме-
ния, Западная Сибирь, Украина, Хабаровский край и др.).
За рубежом эпизоотии токсоплазмоза в популяциях серых крыс уста-
новлены в США, Норвегии, ФРГ (ог 3,2 до 12% зараженных особей, по
данным Д. Н. Засухина и С. П. Васаной, 1955). Циркуляция токсоплазм
обнаружена у серых крыс в Хельсинки (Сгоотооз, Зашиутен, 1955). В Че-
Практическое значение
359
хословакии пораженность серых крыс токсоплазмами колебалась от 1,2
до 20%; возбудитель обнаружен у пих в мозге, сосудистой и лимфатиче-
ской системах. На Тайване токсоплазмы выделены от крыс методом био-
пробы па белых мышах (Уей Уао-СЬше, 1966). Зараженность токсоплаз-
мами серых крыс установлена на маленьких островах в западной части
Тихого океана (2,29% — по: УаПасе еб а[., 4972).
Болеют токсоплазмозом люди, контактирующие с домашними животны-
ми и серыми крысами. Часто случаи заболевания встречаются среди де-
ратизаторов, лаборантов и ветеринарных работников.
Балантидиоз — возбудитель ВаапиФа (паразитические инфу-
зории).
Почти во всех странах мира, преимущественио в районах развитого
свиповодства, отмечаются спорадические случаи балантидиоза среди лю-
дей. Основной резервуар
— свипьи, которые почти поголовпо ипфициро-
ваны балантидиями и постоянно выделяют с фекалиями цистные формы
этих паразитов.
На ряде территорий СССР выявлена зараженность балантидиями серых
крыс. Так, А. А. Авакян (1936) сообщил, что из 27 крыс, отловленных в
Московском зоопарке, 8 оказались ипзвазироваипыми. И. Д. Гвелисиани
(1949) исследовал 10 серых крыс, отловленных в пораженном свинарни-
ке, у одной из них обнаружены бэлантидии. Он не выявил существеппых
различий при изучение балаптидий, выделенных от людей, свиней и
крыс. При исследовании 79 серых крыс, отловленных на территории сви-
нарника в одном из сельских паселенных пунктов Днепропетровской об-
ласти, диагностировали балантидиоз у 4 экз. П. Л. Ианченко и А. Д. Тим-
ченко (1967) установили ипвазироваипость балантидиями 2,6% серых
крыс, отловленных в пригородах Оцессы. В ряде случаев при изучении
спонтанного балаптидиоза у крыс обнаруживали глубокие патологические
изменения стенки кишечника, сходные с поражениями, наблюдаемыми
при балантидиозе у человека. Однако прямых доказательств эпидемиоло-
гической роли ‘крыс при спорадияеском балаптидиозе еще пет.
Гельъминтозы
Энидемиологическая роль серых крысе в распространении тельминтозов
установлена при таких заболеваниях, как трихипеллез, гимеполепидоз,
шистозоматоз, описторхоз и др.
Трихинеллез. Развитие возбудителя трихинеллеза ТисьтеПа
5ригаН$ происходит с участием окончательного и промежуточного хозяев.
Круг хозяев трихиполл очень велик и включает множество домашних и
диких животных, в том числе серых крыс. По данным А. С. Бессонова
(1970). при исследовании 1522 серых крые на различных территориях
СССР было выявлено 323 зараженных особи (2,1%).
Распространение трихинеллеза среди основного носителя
— свиней
связано © распрострапением серой крысы, которая является не только
основным источником заражения свиней, но и основным переносчиком
трихинеллезной инвазии из дикой природы в антроиургические очаги.
12**
360
Глава пятая
Отмечено, что постоянные и интенсивные очаги трихинеллеза находятся
в тех районах, где плотность популяции серых крыс высока и они посто-
янно или в летний период обитают в дикой природе, в которой в основ-
ном заражаются этой инфекцией. К таким районам относятся южные и
западные районы СССР. В то же время времепные очаги трихинеллеза
свипей рогистрируются в северных районах СССР, где серая крыса не
выселяется в дикуо природу или выселяется позпачительная часть нопу-
ляции. Наконец, в районах, неблагоприятных для серой крысы, трихинел-
леза свиней вообще нет.
Трихицнелл у серых крыс находили и при изучении их распростране-
ния в ряде зарубежных стран. Так, на Аляске зараженность серых крыс
достигала 10,7%. Можно полагать, что с развитием промышленного жи-
вотноводетва будет возрастать зпидемиологическая роль инвазий, в под-
держании которых значительная роль принадлежит серым крысам.
Гименоленидоз. Крысы являются окончательным хозяином
крысиного цепия Нушепо]ер!з дпилииа
— одного из возбудителей гиме-
нолепидоза, встречающегося у человска. Заражение зверьков происходит
при случайном проглатывании промежуточных хозяев крысиного цеп-
ня — насекомых, в теле которых локализуются цистицеркоиды. Ото за-
болевание зарегистрировано в различных регионах распространения се-
рых крыс. Н. длийища встречатотся у этих грызунов в течение всего года.
Интенсивность заражения колеблется от 5,8 до 44,8 экз. в различные пе-
риоды года.
За рубежом в тропиках и субтропиках отмечена зараженность серых
крыс возбудителями шисгозомоза. Ряд исследователей отводят крысам
значительную роль в переносе этих инзазий (Яроцкий, 1982).
Таким образом, серые крысы связаны в большей или меньшей стеие-
ни с огромным количеством инфекций и инвазий, описанных для человека
поэтому изучение энизоотий в их популяциях особенно необходимо.
В связи с этим мы еще раз подчеркиваем, что изучепие эпизоотий и
контроль численности этих грызунов приобретают особое значение.
Методы борьбы с серыми крысами
В отечественной литературе первая паиболее подробная и полпая сводка
различпых приемов борьбы с серой крысой содержится в руководстве
Я. Л. Окуневского (1936), где изложен весь накопленный к этому вре-
мени в нашей стране и за рубежом опыт дератизации. Но приводимый
материал не упорядочен и не классифицирован.
Лилть в 1974 г. группа авторов (Вашков и др., 1974) предложила чет-
кую систему классификации дератизации, в которой по средствам воз-
действия выделен ряд способов, объединяемых в три основных метода:
1) биологический, 2) физический и 3) химический.
Таким образом, упорядочепа пе только структура наименований ист-
ребительных мероприятий, но разграничены часто смешиваемые понятия
метода и способа.
—
Практическое значение
361
Из иностранных руководств по борьбе с серой крысой следует отме-
тить книту Штайнигера (З4елисог, 1952) и мопографию Читти (СыМу,
4954).
Биологический метод борьбы
1. Использование хищников. Для уничтожения серых крыс © давних пор
были использованы различные хищники: хорьки, ласки, мангусты, соба-
ки, кошки, птицы, варапы, змеи (Окупевский, 1936; Бештли, 1967; Джек-
сон, 1973).
Па осповании наблюдений Пирсона (Реагзоп, 1964, 1966, цит по:
Воджицкий, 1974) К. Воджицкий делает важный вывод об эффективно-
сти использования хищников для борьбы с грызунами, когда их числен-
ность находится в фазе пика, однако они подавляют ее в тот период, ког-
да какой-либо дополнительный внешний фактор, например, применение
отравленной приманки, способствует снижению се пика. Этот вывод под-
тверждается опытами Элтона в 1958 г. на четырех фермах Англии, где
была проведена обработка территории ратицидами, после чего выпустили
конек. Длительное время на фермах крысы отсутствовали. Но там, где
кошек пе было, числепвость этих грызунов после обработки ратицидами
быстро восстановилась. Можно добиться длительного снижения числен-
ности крысе при условии содержания в помещении хищников, которых
дополнительно подкармливают молоком (Воджицкий, 1974).
Интересны проведенные за последнее время опыты по борьбе с серы-
ми крысами с помощью куницеобразпых (колонки, солонгои; хорьки ока-
зались слишком крупными, чтобы беспрепятственно проникать во все
щели, куда от них прячутся крысы) на различных объектах: в теплицах,
на животноводческих фермах, складах зерна, в яйцехранилищах и др.,
удаленных от населенных пунктов, а также на изолированных объектах,
таких, как корабли. Па искоторых из них может быть достигнуто полное
уничтожение грызунов (Денисов, 1986).
Некоторые авторы придерживаются отрицательного мнепия в вопросе
использования хищников. А. ЦП. Вузякин (19595), например, отмечает бес-
полезность кошек для борьбы с грызунами. Наблюдения многих авторов
подтвердили пезначительпость роли хищников в регуляции численности
серых крыс (Бентли, 1967; Джексон, 1973; СозсхупзКЬ 1974; ЕагВаюз-
Аза, зомВулеЕ, 1979). На полезных или важных в промыеловом отно-
шении видах нередко неблагоприятно сказывается выпуск хищников
(«Втиппег её а|., 1981) вследствие их трудноуправляемой избирательпости
в выборе источников корма (Воджицкий, 1974). По нашему мнению, ис-
пользование хищников вряд ли целесообразно как с точки зрения их
низкой эффективности, так и из-за чрезвычайной трудоемкости всей
процедуры.
2. Бактериальный способ. Главная идея этого способа борьбы с грызу-
пами, в том числе и с серой крысой,— создание эпизоотии в понуляции,
приводящей к ее полной ликвидацин. С этей целью на протяжепии мпо-
гих десятков лет были испытаны знтаммы сальмонелл Данича, Мережков-
360
Глава пятая
Отмечено, что постоянные и интенсивные очаги трихинеллеза находятся
в тех районах, где плотность популяции серых крыс высока и они посто-
янно или в летний период обитают в дикой природе, в которой в основ-
ном заражаются этой инфекцией. К таким районам относятся южные и
западные районы СССР. В то же время времепные очаги трихинеллеза
свипей рогистрируются в северных районах СССР, где серая крыса не
выселяется в дикуо природу или выселяется позпачительная часть нопу-
ляции. Наконец, в районах, неблагоприятных для серой крысы, трихинел-
леза свиней вообще нет.
Трихицнелл у серых крыс находили и при изучении их распростране-
ния в ряде зарубежных стран. Так, на Аляске зараженность серых крыс
достигала 10,7%. Можно полагать, что с развитием промышленного жи-
вотноводетва будет возрастать зпидемиологическая роль инвазий, в под-
держании которых значительная роль принадлежит серым крысам.
Гименоленидоз. Крысы являются окончательным хозяином
крысиного цепия Нушепо]ер!з дпилииа
— одного из возбудителей гиме-
нолепидоза, встречающегося у человска. Заражение зверьков происходит
при случайном проглатывании промежуточных хозяев крысиного цеп-
ня — насекомых, в теле которых локализуются цистицеркоиды. Ото за-
болевание зарегистрировано в различных регионах распространения се-
рых крыс. Н. длийища встречатотся у этих грызунов в течение всего года.
Интенсивность заражения колеблется от 5,8 до 44,8 экз. в различные пе-
риоды года.
За рубежом в тропиках и субтропиках отмечена зараженность серых
крыс возбудителями шисгозомоза. Ряд исследователей отводят крысам
значительную роль в переносе этих инзазий (Яроцкий, 1982).
Таким образом, серые крысы связаны в большей или меньшей стеие-
ни с огромным количеством инфекций и инвазий, описанных для человека
поэтому изучение энизоотий в их популяциях особенно необходимо.
В связи с этим мы еще раз подчеркиваем, что изучепие эпизоотий и
контроль численности этих грызунов приобретают особое значение.
Методы борьбы с серыми крысами
В отечественной литературе первая паиболее подробная и полпая сводка
различпых приемов борьбы с серой крысой содержится в руководстве
Я. Л. Окуневского (1936), где изложен весь накопленный к этому вре-
мени в нашей стране и за рубежом опыт дератизации. Но приводимый
материал не упорядочен и не классифицирован.
Лилть в 1974 г. группа авторов (Вашков и др., 1974) предложила чет-
кую систему классификации дератизации, в которой по средствам воз-
действия выделен ряд способов, объединяемых в три основных метода:
1) биологический, 2) физический и 3) химический.
Таким образом, упорядочепа пе только структура наименований ист-
ребительных мероприятий, но разграничены часто смешиваемые понятия
метода и способа.
—
Практическое значение
361
Из иностранных руководств по борьбе с серой крысой следует отме-
тить книту Штайнигера (З4елисог, 1952) и мопографию Читти (СыМу,
4954).
Биологический метод борьбы
1. Использование хищников. Для уничтожения серых крыс © давних пор
были использованы различные хищники: хорьки, ласки, мангусты, соба-
ки, кошки, птицы, варапы, змеи (Окупевский, 1936; Бештли, 1967; Джек-
сон, 1973).
Па осповании наблюдений Пирсона (Реагзоп, 1964, 1966, цит по:
Воджицкий, 1974) К. Воджицкий делает важный вывод об эффективно-
сти использования хищников для борьбы с грызунами, когда их числен-
ность находится в фазе пика, однако они подавляют ее в тот период, ког-
да какой-либо дополнительный внешний фактор, например, применение
отравленной приманки, способствует снижению се пика. Этот вывод под-
тверждается опытами Элтона в 1958 г. на четырех фермах Англии, где
была проведена обработка территории ратицидами, после чего выпустили
конек. Длительное время на фермах крысы отсутствовали. Но там, где
кошек пе было, числепвость этих грызунов после обработки ратицидами
быстро восстановилась. Можно добиться длительного снижения числен-
ности крысе при условии содержания в помещении хищников, которых
дополнительно подкармливают молоком (Воджицкий, 1974).
Интересны проведенные за последнее время опыты по борьбе с серы-
ми крысами с помощью куницеобразпых (колонки, солонгои; хорьки ока-
зались слишком крупными, чтобы беспрепятственно проникать во все
щели, куда от них прячутся крысы) на различных объектах: в теплицах,
на животноводческих фермах, складах зерна, в яйцехранилищах и др.,
удаленных от населенных пунктов, а также на изолированных объектах,
таких, как корабли. Па искоторых из них может быть достигнуто полное
уничтожение грызунов (Денисов, 1986).
Некоторые авторы придерживаются отрицательного мнепия в вопросе
использования хищников. А. ЦП. Вузякин (19595), например, отмечает бес-
полезность кошек для борьбы с грызунами. Наблюдения многих авторов
подтвердили пезначительпость роли хищников в регуляции численности
серых крыс (Бентли, 1967; Джексон, 1973; СозсхупзКЬ 1974; ЕагВаюз-
Аза, зомВулеЕ, 1979). На полезных или важных в промыеловом отно-
шении видах нередко неблагоприятно сказывается выпуск хищников
(«Втиппег её а|., 1981) вследствие их трудноуправляемой избирательпости
в выборе источников корма (Воджицкий, 1974). По нашему мнению, ис-
пользование хищников вряд ли целесообразно как с точки зрения их
низкой эффективности, так и из-за чрезвычайной трудоемкости всей
процедуры.
2. Бактериальный способ. Главная идея этого способа борьбы с грызу-
пами, в том числе и с серой крысой,— создание эпизоотии в понуляции,
приводящей к ее полной ликвидацин. С этей целью на протяжепии мпо-
гих десятков лет были испытаны знтаммы сальмонелл Данича, Мережков-
362
Тлава пятая
ского, Исаченко, Прохорова (птамм No 5170) и штамм No ВК2С (Вод-
жицкий, 1974).
Несмотря па постоянную работу по усовершенствованию бактериоло-
гического способа, сго эффективпосгь иротив серых крыс сильно колеб-
лется — от 60 до 95% смертности (Прохоров, 1962; Сивнцова, Серебряко-
ва, 1975), а у оставшихея в живых крыс развивается иммупитет и при
повторном применении бактерий эффективность падает (Окупевский,
1936; Соболева и др., 1972).
По критической оценке В. В. Кучерука (1964, с. 17—18), «до настоя-
щего времени пе описано ни одного достоверлого случая коитактпой эпи-
зоотин, вызванпой бактериями, применяемыми для истребления грызу-
пов. Напротив, все инструктивпые указапия строятся в расчете зараже-
ния кайхдого зверька непосредетвенно от примапкн. Исследования, про-
веденпые советскими учепыми, показали, что даже такие высоковирулент-
ные микроорганизмы, как возбудители чумы и туляремии. не приводят к
ноголовному вымирапию грызунов ...Гаким образом, основное предпола-
таемое преимущество бактериального способа борьбы— возмоятиость вы-
взывать естоствонпое развитие энизоотии при использовапии микробов,
употребляемых в настоящое время,— не оправдалось».
Применспие сальмонелл для дератизации приводит к иштепсивному
загрязнению среды и опасно для людей и животных (Воджицкий, 1974).
В связи с этим в ряде стран бактерии для дератизации запрещены и не
примепяются.
В СССР бактериальный ©1060б борьбы с грызунами используется
гланным образом для обработки сельскохозяйетвениых объектов, но сфера
сго применения все более ограничивается ввиду опаспости бактерий для
молодняка животных (Траханов, 1973).
Наилучишм вариалтом применения бактерий оказалось добавлепие к
приманке небольших количеств химических ядов, т. е. объединение хи-
мического и биологического методов. В комбинации © ядами-антикоагу-
дяитами тина варфарипа бактериальные приманки дают смертность
крыс до 96-—400% (Траханов, 1961а, 6; Егоренко, 1963). Тем не ‘менее
комбинированный ©с10соб борьбы пе уменьшает возможности загрязнения
среды сальмонеллами, что ограничивает ого применение.
‹
Физический метод борьбы
1. Механичеекий способ. Среди различных мехапических способов унич-
тожения серых крыс паиболее распространенным является нрименепие
ловушек и капканов, которые. в свото очередь, можно соответственно под-
разделить па два осповных типа: живоловки одноместпые, мпогоместные
(верши) и убивающие — капкапчики Геро и тарелочные дуговые капка-
ны. Отлов крыс дуговыми капканами припципиально отличается от отло-
ва ловушками с приманкой тем, что он основа пе на привлечении крыс,
а на использовании стереотипа их передвижений в наиболее часто иосе-
щаемых ими местах. Высокая эффективность лова крыс дуговыми капка-
нами связана, видимо, с тем, что стереотип движения их часто оказыва-
ется намного сильнее избегания капкана как пового предмета.
Практическое значение
363
Отлов крыс требует спецвальных навыков и постоянной тренировки.
Массовое применепие ловушек и капкапов плохо обученными людьми не
способно оказать существенного воздействия на ход дератизации. Публи-
каций о методике отлова серых крыс таким способом крайне мало, в их
числе наиболее заслуживает внимания работа И. И. Дементьева (1953),
посвященная искусству вылова крыс дуговыми капканами.
Ряд авторов (СВ, 1954; Кузякии, 1963; Лялин, 1974а) выделяют
три осповных категории пасюков по степени их пастороженности к ло-
вушкам: 1) неосторожные, сразу попадающие в настороженные ловупки
с приманкой, 2) осторожные, которые пе трогают насаженпую приманку
отэ—7 до 12—15 дней, и 3 — очевь осторожные.
А. П. Кузякии (1950) предложил пасюков отлавливать по отдельным
кварталам города, а для «дочистки» в пих объектов, где остались единич-
ные крысы, оп рекомендуст использовать сильные яды с охотно избирае-
мой зверьками пищевой или ‘питьевой основой (с. 186). С нашей точки
зрения, вряд ли можно согласиться с тем, что чрезвычайно трудоемкий
метод физического уничтожения поставлен в основу борьбы, а наиболее
эффективпому и экономичному, химическому методу оставлена роль «до-
чистки».
|
В заключение обзора по механическим способам лова хочется сослать-
ся на мноние Д. Друммонда (1974) о том, что :рименение ловушек мало-
пригодно для уничтожения популяции грызунов, но оно годится для лик-
видации пебольшого числа особей, которые не взяли отравленпую ири-
манку. Особенно подходят для этой цели дуговыс капканы.
К физическим методам борьбы относятся и адгезионные ловушки
(ловчий клей), представляющие собой «столики», покрытые липкой мас-
сой, которые расставляют около нор, лазов крыс, вдоль стеи. Но прили-
пают к массе гораздо лучше мыши, чем крысы.
2. Электрический способ. Среди других средств уничтожения пасюков
заслуживает внимания примепепие элтектродератизаторов, но Я. Л. Оку-
невский (1936) пизнет, что опи не всогда убивают крыс, опаспы в по-
жарном отношении, требуют специальных охранительных ностов, что
резко суживает сферу их применения. Тем не менее А. Ц. Леонтьев
(1968) в Узбекистане с помощью электродератизаторов уничтожал в от-
дельных сельскохозяйственных объектах до 2—4 тыс. серых крысе.
По-видимому, для защиты от грызунов объектов, где нет людей и
животных (например, электроподстанций, складов и т. д.), подобные
электроустройства могут быть полезпы,
*х 3. Акустический способ (ультразвуковые установки). Многочисленные
попытки примепять ультразвуковые установки для упичтожения и от-
нугивания крыс не дали положительных результатов (МагзВ, ТасКзоп,
1978; Мах, 1983; Пуцято, 1984).
Несомненным преимуществом применения физического метода дера-
тизации является его высокая степень избирательности в отношении ои-
ределепного вида, и в противоположность химическому методу оп не при-
водит к загрязнепию среды нестицидами (Кио, 1980).
362
Тлава пятая
ского, Исаченко, Прохорова (птамм No 5170) и штамм No ВК2С (Вод-
жицкий, 1974).
Несмотря па постоянную работу по усовершенствованию бактериоло-
гического способа, сго эффективпосгь иротив серых крыс сильно колеб-
лется — от 60 до 95% смертности (Прохоров, 1962; Сивнцова, Серебряко-
ва, 1975), а у оставшихея в живых крыс развивается иммупитет и при
повторном применении бактерий эффективность падает (Окупевский,
1936; Соболева и др., 1972).
По критической оценке В. В. Кучерука (1964, с. 17—18), «до настоя-
щего времени пе описано ни одного достоверлого случая коитактпой эпи-
зоотин, вызванпой бактериями, применяемыми для истребления грызу-
пов. Напротив, все инструктивпые указапия строятся в расчете зараже-
ния кайхдого зверька непосредетвенно от примапкн. Исследования, про-
веденпые советскими учепыми, показали, что даже такие высоковирулент-
ные микроорганизмы, как возбудители чумы и туляремии. не приводят к
ноголовному вымирапию грызунов ...Гаким образом, основное предпола-
таемое преимущество бактериального способа борьбы— возмоятиость вы-
взывать естоствонпое развитие энизоотии при использовапии микробов,
употребляемых в настоящое время,— не оправдалось».
Применспие сальмонелл для дератизации приводит к иштепсивному
загрязнению среды и опасно для людей и животных (Воджицкий, 1974).
В связи с этим в ряде стран бактерии для дератизации запрещены и не
примепяются.
В СССР бактериальный ©1060б борьбы с грызунами используется
гланным образом для обработки сельскохозяйетвениых объектов, но сфера
сго применения все более ограничивается ввиду опаспости бактерий для
молодняка животных (Траханов, 1973).
Наилучишм вариалтом применения бактерий оказалось добавлепие к
приманке небольших количеств химических ядов, т. е. объединение хи-
мического и биологического методов. В комбинации © ядами-антикоагу-
дяитами тина варфарипа бактериальные приманки дают смертность
крыс до 96-—400% (Траханов, 1961а, 6; Егоренко, 1963). Тем не ‘менее
комбинированный ©с10соб борьбы пе уменьшает возможности загрязнения
среды сальмонеллами, что ограничивает ого применение.
‹
Физический метод борьбы
1. Механичеекий способ. Среди различных мехапических способов унич-
тожения серых крыс паиболее распространенным является нрименепие
ловушек и капканов, которые. в свото очередь, можно соответственно под-
разделить па два осповных типа: живоловки одноместпые, мпогоместные
(верши) и убивающие — капкапчики Геро и тарелочные дуговые капка-
ны. Отлов крыс дуговыми капканами припципиально отличается от отло-
ва ловушками с приманкой тем, что он основа пе на привлечении крыс,
а на использовании стереотипа их передвижений в наиболее часто иосе-
щаемых ими местах. Высокая эффективность лова крыс дуговыми капка-
нами связана, видимо, с тем, что стереотип движения их часто оказыва-
ется намного сильнее избегания капкана как пового предмета.
Практическое значение
363
Отлов крыс требует спецвальных навыков и постоянной тренировки.
Массовое применепие ловушек и капкапов плохо обученными людьми не
способно оказать существенного воздействия на ход дератизации. Публи-
каций о методике отлова серых крыс таким способом крайне мало, в их
числе наиболее заслуживает внимания работа И. И. Дементьева (1953),
посвященная искусству вылова крыс дуговыми капканами.
Ряд авторов (СВ, 1954; Кузякии, 1963; Лялин, 1974а) выделяют
три осповных категории пасюков по степени их пастороженности к ло-
вушкам: 1) неосторожные, сразу попадающие в настороженные ловупки
с приманкой, 2) осторожные, которые пе трогают насаженпую приманку
отэ—7 до 12—15 дней, и 3 — очевь осторожные.
А. П. Кузякии (1950) предложил пасюков отлавливать по отдельным
кварталам города, а для «дочистки» в пих объектов, где остались единич-
ные крысы, оп рекомендуст использовать сильные яды с охотно избирае-
мой зверьками пищевой или ‘питьевой основой (с. 186). С нашей точки
зрения, вряд ли можно согласиться с тем, что чрезвычайно трудоемкий
метод физического уничтожения поставлен в основу борьбы, а наиболее
эффективпому и экономичному, химическому методу оставлена роль «до-
чистки».
|
В заключение обзора по механическим способам лова хочется сослать-
ся на мноние Д. Друммонда (1974) о том, что :рименение ловушек мало-
пригодно для уничтожения популяции грызунов, но оно годится для лик-
видации пебольшого числа особей, которые не взяли отравленпую ири-
манку. Особенно подходят для этой цели дуговыс капканы.
К физическим методам борьбы относятся и адгезионные ловушки
(ловчий клей), представляющие собой «столики», покрытые липкой мас-
сой, которые расставляют около нор, лазов крыс, вдоль стеи. Но прили-
пают к массе гораздо лучше мыши, чем крысы.
2. Электрический способ. Среди других средств уничтожения пасюков
заслуживает внимания примепепие элтектродератизаторов, но Я. Л. Оку-
невский (1936) пизнет, что опи не всогда убивают крыс, опаспы в по-
жарном отношении, требуют специальных охранительных ностов, что
резко суживает сферу их применения. Тем не менее А. Ц. Леонтьев
(1968) в Узбекистане с помощью электродератизаторов уничтожал в от-
дельных сельскохозяйственных объектах до 2—4 тыс. серых крысе.
По-видимому, для защиты от грызунов объектов, где нет людей и
животных (например, электроподстанций, складов и т. д.), подобные
электроустройства могут быть полезпы,
*х 3. Акустический способ (ультразвуковые установки). Многочисленные
попытки примепять ультразвуковые установки для упичтожения и от-
нугивания крыс не дали положительных результатов (МагзВ, ТасКзоп,
1978; Мах, 1983; Пуцято, 1984).
Несомненным преимуществом применения физического метода дера-
тизации является его высокая степень избирательности в отношении ои-
ределепного вида, и в противоположность химическому методу оп не при-
водит к загрязнепию среды нестицидами (Кио, 1980).
364
Глава пятая
Химический метод борьбы
1. Удушающие яды (газация). Для борьбы с крысами было испытано
мнозжкество газов: сернистый газ, углекислый газ, окись углерода, хлор,
хлорпикрин, цианистый водород, фосфористый водород, окись этилена,
бромистый метил. Все перечисленные газы вызывали полную гибель этих
грызунов при условии, что они не могли уйти из отравленпой зоны. Вре-
мя их гибели при этом составляло от нескольких минут до нескольких
часов. Но такой же высокой токсичностьо перечисленные газы обладают
по отпошению к другим животным и людям, что требует очепь больших
затрат средств и усилий по обеспечению безопасности ири обработке
ими. Перед газацией строений из них удаляют людей, останавливают
пронзводство и тшательно герметизируют все отверстия. В ряде случаев
обработку газом проводить нельзя, если поблизости есть жилые дома и
предприятия. Недостатком способа газации является и отсутствие в по-
мещениях остаточного действия газа на крыс после окончания обработ-
ки — как только помещения проветрены, они сразу могут быть повторно
заселены крысами. Большим минусом является и высокая стоимость об-
работки. которая тем выше, чем больше объем газируемого помещения.
Некоторые газы химически реагируют с различными поверхпостями, что
резко ограничивает их применение (серпистый газ, хлор), другие адсор-
бируются и с трудом поддаются удалению (хлорпикрип), третьи слиш-
ком токсичпы (окись углерода, бромистый метил, цианистый водород).
В настоящее время газацию применяют только для обработки специ-
альных объектов: судов, вагонов, элеваторов, реже — холодильпиков.
Газация фосфипом применяется для обработки нор серых крыс, рас-
положенных вне построек человека (Ме, 1980). Их обитатели полностью
гибнут при условии заделки нор после газации.
Безусловным достоипством этого метода явллется способность газов
одномоментно уничтожать практически всех грызунов в закрытых поме-
щениях, где примепение других способов невозможно или неэффективно.
2. Кишечные яды и формы их применения
а
Препаративные формы
41. Пищевые отравленные приманки. В пастоящее время
этот способ язляется ввиду своей универсальности основным среди всех
способов дератизации. Отравленные приманки условпо можно разделить
ио содержанито влаги в пищевой оспове на сухие и увлажнепные, причем
последние поедаются значительно лучше (Дукельская, 1948; Рагцей,
Зрепсог, 1953; Друммонд, 1974), но быстрее портятся. Во всох случаях
лучше всего посдаются только свежие, доброкачественные продукты (Со-
колов и др., 1986а).
Поедаемость серыми крысами пищевой основы отравленных приманок
в значительной мере зависит от состава и обилия кормов в привычных
для них условиях обитания. На объектах с однородной кормовой базой
Практическое значение
365
паиболее предночитаемой является пищевая основа, которая восполняет
недостаток отдельных компонентов рациона крыс. На мясокомбинате,
в колбасной мастерской и в холодильнике крысы испытывают явный де-
фицит углеводов. Применение мучных приманок с сахаром (яд зооку-
марин) позволило’ полностью освободить эти объекты от них (Лыскова
и др. 1972; Хамаганов, Шахуров, 1959). По данным 9. А. Гутмана
(1972), на холодильном комбинате серые крысы практически не тронули
хлеб, мясной и рыбпый фарш, зато за сутки они съели 8,4 кг помидоров
© зоокумарином.
В складах зерна, муки, крупы пасюки питаются нысококалорийными
кормами, содержащими большую часть необходимых компопептов, одна-
ко здесь имеется недостаток влаги, поэтому паиболее эффективными ока-
зываются жидкие приманки— молоко, вода с сахаром (Паселхник, 1947;
Айзенштацт, 1956).
В качестве жидких отравленных приманок были также испытаны
растворы этилового спирта (40 капель на 90 мл воды), валерьяны (10—
20% раствор), грушевого сиропа, пиза. У сосудов с водой следы крыс
отмечены в 100%, а у ноилок со спиртовыми растворами—в 55% слу-
чаев. Растворы © 5%-ной концентрацией спирта были более предночитае-
мы, чем со спиртом в иных концентрациях. При сравнении других жид-
ких приманок выяснилось, что крысы их предпочитали в следующей
последовательности: раствор валерьяны, вода, грушевый сироп, пиво.
Зверьки начинали пить эти растворы линь на 2—3-й день оксперимента.
Для того чтобы исключить случаи «подтравливания» крыс, предлагают
использовать в составе жидких приманок сильнодействующие яды доста-
точно высокой концентрации (Эйгелис и др., 1986). Однако пока ие иро-
ведены испытания разных доз этих ядов, не оценены их эффективность,
степень оборонительной реакции, а также пока не прошли практические
испытания, выводы и рекомендации вышеупомянутой работы носят прец-
варительпый характер.
Опыт показывает, что на большинстве объектов, где достаточно влаги,
поилки оказываются малоэффективными. Массовое применение жидких
отравленных приманок в сочетании с зерновыми в г. Новочеркасске
обеспечило снижение численности грызупов на 88,6%, а применение
только зерповых отравленных приманок— на 88,4 (наши данные).
В местах обитания, где рацион кормов содержит все необходимые
компопепты, крысы в качестве приманки предпочитают наиболее привыч-
ный корм; например, в свинарниках
— комбикорм, мясной и рыбный
фар, причем с уменьшением плотности их населения такая выбороч-
сть становится наиболее явной (ЙХареа|, 1962; Спиченко, 1962; Ля-
лин, 1978). На рисовых полях Краснодарского края в июле крысы пред-
почитали рис пшенице и ячменю (Шутова и др., 1981). Таким образом,
состав пищевой основы, правильно сбалансированный с обеспеченностью
серых крыс кормами в местах их обитания, может быть ретающим усло-
нием привлекательности отравленной приманки. Как правило, добавка к
пищевой основе аттрактантов (5—10% сахара или 3% растительного
масла) значительно улучшает ее поедаемость (Вагрей, 1956; Огуси Код:
364
Глава пятая
Химический метод борьбы
1. Удушающие яды (газация). Для борьбы с крысами было испытано
мнозжкество газов: сернистый газ, углекислый газ, окись углерода, хлор,
хлорпикрин, цианистый водород, фосфористый водород, окись этилена,
бромистый метил. Все перечисленные газы вызывали полную гибель этих
грызунов при условии, что они не могли уйти из отравленпой зоны. Вре-
мя их гибели при этом составляло от нескольких минут до нескольких
часов. Но такой же высокой токсичностьо перечисленные газы обладают
по отпошению к другим животным и людям, что требует очепь больших
затрат средств и усилий по обеспечению безопасности ири обработке
ими. Перед газацией строений из них удаляют людей, останавливают
пронзводство и тшательно герметизируют все отверстия. В ряде случаев
обработку газом проводить нельзя, если поблизости есть жилые дома и
предприятия. Недостатком способа газации является и отсутствие в по-
мещениях остаточного действия газа на крыс после окончания обработ-
ки — как только помещения проветрены, они сразу могут быть повторно
заселены крысами. Большим минусом является и высокая стоимость об-
работки. которая тем выше, чем больше объем газируемого помещения.
Некоторые газы химически реагируют с различными поверхпостями, что
резко ограничивает их применение (серпистый газ, хлор), другие адсор-
бируются и с трудом поддаются удалению (хлорпикрип), третьи слиш-
ком токсичпы (окись углерода, бромистый метил, цианистый водород).
В настоящее время газацию применяют только для обработки специ-
альных объектов: судов, вагонов, элеваторов, реже — холодильпиков.
Газация фосфипом применяется для обработки нор серых крыс, рас-
положенных вне построек человека (Ме, 1980). Их обитатели полностью
гибнут при условии заделки нор после газации.
Безусловным достоипством этого метода явллется способность газов
одномоментно уничтожать практически всех грызунов в закрытых поме-
щениях, где примепение других способов невозможно или неэффективно.
2. Кишечные яды и формы их применения
а
Препаративные формы
41. Пищевые отравленные приманки. В пастоящее время
этот способ язляется ввиду своей универсальности основным среди всех
способов дератизации. Отравленные приманки условпо можно разделить
ио содержанито влаги в пищевой оспове на сухие и увлажнепные, причем
последние поедаются значительно лучше (Дукельская, 1948; Рагцей,
Зрепсог, 1953; Друммонд, 1974), но быстрее портятся. Во всох случаях
лучше всего посдаются только свежие, доброкачественные продукты (Со-
колов и др., 1986а).
Поедаемость серыми крысами пищевой основы отравленных приманок
в значительной мере зависит от состава и обилия кормов в привычных
для них условиях обитания. На объектах с однородной кормовой базой
Практическое значение
365
паиболее предночитаемой является пищевая основа, которая восполняет
недостаток отдельных компонентов рациона крыс. На мясокомбинате,
в колбасной мастерской и в холодильнике крысы испытывают явный де-
фицит углеводов. Применение мучных приманок с сахаром (яд зооку-
марин) позволило’ полностью освободить эти объекты от них (Лыскова
и др. 1972; Хамаганов, Шахуров, 1959). По данным 9. А. Гутмана
(1972), на холодильном комбинате серые крысы практически не тронули
хлеб, мясной и рыбпый фарш, зато за сутки они съели 8,4 кг помидоров
© зоокумарином.
В складах зерна, муки, крупы пасюки питаются нысококалорийными
кормами, содержащими большую часть необходимых компопептов, одна-
ко здесь имеется недостаток влаги, поэтому паиболее эффективными ока-
зываются жидкие приманки— молоко, вода с сахаром (Паселхник, 1947;
Айзенштацт, 1956).
В качестве жидких отравленных приманок были также испытаны
растворы этилового спирта (40 капель на 90 мл воды), валерьяны (10—
20% раствор), грушевого сиропа, пиза. У сосудов с водой следы крыс
отмечены в 100%, а у ноилок со спиртовыми растворами—в 55% слу-
чаев. Растворы © 5%-ной концентрацией спирта были более предночитае-
мы, чем со спиртом в иных концентрациях. При сравнении других жид-
ких приманок выяснилось, что крысы их предпочитали в следующей
последовательности: раствор валерьяны, вода, грушевый сироп, пиво.
Зверьки начинали пить эти растворы линь на 2—3-й день оксперимента.
Для того чтобы исключить случаи «подтравливания» крыс, предлагают
использовать в составе жидких приманок сильнодействующие яды доста-
точно высокой концентрации (Эйгелис и др., 1986). Однако пока ие иро-
ведены испытания разных доз этих ядов, не оценены их эффективность,
степень оборонительной реакции, а также пока не прошли практические
испытания, выводы и рекомендации вышеупомянутой работы носят прец-
варительпый характер.
Опыт показывает, что на большинстве объектов, где достаточно влаги,
поилки оказываются малоэффективными. Массовое применение жидких
отравленных приманок в сочетании с зерновыми в г. Новочеркасске
обеспечило снижение численности грызупов на 88,6%, а применение
только зерповых отравленных приманок— на 88,4 (наши данные).
В местах обитания, где рацион кормов содержит все необходимые
компопепты, крысы в качестве приманки предпочитают наиболее привыч-
ный корм; например, в свинарниках
— комбикорм, мясной и рыбный
фар, причем с уменьшением плотности их населения такая выбороч-
сть становится наиболее явной (ЙХареа|, 1962; Спиченко, 1962; Ля-
лин, 1978). На рисовых полях Краснодарского края в июле крысы пред-
почитали рис пшенице и ячменю (Шутова и др., 1981). Таким образом,
состав пищевой основы, правильно сбалансированный с обеспеченностью
серых крыс кормами в местах их обитания, может быть ретающим усло-
нием привлекательности отравленной приманки. Как правило, добавка к
пищевой основе аттрактантов (5—10% сахара или 3% растительного
масла) значительно улучшает ее поедаемость (Вагрей, 1956; Огуси Код:
365
Глава пятая
зи, Токумицу Ивао, 1970; Но\уага Е 81.. 1972; Хамагапов, ПТахуров,
1959), хотя иногда аттрактивное действие добавок не проявляется. а-
пример, на рисовых полях Краснодарского края серые крысы ни летом,
ни зимой не оказывали предпочтения зерну с растительным маслом по
сравнению с чистым зерном (Шутова и др., 1981).
|
Поедаемость приманки зависит не только от ев состава, по и от места
ее раскладки. В пепривычных местах крыса берет корм о мого ра
хуже, чом там, гле она привыкла кормиться (Абелепцев, 19 : м
1952; Таскзоп, 1965; Зыхавауа, 1966; Кирсапов, 1967; Дукельская, 48).
Тот или иной тип приманки подбирают таким образом, чтобы ее поеда-
ние зверьками наиболее благоприятетвовало поступнению в их организм
льной дозы ядов.
м
поприманочные препаративные формы. Сущ-
ность способа заключается в покрытии ядом поверхности пор, подземных
и наземпых ходов сообщения. При этом яд должен быть избирательно
токсичен для крыс и малоопасеп для человека и животных.
Большое значение имеет наполнитель, в составе которого применяется
яд. Наиболее часто такие яды «контактного» действия применяются в
виде дустов, основу которых составляют порошковидные наполиители,
которые по способности приставать к волосяному покрову грызунон рас-
полагаются следующим образом: мука, крахмал, соевая мука, тальк, сили-
кагель (Тапака, 1973). В качестве активио действуютщего начала в соота-
вс дустов Танака рекомендует применять сциллирозид и норбомил, о
рые обладают избирательным действием на крыс (Танака и др.,
;
тко\зю, 1967).
_
Поо
Хентель (Новзсве!, 1955), серая крыса способна
унести на себе 444 мг распыленного порошка с ядом аитикоагулянтом и
после пробега на 1 м теряет примерно четверть прилиниего к ве телу
порошка. Следовательно, при недостаточной эффективности оныливатия
этот снособ приводит к интепсизному загрязнению поверхпостен, к раз-
носу яда зверьками и к возможности попадания сто на иитевые про-
ы.
.
К Зидчительно меньшее загрязнение среды получается при использова-
нии искусственных убежищ — ящиков с отверстиями или трубок, напол-
пенпых тнездозым материалом, пропыленным ядом— соломои, сепом, ва-
той, бумагой. Сами по себе искусствепные убежища пе всегда привлеазт
крыс, поэтому в них целесообразно класть приманку (Спиченко, 962;
Трофимов, 4968). Следовательно, повышение эффективности способа опы-
ливания достигается главным образом за счет использования приманок,
и этот способ следует рассматривать как комбинированный.
Кроме дустов, яды могут применяться в составе липких масс.
в
Одним из первых исследователей, изучавших применимость лиикой
массы © яцом для борьбы с крысами и мышами, был В. П. Никитих
(1955), рекомендовавший для лилкой массы использовать смесь из а:
стей гудрона, 5 частей веретенпого масла, $ частей рыбьего жира, р
касторового масла и 8 частей пищевого растительного масла. Ля работы
при отрицательных температурах (—10°..—-22 С) на те же частей
Практическое значение
367
гудрона берется 8 частей веретенного масла, или 10 частей рыбьего жира,
или 15 частей касторового масла, или, пакопец, 10 частей растительного
масла.
Для определения оптимальной концентрации яда в липкой массе
В. П. Пикитин взвешивал крыс и мышей до контакта с липкой массой,
сразу после такового и спустя некоторое время. Установлено, что среднее
количество прилипшей к их лапам массы равпо для мышей 157 мг, а для
крыс 378 мг. Теряют же с лап линкой массы но выходе с отравленного
вещества: крысы— 60 мг, мыши —7 мг. Следовательно, минимальное
действующее количество вещества, прилишиего к грызуну равно: лля
крыс 318 мг, для мышей — 150 мг. С учетом смертельной дозы яда рас-
считаны его онтимальные дозы: для крысида — 10%, фосфида цинка—
20% (эффективность гибели крые -- 90—98%). Соли мышьяка вызывали
смерть только 70% зверьков. Массу в практических условиях папосили
на листы 20Ж20 см слоем толщиной 3 мм и размещали в наиболее по-
сещаемых крысами местах.
Цак показал опыт дератизации, эта рецептура, предлеженная В. П. Ни-
китиным, широкого применения не напгла. Причин тут несколько, однако
наиболее существеппая из пих — избегание грызупами поверхностей, об-
мазанных липкой массой, которая обладает большой вязкостью, особенно
при низких температурах, и поэтому затрудняет передвижение по ней.
Молодые крысы и домовые мыши гибли зачастую в результате прилипа-
ния к ней. Однако идея применения липкой массы в дополнение к прима-
почпому способу дератизации биологически обоспована.
Практика. дератизации также показывает, что применение наряду с
приманочным бесприманочных способов (раскладка опылеиных ватных
тампонов, которые крысы затаскивают себе в гнезда, фумигация, обра-
ботка их нор и троп пенами и липкими составами) обеспечивает высокий
процент гибели серых крыс (Полежаев и др., 1963а; Волкова, Ткац,
1977; Богачева и др., 1981; Арабов, 1982).
|
Несколько позже были разработаны эффективные липкие композиции
на основе пнепищевых продуктов — солидола, нигрола, вазелина— с до-
бавлением ядов, о которых речь пойдет пиже.
В качестве контактных носителей используют не только технические
смазочные масла, но и полупищевые продукты.
Ипзтереспа модификация липкой массы па оспове смеси низкокало-
рийного маргарипа с ядом, наносимой на поверхность, ио которой пере-
мещались животные. По сравпению с желеподобными и воскоподобпыми
составами маргарин не вызывает у грызунов реакции избегания и более
хохотно поедается ими в процессе чистки шерсти (Оау1з, 1983).
Несмотря на то что крысы © большей охотой слизывают полупищевые
продукты по сравнению с техническими смазками, общим недостатком
первых является относительно более быстрая порча или высыхание. Ста-
бильность лииких масс на вазелине, солидоле несравненно выше.
В заключение сделаем несколько общих замечапий по применению:
контактных носителей ядов.
365
Глава пятая
зи, Токумицу Ивао, 1970; Но\уага Е 81.. 1972; Хамагапов, ПТахуров,
1959), хотя иногда аттрактивное действие добавок не проявляется. а-
пример, на рисовых полях Краснодарского края серые крысы ни летом,
ни зимой не оказывали предпочтения зерну с растительным маслом по
сравнению с чистым зерном (Шутова и др., 1981).
|
Поедаемость приманки зависит не только от ев состава, по и от места
ее раскладки. В пепривычных местах крыса берет корм о мого ра
хуже, чом там, гле она привыкла кормиться (Абелепцев, 19 : м
1952; Таскзоп, 1965; Зыхавауа, 1966; Кирсапов, 1967; Дукельская, 48).
Тот или иной тип приманки подбирают таким образом, чтобы ее поеда-
ние зверьками наиболее благоприятетвовало поступнению в их организм
льной дозы ядов.
м
поприманочные препаративные формы. Сущ-
ность способа заключается в покрытии ядом поверхности пор, подземных
и наземпых ходов сообщения. При этом яд должен быть избирательно
токсичен для крыс и малоопасеп для человека и животных.
Большое значение имеет наполнитель, в составе которого применяется
яд. Наиболее часто такие яды «контактного» действия применяются в
виде дустов, основу которых составляют порошковидные наполиители,
которые по способности приставать к волосяному покрову грызунон рас-
полагаются следующим образом: мука, крахмал, соевая мука, тальк, сили-
кагель (Тапака, 1973). В качестве активио действуютщего начала в соота-
вс дустов Танака рекомендует применять сциллирозид и норбомил, о
рые обладают избирательным действием на крыс (Танака и др.,
;
тко\зю, 1967).
_
Поо
Хентель (Новзсве!, 1955), серая крыса способна
унести на себе 444 мг распыленного порошка с ядом аитикоагулянтом и
после пробега на 1 м теряет примерно четверть прилиниего к ве телу
порошка. Следовательно, при недостаточной эффективности оныливатия
этот снособ приводит к интепсизному загрязнению поверхпостен, к раз-
носу яда зверьками и к возможности попадания сто на иитевые про-
ы.
.
К Зидчительно меньшее загрязнение среды получается при использова-
нии искусственных убежищ — ящиков с отверстиями или трубок, напол-
пенпых тнездозым материалом, пропыленным ядом— соломои, сепом, ва-
той, бумагой. Сами по себе искусствепные убежища пе всегда привлеазт
крыс, поэтому в них целесообразно класть приманку (Спиченко, 962;
Трофимов, 4968). Следовательно, повышение эффективности способа опы-
ливания достигается главным образом за счет использования приманок,
и этот способ следует рассматривать как комбинированный.
Кроме дустов, яды могут применяться в составе липких масс.
в
Одним из первых исследователей, изучавших применимость лиикой
массы © яцом для борьбы с крысами и мышами, был В. П. Никитих
(1955), рекомендовавший для лилкой массы использовать смесь из а:
стей гудрона, 5 частей веретенпого масла, $ частей рыбьего жира, р
касторового масла и 8 частей пищевого растительного масла. Ля работы
при отрицательных температурах (—10°..—-22 С) на те же частей
Практическое значение
367
гудрона берется 8 частей веретенного масла, или 10 частей рыбьего жира,
или 15 частей касторового масла, или, пакопец, 10 частей растительного
масла.
Для определения оптимальной концентрации яда в липкой массе
В. П. Пикитин взвешивал крыс и мышей до контакта с липкой массой,
сразу после такового и спустя некоторое время. Установлено, что среднее
количество прилипшей к их лапам массы равпо для мышей 157 мг, а для
крыс 378 мг. Теряют же с лап линкой массы но выходе с отравленного
вещества: крысы— 60 мг, мыши —7 мг. Следовательно, минимальное
действующее количество вещества, прилишиего к грызуну равно: лля
крыс 318 мг, для мышей — 150 мг. С учетом смертельной дозы яда рас-
считаны его онтимальные дозы: для крысида — 10%, фосфида цинка—
20% (эффективность гибели крые -- 90—98%). Соли мышьяка вызывали
смерть только 70% зверьков. Массу в практических условиях папосили
на листы 20Ж20 см слоем толщиной 3 мм и размещали в наиболее по-
сещаемых крысами местах.
Цак показал опыт дератизации, эта рецептура, предлеженная В. П. Ни-
китиным, широкого применения не напгла. Причин тут несколько, однако
наиболее существеппая из пих — избегание грызупами поверхностей, об-
мазанных липкой массой, которая обладает большой вязкостью, особенно
при низких температурах, и поэтому затрудняет передвижение по ней.
Молодые крысы и домовые мыши гибли зачастую в результате прилипа-
ния к ней. Однако идея применения липкой массы в дополнение к прима-
почпому способу дератизации биологически обоспована.
Практика. дератизации также показывает, что применение наряду с
приманочным бесприманочных способов (раскладка опылеиных ватных
тампонов, которые крысы затаскивают себе в гнезда, фумигация, обра-
ботка их нор и троп пенами и липкими составами) обеспечивает высокий
процент гибели серых крыс (Полежаев и др., 1963а; Волкова, Ткац,
1977; Богачева и др., 1981; Арабов, 1982).
|
Несколько позже были разработаны эффективные липкие композиции
на основе пнепищевых продуктов — солидола, нигрола, вазелина— с до-
бавлением ядов, о которых речь пойдет пиже.
В качестве контактных носителей используют не только технические
смазочные масла, но и полупищевые продукты.
Ипзтереспа модификация липкой массы па оспове смеси низкокало-
рийного маргарипа с ядом, наносимой на поверхность, ио которой пере-
мещались животные. По сравпению с желеподобными и воскоподобпыми
составами маргарин не вызывает у грызунов реакции избегания и более
хохотно поедается ими в процессе чистки шерсти (Оау1з, 1983).
Несмотря на то что крысы © большей охотой слизывают полупищевые
продукты по сравнению с техническими смазками, общим недостатком
первых является относительно более быстрая порча или высыхание. Ста-
бильность лииких масс на вазелине, солидоле несравненно выше.
В заключение сделаем несколько общих замечапий по применению:
контактных носителей ядов.
363
Глава пятая
Прежде всего мы считаем недопустимым примепение ядов острого
действия в составе липких композиций, а также для оныливания, так ках
это представляет прямую угрозу здоровью и жизни людей. Прямое нане-
сение контактных носителей с антикоагулянтами на открытые поверхпо-
сти целиком оправданно на некоторых специфических объектах, нело-
стунпных для человека и домашних животпых
— свалки отходов и капа-
лизация. В помещениях более гигиенично и при этом достаточно эффек-
тивно нанесение контактпых посителей с ядом на дно ящиков, коробок
или просто на подложки из рубероида, толстого полиэтилена или картона,
что предотвразцает стойкое загрязнение полов ратицидами,
Яды острого действия
Подача ядов острого действия с пищевой основой вызывает гибель крыс
после одпократного посдапия ими приманки. К пим относятся кремне-
фтористый натрий, углекислый барий, соединения мышьяка, желтый
фосфор, фосфид цинка, сульфат таллия и другие неорганические соеди-
нения, а также: 1) органические растительные яды — стрихпин, сцилли-
розид (препарат красного морского лука), фторацетат натрия (1080)
и 2) органические синтетические яды — крысид (альфапафтилтиомочеви-
на), тиосемикарбазид, промурит, фторацетамид (1084), фторацетат бария,
монофторип, глифтор, порбомид (шоксин), вакор (ВН — 787) и др. Цры-
сы распознают яды по вкусу, а также при наступлении симптомов отрав-
лепия. Отличительной особенностью большинства перечисленных ядов
является то, что они начинают вызывать симптомы отравления у крысе
с первого же часа после попадания в организм. Особенно хоротпо расло-
знаются зверьками углекислый барий, кремпефтористый патрий, крысил.
Симптомы отравления фторацетамидом и сульфатом талия развиваются
сравнительно медленнее: эти яды хорошо поздаются в составе приманок
на фоне неотравленного корма.
Крысы легко распознают яды острого действия в сухих приманках,
тогда как влажные поедаются ими. Примапки с пизкой копцептрациоей
яда также поедаются лучшие (Огуси НКодзи, Токумицу Ивао, 1970; Ду-
кельская, 1960).
Многие из перечисленных ядов по ряду причин изъяты из примене-
ния в практике здравоохранения ряда стран. Соединения мышьяка, жел-
тый фосфор, сульфат таллия, стрихпип, изъяты, например, из-за высокой
токсичности для домашних животных и человека. Н применению в систе-
ме здравоохрапепия СССР разрешены следующие препараты из числа
ядов острого действия: фосфид цинка, фторацетамид, монофторин. Реаль-
но же используется только фосфид цинка, да и то строго регламентиро-
ванно. Два других из-за очень высокой токсичности применяются лишь
в экстренных случаях и очень локально.
Из многочисленной труппы ядов острого действия наибольшее рас-
пространение получил, как мы уже говорили, фосфид цинка (ЙпзР»)
который, попадая в желудок крысы, реагирует с соляной кислотой и вы-
деляет фосфористый водород (РНз), проникающий в кровь, мозг и дей-
л
ь
л
е
т
ь
и
а
м
ы
Практическое значение 369
ствующий на дыхательный центр. При рекомендуемой инструкцией коп-
центрации его (3%) в прималке этот яд отпосительно менее опасен, чем
мпогие другие, и не вызывает вторичных отравлений. у хищников,
съевптих отравленных крыс (НШ. Сагрепег, 1982; Нееда| её а|., 1984).
Фторацетаты бария, патрия, фторацетамид, сульфат таллия, тиосеми-
карбазид весьма опасны в обращении, вызывают вторичные отравлепия
у хищников, и их применят крайпе редко. Яды, избирательно токсич-
ные для крыс (крысид, норбомид), очень илохо поедаются ими (Никифо-
ров и др., 1963; Трофимов, 1968; Огуси ЦКодзи, Токумицу Ивао, 1970;
Такада Садаото, 1982).
Применение ядов острого действия без прикорма вызывает гибель
50—70% серых крыс, папример фосфид цинка (Спиченко, 1962; Рыль-
ников и др., 1983а; Немченко, 1986), и 87% их в случае фторацетамида
(Рыльников и др., 1983а). Па пекоторых объектах эффективность дейст-
вия фосфида цинка достигает 95—97%, как это имело место в животно-
водческих помещениях Казахстана (Цозлов, 1980а). Это может быть свя-
запо с тем, что приманку с ядом раскладывали в местах, где вместе со
свиньями привыкли питаться и крысы, что апалогично действию предва-
рительного прикорма.
Сравнительные испытания поедаемости серыми крысами приманок с
различными ядами острого действия показали, что чем быстрее зверьки
распознают яд, тем меныне они едят приманку с пим и тем меньше их
погибает (Айзенштадт, 1957; Дукольская, 1948).
В последпее время все чаще появляются работы, в которых указыва-
ется на то, что реакция вторичного избегания отравленной приманки вы-
рабатывается пе столько па яд, сколько на пищевую основу в целом и
на входящие в нее компоненты (сахар, растительное масло и другие ат-
трактанты, внешний вид пищевой основы), па которые у подтравленных
животных вырабатывается условный рефлекс. Это явление показано как
_на серых (Траханов, 1963) и черных крысах (ВВатамау, Ртаказь, 41979:
Таш, Рауде, 1981), так и на другнх видах грызунов (КВап, 9199,
1980; ОПуег её а1., 1982). По мнению этих авторов, наиболее эффектив-
ный с10с0б преодоления реакции вторичпого избегания
— чередование
пищевой основы, аттрактантов, а также ядов. Однако последовательное
чередование компонентов отравленной приманки не всегда приносит по-
ложительные результаты, так как часто оказывается невозможным вы-
яенить характер и причины избегания приманок (неофобия, или
аверсия). Наряду с совершенствовапием тактики, осповапной на чере-
довании приманок (Быковский, Николаева, 1986), более перспективным
может оказаться прием, когда крысам предлагают параллельно несколь-
`ко приманок с ядами острого действия. Например, Штайнигер (Зем:
хег,. 1957) уничтожил остаточную популяцию серых крыс, применив
. 15-20 различпых сочетаний приманок и ядов острого действия, вызы-
вающих наименьшую реакцию настороженности, а именно фторацетаты
натрия и бария, сульфат таллия. Пе исключено, что серые крысы в ус-
ловиях широкого выбора приманок оказываются неспособными диффе-
ренцировать их по степени опасности.
363
Глава пятая
Прежде всего мы считаем недопустимым примепение ядов острого
действия в составе липких композиций, а также для оныливания, так ках
это представляет прямую угрозу здоровью и жизни людей. Прямое нане-
сение контактных носителей с антикоагулянтами на открытые поверхпо-
сти целиком оправданно на некоторых специфических объектах, нело-
стунпных для человека и домашних животпых
— свалки отходов и капа-
лизация. В помещениях более гигиенично и при этом достаточно эффек-
тивно нанесение контактпых посителей с ядом на дно ящиков, коробок
или просто на подложки из рубероида, толстого полиэтилена или картона,
что предотвразцает стойкое загрязнение полов ратицидами,
Яды острого действия
Подача ядов острого действия с пищевой основой вызывает гибель крыс
после одпократного посдапия ими приманки. К пим относятся кремне-
фтористый натрий, углекислый барий, соединения мышьяка, желтый
фосфор, фосфид цинка, сульфат таллия и другие неорганические соеди-
нения, а также: 1) органические растительные яды — стрихпин, сцилли-
розид (препарат красного морского лука), фторацетат натрия (1080)
и 2) органические синтетические яды — крысид (альфапафтилтиомочеви-
на), тиосемикарбазид, промурит, фторацетамид (1084), фторацетат бария,
монофторип, глифтор, порбомид (шоксин), вакор (ВН — 787) и др. Цры-
сы распознают яды по вкусу, а также при наступлении симптомов отрав-
лепия. Отличительной особенностью большинства перечисленных ядов
является то, что они начинают вызывать симптомы отравления у крысе
с первого же часа после попадания в организм. Особенно хоротпо расло-
знаются зверьками углекислый барий, кремпефтористый патрий, крысил.
Симптомы отравления фторацетамидом и сульфатом талия развиваются
сравнительно медленнее: эти яды хорошо поздаются в составе приманок
на фоне неотравленного корма.
Крысы легко распознают яды острого действия в сухих приманках,
тогда как влажные поедаются ими. Примапки с пизкой копцептрациоей
яда также поедаются лучшие (Огуси НКодзи, Токумицу Ивао, 1970; Ду-
кельская, 1960).
Многие из перечисленных ядов по ряду причин изъяты из примене-
ния в практике здравоохранения ряда стран. Соединения мышьяка, жел-
тый фосфор, сульфат таллия, стрихпип, изъяты, например, из-за высокой
токсичности для домашних животных и человека. Н применению в систе-
ме здравоохрапепия СССР разрешены следующие препараты из числа
ядов острого действия: фосфид цинка, фторацетамид, монофторин. Реаль-
но же используется только фосфид цинка, да и то строго регламентиро-
ванно. Два других из-за очень высокой токсичности применяются лишь
в экстренных случаях и очень локально.
Из многочисленной труппы ядов острого действия наибольшее рас-
пространение получил, как мы уже говорили, фосфид цинка (ЙпзР»)
который, попадая в желудок крысы, реагирует с соляной кислотой и вы-
деляет фосфористый водород (РНз), проникающий в кровь, мозг и дей-
л
ь
л
е
т
ь
и
а
м
ы
Практическое значение 369
ствующий на дыхательный центр. При рекомендуемой инструкцией коп-
центрации его (3%) в прималке этот яд отпосительно менее опасен, чем
мпогие другие, и не вызывает вторичных отравлений. у хищников,
съевптих отравленных крыс (НШ. Сагрепег, 1982; Нееда| её а|., 1984).
Фторацетаты бария, патрия, фторацетамид, сульфат таллия, тиосеми-
карбазид весьма опасны в обращении, вызывают вторичные отравлепия
у хищников, и их применят крайпе редко. Яды, избирательно токсич-
ные для крыс (крысид, норбомид), очень илохо поедаются ими (Никифо-
ров и др., 1963; Трофимов, 1968; Огуси ЦКодзи, Токумицу Ивао, 1970;
Такада Садаото, 1982).
Применение ядов острого действия без прикорма вызывает гибель
50—70% серых крыс, папример фосфид цинка (Спиченко, 1962; Рыль-
ников и др., 1983а; Немченко, 1986), и 87% их в случае фторацетамида
(Рыльников и др., 1983а). Па пекоторых объектах эффективность дейст-
вия фосфида цинка достигает 95—97%, как это имело место в животно-
водческих помещениях Казахстана (Цозлов, 1980а). Это может быть свя-
запо с тем, что приманку с ядом раскладывали в местах, где вместе со
свиньями привыкли питаться и крысы, что апалогично действию предва-
рительного прикорма.
Сравнительные испытания поедаемости серыми крысами приманок с
различными ядами острого действия показали, что чем быстрее зверьки
распознают яд, тем меныне они едят приманку с пим и тем меньше их
погибает (Айзенштадт, 1957; Дукольская, 1948).
В последпее время все чаще появляются работы, в которых указыва-
ется на то, что реакция вторичного избегания отравленной приманки вы-
рабатывается пе столько па яд, сколько на пищевую основу в целом и
на входящие в нее компоненты (сахар, растительное масло и другие ат-
трактанты, внешний вид пищевой основы), па которые у подтравленных
животных вырабатывается условный рефлекс. Это явление показано как
_на серых (Траханов, 1963) и черных крысах (ВВатамау, Ртаказь, 41979:
Таш, Рауде, 1981), так и на другнх видах грызунов (КВап, 9199,
1980; ОПуег её а1., 1982). По мнению этих авторов, наиболее эффектив-
ный с10с0б преодоления реакции вторичпого избегания
— чередование
пищевой основы, аттрактантов, а также ядов. Однако последовательное
чередование компонентов отравленной приманки не всегда приносит по-
ложительные результаты, так как часто оказывается невозможным вы-
яенить характер и причины избегания приманок (неофобия, или
аверсия). Наряду с совершенствовапием тактики, осповапной на чере-
довании приманок (Быковский, Николаева, 1986), более перспективным
может оказаться прием, когда крысам предлагают параллельно несколь-
`ко приманок с ядами острого действия. Например, Штайнигер (Зем:
хег,. 1957) уничтожил остаточную популяцию серых крыс, применив
. 15-20 различпых сочетаний приманок и ядов острого действия, вызы-
вающих наименьшую реакцию настороженности, а именно фторацетаты
натрия и бария, сульфат таллия. Пе исключено, что серые крысы в ус-
ловиях широкого выбора приманок оказываются неспособными диффе-
ренцировать их по степени опасности.
370
Глава пятая
Наилучший результат приманки с ядами острого действия дают в тех
случаях, когда крысам сначала некоторое время предлагают корм без
яда, а затем точпо такой же корм с ядом. Этот прием получил назвапие
предварительного прикорма. Длительность предварительного прикорма
может составлять до двух недель и более в зависимости от осторожности
крыс и привлекательпости предложенной приманки (СВ\у, 1954).
Фосфид цинка не рекомендуют примепять в липких массах или в виде
дуста из-за его высокой токсичности но отношению к другим видам жи-
вотных (Траханов, 1964). Но известпы случаи его применения в составе
липких композиций. В. О. Берулава (1972), например, прибавлял 10%
этого яла к массе солидола, а С. В. Вишняков и Н. М. Дукельская
(1957) брали 20% к массе патоки. Названные составы применяли для
обмазки входов нор и троп.
Яды кумулятивного действия
1. Антикоагулянпты. Наибольшую долю среди ядов кумулятивного
действия составляют антикоагулянты крови из группы кумарина (варфа-
рин или зоокумарин, кумахлор, дикумарол и др.) и индадиона (дифена-
цин, фентолацин и др.).
Открытые в 1944 г. дикумарин (Дукельская, 1960), в 1942 г. варфа-
рин (Негтани, 1969), позже ипдадион совертили настоящую революцию
в дератизации. При однократном попадании в оргапиам крыс небольитих
количеств отих ядов симптомы отравления практически не проявляются,
однако при многократном потреблении аптикоагулянтов их токсичность
значительно возрастает не столько в результате накопления яда в орга-
низме, сколько снижения уровпя протромбина, вызывающего нарушения
свертывающей системы крови, что сопровождается увеличением пропи-
цаемости сосудов, кровоизлияпиями во многих внутренних органах и
кожных иокровах, вызывая последующую гибель (Туды Клазапо, 1960).
Кумулятивпая смертельная доза варфарина для серой крысы со-
ставляет 4—8 мг/кг при подаче в течепие 4 дпей (Козакевич и др.
1958; Дукельская, 1960), что близко к токсичности фторацетатов и при-
мерно в 10 раз токсичнее фосфида ципка. В связи с низкой оборонитель-
ной реакцией на антикоагулянты количество поедаемой грызупами при-
манки с этими ядами спижается более медленно по сравнению © ядами
острого действия (ТеПе, 1963; Рароацег, МеСаип, 1976}.
Для упичтожения серой крысы достаточна 0,0054 -ная концентрация
зарфарина, не вызывающая вкусового отвращения. Повышение концент-
рации его до 0,01% в приманке понижает ее поедаемость в 2 раза,
а до 0,054 —в 4 раза (ТеПе, 1963). Н. М. Дукельская (1957) в лабора-
торных опытах показала, что сокращение дачи приманки до 2—8 раз,
так же как и уменьшение интервалов между завтраками до 6—12 ч или
их увеличение до 48 и 72 ч, снижает эффект препарата. Таким образом,
эффективпость варфарина зависит как от числа повторных введений, так
и от их интервалов.
Нрактическое значение
371
Несмотря на большую токсичность варфарина по сравнению со мно-
тими ядами острого действия, опасность отравления им человека и до-
маптних животных значительно ниже, во-первых, ввиду пеобходимых для
этого более высоких пороговых доз яда, во-вторых, из-за доступности и эф-
фективности антидота — витамина К (Дукельская, 1960).
Кроме варфарина, в СССР налажено промызиленное производство еще
двух антикоагулянтов: дифенацина (ратиндан 1 или дифенилацетил-инда-
дион — 1,3) и фентолацина (ратиндан 2 или 2-фепилтолилацетил-индади-
оп— 1,3). При разовом пероральном введении минимальная летальная
доза фентолацина для серых крыс составляет 4 мг/кг, а дифепаципа—
5 мг/кг. Кумулятивная ЛД. для обоих препаратов составляет в сред-
нем 2 мг/кг в течение 4—8 суток. Оборонительная реакция отсутствует
(Фалькенштейн, Егорова, 196/). Безусловным преимуществом этих ядов
является то, что они в течение нескольких лет не теряют своих токсиче-
ских свойств при хранении (Туманов, 1964).
Давая характеристику ядов антикоагулянтов отечественного ироизвод-
ства, И. Ц. Туров (1970) обращает внимание на сравнительно более быст-
рый отказ от поедания примапки с обоими видами ратиндана и на более
короткие сроки гибели от него грызунов по сравнению с зоокумарином.
В литературе появляются попытки (Немчепко, 1986) использовать
дифенацин в расчете на одноразовое поедание за счет увеличения концен-
трации с 0,015 до 0,5%. Однократное поедание 2 г зерна с 0,5% дифена-
ципа приводит к гибели 100% особей в течение 5—7 дней. Как считает
автор этих материалов, такая высокая концентрация яда может оказать-
сл особенно пригодной па объектах с хорошей кормовой базой.
Мы намеренно ограничиваемся рассмотрением только антикоагулянтов
отечественного производства, так как применение зарубежных ядов про-
тивосвертыватющего действия как против серых и черных крыс, так и
против многих других грызунов подробно рассмотрено Бентли (Веп@еу,
1972) и Лупдом (Тлиа, 1977).
Преимущества ядов антикоагулянтов реализуются не.всегда, а только
при соблюдении определенных условий. Прежде всего это зависит от ко-
личества приманки в местах раскладки. Шри высокой численности крыс
приманку © антикоагулянтом рекомендуется класть в количествах от |
до 5 кг па одпо место (Заепиеог, 1955, 1956, 1957). В тех случаях, когда
численность крыс определить трудно, папример при обработке канализа-
ционных колодцев, порции приманки должиы составлять не менее 1 кг
{ КгеВБего, 1963). Многие исследователи рекомендуют класть приманку
з избытке дозами не менее 250 г (ЗшИВ, 1963). Подобная раскладка
обеспечивает ее поедание крысами неоднократно, что необходимо для на-
копления ими летальной дозы.
Количество мест раскладки таклке играет роль. При раскладке при-
манки во множестве мест вероятность ее поедания крысами возрастает и
эффективность борьбы повышается. Если каждая порция рассредоточена
в нескольких рядом стоящих лотках, то это облегчает одновременное по-
требление корма зверьками разного ранга, устраняя внутривидовую кон-
куренцию у мест кормежек (\Уез, ГаП, 1975). Уменьшение числа мест
370
Глава пятая
Наилучший результат приманки с ядами острого действия дают в тех
случаях, когда крысам сначала некоторое время предлагают корм без
яда, а затем точпо такой же корм с ядом. Этот прием получил назвапие
предварительного прикорма. Длительность предварительного прикорма
может составлять до двух недель и более в зависимости от осторожности
крыс и привлекательпости предложенной приманки (СВ\у, 1954).
Фосфид цинка не рекомендуют примепять в липких массах или в виде
дуста из-за его высокой токсичности но отношению к другим видам жи-
вотных (Траханов, 1964). Но известпы случаи его применения в составе
липких композиций. В. О. Берулава (1972), например, прибавлял 10%
этого яла к массе солидола, а С. В. Вишняков и Н. М. Дукельская
(1957) брали 20% к массе патоки. Названные составы применяли для
обмазки входов нор и троп.
Яды кумулятивного действия
1. Антикоагулянпты. Наибольшую долю среди ядов кумулятивного
действия составляют антикоагулянты крови из группы кумарина (варфа-
рин или зоокумарин, кумахлор, дикумарол и др.) и индадиона (дифена-
цин, фентолацин и др.).
Открытые в 1944 г. дикумарин (Дукельская, 1960), в 1942 г. варфа-
рин (Негтани, 1969), позже ипдадион совертили настоящую революцию
в дератизации. При однократном попадании в оргапиам крыс небольитих
количеств отих ядов симптомы отравления практически не проявляются,
однако при многократном потреблении аптикоагулянтов их токсичность
значительно возрастает не столько в результате накопления яда в орга-
низме, сколько снижения уровпя протромбина, вызывающего нарушения
свертывающей системы крови, что сопровождается увеличением пропи-
цаемости сосудов, кровоизлияпиями во многих внутренних органах и
кожных иокровах, вызывая последующую гибель (Туды Клазапо, 1960).
Кумулятивпая смертельная доза варфарина для серой крысы со-
ставляет 4—8 мг/кг при подаче в течепие 4 дпей (Козакевич и др.
1958; Дукельская, 1960), что близко к токсичности фторацетатов и при-
мерно в 10 раз токсичнее фосфида ципка. В связи с низкой оборонитель-
ной реакцией на антикоагулянты количество поедаемой грызупами при-
манки с этими ядами спижается более медленно по сравнению © ядами
острого действия (ТеПе, 1963; Рароацег, МеСаип, 1976}.
Для упичтожения серой крысы достаточна 0,0054 -ная концентрация
зарфарина, не вызывающая вкусового отвращения. Повышение концент-
рации его до 0,01% в приманке понижает ее поедаемость в 2 раза,
а до 0,054 —в 4 раза (ТеПе, 1963). Н. М. Дукельская (1957) в лабора-
торных опытах показала, что сокращение дачи приманки до 2—8 раз,
так же как и уменьшение интервалов между завтраками до 6—12 ч или
их увеличение до 48 и 72 ч, снижает эффект препарата. Таким образом,
эффективпость варфарина зависит как от числа повторных введений, так
и от их интервалов.
Нрактическое значение
371
Несмотря на большую токсичность варфарина по сравнению со мно-
тими ядами острого действия, опасность отравления им человека и до-
маптних животных значительно ниже, во-первых, ввиду пеобходимых для
этого более высоких пороговых доз яда, во-вторых, из-за доступности и эф-
фективности антидота — витамина К (Дукельская, 1960).
Кроме варфарина, в СССР налажено промызиленное производство еще
двух антикоагулянтов: дифенацина (ратиндан 1 или дифенилацетил-инда-
дион — 1,3) и фентолацина (ратиндан 2 или 2-фепилтолилацетил-индади-
оп— 1,3). При разовом пероральном введении минимальная летальная
доза фентолацина для серых крыс составляет 4 мг/кг, а дифепаципа—
5 мг/кг. Кумулятивная ЛД. для обоих препаратов составляет в сред-
нем 2 мг/кг в течение 4—8 суток. Оборонительная реакция отсутствует
(Фалькенштейн, Егорова, 196/). Безусловным преимуществом этих ядов
является то, что они в течение нескольких лет не теряют своих токсиче-
ских свойств при хранении (Туманов, 1964).
Давая характеристику ядов антикоагулянтов отечественного ироизвод-
ства, И. Ц. Туров (1970) обращает внимание на сравнительно более быст-
рый отказ от поедания примапки с обоими видами ратиндана и на более
короткие сроки гибели от него грызунов по сравнению с зоокумарином.
В литературе появляются попытки (Немчепко, 1986) использовать
дифенацин в расчете на одноразовое поедание за счет увеличения концен-
трации с 0,015 до 0,5%. Однократное поедание 2 г зерна с 0,5% дифена-
ципа приводит к гибели 100% особей в течение 5—7 дней. Как считает
автор этих материалов, такая высокая концентрация яда может оказать-
сл особенно пригодной па объектах с хорошей кормовой базой.
Мы намеренно ограничиваемся рассмотрением только антикоагулянтов
отечественного производства, так как применение зарубежных ядов про-
тивосвертыватющего действия как против серых и черных крыс, так и
против многих других грызунов подробно рассмотрено Бентли (Веп@еу,
1972) и Лупдом (Тлиа, 1977).
Преимущества ядов антикоагулянтов реализуются не.всегда, а только
при соблюдении определенных условий. Прежде всего это зависит от ко-
личества приманки в местах раскладки. Шри высокой численности крыс
приманку © антикоагулянтом рекомендуется класть в количествах от |
до 5 кг па одпо место (Заепиеог, 1955, 1956, 1957). В тех случаях, когда
численность крыс определить трудно, папример при обработке канализа-
ционных колодцев, порции приманки должиы составлять не менее 1 кг
{ КгеВБего, 1963). Многие исследователи рекомендуют класть приманку
з избытке дозами не менее 250 г (ЗшИВ, 1963). Подобная раскладка
обеспечивает ее поедание крысами неоднократно, что необходимо для на-
копления ими летальной дозы.
Количество мест раскладки таклке играет роль. При раскладке при-
манки во множестве мест вероятность ее поедания крысами возрастает и
эффективность борьбы повышается. Если каждая порция рассредоточена
в нескольких рядом стоящих лотках, то это облегчает одновременное по-
требление корма зверьками разного ранга, устраняя внутривидовую кон-
куренцию у мест кормежек (\Уез, ГаП, 1975). Уменьшение числа мест
372
Глава пятая
раскладки и порций приманки с антикоагулянтами может быть серьезной
ошибкой.
По сообщению Заплеталя (Йарейа!, 1962), в процессе дератизации
серые крысы потребляют только наиболее обильный на объекте корм.
Тем самым они избегают новых, непривычных приманок, что позноляет
им выжить. Таким образом, раскладка крулных порций примапки с анти-
коагулянтами позволяет уменьшить пастороженноеть зверьков за счет
создания у них представления об отравленной приманке как об основном
корме.
.
РА тикоагулянты в больших дозах (более 0,05% к массе пищевой ос-
новы) вызызают избегание приманок. По сообщению Телле (ТеПе,
4967), ло 10% крыс могут вообще не брать приманку из-за того, что они
ощущают в ней привкус яда. Если в таких случаях использовать при-
манки, в которых одновременно имеется два различных яда, то количе-
ство зверьков, не берущих приманку, сокращается до 1—5 %. При истреб-
лении крыс с помощью приманок, содержащих не два, а три различных
антикоагулянта, доля крыс, взявших ее, повышается до 100 ф, т. е. про-
исходит полное истребление популяции. Эти данные согласуются с дап-
ными Птайнигера (З\епиоет, 1957) о том, что в условиях разнообразия
отравленных рецептур крысы оказываются неспособными дифферепциро-
вать их по степени опасности.
В тех случаях, когда у крыс имеется настороженность по отпошению
ко вновь предложенному корму, очень большую роль играет длительная
сохранность приманки. Не случайно массоное истребление отих грызунов
в населенных пунктах ФРГ и других региопах осуществляется с помощью
долго сохраняющихся приманок из сухих мучных или зерновых продук-
тов (кукурузной дерти, овсяпых хлопьев, муки). Высокая их сохрап-
ность оказывается более важным фактором, чем повышенная вкусовая
привлекательность влажных кормов.
Использование сухих приманок с ядами-антикоагулянтами позволило
довести уничтожение крыс в популяции до 98—99%, а в некоторых слу-
чаях и полностью. Если осторожные зверьки не брали приманку с анти-
коагулянтами, то эти яды применяли для опыливания нор, троп и поверх-
постей, по которым они перемещаются. Именно благодаря ядам-антико-
агулянтам были пачаты первые опыты по практически полному
уничтожению серых крыс в масштабах целото населенного пункта или
торода и нотом проводились во множестве городов (Бентли, 1967).
Хорошие результаты были получены при глубоком пропыливании нор
серых крыс дустом кумарина вне построек человека (ТеПе, 1962), а так-
же 1%-ным дустом варфарина в животноводческих помещениях. В то же
время 0,5%-ный дуст варфарина был малоэффективен при пронылива-
нии земляных нор и несколько лучше действовал при наносении па глад-
кие поверхности (Траханов, 1964). После многократного опыливания
0,5%-ным дустом зоокумарина удалось из 146 объектов населенного
пункта освободить только 28 (20,6%). На 214 объекте численность серых
крыс зпачительно снизилась, но на большей части объектов— 97— она
осталась без изменения. При использовании 1,0$-ного дуста зоокумарина
Практическое значение 373
бло полпостьо освобождено не более 30% объектов. По мнению
В. Г. Полежаева с соавторами (1963), опыливание может быть лишь до-
полнительным, а не основпым способом борьбы с крысами (Полежаев
и др., 1963а). В качестве контактного носителя использовали солидол
(тавот) с добавлением 0,5%-ных дустов зоокумарина и дифенацина в
количестве 25—30% к массе солидола. Чаще использовали солидол ©
зоокумарином. Солидоловую пасту с ратицидом наносили слоем в 2—
3 мм на поверхности троп, ходов, а при необходимости и на материалы,
которыми тампонировали норы. Обработку повторяли через 4—5 дпей,
после чего норы и ходы заделызвали. Расход солидоловой пасты составил
30 г на 1 крысиную и 5—7 г на 1 мышиную нору. В течение 4 месяцев
из 350 объектов, заселенных крысами, было освобождено 294 (эффектив-
пость 84%) (Берулава, 1972). Пригодными для контактного нанесения
оказались пены, однако они сохраняются только в течение двух недель
(Злешижег, 1953; Траханов, 1963) и крысы способны распознавать поверх-
ности, обработанные ими (Пгапипоп@, 1960; Трахапов, 1963).
Каждая из описапиых липких композиций имеет свои плюсы и мину-
сы, которые, как правило, выяснялись уже в процессе практических ис-
пытаний. В. С. Пуцято (1986, 1987) впервые была проведена лаборатор-
ная оценка покрытий из разных ядовитых композиций. Наибольнтая ги-
бель крыс наблюдалась от пенокумарипа и в мепьшей степени от
солидола или пигрола с 0,5%-ным зоокумарином (табл. 35). Более позд-
ние исследования В. С. Пуцято (личное сообщение) позволили выделить
в число наилучиих «посителей» вазелин с 0,5% зоокумарина и 49,5%
талька (талькомагнезита). Исследования на практике показали, что эф-
фективность этой композиции при условии нанесения ее вокруг кормуш-
ки с неотравленным кормом (служащим только для привлечения крыс)
полосой в ширину 25—30 см составила 80—90%.
Добавление 5% тотрациклина в вазелиновую пасту с 0,5% зоокума-
рина и последующий поиск маркера на поперечных шлифах верхних рез-
цов серых крыс (определепие количества и интенсивности свечения в
УФ-лучах меток тетрациклина) позволили проанализировать причины
их выживаемости и гибели. Установлено, что подавляющее большинство
крыс (и самцов, и самок) контактировали с поверхностью, намазанпой
вазелиновой пастой. В течение 2 недель с момента ее нанесения были
уничтожепы практически все крысы. Молодые самцы чаще вступали в
контакт © вазелиновой пастой, чем взрослые, одпако получали более
низкие дозы яда, в результате чего медианы суммарных доз зоокумарина
У молодых и взрослых самцов из числа погибших примерно равпы. Мо-
модые самки более активпо контактировали с вазелиновой пастой в лер-
вые б дней после ее нанесения, доля погибитих молодых была несколько
вылте, чем взрослых. В целом же среди молодых и взрослых обоих полов
выживаемость примерно одинакова — 10,47-4,00 и 7,02=3,41% соответст-
венно. Сопоставление этих данных с материалами, получеппыми при ана-
лизе поедаемости отравленной приманки с 0,025$ зоокумарина, полу-
ченными в аналогичных условиях подвалов многоэтажных коммунальных
зданий, позволило сделать следующие выводы: 1) эффективность вазели-
372
Глава пятая
раскладки и порций приманки с антикоагулянтами может быть серьезной
ошибкой.
По сообщению Заплеталя (Йарейа!, 1962), в процессе дератизации
серые крысы потребляют только наиболее обильный на объекте корм.
Тем самым они избегают новых, непривычных приманок, что позноляет
им выжить. Таким образом, раскладка крулных порций примапки с анти-
коагулянтами позволяет уменьшить пастороженноеть зверьков за счет
создания у них представления об отравленной приманке как об основном
корме.
.
РА тикоагулянты в больших дозах (более 0,05% к массе пищевой ос-
новы) вызызают избегание приманок. По сообщению Телле (ТеПе,
4967), ло 10% крыс могут вообще не брать приманку из-за того, что они
ощущают в ней привкус яда. Если в таких случаях использовать при-
манки, в которых одновременно имеется два различных яда, то количе-
ство зверьков, не берущих приманку, сокращается до 1—5 %. При истреб-
лении крыс с помощью приманок, содержащих не два, а три различных
антикоагулянта, доля крыс, взявших ее, повышается до 100 ф, т. е. про-
исходит полное истребление популяции. Эти данные согласуются с дап-
ными Птайнигера (З\епиоет, 1957) о том, что в условиях разнообразия
отравленных рецептур крысы оказываются неспособными дифферепциро-
вать их по степени опасности.
В тех случаях, когда у крыс имеется настороженность по отпошению
ко вновь предложенному корму, очень большую роль играет длительная
сохранность приманки. Не случайно массоное истребление отих грызунов
в населенных пунктах ФРГ и других региопах осуществляется с помощью
долго сохраняющихся приманок из сухих мучных или зерновых продук-
тов (кукурузной дерти, овсяпых хлопьев, муки). Высокая их сохрап-
ность оказывается более важным фактором, чем повышенная вкусовая
привлекательность влажных кормов.
Использование сухих приманок с ядами-антикоагулянтами позволило
довести уничтожение крыс в популяции до 98—99%, а в некоторых слу-
чаях и полностью. Если осторожные зверьки не брали приманку с анти-
коагулянтами, то эти яды применяли для опыливания нор, троп и поверх-
постей, по которым они перемещаются. Именно благодаря ядам-антико-
агулянтам были пачаты первые опыты по практически полному
уничтожению серых крыс в масштабах целото населенного пункта или
торода и нотом проводились во множестве городов (Бентли, 1967).
Хорошие результаты были получены при глубоком пропыливании нор
серых крыс дустом кумарина вне построек человека (ТеПе, 1962), а так-
же 1%-ным дустом варфарина в животноводческих помещениях. В то же
время 0,5%-ный дуст варфарина был малоэффективен при пронылива-
нии земляных нор и несколько лучше действовал при наносении па глад-
кие поверхности (Траханов, 1964). После многократного опыливания
0,5%-ным дустом зоокумарина удалось из 146 объектов населенного
пункта освободить только 28 (20,6%). На 214 объекте численность серых
крыс зпачительно снизилась, но на большей части объектов— 97— она
осталась без изменения. При использовании 1,0$-ного дуста зоокумарина
Практическое значение 373
бло полпостьо освобождено не более 30% объектов. По мнению
В. Г. Полежаева с соавторами (1963), опыливание может быть лишь до-
полнительным, а не основпым способом борьбы с крысами (Полежаев
и др., 1963а). В качестве контактного носителя использовали солидол
(тавот) с добавлением 0,5%-ных дустов зоокумарина и дифенацина в
количестве 25—30% к массе солидола. Чаще использовали солидол ©
зоокумарином. Солидоловую пасту с ратицидом наносили слоем в 2—
3 мм на поверхности троп, ходов, а при необходимости и на материалы,
которыми тампонировали норы. Обработку повторяли через 4—5 дпей,
после чего норы и ходы заделызвали. Расход солидоловой пасты составил
30 г на 1 крысиную и 5—7 г на 1 мышиную нору. В течение 4 месяцев
из 350 объектов, заселенных крысами, было освобождено 294 (эффектив-
пость 84%) (Берулава, 1972). Пригодными для контактного нанесения
оказались пены, однако они сохраняются только в течение двух недель
(Злешижег, 1953; Траханов, 1963) и крысы способны распознавать поверх-
ности, обработанные ими (Пгапипоп@, 1960; Трахапов, 1963).
Каждая из описапиых липких композиций имеет свои плюсы и мину-
сы, которые, как правило, выяснялись уже в процессе практических ис-
пытаний. В. С. Пуцято (1986, 1987) впервые была проведена лаборатор-
ная оценка покрытий из разных ядовитых композиций. Наибольнтая ги-
бель крыс наблюдалась от пенокумарипа и в мепьшей степени от
солидола или пигрола с 0,5%-ным зоокумарином (табл. 35). Более позд-
ние исследования В. С. Пуцято (личное сообщение) позволили выделить
в число наилучиих «посителей» вазелин с 0,5% зоокумарина и 49,5%
талька (талькомагнезита). Исследования на практике показали, что эф-
фективность этой композиции при условии нанесения ее вокруг кормуш-
ки с неотравленным кормом (служащим только для привлечения крыс)
полосой в ширину 25—30 см составила 80—90%.
Добавление 5% тотрациклина в вазелиновую пасту с 0,5% зоокума-
рина и последующий поиск маркера на поперечных шлифах верхних рез-
цов серых крыс (определепие количества и интенсивности свечения в
УФ-лучах меток тетрациклина) позволили проанализировать причины
их выживаемости и гибели. Установлено, что подавляющее большинство
крыс (и самцов, и самок) контактировали с поверхностью, намазанпой
вазелиновой пастой. В течение 2 недель с момента ее нанесения были
уничтожепы практически все крысы. Молодые самцы чаще вступали в
контакт © вазелиновой пастой, чем взрослые, одпако получали более
низкие дозы яда, в результате чего медианы суммарных доз зоокумарина
У молодых и взрослых самцов из числа погибших примерно равпы. Мо-
модые самки более активпо контактировали с вазелиновой пастой в лер-
вые б дней после ее нанесения, доля погибитих молодых была несколько
вылте, чем взрослых. В целом же среди молодых и взрослых обоих полов
выживаемость примерно одинакова — 10,47-4,00 и 7,02=3,41% соответст-
венно. Сопоставление этих данных с материалами, получеппыми при ана-
лизе поедаемости отравленной приманки с 0,025$ зоокумарина, полу-
ченными в аналогичных условиях подвалов многоэтажных коммунальных
зданий, позволило сделать следующие выводы: 1) эффективность вазели-
374
Глава пятая
Таблица 35. Эффективность ядовитых покрытий
против серых крые (®—30) в условиях лабораторного
экеперимента (по: Нуцято, 1986)
Кратность
ежедневно- | Доля погиб-
то прохо-
ших зверь-
Вид покрытия
ждения
ков, %
покрытия
Дуст с 0,5% зоокумарина
:
В
{
:
+16
Дуст с 1% зоокумарина
:
в
; 0,59
И
40-=18
Солидол с 0,5% зоокумарина
Е
ия
Нигрол с 0,5$ зоокумарина
1
818
Пенокумарин © 2$ натриевой
4
Иа 18
соли зоокумарина
3
=
новой пасты больше в более короткие сроки; 2) ее действие более поиз-
бирательно в отношении разных половых и возрастных групи. ожно
ожидать, что при параллельном применепии этих Двух препаративн х
форм будет цостигнута более высокая эффективность истребления крые
и мепее выраженная избирательность, чем при применении одной из этиз
форм. Этот вывод имеет важное значение при реализации стратегии, а
правленной на закрепление «среднего» фенотина, более вероятпое уни
тожение уклоняющихся вариантов и снижение вероятности укоренения
той или иной формы адаптации (наши совместно с В. В. Мелковой дан-
ные). Показано, что эффективность дератизации, где применялись " ва-
зелиновая паста, и приманки с зоокумарином, составила 87 ф, что было
достоверно (Р<0,05) выше, р где истребление проводилось одпими
ами с зоокумарином
ь).
И
ПР", ффоктивность У аволиновой пасты © 0,5 4 дифенацина и 49,5 таль-
ка (талькомагпезита) оказалась выше 90 }) (наши данные), чем анало-
й
ы с зоокумарином.
м облвльвие й рецептуру чево талька снижает избегание пасты гры-
зунами (личное сообщение В. С. Пуцято), однако, как показал опыт ири-
мепения ев в условиях отрицательных температур, она загустевает ниже
нуля градусов, что снижает ее адгезию и в результате эффективность
действия на грызунов. В связи © этим оказалось целесообразным разра-
ботать пасту, сохраняющую пужный уровень адгезии в условиях, отри-
цательных температур. Такая рецептура была предложена А. Н. а.
вым, Д. Ф. Трахановым и В. С. Пуцято (неопубликованные данные),
а именно смесь вазелина с 2-ным раствором дифенацииа в соотнопте-
нии 3 но мостерилянты. Метод стерилизации в борьбе с грынуна-
ми признан специалистами (Но\ат4, 1967; Матзь, Но\аг@, 1976; Чп1з1,
Практическое значение
375
Атота, 1976), но большинство хемостерилянтов для грызунов все еще
проверяется в эксперименте и в настоящее время в повседпевпой прак-
тике дератизации не используется. Получены многочисленные данные о
веществах, оказывающих отрицательное действие на различпых стадиях
процесса размножения млекопитающих, которые служат источником све-
дений в плане отбора хемостерилянтов, перспективных для борьбы и с
грызунами.
Помимо специфических ядов, на размножение грызунов влияют и
обычные яды (фентолацин, фторацетамид, глифтор), но действие их
кратковременно и не оказывает существенного влияния на последующее
состояние популяции (Левин, 4971; Токарева и др., 1974; Быковский,
Пегельман, 1980).
Наиболее целесообразно применять хемостерилянты в сухих приман-
ках, а также в виде дустов, напыляемых в местах передвижений серых
крыс (Марш, Говард, 1974). Влажные приманки быстро портятся и по-
этому не могут обеспечить получения кумулятивной дозы.
Полная ликвидация популяций возможна только при достаточно дли-
тельпом периоде действия стерилянта, охватывающем по меньшей мере
три поколения (КирНос, Мебане, 1972).
Наиболее эффективна стерилизация обоих полов, стерилизация только
самцов оказалась эффективна лишь в тех случаях, когда популяция гры-
зунов малочисленна. Относительная многочисленность популяции поли-
гамных грызунов, т. е. серых крыс, дающих в год несколько приплодов,
создает математическую вероятность того, что сравнительно небольшое
число нестерильных самцов может услешио конкурировать с преобладаю-
щим числом стерильных самцов (КеппеПу © а1., 1972 — цит. по: Марш,
Говард, 1974). При невозможности стерилизации обоих полов было бы
эффективным применение препарата, стерилизующего по крайней мере
самок. Таким веществом мог бы быть местрапол, по он очень плохо по-
., едается крысами (МеПау@, Моод, 1972).
Опыт стерилизации эстрогеном (ВДН-10134) показал, что серые
крысы едят его в приманках лучше, чем местранол, и он действует более
длительно. При подаче приманки с 0,05% эстрогена через 56 недель пона-
даемость крыс в ловушки сократилась с 66 до 0, в контроле она не сни-
зилась, и беремеппых самок отмечали нсе время (Кеп@е её а1., 1973).
Следует, однако, подчеркнуть, что стероидные стерилянты, несмотря
на их эффективность при применении в приманках, оказываются в ряде
случаев очень дорогостоящими ‘для использования их в практической
дератизации.
*Экопомически более выгодными могут быть нестероидные соединения,
в частности алкирующие агенты (антисперматогенные препараты: этило-
нимин, метапсульфонат, 3-хлор-1,2-пропандиол, а-хлоргидрин и колхи-
цин — препарат, стерилизующий самок.
Перечисленные препараты имеют ряд недостатков: колхицин, напри-
мер, плохо усваивается с пищей. о-хлоргидрин через 2 недели после
скармлизания приманки с 1ф-ным его содержапием вызвал стерилизацию
18 самцов (78,2%) из 23 проверенных, но через 17 недель количество
374
Глава пятая
Таблица 35. Эффективность ядовитых покрытий
против серых крые (®—30) в условиях лабораторного
экеперимента (по: Нуцято, 1986)
Кратность
ежедневно- | Доля погиб-
то прохо-
ших зверь-
Вид покрытия
ждения
ков, %
покрытия
Дуст с 0,5% зоокумарина
:
В
{
:
+16
Дуст с 1% зоокумарина
:
в
; 0,59
И
40-=18
Солидол с 0,5% зоокумарина
Е
ия
Нигрол с 0,5$ зоокумарина
1
818
Пенокумарин © 2$ натриевой
4
Иа 18
соли зоокумарина
3
=
новой пасты больше в более короткие сроки; 2) ее действие более поиз-
бирательно в отношении разных половых и возрастных групи. ожно
ожидать, что при параллельном применепии этих Двух препаративн х
форм будет цостигнута более высокая эффективность истребления крые
и мепее выраженная избирательность, чем при применении одной из этиз
форм. Этот вывод имеет важное значение при реализации стратегии, а
правленной на закрепление «среднего» фенотина, более вероятпое уни
тожение уклоняющихся вариантов и снижение вероятности укоренения
той или иной формы адаптации (наши совместно с В. В. Мелковой дан-
ные). Показано, что эффективность дератизации, где применялись " ва-
зелиновая паста, и приманки с зоокумарином, составила 87 ф, что было
достоверно (Р<0,05) выше, р где истребление проводилось одпими
ами с зоокумарином
ь).
И
ПР", ффоктивность У аволиновой пасты © 0,5 4 дифенацина и 49,5 таль-
ка (талькомагпезита) оказалась выше 90 }) (наши данные), чем анало-
й
ы с зоокумарином.
м облвльвие й рецептуру чево талька снижает избегание пасты гры-
зунами (личное сообщение В. С. Пуцято), однако, как показал опыт ири-
мепения ев в условиях отрицательных температур, она загустевает ниже
нуля градусов, что снижает ее адгезию и в результате эффективность
действия на грызунов. В связи © этим оказалось целесообразным разра-
ботать пасту, сохраняющую пужный уровень адгезии в условиях, отри-
цательных температур. Такая рецептура была предложена А. Н. а.
вым, Д. Ф. Трахановым и В. С. Пуцято (неопубликованные данные),
а именно смесь вазелина с 2-ным раствором дифенацииа в соотнопте-
нии 3 но мостерилянты. Метод стерилизации в борьбе с грынуна-
ми признан специалистами (Но\ат4, 1967; Матзь, Но\аг@, 1976; Чп1з1,
Практическое значение
375
Атота, 1976), но большинство хемостерилянтов для грызунов все еще
проверяется в эксперименте и в настоящее время в повседпевпой прак-
тике дератизации не используется. Получены многочисленные данные о
веществах, оказывающих отрицательное действие на различпых стадиях
процесса размножения млекопитающих, которые служат источником све-
дений в плане отбора хемостерилянтов, перспективных для борьбы и с
грызунами.
Помимо специфических ядов, на размножение грызунов влияют и
обычные яды (фентолацин, фторацетамид, глифтор), но действие их
кратковременно и не оказывает существенного влияния на последующее
состояние популяции (Левин, 4971; Токарева и др., 1974; Быковский,
Пегельман, 1980).
Наиболее целесообразно применять хемостерилянты в сухих приман-
ках, а также в виде дустов, напыляемых в местах передвижений серых
крыс (Марш, Говард, 1974). Влажные приманки быстро портятся и по-
этому не могут обеспечить получения кумулятивной дозы.
Полная ликвидация популяций возможна только при достаточно дли-
тельпом периоде действия стерилянта, охватывающем по меньшей мере
три поколения (КирНос, Мебане, 1972).
Наиболее эффективна стерилизация обоих полов, стерилизация только
самцов оказалась эффективна лишь в тех случаях, когда популяция гры-
зунов малочисленна. Относительная многочисленность популяции поли-
гамных грызунов, т. е. серых крыс, дающих в год несколько приплодов,
создает математическую вероятность того, что сравнительно небольшое
число нестерильных самцов может услешио конкурировать с преобладаю-
щим числом стерильных самцов (КеппеПу © а1., 1972 — цит. по: Марш,
Говард, 1974). При невозможности стерилизации обоих полов было бы
эффективным применение препарата, стерилизующего по крайней мере
самок. Таким веществом мог бы быть местрапол, по он очень плохо по-
., едается крысами (МеПау@, Моод, 1972).
Опыт стерилизации эстрогеном (ВДН-10134) показал, что серые
крысы едят его в приманках лучше, чем местранол, и он действует более
длительно. При подаче приманки с 0,05% эстрогена через 56 недель пона-
даемость крыс в ловушки сократилась с 66 до 0, в контроле она не сни-
зилась, и беремеппых самок отмечали нсе время (Кеп@е её а1., 1973).
Следует, однако, подчеркнуть, что стероидные стерилянты, несмотря
на их эффективность при применении в приманках, оказываются в ряде
случаев очень дорогостоящими ‘для использования их в практической
дератизации.
*Экопомически более выгодными могут быть нестероидные соединения,
в частности алкирующие агенты (антисперматогенные препараты: этило-
нимин, метапсульфонат, 3-хлор-1,2-пропандиол, а-хлоргидрин и колхи-
цин — препарат, стерилизующий самок.
Перечисленные препараты имеют ряд недостатков: колхицин, напри-
мер, плохо усваивается с пищей. о-хлоргидрин через 2 недели после
скармлизания приманки с 1ф-ным его содержапием вызвал стерилизацию
18 самцов (78,2%) из 23 проверенных, но через 17 недель количество
376
Глава пятая
стерильных самцов уменьшилось до 36 %, а из 84 самок 46 (54,71%) были
беремеппыми или кормящими. На контрольном участке из 73 самок в
размножении участвовало 34 (46,64), т. е. опыт дал отрицательные ре-
зультаты (Во\’егтап, Вгоокз, 1974).
аа
Другой опыт с тем же веществом в концентрации 6—8 мг на 11
приманки привел к спижению численности серых крыс на земляных па-
сыпях с 50% примерно до 20%. Из 3000—4000 крыс осталось около
1000 штук и численность популяции оставалась сниженной в течение
лета (Апдтемв её а1., 1974). Судя чо количеству оставитихся зверьков,
результаты опыта неудовлетворительны, ибо при 204$ остаточной но-
пуляции численность се очепь быстро может восстановиться.
По мнению Джексона (1973), эффективные результаты дает только
стерилизация почти 100% популяции. Кроме того, он считает, что хе-
мостерилянты не могут быть широко использованы из-за отсутствия
специфического действия и тем самым опаспы для других видов. С этим
мнением пельзя не согласиться.
По мнению Марша и Говарда (1974), хемостерилянты должны играть
основную роль при упичтожении остаточных популяций, я также лля
снижения числепности крыс в прилегающих ряйонах и для предупрежде-
ния повторного заселения освобожденных от пих районов. Хемостерилян-
ты могут сыграть решающую роль в истреблении крыс, устойчивых к
аптикоагулянтам.
Особое внимание при применении хемостериляитов необходимо обра-
щать на их опаспость для полезных животных (для травоядных и хиш-
ных видов) нри прямых и вторичных отравлениях, а также на степень
загрязнения ими среды с учетом их мутагенного и канцерогепного свой-
ства.
Возможноеть практически полного уничтожения
серой крысы в пределах населенного пункта
Борьба с крысами в объектах разных категорий
Термин «практически полное уничтожение» серой крысы, сформулиро-
ванный Телле (ТеПе, 1968, 1971), означает, что на территории, где пре-
ведепо истребление крыс, возможно существование лишь небольшого чис-
ла их поселений.
Наземные сооружения. Стратегия и тактика борьбы с серыми крыха-
ми в малых, средних, круппых городах, а также в городах-гигантах,
по всей видимости, должны отличаться учетом типа их застройки, при
этом необходимо иметь в виду: а) инфраструктуру популяций крыс,
б) характер их пространственного распределения по отдельным строепи-
ям, впутри помегений, а также по участкам открытых стаций, в) под-
вижность этих грызунов в зависимости от стабильности убежищ и кор-
мовой базы, а также от наличия системы подземных и наземных комму-
никаций.
Практическое значение
ВУ
На основе изучения неравномерпости распределения серых крыс по
строепиям различпых категорий рядом авторов (Кузякин, 1950; 1951;
1963; Траут, 1950; Дукельская, 1959), сообщения которых были пеодно-
кратно подтверждены позднее (Денисов, 1986; Страупманис, 1986), сде-
ланы важные заключения о необходимости концентрации усилий по ис-
треблению крыс с учетом их размещения по объектам и сезонной дина-
мики пространственной структуры. Вместе с тем в силу недостаточной
разработанности вопроса о передвижениях серых крыс в условиях города
эти авторы сделали излишне категоричные выводы об их низкой подвизк-
ности и привязанности к определенным объектам. Основываясь па таких
представлениях, А. П. Кузякип, в свою очередь, сделал неверные орга-
низациопно-методические выводы о возможности «поэтапного» (крартал
за кварталом) уничтожения серых крыс па территории целого города.
Но последующими работами экологического и прикладного характера эта
точка зрения была опровергнута.
Тем пе менее безусловно верен вывод А. П. Кузякина (1963) о том,
что частичное (неполное) истребление пасюков в постройках ведот к
улучшепию жизни зверьков, оставшихся в живых, к повышению их фак-
тической плодовитости и к очень быстрому восстановлению их числепно-
сти до исходного уровия.
Полное истребление крыс во всех постройках невозможно, учитывая,
что средства уничтожения применяются далеко пе так квалифицирован-
но, как слодовало бы. В результате истребление зверьков в болыпинстве
случаев бывает только частичным. Эта объективцая реальпость служит
еще одним препятствием для реализации «поэтапного» освобождения го-
ролов от крысе — к однажды обработанным участкам приходится возвра-
щаться не один раз. Отсюда вытекает пеобходимость систематического
обследования объектов на наличие крыс и в случае необходимости про-
ведения там истребительных мероприятий.
В 60—70-е годы появилея целый ряд работ, свидетельствующих о
`высокой подвижности пасюков в городах (Полежаев и др., 1959; Шура-
Бура и др., 1962; Судейкии и др., 1962; Судейкин, 1976). Эти данные
легли в основу принципа еплопиной систематической дератизации, фор-
мулировка которого стала одним из паиболес важпых этапов в развитии
дератизации. Вряд ли мы ошибемся, если скажем, что стремление прак-
тических работников к увеличению обрабатываемых площадей слижает
внимание к учету особеппостей пространственного распределения серых
крысе в городе. Тем не менее практика дератизации показала, что ве эф-
фективность целиком и полпостью определяется полнотой выявления
всех поселений пасюков, иначе говоря, качеством обследования террито-
‘рии на заселенность ими. В связи с этим 0. В. Тощигипым с соавторами
(1969, 1976, 1977) были предложены контрольно-истребительные площад-
.ки в виде лотков с мукой, содержащей 0,0254 зоокумарина, которые
рекомендовалось расставлять через 4—5 м вдоль стен, причем. отравлен-
ную приманку добавляли лить в тех местах, где наблюдали ее поедапие.
На этом этапе реализации программы сплошной систематической дерати-
зации, когда рост знаний о биологии серых крыс в условиях урбаноцено-
376
Глава пятая
стерильных самцов уменьшилось до 36 %, а из 84 самок 46 (54,71%) были
беремеппыми или кормящими. На контрольном участке из 73 самок в
размножении участвовало 34 (46,64), т. е. опыт дал отрицательные ре-
зультаты (Во\’егтап, Вгоокз, 1974).
аа
Другой опыт с тем же веществом в концентрации 6—8 мг на 11
приманки привел к спижению численности серых крыс на земляных па-
сыпях с 50% примерно до 20%. Из 3000—4000 крыс осталось около
1000 штук и численность популяции оставалась сниженной в течение
лета (Апдтемв её а1., 1974). Судя чо количеству оставитихся зверьков,
результаты опыта неудовлетворительны, ибо при 204$ остаточной но-
пуляции численность се очепь быстро может восстановиться.
По мнению Джексона (1973), эффективные результаты дает только
стерилизация почти 100% популяции. Кроме того, он считает, что хе-
мостерилянты не могут быть широко использованы из-за отсутствия
специфического действия и тем самым опаспы для других видов. С этим
мнением пельзя не согласиться.
По мнению Марша и Говарда (1974), хемостерилянты должны играть
основную роль при упичтожении остаточных популяций, я также лля
снижения числепности крыс в прилегающих ряйонах и для предупрежде-
ния повторного заселения освобожденных от пих районов. Хемостерилян-
ты могут сыграть решающую роль в истреблении крыс, устойчивых к
аптикоагулянтам.
Особое внимание при применении хемостериляитов необходимо обра-
щать на их опаспость для полезных животных (для травоядных и хиш-
ных видов) нри прямых и вторичных отравлениях, а также на степень
загрязнения ими среды с учетом их мутагенного и канцерогепного свой-
ства.
Возможноеть практически полного уничтожения
серой крысы в пределах населенного пункта
Борьба с крысами в объектах разных категорий
Термин «практически полное уничтожение» серой крысы, сформулиро-
ванный Телле (ТеПе, 1968, 1971), означает, что на территории, где пре-
ведепо истребление крыс, возможно существование лишь небольшого чис-
ла их поселений.
Наземные сооружения. Стратегия и тактика борьбы с серыми крыха-
ми в малых, средних, круппых городах, а также в городах-гигантах,
по всей видимости, должны отличаться учетом типа их застройки, при
этом необходимо иметь в виду: а) инфраструктуру популяций крыс,
б) характер их пространственного распределения по отдельным строепи-
ям, впутри помегений, а также по участкам открытых стаций, в) под-
вижность этих грызунов в зависимости от стабильности убежищ и кор-
мовой базы, а также от наличия системы подземных и наземных комму-
никаций.
Практическое значение
ВУ
На основе изучения неравномерпости распределения серых крыс по
строепиям различпых категорий рядом авторов (Кузякин, 1950; 1951;
1963; Траут, 1950; Дукельская, 1959), сообщения которых были пеодно-
кратно подтверждены позднее (Денисов, 1986; Страупманис, 1986), сде-
ланы важные заключения о необходимости концентрации усилий по ис-
треблению крыс с учетом их размещения по объектам и сезонной дина-
мики пространственной структуры. Вместе с тем в силу недостаточной
разработанности вопроса о передвижениях серых крыс в условиях города
эти авторы сделали излишне категоричные выводы об их низкой подвизк-
ности и привязанности к определенным объектам. Основываясь па таких
представлениях, А. П. Кузякип, в свою очередь, сделал неверные орга-
низациопно-методические выводы о возможности «поэтапного» (крартал
за кварталом) уничтожения серых крыс па территории целого города.
Но последующими работами экологического и прикладного характера эта
точка зрения была опровергнута.
Тем пе менее безусловно верен вывод А. П. Кузякина (1963) о том,
что частичное (неполное) истребление пасюков в постройках ведот к
улучшепию жизни зверьков, оставшихся в живых, к повышению их фак-
тической плодовитости и к очень быстрому восстановлению их числепно-
сти до исходного уровия.
Полное истребление крыс во всех постройках невозможно, учитывая,
что средства уничтожения применяются далеко пе так квалифицирован-
но, как слодовало бы. В результате истребление зверьков в болыпинстве
случаев бывает только частичным. Эта объективцая реальпость служит
еще одним препятствием для реализации «поэтапного» освобождения го-
ролов от крысе — к однажды обработанным участкам приходится возвра-
щаться не один раз. Отсюда вытекает пеобходимость систематического
обследования объектов на наличие крыс и в случае необходимости про-
ведения там истребительных мероприятий.
В 60—70-е годы появилея целый ряд работ, свидетельствующих о
`высокой подвижности пасюков в городах (Полежаев и др., 1959; Шура-
Бура и др., 1962; Судейкии и др., 1962; Судейкин, 1976). Эти данные
легли в основу принципа еплопиной систематической дератизации, фор-
мулировка которого стала одним из паиболес важпых этапов в развитии
дератизации. Вряд ли мы ошибемся, если скажем, что стремление прак-
тических работников к увеличению обрабатываемых площадей слижает
внимание к учету особеппостей пространственного распределения серых
крысе в городе. Тем не менее практика дератизации показала, что ве эф-
фективность целиком и полпостью определяется полнотой выявления
всех поселений пасюков, иначе говоря, качеством обследования террито-
‘рии на заселенность ими. В связи с этим 0. В. Тощигипым с соавторами
(1969, 1976, 1977) были предложены контрольно-истребительные площад-
.ки в виде лотков с мукой, содержащей 0,0254 зоокумарина, которые
рекомендовалось расставлять через 4—5 м вдоль стен, причем. отравлен-
ную приманку добавляли лить в тех местах, где наблюдали ее поедапие.
На этом этапе реализации программы сплошной систематической дерати-
зации, когда рост знаний о биологии серых крыс в условиях урбаноцено-
378
Глава пятая
за значительно отставал от происходящих в нем изменений, такой уни-
версальный с©10с0б, соединяющий одповременно обнаружение крыс и их
истребление, оказался весьма эффективным.
Особенно важным для повышения эффективности истребления крыс
является знание тонкой структуры их прострапствеппого распределения
и ее динамики. Такие работы появились только в 80-е годы и касаются
преимущественно жилых домов, которые, согласно В. А. Судейкину
(1986), являтотся категорией строений, наиболее заселенных крысами,
и занимают наибольшую площадь застройки. В. К. Мелкова с соавтора-
ми (1985) выявила причины низкой эффективпости истребления крыс
отравленными приманками при равномерном распределении последних
по территории подвала многоэтажного жилого дома. В. А. Рыльниковым
и А. В. Звопаревым (1985, 1986) показано, что равномерная раскладка
мелких порций приманки © зоокумарипом по всей площади подвала вы-
зывает гибель лишь незначительной части крыс: за 1 месяц— 5 трупов.
Концентрация этой же приманки под мусорокамерами тото же подвала
в течепие последующего месяца привела к гибели 148 крыс. Цак пока-
зал опыт, около каждой камеры должно быть не менее 4 коробок или
ящиков с отравленной приманкой (пе мепышо 1 кг в каждой емкости),
расставленных на расстоянии 4—5 м друг от друга. Такая раскладка
приманки наилучшим образом способствует поеданию крысами. Концен-
трируя отравлеппую приманку под мусорокамерами, мы как бы «при-
зязываем» ее к зоне наибольшей активности крыс и в то же время,
рассредоточивая основную порцию по четырем кормушкам, создаем ус-
ловия, при которых спижается внутривидовая конкуренция около при-
мапки (Рыльников, Звонарев, 1986; Рыльшиков, Сиротинина, 1986). По-
добные исследования в промышленных преднриятиях— категории
строений, которая по заселенности крысами занимает второе место после
жилого соктора, пе проводились, однако по аналогии можно предполагать,
что такая расстановка долговременных точек отравления около мест пн-
тания работников предприятий, расположения пищевых отходов, складн-
рования запасов продовольствия, там, где имеются постоянные и времен-
ные группировки серых крыс, будет наиболее оптимальна. Чем шире зона
активности данпой группировки, чем она болыше по числу входящих в
пее крыс, тем больше должно быть отдельных кормушек. Основное усло-
вие — обеспечение необходимого (для гибели) уровия потреблепия от-
равленной приманки каждым членом группировки, что достигается ее
избытком и спижением внутривидовой конкурепции около мест ее рас-
кладки.
В отношении дератизации объекты ветерипарного надзора могут быть
разделены на две групны. Первую составляют собственно животнозвод-
ческие объекты: свинарники, птичники, коровники, конюлтни, зверофер-
мы ит. д. а во вторую входят те объекты, где ведется переработка
животноводческого сырья и продуктов: мясокомбинаты, заводы по пройз-
водству желатипы, костной муки, по утилизации отходов.
Объекты первой группы наиболее трудны для проведения истреби-
тельных работ из-за опасности отравления полезных животных. Успех
м
с
о
л
и
Практичеекое значение
379
дератизации в этом случае определяется наличием достаточного количе-
ства мест, где можно проводить обработку. С этой целью приходится обо-
рудовать ящики, лотки, трубки (по 20—30 шт. на 1000 м? площади пола)
с одновременной раскладкой в них приманок с ядом: а) из пшеницы;
6) подсолнуха; в) комбикорма или муки (Траханов, 1981). Вторую труд-
ность создает обилие и разнообразие корма для животных, при наличии
которого приманки с ядами хуже поедаются крысами. Для преодоления
этого можно использовать аттрактанты
— сахар (10%) и растительное
масло (3%), добавляемые к пищевой основе приманки (Тощигин и др.,
1977; Хамагапов и др., 1983). Третью трудность представляют крупные
размеры животноводческих объектов (комилексов), в которых содержатся
десятки тысяч голов скота и где популяции крыс пасчитывают по не-
сколько тысяч особей, что заставляет тратить на их истребление большие
количества приманки. В 1961 г. за одни сутки в четырех райопах Тю-
менской области крысы съели более 7 т приманок (Жаров и др., 1974).
Именно по этой причине были разработаны более экономичные способы
борьбы
— были примепены 1,0%-пый дуст зоокумарина для опылепия
поверхностей, по которым перемещаются крысы, ядовитые пены для за-
купорки нор и ядовитые липкие композиции, наносимые на доски, картон
и пачкающие пробегающих по ним крысе (Зацепин, 1969; Пуцято,
1986).
Расположенные па путях перемещений крыс контактные яды позво-
ляют, не тратя пищевых продуктов, снизить заселенность популяции в
несколько раз, после чего обработка отравленными приманками становит-
более рентабельной.
Помимо состава, эффективность применепия отравлеппых приманок,
как мы уже говорили, зависит и от выбора мест раскладки. Там, где
крысы привыкли питаться (например, в кормушках животных), разло-
жонная рядом приманка посдается ими хорошо, а в норах или на путях
. передвижения — хуже (Абеленцев, 1959; Козлов, 1980).
Состав примепяемых ядов имеет очень большое значение. В свинар-
никах наиболее целесообразно применение фентолацина (30—40 мг/кг),
который значительно менее токсичен для свиней, чем зоокумариц
(1—2 мг/кг). В птичниках лучше использовать зоокумарин (200—
300 мг/кг), который малотоксичен для птиц даже при многократном вве-
депии (Тажибаев, 1979).
Мы хотим подчеркнуть еще раз, что яды острого действия (фосфид
цинка, фторацетаты и др.) не могут быть рекомендованы из-за опасности
отравления полезных животных как при случайном контакте с приман-
кой, так и при поедании ими павших крыс (Траханов, Цадиров, 1984).
По мнепию Н. И. Пикифорова © соавторами (1977), исключительно
большое значение для дератизации животноводческих объектов имеют
сухие долго сохраняющиеся примапки, но в последнее время приоритет
отдается бесприманочному способу применения антикоагулянтов (Тра-
ханов, 1985)...
Кроме мероприятий по уничтожению, все большую роль начинают
отводить санитарно-техническим мерам и строительству объектов из пе-
378
Глава пятая
за значительно отставал от происходящих в нем изменений, такой уни-
версальный с©10с0б, соединяющий одповременно обнаружение крыс и их
истребление, оказался весьма эффективным.
Особенно важным для повышения эффективности истребления крыс
является знание тонкой структуры их прострапствеппого распределения
и ее динамики. Такие работы появились только в 80-е годы и касаются
преимущественно жилых домов, которые, согласно В. А. Судейкину
(1986), являтотся категорией строений, наиболее заселенных крысами,
и занимают наибольшую площадь застройки. В. К. Мелкова с соавтора-
ми (1985) выявила причины низкой эффективпости истребления крыс
отравленными приманками при равномерном распределении последних
по территории подвала многоэтажного жилого дома. В. А. Рыльниковым
и А. В. Звопаревым (1985, 1986) показано, что равномерная раскладка
мелких порций приманки © зоокумарипом по всей площади подвала вы-
зывает гибель лишь незначительной части крыс: за 1 месяц— 5 трупов.
Концентрация этой же приманки под мусорокамерами тото же подвала
в течепие последующего месяца привела к гибели 148 крыс. Цак пока-
зал опыт, около каждой камеры должно быть не менее 4 коробок или
ящиков с отравленной приманкой (пе мепышо 1 кг в каждой емкости),
расставленных на расстоянии 4—5 м друг от друга. Такая раскладка
приманки наилучшим образом способствует поеданию крысами. Концен-
трируя отравлеппую приманку под мусорокамерами, мы как бы «при-
зязываем» ее к зоне наибольшей активности крыс и в то же время,
рассредоточивая основную порцию по четырем кормушкам, создаем ус-
ловия, при которых спижается внутривидовая конкуренция около при-
мапки (Рыльников, Звонарев, 1986; Рыльшиков, Сиротинина, 1986). По-
добные исследования в промышленных преднриятиях— категории
строений, которая по заселенности крысами занимает второе место после
жилого соктора, пе проводились, однако по аналогии можно предполагать,
что такая расстановка долговременных точек отравления около мест пн-
тания работников предприятий, расположения пищевых отходов, складн-
рования запасов продовольствия, там, где имеются постоянные и времен-
ные группировки серых крыс, будет наиболее оптимальна. Чем шире зона
активности данпой группировки, чем она болыше по числу входящих в
пее крыс, тем больше должно быть отдельных кормушек. Основное усло-
вие — обеспечение необходимого (для гибели) уровия потреблепия от-
равленной приманки каждым членом группировки, что достигается ее
избытком и спижением внутривидовой конкурепции около мест ее рас-
кладки.
В отношении дератизации объекты ветерипарного надзора могут быть
разделены на две групны. Первую составляют собственно животнозвод-
ческие объекты: свинарники, птичники, коровники, конюлтни, зверофер-
мы ит. д. а во вторую входят те объекты, где ведется переработка
животноводческого сырья и продуктов: мясокомбинаты, заводы по пройз-
водству желатипы, костной муки, по утилизации отходов.
Объекты первой группы наиболее трудны для проведения истреби-
тельных работ из-за опасности отравления полезных животных. Успех
м
с
о
л
и
Практичеекое значение
379
дератизации в этом случае определяется наличием достаточного количе-
ства мест, где можно проводить обработку. С этой целью приходится обо-
рудовать ящики, лотки, трубки (по 20—30 шт. на 1000 м? площади пола)
с одновременной раскладкой в них приманок с ядом: а) из пшеницы;
6) подсолнуха; в) комбикорма или муки (Траханов, 1981). Вторую труд-
ность создает обилие и разнообразие корма для животных, при наличии
которого приманки с ядами хуже поедаются крысами. Для преодоления
этого можно использовать аттрактанты
— сахар (10%) и растительное
масло (3%), добавляемые к пищевой основе приманки (Тощигин и др.,
1977; Хамагапов и др., 1983). Третью трудность представляют крупные
размеры животноводческих объектов (комилексов), в которых содержатся
десятки тысяч голов скота и где популяции крыс пасчитывают по не-
сколько тысяч особей, что заставляет тратить на их истребление большие
количества приманки. В 1961 г. за одни сутки в четырех райопах Тю-
менской области крысы съели более 7 т приманок (Жаров и др., 1974).
Именно по этой причине были разработаны более экономичные способы
борьбы
— были примепены 1,0%-пый дуст зоокумарина для опылепия
поверхностей, по которым перемещаются крысы, ядовитые пены для за-
купорки нор и ядовитые липкие композиции, наносимые на доски, картон
и пачкающие пробегающих по ним крысе (Зацепин, 1969; Пуцято,
1986).
Расположенные па путях перемещений крыс контактные яды позво-
ляют, не тратя пищевых продуктов, снизить заселенность популяции в
несколько раз, после чего обработка отравленными приманками становит-
более рентабельной.
Помимо состава, эффективность применепия отравлеппых приманок,
как мы уже говорили, зависит и от выбора мест раскладки. Там, где
крысы привыкли питаться (например, в кормушках животных), разло-
жонная рядом приманка посдается ими хорошо, а в норах или на путях
. передвижения — хуже (Абеленцев, 1959; Козлов, 1980).
Состав примепяемых ядов имеет очень большое значение. В свинар-
никах наиболее целесообразно применение фентолацина (30—40 мг/кг),
который значительно менее токсичен для свиней, чем зоокумариц
(1—2 мг/кг). В птичниках лучше использовать зоокумарин (200—
300 мг/кг), который малотоксичен для птиц даже при многократном вве-
депии (Тажибаев, 1979).
Мы хотим подчеркнуть еще раз, что яды острого действия (фосфид
цинка, фторацетаты и др.) не могут быть рекомендованы из-за опасности
отравления полезных животных как при случайном контакте с приман-
кой, так и при поедании ими павших крыс (Траханов, Цадиров, 1984).
По мнепию Н. И. Пикифорова © соавторами (1977), исключительно
большое значение для дератизации животноводческих объектов имеют
сухие долго сохраняющиеся примапки, но в последнее время приоритет
отдается бесприманочному способу применения антикоагулянтов (Тра-
ханов, 1985)...
Кроме мероприятий по уничтожению, все большую роль начинают
отводить санитарно-техническим мерам и строительству объектов из пе-
380
Глава пятая
доступных для крыс материалов (Траханов, 1981; Поляков и др., 1983;
Тур, 1080) ‘второй труппы могут применяться все способы борьбы [и
соблюдением обычных мер предосторожности, направлеппых на защиту
от отравления людей, продуктов и внешней среды. Состав приманок в
этих условиях определяется выбором тех компонентов пищи, которые де-
фицитны в данном месте. В условиях мясокомбинатов и заводов то про-
изводетву желатины, мясокостной муки крысы имеют избыток елко,
жиров и практически не имеют углеводов. В связи © этим на мясокомби-
натах пами в течение ряда лет были исиытаны углеводные примапки и
тие результаты.
о акиь ка обработки с аналогичным результатом были проведены под
нашим руководством в Хабаровске, Свердловске, Смоленске, Вязьмс.
Особого внимания заслуживает то обстоятельство, что при учете по-
сещенпых крысами площадок оценка эффективности дератизации прово-
дилась ежемесячно на всей площади объектов без дополпительных меро-
ИР" таким образом, широкое использование мучных приманок с антико-
агулянтами в практике дератизации животноводческих объектов позво-
ляет решить одну из труднейших задач
— оценку эффективности дера-
тизации на больших площадях без повыштепия трудоемкости работ.
Подземные вооружения. Основную трудность при истреблении грызу-
нов в канализационных сооружениях составляет отсутствие там мест,
пригодпых дия раскладки отравленной ириманки. Наиболее оффоктивной
препаративпой формой оказались парафиновые брикеты С No т
рина (80,8% освобожденных колодцев) или с 1% фторацетамида (85,7%
освобожденных колодцев). Не менее эффективным оказалось развепива-
ние фанерок размером 10Ж15 см с нанесепной на них пастой, обтоящей
из 45% вазелина, 40 дуста с добавлением 0,5% зоокумарина и о
растительного масла (87,5—90,5 освобожденных колодцев). В канали-
зации возможна раздельная обработка разрозненных поселений пасюков
гин, 1986).
.
ах У рые крысы концентрируются там, гле работатот люди
(откаточные штреки, «лавы» — места выемки угля, камеры ожидатия,
подстанции, склады взрывчатых вещоств, места трацезы шахтеров).
В этих местах рекомендовано расставлять ящики, в которые раскладыва-
стся в течение 12—24 дней неотравленный корм, затем ето заменит на
приманки с 1% крысида или 5% фосфида цинка. Однократная о работка
по такому плану приводит к полному освобождению шахты от грызунов.
При обработке шахт хлебной крошкой с 5% зоокумарина или пара-
финироваппыми брикетами с 10% зоокумарина предварительного прикор-
ма не проводят, но 4—5-кратная раскладка такой приманки в течени
45—20 дпей полностью освобождает тахту от грызунов. Такая высокая
эффективность дератизации оказалась возможной только при условии
обработки всех мест, заселенных грызунами (Тоцигин, }). ффект ",-
пость парафипированных брикетов с зоокумарином против серых кр
подтверждают и другие авторы (Гажиев и др., 1986а, 1% }.
Практическое значение
381
Наряду с приманочным весьма эффективны комбинированные спосо-
бы: 1) искусственные гнездовья с материалом, пропылеппым 0,5% -пым
дустом дифенацина (ратиндана), около которых раскладывают для при-
влечения крыс прикорм, 2) трубки из типографских отходов с вложенной
внутрь приманкой и по концам изнутри (на ширину 10—15 см) обмазан-
ные вазелиновой пастой с 0,5%-ным зоокумарином или дифенацином.
Эффективность этого способа составляет 78—90 и 84—93% соответствеп-
но, и имеется то преимущество, что даже в условиях повытпенной влаж-
пости, когда примапка быстро плесневеет, здесь опа длительно сохраняет
свою эффективность.
Открытые етации. Для поддержания в городах численности серых
крыс на возможно более низком уровне необходимо их истреблять не
только в наземных постройках и подземных сооружениях, но и в откры-
тых стациях города (свалки отходов, берега речек и прудов, пустыря,
парковая зона). Очевидно, что наиболее трудна эта задача в местах крут-
логодичного обитания пасюков вне построек человека, где опи заселятот
биотспы (обычно около источников воды) природного и антропогенного
происхождения. Истребление крыс там следует проводить на всей пло-
щади открытых стаций в течепие всего года.
Раскладка, например, отравленной приманки не только в постройках
человека, по и па открытой территории г. Риги обеспечила более полное
снижение численности пасюка при меньшей норме расхода, чем при рас-
кладке приманок только в постройках (Тощигин и др., 1980; Шмате,
1983). Круглогодичные поселения этого грызуна более локально приуро-
чены к свалкам и незамерзающим водоемам (Формозов, 1945; Цузякин,
1951; Полежаев и др., 1959; Попов, 1970; и др.). Такие места могут быть
сравнительно легко обнаружены, а потому вопрос уничтожения там крыс
зависит от качества обработки. Однако осенью большая часть мигрирует
в отапливаемые помещения. В таких местах наибольшее значение ириоб-
``ретают так называемые «барьерные» обработки, когда истребление крыс
ведут на путях их миграции в строения человека. Важно подчеркнуть,
что во всех климатических зонах пространственная локализация популя-
ции крыс усиливается в зимний период. Даже в южных районах с от-
носительно мягкой зимой, гие большая часть зверьков зимует в природ-
ных биотопах, наблюдается их пространственная локализация около
наиболсе «кормных» мест; на рисовых полях, например, пасюки концен-
трируются зимой на неперепаханных чеках с большим количеством про-
сыпей зерна, около пезамерзающих водоемов с обилием беспозвоночных,
ляцушек. Истребление крыс на таких участках в зимний период приводит
к длительному снижению их численности на территории, значительно
превьпнающей обработанную.
Объем работ по борьбе с крысами в открытых стациях, а особеппо в
`° природных очагах иктерогеморрагического лептоспироза, должен опре-
деляться также социальным фактором — наиболее важно истреблять этих
грызунов в местах массового отдыха, охоты и рыбной ловли. Эти два ус-
ловия придают борьбе с крысами в открытых стациях целенаправленный
характер, концентрируя силы и средства на наиболее важпых участках
380
Глава пятая
доступных для крыс материалов (Траханов, 1981; Поляков и др., 1983;
Тур, 1080) ‘второй труппы могут применяться все способы борьбы [и
соблюдением обычных мер предосторожности, направлеппых на защиту
от отравления людей, продуктов и внешней среды. Состав приманок в
этих условиях определяется выбором тех компонентов пищи, которые де-
фицитны в данном месте. В условиях мясокомбинатов и заводов то про-
изводетву желатины, мясокостной муки крысы имеют избыток елко,
жиров и практически не имеют углеводов. В связи © этим на мясокомби-
натах пами в течение ряда лет были исиытаны углеводные примапки и
тие результаты.
о акиь ка обработки с аналогичным результатом были проведены под
нашим руководством в Хабаровске, Свердловске, Смоленске, Вязьмс.
Особого внимания заслуживает то обстоятельство, что при учете по-
сещенпых крысами площадок оценка эффективности дератизации прово-
дилась ежемесячно на всей площади объектов без дополпительных меро-
ИР" таким образом, широкое использование мучных приманок с антико-
агулянтами в практике дератизации животноводческих объектов позво-
ляет решить одну из труднейших задач
— оценку эффективности дера-
тизации на больших площадях без повыштепия трудоемкости работ.
Подземные вооружения. Основную трудность при истреблении грызу-
нов в канализационных сооружениях составляет отсутствие там мест,
пригодпых дия раскладки отравленной ириманки. Наиболее оффоктивной
препаративпой формой оказались парафиновые брикеты С No т
рина (80,8% освобожденных колодцев) или с 1% фторацетамида (85,7%
освобожденных колодцев). Не менее эффективным оказалось развепива-
ние фанерок размером 10Ж15 см с нанесепной на них пастой, обтоящей
из 45% вазелина, 40 дуста с добавлением 0,5% зоокумарина и о
растительного масла (87,5—90,5 освобожденных колодцев). В канали-
зации возможна раздельная обработка разрозненных поселений пасюков
гин, 1986).
.
ах У рые крысы концентрируются там, гле работатот люди
(откаточные штреки, «лавы» — места выемки угля, камеры ожидатия,
подстанции, склады взрывчатых вещоств, места трацезы шахтеров).
В этих местах рекомендовано расставлять ящики, в которые раскладыва-
стся в течение 12—24 дней неотравленный корм, затем ето заменит на
приманки с 1% крысида или 5% фосфида цинка. Однократная о работка
по такому плану приводит к полному освобождению шахты от грызунов.
При обработке шахт хлебной крошкой с 5% зоокумарина или пара-
финироваппыми брикетами с 10% зоокумарина предварительного прикор-
ма не проводят, но 4—5-кратная раскладка такой приманки в течени
45—20 дпей полностью освобождает тахту от грызунов. Такая высокая
эффективность дератизации оказалась возможной только при условии
обработки всех мест, заселенных грызунами (Тоцигин, }). ффект ",-
пость парафипированных брикетов с зоокумарином против серых кр
подтверждают и другие авторы (Гажиев и др., 1986а, 1% }.
Практическое значение
381
Наряду с приманочным весьма эффективны комбинированные спосо-
бы: 1) искусственные гнездовья с материалом, пропылеппым 0,5% -пым
дустом дифенацина (ратиндана), около которых раскладывают для при-
влечения крыс прикорм, 2) трубки из типографских отходов с вложенной
внутрь приманкой и по концам изнутри (на ширину 10—15 см) обмазан-
ные вазелиновой пастой с 0,5%-ным зоокумарином или дифенацином.
Эффективность этого способа составляет 78—90 и 84—93% соответствеп-
но, и имеется то преимущество, что даже в условиях повытпенной влаж-
пости, когда примапка быстро плесневеет, здесь опа длительно сохраняет
свою эффективность.
Открытые етации. Для поддержания в городах численности серых
крыс на возможно более низком уровне необходимо их истреблять не
только в наземных постройках и подземных сооружениях, но и в откры-
тых стациях города (свалки отходов, берега речек и прудов, пустыря,
парковая зона). Очевидно, что наиболее трудна эта задача в местах крут-
логодичного обитания пасюков вне построек человека, где опи заселятот
биотспы (обычно около источников воды) природного и антропогенного
происхождения. Истребление крыс там следует проводить на всей пло-
щади открытых стаций в течепие всего года.
Раскладка, например, отравленной приманки не только в постройках
человека, по и па открытой территории г. Риги обеспечила более полное
снижение численности пасюка при меньшей норме расхода, чем при рас-
кладке приманок только в постройках (Тощигин и др., 1980; Шмате,
1983). Круглогодичные поселения этого грызуна более локально приуро-
чены к свалкам и незамерзающим водоемам (Формозов, 1945; Цузякин,
1951; Полежаев и др., 1959; Попов, 1970; и др.). Такие места могут быть
сравнительно легко обнаружены, а потому вопрос уничтожения там крыс
зависит от качества обработки. Однако осенью большая часть мигрирует
в отапливаемые помещения. В таких местах наибольшее значение ириоб-
``ретают так называемые «барьерные» обработки, когда истребление крыс
ведут на путях их миграции в строения человека. Важно подчеркнуть,
что во всех климатических зонах пространственная локализация популя-
ции крыс усиливается в зимний период. Даже в южных районах с от-
носительно мягкой зимой, гие большая часть зверьков зимует в природ-
ных биотопах, наблюдается их пространственная локализация около
наиболсе «кормных» мест; на рисовых полях, например, пасюки концен-
трируются зимой на неперепаханных чеках с большим количеством про-
сыпей зерна, около пезамерзающих водоемов с обилием беспозвоночных,
ляцушек. Истребление крыс на таких участках в зимний период приводит
к длительному снижению их численности на территории, значительно
превьпнающей обработанную.
Объем работ по борьбе с крысами в открытых стациях, а особеппо в
`° природных очагах иктерогеморрагического лептоспироза, должен опре-
деляться также социальным фактором — наиболее важно истреблять этих
грызунов в местах массового отдыха, охоты и рыбной ловли. Эти два ус-
ловия придают борьбе с крысами в открытых стациях целенаправленный
характер, концентрируя силы и средства на наиболее важпых участках
382 Глава пятая
и в тот период года, когда популяция паиболее уязвима для истребитель-
ных мероприятий (Рыльников, Карасева, 1985).
"Гактика применения средств истребления обычно зависит от времопи,
места обработок и тех целой, для которых они предназначены. Так, об-
работка открытой территории вокруг построек в средней полосе ведется
© помощью долговременных отравляющих точек, куда раскладывают ири-
манку с аитикоагулянтами (Тощитин, и др. 1980; Низот, Вептвот,
1981). При обработках открытых стации наиболее трудпая задача о ес-
поечение сохранности ириманок, поэтому рекомендуют применепие их
расфасованными в полиэтиленовые пакетики, а также в виде парафипи-
ровапных брикетов. Готовую отравленную приманку раскладывают С
норы грызунов и другие естественные укрытия или закладывают в труб-
ки из рубероида, картонные трубки из-под типографских отходов, ящич-
ки ит. п.
ИПриманочный способ борьбы с крысами в открытых стациях следует
признать основпым. Другие способы, в том числе бесприманочные (газа-
ция, дустация), менее приемлемы, хотя существует опыт их применепия,
например глубокое пропыливание пор серых крыс дустами аптикоагу-
лянтов (ТеПе, 1962), опыливание троп и закладка в поры проныленных
антикоагуляптами ватных тампонов. (Тощигин и др., 1976; Богачева
и др. 1984). Эффективпость примепения этого способа будет определять-
ся насыщенностью почвы влагой -—- чем почва суше, тем сильнее «пыля-
щий» эффект.
влажным почвам дуст прилипает и смешивастея с
грязью.
Возможна газация фосфипом при условии плотного закрывания всех
выходов крысиной поры (Ме, 1980). Фумигация нор хлорпикрином в
зимний пернод неприемлема, так как при температуре воздуха ниже
12°С он малоэффективен. Применение цианплава и циклона (препара-
тивные формы КСМ) вряд ли возможно па культурах зерповых, учиты-
вая хорошую растворимость солей синильной кислоты в воде и вороят-
ность проникновения их в культуру. Эти срелетва опаспо также’ приме-
нять около строений человека. Истребление легче проводить зимой, когда
поры крыс можно обнаружить среди ножухлой травы и когда отравлеп-
пые приманки поедаются ими более охотно, чем летом — в условиях изо-
билия конкурирующих кормов.
|
.
Опыты по практически полному уничтожению крыс
Первый успелный опыт практически полного уничтожения крыс был
проведен в г. Куксхафене (ФРГ) по методике, предложенной Штейниге-
ром в 1955 г. (Бецивег, 1955, 1957, 1958). Истребительным работам
предшествовали тптательная организационная подготовка, ныявленио всех
мест, где имеются крысы. В течение 20 дней уничтожили 95% зверьков,
ликвидация оставшихся 5% потребовала вдвое больше времени. Главным
средством уничтожения были приманки из муки с ядом варфарином, ко
торые раскладывали в приманочные ящики крупными порциями (1—
5 кг на ящик). Вокруг обработанной зоны также были установлены ящи-
Практическое значение
383
ки с приманкой для упичтожения вновь вселяющихся крыс. Кроме при-
манок, применили варфарин и для опыления нор н трои. Проведенный
опыт показал, что крысы в обработанной зопе упичтожены полпостью и
повторное заселение было возможно только со стороны.
Город Оберхаузен (ФРГ) с 250 тыс. жителей подвергли обработке по
примеру г. Куксхвафепа. В целом там была сильшо заселена канализа-
ция, а также 274 кварталов, в которых и проводили обработку. В канали-
зации на 6 тыс. смотровых колодцев израсходовали 7,2т отравлеппой при-
манки. Примерно через 40 дней после начала обработки было отмечено
полное уничтожение крыс, предусмотренное в договоре города с фирмой,
обязавитейся освободить от них город за 12 недель. Исчезновение зверь-
ков было установлено но прекратцению поедания приманки на обработан-
пых участках. Особое впимапие обращали на защиту границ города от
повторного заселения путем раскладывания резервов отравленной при-
манки (всего было израсходовано 15,5 т па все пазомные объекты и от-
крытую территорию города} (Ки-оБего, 1963, 1969).
Аналогичная обработка в южном районе Гамбурга (ФРГ) позволила
снизить заселенность домовладений с 45 до 1% (Непзсте[, 1965).
Специалистами ФРГ с 1961—1968 гг. в 485 городах и населеппых
пунктах Нижней Саксонии © большим числом жителей (в 1,5 млн чело-
век) лля борьбы © крысами была использована упомянутая выше мето-
дика, предложенпая Штейпигером. Среди обработаппых против зверьков
городов не было таких, численность которых превышала бы 5 тыс. чело-
век, и, по мнению Телле (ТеПе, 1974), эффективпость подобной борьбы
в крупных городах еще неясна и требует проверки.
Заселенпость домовладений крысами увеличивается в осепние месяцы
и связапа с переселепием зверьков осенью из открытых участков города
в строения, что свидетельствует о необходимости интенсификации там
обработок в осенний период. Попытка полностью освободить город от
“крыс на длительное время была предприята в г. Лахти (Финляндия;
МуПушак, 1969), где они заселяли более 40% земельных участков горо-
да. После того как в течение трех месяцев их популяция была уничто-
жена, дальнейшие обработки проводили по заявкам населения. В период
с 1957 по 1968 г. заселеппость крысами колебалась от минимальной в
феврале (0,3%) до максимальной в октябре (2,54%).
В Венгрии попытки полного упичтожения крыс были предприняты в
г. Будапоште (Егд5з, 1972). В течение нескольких последующих лет в
наземных объектах города зверьки были уничтожены, по остались они
в канализационпых сооружениях, где после обработки парафинировавными
брикетами с ракумином-57 заселенность их снизилась с 56 до 234%,
а через 6 месяцев после обработки возросла до 37,5% (Ва]опы её а|.,
1976).
` Результаты дератизации, проведенной в Будапеште, а также в Обер-
хаузепе и других городах, показали, что проблема уничтожения крыс в
крупных городах зависит от эффективности борьбы с ними в канализа-
ционных сооружениях (КатсЬфего, 1963; Негтаип, НотаЪгесВег, 1962;
Непзсве!, 1965).
382 Глава пятая
и в тот период года, когда популяция паиболее уязвима для истребитель-
ных мероприятий (Рыльников, Карасева, 1985).
"Гактика применения средств истребления обычно зависит от времопи,
места обработок и тех целой, для которых они предназначены. Так, об-
работка открытой территории вокруг построек в средней полосе ведется
© помощью долговременных отравляющих точек, куда раскладывают ири-
манку с аитикоагулянтами (Тощитин, и др. 1980; Низот, Вептвот,
1981). При обработках открытых стации наиболее трудпая задача о ес-
поечение сохранности ириманок, поэтому рекомендуют применепие их
расфасованными в полиэтиленовые пакетики, а также в виде парафипи-
ровапных брикетов. Готовую отравленную приманку раскладывают С
норы грызунов и другие естественные укрытия или закладывают в труб-
ки из рубероида, картонные трубки из-под типографских отходов, ящич-
ки ит. п.
ИПриманочный способ борьбы с крысами в открытых стациях следует
признать основпым. Другие способы, в том числе бесприманочные (газа-
ция, дустация), менее приемлемы, хотя существует опыт их применепия,
например глубокое пропыливание пор серых крыс дустами аптикоагу-
лянтов (ТеПе, 1962), опыливание троп и закладка в поры проныленных
антикоагуляптами ватных тампонов. (Тощигин и др., 1976; Богачева
и др. 1984). Эффективпость примепения этого способа будет определять-
ся насыщенностью почвы влагой -—- чем почва суше, тем сильнее «пыля-
щий» эффект.
влажным почвам дуст прилипает и смешивастея с
грязью.
Возможна газация фосфипом при условии плотного закрывания всех
выходов крысиной поры (Ме, 1980). Фумигация нор хлорпикрином в
зимний пернод неприемлема, так как при температуре воздуха ниже
12°С он малоэффективен. Применение цианплава и циклона (препара-
тивные формы КСМ) вряд ли возможно па культурах зерповых, учиты-
вая хорошую растворимость солей синильной кислоты в воде и вороят-
ность проникновения их в культуру. Эти срелетва опаспо также’ приме-
нять около строений человека. Истребление легче проводить зимой, когда
поры крыс можно обнаружить среди ножухлой травы и когда отравлеп-
пые приманки поедаются ими более охотно, чем летом — в условиях изо-
билия конкурирующих кормов.
|
.
Опыты по практически полному уничтожению крыс
Первый успелный опыт практически полного уничтожения крыс был
проведен в г. Куксхафене (ФРГ) по методике, предложенной Штейниге-
ром в 1955 г. (Бецивег, 1955, 1957, 1958). Истребительным работам
предшествовали тптательная организационная подготовка, ныявленио всех
мест, где имеются крысы. В течение 20 дней уничтожили 95% зверьков,
ликвидация оставшихся 5% потребовала вдвое больше времени. Главным
средством уничтожения были приманки из муки с ядом варфарином, ко
торые раскладывали в приманочные ящики крупными порциями (1—
5 кг на ящик). Вокруг обработанной зоны также были установлены ящи-
Практическое значение
383
ки с приманкой для упичтожения вновь вселяющихся крыс. Кроме при-
манок, применили варфарин и для опыления нор н трои. Проведенный
опыт показал, что крысы в обработанной зопе упичтожены полпостью и
повторное заселение было возможно только со стороны.
Город Оберхаузен (ФРГ) с 250 тыс. жителей подвергли обработке по
примеру г. Куксхвафепа. В целом там была сильшо заселена канализа-
ция, а также 274 кварталов, в которых и проводили обработку. В канали-
зации на 6 тыс. смотровых колодцев израсходовали 7,2т отравлеппой при-
манки. Примерно через 40 дней после начала обработки было отмечено
полное уничтожение крыс, предусмотренное в договоре города с фирмой,
обязавитейся освободить от них город за 12 недель. Исчезновение зверь-
ков было установлено но прекратцению поедания приманки на обработан-
пых участках. Особое впимапие обращали на защиту границ города от
повторного заселения путем раскладывания резервов отравленной при-
манки (всего было израсходовано 15,5 т па все пазомные объекты и от-
крытую территорию города} (Ки-оБего, 1963, 1969).
Аналогичная обработка в южном районе Гамбурга (ФРГ) позволила
снизить заселенность домовладений с 45 до 1% (Непзсте[, 1965).
Специалистами ФРГ с 1961—1968 гг. в 485 городах и населеппых
пунктах Нижней Саксонии © большим числом жителей (в 1,5 млн чело-
век) лля борьбы © крысами была использована упомянутая выше мето-
дика, предложенпая Штейпигером. Среди обработаппых против зверьков
городов не было таких, численность которых превышала бы 5 тыс. чело-
век, и, по мнению Телле (ТеПе, 1974), эффективпость подобной борьбы
в крупных городах еще неясна и требует проверки.
Заселенпость домовладений крысами увеличивается в осепние месяцы
и связапа с переселепием зверьков осенью из открытых участков города
в строения, что свидетельствует о необходимости интенсификации там
обработок в осенний период. Попытка полностью освободить город от
“крыс на длительное время была предприята в г. Лахти (Финляндия;
МуПушак, 1969), где они заселяли более 40% земельных участков горо-
да. После того как в течение трех месяцев их популяция была уничто-
жена, дальнейшие обработки проводили по заявкам населения. В период
с 1957 по 1968 г. заселеппость крысами колебалась от минимальной в
феврале (0,3%) до максимальной в октябре (2,54%).
В Венгрии попытки полного упичтожения крыс были предприняты в
г. Будапоште (Егд5з, 1972). В течение нескольких последующих лет в
наземных объектах города зверьки были уничтожены, по остались они
в канализационпых сооружениях, где после обработки парафинировавными
брикетами с ракумином-57 заселенность их снизилась с 56 до 234%,
а через 6 месяцев после обработки возросла до 37,5% (Ва]опы её а|.,
1976).
` Результаты дератизации, проведенной в Будапеште, а также в Обер-
хаузепе и других городах, показали, что проблема уничтожения крыс в
крупных городах зависит от эффективности борьбы с ними в канализа-
ционных сооружениях (КатсЬфего, 1963; Негтаип, НотаЪгесВег, 1962;
Непзсве!, 1965).
384
Глава пятая
В СССР попытка практически полного уничтожения серых крыс была
проведена в крупных населенных пунктах. В ходе экспериментапол-
ностью подтвердились все основпые рекомендации Штайнитера (‘Тощи-
гип, и др., 1969, 1976).
Иногда, как это имело место в т. Волгограде, отдельные поселепия
крыс разобщены и тогда их можно уничтожать поочередио (Шепотьев,
Попов, 1971). Гораздо чаще животные заселяют такие участки и комму-
никации, которые позволяют им образовывать очень крупную популяцию,
состоящую из ряда поселепий, имеющих друг с другом связь.
По мнению Друммонда (1974), еще не доказана ценность метода,
основанного на истреблепии крыс и профилактике повторного заселения
ими объектов с помощью резервов долго сохраняющейся отравленной
приманки, разработанного Штайнигером для обширных сельских райо-
нов, но примеры такой обработки имеются— крысы практически отсут.
ствуют в провинции Альберта (Канада) на площади _661 тыс. км
(Зиопыег, 1964; Вагге&, 1967). Аналогичпая успеиная работа проведена
на островах Ванкувер (ЗшИВ, 1963) и Гельголапд (ТеПе, Уашк, 1963).
Прянции сплошной систематической дератизации был экологически
обоснован и внедрен в СССР В. Г. Полежаевым и др- (1959, 1962). Упо-
мянутые выше работы Телле, Тощигина и Баррета — розультат творче-
ского развития иринцина сплошной систематической дератизации.
На основании анализа материалов по пространствеппому размещению
серых крыс в разных объектах и разных местообитаниях, характера под-
зижности и особенностей их размножения можно выделить некоторые
общие принципы ограпичения численпости грызунов, часть из которых
сформулирована А. П. Кузякиным (1951), подтверждена многими дру-
тими авторами и сводится к следующему: 1) выявление наиболее плотно
васелеппых участков города, района, объекта, их картирование и распро-
деление в соответствии с этим сил и средств истребления; 2) преимущест-
венное использование для борьбы © крысами периодов наибольшей неод-
нородности в динамике пространственной структуры их популяции;
3) борьба в «стациях переживания» в зимнее или ранневесенное время,
особенно в средних и более севервых широтах СССР.
.
Следует заметить, что эти предложения могут быть длополнепы. К ар-
гументации «зимних» обработок можно добавить, что в этот период жиз-
ни популяции наблюдаются паиболее низкие показатели плодовитости,
наиболее высокая смертность и однородпость возрастного состава попу-
ляции, ес фено- и генотипической структуры. Эти закопомерности отчет-
ливо проявляются в умеренных и северных пиротах в популяциях крыс,
обитающих в черте пасоленных пунктов, а также в популяциях крыс,
круглогодично обитающих в природных биотопах в условиях слабоконти-
пентального и субтронического климата па территории СССР и прилежа-
щих стран.
Практическое значение
385
Оценка качества истребительных работ
Учет численности серых крые до и носле проведения истребительных ме-
роприятий позволяет оценить конечные результаты и в тех случаях, кот-
да они оказываются неудовлетворительными; важно установить причины
этого (Айзенштадт, 1949).
В 1968 г. в комеппоугольных шахтах и канализациопных сооружениях
была предпринята попытка определить качество проведенной дератиза-
ции по числу мест, где в дальнейшем были обнаружены крысы, а также
следы их жизнедеятельности (Тотцигип, 1968). Если до истребления
зверьки заселяли 50 капализационных колодцев, а после него 5, то про-
странство, запятое популяцией, считалось соответственно в 10 раз мень-
шим, Способы учета мест обитания крыс по количеству заслеженных
пробных площадок предложили В. А. Вилькович с соавторами (1969),
ТГрэвс с соавторами (Стеауез её а|., 1982) и др. Авторы основывались
на предноложении, что размер площади, занятой популяцией, соответст-
вует численности зворьков и число мест, где они были обнаружены, от-
ражает изменения в численности их популяции. По-видимому, для учета
крыс в условиях канализации, а также для других типов местообитаний,
где изменения численности происходят в основном за счет флуктуаций
в пространетве, этот способ пригоден. Показатель процента освобождеп-
ной от грызунов площади строений широко используется практическими
работниками для оценки качества дератизационных работ (Судейкин,
1975). Предполагается, что изменение грапиц обитания крупных понуля-
ций грызунов приводит к столь существенным изменениям их численно-
сти, что разницей в плотности обитания крыс внутри этих грапиц, на
разных объектах, можно пренебречь. Следует обратить внимание на то,
что критерии оценки площади, свободной от грызунов, в том числе и
‚ крыс, формируются на оспове оцепок их относительной численности в
‘каждом конкретном местообитаний и зачастую основаны не на действи-
тельном отсутствии грызунов, о чем можно судить только по комплексу
таких объективных показателей, как полное отсутствие следов их жизне-
деятельности (помета, следов, жилых входных отверстий, полной сохрап-
ности контрольной приманки, не говоря уже о полном отсутствии вылова
их механическими орудиями лова), а лишь на вылове зверьков ловупка-
ми «Геро» в количестве, меньшем определеппого зпачения. Этот «крите-
рий», по которому судят о качестве труда практических работников,
далек от объективности.
Хочется подчеркпуть, что объективную информацию об относительной
численности серых крыс в каждом местообитании можно собрать, основы-
ваясь только ‘на комплексе показателей под обтиим названием «следы
казнедеятельности», среди которых есть более динамичные например
отпечатки лап, число мест со свежим пометом, и более копсервативные,
такие, как тропы, входпые отверстия. Информативность каждого из этих
показателей меняется от местообитания к местообитанию, поэтому для
получения сравнимых результатов вряд ли правильпо ограничиваться ка-
КИМ-ТО ОДНИМ ИЗ НИХ.
384
Глава пятая
В СССР попытка практически полного уничтожения серых крыс была
проведена в крупных населенных пунктах. В ходе экспериментапол-
ностью подтвердились все основпые рекомендации Штайнитера (‘Тощи-
гип, и др., 1969, 1976).
Иногда, как это имело место в т. Волгограде, отдельные поселепия
крыс разобщены и тогда их можно уничтожать поочередио (Шепотьев,
Попов, 1971). Гораздо чаще животные заселяют такие участки и комму-
никации, которые позволяют им образовывать очень крупную популяцию,
состоящую из ряда поселепий, имеющих друг с другом связь.
По мнению Друммонда (1974), еще не доказана ценность метода,
основанного на истреблепии крыс и профилактике повторного заселения
ими объектов с помощью резервов долго сохраняющейся отравленной
приманки, разработанного Штайнигером для обширных сельских райо-
нов, но примеры такой обработки имеются— крысы практически отсут.
ствуют в провинции Альберта (Канада) на площади _661 тыс. км
(Зиопыег, 1964; Вагге&, 1967). Аналогичпая успеиная работа проведена
на островах Ванкувер (ЗшИВ, 1963) и Гельголапд (ТеПе, Уашк, 1963).
Прянции сплошной систематической дератизации был экологически
обоснован и внедрен в СССР В. Г. Полежаевым и др- (1959, 1962). Упо-
мянутые выше работы Телле, Тощигина и Баррета — розультат творче-
ского развития иринцина сплошной систематической дератизации.
На основании анализа материалов по пространствеппому размещению
серых крыс в разных объектах и разных местообитаниях, характера под-
зижности и особенностей их размножения можно выделить некоторые
общие принципы ограпичения численпости грызунов, часть из которых
сформулирована А. П. Кузякиным (1951), подтверждена многими дру-
тими авторами и сводится к следующему: 1) выявление наиболее плотно
васелеппых участков города, района, объекта, их картирование и распро-
деление в соответствии с этим сил и средств истребления; 2) преимущест-
венное использование для борьбы © крысами периодов наибольшей неод-
нородности в динамике пространственной структуры их популяции;
3) борьба в «стациях переживания» в зимнее или ранневесенное время,
особенно в средних и более севервых широтах СССР.
.
Следует заметить, что эти предложения могут быть длополнепы. К ар-
гументации «зимних» обработок можно добавить, что в этот период жиз-
ни популяции наблюдаются паиболее низкие показатели плодовитости,
наиболее высокая смертность и однородпость возрастного состава попу-
ляции, ес фено- и генотипической структуры. Эти закопомерности отчет-
ливо проявляются в умеренных и северных пиротах в популяциях крыс,
обитающих в черте пасоленных пунктов, а также в популяциях крыс,
круглогодично обитающих в природных биотопах в условиях слабоконти-
пентального и субтронического климата па территории СССР и прилежа-
щих стран.
Практическое значение
385
Оценка качества истребительных работ
Учет численности серых крые до и носле проведения истребительных ме-
роприятий позволяет оценить конечные результаты и в тех случаях, кот-
да они оказываются неудовлетворительными; важно установить причины
этого (Айзенштадт, 1949).
В 1968 г. в комеппоугольных шахтах и канализациопных сооружениях
была предпринята попытка определить качество проведенной дератиза-
ции по числу мест, где в дальнейшем были обнаружены крысы, а также
следы их жизнедеятельности (Тотцигип, 1968). Если до истребления
зверьки заселяли 50 капализационных колодцев, а после него 5, то про-
странство, запятое популяцией, считалось соответственно в 10 раз мень-
шим, Способы учета мест обитания крыс по количеству заслеженных
пробных площадок предложили В. А. Вилькович с соавторами (1969),
ТГрэвс с соавторами (Стеауез её а|., 1982) и др. Авторы основывались
на предноложении, что размер площади, занятой популяцией, соответст-
вует численности зворьков и число мест, где они были обнаружены, от-
ражает изменения в численности их популяции. По-видимому, для учета
крыс в условиях канализации, а также для других типов местообитаний,
где изменения численности происходят в основном за счет флуктуаций
в пространетве, этот способ пригоден. Показатель процента освобождеп-
ной от грызунов площади строений широко используется практическими
работниками для оценки качества дератизационных работ (Судейкин,
1975). Предполагается, что изменение грапиц обитания крупных понуля-
ций грызунов приводит к столь существенным изменениям их численно-
сти, что разницей в плотности обитания крыс внутри этих грапиц, на
разных объектах, можно пренебречь. Следует обратить внимание на то,
что критерии оценки площади, свободной от грызунов, в том числе и
‚ крыс, формируются на оспове оцепок их относительной численности в
‘каждом конкретном местообитаний и зачастую основаны не на действи-
тельном отсутствии грызунов, о чем можно судить только по комплексу
таких объективных показателей, как полное отсутствие следов их жизне-
деятельности (помета, следов, жилых входных отверстий, полной сохрап-
ности контрольной приманки, не говоря уже о полном отсутствии вылова
их механическими орудиями лова), а лишь на вылове зверьков ловупка-
ми «Геро» в количестве, меньшем определеппого зпачения. Этот «крите-
рий», по которому судят о качестве труда практических работников,
далек от объективности.
Хочется подчеркпуть, что объективную информацию об относительной
численности серых крыс в каждом местообитании можно собрать, основы-
ваясь только ‘на комплексе показателей под обтиим названием «следы
казнедеятельности», среди которых есть более динамичные например
отпечатки лап, число мест со свежим пометом, и более копсервативные,
такие, как тропы, входпые отверстия. Информативность каждого из этих
показателей меняется от местообитания к местообитанию, поэтому для
получения сравнимых результатов вряд ли правильпо ограничиваться ка-
КИМ-ТО ОДНИМ ИЗ НИХ.
386
Глава пятая
Друммонд и Реннисон (4974) и Рениисон (Вептузоп, 1977) предложи-
ли опенивать эффективность родентицидов с помощью регулярного (каж-
дые 2—3 дня в течепие одного месяца) взвептивания отравленнойпри-
манки, на осповании чего строится трафик динамики ее поедания с
указанием средпих значений, а также верхнего и нижнего пределов их
варьирования.
Роль профилактических мероприятий в ограничении
числентости серых крыс
Профилактические мероприятия можно разделить на технические и са-
хи ические мероприятия включают: обеспечение плотно-
го закрывания входных дверей, заделку щелей, бетонирование и цемен-
тирование полов, устаповку металлических сеток в окоппых и других
проемах, через которые могут проникать крысы, т. е. обесиечение «кры-
сонепроницаемости». Эти требования должны быть учтены при проекти-
ровании отдельных зланий и целых городов (Полежаев, РИ ; опиает,
1952; Свуйу, 1954; Кузякин, 1955; Матусевич, 1959; Вашков и др.,
1974; Клофе, 1980).
—
Для хранепия фуража и пищевых пролуктов в домах сельской за.
стройки целесообразно использование плотно закрывающихся ларен, 00-
шитых жестью, гладким пластиком, стеклотекстолитом, которые, соглас-
но И. А. Емельяновой (1986), в наименьшей степени прогрызаются кры-
сами. По мнению А. ПН. Кузякина (сообщепие в порядке дополнения),
встроенные лари и отсеки с грызупонепроницаемой обшивкой необходи-
мо включать в строительные проекты усадебных комплексов, животновод-
ческ
спий.
,
"“ Пекоторые авторы считают, что техпические мероприятия дороги и не
обеспечивают полной защиты от крыс (Зоо, 1963; Себе, 1980). В ряде
случаев они и неприменимы. Например, вряд ли возможна кардинальная
перестройка старых городов, традиционпо сохраннющих свой историче
й облик.
и пение мероприятия наряду с санитарными входят в состав
«управлепия внешлей средой обитания крыс», которое, по мнению Дэкек-,
сона (1973), Дэвиса и Кристиана (1977), является наиболее кардиналь-
ным средством регулирования численности грызунов-вредителей. '109т0-
му эти мероприятия должны быть применены там, где они экономически
авны и возможны.
пор арные мероприятия включают в себя очистку строе-
ний, дворов и других территорий от мусора, который служит либо источ
орма. либо убежищем для крыс.
и оличестьо мусора в городах в период с середины семидесятых до
конца восьмидесятых тодов нашего столетия, по подечетам, должно уве-
личиться втрое (Дударев и др., 1974). Пищевые отходы (от 28 до 47% )
в составе мусора пригодны для питания крыс (Когап, 1978), заселяющих
не только неорганизоватитые, но и организованные свалки, где санитарная
Практическое значение
387
служба ие справляется со своими задачами (Соболева и др., 1957; Хама-
ганов, 1965). Численность этих грызунов па свалках достигает 120 экз.
па 1500 м? (Вапищег, 1974).
Освобождение районов от крыс оказывается возможным, когда техни-
ческие и санитарные мероприятия сочетаются с истреблением их химиче-
ским методом и с последующей расстановкой долговременных отравляю-
щих точек, обеспечивающих профилактику заселения грызунами. Как мы
уже упомипали, для этих целей наиболее пригодны сухие долго сохра-
няющиеся приманки.
Подобная активная профилактика с помощью резервов отравленной
приманки широко применяется зарубежными и советскими специалиста-
ми (Кондратенко, 1977; ЗшИм, 1963; Гг@бз, 1972; Джексон, 1973; Про-
кофьева, Ивапов, 1974; Тащигин и лр., 1976, 1980; Николаева и др.,
1977).
Тути адаптации серых крыс при пров?дении длительных
истребительных мероприятий аптикоагу зянтами
В связи с тем что дератизация стала хорошо организованной системой
мероприятий, проводимых в течение длительного срока па большой ча-
сти территории населеппых пупктов, встала проблема профилактики и
преодоления адаптаций грызунов к ядам, Эти адаптации возпикают не-
избежно как ответ на непрерывное действие химических факторов ист-
ребления, стаповящихся факторами отбора экстремального характера.
К числу таких адаптаций можно отнести физнолого-генетическую устой-
чивость к антикоагуляптам и две основные формы оборонительной реак-
ции: неофобию и аверсию (вторичное избегание).
Для изучения оборонительной реакции и физиологической устойчиво-
сти грызунов в условиях лабораторного содержания используют соот-
”-, ветственио кормление животных в условиях выбора между контрольной
и отравленной пищей, а также кормление отравленным кормом (с опре-
доленным процентным содержанием яда) в условиях отсутствия выбора.
Например, процентное содержапие зоокумарина в приманке в таких
случаях составляет 0,005%. 100% чувствительных к зоокумарину серых
крые гибнет при поедании такой приманки в точение 6 дней, причем сни-
жение концентрации яда или сроков кормления, порознь или вместе. ве-
дет к снижению числа погибних особей (ТеПе, 1967; Огаттова, У/Йзов,
1968). Эта методика легла в оспову определения резистентности к вар-
фарину (зоокумарину). Крысы, выживитие после 21 дня наблодепия,
считаются резистентными (\НО, 1975).
Прижизнепное определение активности свертывающей системы крови
у грызунов проводят ио величине протромбипового времепи. Величина
естественной изменчивости протромбинового времени очень велика (Роо]
её а[., 1968), что, по мнению Мартина с сотрудниками (Маги 0% а|.,
1979), затрудняет выделение точного критерия для определения при-
надлежности крысы к той или иной категории по чувствительности к
ядам.
386
Глава пятая
Друммонд и Реннисон (4974) и Рениисон (Вептузоп, 1977) предложи-
ли опенивать эффективность родентицидов с помощью регулярного (каж-
дые 2—3 дня в течепие одного месяца) взвептивания отравленной
при-
манки, на осповании чего строится трафик динамики ее поедания с
указанием средпих значений, а также верхнего и нижнего пределов их
варьирования.
Роль профилактических мероприятий в ограничении
числентости серых крыс
Профилактические мероприятия можно разделить на технические и са-
хи ические мероприятия включают: обеспечение плотно-
го закрывания входных дверей, заделку щелей, бетонирование и цемен-
тирование полов, устаповку металлических сеток в окоппых и других
проемах, через которые могут проникать крысы, т. е. обесиечение «кры-
сонепроницаемости». Эти требования должны быть учтены при проекти-
ровании отдельных зланий и целых городов (Полежаев, РИ ; опиает,
1952; Свуйу, 1954; Кузякин, 1955; Матусевич, 1959; Вашков и др.,
1974; Клофе, 1980).
—
Для хранепия фуража и пищевых пролуктов в домах сельской за.
стройки целесообразно использование плотно закрывающихся ларен, 00-
шитых жестью, гладким пластиком, стеклотекстолитом, которые, соглас-
но И. А. Емельяновой (1986), в наименьшей степени прогрызаются кры-
сами. По мнению А. ПН. Кузякина (сообщепие в порядке дополнения),
встроенные лари и отсеки с грызупонепроницаемой обшивкой необходи-
мо включать в строительные проекты усадебных комплексов, животновод-
ческ
спий.
,
"“ Пекоторые авторы считают, что техпические мероприятия дороги и не
обеспечивают полной защиты от крыс (Зоо, 1963; Себе, 1980). В ряде
случаев они и неприменимы. Например, вряд ли возможна кардинальная
перестройка старых городов, традиционпо сохраннющих свой историче
й облик.
и
пение мероприятия наряду с санитарными входят в состав
«управлепия внешлей средой обитания крыс», которое, по мнению Дэкек-,
сона (1973), Дэвиса и Кристиана (1977), является наиболее кардиналь-
ным средством регулирования численности грызунов-вредителей. '109т0-
му эти мероприятия должны быть применены там, где они экономически
авны и возможны.
пор арные мероприятия включают в себя очистку строе-
ний, дворов и других территорий от мусора, который служит либо источ
орма. либо убежищем для крыс.
и оличестьо мусора в городах в период с середины семидесятых до
конца восьмидесятых тодов нашего столетия, по подечетам, должно уве-
личиться втрое (Дударев и др., 1974). Пищевые отходы (от 28 до 47% )
в составе мусора пригодны для питания крыс (Когап, 1978), заселяющих
не только неорганизоватитые, но и организованные свалки, где санитарная
Практическое значение
387
служба ие справляется со своими задачами (Соболева и др., 1957; Хама-
ганов, 1965). Численность этих грызунов па свалках достигает 120 экз.
па 1500 м? (Вапищег, 1974).
Освобождение районов от крыс оказывается возможным, когда техни-
ческие и санитарные мероприятия сочетаются с истреблением их химиче-
ским методом и с последующей расстановкой долговременных отравляю-
щих точек, обеспечивающих профилактику заселения грызунами. Как мы
уже упомипали, для этих целей наиболее пригодны сухие долго сохра-
няющиеся приманки.
Подобная активная профилактика с помощью резервов отравленной
приманки широко применяется зарубежными и советскими специалиста-
ми (Кондратенко, 1977; ЗшИм, 1963; Гг@бз, 1972; Джексон, 1973; Про-
кофьева, Ивапов, 1974; Тащигин и лр., 1976, 1980; Николаева и др.,
1977).
Тути адаптации серых крыс при пров?дении длительных
истребительных мероприятий аптикоагу зянтами
В связи с тем что дератизация стала хорошо организованной системой
мероприятий, проводимых в течение длительного срока па большой ча-
сти территории населеппых пупктов, встала проблема профилактики и
преодоления адаптаций грызунов к ядам, Эти адаптации возпикают не-
избежно как ответ на непрерывное действие химических факторов ист-
ребления, стаповящихся факторами отбора экстремального характера.
К числу таких адаптаций можно отнести физнолого-генетическую устой-
чивость к антикоагуляптам и две основные формы оборонительной реак-
ции: неофобию и аверсию (вторичное избегание).
Для изучения оборонительной реакции и физиологической устойчиво-
сти грызунов в условиях лабораторного содержания используют соот-
”-, ветственио кормление животных в условиях выбора между контрольной
и отравленной пищей, а также кормление отравленным кормом (с опре-
доленным процентным содержанием яда) в условиях отсутствия выбора.
Например, процентное содержапие зоокумарина в приманке в таких
случаях составляет 0,005%. 100% чувствительных к зоокумарину серых
крые гибнет при поедании такой приманки в точение 6 дней, причем сни-
жение концентрации яда или сроков кормления, порознь или вместе. ве-
дет к снижению числа погибних особей (ТеПе, 1967; Огаттова, У/Йзов,
1968). Эта методика легла в оспову определения резистентности к вар-
фарину (зоокумарину). Крысы, выживитие после 21 дня наблодепия,
считаются резистентными (\НО, 1975).
Прижизнепное определение активности свертывающей системы крови
у грызунов проводят ио величине протромбипового времепи. Величина
естественной изменчивости протромбинового времени очень велика (Роо]
её а[., 1968), что, по мнению Мартина с сотрудниками (Маги 0% а|.,
1979), затрудняет выделение точного критерия для определения при-
надлежности крысы к той или иной категории по чувствительности к
ядам.
338
Глава пятая
Механизм резистентности серых крыс к антикоатулянтам подробно
описан в ряде оригинальных работ, а также публикаций обзорного плана,
среди которых наиболее полной является сводка Шаде (Эепафе, 14974).
По данным большинства опубликовапных работ, существование рези-
стентности определяется аутосомальным доминантным геном. В отноше-
нии биохимического механизма резистентности существуют две гипотезы,
согласно первой из которых в мутантном организме происходит структур-
ное изменение места действия кофермента — витамина К в белковой мо-
лекуле предшественника протромбина, согласно второй — мутация изме-
пяет энзим или энзимную систему, которая превращает 2,3-эпоксид
витамина К в витамин К. В первом случае возрастает конкурентоспособ-
ность витамина К в сравнении с варфарином при соединении с белковой
молекулой. Во втором случае, витамин К образуется из эпоксида в при-
сутствии варфарипа, который в нормальном организме блокирует такое
превращение (О’Веуу её а1., 1968; Ве| е\ а1., 1972).
Резистентность в популяциях серых крыс, иптепсивно обрабатывае-
мых антикоатулянтами, имеет тепденцию к распространению центробеж-
но со скоростью около 3 км в гол. Так, в Великобритании антикоагулян-
ты стали применять в 1953 г. (Стеауез, 1974), в 1958 г. па одной из
ферм в Западной ПГотландии, около г. Глазго, впервые была обнаружена
устойчивость серых крыс к ним; впоследствии территория, занятая ре-
вистентными крысами в Шотландии, расширилась до 2100 км”. Анало-
гичная картина наблюдалась в Англии, Уэльсе. Вскоре после обпаруже-
ния резистентных крыс в Шотландии стали поступать сообщепия об их
наличии в 1962 г. в Ютландии (Дапия), а в 1966 г.— в Нидерландах,
Гвиане, в 70-х годах — в ФРГ и Бельгии, причем в осповпом в сельской
местности (Хроника ВОЗ, 1966; ОрвоЁ, ГБапреуе!8, 1969; Гл, 1969;
Стеауез, 1974; ТеПе, 1972).
В 1971 г. устойчивые к варфарину серые крысы были впервые обна-
ружоны в СТА в сельской местности штата Северная Каролина. К 1975 г.
резистентные крысы были обнаружены в 18 городах и А агропромышлен-
ных пунктах (ТаеКзоп, Каикешет, 1976; ТасКзоп её а1., 1978; Аррегзоп
ей а|., 1981). В СССР к настоящему времени случаи резистентности се-
рых крые к зоокумарину были отмечены только в Ленинграде (Быков-
ский, Пиколаева, 1986) и в отдельных точках Москвы (наши данные),
Для преодоления защитных физиологических и генетических меха-
низмов устойчиности к варфарину применяли более токсичные антикоагу-
лянты: куматстралил, дифенакум и дифепацин, которые на первых порах
вызывали гибель большинства резистентных крыс. Однако успех оказал-
ся временным, что связывают с перекрестной устойчивостью. Например,
в Англии в пределах распространения серых крыс, устойчивых к варфа-
рипу, обнаружены места, где обиталот крысы, устойчивые к куматетра-
лилу и дифепакуму. Постунают сведения об устойчивости даже к анти-
коагулянтам второго поколения: бромадиолону и бродифакуму (ВепЧеу,
1969; Гаю, 4969; Стеауез, 4974; Набег, 1975; Суеауез об а]., 1982; Мас-
пЙюп, СШ, 1987). В США обнаружены крысы, устойчивые к дифепаци-
пу (дифасинону) (Тлапд, 1969, 1977).
Практическое значение
389
Гривс (Стедуез её а|., 1977) и Джексоп с Каукейненом (ТасКзов,
Капкешев, 1976) считают, что применение любых антикоагулянтов не-
целесообразно там, где хотя бы к одному из них обнаружена резистент-
ность.
.
Возможный путь усиления действия антикоагулянтов— добавление
синергистов (Тощигин, Рыльников, 4985).
:
К антикоагулянтам рекомендуют также добавлять стерилянты, кото-
рые предотвращают появление резистентности и ее распространение
{Еитмуаег, 1980; МатзВ, Но\гага, 1976).
Одпако наиболее доступным и эффективным в настоящее время спо-
<собом борьбы с резистентными к антикоагуляптам крысами является ис-
пользование ядов острого действия: 1) фторацетата натрия (ОрВоф, Гап-
е\уе]4, 1969); 2) фоефида цинка (УасКзоп её а|., 1978); 3) вакора (ТасК-
зоп, КацКетет, 1976; Гип@, 1977); 4) сциллирозида (Таша, 1977);
5) норбомида (Гиша, 1977). Перечисленные яды особенно эффективны
при принудительных формах подачи (обмазка, опыливание нор, исполь-
зование пылевых площадок, ящиков, трубок и пр.), поскольку резистент-
ность к некоторым антикоагулянтам (куматетралил) сочетается с реак-
цией отказа от приманки (лапа, 1969).
Можно ‘считать, что профилактика устойчивости должна строиться с
учетом трех осповпых условий, определяющих долю резистентных особей
в популяции: 1) интенсивность отбора с помощью антикоагулянтов;
2) величина притока мигрантов извпе и их генотинический состав по ре-
зистентности; 3) количество витамина К, поступающего в организм с пи-
щей и продуцируемого бактериями кишечника.
жж+
Прослеживая возпикновепие, развитие и копечные результаты примене-
ния различных методов и способов борьбы с серой крысой, мы видим,
‘что наилучшие результаты получаются при соблюдении следующих ус-
ловий: 1} полнота истребления популяции; 2) длительное остаточное
действие яда; 3) безопасность применения его для людей и домашних
животных; 4) высокая экономичность. В паиболышней степени этим тре-
бованиям удовлетворяют яды-антикоагулянты в сухих приманках, в с0-
ставе липких композиций, дустов, нанесенных на поверхпость листов,
трубок; ящиков и т. п., при параллельном с ними использовании всех
возможных технических и санитарных мероприятий.
В организации истребительных и профилактических работ решающую
роль. играет тщательность обследования и выявления поселений крыс.
Если известны все поселения зверьков, их плотность и примерные грани-
цы обитания,то уничтожение популяции этих грызунов вполне реально.
Наиболее благоприятным периодом при проведении истребительных работ
против серых крыс является период до начала весеннего (март—май)
размножения, когда их популяция наиболее однородна по возрастному
составу и, как следствие этого, более одпородна по стереотипам пищевого
поведения отдельных особей. Однако разнокачественность состава попу-
ляции — черта, присущая им даже в зимний период. Поэтому целесооб-
338
Глава пятая
Механизм резистентности серых крыс к антикоатулянтам подробно
описан в ряде оригинальных работ, а также публикаций обзорного плана,
среди которых наиболее полной является сводка Шаде (Эепафе, 14974).
По данным большинства опубликовапных работ, существование рези-
стентности определяется аутосомальным доминантным геном. В отноше-
нии биохимического механизма резистентности существуют две гипотезы,
согласно первой из которых в мутантном организме происходит структур-
ное изменение места действия кофермента — витамина К в белковой мо-
лекуле предшественника протромбина, согласно второй — мутация изме-
пяет энзим или энзимную систему, которая превращает 2,3-эпоксид
витамина К в витамин К. В первом случае возрастает конкурентоспособ-
ность витамина К в сравнении с варфарином при соединении с белковой
молекулой. Во втором случае, витамин К образуется из эпоксида в при-
сутствии варфарипа, который в нормальном организме блокирует такое
превращение (О’Веуу её а1., 1968; Ве| е\ а1., 1972).
Резистентность в популяциях серых крыс, иптепсивно обрабатывае-
мых антикоатулянтами, имеет тепденцию к распространению центробеж-
но со скоростью около 3 км в гол. Так, в Великобритании антикоагулян-
ты стали применять в 1953 г. (Стеауез, 1974), в 1958 г. па одной из
ферм в Западной ПГотландии, около г. Глазго, впервые была обнаружена
устойчивость серых крыс к ним; впоследствии территория, занятая ре-
вистентными крысами в Шотландии, расширилась до 2100 км”. Анало-
гичная картина наблюдалась в Англии, Уэльсе. Вскоре после обпаруже-
ния резистентных крыс в Шотландии стали поступать сообщепия об их
наличии в 1962 г. в Ютландии (Дапия), а в 1966 г.— в Нидерландах,
Гвиане, в 70-х годах — в ФРГ и Бельгии, причем в осповпом в сельской
местности (Хроника ВОЗ, 1966; ОрвоЁ, ГБапреуе!8, 1969; Гл, 1969;
Стеауез, 1974; ТеПе, 1972).
В 1971 г. устойчивые к варфарину серые крысы были впервые обна-
ружоны в СТА в сельской местности штата Северная Каролина. К 1975 г.
резистентные крысы были обнаружены в 18 городах и А агропромышлен-
ных пунктах (ТаеКзоп, Каикешет, 1976; ТасКзоп её а1., 1978; Аррегзоп
ей а|., 1981). В СССР к настоящему времени случаи резистентности се-
рых крые к зоокумарину были отмечены только в Ленинграде (Быков-
ский, Пиколаева, 1986) и в отдельных точках Москвы (наши данные),
Для преодоления защитных физиологических и генетических меха-
низмов устойчиности к варфарину применяли более токсичные антикоагу-
лянты: куматстралил, дифенакум и дифепацин, которые на первых порах
вызывали гибель большинства резистентных крыс. Однако успех оказал-
ся временным, что связывают с перекрестной устойчивостью. Например,
в Англии в пределах распространения серых крыс, устойчивых к варфа-
рипу, обнаружены места, где обиталот крысы, устойчивые к куматетра-
лилу и дифепакуму. Постунают сведения об устойчивости даже к анти-
коагулянтам второго поколения: бромадиолону и бродифакуму (ВепЧеу,
1969; Гаю, 4969; Стеауез, 4974; Набег, 1975; Суеауез об а]., 1982; Мас-
пЙюп, СШ, 1987). В США обнаружены крысы, устойчивые к дифепаци-
пу (дифасинону) (Тлапд, 1969, 1977).
Практическое значение
389
Гривс (Стедуез её а|., 1977) и Джексоп с Каукейненом (ТасКзов,
Капкешев, 1976) считают, что применение любых антикоагулянтов не-
целесообразно там, где хотя бы к одному из них обнаружена резистент-
ность.
.
Возможный путь усиления действия антикоагулянтов— добавление
синергистов (Тощигин, Рыльников, 4985).
:
К антикоагулянтам рекомендуют также добавлять стерилянты, кото-
рые предотвращают появление резистентности и ее распространение
{Еитмуаег, 1980; МатзВ, Но\гага, 1976).
Одпако наиболее доступным и эффективным в настоящее время спо-
<собом борьбы с резистентными к антикоагуляптам крысами является ис-
пользование ядов острого действия: 1) фторацетата натрия (ОрВоф, Гап-
е\уе]4, 1969); 2) фоефида цинка (УасКзоп её а|., 1978); 3) вакора (ТасК-
зоп, КацКетет, 1976; Гип@, 1977); 4) сциллирозида (Таша, 1977);
5) норбомида (Гиша, 1977). Перечисленные яды особенно эффективны
при принудительных формах подачи (обмазка, опыливание нор, исполь-
зование пылевых площадок, ящиков, трубок и пр.), поскольку резистент-
ность к некоторым антикоагулянтам (куматетралил) сочетается с реак-
цией отказа от приманки (лапа, 1969).
Можно ‘считать, что профилактика устойчивости должна строиться с
учетом трех осповпых условий, определяющих долю резистентных особей
в популяции: 1) интенсивность отбора с помощью антикоагулянтов;
2) величина притока мигрантов извпе и их генотинический состав по ре-
зистентности; 3) количество витамина К, поступающего в организм с пи-
щей и продуцируемого бактериями кишечника.
жж+
Прослеживая возпикновепие, развитие и копечные результаты примене-
ния различных методов и способов борьбы с серой крысой, мы видим,
‘что наилучшие результаты получаются при соблюдении следующих ус-
ловий: 1} полнота истребления популяции; 2) длительное остаточное
действие яда; 3) безопасность применения его для людей и домашних
животных; 4) высокая экономичность. В паиболышней степени этим тре-
бованиям удовлетворяют яды-антикоагулянты в сухих приманках, в с0-
ставе липких композиций, дустов, нанесенных на поверхпость листов,
трубок; ящиков и т. п., при параллельном с ними использовании всех
возможных технических и санитарных мероприятий.
В организации истребительных и профилактических работ решающую
роль. играет тщательность обследования и выявления поселений крыс.
Если известны все поселения зверьков, их плотность и примерные грани-
цы обитания,то уничтожение популяции этих грызунов вполне реально.
Наиболее благоприятным периодом при проведении истребительных работ
против серых крыс является период до начала весеннего (март—май)
размножения, когда их популяция наиболее однородна по возрастному
составу и, как следствие этого, более одпородна по стереотипам пищевого
поведения отдельных особей. Однако разнокачественность состава попу-
ляции — черта, присущая им даже в зимний период. Поэтому целесооб-
390
Глава пятая
разно применять параллельно две-три отравлеппых приманки на различ-
ной пищевой основе, комбинируя их с формами контактпого применения
антикоагуляитов, как, например, липкие композиции, дусты. Это позволя-
ет приблизиться к возможности воздействия па крыс всех возрастов с
учетом стереотипа их пищевого поведения, в том числе и па крыс, отка-
зывающцихся поедать отравлеппый корм.
:
Как показали нап исследования, повьипение концентрации антикоа-
гулянтов в препаративных формах может.привести к ускоренному раз-
витию оборонительной реакции в популяции крыс с нормальным уровнем
физиологической устойчивости или К дальнейшему повьшпению рези-
стентности крыс в тех популяциях, где уровень. физиологической устой-
чивости выше нормального.
:
Ввиду быстрого образования у зверьков оборенительной` реакции на
яды острого действия их применение лучгие ограничить одним-двумя ту-
рами в году. Такое чередование не только желательно, по и необходимо
для профилактики возникновепия резистентпости к антикоагулянтам.
Репеллентные методы защиты материалов от крыс
Истребительные мероприятия не всегда приемлемы и эффективны—
применение отравленных приманок рассчитано на то, чтобы зверек вотел
в контакт с иселедуемым им предметом, в то время как ренел-
лент, наоборот, должен отпугнуть его. Известно несколько репеллентных
методов защиты материалов и технических изделий от повреждений, при-
чиняемых пасюком,— химический репеллентный метод, физический и
биологический.
+
Химические` репелленты
Химические репеллепты
— это соединения, которыми обрабатывают
объекты, придавая им отпугивающие свойства. Проблема поиска эффек-
тивных репеллентов очень сложна. Вещества, предлагаемые в качестве
репеллентов, должны отвечать многам требованиям: помимо того, что
они обладают собственно репеллептным действием, они не могут быть.
токсичными, пе должны изменять свойств материалов и не терять своих
репеллептных свойств при введении н них, не должны изменяться под
действием впешней среды (Емельянова, Данилкипа, 1983). Поиск репел-
дентов затруднен еще тем, что пока не выяснен мехапизм их действия,
отсутствуют данные о влиянии различных веществ на рецепторы органов
чувств грызунов, не установлены закопомерности отпугивающего дейст-
вия как сложных составов, так и отдельных ингредиентов химических
соединений (Вилькович, Машерук, 1964).
Методы оценки репеллентных свойетв веществ. В разное время было
предложено несколько методов испытаний и оценки репеллентных
свойств различных химических соединений. Беллак и Де Витт (ВеПаскК,
Ре УМ, 1949) вводили исследуемый препарат в корм и затем предлага-
ли одинаковое количество корма с препаратом и без него отдельно сидя-
Практическое значение
391
щим (по одной в клетке) серым крысам. Эту еду оставляли на нескольк
дней. Каждые сутки остатки несъеденного корма взвешивали и возв м
щали зверькам без добавления. На осповании показателей с еднес\ т -
ного поедания корма был выведен коэффициент отпугивания ость про.
Уйи,
сса подопытного животного в кг; Т — суточ-
дание корма с препаратом; У — суточное поедание корма без нре-
парата; Х — остаток корма без препарата.
р
м По эффективиости препараты разделяют на три группы: К =90--100 —
уно;
ЕЁ =80-90
— препараты, дающие удов-
м
пугивания; А =80 — малоэффективные пре-
лее
епеллентных свойоть имеются еще методы японских ис-
л
й и\арага её а1., 1970) и два метода Шефе
Б
(ЭсВаеЁег, Во\з, 198: ` }
$
ОВ
И
ме
(5 аетет,
‚ 1985), которые они предложили для хомячков и домо-
Применяемые репелленты. В качестве репеллентов испытывались
представители различных классов химических соединений, технические
<меси, продукты растительного происхождения и др. Пока что поиск ве-
ществ для выявления репеллептной активности против серой крысы, как
и против других грызунов, осуществляется эмпирически. Нет теоретиче-
ских предпосылок или общих припципиальных подходов, связывающих
‹троение химических соединений с репеллентной активностью, пе уче.
ны механизмы действия химических веществ на рецепторы отанов
чувств. Вместе с тем очевидно, что репеллентное действие должно быть
<вязапо с созданием дискомфортных условий при действии на органы
вкуса, обоняния, осязания, при действии на рецепторы дыхательны
тей (Фалькенштейн, Осипова, 1976).
У
Задача поиска репеллентов против грызунов имеет несколько аспек-
тов. С одпой стороны, бывает необходимо найти препараты обладающие
отнугивающим действием на расстояпии и предотвращающие таким об-
разом доступ животных в определенные места или районы. Среди хими-
ческих соединений такими должны быть летучие вещества, действующие
раздражающе прежде всего па органы обоняния.
Вместе с тем такие соединения не должны обладать токсичным или
раздражающим действием на человека, находящегося в зоне действия ре-
пеллента. Поскольку действие летучих соединений связано с их испаре-
нием, срок их влияния ограничен, поэтому © их помощью невозможно
обеспечить мпотолетпюю защиту сооружений и технических изделий.
„Пеэтому наряду с поиском репеллентов, обладающих дистанционным
действием, необходимо вести поиск репеллентов контактного действия
К этой группе относятся вещества, действующие на тактильные рецепто-
Ты, например липкие и клейкие ленты и пасты, получившие примепение
для защиты некоторых объектов от птиц. Контактным действием облада-
ют и вещества, раздражающие органы вкуса, и их включают в группу
антифидантов. Сюда же входят и вещества, сами по себе не обладающие
раздражающим или отталкивающим действием, но снижающие обоня-
390
Глава пятая
разно применять параллельно две-три отравлеппых приманки на различ-
ной пищевой основе, комбинируя их с формами контактпого применения
антикоагуляитов, как, например, липкие композиции, дусты. Это позволя-
ет приблизиться к возможности воздействия па крыс всех возрастов с
учетом стереотипа их пищевого поведения, в том числе и па крыс, отка-
зывающцихся поедать отравлеппый корм.
:
Как показали нап исследования, повьипение концентрации антикоа-
гулянтов в препаративных формах может.привести к ускоренному раз-
витию оборонительной реакции в популяции крыс с нормальным уровнем
физиологической устойчивости или К дальнейшему повьшпению рези-
стентности крыс в тех популяциях, где уровень. физиологической устой-
чивости выше нормального.
:
Ввиду быстрого образования у зверьков оборенительной` реакции на
яды острого действия их применение лучгие ограничить одним-двумя ту-
рами в году. Такое чередование не только желательно, по и необходимо
для профилактики возникновепия резистентпости к антикоагулянтам.
Репеллентные методы защиты материалов от крыс
Истребительные мероприятия не всегда приемлемы и эффективны—
применение отравленных приманок рассчитано на то, чтобы зверек вотел
в контакт с иселедуемым им предметом, в то время как ренел-
лент, наоборот, должен отпугнуть его. Известно несколько репеллентных
методов защиты материалов и технических изделий от повреждений, при-
чиняемых пасюком,— химический репеллентный метод, физический и
биологический.
+
Химические` репелленты
Химические репеллепты
— это соединения, которыми обрабатывают
объекты, придавая им отпугивающие свойства. Проблема поиска эффек-
тивных репеллентов очень сложна. Вещества, предлагаемые в качестве
репеллентов, должны отвечать многам требованиям: помимо того, что
они обладают собственно репеллептным действием, они не могут быть.
токсичными, пе должны изменять свойств материалов и не терять своих
репеллептных свойств при введении н них, не должны изменяться под
действием впешней среды (Емельянова, Данилкипа, 1983). Поиск репел-
дентов затруднен еще тем, что пока не выяснен мехапизм их действия,
отсутствуют данные о влиянии различных веществ на рецепторы органов
чувств грызунов, не установлены закопомерности отпугивающего дейст-
вия как сложных составов, так и отдельных ингредиентов химических
соединений (Вилькович, Машерук, 1964).
Методы оценки репеллентных свойетв веществ. В разное время было
предложено несколько методов испытаний и оценки репеллентных
свойств различных химических соединений. Беллак и Де Витт (ВеПаскК,
Ре УМ, 1949) вводили исследуемый препарат в корм и затем предлага-
ли одинаковое количество корма с препаратом и без него отдельно сидя-
Практическое значение
391
щим (по одной в клетке) серым крысам. Эту еду оставляли на нескольк
дней. Каждые сутки остатки несъеденного корма взвешивали и возв м
щали зверькам без добавления. На осповании показателей с еднес\ т -
ного поедания корма был выведен коэффициент отпугивания ость про.
Уйи,
сса подопытного животного в кг; Т — суточ-
дание корма с препаратом; У — суточное поедание корма без нре-
парата; Х — остаток корма без препарата.
р
м По эффективиости препараты разделяют на три группы: К =90--100 —
уно;
ЕЁ =80-90
— препараты, дающие удов-
м
пугивания; А =80 — малоэффективные пре-
лее епеллентных свойоть имеются еще методы японских ис-
л
й и\арага её а1., 1970) и два метода Шефе
Б
(ЭсВаеЁег, Во\з, 198: ` }
$
ОВ
И
ме
(5 аетет,
‚ 1985), которые они предложили для хомячков и домо-
Применяемые репелленты. В качестве репеллентов испытывались
представители различных классов химических соединений, технические
<меси, продукты растительного происхождения и др. Пока что поиск ве-
ществ для выявления репеллептной активности против серой крысы, как
и против других грызунов, осуществляется эмпирически. Нет теоретиче-
ских предпосылок или общих припципиальных подходов, связывающих
‹троение химических соединений с репеллентной активностью, пе уче.
ны механизмы действия химических веществ на рецепторы отанов
чувств. Вместе с тем очевидно, что репеллентное действие должно быть
<вязапо с созданием дискомфортных условий при действии на органы
вкуса, обоняния, осязания, при действии на рецепторы дыхательны
тей (Фалькенштейн, Осипова, 1976).
У
Задача поиска репеллентов против грызунов имеет несколько аспек-
тов. С одпой стороны, бывает необходимо найти препараты обладающие
отнугивающим действием на расстояпии и предотвращающие таким об-
разом доступ животных в определенные места или районы. Среди хими-
ческих соединений такими должны быть летучие вещества, действующие
раздражающе прежде всего па органы обоняния.
Вместе с тем такие соединения не должны обладать токсичным или
раздражающим действием на человека, находящегося в зоне действия ре-
пеллента. Поскольку действие летучих соединений связано с их испаре-
нием, срок их влияния ограничен, поэтому © их помощью невозможно
обеспечить мпотолетпюю защиту сооружений и технических изделий.
„Пеэтому наряду с поиском репеллентов, обладающих дистанционным
действием, необходимо вести поиск репеллентов контактного действия
К этой группе относятся вещества, действующие на тактильные рецепто-
Ты, например липкие и клейкие ленты и пасты, получившие примепение
для защиты некоторых объектов от птиц. Контактным действием облада-
ют и вещества, раздражающие органы вкуса, и их включают в группу
антифидантов. Сюда же входят и вещества, сами по себе не обладающие
раздражающим или отталкивающим действием, но снижающие обоня-
392
Глава пятая
тельпую или вкусовую привлекательность продукта или объекта, который
должен быть защищен от грызуна.
Р связи с этим з группу репеллентов могут входить не только в пря-
мом смысле отпугивающие вещества, но и вещества, делающие защищае-
мый объект непривлекательным для пищевой, исследовательской, гнездо-
строительной или другой деятельности животного. В делом поиск репел-
лентов с теми или иными свойствами должен определяться конкретными
задачами, которые должиы решаться с его помощью, и условиями при-
менения,
Несмотря на отсутствие теоретических предпосылок, поиск репеллен-
тов против грызупов ведется давно. Большой опыт накоплен в сельском
хозяйстве и в быту по использованию различных растепий или расти-
тельных препаратов для защиты пищевых продуктов, прежде всего зер-
новых запасов, садовых деревьев и других полезных растений от повреж-
дений грызупами. К сожалению, попытки применения этого опыта в
промышленных масштабах пока не получили широкого распространения.
Направленные поисковые исследования химиков и зоологов позволили
выявить болыное число химических соедипений, обладающих репеллент-
ной активностью. Наиболее полный обзор химических ренпеллентов против
трызунов. предложенных советскими и зарубежными исследователями,
сделан И. А. Емельяповой и Л. Л. Данилкиной (1983). В обзор вошли
материалы, опубликованные до 1978 г. В нем наиболее полно собрапа и
обобщена патептпая и периодическая литература, впервые проведена
классификация ренеллентов по их принадлежности к тем или иным
классам химических соединений.
В. Г. Полежаев (1959) предложил использовать в качестве репеллен-
та слапцевое масло и альбихтол. За рубежом для отпугивания крыс и
других грызунов применяли препарат ‘© фирменным названием «курб»,
содержащий аммониевые и алюминиевые квасцы (Золе, 1977). Этот же
препарат был рекомендеван в качестве»репеллента для отпугивания птиц
от посевов. Для защиты от крыс в бытовых условиях запатентована
смесь креозота с нафталином в-соотпошении 8 :2, которую готовят пере-
мешиванием при пагревании до температуры 80° и потом обрабатывают
сю паружные поверхности, предотвращая таким образом пропикновение
в дома крыс и муравьев (Такада, 1982). .
Для увеличения продолжительности действия летучих репеллентов
используют различные загустители, порошкообразные наполнители, по-
ристые адсорбенты, пленкообразующие полимерпые связующие (Огита,
1980: Мотиги и др., 1983).
Репеллентные свойства различных полимеров были испытаны Л. Хри-
стовым и М. Матевой (1976).
Таким образом, из рассмотрения представленного выте круга хими-
ческих соединений, показавнгих репеллентную активность, можно заклю-
чить, что это направление является перспективным в комнлексе. меро-
приятий по защите от грызунов. В тех случаях, когда истребительные
меры невозможны или пежелательны, необходимо прибегать к использо-
вапию репеллентов.
Практическое значение
393
Физические репелленты
Известно, что ультразвук с определенным уровнем интенсивности в кило-
терцевом диапазоне‘ частот может оказывать биологическое воздействие
па животных. Под биологическим эффектом воздействия ультразвука в
проблеме защиты от повреждений материалов ионимается возможность
направленного изменения поведения живых организмов (управление по-
ведением) с целью предотвращения или ослабления повреждений.
За рубежом опубликован ряд сообщений о создании приборов и
устройств
— генераторов Ультразвука, предпазпаченных для защиты от
повреждений, причиняемых грызунами. Ультразвуковые генераторы си-
стемы «Пекиг» (ФРГ) рекомендовались для защиты от грызупов мате-
риалов в складских помещениях площадью 1000—3000 м? (Вектра
уоп.., 1982). Сообщалось о разработке ультразвуковых приборов в Анг-
лип марки «Ва Ваш», которые якобы создавали ультразвуковой «барьер»,
препятствующий проникновению крыс в помещения (Те1, 1969).
Представляет опоеделенный интерес краткое сообщение об обратном
ренеллентному действии ультразвука — аттрактантном. Это действие ис-
пользовано в конструкции своеобразной крысоловки, в которой ультра-.
звуковой генератор, работавший на частоте 24 кРц, выполнял роль при-
манки (Дмитриев, 1985).
Устройства и приборы с генераторами ультразвука для борьбы с серой
крысой рекламируются и выпускаются многими зарубежными фирмами
уже па протяжении ряда лет. Однако экспериментально обоспованных
достоверных доказательств, свидетельствующих об эффективности этого
метода, насколько можно судить по публикациям, недостаточно (Ноузата,
МатзН, 1985).
Так, генераторы ультразвука, установленные в экспериментальном
. хранилище и работавшие на частотах 41 и 22 кГц, вызвали кратковре-
менное подавление пищевой активности серых крыс: в озвучиваемом по-
мещенин опа спизилась в течепие первой педели испытапий, одпако за-
тем опять восстановилась (Га Усе Саба, 1977). Кратковременную за-
щитную эффективность показали испытания ультразвуковых приборов
марки «ЗоиЙтой» на серых крысах и домовых мышах, обитавших в раз-
личных сельскохозяйственных помещения (УасКзоп, 1980).
Представляет интерес возможность снижения расхода родентицидов
при использовании их в сочетании с ультразвуком. Было установлено,
что в результате угнетения ультразвуком эпдокрипной системы крысе
можно сократить расход зоокуммарина в два раза и существенно повы-
‘сить эффективность дератизациопных мероприятий (Козорез, 1984).
В связи с разнообразием защищаемых от грызунов объектов, условий
. экспериментов и параметров ультразвуковых генераторов исследования и
пепытания ультразвука, проведенные разными авторами, трудно сравни-
вать между собой. В этих работах не приводится надежных сведений о
регистрации биологических изменений, происходящих в организме живот-
пых под действием ультразвука,
13 Серая крыса
392
Глава пятая
тельпую или вкусовую привлекательность продукта или объекта, который
должен быть защищен от грызуна.
Р связи с этим з группу репеллентов могут входить не только в пря-
мом смысле отпугивающие вещества, но и вещества, делающие защищае-
мый объект непривлекательным для пищевой, исследовательской, гнездо-
строительной или другой деятельности животного. В делом поиск репел-
лентов с теми или иными свойствами должен определяться конкретными
задачами, которые должиы решаться с его помощью, и условиями при-
менения,
Несмотря на отсутствие теоретических предпосылок, поиск репеллен-
тов против грызупов ведется давно. Большой опыт накоплен в сельском
хозяйстве и в быту по использованию различных растепий или расти-
тельных препаратов для защиты пищевых продуктов, прежде всего зер-
новых запасов, садовых деревьев и других полезных растений от повреж-
дений грызупами. К сожалению, попытки применения этого опыта в
промышленных масштабах пока не получили широкого распространения.
Направленные поисковые исследования химиков и зоологов позволили
выявить болыное число химических соедипений, обладающих репеллент-
ной активностью. Наиболее полный обзор химических ренпеллентов против
трызунов. предложенных советскими и зарубежными исследователями,
сделан И. А. Емельяповой и Л. Л. Данилкиной (1983). В обзор вошли
материалы, опубликованные до 1978 г. В нем наиболее полно собрапа и
обобщена патептпая и периодическая литература, впервые проведена
классификация ренеллентов по их принадлежности к тем или иным
классам химических соединений.
В. Г. Полежаев (1959) предложил использовать в качестве репеллен-
та слапцевое масло и альбихтол. За рубежом для отпугивания крыс и
других грызунов применяли препарат ‘© фирменным названием «курб»,
содержащий аммониевые и алюминиевые квасцы (Золе, 1977). Этот же
препарат был рекомендеван в качестве»репеллента для отпугивания птиц
от посевов. Для защиты от крыс в бытовых условиях запатентована
смесь креозота с нафталином в-соотпошении 8 :2, которую готовят пере-
мешиванием при пагревании до температуры 80° и потом обрабатывают
сю паружные поверхности, предотвращая таким образом пропикновение
в дома крыс и муравьев (Такада, 1982). .
Для увеличения продолжительности действия летучих репеллентов
используют различные загустители, порошкообразные наполнители, по-
ристые адсорбенты, пленкообразующие полимерпые связующие (Огита,
1980: Мотиги и др., 1983).
Репеллентные свойства различных полимеров были испытаны Л. Хри-
стовым и М. Матевой (1976).
Таким образом, из рассмотрения представленного выте круга хими-
ческих соединений, показавнгих репеллентную активность, можно заклю-
чить, что это направление является перспективным в комнлексе. меро-
приятий по защите от грызунов. В тех случаях, когда истребительные
меры невозможны или пежелательны, необходимо прибегать к использо-
вапию репеллентов.
Практическое значение
393
Физические репелленты
Известно, что ультразвук с определенным уровнем интенсивности в кило-
терцевом диапазоне‘ частот может оказывать биологическое воздействие
па животных. Под биологическим эффектом воздействия ультразвука в
проблеме защиты от повреждений материалов ионимается возможность
направленного изменения поведения живых организмов (управление по-
ведением) с целью предотвращения или ослабления повреждений.
За рубежом опубликован ряд сообщений о создании приборов и
устройств
— генераторов Ультразвука, предпазпаченных для защиты от
повреждений, причиняемых грызунами. Ультразвуковые генераторы си-
стемы «Пекиг» (ФРГ) рекомендовались для защиты от грызупов мате-
риалов в складских помещениях площадью 1000—3000 м? (Вектра
уоп.., 1982). Сообщалось о разработке ультразвуковых приборов в Анг-
лип марки «Ва Ваш», которые якобы создавали ультразвуковой «барьер»,
препятствующий проникновению крыс в помещения (Те1, 1969).
Представляет опоеделенный интерес краткое сообщение об обратном
ренеллентному действии ультразвука — аттрактантном. Это действие ис-
пользовано в конструкции своеобразной крысоловки, в которой ультра-.
звуковой генератор, работавший на частоте 24 кРц, выполнял роль при-
манки (Дмитриев, 1985).
Устройства и приборы с генераторами ультразвука для борьбы с серой
крысой рекламируются и выпускаются многими зарубежными фирмами
уже па протяжении ряда лет. Однако экспериментально обоспованных
достоверных доказательств, свидетельствующих об эффективности этого
метода, насколько можно судить по публикациям, недостаточно (Ноузата,
МатзН, 1985).
Так, генераторы ультразвука, установленные в экспериментальном
. хранилище и работавшие на частотах 41 и 22 кГц, вызвали кратковре-
менное подавление пищевой активности серых крыс: в озвучиваемом по-
мещенин опа спизилась в течепие первой педели испытапий, одпако за-
тем опять восстановилась (Га Усе Саба, 1977). Кратковременную за-
щитную эффективность показали испытания ультразвуковых приборов
марки «ЗоиЙтой» на серых крысах и домовых мышах, обитавших в раз-
личных сельскохозяйственных помещения (УасКзоп, 1980).
Представляет интерес возможность снижения расхода родентицидов
при использовании их в сочетании с ультразвуком. Было установлено,
что в результате угнетения ультразвуком эпдокрипной системы крысе
можно сократить расход зоокуммарина в два раза и существенно повы-
‘сить эффективность дератизациопных мероприятий (Козорез, 1984).
В связи с разнообразием защищаемых от грызунов объектов, условий
. экспериментов и параметров ультразвуковых генераторов исследования и
пепытания ультразвука, проведенные разными авторами, трудно сравни-
вать между собой. В этих работах не приводится надежных сведений о
регистрации биологических изменений, происходящих в организме живот-
пых под действием ультразвука,
13 Серая крыса
394
Глава пятая
В. Е. Ховард и Р. Е. Марш (Но\аг@, Матзи, 1985) на основании ана-
лиза проведенных работ по испытаниям ультразвуковых приборов для
борьбы с трызупами полагают, что слышимый звук является более эд-
фективным для крыс, чем ультразвук. Записанный на магнитпую пленку
стрессовый крик крысы, убиваемой хорьком, наиболее сильно действует
на зверьков из всех опробованных звуков. Прерывистые звуки, изменяю-
щиеся в пределах целой октавы (6—12кГц), более эффективны, чем в
пределах частот для нолуоктав (6—9; 9—12; 12—46кГщц).
В лабораторных условиях интенсивные звуки вызывали у крыс стрес-
совое состояпие, судороги и смерть.
Исхоля из имеющихся данных (Константинов, Мовчан, 1985) по зву-
ковой коммуникации грызунов, крысы и мыши мотут слышать звуки в
диаиазоне частот от 0,5 до 70 кГц и от 1 до 80 кГц соответетвенно.
У них имеются два участка повышенной чувствительности: первый уча-
сток (диапазон) у крыс в диапазоне 20—25 кГц, у мышей— 10—
20 кГц; второй участок (дианазон) У крыс
— 40—50 кГц, у мышей—
40—60 кГц.
Все исследования, рассмотрепные вылте, проводились в первом диапа-
зопе частот повышепиой чувствительности и не проводились во втором
диапазоне. Порог слышимости ультразвуковых колебаний во втором диа-
’пазоне по интенсивности гораздо ниже всех остальных частот, следова-
тельно, для достижения репеллентного действия ультразвука требуется
меньтий уровепь интенсивности, чем в других областях слухового диа-
пазона,
Варьируя в период продолжительноети излучаемого импульса, ча-
стотой в диапазоне 40—50 кГц и интенсивностью, одновремеппо изменяю-
шимися в хаотическом режиме, чтобы исключить адаптацию крыс, мояс
но добиться паправленного измепепия их поведения,
Вероятно, более тщательный подбор параметров ультразвуковых ге-
нераторов с учетом биологических особенностей и поведения серой крысы
позволит найти оптимальные условия для эффективного репеллентного
воздействия ультразвука на серую крысу.
Биологические репелленты
Возможность применения биологических репеллентов для отпугивания
крыс пока изучена очень слабо, данные, приводимые в литературе, весь-
ма противоречивы. Издавна в народе было замечено, что некоторые ра-
стения отпугивают грызунов. Ренеллептные свойства чернокорня (Супо-
Фоззит ОН стае) изучались многими исследователями (Траханов,
1960, и др.), которые указывали на отвращение зверьков к этому расте-
нию. Однако В. А. Вильковичем и Н. Н. Гуревичем (1968) были проведе-
пы опыты на поедание и погрызы крысами листьев чернокорня, в ре-
зультате которых это растение не проявило отпугивающих свойств.
Д. Ф. Траханов (1961а, б), специально занимавшийся изучением ренел-
лентных свойств чернокорня, также пе получил положительного резуль-
тата. По его мнению, возможно, ренеллентные свойства проявляются в
Практическое значение
395
определенные сезоны года, в определенных природпых условиях и зонах.
Отмечали также репеллентные свойства бузины (ЗатБислз и1ега), лав-
ровишни (Таптосегазиз оЁета]з) (Емельянова, Данилкина, 1983), зе-
ленчука (Гапчазхгилм га!еоЪ9оют) (Артамонов, 1986), черкеза (Заза
Мег) (В. С. Соколов, 1952). Необходимо полное и обстоятельное ис-
следование этого вопроса, так как применение растений для отпугивания
грызунов может принести немалую экономию химических и других
средств защиты.
Предлагались и некоторые препараты растительного происхождения,
например альгопаста. Она представляет собой экстракт из синезеленых
водорослей и выпускается в виде пасты темпо-зеленого цвета. Однако ла-
бораторные и полевые испытания (Емельянова, Жарикова, 1984) не под-
твердили репеллентных свойств препарата. Предлагалось также примене-
ние смолистых веществ хвои сосны или ели (Семаков и др., 1983)
и хвойного натурального экстракта (Семаков и др., 1984) в качестве ре-
пеллепта для грызунов. Но необходимы дальнейшие исследования этого
вопроса.
13*
394
Глава пятая
В. Е. Ховард и Р. Е. Марш (Но\аг@, Матзи, 1985) на основании ана-
лиза проведенных работ по испытаниям ультразвуковых приборов для
борьбы с трызупами полагают, что слышимый звук является более эд-
фективным для крыс, чем ультразвук. Записанный на магнитпую пленку
стрессовый крик крысы, убиваемой хорьком, наиболее сильно действует
на зверьков из всех опробованных звуков. Прерывистые звуки, изменяю-
щиеся в пределах целой октавы (6—12кГц), более эффективны, чем в
пределах частот для нолуоктав (6—9; 9—12; 12—46кГщц).
В лабораторных условиях интенсивные звуки вызывали у крыс стрес-
совое состояпие, судороги и смерть.
Исхоля из имеющихся данных (Константинов, Мовчан, 1985) по зву-
ковой коммуникации грызунов, крысы и мыши мотут слышать звуки в
диаиазоне частот от 0,5 до 70 кГц и от 1 до 80 кГц соответетвенно.
У них имеются два участка повышенной чувствительности: первый уча-
сток (диапазон) у крыс в диапазоне 20—25 кГц, у мышей— 10—
20 кГц; второй участок (дианазон) У крыс
— 40—50 кГц, у мышей—
40—60 кГц.
Все исследования, рассмотрепные вылте, проводились в первом диапа-
зопе частот повышепиой чувствительности и не проводились во втором
диапазоне. Порог слышимости ультразвуковых колебаний во втором диа-
’пазоне по интенсивности гораздо ниже всех остальных частот, следова-
тельно, для достижения репеллентного действия ультразвука требуется
меньтий уровепь интенсивности, чем в других областях слухового диа-
пазона,
Варьируя в период продолжительноети излучаемого импульса, ча-
стотой в диапазоне 40—50 кГц и интенсивностью, одновремеппо изменяю-
шимися в хаотическом режиме, чтобы исключить адаптацию крыс, мояс
но добиться паправленного измепепия их поведения,
Вероятно, более тщательный подбор параметров ультразвуковых ге-
нераторов с учетом биологических особенностей и поведения серой крысы
позволит найти оптимальные условия для эффективного репеллентного
воздействия ультразвука на серую крысу.
Биологические репелленты
Возможность применения биологических репеллентов для отпугивания
крыс пока изучена очень слабо, данные, приводимые в литературе, весь-
ма противоречивы. Издавна в народе было замечено, что некоторые ра-
стения отпугивают грызунов. Ренеллептные свойства чернокорня (Супо-
Фоззит ОН стае) изучались многими исследователями (Траханов,
1960, и др.), которые указывали на отвращение зверьков к этому расте-
нию. Однако В. А. Вильковичем и Н. Н. Гуревичем (1968) были проведе-
пы опыты на поедание и погрызы крысами листьев чернокорня, в ре-
зультате которых это растение не проявило отпугивающих свойств.
Д. Ф. Траханов (1961а, б), специально занимавшийся изучением ренел-
лентных свойств чернокорня, также пе получил положительного резуль-
тата. По его мнению, возможно, ренеллентные свойства проявляются в
Практическое значение
395
определенные сезоны года, в определенных природпых условиях и зонах.
Отмечали также репеллентные свойства бузины (ЗатБислз и1ега), лав-
ровишни (Таптосегазиз оЁета]з) (Емельянова, Данилкина, 1983), зе-
ленчука (Гапчазхгилм га!еоЪ9оют) (Артамонов, 1986), черкеза (Заза
Мег) (В. С. Соколов, 1952). Необходимо полное и обстоятельное ис-
следование этого вопроса, так как применение растений для отпугивания
грызунов может принести немалую экономию химических и других
средств защиты.
Предлагались и некоторые препараты растительного происхождения,
например альгопаста. Она представляет собой экстракт из синезеленых
водорослей и выпускается в виде пасты темпо-зеленого цвета. Однако ла-
бораторные и полевые испытания (Емельянова, Жарикова, 1984) не под-
твердили репеллентных свойств препарата. Предлагалось также примене-
ние смолистых веществ хвои сосны или ели (Семаков и др., 1983)
и хвойного натурального экстракта (Семаков и др., 1984) в качестве ре-
пеллепта для грызунов. Но необходимы дальнейшие исследования этого
вопроса.
13*
Литература
Абеленцев В. И. Особенности распределе-
ния серых крыс на животноводческих
фермах Украины и организация борь-
бы с ними // Проблемы паразитологии:
Тр. И науч. конф. паразитологов УССР.
Киев: Изд-во АН УССР, 1956. С. 319-
322.
Абеленцев В. И. О приманках для борь-
бы © крысами на животноводческих
фермах // Ветеринария. 1959. No3.
С. 65-66.
|
Абеленцев В. И. Экологические особен-
ности размещения крыс на животно-
водческих фермах Украины// Тр. Ин-
та зоологии АН УССР. 1960. Т. 16.
С. 43—49.
,
Авакян А. А. Крысы-носители (Ва]ап та
с01:) // Вестн. микробиологии, эпидемио-
логии, паразитологии. 1936. Т. 15, No 1.
С. 25.
Айзенштадт Д. С. Некоторые данные по
распространению и образу жизни се-
рой крысы (КаИлз погуер1сиз Ветк)
лесного района северо-западной части
РСФСР 1 Зоол, журн. 1945. Т. 24, No 3.
С. 182—189.
Айзенштадт Д, С. Методика обследования
и учет эффективности дератизации И
Воен.-мед. журн. 1949. No 14. С. 45.
Айзенштадт Д. С. Расселение серых крые
влоль железнодорожного полотна //При-
рода. 1950а. No 4. С. 62—63.
Айзенштадт Д. С. Гнезда серых. крыс,
устраиваемые во время половодья //
Природа. 19506. No 6. С. 81—82.
Айзенштадт Д. С. К вопросу об истории
и путях расселения пастоков (ВаМаз
погуезсив Вегк) в пределах СССР//
Зоол. журн. 1955. Т. 34, No 5. С. 1145—
1152.
Айзенштадт Д. С. Выбор, чередование
ядов и способов их применения для
истребления серых крыс // Труды Азер-
байджанской противочумной станции:
(Посвящены 40-летию Великой Ок-
тябрьской социалистической револю-
ции). Баку, 1956. Т. 11. С. 145—151.
Айзенштадт Д. С. Пути повышения эф-
фективности истребления серых крыс
(Вамиз потуевсиз ВегК) химическим
методом // Вопр. экологии. Киев: Изд-
во Киев. ун-та, 1957. Т. 2. С. 254-
261.
Айаенштадт Д. С. Характер взаимоотно-
шений между черной крысой (Вайлз
газ Г.) и серой крысой (Ва лз пог-
уер1сиз ВегК) // Зоол. журн. 1959. Т. 38,
No 9. С. 1396—1405.
Айзенштадт Д. С., Арсеньев А. А. Истреб-
ление серых крыс в канализации // Ги-
гиена и санитария. 1954, No 8. С. 2
28.
Айсели А. А. Экспериментальная туля-
ремия у серой крысы // Изв. Иркут.
науч.-исслед. нротивочум. ин-та Сиби-
и и Дал. Востока. 1951. Т. 9. С. 5-
8.
Алания И. И., Бугорская А. К., Читиаш-
вили В. К. и др. Некоторые материалы
по оценке значения серых крыс как
носителей инфекции в Грузии//
ГУ съезд Веесоюз. териол. о-ва: Тез.
докл. рабоч. засед., 27—31 янв. 1986 г.,
Москва. М. АН СССР, 1986. Т. 3.
С. 307—308.
Алания И. И., Ростигаев Б. А., Ширано-
вич П. И., Дзнеладзе М. Т. Материалы
к фауне блох Аджарии. Сообщ. 1. Ви-
довой состав и зональное размещение
блох // Тр. Арм. противочум. станцил.
1964. Вып. 3. С. 407—435,
Алекперов Ф. П. Серая крыса
— носитель
лептоспир серогругаты Сат1соЙ]а на Ап-
шеронском полуострове // Экология и
медицинское значение серой крысы
{Вабаз погуерсиз ВегК). М.: Наука,
41988. С. 15-76.
.
Алекперов Ф. П. Серые крысы - носители
леитоепир серогруппы Рошова на тер-
ритории Азербайджанской СОРИ
ТУ съезд Всесоюз. териол. о-ва: Тез.
докл. М. 1986. Т. 3. С. 836—388.
".
Алексеева 9. В. Численность ископаемой
серой крысы в Южном Приморье // Там
же. 1986. Т. 3. С. 255—256.
Али-Заде Ф. М. Случаи бешенства у лю-
дей после укуса крысы и ослюнения
травоядными // Журн. микробиологии,
эпидемиологии и иммунобиологии. 1958.
No2.С.65.
Алиев М. Н., Булах О. С., Балыбов А. Н.,
Рябцева Н. Л. Реактивность серых крыс
в ответ на введение возбудителя чумы //
Экология и медицинское значение се-
рой крысы (ВаМлз погуереаз ВегК).
М.: АН СССР, 1983. С. 64—66.
Литература 397
Алтарева Н. Д. О восприимчивости белых
крыс к бруцеллам // Журн. микробиоло-
гии, эпидемиологии и ‘иммунобиологии.
1952. No 6. С. 29-30.
Амосенкова Н. И., Попова Е. М. Наблю-
дения над основным резервуаром икте-
ро-геморрагических лептоснир // Там же,
1954. No 12. С. 67—70.
Ананьин В. В. К вопросу о ‘носительстве
леитосиир среди диких крыс // Там же.
1941. No 12. С. 85-81.
Ананъин В. В., Карасева Е. В. Лептоспи-
роз у обезьян // Вопросы краевой общей
и экспериментальной паразитологии.
М.: Медгиз, 1955. Т. 9. С. 45—50.
Ананъин В. В., Карасева Е. В. Природная
очаговость лептоспирозов. М.: Медгиз,
1961. 289 с.
Ананьин В. В., Карасева Е. В. Географи-
ческое распространение лептоспиро-
зов // Лептоспирозы людей и животных.
М.: Медицина, 1971..С. 269—313.
Ананьина Ю. В., Чернуха Ю. Г., Карасе-
ва Е. В. и др. Идентификация штам-
мов лептоспир серотрунпы 1чеговае-
поггназае, выделенных на территории
СССР от серых крыс и из других источ-
ников // Материалы по экологии и ме-
‚ тодам ограничения численности серой
крысы. М.; АН СССР, 1987. С. 231—243.
Андреева Г. С., Сахно Е. И., Соловьев В.И.
Циркуляция возбудителя кишечного
нерсиниоза среди серых крыс в г. Одес-
се// Экология и медицинское значение
серой крысы: Материалы Т рабоч. со-
вещ. по серой ‘крысе (провед. 31.1—3.1.
1983). М.: АН СССР, 1983. С. 83—84.
Анисимова Т. И. Сезонная восириимчи-
вость к чуме серых крыс (ВаМмз пог-
уе1сиз ВегК) в умеренных широтах и
ее возможное значение в эпизоотологии
чумы // Науч, конф. по наразитологии,
эпизоотологии, опидемиологии и др.
вонр. природ. очаговости чумы: Тез.
докл. Ставрополь: «Микроб», 1957. С, 5.
Анисимова Т. И. К.вопросу о существо-
вании очагов элизоотии чумы крыси-
ного тина // Тр. Среднеаз. противочум.
* ин-та. 1959. С. 24—26, .
Анисимова Т. И. Экспериментальная чума
у серых крыс: Автореф. дис ... канд.
‚мед. наук. Саратов: «Микроб», 1962.
З1 с.
у
Антипина В. Е., Маслов С. П. История
формирования экосистем Крыма в-позд-
нем голоцене // УТ Всесоюз. зоогеогр.
конф., Ленинград, 6-8 февр. 1984 г.:
Тез. докл. Л.. 1984. С. 164—165.
Антонова Т. П., Муха Ю. Г., Климчен-
ко Л, Ф. К вопросу о заболеваемости
лептоспирозом в г. Киеве // Лептоепиро-
зы.: Тез. докл. Веесоюз. конф. по лепто-
спирозам. Киев: Наук. думка, 1979.
С. 81—82.
Антонюк В. Я., Олькова Н. В., Шахно-
вич Г. Ф. и др. Задачи ограничения
численности серой крысы в порту Осет-
рово и мероприятия против ее завоза
в Якутскую АССР // Вопросы региональ-
ной гигиены, санитарии и эпидемиоло-
гии. Якутск: Минздрав, 1987. Вып. 2.
С. 184—186.
Арабов 9. Б. Уничтожение грызунов //
Кролиководотво и звероводство. 1984.
No5.С.37-38. |
Аргиропуло А. И. Семейство Маг ае —
мыши. М.; Л.: Изд-во АН СССР, 1940.
170 с. (Фауна СССР. Млекопитающие;
Т. 3, вын. 5).
Аристов А. А., Гамбарян П. П., Мамо-
нов С. О. Адаптация к лазанию в роде
(Мог ае, Кодепиа) // Грызуны: Мате-
риалы УТ Всесоюз. совещ., Ленинград,
25—28 янв. 1983 г. Л.: Наука, 1984.
С. 140—141.
Артамонов В. И. Растение и чистота при-
родной среды. М.: Наука, 1986. 176 с.
Афанасьев С. Ф. Серая крыса — обитатель
плавней реки Таловки // Изв. Грозн. обл.
краевед. музея. 1956. Вын. 7—8. С. 238-—
240
Бабалова Е. Г, Шарашидзе Т. Г., НКуха-
лейшвили И. М. и пр. К распростране-
нию крысиного риккетсиоза в ГССР.
‚
Сообщ..2 / Тр. ГНИЙ вакцин и сыворо-
ток. 1958. Т. 3. С. 155—151.
Базилевский Б, Г. Этюды но изучению
этологии и эпидемиологии инфокциоп-
ной желтухи // Журн. микробиологии,
патологии и инфекц. болезней. 1929.
Т. 6, No З/4. С. 32—45.
Бакеев Н. П. О зимнем питании серой
крысы (В. погуер1саз) в Приазовских
плавнях // Бюл. МОИЦ. Отд. биол. 1967.
Т. 72, No 3. С. 124-125.
Бакутис В. 9. Санитарное благоустройст-
во городов. М.: Мин-во коммун. хоз-ва
РСФСР, 1956. 3144 с.
Баландин Г. А. Бруцеллез. Ростов н/Д,
1958. 42 с,
Банников А. Г. Млекопитающие Мон-
гольской Народной Республики. М.:
АН СССР, 1954. 420 с.
Бароян О. В. Судьба конвекционных бо-
лезпей (проптлое, настоящее, будущее).
М.: Медицина, 1971. 326 с.
Башенина И. В. Пути адаптации мыне-
видных грызунов. М.: Наука, 1977. 352 с.
Литература
Абеленцев В. И. Особенности распределе-
ния серых крыс на животноводческих
фермах Украины и организация борь-
бы с ними // Проблемы паразитологии:
Тр. И науч. конф. паразитологов УССР.
Киев: Изд-во АН УССР, 1956. С. 319-
322.
Абеленцев В. И. О приманках для борь-
бы © крысами на животноводческих
фермах // Ветеринария. 1959. No3.
С. 65-66.
|
Абеленцев В. И. Экологические особен-
ности размещения крыс на животно-
водческих фермах Украины
// Тр. Ин-
та зоологии АН УССР. 1960. Т. 16.
С. 43—49.
,
Авакян А. А. Крысы-носители (Ва]ап та
с01:) // Вестн. микробиологии, эпидемио-
логии, паразитологии. 1936. Т. 15, No 1.
С. 25.
Айзенштадт Д. С. Некоторые данные по
распространению и образу жизни се-
рой крысы (КаИлз погуер1сиз Ветк)
лесного района северо-западной части
РСФСР 1 Зоол, журн. 1945. Т. 24, No 3.
С. 182—189.
Айзенштадт Д, С. Методика обследования
и учет эффективности дератизации И
Воен.-мед. журн. 1949. No 14. С. 45.
Айзенштадт Д. С. Расселение серых крые
влоль железнодорожного полотна //При-
рода. 1950а. No 4. С. 62—63.
Айзенштадт Д. С. Гнезда серых. крыс,
устраиваемые во время половодья //
Природа. 19506. No 6. С. 81—82.
Айзенштадт Д. С. К вопросу об истории
и путях расселения пастоков (ВаМаз
погуезсив Вегк) в пределах СССР//
Зоол. журн. 1955. Т. 34, No 5. С. 1145—
1152.
Айзенштадт Д. С. Выбор, чередование
ядов и способов их применения для
истребления серых крыс // Труды Азер-
байджанской противочумной станции:
(Посвящены 40-летию Великой Ок-
тябрьской социалистической револю-
ции). Баку, 1956. Т. 11. С. 145—151.
Айзенштадт Д. С. Пути повышения эф-
фективности истребления серых крыс
(Вамиз потуевсиз ВегК) химическим
методом // Вопр. экологии. Киев: Изд-
во Киев. ун-та, 1957. Т. 2. С. 254-
261.
Айаенштадт Д. С. Характер взаимоотно-
шений между черной крысой (Вайлз
газ Г.) и серой крысой (Ва лз пог-
уер1сиз ВегК) // Зоол. журн. 1959. Т. 38,
No 9. С. 1396—1405.
Айзенштадт Д. С., Арсеньев А. А. Истреб-
ление серых крыс в канализации // Ги-
гиена и санитария. 1954, No 8. С. 2
28.
Айсели А. А. Экспериментальная туля-
ремия у серой крысы // Изв. Иркут.
науч.-исслед. нротивочум. ин-та Сиби-
и и Дал. Востока. 1951. Т. 9. С. 5-
8.
Алания И. И., Бугорская А. К., Читиаш-
вили В. К. и др. Некоторые материалы
по оценке значения серых крыс как
носителей инфекции в Грузии//
ГУ съезд Веесоюз. териол. о-ва: Тез.
докл. рабоч. засед., 27—31 янв. 1986 г.,
Москва. М. АН СССР, 1986. Т. 3.
С. 307—308.
Алания И. И., Ростигаев Б. А., Ширано-
вич П. И., Дзнеладзе М. Т. Материалы
к фауне блох Аджарии. Сообщ. 1. Ви-
довой состав и зональное размещение
блох // Тр. Арм. противочум. станцил.
1964. Вып. 3. С. 407—435,
Алекперов Ф. П. Серая крыса
— носитель
лептоспир серогругаты Сат1соЙ]а на Ап-
шеронском полуострове // Экология и
медицинское значение серой крысы
{Вабаз погуерсиз ВегК). М.: Наука,
41988. С. 15-76.
.
Алекперов Ф. П. Серые крысы - носители
леитоепир серогруппы Рошова на тер-
ритории Азербайджанской СОРИ
ТУ съезд Всесоюз. териол. о-ва: Тез.
докл. М. 1986. Т. 3. С. 836—388.
".
Алексеева 9. В. Численность ископаемой
серой крысы в Южном Приморье // Там
же. 1986. Т. 3. С. 255—256.
Али-Заде Ф. М. Случаи бешенства у лю-
дей после укуса крысы и ослюнения
травоядными // Журн. микробиологии,
эпидемиологии и иммунобиологии. 1958.
No2.С.65.
Алиев М. Н., Булах О. С., Балыбов А. Н.,
Рябцева Н. Л. Реактивность серых крыс
в ответ на введение возбудителя чумы //
Экология и медицинское значение се-
рой крысы (ВаМлз погуереаз ВегК).
М.: АН СССР, 1983. С. 64—66.
Литература 397
Алтарева Н. Д. О восприимчивости белых
крыс к бруцеллам // Журн. микробиоло-
гии, эпидемиологии и ‘иммунобиологии.
1952. No 6. С. 29-30.
Амосенкова Н. И., Попова Е. М. Наблю-
дения над основным резервуаром икте-
ро-геморрагических лептоснир // Там же,
1954. No 12. С. 67—70.
Ананьин В. В. К вопросу о ‘носительстве
леитосиир среди диких крыс // Там же.
1941. No 12. С. 85-81.
Ананъин В. В., Карасева Е. В. Лептоспи-
роз у обезьян // Вопросы краевой общей
и экспериментальной паразитологии.
М.: Медгиз, 1955. Т. 9. С. 45—50.
Ананъин В. В., Карасева Е. В. Природная
очаговость лептоспирозов. М.: Медгиз,
1961. 289 с.
Ананьин В. В., Карасева Е. В. Географи-
ческое распространение лептоспиро-
зов // Лептоспирозы людей и животных.
М.: Медицина, 1971..С. 269—313.
Ананьина Ю. В., Чернуха Ю. Г., Карасе-
ва Е. В. и др. Идентификация штам-
мов лептоспир серотрунпы 1чеговае-
поггназае, выделенных на территории
СССР от серых крыс и из других источ-
ников // Материалы по экологии и ме-
‚ тодам ограничения численности серой
крысы. М.; АН СССР, 1987. С. 231—243.
Андреева Г. С., Сахно Е. И., Соловьев В.И.
Циркуляция возбудителя кишечного
нерсиниоза среди серых крыс в г. Одес-
се// Экология и медицинское значение
серой крысы: Материалы Т рабоч. со-
вещ. по серой ‘крысе (провед. 31.1—3.1.
1983). М.: АН СССР, 1983. С. 83—84.
Анисимова Т. И. Сезонная восириимчи-
вость к чуме серых крыс (ВаМмз пог-
уе1сиз ВегК) в умеренных широтах и
ее возможное значение в эпизоотологии
чумы // Науч, конф. по наразитологии,
эпизоотологии, опидемиологии и др.
вонр. природ. очаговости чумы: Тез.
докл. Ставрополь: «Микроб», 1957. С, 5.
Анисимова Т. И. К.вопросу о существо-
вании очагов элизоотии чумы крыси-
ного тина // Тр. Среднеаз. противочум.
* ин-та. 1959. С. 24—26, .
Анисимова Т. И. Экспериментальная чума
у серых крыс: Автореф. дис ... канд.
‚мед. наук. Саратов: «Микроб», 1962.
З1 с.
у
Антипина В. Е., Маслов С. П. История
формирования экосистем Крыма в-позд-
нем голоцене // УТ Всесоюз. зоогеогр.
конф., Ленинград, 6-8 февр. 1984 г.:
Тез. докл. Л.. 1984. С. 164—165.
Антонова Т. П., Муха Ю. Г., Климчен-
ко Л, Ф. К вопросу о заболеваемости
лептоспирозом в г. Киеве // Лептоепиро-
зы.: Тез. докл. Веесоюз. конф. по лепто-
спирозам. Киев: Наук. думка, 1979.
С. 81—82.
Антонюк В. Я., Олькова Н. В., Шахно-
вич Г. Ф. и др. Задачи ограничения
численности серой крысы в порту Осет-
рово и мероприятия против ее завоза
в Якутскую АССР // Вопросы региональ-
ной гигиены, санитарии и эпидемиоло-
гии. Якутск: Минздрав, 1987. Вып. 2.
С. 184—186.
Арабов 9. Б. Уничтожение грызунов //
Кролиководотво и звероводство. 1984.
No5.С.37-38. |
Аргиропуло А. И. Семейство Маг ае —
мыши. М.; Л.: Изд-во АН СССР, 1940.
170 с. (Фауна СССР. Млекопитающие;
Т. 3, вын. 5).
Аристов А. А., Гамбарян П. П., Мамо-
нов С. О. Адаптация к лазанию в роде
(Мог ае, Кодепиа) // Грызуны: Мате-
риалы УТ Всесоюз. совещ., Ленинград,
25—28 янв. 1983 г. Л.: Наука, 1984.
С. 140—141.
Артамонов В. И. Растение и чистота при-
родной среды. М.: Наука, 1986. 176 с.
Афанасьев С. Ф. Серая крыса — обитатель
плавней реки Таловки // Изв. Грозн. обл.
краевед. музея. 1956. Вын. 7—8. С. 238-—
240
Бабалова Е. Г, Шарашидзе Т. Г., НКуха-
лейшвили И. М. и пр. К распростране-
нию крысиного риккетсиоза в ГССР.
‚
Сообщ..2 / Тр. ГНИЙ вакцин и сыворо-
ток. 1958. Т. 3. С. 155—151.
Базилевский Б, Г. Этюды но изучению
этологии и эпидемиологии инфокциоп-
ной желтухи // Журн. микробиологии,
патологии и инфекц. болезней. 1929.
Т. 6, No З/4. С. 32—45.
Бакеев Н. П. О зимнем питании серой
крысы (В. погуер1саз) в Приазовских
плавнях // Бюл. МОИЦ. Отд. биол. 1967.
Т. 72, No 3. С. 124-125.
Бакутис В. 9. Санитарное благоустройст-
во городов. М.: Мин-во коммун. хоз-ва
РСФСР, 1956. 3144 с.
Баландин Г. А. Бруцеллез. Ростов н/Д,
1958. 42 с,
Банников А. Г. Млекопитающие Мон-
гольской Народной Республики. М.:
АН СССР, 1954. 420 с.
Бароян О. В. Судьба конвекционных бо-
лезпей (проптлое, настоящее, будущее).
М.: Медицина, 1971. 326 с.
Башенина И. В. Пути адаптации мыне-
видных грызунов. М.: Наука, 1977. 352 с.
398
Литература
Безденежных И. С. Шаферштейн Д. Л.
Носители лептоспир типа Сапсоа у
диких крыс на острове Сахалине //
Журн. микробиологии, эпидемиологии
и иммунобиологии. 1954, No 12. С. 71.
Бейшебаев К. К. Туркестанская крыса //
Тр. Ип-та зоологии и паразитологии
АН КиреССР. 1957. Вып. 4. С. 101-103.
Бейшебаев К. ПК. О размножении турке-
станской крысы// Изв. АН КиргССР.
Сер. биол. 1962. Т. 4, No 1. С. 27-39.
Бекасова Т. С. Эволюция кариотица крыс
подрода Наив (Водепиа. Митае) //
Эволюционные исследования. Макро-
эволюция. Владивосток: Изд-во Даль-
невост. ун-та, 1984. С. 140--158.
Белова М. А. Эризипелоид в Тульской
обл.: Автореф. дис .. канд. мед. наук.
Тула: Мед. ин-т, 1972. 16 с.
Бёме Л. Б. Дикие звери Северо-Кавказ-
ского края. Пятигорек: Госиздат, 1936.
435 с.
Бентли Е. В. Борьба с грызунами // Хро-
ника ВОЗ. 1967. Т. 21, No9. С. 359-
364.
Бентли Е. В. Обзор ратидидон-антикоагу-
лянтов, применяемых х настоящее вре-
мя // Бюл. ВОЗ. 1972. Т. 47. С. 275-280.
Беньковский Л. М. Паеюк на острове
Итуруц //ТУ съезд Всесоюз. териол.
о-ва: Тез. докл, М., 1986. Т. 3. С. 249-
250.
Березанцев Ю. А. Трихинеллез. М. Ме-
Ддицина. 1974. 160 с.
Бернар Ж. Период размножения и пло-
довитость грызунов в Тунисе // Реф.
докл. Г Междунар. конгр. по млекопи-
тающим. М.: Наука, 1974. С. 59—60.
Бернасовская Е. П., Компанцев Н. Ф.,
Моисеева А. В., Клименко Л. Ф. Серые
крысы
— источник заражения людей
иктерогеморрагическим лептоснирозом
в Украинской ССР // Экология и меди-.
цинское значение серой крысы (ВаЦлз
поте оц ВегК). М.: АН СССР, 1983.
‚ 76.
Бернасовская Е. П. Кондратенко В. Н.
Эпидемиология иктерогеморрагического
лептоспироза: (Экол. аспекты) // Журн.
микробиологии, эпидемиологии и им-
мунобиологии. 1987. No 10. С. 43—49.
Бернасовская Е. П., Моисеева А. В., Кож-
панцев Н. Ф. и др. Серые крысы
- ис-
точник заражения людей иктерогемор-
рагическим септоспировом в Украин-
ской ССР// Серая ‚крыса. М.: АН СССР,
1986. Т. 2. С. 19-25
Бернштейн А. Д. Некоторыс особенности
биологии черной крысы в Абхазии //
Бюл. МОИЦ. Отд. биол. 1959. Т. 64, No 1
С. 5-14. .
Берулава` В. С. К методике проведения
дератизации // Фельдшер и акушерка.
1972. No 3. С. 14-—16..
Бессонов А, С. Эпизоотология (эзпидемио-
логия), диагностика и профилактика
трихинеллеза: Автореф. дис. ... д-ра мед.
паук. М., 1970. 32 в.
Бибиков Д. И., Карасева Е. В., Константи-
нов В. М. 06 оеновных подходах
к управлению популяциями проблем-
ных животных // Влияние антропоген-
ной трансформации ландшафта на на-
селение наземных позвоночных живот-
ных: 'Гез. Всесоюз. совещ. М.: Агропро-
миздат, 1987. Ч. 1. С. 43—48.
Бибикова В. А., Класовский Л. Н. Переда-
ча чумы блохами. М. Медицина, 1975.
188 с.
Бич Ф. А. Инстинктивное поведение. Раз-
множение // Экспериментальная психо-
логия. М.: Изд-во иностр. лит. 1960.
С. 513—576.
Бобринский П. А. ‚ Кузнецов Б. А., Кузя-
кин А. П. Определитель млекопитаю-
щих СССР. М.: Изд-во МГУ, 1965. 382 с.
Богачева М. А., Гудкова 9. И., Сидорен-
ков В. Н., Киреева Н. И. Опыт работы
по освобождению туркомплекса «Ияз-
майлово» от грызунов // Современные
направления медицинской дезинсекции
и дератизации: Тез. докл. Москва, 17—
18 окт. 1984. М.: Минздрав СССР, 1981.
С. 185—186.
Боженко В. ПН. Дельтовый природный очаг
туляремии юга РСФСР //Тр. Рост. гос.
противочум. ин-та. 1957. Т. 12. С. 326—
ЗАЗ.
Большанов В. Н., Кубанцев Б. С. Поло-
вая структура иопуляций млекопитаю-
щих и. ее динамика. М.: Наука, 1984.
232 с.
.
Бондарь Е. Н. Грызуны насоленных
пупктов Туркменистана // Изв. Туркм.
фил. АН СССР. 1946. No 1. С. 67—71.
Борисенко В. А. Распространение серой
крысы в Кустанайской обл. // Изв. АН
КазССР. Сер. биол. 1969. Вып. 2. С. 35—
42,
Борисенко В. А. Серая, или амбарная,
крыса, пасюк// Млекопитающие Ка-
захстана. Алма-Ата: Наука, 1977. Т. 1,
ч. 2. С, 369—383.
Бояркин И. В. Сравнительная экология
зайцеобразных и трызунов западной
части Тувинской АССР. Иркутск, 1984.
170 с.
Бродский А. С., Ножквина Л. Б. Новерх-
Литература
399
ностные воды. Атлас Сахалинской обл.
М.: ГУГН, 1967. С. 84-89.
Буров А. И., Крицкая 3. Ф. Лептоспиро-
носительство у серых крыс в г. Одессе //
Журн. микробиологии, эпидемиологии
и иммунобиологии. 1964, No 10. С. 134-
136.
Бухникашвили А. Крученкова Е. Онто-
генез социального поведения большой
песчанки // Групповое поведение живот-
ных. М.: Наука, 1976. С. 37—40.
Быковский В. А., Николаева Н. Н. Ис-
следования серых крыс на устойчивость
к традиционным ратицидам и испыта-
ния новых препаратов //ТУ съезд Все-
союз. териол. о-ва: Тез. докл. М., 1986
Т. 3. С. 367—368.
Быковский В. А., Пегельман С. Г. Влия-
пие тонсикапта и патогепа на плодо-
витость грызунов // Грызуны; Материа-
лы У Всесоюз. совещ. М.: Наука, 1980.
С. 388—390.
Варшавский Б. С, Буркин В. С.; Илю-
хин А. А. и др. Материалы по распрост-
ранению и численности серой крысы в
Астраханской области // Экология и ме-
дицинское значение серой крысы (Вас
{3 погуевсиз ВегК). М: АН СССР,
1983. С. 28—30.
Варшавский С. Н., Шилов М. П., По-
пов Н. В. и др. Обзор совремепного
распространения серой крысы в онзо-
отичных по чуме районах на Европей-
ском Юго-Востоке, Кавказе, Казахстане
и Средней Азии и некоторые задачи
дальнейших исследований // Материалы
по экологии и методам ограничения
численности серой крысы. М.: Наука,
1987. С. 32—68.
Варшавский С. Н., Шилов М. Н., Сурвил-
10 и др. Расширение ареала и совре-
менное распрострапепие серой крысы
в северо-западном Прикаспийском оча-
те чумы // Серая крыса. М.: Наука, 1986.
Т. 2. С. 32—4/.
Вашков В. И., Вишняков С. В., Полежа-
ев В. Г. и др. Борьба с грызунами в
городах и населенных пунктах сель-
ской местности. М.: Медицина, 1974,
*255 с.
Ващенок Г. И. Андрейчик 9. П., Ваще-
нок В. С. и др. Иерсиниозы в Ленин-
граде: Болезни © природной очаго-
востью // Тр. Ин-та эпидемиологии и
микробиологии им. Пастера. 1983. Т. 60.
С. 82—87.
Верещагин Н. К. Млекопитающие Кав-
каза: История формирования фауны.
М.: Л.: Изд-во АН СССР, 1959. 70А с.
Вершилова ПН. А., Нетрищева П. А., Кот-
лярова Х. С., Тер-Вартанов В. Н. К воп-
росу о возможности природной очаго-
вости бруцеллеза // Природная очаго-
вость и эпидемиология 060бо опасных
инфекционных заболеваний. Саратов:
«Микроб», 1959. С. 466—475.
Вигоров Ю. Л. Морфологическая измен-
чивость пасюка в северной части арса-
ла Евразии // Исследование мелких млс-
копитающих на Урале: (Пробл. терио-
логии на Урале): Препринт. Сверд-
ловск: УНЦ АН СССР, 1985. С. 9—12.
Вилькович В. А., Гуревич Н. Н. Испыта-
ние средств, отпугивающих грызунов
от морских судов // Тр. НИИ гигиены
и вод. транен. 1968. Выи. 1. С. 348—
354
Вильковии В. А. Машерук С. В. Поиск
репеллептов для защиты морских су-
дов от грызунов // Материалы науч.
конф. по вопр. гигиены вод. трансп.:
{По итогам за 1961—1963 гг.). М., 1964,
С. 80-842.
Вилькович В. А., Сахарова Т. В., Дадо-
нов Ю. В. и др. Разработка методики
определения численности грызунов в
портах // Материалы Всесоюз. конф. по
вопр. дезинфекции и стерилизации
(29 сент—2 окт. 1969 г., Москва). М.:
Минздрав СССР, 1969. С. 158—159.
Виноградов Б. С., Аргиропуло А. И. Фзу-
на СССР. Млекопитающие: Определи-
тель грызунов. М.; Л.: Изд-во АН СССР,
1941. 241 с.
Виноградов Б. С., Громов И. М. Грызуны
фауны СССР. М.; Л. Изд-во АН СССР,
1952. 297 с.
.
Вишняков С. В. Размножение и плодови-
тость городских грызунов // Тр. Цептр.
пауч.-исслед. дезинфекц. ип-та. 1970,
Т. 19. С. 422—430.
Вишняков С. В., Дукельская Н. М., Ива-
нова В. В. Относительный учет числен-
ности грызунов в городских условиях //
Зоол. журн. 1955. Т. 34, No4. 0. 902-
914.
Вишняков С. В., Дукельская Н. М. Но-
вый вариант химического метода борь-
бы с грызунами в городских условиях //
Тр. Центр. науч-иселед. дезинфекц.
ин-та. 1956. Т. 9. С. 254—258.
Воджицкий К. Перспективы биологиче-
ской борьбы с популяциями грызунов /1
Бюл. ВОЗ. 1974. Т. 48, No4. С. 461-
174.
Волкова К.С., Ткац И. Н. Об организации
и проведении дератизационных меро-
приятий в Хабаровском крае // Пробле-
398
Литература
Безденежных И. С. Шаферштейн Д. Л.
Носители лептоспир типа Сапсоа у
диких крыс на острове Сахалине //
Журн. микробиологии, эпидемиологии
и иммунобиологии. 1954, No 12. С. 71.
Бейшебаев К. К. Туркестанская крыса //
Тр. Ип-та зоологии и паразитологии
АН КиреССР. 1957. Вып. 4. С. 101-103.
Бейшебаев К. ПК. О размножении турке-
станской крысы// Изв. АН КиргССР.
Сер. биол. 1962. Т. 4, No 1. С. 27-39.
Бекасова Т. С. Эволюция кариотица крыс
подрода Наив (Водепиа. Митае) //
Эволюционные исследования. Макро-
эволюция. Владивосток: Изд-во Даль-
невост. ун-та, 1984. С. 140--158.
Белова М. А. Эризипелоид в Тульской
обл.: Автореф. дис .. канд. мед. наук.
Тула: Мед. ин-т, 1972. 16 с.
Бёме Л. Б. Дикие звери Северо-Кавказ-
ского края. Пятигорек: Госиздат, 1936.
435 с.
Бентли Е. В. Борьба с грызунами // Хро-
ника ВОЗ. 1967. Т. 21, No9. С. 359-
364.
Бентли Е. В. Обзор ратидидон-антикоагу-
лянтов, применяемых х настоящее вре-
мя // Бюл. ВОЗ. 1972. Т. 47. С. 275-280.
Беньковский Л. М. Паеюк на острове
Итуруц //ТУ съезд Всесоюз. териол.
о-ва: Тез. докл, М., 1986. Т. 3. С. 249-
250.
Березанцев Ю. А. Трихинеллез. М. Ме-
Ддицина. 1974. 160 с.
Бернар Ж. Период размножения и пло-
довитость грызунов в Тунисе // Реф.
докл. Г Междунар. конгр. по млекопи-
тающим. М.: Наука, 1974. С. 59—60.
Бернасовская Е. П., Компанцев Н. Ф.,
Моисеева А. В., Клименко Л. Ф. Серые
крысы
— источник заражения людей
иктерогеморрагическим лептоснирозом
в Украинской ССР // Экология и меди-.
цинское значение серой крысы (ВаЦлз
поте оц ВегК). М.: АН СССР, 1983.
‚ 76.
Бернасовская Е. П. Кондратенко В. Н.
Эпидемиология иктерогеморрагического
лептоспироза: (Экол. аспекты) // Журн.
микробиологии, эпидемиологии и им-
мунобиологии. 1987. No 10. С. 43—49.
Бернасовская Е. П., Моисеева А. В., Кож-
панцев Н. Ф. и др. Серые крысы
- ис-
точник заражения людей иктерогемор-
рагическим септоспировом в Украин-
ской ССР// Серая ‚крыса. М.: АН СССР,
1986. Т. 2. С. 19-25
Бернштейн А. Д. Некоторыс особенности
биологии черной крысы в Абхазии //
Бюл. МОИЦ. Отд. биол. 1959. Т. 64, No 1
С. 5-14. .
Берулава` В. С. К методике проведения
дератизации // Фельдшер и акушерка.
1972. No 3. С. 14-—16..
Бессонов А, С. Эпизоотология (эзпидемио-
логия), диагностика и профилактика
трихинеллеза: Автореф. дис. ... д-ра мед.
паук. М., 1970. 32 в.
Бибиков Д. И., Карасева Е. В., Константи-
нов В. М. 06 оеновных подходах
к управлению популяциями проблем-
ных животных // Влияние антропоген-
ной трансформации ландшафта на на-
селение наземных позвоночных живот-
ных: 'Гез. Всесоюз. совещ. М.: Агропро-
миздат, 1987. Ч. 1. С. 43—48.
Бибикова В. А., Класовский Л. Н. Переда-
ча чумы блохами. М. Медицина, 1975.
188 с.
Бич Ф. А. Инстинктивное поведение. Раз-
множение // Экспериментальная психо-
логия. М.: Изд-во иностр. лит. 1960.
С. 513—576.
Бобринский П. А. ‚ Кузнецов Б. А., Кузя-
кин А. П. Определитель млекопитаю-
щих СССР. М.: Изд-во МГУ, 1965. 382 с.
Богачева М. А., Гудкова 9. И., Сидорен-
ков В. Н., Киреева Н. И. Опыт работы
по освобождению туркомплекса «Ияз-
майлово» от грызунов // Современные
направления медицинской дезинсекции
и дератизации: Тез. докл. Москва, 17—
18 окт. 1984. М.: Минздрав СССР, 1981.
С. 185—186.
Боженко В. ПН. Дельтовый природный очаг
туляремии юга РСФСР //Тр. Рост. гос.
противочум. ин-та. 1957. Т. 12. С. 326—
ЗАЗ.
Большанов В. Н., Кубанцев Б. С. Поло-
вая структура иопуляций млекопитаю-
щих и. ее динамика. М.: Наука, 1984.
232 с.
.
Бондарь Е. Н. Грызуны насоленных
пупктов Туркменистана // Изв. Туркм.
фил. АН СССР. 1946. No 1. С. 67—71.
Борисенко В. А. Распространение серой
крысы в Кустанайской обл. // Изв. АН
КазССР. Сер. биол. 1969. Вып. 2. С. 35—
42,
Борисенко В. А. Серая, или амбарная,
крыса, пасюк// Млекопитающие Ка-
захстана. Алма-Ата: Наука, 1977. Т. 1,
ч. 2. С, 369—383.
Бояркин И. В. Сравнительная экология
зайцеобразных и трызунов западной
части Тувинской АССР. Иркутск, 1984.
170 с.
Бродский А. С., Ножквина Л. Б. Новерх-
Литература
399
ностные воды. Атлас Сахалинской обл.
М.: ГУГН, 1967. С. 84-89.
Буров А. И., Крицкая 3. Ф. Лептоспиро-
носительство у серых крыс в г. Одессе //
Журн. микробиологии, эпидемиологии
и иммунобиологии. 1964, No 10. С. 134-
136.
Бухникашвили А. Крученкова Е. Онто-
генез социального поведения большой
песчанки // Групповое поведение живот-
ных. М.: Наука, 1976. С. 37—40.
Быковский В. А., Николаева Н. Н. Ис-
следования серых крыс на устойчивость
к традиционным ратицидам и испыта-
ния новых препаратов //ТУ съезд Все-
союз. териол. о-ва: Тез. докл. М., 1986
Т. 3. С. 367—368.
Быковский В. А., Пегельман С. Г. Влия-
пие тонсикапта и патогепа на плодо-
витость грызунов // Грызуны; Материа-
лы У Всесоюз. совещ. М.: Наука, 1980.
С. 388—390.
Варшавский Б. С, Буркин В. С.; Илю-
хин А. А. и др. Материалы по распрост-
ранению и численности серой крысы в
Астраханской области // Экология и ме-
дицинское значение серой крысы (Вас
{3 погуевсиз ВегК). М: АН СССР,
1983. С. 28—30.
Варшавский С. Н., Шилов М. П., По-
пов Н. В. и др. Обзор совремепного
распространения серой крысы в онзо-
отичных по чуме районах на Европей-
ском Юго-Востоке, Кавказе, Казахстане
и Средней Азии и некоторые задачи
дальнейших исследований // Материалы
по экологии и методам ограничения
численности серой крысы. М.: Наука,
1987. С. 32—68.
Варшавский С. Н., Шилов М. Н., Сурвил-
10 и др. Расширение ареала и совре-
менное распрострапепие серой крысы
в северо-западном Прикаспийском оча-
те чумы // Серая крыса. М.: Наука, 1986.
Т. 2. С. 32—4/.
Вашков В. И., Вишняков С. В., Полежа-
ев В. Г. и др. Борьба с грызунами в
городах и населенных пунктах сель-
ской местности. М.: Медицина, 1974,
*255 с.
Ващенок Г. И. Андрейчик 9. П., Ваще-
нок В. С. и др. Иерсиниозы в Ленин-
граде: Болезни © природной очаго-
востью // Тр. Ин-та эпидемиологии и
микробиологии им. Пастера. 1983. Т. 60.
С. 82—87.
Верещагин Н. К. Млекопитающие Кав-
каза: История формирования фауны.
М.: Л.: Изд-во АН СССР, 1959. 70А с.
Вершилова ПН. А., Нетрищева П. А., Кот-
лярова Х. С., Тер-Вартанов В. Н. К воп-
росу о возможности природной очаго-
вости бруцеллеза // Природная очаго-
вость и эпидемиология 060бо опасных
инфекционных заболеваний. Саратов:
«Микроб», 1959. С. 466—475.
Вигоров Ю. Л. Морфологическая измен-
чивость пасюка в северной части арса-
ла Евразии // Исследование мелких млс-
копитающих на Урале: (Пробл. терио-
логии на Урале): Препринт. Сверд-
ловск: УНЦ АН СССР, 1985. С. 9—12.
Вилькович В. А., Гуревич Н. Н. Испыта-
ние средств, отпугивающих грызунов
от морских судов // Тр. НИИ гигиены
и вод. транен. 1968. Выи. 1. С. 348—
354
Вильковии В. А. Машерук С. В. Поиск
репеллептов для защиты морских су-
дов от грызунов // Материалы науч.
конф. по вопр. гигиены вод. трансп.:
{По итогам за 1961—1963 гг.). М., 1964,
С. 80-842.
Вилькович В. А., Сахарова Т. В., Дадо-
нов Ю. В. и др. Разработка методики
определения численности грызунов в
портах // Материалы Всесоюз. конф. по
вопр. дезинфекции и стерилизации
(29 сент—2 окт. 1969 г., Москва). М.:
Минздрав СССР, 1969. С. 158—159.
Виноградов Б. С., Аргиропуло А. И. Фзу-
на СССР. Млекопитающие: Определи-
тель грызунов. М.; Л.: Изд-во АН СССР,
1941. 241 с.
Виноградов Б. С., Громов И. М. Грызуны
фауны СССР. М.; Л. Изд-во АН СССР,
1952. 297 с.
.
Вишняков С. В. Размножение и плодови-
тость городских грызунов // Тр. Цептр.
пауч.-исслед. дезинфекц. ип-та. 1970,
Т. 19. С. 422—430.
Вишняков С. В., Дукельская Н. М., Ива-
нова В. В. Относительный учет числен-
ности грызунов в городских условиях //
Зоол. журн. 1955. Т. 34, No4. 0. 902-
914.
Вишняков С. В., Дукельская Н. М. Но-
вый вариант химического метода борь-
бы с грызунами в городских условиях //
Тр. Центр. науч-иселед. дезинфекц.
ин-та. 1956. Т. 9. С. 254—258.
Воджицкий К. Перспективы биологиче-
ской борьбы с популяциями грызунов /1
Бюл. ВОЗ. 1974. Т. 48, No4. С. 461-
174.
Волкова К.С., Ткац И. Н. Об организации
и проведении дератизационных меро-
приятий в Хабаровском крае // Пробле-
400
Литература
мы дезинфекции и стерилизации: (Ма-
риал ции 23—25 ноябр. 1977 г.).
М.: Минздрав СССР, 1977. Ч. 2. С. 76.
Волянский Б. Зазйтки про звцЯ в Кер-
ченського пвоетрова (Крим) // Тр. Ф!з.-
мат. выд. Всеукр. АН. 1929. Т. 43, вип. 1.
С. 29—38.
Воронов В. Г. Млекопитающие Курильских
островов. Л.: Наука, 1974. 157 с.
Воронцов Н. Н. Эволюция пищеваритель-
ной системы грызунов: (Мыптеобраз-
ные). Новосибирск: Наука, 1967а. 289 с.
Воронцов Н. Н. Историческая зоогеогра-
фия мышеобразных (МиЧае) грызу-
нов // Проблемы эволюции. Новоси-
бирск: Наука, 19676. С. 116—141.
Гавриловская И. П., Бойко В. А. Гемор-
рагическая лихорадка с почечным син-
дромом: Науч. обзор ВНИИМИ. М., 1985.
74 с.
Гажиев В. В., Сергеева Л. А., Степинец
В. И. и др. Некоторые данные о меди-
цинском значении серой крысы в Донец-
кой области // ТУ съезд Всесоюз. териол.
о-ва: Тез. докл. М.: АН СССР, 1986. Т. 3.
С. 305—306.
Гажиев В. В., Степанец В. И., Анищенко
Г. А. и др. Борьба с синантропными
трызунами в шахтах Донбасса // Там
же. 198ба. Т. 3. С. 389—390.
Гажиев В. В., Степанец В. И., Грянов
В. И. и др. Борьба с серой крысой в
шахтах Донецкой обл. // Материалы по
экологии и методам ограничения чис-
ленности серой крысы. М.: Наука, 1987.
С. 321-322.
Гамалея Н. Ф. Эпидемиология Одесской
чумы 1902 г.//Чума в Одессе, истори-
ческое и эпидемиологическое исследо-
вапие. Одесса, 1908. 'Г. 4. С. 131-172.
Гамбарян П. Н., Дукельская Н. М. Крыса,
М.: Сов. наука, 1955. 254 с.
Гзелисиани И. Д. Крысы как резервуар
Ва!апи ут сой// Журн. мед, парази-
тологии паразитар. болезней. 1949.
Выи. 6. С. 499—500.
Гиренко Л. Л. К вопросу о распростра-
нении черной крысы в УССР // Тр. Зоол.
музея Киев. ун-та. 1950. Т. 2. С. 75-
95.
Голещилин А. К. Оргапизация и прояе-
дение одномоментпой сплошной дера-
тизации в условиях населенного пунк-
та: Автореф. дис. ... канд. биол. наук.
Кишинев, 1955. 20 с.
Гольденштейн 3. А. Заболеваемость лепто-
спирозом в Краснодарском крае, его
лабораторная диагностика и осно-
вы предупреждения лептоспироза // Тез.
докл. УТ Веесозоз. науч. конф. по лепто-
спирозу. Баку, 1976. С. 67-68.
Гольцман М. Е., Наумов Н. Н., Николь-
ский А. А. и др. Социальное поведение
болыпой песчанки (НВомБотуз орз
Тя.) И Поведение млекопитающих. М.:
Наука, 1977. С. 5-69.
Горшанова Е. Н. Носительство [.. 1ага5з0-
у: среди синантронных грызунов //
Журн. микробиологии, эпидемиологии
и иммунобиологии. 1962. No4. С. 53—
54.
Гребенчук А. Н., Бакулина Л. И., Ваще-
нок Г. И. и др. Сальмонеллез у грызу-
нов в Ленинграде // Там же. 1965. No 6.
С. 43—46.
Громов И. М. Отряд Грызуны Водепма //
Каталог млекопитающих СССР: Плио-
цен-современность. Л.: Наука, 1981.
С. 75-247.
Громов И. М., Гуреев А. А., Новиков Г. А.
и др. Млекопитающие фауны СССР. М.;
Л.: Наука; 1963. Ч. 1. 639 с.
Груздев В. В. Зоны наибольшей вредности
серых крыс в сельской. местности //
Биол. науки. 1959. No 2. С. 39—44.
Груздев В. В. Характеристика численно-
сти и вредности грызунов при комп-
лексном районировании для сельского
хозяйства // Материалы ГУ Межвуз. со-
вещ. по районированию для сел. хоз-ва.
М.: Колос, 1963. С. 21—24,
Губарев Л. Д. Распространение серой
крысы (ВаМиз погусе1сяз ВегК) в вос-
точных районах Ростовской области //
Тр. Рост. противочум. ин-та. 1941. Т. 2.
С. 133—144.
Гутман 9. А. Приманка из свежих поми-
доров с зоокумарилом для уничтоже-
ния грызунов // Ветеринария. 1972.7No 5.
[9
Гуцель Ю. Н., Рожков Ф. Ф., Даданов
В. и др..Некоторые вопросы дера-
тизации в морском порту //'Экология и,
медицинское значение серой крысы
(Ва 1$ потуессив ВетК). М.: Наука,
1983. С. 28—31.
|
|
Давыдов Г. С. Грызуны Северного 'Гад-
жикистана. Душанбе: Тадж. ун-т, 1964.
272 с.
Давыдов Г. С. Грызуны
— вредители по-
ливных культур и пастбищ Таджикис-
тана и борьба с ними. Душанбе, 1978.
55 с.
р
Давыдов Г. С. Распространение и дина-
мика численности серой крысы в Се-
верном Таджикистане // ТУ съезд Вес-
союз. териол. о-ва: Тез. докл. М., 1986.
Т. 3. С. 281-282,
Литература
4041
Дадонов Ю. В., Арканников А. Н., Го-
лубков Н.Е. и др. Испытания на устой-
чивость к повреждению грызунами раз-
личных материалов, применяемых в
электронной радиопромышленности //
Насекомые и грызуны
— разрушители
материалов и технических изделий. М.:
Наука, 1983. С. 206—208.
Даль С. К. Животлый мир Армянской
ССР. Т. 1. Позвопочпые животные. Ере-
ван, 1954. 415 с.
Данилкина Л. Л. Грызущая деятельность
серой крысы // Грызуны: Материалы УТ
Всесоюз. совещ. (Ленинград, 25—28 янв.
1983 г.). Л.: Наука, 1984. С. 254—255.
Дао Ван Тьен. Материалы по фауне поз-
воночных Вьетнама // Зоол. журн. 1962.
Т. 41, No 5, С. 724-734.
Дао Ван Тьен. Опыт зоогеографического
районирования Вьетнама // Там же. 1978.
Т. 51, No 4. С. 582—586.
Девяткина М. С. Некоторые данные о
серой крысе в природных биотопах но-
бережья Японского моря // Докл. Иркут.
НИИ. 1966. Т. 7. С. 213-215.
Демберил Ж. Распространение серой кры-
сы в МНР// Изв. Иркут. науч. иселед.
противочум. ин-та Сибири. и Дал. Вос-
тока. 1962. Т. 24. С. 332-337.
Дементьев П. И. Способы ловли пасюков
капканами // Рационализация техники
охотничьего промысла, М., 1953. Вып. 2.
С. 47-52.
Денисов Е. И. О применении куницеоб-
разных в борьбе с серыми крысами//
Серая крыса. М.: АН СССР, 1986. Т. 2.
С. 140—147.
Джексон В. Изучение биологии и пове-
дения грызунов как основа разработки
мер борьбы с ними
// Бюл. ВОЗ. 1973.
Т. 41, No 3. С. 283—288.
Дзюбак В. Ф., Погребко А. Г., Сурков
В. С. Значение серой крысы в распро-
странении псездотуберкулезной инфек-
ции на Сахалине и Курильских остро-
зах // Экология и медицинское значение
серой крысы (Ваз погуес1еиз ВегК}.
М.: АН СССР, 1983. С. 78—19.
Дмитриев А. Крысоловка электронного
“века // Химия и жизнь. 41985. No4.
С. 54.
Дмитриева Т. В. Закономерности размс-
жения мышевидных грызунов в насс-
ленных пунктах городского типа // Тр.
Воронеж. ун-та. 1958. Т. 56, вып. 2.
С. 25—80,
Дмитриева Т. В. К вопросу о заселении
Воронежской области серой крысой //
Некоторые проблемы биологии и почно-
ведения: (Материалы отчет. науч. копф.
биол.-почв. фак. ВГУ, 1969 г.). Воронеж,
1970. Выц. 4. С. 59—62.
Днитриева Т.Н. Некоторые особепности
размещения серой крысы по террито-
рии областей Черноземного центра //
Тр. Воронеж. ун-та. 1971. Т. 93, выш. 4.
С. 71-79.
Дмитриева Т. В. Динамика размещения
ссрой крысы в юго-восточной части
Черноземного центра // Тр. П Веесоюз.
совещ. но млекопитающим. М.: АН
СССР, 1975. С. 239—240.
Дмитриева Т. В., Ковалев М, И. Разме-
щение серой крысы на территории Бел-
городской области // Проблемы изуче-
ния и охраны ландшафтов, Воронеж,
1974. С. 320.
Дмитриева Т. В., Филин А. Д. Размеще-
ние крыс по территории Липецкой об-
ласти // Охрана природы и рациональ-
ное использование природных ресурсов
Центрально-Черноземной полосы. Воро-
неж, 1972. С. 109—444.
Домбровский В. В. Серая крыса и другие
виды млекопитающих в населенных
пунктах Московской области// Эколо-
гия и медицинское значение серой кры-
сы (Ваз потуееаз ВетКк). М.: АН
СССР, 1983. С. 10—11.
Друммонд Д. Биология домашних гры-
зунов и борьба с ними // Борьба с пе-
реносчиками болезней и международ-
ное здравоохранение. Женева: Изд-во
ВОЗ, 1974. С. 54—16.
Друммонд Д. Реннисон Б. Выявление
резистентности грызунов к антикоагу-
лянтам // Бюл. ВОЗ. 1974. Т. 48, No 2.
С. 254—258.
Дубровский В. Ю. Серая крыса на же-
лезнодорожных вокзалах // Материалы
но экологии и методам ограничения
численности серой крысы. М.: Наука,
1987. С. 207—244.
Дударев А. Я., Новиков Г. В., Разнощик
В. В. Ольшанский В. А. Санитарная
очистка городов. Л.: Медицина, 1974.
с.
Дукельская Н. М. Распространепие и био-
логия серой крысы // Тр. Центр. науч.-
исслед. дезинфекц. ин-та. 1947. Т. 3.
С. 223—233.
Дукельская Н. М. Подбор нищевых про-
дуктов для изготовления отравленных
приманок для крыс // Там же. 1948. Т.4.
С. 145—150.
Дукельская Н. М. Новое средство борьбы
с крысами // Защита растений от вре-
дителей и болезней. 1957. No 4. С. 60.
400
Литература
мы дезинфекции и стерилизации: (Ма-
риал ции 23—25 ноябр. 1977 г.).
М.: Минздрав СССР, 1977. Ч. 2. С. 76.
Волянский Б. Зазйтки про звцЯ в Кер-
ченського пвоетрова (Крим) // Тр. Ф!з.-
мат. выд. Всеукр. АН. 1929. Т. 43, вип. 1.
С. 29—38.
Воронов В. Г. Млекопитающие Курильских
островов. Л.: Наука, 1974. 157 с.
Воронцов Н. Н. Эволюция пищеваритель-
ной системы грызунов: (Мыптеобраз-
ные). Новосибирск: Наука, 1967а. 289 с.
Воронцов Н. Н. Историческая зоогеогра-
фия мышеобразных (МиЧае) грызу-
нов // Проблемы эволюции. Новоси-
бирск: Наука, 19676. С. 116—141.
Гавриловская И. П., Бойко В. А. Гемор-
рагическая лихорадка с почечным син-
дромом: Науч. обзор ВНИИМИ. М., 1985.
74 с.
Гажиев В. В., Сергеева Л. А., Степинец
В. И. и др. Некоторые данные о меди-
цинском значении серой крысы в Донец-
кой области // ТУ съезд Всесоюз. териол.
о-ва: Тез. докл. М.: АН СССР, 1986. Т. 3.
С. 305—306.
Гажиев В. В., Степанец В. И., Анищенко
Г. А. и др. Борьба с синантропными
трызунами в шахтах Донбасса // Там
же. 198ба. Т. 3. С. 389—390.
Гажиев В. В., Степанец В. И., Грянов
В. И. и др. Борьба с серой крысой в
шахтах Донецкой обл. // Материалы по
экологии и методам ограничения чис-
ленности серой крысы. М.: Наука, 1987.
С. 321-322.
Гамалея Н. Ф. Эпидемиология Одесской
чумы 1902 г.//Чума в Одессе, истори-
ческое и эпидемиологическое исследо-
вапие. Одесса, 1908. 'Г. 4. С. 131-172.
Гамбарян П. Н., Дукельская Н. М. Крыса,
М.: Сов. наука, 1955. 254 с.
Гзелисиани И. Д. Крысы как резервуар
Ва!апи ут сой// Журн. мед, парази-
тологии паразитар. болезней. 1949.
Выи. 6. С. 499—500.
Гиренко Л. Л. К вопросу о распростра-
нении черной крысы в УССР // Тр. Зоол.
музея Киев. ун-та. 1950. Т. 2. С. 75-
95.
Голещилин А. К. Оргапизация и прояе-
дение одномоментпой сплошной дера-
тизации в условиях населенного пунк-
та: Автореф. дис. ... канд. биол. наук.
Кишинев, 1955. 20 с.
Гольденштейн 3. А. Заболеваемость лепто-
спирозом в Краснодарском крае, его
лабораторная диагностика и осно-
вы предупреждения лептоспироза // Тез.
докл. УТ Веесозоз. науч. конф. по лепто-
спирозу. Баку, 1976. С. 67-68.
Гольцман М. Е., Наумов Н. Н., Николь-
ский А. А. и др. Социальное поведение
болыпой песчанки (НВомБотуз орз
Тя.) И Поведение млекопитающих. М.:
Наука, 1977. С. 5-69.
Горшанова Е. Н. Носительство [.. 1ага5з0-
у: среди синантронных грызунов //
Журн. микробиологии, эпидемиологии
и иммунобиологии. 1962. No4. С. 53—
54.
Гребенчук А. Н., Бакулина Л. И., Ваще-
нок Г. И. и др. Сальмонеллез у грызу-
нов в Ленинграде // Там же. 1965. No 6.
С. 43—46.
Громов И. М. Отряд Грызуны Водепма //
Каталог млекопитающих СССР: Плио-
цен-современность. Л.: Наука, 1981.
С. 75-247.
Громов И. М., Гуреев А. А., Новиков Г. А.
и др. Млекопитающие фауны СССР. М.;
Л.: Наука; 1963. Ч. 1. 639 с.
Груздев В. В. Зоны наибольшей вредности
серых крыс в сельской. местности //
Биол. науки. 1959. No 2. С. 39—44.
Груздев В. В. Характеристика численно-
сти и вредности грызунов при комп-
лексном районировании для сельского
хозяйства // Материалы ГУ Межвуз. со-
вещ. по районированию для сел. хоз-ва.
М.: Колос, 1963. С. 21—24,
Губарев Л. Д. Распространение серой
крысы (ВаМиз погусе1сяз ВегК) в вос-
точных районах Ростовской области //
Тр. Рост. противочум. ин-та. 1941. Т. 2.
С. 133—144.
Гутман 9. А. Приманка из свежих поми-
доров с зоокумарилом для уничтоже-
ния грызунов // Ветеринария. 1972.7No 5.
[9
Гуцель Ю. Н., Рожков Ф. Ф., Даданов
В. и др..Некоторые вопросы дера-
тизации в морском порту //'Экология и,
медицинское значение серой крысы
(Ва 1$ потуессив ВетК). М.: Наука,
1983. С. 28—31.
|
|
Давыдов Г. С. Грызуны Северного 'Гад-
жикистана. Душанбе: Тадж. ун-т, 1964.
272 с.
Давыдов Г. С. Грызуны
— вредители по-
ливных культур и пастбищ Таджикис-
тана и борьба с ними. Душанбе, 1978.
55 с.
р
Давыдов Г. С. Распространение и дина-
мика численности серой крысы в Се-
верном Таджикистане // ТУ съезд Вес-
союз. териол. о-ва: Тез. докл. М., 1986.
Т. 3. С. 281-282,
Литература
4041
Дадонов Ю. В., Арканников А. Н., Го-
лубков Н.Е. и др. Испытания на устой-
чивость к повреждению грызунами раз-
личных материалов, применяемых в
электронной радиопромышленности //
Насекомые и грызуны
— разрушители
материалов и технических изделий. М.:
Наука, 1983. С. 206—208.
Даль С. К. Животлый мир Армянской
ССР. Т. 1. Позвопочпые животные. Ере-
ван, 1954. 415 с.
Данилкина Л. Л. Грызущая деятельность
серой крысы // Грызуны: Материалы УТ
Всесоюз. совещ. (Ленинград, 25—28 янв.
1983 г.). Л.: Наука, 1984. С. 254—255.
Дао Ван Тьен. Материалы по фауне поз-
воночных Вьетнама // Зоол. журн. 1962.
Т. 41, No 5, С. 724-734.
Дао Ван Тьен. Опыт зоогеографического
районирования Вьетнама // Там же. 1978.
Т. 51, No 4. С. 582—586.
Девяткина М. С. Некоторые данные о
серой крысе в природных биотопах но-
бережья Японского моря // Докл. Иркут.
НИИ. 1966. Т. 7. С. 213-215.
Демберил Ж. Распространение серой кры-
сы в МНР// Изв. Иркут. науч. иселед.
противочум. ин-та Сибири. и Дал. Вос-
тока. 1962. Т. 24. С. 332-337.
Дементьев П. И. Способы ловли пасюков
капканами // Рационализация техники
охотничьего промысла, М., 1953. Вып. 2.
С. 47-52.
Денисов Е. И. О применении куницеоб-
разных в борьбе с серыми крысами//
Серая крыса. М.: АН СССР, 1986. Т. 2.
С. 140—147.
Джексон В. Изучение биологии и пове-
дения грызунов как основа разработки
мер борьбы с ними
// Бюл. ВОЗ. 1973.
Т. 41, No 3. С. 283—288.
Дзюбак В. Ф., Погребко А. Г., Сурков
В. С. Значение серой крысы в распро-
странении псездотуберкулезной инфек-
ции на Сахалине и Курильских остро-
зах // Экология и медицинское значение
серой крысы (Ваз погуес1еиз ВегК}.
М.: АН СССР, 1983. С. 78—19.
Дмитриев А. Крысоловка электронного
“века // Химия и жизнь. 41985. No4.
С. 54.
Дмитриева Т. В. Закономерности размс-
жения мышевидных грызунов в насс-
ленных пунктах городского типа // Тр.
Воронеж. ун-та. 1958. Т. 56, вып. 2.
С. 25—80,
Дмитриева Т. В. К вопросу о заселении
Воронежской области серой крысой //
Некоторые проблемы биологии и почно-
ведения: (Материалы отчет. науч. копф.
биол.-почв. фак. ВГУ, 1969 г.). Воронеж,
1970. Выц. 4. С. 59—62.
Днитриева Т.Н. Некоторые особепности
размещения серой крысы по террито-
рии областей Черноземного центра //
Тр. Воронеж. ун-та. 1971. Т. 93, выш. 4.
С. 71-79.
Дмитриева Т. В. Динамика размещения
ссрой крысы в юго-восточной части
Черноземного центра // Тр. П Веесоюз.
совещ. но млекопитающим. М.: АН
СССР, 1975. С. 239—240.
Дмитриева Т. В., Ковалев М, И. Разме-
щение серой крысы на территории Бел-
городской области // Проблемы изуче-
ния и охраны ландшафтов, Воронеж,
1974. С. 320.
Дмитриева Т. В., Филин А. Д. Размеще-
ние крыс по территории Липецкой об-
ласти // Охрана природы и рациональ-
ное использование природных ресурсов
Центрально-Черноземной полосы. Воро-
неж, 1972. С. 109—444.
Домбровский В. В. Серая крыса и другие
виды млекопитающих в населенных
пунктах Московской области// Эколо-
гия и медицинское значение серой кры-
сы (Ваз потуееаз ВетКк). М.: АН
СССР, 1983. С. 10—11.
Друммонд Д. Биология домашних гры-
зунов и борьба с ними // Борьба с пе-
реносчиками болезней и международ-
ное здравоохранение. Женева: Изд-во
ВОЗ, 1974. С. 54—16.
Друммонд Д. Реннисон Б. Выявление
резистентности грызунов к антикоагу-
лянтам // Бюл. ВОЗ. 1974. Т. 48, No 2.
С. 254—258.
Дубровский В. Ю. Серая крыса на же-
лезнодорожных вокзалах // Материалы
но экологии и методам ограничения
численности серой крысы. М.: Наука,
1987. С. 207—244.
Дударев А. Я., Новиков Г. В., Разнощик
В. В. Ольшанский В. А. Санитарная
очистка городов. Л.: Медицина, 1974.
с.
Дукельская Н. М. Распространепие и био-
логия серой крысы // Тр. Центр. науч.-
исслед. дезинфекц. ин-та. 1947. Т. 3.
С. 223—233.
Дукельская Н. М. Подбор нищевых про-
дуктов для изготовления отравленных
приманок для крыс // Там же. 1948. Т.4.
С. 145—150.
Дукельская Н. М. Новое средство борьбы
с крысами // Защита растений от вре-
дителей и болезней. 1957. No 4. С. 60.
405
Литература
Дукельская Н. М. Подвижность серых
крые в населенных пунктах и влияние
деятельности человека на их расселе-
ние // Проблемы зоогеотрафии суши.
Львов. ун-та, 4959. С. 90—94.
Дукельская Н. М. Повые средства борь-
бы с крысами // Фауна и экология гры-
зунов. 1960. Вып. 6. С. 208—230.
Дьюсбери Д. Поведение животных. Срав-
нительные аспекты. М; Мир, 1981.
480 с.
Дэвис Д. Е., Кристиан Д. Д. Регуляция
популяций — млекопитающих/// Успехи
современной териологии. М.: Наука,
1977. С. 46—54.
Евдокимова 0. А., Чернуха Ю. Г., Анань-
ина Ю. В., Зайцев С. В. Источники ин-
фекции в очагах иктерогеморрагическо-
го лептосцироза // Материалы Всесоюз.
конф. по зоопозам. М.: Минздрав СССР,
1988. С. 44—45.
Егоренко Г. Г. Комбинированное приме-
ненис бактерийных препаратов и ра-
тиндана // Тр. ВНИИ вет. санитарии.
1963. Т. 22. С. 173—177.
Егоров П. Г., Хмелева А. С. Новые пунк-
ты обпаружения серой крысы в Яку-
тии // Вопр. регион. гигиены, санитарии
и эпидемиологии: ('Гез. докл. науч.
практ. конф., 18—19 дек. 1987 г.).
Якутск, 1987. Выш. 2. С. 186—187.
Егоров (). В. Материалы но питанию фи-
лина в Якутии // Тр. проблем. и темат.
совещ. Зоол. ин-та АН СССР. 1960.
Вып. 9. С. 191—204.
Ежова Н. Г. Лептоспиропосительство у
серых крыс // Журн. микробиологии,
эпидемиологии и иммунобиологии. 1955.
No4. С. 101-108.
Ельшин С. Б. Распространение и числен-
ность домовой мыши и серой крысы на
севере Западной Сибири // Грызуны:
Материалы УТ Всесоюз. совещ. Л.: Нау-
ка, 1983. С. 378—379.
й
Емельянова И. А. Способность серой кры-
сы повреждать сооружения и материа-
лы // Экология и медицинское значение
серой крысы (Ваиз погуезсиз ВегК).
М.: Паука, 1983. С. 89-90.
Емельянова И. А. Способность серой кры-
сы повреждать сооружения и материа-
лы // Серая крыса. М.: АН СССР, 1986.
Т. 2. С. 104-145.
Емельянова И. А., Данилвина Л. Л. Ре-
пелленты, предлагаемые для защиты
материалов от повреждения грызуна-
ми // Насекомые и грызуны — разруши-
тели материалов и технических изде-
лий. М.: Наука, 1983. С. 175—190.
Емельянова И. А., Жарикова Е. Н. Изу-
чение репеллентных свойств препарата
«Альгонаста» в полевых и лаборатор-
ных условиях// Защита материалов и
техники от повреждений, причиняемых
насекомыми и грызупами: Тез. Всесоюз.
симпоз. М.: Наука, 1984. С. 139—141.
Емельянова И. А., Малыгин В. М. Испы-
тание устойчивости синтетических ма-
териалов к повреждению грызунами //
Биологические повреждения строитель-
ных и промышленных материалов. М.,
1973. С. 194—197.
Ефимов В. И. Распространение, числен-
ность и некоторые черты экологии се-
рой крысы в северо-западной Туркме-
нии // Материалы по экологии и мето-
дам ограничения численности серой
крысы. М.: Наука, 1987. С. 69—75.
Жарков И. В. Колебания численности мы-
птевидных грызунов в Кавказском за-
поведнике // Тр. Вавк. гос. заповедника,
1949. Выш. 3. С. 65-80.
`
Жаров В. Г., Ямов В. 3., Паленов А. К.
Новое в организации дератизации жи-
вотноводческих объектов: Рекоменда-
ции/Гюмен. обл. правление науч.-техн.
о-ва сел. хоззва; ВНИИ вет. энтомоло-
тии и арахнологии. Гюмень, 1974.
Жидбан Ж. Оценка вредоносной деятель-
ности млекопитающих в регионах с
развитым сельским и лесным хозяйст-
вами // Успехи современной териологии.
М.: Наука, 1977. С. 55—61.
Жирнов А. Е. Особенности размножения
серых крыс в Ленинграде // Грызуны:
Материалы У Всесоюз. совещ. М.: Нау-
ка, 1980. С. 191—192.
Жирнов А. Е. Н характеристике серых
крыс Ленинграда
// Экология и меди-
нинское значение серой крысы (ВаИиз
погузтсиз ВегК). М.: Наука, 1983. С. 12—
в..
Жиров А. Я., Дурас Т.И., Карманова Е. В.,
Фищенко И. Г. О выделении возбуди-
теля псевдотуберкулеза в районе г. На-
ходки // Докл. Иркут. противочум. ин-та
Сибири и Дал. Востока. 1983. Выш. 5.
С. 23—25.
Житков Б. М. Материалы по фауне мле-
копитающих Симбирской губернии//
Дневник зоол. отд-ния и 300л. музея.
1898. Т. 2, No 8. С. 1-27.
Житков Б. М. Замечания о крысах и не-
которые условия их исследования //
Зоол. журн. 1944. Т. 23, No 2/3. С. 91-
97.
Литература
403
Заболотный Д. К. Характерные особен-
ности эпидемии 1910 г. // Чума в Одес-
се в 1910 г. СИб., 1912. С. 18—22,
Зарудный Н. Заметки по фауне млеко-
нитающих Оренбургского края // Мате-
риалы к познанию фауны и флоры Рос-
сийской империи. Отд. зоол. М.: МОИП,
1897. Вын. 3. С. 4-42.
Засухин Д. Н. Проблема токсоплазмоза //
Борьба с болезнями, общими для че-
ловека и животных. Зоонозы. М.: М-во
сел. хоз-ва СССР, 1961. С. 98—108.
Засутин Д. Н. Природная очаговость ток-
соплазмоза // Вопр. эпидемиологии мик-
робиологии и иммунологии. 1965. Т. 16.
С. 144—145.
Засухин Д. Н., Васина С. Г. Токсоплаз-
моз // Природная очаговость болезней
человека и краевая эпидемиология. М.:
Медгиз, 1955. С. 299—316.
Зацепин В. Г. Испытания ядовитых пен
для дератизации // Проблемы встери-
парной санитарии. М.: Минздрав СССР,
1969. С. 630—635.
Зверев М. Д. К изучению серых крыс в
Сибири и методика борьбы с ними//
Изв. Сиб. краев. станции защиты рас-
тений. 1930. No 4/7. С. 61-68.
Зинневич 9. П., Курятов Н. С. Состав ле-
тучих кислот и оснований мочи неко-
торых видов грызунов // Веесоюз. совещ,
по хим. коммуникации животных. М.:
АН СССР, 1979. С. 86-87.
Златогоров С. И. О борьбе с крысами
в Северо-Американских Соединенных
Штатах // Профилакт. медицина. 1996.
Т. 48. No 4. С. 475—484.
Змеев Г. Я. Анкилостомоз, ангвиллулез
и другие гельминтозы среди рабочих
каменноугольных рудников ДВК // Тр.
Дальневост. краев. бактериол. ин-та.
1931. Выш. 1. С. 101-105.
Зубко В. И., Соловьев В. И., Браверман
Г. К. К вопросу эпидемиологии ленто-
спироза в Одесской области // Лептоспи-
розы: Тез. докл. УПЕ Всесоюз. конф.
по лептоспирозам. Тбилиси: Сабчота
Сакартвело, 1983. С. 103—104.
Иванидзе 5). А. Обнаружение природных
« очагов ГЛИС в Грузинской ССР // Вопр.
вирусологии. 1987. No 5. С. 607—610.
Иванкина Е. В. Динамика численности и
‚структура населения рыжей полевки в
`Подмосковье: Автореф. дис. .. канд.
биол. наук. М., 1987. 22 в.
Иванова В. В. Состояние дератизацион-
ных работ, проводимых в городах Со-
ветского Союза дезинфекционными
станциями в 1955 и 1956 г.. // Тр. Центр.
науч.-исслед. дезинфекц. ин-та. 1959.
Т. 12. С. 259—263.
Ивантер 9. В. Популяционная экология
мелких млекопитающих таежного севе-
ро-запада СССР. Л.: Изд-во ЛГУ, 1975.
246 с
Иванченко П. Л., Тимченко А. Д. К рас-
пространению балантидиоза и гельми-
нелепидоза среди серых крыс // Проб-
лемы паразитологии: (Тез, докл. У науч.
конф. паразитологов УССР). Киев: Наук.
думка, 1967, С. 45—46.
Ильенко А. И., Зубчанинова Е. В. Кругло-
годичное наблюдение за мечеными ры-
жими полевками и лесными мышами
в Подмосковье // Зоол. журн. 1963. Т. 42,
No 4. С. 609—616.
Инетрукция по учету численности гры-
зунов для противочумных станций Со-
ветского Союза. Саратов: Минздрав
СССР, 1978. 79 с.
Иоффе Г. Д. Опыт работы по дератиза-
ции в угольных шахтах Луганекой об-
ласти // Гигиена и санитария. 1959. No 9.
С. 45—47.
Исмаилов А. Ш., Лобанова Т. И., Раби-
нович Б. К. и др. Некоторые данные
по распространению черных и серых
крыс в Азербайджане // ГУ съезд Все-
союз. териол. о-ва: Тез. докл. М., 1986.
Т. 3, С. 276—277.
Кавтарадзе К. Н., Бернштейн А. Д., Ква-
рухелия Г. Я. К вопросу об источниках
лептоспироза в Абхазской АССР //
Журн. микробиологии, эпидемиологии
и иммунобиологии. 1957. No 9. С. 60—63.
Кадацкий Н. Г. Грызупы Талыша и Лен-
коранской низменности и их распрост-
ранение по ландшафтно-географиче-
ским районам // Зоол. журн. 1964. Т. 43,
No 11. С. 1693—1707.
Кайзер Г. А. Итоги работы но службе
учета численности грызунов на юто-
востоке СССР в 1939 г.//Грызуны и
борьба с ними. Алма-Ата, 1944. Вып. 1.
С. 225—234.
Калабутов Н. И., Раевский В. В. Методи-
ка изучения покоторых вопросов эко-
логии мышевидных грызунов // Вестн.
микробиологии, энидемиологии и пара-
зитологии. 1933. Т. 42, выш. 1. С. 128-—
Каменов Д. А. Экопого-физиологические
механизмы формирования и поддержа-
ния структуры популяции диких домо-
вых мышей (Миз шизсиа$): Автореф.
дис. .. канд. биол. наук. М.: МГУ, 1973.
18 с.
405
Литература
Дукельская Н. М. Подвижность серых
крые в населенных пунктах и влияние
деятельности человека на их расселе-
ние // Проблемы зоогеотрафии суши.
Львов. ун-та, 4959. С. 90—94.
Дукельская Н. М. Повые средства борь-
бы с крысами // Фауна и экология гры-
зунов. 1960. Вып. 6. С. 208—230.
Дьюсбери Д. Поведение животных. Срав-
нительные аспекты. М; Мир, 1981.
480 с.
Дэвис Д. Е., Кристиан Д. Д. Регуляция
популяций — млекопитающих/// Успехи
современной териологии. М.: Наука,
1977. С. 46—54.
Евдокимова 0. А., Чернуха Ю. Г., Анань-
ина Ю. В., Зайцев С. В. Источники ин-
фекции в очагах иктерогеморрагическо-
го лептосцироза // Материалы Всесоюз.
конф. по зоопозам. М.: Минздрав СССР,
1988. С. 44—45.
Егоренко Г. Г. Комбинированное приме-
ненис бактерийных препаратов и ра-
тиндана // Тр. ВНИИ вет. санитарии.
1963. Т. 22. С. 173—177.
Егоров П. Г., Хмелева А. С. Новые пунк-
ты обпаружения серой крысы в Яку-
тии // Вопр. регион. гигиены, санитарии
и эпидемиологии: ('Гез. докл. науч.
практ. конф., 18—19 дек. 1987 г.).
Якутск, 1987. Выш. 2. С. 186—187.
Егоров (). В. Материалы но питанию фи-
лина в Якутии // Тр. проблем. и темат.
совещ. Зоол. ин-та АН СССР. 1960.
Вып. 9. С. 191—204.
Ежова Н. Г. Лептоспиропосительство у
серых крыс // Журн. микробиологии,
эпидемиологии и иммунобиологии. 1955.
No4. С. 101-108.
Ельшин С. Б. Распространение и числен-
ность домовой мыши и серой крысы на
севере Западной Сибири // Грызуны:
Материалы УТ Всесоюз. совещ. Л.: Нау-
ка, 1983. С. 378—379.
й
Емельянова И. А. Способность серой кры-
сы повреждать сооружения и материа-
лы // Экология и медицинское значение
серой крысы (Ваиз погуезсиз ВегК).
М.: Паука, 1983. С. 89-90.
Емельянова И. А. Способность серой кры-
сы повреждать сооружения и материа-
лы // Серая крыса. М.: АН СССР, 1986.
Т. 2. С. 104-145.
Емельянова И. А., Данилвина Л. Л. Ре-
пелленты, предлагаемые для защиты
материалов от повреждения грызуна-
ми // Насекомые и грызуны — разруши-
тели материалов и технических изде-
лий. М.: Наука, 1983. С. 175—190.
Емельянова И. А., Жарикова Е. Н. Изу-
чение репеллентных свойств препарата
«Альгонаста» в полевых и лаборатор-
ных условиях// Защита материалов и
техники от повреждений, причиняемых
насекомыми и грызупами: Тез. Всесоюз.
симпоз. М.: Наука, 1984. С. 139—141.
Емельянова И. А., Малыгин В. М. Испы-
тание устойчивости синтетических ма-
териалов к повреждению грызунами //
Биологические повреждения строитель-
ных и промышленных материалов. М.,
1973. С. 194—197.
Ефимов В. И. Распространение, числен-
ность и некоторые черты экологии се-
рой крысы в северо-западной Туркме-
нии // Материалы по экологии и мето-
дам ограничения численности серой
крысы. М.: Наука, 1987. С. 69—75.
Жарков И. В. Колебания численности мы-
птевидных грызунов в Кавказском за-
поведнике // Тр. Вавк. гос. заповедника,
1949. Выш. 3. С. 65-80.
`
Жаров В. Г., Ямов В. 3., Паленов А. К.
Новое в организации дератизации жи-
вотноводческих объектов: Рекоменда-
ции/Гюмен. обл. правление науч.-техн.
о-ва сел. хоззва; ВНИИ вет. энтомоло-
тии и арахнологии. Гюмень, 1974.
Жидбан Ж. Оценка вредоносной деятель-
ности млекопитающих в регионах с
развитым сельским и лесным хозяйст-
вами // Успехи современной териологии.
М.: Наука, 1977. С. 55—61.
Жирнов А. Е. Особенности размножения
серых крыс в Ленинграде // Грызуны:
Материалы У Всесоюз. совещ. М.: Нау-
ка, 1980. С. 191—192.
Жирнов А. Е. Н характеристике серых
крыс Ленинграда
// Экология и меди-
нинское значение серой крысы (ВаИиз
погузтсиз ВегК). М.: Наука, 1983. С. 12—
в..
Жиров А. Я., Дурас Т.И., Карманова Е. В.,
Фищенко И. Г. О выделении возбуди-
теля псевдотуберкулеза в районе г. На-
ходки // Докл. Иркут. противочум. ин-та
Сибири и Дал. Востока. 1983. Выш. 5.
С. 23—25.
Житков Б. М. Материалы по фауне мле-
копитающих Симбирской губернии//
Дневник зоол. отд-ния и 300л. музея.
1898. Т. 2, No 8. С. 1-27.
Житков Б. М. Замечания о крысах и не-
которые условия их исследования //
Зоол. журн. 1944. Т. 23, No 2/3. С. 91-
97.
Литература
403
Заболотный Д. К. Характерные особен-
ности эпидемии 1910 г. // Чума в Одес-
се в 1910 г. СИб., 1912. С. 18—22,
Зарудный Н. Заметки по фауне млеко-
нитающих Оренбургского края // Мате-
риалы к познанию фауны и флоры Рос-
сийской империи. Отд. зоол. М.: МОИП,
1897. Вын. 3. С. 4-42.
Засухин Д. Н. Проблема токсоплазмоза //
Борьба с болезнями, общими для че-
ловека и животных. Зоонозы. М.: М-во
сел. хоз-ва СССР, 1961. С. 98—108.
Засутин Д. Н. Природная очаговость ток-
соплазмоза // Вопр. эпидемиологии мик-
робиологии и иммунологии. 1965. Т. 16.
С. 144—145.
Засухин Д. Н., Васина С. Г. Токсоплаз-
моз // Природная очаговость болезней
человека и краевая эпидемиология. М.:
Медгиз, 1955. С. 299—316.
Зацепин В. Г. Испытания ядовитых пен
для дератизации // Проблемы встери-
парной санитарии. М.: Минздрав СССР,
1969. С. 630—635.
Зверев М. Д. К изучению серых крыс в
Сибири и методика борьбы с ними//
Изв. Сиб. краев. станции защиты рас-
тений. 1930. No 4/7. С. 61-68.
Зинневич 9. П., Курятов Н. С. Состав ле-
тучих кислот и оснований мочи неко-
торых видов грызунов // Веесоюз. совещ,
по хим. коммуникации животных. М.:
АН СССР, 1979. С. 86-87.
Златогоров С. И. О борьбе с крысами
в Северо-Американских Соединенных
Штатах // Профилакт. медицина. 1996.
Т. 48. No 4. С. 475—484.
Змеев Г. Я. Анкилостомоз, ангвиллулез
и другие гельминтозы среди рабочих
каменноугольных рудников ДВК // Тр.
Дальневост. краев. бактериол. ин-та.
1931. Выш. 1. С. 101-105.
Зубко В. И., Соловьев В. И., Браверман
Г. К. К вопросу эпидемиологии ленто-
спироза в Одесской области // Лептоспи-
розы: Тез. докл. УПЕ Всесоюз. конф.
по лептоспирозам. Тбилиси: Сабчота
Сакартвело, 1983. С. 103—104.
Иванидзе 5). А. Обнаружение природных
« очагов ГЛИС в Грузинской ССР // Вопр.
вирусологии. 1987. No 5. С. 607—610.
Иванкина Е. В. Динамика численности и
‚структура населения рыжей полевки в
`Подмосковье: Автореф. дис. .. канд.
биол. наук. М., 1987. 22 в.
Иванова В. В. Состояние дератизацион-
ных работ, проводимых в городах Со-
ветского Союза дезинфекционными
станциями в 1955 и 1956 г.. // Тр. Центр.
науч.-исслед. дезинфекц. ин-та. 1959.
Т. 12. С. 259—263.
Ивантер 9. В. Популяционная экология
мелких млекопитающих таежного севе-
ро-запада СССР. Л.: Изд-во ЛГУ, 1975.
246 с
Иванченко П. Л., Тимченко А. Д. К рас-
пространению балантидиоза и гельми-
нелепидоза среди серых крыс // Проб-
лемы паразитологии: (Тез, докл. У науч.
конф. паразитологов УССР). Киев: Наук.
думка, 1967, С. 45—46.
Ильенко А. И., Зубчанинова Е. В. Кругло-
годичное наблюдение за мечеными ры-
жими полевками и лесными мышами
в Подмосковье // Зоол. журн. 1963. Т. 42,
No 4. С. 609—616.
Инетрукция по учету численности гры-
зунов для противочумных станций Со-
ветского Союза. Саратов: Минздрав
СССР, 1978. 79 с.
Иоффе Г. Д. Опыт работы по дератиза-
ции в угольных шахтах Луганекой об-
ласти // Гигиена и санитария. 1959. No 9.
С. 45—47.
Исмаилов А. Ш., Лобанова Т. И., Раби-
нович Б. К. и др. Некоторые данные
по распространению черных и серых
крыс в Азербайджане // ГУ съезд Все-
союз. териол. о-ва: Тез. докл. М., 1986.
Т. 3, С. 276—277.
Кавтарадзе К. Н., Бернштейн А. Д., Ква-
рухелия Г. Я. К вопросу об источниках
лептоспироза в Абхазской АССР //
Журн. микробиологии, эпидемиологии
и иммунобиологии. 1957. No 9. С. 60—63.
Кадацкий Н. Г. Грызупы Талыша и Лен-
коранской низменности и их распрост-
ранение по ландшафтно-географиче-
ским районам // Зоол. журн. 1964. Т. 43,
No 11. С. 1693—1707.
Кайзер Г. А. Итоги работы но службе
учета численности грызунов на юто-
востоке СССР в 1939 г.//Грызуны и
борьба с ними. Алма-Ата, 1944. Вып. 1.
С. 225—234.
Калабутов Н. И., Раевский В. В. Методи-
ка изучения покоторых вопросов эко-
логии мышевидных грызунов // Вестн.
микробиологии, энидемиологии и пара-
зитологии. 1933. Т. 42, выш. 1. С. 128-—
Каменов Д. А. Экопого-физиологические
механизмы формирования и поддержа-
ния структуры популяции диких домо-
вых мышей (Миз шизсиа$): Автореф.
дис. .. канд. биол. наук. М.: МГУ, 1973.
18 с.
404
Литература
Као Ван Шунг, Фам Дык Тьен, Чан Ван
Минь. К экологии зайцеобразных и
грызунов плато Тай-Нгуен // Животный
мир Вьетнама. М.: Наука, 1982. С. 52—
ева Е. В. Изучение ве помощью ме-
чения особенностей использования тер-
ритории обыкновенным хомяком в Ал-
тайском крае // Зоол. журн. 1962. Т. 44,
No 2. С. 275-285.
й
Карасева Е. В. Значение особенностей по-
ведения серых крыс в передаче инфек-
ции при лептоспирозе // Лептоспирозы:
Тез. докл. УПТ Всесоюз. конф. по ленто-
спирозам. Тбилиси: Сабчота Сакартве-
ло, 1983. С. 107—109.
Карасева Е. В., Ильченко А. И. Некоторые
особенности биологии полевкиэконом-
ки, изученные методом мечи ве:
ков // Фауна и экология грыз ‚ М.:
ра МГУ, 1957. Вып. 6. С. 171-185.
Карасева Е. В., Козлов А. Н, Мерзликин
И. Р. Поверхностпые гнезда, норы и
другие убежища серой крысы [1 Рас-
пространение и экология серой крысы
и мотоды ограничения ее численности.
М.: Наука, 4985. С. 53—71.
Карасева Е. В., Рыльников В. А., Белень-
кий В. А., Беленькая Е. М. Некоторые
черты экологии серых крыс средней по-
лосы Росеии в котловине озера Неро
(Ярославекая обл.} и их роль в при
родном очаге лептоснироза // Серая кры-
са.М.:АНСССЬ,1986а.Т.2.С.3—
18.
Карасева Е. В. Рыльников В. А., Дуби-
нина Н. В. и др. Влияние рисоссяния
на природные очаги лептоспироза н
Кубани Бюл. МОИП. Отд. биол. 1986.
Т, 91, No 5. С. 29—40.
Карасева Е. В., Свешникова Н. Н. Дикие
позвоночные животные — носители леп-
тоепир в природе и характер эпизоотии
в их популяциях // Лептоспирозы лю“
дей и животных. М.: Медицина, 1971.
. 163—207.
сасева Е. В. Свешникова Н. П. Роль
серых крыс в очагах лептоспирозов й
ТУ съезд Всеесоюз. териол. о-ва: Тез.
докл. М., 1986. Т. 3. С. 320-324.
Карасева Е. В., Свешникова Н. П. Серая
крыса в природных и антропургических
очатах лептоспироза // Материалы о
экологии и методам ограничения чис-
ленности серой крысы. М.: АН СССР,
987. С. 215-237.
абисва Е. В., Семенова Л. П., Солошен-
ко И, 3. и др. Природные очаги л -
спироза еверо-Осетинской АССР //
Журн. микробиологии, эпидемиологии и
и упобмологии. 1963. No 15. С. 53-
55.
:
Каримова 3. Х. Лептоспирозная желтуха
в Казани // Тез. докл. науч.-практ. конф
но лептоспирозу. М.: Медгиз, 1954.
С. 12—14.
Каркадиновская И. А. 9 бруцеллезе. ди-
ких серых крыс // Журн. микробиоло-
гии, эпидемиологии и иммунобиоло-
гии. 1936. Т. 16, вым. 3. С. 388.
м
Карноихова Н. Г. Определение возрас
рых и черных крысе // Экология. 4974.
No 2. С. 71-76.
Кащенко Н. Ф. Крысы и заместители их
в Западной Сибири и Туркестане // Еже-
тодник Зоол. музея АН. 1912. Т. 47.
С. 370-390.
й
Квашнин С. А. Наблюдения за группои
серых крыс в искусственных услови-
ях // П Всесоюз. конф. молодых ученых.
.М.: Наука, 1975. С. 147—148.
Квашнин С. А. Взаимоотнотения самцов
диких серых крыс в эксперименталь-
. ных группах // Конф. молодых ученых.
Кисв; Наук. думка, 1978. С. 18—22.
Квашнин С. А. Пространственно-этоло-
гическая организация колонии серых
крыс // Материалы ПТ съезда Всесоюз.
ре ва, М. 1982. Т. 2. С. 139-140.
Квашнин С. А. Игровое поведение у ди
ких серых крыс’ (НаМлз погуерлсиз
ВегК) // Материалы 1 Всесоюз. конф.
по поведению животных. М.: Наука,
198За. Т. 4. С. 144-145.
Квашнин? С. 4. Поведение серых крысе
(ВаНиз погуерсиз Вегк) в естествен-
вых и искусственных условиях: Авто-
реф. дис. ... канд. биол. наук. М., 19836.
27 с.
.
Квашнин ©. А. Некоторые наблюдения
за восстановлением популяции серых
крыс после дератизации |/ ГУ съезд Вее-
союз. териол. о-ва: Тез. докл. М., 19886.
Т. 3. С. 149.
Квашнии С. А., Карасева Е. В. Простран-
ствоенно-этологическая структура и 060-
бенности поведения серых крыс в ес-
тественных условиях // Грызуны: Ма-
териалы УЕ Вессоюз, совещ. Л.: Наука,
1983. С. 388—390.
Квашнин С. .А., Карасева Е. В. К изуче-
нию прострапственно-этологической
структуры поселений серых крыс и
особенностей их поведения в открытых
биотопах // Распространение и экология
серой крысы и методы ограпичения ее
Числонности. М.: Наука, 1985. С. 129-
146.
`
Литература
405
Квашнин С. А., Назарова А. В., Сироти-
нина Е. В., Телицина А. Ю. К изучению
суточной вненоровой активности серых
крые в Узбекистане // Сорая крыса. М.:
Наука, 1986. Т. 1. С. 169—177.
Ивашник С. А., Суров А. В. Групповое
поведение диких серых крыс (КВаМлз
погуея1ств ВегЕ) в условиях неволи //
Групповое поведение животных: (И
Всесоюз. конф. по поведению живот-
ных). М.: АН СССР, 1976. С. 169—172.
Кесслер К. Ф. Некоторые заметки о во-
дящихся в России крысах // Натура-
лист. 1866. Т. 3, No 17. С. 15-19.
Ким Т. А., Лихашерстов В. М. О размно-
жении пасюка в г, Красноярске // Учен.
зап. Краенояр. пед. ин-та. 1964. Т. 20,
вып. 2. С. 81—81.
Кирис И. Д. Методика и техника опре-
деления возраста и анализ возрастного
состава популяции белки // Бюл. МОИП.
Отд. биол. 1937. Т. 46, No1, С. 36-
42,
Кирсанов М. ПН. Опыт повышения эффек-
тивности истребления серых крыс от-
равленными приманками // Грызуны и
их эктопаразиты Саратов: Изд-во Са-
рат. ун-та, 1967. С. 364—368.
„Кирьянов Г. И. Серая крыса (пасюк) в
Алтайском крае й Докл. Иркут. науч.-
исслед. противочум. ин-та Сибири и
Дал. Востока. 1962. Вып. 4, С. 148-
152.
„Класовский И. Л., Плотников В. А. Эпи-
зоотические проявления лептоспироз-
ной инфекции у серых крыс в живот-
новодческих объектах Краснодарского
края //ТУ съезд Всесоюз. териол. о-ва:
Тез. докл. М., 1986, Т. 3. С, 334-335.
Клевезаль Г. А. Рост наземных позвоноч-
ных
// Рост животных/Под ред. Л. П.
Позпапипа. М.: Наука, 1973а, С. 116—
Клевезаль Г. А. Об ограничениях и но-
вых возможностях использования сло-
ев в тканях зуба и кости для опреде-
ления возраста млекопитающих // Зоол.
журн. 19756. Т. 52, No 5. С. 757-765.
Клевезаль Г. А. Некоторые методические
аспекты изучения возраста и роста мле-
копитающих // Междунар. териол.
конгр.: (Реф. докл. 6-12 июня 1974 г.,
Москва). М.: Наука, 1974. Т. 4. С. 270.
Клевезаль Г. А. Метод изучения запаса-
ния корма и потребления запасов у
грызунов в природе // Зоол. журн. 1984.
Т. 63, No 2. С. 276—283.
Клевезаль Г. А., Клейненберг С. Е. Оп-
ределение возраста животных по сло-
истым структурам зубов и кости. М.:
Наука, 1967. 144 с.
Клименко А. 3., Клименко 0. И. Патоген-
нал микрофлора грызунов южной час-
ти Сахалина // Природа Сахалина и здо-
ровье человека. Южно-Сахалинск: Ин.
изд-во, 1962. С. 178—182.
Клодницкий Н. И,, Кац Л. Я. Роль крыс
в эпидемиологии китечных инфекций.
Сообщ. 1// Журн. микробиологии, эпи-
демиологии и иммунобиологии. 1936.
Т. 17, вып. 4. С. 24.
Ковалевский К. Л. Лабораторное живот-
новодство. 2-е изд., перераб. и доп. М.:
Медгиз, 1958. 324 с.
Ковалевский Ю. В., Коренберг 9. И. Фор-
мирование фауны синантронных гры-
зунов в зоне Байкало-Амурской желез-
нодорожной магистрали // Бюл. МОИП.
Отд. биол. 1983. Т. 88, No4. С. 89-77.
Коган Н. И. Мусор
- проблема физико-
химическая // Наука и жизнь. 1978. No 7.
С. 33—38.
Козакевим В. П., Домарадский И. В., Бах-
рал Е. Э. Яды, применяемые для борьбы
с хранителями и. переносчиками: 0с0бо
опасных инфекций. М.: Медгиз, 1958.
156 с.
у
Козлов А. Н. Подвижность и сезонные
миграции серой крысы в сельской мест-
ности Северного Казахстана// Вестн.
зоологии. 1978. No 1. С. 53—57.
Козлов А. Н. Заселение Северного На-
захстана серой крысой в результате
освоения целинных земель // Экология.
1979. No 6. С. 89—91.
Козлов А. Н. Особенности изменения чис-
ленности серой крысы в животновод-
ческих помещениях Северного Казах-
стана // Грызуны: Материалы У Все-
союз. совещ. М.: Наука, 1980а, С. 208—
Козлов А. П. Биология серой крысы в
районах освоения целинных земель Се-
верного Казахстана: Автореф. дис. ...
кан, биол. наук. Свердловек, 19806.
с.
Козлов А. Н. Размножение серой крысы
(В. потуес1сив) в животноводческих по-
мещениях Северного Казахстана // Зоол.
журн. 1981. Т. 60, No 4. С. 587—594.
Козлов А. Н. Зависимость параметров
размножения от плотности популяции
на нримере крысы // Млекопитающие
СССР: ПТ съезд Всесоюз. териол. о-ва:
Тез. докл. М., 1982. Т. 1. С. 249.
Козлов А. Н. Современное распростране-
ние серой крывы на территории Черно-
земного центра // Экология и медицин-
404
Литература
Као Ван Шунг, Фам Дык Тьен, Чан Ван
Минь. К экологии зайцеобразных и
грызунов плато Тай-Нгуен // Животный
мир Вьетнама. М.: Наука, 1982. С. 52—
ева Е. В. Изучение ве помощью ме-
чения особенностей использования тер-
ритории обыкновенным хомяком в Ал-
тайском крае // Зоол. журн. 1962. Т. 44,
No 2. С. 275-285.
й
Карасева Е. В. Значение особенностей по-
ведения серых крыс в передаче инфек-
ции при лептоспирозе // Лептоспирозы:
Тез. докл. УПТ Всесоюз. конф. по ленто-
спирозам. Тбилиси: Сабчота Сакартве-
ло, 1983. С. 107—109.
Карасева Е. В., Ильченко А. И. Некоторые
особенности биологии полевкиэконом-
ки, изученные методом мечи ве:
ков // Фауна и экология грыз ‚ М.:
ра МГУ, 1957. Вып. 6. С. 171-185.
Карасева Е. В., Козлов А. Н, Мерзликин
И. Р. Поверхностпые гнезда, норы и
другие убежища серой крысы [1 Рас-
пространение и экология серой крысы
и мотоды ограничения ее численности.
М.: Наука, 4985. С. 53—71.
Карасева Е. В., Рыльников В. А., Белень-
кий В. А., Беленькая Е. М. Некоторые
черты экологии серых крыс средней по-
лосы Росеии в котловине озера Неро
(Ярославекая обл.} и их роль в при
родном очаге лептоснироза // Серая кры-
са.М.:АНСССЬ,1986а.Т.2.С.3—
18.
Карасева Е. В. Рыльников В. А., Дуби-
нина Н. В. и др. Влияние рисоссяния
на природные очаги лептоспироза н
Кубани Бюл. МОИП. Отд. биол. 1986.
Т, 91, No 5. С. 29—40.
Карасева Е. В., Свешникова Н. Н. Дикие
позвоночные животные — носители леп-
тоепир в природе и характер эпизоотии
в их популяциях // Лептоспирозы лю“
дей и животных. М.: Медицина, 1971.
. 163—207.
сасева Е. В. Свешникова Н. П. Роль
серых крыс в очагах лептоспирозов й
ТУ съезд Всеесоюз. териол. о-ва: Тез.
докл. М., 1986. Т. 3. С. 320-324.
Карасева Е. В., Свешникова Н. П. Серая
крыса в природных и антропургических
очатах лептоспироза // Материалы о
экологии и методам ограничения чис-
ленности серой крысы. М.: АН СССР,
987. С. 215-237.
абисва Е. В., Семенова Л. П., Солошен-
ко И, 3. и др. Природные очаги л -
спироза еверо-Осетинской АССР //
Журн. микробиологии, эпидемиологии и
и упобмологии. 1963. No 15. С. 53-
55.
:
Каримова 3. Х. Лептоспирозная желтуха
в Казани // Тез. докл. науч.-практ. конф
но лептоспирозу. М.: Медгиз, 1954.
С. 12—14.
Каркадиновская И. А. 9 бруцеллезе. ди-
ких серых крыс // Журн. микробиоло-
гии, эпидемиологии и иммунобиоло-
гии. 1936. Т. 16, вым. 3. С. 388.
м
Карноихова Н. Г. Определение возрас
рых и черных крысе // Экология. 4974.
No 2. С. 71-76.
Кащенко Н. Ф. Крысы и заместители их
в Западной Сибири и Туркестане // Еже-
тодник Зоол. музея АН. 1912. Т. 47.
С. 370-390.
й
Квашнин С. А. Наблюдения за группои
серых крыс в искусственных услови-
ях // П Всесоюз. конф. молодых ученых.
.М.: Наука, 1975. С. 147—148.
Квашнин С. А. Взаимоотнотения самцов
диких серых крыс в эксперименталь-
. ных группах // Конф. молодых ученых.
Кисв; Наук. думка, 1978. С. 18—22.
Квашнин С. А. Пространственно-этоло-
гическая организация колонии серых
крыс // Материалы ПТ съезда Всесоюз.
ре ва, М. 1982. Т. 2. С. 139-140.
Квашнин С. А. Игровое поведение у ди
ких серых крыс’ (НаМлз погуерлсиз
ВегК) // Материалы 1 Всесоюз. конф.
по поведению животных. М.: Наука,
198За. Т. 4. С. 144-145.
Квашнин? С. 4. Поведение серых крысе
(ВаНиз погуерсиз Вегк) в естествен-
вых и искусственных условиях: Авто-
реф. дис. ... канд. биол. наук. М., 19836.
27 с.
.
Квашнин ©. А. Некоторые наблюдения
за восстановлением популяции серых
крыс после дератизации |/ ГУ съезд Вее-
союз. териол. о-ва: Тез. докл. М., 19886.
Т. 3. С. 149.
Квашнии С. А., Карасева Е. В. Простран-
ствоенно-этологическая структура и 060-
бенности поведения серых крыс в ес-
тественных условиях // Грызуны: Ма-
териалы УЕ Вессоюз, совещ. Л.: Наука,
1983. С. 388—390.
Квашнин С. .А., Карасева Е. В. К изуче-
нию прострапственно-этологической
структуры поселений серых крыс и
особенностей их поведения в открытых
биотопах // Распространение и экология
серой крысы и методы ограпичения ее
Числонности. М.: Наука, 1985. С. 129-
146.
`
Литература
405
Квашнин С. А., Назарова А. В., Сироти-
нина Е. В., Телицина А. Ю. К изучению
суточной вненоровой активности серых
крые в Узбекистане // Сорая крыса. М.:
Наука, 1986. Т. 1. С. 169—177.
Ивашник С. А., Суров А. В. Групповое
поведение диких серых крыс (КВаМлз
погуея1ств ВегЕ) в условиях неволи //
Групповое поведение животных: (И
Всесоюз. конф. по поведению живот-
ных). М.: АН СССР, 1976. С. 169—172.
Кесслер К. Ф. Некоторые заметки о во-
дящихся в России крысах // Натура-
лист. 1866. Т. 3, No 17. С. 15-19.
Ким Т. А., Лихашерстов В. М. О размно-
жении пасюка в г, Красноярске // Учен.
зап. Краенояр. пед. ин-та. 1964. Т. 20,
вып. 2. С. 81—81.
Кирис И. Д. Методика и техника опре-
деления возраста и анализ возрастного
состава популяции белки // Бюл. МОИП.
Отд. биол. 1937. Т. 46, No1, С. 36-
42,
Кирсанов М. ПН. Опыт повышения эффек-
тивности истребления серых крыс от-
равленными приманками // Грызуны и
их эктопаразиты Саратов: Изд-во Са-
рат. ун-та, 1967. С. 364—368.
„Кирьянов Г. И. Серая крыса (пасюк) в
Алтайском крае й Докл. Иркут. науч.-
исслед. противочум. ин-та Сибири и
Дал. Востока. 1962. Вып. 4, С. 148-
152.
„Класовский И. Л., Плотников В. А. Эпи-
зоотические проявления лептоспироз-
ной инфекции у серых крыс в живот-
новодческих объектах Краснодарского
края //ТУ съезд Всесоюз. териол. о-ва:
Тез. докл. М., 1986, Т. 3. С, 334-335.
Клевезаль Г. А. Рост наземных позвоноч-
ных
// Рост животных/Под ред. Л. П.
Позпапипа. М.: Наука, 1973а, С. 116—
Клевезаль Г. А. Об ограничениях и но-
вых возможностях использования сло-
ев в тканях зуба и кости для опреде-
ления возраста млекопитающих // Зоол.
журн. 19756. Т. 52, No 5. С. 757-765.
Клевезаль Г. А. Некоторые методические
аспекты изучения возраста и роста мле-
копитающих // Междунар. териол.
конгр.: (Реф. докл. 6-12 июня 1974 г.,
Москва). М.: Наука, 1974. Т. 4. С. 270.
Клевезаль Г. А. Метод изучения запаса-
ния корма и потребления запасов у
грызунов в природе // Зоол. журн. 1984.
Т. 63, No 2. С. 276—283.
Клевезаль Г. А., Клейненберг С. Е. Оп-
ределение возраста животных по сло-
истым структурам зубов и кости. М.:
Наука, 1967. 144 с.
Клименко А. 3., Клименко 0. И. Патоген-
нал микрофлора грызунов южной час-
ти Сахалина // Природа Сахалина и здо-
ровье человека. Южно-Сахалинск: Ин.
изд-во, 1962. С. 178—182.
Клодницкий Н. И,, Кац Л. Я. Роль крыс
в эпидемиологии китечных инфекций.
Сообщ. 1// Журн. микробиологии, эпи-
демиологии и иммунобиологии. 1936.
Т. 17, вып. 4. С. 24.
Ковалевский К. Л. Лабораторное живот-
новодство. 2-е изд., перераб. и доп. М.:
Медгиз, 1958. 324 с.
Ковалевский Ю. В., Коренберг 9. И. Фор-
мирование фауны синантронных гры-
зунов в зоне Байкало-Амурской желез-
нодорожной магистрали // Бюл. МОИП.
Отд. биол. 1983. Т. 88, No4. С. 89-77.
Коган Н. И. Мусор
- проблема физико-
химическая // Наука и жизнь. 1978. No 7.
С. 33—38.
Козакевим В. П., Домарадский И. В., Бах-
рал Е. Э. Яды, применяемые для борьбы
с хранителями и. переносчиками: 0с0бо
опасных инфекций. М.: Медгиз, 1958.
156 с.
у
Козлов А. Н. Подвижность и сезонные
миграции серой крысы в сельской мест-
ности Северного Казахстана// Вестн.
зоологии. 1978. No 1. С. 53—57.
Козлов А. Н. Заселение Северного На-
захстана серой крысой в результате
освоения целинных земель // Экология.
1979. No 6. С. 89—91.
Козлов А. Н. Особенности изменения чис-
ленности серой крысы в животновод-
ческих помещениях Северного Казах-
стана // Грызуны: Материалы У Все-
союз. совещ. М.: Наука, 1980а, С. 208—
Козлов А. П. Биология серой крысы в
районах освоения целинных земель Се-
верного Казахстана: Автореф. дис. ...
кан, биол. наук. Свердловек, 19806.
с.
Козлов А. Н. Размножение серой крысы
(В. потуес1сив) в животноводческих по-
мещениях Северного Казахстана // Зоол.
журн. 1981. Т. 60, No 4. С. 587—594.
Козлов А. Н. Зависимость параметров
размножения от плотности популяции
на нримере крысы // Млекопитающие
СССР: ПТ съезд Всесоюз. териол. о-ва:
Тез. докл. М., 1982. Т. 1. С. 249.
Козлов А. Н. Современное распростране-
ние серой крывы на территории Черно-
земного центра // Экология и медицин-
406
Литература
ское значение серой крысы (ВаМлз пот-
уез!сиз ВсгЕК). М.; Наука, 1983а. С. 854—
35
Козлов А. Н. Пространственная и иерар-
хическая структура поселений серой
крысы в животноводческих помещени-
ях // Прикладная этология. М.: Наука,
19836. С. 122-125.
Козлов А. Н. Распространение и места
обитания серых крыс (Вайаз погуез1-
си ВегК) в сравнении е черными кры-
сами (КаЙиз таНиз Г.) в Центральпо-
Черноземном районе РСФСР // Серая
крыса. М.: Наука, 1986а. Т. 1. С. 58—82.
Козлов А. Н. Вев и размеры эмбрионов
у серой крысы //4-ый съезд Всесоюз.
териол. о-ва (тез. докл. раб. заседаний,
27—31 января 1986 г., Москва). М.: АП
СССР, 19866. Т. 3. С. 254—255.
Козлов А. Н. Иптенсивность размножения
серой крысы в поселениях различной
плотности // Журн. общ. биологии. 1987.
Т. 48, No 1. С. 115-123.
Козлов Н. И. Вопросы экологии серой
крысы в животноводческих помещени-
ях Северного Казахстана// РТ Всесоюз.
совещ. по млекопитающим. М.: Изд-во
МГУ, 1961. С. 37-88.
Козлов Н. П., Козлов А. Н. Распроетра-
нение и обитание серых крые в Север-
ном Казахстане // Вестн. с.-х. науки Ка-
захстана. 1970. No 6. С. 103—105.
Козлов Н. П., Козлов А. Н. К вопросу
роющей деятельности ссрой крысы в
Северном Казахстане // Тр. [ науч. конф.
Целиногр. вет. станции. Целиноград,
1971. С. 132—135.
Козлов Н. Н., Козлов А. НП. Суточная на-
земная активность серых крыс в жи-
вотноводческих помещениях Северного
Казахстана // Вестн. зоологии. 1977. No 3.
С. 87-88.
Козорез В. Ф. Влияние ультразвука на
крыс // Использование новых лекарет-
венных средств в лечении и профилак-
тике заболеваний и отравлений живот-
ных на промышленных комплексах и
в хозяйствах
— важнейший фактор по-
вышения продуктивности животновод-
ства: Тез. докл. Челябинск, 1984. С. 59—
60
Коковин И. П., Свешиикова П. П., Ме-
жидова М. М. и др. Роль сельскохозяй-
ственных и диких животных в заболе-
ваниях лептоспирозом людей в Чечено-
Ингушекой АССР // Лептоспирозы: Тр.
ПГ Всесоюз. конф. по лептоспирозам че-
ловека и животных. М.: Минздрав
СССР, 1962. С. 227—230,
Коковин И. Л., Чернуха Ю. Г., Карасева
Е. В. и пр. Особенноети зараженности
лептоспирозом серых крыс на Север-
ном Кавказе //Т Всевоюз. съезд пара-
зитологов: Тез. докл. Киев: Наук. дум-
ка, 1978. С. 64—66.
Колесников И. И. О появлении серой кры-
сы в Ташкенте // Тр, Среднеаз. ун-та,
Биол. науки. 1952. Вып. 32, кн. 2, С. 18—
25.
Коли Г. Анализ популяций позвоночных.
М.: Мир, 1979. 362 с.
Колосов А. М. Распространение, биоло-
гия и эпидемиологическое значение
крыс на Дальнем Востоке. Владивосток,
1945. 62 с. (Тр. Дальневост. базы им.
акад. В. Л. Комарова. Сер. зоол.
Вып. !).
Колосов А. М. Фауна Дальнего Востока
и ее охрана в зоне БАМ. М.: Россель-
хозиздат, 1978. 222 с.
Кондратенко Е. П. Опыт проведения де-
ратизации с оборудованием долгодей-
слвующих точек отравления грызунов
в г. Кемерово за период 1969—1974 гг. //
Проблемы дезинфекции и стерилиза-
ции: (Материалы симпоз., 23—25 нояб.
1977 г., Москва). М. Минздрав СССР,
1977. Ч. 2. С. 81—82.
Кондратенко В. Ф., Мзовкин 9. А., Бунин
И. К. и др. Эпизоотия иктерогеморра-
гического лептоспироза среди серых
крыс на рыборазводческих прудах Рос-
товской области //1У съезд Всесоюз.
териол. о-ва: Тез. докл. М., 1986. Т. 8.
С. 326—828.
Кондрашкин Г. А. О серых крысах (КаНиз
погуес1сиз ВегЕ) дельты Волги
// Бюл.
МОИП. Отд. биол. 1949. Т. 54, No1.
С. 29-33.
Кондрашкин Г. А, О серых крысах дель-
ты Волги // Грызуны и борьба с ними.
Саратов, 1950. Вын, 3. С. 189—200.
Константинов А. И., Мовчан В. Н. Звуки
в жизни зворей. Л.: Изд-во ЛГУ, 1985,
304 с.
Кормилицина В. В. Изменения численно-
сти и видового состава грызунов Кав-
казского заповедника за период 1935—
1977 гг. // Грызуны: Материалы У Все-
союз. совещ. М.: Наука, 1980. С. 212—
213.
Корытин С. А. Влияние различных запа-
хов на поведение зверей // Тр. ВНИИОЗ.
1976. Вып. 286. С. 174-364.
Костенко В. А. Закономерности биотопи-
ческого размещения и распределения
грызунов на Дальнем Востоке СССР //
Наземные млекопитающие Дальнего
Литература
407
Востока СССР. Владивосток, 1976. С. 3—
62.
Красовский Д. В. Материалы к познапию
млекопитающих Горпой Ипгушетии.
Владикавказ: Ингуш. НИИ Краеведе-
ния, 1929. С. 66—92.
Крымов А. А. Игра и развитие мотивации
поведения у крыс // Вестн. МГУ. Сер.
14, Психология. 1981. No 4. С. 39—47.
Крымов А. А. Развитие психической дея-
тельпости млеконитающих в игровом
периоде оптогенеза: Автореф. дис.
канд. психол. наук. М., 1983. 21 с.
Ксенц А. С., Лялин В. Г. Серая крыса в
естественных биотопах таежной зоны
Западной Сибири// ГУ съезд Всесоюз.
териол. о-ва: Тез. докл. М., 1986. Т. 3.
С. 236—237.
Кузиков И. В. Ареал серой крысы в
СССР в прошлом и настоящем // Эко-
логия и медицинское значение серой
крысы (ВаНиз погуейсиз ВегК). М.
Наука, 1983. С. 22—23.
|
Кузьмичев В. Я., Федорова Е. И., Наси-
булина А. С. и др. К вопросу о возмож-
ности развития профессионального ан-
килостомоза и других гельминтозов на
шахтах Карагандинского угольного бас-
ссйна // Изв. АН КазССР. Медицина и
физиология. 1958. Вын. 1 (а}. С. 67—
70.
Кузякин А. ПН. К методике учета и вы-
лова серых крыс в городских объек-
тах // Грызуны и борьба с ними. Сара-
тов: «Микроб», 1950. Выш. 3. ©. 127-
138.
Кузявин А. П. История расселения, сов-
“. ременное распространение и места оби-
тания пасюка в СССР // Фауна и эколо-
гия грызунов. Н. С. Отд. зоол. 1951.
Выц. 4. С. 23—81.
Кузякин А. П. О размножении пасюков
в городах // Бюл. МОИП. Отд. биол.
4952. Т. 57, No 3. С. 15-26.
Кузякин А. П. Истребление крыс и мы-
тей в жилищах и надворных построй-
ках. М.: Медгиз, 1955. 46 с.
Кузякин А. Н. Биологические основы де-
ратизации // Учен. зац. Моск. обл. пед.
ин-та им. Н. К. Крупской. 1963. Т. 126,
вып. 6. С. 89—53.
Кузякин А. П., Второв П.П., Дроздов Н.Н.
Отряд Грызуны (Водепйа) // Жизнь жи-
вотных. М.: Знание, 1971. Т. 6: Млеко-
питающие или звери. С. 152-251.
Кулагин С. М., Земская А. А. Гамазовый
клещ
(АПодегиапу5з — запойтеи$
Низ) как переносчик везикулезного
риккетсиоза // Вопр.
1953. No 8. С. 15—16.
Кулагин С. М., Имамалиев С. А. Эидеми-
ческий крысиный риккетсиоз // Журн.
микробиологии, эпидемиологии и им-
мунол. 1952. No 12. С. 10—45.
Кулагин С. М., Тарасевич И. В. Лихорад-
ка цуцугамуши. М.: Медицина, 1972.
80е
паразитологии.
Кулов А. Б. Распространение лептоспи-
розов на территории Грузинской ССР //
Материалы 1 науч. конф. молодых спе-
циалистов противочум. учреждений
Кавказа. Ставрополь, 1987. С. 115—124.
Кунцевич М. В. Различение лаборатор-
ными крысами запаха мочи стресси-
рованных и нестрессированных сороди-
чей // Химическая коммуникация жи-
вотных! 'Геория и практика. М.: Наука,
1986. С. 315-320.
Купресова О. К., Певцов Н. П. О воспри-
имчивости серой крысы к тудяремии
и ее возможной роли как источника
распространения инфекции // Журп.
микробиологии, эпидемиологии и им-
мунобиологии. 1951. No 11. С. 15—20.
Курятников Н. Н. Серая крыса в горах
Центрального Кавказа // Серая крыса.
М.; Наука, 1986. Т: 1. С. 92-102.
Кучерова П. Т. О бруцеллезе серых крыс //
Журн. микробиологии, эпидемиологии
и иммунобиологии. 1957. No 9. С. 25-
Кучерова Н. Т. К вопросу о зараженности
бруцеллезом диких серых крыс в оча-
гах бруцеллеза сельскохозяйственных
животных // Зоонозные инфекции. Киев:
Наук. думка, 1959. С. 85—88.
Кучерук В. В. Грызуны
— обитатели жи-
лин человека в Восточной Монголии //
Зоол. журн. 1946. Т. 25, No2. (С. 175-
183,
Кучерук В. В. Борьба с грызунами
— по-
сителями болезней. М.: Медицина, 4964.
7с.
Кучерук В. В. Вопросы палеогепезиса
природных очагов чумы в связи с ис-
торией фауны грызунов // Фауна и эко-
логия грызунов. М.: Изд-во МТУ, 1965.
Вып. 7. С. 5-86.
,
Кучерук В. В. Норы млекопитающих—
их строение, использование и типоло-
гия // Фауна и экология грызунов. М.:
Изд-во МГУ, 1983. Вып. 45. С. 5—54.
Кучерук В. В., Коренберг 5. И. Количест-
венный учет важнейших теплокровных
носителей болезней // Методы изучения
природных очагов болезней человека.
М.: Медицина, 1964. С. 129—154.
406
Литература
ское значение серой крысы (ВаМлз пот-
уез!сиз ВсгЕК). М.; Наука, 1983а. С. 854—
35
Козлов А. Н. Пространственная и иерар-
хическая структура поселений серой
крысы в животноводческих помещени-
ях // Прикладная этология. М.: Наука,
19836. С. 122-125.
Козлов А. Н. Распространение и места
обитания серых крыс (Вайаз погуез1-
си ВегК) в сравнении е черными кры-
сами (КаЙиз таНиз Г.) в Центральпо-
Черноземном районе РСФСР // Серая
крыса. М.: Наука, 1986а. Т. 1. С. 58—82.
Козлов А. Н. Вев и размеры эмбрионов
у серой крысы //4-ый съезд Всесоюз.
териол. о-ва (тез. докл. раб. заседаний,
27—31 января 1986 г., Москва). М.: АП
СССР, 19866. Т. 3. С. 254—255.
Козлов А. Н. Иптенсивность размножения
серой крысы в поселениях различной
плотности // Журн. общ. биологии. 1987.
Т. 48, No 1. С. 115-123.
Козлов Н. И. Вопросы экологии серой
крысы в животноводческих помещени-
ях Северного Казахстана// РТ Всесоюз.
совещ. по млекопитающим. М.: Изд-во
МГУ, 1961. С. 37-88.
Козлов Н. П., Козлов А. Н. Распроетра-
нение и обитание серых крые в Север-
ном Казахстане // Вестн. с.-х. науки Ка-
захстана. 1970. No 6. С. 103—105.
Козлов Н. П., Козлов А. Н. К вопросу
роющей деятельности ссрой крысы в
Северном Казахстане // Тр. [ науч. конф.
Целиногр. вет. станции. Целиноград,
1971. С. 132—135.
Козлов Н. Н., Козлов А. НП. Суточная на-
земная активность серых крыс в жи-
вотноводческих помещениях Северного
Казахстана // Вестн. зоологии. 1977. No 3.
С. 87-88.
Козорез В. Ф. Влияние ультразвука на
крыс // Использование новых лекарет-
венных средств в лечении и профилак-
тике заболеваний и отравлений живот-
ных на промышленных комплексах и
в хозяйствах
— важнейший фактор по-
вышения продуктивности животновод-
ства: Тез. докл. Челябинск, 1984. С. 59—
60
Коковин И. П., Свешиикова П. П., Ме-
жидова М. М. и др. Роль сельскохозяй-
ственных и диких животных в заболе-
ваниях лептоспирозом людей в Чечено-
Ингушекой АССР // Лептоспирозы: Тр.
ПГ Всесоюз. конф. по лептоспирозам че-
ловека и животных. М.: Минздрав
СССР, 1962. С. 227—230,
Коковин И. Л., Чернуха Ю. Г., Карасева
Е. В. и пр. Особенноети зараженности
лептоспирозом серых крыс на Север-
ном Кавказе //Т Всевоюз. съезд пара-
зитологов: Тез. докл. Киев: Наук. дум-
ка, 1978. С. 64—66.
Колесников И. И. О появлении серой кры-
сы в Ташкенте // Тр, Среднеаз. ун-та,
Биол. науки. 1952. Вып. 32, кн. 2, С. 18—
25.
Коли Г. Анализ популяций позвоночных.
М.: Мир, 1979. 362 с.
Колосов А. М. Распространение, биоло-
гия и эпидемиологическое значение
крыс на Дальнем Востоке. Владивосток,
1945. 62 с. (Тр. Дальневост. базы им.
акад. В. Л. Комарова. Сер. зоол.
Вып. !).
Колосов А. М. Фауна Дальнего Востока
и ее охрана в зоне БАМ. М.: Россель-
хозиздат, 1978. 222 с.
Кондратенко Е. П. Опыт проведения де-
ратизации с оборудованием долгодей-
слвующих точек отравления грызунов
в г. Кемерово за период 1969—1974 гг. //
Проблемы дезинфекции и стерилиза-
ции: (Материалы симпоз., 23—25 нояб.
1977 г., Москва). М. Минздрав СССР,
1977. Ч. 2. С. 81—82.
Кондратенко В. Ф., Мзовкин 9. А., Бунин
И. К. и др. Эпизоотия иктерогеморра-
гического лептоспироза среди серых
крыс на рыборазводческих прудах Рос-
товской области //1У съезд Всесоюз.
териол. о-ва: Тез. докл. М., 1986. Т. 8.
С. 326—828.
Кондрашкин Г. А. О серых крысах (КаНиз
погуес1сиз ВегЕ) дельты Волги
// Бюл.
МОИП. Отд. биол. 1949. Т. 54, No1.
С. 29-33.
Кондрашкин Г. А, О серых крысах дель-
ты Волги // Грызуны и борьба с ними.
Саратов, 1950. Вын, 3. С. 189—200.
Константинов А. И., Мовчан В. Н. Звуки
в жизни зворей. Л.: Изд-во ЛГУ, 1985,
304 с.
Кормилицина В. В. Изменения численно-
сти и видового состава грызунов Кав-
казского заповедника за период 1935—
1977 гг. // Грызуны: Материалы У Все-
союз. совещ. М.: Наука, 1980. С. 212—
213.
Корытин С. А. Влияние различных запа-
хов на поведение зверей // Тр. ВНИИОЗ.
1976. Вып. 286. С. 174-364.
Костенко В. А. Закономерности биотопи-
ческого размещения и распределения
грызунов на Дальнем Востоке СССР //
Наземные млекопитающие Дальнего
Литература
407
Востока СССР. Владивосток, 1976. С. 3—
62.
Красовский Д. В. Материалы к познапию
млекопитающих Горпой Ипгушетии.
Владикавказ: Ингуш. НИИ Краеведе-
ния, 1929. С. 66—92.
Крымов А. А. Игра и развитие мотивации
поведения у крыс // Вестн. МГУ. Сер.
14, Психология. 1981. No 4. С. 39—47.
Крымов А. А. Развитие психической дея-
тельпости млеконитающих в игровом
периоде оптогенеза: Автореф. дис.
канд. психол. наук. М., 1983. 21 с.
Ксенц А. С., Лялин В. Г. Серая крыса в
естественных биотопах таежной зоны
Западной Сибири// ГУ съезд Всесоюз.
териол. о-ва: Тез. докл. М., 1986. Т. 3.
С. 236—237.
Кузиков И. В. Ареал серой крысы в
СССР в прошлом и настоящем // Эко-
логия и медицинское значение серой
крысы (ВаНиз погуейсиз ВегК). М.
Наука, 1983. С. 22—23.
|
Кузьмичев В. Я., Федорова Е. И., Наси-
булина А. С. и др. К вопросу о возмож-
ности развития профессионального ан-
килостомоза и других гельминтозов на
шахтах Карагандинского угольного бас-
ссйна // Изв. АН КазССР. Медицина и
физиология. 1958. Вын. 1 (а}. С. 67—
70.
Кузякин А. ПН. К методике учета и вы-
лова серых крыс в городских объек-
тах // Грызуны и борьба с ними. Сара-
тов: «Микроб», 1950. Выш. 3. ©. 127-
138.
Кузявин А. П. История расселения, сов-
“. ременное распространение и места оби-
тания пасюка в СССР // Фауна и эколо-
гия грызунов. Н. С. Отд. зоол. 1951.
Выц. 4. С. 23—81.
Кузякин А. П. О размножении пасюков
в городах // Бюл. МОИП. Отд. биол.
4952. Т. 57, No 3. С. 15-26.
Кузякин А. П. Истребление крыс и мы-
тей в жилищах и надворных построй-
ках. М.: Медгиз, 1955. 46 с.
Кузякин А. Н. Биологические основы де-
ратизации // Учен. зац. Моск. обл. пед.
ин-та им. Н. К. Крупской. 1963. Т. 126,
вып. 6. С. 89—53.
Кузякин А. П., Второв П.П., Дроздов Н.Н.
Отряд Грызуны (Водепйа) // Жизнь жи-
вотных. М.: Знание, 1971. Т. 6: Млеко-
питающие или звери. С. 152-251.
Кулагин С. М., Земская А. А. Гамазовый
клещ
(АПодегиапу5з — запойтеи$
Низ) как переносчик везикулезного
риккетсиоза // Вопр.
1953. No 8. С. 15—16.
Кулагин С. М., Имамалиев С. А. Эидеми-
ческий крысиный риккетсиоз // Журн.
микробиологии, эпидемиологии и им-
мунол. 1952. No 12. С. 10—45.
Кулагин С. М., Тарасевич И. В. Лихорад-
ка цуцугамуши. М.: Медицина, 1972.
80е
паразитологии.
Кулов А. Б. Распространение лептоспи-
розов на территории Грузинской ССР //
Материалы 1 науч. конф. молодых спе-
циалистов противочум. учреждений
Кавказа. Ставрополь, 1987. С. 115—124.
Кунцевич М. В. Различение лаборатор-
ными крысами запаха мочи стресси-
рованных и нестрессированных сороди-
чей // Химическая коммуникация жи-
вотных! 'Геория и практика. М.: Наука,
1986. С. 315-320.
Купресова О. К., Певцов Н. П. О воспри-
имчивости серой крысы к тудяремии
и ее возможной роли как источника
распространения инфекции // Журп.
микробиологии, эпидемиологии и им-
мунобиологии. 1951. No 11. С. 15—20.
Курятников Н. Н. Серая крыса в горах
Центрального Кавказа // Серая крыса.
М.; Наука, 1986. Т: 1. С. 92-102.
Кучерова П. Т. О бруцеллезе серых крыс //
Журн. микробиологии, эпидемиологии
и иммунобиологии. 1957. No 9. С. 25-
Кучерова Н. Т. К вопросу о зараженности
бруцеллезом диких серых крыс в оча-
гах бруцеллеза сельскохозяйственных
животных // Зоонозные инфекции. Киев:
Наук. думка, 1959. С. 85—88.
Кучерук В. В. Грызуны
— обитатели жи-
лин человека в Восточной Монголии //
Зоол. журн. 1946. Т. 25, No2. (С. 175-
183,
Кучерук В. В. Борьба с грызунами
— по-
сителями болезней. М.: Медицина, 4964.
7с.
Кучерук В. В. Вопросы палеогепезиса
природных очагов чумы в связи с ис-
торией фауны грызунов // Фауна и эко-
логия грызунов. М.: Изд-во МТУ, 1965.
Вып. 7. С. 5-86.
,
Кучерук В. В. Норы млекопитающих—
их строение, использование и типоло-
гия // Фауна и экология грызунов. М.:
Изд-во МГУ, 1983. Вып. 45. С. 5—54.
Кучерук В. В., Коренберг 5. И. Количест-
венный учет важнейших теплокровных
носителей болезней // Методы изучения
природных очагов болезней человека.
М.: Медицина, 1964. С. 129—154.
408
Литература
Кучерук В. В., Кузиков И. В. Современ-
ный ареал серой крысы // Распростра-
нение и экология серой крысы и мето-
ды ограпичения ее численности, М.:
Наука, 1985. С. 17—52.
Кучерук В. В., Лапшов В. А. Серая кры-
са (Вайиз потуессиз ВегК) и другие
синантропные грызуны океапических
островов // Материалы по экологии и
методам ограничения численности се-
рой крысы. М.: Наука, 1987. С. 5-31.
Кучерук В. В., Никитина Н. А. Основные
задачи и итоги мечения млекопитаю-
щих в СССР//Вопросы териологии:
Итоги мечения млекопитающих.
Наука, 1980. С. 3—9.
Лаврова М. Я. О роли грызунов в лепто-
спирозных очагах низовий Кубани //
Зоол. журн. 1962. Т. 41, No 7. ©. 1067-—
1074.
Лаврова М. Я., Минина Т. С. К биологии
и эпизоотологическому значению серых
крыс в лептоспирозных очагах низовий
Кубани // Материалы докл. науч. конф.
по лентоспирозам зон Северного Кав-
каза и Поволжья, 17—18 дек. 1968. Крас-
нодар, 1968. С. 147—155.
Лавровский А. А., Козакевич В. П., Вар-
шавский С. Н. Природные очаги чумы
в Передней и Юто-Западной частях
Азии // Проблемы особо опаспых инфек-
ций. Саратов: «Микроб», 1973. Вып. 2.
С. 9-22.
.
Лавровский А. А., Колесников И. М. Ма-
териалы к познанию грызунов Дагес-
танской АССР //Тр. Противочум. ин-та
Кавказа и Закавказья. 1956. Вы. 1.
С. 253—277.
й
Лаптев И. П. Млекопитающие таежной
зоны Западной Сибири. Томск: Изд-во
Том. ун-та, 1958. 258 с.
Леви М. И., Судейкин В. А. О методике
учета численности серых крыс (ВаМлз
погуе21с1$) в жилых и производствеп-
ных помещениях города Москвы // Зоол.
журн. 1977. Т. 61. Выш. 4. С. 1067—1070.
Левин В. Г. Динамика некоторых морфо-
физиологических изменений в организ-
ме обыкновенных полевок, затравлен-
ных фентолацином // Зап. Ленингр. с.-х.
ин-та. 1974. Т. 156. С. 125—130.
Леонтьев А. К. О распространении серой
крысы в Узбекистане // Тр. ВНИИ вет.
санитарии. 1968. Т. 30. С. 420—424.
Ливявичене П. М. Мытевидные грызуны
Литовской ССР: Автореф. дис. ... канд.
биол. наук. Вильнюс, 1960. 21 в.
Липаев В. М. Новые данные о распроет-
ранении трызунов северо-восточной
Монголии// Изв. Иркут. науч.-иселед.
противочум. ин-та Сибири и Дал, Вос-
тока. 1963. Т. 25. С. 273—280.
Липаезв В. М., Бусоедова Н. М. Серая
крыса— основной носитель зоонозов в
антропургических очагах Нижнего При-
амурья // Экология и медицинское зна-
чение серой крысы (КаИл$ погуес!ел$
ВетКк). (Мат. 1-го рабоч. совещ. но се-
рой крысе, 31.1-8.11.1983 г., Москва).
М.: АН СССР, 1983. (. 61—62.
Лобанова Т. И., Петросян 9. А. Грызу-
ны Азербайджана и их распростране-
ние // Тр. юбил. науч. конф. Азерб. про-
тивочум. станции. Баку, 1959. Т. 2.
С. 94-105.
Лозан М. Н. История становления фзу-
ны и экологии рецентных видов // Гры-
зуны Молдавии. Кишипев: Штиинца,
1971. Т. 2. С. 83—13.
Лозан М. Н., Кучук А. П. Популяции ди-
ких и сивантропных серых крыс в Мол-
давии и некоторые вопросы микроэво-
люции // Вопросы экологии и практи-
ческого значения птиц и млекопитаю-
щих Молдавии. Киптинев: Штиинца,
1969. No 4. С. 85—98.
Лозан М. Н., Нивул В. С. Социальная
структура популяции синантроцных се-
рых крыс // Экология птиц и млекопи-
тающих Молдавии. Кишинев: Штиин-
ца, 1984. С. 44—58.
Лозан М. Н., Никул В. С. Характер ис-
пользования участка обитания синан-
тропными крысами // Прякладная этоло-
гия. М.: Наука, 1983. С. 131—133.
Лозан М. Н., Никул В. С., Орлов А. С.
Изучение суточного ритма общей ак-
тивности и основных форм поведения
во внутрипопуляционной группировке
синантропных серых крыс // Фауна, эко-
логия и физиология животных. Киши-
нев: Штиинца, 1980. С. 23—29.
Лыскова В. С., Петрова Н. Н., Цивиле-
ва И. Я. и др. Эффективность дерати-
зации при применении долгодействую-
щих мучных приманок // Материалы
6-й науч.-практ. конф. сан.-эпид. служ-
бы Свердл. обл. (20 дек. 1972 т.). Сверд-
ловск, 1972. С. 194—197.
Лысов Е. С. Опыт освобождения от гры-
зунов гостиницы // Проблемы дезинфек-
ционного обслуживания крупных тгра-
достроительных объектов: (Тез. Все-
союз. конф., 28—29 февр. 1984 г.). М.:
Минздрав СССР, 1984. С. 80—81.
Львов Г. Н. Колхидекий пасюк// Учен.
зап. Горьк. ун-та. 1949. Вып. 14. С. 39—
51.
Литература
409
Лютов Ю. Г., Ващенок В. С., Шибалов
В. А. и др. Патогенная микрофлора
грызунов и насекомоядных Ленинграда
и его пригородов. Эпидемиология и про-
филактика природно-очаговых инфек-
ций// Тр. Ин-та Пастера. 1980. Т. 55.
С. 77-82.
Лютов Ю. Г., Шибалов В. А., Богоявлен-
ский Б. В. и др. Серая крыса
— резер-
вуар и носитель возбудителей зооноз-
ных болезней в Ленинграде // Экология
и медицинское значение серой крысы
(Ваз погуер1сиз ВегК). М.: Наука,
1983. С. 68—64.
Ляйстер А. Ф. Млекопитающие Эриван-
ской губернии //К фауне Эриванской
губ. Т. ГУ. Эривань, 1912. С. 21-1415.
Лялин В. Г. К экологии серой крысы в
Западной Сибири // Зоологические проб-
лемы Сибири. Новосибирск: Наука,
1972. С. 245.
Лялин В. Г. Изучение передвижения се-
рых крыс по территории сельских на-
селенных пунктов Томской области //
Вопросы ботаники, зоологии и почво-
ведения. Томск: Изд-во Том. ун-та, 1973.
С. 225—232,
Лялин В. Г. Некоторые особенности по-
ведения серых крыс //Тр. НИИ биоло-
гии и биофизики при Том. ин-те. 4974а.
Т. 4. С. 38—44.
Лялин В. Г. Современное распростране-
ние серой крысы в Западной Сибири //
Там же, 19746. ТГ. 4. С. 45-50.
Лялин В. Г. Распространение серой кры-
сы в Томекой области
// Природное и
сельскохозяйственное районирование
”^. СССР. М.: Изд-во МГУ, 1975, С, 422—
424.
Лялин В. Г. Некоторые особенности пи-
тания серых крыс //Вопр. биологии.
томок: Изд-во Том. ун-та, 1978. С. 3—
Лялин В. Г. Оценка экономического ущер-
ба, наносимого серой крысой в Запад-
ной Сибири // Экология и медицинское
значение серой крысы (Ваз потуес|-
си5 ВегК). М.: Наука, 1983. С. 86—87.
Ляфрунова К. Л. Социальное новедение и
химическая коммуникация серых крыс
(Вабиаз погуешеиз): торе, дис. ...
канд. биол. наук. М.: Ин-т эвол. морфо-
логии и экологии животных АН СССР,
1980. 30 с.
Мажникова Г. И. Эпизоотология ленто-
спироза в Красподарском крае // Лепто-
спирозы: Тез. докл. УП Всесоюз. конф.
10 лептоспирозам. Киев: Минздрав
СССР, 1979. С. 124—122.
Мазин Л. П., Судейкин В. А. Циклы од-
новременного истребления серых крыс
в постройках болыпотго города// Мате-
риалы по экологии и методам ограни-
чения численности серой крысы. М.:
Наука, 1987. С. 335-364.
Максимов А. А. Материалы к фауне мы-
шевидных грызунов Прибайкальской
части Восточно-Сибирекой области //
Труды по защите растений Восточной
Сибири. Иркутск, 1937. С. 126—147.
Малахов Ю. А. О носительстве листерий
} крыс // Ветеринария. 1962. No 2. С. 41-
2
Малахов Ю. А. Крысы— источник листе-
риозной инфекции // Тр. ВНИИ вет. са-
нитарии. 1964. Т. 24. С. 343—353.
Малиновский Л. Н., Заболотный Д. К.//
Чума в Одессе в 1910 г. СИб., 1912.
С. 15—81.
Малолетков И. С. Эпидемиология чумы в
портовых городах и меры по преду-
нреждению ее завоза: Автореф. дис. ...
канд. мед. наук. Одесса, 1963. 22 с.
Мальцев В. П. Возрастные изменения б0-
левых акустических сигналов, издавае-
мых крысятами // Журн. эволюц. био-
химии и физиологии. 1974. Т. 10, No 4.
С. 386—394.
Мамед-Заде У. А., Петросян 9. А., Над-
жафов А. Ф. Сезонные изменения чув-
ствительности к чуме серых крыс в
связи с их некоторыми эколого-физио-
логическими особенностями в условиях
Апшерона // Гр. Азерб. противочум.
станции. 1962. Т. 3. С. 37—46.
Марш Р. Е., Говард В. Е. Перспективы
применения хемостерилянтов и гене-
тических методов для борьбы с грызу-
нами // Бюл. ВОЗ. 1974. Т. 48, No3.
С. 381-389.
Маслов С. П., Шилов И. А., Каменов Д.А.
Роль уровня различий животных по
подвижности нервной системы в воз-
никновении «несовместимых» группиро-
вок мышевидных грызунов // Докл. АН
СССР. 1974. Т. 216, No 9. С. 670-673.
Матева М. Х., Христов Л. Динамика на
численоста на синантропните гризачи
р ‚България // Екология. 1986. Т. 18.
. 8—Ю.
Матусевич В. Ф. Опыт сплошной дерати-
зации на животноводческих фермах
Хмельницкой — области // Ветеринария.
1959. No 4. С. 69—74.
Маяков А. А., Шепель А. И. Определение
вида и пола некоторых млекопитающих
о костям таза, голени и бедра // Зоол.
журн. 1987, Т. 46, No 2. С. 288—294.
408
Литература
Кучерук В. В., Кузиков И. В. Современ-
ный ареал серой крысы // Распростра-
нение и экология серой крысы и мето-
ды ограпичения ее численности, М.:
Наука, 1985. С. 17—52.
Кучерук В. В., Лапшов В. А. Серая кры-
са (Вайиз потуессиз ВегК) и другие
синантропные грызуны океапических
островов // Материалы по экологии и
методам ограничения численности се-
рой крысы. М.: Наука, 1987. С. 5-31.
Кучерук В. В., Никитина Н. А. Основные
задачи и итоги мечения млекопитаю-
щих в СССР//Вопросы териологии:
Итоги мечения млекопитающих.
Наука, 1980. С. 3—9.
Лаврова М. Я. О роли грызунов в лепто-
спирозных очагах низовий Кубани //
Зоол. журн. 1962. Т. 41, No 7. ©. 1067-—
1074.
Лаврова М. Я., Минина Т. С. К биологии
и эпизоотологическому значению серых
крыс в лептоспирозных очагах низовий
Кубани // Материалы докл. науч. конф.
по лентоспирозам зон Северного Кав-
каза и Поволжья, 17—18 дек. 1968. Крас-
нодар, 1968. С. 147—155.
Лавровский А. А., Козакевич В. П., Вар-
шавский С. Н. Природные очаги чумы
в Передней и Юто-Западной частях
Азии // Проблемы особо опаспых инфек-
ций. Саратов: «Микроб», 1973. Вып. 2.
С. 9-22.
.
Лавровский А. А., Колесников И. М. Ма-
териалы к познанию грызунов Дагес-
танской АССР //Тр. Противочум. ин-та
Кавказа и Закавказья. 1956. Вы. 1.
С. 253—277.
й
Лаптев И. П. Млекопитающие таежной
зоны Западной Сибири. Томск: Изд-во
Том. ун-та, 1958. 258 с.
Леви М. И., Судейкин В. А. О методике
учета численности серых крыс (ВаМлз
погуе21с1$) в жилых и производствеп-
ных помещениях города Москвы // Зоол.
журн. 1977. Т. 61. Выш. 4. С. 1067—1070.
Левин В. Г. Динамика некоторых морфо-
физиологических изменений в организ-
ме обыкновенных полевок, затравлен-
ных фентолацином // Зап. Ленингр. с.-х.
ин-та. 1974. Т. 156. С. 125—130.
Леонтьев А. К. О распространении серой
крысы в Узбекистане // Тр. ВНИИ вет.
санитарии. 1968. Т. 30. С. 420—424.
Ливявичене П. М. Мытевидные грызуны
Литовской ССР: Автореф. дис. ... канд.
биол. наук. Вильнюс, 1960. 21 в.
Липаев В. М. Новые данные о распроет-
ранении трызунов северо-восточной
Монголии// Изв. Иркут. науч.-иселед.
противочум. ин-та Сибири и Дал, Вос-
тока. 1963. Т. 25. С. 273—280.
Липаезв В. М., Бусоедова Н. М. Серая
крыса— основной носитель зоонозов в
антропургических очагах Нижнего При-
амурья // Экология и медицинское зна-
чение серой крысы (КаИл$ погуес!ел$
ВетКк). (Мат. 1-го рабоч. совещ. но се-
рой крысе, 31.1-8.11.1983 г., Москва).
М.: АН СССР, 1983. (. 61—62.
Лобанова Т. И., Петросян 9. А. Грызу-
ны Азербайджана и их распростране-
ние // Тр. юбил. науч. конф. Азерб. про-
тивочум. станции. Баку, 1959. Т. 2.
С. 94-105.
Лозан М. Н. История становления фзу-
ны и экологии рецентных видов // Гры-
зуны Молдавии. Кишипев: Штиинца,
1971. Т. 2. С. 83—13.
Лозан М. Н., Кучук А. П. Популяции ди-
ких и сивантропных серых крыс в Мол-
давии и некоторые вопросы микроэво-
люции // Вопросы экологии и практи-
ческого значения птиц и млекопитаю-
щих Молдавии. Киптинев: Штиинца,
1969. No 4. С. 85—98.
Лозан М. Н., Нивул В. С. Социальная
структура популяции синантроцных се-
рых крыс // Экология птиц и млекопи-
тающих Молдавии. Кишинев: Штиин-
ца, 1984. С. 44—58.
Лозан М. Н., Никул В. С. Характер ис-
пользования участка обитания синан-
тропными крысами // Прякладная этоло-
гия. М.: Наука, 1983. С. 131—133.
Лозан М. Н., Никул В. С., Орлов А. С.
Изучение суточного ритма общей ак-
тивности и основных форм поведения
во внутрипопуляционной группировке
синантропных серых крыс // Фауна, эко-
логия и физиология животных. Киши-
нев: Штиинца, 1980. С. 23—29.
Лыскова В. С., Петрова Н. Н., Цивиле-
ва И. Я. и др. Эффективность дерати-
зации при применении долгодействую-
щих мучных приманок // Материалы
6-й науч.-практ. конф. сан.-эпид. служ-
бы Свердл. обл. (20 дек. 1972 т.). Сверд-
ловск, 1972. С. 194—197.
Лысов Е. С. Опыт освобождения от гры-
зунов гостиницы // Проблемы дезинфек-
ционного обслуживания крупных тгра-
достроительных объектов: (Тез. Все-
союз. конф., 28—29 февр. 1984 г.). М.:
Минздрав СССР, 1984. С. 80—81.
Львов Г. Н. Колхидекий пасюк// Учен.
зап. Горьк. ун-та. 1949. Вып. 14. С. 39—
51.
Литература
409
Лютов Ю. Г., Ващенок В. С., Шибалов
В. А. и др. Патогенная микрофлора
грызунов и насекомоядных Ленинграда
и его пригородов. Эпидемиология и про-
филактика природно-очаговых инфек-
ций// Тр. Ин-та Пастера. 1980. Т. 55.
С. 77-82.
Лютов Ю. Г., Шибалов В. А., Богоявлен-
ский Б. В. и др. Серая крыса
— резер-
вуар и носитель возбудителей зооноз-
ных болезней в Ленинграде // Экология
и медицинское значение серой крысы
(Ваз погуер1сиз ВегК). М.: Наука,
1983. С. 68—64.
Ляйстер А. Ф. Млекопитающие Эриван-
ской губернии //К фауне Эриванской
губ. Т. ГУ. Эривань, 1912. С. 21-1415.
Лялин В. Г. К экологии серой крысы в
Западной Сибири // Зоологические проб-
лемы Сибири. Новосибирск: Наука,
1972. С. 245.
Лялин В. Г. Изучение передвижения се-
рых крыс по территории сельских на-
селенных пунктов Томской области //
Вопросы ботаники, зоологии и почво-
ведения. Томск: Изд-во Том. ун-та, 1973.
С. 225—232,
Лялин В. Г. Некоторые особенности по-
ведения серых крыс //Тр. НИИ биоло-
гии и биофизики при Том. ин-те. 4974а.
Т. 4. С. 38—44.
Лялин В. Г. Современное распростране-
ние серой крысы в Западной Сибири //
Там же, 19746. ТГ. 4. С. 45-50.
Лялин В. Г. Распространение серой кры-
сы в Томекой области
// Природное и
сельскохозяйственное районирование
”^. СССР. М.: Изд-во МГУ, 1975, С, 422—
424.
Лялин В. Г. Некоторые особенности пи-
тания серых крыс //Вопр. биологии.
томок: Изд-во Том. ун-та, 1978. С. 3—
Лялин В. Г. Оценка экономического ущер-
ба, наносимого серой крысой в Запад-
ной Сибири // Экология и медицинское
значение серой крысы (Ваз потуес|-
си5 ВегК). М.: Наука, 1983. С. 86—87.
Ляфрунова К. Л. Социальное новедение и
химическая коммуникация серых крыс
(Вабиаз погуешеиз): торе, дис. ...
канд. биол. наук. М.: Ин-т эвол. морфо-
логии и экологии животных АН СССР,
1980. 30 с.
Мажникова Г. И. Эпизоотология ленто-
спироза в Красподарском крае // Лепто-
спирозы: Тез. докл. УП Всесоюз. конф.
10 лептоспирозам. Киев: Минздрав
СССР, 1979. С. 124—122.
Мазин Л. П., Судейкин В. А. Циклы од-
новременного истребления серых крыс
в постройках болыпотго города// Мате-
риалы по экологии и методам ограни-
чения численности серой крысы. М.:
Наука, 1987. С. 335-364.
Максимов А. А. Материалы к фауне мы-
шевидных грызунов Прибайкальской
части Восточно-Сибирекой области //
Труды по защите растений Восточной
Сибири. Иркутск, 1937. С. 126—147.
Малахов Ю. А. О носительстве листерий
} крыс // Ветеринария. 1962. No 2. С. 41-
2
Малахов Ю. А. Крысы— источник листе-
риозной инфекции // Тр. ВНИИ вет. са-
нитарии. 1964. Т. 24. С. 343—353.
Малиновский Л. Н., Заболотный Д. К.//
Чума в Одессе в 1910 г. СИб., 1912.
С. 15—81.
Малолетков И. С. Эпидемиология чумы в
портовых городах и меры по преду-
нреждению ее завоза: Автореф. дис. ...
канд. мед. наук. Одесса, 1963. 22 с.
Мальцев В. П. Возрастные изменения б0-
левых акустических сигналов, издавае-
мых крысятами // Журн. эволюц. био-
химии и физиологии. 1974. Т. 10, No 4.
С. 386—394.
Мамед-Заде У. А., Петросян 9. А., Над-
жафов А. Ф. Сезонные изменения чув-
ствительности к чуме серых крыс в
связи с их некоторыми эколого-физио-
логическими особенностями в условиях
Апшерона // Гр. Азерб. противочум.
станции. 1962. Т. 3. С. 37—46.
Марш Р. Е., Говард В. Е. Перспективы
применения хемостерилянтов и гене-
тических методов для борьбы с грызу-
нами // Бюл. ВОЗ. 1974. Т. 48, No3.
С. 381-389.
Маслов С. П., Шилов И. А., Каменов Д.А.
Роль уровня различий животных по
подвижности нервной системы в воз-
никновении «несовместимых» группиро-
вок мышевидных грызунов // Докл. АН
СССР. 1974. Т. 216, No 9. С. 670-673.
Матева М. Х., Христов Л. Динамика на
численоста на синантропните гризачи
р ‚България // Екология. 1986. Т. 18.
. 8—Ю.
Матусевич В. Ф. Опыт сплошной дерати-
зации на животноводческих фермах
Хмельницкой — области // Ветеринария.
1959. No 4. С. 69—74.
Маяков А. А., Шепель А. И. Определение
вида и пола некоторых млекопитающих
о костям таза, голени и бедра // Зоол.
журн. 1987, Т. 46, No 2. С. 288—294.
410
Литература
Мелкова В. К. Особенности обитания се-
рых крыс в многоэтажных жилых до”
мах // Материалы по экологии и мето-
дам ограничения численности серой
крысы. М.: Наука, 1987. С. 179—207.
Мелкова В. К., Кунашев М. В. О сезон-
ном заселении серыми крысами неко-
торых строений // Экология и медицин-
ское значение серой крысы (КаЙлз пот-
уегсиз ВегЕ). М.: Наука, 1983. С. 99-
100.
Мелкова В. К., Туров И. С. Мозаичный
характер распределения серых крыс в
городе как основа целенаправленной
борьбы с ними // ТУ сЪеЗД Всесозюз,
териол. о-ва: Тез. докл. М., 1986. Т. 3.
С. 377-878.
Медкова В. К., Туров И. С., Одинец А. А.
и др. Характер размещения серых крыс
в подвалах жилых домов и его измене-
ния под влиянием дератизационных
мероприятий // Проблемы дезинфекции
и стерилизации: Сб. науч. тр. М. Мин-
здрав СССР, 1985. С. 168—170.
Мерзликин И. Р. Некоторые аспекты кор-
модобывающей деятельности серых
крыс в природных биотопах // ТУ съезд
Всесоюз. териол. о-ва: Тез. докл. М.,
1986. Т. 3. С. 242—244.
Мерзликин И. Р, Материалы по размно-
жению серой крысы в условиях северо-
восточной Украины // Материалы по
экологии и методам ограничения чис-
ленности серой крысы. М.: АН ссср,
1987а. С. 159—178.
Мераликин И. Р. Некоторые наблюдения
за характером использования участка
обитания ссрой крысой // Материалы по
экологии и методам ограничения чис-
ленности серой крысы. М. Наука,
49876. С. 113—128.
Мерзликин И. Р. Разновидность назем-
ных мост отдыха серой крысы в при-
роде // Там же. 19876. С. 143—158.
Меркова М. А. Некоторые данные по эко_
логии рыжей полевки и желтогорлой
мыши юга Московской обл. Телерма-
новской роши // Бюл. МОИП. Отд. биол.
41955. Т. 60, No 1. 0. 21-31.
Методические указапия по борьбе с се-
рой крысой (Вайлз погуез1сиз) в при-
родных очагах иктерогеморрагического
‘лептоспироза. М.: Минздрав СССР, 1984.
12 с.
Мешкова Н. ПН. Ориентировочно-исследо-
вательская деятельность серой крысы
в «откоытом поле»: (Зоопеихол. ана-
лиз) // Бюл. МОИП. Отд. биол. 1981.
Т. 86, No В. С. 22—29.
Мигулин А. А. Звери Украинской ССР.
Киев: Наук. думка, 1938. 426 с. На укр.
яз.
.
Милютин А. И. Диморфизм окраски в
естественных понуляциях пасюков (Ва
{из потуевсл$) // Науч
с.-х, ин-та. 1981. Т. 405. С. 50-54.
Милютин А. И. Изменчивость окраски
меха в естественных полуляциях серой
крысы // ПТ съезд Всесоюз. териол. о-ва:
Тез. докл. М., 1982. Т. 2. С. 55—56.
Милютин А. И. О времени появления се-
рой крысы в Европе // Экология и ме-
дицинское значение серой крысы (Ван
из потуедсие ВетК). М.: Наука, 19832.
С. 19—20.
Милютин А. И. К вопросу о времени
появления серой крысы в Европе //
Резн шик. ТаШит, 19836. Вын. 2.
С. 51—62.
Милютин А. И. Серая (ВаЙмз погуе21сл1$)
и черная (Вали таМиз) крысы в При-
балтике: распространение и межвидо-
вые отношения
// Серая крыса (эколо-
тия и распространение). ТГ. 1. М.: АН
СССР, 1986.
Милютин Л. И., Езерскене 9. Н. Распро-
странение крыс (род Ваз) в При-
балтике. Сообщ. 1// Еезй и\И9. Тал-
линн, 1984. Выш. 3. С. 39—50.
Милютин А. И., Езерснене Э. П. Распрост-
ранение крысе (род Ваз) в Прибал-
тике. Сообщ. 2 // ЕезН ак. Таллинн,
1986. Вып. 4. С. 35—45.
Мирзоев Ш. М. Организация работ по
дератизации // Ветеринария. 1974. No 10.
С. 28—30.
Мирзоев Ш. М. Защита свиноводческих
комплексов от грызунов // Там же. 1978.
No3.С.16-18. °
|
Мирзоев Ш. М. Зазщита крупных живот-
новодческих хозяйств и комплексов от
синантропных грызунов: Автореф. дис.
... дера биол. наук. М., 1984. 20 с.
Миронов Н. П. К вопросу об эволюции и
природной очаговости бруцеллеза //
Журп. микробиологии, эпидемиологии
и иммунологии. 1964. No 12. С. 60—65.
Миронов Н. П., Карпузиди К. С. Климен-
ко И. 3. и др. Источники и переносчи-
ки чумы и туляремии. М., 1965. 145 с.
Мипонов Н. Н., Рожков В. С., Денисов
П. С., Некрасов П. А. Изменение гра-
ницы ареала серой крысы в восточных
районах Ростовской области и запад-
ных районах Калмыцкой АССР // Науч.
работы Элистинской противочум. ст.
4961. Вып. 2. С. 215-286.
тр. Ленингр.
Литература
414
Митропольский О. В., Лобызова В. П., Се-
дин В. И. Размножение серой крысы в
Узбекистане // ГУ съезд Всесоюз. те-
риол. о-ва: Тез. докл. М.
.т.
С. 250—252. докт. М» 1986. т.
Михайленко А. Г. Пространственное рас-
пределение серых крыс на свинофер-
ме // Млекопитающие СССР: Тез. докл.
3-й Всесоюз. конф. териол. о-ва. М.;: АН
СССР, 1982. Т. 4. С. 250-251.
Михайленко А. Г. Основные черты со-
циального поведения синантропных се-
рых крыс на животноводческой ферме //
дктувльные Вопросы дезинфекции и
терилизации: (Тез. науч.-практ, конф.).
М.: АН СССР, 1987. С. Я
о=ф
Михайлова О. А., Якунина Т. И. Обнару-
жение возбудителя листериоза у тгры-
зунов города Владивостока// Изв. Ир-
кут. паче послед. противочум. ин-та
ибири и Дал. Востока.
:
24
Сибири д
а. 1962. Т. 24.
Моргилевская И. Е., Цителова 3. К., Ми-
и А. Грызуны города Тбилиси //
. Зоол. ин-та
о,
ССР. 1974. Т. 7.
Мороз А. Н, Хижинская 0. П. Поражен-
ность листереллами грызунов г. Киева
и его окрестностей // Зоонозные инфек-
ции. Киев: Наук. думка, 1959. С. 254—
Москва в цифрах: Стат. ежегодник. М.:
Финансы и статистика, 1982. 222 с,
Мотиги К., Нагаи Х., Хасэбэ Я., Осэк
,
.
.
иТ.
Пат,МКИС09Ь5/4,АОМ57/4
Япония. Покрытие для отпугивания
крые. Заявл. 7.06.82; Опубл. 14.12.83.
“Мясников Ю. А. Эризипелоид
// Общ. и
частн. эпидемиол. М.: М
Т. 2. С. 423—486.
‘дищина, 1973.
Мясников Ю. А. Распространение и ко-
лебания численности грызунов, зайце-
образных и насекомоядных Тульской
области // Фауна и экология грызунов.
М.: Паука, 1976. Выш. 13. С. 164—236.
Мясников Ю. А., Белова М. А. О типах
природных и антропургических очагов
офизиполоида // Вопросы природного
с ата болезней, Алма-Ата, 1978. Вып. 9.
Набенов К. Н. Эффективность дератиза-
цибнных мероприятий в синантронных
нопуляциях домовой мыши // Современ-
ные направления медицинской дезин-
секции и дератизации: (Тез. докл.). М.:
Медицина, 1981. С. 105—106.
Наумов Н. П. Определение возраста бел-
ки // Учен. зап.
С. 15-20.
Наумов Н. Н. определение возраста ма-
лого суслика // Защита ре
й
его суслика, щита растений. 1955.
Наумов Н. П. Типы поселений грызу-
нов и их экологическое значение //
Воол. журн. 1954. Т. 33, No2. С. 268—
МГУ. 1934. Вын. 2.
Наумов Н. П. Структура популяций и
ДараЖИКА ПИСлОННОСТ наземных поз-
воночных ам же. 1967. Т. 46, No
С. 1470—1486.
Юм
Море Н. Н. Пространственные струк-
ы вида млекопитающих // Там .
1971. Т. 50, No 7. С. 351—362. !
вы
Наумов Н. П., Гольцман М. Е. Новеден-
ческие и физиологические реакции мле-
Поитющих на запахи сородичей по
виду спехи соврем. биологии. 1972.
Т. 73, No 3. С. 427-444.
9
Наумов Н. И., Емельянова И. А. Грызу-
ны — разрушители материалов, изделий,
сооружений // Проблемы защиты от
иоповреждений. М.: Знание, 1979.
С. 33—47. (Сер. биол.; No 1).
Нацкий К. В., Игнатович В. 0. К вопросу
о значении грызунов в очагах дальне-
восточной скарлатиноподобной лихорад-
ки (псевдотуберкулеза) во Владивосто-
ке // 1 Межобл. науч.-практ. конф. по
псендотуберкулезу ’ (дальневосточной
скарлатиноподобной лихорадке): Тез.
докл. Владивосток, 1970. С. 62—63.
Некипелов ПН. В. Улучшенный способ
отлова крыс в верши// Изв. Иркут.
науч.-исслед. противочум, ин-та Сибири
Н и Дал. Востока. 1957. Т. 16. С. 247—248.
‘пипелов Н. 2 домовые грызуны в Юто-
осточном Забайкалье // Там же.
Т. 19. С. 72-82.
17 Там ко. 1958,
Некипелов Н. В. Эпизоотология чумы в
Монгольской Народной Республике //
Там же. 1959. Т. 12. С. 108—243.
Немченко Л. С. Меры борьбы с серой
крысой при экстренной профилактике //
ГУ съезд Всесоюз. териол. о-ва: Тез.
докл. рабочих засед., Москва, 27—31 ян-
варя 1986 г. М.: АН СССР.
Варя 1986.
‚ 1986. Т. 3.
Немченко Л. С., Бытов Г. Г. Обитание сс-
рои крысы в нриродных биотопах Муй-
ско-Куандинской котловины // Экология
едищинское зпачение серой крысы
аИяз потуеб!сив ВегК). М.:
1983. С. 15—46,
}. М. Наука
Пестеренко В. А. Обоснование мер борь-
бы с грызунами
— вредителями риса в
сельском хозяйстве. Динамика числен-
410
Литература
Мелкова В. К. Особенности обитания се-
рых крыс в многоэтажных жилых до”
мах // Материалы по экологии и мето-
дам ограничения численности серой
крысы. М.: Наука, 1987. С. 179—207.
Мелкова В. К., Кунашев М. В. О сезон-
ном заселении серыми крысами неко-
торых строений // Экология и медицин-
ское значение серой крысы (КаЙлз пот-
уегсиз ВегЕ). М.: Наука, 1983. С. 99-
100.
Мелкова В. К., Туров И. С. Мозаичный
характер распределения серых крыс в
городе как основа целенаправленной
борьбы с ними // ТУ сЪеЗД Всесозюз,
териол. о-ва: Тез. докл. М., 1986. Т. 3.
С. 377-878.
Медкова В. К., Туров И. С., Одинец А. А.
и др. Характер размещения серых крыс
в подвалах жилых домов и его измене-
ния под влиянием дератизационных
мероприятий // Проблемы дезинфекции
и стерилизации: Сб. науч. тр. М. Мин-
здрав СССР, 1985. С. 168—170.
Мерзликин И. Р. Некоторые аспекты кор-
модобывающей деятельности серых
крыс в природных биотопах // ТУ съезд
Всесоюз. териол. о-ва: Тез. докл. М.,
1986. Т. 3. С. 242—244.
Мерзликин И. Р, Материалы по размно-
жению серой крысы в условиях северо-
восточной Украины // Материалы по
экологии и методам ограничения чис-
ленности серой крысы. М.: АН ссср,
1987а. С. 159—178.
Мераликин И. Р. Некоторые наблюдения
за характером использования участка
обитания ссрой крысой // Материалы по
экологии и методам ограничения чис-
ленности серой крысы. М. Наука,
49876. С. 113—128.
Мерзликин И. Р. Разновидность назем-
ных мост отдыха серой крысы в при-
роде // Там же. 19876. С. 143—158.
Меркова М. А. Некоторые данные по эко_
логии рыжей полевки и желтогорлой
мыши юга Московской обл. Телерма-
новской роши // Бюл. МОИП. Отд. биол.
41955. Т. 60, No 1. 0. 21-31.
Методические указапия по борьбе с се-
рой крысой (Вайлз погуез1сиз) в при-
родных очагах иктерогеморрагического
‘лептоспироза. М.: Минздрав СССР, 1984.
12 с.
Мешкова Н. ПН. Ориентировочно-исследо-
вательская деятельность серой крысы
в «откоытом поле»: (Зоопеихол. ана-
лиз) // Бюл. МОИП. Отд. биол. 1981.
Т. 86, No В. С. 22—29.
Мигулин А. А. Звери Украинской ССР.
Киев: Наук. думка, 1938. 426 с. На укр.
яз.
.
Милютин А. И. Диморфизм окраски в
естественных понуляциях пасюков (Ва
{из потуевсл$) // Науч
с.-х, ин-та. 1981. Т. 405. С. 50-54.
Милютин А. И. Изменчивость окраски
меха в естественных полуляциях серой
крысы // ПТ съезд Всесоюз. териол. о-ва:
Тез. докл. М., 1982. Т. 2. С. 55—56.
Милютин А. И. О времени появления се-
рой крысы в Европе // Экология и ме-
дицинское значение серой крысы (Ван
из потуедсие ВетК). М.: Наука, 19832.
С. 19—20.
Милютин А. И. К вопросу о времени
появления серой крысы в Европе //
Резн шик. ТаШит, 19836. Вын. 2.
С. 51—62.
Милютин А. И. Серая (ВаЙмз погуе21сл1$)
и черная (Вали таМиз) крысы в При-
балтике: распространение и межвидо-
вые отношения
// Серая крыса (эколо-
тия и распространение). ТГ. 1. М.: АН
СССР, 1986.
Милютин Л. И., Езерскене 9. Н. Распро-
странение крыс (род Ваз) в При-
балтике. Сообщ. 1// Еезй и\И9. Тал-
линн, 1984. Выш. 3. С. 39—50.
Милютин А. И., Езерснене Э. П. Распрост-
ранение крысе (род Ваз) в Прибал-
тике. Сообщ. 2 // ЕезН ак. Таллинн,
1986. Вып. 4. С. 35—45.
Мирзоев Ш. М. Организация работ по
дератизации // Ветеринария. 1974. No 10.
С. 28—30.
Мирзоев Ш. М. Защита свиноводческих
комплексов от грызунов // Там же. 1978.
No3.С.16-18. °
|
Мирзоев Ш. М. Зазщита крупных живот-
новодческих хозяйств и комплексов от
синантропных грызунов: Автореф. дис.
... дера биол. наук. М., 1984. 20 с.
Миронов Н. П. К вопросу об эволюции и
природной очаговости бруцеллеза //
Журп. микробиологии, эпидемиологии
и иммунологии. 1964. No 12. С. 60—65.
Миронов Н. П., Карпузиди К. С. Климен-
ко И. 3. и др. Источники и переносчи-
ки чумы и туляремии. М., 1965. 145 с.
Мипонов Н. Н., Рожков В. С., Денисов
П. С., Некрасов П. А. Изменение гра-
ницы ареала серой крысы в восточных
районах Ростовской области и запад-
ных районах Калмыцкой АССР // Науч.
работы Элистинской противочум. ст.
4961. Вып. 2. С. 215-286.
тр. Ленингр.
Литература
414
Митропольский О. В., Лобызова В. П., Се-
дин В. И. Размножение серой крысы в
Узбекистане // ГУ съезд Всесоюз. те-
риол. о-ва: Тез. докл. М.
.т.
С. 250—252. докт. М» 1986. т.
Михайленко А. Г. Пространственное рас-
пределение серых крыс на свинофер-
ме // Млекопитающие СССР: Тез. докл.
3-й Всесоюз. конф. териол. о-ва. М.;: АН
СССР, 1982. Т. 4. С. 250-251.
Михайленко А. Г. Основные черты со-
циального поведения синантропных се-
рых крыс на животноводческой ферме //
дктувльные Вопросы дезинфекции и
терилизации: (Тез. науч.-практ, конф.).
М.: АН СССР, 1987. С. Я
о=ф
Михайлова О. А., Якунина Т. И. Обнару-
жение возбудителя листериоза у тгры-
зунов города Владивостока// Изв. Ир-
кут. паче послед. противочум. ин-та
ибири и Дал. Востока.
:
24
Сибири д
а. 1962. Т. 24.
Моргилевская И. Е., Цителова 3. К., Ми-
и А. Грызуны города Тбилиси //
. Зоол. ин-та
о,
ССР. 1974. Т. 7.
Мороз А. Н, Хижинская 0. П. Поражен-
ность листереллами грызунов г. Киева
и его окрестностей // Зоонозные инфек-
ции. Киев: Наук. думка, 1959. С. 254—
Москва в цифрах: Стат. ежегодник. М.:
Финансы и статистика, 1982. 222 с,
Мотиги К., Нагаи Х., Хасэбэ Я., Осэк
,
.
.
иТ.
Пат,МКИС09Ь5/4,АОМ57/4
Япония. Покрытие для отпугивания
крые. Заявл. 7.06.82; Опубл. 14.12.83.
“Мясников Ю. А. Эризипелоид
// Общ. и
частн. эпидемиол. М.: М
Т. 2. С. 423—486.
‘дищина, 1973.
Мясников Ю. А. Распространение и ко-
лебания численности грызунов, зайце-
образных и насекомоядных Тульской
области // Фауна и экология грызунов.
М.: Паука, 1976. Выш. 13. С. 164—236.
Мясников Ю. А., Белова М. А. О типах
природных и антропургических очагов
офизиполоида // Вопросы природного
с ата болезней, Алма-Ата, 1978. Вып. 9.
Набенов К. Н. Эффективность дератиза-
цибнных мероприятий в синантронных
нопуляциях домовой мыши // Современ-
ные направления медицинской дезин-
секции и дератизации: (Тез. докл.). М.:
Медицина, 1981. С. 105—106.
Наумов Н. П. Определение возраста бел-
ки // Учен. зап.
С. 15-20.
Наумов Н. Н. определение возраста ма-
лого суслика // Защита ре
й
его суслика, щита растений. 1955.
Наумов Н. П. Типы поселений грызу-
нов и их экологическое значение //
Воол. журн. 1954. Т. 33, No2. С. 268—
МГУ. 1934. Вын. 2.
Наумов Н. П. Структура популяций и
ДараЖИКА ПИСлОННОСТ наземных поз-
воночных ам же. 1967. Т. 46, No
С. 1470—1486.
Юм
Море Н. Н. Пространственные струк-
ы вида млекопитающих // Там .
1971. Т. 50, No 7. С. 351—362. !
вы
Наумов Н. П., Гольцман М. Е. Новеден-
ческие и физиологические реакции мле-
Поитющих на запахи сородичей по
виду спехи соврем. биологии. 1972.
Т. 73, No 3. С. 427-444.
9
Наумов Н. И., Емельянова И. А. Грызу-
ны — разрушители материалов, изделий,
сооружений // Проблемы защиты от
иоповреждений. М.: Знание, 1979.
С. 33—47. (Сер. биол.; No 1).
Нацкий К. В., Игнатович В. 0. К вопросу
о значении грызунов в очагах дальне-
восточной скарлатиноподобной лихорад-
ки (псевдотуберкулеза) во Владивосто-
ке // 1 Межобл. науч.-практ. конф. по
псендотуберкулезу ’ (дальневосточной
скарлатиноподобной лихорадке): Тез.
докл. Владивосток, 1970. С. 62—63.
Некипелов ПН. В. Улучшенный способ
отлова крыс в верши// Изв. Иркут.
науч.-исслед. противочум, ин-та Сибири
Н и Дал. Востока. 1957. Т. 16. С. 247—248.
‘пипелов Н. 2 домовые грызуны в Юто-
осточном Забайкалье // Там же.
Т. 19. С. 72-82.
17 Там ко. 1958,
Некипелов Н. В. Эпизоотология чумы в
Монгольской Народной Республике //
Там же. 1959. Т. 12. С. 108—243.
Немченко Л. С. Меры борьбы с серой
крысой при экстренной профилактике //
ГУ съезд Всесоюз. териол. о-ва: Тез.
докл. рабочих засед., Москва, 27—31 ян-
варя 1986 г. М.: АН СССР.
Варя 1986.
‚ 1986. Т. 3.
Немченко Л. С., Бытов Г. Г. Обитание сс-
рои крысы в нриродных биотопах Муй-
ско-Куандинской котловины // Экология
едищинское зпачение серой крысы
аИяз потуеб!сив ВегК). М.:
1983. С. 15—46,
}. М. Наука
Пестеренко В. А. Обоснование мер борь-
бы с грызунами
— вредителями риса в
сельском хозяйстве. Динамика числен-
419 Литература
ности грызупов на ДВ СССР и их роль
в естественных сообществах и агроце-
нозах // Тез. докл. П регион. науч. конф.
АН СССР. Владивосток, 1985. С. 40—
41.
Нечаева Н. И. Избирательное отношение
некоторых мышевидных грызунов к
различным кормам // Изв. Иркут. науч.-
исслед. противочум. ин-та Сибири и
Дал. Востока. 1954. Т. 18. С. 120—128.
Пикитин В. Н. К биологии домовой кры-
сы // Природа. 1950. No 3. С. 70-71.
Никитин В. Н. Борьба с домовыми гры-
зунами с помощью отравленной клес-
вой массы // Грызуны и борьба с ними.
Саратов: Ки. изд-во, 1955. Вып. 4.
С. 268—210.
Никитина Н. А. Мытпи // Вопросы терио-
логии: итоги мечения млекопитающих.
М.: Наука, 1980. С. 157—176.
Никитюк П. М. Роль животных и птиц
как источников сальмонеллезпых забо-
леваний человека: (Обзор лит.) // Журн.
микробиологии, эпидемиологии и имму-
нобиологии. 1970. No 10. С. 123-130.
Нивифоров Н. И., Зацепин В. Г., Винову-
ров В. В. Борьба с мышевидными гры-
зунами на фермах. М.: Колос, 1977.
140 с.
Никифоров Н. И., Зотова А. А., Траханов
Д. Ф., Егоренко В. В. Итоги научно-
исследовательской работы на основе
массового истребления мышевидных
грызунов на фермах колхозов и совхо-
зов Тамбовской области //Тр. ВНИИ
вет. санитарии. 1963. Г. 22, С. 153—161.
Никифоров П. И., Тратанов Д. Ф., Заце-
пин .В. Г. Испытание водных приманок
для истребления серых крые // Там же.
1964. Т. 24. С. 354—359.
Никифоров ГО. Н., Галацевич Н. Ф. К воп-
росу о расселении и численности серой
крысы в Тувинской АССР ИТУ съезд
Всесоюз. териол. о-ва: Тез. докл. М.
1986. Г. 3. С. 288—289.
Пикифорова А. С. Роль серых крыс в
эпидемиологии водной лихорадки типа
Моняков // Журн. микробиологии, эпи-
демиологии и иммунобиологии. 1957.
No 7. С. 81-82.
Николаев Н. И. Лептоспироз среди крые
крупного города // Там же. 1945. No 12.
С. 70—71.
Николаева ПН. Н., Мельников Е. К., Швай-
дель Г. В. Опыт применения сухих от-
равленных приманок для дератизации й
Проблемы дезинфекции и стерилиза-
ции. М.: Минздрав СССР, 1977. Ч. 2.
С. 88—89.
Никул В. С. Современное состояние чис-
ленности крыс (Вабиз погуео1е18) в
Молдавии // Фауна, экология и физио-
логия животных. Кишинев, 1978. С. 35—
40.
.
Новиков Г. А. Айрапетянц А. 9., Пувин-
ский 10. В. и др. Звери Лепинтрадской.
области. Л.: Изд-во ЛГУ, 1970. 360 с.
Норман Г-Д. Международпые санитар-
ныс конференции 1851—1938 тг. Жене-
ва: ВОЗ, 1976. 124 с.
Овасапян 0. В. Туляремия в районах Ле-
нинаканского бассейна и ее эпизоото-
логические и эпидемиологические 0со-
бенности: Автореф. дис ... канд. мед.
наук. Ереван, 1957. 39 с.
Овасапян О. В., Есадисанян М. М., Гало-
ян В. О. О возможности носительства
эризипелотриксов у серых крыс // Журн.
микробиологии, эпидемиологии и имму-
нологии. 1964. No 12. С. 35-38.
Огита К. Пат. В ОП 7/00, В ОП 13/00,
No 55-23098 Япония. Препараты, сенси-
билизированные гелем полиакрилами-
да. Заявл. 25.04.74; Онубл. 20.06.80.
Огнева Н. С. Особенности распростране-
ния листериоза среди грызунов в усло-
виях больптого города // Журн. микро-
биологии, эпидемиологии и иммуноло-
гии. 1961. No 5. С. 69—74.
Огнева Н. С. Об эпизоотологии листериоза
трызунов // Зоол. журн. 1964. Т. 43, No 9.
С. 1873—1380.
Огуси К., Товумицу И. Изучение роден-
тицидов. Ч. 2. Предпочитаемость серой
крысой приманок с острыми ядами и
их эффективность // Тар. 7. Защ. 2001.
41970. Уа1. 21, No3. Р. 181-185.
Окладников А. П. Следы палеолита в до-
лине р. Лены // Палеолит и пеолит
СССР. М.; Л., 1953. С. 227—265.
Окуневский Я. Л. Практическое руковод-
ство по дезинфекции. Т. 4. Дезинсек-
ция, дератизация. М.; Л.: Биомедгиз,
41986. 755 с.
-
у
Олсуфьев Н. Г., Дунаева Т. Н. Экспери-
ментальная туляремия у лабораторных
животных
// Туляремия. М.: Медгиз,
1960. С. 96—135.
Олькова Н. В. Антонюк В. Я., Солдатов
Г. М. и др. Современное состояние чис-
ленности есрой крысы в населенных
пунктах Сибири и Дальнего Востока //
Материалы по экологии и методам огра-
ничения численности серой крысы. М.:
Наука, 1987. С. 76-98.
Опарина 0. С. Распределение и числен-
ность серой крысы в условиях аптро-
погенно преобразованного ландшафта
Литературы
413
Волго-Ахтубинской поймы // Природная
очаговость и профилактика зоонозов.
Саратов: Минздрав СССР, 1987. С. 71-—
Опарина О. С., Опарин М. Л., Самарин
Е. Г. К экологии серой. крысы в Северо-
Восточном Прикаспии // Эпизоотология
природно-очаговых инфекций. Саратов,
_ 1985. С. 54—60.
Орлов В. Н., Булатова Н. Ш. Сравпитель-
ная цитогенетика и кариосистематика
млекопитающих. М.:; Наука, 1983. 405 с.
Осташев С. Н. Распространение, места
обитания серых крыс и экономический
ущерб, наносимый ими в Кировской об-
ласти // Тр. Киров. с.-х. ин-та. 1967.
Т. 20, вып. 39. С. 73—78.
Очиров Ю. Д. К распространению и чис-
лепности некоторых млекопитающих
северпых районов Читинской области //
Изв. Иркут. науч-исслед. противочум.
ин-та Сибири и Дал. Востока. 1968а.
Т. 27, С. 69—74.
ОЭчиров Ю. Д. Распространение некото-
рых видов млекопитающих в Северо-
Восточном Забайкалье // Зоол.. журн.
19686. Т. 47. No 12. С. 1065—1070.
Эчиров Ю. Д., Башанов К. А. Млекопи-
тающие Тувы. Кызыл, 1975. 452 с,
Очиров Ю. Д., Бондарчук А. С. Грызуны
населенных пунктов Витимо-Олекмин-
ской горной страны // Изв. Иркут. науч.-
исслед. протиночум. ин-та Сибири и
Дал. Востока. 1963. Т. 25. С. 243—247.
Павлов М. П., Кирис И. Б. Питание. ли-
сицы (Ушрез ушрез Г.) в приазовских
плавнях Кубани, заселенных нутрией
”-, (Муосавёот соуриз Ме!:) //Зоол. журн.
1956. Т. 35, No 6. С. 879-906.
Панина Т. В. Расширение ареала серой
крысы в Тульской области и взаимо-
отноптения между серой и черной кры-
сой // Серая крыса. М.: Наука, 1986а.
"Г. +. С. 178—246.
Панина Т. В. Расширение ареала серой
крыем в Тульской области и заселение
ето г. Тулы // ГУ съезд Веесоюз. териол.
о-ва: Тез. докл. М., 19866. Т. 3. С. 2688—
Панкратов А. В. О бруцеллезе у крыс //
Арх. биол. наук. 1939. Т. 53, вып. 1.
С. 28—30.
Панов Е. Н. Поведение животных и это-
логическая структура популяций. М.:
Наука, 1983. 423. с.
у
Пантелеев П. А. Популяционная эколо-
гия водяной полевки и меры борьбы с
ней, М.: Наука, 1968. 255 с.
Пасешник А. А. К вопросу размножае-
мости крыс в г. Москве // Тр. Моск. де-
зинфекц. ин-та. 1936. Вып. (. С. 70-76.
Пасешник А. А. Пищезые приманки и не-
пищевое отравление серых крыс (Ва(-
{13 погуер1ец$ ВегЕ) // Журн. микробио-
логии, эпидемиолотии и иммунобиоло-
гии. 1947. No 10. С. 22—26.
Петров В. С., Леонтьев М. Н. Распростра-
нение и экология свободно живущих
понуляций в условиях большого горо-
да и его окрестностей // Наземные и
водные экосистемы. Горький: Горьк.
ун-т, 1977. Выш. 1. С. 171—174.
Петров В. С, Леонтьев М. Н., Соловьев
. К. и др. К изучению фауны и эко-
логии грызунов большого города // Гры-
зуны: Материалы У Всесотоз. совеще М.:
Наука, 1980. С. 434—435.
Петров Е. М. Результаты серогрупповой
идентификации ттаммов лептоспир из
Социалистической Республики Вьет-
нам // Лептоспирозы. Тез. докл. 8-й `Все-
союз. конф. по лептоспирозам. Тбилиси:
Сабчота Сакартвело, 1983, С. 133—135.
Петров Е. М., Нгуен Ван Нье. Крысы —
лентоспироносители в СРВ // Лептосии-
розы: УП Всесоюз. конф. по лентоспи-
розам: Тез. докл. Тбилиси: Сабчота Са-
картвело, 1983. С. 134—135.
Петров П. А., Адамян А. 0. Грызуны па-
селенных пунктов высокогорных райо-
нов востока Армении // Особо опас-
ные ‘инфекции на Кавказе. Ста
р 1966. С. 435-437.
протоль,
етросян 9. А., Лобанова Т. И., Некрасов
П. А., Исаева 5. В. Численность ры.
нов и их эктопаразитов в населенных
пунктах Апшеронского полуострова в
1955—1959 гг.//Тр. Азерб. противочум.
станции. 1962. 'Г. 3. С. 432-140.
Плохинский Н. А. Алгоритмы биометрии
М.: Изд-во МГУ, 1980. 150 с.
р
Наятер-Плохоцкий К. Грызуны — вредите-
Ли риса Дальневосточного края. Хаба-
ровск, 1931. 48 с.
Паятер-Плохоцкий К. А. Вредные и по-
лезные млекопитающие в сельском хо-
зяйстве Дальневосточного края. Хаба-
ровек: Дальненост. краев. изд-во, 1936.
180 с.
'
Повалишина Т. П. Картографический ана-
лиз результатов обследования мелких
млекопитающих па антиген вируса
глис. на территории СССР // Актуаль-
ные проблемы медицилской вирусоло-
гии. М.: Ин-т полиомиелита АМН СССР
1985. С. 126—128.
у
419 Литература
ности грызупов на ДВ СССР и их роль
в естественных сообществах и агроце-
нозах // Тез. докл. П регион. науч. конф.
АН СССР. Владивосток, 1985. С. 40—
41.
Нечаева Н. И. Избирательное отношение
некоторых мышевидных грызунов к
различным кормам // Изв. Иркут. науч.-
исслед. противочум. ин-та Сибири и
Дал. Востока. 1954. Т. 18. С. 120—128.
Пикитин В. Н. К биологии домовой кры-
сы // Природа. 1950. No 3. С. 70-71.
Никитин В. Н. Борьба с домовыми гры-
зунами с помощью отравленной клес-
вой массы // Грызуны и борьба с ними.
Саратов: Ки. изд-во, 1955. Вып. 4.
С. 268—210.
Никитина Н. А. Мытпи // Вопросы терио-
логии: итоги мечения млекопитающих.
М.: Наука, 1980. С. 157—176.
Никитюк П. М. Роль животных и птиц
как источников сальмонеллезпых забо-
леваний человека: (Обзор лит.) // Журн.
микробиологии, эпидемиологии и имму-
нобиологии. 1970. No 10. С. 123-130.
Нивифоров Н. И., Зацепин В. Г., Винову-
ров В. В. Борьба с мышевидными гры-
зунами на фермах. М.: Колос, 1977.
140 с.
Никифоров Н. И., Зотова А. А., Траханов
Д. Ф., Егоренко В. В. Итоги научно-
исследовательской работы на основе
массового истребления мышевидных
грызунов на фермах колхозов и совхо-
зов Тамбовской области //Тр. ВНИИ
вет. санитарии. 1963. Г. 22, С. 153—161.
Никифоров П. И., Тратанов Д. Ф., Заце-
пин .В. Г. Испытание водных приманок
для истребления серых крые // Там же.
1964. Т. 24. С. 354—359.
Никифоров ГО. Н., Галацевич Н. Ф. К воп-
росу о расселении и численности серой
крысы в Тувинской АССР ИТУ съезд
Всесоюз. териол. о-ва: Тез. докл. М.
1986. Г. 3. С. 288—289.
Пикифорова А. С. Роль серых крыс в
эпидемиологии водной лихорадки типа
Моняков // Журн. микробиологии, эпи-
демиологии и иммунобиологии. 1957.
No 7. С. 81-82.
Николаев Н. И. Лептоспироз среди крые
крупного города // Там же. 1945. No 12.
С. 70—71.
Николаева ПН. Н., Мельников Е. К., Швай-
дель Г. В. Опыт применения сухих от-
равленных приманок для дератизации й
Проблемы дезинфекции и стерилиза-
ции. М.: Минздрав СССР, 1977. Ч. 2.
С. 88—89.
Никул В. С. Современное состояние чис-
ленности крыс (Вабиз погуео1е18) в
Молдавии // Фауна, экология и физио-
логия животных. Кишинев, 1978. С. 35—
40.
.
Новиков Г. А. Айрапетянц А. 9., Пувин-
ский 10. В. и др. Звери Лепинтрадской.
области. Л.: Изд-во ЛГУ, 1970. 360 с.
Норман Г-Д. Международпые санитар-
ныс конференции 1851—1938 тг. Жене-
ва: ВОЗ, 1976. 124 с.
Овасапян 0. В. Туляремия в районах Ле-
нинаканского бассейна и ее эпизоото-
логические и эпидемиологические 0со-
бенности: Автореф. дис ... канд. мед.
наук. Ереван, 1957. 39 с.
Овасапян О. В., Есадисанян М. М., Гало-
ян В. О. О возможности носительства
эризипелотриксов у серых крыс // Журн.
микробиологии, эпидемиологии и имму-
нологии. 1964. No 12. С. 35-38.
Огита К. Пат. В ОП 7/00, В ОП 13/00,
No 55-23098 Япония. Препараты, сенси-
билизированные гелем полиакрилами-
да. Заявл. 25.04.74; Онубл. 20.06.80.
Огнева Н. С. Особенности распростране-
ния листериоза среди грызунов в усло-
виях больптого города // Журн. микро-
биологии, эпидемиологии и иммуноло-
гии. 1961. No 5. С. 69—74.
Огнева Н. С. Об эпизоотологии листериоза
трызунов // Зоол. журн. 1964. Т. 43, No 9.
С. 1873—1380.
Огуси К., Товумицу И. Изучение роден-
тицидов. Ч. 2. Предпочитаемость серой
крысой приманок с острыми ядами и
их эффективность // Тар. 7. Защ. 2001.
41970. Уа1. 21, No3. Р. 181-185.
Окладников А. П. Следы палеолита в до-
лине р. Лены // Палеолит и пеолит
СССР. М.; Л., 1953. С. 227—265.
Окуневский Я. Л. Практическое руковод-
ство по дезинфекции. Т. 4. Дезинсек-
ция, дератизация. М.; Л.: Биомедгиз,
41986. 755 с.
-
у
Олсуфьев Н. Г., Дунаева Т. Н. Экспери-
ментальная туляремия у лабораторных
животных
// Туляремия. М.: Медгиз,
1960. С. 96—135.
Олькова Н. В. Антонюк В. Я., Солдатов
Г. М. и др. Современное состояние чис-
ленности есрой крысы в населенных
пунктах Сибири и Дальнего Востока //
Материалы по экологии и методам огра-
ничения численности серой крысы. М.:
Наука, 1987. С. 76-98.
Опарина 0. С. Распределение и числен-
ность серой крысы в условиях аптро-
погенно преобразованного ландшафта
Литературы
413
Волго-Ахтубинской поймы // Природная
очаговость и профилактика зоонозов.
Саратов: Минздрав СССР, 1987. С. 71-—
Опарина О. С., Опарин М. Л., Самарин
Е. Г. К экологии серой. крысы в Северо-
Восточном Прикаспии // Эпизоотология
природно-очаговых инфекций. Саратов,
_ 1985. С. 54—60.
Орлов В. Н., Булатова Н. Ш. Сравпитель-
ная цитогенетика и кариосистематика
млекопитающих. М.:; Наука, 1983. 405 с.
Осташев С. Н. Распространение, места
обитания серых крыс и экономический
ущерб, наносимый ими в Кировской об-
ласти // Тр. Киров. с.-х. ин-та. 1967.
Т. 20, вып. 39. С. 73—78.
Очиров Ю. Д. К распространению и чис-
лепности некоторых млекопитающих
северпых районов Читинской области //
Изв. Иркут. науч-исслед. противочум.
ин-та Сибири и Дал. Востока. 1968а.
Т. 27, С. 69—74.
ОЭчиров Ю. Д. Распространение некото-
рых видов млекопитающих в Северо-
Восточном Забайкалье // Зоол.. журн.
19686. Т. 47. No 12. С. 1065—1070.
Эчиров Ю. Д., Башанов К. А. Млекопи-
тающие Тувы. Кызыл, 1975. 452 с,
Очиров Ю. Д., Бондарчук А. С. Грызуны
населенных пунктов Витимо-Олекмин-
ской горной страны // Изв. Иркут. науч.-
исслед. протиночум. ин-та Сибири и
Дал. Востока. 1963. Т. 25. С. 243—247.
Павлов М. П., Кирис И. Б. Питание. ли-
сицы (Ушрез ушрез Г.) в приазовских
плавнях Кубани, заселенных нутрией
”-, (Муосавёот соуриз Ме!:) //Зоол. журн.
1956. Т. 35, No 6. С. 879-906.
Панина Т. В. Расширение ареала серой
крысы в Тульской области и взаимо-
отноптения между серой и черной кры-
сой // Серая крыса. М.: Наука, 1986а.
"Г. +. С. 178—246.
Панина Т. В. Расширение ареала серой
крыем в Тульской области и заселение
ето г. Тулы // ГУ съезд Веесоюз. териол.
о-ва: Тез. докл. М., 19866. Т. 3. С. 2688—
Панкратов А. В. О бруцеллезе у крыс //
Арх. биол. наук. 1939. Т. 53, вып. 1.
С. 28—30.
Панов Е. Н. Поведение животных и это-
логическая структура популяций. М.:
Наука, 1983. 423. с.
у
Пантелеев П. А. Популяционная эколо-
гия водяной полевки и меры борьбы с
ней, М.: Наука, 1968. 255 с.
Пасешник А. А. К вопросу размножае-
мости крыс в г. Москве // Тр. Моск. де-
зинфекц. ин-та. 1936. Вып. (. С. 70-76.
Пасешник А. А. Пищезые приманки и не-
пищевое отравление серых крыс (Ва(-
{13 погуер1ец$ ВегЕ) // Журн. микробио-
логии, эпидемиолотии и иммунобиоло-
гии. 1947. No 10. С. 22—26.
Петров В. С., Леонтьев М. Н. Распростра-
нение и экология свободно живущих
понуляций в условиях большого горо-
да и его окрестностей // Наземные и
водные экосистемы. Горький: Горьк.
ун-т, 1977. Выш. 1. С. 171—174.
Петров В. С, Леонтьев М. Н., Соловьев
. К. и др. К изучению фауны и эко-
логии грызунов большого города // Гры-
зуны: Материалы У Всесотоз. совеще М.:
Наука, 1980. С. 434—435.
Петров Е. М. Результаты серогрупповой
идентификации ттаммов лептоспир из
Социалистической Республики Вьет-
нам // Лептоспирозы. Тез. докл. 8-й `Все-
союз. конф. по лептоспирозам. Тбилиси:
Сабчота Сакартвело, 1983, С. 133—135.
Петров Е. М., Нгуен Ван Нье. Крысы —
лентоспироносители в СРВ // Лептосии-
розы: УП Всесоюз. конф. по лентоспи-
розам: Тез. докл. Тбилиси: Сабчота Са-
картвело, 1983. С. 134—135.
Петров П. А., Адамян А. 0. Грызуны па-
селенных пунктов высокогорных райо-
нов востока Армении // Особо опас-
ные ‘инфекции на Кавказе. Ста
р 1966. С. 435-437.
протоль,
етросян 9. А., Лобанова Т. И., Некрасов
П. А., Исаева 5. В. Численность ры.
нов и их эктопаразитов в населенных
пунктах Апшеронского полуострова в
1955—1959 гг.//Тр. Азерб. противочум.
станции. 1962. 'Г. 3. С. 432-140.
Плохинский Н. А. Алгоритмы биометрии
М.: Изд-во МГУ, 1980. 150 с.
р
Наятер-Плохоцкий К. Грызуны — вредите-
Ли риса Дальневосточного края. Хаба-
ровск, 1931. 48 с.
Паятер-Плохоцкий К. А. Вредные и по-
лезные млекопитающие в сельском хо-
зяйстве Дальневосточного края. Хаба-
ровек: Дальненост. краев. изд-во, 1936.
180 с.
'
Повалишина Т. П. Картографический ана-
лиз результатов обследования мелких
млекопитающих па антиген вируса
глис. на территории СССР // Актуаль-
ные проблемы медицилской вирусоло-
гии. М.: Ин-т полиомиелита АМН СССР
1985. С. 126—128.
у
414
Литература
Подболотов К. В., Сурков В. С. К эпизо-
отологической оценке кишечного иер-
синиоза в Сахалинской области // Журн.
микробиологии, эпидемиологии и имму-
нобиологии. 1984. No 2. С. 71-73.
Подкорытов Ю. Н., Захаров Т. Ф., Банду-
раС.А.идр.Сераякрысавсвязис
заболеваниями иктерогеморрагическим
лептоспирозом в Крыму //1У съезд
Всесоюз. териол. о-ва: Тез. докл. М.,
4986. Т. 3. С. 383—834.
Покровская В. И. Псевпотуберкулез в
Новгородской области // Природно-оча-
товые инфекции. Л., 1980. С. 162—166.
(Тр. Ленингр. ИЭМ; Т. 44).
Полежаев В. Г. Крысонеироницаемость
и роль подвальных помещений // Тр.
Центр. науч.-исслед. дезинфекц. ин-та.
4947. Т. 3. С. 217—222.
Полежаев В. Г. Изучение поведепия се-
рой крысы в зависимости от факторов
внешней среды
// Там же. 1950. Т. 6.
С. 237—243.
Полежаев В. Г. Препараты, отпугиваю-
щие грызунов // Там же. 1959. Т. 141.
С. 267—278.
Полежаев В. Г., Земит И. И., Раценис
Э.К. и др. Изучение передвижений се-
рых крыс в условиях города для обос-
нования методики дератизационных ра-
бот // Там же. 1959. Т. 12. С. 245-254.
Полежаев В. Г., Кирин Л. А., Залежский
Г. В. и др. Эффективность дератизации,
проводимой в населенных пунктах
только методом опыливания // Там же.
1963. Т. 16. С. 302—809.
Полежаев В. Г., Кирин Л. А., Меерсон
Г. М. и др. Состояние дератизационных
работ в республиканских, краевых и
областных центрах // Тез. докл. науч.
конф. Центр. науч.-исслед. дезинфекц.
ин-та. М.: Минздрав СССР, 1963а. С. 56-—
57.
Полежаев В. Г., Тощизин Ю. В., Кирин
А. и др. Изучение миграции грызу-
нов путем мечепия их радиоактивным
изотопом // Тр. Центр. науч.-исслед. де-
зинфекц. ин-та. 1962. Т. 15. С. 295-299.
Поляков А. А., Бочаров Д. А., Шептунин
В. П. Ветеринарная санитария и гигие-
па предприятий мясной и молочной
промыптленности. М.: Лег. и пищ. пром-
сть, 1983. 232 с.
Пономарева Т. Н., Родкевич Л. В. Пасте-
реллез у грызунов большого города //
Журн. микробиологии, эпидемиологии и
иммунологии. 1964. No 5. С. 144—145.
Попов В. А. Млекопитающие Волжеко-
Камского края (насекомоядные, руко-
крылые, грызуны). Казань: Изд-во Ка-
зан. ун-та, 1970. 282 с. -
Нопов В. П., Тесленко Е. Б. Чума у се-
рых крыс // Современные аспекты про-
филактики зоонозных инфекций. Ир-
кутск, 1984. Ч. 1. С, 42—44.
Ноярков Д. В. Новые данные о серой
крысе в природных биотопах европей-
ской части СССР
// Зоол. журн. 1961.
Т. 40, No 10. С. 1552—1556.
Поярков Д. В., Пояркова Н. Н. В изуче-
нию антропургических очагов сальмо-
неллезов млекопитающих и птиц в
г. Москве // Животное население Моск-
вы и Подмосковья, его изучение, охра-
на и направленное преобразование. М.:
Меджцина, 1967. С. 98—100.
Прокофьева 3. В., Иванов М. А. Расселе-
"ние серой крысы и борьба с ней на тер-
ритории Ставропольского края // Особо
опасные инфекции на Кавказе. Ставро-
поль, 1974. Вып. {. С. 174-176.
Промптов В. П. Пасюк в Татикентском
оазисе // Узб. биол. журн. 1962. No 3.
С. 66—69.
Промптов В. Н., Устименко С. Ф., Ефи-
мов А, Ю., Каштанкин М. Н. Измене-
ние границ ареала и некоторых морфо-
логических признаков у серой крысы
в Средней Азии//Там же. 19841. No 4.
С. 52—54.
Промптов В. П., Шестакова Л. Н., Каш-
танкин М. Н., Ефимов А. Ю. Современ-
пос состояние и тенденция изменения
ареала серой крысы в Узбекистане и
на смежных © ним территориях // Эко-
логия и медицинское значение серой
крысы (ВаЙиз погуессиз ВегК): Ма-
териалы 1-го рабоч. совещ. по серой
крысе. М.: АН СССР, 1983. С. 26-28.
Иромптов В. П., Шеставова Л. И., Каш-
танкиц М. Н., Ефимов А. Ю. Эпидеми-
ческое и хозлиственное значение серой
крысы в связи с дальлейшим раеселе-
нием ее по территории республик Сред:
ней Азии и того-запада Казахетана //
Серая крыса. М.: АН СССР, 1986. Т. 2.
С. 62-68.
Прохоров М. И. Микробиологический ме-
тод борьбы с вредными грызунами. П.;
М.: Сельхозиздат, 1962. 133 с.
Пузанов И. И. Предварительные итоги
изучения фауны позвоночных Крым-
ского зановедника // Работы по изуче-
нию фауны Крымского гос. заповедни-
ка. М; Л. 1981. С. 5-38.
Пуцято В. С. Применение звука в дера-
тизации // Вопросы ветеринарной дезин-
з
н
а
О
В
Ш
р
а
р
о
л
к
н
ы
ы
а
Литература 415
фекции на совремеппом этапе. М. Агро-
промиздат, 1984. С. 140-146.
Пуцито В. С. Ндовитые покрытия в борь-
бе с крысами //ТУ съезд Всесоюз. те-
риол. о-ва: Тез. докл. М., 1986. Т. 3.
С. 394—398.
Пуцято В. С. Эффективность ядовитых
покрытий из дустов линких масс и пен
в борьбе с крысами// Материалы по
экологии и методам ограничения чис-
ленности серой крысы. М.: Наука, 1987.
С. 281—307.
Пушница Ф. А., Харченко А, П., Дзнелад-
зе М. Т., Михеева К. Е. Видовой состав.
размещение и числепность синантроп-
пых грызунов в нортах и портовых го-
родах европейской части СССР. Сообщ. 1.
Грызуны портовых городов Черно-
морского побережья // Тр. Рост. проти-
вочум. ин-та. 1960. Т. 47, С. 48-56,
Нушница Ф. А., Ширанович П. И. Рас-
прострапение крысиных блох в пре-
делах европейской части СССР // Там
же. 1960. Т. 17. С. 64-71.
Райхлин М. И. Материалы о природных
и антропургических очагах лептоспи-
розов в Таймырском национальном ок-
руге // Природно-очаговые — антропозо-
онозы: (Тез. докл. к 1Х Всесоюз. конф.
по природ. очаговости болезней чело-
вска и животных, 18—24 мая, 1976 г.).
Омск, 1976. С. 101-103.
Ревин Ю. В., Шадрина Е. Г. Репродук-
тивпые параметры популяции серых
крыс в Центральной Якутии // ГУ съезд
Веесоюз. териол. о-ва: Тез. докл. М.,
„ 1986. Т. 3. С. 252—254.
Реймерс Н. Ф., Воронов Г. А., Загород-
ский В. Е. Алина’А. В. Насекомояд-
ные и трызуны Сахалина и Куриль-
ских островов
// Сборник по экологии
и териологии. Пермь, 1968. Вып. 3.
С. 35-99.
Риверс Т. Вирусные и риккетсиозные
инфекции человека. М.: Изд-во иностр.
лит-ры, 1955. С. 670—682.
Ройхель М. М. Совремепные методы де-
ратизации в Западной Европе и Амери-
кеу/ Гигиена и эпидемиология. 1957.
No 4. С. 95-105.
Романов Г. В. О хищных наклонностях
серой крысы // Зоол. журн. 1963. Т. 43,
No 3. С. 470—474.
Романова Г. А. Грызуны населенных
пунктов Якутии // Биологические проб-
лемы севера. Якутск: Якут. фил. СО
АН СССР, 1974. Вы. 4. С. 93-98.
Романова Г. А. Экология серой крысы в
‚
Якутии // Экология и медицинское зна-
чение серой крысы (ВаМиз погуер1сиз
ВегК). М.: Паука, 1983. С. 18—19.
Романова Г. А. Типы окраски серой кры-
сы (Каз погуер!сиз) в Якутии // Фе-
нотика понуляций: Материалы ТИ Все-
юз. совещ. М.: Наука, 1985. С. 235—
Романова Г. А. Некоторые черты эко-
логии серой крысы в Якутии// Серая
крыса. М.: Наука, 1988. Т. 1. С. 103—
Романова Г. Л., Верещагина С. В. Уш-
ницкий Г. Н. Пути распространения
и характер расселения серой крысы
в Якутии / ГУ съезд Всесоюз. териол.
си: Тез. докл. М., 1986. Т. 3, С. 285—
Росицкий Б., Кратожвил И. Синантропия
млекопитающих и роль синантропи-
ческих и экзоантропических грызунов
в природных очагах болезней // Чехосл.
биология. 1953. Т. 2, вып. 5. С. 283—
295.
Россиков К. Н. Обзор млекопитающих
животных долины р. Малки // Зап. Акад.
наук. Физ.-мат. отд. 1887. Т. 54. С.1-
Рубакин А. Международная конференция
по борьбе с крысами в Париже // Ги-
тиена и эпидемиология. 1928. No4.
С. 95—104.
Русев И. Т., Бабенко Н. П., Соловьев
В. И., Харченко А. П. Характер раз-
множения серых крыс на территории
города и порта Одессы
// Экология и
медицинское значение серой крысы
(КаМиз погуессиз ВегЕ). М.: Наука
1983. С. 12—13.
'
Русев И. Т., Соловьев В. И. Некоторые
особенности серой крысы в пойме Ниж-
него Днестра
// Материалы по зэколо-
гии и методам ограничения численно-
сти серой крысы, М.: Наука, 1987.
С. 429—143.
Рыльников В. А. Экология серых крыс
и меры ограничения их численности
в природных очагах лептоспирова: Ав-
тореф. дис. канд. биол. наук. М.
1988. 27 с.
ь
зук. М,
Рыльников В. А., Беленький В. А., Во-
лынская Е. В. Метод идентификации
красителей в экскрементах грызунов
и ого применение для изучения под-
вижности серых крыс в природе // Бол.
МОИП. Отд. биол. 108 те 4.
С. 77-84.
'
Рыльников В. А., Звонарев А. В. Способ
количественной оценки контакта тры-
зунов с ратицидами // Проблемы дезин-
414
Литература
Подболотов К. В., Сурков В. С. К эпизо-
отологической оценке кишечного иер-
синиоза в Сахалинской области // Журн.
микробиологии, эпидемиологии и имму-
нобиологии. 1984. No 2. С. 71-73.
Подкорытов Ю. Н., Захаров Т. Ф., Банду-
раС.А.идр.Сераякрысавсвязис
заболеваниями иктерогеморрагическим
лептоспирозом в Крыму //1У съезд
Всесоюз. териол. о-ва: Тез. докл. М.,
4986. Т. 3. С. 383—834.
Покровская В. И. Псевпотуберкулез в
Новгородской области // Природно-оча-
товые инфекции. Л., 1980. С. 162—166.
(Тр. Ленингр. ИЭМ; Т. 44).
Полежаев В. Г. Крысонеироницаемость
и роль подвальных помещений // Тр.
Центр. науч.-исслед. дезинфекц. ин-та.
4947. Т. 3. С. 217—222.
Полежаев В. Г. Изучение поведепия се-
рой крысы в зависимости от факторов
внешней среды
// Там же. 1950. Т. 6.
С. 237—243.
Полежаев В. Г. Препараты, отпугиваю-
щие грызунов // Там же. 1959. Т. 141.
С. 267—278.
Полежаев В. Г., Земит И. И., Раценис
Э.К. и др. Изучение передвижений се-
рых крыс в условиях города для обос-
нования методики дератизационных ра-
бот // Там же. 1959. Т. 12. С. 245-254.
Полежаев В. Г., Кирин Л. А., Залежский
Г. В. и др. Эффективность дератизации,
проводимой в населенных пунктах
только методом опыливания // Там же.
1963. Т. 16. С. 302—809.
Полежаев В. Г., Кирин Л. А., Меерсон
Г. М. и др. Состояние дератизационных
работ в республиканских, краевых и
областных центрах // Тез. докл. науч.
конф. Центр. науч.-исслед. дезинфекц.
ин-та. М.: Минздрав СССР, 1963а. С. 56-—
57.
Полежаев В. Г., Тощизин Ю. В., Кирин
А. и др. Изучение миграции грызу-
нов путем мечепия их радиоактивным
изотопом // Тр. Центр. науч.-исслед. де-
зинфекц. ин-та. 1962. Т. 15. С. 295-299.
Поляков А. А., Бочаров Д. А., Шептунин
В. П. Ветеринарная санитария и гигие-
па предприятий мясной и молочной
промыптленности. М.: Лег. и пищ. пром-
сть, 1983. 232 с.
Пономарева Т. Н., Родкевич Л. В. Пасте-
реллез у грызунов большого города //
Журн. микробиологии, эпидемиологии и
иммунологии. 1964. No 5. С. 144—145.
Попов В. А. Млекопитающие Волжеко-
Камского края (насекомоядные, руко-
крылые, грызуны). Казань: Изд-во Ка-
зан. ун-та, 1970. 282 с. -
Нопов В. П., Тесленко Е. Б. Чума у се-
рых крыс // Современные аспекты про-
филактики зоонозных инфекций. Ир-
кутск, 1984. Ч. 1. С, 42—44.
Ноярков Д. В. Новые данные о серой
крысе в природных биотопах европей-
ской части СССР
// Зоол. журн. 1961.
Т. 40, No 10. С. 1552—1556.
Поярков Д. В., Пояркова Н. Н. В изуче-
нию антропургических очагов сальмо-
неллезов млекопитающих и птиц в
г. Москве // Животное население Моск-
вы и Подмосковья, его изучение, охра-
на и направленное преобразование. М.:
Меджцина, 1967. С. 98—100.
Прокофьева 3. В., Иванов М. А. Расселе-
"ние серой крысы и борьба с ней на тер-
ритории Ставропольского края // Особо
опасные инфекции на Кавказе. Ставро-
поль, 1974. Вып. {. С. 174-176.
Промптов В. П. Пасюк в Татикентском
оазисе // Узб. биол. журн. 1962. No 3.
С. 66—69.
Промптов В. Н., Устименко С. Ф., Ефи-
мов А, Ю., Каштанкин М. Н. Измене-
ние границ ареала и некоторых морфо-
логических признаков у серой крысы
в Средней Азии//Там же. 19841. No 4.
С. 52—54.
Промптов В. П., Шестакова Л. Н., Каш-
танкин М. Н., Ефимов А. Ю. Современ-
пос состояние и тенденция изменения
ареала серой крысы в Узбекистане и
на смежных © ним территориях // Эко-
логия и медицинское значение серой
крысы (ВаЙиз погуессиз ВегК): Ма-
териалы 1-го рабоч. совещ. по серой
крысе. М.: АН СССР, 1983. С. 26-28.
Иромптов В. П., Шеставова Л. И., Каш-
танкиц М. Н., Ефимов А. Ю. Эпидеми-
ческое и хозлиственное значение серой
крысы в связи с дальлейшим раеселе-
нием ее по территории республик Сред:
ней Азии и того-запада Казахетана //
Серая крыса. М.: АН СССР, 1986. Т. 2.
С. 62-68.
Прохоров М. И. Микробиологический ме-
тод борьбы с вредными грызунами. П.;
М.: Сельхозиздат, 1962. 133 с.
Пузанов И. И. Предварительные итоги
изучения фауны позвоночных Крым-
ского зановедника // Работы по изуче-
нию фауны Крымского гос. заповедни-
ка. М; Л. 1981. С. 5-38.
Пуцято В. С. Применение звука в дера-
тизации // Вопросы ветеринарной дезин-
з
н
а
О
В
Ш
р
а
р
о
л
к
н
ы
ы
а
Литература 415
фекции на совремеппом этапе. М. Агро-
промиздат, 1984. С. 140-146.
Пуцито В. С. Ндовитые покрытия в борь-
бе с крысами //ТУ съезд Всесоюз. те-
риол. о-ва: Тез. докл. М., 1986. Т. 3.
С. 394—398.
Пуцято В. С. Эффективность ядовитых
покрытий из дустов линких масс и пен
в борьбе с крысами// Материалы по
экологии и методам ограничения чис-
ленности серой крысы. М.: Наука, 1987.
С. 281—307.
Пушница Ф. А., Харченко А, П., Дзнелад-
зе М. Т., Михеева К. Е. Видовой состав.
размещение и числепность синантроп-
пых грызунов в нортах и портовых го-
родах европейской части СССР. Сообщ. 1.
Грызуны портовых городов Черно-
морского побережья // Тр. Рост. проти-
вочум. ин-та. 1960. Т. 47, С. 48-56,
Нушница Ф. А., Ширанович П. И. Рас-
прострапение крысиных блох в пре-
делах европейской части СССР // Там
же. 1960. Т. 17. С. 64-71.
Райхлин М. И. Материалы о природных
и антропургических очагах лептоспи-
розов в Таймырском национальном ок-
руге // Природно-очаговые — антропозо-
онозы: (Тез. докл. к 1Х Всесоюз. конф.
по природ. очаговости болезней чело-
вска и животных, 18—24 мая, 1976 г.).
Омск, 1976. С. 101-103.
Ревин Ю. В., Шадрина Е. Г. Репродук-
тивпые параметры популяции серых
крыс в Центральной Якутии // ГУ съезд
Веесоюз. териол. о-ва: Тез. докл. М.,
„ 1986. Т. 3. С. 252—254.
Реймерс Н. Ф., Воронов Г. А., Загород-
ский В. Е. Алина’А. В. Насекомояд-
ные и трызуны Сахалина и Куриль-
ских островов
// Сборник по экологии
и териологии. Пермь, 1968. Вып. 3.
С. 35-99.
Риверс Т. Вирусные и риккетсиозные
инфекции человека. М.: Изд-во иностр.
лит-ры, 1955. С. 670—682.
Ройхель М. М. Совремепные методы де-
ратизации в Западной Европе и Амери-
кеу/ Гигиена и эпидемиология. 1957.
No 4. С. 95-105.
Романов Г. В. О хищных наклонностях
серой крысы // Зоол. журн. 1963. Т. 43,
No 3. С. 470—474.
Романова Г. А. Грызуны населенных
пунктов Якутии // Биологические проб-
лемы севера. Якутск: Якут. фил. СО
АН СССР, 1974. Вы. 4. С. 93-98.
Романова Г. А. Экология серой крысы в
‚
Якутии // Экология и медицинское зна-
чение серой крысы (ВаМиз погуер1сиз
ВегК). М.: Паука, 1983. С. 18—19.
Романова Г. А. Типы окраски серой кры-
сы (Каз погуер!сиз) в Якутии // Фе-
нотика понуляций: Материалы ТИ Все-
юз. совещ. М.: Наука, 1985. С. 235—
Романова Г. А. Некоторые черты эко-
логии серой крысы в Якутии// Серая
крыса. М.: Наука, 1988. Т. 1. С. 103—
Романова Г. Л., Верещагина С. В. Уш-
ницкий Г. Н. Пути распространения
и характер расселения серой крысы
в Якутии / ГУ съезд Всесоюз. териол.
си: Тез. докл. М., 1986. Т. 3, С. 285—
Росицкий Б., Кратожвил И. Синантропия
млекопитающих и роль синантропи-
ческих и экзоантропических грызунов
в природных очагах болезней // Чехосл.
биология. 1953. Т. 2, вып. 5. С. 283—
295.
Россиков К. Н. Обзор млекопитающих
животных долины р. Малки // Зап. Акад.
наук. Физ.-мат. отд. 1887. Т. 54. С.1-
Рубакин А. Международная конференция
по борьбе с крысами в Париже // Ги-
тиена и эпидемиология. 1928. No4.
С. 95—104.
Русев И. Т., Бабенко Н. П., Соловьев
В. И., Харченко А. П. Характер раз-
множения серых крыс на территории
города и порта Одессы
// Экология и
медицинское значение серой крысы
(КаМиз погуессиз ВегЕ). М.: Наука
1983. С. 12—13.
'
Русев И. Т., Соловьев В. И. Некоторые
особенности серой крысы в пойме Ниж-
него Днестра
// Материалы по зэколо-
гии и методам ограничения численно-
сти серой крысы, М.: Наука, 1987.
С. 429—143.
Рыльников В. А. Экология серых крыс
и меры ограничения их численности
в природных очагах лептоспирова: Ав-
тореф. дис. канд. биол. наук. М.
1988. 27 с.
ь
зук. М,
Рыльников В. А., Беленький В. А., Во-
лынская Е. В. Метод идентификации
красителей в экскрементах грызунов
и ого применение для изучения под-
вижности серых крыс в природе // Бол.
МОИП. Отд. биол. 108 те 4.
С. 77-84.
'
Рыльников В. А., Звонарев А. В. Способ
количественной оценки контакта тры-
зунов с ратицидами // Проблемы дезин-
416
Литература
фекции и стерилизации. М.: Минздрав
СССР, 1985. С. 148—153.
Рыльников В. А., Звонарев А. В. Анализ
популяции серых крыс в процессе их
истребления антикоагулянтами // Серая
крыса. М.: АН СССР, 1986. Т. 2. С. 416—
139.
Рыльников В. А., Карасева Е. В. Особен-
ности экологии серых крыс на рисо-
вых полях Кубани и меры ограничения
их численности// Распространение и
экология серой крысы и методы огра-
ничения се численности. М.: Наука,
1985. С. 71—112.
Рыльников В. А., Карасева Е. В., Дуби-
нина Н. В. Применение гистологиче-
ских красителей и тетрациклина для
изучения подвижности серых крыс на
рисовых полях Краснодарского края Й
Зоол. журн. 1984. Т. 60. No 6. С. 264-
265.
Рыльников В. А., Карасева Е. В., Дуби-
нина Н. В. Стратегия борьбы с серой
крысой в природных очагах лептосии-
роза на рисовых полях // Экология и
медицинское значение серой крысы
(Пабиз погусй1сл8 ВегК). (материалы
1-го рабоч. совещ. по серой крысе,31 ян-
варя —3 февраля 1983 г. Москва). М.:
АН СССР, 1983. С. 71-73.
Рыльников В. А., Сиротинина Е. В. Пи-
щевая активность и взаимоотношения
серых крыс около кормушек с неотрав-
ленной пищей в условиях их естест-
венного обитания // Вопросы дезинфек-
ции и стерилизации: Сб. науч. тр. М.:
Минздрав СССР, 4986. С. 162—170.
Рябцева Н. Л. Грызуны-синантропы Вос-
точного Закавказья и особенности про-
филактики чумы в населенных пунк-
тах // Эпидемиология и эпизоотология
чумы. Саратов, 1980. С. 59—64.
Сакаль Н. Н. Псевдотуберкулез и неко-
торые другие природно-очатовые ин-
фекции в Закарпатье: Автореф. дис ...
канд. биол. наук. Львов, 1976. 14 с.
Сакварелидзе Л. А., Боцвадзе 9. Ш., Нер-
сенов В. А. и др. О первых случаях
выделения псендотуберкулеза от лю-
дей в Грузии// Журн. микробиологии,
эпидемиологии и иммунологии. 1983.
No 7. С. 117-118.
Самарский С. Л., Картель М. В. Размно-
жение серой крысы на экивотновод-
чоских комплексах центральной лесо-
степи УССР // Серая крыса. М.: Наука,
1986. Т. 1. С. 129-138.
Сатунин К. А. О млекопитающих степей
северо-восточного Кавказа // Изв. Кавк.
музея. 1904. Т. 1, вып. 4. С. 1-10.
Сахаров П. П. Лабораторные животные.
М.; Л.: Издво ЛГУ, 1937. 228 с.
Свириденко П. А. Питание мышевидных
грызунов и их значение в проблеме
возобновления леса // Зоол. журн. 1940.
Т. 19, No4. С. 681-708.
Север Т. А. Грызуны города Красновод-
ска // Тр. Туркм. противочум. станции.
1958. Т. 1. С. 164—164.
Селимов М. А. Бешенство. М.; Медицина,
4978. 336 с.
Семаков В. В., Семивина В. П., Выро-
дов В. А. и др. А. е. 904209 СССР, МКИ
А ОГМ 31/00. Репеллент против гры-
зунов. Заявл. 21.07.80; Опубл. 12/У. 1983,
Бюл. No 3.
Семаков В. В., Семикина В. Н., Выробов
В.И.идр.А.с.914049СССР,МКИА
ОХ 61/00. Репеллент против мышевид-
ных трызунов. Заявл. 21.07.80; Опубл.
5ЛИ 1984, Бюл. No 10.
Сержанин И. ПН. Млекопитающие Бело-
руссии. Минск, 1961. 348 с.
Сильвестров В. С., Стариков А. Е., Се-
менов В. Е. и др. Особенности распро-
странения серой крысы в открытых
местообитаниях г. Москвы// ТУ съезд.
Всесоюз. териол. о-ва: Тез. докл. М.
1986. Т. 3. С. 232.
Синичвина А. А. Сезонные и возрастные
изменения реакции серых крыс на
влияние температуры среды // Науч-
конф. по природ. очаговости и эпиде-
миям 06060 опасных инфекций. М.:
Медгиз, 1957. С. 18—20.
Синцова ПЛ. Я., Серебрякова Л. К. Ис-
пользование бактородентицидов в борь-
бе с грызунами-Исполуване на бак-
тороденцида борбота с грызачите /
Биол. средства за раст. защита. С.:
Земиздат, 1975. С. 457-464. На болг.’
яз.
Скалон В. Н. К изучению фауны грызу-
нов Восточно-Сибирского края // Труды
по защите растений Восточной Сибири.
Иркутск, 1933. С. 45—50.
Скалов В. Н. К фаупе млекопитающих
Кентейского аймака Монгольской На-
родной Республики // Бюл. МОИП. Отд.
биол. 1949. Т. 54, No 3. С. 83—45.
Скурат Л. Н., Ляпунова К. Л., Дадикян
К. М. Поведенческие реакции серых
крыс (В. погуезсиз. погу.) на запах
специфических желез самцов и самок
своего вида // ТГ Всесоюз. совещ. по хим.
Литература
447
коммуникации. М.: ВАСХНИЛ, 1979.
С. 161—163.
Славин Н. И., Мединский Г. Н. К воп-
росу о носительстве серыми крысами
лецтоспир типа моняков в Эстонской
ССР // Журн. микробиологии, эпидемио-
логии и иммунобиологии. 1956. No 9.
С. 57—60.
Слоним А. Д. Среда и поведение: Фор-
мирование адаптивного поведения. Л.:
Наука, 1976. 270 с.
Слудский А. А., Бекенов А., Борисенко
В. А. и др. Млекопитающие Казахста-
на. Алма-Ата, 1977. Т. 1, ч. 2. 536 с.
Смирин В. М. Позы и выразительные
движения серой крысы // Серая крыса.
М.: АН СССР, 1986. Т. 2. С. 242—254.
Смирнов Е. Н. К методике определения
возраста серых крыс // Зоол. журн. 1968.
Т. 41, No 1. С. 922-924.
Соболева Г. В., Синцова Л. Н., Свободо-
ва А. А., Карандаева В. В. Изучение
причины невосприимчивости некоторых
видов грызунов к ЗавпопеПа епфег!13 //
Бактериальные средства и методы
борьбы с насекомыми и грызунами.
Л.: Колос, 1972. С. 147-120.
Соболева Г. В., Смирнова К. В., Залеж-
ский Г. В. Материалы по заселенности
грызунами и методике дератизации го-
родских свалок// Центр. пауч.-исслед.
дезинфекц. ин-т. Тез. докл. науч. конф.
ин-та, 1957 г. М.: Минздрав СССР, 1957.
С. 445-146.
Соколов В. Е. Систематика млекопитаю-
щих. М.: Изд-во МГУ, 1977. 494 с.
Соколов В. Е., Анисимова О. Ю., Семио-
`` ина Е. И., Зинкевич 9. Н. Обонятель-
ная коммуникация у серых крыс //
Распространение и экология серой кры-
сы и методы ограничения ее числен-
ноети. М.: Наука, 1985. С. 208—218.
Соколов В. Е., Граевская Б. М., Золота-
рева Н. Н. и др. Метаболические и по-
веденческие реакции крыс на обоня-
тельный сигпал: Зависимость от поло-
жения 0с0би в иерархической струк-
туре группы
// Журн. общ. биологии.
1980. Т. 41, No 2. С. 219-225.
Соколов В. Е., Заумыслова 0. Ю., Зин-
`кевич Э. И. Распознавание самцами ее-
рых крыс пола особей своего и чужо-
го вида по обонятельным сигналам //
` Экология и медицинское значение сс-
рой крысы (Ва Ми погуезеаз ВегК).
М.: Наука, 1983а. С. 50—56.
Соколов В. Е, Заумыслова 0. Ю., Зинке-
вич 9. П. Экспериментальная модель
для изучения влияния обонятельных
сигналов на агрессивное поведение
самцов серой крысы (В. погусб1си8
ВегК) // Химическая коммуникация жи-
вотных: Теория и практика. М.: Нау-
ка, 1986. С. 345-349.
Соколов В. Е., Карасева Ё. В. Серая кры-
са —- жизненная форма тгрызуна-синан-
тропа // Экология и медицинское зна-
чение серой крысы (ПаИаз погуер1с0$
ВегК). М.: АН СССР, 1983. С. 4-8.
Соколов В. Е., Карасева Е. В. Серая кры-
са — жизненная форма грызуна-синан-
тропа // Распространение и экология се-
рой крысы и методы ограничения ее
численности. М.: Наука, 1985. С. 6-
47
Соколов В. Е., Квашнин С. А. Влияние
родителей на развитие, поведение и
взаимоотношения молодых серых крыс
(ВаНиз погуез1сиз ВегК) // Материалы
1 Всесоюз. конф. по поведению жи-
вотных. М.: Наука, 1983а. Т. 1. С. 143—
1Ал.
. Соколов В. Е., Квашнин С. А. Онтогенез
поведения серых крыс в послегнездо-
вой период. Сообщ. 1, 2// Экология и
медицинское значение серой ысы
(Ваз потуезсив ВегК). М.: АН СССР,
-” 19886. С. 47-50.
Соколов В. Е., Квашнин С. А. Оптогепез
поведения диких серых крыс в после-
гисздовой период // Распростраиение и
экология серой крысы и методы отра-
ничения се численности. М.; Наука,
1985. С. 147—470.
Соколов В. Е., Ляпунова К. Л. Реакция
серых крыс на обонятельные сигналы
особей своего вида в зависимостиот
пола и социальпого ранга // Зоол. журн.
1979. Т. 58, No 2. С. 218—229.
Соволов В. Е., Ляпунова К. Л. Особен-
ности иерархической структуры сооб-
ществ серых крыс (Ваз погуоси$
погуев1сиз Вегк) в условиях вольерно-
го содержания
// Ферономы и поведе-
ние, М.: Наука, 1982. С. 162—179.
Соколов В. Е. Ляпунова К. Л., Зинке-
вич 9. П. Химическая коммуникация
серой крысы (ВаИлз погуес!1е18 погуе-
21с18 ВсгЕ) // Там же. 1982. С. 179-195.
Соколов В. Е, Ляпунова К. Л., Зинке-
вич 9. П., Судейкин В. А. А. с. 877807
СССР. Феромон серых крые для их от-
лова. Опубл. 1.07.81.
Соколов В. Е., Ляпунова К. Л., Котенко-
ва Е. В. и др. Распознавание принад-
лежности особи к своему виду по обо-
нятельным сигналам у серых крыс, до-
мовых мышей и больших песчанок //
416
Литература
фекции и стерилизации. М.: Минздрав
СССР, 1985. С. 148—153.
Рыльников В. А., Звонарев А. В. Анализ
популяции серых крыс в процессе их
истребления антикоагулянтами // Серая
крыса. М.: АН СССР, 1986. Т. 2. С. 416—
139.
Рыльников В. А., Карасева Е. В. Особен-
ности экологии серых крыс на рисо-
вых полях Кубани и меры ограничения
их численности// Распространение и
экология серой крысы и методы огра-
ничения се численности. М.: Наука,
1985. С. 71—112.
Рыльников В. А., Карасева Е. В., Дуби-
нина Н. В. Применение гистологиче-
ских красителей и тетрациклина для
изучения подвижности серых крыс на
рисовых полях Краснодарского края Й
Зоол. журн. 1984. Т. 60. No 6. С. 264-
265.
Рыльников В. А., Карасева Е. В., Дуби-
нина Н. В. Стратегия борьбы с серой
крысой в природных очагах лептосии-
роза на рисовых полях // Экология и
медицинское значение серой крысы
(Пабиз погусй1сл8 ВегК). (материалы
1-го рабоч. совещ. по серой крысе,31 ян-
варя —3 февраля 1983 г. Москва). М.:
АН СССР, 1983. С. 71-73.
Рыльников В. А., Сиротинина Е. В. Пи-
щевая активность и взаимоотношения
серых крыс около кормушек с неотрав-
ленной пищей в условиях их естест-
венного обитания // Вопросы дезинфек-
ции и стерилизации: Сб. науч. тр. М.:
Минздрав СССР, 4986. С. 162—170.
Рябцева Н. Л. Грызуны-синантропы Вос-
точного Закавказья и особенности про-
филактики чумы в населенных пунк-
тах // Эпидемиология и эпизоотология
чумы. Саратов, 1980. С. 59—64.
Сакаль Н. Н. Псевдотуберкулез и неко-
торые другие природно-очатовые ин-
фекции в Закарпатье: Автореф. дис ...
канд. биол. наук. Львов, 1976. 14 с.
Сакварелидзе Л. А., Боцвадзе 9. Ш., Нер-
сенов В. А. и др. О первых случаях
выделения псендотуберкулеза от лю-
дей в Грузии// Журн. микробиологии,
эпидемиологии и иммунологии. 1983.
No 7. С. 117-118.
Самарский С. Л., Картель М. В. Размно-
жение серой крысы на экивотновод-
чоских комплексах центральной лесо-
степи УССР // Серая крыса. М.: Наука,
1986. Т. 1. С. 129-138.
Сатунин К. А. О млекопитающих степей
северо-восточного Кавказа // Изв. Кавк.
музея. 1904. Т. 1, вып. 4. С. 1-10.
Сахаров П. П. Лабораторные животные.
М.; Л.: Издво ЛГУ, 1937. 228 с.
Свириденко П. А. Питание мышевидных
грызунов и их значение в проблеме
возобновления леса // Зоол. журн. 1940.
Т. 19, No4. С. 681-708.
Север Т. А. Грызуны города Красновод-
ска // Тр. Туркм. противочум. станции.
1958. Т. 1. С. 164—164.
Селимов М. А. Бешенство. М.; Медицина,
4978. 336 с.
Семаков В. В., Семивина В. П., Выро-
дов В. А. и др. А. е. 904209 СССР, МКИ
А ОГМ 31/00. Репеллент против гры-
зунов. Заявл. 21.07.80; Опубл. 12/У. 1983,
Бюл. No 3.
Семаков В. В., Семикина В. Н., Выробов
В.И.идр.А.с.914049СССР,МКИА
ОХ 61/00. Репеллент против мышевид-
ных трызунов. Заявл. 21.07.80; Опубл.
5ЛИ 1984, Бюл. No 10.
Сержанин И. ПН. Млекопитающие Бело-
руссии. Минск, 1961. 348 с.
Сильвестров В. С., Стариков А. Е., Се-
менов В. Е. и др. Особенности распро-
странения серой крысы в открытых
местообитаниях г. Москвы// ТУ съезд.
Всесоюз. териол. о-ва: Тез. докл. М.
1986. Т. 3. С. 232.
Синичвина А. А. Сезонные и возрастные
изменения реакции серых крыс на
влияние температуры среды // Науч-
конф. по природ. очаговости и эпиде-
миям 06060 опасных инфекций. М.:
Медгиз, 1957. С. 18—20.
Синцова ПЛ. Я., Серебрякова Л. К. Ис-
пользование бактородентицидов в борь-
бе с грызунами-Исполуване на бак-
тороденцида борбота с грызачите /
Биол. средства за раст. защита. С.:
Земиздат, 1975. С. 457-464. На болг.’
яз.
Скалон В. Н. К изучению фауны грызу-
нов Восточно-Сибирского края // Труды
по защите растений Восточной Сибири.
Иркутск, 1933. С. 45—50.
Скалов В. Н. К фаупе млекопитающих
Кентейского аймака Монгольской На-
родной Республики // Бюл. МОИП. Отд.
биол. 1949. Т. 54, No 3. С. 83—45.
Скурат Л. Н., Ляпунова К. Л., Дадикян
К. М. Поведенческие реакции серых
крыс (В. погуезсиз. погу.) на запах
специфических желез самцов и самок
своего вида // ТГ Всесоюз. совещ. по хим.
Литература
447
коммуникации. М.: ВАСХНИЛ, 1979.
С. 161—163.
Славин Н. И., Мединский Г. Н. К воп-
росу о носительстве серыми крысами
лецтоспир типа моняков в Эстонской
ССР // Журн. микробиологии, эпидемио-
логии и иммунобиологии. 1956. No 9.
С. 57—60.
Слоним А. Д. Среда и поведение: Фор-
мирование адаптивного поведения. Л.:
Наука, 1976. 270 с.
Слудский А. А., Бекенов А., Борисенко
В. А. и др. Млекопитающие Казахста-
на. Алма-Ата, 1977. Т. 1, ч. 2. 536 с.
Смирин В. М. Позы и выразительные
движения серой крысы // Серая крыса.
М.: АН СССР, 1986. Т. 2. С. 242—254.
Смирнов Е. Н. К методике определения
возраста серых крыс // Зоол. журн. 1968.
Т. 41, No 1. С. 922-924.
Соболева Г. В., Синцова Л. Н., Свободо-
ва А. А., Карандаева В. В. Изучение
причины невосприимчивости некоторых
видов грызунов к ЗавпопеПа епфег!13 //
Бактериальные средства и методы
борьбы с насекомыми и грызунами.
Л.: Колос, 1972. С. 147-120.
Соболева Г. В., Смирнова К. В., Залеж-
ский Г. В. Материалы по заселенности
грызунами и методике дератизации го-
родских свалок// Центр. пауч.-исслед.
дезинфекц. ин-т. Тез. докл. науч. конф.
ин-та, 1957 г. М.: Минздрав СССР, 1957.
С. 445-146.
Соколов В. Е. Систематика млекопитаю-
щих. М.: Изд-во МГУ, 1977. 494 с.
Соколов В. Е., Анисимова О. Ю., Семио-
`` ина Е. И., Зинкевич 9. Н. Обонятель-
ная коммуникация у серых крыс //
Распространение и экология серой кры-
сы и методы ограничения ее числен-
ноети. М.: Наука, 1985. С. 208—218.
Соколов В. Е., Граевская Б. М., Золота-
рева Н. Н. и др. Метаболические и по-
веденческие реакции крыс на обоня-
тельный сигпал: Зависимость от поло-
жения 0с0би в иерархической струк-
туре группы
// Журн. общ. биологии.
1980. Т. 41, No 2. С. 219-225.
Соколов В. Е., Заумыслова 0. Ю., Зин-
`кевич Э. И. Распознавание самцами ее-
рых крыс пола особей своего и чужо-
го вида по обонятельным сигналам //
` Экология и медицинское значение сс-
рой крысы (Ва Ми погуезеаз ВегК).
М.: Наука, 1983а. С. 50—56.
Соколов В. Е, Заумыслова 0. Ю., Зинке-
вич 9. П. Экспериментальная модель
для изучения влияния обонятельных
сигналов на агрессивное поведение
самцов серой крысы (В. погусб1си8
ВегК) // Химическая коммуникация жи-
вотных: Теория и практика. М.: Нау-
ка, 1986. С. 345-349.
Соколов В. Е., Карасева Ё. В. Серая кры-
са —- жизненная форма тгрызуна-синан-
тропа // Экология и медицинское зна-
чение серой крысы (ПаИаз погуер1с0$
ВегК). М.: АН СССР, 1983. С. 4-8.
Соколов В. Е., Карасева Е. В. Серая кры-
са — жизненная форма грызуна-синан-
тропа // Распространение и экология се-
рой крысы и методы ограничения ее
численности. М.: Наука, 1985. С. 6-
47
Соколов В. Е., Квашнин С. А. Влияние
родителей на развитие, поведение и
взаимоотношения молодых серых крыс
(ВаНиз погуез1сиз ВегК) // Материалы
1 Всесоюз. конф. по поведению жи-
вотных. М.: Наука, 1983а. Т. 1. С. 143—
1Ал.
. Соколов В. Е., Квашнин С. А. Онтогенез
поведения серых крыс в послегнездо-
вой период. Сообщ. 1, 2// Экология и
медицинское значение серой ысы
(Ваз потуезсив ВегК). М.: АН СССР,
-” 19886. С. 47-50.
Соколов В. Е., Квашнин С. А. Оптогепез
поведения диких серых крыс в после-
гисздовой период // Распростраиение и
экология серой крысы и методы отра-
ничения се численности. М.; Наука,
1985. С. 147—470.
Соколов В. Е., Ляпунова К. Л. Реакция
серых крыс на обонятельные сигналы
особей своего вида в зависимостиот
пола и социальпого ранга // Зоол. журн.
1979. Т. 58, No 2. С. 218—229.
Соволов В. Е., Ляпунова К. Л. Особен-
ности иерархической структуры сооб-
ществ серых крыс (Ваз погуоси$
погуев1сиз Вегк) в условиях вольерно-
го содержания
// Ферономы и поведе-
ние, М.: Наука, 1982. С. 162—179.
Соколов В. Е. Ляпунова К. Л., Зинке-
вич 9. П. Химическая коммуникация
серой крысы (ВаИлз погуес!1е18 погуе-
21с18 ВсгЕ) // Там же. 1982. С. 179-195.
Соколов В. Е, Ляпунова К. Л., Зинке-
вич 9. П., Судейкин В. А. А. с. 877807
СССР. Феромон серых крые для их от-
лова. Опубл. 1.07.81.
Соколов В. Е., Ляпунова К. Л., Котенко-
ва Е. В. и др. Распознавание принад-
лежности особи к своему виду по обо-
нятельным сигналам у серых крыс, до-
мовых мышей и больших песчанок //
418
Литература
1 Всесоюз. совещ. по хим. коммуника-
ции животных: Тезисы. М. 1979а. С. 167.
Соколов В. Е, Ляпунова К. Л., Суров
В. А., Зинкевич 9. П. Феромон припад-
лежности к своему виду у сорых крыс
(ПаНиз погуее1си8 погу.): «Обонятель-
ный образ//П Междунар. териол.
конгр. Брно, 1978. с. 383.
Соколов В. Е., Ляпунова К. Л., Суров А.В.
и др. Феромон принадлежности особи
к своему виду у серых крыс {Ваз
погуса1ств // Докл. АН СОСР. 19796.
Т. 248, No 6. С. 1506—1509.
Соколов В. Е., Ляпунова К. Л., Хорли-
на И. М. Особенности поведения и фор-
мирования социальной структуры труп-
пы серых крыс (В. погуес1сиз ВегК) в
искусственных условиях // Поведение
млекопитающих. М.: Наука, 1977. С. 84—
106.
Соколов В. Е, Пронъкова 0. Ю., Зинке-
вич 9. ПН. Распознавание самками се-
рых крыс (НаЙмз погуса1е0з) пола и
физического состояния сородичеи по
обопятельным сигналам // П Всесоюз.
совещ. по хим. коммуникации живот-
ных. М.: АН СССР, 19836. С. 126.
Соколов Б. Е, Шилова С. А, Шутова М.И,,
Карасева Е. В. Экономический ущерб,
наносимый серой крысой // Серая кры-
са. М.. АН СССР, 1986ба. Т. 1. С. 5-16.
Соколов В. С. Алкалоидные растения
СССР. М.: Минздрав СССР, 1952. 815 с.
Сокур И. Т. Млекопитающие фауны Ук
раины и их хозяйственное значение. Ви-
ев: Рад. тик. 1960. 241 с. На укр. яз.
Солдатов Г. М., Шереметьев С. А., Лоши-
ков В. Е. Распрострапение и динамика
численности серой крысы (НаНаз пог-
уес1сив ВегК) // Материалы | рабочего
совещ. по серой крысе. М.: Наука, 1983.
С. 43—44,
Солитерман Н. Л. Экспериментальные
наблюдения по эондемическому сыпному
тифу //Риккетсии и риккетсиозы. М.:
АМН СССР, 1948. 205 с.
Соловьев В. И., Бабенко Н. П., Бондарь
Л. Н. и др. Распространение серой кры-
сы на Правобережной Украине //
ТУ съезд Всесоюз. териол. о-ва; Тез.
докл. М., 1986. Т. 3. С. 272—273.
Соломкин Н. С. Листереллез сельскохо-
зяйственных животных. М.: Сельхозгиз,
4959. 62 с.
Сомов Г. П. Дальневосточная скарлатино-
подобная лихорадка. М.: Медицина,
1979. 184 с.
Сомов Г. Н., Шубин Ф. Н., Топаченко И.М.
и др. Изучение зооантропонозных ин-
фекций на Чукотке // Природно-очаго-
вые заболевания в зонах освоения Си-
бири и Крайнего Севера. Новосибирск,
1977. С. 16—21. (Науч. тр. Новосиб. фил.
АН СССР),
:
Сомова Н. Г., Герасюк Л. Г., Афанасьева
М. К. и др. К вопросу об эндемическом.
крысином сыпном тифе на Черномор-
ском побережье // Журн. микробиоло-
гии, эпидемиологии и иммупобиологии.
1960. No 2. С. 51—56.
Сорокина 3. С., Бацин В. Т., Коржов НП. Н.
Размещение и числепность серой кры-
сы на территории Дагестанской, Че-
чено-Ингушской, Северо-Осетинской
АССР и восточных районов Ставроцоль-
ского края // Серая крыса. М.: Наука,
1986. Т. 1. С. 83—91.
Сорокина 3. С., Потапова Е. А. Грызуны
населенных пунктов Дагестанского гор-
ного очага чумы // Особо опасные ин-
фекции на Кавказе. Ставрополь, 1987.
С. 307—309.
Соснихина Т. М. Грызуны города Кирова-
кана // Изв. АН АрыССР. Естеств. нау-
ки. 1946. No 3. С. 67—78.
Соснихина Т. М. Серый хомячок Сиее-
118 ииогаогназ Ра] в условиях Армян-
ской ССР // Зоологический сборник АН
Армянской ССР. Ереван, 1950. Вып. 7.
С. 55—82.
Сосницкий В. И. Природные очаги ту-
ляремии в Архангельской области. При-
родно-очаговые инфекции Тр. Ин-та
эпидемиологии и микробиологии
им. Пастера. 1973. Т. 44. С. 149—161.
Соспицкий В. И. Источники лептоспироза
в городе Архангельске // Тез. докл.
УП Всесоюз. конф. по лептоспирозам.
Киев: Наук. думка, 1979. С. 158—160.
Сосницкий В. И, Некрасова Л. И. Мате
риалы к эпидемиологии туляремии в
Ненецком национальном округе Архан-
тельской области // Тр. Ип-та эпидеми-
ологии и микробиологии им. Пастера.
1966. Т. 34. С. 83-93.
Сосновский Д. И. Опыт классификации
растительных формаций Грузии // За-
кавк. краевед. сб. Сер. А: Естествозна-
ние. 1930. Вып. 4. С. 128—132.
Спиченко Д. А. К методике борьбы с сс-
рой крысой // Проблемы зоологических
исследований в Сибири. Горно-Алтайск:
Кн. изд-во, 1962. С. 224—226.
Стадутин О. В. Пасюни заселяют новые
территории з Свердловской области // Тр.
П Всесотоз. совещ. по млекопитающим.
М.: Изд-во МГУ, 1975. С. 238.
Литература
4419
Стальмакова В. А. О нахождении черепа
туркестанской крысы (Ваз вигКезёа-
11005 Завиш) в низовьях Амударьи //
Тр. Ил-та биологии АН ТССР. 1956.
Т. 4. С. 55-57.
„Степанов И. В. Материалы по биологии
размножения крыс и мышей в Бату-
ми // Бюл. МОИП. Отд. биол. 1946.
Т. 51, No 6. С. 29-35.
Степанова Н. В., Тимошков В. В., Буви-
на С. Г. и др. Серая крыса
— носитель
возбудителя зоонозных инфекций в
г. Москве //ТУ съезд Всесоюз. териол.
о-ва: Тез. докл. М., 1986. Т. 3. С. 808—
304.
'Степин Е. К. Грызуны низовий Амура //
Изв. Иркут. науч.-исслед. противочум.
ин-та Сибири и Дал. Востока. 41952.
Т. 10. С. 15—47.
`Стержова Н. Н., Ахундова 9. Д. Результа-
ты серологических исследований на кле-
щевой сыпной тиф в Азербайджан-
ской ССР //Тр. НИИ вирусологии, мик-
робиологии и гигиены им. Мусабекова.
1971. Т. 24. С. 62-66.
Стогов В. И. Серая крыса и опыт борьбы
с ней в Алма-Ате //ТУ съезд Всесоюз.
териол. о-ва: Тез. докл. М., 1986. Т. 3.
С. 385—387.
Стогов В. И., Байтанаев О. А. Серая кры-
са в зоне канала Иртыш- Караганда //
Экология. и медицинское значение се-
рой крысы (Ваз погуес1еиз ВегК). М.:
Наука, 1983. С. 14-15.
Страупмание В. Х. Интенсивность засе-
ления серой крысой договорных объек-
тов отдельных городов и сельских райо-
нов Латвийской ССР //ТГУ съезд Все-
союз. териол. о-ва; Тез. докл. М., 1986.
Т. 3. С. 280-231.
'
Судейкин В. А. Итоги борьбы с серой кры-
сой в г. Москве за последние десятиле-
тия // Тр. П Воссотоз. совещ. но млеко-
питаюцщим. М.: зд-во
Г
5.
С. 284-286.
^ МГУ, 1975
Судейкин В. А. Миграции серых крыс в
условиях большого города // Фауна и
экойогия грызунов. М.: Изд-во МГУ
. 1978. Выш. 3. С. 41-85.
у
Судейвин В. А. Опыт оценки экономиче-
ского ущерба, причиняемого серыми
. крысами в Москве // Экология и меди-
цинское значение серой крысы (ВаНлз
погуер1ст8 ВееК). М.: АН СССР, 1983.
С. 87-89.
Судейкин В. А. Некоторые материалы о
вреде, наносимом серыми крысами здо-
ровью людей // Экология и медицин-
ское значение серой крысы (ВаКаз пог-
уер1стз ВегК). М, АН СССР, 1983а.
С. 84-85.
`
Судейкии В. А. Город как среда обитания
серой крысы // Серая крыса. М.: Нау-
ка, 1986. Т. 1. С. 34-53.
Судейкин В. А., Ляпунова К. Л., Зинке-
вич Э. П. Опыт исследования некото-
рых компонентов мочи серых крыс при
их в отлове // Грызуны: Материалы
сесоюз, совещ. Л.: Наука, 1983.
С. 595—596. м
ука, 1983
Судейкин В. А., Харламов В. П., Судейки-
на М. В. Опыт изучения миграции се-
рых крыс в условиях большого города
методом радиоактивной маркировки //
зоол. журн. 1962, Т. 24, No 9, С. 1409-
Сурков В. С. Эпидемиологическое и эко-
номическое значение серой крысы на
Сахалине // Докл. Иркут. науч.-иселед.
противочум. ин-та Сибири и Дал. Вос-
тока. 1971. Выш. 9. С. 205-206.
Сурков В. С. Мышеобразные (Миг!Чае) и
хомякообразные (М1сгоНпае) Сахалина
и их роль как носителей природно-оча-
говых инфекций и гельминтозов: Авто-
реф. дис. ... канд. биол. наук. Владиво-
сток, 1972. 24 с.
Сурков В. С. О некоторых экологических
особенностях серой крысы на Сахали-
не и Курильских островах // Экология
и медицинское значение серой крысы
(Вабиз погуео1ст8 ВегЕ). М.: АН СССР
1983, С. 39—44.
'
Сурков В. С. Некоторые экологические
особенности серой крысы Сахалина и
Южных Курил // Серая крыса. М.: Нау-
ка, 1986а. Т. 4. С. 114-128.
Сурков В. С. К вопросу о питании серой
крысы, обитающей в естественных ланд-
птафтах о-ва Сахалина // ТУ съезд Все-
союз. териол. о-ва: Тез. докл. М., 19866.
Т. 3. С. 244-245.
Сурков В. С., Гайдукова Н. С. К структу-
ре очагов псевдотуберкулеза на Саха-
лине // Проблемы особо опасных инфек-
ций Саратов, 1972. Вып. 3(25). С. 199—
Сурков В. С., Рябов С. В. К вопросу о чис-
ленпости и размножении серой крысы
в портовых городах Сахалина // Эколо-
гия и медицинское значение серой кры-
сы (Вай погус!сз ВегК). М.: Нау-
ка, 1983. С. 44—43.
Тажибаезв А. С. Профилактика болезней,
распространяемых
мышевидными
грызунами. Алма-Ата: Кайнар, 1979.
с.
418
Литература
1 Всесоюз. совещ. по хим. коммуника-
ции животных: Тезисы. М. 1979а. С. 167.
Соколов В. Е, Ляпунова К. Л., Суров
В. А., Зинкевич 9. П. Феромон припад-
лежности к своему виду у сорых крыс
(ПаНиз погуее1си8 погу.): «Обонятель-
ный образ//П Междунар. териол.
конгр. Брно, 1978. с. 383.
Соколов В. Е., Ляпунова К. Л., Суров А.В.
и др. Феромон принадлежности особи
к своему виду у серых крыс {Ваз
погуса1ств // Докл. АН СОСР. 19796.
Т. 248, No 6. С. 1506—1509.
Соколов В. Е., Ляпунова К. Л., Хорли-
на И. М. Особенности поведения и фор-
мирования социальной структуры труп-
пы серых крыс (В. погуес1сиз ВегК) в
искусственных условиях // Поведение
млекопитающих. М.: Наука, 1977. С. 84—
106.
Соколов В. Е, Пронъкова 0. Ю., Зинке-
вич 9. ПН. Распознавание самками се-
рых крыс (НаЙмз погуса1е0з) пола и
физического состояния сородичеи по
обопятельным сигналам // П Всесоюз.
совещ. по хим. коммуникации живот-
ных. М.: АН СССР, 19836. С. 126.
Соколов Б. Е, Шилова С. А, Шутова М.И,,
Карасева Е. В. Экономический ущерб,
наносимый серой крысой // Серая кры-
са. М.. АН СССР, 1986ба. Т. 1. С. 5-16.
Соколов В. С. Алкалоидные растения
СССР. М.: Минздрав СССР, 1952. 815 с.
Сокур И. Т. Млекопитающие фауны Ук
раины и их хозяйственное значение. Ви-
ев: Рад. тик. 1960. 241 с. На укр. яз.
Солдатов Г. М., Шереметьев С. А., Лоши-
ков В. Е. Распрострапение и динамика
численности серой крысы (НаНаз пог-
уес1сив ВегК) // Материалы | рабочего
совещ. по серой крысе. М.: Наука, 1983.
С. 43—44,
Солитерман Н. Л. Экспериментальные
наблюдения по эондемическому сыпному
тифу //Риккетсии и риккетсиозы. М.:
АМН СССР, 1948. 205 с.
Соловьев В. И., Бабенко Н. П., Бондарь
Л. Н. и др. Распространение серой кры-
сы на Правобережной Украине //
ТУ съезд Всесоюз. териол. о-ва; Тез.
докл. М., 1986. Т. 3. С. 272—273.
Соломкин Н. С. Листереллез сельскохо-
зяйственных животных. М.: Сельхозгиз,
4959. 62 с.
Сомов Г. П. Дальневосточная скарлатино-
подобная лихорадка. М.: Медицина,
1979. 184 с.
Сомов Г. Н., Шубин Ф. Н., Топаченко И.М.
и др. Изучение зооантропонозных ин-
фекций на Чукотке // Природно-очаго-
вые заболевания в зонах освоения Си-
бири и Крайнего Севера. Новосибирск,
1977. С. 16—21. (Науч. тр. Новосиб. фил.
АН СССР),
:
Сомова Н. Г., Герасюк Л. Г., Афанасьева
М. К. и др. К вопросу об эндемическом.
крысином сыпном тифе на Черномор-
ском побережье // Журн. микробиоло-
гии, эпидемиологии и иммупобиологии.
1960. No 2. С. 51—56.
Сорокина 3. С., Бацин В. Т., Коржов НП. Н.
Размещение и числепность серой кры-
сы на территории Дагестанской, Че-
чено-Ингушской, Северо-Осетинской
АССР и восточных районов Ставроцоль-
ского края // Серая крыса. М.: Наука,
1986. Т. 1. С. 83—91.
Сорокина 3. С., Потапова Е. А. Грызуны
населенных пунктов Дагестанского гор-
ного очага чумы // Особо опасные ин-
фекции на Кавказе. Ставрополь, 1987.
С. 307—309.
Соснихина Т. М. Грызуны города Кирова-
кана // Изв. АН АрыССР. Естеств. нау-
ки. 1946. No 3. С. 67—78.
Соснихина Т. М. Серый хомячок Сиее-
118 ииогаогназ Ра] в условиях Армян-
ской ССР // Зоологический сборник АН
Армянской ССР. Ереван, 1950. Вып. 7.
С. 55—82.
Сосницкий В. И. Природные очаги ту-
ляремии в Архангельской области. При-
родно-очаговые инфекции Тр. Ин-та
эпидемиологии и микробиологии
им. Пастера. 1973. Т. 44. С. 149—161.
Соспицкий В. И. Источники лептоспироза
в городе Архангельске // Тез. докл.
УП Всесоюз. конф. по лептоспирозам.
Киев: Наук. думка, 1979. С. 158—160.
Сосницкий В. И, Некрасова Л. И. Мате
риалы к эпидемиологии туляремии в
Ненецком национальном округе Архан-
тельской области // Тр. Ип-та эпидеми-
ологии и микробиологии им. Пастера.
1966. Т. 34. С. 83-93.
Сосновский Д. И. Опыт классификации
растительных формаций Грузии // За-
кавк. краевед. сб. Сер. А: Естествозна-
ние. 1930. Вып. 4. С. 128—132.
Спиченко Д. А. К методике борьбы с сс-
рой крысой // Проблемы зоологических
исследований в Сибири. Горно-Алтайск:
Кн. изд-во, 1962. С. 224—226.
Стадутин О. В. Пасюни заселяют новые
территории з Свердловской области // Тр.
П Всесотоз. совещ. по млекопитающим.
М.: Изд-во МГУ, 1975. С. 238.
Литература 4419
Стальмакова В. А. О нахождении черепа
туркестанской крысы (Ваз вигКезёа-
11005 Завиш) в низовьях Амударьи //
Тр. Ил-та биологии АН ТССР. 1956.
Т. 4. С. 55-57.
„Степанов И. В. Материалы по биологии
размножения крыс и мышей в Бату-
ми // Бюл. МОИП. Отд. биол. 1946.
Т. 51, No 6. С. 29-35.
Степанова Н. В., Тимошков В. В., Буви-
на С. Г. и др. Серая крыса
— носитель
возбудителя зоонозных инфекций в
г. Москве //ТУ съезд Всесоюз. териол.
о-ва: Тез. докл. М., 1986. Т. 3. С. 808—
304.
'Степин Е. К. Грызуны низовий Амура //
Изв. Иркут. науч.-исслед. противочум.
ин-та Сибири и Дал. Востока. 41952.
Т. 10. С. 15—47.
`Стержова Н. Н., Ахундова 9. Д. Результа-
ты серологических исследований на кле-
щевой сыпной тиф в Азербайджан-
ской ССР //Тр. НИИ вирусологии, мик-
робиологии и гигиены им. Мусабекова.
1971. Т. 24. С. 62-66.
Стогов В. И. Серая крыса и опыт борьбы
с ней в Алма-Ате //ТУ съезд Всесоюз.
териол. о-ва: Тез. докл. М., 1986. Т. 3.
С. 385—387.
Стогов В. И., Байтанаев О. А. Серая кры-
са в зоне канала Иртыш- Караганда //
Экология. и медицинское значение се-
рой крысы (Ваз погуес1еиз ВегК). М.:
Наука, 1983. С. 14-15.
Страупмание В. Х. Интенсивность засе-
ления серой крысой договорных объек-
тов отдельных городов и сельских райо-
нов Латвийской ССР //ТГУ съезд Все-
союз. териол. о-ва; Тез. докл. М., 1986.
Т. 3. С. 280-231.
'
Судейкин В. А. Итоги борьбы с серой кры-
сой в г. Москве за последние десятиле-
тия // Тр. П Воссотоз. совещ. но млеко-
питаюцщим. М.: зд-во
Г
5.
С. 284-286.
^ МГУ, 1975
Судейкин В. А. Миграции серых крыс в
условиях большого города // Фауна и
экойогия грызунов. М.: Изд-во МГУ
. 1978. Выш. 3. С. 41-85.
у
Судейвин В. А. Опыт оценки экономиче-
ского ущерба, причиняемого серыми
. крысами в Москве // Экология и меди-
цинское значение серой крысы (ВаНлз
погуер1ст8 ВееК). М.: АН СССР, 1983.
С. 87-89.
Судейкин В. А. Некоторые материалы о
вреде, наносимом серыми крысами здо-
ровью людей // Экология и медицин-
ское значение серой крысы (ВаКаз пог-
уер1стз ВегК). М, АН СССР, 1983а.
С. 84-85.
`
Судейкии В. А. Город как среда обитания
серой крысы // Серая крыса. М.: Нау-
ка, 1986. Т. 1. С. 34-53.
Судейкин В. А., Ляпунова К. Л., Зинке-
вич Э. П. Опыт исследования некото-
рых компонентов мочи серых крыс при
их в отлове // Грызуны: Материалы
сесоюз, совещ. Л.: Наука, 1983.
С. 595—596. м
ука, 1983
Судейкин В. А., Харламов В. П., Судейки-
на М. В. Опыт изучения миграции се-
рых крыс в условиях большого города
методом радиоактивной маркировки //
зоол. журн. 1962, Т. 24, No 9, С. 1409-
Сурков В. С. Эпидемиологическое и эко-
номическое значение серой крысы на
Сахалине // Докл. Иркут. науч.-иселед.
противочум. ин-та Сибири и Дал. Вос-
тока. 1971. Выш. 9. С. 205-206.
Сурков В. С. Мышеобразные (Миг!Чае) и
хомякообразные (М1сгоНпае) Сахалина
и их роль как носителей природно-оча-
говых инфекций и гельминтозов: Авто-
реф. дис. ... канд. биол. наук. Владиво-
сток, 1972. 24 с.
Сурков В. С. О некоторых экологических
особенностях серой крысы на Сахали-
не и Курильских островах // Экология
и медицинское значение серой крысы
(Вабиз погуео1ст8 ВегЕ). М.: АН СССР
1983, С. 39—44.
'
Сурков В. С. Некоторые экологические
особенности серой крысы Сахалина и
Южных Курил // Серая крыса. М.: Нау-
ка, 1986а. Т. 4. С. 114-128.
Сурков В. С. К вопросу о питании серой
крысы, обитающей в естественных ланд-
птафтах о-ва Сахалина // ТУ съезд Все-
союз. териол. о-ва: Тез. докл. М., 19866.
Т. 3. С. 244-245.
Сурков В. С., Гайдукова Н. С. К структу-
ре очагов псевдотуберкулеза на Саха-
лине // Проблемы особо опасных инфек-
ций Саратов, 1972. Вып. 3(25). С. 199—
Сурков В. С., Рябов С. В. К вопросу о чис-
ленпости и размножении серой крысы
в портовых городах Сахалина // Эколо-
гия и медицинское значение серой кры-
сы (Вай погус!сз ВегК). М.: Нау-
ка, 1983. С. 44—43.
Тажибаезв А. С. Профилактика болезней,
распространяемых
мышевидными
грызунами. Алма-Ата: Кайнар, 1979.
с.
420
Литература
Такад С. Пат. МКИ А 01М 31/08, А 01%
21/00 Япония. Бытовой антисептик и
инсектицидное масло. Заявл. 16.03.81;
Опубл. 22.09.82.
Танака И. Оценка родентицидов в пыле-
видных препаратах, пачкающих грызу-
нов // Тар. 7. Запй. 2001. 1973. Уо|. 23,
М 3. Р. 219-223.
Танака И., Огата К., Судзуки Т. Некото-
рые опыты определения действия нохо-
то родентицида на крыс и мышей и
1519. 1967. Ус. 18, М 1. Р. 39—43.
Тарасов М. ИП. Границы распространения
некоторых видов нтиц и млекопитаю-
щих на севере Средней Сибири // Зоол.
журн. 1965. Т, 44. No 12. С. 1835-1841. _
Татаринов К. А. Щур звичйний у верх
в'ях Дн!стра//Наук. зап. Львв. наук.
природозн. музею АН УРСР. 1954. Т. 3,
С. 77.
Татаринов К. А. Звери западных обла-
стей Украины. Киев, 1956. 188 с. На
укр. яз.
Темботов А. К. География млекопитазю-
щих Северного Кавказа. Нальчик: Эль-
брус, 1972. 245 с.
.
Теплов В. П., Теплова Е. Н. Млекопитаю-
щие Печеро-Ылычсекого зановедника //
Тр. Печеро-Ылыч. гос. заповедника.
1947. Выш. 5. С. 3—84.
Терещенко М. П. К обнаружению возбу-
дителя листереллеза у грызунов в усло-
виях большого города. Нриродная оча-
говость болезпей человека и краевая
эцидемиология // Работы научной сес-
сии МЗ СССР, АМН СССР, ИЭМ АМН
СССР. М.: Минздрав СССР, 1955. С. 182—
183.
Терещенко М. П., Есаджанян М. М., Ми-
рошниченко М. А. и др. Об эпидемио-
логическом значении овец при туляре-
мии// Журн. микробиологии, эпидемио-
логии и иммунологии, 1956. No 9. С. 34—
86.
Терещенко М. П., Родкевич Л. В., Кова-
лева Р. В. Бактериальные инфекции у
грызунов большого города // Тез. докл,
науч. конф. по природ. очаговости и
эпидемиологии 0с0бо опасных инфек-
ционных заболеваний, Саратов: «Мик-
роб», 1957. С. 383—384.
Тимофеева А. А. Грызуны на судах Са-
халинекой области // Докл. Иркут. про-
тивочум. ин-та. Хабаровск, 1962. Выш. 3.
С. 162—164.
Тимофеева А. А., Сурков В. С. К эпиде-
миологической оценке основных ланд-
шафтов Сахалина в связи с биотопиче-
ским распредолением грызунов // Изв.
Иркут. пауч.-иселед. противочум. ин-та
Сибири и Дол. Востока. 1968. Т. 27.
С. 75-88.
.
Тихенко И. И., Таран И. Ф., Лямкин Г. И.
К изучению механизмов циркуляции
возбудителя бруцеллеза среди мыше-
зилвых- грызунов // Болезни с природ-
ной очаговостью на Кавказе: (Тез. докл.
краев. науч. конф., посвящ. 60-летию
образования СССР). Ставрополь, 1982.
С. 136—137.
Тилвинская М. В. Материалы к экологии
серой крысы (ВаНаз погуер!сиз ВегК)
в природных биотопах // Учен. зап. Чу-
ваш. гое. пед. ин-та. 1964. Вып. 20.
С. 48—60.
Тихвинский В. И. Биология рыжеватого
суслика // Работы Волжеко-Камской зо-
нальной охотничьс-промысловой биоло-
гической станции. Казань: Изд-во Ка-
зан. ин-та, 1932. Вып. 2. С. 28-39.
Тихомиров А. А., Корчагин А. Н. Списки
и описание предметов, находящихся в
Зоологическом музее имп. Моск. ун-та.
Отдел четвертый. Списки и описание
коллекции млекопитающих музея // Изв.
имп. о-ва любителей естествознания,
1889. Т. 56, вып. 4, No 1. С. 1500.
Токалова К. А., Нопова Е. М., Сушкевич
Н. И. Результаты дальнейшего изуче-
ния лептоспирозной инфекции в Кали-
нинградской обл. // Тр. Ин-та эпидемио-
логии и микробиологии им. Пастера.
1963. Т. 25. С. 53—57.
Токарева Т. Г., Туров И. С., Алексеева
А. Н..Действие фторацетамида на пло-
довитость белых мышей: (Предвар. со-
общ.) // Журн. микробиологии, эпиде-
миологии и иммунобиологии. 1974. No 8.
С. 24—26.
Токаревич К. П. Лептоспирозная желтуха.
(болезнь Вейля) в Ленинграде во время,
Великой Отечественной войны и блока-
ды. Л.: Медгиз, 1947. 120 с.
Токаревич К. Н., Гавеман А. К. Возбуди-
тели болезни Боткина—Вейля у диких
крыс в Ленинграде // Сов. врачеб. журн.
1986. No 7. С. 511—516.
Токаревич К. Н., Германова Е. И., Гаве-
ман А. К. О спонтанной зараженноети
крыс бактериями псевдотуберкулеза //
Вестн. микробиологии, эпидемиологии
и паразитологии. 1940. Т. 19, No 2.
С. 226—235.
Тонышева В. С., Шиндяпин А. И. Меро-
приятия по борьбе с грызунами ‘в Мат-
чинском ‘районе // Материалы Ш Респ.
Литература 424
науч.-практ, конф. по дезинфекц. делу.
Дунтанбе, 1968. С. 144—117.
Тощигин Ю. В. Методика и организация
борьбы с серой крысой в канализаци-
онных сооружениях тородон и уголь-
ных шахтах: Автореф. дис. .. канд.
биол. наук. М., 1968. 16 с.
Тощигин Ю. В. Определение численно-
сти серой крысы с помощью пылевых
площадок // Экология и медицинское
значение серой крысы (Ваз погуер1-
смз ВегК). М.: АН СССР, 1983. С. 92—93.
Тошщигин Ю. В. Особенности экологии се-
рой крысы в подземных сооружениях и
методы ограничения ее численности //
Серая крыса, М.: АН СССР, 1986, Т. 2.
С. 69—88.
Тощигин Ю. В., Рыльников В. А. Совре-
менное состояние и перспективы регу-
ляции численности серой крысы// Рас-
пространенив и экология серой крысы и
методы ограничения ее численности. М.:
Наука, 1985. С. 242—273.
Тощигин Ю. В., Жукова Л. Д., Нодъем-
щикова Л. Г. и др. К вопросу об органи-
зации и методике практически полного
уничтожения синантропных грызунов
в населенном пункте// Проблемы дезин-
фекции и стерилизации. М.: Минздрав
СССР, 1976. Вып. 26. С. 240—245.
Тошигин Ю. В., Жукова Л. Д., Сирота
В. И. и др. Об оценке эффективности
дератизации в населенных пунктах //
Проблемы дезинфекции и стерилизации:
(Материалы симпоз. 23—25 нояб.
1977 г.). М.: Минздрав СССР, 1977. Ч. 2.
й
.
”., С. 98-99.
Тощигин Ю. В., Писаренко Г. Я., Кривуля
С. Д. и др. Н вопросу о стратегии борь-
бы с грызунами в крунном городе //
Грызуны: Материалы У Всесоюз. со-
веш. М.: Наука, 1980. С. 456—457.
Тощигин Ю. В., Полежаев В. Г., Грепачев
И. В. и др. Методика и организация
практически полного уничтожения се-
рых крыс на участке города // Матери-
алы Всесо1оз. конф. по вопр. дезинфек-
ции и стерилизации (29—30 селт.—1—
2 ‘окт. 1969 г., Москва). М.: Минздрав
СССР, 4969. С. 444.
Тощигин Ю. В., Харченко А. П. Распро-
странение, гнездование и условия про-
кормления серых крыс в угольных шах-
тах и канализациопных системах го-
родов // Тр. Центр. науч.-исслед. дезин-
фекц. ин-та. 1967. Вып. 18. С. 66.
Траут И. И. Численность и размещение
грызунов в населенном пункте город-
ского типа // Грызуны и борьба с ними.
Саратов, 1950. Вып. 3. С. 243—254.
Траханов Д. Ф. Изучение отпугивающего
действия чернокорня на серых крыс //
Тр. ВНИИ вет, санитарии. 1960. Т. 416.
С. 15—20.
Тратанов Д. Ф. Разработка эффективного
метода дератизации птичников: Азвто-
реф. дис. „. канд. биол. наук. М., 1964а.
с.
Траханов Д. Ф. `Комбинированный метод
борьбы с крысами // Тр. ВНИИ вет. са-
нитарии. 19616. Т. 19. С. 205-211.
Траханов Д. Ф. Защитно-рефлекторные
реакции серых крыс на отравленные
приманки // Проблемы ветеринарной са-
нитарии. М.: Медицина, 1963. Т. 22.
С. 162-172.
Траханов Д. Ф. Материалы к обоснованию
метода опыливания при дератизации //
Проблемы ветеринарной санитарии. М.:
Колос, 1964. С. 189—194.
Траланов Д. Ф. Дератизация в объектах
животноводства СССР: Автореф. дис.
„.ора с.-х. наук. М., 1978. 34 с.
Траханов Д. Ф. Меры борьбы с синантроп-
ными грызунами // Санитария и гигие-
на содержания животных. М.: Колос,
1981. С. 94—94.
Траханов Д. Ф. Пути повышения эффек-
тивности дератизации в животноводст-
ве // Экология и медицинское значение
серой крысы (Вайиз погуес1са$ ВегК).
М.: АН СССР, 1983. С. 98—99.
Траханов Д. Ф. Современное состояние и
перспективы ветеринарных разработок
по дератизации // Проблемы ветеринар-
ной санитарии и зоогигиены в промыш-
ленном животноводстве. М.: Колос,
1985. С. 81—86.
Траханов Д. Ф., Кадиров А. Ф. Проблемы
защиты животноводческих комплексов
‚от грызунов // Современные направле-
ния медицинской дезинфекции и дера-
тизации, М.: Минздрав СССР, 1981.
С. 163.
Триполитова А. А., Борисова Г. В. Лис-
териоз. Томек: Изд-во Том. ин-та, 1965.
257 с.
:
Троп И. Е., Чуловский И. К. Обнаружение
патогенных лептоснир типов иктероге-
моррагиа У серых крыс в г. Омске//
Тр. Омского НИИ эпидемиологии, мик-
робиологии и гигиены. 1959. No 6. С. 21—
23
Троп И. Е. Чуловский И. К. Заражен-
ность патогенными микроорганизмами
кишечной группы серых крыс, отлов-
ленных на пищевых предприятиях и
420
Литература
Такад С. Пат. МКИ А 01М 31/08, А 01%
21/00 Япония. Бытовой антисептик и
инсектицидное масло. Заявл. 16.03.81;
Опубл. 22.09.82.
Танака И. Оценка родентицидов в пыле-
видных препаратах, пачкающих грызу-
нов // Тар. 7. Запй. 2001. 1973. Уо|. 23,
М 3. Р. 219-223.
Танака И., Огата К., Судзуки Т. Некото-
рые опыты определения действия нохо-
то родентицида на крыс и мышей и
1519. 1967. Ус. 18, М 1. Р. 39—43.
Тарасов М. ИП. Границы распространения
некоторых видов нтиц и млекопитаю-
щих на севере Средней Сибири // Зоол.
журн. 1965. Т, 44. No 12. С. 1835-1841. _
Татаринов К. А. Щур звичйний у верх
в'ях Дн!стра//Наук. зап. Львв. наук.
природозн. музею АН УРСР. 1954. Т. 3,
С. 77.
Татаринов К. А. Звери западных обла-
стей Украины. Киев, 1956. 188 с. На
укр. яз.
Темботов А. К. География млекопитазю-
щих Северного Кавказа. Нальчик: Эль-
брус, 1972. 245 с.
.
Теплов В. П., Теплова Е. Н. Млекопитаю-
щие Печеро-Ылычсекого зановедника //
Тр. Печеро-Ылыч. гос. заповедника.
1947. Выш. 5. С. 3—84.
Терещенко М. П. К обнаружению возбу-
дителя листереллеза у грызунов в усло-
виях большого города. Нриродная оча-
говость болезпей человека и краевая
эцидемиология // Работы научной сес-
сии МЗ СССР, АМН СССР, ИЭМ АМН
СССР. М.: Минздрав СССР, 1955. С. 182—
183.
Терещенко М. П., Есаджанян М. М., Ми-
рошниченко М. А. и др. Об эпидемио-
логическом значении овец при туляре-
мии// Журн. микробиологии, эпидемио-
логии и иммунологии, 1956. No 9. С. 34—
86.
Терещенко М. П., Родкевич Л. В., Кова-
лева Р. В. Бактериальные инфекции у
грызунов большого города // Тез. докл,
науч. конф. по природ. очаговости и
эпидемиологии 0с0бо опасных инфек-
ционных заболеваний, Саратов: «Мик-
роб», 1957. С. 383—384.
Тимофеева А. А. Грызуны на судах Са-
халинекой области // Докл. Иркут. про-
тивочум. ин-та. Хабаровск, 1962. Выш. 3.
С. 162—164.
Тимофеева А. А., Сурков В. С. К эпиде-
миологической оценке основных ланд-
шафтов Сахалина в связи с биотопиче-
ским распредолением грызунов // Изв.
Иркут. пауч.-иселед. противочум. ин-та
Сибири и Дол. Востока. 1968. Т. 27.
С. 75-88.
.
Тихенко И. И., Таран И. Ф., Лямкин Г. И.
К изучению механизмов циркуляции
возбудителя бруцеллеза среди мыше-
зилвых- грызунов // Болезни с природ-
ной очаговостью на Кавказе: (Тез. докл.
краев. науч. конф., посвящ. 60-летию
образования СССР). Ставрополь, 1982.
С. 136—137.
Тилвинская М. В. Материалы к экологии
серой крысы (ВаНаз погуер!сиз ВегК)
в природных биотопах // Учен. зап. Чу-
ваш. гое. пед. ин-та. 1964. Вып. 20.
С. 48—60.
Тихвинский В. И. Биология рыжеватого
суслика // Работы Волжеко-Камской зо-
нальной охотничьс-промысловой биоло-
гической станции. Казань: Изд-во Ка-
зан. ин-та, 1932. Вып. 2. С. 28-39.
Тихомиров А. А., Корчагин А. Н. Списки
и описание предметов, находящихся в
Зоологическом музее имп. Моск. ун-та.
Отдел четвертый. Списки и описание
коллекции млекопитающих музея // Изв.
имп. о-ва любителей естествознания,
1889. Т. 56, вып. 4, No 1. С. 1500.
Токалова К. А., Нопова Е. М., Сушкевич
Н. И. Результаты дальнейшего изуче-
ния лептоспирозной инфекции в Кали-
нинградской обл. // Тр. Ин-та эпидемио-
логии и микробиологии им. Пастера.
1963. Т. 25. С. 53—57.
Токарева Т. Г., Туров И. С., Алексеева
А. Н..Действие фторацетамида на пло-
довитость белых мышей: (Предвар. со-
общ.) // Журн. микробиологии, эпиде-
миологии и иммунобиологии. 1974. No 8.
С. 24—26.
Токаревич К. П. Лептоспирозная желтуха.
(болезнь Вейля) в Ленинграде во время,
Великой Отечественной войны и блока-
ды. Л.: Медгиз, 1947. 120 с.
Токаревич К. Н., Гавеман А. К. Возбуди-
тели болезни Боткина—Вейля у диких
крыс в Ленинграде // Сов. врачеб. журн.
1986. No 7. С. 511—516.
Токаревич К. Н., Германова Е. И., Гаве-
ман А. К. О спонтанной зараженноети
крыс бактериями псевдотуберкулеза //
Вестн. микробиологии, эпидемиологии
и паразитологии. 1940. Т. 19, No 2.
С. 226—235.
Тонышева В. С., Шиндяпин А. И. Меро-
приятия по борьбе с грызунами ‘в Мат-
чинском ‘районе // Материалы Ш Респ.
Литература 424
науч.-практ, конф. по дезинфекц. делу.
Дунтанбе, 1968. С. 144—117.
Тощигин Ю. В. Методика и организация
борьбы с серой крысой в канализаци-
онных сооружениях тородон и уголь-
ных шахтах: Автореф. дис. .. канд.
биол. наук. М., 1968. 16 с.
Тощигин Ю. В. Определение численно-
сти серой крысы с помощью пылевых
площадок // Экология и медицинское
значение серой крысы (Ваз погуер1-
смз ВегК). М.: АН СССР, 1983. С. 92—93.
Тошщигин Ю. В. Особенности экологии се-
рой крысы в подземных сооружениях и
методы ограничения ее численности //
Серая крыса, М.: АН СССР, 1986, Т. 2.
С. 69—88.
Тощигин Ю. В., Рыльников В. А. Совре-
менное состояние и перспективы регу-
ляции численности серой крысы// Рас-
пространенив и экология серой крысы и
методы ограничения ее численности. М.:
Наука, 1985. С. 242—273.
Тощигин Ю. В., Жукова Л. Д., Нодъем-
щикова Л. Г. и др. К вопросу об органи-
зации и методике практически полного
уничтожения синантропных грызунов
в населенном пункте// Проблемы дезин-
фекции и стерилизации. М.: Минздрав
СССР, 1976. Вып. 26. С. 240—245.
Тошигин Ю. В., Жукова Л. Д., Сирота
В. И. и др. Об оценке эффективности
дератизации в населенных пунктах //
Проблемы дезинфекции и стерилизации:
(Материалы симпоз. 23—25 нояб.
1977 г.). М.: Минздрав СССР, 1977. Ч. 2.
й
.
”., С. 98-99.
Тощигин Ю. В., Писаренко Г. Я., Кривуля
С. Д. и др. Н вопросу о стратегии борь-
бы с грызунами в крунном городе //
Грызуны: Материалы У Всесоюз. со-
веш. М.: Наука, 1980. С. 456—457.
Тощигин Ю. В., Полежаев В. Г., Грепачев
И. В. и др. Методика и организация
практически полного уничтожения се-
рых крыс на участке города // Матери-
алы Всесо1оз. конф. по вопр. дезинфек-
ции и стерилизации (29—30 селт.—1—
2 ‘окт. 1969 г., Москва). М.: Минздрав
СССР, 4969. С. 444.
Тощигин Ю. В., Харченко А. П. Распро-
странение, гнездование и условия про-
кормления серых крыс в угольных шах-
тах и канализациопных системах го-
родов // Тр. Центр. науч.-исслед. дезин-
фекц. ин-та. 1967. Вып. 18. С. 66.
Траут И. И. Численность и размещение
грызунов в населенном пункте город-
ского типа // Грызуны и борьба с ними.
Саратов, 1950. Вып. 3. С. 243—254.
Траханов Д. Ф. Изучение отпугивающего
действия чернокорня на серых крыс //
Тр. ВНИИ вет, санитарии. 1960. Т. 416.
С. 15—20.
Тратанов Д. Ф. Разработка эффективного
метода дератизации птичников: Азвто-
реф. дис. „. канд. биол. наук. М., 1964а.
с.
Траханов Д. Ф. `Комбинированный метод
борьбы с крысами // Тр. ВНИИ вет. са-
нитарии. 19616. Т. 19. С. 205-211.
Траханов Д. Ф. Защитно-рефлекторные
реакции серых крыс на отравленные
приманки // Проблемы ветеринарной са-
нитарии. М.: Медицина, 1963. Т. 22.
С. 162-172.
Траханов Д. Ф. Материалы к обоснованию
метода опыливания при дератизации //
Проблемы ветеринарной санитарии. М.:
Колос, 1964. С. 189—194.
Траланов Д. Ф. Дератизация в объектах
животноводства СССР: Автореф. дис.
„.ора с.-х. наук. М., 1978. 34 с.
Траханов Д. Ф. Меры борьбы с синантроп-
ными грызунами // Санитария и гигие-
на содержания животных. М.: Колос,
1981. С. 94—94.
Траханов Д. Ф. Пути повышения эффек-
тивности дератизации в животноводст-
ве // Экология и медицинское значение
серой крысы (Вайиз погуес1са$ ВегК).
М.: АН СССР, 1983. С. 98—99.
Траханов Д. Ф. Современное состояние и
перспективы ветеринарных разработок
по дератизации // Проблемы ветеринар-
ной санитарии и зоогигиены в промыш-
ленном животноводстве. М.: Колос,
1985. С. 81—86.
Траханов Д. Ф., Кадиров А. Ф. Проблемы
защиты животноводческих комплексов
‚от грызунов // Современные направле-
ния медицинской дезинфекции и дера-
тизации, М.: Минздрав СССР, 1981.
С. 163.
Триполитова А. А., Борисова Г. В. Лис-
териоз. Томек: Изд-во Том. ин-та, 1965.
257 с.
:
Троп И. Е., Чуловский И. К. Обнаружение
патогенных лептоснир типов иктероге-
моррагиа У серых крыс в г. Омске//
Тр. Омского НИИ эпидемиологии, мик-
робиологии и гигиены. 1959. No 6. С. 21—
23
Троп И. Е. Чуловский И. К. Заражен-
ность патогенными микроорганизмами
кишечной группы серых крыс, отлов-
ленных на пищевых предприятиях и
499 Литература
микробиологии,
свинофермах // Журн.
фермах // Жур
И
эпидемиологии и иммунологии.
No9.С.143.
Трофимов М. П. Эффективный метод де-
ратизации // Материалы науч-произв.
конф. по болезням с.-х. животных и
птиц. Псков, 1968. С. 229—231.
Трофимчук Г. Л. Полигостальность при-
родных очагов ГЛИС в Приморском
крае // Актуальные проблемы медицин-
ской вирусологии. М.: Ин-т нолиомие-
лита АМН СССР, 1985, С. 147.
Туманов И. Л. Уничтожение грызунов ра-
тинданом // Ветеринария. 1964. No 11.
С. 95-97.
Тупикова Н. В. Изучение размножения и
возрастного состава популяции мелких
млекопитающих // Методы изучения
природных очагов болезней челонека/
Под. ред. Л. А. Петрищевой, Н. Г. Ол-
суфьева. М.: Медицина, 1964. С. 154—
191.
Тупикова П. В. Зоологическое картогра-
фирование. М.: Изд-во МГУ, 1969. 250 с.
Тупикова Н. В., Медведева И. М. Определе-
ние возраста эмбрионов как один из
методов изучения размножения гры-
зунов // Зоол. журн. 1956. "Г. 25, No 10.
С. 1574—1582.
Тупикова Н. В., Тесленко Е. Б. Составле-
ние кадастрово-справочных карт рас-
пространения видов // Бюл. МОИП. Отд.
биол. 1967. Т. 72, No 1. С. 5-14.
Тупикова Н. В., Швецов Ю. Г. Размноже-
ние водяной крысы в Волго-Ахтубин-
ской пойме //Зоол. журн. 1956. Т. 35,
No 1. С. 130-140.
Тураев А. О новом местонахождении се-
рой крысы в юго-западном Таджикиста-
не //ГУ съезд Всесоюз. териол. о-ва:
Тез. докл. М., 1986. Т, 3. С. 283—284.
Туров И. С. Материалы к сравнительной
характеристике некоторых дератизаци-
онных препаратов при изучении их в
вольере //Тр. Центр. научгиселед. де-
зинфекц. ин-та. 1970. Вып. 19. С. 411—
АЛТ.
Турянин И. И. О численности и некото-
рых особенностях экологии серой кры-
сы на Карпатах // ГУ съезд Всесоюл.
териол. о-ва: Тез. докл. М., 1986. Т. 3.
С. 256—257.
Уголев М. А. Материалы о деятельности
слюнных желез у грызунов // Опыт изу-
чения регуляции физиологических
функций. М,; Л.: Изд-во АН СССР, 1959.
Т. 4. С. 1973—1983.
Уждавини 9. И. Групповые отношения
животных. Л. Наука, 1980. 144 с.
Усова В. С. Экология и медицинское зна-
чение серой крысы на территории Вла-
димирской области // ТУ съезд Всесоюз.
териол. о-ва; Тез. докл. М., 1986, 'Г. 8.
С. 137—138.
Фабри К. 9. К проблеме игры у живот-
ных // Бюл. МОИП. Отд. биол. 1973.
Т. 15, No 3. С. 137-146.
Фабри К. 9. Основы зоопсихологии. М.:
Изд-во МГУ, 1976. 287 с.
Фабри К. 9. Игры животных и игры де-
тей: (Сравнительно-пеихологические ас-
пекты) // Вопр. пеихологии. 1982. No 3.
С. 26—34.
Фоабри К. Э., Филиппова Г. Г. К проблеме
имитационного научения у животных //
Вестн. МГУ. Сер. 14, Психология. 1982.
No 2, С. 55-61.
Фалькенштейн Б. Ю., Егорова Л. В. Ра-
тиндан // Защита растений от вредите-
лей и болезней. 1964. No 10. С, 33—34.
Фалькенштейн Б. Ю., Осипова В. И. Реф-
лекторные механизмы репеллентного
действия запахов на млекопитающих //
Бюл. ВИЗР. 1976. No 36. С. 50—54.
Фаршатов А. Г. Распространение серой
крысы на территории Коми АССР и
усиление этого процесса в связи © раз-
витием животноводческих хозяйств //
ТУ съезд Всесоюз. териол. о-ва: Тез.
докл. М. 1988. Т. 3. С. 284—285.
Федоров В. Д., Гильманов Т. Г. Экология.
М.: Изд-во МГУ, 41980. 464 с.
Федоров В. Н., Казавевич В. Н. Современ-
ное распространение чумы в зарубеж-
ных странах // Природная очаговость и
эпидемиология особо опасных инфекци-
онных заболеваний. Саратов: «Микроб»,
1959. С. 18—39.
|
Федоров Л. И., Евсеева Т. И., Щербина
Р. Д., Бунин В. Т. Заболевания людей
псевдотуберкулезом в Сахалинской об-
ласти // Первая межобл. науч.-практ.
конф. по псевдотуберкулезу человека
(дальневост. скарлатиноподоб. лихорад-
ке): Тез. докл. Владивосток, 1970.
С. 51—52.
Федорова Н. И. Эпидемиология и-профи-
лактика Ку-риккетсиоза. М.: Медицина,
1968. 230 с.
Федосеева М. Ф., Копытквин С. И., Баже-
нова А. А. Туляремия в Псковской об-
ласти // Тр. Ин-та эпидемиологии и
микробиологии им. Пастера. 1967. Т. 31.
С. 43—56.
Фетисов А. С. Крыса-карако в Восточной
Сибири. Иркутск: Обл. кн. изд-во, 1945.
32 с.
Фетисов А. С. Материалы по сезонной
о
м
в
о
л
и
Литература
493
динамике числеппости грызунов в насе-
ленных пунктах Забайкалья // Изв, Ир-
кут. науч.-исслед. противочум. ин-та
Сибири и Дал. Востока. 1946. Т. 6.
С. 195—204.
Фирстов Н. И. Еще 0б избирательном от-
ношении серых крыс к различным ви-
дам отравленных приманок и сравни-
тельной эффективности некоторых ору-
дий лова // Там же. 1958. Т. 49. С. 161—
16
Флегонтова А. Д., Малафеева Л.С. Актив-
ность в передаче чумы блох рода Сега-
форпуПаз //Тр. Арм. противочум. стан-
ции. 1960. Вып. 4, С. 127—135.
Флеров К. К. Очерк фауны млекопитаю-
щих долины реки Чороха // Ежегодпик
зоол. музея АП СССР. 1927. Т. 28,
С. 63—73.
Формозов А. Н. Несколько дополнений к
статье Б. М. Житкова «Замечания о
крысах и некоторых условиях их ис-
следования» // Зоол. журн. 1945. 1. 24,
No 2. С. 126—134.
Хайрулин Н. Н. Эколого-морфологический
анализ двух популяций серых крыс
(ВаИаз погусотеиз ВегК, 1769}; Авто-
реф. дис. .. канд. биол. наук. Сверд-
ловск, 1972. 37 с.
Хамаганов С. А. Серые крысы в природ-
ных биотопах Хабаровского края // Воп-
росы географии Дальнего Востока. Вла-
дивосток, 1965. Вып. 7. С. 268—271.
Хамаганов С. А. О питании серых крыс
в природных биотопах Среднего При-
амурья // Экология млекопитающих и
птиц. М.: Наука, 1967. С. 206-209.
„. Хамаганов С. А. Экология серой крысы и
вопросы борьбы с нею на Дальнем Вос-
токе: Автореф. дис. „.канд. биол. наук.
Владивосток, 1968. 21 с.
Хамаганов С. А. Распространение серой
крысы на Дальпем Востоке // Фауна и
экология грызунов. М., 1972. Вып. 11.
С. 149—158.
Хамаганов С. А. Норы и другие типы убе-
жищ серой крысы в природных био-
топах Приамурья // Проблемы особо
опасных инфекций. Иркутск: Медицина,
1975. Выш. 4. С, 38—44.
- Хамаганов С. А. О современном ареале
серой крысы в Восточной Сибири и на
Дальпем Востоке // Экология и медицин-
ское значение серой крысы (Валя пог-
усов Вет). М.: Наука, 1983. С. 36-—
Хамаганов С. А. Экология серой крысы в
Восточной Сибири и на Дальнем Восто-
ке // Распространение и экология серой
крысы и методы отраничения ее чис-
ленности. М.: Наука, 1985. С. 112—127.
Хамаганов С. А., Греков В. Г. Распроет-
ранение и численность серых крыс в
природных биотопах Читинской обла-
сти // Природно-очаговые инфекции в
Забайкалье. Чита, 1983. С. 100—102.
Хамаганов С. Д., Денисов Е. Т., Липаев
В. М. и др. Современное распростране-
ние серых крыс в Восточной Сибири и
на Дальнем Востоке // Серая крыса. М.:
Наука, 1986. Т. 1. С. 54—67.
Хамаганов С. А., Рогов А. Г., Горелышев
П. П. К вопросу эффективной дерати-
зации на птицефермах // Экология и ме-
дицинское значение серой крысы (Вас
ш5 погуер1еиз ВегЕ). М.: АН СССР,
1983. С. 106—107.
Хамаганов С. А. Шахуров Д. В. Отрав-
ленные приманки в борьбе с домовыми
грызунами // Изв. Иркут. науч.-исслед.
противочум, ин-та Сибири и Дал. Восто-
ка. 1959. Т. 24. С. 364-369.
Хвещенко С. М. Течение инфекционного
процесса экспериментальной чумы у се-
рых крыс // Докл. Иркут. науч.-иеслед.
противочум. ин-та Сибири и Дал. Вос-
тока. 1962. Вып. 5. С. 3—4.
Хвещенко Е. М. Течение инфекционного
процесса при чуме у серых и черных
крыс в эксперименте // Там же. 1963.
Выи. 6. С. 63—64.
Хвещенко Е. Н., Синцова Н. В., Падалко
3. Ф. Случай выделения листерелл в
г. Ворошилове // Изв. Иркутск. науч.-ис-
след. противочум. ин-та Сибири и Дал.
Востока. 1956. Т. 14. С. 28—29.
Христов Л., Матева М. Изследване на ре-
пелентните свойства на полимери спря-
мо гризачи// Екология. 1976. No2.
С. 61-68.
Цихистави ПТ. Г., Джмухадзе И. М. Мате-
риалы но грызунам Сванетии // Особо
опасные инфекции на Кавказе. Ставро-
ноль, 1984. С. 185-486.
Цилистави Ш. Г., Читиавшвили В. К.
К изучению распространения серой кры-
сы в Грузии //ТУ съезд Всесоюз. тери-
ол. о-ва: Тез. докл. М., 1986. Т. 3.
С. 273—275.
Цуци Ц. Мураками К. Пат. кл. З0Т 342.
(А 01У 17/12) Япония. Репелленты для
грызунов. Заявл. 25.0670; Опубл.
20.11.73.
Чабовский В, И., Даргольц В. Р. Об изме-
нении фауны мелких позвоночных в
зависимости от степени обжитости по-
селкох // Бюл. МОИП. Отд. биол. 1964.
Т. 59, No 4. С. 38-50.
499 Литература
микробиологии,
свинофермах // Журн.
фермах // Жур
И
эпидемиологии и иммунологии.
No9.С.143.
Трофимов М. П. Эффективный метод де-
ратизации // Материалы науч-произв.
конф. по болезням с.-х. животных и
птиц. Псков, 1968. С. 229—231.
Трофимчук Г. Л. Полигостальность при-
родных очагов ГЛИС в Приморском
крае // Актуальные проблемы медицин-
ской вирусологии. М.: Ин-т нолиомие-
лита АМН СССР, 1985, С. 147.
Туманов И. Л. Уничтожение грызунов ра-
тинданом // Ветеринария. 1964. No 11.
С. 95-97.
Тупикова Н. В. Изучение размножения и
возрастного состава популяции мелких
млекопитающих // Методы изучения
природных очагов болезней челонека/
Под. ред. Л. А. Петрищевой, Н. Г. Ол-
суфьева. М.: Медицина, 1964. С. 154—
191.
Тупикова П. В. Зоологическое картогра-
фирование. М.: Изд-во МГУ, 1969. 250 с.
Тупикова Н. В., Медведева И. М. Определе-
ние возраста эмбрионов как один из
методов изучения размножения гры-
зунов // Зоол. журн. 1956. "Г. 25, No 10.
С. 1574—1582.
Тупикова Н. В., Тесленко Е. Б. Составле-
ние кадастрово-справочных карт рас-
пространения видов // Бюл. МОИП. Отд.
биол. 1967. Т. 72, No 1. С. 5-14.
Тупикова Н. В., Швецов Ю. Г. Размноже-
ние водяной крысы в Волго-Ахтубин-
ской пойме //Зоол. журн. 1956. Т. 35,
No 1. С. 130-140.
Тураев А. О новом местонахождении се-
рой крысы в юго-западном Таджикиста-
не //ГУ съезд Всесоюз. териол. о-ва:
Тез. докл. М., 1986. Т, 3. С. 283—284.
Туров И. С. Материалы к сравнительной
характеристике некоторых дератизаци-
онных препаратов при изучении их в
вольере //Тр. Центр. научгиселед. де-
зинфекц. ин-та. 1970. Вып. 19. С. 411—
АЛТ.
Турянин И. И. О численности и некото-
рых особенностях экологии серой кры-
сы на Карпатах // ГУ съезд Всесоюл.
териол. о-ва: Тез. докл. М., 1986. Т. 3.
С. 256—257.
Уголев М. А. Материалы о деятельности
слюнных желез у грызунов // Опыт изу-
чения регуляции физиологических
функций. М,; Л.: Изд-во АН СССР, 1959.
Т. 4. С. 1973—1983.
Уждавини 9. И. Групповые отношения
животных. Л. Наука, 1980. 144 с.
Усова В. С. Экология и медицинское зна-
чение серой крысы на территории Вла-
димирской области // ТУ съезд Всесоюз.
териол. о-ва; Тез. докл. М., 1986, 'Г. 8.
С. 137—138.
Фабри К. 9. К проблеме игры у живот-
ных // Бюл. МОИП. Отд. биол. 1973.
Т. 15, No 3. С. 137-146.
Фабри К. 9. Основы зоопсихологии. М.:
Изд-во МГУ, 1976. 287 с.
Фабри К. 9. Игры животных и игры де-
тей: (Сравнительно-пеихологические ас-
пекты) // Вопр. пеихологии. 1982. No 3.
С. 26—34.
Фоабри К. Э., Филиппова Г. Г. К проблеме
имитационного научения у животных //
Вестн. МГУ. Сер. 14, Психология. 1982.
No 2, С. 55-61.
Фалькенштейн Б. Ю., Егорова Л. В. Ра-
тиндан // Защита растений от вредите-
лей и болезней. 1964. No 10. С, 33—34.
Фалькенштейн Б. Ю., Осипова В. И. Реф-
лекторные механизмы репеллентного
действия запахов на млекопитающих //
Бюл. ВИЗР. 1976. No 36. С. 50—54.
Фаршатов А. Г. Распространение серой
крысы на территории Коми АССР и
усиление этого процесса в связи © раз-
витием животноводческих хозяйств //
ТУ съезд Всесоюз. териол. о-ва: Тез.
докл. М. 1988. Т. 3. С. 284—285.
Федоров В. Д., Гильманов Т. Г. Экология.
М.: Изд-во МГУ, 41980. 464 с.
Федоров В. Н., Казавевич В. Н. Современ-
ное распространение чумы в зарубеж-
ных странах // Природная очаговость и
эпидемиология особо опасных инфекци-
онных заболеваний. Саратов: «Микроб»,
1959. С. 18—39.
|
Федоров Л. И., Евсеева Т. И., Щербина
Р. Д., Бунин В. Т. Заболевания людей
псевдотуберкулезом в Сахалинской об-
ласти // Первая межобл. науч.-практ.
конф. по псевдотуберкулезу человека
(дальневост. скарлатиноподоб. лихорад-
ке): Тез. докл. Владивосток, 1970.
С. 51—52.
Федорова Н. И. Эпидемиология и-профи-
лактика Ку-риккетсиоза. М.: Медицина,
1968. 230 с.
Федосеева М. Ф., Копытквин С. И., Баже-
нова А. А. Туляремия в Псковской об-
ласти // Тр. Ин-та эпидемиологии и
микробиологии им. Пастера. 1967. Т. 31.
С. 43—56.
Фетисов А. С. Крыса-карако в Восточной
Сибири. Иркутск: Обл. кн. изд-во, 1945.
32 с.
Фетисов А. С. Материалы по сезонной
о
м
в
о
л
и
Литература
493
динамике числеппости грызунов в насе-
ленных пунктах Забайкалья // Изв, Ир-
кут. науч.-исслед. противочум. ин-та
Сибири и Дал. Востока. 1946. Т. 6.
С. 195—204.
Фирстов Н. И. Еще 0б избирательном от-
ношении серых крыс к различным ви-
дам отравленных приманок и сравни-
тельной эффективности некоторых ору-
дий лова // Там же. 1958. Т. 49. С. 161—
16
Флегонтова А. Д., Малафеева Л.С. Актив-
ность в передаче чумы блох рода Сега-
форпуПаз //Тр. Арм. противочум. стан-
ции. 1960. Вып. 4, С. 127—135.
Флеров К. К. Очерк фауны млекопитаю-
щих долины реки Чороха // Ежегодпик
зоол. музея АП СССР. 1927. Т. 28,
С. 63—73.
Формозов А. Н. Несколько дополнений к
статье Б. М. Житкова «Замечания о
крысах и некоторых условиях их ис-
следования» // Зоол. журн. 1945. 1. 24,
No 2. С. 126—134.
Хайрулин Н. Н. Эколого-морфологический
анализ двух популяций серых крыс
(ВаИаз погусотеиз ВегК, 1769}; Авто-
реф. дис. .. канд. биол. наук. Сверд-
ловск, 1972. 37 с.
Хамаганов С. А. Серые крысы в природ-
ных биотопах Хабаровского края // Воп-
росы географии Дальнего Востока. Вла-
дивосток, 1965. Вып. 7. С. 268—271.
Хамаганов С. А. О питании серых крыс
в природных биотопах Среднего При-
амурья // Экология млекопитающих и
птиц. М.: Наука, 1967. С. 206-209.
„. Хамаганов С. А. Экология серой крысы и
вопросы борьбы с нею на Дальнем Вос-
токе: Автореф. дис. „.канд. биол. наук.
Владивосток, 1968. 21 с.
Хамаганов С. А. Распространение серой
крысы на Дальпем Востоке // Фауна и
экология грызунов. М., 1972. Вып. 11.
С. 149—158.
Хамаганов С. А. Норы и другие типы убе-
жищ серой крысы в природных био-
топах Приамурья // Проблемы особо
опасных инфекций. Иркутск: Медицина,
1975. Выш. 4. С, 38—44.
- Хамаганов С. А. О современном ареале
серой крысы в Восточной Сибири и на
Дальпем Востоке // Экология и медицин-
ское значение серой крысы (Валя пог-
усов Вет). М.: Наука, 1983. С. 36-—
Хамаганов С. А. Экология серой крысы в
Восточной Сибири и на Дальнем Восто-
ке // Распространение и экология серой
крысы и методы отраничения ее чис-
ленности. М.: Наука, 1985. С. 112—127.
Хамаганов С. А., Греков В. Г. Распроет-
ранение и численность серых крыс в
природных биотопах Читинской обла-
сти // Природно-очаговые инфекции в
Забайкалье. Чита, 1983. С. 100—102.
Хамаганов С. Д., Денисов Е. Т., Липаев
В. М. и др. Современное распростране-
ние серых крыс в Восточной Сибири и
на Дальнем Востоке // Серая крыса. М.:
Наука, 1986. Т. 1. С. 54—67.
Хамаганов С. А., Рогов А. Г., Горелышев
П. П. К вопросу эффективной дерати-
зации на птицефермах // Экология и ме-
дицинское значение серой крысы (Вас
ш5 погуер1еиз ВегЕ). М.: АН СССР,
1983. С. 106—107.
Хамаганов С. А. Шахуров Д. В. Отрав-
ленные приманки в борьбе с домовыми
грызунами // Изв. Иркут. науч.-исслед.
противочум, ин-та Сибири и Дал. Восто-
ка. 1959. Т. 24. С. 364-369.
Хвещенко С. М. Течение инфекционного
процесса экспериментальной чумы у се-
рых крыс // Докл. Иркут. науч.-иеслед.
противочум. ин-та Сибири и Дал. Вос-
тока. 1962. Вып. 5. С. 3—4.
Хвещенко Е. М. Течение инфекционного
процесса при чуме у серых и черных
крыс в эксперименте // Там же. 1963.
Выи. 6. С. 63—64.
Хвещенко Е. Н., Синцова Н. В., Падалко
3. Ф. Случай выделения листерелл в
г. Ворошилове // Изв. Иркутск. науч.-ис-
след. противочум. ин-та Сибири и Дал.
Востока. 1956. Т. 14. С. 28—29.
Христов Л., Матева М. Изследване на ре-
пелентните свойства на полимери спря-
мо гризачи// Екология. 1976. No2.
С. 61-68.
Цихистави ПТ. Г., Джмухадзе И. М. Мате-
риалы но грызунам Сванетии // Особо
опасные инфекции на Кавказе. Ставро-
ноль, 1984. С. 185-486.
Цилистави Ш. Г., Читиавшвили В. К.
К изучению распространения серой кры-
сы в Грузии //ТУ съезд Всесоюз. тери-
ол. о-ва: Тез. докл. М., 1986. Т. 3.
С. 273—275.
Цуци Ц. Мураками К. Пат. кл. З0Т 342.
(А 01У 17/12) Япония. Репелленты для
грызунов. Заявл. 25.0670; Опубл.
20.11.73.
Чабовский В, И., Даргольц В. Р. Об изме-
нении фауны мелких позвоночных в
зависимости от степени обжитости по-
селкох // Бюл. МОИП. Отд. биол. 1964.
Т. 59, No 4. С. 38-50.
494
Литература
Чарушина А. М. Население мелких мле-
копитающих основных ландттафтов Ки-
ровской области // Материалы о фауне
северо-востока европейской части Со-
ветского Союза. Киров, 1973. С. 44—56.
Черномордиков В. В. Некоторые матери-
алы к сравнительной характеристике
питания и обмена серой и черной кры-
сы // Совещ, по экол. физиологии: (Тез.
докл.). М.; Л.: Изд-во АН СССР, 1959.
Т. 2. С. 149—124.
Черномордиков В. В., Горячева М. Г. Су-
точный ритм активности и реакция на
свет у рыжих, черных и туркестанских
крыс //Т Всесоюз. совещ. по млекопи-
тающим. М.; Л.: Издво МТУ, 1961.
Т. 2. С. 195-108.
Чистяков А. А. Обнаружение серой кры-
сы на полуострове Ямал // Сборник на-
учных работ Тюменского НИИ краевой
инфекционной патологии. Тюмень,
1965. No 1.С. 231-282.
Читиашвили В. К., Цихистави В. К., Ку-
лов А. Б. Распространение, некоторые
аспекты экологии серых крые в Гру-
зии и ее медицинское значение // Ма-
териалы по эколотии и методам огра-
ничения численности серой крысы. М.:
АН СССР, 1987. С. 99—413.
Шарашидзе Т. Г. Заразные свойства мочи
диких крыс и лабораторных животных
при крысином риккетсиозе // Тр. ГНИИ
вакцин и сывороток, 1958. Т. 3. С. 159—
160.
Шаферштейн Д. Л., Айзенштадт Д. С.
Изучение зараженноети серых крыс
лептоспирозом на юго-западе Украи-
ны // Журн. микробиологии, эпидемио-
логии и иммунобиологии. 1959. No 11.
С. 116.
Шварц С. С. О возрастной структуре по-
пуляции млекопитающих// Тр. Урал.
отд. МОИП. 1959. Вып. 2. С, 3-22.
Швецов Ю. Г. Мелкие млекопитающие
Байкальской котловины. Новосибирск:
Наука, 1977. 457 с.
Шепелева В. К. Некоторые особенности
нитания пасюка (ВаИиз потуе21с8
Вегк) и черной крысы (В. гаНллз Г.) //
Массовые размножения животных и их
прогнозы: Тез. докл. Ш экол. конф.
Киев: Наук. думка, 1950. Ч. 2. С. 124-
424.
Шервинская А. И. Антигенная структу-
ра иктерогеморрагических ттаммов леп-
тоспир, встречающихся в СССР И Журн.
микробиологии, эпидемиологии и им-
мунологии. 1968. No 8. С. 68—65.
Шидловский М. В. Родентофауна Черно-
морекого побережья Грузии // Тр. Зоол.
ин-та АН ГССР. 1947. Т. 7. С. 97-146.
Шидловский М. В. Млекопитающие фау-
ны высокогорья Большого Кавказа в
Грузии // Фауна высокогорья Большого
Кавказа в пределах Грузии. Тбилиси:
Мецниереба, 1964. С. 175-198.
Пыдловский М. В. Определитель грызу-
нов Закавказья. 2-е изд. Тбилиси: Мец-
ниереба, 1976. 255 с.
Шилов И. А. Эколого-физиологические ос-
новы популяционных отношений у жи-
вотных. М.: Изд-во МГУ, 1977. 262 с.
Шилов И. А. Физиологическая экология
животных. М.: Выси. шк., 1985. 328 с.
Шилов М. Н., Варшавский С. Н., Попов
А. В. и др. Распространение, числен-
ность и опидемиологическое значение
серой крысы в природных очагах чумы
низовьев Волги и Урала // Серая крыса.
М.: АН СССР. 1986. Т. 2. С. 45—62.
Шилов М. Н., Варшавский С. Н., Сурвил-
ло А. В. и др. Расширение ареала и
современное раснространение серой
крысы в Калмыцкой АССР и восточ-
ных районах Ростовской области // Ма-
териалы ! рабочего совещ. по серой кры-
се. М.; АН СССР, 1983. С. 30-31.
Шмитс А. Л. Тактика дератизации и пре-
дунреждения заселения строений се-
рыми крысами в г. Риге // Эколотия и
медицинское значение серой крысы
(Вайлз погуез1сиз ВегЕ). М.: АН СССР,
1983. С. 105—106.
Шубин Ф. Н., Сомов Г. П., Токаченко
И. М. и др. Изучение иерсиниозов на
Чукотке // Природно-очаговые зооантро-
понозы на Крайнем Севере. Новоси-
бирск: Наука, 1978. С. 93—98.
Шура-Бура Б. И., Тарарин Р. А., Ключ-
ник Н. С. Опыт изучения миграций се-
рых крыс методом меченых атомов //
Журн. микробиологии, эпидемиологии
и иммунобиологии. 1962. No 12. С. 76-
81.
Шура-Бура Б. Л., Харламов В. И. Радио-
автография как метод выявления ме-
ченых грызунов и их эктопаразитов
при изучении вопросов миграции //
Зоол. журн. 1966. Т. 40, No 2. С. 15—17.
Шутова М. И., Вигоров Ю. Л., Авдеев
Д. В. Синантропные грызуны поселков
восточной части БАМ // Проблемы де-
зинфекции и стерилизации: Сб, науч.
тр. М.: Минздрав СССР, 1985. С. 153—
161.
Шутова М. И., Рыльников В. А., Иваниц-
кая Е. Г., Левина Д.Е. Принцииы изу-
чения предпочитаемости грызунами раз-
‹
Литература
425
личных приманок // Основные направле-
ния развития науки и практики дезин-
фекционнохо дела: Сб. науч. тр. Моск.
науч.-исслед. ин-та вакцин и сывороток
им. И. И. Мечникова. М.: Минздрав
СССР, 1981. С. 96-99.
Щастный С. М. Отчет по исследованию
крыс на Одесской бактериологической
станции с сентября 4940 г. по декабрь
1941 г.// Чума в Одессе в 1910 г. СИб.,
1912. С. 91-134.
Щепотьев Н. В., Попов Н. А. О путях к
уничтожению серых крыс в условиях
крупного паселепного пункта (на при-
мере г. Волгограда) // Проблемы особо
опасных инфекций. Саратов, 1971.
Вып. 5(24). С. 35—40.
Щепотьев П. В., Гкачева Н. В. Современ-
ное распространение серой крысы в
Пижнем Поволжье и возможные пути
ее расселения // Зоол. журн. 1966. ТГ. 45,
No 3. С. 468—471.
Эйгелие Ю. К. Влияние хозяйственной
деятельности человека на природные
очаги некоторых болезней
// ХГ съезд
Всесоюз. териол. о-ва: (Пленар. докл.,
31 янв.--4 февр. 1978). М.: Всесотоз. те-
риол. 0-во АН СССР, 1980. С. 128—139.
Эйгелис Ю. Е. Грызуны Восточного Закав-
казья и проблема оздоровлепия мест-
ных очагов чумы. Саратов: «Микроб»,
19800. 262 с.
Эйгелис Ю. А., Сурвилило А. В., Варшав-
спий Б. С. Испытание жидких отравлен-
ных приманок при борьбе с серой кры-
сой// ТУ съезд Всесоюз. териол. о-ва:
*Гез. докл. М., 1986. Т. 3. С. 372—374.
Элтон Ч. Экология наптествий животных
и растений. М.; ` Изд-во иностр. лит.,
1960. 230 с.
Эльконин Д. Б. Пеихология игры. М.: Пе-
Дагогика, 1978. 159 с.
Аскегтап 5. Н., Ноет М. А., Тетег Н.
Азе аЁ шаегпа! зерагаНол ал9 вазе
егоз1оп зазсериШИ оЁ бе га // РвуеВо-
зот. Мей. 1975. У]. 37. Р. 180—184.
Адатз О. В. Тве г@ацоп оЁ зеещ-татКтя
оТасё0ту шуезИдаНой ап@ зреейе роз-
Бтез ш 1з9]абоп-шаасеё Вориое м
газ // Вевау!юие. 1976. Уо1. 56, М 3/4.
Ь. 286-297.
Айатз No, Восе В. А 1опенадта за4у
оЁ Чотапсе Ш ап опё4оог союпу оЁ
ФотезЫс газ //7. Сошр. РзуеВо|. 1983.
\о1. 97, М 1.Р. 24-83.
А ег МNo., Ао 7. Те Бевауог оЁ сот-
попса ос: ай апауз!3 оГ Фе 22 ЕН;
14 Серая крыса
Юдин Б. С., Кривошеев В. Г., Беляев В. Г.
Мелкие млекопитающие севера Даль-
вого Востока. Новосибирск: Наука, 1976.
70 с.
Ющенко Г. В. Нсевдотуберкулез у гры-
зунов в условиях большого города //
Журн. микробиологии, эпидемиологии
и иммунологии. 1957. No 1. С. 144—119.
Ющенко Г. В. Псевдотуберкулез челове-
ка: (Обзор) // Там же. 1964. No 2. С. 88—
2.
Ющенко Г. В. Некоторые аспекты эколо-
гии, эпидемиологии и клиники пеевдо-
туберкулеза. Природно-очаговые антро-
позоонозы// Тез. докл. к ТХ Всесоюз.
конф. по природ. очаговости болезней
человека и животных, Омск, 1976.
С. 114—145.
Ющенко Г. В., Терещенко М. П., Нонома-
рева Т. Н. и др. Особенности распрост-
ранения некоторых бактериальных ин-
фекций в популяции городских грызу-
нов // Журл. микробиологии, эпидемио-
логии и иммунологии. 1965. No 3. С. 101-
106.
Яковлев М. Г. Некоторые экологические
особенности серой крысы в низовьях
реки Дон // Тр. Рост. науч.-исслед. про-
тивочум. ин-та. 1949. Т. 8, No 7. С. 89-
98.
Нкунина Т. И., Михайлова О. А,‚, Шереме-
тъев С. А. Псевдотуберкулез синант-
ропных грызунов в г. Владивостоке //
Дальневосточная скарлатиноподобная
лихорадка, Владивосток: Изд-во Дальне-
вост. ун-та. 1974. С. 39—45.
Яроцкий Л. С. Шистозомозы. М.: Меди-
цина, 1982. 279 с.
Яцимирская-Кронтовская М. К. Клещевой
сыпной тиф // Тр. Всесоюз. конф. мик-
робиологии, эпидемиологии, инфекц. б0-
лезней. М.: Медгиз, 1940. С. 234—241.
са] оЁ га (ВаИиз погуертсиз) // Ашег.
2001. 1979. Уч. 19, М 2. Р. 493—508.
Аё5пег В. Ваз ап@ рше сопез // 1зг. Гап@
ап Машге. 1981. Уо1. 6, М 2. Р. 94-97.
Цит. по; РЖ, 1984. No 8. Реф. И(387).
Аззпег В., Тегке У. НаБиа ехройаНоп
{Вгооср саШага! #га0511135101: рте-со-
пе {еед те Бопау1тог ш МасК га&з // Е&Во-
1обу 85: 191 Пиегп. ео1. сопё. Тошлве.
1985. Уо1. 1. Р. 148.
А1бегз 7. В. ОНасоту сопайопа 10 Ъе-
Вамтога| Чеуе!ортеп{ ш годепз // Мащ-
таПап оМасНоп, гергодисНуе ргосеззез
ап9 Берау!оюг. М. У.: Аса@. ргезз, 1976.
Р. 67—94.
АБег 7. В., Сще} В. <. ОЦаедогу слез
494
Литература
Чарушина А. М. Население мелких мле-
копитающих основных ландттафтов Ки-
ровской области // Материалы о фауне
северо-востока европейской части Со-
ветского Союза. Киров, 1973. С. 44—56.
Черномордиков В. В. Некоторые матери-
алы к сравнительной характеристике
питания и обмена серой и черной кры-
сы // Совещ, по экол. физиологии: (Тез.
докл.). М.; Л.: Изд-во АН СССР, 1959.
Т. 2. С. 149—124.
Черномордиков В. В., Горячева М. Г. Су-
точный ритм активности и реакция на
свет у рыжих, черных и туркестанских
крыс //Т Всесоюз. совещ. по млекопи-
тающим. М.; Л.: Издво МТУ, 1961.
Т. 2. С. 195-108.
Чистяков А. А. Обнаружение серой кры-
сы на полуострове Ямал // Сборник на-
учных работ Тюменского НИИ краевой
инфекционной патологии. Тюмень,
1965. No 1.С. 231-282.
Читиашвили В. К., Цихистави В. К., Ку-
лов А. Б. Распространение, некоторые
аспекты экологии серых крые в Гру-
зии и ее медицинское значение // Ма-
териалы по эколотии и методам огра-
ничения численности серой крысы. М.:
АН СССР, 1987. С. 99—413.
Шарашидзе Т. Г. Заразные свойства мочи
диких крыс и лабораторных животных
при крысином риккетсиозе // Тр. ГНИИ
вакцин и сывороток, 1958. Т. 3. С. 159—
160.
Шаферштейн Д. Л., Айзенштадт Д. С.
Изучение зараженноети серых крыс
лептоспирозом на юго-западе Украи-
ны // Журн. микробиологии, эпидемио-
логии и иммунобиологии. 1959. No 11.
С. 116.
Шварц С. С. О возрастной структуре по-
пуляции млекопитающих// Тр. Урал.
отд. МОИП. 1959. Вып. 2. С, 3-22.
Швецов Ю. Г. Мелкие млекопитающие
Байкальской котловины. Новосибирск:
Наука, 1977. 457 с.
Шепелева В. К. Некоторые особенности
нитания пасюка (ВаИиз потуе21с8
Вегк) и черной крысы (В. гаНллз Г.) //
Массовые размножения животных и их
прогнозы: Тез. докл. Ш экол. конф.
Киев: Наук. думка, 1950. Ч. 2. С. 124-
424.
Шервинская А. И. Антигенная структу-
ра иктерогеморрагических ттаммов леп-
тоспир, встречающихся в СССР И Журн.
микробиологии, эпидемиологии и им-
мунологии. 1968. No 8. С. 68—65.
Шидловский М. В. Родентофауна Черно-
морекого побережья Грузии // Тр. Зоол.
ин-та АН ГССР. 1947. Т. 7. С. 97-146.
Шидловский М. В. Млекопитающие фау-
ны высокогорья Большого Кавказа в
Грузии // Фауна высокогорья Большого
Кавказа в пределах Грузии. Тбилиси:
Мецниереба, 1964. С. 175-198.
Пыдловский М. В. Определитель грызу-
нов Закавказья. 2-е изд. Тбилиси: Мец-
ниереба, 1976. 255 с.
Шилов И. А. Эколого-физиологические ос-
новы популяционных отношений у жи-
вотных. М.: Изд-во МГУ, 1977. 262 с.
Шилов И. А. Физиологическая экология
животных. М.: Выси. шк., 1985. 328 с.
Шилов М. Н., Варшавский С. Н., Попов
А. В. и др. Распространение, числен-
ность и опидемиологическое значение
серой крысы в природных очагах чумы
низовьев Волги и Урала // Серая крыса.
М.: АН СССР. 1986. Т. 2. С. 45—62.
Шилов М. Н., Варшавский С. Н., Сурвил-
ло А. В. и др. Расширение ареала и
современное раснространение серой
крысы в Калмыцкой АССР и восточ-
ных районах Ростовской области // Ма-
териалы ! рабочего совещ. по серой кры-
се. М.; АН СССР, 1983. С. 30-31.
Шмитс А. Л. Тактика дератизации и пре-
дунреждения заселения строений се-
рыми крысами в г. Риге // Эколотия и
медицинское значение серой крысы
(Вайлз погуез1сиз ВегЕ). М.: АН СССР,
1983. С. 105—106.
Шубин Ф. Н., Сомов Г. П., Токаченко
И. М. и др. Изучение иерсиниозов на
Чукотке // Природно-очаговые зооантро-
понозы на Крайнем Севере. Новоси-
бирск: Наука, 1978. С. 93—98.
Шура-Бура Б. И., Тарарин Р. А., Ключ-
ник Н. С. Опыт изучения миграций се-
рых крыс методом меченых атомов //
Журн. микробиологии, эпидемиологии
и иммунобиологии. 1962. No 12. С. 76-
81.
Шура-Бура Б. Л., Харламов В. И. Радио-
автография как метод выявления ме-
ченых грызунов и их эктопаразитов
при изучении вопросов миграции //
Зоол. журн. 1966. Т. 40, No 2. С. 15—17.
Шутова М. И., Вигоров Ю. Л., Авдеев
Д. В. Синантропные грызуны поселков
восточной части БАМ // Проблемы де-
зинфекции и стерилизации: Сб, науч.
тр. М.: Минздрав СССР, 1985. С. 153—
161.
Шутова М. И., Рыльников В. А., Иваниц-
кая Е. Г., Левина Д.Е. Принцииы изу-
чения предпочитаемости грызунами раз-
‹
Литература
425
личных приманок // Основные направле-
ния развития науки и практики дезин-
фекционнохо дела: Сб. науч. тр. Моск.
науч.-исслед. ин-та вакцин и сывороток
им. И. И. Мечникова. М.: Минздрав
СССР, 1981. С. 96-99.
Щастный С. М. Отчет по исследованию
крыс на Одесской бактериологической
станции с сентября 4940 г. по декабрь
1941 г.// Чума в Одессе в 1910 г. СИб.,
1912. С. 91-134.
Щепотьев Н. В., Попов Н. А. О путях к
уничтожению серых крыс в условиях
крупного паселепного пункта (на при-
мере г. Волгограда) // Проблемы особо
опасных инфекций. Саратов, 1971.
Вып. 5(24). С. 35—40.
Щепотьев П. В., Гкачева Н. В. Современ-
ное распространение серой крысы в
Пижнем Поволжье и возможные пути
ее расселения // Зоол. журн. 1966. ТГ. 45,
No 3. С. 468—471.
Эйгелие Ю. К. Влияние хозяйственной
деятельности человека на природные
очаги некоторых болезней
// ХГ съезд
Всесоюз. териол. о-ва: (Пленар. докл.,
31 янв.--4 февр. 1978). М.: Всесотоз. те-
риол. 0-во АН СССР, 1980. С. 128—139.
Эйгелис Ю. Е. Грызуны Восточного Закав-
казья и проблема оздоровлепия мест-
ных очагов чумы. Саратов: «Микроб»,
19800. 262 с.
Эйгелис Ю. А., Сурвилило А. В., Варшав-
спий Б. С. Испытание жидких отравлен-
ных приманок при борьбе с серой кры-
сой// ТУ съезд Всесоюз. териол. о-ва:
*Гез. докл. М., 1986. Т. 3. С. 372—374.
Элтон Ч. Экология наптествий животных
и растений. М.; ` Изд-во иностр. лит.,
1960. 230 с.
Эльконин Д. Б. Пеихология игры. М.: Пе-
Дагогика, 1978. 159 с.
Аскегтап 5. Н., Ноет М. А., Тетег Н.
Азе аЁ шаегпа! зерагаНол ал9 вазе
егоз1оп зазсериШИ оЁ бе га // РвуеВо-
зот. Мей. 1975. У]. 37. Р. 180—184.
Адатз О. В. Тве г@ацоп оЁ зеещ-татКтя
оТасё0ту шуезИдаНой ап@ зреейе роз-
Бтез ш 1з9]абоп-шаасеё Вориое м
газ // Вевау!юие. 1976. Уо1. 56, М 3/4.
Ь. 286-297.
Айатз No, Восе В. А 1опенадта за4у
оЁ Чотапсе Ш ап опё4оог союпу оЁ
ФотезЫс газ //7. Сошр. РзуеВо|. 1983.
\о1. 97, М 1.Р. 24-83.
А ег МNo., Ао 7. Те Бевауог оЁ сот-
попса ос: ай апауз!3 оГ Фе 22 ЕН;
14 Серая крыса
Юдин Б. С., Кривошеев В. Г., Беляев В. Г.
Мелкие млекопитающие севера Даль-
вого Востока. Новосибирск: Наука, 1976.
70 с.
Ющенко Г. В. Нсевдотуберкулез у гры-
зунов в условиях большого города //
Журн. микробиологии, эпидемиологии
и иммунологии. 1957. No 1. С. 144—119.
Ющенко Г. В. Псевдотуберкулез челове-
ка: (Обзор) // Там же. 1964. No 2. С. 88—
2.
Ющенко Г. В. Некоторые аспекты эколо-
гии, эпидемиологии и клиники пеевдо-
туберкулеза. Природно-очаговые антро-
позоонозы// Тез. докл. к ТХ Всесоюз.
конф. по природ. очаговости болезней
человека и животных, Омск, 1976.
С. 114—145.
Ющенко Г. В., Терещенко М. П., Нонома-
рева Т. Н. и др. Особенности распрост-
ранения некоторых бактериальных ин-
фекций в популяции городских грызу-
нов // Журл. микробиологии, эпидемио-
логии и иммунологии. 1965. No 3. С. 101-
106.
Яковлев М. Г. Некоторые экологические
особенности серой крысы в низовьях
реки Дон // Тр. Рост. науч.-исслед. про-
тивочум. ин-та. 1949. Т. 8, No 7. С. 89-
98.
Нкунина Т. И., Михайлова О. А,‚, Шереме-
тъев С. А. Псевдотуберкулез синант-
ропных грызунов в г. Владивостоке //
Дальневосточная скарлатиноподобная
лихорадка, Владивосток: Изд-во Дальне-
вост. ун-та. 1974. С. 39—45.
Яроцкий Л. С. Шистозомозы. М.: Меди-
цина, 1982. 279 с.
Яцимирская-Кронтовская М. К. Клещевой
сыпной тиф // Тр. Всесоюз. конф. мик-
робиологии, эпидемиологии, инфекц. б0-
лезней. М.: Медгиз, 1940. С. 234—241.
са] оЁ га (ВаИиз погуертсиз) // Ашег.
2001. 1979. Уч. 19, М 2. Р. 493—508.
Аё5пег В. Ваз ап@ рше сопез // 1зг. Гап@
ап Машге. 1981. Уо1. 6, М 2. Р. 94-97.
Цит. по; РЖ, 1984. No 8. Реф. И(387).
Аззпег В., Тегке У. НаБиа ехройаНоп
{Вгооср саШага! #га0511135101: рте-со-
пе {еед те Бопау1тог ш МасК га&з // Е&Во-
1обу 85: 191 Пиегп. ео1. сопё. Тошлве.
1985. Уо1. 1. Р. 148.
А1бегз 7. В. ОНасоту сопайопа 10 Ъе-
Вамтога| Чеуе!ортеп{ ш годепз // Мащ-
таПап оМасНоп, гергодисНуе ргосеззез
ап9 Берау!оюг. М. У.: Аса@. ргезз, 1976.
Р. 67—94.
АБег 7. В., Сще} В. <. ОЦаедогу слез
126 Литература
ап моусшепи зишаН шефанае ше
тазрееШе асотсвавоп ш \09 Могуау
та // 7. Сор. ап@ Рруз!01. Рвусво1. 1978.
Уо1. 87. Р. 233—242.
А1Бегз 7. В., Мау В. Оехеюрутлеп" оё па-
за] гезргаНоп ап@ зи пе 1т таё // РВу-
5101. ап ВеБау. 1980а. \01. 24, М 5.
Р.. 957—963.
А1Ъетз 7. В., Мау В. Ощосепу оЁ оНасН-
оп: Пеу@орзтейё ог газ зепзуйу ©
игте ао ашу| асебае // No9. 1980.
У}. 24, МБ. Р. 965—970.
АТВет: 7. В., Мау В. Моплаб Шуе, Фегио-
(асе шдасНоп 01 Ва! ВаоаНос 11 га
рирз // Оеуе]ор. РзуеВо]. 1984. \о1. 17,
МА. Р. 161—181.
АПеп С. М. А сресКИз оЁ АНлеап татш-
та! // Ва. Маз. Сор. 2001. Нагуаг@.
1939. Ув]. 83. Р. 763.
АШь 7. Т., Вапкз Е. М. ЕНесз оЁ {етре-
табте оп иНтазопи@ ргодисвов Бу ш-
Гапё ато га\з // Оеуе]ор. РзусвоЫо1.
1971. Уд. 4, М2. Р. 149-156.
АЙ 1. Т., Вапкз Е. М. Ечосйопа] азресз
оЁ иКгазоциб ргобасНоп Бу пШашЕ а
по гаёз // Аппи. Вефау. 1972. Ув. 20.
Р. 115-419.
А10пз0 В. А., УШакотез О. А. Е., бапспег
А., Сапш 7. М. Амаю Кагуовуре (20-42)
ш Ше ЫасК гаё (ВаНлаз табаз) а Ме
х1со // Апи. Сешеё. 1982. Ув. 25, М 3.
Р. 133—135.
А1зюп 7, Вгоот 5. Герозргоз!3 Ш тат
ап апппа?з. ЕдшЬогсй; Г, 1958. 500 р.
Айтапп 5. А. ТВе заду оЁ рениа{е зоса1
сошташсаНоп // Зос1а] сотиозисаНол
атопя рейпа(ез. СЫсасо: Ушу. СЫсабо
ргезз, 1967. Р. 300.
`
А1рагех А., баз Г. оеасюЧазе сБапрез
1 {Ме деуеюрше пиезНла {тасё о 1Ве
га! // Майе. 1971. Ус]. 190. Р. 826—827.
АтогЕ б., Сына М., Сошой Г. Зи го-
ЧИотг (СИ 9Чае, АгусоНдае, Миг!ае)
Ое!’ЦаПа репшзщаге ед лазаге 1 гар-
рофо аНаштыее МосШтансо МедПегга-
пео // АппоаНа, 1984. У]. 14, М 1/3.
Р. 217-269.
Апдегзоп 7. У’. Те ргодасНоп о аЙтазо-
пс Бу ЛаБогафоту гаёз ап@ о ег шаш-
зта1з // Зс1елсе. 1954. \0]. 119. Р. 808.
Апагеш; В. И’, Вейпар В. И’, `Кеспап
Е. /. ОетостарЬ1с ап4 гезропзез оЁ Мог-
уау га ю ап ЁегИШеу сошго] // 7. \УП9-
Ше Мапас. 1974. Уо|. 38, М 4. Р. 868—
874.
Ашзко 1. Т., Зиег 5. Ё., МеСИитосЁ М. К.
её а1. Тье ге]айопзрар Бейуееп 22 КН»
и|(газотс 3101213 ап зос10зехиа| Бера-
\тог м фе гаё//Т. Сошр. ап Рвуз101.
Рзусво|. 1978. \0]. 92. Р. 824—829,
Аррегзоп С. 5., Запаег; О. Т., Каикетеп
р. Е. Гафогаогу ап@ Не]9 (113.оЁ {Ве
годепис!е Бго4Насойта абазё зуага-
1о-гез13ап{ Могмау газ // РезЫс. Зе
1981. Уи. 12, М 6. Р. 662—668.
АгтИаве К. В. $оиа] Бева\юог ап роро-
1апод Фупайся оЁ шагио$ /} ОЦсов. 1975.
\о1. 26. Р. 341—354.
Аттиаве К. В. Зо] уатсву ш с ус|-
1о%-Бе!Ше4д шагто: а роршаНоп-Бенау10-
ха] зузбежт // Аюпт. Верах. 1977. Уо/. 25.
Р. 585—598.
Агоп С., Съиеаи О. Ргезатед туоуететф
оЁ рЬегопопез п шайпях Бепау!ог т, {пе
гаё // Погтопе Верах. 1974. У01. 2.
Р. 315—323.
Агоп С., Нооз 1., Азсь С. ЕНес& о{ гето-
уа| оЁ {Ле оНасвогу Баз оп шайие Ъе-
Ваутог ап оуШайоп шт {Ме га // Меп-
гоепдосгто]озу. 1970. У]. 6. Р. 109—117.
Аззе{ Бег В. Н. Ое уегзрее Че уап 4е
ЮМете 2о0#41егеп 1 Ве]е уап де Вап8
уап ЬгааКБаепапаТузе // ВиП. [13% гоу.
Бет. паг. Ве. 1974. Уд]. 47, М 5. Р. 60.
Анау 7. В. Вед шИез апд фурВиз. Г.: АФ-
Лопе, 1968. 102 р.
Арегу Р. М. Тье 91зрегза| оЁ Ъго\п газ
ВаНа$ погуе21с1$ ап пе\ зресоепз
{тот 19 сепфату Саре Тоуп // Машта-
Па. 1985. У01. 49, М 4. Р. 573—576.
Ваыску А., Очадаотга 9., Райзек Г. её а|,
0зе о{ габто1зоюре (есвонтаез Гог дебег-
шииео Фе \уеаптх рего@ 1ш ехрег!-
теюа! апоа1з // Руз101. БоВетлоз]. 1970.
\Уо1. 19. Р. 457—467.
Вафифет? В. ЕрЧети1оо21е Ч4ез Герозр!-
го5ез ФЦаНо
// 20010565 сопа1ззапсез
Чери фае попуеПез: СоПоч. ОМ5/ЕАО
ве 1е8 70010868. Сбпеуе, 1954. Р. 212.
Вафифет В., Мозсоте: 7. Перозритозе! пе]-
1е г!зае 9е Гао е Ч4еШа баг4деупей //
АН: 136 Зарег. ЗавпНа. 1955. Уч. 18.
Р. 70.
Ваепптаег Г. Р. Тре гейаБШу оЁ дот!
пайсе огфегз т гаёз// Апот. ВеБах.
1966. Уо1. 14. Р. 367—371.
Раептткег Г. Р. Сотраг!зоп о? Бевауоп-
га] деуортеюе 1 зосаПу 1з01ей апд
отопрей га!з // 194. 1967. УоЁ 15.
Р. 312-828.
Ваепптеег В. Зоште сопзедистсе$ 0 аб-
этеззтуе Бера\1ог: А з@есНуе ге\е\м оЁ
{Ве Пстабаге оп о{Вег апта]5 // Арёгез.
Вера. 1974. Уо|. 1. Р. 17-37.
Варота Б., бай Е., Гапе Е., Уатов В.
ЕЁ степсу о{ ега@сайте газ \ИВ рагаЁ-
Литература
427
Но Байз// уеги. РезЕ Соп
о]. 18, М6. Р. 1245.
58.
Вала Р.Р. ТЬе емо йоп 0Ё магтой 30-
е16ез: а репега! {еогу // Зетее. ,
Уо1, 185. 2. 415-420. У! с. 1974.
Ватра! В. 7. Апегрась Р., Сеует Г. А.,
МеПчозй Т. К. ОИтазоше уосаП2анойв
ш_ гаф зехиа| Берлау1ох // Ашег. 7.001.
1979. Уч. 19, No2. Р. 469-480.
Вагпеа В. 71, Сеуег Г. А. Зехиа! Ъева-
мог: о
роедаотаогу опр 9
пе та[е гаё е1епсе.
аа
1972. Уо]. 176.
Вагре@4 В., Сеусг Г. ТВе оНтазове розте-
Дасаюту уосаЙтаноп апа е розе]а-
оао теГгасвогу репой оЁ Ше таГе //
‚ Сошр. ап
3191. Р%
5
\о1. 88. Р. 723-734
енот 1975.
Ватпей 5. А. Сотренноп атопх \Па
газ // Майе. 1955. Уо]. 175. Р. 486—497,
Вагпей 5. А. Вепау1ог сотропетз ш фе
Теефто о! м9 ап4 1аБогатогу та //
Веваутоиг. 1956. Уд]. 9, М 1. Р, 24-43.
Вагпей 5. и Ап ау о{ зоба! реБа-
Утот т 19 га // Ргое. 7001. бое.
Уо1. 130. Р. 107—152.
ос. 195За.
Ватпей 5. А. Гафогаогу ей ойз {ог Ъе-
Вахтог вез о Йа гаЁз //Т. Апип.
Тесвп. Аззос. 19585. Уо|. 9. Р. 6—14.
Ватпей 5. А. А заду о! БеБауюце: Рей
спеё оГ ело]обу апд Бера\оиг рзусво-
108у Ир ауе таиЦу т Фе таё 1:
Мелеп, 1963. 288 р.
Вагпей 5. А. 30621 згезз // Уемропиз
ш Ыо1оду. [.: ВайегуотИлз, 1964. Р. 260.
Вагней 5. А. Ваз / 8“. Азтог. 41967
\Уа1. 246, М1.Р. 79-86.
о
Ватпей 5. А. "Гра гаё: А за4у о Беваутоиг.
Сусаво; Г.: Ому. СЫсаво ргезз, 1975.
р.
у
.
Вате! 5. А. Вит 7. ЕаШу зНииЦаноп
п@ шатра ЪеВау1отг // Ма
ой, 213, В. ЧО
Майе. 1967
Вагпен 8. А., брепсег М. Ехрегипениз оп
о Фоод ргеЁегепсез оЁ \И га (Ва
5 погуео1сиз Вегкепро
1
1953. Уа. Е М.Р м // Вувтете,
Ватоп А., Вгоойзте К. Н. Митап В. А.
ЕНесёз оЁ папе ап адаНз зроскта-
ита проп ]феатишо шт Ме ада жЪие
та // 7. Сошр. ап РВузю]. РзусНо].
1957. Уо1. 50. Р. 530—534.
Ваттей С. Е. Ваз шт ргане аот1саНаге //
т. 1пзё. Вет. 1967. \У0]. 22, М6. Р. 43—
++,
-
Виезоп С. А Шеоту оЁ Рау апё Га
//
РэзусМаЕ. Веу. 1955, Те х Р. р
Вашеу У. М., Зтиь`Р. 1. No. Ваткош 6.1.
Тре ерЧеп!оору о 1ерюзр!г05185 Ш
Мог Оцеспз]ап4. 2. Кагбег оЪзегуа-
от8 оп Ве [035 т бе Моззшаи 9156-
1166 // У. Нуз. 1964. Уд. 62, М4. Р. 485-—
А9л.
Ваиег 7. Н. ЕНесу оЁ зпафегпа] эва(е оп ве
ето го пез оЧогз о{ ВоодеЯ
та(5 У10]. апа Вевау. 1983. . 30.
В Р. 229—232.
Уо. 30
бит ег М. Пъег ]азНе Убре! пла Зааег
а]; КаНатГо]сег ап МавПеррев поё м.
Пенкецеп 2 Штег Бо]ор1зсНеп Векйтр-
Рапз // Аша. Эсв5аНлозК., РНапт. пад
ромео, 4971. Ва. 44, М9. 3. 138—
Ваилиег И., Маптапп-Ецеппе С., боат-
Йо ХТ. её а1. ЭсВаЧИеве Ее] дпасег т Рог-
ще ТЬЯ. 1984. ВВ. 57, МТ. $. 134-
Варетось Р. В. Адатз М., Махзоп Г. В.
Уоз:4а Т. Н. Сепеые аетепцаной
атопа Кагуобур!с Тоглв оЁ (Ле ЫасК га
Ваз гаМлз // Сепенсв. 1983. -УоЁ. 105.
No4. Р. 969-083.
у
ВавегюсЕ Р. В., Райз С. В. 5., Ааатз М.,
Сое 5. Н. Сепенса! геаНотзррз
ашопе АпзгаНап гоЧеп{з (Миг Чае) //
Апзга]. 1. 7001. 1981. Ус. 29, МЗ
Р. 289—303.
ВатегзосК Р. В., Райз С. В. 5., Новамь
7. Т. Кагуоуре Чайа ап Ме зресне
Збайлв ап фахополис аНиез о? Ван
{15 У 03158118 апа В. зог414 105 ИТЫа.
1975. Уо1. 23, М 3. Р. 293—294.
Варетз ось Р. В., Тань С. И. 5., Нова
7. Т. её в]. СБготозоте еуош оп п Алз-
табап годет(з. 2. ТВе ВаНаз отопр //
рготаовотии. 1977. Уо|. 61, No3. Р. 297-
Веасй Г. А. Сошраг!зоп оЁ соршаюгу Ъе-
Вачтошт о ав га га1зе т 13а от,
сопабкайов ап зертегаНои // 7.
.
РзусНо|. 1942. Уч, 60, Р. 124. 7. белее
Веасй Е. А. Сохгепё сопсерз о! Рау т
аппа|
// Атег. Мабог. 1945. Уо| 79.
Р. 523—541.
Веась, Е. А., Таупез 7, Зил91е8 оЁ пабогоа]
тоитеуши Ш гав. 1. ВесосйНов о
уоппе //Т. Матиа!. 41958. Уф, 37.
Р. 177—180.
ВеаНу ТУ. И’. Подее А. М., Войре Г. У.
её а]. Рзуспотоюг зи аи, зоса|
ЧермуаМоп ап рау ш ауепПе газ й
РВагтасо]. В1ослет. апа Вевау. 1982,
\Уо|. 16. Р. 417—422.
Веану У’. И’, РодЕе А. М., Тао" К.
её а Зер! 1езюпз тогеазе р]ау Пов-
Ире ш уепПе гафз // РВузю1. ап@ Ве-
Вау. 1982. Уч. 28. Р. 649-652.
Вевисратр С. В., Магии }. С., Шейти-
126 Литература
ап моусшепи зишаН шефанае ше
тазрееШе асотсвавоп ш \09 Могуау
та // 7. Сор. ап@ Рруз!01. Рвусво1. 1978.
Уо1. 87. Р. 233—242.
А1Бегз 7. В., Мау В. Оехеюрутлеп" оё па-
за] гезргаНоп ап@ зи пе 1т таё // РВу-
5101. ап ВеБау. 1980а. \01. 24, М 5.
Р.. 957—963.
А1Ъетз 7. В., Мау В. Ощосепу оЁ оНасН-
оп: Пеу@орзтейё ог газ зепзуйу ©
игте ао ашу| асебае // No9. 1980.
У}. 24, МБ. Р. 965—970.
АТВет: 7. В., Мау В. Моплаб Шуе, Фегио-
(асе шдасНоп 01 Ва! ВаоаНос 11 га
рирз // Оеуе]ор. РзуеВо]. 1984. \о1. 17,
МА. Р. 161—181.
АПеп С. М. А сресКИз оЁ АНлеап татш-
та! // Ва. Маз. Сор. 2001. Нагуаг@.
1939. Ув]. 83. Р. 763.
АШь 7. Т., Вапкз Е. М. ЕНесз оЁ {етре-
табте оп иНтазопи@ ргодисвов Бу ш-
Гапё ато га\з // Оеуе]ор. РзусвоЫо1.
1971. Уд. 4, М2. Р. 149-156.
АЙ 1. Т., Вапкз Е. М. Ечосйопа] азресз
оЁ иКгазоциб ргобасНоп Бу пШашЕ а
по гаёз // Аппи. Вефау. 1972. Ув. 20.
Р. 115-419.
А10пз0 В. А., УШакотез О. А. Е., бапспег
А., Сапш 7. М. Амаю Кагуовуре (20-42)
ш Ше ЫасК гаё (ВаНлаз табаз) а Ме
х1со // Апи. Сешеё. 1982. Ув. 25, М 3.
Р. 133—135.
А1зюп 7, Вгоот 5. Герозргоз!3 Ш тат
ап апппа?з. ЕдшЬогсй; Г, 1958. 500 р.
Айтапп 5. А. ТВе заду оЁ рениа{е зоса1
сошташсаНоп // Зос1а] сотиозисаНол
атопя рейпа(ез. СЫсасо: Ушу. СЫсабо
ргезз, 1967. Р. 300.
`
А1рагех А., баз Г. оеасюЧазе сБапрез
1 {Ме деуеюрше пиезНла {тасё о 1Ве
га! // Майе. 1971. Ус]. 190. Р. 826—827.
АтогЕ б., Сына М., Сошой Г. Зи го-
ЧИотг (СИ 9Чае, АгусоНдае, Миг!ае)
Ое!’ЦаПа репшзщаге ед лазаге 1 гар-
рофо аНаштыее МосШтансо МедПегга-
пео // АппоаНа, 1984. У]. 14, М 1/3.
Р. 217-269.
Апдегзоп 7. У’. Те ргодасНоп о аЙтазо-
пс Бу ЛаБогафоту гаёз ап@ о ег шаш-
зта1з // Зс1елсе. 1954. \0]. 119. Р. 808.
Апагеш; В. И’, Вейпар В. И’, `Кеспап
Е. /. ОетостарЬ1с ап4 гезропзез оЁ Мог-
уау га ю ап ЁегИШеу сошго] // 7. \УП9-
Ше Мапас. 1974. Уо|. 38, М 4. Р. 868—
874.
Ашзко 1. Т., Зиег 5. Ё., МеСИитосЁ М. К.
её а1. Тье ге]айопзрар Бейуееп 22 КН»
и|(газотс 3101213 ап зос10зехиа| Бера-
\тог м фе гаё//Т. Сошр. ап Рвуз101.
Рзусво|. 1978. \0]. 92. Р. 824—829,
Аррегзоп С. 5., Запаег; О. Т., Каикетеп
р. Е. Гафогаогу ап@ Не]9 (113.оЁ {Ве
годепис!е Бго4Насойта абазё зуага-
1о-гез13ап{ Могмау газ // РезЫс. Зе
1981. Уи. 12, М 6. Р. 662—668.
АгтИаве К. В. $оиа] Бева\юог ап роро-
1апод Фупайся оЁ шагио$ /} ОЦсов. 1975.
\о1. 26. Р. 341—354.
Аттиаве К. В. Зо] уатсву ш с ус|-
1о%-Бе!Ше4д шагто: а роршаНоп-Бенау10-
ха] зузбежт // Аюпт. Верах. 1977. Уо/. 25.
Р. 585—598.
Агоп С., Съиеаи О. Ргезатед туоуететф
оЁ рЬегопопез п шайпях Бепау!ог т, {пе
гаё // Погтопе Верах. 1974. У01. 2.
Р. 315—323.
Агоп С., Нооз 1., Азсь С. ЕНес& о{ гето-
уа| оЁ {Ле оНасвогу Баз оп шайие Ъе-
Ваутог ап оуШайоп шт {Ме га // Меп-
гоепдосгто]озу. 1970. У]. 6. Р. 109—117.
Аззе{ Бег В. Н. Ое уегзрее Че уап 4е
ЮМете 2о0#41егеп 1 Ве]е уап де Вап8
уап ЬгааКБаепапаТузе // ВиП. [13% гоу.
Бет. паг. Ве. 1974. Уд]. 47, М 5. Р. 60.
Анау 7. В. Вед шИез апд фурВиз. Г.: АФ-
Лопе, 1968. 102 р.
Арегу Р. М. Тье 91зрегза| оЁ Ъго\п газ
ВаНа$ погуе21с1$ ап пе\ зресоепз
{тот 19 сепфату Саре Тоуп // Машта-
Па. 1985. У01. 49, М 4. Р. 573—576.
Ваыску А., Очадаотга 9., Райзек Г. её а|,
0зе о{ габто1зоюре (есвонтаез Гог дебег-
шииео Фе \уеаптх рего@ 1ш ехрег!-
теюа! апоа1з // Руз101. БоВетлоз]. 1970.
\Уо1. 19. Р. 457—467.
Вафифет? В. ЕрЧети1оо21е Ч4ез Герозр!-
го5ез ФЦаНо
// 20010565 сопа1ззапсез
Чери фае попуеПез: СоПоч. ОМ5/ЕАО
ве 1е8 70010868. Сбпеуе, 1954. Р. 212.
Вафифет В., Мозсоте: 7. Перозритозе! пе]-
1е г!зае 9е Гао е Ч4еШа баг4деупей //
АН: 136 Зарег. ЗавпНа. 1955. Уч. 18.
Р. 70.
Ваепптаег Г. Р. Тре гейаБШу оЁ дот!
пайсе огфегз т гаёз// Апот. ВеБах.
1966. Уо1. 14. Р. 367—371.
Раептткег Г. Р. Сотраг!зоп о? Бевауоп-
га] деуортеюе 1 зосаПу 1з01ей апд
отопрей га!з // 194. 1967. УоЁ 15.
Р. 312-828.
Ваепптеег В. Зоште сопзедистсе$ 0 аб-
этеззтуе Бера\1ог: А з@есНуе ге\е\м оЁ
{Ве Пстабаге оп о{Вег апта]5 // Арёгез.
Вера. 1974. Уо|. 1. Р. 17-37.
Варота Б., бай Е., Гапе Е., Уатов В.
ЕЁ степсу о{ ега@сайте газ \ИВ рагаЁ-
Литература
427
Но Байз// уеги. РезЕ Соп
о]. 18, М6. Р. 1245.
58.
Вала Р.Р. ТЬе емо йоп 0Ё магтой 30-
е16ез: а репега! {еогу // Зетее. ,
Уо1, 185. 2. 415-420. У! с. 1974.
Ватра! В. 7. Апегрась Р., Сеует Г. А.,
МеПчозй Т. К. ОИтазоше уосаП2анойв
ш_ гаф зехиа| Берлау1ох // Ашег. 7.001.
1979. Уч. 19, No2. Р. 469-480.
Вагпеа В. 71, Сеуег Г. А. Зехиа! Ъева-
мог: о
роедаотаогу опр 9
пе та[е гаё е1епсе.
аа
1972. Уо]. 176.
Вагре@4 В., Сеусг Г. ТВе оНтазове розте-
Дасаюту уосаЙтаноп апа е розе]а-
оао теГгасвогу репой оЁ Ше таГе //
‚ Сошр. ап
3191. Р%
5
\о1. 88. Р. 723-734
енот 1975.
Ватпей 5. А. Сотренноп атопх \Па
газ // Майе. 1955. Уо]. 175. Р. 486—497,
Вагпей 5. А. Вепау1ог сотропетз ш фе
Теефто о! м9 ап4 1аБогатогу та //
Веваутоиг. 1956. Уд]. 9, М 1. Р, 24-43.
Вагпей 5. и Ап ау о{ зоба! реБа-
Утот т 19 га // Ргое. 7001. бое.
Уо1. 130. Р. 107—152.
ос. 195За.
Ватпей 5. А. Гафогаогу ей ойз {ог Ъе-
Вахтог вез о Йа гаЁз //Т. Апип.
Тесвп. Аззос. 19585. Уо|. 9. Р. 6—14.
Ватпей 5. А. А заду о! БеБауюце: Рей
спеё оГ ело]обу апд Бера\оиг рзусво-
108у Ир ауе таиЦу т Фе таё 1:
Мелеп, 1963. 288 р.
Вагпей 5. А. 30621 згезз // Уемропиз
ш Ыо1оду. [.: ВайегуотИлз, 1964. Р. 260.
Вагней 5. А. Ваз / 8“. Азтог. 41967
\Уа1. 246, М1.Р. 79-86.
о
Ватпей 5. А. "Гра гаё: А за4у о Беваутоиг.
Сусаво; Г.: Ому. СЫсаво ргезз, 1975.
р.
у
.
Вате! 5. А. Вит 7. ЕаШу зНииЦаноп
п@ шатра ЪеВау1отг // Ма
ой, 213, В. ЧО
Майе. 1967
Вагпен 8. А., брепсег М. Ехрегипениз оп
о Фоод ргеЁегепсез оЁ \И га (Ва
5 погуео1сиз Вегкепро
1
1953. Уа. Е М.Р м // Вувтете,
Ватоп А., Вгоойзте К. Н. Митап В. А.
ЕНесёз оЁ папе ап адаНз зроскта-
ита проп ]феатишо шт Ме ада жЪие
та // 7. Сошр. ап РВузю]. РзусНо].
1957. Уо1. 50. Р. 530—534.
Ваттей С. Е. Ваз шт ргане аот1саНаге //
т. 1пзё. Вет. 1967. \У0]. 22, М6. Р. 43—
++,
-
Виезоп С. А Шеоту оЁ Рау апё Га
//
РэзусМаЕ. Веу. 1955, Те х Р. р
Вашеу У. М., Зтиь`Р. 1. No. Ваткош 6.1.
Тре ерЧеп!оору о 1ерюзр!г05185 Ш
Мог Оцеспз]ап4. 2. Кагбег оЪзегуа-
от8 оп Ве [035 т бе Моззшаи 9156-
1166 // У. Нуз. 1964. Уд. 62, М4. Р. 485-—
А9л.
Ваиег 7. Н. ЕНесу оЁ зпафегпа] эва(е оп ве
ето го пез оЧогз о{ ВоодеЯ
та(5 У10]. апа Вевау. 1983. . 30.
В Р. 229—232.
Уо. 30
бит ег М. Пъег ]азНе Убре! пла Зааег
а]; КаНатГо]сег ап МавПеррев поё м.
Пенкецеп 2 Штег Бо]ор1зсНеп Векйтр-
Рапз // Аша. Эсв5аНлозК., РНапт. пад
ромео, 4971. Ва. 44, М9. 3. 138—
Ваилиег И., Маптапп-Ецеппе С., боат-
Йо ХТ. её а1. ЭсВаЧИеве Ее] дпасег т Рог-
ще ТЬЯ. 1984. ВВ. 57, МТ. $. 134-
Варетось Р. В. Адатз М., Махзоп Г. В.
Уоз:4а Т. Н. Сепеые аетепцаной
атопа Кагуобур!с Тоглв оЁ (Ле ЫасК га
Ваз гаМлз // Сепенсв. 1983. -УоЁ. 105.
No4. Р. 969-083.
у
ВавегюсЕ Р. В., Райз С. В. 5., Ааатз М.,
Сое 5. Н. Сепенса! геаНотзррз
ашопе АпзгаНап гоЧеп{з (Миг Чае) //
Апзга]. 1. 7001. 1981. Ус. 29, МЗ
Р. 289—303.
ВатегзосК Р. В., Райз С. В. 5., Новамь
7. Т. Кагуоуре Чайа ап Ме зресне
Збайлв ап фахополис аНиез о? Ван
{15 У 03158118 апа В. зог414 105 ИТЫа.
1975. Уо1. 23, М 3. Р. 293—294.
Варетз ось Р. В., Тань С. И. 5., Нова
7. Т. её в]. СБготозоте еуош оп п Алз-
табап годет(з. 2. ТВе ВаНаз отопр //
рготаовотии. 1977. Уо|. 61, No3. Р. 297-
Веасй Г. А. Сошраг!зоп оЁ соршаюгу Ъе-
Вачтошт о ав га га1зе т 13а от,
сопабкайов ап зертегаНои // 7.
.
РзусНо|. 1942. Уч, 60, Р. 124. 7. белее
Веасй Е. А. Сохгепё сопсерз о! Рау т
аппа|
// Атег. Мабог. 1945. Уо| 79.
Р. 523—541.
Веась, Е. А., Таупез 7, Зил91е8 оЁ пабогоа]
тоитеуши Ш гав. 1. ВесосйНов о
уоппе //Т. Матиа!. 41958. Уф, 37.
Р. 177—180.
ВеаНу ТУ. И’. Подее А. М., Войре Г. У.
её а]. Рзуспотоюг зи аи, зоса|
ЧермуаМоп ап рау ш ауепПе газ й
РВагтасо]. В1ослет. апа Вевау. 1982,
\Уо|. 16. Р. 417—422.
Веану У’. И’, РодЕе А. М., Тао" К.
её а Зер! 1езюпз тогеазе р]ау Пов-
Ире ш уепПе гафз // РВузю1. ап@ Ве-
Вау. 1982. Уч. 28. Р. 649-652.
Вевисратр С. В., Магии }. С., Шейти-
498 Литература
1юп 7. Г., МузосЫ С. т. Тве ассеззогу
оНасфогу зузюш: го]е ш шапцепапее
о сВемошуезисаюгу Бераутоиг// СНе-
пса! юпа! ш уегеЪгаюз. М. У.: Ре-
пои ргезз, 1983. Уд. 3. Р. 73—86.
Вескег К. дабато Цайаз Езерег, 1803 //
Нап@Биел 4ег З&аоейеге Епгораз.
УЛезЪадеп: АкаЧ.-Уег|., 1978. Ва. 1:
Марейете. 5. 382—420.
В
,
Векйтрите уоп Памеп пп@ Мёизеп т
УбгазеБаП // МаШе Мазер легбеспи.
1982. Ва. 119, Г 3. $. 34. а лока
Ко! М. зоба ау Ш соу01е$,
Верой 4058 И обыесе, 1974а. Уо|. 24.
Р. 225—230.
В
Веко}! М. оса! р]ау апд р|ау-зоНстя
11 Нап сап1з
// Ашег. 7оо[. 19745.
У. 14. Р. 323—340.
и
еко{! М. Апипа] рау ап@ БеБау1опга
и // Ахтет. Мафат. 1975. Уо|. 109.
Р. 601-603.
,
Веко! М. МаттаНап 91зрегза: ап@ фе
отогепу оЁ шаг/Чца! Бевауоига] рБе-
пофурев
// ТЬ19. 1977. Уо]. 141. Р. 715-
752.
,
Кой М. Зоса|] р1ау Бевауюпг // Вю-
а па 1984. У\о|. 34, М 4. Р. 228-
233.
Вей В. И’. ОNoгазоио98 шт зтаП годетиз;:
АгопзаЕргодисе@ ап@ агодза-ргоди-
сшр // Оеуе]ор. РзусвоШо!. 1974. Уо1. 7,
М1. Р. 39—42.
Вей В. М’, МизейКе И’. Сотгу т. Н.,
Расйтеп У. Папе збплайопап@
иНтазоте зюпаПИу а роз е шеф аог
0# еату Вап@Нле рВепотепе // 119.
1974. Уо1. 4, М 3. Р. 181-191. =
ВеП В. С., Задов: 7. А., Мойзертет Г.А.
Меспап1зт оЁ асмоп о# аа; муаг-
Тат ап9 тебабоНзт ог уНатт К: и
Вюсвепизту. 1972. Ус. 144, М 10.
Р. 1959—1961.
Вейаск В. Е. Ре И’ ТУ. В. А@уапсед
за41ез ш Фе еуашайоп оЁ сПелиса|
гереПешз // Т. Атег. РВаги. Аззос. 1949.
Уо1. 38, М 2.Р. 109—112. ,
ВепЦеу Е. No’. Сошхо[ оЁ газ ш зе\мусгз //
М. Аст. ЕзВ ап@ Роо4. Тесвп. ВиИ.
1960. М 10. Р. 22.
м
еу Е. No. ТАе уаагиь гезжапее
а
шт ода апд \ез // Вас
бепьююе пп@ ВамепБекйтрЁшс:
(УотиавзуегаицаЦиня 4ез Уегешз тг
УТаззег-, Водеп- ип@ ГаЙтучепе ата 28
019 29. Моуешфег 1968, КопотеззВаНе,
ВетНи). За Идагь, 1969. 5. 19—25. ь
ВепНеу Е. И’. А теме\у оЁ апИсоахи]ащ
годепис14ез ш саггеп пзе // ВаН. Уго 9
Неа Отгбан!2. 1972. \У01. 47. Р. 275-
280.
.
ВегЕ О. 5, Ваепткег В. Ен Бу 1а-
Богафогу газ Чигте НобЫте епсойи-
Чегз
// Вевау. В1юоТ. 1973. Уд. 8, М 6.
Р. 733—741.
,
Вегтапф в. Ташог Г. ПиегасИуе еНесз
оЁ ехремепсе ап@ оМасфогу ЫМЬ 1е510п0
Ш ша|е гаё соршаНоп // Рруз!0|. ап@ Ве-
Вах. 1969, Ув]. 4. Р. 13—17.
Вегпага 7, Сойпт С. А ргорогв Фе 4еса{
де загили10 8 (ВаИаз погуер!сив Вегк)
еп отапде саНиге // Рагаз са. 1966.
Уо1. 16. Р. 140—148.
о|
Вегпзфен Г. Тре еНеез о{ уамаНотз т
Бала паров ]еагшис ап геепнол //
. Сотр. ав Р\ЬузоЁ Рэусво|. 1957.
\1. 50. Р. 162.
.
Вйаташа] О., Ртаказь 1. АззобаНоп оЁ
Бай збупезз ИВ Фе сотропеш о
ро1зопей {009 зи Ваббаз гаИлаз // 7бзевг.
аНоет. 7оо{. 1979. Ва. 66, М 3. $. 329-—
335
Бтррат И’. Е. бт [ий ТУ. 7. ТВе еНесь
оЁ @1Негешь еп тотевз диго раЁап-
су оп ада Бевауюог ш Фе таб/
1. Сотр. ап@ Рвузо1. Рзусво|. 1952.
Уо1. 45. Р. 307-3412.
ВгЕе Г. Г. ЗсепелпатЕте апд ре сего
суе оЁ те 1ета!е гаё// Апии. Верах.
1978. Уо|. 26. Р. 1165-1168.
,
БтЁе Г. 1, Меапеу М. 7. допада| юго193
ап@ фе зехиа! 91егепНаНол оЁ р!ау Ъе-
Вамтойг оЁ Ще гаё// Ргос. 48 Пиеги.
ефо[. соп{. ВизЬале, 1983. АЪзт. 28.
Р. 13.
,
ВигЁе Г. Г., байег О. Зееретагктя Бе-
Вау Ш тезропзе ®ю опере
тз Бу Ше гаь ВаИмз потуееи$
па 1984. Уо1, 32, М 2. Р. 493—
500.
Вапсвага В. Г, Гиту Р. Р. Р., | Вап-
срага Р. С. Сопзресе зонда т
Неетапошо Вацаз погуеа1е0з` ош’
зваЫе ап@ ппзаЫе роршаНопз // Рзу.
сво[. Вес. 1985. Уо|. 35, М 3. Р. 329-
336.
..
Напрта И’. Т. Тве Ёзипа оЁ ВчазВ Шш
Ч1а, топе Сеуоп апд Вигта: Мат -
тпаНа. 1.: Тауог ап@ Егапез, 1888-.
891. 647 р.
Вы Е. М. Редегзен Р. Е. Ргепаё| апЯ
розбиайа! Чеёегичтатя оЁ Те 136 засК-
Попе ер1зоде ш &Ыпо гай // Оеуеор.
Рзусво 101. 1982. \01|. 4. Р. 349-3583.
Воке В. Боше Бера\юогаГ {е563 оЁ 90-
езИсаНоп ш Могуау гафз // Вевауюог.
1972. Уд]. 42. Р. 198-231.
.
Восе В. Е. Вштомз оЁ УП ап ато
Литература 429
тай еНесь оЁ ЧотезисаНоп, ом Чоот, асе,
ехренчептсе ап таегпа! чае ИТ. Сошр.
ап Рпузо]. РзусвоГ. 1977, Ув. 91.
Р. 649—664.
ВоЙез В. С., Моой; Р. у Тве опюосепу
оЁ Ъеваутоог шт Фе ато габ// Апиа.
Вевау. 1964. Уо]. 42. Р. 427—441.
Вошегтап А. М., ВтосЁ: }. Е. ЕуащаНоп
0Ё 10-5897 аз а ше спешоз{егНаиь {юг
таб сошто!//Т. УЯШе Мапас. 1974.
Ув]. 35, М 4. Р. 648—624.
Втешайег Ф., Геоп М. ЕасИаНоп оР тацег-
па} (гаозрогё Бу Могуау гае рирз//
7. Сотр. апа Рвузто|. Рзусво!. 1980.
У]. 94. Р. 80-88.
Внавез В. Рататамон: Из гое м Ще
Топрйегт тоепНов оЁ шабегиа] Берау1опт
Ш Че габ// РВузюо]. ап Венах. 1977.
У]. 18. Р. 487—490.
ВнаЕез В. Пеюпиоп о гар! опзеё о
шафетпа! Берау1оиг Читая ргеспавсу
т реширагой$ таз // Вевау. В!01. 1978.
\о1. 24. р. 143—147.
Внавез В. 5., РЕ Возе Н., Гоипаез р. р.
Ргоасив запиМаНон о та{егпа! Ъе-
ат ш ета та // Зеаеспее, 1985.
\0]. 227. Р. 783—784.
Вт4вез В. 5., ВозепЫаи 1. 5., едет Н. Н.
Зегиш рговезюгопе сопсещгаНоп ап
табегпа! Берауойг т газ аНег ргед-
папсу (егиипаНоп: Беваутопга! зеашо-
Лабоп аНег ргорез(егопе УИ гауга! апа
ШНЫбоп Ъу рговезёегопе тшайпиапсе //
опдостиоюду. 1978а. Уд. 102. Р. 258—
67.
Вт4еез В. 5., Нозепан 7. 5., Редег Н. Н.
Зитшайоп оЁ шаегпа! гезропз{уепезз
аНег ргерпапсу ‘етиыпаной т таё/
Ноттове Вевау. 1978Ъ. У]. 10. Р. 225—
5.
Вгть ЕР. Н. вап 4еп. Ге ЗаисеНеге Епго-
раз. НатЬиге; В., 1956. 525 $,
Бтойе 7., КазЁ С. Е., Стаи Е. Могуау
таз, (Ваоз погусй!сиз) оп сете! ваЪ-
Ые//Т. 2001. 1983. Уа, 195, М4
Р. 542—646,
Втопзоп Е. Н., Беззагатз С. В (его! Ног-
%Шопез ап@ аротезайуе Ъевах1отг // ТВе
рНузю1обу о аротеззоп а09 Че!оав,
No У.; 6.: Р]еший ргезз, 1974. Р. 215—
230.
Втопзоп С. И’. Тре Чеуейортепв оЁ {еаг
Ш шап апд_ офег апииа1з
// СЬИа Бе-
уеЮр. 1968а. \о]. 39, Р. 409—430.
Втопзоп С. И’. ТЬе {еаг ОЕ поуеИу // Рэу-
‚ сво]. Ву. 1968. Уо|. 69. Р. 350-358.
Втоой: Г.Е, Вошегтапв А. М. Ргеегепсе
0Ё \Па Могчау гоёз Гог таз, зее43,
ап@ ]ейитез // Резё Сошее. 1973. Уо1. 41,
М 8. Р. 16, 18, 36, 38—39.
Втоойз 7. Е., Ша Мати, ИаШюв РБ. И’.
©ь а!. Р/ахое т вта шаха! ап@
Витая т Вапяооп, Вага // Зои феасЕ
Азап Т. Ттор Мед. Ро. НеаНВ. 1977.
Уо1. 8, No 3. Р. 835-344.
Втоойзфапк В. МУ. г. И’Йзоп РБ. А. А.
СюпЕь 6. Тве ш мо праКе оЁ ЗН-ап-
Чгоза-4, 16-Ч1еп-З-опе {1 Нззиез оЁ Ме
ад ша]е гаф //Т. Епбостизо]. 1973,
Уо1. 57. Р. 1-44.
Втоши В. Е. Оцесе атесей игте-магКто
Ъу ше газ (Вайлз погуей1юлз) /
Вевау. В]. 1975. У0]. 15. Р. 251-254.
Втошт В. Е. Ойопг ртеегепсе ал птше.
штат ше зсаез ш шые ап@ {ета
газ: еНесз о допадесюту ап@ зехиа|
ехречепсе оп гезропзез 1ю сопзрес с
одотв // 7. Сотар. РБусВо1. 19778. \а]. 94.
Р. 1190-2006.
Втошпт В. Е. ОЧопт ртеегепсе 5са]ез т
гавз // ОНасНоп ап тазе. [.: Ош,
ргезз, 1977Ъ. Уо]. 6. Р. 188.
Втошт В. Е. Тве э2-КНа рге-сори]аюгу
уосаНгайот оЁ ме шае га // РВуз01.
ап Вевау. 1979. Уо|. 92. р. 483—489.
Втошп В. Е. Ртеетепсев о[ рге- ап@ розё-
\жеапНие Топа- ЕКуапз газ Юг 065
оЧогз// ГЫ. 1982. Уч, 29. Р. 865—874.
Втгошп В, Е. ТВе годетив: зибог4ег Муо-
тогрВа // ЗосЁа| одогз {п шапта]3. Ох-
Тог4: Сатепдоп ргезз, 1985. Р. 345—457.
Втошп В. 2. Ноювса1 Таскюгз Ш дотезис
тодош соп4"о]. ВоскуШе, 1969. Р. 442—
125.
Втоштей В. Г., Срап #-Сйалп. Витомше
т No19 аз аошезнс Могмау газ И
г. Мапа]. 1977. Уд]. 58, No2. Р. 239—
240.
зан а09 ЛасбаНов- ш Ше га
7. Вергой. Ееге. 1964. Уо1. 2. Р. 17—34.
Втиппег Н., З1еуетз Р. Г, Васклиег 1. В.
Тпигодисед аттоа8 п Уаюна // Узсю-
пап Мабг. 1984. Уд. 98, М 1. Р. 5-17.
Втипо Л. Р., Тейог М. М. Веазе Е. М.
Пепзогу Чести+таю 8 оЁ зисКНас Бева-
ог т эеапНиг гаЁз ИТ. Сошр. апа
РЬузо!. Рвусво]. 1980. \01. 94, М 14.
Р. 115—127.
:
Витиз В. А., Тротаз В. Г., рарз 5. Е.
ТВтгее-Нта|! зедиепсез о{ теугат@ тасп!-
{4ез \И одог слез шахииед // Ва.
т вусвопот. Зос. 1981. \Уо|. 17. Р. 266—
2
Втисе И. М. ОБзегуайоп оп Фе ия
й
ВиНег 5. В, бизыта М. Е, бета.
Бетз 5. М. Мабегпа] БеНах!ог аз хези]а-
ог оЁ роуаште Бозущезз т Баш?
498 Литература
1юп 7. Г., МузосЫ С. т. Тве ассеззогу
оНасфогу зузюш: го]е ш шапцепапее
о сВемошуезисаюгу Бераутоиг// СНе-
пса! юпа! ш уегеЪгаюз. М. У.: Ре-
пои ргезз, 1983. Уд. 3. Р. 73—86.
Вескег К. дабато Цайаз Езерег, 1803 //
Нап@Биел 4ег З&аоейеге Епгораз.
УЛезЪадеп: АкаЧ.-Уег|., 1978. Ва. 1:
Марейете. 5. 382—420.
В
,
Векйтрите уоп Памеп пп@ Мёизеп т
УбгазеБаП // МаШе Мазер легбеспи.
1982. Ва. 119, Г 3. $. 34. а лока
Ко! М. зоба ау Ш соу01е$,
Верой 4058 И обыесе, 1974а. Уо|. 24.
Р. 225—230.
В
Веко}! М. оса! р]ау апд р|ау-зоНстя
11 Нап сап1з
// Ашег. 7оо[. 19745.
У. 14. Р. 323—340.
и
еко{! М. Апипа] рау ап@ БеБау1опга
и // Ахтет. Мафат. 1975. Уо|. 109.
Р. 601-603.
,
Веко! М. МаттаНап 91зрегза: ап@ фе
отогепу оЁ шаг/Чца! Бевауоига] рБе-
пофурев
// ТЬ19. 1977. Уо]. 141. Р. 715-
752.
,
Кой М. Зоса|] р1ау Бевауюпг // Вю-
а па 1984. У\о|. 34, М 4. Р. 228-
233.
Вей В. И’. ОNoгазоио98 шт зтаП годетиз;:
АгопзаЕргодисе@ ап@ агодза-ргоди-
сшр // Оеуе]ор. РзусвоШо!. 1974. Уо1. 7,
М1. Р. 39—42.
Вей В. М’, МизейКе И’. Сотгу т. Н.,
Расйтеп У. Папе збплайопап@
иНтазоте зюпаПИу а роз е шеф аог
0# еату Вап@Нле рВепотепе // 119.
1974. Уо1. 4, М 3. Р. 181-191. =
ВеП В. С., Задов: 7. А., Мойзертет Г.А.
Меспап1зт оЁ асмоп о# аа; муаг-
Тат ап9 тебабоНзт ог уНатт К: и
Вюсвепизту. 1972. Ус. 144, М 10.
Р. 1959—1961.
Вейаск В. Е. Ре И’ ТУ. В. А@уапсед
за41ез ш Фе еуашайоп оЁ сПелиса|
гереПешз // Т. Атег. РВаги. Аззос. 1949.
Уо1. 38, М 2.Р. 109—112. ,
ВепЦеу Е. No’. Сошхо[ оЁ газ ш зе\мусгз //
М. Аст. ЕзВ ап@ Роо4. Тесвп. ВиИ.
1960. М 10. Р. 22.
м
еу Е. No. ТАе уаагиь гезжапее
а
шт ода апд \ез // Вас
бепьююе пп@ ВамепБекйтрЁшс:
(УотиавзуегаицаЦиня 4ез Уегешз тг
УТаззег-, Водеп- ип@ ГаЙтучепе ата 28
019 29. Моуешфег 1968, КопотеззВаНе,
ВетНи). За Идагь, 1969. 5. 19—25. ь
ВепНеу Е. И’. А теме\у оЁ апИсоахи]ащ
годепис14ез ш саггеп пзе // ВаН. Уго 9
Неа Отгбан!2. 1972. \У01. 47. Р. 275-
280.
.
ВегЕ О. 5, Ваепткег В. Ен Бу 1а-
Богафогу газ Чигте НобЫте епсойи-
Чегз
// Вевау. В1юоТ. 1973. Уд. 8, М 6.
Р. 733—741.
,
Вегтапф в. Ташог Г. ПиегасИуе еНесз
оЁ ехремепсе ап@ оМасфогу ЫМЬ 1е510п0
Ш ша|е гаё соршаНоп // Рруз!0|. ап@ Ве-
Вах. 1969, Ув]. 4. Р. 13—17.
Вегпага 7, Сойпт С. А ргорогв Фе 4еса{
де загили10 8 (ВаИаз погуер!сив Вегк)
еп отапде саНиге // Рагаз са. 1966.
Уо1. 16. Р. 140—148.
о|
Вегпзфен Г. Тре еНеез о{ уамаНотз т
Бала паров ]еагшис ап геепнол //
. Сотр. ав Р\ЬузоЁ Рэусво|. 1957.
\1. 50. Р. 162.
.
Вйаташа] О., Ртаказь 1. АззобаНоп оЁ
Бай збупезз ИВ Фе сотропеш о
ро1зопей {009 зи Ваббаз гаИлаз // 7бзевг.
аНоет. 7оо{. 1979. Ва. 66, М 3. $. 329-—
335
Бтррат И’. Е. бт [ий ТУ. 7. ТВе еНесь
оЁ @1Негешь еп тотевз диго раЁап-
су оп ада Бевауюог ш Фе таб/
1. Сотр. ап@ Рвузо1. Рзусво|. 1952.
Уо1. 45. Р. 307-3412.
ВгЕе Г. Г. ЗсепелпатЕте апд ре сего
суе оЁ те 1ета!е гаё// Апии. Верах.
1978. Уо|. 26. Р. 1165-1168.
,
БтЁе Г. 1, Меапеу М. 7. допада| юго193
ап@ фе зехиа! 91егепНаНол оЁ р!ау Ъе-
Вамтойг оЁ Ще гаё// Ргос. 48 Пиеги.
ефо[. соп{. ВизЬале, 1983. АЪзт. 28.
Р. 13.
,
ВигЁе Г. Г., байег О. Зееретагктя Бе-
Вау Ш тезропзе ®ю опере
тз Бу Ше гаь ВаИмз потуееи$
па 1984. Уо1, 32, М 2. Р. 493—
500.
Вапсвага В. Г, Гиту Р. Р. Р., | Вап-
срага Р. С. Сопзресе зонда т
Неетапошо Вацаз погуеа1е0з` ош’
зваЫе ап@ ппзаЫе роршаНопз // Рзу.
сво[. Вес. 1985. Уо|. 35, М 3. Р. 329-
336.
..
Напрта И’. Т. Тве Ёзипа оЁ ВчазВ Шш
Ч1а, топе Сеуоп апд Вигта: Мат -
тпаНа. 1.: Тауог ап@ Егапез, 1888-.
891. 647 р.
Вы Е. М. Редегзен Р. Е. Ргепаё| апЯ
розбиайа! Чеёегичтатя оЁ Те 136 засК-
Попе ер1зоде ш &Ыпо гай // Оеуеор.
Рзусво 101. 1982. \01|. 4. Р. 349-3583.
Воке В. Боше Бера\юогаГ {е563 оЁ 90-
езИсаНоп ш Могуау гафз // Вевауюог.
1972. Уд]. 42. Р. 198-231.
.
Восе В. Е. Вштомз оЁ УП ап ато
Литература 429
тай еНесь оЁ ЧотезисаНоп, ом Чоот, асе,
ехренчептсе ап таегпа! чае ИТ. Сошр.
ап Рпузо]. РзусвоГ. 1977, Ув. 91.
Р. 649—664.
ВоЙез В. С., Моой; Р. у Тве опюосепу
оЁ Ъеваутоог шт Фе ато габ// Апиа.
Вевау. 1964. Уо]. 42. Р. 427—441.
Вошегтап А. М., ВтосЁ: }. Е. ЕуащаНоп
0Ё 10-5897 аз а ше спешоз{егНаиь {юг
таб сошто!//Т. УЯШе Мапас. 1974.
Ув]. 35, М 4. Р. 648—624.
Втешайег Ф., Геоп М. ЕасИаНоп оР тацег-
па} (гаозрогё Бу Могуау гае рирз//
7. Сотр. апа Рвузто|. Рзусво!. 1980.
У]. 94. Р. 80-88.
Внавез В. Рататамон: Из гое м Ще
Топрйегт тоепНов оЁ шабегиа] Берау1опт
Ш Че габ// РВузюо]. ап Венах. 1977.
У]. 18. Р. 487—490.
ВнаЕез В. Пеюпиоп о гар! опзеё о
шафетпа! Берау1оиг Читая ргеспавсу
т реширагой$ таз // Вевау. В!01. 1978.
\о1. 24. р. 143—147.
Внавез В. 5., РЕ Возе Н., Гоипаез р. р.
Ргоасив запиМаНон о та{егпа! Ъе-
ат ш ета та // Зеаеспее, 1985.
\0]. 227. Р. 783—784.
Вт4вез В. 5., ВозепЫаи 1. 5., едет Н. Н.
Зегиш рговезюгопе сопсещгаНоп ап
табегпа! Берауойг т газ аНег ргед-
папсу (егиипаНоп: Беваутопга! зеашо-
Лабоп аНег ргорез(егопе УИ гауга! апа
ШНЫбоп Ъу рговезёегопе тшайпиапсе //
опдостиоюду. 1978а. Уд. 102. Р. 258—
67.
Вт4еез В. 5., Нозепан 7. 5., Редег Н. Н.
Зитшайоп оЁ шаегпа! гезропз{уепезз
аНег ргерпапсу ‘етиыпаной т таё/
Ноттове Вевау. 1978Ъ. У]. 10. Р. 225—
5.
Вгть ЕР. Н. вап 4еп. Ге ЗаисеНеге Епго-
раз. НатЬиге; В., 1956. 525 $,
Бтойе 7., КазЁ С. Е., Стаи Е. Могуау
таз, (Ваоз погусй!сиз) оп сете! ваЪ-
Ые//Т. 2001. 1983. Уа, 195, М4
Р. 542—646,
Втопзоп Е. Н., Беззагатз С. В (его! Ног-
%Шопез ап@ аротезайуе Ъевах1отг // ТВе
рНузю1обу о аротеззоп а09 Че!оав,
No У.; 6.: Р]еший ргезз, 1974. Р. 215—
230.
Втопзоп С. И’. Тре Чеуейортепв оЁ {еаг
Ш шап апд_ офег апииа1з
// СЬИа Бе-
уеЮр. 1968а. \о]. 39, Р. 409—430.
Втопзоп С. И’. ТЬе {еаг ОЕ поуеИу // Рэу-
‚ сво]. Ву. 1968. Уо|. 69. Р. 350-358.
Втоой: Г.Е, Вошегтапв А. М. Ргеегепсе
0Ё \Па Могчау гоёз Гог таз, зее43,
ап@ ]ейитез // Резё Сошее. 1973. Уо1. 41,
М 8. Р. 16, 18, 36, 38—39.
Втоойз 7. Е., Ша Мати, ИаШюв РБ. И’.
©ь а!. Р/ахое т вта шаха! ап@
Витая т Вапяооп, Вага // Зои феасЕ
Азап Т. Ттор Мед. Ро. НеаНВ. 1977.
Уо1. 8, No 3. Р. 835-344.
Втоойзфапк В. МУ. г. И’Йзоп РБ. А. А.
СюпЕь 6. Тве ш мо праКе оЁ ЗН-ап-
Чгоза-4, 16-Ч1еп-З-опе {1 Нззиез оЁ Ме
ад ша]е гаф //Т. Епбостизо]. 1973,
Уо1. 57. Р. 1-44.
Втоши В. Е. Оцесе атесей игте-магКто
Ъу ше газ (Вайлз погуей1юлз) /
Вевау. В]. 1975. У0]. 15. Р. 251-254.
Втошт В. Е. Ойопг ртеегепсе ал птше.
штат ше зсаез ш шые ап@ {ета
газ: еНесз о допадесюту ап@ зехиа|
ехречепсе оп гезропзез 1ю сопзрес с
одотв // 7. Сотар. РБусВо1. 19778. \а]. 94.
Р. 1190-2006.
Втошпт В. Е. ОЧопт ртеегепсе 5са]ез т
гавз // ОНасНоп ап тазе. [.: Ош,
ргезз, 1977Ъ. Уо]. 6. Р. 188.
Втошт В. Е. Тве э2-КНа рге-сори]аюгу
уосаНгайот оЁ ме шае га // РВуз01.
ап Вевау. 1979. Уо|. 92. р. 483—489.
Втошп В. Е. Ртеетепсев о[ рге- ап@ розё-
\жеапНие Топа- ЕКуапз газ Юг 065
оЧогз// ГЫ. 1982. Уч, 29. Р. 865—874.
Втгошп В, Е. ТВе годетив: зибог4ег Муо-
тогрВа // ЗосЁа| одогз {п шапта]3. Ох-
Тог4: Сатепдоп ргезз, 1985. Р. 345—457.
Втошп В. 2. Ноювса1 Таскюгз Ш дотезис
тодош соп4"о]. ВоскуШе, 1969. Р. 442—
125.
Втоштей В. Г., Срап #-Сйалп. Витомше
т No19 аз аошезнс Могмау газ И
г. Мапа]. 1977. Уд]. 58, No2. Р. 239—
240.
зан а09 ЛасбаНов- ш Ше га
7. Вергой. Ееге. 1964. Уо1. 2. Р. 17—34.
Втиппег Н., З1еуетз Р. Г, Васклиег 1. В.
Тпигодисед аттоа8 п Уаюна // Узсю-
пап Мабг. 1984. Уд. 98, М 1. Р. 5-17.
Втипо Л. Р., Тейог М. М. Веазе Е. М.
Пепзогу Чести+таю 8 оЁ зисКНас Бева-
ог т эеапНиг гаЁз ИТ. Сошр. апа
РЬузо!. Рвусво]. 1980. \01. 94, М 14.
Р. 115—127.
:
Витиз В. А., Тротаз В. Г., рарз 5. Е.
ТВтгее-Нта|! зедиепсез о{ теугат@ тасп!-
{4ез \И одог слез шахииед // Ва.
т вусвопот. Зос. 1981. \Уо|. 17. Р. 266—
2
Втисе И. М. ОБзегуайоп оп Фе ия
й
ВиНег 5. В, бизыта М. Е, бета.
Бетз 5. М. Мабегпа] БеНах!ог аз хези]а-
ог оЁ роуаште Бозущезз т Баш?
430
Литература
ап Пеаг 0{ Ме деуейоршя га рир//
Зеепсе. 1978. У]. 199, Р. 445—447.
Саоип 7. В. Те деуе!оршепё ай го
о? зоса1 забав ашопб ПЧ Могуау
таз // Апаь, Вес. 1948а. Уо1. 101. Р. 694.
Сапоип Т. В. МомаШу ва шоуетет о
Ъго\уп гафз (ВаМаз погуео1сив) 1 аг-
инааПу зарегзафагайед рормйаНоп//
7. \МЦаШе Мапае, 19485. УоЁ. 12.
Р. 161—172.
Са!фоип 7. В. а аепсе оЁ зрасе ап Ите
оп фе $061а1 Бервазлойг оЁ Ме гаё//
Апар. Вес. 41949. Уо|. 105. Р. 508.
Сийони У. В. ТВе есообу ап з061010у
о{ Могжау таь Вефезда: 08 Шер.
Неа Н, 1962. 288 р.
Сафопп 7. В. Тве есоюбу ап зос10Ъ10]0-
ху оЁ (Ме Могуау таё. УУазв. (О. С.);
Ве!тезда, 1963. 320 р.
Сайеюп М. О., Миззег @- Сб. Миго9 го-
Детия
// Отдегв ап@ {ап Шез оЁ тесепф
тата! о Те 0119. М. У. УПеу,
1984. Р. 289—879.
Сатг No’. Г. РЬеготопа! зех аИтаса!з т
Ве Могуау гай // Мопуега1 сошзпашса-
Чоп. No У.; [.: Репа ргезз, 1974.
. Р. 103—181.
Сатг No’. Г. Низсй ФХ. Т., Ваз 7. М. Вез-
ропзез о{ мае габз 10 обогз {топ 1а-
тШаг у6т308 поуе| !ета[ев // Верау.
Меига[. В!ю]. 1980. Уо1. 29, М 3. Р. 381-
337.
Сагг И’. 7, Ктатез Г,., Сопяатло В. 7.
Рееуюиз зехиа| ехреепсе ап оНасбо-
гу ргейегенее {ог поуе| уетзи8 отита
зех рагёпегз {т гаёз //Т. Сотр. ап РВу-
° $101. Рзусро/|. 1970. УоТ. 71. Р. 216-222.
Сагг ИУ. У., Гоер Г. 5. [Язятвег М. Г.
Везропзез оЁ Ме гаЁз зех одогз // 119.
1965. Уа1. 59. Р. 370-377.
СазНе И’. Е. ТНе ЧотезИсаНоп оЁ Фе
та // Ртос. Маё. Асад. Зе. 08. 1947.
\Уо1. 33, М 5. Г. 109—117.
у
Санатеш М., УетпеЕМоиту Е. Статей 7.
Сопто] оЁ Ще габ‘оПШасоту Бы Ь аси-
уНу 1адисей Бу Моюз1саНу зюийсат
'’одогз // РНуз10|. ап@ Вевау. 1977. Уо|. 19.
Р. 381—387.
Срави А., бсйаерег С.
Атоп С. Ногтова| сокбто: оЁ 1Ъе рег
серНоп оЁ \Noе оНацюгу 3101а18 УМев
фасПНае 10г405з Берауог ш Ше шае
таф // Па. 1985. У]. 35. Р. 729—735.
Среиеаи П., Поозе 1., Атоп С. Асмоп 4е
Гагте шае оп 1етеПе ргоуепапё де
таз погтаох оп сазтез зиг 1а дотее
суе оезбга|`‘сВе? Noе таМе //С. г. 506.
М о1. 1972. Уо|. 3. Р. 1149-1418.
Свайеаи .Ю., Ноозе 7., Р1аз-Возег 5. её а1.
батта В.
Ногтопа[ шесраз1зтз, шуоуеё ш Фе
соп!то| оЁ осзтгоиз сус1е Чотайоп Ъу
4Ъе обоиг оЁ имие ш фе гаё// Аба еп-
Чосг! ло]. 1976. Уо]. 82. Р. 426—435.
Сыну О. А тайуе сепваз шефо@ Тог
Вго\уп гаёз (Вайлё погуес!сив) // Маш-
ге. 1942. Уо]. 150. Р. 59-60.
Сшину РБ. Сопфто| оЁ газ ап писе. Ох-
1ог4: СТагепдой ртезв, 4954. Уд. 2.
532 р.
СитзНап 7. 7. Зома]! зафот@та@оп, рорч-
1лаНоп 4епзНу апд шатотаНап еуоаН-
оп // Зепсе. 1970. \У01. 149, М 3927.
р. 84—90.
Сауюп В. В., No1301 А. М., Еташа Г. О.
'ГВе зКш з4егоз 0Ё погша! апйб итрата-
10]-(геае гаёз//Т. ар! Вез. 1973.
Уо1. 4. Р. 166—178.
Соиёй С. С. етич апд рорабоп
ргоелз // Азпег. Зс. 41965. Уо|. 53,
М2. Р. 199-212.
Сотроп В. Р., Косй М. О., Агп@ ИУ. 7.
ЕНесз о{ ша{егпа[ одог ов саг@ас гайе
о{ шаогпаПу зерагабе4 ша та //
РЬуз01. ап@ Верау. 1977. У01. 18.
Р. 769—773.
СопЕйп Р., Неерепезз Г. No’. Мабгайоп
о (етрегаате Вотлеоз$а$13 т фе га//
Атег. Г. РЬуз101. 1971. Ув. 220. Р. 333—
336
Сотфеё С. В. ТВе татта18 оЁ Фе Ра-
фаеатсбс тео1оп: А фахопоне теме.
Г.; ИБаса: Сотпе! Ушу. ргезз, 1978.
ЗАА р.
.
Согинап 7. @., Иаппейу К. 7. Огазвоше
хосаНяаНоп о ЧеЁеае тшае газ //
7. Сотшр. ап РЬузо| РзусВо!. 1979.
У\61. 93. Р. 105—115.
Спет 7. К. ТегасусПпез аз а Ниогезсетй
тагкег 1 Бопез ап@ (ее оЁ годен //
7. УПаН. Марас. 1970. У\о]. 34(40).
Р. 829-834.
Стоой 7. Н. Бома! Бепауюиг апд еФо1о-
‘цу // Зоса1 Бера\з1оиг шп Ыт4з ап@ шат-'
ша]з. М. У.: Асад. ргезз, 1970. Р. 21—40.
Сошле А. Т., РоЙеу 5. 7. ТБе шатитагу
2]ап@ а09 ПасбаНоп
// Зех ап ниегаа|
зесгеНопз. ВаNoтоте, 1964. Р. 590—642.
ракхутре-АЦота У. С., Ветоп О. Вера-
уопга| шоп ап Фе асе аб зос1а1
1зоТаНоп фт гаёз //Т. Сотр. ап@ РВуз101.
РусВо!. 1984. Уо|. 36, М 1. Р. 27-38.
Рау М. Еайу зитщаНол оЁ годеюёз:
А смисаЁ гемезху оЁ ргезешёЕ пуетрге-
Нопз
// ВгИ. 7. Рзусво|. 1973. Уо|.: 64.
Р. 435—460.
Раз РБ. Е. ТВе загуа! оЁ Иа Ьго\т
таз оп а Магу!апа {ат // Есо]осу. 1948.
Уо1. 29, М 4. Р. 437—448.
Литература 431
Рарз О. Е. А сотрамзоп о тергодисиуе
роепйа] оЁ 0 га роршаНонз// Есо-
1юсу. 1954а. У&. 32, М З.Р. 469-475.
Раз Ш. Е. Те геаИоп Бебуееп ее!
о рораНоп ап 312е ап@ зех оЁ Мог-
уау гаёз
// No19. 1954. Уо]. 32, М 3.
Р. 462—464.
Раз О. Е. Тье теабоп ребувеп 1вуе@] о!
рорШайоп ап ргебпапсу оЁ Могхау
К // Па. 1951с. У01. 32, М 3. Р. 459-
Раз В. Е. ТВе спагаскег1зЫс оЁ гаё ро-
ршайоп // Опаг. Веу. В10|. — 1953а.
\о1. 28, М 4. Р. 373-401.
Рав 2. Е. Апа1узз о{ Воте гапае ош
гесариге 9@//1. Матша. 1953.
\Уо1. 34, No 3. Р. 352—358.
Раз 0. Е, Етеп /. Т. ПШегепца| 4та-
раБ у оЁ гаёз ассот@тое 140 зе апд
зех //Т. УУПаШе Мапар. 1956. Уо1. 20,
М 3. Р. 41—43.
Раыз БР. Е., Еп4еп УХ. Т., бюйез А. М’.
ЗаФез оЁ Воше гапае т Фе Бом
гаф//Т. Мата]. 1948. Уо|. 29, М 2.
Р. 207—225.
Раз р. Е., ХоПу Р. Рипеез шп шаитта-
юву. К. У.: Вашво]4, 1963. 243 р.
Раз В. А. `ЕФЫс {81 аз а сагег бог
сотбасё годепнс1Чев // Пиеги, Резё Сотит.
1983. Уо|. 25, М 2. Р. 46-47.
Рекеузег Р. Г. Гез шатиШегез де ГА-
Чие Моте Егапса1зе. аКег: 1ЕАМ, 1955.
426 р.
Реапое Р. Пе Па гагее да Зоагити|0 а
Матасап// ВиП. 506. ра о|. ехоё. 1925,
М 18. Р. 152-1583.
ДРеппепьеги У. No Ап амер No0 {зо]е
ст ев ремо@з о{ деуеортешь 18 Ще
га //1. Сошр. ап РВузоГ. Рзусва,
1962. Уо|. 55, М Б.Р. 813—845.
ДеппепфегЕ У. No. А сопядегаНоп оЁ изс-
Г ле55 оЁ Ве ст са] рето9 пуроезз
аз аррНеЯ © фе зНииаНоп оЁ годен
т ПЫапеу// Еау ехремепсе ап@ Ье-
Ва\1ог: ТЬе рзусвоБоюру оЁ еуеор-
теле. Эрго е19 (11.): Твотшаз, 1968,
Р. 142—167,
Деппепреги. У. М. бтыа Г, 7. Зоча-
« зееКло ап поуебу-зееюио ЪеВах!отг
аз а поЧоп о аШогевиа] геагте ЬВ1-
36015 // 7. АБпогт. $06. РзусВо]. 1964.
‚
Уо]. 69. Р. 453.
Беппепфетя ТУ. No., НоПошау И’. К., Ро1-
Ипдег М. 1. Уае ваш аз сопзедиепсе
оЁ шацегпа] Бела\оиг т фе гаё// Ве-
Вау. В10]. 1976. Уо1. 47. Р. 51—60.
ДеппепфетЕ У. М., Ниакепз С. А., Пат
тор М. 2. М!ее геатёед \%Иф газ шод-
ВсаНоп оЁ Ъерау1ютг Бу еатЁу ехрегеп-
се \”ИЙ апобег зревез // Заепсе. 1964.
\Уо]. 143. Р. 380—381.
Деппепфеги Г. МNo., Кагаз С. В. Пщегаснуе
сНебз оЁ аде ап доганоп о! шШап6
схрейепее оп адоН, ]еагиао // РэусВо1.
Вер. 1960. \о1. 7. Р. 313.
Реппепфети Г. No., Кагаз @. С. ПиегасН-
ус с есбз оЁ ИМапе ап ада ехре-
епсе проп уешВё гаш ава пораШу
т Ше гаф//Т. Сотр. ап Рвуз1о]. Рзу-
сВо|. 1961. Уч]. 54. Р. 170.
.
ДРеппепьегЕ У. No. Момов 1. В. С., К!-
пе No. Т., Стош Г. 7. ЕНесё8 о дотаНоп
о Чаше зИлиаНоп ироп етоНопа-
Шу] Сапа. 7. Рзусво|. 1962а. Уо. 16.
Деппепфетк ТУ. No, ОНиег О. Н., 5ер-
йеп; М. М’. ЕНесь оЁ шафегпа| Тасюгз
про сто\ В ап@ Ъерау!оиг оЁ Ч1е га //
СЬИ9 Оеуеюр. 4962Ъ. У]. 33. Р. 65.
РеппепфегЕ У. М., Позепфеги К. М. Моп-
бепейе тапзти5ю0 оЁ пуюгтаНоп //
Мабите. 1967. Уо1. 246. Р. 549—550.
РеппепфегЕ У. No, Позепфеги К. М., 2аг-
тош М. Х. М1се геагеЯ у газ апп:
еНеср ш адаЙРоод проп р]азта согН-
созбегопе ап@ орепйе!4 аснуйу // Рву-
т ап@ Вевау. 1969. Уо1. 4. Р. 705—
"7.
ДеппепьегЕ У. No., бтий 7. ЕНовз о т-
ТатеПе эишабой ап асе проп Бева-
У1юит// 7. Сошр. ап Рьуз10|. РэусВо].
1963. \о1. 56. Р. 307-312.
Веогаз Р. Т. Ваз ап@ тет сопо //
119. Г. Еююшо]. 1964. ЗИуег УоЪЙес
Митьет. Р. 407—448.
Деотаз В. К. Ва шапаре ап@ Вох ю
Навь #// Резис4ез. 1975. У]. 9, М 8.
Р. 38—43.
Регис® Е. Н., Роре 7. Н. Мигте пурриз,
И\се, газ ап@ Неаз оп Ме ПагНие
Роуз
// Мед. Т. Амзиа[. 1960. Уо1. 2,
М 24. Р. 924-928.
Ре бапиз П., Зевтайз Г. И’, ТВе шо#фет-
Пиег те]абопзЫр ш асхеюршеюа! га
зи ез: сапиаНзи // Оеуеюр. Рзусво-
Ыо1, 1984. Уо|. 47. Р. 255—262.
Дезз-Рывфен Р., Газой В. Зоте оь-
зегуаНоп оп Ме ошюобенез!з оЁ. в0с1а]
пиегасИопз Ш Фе га //МопИ. 2001,
Ца]. 1979. Уо1. 13, М 23. Р. 204—202.
Пе М Р. Р. Зоип@ аз а годере деегтель //
Резр Сопг. 1969. Уо1. 37, М. В. Р. 36,
38, 40—42, 44.
Гез В. Р. Те еНесё оЁ ап ежегосерйуе
зи оп мк едесбоп т Ласбайия
та /Т. Рзусво]. 1968. Ув]. 197. Р. 37—
+0.
[Иетег Г. У., Въоаез В. Тшагена ш 14
430
Литература
ап Пеаг 0{ Ме деуейоршя га рир//
Зеепсе. 1978. У]. 199, Р. 445—447.
Саоип 7. В. Те деуе!оршепё ай го
о? зоса1 забав ашопб ПЧ Могуау
таз // Апаь, Вес. 1948а. Уо1. 101. Р. 694.
Сапоип Т. В. МомаШу ва шоуетет о
Ъго\уп гафз (ВаМаз погуео1сив) 1 аг-
инааПу зарегзафагайед рормйаНоп//
7. \МЦаШе Мапае, 19485. УоЁ. 12.
Р. 161—172.
Са!фоип 7. В. а аепсе оЁ зрасе ап Ите
оп фе $061а1 Бервазлойг оЁ Ме гаё//
Апар. Вес. 41949. Уо|. 105. Р. 508.
Сийони У. В. ТВе есообу ап з061010у
о{ Могжау таь Вефезда: 08 Шер.
НеаН, 1962. 288 р.
Сафопп 7. В. Тве есоюбу ап зос10Ъ10]0-
ху оЁ (Ме Могуау таё. УУазв. (О. С.);
Ве!тезда, 1963. 320 р.
Сайеюп М. О., Миззег @- Сб. Миго9 го-
Детия
// Отдегв ап@ {ап Шез оЁ тесепф
тата! о Те 0119. М. У. УПеу,
1984. Р. 289—879.
Сатг No’. Г. РЬеготопа! зех аИтаса!з т
Ве Могуау гай // Мопуега1 сошзпашса-
Чоп. No У.; [.: Репа ргезз, 1974.
. Р. 103—181.
Сатг No’. Г. Низсй ФХ. Т., Ваз 7. М. Вез-
ропзез о{ мае габз 10 обогз {топ 1а-
тШаг у6т308 поуе| !ета[ев // Верау.
Меига[. В!ю]. 1980. Уо1. 29, М 3. Р. 381-
337.
Сагг И’. 7, Ктатез Г,., Сопяатло В. 7.
Рееуюиз зехиа| ехреепсе ап оНасбо-
гу ргейегенее {ог поуе| уетзи8 отита
зех рагёпегз {т гаёз //Т. Сотр. ап РВу-
° $101. Рзусро/|. 1970. УоТ. 71. Р. 216-222.
Сагг ИУ. У., Гоер Г. 5. [Язятвег М. Г.
Везропзез оЁ Ме гаЁз зех одогз // 119.
1965. Уа1. 59. Р. 370-377.
СазНе И’. Е. ТНе ЧотезИсаНоп оЁ Фе
та // Ртос. Маё. Асад. Зе. 08. 1947.
\Уо1. 33, М 5. Г. 109—117.
у
Санатеш М., УетпеЕМоиту Е. Статей 7.
Сопто] оЁ Ще габ‘оПШасоту Бы Ь аси-
уНу 1адисей Бу Моюз1саНу зюийсат
'’одогз // РНуз10|. ап@ Вевау. 1977. Уо|. 19.
Р. 381—387.
Срави А., бсйаерег С.
Атоп С. Ногтова| сокбто: оЁ 1Ъе рег
серНоп оЁ \Noе оНацюгу 3101а18 УМев
фасПНае 10г405з Берауог ш Ше шае
таф // Па. 1985. У]. 35. Р. 729—735.
Среиеаи П., Поозе 1., Атоп С. Асмоп 4е
Гагте шае оп 1етеПе ргоуепапё де
таз погтаох оп сазтез зиг 1а дотее
суе оезбга|`‘сВе? Noе таМе //С. г. 506.
М о1. 1972. Уо|. 3. Р. 1149-1418.
Свайеаи .Ю., Ноозе 7., Р1аз-Возег 5. её а1.
батта В.
Ногтопа[ шесраз1зтз, шуоуеё ш Фе
соп!то| оЁ осзтгоиз сус1е Чотайоп Ъу
4Ъе обоиг оЁ имие ш фе гаё// Аба еп-
Чосг! ло]. 1976. Уо]. 82. Р. 426—435.
Сыну О. А тайуе сепваз шефо@ Тог
Вго\уп гаёз (Вайлё погуес!сив) // Маш-
ге. 1942. Уо]. 150. Р. 59-60.
Сшину РБ. Сопфто| оЁ газ ап писе. Ох-
1ог4: СТагепдой ртезв, 4954. Уд. 2.
532 р.
СитзНап 7. 7. Зома]! зафот@та@оп, рорч-
1лаНоп 4епзНу апд шатотаНап еуоаН-
оп // Зепсе. 1970. \У01. 149, М 3927.
р. 84—90.
Сауюп В. В., No1301 А. М., Еташа Г. О.
'ГВе зКш з4егоз 0Ё погша! апйб итрата-
10]-(геае гаёз//Т. ар! Вез. 1973.
Уо1. 4. Р. 166—178.
Соиёй С. С. етич апд рорабоп
ргоелз // Азпег. Зс. 41965. Уо|. 53,
М2. Р. 199-212.
Сотроп В. Р., Косй М. О., Агп@ ИУ. 7.
ЕНесз о{ ша{егпа[ одог ов саг@ас гайе
о{ шаогпаПу зерагабе4 ша та //
РЬуз01. ап@ Верау. 1977. У01. 18.
Р. 769—773.
СопЕйп Р., Неерепезз Г. No’. Мабгайоп
о (етрегаате Вотлеоз$а$13 т фе га//
Атег. Г. РЬуз101. 1971. Ув. 220. Р. 333—
336
Сотфеё С. В. ТВе татта18 оЁ Фе Ра-
фаеатсбс тео1оп: А фахопоне теме.
Г.; ИБаса: Сотпе! Ушу. ргезз, 1978.
ЗАА р.
.
Согинап 7. @., Иаппейу К. 7. Огазвоше
хосаНяаНоп о ЧеЁеае тшае газ //
7. Сотшр. ап РЬузо| РзусВо!. 1979.
У\61. 93. Р. 105—115.
Спет 7. К. ТегасусПпез аз а Ниогезсетй
тагкег 1 Бопез ап@ (ее оЁ годен //
7. УПаН. Марас. 1970. У\о]. 34(40).
Р. 829-834.
Стоой 7. Н. Бома! Бепауюиг апд еФо1о-
‘цу // Зоса1 Бера\з1оиг шп Ыт4з ап@ шат-'
ша]з. М. У.: Асад. ргезз, 1970. Р. 21—40.
Сошле А. Т., РоЙеу 5. 7. ТБе шатитагу
2]ап@ а09 ПасбаНоп
// Зех ап ниегаа|
зесгеНопз. ВаNoтоте, 1964. Р. 590—642.
ракхутре-АЦота У. С., Ветоп О. Вера-
уопга| шоп ап Фе асе аб зос1а1
1зоТаНоп фт гаёз //Т. Сотр. ап@ РВуз101.
РусВо!. 1984. Уо|. 36, М 1. Р. 27-38.
Рау М. Еайу зитщаНол оЁ годеюёз:
А смисаЁ гемезху оЁ ргезешёЕ пуетрге-
Нопз
// ВгИ. 7. Рзусво|. 1973. Уо|.: 64.
Р. 435—460.
Раз РБ. Е. ТВе загуа! оЁ Иа Ьго\т
таз оп а Магу!апа {ат // Есо]осу. 1948.
Уо1. 29, М 4. Р. 437—448.
Литература 431
Рарз О. Е. А сотрамзоп о тергодисиуе
роепйа] оЁ 0 га роршаНонз// Есо-
1юсу. 1954а. У&. 32, М З.Р. 469-475.
Раз Ш. Е. Те геаИоп Бебуееп ее!
о рораНоп ап 312е ап@ зех оЁ Мог-
уау гаёз
// No19. 1954. Уо]. 32, М 3.
Р. 462—464.
Раз О. Е. Тье теабоп ребувеп 1вуе@] о!
рорШайоп ап ргебпапсу оЁ Могхау
К // Па. 1951с. У01. 32, М 3. Р. 459-
Раз В. Е. ТВе спагаскег1зЫс оЁ гаё ро-
ршайоп // Опаг. Веу. В10|. — 1953а.
\о1. 28, М 4. Р. 373-401.
Рав 2. Е. Апа1узз о{ Воте гапае ош
гесариге 9@//1. Матша. 1953.
\Уо1. 34, No 3. Р. 352—358.
Раз 0. Е, Етеп /. Т. ПШегепца| 4та-
раБ у оЁ гаёз ассот@тое 140 зе апд
зех //Т. УУПаШе Мапар. 1956. Уо1. 20,
М 3. Р. 41—43.
Раыз БР. Е., Еп4еп УХ. Т., бюйез А. М’.
ЗаФез оЁ Воше гапае т Фе Бом
гаф//Т. Мата]. 1948. Уо|. 29, М 2.
Р. 207—225.
Раз р. Е., ХоПу Р. Рипеез шп шаитта-
юву. К. У.: Вашво]4, 1963. 243 р.
Раз В. А. `ЕФЫс {81 аз а сагег бог
сотбасё годепнс1Чев // Пиеги, Резё Сотит.
1983. Уо|. 25, М 2. Р. 46-47.
Рекеузег Р. Г. Гез шатиШегез де ГА-
Чие Моте Егапса1зе. аКег: 1ЕАМ, 1955.
426 р.
Реапое Р. Пе Па гагее да Зоагити|0 а
Матасап// ВиП. 506. ра о|. ехоё. 1925,
М 18. Р. 152-1583.
ДРеппепьеги У. No Ап амер No0 {зо]е
ст ев ремо@з о{ деуеортешь 18 Ще
га //1. Сошр. ап РВузоГ. Рзусва,
1962. Уо|. 55, М Б.Р. 813—845.
ДеппепфегЕ У. No. А сопядегаНоп оЁ изс-
Г ле55 оЁ Ве ст са] рето9 пуроезз
аз аррНеЯ © фе зНииаНоп оЁ годен
т ПЫапеу// Еау ехремепсе ап@ Ье-
Ва\1ог: ТЬе рзусвоБоюру оЁ еуеор-
теле. Эрго е19 (11.): Твотшаз, 1968,
Р. 142—167,
Деппепреги. У. М. бтыа Г, 7. Зоча-
« зееКло ап поуебу-зееюио ЪеВах!отг
аз а поЧоп о аШогевиа] геагте ЬВ1-
36015 // 7. АБпогт. $06. РзусВо]. 1964.
‚
Уо]. 69. Р. 453.
Беппепфетя ТУ. No., НоПошау И’. К., Ро1-
Ипдег М. 1. Уае ваш аз сопзедиепсе
оЁ шацегпа] Бела\оиг т фе гаё// Ве-
Вау. В10]. 1976. Уо1. 47. Р. 51—60.
ДеппепфетЕ У. М., Ниакепз С. А., Пат
тор М. 2. М!ее геатёед \%Иф газ шод-
ВсаНоп оЁ Ъерау1ютг Бу еатЁу ехрегеп-
се \”ИЙ апобег зревез // Заепсе. 1964.
\Уо]. 143. Р. 380—381.
Деппепфеги Г. МNo., Кагаз С. В. Пщегаснуе
сНебз оЁ аде ап доганоп о! шШап6
схрейепее оп адоН, ]еагиао // РэусВо1.
Вер. 1960. \о1. 7. Р. 313.
Реппепфети Г. No., Кагаз @. С. ПиегасН-
ус с есбз оЁ ИМапе ап ада ехре-
епсе проп уешВё гаш ава пораШу
т Ше гаф//Т. Сотр. ап Рвуз1о]. Рзу-
сВо|. 1961. Уч]. 54. Р. 170.
.
ДРеппепьегЕ У. No. Момов 1. В. С., К!-
пе No. Т., Стош Г. 7. ЕНесё8 о дотаНоп
о Чаше зИлиаНоп ироп етоНопа-
Шу] Сапа. 7. Рзусво|. 1962а. Уо. 16.
Деппепфетк ТУ. No, ОНиег О. Н., 5ер-
йеп; М. М’. ЕНесь оЁ шафегпа| Тасюгз
про сто\ В ап@ Ъерау!оиг оЁ Ч1е га //
СЬИ9 Оеуеюр. 4962Ъ. У]. 33. Р. 65.
РеппепфегЕ У. М., Позепфеги К. М. Моп-
бепейе тапзти5ю0 оЁ пуюгтаНоп //
Мабите. 1967. Уо1. 246. Р. 549—550.
РеппепфегЕ У. No, Позепфеги К. М., 2аг-
тош М. Х. М1се геагеЯ у газ апп:
еНеср ш адаЙРоод проп р]азта согН-
созбегопе ап@ орепйе!4 аснуйу // Рву-
т ап@ Вевау. 1969. Уо1. 4. Р. 705—
"7.
ДеппепьегЕ У. No., бтий 7. ЕНовз о т-
ТатеПе эишабой ап асе проп Бева-
У1юит// 7. Сошр. ап Рьуз10|. РэусВо].
1963. \о1. 56. Р. 307-312.
Веогаз Р. Т. Ваз ап@ тет сопо //
119. Г. Еююшо]. 1964. ЗИуег УоЪЙес
Митьет. Р. 407—448.
Деотаз В. К. Ва шапаре ап@ Вох ю
Навь #// Резис4ез. 1975. У]. 9, М 8.
Р. 38—43.
Регис® Е. Н., Роре 7. Н. Мигте пурриз,
И\се, газ ап@ Неаз оп Ме ПагНие
Роуз
// Мед. Т. Амзиа[. 1960. Уо1. 2,
М 24. Р. 924-928.
Ре бапиз П., Зевтайз Г. И’, ТВе шо#фет-
Пиег те]абопзЫр ш асхеюршеюа! га
зи ез: сапиаНзи // Оеуеюр. Рзусво-
Ыо1, 1984. Уо|. 47. Р. 255—262.
Дезз-Рывфен Р., Газой В. Зоте оь-
зегуаНоп оп Ме ошюобенез!з оЁ. в0с1а]
пиегасИопз Ш Фе га //МопИ. 2001,
Ца]. 1979. Уо1. 13, М 23. Р. 204—202.
Пе М Р. Р. Зоип@ аз а годере деегтель //
Резр Сопг. 1969. Уо1. 37, М. В. Р. 36,
38, 40—42, 44.
Гез В. Р. Те еНесё оЁ ап ежегосерйуе
зи оп мк едесбоп т Ласбайия
та /Т. Рзусво]. 1968. Ув]. 197. Р. 37—
+0.
[Иетег Г. У., Въоаез В. Тшагена ш 14
432 Литература
таз //Т. ТиЁесё. П15еазез. 1926. Уо1. 38.
Р. 541.
Р:аейеп Е, Мои: С. Тат Эйпиейег-
{зопа
бадап
— уеНеге — Егзпасв\уе1зе
и09 Бетегкепзуегюе ГКопде// Бёпве-
Негк. МИ. 1984-1985. Ва. 32, М 2.
$3. 205—209.
Докетот 1, Уап Иптетвта @. бша-
аь 1. Е. ЕуЩетсе Тог т@еазе оЁ ча-
зоргеззт ап@ охуюсш 1ш сегебгозрта]
Пиза // Меогово4остто]осу. 1977. Уи]. 24.
Р. 108—118.
Дойтзег М. Х., НоНошау И’. В., Эеппеп-
фегг Т. Н. ТЬе дехеюртеоё оЁ Бера-
у1оига! сотреепсе ш Ше гаё// Мшег-
па! пЯиепее ап вау БеВзу!оиг. 1980.
Р. 27—55.
Доп 4зоп Н. ТНе га. РЫПаас]рШа, 1924.
Рё2,211р-
РоВ.А,БыуГ.А.ЕЙесзоЁБап@Вис
аб уат0из абез оп ег ореп-Не@ Бе-
Ваутосг // Сапа4. Т. Рэзусво|. 1967.
Уо/. 21. Р. 463—470.
Розу В. Г. Одог-ро14е4 Беваз1оиг ш шат-
ипа]5 // Ехремепа. 1986. Уо|. 42, М 3.
Р. 257-270.
Доре И’. Е., бвейтите В. Тгорса! га& и-
4ез, Мропуззиз Басон Н!тз, уесютв о#
епдеп!с фурВиз // 7. Атмег. Мед. Аззос.
1931. Уо|. 97. Р. 1506-1510.
Птешз 0. В. А дезстрауе зу ой заса!
Чеуе1ортаетр 3т ФатПу ртопрз о{Т газ
(Пафа$ погуертстз) // ВыН. РзусВопот.
бос. 1982. УоТ. 19, М 3. Р. 177-180.
[титтопа В. С. РатИа] ахо!Чапсе годеп-
91а] Чазё Бу АаМиз гайаз Г. // Ра-
таз са. 1960а. \Уо|. 16, М 1. Р. 1-6.
Ртиттопа ВБ. С. ТЪе фооа о Вайлаз пог-
у601603 Ш ап агеа оЁ зеа \аП, зай
тагзВ ап@ ши@Йаб// У. Аппа. Есо].
4960. Уо|, 29, М 2. Р. 341-347.
Ргиттопа В. С., И’Изоп Е. Г. ТаЪогайо-
гу шуезНвайопз оЁ тезапсе 10 угаг-
аш оГВамаз потуер1еив Век. Ш
Мопвотесгузте апа ЗевгорзNoте // Апи.
Арр!. Во. 1968. У\Уо/. 61. Р. 303-3412.
Евюоп Р., итей С. 5. ВергодасНуе гме
шт \19 габ// баелсе. 1928. УоГ. 67.
Р. 555-556.
Есфапфеа Е. Г. В., Козсбо М. Втой-
тап 5. РгеетепИа|] пезипе ш ]етоп-
зсетёед еп\топтпет 1ш гаёз теагед оп
1етоп-зсет{ед Бедашо Йош ЫФ ©
уеаптше // Рруз!01. апа ВерБау. 1982.
Уа1. 3, М 1.Р. 47—49.
Кске О. Н. Ап шуаз1юоп оЁ Могуау газ ш
зо ез6 Сеогойа// 7. Маттоа]. 1954.
Уи]. 35. Р, 524—525.
ЕЦегтап У. В. ЕатаШез ап@ вепега оЁ Н-
УПЯ годе. Г: Вы. Миз. (Май.
Н13Ё.), 1940. Уо|. 1: Водетёз оМег Тай
Митае. ХХУЬ 689 р.
ЕЦегтап 7. В. ЕашШез ап@ депега оЁ \-
Ушс тодепз. 1: ВгИ. Маз. (Маг.
Н!.), 1944. Уо|. 2: ЕатПу МианФае.
ХИ, 690 р.
ЕЦегтап 7. В. Еаш Иез вп сепега о
Пуше годе. Г. Вр. Миав. (Маг.
Н!3.), 1949. Ус. 3, ре 1. Р. У. 240 р.
ЕЦегтап 7. В. ТЬе !аапа о Та шеГл-
4ше Ра&${ап, Вигша ап@ Сеу]оп //
Матта[Па. СасаНа, 1961. Ув. 3: Во-
ЧепНа, рё 2: Миг Чае. Р. 401.
ЕЙегтап ФУ. В., Могизоп-5сой Т. С. 5.
СвескИ55 ог РааеатсИс ав@ Тадап
талта1$, 1758—1946. 1.: Втё. Муз.
(Маг. Н1з%), 1951. 810 р.
ЕпЦеп 7. Т., АЦеп 7Т., Бювез И. Ит-
з0т С. Р. Те тафе оЁ гесоуегу оЁ 4е-
стае роршШаНор8 оЁ Бтоми газ Ш
пабите
// Есо!осу. 1948. У01. 29, М 2.
Р. 133—145.
Епцеп 7. Г, Бамз Р. к. ОщегийтаНоп оЁ
тергодисйуе га{ез ш га роработз Бу
ехашшаНоп о{ сагсаззез // РВуз101. 7001.
1948. У0]. 21. Р. 59—65.
Етабз Су. РгоМете ипд МёоНерКенеп 4ег
Айзгойиис ег Ваеп // Азевг. 5ез.
Нус. 1972. ВЧ. 18, М Т. 8. 499—502.
Егриф ФТ, быфе М. Пе Агезе аизсе-
\аВИег Кешпзйаоегагюп ш 4ег ООВ //
Негсуша. 1986. Ва. 23, М 3. 8. 257-
304.
ЕтзЁЕт М. 5. ОБеппепЬеге И. Н.. бо&4-
тап В. р. Автезюп ш Фе Лабайпя
та: еМесё оЁ 1пётадег асе апй {е$# аге-
пе // Верау. В1оЁ. 1978. У. 23. Р. 52-—
66.
Етеьев С. Р. Зучета Нет! АпиваНз
рег С1аззез, От@ тез, Сепега, бресез,
Уатеаез спт бупопупиа её Ноа
Апиа ит. (125313 1. МатштшаНа. Но|-
тае, 1777. 381 р.
Ез|трег Р. С., Гафуизоп Н. ТУ. Ате фе‘
сопзёгатз оп Пеагие@ гезропзез 10 040т5
{гот ге\уагдей ап поп-геуагдей газ? //
Апию. Беаги. Верау. 1980. Уд. 8.
Р. 452—456.
Ешег В. Е. Тве Ы010ху ап4 ЪБеНауойг 01
а Гтее-Нуше рорайор оЁ ШасК гай
(ВаИлмлз газ) // Авпи. Верау. Мопост.
1974, Уо1. 4, М З.Р. 127-114.
Равеп В. М. Зёесиуе ап еуомЯопагу
а5рес4з оЁ апниа! р]ау // Атег. Май.
1974. Уо|. 108. Р. 850-858.
Рареп В. М. Ашта| р1ау Бераутопг. М. У.:
Охюга Чшу. ргезз, 1981. 216 р.
Ралгьа О. У. Вевахмюг оё Фзрегзшо деег
Литература
435
июе (Регоузсиз шашешабаз) // Ве-
Вау. Есо!., 3061010]. 1978. Уо1. 3, М3.
Р. 265—282.
ЕКаграпз-Азаа А. бошйисЕ С. Н. Рори-
1аНоп есооху оЁ Могуау таз шт Фе
ВаНвпоте 200 апё Ога НШ Рагк Ва
итоге, МагуУЛап9 // Апп. 00|. 1979.
Уо1. 15, М 1.Р. 1-42.
Рава 7. УеПо\у пийап ше Ъгодуп таё,
Вайаз погуес1сиз (Веепрот6) // Ава
306. 2001]. Боветоз. 1963. \9|. 27.
Р. 209—240.
РИиайа 7. П1Шегепсез ш тВуфт оЁ асы-
УНу ш ЫасК гаёз (ВаИл8 таМиз) апа
Бтоуп газ (Вабаз потуер!сиз) // Усэт.
СезКоз1. зроес. 200]. 1965. Зу. 29, М 1.
$. 70-84.
Евзсйег И’..@. воп. Раз Мабопатизейт
Чег МабигаезсЫ сре га Рашз. Рат!з, 1803.
Ва. 2. 422 $.
Ризшщег И’. П. Водепйеев, ВойеНс!-
4ез// Сетеа1 Гоодз УМоа. 1980. У@. 25,
М 12. Р. 751-753.
Рапаета Р., МораКова У. ЕМесё ой то-
Ффег оп Ше 4еу@оршепе оЁ асотезыуе
Берауоиг — гайз // Пеуе]ор. Рзусвоо1.
1974. Уч]. 8, М 1. Р. 49-54.
Паппёу К., Готе В. Те шИоспее Ёеша-
1е5 проп асатеззют т дошезисаю8
тша]е газ (Вабаз погуес1сиз) // Апиа.
Ворау. 1977. Ус]. 25, М 3. Р. 654—
Юетити А. 5. Сопёто] оЁ 100 пиаКе Ш
ле ]асаыта тай: го]е оЁ зисКНие ап
Воттолез // РЬуз10]. ап Пефау. 1976.
‚Мот. 17. Р. 841—848.
ВЧетитЕ А. 5. Возепан 7. 5. Маегпа!
Бева\опг 1 Ше упет апд Лабанис
гафз // 7. Сошр. ап РВуз1о1. РзусвоГ.
1974а. УоТ. 86, М5.Р. 957-972.
Непипя А. 5., Нозеп ши 7. 5. ОНасоту
теаНоп 0{ шафегпа|1 Ъева\отт Ш
гаёз. 1. ЕНесёз оЁ оМасфогу БиЬ гето-
уа] т ехремепсеё ап шехремелсей
Табациа ап@ суше етаез // ИЫ8.
1974Ъ, Уч. 86, М 3. Р. 221-232.
Яепипя А. 5., Возепьиие 1. 5. ОПасдюогу
« товфаНоп оЁ шаегпа! Ъевауюц Ш
газ. 2, ЕНесё оЁ рерВегаПу шдисед
апо5111с ап@ 1ез1оп оЁ Фе 1а6ега] оШас-
фогу цтав6 т рир-шдисейд утеаз // Па.
`1974с. Ус. 86, М 3. Р. 233-246.
РепитЕ А. 5., Гассатто Ё., ТатЪоззо Г.
Уотегопаза! ап оНасёоту зучет то-
ОщаНоп 0{ шабегоа! Ъерауоиг ш Фе
"а! Зо1епос, 1979. Уб|. 203. Р; 372—
Еетте С. А. Ваз ап@ тшоа ехНаеНоп И
Моюгив. 1969. Уа1. 46, М 3. Р. 210-241.
ЕЯошет 5. 5. No0498 оп фе гссепё шапиа]
0 Есурь // Ргос. 2001. Зое. Бопдоп. 1982.
У]. 12. Р. 369—450.
Рох М. И’. Мепто-Бевауюга! ошюбепу:
А зувезз оЁ ео]орса! ап пеигор-
Вуз] оса] сопсерёз // Вгат Вез. 1966.
\о1. 2. Р. 3—20.
Ртапсз В. Г. 22-КНа саНз Бу 1зо1щеЯ
218 // Мавше. 1977. \о|. 265. Р. 2368-
Итезв ФТ. М’, Тай Све-Свипя, Гей СЪиап-
Ниапк, СфапЕ Сфе-Топв. Г9р10зрто8 т
шап ап годеп{з ш Тагмап // Атег.
7. Тгор. Ме4. апд Нуе. 1968. Уа. 17.
М5.Р. 17-18.
Рнейтап М. Т., Втито 1. Р., АЪемз }. В.
РВуз!01ор1са] ап Борау!10ига! сопзеди-
епсез шт габ оЁ уужег госусПпа дагше
ЛасбаНоп //7. Сошр. ава Р|Вузю] Рзу-
сво]. 1981. Уч. 95. Р. 25-35.
.
Сие] В. С. (/1т.). Зома|. гапзизюп оЁ
асфие4 Бевауютг // Аду. За4у.. Ъевах..
1976. Уо]. 6. Р. 77-100.
Сие} В. С. (1е.). Тре вос мапзпиззют:
07 {004-ртеегепсез: ап адаранов Юг
уеашр: ш габз //Т. Сотр. ап. Рьуз!о].
Рзусво]. 1977. Уд]. 91. Р. 4436-1140.
Се} В. С. (1т.). Птуте Чог 1004 апауз
оЁ а розз!е сазе о{ зоста| |еагшиа т
а таёз // П54. 1980. Уо{. 94. Р. 416=
Саше} В. С. (Ть.). Тье есо]обу оЁ зуеашиа //
Рагетца] саге ш изанипа1/Е4..П. 7. Си-
Ъегшк, Р. Н. КюрЁег. М. У.; 6.: Репа
ргезз, 1981. Р. 214—243.
Сие] В. С. (1т.). ЧиПеаноп Ъу Могуау
газ (В. погуерюиз) оЁ шие шез-
завез сопсегита 915ап4 {0043 //Т. Сошр.
204 РВуз!01. Рзусвой. 1983. Уа. 97.
Р. 364—371.
Са1е] В. С. (7г.). ОМасвюогу сошлтиеаНов
атпоп га{з: п\огтайов сопсегите 4-
3206 Чейз
// СБетуса1 151813 1 уег-
фебгавез. М. У.; Ё.: Р]епат ргезз, 1986.
Уо1. 4. Р. 487—505.
Сие} В. С. (1т.}, ВесЁЕ М. Адуегяхе апа
аМгасНуе шагКто 0 1юхе ап за
10048 Ъу Могуау га!з // ВеВау. Меита|.
В1о|. 1985. Уо|. 43. Р. 298—310.
Сие} В. С. (7е.), Сагк М. М. Зое! Гас-
{от8 т Ше ро!1з0п ауо!Чапсе ап@ гей
Берау1опг о? уИЧ ап@ дотезнсаю@ таё
рирз //Т. Сотр. ип@ РНузюо]. РвусВо|.
19 4а. Уд]. 75. Р. 341-357.
Сие} В. С. (1), СатЬ М. М. Рагепео!-
зрешя НиегасНой Чеюогише Ите ап@
Расе оЁ Нтёв тоезНоп оЁ зоНа Тоой Бу
No14 таб рирз// Рзуспо]. 561. 1974Ъ.
Уо]. 25. Р. 15-46.
432 Литература
таз //Т. ТиЁесё. П15еазез. 1926. Уо1. 38.
Р. 541.
Р:аейеп Е, Мои: С. Тат Эйпиейег-
{зопа
бадап
— уеНеге — Егзпасв\уе1зе
и09 Бетегкепзуегюе ГКопде// Бёпве-
Негк. МИ. 1984-1985. Ва. 32, М 2.
$3. 205—209.
Докетот 1, Уап Иптетвта @. бша-
аь 1. Е. ЕуЩетсе Тог т@еазе оЁ ча-
зоргеззт ап@ охуюсш 1ш сегебгозрта]
Пиза // Меогово4остто]осу. 1977. Уи]. 24.
Р. 108—118.
Дойтзег М. Х., НоНошау И’. В., Эеппеп-
фегг Т. Н. ТЬе дехеюртеоё оЁ Бера-
у1оига! сотреепсе ш Ше гаё// Мшег-
па! пЯиепее ап вау БеВзу!оиг. 1980.
Р. 27—55.
Доп 4зоп Н. ТНе га. РЫПаас]рШа, 1924.
Рё2,211р-
РоВ.А,БыуГ.А.ЕЙесзоЁБап@Вис
аб уат0из абез оп ег ореп-Не@ Бе-
Ваутосг // Сапа4. Т. Рэзусво|. 1967.
Уо/. 21. Р. 463—470.
Розу В. Г. Одог-ро14е4 Беваз1оиг ш шат-
ипа]5 // Ехремепа. 1986. Уо|. 42, М 3.
Р. 257-270.
Доре И’. Е., бвейтите В. Тгорса! га& и-
4ез, Мропуззиз Басон Н!тз, уесютв о#
епдеп!с фурВиз // 7. Атмег. Мед. Аззос.
1931. Уо|. 97. Р. 1506-1510.
Птешз 0. В. А дезстрауе зу ой заса!
Чеуе1ортаетр 3т ФатПу ртопрз о{Т газ
(Пафа$ погуертстз) // ВыН. РзусВопот.
бос. 1982. УоТ. 19, М 3. Р. 177-180.
[титтопа В. С. РатИа] ахо!Чапсе годеп-
91а] Чазё Бу АаМиз гайаз Г. // Ра-
таз са. 1960а. \Уо|. 16, М 1. Р. 1-6.
Ртиттопа ВБ. С. ТЪе фооа о Вайлаз пог-
у601603 Ш ап агеа оЁ зеа \аП, зай
тагзВ ап@ ши@Йаб// У. Аппа. Есо].
4960. Уо|, 29, М 2. Р. 341-347.
Ргиттопа В. С., И’Изоп Е. Г. ТаЪогайо-
гу шуезНвайопз оЁ тезапсе 10 угаг-
аш оГВамаз потуер1еив Век. Ш
Мопвотесгузте апа ЗевгорзNoте // Апи.
Арр!. Во. 1968. У\Уо/. 61. Р. 303-3412.
Евюоп Р., итей С. 5. ВергодасНуе гме
шт \19 габ// баелсе. 1928. УоГ. 67.
Р. 555-556.
Есфапфеа Е. Г. В., Козсбо М. Втой-
тап 5. РгеетепИа|] пезипе ш ]етоп-
зсетёед еп\топтпет 1ш гаёз теагед оп
1етоп-зсет{ед Бедашо Йош ЫФ ©
уеаптше // Рруз!01. апа ВерБау. 1982.
Уа1. 3, М 1.Р. 47—49.
Кске О. Н. Ап шуаз1юоп оЁ Могуау газ ш
зо ез6 Сеогойа// 7. Маттоа]. 1954.
Уи]. 35. Р, 524—525.
ЕЦегтап У. В. ЕатаШез ап@ вепега оЁ Н-
УПЯ годе. Г: Вы. Миз. (Май.
Н13Ё.), 1940. Уо|. 1: Водетёз оМег Тай
Митае. ХХУЬ 689 р.
ЕЦегтап 7. В. ЕашШез ап@ депега оЁ \-
Ушс тодепз. 1: ВгИ. Маз. (Маг.
Н!.), 1944. Уо|. 2: ЕатПу МианФае.
ХИ, 690 р.
ЕЦегтап 7. В. Еаш Иез вп сепега о
Пуше годе. Г. Вр. Миав. (Маг.
Н!3.), 1949. Ус. 3, ре 1. Р. У. 240 р.
ЕЦегтап 7. В. ТЬе !аапа о Та шеГл-
4ше Ра&${ап, Вигша ап@ Сеу]оп //
Матта[Па. СасаНа, 1961. Ув. 3: Во-
ЧепНа, рё 2: Миг Чае. Р. 401.
ЕЙегтап ФУ. В., Могизоп-5сой Т. С. 5.
СвескИ55 ог РааеатсИс ав@ Тадап
талта1$, 1758—1946. 1.: Втё. Муз.
(Маг. Н1з%), 1951. 810 р.
ЕпЦеп 7. Т., АЦеп 7Т., Бювез И. Ит-
з0т С. Р. Те тафе оЁ гесоуегу оЁ 4е-
стае роршШаНор8 оЁ Бтоми газ Ш
пабите
// Есо!осу. 1948. У01. 29, М 2.
Р. 133—145.
Епцеп 7. Г, Бамз Р. к. ОщегийтаНоп оЁ
тергодисйуе га{ез ш га роработз Бу
ехашшаНоп о{ сагсаззез // РВуз101. 7001.
1948. У0]. 21. Р. 59—65.
Етабз Су. РгоМете ипд МёоНерКенеп 4ег
Айзгойиис ег Ваеп // Азевг. 5ез.
Нус. 1972. ВЧ. 18, М Т. 8. 499—502.
Егриф ФТ, быфе М. Пе Агезе аизсе-
\аВИег Кешпзйаоегагюп ш 4ег ООВ //
Негсуша. 1986. Ва. 23, М 3. 8. 257-
304.
ЕтзЁЕт М. 5. ОБеппепЬеге И. Н.. бо&4-
тап В. р. Автезюп ш Фе Лабайпя
та: еМесё оЁ 1пётадег асе апй {е$# аге-
пе // Верау. В1оЁ. 1978. У. 23. Р. 52-—
66.
Етеьев С. Р. Зучета Нет! АпиваНз
рег С1аззез, От@ тез, Сепега, бресез,
Уатеаез спт бупопупиа её Ноа
Апиа ит. (125313 1. МатштшаНа. Но|-
тае, 1777. 381 р.
Ез|трег Р. С., Гафуизоп Н. ТУ. Ате фе‘
сопзёгатз оп Пеагие@ гезропзез 10 040т5
{гот ге\уагдей ап поп-геуагдей газ? //
Апию. Беаги. Верау. 1980. Уд. 8.
Р. 452—456.
Ешег В. Е. Тве Ы010ху ап4 ЪБеНауойг 01
а Гтее-Нуше рорайор оЁ ШасК гай
(ВаИлмлз газ) // Авпи. Верау. Мопост.
1974, Уо1. 4, М З.Р. 127-114.
Равеп В. М. Зёесиуе ап еуомЯопагу
а5рес4з оЁ апниа! р]ау // Атег. Май.
1974. Уо|. 108. Р. 850-858.
Рареп В. М. Ашта| р1ау Бераутопг. М. У.:
Охюга Чшу. ргезз, 1981. 216 р.
Ралгьа О. У. Вевахмюг оё Фзрегзшо деег
Литература
435
июе (Регоузсиз шашешабаз) // Ве-
Вау. Есо!., 3061010]. 1978. Уо1. 3, М3.
Р. 265—282.
ЕКаграпз-Азаа А. бошйисЕ С. Н. Рори-
1аНоп есооху оЁ Могуау таз шт Фе
ВаНвпоте 200 апё Ога НШ Рагк Ва
итоге, МагуУЛап9 // Апп. 00|. 1979.
Уо1. 15, М 1.Р. 1-42.
Рава 7. УеПо\у пийап ше Ъгодуп таё,
Вайаз погуес1сиз (Веепрот6) // Ава
306. 2001]. Боветоз. 1963. \9|. 27.
Р. 209—240.
РИиайа 7. П1Шегепсез ш тВуфт оЁ асы-
УНу ш ЫасК гаёз (ВаИл8 таМиз) апа
Бтоуп газ (Вабаз потуер!сиз) // Усэт.
СезКоз1. зроес. 200]. 1965. Зу. 29, М 1.
$. 70-84.
Евзсйег И’..@. воп. Раз Мабопатизейт
Чег МабигаезсЫ сре га Рашз. Рат!з, 1803.
Ва. 2. 422 $.
Ризшщег И’. П. Водепйеев, ВойеНс!-
4ез// Сетеа1 Гоодз УМоа. 1980. У@. 25,
М 12. Р. 751-753.
Рапаета Р., МораКова У. ЕМесё ой то-
Ффег оп Ше 4еу@оршепе оЁ асотезыуе
Берауоиг — гайз // Пеуе]ор. Рзусвоо1.
1974. Уч]. 8, М 1. Р. 49-54.
Паппёу К., Готе В. Те шИоспее Ёеша-
1е5 проп асатеззют т дошезисаю8
тша]е газ (Вабаз погуес1сиз) // Апиа.
Ворау. 1977. Ус]. 25, М 3. Р. 654—
Юетити А. 5. Сопёто] оЁ 100 пиаКе Ш
ле ]асаыта тай: го]е оЁ зисКНие ап
Воттолез // РЬуз10]. ап Пефау. 1976.
‚Мот. 17. Р. 841—848.
ВЧетитЕ А. 5. Возепан 7. 5. Маегпа!
Бева\опг 1 Ше упет апд Лабанис
гафз // 7. Сошр. ап РВуз1о1. РзусвоГ.
1974а. УоТ. 86, М5.Р. 957-972.
Непипя А. 5., Нозеп ши 7. 5. ОНасоту
теаНоп 0{ шафегпа|1 Ъева\отт Ш
гаёз. 1. ЕНесёз оЁ оМасфогу БиЬ гето-
уа] т ехремепсеё ап шехремелсей
Табациа ап@ суше етаез // ИЫ8.
1974Ъ, Уч. 86, М 3. Р. 221-232.
Яепипя А. 5., Возепьиие 1. 5. ОПасдюогу
« товфаНоп оЁ шаегпа! Ъевауюц Ш
газ. 2, ЕНесё оЁ рерВегаПу шдисед
апо5111с ап@ 1ез1оп оЁ Фе 1а6ега] оШас-
фогу цтав6 т рир-шдисейд утеаз // Па.
`1974с. Ус. 86, М 3. Р. 233-246.
РепитЕ А. 5., Гассатто Ё., ТатЪоззо Г.
Уотегопаза! ап оНасёоту зучет то-
ОщаНоп 0{ шабегоа! Ъерауоиг ш Фе
"а! Зо1епос, 1979. Уб|. 203. Р; 372—
Еетте С. А. Ваз ап@ тшоа ехНаеНоп И
Моюгив. 1969. Уа1. 46, М 3. Р. 210-241.
ЕЯошет 5. 5. No0498 оп фе гссепё шапиа]
0 Есурь // Ргос. 2001. Зое. Бопдоп. 1982.
У]. 12. Р. 369—450.
Рох М. И’. Мепто-Бевауюга! ошюбепу:
А зувезз оЁ ео]орса! ап пеигор-
Вуз] оса] сопсерёз // Вгат Вез. 1966.
\о1. 2. Р. 3—20.
Ртапсз В. Г. 22-КНа саНз Бу 1зо1щеЯ
218 // Мавше. 1977. \о|. 265. Р. 2368-
Итезв ФТ. М’, Тай Све-Свипя, Гей СЪиап-
Ниапк, СфапЕ Сфе-Топв. Г9р10зрто8 т
шап ап годеп{з ш Тагмап // Атег.
7. Тгор. Ме4. апд Нуе. 1968. Уа. 17.
М5.Р. 17-18.
Рнейтап М. Т., Втито 1. Р., АЪемз }. В.
РВуз!01ор1са] ап Борау!10ига! сопзеди-
епсез шт габ оЁ уужег госусПпа дагше
ЛасбаНоп //7. Сошр. ава Р|Вузю] Рзу-
сво]. 1981. Уч. 95. Р. 25-35.
.
Сие] В. С. (/1т.). Зома|. гапзизюп оЁ
асфие4 Бевауютг // Аду. За4у.. Ъевах..
1976. Уо]. 6. Р. 77-100.
Сие} В. С. (1е.). Тре вос мапзпиззют:
07 {004-ртеегепсез: ап адаранов Юг
уеашр: ш габз //Т. Сотр. ап. Рьуз!о].
Рзусво]. 1977. Уд]. 91. Р. 4436-1140.
Се} В. С. (1т.). Птуте Чог 1004 апауз
оЁ а розз!е сазе о{ зоста| |еагшиа т
а таёз // П54. 1980. Уо{. 94. Р. 416=
Саше} В. С. (Ть.). Тье есо]обу оЁ зуеашиа //
Рагетца] саге ш изанипа1/Е4..П. 7. Си-
Ъегшк, Р. Н. КюрЁег. М. У.; 6.: Репа
ргезз, 1981. Р. 214—243.
Сие] В. С. (1т.). ЧиПеаноп Ъу Могуау
газ (В. погуерюиз) оЁ шие шез-
завез сопсегита 915ап4 {0043 //Т. Сошр.
204 РВуз!01. Рзусвой. 1983. Уа. 97.
Р. 364—371.
Са1е] В. С. (7г.). ОМасвюогу сошлтиеаНов
атпоп га{з: п\огтайов сопсегите 4-
3206 Чейз
// СБетуса1 151813 1 уег-
фебгавез. М. У.; Ё.: Р]епат ргезз, 1986.
Уо1. 4. Р. 487—505.
Сие} В. С. (1т.}, ВесЁЕ М. Адуегяхе апа
аМгасНуе шагКто 0 1юхе ап за
10048 Ъу Могуау га!з // ВеВау. Меита|.
В1о|. 1985. Уо|. 43. Р. 298—310.
Сие} В. С. (7е.), Сагк М. М. Зое! Гас-
{от8 т Ше ро!1з0п ауо!Чапсе ап@ гей
Берау1опг о? уИЧ ап@ дотезнсаю@ таё
рирз //Т. Сотр. ип@ РНузюо]. РвусВо|.
19 4а. Уд]. 75. Р. 341-357.
Сие} В. С. (1), СатЬ М. М. Рагепео!-
зрешя НиегасНой Чеюогише Ите ап@
Расе оЁ Нтёв тоезНоп оЁ зоНа Тоой Бу
No14 таб рирз// Рзуспо]. 561. 1974Ъ.
Уо]. 25. Р. 15-46.
4ЗА Литература
бе! В. 6. (1е.), Нефег 1. Тье тойе оЁ
се 4иа1 оНасфогу сиез ш Ме даег-
пуптаоб ой {еедше зИе з@есйоп оЁ 40-
тезЫс газ //Т. Сотр. ав@ Рруз1о]. Рзу-
СВО. 1976. Уо1. 90. Р. 727-789.
сще} В. 6. (1.), Кеппей Б. 6. Я ет М.
Решпопзтаюг шЯчепсе оп обзегуег 91с+
ргеетспее: ЕИесёз оЁ эпр!е ехрозите
ап ФНе ргезепее 0 а дотопзигаог /
Апиа, Теагп. Верау. 1985. Уо1. 13, М 1.
Р. 25—30.
бер 6. В. (1т.), Мизсиз Р. А. ОНасюгу
тефаноп оЁ лофег — уобпв сошасё т
Гопо-— Ехапз газ // 7. Сор. зп4 РВуз101.
Рзусво!. 1979. \о|. 93. Р. 708—710.
бие! ВН. ©. (1°.), бйеггу ШБ. Е. Мотег’з
тЙК а шей Рог тапзииззюл 0 сиез
теНеснис Ше Йауот оЁ тошег Фе
ТЫ. 1973. Ув. 83. Р. 374-378.
Сие В. С. (7т.}, Ижтоте 5. М. Тгапзег
оЁ ПРогтаноп сопсегишя 41зваюё 10005:
а 1аЪогайогу шуезЫраноп оф Ме «т-
тоттаНоп сехите» пуро#ез$ // Апит. Ве-
вау. 1983. Уо|. 31. Р. 748—758.
бойево А., бапсйез-Счайо У. Е. Раегото-
па| тедмайоп оЁ Фе езгоц5 суше ш
{Ве {ела|с гаф Бу Noе ассеззогу оШасо-
гу зузет // АМа епбостто|. — 1979.
Уо1. 225, варр!. Р. 256.
СаПево А.. Запсвез-Сцадо Т. Е. Мота 0.
Вос оЁ Ше уошегопаза! зузет 1 рВе
готаопа! тесербов 11 ‘Фе гаЁ//Омег-
тнипе оЁ Бераугоог апд снеписа] вИ-
той. 1.: ТВГ ргезз, 1982. Р. 57-64.
Сагаа 7. КоеИти В. А. ВааНоп оЁ смез
{0 сопвециепее и ауоансе 1еагита //
РаусНо!. Зе1. 1966. Уо1. 4. Р. 123—124.
Сашлепошз 1 А. М. Светиса| айтасат{$ о!
Ве гаё ргерама! Зап // СВеписа] 512-
па!5 ш уееБгавез. М. У.: Р/с `ргезз.
1977. Р. 45—59.
Санлепошзы Л. М. Бе Мсош О. В., 5а-
сешсл-ЗариллаЯ: М. Апдговеп 4ереп-
Чепсе оЁ а шаге рЬеготове ш га
итше // Ногтопе Вевах. 1976. \01. 7,
М 4, Р. 401—405.
Саишепош:м А. М. ОтзщаЕ Р. ХТ, 5 шсе-
идст-Заритйзааз М. Тозерв В. М. Руе-
зепз оф зех р|еготопе т ргераиа|
5]1ап9з ой шае га 7. Епдосгио!. 1975.
Уо1. 67. Р. 283—288.
СеттеЕе Н., Метаттег 7. Рог Тахопо-
пие Чег Сабишо ВаМиз (Водепиа, Ми-
т14ае) // Знпремегк. МИ. 1984. Ва. 49,
No2. $. 104—116.
|
Сегай Н. Б. Тата Т. [., СетаЙ А. А.
Пузгарноп оЁ Фе: мае газ зохиа! Бе-
‚ Бамопг 1ш@исеё Ъу зоба 1з91абоп//
Апим. Веряу. 1967. Ус]. 15. Р. 54-58.
Сецег Г. ОЦасюту ап Шегта] а чепсез
оп аИгазот!с уосайтаНоп Фиша Чеуе-
1ортепё 11 гофетив // Ашег. 700[. 1979.
\Уо1. 19. Р. 420—431.
.
Сеуег Г. А. Вагнеа В. Г. ГзИлепсе о
гопада! вогтопез ап4 зехиа1 Берау1оиг
оп аЦтазоме уосаНхаМоп 11 га. 1.
Тееациене оЁ 1ета]е //Т. Сошр. ап РВу-
$101. Рэусрвой. 1978. Уи. 92. Р. 438—446.
Сеуег Г. А., Ватра В. 7. Мегиовзй Т. К.
ТаЙаетсе о! гопада! Берау!оот оп ига
Зое уосаН2аНоп No1 газ. 2. Ттеавпец
оЁ мае // ТЫ. 1978а. УоГ. 92. Р. 447 —
456.
Сеуег Г. А., МеГиозь Т. К., Вагнаа В. Л.
ЕНесз оЁ иЙтазоте уосаНйтаНопз апЯ
тае’з игше оп Гетоа]е гаё геа@ 11055 {0
тра / 14. 19785. Уо1. 92. Р. 457—462.
Сиеке В. ТНе пес {ог Имедтайоп оЁ есо-
1од1са1, рВузю]озеа! ап е{НоТоб1са! ге-
збагсл т 10060 сопёто| // РАБ. ОЕРР,
1970. Уо]. 58. Р. 49-54.
СШен А. No, Воитвооп Б. А., чи
рап К, А., Ааег М. Т. Мофог-уеатия
пиегасНор ша Могмау га! шт ЧВе рге-
зепсе оЁ а зиссезыуе НИег ргодис1е4 Бу
розёрагьатт шания // РВуз1!. ав Вевау.
1983. Ув]. 30. Р. 267—274.
СизВ С. К, Агога К. К. 549168 оп го-
деп{з ава Мет сопито1. 13. СвешояегЕ-
]ап4з ‘есри1чиез юг годе сопто] //
Вай. Сташ ТесВио, 1976. Уо1. 14, М 3.
Р. 219-228.
боЧтап 1. В. Те еНесёз о! Тапа Нае апа
звосКте т НУапеу проп адиз ЪеВа-
утоиг т фе аЫшо га //Т. Селе. Рзу-
сро!. 1964. Ус. 104. Р. 301-310.
“Согаоп тив С. Е., Титпег Ё. Н. Нате
зон 7. Г., Втоот 7. С. Апиа 1ербюозр-
0515 т Майауа. 2. ГосаНЦез затр!е4 //
ВоЙ. Огргаше. топ. заше. 1961а.
\Уо1. 24, М 1. Р. 23—34.
Согаоп бтив С. Е. Титпег Ё. Н., Натп-,
зоп 7. Г., Втоот 7. С. Апиа! 1ербозрто-
815 ш Маауа. 1. Ме о43. зоопсовтар[1-
са! БасКкотоиЯ ап@ Ъгоаф апа|уз$ оЁ
тези!(з // ТЫ. 1961. Уо1. 24, М 4. Р. 5—
21.
Созафел Х., Горел-ГРиег М. Т. Тве пам-
га} сот!Иез оЁ зта] хлалта8 (-
зесыуотез ап годе!) о Сайаюша
(Зраш) // М!зе. 2001. 1985. М 9. Р. 375-—
387
Созсзутз 7. ММу\м Чгареттусв ззако\
риаКо\ роршаее оНаг// \УЛад. еКо1.
1974. Уо|. 20, М 3. $. 207-289.
Соигевисв @. НасЁ М. М. АТЯЬЙЦУ Ш
Ве гаф//У. Сор. ава Рвузо!. Рзусво|.
1966. Уо1. 62. Р. 289.
„
Литература
135
Стат Е. С. Ап апа]уз1з ой {Не зола! Ъе-
Пауюиг оЁ Ме шае Пафотаюгу гаЁ//
Верауюиг. 1963. Уо|. 24. Р. 260-281.
ата Е. С., МасЕйщозй 7. No. А сотраг-
50п о те зоста! розшоз 0{ зоше сош-
шоп |аБботаюту го4етз 1
Уо1. 21, Р. 246-259. та. 1983.
Стая No. С. ОТап тодеоеБогие 491зеазе
ап тоделё тай Ш [згае] апа
пеовБотте сопптез // Тэг. 7. Мед. 31.
1973. \Уо1. 9(8). Р. 969-979.
Стау Р., СпеЯеу 5. Оечортетё оЁ та-
тра] рерауюиг т пИНрагопз габв: е#-
Рос а ап9 вау шаетпа| ехре-
иепее // 7. Сошр. ап@ ГNoуз01. Рзу
1984. У. 08 В. 9199. © ое Реут
баров 1. ие 10 ап/соаРШат
11 годен // РезИе. 31. 1974. Уд. :
си 8. Р. 276-279.
Бу
тевуез 7. Н., Ве4{еги В., Аутез Р. В,
СШ7.Е.Маагтозанее:аны
се ро!утотНзт 11 Могмау гаё// Сепе-
Неа ВезеагеВ. 1977. \Уо]. 30. Р. 257—
Стеарез 7. Н. Арерйета р. 5. и
Ке1@ 91218 о ОЕ
а
соваиатив аватя аЁепасойт-геззать
оту’ау гаф рораНотз //Т. Нуе.
\Уо1. 89, No2. ЗО
УВ. 1982,
Сирия И’. Л, батткег 17. Р. Тре сНесз
оЁ пиепзе зЫпиаНоп ехрегелсеф 4-
ти иуапсу оп ад Бела\топг 11 Фе
га . отр. ап РНузю1.
1052. от. 45. В, 304-306 Русво
Стоптооз Р., бттеп А. Тохор]азт0$! т
‚ Могуау газ па Не]зшЮ
// Апиа. Мед.
ЕхрН. В10]. Еегииеа. 1955. Уо|. 33
с. Р. 44,
|
гозкеггу Р. С., 5тИВ С., Геоп М., Ми-
сле Ц Е. 4. Тпегшогев Иа Чо апа а.
егра. спауюг о ег
о
99080 Чаи
бтоззеё ПО., Ройпе А. Тве машз оЁ Ван
105: газ аз зиб]есз ш агИсез ри
$Нед т Топгпа| о? Созпрагайуе Рвуз1010-
21са! Рзусро]огу ош 1969 урголеВ
1984 // Рвуспо]. Вер. 1982. Уч. 54, М 3.
ре 1. Р. 989-970.
'
Стозрепог С. Е. Майрек Н., Мепа Е. А.
А зу о! асёог8 туоуеЯ п Те 4е-
у@юоршетё о! Фе ехёегосерыуе гееазе
оЁ ргоасйп ш Фе Тасбайпе гаф// Ног-
топе Верау. 1970. Уо]. 1. Р. 144-120.
Стозуепог С. Е., Мепа РЁ. Мепта| ап@ дот-
това] оо о? шПК зестейоп апа
тИЕ е]ес@оп // ГаббаНоп.
.
то
у
1974. Уд. 1.
Стош Г. 7. ЕНесё о? НИег эле, асе о
Усйп ан@ рагбу оп фозег клоет Ъе-
’
Вауоцг 1 ВаМаз погуеслсиз // Ат
аТЗ
Стоа Г. Г, А4ег В. Сопйпаоиз тесотфшт
0# тафетра! Бераутоиг 1 Вабиз погуе-
21св
// 9. 1969. Уо|. 17. Р. 722-729.
Стиепае! А. 0., Атпоа 1’. У. ЕНесз о
вату зоба! дерМуаНор оп гергофасмуе
Беваутог о{ ша! та{//Т. Сошр. апй
Ррузо. РзусВо!. 1969. Уо1. 67. Р. 123—
Сифетиск Ш. Т, Аве; /. В. Ммегпя
Е 9 учи: ого ехсНапое ап@
тохипа{е соло // РВуз10].
пера,
СпрегисЁ р. Г, АВетз ХТ. В. Мабетра!
Пони Ву мы ара 1абаНай таб:
хаег 1тапег ош ри
19.
УоЕ 3. В. 501-506 та. 1985.
Нааз С. Е., И’Изоп No, Вмимоп В. В.
ОБзетуа 00$ оп Могмуау газ, Вайлв.
погуес1сиз, м Кофак, АазКа // Сапа.
Ее 9-Манаг. 1986. Уо. 100, М 3. Р. 316.
Нает м. РА \меароп аратзь Фе гез1-
апф та ес Чез. 1975.
у
За та
1975. У. 11, М9.
Найп Р. Те ЧеуеТорхлете оЁ (Пегтотеси-
1аНоп. 2. ТВе рагсранов оЁ те Фу-
го ввод ш Ше деу@ортете оЁ фег-
погезиа оп т габ // РВуз1о1
--
1103]. 1958. Уо/. 5. Р. и ое
Нар Р.,. Ко 4орзку О., Ктесейк 1. её &
Еп4оске ап@ шеаЪоНс азресаз оЁ с
Чеус]оршеш оЁ Вотеотетту ш Ше
га // Зошанс заБИиу о Ме пез
Боги. Возбюш, 1964. Р. 131-155.
НаП Е. В. ТЬс шашта!з оГ Мог Аше-
пса. No. У.: УУЦеу, 1981. У. 2. 2000 р.
Ной 8. в. Ке!зоп К. В. ТВе аашта]8 о
тот
шегка. М. У.:
1959. Уо1. 2. 538 р. Копа ртов,
НаЙ И’. 6., Статег С. Р., В!азз Е. М.
етот свапосз 1 зисКЦио ой
га
и1рз абаге.
та НН
1975. У. 258.
На! ИУ. 6., Нозеп Май 7. 5. ЗаеКИаи Ъе-
Вачлош аа о сопто| ш {Фе 4еуе-
оршс гаё рирз // Т. Созар. Р$
ар,
РЗУСЛО, 197.
Натаг М. Те рдеодтарМзеве УегЬтеНаис
19 бкоюс1зсВе Туреп 4ег \Уапдегга!-
4е (Вайиз погуеб!си;) ш Вишёшнен //
вет. апсем. 2001. 1973. М 3. $8. 219—
Натаг. М. МесВатзей сот4го] о# Миг9ае
шт 139аю@ Го //ЕРРО Вой. 1977.
\о1. 7(2). Р. 541-550.
Патаг М. 50а М. Та А. Терса
ега2егй ш опНейе аог!еде. С
4ос. аот. Вие., 1968а. 86 р.
а
4ЗА Литература
бе! В. 6. (1е.), Нефег 1. Тье тойе оЁ
се 4иа1 оНасфогу сиез ш Ме даег-
пуптаоб ой {еедше зИе з@есйоп оЁ 40-
тезЫс газ //Т. Сотр. ав@ Рруз1о]. Рзу-
СВО. 1976. Уо1. 90. Р. 727-789.
сще} В. 6. (1.), Кеппей Б. 6. Я ет М.
Решпопзтаюг шЯчепсе оп обзегуег 91с+
ргеетспее: ЕИесёз оЁ эпр!е ехрозите
ап ФНе ргезепее 0 а дотопзигаог /
Апиа, Теагп. Верау. 1985. Уо1. 13, М 1.
Р. 25—30.
бер 6. В. (1т.), Мизсиз Р. А. ОНасюгу
тефаноп оЁ лофег — уобпв сошасё т
Гопо-— Ехапз газ // 7. Сор. зп4 РВуз101.
Рзусво!. 1979. \о|. 93. Р. 708—710.
бие! ВН. ©. (1°.), бйеггу ШБ. Е. Мотег’з
тЙК а шей Рог тапзииззюл 0 сиез
теНеснис Ше Йауот оЁ тошег Фе
ТЫ. 1973. Ув. 83. Р. 374-378.
Сие В. С. (7т.}, Ижтоте 5. М. Тгапзег
оЁ ПРогтаноп сопсегишя 41зваюё 10005:
а 1аЪогайогу шуезЫраноп оф Ме «т-
тоттаНоп сехите» пуро#ез$ // Апит. Ве-
вау. 1983. Уо|. 31. Р. 748—758.
бойево А., бапсйез-Счайо У. Е. Раегото-
па| тедмайоп оЁ Фе езгоц5 суше ш
{Ве {ела|с гаф Бу Noе ассеззогу оШасо-
гу зузет // АМа епбостто|. — 1979.
Уо1. 225, варр!. Р. 256.
СаПево А.. Запсвез-Сцадо Т. Е. Мота 0.
Вос оЁ Ше уошегопаза! зузет 1 рВе
готаопа! тесербов 11 ‘Фе гаЁ//Омег-
тнипе оЁ Бераугоог апд снеписа] вИ-
той. 1.: ТВГ ргезз, 1982. Р. 57-64.
Сагаа 7. КоеИти В. А. ВааНоп оЁ смез
{0 сопвециепее и ауоансе 1еагита //
РаусНо!. Зе1. 1966. Уо1. 4. Р. 123—124.
Сашлепошз 1 А. М. Светиса| айтасат{$ о!
Ве гаё ргерама! Зап // СВеписа] 512-
па!5 ш уееБгавез. М. У.: Р/с `ргезз.
1977. Р. 45—59.
Санлепошзы Л. М. Бе Мсош О. В., 5а-
сешсл-ЗариллаЯ: М. Апдговеп 4ереп-
Чепсе оЁ а шаге рЬеготове ш га
итше // Ногтопе Вевах. 1976. \01. 7,
М 4, Р. 401—405.
Саишепош:м А. М. ОтзщаЕ Р. ХТ, 5 шсе-
идст-Заритйзааз М. Тозерв В. М. Руе-
зепз оф зех р|еготопе т ргераиа|
5]1ап9з ой шае га 7. Епдосгио!. 1975.
Уо1. 67. Р. 283—288.
СеттеЕе Н., Метаттег 7. Рог Тахопо-
пие Чег Сабишо ВаМиз (Водепиа, Ми-
т14ае) // Знпремегк. МИ. 1984. Ва. 49,
No2. $. 104—116.
|
Сегай Н. Б. Тата Т. [., СетаЙ А. А.
Пузгарноп оЁ Фе: мае газ зохиа! Бе-
‚ Бамопг 1ш@исеё Ъу зоба 1з91абоп//
Апим. Веряу. 1967. Ус]. 15. Р. 54-58.
Сецег Г. ОЦасюту ап Шегта] а чепсез
оп аИгазот!с уосайтаНоп Фиша Чеуе-
1ортепё 11 гофетив // Ашег. 700[. 1979.
\Уо1. 19. Р. 420—431.
.
Сеуег Г. А. Вагнеа В. Г. ГзИлепсе о
гопада! вогтопез ап4 зехиа1 Берау1оиг
оп аЦтазоме уосаНхаМоп 11 га. 1.
Тееациене оЁ 1ета]е //Т. Сошр. ап РВу-
$101. Рэусрвой. 1978. Уи. 92. Р. 438—446.
Сеуег Г. А., Ватра В. 7. Мегиовзй Т. К.
ТаЙаетсе о! гопада! Берау!оот оп ига
Зое уосаН2аНоп No1 газ. 2. Ттеавпец
оЁ мае // ТЫ. 1978а. УоГ. 92. Р. 447 —
456.
Сеуег Г. А., МеГиозь Т. К., Вагнаа В. Л.
ЕНесз оЁ иЙтазоте уосаНйтаНопз апЯ
тае’з игше оп Гетоа]е гаё геа@ 11055 {0
тра / 14. 19785. Уо1. 92. Р. 457—462.
Сиеке В. ТНе пес {ог Имедтайоп оЁ есо-
1од1са1, рВузю]озеа! ап е{НоТоб1са! ге-
збагсл т 10060 сопёто| // РАБ. ОЕРР,
1970. Уо]. 58. Р. 49-54.
СШен А. No, Воитвооп Б. А., чи
рап К, А., Ааег М. Т. Мофог-уеатия
пиегасНор ша Могмау га! шт ЧВе рге-
зепсе оЁ а зиссезыуе НИег ргодис1е4 Бу
розёрагьатт шания // РВуз1!. ав Вевау.
1983. Ув]. 30. Р. 267—274.
СизВ С. К, Агога К. К. 549168 оп го-
деп{з ава Мет сопито1. 13. СвешояегЕ-
]ап4з ‘есри1чиез юг годе сопто] //
Вай. Сташ ТесВио, 1976. Уо1. 14, М 3.
Р. 219-228.
боЧтап 1. В. Те еНесёз о! Тапа Нае апа
звосКте т НУапеу проп адиз ЪеВа-
утоиг т фе аЫшо га //Т. Селе. Рзу-
сро!. 1964. Ус. 104. Р. 301-310.
“Согаоп тив С. Е., Титпег Ё. Н. Нате
зон 7. Г., Втоот 7. С. Апиа 1ербюозр-
0515 т Майауа. 2. ГосаНЦез затр!е4 //
ВоЙ. Огргаше. топ. заше. 1961а.
\Уо1. 24, М 1. Р. 23—34.
Согаоп бтив С. Е. Титпег Ё. Н., Натп-,
зоп 7. Г., Втоот 7. С. Апиа! 1ербозрто-
815 ш Маауа. 1. Ме о43. зоопсовтар[1-
са! БасКкотоиЯ ап@ Ъгоаф апа|уз$ оЁ
тези!(з // ТЫ. 1961. Уо1. 24, М 4. Р. 5—
21.
Созафел Х., Горел-ГРиег М. Т. Тве пам-
га} сот!Иез оЁ зта] хлалта8 (-
зесыуотез ап годе!) о Сайаюша
(Зраш) // М!зе. 2001. 1985. М 9. Р. 375-—
387
Созсзутз 7. ММу\м Чгареттусв ззако\
риаКо\ роршаее оНаг// \УЛад. еКо1.
1974. Уо|. 20, М 3. $. 207-289.
Соигевисв @. НасЁ М. М. АТЯЬЙЦУ Ш
Ве гаф//У. Сор. ава Рвузо!. Рзусво|.
1966. Уо1. 62. Р. 289.
„
Литература
135
Стат Е. С. Ап апа]уз1з ой {Не зола! Ъе-
Пауюиг оЁ Ме шае Пафотаюгу гаЁ//
Верауюиг. 1963. Уо|. 24. Р. 260-281.
ата Е. С., МасЕйщозй 7. No. А сотраг-
50п о те зоста! розшоз 0{ зоше сош-
шоп |аБботаюту го4етз 1
Уо1. 21, Р. 246-259. та. 1983.
Стая No. С. ОТап тодеоеБогие 491зеазе
ап тоделё тай Ш [згае] апа
пеовБотте сопптез // Тэг. 7. Мед. 31.
1973. \Уо1. 9(8). Р. 969-979.
Стау Р., СпеЯеу 5. Оечортетё оЁ та-
тра] рерауюиг т пИНрагопз габв: е#-
Рос а ап9 вау шаетпа| ехре-
иепее // 7. Сошр. ап@ ГNoуз01. Рзу
1984. У. 08 В. 9199. © ое Реут
баров 1. ие 10 ап/соаРШат
11 годен // РезИе. 31. 1974. Уд. :
си 8. Р. 276-279.
Бу
тевуез 7. Н., Ве4{еги В., Аутез Р. В,
СШ7.Е.Маагтозанее:аны
се ро!утотНзт 11 Могмау гаё// Сепе-
Неа ВезеагеВ. 1977. \Уо]. 30. Р. 257—
Стеарез 7. Н. Арерйета р. 5. и
Ке1@ 91218 о ОЕ
а
соваиатив аватя аЁепасойт-геззать
оту’ау гаф рораНотз //Т. Нуе.
\Уо1. 89, No2. ЗО
УВ. 1982,
Сирия И’. Л, батткег 17. Р. Тре сНесз
оЁ пиепзе зЫпиаНоп ехрегелсеф 4-
ти иуапсу оп ад Бела\топг 11 Фе
га . отр. ап РНузю1.
1052. от. 45. В, 304-306 Русво
Стоптооз Р., бттеп А. Тохор]азт0$! т
‚ Могуау газ па Не]зшЮ
// Апиа. Мед.
ЕхрН. В10]. Еегииеа. 1955. Уо|. 33
с. Р. 44,
|
гозкеггу Р. С., 5тИВ С., Геоп М., Ми-
сле Ц Е. 4. Тпегшогев Иа Чо апа а.
егра. спауюг о ег
о
99080 Чаи
бтоззеё ПО., Ройпе А. Тве машз оЁ Ван
105: газ аз зиб]есз ш агИсез ри
$Нед т Топгпа| о? Созпрагайуе Рвуз1010-
21са! Рзусро]огу ош 1969 урголеВ
1984 // Рвуспо]. Вер. 1982. Уч. 54, М 3.
ре 1. Р. 989-970.
'
Стозрепог С. Е. Майрек Н., Мепа Е. А.
А зу о! асёог8 туоуеЯ п Те 4е-
у@юоршетё о! Фе ехёегосерыуе гееазе
оЁ ргоасйп ш Фе Тасбайпе гаф// Ног-
топе Верау. 1970. Уо]. 1. Р. 144-120.
Стозуепог С. Е., Мепа РЁ. Мепта| ап@ дот-
това] оо о? шПК зестейоп апа
тИЕ е]ес@оп // ГаббаНоп.
.
то
у
1974. Уд. 1.
Стош Г. 7. ЕНесё о? НИег эле, асе о
Усйп ан@ рагбу оп фозег клоет Ъе-
’
Вауоцг 1 ВаМаз погуеслсиз // Ат
аТЗ
Стоа Г. Г, А4ег В. Сопйпаоиз тесотфшт
0# тафетра! Бераутоиг 1 Вабиз погуе-
21св
// 9. 1969. Уо|. 17. Р. 722-729.
Стиепае! А. 0., Атпоа 1’. У. ЕНесз о
вату зоба! дерМуаНор оп гергофасмуе
Беваутог о{ ша! та{//Т. Сошр. апй
Ррузо. РзусВо!. 1969. Уо1. 67. Р. 123—
Сифетиск Ш. Т, Аве; /. В. Ммегпя
Е 9 учи: ого ехсНапое ап@
тохипа{е соло // РВуз10].
пера,
СпрегисЁ р. Г, АВетз ХТ. В. Мабетра!
Пони Ву мы ара 1абаНай таб:
хаег 1тапег ош ри
19.
УоЕ 3. В. 501-506 та. 1985.
Нааз С. Е., И’Изоп No, Вмимоп В. В.
ОБзетуа 00$ оп Могмуау газ, Вайлв.
погуес1сиз, м Кофак, АазКа // Сапа.
Ее 9-Манаг. 1986. Уо. 100, М 3. Р. 316.
Нает м. РА \меароп аратзь Фе гез1-
апф та ес Чез. 1975.
у
За та
1975. У. 11, М9.
Найп Р. Те ЧеуеТорхлете оЁ (Пегтотеси-
1аНоп. 2. ТВе рагсранов оЁ те Фу-
го ввод ш Ше деу@ортете оЁ фег-
погезиа оп т габ // РВуз1о1
--
1103]. 1958. Уо/. 5. Р. и ое
Нар Р.,. Ко 4орзку О., Ктесейк 1. её &
Еп4оске ап@ шеаЪоНс азресаз оЁ с
Чеус]оршеш оЁ Вотеотетту ш Ше
га // Зошанс заБИиу о Ме пез
Боги. Возбюш, 1964. Р. 131-155.
НаП Е. В. ТЬс шашта!з оГ Мог Аше-
пса. No. У.: УУЦеу, 1981. У. 2. 2000 р.
Ной 8. в. Ке!зоп К. В. ТВе аашта]8 о
тот
шегка. М. У.:
1959. Уо1. 2. 538 р. Копа ртов,
НаЙ И’. 6., Статег С. Р., В!азз Е. М.
етот свапосз 1 зисКЦио ой
га
и1рз абаге.
та НН
1975. У. 258.
На! ИУ. 6., Нозеп Май 7. 5. ЗаеКИаи Ъе-
Вачлош аа о сопто| ш {Фе 4еуе-
оршс гаё рирз // Т. Созар. Р$
ар,
РЗУСЛО, 197.
Натаг М. Те рдеодтарМзеве УегЬтеНаис
19 бкоюс1зсВе Туреп 4ег \Уапдегга!-
4е (Вайиз погуеб!си;) ш Вишёшнен //
вет. апсем. 2001. 1973. М 3. $8. 219—
Натаг. М. МесВатзей сот4го] о# Миг9ае
шт 139аю@ Го //ЕРРО Вой. 1977.
\о1. 7(2). Р. 541-550.
Патаг М. 50а М. Та А. Терса
ега2егй ш опНейе аог!еде. С
4ос. аот. Вие., 1968а. 86 р.
а
436 Литература
Натаг М., бота М., Та АТ. Возеюаге-
1е 1 сошЬайетеа ]от// Сетит. 4ес. аот.
Вис., 1968. 71 р.
Напзеп 5. Мопиипе Бера\!ог ап тесери-
уе Бевау!очг ш Чеуеорття Фета!е га
ап Фе еНесё 01 востаЁ 1801айоп // РВу-
510]. апа Вевау. 1977. Уо]. 19, No 6.
Р. 149-752.
Натош П. Г. Аве-та!е ог реег аЙесНопаТ
зузбетв // Аду. $19. ВеВау. 1969. Уо1. 2.
Р. 333—383.
Пагтяюв No. С, неё! С. Т. ГЯзифайов
апд сопЁто] о{ гаёз ш Нуе ВосКу Моип-
бат завез //РаЫ. НеаЙн Вер. 1960.
УоТ. 75, М 11. Р. 4077—1084.
Нагтзоп У. Г. НаБИаф о{ зоше Маауай
газ // Ртос. 200]. 806. Гопдой. 1957.
Уо1. 128, М 1.Р. 1-24,
Нагизоп 0). Г. Тье тайита!5 оЁ Агава.
Уо]1. 3. Габвошогрва, Водепйа. Г.: Е.
Вепи. 1972. Р. 670.
Нат М. Ваз. Ь.: АШзоп, ВазБу, 1982.
172 р.
НагНеу 0. 7Т., Взфор Т. Л. Поше гапсе
ап@ шоусштелё 1 роршШаНопз оЁ ПаНи$
погуев1еиз ро]утотрЬе юг хагфагтпге-
звиапее // В10]. 7. Глппеап 306. 1979.
\Уо]. 12. Р. 19—43.
Насй А. М., ИТЪегЕ С. 5., Ваазе Т., бт
се Н. С. Гопё 1ета 1воЙаМоп тез т
га{з // Заепсе. 4963. Ув]. 142. Р. 507.
Нафашеиу 5. С, ВшсЁтоге О. К. Есо]0-
о1са| азрес!ё о Ме еретш10]оду оЁ ш-
Геснов \ИВ ПЛерюзритез оЁ Фе ВаПат
вегостопр т Ме ЫасЕ гаё (ВаМиз га[-
415) ап@ {Те Бго\хп га (ВаИаз потуе-
0113) т Мем 2еяапа//Т. Нуя. 1984.
Уа1. 87, М 3. Р. 427—436.
Науазм 1. Г, Уопекаша Н., боюой 0.
её а!. ЕуоаНопа| азресёз оЁ уаматё $у-
рез 0 гаф шИосвопдта! ОМАз // В!о-
сВНо. её Борвуз. аба. 1979. Уо|. 564.
Р. 202—211.
Неапеу Г. В., Нафог 2. 5. Мата! 9
П}пасаф ап З1атоао 13}ап4з, РЫПррЕ
пез // Оссаз. Рар. Миз. 7001. Пзиу. М1ев.
1982. М 669. Р. 1-30.
Некао! Р. Г., биз Т. А., ие Е. С. 5е-
сопдагу сНесёз оЁ годеписез оп тат-
паба ргедафогв // УГо 9х 9е Китгреагег
сопЁ, Егозфиго, 3—14 Арг. 1981: Ргос.
Егозфиге, 1984. Уо1. 3. Р. 1781—1793.
Непзесйе! 7. Ощегзислилеет Бег 91е уоп
Наоз- 009 УУ’апдеггайеп муегзеШеррее
ЭЗнепритегтепее // Ап. ЗевадНаозкК.
1955. В@. 28, No 7. 5. 97-101.
'Непзсйе 7. ВаметЪеаП пи НатЬигаег
‚ Збаааесееё <паНсН ег Мог4епЬепе уог
и19 пасВ дег ВаНетЪек&тр!апо, 1963 //
РИЙаптепзср7-МасЬт. «Вауег». 1965.
Ва. 18, М4. 5. 151-164.
Неррег Р. 6. УЪШшв тесопи оп ш Фе
так // Апиа. Вевау. 1983. Уд. 31.
р. 1177—1191.
Негтапп С. ГЛе веъгааеВИсВей Воден\1-
с9еп опа {ге Армердиюя ш опис
Тап4еги 03- ир@ \Уечеогорав
// Вае
{епЬ1о1021е пир ВайепЬеКал ато: (УогЕ-
тасзусгапзаатс сз Уегешз Гаг \Уаб-
зег, Во4еп- ип4 ГаНрубепе ат 28.
па 29. МоуетБег 1968, КопотеззваПе,
ВегНа). ЗбиИрать 1969. $. 179—197.
Негтапи С., Потфтгесрег 5. Вайе- ип9
Маизерек&трйлир п Васитй1-57 Ргй-
рага{еп // РНалтепвср
и -Масйг. «Вауег».
1962. ВЧ. 15, М 2. $. 93—112.
ИНегто4з0в М. Р., Зише 7. И’, ив К.Р.
МТататт тебаБоНзт ап уМапит К те-
фитетет ш Фе магРагт гез1зват
таб
// Атег. Г. РБуз1ю1. 1969. Уо1. 217,
М 5.Р. 1346-1319.
Негпапае: А. ОЭтиттопа О. С. А заду
оЁ годеп+ датаре {0 No004 ш зоше сифав
\затевойзез апа Ше 03 о! ргеуепиое
и //Т. %юте@ Ргод. Вез. 1984. У]. 29,
М2.Р. 83—86.
Нега У’. ОЪег @е Наше асг Масазавне
Чег У’ап4еггаИе из спыдег апдетег
Мазег // Апиа, ЗеваЯНиезК. 1950. ВЯ. 23,
М 10. 8. 150.
НепепкоШ Г. В., ГазЁ В. Б. ЕНесё оЁ 1ас-
ц4аНоп оЁ рговечегопе ]есНопз а@пз-
013 {его Бефоге ап аЙетг рагатаНот
ш Фе гаё// Ргос. ос. Ехр. ВТ. ата
Мед. 1973. Уо|. 142. Р. 506-510.
Неуегда М Т. Еам-ЮЩуа: ПВасК 10 пайте.
К. У, 1974. 292 р.
ПШ р. А., Поетзоп Н. А, битег-
1апа 1’. Г. Втоуй таёё (ВаМиз пог-
уе21с05) сИиЬше No оБаш $оез ап4
МасКЪегтез//Т. 2001. 1983. \в|. 200,
М2.Р.302.
НШ Е. Е., Сагремег Г. 17. Везропзез о
З1Бетап {егге{з No зесопаагу 2116 рро-
рые ро1зопшо //Т. УПаШе Маваз.
1982. Уо1. 46, М 3. Р. 678—685.
Ната РО. М., МNoМазз В. ). Стоу®ф апа
зехиа! таштайоп оЁ ]аБога(огу-гсагей
\1а КВаИаз погуса!сиз, В. гаМаз ап@
В. ехи]апз ш Назан //У. Машта!. 1974.
Уо!. 55, М 2. Р. 472—474.
НигзЕ Е. Везеатевез оп \Noе рагазИоюсу
о{ Раис // Сеуюп Т. 3с1. Ме4. 1926-
1927. Уо|. 1, рф 4/5. Р. 157-267, 279-
451.
Ни ?. С., РЬИИрз Г., Азало Н. Ногтопа|
Чести {таз оЁ ргеЁегепее {ог етае
з6х одог 11 габз: Рар. ргезетеЯ ав ле
Литература 437
тееь, оЁ 1Ие Рзуспопот, 506. Зап Апю-
а ('Гех.), 1970.
Носктап С. Н. Ргепа] табегпа| з5'е3з
т Ше таб Из еНесз оп ешоНопа! Бе-
Вау1оиг ш Ше оН рита // 7. Сотр. апа
Влучо. РзусВо]. 1964. У0]. 54. Р. 679—
'|.
Ноедеп 7. вап 4ег. Тре рафовешсну о!
1ерюзртае No Йе9 тодегз 11 згае| //
7. ПЧесё. О1зеазез. 1964. Ус]. 95. Р. 243.
Ноедет 1. рап 4ет., бтепьети Е. Герюзрта
П\есНо0з т гаёз ш Тзгае] // Тгор. Сеост,
Мед. 1967. Уо]. 16, М4. Р. 377—384.
Нофег М.А. Тре еНесь оф Бей та(егпа1
зерагайоп оп Бервауютг ап@ ВеагЁ гае
оЁ бхо \ееК 01 гаё рире // РНуз101. ап
ВеВах. 1973. \о]. 10. Р. 423-437.
Нофег М. А. Зигууа апд тесоусту о
рВузТово ТопеНопз аМег еат!у шацег-
па! зерагамоп 11 газ // ТЫа.
Уо1. 45. Р. 475-480. и 197.
Ноег М. А. Рагепа] сопыБоНопз 40 Фе
Чеусюркепй оЁ Тейт оЙзогша // Рагсп-
121 Бевау1опг т ташта13/Е9. О. 7. бо-
Бегшек, Р. Н. Кюор[ег. М. У.; Ь.: Р]е-
пи ргезз, 1984. Р. 77—145.
Нофег М. А., брт НП. Согиго| о еер-
жаке збаез ш Фе ш!апе га Бу Геаб-
тез оЁ фе шоШег-иРат те]аНопзВр //
Оеуеюр. Рвусвомо1. 1982. \01. 45.
Р. 229—243.
Нофег М. А., брат Н, Этий Р. Е\1депсе
4Ваф таегпа] уешта! эт зибзапсез
рготофе зисКИие Ш ап габз // РВу-
о. ап Верах. 1976. У]. 47. Р. 431—
Нофег М. А., У’етег В. Т\е деус]орштеп+
ап тесрап!зтз оЁ сага1отезртаогу
роте ю оаетиа Чериуавот 11 га
рирз // Рзуспозота. Ме. 1974.
Р. 353—362.
КУ38
Нойптка С. Е., Сатзоп А. Б. Рир аИгаси-
оп о Л1абайие бргазпе—Паз]еу газ //
Вейау. В!101. 1976. Уо|. 16. Г. 489-505.
Нозег М. 'ГНе вепегс патез Ерниуз
ап@ ВаИаз // Ргос. В101. $ос. УТазв.
ива. Уо1. 29. Р. 125-128.
оШзег No ТЬе Фуре зрес1ез о Ван
«_ Па. 1916Ь. Уо1. 9, р. 205—208. ий
`НоНошау. И’. (7ь.}, РоШптвег М. 7. Беп-
пепрегеУ. Н. Рагтабов ше гаё
| Чезсеруоп 804 — аззезтень // Ма1егпа!
ШНоепсез апа вау — Бепауоог/Ед.
УМ. Р. Зтофегтап, В. No. Вей. 1... 4980.
Р. 1-26.
у
Нотова М. Оп фе ргоМет оЁ ехоатито-
ре оссигепсе оЁ ВаМаз погуессиз//
ЕоПа 200]. 1983. No 3. Р. 203—244.
Ноорзтай Н. А БчеЁ гемем оЁ фе соп-
1етрогагу ап татта!з о Едурё (тп-
сте ЭЗша!). 2. Таготогрва ‘ап9 Во-
Чепыа //7. Есурь Ри. НеаЙNo Аззос.
1963. Уа1. 38, М 1. Р. 1-35.
НотифсNo В. Н. ТЬе ошобепу оЁ зоба
те
ту зЧате! (Зегагиз
сагоНпепз15
Чзепг. Тегрз
.
.
ое
рзусво]. 1972
Ношата No. Е. Вйосошио] ап@ сВетозе-
гаи // Резё сопуго]. М. У.; [,: Асад.
ргезз, 1967. Р. 343—386.
Ношата No. Е., Могзр В. Е. Отазошез
204 е]естотаспейс сотой оЁ годеш //
пса 200], {епп. 1985. Ус]. 173, Р. 167-
Ношага И. Е., Магзь В. Е., Ратицег 5. Р.
Ваф ассерапсе оЁ ЧИЧегош зираг соп-
сеттаНопз ш мафег БаНз // иегп. Резь
Сопг. 1972. Уо1, 14, М 6. Р. 17, 19-20.
Нивег С. С. ТЪе Чвуе!ортепь оЁ Фе а.
по гаф, Миз погуер!сиз ашаз // 7. Мог-
рВо1. 1945. Уо1. 26, М 10. Р. 247—958.
Нисё ИП. Г, Га Выт 7. Вше епиасе
Мепо4е гг Везитлите ог МасеНег-
гез15фет? уоп {есрзелеп Мапа //
Ап2. ЭерёаНаязК. 1959. В4. 32, М 1.
$. 9—10.
Низоп И’.. Пептизоп В. О. Зеазопа] уата-
ыЫШу 0# Могуау гар (Ваз погус1с03)
печаНов оЁ авгеаата| ргети1ззез //
9 1981. \Уо|. 94 М 2.Р. 957-
ит Т. Хоню добуцучаку дзасси И
7. Мапита|. 506. ар. 1979. Ус. 7 М
Р. 233—239, р
о
У
58
сноп и В. ИЯ райегиз Ш 9о-
тезие годел{з // Резь Сопи:. 1965. Уо1. 33.
Р. 11—13, 50, 52.
оз
Таскзоп И’. В. ТЛбгазощев ргофесё е
ати ТЫЯ. 1980. Ув. 48, М $. Р. 28
ТасЁзоп No’. В., Втошп М. Г., Азрюп А. В.
СопбтоШиа& теззапф газ ш СЫсаро //
1Ы9. 1978. У]. 46, М 8. Р. 47, 19,20.
УасЁзоп И’. В., Капкетепв Р. Е. 'АпЧсоа-
Шарф геззапсе ап@ годе сопуго| /
Ртос. 3:4 Пцеги. Модестаа. зУушр.,
Клип, 1975. 1, 1976. Р. 303-308,
7асобз Г. Г. ЕоззЙ годе (В:2отуЧае
ап@ Ми Чае) ош Меосепе ЗмаЦК 4е-
розИз, Ракбар. 1978. ХТ, 103 р. (Маз.
Мог Ап. Ргезз. Ви; М 52).
ип А. Р., Рашае У. А. Везропзез оЁ а
фгее-NoУте роршаНотз оЁ ВаНаз гайиз
(ГЛол.} ю\%аг@з ро1зопе@ БаН апф пех
1004 // 114. 1. Ехр. Во. 1981. Уо. 19.
Р. 678—679.
Зактьошз М., Тегье! Г. Рир апа оБзе
сагтуша Бу тмегра!у ап@ поптабег-
436 Литература
Натаг М., бота М., Та АТ. Возеюаге-
1е 1 сошЬайетеа ]от// Сетит. 4ес. аот.
Вис., 1968. 71 р.
Напзеп 5. Мопиипе Бера\!ог ап тесери-
уе Бевау!очг ш Чеуеорття Фета!е га
ап Фе еНесё 01 востаЁ 1801айоп // РВу-
510]. апа Вевау. 1977. Уо]. 19, No 6.
Р. 149-752.
Натош П. Г. Аве-та!е ог реег аЙесНопаТ
зузбетв // Аду. $19. ВеВау. 1969. Уо1. 2.
Р. 333—383.
Пагтяюв No. С, неё! С. Т. ГЯзифайов
апд сопЁто] о{ гаёз ш Нуе ВосКу Моип-
бат завез //РаЫ. НеаЙн Вер. 1960.
УоТ. 75, М 11. Р. 4077—1084.
Нагтзоп У. Г. НаБИаф о{ зоше Маауай
газ // Ртос. 200]. 806. Гопдой. 1957.
Уо1. 128, М 1.Р. 1-24,
Нагизоп 0). Г. Тье тайита!5 оЁ Агава.
Уо]1. 3. Габвошогрва, Водепйа. Г.: Е.
Вепи. 1972. Р. 670.
Нат М. Ваз. Ь.: АШзоп, ВазБу, 1982.
172 р.
НагНеу 0. 7Т., Взфор Т. Л. Поше гапсе
ап@ шоусштелё 1 роршШаНопз оЁ ПаНи$
погуев1еиз ро]утотрЬе юг хагфагтпге-
звиапее // В10]. 7. Глппеап 306. 1979.
\Уо]. 12. Р. 19—43.
Насй А. М., ИТЪегЕ С. 5., Ваазе Т., бт
се Н. С. Гопё 1ета 1воЙаМоп тез т
га{з // Заепсе. 4963. Ув]. 142. Р. 507.
Нафашеиу 5. С, ВшсЁтоге О. К. Есо]0-
о1са| азрес!ё о Ме еретш10]оду оЁ ш-
Геснов \ИВ ПЛерюзритез оЁ Фе ВаПат
вегостопр т Ме ЫасЕ гаё (ВаМиз га[-
415) ап@ {Те Бго\хп га (ВаИаз потуе-
0113) т Мем 2еяапа//Т. Нуя. 1984.
Уа1. 87, М 3. Р. 427—436.
Науазм 1. Г, Уопекаша Н., боюой 0.
её а!. ЕуоаНопа| азресёз оЁ уаматё $у-
рез 0 гаф шИосвопдта! ОМАз // В!о-
сВНо. её Борвуз. аба. 1979. Уо|. 564.
Р. 202—211.
Неапеу Г. В., Нафог 2. 5. Мата! 9
П}пасаф ап З1атоао 13}ап4з, РЫПррЕ
пез // Оссаз. Рар. Миз. 7001. Пзиу. М1ев.
1982. М 669. Р. 1-30.
Некао! Р. Г., биз Т. А., ие Е. С. 5е-
сопдагу сНесёз оЁ годеписез оп тат-
паба ргедафогв // УГо 9х 9е Китгреагег
сопЁ, Егозфиго, 3—14 Арг. 1981: Ргос.
Егозфиге, 1984. Уо1. 3. Р. 1781—1793.
Непзесйе! 7. Ощегзислилеет Бег 91е уоп
Наоз- 009 УУ’апдеггайеп муегзеШеррее
ЭЗнепритегтепее // Ап. ЗевадНаозкК.
1955. В@. 28, No 7. 5. 97-101.
'Непзсйе 7. ВаметЪеаП пи НатЬигаег
‚ Збаааесееё <паНсН ег Мог4епЬепе уог
и19 пасВ дег ВаНетЪек&тр!апо, 1963 //
РИЙаптепзср7-МасЬт. «Вауег». 1965.
Ва. 18, М4. 5. 151-164.
Неррег Р. 6. УЪШшв тесопи оп ш Фе
так // Апиа. Вевау. 1983. Уд. 31.
р. 1177—1191.
Негтапп С. ГЛе веъгааеВИсВей Воден\1-
с9еп опа {ге Армердиюя ш опис
Тап4еги 03- ир@ \Уечеогорав
// Вае
{епЬ1о1021е пир ВайепЬеКал ато: (УогЕ-
тасзусгапзаатс сз Уегешз Гаг \Уаб-
зег, Во4еп- ип4 ГаНрубепе ат 28.
па 29. МоуетБег 1968, КопотеззваПе,
ВегНа). ЗбиИрать 1969. $. 179—197.
Негтапи С., Потфтгесрег 5. Вайе- ип9
Маизерек&трйлир п Васитй1-57 Ргй-
рага{еп // РНалтепвср
и -Масйг. «Вауег».
1962. ВЧ. 15, М 2. $. 93—112.
ИНегто4з0в М. Р., Зише 7. И’, ив К.Р.
МТататт тебаБоНзт ап уМапит К те-
фитетет ш Фе магРагт гез1зват
таб
// Атег. Г. РБуз1ю1. 1969. Уо1. 217,
М 5.Р. 1346-1319.
Негпапае: А. ОЭтиттопа О. С. А заду
оЁ годеп+ датаре {0 No004 ш зоше сифав
\затевойзез апа Ше 03 о! ргеуепиое
и //Т. %юте@ Ргод. Вез. 1984. У]. 29,
М2.Р. 83—86.
Нега У’. ОЪег @е Наше асг Масазавне
Чег У’ап4еггаИе из спыдег апдетег
Мазег // Апиа, ЗеваЯНиезК. 1950. ВЯ. 23,
М 10. 8. 150.
НепепкоШ Г. В., ГазЁ В. Б. ЕНесё оЁ 1ас-
ц4аНоп оЁ рговечегопе ]есНопз а@пз-
013 {его Бефоге ап аЙетг рагатаНот
ш Фе гаё// Ргос. ос. Ехр. ВТ. ата
Мед. 1973. Уо|. 142. Р. 506-510.
Неуегда М Т. Еам-ЮЩуа: ПВасК 10 пайте.
К. У, 1974. 292 р.
ПШ р. А., Поетзоп Н. А, битег-
1апа 1’. Г. Втоуй таёё (ВаМиз пог-
уе21с05) сИиЬше No оБаш $оез ап4
МасКЪегтез//Т. 2001. 1983. \в|. 200,
М2.Р.302.
НШ Е. Е., Сагремег Г. 17. Везропзез о
З1Бетап {егге{з No зесопаагу 2116 рро-
рые ро1зопшо //Т. УПаШе Маваз.
1982. Уо1. 46, М 3. Р. 678—685.
Ната РО. М., МNoМазз В. ). Стоу®ф апа
зехиа! таштайоп оЁ ]аБога(огу-гсагей
\1а КВаИаз погуса!сиз, В. гаМаз ап@
В. ехи]апз ш Назан //У. Машта!. 1974.
Уо!. 55, М 2. Р. 472—474.
НигзЕ Е. Везеатевез оп \Noе рагазИоюсу
о{ Раис // Сеуюп Т. 3с1. Ме4. 1926-
1927. Уо|. 1, рф 4/5. Р. 157-267, 279-
451.
Ни ?. С., РЬИИрз Г., Азало Н. Ногтопа|
Чести {таз оЁ ргеЁегепее {ог етае
з6х одог 11 габз: Рар. ргезетеЯ ав ле
Литература 437
тееь, оЁ 1Ие Рзуспопот, 506. Зап Апю-
а ('Гех.), 1970.
Носктап С. Н. Ргепа] табегпа| з5'е3з
т Ше таб Из еНесз оп ешоНопа! Бе-
Вау1оиг ш Ше оН рита // 7. Сотр. апа
Влучо. РзусВо]. 1964. У0]. 54. Р. 679—
'|.
Ноедеп 7. вап 4ег. Тре рафовешсну о!
1ерюзртае No Йе9 тодегз 11 згае| //
7. ПЧесё. О1зеазез. 1964. Ус]. 95. Р. 243.
Ноедет 1. рап 4ет., бтепьети Е. Герюзрта
П\есНо0з т гаёз ш Тзгае] // Тгор. Сеост,
Мед. 1967. Уо]. 16, М4. Р. 377—384.
Нофег М.А. Тре еНесь оф Бей та(егпа1
зерагайоп оп Бервауютг ап@ ВеагЁ гае
оЁ бхо \ееК 01 гаё рире // РНуз101. ап
ВеВах. 1973. \о]. 10. Р. 423-437.
Нофег М. А. Зигууа апд тесоусту о
рВузТово ТопеНопз аМег еат!у шацег-
па! зерагамоп 11 газ // ТЫа.
Уо1. 45. Р. 475-480. и 197.
Ноег М. А. Рагепа] сопыБоНопз 40 Фе
Чеусюркепй оЁ Тейт оЙзогша // Рагсп-
121 Бевау1опг т ташта13/Е9. О. 7. бо-
Бегшек, Р. Н. Кюор[ег. М. У.; Ь.: Р]е-
пи ргезз, 1984. Р. 77—145.
Нофег М. А., брт НП. Согиго| о еер-
жаке збаез ш Фе ш!апе га Бу Геаб-
тез оЁ фе шоШег-иРат те]аНопзВр //
Оеуеюр. Рвусвомо1. 1982. \01. 45.
Р. 229—243.
Нофег М. А., брат Н, Этий Р. Е\1депсе
4Ваф таегпа] уешта! эт зибзапсез
рготофе зисКИие Ш ап габз // РВу-
о. ап Верах. 1976. У]. 47. Р. 431—
Нофег М. А., У’етег В. Т\е деус]орштеп+
ап тесрап!зтз оЁ сага1отезртаогу
роте ю оаетиа Чериуавот 11 га
рирз // Рзуспозота. Ме. 1974.
Р. 353—362.
КУ38
Нойптка С. Е., Сатзоп А. Б. Рир аИгаси-
оп о Л1абайие бргазпе—Паз]еу газ //
Вейау. В!101. 1976. Уо|. 16. Г. 489-505.
Нозег М. 'ГНе вепегс патез Ерниуз
ап@ ВаИаз // Ргос. В101. $ос. УТазв.
ива. Уо1. 29. Р. 125-128.
оШзег No ТЬе Фуре зрес1ез о Ван
«_ Па. 1916Ь. Уо1. 9, р. 205—208. ий
`НоНошау. И’. (7ь.}, РоШптвег М. 7. Беп-
пепрегеУ. Н. Рагтабов ше гаё
| Чезсеруоп 804 — аззезтень // Ма1егпа!
ШНоепсез апа вау — Бепауоог/Ед.
УМ. Р. Зтофегтап, В. No. Вей. 1... 4980.
Р. 1-26.
у
Нотова М. Оп фе ргоМет оЁ ехоатито-
ре оссигепсе оЁ ВаМаз погуессиз//
ЕоПа 200]. 1983. No 3. Р. 203—244.
Ноорзтай Н. А БчеЁ гемем оЁ фе соп-
1етрогагу ап татта!з о Едурё (тп-
сте ЭЗша!). 2. Таготогрва ‘ап9 Во-
Чепыа //7. Есурь Ри. НеаЙNo Аззос.
1963. Уа1. 38, М 1. Р. 1-35.
НотифсNo В. Н. ТЬе ошобепу оЁ зоба
те
ту зЧате! (Зегагиз
сагоНпепз15
Чзепг. Тегрз
.
.
ое
рзусво]. 1972
Ношата No. Е. Вйосошио] ап@ сВетозе-
гаи // Резё сопуго]. М. У.; [,: Асад.
ргезз, 1967. Р. 343—386.
Ношата No. Е., Могзр В. Е. Отазошез
204 е]естотаспейс сотой оЁ годеш //
пса 200], {епп. 1985. Ус]. 173, Р. 167-
Ношага И. Е., Магзь В. Е., Ратицег 5. Р.
Ваф ассерапсе оЁ ЧИЧегош зираг соп-
сеттаНопз ш мафег БаНз // иегп. Резь
Сопг. 1972. Уо1, 14, М 6. Р. 17, 19-20.
Нивег С. С. ТЪе Чвуе!ортепь оЁ Фе а.
по гаф, Миз погуер!сиз ашаз // 7. Мог-
рВо1. 1945. Уо1. 26, М 10. Р. 247—958.
Нисё ИП. Г, Га Выт 7. Вше епиасе
Мепо4е гг Везитлите ог МасеНег-
гез15фет? уоп {есрзелеп Мапа //
Ап2. ЭерёаНаязК. 1959. В4. 32, М 1.
$. 9—10.
Низоп И’.. Пептизоп В. О. Зеазопа] уата-
ыЫШу 0# Могуау гар (Ваз погус1с03)
печаНов оЁ авгеаата| ргети1ззез //
9 1981. \Уо|. 94 М 2.Р. 957-
ит Т. Хоню добуцучаку дзасси И
7. Мапита|. 506. ар. 1979. Ус. 7 М
Р. 233—239, р
о
У
58
сноп и В. ИЯ райегиз Ш 9о-
тезие годел{з // Резь Сопи:. 1965. Уо1. 33.
Р. 11—13, 50, 52.
оз
Таскзоп И’. В. ТЛбгазощев ргофесё е
ати ТЫЯ. 1980. Ув. 48, М $. Р. 28
ТасЁзоп No’. В., Втошп М. Г., Азрюп А. В.
СопбтоШиа& теззапф газ ш СЫсаро //
1Ы9. 1978. У]. 46, М 8. Р. 47, 19,20.
УасЁзоп И’. В., Капкетепв Р. Е. 'АпЧсоа-
Шарф геззапсе ап@ годе сопуго| /
Ртос. 3:4 Пцеги. Модестаа. зУушр.,
Клип, 1975. 1, 1976. Р. 303-308,
7асобз Г. Г. ЕоззЙ годе (В:2отуЧае
ап@ Ми Чае) ош Меосепе ЗмаЦК 4е-
розИз, Ракбар. 1978. ХТ, 103 р. (Маз.
Мог Ап. Ргезз. Ви; М 52).
ип А. Р., Рашае У. А. Везропзез оЁ а
фгее-NoУте роршаНотз оЁ ВаНаз гайиз
(ГЛол.} ю\%аг@з ро1зопе@ БаН апф пех
1004 // 114. 1. Ехр. Во. 1981. Уо. 19.
Р. 678—679.
Зактьошз М., Тегье! Г. Рир апа оБзе
сагтуша Бу тмегра!у ап@ поптабег-
438
Литература
паПу февауше {етае ато гаёз (Ва
{из погуер1ел5) //Т. Сотар. Рзусво]. 1984.
Уч]. 98. Р. ЗМ- 347.
Уапез 7. Геоп М. Реетпитатйз ой таоЩег-
убий сопфасе т Могуау газЛ РВу-
310]. ап@ Вепау. 1983. Ув. 30. Р. 919-
935.
о|е 7. М. Сепобуре ап ргепайёе] ала
ргетайтя зтезз Ицегась 0 аМесф ада
Белауюгт Ш 1газ//Заепсе. 1965.
Уо1. 150. Р. 1844.
Товпзоп О. Лъе АизтаНап 1ерфозр!гоз$ //
Мед. Т. Аязга|. 1950. \Уо1. 20. Р. 724.
Топез 7. К., Тойпзоп Р. Н. Зупоряз 9
Ие ]асотогрЬз ап годет{5 оЁ Котеа //
Ом. Капз. Ра. Миз. Мабг. НИ.
1965. Ус. 16, М 2. Р. 357-407.
Татутег В. АвеТайиез апй-Вскейыа
сВе; ВаНчз потуео1сиз а Тип!8 еп аш,
а Шеф е! ао 1959 // ВыП. 506. рабо].
ехоё. 1960. Ус. 53. Р. 85-89.
Кайт А-Я. Н., Мизыа А. М. Сеобгар-
са Чзирайой 0 годез 1 зоше
ргоупсез оЁ Штад ИВ геетепсее ю
Фет Ыоюрез // 8101. $1. Вез. 1983.
У. 14. М2 Р. 1-24.
Кесре С. Атоп С. ОМасюгу сиез ап@
ассезвогу оМасфоту БШЪ 1езоп: еЁесф5
оп зехна! ревау1оиг 1 Фе суеНе ета
га\з // Рнуз101. ап Вепау. 1984. Уо/1. 33,
М 1.Р. 45—48.
КепааИ Р. В. Зеазопа] свапдез оЁ зех
таНо т Могуау гаф (ВаНаз погуер1-
сз) рорщаНопз ш \Уа1ез// Г, 209.
1984. У]. 203, М 2. Р. 284—291.
Кепйе К. Е.„ Газагиз А. Воше Е. Р.
е а1. Зе Шзайоп оЁ годеп ап оШег
реэз чзше а зуп Мейс оезбтореп // Ма-
фиге. 1973. Уо|. 244. Р. 105—108.
Кеппеду С. У. Наг4егоротриутт: а пех
ротрвугйз Рот Фе Наг4емап 2]апзо#
бе гаф// Сотар. В!юсВешт. ап Р\уз.
1970. Уо1. 36. Р. 21-36.
Кемег С. В. Роупемап га ргедаМоп оп
пезйпи Гаузап аNoайгоззез ап@ оШег
Рас с зеаЫтаз
// АдЕ. 1967. Уо|. 84,
МЗ.Р. 426—630.
Крап 2, ааа У. 1. Ро1зоп- ава рые
зНупезз 07 Мог Теги Рана Зфанте! Еи-
патро!аз реппапй реппапи УУтопе\-
оп // АррЁ. Епюошо]. ава 2001. 1980.
Уо]. 15, М2. Р. 175—177.
КИрачей 5. Гее Т. Мойз Н. Тве тша-
1егпа! рВегошопе о Фе таб 1е5Ы пя
зоте аззитриопв опде{уше а Вурое-
$13 // РВуз10]. ава Вевау. 1983. \У01. 30.
Р. 539—543.
No.
Ктр О. Г. А теме\ ап4 пуегргеаоп. оЁ
зоше азресёз оЁ Фе. пфапетоег ге-
Тайопзр 1 шаттаа]з ап@ No48 // Рзу-
с1о]. Ву. 1966. Уч. 65. Р. 143-155.
Кит Н. О. Тве отомАй ап4 хата ИИу т
Фе Боду ме! оф Че Мт\хау та
(Миз погуезтсяз) // Апаё. Пес.‘ 1923.
Уо|. 25. Р. 15—18.
Кте Н. 1. ТАМет ртодасНоп ап4 зех та-
Нот газ/ГЫ. 1924.У. 27.Р.337-
366.
Кти Н. Ь. ГМе ртосеззез оЁ \\е огау
Мот\уау гаф Чагше 1еп репегайопз ш
сарНУуЦу // Ашег. Апаё. Меш. 1929. М 14.
Р. 11-10.
КтвН.Ь.ВИмешВшФевтеуNoг-
мау гаф ап@ Ще Гасютз Ша шЙПчепсе-
1 // Ата. Вес. 1935. У]. 63. Р. 325-—
354.
Ктз Н. О. ТМе рхосеззез ш втау Могуау
газ Чагша Точщееп уеатз Ш сариу-
$у // Атег. Апвак Мет. 41939. М 17.
Р. 17-32.
Ктё Н. О., Бопа Азот Н. Н. Те ргосез-
зез ап@ э12е оЁ &Ве Бофу ап отрапв оЁ
Ще отау Могуау таб Фагто {еп сепе-
гаМопз ш сарНуйу // Та. 1929. М 14.
Р. 1—106.
КтЕ М. С. Зита вепегаНзаНот оГ
соп@Нопей {еа’ т гаёз оуег Ише: о]-
{аогу слез ап айдтепаЁ асйуЦу//
1. Сотр. апа Р\у1о]. РэзусВоГ. 1969.
\Уо1. 69. Р. 590-600.
Ктя М. @., Рияег Н. Р., Гивизю Е. Г...
РНетепНа! ргеегепсез Фог ап асИуа-
Ноп Бу Ще одотМегочз сотарагииепь о#
а зПа Ме ох ш {еаг сопашопей апд
па!уе газ // Везам. В1о]. 1975. Уч. 13.
Р. 175—181.
Ктздоп 7. Еа АЙЯсап таапипа]з // Ап
аЙаз о! еуофайов ш Аа. Г.; М. У.,
1974. Уо1. 28. Р. 761.
`
Кигсрфега Е. Весов ИБег @е ВаМепЪе-
КАтарЕапо 31 4ег За ОЪегВаязеп пп
ТаВге 1962 // РНаптепзсии2-МасВг. «Вау-
ег›. 1963. Ва. 16, М 1. $. 37—48.
КатсйВетя Е. ТИеипе 4ез ептор&!зсВеп
Ванепрез{ап4ез
— Чюр!е одег МбеПс\-
Кей? // Найепо1од1е чп 4 Вабепфек&тр-
Гал: (Уоттарзуегат{аШтя 4ез Уе-
телз г Уаязег-, Водеп- ипа ГлаЪу-
о1епе ат 28. ип@ 29. Моуетьег 1968,
КопотгеззпаПе, ВегНп). ЭбаИхжагь, 1969.
8. 167-177.
КирИив Е. Е., Мебитме 7. 0. РобепИа!
го]е оЁ зегШлайой ог зарргезашя гай
рораНопз: А ШМеотейса|! аррга!за! //
ТесNoп. ВаН. 0$ Бер. Аст. 1972. М 1455.
Р. 27.
Кпоггр. И’., Уапта-Реша Т.,Гирзеи М.А.
Эагасфике ап@ Рапсоп оЁ 433 Игоив
Литература
439
риЪезсепзе // Еп4остоюзу. 1970. Уо1. 86.
Р, 1298—1304.
Кпое С. Е. Рвузса[ сотиго! оЁ годете //
Сегеа1 Еоойз Уст. 1980. Уо]. 25, М 12.
Р. 748—749.
Когапуё Г. Лл5зак, Татазу ТУ. Кате-
таз Г. Вевам!опта! ап весихорвуз!0]0-
81са1 аМетр!з No0 ешеаю сештаЁ пег-
уоиз зубеш теспап!5тз гезропз]е
Гот шаегпа| Ъеваутотг // АгсВ. Зех. Ве-
Вау. 1976. Уо|. 5. Р. 503-510.
Ктефз С. 1. А темеу оГ Не СЫМу Вурон-
10518 ог рорШайоп гершаНоп // Сапад.
1. 7001, 1978. Ув]. 56, М 12. Р. 2463—
2480.
Ктесей 7. ТВе \уеашши ремо4 аз а ст
са! ремо@ оЁ аехеюртен& // Розвайа1
4еуе!ортеп& оЁ рпепойуре. Ргарие: Аса-
Четита, 1970. Р. 33.
Ктесйеьзву 7. НегейИагу парате оф «Ву-
роезез» //Т. Сошр. РзусВо]. 1933.
\Уо1, 16. Р. 148—160.
Ктей Ме! В. Н. ТЬе еНесёз оЁ ]асфаНоп оп
бе Пиратаноп оЁ оуа а сопслитен
ргезпапеу ш 1е гаф// Апаё. Вес. 1944.
Уч. 81. Р. 48—65.
„Кисйетий У. У. Зупап горю годен! апа
Фе эшшЙсапсе т {Ме 1тапзти! 33100 0
иМесопз // ТВеотейса! диезНопз о? па-
бота! 1061 91звазев. Ргавие, 1965.
Р. 353—866.
Кийп С. М., Вийег 5. В., УрапьегЕ 5. М.
З@есНуе Чергеззюн оЁ зегита ото
Воттопе догшя таафегпа! дергуайов т
тай рирз// Заепее. 1978. УоЁ 201.
Р. 1034-1036.
Кишабата М. ЭмтизМ А. Тиеаа ПН.
М ему тео Тог {езНпя габ тереПети //
Ар. Ет\отпой. ава 2001. 1970. Уо1. 5,
М 2. Р. 84-94.
Тапаааег М. В., Ипезе В. Е., Сагг 1’. 1.
Везропзез оф зехцаПу ехремепсей апа
паНуе газ {0 с1ез гот гесермуе уз.
попгссерЫуе {ета1ез // Апиа. Геагп. Ве-
‚
Вау. 1977. Уо]. 5, М4. Р. 398—402.
Татзоп С. А ргоееНоп 109% ш писе {ог
1ЧепыНсайов оф 1ерюзр!тозев 1с4его-
ВаетогтВаятае: (\Уе5 1звазе) // Ри.
` НеаМЬ Вер. 1944. Уч]. 56. Р. 1593.
Таитетё Р. `Опе тесыйсаНоп: 1е Загтиа10
ВаИиз погуер1стз ВегК, а 616 101216 А
фот А Вей ОчоН (304 огапа1е) // Вай.
Зос. 2001. Егапсе. 1937. Ув]. 67. Р. 344-—
ТапчеЕ.М.0.,НШТУТ.Е.ЫзоПапа
татта]з о? Мех Сишеа, С@ефез ап4
а]асешф 13]ап4з, 1758—1952. 1: Вти.
Миз. (Маг. Н!5.), 1954. 175 р.
Та Ге С. бцайь ПР. Лазоци@ аз а
Чеететь No ВаНиз погуерсиз // У. 9ю-
ге Рго4. Вез. 1977. Уо|. 13. Р. 23—28.
Ге Рис 7. И’., ЭтИЬ С. А., Тойтзоп К. М.
Папаап-НЕе угизез тот ЧотпезЫс га{з
сарагед п бе ОпИеЯ $+ез
// Атег,
3. Тгор. Мей. ав@ Нур. 1984. Уд. 33,
КNo5.Р. 992-998.
Ге Вис 7. И’, утий С. А., Ртето Е.Р. в
1. [зоаНоп о? а Нащаац-ге]а1е уз
гот ВтаШап гафз ап зегоос1е е%-
Чепсе о{ Из \1Че зргеаЯ д13раноп т
Зои Ашегса // Л. 1985. Уо1. 27, М 4.
Р. 810—815.
Гее Р. No’. УапаЕтата В. 616 С. 7. (Т'.),
Саазек О. С. Раорепез!8 оЁ схрог-
пела! Нашаап Утиз Несов 11 1а-
рогафюгу гайз // АтеВ. У1го]. 1986. Уо1. 88.
Р. 57-66.
ГееТ.М.,ГееС.МойзН.Расат
Пуег су050! ап р|еготопа| ети!510п
т ЧФе гаф// РЬуз10]. ава ВеБау. 41982.
Уо1. 26. Р. 1429-1131.
ее Т. М., Мой: Н. ТВе шамегпа! р\е-
гошопе ап@ 4еохуспоНе ас14 ш г@аноп
40 гаш шуеНп т {Ве рголусапНие габ//
14. 1984а. Уо|. 33. Р. 394-395.
Гее Т. М., Мой: Н. Тье таегпа| рАего-
шопе ап ЧеохусвоНе аа 11 Ме за
У1ма[ оЁ ргемуеапНир газ
// Па. 1984Ъ.
Уо1. 33. Р. 931-935.
Гевепаге В. 1е репр1ешети 4е Ре Еюго-
ра // Мет. Мих. па. No15 пабит. 1966.
Уо1. 41. Р. 213—220.
Те Мазпеп У. 18 рИепотепез оНасдо-
зехие!з све? 1е гаф Ыапс// АгеВ. Зе.
РNoуз!10]. 1952. Уа1. 6. Р. 295-332.
Теоп М. Мыегпа! рИеготопе // РВуз101.
ап Вевау. 1974. У]. 13. Р. 441-453.
Геоп М. Гагу сопбто| 07 тафегпа! рйе-
тошопе ш Пабаной гаф// Та. 1975.
Уо1. 14. Р. 314—319.
Геоп М. Аде5 Г, Соорегзтаёр В. ТЬег-
ша] Итйабоп оЁ шо Тег-уовпар сопбас
т Могуау га // Беуе1ор. РзусвоБо1.
1985. Уо1. 18. Р. 85-105.
Геоп М., СтозКетту Р. С., бтий Ц. Е.
ТВегиаа1 сопго] 9 тоег-уопио сопбас
Ш таз // РВузю0|]. ап Верау. 1978.
Уо1. 21. Р. 798—811.
Геоп М., Се} В. С., Вейзе С. Н. Еза-
Ъ1 35 тепё оЁ рвеготопа] Ъоп4$ апа 41%
сВо1зе 1 уоппе гай Бу ойойг ргеехро-
зе /ТЬ4. 4977. Уо1. 18. Р. 887-
Теоп М. Мой; Н. Маегпа! рВеготопе:
А1стнитайоп Фу ргеувапНайя ато
газ // 1514. 1974. Уд. 7. Р. 265-267.
Геоп М., Мойз ВН. Те деуортеть о
438
Литература
паПу февауше {етае ато гаёз (Ва
{из погуер1ел5) //Т. Сотар. Рзусво]. 1984.
Уч]. 98. Р. ЗМ- 347.
Уапез 7. Геоп М. Реетпитатйз ой таоЩег-
убий сопфасе т Могуау газЛ РВу-
310]. ап@ Вепау. 1983. Ув. 30. Р. 919-
935.
о|е 7. М. Сепобуре ап ргепайёе] ала
ргетайтя зтезз Ицегась 0 аМесф ада
Белауюгт Ш 1газ//Заепсе. 1965.
Уо1. 150. Р. 1844.
Товпзоп О. Лъе АизтаНап 1ерфозр!гоз$ //
Мед. Т. Аязга|. 1950. \Уо1. 20. Р. 724.
Топез 7. К., Тойпзоп Р. Н. Зупоряз 9
Ие ]асотогрЬз ап годет{5 оЁ Котеа //
Ом. Капз. Ра. Миз. Мабг. НИ.
1965. Ус. 16, М 2. Р. 357-407.
Татутег В. АвеТайиез апй-Вскейыа
сВе; ВаНчз потуео1сиз а Тип!8 еп аш,
а Шеф е! ао 1959 // ВыП. 506. рабо].
ехоё. 1960. Ус. 53. Р. 85-89.
Кайт А-Я. Н., Мизыа А. М. Сеобгар-
са Чзирайой 0 годез 1 зоше
ргоупсез оЁ Штад ИВ геетепсее ю
Фет Ыоюрез // 8101. $1. Вез. 1983.
У. 14. М2 Р. 1-24.
Кесре С. Атоп С. ОМасюгу сиез ап@
ассезвогу оМасфоту БШ Ъ 1езоп: еЁесф5
оп зехна! ревау1оиг 1 Фе суеНе ета
га\з // Рнуз101. ап Вепау. 1984. Уо/1. 33,
М 1.Р. 45—48.
КепааИ Р. В. Зеазопа] свапдез оЁ зех
таНо т Могуау гаф (ВаНаз погуер1-
сз) рорщаНопз ш \Уа1ез// Г, 209.
1984. У]. 203, М 2. Р. 284—291.
Кепйе К. Е.„ Газагиз А. Воше Е. Р.
е а1. Зе Шзайоп оЁ годеп ап оШег
реэз чзше а зуп Мейс оезбтореп // Ма-
фиге. 1973. Уо|. 244. Р. 105—108.
Кеппеду С. У. Наг4егоротриутт: а пех
ротрвугйз Рот Фе Наг4емап 2]апзо#
бе гаф// Сотар. В!юсВешт. ап Р\уз.
1970. Уо1. 36. Р. 21-36.
Кемег С. В. Роупемап га ргедаМоп оп
пезйпи Гаузап аNoайгоззез ап@ оШег
Рас с зеаЫтаз
// АдЕ. 1967. Уо|. 84,
МЗ.Р. 426—630.
Крап 2, ааа У. 1. Ро1зоп- ава рые
зНупезз 07 Мог Теги Рана Зфанте! Еи-
патро!аз реппапй реппапи УУтопе\-
оп // АррЁ. Епюошо]. ава 2001. 1980.
Уо]. 15, М2. Р. 175—177.
КИрачей 5. Гее Т. Мойз Н. Тве тша-
1егпа! рВегошопе о Фе таб 1е5Ы пя
зоте аззитриопв опде{уше а Вурое-
$13 // РВуз10]. ава Вевау. 1983. \У01. 30.
Р. 539—543.
No.
Ктр О. Г. А теме\ ап4 пуегргеаоп. оЁ
зоше азресёз оЁ Фе. пфапетоег ге-
Тайопзр 1 шаттаа]з ап@ No48 // Рзу-
с1о]. Ву. 1966. Уч. 65. Р. 143-155.
Кит Н. О. Тве отомАй ап4 хата ИИу т
Фе Боду ме! оф Че Мт\хау та
(Миз погуезтсяз) // Апаё. Пес.‘ 1923.
Уо|. 25. Р. 15—18.
Кте Н. 1. ТАМет ртодасНоп ап4 зех та-
Нотгаз/ГЫ. 1924.У. 27.Р.337-
366.
Кти Н. Ь. ГМе ртосеззез оЁ \\е огау
Мот\уау гаф Чагше 1еп репегайопз ш
сарНУуЦу // Ашег. Апаё. Меш. 1929. М 14.
Р. 11-10.
КтвН.Ь.ВИмешВшФевтеуNoг-
мау гаф ап@ Ще Гасютз Ша шЙПчепсе-
1 // Ата. Вес. 1935. У]. 63. Р. 325-—
354.
Ктз Н. О. ТМе рхосеззез ш втау Могуау
газ Чагша Точщееп уеатз Ш сариу-
$у // Атег. Апвак Мет. 41939. М 17.
Р. 17-32.
Ктё Н. О., Бопа Азот Н. Н. Те ргосез-
зез ап@ э12е оЁ &Ве Бофу ап отрапв оЁ
Ще отау Могуау таб Фагто {еп сепе-
гаМопз ш сарНуйу // Та. 1929. М 14.
Р. 1—106.
КтЕ М. С. Зита вепегаНзаНот оГ
соп@Нопей {еа’ т гаёз оуег Ише: о]-
{аогу слез ап айдтепаЁ асйуЦу//
1. Сотр. апа Р\у1о]. РэзусВоГ. 1969.
\Уо1. 69. Р. 590-600.
Ктя М. @., Рияег Н. Р., Гивизю Е. Г...
РНетепНа! ргеегепсез Фог ап асИуа-
Ноп Бу Ще одотМегочз сотарагииепь о#
а зПа Ме ох ш {еаг сопашопей апд
па!уе газ // Везам. В1о]. 1975. Уч. 13.
Р. 175—181.
Ктздоп 7. Еа АЙЯсап таапипа]з // Ап
аЙаз о! еуофайов ш Аа. Г.; М. У.,
1974. Уо1. 28. Р. 761.
`
Кигсрфега Е. Весов ИБег @е ВаМепЪе-
КАтарЕапо 31 4ег За ОЪегВаязеп пп
ТаВге 1962 // РНаптепзсии2-МасВг. «Вау-
ег›. 1963. Ва. 16, М 1. $. 37—48.
КатсйВетя Е. ТИеипе 4ез ептор&!зсВеп
Ванепрез{ап4ез
— Чюр!е одег МбеПс\-
Кей? // Найепо1од1е чп 4 Вабепфек&тр-
Гал: (Уоттарзуегат{аШтя 4ез Уе-
телз г Уаязег-, Водеп- ипа ГлаЪу-
о1епе ат 28. ип@ 29. Моуетьег 1968,
КопотгеззпаПе, ВегНп). ЭбаИхжагь, 1969.
8. 167-177.
КирИив Е. Е., Мебитме 7. 0. РобепИа!
го]е оЁ зегШлайой ог зарргезашя гай
рораНопз: А ШМеотейса|! аррга!за! //
ТесNoп. ВаН. 0$ Бер. Аст. 1972. М 1455.
Р. 27.
Кпоггр. И’., Уапта-Реша Т.,Гирзеи М.А.
Эагасфике ап@ Рапсоп оЁ 433 Игоив
Литература
439
риЪезсепзе // Еп4остоюзу. 1970. Уо1. 86.
Р, 1298—1304.
Кпое С. Е. Рвузса[ сотиго! оЁ годете //
Сегеа1 Еоойз Уст. 1980. Уо]. 25, М 12.
Р. 748—749.
Когапуё Г. Лл5зак, Татазу ТУ. Кате-
таз Г. Вевам!опта! ап весихорвуз!0]0-
81са1 аМетр!з No0 ешеаю сештаЁ пег-
уоиз зубеш теспап!5тз гезропз]е
Гот шаегпа| Ъеваутотг // АгсВ. Зех. Ве-
Вау. 1976. Уо|. 5. Р. 503-510.
Ктефз С. 1. А темеу оГ Не СЫМу Вурон-
10518 ог рорШайоп гершаНоп // Сапад.
1. 7001, 1978. Ув]. 56, М 12. Р. 2463—
2480.
Ктесей 7. ТВе \уеашши ремо4 аз а ст
са! ремо@ оЁ аехеюртен& // Розвайа1
4еуе!ортеп& оЁ рпепойуре. Ргарие: Аса-
Четита, 1970. Р. 33.
Ктесйеьзву 7. НегейИагу парате оф «Ву-
роезез» //Т. Сошр. РзусВо]. 1933.
\Уо1, 16. Р. 148—160.
Ктей Ме! В. Н. ТЬе еНесёз оЁ ]асфаНоп оп
бе Пиратаноп оЁ оуа а сопслитен
ргезпапеу ш 1е гаф// Апаё. Вес. 1944.
Уч. 81. Р. 48—65.
„Кисйетий У. У. Зупап горю годен! апа
Фе эшшЙсапсе т {Ме 1тапзти! 33100 0
иМесопз // ТВеотейса! диезНопз о? па-
бота! 1061 91звазев. Ргавие, 1965.
Р. 353—866.
Кийп С. М., Вийег 5. В., УрапьегЕ 5. М.
З@есНуе Чергеззюн оЁ зегита ото
Воттопе догшя таафегпа! дергуайов т
тай рирз// Заепее. 1978. УоЁ 201.
Р. 1034-1036.
Кишабата М. ЭмтизМ А. Тиеаа ПН.
М ему тео Тог {езНпя габ тереПети //
Ар. Ет\отпой. ава 2001. 1970. Уо1. 5,
М 2. Р. 84-94.
Тапаааег М. В., Ипезе В. Е., Сагг 1’. 1.
Везропзез оф зехцаПу ехремепсей апа
паНуе газ {0 с1ез гот гесермуе уз.
попгссерЫуе {ета1ез // Апиа. Геагп. Ве-
‚
Вау. 1977. Уо]. 5, М4. Р. 398—402.
Татзоп С. А ргоееНоп 109% ш писе {ог
1ЧепыНсайов оф 1ерюзр!тозев 1с4его-
ВаетогтВаятае: (\Уе5 1звазе) // Ри.
` НеаМЬ Вер. 1944. Уч]. 56. Р. 1593.
Таитетё Р. `Опе тесыйсаНоп: 1е Загтиа10
ВаИиз погуер1стз ВегК, а 616 101216 А
фот А Вей ОчоН (304 огапа1е) // Вай.
Зос. 2001. Егапсе. 1937. Ув]. 67. Р. 344-—
ТапчеЕ.М.0.,НШТУТ.Е.ЫзоПапа
татта]з о? Мех Сишеа, С@ефез ап4
а]асешф 13]ап4з, 1758—1952. 1: Вти.
Миз. (Маг. Н!5.), 1954. 175 р.
Та Ге С. бцайь ПР. Лазоци@ аз а
Чеететь No ВаНиз погуерсиз // У. 9ю-
ге Рго4. Вез. 1977. Уо|. 13. Р. 23—28.
Ге Рис 7. И’., ЭтИЬ С. А., Тойтзоп К. М.
Папаап-НЕе угизез тот ЧотпезЫс га{з
сарагед п бе ОпИеЯ $+ез
// Атег,
3. Тгор. Мей. ав@ Нур. 1984. Уд. 33,
КNo5.Р. 992-998.
Ге Вис 7. И’, утий С. А., Ртето Е.Р. в
1. [зоаНоп о? а Нащаац-ге]а1е уз
гот ВтаШап гафз ап зегоос1е е%-
Чепсе о{ Из \1Че зргеаЯ д13раноп т
Зои Ашегса // Л. 1985. Уо1. 27, М 4.
Р. 810—815.
Гее Р. No’. УапаЕтата В. 616 С. 7. (Т'.),
Саазек О. С. Раорепез!8 оЁ схрог-
пела! Нашаап Утиз Несов 11 1а-
рогафюгу гайз // АтеВ. У1го]. 1986. Уо1. 88.
Р. 57-66.
ГееТ.М.,ГееС.МойзН.Расат
Пуег су050! ап р|еготопа| ети!510п
т ЧФе гаф// РЬуз10]. ава ВеБау. 41982.
Уо1. 26. Р. 1429-1131.
ее Т. М., Мой: Н. ТВе шамегпа! р\е-
гошопе ап@ 4еохуспоНе ас14 ш г@аноп
40 гаш шуеНп т {Ве рголусапНие габ//
14. 1984а. Уо|. 33. Р. 394-395.
Гее Т. М., Мой: Н. Тье таегпа| рАего-
шопе ап ЧеохусвоНе аа 11 Ме за
У1ма[ оЁ ргемуеапНир газ
// Па. 1984Ъ.
Уо1. 33. Р. 931-935.
Гевепаге В. 1е репр1ешети 4е Ре Еюго-
ра // Мет. Мих. па. No15 пабит. 1966.
Уо1. 41. Р. 213—220.
Те Мазпеп У. 18 рИепотепез оНасдо-
зехие!з све? 1е гаф Ыапс// АгеВ. Зе.
РNoуз!10]. 1952. Уа1. 6. Р. 295-332.
Теоп М. Мыегпа! рИеготопе // РВуз101.
ап Вевау. 1974. У]. 13. Р. 441-453.
Геоп М. Гагу сопбто| 07 тафегпа! рйе-
тошопе ш Пабаной гаф// Та. 1975.
Уо1. 14. Р. 314—319.
Геоп М. Аде5 Г, Соорегзтаёр В. ТЬег-
ша] Итйабоп оЁ шо Тег-уовпар сопбас
т Могуау га // Беуе1ор. РзусвоБо1.
1985. Уо1. 18. Р. 85-105.
Геоп М., СтозКетту Р. С., бтий Ц. Е.
ТВегиаа1 сопго] 9 тоег-уопио сопбас
Ш таз // РВузю0|]. ап Верау. 1978.
Уо1. 21. Р. 798—811.
Геоп М., Се} В. С., Вейзе С. Н. Еза-
Ъ1 35 тепё оЁ рвеготопа] Ъоп4$ апа 41%
сВо1зе 1 уоппе гай Бу ойойг ргеехро-
зе /ТЬ4. 4977. Уо1. 18. Р. 887-
Теоп М. Мой; Н. Маегпа! рВеготопе:
А1стнитайоп Фу ргеувапНайя ато
газ // 1514. 1974. Уд. 7. Р. 265-267.
Геоп М., Мойз ВН. Те деуортеть о
440
Литература
{Вс рВеготопа! Бопа п йе ато гаф//
Ты4. 1972. Уо1. 8. Р. 684-688.
Геоп М., Мой; Н. ТЪе деуеюршепь оё
{Те ретотопа] Ъоп4 {1 {Ве ато га% //
ТЫ. 1973. Уд]. 10. Р. 65—67.
Геоп М. Мой: Н. Епаосгше сопио] 9
{Ве тафегпа] рВегошопе ш Фе роз
рагина {ешае таб //РЬузю]. Вевам.
1975. Уд. 10. Р. 65—67.
ГезЦе Р. Н., Уепа Ще; 0.М., Уепаез Г,. 59.
Тие 1е Шу ап@ роруй або эгиасбаке оЁ
Ще гоу габ (Ваз погуе?1си8) т
сотп ГЮКз ап@ зоте о ег ВаБИате //
Ргос. 2001. б0с. Гоп4оп. 1952. У@1. 122,
р61.Р. 187-239.
Герт В., Утетп Е. Маегпа| шиепсез оп
опбюсепу оЁ зисКир ап@ {ее тр гВуИниз
{1 Ве таё// У. Сотр. апа Руузю]? Рзу-
сво]. 1975. Уо1. 89. Р. 7141-7241.
Тепте 5. ЕНесь о# вату Чермуайоп ап@
4е]ауе \еапше оп ауоапсе |еагишя
т Ше аПло гаё// АгеВ. Мечго]. Рзу-
сБ1а'. 1958. Уо]. 79. Р. 241.
Гете 5. Ап епдостте Шеогу о пфап-
Не зишанон // Зиниайоп ш сайу
ИМапсу. No. У.: АшЬгозе, 1969. Р. 45—
63.
Герте 5., МёИлпз В. Е. Ногтопа1 1аЙиеп-
се оп га отбатй2аНоп ш И{аоф габз //
Зс1епсе. 1966. Уо!. 152. Р. 1585.
Гете 5. Онз Г. 5. Тве еМесз оЁ Вапа-
Низ Бе{оге ап4 аНег \уеаптя оп Не те-
3 апсе оё ато таз 10 1Вет 4ергуа-
оп // Сапад. Т. Рзусво|. 1958. Уо/1. 12.
Р. 103—108.
Теияя Р. В, бейчерт 7. А. О\тазоипа
ргодисНоп Ъу ргебпап газ // Верам.
апа МNo ига]. В101. 1982. У\о1. 35, М 4.
Р. 422—425.
фегтап М., Глбегтап 0. Гацазе 4ей-
с1епсу: а сепене шесВазизиа \МеВ ге-
пи|а{ез (Ве Ише о{ уеашия // Атег. Ма-
фиг. 1978. Ув]. 112. Р. 625-627.
Шппаеиз С. Зумеша Мабагае. 10 е4.
Ношнае, 1758. Т. 1. 824 р.
Госкага ПВ. В., Наеге Н. А сотарагзоп
ОЁ гаф ап@ Батзег регод1еИез // Сот-
шип. Вевау. В101. 1969. — Аъа.
М 1169005. Р. 105-108.
Гозоз К. Рау т татита1$ // Зушр. 2001.
Зос. Гопаоп. 1966. М 18. Р. 1-9.
Топе Т. А., Етапз Н. М. ТЬе оезтойз
суше т Ве гаф ап Из аззочафей рНе-
потепз // Мет. Ошху. СаШ. 1922, Уо1. 6.
Р. 1-148.
`
Тоге В. Еогарше ап о Воаг4ши о{
\НЯ Вайоз. погуес1еиз.ш Фе шфапв
епугоптети // Венау. ап Мепга]. В10].
1980. Ус]. 29, М 4. Р. 527-531.
.
Готе В, Раппейу К. Еагта Р. ОЙтазво-
ап4з ргодисе@ Ъу газ ассотшрапу 4е-
стеазез ш питазреыс Невое // Ао
отез. Верау. 1976. Уо|. 2. Р. 175—181.
Гоге В. К. РаппеПу К. Г. Сотрагайуе
ба ез оЁ УПА ап4 дотлезЫс гафз: зоте
АНЙсь ев п 1з9аНио фе еНесфз оЁ
сепофуре ап еп\Чгоптен+
// 1514. 1981.
Уо1. 7. Р. 253—270.
Гоге В. К., 9Иро-Юайети, А. Ровбуеатте
$06121 ехрешепсе ап а@оМ аертеззоп
т газ ВУ, ап Вевау. 1984.
\01. 33, М 4.Р. 571—514.
Готепл К. ТВе сошрагаНуе шефой п за
душа шпае БеВауюнг райегиз // Зушр.
Зое. Ехр. В10|. 1950. Уд. 4. Р. 221-
268.
Готепз К. Р]ауз ап@ уасчит аси ев /!
ГуазИюкь Чапз 1е сотромешет 4ез
апппаох её 4с Готше. Р.: Маззоп,
1956. Р. 633—645.
Готепа К. Оп авотезаюоп. Г.: Мефчеп,
1966. 273 р.
Гогепт К. Оп аратеззюп. М. У. Ващап,
1967. 306 р.
Тон ЛР. Е., Возев ай У. 5. Беуортептё
о тафегла! тезропзУетезз дигте
ртевпапсу шт Фе гаё// Рееги{тап $ оЁ
фапе Берамтог/Е4. В. М. Розе. [., 1969.
Уо1. 4. Р. 61—61.
Гпобапо 0., Готе В. Атитезяюн ап з0-
са] ехреценсе ш Чотшезисаце газ //
7. Сошр. ап РЬузо]. Рэзусво]. 1975.
У]. 88. Р. 917-9283.
Гита М. Веззаптсе No апйсоайи]апт8 т
РептагК // ВаЙев101021е ип ВаИеп-
ЪекАтарРтое: (Уогтавзуетатв
ате4ез
Уегепз т УУаззег, Водеп- ип Бай-
пуйеле ат 28, ип4 29. Моуетфег 1968,
КопртеззваПе, ВегПп). ЭбиИхагь 1969.
5. 27—38.
Гала М. Зома] тесвВап!зт8 ап осла]
вртисфигеЧп газ ап@ ти1се // Псой. Вой.
1975. М 19.Р. 255-260.
-
Гата М. Ме\у тодепис@Чез ата апН-
соарпап-гез1{апсе газ ап писе //
ВиП. ОЕРР. 4977. Уч]. 7, М2. Р. 503—
508.
Тудей К. Бу В. Г. Ма!е гаф одог рге-
Тегепсез {ог Ёета!е итше аз а апсНоп
0 зехиа] ехреепсе, игпе азе ап4 111-
пе з01г8 // Ногтопе Вервах. 1972. У]. 3.
Р. 205-242.
Маскау-бт А., Гат О. в. Рзеглита-
Чоп о{ офоитз ош этеззей газ Ъу
поп-3теззе газ // РВуз10]. апа Вевау.
1980. Уч. 24. Р. 669—704.
МасКкау-5ит А., Гате РБ. 6. ТЬе зоотсе8
„
Литература
444
0# ойогз 1тош этеззей газе // П14. 1984.
Уо1. 27. Р. 541—513.
Мас Поп А. В., СШ С. Е. Те оссогапсе
эп си Нсапсе оЁ годепие!Че-гезэ5ап-
св м \Ше У. 0. К.// Зюогаве Рго4. Рез
Сопуг. 1987. М 37. Р. 89—95.
МаШопх М. Зиг 1ез зоцгсез 4е сотфапи-
паНо0$ раг 1ез 1ерюзриез еп АЯ1аие
Ча МNo ога // 7лзеВт. Тторепшей. ип4 Рата-
зо. 1965. Ва. 16, М 3. $. 291-298.
МаШоих М., Вепгапе М. Азремз ер4е-
11010914063 4апз Та гертоп 4е Мекпез //
Магое 164. 1970. Ус. 50, М 537.
Р. 442—444.
Маттаге Г. Т., Мой: Н. Зеглм ргоасип
1еуе!5 ап уайта| сусу ш сопсауеа-
%е4 апа ]абацие {ета]е гай // Рвуз1ю1.
ап@ Верах. 1978. У]. 24. Р. 525-528.
Матг 1. No, ГИйяюоп Г. 6. Зоя] аНасЬ-
пепф М газ Бу офог ап@ аое// Ве-
Вау!юг. 1969, Уа. 33. Р. 277—282.
Мать В. Е., Ношага У’. Е. Мех регзреси-
уез т тодепф аю4 шапциа! сопго] //
Ргос. Зг4 Пцеги. Ыо4еста4. зушр., Ктр-
звоп, 1975. Т., 1976. Р. 317—329.
Матзй В. Е., Хаскзоп 7. В. Епугоптеп-
{а1 сопло] оР таз. Рё 2//Резь Сом.
1978. Ув. 46, No 9. Р. 26—29, 40.
МагзйаЙ 7. Т. ЕатЙу Магае: газ апа
писе // Мапа! оЁ ТваПапа. ВапоКок:
дос. Сопзегу. УУ/ПаШе, 1977. Р. 396—
На.
Магип Г. Т., АNoетз 7. В. Тазце аустаоп
$0 тофегз шИЕ; Ще аве теме го]е
0 поташа 11 аси5 НЮп ап@ ехргезаоп
ОТ а 1еагпей аззосайоп //Т. Сотр. ав4
: Ру. РзусВо1. 1979. Уд]. 93. Р. 430-—
МагитеЕ Г., Очаги В., Соитоф М. Звазо-
па] Ътеедтр: сПапоез 11 сопада! ас-
УНу // Аба 700]. {епп. 1984. М 174.
Р. 157—163.
Мауег А. Б., Позепой 7. 5. Опюрепу
07 тафегпа] ЪеНа\у!оиг 1 Ве 1афогайогу
га:еа[уорит18No0274аузо1а
уония // Реуеор. — РэусвоЫо1. 1979.
У]. 12. Р. 407-424.
Мауег А. Р., Возеп Ман 7. 5. Ногтопа!
бегасНоп УИ зЫпиИ из ап зИлаНо-
па| Гасбогз 11 Ше п аНоп оЁ таегпа|
Бева\1опг. п попртеспатф газ // {. Сотр.
ап@ Рвузо]. Рэзусро]. 1980. Уо|. 94.
Р. 1049—1059.
Мауег А. Р., ВозепМан 1. 5. Ртерагилт
спаппез м шаегпа| гезропз{уепез$ апа
пезф Чеепзе 1т Ме гаё// У. Сотр. Рзу-
сво]. 1984. Уч. 98. Р. 177—188.
Ма У: Сытя. Еаита НеЙопоуапеа:
МаштиаНа. Нагро, 1986. 520 р.
МсВмае С. А вепега] ЧПеогу оЁ зос1а1
отраштайоп ап4 фераз!1отг // Ошу. Оце-
01851. Рар. Уеё. Зе. 1964. Уд]. 1, М 2,
Р. 75-110.
ИсСиюоск М. Зосла] сопио! 07 Ве оча-
Нап суе ап Фе РапсНоп оЁ оезиав
зупевгопу // Атпег. 7001. 1984. У0]. 21,
М 1.Р. 243-256.
МеСитоск М. Модацоп оЁ Пе оезтопз
су4е Бу рвеготопез {тот ртецпапь апа
асан Гета]ез // В10]. Вергод. 1983а.
Уо1. 28. Р. 823—829.
МеСТимоск М. К. Зупсвтотия очамап
ав4 048 суШез Бу 1етае рВегошо-
пез // СВешуса|] 3101а]3 1 уемергаез,
М. У. Г: Асай. ртезз, 1983. Уо|. 3.
Р. 159-178.
МсСИтоск М. К. Реготопа! гераНоп
о ФНе оуатап суфе: епеВапсетень зир-
ргеззюп ап@ зупейгопу // РНеготопех
ап гергодисНоп ш шашша. М: У.
Асад. ргезз, 1983с. Р. 143-143.
Мераяа С. С., Мооа Е. М. Воортса[
азресёз о! пгЪап га сошто] // «НЗМНА»
Неана Вер. 1972. Ус. 87, No1. Р. 47-
Меронвай ., Мероивай К. Миез оп
т3Нпсё ап пиеШоепсее Ш газ ао
са{з//Т. Сотр. Рзусво]. 1972. Уи, 7.
Р. 45-175.
Мепиозр Т. К., Вагна@а Н. 7., Сеуег Г. А.
(4тазош1е уосаНзаНотв ТасИНаю зе-
хпа|! Берах1оцг оЁ {ета!е гафз // Мафате.
1978. Уо1. 272. Р. 163—164.
Меапеу М. 7, Звшат 7. А 4езст!риуе
ЗйзЧу оф з0с!а1 Чеуеоршешь шт Ше га
(Ваз — потуес!сиз) // Атиш. Вевау.
198 1а. Ув]. 29, М 1.Р. 34—45.
Меапеу М. 1., пешатм Т. Меопайа1 апато-
вепз п иепсе {Ве зоба! рау оЁ рге-
рафезсеш, гай // Ногтопе Верау. 1981Ъ.
\Уо1. 15. Р. 197-243.
Меапеу М. 1. У1ешам Т., Веану ТУ. У.
Тве шИпепсе о? 2Тасосогысоез дигио
Пе пеопабаГ рег!о4 оп \Noе 4еуеортел+
< рау-ИеВЫте Ш Могуау гаф рирз //
П14. 1982. Уо]. 16. Р. 475.
Меапеу М. 71, 5б1ешат УТ, Рошт Р.,
МсЕшеп В. 5. Зехиа! аНегеннанов о!
306181 р1ау 11 таф рирз 13 шеджед
Бу ШШе пеопаа! апдгосеп-гесерюг зуз-
Фет // Мецгоеп4ост!по]ору. 1983. Уо1. 37.
Р. 85—90.
Меейап А. Р. Ваз ап@ пысе; Твеш Бо-
Ту ап сопбго|. Еаёф СтшяеаЯ, 1984.
р.
Мепа Е. В@еазе оГ ргоасНи Ш гафз Бу
ехфегогесерИуе зНтшаМоп зепзогу`зН-
440
Литература
{Вс рВеготопа! Бопа п йе ато гаф//
Ты4. 1972. Уо1. 8. Р. 684-688.
Геоп М., Мой; Н. ТЪе деуеюршепь оё
{Те ретотопа] Ъоп4 {1 {Ве ато га% //
ТЫ. 1973. Уд]. 10. Р. 65—67.
Геоп М. Мой: Н. Епаосгше сопио] 9
{Ве тафегпа] рВегошопе ш Фе роз
рагина {ешае таб //РЬузю]. Вевам.
1975. Уд. 10. Р. 65—67.
ГезЦе Р. Н., Уепа Ще; 0.М., Уепаез Г,. 59.
Тие 1е Шу ап@ роруй або эгиасбаке оЁ
Ще гоу габ (Ваз погуе?1си8) т
сотп ГЮКз ап@ зоте о ег ВаБИате //
Ргос. 2001. б0с. Гоп4оп. 1952. У@1. 122,
р61.Р. 187-239.
Герт В., Утетп Е. Маегпа| шиепсез оп
опбюсепу оЁ зисКир ап@ {ее тр гВуИниз
{1 Ве таё// У. Сотр. апа Руузю]? Рзу-
сво]. 1975. Уо1. 89. Р. 7141-7241.
Тепте 5. ЕНесь о# вату Чермуайоп ап@
4е]ауе \еапше оп ауоапсе |еагишя
т Ше аПло гаё// АгеВ. Мечго]. Рзу-
сБ1а'. 1958. Уо]. 79. Р. 241.
Гете 5. Ап епдостте Шеогу о пфап-
Не зишанон // Зиниайоп ш сайу
ИМапсу. No. У.: АшЬгозе, 1969. Р. 45—
63.
Герте 5., МёИлпз В. Е. Ногтопа1 1аЙиеп-
се оп га отбатй2аНоп ш И{аоф габз //
Зс1епсе. 1966. Уо!. 152. Р. 1585.
Гете 5. Онз Г. 5. Тве еМесз оЁ Вапа-
Низ Бе{оге ап4 аНег \уеаптя оп Не те-
3 апсе оё ато таз 10 1Вет 4ергуа-
оп // Сапад. Т. Рзусво|. 1958. Уо/1. 12.
Р. 103—108.
Теияя Р. В, бейчерт 7. А. О\тазоипа
ргодисНоп Ъу ргебпап газ // Верам.
апа МNo ига]. В101. 1982. У\о1. 35, М 4.
Р. 422—425.
фегтап М., Глбегтап 0. Гацазе 4ей-
с1епсу: а сепене шесВазизиа \МеВ ге-
пи|а{ез (Ве Ише о{ уеашия // Атег. Ма-
фиг. 1978. Ув]. 112. Р. 625-627.
Шппаеиз С. Зумеша Мабагае. 10 е4.
Ношнае, 1758. Т. 1. 824 р.
Госкага ПВ. В., Наеге Н. А сотарагзоп
ОЁ гаф ап@ Батзег регод1еИез // Сот-
шип. Вевау. В101. 1969. — Аъа.
М 1169005. Р. 105-108.
Гозоз К. Рау т татита1$ // Зушр. 2001.
Зос. Гопаоп. 1966. М 18. Р. 1-9.
Топе Т. А., Етапз Н. М. ТЬе оезтойз
суше т Ве гаф ап Из аззочафей рНе-
потепз // Мет. Ошху. СаШ. 1922, Уо1. 6.
Р. 1-148.
`
Тоге В. Еогарше ап о Воаг4ши о{
\НЯ Вайоз. погуес1еиз.ш Фе шфапв
епугоптети // Венау. ап Мепга]. В10].
1980. Ус]. 29, М 4. Р. 527-531.
.
Готе В, Раппейу К. Еагта Р. ОЙтазво-
ап4з ргодисе@ Ъу газ ассотшрапу 4е-
стеазез ш питазреыс Невое // Ао
отез. Верау. 1976. Уо|. 2. Р. 175—181.
Гоге В. К. РаппеПу К. Г. Сотрагайуе
ба ез оЁ УПА ап4 дотлезЫс гафз: зоте
АНЙсь ев п 1з9аНио фе еНесфз оЁ
сепофуре ап еп\Чгоптен+
// 1514. 1981.
Уо1. 7. Р. 253—270.
Гоге В. К., 9Иро-Юайети, А. Ровбуеатте
$06121 ехрешепсе ап а@оМ аертеззоп
т газ ВУ, ап Вевау. 1984.
\01. 33, М 4.Р. 571—514.
Готепл К. ТВе сошрагаНуе шефой п за
душа шпае БеВауюнг райегиз // Зушр.
Зое. Ехр. В10|. 1950. Уд. 4. Р. 221-
268.
Готепз К. Р]ауз ап@ уасчит аси ев /!
ГуазИюкь Чапз 1е сотромешет 4ез
апппаох её 4с Готше. Р.: Маззоп,
1956. Р. 633—645.
Готепа К. Оп авотезаюоп. Г.: Мефчеп,
1966. 273 р.
Гогепт К. Оп аратеззюп. М. У. Ващап,
1967. 306 р.
Тон ЛР. Е., Возев ай У. 5. Беуортептё
о тафегла! тезропзУетезз дигте
ртевпапсу шт Фе гаё// Рееги{тап $ оЁ
фапе Берамтог/Е4. В. М. Розе. [., 1969.
Уо1. 4. Р. 61—61.
Гпобапо 0., Готе В. Атитезяюн ап з0-
са] ехреценсе ш Чотшезисаце газ //
7. Сошр. ап РЬузо]. Рэзусво]. 1975.
У]. 88. Р. 917-9283.
Гита М. Веззаптсе No апйсоайи]апт8 т
РептагК // ВаЙев101021е ип ВаИеп-
ЪекАтарРтое: (Уогтавзуетатв
ате4ез
Уегепз т УУаззег, Водеп- ип Бай-
пуйеле ат 28, ип4 29. Моуетфег 1968,
КопртеззваПе, ВегПп). ЭбиИхагь 1969.
5. 27—38.
Гала М. Зома] тесвВап!зт8 ап осла]
вртисфигеЧп газ ап@ ти1се // Псой. Вой.
1975. М 19.Р. 255-260.
-
Гата М. Ме\у тодепис@Чез ата апН-
соарпап-гез1{апсе газ ап писе //
ВиП. ОЕРР. 4977. Уч]. 7, М2. Р. 503—
508.
Тудей К. Бу В. Г. Ма!е гаф одог рге-
Тегепсез {ог Ёета!е итше аз а апсНоп
0 зехиа] ехреепсе, игпе азе ап4 111-
пе з01г8 // Ногтопе Вервах. 1972. У]. 3.
Р. 205-242.
Маскау-бт А., Гат О. в. Рзеглита-
Чоп о{ офоитз ош этеззей газ Ъу
поп-3теззе газ // РВуз10]. апа Вевау.
1980. Уч. 24. Р. 669—704.
МасКкау-5ит А., Гате РБ. 6. ТЬе зоотсе8
„
Литература
444
0# ойогз 1тош этеззей газе // П14. 1984.
Уо1. 27. Р. 541—513.
Мас Поп А. В., СШ С. Е. Те оссогапсе
эп си Нсапсе оЁ годепие!Че-гезэ5ап-
св м \Ше У. 0. К.// Зюогаве Рго4. Рез
Сопуг. 1987. М 37. Р. 89—95.
МаШопх М. Зиг 1ез зоцгсез 4е сотфапи-
паНо0$ раг 1ез 1ерюзриез еп АЯ1аие
Ча МNo ога // 7лзеВт. Тторепшей. ип4 Рата-
зо. 1965. Ва. 16, М 3. $. 291-298.
МаШоих М., Вепгапе М. Азремз ер4е-
11010914063 4апз Та гертоп 4е Мекпез //
Магое 164. 1970. Ус. 50, М 537.
Р. 442—444.
Маттаге Г. Т., Мой: Н. Зеглм ргоасип
1еуе!5 ап уайта| сусу ш сопсауеа-
%е4 апа ]абацие {ета]е гай // Рвуз1ю1.
ап@ Верах. 1978. У]. 24. Р. 525-528.
Матг 1. No, ГИйяюоп Г. 6. Зоя] аНасЬ-
пепф М газ Бу офог ап@ аое// Ве-
Вау!юг. 1969, Уа. 33. Р. 277—282.
Мать В. Е., Ношага У’. Е. Мех регзреси-
уез т тодепф аю4 шапциа! сопго] //
Ргос. Зг4 Пцеги. Ыо4еста4. зушр., Ктр-
звоп, 1975. Т., 1976. Р. 317—329.
Матзй В. Е., Хаскзоп 7. В. Епугоптеп-
{а1 сопло] оР таз. Рё 2//Резь Сом.
1978. Ув. 46, No 9. Р. 26—29, 40.
МагзйаЙ 7. Т. ЕатЙу Магае: газ апа
писе // Мапа! оЁ ТваПапа. ВапоКок:
дос. Сопзегу. УУ/ПаШе, 1977. Р. 396—
На.
Магип Г. Т., АNoетз 7. В. Тазце аустаоп
$0 тофегз шИЕ; Ще аве теме го]е
0 поташа 11 аси5 НЮп ап@ ехргезаоп
ОТ а 1еагпей аззосайоп //Т. Сотр. ав4
: Ру. РзусВо1. 1979. Уд]. 93. Р. 430-—
МагитеЕ Г., Очаги В., Соитоф М. Звазо-
па] Ътеедтр: сПапоез 11 сопада! ас-
УНу // Аба 700]. {епп. 1984. М 174.
Р. 157—163.
Мауег А. Б., Позепой 7. 5. Опюрепу
07 тафегпа] ЪеНа\у!оиг 1 Ве 1афогайогу
га:еа[уорит18No0274аузо1а
уония // Реуеор. — РэусвоЫо1. 1979.
У]. 12. Р. 407-424.
Мауег А. Р., Возеп Ман 7. 5. Ногтопа!
бегасНоп УИ зЫпиИ из ап зИлаНо-
па| Гасбогз 11 Ше п аНоп оЁ таегпа|
Бева\1опг. п попртеспатф газ // {. Сотр.
ап@ Рвузо]. Рэзусро]. 1980. Уо|. 94.
Р. 1049—1059.
Мауег А. Р., ВозепМан 1. 5. Ртерагилт
спаппез м шаегпа| гезропз{уепез$ апа
пезф Чеепзе 1т Ме гаё// У. Сотр. Рзу-
сво]. 1984. Уч. 98. Р. 177—188.
Ма У: Сытя. Еаита НеЙопоуапеа:
МаштиаНа. Нагро, 1986. 520 р.
МсВмае С. А вепега] ЧПеогу оЁ зос1а1
отраштайоп ап4 фераз!1отг // Ошу. Оце-
01851. Рар. Уеё. Зе. 1964. Уд]. 1, М 2,
Р. 75-110.
ИсСиюоск М. Зосла] сопио! 07 Ве оча-
Нап суе ап Фе РапсНоп оЁ оезиав
зупевгопу // Атпег. 7001. 1984. У0]. 21,
М 1.Р. 243-256.
МеСитоск М. Модацоп оЁ Пе оезтопз
су4е Бу рвеготопез {тот ртецпапь апа
асан Гета]ез // В10]. Вергод. 1983а.
Уо1. 28. Р. 823—829.
МеСТимоск М. К. Зупсвтотия очамап
ав4 048 суШез Бу 1етае рВегошо-
пез // СВешуса|] 3101а]3 1 уемергаез,
М. У. Г: Асай. ртезз, 1983. Уо|. 3.
Р. 159-178.
МсСИтоск М. К. Реготопа! гераНоп
о ФНе оуатап суфе: епеВапсетень зир-
ргеззюп ап@ зупейгопу // РНеготопех
ап гергодисНоп ш шашша. М: У.
Асад. ргезз, 1983с. Р. 143-143.
Мераяа С. С., Мооа Е. М. Воортса[
азресёз о! пгЪап га сошто] // «НЗМНА»
Неана Вер. 1972. Ус. 87, No1. Р. 47-
Меронвай ., Мероивай К. Миез оп
т3Нпсё ап пиеШоепсее Ш газ ао
са{з//Т. Сотр. Рзусво]. 1972. Уи, 7.
Р. 45-175.
Мепиозр Т. К., Вагна@а Н. 7., Сеуег Г. А.
(4тазош1е уосаНзаНотв ТасИНаю зе-
хпа|! Берах1оцг оЁ {ета!е гафз // Мафате.
1978. Уо1. 272. Р. 163—164.
Меапеу М. 7, Звшат 7. А 4езст!риуе
ЗйзЧу оф з0с!а1 Чеуеоршешь шт Ше га
(Ваз — потуес!сиз) // Атиш. Вевау.
198 1а. Ув]. 29, М 1.Р. 34—45.
Меапеу М. 1., пешатм Т. Меопайа1 апато-
вепз п иепсе {Ве зоба! рау оЁ рге-
рафезсеш, гай // Ногтопе Верау. 1981Ъ.
\Уо1. 15. Р. 197-243.
Меапеу М. 1. У1ешам Т., Веану ТУ. У.
Тве шИпепсе о? 2Тасосогысоез дигио
Пе пеопабаГ рег!о4 оп \Noе 4еуеортел+
< рау-ИеВЫте Ш Могуау гаф рирз //
П14. 1982. Уо]. 16. Р. 475.
Меапеу М. 71, 5б1ешат УТ, Рошт Р.,
МсЕшеп В. 5. Зехиа! аНегеннанов о!
306181 р1ау 11 таф рирз 13 шеджед
Бу ШШе пеопаа! апдгосеп-гесерюг зуз-
Фет // Мецгоеп4ост!по]ору. 1983. Уо1. 37.
Р. 85—90.
Меейап А. Р. Ваз ап@ пысе; Твеш Бо-
Ту ап сопбго|. Еаёф СтшяеаЯ, 1984.
р.
Мепа Е. В@еазе оГ ргоасНи Ш гафз Бу
ехфегогесерИуе зНтшаМоп зепзогу`зН-
442
Литература
ти шуо]уе4
// Ногиопе ВеЪау. 1971.
Уи. 2. Р. 107-116.
Мепа Т., Стозрепог С. Е. ЕНесв оЁ пит-
Ъег оЁ рирз ироп засКЫпрутаисе@ Та|
ш римйату ргоасЫт сопсептаНоп ап4
шИЕ еесйоп т \е га+ // Еп4ост!пло]оду.
1968. Уо1. 87. Р. 623—626.
Мепдоза 5. Р., бое С. Г, Этотет-
тап 7. Р. её а]. Еипсопа] сопзефаеп-
сез оЁ аМасВтаеть а сошратзоп о? мо
зрес1ез
// Майегпа! чепсез оп еау
Ъеваутоит/Е4. У. Р. Зшоегктап,
В. \У. Вей. Т.., 4980. Р. 201—210.
Мепжез 7. Г., )епиз Е. Нап@оок оё МNo%
Сишеа годетз. Уаи, 1979. 68 р.
Мегсег О., Мегаег М., Вопва Т., ЁКотз-
ег Р. Бееиштайот да 4еБиё 4е 1а
рифеме сБез 1е гаф та!е её ЁещеНе
(Вамаз погуер1сиз): АррНсаймот & Т6-
{п4е 4ез зомеВез Зргарие— Ба\еу, У
маг еь Гопо—Еуапя // $с1. 1еасВ. апп.
]ар. 1982. Т. 7, М 3. Р. 205—215.
Мезе; Т. С., Возеп 7. Бопыпапсе БеВа-
уг аз а леноп оЁ има Ше зИти-
1айоп Чп Ме гаЁ// Агсв. Сеп. РвусВо1.
1960. Уч. 3. Г. 58—56.
МШег С. 5. ТЬе пепее паше оЁ Ще
Вопзе гаё// Ргос. В. 80е. УМазй. 1910.
Уи1. 23. Р. 57—60.
Мшег 6. 5. 14% оф Мой Атетсап те-
сепф шапииа1$// 0$ Маё Миз. Ва.
1923. М 128. Р. 32.
МШег М. ВергодосНоп т ие Вго\уп газ //
Ашег. Мабаг. 1911. Уч. 45, М 3. Р. 623--
635.
Мтейе Н. Р. Тербозртомз ш Воде
ап@ Мопго0$ оп е 13ап@ оЁ Нажуай //
Ат. 7. Тторр. Мед. Нуз. 1964. Уо1. 13.
Р. 821-832.
Мзоппе Х. ВерагНМов веостар еле ас-
{чиеПе 4е ВаНиз тайлз Ыпие, 1758 ©
4е ЦПамаз погуес!сиз Вегкеппосце. 1769
вп Ттап // ВаЙ. Газ. В. За. Маё Ве].
1956. Ус. 32, М 49. Р. 4М.
Мзоппе Х. Боппесз зиг 1е5 топвеогз
Чип Гоуег 4е резе еп // Вы. 19,
В. $1. Ма. Вею. 1959. Уо1. 35, М 17.
Р. 1-32.
М1зоппе Х. Апаузе гообеортарЫдие 4е$
тапниИегез 4е ГТтап // Мет. 115. гоу.
361. пафмг. Ве. бег. 2. 1960. Газе. 59.
Р. 157.
М:5оппе Х. Ай\сав ава 14о-АлэбгаНап
Мигае: Еуошопагу 1теп4з // Зе1. 2001.
1969. М 172. Р, 219.
Мисье 2. Ехрегииепз оп пеорцоМа т
\Па ап@ 1арогафогу габз: А говуаNoаа-
Чоп //Т. Сотр. ава Рпузю РзусНо].
1976. Уо. 90. Р. 190—197.
Морату В. Мзйта 5. ЕНесфз оЁ в0с1а|
1зоайоп ап ада Вап@ те оп ореп
Бе! етонопаШу о аПтпо газ //
Т. Рэусво!. Вез. 1984. Ус. 28, М 1.
Р. 52-58.
у
Мой Н. Парги ти: Ешритса: Базз
ап@ ‘Меотейса! ап сапсе
// РзусВо1.
Ви|. 1960. Уо1. 57. Р. 291.
Мой Н., СеЙег Б., Герт В. Мабегра]
рекам!отг ш бе ю!аПу шаттесфот!-
тей гаф//У. Сошр. ав@ Рвуз101. РзусВо!.
1967. Уо]. 64. Р. 225—229.
Мой» Н. ГещаМ Г,., Воифапа Б. Рго]оп-
РаНоп оЁ рЬегопзова} еп оп т Ме
таегпа| га // РВуз101. ап@ Вевах. 1974.
У\1. 12. Р. 409—412.
Мой: Н., Геоп М. Зитиа$ сопо! 9
{Ве шабгпа| рВеготопе 1 фе 1асба-
Моб гаф// ТЯ. 1973. Уд. 10. Р. 69-
71.
Мойз Н. Гат М., Геоп М. Митап М.
Ногтопва} тачсНоп оЁ шаегиа! БеБа-
уоот а Че оуапесюшией паШрагоп5
газ // Па. 1970. Ув. 5. Р. 1378-1375.
Мооге С. Г. Ап оМабюгу Базз Бог табог-
па! Чзеглитаноп 0{ зех оЁ оЁзрета
{1 таф// Апиа. Вефау. 1981. Уой. 29.
р. 383—386.
Мооте С. Г. Майегпа! сопйрийот$ ю Фе
Чеуе]ортетв оЁ тазсиНие зехиа! Бева-
мог ш \\е ГаБогафогу газ // Реуор.
РзусвоБ1о1. 1984. Ув. 17. Р. 347-356.
Мооге С. Г. Зех @егепсез ш игтагу
офогз ргодисе4 Ъу уопля 1афогацюгу газ,
Вайиз потуер1сиз // 7. Сотшр. Рзусво|.
1985. Уо]. 99, М 3. Р. 336--344.
Мотца С. Н., Мазнига У., Мопкаша 5. Н.,
Киатита Т. Апе Череп4етф 1таптзти1$-
оп 0 Нетогтвасе Геуег \ИВ гепа|
зупагоше (НЕВ$) уаз ш газ // АтеЙ.
У!го|. 1985. Уо|. 85. Р. 145—149.
Моншей К., 5ю Т., Тзистуа К. ТЯНе-
тепсе ш апушо ас сотроз оп о? ве-
ги 1тапзегыт атопе уамопя $рес!ез
оЁ{ ВаИлз // Аппи. Вер. Маб. зе. бепев. '
Тар. 1971. Ув]. 24. Р. 34-86.
Мотта М. Теуе! о? тафегиа| згезз диття
1х0 ргеспапсу рег!0з оп гаф оНзргите
Берауточт // Рвуспопош. 561. 1965. Уо|. 3.
Р. 7.
Могтзов В. В, Гидыязоп Н. И’. Плзете-
штайоп Бу газ оЁ сопзресШе одогз
оЁ тезгаг@ ап4 повтемат@ // Заепсе. 1970.
Уо1. 167. Р. 904—905.
Миг А. С. Уохтаизземаитееп Е ете
хиКзате Мавсйегьекйтрие ип Ье-
Бепзиние/Ъейчеь // 3\%155 Роо4. 1983.
\Уо1. 3, М 11. Р. 21-27.
Мигашы Е. Г, 08 И. Опзатайед мах
„
Литература
443
вет 11 Ще ПВаг4егап о]ап@ оЁ Ще
Раф// (кет. ап@ Р|уз. ТА Ча. 1974.
\о1. 18. Р. 155-158.
Миззег С. ©. Ертуз Бепбце(епз13, а сот-
розЦе, ап4 опе тоореоетарме хех о!
га\ ап поизе Гааптаз ш Фе РЫШрр:-
пез ап@ СееЪез // Атег. Миз. Мочи.
1977. М 2624. Р. 1-15.
Миззег С. С. А пем оепиз оЁ аогеа!
га Нош \мезр Фауа, Ттдопеза// 7001.
уегВ. 1981а. М 189. Р. 1-35.
М5ег С. С. ВезаЦз оЁ {\е Агено4 сх-
ред! оп, М 105: МNo1ез оп зучетаНсь
о т4о-Ма]ауап ши гойепз, апа
913 оп5 0 пе\ вепега ап@ зрес1ез
Пот Сеуоп, Зшазуезр ап 4Ве РЫШр-
р1тез
// Ва. Ашег. Миз. Маг. [15
1981Ъ. Уо1. 168, агё. 3. Р. 225—834.
Миззег С. С. Тпе лапе хаб оГ Еогез апа
Из геауез еазё оГ Вотопео апа Вай //
ТЯ. 1981с. Уо|. 169, ак. 2. Р. 617—176.
Миззег (. С. ТЬе Тнлй газ // Модеги
Опабегпагу тезеагсВ т бои еазь Аза.
1982. Уч. 7. Р. 65-85.
Миззег С. С., Воеай М. А пе\ репиз о!
шим гофете {го Ме Котойдо 15апаз
ш Мизацеперага//7. Матша|. 1980.
Уо1. 61. Р. 395—413.
Миззег ((. С., Сайра О. Везиз оф Атей-
Ъо14 ехре@ #015, М. 106: ТаепнНез оЁ
таз Нош РШаи Магата апа ощфег
13ап9з ой Еа$ Вотопео // Атег. Мив.
МNo чи. 1982. М 2726. Р. 30.
Миззег (. (., Неапеу 1. В. Риррше
Ваз: А пех. зрес!е5 ош Ще Чиа
дтомрааво ТЫ. 1985. No 2818. Р. 1-
Миззег С. С. МешсотЬ С. Мау!ап пт-
г143 ап Фе п1ап6 таф о Зишайта //
ВоН. Ашег. Миз. Мабт. Не 1983.
У\о1. 174, ат 4. Р. 327-598.
Миззег С. б., Мешсоть С. Бейт юпз 9
Госьтезо Вашиз 1озеа апд а печ зре-
©1ез {том Уюсбташ
// Ашег. Миз. Моуи.
1985. М 28144. Р. 1-32.
Муйутам А. Ап вау арртоасЬ No0 а
га-тее юут ш Ешщана // Уетет Уаз
зег-, Во@еп- ипа Тл фур. 1969. М 32.
_ 8 161—162.
Мезрте А. Тагра А., Авозт М., Све-
еп В. пасе ае иЧезамой рог Терю-
зргаз еп 1а$ габаз (Вале погуе01с11$}
461 та{а4дего тип!сра! 4е ЗапНаго,
СВНе // Во]. Погм. рагазй. Се. 1951.
Уо1. 6. Р. 6-7.
МNoМею С. 4е. Ч поуо тефюйо рага сот-
Бафе а0з гоедотез; паз Исасао сош [аз-
Гео 4е аТатито // Вююг1со. 1980. Уо1. 46,
М 1/2. Р. 3-9.
Мепраиз И’. Птзаснеп ип@ Стео2еп уоп
Карела! еипоеп апесЬ ВаИеп // Апя.
Таг ЭсвааЙтозк. 1957. Ва. 30, М 6,
5. 81—88.
Мсйо[30п А. 7Т., ИУ’атгпег Б. ИУ. ТЬе го-
4ет1з 0! Мех Сяедоша //Т. Маптаа|.
1953. У. 34, М2. Р. 168-179.
Мсоа14е; No. ЭЕт Пр. 2. ТЛрй9 с1авз
сошрозоп о{ затр тот уатопз зре-
с1ез ап апаютиса! зНез //Т. Атег. ОП
Сет. З0с. 1965. \0]. 42. Р. 694-702.
МЁкагт Т. Сотрагамуе спепузгу оЁ з6-
из // Т. Тауезё. Бегшафю]. 1974. Уд]. 62.
Р. 257—267.
Морис Н. ЗриосВаеа 1еетопаетоггВа-
ае ш Атегсап \%14 газ апа Из ге-
1айоп \Noю Ше Тарапезе ап@ Еигореап.
тат /1. Ехр. Мей. 1947. Уо1. 25.
Мото Е. ОИтазоии@ Ш уопие тодетив.
2. Спапвез И аве ш ато газ//
длит. Вевау. 1968. Ув]. 16, М 1. Р. 129-
134.
Мото Е. Те опзеф ап@ Авуеортене о{
таетпаР Фепауютг 1 га, Башеег
ап писе// Аду. За4у Ъевау. 1972а.
\Уо1. 4. р. 107—145.
NoМитоё Е. ОНгазочидз ап та{егпа] }е-
Ва\1оие ш зшаП годелз // Вечер. Рзу-
спорю. 1972. Уч. 5, МА. ТР. 371-
387.
Моилек: 9, Агтиаве К. В. Верау1оиг оЁ
уауепПе уеПо\у-БеШей шпагой: р]ау
ап@ зоба] иместацоп // 2АзеВг. Ттегрзу-
спо]. 1979. Ва. 51. 5. 85-105.
МNoитап М, Вгаш тесвВат!з1$ 0Ё тпафетпа!
февамоцг ш Ие гаф// Ногиопез ап4
репауог ш ШоВег уемергаез. М. У.
Токуо: ВаНВагаг, 1983. Р. 69-85.
Noитап М., Геоп М., Мой; Н. Пцегегеп-
се УИ рго]асйп ге]еазе ап Ше ша-
1етпа! Бевах1оиг оЁ Гема]е газ
// Нот-
топе. Верау. 1972. Уч]. 3. Р. 29-38.
Оков Л. Е. Те ‘етрегаваге ге]аНон оЁ
уосаН»аНоп ш Нап со4еп Батзетгз
апа УЙжаг га{з // У. Хоо[. 1974. Ус]. 164.
р. 221-231.
Окоп Е. Е. ЕКабютз аНесИля иИхазоппа
ргодасйоп ш шРапф годету$ // Па. 1972.
уч. 168. Р, 139—148.
Обераг Е. Меююте еКооое рогпатКку о
ро{КалосВ бъусатусв ВаЙлз погуеб1сив
Я]теюй па уусбодпот ЗШюуетзКи // 2Ъ.
ред. Так. Ргезоуе ппу. Ко&се. 1980.
Вос. 18, М 1. 5. 209-243.
Обетог Г. Гатогепозь оБекюу рожапот
бЪусатуш (Ва баз погуер1еи8 Вегк.
1769) па Чефтасй ЗЛюоуепза
// Во 61а
442
Литература
ти шуо]уе4
// Ногиопе ВеЪау. 1971.
Уи. 2. Р. 107-116.
Мепа Т., Стозрепог С. Е. ЕНесв оЁ пит-
Ъег оЁ рирз ироп засКЫпрутаисе@ Та|
ш римйату ргоасЫт сопсептаНоп ап4
шИЕ еесйоп т \е га+ // Еп4ост!пло]оду.
1968. Уо1. 87. Р. 623—626.
Мепдоза 5. Р., бое С. Г, Этотет-
тап 7. Р. её а]. Еипсопа] сопзефаеп-
сез оЁ аМасВтаеть а сошратзоп о? мо
зрес1ез
// Майегпа! чепсез оп еау
Ъеваутоит/Е4. У. Р. Зшоегктап,
В. \У. Вей. Т.., 4980. Р. 201—210.
Мепжез 7. Г., )епиз Е. Нап@оок оё МNo%
Сишеа годетз. Уаи, 1979. 68 р.
Мегсег О., Мегаег М., Вопва Т., ЁКотз-
ег Р. Бееиштайот да 4еБиё 4е 1а
рифеме сБез 1е гаф та!е её ЁещеНе
(Вамаз погуер1сиз): АррНсаймот & Т6-
{п4е 4ез зомеВез Зргарие— Ба\еу, У
маг еь Гопо—Еуапя // $с1. 1еасВ. апп.
]ар. 1982. Т. 7, М 3. Р. 205—215.
Мезе; Т. С., Возеп 7. Бопыпапсе БеВа-
уг аз а леноп оЁ има Ше зИти-
1айоп Чп Ме гаЁ// Агсв. Сеп. РвусВо1.
1960. Уч. 3. Г. 58—56.
МШег С. 5. ТЬе пепее паше оЁ Ще
Вопзе гаё// Ргос. В. 80е. УМазй. 1910.
Уи1. 23. Р. 57—60.
Мшег 6. 5. 14% оф Мой Атетсап те-
сепф шапииа1$
// 0$ Маё Миз. Ва.
1923. М 128. Р. 32.
МШег М. ВергодосНоп т ие Вго\уп газ //
Ашег. Мабаг. 1911. Уч. 45, М 3. Р. 623--
635.
Мтейе Н. Р. Тербозртомз ш Воде
ап@ Мопго0$ оп е 13ап@ оЁ Нажуай //
Ат. 7. Тторр. Мед. Нуз. 1964. Уо1. 13.
Р. 821-832.
Мзоппе Х. ВерагНМов веостар еле ас-
{чиеПе 4е ВаНиз тайлз Ыпие, 1758 ©
4е ЦПамаз погуес!сиз Вегкеппосце. 1769
вп Ттап // ВаЙ. Газ. В. За. Маё Ве].
1956. Ус. 32, М 49. Р. 4М.
Мзоппе Х. Боппесз зиг 1е5 топвеогз
Чип Гоуег 4е резе еп // Вы. 19,
В. $1. Ма. Вею. 1959. Уо1. 35, М 17.
Р. 1-32.
М1зоппе Х. Апаузе гообеортарЫдие 4е$
тапниИегез 4е ГТтап // Мет. 115. гоу.
361. пафмг. Ве. бег. 2. 1960. Газе. 59.
Р. 157.
М:5оппе Х. Ай\сав ава 14о-АлэбгаНап
Мигае: Еуошопагу 1теп4з // Зе1. 2001.
1969. М 172. Р, 219.
Мисье 2. Ехрегииепз оп пеорцоМа т
\Па ап@ 1арогафогу габз: А говуаNoаа-
Чоп //Т. Сотр. ава Рпузю РзусНо].
1976. Уо. 90. Р. 190—197.
Морату В. Мзйта 5. ЕНесфз оЁ в0с1а|
1зоайоп ап ада Вап@ те оп ореп
Бе! етонопаШу о аПтпо газ //
Т. Рэусво!. Вез. 1984. Ус. 28, М 1.
Р. 52-58.
у
Мой Н. Парги ти: Ешритса: Базз
ап@ ‘Меотейса! ап сапсе
// РзусВо1.
Ви|. 1960. Уо1. 57. Р. 291.
Мой Н., СеЙег Б., Герт В. Мабегра]
рекам!отг ш бе ю!аПу шаттесфот!-
тей гаф//У. Сошр. ав@ Рвуз101. РзусВо!.
1967. Уо]. 64. Р. 225—229.
Мой» Н. ГещаМ Г,., Воифапа Б. Рго]оп-
РаНоп оЁ рЬегопзова} еп оп т Ме
таегпа| га // РВуз101. ап@ Вевах. 1974.
У\1. 12. Р. 409—412.
Мой: Н., Геоп М. Зитиа$ сопо! 9
{Ве шабгпа| рВеготопе 1 фе 1асба-
Моб гаф// ТЯ. 1973. Уд. 10. Р. 69-
71.
Мойз Н. Гат М., Геоп М. Митап М.
Ногтопва} тачсНоп оЁ шаегиа! БеБа-
уоот а Че оуапесюшией паШрагоп5
газ // Па. 1970. Ув. 5. Р. 1378-1375.
Мооге С. Г. Ап оМабюгу Базз Бог табог-
па! Чзеглитаноп 0{ зех оЁ оЁзрета
{1 таф// Апиа. Вефау. 1981. Уой. 29.
р. 383—386.
Мооте С. Г. Майегпа! сопйрийот$ ю Фе
Чеуе]ортетв оЁ тазсиНие зехиа! Бева-
мог ш \\е ГаБогафогу газ // Реуор.
РзусвоБ1о1. 1984. Ув. 17. Р. 347-356.
Мооге С. Г. Зех @егепсез ш игтагу
офогз ргодисе4 Ъу уопля 1афогацюгу газ,
Вайиз потуер1сиз // 7. Сотшр. Рзусво|.
1985. Уо]. 99, М 3. Р. 336--344.
Мотца С. Н., Мазнига У., Мопкаша 5. Н.,
Киатита Т. Апе Череп4етф 1таптзти1$-
оп 0 Нетогтвасе Геуег \ИВ гепа|
зупагоше (НЕВ$) уаз ш газ // АтеЙ.
У!го|. 1985. Уо|. 85. Р. 145—149.
Моншей К., 5ю Т., Тзистуа К. ТЯНе-
тепсе ш апушо ас сотроз оп о? ве-
ги 1тапзегыт атопе уамопя $рес!ез
оЁ{ ВаИлз // Аппи. Вер. Маб. зе. бепев. '
Тар. 1971. Ув]. 24. Р. 34-86.
Мотта М. Теуе! о? тафегиа| згезз диття
1х0 ргеспапсу рег!0з оп гаф оНзргите
Берауточт // Рвуспопош. 561. 1965. Уо|. 3.
Р. 7.
Могтзов В. В, Гидыязоп Н. И’. Плзете-
штайоп Бу газ оЁ сопзресШе одогз
оЁ тезгаг@ ап4 повтемат@ // Заепсе. 1970.
Уо1. 167. Р. 904—905.
Миг А. С. Уохтаизземаитееп Е ете
хиКзате Мавсйегьекйтрие ип Ье-
Бепзиние/Ъейчеь // 3\%155 Роо4. 1983.
\Уо1. 3, М 11. Р. 21-27.
Мигашы Е. Г, 08 И. Опзатайед мах
„
Литература
443
вет 11 Ще ПВаг4егап о]ап@ оЁ Ще
Раф// (кет. ап@ Р|уз. ТА Ча. 1974.
\о1. 18. Р. 155-158.
Миззег С. ©. Ертуз Бепбце(епз13, а сот-
розЦе, ап4 опе тоореоетарме хех о!
га\ ап поизе Гааптаз ш Фе РЫШрр:-
пез ап@ СееЪез // Атег. Миз. Мочи.
1977. М 2624. Р. 1-15.
Миззег С. С. А пем оепиз оЁ аогеа!
га Нош \мезр Фауа, Ттдопеза// 7001.
уегВ. 1981а. М 189. Р. 1-35.
М5ег С. С. ВезаЦз оЁ {\е Агено4 сх-
ред! оп, М 105: МNo1ез оп зучетаНсь
о т4о-Ма]ауап ши гойепз, апа
913 оп5 0 пе\ вепега ап@ зрес1ез
Пот Сеуоп, Зшазуезр ап 4Ве РЫШр-
р1тез
// Ва. Ашег. Миз. Маг. [15
1981Ъ. Уо1. 168, агё. 3. Р. 225—834.
Миззег С. С. Тпе лапе хаб оГ Еогез апа
Из геауез еазё оГ Вотопео апа Вай //
ТЯ. 1981с. Уо|. 169, ак. 2. Р. 617—176.
Миззег (. С. ТЬе Тнлй газ // Модеги
Опабегпагу тезеагсВ т бои еазь Аза.
1982. Уч. 7. Р. 65-85.
Миззег С. С., Воеай М. А пе\ репиз о!
шим гофете {го Ме Котойдо 15апаз
ш Мизацеперага//7. Матша|. 1980.
Уо1. 61. Р. 395—413.
Миззег ((. С., Сайра О. Везиз оф Атей-
Ъо14 ехре@ #015, М. 106: ТаепнНез оЁ
таз Нош РШаи Магата апа ощфег
13ап9з ой Еа$ Вотопео // Атег. Мив.
МNo чи. 1982. М 2726. Р. 30.
Миззег (. (., Неапеу 1. В. Риррше
Ваз: А пех. зрес!е5 ош Ще Чиа
дтомрааво ТЫ. 1985. No 2818. Р. 1-
Миззег С. С. МешсотЬ С. Мау!ап пт-
г143 ап Фе п1ап6 таф о Зишайта //
ВоН. Ашег. Миз. Мабт. Не 1983.
У\о1. 174, ат 4. Р. 327-598.
Миззег С. б., Мешсоть С. Бейт юпз 9
Госьтезо Вашиз 1озеа апд а печ зре-
©1ез {том Уюсбташ
// Ашег. Миз. Моуи.
1985. М 28144. Р. 1-32.
Муйутам А. Ап вау арртоасЬ No0 а
га-тее юут ш Ешщана // Уетет Уаз
зег-, Во@еп- ипа Тл фур. 1969. М 32.
_ 8 161—162.
Мезрте А. Тагра А., Авозт М., Све-
еп В. пасе ае иЧезамой рог Терю-
зргаз еп 1а$ габаз (Вале погуе01с11$}
461 та{а4дего тип!сра! 4е ЗапНаго,
СВНе // Во]. Погм. рагазй. Се. 1951.
Уо1. 6. Р. 6-7.
МNoМею С. 4е. Ч поуо тефюйо рага сот-
Бафе а0з гоедотез; паз Исасао сош [аз-
Гео 4е аТатито // Вююг1со. 1980. Уо1. 46,
М 1/2. Р. 3-9.
Мепраиз И’. Птзаснеп ип@ Стео2еп уоп
Карела! еипоеп апесЬ ВаИеп // Апя.
Таг ЭсвааЙтозк. 1957. Ва. 30, М 6,
5. 81—88.
Мсйо[30п А. 7Т., ИУ’атгпег Б. ИУ. ТЬе го-
4ет1з 0! Мех Сяедоша //Т. Маптаа|.
1953. У. 34, М2. Р. 168-179.
Мсоа14е; No. ЭЕт Пр. 2. ТЛрй9 с1авз
сошрозоп о{ затр тот уатопз зре-
с1ез ап апаютиса! зНез //Т. Атег. ОП
Сет. З0с. 1965. \0]. 42. Р. 694-702.
МЁкагт Т. Сотрагамуе спепузгу оЁ з6-
из // Т. Тауезё. Бегшафю]. 1974. Уд]. 62.
Р. 257—267.
Морис Н. ЗриосВаеа 1еетопаетоггВа-
ае ш Атегсап \%14 газ апа Из ге-
1айоп \Noю Ше Тарапезе ап@ Еигореап.
тат /1. Ехр. Мей. 1947. Уо1. 25.
Мото Е. ОИтазоии@ Ш уопие тодетив.
2. Спапвез И аве ш ато газ//
длит. Вевау. 1968. Ув]. 16, М 1. Р. 129-
134.
Мото Е. Те опзеф ап@ Авуеортене о{
таетпаР Фепауютг 1 га, Башеег
ап писе// Аду. За4у Ъевау. 1972а.
\Уо1. 4. р. 107—145.
NoМитоё Е. ОНгазочидз ап та{егпа] }е-
Ва\1оие ш зшаП годелз // Вечер. Рзу-
спорю. 1972. Уч. 5, МА. ТР. 371-
387.
Моилек: 9, Агтиаве К. В. Верау1оиг оЁ
уауепПе уеПо\у-БеШей шпагой: р]ау
ап@ зоба] иместацоп // 2АзеВг. Ттегрзу-
спо]. 1979. Ва. 51. 5. 85-105.
МNoитап М, Вгаш тесвВат!з1$ 0Ё тпафетпа!
февамоцг ш Ие гаф// Ногиопез ап4
репауог ш ШоВег уемергаез. М. У.
Токуо: ВаНВагаг, 1983. Р. 69-85.
Noитап М., Геоп М., Мой; Н. Пцегегеп-
се УИ рго]асйп ге]еазе ап Ше ша-
1етпа! Бевах1оиг оЁ Гема]е газ
// Нот-
топе. Верау. 1972. Уч]. 3. Р. 29-38.
Оков Л. Е. Те ‘етрегаваге ге]аНон оЁ
уосаН»аНоп ш Нап со4еп Батзетгз
апа УЙжаг га{з // У. Хоо[. 1974. Ус]. 164.
р. 221-231.
Окоп Е. Е. ЕКабютз аНесИля иИхазоппа
ргодасйоп ш шРапф годету$ // Па. 1972.
уч. 168. Р, 139—148.
Обераг Е. Меююте еКооое рогпатКку о
ро{КалосВ бъусатусв ВаЙлз погуеб1сив
Я]теюй па уусбодпот ЗШюуетзКи // 2Ъ.
ред. Так. Ргезоуе ппу. Ко&се. 1980.
Вос. 18, М 1. 5. 209-243.
Обетог Г. Гатогепозь оБекюу рожапот
бЪусатуш (Ва баз погуер1еи8 Вегк.
1769) па Чефтасй ЗЛюоуепза
// Во 61а
ДАЛ
Литература
(С558). 1986. Вос. 41, М 2. $8. 173-
180
Оно! М., б1ешрат 7. Зех ЧШегепсез п Ме
р!ау Ъевауючг оЁ ртериарезсетв газ //
Рьуз101. ап Вевау. 1978. У. 20.
Р. 113—145.
Опоег А. Т., ИВешег 5. Н., боо@дтр С. Б.
Е! суашамоп сопёго| о Те табы
Огусоавиз сатеив // Аз га]. УУПаН-
Ре Вев. 1982. М 9. Р. 125—134.
ОПиет В., Моз Т., Оотзейо В. Г. Маегпа!
арот15Не Бевамошг 11 гайз // ЕФо]о-
су 85: 19% Пиегп. ео]. сопф. Тоиюи-
зе, 1985. \01. 1. Р. 235.
Орйо| А. 7, Гапвегёа р. \. Убамагие
тез1{апсе ш Фе МеФегапаз // ВаМеп-
Боюе ипа ВаИепЪеКатарЁлае: (Уог{-
гарзуегап(аЦипе 4ез Усгешз Ёаг УМаз-
зег-, Войеп-, ип Та Фубеве аш 28.
ип@ 29. МоуетЪег 1968, ГопогеззваПе,
ВегИп). Зи Исагц, 1969. 3. 19—25.
О’Вепу В. А. Рой У. С, Авефег Р. М.
НегедЦагу гез1ебапсе No0 соптатт апй-
соабапв агиоз ш тап ап га// Апп.
No. У. Асай. 51. 1968. Уо]1. 151. Р. 913-
981.
Озрогп Б. 1, Нёту Г. ТNoе сошетрогагу
Тат тата! оЁ Ерурё (тоадше 8
па!) // Е1е141апа: 2001. No. $. 1980. М 5.
Р. 579.
Озшай (4. Г., Маег Ц. ТУ. ОПабюту, Мет-
ша], ап 1асИа! шНиепсез оп папе
иЦгазоп!е уосаНзайоп 1 гаё// Беуе]ор.
Рзусво101. 1975. Уо1. 7, М 2.Р. 129-
135.
Ота К., Уокоуата А. РхезатрИоп о? 1асфа-
Чоп Бу засКИоо ш 1абфайпе гаёз аНег
тетоуа1 оЁ Иегз //Т. Епдостто1. 4965.
Уо1. 2. Р. 185-194.
РаГтийет 5. 9., Мсбанп У. А. В@аНот-
ЗЬТр о# ассер{апсе ап тотбаШу о{ ап-
Нсоари]ат® раН$ 10 габз // Ва. Епутоп.
Сот{ат. ап@ Тох!со1. 1976. М 6. Р. 750-—
755.
Ра]тег В. 5. ТЬе шашша сие, таш-
11а] 01 No Ашенса пог оЁ Мех!со.
М, У.: Бои Медау, 1954. 384 р.
Рапрзерр 7. Тве опюсепу о мау ш
гафз // Оеуе!ор. РзусБоБ101. 1981. Уо1. 14.
Р. 327—332.
Рапкзерр 7, Веану ТУ. И’. Зосэ1 Черт
уаЯоп апа р]ау т га // ВеВау. Мепга|.
Вто]. 1980. Уч. 30. Р. 197-206.
РапЁзерр 7, билу $. Могтапзей Г. ТВе
реусвооюгу о0Ё Рау: ТВеогейса! апа
те{Подою21са1 регзресНуез // Мепгозс1.
В1оБеВау. Вех. 1984. УоЁ 8, М 4.
Р. 463—492.
Ратгой В. Г. ЕМесь о{ сазтайв ош зе-
хив| агочза| 11 Ще гаф, дефегиштей Гот
гесог4з 0 роз-е]асаогу ийтавотс
уосаНтаНопз
// РВузю. ВеВах.. 1976.
Уо|. 16. Р. 689—692.
Разеит No, Тогтз 7У., Тора М., ГзЕап-
4аг ПР. Сепенс @1НетепНайот т 1190-
пезап апа ЕгепсВ гафз о# Фе заЪхепиз
ВаНиз // Р1освеш. Зуз. Есо|. 1982,
Ус]. 10, М 2. Р. 191—196.
РалаЕ No. А заду оп Фе ргохпиайе сош-
розой ап пийуе уаше оЁ фатфоо
(ВаЪЪизза учат) огашз Фог писе//
104. 7. Noи. Ре. 1979. Уч. 16, М 2.
Р. 52-53.
Реай В. ТВе Шоюсу оЁ бе стом оЁ
рориаНоп. М. У., 1925. 300 р.
Рейегзеп С. А., Ргапве А. Ф. (7т.). Тодлс-
Яоп 0 шаегпа! Бева\опг ш уШешт
гафз аНег ТитасегеБтоуе
те ]аг ати!-
итаНоп о охуюст// Ргос. Маф. Асад.
31. 05. 1979. Уо. 79. Р. 6661—6665.
РееЁе Н. У. 5., бетег А. В. НаБ\тлайой
40 епутоптетёа! ап@ зребе апаИогу
зНтоН п Фе габ// Вевау. Сототаол.
В1о|. А. 1970. Уч, 5, М 1. Р. 29-39.
Реггу 7. 5. ТВе тергодисИоп оЁ Noе *На
Вгохп гаё (ВаЙмз потуеелз Бехе-
Ъеп) //Ргос. 2001. Зое. Гопаоп. 1945.
Ус]. 115, рё 1. Р. 19—46.
Рецег Р., Зап Слтопз М-С. Без топееотз
4и Магос // Тгау. 15%. вс1. сВегШ, З6г.
2001. 1965. Уо1. 31. Р. 1-58.
Рта}! О. И’., Р}арртап С. Вевам1опга! эва
еесгорвузо
о 1са[ гезропзез оЁ шае
таз Ф шэе птте одогз
// ОНасНол
ап фаз. М. У.: ВоскКеПег От!\. ргезз,
1969. Уо1. 3. Р. 258—267.
Рняег Н. Р., Гут; А. Ргепайа! рзусБо-
1ор1са| тез еНесвз оп оНзргше Ъева-
Уопг Ш газ // Апзта|. Т. РзусВо]. 1983.
Уо1. 35, М 1. Р. 21-36.
РЧегсе 7. Т., Мшай В. М. ЗеН-расе4 зехиа|
Ъезамойг ш Ше {етае га //Т. Сошр.
ап@ Рвуз10]. РзусВо]. 1961. У. 54.
Р. 310—313.
Рзапо Х., /!огег Т. С. Вигго\з ап@ Гее то
оЁ \Noе Мог\ау габ//Т. Мата. 1948.
Уо1. 23, М 4.Р. 374-383.
Равие ш Ашегсаз. \МазВ. (Ъ. С.), 1945.
280 р. (Рап Ашег. Зап. Виг. РаБ].;
М 225).
Ройчег Е. Е., Эта Е. О. ЗоааЙтте Гапс-
Н008 0{ ргипа{е рау // Атег. 7ооЁ. 1974.
Уо1. 14. Р. 275—287.
РоПасЁ Е. Г., басйз В. О. Мае соршаюогу
БеБауюог ап@ {ее табегоа|1 Ъевз-
у1очг ш реопабаПу рафесюш1тей гай //
РЬуз10]. ап@ ВеБау. 4975. Уо|. 14.
Р. 337-343.
Литература
445
Род 7. С., О’Вейу В. А. 5ейпадег-
тап Г. Т, Мехапдег М. Уафант те-
я запсе ш \Ше гаф// Ашех. Т. РЬузо1.
1968. Уо1. 215, М 3. Р. 627—631.
Рофе Т. В. Аботезыче р]ау ш ро]е-
саёз //Зутр. 200|. $0с. Гопфоп. 1966.
\Уо1. 18. Р. 28—44.
Робе Т. В., Юз ФТ. Ап ТтуезиеаНоп ой
р1ауй] Ъерачмюит ш ВаНиз погуео1сиз
ап@ Миз тшиазса!аз // Ргос. 2001. $06.
Гопдоп. 1975. УоТ. 175, No 1.Р. 61-72.
Рофе Т. В., Ев 7. Ап шуезываноп о#
шагядиа], асе ап зехоа! @етепсез
1 Фе разу оЁ Ваз погуевеиз (Матш-
пзаПа, Нодепйа) //Т. 2001. 1976. Уд. 179,
М 2.Р. 249-260.
Рорр 1. Еще Её аЙеБегер!Чете Бег Ет-
5епрИйскКего // 24зсВг. Нур. ип Тыек,
1950. Ва. 431, М 6. 3. 575.
Розадаз-Апагешз А., Рорег Т. @. бов
1гапз11 5101 0Ё {004 ргеегепсоз п ад
га! // Апит. Вевау. 1983. Ус 34.
р. 265—271.
Рысе Е. С. Ногтопа] сопёто! оЁ игтпе-тахг-
Юше Ш %1а ап аотезие Могуау
га{з // Ногтопе Верах. 1975. Уч. 6.
Р. 393—397.
Руе Т., Воппег И’. Еега] Бго\уп газ, Ва
105 погуея1с1з. ш Зои Сеогоа (Зои
АЧапвйс Осеап) // 5. 2001. 1980. Уа1. 192,
М2.Р. 237-255.
Патап В., Эрагта Т. Кагуогуре еуиНоп
апа зреНаНоп ш сепиз Ваз Е1зеПег //
}. 59. апд Тадиз. Вез. 1977. Уо1. 36,
No 8. Р. 385—404.
Ватраиа М. Турообу оЁ \уПА гаф рори-
’Лаботз // АСЁа 2001. Ёепп. 1984. Уд]. 174.
Р. 233—236.
ВапЕ С, Г. ТЬв годепб$ оЁ ТГлБуа // ВаИ.
03 Маё Миз. 1968. М 275. Р. 264.
Кети Е. Мае Кодитаа Птпа4е то
$4086 }а Ы14ез // Тоодиз. 1923. М Т.
Р. 429—435.
ПЯепп1з0оп В. О. Мефойз о[ ЧезИпо то-
Чепйс4ез 11 Ме Пе! асашзь таз //
Резиес. 31. 1977. \01. 8. Р. 405—413
Всратаз П. В., Э1ерепз р. А. Еуепсе
Гог тагктя НВ игше Бу га // Ведах.
` Во]. 1977. У]. 12. Р. 547-528.
Е спагаз М. Р. М. П4аобе Вап4Ние т
тойепёз: а г6азвеззтепф т Ме Порё о{
‚ тесеп з611ев о{ тафегпа!у Ъепамточг //
Апни. Вевау. 1966. Уо1. 14. Р. 582.
Всртопа (., Засйз В. 0. Маюгпа! 415ст1-
индаНоп 0 рир зех ш га // Беуеюр.
РзусВо]. 1984. \о1. 47, М 1. Р. 87-89.
Аца М. А., Мойаттей А. Н. Н., 5ра-
шатКу А. А., Атауа М. 5. те @епсе апа
Чтиво оГ дошезыс ап@ соттаепва]
тодет1в ш САВ///. Геуре. Ра. НеаНЬ
Аззос. 1969. У]. 44, М 2. Р. 427—146.
Ншез В. И’, Апфет М. А., Гапя И. 9-
зопепко Р. С. Рифегбу ш \\е гаё: то-
ааНоп зу шеаюпш ап@ Но // 7. Мен-
и Ттапзи1з3. 1986. Зирр1. 21. Р. 81-
108.
Йофегз Т. Т. Тье шага оЁ Ра зап.
Г.: Вепп, 1977. 361 р.
Нойтзоп В. СепеНсз оЁ Ще Могмуау гаё
No. У.: Регеапаоп ргозз, 1965. 804 р.
ВофцаШе У. А. Вокеё 7. Ее оъзегуай-
опз оп 6 зоба! Бева\м1опг оЁ Фе М от-
мау габз
// Во]. Вевау. 1976. Уо|. 4.
Р. 289-308.
Восс С., Рвапо УТ, Угату У., Эюгег Г.
Вигтомз ап Теедтез оЁ фе Могмуау
гай //Т. Мата]. 1948. Уд. 29, М 4.
Р. 374-383.
Позепретге К., Беппепвега У. Н., Каг-
гош# М. Х. Ме теагед \мИН гаф апп:
{фе го]е оЁ габ-томзе сошасв ш тей
Тай Ъева\1опга! ап@ рВузоорса|
сВапоез5 ш Ще шоизе // Ашт. Верах.
1970. Уо1. 18. Р. 188—143.
Аозеп ай 7. 5. Моппогтопа! }аз13 о
та4егпа! Берауюопг ш Фе газ //Заеп-
се. 1967. Ус. 156. Р. 1512—1514,
ПНозепай 7. 5. Тце 4еуоршеть о! та-
1етпа] гезропуУепез$ 1 Ве каф // Аюег.
Г. ОтпорзусВоГ. 1969. Ус. 39. Р. 36-—
50
Возепра! 7. 5., Гейттап РБ. В. Маегра!
феВау1оиг ш Те 1аБогабогу гаф// Мабег-
па| ревауюог ш шашита]. М. У: МЕ
1еу. 1963. Р. 8—57.
Возепай У. 5., Мауег А. р., беде Н. 7.
Маегпа! Бевау!1ог атопа е попрерпа-
1% шанипа|з // Нап4Ъ. Вевах. Меиго 01.
1985. Ус]. 7. Р. 229-298.
Позеп ай 7. 5., безе Н. У. Нузегесо-
шу-шдисе тпабегпа! Бевамочг фагии
ргеопапсу Шш \ШМе таф//7Т. Сошр. ана
Рвуз1о1. Рвусво1. 1975. Ув. 89. Р. 685—
700.
Нозеп Мей У. 5., ере! Н. 7. Мабюгпа[ Ъе-
Вауюпт шт Пе Таотаюгу гаф// Маёег-
па| Ш иерсез оп еаТу Бепаутоит/ВА.
УУ. Р. Зтофегтат, В. У. Ве. Ё., 4980.
Р. 155—200.
Возепан 7. 5., Чезе! Н. 7, Мауег А. В.
Ргобтезз ш {Ве зу оЁ табегоа! Ъе-
Вауоцг Ш Фе таб
// Аду. Зю@у Вевам.
1979. \о1. 10. Р. 225-3141.
Позепсшев М. В., Веппеё Е. Г., Бы-
топа М. С. Вгайт сВапеез ш тезропзе
$0 ехрешепсе // 3с1, Атег, 1972. Уч]. 226,
М2.Р. 15-18.
Аоз=КошзNo А. Р. СотрагаИуе юхеЦу оЁ
ДАЛ
Литература
(С558). 1986. Вос. 41, М 2. $8. 173-
180
Оно! М., б1ешрат 7. Зех ЧШегепсез п Ме
р!ау Ъевауючг оЁ ртериарезсетв газ //
Рьуз101. ап Вевау. 1978. У. 20.
Р. 113—145.
Опоег А. Т., ИВешег 5. Н., боо@дтр С. Б.
Е! суашамоп сопёго| о Те табы
Огусоавиз сатеив // Аз га]. УУПаН-
Ре Вев. 1982. М 9. Р. 125—134.
ОПиет В., Моз Т., Оотзейо В. Г. Маегпа!
арот15Не Бевамошг 11 гайз // ЕФо]о-
су 85: 19% Пиегп. ео]. сопф. Тоиюи-
зе, 1985. \01. 1. Р. 235.
Орйо| А. 7, Гапвегёа р. \. Убамагие
тез1{апсе ш Фе МеФегапаз // ВаМеп-
Боюе ипа ВаИепЪеКатарЁлае: (Уог{-
гарзуегап(аЦипе 4ез Усгешз Ёаг УМаз-
зег-, Войеп-, ип Та Фубеве аш 28.
ип@ 29. МоуетЪег 1968, ГопогеззваПе,
ВегИп). Зи Исагц, 1969. 3. 19—25.
О’Вепу В. А. Рой У. С, Авефег Р. М.
НегедЦагу гез1ебапсе No0 соптатт апй-
соабапв агиоз ш тап ап га// Апп.
No. У. Асай. 51. 1968. Уо]1. 151. Р. 913-
981.
Озрогп Б. 1, Нёту Г. ТNoе сошетрогагу
Тат тата! оЁ Ерурё (тоадше 8
па!) // Е1е141апа: 2001. No. $. 1980. М 5.
Р. 579.
Озшай (4. Г., Маег Ц. ТУ. ОПабюту, Мет-
ша], ап 1асИа! шНиепсез оп папе
иЦгазоп!е уосаНзайоп 1 гаё// Беуе]ор.
Рзусво101. 1975. Уо1. 7, М 2.Р. 129-
135.
Ота К., Уокоуата А. РхезатрИоп о? 1асфа-
Чоп Бу засКИоо ш 1абфайпе гаёз аНег
тетоуа1 оЁ Иегз //Т. Епдостто1. 4965.
Уо1. 2. Р. 185-194.
РаГтийет 5. 9., Мсбанп У. А. В@аНот-
ЗЬТр о# ассер{апсе ап тотбаШу о{ ап-
Нсоари]ат® раН$ 10 габз // Ва. Епутоп.
Сот{ат. ап@ Тох!со1. 1976. М 6. Р. 750-—
755.
Ра]тег В. 5. ТЬе шашша сие, таш-
11а] 01 No Ашенса пог оЁ Мех!со.
М, У.: Бои Медау, 1954. 384 р.
Рапрзерр 7. Тве опюсепу о мау ш
гафз // Оеуе!ор. РзусБоБ101. 1981. Уо1. 14.
Р. 327—332.
Рапкзерр 7, Веану ТУ. И’. Зосэ1 Черт
уаЯоп апа р]ау т га // ВеВау. Мепга|.
Вто]. 1980. Уч. 30. Р. 197-206.
РапЁзерр 7, билу $. Могтапзей Г. ТВе
реусвооюгу о0Ё Рау: ТВеогейса! апа
те{Подою21са1 регзресНуез // Мепгозс1.
В1оБеВау. Вех. 1984. УоЁ 8, М 4.
Р. 463—492.
Ратгой В. Г. ЕМесь о{ сазтайв ош зе-
хив| агочза| 11 Ще гаф, дефегиштей Гот
гесог4з 0 роз-е]асаогу ийтавотс
уосаНтаНопз
// РВузю. ВеВах.. 1976.
Уо|. 16. Р. 689—692.
Разеит No, Тогтз 7У., Тора М., ГзЕап-
4аг ПР. Сепенс @1НетепНайот т 1190-
пезап апа ЕгепсВ гафз о# Фе заЪхепиз
ВаНиз // Р1освеш. Зуз. Есо|. 1982,
Ус]. 10, М 2. Р. 191—196.
РалаЕ No. А заду оп Фе ргохпиайе сош-
розой ап пийуе уаше оЁ фатфоо
(ВаЪЪизза учат) огашз Фог писе//
104. 7. Noи. Ре. 1979. Уч. 16, М 2.
Р. 52-53.
Реай В. ТВе Шоюсу оЁ бе стом оЁ
рориаНоп. М. У., 1925. 300 р.
Рейегзеп С. А., Ргапве А. Ф. (7т.). Тодлс-
Яоп 0 шаегпа! Бева\опг ш уШешт
гафз аНег ТитасегеБтоуе
те ]аг ати!-
итаНоп о охуюст// Ргос. Маф. Асад.
31. 05. 1979. Уо. 79. Р. 6661—6665.
РееЁе Н. У. 5., бетег А. В. НаБ\тлайой
40 епутоптетёа! ап@ зребе апаИогу
зНтоН п Фе габ// Вевау. Сототаол.
В1о|. А. 1970. Уч, 5, М 1. Р. 29-39.
Реггу 7. 5. ТВе тергодисИоп оЁ Noе *На
Вгохп гаё (ВаЙмз потуеелз Бехе-
Ъеп) //Ргос. 2001. Зое. Гопаоп. 1945.
Ус]. 115, рё 1. Р. 19—46.
Рецег Р., Зап Слтопз М-С. Без топееотз
4и Магос // Тгау. 15%. вс1. сВегШ, З6г.
2001. 1965. Уо1. 31. Р. 1-58.
Рта}! О. И’., Р}арртап С. Вевам1опга! эва
еесгорвузо
о 1са[ гезропзез оЁ шае
таз Ф шэе птте одогз
// ОНасНол
ап фаз. М. У.: ВоскКеПег От!\. ргезз,
1969. Уо1. 3. Р. 258—267.
Рняег Н. Р., Гут; А. Ргепайа! рзусБо-
1ор1са| тез еНесвз оп оНзргше Ъева-
Уопг Ш газ // Апзта|. Т. РзусВо]. 1983.
Уо1. 35, М 1. Р. 21-36.
РЧегсе 7. Т., Мшай В. М. ЗеН-расе4 зехиа|
Ъезамойг ш Ше {етае га //Т. Сошр.
ап@ Рвуз10]. РзусВо]. 1961. У. 54.
Р. 310—313.
Рзапо Х., /!огег Т. С. Вигго\з ап@ Гее то
оЁ \Noе Мог\ау габ//Т. Мата. 1948.
Уо1. 23, М 4.Р. 374-383.
Равие ш Ашегсаз. \МазВ. (Ъ. С.), 1945.
280 р. (Рап Ашег. Зап. Виг. РаБ].;
М 225).
Ройчег Е. Е., Эта Е. О. ЗоааЙтте Гапс-
Н008 0{ ргипа{е рау // Атег. 7ооЁ. 1974.
Уо1. 14. Р. 275—287.
РоПасЁ Е. Г., басйз В. О. Мае соршаюогу
БеБауюог ап@ {ее табегоа|1 Ъевз-
у1очг ш реопабаПу рафесюш1тей гай //
РЬуз10]. ап@ ВеБау. 4975. Уо|. 14.
Р. 337-343.
Литература
445
Род 7. С., О’Вейу В. А. 5ейпадег-
тап Г. Т, Мехапдег М. Уафант те-
я запсе ш \Ше гаф// Ашех. Т. РЬузо1.
1968. Уо1. 215, М 3. Р. 627—631.
Рофе Т. В. Аботезыче р]ау ш ро]е-
саёз //Зутр. 200|. $0с. Гопфоп. 1966.
\Уо1. 18. Р. 28—44.
Робе Т. В., Юз ФТ. Ап ТтуезиеаНоп ой
р1ауй] Ъерачмюит ш ВаНиз погуео1сиз
ап@ Миз тшиазса!аз // Ргос. 2001. $06.
Гопдоп. 1975. УоТ. 175, No 1.Р. 61-72.
Рофе Т. В., Ев 7. Ап шуезываноп о#
шагядиа], асе ап зехоа! @етепсез
1 Фе разу оЁ Ваз погуевеиз (Матш-
пзаПа, Нодепйа) //Т. 2001. 1976. Уд. 179,
М 2.Р. 249-260.
Рорр 1. Еще Её аЙеБегер!Чете Бег Ет-
5епрИйскКего // 24зсВг. Нур. ип Тыек,
1950. Ва. 431, М 6. 3. 575.
Розадаз-Апагешз А., Рорег Т. @. бов
1гапз11 5101 0Ё {004 ргеегепсоз п ад
га! // Апит. Вевау. 1983. Ус 34.
р. 265—271.
Рысе Е. С. Ногтопа] сопёто! оЁ игтпе-тахг-
Юше Ш %1а ап аотезие Могуау
га{з // Ногтопе Верах. 1975. Уч. 6.
Р. 393—397.
Руе Т., Воппег И’. Еега] Бго\уп газ, Ва
105 погуея1с1з. ш Зои Сеогоа (Зои
АЧапвйс Осеап) // 5. 2001. 1980. Уа1. 192,
М2.Р. 237-255.
Патап В., Эрагта Т. Кагуогуре еуиНоп
апа зреНаНоп ш сепиз Ваз Е1зеПег //
}. 59. апд Тадиз. Вез. 1977. Уо1. 36,
No 8. Р. 385—404.
Ватраиа М. Турообу оЁ \уПА гаф рори-
’Лаботз // АСЁа 2001. Ёепп. 1984. Уд]. 174.
Р. 233—236.
ВапЕ С, Г. ТЬв годепб$ оЁ ТГлБуа // ВаИ.
03 Маё Миз. 1968. М 275. Р. 264.
Кети Е. Мае Кодитаа Птпа4е то
$4086 }а Ы14ез // Тоодиз. 1923. М Т.
Р. 429—435.
ПЯепп1з0оп В. О. Мефойз о[ ЧезИпо то-
Чепйс4ез 11 Ме Пе! асашзь таз //
Резиес. 31. 1977. \01. 8. Р. 405—413
Всратаз П. В., Э1ерепз р. А. Еуепсе
Гог тагктя НВ игше Бу га // Ведах.
` Во]. 1977. У]. 12. Р. 547-528.
Е спагаз М. Р. М. П4аобе Вап4Ние т
тойепёз: а г6азвеззтепф т Ме Порё о{
‚ тесеп з611ев о{ тафегпа!у Ъепамточг //
Апни. Вевау. 1966. Уо1. 14. Р. 582.
Всртопа (., Засйз В. 0. Маюгпа! 415ст1-
индаНоп 0 рир зех ш га // Беуеюр.
РзусВо]. 1984. \о1. 47, М 1. Р. 87-89.
Аца М. А., Мойаттей А. Н. Н., 5ра-
шатКу А. А., Атауа М. 5. те @епсе апа
Чтиво оГ дошезыс ап@ соттаепва]
тодет1в ш САВ///. Геуре. Ра. НеаНЬ
Аззос. 1969. У]. 44, М 2. Р. 427—146.
Ншез В. И’, Апфет М. А., Гапя И. 9-
зопепко Р. С. Рифегбу ш \\е гаё: то-
ааНоп зу шеаюпш ап@ Но // 7. Мен-
и Ттапзи1з3. 1986. Зирр1. 21. Р. 81-
108.
Йофегз Т. Т. Тье шага оЁ Ра зап.
Г.: Вепп, 1977. 361 р.
Нойтзоп В. СепеНсз оЁ Ще Могмуау гаё
No. У.: Регеапаоп ргозз, 1965. 804 р.
ВофцаШе У. А. Вокеё 7. Ее оъзегуай-
опз оп 6 зоба! Бева\м1опг оЁ Фе М от-
мау габз
// Во]. Вевау. 1976. Уо|. 4.
Р. 289-308.
Восс С., Рвапо УТ, Угату У., Эюгег Г.
Вигтомз ап Теедтез оЁ фе Могмуау
гай //Т. Мата]. 1948. Уд. 29, М 4.
Р. 374-383.
Позепретге К., Беппепвега У. Н., Каг-
гош# М. Х. Ме теагед \мИН гаф апп:
{фе го]е оЁ габ-томзе сошасв ш тей
Тай Ъева\1опга! ап@ рВузоорса|
сВапоез5 ш Ще шоизе // Ашт. Верах.
1970. Уо1. 18. Р. 188—143.
Аозеп ай 7. 5. Моппогтопа! }аз13 о
та4егпа! Берауюопг ш Фе газ //Заеп-
се. 1967. Ус. 156. Р. 1512—1514,
ПНозепай 7. 5. Тце 4еуоршеть о! та-
1етпа] гезропуУепез$ 1 Ве каф // Аюег.
Г. ОтпорзусВоГ. 1969. Ус. 39. Р. 36-—
50
Возепра! 7. 5., Гейттап РБ. В. Маегра!
феВау1оиг ш Те 1аБогабогу гаф// Мабег-
па| ревауюог ш шашита]. М. У: МЕ
1еу. 1963. Р. 8—57.
Возепай У. 5., Мауег А. р., беде Н. 7.
Маегпа! Бевау!1ог атопа е попрерпа-
1% шанипа|з // Нап4Ъ. Вевах. Меиго 01.
1985. Ус]. 7. Р. 229-298.
Позеп ай 7. 5., безе Н. У. Нузегесо-
шу-шдисе тпабегпа! Бевамочг фагии
ргеопапсу Шш \ШМе таф//7Т. Сошр. ана
Рвуз1о1. Рвусво1. 1975. Ув. 89. Р. 685—
700.
Нозеп Мей У. 5., ере! Н. 7. Мабюгпа[ Ъе-
Вауюпт шт Пе Таотаюгу гаф// Маёег-
па| Ш иерсез оп еаТу Бепаутоит/ВА.
УУ. Р. Зтофегтат, В. У. Ве. Ё., 4980.
Р. 155—200.
Возепан 7. 5., Чезе! Н. 7, Мауег А. В.
Ргобтезз ш {Ве зу оЁ табегоа! Ъе-
Вауоцг Ш Фе таб
// Аду. Зю@у Вевам.
1979. \о1. 10. Р. 225-3141.
Позепсшев М. В., Веппеё Е. Г., Бы-
топа М. С. Вгайт сВапеез ш тезропзе
$0 ехрешепсе // 3с1, Атег, 1972. Уч]. 226,
М2.Р. 15-18.
Аоз=КошзNo А. Р. СотрагаИуе юхеЦу оЁ
446
Литература
тодепис!
дез // Ес4. Ргос. 1967. Уо?. 26,
No4.Р. 1082—1088.
ВоферИа 7. Тце согриз Пиечт — рНаНа-
ту теаНопзВрз: Тие азвосмаНоп Ъе!-
муееп Ще сапзе оГ Пнесотор!а зестеЙоп
ап@ \Ше сапзе о? ГоШеШаг дфиезепсе
Читте Табаоп // Епдосео]ову. 1960.
Уо1. 67. Р. 9—41.
Воше Г. Р. Ваз ап@ писе ш меКз/
Аве Щите. 1962. У]. 7. Р. 28—30.
Виду 7. И’., СвеаНе М. В. ОМасюту зИ-
ша|Н ап@ Ие рзеадосхипсйон еНесв //
З1епсе. 1977. Уо|. 198. Р. 845—846.
Виззе Р. А. П4апе зишШайоп ш го-
Чеп{з: А сопы4егайот оЁ роззИе те-
сВап1эта // Рзусво]. Ви. 1971. М 3.
Р. 192—202.
Засйз В. Р.. Мезй П. Ь. Рафема! деуе-
1ортепё оЁ репЙе геЙехез ап@ сорийа-
Нов ш мае га // Рзуспопеигоетдо-
стпо]осу. 1979. \01. 4. Р. 287—296.
бафег В. М. Е. Подовлса] сопзедцепсез
0 Пасбамоп // Асба 2001. [епп. 1984.
\о1. 171.Р. 179—182.
бат: Стопз М.С. Тез шашиШегез @е
Егапсе её 4и Вепашх. Р.: РОТХ, 1973.
481 р.
без С. 2. ЧЦтазовтЯ ап@ тшайое Бе-
Вауопг ш годепёз УИ зоте офзегуа-
900$ оп офег Бевауоига| зИиайот$//
7. 2001. 1972а. Ус]. 168, М 2. Р. 149-
164.
5е @. О. Ойтазопи@ апд аротезяуе
Бевау1оиг т габз апд офег зта|Н пзат-
та]// Арпи. Вевау. 1972Ъ. Уо]. 20,
Р. 88—100.
51 ©. Р., Руе Р. ОНгазотие сопииит-
сайоп Ъу апиа1з. Т.: СВаршап ап4д
НаН, 1974. 281 р.
5а(е; С. О., 5тий У. С. Сотрагайуе з1-
41е$ о Ше иКтазот1с саНз о шащ
тие — годен» // Оеус]ор. — Рзусво о.
1978. Ус. 11, М 6. Р. 595—619.
бапсйез-Спа4о 7. Е. туд метеп® оф Ме
уотегопаза| зузешт т ЧМе тергодисй-
уе рвуз101юсу оЁ Ме га*// ОНасИоп ап&
епдосгле геоц]айоп. Т.: ТАБ ргезз,
1982. Р. 209-218.
бапслез-Стадо ХФ. Р., ааЦево А. Мае ш-
Чисей ргесостоптз-раЪегу ш Фе Гота
тай Вое оЁ Ме уотетопаза] зузет //
Ас+а епдосгло|. 1979. Уо|. 91, заррЁ.
225. Р. 254,
бапказишап Т!., Ропвргади Р., Водфаае-
{а Р. её а1. Мите 1урВиз ш ТВаПазЯ //
Ттапз. Воу. 306. Тгор. Мед. апа Нур.
1969. Ус]. 63, No5.Р. 639—643.
баю МNo., НаПег Е. М’. Рошей В. О., Неп-
т Р. Т. Зехиа! шашгайов т БаЪес®-
111269 Тета!е га{з//Т. Вергод. Еег%.
1984. Уо1. 36. Р. 301-309.
5сйаае В. Гиегаитетсь& гаш Ве$13еп7-
тесзап1зшиз 4ег УУвидеггайе петепй-
ег шапеЖеп Ап соави]атщеп // Апоех.
Рагазио. 1974. Ва. 15, М4. 8. 201-
Зерие [ег Е. И’., ВоиДез И’. Д. Асще ога]
10х16у ап@ гереНепсу оЁ 933 сВели-
са! 10 Копзе ап 4еег писе // АгсВ.
Елутоп. Соп{ат. ап@ Тох!с01. 1985.
У]. 14. Р. 411-129.
Зераерег С. За М. А, Кеюйе С.
Агоп С. ОНасюту шу/топтет ап@ 1ог-
90518 Бевауюиг ш фе Гете ап шаГе
гаф // ОПасИоп ап@ епфосгте геощаНол.
Г.: ТВГ ртезз, 1982. Р. 115—125.
бейШег Е. Г. Есоюру ап пеаМ оЁ Ван
{0$ аб Моше, АЙазка //Т. Матита|. 1956.
У\о1. 37. Г. 181-188.
бевтапойт С. 5., Нази 9. Е, Битут-
ре ФТ. М. её а1. Апйвегие ап депейс
ргорегНез оЁ утизоз Пике 10 Нетог-
Грар1с Теуег УИ гепа| зупакоше // 8с1-
епее. 1985. Уо|. 227. Р. 1041-1044.
Эсйтиа Е. Вотаегее сте ЕщепеембПе
1 Апр (ВТ) // Отл Во]. ВеоЪ. 1984.
Ва. 81, No 3. 5. 249-254.
Эсйпиттепьегиег Р. Н., Напзоп ЕЁ. В., Маг-
ни В. 7. Терювртов8: Топе-бетий 8ит-
уеШапсе оп ап ПШпо1$ Гага
// Атаег.
2. ЕрЯ. 1970. Уд]. 92, М 4. Р. 223—
239.
успиртег И’., Киепеп И’. А. Не моогВо-
шеп 4ег зроспаеюет уап Ве {уре 4ет
Эр. 1юбегопаетоттваятае Ъ1] гаМел, т
АтяетЧат // Ме. ТазсВг. у. Сепеезк.
1923. Уо1. 67. Р. 20—22.
Эспитасйег 5. К., Мойх Н. ЛЪе таегпа!
рНеготопе оЁ {Ве гаф аз ап шла 38-
шиаз 1ог ргезуеапиих уоцпо // РВузю|-
ап@ Вевау. 1982. Уч]. 3, М 1. Р. 67-71.
ЭейшагЕ Е. СазуИсаЙол, оош ап Че
ионов оЁ сопилептза! газ
// Вай. *
УУо4 НеаМ\ Отгалих. 1960. Уо|.. 2/3.
Р. 411-416.
5сон Н. ВодепеЪогпе Фзеазе сопио]
Игоцей тгодепё эюрраве. Рь 1, 2// Рез
Сопи. 1963. \01. 31, М 8. Р. 30-46;
М Э.Р. 18—28.
5сой 7. Р. Ст са] рег1о48 ш Ще 4еуеор-
еп 0{ зос1а1 Бева\мюочг 11 рирруез //
Русвозот. Мей. 1958. Уо1. 20. Р. 42-
5.
.
5сон 7. Р. Авог1зИс Бевазюцг оЁ гл1се
ап га: А теме\ // Атег. 7001. 1966.
М 6. Р. 683—701.
5со ФТ. Р., Етедемсвоп Е. Те салзе 9
„
Литература ___ 447
Невыпо 11 пусе ап@ газ // Рнуз10]. 2001.
1951. Уо1. 24, М2.Р. 273-309.
5сой ХТ. Р., Гайег Г. Ё. Сепенсз ап@ з0-
с1а! Бевау1отт оё Фе дор. Сы саро: Ошу,
С сабо ргезз, 1965. 215 р.
5еа1 5. С. Пр!аептоотса! за ез оп р!а-
пе ш Са]сойа: ТЬе гое оЁ Чотезис
ап реготезЫс годеп(з 1 Ще тат-
1епапее о? расе пуесйоп // То4. 7. Мед.
Вез. 1964. Уд1. 49, М 6.Р. 1019—1038.
5еайе А. С. Сошрагайуе вопоМс5 оЁ соа+
со]оцг а пататаа]з. Г., 1968. 400 р.
бей Н. Тве тшоабегпа] шзИпсё 1 апрпа!
зиЪ]есёз // РзусВозоти. Мед. 1954а.
Ус}. 21. Р. 285.
бей; Р. Р. р. Тве еНеосз о пап
ехремепоез проп ада Беваутоит п
апниа| 5050645. 1. ЕНесё оё ИМег яме
Чате шРапсу проп ада Ъевахтойг
1 Ме гаф// Атег. 9. РзусШае. 1954.
Уо1. 1410. Р. 916.
бешей 4. 2. ЧИтазоитдз ш ад го-
Чеп+з
// Мате. 1967. УоЁ. 215. Р. 512.
бешей @. 2. ОНгазоме зи’па1з ош го-
Чегиз // ОЦгазотиез. 1970. Уа1. 8, М 1.
Р. 26—30.
брарто 5., ЭЧаз М. Вевамопта| гезроп-
зе оЁ пап газ No шаегла[ одох//
Рьуз101. ап Вевау. 1970. Уо. 5.
Р. 815-817.
брепкег А. От Тюиг-|еясей епепиез // За-
фофу бег. 1980. Уо|. 8, М 4. Р. 5-6.
Я еве Н. 7. Позепан Т. 5. БигаНоп о
озговеп энтамоп ап ргорежегопе
2ЫЫЯоп оЁ шабегпа! феВау1юог ш ргез-
‘ папсе- тетита{е4 га // Ногшопе Вейау.
1975. Уо1. 11. Р. 12—19.
51 трзов С. 6. ТВе рише]ез оГ сЛазз-
саНоп ап а «ЛаззШсаноп оЁ тшаш-
11815 // ВяП. Атшег, Маз. Маг. Н136.
1945. Уо1. 85. Р. 1-350.
Этнег 7. 7. Аповепиа! ехрогаНоп ап@
1езбозбегопе-ргор1опа{е-пдисей — зехоа|
агоиза! ш гафз // Верау. В1ю01. 1972. \в1. 7.
Р. 743—741.
ЭтЕР Р. 7Т., Горась Е. ОНаслогу Баесфо-
ту ап4 питта репаутоиг т гаф рире //
Реуе1ор. Рзусвоью01. 1975. Уд. 8.
Р. 151-154.
Этер Р. Т., Гискег А. М.. Нофег М. А.
3 Нес! 0 паза! 7150. шыпамоп ап
оасфогу Быесоту оп гаё рирз // РВу-
$101. ап@ Вевау. 1976. \Уо1. 17. Р. 373—
382.
ЗТописк В. М., Сагрещег М. Г., Риазсо В.
Пучаноп 0[ шаегва! Ъовамюиг т
рута шиНрагойз газ // Ногизопе Ве-
ау. 1973. Уо1. 4. Р. 53—59.
бтаЙ У’. 5. Noмез оп Ще рзусЬ1е дече-
1оршепЕ оп Ше уоцие \ВЦе га\ // Аюег.
7. Рзусво]. 1899, Уо1. 11. Р. 80—100.
тив Г. Ап ехрегилеша] га ега@саНов
рговгаш 11 ап игБап агеа // Ра. Не-
ай} Вер. 1963. Уо|. 78. М 9. Р. 807-
811.
5тий 17. 7. Тне Бева\1опг ог соттилг-
саНия. Сатфт! се: Нагуага От. ргезз,
1977. 242 р.
Утойегтап М. Р. Каа| еагуле 1 е-
то // ЕЕТоюсу 85: 19 Пиегп. паб. еФо1.
сот: АБзёгасе. Тоцючизе, 1985. Уо|. 1.
Р. 85.
Утотегтап И’. Р., ВеЙ В. И’., Загаес 7.
е{ а1. Мафегпа| гезропзез ю паи у0-
са|НзаНопз вп оМасюгу сиез т га
ап@ пусе // ВеВау. Во]. 1974. Уо|. 12,
МNo1.Р. 55—66.
бтотегтап No. Р., ИПепег 5. А., Мепдо-
за 5. Р., Герте 8. РИиаЦагу
— айгева]
тезропз{уетезз о{ гаф то{Тегз 10 пох10и5
эра ав зЫшоН ргодисей Ъу рирз //
СЪа Еопл@. Зушр. 1976. Уо1. 45. Р. 5- °
25.
бопЕ Сап, Напе СЪапазВон, Глаа Ниамт
© а1. ТзоаНоп от ЕНЕ-те]аед асе
Кот ВаМиз погуез1сия сарытед Кош
рабетз Воте ш епфешис агеа оЁ Фе
шИа ‘уре оЁ ВетогтВаяе 1еуег
// Аца
тасгорю!. зпса. 1982. Уо]. 22, М 4.
Р. 373-3177.
Эрве! 7. М., Етазег В. «Ропизапсе» т
ЧВе Тафогаюгу гас Те етегоепсе о
отоонциа // 74зсВг. ТиегрзусВо!. 1974.
Ва. 4. 8. 59-69.
.
Умраата А. 5. Ваз ап@ Фет сопго1
Гау4еу. Т. Зе. Тесрпо]. 1966. \01. 4,
МА. Р. 207-240.
|
Э1езег В. И’., Ниапе Н. Н. Н. ТВе герго-
Чисиуе ЧесИпе ш тайе га{$ // Мепгоет-
Чостто бу. Аста. М. У; ТГ. 1983.
Р. 123—142.
Яетег А. Ё. МомаШу тез те Бош Ш-
{тазреНос Нойбое п зоше стоип@ зааи-
те] роршаНоп //7Т. Маша]. 1972.
Уч. 53, М 3,Р. 15—16.
ейивег Е. Вейтёсе гаг Зомлооре ипа
зопзНоеп В10о1е 4ег \Мапаегтайе //
ЯАзевг. ТлегрзусВо|. 1950. Ва. 7. $. 356—
3 79.
бЧетищег К. Вамепьююе ип ВаНептре-
Кипра етзсВЦеззНсВ т ег Тохо-
1овле вартёчеВНсвеп ВаМепре. Зиц-
1рагы Ешке, 1952. 149 5.
бтепивег РЁ. Кобзон е ег ВаНептфе-
КАтарЕ ие // 25]. Васбегю|., РагазЦецк.,
ТоеЕНопзКгаткВ. ип@ Ну. 1958.
Ва. 160, М 1/5. 5. 300-304.
б1епичег Е. Ге уоПаодше Везейриия
446
Литература
тодепис!
дез // Ес4. Ргос. 1967. Уо?. 26,
No4.Р. 1082—1088.
ВоферИа 7. Тце согриз Пиечт — рНаНа-
ту теаНопзВрз: Тие азвосмаНоп Ъе!-
муееп Ще сапзе оГ Пнесотор!а зестеЙоп
ап@ \Ше сапзе о? ГоШеШаг дфиезепсе
Читте Табаоп // Епдосео]ову. 1960.
Уо1. 67. Р. 9—41.
Воше Г. Р. Ваз ап@ писе ш меКз/
Аве Щите. 1962. У]. 7. Р. 28—30.
Виду 7. И’., СвеаНе М. В. ОМасюту зИ-
ша|Н ап@ Ие рзеадосхипсйон еНесв //
З1епсе. 1977. Уо|. 198. Р. 845—846.
Виззе Р. А. П4апе зишШайоп ш го-
Чеп{з: А сопы4егайот оЁ роззИе те-
сВап1эта // Рзусво]. Ви. 1971. М 3.
Р. 192—202.
Засйз В. Р.. Мезй П. Ь. Рафема! деуе-
1ортепё оЁ репЙе геЙехез ап@ сорийа-
Нов ш мае га // Рзуспопеигоетдо-
стпо]осу. 1979. \01. 4. Р. 287—296.
бафег В. М. Е. Подовлса] сопзедцепсез
0 Пасбамоп // Асба 2001. [епп. 1984.
\о1. 171.Р. 179—182.
бат: Стопз М.С. Тез шашиШегез @е
Егапсе её 4и Вепашх. Р.: РОТХ, 1973.
481 р.
без С. 2. ЧЦтазовтЯ ап@ тшайое Бе-
Вауопг ш годепёз УИ зоте офзегуа-
900$ оп офег Бевауоига| зИиайот$//
7. 2001. 1972а. Ус]. 168, М 2. Р. 149-
164.
5е @. О. Ойтазопи@ апд аротезяуе
Бевау1оиг т габз апд офег зта|Н пзат-
та]// Арпи. Вевау. 1972Ъ. Уо]. 20,
Р. 88—100.
51 ©. Р., Руе Р. ОНгазотие сопииит-
сайоп Ъу апиа1з. Т.: СВаршап ап4д
НаН, 1974. 281 р.
5а(е; С. О., 5тий У. С. Сотрагайуе з1-
41е$ о Ше иКтазот1с саНз о шащ
тие — годен» // Оеус]ор. — Рзусво о.
1978. Ус. 11, М 6. Р. 595—619.
бапсйез-Спа4о 7. Е. туд метеп® оф Ме
уотегопаза| зузешт т ЧМе тергодисй-
уе рвуз101юсу оЁ Ме га*// ОНасИоп ап&
епдосгле геоц]айоп. Т.: ТАБ ргезз,
1982. Р. 209-218.
бапслез-Стадо ХФ. Р., ааЦево А. Мае ш-
Чисей ргесостоптз-раЪегу ш Фе Гота
тай Вое оЁ Ме уотетопаза] зузет //
Ас+а епдосгло|. 1979. Уо|. 91, заррЁ.
225. Р. 254,
бапказишап Т!., Ропвргади Р., Водфаае-
{а Р. её а1. Мите 1урВиз ш ТВаПазЯ //
Ттапз. Воу. 306. Тгор. Мед. апа Нур.
1969. Ус]. 63, No5.Р. 639—643.
баю МNo., НаПег Е. М’. Рошей В. О., Неп-
т Р. Т. Зехиа! шашгайов т БаЪес®-
111269 Тета!е га{з//Т. Вергод. Еег%.
1984. Уо1. 36. Р. 301-309.
5сйаае В. Гиегаитетсь& гаш Ве$13еп7-
тесзап1зшиз 4ег УУвидеггайе петепй-
ег шапеЖеп Ап соави]атщеп // Апоех.
Рагазио. 1974. Ва. 15, М4. 8. 201-
Зерие [ег Е. И’., ВоиДез И’. Д. Асще ога]
10х16у ап@ гереНепсу оЁ 933 сВели-
са! 10 Копзе ап 4еег писе // АгсВ.
Елутоп. Соп{ат. ап@ Тох!с01. 1985.
У]. 14. Р. 411-129.
Зераерег С. За М. А, Кеюйе С.
Агоп С. ОНасюту шу/топтет ап@ 1ог-
90518 Бевауюиг ш фе Гете ап шаГе
гаф // ОПасИоп ап@ епфосгте геощаНол.
Г.: ТВГ ртезз, 1982. Р. 115—125.
бейШег Е. Г. Есоюру ап пеаМ оЁ Ван
{0$ аб Моше, АЙазка //Т. Матита|. 1956.
У\о1. 37. Г. 181-188.
бевтапойт С. 5., Нази 9. Е, Битут-
ре ФТ. М. её а1. Апйвегие ап депейс
ргорегНез оЁ утизоз Пике 10 Нетог-
Грар1с Теуег УИ гепа| зупакоше // 8с1-
епее. 1985. Уо|. 227. Р. 1041-1044.
Эсйтиа Е. Вотаегее сте ЕщепеембПе
1 Апр (ВТ) // Отл Во]. ВеоЪ. 1984.
Ва. 81, No 3. 5. 249-254.
Эсйпиттепьегиег Р. Н., Напзоп ЕЁ. В., Маг-
ни В. 7. Терювртов8: Топе-бетий 8ит-
уеШапсе оп ап ПШпо1$ Гага
// Атаег.
2. ЕрЯ. 1970. Уд]. 92, М 4. Р. 223—
239.
успиртег И’., Киепеп И’. А. Не моогВо-
шеп 4ег зроспаеюет уап Ве {уре 4ет
Эр. 1юбегопаетоттваятае Ъ1] гаМел, т
АтяетЧат // Ме. ТазсВг. у. Сепеезк.
1923. Уо1. 67. Р. 20—22.
Эспитасйег 5. К., Мойх Н. ЛЪе таегпа!
рНеготопе оЁ {Ве гаф аз ап шла 38-
шиаз 1ог ргезуеапиих уоцпо // РВузю|-
ап@ Вевау. 1982. Уч]. 3, М 1. Р. 67-71.
ЭейшагЕ Е. СазуИсаЙол, оош ап Че
ионов оЁ сопилептза! газ
// Вай. *
УУо4 НеаМ\ Отгалих. 1960. Уо|.. 2/3.
Р. 411-416.
5сон Н. ВодепеЪогпе Фзеазе сопио]
Игоцей тгодепё эюрраве. Рь 1, 2// Рез
Сопи. 1963. \01. 31, М 8. Р. 30-46;
М Э.Р. 18—28.
5сой 7. Р. Ст са] рег1о48 ш Ще 4еуеор-
еп 0{ зос1а1 Бева\мюочг 11 рирруез //
Русвозот. Мей. 1958. Уо1. 20. Р. 42-
5.
.
5сон 7. Р. Авог1зИс Бевазюцг оЁ гл1се
ап га: А теме\ // Атег. 7001. 1966.
М 6. Р. 683—701.
5со ФТ. Р., Етедемсвоп Е. Те салзе 9
„
Литература ___ 447
Невыпо 11 пусе ап@ газ // Рнуз10]. 2001.
1951. Уо1. 24, М2.Р. 273-309.
5сой ХТ. Р., Гайег Г. Ё. Сепенсз ап@ з0-
с1а! Бевау1отт оё Фе дор. Сы саро: Ошу,
С сабо ргезз, 1965. 215 р.
5еа1 5. С. Пр!аептоотса! за ез оп р!а-
пе ш Са]сойа: ТЬе гое оЁ Чотезис
ап реготезЫс годеп(з 1 Ще тат-
1епапее о? расе пуесйоп // То4. 7. Мед.
Вез. 1964. Уд1. 49, М 6.Р. 1019—1038.
5еайе А. С. Сошрагайуе вопоМс5 оЁ соа+
со]оцг а пататаа]з. Г., 1968. 400 р.
бей Н. Тве тшоабегпа] шзИпсё 1 апрпа!
зиЪ]есёз // РзусВозоти. Мед. 1954а.
Ус}. 21. Р. 285.
бей; Р. Р. р. Тве еНеосз о пап
ехремепоез проп ада Беваутоит п
апниа| 5050645. 1. ЕНесё оё ИМег яме
Чате шРапсу проп ада Ъевахтойг
1 Ме гаф// Атег. 9. РзусШае. 1954.
Уо1. 1410. Р. 916.
бешей 4. 2. ЧИтазоитдз ш ад го-
Чеп+з
// Мате. 1967. УоЁ. 215. Р. 512.
бешей @. 2. ОНгазоме зи’па1з ош го-
Чегиз // ОЦгазотиез. 1970. Уа1. 8, М 1.
Р. 26—30.
брарто 5., ЭЧаз М. Вевамопта| гезроп-
зе оЁ пап газ No шаегла[ одох//
Рьуз101. ап Вевау. 1970. Уо. 5.
Р. 815-817.
брепкег А. От Тюиг-|еясей епепиез // За-
фофу бег. 1980. Уо|. 8, М 4. Р. 5-6.
Я еве Н. 7. Позепан Т. 5. БигаНоп о
озговеп энтамоп ап ргорежегопе
2ЫЫЯоп оЁ шабегпа! феВау1юог ш ргез-
‘ папсе- тетита{е4 га // Ногшопе Вейау.
1975. Уо1. 11. Р. 12—19.
51 трзов С. 6. ТВе рише]ез оГ сЛазз-
саНоп ап а «ЛаззШсаноп оЁ тшаш-
11815 // ВяП. Атшег, Маз. Маг. Н136.
1945. Уо1. 85. Р. 1-350.
Этнег 7. 7. Аповепиа! ехрогаНоп ап@
1езбозбегопе-ргор1опа{е-пдисей — зехоа|
агоиза! ш гафз // Верау. В1ю01. 1972. \в1. 7.
Р. 743—741.
ЭтЕР Р. 7Т., Горась Е. ОНаслогу Баесфо-
ту ап4 питта репаутоиг т гаф рире //
Реуе1ор. Рзусвоью01. 1975. Уд. 8.
Р. 151-154.
Этер Р. Т., Гискег А. М.. Нофег М. А.
3 Нес! 0 паза! 7150. шыпамоп ап
оасфогу Быесоту оп гаё рирз // РВу-
$101. ап@ Вевау. 1976. \Уо1. 17. Р. 373—
382.
ЗТописк В. М., Сагрещег М. Г., Риазсо В.
Пучаноп 0[ шаегва! Ъовамюиг т
рута шиНрагойз газ // Ногизопе Ве-
ау. 1973. Уо1. 4. Р. 53—59.
бтаЙ У’. 5. Noмез оп Ще рзусЬ1е дече-
1оршепЕ оп Ше уоцие \ВЦе га\ // Аюег.
7. Рзусво]. 1899, Уо1. 11. Р. 80—100.
тив Г. Ап ехрегилеша] га ега@саНов
рговгаш 11 ап игБап агеа // Ра. Не-
ай} Вер. 1963. Уо|. 78. М 9. Р. 807-
811.
5тий 17. 7. Тне Бева\1опг ог соттилг-
саНия. Сатфт! се: Нагуага От. ргезз,
1977. 242 р.
Утойегтап М. Р. Каа| еагуле 1 е-
то // ЕЕТоюсу 85: 19 Пиегп. паб. еФо1.
сот: АБзёгасе. Тоцючизе, 1985. Уо|. 1.
Р. 85.
Утотегтап И’. Р., ВеЙ В. И’., Загаес 7.
е{ а1. Мафегпа| гезропзез ю паи у0-
са|НзаНопз вп оМасюгу сиез т га
ап@ пусе // ВеВау. Во]. 1974. Уо|. 12,
МNo1.Р. 55—66.
бтотегтап No. Р., ИПепег 5. А., Мепдо-
за 5. Р., Герте 8. РИиаЦагу
— айгева]
тезропз{уетезз о{ гаф то{Тегз 10 пох10и5
эра ав зЫшоН ргодисей Ъу рирз //
СЪа Еопл@. Зушр. 1976. Уо1. 45. Р. 5- °
25.
бопЕ Сап, Напе СЪапазВон, Глаа Ниамт
© а1. ТзоаНоп от ЕНЕ-те]аед асе
Кот ВаМиз погуез1сия сарытед Кош
рабетз Воте ш епфешис агеа оЁ Фе
шИа ‘уре оЁ ВетогтВаяе 1еуег
// Аца
тасгорю!. зпса. 1982. Уо]. 22, М 4.
Р. 373-3177.
Эрве! 7. М., Етазег В. «Ропизапсе» т
ЧВе Тафогаюгу гас Те етегоепсе о
отоонциа // 74зсВг. ТиегрзусВо!. 1974.
Ва. 4. 8. 59-69.
.
Умраата А. 5. Ваз ап@ Фет сопго1
Гау4еу. Т. Зе. Тесрпо]. 1966. \01. 4,
МА. Р. 207-240.
|
Э1езег В. И’., Ниапе Н. Н. Н. ТВе герго-
Чисиуе ЧесИпе ш тайе га{$ // Мепгоет-
Чостто бу. Аста. М. У; ТГ. 1983.
Р. 123—142.
Яетег А. Ё. МомаШу тез те Бош Ш-
{тазреНос Нойбое п зоше стоип@ зааи-
те] роршаНоп //7Т. Маша]. 1972.
Уч. 53, М 3,Р. 15—16.
ейивег Е. Вейтёсе гаг Зомлооре ипа
зопзНоеп В10о1е 4ег \Мапаегтайе //
ЯАзевг. ТлегрзусВо|. 1950. Ва. 7. $. 356—
3 79.
бЧетищег К. Вамепьююе ип ВаНептре-
Кипра етзсВЦеззНсВ т ег Тохо-
1овле вартёчеВНсвеп ВаМепре. Зиц-
1рагы Ешке, 1952. 149 5.
бтепивег РЁ. Кобзон е ег ВаНептфе-
КАтарЕ ие // 25]. Васбегю|., РагазЦецк.,
ТоеЕНопзКгаткВ. ип@ Ну. 1958.
Ва. 160, М 1/5. 5. 300-304.
б1епичег Е. Ге уоПаодше Везейриия
448
Литература
ег ВаИеп пп Наеп- пла Еземта9и-
зачесе 1е уоп СихВауер // Мепез АгеВ.
NoедегзасВзеп. 1955. В4. 8/43, Н. 5.
5. 395—397.
Зфепивег РЕ. Вайег!тыег НаРеп пл га@-
фептеез ЕАзе ти -чесе ев {п СихВа-
уеп // РезшеК, ипа Сезипанеу. 1956.
В4. 48. Н. 11. $, 30—31.
Блеривег Г. п НаГеп овпе НаНеп // Коз-
оз. 1957. Н. 6. 8. 295—299.
Э1еинег Г. 2 Тарте гаценеез Е зсйе-
теша еп ива ЕзоБтдизьчетеек т
Сихпауел // РезоК. пой Сезап@ейзуу.
1958. Ва. 50. Н. 6. $. 86-88.
буейияег Е. ПаМеп Котттеп аиз дет
3УС
// ПЫа. 1960. Ва. 52, Н. 12. $. 473—
475.
Чепикег Р. Ее таметйтее Ргоуше //
ПЫ4. 1961. Ва. 53. И. 7. 8. 105-107.
Зетп 7. 7. Везропзез ор та гаёз 10 зех
офогз // Рвуз101. ав Вевах. 1970.
Уч. 5.Р. 519-524.
51егп 7. М. ЕНесф оЁ огоосгурыте оп
розбрагиии табегпа! Ъепа\1оиг, оуаап
сусШу ап4 [004 пуаке 11 га\з // Вевау.
В101. 1977. \о1. 21. Р. 134-140.
51етп 7. М., МеКтпоп Г. А. Зепвоту теоп-
Лайоп оЁ табегпа] Бевам1оие п гафз: Е
1066 оЁ рир аве// Беуе]ор. РзусВоЫо1.
1978. Уо|. 11. Р. 579-587.
5фети 7. М., Пека Н. Г. Реоасип гееаве
т Лафайле риширагоиз ап@ тшира-
гопз {Пеесфоти12е ап шафегпа! утот
тайез ехрозей No0 рир зЫшоН // Вевах.
В191. 1978. Уо1. 19. Р. 177-182.
ети 7. М. Твотаз р. А., Ваьи У. Ваг-
реа В. 1. Оо рир аНтазоте стез ргоуо-
се ргоасйп зесгемоп т ЛасфаНоп габз? //
Ногиопе Вевау. 1984. \0|. 13, М 1,
Р. 86—94.
5{етенз О. А., Сегтов-Тпотаз 0. А. ЕН
теасйоп 1 газ Noю сопзресНе Язвче //
Рвуз1о] Вебау. 1977. Уо|. 18, М 1.
Р. 47-54.
Берет; О. Л. Коцег Е. Р. Ореп-Неа
гезропзе$ о{ гафз офогз {тот зтеззе@ ап@
попзгеззей ргедесеззогз // Верау. В101.
1972. Ув]. 7, М 4.Р. 519-525,
Зюа4ат 1. М. Зеазоп уамаБИиу оф Мог-
мау гаё (ВаМиз погуед1стз) шШезаНоп
0Ё авмеш а] ртетизе //Т. 200]. 1984.
Уо]. 194. Р. 257—289.
Эюпе Е., ВоппеЁ К., Но]ег М. А. Зикита
ап 4еувортепе оЁ{ шабегпа]у Дертуед
гафз: Во]е оЁ Ъоду 1ютрегабиге // Рзу-
сВозот. Мед. 1976. Уз. 38. Р. 242—
249.
510пе В. 7. Сопыо] оЁ Ыта ап@ апира1
Фатаре 10 сторз Ъу Ме сНешиса] ге-
реет «СИВВ» // \У. Ат. Тесвп. Веу.
1977. Топе. Р. 55—56, 58.
|
таюоп С., Вегсоле С., бтаюп А. её а.
За гозегуоги пабига! Че Тервозр!-
те ат 4еНа Бопаеу:
// Веу. те4. св.
{Н5В). 1977. Уо1. 81, М 2.Р. 233—237.
51топа Р. С. РорМаНоп дупапиез 9 Вайия
гаМиз ап4 В. погуедеаз ш гратап Ва-
Ъапё//Т. Магииа!. 1982. Уо|. 63, М 1.
Р. 151-154.
Этирр В. 1, Геризву Ш. А. Зоба| гапз-
пуззюй оф {004 рге{егепсез 11 ада
Воо4е гаёз (ВаМаз погуев1еив) /!
7. Сошр. РзусвоЁ. 1984. Уо|. 98. Р. 257—
266.
Такавазя Г. К. Готе В. К. Рау Новиие
ап Фе 4еуоршептф о або Ыс Ъе-
Вамоиг ш шае апд 1ета|е га1{$ // Аб-
огез. Ведах. 1983. Уд]. 9, М 3. Р. 217-
227
у
Такайазм Г. К., Твотаз 0. А., Ваг|-
е14 В. 7. Апа]уз13 9 иИтазоте уосаНта-
Нопз епыймей Ъу тея4еАз Чигше а9-
втезыуе епсоитиегз атопо га1з (Вацие
погуе1сиз) //Т. Сошр. РзусПо|. 1983.
Уо1. 97, М 3. Р. 207-212.
Тапиуа Т. Весеп6 адуапсез 1 збабез оЁ
Тя\зиратизН1 41зеазе т Фарап. То-
Куо: Мед. си. 1пс., 1962. 302 р.
Тапака В. Оп ЧШМетепНа| гезропзе 0 Ше
{тарз 0Р тагКей ап питагкей зта|.
шатагаа]з // Аппо{. 2001. Тар. 1956.
Уч]. 29, М1. Р. 44-51.
Тапя С., бип Г, мп М. ГНесз ОР зо-
с1а]1 1зо1аНоп оп Фе Бепа\1отг о? тот:
ар габз Чагииу зов] згезз // Асфа рзу-
ера, зныса. 1984. Уо1. 16, М 4. Р. 435—
440
Тие С. И. Н. Зоте Мимаае оГ фе Тодо-
АпзбгаНап геотоп // ВЦ. Ашег. Миз.
Маг. Шзё. 1936. Уч]. 72, ам, 6. Р. 504—
728
Ташог @. Т. Еойуоя т ауепПе газ ао
{пе ответу оЁ абоп1зМе Бевамютх //
7. Сошр. апа Рвуз101. Рзусво|. 1980.
Уо]. 94. Р. 953—961.
ТГауог в. Г, НаЦег 7., Ват Ко @., И’еззз 1.
Сопзрес!Не птше таткте шт та1е-Ре-
ша!е раз о{ Лафогаботу газ // Рвуз101..
ап@ Верау. 1984. Уо]. 32. Р. 544-546.
Тауог С. Т., НаПег Т., Вевап ПО. Еетае
гаёз ргеег ап агеа уасайей Бу а ШВ
чозючщегопе шае // 114. 1982. У\о|. 28.
Р. 953—958.
Тауог ФТ. М., Ногпег В. Е. ВезаЦз оЁ 1е:
АтсЬЪо ехре@Нолз, М 98: ЗужетаНся:
09 памуе АчэтаНап ВаИаз (Водепца,
Мин Чае) // Ви. Ашег. Миз. Мабиг. Н!з.
1973. Уо|. 150, агб. 1. Р. 4—180.:
„
Литература
449
Тают 7. М., Саафу У. Н., 5тиВ 5. С.
Мануе Цабиз, ]ап4 }г19сез, ап@ Апяга-
Нап гезтоп //Т Машшз[. 1983. У. 64,
М 3. Р. 463—475.
Талог ФУ. М., Сщафу У. Н., Уап Беи-
зепН.М.Атем!100оЩесепизВае-
паз (Водепыа, МитЧае) ше Мех
‘итеап гед1оп // ВоП. Ашег. Маз. Ма-
Ш. НИЗ 1982. Уо|. 173, агё. 3. Р. 177-
36
Гайог К. О. бепаз Вамиаз // ПапаъосКк о!
ВгИЯзВ тпапила13, 210 е4. Охгога: Их.
ргезз, 1977. Р. 234-247.
Тауог К. Б. Вапее о шоуешеп ап@ асё-
УНу оЁ соттоп габз (Ваз погуер!сл$)
оп авттсаНате Тапа //Т. Арр1. Есо|. 1978.
Уо1. 45. Р. 663—677.
Ташог К. В, Паттоп Г. Е., Оиу В. $.
Тре геасИопз оЁ соттроп газ 10 Гог &у-
рез оЁ Пуе-сарыше тар // 19. 1974.
Уа1, 41. Р. 453—459.
Тауог К. О. Опу В. Г. Мова зузет
о{ Фе зу оЁ гар тоуетшеш, // Мат-
а. Веу. 1973. Ус. 3 М2.Р. 80-
Тауог К. Р., Оу В. 7. Гопо Я запасе по-
уетет{з оЁ а сошиюоп гаф (ВаЙиз пог-
Уе51спз) теувей фу гаобтасКшо //
МаттаНа. 1978. Ув|. 42, М 1.Р. 63—
71.
Тауог К. П., Опу В. Т, Сигпей У. Соп-
рачзоп о{ Штее те оз {ог езбтаНио
бе пшофегз оЁ сотиоп га1в (Вабаз
погуес1с13) // Па. 1981. Уо1. 45, М 4.
Р. 403--443,
Тесрег М. Н., Ваз; Е. М. ЗасЕНие Ш
° пезфоги гаёз: еПпитаеа Бу пре 1э-
уаре гезме@ Ъу рир заа // Зфепсе.
1977а. Уо|. 193. Р. 422—425.
Тесйег М. Н. Ваз; Е. М. Ен васКИая
гезропзе оЁ Ще пежфоги ато гаё: \е
го]е оЁ ЛафаНоп ап@ атиюнс-Ниа//
14. 1977Ъ. Уо1. 198. Р. 635—637.
Тесйег М. Н., апт Г. Е., ТИНатз М.
еф а. биту! уа], отомИб. заеКНаи Бева-
Уоог о{ пеопабаНу БиШБесюнизе габз //
Р|Вузю1. ап Вевау. 1978. У9. 2.
Р. 553—561.
Теце Н. 7. Уапаегганеп по ЕтеЙапа—
ош пецез ВекатрТиатозргоет? // Аля.
ЗевааНпязКк. 1962. Ва. 35, М 4. $. 53—
54
“Гейе Н. 1. Бе позсЬиаскзаБ\уе!вепае
УИткил& ешшшег СитагтаетРуаце // Ма-
ситюетЪ]. 0%. РНаптепзетияел8.
1963. Ва. 15, No 5. 8. 70—73.
ТеЦе Н. 7. Вейгая гиг Кепиниз 4ег Ует-
ВаНепзлуе!зеп уоп ВаЦеп // Аппе\. 700].
1966. М 53. 3. 129—196.
ТеНе Н.Т. Ге АпзуаШ уоп Водепы214еп
Гаг Вайепуег юппоев ипа 16г фе Ве
Берайиар ешез таМешгееп Япзап-
4ез
// Ап. ЗевёаНирзк. 1967. Ва. 40,
М 11. $. 161-166.
Тейе ПН. 7. ?мг Ргоетайк.4ез ВергИ-
Рез чтайеш
тер» // ПЧ. 1968. Ва. 441,
М 9. 5. 149-125.
Те|е Н. 7. КогётоЦе ива Вемегнтиезтад-
&ПеВКейеп уоп отоззгёиииюет Векёит-
Рапозмаопеп седеп \апдегтаНет
({ВаИмз — погуее1си5) // 7АзеВг. апеех.
Соо1. 1971. Ва. 58, No2. 8. 141-167.
ТеЦе Н. 7. Вемзепт? уоп \У/апдегтанеп
девепйрег \УУагЁатт 11 ВипдезтераЬИЕ
Бешзе ана // Ап. Зев5аНлозк. РЙал-
попвовия. 1972. 34. 45, No2. $. 17-
2
Тейе_Н. 7, Уаик С. ОЪъег баз УегнаНеп
914 4е УетосВишо 4ег УУапдетгаце
(Вайаз погуешеиз ВегКк.}) пи Табге
1962, ащЁ Неюофапа // Ап? ЗевааНиозК.
1963. В4. 36, М 5. $. 70-73.
Тегке /., Ратазза О. А., башег СЪ. Н.
О газо е сыез Кош шШфап та8 зН-
„, пиае рговеци гееазе т Лабайне
тпоПотз // Ногшопе ВеВау. 1979. У01. 12,
М1.Р. 95-102.
Тегке! Х., Возеп Ман У. 5. Маегпа] Ъо-
Ба\1опг Га@исей Бу шаегпа]-Зо04 р1аз-
па И1]е0еа пцо упрш гав// 7. Сотр.
ап РВуз0]. РэзусВо]. 1968. Уо|. 65.
Р. 475—482.
Тегке 7., Возепой У. 5. Азрес1$ о
поппогшопа! таёегпа| февау:оиг п Фе
га// Ногшопе Вевау. 1971. Уд. 2,
М2.Р. 161-171.
Тегке! 7, Возепа 7. 5. Ногтопа! #ас-
фот ипаеутя шабегпа! БеБау!оцг а
раготаМоп: сгоззгапз азот Ъегуееп
Небу шоушь га{з//). Сотр. ава РВу-
8101. РзусВоГ. 1972. Уо]. 80. Р. 365—
371.
Тщегтатт А. В. ТщеЧепсе о ]ерюзриго-
313 т Ме Бетой, гаф рорщаНоп // Атпег,
7. Тгор. Мей. ап4 Ну. 1977. \о]. 96.
Р. 970—974.
Тмегтапи А. В. ТЬе Могуау 15 аз а
зе]есИуе сВголе сагмег оЁ Герюзрига
1с4еговаетоггваотае //Т. \УПаШМе Гузеа-
зез. 1981. УоТ. 47, М 1. Р. 39—43.
Твотаз Р. А, Ношата 5. В., Ваграа В.7,
Мае ргодисей иИтазот:е уосаНгайопв
эп@ тамир райегиз ш Тета!е гайз//
7. Сошр. ап Рвузо]. Рэзусво!. 1982.
Уо1. 96. Р. 807-815.
Тротаз 0. А., Таровазт Г. К. Вагй-
214 В. 7. Апа]узз оЁ иНтазопе уоса|.
таНопз еп1Ме4 Ъу Иицтиегз Читтр ар-
448
Литература
ег ВаИеп пп Наеп- пла Еземта9и-
зачесе 1е уоп СихВауер // Мепез АгеВ.
NoедегзасВзеп. 1955. В4. 8/43, Н. 5.
5. 395—397.
Зфепивег РЕ. Вайег!тыег НаРеп пл га@-
фептеез ЕАзе ти -чесе ев {п СихВа-
уеп // РезшеК, ипа Сезипанеу. 1956.
В4. 48. Н. 11. $, 30—31.
Блеривег Г. п НаГеп овпе НаНеп // Коз-
оз. 1957. Н. 6. 8. 295—299.
Э1еинег Г. 2 Тарте гаценеез Е зсйе-
теша еп ива ЕзоБтдизьчетеек т
Сихпауел // РезоК. пой Сезап@ейзуу.
1958. Ва. 50. Н. 6. $. 86-88.
буейияег Е. ПаМеп Котттеп аиз дет
3УС
// ПЫа. 1960. Ва. 52, Н. 12. $. 473—
475.
Чепикег Р. Ее таметйтее Ргоуше //
ПЫ4. 1961. Ва. 53. И. 7. 8. 105-107.
Зетп 7. 7. Везропзез ор та гаёз 10 зех
офогз // Рвуз101. ав Вевах. 1970.
Уч. 5.Р. 519-524.
51егп 7. М. ЕНесф оЁ огоосгурыте оп
розбрагиии табегпа! Ъепа\1оиг, оуаап
сусШу ап4 [004 пуаке 11 га\з // Вевау.
В101. 1977. \о1. 21. Р. 134-140.
51етп 7. М., МеКтпоп Г. А. Зепвоту теоп-
Лайоп оЁ табегпа] Бевам1оие п гафз: Е
1066 оЁ рир аве// Беуе]ор. РзусВоЫо1.
1978. Уо|. 11. Р. 579-587.
5фети 7. М., Пека Н. Г. Реоасип гееаве
т Лафайле риширагоиз ап@ тшира-
гопз {Пеесфоти12е ап шафегпа! утот
тайез ехрозей No0 рир зЫшоН // Вевах.
В191. 1978. Уо1. 19. Р. 177-182.
ети 7. М. Твотаз р. А., Ваьи У. Ваг-
реа В. 1. Оо рир аНтазоте стез ргоуо-
се ргоасйп зесгемоп т ЛасфаНоп габз? //
Ногиопе Вевау. 1984. \0|. 13, М 1,
Р. 86—94.
5{етенз О. А., Сегтов-Тпотаз 0. А. ЕН
теасйоп 1 газ Noю сопзресНе Язвче //
Рвуз1о] Вебау. 1977. Уо|. 18, М 1.
Р. 47-54.
Берет; О. Л. Коцег Е. Р. Ореп-Неа
гезропзе$ о{ гафз офогз {тот зтеззе@ ап@
попзгеззей ргедесеззогз // Верау. В101.
1972. Ув]. 7, М 4.Р. 519-525,
Зюа4ат 1. М. Зеазоп уамаБИиу оф Мог-
мау гаё (ВаМиз погуед1стз) шШезаНоп
0Ё авмеш а] ртетизе //Т. 200]. 1984.
Уо]. 194. Р. 257—289.
Эюпе Е., ВоппеЁ К., Но]ег М. А. Зикита
ап 4еувортепе оЁ{ шабегпа]у Дертуед
гафз: Во]е оЁ Ъоду 1ютрегабиге // Рзу-
сВозот. Мед. 1976. Уз. 38. Р. 242—
249.
510пе В. 7. Сопыо] оЁ Ыта ап@ апира1
Фатаре 10 сторз Ъу Ме сНешиса] ге-
реет «СИВВ» // \У. Ат. Тесвп. Веу.
1977. Топе. Р. 55—56, 58.
|
таюоп С., Вегсоле С., бтаюп А. её а.
За гозегуоги пабига! Че Тервозр!-
те ат 4еНа Бопаеу:
// Веу. те4. св.
{Н5В). 1977. Уо1. 81, М 2.Р. 233—237.
51топа Р. С. РорМаНоп дупапиез 9 Вайия
гаМиз ап4 В. погуедеаз ш гратап Ва-
Ъапё//Т. Магииа!. 1982. Уо|. 63, М 1.
Р. 151-154.
Этирр В. 1, Геризву Ш. А. Зоба| гапз-
пуззюй оф {004 рге{егепсез 11 ада
Воо4е гаёз (ВаМаз погуев1еив) /!
7. Сошр. РзусвоЁ. 1984. Уо|. 98. Р. 257—
266.
Такавазя Г. К. Готе В. К. Рау Новиие
ап Фе 4еуоршептф о або Ыс Ъе-
Вамоиг ш шае апд 1ета|е га1{$ // Аб-
огез. Ведах. 1983. Уд]. 9, М 3. Р. 217-
227
у
Такайазм Г. К., Твотаз 0. А., Ваг|-
е14 В. 7. Апа]уз13 9 иИтазоте уосаНта-
Нопз епыймей Ъу тея4еАз Чигше а9-
втезыуе епсоитиегз атопо га1з (Вацие
погуе1сиз) //Т. Сошр. РзусПо|. 1983.
Уо1. 97, М 3. Р. 207-212.
Тапиуа Т. Весеп6 адуапсез 1 збабез оЁ
Тя\зиратизН1 41зеазе т Фарап. То-
Куо: Мед. си. 1пс., 1962. 302 р.
Тапака В. Оп ЧШМетепНа| гезропзе 0 Ше
{тарз 0Р тагКей ап питагкей зта|.
шатагаа]з // Аппо{. 2001. Тар. 1956.
Уч]. 29, М1. Р. 44-51.
Тапя С., бип Г, мп М. ГНесз ОР зо-
с1а]1 1зо1аНоп оп Фе Бепа\1отг о? тот:
ар габз Чагииу зов] згезз // Асфа рзу-
ера, зныса. 1984. Уо1. 16, М 4. Р. 435—
440
Тие С. И. Н. Зоте Мимаае оГ фе Тодо-
АпзбгаНап геотоп // ВЦ. Ашег. Миз.
Маг. Шзё. 1936. Уч]. 72, ам, 6. Р. 504—
728
Ташог @. Т. Еойуоя т ауепПе газ ао
{пе ответу оЁ абоп1зМе Бевамютх //
7. Сошр. апа Рвуз101. Рзусво|. 1980.
Уо]. 94. Р. 953—961.
ТГауог в. Г, НаЦег 7., Ват Ко @., И’еззз 1.
Сопзрес!Не птше таткте шт та1е-Ре-
ша!е раз о{ Лафогаботу газ // Рвуз101..
ап@ Верау. 1984. Уо]. 32. Р. 544-546.
Тауог С. Т., НаПег Т., Вевап ПО. Еетае
гаёз ргеег ап агеа уасайей Бу а ШВ
чозючщегопе шае // 114. 1982. У\о|. 28.
Р. 953—958.
Тауог ФТ. М., Ногпег В. Е. ВезаЦз оЁ 1е:
АтсЬЪо ехре@Нолз, М 98: ЗужетаНся:
09 памуе АчэтаНап ВаИаз (Водепца,
Мин Чае) // Ви. Ашег. Миз. Мабиг. Н!з.
1973. Уо|. 150, агб. 1. Р. 4—180.:
„
Литература
449
Тают 7. М., Саафу У. Н., 5тиВ 5. С.
Мануе Цабиз, ]ап4 }г19сез, ап@ Апяга-
Нап гезтоп //Т Машшз[. 1983. У. 64,
М 3. Р. 463—475.
Талог ФУ. М., Сщафу У. Н., Уап Беи-
зепН.М.Атем!100оЩесепизВае-
паз (Водепыа, МитЧае) ше Мех
‘итеап гед1оп // ВоП. Ашег. Маз. Ма-
Ш. НИЗ 1982. Уо|. 173, агё. 3. Р. 177-
36
Гайог К. О. бепаз Вамиаз // ПапаъосКк о!
ВгИЯзВ тпапила13, 210 е4. Охгога: Их.
ргезз, 1977. Р. 234-247.
Тауог К. Б. Вапее о шоуешеп ап@ асё-
УНу оЁ соттоп габз (Ваз погуер!сл$)
оп авттсаНате Тапа //Т. Арр1. Есо|. 1978.
Уо1. 45. Р. 663—677.
Ташог К. В, Паттоп Г. Е., Оиу В. $.
Тре геасИопз оЁ соттроп газ 10 Гог &у-
рез оЁ Пуе-сарыше тар // 19. 1974.
Уа1, 41. Р. 453—459.
Тауог К. О. Опу В. Г. Мова зузет
о{ Фе зу оЁ гар тоуетшеш, // Мат-
а. Веу. 1973. Ус. 3 М2.Р. 80-
Тауог К. Р., Оу В. 7. Гопо Я запасе по-
уетет{з оЁ а сошиюоп гаф (ВаЙиз пог-
Уе51спз) теувей фу гаобтасКшо //
МаттаНа. 1978. Ув|. 42, М 1.Р. 63—
71.
Тауог К. П., Опу В. Т, Сигпей У. Соп-
рачзоп о{ Штее те оз {ог езбтаНио
бе пшофегз оЁ сотиоп га1в (Вабаз
погуес1с13) // Па. 1981. Уо1. 45, М 4.
Р. 403--443,
Тесрег М. Н., Ваз; Е. М. ЗасЕНие Ш
° пезфоги гаёз: еПпитаеа Бу пре 1э-
уаре гезме@ Ъу рир заа // Зфепсе.
1977а. Уо|. 193. Р. 422—425.
Тесйег М. Н. Ваз; Е. М. Ен васКИая
гезропзе оЁ Ще пежфоги ато гаё: \е
го]е оЁ ЛафаНоп ап@ атиюнс-Ниа//
14. 1977Ъ. Уо1. 198. Р. 635—637.
Тесйег М. Н., апт Г. Е., ТИНатз М.
еф а. биту! уа], отомИб. заеКНаи Бева-
Уоог о{ пеопабаНу БиШБесюнизе габз //
Р|Вузю1. ап Вевау. 1978. У9. 2.
Р. 553—561.
Теце Н. 7. Уапаегганеп по ЕтеЙапа—
ош пецез ВекатрТиатозргоет? // Аля.
ЗевааНпязКк. 1962. Ва. 35, М 4. $. 53—
54
“Гейе Н. 1. Бе позсЬиаскзаБ\уе!вепае
УИткил& ешшшег СитагтаетРуаце // Ма-
ситюетЪ]. 0%. РНаптепзетияел8.
1963. Ва. 15, No 5. 8. 70—73.
ТеЦе Н. 7. Вейгая гиг Кепиниз 4ег Ует-
ВаНепзлуе!зеп уоп ВаЦеп // Аппе\. 700].
1966. М 53. 3. 129—196.
ТеНе Н.Т. Ге АпзуаШ уоп Водепы214еп
Гаг Вайепуег юппоев ипа 16г фе Ве
Берайиар ешез таМешгееп Япзап-
4ез
// Ап. ЗевёаНирзк. 1967. Ва. 40,
М 11. $. 161-166.
Тейе ПН. 7. ?мг Ргоетайк.4ез ВергИ-
Рез чтайеш
тер» // ПЧ. 1968. Ва. 441,
М 9. 5. 149-125.
Те|е Н. 7. КогётоЦе ива Вемегнтиезтад-
&ПеВКейеп уоп отоззгёиииюет Векёит-
Рапозмаопеп седеп \апдегтаНет
({ВаИмз — погуее1си5) // 7АзеВг. апеех.
Соо1. 1971. Ва. 58, No2. 8. 141-167.
ТеЦе Н. 7. Вемзепт? уоп \У/апдегтанеп
девепйрег \УУагЁатт 11 ВипдезтераЬИЕ
Бешзе ана // Ап. Зев5аНлозк. РЙал-
попвовия. 1972. 34. 45, No2. $. 17-
2
Тейе_Н. 7, Уаик С. ОЪъег баз УегнаНеп
914 4е УетосВишо 4ег УУапдетгаце
(Вайаз погуешеиз ВегКк.}) пи Табге
1962, ащЁ Неюофапа // Ап? ЗевааНиозК.
1963. В4. 36, М 5. $. 70-73.
Тегке /., Ратазза О. А., башег СЪ. Н.
О газо е сыез Кош шШфап та8 зН-
„, пиае рговеци гееазе т Лабайне
тпоПотз // Ногшопе ВеВау. 1979. У01. 12,
М1.Р. 95-102.
Тегке! Х., Возеп Ман У. 5. Маегпа] Ъо-
Ба\1опг Га@исей Бу шаегпа]-Зо04 р1аз-
па И1]е0еа пцо упрш гав// 7. Сотр.
ап РВуз0]. РэзусВо]. 1968. Уо|. 65.
Р. 475—482.
Тегке 7., Возепой У. 5. Азрес1$ о
поппогшопа! таёегпа| февау:оиг п Фе
га// Ногшопе Вевау. 1971. Уд. 2,
М2.Р. 161-171.
Тегке! 7, Возепа 7. 5. Ногтопа! #ас-
фот ипаеутя шабегпа! БеБау!оцг а
раготаМоп: сгоззгапз азот Ъегуееп
Небу шоушь га{з//). Сотр. ава РВу-
8101. РзусВоГ. 1972. Уо]. 80. Р. 365—
371.
Тщегтатт А. В. ТщеЧепсе о ]ерюзриго-
313 т Ме Бетой, гаф рорщаНоп // Атпег,
7. Тгор. Мей. ап4 Ну. 1977. \о]. 96.
Р. 970—974.
Тмегтапи А. В. ТЬе Могуау 15 аз а
зе]есИуе сВголе сагмег оЁ Герюзрига
1с4еговаетоггваотае //Т. \УПаШМе Гузеа-
зез. 1981. УоТ. 47, М 1. Р. 39—43.
Твотаз Р. А, Ношата 5. В., Ваграа В.7,
Мае ргодисей иИтазот:е уосаНгайопв
эп@ тамир райегиз ш Тета!е гайз//
7. Сошр. ап Рвузо]. Рэзусво!. 1982.
Уо1. 96. Р. 807-815.
Тротаз 0. А., Таровазт Г. К. Вагй-
214 В. 7. Апа]узз оЁ иНтазопе уоса|.
таНопз еп1Ме4 Ъу Иицтиегз Читтр ар-
450
Литература
дтезыуе есоитиегв ашопя га (ВаНи$
потуер1сив) // Па. 1983. Уо1. 97. Р. 204—
206.
Тротаз О. А, Таша: Г., Ватнаа В. Ф.
ЕНесё5 оЁ деуосаПеаНот 0# 11е таа]е оп
тшайие ревахюйг т та+з
// 114. 1984.
\о1. 95. Р. 630—637.
Тротаз 0. ТЬе репегс аггапаетепё оЁ
Те АизраНап шигтез ВИНего гегогей
10 «Миз»//Апп. ап@ Мас. Мабат. НУ.
бег, 8, 1910. У. 6. Р. 603—607.
Твотрзоп И’. П. Ищвоп У. Срагиз-
гот ТУ. В. Те еес4з оЁ ргепа{а! та-
{егпа| этезз оп озреше Бевау!опг ш
га(з
// Рвусво]. Моповт. 1962. Уо|. 76,
М 38.Р. 12-15.
Трог О. Н., Нойошау И. В. (1т.). Апозиа
ап рау НабИие фова\мюоит Ш ргери-
Ъезсетё ша ап@ Фета!е газ// Рву-
$10]. ‘ап Вевах. 1982. Уо|. 29. Р. 281-
285.
ТРог 0. Н., НоЙошву И’. В. (1'".). Зеоро-
ауте МосКз р]ау-Невыля ш ауеше
та{з // ТЯ. 1983а. Уо|. 30. Р. 545—549.
Трог р. Н. НоПошау И’. В. (1т.). Рву-
зоНеНамов Бева\отг Ш ГауепНИе та|е
апа Геша е га // Апиа. ЁБеаги. Вевах.
1983Ъ, Ув. 11. Р. 178—178.
Трог Р. Н., НоЦошау ТУ. В. (7г.). Зех
ап зобаЁ р!ау 11 фиуевПе гафз (На Низ
погуео1стз) //7. Сошр. РэусНо|. 1984.
\Уо1. 98, М 3. Р. 276-284.
Тпог Р. Н., НоПошау 1’. В. (".). Бехе-
1оршепа! апа!узез оЁ зоса] ау Ъе-
Вачотг т ромедИе гаёз // ВиП. РзуеВо-
пог. бое. 19845. Ув. 22, М 6. Р. 587.
ТРогре И’. Н. ВииаШтаНоп т ощобепу.
4. Апниа! р1ау // РЬПоз. Тгапз. Воу. 306.
Гопдоп. В. 1966. Уо1. 772, М 154.
Р. 311-319.
ТИпеу Е. Вевауюпг т Из геабоп ю Шь
деуе!ортлевь оЁ Фе гаш // ВаН. Мета!
Таз. 1933. М 11.Р. 252-358.
Топись Р. 0. Матта8 ш Намуай: А зу-
порз1з ап побаМопар ЫЪНортарйу [1
В1зрор. Миз. Зрес. Ри. 1969. М 51.
°Р. 238.
Топег 7. Р., А@ег No. Т. Роепсу оЁ тай
ефастаНопз уагез УИВ пеш отаег впа
УИ} ша аче // Рьуз10]. ап@ Вевах.
1985. \а1, 35, No 1.Р. 113—145.
Торзт У. Г., рта 7, Ттепи Напв Оиу.
ТВе годеп гезогуой оЁ 1ерфюзритае т
Могв Уешат // АтеН. гоши. . рао!.
ехр. её пустоБоЁ 1970. \о1. 29, No 1/2.
Р. 331—338.
Ттоиеззат Е. Г. Сааовие дез ташш!е-
тез // Ви]. Зое. Гфадез ЗеЁ. Ап4егв. 1881.
Уи. 10. Р. 18-21.
Ттопеззат Е. Г. Водепиа
// Са{а105 05
татштаНит бат ууепили дааш #0581
Паш. В. 1897. Еазс. 3. Р. 458—664.
Тгонеззат Е. Ё. Сщаювиз таттаНат
{ат уепаниа дааш ТоззЙи. Зирр!е-
тепиит. В. 1904-1905. УП, 929 р.
Тискег Н. А, Тфаейег И. Рийату
ргохИВ Ногшопе апд Нцетииялтя Вогизо-
пе солфешб аНег уагоиз пигяше пиеп-
&1ез
// Ргое. бое. Ехр. ПВ. ав@ Мед.
1968. Уо|. 129. Р. 578—580.
Тише в. Г. Пуечайоп оЁ Пе топ га
(Вайиз погуер1еия) 1 г пишез о
Ме Выизь Т&апдз//7. Нув. 1964.
\о1. 59, М 3. Р. 271-284.
ТьшЕ С. Г. А темех о{ Фе оссигепсе
ш ВизН тпатитаа]з оЁ Ще таа]от огба-
п1зтз 0{ 2оопоНе пароепз // Магата|.
Веу. 1980. У]. 140. М 4. Р. 139-149.
Туии К. Пметгеайов фефбуееп Бай соп-
зирИоп, ргоюпзаНой ог робот
те ап Весгеазе соЁ 004 сарШату гв-
зфапсе Фу соптаага годепйс1Че, \уаг-
{ага // Noрроп. Оуо ВоБифви ба 2аз-
&В1. 1960, М 4. Р. 31-37.
.
Ущена ХТ. В., Ву М. К. Ызстоитаой
о Ме одог оЁ 56836 гафз // З4епсе.
1968. Уоф. 161. Р. 599-601.
ТапаепфегЕй 7. С. Ассегайоп о! зохиа]
шаиганоп т ета гафз Бу шае $8.
ш\ШаНоп // Вергод. Кег. 1976. Ус]. 46.
Р. 451-453.
Тип Ве! 1. Зетепуа Мена оуегсВ& уап 4е
1орюзр!тояз Ш Веюе
// ВЮ. вепеез.
копае. 1958. Уд], 14, М 18. Р. 20.
УапВер1,ВатеВ.УапNoМ.Еп-
гие пистомооае вегооаие ©
ер!Ченио\ов1Чие зит 1а !ербюзр!гозе аи
Зепера!// М6а. Айг. поте. 1969. Уо1, 16.
No2. Г. 165-171.
.
Уазетиз Г. СепеНсв ой Ше уНИе реШед
авоииеооиг 11 Фе МNoтуау гай // РАзеВт.
\егзисйзНогк. 1972. В4. 14, Н. 3.
$. 142-147.
Уаих С. Ешше Ветегкипоен зала Уог-
Коштеп пп УетВаЦеп 4ег У\ап4егга{-
(е (В. погуевсиз Вегк. 1769) ай ег
[1361 Не]еоо1апд // ЗАивеНегк. М1. 1958.
Ва. 6, М2. 8. 74-76.
Уев Уао-Сатя. Тзоайоп оп фохоразше
{тот ИЯ газ 1 Тагуап // Еадепис
Пузеазез Ви. Масазак ых. 1966.
\Уо1. 8, М 3. Р. 153-157.
Регпе-Маиту Е. Тепаему|-Иматойпе Ш
тох Рнесез: а пабта| айагиние забЗап-
се {юг Ше гаф // ОМасйопт ап@ фазе. Г:
ТВТ, ргезз, 1980. Ус. 7. Р. 407.
Тегие!-Маигу Е. Втопене 7. СВагвобюг-
хайоп ап@ 1ЧепыНсаНоп оЁ Ще рВего-
Литература
451
шопе о! Ме гаф рирз ргероба! 21апа/
ЕЛоову 85: 19% нет НОЕ и
Тошоизе, 1985. Уо1. 1. Р, 90.
УегпеН-Маигу Е, Рок Е. Н., Ретае А.
Обгастате-асцуйу геанмопзр о? згезё-
шдисшо одогап{з 1 Фе хаб //Т. Свет,
ЕсоГ. 1984. Ув]. 10, М 7.Р. 1007—1018.
Тов ске! У. Га гератайоп 4ез езресез
В. погуесеив еф В. гаНаз а Почцайа //
мед. (тор. 1958. Уо1. 18, М4. Р. 677-
Товске!? /, Тащегаз (+. Га гератИНоп 4ез
езресез В. погуей1с0$ её В. таНмз а
ола // Пула. 1960. Уо1. 20, М 5. Р. 625-
Тоогнеез ФТ. И’., Ветдеу No. В. МИга| сей
гезропзез ю \Ше ойогз о{ гечаг@ ап@
поптеууаг@ // РЬуз101. — Рзусво. — 1981.
Уо1. 9. Р. 164-170.
УУасег А. Пешзсве Еогз-ЗоНии-Шега-
фи, 1969—1974, — Мавейете // дзейг.
РНаптетзена. 1977. Ва. 84, М 71.
$. 475-479.
УаШятава К., Кпизов 7. Т., Ущеп В. 7.
ЕНесф8 о{ 13 ]айоп алгше деуе]ортене
оп теасыуНу ап Вотесайе агот15Ыс
Ъера\уопг ш гай // Аротез. Венау. 1983.
\о]. 9, М1. Р. 29—40.
ИчИатз 5. Л. ТЦе ргезешь збафе оЁ 1ер-
(03р1г05:3 41801093 ап сощто]. Ё.:
МЛоН, 1986. 262 р.
УТаПасе С. Р., МатзваИ Г., МатзРай М.
Саёз, гафз ап 1юхор1аз110513 оп а зтаП
Расе 14апа // Атег. 7. Ер Чего.
1972. Уч. 5.Р. 475—482.
ЧаПег 7., Иерег М. Р15еазев тапзпией
Бу гаёз ап писе. М. У., 1982. 182 р.
И’ата Т. Е. Ргепа&а] тез {е11120$ ап
ЧетазсиНиитез Ше Бепауопг оЁ таТез //
ЭЛепсе. 1972. \01. 175. Р. 82-84.
Уата Р.„ Фарил А, Ле ппромапсе ой
сена аззетЫаре оЁ }г4$ аз «иог-
шайоп-сешгез» Гог {004 Ниша // Пиз.
‚1973. УоЁ. 145. Р. 517-534.
Усов Т. 5., Реггу 7. 5. Ехрегитеп{: оп
га соо] ш Раезуше ав@ Фе $0-
Дал // Сотуто| 07 гафз ап пес. Охота:
ОУ. ргезв, 1954. Р. 500-521.
\ Май: С. И. 5. Тве 10048 езеп Бу зоше
АпзтаНап го4ет{ (Мом Аае) // Апзага!.
\МПаШе Вез. 1977. Уо1. 4. Р. 151-157.
Жан С. Н. 5. УосаПзаНотз оё пте зре-
с1ез оЁ таб (Ваз; Миндае) И. Тод.
1980. Уо|. 191, МА. Р. 531—555.
ЖТекяет О. В. ЗутраФенс ГапсНоп апй
ЧемеоршетЕ оЁ (етрегайиге теощай-
оп // Ашег. 7. РNoуз1о]. 1964. Уо|. 206.
Р. 823—826.
Иез В. В., Ра М. ИУ. Водисате Ищегас-
Нопв атопе газ 10 паргоуе Бай, асеер-
фапсе // РЫЙрр. Аог. 1975. Уч. 59,
М 1/2. Р. 34-36.
У/йеаЦеу У. В., Затез А. Т. Тве сотро-
ЭНоп о Ме зерит оЁ зоте соттоп
тоЧепз // Взосвеш. т. 1957. У. 65
Р. 36—42.
Илезпег В. Р., 5йеага No М. Маегпа| Ъе-
Вауоог ш Фе га|, Т.; ОНуег ап4 Воу4,
1933. 150 р.
ТИиатз Е., сои 7. Р. Тве деуеюртети
оЁ зочаГ репамюоик раМегиз т Ше топ-
зе 11 геЙайоп No пабга| регто43з // Вен э-
Уют. 1953. Уо|. 47. Р. 38—63.
Й’Изоп Г. 0. Зомотоюру: ТЬе пе\у зуп-
{Не513. 74 е4. Сатьг ее, 1982. 697 р.
Тан Р. Негетя-блтазсЬИ В., Вапег В.
ВаЦиз гаймз ип@ Вайаз погуер1сиз шт
Озетгесв ип Чегеп Опегзсвеипо
ап ЗевёдеЁ ип@ розбсгап1а]!ет Зкее& //
Ми. АБ. 2001, Гардезтиз. Уоаппеит.
1980. Ва. 9, М 3. $. 141—188.
ТопР.0О.,ТооН.С.ОпЩетооЁга
оЁ Рарееё ТЗапд
// АррЁ. 2оо]. 1960.
Уча. 15. МЗ. Р. 49—52.
УТоойз4е В., Геоп М. ТАегтоепдостие
о цепсев оц таайегпа! пез ше фева-
Уюит Чт гаёз//Т. Сор. апа Р\узю1.
Рзусло1. 1980. Уо1. 94. Р. 61-68.
Иоойз4е В., Геоп М., Аната М. еб а1.
Рго|асит-зето! тЯиепсе$ оп {Ве Жег-
шаЁ Ъаз1з {ог поПег-уопте сотфас шп
Могуау гайз // НИЧ. 4981. УоТ. 95.
Р. 771-780.
1Т’и Р. Оп зифзресШе ЧШегепнаНоп оЁ
Ъгохи га (ВаМаз погуе1сиз ВегКеп-
Во) ш СЫта// Аба его]. О зпыса.
1982. Уч. 2, М 1.Р. 107-142.
Й’уппе-Едшагаз Т. С. Зо] ограпаНоп
аз а роршаНоп тесшаНоп // Зугр. 2001.
ос. ТГопаоп. 1965. Уо. 14. Р. 173-
78.
Уаеп Ь. И’., Гагреп М. Г., КосК Ш. Са-
{а1одие оф Фе ташта о? ЕЧюра.
2. Тазесиуога ап тодепНа
// Моп{оге
1001. На. No. 5. 1976. М 1, зарр. 8.
Р. 1148.
Уокоуата А., О К. ЕНесь оЁ охуост
херйасетеф оп 1абаНоп т габз ]еаг-
пише Вурофа]ание 1ез1опз // Епдостио].
Тар. 1959. Уч]. 6. Р. 286—274.
Уо4а Т. П. Суюсепейсв о! Ме ШасКк
га": Кагуобуре еуошИоп ап зресез
1 НегелНаМой. ТоКуо; Ва тоге. 1980.
256 р.
Уояа Т. Н. Ситотловота{ ап@ 1 осВепи-
са] еуооп Ш Фе сепиз Вайлз // Ас4а
200]. Тепп. 1985. Уо1. 170. Р. 7-14.
Уонпе И’. С., Вотз в. Г, Вапаев В. С.
450
Литература
дтезыуе есоитиегв ашопя га (ВаНи$
потуер1сив) // Па. 1983. Уо1. 97. Р. 204—
206.
Тротаз О. А, Таша: Г., Ватнаа В. Ф.
ЕНесё5 оЁ деуосаПеаНот 0# 11е таа]е оп
тшайие ревахюйг т та+з
// 114. 1984.
\о1. 95. Р. 630—637.
Тротаз 0. ТЬе репегс аггапаетепё оЁ
Те АизраНап шигтез ВИНего гегогей
10 «Миз»//Апп. ап@ Мас. Мабат. НУ.
бег, 8, 1910. У. 6. Р. 603—607.
Твотрзоп И’. П. Ищвоп У. Срагиз-
гот ТУ. В. Те еес4з оЁ ргепа{а! та-
{егпа| этезз оп озреше Бевау!опг ш
га(з
// Рвусво]. Моповт. 1962. Уо|. 76,
М 38.Р. 12-15.
Трог О. Н., Нойошау И. В. (1т.). Апозиа
ап рау НабИие фова\мюоит Ш ргери-
Ъезсетё ша ап@ Фета!е газ// Рву-
$10]. ‘ап Вевах. 1982. Уо|. 29. Р. 281-
285.
ТРог 0. Н., НоЙошву И’. В. (1'".). Зеоро-
ауте МосКз р]ау-Невыля ш ауеше
та{з // ТЯ. 1983а. Уо|. 30. Р. 545—549.
Трог р. Н. НоПошау И’. В. (1т.). Рву-
зоНеНамов Бева\отг Ш ГауепНИе та|е
апа Геша е га // Апиа. ЁБеаги. Вевах.
1983Ъ, Ув. 11. Р. 178—178.
Трог Р. Н., НоЦошау ТУ. В. (7г.). Зех
ап зобаЁ р!ау 11 фиуевПе гафз (На Низ
погуео1стз) //7. Сошр. РэусНо|. 1984.
\Уо1. 98, М 3. Р. 276-284.
Тпог Р. Н., НоПошау 1’. В. (".). Бехе-
1оршепа! апа!узез оЁ зоса] ау Ъе-
Вачотг т ромедИе гаёз // ВиП. РзуеВо-
пог. бое. 19845. Ув. 22, М 6. Р. 587.
ТРогре И’. Н. ВииаШтаНоп т ощобепу.
4. Апниа! р1ау // РЬПоз. Тгапз. Воу. 306.
Гопдоп. В. 1966. Уо1. 772, М 154.
Р. 311-319.
ТИпеу Е. Вевауюпг т Из геабоп ю Шь
деуе!ортлевь оЁ Фе гаш // ВаН. Мета!
Таз. 1933. М 11.Р. 252-358.
Топись Р. 0. Матта8 ш Намуай: А зу-
порз1з ап побаМопар ЫЪНортарйу [1
В1зрор. Миз. Зрес. Ри. 1969. М 51.
°Р. 238.
Топег 7. Р., А@ег No. Т. Роепсу оЁ тай
ефастаНопз уагез УИВ пеш отаег впа
УИ} ша аче // Рьуз10]. ап@ Вевах.
1985. \а1, 35, No 1.Р. 113—145.
Торзт У. Г., рта 7, Ттепи Напв Оиу.
ТВе годеп гезогуой оЁ 1ерфюзритае т
Могв Уешат // АтеН. гоши. . рао!.
ехр. её пустоБоЁ 1970. \о1. 29, No 1/2.
Р. 331—338.
Ттоиеззат Е. Г. Сааовие дез ташш!е-
тез // Ви]. Зое. Гфадез ЗеЁ. Ап4егв. 1881.
Уи. 10. Р. 18-21.
Ттопеззат Е. Г. Водепиа
// Са{а105 05
татштаНит бат ууепили дааш #0581
Паш. В. 1897. Еазс. 3. Р. 458—664.
Тгонеззат Е. Ё. Сщаювиз таттаНат
{ат уепаниа дааш ТоззЙи. Зирр!е-
тепиит. В. 1904-1905. УП, 929 р.
Тискег Н. А, Тфаейег И. Рийату
ргохИВ Ногшопе апд Нцетииялтя Вогизо-
пе солфешб аНег уагоиз пигяше пиеп-
&1ез
// Ргое. бое. Ехр. ПВ. ав@ Мед.
1968. Уо|. 129. Р. 578—580.
Тише в. Г. Пуечайоп оЁ Пе топ га
(Вайиз погуер1еия) 1 г пишез о
Ме Выизь Т&апдз//7. Нув. 1964.
\о1. 59, М 3. Р. 271-284.
ТьшЕ С. Г. А темех о{ Фе оссигепсе
ш ВизН тпатитаа]з оЁ Ще таа]от огба-
п1зтз 0{ 2оопоНе пароепз // Магата|.
Веу. 1980. У]. 140. М 4. Р. 139-149.
Туии К. Пметгеайов фефбуееп Бай соп-
зирИоп, ргоюпзаНой ог робот
те ап Весгеазе соЁ 004 сарШату гв-
зфапсе Фу соптаага годепйс1Че, \уаг-
{ага // Noрроп. Оуо ВоБифви ба 2аз-
&В1. 1960, М 4. Р. 31-37.
.
Ущена ХТ. В., Ву М. К. Ызстоитаой
о Ме одог оЁ 56836 гафз // З4епсе.
1968. Уоф. 161. Р. 599-601.
ТапаепфегЕй 7. С. Ассегайоп о! зохиа]
шаиганоп т ета гафз Бу шае $8.
ш\ШаНоп // Вергод. Кег. 1976. Ус]. 46.
Р. 451-453.
Тип Ве! 1. Зетепуа Мена оуегсВ& уап 4е
1орюзр!тояз Ш Веюе
// ВЮ. вепеез.
копае. 1958. Уд], 14, М 18. Р. 20.
УапВер1,ВатеВ.УапNoМ.Еп-
гие пистомооае вегооаие ©
ер!Ченио\ов1Чие зит 1а !ербюзр!гозе аи
Зепера!// М6а. Айг. поте. 1969. Уо1, 16.
No2. Г. 165-171.
.
Уазетиз Г. СепеНсв ой Ше уНИе реШед
авоииеооиг 11 Фе МNoтуау гай // РАзеВт.
\егзисйзНогк. 1972. В4. 14, Н. 3.
$. 142-147.
Уаих С. Ешше Ветегкипоен зала Уог-
Коштеп пп УетВаЦеп 4ег У\ап4егга{-
(е (В. погуевсиз Вегк. 1769) ай ег
[1361 Не]еоо1апд // ЗАивеНегк. М1. 1958.
Ва. 6, М2. 8. 74-76.
Уев Уао-Сатя. Тзоайоп оп фохоразше
{тот ИЯ газ 1 Тагуап // Еадепис
Пузеазез Ви. Масазак ых. 1966.
\Уо1. 8, М 3. Р. 153-157.
Регпе-Маиту Е. Тепаему|-Иматойпе Ш
тох Рнесез: а пабта| айагиние забЗап-
се {юг Ше гаф // ОМасйопт ап@ фазе. Г:
ТВТ, ргезз, 1980. Ус. 7. Р. 407.
Тегие!-Маигу Е. Втопене 7. СВагвобюг-
хайоп ап@ 1ЧепыНсаНоп оЁ Ще рВего-
Литература
451
шопе о! Ме гаф рирз ргероба! 21апа/
ЕЛоову 85: 19% нет НОЕ и
Тошоизе, 1985. Уо1. 1. Р, 90.
УегпеН-Маигу Е, Рок Е. Н., Ретае А.
Обгастате-асцуйу геанмопзр о? згезё-
шдисшо одогап{з 1 Фе хаб //Т. Свет,
ЕсоГ. 1984. Ув]. 10, М 7.Р. 1007—1018.
Тов ске! У. Га гератайоп 4ез езресез
В. погуесеив еф В. гаНаз а Почцайа //
мед. (тор. 1958. Уо1. 18, М4. Р. 677-
Товске!? /, Тащегаз (+. Га гератИНоп 4ез
езресез В. погуей1с0$ её В. таНмз а
ола // Пула. 1960. Уо1. 20, М 5. Р. 625-
Тоогнеез ФТ. И’., Ветдеу No. В. МИга| сей
гезропзез ю \Ше ойогз о{ гечаг@ ап@
поптеууаг@ // РЬуз101. — Рзусво. — 1981.
Уо1. 9. Р. 164-170.
УУасег А. Пешзсве Еогз-ЗоНии-Шега-
фи, 1969—1974, — Мавейете // дзейг.
РНаптетзена. 1977. Ва. 84, М 71.
$. 475-479.
УаШятава К., Кпизов 7. Т., Ущеп В. 7.
ЕНесф8 о{ 13 ]айоп алгше деуе]ортене
оп теасыуНу ап Вотесайе агот15Ыс
Ъера\уопг ш гай // Аротез. Венау. 1983.
\о]. 9, М1. Р. 29—40.
ИчИатз 5. Л. ТЦе ргезешь збафе оЁ 1ер-
(03р1г05:3 41801093 ап сощто]. Ё.:
МЛоН, 1986. 262 р.
УТаПасе С. Р., МатзваИ Г., МатзРай М.
Саёз, гафз ап 1юхор1аз110513 оп а зтаП
Расе 14апа // Атег. 7. Ер Чего.
1972. Уч. 5.Р. 475—482.
ЧаПег 7., Иерег М. Р15еазев тапзпией
Бу гаёз ап писе. М. У., 1982. 182 р.
И’ата Т. Е. Ргепа&а] тез {е11120$ ап
ЧетазсиНиитез Ше Бепауопг оЁ таТез //
ЭЛепсе. 1972. \01. 175. Р. 82-84.
Уата Р.„ Фарил А, Ле ппромапсе ой
сена аззетЫаре оЁ }г4$ аз «иог-
шайоп-сешгез» Гог {004 Ниша // Пиз.
‚1973. УоЁ. 145. Р. 517-534.
Усов Т. 5., Реггу 7. 5. Ехрегитеп{: оп
га соо] ш Раезуше ав@ Фе $0-
Дал // Сотуто| 07 гафз ап пес. Охота:
ОУ. ргезв, 1954. Р. 500-521.
\ Май: С. И. 5. Тве 10048 езеп Бу зоше
АпзтаНап го4ет{ (Мом Аае) // Апзага!.
\МПаШе Вез. 1977. Уо1. 4. Р. 151-157.
Жан С. Н. 5. УосаПзаНотз оё пте зре-
с1ез оЁ таб (Ваз; Миндае) И. Тод.
1980. Уо|. 191, МА. Р. 531—555.
ЖТекяет О. В. ЗутраФенс ГапсНоп апй
ЧемеоршетЕ оЁ (етрегайиге теощай-
оп // Ашег. 7. РNoуз1о]. 1964. Уо|. 206.
Р. 823—826.
Иез В. В., Ра М. ИУ. Водисате Ищегас-
Нопв атопе газ 10 паргоуе Бай, асеер-
фапсе // РЫЙрр. Аог. 1975. Уч. 59,
М 1/2. Р. 34-36.
У/йеаЦеу У. В., Затез А. Т. Тве сотро-
ЭНоп о Ме зерит оЁ зоте соттоп
тоЧепз // Взосвеш. т. 1957. У. 65
Р. 36—42.
Илезпег В. Р., 5йеага No М. Маегпа| Ъе-
Вауоог ш Фе га|, Т.; ОНуег ап4 Воу4,
1933. 150 р.
ТИиатз Е., сои 7. Р. Тве деуеюртети
оЁ зочаГ репамюоик раМегиз т Ше топ-
зе 11 геЙайоп No пабга| регто43з // Вен э-
Уют. 1953. Уо|. 47. Р. 38—63.
Й’Изоп Г. 0. Зомотоюру: ТЬе пе\у зуп-
{Не513. 74 е4. Сатьг ее, 1982. 697 р.
Тан Р. Негетя-блтазсЬИ В., Вапег В.
ВаЦиз гаймз ип@ Вайаз погуер1сиз шт
Озетгесв ип Чегеп Опегзсвеипо
ап ЗевёдеЁ ип@ розбсгап1а]!ет Зкее& //
Ми. АБ. 2001, Гардезтиз. Уоаппеит.
1980. Ва. 9, М 3. $. 141—188.
ТопР.0О.,ТооН.С.ОпЩетооЁга
оЁ Рарееё ТЗапд
// АррЁ. 2оо]. 1960.
Уча. 15. МЗ. Р. 49—52.
УТоойз4е В., Геоп М. ТАегтоепдостие
о цепсев оц таайегпа! пез ше фева-
Уюит Чт гаёз//Т. Сор. апа Р\узю1.
Рзусло1. 1980. Уо1. 94. Р. 61-68.
Иоойз4е В., Геоп М., Аната М. еб а1.
Рго|асит-зето! тЯиепсе$ оп {Ве Жег-
шаЁ Ъаз1з {ог поПег-уопте сотфас шп
Могуау гайз // НИЧ. 4981. УоТ. 95.
Р. 771-780.
1Т’и Р. Оп зифзресШе ЧШегепнаНоп оЁ
Ъгохи га (ВаМаз погуе1сиз ВегКеп-
Во) ш СЫта// Аба его]. О зпыса.
1982. Уч. 2, М 1.Р. 107-142.
Й’уппе-Едшагаз Т. С. Зо] ограпаНоп
аз а роршаНоп тесшаНоп // Зугр. 2001.
ос. ТГопаоп. 1965. Уо. 14. Р. 173-
78.
Уаеп Ь. И’., Гагреп М. Г., КосК Ш. Са-
{а1одие оф Фе ташта о? ЕЧюра.
2. Тазесиуога ап тодепНа
// Моп{оге
1001. На. No. 5. 1976. М 1, зарр. 8.
Р. 1148.
Уокоуата А., О К. ЕНесь оЁ охуост
херйасетеф оп 1абаНоп т габз ]еаг-
пише Вурофа]ание 1ез1опз // Епдостио].
Тар. 1959. Уч]. 6. Р. 286—274.
Уо4а Т. П. Суюсепейсв о! Ме ШасКк
га": Кагуобуре еуошИоп ап зресез
1 НегелНаМой. ТоКуо; Ва тоге. 1980.
256 р.
Уояа Т. Н. Ситотловота{ ап@ 1 осВепи-
са] еуооп Ш Фе сепиз Вайлз // Ас4а
200]. Тепп. 1985. Уо1. 170. Р. 7-14.
Уонпе И’. С., Вотз в. Г, Вапаев В. С.
452 Литература
ТЬе уаята| зтеаг расе, зехиаЁ ге-
серыуйу ап Ите оЁ оуШаНоп 1 Ше
ато гаф// Апаф. Вес. 1941. Уо|. 80.
Р. 37-45.
.
.
Харекй М. О уоЪе паупа@у Ъ0}е ргой
рожапе (В. погуемеиз) у родпипкасЬ
зете4де]зкусв оБека // 2001. Избу. 1962.
у. 11, М1. $. 81-92.
Зале] М. Оп Ше осситепсе оё Ме
Бго\и га!з В. погуео1сиз ВегК, ипдег
паба! сопНоп м С2еспозочхаКта//
7001. Тау (ЕоПа тооов а). 1964.
Уа1. 13, М З.Р. 125-134.
2арени М., баяет 7. Роршасе Ктузу Ван
{13 погуегсиз ВетК. у ргозтей аза-
пасбей дат и Ораху// Па. 1983.
Уо1. 32, М 4. 5. 293—300.
Фаттош М. Х., Сапдатап В., Деппеп-
Бег Г. Н. Ргасйт: 13 Ц аш еззепИа1
Вогиопе {юг шаегпа| Бейа\ойг т
тпаттла3? // Ногтопе Вевау.
—
1974.
Ув1. 2. Р. 343-354.
7етег А. В., РееЁ Н. Г. 5. НаЪйваНоп
0Ё гезропзе 40 Ч915те5$ стез шт фе таб
Меазигетеле Бу ап шпае зарргезз оп
феспт1ще // Соштаип. Вевау. В10]. А.
1969. Уа1. 3, М 5/6. Р. 249—252.
2еп Уой Н., бака! 5., Магауапа Т., Уа-
павага У. Тзайоп оЁ Уегзима стйего-
со|иса ап Уегзииа рзезаофаЪогси10515
{тот элуше, сейее ап гаёз а ап аБа{-
{от
// Зар. У. Мистоыой. 1974. Уо|Ё. 18,
М1. Р. 103—105.
та 7, Кг В. Кагуобурез о! Глгореап
ташита]з. 2//Ама 561. Мант. Асад.
Зс1. Вобетозюотуасае. Втпо, 1984. Уо1. 18,
М 8.Р. 1-62.
Зиттагу
Тне шопосгарВ «ТВе Могуау га сошашз 44а оп фахопоту, @з'Пиачоп, есоюбу,
епоТору, ргасЫса] за Исапсе ап депеЙу сопго! оЁ 8 зрес!ез. А Ызюгса1 теде\
0Ё бахопописа! зе п Ме вепиз Ваз ава т Ще зр/с1ез оЁ М5 пепиз 13 р1уеп.
Тиле таогрвоюзса| Кагуоурса! ап о'Вег езстрЫопз оЁ 1Тезе {аха аго ргоу!4ей.
Тве @зитоНот о? Фе Мог\уау гаф оп аН оЁ 1Noе сопИпенйз 18 езёииа(ед. Тке статеш
агеа ш а] рагёз оЁ Ще ог! 13 @езстфе4. Тве аз фиНов оЁ Ве зремез т Че
О$$8 15 соуегей ш 4еай, ав Noе гезйем оЁ Ве сВапоез ш 13 агеа ш Ще 1а%
50 уеагз 13 ргоу!4е4. ЭИТегев& те оёз ог суашайов о{ диапу аге с" саПу апаЁу-
зе4. Рагислагсз оф Ще Могузу каф ПаЪНайз ш пабаге, 1 Ве забигЬз ап@ ш е4ез
аге ргезещей. Тре есо]ов1са! р]азИеНу оЁ Не зрес1сз 13 Чезстфейд: фе аБИИу ог
зитутуа! 1 уамомз сот! опз шеш@шя Ше ехтеше опез. ТВе зиасёте оЁ Богтолуз
апб офег геЁикез 13 41зсиззей, (овеПег НИ рагНси]атНез о{ Бееййав, изе оЁ 1еггИогу,
шоуау, Бтеедте апф Чеай. Раба оп 4Ве ошюпепез!8 0Ё фепазлог аге заррНей.
Тне заше БеВау!ог ап шесвашзтаз 0Ё пуеггеайотз фебусей шоШег ап оНзрише
13 апазед. ТЬе дупат$ т зоба! пиегаеНотз Бегуееп Ше апйпа|з 15 тезмед ш
гебресё оЁ Черепфепсе Фот Шей иде, зех ап т апа! доабЫез. ТВе реоЫетз шт
сВеписа] ап@ аспзЫса! сотштиииеаНон {т газ, реги! те сегмайш адариуе роззШиез
0Ё 116 зрес1ез, ате апаГузей ш 4е{аП. ТВе гое оЁ \е Могуау га ш зргеадте уйгаз,
Басета! ап@ ргоюхоаш ПМесйопз, Чапкегоиз {ог тшеп, {5 зВо\и. ТЬе Вата сапзей
Бу Ме Мгуау гаё ш Че зрЬеге оЁ есопопусз 13 гезле\ей. Те ситгеп& тао обв ой
даа Ну годаноп ш 113 зрес1ез ап@ Ше адарбаНот$ оЁ Ме ЛаНег 0 Тюозе шИшепсез
аге 91зстззе4д.
452 Литература
ТЬе уаята| зтеаг расе, зехиаЁ ге-
серыуйу ап Ите оЁ оуШаНоп 1 Ше
ато гаф// Апаф. Вес. 1941. Уо|. 80.
Р. 37-45.
.
.
Харекй М. О уоЪе паупа@у Ъ0}е ргой
рожапе (В. погуемеиз) у родпипкасЬ
зете4де]зкусв оБека // 2001. Избу. 1962.
у. 11, М1. $. 81-92.
Зале] М. Оп Ше осситепсе оё Ме
Бго\и га!з В. погуео1сиз ВегК, ипдег
паба! сопНоп м С2еспозочхаКта//
7001. Тау (ЕоПа тооов а). 1964.
Уа1. 13, М З.Р. 125-134.
2арени М., баяет 7. Роршасе Ктузу Ван
{13 погуегсиз ВетК. у ргозтей аза-
пасбей дат и Ораху// Па. 1983.
Уо1. 32, М 4. 5. 293—300.
Фаттош М. Х., Сапдатап В., Деппеп-
Бег Г. Н. Ргасйт: 13 Ц аш еззепИа1
Вогиопе {юг шаегпа| Бейа\ойг т
тпаттла3? // Ногтопе Вевау.
—
1974.
Ув1. 2. Р. 343-354.
7етег А. В., РееЁ Н. Г. 5. НаЪйваНоп
0Ё гезропзе 40 Ч915те5$ стез шт фе таб
Меазигетеле Бу ап шпае зарргезз оп
феспт1ще // Соштаип. Вевау. В10]. А.
1969. Уа1. 3, М 5/6. Р. 249—252.
2еп Уой Н., бака! 5., Магауапа Т., Уа-
павага У. Тзайоп оЁ Уегзима стйего-
со|иса ап Уегзииа рзезаофаЪогси10515
{тот элуше, сейее ап гаёз а ап аБа{-
{от
// Зар. У. Мистоыой. 1974. Уо|Ё. 18,
М1. Р. 103—105.
та 7, Кг В. Кагуобурез о! Глгореап
ташита]з. 2//Ама 561. Мант. Асад.
Зс1. Вобетозюотуасае. Втпо, 1984. Уо1. 18,
М 8.Р. 1-62.
Зиттагу
Тне шопосгарВ «ТВе Могуау га сошашз 44а оп фахопоту, @з'Пиачоп, есоюбу,
епоТору, ргасЫса] за Исапсе ап депеЙу сопго! оЁ 8 зрес!ез. А Ызюгса1 теде\
0Ё бахопописа! зе п Ме вепиз Ваз ава т Ще зр/с1ез оЁ М5 пепиз 13 р1уеп.
Тиле таогрвоюзса| Кагуоурса! ап о'Вег езстрЫопз оЁ 1Тезе {аха аго ргоу!4ей.
Тве @зитоНот о? Фе Мог\уау гаф оп аН оЁ 1Noе сопИпенйз 18 езёииа(ед. Тке статеш
агеа ш а] рагёз оЁ Ще ог! 13 @езстфе4. Тве аз фиНов оЁ Ве зремез т Че
О$$8 15 соуегей ш 4еай, ав Noе гезйем оЁ Ве сВапоез ш 13 агеа ш Ще 1а%
50 уеагз 13 ргоу!4е4. ЭИТегев& те оёз ог суашайов о{ диапу аге с" саПу апаЁу-
зе4. Рагислагсз оф Ще Могузу каф ПаЪНайз ш пабаге, 1 Ве забигЬз ап@ ш е4ез
аге ргезещей. Тре есо]ов1са! р]азИеНу оЁ Не зрес1сз 13 Чезстфейд: фе аБИИу ог
зитутуа! 1 уамомз сот! опз шеш@шя Ше ехтеше опез. ТВе зиасёте оЁ Богтолуз
апб офег геЁикез 13 41зсиззей, (овеПег НИ рагНси]атНез о{ Бееййав, изе оЁ 1еггИогу,
шоуау, Бтеедте апф Чеай. Раба оп 4Ве ошюпепез!8 0Ё фепазлог аге заррНей.
Тне заше БеВау!ог ап шесвашзтаз 0Ё пуеггеайотз фебусей шоШег ап оНзрише
13 апазед. ТЬе дупат$ т зоба! пиегаеНотз Бегуееп Ше апйпа|з 15 тезмед ш
гебресё оЁ Черепфепсе Фот Шей иде, зех ап т апа! доабЫез. ТВе реоЫетз шт
сВеписа] ап@ аспзЫса! сотштиииеаНон {т газ, реги! те сегмайш адариуе роззШиез
0Ё 116 зрес1ез, ате апаГузей ш 4е{аП. ТВе гое оЁ \е Могуау га ш зргеадте уйгаз,
Басета! ап@ ргоюхоаш ПМесйопз, Чапкегоиз {ог тшеп, {5 зВо\и. ТЬе Вата сапзей
Бу Ме Мгуау гаё ш Че зрЬеге оЁ есопопусз 13 гезле\ей. Те ситгеп& тао обв ой
даа Ну годаноп ш 113 зрес1ез ап@ Ше адарбаНот$ оЁ Ме ЛаНег 0 Тюозе шИшепсез
аге 91зстззе4д.
Оглавление
Предисловие
Глава первая
Систематика (А. И. Милютин)
Систематика рода Ва из
Систематика ВаМиз погуес!си$
Глава вторая
Ареал (В. В. Кучерук)
Глава третья
Экология
Места обитания (©. в. ` Карасева, д. Н. Козлов, в. К. Мелко, д. `. ‚Тре
ханов, И. С. Туров, И. Новатный)
Убежища (Е. В. Карасева)
Питание (М. И. Шутова)
Подвижность (Е. В. Карасева)
Размножение, возрастной состав и смертность (В. ГА. Рыльников)
Глава четвертая
Поведение
.
Онтогенез поведения (в. Е. боколов, 6. А. ` Квашнин)
Взаимоотношения между матерью и детепитами (. ИА рученкова,
М. Е. Гольцман)
Социальная оргацизация (в. Е. боков, с. А. ` Квашнин)
Обонятельная коммуникация (В. Е. Соколов, В. Д. Вознесенская,
Е. Ю. Павлова, 9. П. Зинкевич)
ЗА
Акустическая коммуникация (А. А. Никольский)
Позы и выразительные движения (С. А. Квашнин)
Глава пятая
Практическое значение
Экономический ущерб (Б. Е. ополов, В. В. ` Бобров)
Медицинское значение (Ё. В. Карасева, В. И. Соловьев, И. 1. ` Гаврилов.
ская, И. В. Тарасевич, Ю. В. Ананьина, Т. Н. Дунаева, Ю. А. Мясников)
Методы борьбы с серыми крысами (В. А. Рыльников, Ю. В. Тощигин)
Репеллентные методы защиты Материалов от крыс (Е. В. Бочаров,
7. П. Ирючков)
..
уни
Литература
Зиштагу
25
34
85
128
144
161
181
230
230
241
275
289
306
325
332
332
338
360
390
396
453
Сошеп5
Ротезгота
Сварет 1
бумештаНез (А. Г. Мйуинт)
БуетаИс$ оЁ пепиз НаМи8
ЗузетаНс$ ВаЙиз погуесиз
СБарег 2
Валае (УТ. Т. Кисйегай)
СВарег 3
Есо]юсу
еее
НаБбайв (Е. У. Капазета. А. К. Коль, у. К. МеЦора, Р. Р. Ттафйапоь,
Т. 5. Титор, Г. Моъату)
(.
ЗО
Соуетз (Е. У. Кагазера)
Еее шр (М. Г. УВьдова)
МоБИиу (Е. Г. Кагазега)
.
ВгееЯ тя, апе сошрозоп апд томайву, (и. ГА. `Вувыфоь)
СВар1ег 4
Веъау1юие
Вепауюот поел (у. я. "борооь, 5. и. "Коазтит)
ГоцеггаНопз5рз Фебуееп шошег ап@ уоцпр (Е. Р. Кгисйепкова.
М. Е. Сойзтап)
.
Зоеа] огоапмаНов (ТГ. Е. `бокооь, 5. А. `Кразвит)
ОНасюту сопитлимсаНот (У. Я. бокоюь, У. РБ. Уозпеветзкауа, Е. Уи. Рав.
{ора, В. Р. Итьенсь)
.
АсовзИс сопиаищеаной (Д. А. Мо Ьу)
Розфигез ап ехргеззуе шо\ететйз (5. А. Къаввит)
(Парт 5
РгаеНеа! ппробапее
.
Есопоти1с датаяе (Т. Л. бовооь, у. т. Вотоь)
Мей!са! 1трогапсе (Л. У. Кагазега, У. Г. баюовуеь, [. ТА. бавтиогзКаца,
Г. И. Татазелей, Уи. У. Апапута, Т. М. Бипаера, Ув. А. Муазт вос}
Ме043 о{Ё сошто|! (ТУ. А. Ву ов, Уи. У. Гозсмит)
ВереПеп+ пеВо4$ 0#Е ргуесИоп 0{ тафет!а]$ и. У. Восвагоь, Т. ‘р. Кери:
Ков)
Веегепсез
Эиитагу
25
З4
85.
85
128
144
161
184
230:
230
247
275.
289.
306
325
332
332
338
360
390
396
458
Оглавление
Предисловие
Глава первая
Систематика (А. И. Милютин)
Систематика рода Ва из
Систематика ВаМиз погуес!си$
Глава вторая
Ареал (В. В. Кучерук)
Глава третья
Экология
Места обитания (©. в. ` Карасева, д. Н. Козлов, в. К. Мелко, д. `. ‚Тре
ханов, И. С. Туров, И. Новатный)
Убежища (Е. В. Карасева)
Питание (М. И. Шутова)
Подвижность (Е. В. Карасева)
Размножение, возрастной состав и смертность (В. ГА. Рыльников)
Глава четвертая
Поведение
.
Онтогенез поведения (в. Е. боколов, 6. А. ` Квашнин)
Взаимоотношения между матерью и детепитами (. ИА рученкова,
М. Е. Гольцман)
Социальная оргацизация (в. Е. боков, с. А. ` Квашнин)
Обонятельная коммуникация (В. Е. Соколов, В. Д. Вознесенская,
Е. Ю. Павлова, 9. П. Зинкевич)
ЗА
Акустическая коммуникация (А. А. Никольский)
Позы и выразительные движения (С. А. Квашнин)
Глава пятая
Практическое значение
Экономический ущерб (Б. Е. ополов, В. В. ` Бобров)
Медицинское значение (Ё. В. Карасева, В. И. Соловьев, И. 1. ` Гаврилов.
ская, И. В. Тарасевич, Ю. В. Ананьина, Т. Н. Дунаева, Ю. А. Мясников)
Методы борьбы с серыми крысами (В. А. Рыльников, Ю. В. Тощигин)
Репеллентные методы защиты Материалов от крыс (Е. В. Бочаров,
7. П. Ирючков)
..
уни
Литература
Зиштагу
25
34
85
128
144
161
181
230
230
241
275
289
306
325
332
332
338
360
390
396
453
Сошеп5
Ротезгота
Сварет 1
бумештаНез (А. Г. Мйуинт)
БуетаИс$ оЁ пепиз НаМи8
ЗузетаНс$ ВаЙиз погуесиз
СБарег 2
Валае (УТ. Т. Кисйегай)
СВарег 3
Есо]юсу
еее
НаБбайв (Е. У. Капазета. А. К. Коль, у. К. МеЦора, Р. Р. Ттафйапоь,
Т. 5. Титор, Г. Моъату)
(.
ЗО
Соуетз (Е. У. Кагазера)
Еее шр (М. Г. УВьдова)
МоБИиу (Е. Г. Кагазега) .
ВгееЯ тя, апе сошрозоп апд томайву, (и. ГА. `Вувыфоь)
СВар1ег 4
Веъау1юие
Вепауюот поел (у. я. "борооь, 5. и. "Коазтит)
ГоцеггаНопз5рз Фебуееп шошег ап@ уоцпр (Е. Р. Кгисйепкова.
М. Е. Сойзтап)
.
Зоеа] огоапмаНов (ТГ. Е. `бокооь, 5. А. `Кразвит)
ОНасюту сопитлимсаНот (У. Я. бокоюь, У. РБ. Уозпеветзкауа, Е. Уи. Рав.
{ора, В. Р. Итьенсь)
.
АсовзИс сопиаищеаной (Д. А. Мо Ьу)
Розфигез ап ехргеззуе шо\ететйз (5. А. Къаввит)
(Парт 5
РгаеНеа! ппробапее
.
Есопоти1с датаяе (Т. Л. бовооь, у. т. Вотоь)
Мей!са! 1трогапсе (Л. У. Кагазега, У. Г. баюовуеь, [. ТА. бавтиогзКаца,
Г. И. Татазелей, Уи. У. Апапута, Т. М. Бипаера, Ув. А. Муазт вос}
Ме043 о{Ё сошто|! (ТУ. А. Ву ов, Уи. У. Гозсмит)
ВереПеп+ пеВо4$ 0#Е ргуесИоп 0{ тафет!а]$ и. У. Восвагоь, Т. ‘р. Кери:
Ков)
Веегепсез
Эиитагу
25
З4
85.
85
128
144
161
184
230:
230
247
275.
289.
306
325
332
332
338
360
390
396
458
Научное издание
Серая крыса
Систематика, экология,
регуляция численности
Утверждено в печати
Институтом эволюционной морфологии
и экологии животных им. А. Н. Северцова
АН СССР
Редактор
Г. М. Орлова
Художник
С. А. Литвак
Художественный редактор
Н. Н. Михайлова
Технический редактор
Л. И. Куприянова
Корректоры |
А. Б. Васильев, К. П, Лосева
ИБ No 39574
Сдано в набор 4.07.89.
Подписано к печати 25.12.89
7-18 956. Форма 70 х 90: лв
Бумага типографская No 1
Гарнитура обыкновенная
Печать высокая
Усл. печ. л. 33,34 Усл. кр. отт. 83,63 Уч.-изд. д. 86,9
Тираж 41800 экз. Тип. зак. 8464
Цена6р.80к.
Ордена Трудового Красного Знамени
издательства «Наука»
117864, ГСП-7, Москва, В-485,
Профсоюзная ул., 90
2-я типография издательства «Наука»
121099, Москва, Г-99, Шубинский пер., 6
Научное издание
Серая крыса
Систематика, экология,
регуляция численности
Утверждено в печати
Институтом эволюционной морфологии
и экологии животных им. А. Н. Северцова
АН СССР
Редактор
Г. М. Орлова
Художник
С. А. Литвак
Художественный редактор
Н. Н. Михайлова
Технический редактор
Л. И. Куприянова
Корректоры |
А. Б. Васильев, К. П, Лосева
ИБ No 39574
Сдано в набор 4.07.89.
Подписано к печати 25.12.89
7-18 956. Форма 70 х 90: лв
Бумага типографская No 1
Гарнитура обыкновенная
Печать высокая
Усл. печ. л. 33,34 Усл. кр. отт. 83,63 Уч.-изд. д. 86,9
Тираж 41800 экз. Тип. зак. 8464
Цена6р.80к.
Ордена Трудового Красного Знамени
издательства «Наука»
117864, ГСП-7, Москва, В-485,
Профсоюзная ул., 90
2-я типография издательства «Наука»
121099, Москва, Г-99, Шубинский пер., 6
$58 Асадету оЁ Зстепсе$
А. М. Зеуей20у пзицие
о Еуо!инопагу 'Апита!
Могрроову апд Есоову
Заепийс Соипсй
Юг Вююрка! Сита
о Мапаретет, Кесоп$игисноп
апа Сопзегуацоп ог Апипа!5
6р.80к.
Зрес1ез
оЁ пе Раипа оГ {Пе 0558.
апа {Ре Сопивцои$
Соипез
Зоме! СотпиШее
Гог Ше ИМЕЗСО Рговгатте
„Мап апд ВюзрНеге”
Noгуау Ваё
дуяетанс$
Есо]ову
Роршапоп сотой
Маика Риыкйег$