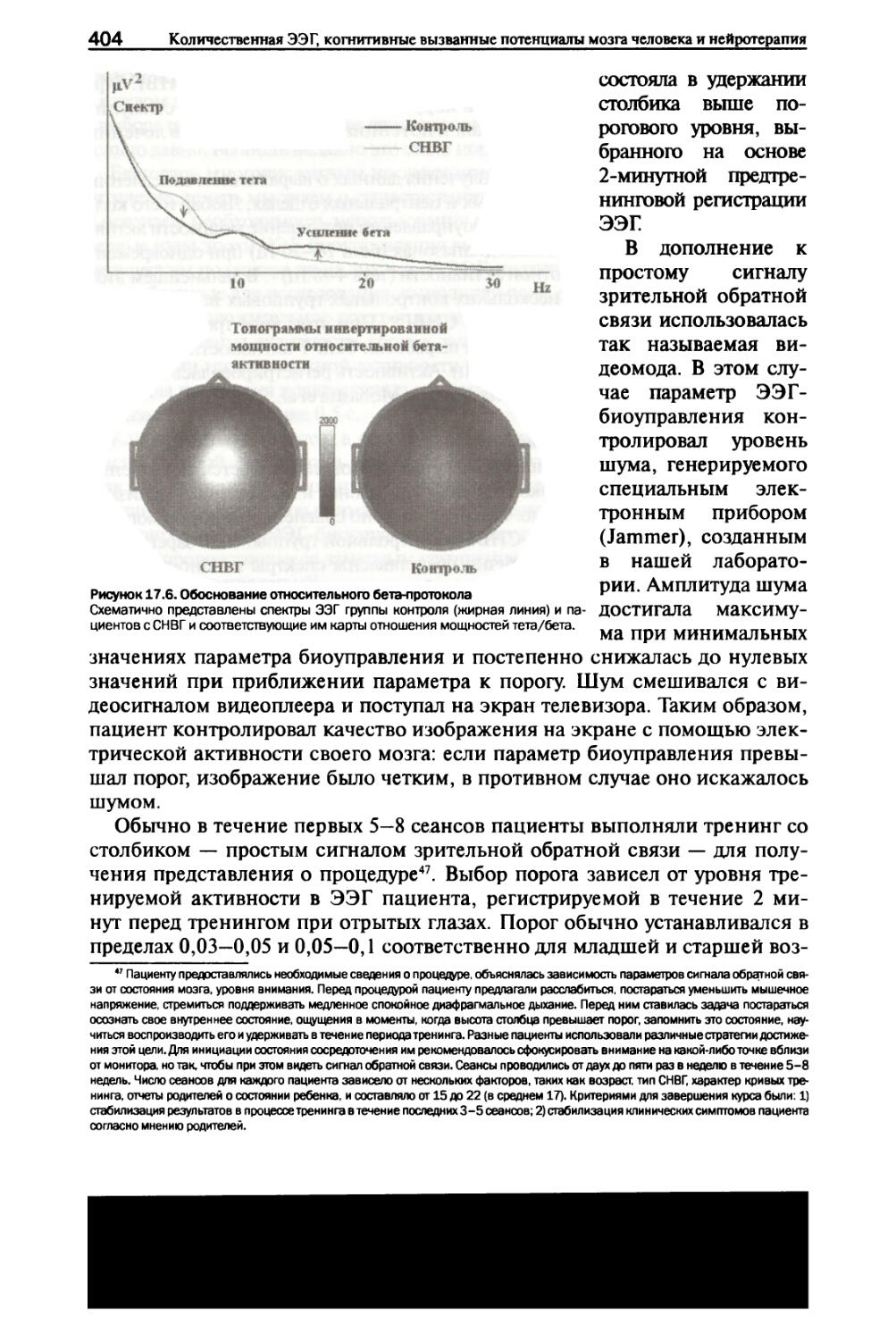
/












































































































































































































































































































































































































































































































Author: Кропотов Ю.Д.
Tags: нервная система органы чувств невропатология алкоголизм как болезнь нейрохирургия психиатрия медицина здоровье
ISBN: 978-617-7001-39-2
Year: 2010




Text
Ю.Д. Кропотов
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Перевод с английского к.м.н. В.Н. Гринь-Яценко, к.б.н. Н.В. Шемякиной, к.б.н. Ж.В. Нагорновой, к.б.н. М.В. Киреева.
Редакция русского текста к.б.н. В.А. Пономарева
Донецк
Издатель Заславский А.Ю.
2010
УДК 612.82-073.97-08
БЕК 56.1
К83
Книга издается при содействии Общественной организации «Всеукраинская Ассоциация клинических нейрофизиологов».
Ответственность за содержание, достоверность и орфографию рекламных материалов несет рекламодатель.
Кропотов Ю.Д.
К83 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия / Перевод с англ, под ред. В.А. Пономарева. — Донецк: Издатель Заславский А.Ю., 2010. — 512 с.
ISBN 978-617-7001-39-2
Книга дает общее представление о двух электрофизиологических параметрах жизнедеятельности мозга: статистических спектральных характеристиках спонтанной ЭЭГ и когнитивных вызванных потенциалах. Книга описывает методологию исследования этих показателей мозга, разработанную в Институге мозга человека РАН. Эта методология включает в себя такие современные методы анализа, как пространственная фильтрация при коррекции артефактов глазных движений, метод независимых компонент для выделения составляющих спонтанной ЭЭГ и вызванных потенциалов, электромагнитная томография низкого разрешения, волновой анализ, вызванная деситгхронизация и некоторые другие. Книга также является первой попыткой выявления эндофенотипов (биологических маркеров) некоторых заболеваний мозга, таких как синдром нарушения внимания с гиперактивностью, шизофрения, синдром навязчивых состояний, депрессия... В основе этого диагностического подхода лежит нормативная база данных, включающая характеристики спонтанной и вызванной активности мозга более 1000 здоровых испытуемых, записанных в состоянии спокойного бодрствования с открытыми и закрытыми глазами и в пяти тестах на внимание, чтение, математические операции, слуховое и зрительное восприятие. Книга снабжена обучающим программным обеспечением, позволяющим читателю овладеть основными методами количественной ЭЭГ и когнитивных вызванных потенциалов. Книга рассчитана на нейрофизиологов, психофизиологов, психологов, неврологов и психиатров.
УДК 612.82-073.97-08
БЕК 56.1
ISBN 978-617-7001-39-2
© Кропотов Ю.Д., 2010
© Видавець Заславський О.Ю., 2010
Содержание
Предисловие к русскому изданию.....................................8
Введение: теоретические основы количественной ЭЭГ и нейротерапии...11
1. Место ЭЭГ в нейронауке и медицине...........................14
2. От нейронных спайков — к локальным корковым потенциалам, и далее — к скальповой ЭЭГ.....................................20
3. Эндофенотипы и индивидуальные различия......................24
4. Фармако-ЭЭГ.................................................29
5. Предпосылки нейротерапии....................................30
ЧАСТЫ. Ритмы ЭЭГ
Глава 1. Мед ленные, сверхмед ленные потенциалы и дельта-ритмы
I. Происхождение скальповых потенциалов........................42
II. Сверхмедленные колебания...................................49
III. Медленные волны глубокого сна.............................52
IV. Дельта-колебания...........................................54
V. Заключение..................................................58
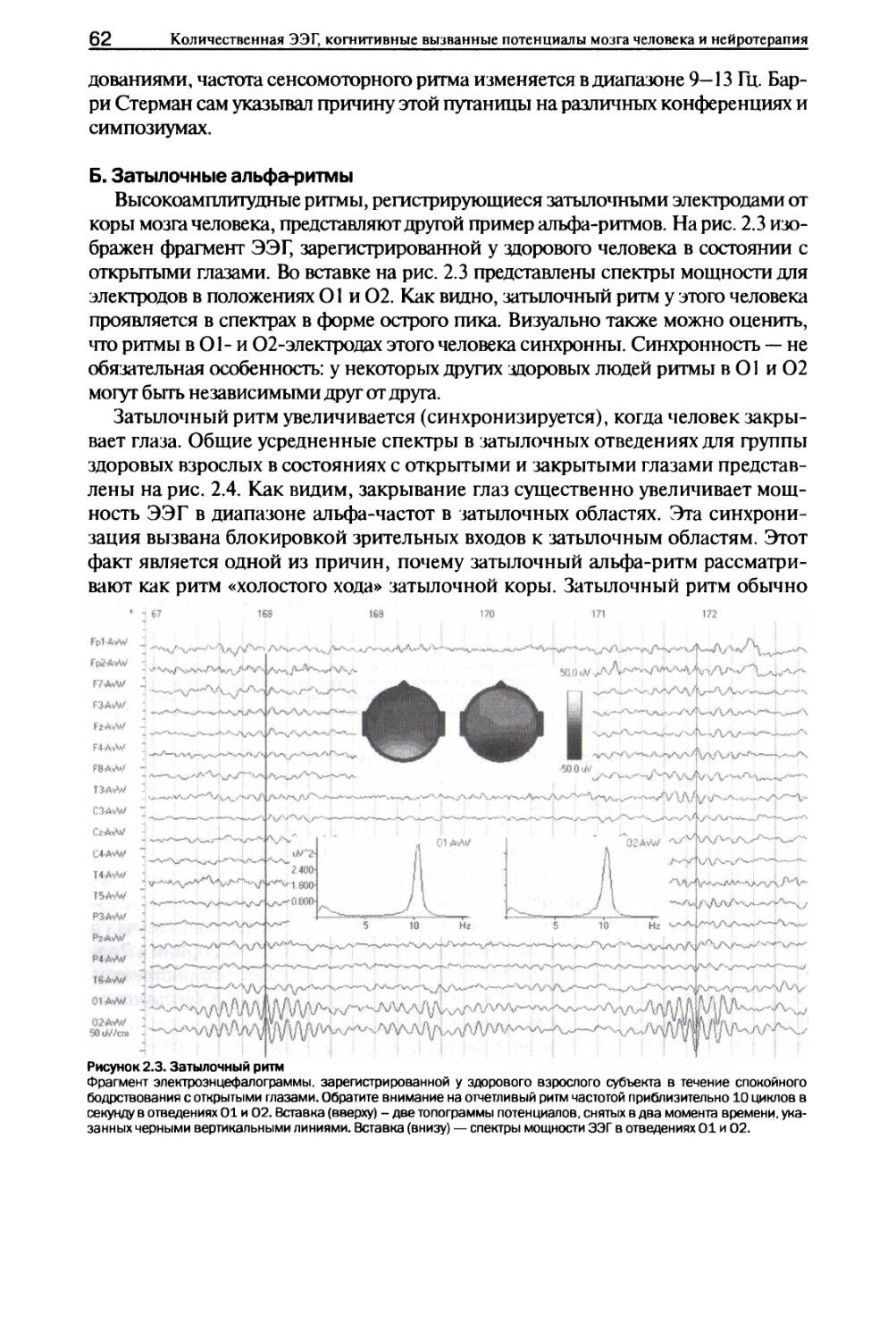
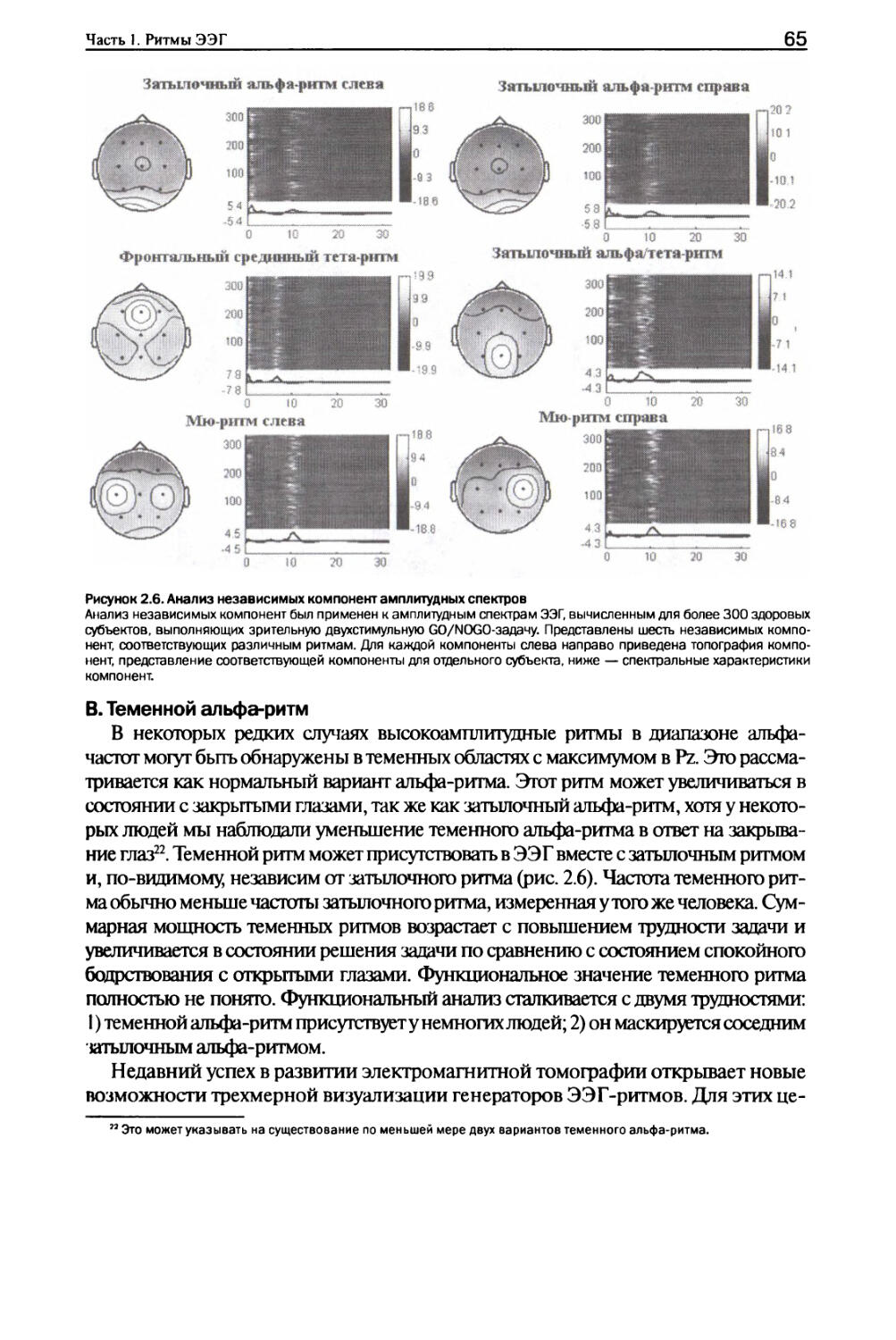
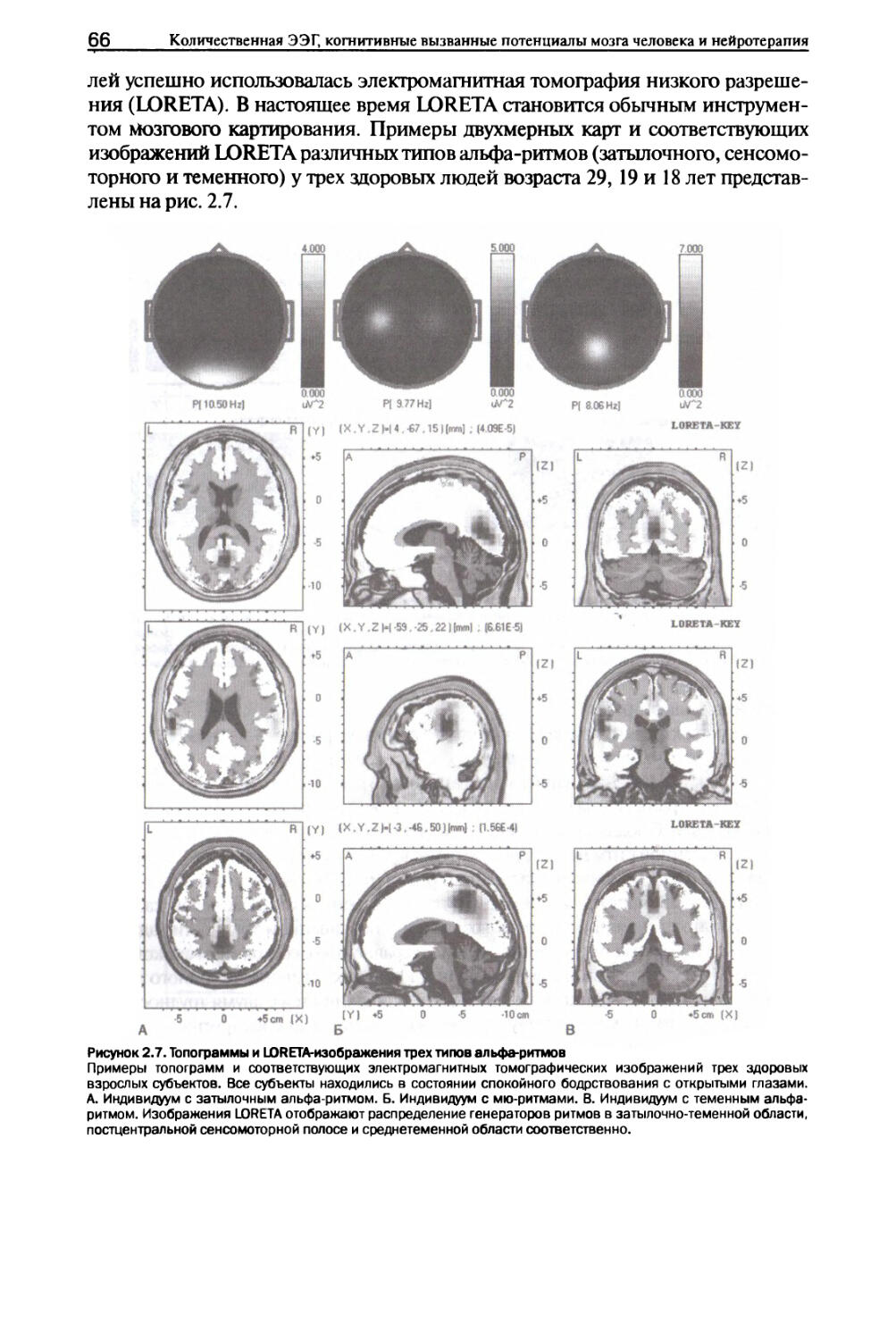
Глава 2. Альфа-ритмы I. Типы альфа-ритмов...........................................59
II. Нейронные механизмы........................................67
III. Изменения в ответ на решение задач........................75
IV. Функциональное значение....................................78
V. Аномальные альфа-ритмы......................................80
VI. Заключение.................................................84
Глава 3. Бета-ритмы
I. Типы бета-ритмов............................................86
II. Нейронные механизмы........................................91
III. Гамма-активность..........................................94
IV. Функциональное значение................................... 96
V. Аномальные бета-ритмы...................................... 99
VI. Заключение...............................................100
Глава 4. Среднелобный тета-ритм I. Характеристики.............................................101
II. Нейронные механизмы.......................................104
III. Изменения в ответ на решение задач.......................111
IV Функциональное значение...................................114
V Аномальные тета-ритмы......................................115
VI. Заключение................................................116
Глава 5. Пароксизмальная активность I. Спайки.....................................................118
II. Нейронные механизмы.......................................121
III. Заключение...............................................123
Глава 6. Эндофенотипы количественной ЭЭГ
I. Воспроизводимость результатов от теста к тесту.............125
II. Отражение функционирования мозговых систем................129
III. Наследуемость............................................129
IV Заключение................................................130
Содержание 5
Глава 7. QEEG во время сна I. Анатомические основы.........................................131
II. ЭЭГ-корреляты сна..........................................132
III. Функциональное значение сна...............................133
IV. Биспектральный индекс......................................134
Глава 8. Методы анализа фоновой ЭЭГ
I. Анатомическая локализация...................................137
II. Поля Бродмана..............................................137
III. Международная система размещения электродов 10—20.........139
IV Электроды..................................................139
V. Усилители..................................................141
VI. Переведение ЭЭГ в цифровую форму..........................142
VII. Монтаж....................................................142
VIII. Анализ Фурье.............................................145
IX. Картирование ЭЭГ...........................................148
X. Фильтрация.................................................148
XI. Биспектры.................................................150
XII. Когерентность.............................................151
XIII. Вызванная десинхронизация................................153
XIV Волновое (вейвлет) преобразование........................154
XV. Слепое разделение источников и анализ независимых компонент (ICA).155
XVI. Коррекция артефактов пространственной фильтрацией........160
XVII. Другие типы артефактов..................................161
XVIII. Обратная задача и оценка диполя.........................164
XIX. Электромагнитная томография низкого разрешения (LORETA)..165
XX. Зависимая от уровня оксигенации крови фМРТ (BOLD fMRI)....169
XXI. Корданс..................................................170
XXII. Нормальные распределения и отклонение от нормы..........171
XXIII. Доступные в настоящее время базы данных.................173
Глава 9. Практика I. Введение.....................................................177
II. Программное обеспечение Ed-EEG.............................180
III. Упражнения................................................191
ЧАСТЬ 2. Вызванные потенциалы
Глава 10. Сенсорные системы I. Анатомия....................................................201
II. Обработка зрительной информации...........................204
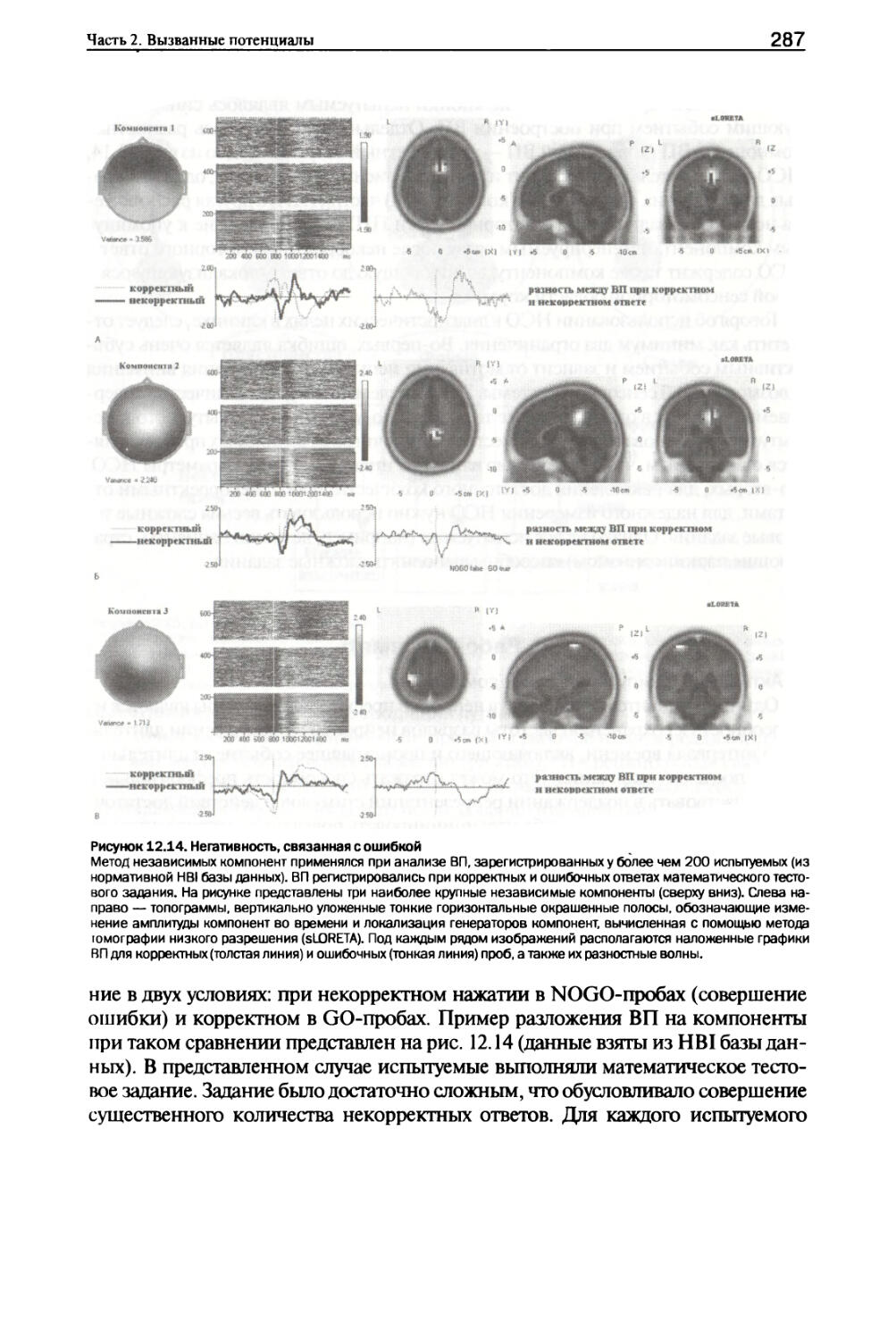
III. Разложение ВП одной пробы на независимые компоненты......211
IV. Разложение усредненного ВП на отдельные компоненты........214
V. Информационные потоки слуховой информации..................218
VI. Соматосенсорная модальность..............................222
VII. Детекция изменений.......................................223
VI11. Типы сенсорных систем...................................233
IX. Диагностическая ценность сенсорных ВП.....................235
X. Заключение................................................237
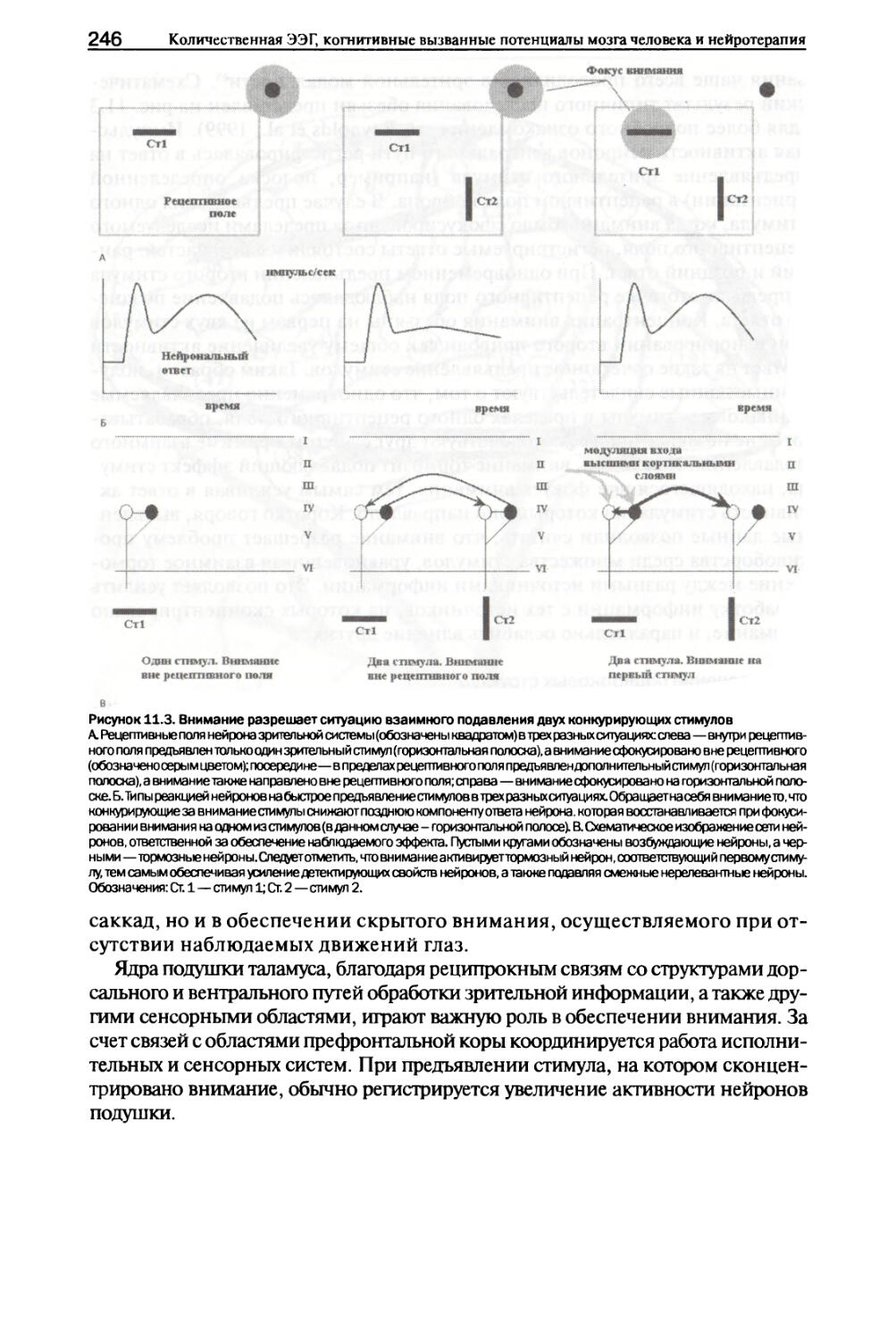
I лава 11. Система внимания I. Психология..................................................239
II. Анатомия..................................................243
6 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
III. Модуляция процессов обработки сенсорной информации.......244
IV Нейрофизиология...........................................248
V. Нейрональные сети.........................................249
VI. Поздние позитивные компоненты ВП.........................252
VII. Заключение...............................................259
Глава 12. Исполнительные системы I. Психология.................................................260
II. Базальные ганглии как «темный подвал» мозга...............261
III. Префронтальная кора и исполнительный контроль............273
IV Операции вовлечения и отвлечения..........................276
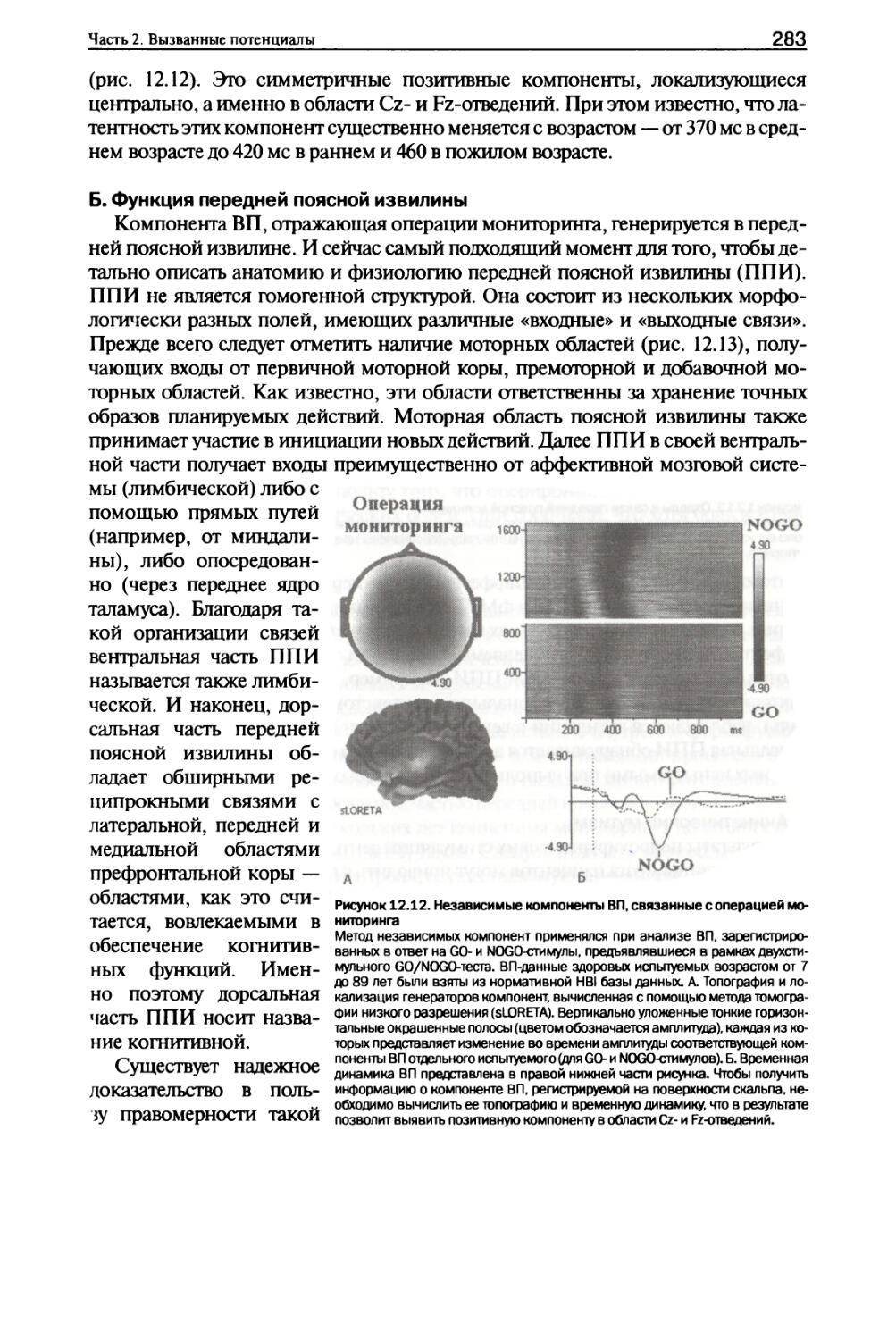
V. Операции мониторинга......................................282
VI. Рабочая память............................................288
VII. Дофамин как медиатор исполнительных систем...............291
VIII. Заключение..............................................295
Глава 13. Аффективная система I. Психология.................................................297
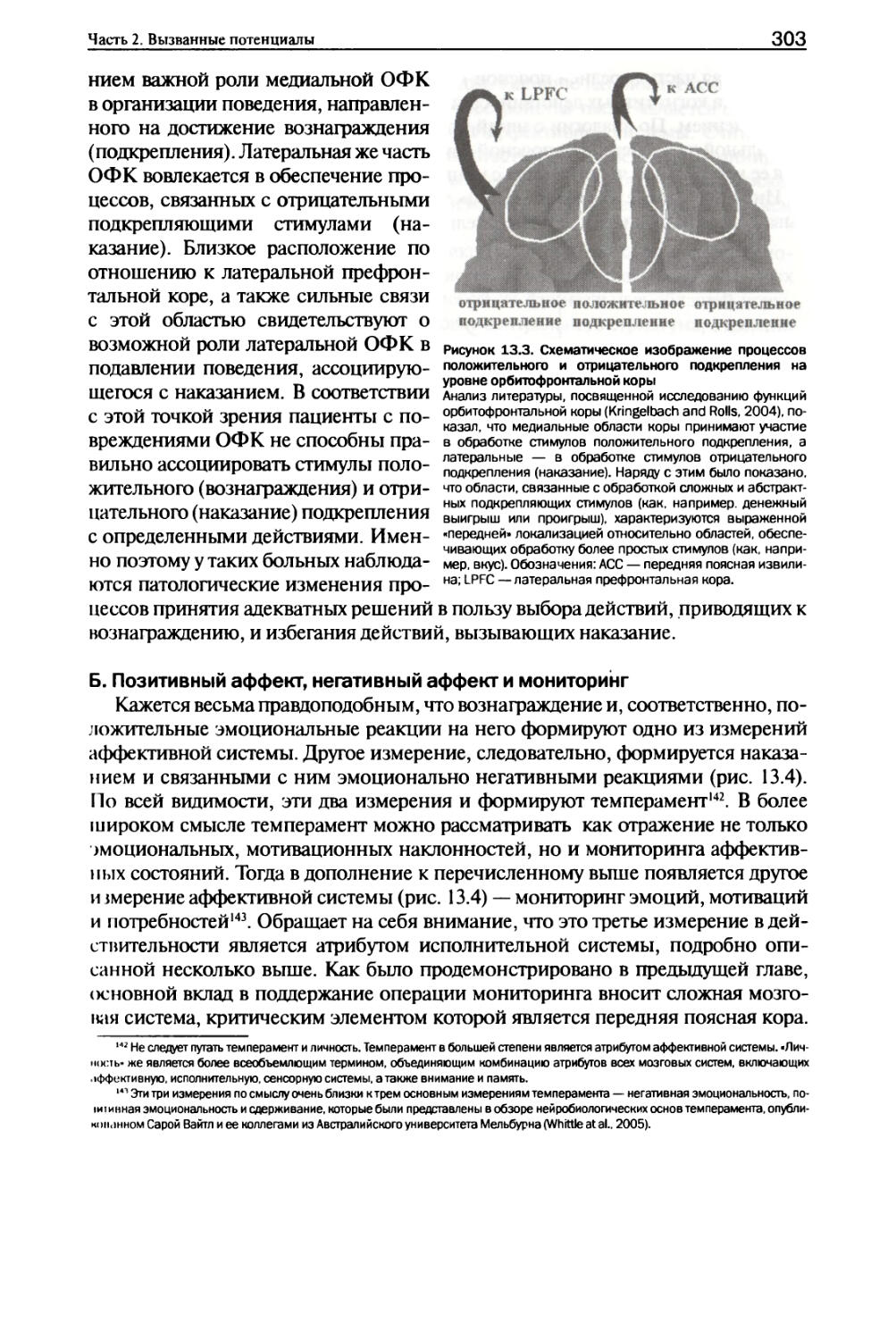
И. Анатомия...................................................299
III. Физиология...............................................301
IV Этапы реакций аффективной системы.........................309
V. Серотонин как основной медиатор аффективной системы.......311
VI. Заключение................................................312
Глава 14. Системы памяти I. Психология.................................................314
II. Декларативная память......................................315
III. Ацетилхолин как основной медиатор системы обеспечения декларативной памяти..........................................319
IV. ВП-показатели эпизодической памяти........................321
V. Система процедурной памятиЗ...............................322
VI. Основные медиаторы мозговой системы процедурной памяти....326
VII. Заключение...............................................327
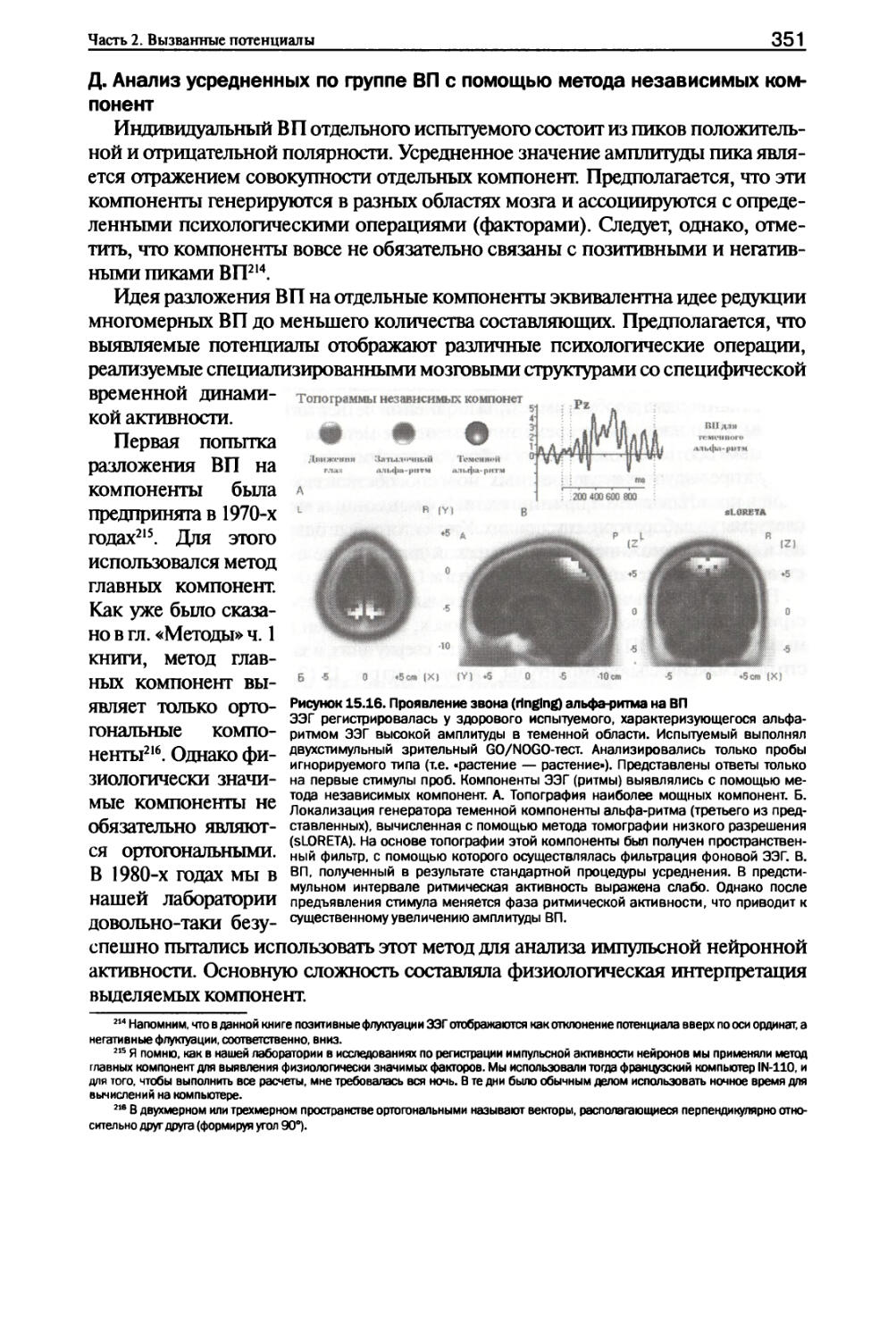
Глава 15. Методы: нейронные сети и вызванные потенциалы I. Информационные процессы нейронных сетей....................329
II. Нейротрансмиттеры и нейромодуляторы.......................341
III. Методы анализа вызванных потенциалов.....................346
FV Вызванные потенциалы в фармакологических исследованиях.................................................353
V Поведенческие парадигмы.....................................356
Глава 16. Практика I. Введение...................................................366
II. Формирование тестового задания............................368
III. Программа EdEEG..........................................370
IV Упражнения................................................377
ЧАСТЬ 3. Расстройства систем мозга
Глава 17. Сшщром нарушения внимания с гиперактивностью I. Клинические симптомы.......................................386
II. Генетические и внешние факторы............................388
III. Структурные и физиологические корреляты..................389
IV Корреляты когнитивных вызванных потенциалов...............392
Содержание 7
V Дофаминовая гипотеза СНВГ..................................399
VI. Лечение...................................................400
VII. Заключение................................................408
Глава 18. Шизофрения I. Клинические симптомы........................................410
II. Генетические и внешние факторы.............................411
III. Структурные и физиологические корреляты...................411
IV Дофаминовая гипотеза шизофрении............................415
V. Лечение....................................................417
VI. Заключение................................................419
Глава 19. Аддиктивные расстройства I. Описание поведения..........................................421
II. Структурные и физиологические корреляты....................422
III. Этапы аддиктивного процесса...............................423
IV Лечение....................................................427
V. Заключение.................................................429
Глава 20. Обсессивно-компульсивное расстройство I. Описание поведения..........................................431
II. Генетические факторы и коморбидность.......................432
III. Структурные и физиологические корреляты...................432
IV Медиаторы..................................................434
V Лечение....................................................434
VI. Заключение................................................436
Глава 21. Депрессия I. Клиническая картина.........................................438
II. Структурные и физиологические корреляты....................439
III. Нейрональная модель.......................................444
IV Лечение....................................................446
V. Заключение.................................................448
Глава 22. Болезнь Альцгеймера I. Описание поведения..........................................450
II. Медиаторы..................................................450
III. Модель нейронной сети.....................................451
IV. Структурные и физиологические корреляты....................452
V. Лечение....................................................453
VI. Заключение................................................454
Глава 23. Методы нейротерапии I. Плацебо.....................................................455
II . ЭЭГ-биоуправление.........................................457
111 . Глубинная стимуляция мозга...............................477
IV Транскраниальная магнитная стимуляция......................479
V Транскраниальная микрополяризация..........................480
Заключение I. Общие принципы анализа ЭЭГ и нейротерапии...................487
11.11 редметы дальнейших исследований..........................492
Литература........................................................496
Предисловие к русскому изданию
Идея написать эту книгу появилась в декабре 2002 года на живописном приморском курорте около Лиссабона, где Джей Гункельман и я проводили школу по количественной ЭЭГ и нейротерапии. Эта школа была одним из первых мероприятий, организованных и спонсируемых Европейским отделением Международного общества нейрорегуляции. Дело в том, что в Америке в 90-е годы XX столетия большую популярность получило направление «нейротерапия». Нейротерапия как методология модуляции физиологии мозга основывается на регистрации ЭЭГ и на биологической обратной связи, осуществляемой текущими параметрами этого электрофизиологического параметра деятельности мозга человека. Следствием популярности нейротерапии стало создание сначала американского, а потом и международного общества по нейрорегуляции. Это общество в настоящее время насчитывает более тысячи членов, которые ежегодно собираются на конференциях, проводимых в различных штатах Америки.
Следует сразу сказать, что методика биологической обратной связи была разработана в конце 60-х годов в Институте экспериментальной медицины в отделе экологической физиологии под руководством профессора Николая Николаевича Василевского. Эта методика одновременно с экспериментальными работами на животных стала использоваться в клинике для коррекции некоторых устойчивых патологических состояний мозга человека. В частности, клинические испытания метода проводились профессором Наталией Васильевной Черниговской в отделе нейрофизиологии человека, возглавляемом академиком Наталией Петровной Бехтеревой. В 90-е годы в нашей лаборатории Института мозга человека РАН мы стали использовать биологическую обратную связь для коррекции поведения детей с синдромом нарушения внимания с гиперактивностью. Поскольку все эти отечественные работы проводились параллельно с американскими исследованиями, очень трудно отдать приоритет той или другой стороне. Да это и не важно. Главное то, что в конце 90-х годов было фактически два центра научных исследований в области биологической обратной связи — один в России, другой в США. Поэтому неудивительно, что на одной европейской школе по нейротерапии в 2002 году занятия проводились американцем Джеем Гункельманом и русским, автором настоящей книги Юрием Дмитриевичем Кропотовым.
На этой школе наши ученики задавали один и тот же вопрос: «Где все это можно прочитать?» И надо сказать, это был один из немногих вопросов, на
Предисловие к русскому изданию
9
которые мы не могли дать вразумительного ответа. Все дело в том, что не существовало такого учебника. Вернее, были книги по практическому использованию биологической обратной связи, но теоретическое обоснование метода полностью отсутствовало. Поэтому практиков нейротерапии часто критиковали за отсутствие теории. Потребность в такой книге становилась все более и более очевидной.
В 1995 году в журнале Международного общества по психофизиологии была опубликована наша статья по применению когнитивных вызванных потенциалов для оценки успешности нейротерапии у детей с синдромом нарушения внимания. Эта статья получила огромный резонанс в научных кругах и стала наиболее цитируемой статьей в области психофизиологии. Указанный журнал издается международным издательством «Элзавир», которое также специализируется на выпуске академических изданий и постоянно отслеживает точки роста в различных областях науки. Сара Пурди — главный редактор секции нейронаук «Элзавира» — обратилась ко мне с предложением написать научную монографию на тему «Количественная ЭЭГ и нейротерапия». Естественно, книга должна была быть на английском языке. Я принял это предложение.
В те годы (и по настоящее время) я был приглашен читать лекции по этой области нейронаук в Норвежский университет науки и технологии в Тронд-хейме. Будучи профессором этого крупнейшего в Норвегии университета, я имел прямой доступ ко всем журналам, что значительно облегчило анализ литературы. Большая часть книги была написана во время долгих (с двумя остановками) перелетов из Санкт-Петербурга в Трондхейм.
В нашей лаборатории старшим научным сотрудником, кандидатом биологических наук Пономаревым Валерием Александровичем было разработано программное обеспечение для анализа количественной ЭЭГ и когнитивных вызванных потенциалов. Эта работа фактически началась в 80-х годах, когда, получив французский мини-компьютер ИН-110 для исследований импульсной активности нейронов, Юрий Львович Гоголицын, Сергей Викторович Пахомов и автор этой книги создали первую аппаратнокомпьютерную систему обработки физиологических сигналов. Далее работа над программным обеспечением продолжалась Валерием Александровичем Пономаревым и была доведена до совершенства. Имея, таким образом, в руках одну из лучших в мире программ, я получил уникальную возможность проверить многие теоретические идеи на практике.
И наконец, к тому времени, когда я начал работу над рукописью книги, была собрана достаточно представительная референтная база данных. Эта работа была выполнена в нескольких научных центрах Европы к рамках частных проектов и проектов Европейского сообщества. Она координировалась ученым и практиком из Швейцарии доктором Андреасом Мюллером. Референтная база, получившая название «Человеческий мозговой индекс» — ЧМИ (Human Brain Index — HBI), включала в себя записи ЭЭГ более тысячи здоровых испытуемых и большого числа
10 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
больных с различными психическими заболеваниями, в том числе нарушением внимания, шизофренией, депрессией, навязчивыми состояниями. 19-канальная ЭЭГ регистрировалась у здоровых испытуемых и больных в состоянии спокойного бодрствования с открытыми и закрытыми глазами, а также во время выполнения психологических тестов на внимание, память, зрительное и слуховое восприятие, математические операции, распознавание письменной и устной речи, оценку эмоционального состояния людей по фотографиям. Таким образом, я получил доступ к огромному количеству данных, которые можно было обработать и проанализировать.
После того как эта моя книга — «Quantitative EEG, event related potentials and neurotherapy» была написана и сдана в редакцию, я решил организовать Всероссийскую конференцию с практически тем же названием. Эта конференция впервые была проведена в Санкт-Петербурге в конце октября 2007 года. Организацию конференции спонсировали несколько российских фирм по производству медицинской техники, среди которых ключевую роль сыграла Санкт-Петербургская фирма «Ми-цар» во главе с ее директором Сергеем Васильевичем Васильевым. Конференция, в организации которой активное участие приняли ученые из Москвы и Санкт-Петербурга, привлекла большое число участников, единодушно приветствовавших организацию ежегодных таких конференций. Следующая конференция была проведена в 2009-м. На круглом столе, посвященном перспективам развития отрасли, наши украинские коллеги во главе с Зубовым Павлом Геннадиевичем предложили опубликовать эту книгу в Украине. Перевод с английского на русский был сделан сотрудниками Института мозга человека В.Н. Гринь-Яценко, Н.В. Шемякиной, Ж.В. Нагорновой, М.В. Киреевой под редакцией В.А. Пономарева.
Эта книга не была бы возможной без труда огромного количества людей, помогавших мне в проведении исследований и поддерживающих меня во время ее написания. Невозможно всех перечислить. Хочу поклониться своему учителю — Наталье Петровне Бехтеревой, сказать слова признательности директору нашего института Святославу Всеволодовичу Медведеву. Особую благодарность хочу выразить сотрудникам моей лаборатории В.А. Пономареву, В.Н. Гринь-Яценко, Ю.И. Полякову, Л.С. Чутко, Е.П. Терещенко, Е.А. Яковенко, Н.С. Никишеной, А.В. Севостьянову, О.В. Кара, И.З. Кремень, В.Н. Нечаеву Из зарубежных коллег хотелось бы выделить Дага Ричардсона и Кортиса Крайпа за финансовую поддержку, Джея Гункельмана, Андреаса Мюллера, Зигфрида Отмара, Джоела Любара, Тони и Беверли Стеффертов, Джона Грузилье, Джонел Виллар, Вице ван дер Цваага, Кнута Хестада за плодотворные научные обсуждения.
Февраль 2010, Санкт-Петербург
Понять — значит упростить.
Аркадий и Борис Стругацкие.
Волны гасят ветер
Введение: теоретические основы количественной ЭЭГ и нейротерапии
Мозг человека — это наисложнейшее образование на земле. Функционирование этой анатомической структуры определяет все многообразие поведения человека. В течение последних 80 лет фундаментальная наука разработала несколько методов изучения работы человеческого мозга. Эти методы используют как инвазивные, так и неинвазивные средства. Инвазивные методы, такие как регистрация импульсной активности одиночных нейронов и нейронных популяций, запись локальных электрических потенциалов и внутричерепных когнитивных вызванных потенциалов, полярографическая регистрация уровня оксигенации ткани мозга, требуют имплантации электродов в ткани мозга человека (в клинических целях) или животных (в целях эксперимента). К числу неинвазивных методов относят магнитно-резонансную томографию, позитронно-эмиссионную томографию, магнитоэнцефалографию и электроэнцефалографию и некоторые другие. Как инвазивные, так и неинвазивные методы открывают для нас несколько перекрывающихся, но все же разных «окон», позволяющих смотреть на живой мозг с разных точек зрения. ЭЭГ — лишь один из этих методов!
Во введении мы намерены: 1) представить читателю общий обзор методов оценки работы мозга; 2) показать сходство и различия между ЭЭГ и другими методами; 3) кратко обозначить потенциальные возможности ЭЭГ в раскрытии индивидуальных особенностей здорового мозга и определении эндофенотипов больного мозга; 4) изложить основную идею о том, что предъявление информации об основных электрофизиологических и метаболических параметрах работы мозга в виде сигнала обратной связи дает возможность здоровым людям и пациентам произвольно (если параметр представлен в сенсорной модальности) или непроизвольно (если мозг подвергается воздействию электрического тока) влиять на работу своего мозга.
Глоссарий
Потенциал действия нейрона (иногда называемый нейронным спайком) — это нейронный разряд, вызванный быстрым (порядка миллисекунд) открытием и закрытием Na+- и К+- ионных каналов мембраны нейрона. Разряд возникает и случае, если деполяризация мембраны достигает пороговых значений. Потенциал действия — самое элементарное событие в процессе обработки ин-
12 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
формации в нейронных сетях, связанное с передачей «байта» информации от одного нейрона к другим.
Биохимия — наука, изучающая химическую природу веществ, входящих в состав живых организмов, и химические процессы, лежащие в основе их жизнедеятельности.
Картирование мозга — это дисциплина, возникшая недавно на стыке медицины и нейронауки. Картирование мозга разделяется на две основные категории — структурное картирование и функциональное картирование. Структурное картирование занимается 3-мерными пространственными параметрами, характеризующими анатомическую и биохимическую структуры мозга. Функциональное картирование имеет отношение к динамическим параметрам работы мозга. МРТ, ПЭТ и ЭЭГ — примеры методов функционального картирования.
Глубокая стимуляция мозга — длительное применение коротких импульсов электрического тока с помощью имплантированных электродов, ведущее, как предполагается, к функциональной блокаде стимулируемых нейронных сетей.
Электроэнцефалограмма (ЭЭГ) — запись биоэлектрических потенциалов мозга с помощью электродов, расположенных на голове.
Электрофизиология — раздел физиологии, изучающий электрические явления в организме при различных видах его деятельности.
Эндофенотипы — количественные характеристики мозга или организма (например, показатели мощности ЭЭГ в конкретных частотных диапазонах или компоненты когнитивных ВП), определяющие индивидуальную предрасположенность к развитию или манифестации определенных заболеваний или особенностей поведения.
Когнитивные вызванные потенциалы — локальные электрические потенциалы, зарегистрированные в процессе выполнения психологических тестов (например, oddball или GO/NOGO) и усредненные для каждого вида проб. В процессе усреднения спонтанные положительные и отрицательные колебания взаимно погашают друг друга, оставляя усредненные потенциалы, связанные с различными стадиями обработки информации в мозге.
Функциональная МРТ — разновидность магнитно-резонансной томографии, позволяющая изучать изменение локального кровотока и метаболических реакций мозга, связанных с выполнением какой-либо деятельности. Метод основан на измерении неоднородности атомов кислорода, обусловленных сложными биохимическими процессами оксигенации крови, потребления кислорода и активации нервных клеток.
Ген — функционально является единицей наследственности и структурно заключен в нить ДНК. Ген содержит промотор, контролирующий активность гена и кодирующий последовательность нуклеотидов, определяющую его свойства.
Генотип — совокупность всех генов, локализованных в хромосомах данного организма. Термины «генотип» и «фенотип» отражают два полярных уровня организации — молекулярный уровень ДНК и целостное поведение человека. Генотип и фенотип не связаны напрямую: некоторые гены могут реализовать
Введение: теоретические основы количественной ЭЭГ и нейротерапии
13
конкретный фенотип только при определенных условиях окружающей среды, в то время как некоторые фенотипы — результат влияния множества генов.
Локальные потенциалы — потенциалы, зарегистрированные с помощью микро-и макро-электродов, помещенных в ткань мозга. Эти потенциалы генерируются мембранами локальных нейронов и глии.
Магнитно-резонансная томография (МРТ) — метод измерения плотности распределения некоторых элементарных магнитов (таких как атомы водорода), помещенных в магнитное поле, с помощью регистрации ответа этих магнитов на радиосигналы. Этот ответ называется магнитным резонансом.
Магнитоэнцефалография (МЭГ) — неинвазивный метод исследования электрической активности мозга путем регистрации магнитных полей, связанных с мозговой активностью. Поскольку мозг излучает крайне слабые магнитные поля, для регистрации сигнала используются чрезвычайно чувствительные датчики магнитного поля, работающие при очень низких температурах, обеспечивающих сверхпроводимость.
Молекулярная генетика — отрасль науки, изучающая молекулярные основы ДНК, РНК и связанные с ними молекулярные структуры клеток живого организма.
Активность нейронной популяции — это последовательность потенциалов действия группы нейронов, окружающих электрод, помещенный в ткань мозга. Активность нейронной популяции регистрируется с помощью внеклеточных микро- (диаметром приблизительно несколько микрон) или макроэлектродов (до 100 микрон).
Нейронаука — это отрасль науки, изучающая мозг и его связь с поведением человека и животных.
Нейротерапия — это ряд основанных на нейрофизиологии методов, используемых для изменения мозговых функций. Эти методы включают ЭЭГ-БОС, транскраниальную микрополяризацию и транскраниальную магнитную стимуляцию.
Фенотип — это совокупность всех признаков и свойств организма, формирующихся при взаимодействии его генотипа с окружающей средой. Поскольку фенотипы доступны наблюдению в гораздо большей степени, чем генотипы, такие отрасли современной медицины, как неврология и психиатрия, используют поведенческие фенотипы для классификации и диагностики заболеваний мозга.
Полярографический метод измерения уровня концентрации кислорода основан на том факте, что напряжение —0,63 V, подаваемое на поляризующий электрод (например, золотой проволочный электрод, помещенный в ткань мозга), вызывает ток, пропорциональный по величине концентрации кислорода в ткани мозга.
Психология — наука о закономерностях, механизме и фактах психической жизни и поведения человека и животных.
Количественная ЭЭГ— совокупность методов количественной обработки ЭЭГ-сигналов. Количественная ЭЭГ включает спектральный анализ и частотновременное представление мощности ЭЭГ (вейвлет-анализ).
14 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Активность одиночного нейрона — последовательность потенциалов действия отдельного нейрона, зарегистрированная с помощью электрода, введенного в нейрон (внутриклеточная регистрация) или расположенного рядом с нейроном (внеклеточная регистрация).
Стереотаксическая нейрохирургия — это микрохирургическое вмешательство в глубокие структуры мозга в целях их разрушения, биопсии или имплантации электродов, основанное на нейрокартировании с использованием трехмерной системы внутримозговых координат.
1. Место ЭЭГ в нейронауке и медицине
А. Цели нейронауки
Поведение человека обычно подразделяют на две основные категории: нормальное, т.е. соответствующее поведенческому паттерну здоровых индивидов, и аномальное, обусловленное нарушениями психической деятельности. Нормальное поведение изучается главным образом фундаментальными науками, обычно объединенными под названием «нейронаука», в то время как аномальное поведение — предмет исследования медицинских наук. Нейронаука исследует мозг и его связь с психической деятельностью (рис. В. 1). Изучение мозга человека, помимо удовлетворения чистого любопытства, преследует две основные цели. Первая из них базируется на неврологии, психиатрии, психотерапии и медицинской психологии и направлена: 1) на определение анатомических и физиологических маркеров заболеваний мозга; 2) лечение расстройств психической деятельности. Другая задача нейронауки — имитация работы мозга в различных прикладных целях. Реализация этой цели связана с математическим моделированием нейронных сетей и созданием электронных приборов, называемых ней-
Фундаментальная наука Нормальное поведение Исследование Медицина Ненормальное поведение Исследование Диагностика Лечение
Психология Нейрофизиология МКБ-10 Психохирургия Психотерапия Фенотнн ж X Д
Нейрофизиология 11атофизио7101 ия МРТ,фМРТ,ПЭТ Количественная ЭЭГ ээг,вп Нейротержтня м Я К g
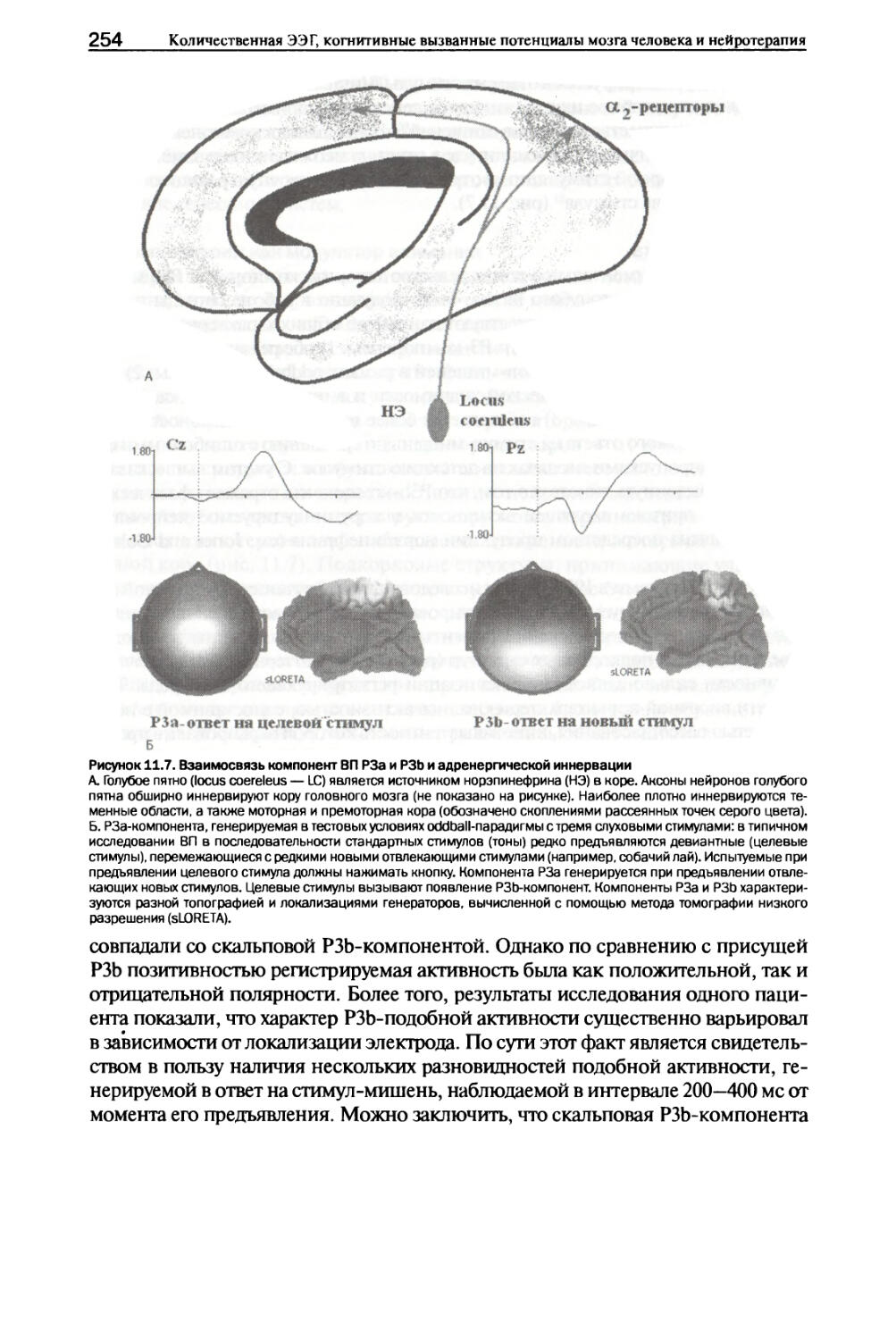
Эндофенотнп
Биохиьшя Фармакология Рецепторы, ионные канал*! ИТ.Д. Фармакотерапия № О CQ
Молекулярная генетика Мсдвдюккая генетика ДНК, гены Генная юокенерия Геногни
Рисунок В.1. Место количественной ЭЭГ и нейротерапии в фундаментальных и медицинских науках
Изучение нормального поведения — предмет изучения психологии, физиологии, биохимии и молекулярной генетики. Исследование аномального поведения относится к сфере нейрофизиологии, патофизиологии, фармакологии и меди цинской генетики. Постановка диагноза осуществляется на основе описания поведения согласно руководствам DSM-IV или ICD-10. В современной медицине четко прослеживается тенденция к поиску так называемых эндофенотипов — объективных диагностических параметров, специфичных для данного заболевания. Лечение проводится в соответ ствии с четырьмя различными уровнями организации мозга/поведения с помощью психотерапии, психохирургии, ней ротерапии и фармакотерапии.
Введение: теоретические основы количественной ЭЭГ и нейротерапии 15
рокомпьютерами, основанных на принципах обработки информации в мозге. Объем этой книги ограничен первой целью. Мы собираемся представить экспериментальные данные, теоретические сведения и методологические инструменты, которые могут помочь неврологам, психиатрам, психологам, психотерапевтам и работникам здравоохранения в оценке мозговых дисфункций и на основе этой оценки обеспечить оптимальное лечение их пациентов.
Б. Цели психиатрии и неврологии
Одна из главных задач психиатрии и неврологии связана с объективной оценкой дисфункций мозговой деятельности. В настоящее время единственным способом диагностики психических расстройств является использование описания отклонений в поведении с помощью специфической терминологии, данной в таких руководствах, как Diagnostic Statistical Manual в США (DSM —IV) или International Classification of Disorders в Европе (ICD-10) (рис. В. 1).
Согласно этим руководствам все мозговые дисфункции разделяются на отдельные категории, называемые заболеваниями. Каждое заболевание имеет свое название (например, болезнь Паркинсона или шизофрения) и определяется входящими в его состав симптомами (основными аномальными поведенческими паттернами) и/или синдромами (группами симптомов). Пример описания симптомов отдельного психического расс+ройства представлен в перечне поведенческих паттернов при СНВГ* 1 (синдром нарушения внимания и гиперактивности).
В. Фенотип и генотип
Важная роль в функционировании нейронных сетей отводится медиаторам и рецепторам, определяющим динамические свойства синаптических связей в нейронных сетях. Изучение субклеточных процессов и молекулярных механизмов работы синапсов относится к разделу биохимии в сфере фундаментальных наук и фармакологии2 в сфере медицины.
Синтез нейромедиаторов, работа ионных каналов и рецепторов контролируются генами. Гены определяют генотип индивида. Термин «фенотип» отражает подлинные анатомические и физиологические черты индивида и как следствие этих черт — его поведение. Генотип — важнейший, но не единственный фактор, влияющий на формирование фенотипа. Как нормальные, так и аномальные поведенческие паттерны человека возникают на основе сложного взаимодействия генетических факторов и влияния окружающей среды. Механизмы этого взаи-модействия пока недостаточно изучены3.
* См., например, проект «Википедия», многоязычную энциклопедию со свободно распространяемым в Интернете содержимым. Википедия создается совместными усилиями добровольцев всего мира. За редким исключением, ее статьи могут редактироваться любым пользователем Интернета. Википедия — слово-гибрид составленное из слов «вики» (тип совместного сайта в Интернете) и •энциклопедия». Со времени ее создания в 2001 г. Википедия быстро выросла и превратилась в один из самых больших справочных (мигов. Описание СНВГ и его симптомов вы можете найти на: http://en.wikipedia.org/wiki/Attention-Deficit_Hyperactivity_Disonder.
’ Хотя известно, что психоактивные средства использовались еще тысячелетия назад психофармакология как наука возникла в 1 МЬО-х, когда случайно была обнаружена эффективность этих средств при шизофрении и депрессии
1 Сложная структура организации мозга — основной фактор, затрудняющий понимание взаимосвязи генотипа и феноти-
па. Дело в том, что клетки мозга проявляют разные свойства в составе различных мозговых систем, и в то же время их взаимодействия подвержены изменениям под влиянием факторов внешней среды. Как следствие, комплексное взаимодействие
юное и внешней среды определяет повреждение нейронных систем и выполняемых ими функций.
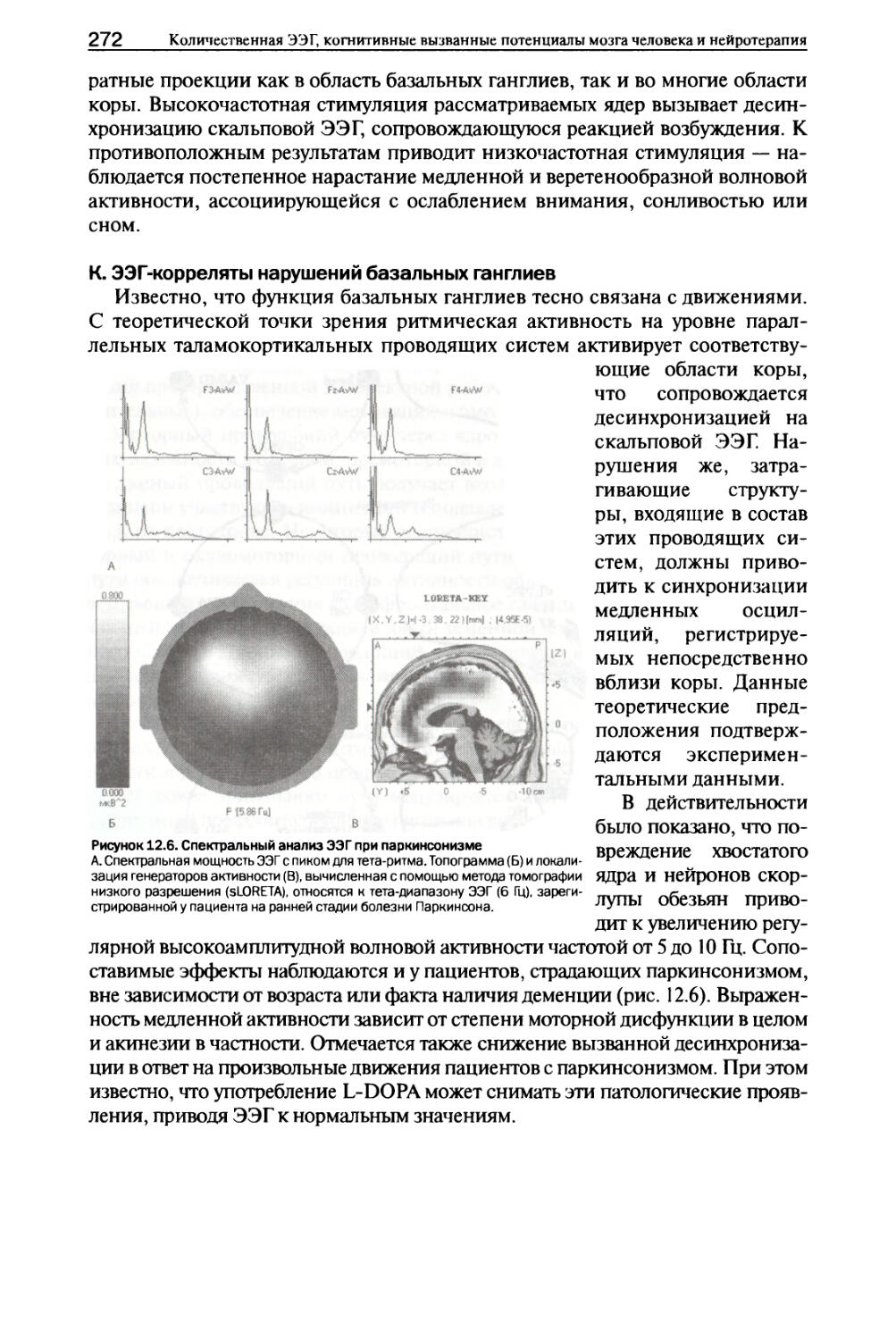
16 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Число исследований в нейронауке экспоненциально увеличивается, и знания, полученные за последние 10—20 лет, значительно изменили наши представления о мозге. Эти изменения определили новый подход к клиническому применению электроэнцефалографии (ЭЭГ). Однако ЭЭГ — не единственный метод, предоставляемый нейронаукой в распоряжение клиницистов. Из других методов наиболее известны магнитоэнцефалография (МЭГ), позитронно-эмиссионная томография (ПЭТ) и магнитно-резонансная томография (МРТ). Все они обычно объединяются под общим названием «нейрокартирование», и каждый занимается исследованием специфических нейроанатомических и нейрофизиологических параметров, открывая, таким образом, маленькие «окна в мозг». Ниже представлена взаимосвязь этих методов и электроэнцефалографии.
Г. Магнитоэнцефалография — метод, дополняющий ЭЭГ
ЭЭГ как метод отражения активности мозга человека была открыта немецким ученым Гансом Бергером около 80 лет назад. С тех пор периоды процветания этого метода и энтузиазма в отношение его чередовались с периодами его упадка и забвения. Периоды процветания были связаны с появлением новых технологий регистрации и анализа данных ЭЭГ, в то время как периоды забвения были обусловлены появлением альтернативных методов нейрокартирования. В настоящее время известно, что ЭЭГ в основной своей части представляет результат суммации микродиполей, образованных пирамидными клетками, ориентированными перпендикулярно к поверхности головы. Такая ориентация электрических диполей означает, что ЭЭГ наиболее чувствительна к радиально расположенным источникам. Помимо электрических полей нейроны генерируют магнитные поля4. МЭГ — это сумма магнитных полей, возникающих в сети электрических диполей. Эти диполи соответствуют электрическим потенциалам групп пирамидных нейронов, расположенных параллельно поверхности головы, т.е. в извилинах коры. Таким образом, МЭГ и ЭЭГ могут рассматриваться как дополняющие друг друга методы: максимальный эффект в МЭГ вызывается тангенциальными токами, т.е. токами в извилинах, в то время как максимальный эффект в ЭЭГ обусловлен радиальными токами, в частности токами в бороздах, расположенных близко к поверхности (рис. В.2). В отличие от электрических сигналов ЭЭГ магнитные поля не искажаются при прохождении через оболочки мозга, кости черепа и мягкие ткани головы5.
Д. Магнитно-резонансная томография
Метод МРТ базируется на физическом феномене, известном как магнитный резонанс. Многие химические элементы, например такие как атом водорода, являются своеобразными элементарными магнитами. В обычном состоянии ориентация этих крошечных магнитов случайна. Однако если приложить внеш-
4 Магнитные поля в действительности генерируются ионными токами в нейронах. Ионные мембранные токи отдельных нейронов могут рассматриваться как токовые диполи.
5 МЭГ возникла в 1970-х, и ее становление было в значительной степени поддержано недавними достижениями в сфере компьютерного анализа и технических разработок. По сравнению с ЭЭГ для МЭГ характерно относительно низкое пространственное и чрезвычайно высокое временное разрешение (около 1 мс). Однако высокая стоимость оборудования для МЭГ и низкое отношение «сигнал/шум» сужает возможности клинического ее использования. В настоящее время применение МЭГ в клинике ограничено локализацией спайковой эпилептиформной активности.
Введение: теоретические основы количественной ЭЭГ и нейротерапии
17
Ответы на радиальные нейрональные источники
Нет Ответа
о
Ответы на тангенциальные нейрональные источники
ЭЭГ Б
МЭГ ПЭТ
А
Рисунок В.2. Методы ЭЭГ, МЭГ и ПЭТ
Слева схематично представлена кора извилин мозга с помощью двух электрических диполей, изображенных стрелками. Кровеносные сосуды вокруг нейронных генераторов помечены темно-серым цветом. Три типа датчиков схематически приведены справа посередине: металлический электрод для регистрации ЭЭГ, СКВИД (сверхпроводящий квантовый интерферометр) для регистрации МЭГ и фотоумножитель для ПЭТ, предназначенный для детекции вспышки света, излучаемой сцинтиллятором при достижении датчика гамма-фотоном. Изменения данных ЭЭГ, МЭГ и ПЭТ в ответ на кратковременное повышение нейронной активности показаны для тангенциального диполя (справа внизу) и для радиального диполя (справа вверху). ЭЭГ оценивает радиально ориентированный диполь, в то время как МЭГ — тангенциально ориентированный диполь. Задержка сигнала между событиями на уровне нейрона по данным ЭЭГ и МЭГ почти отсутствует. При использовании радиоактивной воды сигнал ПЭТ позволяет оценить медленные изменения локального кровотока, сопутствующие событиям на уровне нейрона с длительной (около 6 с) задержкой.
нее магнитное поле, эти магнитики выстроятся вдоль поля, как стрелка компаса ориентируется вдоль магнитного поля Земли. Внешние магнитные волны в радиодиапазоне, проходя через намагниченные таким образом области, заставляют эти элементарные магниты вращаться в определенном направлении. После прекращения потока радиоволн атомы возвращаются в исходное положение и излучают волны отдачи, которые регистрируются магнитными детекторами томографа. Амплитуды последних радиоволн являются исходными данными для МРТ При этом мощность сигнала определяется плотностью и магнитными свойствами элементарных магнитов. Для того чтобы восстановить плотность распределения этих магнитов в трехмерном пространстве, используются специальные математические методы реконструкции изображения6.
Е. Позитронно-эмиссионная томография
ПЭТ основывается на использовании физических свойств изотопов — радиоактивных форм простых атомов (таких как водород, кислород, фтор), которые, распадаясь, испускают позитроны. Радиоактивные атомы объединяются в более сложные молекулы, такие как кислород, вода или глюкоза. При проведении ПЭТ-исследования радиоактивные вещества вводятся в кровь пациентов и по сосудам достигают мозга. Эти вещества потребляются клетками определенных областей мозга. Радиоактивное вещество испускает позитроны. Позитроны, сталкива
* Многие неврологи и нейрохирурги, рассматривая снимки МРТ, даже не подозревают, что исходные данные выглядят совсем не так, как •н» МРТ-снимках. Исходные данные обрабатываются путем сложных математических опереций, требующих огромных (даже по современным понятиям) вычислительных мощностей. Пространственное разрешение, достигаемое с помощью МРТ, поистине изумляет. На MPT-снимках видны отдельные извилины, мозолистое тело, хвостатое ядро и даже более мелкие структуры, такие как мамиллярные тела и ядра таламуса.
18 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
ясь с электронами, аннигилируют с излучением двух гамма-квантов на каждое столкновение. Эти гамма-кванты регистрируются специальными датчиками, расположенными вокруг головы испытуемого, причем число столкновений прямо пропорционально метаболической активности соответствующего участка мозга. Иными словами, чем более активна область мозга, тем больше радиоизотопов она поглощает и тем больший уровень гамма-излучения будет зарегистрирован из этой области7. Для того чтобы восстановить распределение плотности радиоактивного вещества в трехмерном пространстве, используются специальные математические методы реконструкции, подобные тем, которые применяются в магнитно-резонансной томографии (МРТ). Однако пространственная разрешающая способность у ПЭТ значительно ниже, чем у МРТ, что не позволяет на снимках ПЭТ различать малые структуры мозга, например ядра таламуса и мамиллярные тела8.
Ж. Функциональная МРТ
Недавно была разработана модификация МРТ, так называемая функциональная МРТ, направленная на изучение изменений кровотока/метаболизма в тканях мозга при функциональных нагрузках. Этот метод основан на том факте, что любая локальная активность нейронов мозга вызывает изменение гемодинамики в этой области мозга. Сигнал на основе BOLD-контраста (bloodoxygenation-level-dependent contrast — контраст, зависящий от степени насыщения крови кислородом) в функциональной МРТ поступает от атомов водорода, входящих в состав молекул воды, в изобилии присутствующей в тканях мозга. Сигнал, регистрируемый в радиодиапазоне, с течением времени ослабевает под влиянием различных факторов, включая наличие неоднородностей в магнитном поле. Неоднородность отчасти связана с изменениями в насыщении крови кислородом. Диокси- и оксигемоглобин обладают различными магнитными свойствами, причем большая неоднородность присуща диоксигемоглобину. В связи с этим повышение концентрации диоксигемоглобина приводит к снижению интенсивности изображения.
3. Полиграфический метод исследования оксигенации мозга
Взаимосвязь между тремя процессами: 1) импульсной активностью нейронов, 2) потреблением кислорода и глюкозы нервными клетками и 3) локальным кровотоком довольно сложна и недостаточно изучена. Любое быстрое изменение импульсной активности нейрона ведет к медленным (с задержкой приблизительно на 6—10 секунд) изменениям локального кровотока и содержания кислорода во внеклеточной жидкости. В 1970-х в нашей
7 Исходным параметром для ПЭТ служит число столкновений — моментов излучения двух гамма-квантов. В ПЭТ-камере находится довольно большое число датчиков, регистрирующих гамма-кванты, и они окружают голову своеобразным кольцом, а точнее несколькими кольцами или цилиндрами.
8 ПЭТ — это довольно дорогостоящий и небезопасный метод для его работы необходимо иметь циклотрон и специальную радиохимическую лабораторию. Для того чтобы уменьшить дозу радиации, получаемую испытуемым, ему вводят сравнительно небольшие дозы изотопов, что, в свою очередь, не может не отразиться на качестве получаемых ПЭТ-изображений. Однако для решения некоторых научных задач (таких как изучение плотности рецепторов обратного захвата дофамина) ПЭТ, по-видимому, является единственным методом, доступным на настоящий момент.
Введение: теоретические основы количественной ЭЭГ и нейротерапии
19
лаборатории мы совместно с В.Б. Гречиным 9 использовали полярографический метод для изучения уровня концентрации внеклеточного кислорода у неврологических пациентов. Полярографический метод измерения уровня концентрации кислорода основан на том факте, что напряжение —0,63 V, подаваемое на поляризующий электрод (например, золотой проволочный электрод, помещенный в ткань мозга), вызывает ток, прямо пропорциональный по величине концентрации кислорода в местной ткани. В наших исследованиях мы показали, что локальный уровень концентрации кислорода в мозговой ткани непостоянен и колеблется с очень низкой
Спонтанные колебания уровня напряжешш кислорода рО>
10 23 ЗЙ
вызванная импульсная активность нейронов
О 10 28 30 еек
вызванные колебания
В
Рисунок В.З. Сверхмедленные колебания локального уровня концентрации кислорода в ткани мозга
Вверху—спонтанные колебания рО2 в хвостатом ядре и в двух ядрах таламуса. зарегистрированные с помощью полярографического метода у пациентов с болезнью Паркинсона, которым с диагностической и терапевтической целью были имплантированы золотые электроды. Внизу — импульсная активность нейронной популяции, усредненная для 10 одновременно зарегистрированных проб (слева) и рО2 (справа) в вентральном таламусе мозга человека. Импульсная активность нейронов и рО, измерялись одними и теми же электродами. Каждая проба состояла в арифметической операции (сложение или вычитание) с двумя числами, предъявленными в начале пробы.
периодичностью — приблизительно 6—10 циклов в минуту. Эти колебания отражают комплексные метаболические процессы в мозге, связанные с потреблением кислорода в нейронных сетях и регуляцией локального кровотока. Наиболее поразительным свойством этих колебаний оказалось то, что они согласовывались с медленными колебаниями электрических потенциалов коры, регистрируемых со скальпа. Эти десятисекундные колебания могли появляться при определенных заданиях, например при движениях руки или выполнении арифметических действий. Пример такой реакции локального уровня концентрации кислорода в глубокой структуре мозга показан на рис. В.З. Резкое повышение региональной импульсной активности вызывает медленные изменения напряжения внеклеточного кислорода длительностью приблизительно 12 секунд, таким образом, первый максимум ответа возникает с задержкой 6 секунд10. Этот пример ясно показывает, что для исследования процессов обработки информации
* Это были пациенты с болезнью Паркинсона, не поддававшиеся ни одному из традиционных методов лечения и согласив ши«юя на проведение стереотаксической операции. Золотые электроды толщиной 100 микрон были имплантированы в такие ( Ц)уктуры базальных ганглиев, как бледный шар и скорлупа, а также в вентральные ядра таламуса. Результаты этих исследований приведены в нашем труде, опубликованном издательством Академии наук СССР в 1979 году (Гречин. Кропотов. 1979).
|и В некоторых областях мозга отсроченному повышению уровня р03 предшествовало кратковременное (около 1-3 секунд) снижение локального уровня концентрации внеклеточного кислорода.
20
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
в нейронных сетях и изменений метаболической активности необходимы разные временные шкалы.
Около 30 лет спустя подобные десятисекундные спонтанные колебания электрической активности мозга наблюдались в blood oxygen level dependent сигнале при проведении функциональной магнитно-резонансной томографии (фМРТ)11. Эти колебания отражают последовательное вовлечение различных регионов, входящих в состав определенных систем, таких как соматосенсорная, зрительная или слуховая системы. Функциональное значение этих колебаний не установлено. В одном из наших исследований (Кропотов, Гречин, 1979) мы показали, что фазы убывания уровня кислорода могут быть связаны с консолидацией памяти при переводе кратковременных электрических сигналов в нейронных сетях в долговременные метаболические изменения.
2. От нейронных спайное — к локальным корковым потенциалам, и далее — к скальповой ЭЭГ
А. Импульсная нейронная активность
В настоящее время известно, что работа мозга определяется совместным функционированием нейронных ансамблей, а не активностью одиночных нейронов. Но 40 лет назад ученые придерживались противоположной точки зрения, предполагая, что только исследование поведения отдельных нейронов сможет дать ключ к пониманию функций всего мозга. В те дни возник новый метод — регистрация активности одиночного нейрона12 (рис. В.4). Большинство нейрофизиологов покинуло область классической ЭЭГ (единственного доступного в те дни электрофизиологического метода) и приступило к исследованию активности одиночных нейронов13. Это были годы эйфории.
В некоторой степени ожидания были оправданы. Действительно, открытие свойств простых и сложных клеток зрительной коры Хюбелем и Визелем (награжденными Нобелевской премией в 1981 г.) казалось, подтвердило предположение о том, что изучение поведения одиночных нейронов вносит новый вклад в понимание процессов обработки информации в мозге животных14.
Б. Профили нейрональных реакций
В 1970-х, изучая реакции одиночных нейронов и нейронных популяций мозга человека с помощью крошечных электродов, имплантированных в различные
u См. обзор Fox. Raichel, 2007.
u Техника предполагала использование микроэлектродов (диаметром в несколько микрон) и специальных усилителей с полосой пропускания от 100 Гц д о 10 кГц. Самым интересным в нашей работе в те дни было то. что в некоторых редких (увы) случаях мы могли регистрировать импульсную активность одиночных нейронов с помощью макроэлектрсдов, имплантированных в мозг неврологических пациентов.
13 Мой научный руководитель, профессор Н.П. Бехтерева, первой решила регистрировать импульсную активность нейронов у пациентов. Она считала, что ЭЭГ зачастую зашумлена и неспецифична, и верила, что исследование импульсной активности нейронов — лучший путь изучения нейронных механизмов познания. Электроды имплантировались пациентам в глубокие структуры мозга в диагностических и лечебных целях.
14 В области исследований зрения регистрация активности одиночных нейронов выявила существование нескольких классов нейронов в первичной и вторичной зрительной коре. Эти классы по-разному отвечали на предъявление специфических зрительных паттернов, таких как стационарные или движущиеся пространственные решетки, цветовые паттерны и другие, более сложные зрительные стимулы. Более того, некоторые нейроны сохраняли свою активность после прекращения подачи стимулов, обнаруживая способность к запоминанию, в то время как активность других в этой ситуации угасала.
Введение: теоретические основы количественной ЭЭГ и нейротерапии
21
Потенциалы действия
3*10'3 см
Нейрон
Локальные потенциалы коры
500 мкВ
Кожа
Череп
твердая оболочка паутинная оболочка мягкая оболочка
кора мозга
ЭЭГ
50 мкВ
В
Рисунок В.4. Три уровня электрических событий в мозге
Уровень одиночного нейрона соответствует 30 микронам пространственной шкалы и длительности спайка, равной 1 мс. Локальные потенциалы измеряются с помощью макроэлектродов, помещенных в ткань мозга, и различимы при использовании масштаба в 3 мм. ЭЭГ, зарегистрированная со скальпа, имеет пространственное разрешение, равное нескольким сантиметрам. Обратите внимание на 10-кратное различие масштабов амплитуды потенциалов, зарегистрированных интракортикально и с поверхности коры.
области мозга неврологических и психиатрических пациентов, мы случайно обнаружили сходство нейрональных реакций в некоторых отделах мозга15 (рис. В.5). Оно выражалось в подобии так называемых профилей нейрональных реакций — усредненных изображений ответов всех нейронов, зарегистрированных в определенной анатомической структуре. Сравнение профилей реакций с локальными потенциалами, зарегистрированными теми же имплантированными электродами, показало, что коллективное поведение нейронов находит довольно точное отражение в локальных потенциалах (рис. В.6).
В. Локальные потенциалы
Помня об этих результатах, в начале 90-х мы стали использовать регистрацию локальных потенциалов для анализа этапов обработки информации в
15 Инвазивные регистрации электрической активности мозга во время стереотаксических операций необходимы для локализа-||ии эон коры, предназначенных для последующего разрушения или стимуляции. Эти исследования включают измерение локальных потенциалов, отражающих согласованную активность небольших клеточных ансамблей, или оценку активности одиночных клеток с помощью микроэлектродов. Поскольку для проведения инвазивных исследований требуются четкие клинические показания, они не могут дать информации о работе нейронных сетей в норме. Тем не менее эти данные ценны для понимания патофизиологии нейропатических расстройств и для сопоставления экспериментов на животных с клиническими ситуациями. Помимо этого, результаты этим работ способны помочь в понимании базовых механизмов мозговой деятельности, таких как восприятие, двигательный контроль, формирование памяти и речи и даже сознания.
22
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Импульсная активность нейронов
Ignore NOGO GO
Рисунок В.5. Профили нейрональных реакций в подкорковых структурах
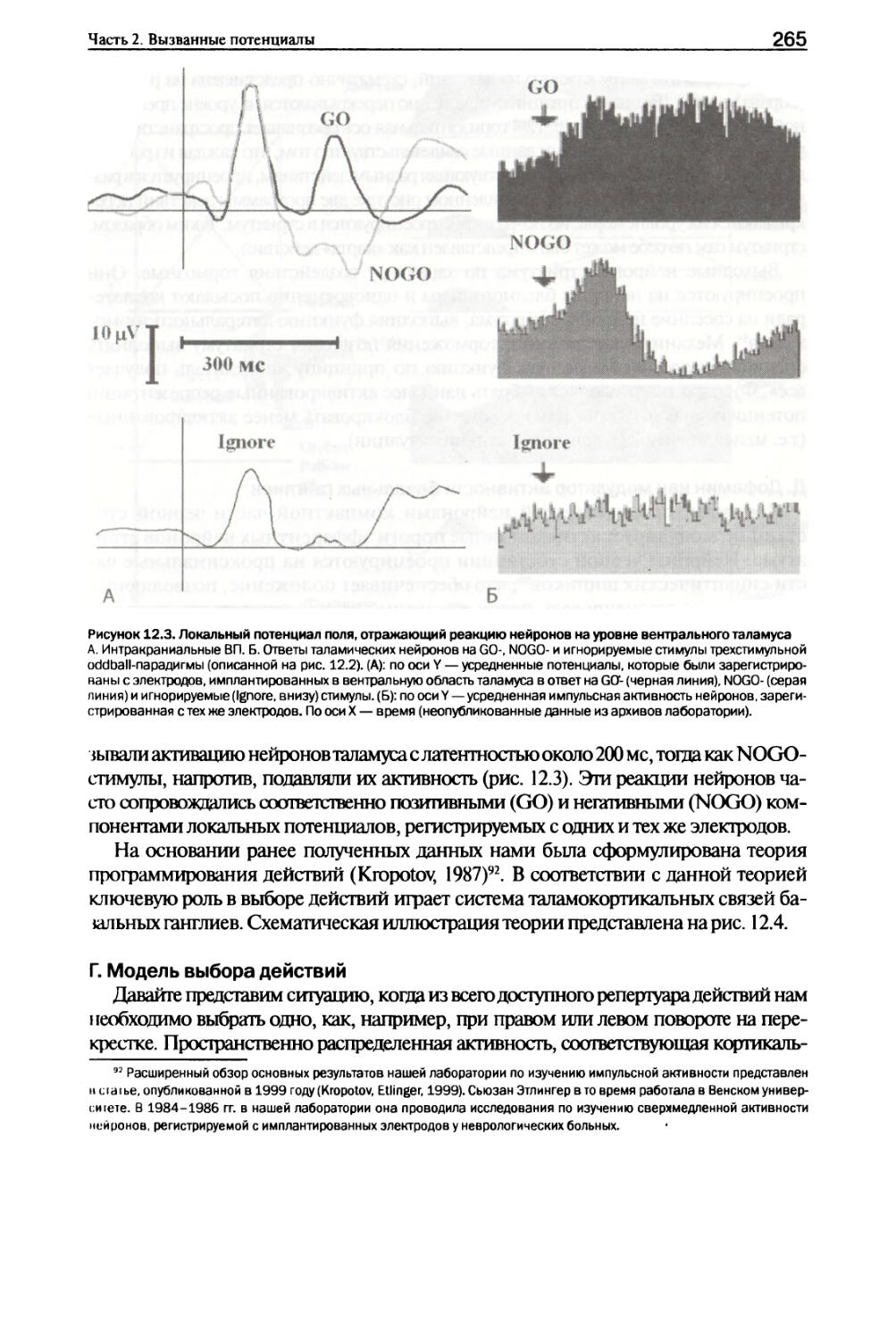
Активность нейронной популяции (вверху) при выполнении двухстимульного GO/NOGO-теста оценивалась с помощью электродрр, имплантированных в базальные ганглии и таламус пациентов. Каждая горизонтальная линия (посередине) соответствует отдельной нейронной популяции. Черные (серые) клетки: значимое (р < 0,01) увеличение (уменьшение) частоты разрядов по отношению к фоновой частоте разрядов. Бинарные постстимульные гистограммы были усреднены для получения профилей реакций (внизу). Вертикальные оси: число нейронных популяций, значимо активированных (выше) или инактивированных (ниже); пунктирные горизонтальные линии: уровни значимости р < 0,01. Горизонтальные оси: время в 100 мс bins. Слева направо представлены типы проб: игнорируемые, NOGO- и GO-пробы. SI, S2, Т — предъявления первого, второго и третьего стимулов в пробах. Заимствовано из Kropotov et al. (1997).
мозге человека. Работа проводилась совместно с Р. Наатаненом и его коллегами из Отдела исследований головного мозга (Cerebral Brain Research Unit (CBRU)) Университета г. Хельсинки16. Во время этих исследований мы обнаружили, что определенные области слуховой коры генерируют различные вызванные потенциалы, что дало нам возможность изучать детали обработки информации в слуховой коре человека17.
16 Задолго до этого, в 1978 г., Р. Наатанен обнаружил компонент когнитивных вызванных потенциалов, названный mismatch negativity (MMN). Этот компонент не был связан с вниманием и появлялся в ответ на предъявление редких девиантных стимулов, в отличие от когнитивных ВП, возникающих при предъявлении стандартных стимулов в oddball-парадигме.
17 Действительно, первичное слуховое поле 41 -кодировало в своих вызванных локальных потенциалах частоты формант акустических стимулов в диапазоне ранних (до 50 мс) временных интервалов, но не обнаруживало никаких эффектов -памяти», т.е. не проявляло привыкания к повторяющимся слуховым стимулам. В противоположность этому для ответов вторичного слухового поля 42 было характерно существенное привыкание, что доказывало наличие выраженного эффекта -памяти» в этой области. Самые впечатляющие ответы генерировались в слуховом ассоциативном поле 22, где возникал дополнительный потенциал в случае, если стимул не соответствовал ряду предшествовавших монотонно повторяющихся стимулов. Комбинация этих трех типов локальных потенциалов, по-видимому, находит отражение в регистрируемом с поверхности головы негативном колебании, названном Mismatch Negativity
Введение: теоретические основы количественной ЭЭГ и нейротерапии
23
Рисунок В.6. Когнитивные вызванные потенциалы, зарегистрированные со скальпа и с внутрикорковых электродов В тесте стандартные акустические стимулы (тон частотой 1000 Гц и длительностью 100 мс) предъявлялись последовательно с интервалами в 800 мс и в случайном порядке смешивались с редкими (вероятность — 15 %) девиантными (тон частотой 1300 Гц) стимулами. Испытуемые читали книгу. Обратите внимание на 10-кратное различие амплитуды сигналов, зарегистрированных со скальпа и интракортикально. Заметьте также, что паттерны ответов различны в трех разных слуховых зонах: поле Бродмана 41 — первичная слуховая кора, поле Бродмана 42 — вторичная слуховая кора, поле Бродмана 22 — ассоциативная слуховая зона.
Г. Взаимосвязь локальных потенциалов и скальповой ЭЭГ
Результаты исследований представлены на рис. В.6 и отражают два явления: I) усредненные по всем пробам локальные потенциалы дают детальную информацию о временном и пространственном распределении локальных операций в нейронных сетях коры; 2) локальные потенциалы находят отражение в когнитивных вызванных потенциалах, регистрируемых с поверхности головы, со значительно меньшими амплитудными значениями. Сходные выводы могут быть сделаны относительно взаимосвязи спонтанной ЭЭГ и интракор-тикальных локальных потенциалов. В нашей лаборатории при изучении спонтанных локальных потенциалов, зарегистрированных с имплантированных электродов, мы столкнулись с тем фактом, что в различных полях Бродмана выяляются довольно отличающиеся паттерны колебаний. Пример такой записи у пациента с вживленными электродами представлен на рис. В.7.
Д. Возрождение ЭЭГ в наши дни
Несмотря на то что метод ЭЭГ возник почти 80 лет назад, самые быстрые темпы его развития наблюдаются только в последнее время. В наши дни мы видим возрождение ЭЭГ Существует по меньшей мере четыре причины этого явления. I крвая связана с недавним появлением новых методов анализа ЭЭГ, таких как техника пространственной фильтрации при коррекции артефактов, анализ независи-
24 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
; а j X i Я; за j » | я i Ж j %
г*
rc« /'/ V\Aai'';JW’/W/-’4Jw ЫЧ*''1 rc«Ц^/ у Шф/у И^-фп ''^"'if(№л чУтЧ
J°c -" ' J\' f- <' * Ф 0’' ’!'? t W/ v
ex Lд___________Ly^
Рисунок B.7. Спонтанные локальные потенциалы и ЭЭГ
Спонтанные локальные потенциалы, зарегистрированные с помощью интракортикальных электродов в различных областях мозга (ПБ 41,46,42,22, гиппокамп, миндалина, передняя цингулярная извилина) по сравнению с ЭЭГ, зарегистрированной со скальпа в точке Cz. Обратите внимание на то, что в различных зонах генерируются различные паттерны локальных потенциалов. В среднем они в 10 раз превосходят по величине скальповую ЭЭГ, довольно независимы и отличаются по доминирующей частоте.
мых компонент когнитивных ВП, вэйвлет-анализ, электромагнитная томография и некоторые другие методы. Вторая причина заключается в относительной дешевизне современных элекгроэнцефалографов. Действительно, в наши дни приборы для регистрации ЭЭГ стоят от нескольких тысяч до нескольких десятков тысяч долларов США, что недорого по сравнению с многомиллионной стоимостью оборудования для МРТ и ПЭТ Третья причина — значительный рост наших познаний о механизмах генерации волн спонтанной ЭЭГ и функционального значения компонентов когнитивных ВП. И, наконец, четвертая — высокое временное разрешение сигналов ЭЭГ и когнитивных ВП, что принципиально не может быть достигнуто другими техниками нейрокартирования. Методы ЭЭГ и когнитивных ВП обеспечивают временное разрешение сигналов в несколько миллисекунд, в то время как ПЭТ и МРТ ограничены разрешением в несколько секунд (рис. В. 8).
3. Эндофенотипы и индивидуальные различия
А. Биологические маркеры заболеваний
Поведение индивидуума, по сути, может рассматриваться как набор всевозможных действий, которые он способен выполнить, — репертуар поведения. Как нам известно из нейронауки, поведение определяется сложным взаимодействием различных систем мозга, играющих разные роли в планировании, выполнении и запоминании действий. Функционирование этих мозговых систем, в свою очередь, обусловлено влиянием генов, их сложными взаимодействиями друг с другом и внешней средой. Таким образом, поведение можно рассматривать как
Введение: теоретические основы количественной ЭЭГ и нейротерапии
25
фенотип индивида. В психиатрии биологические маркеры заболеваний известны как эндофенотипы. Этот термин происходит от греческого слова «эндос», что означает «внутренний», «внутри». Иными словами, эндофенотип представляет собой измеряемый параметр живого организма или мозга, занимающий промежуточное положение между фенотипом и генотипом (рис. В.9). Эндофенотип становится важным понятием при изучении комплекса психиатрических расстройств18.
Б. Взаимосвязь с работой мозговых систем
Природа эндофенотипа может быть нейрофизиологической, биохимической,
Рисунок В.8. Пространственное и временное разрешение ЭЭГ и других методов
Пространственное (ось Y) и временное (ось X) разрешения изображены в виде прямоугольников. Обратите внимание на отсутствие временного разрешения в техниках нейроанатомии. Аббревиатуры: компьютерная томография (КТ), позитронная эмиссионная томография, магнитно-резонансная томография. Нейроанатомия связана с различными пространственными шкалами. Пространственное разрешение микроскопа меньше размера клетки (около 30 микрон — 3 х IO-3 см). ПЭТ и МРТ дают гораздо меньшее пространственное разрешение. Нейрофизиологические параметры включают электрические и электромагнитные свойства нейронов и нейронных сетей и служат предметом изучения клеточной электрофизиологии, скальповой ЭЭГ и когнитивных ВП.
эндокринологической, нейроанатомической или нейропсихологической. Эндо-фенотипы являются более простыми ключами к пониманию генетических механизмов по сравнению с поведенческими симптомами. Основная идея этой кон-
цепции связана с предположением, что более точный психиатрический диагноз можно поставить на основании знаний о системах мозга (таких как система управления) и психических операциях (таких как двигательная установка и контроль поведения), связанных с определенными психологическими процессами. Например, нарушение процессов селекции действий может лежать в основе некоторых форм шизофрении. Следовательно, в данном случае основной задачей современной психиатрии, ориентированной на знание биологии, мог бы стать поиск биологических маркеров этих процессов и выявление их избирательного повреждения при шизофрении. В последнее время для создания гипотез о нейронных сетях, вовлеченных при таких заболеваниях, как депрессия, шизофрения и СНВГ, стали использоваться методы функциональной МРТ, ЭЭГ и когнитивных ВП.
В. Закон перевернутой U-образной кривой
В одной и той же ситуации разные люди ведут себя по-разному Некоторые из нас почти всегда довольны жизнью, другие же склонны к депрессивному настроению; одни успешнее воспринимают зрительную информацию, а другие —
“ Термин возник в 1966 г. и был использован в психиатрии Готтесманом и Шиледсом в 1972 г. (см. Gottesman, Gould, 2003). 1прмин эндофенотип" постепенно вытесняет другие семантически сходные термины, такие как «биологический маркер», «маркер уязвимости», «скрытая акцентуация».
26
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
СИШГГОМ1 Симптом! Симптом 3 Симптом 4
Поведение
Рисунок В.9. Эндофенотипы как биологические маркеры проявления генетических свойств в поведенческом паттерне
Гены отвечают за выработку специфических пептидов, определяющих функционирование определенных систем мозга. Дисфункция этих систем связана с характерными симптомами. Биологический маркер или эндофенотип — это параметр, отражающий работу системы мозга.
слуховую; одни никогда ничего не забывают, а другие не могут вспомнить, что ели вчера на обед19... Согласно положениям этой книги, мозг разделяется на несколько функциональных систем, играющих различные роли в организации поведения: сенсорная система, эмоциональная система, система управления, системы памяти и внимания. Каждая из этих систем может рассматриваться как комплексная нейронная сеть. Нейронные элементы этой струкзуры получают многочисленные входные сигналы и трансформируют их в аксонные потенциалы (спайки). Операция перевода данных на входе в данные на выходе представляет собой нелинейную взаимосвязь, описанную сигмовидной функцией. Подобно этому, передаточная функция в нейронной сети в целом также может быть описана сигмовидной функцией, представленной на рис. В. 10. Форма этой функции в действительности означает, что: 1) активация нейронной сети снижена при низком уровне информации на входе, когда входные сигналы большинства нейронов не превышают пороговых значений; 2) изменение активности нейронной сети находится в почти линейной зависимости от средних данных на входе; 3) активация нейронной сети достигает плато при наивысшем уровне информации на входе — так называемый «эффект потолка». Исходя из этого, мы можем предположить, что эффективность системы определяется ее возможностью реагировать на небольшие изменения входных сигналов. Математически эффективность системы определяется первой производной dO/dl (рис. В. 10). Первая произ
19 Химией были открыты различия между химическими элементами, и в XIX веке были затрачены огромные усилия на создание периодической системы Менделеева — системы классификации элементов. В психологии основы классификации личностей были заложены Павловым в начале XX столетия и позднее, в 1950 г. — Айзенком и его последователями.
Введение: теоретические основы количественной ЭЭГ и нейротерапии
27
водная представлена так называемой перевернутой U-образной кривой. В психофизиологии это явление известно как закон Йеркса—Додсона20.
Как показано на рис. В. 10, нейронные сети характеризуются двумя параметрами: уровнем активации, т.е. амплитудой входного сигнала, управляющего системой, и реактивностью системы, т.е. ее способностью реагировать на небольшие изменения потока информации на входе. В нейрофизиологических исследованиях эти два параметра обычно называются тонической и фазической активацией. Мы предполагаем, что тоническая и фазическая активация мозга имеют разное функциональное значе-
Рисунок В.10. Закон перевернутой U-образной кривой
Вверху схематичное изображение зависимости общей активности гипотетической нейронной сети от входного сигнала, управляющего системой. Внизу: схематичное изображение зависимости ответа системы от сигнала на входе. Ответ определяется как реакция системы на небольшое начальное повышение сигнала на входе.
ние: первая ассоциируется с состоянием, а вторая — с ответом. Например, для системы внимания тоническая активация может быть связана с неспецифическим бодрствованием, в то время как фазическая активация — с селективным вниманием21.
Г. Теории типологии личности Павлова, Айзенка и современных исследователей
Межличностные вариации, по-видимому, определяются различиями в уровне общей активации мозговых систем. На рис. В.10 видно, что «положение» мозговой системы на кривой вход/выход определяет уровень ее тонической активации (как, например, бодрствование для системы внимания) и реактивности (например, привлечение внимания). Для каждой конкретной мозговой системы все субъекты могут быть представлены в виде точек на пе
20 Закон Йеркса — Додсона был сформулирован психологами Р.М. Йерксом и ДД Додсоном в 1908 г. Закон описывает эмпирическую взаимосвязь между уровнем бодрствования и эффективностью деятельности. Он гласит, что эффективность деятельности возрастает с увеличением уровня бодрствования только до определенного момента: при слишком выраженном увеличении уровня бодрствования она снижается.
’* Как будет показано ниже, общий тонический сигнал на входе системы внимания регулируется уровнем норадреналина — пеп-1идв синтезируемого в голубом пятне ствола мозга, в то время как фазическая активация определяется сенсорными стимулами, привлекающими внимание.
28 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
ревернутой U-образной кривой и разделены на три группы: нижнюю, среднюю и верхнюю, в зависимости от положения на этой кривой. Субъекты в этих трех группах по-разному реагируют на небольшое повышение уровня сигнала на входе. Наиболее выраженные, оптимальные реакции отмечаются у «средней» группы. «Нижняя» и «верхняя» группы реагируют сходно, с меньшей (не оптимальной) выраженностью, однако ведут себя по-разному при повышении уровня входного сигнала. «Нижняя» группа отвечает лучше при воздействии факторов, увеличивающих уровень входных сигналов, в то время как «верхняя» группа в этой ситуации снижает свою эффективность. Факторами, повышающими уровень входного сигнала, могут быть стрессовые влияния внешней среды или изменение концентрации медиаторов восходящей активирующей системы ствола мозга.
Во времена Павлова и Айзенка физиологические основы межличностных различий были недостаточно поняты. Павлов применял условные рефлексы в качестве физиологического параметра для распознавания типов нервной системы, в то время как Айзенк использовал время реакции как индикатор скорости обработки информации. В 1960-х введение когнитивных ВП в качестве параметров, отражающих этапы обработки информации, открыло новые горизонты. Первое указание на то, что все индивиды могут быть объективно разделены на две группы, возникло при исследовании взаимосвязи между амплитудой комплекса N1/Р2 и громкостью слухового стимула в слуховых вызванных потенциалах. Было выделено две группы испытуемых: к первой группе были отнесены те, кто увеличивал амплитуду вызванных потенциалов при увеличении громкости слухового стимула, а во второй группе зависимость от громкости сигналов была незначительной.
Наиболее влиятельная теория о межличностных различиях разделяет людей на экстравертов и интровертов22. Эта теория (впервые предложенная Айзенком) основывает такое разделение на уровне бодрствования в системе внимания: у экстравертов уровень бодрствования ниже23. Это положение недавно была протестировано с помощью метода вызванных потенциалов. Поздний положительный компонент, называемый Р300, служил индикатором реактивности коры в ответ на значимые стимулы. В соответствии с теорией экстраверсии/интровер
22 Понятия «экстраверсия» и «интроверсия» использовались К.Г. Юнгом для объяснения различного способа управления своей внутренней энергией» у различных людей. Если субъект предпочитает проводить время во внешнем мире людей и вещей, он относится ктипу экстравертов. Он «заряжается» энергией, активно вовлекаясь в события и участвуя во многих видах деятельности. Он получает энергию от окружающих и с радостью делится ею с другими. Он часто лучше понимает проблему при ее открытом обсуждении и при возможности слышать мнения других людей. Если же субъект склонен к погружению во внутренний мир идей и воображения, он принадлежит к типу интровертов. Согласно Г. Айзенку, тип реагирования индивида определяется восходящей активирующей системой. Если активность этой системы снижена, индивид нуждается в большей стимуляции и относится ктипу экстравертов. При повышенной ее активности стимуляция коры избыточна, и индивид относится к типу интровертов. Таким образом, интроверты демонстрируют избегание стимуляции, а экстраверты — повышенную потребность в ней.
23 Другая идея классификации межличностных различий связана с эмоциональной системой. Она предполагает наличие трех компонентов, свойственных темпераменту, таких как Негативная эмоциональность (НЭ), Позитивная эмоциональность (ПЭ), и Сдержанность (см. Whittle et al., 2005). Индивидам с НЭ свойственна повышенная чувствительность к подавлению, избеганию и наказанию. Они склонны к переживанию разнообразных негативных эмоций, таких как страх, тревога, печаль, чувство вины. Индивиды с высокой ПЭ активно взаимодействуют с внешним миром, отличаясь повышенной чувствительностью к контактам и поощрению и склонностью к переживанию разнообразных позитивных эмоций, таких как радость, счастье, энтузиазм и гордость. К сдержанным относятся индивиды с высоким уровнем контроля импульсивности и эмоций, способностью концентрировать внимание и воздерживаться от удовольствий. Индивидуумы с выраженными проявлениями этого компонента отличаются старательностью, упорством, надежностью и ответственностью.
Введение: теоретические основы количественной ЭЭГ и нейротерапии
29
сии, основанной на показателях бодрствования, у интровертов по сравнению с экстравертами выявлялся более выраженный компонент Р300 при предъявлении значимых стимулов (Beauducel et al, 2005).
При исследовании ЭЭГ и вызванных потенциалов к индивидуальным различиям традиционно относились как к нежелательному статистическому шуму
Но индивидуальные отличия проявляют заметную стабильность у одного и того же индивида, что позволяет предположить, что они являются не случайными колебаниями, а скорее чертой личности. Для дифференциации межличностных различий и отклонений от нормы традиционно используются нормативные базы данных ЭЭГ и когнитивных ВП.
4. Фармако-ЭЭГ
А. Цели
Идея об использовании количественной ЭЭГ в фармакологии в качестве показателя реакции мозга в ответ на лекарственные вещества родилась в начале 1960-х. Сначала ставились скромные задачи: найти новый функционально-ориентированный метод классификации лекарственных эффектов в качестве альтернативы структурным методам, основанным на изучении сходства химических веществ. В течение последних 50 лет мы столкнулись с быстрым ростом числа исследований в этой области. В 1980 г. было основано Международное общество фармако-ЭЭГ (International Pharmaco-EEG society, I PEG)24, и с тех пор ежегодно проводятся собрания этого общества. Было показано, что различные классы лекарственных веществ по-разному влияют на пространственно-временные параметры (в виде спектрограмм) фоновой ЭЭГ25. Однако нередко индивидуальные профили ЭЭГ для разных классов лекарственных веществ могут частично совпадать. Вероятно, это происходит по трем причинам: 1) некорректное определение параметров количественной ЭЭГ с последующим приданием им неадекватного функционального значения; 2) выраженные межиндивидуальные различия, представленные несколькими классами паттернов ЭЭГ; 3) сходство эффектов отдельных классов лекарств.
Было показано, что некоторые психотропные средства оказывают на ЭЭГ влияние, противоположное изменениям в ЭЭГ, ассоциируемым с соответствующими психическими расстройствами (принцип «ключ — замок»). Для предсказания реакции пациентов на антидепрессанты был изобретен26 параметр приближенной оценки локальной метаболической активности. Для контроля эффективности лечения пациентов с шизофренией и деменцией также использовались компоненты когнитивных ВП, например РЗОО.
24IPEG — сообщество ученых, участвующих в исследованиях электрофизиологии мозга в сфере предклинической и клинической фармакологии, нейротоксикологии и в смежных областях. Цели IPEG: 1) стать платформой для дискуссий как ученых, участвующих в академических и промышленных исследованиях, так и клиницистов, занятых изучением фармако-ЭЭГ; 2) содействовать использо-панию электроэнцефалографии в изучении работы мозга в сфере предклинической и клинической фармакологии и смежных наук; 3) обеспечивать руководство исследованиями важных аспектов фармако-ЭЭГ.
л Эффект измерялся как изменение параметров количественной ЭЭГ по сравнению с плацебо.
м Исследования были проведены в UCLA.
30
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Б. Ограничения
Однако, несмотря на прогресс, до недавнего времени фармако-ЭЭГ оставалась эмпирическим методом со слабой теоретической базой, не оказывавшим существенного влияния на фармакологическую индустрию и не находившим широкого применения в клинике. Препятствия в использовании фармако-ЭЭГ были связаны с недостаточным пониманием механизмов возникновения ритмов ЭЭГ и компонентов ВП и ограниченностью методов извлечения адекватной информации из доступных данных. Одна из задач этой книги — уменьшить эти препятствия: 1) изложить новые концепции касательно механизмов генерации ритмов ЭЭГ; 2) представить недавно разработанные методы количественной оценки ЭЭГ, такие как sLORETA, пространственная фильтрация артефактов, нейрометрика; 3) показать физиологическое значение независимых компонент когнитивных ВП, полученных с помощью недавно разработанного метода анализа независимых компонент; 4) представить методы, позволяющие оценивать достоверные27 параметры электрофизиологии мозга.
В. Новые горизонты
Помимо спектрограмм ЭЭГ в оценке эффектов лекарственных веществ также использовались компоненты когнитивных ВП. Их применение было довольно лимитированным и ограничивалось главным образом компонентами N1/P2 и РЗ слуховых ВП28. В рамках этого исследования работа серотонинергической системы при депрессии оценивалась с помощью определения зависимости слухового N1 /Р2 компонента от громкости звука, а мониторинг холинергической системы проводился с использованием компонента РЗЬ. Результаты этого исследования обнадеживают. Новый метод разложения исходных ВП на независимые компоненты увеличивает значение этого подхода и открывает новые горизонты для фармако-ЭЭГ. Дело в том, что амплитуда и латентные периоды компонентов вызванных потенциалов имеет большее диагностическое значение по сравнению с амплитудой мощности спектров ЭЭГ Методология разработана на основе базы данных Института мозга человека, описана в последующих главах и представляет собой инструмент для будущих исследований (рис. B.l 1).
5. Предпосылки нейротерапии
А. ЭЭГ-биоуправление (биологическая обратная связь по ЭЭГ — ЭЭГ-БОС)
Использование спектрального анализа ЭЭГ позволило выявить существенные отличия амплитудных значений ЭЭГ в некоторых частотных диапазонах при ряде мозговых дисфункций от этих показателей у здоровых лиц29. ЭЭГ-биоуправление представляет собой инструмент для коррекции этих от
27 Надежность метода является серьезной проблемой, ограничивающей применение ЭЭГ. Дело в том, что мозг — это «шумящий компьютер», извлекающий необходимую информацию из фонового шума. Шумящий процессор генерирует зашумленные электрические сигналы. Для достижения приемлемого отношения сигнал/шум при регистрации ЭЭГ и ВП было предложено несколько методов.
28 См. обзор по когнитивным ВП в нейропсихофармакологии Pogarell и др.. 2006.
29 Например, у довольно большого числа детей с СНВГ выявлено увеличение отношения «тета/бета» в лобно-центральных областях, что связано с гипоактивацией лобных долей.
Введение: теоретические основы количественной ЭЭГ и нейротерапии
31
клонений от нормы. ЭЭГ-БОС основана на трех научных фактах. Во-первых, параметры ЭЭГ отражают мозговую дисфункцию при определенных заболеваниях. Во-вторых, человек может произвольно изменять свое состояние, и эти изменения могут быть связаны с увеличением или снижением этих параметров (рис. В. 12 слева). В-третьих, мозг способен запоминать это новое состояние и долгое время сохранять его не только в лабораторных условиях, но и в другой обстановке, например дома или в школе. Исторически ЭЭГ-БОС
Генераторы двух компонент ВП
А Б
Рисунок В.11. Основа для применения когнитивных ВП в фармакотерапии (пример)
Слева — распределение дофамина в коре, ассоциированной с вентральным отделом покрышки ствола мозга. Наиболее плотное распределение дофамина сосредоточено в передней цингулярной коре. Справа — две из многочисленных компонент когнитивных ВП, зарегистрированных в ответ на NOGO-стимулы в двустимульном GO/NOGO-тесте: вверху — компонента мониторинга Р400, генерируемая в передней цингулярной коре (sLDRETA изображение динамики во времени), внизу - компонента сравнения Р200, генерируемая в височно-затылочной коре. Компонента Р400 может найти применение в нейропсихофармакологии в качестве параметра функционирования передней цингулярной коры.
впервые была использована в клинической практике для лечения пациентов с эпилепсией. Этому предшествовали серьезные лабораторные исследования на кошках. Позднее ЭЭГ-биоуправление стало применяться для коррекции поведения у детей с СНВГ. В настоящее время этот метод используется при разнообразных мозговых дисфункциях.
Следует отметить, что параметры спектров ЭЭГ — не единственные физиологические показатели, отражающие работу мозга. По существу, такими параметрами могли бы быть: вызванная десинхронизация/синхронизация как мера реактивности ритмов ЭЭГ, компоненты когнитивных ВП как показатели этапов обработки информации в нейронных сетях мозга, сверхмедленные потенциалы мозга, сигналы фМРТ и другие параметры гемо-динамической/метаболической активности мозга. Информация об этих
32
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
данных может быть «возвращена» индивидууму в виде сигналов обратной связи и, следовательно, использована для произвольного контроля над мозговыми функциями.
Б. Интерфейс «мозг — компьютер»
Биологическая обратная связь по электрофизиологическим параметрам может использоваться не только для саморегуляции работы мозга. Она может применяться ддя обмена информацией с внешним миром и для управления техническими приборами, например протезами и микропроцессорами. Этот тип использования биологической обратной связи назван «интерфейс «мозг-компьютер», Brain-Computer Interface (BCI). Обычно в процессе освоения BCI человек учится контролировать физиологические параметры работы своего мозга (такие как ЭЭГ или когнитивные ВП).
В. Транскраниальная микрополяризация (ТКМП)
Наши знания о работе мозга могут быть объединены в два постулата: обработка информации представлена быстрыми аксонными потенциалами нейронов30, в то время как модуляция этой обработки выражена в более медленных колебаниях потенциалов. Нам также известно, что некоторые физиологические процессы, такие как внимание или подготовка к действию, связаны со сверхмедленными сдвигами электрических потенциалов, зарегистрированными со скальпа. Логично предположить, что непосредственное воздействие электрического тока на нейронные сети будет оказывать модулирующее влияние на работу мозга. Существует несколько методов такого воздействия. Один из них состоит в применении постоянного тока для изменения поляризации кортикальных нейронов. Применение анодного или катодного тока, направленного вдоль апикальных дендритов пирамидных клеток коры, может изменить их мембранные потенциалы вблизи аксонных холмиков и, следовательно, изменить вероятность возникновения нейронных разрядов в ответ на некоторые внешние стимулы31 (рис. В. 12 справа). Многие исследования показали, что анодная ТКМП является активирующей процедурой, в то время как катодная ТКМП оказывает противоположный эффект32. Стимуляция постоянным током стала изучаться в лабораторных условиях в 1960-х. Однако успехи вскоре зародившейся психофармакологии затмили первые положительные результаты электростимуляции. Финансовая поддержка исследований стимуляций постоянным током на Западе была сокращена, и в течение двух десятилетий СССР33 являлся единственной страной, где продолжались эти работы. Теперь мы наблюдаем возрождение этого метода. Недавно он был использован в реабилитации пациентов, перенесших инсульт. В Центре нейротерапии Института мозга человека Российской академии наук мы применяем этот метод у пациентов с СНВГ и задержками речи.
30 Вспомните, что длительность каждого потенциала действия составляет всего лишь 1 мс.
31 Эти изменения связаны с повышением или снижением возбудимости нейронов коры.
32 В 1970-х мы искали простую методику для временной активации или подавления импульсной активности нейронов вблизи имплантированных электродов. Было обнаружено, что этот эффект мог быть достигнут при применении постоянного тока. Профессор Бехтерева первой предложила эту идею.
33 Одной иэ положительных сторон системы науки в СССР было то, что работа ученых оплачивалась вне зависимости от того, чем они занимались, и большинство из них проводили исследования в сфере своих интересов.
Введение: теоретические основы количественной ЭЭГ и нейротерапии
33
Б
Рисунок В.12. Схема ЭЭГ-биоуправления и ТЕМП
Вверху: пример ЭЭГ, зарегистрированной со скальпа, у ребенка с СНВГ. Спектры мощности рассчитаны и сопоставлены С базой данных (не показано). Параметр ЭЭГ-БОС вычисляется и предъявляется пациенту на экране. Пациенту удается подавлять тета-активность и увеличивать бета-активность (внизу). Такой же результат может быть достигнут при деполяризации пирамидных нейронов путем предъявления постоянного тока с помощью металлического электрода, расположенного на коже головы.
Г. ТМС, ТЭС и другие виды стимуляции
Следует подчеркнуть, что основанные на электрофизиологии способы модуляции работы мозга не ограничиваются транскраниальной микрополяризацией и ЭЭГ-БОС. В число методов нейротерапии входят также транскраниальная магнитная стимуляция (ТМС), стимуляция переменным током, электроконвульсивная терапия (ЭКТ), глубинная стимуляция мозга (deep brain stimulation — DBS) и некоторые другие. Основная идея всех этих методов заключается в стимуляции (или подавлении) активности нейронов определенных зон коры с помощью электромагнитных полей.
ЧАСТЬ 1. Ритмы ЭЭГ
Один из самых основных законов вселенной — закон периодичности.
Георгий Будзаки. Ритмы мозга
Введение
Новые открытия в области электроэнцефалографии (ЭЭГ), сделанные в течение нескольких прошлых десятилетий, существенно изменили классические представления, на которых базировалось обучение психиатров и неврологов в медицинских школах. Согласно классическим представлениям, обработка информации в человеческом мозгу реализуется за счет импульсной (спайковой) активности отдельных нейронов. Осцилляторная электрическая активность (например, ритмы ЭЭГ) в худшем случае отвергалась и игнорировалась, в лучшем — считалась фоновой активностью. В качестве примера может быть приведена книга Principles of neuroscience под редакцией Е. Kandel, J. Schwartz и Т. Jessel, которая считается «библией» ученых, занимающихся исследованием деятельности мозга. В четвертом издании глава про электрические колебания ЭЭГ отсутствовала, а глава Джона Мартина (John Martin) «Коллективная электрическая активность нейронов: электроэнцефалограмма и механизмы эпилепсии», представленная в третьем издании, была отвергнута.
В течение последних нескольких лет ситуация медленно изменяется. Теперь мы сталкиваемся с ренессансом ЭЭГ, что связано с появлением новых методов оценки человеческой ЭЭГ и новых экспериментальных результатов в исследованиях на животных, которые позволили электрофизиологам обнаружить, что изменения в колебательных паттернах ЭЭГ играют определяющую роль в поддержании мозговых функций и, следовательно, могут использоваться как мощный инструмент для диагноза мозговых дисфункций.
С общей точки зрения, колебания присутствуют во всех физических и биологических системах, стремящихся достигнуть равновесия. Почти во всех случаях колебания появляются, когда система управляется дву-
Часть 1. Ритмы ЭЭГ
35
мя противоположными процессами: выводящим систему из равновесия и возвращающим ее обратно к равновесию. В этом отношении колебания ЭЭГ не отличаются от колебаний в других биологических системах. В случае любого наблюдаемого ритма ЭЭГ (такого как альфа, бета или тета) мы всегда можем обнаружить силу, которая выводит нейрон или нейронную сеть из состояния равновесия, и силу, которая возвращает их назад.
Однако колебания могут быть не только отражением действия двух противоположных сил в нейронных сетях, но гипотетически также могут служить источником объединяющего начала в организации нейронных сетей. Например, изменения суммарного локального электрического потенциала, созданного нейронами — генераторами данного ритма, могут вовлечь в этот процесс другие нейроны, которые непосредственно не участвуют в генерации ритма. Это вовлечение синхронизирует активность всех нейронов нейронной сети с генераторами ритма. Несмотря на попытки доказать это предположение, мы все еще не знаем, правильно оно или нет.
DC сверхмедленные медленные
Рисунок 4.1, В.1. Частотные диапазоны в спектре ЭЭГ
Границы частот строго не определены. Однако полноценность частотной классификации доказывается всей историей ЭЭГ.
В этой части книги описываются ритмы ЭЭГ в диапазоне частот от О до 70 Гц. Данная частотная полоса охватывает несколько категорий электрических явлений, регистрирующихся от скальпа. Эти биоэлектрические явления традиционно разделены на следующие типы: сдвиги DC-потенциалов, декасекундные колебания и медленные волны, дельта-, тета-, альфа- и бета-ритмы ЭЭГ (рис. 4.1, В.1). Здесь необходимо подчеркнуть, что понятие ритма предполагает, что ритм представляет собой регулярные изменения электрического потенциала, измеренного электродами от скальпа. Если к записи ЭЭГ, содержащей ритмы, применяется преобразование Фурье или волновое (wavelet) преобразование, то эти ритмы выявляются в соответствующих спектрах в форме пиков.
Для регистрации декасекундных колебаний необходимы специальные усилители. Декасекундные колебания, как правило, не рассматриваются
36 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
в обычной ЭЭГ. Дельта-ритмы охватывают диапазон частот от 1 до 4 Гц, тета-ритмы — 4—8 Гц, альфа-ритмы — 8—13 Гц и бета-ритмы имеют частоту выше 13 Гц. Тета-, альфа- и бета-ритмы присутствуют в нормальной ЭЭГ, регистрируются в состоянии покоя (с закрытыми или открытыми глазами) и в условиях решения различных задач. Дельта-ритмы в нормальном мозге выражены на спектрограммах в форме пиков только во время состояния глубокого сна. Хотя основные ритмы ЭЭГ известны со времени Бергера (конец 1920-х), их нейрофизиологические основы начинают выясняться только недавно, начиная приблизительно с 1980-х годов.
Надо подчеркнуть, что ЭЭГ — это чувствительный маркер состояния человека: ритмы ЭЭГ существенно изменяются, когда человек засыпает и переходит от одной стадии сна к другой. Например, во II стадии сна появляется определенные колебания, названные сонными веретенами. Веретена сна исчезают в то время, как в дальнейших стадиях сна развиваются тета- и дельта-ритмы. Во время бодрствования ритмы могут определять меру реакции мозга на различные психологические задачи. Например, затылочные альфа-ритмы подавляются (реакция десинхронизации) в то время, как лобные бета-ритмы увеличиваются (реакция синхронизации) в ответ на поведенчески значащие зрительные стимулы.
В поврежденном мозге нормальные механизмы генерации ритмов ЭЭГ могут нарушаться, и ритмы могут: 1) становиться медленнее по частоте (замедление ЭЭГ); 2) иметь необычную локализацию (например, альфа-ритмы в височных областях); 3) стать выше по амплитуде (гиперсинхронизация) и в большей синхронности с другими областями (гиперкогерентность). В некоторых серьезных случаях (характеризующихся, например, разобщением корковых областей и подкорковых структур, в результате травмы или опухоли) может появиться медленный ритм с частотой дельта-диапазона (1—3 Гц). Также нормальные механизмы синхронизации могут быть усилены, тогда на ЭЭГ появляются спайки или паттерны «спайк/медленная волна», указывающие на очаг эпилептиформной активности в человеческом мозгу, который иногда может привести к эпилептическому приступу. Нормативные базы данных могут помочь электроэнцефалографисту выявить перечисленные аномальные паттерны и оценить уровень статистической значимости отклонения от нормы для каждого исследуемого человека. Пространственная локализация генераторов ритмов ЭЭГ может быть оценена различными методами, такими как дипольная аппроксимация или электромагнитная томография низкого разрешения (LORETA).
Пюссарий
Альфа-ритм — ритмическая активность ЭЭГ, регистрирующейся в области первичных или вторичных зон сенсорной коры при открытых или закрытых глазах
Часть I. Ритмы ЭЭГ
37
в состоянии покоя. При активации сенсорных областей коры альфа-ритм угнетается. У здорового человека альфа-ритм регистрируется в области теменной, затылочной и сенсомоторной коры (mu или сенсомоторные ритмы). Частота альфа-ритма варьирует в пределах от 8 до 13 Гц.
Барбитураты — лекарственные препараты, которые связываются с седативногипнотическими областями ГАМК А-рецепторов и обеспечивают открытие Cl-каналов. Барбитураты принадлежат к классу агонистов ГАМК и применяются в качестве снотворного, например, как для предоперационной анестезии, так и для снятия тревожности.
Бета-ритм — ритмическая ЭЭГ и МЭГ-активность более 13 Гц. Иногда бета-ритм подразделяется на категории: низкий бета-ритм (бета-1) — от 13 до 21 Гц; высокий бета-ритм (бета-2) — от 21 до 30 Гц и гамма-ритм — активность с частотой более 30 Гц.
Биспектральный индекс (BIS) ЭЭГ— эмпирическая, статически вычисляемая переменная, которая позволяет получить информацию о взаимодействии корковых и подкорковых областей мозга. BIS, который выражается в единицах от 0 до 100, надежно и адекватно характеризует состояние сознания в норме. При значениях BIS, равных 95 и более, констатируется состояние полного сознания.
Вертикальная организация коры — отражает тот факт, что пирамидные клетки коры со своими длинными апикальными дендритами ориентированы перпендикулярно поверхности. При этом разные слои играют разные функциональные роли в обработке входной информации и передаче выходной.
Возбуждающие нейроны — нейроны, генерирующие так называемые возбуждающие постсинаптические потенциалы, которые деполяризуют постсинаптическую мембрану и, следовательно, смещают мембранный потенциал в направлении порога генерации потенциала. В частности, глутамат считается быстрым активирующим медиатором для множества нейронов коры головного мозга.
Вызванная десинхронизация/синхронизация (ERD/ERS) — параметр, который определяет меру процентного снижения/увеличения спектральной мощности ЭЭГ в заданной частотной полосе и интервале времени в ответ на предъявление стимула.
Гйперполяризация — увеличение мембранного потенциала нейрона, обычно вызывается диффузией анионов (таких как СГ) в клетку посредством открытия соответствующих мембранных каналов. Изменение потенциала, вызванное током анионов, называется тормозным постсинаптическим потенциалом (ТПСП).
1йппокампальный тета-ритм — ритмическая активность в частотном диапазоне (И 3 до 10 Гц, обнаруживаемая в гиппокампе млекопитающих и взаимосвязанных с ним анатомических структурах мозга.
Дельта-ритм кортикальный — ЭЭГ-ритм, генерируемый при участии таламо-ко|л икального механизма. Преобладает в ЭЭГ, когда области коры отключены о г соответствующих ядер таламуса.
Дельта-ритм таламический — ЭЭГ-ритм, регистрируемый с поверхности скальпа и генерируемый в таламусе при взаимодействии двух ионных токов таламо-коргикальных нейронов (ток катионов, деполяризующий мембранный потен
38 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
циал, и кратковременный низкопороговый Са++-ток), ответственных за генерацию кальциевых спайков.
Деполяризация — снижение мембранного потенциала, обычно вызываемого током катионов внутрь клетки (Na+ и Са++), обусловленного открытием соответствующих ионных каналов синаптической мембраны. Изменения потенциала, вызываемые этими катионными токами, называются вызванными постсинаптическим потенциалами (ВПСП).
Дифференциальный усилитель — усиливает разницу между двумя входящими потенциалами VI и V2.
Долговременная потенциация — увеличение синаптической эффективности между двумя нейронами, вызванное высокочастотной стимуляцией пресинаптиче-ской терминали.
Кальциевый спайк — восстановительная деполяризация таламокортикальных клеток, которая следует за сильной гиперполяризацией нейронов. Деполяризация обеспечивается кратковременным низкопороговым Са' -током (Q, который инактивируется, когда нейрон деполяризован, и деинактивируется при гиперполяризации.
Когерентность — мера синхронизации ЭЭГ-активности, зарегистрированной с разных областей скальпа. Когерентность отражает корреляцию спектральной мощности ЭЭГ, вычисленной для разных электродов в одном и том же частотном диапазоне.
Локальный усредненный референтный монтаж — способ расчета монтажа, при котором из потенциала анализируемого электрода вычитается локальный усредненный потенциал, получаемый путем усреднения значений потенциалов небольшого количества электродов, располагающихся в непосредственной от него близости. Существует несколько видов усредненных монтажей (Лапласиана, Лемоса, Хьерта).
Медленные волны сна — ЭЭГ-активность частотой от 0,3 до 1 Гц, доминирующая в ЭЭГ на стадиях глубокого сна. Данный тип активности генерируется благодаря кооперированным механизмам рекуррентных корковых цепей и связан с периодическими флуктуациями состояний покоя и бодрствования.
Международная система расположения электродов «10—20» — принята в 1959 году, свое название получила благодаря тому, что расстояние от любого электрода до другого определяется как 10 или 20 % индивидуально измеренных размеров головы.
Метод независимых компонент (ICA) — метод разрешения проблемы слепого разделения источников. Применительно ЭЭГ проблема может быть сформулирована как нахождение независимых кортикальных генераторов потенциала, регистрируемого с поверхности скальпа.
Монтаж — метод, в соответствии с которым вычисляются ЭЭГ-потенциалы. Наиболее простым монтажом (монтаж объединенных ушей) является вычисление электродного потенциала относительно двух объединенных электродов, располагающихся на мочках правого и левого уха.
Часть I. Ритмы ЭЭГ
39
Неполяризующийся электрод — электрод, который не поляризуется. Это означает, что если через электрод пропустить ток, то его потенциал значимо не изменится относительно равновесного потенциала. Хлорсеребряный электрод — пример неполяризующегося электрода.
Область Бродмана — область коры головного мозга человека, определяемая на основе данных спектроскопии при гистохимическом окрашивании клеток ткани мозга. Области Бродмана были выявлены в 1909 году немецким неврологом Корбинианом Бродманом и обозначены числами от 1 до 52.
Обратная задача — проблема нахождения множества элементарных диполей в коре головного мозга (иногда называемых плотностью нейронных генераторов), которые наилучшим образом аппроксимируют потенциалы, зарегистрированные с множества электродов на скальпе.
Общий усредненный референтный монтаж — монтаж, при котором потенциалы электродов измеряются относительно общего усредненного референта, т.е. потенциала, полученного при усреднении значений, зарегистрированных со всех электродов.
Пачка спайков — два и более разряда нейронов, сменяемых периодом покоя. Такой режим работы нейронов таламуса устанавливается в ответ на длительную гиперполяризацию, благодаря механизму генерации Са-зависимых спайков. Вспышки активности нейронов коры мозга индуцируются в ответ на деполяризующие токи.
Подавление синфазного сигнала (CMR) — определяет возможность дифференциального усилителя быть как можно более близким к идеальному (выходной сигнал равен нулю, когда VI = V2). Выражается как отношение выходного сигнала при VI = V2 (они соединены с одним источником) к выходному сигналу, когда только один вход не равен нулю. CMR измеряется в децибелах (дБ).
Поляризующийся электрод — электрод, который легко поляризуется. Это означает, что если через электрод пропустить ток, то его потенциал изменится значимо относительно равновесного потенциала. Золотой электрод является примером поляризующегося электрода.
Полярографический метод измерения внеклеточного кислорода — метод измерения кислорода в живых тканях, при котором индуцируемый электрический ток при подаваемом напряжении в —0,63 V пропорционален концентрации внеклеточного кислорода.
Потенциал постоянного тока — потенциал, регистрирующийся с поверхности скальпа при помощи неполяризующегося электрода (например, хлорсеребря-ные электроды) в частотном диапазоне от 0,04 до 0,16 Гц.
Правило Хебба — правило, сформулированное в 1949 году Дональдом Хеббом, постулирующее возможный механизм запоминания. В соответствии с законом Хебба, если пресинаптический нейрон А активен и постсинаптический нейрон В также находится в активном состоянии, тогда синапс АВ будет усиленным.
40
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Прямая задача — проблема вычисления потенциала, зарегистрированного с поверхности скальпа, отражающего активность одного или набора диполей, локализованных в коре головного мозга. Решение прямой задачи может быть выражено с помощью физической формулы и численно разрешено компьютерными средствами.
Реакция десинхронизации — соответствует подавлению соответствующей ритмической активности ЭЭГ.
Режекторный фильтр — очень точный фильтр, который уменьшает сигнал определенной частоты. В ЭЭГ используется режекторный фильтр 50 (60) Гц для устранения шума электрической сети комнаты.
Референтная запись — запись ЭЭГ-сигнала со вторым (референтным) электродом, обычно располагаемым на мочках ушей, сосцевидных отростках или кончике носа, т.е. удаленно от источника нейронной активности. Такая запись отличается от той, при которой используется последовательное подключение электродов, когда два электрода (один из них референтный) расположены близко от источника регистрируемой активности.
Сверхмедленная активность — тип ЭЭГ-активности, который может быть зарегистрирован только при помощи специальных усилителей (усилителей постоянного тока), включает колебания с периодом от нескольких до десятков секунд. Механизм генерации этих колебаний не известен, но предполагается, что это связано с медленными метаболическими процессами мозга.
Спектр — рассчитывается с помощью метода быстрого преобразования Фурье, который позволяет разложить сигнал ЭЭГ на множество синусоидальных функций разных частот, амплитуд и фаз. Спектр демонстрирует, каким образом амплитуда, мощность или фаза определенной синусоидальной гармоники зависят от синусоидальной частоты ЭЭГ-сигнала.
Таламокортикальные нейроны — нейроны таламуса, проецирующие связи в соответствующие области коры и получающие обратно активирующие связи от этих областей.
Таламус — подкорковая структура мозга, которая через соответствующие сенсорные ядра контролирует поток сенсорной информации, поступающий в кору головного мозга, а также регулирует активность областей префронтальной коры при помощи различных наборов ядер.
Тета-ритм — ЭЭГ-активность от 4 до 8 Гц.
Тормозные нейроны — нейроны, активность которых обеспечивает генерацию тормозных постсинаптических потенциалов, вызывающих гиперполяризацию постсинаптический мембраны, и вследствие этого смещающих мембранный потенциал от порога генерации потенциала действия и снижающих вероятность его появления.
Усилитель — основная компонента любого элеткроэнцефалографа, который позволяет усиливать слабый (30—100 мкВ) ЭЭГ-сигнал. 1 мкВ = 0,000001 В.
Условное негативное отклонение (УНО) — медленный негативный сдвиг потенциала, связанный с подготовительной активностью испытуемого, как, например, подготовкой к предъявлению стимула или реализации моторного ответа.
Часть I. Ритмы ЭЭГ 41
Фильтр высоких частот — подавляет низкие частоты ЭЭГ-сигнала, пропуская высокие. Фильтр характеризуется нижней граничной частотой, которую принято обозначать в виде эквивалентной постоянной времени (в секундах).
Фильтр низких частот — аналоговый или цифровой фильтр, который подавляет высокие частоты ЭЭГ-сигнала и пропускает низкие.
Фронтальный тета-ритм средней линии — спонтанные или индуцированные выполнением задания кратковременные вспышки ритмической (от 5,5 до 8,5 Гц) активности, регистрируемые в области фронтальных отведений с максимумом в Fz. Наблюдается у здоровых взрослых испытуемых. Этот тета-ритм синхронизирован с реализацией действий в ответ на значимые события окружающей среды и ассоциируется с процессами вспоминания и кодирования следов памяти.
Частота дискретизации — частота, с которой регистрируется ЭЭГ В соответствии с теоремой Найквиста, частота дискретизации должна быть вдвое больше самой высокой из возможных частот регистрируемого ЭЭГ-сигнала.
42
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Глава 1.
Медленные, сверхмедленные потенциалы и дельта-ритмы
I. Происхождение скальповых потенциалов
А. Внутрикорковая организация
Электроды, расположенные на скальпе, регистрируют потенциалы главным образом от корковых областей, расположенных около поверхности головы. Ниже будет детально описана анатомия и физиология корковых генераторов, которые производят потенциалы, распространяющиеся к поверхности головы и измеряющиеся скальповыми электродами в форме электроэнцефалограммы. Нейроны с их аксонами, сомами и дендритами — первичные кандидаты на роль таких генераторов потенциалов. Один из основных принципов корковой организации — существование в коре возбуждающих и тормозных нейронов (рис. 1.1). Эти типы нейронов предназначены природой для формирования сложных действий в корковых нейронных сетях: 1) обнаружение совпадений1; 2) извлечение пространственных особенностей сигнала2; 3) реверберация нейронной активности3; 4) сравнение входов4 и некоторые другие определенные и гипотетические действия.
В общих чертах процессы возбуждения и торможения в коре сбалансированы. Как возбуждающие, так и тормозные клетки коры получают возбуждающие входы от таламуса. Таламус, в свою очередь, рассматривается как корковые ворота, так как все сенсорные пути, берущие начало в сенсорных органах (например, сетчатка, кохлеарные ядра), проходят через релейные ядра таламуса. Нерелей-
1 Обнаружение совпадения необходимо для того, чтобы увеличить отношение •сигнал/шум», обрабатывающегося нейронной се тью. Когда входы к ассоциативной коре от первичных и вторичных сенсорных областей, извлекающих различные особенности стимула. совпадают во времени, датчик совпадения сигнализирует, что эти входы не случайны, а принадлежат тому же самому стимулу.
2 Пространственная фильтрация выполняет ту же функцию (увеличение отношения «сигнал/шум»), что и обнаружение совпадения, но имеет другой механизм. Этот механизм основывается на пространственной (а не временной, как в случае обнаружения совпадения) интеграции раздражений, прибывающих в определенный нейрон от пространственно распределенной сети. Возбуждающие и тормозные входы формируют сложный пространственный фильтр. Например, рецепторные поля сложных клеток в первичной зрительной коре можно было бы рассматривать как пространственные фильтры Габора, в то время как соответствующее действие в зрительной системе рассматривают как пространственную фильтрацию.
3 Реверберация сигнала в нейронных сетях необходима, чтобы сохранять сигнал в краткосрочной памяти. Теоретическая концепция реверберации была известна с 1940-х, но только недавно было получено прямое нейрофизиологическое доказательство. Критическая структурная предпосылка для реверберации — существование рекуррентных цепей в нейронных сетях мозга.
4 Сравнение двух входов — произведенного ожидаемой моделью поведения и произведенного текущей ситуацией — играет критическую роль в сенсорных и исполнительных системах. В сенсорной системе это необходимо для обнаружения изменений, то есть для обнаружения любого изменения в монотонном сенсорном возбуждении. В исполнительной системе действие сравнения необходимо для обнаружения несоответствия между запланированным действием и действием, которое было фактически выполнено субъектом. Обнаружение этого изменения, в свою очередь, служит для того, чтобы контролировать поведение в общем, и для тою, чтобы исправлять ошибки в частности.
Часть I. Ритмы ЭЭГ
43
Рисунок 1.1. Организация корковых связей
А. Связи возбуждающих нейронов (изображенных пустыми кругами) и тормозных нейронов (изображенных черными кругами) в поверхностных, средних и глубоких слоях коры. Б. Связи между двумя корковыми областями: расположенной на более низком уровне (изображена слева) и на более высоком уровне (изображена справа) корковой иерархии. В. Пирамидная и звездчатая клетки коры.
ные ядра таламуса получают информацию от базальных ганглиев, так же как и си первичных сенсорных корковых областей, и посылают проекции к остальной части коры. Наибольшее количество входов от таламуса в кору распределено в пределах срединных корковых областей. Кроме того, возбуждающие и тормозные нейроны получают прямые проекции от корковых областей, расположенных на более низком уровне корковой иерархии, и обратные проекции от корковых областей, расположенных на более высоком уровне корковой иерархии5.
Возбуждающие нейроны в пределах локального коркового модуля связаны друге другом, формируя цепь реверберации, которая оказывается ответственной за поддержа-11ис локальной активации на какое-то время дольше, чем длительность действия сти-
* Эти таламо-кортмкальные взаимосвязи первого и более высокого порядка, прямые и обратные межкорковые связи так же, как местные виутрикорковые связи, формируют сложные нейронные цепи, представляют основу для сложного человеческого поведения.
44 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия мула. Этот процесс функционально основывается на следующих процессах локальных прямой и обратной передачи: когда группа возбужденных нейронов посылает возбуждающие потенциалы соседним нейронам, они разряжаются с некоторой задержкой и посылают возбуждающие потенциалы назад к первоначально задействованной группе клеток, активизируя их снова (рекуррентно). Эта последовательность прямой и обратной передачи возбуждения ведет к лавинообразному возбуждению и ответственна за поддержание активации в пределах локальной цепи.
Тормозные нейроны получают возбуждающие импульсы непосредственно от таламуса и через окружающие возбуждающие нейроны. Тормозные нейроны имеют обратные проекции на локальные возбуждающие нейроны. Тормозные нейроны выполняют по крайней мере три важных функции. Во-первых, текущее торможение не позволяет «перевозбуждаться» локальным корковым цепям. Во-вторых, пространственный паттерн распределения тормозных клеток формирует сложные воспринимающие поля нейронов в коре. В-третьих, комбинация торможения и возбуждения может формировать различные типы операций сравнения в пределах коры. Возбуждающие нейроны в коре разделены на два типа: пирамидные клетки и звездчатые клетки6. Примеры этих двух типов возбуждающих нейронов представлены на рис. 1.1. Короткая длина дендритов позволяет звездчатым нейронам обрабатывать поступающие сигналы быстро и с высокой временной точностью. Пирамидный нейрон отличается от звездчатого нейрона наличием очень длинного, так называемого апикального дендрита, который в 6 раз длиннее, чем базальные дендриты. Апикальный дендрит ориентирован перпендикулярно к корковой поверхности.
Этот тип ориентации дендритов называют вертикальной организацией в отличие от горизонтальной (вдоль корковой поверхности). Короткие дендриты пирамидных клеток простираются от тела нейрона практически во всех направлениях и называются базальными дендритами. Возможно, апикальные дендриты играют особую роль в обработке информации в пределах корковых нейронных сетей. Эта функция все еще требует объяснения, но мы знаем наверняка, что длинные апикальные дендриты пирамидных нейронов коры играют критическую роль в генерации ЭЭГ, регистрируемой со скальпа.
Разнообразие функций коркового торможения достигается разнообразными классами тормозных интернейронов: корзинчатыми, клеткими-канделябрами, нейронами двойного букета, нейроглиаформными клетками. Аксоны этих интернейронов имеют высокоспецифичную геометрию и паттерны иннервации. Клеточные механизмы, которые определяют тормозные связи, только начинают становиться быть понятными7.
в Пирамидные клетки имеют сому, внешне похожую на пирамиду от которой отходят дендриты, покрытые шипиками. Звездчатые клетки в основном меньше, и их клеточные тела больше напоминают форму звезды. Они также имеют дендриты, которые покрыты многочисленными шипиками с глутаматными рецепторами. Интернейроны (другой тип корковых клеток) имеют более округлые клеточные тела и на их дендритах мало или совсем нет шипиков. За последние годы анатомы дали описания множеству уникальных подтипов иктер-нейроных нейронов. Среди них клетки-канделябры (chandelier cell)—тормозные клетки с аксонами, напоминающими канделябры; ней-роглиаформные клетки — тормозные, напоминающие глиальные клетки; двубукетные клетки — тормозные с аксональными коллатералями и дендритами, простирающимися вертикально в тесных пучках; корзинчатые клетки — тормозные с аксональными ветвлениями, заканчивающимися на пирамидных клетках коры в манере, напоминающей корзину, и несколько других типов.
7 Для современного обзора по этой теме см. статью Huang, Di Cristo and Ango (2007).
Часть 1. Ритмы ЭЭГ
45
Б. Мембранные потенциалы
Чтобы описать механизмы возникновения ЭЭГ, необходимо начать с описания мембранных потенциалов. Как мы знаем из электрофизиологии, мембрана нейронов — место, где возникают электрические потенциалы. В состоянии покоя мембрана нейронов электрически поляризована. Поляризация мембраны достигается активным процессом, названным Na+-K+-nacocoM (рис. 1.2). Помимо этого насоса, мембранный потенциал определяется функционированием ионных каналов. Ионные каналы — пути движения ионов через плазматическую мембрану Потоки ионов через мембрану от положительно и отрицательно заряженных поверхностей формируют мембранный потенциал8.
Рисунок 1.2. (лавные элементы нейронной мембраны
А. Схематическое представление мембраны с двойным липидным слоем, формирующим мембрану, с ионным каналом и К- Na-насосом. Внешняя поверхность мембраны заряжена положительно, а внутренняя поверхность — отрицательно. Б. Схема пирамидного нейрона с обозначением основных частей.
В состоянии покоя внутренняя поверхность мембраны заряжена отрицательно по отношению к внешней поверхности. Так, мембранный потенциал нейронов определяется как V _ = V — V , где V . — потенциал к г мембр внутр наруж’ внутр (наруж)
во внутри- (вне-) клеточной поверхности мембраны. В большинстве нейронов VMeM6p изменяется в пределах от —65 до —75 мВ.
Ионные каналы состоят из нескольких различных белков, субъединиц, (х-субьединицд, представляющая собой проводящую ионы пору, также как сайты для связывания с эндогенными лигандами типа синаптических нейротрансмиттеров (или I icI триродными молекулами, такими как некоторые фармацевтические препараты).
* (ок ионов через открытый ионный канал пассивен, не требует никаких затрат метаболической энергии. Ионный ток определяем дпл силы: химический градиент и электрический потенциал по разные стороны мембраны. Химический градиент вынуждает ионы диффундировать из мест с более высокими концентрациями к местам с более низкими концентрациями соответствующего иона.
Мантрический потенциал мембраны формируется положительно и отрицательно заряженными поверхностями. Положительно за-|жжлнная поверхность мембраны притягивает отрицательно заряженные ионы (анионы) и отталкивает положительно заряженные ионы (катионы), и наоборот. Na*, К*, Са**-ионы — примеры катионов, С1~ — пример анионов.
46 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Ионный канал как белковая молекула имеет два и больше конформационных состояния, которые являются относительно устойчивыми. Одно из состояний — закрытое, другое названо инактивированным. В обоих состояниях каналы не проводят ионы. Существует несколько способов открыть ионные каналы, и каждый канал имеет свои собственные условия открывания. Согласно этому качеству ионные каналы разделены на несколько групп. Две из них — лиганд-зависимые и потенциалзависимые каналы (рис. 1.3). Лиганд-зависимые ионные каналы открываются в ответ на присоединение лиганда (нейромедиатора или гормона) к рецептору ионного канала. Энергия от присоединения лиганда открывает канал. Потенциалзависимые ионные каналы открываются в ответ на изменения мембранного потенциала. Лиганд-зависимые ионные каналы играют критическую роль в эффектах, производимых фармакологическими агентами при лечении различных мозговых дисфункций. Потенциалзависимые каналы играют критическую роль в эффектах транскраниальной магнитной сти
Ме дня гор
Канал закрыт
Медиатор еиязываекя е рецептором
Kaitan открыт
Канал зякрыг Канал открыт
Рисунок 1.3. Лиганд* и потенциалзависимые ионные каналы
Лигандзависимый канал открывается, когда с рецепторной частью канала связывается молекула посредника (изображена в виде квадрата). Потенциалзависимый канал открывается, когда мембрана нейрона деполяризуется.
муляции, микрополяризации (transcranial Direct Current Stimulation) и других электрофизиологических терапевтических методах.
Известно больше дюжины основных типов каналов. Каждый основной тип имеет несколько подтипов (тесно связанных изоформ), которые отличаются по скорости открывания и закрытия ворот и их чувствительности к различным факторам. Богатое разнообразие подтипов ионных каналов формирует основу для разнообразия функций нейронных сетей.
В. Синаптическая передача
Когда нейрон разряжается, он посылает потенциал действия через аксон к другой клетке, названной постсинаптическим нейроном. В синапсе, контакте между этими двумя нейронами, потенциал действия высвобождает ме-
Часть I. Ритмы ЭЭГ
47
диатор. Медиатор распространяется в синаптическую щель и связывается с лиганд-зависимыми ионными каналами на постсинаптической мембране. Канал открывается и позволяет току катионов или анионов (в зависимости от типа нейронов — возбуждающего или тормозного) воздействовать на дендрит. Так как суммарный заряд дендрита равен нулю, ионный ток, направленный внутрь, на его постсинаптической мембране компенсируется ионным током, направленным наружу в противоположной части дендрита. Этот поток ионов внутрь дендрита и наружу производит маленький электрический ток (рис. 1.4).
Внеклеточное пространство
Na+ канал Внутриклеточное
закрыт пространстве
Внекяето'шое пространство
Na -канал Внутриклеточное открыт пространство
Рисунок 1.4. Ионные токи через мембрану
А. Схематическое представление закрытого и открытого дендритного Na-канала. Б. Стрелки показывают ионные токи во внеклеточном и внутриклеточном пространстве. Обратите внимание, что поток ионов (электрический ток) образует электрический диполь с положительным полюсом у основания дендрита.
Г. Пирамидные клетки как простейшие электрические диполи
Маленькие электрические токи всех дендритов одного нейрона суммируются по-разному в различных типах нейронов. Если нейрон — звездчатая клетка и возбуждающие синапсы покрывают множество дендритов, ориентированных в различных направлениях, суммарный ток на уровне клетки в целом будет близок к нулю, и суммарный потенциал, произведенный звездчатой клеткой, будет незначителен. Но если нейрон — пирамидная клетка и возбуждающие синапсы расположены на апикальных дендритах, суммарный ток пирамидной клетки будет направлен вдоль апикального дендрита, и нейрон может быть представлен как маленький диполь с отрицательным полюсом в апикальном дендрите и с поло
48
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
жительным полюсом в базальной части нейрона (рис. 1.5). Также, если возбуждающие синапсы активизированы на соме и базальных дендритах нейрона, нейрон становится диполем, который заряжен отрицательно в соме и положительно — в апикальном дендрите.
ЭЭГ — это результат общей электрической активности пирамидных нейронов, то есть суммирования сотен тысяч маленьких диполей, соответствующих этим пирамидным нейронам. Диполи образованы локальными токами, которые связаны с возбуждающими и тормозными постсинаптическими потенциалами. Возбуждающие постсинаптические потенциалы деполяризуют мембрану, приближая ее к порогу возникновения спайка, и делают выше вероятность овершута (перезарядки мембраны при возникновении ПД). Тормозные постсинаптические потенциалы гиперполяризуют мембрану, уводя
I_____I
200 мкм
Рисунок 1.5. Схематическое представление электрических токов в пирамидных и звездчатых клетках
А. Пирамидная клетка с длинным апикальным дендритом. Б. Звездчатая клетка. На этом примере поток Na* внутрь клетки вызван связыванием молекул возбуждающего медиатора глутамата с рецепторами дендрита. Обратите внимание, что ионные токи в пирамидной клетке формируют отдельный диполь, в то время как ионные токи в звездчатой клетке формируют вне клетки нулевое поле.
Часть 1. Ритмы ЭЭГ
49
ее от порога генерации спайка, и таким образом уменьшают вероятность перезарядки.
Продолжительность постсинаптических потенциалов в десять раз больше продолжительности отдельного спайка. Как мы знаем, продолжительность постсинаптического потенциала зависит от типа соединения рецептора-медиатора, но длится по крайней мере 20—30 мс, что соответствует частоте 30—50 Гц. Продолжительность спайка — приблизительно 1 мс, что соответствует частоте 1000 Гц.
Кроме того, пространство, занятое синапсами, где производятся постсинаптические потенциалы, намного больше, чем пространство, занятое аксональным бугорком, где возникают спайки. Из этих двух факторов следует, что постсинаптические потенциалы играют критическую роль в генерации электрических потенциалов, регистрируемых со скальпа в диапазоне частот 0-70 Гц и известных как электроэнцефалограмма.
Дипольная структура внутр икорковых потенциалов экспериментально демонстрируется, когда локальные электрические потенциалы, измеренные в различных корковых слоях посредством сетки микроэлектродов, помещенных в кору, показывают явную смену полярности потенциала в средних слоях коры. Эти внутрикорковые диполи производят скальповые потенциалы, которые могут быть вычислены согласно законам физики. Проблема вычисления скальповых потенциалов от известных внутрикорковых диполей известна как прямая задача.
Синхронизация нейронных элементов — другой фактор, который определяет скальповый электрический потенциал. Интуитивно понятно, что для того чтобы возник существенный потенциал на относительно отдаленном расстоянии от корковых слоев (на поверхности скальпа), нейроны должны быть не только должным образом ориентированы, но они должны работать вместе, синхронно. Например, область источника эпилептического спайка, который внезапно перекрывает фоновую активность, была экспериментально оценена приблизительно в 2000 мм2, в то время как для других элементов ЭЭГ может быть достаточно кластера в 40—200 мм2. Так или иначе синхронизация нейронных элементов на расстоянии десятков миллиметров — необходимый фактор для возможности регистрации элементов ЭЭГ со скальпа.
II. Сверхмедленные колебания
А. Спонтанная активность
Сверхмедленные потенциалы регистрируются специальными усилителями с постоянной времени больше 10 секунд9 и представляют собой периодические колебания скальповых потенциалов с периодами от нескольких секунд до нескольких десятков секунд (рис. 1.6). Хотя сверхмедленные потен-
• Иногда усилители для измерения сверхмедленных потенциалов называют DC-усилителями (усилителями постоянного 1ОИЛ).
50
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Б
Рисунок 1.6. Сверхмедленные, медленные и более быстрые ритмы ЭЭГ
Пример ЭЭГ, зарегистрированной в отведении FT7 у здорового 19-летнего волонтера в состоянии с закрытыми глазами. А. ЭЭГ в широкой частотной полосе от 0,016 Гц (соответствует постоянной времени 10 с) до 150 Гц. Б. ЭЭГ, зарегистрированная в «обычной» полосе частот ЭЭГ от 0,16 до 30 Гц. Обратите внимание на различие масштабов времени — 7,5 мм/с для верхней записи и 30 мм/с —для нижней записи.
циалы были обнаружены в начале 1960-х, их физиологические механизмы и функциональное значение остаются неясными до сих пор.
Существуют веские эмпирические доказательства, указывающие на то, что декасекундные колебания в мозге человека могут быть связаны с периодическими колебаниями метаболической активности мозговой ткани, которая характеризуется медленными колебаниями внеклеточного кислорода и локального кровотока. В исследованиях, выполненных в нашей лаборатории в 1970-х10, была измерена концентрация внеклеточного кислорода от поляризующихся (золотых) электродов полярографическим методом. В этом методе слабое отрицательное напряжение (—0,63 V), прикладываемое к золотому электроду, вызывает электрический ток, который пропорционален концентрации внеклеточного кислорода. Примеры спонтанных колебаний рО2 (напряжения кислорода) представлены на рис. В.З раздела «Введение»11 12. Подобные колебания также были зарегистрированы у человека от неполяризующихся скальповых электродов |2.
10 В те годы я работал с пациентами с паркинсонизмом, которым были вживлены электроды в диагностических и лечебных целях. Эти пациенты были не восприимчивы ко всем обычным формам лечения и подвергались стереотактическим воздействиям. Им им плантировались золотые электроды в базальные ганглии, таламус и премоторную кору.
11 Напомним, что эти колебания имеют относительно высокую амплитуде с периодами колебаний 30 секунд (0,03 Гц), различные паттерны в различных мозговых областях и изменяются в ответ на выполнение субъектом различных задач.
12 Сверхмедленные колебания стали предметом интенсивного изучения в 1970-1980-х в Советском Союзе. Пионеры в этой области — Нина Аладжалова из Москвы и Валентина Илюхина из Института экспериментальной медицины в Санкт-Петербурге. В Австрии подобные исследования выполнялись Гербертом Боуером и Джезилэром Гатгманном (Herbert Bouer Gezilher Guttmann) из Венского уни верситета. Исследование в эти ранние годы было главным образом связано с эмпирическими вопросами о том, как сверхмедленнып колебания изменяются при выполнении психологических задач и как они моделируют качество выполнения этих задач. В частности, ав
Часть 1. Ритмы ЭЭГ
51
В недавнем объединенном исследовании университетов Хельсинки (Финляндия) и Вашингтона (США) (S. Vanhatalo et al., 2004) было обнаружено, что сверхмедленные колебания тесно связаны с более быстрой активностью ЭЭГ. Например, отрицательные фазы сверхмедленных колебаний были связаны с увеличением вероятности возникновения К-комплексов и межприступных спайков у пациентов. Эти новые результаты подтверждают ранние представления относительно сверхмедленных колебаний как медленных, циклических модуляций общей корковой возбудимости.
Б. Подготовительная активность
Помимо сверхмедленных спонтанных (то есть не связанных с заметными изменениями поведения) колебаний в корковых потенциалах существует специфическая медленная мозговая активность, связанная с когнитивными функциями. Хороший пример медленной когнитивной активности — группа различных подготовительных операций. Эта активность появляется, когда в отсутствие каких-либо поведенческих актов субъект готовится к движению или к предъявлению стимула. Такая подготовительная активность может длиться до нескольких секунд и дальше. Электрофизиологический показатель такой подготовительной активности (ЭЭГ-феномен) назван «условным отрицательным отклонением», оно впервые было обнаружено в 1950-х годах известным английским невропатологом Греем Уолтером (Grey Walter).
В 1970—1980-х подготовительная активность в человеческом мозгу исследовалась посредством электрофизиологических методов в Тубенгенском университете в Германии (R. Rockstroh, Niels Birbaumer). Тубенгенская теория подготовительной активности может быть кратко сформулирована следующим образом: когда мозг готовится получить стимул или совершить моторное действие, апикальные (наиболее поверхностные) дендриты пирамидных клеток получают возбуждающие импульсы от более высоких корковых областей. Пирамидные клетки, таким образом, подготовлены так, чтобы была облегчена обработка информации в корковых сетях. Эго облегчение было продемонстрировано улучшением качества выполнения заданий (например, способностью узнать стимул, предъявляемый почти на пороге восприятия) во время отрицательных фаз декасекуцдных колебаний.
Если теория верна, отрицательная фаза постоянных потенциалов должна облегчать судорожную активность, типа спайков и комплексов «спайк — медленная волна», в соответствующих корковых областях, в то время как подавление корко-1ЮЙ негативности и увеличение позитивности должно вызывать противоположный эффект — подавление судорожной деятельности.
В исследованиях Тубенгенской группы была сделана попытка использовать эту |к*гуляторную функцию постоянных потенциалов для лечения пациентов с эпилепсией. Пациенты с эпилепсией с помощью технологий биообратной связи были обучены регулировать медленные корковые потенциалы. Самое важное открытие состояло в том, что пациенты со сложными парциальными и вторично генерализованными припадками с большей вероятностью достигали сокращения приступов, мрийсквя группа показала, что отрицательные сдвиги постоянных потенциалов связаны с улучшением выполнения задач (увеличивают чиоло правильных ответов), в то время как положительные сдвиги корковых потенциалов были связаны со снижением эффективности выполнения задач.
52 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
если они демонстрировали хорошее управление медленными корковыми потенциалами в конце обучения.
Другой способ исследовать регуляторную функцию постоянных потенциалов выражен в методе транскраниальной электростимуляции. Метод состоит из воздействия постоянного электрического тока очень низкой интенсивности (ниже 1 мА) через скальповые электроды (анод и катод). Систематические исследования метода начались в 1960-х, но сократились из-за обширного применения фармакологических методов лечения мозговых дисфункций. Теперь мы сталкиваемся с ренессансом этого метода, который может использоваться как альтернативный метод для активации (или торможения) локальной корковой активности при лечении традиционных мозговых нарушений (таких как синдром навязчивых состояний, депрессии) или в ходе реабилитации после инсульта или мозговой травмы.
III. Медленные волны глубокого сна
A. Up- и down-состояния
Медленные колебания — доминирующий паттерн во время медленноволнового сна и некоторых форм анастезии. Частота медленных колебаний — в пределах диапазона 0,3— 1 Гц. Активность в этой полосе частот также присутствует во время других состояний, таких как состояние с открытыми глазами, закрытыми глазами и во время решения различных задач. Однако вопрос, остается ли механизм возникновения этой активности во время бодрствования тем же, что и во время сна, еще требует ответа.
Наличие медленных колебаний после обширных таламических повреждений указывает, что медленные волны генерируются в пределах коры. Медленная волна представляет последовательность up- и down-состояний корковой нейронной сети. Up-состояние характеризуется активацией (увеличением частоты разрядов) корковых нейронов, связанной с деполяризацией этих клеток (рис. 1.7). Активность в этом состоянии постоянна в течение приблизительно половины секунды из-за четкого баланса между возбуждением и торможением в пределах текущей нейронной сети. Down-состояние характеризуется подавлением импульсной активности нейронов вследствие гиперполяризации корковых нейронов. Детальные нейронные механизмы генерации медленных волн остаются неизвестными. Один из гипотетических механизмов (Баженов и другие, 2002) состоит в том, что случайная суммация маленьких возбуждающих постсинаптиче-ских потенциалов в down-состоянии может активизировать постоянный ток натрия и деполяризовать мембрану выше порога генерации электрических спайков (разрядов).
Хотя медленные волны генерируются внутрикорковыми механизмами, периодические изменения между up- и down-состояниями упорядочивают таламические электрические паттерны (типа дельта-ритмов и сонных веретен) посредством
Часть 1. Ритмы ЭЭГ
53
кортико-таламических путей. Вследствие их синхронизирующего влияния на нейронную активность в пределах коры головного мозга и во взаимодействии с таламическими и гиппокампальными кругами, медленные колебания, как предполагается, лежат в основе консолидации памяти в течение сна.
Б. Транскраниальная индукция медленных волн
Неясно, имеют ли медленные корковые потенциалы и их внеклеточный эквивалент какое-либо физиологическое значение сами по себе. Попытка проверить эту гипотезу была недавно предпринята применением внеклеточных медленных переменных токов (Marshal et al., 2006). Током воздействовали на молодых здоровых добровольцев через стимулирующие электроды, прикладываемые с двух сторон в лобно-латеральных положениях (участки анодной, положительной поляризации) и на мастоиды (рис. 1.8 вверху)13. Показано, что стимулирование медлен-
Отведение с
поверхности коры
--mV
Рисунок 1.7. Внеклеточная и внутриклеточная регистрация потенциалов
А. Медленные колебания локального внеклеточного потенциала. Б. Данные внутриклеточной регистрации. Обратите внимание, что отрицательные изменения внеклеточного потенциала совпадают с увеличением импульсной активности нейронов — up-состояние, в то время как положительные изменения внеклеточного потенциала связаны с подавлением нейронных разрядов — down-состояние. Обратите внимание также, что медленные колебания мембранного потенциала нейрона обратны по отношению к локальным полевым потенциалам. Адаптировано из Haider et al. (2006).
ными токами с помощью транскраниального воздействия в течение раннего ночного сна увеличивает сохранение воспоминаний в памяти. Стимуляция вызвала увеличение продолжительности следующего промежутка медленноволнового сна и увеличение корковых медленноволновых колебаний и медленной веретеннообразной активности в коре (рис. 1.8 внизу).
u Транскраниально применялись колебания тока в частоте 0,75 Гц с максимальной плотностью тика 0,5 мА/см2. Стимуляция начиналась через 4 минуты после того, как субъекты вступали во вторую стадию сна NREM (впервые после засыпания), во время, когда сон должен прогрессировать в стадию с медленными волнами. Субъекты были проверены дважды — в состоянии стимуляции и в состоянии ложной стимуляции. В каждом условии вечером перед сном субъекты разучивали различную информацию. Качество запоминания проверялось слодющим утром. Декларативная память измерялась запоминанием ассоциативн ых пар (pained-associate leamingtask). Предъявлялся список из 46 пар слов, необходимо было правильно воспроизвести не менее 60 % (при немедленном предъявлении первого слова из пары («указывающего' слова) была поставлена задача назвать второе слово). При проверке воспроизведения утром после сна указывающие слова показывались снова, и испытуемые должны были вспомнить соответствующее ответное слово.
54 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
IV. Дельта-колебания
В ЭЭГ человека были выделены 2 типа дельта-колебаний: первый имеет корковое происхождение, в то время как второй генерируется в таламусе. Нейронные механизмы корковой генерации дельта-ритма неизвестны. Единственно, что мы знаем: хирургическое удаление таламуса, равно как и разъединения коры и таламуса, приводят к существенному повышению дельта-активности, регистрирующейся от скальпа. Эти факты указывают, что тип дельта-ритмов, генерируемый за счет внутрикор-
Бодрствование ---
БДГ
Стадии сна 1 2 3
4
Стимуляция 0,75 Hz
Рисунок 1.8. Модулированная транскраниальная микрополяризация мозга улучшает декларативную память и увеличивает медленноволновую активность
А. Временная диаграмма эксперимента. Обозначено время запоминания и извлечения информации из памяти, интервалы стимуляции, период выключения света (горизонтальная серая полоса) и сон, представленный гипнограммой. Б. Выполнение задач на декларативную память при условии стимуляций во время ночного сна и при условии ложных стимуляций. Эффективность выполнения выражается в виде разницы между числом правильно названных слов и общим числом слов, предложенных для запоминания. В. Диаграммы усредненной мощности спектров ЭЭГ в течение периодов с электрическими стимуляциями (серый столбик) и в периодов с ложными стимуляциями (черный столбик) в среднелобном отведении. Заштрихованные области указывают диапазоны частот медленных колебаний (0,5-1 Гц) и медленной лобной веретенообразной активности (верхний ряд, 8-12 Гц), адаптировано из Marshal et al. (2006). Обозначения: БДГ — фаза быстрых движений глаз.
Часть 1. Ритмы ЭЭГ
55
Рисунок 1.9. Генерация дельта-ритма в таламических нейронах
А. Таламокортикальная нейронная сеть. Возбуждающие нейроны и возбуждающие связи обозначены черным. Тормозные нейроны и тормозные связи обозначены серым. Б. Вверху - сдвиг от деполяризации к гиперполяризации, приводящий к переходу от режима отдельных спайков к режиму пачечной импульсации. В пачечном режиме потенциал мембраны колеблется с частотой дельта- активности, которая в свою очередь является результатом взаимодействия двух потоков ионов: катионного тока lh, который активируется, когда нейрон гиперполяризован, и переходного низкопорогового тока Ca++-lt, который ответственен за возникновение Са*+-спайков. С. Открытие мембранных каналов в течение первых двух стадий возникновения Са++-спайков.
ковых механизмов, вероятно, связан с некоторыми медленными процессами в пределах коры14.
А. Дельта-ритм сна
В отличие от дельта-акгивности, производимой в коре, механизмы таламической нейрональной генерации дельта-ритмов хорошо известны. При внутриклеточной регистрации активности в животных препаратах показано, что дельта-ритм может генерироваться отдельной таламокортикальной кдеткой, то есть таламической клеткой, имеющей проекции к соответствующей корковой области. Схема таламокортикальной цепи, вовлеченной в генерацию дельта-ритма, изображена на рис. 1.9.
Здесь надо подчеркнуть, что та же цепь вовлечена в генерацию веретен сна (периодическая активность с частотой около 13 Гц) и альфа-ритмов (периодическая активность с частотой около 10 Гц) во время бодрствования. Эти ритмические активности проявляются на различных уровнях поляризации таламокорковых клеток: альфа-ритмы появляются, когда таламокорковые нейроны относительно деполяризованы, сонные веретена — когда эти клетки относительно гиперполяризованы, дельта-ритмы появляются на самом глубоком уровне гиперполяризации таламокорковых нейронов.
Для генерации веретен сна и альфа-ритмов необходимы свойства сетей, такие как синхронизированное торможение ретикулярными нейронами таламуса. В отличие от сонных веретен и альфа-ритмов дельта-волны генерируются отдельными клетками. Ритм генерируется взаимодействием двух токов (и сле
,4Дельта-колебания могут быть связаны с медленно развивающейся синаптической депрессией. Мы знаем, что постсинаптическая депрессия может длиться сотни миллисекунд что соответствует дельта-диапазону частот. Положительные и отрицательные стадии дельта-колебаний, генерируемых в коре, могут быть связаны с синаптической депрессией, которая следует за текущей активацией.
56 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
довательно — двумя типами ионных каналов) в таламо-кортикальных нейронах (рис. 1.10).
Первый ток называют Ih — это катионный ток, который активизирован, когда нейрон гиперполяризован, и, как следствие, он возвращает мембранный потенциал из гиперполяризованного состояния к менее поляризованному. Второй поток называют It — это низкопороговый ток кальция15. Во время деполяризованного состояния нейрона (состояние бодрствования) этот ток отсутствует. Он появляется только при гиперполяризации нейрона, во время глубокого сна из-за подавления сигналов от восходящей активационной системы ствола мозга, так же как из-за подавления входов от других мозговых систем.
It-поток отвечает за возникновение Са++-спайков. На вершине фазы деполяризации Са++-спайка нейрон генерирует вспышку обычных K ‘-Na++-спайков. Эти спайки через таламокорковые пути передаются соответствующим корковым областям и генерируют постсинаптические потенциалы, которые, в свою очередь, регистрируются от скальпа в форме дельта-ритма.
Генерация низкопороговых кальциевых спайков посредством 1(-тока впервые была продемонстрирована в таламических нейронах Rodolfo Llinas и Jahnsen в 1984 г., а также была зарегистрирована in vivo Marcelo Steriade и коллегами. Безусловно, можно отметить, что большое влияние на наше понимание клеточных механизмов генерации ЭЭГ было оказано McCormic и его сотрудниками.
Б. Низкопороговый пачечный режим пульсаций таламических нейронов
Дельта-колебания связаны с так называемым пачечным режимом пульсаций таламо-кортикальных нейронов, который появляется только тогда, когда эти нейроны сильно гиперполяризованы. В этом состоянии отношения между входным и результирующим импульсом отдельной клетки представляют собой ступенчатообразную функцию, отражающую тот факт, что выходная частота разрядов таламо-кортикальных нейронов не зависит от входящего тока (внизу рис. 1.10). Этот режим пульсаций нейронов отличается от тонического состояния (или единичных разрядов), в котором таламо-кортикальные нейроны отвечают на входящие деполяризационные токи линейной зависимостью: чем больше входящий ток, тем больше разрядов на выходе.
Кальциевые спайки отдачи — не единственный способ вызвать пачечный режим разрядов. В коре существует целый особый набор нейронов, которые генерирую спайки в пачечном режиме. Этот режим может определяться внутренними особенностями нейронов или нейронной сети, к которой принадлежит нейрон. Пачечный режим пульсаций корковых клеток, вызванный деполяризующими токами, отличается от состояния пачечного режима пульсаций таламических нейронов (релейных и ретикулярных), вызванного гиперполяризующими токами.
151(-поток ответственен за генерацию так называемых Са”-спайков. Са”-спайк представляет собой ток ионов Са” внутрь, который деполяризует мембрану »иламонор1иклльиых нейронов. I, управляется двумя воротами: активационными и инактивационными, It активируется при - 70 мН, при относительно низком порою. поэтому он называется низкопороговым и приводит к возникновению низкопорогового кальциппою (.ннйнл, Н этом кто отличие oi обычною Na* спайка, который имеет более высокий порог активации, соответствующий приблити1плыю 'ИТ мН Д|тутоп отличил млжду обычными Ма'-спайками и низкопороговыми Са"-спайками — пппллпжительностьспникоп 1.... Nii iiuhiuiih i м< н н>прнмн млн п|м»/у)лжительностьСа“-спайков — 50мс.
Часть 1. Ритмы ЭЭГ
57
Спаики/с
Режим одиночных спайков
А
Г ток (рА)
Пачечный режим
Рисунок 1.10. Тонический и пачечный режимы пульсаций таламокортикальных нейронов
Схематическое представление тонического (А) и пачечного (Б) режима импульсаций таламокортикальных клеток. В, Г. Схематическое представление отношений ввода — вывода для отдельной клетки в этих двух режимах. Входная переменная — амплитуда деполяризующего импульса тока. Результирующая переменная — частота разрядов клетки. Исходные потенциалы 50 мВ для тонического режима и -80 мВ для пачечного режима.
В. Патологические ритмы дельты
Феноменология дельта-активности, появляющаяся при некоторых серьезных мозговых отклонениях, очень сложна. В главе известной книги Electroencephalography под редакцией Ernst Niedermeyer и Fernando Lopes Da Silva, названной Nonspecific abnormal EEG patterns, Frank Sharbrough суммировал все известные феномены, связанные с дельта-активностью в несколько категорий. Эти категории: неустойчивая ритмичная дельта-активность (IRDA), лобная неустойчивая ритмичная дельта-активность (FIRDA), постоянная неритмичная дельта-активность (PNDA). Например, центральную PNDA рассматривают как один из самых надежных признаков серьезного локального мозгового нарушения типа инсульта или мозговой травмы. Необходимо подчеркнуть, что ЭЭГ отражает главным образом корковые явления, в то время как подкорковые структуры имеют очень небольшое отражение в ЭЭГ. Таким образом, обнаруженные зоны повреждения мозга по данным ЭЭГ и трехмерной локализации пораженного участка мозга, например, с использованием возможностей компьютерной и магнитно-резонансной
58 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
томографии, не обязательно будут совпадать. Более того, иногда видимые на ЭЭГ нарушения не картируются более высокотехнологичными КТ- и МРТ-методами и наоборот существенные нарушения по данным перечисленных методик в ЭЭГ могут характеризоваться незначительными изменениями. Для того чтобы иметь объективное представление о нарушениях в мозге человека при использовании метода ЭЭГ, необходимо визуальный анализ ЭЭГ подкреплять данными спектрального анализа и результатами исследований вызванных потенциалов.
V. Заключение
Частотный диапазон ниже 0,1 Гц соответствует декасекундным (с периодами в десятки секунд) колебаниям. Сверхмедленные потенциалы появляются в связи с медленными метаболическими процессами. Когда эти процессы измеряются при регистрации локального кровотока и/или концентрации кислорода, самая поразительная особенность динамики этих двух метаболических параметров — появление сверхмедленных квазипериоди-ческих колебаний. В настоящее время сверхмедленные колебания обнаружены также в исследованиях фМРТ и взаимосвязаны с колебаниями локального кровотока. Медленные волны находятся в диапазоне 0,1 — 1 Гц. Медленные волны присутствуют в электроэнцефалограмме во всех состояниях от глубокого сна до состояния сосредоточенного внимания. Они доминируют в ЭЭГ в глубоком сне, и одна из его стадий имеет название «медленноволновый сон». Медленные колебания характеризуются ритмичными циклами деполяризации мембран корковых нейронов (UP-состояния), сменяющиеся их гиперполяризацией (DOWN-состояния). UP-состояния связаны с увеличением электрической активности (частоты спайков) групп корковых нейронов, в то время как DOWN-состояния связаны с уменьшением генерации разрядов нейронами.
Дельта-колебания сна (в пределах диапазона от 1 до 4 Гц) генерируются при взаимодействии двух потоков ионов (и следовательно — работы двух типов ионных каналов) при передаче таламическими нейронами импульсов к соответствующим корковым областям. Взаимодействие «возбуждающих» и «тормозных» токов ионов в таламической мембране, ответственной за генерацию кальциевых разрядов, вызывает деполяризацию в ответ на длительную гиперполяризацию.
Часть 1. Ритмы ЭЭГ
59
Глава 2.
Альфа-ритмы
I. Тйпы альфа-ритмов
Немецкий психолог Ганс Бергер (Gans Beiger) был первым, кто наблюдал альфа-ритмы, зарегистрированные от поверхности головы человека. Он сделал запись этого типа ритмов с поверхности головы субъектов, сидевших спокойно с закрытыми глазами. Ученый опубликовал свое открытие в 1929 году в статье Uber das Elektrenkephalogramm das Menschen — «Поверхностная электроэнцефалограмма человека» — и назвал эти электрические феномены «волнами первого порядка», или «а (алъфа)-волнами».
Бергер далее показал, что альфа-ритмы блокируются после открывания глаз или во время некоторых типов умственной активности, ведя к появлению «волн второго порядка», или «р (бета)-волн». Результаты Бергера были позже подтверждены несколькими другими исследователями, в частности Adrian and Matthews, которые ввели блестящую концепцию (которая остается верной до настоящего времени), что различные сенсорные области мозга имеют собственные альфа-ритмы (ритмы «холостого хода»), которые, по их предположениям, представляют состояние «отдыха», или «праздности» этой области мозга. Несмотря на то что со времен Бергера в электрофизиологии накопилась масса знаний относительно феноменологии альфа-ритмов, множество гипотез относительно механизмов и функционального значения этих ритмов не сходятся в общую теорию. Эта двусмысленность, вероятно, отражает разнородность альфа-ритмов.
Обычно альфа-ритмы представляют как ритмы нормальной человеческой ЭЭГ с частотами в диапазоне от 8 до 13 Гц. Определение альфа-ритмов как отдельной категории мозговой ритмики также предполагает особое пространственное распределение, мощный ответ на блокирование соответствующего сенсорного входа16 и особый механизм генерации. Существует не один, а несколько вариантов альфа-ритмов. Этот факт был известен с 1950-х (упоминается известным английским электрофизиологом Грэем Уолтером), однако достаточно описан он только недавно, когда стали обычной процедурой методы картирования мозга человека.
А. Мю-ритмы
Мы начнем наше описание с сенсомоторных альфа-ритмов17. На рис. 2.1 изображен фрагмент ЭЭГ здорового взрослого человека в состоянии с открытыми глазами.
“ Например, закрывание глаз, то есть уменьшение сенсорного входа к затылочно-теменным областям, ведет к увеличению мощности задних (затылочного и теменного) альфа-ритмов.
17 Сенсомоторные ритмы имеют немного более высокую частоту, чем затылочные альфа-ритмы.
60
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
FplAvW FpZAvW Г7АЫ
F3AA-/
FlAvW
F4AMW
F8AAV
T3AvW
C3AAV
C4AvW
T4AvW
T5AvW
P3AvW
PjAvW
P4AvW
T6AvW
OlAvW
02AvW
W«AMcm
33 34 X X 37 X X
Рисунок 2.1. Мю-ритмы
Фрагмент электроэнцефалограммы, зарегистрированной у здорового взрослого субъекта в течение спокойного бодрствования с открытыми глазами. Обратите внимание на различные ритмы с частотой приблизительно 10 циклов в секунду в отведениях 03 и 04. Вставка — две топограммы потенциалов, снятых в два момента времени, указанных черными вертикальными линиями.
Можно ясно видеть два различных ритмичных паттерна ЭЭГ в отведениях СЗ и С4. Эти колебания, по-вцдимому, независимы друг от друга и имеют специфическую форму с острыми отрицательными пиками, которые напоминают греческую букву р (мю). Эта определенная форма18 дает одно из названий этого ЭЭГ-паттерна — мю-ритма. Другое название — сенсомоторный ритм — дано благодаря тому, что ритм локализован над сенсомоторными участками коры (см. также карты потенциалов, построенные для двух различных моментов времени). В некоторых статьях этот ритм также называют роландическим, потому что он обнаруживается над роландической бороздой. Сенсомоторные ритмы могут блокироваться движением соответствующей руки или прикосновением к руке. И наоборот, расслабление мускулов увеличивает сенсомоторные ритмы19.
Здесь надо подчеркнуть, что хотя мю-ритмы обнаружены и в левом, и в правом полушариях (и во многих случаях выглядят весьма симметричными в спектрах ЭЭГ), они обычно независимы, то есть производятся независимыми генераторами. Эго продемонстрировано на рис. 2.2 при помощи когерентного анализа.
На рис. 2.2 представлены спектры, вычисленные для здорового человека, во всех 19 каналах в состоянии спокойного бодрствования с открытыми глазами. Как выявляется на спектрах ЭЭГ, в ЭЭГ доминируют два различных ритма: один с максимумом мощности ЭЭГ в отведении СЗ с пиковой частотой 10 Гц и другой с максимумом мощности ЭЭГ в отведении С4 с пиковой частотой 9,5 Гц. Карты когерентности, вычисленной отдельно для СЗ и С4, представлены внизу справа. Как видно, сенсомоторный ритм с максимумом в СЗ имеет самую высокую когерентность на частоте ле-
18 Напомним, что мю-ритм стоит за движениями.
19 Подавление сенсомоторных ритмов реальными или воображаемыми движениями, по-видимому, вызывается увеличением со матосенсорного входа от рецепторов тела к сенсомоторной полосе. Этот вид реактивности похож на реактивность затылочных альфа-ритмов в ответ на открывание глаз.
Часть 1. Ритмы ЭЭГ
61
Рисунок 2.2. Независимость лево- и правосторонних мю-ритмов
Пример спектров мощности здорового взрослого субъекта. А. Топограммы распределения мощности левого и правого мю-ритмов в максимуме их амплитуды. Обратите внимание, что эти мю-ритмы немного отличаются по частоте. Б. Топограммы когерентности отведений СЗ и 04 соответственно. Обратите внимание, что положительный пик когерентности в одной стороне сопровождается отрицательным пиком в противоположной стороне, что указывает на независимость левых и правых мю-ритмов.
вого ритма (10 Гц) с близлежащими электродами левого полушария и самую низкую когерентность с С4 электродом и, наоборот, ритм в С4 не работает синхронно с ритмом в отведении СЗ на своей пиковой частоте (9,5 Гц)20.
В литературе, посвященной нейробиообратной связи, была небольшая путаница при определении этого ритма. В некоторых статьях можно найти, что этот ритм определяется в диапазоне частот от 12 до 15 Гц. Это типичный пример несоответствующего применения к людям результатов, полученных в исследовании на животных. Дело в том, что сенсомоторный ритм интенсивно изучал в 1960-х годах Барри Стерман (Barry Sterman) из университета Калифорнии в Лос-Анджелесе (США). В экспериментах на кошках он обнаружил, что инструментально поощряемая генерация ритма ЭЭГ, регистрирующегося от сенсомоторной коры кошек, приводит к большей устойчивости к развитию приступов. Он обозначил этот ритм как сенсомоторный ритм. У кошек этот ритм находился в диапазоне 12—15 Гц. У людей, что недвусмысленно продемонстрировано иссле-
30 Мы не нашли в литературе систематических исследований, показывающих, что частота сенсомоторного ритма значительно изменяется с возрастом. Этот ритм в пределах того же самого диапазона частот может быть обнаружен и у младенцев, и у пожилых людей, вероятно, это отражает раннее созревание сенсомоторного ритма и умеренное старение.
62 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
даваниями, частота сенсомоторного ритма изменяется в диапазоне 9—13 Гц. Барри Стерман сам указывал причину этой путаницы на различных конференциях и симпозиумах.
Б. Затылочные альфа-ритмы
Высокоамплитудные ритмы, регистрирующиеся затылочными электродами от коры мозга человека, представляют другой пример альфа-ритмов. На рис. 2.3 изображен фрагмент ЭЭГ, зарегистрированной у здорового человека в состоянии с открытыми глазами. Во вставке на рис. 2.3 представлены спектры мощности для электродов в положениях 01 и 02. Как видно, затылочный ритм у этого человека проявляется в спектрах в форме острого пика. Визуально также можно оценить, что ритмы в 01- и 02-электродах этого человека синхронны. Синхронность — не обязательная особенность: у некоторых других здоровых людей ритмы в 01 и 02 могут быть независимыми друг от друга.
Затылочный ритм увеличивается (синхронизируется), когда человек закрывает глаза. Общие усредненные спектры в затылочных отведениях для группы здоровых взрослых в состояниях с открытыми и закрытыми глазами представлены на рис. 2.4. Как видим, закрывание глаз существенно увеличивает мощность ЭЭГ в диапазоне альфа-частот в затылочных областях. Эта синхронизация вызвана блокировкой зрительных входов к затылочным областям. Этот факт является одной из причин, почему затылочный альфа-ритм рассматривают как ритм «холостого хода» затылочной коры. Затылочный ритм обычно
FplAvW
Fp2-AvW FZAvW
F3AvW
FzAvW
F4AyW
FOAyW
И-AvW
C3-AvW
Гл-MW
C4AvW
T4-AyW
T5AvW
P3AvW
PzMW
P4A'AV
TSAvW
OVAyW
SOtA/.-'cm
170
Рисунок 2.3. Затылочный ритм
Фрагмент электроэнцефалограммы, зарегистрированной у здорового взрослого субъекта в течение спокойного бодрствования с открытыми глазами. Обратите внимание на отчетливый ритм частотой приблизительно 10 циклов в секунду в отведениях 01 и 02. Вставка (вверху) - две топограммы потенциалов, снятых в два момента времени, указанных черными вертикальными линиями. Вставка (внизу) — спектры мощности ЭЭГ в отведениях 01 и 02.
Часть 1. Ритмы ЭЭГ
63
Р[ 10.01 Hz] uV'2
Топограмма спектральной мощности
Б при закрытых глазах
Рисунок 2.4. Синхронизация затылочного альфа-ритма в состоянии с закрытыми глазами
Усредненные спектры для группы здоровых взрослых субъектов в состоянии спокойного бодрствования с закрытыми глазами по сравнению с состоянием с открытыми глазами. Внизу топограмма разностного спектра для этих двух состояний.
доминирует в записи ЭЭГ (также как и в спектрах) во время состояния спокойного бодрствования с закрытыми глазами. В некоторых редких случаях существуют не один, а два затылочных альфа-ритйа. Они могут генерироваться в различных областях затылочной доли и могут иметь различные пиковые частоты.
Согласно литературе, частота затылочных ритмов изменяется с возрастом. На рис. 2.5 изображены спектры ЭЭГ, усредненные по группам здоровых людей в возрасте от 7 до 89 лет. Наблюдается небольшое увеличение средней частоты с 7 до 20 лет и небольшое уменьшение в последующие годы. Однако разница спектров ЭЭГ, представленная шириной пика, является большей, чем изменения с возрастом. Частота доминирующего альфа-ритма у здоровых пожилых людей приблизительно 10 Гц, так что частота затылочного альфа-ритма меньше чем 7,5 Гц обычно расценивается как умеренное отклонение21.
21 Из-за снижения альфа-частоты с возрастом часто предполагалось, что частота альфа-активности может отражать более «острый» ум, те. мозг «более умных» людей характеризуется более высокими частотами альфа-ритмов. Группа исследователей из Vnje (свободного) университета Амстердама провела исследование 271 близнецового семейства. Показатель интеллекта оценивался голландской версией опросника Векслера (Wechsler Adult Intelligence Scale, WAIS), из которого рассчитывались четыре параметра (устное понимание, рабочая память, перцепционная организация и скорость обработки). Индивидуальные пиковые частоты оценивались согласно методу, описанному Klimesch (1999). Моделирование структурного уравнения показало, что пиковая частота и измеренные параметры интеллекта имеют вышние показатели наследуемости (диапазон 66-83 %), но не было показано никакой корреляции межде пиковой частотой альфа-ритма и любым из четырех параметров WAIS. Однако наш опыт показывает, что замедление доминирующего ритма ниже нормативного диапазона ж) крайней мере на два стандартных отклонения нужно рассматривать как отклонение от нормы.
64
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
uV> 1.200-0 800-0400-
РИСУНОК 2.5. Зависимость затылочного альфа-ритма и мю-ритмов от возраста
Обратите внимание, что усредненная мощность ЭЭГ в альфа-диапазоне заметно уменьшается с возрастом, в то время как частота максимума мощности усредненных спектров (частота ритмов) изменяется незначительно.
Часть 1. Ритмы ЭЭГ
65
Затьшо'шьш альфа-ритм слева
Затылочный альфа-ритм справа
Рисунок 2.6. Анализ независимых компонент амплитудных спектров
Анализ независимых компонент был применен к амплитудным спектрам ЭЭГ, вычисленным для более 300 здоровых субъектов, выполняющих зрительную двухстимульную GO/NOGO-задачу. Представлены шесть независимых компонент, соответствующих различным ритмам. Для каждой компоненты слева направо приведена топография компонент, представление соответствующей компоненты для отдельного субъекта, ниже — спектральные характеристики компонент.
В. Теменной альфа-ритм
В некоторых редких случаях высокоамплитудные ритмы в диапазоне альфа-частот могут быть обнаружены в теменных областях с максимумом в Pz. Это рассматривается как нормальный вариант альфа-ритма. Этот ритм может увеличиваться в состоянии с закрытыми глазами, так же как затылочный альфа-ритм, хотя у некоторых людей мы наблюдали уменьшение теменного альфа-ритма в ответ на закрывание глаз22. Теменной ритм может присутствовать в ЭЭГ вместе с затылочным ритмом и, по-видимому, независим от затылочного ритма (рис. 2.6). Частота теменного ритма обычно меньше частоты затылочного ритма, измеренная у того же человека. Суммарная мощность теменных ритмов возрастает с повышением трудности задачи и увеличивается в состоянии решения задачи по сравнению с состоянием спокойного бодрствования с открытыми глазами. Функциональное значение теменного ритма полностью не понято. Функциональный анализ сталкивается с двумя трудностями: 1) теменной альфа-ритм присутствуету немногих людей; 2) он маскируется соседним затылочным альфа-ритмом.
Недавний успех в развитии электромагнитной томографии открывает новые возможности трехмерной визуализации генераторов ЭЭГ-ритмов. Для этих це-
73 Это может указывать на существование по меньшей мере двух вариантов теменного альфа-ритма.
66 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
лей успешно использовалась электромагнитная томография низкого разрешения (LORETA). В настоящее время LORETA становится обычным инструментом Мозгового картирования. Примеры двухмерных карт и соответствующих изображений LORETA различных типов альфа-ритмов (затылочного, сенсомоторного и теменного) у трех здоровых людей возраста 29, 19 и 18 лет представлены на рис. 2.7.
Рисунок 2.7. Топограммы и LORETA-изображения трех типов альфа-ритмов
Примеры топограмм и соответствующих электромагнитных томографических изображений трех здоровых взрослых субъектов. Все субъекты находились в состоянии спокойного бодрствования с открытыми глазами. А. Индивидуум с затылочным альфа-ритмом. Б. Индивидуум с мю-ритмами. В. Индивидуум с теменным альфа-ритмом. Изображения LORETA отображают распределение генераторов ритмов в затылочно-теменной области, постцентральной сенсомоторной полосе и среднетеменной области соответственно.
Часть I. Ритмы ЭЭГ
67
Три ритма, представленные на рис. 2.7, обнаружены у трех различных людей. Однако в некоторых случаях все три ритма могут быть найдены у одного нормального (здорового) человека. Более того, даже альфа-ритм с определенной локализацией (такой как затылочный) может иметь два и больше подритма с немного различной локализацией и с различными частотами.
II. Нейронные механизмы
А. Взаимосвязь с корковой дезактивацией
Электрическая стимуляция внутриламинарных ядер таламуса подавляет альфа-ритмы (реакция десинхронизации). Высокочастотная стимуляция таламических ядер может увеличивать суммарную активность коры, действуя как при реакции пробуждения (arousal). В соответствии с этими наблюдениями, повреждения в таламусе ведут к отчетливой дезорганизации или даже полному подавлению альфа-активности в ЭЭГ Эти данные подтверждают ранние предположения, что альфа-ритмы контролируются регулярной таламической активностью.
Исследования корреляции между позитронно-эмисмионно-томографическими, ЭЭГ- или фМРТ-сигналами поддерживают гипотезу о том, что альфа-активность представляет собой ритмы состояния «холостого хода» соответствующих корковых областей. Для затылочного альфа-ритма это показано во многих независимых статьях. Поведенческая парадигма, используемая в этих исследованиях, состоит из последовательного открывания и закрывания глаз, которые, соответственно, ведут к подавлению и повышению затылочной альфа-активности. В большинстве современных исследований показано, что во время увеличения затылочных альфа-ритмов наблюдается уменьшение активации затылочных областей, измеряемой методами оценки насыщенности крови кислородом, кровотока или метаболизма глюкозы23. Связи метаболических параметров подкорковых (включая таламические) структур с амплитудой альфа-активности ЭЭГ менее хорошо изучены и противоречивы.
Б. Таламокортикальные пути
Одна из первых идей в 1960-х заключалась в том, что альфа-ритмы и сонные веретена естественного сна имеют подобные нейронные механизмы24. В те годы Andersen и коллеги выполнили ряд изящных экспериментов, показывающих, что веретена бирбитуратного сна генерируются на уровне таламуса. Ключевую роль в генерации иерстен барбитуратного сна играли фазически разряжающиеся тормозные нейроны, приводящие к вспышкам потенциалов действия в таламических нейронах (по-ci ингибиторный обратный эффект), имеющих проекции к коре. Сначала считалось, •пи эги тормозные нейроны являются интернейронами локальных цепей, но позже
"Например, в недавнем исследовании (Feige et al., 2005) альфа-компонент были извлечен посредством анализа независимых homhoiwntt, в то время как метаболическая активность оценена при помощи параметра насыщения крови кислородом (blood-oxygen-Имй (bpnndent. BOLD), измеренного в парадигме исследований «глаза открытые/закрытые». Таламус, как было показано, имеет по-ложи1вльную корреляцию с амплитудой затылочной альфа-активности, в то время как широко распределенные области в затылочно-икианным корковых отведениях имели отрицательную корреляцию с BOLD-сигналом.
° II исследованиях на животных сонные веретена, подобные сонным веретенам человека, моделировались сном под дей-I Iинам барбитуратов.
68 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
было показано, что в действительности эти клетки — тормозные ГАМКергические нейроны, расположенные в ретикулярном ядре таламуса.
Необходимо более подробно описать схему и принципы информационного потока в таламусе. Таламус — парные ядра, расположенные около центра мозга, — играет определяющую роль в управлении информационными потоками от рецепторов к сенсорным областям коры. За эту функцию он и получил свое название «ворота к коре». Действительно, почти каждый бит сенсорной информации попадает в кору через таламус. Помимо этой функции, связанной с сенсорными вхо
LGB
Рисунок 2.8. Главные ядра таламуса
Главные сенсорные релейные ядра — LGB (латеральное коленчатое тело), MGB (медиальное коленчатое тело) и VP (заднее вентральное ядро). Большое ядро второго порядка — Pul (подушка таламуса). Все ядра (кроме переднего ядра) покрыты ретикулярным ядром таламуса (Rt).
дами, таламус также играет важную роль в модуляции потока информации в ассоциативные, моторные и эмоциональные корковые области25.
Существуют две главные компоненты таламуса: дорзальный таламус (мы будем называть его таламус), состоящий примерно из 15 ядер, и вентральный таламус, главной частью которого является ретикулярное ядро. Ретикулярное ядро формирует своего рода щит вокруг дорзального таламуса (рис. 2.8). Оно состоит из тормозных ГАМКергических нейронов, которые имеют обратные проекции к таламусу Ретикулярные нейроны получают прямые возбуждающие проекции от таламуса, возбуждающие проекции об
ратной связи от коры и тормозные проекции восходящих активирующих систем ствола мозга (рис. 2.9).
Связанные с сенсорными потоками ядра таламуса могут быть разделены на релейные ядра первого порядка и ядра высшего порядка. Релейные ядра первого порядка получают сенсорную информацию непосредственно от рецепторов,
в то время как переключающие ядра высшего порядка получают сенсорную информацию, обработанную на более высоких иерархических слоях: 1) от корковых сенсорных областей посредством кортико-таламических проекций; 2) от ядер, вовлеченных в моторные действия (такие как ориентация тела, головы или глаз). Примеры ядер первого порядка — латеральное коленчатое тело для зрительной модальности, медиальное коленчатое ядро для слуховой модальности и заднее вентральное ядро для соматосенсорной модальности (рис. 2.8). Примеры ядер высшего порядка — подушка таламуса для зрительной модальности, заднее ядро для соматосенсорной модальности (рис. 2.8).
35 Различные взгляды на функции таламуса представлены в книге двух экспертов в исследованиях таламуса Murray Sherman and R.W. Giillery «Исследование таламуса и его роли в корковой деятельности». Согласно их мнению, у каждого ядра свои отдельные водители (афференты) и модуляторы. Классические сенсорные афференты — зрительные, слуховые, соматосенсорные, вкусовые — рассматриваются как водители к таламическим ядрам первого порядка. Другие типы водителей — мозжечок и мамиллярные афференты — для вентралатерального и переднего ядер соответственно. Третий тип водителей — 5-й слой кортико-таламических аксонов — для ядер более высокого порядка. Книга включает хороший обзор современных исследований относительно таламокортикальных связей и их функциональной роли.
Часть I. Ритмы ЭЭГ
69
Простейший взгляд на таламус подчеркивает его релейную функцию, то есть способность передавать сенсорную информацию к коре. Однако анатомические данные указывают, что даже для таламических ядер первого порядка первичные сенсорные входы составляют меньше чем 20 % всех синаптических входов. Этот факт поддерживает другой (и более реа-
Нейроны:
Кора
Таламус
кортикальные
Восходящая
ак т ив анионная
пир амид яльньк звездчатые
200 ms
Рисунок 2.9. Таламокортикальные цепи генерации сонных веретен
А. То же, что на рис. 1.9. Б. Импульс ретикулярного нейрона вызывает сильное торможение таламокортикальной клетки, котороедеинактивируетСа+*-каналы; токСа++ приводит к деполяризации таламокортикального нейрона. На вершине этой деполяризации нейроны производят разряды №+-К+-спайков. Обозначения: RE — нейроны ретикулярной формации; ТС — таламокортикальные нейроны.
листичный) взгляд на таламус, подчеркивающий значение несенсорных входов таламуса и, следовательно, его модулирующую функцию. Сложные цепи, вклю-
чающие таламические, ретикулярные и корковые узлы, играют важную роль в модуляции сенсорного информационного потока к коре. Способ сенсорной модуляции, по-видимому, существенно зависит от состояния мозга и отличается в разных стадиях сна и в различных стадиях информационного потока во время бодрство
вания.
В. Сонные веретена
Описание модуляторных функций таламуса мы начнем с описания сонных веретен. Веретена сна появляются в ЭЭГ только во время легкой дремоты (стадия 2) или медленноволнового сна (стадии 3 и 4). Это ритмичные синусоидальные волны с частотами от 10 до 14 Гц. Волны сгруппированы в циклы (вспышки) продолжительностью 0,5—3 секунды, которые могут повторяться с интервалами 3-10 секунд. Веретена сна проявляются в очень раннем возрасте (6—8 недель), наиболее выражены во время детства и юности и снижаются по амплитуде с возящим. Примеры веретен сна одного здорового человека в возрасте 23 лет представлены на рис. 2.1026.
Упрощенная схема генерации веретен сна представлена на рис. 2.9. Согласно иий схеме, для генерации веретен необходима сеть, состоящая по меньшей мере и»двух цементов — таламического ретикулярного нейрона и таламокортикаль
м Практически существуют два типа веретен сна, которые различаются по распределению и частоте. Высокоча-кнннып веретена имеют максимум пиковой частоты в 14 Гц. Максимальная амплитуда их распределения расположена Niihpyf С/. Они первые появляются в ЭЭГ в течение легкой дремоты. Более медленные (с максимумом около 12 Гц) вари тине появляются во 2-й стадии сна. когда сон углубляется. В течение перехода от 2-й стадии к 3-й появляются еще П|Н1яо медленные веретена (около 10 Гц). Максимум распределения более медленного типа веретен сна расположен НННИИ I i
70 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
ного нейрона, и взаимодействие между ними. Согласно этой схеме, вспышка импульсов тормозных нейронов ретикулярного таламического ядра вызывает сильные тормозные постсинаптические потенциалы в таламокортикальных нейронах. Эта гиперполяризация деинактивирует низкопороговый поток Са++. Поток Са++ вовнутрь вызывает обратную деполяризацию (обратный эффект) таламокортикальных клеток (Са++-спайк) и возникновение вспышки. Разряды таламокортикальных клеток возбуждают ретикулярные нейроны посредством прямых связей, позволяя циклу начаться снова.
Спонтанные веретена синхронизованы на больших корковых областях. Кортико-таламические пути играют важную роль в этой синхронизации. Действительно, перерезка этих путей вызывает десинхронизацию таламической активности, которая изменяется в дезорганизованный паттерн с низкой пространственно-временной когерентностью.
Г. Альфа-ритмы в состоянии бодрствования
Необходимо подчеркнуть, что нейронная схема, описанная выше, базируется на исследованиях, проведенных на анестезированных животных, и объясняет механизм генерации особого типа альфаподобных колебаний — сонных веретен. Веретена сна состоят из растущих и уменьшающихся колебаний потенциалов в диапазоне частот 10—14 Гц. В отличие от альфа-вспышек (которые могут длиться долго) веретена сна обычно короче и длятся только 1—3 секунды, снова возникая каждые 5—15 секунд27. Хотя некоторые авторы попытались перенести механизмы генерации веретен сна для объяснения альфа-ритмичности в состоянии бодрствования, недавние исследования привнесли сомнения в это простое объяснение. Действительно, более детальное рассмотрение показывает, что веретена сна и альфа-ритмы — различные явления.
Во-первых, альфа-ритмы и веретена сна проявляются в двух очень разных поведенческих состояниях (в состоянии расслабленного бодрствования по сравнению с состоянием сна или анастезии). Во вторых, альфа-ритмы случайно увеличиваются и уменьшаются в течение нескольких секунд, тогда как барбитуратные веретена имеют более короткую продолжительность. В-третьих, альфа-ритмы у людей имеют локальное корковое распределение, ограниченное первичными сенсорными областями, тогда как веретена сна более широко распространены с максимумом в центральных областях. И наконец, альфа-ритмы имеют более низкую частоту, чем сонные веретена.
Новая стадия в поисках нейронных механизмов генерации альфа-ритмов наступила совсем недавно с открытием того, что колебания в диапазоне альфа-частот можно смоделировать in vitro на препарате среза латерального коленчатого тела кошачьего мозга — части таламуса, передающей информацию зрительной коре (Hughes et al., 2004). Напомним, что в этом препарате таламуса корковая часть отсутствует и, следовательно, нет прямого
27 Хотя затылочные альфа-ритмы могут показать более выраженную веретеноподобную форму (см. рис. 2.10), термин «веретена сна» широко используется для вышеупомянутого паттерна ЭЭГ, чтобы избежать путаницы.
Часть I. Ритмы ЭЭГ
71
К-комплекс Веретена 14 Гц
Рисунок 2.10. Веретена сна
;КМ Оыла зарегистрирована у здорового субъекта 23 лет в течение трех стадий засыпания. В состоянии расслаблении (слева) в ЭЭГ присутствуют два альфа-ритма: затылочный альфа-ритм (запись с отведения 01) и мю-ритм (запись с отедения СЗ). Спектры ЭЭГ в этих отведениях и топограммы ритмов представлены выше. Стадия сонливости (в цмнгре) связана с подавлением этих ритмов. В течение следующей стадии сна появляются К-комплексы, состоящие и । нысокоамплитудной негативной волны, за которой следует меньшая позитивная волна, за ними следуют веретена |»«л (14 ПО с максимумом амплитуды в центральном отведении (см. запись с отведения Cz). Кроме того, присутствует I in iM с частотой 12 Гц и максимумом амплитуды в лобном отведении.
способа смоделировать таламокорковые взаимодействия. Отсутствующие модулирующие обратные кортико-таламические взаимодействия можно смоделировать искусственно в экспериментальных условиях, фармакологически активизируя глутаматные рецепторы в местах контактов кортикально-ыламических волокон и таламических нейронов.
Fp‘
Веретена 12 Гц
Д. Панковой режим высокопороговой пульсации таламических нейронов
Внутриклеточная регистрация активности таламокорковых нейронов и препаратах срезов таламуса показывает новую форму вспышек (нейронных пачек), которые происходят при относительной деполяризации
72.Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
(> —55 мВ) мембранных потенциалов и названы высокопороговыми вспышками28 (Hughes et al., 2002; Hughes et al., 2004) (рис. 2.11). Наиболее существенное свойство высокопороговых вспышек заключается в том, что они происходят периодично в широком диапазоне частот, который охватывает альфа- и тета-ритмы (то есть 4—13 Гц), со специфической частотой в любой момент определяемой степени нейронной деполяризации. Было обнаружено, что более сильная активация глутаматных рецепторов ведет к появлению ритмов с альфа-частотами, тогда как уменьшение уровня активации вызывает тета-волны. Таким образом, на таламическом уровне, альфа- и тета-волны формируют непрерывную активность, которая генерируется одними и теми же нейронными механизмами. Можно допустить, что уменьшение in vitro уровня таламической глютаматергической активации соответствует снижению возбуждения (arousal) in vivo. Следовательно, наблюдаемое у отдельного человека замедление альфа-колебаний (например, ниже 7,5 Гц для затылочного альфа-ритма в состоянии бодрствования) может быть связано с уменьшением уровня активации (arousal) у этого человека.
Возникает вопрос: что заставляет таламокортикальные клетки генерировать колебания синхронно? Эксперименты in vitro продемонстрировали, что синхронизация высокопороговых разрядов, вызывающих ритмичные альфа- и тета-колебания, зависят от наличия щелевидных контактов29 между таламическими нейронами. Hughes et al. (2004) показали, что альфа- и тета-ритмы в препаратах срезов были восприимчивы к большому диапазону веществ, которые действуют на функции щелевидных контактов, но сохраняются после блокады обычной химической синаптической передачи.
Ритмичные вспышки таламокорковых нейронов в альфа-диапазоне передаются на корковые нейроны через таламокорковые пути. Входящие вспышки создают постсинаптические потенциалы, которые измеряются со скальпа в форме альфа-ритмов. Пространственное распределение корковых генераторов различных типов альфа-ритмов (полученные методом LORETA) показывает три различных таламических источника альфа-ритмов, соответствующих трем типам альфа-ритмов.
Первый таламический генератор расположен в заднем вентральном ядре и соотносится с сенсомоторным ритмом (рис. 2.12). Этот ритм генерируется в постцентральной борозде теменной доли, которая соответствует первичной соматосенсорной коре. Как мы знаем из анатомии и физиологии, эта часть человеческой коры получает проекции от заднего вентрального ядра таламуса — таламического ядра первого порядка соматосенсорной модальности.
28 При рассмотрении ионной проводимости высокопороговые разряды, по-видимому, критически зависят от активации дендрит ных 1(-токов, переходных Са~-токов. Однако здесь надо подчеркнуть, что высокопороговые вспышки очень отличаются от низкопороговых вспышек во время сна временным паттерном и уровнем гиперполяризации таламокорковых нейронов.
29 Щелевидые контакты — морфологическая структура электрического синапса — весьма редкий тип соединения между нейронами в мозгу. В отличие от обычных химических синапсов, где передача сигналов вовлекает выпуск и распространение химического вещества — медиатора — в электрическом синапсе проводимость электрического сигнала от одного нейрона к другому происходит через пассивные электрические токи между пре- и постсинаптическими мембранами.
Часть 1. Ритмы ЭЭГ
73
Срез in vitro
Актив ацня глутаматных рецепторов
Рисунок 2.11. Высокопороговая таламическая пульсация
А. Схема среза таламического препарата (латеральное коленчатое тело). При гиперполяризованной мембране таламический нейрон показывает низкопороговые Са++-разряды (Б) и соответствующий пачечный режим пульсаций. Индивидуальные разряды обладают характерным паттерном межспайковых интервалов, состоящим из коротких межспайковых интервалов в начале разрядов (~ 2-5 миллисекунд), градуально увеличивающихся в процессе. Когда в таламическом препарате смоделирована кортико-таламическая обратная связь, путем экспериментальной активации подтипа глутаматных рецепторов (В), таламические нейроны подвергаются продолжительной деполяризации. В этом деполяризованном состоянии они показывают высокопороговую пульсацию (Г). Индивидуальные высокопороговые спайки имеют временный паттерн, отличающийся от низкопороговых Са++-спайков относительно большими межспайковыми интервалами. Адаптировано из Hughes et al. (2004).
Два других таламических генератора соотносятся с затылочным и теменным альфа-ритмами (рис. 2.13). Затылочный альфа-ритм генерируется около шпорной борозды, которая относится к первичной зрительной коре. Эта область затылочной коры получает проекции от латерального коленчатого тела — релейного ядра первого порядка (зрительная модальность). Теменной альфа-ритм генерируется в 7-м поле Бродмана (ПБ 7). Эта область получает проекции от пульвинарного ядра таламуса (подушки) — релейного ядра второго порядка (зрительная модальность).
Как видно на рис. 2.12 и 2.13, только расположенные в задней части таламуса ядра (заднее вентральное, подушка, латеральное коленчатое ядро) вовлечены в генерацию альфа-колебаний в здоровом человеческом мозгу. Мы нс знаем, отражает ли этот факт относительно более низкий уровень деполяризации таламокорковых нейронов в этих ядрах или некоторые другие присущие клеткам этих ядер свойства или особенности нейронных сетей задних ядер таламуса30.
* I tepeoe предложение выглядит более реалистическим. Если это верно, тогда более низкий уровень деполяризации можно Амлп Аы объяснить меньшим количеством возбуждающих входов к таламокорковым нейронам первого порядка и таламическим |тпяАным ядрам второго порядка. В соответствии с этим предложением любое поведение, направленное на уменьшение сенсор-1ННП яходв (например, закрывание глаз для зрительной модальности или расслабление мышц для соматосенсорной модальности) увеличивает соответствующий альфа-ритм. Другой способ уменьшить деполяризацию в таламокорковых клетках, возможно, связан । уменьшением общего возбуждения корковых нейронов, имеющих обратные проекции к таламусу.
74
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
(LORETA)
Рисунок 2.12. Генераторы мю-ритма
Мю-ритм регистрируется над сенсомоторной извилиной с максимумом амплитуды в отведениях СЗ, С4 (см. ЭЭГ). Электромагнитная томография (UDRETA) указывает генераторы в соматосенсорной коре. Эта кора получает входные сигналы от вентрального заднего ядра таламуса. Обозначения: VP (заднее вентральное ядро).
Рисунок 2.13. Генераторы затылочных и теменных альфа-ритмов
Затылочный альфа-ритм регистрируется в затылочных областях с максимумом амплитуды в 01,02 (см. запись ЭЭГ), в то время как теменной альфа-ритм регистрируется в теменной области с максимумом амплитуды в Pz (см. запись ЭЭГ). Электромагнитная томография (LORETA) указывает генераторы в шпорной борозде для затылочного альфа-ритма и в среднетеменной коре для теменного альфа-ритма. Затылочные и теменные области получают входные сиг налы от латерального коленчатого тела и подушки таламуса соответственно.
Часть 1. Ритмы ЭЭГ
75
III. Изменения в ответ на решение задач
Согласно теории генерации альфа-волн, представленной выше, любое увеличение сигнала на сенсорном входе должно подавить соответствующий альфа-ритм. Реакцию подавления называют реакцией десинхронизации, потому что высокоамплитудные альфа-волны можно рассмотривать как результат синхронизации в нейронных сетях. Реакция десинхронизации альфа-ритмов в ответ на сенсорное возбуждение действительно наблюдалась во многих исследованиях. Эти исследования также внесли некоторую ясность относительно механизмов генерации альфа-ритмов и их функционального значения.
А. Вызванная десинхронизация мю-ритмов в ответ на двигательную активность
Ведущую роль в этих исследованиях сыграла группа исследователей из технического университета в Граце (Швейцария), возглавляемая Gert Pfurtscheller. В 1979 году он впервые опубликовал методологию параметрической оценки ЭЭГ-десинхронизации. Проанализированный параметр был назван вызванной десинхронизацией (ERD — event related desynchronization). В некоторых исследованиях вместо подавления альфа-активности при двигательных актах наблюдалось повышение мощности альфа-ритма. Эту реакцию назвали синхронизацией, а параметр, этому соответствующий, — вызванной синхронизацией (ERS)31.
Для пространственного картирования ERD/ERS обычно применяются монтаж локального среднего или монтажи подобные преобразованию Лапласа. Пример вычисления и картирования ERD показан на рис. 2.14, где представлены данные здорового взрослого человека. В спектрах ЭЭГ этого человека наблюдается отчетливый мю-ритм как в состоянии с открытыми глазами, так и в состоянии с закрытыми глазами. Этот испытуемый также выполнял двухстимульную GO/NOGO и слуховую oddball-задачи. Для оценки вызванной Десинхронизации в ответ на стимулы, предполагавшие двигательный ответ (нажатие на кнопку), анализ был проведен в соответствии с разработанным G. Pfurtscheller методом. В верхней части рис. 2.14 представлены вызванная десинхро-низация/синхронизация между первым предупреждающим стимулом32 и вторым, который может быть или GO-стимулом (запускающим двигательную реакцию), или NOGO-стимулом (не требующим двигательного ответа). Можно заметить, что подготовительный период сопровождается десинхронизацией мю-ритма в контралатеральном полушарии и синхронизацией в ипсилатеральном полушарии. Внизу на рис. 2.14
11 Классический метод вычисления ERD/ERS предполагает следующие стадии. Во-первых, на ЭЭГ, зарегистрированной во время выполнения предложенных испытуемым заданий, должна быть осуществлена частотная фильтрация. Эта процедера позволяет исследова-frviK) выделить альфа-ритм. В настоящее время наряду с временной фильтрацией иногда применяется пространственная фильтрация, пойманная на анализе независимых компонент сырой ЭЭГ. Вторая стадия состоит из возведения в квадрат значений ЭЭГ-сигналов. В Iт.ту>|ыате этой процедуры вычисляется мощность сигналов ЭЭГ. Третья стадия заключается в усреднении мощности ЭЭГ для всех проб, ‘кнмртэя стадия необходима для того, чтобы сгладить данные и уменьшить их вариативность. И наконец, вычисляются относительные «шчания ERD/ERS. Они могут быть рассчитаны в процентах (для более подробного рассмотрения см. «Методы» главы 1).
” Испытуемый должен был ждать предъявления второго стимула, в то время как от него не требовалось никакой двигательной |ммкции (нажатия правым пальцем).
76
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Рисунок 2.14. Вызванная десинхронизация во время подготовки к движению и во время самого движения А. Вызванная десинхронизация, вычисленная для здорового взрослого субъекта, выполняющего двухстимульную GO/NOGO-задачу. Предъявляются пары изображений животных и растений в различных комбинациях, задача — нажать кнопку в ответ на два предъявленные подряд изображения животных. Обратите внимание на десинхронизацию мю-ритма с левой стороны, сопровождаемую вызванной синхронизацией в правой стороне во время подготовки к движению. Б. Вызванная десинхронизация, вычисленная для того же самого субъекта, выполняющего слуховую oddball-задачу. Значительная вызванная десинхронизация наблюдается при движениях в ответ на целевой стимул.
Обратите внимание, что десинхронизация в одной области сопровождается противоположной реакцией — синхронизацией — в смежной области.
представлены вызванная десинхронизация/синхронизация в ответ на слуховые стимулы: редкий стимул-мишень, предъявление которого требовало нажатия правым пальцем на кнопку, и стимул «не мишень», в ответ на который нажимать на кнопку не требовалось. Можно видеть, что предъявление только стимула-мишени вызывает сильную десинхронизацию мю-ритма.
Используя методологию вызванной десинхронизации/синхронизации в поведенческих исследованиях, связанных с сознательными движениями, группа из Граца сообщила о существовании нескольких подтипов роландических ритмов. Они показали, что мю-ритмы могут быть дифференцированы на основе их пространственной локализации. Например, области лица и ноги в сенсомоторной коре производят различные мю-ритмы. Группа Граца также показала, что мю-ритмы могут разделяться по частотам. Более низкочастотный (8—10 Гц) мю-ритм показывает широко распространенный неспецифический к типу движений паттерн вызванной десинхронизации, сходный для движений пальцев или ступней, тогда как более высокочастотный (10—13 Гц) мю-ритм демонстрирует более сфокусированный, специфический для типов движения паттерн вызванной десинхронизации, отличающийся для движений ступней и пальцев. Оба эти ритма блокируются перед и
Часть 1. Ритмы ЭЭГ
77
во время движений, и поэтому их можно рассматривать как мю-ритмы, хотя их реактивные компоненты различны33.
Согласно данным, полученным группой из Граца, десинхронизация высокочастотного мю-ритма соматотопически специфична и топографически ограничена. Десинхронизация этого ритма начинается в области контралатеральной руки за две секунды перед началом сознательного движения ипсилатеральной руки. Десинхронизация часто сопровождается синхронизацией в центрально-теменных областях, и была названа «очаговой (локальной) десинхронизацией, окруженной зоной синхронизации». В целом этот термин описывает наблюдение, что десинхронизация ритмичной альфа-активности происходит не в изоляции, а может сопровождаться синхронизацией в соседних корковых областях. Области латеральной синхронизации могут соответствовать той же самой сенсорной модальности или другой модальности.
Чтобы объяснять явление «локальной десинхронизации, окруженной зоной синхронизации», G. Pfurtscheller предложил таламический селективный, воротный механизм. Он утверждал, что та же самая таламическая модель, которая использовалась для объяснения возникновения сонных веретен, может быть применена для объяснения генерации мю-ритма. Определяющую роль в его схеме играют взаимодействия между таламокорковыми клетками и тормозными нейронами таламического ретикулярного ядра. Последнее формирует топографически организованный тормозной механизм обратной связи, который способен управлять информационным потоком через таламус. Эту функцию управления таламических ретикулярных тормозных нейронов называют «селекцией, отбором,, или воротным механизмом». Когда ворота закрыты, таламические нейроны разряжаются в виде пачек спайков, которые проявляются в корковом мю-ритме. Когда ворота открыты, таламические нейроны разряжаются в тоновом режиме, который проявляется в десинхронизации мю-ритма. Согласно предложенному механизму, латеральное торможение, которое существует между ретикулярными нейронами, отвечает за явление «локальной десинхронизации, окруженной зоной синхронизации»34. Исходя из всего вышесказанного, можно заключить, что роландические мю-ритмы подавляются в ответ на реальные или воображаемые движения.
м В отличие от очаговой (фокальной) и соматотопически специфической десинхронизации в высоких частотах неспецифическая десинхронизация более низкчастотного мю-ритма, обнаруженная в лобных, центральных, и теменных областях, распространена широко. Она может указывать на существование распределенной неспецифической нейронной сети в сенсомоторной коре. Эта сеть активизируется различными типами двигательного поведения, но не обязательно необходима для поддержки определенного движения. Одна гипотеза предполагает, что эта система может действовать как относительно неспецифический механизм, который подготавливает соматосенсорные и моторные нейроны перед определенным двигательным актом. Другая интерпретация этой неспецифической системы, связанной с низкочастотным мю-ритмом, такова, что эта система, возможно, служит нейрофизиологическим механизмом для общей двигательной установки, включая активацию премоторных и теменных областей в дополнение к первичным оенсомоторным. Действительно, десинхронизация низкочастотной альфа-активности была получена в ответ на различные неспецифические для определенной задачи факторы, которые могут быть объединены под термином «внимание». Такая вызванная десинхро-нимция топографически широко распространена по скальпу и, возможно, отражает активацию общих ресурсов для решения задачи, общие требования, обусловленные заданием, и процессы внимания.
м Здесь необходимо подчеркнуть, что ретикулярное таламическое ядро—не единственное место в мозге, связанное с латеральным торможением. Подобный эффект может возникать при латеральном торможением в стриатуме — части базальных ганглиев, — который управляет информационным потоком от таламуса к корковым областям контроля и принятия решений — областям коры, которые вовлечены в программирование действий.
78
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Б. Вызванная десинхронизация затылочных ритмов в ответ на зрительные стимулы
Похожие результаты показаны и для затылочных альфа-ритмов: они подавляются в ответ на зрительные стимулы. Кроме того, реакции в альфа-диапазоне отражают умственное напряжение, связанное с обработкой этих стимулов. Этот факт проиллюстрирован на рис. 2.15, где представлены данные, взятые из HBI базы данных. Вызванная десинхронизация/синхрони-зация для альфа-диапазона частот (8—12 Гц) вычислялась для группы здоровых людей в возрасте 12—13 лет. Они выполнили двухстимульное зрительное задание. Вызванная десинхронизация в затылочных областях в ответ на релевантные и нерелевантные стимулы представлена слева. Предъявление в паре стимулов первым релевантного стимула указывало на то, что испытуемые должны были ждать второго стимула (после которого требовался ответ GO или NOGO), в то время как предъявление нерелевантного стимула указывало, что вся проба должна игнорироваться. Как можно видеть, на первом этапе обработки информации (до 240 мс после стимула) оба типа стимулов выявляли одну и ту же десинхронизацию затылочной альфа-активности. Однако более поздние этапы обработки информации связаны с большей десинхронизацией затылочной альфа-активности для релевантного стимула по сравнению с нерелевантным35.
IV. Функциональное значение
А. Альфа-ритмы как ритмы «холостого хода» в ЭЭГ-активности
Как показывают фМРТ-исследования, величина синхронизации альфа-ритмов коррелирует отрицательно с метаболической активностью. Следовательно, альфа-ритмы рассматриваются как ритмы работы на «холостом ходу». Согласно этому представлению, мы могли бы ожидать, что появление альфа-ритмов в определенных сенсорных областях тормозит в них обработку сенсорной информации36. Альфа-ритмы являются эпифеноменами, отражающими ритмические таламические разряды, передающиеся коре. Эти разряды отражают уровень нейронной деполяризации, которая, в свою очередь, отражает дисбаланс между возбуждающими и тормозными постсинаптическими потенциалами, приходящими в тала-мокоркальные нейроны (рис. 2.16). Если возбуждение достаточно сильно для того, чтобы деполяризовать таламо-кортикальные нейроны, то эти нейроны перестают генерировать пачки импульсов и переходят в другой
35 Также обратите внимание на то, что подготовка ко второму стимулу, за которым могло следовать нажатие правым пальцем, связано с вызванной десинхронизацией мю-ритма в отведении СЗ.
36 Эта гипотеза была проверена в недавнем исследовании Ergenoglu с коллегами (2004). В этом исследовании стимулы предъявлялись на пороге восприятия так, чтобы приблизительно в 50 % проб человек не мог их опознать. Интенсивность стимулов была подобрана для каждого человека согласно его или ее способностям восприятия. Пробы были разделены на две группы: пробы с обнаруженными и необнаруженными стимулами. Между этими двумя условиями было получено достоверное различие относительной мощности альфа-ритма, которая была значительно ниже перед обнаруженными стимулами, вместе с достоверно (р < 0,001) более высокими амплитудами вызванных потенциалов (ERP). Эти результаты показывают, что коротколатентные изменения состояния возбудимости мозга отражаются в альфа-ритмах ЭЭГ.
Часть 1. Ритмы ЭЭГ
79
режим, в котором информация от воспринимающих органов передается коре без всякого ограничения. Мы называем такой сенсорный путь активным каналом.
Б. Латеральное торможение в активации альфа-ритмов
Если избыток торможения или недостаток возбуждения приводит мембранный потенциал в состояние гиперполяризации (то есть уровень возбудимости клетки падает), нейроны начинают разряжаться вспышками. Недостаток возбуждения может быть достигнут подавлением сенсорного входа на релейных (переключающих) ядрах таламуса, например, закрыванием глаз (для зрительной модальности). Избыток торможения может быть достигнут посредством латерального торможения, которое создается ретикулярными нейронами в близлежащем (активном) канале, возбуждающимися через связи от нейронов в активном канале как на корковом, так и на таламическом уровнях. Таламокортикальные нейроны в таком канале, который мы называем подавляемым каналом, производят разряды в частотах альфа-ритмов. Чтобы альфа-ритмы были зарегистрированы от скальпа, нейроны в подавляемом канале должны быть синхронизированы. При этом режиме синхронизации передача сенсорной информации к коре ограничена. Она полностью не блокируется как в течение ранних стадий сна, когда таламические нейроны генерируют сонные веретена, однако ограничена до некоторого уровня, такого что опознавание (обработка) стимула при этом режиме оказывается неполной.
И наконец, частота вспышек, произведенных в состоянии бодрствования в сетях подавляемого канала, определена уровнем поляризации таламокортикальных нейронов: чем больше они заторможены, тем медленней частота. В некоторых патологических случаях у взрослых эта частота может быть ниже 8 Гц.
Рисунок 2.15. Вызванная десинхронизация затылочных альфа-ритмов в ответ на зрительное возбуждение
11редстаалена усредненная вызванная десинхронизация, рассчитанная в диапазоне частот 8-12 Гцдля релевантных и нерелевантных стимулов в двухстимульном GO/NOGO-тесте. Для примера была выбрана группа здоровых субъектов 12-13 лет. Обратите внимание, что релевантный стимул вызывает более длинный ответ, но с такой же амплитудой на начальных С1ндиях обработки, что и нерелевантный стимул. Обозначения: т1, т2—топограммы вызванной десинхронизации.
80
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
ЭЭГ
Пирамидный нейрон коры
Тормозным нейрон ретикулярного ядра
Таламокортикальный j
нейрон /
Активный канал
Подав ляемый канал
Сенсорные рецепторы
Рисунок 2.16. Латеральное торможение в таламокортикальных цепях
Схематично представлены два канала: активный (слева) и подавляемый (справа). Тормозные нейроны ретикулярного ядра в активном канале активируются и тормозят таламокортикальные нейроны в смежном канале. Нейроны в подавляемом ijanane гиперполяризованы и генерируют альфа-ритмы.
V. Аномальные альфа-ритмы
А. Полное отсутствие альфа-ритмов
В 1949 г. Moizzi и Magoun впервые показали, что стимуляция ретикулярной формации ствола мозга активизирует кору и переводит высокоамплитудную, низкочастотную ЭЭГ (т.е. ЭЭГ с заметными альфа-ритмами) в низкоамплитудную, высокочастотную ЭЭГ37 В нормальном мозге альфа-ритмы ЭЭГ имеют наибольшую амплитуду. Когда альфа-ритмы отсутствуют, запись ЭЭГ выглядит и называется низкоамплитудной. Если более высокочастотные ритмы не подавлены по сравнению с нормой, запись ЭЭГ называют низкоамплитудной высокочастотной ЭЭГ. На спектрах мощности таких ЭЭГ отсутствуют существенные пики альфа-частот в любых состояниях (и с открытыми, и с закрытыми глазами). Эмпирически низкоамплитудная запись, определяемая как запись в состояниях с открытыми и с закрытыми глазами с амплитудой ЭЭГ в монтаже относительно объединенных ушных электродов, не больше чем 20 мкВ во всех корковых областях. Как можно видеть, это определение весьма субъективно и очень пространно.
Низкоамплитудные ЭЭГ обнаруживаются у 4— 10 % здоровых взрослых субъектов в зависимости от критериев оценки и едва ли могут рассматриваться как откло
37 Этот эффект частично обусловлен средними таламическими ядрами. Электрическая стимуляция внутриламинарных ядер тала муса по отдельности вызывает тот же самый эффект. Фактически реакция активации состоит из подавления альфа-ритмов (реакция альфа-десинхронизации) и увеличения высокочастотной активности (реакция бета-синхронизации).
Часть 1. Ритмы ЭЭГ
81
нение от нормы. Скорее низкоамплитудные ЭЭГ могут быть связаны с некоторой индивидуальной особенностью. Низкоамплитудные ЭЭГ найдены в семьях алкоголиков и наркоманов. Потребление алкоголя, похоже, работает как прием «самолечения» у этой части населения с целью уменьшить гиперактивацию коры.
Б. Альфа-ритмы в необычных участках
У детей и молодых взрослых альфа-активность ЭЭГ может быть обнаружена в затылочных областях (01, 02), теменной области (Pz) и в сенсомоторной зоне (СЗ, С4). Однако скальповое распределение альфа-активности (особенно в состоянии с открытыми глазами) изменяется с возрастом, и у пожилых людей становятся заметными височные альфа-ритмы. Niedermeyer (1997) описал височный альфоподобный ритм, обнаруженный главным образом в передне- и средневисочных областях, характеризующийся умеренной анормальностью, и предположил, что он может быть признаком начальных цереброваскулярных нарушений. Он также упоминает, что в период полового созревания этот паттерн, обнаруживаемый в височной доле, может скрывать очаг эпилептогенеза. Ритмичная альфа-активность также может маскировать ритмичные пароксизмальные вспышки, без проявления каких-либо острых компонент.
Наш опыт работы со здоровыми испытуемыми и пациентами позволяет сделать следующее заключение: если у отдельного пациента: 1) максимум ритмичной активности в пределах диапазона 7— 13 Гц локализован в отведениях, отличных от упомянутых для нормы, 2) сам ритм заметен настолько, что наблюдается существенное отклонение от нормы и в абсолютной, и в относительной мощности, — тогда этот ритм можно считать анормальным. В нашей практике максимумы распределений анормальных альфа-ритмов были обнаружены в задневисочных областях (например, в связи со звоном в ушах или травмой позвоночника), в теменных областях левого полушария (в связи с дислексией), в средних и в передних височных областях (в связи с возрастными цереброваскулярными нарушениями). Только в немногих случаях анормальные альфа-ритмы были обнаружены в лобных областях.
Пример спектральных характеристик ЭЭГ пациента38 с аномальными альфа-ритмами представлен на рис. 2.17. На сырой записи ЭЭГ проявляются два типа ритмов в пределах альфа-частот: первый с частотой 9,5 Гц, расположенный в левой средней височной области, и второй с частотой 7,3 Гц, расположенный в зоне отведения Fz. Результаты сравнения с нормативной базой данных и изображениями sLORETA представлены на рис. 2.17 справа сверху.
В целом изменения в нормальном функционировании таламокортикальных путей могут заканчиваться рядом неврологических расстройств, например, эпи
м Это пациент 13 лет с показателем интеллекта 88, посещает специальную школу. У него синдром Блоха — Сульцбергера — редкая болезнь из группы генетических заболеваний, которые ведут к появлению характерных зон беспигментной кожи и также заграждают мозг, глаза, ногти и волосы. В большинстве случаев синдром Блоха — Сульцбергера вызван мутациями в гене, называемом NI МО. Изменения цвета кожи вызваны чрезмерным отложением меланина (нормального пигмента кожи). Неврологические проблемы включают мозговую атрофию, формирование маленьких полостей в центральном белом веществе мозга. Приблизительно Л) Н детей с синдромом Блоха — Сульцбергера имеют замедленное двигательное развитие, слабые мышцы в одной или обеих частях fw/ш, задержки умственного развития и приступы. У них также с большой вероятностью возникают проблемы с органами зрения (например, косоглазие, катаракта и серьезное ухудшение зрения).
82 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
лептическими припадками и тремором при болезни Паркинсона, оба эти расстройства имеют ритмичные компоненты. Стимуляция или разрушение соответствующей части таламуса (например, зон вентральных ядер) — один из принятых методов облегчения тремора, по всей видимости, связанный с разрушением ритмической активности таламокортикальных сетей. Аномальная ритмическая активность таламических клеток (как показано на пациентах со вживленными электродами) может наблюдаться при некоторых неврологических расстройствах, связанных с нарушением поведения, которые не обязательно имеют ритмическую природу. Например, регистрация активности таламических нейронов у пациентов, страдающих хронической болью, как результата сенсорной деафферентации (так называемые фантомные боли) демонстрирует присутствие аномальных ритмических вспышек потенциалов действия. В этом случае стереотактические лизисы таламических ядер ведут к уменьшению фантомных болей39.
В. Лобная альфа-асимметрия
Другая важная характеристика анормальности альфа-ритма — его асимметрия. Как было указано выше, несмотря на то что усредненные спектры являются весьма симметричными, для здорового человека возможна асимметрия в диапазоне от 10 до 50 %. Однако если асимметрия больше 50 %, это может быть следствием какой-то патологии. В таких случаях сравнение с базой данных может показать, насколько наблюдаемая асимметрия отклоняется от нормальности. Пример патологической альфа-асимметрии на рис. 2.18. Здесь представлены спектральные характеристики ЭЭГ пациента с сенсорным игнорированием, вызванным повреждением правой затылочно-теменной области. Обратите внимание на большую (почти 100 %) асимметрию затылочного альфа-ритма в состояниях и с открытыми, и с закрытыми глазами.
Усредненные спектры мощности ЭЭГ в альфа-диапазоне для различных возрастных групп выглядят весьма симметричными (рис. 2.19). Другими словами, левое и правое полушария производят в среднем ЭЭГ-паттерны со схожей амплитудой и частотой. Но при индивидуальном измерении асимметрия мощности может быть достаточно большой. Возникает вопрос, не может ли эта асимметрия быть мерой некоторых индивидуальных черт, таких как интраверсия/экстраверсия или превалирование положительных/отрицательных эмоциональных состояний.
Больше двух десятилетий назад ряд исследователей занялись проблемой электроэнцефалографической асимметрии при эмоциональных переживаниях. Исследование было вдохновлено теорией Ричарда Дэвидсона об относительно асимметричном вовлечении орбито-фронтальной коры в эмоциональные реакции. Если говорить проще, согласно его теории, левое полушарие склоняется к обработке
39 Состояние, при котором таламокортикальные сети производят несоответствующую ритмичную активность (например, медленные ритмичные вспышки при бодрствующем мозге), было названо Uinaas и коллегами таламической дизритмией*. Дизритмия в этом смысле присутствует в некоторых формах эпилепсии и синдромов, сопровождающихся тремором. Оба этик типа нарушений представляют собой аномальные состояния таламокортикальной сети, сопровождаемые стереотипным по ведением, таким как ритмичные движения руками при треморе или колебаниями спайк/волна» с частотой 3 Гц на ЭЭГ но время генерализованных абсансов (припадков). Есть предположение, что подобная таламокортикальная дизритмия можп! лежать в основе других расстройств, включая психоневрологические. В настоящее время это интересное и провокационной предложение Uinbs и коллег нужно считать строго гипотетическим предположением, нуждающимся в большом количеств исследований.
Часть I. Ритмы ЭЭГ
83
Рисунок 2.17. Случай аномальной локализации альфаподобных ритмов
Вверху слева — фрагмент ЭЭГ в состоянии с открытыми глазами. Представлены отклонения от нормы спектров ЭЭГ и соответствующие топограммы. Внизу — sLORETA-изображения генераторов аномальных ритмов.
эмоционально-положительных стимулов, в то время как правое полушарие обрабатывает главным образом отрицательные эмоциональные стимулы. Это исследование привело к предположению о том, что лобная асимметрия ЭЭГ в состоянии спокойного бодрствования отражает асимметрию переднекорковой активности, которая действует как «предупреждающий сигнал» для возникновения эмоций и является фактором для индивидуальных различий в эмоциональном/мотивацион-ном поведении40.
40 В обычном исследовании шаги экспериментальной процедуры должны быть следующими. Спонтанная ЭЭГ регистрируется в со-г|(>янии спокойного бодрствования у здоровых добровольцев. Участки асимметрии мощности (Р) альфа-ритмов вычисляются как отношение: Р (L) - Р (R)/P (L) + Р (R). где Р (L) и Р (R) — мощность ЭЭГ в альфа-диапазоне в левых (L) и в правых (R) гомологичных электродах, .ho отношение служит мерой передней асимметрии корковой деятельности. Принимая во внимание то, что альфа ритм является ритмом холостого хода, положительная асимметрия Р (L) > Р (R) служит индикатором большей корковой активации в правом полушарии, и наоборот, отрицательная асимметрия Р (L) < Р (R) служит индикатором большей корковой активации в левом полушарии. Аспекты «национального стиля поведения испытуемых измеряются экспериментально или с помощью психологичеких опросников. Статистический анализ сосредоточен на соотношении между асимметрией и эмоционально-когнитивным аспектом деятельности (аффектом).
84
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Рисунок 2.18. Случай аномальной асимметрии затылочного альфа-ритма
А. Спектры в отведениях 01,02 (для состояний с открытыми и закрытыми глазами) и голограмма спектров ЭЭГ (для состояния с закрытыми глазами),
Результаты многих исследовании поддерживают теорию Дэвидсона, но некоторые авторы отмечают существенные противоречия в данных. Не так давно подобная ситуация стимулировала поиск методологических факторов, которые могли внести вклад в расхождение результатов. Для обзора методологических проблем см. Hagemann и др. (2005).
Согласно нашему
опыту, противоречие в данных возможно по двум причинам. Во-первых, индекс
асимметрии — это отношение двух случайных переменных, одна из которых — различие между двумя переменными. Следовательно, стандартное отклонение альфа-асимметрии как отношения пропорционально сумме двух стандартных отклонений обеих частей этого отношения. Различие между мощностью ЭЭГ с правой и левой стороны у большинства здоровых субъектов очень маленькое, следовательно, ошибка в вычислении индекса асимметрии весьма большая. Далее регистрация проводится с той стороны, где присутствует много мышечных артефактов и артефактов движений глаз. Второй фактор состоит в фактическом отсутствии заметной альфа-активности в лобных областях (рис. 2.19). Обратите внимание на отсутствие какого-либо пика в диапазоне альфа-частот в спектрах ЭЭГ в отведениях F3, F4, что недвусмысленно указывает на отсутствие генераторов альфа-ритма в лобных областях.
VI. Заключение
В состоянии спокойного расслабленного бодрствования с закрытыми глазами мозг человека продуцирует несколько типов ритмичной электрической активности в диапазоне альфа-частот (8—13 Гц). Обычно выделяют два основных типа альфа-ритмов, регистрируемых со скальпа: 1) затылочные альфа-ритмы, регистрирующиеся в затылочных или затылочно-теменных областях, и 2) роландический, или мю-ритм, регистрирующийся с сенсомоторной коры. Альфа-ритмы возникают в результате ритмичной активности таламических ядер: каждый ритм возникает в соответствующем таламическим ядре. Частота затылочного альфа-ритма немного изменяется с возрастом, достигая своего максимума в возрасте 15—20 лет.
Часть I. Ритмы ЭЭГ
85
11-12 15-14 15-16 Возраст
Рисунок 2.19. Асимметрия альфа-активности
Усредненные спектры для пяти групп здоровых субъектов различного возраста наложены друг на друга в парах F3-F4, СЗ-С4, РЗ-Р4, 01-02. Отрицательные различия (например, при СЗ < С4) показаны черным цветом. Обратите внимание, что нет асимметрий, устойчивых во всех возрастных группах.
Затылочные альфа-ритмы подавляются в ответ на зрительное возбуждение, в то время как роландические ритмы отвечают реакцией десинхронизации (уменьшения амплитуды) на реальные или воображаемые действия. Альфа-ритмы необходимо отделять и отличать от сонных веретен. Эти две различные категории ритмов наблюдаются в различных состояниях (во сне по сравнению с бодрствованием), имеют различное пространственное распределение (веретена сна имеют широкое центральное распределение, в то время как альфа-ритмы расположены около первичных сенсорных корковых областей), различные частоты (веретена сна — приблизительно 13—14 Гц в отличие от альфа-частот, которые изменяются между 8 и 13 Гц). Механизмы генерации сонных веретен подробно исследовались на модели животных. Что касается альфа-ритмов, механизмы их генерации известны не полностью. Мощность альфа-активности обратно пропорционально коррелирует с метаболической деятельностью в соответствующей корковой области, приводя к функциональному объяснению альфа-ритмов как нерабочих ритмов коры (ритмов расслабления и «холостого хода»).
86
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Глава 3. Бета-ритмы
1.Тйпы бета-ритмов
Бета-ритмы, проявляющиеся в отдельных пиках на спектрограммах, у здоровых людей обнаруживаются в различных областях коры. Бета-ритмы с частотой 13—30 Гц наиболее часто выявляются в лобных или центральных областях по сравнению с задними отделами коры. Могут быть выделены по крайней мере два различных бета-ритма: бета-ритмы с максимумами в спектрах ЭЭГ, расположенных в сенсомоторной зоне, — роландические бета-ритмы, и бета-ритмы, локализованные более фронтально, — лобные бета-ритмы. Они проявляются в форме пиков на индивидуальных спектрах. Однако только у немногих здоровых испытуемых выявляется отчетливый бета-ритм, и, как следствие, усредненные спектры в состояниях и с открытыми, и с закрытыми глазами имеют отчетливые максимумы в
f₽i
Мим
F8
J Л
,ЛА vvy Л W^-Maa*wv'
rs
01
02
Рисунок 3.1. Срединный бета-ритм
А. Фрагмент 19-канальной ЭЭГ здорового субъекта 48 лет в состоянии с открытыми глазами. Б. Спектры в альфа- и бета-диапазонах частот и соответствующие топограммы. Обратите внимание, что вместе с мю-ритмами, зарегистрированными в СЗ, С4, ЭЭГ имеет выраженный веретенообразный бета-ритм, регистрируемый в отведении Cz.
Часть I. Ритмы ЭЭГ
87
диапазоне бета-частот. Амплитуда бета-ритмов при измерении с объединенными ушными референтами меньше 20 мкВ41.
А. Роландические бета-ритмы
Роландические бета-ритмы наблюдаются как спонтанная активность в состояниях с открытыми й закрытыми глазами у здоровых испытуемых в сенсомоторных зонах (СЗ, Cz, С4). Хотя частота роландических бета-ритмов приблизительно 20 Гц (то есть в два раза чаще мю-ритма), их нельзя рассматривать как субгармонику низкочастотных колебаний мю-ритма. Действительно, как показали МЭГ-исследования, мю- и бета-ритмы имеют различные источники в первичной соматосенсорной и моторной коре соответственно (Хари и Salmelin, 1997, для обзора). Роландические бета-ритмы могут бьпъ обнаружены в ЭЭГ при отсутствии роландических мю-ритмов, а также вместе с роландическими мю-ритмами, но в различных отведениях (рис. 3.1). Частота роландического бета-ритма (максимум пика в спектрах) у разных людей может различаться, в широком диапазоне от 14 до 30 Гц. Кроме того, частота роландических бета-ритмов зависит от возраста.
Б. Паттерны десинхронизации/синхронизации в ответ на двигательную активность
Роландический бета-ритм модулируется при выполнении различных двигательных и когнитивных задач. Пример динамики роландического бета-ритма в течение выполнения GO/NOGO-задачи в группе 15—16-летних подростков представлен на рис. 3.2. Как видно на рис. 3.2, роландический бета-ритм, зарегистрированный в СЗ, отчетливо подавляется во время нажатий пальцем после предъявления второго стимула в GO-пробах, так же как во время периодов подготовки, когда испытуемые готовились сделать движение, в пробах GO и NOGO, но не в игнорируемых пробах, когда испытуемому не надо было нажимать на кнопку. Обратите внимание, что уменьшение роландического бета-ритма (названное десинхронизацией) сопровождается уменьшением мощности ЭЭГ в альфа-диапазоне частот — десинхронизацией мю-ритма. За подавлением роландического бета-ритма, связанного с нажатием пальцем, следует обратная бета-синхронизация. Эта синхронизация имеет место во время продолжающейся десинхронизации мю-ритма, указывая на то, что роландические бета- и мю-ритмы имеют различную динамику42 43.
Поскольку движение связано с глобальной активацией нейронов в сенсомоторной коре, мы можем предполагать, что роландическая бета-активность появляется, когда соответствующая нейронная система в сенсомоторной коре расслабляется после фазы сильной активации. Другими словами, увеличение роланди-
41 Асимметрия бета-ритмов обычно не превышает 35-40 %. Эмпирически любая асимметрия больше 50 % может считаться от-
клонением и заслуживает дополнительного внимания.
43 Необходимо подчеркнуть, что в исследованиях, представленных выше, предъявление стимулов служит точкой отсчета синхронизации. Однако так как роландические бета-ритмы связаны с движениями, разумно использовать как точки отсчета для вычисления вызванной десинхронизации сами движения, а не стимулы, вызывающие движения. В исследованиях, выполненных группой исследователей из Граца, возглавляемой Gert Pfurtscheller, испытуемых с множеством электродов, расположенных в зоне сенсомоторной коры, просили нажимать на кнопку произвольно и в удобном темпе. Исследования показали, что само движение сопровождается подавлением роландической бета-активности. Вслед за подавлением наблюдался сильный обратный эффект — бета-синхронизация.
88
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Рисунок 3.2. Подавление роландических бета-ритмов моторной активностью
Представлены усредненные данные для группы 15-16-летних здоровых детей, выполняющих двухстимульную GO/ NOGO-задачу. А. Мощностно-частотно-временное представление волнового (вейвлет) анализа ЭЭГ-реакций в GO, NOGO и игнорируемых пробах (Ignore). Расположение в колонке — сверху вниз. Приведены данные для отведения СЗ. Б. Топограммы ЭЭГ-реакций в частотной полосе 18-24 Гц в GO-пробах, моменты времени, соответствующие то-пограммам, показаны стрелками.
ческой бета-активности — явление «отдачи», которое можно рассматривать как след постактивации.
В. Лобные бета-ритмы
Второй тип бета-ритмов — лобные, которые обычно проявляются в спектрограммах лобных отведений как широкий пик с маленькой амплитудой. Такая форма спектра отражает низкую амплитуду этой ритмической активности и их нерегулярный паттерн. Бета-волны в монтаже с объединенным референтом редко, приблизительно у 2 % здорового населения, превышают 20 мкВ. Эти ритмы отличаются от альфа-ритмов центральных и задних отделов, которые обычно имеют более регулярные паттерны и выражаются в узком пике спектрограммы (рис. 3.3).
В F3-, Fz- и Р4-отведениях ЭЭГ спектры имеют отдельные максимумы в пределах 19 Гц.
Г. Паттерны десинхронизации/синхронизации в ответ на когнитивные задачи
В отличие от роландических бета-ритмов (которые появляются при двигательных задачах) лобные появляются при решении когнитивных задач, связанных с оценкой стимула и принятием решения. У некоторых людей перед лобной бета-синхронизацией может возникать бета-десинхронизация. Но во всех случаях для развития лобной бета-активности требуется постстимульный интервал в несколько сотен миллисекунд. Лобный бета-ритм может присутствовать у человека вместе с роландическим (рис. 3.4). Первое предъявление стимула в GO- и NOGO-пробах вызывает начальную де-
Часть I. Ритмы ЭЭГ
89
108 109 110 111 112
F₽1 ‘ /V^W4^ /W^MWtf\KW\
F₽a: V*
* ' H*
Fl:
W • YT^'K^MaWV^
re J/W/vjj4^/lv^^^
C4
T5
: Vq^v^W ^MAaXi/лМ/ V
T6
01
02AvW 50JZ/cm
Рисунок 3.3. Лобный бета-ритм
А. Фрагмент записи 19-канальной ЭЭГ здорового субъекта 53 лет в состоянии с открытыми глазами. Б. Спектры в отведениях F4 и Pz, топограммы в альфа- и бета-частотных диапазонах. Обратите внимание, что вместе с затылочным альфа-ритмом, регистрирующимся в отведениях 01,02, на ЭЭГ выявляется заметный веретенообразный бета-ритм в лобных областях с максимумами в отведениях F3, F4. Обратите внимание также, что мощность бета-ритма в три раза меньше мощности альфа-ритма.
синхронизацию с последующей синхронизацией отдачи в лобных областях (F3, F4). Напротив, бета-ритм в сенсомоторной зоне (СЗ) десинхронизируется во время подготовки к движению по отношению к синхронизации бета-ритма сразу после движения.
Мы выбрали для вышеупомянутого примера синхронизацию лобной бета-активности у одного здорового взрослого человека с отчетливым лобным бета-ритмом. Подобные данные могут быть получены на группах здоровых людей. На рис. 3.5, где представлены усредненные частотновременные реакции ЭЭГ-ответов при выполнении GO/NOGO-заданий группой 15—16-летних подростков, обратите внимание на то, что бета-синхронизация в ответ на первые стимулы в парах GO и NOGO начинается со значительной задержкой и достигает максимума в пределах 800 мс. Лобная бета-синхронизация не наблюдается в игнорируемых пробах (то есть в пробах, где предъявление первого стимула указывало на то, что вся проба должна игнорироваться).
Синхронизация лобного бета-ритма, вызванная каким-либо заданием, может быть измерена как различие между суммарной мощностью ЭЭГ
90
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Рисунок 3.4. Синхронизация лобного бета-ритма (отдельный случай)
Данные, зарегистрированные у здорового 45-летнего субъекта, выполняющего двухстимульную GO/NOGO-задачу. А. Мощностно-частотно-временное представление (волновой (вейвлет) анализ) ЭЭГ-реакций в GO- и NOGO-пробах в отведении F3, а также в отведении СЗ в GO-пробах. Б. Динамика относительной мощности ЭЭГ в диапазоне 18-24 Гц и топограммы, снятые в моменты, отмеченные стрелками. Обратите внимание, что предъявление стимула (требующего принятия решения, но не вызывающего движение) вызывало незначительное подавление лобной бета-активности, сопровождаемое синхронизацией через 500 мс от включения стимула. Обозначения: Стим. 1, 2 — время предъявления 1-го и 2-го стимулов проб; 11, 2 — соответствующие моменты времени, в которых вычислялись топограммы.
в бета-диапазоне при решении поставленной задачи43 и соответствующим параметром в состоянии спокойного бодрствования с открытыми глазами. Различия между спектрами в бета-диапазоне ЭЭГ при выполнении какой-либо когнитивной задачи могут являться мерой активации лобных долей при решении поставленных задач. Это отображено на рис. 3.6, где спектры ЭЭГ для двух различных задач (математической и GO/NOGO) наложены на спектры, вычисленные в состоянии спокойного бодрствования с открытыми глазами. Карта различий в бета-диапазоне частот показывает, что вызванная решением поставленной задачи бета-активность распределена в лобных зонах. Кроме того, степень увеличения лобного бета-ритма зависит от трудности задачи44.
43 Спектры вычисляются для всей записи ЭЭГ, зарегистрированной во время выполнения поставленного задания.
44 Для сравнения изменения бета-активности, связанные с решением двух психологических задач, изображены на рис. 1.3.6. Первая задача — трудная математическая задача, которая требует интенсивного включения рабочей памяти. Вторая — более легкая GO/NOGO-задача. В ней волонтер должен реагировать только в 25 % проб во время выполнения простой задачи на различение. Трудность задач выражается в количестве ошибок, которые намного больше для математической задачи, чем для задачи GO/NOGO. Можно видеть, что трудная задача ведет к увеличению активности ЭЭГ в диапазоне частот от 14 до 20 Гц, вдвое больше, чем увеличение бета-активности при выполнении простой задачи.
Часть 1. Ритмы ЭЭГ
91
Рисунок 3.5. Синхронизация лобной бета-активности (усреднение по группе)
Данные, усредненные по группе 15-16-летних здоровых субъектов. Подписи те же, что и на рис. 3.2.
II. Нейронные механизмы
А. Связь с корковой активацией
Существуют тесные отношения между, мощностью ЭЭГ в бета-диапазоне частот и метаболической деятельностью в соответствующей корковой области мозга человека. Это показано и для спонтанной ЭЭГ, и для вызванных ответов.
Одно из наиболее широкоизвестных исследований в этом направлении было сделано группой ученых из UCLA (Jan Cook и др., 1998). Они выполнили одновременную регистрацию многоканальной ЭЭГ и ПЭТ-исследование (с использованием радиоактивной воды) у здоровых взрослых людей в состоянии покоя и при выполнении простой двигательной задачи45. На основе своих исследований авторы заключили, что: 1) топографическое картирование ЭЭГ может точно отразить локальное мозговую функцию способом, сопоставимым с другим методам; 2) выбор ЭЭГ-измерений и монтажа существенно влияет на степень, с которой ЭЭГ отражает локальную метаболическую активность. Результаты исследования продемонстрировали, что мощность ЭЭГ в бета-диапазоне частот,
4В Данные ЭЭГ были обработаны с использованием 3 различных монтажей, оценивались значения как относительной, так и абсолютной мощности ЭЭГ. Результаты исследования показали, что относительная мощность ЭЭГ более сильно коррелирует с перфузией, чем абсолютная мощность. Кроме того, вычисление мощности для пар биполярных электродов и усреднение мощности между электродными парами, разделяющими общий электрод привели к более сильной корреляции с перфузией, чем данные, вычисленные для монтажа с одним референтным электродом.
92
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
бета =14-16 Hz
Рисунок 3.6. Вызванная бетэсинхронизация на спектрах мощности
А и Б. Усредненные данные по группе 13-14-летних здоровых субъектов. Вверху— наложенные спектры мощности ЭЭГ, вычисленные в состоянии решения математической (трудной) и двухстимульной GO/NOGO (легкой) задач по сравнению с состоянием спокойного бодрствования с открытыми глазами. В середине — разностные спектры с вертикальными полосами, указывающими уровень доверительной вероятности различий (р < 0,05). В. Разностные топограммы мощности для диапазона частот 14-16 Гц. Обозначения: тест — математическое тестовое задание; ГО — состояние с открытыми глазами.
вычисленная с использованием локального среднего монтажа, положительно коррелирует с метаболической активностью в соответствующей локальной корковой области (рис. 3.7)46.
Тесные отношения между мощностью ЭЭГ и метаболической активностью мозга человека дают основание предполагать, что абсолютные значения изменений во всех диапазонах ЭЭГ могут служить индексом метаболической активации коры при решении определенных задач. Сравнение между fMRI и вызванной электромагнитной активностью было сделано в исследовании Singh (2002). Они сделали запись МЭГ- и fMRI-параметров при выполнении задачи на скорость распознавания букв при зашумлении сигнала. Изменения мощности МЭГ были нор-
мализованы и усреднены между волонтерами. Результаты показывают, что вызванные решением поставленной задачи частотно-специфические изменения корковой синхронизации, обнаруженной на МЭГ, соответствуют тем областям мозга, в которых наблюдается вызванный корковый гемодинамический fMRI-ответ. Большинство этих изменений на МЭГ являлось вызванной десинхронизацией (ERDs) в бета- и альфа-диапазонах частот ЭЭГ.
Б. Чувствительность к ГАМК-агонистам
Самая поразительная особенность бета-ритмов — их чувствительность к ГАМК-агонистам. Действительно, мощность бета-активности увеличивается после приема барбитуратов, некоторых небарбитуратных успокоительных средств и легких транквилизаторов47. На рис. 3.8 схематично объясняется действие вышеупомянутых средств. Показан СГ-лиганд-зависимый канал. Когда канал открыт, наблюдается инфузия (ток вовнутрь) в нейрон ионов СГ. Этот приток гиперполяризует мембрану и уменьшает вероятность возникновения нейронного разряда. Канал — сложная молекула, которая имеет несколько участков связывания: участок связывания ГАМК, зоны связывания седатив-
46 Надо отметить, что мощность ЭЭГ в других диапазонах частот также коррелирует с метаболической активностью, но различными способами: альфа-активность, например, отрицательно коррелируете перфузией.
47 Обратите внимание, что по контрасту с увеличением мощности бета-ритма средняя частота роландического бета-ритма после приема барбитуратов уменьшается (Jensen и др., 2005).
Часть 1. Ритмы ЭЭГ
93
Частная
Рисунок 3.7. Взаимосвязи ЭЭГ и метаболизма
Соотношения между значениями локальной перфузии (ПЭТ) и относительной мощности ЭЭГ (при использовании локального взвешенного среднего монтажа). Статистическая значимость обозначена горизонтальными штриховыми линиями, представляющими величину, в которой коэффициент корреляции достигает значения р = 0,001. Адаптировано из Cook et al. (1998).
ных и анксиолитических веществ. Участок связывания ГАМК имеет сродство к молекуле самой ГАМК. Участок связывания седативных веществ имеет сродство к алкоголю или барбитуратам, таким как фенобарбитал. Участок связыва-
СГ канал
я ГАМК
0 бензодиазепин
ж барбитурат
НИЯ анксиолитических Рисунок 3.8. Схема ГАМК-рецептора
Показан лиганд-зависимый СТ -канал, который имеет три различных сайта (седа-ВещеСТВ связывается С тивный, антианксиолитический сайт, ГАМК-рецептор) со сродством к различным беНЗОД ИазеПИНаМИ та- схематично изображенным в виде различных геометрических форм,
кими как валиум, либриум48. Тормозные и возбуждающие синапсы отличаются по положению на мембране. Возбуждающие синапсы обычно располагаются на дендритах, в то время как тормозные синапсы расположены на теле клетки.
Идея о том, что колебания генерируются в результате коллективного поведения нейронов в сетях благодаря возвратному (циклическому) взаимодействию между ними, появилась давным-давно. Было предпринято много попыток смоделировать такие сети. Эти попытки находятся в рамках отдельной теоретической отрасли нейронаук, названной нейронными
сетями._______________
48 Из-за их различных действий на ГАМК-рецепторы седативные и анксиолитические лекарственные средства никогда не должны приниматься вместе. Объединенные дозы двух типов лекарств могут произвести к коме или смерти.
94
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
В. Торможение в корковых цепях
Исследование в смоделированных нейронных сетях показало, что тормозные
нейроны могут играть определяющую роль в генерации колебаний. Общие сети, генерирующие колебания, состоят из двух типов реципрокно связанных нейронных цепей (сетей): тормозной и возбуждающей. Представьте теперь себе, что в такой сети заблокированы тормозные процессы. Благодаря рекуррентному возбуждению такая сеть будет лавинообразно самовозбуждаться по принципу положительной обратной связи. При такой обратной связи чем больше нейронов
активно, тем больше они активируют другие нейроны через прямые связи, и тем больше они получают возбуждение через обратные связи. Положительная обратная связь может вести к сверхактивации сети. В действительности (в реальных
Рисунок 3.9. Нейронная модель бета-колебаний
Упрощенная модель колебаний в сети с возбуждающими и тормозными нейронами. Обратите внимание, что активность тормозных и возбуждающих нейронов приблизительно противофазна.
условиях) такая сверхактивация ограничивается торможением. Тормозные нейроны оказывают регулирование действия на эти лавинообразные реакции: они периодически прерывают сверхактивацию сети. Частоты этих периодических прерываний сильно зависят от продолжительности тормозных постсинаптических потенциалов. Последовательность событий в такой нейронной сети схематично представлена на рис. 3.9. Сеть содержит возбуждающие и тор
мозные клетки с взаимосвязями между ними и входами от внешних источников. Внешние входы служат источниками активации сети.
Выше приведена гипотетическая и упрощенная схема совокупных (групповых) нейронных осцилляций48 49. Такое поведение нейронов возможно, если: 1) внешнее возбуждение достаточно сильно, чтобы вызвать лавинообразную реакцию; 2) возбуждающие связи достаточно сильны, чтобы удерживать возбуждение; 3) тормозные связи достаточно сильны для того, чтобы остановить лавину50.
III. Пима-активность
Гамма-ритмы (> 30 Гц) имеют низкую энергию и поэтому трудны для записи. Действительно, для удаления 50 (60) Гц артефактов наводки электросети необходимы специальные так называемые режекторные фильтры. Режекторные фильтры должны быть очень точны, чтобы позволить анализ в гамма-частотах ЭЭГ, а
48 Однако она достаточно хорошо отражает внутренние связи в стандартной (эталон ной) корковой цепи, которая может считаться
примером близким к реальным корковым сетям.
50 Новое подтверждение причастности ГАМК-рецепторов в генерации бета-активности недавно было получено в совместных электроэнцефалографических/молекулярно-генетических исследованиях. Самая сильная связь с мощностью ЭЭГ в бета-частотах наблюдалась у короткого плеча хромосомы 4. Эта область содержит группу генов ГАМК А-рецептора (Porjesz и др., 2005).
Часть 1. Ритмы ЭЭГ
95
не в частотах сетевой наводки в 50 (60) Гц. Гамма-колебания могут быть проанализированы при записи ЭЭГ в диапазоне выше 30 Гц, в случае если будут использованы упомянутые ранее режекторные фильтры 50 (60) Гц.
А. Временная связь
В экспериментах на животных было сделано предположение, что синхронизация между нейронными элементами в частоте 40 Гц может быть специальным механизмом нейронной кооперации, который назван временной связью. Эта временная координация в режиме пульсаций пространственно распределенных нейронов, кодирующих различные особенности одного и того же образа, необходима, чтобы интегрировать вместе представление об этих различных особенностях в определенный объект перцепции. Большая часть работ в этой области была сделана в Max-Planck-Institute for Brain Research во Франкфурте (Германия), в группе, возглавляемой Вольфом Зингером (Vvblf Singer). Недавно было сделано предположение, что нейронная синхронизация также может играть важную роль в процессе сознательной обработки информации (см., например, Engel, Singer, 2001).
Регистрация скальповой ЭЭГ неизменно обнаруживает существование синхронизированной колебательной активности в гамма-диапазоне, когда люди воспринимают целостный зрительный образ. Эго было продемонстрировано Tallon-Baudry и Bertrand в 1999. В этом исследовании авторы предъявляли субъектам квазислучайные пунктирные изображения. Первоначально субъекты воспринимали такие стимулы как бессмысленные капли. Однако когда испытуемых обучили находить изображение собаки-далматинца, которое было скрыто на картинах, предъявляемые зрительные стимулы стали для испытуемых значащими и вызвали появление гамма-акгивности в затылочных зонах коры на 280 мс после предъявления стимула. Вероятная интерпретация этих результатов заключается в том, что объекты, вызывающие восприятие целостного зрительного образа, активируют зрительные области, которые синхронизируются в гамма-диапазоне.
Интракраниальная регистрация со вживленных электродов, сделанная теми же самыми авторами (2005), показала, что вызванная гамма-синхронизация может быть обнаружена в латеральной затылочной и затылочно-височной (веретенообразной извилине) зонах коре. Эти области неизменно демонстрировали большие гамма-колебания при кодировании визуального стимула, в то время как другие экстрастриарные области оставались систематически молчащими51.
В имеющихся у нас интракраниальных записях электрической активности мозга локальных полевых потенциалов у пациентов с эпилепсией значительная гамма-синхронизация наблюдалась в поле Бродманна 41 — первичной слуховой коре — в ответ на новые стимулы в слуховой oddball-парадигме. Это продемонстрировано на рис. 3.10 с помощью усредненных wavelet-трансформаций ответов локальных потенциалов поля. Как видим, только новые стимулы независимо от того, было ли состояние выполнения задачи пассивным (пациент только смотрел телевизор, в то время как предъявлялись слуховые стимулы) или активным (паци-
51 Одновременно с wavelet-анализом вызванной ЭЭГ авторы также вычислили вызванные потенциалы (ERPs), подробно описанные во 2-й главе книги. Они показали, что вызванная гамма-активность и ВП — различные сущности, т.н. они могут присутствовать или отсутствовать в различных интракраниальных областях независимо друг от друга.
96
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Акптное условие
Пассивное условие
Рисунок 3.10. Синхронизация гамма-активности при внутричерепных регистрациях
У пациента с эпилепсией, которому для диагностических целей и лечения были имплантированы электроды, проводилась регистрация локальных потенциалов в различных областях левой латеральной височной коры (области 41,22) и структурах левой средневисочной области (гиппокамп, миндалина). Самая сильная синхронизация гамма-активности в первичной слуховой коре обнаруживалась в ответ на новые стимулы. Обратите внимание на то, что параметры локального потенциала, ВП и вейвлет-разложения не различаются между пассивным и активным состояниями, указывая, что обработка информации в первичной слуховой коре является независимой от внимания. Стимулы продолжительностью 100 мс включались в момент времени, обозначенный как 0 мс. Обозначения: st — стандартный стмимул, dev — девиантный стимул, nov — новый стимул.
ент должен был нажать кнопку в ответ на девиантные стимулы и игнорировать девиантные и новые стимулы, когда они предъявляются вместе), выявляют сильное (приблизительно 100 %) увеличение интракраниальной мощности ЭЭГ в пределах диапазона частот от 40 до 55 Гц и латентностью 120-250 мс. Функциональное значение этого ответа не ясно. Robert Knight нашел подобные ответы в интракраниальных исследованиях и связал этот тип гамма-синхронизации с кодированием существенных признаков слуховых стимулов.
IV. Функциональное значение
Бета-синхронизация в задачах, связанных со зрительной модальностью, может наблюдаться не только в соматосенсорных и лобных, но также и в затылочных областях коры (рис. 3.11). В этих областях бета-синхронизация происходит в ответ на игнорируемые или NOGO-стимулы, то есть на стимулы, которые сопровождаются «расслаблением» субъекта, но не на стимулы (типа GO-стимулы), которые нуждаются в дальнейшей обработке или реакциях. На рис. 3.11 показаны усредненные значения для wavelet-преобразования и вызванной де/синхронизации (ERD/ERS) в диапазоне бета-частот для левой затылочной области (электрод 01). Как видно на рисунке, бета-синхронизация в затылочной области сопровождается предшествующей ей значительной десинхронизацией и в альфа-, и в бета-диапазонах частот.
Часть 1. Ритмы ЭЭГ
97
А. Бета-ритмы как следы постактивации
На рис. 3.11 самым важным фактом в отношении нейронных механизмов является наблюдение, что перед синхронизацией в бета-диапазоне наблюдалось увеличение отрицательных колебаний в вызванных потенциалах, зарегистрированных одновременно от того же самого электрода52. Как будет показано во второй главе этой книги, положительные компоненты ВП (по крайней мере в затылоч
ных областях в ответ на визуальные стимулы и для определенного возраста) связаны с возбуждающими постсинаптическими потенциалами, в то время как отрицательные компоненты ВП, по крайней мере частично, связаны с тормозными постсинаптическими потенциалами53. Принимая это во внимание, мы можем заключить, что бета-синхронизация — след постактивации, которая следует за сильной активацией затылочной коры. Этот след постактивации возни-
в
Рисунок 3.11. Бета-синхронизация в затылочных областях
Приведены данные, усредненные по группе здоровых субъектов 14-15 лет. А. Усредненные мощностно-частотно-временные представления (волновой (вейвлет) анализ) ЭЭГ-реакций в отведении 01 для проб NOGO и игнорируемых проб. Б. Вызванная де/синхронизация (ERD/ERS) в бета-диапазоне частот (16-21 Гц) в NOGO и игнорируемых пробах, наложенные друг на друга. В. ВП, вычисленный для того же электрода и тех же условий выполнения задачи.
кает в результате включения тормозных взаимодействий в корковой нейронной сети (рис. 3.12).
Это заключение соответствует ряду наблюдений о синхронизации роландической и фронтальной бета-активности. Вспомним, что мы связали вызванную бета-синхронизацию с постактивационными (возвратными), эффектами в соматосенсорной и в лобной коре. Мы допускаем, что различные типы бета-синхронизации (в полосе от 14 до 30 Гц), возникающие при решении задач, в ответ на стимул или движение можно рассматривать как единичный феномен — след постактивации.
Б. Перезагрузка обработки информации
Функциональное значение этого явления не ясно. Это может быть след памяти, который сохраняется в нервной системе, после ее активации. Однако согласно такому предположению, след должен быть сильнее для ре-
“ Различия в негативных компонентах ВП в ответ на вторые стимулы между NOGO и игнорируемыми условиями возникают за 200 мс перед любыми различиями в бета-синхронизации. Это наблюдение указывает на то, что бета-синхронизация следует за изменениями в ВП со значимой задержкой.
“ Обратите внимание, это утверждение — сильное упрощение и может быть истинным только для этого особого случая.
98
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Рисунок 3.12. Бета-синхронизация как след постактивации
Схематически изображены синаптические события в корковой цепи в ответ на сенсорный стимул. А. Временное течение постсинаптических возбуждающих и тормозных потенциалов в ответ на стимул. Б. Основные события двух последовательных стадий обработки информации. Первая стадия связана с активацией нейронов через возвратные возбуждающие синаптические связи. В течение этой стадии бета-ритм подавляется и информация обрабатывается активно. Вторая стадия связана с включением тормозных нейронов и подавлением возбуждающих нейронов. Увеличение торможения ведет к выраженной синхронизации бета-ритма (в центре). Функционально этот постактивационный след возвращает нейронную сеть в исходное состояние для дальнейшей обработки новой информации. Обозначения: ПСП — постсинаптический потенциал.
левантных стимулов по сравнению с нерелевантными. Но мы наблюдаем прямо противоположное явление — след намного сильнее для нерелевантных стимулов, когда больше нет необходимости в использовании памяти. Самое вероятное объяснение наблюдаемых эффектов: бета-синхронизация как отдельное явление представляет собой процесс перезагрузки. Этот процесс необходим, чтобы стереть результаты предыдущей активации и соответствующих «вычислений» и подготовить систему к новым действиям.
Так или иначе бета-активность нужно рассматривать как отсроченный индекс корковой активации. Это происходит как реакция на сильную активацию корковых нейронных сетей, когда тормозные нейроны получают достаточное возбуждение от внешних входов и рядом расположенных возбуждающих нейронов. В течение этой стадии обработки сигнала тормозные нейроны начинают давать разряды, чтобы подавить сильную активацию.
Это торможение происходит циклично, и каждый цикл — цикл бета-акгивности, зарегистрированной со скальпа. В здоровом мозгу бета-активность может проявляться как «акт перезагрузки», который «стирает» последствия сильной активации в нейронных сетях, и это позволяет сетям обрабатывать информацию снова и снова.
При отклонении от нормы, когда это происходит почти постоянно и отражается в слишком высоком уровне бета-активности, она может рассматриваться как показатель гиперактивации и раздражения соответствующей корковой области. Синхронизация ритма в бета-диапазоне ЭЭГ также может увеличиваться, если облегчаются (по-тенциируются) тормозные взаимодействия, например, после употребления седативных лекарств и антидепрессантов.
Часть 1. Ритмы ЭЭГ
99
V. Аномальные бета-ритмы
А. Потребность в нормативных базах данных
Как мы уже упоминали, генерация бета-ритмов связана с некоторыми специфическими состояниями коры. Эти состояния характеризуются высоким уровнем внешнего возбуждающего входа и относительно сильным внутрикорковым торможением. Эти состояния возникают во время постактивационной обработки информации в ответ на короткое предъявление стимула, когда тормозные нейроны включаются в работу, чтобы подавить деятельность нейронов, активных на первой стадии сенсорной обработки информации. Чтобы понять, является ли бета-ритм у отдельного человека слишком большим или слишком низким, необходимо сравнение данных по этому человеку с общей нормативной базой.
Когда у человека обнаруживается избыток или, наоборот, недостаток бета-активности, этот факт необходимо интерпретировать с осторожностью. Во-первых, для начала нужно оценить распределение бета-активности по поверхности головы. Нетрадиционному распределению бета-ритма, например в теменных или височных отведениях, должно быть уделено больше внимания, чем его распределению в сенсомоторных или лобных отведениях. Во-вторых, асимметрию бета-активности (выше 50 %) нужно рассматривать как дополнительный признак отклонения от нормы. В-третьих, реактивность наблюдаемого чрезмерного бета-ритма — важный показатель его функционирования. Если чрезмерная бета-активность у отдельного человека не синхронизируется в ответ на условие какого-либо задания так же, как в нормативной группе, то это наблюдение служит дополнительным индикатором отклонения от нормы. В-четвертых, необходимо принимать во внимание жалобы пациента, для чего необходимо вести протокол проводимой с ним текущей терапии и медикаментозного лечения.
Б. Корковая ирритация (раздражение)
Увеличение бета-активности и соответствующей сверхактивации может происходить в областях, связанных с фокусом эпилептической активности, например во время предэпилептических аур. Другой пример: корковая гиперактивация может возникать при галлюцинациях. Слуховые галлюцинации — один из позитивных симптомов, связанных с шизофренией. ПЭТ-исследования показывают, что галлюцинации связаны с гиперметаболизмом в слуховых корковых областях. В соответствии с исследованиями, связывающими бета-активность с изменениями локального мозгового метаболизма, в МЭГ-исследованиях также было получено увеличение активности в бета-диапазоне ЭЭГ (12,5-30 Гц) в левой слуховой коре, сопровождавшее галлюцинации (Ropohl и др., 2003).
Иногда увеличение бета-акгивности может визуализироваться как веретенообразный бета-ритм. Этот паттерн, как известно, связан с «корковым раздражением», вирусными или лекарственными энцефалопатиями и эпилепсией. Такая неправильная бета-активность выглядит как увеличивающиеся и уменьшающиеся веретена. Избыток бета-активности наблюдается меньше чем у 10 % населения, страдающего синдромом нарушения внимания и гиперактивности (СНВГ).
100 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
VI. Заключение
Полоса бета-ритма — это частоты ЭЭГ выше 13 Гц. Первым бета-ритм описал Ганс Бергер, эту мозговую электрическую активность связывали со сфокусированным вниманием. В полосе бета-ритма ЭЭГ можно выделить несколько его типов. Гетерогенность бета-ритма, разнородность подразумевает его традиционное деление на следующие поддиапазоны: низкочастотный бета-ритм — от 13 до 20 Гц, высокочастотный бета-ритм — от 21 до 30 Гц, гамма-активность — от 31 Гц и выше. Иногда выделяется отдельно особый тип «40 Гц активность». Этот тип бета-активности привлек большой интерес ученых в 1980-х. Исследования были сосредоточены на проблеме связанности восприятия. Было показано (и теоретически, и экспериментально), что сети тормозных промежуточных нейронов вовлечены в генерацию бета-ритмов. Причастность тормозных нейронов к генерации бета-ритма поддерживается идеей о чувствительности бета-ритмов к ГАМКергическим агонистам — фармацевтическим препаратам, которые подражают (имитируют) действию ГАМК, — основного тормозного медиатора в центральной нервной системе. ГАМК-агонисты, такие, например, как барбитураты и бензодиазепины, увеличивают мощность высокочастотных ритмов. В здоровом мозгу было показано, что бета-активность положительно коррелирует с метаболической активностью в корковой зоне, находящейся под записывающим электродом.
Часть 1. Ритмы ЭЭГ
101
Глава 4.
Среднелобный тета-ритм
I. Характеристики
А. Пространственное распределение
Согласно международной номенклатуре, частотный диапазон тета-ритма — 4—8 Гц (IFSECN, 1974). В 1950 году Arellano и Schwab наблюдали ритм с частотой 4—7 колебаний в секунду на центральной линии спереди от вертекса во время решения человеком мыслительной задачи. Ishihara и Yoshii
Стам. 1
Стам 2
Рисунок 4.1. Среднелобный тета-ритм у здорового 19-летнего субъекта
А. Нативная ЭЭГ, зарегистрированная в трех пробах в точке Fz во время решения математической задачи, в которой зрительные стимулы предъявлялись парами. Первый стимул представлял собой арифметическое выражение (например, 2 + 2), второй стимул — число, либо равнявшееся результату арифметической формулы (4), либо нет (5). Испытуемый нажимал на кнопку в случае совпадения. Обратите внимание — вспышки тета-ритма не синхронизированы по фазе с предъявлением стимулов. Б. sLORETA-изображения распределения источников генераторов данного тета-ритма. В. Усредненные по пробам результаты вейвлет-разложения нативной ЭЭГ: ось X — время, ось Y—частота, цветом кодируется величина изменения мощности сигналов. Г. Топограмма распределения синхронизации тета-ритма в момент времени 700 мс после предъявления первого стимула. Обозначения: стим. 1,2 — соответствующие стимулы проб; т — синхронизация ЭЭГ в тета-диапазоне.
102
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Рисунок 4.2. Среднелобная тета-активность как отдельная ритмичная компонента ЭЭГ
ЭЭГ, зарегистрированная у здорового 20-летнего субъекта во время решения математической задачи. А, Б, В, Г, Д. Результаты разложения ЭЭГ на независимые компоненты. Для каждой компоненты: слева — ее топография, внизу— усредненный спектр мощности, справа — кодированные цветом спектры мощности компоненты для всех последовательных 4-секундных эпох, нижняя цветная линия соответствует началу решения задачи. Е. Топограммы мощности ЭЭГ, вычисленные для трех различных частот.
из Японии индуцировали в ЭЭГ человека возникновение тета-ритма, дав ему задание на непрерывное арифметическое сложение. Эта тета-активность была представлена последовательными ритмическими волнами с частотой 6—7 Гц и с максимальной амплитудой в среднелобной зоне. Авторы назвали этот ритм среднелобным (frontal midline theta).
Пример среднелобной тета-активности, зарегистрированной у одного испытуемого из нашей нормативной базы, представлен на рис. 4.1. 19-летняя студентка выполняла задание, в котором она должна была сделать простое арифметическое действие — сложить две цифры, предъявленные на экране
Часть 1. Ритмы ЭЭГ
103
на 100 мс, и затем сравнить результат с числом, предъявленным на экране на секунду позже. Усилие, связанное с математическим действием, вызывает вспышку из 2—3 колебаний в полосе тета-ритма. Эти вспышки не были синхронизированы (не связаны во времени) с предъявлением стимулов и, следовательно, могли быть выявлены только методом вызванной синхрониза-ции/десинхронизации (см. «Методы»). Вспышки тета-ритма наблюдались не в каждой пробе, а случайно каждые 3 или 5 проб с периодом времени между отдельными вспышками приблизительно 20 секунд. sLORETA-образы указывают на то, что эти вспышки были произведены в средней части префронтальной коры (включая зоны передней поясной извилины).
Среднелобный тета-ритм принадлежит к отдельной категории мозговых ритмов и, следовательно, независим от альфа-ритмов (рис. 4.2). Анализ независимых компонент был применен к сырой ЭЭГ здорового испытуемого, выполняющего математическую задачу.
Как видно в нижней части рисунка, спектральная карта ЭЭГ для частоты 6,8 Гц (средняя частота тета-ритма, зарегистрированного у испытуемого) перекрывается с картой альфа-ритмов (затылочный и теменноцентральный), зарегистрированных у испытуемого. Однако применение метода независимых компонент (ICA-метод) к сырой ЭЭГ позволило выделить пять различных независимых компонент, первый из которых соответствует среднелобному тета-ритму.
В исследовании Hiroshi Asada и коллег, чтобы выяснить, где располагается источник ритмичной активности, были использованы методы МЭГ и ЭЭГ. Они одновременно регистрировали физиологические параметры у здоровых испытуемых при выполнении ими различных умственных задач (использовалась система МЭГ для всей головы).
Параметры МЭГ усреднялись во время каждого цикла среднелобного тета-ритма, в качестве начала цикла использовались индивидуальные позитивные пики среднелобных тета-волн в лобно-центральном отведении. Далее усредненные тета-компоненты МЭГ-сигналов были проанализированы с использованием мульти-дипольной модели. Были выделены два источника — в префронтальной средней поверхности коры и передней поясной коре (АСС). Эти области были «альтернативно» по очереди активированы приблизительно в 40—120 градусных фазовых сдвигах в течение одного цикла среднелобной тега-активности54.
Б. Индивидуальные особенности людей с выраженным тета-ритмом
Во время состояния спокойного бодрствования (REST) с открытыми или закрытыми глазами среднелобный тета-ритм в виде заметного пика прояв
м В работах другой японской группы (Sasaki и др.) срединная локализация генератора среднелобного тета-ритма была подвергнута сомнению. Эта группа продемонстрировала, что электрические диполи, ответственные за генерацию вспышек среднелобной тета-активности (5-7 Гц) рассредоточены в широкой зоне лобной коры обоих полушарий во время выполнения мысленных вычислений и заданий, связанных с музыкой. Они дали интерпретацию для этого противоречия следующим обрезом: тета-активность генерируется в областях лобной коры обоих полушарий, а максимальная активность тета-ритма ЭЭГ регистрируется на средней линии, потому что внеклеточные электрические токи, генерируемые пирамидными нейронами лобной коры обоих полушарий, суммируются, проходя через объемный проводник, имеющий несколько слоев с различным электрическим сопротивлением. Направление течения токов вдоль средней линии проявляется главным образом благодаря эффекту шунтирования, вызванного низким сопротивлением спинномозговой жидкости.
104 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
ляется в спектрах ЭЭГ только у незначительного количества испытуемых. У остальных испытуемых не выявляется никаких признаков такой ритмической активности в спокойном состоянии. В качестве объяснения можно предположить, что из-за глубокого расположения мозговых ритмических генераторов среднелобный тета-ритм проявляется и регистрируется с поверхности скальпа только в редких случаях. Необходимо принимать во внимание, что для каждого коркового ритма, записываемого со скальпа, должны выполняться следующие требования. Во-первых, источники генерируемого ритма должны быть около поверхности, а не слишком глубоко в мозгу. Во-вторых, работа источников должна быть синхронизирована на относительно большом пространстве мозга. В-третьих, источники должны быть достаточно сильны для того, чтобы генерируемая ими электрическая активность могла быть записана с поверхности головы55.
Согласно данным японских исследователей (обзор Inanaga, 1998), среднелобный тета-ритм встречается у 43 % взрослых испытуемых (18—28 лет) и у 11 % испытуемых старше этого возраста. В исследованиях японских коллег здоровые испытуемые были разделены на 3 группы по выраженности среднелобного тета в зоне лобного отведения (Fz) во время решения ими арифметических задач: с низким, средним и высоким уровнем выраженности среднелобной тета-активности56. Группа с высоким уровнем выраженности среднелобного тета-ритма продемонстрировала самый низкий уровень тревожности и невротизации и самый высокий уровень экстраверсии. С другой стороны, группа с низкой выраженностью среднелобного тета-ритма показала противоположные результаты: для них был характерен самый высокий уровень тревожности и невротизма и самый низкий уровень экстраверсии. В своем исследовании японские коллеги не выявили достоверных различий между группами испытуемых с разной выраженностью среднелобного тета-ритма в отношении решаемой задачи. Таким образом, было сделано заключение, что выраженность среднелобного тета-ритма связана в основном с личностными характеристиками испытуемых и уровнем их тревожности.
II. Нейронные механизмы
А. Связь с корковой активацией
В отличие от альфа-ритмов среднелобный тета-ритм может быть связан с активацией и увеличением метаболической активности в средней лобной области и передней поясной извилине. У пациентов с диагнозом эпилепсии электрическая стимуляция зон передней поясной извилины через интрацеребральные электроды приводила к возникновению 3—8 Гц ритма, регистрируемого в среднелобных зонах. Такая активность наблюдалась в среднелобных зонах у пациентов и без
“ Другое объяснение может состоять в том, что генерация среднелобного тета зависит от генетических факторов и может проявляться только у особой группы испытуемых.
“Для оценки личностных характеристик и уровня тревожности испытуемых были использованы тесты Maudsley Personality Inventory и Taylor's Manifest Anxiety Scale, выполнявшиеся испытуемыми после проведения ЭЭГ-исследований.
Часть 1. Ритмы ЭЭГ
105
электрической стимуляции (Talairach и др., 1973). Исследовательская группа Гарвардского университета и университета Висконсина (США) (Pizzagalli и др., 2003) была первой, которая изучила связь между среднелобной тета-активностью и метаболизмом в человеческом мозгу. Одновременная регистрация электрической мозговой активности (ЭЭГ) и метаболизма глюкозы (ПЭТ-исследование) была выполнена на группе здоровых испытуемых. Данные ЭЭГ были проанализированы с позиций выявления мозгового источника генерации указанной активности, что позволило сопоставить ЭЭГ- и ПЭТ-данные на уровне оценки активации в векселях. Именно для тета-ритма был выявлен самый большой кластер в передней поясной извилине (Поля Бродмана — 24/32), демонстрирующий положительную корреляцию между плотностью тока и метаболизмом глюкозы.
Б. Связь с гиппокампальными тета-ритмами
Среднелобный тета-ритм в ЭЭГ человека часто связывается с гиппокампальной тета-активностью в мозге млекопитающих. Причина такого связывания очень проста: гиппокамп млекопитающих обладает поразительной способностью к генерации тета-ритма. Эта ритмическая активность может быть зарегистрирована макроэлектродами во всех частях гиппокампа и в большинстве связанных с ним анатомических структур. Этот тип общего внеклеточно регистрируемого явления отражает кооперативное поведение большого количества гиппокампальных пирамидных клеток. Гиппокампальные пирамидные клетки, имеющие одинаковую ориентацию в пространстве, генерируют синхронную и периодическую электрическую активность в частоте тета-ритма. Частоты этих колебаний изменяются в весьма широком диапазоне от 4 до 10 Гц, имеют свои особенности и зависят от функционального состояния. Эти типы колебаний обычно объединяются под общим названием «гиппокампальные тета-ритмы» или, в более широком смысле, «лимбические тета-ритмы».
В. Лимбическая система гиппокампальных тета-ритмов
Мозговые структуры, в которых возможна регистрация тета-ритма, изображены на рис. 4.3. Эти структуры фактически представляют собой петлю (круг)57. Информационный поток в этом круге может быть сведен к следующему: 1) гиппокамп получает полисенсорный сигнал через обонятельную кору средней височной доли от различных височных и теменных сенсорных областей и сигнал от передней части поясной извилины; 2) гиппокамп интегрирует эту информацию и посылает результаты обработки к мамиллярным телам через гипоталамус, через пучок аксонов, называемый сводом (fornix); 3) мамиллярные тела посылают информацию дальше к переднему ядру таламуса через мамиллярноталамический пучок; 4) переднее ядро таламуса перенаправляет информацию к передней поясной извилине, а ее обратные связи с гиппокампом замыкают петлю Папеца.
57 Эту петлю также называют кругом Папеца. В 1937 гиду американский нейроанатом Джон Папец обозначил мозговую петлю, которая должна была начинаться и заканчиваться в гиппокампе. Он предположил, что этот круг структур играет определяющую роль п обеспечении эмоциональных процессов. Первоначальные представления о том, что круг Папеца связан только с эмоциями, были отставлены и заменены предположением, что этот круг структур главным образом вовлечен в процессы памяти. Это предположение было основано на результатах исследований, показывавших, что повреждения отдельных структур из круга Папеца в первую очередь нарушали эпизодическую память, а не эмоциональные функции.
106 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Г. Классическая модель возникновения гиппокампального тета-ритма
Согласно клиническим данным, гиппокамп — это структура мозга, необходимая для консолидации эпизодической памяти. Несмотря на интенсивные экспе
риментальные исследования, связь между молекулярными механизмами памяти, с одной стороны, и функционированием на уровне нейронной сети, с другой стороны, остается неизвестной. Одна из ключевых проблем для современных исследований — понять, как множеством отдельных нейронов генерируется тета-ритм и какую роль эта ритмическая активность играет в формировании памяти. С этой целью гиппокампальные тета-ритмы особенно активно изучались на животных, особенно у крыс. Эти исследования позволили сформировать классическую модель генерации тета-ритма в гиппокампе млекопитающих (рис. 4.4).
Согласно этой модели, тета-ритм возникает в стволе головного мозга как неритмичная нейронная активация (обозначено стрелкой на рис. 4.4). Нейронные элементы ствола мозга включают норадренергические клетки, расположенные в голубом пятне, серотонинергические клетки — в ядрах шва, допаминергические клетки — в передней части покрышки и компактной части черной субстанции. Эти клетки активируются напрямую через чувствительные коллатерали нейронов или косвенно через обратные проекции от коры, что ведет к активации клеток в ядра перегородки, расположенных в основании переднего мозга. Активация кле
ток в ядрах перегородки вызывает вспышки тета-ритма.
Важную роль в генерации тета-ритма играют реципрокные взаимодействия между нейронами перегородки58 и гиппокампа. В состоянии спокойного бодрствования фоновая активность тормозных нейронов в гиппокампе подавляет гиппокампальные пирамидные клетки так, чтобы никакая сенсорная информация не кодировалась в след памяти. В случае же если появляется значимый стимул (например, эмоционально-значимый), этот стимул через активирующую си
Рисунок 4.3. Анатомические структуры, в которых наблюдаются тета-ритмы
Гиппокамп (Hip) получает мультимодальные сенсорные входы, смешанные со входами от передней поясной коры, обрабатывает их и посылает результаты сосцевидным (мамиллярным) телам (МВ) гипоталамуса через путь, названный Сводом (Fornix), где они объединяются с другими входами и посылаются к переднему ядру (А) таламуса. Переднее ядро служит релейным ядром, посылающим свои выходы к поясной извилине (GC).
стему ствола мозга увеличивает активность возбуждающих и тормозных нейронов перегородки. Тормозные нейроны перегородки снижают активность тормозных клеток гиппокампа. Эго торможение активности тормозных клеток гиппокампа (растормаживание) удаляет фоновое торможение пира-
58 Область включает среднюю перегородку и диагональную зону Вгоса.
Часть 1. Ритмы ЭЭГ
107
Перегородка
Гиппокамп
Рисунок 4.4. Нейронная сеть генерации гиппокампального тета-ритма
А. Сеть, по-видимому, предназначена природой для преобразования сильной временной входной активации от ствола мозга и мультимодальных корковых областей во вспышки тета-ритма. Сеть состоит из возбуждающих и тормозных нейронов, расположенных в области перегородки и в гиппокампе. Нейроны перегородки посредством торможения тормозных гиппокампальных нейронов позволяют сенсорному информационному потоку проходить через гиппокамп. Взаимные тормозные связи между перегородкой и гиппокампом производят вспышки тета-ритма. Как на всех других рисунках, возбуждающие нейроны и возбуждающие связи отмечены черным, в то время как тормозные связи — серым. Б. Вспышки потенциалов действий, генерируемые различными частями сети.
Тормозные нейроны
перегородки --------
I ормозные нейроны гиппокампа
Пирамидные нейроны
Вспышки тета рнгма
мидных клеток и в конечном счете способствует их работе. Такая последовательность нейронных событий запускает цикл тета-ритма.
Далее активация пирамидных клеток гиппокампа тормозит тормозные клетки в ядрах перегородки, которые, в свою очередь, снимают торможение с гиппокампальных тормозных нейронов, а те далее тормозят пирамидные клетки. Так завершается цикл. В этой схеме зону перегородки рассматривают как «узловую точку», куда поступают неритмические колебания от ретикулярной системы ствола мозга и где они преобразуются в ритмичные сигналы и передаются в гиппокамп. Гиппокампальный тета-ритм зависит от силы входящих сигналов от нейронов перегородки: когда достигается некоторый критический уровень активации нейронов перегородки — возникает гиппокампальный тета-ритм59.
А Вовлечение в процессы памяти
Существуют данные, которые поддерживают концепцию о том, что тета-ритмы играют важную роль в обеспечении процессов памяти. Во-первых, долгосрочная потенциация (long-term potentiation, LTP) как механизм синаптической модификации60, чувствительный к стадии тета-ритма, улучшается на пике цикла и ухудшается на спаде цикла (Huerta и Lisman, 1993). Во-вторых, в рядк исследований in vitro и in vivo продемонстрировано что индукция долгосрочной потенци-
“ В гиппокампе животных существуют не один, а по крайней мере д ва типа ритмической тета-активности. Первый тип связан с произвольными движениями и не изменяется поддействием атропина или холинергического истощения. Второй тип тета-активности иногда возникает в состоянии неподвижности, снимается атропином и может спонтанно возникать при уретановой анестезии (Kramis и др., 1975).
“ Долгосрочная потенциация (LTP) была открыта в гиппокампе млекопитающих Terje Lamo в 1966 году. В экспериментах in vitro, in vivo было показано, что серия коротких высокочастотных электрических стимуляций усиливала работу синапса (или облегчала) на несколько минут, иногда часов и дней. В связи с этим LTP рассматривается как один из главных механизмов формирования памяти.
108 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
ации оптимальна, когда временной интервал между стимулами приближается к 200 мс, т.е. при частоте тета-ритма. В-третьих, разрушения перегородки не только блокируют возникновение тета-ритма в гиппокампе, но и приводят к ухудшению памяти. Эти наблюдения предполагают, что этот тета-ритм является своего рода управляющим механизмом для синаптической пластичности61.
Следует подчеркнуть, что гиппокамп — не единственная область в мозгу, генерирующая тета-ритмы. Этот ритм был найден в большом количестве мозговых структур: парагиппокампальная кора, передняя поясная извилина, мамиллярные тела гипоталамуса, медиадорсальное ядро таламуса (рис. 4.3). Все эти структуры принадлежат лимбической системе. Несмотря на то что эти структуры мозга самостоятельно не способны генерировать тета-ритм, они связаны друг с другом и обеспечивают сложную нейронную сеть, выполняющую различные функции, и, следовательно, играют несколько возможных ролей для реализации тета-ритма. Gyoigy Buzaky, мировой эксперт в этой области, предложил использование термина «лимбические тета-осцилляции» вместо гиппокампального тета-ритма, что отражает сложность и распределенный характер природы возникновения тета-колебаний.
Е. Тета-активность как отражение кванта-информации
Этому взгляду на функциональное значение лимбической системы соответствует открытие долгосрочной потенциации и ее взаимоотношений с гиппокампальным тета-ритмом. Действительно, как мы упоминали выше, индукция долгосрочной потенциации оптимальна, когда стимулы повторяются с периодичностью в диапазоне тета-ритма. Чтобы вызывать долгосрочную потенциацию, ритмичное возбуждение в частоте тета-ритма не обязательно: достаточно даже двух высокочастотных разрядов с интервалом в 200 мс, чтобы вызвать долговременные изменения в синаптической работе. Эти данные явно показывают, что для того, чтобы запоминать, мозг не нуждается в постоянном тета-ритме — достаточно и двух неполных циклов. Именно поэтому цикл тета-ритма можно рассматривать как информационный квант. Этот квант, похоже, служит для функциональной связи различных лимбических структур, позволяющих кодирование некоторого эпизода нашей ежедневной жизни в воспоминание (паттерн памяти).
Ж. Гиппокамп как карта эпизодов памяти
Самое распространенное представление о функциях гиппокампа — это то, что гиппокамп кодирует в сжатой форме и временно хранит возникшие на уровне неокортекса эпизоды памяти (McNaughton, 1989). Фактически есть две части любого эпизодического события: сенсорная составляющая (включающая нейронные элементы, кодирующие различные свойства зрительных стимулов, слуховых сигналов, телесных ощущений) и связанная с действием
61 Как показано, тета-ритм играет роль в нейронном кодировании места (O’Keefe и Recce, 1993). Когда крыса пересекает определенное место, гиппокампальные «клетки места» дают разряды на самой ранней стадии тета-осцилляции. Эта информация может значительно улучшить точность определения положения животного в пространстве. Другая точка зрения (Bland, Oddie, 2001) предполагает, что тета-ритм играет роль в сенсомоторной интеграции. В соответствии с этой гипотезой человеческие тета-осцилляции происходят во время эвристического, исследовательского поиска и целенаправленного поведения, а также при виртуальных движениях, когда сенсорная информация и планирование движения находятся в состоянии непрерывного изменения; по сравнению с периодами самоконтролируемого состояния неподвижности.
Часть 1. Ритмы ЭЭГ
109
составляющая (включающая нейронные элементы, кодирующие выполнение действия, эмоциональное состояние, побуждение). Эти две составляющие события кодируются в различных корковых областях: в париетально-височных зонах — сенсорная компонента, и в областях лобной доли — связанные с событием действия. Передняя поясная извилина служит промежуточной интегративной областью в префронтальной коре. Она интегрирует всю информацию от различных префронтальных областей и посылает результаты обработки информации к моторным областям и в гиппокамп. Хотя задняя и передняя части поясной извилины связаны друг с другом, они кодируют совершенно
Время
Рисунок 4.5. Гйппокамп как устройство картирования памяти
Сенсорная составляющая индивидульных эпизодов картируется в модально-специфических задних отделах коры, в то время как двигательная составляющая эпизодов картируется в префронтальных областях коры. Обе репрезентативные схемы, в свою очередь, картируются в гиппокампе. Следы, существующие за счет реверберации в префронтальных и задних отделах коры, сохраняются только в течение короткого времени (изображены прерывистой линией), в го время как следы, основанные на долгосрочной потенциации в гиппокампальных клетках, сохраняются в течение более длительного времени (обозначены серым), прежде чем они объединяются и сохраняются в долгосрочной памяти (обозначены черным).
110
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
различные параметры события. Гиппокамп же — это зона мозга, где все различные и пространственно распределенные представления о событии сходятся в единый активационный паттерн или сегмент62.
3. Тета-ритм и консолидация следов памяти
Без гиппокампа и связанных с ним структур63 след памяти в этих областях может быть сохранен только в рабочей памяти в форме реверберации нейронных импульсов в рекуррентных нейронных сетях. Этот след рабочей памяти существует несколько секунд и очень чувствителен к помехам — любой новый эпизод вызывает распространение следа памяти, новый след накладывается на след от предыдущего события и уничтожает этот.
Наша гипотеза состоит в том, что гиппокампальный след сохраняется на более длительное время. Продолжительность гиппокампального следа, возможно, определяется долгосрочным облегчением, вызванным высокочастотными вспышками разрядов, которые следуют за друг другом через оптимальные (в диапазоне тета-ритма) временные интервалы. Такой ход вещей позволяет гиппокампальной системе быстро воспринимать новую информацию, не разрушая ранее сформированные следы памяти, сохраненные в коре головного мозга, и в то же самое время позволяет постепенно интегрировать новую информацию с имеющейся в памяти и структурировать ее.
Вышеупомянутая модель фактически предполагает, что гиппокамп служит временным хранилищем событий. Важное событие вызывает вспышку тета-активности, которая отражает синхронную активацию гиппокампальных клеток, связанных со «сжатым» следом. Если событие не достаточно важно, то оно не вызывает вспышки тета-активности и консолидации следа не будет.
Следы в гиппокампе становятся активными или при эксплицитном воспроизведении, или при имплицвдных процессах, например, во сне. Это вызывает восстановление соответствующей корковой памяти, проявляющееся в возрастании корковых взаимодействий, вероятно, вовлекающих местную синаптическую консолидацию.
Таким образом, гиппокамп использует «репрезентации» событий, чтобы неоднократно обновлять репрезентации на уровне корковых связей, до тех пор пока они не станут консолидированными и независимыми от гиппокампа64.
62 С позиции анатомии гиппокамп идеально подходит для картирования. Он состоит из большого количества долей (сегментов), выровненных вдоль одной борозды. Активация ограниченного числа сегментов гиппокампа будет успешно картировать корковые представительства произошедшего эпизода в пространственный паттерн активности гиппокампа.
63 Мамиллярные тела гипоталамуса и переднее ядро таламуса.
64 Существует много свидетельств в пользу того, что сон играет важную роль в синаптической пластичности и памяти. Было показано, что сон важен после зрительного обучения для консолидации некоторых форм процедурной памяти, например, зрительно дискриминируемых навыков. Однако облегчающий запоминание эффект сна не ограничен наблюдениями только для процедурной памяти. Так, например, известно, что лишение сна ухудшает эпизодическую память. На основе наблюдаемых явлений было сделано заключение, что эффект облегчения консолидации памяти во сне может зависеть от синхронизированной медленноволновой нейронной активности. В качестве примера — модель эпизодической памяти (Buzsaki G., 1989) постулирует, что в состоянии бодрствования информация первоначально сохраняется в САЗ-области гиппокампа, через изменения силы взаимодействий между пирамидными нейронами. Позже, во время медленного сна, синхронизированные популяционные разряды САЗ-нейронов будут •перезапускать, перепрезентовать» следы, сохраненные в САЗ-нейронной сети, и через обонятельную кору реактивировать ассоциативные корковые нейроны, воспроизводя «основные характеристики» интересующего события. В конечном счете этот перезапуск сохраненных следов памяти ведет к долгосрочным синаптическим изменениям в ассоциативных корковых сетях.
Часть 1. Ритмы ЭЭГ
111
Это временное воспроизведение события (следа) в гиппокампе оказывается относительно нечувствительным к помехам, т.к. следы отдельных событий фиксируются в отдельных частях пространства гиппокампа.
III. Изменения в ответ на решение задач
А. Увеличение нагрузки на память
Основываясь на успехах исследований тета-ритма на животных, тета-ритм человека интенсивно начали изучать в последние два десятилетия. В одном из первых исследований, выполненных Alan Gevins (EEG systems Laboratory and SAM technology in San Francisco), изменения корковой активности при решении задач на память были исследованы с помощью электроэнцефалограмм, регистрируемых от 115 каналов. Испытуемые выполняли задания, требующие сравнения каждого нового стимула с предыдущим на основании его вербальных или пространственных признаков. В этом исследовании было получено увеличение среднелобного тета-ритма с увеличением нагрузки на память. В исследовании наблюдалось увеличение сигналов тета-ритма и улучшение выполнения задания после тренировки.
Б. Два типа ответов в тета-диапазоне
Ведущая роль в изучении тета-ритма людей принадлежит группе Wblfgang Klimesch из Зальцбурга (Австрия). Группа получила свидетельства того, что существует несколько типов тета-активности, по-разному соотносящихся с заданиями на память и внимание. Первый тип может быть связан с направленным вниманием или рабочей памятью. Этот тип обусловленного вниманием тета-ритма выражен в длительных тонических колебаниях в диапазоне частоты тета-ритма. Фазические вспышки тета-активности рассматриваются как основа для двух переходных типов тета-ответов: теменной тета-активности, которая связана с кодированием эпизодической памяти, и лобной активности, которая связана с извлечением из памяти. Эго заключение поддерживается многочисленными исследованиями группы Климента, которая обнаружила, что степень синхронизации тета-ритма при кодировании определяет эффективность извлечения из памяти и что вызванная тета-синхронизация во время опознания старых слов оказывается больше, чем для новых.
Подтверждение существования двух фазических ответов в тета-диапазоне обнаруживается в работах Nobuaki Nishiyama и Yoko Yamaguchi в Brain Science Institute (BSI), RIKEN (Япония). Они записывали ЭЭГ от 64 каналов во время виртуального прохождения лабиринта. Колебания в тета-диапазоне наблюдались в виде вспышек, состоящих из нескольких циклов. Эти вспышки обычно были локализованы в двух зонах коры: лобной и теменно-височной. Первый тип вспышек был определен как среднелобный тета-ритм. Активность в теменновисочных зонах сопровождалась вспышками среднелобного тета-ритма, в то время как фазовые различия активности между этими двумя областями время от времени изменялись.
112 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Два типа тетаподобной активности также наблюдались в исследованиях, составляющих нормативную HBI базу данных. Усредненные значения воспроизводимых ЭЭГ-ответов при выполнении двухстимульных GO/NOGO-задач представлены на рис. 4.6. Стимулы разделены на две категории: значащие и незначащие65. Можно видеть, что эти стимулы вызывают синхронизацию тета-подобной активности в лобных и теменных областях. Эти тета-подобные изменения отличаются не только в отношении пространственного распределения (лобная область в сравнении с теменной), но и в отношении временной динамики, внимания и характера привыкания к повторным пробам.
Fz
Значимый
Незначнмын
Тета « 5-7 Гц
200 400 600 800 ms
Рисунок 4.6. Два типа тетаподобной активности (волновой анализ)
Представлены усредненные вейвлет-преобразования ЭЭГ-реакций при выполнении двухстимульной GO/NOGO-задачи. Значащие (животное на первом месте пары) и незначащие (растение на первом месте) стимулы вызывают тета подобную активность в лобных и теменных областях. Эти вызванные ритмы имеют различный временный паттерн: теменная тета-активность отличается более короткой латентностью: 350 мс по сравнению с 500 медля лобного тета-ритма. Лобная тета-активность критически зависит от поведенческого значения стимула, в то время как теменная тета-активность не зависит от значения стимула.
Действительно, во-первых, пик латентности ответов в теменных областях меньше, чем в лобных областях. Во-вторых, лобная тета-синхронизация в сравнении с теменной тета-синхронизацией выше на значимый стимул по сравнению с ответом на незначимый. И, в-третьих, лобный тета-ритм демонстрировал привыкание к повторяющимся пробам, в то время как теменной тета-ритм этого не показал (рис. 4.6).
Два типа тета-подобной активности могут быть выделены простым вычитанием спектров, измеренных в состояниях с открытыми глазами, из спектров, измеренных в условиях решения задачи (в качестве примера может быть выбрано математическое задание). Результаты вычитания для группы здоровых 13—14-лет-них испытуемых представлены на рис. 4.7. Обратите внимание, что выполнение задания связано с большей мощностью ЭЭГ в тета-диапазоне (4—8 Гц) в лобной области Fz и с большей мощностью ЭЭГ в диапазоне частоты 5,5—9,5 Гц в темен
65 Значащие стимулы — изображения животных на первом месте в парах стимулов. После предъявления этих стимулов испытуемые должны были подготовиться к следующему предъявлению стимула и нажимать кнопку, если второй стимул соответствовал первому. Незначимыми являлись изображения растений на первом месте в пробе. После предъявления этого стимула испытуемые могли расслабиться и проигнорировать всю пробу.
Часть 1. Ритмы ЭЭГ
113
ной области Pz66. Также обращает на себя внимание подавление мощности ЭЭГ в сенсомоторной коре из-за постоянных нажатий на кнопку, которые, как мы уже знаем, вызывают подавление мю-ритма в сенсомоторной коре.
Задачи, в которых среднелобный тета-ритм является наиболее распространенным, — те, в которых испытуемый находится как в состоянии концентрации, так и расслабления. В примерах, представленных выше, были двухстимульный GO/ NOGO-тест и математическое задание. Другие примеры заданий, связанных с индукцией среднелобного тета-ритма, — это игры, такие как тетрис, задания на во-
Фронтальный срединный тетя-ритм
Теменной альфа-ритм
Мю-ритм
Рисунок 4.7. Два типа тетаподобной активности (спектральный анализ)
А. Результат вычитания спектров мощности, вычисленных для состояния с открытыми глазами, из спектров, полученных во время решения математической задачи для группы здоровых 13-14-летних детей. Б. Топограммы распределения разностных спектров мощности для различных частот.
ждение и др. В одном исследовании (Laukka и др., 1995) испытуемые были заняты в задаче на вождение на тренажере, в которой они должны были научиться ориентироваться и двигаться через ряд улиц, предъявляемых в виде анимации на компьютерном экране. В этом исследовании было выяснено, что процент среднелобного тета-ритма увеличился, по мере того как испытуемые учились лучше справляться с поставленной задачей и делали больше правильных решений.
В. Проявление в гипнозе
Состояние, которое также связывают с появлением среднелобного тета-ритма, — это гипноз. Чтобы быть загипнотизированными, люди должны со
•• Этот тип ритмической активности занимает диапазон альфа- и тета-ритмов. Именно поэтому мы называем его альфа-тета-ритм.
114 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
средоточиться на себе и на человеке, который их гипнотизирует. Если они отвлечены, состояние транса будет разрушено. Высокогипнабельные люди показывают достоверно больше тета-ритма до и во время гипноза, чем низко-гипнабельные (Crawford, 1994). Другое уникальное человеческое состояние, связывающее тета-активность с вниманием, — это медитация. Есть данные о регистрации ЭЭГ во время дзен-медитаций. По мере того, как практикующие дзэн-медитации люди достигали все более и более глубоких медитативных состояний или трансов, типичные образцы активности ЭЭГ, начинаясь с альфа-активности (8—10 Гц), постепенно сдвигались в диапазон тета-активности ЭЭГ
IV. Функциональное значение
А. Взаимосвязь между двумя типами тета-ответов человека и двумя типами тета-ответов у животных
Детальный анализ wavelet-преобразования нейронных ответов при выполнении различных заданий (полученных из нормативной HBI базы данных) показывает, что есть по меньшей мере два типа тета-подобной активности. Первый — среднелобный тета-ритм. Он проявляется в коротких вспышках в пределах диапазона 5—8 Гц. Этот ритм становится более выраженным с увеличением сложности задачи, оказывается выше при восприятии значащих стимулов и снижается, если тот же самый стимул представлен несколько раз. Второй — теменной альфа-тета-ритм может быть трудно различим в спектрах, вычисленных для состояний с открытыми или закрытыми глазами, и появляется только с возникновением мыслительной задачи. Теменной альфа-тета-ритм может появляться даже в виде более коротких вспышек по сравнению со среднелобным тета. В среднем этот ритм более низкой амплитуды, чем среднелобный тета, он не зависит от значения стимула и не подвержен привыканию к повторяющимся стимулам. Этот ритм не так давно был выявлен и еще не исследован подробно.
Механизмы генерации этих ритмов неизвестны. Их взаимоотношения с гиппокампальными тета-ритмами не были доказаны. Однако есть некоторые аналогии между этими человеческими ритмами и двумя типами тета-активности, обнаруженной в гиппокампальной системе в исследованиях на животных, упоминание о которых не может быть опущено в этом контексте. В исследованиях на животных два типа гиппокампальных тета-ритмов отличаются друг от друга на основе фармакологической чувствительности, частоты и поведения. Первый тип не блокируется ацетилхолиновыми антагонистами, но чувствителен к анастезирующим агентам типа уретана, этилового эфира и алкоголя, имеет частоту 6—10 Гц и возникает при произвольных движениях. Он не связан с привыканием (не адаптируется). В человеческой ЭЭГ-записи этот высокочастотный тета может быть соотнесен
Часть 1. Ритмы ЭЭГ
115
с теменным тета-ритмом. Он вызывается короткими вспышками, не угнетается, может быть связан с сенсорным кодированием.
Второй тип тета у животных угнетается антагонистами мускариновых (холинергических) рецепторов типа атропина. Пиковая частота этого ритма обычно 4—6 Гц (ниже, чем у первого типа). Этот тип тета-колебаний возникает при передвижениях, неподвижности и уретановой анастезии. Второй тип тета-ритма по сравнению с первым типом тренируется у интактного животного, появляется в ответ на сенсорные стимулы, когда животное находится в возбужденном состоянии. Спонтанное возникновение тета-ритма второго типа у крыс исключительно редко и имеет очень короткую продолжительность. Второй тип тета-ритма может возникать при тренировках и, вероятно, связан с научением. Возникновение второго типа тета-ритма опосредовано наличием холинергических входов в гиппокампальную формацию от центральной перегородки67. В ЭЭГ человека при регистрации со скальпа аналог этого ритма может быть соотнесен со среднелобным тета-ритмом, который сильно тренируется и, вероятно, связан с эпизодической памятью.
Среднелобный тета-ритм в состоянии спокойного бодрствования может быть обнаружен в сырой ЭЭГ только в небольшой группе нормальной выборки испытуемых и соотнесен с генетически детерминированными поведенческими чертами: низким уровнем тревожности и нейротизации, высоким уровнем экстраверсии. Есть также исследования, рассматривающие среднелобный тета-ритм как признак освобождения от чувства беспокойства68. Мы теперь знаем по крайней мере три различных способа увеличения среднелобной тета-активности — медитация, биообратная связь, бинауральная техника69.
V. Аномальные тета-ритмы
Клиническое значение нормального среднелобного тета-ритма и теменного альфа-тета-ритмов ограничено по нескольким причинам. Во-первых, эти ритмы имеют короткую длительность и появляются только в состояниях выполнения каких-либо заданий в зависимости от трудности задачи. Во-вторых, даже при выполнении заданий они могут быть замечены в спектрах ЭЭГ в форме пиков только у части здорового населения. В-третьих, трудность регистрации этих ритмов создает проблемы для систематических исследований их нейронных механизмов.
87 Наиболее интригующая часть этой истории исходит от наблюдений, что те же самые пирамидные нейроны могут участвовать в генерации первого и второго типа тета-колебаний, т.е. те же самые анатомические структуры могут быть вовлечены в производство обоих ритмов.
88 Например, в исследованиях японской группы ЭЭГ была зарегистрирована на пациентах с общим нарушением уровня тревожности, уровень определялся шкалой Гамильтона (Hamilton Rating Scale for Anxiety) и опросником Спильергера (Spielberger’s State-Trait Anxiety Inventory) один раз в неделю. Согласно их данным, появление среднелобной тета-активности могло быть сопряжено с улучшением состояния пациентов и снижением уровня тревожности (Suetsugi и др., 2000).
88 Бинауральная стимуляция — важный элемент запатентованной акустической системы, разработанной Робертом А. Монро. В этом методе индивидуумы выполняют задания, включающие дыхательные упражнения, контролируемое расслабление, визуализацию и акустическую стимуляцию. Последнее задание базируется на способности мозга обнаруживать различия в частотах слуховых тонов, предъявленных в левое и правое уши. Восприятие бинаурального сигнала происходит на частоте различия между двумя слуховыми входами. Бинауральные сигналы можно легко различать в низких частотах (< 30 Гц), которые являются характерными для спектра ЭЭГ. Восприятие бинауральных тонов связано с последующим частотным ответом ЭЭГ. Это явление описано Atwater (1997).
116 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
А. Подтип среднелобного тета-ритма при синдроме нарушения внимания и гиперактивности
На текущей стадии исследований мы имеем представления о том, что существует условие, связанное с отклонениями от нормы при генерации среднелобного тета-ритма. Это редкий подтип — часть населения с синдромом нарушения внимания и гиперактивности. Особыми признаками такого населения являются сложные социальные отношения и неспособность корректировать свое поведение. С электрофизиологической точки зрения этот СНВГ-подтип характеризуется необычной выраженностью среднелобного тета-ритма. Присутствие ритма выражено в чрезвычайно высоком и остром пике на спектрах ЭЭГ в Fz-отведении в диапазоне частот от 5,5 до 8 Гц и чрезвычайно низкой синхронизацией тета-ритма в ответ на значащие стимулы при выполнении двухстимульной GO/NOGO и математических задач.
Б. Тета-ритмы в нелобных областях
Появление тета-ритмов в других областях кроме отведения Fz (и до некоторой степени в Pz) следует считать ненормальным. В таких случаях необходимо сравнение с нормативной базой данных для того, чтобы определить уровень достоверности наблюдаемого отклонения от нормы (рис. 4.8). ЭЭГ была зарегистрирована на пациенте через нескольких дней после закрытой черепно-мозговой травмы. Можно видеть высокий по амплитуде тета-ритм в F3-, Р4-отведениях, мощность которых значительно (р < 0,001) отличается от нормативных данных. Обратите внимание также на то, что отклонения в диапазоне тета-ритма сопровождаются чрезвычайно низкой мощностью бета-активности, широко распределенной по коре.
VI. Заключение
Тета-ритм — ритм с частотой больше 4 и меньше 8 Гц. Японские ученые впервые индуцировали проявление тета-активности ЭЭГ, предложив испытуемым умственную задачу на непрерывное арифметическое сложение. Эта тета-активность проявилась в виде последовательности ритмичных волн с частотой 6—7 Гц с максимальной амплитудой вокруг лобной средней линии и была названа среднелобным тета-ритмом. Среднелобный тета-ритм часто связывается с гиппокампальными тета-ритмами. Существуют гипотезы, согласно которым гиппокампальные тета-колебания вовлечены в кодирование следов памяти и извлечение из памяти. Регистрация электрической активности в гиппокампе человека доступна только в редких случаях сте-реотактических операций у пациентов с эпилепсией со вживленными для диагностических и лечебных целей электродами. Среднелобный тета-ритм указывает на индивидуальные различия и связан с некоторыми чертами индивидуальности: выраженность среднелобного тета-ритма отрицательно
Часть 1. Ритмы ЭЭГ
117
Рисунок 4.8. Аномальный тета-ритм
А. Фрагмент ЭЭГ, зарегистрированной у 32-летнего пациента через 5 дней после внутричерепной мозговой травмы. Б. Спектр мощности сигнала в отведении F3 и отклонение от нормы в отведении F3. С. Карты отклонений от нормы в тета- и бета-частотах и изображения sLORETA эквивалентных генераторов тета-ритма.
коррелирует с уровнем тревожности и невротизма личности, положительно — с уровнем экстраверсии. Среднелобный тета-ритм также коррелирует с изменениями уровня тревожности, вызванными успокаивающими лекарственными препаратами.
118
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Глава 5.
Пароксизмальная активность
Рисунок 5.1. Временные и амплитудные параметры формы волнового сигнала, используемые для автоматического обнаружения спайка Объяснения см. в тексте.
I. Спайки
При визуальном просмотре ЭЭГ аномальные паттерны в основном разделяются на две группы: медленные волны70 и спайки. Спайк — пароксизмальный (т.е. внезапно появляющийся) электрический разряд, который превышает фоновую активность и напоминает длинный шип. Напомним, что в нормальном мозгу процессы коркового возбуждения и торможения должны быть сбалансированы. В этом случае кора во время бодрствования генерирует нормальную регулярную электрическую активность, такую как альфа-ритмы, бета-ритмы и среднелобный тета-ритм, которые регистрируются со скальпа благодаря синхронной активации миллионов нейронов. Если баланс «торможение — возбуждение» нарушается так, что процесс возбуждения начинает превалировать над торможением и нейронные сети гиперсинхронизируются, кора начинает генерировать аномальную электрическую активность, названную пароксизмами71. Эти отклонения в большинстве случаев могут быть зарегистрированы со скальпа методом обычной ЭЭГ в виде определенных электрографиче
ских образцов. Есть несколько типов пароксизмальной активности, наиболее распространенные из которых — спайки, острые волны, комплексы «спайк — медленная волна». В экстремальных случаях корковая гиперсинхронизация в эпилептогенном очаге может привести к судорожному припадку — комплексу стереотипных и неконтролируемых судорожных изменений поведения, таких как подергивания и конвульсии, сопровождающиеся проходящей потерей сознания. Тяжесть припадков может быть разной: некоторые пациенты могут трястись и падать, у других припадок может быть замечен только самыми внимательными членами семейства.
70 Аномальные медленные волны были описаны в предыдущих главах при описании дельта-, тета- и альфа-ритмов.
71 Пароксизмы в ЭЭГ — это аномальная электрическая активность чрезмерной синхронизации.
Часть 1. Ритмы ЭЭГ
119
А. Пространственно-временные характеристики
Есть три характеристики, по которым можно выделить спайк или острую волну в ЭЭГ: пароксизмальный характер, высокая степень заостренности сигнала и короткая длительность (рис. 5.1)72.
Пароксизмальный характер спайка частично связан с его амплитудой: спайк выделяется из фоновой активности. Амплитуда спайка, в свою очередь, зависит от нескольких факторов: 1) положения фокуса эпилептиформной активности — чем он глубже в мозгу, тем меньше амплитуда спайка; 2) размера эпилептогенного фокуса: спайки с более низкой амплитудой генерируются более мелкими фокусами; 3) ориентации электрического диполя, соответствующего спайку: тангенциальные диполи производят спайки меньших амплитуд по сравнению с радиально ориентированными диполями. Амплитудные параметры, которые мы используем в нормативной HBI базе данных для автоматического обнаружения спайков, представлены на рис. 5.1. Степень остроты спайка может быть оценена как вторая производная сигнала ЭЭГ. Вторая производная, вычисленная в максимуме (или минимуме) для спайка, устанавливается больше чем 0,3 мкВ/мс2.
Другой параметр заостренности сигнала — наклон, определяемый как максимальная амплитуда первой временной производной. Этот параметр для спайка устанавливался большим, чем 1 мкВ/мс. По определению, длительность спайка должна быть меньше чем 70 мс, в то время как длительность острой волны — обычно от 70 до 200 мс, длительности некоторых временных параметров спайка представлены на рис. 5.1. Например, продолжительность интервала между двумя последовательными максимумами или минимумами — меньше 150 мс (но больше 50 мс).
Наш опыт показывает, что выбранные параметры позволяют надежно выявлять большинство спайков, которые могут быть визуально обнаружены опытным электроэнцефалографистом.
Другой тип пароксизмальной активности — комплексы спайк-волна. Это определение относится к спайку, который сопровождается явной волной. Если спайк связан с синхронным возбуждением многих нейронов, следующая за ним волна связана с тормозными постсинаптическими потенциалами, которые следуют за возбуждением. Здесь надо подчеркнуть, что появление волны после спайка не переводит эту пароксизмальную активность в другую категорию. Оба паттерна принадлежат к пароксизмальным образцам и отражают синхронную активность эпилептогенного фокуса.
Пространственное положение эпилептогенного фокуса в коре точно определяет характер припадков. Локализация эпилептогенных фокусов у разных пациентов может быть разной. Эпилептогенные фокусы могут быть лобными (т.е. премоторными, срединными или орби
72 Параметры взяты из статьи Ktonas RY. Автоматическое обнаружение спайка и острой волны // Методы анализа электрических и магнитных мозговых сигналов. Руководство ЭЭГ (revised series, Vol 1) / A.S. Gevins and A. Remond (Eds). Elsevier Science Publishefs B.V.,1987 — R 211-24L
120
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
тальными), височными (передними, медиобазальными, задними), теменными, затылочными. В некоторых случаях эпилептогенный фокус располагается в гиппокампе или связанных с ним структурах. В таких случаях поверхностное обнаружение спайков затруднено из-за низкой амплитуды спайков, регистрируемых от скальпа.
Б. Автоматическое обнаружение спайков
Пример автоматического обнаружения спайков представлен на рис. 5.2. ЭЭГ была зарегистрирована у пациента с эпилепсией в возрасте 7 лет. Так как эти спайки были обнаружены в период между припадками, они называются промежуточными (interictal) спайками73. Преимущество автоматического обнаружения спайков состоит в том, что процедура позволяет нам усреднить обнаруженные спайки и получить надежную картину их распределения, пространствен
02Av ~
А
Рисунок 5.2. Автоматическое обнаружение спайков
А. Фрагмент электроэнцефалограммы, зарегистрированной у 7-летнего пациента, страдающего эпилепсией. Спайк, отмеченный вертикальной линией и полосой с номером канала максимальной амплитуды (внизу), был обнаружен компьютерным алгоритмом, реализованном в программном обеспечении института мозга человека, на основе амплитудно-временных параметров, представленных на рис. 5.1. Б. Усредненный спайк, его двухмерная топограм-ма, оценка эквивалентного диполя и sLORETA-изображение генераторов спайка. Полосы в основании графиков ВП показывают уровни значимости отклонения потенциала фона от среднего фонового.
ного распределение генераторов при помощи sLORETA и локализации эквивалентного диполя в пределах мозга. Результаты усреднения спайков представлены на рис. 5.274. Как видно на рис. 5.2, спайки с максимумом амплитуды в отведении F8 имеют отрицательные значе-
73 Параметры обнаружения спайков соответствуют всем выше перечисленным параметрам по амплитуде и временным характеристикам. Например, Да = 130 мкВ, Т2 = 40 мс, программа автоматически обнаруживает спайки, которые могут быть выделены визуально.
74 Согласно алгоритму, представленному выше, программы автоматически обнаруживает спайки, бегущие отдельно через все каналы. Когда спайк в канале усреднен, его потенциальное распределение по голове во время пика приближено единственным(отдельным) диполем; squired-различие между реальным потенциалом и его приближением усреднено по всем каналам и разделено на полную энергию. Вычисленный параметр называют относительной остаточной энергией, которая определяет, как хорошо спайк может быть приближен диполем. В нашей лаборатории мы устанавливаем этот параметр на 0,2.
Часть 1. Ритмы ЭЭГ
121
Рисунок 5.3. Спайки при внутричерепной регистрации
Локальные полевые потенциалы, зарегистрированные в гиппокампе и смежных структурах у пациента, страдающего эпилепсией, которому для диагностических целей были имплантированы электроды. Обратите внимание на два различных типа спайков, произведенных различными структурами. Также обратите внимание, что на записи со скальповых электродов (для сравнения представлен электрод ТЗ) не выявляется никаких значимых спайков.
ния. Эта негативность (отрицательность) — отражение сильной деполяризации, которая имеет место в эпилептогенном фокусе. Однако так как диполь ориентирован тангенциально к поверхности, одновременно может быть обнаружен небольшой положительный пик в отведении Fz.
В. Интракраниально регистрируемые спайки
Необходимо подчеркнуть, что нет прямых отношений между эпилепсией и пароксизмальными образцами. Иногда спайки могут не обнаруживаться при скальповой регистрации и проявляются только на записи с интракраниальных электродов (рис. 5.3).
Иногда, наоборот, на спонтанной ЭЭГ выявляются спайки, однако эта эпилептиформная активность не выражается в эпилептических припадках, и человек может вообще никогда их не иметь. Последний случай — это ситуация, когда мы должны быть очень осторожны. Дело в том, что наличие эпилептогенного фокуса в коре может не вызывать заметных приступов и припадков 75, но гиперсинхронизация в фокусе может нарушать нормальный поток информации в коре и может быть связана с мозговыми дисфункциями, специфическими для отдельных локализаций (рис. 5.4).
Или приступы являются настолько легкими, что могут оставаться незаметными для самого пациента и окружающих.
122
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
SOOuV
Рисунок 5.4. Спайки в ЭЭГ 6-летнего пациента без эпилептических припадков
Девочка имела трудности с речью. Слева направо — усредненный спайк (А), двухмерная топограмма (Б), трехмерное sUORETA-изображение генератора спайка (В), оценка эквивалентного диполя (Г). Д. Изображение генератора спайков на срезах мозга (sLORETA).
II. Нейронные механизмы
А. Недостаточное торможение
Наши знания относительно механизмов эпилепсии в основном получены из экспериментов на животных. «Межприступные» спайки отражают синхронную деятельность многих сотен тысяч (или миллионов) нейронов. Избыток синхронизации может достигаться изменением баланса между возбуждением и торможением возбуждения. Это может происходить в результате увеличения возбуждения или уменьшения торможения или в результате обоих этих процессов. Внутриклеточные регистрации в корковых нейронах экспериментального эпилептического фокуса показали, что начальная компонента спайка сопровождается так называемым «пароксизмальным» сдвигом уровня деполяризации. Изменение уровня деполяризации создается возбуждающими постсинаптическими потенциалами, которые увеличиваются и последовательно усиливаются собственными потенциалзависимыми мембранными ответами. Увеличение может быть результатом разнообразных механизмов. Один из них — уменьшение внутрикоркового торможения. Это было недавно обнаружено при фокальной корковой дисплазии76, которая с медицинской точки зрения является распространенной причиной тяжелой эпилепсии (Calcagnotto и др., 2005).
Б. Биообратные связи
Наличие повторных (рекуррентных) неспровоцируемых приступов является признаком эпилепсии как общего хронического неврологи
76 Этот термин относится к состоянию, при котором кора в развивающемся зародыше не формируется должным образом. Дисплазия по-гречески означает «плохое формирование».
Часть 1. Ритмы ЭЭГ
123
ческого заболевания77. Эпилепсия — первое нарушение, для которого применялась нейротерапия. Было показано, что при некоторых типах эпилепсии, когда лечение не помогало или имело нежелательные побочные действия, БОС-тренинги 12—14 Гц активности сенсомоторной полосы достоверно уменьшали количество судорожных припадков. Барри Стерман из Калифорнийского университета в Лос-Анджелесе был первым, кто начал вводить этот метод в клиническую практику.
В. Эпилептология
Существует много различных типов эпилепсии, каждый из которых отличается собственной уникальной комбинацией форм приступов, возраста возникновения, ЭЭГ-коррелят, методов лечения и прогнозов. Эпилепсия как предмет отдельной отрасли медицины — эпилептологии — не может быть подробно описана в этой книге. Здесь мы только хотим подчеркнуть, что регистрация ЭЭГ в случаях подозрений на эпилепсию сопровождается провоцирующими приступы процедурами, например, гипервентиляцией78, фотостимуляцией и депривацией сна, пока судорожная активность не станет обнаруживаться визуально при просмотре ЭЭГ квалифицированными клиницистами. Метод же количественной ЭЭГ, который главным образом сосредоточен на оценке количественных параметров ЭЭГ в различных условиях, таких как спокойное бодрствование с открытыми и закрытыми глазами или во время решения различных задач, которые специально разработаны для исследования сенсорных и высших когнитивных функций мозга, таких как контроль и принятие решений, эмоции и память. Таким образом, анализ QEEG, главным образом, используется для исследования таких мозговых нарушений, как СНВГ, депрессии, шизофрения и др. Однако наш опыт работы с QEEG показывает, что некоторые проблемы поведения (например, СНВГ) в редких случаях могут быть последствием наличия в коре эпилептогенного фокуса79.
III. Заключение
В нормальном мозгу процессы коркового возбуждения и торможения хорошо сбалансированы. Кора со сбалансированными процессами торможения и возбуждения в состоянии спокойного бодрствования генерирует нормальные регулярные электрические колебания: альфа-ритмы,
77 Как можно видеть из этого определения, просто наличие судорог не является диагностическим признаком эпилепсии, т.к. судороги могут быть вызваны травматическими событиями: травмой, недостатком кислорода, опухолями, отравлением ядовитыми химикатами или высокой температурой. В прошлом эпилепсию часто связывали с религиозными переживаниями или даже с демоническим воздействием. В настоящее время эпилептические приступы связывают с чрезмерной синхронной активностью нейронов мозга. Приблизительно 0,5-1 % населения страдает эпилепсией. Эпилепсия обычно контролируется медикаментозно, в трудных случаях в качестве альтернативного способа лечения может применяться хирургическое вмешательство.
78 Например, в недавнем исследовании с записью ЭЭГ и видеомониторингом 97 пациентов у 24 пациентов были обнаружены клинические приступы, связанные с изменениями ЭЭГ во время гипервентиляции, главным образом в первые 4 минуты (Guaranha и др., 2005).
79 Если судорожные паттерны наблюдаются в ЭЭГ в любом из условий (с открытыми/закрытыми глазами или во время решения задач), пациенту рекомендуют обратиться к эпилептологу для более тщательных исследований.
124 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
бета-ритмы и среднелобный тета-ритм. Если указанный баланс нарушен так, что возбуждение превышает торможение, кора начинает производить аномальные паттерны электрической активности, которые называются пароксизмами. В большинстве случаев они могут быть зарегистрированы с помощью обычной скальповой ЭЭГ в форме определенных электрографических паттернов. Есть несколько типов эпилептиформной активности, наиболее распространенные из которых — спайки, острые волны, комплексы «спайк — медленная волна». Спайки обычно генерируются локальной корковой областью, называемой фокусом. С использованием таких современных методов, как электромагнитная томография, LORETA или анализ диполей, эпилептогенные фокусы могут быть локализованы в коре с хорошей точностью. С 1950-х годов долгие годы обнаружение спайков визуальным просмотром ЭЭГ оставалось единственным методом. За прошлые 10—20 лет были разработаны весьма надежные методы для автоматического обнаружения спайков. В настоящее время большинство современных QEEG-систем имеет дополнительные инструменты для автоматического обнаружения спайков, которые помогают практикам обнаружить и проанализировать пароксизмальную активность в мозгу.
Часть 1. Ритмы ЭЭГ
125
Глава 6.
Эндофенотипы количественной ЭЭГ
Во «Введении» книги мы представили концепцию эндофенотипов как биологических маркеров заболеваний мозга. Эндофенотип должен отвечать следующим требованиям: 1) быть устойчивым и восстанавливаемым в интервалах времени, в течение которых поведенческие паттерны, связанные с определенным состоянием мозга, остаются неизменными; 2) отражать функцию некоторой мозговой системы, которая определенным способом управляет человеческим поведением; 3) быть наследуемым, то есть у гомозиготных близнецов он должен быть весьма похожим.
В этой главе мы собираемся показать, что спектры, или, лучше сказать амплитуды, фоновой ЭЭГ в некоторых диапазонах частот отвечают этим требованиям и, следовательно, могут рассматриваться как эндофенотипы.
I. Воспроизводимость результатов от теста к тесту
Основные ЭЭГ-колебания (такие как альфа-ритмы и среднелобный тета-ритм) периодически уменьшаются и увеличиваются с течением времени. Степень изменчивости основных колебаний зависит от диапазона частот и состояния (глаза открытые/закрытые, решение задач) человека. Например, альфа-веретена в затылочных областях изменяются с периодами в несколько секунд, вспышки среднелобного тета-ритма появляются с периодами в несколько декасекунд. Однако при усреднении достаточно длительных интервалов записи ЭЭГ результирующие спектры становятся весьма устойчивыми характеристиками мозга. Возникает вопрос, какая минимальная длина записи ЭЭГ необходима для спектрального анализа80, для стабильной и надежной оценки.
Чтобы создать у читателя ощущение нестационарности (изменчивости) ЭЭГ, на рис. 6.1 слева сверху представлена необработанная ЭЭГ, зарегистрированная у здорового субъекта в отведении 02 в условиях с закрытыми глазами. Спектры ЭЭГ, вычисленные для последовательных временных интервалов в 4, 40 и 120 секунд, представлены ниже. Можно видеть, что спектры для 4-секундных отрезков ЭЭГ сильно отличаются, тогда как для 120-секундных отрезков ЭЭГ становятся почти идентичными.
80 Не путайте длину интервала записи ЭЭГ, взятого для анализа, с эпохой анализа для быстрого преобразования Фурье. Эпоха определяет частотное разрешение спектров: чем длиннее эпоха анализа, тем выше частотное разрешение. Длина интервала анализа (включающая некоторое количество эпох анализа) определяет надежность оценки спектров.
126
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Рисунок 6.1. Изменчивость ЭЭГ
А. Затылочные альфа-ритмы уменьшаются и увеличиваются в пределах интервала в одну секунду. Б. Спектры Фурье, вычисленные для 4-секундных последовательных эпох анализа, в течение 40- и 120- секундных периодов анализа. Весь период анализа был разделен на 4-секундные эпохи анализа. Преобразование Фурье (слева внизу) применялось отдельно к каждой эпохе анализа, затем результирующие спектры были усреднены. Можно заметить, что изменчивость спектров снижается с увеличением длины анализируемого интервала. В. Зависимость вариабельности спектров ЭЭГ от длины анализируемого интервала. Значения Y — коэффициент изменчивости (КИ) для мощности в альфа-диапазоне частот при увеличении размера анализируемого интервала (ось X) от 4 до 400 с. Для каждой индивидуальной записи ЭЭГ была построена такая кривая зависимости вычислением коэффициентов изменчивости для 100 случайно отобранных длин анализируемых интервалов, далее значения были усреднены по группе. Адаптировано из Maltez et al. (2004).
Существуют работы, посвященные изучению изменчивости различных спектральных параметров ЭЭГ, использующие вариабельность, коэффициенты вариативности (КВ)81 и воспроизводимости, измеряемой как коэффициент корреляции между последовательными измерениями. Группой ученых из Швеции (Karolinska Hospital in Stockholm) недавно было выполнено подобное исследование. У здоровых испытуемых регистрировалась ЭЭГ в течение 15 минут, при этом испытуемые были проинструктированы открывать глаза на 5 секунд каждые 60 секунд, а затем снова закрывать их82. Для анализа методом быстрого преобразования Фурье была выбрана 4-секундная эпоха. Коэффициент вариативности изменялся обратно пропорционально квадратному корню времени (справа на рис. 6.1). По данным этого исследования, альфа-ритм имел самый высокий коэффициент вариативности, а тета-ритм — самый низкий.
На основе графика, приведенного на рис. 6.1, мы можем оценить, что для того, чтобы коэффициент вариативности был меньше 0,05, мы долж-
81 Коэффициент вариативности определяется как стандартное отклонение, разделенное на среднее.
83 Воспроизводимость результатов в записи оценивалась при разделении каждой записи на два набора данных. При сравнении первой половины данных со второй половиной рассчитывался коэффициент корреляции Пирсона. Он был высоким: в пределах от 0,93 до 0,98 для мощности альфа-, бета- и тета-активности. При сравнении нечетных периодов регистрации ЭЭГ с закрытыми глазами с четными периодами регистрации ЭЭГ с закрытыми глазами коэффициенты корреляции Пирсона были 0,97-0,99 для мощности всех частотных диапазонов. Также было исследовано влияние размера анализируемого интервала ЭЭГ на вариативность спектров при помощи вычисления коэффициентов вариативности при изменении длительности анализируемого интервала (числа анализируемых 4-х секундных эпох), выбираемого случайно для каждой записи, от минимального в 4 секунды (1 эпоха) до максимального в 400 секунд (100 эпох). Для каждой длины анализируемого интервала были рассчитаны коэффициенты вариативности с усреднением ста выборов, полученные значения затем были усреднены между людьми.
Рисунок 6.2. Длительность записи ЭЭГ в три минуты обеспечивает достоверные и воспроизводимые спектры ЭЭГ
Спектры мощности и топограммы ЭЭГ, зарегистрированных у одного субъекта в течение 3 минут, с интервалом между регистрациями 7 дней.
1. Ритмы ЭЭГ_____________________________________________127
Рисунок 6.3. Наследуемость спектров ЭЭГ
А. Спектры мощности в состоянии с открытыми глазами, вычисленные для двух пар близнецов: монозиготных близнецов (М3) и дизиготных близнецов (ДЗ). Б. Разностные спектры, полученные вычитанием усредненных спектров нормативной группы из индивидуальных спектров. Обратите внимание, что трое из 4 близнецов имеют отклонение от нормы в альфа-диапазоне ЭЭГ в отведении ТЗ. В. Схематичное представление рефлектограмм для двух групп близнецов, где Pl (Р2) — спектры мощности данного диапазона частот для первого (второго) близнеца в паре. Г. Наследуемость, вычисленная согласно формуле, приведенной в тексте, как функция частоты. Адаптировано из Smit etal. (2005).
Часть 1. Ритмы ЭЭГ
129
ны иметь запись ЭЭГ длиной по крайней мере 3 минуты. Пример изменчивости спектров, проанализированных на 3-минутной записи ЭЭГ, показан на рис. 6.2. Как видим, спектры, полученные в записи ЭЭГ, сделанной во время состояния с открытыми глазами в течение 3 минут с интервалом между регистрациями в 7 дней, очень похожи и могут рассматриваться как надежная и стабильная оценка функционального состояния мозга.
II. Отражение функционирования мозговых систем
ЭЭГ — сложная комбинация ритмов. Например, спектры ЭЭГ пациента, представленные на рис. 6.2, включают лобный тета-ритм, затылочный альфа-ритм, задневисочную низкочастотную бета-активность и центральную высокочастотную бета-активность. При сравнении с нормативной базой данных лобная тета-активность и задневисочная низкочастотная бета-активность оказались вне диапазона нормы.
Различные ритмы ЭЭГ отражают различные механизмы. Альфа-ритмы отражают состояние таламокорковых путей. Среднелобный тета-ритм отражает функционирование лимбической системы. Бета-ритмы более локальны и отражают состояние локальных корковых областей. В предыдущих главах мы пытались показать взаимосвязь между ритмами ЭЭГ и мозговыми системами и, соответственно, мозговыми дисфункциями. Таким образом, определение аномальной ритмической активности ЭЭГ и связь этих отклонений с различными мозговыми системами соответствует второму критерию эндофенотипов как биологических маркеров болезни.
III. Наследуемость
Есть надежные экспериментальное данные, указывающие, что спектральные характеристики ЭЭГ наследуются. Меры наследуемости спектров ЭЭГ схематично представлены на рис. 6.3. Кривые слева взяты из базы данных Института мозга человека, в то время как схематическое представление наследуемости спектров ЭЭГ основывается на результатах недавнего исследования группы (Smit и др., 2005) из Vrije Universiteit (Амстердам, Нидерланды)83. Результаты показали, что в большинстве частотных спектров индивидуальные различия взрослой ЭЭГ в значительной степени определяются генетическими факторами.
83 Это было близнецовое исследование, в котором сравнивалось внутрипарное сходство генетически идентичных (монозиготных близнецов) и генетически неидентичных близнецов (дизиготных близнецов). Если сходство М3 (г) для мощности ЭЭГ выше, чем сходство ДЗ, это свидетельствует о генетических влияниях на ЭЭГ. Простая формула вычисления вклада генетических влияний (наследуемости — h2) на общую изменчивость: h2 = 2 (гМЗ - гДЗ), где гМЗ и гДЗ определяют значение внутрипарного сходства для близнецов ДЗ и МЭ. ЭЭГ регистрировалась от 19 отведений в состоянии с закрытыми глазами у 142 монозиготных и 167 дизиготных пар близнецов и их потомстве, в общей сложности у 760 субъектов.
130 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Эндофенотип становится важной концепцией в исследовании сложных психических болезней. Термин был введен в 1966 году и применялся в психиатрии Gottesman и Shileds в 1972 (Gottesman, Gould, 2003). Он постепенно заменяет другие семантически подобные термины, такие как «биологический маркер», «маркер уязвимости», «субклиническая характеристика» и даже «переходный фенотип»84. Например, согласно поиску в MEDLINE, термин использовался в 16 статьях за 30-летний период до 2000, но в 84 статьях только за один 2006 год. Понятие «эндофенотип» основывается на предположении, что более правильный психиатрический диагноз может быть сделан только на основании представлений о состоянии мозговых систем пациента. В ряде работ этот принцип реализуется, и исследователи использовали фМРТ для визуализации нейронных цепей, активных при таких заболеваниях, как депрессия, шизофрения и СНВГ В этой главе мы показали, что спектральные характеристики фоновой ЭЭГ могут также использоваться в качестве эндофенотипов.
IV. Заключение
В психиатрии эндофенотип обозначает измеримый индекс мозгового функционирования. Эндофенотип отвечает следующим требованиям:
1. Он должен быть устойчивым и восстанавливаемым в интервалах времени, в течение которых поведенческие паттерны, связанные с определенным состоянием мозга, остаются неизменными.
2. Он должен отражать функцию некоторой мозговой системы, которая определенным способом управляет человеческим поведением.
3. Он должен быть наследуемым, то есть у гомозиготных близнецов быть весьма похожим.
В этой главе мы показали, что спектры ЭЭГ отвечают этим требованиям.
* Эндофенотип может быть нейрофизиологическим, биохимическим, эндокринным, нейроанатомическим или нейропсихологическим по характеру. Эндофенотипы дают более простое указание на генетические механизмы, чем поведенческие характеристики.
Часть 1. Ритмы ЭЭГ
131
Глава 7.
QEEG во время сна
I. Анатомические основы
А. Мозговые структуры, задействованные в регуляции цикла сна и бодрствования
В течение дня мозг не остается тем же самым, что может быть продемонстрировано на примере сверхмедленных ритмов, изменения которых следуют за сменой дня и ночи, производимыми вращением Земли вокруг Солнца. Этот ритм называют циркадным ритмом (от латинского circa — вокруг и dies — ежедневно). Циркадный ритм грубо может быть разделен на две стадии: бодрствование и сон. В среднем мы проводим 30 % нашей жизни во сне. Продолжительность сна у людей имеет колоколообразное распределение со средней продолжительностью сна 7,0—7,9 ч. Поведенчески сон определяется четырьмя критериями: I) уменьшенная моторная активность; 2) уменьшенный ответ на внешние стимулы; 3) стереотипия положения; 4) легкая обратимость (в отличие от комы).
Во время Первой мировой войны мир был охвачен пандемией encephalitis lethargica — вирусной инфекции мозга. Инфекция у большинства индивидуумов была причиной глубокого и длительного состояния сонливости. Венский невропатолог барон Константин фон Экономо (Constantin von Economo) сообщил, что это состояние длительной сонливости происходит из-за повреждений зон заднего гипоталамуса и передних отделов среднего мозга. Он также сообщил, что у некоторой группы пациентов существовала противоположная проблема — длительное состояние бессонницы. Это было связано с повреждениями предоптической области и основания мозга. На основании этих наблюдений von Economo предположил, что область гипоталамуса около зрительной хиазмы содержит нейроны сна (sleep-promoting neurons), тогда как задний гипоталамус содержит нейроны бодрствования (neurons that promote wakefulness), повреждение которых приводит к нарколепсии85.
В последующие годы его наблюдения относительно возникающих сонных эффектов при повреждении заднего бокового гипоталамуса были воспроизведены при повреждениях мозга у обезьян, крыс и кошек. Однако основные нейронные цепи поддержания бодрствования были точно определены только в 1980—1990-х, а пути, ответственные за гипоталамическое регулирование сна, начали проясняться только в последние пять лет (рис. 7.1). Как показывают нейрофизиологические данные, вентролатеральное предоптическое ядро
85 Болезнь, при которой индивиды могут заснуть в несоответствующее (неподходящее) время.
132_______Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
(VPO) содержит ГАМКергические и Таланинергические нейроны, которые активны во время сна и тормозят моноаминергические группы клеток в восходящей активирующей системе (таких как ядра шва и голубого пятна) и, следовательно, необходимы для нормального сна. Задний латеральный гипоталамус (PLH) содержит нейроны, вырабатывающие орексин/гипокретин, необходимые
Рисунок 7.1. Мозговые ядра, вовлеченные в регулирова- ДЛЯ поддержания НОрмаЛЬНОГО бодр-ние цикла «бодрствование — сон» СТВОВаНИЯ, В ТО Время Как ИХ ПОВреЖ-
VL РО — вентролатеральное преоптическое ядро, PL Н — за-
дний латеральный гипоталамус, Raphe — ядра шва ствола ДеНИе Ведет К НарКОЛеПСИИ. Суще-мозга, LC — голубое пятно. СТВуеТ МОДеЛЬ, В КОТОРОЙ НвЙрОНЫ
бодрствования и сна тормозят друг друга (на рис. 7.1 не показана), обеспечивая стабильное бодрствование и сон.
II. ЭЭГ-корреляты сна
A. REM- и NREM-сон (парадоксальный и непарадоксальный сон)
В течение многих столетий сон расценивался как простая остановка поведенческой активности. Сегодня мы знаем, что сон — комплексное и высокоорганизованное общее состояние, которое, в свою очередь, может быть разделено на отдельные стадии. Во время сна корковые нейроны подвергаются медленным колебаниям мембранного потенциала, которые проявляются в ЭЭГ как медленноволновая активность < 4 Гц (slow wave activity — SWA). Количество медленноволновой активности регулируется гомеостатически, увеличиваясь после бодрствования и постепенно уменьшаясь во время сна. Предполагается, что гомеостаз медленноволновых ритмов может отражать изменения в синапсах, лежащие в основе клеточной потребности во сне.
У млекопитающих есть два типа сна: сон с быстрыми движениями глаз, быстрый сон (rapid eye movement — REM) и медленный сон (non-REM — NREM). Они определяются в терминах электрофизиологических признаков, которые обнаруживаются при совмещении методов электроэнцефалографии, электро-окулографии и электромиографии, совместное измерение которых называется полисомнографией. REM-сон (также известный как парадоксальный, активный или десинхронизованный сон) характеризуется следующими особенностями: активность ЭЭГ похожа на бодрствование и активацию (высокочастотная, низкоамплитудная, или десинхронизованная активность ЭЭГ); одиночные и множественные быстрые движения глаз в ЭОГ; и очень низкий мышечный тон (атония) в электромиограмме.
Часть 1. Ритмы ЭЭГ
133
Б. Стадии NREM-сна
NREM-сон можно разделить на четыре стадии, соответствующие увеличению глубины сна, что проявляется на ЭЭГ увеличением доминирования высокоамплитудной, низкочастотной (синхронизированной) волновой активности. Такие низкочастотные волны доминируют при самых глубоких стадиях NREM-сна (стадии III и IV, также называемые медленноволновым сном)86. Первые шаги засыпания представляют собой сложную последовательность событий. Состояние расслабления характеризуется высоким индексом альфа-ритмов в затылочных или центральных областях. Периоды сонливости (первая стадия сна) связаны с подавлением этих ритмов. Во второй стадии сна появляются новые ЭЭГ-феномены, названные сонными веретенами. Стадия II NREM-сна также характеризуется медленными (< 1 Гц) колебаниями, которые ассоциируются с появлением К-комплексов. NREM- и RЕМ-сон чередуются в каждом из 4—5 циклов, проходящих в ночном сне взрослого человека. Рано ночью NREM сон более глубок и занимает непропорционально большое время, особенно в первом цикле, в то время как REM-эпоха может быть короткой или прерывистой. Позже ночью стадия NREM-сна становится более короткой, и все большее время каждого цикла занимает REM-coh.
III. Функциональное значение сна
Функциональное значение сна все еще остается неизвестным, несмотря на множество фактов и еще большее число теорий. В этом невежестве, вероятно, кроется главная причина, почему наше общество так неуважительно относится ко сну, предполагая, что сон просто забирает драгоценные мгновения нашей жизни. Теперь мы знаем, что сон влияет на все: нашу иммунную систему, когнитивные способности87 и психическое здоровье. Нарушение цикла сна/бодрствования приводит к широкому диапазону нарушений, включая ухудшение внимания и памяти, депрессии, уменьшение мотиваций, нарушения метаболизма, ослабление иммунитета и увеличение риска развития рака.
А. Консолидация памяти
Открытия, сделанные в исследованиях на животных, также как в исследованиях с участием людей, показали, что существует тесная взаимосвязь между сном и так называемой «зависимой от сна памятью». Во многих исследованиях на животных показано, что лишение сна после научения определенным действиям нарушает выполнение этих действий в последующих
86 Циклическая организация сна варьирует внутри и между видами. Длина периода каждой REM/NREM-фаз растет с увеличением размеров мозга между видами, а глубина и пропорции NREM-стадий в каждом цикле увеличивается с процессом созревания мозга внутри вида. Комплексность NREM-сна — функция таких мозговых систем, как таламокортикальные цепи, которые достигают максимального развития только у зрелых людей и постепенно деградируют в пожилом возрасте. Поэтому можно заключить, что дифференциация сна — функция мозговой дифференциации, принцип которой указывает и механистические, и функциональные связи между сном и другими функциями мозга.
87 Когда люди спят ограниченное время (3 или 5 часов в день) более чем 7 дней подряд, наблюдаются дозозависимые снижения бдительности и эффективности работы. Точно так же, когда длительность периодов сна постепенно увеличивается после длительного ограничения сна, эффективность работы увеличивается. Интересно, что первые несколько часов сна, по-видимому, особенно важны для восстановления. Это может объяснять, почему сон даже продолжительностью в 10 минут, как было показано, улучшает состояние.
134 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
пробах. У людей выполнение различных задач значительно улучшается после ночи сна. Кроме того, выборочное нарушение REM, но не NREM-сна «сводит на нет» это повышение эффективности выполняемой деятельности. В более детальных исследованиях испытуемые были выборочно лишены медленноволнового сна, стадий 3 и 4 NREM и REM-сна. Были сделано заключение, что консолидация памяти начинается в стадии медленноволнового сна и затем увеличивается в течение REM-сна. Если верна гипотеза о зависимости от сна качества обработки следов памяти, тогда одним из аспектов нарушения сна будет ухудшение механизмов пластичности мозга.
Б. Иммунная система
У людей активность клеток-киллеров может понижаться на целых 30 % после только одной бессонной ночи. Потеря сна также вредит многим другим параметрам иммунной системы, включая свободно циркулирующие иммунные комплексы, вторичный антигенный ответ и выработку антигенов. Нарушение сна и постоянное психическое напряжение увеличивают концентрации кортизола в крови. Одна бессонная ночь может поднять концентрацию кортизола к следующему вечеру почти на 50 %. Высокий уровень кортизола подавляет иммунную систему, так что при чрезмерном утомлении люди белее восприимчивы к болезни.
В. Психические нарушения
Психические дисфункции почти всегда связаны с нарушениями сна. Бессонница и ранние утренние пробуждения — основные признаки депрессии. Кроме того, переменчивость (нестабильность) сна — точный индикатор повторения депрессии у пациентов, проходящих лечение. Депрессия часто сопровождается тревожными расстройствами, которые также тесно взаимосвязаны с хроническими нарушениями сна. Когда мы испытываем чрезмерное беспокойство, страдают сон, работа, чувства удовольствия и радости, отношения с окружающими. Панические, посттравматические, общие тревожные расстройства и социальные фобии — все они связаны с нарушением сна. Исследование населения в нескольких европейских странах показало, что тревожность связана с бессонницей у 50 % индивидуумов, имеющих в анамнезе психические расстройства.
IV. Биспектральный индекс
Биспекгры88 — хороший показатель мозговых изменений в цикле бодрство-вание/сон. Биспектральный анализ был впервые разработан в начале 1960-х для характеристики фазовых отношений и нелинейных свойств волновых движений во время изменения атмосферного давления. Применение биспектров
" Биспектральный анализ — следующий уровень после спектрального. Принимая во внимание, что фазовый спектр, получаемый в результате анализа Фурье, измеряет фазу частотных компонент относительно начала эпохи, биспектры измеряют корреляцию фазы между различными частотными компонентами (детальнее см. «Методы»).
Часть 1. Ритмы ЭЭГ
135
к анализу ЭЭГ произошло с развитием высокоскоростной микрокомпьютерной технологии. Биспектральный анализ был предложен как инструмент обработки ЭЭГ в конце 1980 — начале 1990-х, при том что еще даже теоретическая связь между физиологией нейронных сетей коры мозга и частотной синхронностью не была предложена. Современные теории выдвигают гипотезу, что тесные (сильные) фазовые отношения ЭЭГ обратно пропорциональны числу независимых ЭЭГ-пейсмейкеров в мозгу.
А. Взаимосвязь с глубиной анестезии
Биспектральный индекс (BIS, Aspect Medical Systems, Natick, MA) яв-
Биспектральный Состояние
индекс
100 г-Спокойное бодрствование
90
80
70
60
50
40
Умеренный седативный эффект
I лгубокий — седативный эффект
Умеренное — гипотическое состояние
Потеря способности реагировать на болевые стимулы
(лубокое гипнотическое состояние
Рисунок 7.2. Значения BIS и взаимосвязь с различными степенями наркоза Потеря способности к реагированию на вредные стимулы может встречаться в диапазоне значений BIS от 0 до 65. Значения больше 65 связаны с увеличением способности к реагированию во время операции и способности послеоперационного воспоминания операционных вмешательств. Для различных седативных шкал, используемых в исследовании, значения BIS были обратно пропорциональны увеличению глубины наркоза. Значения 90-100 связаны с паттерном ЭЭГ при сосредоточенном внимании, а значение 0 отражает полностью подавленную ЭЭГ (изоэлектрический паттерн).
ляется комплексным параметром, составленным из комбинации временной области, частотной и спектральных субпараметров высокого порядка89. Это уникальный среди параметров QEEG, потому что он объединяет несколько независимых дескрипторов ЭЭГ в единственную переменную. BIS — безразмерный параметр ЭЭГ в пределах от 0 до 100. Он определяется в результате анализа Фурье и биспектрального анализа безартефактных участков записи ЭЭГ BIS рассчитывается алгоритмом, который объединяет три фактора: 1) степень совпадения фаз волновых сигналов ЭЭГ (бикогерентность); 2) мощность ЭЭГ в дельта- (1 —4 Гц) по отношению к бета-диапазону (13—30 Гц) (спектры мощности); 3) пропорция ЭЭГ, которая является изоэлектрической. Степень бикогерентности ЭЭГ увеличивается с увеличением глубины анестезии (рис. 7.2) и обратно пропорциональна результирующему индексу BIS.
89 Особая смесь субпараметров индексов BIS была эмпирически получена из специально собранной базы данных ЭЭГ и поведенческих опросников, представляющей приблизительно 1500 случаев анестезии (почти 5000 часов записей), с использованием различных доз и лекарственных препаратов. Затем индексы BIS были проверены на другой популяции. Дополнительное значение (мощность) в каждом отведении ЭЭГ изменяется с глубиной анестезии. Устройства, которые осуществляют BIS, — в настоящее время единственные, одобренные Food and Drug Administration для продажи в качестве мониторов эффекта анестезии на мозг. Хотя биспектральный анализ вовлекает сложные математические преобразования, современные компьютеры достаточно мощны для их вычислений в реальном времени. Для регистрации ЭЭГ пациента требуется BIS-датчик. Датчик—обычно сенсорная полоска, которая имеет маркеры локализации для упрощения размещения датчика.
136
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
V. Заключение
Сон — общее состояние, отличается от бодрствования поведенчески и нейрофизиологически. Сон определяется уменьшением моторной активности; уменьшением ответа на внешние стимулы, стереотипией положения; легкой обратимостью (отличие сна от комы). Бодрствование и сон далее можно разделить на отдельные периоды с различными поведенческими паттернами. Сон состоит из пяти стадий (от сонливости до глубокого медленного и пародоксального сна). Каждый из этих периодов характеризуется различными нейрофизиологическими механизмами, генерирующими различные паттерны ЭЭГ. Как показывают нейрофизиологические данные, передняя часть гипоталамуса содержит GABA-эргические нейроны, которые активны в течение сна и тормозят моноаминерги-ческие клетки восходящей активирующей системы мозга и, следовательно, необходимы для нормального сна. Задний гипоталамус содержит нейроны, вырабатывающие орексин/гипокретин, необходимые для поддержания нормального бодрствования, их повреждение ведет к нарколепсии. Модель сна предполагает, что нейроны бодрствования и сна тормозят друг друга, что приводит к стабильному циклу сна и бодрствования. ЭЭГ можно рассматривать как лучший из известных методов для определения состояний сна. Биспектральный индекс — сложный параметр, получаемый при анализе с использованием преобразования Фурье и би-спектрального анализа безартефактных участков записи ЭЭГ BIS рассчитывается алгоритмом, который объединяет три фактора: I) степень совпадения фаз волновых сигналов ЭЭГ (бикогерентность); 2) мощность ЭЭГ в дельта- (1—4 Гц) по отношению к бета-диапазону (13—30 Гц) (спектры мощности); 3) пропорция ЭЭГ, которая является изоэлектрической. Степень бикогерентности ЭЭГ повышается с увеличением глубины анестезии и обратно пропорциональна результирующему индексу BIS.
Часть 1. Ритмы ЭЭГ
137
Глава 8.
Методы анализа фоновой ЭЭГ
I. Анатомическая локализация
Для описания локализации различных анатомических структур мозга используется специальная терминология (рис. 8.1). Направления для анатомической локализации называются: латерально-медиальное (боковое — срединное); переднезаднее (рострально-каудальное); дорзально-вентральное (спинно-брюшное). Части мозга, образованные различными плоскостями, называются коронарной, горизонтальной и сагиттальной частями.
II. Поля Бродмана
Термин «кортекс» означает «кора»: как и кора у дерева, он содержит много складок. В коре выделяют борозды — складки (углубления) и извилины (выпу-
Схема мозга рассечена на горизонтальный, сагиттальный и коронарный планы. Направления (медиальное — латеральное, дорсальное — вентральное, ростральное — каудальное) изображены стрелками.
138 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
клые гребни свернутой ткани). Около 2/3 коры заключено в глубинах борозд. В развернутом виде человеческая кора закрывает область приблизительно в 2 м2.
Вся кора в каждом полушарии разделена на четыре доли: затылочную, височную, теменную и лобную. Кора состоит из 52 анатомически различных областей, описанных немецким анатомом Корбинианом Бродманом (Korbinian Brodmann) в 1909 году (рис. 8.2). Эта классификация все еще используется. Надо отметить,
Рисунок 8.2. Поля Бродмана
Схема человеческой коры с числами, соответствующими номерам полей Бродмана (определенных Бродманом в 1909 г. на основе цитоархитектонической организации коры).
Часть I. Ритмы ЭЭГ
139
что нумерация, использованная Бродманом, отражает только порядок, в котором Бродман брал образцы мозга, и не имеет другого систематического значения. В дополнение к номеру поля по Бродману (например, поле Бродмана 17) корковая область может быть соотнесена с ее цитоархитектоническим строением (например, полосатая (стриарная) кора называется так благодаря ее внешнему виду в микроскопе), анатомическим положением относительно борозд и извилин (например, шпорная) и ее функциональным названием, согласно предполагаемой функциональной области (например, первичная зрительная кора).
III. Международная система размещения электродов 10-20
В 1959 году были установлены первые стандарты регистрации ЭЭГ Среди них была система размещения электродов, которую называют международной системой размещения электродов 10—20, или просто системой 10-20 (рис. 8.3)90.
Названия электродов включают первую букву латинского названия области, на которую ставится электрод, и номер, указывающий сторону и расположение электрода в пределах этой области. Fpl, Fp2 — переднелобные (prefrontal), F3, F4 — лобные (frontal), Fz — среднелобный, СЗ, С4 — центральные (central), Cz — центральный вертексный, РЗ, Р4 — теменные (parietal), Pz — центральнотемен-
Рисунок 8.3. Международная система 10-20
Каждое положение электрода (отведение) обозначается буквой (для определения доли мозга) и номером (или другой буквой) для определения локализации электрода в пределах доли. Любой электрод располагается от других электродов на расстоянии, равном 10 или 20 % некоторых измеренных индивидуально расстояний.
ной, F7, F8 — передневисочные, ТЗ, Т4 — средневисочные (temporal), Т5, Тб — задневисочные, 01,02 — затылочная (occipital), Al, А2 — ушные. Нечетные но-
мера указывают на левое полушарие, четные — на правое.
IV. Электроды
Электроды в почти всех коммерчески доступных наборах сделаны из металла. Теоретически оптимальные электроды для того, чтобы измерять медленные изменения потенциалов, — неполяризующиеся электроды, такие как серебряные
90 Название вытекает из того факта, что любой электрод располагается от другого на расстоянии 10 или 20 % от некоторых измеренных расстояний. В направлении от носа до затылка это расстояние определяется окружностью головы, измеренной от переносицы (nasion) до зрительного бугра (inion). В направлении от одного уха до другого расстояние определяется окружностью головы, измеренной от ушной ямки одного уха через вертекс до противоположного уха. Расстояние между окружными электродами определяется как 10 % окружности головы, измеренной сразу над бровями и ушами.
140
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
V _ I Укоры \ R
+ кусил
Б
Рисунок 8.4. Упрощенная электрическая схема регистрации электроэнцефалограммы
А. Металлические электроды присоединены к коже посредством проводящего геля, вольтметр измеряет разницу потенциалов между двумя электродами. Б. Потенциал Укоры усиливается усилителем. Укоры — потенциал, произведенный корой, R1 и R2 — сопротивления электродов или, лучше сказать, сопротивления контактов электродов с головой, главным образом определяемые проводящим гелем и качеством контакта между электродами и кожей. Rycnn — сопротивление усилителя.
(хлор-серебряные электроды, то есть электроды, сделанные из серебра (Ag)) и покрытые слоем хлорида серебра (AgCl). Фактически для того, чтобы зарегистрировать обычную ЭЭГ (например, в пределах диапазона от 0,1 до 70 Гц), могут использоваться и другие металлы91.
Любой металлический электрод, помещенный в проводящий раствор, формирует так называемый двойной электрический слой между раствором и металлом. Этот электрический слой формируется благодаря потоку ионов от металлической поверхности к раствору и обратно. Двойной электрический слой — источник разницы потенциалов между металлическим электродом и раствором. Значение это
91 Такие электроды могут быть сделаны из золота (например, Grass-Telefactor, США), олова (Electro-Cap International. США), серебра (Technomed Europe, Нидерланды), платины (Ad-Tech Medical Insrument Corporation, США) или нержавеющей стали (Ad-Tech Medical . Insrument Corporation, США).
Часть 1. Ритмы ЭЭГ
141
го потенциала — сложная функция материала электрода, состава электролита и температуры. В Ag/AgCl-электродах обмен ионами в двойном электрическом слое очень высок, так что любой ток, приложенный к электроду, не может изменить потенциал двойного слоя, и такие электроды называются неполяризующимися электродами92.
В элекгроэнцефалографах усиливается потенциал между электроэнцефалографическим и референтным электродом. Вспомним, что каждый металл производит свой собственный потенциал двойного слоя между электродом и гелем. Когда один металл используется для референтного и записывающего электродов, потенциалы двойного слоя нейтрализуют друг друга. Однако использование различных металлов для записывающих и референтных электродов может привести к существенному дисбалансу между потенциалами и может изменить нормальные условия записи ЭЭГ В этом причина, почему все электроды для регистрации ЭЭГ должны быть сделаны из одного металла.
Электрическая схема измерения коркового потенциала между двумя электродами показана на рис. 8.4. Обратите внимание, что часть измеренного потенциала падает на электродных резисторах согласно закону Ома93. В этом причина, почему сопротивление электродов (иногда упоминается под общим названием «импеданс») обычно удерживается настолько низким, насколько возможно. Для научных публикаций обычно требуется импеданс электродов для регистрации ЭЭГ ниже 5 кОм.
V. Усилители
Основной компонент любого элекгроэнцефалографа (прибора для регистрации ЭЭГ) — усилитель. Он усиливает сигнал ЭЭГ В записи ЭЭГ взрослого человека амплитуда сигналов должна быть приблизительно 30—50 мкВ. Современные элекгроэнцефалографы включают от 19 до 256 усилителей или каналов и используют дифференциальные усилители (рис. 8.5).
Дифференциальные усилители просто уве-
' вых ~K*(V1~V2)
Рисунок 8.5. Дифференциальный усилитель Упрощенная схема дифференциального усилителя. Он усиливает разницу между двумя потенциалами VI и V2. К — коэффициент увеличения.
личивают разницу потенциалов между двумя входными сигналами. Дифференциальные усилители имеют преимущество, состоящее в том, что они позволяют снизить (подавить) синфаз-
ные помехи, артефакты (затрагивающий V1 и V2), такие как наведенное напряжение сети, и другие внешние источники электрических сигналов. В идеале, когда
два входа имеют один и тот же общий входной сигнал, результирующий сигнал
92 «Неполяризующиеся» буквально означает, что при действии на электрод электрического тока потенциал электрода значительно не смещается от равновесия.
93 Закон Ома определяет отношения между напряжением и током в проводнике. Это положение: разница потенциалов (напряжение) в проводнике пропорциональна электрическому току через него. Коэффициент пропорциональности называется сопротивлением (R).
142 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
равен нулю. На практике результирующий сигнал очень мал, но не равен нулю. Способность усилителей быть так близко к идеалу, насколько возможно, отражается в параметре, названном ослаблением синфазного сигнала (Common Mode Rejection, CMR), выражаемом как отношение результирующего сигнала, когда VI = V2 (они связаны с одним и тем же источником) к результирующему сигналу, когда только один входной сигнал отличается от нуля. CMR измеряется в децибелах. Например, CMR = —100 децибелов при 50 Гц.
VI. Переведение ЭЭГ в цифровую форму
В настоящее время большинство элекгроэнцефалографов цифровые. Они переводят непрерывные (так называемый аналоговые) сигналы ЭЭГ в цифровую форму (набор чисел). Числа представляют электродные потенциалы, измеренные в дискретных временных точках. Количество измеренных точек в секунду называют частотой отсчетов элекгроэнцефалографа (или частотой дискретизации, оцифровки). Например, частота отсчетов 250 Гц означает, что сигнал ЭЭГ от одного электрода в течение 1 секунды представлен в компьютере как набор из 250 чисел. При такой частоте отсчетов интервал между двумя последовательными измерениями 4 мс. Существенно, чтобы частота отсчетов была более чем в два раза больше, чем самая высокая анализируемая частота поступающего сигнала (теорема о дискретном представлении Шеннона — Котельникова)94. Числа сохраняются в компьютере в двоичной системе, то есть в виде набора битов95. Если элекгроэнцефалограф использует 14-битный аналогово-цифровой (А/D) конвертер, его разрешение 1/8192, или приблизительно 0,0001 максимальной производительности. Так, если элекгроэнцефалограф предназначен для измерения сигнала в пределах диапазона +1 мВ до — 1 мВ, размер шага в его 14-битном преобразовании равен 0,25 мкВ.
VII. Монтаж
А. Объединенный ушной референт
В элекгроэнцефалографах с дифференциальными усилителями мы измеряем потенциалы электрода всегда по отношению к некоторому другому потенциалу или потенциалам. Однако традиционно один вид измерения, когда референтный электрод расположен на части головы, по возможности собирающей меньшее количество сигналов от корковых нейронов, называют референтным. В референтной записи референтный электрод обычно расположен на мочке уха, мастоидах или кончике носа. Этот тип регистрации также называют моно-полярной регистрацией. Когда же оба электрода расположены на скальпе над корковыми ЭЭГ-генераторами и значение мощности электрических процессов
94 Консервативная модель фактически призывает к тому, чтобы частота отсчетов была в 4-10 раз выше, чем самая высокая анализируемая частота сигнала.
95 Бит (наименьшая единица информации), может быть равным 0 или 1. Так, число, которое включает 14 битов. — это число между -213 и +213, или -8192 и +8192.
Часть 1. Ритмы ЭЭГ
143
в одном электроде рассматривается относительно другого, этот тип регистрации называют биполярным.
Есть много схем ЭЭГ-измерений. Все они объединены под общим названием монтажи96. Монтаж — это принцип, согласно которому измеряются потенциалы ЭЭГ. Самый простой принцип (так называемый монтаж с объединенным ушным электродом) измеряет потенциалы электродов по отношению к двум объединенным электродам, расположенным на мочках левого и правого уха. Объединенные мастоидные электроды, электрод на кончике носа или подбородке могут также использоваться как референтные электроды97. В идеале референтный электрод должен быть электрически нейтрален, то есть его потенциал не должен изменяться в течение регистрации. К сожалению, из-за проводимости скальпа все отведения на голове в некотором смысле электрически активны и нет никакого места, на которое можно было бы поместить референтный электрод для выполнения критериев идеального референта. Это означает, что электрический потенциал, зарегистрированный от любого отдельного электрода, будет зависеть от референта.
Б. Общий средний монтаж
Один из методов, который касается этой проблемы, — это общий средний референтный монтаж. Он был разработан в начале 1950-х. Процедура проста: потенциал усредняется по всем электродам (назван общим средним потенциалом) и вычитается из любого отдельного потенциала. Процедура выражается в следующей формуле:
V.^-EV/n,
где Е — сумма по всем электродам.
Если бы голова была сферой и электроды могли бы быть помещены во все точки этой сферы, то общим средним числом был бы ноль и общий средний монтаж был бы совершенным решением. Однако в действительности дело обстоит не так: голова не сфера и электроды приложены только к верхним и боковым поверхностям головы.
В. Локальный средний монтаж
Его вариант — локальный средний референтный монтаж. Термин «локальный средний референт» относится к созданию уникального референта для каждого электрода. В этом случае для вычисления комплексного референта используется небольшое количество электродов около целевого электрода.
96 Преимущество современных цифровых ЭЭГ-систем (по сравнению со старыми аналоговыми системами) состоит в том, что цифровая ЭЭГ-система позволяет определять и изменять монтаж и установки фильтров во время просмотра ЭЭГ, а не во время регистрации. ЭЭГ обычно регистрируется с минимально ограничительными аналоговыми установками фильтров и с потенциалами во всех электродах, измеренными относительно единственного референта (такого как, например, референт на кончике носа или объединенный ушной референт). Компьютерная программа может затем обработать данные так, как они будут показаны на экране или выведены на принтер, трансформируя первоначальный референтный монтаж на любой желаемый референтный или биполярный монтажи, и, если необходимо, в цифровой форме фильтруя данные. Если необходимо, один и тот же фрагмент ЭЭГ может быть показан неоднократно, с использованием различных монтажей и фильтров. Исторически в электроэнцефалографии использовалось много различных монтажей. Разнообразие этих монтажей определялось техническими возможностями, доступными исследователям в прошлом.
97 Обратите внимание, что в книге, если не упомянуто иначе, ЭЭГ была зарегистрирована по отношению к объединенному ушному электроду.
144
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Объединенный ушной
Общий средний
Относительные потенциалы
Локальный средний
Рисунок 8.6. Три монтажа
А. В монтаже с объединенным ушным референтным электродом потенциал активного электрода измеряется по отношению к потенциалу двух физически объединенных ушных электродов. Б. В общем среднем монтаже потенциал активного электрода измеряется по отношению к потенциалу, усредненному по всем электродам. В. В локальном среднем монтаже потенциал активного электрода (например, Cz) измеряется по отношению к потенциалу, усредненному по смежным (соседним) электродам. Референтные электроды изображены черным.
В формуле, представленной выше, Е теперь — сумма потенциалов по электродам, окружающим i-й электрод98. В общих чертах любой монтаж может быть определен формулой:
У=У-^хУг
где w — веса. Следовательно, любой монтаж характеризуется его весами w... Значения весов могут быть найдены в настройках любого современного программного обеспечения.
Пример локального среднего референтного монтажа — лапласиановский монтаж. Создание лапласиановского монтажа вовлекает оценку оператора Лапласа для каждого электрода. Математически оператор Лапласа — вторая частная пространственная производная от потенциального поля. С точки зрения физики, вторая пространственная производная пропорциональна электрическому току в соответствующей точке поверхности головы. В свою очередь этот ток позволяет нам оценить силу лежащего в его основе нейронного генератора.
Первоначально описанный Pitts в 1952 году метод требовал 3-мерного множества близко расположенных электродов, те. ситуации, при которой необходимо использование внутричерепных глубинных электродов. Модификации для применения поверхностных потенциалов были разработаны Hjorth в 1975 г Hjorth показал, что вычисления могут быть упрощены, сделав множество предположений о структуре коры, и допустил, что в пределах специфического слоя коры существует однородная проводимость. В результате потенциальные поля, ориентированные
98 Например, если мы хотим определить локальный референт для, скажем, Cz, мы должны вычислить среднее значение по электродам, окружающим Cz, в этом случае: Fz, СЗ, С4, Pz. Для нашего электрода Cz мы имели бы следующее выражение для локального усреднения: - (Vfi + VC3 + VM + VJ/4.
Часть 1. Ритмы ЭЭГ
145
Относительно объединенного ушного референта
Относительно локального среднего референта
(по Лемосу)
Б
Рисунок 8.7. Сравнение двух монтажей
Топограммы спектров мощности здорового субъекта в состоянии с закрытыми глазами при использовании монтажа с объединенным ушным электродом (А) и локального среднего монтажа (Б). Обратите внимание, что локальный средний монтаж обеспечивает более локализованное распределение источников потенциала.
параллельно каждому слою, гомогенны. Поэтому одномерные отведения являются единственно подходящими, т.к. их поля перпендикулярно ориентированы к слою костной ткани. В этом методе вторая пространственная производная потенциального поля ограничена электрическим током, перпендикулярным к корковой поверхности". Метод Hjorth был позже изменен Лемосом.
На рис. 8.7 представлены карты спектров ЭЭГ, вычисленные для двух частот для указанных монтажей: монополярного и локального среднего (Lemos) монтажа. В этой редкой ЭЭГ, зарегистрированной у здорового субъекта, взятой из нормативной HBI базы данных, есть два четких альфа-ритма с перекрывающимися частотами: мю-ритм и теменной альфа-ритм. Обратите внимание, что лучшее пространственное разрешение и четкое «отделение» сенсомоторного ритма от теменного ритма обеспечивается локальным средним монтажом.
VIII. Анализ Фурье
А. Спектры
Важным свойством ЭЭГ является ее ритмичность. Как мы узнали из этой части книги, существует несколько ритмов ЭЭГ, которые отличаются частотой, местопо- *
99 С лапласиановским монтажом связано множество ограничений. Во-первых, пространственная точность сильно зависит от числа электродов и межэлектродного расстояния. Во-вторых, потенциалы в периферии области измерения не могут быть надежно оценены, так как электроды в этих точках не полностью окружены электродами. В-третьих, локализация диффузных источников может быть значительно искажена или даже пренебрегаться. Ввиду этих ограничений ясно, что Лапласиановский монтаж более пригоден для локализации относительно центральных источников.
146 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
ложением, механизмами генерации и функциональным значением. Ритмы развиваются во времени, они растут и убывают. В качестве репрезентативного и надежного обычно рассматривается 3-минутный интервал ЭЭГ или более длительная запись, т.к. весьма трудно оценить всю информацию о ритмах при рассмотрении каждый раз только нескольких секунд регистрации ЭЭГ Мощный метод сокращения информации о ритмичной активности во времени создается анализом Фурье.
Теорема Фурье утверждает, что любой сигнал X (t) в пределах времени от 0 до Т100 может быть расчленен на набор простых синусоидальных функций, названных рядом Фурье (рис. 8.8), то есть:
X(t) = А,, + + (рп),
Сигнал
А
б
Разложение
Фурье
WWVW 13
wwvwwvw +1/5
где L — сумма по всем частотам (гармоника), f — собственная частота сигнала f= 1/Т, ф — фаза, n = 1,2,3, Ап — амплитуда, фп — фаза синусоидальной компоненты (п — порядок гармоники).
Цель анализа Фурье состоит в том, чтобы найти эти параметры. Они могут быть представлены в форме спектров амплитуд (Ап в зависимости от частоты nf), спектров мощности (квадрат амплитуды А2п) и фазовых спектров (фп в зависимости от частоты nf). Подход на основе прямого суммирования в вычислении Ап и фп требует большого времени для расчетов. В 1965 году Cooly и Tukey создали более эффективный алгоритм для вычисления ряда Фурье, названный алгоритмом быстрого преобразования Фурье (fast Fourier transformation — FFT).
Временной интервал регистрации ЭЭГ, например, в состоянии с открытыми глазами, обычно равен 3 минутам. Это означает, что он может быть разделен на 45 четырехсекундных эпох. Разделение непрерывного потока сигнала на конечные эпохи может внести искажение спектров артефактными частотами, созданными резкими переходами на концах эпохи. Этот тип искажения минимизируется умножением потенциала в пределах эпохи анализа на соответствующие значения временного окна. Временное окно — число
вой ряд, содержащий то же самое число элементов, как число отсчетов сигнала в эпохе. Значения элементов временного окна стремятся к нулю на обоих концах ряда и к единице в середине. Было предложено множество разнообразных временных окон, включая прямоугольное, треугольное, окно Ханна и Blackman-функции.
100 Т — эпоха для анализа Фурье, или эпоха анализа. Чем больше эпоха, тем больше число гармоник и выше частотное разрешение для спектров. Например, для эпохи анализа в 4 секунды частотное разрешение 1/4 = 0,25 Гц.
WVWAftWWVWW “ 1/
Гармоники в
Рисунок 8.8. Разложение прямоугольного сигнала на синусоидальные гармоники
Сигнал (А) раскладывается на гармоники (Б) с различными частотами и амплитудами (числа справа). Чем больше добавляется гармоник, тем лучше соответствует результат суммации (С) исходному сигналу.
Часть 1. Ритмы ЭЭГ
147
Применяющиеся различные временные окна создают другую проблему: отсчеты около начала и конца эпох анализа дают меньший вклад в спектры по сравнению со средними отсчетами. Чтобы компенсировать это искажение, выбираются эпохи с перекрыванием. Перекрывание обычно составляет половину эпохи10’.
Б. Динамика спектров и усредненные спектры
Спектры могут быть представлены различными способами. Один из них представляет динамику спектров, вычисленных для каждой эпохи (или множества эпох) последовательно (рис. 8.9, вверху). Как видим, пики в спектрах, соответствующие мю-ритмам, в последовательных временных интервалах убывают и возрастают. Другой способ представления спектров — усреднение спектров по всему интервалу регистрации (рис. 8.9, внизу)101 102. Усредненные спектры более стабильны и могут надежно характеризовать состояние человека. Три минуты — минимальная продолжительность регистрации ЭЭГ для получения относительно устойчивых спектров.
В. Относительные спектры
Амплитуда ЭЭГ, зарегистрированной у отдельного субъекта, зависит от многих факторов, включая нейрофизиологические, анатомические и физические свойства мозга и окружающих тканей (кожи, кости, твердых и мягких мозговых оболочек). Эти параметры изменяются от одного человека к другому и в основном неизвестны. Указанные различия приводят к большой вариабельности абсолютных спектров ЭЭГ (рис. 8.10, вверху). Чтобы скомпенсировать эту вариабельность, иногда вычисляется относительная мощность ЭЭГ (рис. 8.10, внизу). Как видно, изменчивость абсолютной мощности выше, чем изменчивость относительной мощности. Так, когда вычисляются Z-показатели103 для оценки уровня статистической значимости отклонения индивидуального спектра от среднего по группе здоровых субъектов, большие колебания абсолютных спектров ЭЭГ в нормальной группе понизили бы уровень статистической значимости отклонения от нормальности.
Формула для вычисления относительных спектров проста:
Pr(f) = Pa(f)/(LPa(Q),
где Pr(f) — относительная мощность ЭЭГ на частоте f, Pa(f) — абсолютная мощность ЭЭГ на той же самой частоте, L — сумма мощностей ЭЭГ всего спектра или его части.
101 Параметры анализа Фурье могут быть приспособлены к целям определенной задачи: длина эпохи (обычно от 1 до 8 секунд), тип перекрывания между эпохами (обычно выбирается 50 % перекрывания), тип временного окна (лучшие результаты достигаются временным окном Ханна). Почти во всех коммерчески доступных программных обеспечениях эти параметры могут быть настроены пользователем.
102 Спектры мощности, представленные на рис. 8.9, рассчитаны так: 1) длительность эпохи установлена равной 4 секундам; 2) интервал всей записи ЭЭГ (беэ интервалов, содержащих артефакты) разделен на равные эпохи; 3) перекрывание эпох установлено равным 50 %, начиная со второй эпохи первые 50 % каждой эпохи перекрываются с последними 50 % предыдущей эпохи; 4) выбрано временное окно Ханна; 5) спектр мощности вычислен посредством быстрого преобразования Фурье; 6) усредненный спектр рассчитан усреднением по всем эпохам записи ЭЭГ для каждого отдельного канала.
103 Z-показатель для статистического распределения — отношение отклонения от среднего значения к стандартному отклонению (вариабельности).
148
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Рисунок 8.9. Два способа представления спектров мощности ЭЭГ
А. Динамика спектров мощности, вычисленных для 3 каналов (СЗ, Cz и С4). Направление времени — сверху вниз. Б Спектры ЭЭГ, усредненные по всем эпохам. Монтаж — локальный средний монтаж (Lemos). Эпоха анализа 4 секунды. В динамике спектров каждый спектр — усреднение по четырем последовательным эпохам.
DC Картирование ЭЭГ
Когда ЭЭГ зарегистрирована от многих электродов, которые охватывают всю кору, можно вычислить двухмерные (2D) изображения измеренных характеристик ЭЭГ. Характеристиками могут быть или изменения потенциалов, или усредненная мощность (амплитуда, фаза) в определенной частоте104. Два типа карт представлены на рис. 8.11.
X. Фильтрация
А. Фильтры низких и высоких частот
Цифровые фильтры играют важную роль в анализе ЭЭГ. Функция фильтров состоит в подавлении данной частотной компоненты в сигнале и пропускании первоначального сигнала за исключением этих подавленных компонент. В отличие от аналоговых фильтров работа цифровых выполняется математическими действиями на сигнал.
104 Идея картирования мозга с помощью ЭЭГ была предложена еще Grey Walter в 1936 г. Он использовал эту технику для идентификации аномальной электрической активности мозговых областей вокруг опухоли. В наше время картирование ЭЭГ-характеристик стало обычной процедурой.
Часть 1. Ритмы ЭЭГ
149
Рисунок 8.10. Спектры абсолютной мощности по сравнению со спектрами относительной мощности
Спектры абсолютной (А) и относительной (Б) мощности ЭЭГ 28 здоровых субъектов возрастом 15 лет наложены друг на друга. Обратите внимание, что отклонения меньше для относительной мощности по сравнению с абсолютной мощностью.
В функциональном отношении есть много типов фильтров, таких как фильтр низких частот, фильтр высоких частот и режекторный. Фильтр низких частот пропускает только низкочастотные сигналы (сигналы ниже некоторой указанной частоты). Фильтр высоких частот работает так же, только наоборот, отклоняя частотные компоненты ниже некоторого указанного порога и пропуская более высокочастотные составляющие. Фильтры обычно определяются их реакциями на индивидуальные частотные компоненты, составляющие входной сигнал.
Б. КИХ и БИХ-фильтры
В зависимости от алгоритмов расчета существуют два типа фильтров: фильтры с конечной импульсной характеристикой (КИХ) (finite impulse response — FIR) и фильтры с бесконечной импульсной характеристикой (БИХ) (infinite impulse response — HR). Последние также называются рекурсивными фильтрами105. Для фильтров КИХ текущий отсчет (уп) рассчитывается исключительно от текущих и предыдущих входных значений (хп, х , хп2). Напротив, в рекурсивных фильтрах в дополнение к значению входного сигнала также используют значения предыдущих результирующих сигналов. Слово «рекурсивное» буквально означает «бегущий назад» и относится к факту, что предварительно расчетные значения выходного сигнала возвращаются при вычислении последующих выходных сигналов. Поэтому выражение рекурсивного фильтра содержит не только входные значения (хп, хп|, хп_2), но и выходные (уп_р уп2). Для достижения требуемой амплитудно-частотной характеристики рекурсивные фильтры имеют более низкий порядок по сравнению с аналогичными нерекурсивными, и, таким образом, используется меньше элементарных вычислительных операций. Чтобы отфильтровать 50 (60) Гц шум сетевой наводки, применяется режекторный фильтр.
105 Импульсная характеристика цифрового фильтра — пропускание фильтра, когда на его входе есть единичный сигнал (единичная величина 1 во время t = 0). Ответ КИХ-фильтра имеет конечную продолжительность. Ответ БИХ-фильтра теоретически (но не фактически) продолжается бесконечно из-за рекурсивной природы фильтра.
150
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Рисунок 8.11. Два типа картирования ЭЭГ (
к. Фрагмент ЭЭГ с двумя вертикальными линиями, отмечающими два момента времени, — tin t2. Б. Карты потенциалов, сделанных во время первого и второго моментов. В. "Голограмма спектра мощности на частоте 11 Гц, усредненного на 3-минутном отрезке ЭЭГ.
XI. Биспектры
Спектры представляют собой статистические оценки параметров второго порядка для сигналов ЭЭГ. Они не обеспечивают информацию о том, как колебания в различных частотах взаимодействуют друг с другом. Статистические данные высшего порядка включают биспектры. Биспектральный анализ исследует отношения между синусоидами в двух первичных частотах, и f2, и модулирующей компонентой в частоте f, + f2. Этот набор трех частотных компонент известен как триплет (fp f2 и f( + f2). Для каждого триплета может быть рассчитан биспектр В (fp f2), количественно включающий информацию о фазе и мощности. Биспектр может быть разложен: 1) на информацию о фазе, или бикогерентность, BIC (fp f2); 2) общую величину членов триплета, или фактический триплетный продукт, RTP (fl, f2). Высокое значение бикогерентности в fp f2 указывает, что есть фазовое сцепление (взаимосвязь) в пределах тройки частот fp f2, и f( + f2. Жесткое фазовое
Часть 1. Ритмы ЭЭГ
151
сцепление может подразумевать, что синусоидальные компоненты в ft и f2 имеют общий генератор. Так, функциональное значение бикогерентности более просто: фазовый спектр при обычном преобразовании Фурье измеряет фазу частотных компонент относительно начала эпохи, тогда как бикогерентность измеряет корреляцию фаз между различными частотными компонентами. Бикогерентность, BIC (fp f2) — это число, которое изменяется от 0 до 1 пропорционально степени фазового сцепления в триплете частот. Мы еще сомневаемся, какое конкретно физиологическое значение имеют эти фазовые отношения, однако самая простая модель предполагает, что тесные фазовые отношения имеют обратно пропорциональное отношение к числу независимых пейсмейкерных элементов ЭЭГ.
XII. Когерентность
А. Физиологическое значение
С анатомической точки зрения самая поразительная особенность мозга — обилие возможностей взаимосвязи между нейронами. С функциональной точки зрения эта способность к взаимодействию отражается в синхронизации активности в пределах мозга: нейроны в анатомически связанных структурах имеют тенденцию генерировать разряды синхронно. Электрофизиологические данные показывают, что эта синхронность в разрядах повторяется для различных частот. Частота, на которой происходит синхронизация, по-видимому, определяет функциональное значение взаимосвязей. Например, альфа-частоты — это частоты'покоя сенсорных систем, и синхронизация в частоте 10 Гц указывает на состояние сенсорной системы, когда нейроны не передают сенсорную информацию, но готовы начать, кем только появится релевантный стимул. Предполагается, что синхронизация колебаний в гамма-диапазоне может быть обязательным механизмом объединения различных особенностей объекта в целостный объект перцепции. Синхронизация в частоте 40 Гц указывает на синхронную активацию нейронов, ответственных за обнаружение различных особенностей одного и того же стимула. Нарушение нормальной синхронизации может быть признаком неврологической или психической дисфункции. Например, аномальный паттерн синхронизаций между различными частями базальных ганглиев может быть связан с возникновением тремора и дискинезии при болезни Паркинсона.
Б. Представление отклонений от нормальности
В ЭЭГ мерой синхронизации между двумя различными скалъповыми отведениями является параметр, названный когерентность106. С функциональной
106 Когерентность вычисляется для двух каналов, т.е. если вы имеете 19 электродов, общее количество пар 19 х 18 / 2 = 171. Это количество значений невозможно нанести на одну голограмму. Было предложено несколько способов представления когерентности. На рис. 8.12 мы предлагаем один из них.
152
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
ооз ооб --------- -аоз — 0.06
Рисунок 8.12. Недостаточная межполушарная когерентность у субъекта с атрофией мозолистого тела
А. Диаграммы связей для трех диапазонов частот и трех условий регистрации. Обозначены только межполушарные связи, в которых когерентность ниже или выше нормальной когерентности с порогами, определенными внизу. Б. Примеры отклонений когерентности от нормы для двух пар электродов.
точки зрения, когерентность — мера корреляции между мощностями ЭЭГ, вычисленными в одном и том же диапазоне частот, но в различных отведениях107. Математически когерентность ЭЭГ вычисляется следующим образом:
где (f) — значение кросс-спектра на данной частоте f, (f) и (f) — значения автоспектров х и у в той же частоте.
На рис. 8.12 показаны результаты сравнения когерентности ЭЭГ, вычисленной у 45-летнего субъекта с нормативными данными. Это человек с нормальным адаптивным поведением, без психиатрических и неврологических жалоб, но с частичной атрофией мозолистого тела108. Справа изображены результаты вычитания нормативной когерентности (из базы данных) из когерентности, вычисленной для субъекта. Когерентность вычислена для двух пар электродов: СЗ-С4 и F3—F4. ЭЭГ была зарегистрирована в трех различных условиях: с открытыми глазами, при решении GO/NOGO и математической задач. На этой
107 Коэффициент корреляции определяется как R = (Z ху) / (>/х2 >/у2). где х и у — отклонения от средних двух переменных. В нашем конкретном случае х и у соответствуют мощности ЭЭГ, измеренной в двух отведениях в некотором диапазоне частот. R изменяется между -1 и +1. Так, если две переменные х и у изменяются синхронно и в одном направлении во времени, коэффициент корреляции между этими двумя переменными стремится к 1. Если две переменные изменяются не в фазе, коэффициент корреляции между этими переменными -I.
1М Волокна аксонов, соединяющие два полушария.
Часть I. Ритмы ЭЭГ
153
картине изображены только симметричные межполушарные связи. Полосы пропускания ЭЭГ были отрегулированы согласно индивидуальным спектральным характеристикам ЭЭГ Этот способ представления данных ясно показывает, что когерентность между двумя полушариями у этого субъекта всегда меньше, чем в нормальной группе.
XIII. Вызванная десинхронизация
Термин «десинхронизация» касается общего наблюдения, что предъявление релевантного зрительного стимула десинхронизирует затылочный альфа-ритм. На рис. 8.13 представлен фрагмент ЭЭГ во время пробы теста GO/NOGO. Как видим, зрительные стимулы, сопровождаемые ответом субъекта, вызывают уменьшение альфа активности, регистрирующейся в О1,02.
Gert Pfurtsheller в 1977 году для вычисления индекса десинхронизации предложил следующий метод: сначала ЭЭГ фильтруется в некотором диапазоне частот (в случае, представленном на рис. 8.13, был выбран альфа-диапазон от 9 до 11 Гц), затем отфильтрованная ЭЭГ возводится в квадрат для получения мощности ЭЭГ и сгла-
Рисунок 8.13. Вычисление вызванной десинхронизации/синхронизации
А. Фрагмент ЭЭГ, зарегистрированной при выполнении GO/NOGO-задачи и отфильтрованной в диапазоне частот альфа-ритма. Б. Стадии вычисления вызванной десинхронизации. Описание см. в тексте.
живается (фактически в варианте, предложенном Pfurtsheller, сглаживание выполняется на последующих стадиях), и наконец, паттерны мощности ЭЭГ усредняются между всеми пробами. Для получения относительных значений индекса десинхронизации рассчитывается Pbg — средняя мощность на фоновом интервале ЭЭГ (то есть в интервале, предшествующем стимулу), тогда относительное уменьшение (или увеличение) ЭЭГ после предъявления стимула рассчитывается по формуле:
154
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
ERD-KP.-P^/PJ.
Этот параметр был назван вызванной десинхронизацией. Однако в других частотных диапазонах, также как и в альфа-диапазоне, но в иных временных интервалах, наблюдалось увеличение мощности ЭЭГ вместо десинхронизации. Увеличение было отмечено как синхронизация, и соответствующий параметр назвали вызванной синхронизацией.
XIV. Волновое (вейвлет) преобразование
В анализе Фурье сигнал ЭЭГ преобразуется в набор синусоидальных функций с различными частотами, амплитудами и фазой. Эти функции во временном промежутке стремятся к бесконечности. Следовательно, преобразование Фурье не дает четкого представления о временной динамике. Это преобразование не работает, когда мы хотим изучить, как различные ритмы ЭЭГ отвечают на краткие предъявления стимулов или на быстрые движе
Рисунок 8.14. Мощностно-частотно-временные представления (вейвлет-анализ)
А. Сигнал ЭЭГ — s (t). Б. Вейвлеты Морле (Morlet's), также называемые волновыми сигналами. В. Схематичное трехмерное представление разложения сигнала на волновые функции Морле. Г. Мощностно-часто шо-временное представление сигнала, усредненного по пробам.
Часть I. Ритмы ЭЭГ
155
ния. В таких случаях естественно расчленять ЭЭГ не на непрерывные синусоидальные функции, а на колебательные волновые сигналы конечной длины (рис. 8.14, вверху справа). Такие формы волн, представленные на рис. 8.14, называют вейвлетами (wavelets). В математике вейвлет-преобразование относится к представлению сигнала в терминах колебания сигналов конечной длины. Эта форма волны измеряется и переводится в соответствие со входным сигналом.
Используемые главным образом функции — набор вейвлетов Морле (Morlet), определенные уравнением:
W(t, f) = A ехр(—(t-t^ / 2s2) exp(2brft).
W(t,f) имеют Гауссовскую форму как во временной шкале (относительно времени t0), так и в частотной области (относительно частоты fV вейвлеты имеют одно и то же число циклов для различных частотных диапазонов, что приводит к их различной длительности.
Анализируемый сигнал s(t) преобразуется волновым анализом подобно преобразованию с помощью синусоидальных функций:
P(t,f) = I w(t,f) X s(t) I2
Результат возводится в квадрат и дает зависимую от времени мощность ЭЭГ около частоты f. Повторение этого вычисления для ряда волновых сигналов, имеющих различные частоты f, обеспечивает временно-частотное представление мощности компонент сигнала (рис. 8.14, внизу).
XV. Слепое разделение источников и анализ независимых компонент (ЮА)
ЭЭГ — сумма различных ритмов, произведенных в различных частотах генераторами, расположенными в различных корковых областях. Например, генераторы мю-ритма 9— 13 Гц расположены в сенсомоторной коре', в то время как генераторы среднелобного тета-ритма расположены в средней предлобной коре и передней поясной извилине. Любой электрод, помещенный в лобноцентральную область, регистрирует сумму этих двух ритмов. Подобным образом вызванные потенциалы (см. «Часть 2» книги) представляют сумму различных компонент, произведенных различными корковыми генераторами в различных временных интервалах. Например, генераторы в ранней (от 80 до 200 мс) стадии зрительной обработки информации расположены в различных областях задней коры, таких как стриарная, ггрестриарная кора, передний зрительный поток (нижняя —височная кора) и задний поток (теменная кора). В этих двух примерах, если мы хотим найти независимые генераторы, которые производят смешанные потенциалы в коре, мы должны решить так называемую проблему слепого разделения.
156 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
А. Математическая формулировка
Математически эта проблема может быть сформулирована следующим образом. Предположим, что мы имеем N исходных сигналов или переменных (типа нейронных диполей, которые находятся в специфической корковой области и производят специфический ритм) s = {s,, s2, ...,sN}, где si-вектор109 — функция времени110. Если мы далее рассматриваем время как последовательность дискретных временных точек t, тогда динамику времени si можно рассматривать как матрицу S с N-рядами (источники) и Т-колонками (временные отсчеты). Предположим далее, что согласно законам физики (таким как проведение электрических потенциалов в объеме) эти сигналы почти немедленно проектируются к поверхности головы и линейно смешиваются в данном отведении. Если общее количество электродов N, тогда:
P = MS,
где Р = {Pl, Р2,..., PL} — матрица временной динамики потенциала (состоящий из N-рядов и Т-колонок), М — так называемая матрица смешивания, которая смешивает независимые источники в скальповые потенциалы, зарегистрированные отдельными электродами:
М21,MN1 м _ М12 м22,MN2
M1N mnn
Цель проблемы слепого разделения источников математически может быть сформулирована как вычисление значений вектора S, нахождение матрицы W так, чтобы:
U = WP,
где U — оценка S.
Обратите внимание, что матрицу W называют несмешанной матрицей, потому что она преобразует смешанные потенциалы в их источники.
Согласно определениям линейной алгебры:
P = W’ U,
где W'1 — обратная матрица W (вспомните, что W’1 — оценка матрицы смешивания М).
109 Векторы — предмет линейной алгебры, отрасли математики, занятой изучением векторов, векторного пространства, линейных преобразований и систем линейных уравнений.
110 В особом случае, когда сигнал преобразован преобразованием Фурье или волновым преобразованием в частотную область, время может быть заменено частотой.
Часть I. Ритмы ЭЭГ
157
Б. Пространственные фильтры разложения на независимые компоненты
Обратите внимание, что i-ряд матрицы W называют пространственным фильтром соответствующей компоненты, потому что он извлекает (отфильтровывает) независимую компоненту из смешанного сочетания их всех. Эти фильтры можно рассматривать как топографии соответствующих компонент и визуализировать в форме карт (см. карты справа рис. 8.15).
Предполагается, что локализация источников независимых компонент посто
янна, то есть не меняет местоположение в течение времени регистрации. Активации соответствующих источников (Ц) можно рассматривать как волновые сигналы независимой компоненты. Обратите внимание, что получение фактической амплитуды независимого источника на скальпе требует перемножения волнового сигнала и соответствующей топографии. Так как единственная значащая величина — фактическая амплитуда источника на поверхности головы, мы можем «умножить» как сам сигнал, так и топографию на «—1», не изменяя этой величины. Это создает двусмысленность в определении полярности волнового сигнала и его положения. Так, ни признак положения, ни признак волнового сигнала не является значащим сам по себе.
Рисунок 8.15. Слепое разделение источников ЭЭГ
А. Нативная ЭЭГ. Ь. Временная динамика независимых компонент (U). В. Топографии независимых компонент (Wr1). Нативная ЭЭГ равна произведению матриц W^U.
Телеграммы
Представим проблему слепого разделения источников в графической форме. Здесь мы должны подчеркнуть, что трудно вообразить многомерное пространство и еще более трудно вообразить линейные преобразования в этом пространстве. Так, для простоты мы рассматриваем двухмерное пространство (рис. 8.16). Предположим, мы имеем два независимых источника (переменные), которые определяют два параметра данной системы, такие как X, изображенную в горизонтальной оси, и Y, изображенную в вертикальной оси. Параметры X и Y могут быть измерены, в то время как задача состоит в том, чтобы найти независимые источники. Если переменные независимы и они изменяются во времени, набор состояний системы может напоминать то, что представлено на рис. 8.16, где каж
дая точка соответствует состоянию системы в последовательных временных интервалах111. Обратите внимание также, что на рис. 8.16 переменные не являются
111 Обратите внимание, что в этом примере информация о временной динамике системы не принимается во внимание.
158 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
гауссовскими, то есть они очень разбросаны. Таким образом, проблема слепого разделения источников должна состоять в том, чтобы выделить векторы, которые соотносятся с независимыми переменными (или источниками), зная только измеренные параметры системы X и У.
В. Анализ независимых компонент (ICA) в сравнении с анализом главных компонент (РСА)
Существует несколько попыток решить задачу слепого разделения источников. Наиболее мощный из них — анализ независимых компонент (Independent Component Analysis, ICA). ICA был впервые представлен в начале 1980-х, а для ЭЭГ был применен в 1995 году для коррекции артефактов. В общих чертах ICA — специальная область математики, и для выполнения анализа независимых компонент используются численные алгоритмы. Эти алгоритмы базируются на оптимизации (нахождении минимумов или максимумов) некоторых функционалов. ICA внешне связан с классическими методами, такими как анализ главных компонент и факторный анализ. Но ICA намного более мощная техника, способная к обнаружению переменных или источников, лежащих в основе, тогда когда классические методы полностью терпят неудачу. Последнее утверждение продемонстрировано на рис. 8.16. Рисунок представляет случай двух независимых, неорто-
Рисунок 8.16. Сравнение методов анализа независимых и главных компонент
Неортогональная смесь двух распределений в плоскости X-Y, полученных двумя методами. Анализ главных компонент, РСА (Б), дает ортогональные оси в направлениях максимальной дисперсии. Анализ независимых компонент, ICA (А), отражает характер распределений.
тональных и негауссовских переменных, которые смешаны в две измеряемые переменные (ось X и ось Y).
Решения, полученные методом анализа главных компонент и методом анализа независимых компонент, схематично представлены векторами из тонких линий. Явно, что оси ICA лучше описывают структуру этих данных, чем РСА.
Ключевое предположение, используемое в ICA, — то, что источники являются статистически независимыми. Это предположение, по-видимому, очень точно соответствует данным ЭЭГ Действительно, все экспериментальные свидетельства,
Часть I. Ритмы ЭЭГ
159
представленные в этой главе книги, указывают, что ЭЭГ формируется потенциалами, произведенными различными источниками, расположенными в различных корковых областях. Статистическая независимость означает, что измерение потенциала, произведенного в отдельном источнике (скажем, в левой сенсомоторной полосе, что производит мю-ритм) в данный момент, не позволяет сделать никаких предположений относительно источника в другой области мозга (скажем, в правой сенсомоторной полосе, или затылочной области, или передней
Рисунок 8.17. Разложение нативной ЭЭГ на независимые компоненты
А. Независимые компоненты с соответствующими LORETA-изображениями источников. Б. Фрагмент ЭЭГ-сигнала здорового 19-летнего субъекта во время выполнения GO/NOGO-задачи с топограммами трех независимых компонент, изображенными ниже. В. Спектр мощности с топограммой спектра на частоте 9,5 Гц.
160 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
цингулярной извилине). Предположение о статистической независимости, по-видимому, также верно для компонент, наблюдаемых в вызванных потенциалах (для большего количества деталей см. часть 2 этой книги).
Большинство методов ICA выполняется с использованием информацион-ноых алгоритмов обучения. Различные лаборатории используют различные варианты алгоритмов обучения, такие как JADE, InfoMax ICA и FastICA. Когда эти алгоритмы были сравнены, они, в общем, работают одинаково хорошо. В референтной базе данных, описываемой в этой книге, мы используем алгоритм градиентного спуска, разработанный Скоттом Макейгом с соавторами (http://www.cnl.salk.edu/ -scott/ica.html). Надо подчеркнуть, что эти алгоритмы ICA рассматривают только статистику карт данных, зарегистрированных в различные моменты, не учитывая временной порядок этих карт112.
Пример применения ICA к сырым ЭЭГ-данным представлен на рис. 8.17. Как видим из временной динамики ЭЭГ и из сжатых частотных представлений (спектров), данная ЭЭГ генерирована тремя различными источниками, расположенными соответственно около каналов СЗ, С4 и 01,02. Эти источники соответствуют левому мю-ритму, правому мю-ритму и затылочному альфа-ритму. Средние частоты, соответствующие этим ритмам, очень близки друг к другу, составляя 9,5; 9,3 и 10,2 Гц соответственно. Мы применяли метод ICA к трехминутной записи ЭЭГ. Положения трех независимых компонент представлены ниже. Эти положения можно рассматривать как пространственные фильтры. На рис. 8.17 слева представлены результаты применения таких фильтров к сырой ЭЭГ; в середине — изображения, полученные методом LORETA для выбранных компонент.
XVI. Коррекция артефактов пространственной фильтрацией
А. Движения глаз
Электроэнцефалограмма загрязняется различными артефактами. Артефакты движения глаз зачастую самые большие в записи ЭЭГ. Они производятся вертикальными и горизонтальными движениями глаз. Главный источник артефактов — потенциал глазного яблока. Глазное яблоко действует как электрический диполь с положительным полюсом, ориентированным вперед. Моргание как результат возвратного восходящего вертикального движения глаз производит положительное отклонение (прогиб) в лобных областях с максимумом в электродах Fpl, Fp2. Закрывание глаз связано с похожим артефактом, в то время как открывание глаз как результат нисходящего вертикального движения глаз приводит к отрицательному отклонению в электродах Fpl, Fp2. Горизонтальные движения глаз (также названные саккадами) производят противоположные изменения потенциалов в электродах F7, F8. На рис. 8.18 представлена последовательность горизонтальных движений глаз (саккад) и миганий.
112 Так называемый подход слепой идентификации второго порядка (SOBI) был разработан в 1994 году.
Часть I. Ритмы ЭЭГ
161
Рисунок 8.18. Артефакты горизонтальных и вертикальных движений глаз
А. Фрагмент ЭЭГ, зарегистрированной в условии решения задачи у здорового субъекта 17 лет. Горизонтальное движение глаз (елевой стороны регистрации) с соответствующей топограммой распределения потенциалов (негативность в отведении F7 и позитивность в отведении F8). Моргание (справа) с соответствующей топограммой распределения потенциалов (позитивность в отведениях Fpl, Fp2). Б. Модель глаза как электрического диполя с положительным полюсом, ориентированным вперед.
Б. Коррекция артефактов движений глаз
Один из способов коррекции движении глаз состоит в записи окулограммы и вычислении весов влияния глазных артефактов для каждого канала ЭЭГ Далее значения сигнала окулограммы (EOG) вычитаются из первоначальных сигналов ЭЭГ в каждом канале с учетом этих весов.
Недавно был предложен новый метод, который базируется на подходе ICA. Основная идея — разложение сигнала ЭЭГ на две компоненты: ту, которая связана с нейронной электрической активностью, и другую, которая соответствует артефактам. Каждая компонента состоит из волнового сигнала, описывающего временное течение моделируемой активности, и вектора топографии, описывающего, как волновой сигнал вносит вклад в каждый зарегистрированный сигнал. Артефактная активность может быть восстановлена как результат топографии артефакта и волнового сигнала.
На рис. 8.19 представлен пример применения метода ICA для коррекции артефактов.
XVII. Другие типы артефактов
ЭЭГ может быть загрязнена артефактами различных некорковых источников. Помимо медленных изменений потенциала, связанных с вертикальными и горизонтальными движениями глаз, есть много других типов артефактов. Мы перечислим здесь только самые распространенные артефакты, такие как мышечные, кардиографические и артефакты пульса.
162
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Исходная ЭЭГ
ЛЬ Топограммы основных
А компонентов
Г7-АЛ/
Fz-AvW
П-AvW
FSAvW
T3AvW
СЗ-АЛ/ X>-KA/-vy\/yyy/V''--^^
CzAvW VvAnVv^'^'^WVV
Корректированная ЭЭГ
I” , I* ______________I
г »w i .;;
Fp1-AvW
х4мл
O1-A-Ay
02Av\v
ECG
Т5-АЛ/
РЗ-А'Л/ X
Рисунок 8.19. Коррекция артефактов посредством пространственной фильтрации
А. Фрагмент «сырой» ЭЭГ с артефактом моргания, топографии независимых компонент представлены ниже. Б. Результат фильтрации исходной ЭЭГ пространственным фильтром, вычисленным по топографиям вертикальных движений глаз. Карты внизу показывают топографии первых четырех независимых компонент наибольшая компонента соответствует вертикальным движениям глаз и наименьшая — горизонтальным движениям глаз.
А. Мышечные артефакты
Мышечные артефакты являются результатом электрической активности мускулов. В частности, лобные и височные мышцы — самый распространенный источник миогенной активности соответственно в лобных отведениях (главным образом Fpl и Fp2) и в височных отведениях (главным образом ТЗ, Т4). Пример активности височных мышц представлен на рис. 8.20. Обычно не трудно отделить мышечную активность от корковой активности в бета-диапазоне. Действительно, в спектрах диапазон мышечных артефактов обычно более широк, чем диапазон бета-активности. Из-за этого при регистрации мускульная активность выглядит как более толстая линия по сравнению с подлинной ЭЭГ113. Отдельные мышечные разряды могут напоминать эпилептические спайки, но мышечные спайки короче по продолжительности и ограничены только одним электродом.
Б. Артефакт кардиограммы
У людей с короткой шеей и большим сердцем электрические поля сердца могут регистрироваться ушными или другими базальными электродами. Трудно спутать артефакты кардиограммы с эпилептиче-
113 Нужно помнить, что центральные электроды, например Fz. Cz и Pz, расположены в местах, где нет сильных мускулов, поэтому они не регистрируют мускульную активность.
Часть 1. Ритмы ЭЭГ
Т63
А Б
Рисунок 8.20. Мышечные артефакты
А. Фрагмент ЭЭГ, загрязненный миогенными артефактами в отведениях ТЗ, Т4. Б. Спектры мощности мышечной активности (ТЗ) по сравнению с бета-активностью (Р4).
скими спайками, т.к. эти артефакты регулярны и обычно проявляются с одной полярностью во многих электродах (рис. 8.20). Одновременная запись кардиограммы обычно помогает дифференцировать эти артефакты (помечены как артефакты кардиограммы), но опытный электроэнцефалографист легко может сделать это и без такой регистрации.
В. Кардиобаллистический артефакт
Другой распространенный тип не имеющих отношения к мозгу изменений потенциалов называют вазопрессорными (кардио-баллистическими) артефактами. Этот тип артефактов вызван периодическим (с периодом биения сердца) движением электрода, расположенного сразу над кровеносным сосудом головы. Пульсация сосуда перемещает электрод, вызывая периодический артефакт. Вазопрессорный артефакт обычно наблюдается под одним электродом. Поэтому данный артефакт может быть лучше замечен при применении локального среднего монтажа (сравните линии ЭЭГ на рис. 8.22, сделанные при двух монтажах: референтном и локальном среднем монтаже). Такой артефакт хорошо заметен на карте спектров ЭЭГ как локальный пик на частоте приблизительно в 1 Гц.
164
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
XVIII. Обратная задача и оценка диполя
Как мы узнали из этой части книги, элемент ЭЭГ-генератора можно рассматривать как диполь, расположенный в пределах коры и ориентированный перпендикулярно к поверхности коры. В некоторых случаях паттернов ЭЭГ, типа эпилептиформных спайков, модель одиночного диполя может бьпь хорошим приближением нейронного генератора ЭЭГ-паттерна. Для случаев, когда генератор ЭЭГ-патгернов ограничен и может быть описан одиночным диполем, скаль-повое распределение потенциала можно выразить в физическом уравнении и ма
тематически вычислить.
В физике это называют прямой проблемой, и ее решение называют прямым решением. Чтобы решить прямую задачу, мы должны знать: 1) местоположение ди-
поля на корковой поверхности; 2) ориентацию диполя; 3) проводимость оболочек (таких как кость, кожа), которые окружают диполь. Модели трех-четырех кон-
***
P3Rd
p4Rd
6R”
центрических оболочек могут дать хорошее решение. Недавно были представлены реалистические модели головы. Они принимают во внимание такие параметры, как форма и анизотропия головы.
Если дипольная модель правильна, мы можем решить обратную задачу, то есть мы можем найти положение, ориентацию и силу диполя, который более всего соответствует наблюдаемому потенциалу. С математической точки зрения процедура представляет собой нахождение минимума функции, которая выражает раз-
Рисунок 8.21. ЭЭГ-артефакты
Фрагмент ЭЭГ с периодическим(приблизительно одно колебание в секунду) артефактом ЭКГ. Артефакт более отчетливо выявляется в отведении ТЗ, благодаря более низкоамплитудной фоновой активности ЭЭ1 в этом отведении.
личие между реальным и моделируемым распределением. Процедура может совершаться
с помощью нескольких алгоритмов. Почти все из них дают весьма хорошие и по
хожие решения.
В программном обеспечении Института мозга человека мы используем сферическую модель головы как объемного проводника с четырьмя оболочками. Эта модель, по-видимому, есть самое простое приближение, которое рассматривает мозг как состоящий из четырех концентрических сфер, представляющие соответственно кору, спинномозговую жидкость, череп и кожу головы. Решение обратной проблемы методом наименьших квадратов выполняется методом сопряженных градиентов с дополнительными ограниче
ниями.
Часть 1. Ритмы ЭЭГ 165
XIX. Электромагнитная томография низкого разрешения (LORETA)
А. Неопределенность обратной задачи
Спайки — весьма редкие паттерны ЭЭГ. В большинстве случаев паттерны ЭЭГ генерируются распределенными источниками, то есть корковыми генераторами, распределенными в удаленных областях коры.
Рисунок 8.22. Сердечно-сосудистые артефакты
Фрагмент ЭЭГ с периодическим сердечно-сосудистым артефактом. Артефакт выявляется более четко, когда используется локальный средний монтаж. Артефакт выявляется и на топограмме спектров, вычисленной для частоты пульса.
166 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
В этих случаях потенциал ЭЭГ, зарегистрированный от данного электрода — сумма многочисленных элементарных диполей, расположенных не только под электродом, но также и в отдаленных частях коры. Проблему обнаружения этих множественных диполей, зная только потенциалы, зарегистрированные множественными скальповыми электродами, называют обратной проблемой.
Теоретически обратное решение неоднозначно, т.е. есть неопределенное число различных исходных конфигураций, могут соответствовать измеренному электрическому полю114. В 1984 году Hamalainen и Ilmoniemi были первыми, кто ввел томографическое решение обратной проблемы ЭЭГ/МЭГ, хорошо известное как решение наименьшей нормы. Однако решение наименьшей нормы ошибочно располагает существующие глубокие источники на наиболее удаленную от центра кору и, таким образом, порождает чрезмерно большие ошибки в локализации корковых источников.
Лучшая точность в локализации тестовых источников дается методологическим подходом, именуемым томографией низкого разрешения (Low Resolution Tomography) или кратко LORETA. Метод LORETA был сначала представлен в 1994 году Roberto Pascual-Marqui и коллегами. LORETA базируется на некотором электрофизиологическом и нейроанатомиче-ском ограничениях. Это предположение, что соседние115 корковые области производят похожие локальные полевые потенциалы, которые электрофизиологически могут быть результатом синхронной активации соседних нейронов. С математической точки зрения эти физиологические ограничения подразумевают гладкость пространственно распределенного потенциала. В сущности, LORETA дает наиболее сглаженное из всех возможных обратных решений. Обратное решение этого метода соответствует трехмерному распределению нейронной электрической активности, которая обнаруживает максимум синхронизации (в терминах ориентации и силы) между соседними нейронными популяциями (представленными смежными вокселями).
Б. Матрица для решения обратной задачи
В подходе LORETA кора моделируется как плотная сетка объемных элементов (вокселей) в переведенном в цифровую форму атласе Талерака (Brain Imaging Center, Montreal Neurological Institute). В старой версии LORETA число вокселей было 2394, и соответствовало пространственному разрешению 7 мм.
Плотность электрического тока J в вокселях определяет потенциал Ф116 * 118. Согласно физическому Закону Лапласа, т.е. зная плотность электрического
114 Неоднозначность обратной проблемы впервые была обозначена выдающимся немецким врачом и физиком Гельмгольцем (Hermann von Helmholtz) почти 150 лет назад.
115 Соседние в этом контексте означают те, которые имеют пространственное разрешение порядка 7 мм. В наших собственных
исследованиях со вживленными электродами мы использовали пучки электродов с межэлектродным расстоянием 3 мм. Золотые электроды были стереотаксически внедрены в мозг пациентов для диагностических целей. В этих исследованиях локальные потен-
циалы, зарегистрированные от соседних электродов, в большинстве случаев выглядели весьма похожими.
118 Обратите внимание, что Ф — вектор, то есть не одно, а несколько значений, соответствующих потенциалам, зарегистрированным в различных отведениях.
Часть 1. Ритмы ЭЭГ
167
тока в коре, мы можем определить потенциалы в любой точке на скальпе по формуле117:
0=tfxJ + C,
где К — известный оператор118.
В. Минимизация функции
Математически обратное решение состоит в том, чтобы найти J, зная Ф. Один способ нахождения обратного решения может быть получен путем нахождения минимума следующей функцией:
Г=||Ф-Лху_с||2+а||ВИ</||2,
где а — параметр регуляризации, В — оператор Лапласа.
Эта функция должна быть минимизирована относительно J и с, для данных К, Ф и а. Решение в явном виде этой проблемы минимизации получено:
Г=ТФ,
где Т- (WBrBW)‘KrH [НК(WB'BWyWH + аН] ,Н = 1- 11Т/1Т1.
Как можно видеть, Т — матрица, которая может быть вычислена в явном виде для данного положения электродов, и параметр регуляризации. Зная потенциалы Ф, зарегистрированные от данного набора скальповых электродов в данном временном интервале, мы можем вычислить плотность электрического тока' источников, производящих эти потенциалы, просто умножением вектора Ф на матрицу Т Эти вычисления заранее сформированы программным обеспечением LORETA117 118 119.
Примеры применения LORETA для картирования N1-компоненты вызванных потенциалов в ответ на зрительные и слуховые стимулы представлены на рис. 8.23. Обратите внимание, что генераторы расположены в затылочной области для зрительного стимула и в височной — для слухового стимула. Эти локализации очень точно соответствуют нашим знаниям об обработке информации в мозге человека.
Здесь мы должны подчеркнуть, что все в настоящее время доступные обратные томографические представления — не точные решения вопроса, а скорее приближения к действительности. Способность корректно локализовать нейронные источники потенциалов — единственный критерий для валидации любого метода в электромагнитной томографии. Практика — критерий истинности. В течение прошлого десятилетия во многих иссле-
117 Это математическое выражение прямого решения.
118 Матрица К вычисляется согласно физической формуле, которая аналитически выражает потенциал, произведенный электрическим диполем.
119 LORETA-KEY — некоммерческое научное программное обеспечение, не связанное ни с какими коммерческими предприятиями. Программный пакет, который мы используем в базе данных Института мозга человека, тот же самый, который Roberto Pacual-Marqui и его группа используют в Key-Institute for Brain-Mind research in Zurich.
168 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Рисунок 8.23. Электромагнитная томография низкого разрешения (LORETA)
А и В. LORETA-изображения зрительных и слуховых Nl-компонент здорового субъекта. Слуховой стимул — тон 1000 Гц продолжительностью 100 мс, зрительный стимул — цифра 9, предъявляемая на 100 мс. Б. и Г. Соответствующие ВП и двухмерные топограммы.
дованиях с регистрацией ЭЭГ и ВП было показано хорошее соответствие между изображениями LORETA и ожидающимися, которые могут быть сделаны на основании различных нейрофизиологических исследований120.
Г. sLORETA — ошибки нулевой локализации
Несмотря на все предыдущие усилия, LORETA-решения производят изображения с систематическими ошибками ненулевой локализации. Одна из проблем, которые создают ошибки локализации, — изменчивость плотности корковых генераторов в корковом слое. Один из способов избегания систематической ошибки локализации состоит в вычислении и изображении стандартизи
120 Список этих публикаций может быть найден на вебсайте Key Institute: www.unizh.ch/keyinst/NewLORETA
Часть 1. Ритмы ЭЭГ
169
рованной плотности электрического тока. Роберто Паскуаль-Марки (Roberto Rascual-Marqui) в 2002 году предложил метод, в котором заключение о локализации базируется на стандартизации оценок плотности электрического тока. Свободное программное обеспечение может быть найдено на его вебсайте. На большинстве рисунков, представленных ниже в книге, мы используем изображения sLORETA.
XX. Зависимая от уровня оксигенации крови фМРТ (BOLD fMRI)
Хотя эта книга касается электрофизиологии, мы не можем избегнуть обсуждения метода функциональной магнитно-резонансной томографии. Этот метод широко используется в многочисленных лабораториях во всем мире, и в течение прошлого’десятилетия являлся главным источником наших знаний относительно мозговых функций. Параметр, который выявляется в фМРТ-исследованиях, — так называемый «зависимый от уровня оксигенации крови» (blood oxygen level dependent) или кратко BOLD. Для вычисления этого параметра сигнал идет от водородных атомов, которыми изобилуют молекулы воды в мозге121.
После воздействия радиочастотами водородные атомы испускают энергию той же самой высокой частоты, пока постепенно не возвращаются к состоянию равновесия. MPT-сканер измеряет общую сумму испускаемой энергии. Этот сигнал угасает через какое-то время вследствие различных факторов, включая присутствие неоднородностей в магнитном поле (рис. 8.24).
Большая неоднородность приводит к уменьшению интенсивности изображения, потому что каждый водородный атом испытывает немного различную силу магнитного поля, и после короткого времени (обычно называемого Т2*) испускаемые ими волны гасят друг друга. Метод BOLD фМРТ предназначен прежде всего для измерения изменений неоднородности магнитного поля в пределах каждого маленького объема ткани, которые происходят вследствие изменений в насыщении кислородом крови122.
А. Трансформационная модель фМРТ-ответа
В исследованиях корреляции ЭЭГ-фМРТ мы должны принимать во внимание, что изменения потока крови весьма медленны и следуют за быстрыми изменениями электрической нейронной активности с длительной (приблизительно в несколько секунд) задержкой. Модель гемодинамического ответа предполагает, что есть три фазы BOLD фМРТ-ответа на временное увеличение электрической нейронной активности (рис. 8.25): начальное, небольшое уменьшение ниже ба-
121 Как мы узнали во введении, метод МРТ основывается на факте, что в присутствии магнитного поля водородные атомы поглощают направленную энергию в характерных радиочастотах (-64 МГц для стандартного, клинического MPT-сканера на 1,5 Тесл).
122 Деокси- и оксигемоглобин имеют различные магнитные свойства; деоксигемоглобин является парамагнитным и вводит неоднородность в близлежащее магнитное поле, тогда как оксигемоглобин является слабо диамагнитным и имеет небольшой эффект. Следовательно, увеличение в концентрации деоксигемоглобина (из-за потребления кислорода близлежащими нейронами) может являться причиной уменьшения четкости фМРТ-изображения из-за снижения интенсивности его сигнала.
170
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Рисунок 8.24. Измерение метаболической активности методом фМРТ
Поток крови в сосуде приносит кислород к нейронам посредством оксигемоглобина. По мере того как кислород потребляется нейронами, оксигемоглобин превращается в деоксигемоглобин. Деоксигемоглобин как парамагнетик вызывает неоднородность магнитного поля, что может быть измерено падением четкости сигнала МРТ.
зовой линии вследствие начального периода потребления кислорода, которое сменяется большим увеличением выше базовой линии из-за переизбытка окисленной крови, и затем обратное уменьшение снова ниже базовой линии.
Учитывая измеренное временное течение нейронной активности, можно вычислить временное течение фМРТ-ответа. В исследованиях корреляции ЭЭГ-фМРТ это способ найти взаимосвязь между быстро изменяющимися паттернами ЭЭГ и медленными метаболическими колебаниями, измеряемыми фМРТ Надо
отметить, что большинство исследований фМРТ построено наоборот: они делают выводы о нейронной активности из фМРТ-ответа.
XXI. Корданс
Корданс (cordance) как сложная характеристика ЭЭГ была изначально предложена группой ученых из университета Калифорнии в Лос-Анджелесе (Cook и др., 1998). Корданс вычисляется нормализацией (с получением Z-множества) абсолютных и относительных значений мощности ЭЭГ и сложением этих значений. Значения корданса рассчитываются в три шага: сначала вычисляются значения мощности ЭЭГ с помощью так называемого специального монтажа, в котором значения мощности пар электродов, разделяющих общий электрод, усредняются вместе, приводя к измерению относительной мощности123. Относительная мощность рассчитывается как процент мощности в каждом диапазоне частот относительно всего спектра. Во-вторых, эти абсолютные и относительные значения мощности для каждой индивидуальной записи ЭЭГ нормализуются для электродных участков с использованием статистического Z-преобразования для каждого электродного участка — sb каждом диапазоне частот f, получая Anonn(s,f) и Rnonn(s,f) соответственно. Надо отметить, что эти Z-показатели базируются на усредненной
123 Это похоже на локальный средний монтаж, но корданс рекомбинирует значения мощности, тогда как метод локального усреднения рекомбинирует сигналы разности потенциалов.
Часть 1. Ритмы ЭЭГ
171
мощности в каждом диапазоне частот для всех электродов в пределах данной записи количественной ЭЭГ, и нет Z-показателей, относящихся к нормативной популяции (как в нейрометрическом подходе, см. ниже). Процесс нормализации приводит абсолютные и относительные значения мощности к общим единицам (единицам стандартного отклонения или Z-показателям), которые позволяют их комбинировать. В-третьих, значения корданса формируются суммированием Z-показателей для нормализованных абсолютной и относительной мощностей для каждого электродного участка и в каждом диапазоне частот:
Корданс имеет более высокие корреляции с региональным мозговым кровотоком, чем абсолютная или относительная мощность по отдельности (Leuchter и др., 1999). Эта мера использовалась для предсказания реакции на лечение антидепрессантами в группе пациентов с депрессией.
XXII. Нормальные распределения и отклонение от нормы
А. Нормативная база данных
Регистрация ЭЭГ и вычисление спектральных характеристик, таких как мощность и когерентность, — только начальные стадии анализа количественной ЭЭГ (QEEG). Самая важная стадия — сравнение вычисленных параметров с нормативными данными124. Определение нормативной базы данных как представления диапазона «нормальности» в пределах всего населения поднимает проблему того, что подразумевают под нормальным. Предполагается, что нормативная база данных должна состоять из многих людей, для которых точно установлено, что они не страдают психоневрологическими нарушениями, повреждениями мозга, не употребляют наркотики или лекарственные препараты, в анамнезе которых нет неврологических и психиатрических нарушений.
В случае если какая-то из перечисленных характеристик все-таки присуща человеку, он не может быть внесен в нормативную базу. Здоровые (нормальные) индивидуумы должны иметь хороший контакт в семье, школе или на работе и
Рисунок 8.25. Трансформационная модель fMRI-ответов
А. Нейронный импульсный ответ. Б. Гипотетическая гемодинамическая функция импульсного ответа (haemodynamic impulse response function — HIRF), оцениваемая как гемодинамический ответ на краткий всплеск нейронной активности.
124 Здесь надо подчеркнуть, что мы говорим о количественной ЭЭГ, а не о клинической (медицинской) оценке ЭЭГ. Последняя основывается на визуальном просмотре и поиске значащих паттернов ЭЭГ. таких как спайки, медленные волны, и различных типов пароксизмов. Анализ количественной ЭЭГ не исключает, а фактически подразумевает клиническую оценку ЭЭГ как один из необходимых шагов.
172
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
в обществе. База данных должна представлять различные возраста, пол, этническую принадлежность, социально-экономические статусы и другие демографические факторы, представленные в целостном обществе. Это критерии включения человека в нормативные базы данных. Особое отношение к педиатрическим базам данных, где коренные изменения, связанные с развитием, происходят в течение относительно коротких временных интервалов. Для создания таких баз данных необходимы ЭЭГ очень многих детей каждого года возраста125.
Б. Нормальные и логарифмически нормальные распределения
Известный факт, что любой параметр в нормальной популяции, если он зависит от многих факторов, соответствует нормальному (гауссовскому)126 или
-2сг < z < +2 а Р < 0-05
-2.6g < z<+2.6<т р<0 01
-3g<z<+3g р< 0.003
Рисунок 8.26. Нормальное распределение См. текст.
логарифмически нормальному распределению127. В случае логарифмически нормального распределения применяется так называемая процедура нормализации, которая заключается в логарифмировании значений параметра.
Гауссовское (нормальное) распределение данной случайной величины х характеризуют только два параметра — среднее (X) и стандартное отклонение (о). Среднее отражает усредненное значение параметра, в то время как стандартное отклонение отражает, насколько индивидуальные параметры отличаются от этого среднего значения.
В. Z- показатель
Если для индивидуального параметра х (например, мощности ЭЭГ в диапазоне частот 9—10 Гц, вычисленной для отдельного индивидуума) разделить отклонение от среднего этой величины на стандартное отклонение, то мы получим меру отклонения от средней нормы для индивидуального значения х. В статистике эту меру называют Z-показателем:
Z=(x-X)/<r.
Распределение Z-показателя представлено на рис. 8.26, где ось У представляет плотность вероятности. Так, например, вероятность Z-показателя
125 Фактически никакая база данных не соответствует этим строгим критериям. Следовательно, в настоящее время доступные базы данных более подходяще рассматривать как референтные, а не нормативные базы данных. Однако для практических целей базы данных обеспечивают чрезвычайно существенную информацию и они позволяют практику весьма точно оценить отклонения от нормальности конкретного пациента.
126 В самой простой форме центральная предельная теорема утверждает, что сумма большого числа независимых наблюдений из одного и того же распределения имеет при некоторых общих условиях нормальное распределение. Опытным путем замечено, что различные природные признаки, такие как высота индивидуумов, имеют приблизительно нормальное распределение. Предложено объяснение, что эти признаки являются суммами большого количества независимых случайных эффектов и. следовательно, приблизительно нормально распределены согласно центральной предельной теореме.
127 Переменная может быть смоделирована как имеющая логарифмическое нормальное распределение, если ее можно считать мультипликативным продуктом (а не суммой, как в нормальном распределении) многих независимых факторов.
Часть I. Ритмы ЭЭГ
173
меньше чем 2 и больше чем —2 определяется областью под соответствующей кривой. Для |Z| < 2 эти области очень малы и составляют приблизительно 0,05. Это определяет вероятность, с которой Z-показатель у нормального индивидуума будет меньше чем 2. В клинической практике уровень достоверности 0,05 рассматривается как надежный показатель отклонения от нормы.
XXIII. Доступные в настоящее время базы данных
A. NxLink
Первая нормативная база данных была разработана группой исследователей из Нью-Йоркского университета, возглавляемой Роем Джоном, в конце 1980-х. Термин «нейрометрика» (neurometries) сначала применялся этой группой для описания аналогии с психометрической оценкой, обычно используемой в клинической психологии. В нейрометрической базе данных присутствуют следующие параметры ЭЭГ: абсолютная мощность, относительная мощность, когерентность, средняя частота в пределах диапазона ритма и симметрии (левая — правая и передняя — задняя), рассчитанные приблизительно с двухминутной записи ЭЭГ (без удаления артефактов) с закрытыми глазами. Проанализированный диапазон частот ЭЭГ от 0,5 до 25 Гц. Извлеченные параметры преобразованы до гауссовского (нормального) распределения. Две тысячи восемьдесят четыре (2084) переменные вычислены для каждого члена базы данных. Была отмечена корреляция особенностей ЭЭГ с возрастом и были получены уравнения регрессии для учета эффектов возраста. Одномерные и многомерные Z-параметры (Z-scores) были вычислены для характеристики отклонений индивидуума от среднего по популяции. База данных NxLink включает измерения приблизительно от 782 нормальных индивидуумов. Из этого общего количества 356 — в возрасте 6— 16 лет и 426 — в возрасте 16—90. Более чем 4000 клинических наблюдений использовались в дискриминантной части программного обеспечения. Ограничения базы данных следующие: 1) были проанализированы и нормализованы записи ЭЭГ только в состоянии покоя с закрытыми глазами128; 2) сравнения сделаны только с частотными полосами ЭЭГ, такими как дельта-, тета-, альфа- и низкочастотный бета-диапазоны129.
Б. Neuroguide
База данных, разработанная Робертом В. Тэтчером (Thatcher Lifespan Normative EEG Database — Neuroguide), в настоящее время содержит информацию от 625 индивидуумов, охватывая возрастной диапазон от 2 месяцев до 82,6 года. Используются передовые методы, включающие: более обширную перекрестную проверку и
128 Недавние исследования показывают, что диагностическое значение ЭЭГ в состоянии с закрытыми глазами ниже, чем ЭЭГ в состоянии с открытыми глазами и в условиях решения задачи. Действительно, доминирующий затылочный альфа-ритм. представленный в большинстве записей в состоянии с закрытыми глазами, маскирует другие ритмы, представленные в ЭЭГ.
129 Анализ тонкой структуры ритмов ЭЭГ невозможен при таком подходе. Однако, как мы видели выше, идентификация нескольких независимых ритмов в пределах широкого диапазона может иметь клиническое значение.
174 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
проверку гауссовских распределений для среднего взвешенного монтажа, для монтажа с объединенными ушными электродами, Лапласиана в состоянии с открытыми и закрытыми глазами. База данных NeuroGuide была проверена и перепроверена, для каждого монтажа и состояния была рассчитана чувствительность статистических распределений. Девятьсот сорок три (943) переменные вычислены для каждого субъекта, включая значения абсолютной и относительной мощности, когерентности, фазы, асимметрии и отношений мощности. Значения Z-параметров доступны в отдельных одночастотных интервалах. Скользящее среднее используется для вычисления нормы, соответствующей возрастам. Результаты проверены на гауссовское распределение. Главное ограничение этой базы данных — отсутствие регистрации ЭЭГ в условиях выполнения задач.
B.SKIL
База данных SKIL, разработанная Барри Стерманом и Дэвидом Кэйзером, в настоящее время включает 135 взрослых в возрасте 18—55 лет. Референтная популяция состоит из студентов и лабораторного персонала (50 %), волонтеров, набранных из населения (25 %), и персонала американских воздушных сил (25 %). База данных SKIL включает записи ЭЭГ в покое (при закрытых и открытых глазах) и во время состояния решения задач, включая чтение, математические задачи и некоторые другие. На данный момент доступна коррекция записей, которая базируется на объединенных перекрестной и лонгитюдной информации, а не предпочтительном методе отслеживания внугрисубъектных изменений с течением времени. База данных SKIL охватывает ограниченный диапазон частот от 2 до 25 Гц. База данных содержит нормы для каждого единичного приращения в Гц в этом диапазоне частот. В базе данных SKIL используется исключительно объединенный ушной референт. Ограничения базы данных: 1) никакая нормативная информация, касающаяся детей или молодых взрослых, в настоящее время не доступна; 2) задачи не были стандартизированы, чтобы рассчитать вызванные потенциалы.
Г. Neurorep
Билл Хадспет (Bill Hudspeth) предлагает Neurorep AQR (база эталонных ЭЭГ-данных для взрослых130). Одна из самых полезных особенностей работы Хадспета — акцент на надежности измерений, полученных от отдельных пациентов, и важности изменчивости ЭЭГ во времени как клинического индекса. Данные ЭЭГ доступны для состояний с открытыми и с закрытыми глазами. База данных включает значения абсолютной и относительной мощности для 19 скальповых электродов и всех комбинаций пар электродов для когерентности, фазы, асимметрии и индексов корреляции. Общее количество индивидуумов в AQR в настоящее время довольно незначительное (< 50), но собираются дополнительные данные.
Д. Франк X. Даффай, MD
Франк X. Даффай, MD, также разработал базу данных ЭЭГ, которая включает состояния с открытыми и с закрытыми глазами и охватывает широкий
130 Можно найти на www.neurorep.com
Часть 1. Ритмы ЭЭГ
175
возрастной диапазон, в том числе детей и взрослых. Доступны данные ЭЭГ для 19 электродов, включены также значения слуховых и зрительных ВП. Эта база данных прежде использовалась в нескольких коммерческих нейродиаг-ностических инструментах (Nicolet, QSI), но, как нам известно, в настоящее время недоступна.
Е. База данных Novatech LORETA
Сначала база данных LORETA была разработана Марко Конхедо и Лесли Шер-лином из NovaTech. В базе данных в настоящее время имеются 84 наблюдения и активно добавляются новые. Эта ЭЭГ-технология визуализации дает возможность томографического представления источников ЭЭГ в 3-мерном пространстве. Эта база данных полезна не только для выявления отклонений, но и для приблизительной оценки локализации областей мозга, вовлеченных в заданную деятельность.
Ж. База данных BRC
Одно из самых захватывающих событий в области создания баз данных ЭЭГ в течение прошлых нескольких лет — разработка первой стандартизированной Международной базы данных мозга. В ней преодолеваются проблемы баз данных, представленных выше. Проводились консультации с консорциумом ведущих нейрофизиологов и неврологов для оптимального выбора тестов, которые задействуют главные мозговые сети и процессы и позволят их оценить в течение короткого времени. Были устроены шесть лабораторий с идентичным оборудованием и программным обеспечением (в Нью-Йорке, Род-Айленде, Лондоне, Голландии, Аделаиде и Сиднее) под эгидой общественной компании (The Brain Resource Company131), с новыми лабораториями, которые будут добавляться в дальнейшем. Были зарегистрированы сотни нормальных испытуемых, также недавно началась оценка групп клинических пациентов.
База данных BRC включает данные не только ЭЭГ- и ERP-параметров при выпол нении батареи простых задач (такие как слуховые и зрительные oddball -задачи), но также при выполнении батареи разносторонних психологических тестов, регистрация при этом проводилась при использовании сенсорных мониторов132. Также для многих отдельных индивидуумов получены структурные и функциональные MRI-данные. В дальнейшем будет систематически собираться генетическая информация для сравнения с нейроанатомическими, нейрофизиологическими и психометрическими измерениями.
3. База данных HBI
База данных HBI была недавно разработана как совместное научно-практическое предприятие трех человек: программиста Валерия Пономарева, психолога Андреаса Мюллера и автора этой книги. Эта база данных базируется на методологии, разработанной в лаборатории нейробиологии
131 Информация об этой компании может быть найдена на www.brainresource.com
132 Батарея психологических тестов включает тесты на время реакции, на оценку времени, на краткосрочную память, на воспроизведение из памяти, на оценку беглости речи, на переключение внимания и др.
176 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
программирования действия Института мозга человека Российской академии наук (зав. лаб. Ю.Д. Кропотов). Методология представлена в настоящей книге, а история создания базы данных описана во «Введении» к книге. База данных включает ЭЭГ, зарегистрированную в состоянии с открытыми и закрытыми глазами в состоянии спокойного бодрствования и в условиях выполнения пяти различных задач (двухстимульной GO/NOGO-задачи, математической задачи, чтения, слуховой задачи на воспроизведение и слуховых oddball-задач) у более чем 1000 относительно здоровых испытуемых в возрасте от 7 до 98 лет. База данных также включает записи больше чем 1000 детей и подростков с диагнозом СДВГ, также как многочисленные записи ЭЭГ пациентов с другими диагнозами (пациенты с эпилепсией, обсессивно-компульсивным синдромом, склонностью к депрессиям, травмами шеи и т.д.)
Чтобы уменьшить время для предварительной обработки данных, были автоматизированы некоторые процедуры (такие как коррекция артефактов и обнаружения спайков). Для оценки общих и локальных характеристик ЭЭГ используется три монтажа (монтаж с объединенным ушным референтом, средний и взвешенный средний референт). Абсолютная и относительная амплитуда и спектры мощности, средняя и парная когерентность, волновой (вейвлет) анализ и ВП-параметры вычислены и нанесены на карту в двухмерные или в трехмерные изображения, с использованием технологий LORETA и sLORETA. Также доступны методы оценки диполя. Кроме того, ВП подвергнуты анализу независимых компонент. С использованием этого метода выявлены отдельные компоненты, связанные с отдельными психическими операциями. Каждая компонента характеризуется временной динамикой и распределением. На основе этих распределений созданы пространственные фильтры, которые позволяют пользователям выявить амплитуду и латентность каждой компоненты индивидуального ВП. Сравнение этих параметров с нормативными данными создает понимание различных стадий обработки информации у людей.
Глава 9. Практика
I. Введение
Книга снабжена обучающим программным обеспечением. Программное обеспечение — компьютерная программа (Ed-EEG.exe), которая использует основные алгоритмы обработки ЭЭГ, описанные в книге133. Программное обеспечение можно загрузить с сайта http://www.meddia.com.ua. Постоянная страница программы: http://www.meddia.com.ua/edeeg. Кроме программного обеспечения с этой страницы можно загрузить файлы ЭЭГ, зарегистрированные у нормальных субъектов и пациентов Для этой части книги мы выбрали регистрацию ЭЭГ в состояниях спокойного бодрствования с открытыми и закрытыми глазами. Надо подчеркнуть, что мы не собираемся представлять полный атлас различных паттернов ЭЭГ134. Обучающая цель программного обеспечения скорее состоит в том, чтобы помочь читателю попробовать использовать основные методологические принципы анализа фоновой ЭЭГ.
А. Виды (категории) обработки ЭЭГ
Целый набор методов анализа фоновой ЭЭГ может быть разделен на следующие категории: 1) управление данными в ЭЭГ-формате; 2) предварительная обработка ЭЭГ, включающая коррекцию артефактов; 3) представление электрических потенциалов на голове в форме корковых генераторов посредством электромагнитной томографии или определения параметров эквивалентного диполя; 4) преобразование Фурье, которое включает вычисление абсолютных и относительных спектров ЭЭГ для отдельных каналов, когерентности для пар каналов; 5) вейвлет-анализ, включающий вычисление вызванной де-/синхро-низации (ERD/ERS); 6) сравнение с нормативной базой данных, включающее вычисление Z-значений как меры статистической значимости отклонения от нормального распределения, вычисленного для здоровых субъектов соответствующего возраста; 7) управление файлами, в которых сохраняются результаты анализа ЭЭГ; 8) компиляция заключения, т.е. представление результатов обработки в компактной осмысленной форме с заключениями и рекомендациями для терапии.
133 Программное обеспечение было написано Валерием Пономаревым — старшим научным сотрудником в лаборатории автора книги из Института мозга человека Российской академии наук в Санкт-Петербурге. Оно базируется на методологических принципах, развиваемых в лаборатории. Расширенная версия этого программного обеспечения коммерчески доступна.
134 Для современного атласа паттернов ЭЭГ и соответствующих изображений LORETA см. Zumsteg и др. (2004).
178
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Экспорт файлов
Импорт файлов
Конвертирование файлов в другой формат
Изменение монтажей
Коррекция , артефактов V
Двумерное картирование
Электромагнитная томография
Дипольное /
приб.пижение V
Абсолютный и относительный . / спектр v
Вейвлет анализ
Управление данными
Сравнение с / нормативной v базой данных
Составление заключения
Когерентность
Вызванная де/синхрон» гз алия
Вычисление средних групповых
Рекомендации
для терапии
Би спектр
Когерентность связанная с событием
Статистический анализ
Рисунок 9.1. Методы обработки ЭЭГ
Методы разделены на категории (представлены сверху вниз в рядах). Отмечены методы, доступные в программе Ed-EEG. А. Управление форматами ЭЭГ-данных. Б. Предварительная обработка ЭЭГ. В. Двухмерное картирование ЭЭГ и LORETA. Г. Анализ Фурье. Д. Волновое (вейвлет) преобразование. Е. Управление файлами. Ж. Сравнение с нормативной базой данных. 3. Автоматизированное составление заключения.
Эти виды обработки ЭЭГ схематично представлены на рис. 9.1. Для образовательного программного обеспечения мы выбрали только несколько видов анализа ЭЭГ. Те методы, которые не включены в программу Ed-EEG, кратко упомянуты ниже, чтобы дать читателю представление о том, что данные методы могли бы предложить практику.
Б. Форматы данных ЭЭГ
Как мы узнали из этой части книги, ЭЭГ регистрируется посредством усилителей. Усилители увеличивают низкоамплитудные потенциалы с человеческой головы в достаточно большие сигналы, которые далее преобразовываются в цифровой формат посредством аналогово-цифрового преобразователя (АЦП). Чтобы прочитать выходную информацию с АЦП, программное обеспечение, в свою очередь, нуждается в специальной стандартной программе, называемой
Часть 1. Ритмы ЭЭГ
179
драйвером135. Драйвер позволяет программному обеспечению «читать» значения мозговых потенциалов от АЦП и хранить их на диске. Есть несколько стандартных протоколов (форматов) хранения ЭЭГ-данных надиске, такие как Европейский формат данных (с расширением .edf), Универсальный формат данных (с расширением .udf) и EEG ASCII-формат (с расширением .txt). Файлы данных хранят числа, соответствующие потенциалам, измеренные в последовательных интервалах времени в каждом из ЭЭГ-каналов в форме таблиц (каналы — время). Кроме того, файлы данных хранят дополнительную информацию, такую как имя пациента, дата рождения, число каналов, частота отсчетов. В дополнение к стандартным форматам данных некоторые компании создают собственные форматы данных, специальные для их систем регистрации/обра-ботки ЭЭГ136.
В. Управление данными
Когда число записей, которое делает пользователь в своей практике, превышает несколько сотен, становится трудно управлять данными137. Специальные инструменты позволяют пользователю манипулировать данными. Один из них — индивидуальная база данных138. Индивидуальная база данных ЭЭГ — это структурированное собрание записей ЭЭГ, полученных пользователем. Записи сохранены в компьютере в специальном порядке так, чтобы управляющая программа могла обращаться к ним, чтобы ответить на вопросы. Например, нативные ЭЭГ-записи и результаты их обработки (такие как спектры, вейвлет-преобразования или вызванные потенциалы) обычно хранятся в различных папках. Эти различные категории файлов описываются в соответствии с определенной схемой. Например, схема описывает объекты, которые представлены в базе данных именем пациента, датой рождения, датой регистрации, пола, диагноза, вычисленных параметров и т.д. Схема становится важной, когда мы хотим получить усредненные значения для данного возраста, пола или диагноза. Эго также помогает искать отдельного пациента или отдельную дату регистрации. Компьютерная программа, используемая для управления и запроса информации из базы данных, известна как система управления базой данных. Первые системы управления базы данных появились в 1960-х. В области ЭЭГ они стали стандартным инструментом.
Г. Редактирование и компилирование QEEG-заключения
Просмотр и анализ ЭЭГ-данных обычно сопровождаются компиляцией заключения относительно ЭЭГ пациента. Для этого используются специаль
135 В общих чертах драйвер — это программа, которая управляет устройством. Каждое устройство типа принтера или усилителя ЭЭГ должно иметь драйвер. Обычные общие драйверы входят в операционную систему. Для ЭЭГ-усилителя вам необходимо загрузить новый драйвер, когда вы соединяете усилитель с вашим компьютером. В среде Windows драйверы часто имеют расширение SYS. Драйвер работает подобно переводчику между устройством и программой, которая использует устройство, принимая команды из программы и переводя их в специализированные команды для устройства.
136 Например, компания Lexicor имеет специальный формат данных с расширением *.dat, в то время как ООО Mitsar использует специальный формате расширением *.EEG.
137 Например, пользователь хочет проверить предположение, что во многих ЭЭГ-записях пациентов с травмами шеи, которые посетили его офис в течение прошлого года, наблюдается превышение альфа-активности в височных областях. Чтобы проверить эту гипотезу, пользователь должен выбрать данные всех пациентов с травмами шеи определенного возраста (например, от 20 до 30 лет), усреднить данные для определенного состояния (например, получить усредненные спектры в состоянии с открытыми глазами) и сравнить результате нормативными данными.
138 Не путайте с нормативной базой данных. Индивидуальная база данных включает данные, полученные индивидуальным практиком.
180 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
ные подпрограммы, позволяющие практику вносить в заключение всю доступную информацию относительно нативных данных (например, фрагменты ЭЭГ в различных состояниях) и результатов обработки (такую как мощность, амплитуда или относительные спектры ЭЭГ, вызванная десинхронизация и т.д.). Компиляция заключения также включает интерпретацию наблюдаемых результатов и терапевтические рекомендации, в том числе рекомендацию для протоколов процедур нейробиообратной связи и размещения электродов при транскраниальной магнитной стимуляции (Direct Current Stimulation).
II. Программное обеспечение Ed-EEG
А. Установка
Перед началом установки загрузите со страницы http://www.meddia.com.ua/ edeeg архив дистрибутива программы: файл EDEEG_SETUP.ZIP. Извлеките из архива папку EdEEG. В папке EdEEG должен быть файл SETUP.EXE. Запустите его и следуйте инструкциям программы установки. Нажмите кнопку Finish, чтобы закончить установку Программа будет установлена на жестком диске вашего компьютера в папку Ed-EEG.
Б. Папки для обработки данных
В подпапке DATA (рис. 9.2) вы найдете файлы, в которых хранятся цифровые записи ЭЭГ, зарегистрированные у здоровых субъектов и у пациентов. Файлы имеют расширение .eeg139. В папке находятся четыре файла ЭЭГ: S1EO ЕС, S2 E0 ЕС, S3_VCPT, S4 VCPT, зарегистрированные у здоровых субъектов и у пациентов соответственно «Глаза открытые/Глаза закрытые» (ЕО ЕС) в состояниях спокойного бодрствования и во время выполнения двухстимульной GO/NOGO-задачи. Помимо подпапки DATA, главная пап-
Ed-EEG
СПЕКТРЫ ВЫЗВАННАЯ i ВЫЗВАННЫЕ
: НРИМ4РЫ | СТШПГЫ Н дасиюфОНИЗАЦИЯ J ПОТИЩИАЛЫ!
/ ' **.spc **.erd **.erp
I ЗДОРОВЫЙ ; | ПАЦИЕНТ "1
ИСПЫТУЕМЫЙ ; '
**.eeg **.eeg
Рисунок 9.2. Организация файлов в папке Ed-EEG
На схеме: внутри прямоугольников — имена папок, под прямоугольниками — расширение файлов, которые хранятся в соответствующей папке.
139 Файлы включают 19-канальную ЭЭГ + 1 канал меток нажатия кнопки, зарегистрированную усилителями Mitsar (ООО Mftsar, Санкт-Петербург, Россия).
Часть 1. Ритмы ЭЭГ
181
Не View Analysis Setup
Рисунок 9.3. Главное окно, окно диполя и окно топограмм программы Ed-EEG
Вид компьютерного экрана после старта EdEEG.exe. Большое окно — главное (ЭЭГ) окно, которое позволяет пользователю просматривать записи ЭЭГ и анализировать ЭЭГ различными методами. Маленькое левое окно — окно диполя, которое изображает положение и ориентацию электрического диполя, соответствующего выбранному распределению потенциалов скальпа. Маленькое правое окно — окно топограмм, которое изображает двухмерные карты скальповых потенциалов, соответствующие моментам времени, выбранным левым и правым маркерами.
ка Ed-EEG включает три подпапки по имени SPEC, WAVELET и ERP. Эти подпапки будут хранить результаты анализа ЭЭГ, который вы выполните во время практики с программным обеспечением. Подпапки, как предполагается, будут содержать файлы с соответствующими расширениями: **.spc для спектров, **.erd для вызванной де-/синхронизации и вейвлет-преобразования, **.егр для вызванных потенциалов.
Для освоения основных принципов анализа фоновой ЭЭГ следуйте шагами, описанными ниже. Обратите внимание, что в развитии программного обеспечения мы следовали главным принципам продуктов Microsoft. Так, для тех, кто использует платформу Windows MS, функциональное значение и положение главных меню будут казаться знакомыми.
В. Шаг 1. Открытие ЭЭГ-файла
Запустите EdEEG.exe программу из папки Ed-EEG. Будут открыты следующие три окна (рис. 9.3): главное окно, окно диполя (слева) и окно карты
(справа).
В главном окне вы можете видеть следующие меню: файл, вид, анализ, установки. Каждое меню имеет свои собственные команды (рис. 9.4). Некоторые из самых распространенных представле-
Decrease Gain NUM -increase Gain NUM ♦
SE Decrease Speed /
55 increase Speed NUM ♦
Select Mortage...
S3 Man Arif sets. d Artifacts correction. .
Bsndiarqs filter
Comparison of resets.,
ны на панели инструментов (Toolbars).
Чтобы просмотреть запись
Рисунок 9.4. Команды меню «Файл», «Вид» и «Анализ»
Ниже приведены основные команды и соответствующие инструменты меню «Файл», «Вид» и «Анализ».
ЭЭГ, сохраненную в подпапке DATA, перейдите в меню «Файл» и щелкните ко-
манду «Открыть файл». Выберите любой файл и откройте его. Откроется окно
ЭЭГ, подобное представленному на (рис. 9.5).
182
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Sp<»dj30inm/t й*п 3 В****! ООО | Low Cut |*]|'оз (O.53Hj)jj High Cut |H»| [эО 3 N<*ch I <5®
"3 <. Панель
Fpl Av 50^/ст Fp2Av
F7Av 50iA7cm F3Av
F4Av 50<Л>/ст
F8Av 50tV/cm
фильтров Панель времени
Выбранный интервал
T5Av
P3Av
РгА.
01 Av SQ.^.
02Av 50t*7em
!
Панель
названий каналов
Окно диполя
Окно топограммы
Панель
3t.5uM T[R-ty»^.2«)s Я I- 37.6UV А“ <8.1 uv <---- СТЯТуСв
Левый вертикальный маркер Правый вертикальный маркер
Рисунок 9.5. Просмотр фрагмента ЭЭГ
Изображены основные компоненты, необходимые для просмотра и простой предварительной обработки. Панель фильтров — параметры цифровой фильтрации для установки параметров фильтра низких частот, фильтра высоких частот и режекторного фильтра. Панель времени показывает текущее время регистрации в секундах. Левые и правые вертикальные маркеры времени активны в пределах этой панели. Панель каналов содержит имена каналов («Активный» электрод — Референт) и чувствительность в мкВ/см. Строка текущего состояния — потенциал под левым (L) и правым (R) маркерами выбранного канала. Т [R-L] — временной интервал между правыми и левыми маркерами. А — амплитуда ЭЭГ (от пика до пика) для выбранного интервала. F — средняя частота в выбранном интервале. Окно топограмм - двухмерные топограммы потенциалов в левом и правом маркерах времени. Окно диполя — два эквивалентных диполя, которые соответствуют распределению корковых потенциалов под левым и правым вертикальным маркерами.
Г. Шаг 2. Просмотр записи ЭЭГ
Главное (ЭЭГ) окно используется, чтобы просмотреть зарегистрированную ЭЭГ. Оно имеет: панель имен каналов (размещенную в левой стороне компьютерного экрана), строку текущего состояния (размещенную в основании) и панель фильтров (размещенную наверху). Панель имен каналов используется для определения и выбора каналов.
Выбор канала выполняется просто, щелчком на прямоугольнике с соответствующим названием канала в панели имен каналов (например, Fz). Прямоугольник с названием канала подсвечивается и считается выделенным. Выбор фрагмента времени, необходимого для дальнейшего анализа производится двумя вертикальными маркерами, в пределах панели времени, размещенной сразу над записью ЭЭГ. Числа в панели времени соответствуют времени регистрации в секундах. Левый вертикальный маркер времени устанавливается щелчком левой кнопкой мыши в пределах панели времени. Правый вертикальный маркер устанавливается щелчком правой кнопки мыши. Выбранный интервал подкрашивается наверху.
Часть 1. Ритмы ЭЭГ
183
Двухмерные (2D) карты потенциалов в левом и правом маркерах времени представлены в окне карт соответственно на левой и правой сторонах окна. В дополнение к 2В-картам программное обеспечение вычисляет параметры двух эквивалентных диполей, которые соответствуют распределению корковых потенциалов, измеренных в «левом» и «правом» моментах. Эти диполи представлены в окне диполя в красном цвете для «левого» и в синем цвете для «правого» момента времени. И наконец, программное обеспечение позволяет пользователю выполнить электромагнитную томографию (LORETA) как для мгновенных потенциалов, соответствующих левому и правому маркерам, или на кросс-спектрах ЭЭГ, вычисленных для интервала между маркерами140. Чтобы выполнить LORETA на потенциалах ЭЭГ, определенных двумя временными маркерами, выполните «Анализ» —► «LORETA» для потенциалов ЭЭГ. Чтобы выполнить LORETA на кроссспектрах ЭЭГ между этими двумя маркерами, выполните «Анализ» —> «LORETA» для мощности ЭЭГ. При выполнении в первый раз программа запросит путь к главной папке программного обеспечения LORETA, которое должно быть уже установлено на компьютере пользователя.
Строка текущего состояния окна ЭЭГ, представленная у нижней границы окна ЭЭГ, показывает параметры для выбранного канала, такие как L — амплитуда потенциала в выбранном канале в левом маркере, R — в выбранном канале в правом маркере, Т (R—L) — временной интервал между маркерами, R—L — различие потенциалов между маркерами, А — амплитуда ЭЭГ для выбранного интервала (различие между максимальным и минимальным значениями), F — доминирующая частота фрагмента ЭЭГ в отобранном канале.
Д. ШагЗ. Предварительная обработка записи ЭЭГ — фильтрование
Панель фильтров — это орган управления, который позволяет пользователю установить параметры цифровых фильтра низкой частоты и фильтра высокой частоты, также как изменять масштабы времени (скорость) и амплитуды (чувствительность). Выпадающее меню «Скорость» используется, чтобы выбрать горизонтальный (время) масштаб. Выпадающее меню «Чувствительность» используется, чтобы выбрать вертикальный (амплитуда) масштаб. Чтобы изменить чувствительность только для отобранного канала, держите нажатой клавишу Ctrl. Выпадающее меню ФНЧ (Гц) используется, чтобы выбрать полосу пропускания ЭЭГ и отсечь высокие частоты. Для изменения фильтра высоких частот только для выбранного канала нажмите и удерживайте клавишу Ctrl. Выпадающее меню ФВЧ (с) используется, чтобы выбрать полосу пропускания ЭЭГ и отсечь низкие частоты. Чтобы изменить полосу пропускания низких частот только для выбранного канала, нажмите и удерживайте клавишу Ctrl, иначе фильтр низких частот будет изменен для всех каналов, видимых в монтаже. Выпадающее меню «50 (60) Гц» используется для включения (выключения) режекторного фильтра 50 (60) Гц. Чтобы изменить фильтр только для выбранного канала, удерживайте клавишу Ctrl. Поле изолинии
140 Чтобы имелась возможность выполнить эту опцию, пользователь должен установить некоммерческое программное обеспечение LORETA. Программное обеспечение находится на сайте KEY Institute for Brain-Mind Research in Zurich, Switzerland http://www.uzh.ch/keyinst/NewLORETA/. Программа была разработана Roberto D. Pascual-Marqui. При установке программы LORETA следуйте за инструкциями на сайте.
184 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
используется, чтобы изменять изолинию регистрации ЭЭГ. Чтобы изменить изолинию только для выбранного канала, держите нажатой клавишу Ctrl.
В некоторых случаях141 необходимо отфильтровать записи ЭЭГ-фильтра в узком диапазоне частот. Эго производится полосовым фильтром в меню «Анализ» (рис. 9.6). Но перед выполнением этой операции пользователь должен определить параметры фильтра. Это можно сделать в меню «Настройка» —> «Стандарт-
Ц........t............И...,....г...1
Fpl-AvW”
Частотные диапазоны ЭЭГ
Параметры частотной фильтрации
Рисунок 9.6. Цифровая фильтрация ЭЭГ
А. Нативная ЭЭГ. Б. Отфильтрованная ЭЭГ. В. ЭЭГ-диапазоны, обычно используемые для фильтрации. Диапазоны могут быть изменены пользователем, также могут быть добавлены новые диапазоны. Г. Параметры фильтра.
141 Например, при вычислении вызванной де-/синхронизации (ERD/ERS) или выполнении электромагнитной томографии (LORETA).
Часть 1. Ритмы ЭЭГ
185
ные диапазоны» ЭЭГ. В примере, представленном на рис. 9.6, мы выбрали цифровой фильтр Кайзера порядка 512 в пределах диапазона частот 9,00-11,00 Гц. Можно также использовать параметры по умолчанию, такие как стандартные диапазоны ЭЭГ или диапазоны в 1 Гц.
Е. Шаг 4. Предварительная обработка записи ЭЭГ — установка монтажа
ЭЭГ записывается на диске в монтаже с объединенными ушными электродами в пределах всей полосы пропускания ЭЭГ-усилителя. Для просмотра и анализа данных может быть выбран другой монтаж. Это можно сделать нажатием команды «Выбор монтажа» в меню «Вид». Появится окно «Задание монтажа». В нем есть пять закладок: каналы, референты, цвета, вид, электроды. Обратите внимание на различие между электродами и каналами. Электроды — физические места, где
расположены металлические электроды. Каждое положение электрода определяется двумя сферическими координатами на голове142. Они представлены на рис. 9.7 как значения тета и фи.
Канал по определению — относительная запись, которая измеряет разницу потенциалов между физическим электродом и референтом. Запись канала изо-
Channels | Refetenls | Colots | View ВЬ&йЙОГ |
Montage name [DataBase jrj
Theta Ph Theta Phi Theta Phi Theta Phi Theta Phi
ТРз’|?2~*Г^ Fhpz|72~|9O“ P [тГ* pT” F
Fh7|72~|U4 H рТрЗ? P s [зГрЙГ F И [ЙГ [ЙГ P Тё |72~[36~
P npFfwo p сз|зб~[т80 Fc,|F"|iT~ РС4[зГ'рГ~ Р^|72~рГ“
F T5|F“|zie P P3 pe“|225 F P? [з(Г|270 Рм|ЙГ[315 РГ&|тГ'|323Г
PaI PUipF|^ Р=ъ[^“|270 pQzpFpSB PAz
Г
F F 8=o2 F Eeg F Bic4 F 8ю5 F 8»E F S»7 F
P Connect channels A1 andA2
Рисунок 9.7. Координаты электродов в системе 10-20
Каждое положение электрода определяется двумя сферическими координатами на голове — значениями Theta и Phi.
бражена в главном (ЭЭГ) окне как амплитудно-временной график. Несколько типов референтов представлены в диалоговом боксе «Вид» (рис. 9.8). Слева направо: объединенный ушной электрод (Ref), средний (Av) и взвешенный средний (AvW).
Чтобы определить канал, необходимо определить параметры регистрации, представленные на рис. 9.9: электрод (для активного участка при регистрации) и референт (для референтного участка при регистрации). Электрод и референт
устанавливаются для определенного канала просто нажатием соответствующих имен на электроде и референте. Обратите внимание, что пользователь может проектировать свой собственный монтаж, используя эту простую процедуру. Для добавления канала используется кнопка «Добавить», чтобы удалить канал, используется кнопка «Удалить». В монтаже по умолчанию все эти параметры определены, и пользователь может не беспокоиться о них. Но в некоторых случаях параметры должны быть определены конкретным пользователем.
Мы рекомендуем начать с монтажа «База данных». Для того чтобы выбрать этот монтаж, вы должны только выбрать «База данных» в списке монтажа в меню «Референты» (рис. 9.10).
142 Система координат следующая ось X идет от левого уха до правого, ось Y — от затылка до лба, ось Z направлена вверх.
186
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Channels | Referents ; Colon View j Electrodes ;
Montage name СЙЗПЖ
Рисунок 9.8. Вид монтажа электродов
Электроды схематично представлены в прямоугольниках. Прямоугольники связаны друг с другом или с референтным электродом, согласно монтажу. В примере представлен локальный средний монтаж.
Ж. Шаг 5. Удаление артефактов
Удаление артефактов обозначает методы устранения и исправления артефактов.
Мы предлагаем три метода: ручное удаление (удаление), автоматическая разметка
Cbanr*fe- | Refetenu | Cotets | View | Electrodes |
No | Ratre
1 Fpi-ал/ g] 2 F₽2-AvW Я 3 F7AvW *3 4. F2A<AV v] 5. Fz-AvW g fi F4AvW
7. F8-A-A* g 8 T3AvW Й 2 C3AvW ivj 10 Cz-AvW 53 11 C4-AvW 53 T4-Av',V 53 13. TSAw/ gj14 P&W F3] 15 Pz-AvW M IS P4AvW S3 17. Tfc-AvW 53 18- O1-AV-.V 3 is ozavw
Low Cut j High Ctrl | Notch." | ?G53Hz) 30 4555
(053 Hz) 30 4585
{053 Hz) 30 4555
=0.53 Hz) 30 45-55
(0.53 Hz) 30 45-58
(Й53Нг) 30 4555
(053 Нг) 30 45-55
(053 Hz) 30 4555
(053 Hz) 30 4585
(0.53 Hz) 30 45-55
=0.53 Hz) 30 45-55
{053 Hz) 30 45-58
{053 Нг) 30 4555
(053 Hz) 30 4555
(053 Hz) » 4585
(0.53 Hz) 30 4555
(0.53 Hz) 30 45-55
{0.53 Hz) 30 45-55
{053 Hz) 30 45-58
Mcntage r=«r>e |Йаёд$е
Etectisde Нйегегй
LJ „.J cJ J Ж1 Ш
Gain: ВВМ.Г
Low cut I s): |05 (032 Hz) 3
H^i cut fHzt { ЭО Н? 3
Notch 50 (60) H£
Add_____j --___________j
Stows* | Hite aS |
| OK | ; Cancel!: | Apply j Help
Рисунок 9.9. Диалоговое окно «Задание монтажа, каналы».
Параметры каналы определяются электродом, референтом, чувствительностью, фильтром высоких частот, фильтром низких частот, и режекторным фильтром. Эти параметры могут быть изменены в правой части и изображены в ле вой части окна. Название (имя) монтажа показывается сверху справа.
артефактов и коррекция артефактов. Эти методы представляют различные способы работы с артефактами.
Есть специальный инструмент («Вырезать») в меню «Правка», чтобы осуществить функцию исключения артефактных интервалов записи из последующего анализа. Перед нажатием инструмента необходимо найти артефакт визуальным осмотром143, выбрать его посредством двух вертикальных линий. После нажатия бокса «Вырезать» вы будете видеть отмеченный синей полосой интервал.
143 Обнаружение артефакта визуальным просмотром — искусство. Методология описана в книге Д. Коридон Хаммонд и Джей (ункелман «Искусство удаления артефактов» (Hammond and Jay Gunkelman. The art of artifactmg / Published by Society for Neuronal Regulation in 2001).
Часть 1. Ритмы ЭЭГ
187
Визуальный инспекционный способ удаления артефактов является трудоемким, субъективным и ведет к различным результатам в зависимости от опыта и предубеждения пользователя. Мы предлагаем метод, который выполняет автоматический поиск артефактов. Автоматическая разметка артефактов производится командой «Отметить артефакты» меню «Анализ». Поиск артефактов производится на основе заданных параметров. Пример установок параметров представлен на рис. 9.11. Параметры были определены экспериментально. Фрагменты ЭЭГ, которые соответствуют следующим критериям, рассматриваются как артефакты: 1) амплитуда сигнала превышает 100 мкВ; 2) амплитуда медленных волн в диапазоне частот от 0 до 1 Гц выше 50 мкВ; 3) высокочастотная активность в диапазоне частот от 20 до 35 Гц выше 35 мкВ. Чтобы управлять этой процедурой, нажмите команду «Отметить артефакты» в меню «Анализ». Программа отметит артефакты синего цвета метками и не будет принимать отмеченные фрагменты в расчетах
при дальнейшем анализе.
И наконец, третий метод удаления артефактов — метод пространственной фильтрации (обратите внимание, что для корректного анализа ЭЭГ необходимо сначала выполнить пространственную фильтрацию артефактов, а затем разметку артефактов). Хорошие результаты пространственного метода фильтрации получены для артефактов движения глаз. Процедура начинается с определения эпохи ЭЭГ, содержащей артефакты. Эта эпоха используется как шаблон для создания фильтров артефактов. После определения эпохи ЭЭГ можно начать процедуру коррекции артефактов нажатием команды «Коррекция артефактов» в меню «Анализ». Программное обеспечение разлагает выбранный фрагмент ЭЭГ на компоненты, используя анализ главных компонент или анализ независимых компонент. Временная динамика и топограммы РСА-компонент представлены справа: окно, которое появляется после нажатия команды «Коррекция артефактов» (рис. 9.12).
Некоторые из компонент соответствуют сигналу ЭЭГ, в то время как другие
Chennds Cofers | View | Efectiotfes j
Montage name- pgjjjWj
...........Av«ege <av,........ ............Weighted average WvW) Г : P TFpa P'Ftf Г^:
P F7 P F3 & Ft S? Г4 P F8 : -------------------------
P ГЗ P СЗ P Сг P C4 P U |W«^eve.agew Lerner
P T5 P РЗ P Рг P P4 P T6
P 04 Г Ог P 03 М<хйу Usmfaroeto rrxfeis:
Г Stt
Г- Г.::, Г. Г:
Рисунок 9.10. Диалоговое окно "Задание монтажа, референты» Тип локального (взвешенного) среднего монтажа можно определить щелчком справа. Электроды, которые будут усреднены для общего среднего монтажа, определяются слева.
Charnels
^FpZAvW
gF7AvW
$F3AvW
SSFrAvW
£F4-AvW
5gF&AvW
£T3AvW
vC3-AvW
j£C4-AvW
^T4AvW
^T5AvW
£P3AvW
^Ps-AvW
^P4-AvW
PjTS-AvW gjGlAVW ^02-AvW
lOnVEEG 3
& Voltage Threshold Condition Level (uV): fiOO
: P Slow Waves Ampftude(uV):pCl
Frequency from: |0 to jl
[P Fast Waves AmpBtodemf»™
Frequency from p® to [35
(ndude val before (ms): роб""""" Includeintetvalafter(ms): p200~~"™
P Cleat previous set ot artifact intervals
Q% 100*
OK I Cancel j Cleat ail ! I Load from database 1
Рисунок 9.11. Параметры для автоматического поиска и отметки артефактов
Слева — каналы, которые вы собираетесь обработать при поиске артефактов. Справа — порог амплитуды сигналов в мкВ, пороговые параметры для медленных и высокочастотных артефактов.
соответствуют артефактам, таким как вертикальные или горизонтальные дви-
188 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
жения глаз (рис. 9.12).
Мы рекомендуем читателю для коррекции артефактов использовать метод ICA — анализ независимых компонент. Чтобы использовать эту функцию, щелкните по соответствующему изображению (независимые компоненты) в панели «Метод» окна «Коррекция артефактов». Обратите внимание, что для использования метода ICA необходим длительный интервал времени для анализа. Мы рекомендуем выбор целого фрагмента ЭЭГ нажатием левой кнопки мыши в пределах окна времени в начале записи и правой кнопки мыши в конце записи. После щелчка на кнопке ICA программа будет раскладывать нативную ЭЭГ на независимые компоненты. Обратите внимание, что вычисления могут занять несколько минут! Временная динамика и топограммы компонент ICA появятся в правой стороне окна.
Пользователь должен выбрать топограмму артефакта, который нужно исправить. Топограммы вертикальных и горизонтальных движений глаза отмечены на рис. 9.12 стрелками. Выбор необходимой компоненты можно сделать просто щелчком на соответствующей топограмме у правой границы окна, и соответствующая топограмма будет отмечена красным. Пространственный фильтр вычисляется на основе выбранных топограмм. Этот пространственный фильтр применя-
Рисунок 9.12. Окно коррекции артефактов
Внизу — полоса прокрутки, кнопки скорости, чувствительности и метода пространственной фильтрации (главные компоненты или независимые компоненты). Справа — топограммы компонент, полученных с помощью РСА или ICA. Сверху (слева направо) — нативная ЭЭГ, исправленная ЭЭГ, исключенные артефакты и временная динамика полученных главных или независимых компонент ЭЭГ.
Часть 1. Ритмы ЭЭГ
189
ется ко всей записи ЭЭГ после нажатия на кнопку ОК в окне «Коррекции артефактов».
3. Шаг 6. Преобразование Фурье
После устранения и коррекции артефактов мы можем выполнить преобразование Фурье ЭЭГ-сигнала. Это можно сделать с помощью команды «Спектры ЭЭГ» в меню «Анализ». Появится окно «Расчет спектров ЭЭГ» (рис. 9.13). Параметры окна описывают: 1) часть записи ЭЭГ, которую мы собираемся анализировать: выбранный между маркерами (временной интервал, выделенный левыми и правыми вертикальными маркерами времени), фрагмент (фрагмент, который был определен в течение регистрации, например Глаза открытые/Тлаза закрытые) или весь файл ЭЭГ; 2) параметры быстрого преобразования Фурье, такие как длительность эпохи, способ перекрывания эпох и временное окно; 3) виды дополнительной обработки, такие как вычисление динамики спектров, вычисление когерентности и фазовых спектров; 4) числа эпох для усреднения при вычислении динамики спек
тров.
Мы рекомендуем загружать параметры спектрального анализа из базы данных щелчком на соответствующей кнопке окна «Загрузить из базы данных». После нажатия кнопки ОК появятся спектры мощности в виде, представленном на рис. 9.14. На этом рисунке каждый электрод представлен как отдельный график: мощность (квадрат амплитуды) ЭЭГ — по оси Y, частоты ЭЭГ — по оси X. Мощность ЭЭГ в выбранной частоте может быть изображена на двухмерной голограмме. Чтобы добавить голограмму распределения мощности, необходимо выбрать некоторую частоту, активируя курсор в любом из графиков. Это можно сделать нажатием
г Т йпе inter
Fiagnwnt J
Г MEEGSe
Channels. |eeG сф 3
Epoch tentgh
i С 1 second
i 2 second?
i 4 seconds
i Г 8 seconds
C 16 seconds
C 32 seconds
64 seconds
-Overlaying
; 50*
i C Hone
Гт window
О Square
Г Bartlett
Hanning Г Welch
Additional processing...................•;.................
C None
C Spectra dj*»nws Averaging epoch
Cefcutete coherence
C C<dcuaie coherence and phase spectra
C Keep raw spectra
load |
Save |
Ioad from database
Рисунок 9.13. Окно параметров для спектрального анализа
Сверху вниз — интервал анализа, параметры быстрого преобразования Фурье, параметры дополнительной обработки с числом усредненных эпох, используемым для расчета динамики спектров.
правой кнопки мыши в пределах любого графика. После того как будет отпущена эта кнопка, на экране появится всплывающее меню. Чтобы добавить голограмму для выбранной частоты, используйте команду «Добавить голограмму». Спектры могут быть представлены в абсолютной амплитуде, относительной мощности и в логарифмических значениях. Используйте соответствующие единицы, чтобы выбрать, какие параметры вы хотите использовать для изображения на графиках и голограммах: на рис. 9.13 представлены спектры мощности 19-канальной ЭЭГ. Каждый спектр усреднен за 3 мин в состоянии с открытыми глазами. Три топограммы, сделанные для трех различных частот, показаны в низу рис. 9.13. Частоты
190
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Рисунок 9.14. Пример результатов спектрального анализа
А. Спектры 19 электродов, размещенные в окне подобно локализации электродов на скальпе. Ось Y — мощность ЭЭГ в мкВ2, ось X — частота ЭЭГ в Гц. Б. Топограммы мощности ЭЭГ в трех различных частотах: 10,5 Гц, 12,9 Гц, 5,6 Гц. Масштабы представлены справа от каждой карты.
определены пиками спектров ЭЭГ и соответствуют мю-ритму (с пиками в СЗ, С4), теменному альфа-ритму (с пиком в Pz) и центральному тета-ритму (Cz).
И. Шаг 7. Сравнение с нормативной базой данных
После обнаружения пиков на спектрах и их картографии нам необходимо узнать, отличается ли достоверно мощность ЭЭГ индивидуальных спектров от нормативных данных или нет. В этом контексте «достоверно» означает доверительный уровень отклонения специфической особенности индивидуальных спектров (абсолютной мощности или амплитуды, относительной мощности, когерентности) от среднего значения, вычисленного для группы здоровых субъектов того же возраста. Для этого в меню «Анализ» щелкните командой «Сравнение результатов». В следующем диалоговом окне нажмите «Загрузить из базы данных», и вы увидите окно, представленное на рис. 9.15. Слева в этом окне можно видеть имя пациента, ЭЭГ которого вы анализируете, и дату регистрации. Справа в этом окне видны различные варианты для сравнения. Если вы анализируете спектры ЭЭГ в состоянии с открытыми глазами, щелкните EEG spectra Eyes Open и затем нажмите ОК.
Программа вычислит различия между индивидуальными спектрами и сред-
Часть I. Ритмы ЭЭГ
191
ними значениями нормативной группы. Разностные спектры (пациент-норма) будут показаны для каждого канала. Ниже разностных спектров вы можете увидеть вертикальные полосы, которые соответствуют уровню достоверности отклонения от нормальности. В зависимости от длины изображаются следующие
уровни достоверности: маленькие полосы — р < 0,05, средние полосы — р < 0,01, длинные полосы — р < 0,001. Кроме того, точное значение уровня достоверности показано в правой стороне окна, когда вы помещаете курсор (нажимая правую кнопку мыши) на определенную частоту на спектрах, соответствующих интересующему вас каналу (например, Cz в случае на рис. 9.16). Вместе с уровнем достоверности программа показывает частоту и амплитуду анализируемого параметра (рис. 9.16 справа, внизу).
gEEG Spectra Patient X118/05729071 ф EEG Spectra Eyes Dosed ЛдхЙогу Mathematical Reading VCPT Й Event Related Potentials Auditory Mathematical Reading Visual CPI В Event Related Potentials Components Visual CPI
Load Database
Рисунок 9.15. Окно сравнения с базой данных
Слева — имя пациента и дата регистрации. Справа — спектры ЭЭГ, ВП и независимые компоненты для ВП. Категории справа загружаются из нормативной базы данных, щелчком мыши на правой нижней кнопке — «Загрузить из базы данных».
III. Упражнения
Программное обеспечение Ed-EEG снабжено двумя ЭЭГ-файлами: один файл здорового субъекта, другой файл пациента. Оба субъекта принадлежат одной возрастной группе 13—14 лет. ЭЭГ были зарегистрированы в двух состояниях спокойного бодрствования: с открытыми и закрытыми глазами. Соответствующие фрагменты помечены как «Глаза открытые» (Eyes open) и «Глаза закрытые» (Eyes closed).
Мы предлагаем читателю потренироваться на этих двух файлах и ответить на следующие вопросы:
7. Какой файл соответствует здоровому субъекту, какой соответствует пациенту?
2. Какие нормальные ритмы могут быть обнаружены в записях ЭЭГ?
3. Какая мозговая дисфункция обнаруживается в записи ЭЭГпациента ?
4. Какое нарушение (ADHD, OCD или дислексия) может быть связано с наблюдаемой мозговой дисфункцией ?
Для того чтобы ответить на эти вопросы, выполните шаги ЭЭГ-анализа, описанные выше. Кратко действия следующие:
1. Запустите программу EdEEG.exe.
2. В меню «Файл» выполните команду «Открыть файл».
3. Откройте файл из подпапки Data папки EdEEG (если установки Typical расположены надиске С). Там находятся два файла: S 1_ЕОЕС и S2_EOEC. Анализируйте каждый из файлов отдельно.
4. Используйте кнопки линейки инструментов ( « ◄ ► ►►), чтобы рассмотреть записи. Эти кнопки соответствуют командам «Остановить», «Быстро пере-
192
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Рисунок 9.16. Результаты сравнения спектров с нормативной HBI базой данных
Графики — различия между индивидуальными спектрами и нормативной базой данных для 19 электродов. Ниже каждого графика расположены вертикальные полосы, соответствующие уровню значимости отклонения от нормы: маленькие полосы — р < 0,05, средние полосы — р < 0,01. Внизу справа — значение расхождения, частота и уровни значимости для выбранной частоты. Внизу слева — карта разностных спектров на частоте 5,9 Гц.
мотать назад», «Медленно перемотать назад», «Медленно воспроизвести», «Быстро воспроизвести». Вы можете также просмотреть ЭЭГ в различных временных интервалах с помощью полосы прокрутки, расположенной ниже записи ЭЭГ.
5. В меню «Вид» выполните «Выбор монтажа» и из окна «Имя монтажа» выберите «монтаж Версии 1 базы данных (DataBase Version 1 montage)». Это имя монтажа последнее в списке монтажа. Щелкнув в диалоговом боксе «Референтов» в окне «Параметры монтажа», удостоверьтесь, что это монтаж «Взвешенное среднее Lemos». Тогда нажмите ОК. Запись ЭЭГ появится в отобранном монтаже.
6. Визуальным просмотром найдите артефакты вертикальных и горизонтальных движений глаз.
7. Выберите всю запись. Это можно сделать, щелкнув левой кнопкой мыши в начале записи и правой кнопкой — в конце записи. Щелчок должен быть сделан в пределах временной шкалы, расположенной выше записи. После щелчка правой кнопкой временной шкалы станет желтым.
8. В меню «Анализ» выполните команду «Коррекция артефактов». В топограм-мах, представленных справа проявлявшегося окна, первая (сверху) топограмма выделена красным кругом. Эта компонента самая сильная и соответствует обычно вертикальным артефактам движений глаза. Удостоверьтесь, что это именно так, и щелкните кнопкой «Анализ независимых компонент» (ICA) в «Методах». Нажмите «ОК». Программное обеспечение отфильтрует этот тип артефактов из всей
Часть 1. Ритмы ЭЭГ
193
записи ЭЭГ. Визуальным просмотром записи удостоверьтесь, что соответствующий артефакт исправлен.
9. В меню «Анализ» выполните команду «Отметить артефакты». Загрузите параметры отклонения артефакта от базы данных, щелкнув команду «Загрузить из базы данных». После нажатия «ОК» программное обеспечение отметит артефакты, которые соответствуют критериям.
10. В меню «Анализ» выполните командой «Спектры ЭЭГ». Загрузите параметры вычисления спектров ЭЭГ из базы данных, щелкнув команду «Загрузить из базы данных». Выберите временной интервал, щелкнув «Фрагмент» и в дальнейшем выбрав «Глаза открытые» (Eyes Open) (то же самое можно проделать для состояния «Глаза закрытые». Программное обеспечение вычислит спектры для состояния «Глаза открытые» (Eyes Open) для каждого электрода отдельно и представит их в форме 19 графиков, расположенных в окне способом, подобном тому, как электроды расположены на скальпе.
11. В меню «Анализ» выполните команду «Сравнение результатов». Появится окно с именем «Сравнение результатов обработки». Для загрузки спектров ЭЭГ из нормативной базы данных щелкните кнопку «Загрузить базу данных». Слева от окна показывается имя спектров ЭЭГ данного субъекта, в то время как справа показываются имена спектров ЭЭГ из нормативной базы данных. Щелкните на спектрах субъекта слева и «EEG spectra Eyes Open» состояние справа. Нажмите «ОК».
12. Спектры различия (база данных субъекта) появятся в окне сравнения (рис. 9.16). Обратите внимание, что статический уровень значимости отклонения от нормальности представлен в форме маленьких вертикальных полос ниже спектров.
Сделайте тот же самый анализ со вторым файлом и попробуйте ответить на вопросы, представленные выше.
ЧАСТЬ 2.
Вызванные потенциалы
В первой части книги мы уже говорили о том, что генерация разных ритмов ЭЭГ осуществляется за счет разных нейронных механизмов. Считается, что эти ритмы, регистрирующиеся в состоянии спокойного бодрствования при открытых и закрытых глазах, отражают фоновую или спонтанную активность мозга. Однако с появлением стимула или при выполнении движения в течение первых 200 мс наблюдается подавление (десинхронизация) активности в альфа- и бета-частотных диапазонах ЭЭГ. Динамические характеристики этих ритмов свидетельствуют о том, что сами по себе они отражают не процессы обработки информации, а, скорее, модуляцию информационных потоков в мозге. Существуют две разные функции информационных потоков мозга: 1) функция, ассоциирующаяся с информационными потоками, связанными с обработкой сенсорной информации или действий; 2) функция модуляции информационных потоков. Как было продемонстрировано в первой части книги, модуляция информационных потоков проявляется в виде синхронизации или десинхронизации ЭЭГ-активности.
Этапы обработки информации анализируются с помощью вызванных потенциалов (ВП). В отличие от ЭЭГ необходимым условием для регистрации ВП является четкая временная связь регистрируемой активности с конкретным событием, будь то движение или появление стимула. Задачи, используемые в исследованиях с регистрацией ВП, широко охватывают разнообразные сенсорные, когнитивные и моторные функции. Они включают различные типы тестовых заданий по детекции и распознаванию стимулов разных сенсорных модальностей, задания с отсроченной реакцией для изучения рабочей памяти, GO/NOGO-тесты для оценки функций контроля исполнения и многие другие. Каждое конкретное задание ассоциируется с определенным набором психологических операций, например, таких как детекция и распознавание стимулов, обновление рабочей памяти, инициация или подавление действия, мониторинг результата действия и т.д. В свою очередь, каждая из психологических операций вызывает временное появление паттернов акти-вации/деактивации нейронов в определенных областях мозга. Сумма синхронно генерируемых и связанных с событием постсинаптических потенциалов регистрируется с поверхности скальпа в виде компоненты вызванных потенциалов — отклонения потенциала, которое ограничено во времени и пространственно локализовано.
Часть 2. Вызванные потенциалы
195
В ВП-исследованиях первых лет анализируемые компоненты определялись как позитивные или негативные пики, наблюдавшиеся либо на самих ВП, либо на разностных ВП. Разностные ВП получались в результате вычитания ВП, зарегистрированного в задаче, не требующей вовлечения определенных психологических операций, из ВП при другой задаче, где эти операции предположительно должны были протекать1. Отклонения потенциала, видимые на разностных ВП, могут быть подразделены на разные классы в зависимости от их латентности и полярности (позитивные или негативные), как, например: Р100, N100, N200, Р200, РЗОО, N400, где Р используется для обозначения позитивности, a N — негативности, а число обозначает латентный период (ЛП) пика в миллисекундах. Однако пиковая латентность не является исчерпывающей характеристикой описания компонент ВП. В частности, пиковая латентность так называемой РЗЬ-компоненты, в зависимости от сложности задачи по дискриминации целевого/нецелевого стимула, может варьировать в пределах нескольких сотен миллисекунд. Даже полярность определенной компоненты может зависеть от условий регистрации ВП. Так, компонента С1, которая генерируется в VI области зрительной коры, негативна для стимулов, предъявляемых в верхней части зрительного поля, и позитивна для стимулов из нижней его области. Причиной этого является особая анатомическая пространственная организация 17-го поля Бродмана (ПБ) коры головного мозга.
Другой подход в классификации ВП подразумевает рассмотрение компонент с позиций их функциональной значимости. Существует несколько компонент ВП, которые регистрируются только в определенных поведенческих парадигмах и носят специальные обозначения в соответствии с их предполагаемой функцией2 * * * * 7.
Метод ВП получил широкое распространение в области когнитивных нейронаук более 40 лет назад — в 60-х годах прошлого столетия. Первая попытка разделения ВП на компоненты была предпринята в 70-х годах с помощью факторного анализа и метода главных компонент. Однако эти методики позволяли выявить только ортогональные (в строго математическом смысле) компоненты ВП, что явно является ограничением, поскольку ясно, что компоненты не обязательно являются ортогональными. Развитие новых методов объективного разделения компонент (как, например, метод независимых компонент) преодолели недостатки старых подходов и открывают новые возможности исследований в этом направлении. Накапливаемое знание демонстрирует высокую эффективность и диагностическую мощность метода оценки независимых компонент ВП как эндофенотипов дисфункций мозга.
1 Простым примером такого типа заданий может являться тест, в котором зрительные стимулы (например, цифры) предъяв-
ляются на пороге опознания. Испытуемые инструктировались называть цифру или говорить «нет» (если не распознавали) после
появления разрешающего стимула, предъявлявшегося секунду спустя после цифры. Логично предполагать, что две категории
стимулов (распознанные» и «нераспознанные») отличаются одной из психологических операций (распознавание объекта), а раз-
ностная волна между ВП-ответами на эти стимулы позволит получить нам пространственно-временные корреляты этой психоло-
гической операции.
7 Одними из наиболее исследованных компонент ВП являются: «негативность рассогласования» (HP) как показатель детекции изменений повторяющейся слуховой стимуляции; «процессная негативность» (ПН) как показатель фокусирования внимания на одном из сенсорных каналов; «негативность ошибки» (НО) как показатель некорректных, ошибочных действий в длительном выполнении тестового задания; N2 NOGO компонента как показатель подавления моторной реакции, РЗЬ-компоненты как показатель обновления рабочей памяти; РЗа-компоненты как показатель непроизвольного переключения внимания.
196 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Данная часть книги посвящена компонентам ВП, их генерации, функциональной значимости и диагностической ценности3. Мы классифицируем ВП, исходя из их функциональной значимости. Мы предполагаем, что компоненты ВП отражают разные психологические процессы, которые обеспечиваются различными мозговыми системами.
Пюссарий
Аффективное состояние — состояние, характеризуемое побуждениями, эмоциями и мотивациями.
Агнозия — состояние, при котором пациент не может дать название рассматриваемому объекту, но демонстрирует знание правильного его использования и может классифицировать в группу морфологически разных, но функционально однородных предметов.
Амигдала — группа ядер, получающих входы от полимодальных областей височной и теменной коры через гиппокамп и посылающих информацию в области префронтальной коры через таламус. Основной функцией амигдалы считается участие в обеспечении эмоции страха и реализации поведенческих реакций, связанных со страхом.
Модель окружающей среды (Anticipating schemata) — когнитивные структуры, которые обеспечивают высокую степень готовности субъекта к перцепции какого-либо определенного аспекта информации на фоне остальных.
Внимание — с психологической точки зрения является когнитивным механизмом, позволяющим посредством перераспределения ограниченных ресурсов обрабатывать сенсорную информацию выбранного источника более детально по сравнению с игнорируемым. Внимание может быть также определено как готовность воспринимать определенный тип стимулов, т.е. состояние ожидания сенсорного события. С учетом этого определения следует отличать от процессов подготовки моторного ответа как состояния готовности сделать движение.
Восходящая обработка информации — передача сенсорной информации от иерархически низших центров к высшим. Поскольку обычно в нейронных сетях мозга такие восходящие процессы сопровождаются соответствующими нисходящими процессами, их условное разделение несет сугубо теоретический характер.
Канонические кортикальные цепи — гипотетические нейронные сети коры головного мозга, обеспечивающие сложно организованную обработку входящей информации. Термин «цепь» был введен в литературу Родни Дугласом и Ке-ваном Мартином в 1989 году. Основная идея теоретической модели авторов базируется на представлении о существовании кортикальных цепей, образованных локальными возбудительными и тормозными путями проведения информации. Именно такая организация обеспечивает ряд важных свойств кортикальных цепей.
3 Для первоначального ознакомления с техникой ВП я бы рекомендовал книгу Стивена Лака (Steven Luck 2005), которая написана ясным . языком и содержит детальное описание методов ВП.
Часть 2. Вызванные потенциалы
197
Операции сравнения — гипотетические операции, протекающие в сенсорных областях коры, основной целью которых является выявление любого отклонения от ожидаемой модели окружающей среды (anticipatory schemata) и модулирование поведения с учетом этих отклонений. Примером такой операции детекции в слуховой модальности является известный эффект «негативности рассогласования» — компонента вызванных потенциалов, регистрируемая в ответ на девиантный стимул, предъявляемый в ряду стандартных.
Операция отвлечения (Disengagement) или подавления ответа — процесс, противоположный операции вовлечения, при котором наблюдается подавление ресурсов, необходимых для реализации подготовленного действия.
Дорсальный и вентральный потоки проведения зрительной информации берут начало от отдельных областей первичной зрительной коры и посылают входы в височные и теменные зоны коры. Вентральный путь обработки зрительной информации вовлекается в обеспечение распознавания разных объектов (преимущественно отличающихся по форме и цвету). Дорсальный путь обеспечивает кодирование пространственных характеристик объектов и контроль процессов манипуляций и ориентировки по отношению к ним.
Эмоция — поведенческая реакция (изменение частоты сердечных сокращений, мимики, речи) на вознаграждение (положительная эмоция) или наказание (негативная эмоция), а также на представление этих поведенческих событий. Эмоции как психологические сущности могут быть разделены на реакции и ощущения.
Эмоциональный ответ — соматосенсорная (соматическая или лицевая) или эндокринная реакция на эмоционально значимый стимул — наказания или вознаграждения.
Операция вовлечения — операция, реализуемая исполнительной системой, вызывающая активацию корко-подкорковых ресурсов, необходимых для реализации действия. Операция вовлечения подразумевает существование активного мозгового механизма, обеспечивающего растормаживание нейронов коры, что является необходимой предпосылкой для реализации определенного моторного или когнитивного действия.
Эпизодическая память — память на специфические события, датированные определенным временем. Эпизодическая память также обеспечивает формирование связи между разными событиями.
ПСС (потенциалы, связанные с событиями) — потенциалы мозга, связанные с информационными потоками в коре головного мозга, вызываемыми событиями окружающей среды (например, последовательно предъявляемые повторяющиеся стимулы или повторяющееся сгибание пальца руки). В основе техники получения ПСС лежит принцип усреднения потенциалов, позволяющий увеличить отношение сигнала к шуму.
Исполнительные функции — относятся к типу процессов контроля или мониторинга моторных, сенсорных или когнитивных действий при целенаправленном поведении. Основной вклад в обеспечение исполнительных функций вносят лобные области коры, базальные ганглии, а также другие подкорковые структуры, необходимые для реализации контроля.
198 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Эксплицитная память (сознательная) — вид памяти, при котором сохранение и воспроизведение информации контролируется сознанием. Событие, запоминаемое эксплицитно, впоследствии может произвольно вспоминаться испытуемым. Эксплицитную память также называют декларативной.
Чувство (эмоциональное чувство) — субъективное эмоциональное состояние, вызванное эмоционально значимым стимулом. Примерами таких состояний являются радость и грусть, смелость и страх, горе и счастье, любовь и ненависть и т.д. Основной вклад в обеспечение эмоциональных состояний вносит орбитофронтальная кора.
Имплицитная память — приобретенные навыки или знания, которые могут демонстрироваться субъектом бессознательно. Часто имплицитную память называют недекларативной или процедурной.
Обработка информации в мозге представляет широкий класс явлений трансформации импульсной активности эфферентных нейронов рецептивных органов (как, например, сетчатка) в импульсную активность и снижение мембранного потенциала нейронов, входящих в состав различных мозговых нейронных сетей мозга. В свою очередь, информационные процессы могут функционально подразделяться на информационный поток и информационную модуляцию. Эти две операции могут обеспечиваться разными классами нейромедиаторов: быстродействующими медиаторами (например, гамма-аминомасляная кислота — ГАМК) и медленнодействующими медиаторами (такие как дофамин, норадреналин, серотонин и ацетилхолин).
Латеральное торможение — разновидность связи нейронов в нейронной сети, при которой нейроны оказывают тормозное влияние на нейроны ближайшего окружения и тем самым препятствуют распространению возбуждения в латеральном направлении. Впервые явление латерального торможения было описано при изучении сетчатки глаза в 50-х годах прошлого столетия. За счет латерального торможения в зрительной системе обеспечивается выделение высокого градиента яркости видимых объектов (таких как полосы Маха).
Лейкотомия — психохирургическая процедура, во время которой при помощи специального ножа через глазницу производилось отсечение нижневентральной части префронтальной коры. Начало этим операциям, которые были популярны в 1950-х годах, до открытия психотропных лекарств, положил португальский невролог Эгас Мониз.
Лимбическая система — термин, введенный в литературу Полем Брока, под которым понимают набор структур мозга, выделяемых в единую группу, образующих граничную область около ствола мозга. Лимбическая система играет важную роль в обеспечении эмоциональных реакций.
Долгосрочная потенциация (LTP) — устойчивое увеличение амплитуды возбуждающего постсинаптического потенциала, возникающее в результате высокочастотной стимуляции (тетанической) афферентных проводящих путей. LTP рассматривается в качестве клеточной модели научения и памяти.
Консолидация памяти — процесс развития необратимых изменений в синаптической передаче, происходящий на нейронном уровне. На психологическом
Часть 2. Вызванные потенциалы
199
уровне консолидация памяти ассоциируется с процессами формирования долгосрочной памяти, которая практически не распадается в течение жизни.
Мониторинг — является новой концепцией, в соответствии с которой существует психологический механизм контроля качества реализуемой деятельности и модуляции процессов когнитивного контроля, обеспечивающих распределение ресурсов, компенсирующий конфликт между ожидаемым и совершенным действием.
Мотивация — желание или потребность, управляющая целенаправленным поведением.
Моторная подготовительная установка — когнитивный механизм, позволяющий концентрироваться на выполнении конкретного моторного акта и подавлять другие, не имеющие отношения к ситуации действия. Она проявляется в готовности выполнить действие. Внимание и моторная подготовительная установка являются элементами рабочей памяти.
Отрицательное подкрепление (наказание) — стимул, появление которого приводит к оптимизации поведения, направленной на избегание наказания.
Сенсорный дефицит (Neglect) — неврологический синдром, при котором в результате повреждения области мозга у пациентов наблюдается выраженная неспособность воспринимать сенсорную информацию, предъявляемую в контралатеральном сенсорном поле относительно локализации повреждения. Сенсорный дефицит часто связан с повреждением правой теменной коры, однако повреждения в подкорковых структурах (таких как таламус, базальные ганглии и верхние бугры четверохолмия) также могут приводить к сенсорному дефициту.
Нейромодулятор — нейромедиатор медленного действия, модулирующий информационные процессы.
Нейронная сеть — сеть нейронов мозга, обеспечивающая различные функции.
Нейротрансмиттер — нейромедиатор быстрого действия, обеспечивающий распространение информации в пределах нейронной сети.
Орбитофронтальная кора — часть области префронтальной коры, получающая большое количество входов от всех сенсорных систем мозга. Для орбитофронтальной коры характерны разные паттерны активации при предъявлении положительных (вознаграждение) и отрицательных (наказание) подкрепляющих стимулов. Пациенты с нарушениями в области орбитофронтальной коры (как в известном случае Финеаса Кэйджа) теряют способность эффективно дифференцировать положительные и отрицательные подкрепляющие стимулы и тем самым адекватно управлять своим поведением (избегать ситуаций с высокой степенью получения наказания).
Положительное подкрепление — стимул, который проводит к более частому проявлению определенного поведения, позволяющего достигнуть вознаграждения.
Прайминг — усиление процессов распознавания, воспроизведения или склонностей к определенному типу стимула при повторном его восприятии.
Рецептивное поле клетки сенсорной системы — обособленная область внешнего (например, зрение или слух) или внутреннего (например, соматосенсор
200 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
ная модальность) по отношению к субъекту пространства, появление стимула в котором приводит к активации или торможению данной клетки. Стимулы, предъявляемые вне пределов такого рецептивного поля, не влияют на активность нейрона.
Рецептивное поле нейрона — область пространства (внешнего или внутреннего по отношению к субъекту), в которой появление стимула изменяет паттерны активности этого нейрона. Понятие рецептивного поля употребляется для нейронов сенсорных систем. В отношении нейронов лобных областей коры и базальных ганглиев использование термина «рецептивное поле» ограничено, поскольку они в основном реагируют на действия (а не на сенсорные стимулы как таковые).
Репрезентация (представительство) — локализованная или распределенная нейронная сеть, которая обеспечивает хранение информации (память) или, в случае активации сети, доступ к информации. Репрезентации могут быть генетически детерминированы или сформированы в процессе научения.
Скотома — локальная утрата зрительного восприятия в определенной части зрительного поля, вызванная локализованным повреждением первичной зрительной коры мозга.
Операция выбора — процесс активации репрезентации. Считается, что операция выбора сопровождается синхронными осцилляциями в частотных диапазонах тета (4—8 Гц) и гамма- (40 Гц) ритмов. Эти осцилляции формируют оптимальный режим активности, позволяющий извлекать информацию из памяти.
Семантическая память — вид памяти о фактической информации об окружающем мире, понятиях и значениях слов.
Сенсорная модальность — понятие, относящееся к сенсорным системам, обеспечивающим обработку информации, поступающей с рецепторов определенного типа. Наиболее полно изученными сенсорными системами являются: зрительная (способность различать формы, цвет, интенсивность, движение и пространственные отношения зрительных объектов), слуховая (способность слышать и определять источник звуков), соматическая сенсорная, или соматосенсорная (способность ощущать прикосновение, боль, изменения температуры, механическое смещение мышц и суставов).
Сенсорные системы — включают рецепторные органы, подкорковые и корковые нейронные сети, которые специфически реагируют на активацию соответствующих рецепторов.
Нисходящие процессы — распространение информации от высших мозговых центров к низшим на уровне иерархически организованных сенсорных систем. При этом модулируются информационные процессы иерархически более низких сенсорных областей с помощью информации, извлекаемой из памяти высшими центрами.
Рабочая память — определяется как процессы активной манипуляции временно сохраненной информацией для осуществления сенсомоторных или когнитивных действий, таких как речь, планирование, принятие решения и т.д.
Часть 2. Вызванные потенциалы
201
Глава 10. Сенсорные системы
Знания приходят к человеку через дверь чувств.
Гераклит, VI в. до н.э.
Мы воспринимаем изменения во внутреннем и внешнем мире посредством рецепторов разных видов4. Рецепторы можно подразделить на два больших класса экстерорецепторов (внешние) и интерорецепторов (внутренние). Эксте-рорецепторы позволяют получать информацию о событиях окружающего мира, а интерорецепторы — о том, что происходит в организме. Активность некоторых интерорецепторов никогда не достигает нашего сознания. Сенсорные элементы системы восприятия внешнего мира крайне специализированы5. Существуют разные виды сенсорных модальностей, которые позволяют нам воспринимать изображения, звуки, движения тела, а также продуцируют боль, вкус, запах, осязание.
Важно отметить, что мы воспринимаем не объект как таковой, а импульсную активность выходных нейронов рецепторных органов. В свою очередь, эта активность нейронов возникает в результате взаимодействия рецептора и объекта. Области мозга, которые реагируют на стимуляцию определенных рецепторов, обычно рассматриваются как соответствующие сенсорные системы (зрительная, слуховая). Среди всех сенсорных систем наиболее изученной является зрительная. В настоящей главе мы уделим внимание рассмотрению зрительной, слуховой и соматосенсорной модальностей, поскольку данные системы доставляют в мозг наибольшее количество сенсорной информации и лучше всего исследованы.
I. Анатомия
А. Поля Бродмана и ядра таламуса
Сенсорные сигналы от рецепторных органов достигают первичных сенсорных областей коры через релейные ядра таламуса. Основные сенсорные модальности локализуются в следующих областях: поле Бродмана 17 — первичная зрительная кора или область VI, ПБ 41 — первичная слуховая кора или область А1,
4 Специализированные рецепторные клетки преобразуют стимулы в частоту разрядов (импульсную активность) эфферентных клеток рецепторных органов. В частности, колбочки сетчатки, чувствительные к свету и не чувствительные к звуку, передают свои реакции на свет к ганглиозным клеткам глаза.
5 Млекопитающие, например, не могут видеть в инфракрасном диапазоне или воспринимать магнитное поле. Строение и функция зрения хищников отличается от организации зрительной системы травоядных. Кроме того, способность восприятия сильно зависит от приобретаемого опыта. Например, эскимосы, проводящие всю свою жизнь в снегах, могут различать множество его оттенков. Хороший музыкант может различить отдельные ноты звуков.
202
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Рисунок 10.1. Упорядоченные кортикальные проекции сенсорных систем
Сенсорные ядра таламуса (Б) выполняют воротную функцию, контролируя активность первичных сенсорных областей коры (А). На рисунке представлено три модальности — зрительная, слуховая и соматосенсорная. Схематическое изображение вентрального и дорсального зрительных проводящих путей (A). LGB — латеральное коленчатое тело, MGB — медиальное коленчатое тело, LP — латеральное заднее ядро таламуса.
ПБ 1 — первичная соматосенсорная кора или область S1. Латеральное коленчатое тело (Л КТ) является релейным ядром для зрительной модальности сигналов, медиальное коленчатое тело (МКТ) — для слуховой, а заднее латеральное ядро — для соматосенсорной.
Б. Топографическая организация
Каждая из первичных сенсорных областей коры характеризуется топографической организацией проекций6. Нейроны первичной сенсорной коры и таламические ядра формируют сильно взаимосвязанную нейронную сеть. Важной особенностью этой сети является способность генерировать ритмическую активность в частотном диапазоне альфа-ритма (более детально см. гл. 2). Как уже упоминалось в первой части книги, альфа-ритм отражает неактивное состояние сенсорной системы — состояние, при котором сенсорная кора не участвует в активной обработке сенсорной информации, но за долю секунды может переключиться в активный режим.
В. Параллельные пути проведения информации
В качестве примера рассмотрим пути проведения информации в зрительной системе7. С механической точки зрения окружающий нас мир состоит из раз
6 Термин «топографическая репрезентация» буквально означает следующее: смежные клетки рецептивного органа проецируют свои связи на смежные нейроны коры, в результате на уровне коры мозга формируется «карта» этого рецептивного органа.
7 В зрительной модальности информация от сетчатки глаза до коры передается пучком, содержащим около 1 000 000 аксонов. В то же время в слуховой модальности передача информации от улитки до слуховой коры обеспечивается только 30000 аксонов.
Часть 2. Вызванные потенциалы
203
личных объектов различной локализации в пространстве, двигающихся с разной скоростью. Зрение обеспечивает нас полезным инструментом, позволяющим знать: 1) что располагается в пространстве; 2) где именно в этом пространстве это «что» располагается; 3) как субъект может манипулировать этим «что» или сделать направленное движение в его сторону.
Для решения этих задач природа развила мозговые пути проведения информации (или потоки), передающие зрительную информацию разного вида. Эти пути берут свое начало от коленчатых тел таламуса, релейных ядер, состоящих из двух видов клеток, формирующих разные слои ядер: маленькие (парвоцеллюлярные) и крупные (магноцеллюлярные)8. Эти две системы (парвоцеллюлярная и магноцел-люлярная) остаются разделенными на уровне первичной зрительной коры (VI) и дают начало двум мозговым путям обработки зрительной информации.
Один из путей получил название вентрального, или «что»-системы (рис. 10.1) и включает вентролатеральную престриарную кору (части 18 и 19 ПБ) и нижневисочную кору. Стриарная, престриарная и нижневисочная кора составляют иерархически организованную систему, в которой сигналы передаются от первого уровня ко второму, от второго к третьему и так далее. Основной целью такой организации данного зрительного пути является разбиение первоначального образа на отдельные компоненты9, характеризующиеся ориентацией, пространственной частотой и цветом.
Скорость и позиция зрительного объекта кодируется так называемой дорсальной системой обработки зрительной информации или дорсальным путем, включающим область теменной коры. Важной особенностью этого пути проведения зрительной информации является не только кодирование пространственных отношений зрительной сцены (т.е. «где» располагается объект), но и организация зрительно управляемых моторных программ (функция «как»). Задние области теменной коры имеют связи с премоторными центрами лобной доли, что позволяет обеспечивать реализацию целого класса ориентировочных и манипуляционных действий. Например, таких как фиксация взгляда на выбранном объекте, схватывание объекта и манипуляции с ним. В этом отношении дорсальная система тесно связана с процедурной памятью (см. гл. 14).
Вентральный и дорсальный пути обработки зрительной информации не являются строго параллельными, поскольку взаимодействуют на разных иерархических уровнях. Кроме того, в обеих системах существуют обратные связи, обеспечивающие возвращение информации от более высших уровней к низшим. Операции, связанные с вниманием и рабочей памятью, реализуются при помощи этих обратных связей (нисходящие потоки информации). В то же время связи между вентральной и дорсальной проводящими системами позволяют координировать обработку информации и формировать из различных сенсорных характеристик единый перцептивный образ.
8 Наименьшие по размеру клетки располагаются в 4 верхних слоях латерального коленчатого тела и составляют 80 % от общего числа релейных клеток коленчатого тела. Клетки наибольшего размера локализуются в двух нижних слоях и соответственно составляют 20 % от общего числа нейронов.
9 Говоря о первоначальном образе, имеют в виду активность ганглиозных клеток глаза, кодирующих попадание света на разные области сетчатки в генерацию потенциалов действия.
204 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Ядра подушки как координаторы информационных потоков. Обработка информации, осуществляемая в дорсальном и вентральном путях при участии престри-арной, височной и теменной коры, не ограничивается внутренними связями между этими областями. Ядра подушки играют важную роль в координации процессов обработки зрительной информации (рис. 10.1). Недавние исследования показывают, что нейроны подушки увеличивают активность при концентрации внимания на релевантном объекте и, напротив, подавляют свою активность в ответ на предъявление иррелевантного стимула10. Тем самым обнаруженный эффект указывает на вовлечение этих нейронов в обеспечение контроля обработки зрительной информации в зрительной коре, что наиболее выражено при операциях, связанных с вниманием. Некоторые нейроны подушки, например, реагируют на движущиеся объекты в период фиксации взгляда, но не активны в период саккадических движений глаз, таким образом различая движение объекта от движений глаза. При этом участие ядер подушки в контроле процессов обработки информации соответствующими областями коры не ограничивается только зрительной модальностью: существуют данные, свидетельствующие о реакции нейронов подушки на соматосенсорные и слуховые стимулы.
После обработки сенсорная информация: 1) перенаправляется в области префронтальной коры через последовательные (заднефронтальные) пути; 2) распространяется к гиппокампу через ринальную кору. Дополнительно существуют проекции «обратной связи» к дорсальной и вентральной системам обработки зрительной информации от префронтальной коры и гиппокампа.
II. Обработка зрительной информации
A. ON- и OFF-рецептивные поля
Основные знания о нейронных механизмах обработки зрительной информации были получены в работах по исследованию одиночных нейронов животных. Обычно животному предъявляли разные зрительные стимулы и регистрировали импульсную активность отдельного нейрона. В качестве стимулов использовались светлые пятна, определенным образом ориентированные полоски или пространственные решетки. Такие исследования помогали выявлять рецептивные поля конкретного нейрона. На основе реагирования на освещенность или темноту выделяют два класса клеток: ON- и OFF-нейроны. Эти клетки в наибольшем количестве представлены на самых низших уровнях системы обработки зрительных образов11. Рецептивное поле нейронов может быть аппроксимировано разницей гауссовых функций (так называемые DOG-функции, рис. 10.2), характеризуемых соответственно малым и большим пространственным распределением: первое соответствует входу возбуждения, а второе — тормозному воздействию на нейрон. Проведенные расчеты показали, что латеральное торможение играет ключевую роль в формировании таких рецептивных полей (рис. 10.3).
10 Для обзора см. Robinson and Petersen, 1992.
11 Низшие уровни включают ганглиозные клетки сетчатки и нейроны латерального коленчатого тела таламуса.
Часть 2. Вызванные потенциалы
205
Б
Рисунок 10.2. ON- и OFF-нейроны зрительной системы
В верхней части рисунка изображены рецептивные поля ON- (справа) и OFF-клеток (слева). Серым (черным) цветом отображены части рецептивного поля, которые активируются (тормозятся) в ответ на предъявление небольшого пятна света. Внизу представлены ответы двух типов нейронов на свет и темноту (отображены справа).
Б. Пространственная фильтрация на уровне таламуса и коры
Нейроны таламуса и первичной зрительной коры характеризуются существенно различающимися рецептивными полями (сравнение двух фильтров зрительной информации, соответствующих рецептивным полям нейронов Л КТ и зрительной коры, представлено на рис. 10.3). Рецептивное поле нейрона первичной зрительной коры может быть выражено функции Габора12, а нейронов Л КТ и сетчатки — при помощи DOG-функций.
В 1990-х годах в нашей лаборатории была разработана модель канонического кортикального модуля, которая имитировала структуру и функцию зрительной коры (более детально описана в гл. «Методы» части 2). За основу данной модели было взято пространственное (анатомическое) распределение ON- и OFF-клеток тормозного слоя зрительной коры. В соответствии с нашими предположениями, ON- и OFF-клетки были организованы в модули по принципу спиц в колесе. Вследствие такого пространственного расположения и тормозной природы нейронов, влияющих на активность эфферентных нейронов, рецептивные поля клеток, входящих в состав отдельного модуля, можно выразить с помощью функции Габора.
12 Функция Габора является линейным фильтром, импульсный ответ которого определяется гармонической функцией, помноженной на гауссовскую функцию. В результате свертки фильтра с сигналом формируется так называемое пространство Габора, которое успешно применяется в техническом зрении.
206 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Необходимым условием выявления специфических характеристик сенсорного входа зрительной модальности является вовлечение тормозных нейронов. В действительности функции (Габора и DOG), привлекаемые для описания рецептивных полей нейронов зрительной системы, включают как позитивные (активирующие), так и негативные (тормозящие) участки. При регистрации электрической активности с поверхности скальпа возбудительная и тормозная активность отражается противоположными флуктуациями потенциала.
В. Вентральный и дорсальный пути обработки зрительной информации
Зрительная кора макак составляет около 60 % неокортекса. Экспериментальные данные показывают, что изображения, отображающиеся на сетчатке, приводят к активации множества областей коры. Установлено, что у обезьян в обеспечении обработки зрительной информации участвует более двух десятков областей мозга13. В силу анатомической и функциональной организации мозговой системы зрения обработка информации протекает в двух направлениях, по так называемым дорсальному
Рисунок 10.3. Рецептивные поля нейронов таламуса и зрительной коры Под DOG-фил ыром подразумевается разница двух гауссианов (Difference of two Gaussians), который аппроксимирует рецептивные поля на уровне латерального коленчатого тела. Фильтр (абора является продуктом гармонической и гауссовской функций и аппроксимирует рецептивные поля простых клеток нейронов первичной зрительной коры. Обозначения: LGB — латеральное коленчатое тело.
и вентральному зрительным путям, которые впервые были описаны Ангер-лейдером и Мишкиным в 1980 году. Они предположили, что эти пути характеризуются разными возможностями в обработке зрительной информации: 1) теменной путь обеспечивает восприятие пространственных отношений; 2) височный связан со зрительным распознаванием объектов. Дальнейшие исследования показали, что каждый из потоков обработки зрительной информации может быть подразделен и на более мелкие специализированные пути. В свою очередь, эти пути берут начало от так называемых капель
13 В теории определение специализации области коры должно опираться на ряд критериев, среди которых применительно, например, к зрительной коре можно выделить наличие отдельной топографической репрезентации видимого слова, специфических анатомических связей и специфических (отличные от остальных) свойств реагирования нейронов, архитектоники, а также определенного характера поведенческих нарушений, вызываемых повреждением этой области. Однако на практике использование разных технических подходов для проверки всех этих критериев крайне затруднено, и поэтому до сих пор окончательная идентификация и локализация специфических областей не проведена.
Часть 2. Вызванные потенциалы
207
(blobs) и межкапель (interblobs) первичной зрительной коры14. В то время как «капли» принимают участие в цветовом восприятии, «межкапли» обеспечивают восприятие формы и движения объектов. По такому же принципу организована и вторичная зрительная кора (ПБ 18), но способом, отличным от первичной15.
В пределах дорсального и вентрального путей различные характеристики видимого мира (форма, цвет, движение и пространственные отношения) обрабатываются в отдельных потоках, которые связаны с соответствующими функциями: распознавание отдельных объектов, выявление пространственных отношений между объектами, а также определение способов манипуляции и их использования. Например, нейроны вентрального пути проявляют избирательную чувствительность к форме, цвету и текстуре объектов. Нейроны дорсального пути, напротив, не реагируют на данные атрибуты, но демонстрируют селективную активность в отношении скорости и направления движения, а также избирательно реагируют на стимулы, предъявляемые в области пространства, на котором сконцентрировано внимание.
Магноцеллюлярная (М) система, формируемая при участии больших клеток Л КТ, в основном ассоциируется с дорсальным путем и определяет функциональные характеристики его нейронов. В ряде последних исследований было показано, что нейроны дорсального пути быстрее реагируют на стимулы по сравнению с нейронами вентрального пути (парвоцеллюлярная (П) система)16. В частности, у обезьян латентный период нейронов таламуса М-системы в среднем составляет 30 мс, тогда как Л П нейронов П-системы — на 20 больше. На уровне коры эта разница увеличивается до 40 мс.
Г. Иерархическая организация
Обработка зрительной информации в пределах каждого из каналов иерархически организована (рис 10.1). В частности, обработка формы объекта на уровне первичной зрительной коры (ПБ 17) начинается с фильтрации на основе функции Табора, а нейроны нижней височной коры уже селективно реагируют на глобальные свойства объекта, например такие как специфическая форма руки, лица и т.д. Таким образом, средний размер рецептивного поля увеличивается на протяжении пути от затылочной коры до височной17. Следовательно, нейроны первичной зрительной коры ответственны за обработку локальных характеристик видимых объектов, в то время как нижневисочные области коры, по всей видимости, вовлечены в восприятие объекта как целого — термин, известный как гештальт. В действительности пациенты, перенесшие операцию право-передней височной лоботомии, существенно утрачивают способность восприятия сложных зрительных объектов (лица или непривычные, абстрактные изображения). Изначально для описания этого синдрома
14 Эти подразделения зрительной коры могут быть визуализированы при окрашивании коры энзимом цитохромоксидазой, который играет важную роль в метаболизме клетки. При окрашивании под микроскопом можно увидеть прилегающие друг к другу отдельные участки первичной зрительной коры в виде случайных «капель» (blob) (Wong-Riley et al., 1993).
15 При окрашивании цитохромоксидазой ПБ 18 появляется паттерн перемешанных толстых и тонких полосок с пустотами.
16 Это исследование было проведено Мэтью Смоленски в 1998 г. (Schmolesky et al., 1998), который использовал одну и ту же экспериментальную и аналитическую технику для исследования всех областей мозга.
17 На уровне парафовеального окружения рецептивные поля нейронов составляют около 1,58 градусов в ПБ 17 и около 48 в ПБ 37, тогда как средний размер рецептивного поля нейронов 38 ПБ — 26 х 268 градусов. Таким образом, оказывается, что большие по размеру рецептивные поля иерархически высших структур формируются из небольших рецептивных полей структур коры низшего уровня.
208 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
использовался термин «психическая слепота». Позднее Фрейд ввел понятие агнозии. Визуальной агнозией страдают пациенты, которые не могут назвать предмет, показать, как он используется, или сортировать морфологически разные объекты одинакового функционального значения. В то же время у таких пациентов сохраняется острота зрения как в центре, так и на периферии зрительного поля18.
Д. Вычислительные карты (computational maps)
На каждом иерархическом уровне коры представлена упорядоченная карта различных характеристик видимых объектов. Например, одна клетка первичной зрительной коры может селективно отвечать на предъявление световой полоски определенных ориентаций19. Нейроны, проявляющие такие свойства, были названы детекторами линий. Форма организации детекторов линий в зрительной коре такова, что параметр ориентации на протяжении зрительной коры изменяется медленно и имеет повторяющийся паттерн. На основе представлений о регулярности организации зрительной коры было разработано понятие повторяющихся вычислительных карт. На сегодняшний день под вычислительными картами понимают ключевые строительные блоки инфраструктуры систем обработки информации в мозге. Впоследствии экспериментально были обнаружены данные, свидетельствующие о существовании нейронных карт на высших уровнях обработки зрительной информации, избирательно реагирующие на направление движения и пространственную частоту объекта20.
Е. Модель окружающей среды (Anticipating schemata)
Давайте представим человека с катарактой — заболеванием, при котором хрусталик глаза или окружающая прозрачная мембрана становится непроницаемой и свет не может попасть на сетчатку. Далее, если с помощью операции такой хрусталик заменить на искусственный, то сигналы от сетчатки начнут попадать в мозг. Сможет ли такой пациент видеть мир таким, каким видят его обычные, здоровые люди? Как показывают психологические наблюдения, если данное заболевание глаз происходит в раннем возрасте, то в результате таких операций люди не могут воспринимать зрительный мир полноценно. Приведенный пример указывает на то, что для нормального процесса восприятия необходима определенная «инструкция», направляющая сенсорные информационные потоки подобающим способом. Как кажется, эти «инструкции» развиваются в процессе взросления в соответствии с генетически детерминированной нейронной организацией. С самого раннего детства, шаг за шагом, мы активно учимся взаимодействовать с объектами окружающего мира. Мы исследуем мир, поворачивая голову и глаза, манипулируем объектами при помощи рук и пальцев.
18 Нарушения зрительно специфичны, т.е. поведение остается без изменений, когда пациенты слышат характерный звук или касаются предмета. В свою очередь, зрительная агнозия делится на более мелкие категории, такие как двигательная или цветовая агнозия, в зависимости от локализации зоны повреждения.
19 За эти исследования Хыобел и Визель были награждены Нобелевской премией 1981 года.
20 Было показано, что локальная область коры размером 500 х 500 микрон включает в себя все возможные ориентации и пространственные частоты. В наших исследованиях такая область коры получила название канонического кортикального модуля (см. «Методы» ч. 2).
Часть 2, Вызванные потенциалы
209
Идея модели окружающей среды как организующего фактора зрительного восприятия была предложена Найссером (Neisser). В соответствии с его точкой зрения антиципирующая схема, или внутренняя модель мира, является предвосхищающей когнитивной структурой, обеспечивающей готовность воспринимать определенные виды сенсорной информации по отношению к другим. Модель окружающей среды не только управляет перцептивной активностью, но и сама видоизменяется в процессе восприятия (Neisser, 1976). В последнее время в литературе схемы Найссера модифицировались в понятие восходящих и нисходящих потоков обработки информации. При этом концепция была заменена концепцией репрезентаций (памяти) объектов и их пространственных отношений.
Ж. Распознавание лица
Обработка изображений лиц является одной из удивительных способностей человека. Нам достаточно беглого взгляда, чтобы определить, знакомо или нет данное лицо, на основании анализа выражения лица определять эмоциональное состояние, а в некоторых случаях и судить, о чем думает человек. Многими лабораториями мира делались попытки обнаружить так называемый «бабушкин» нейрон, избирательно активирующийся на предъявление определенного лица. Несмотря на то что такие нейроны были обнаружены в нижней височной коре мозга, крайне сложно непротиворечиво доказать, что эти нейроны кодируют информацию, действительно относящуюся к конкретному лицу. Вполне вероятно, что эти нейроны активируются в ответ на более широкий класс сложных объектов или простой паттерн, как, например, топография, являющийся свойственной характеристикой лица.
3. Множественные компоненты ВП
В недавнем исследовании, проведенном учеными Джоном Фоксом и Грегори Симпсоном из отдела нейронаук Медицинского колледжа им. Альберта Эйнштейна, проводилась регистрация ВП высокого пространственного разрешения (с помощью 128 электродов) при зрительном предъявлении красных билатеральных дисков, время предъявления которых составляло 280 мс. При обработке ВП для более тщательного анализа структуры С1 компоненты ВП, которая, как считается, генерируется в области стриарной коры, использовалось преобразование Лапласа. Полученные в этом исследовании данные свидетельствуют о наличии множества источников генерации компонент, диапазон латентных периодов которых традиционен для С1. При этом дорсальные генераторы по сравнению с вентральными характеризуются меньшими значениями ЛП. Удивительным результатом исследования явилось и то, что дорсолатеральная префронтальная кора вовлекается в обработку зрительной информации уже спустя 80 мс с момента появления стимула. Если учесть, что затылочная кора активируется за 56 мс от начала предъявления стимула, то широко распределенная система обработки зрительной информации, включающая сенсорные, теменную и лобные области коры, активируется в течение 30 мс. Данные факты свидетельствует о более высокой скорости работы сенсорных систем мозга по сравнению с более ранними, традиционными представлениями.
210
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
И. Топография коры
Топографическая организация коры является одним из фундаментальных принципов строения мозга21. Топография характерна не только для иерархически низких уровней информационного потока, но также обнаруживается и на более высоких уровнях обработки информации. На уровне зрительной коры различные характеристики объектов активируют специфические области, ответственные за их обработку. В частности, оказывается, что лица, буквы и слова ассоциируются с областями, ответственными за обработку информации центра зрительного поля, а изображения зданий — с периферическими. Например, в исследовании Леви и др. (2001) было четко показано, что изображение лица активирует области зрительной коры, которые накладываются на зоны, топографически соответствующие корковому представительству фове-ального зрения. В то же время распознавание изображения зданий харакге-
5
Рисунок 10.4. ВП на разные категории зрительных стимулов
Всего использовалось 9 категорий стимулов (А.) Усредненные по группе испытуемых ВП, соответствующие стимулам двух категорий и зарегистрированные в отведении Р8 в интервале от 0 до 400 мс с момента предъявления стимула. Характерно, что предъявлению изображения лица соответствует большая амплитуда компонент Р100 и N170 (см. Itier и Taylor, 2004).
ризовалось сложно организованным паттерном целого набора активирующихся структур, в состав которого входили и области зрительной коры, вовлекаемые в обеспечение восприятия периферической зрительной информации.
К. Усиление N170 компоненты ВП в ответ на предъявление изображения лица
ВП-исследования воспроизводят результаты, полученные в фМРТ-исследо-ваниях. Роксана Итер (Roxane Itier) из Ротманского исследовательского института (Торонто, США) и Пол Саба-тир (Paul Sabatier) из Университета Тулузы (Франция) регистрировали ВП в ответ на предъявление разных категорий стимулов — лиц, домов, различных текстур и т.д. В соответствии с данными, полученными в этих исследованиях, предъявление лиц (даже инвертированных) приводило к увеличению амплитуды компоненты N170 (рис. 10.4).
21 Имеются клинические доказательства топографической организации стриарной и престриарной коры. Пациенты с небольшими нарушениями коры (вызванными небольшими инсультами или опухолями) не способны видеть объекты, расположенные в соответствующей области зрительной сцены. Такой вид функциональных нарушений носит название -слепота зрительной коры» (подчеркивая вовлечение коры в сознательное зрительное восприятие), а дефект зрительного восприятия называется скотомой. Примечательно, что любой из нас имеет этот дефект. Это так называемое слепое пятно нормальной сетчатки — небольшая (около 2 миллиметров) область выхода зрительного нерва без рецепторов сетчатки. Так как оптический диск расположен медиально относительно центра зрения обоих глаз, то свет, приходящий из единственной точки бинокулярной зоны, никогда не попадает на оба диска, так что мы обычно не замечаем это слепое пятно.
Часть 2, Вызванные потенциалы 211
Несмотря на то что авторы не применяли метод LORETA при обработке своих данных, динамика двумерных карт (топограмм) свидетельствует о разной топографии компоненты при предъявлении лиц по сравнению с пространственным распределением N170 для других стимулов. Полученные этими авторами данные также отражают одно из преимуществ метода ВП перед другими методами регистрации мозговой активности — высокую временную разрешающую способность. В действительности, регистрация ВП позволяет оценить изменения электрической активности с точностью до нескольких миллисекунд, в то время как фМТР- и ПЭТ-методы оценивают состояние в секундном диапазоне. Эта отличительная особенность ВП позволяет ученым изучать этапы обработки информации мозгом человека.
III. Разложение ВП одной пробы на независимые компоненты
Аналогично тому, как в ЭЭГ выделяется несколько ритмических компонент, вызванный потенциал представляет собой совокупность (сумму) нескольких компонент, генерируемых разными областями коры в разные моменты времени. На рис. 10.5 изображены ВП, зарегистрированные у 26-летнего испытуемого в ответ на предъявление одинаковых зрительных стимулов и комбинацию слуховых и зрительных стимулов, принадлежащих разным категориям. В качестве зрительных стимулов использовались кратковременные (100 мс) предъявления изображений людей и растений. Слуховые стимулы — случайная последовательность тонов разной частоты, каждый из которых предъявлялся на 20 мс. В разных пробах тоны предъявлялись в разных комбинациях и тем самым при каждом предъявлении звучали по-новому. Предполагалось, что испытуемый просто воспринимает зрительную и слуховую стимуляцию, без какой-либо определенной реакции на предъявление.
Как видно на усредненном по всем пробам вызванном потенциале (рис. 10.5), предъявление зрительного стимула вызывало негативное отклонение потенциала с латентным периодом 160 мс в затылочных областях. Одновременное предъявление зрительных и «новых» слуховых стимулов помимо негативности в затылочной области характеризовалось позитивными отклонениям в левой височной (Л П120 мс) и центрально-фронтальной (ЛП 200 мс) области с максимумом в Cz-отведении. Следовательно, можно предположить, что вызванный потенциал при одновременной слуховой и зрительной стимуляции состоит по крайней мере из трех разных компонент, распространенных в лобно-височно-затылочных областях и достигающих своих максимальных значений в разные моменты времени.
На примере рассмотренного ВП можно рассуждать о том, что усредненный вызванный потенциал представляет собой совокупность по меньшей мере трех гипотетических компонент. Для доказательства данного предположения необходимо использовать информацию каждого отрезка ЭЭГ, относящегося к каждой отдельной пробе. Для этого необходимо выяснить, как коррелируют между собой потенциалы различных электродов. В действительности мы
212
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
предполагаем, что потенциалы электродов, регистрирующих активность генераторов одной компоненты, коррелируют между собой в большей степени, чем потенциалы электродов, записывающие активность генераторов двух разных компонент. Информация, касающаяся корреляции активности электродов, содержится в так называемой ковариационной матрице. В течение многих лет ученые пытались решить проблему выделения компонент из ковариационной матрицы. Так называемый метод главных компонент (РСА — principle component analysis), предложенный в 60-х годах прошлого столетия, является одним из подходов к разрешению этой проблемы. С математической точки зрения данный метод позволяет редуцировать многомерные наборы данных (в нашем случае 19-канальная ЭЭГ) до меньшего количества значений (для ЭЭГ, 3 или 4 компонент). К сожалению, в методе главных компонент компоненты ортогональны, в то время как физиологически значимые компоненты не обязательно ортогональны22. Именно поэтому метод главных компонент весьма ограниченно используется в анализе биологических данных в целом и в ЭЭГ/ВП данных в частности.
Несколько лет назад появился новый метод, получивший название метода независимых компонент (ICA — independent component analysis). В основе метода лежит предположение о том, что источники компонент статистически независимы (т.е. не обязательно ортогональны, как при методе главных компонент). По-видимому, это предположение соответствует природе генераторов компонент вызванных потенциалов. В действительности информационный поток, вызываемый предъявлением стимула, последовательно активирует различные, иерархически организованные области коры. В результате генераторы разных компонент активируются в разные интервалы времени в разных областях коры.
На рис. 10.5 (справа) представлен результат применения метода независимых компонент для анализа вызванного потенциала одной пробы, зарегистрированного с нескольких электродов. Следует отметить, что ответы, полученные в одной пробе, разбиваются на три компоненты, отличающиеся по локализации генераторов и латентности.
Необходимо подчеркнуть, что компоненты ВП для одной пробы выглядят весьма зашумленными. На самом деле максимальные значения амплитуд усредненных ВП варьируют от 6 до 8 мкВ. Аналогичные показатели ЭЭГ колеблются около 10 мкВ. Эти параметры соответствуют соотношению сигнала к шуму, равному 1. В то же время усреднение около 100 участков ЭЭГ увеличивает это соотношение в 10 раз. Таким образом, лучшие результаты могут быть получены, если работать с усредненными вызванными потенциалами. Однако в этой ситуации необходимо иметь много испытуемых (100—200 человек, в зависимости от исследовательских условий).
22 В 1980-х годах в нашей лаборатории на основе метода главных компонент был разработан подход к анализу активности нейронов мозга, регистрируемой у пациентов с имплантированными электродами. В ответ на повторяющиеся психологические пробы вычислялась частота разрядки одного или множества нейронов. Анализировалась ковариационная матрица и ее собственные векторы. При предъявлении стимулов наблюдались сильные изменения весовых коэффициентов компонентов, однако интерпретация этих изменений была столь неоднозначной, что после нескольких лет поиска надежного физиологического объяснения этих реакций мы перестали использовать метод главных компонент.
ВП на зрительные я зрительные+слуговые стимулы
Рисунок 10.5. Разложение отдельных проб ЭЭГ на независимые компоненты
19-канальная запись ЭЭГ проводилась на 26-летнем испытуемом, который выполнял двухстимульный вариант GO/NOGO-теста. 50 % проб были неревантными (т.е. не требовали ответа) и представляли последовательное предъявление двух стимулов типа «растение — растение» (соотв. изображение) и «растение — человек + новый акустический стимул». А. ВП, усредненный по группе стимулов типа «растение —человек + новый акустический стимул», который регистрировался в ответ на второй стимул пробы. Там же изображены топограммы потенциалов, усредненных для разных временных значений. Б. Пример использования метода независимых компонент к анализу отрезков ЭЭГ, соответствующих отдельным (единичным) пробам, в которых одновременно предъявлялись и слуховые, и зрительные стимулы. Представлены топография, динамика отдельной пробы ЭЭГ, а также изображения локализации генераторов компонент, вычисленные с помощью метода томографии низкого разрешения (sLORETA). Следует обратить внимание, что выделяемые компоненты характеризуются разными расположениями мест генераций и временной динамикой.
Часть 2. Вызванные потенциалы___________________________________________________________213
214
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
IV. Разложение усредненного ВП на отдельные компоненты
Метод независимых компонент может быть применен к набору индивидуальных ВП. В этом случае выделяемые компоненты ВП будут отражать активность генераторов, являющихся общими для выбранной группы испытуемых. Ниже будет представлен пример использования этого метода. Индивидуальные ВП были выбраны из нормативной базы данных (HBI Database), содержащей ВП-данные около 1000 здоровых человек23.
При анализе ВП-данных целью метода независимых компонент является использование разницы в распределении по скальпу генераторов ВП, общей для всей популяции здоровых испытуемых, для выделения времени их активации. Мы исходим из предположения, что разные этапы обработки информации ассоциируются с различными корковыми структурами и разными (а до некоторого предела — и независимыми) временными паттернами. Следует напомнить, что построение компонент ВП связано с оптимизацией взаимной независимости всех кривых времени активации. Говоря более детально, в основе метода независимых компонент, используемого в анализе ВП, лежат три основных допущения (предположения): 1) генераторы, локализующиеся в разных областях коры, продуцируют потенциалы, которые линейно суммируются на регистрирующих скальповых электродах; 2) ВП представляют собой сумму активаций специфического набора пространственно стабильных мозговых нейронных сетей; 3) временные характеристики этих активаций у разных испытуемых независимы24.
В качестве примера мы собираемся использовать двухстимульный GO/NOGO-тест, специально разработанный для нормативной базы данных. Тестовое задание состоит из 400 стимулов, предъявляемых испытуемым каждые 3 секунды (рис. 10.6 ). Стимулы разделяются на следующие 3 категории: 1) 20 разных изображений животных — обозначаются буквой А; 2) 20 разных изображений растений — Р; 3) 20 разных изображений людей, предъявляемых совместно с искусственным «неожиданным» звуком — HS25. Пробы включали предъявление пары стимулов с межстимульным интервалом 1,1с. Для анализа были выбраны 4 типа пробы: А-А, А-P, Р-Р иР-HS. Все пробы сгруппированы в 4 сессии по 100 в каждой. В каждой сессии использовался уникальный набор из 5 стимулов типа А, Р и HS. Каждая сессия состояла из псевдорандомизирован-ного предъявления 100 пар стимулов с равной вероятностью для каждого типа пробы и стимула.
После преобразования зарегистрированной активности относительно общего усредненного референта для разложения компонент ВП был использован метод независимых компонент, подробно описанный в работе Makage (1999). На рис. 10.7 представлены результаты разложения всего набора ВП, зарегистриро-
23 В первой части настоящей книги я уже уделял внимание основам метода независимых компонент на примере нахождения независимых генераторов ритмов фоновой ЭЭГ-активности. Метод применялся для анализа индивидуальных ЭЭГ-данных и значений спектральной мощности. При одном подходе удавалось выделить индивидуальные источники, а при втором — выделить основные источники, общие для всех здоровых испытуемых. Теоретически такая же процедура может быть выполнена и для ВП, полученных в ответ на стимул или движение.
24 Предположения 2 и 3 основаны на многочисленных экспериментальных данных, полученных в исследованиях животных, которые свидетельствуют об активациях нейронов различных областей мозга на разных этапах обработки информации. Эти этапы характеризуются кодированием свойств стимулов и паттернами времени активации.
25 Случайно варьируемые новые звуки составлялись из пяти 20-миллисекундных фрагментов тонов разной частоты (500,1000, 1500,2000, и 2500 Гц). Интенсивность стимулов была около 75 Дб.
Часть 2. Вызванные потенциалы
215
Рисунок 10.6. Типы стимулов, используемых в GO/NOGO-тесте Пояснения см. в тексте.
ванных у группы здоровых испытуемых в возрасте от 7 до 89 лет. Анализировались ВП в ответ на второй стимул пробы типа Р-Р. В дальнейшем мы будет рассматривать это условие как «нерелевантное». Как видно на рис. 10.7, в ответ на «нерелевантные» зрительные стимулы (т.е. стимулы, не изменяющие поведение) выделяется ряд основных независимых компонент.
Рисунок 10.7. Независимые компоненты, выделяемые при анализе ВП для нерелеватных зрительных стимулов двухстимульного GO/NOGO-теста
Слева направо: 1) топография компонент и значения их вариабельности (под топограммами); 2) вертикально уложенные тонкие горизонтальные окрашенные полосы (цвет обозначает амплитуду), каждая из которых представляет изменение во времени амплитуды соответствующей компоненты индивидуальных ВП. Всего представлены компоненты около 800 здоровых испытуемых, возраст которых колеблется в пределах от 7 (расположенные внизу) до 89 лет (расположены вверху); 3) временная шкала компонент со стрелками и числами, обозначающими латентность первого позитивного пика; 4) локализация генераторов компонент, вычисленная с помощью метода томографии низкого разрешения (sLORETA). Шкалы представлены вблизи соответствующих изображений.
216____Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Часть 2, Вызванные потенциалы 217
Первая компонента генерируется в теменно-затылочной области. Временная динамика этой компоненты сильно зависит от возраста. В возрасте от 7 до 13 лет первое отклонение этой компоненты характеризуется позитивными значениями амплитуды и латентностью 116 мс. После 13 лет изначально позитивное отклонение превращается в негативное26.
Пиковая латентность второй компоненты — 132 мс. Удивителен тот факт, что в отличие от первой компоненты с возрастом позитивность второй не меняется. Эта компонента независимо генерируется в теменно-затылочной области правого и левого полушария.
Третья компонента появляется позже первых двух. Ранняя часть компоненты, характеризующаяся позитивным отклонением с пиковой латентностью 148 мс, сменяется последовательностью негативных и позитивных волн.
Полученные данные можно интерпретировать как представление последовательных этапов обработки информации на уровне дорсального (первый и второй компоненты, система «где») и вентрального пути проведения зрительной информации (вторая компонента, система «что»).
А. Компоненты ВП как последовательность тормозных и активирующих событий (модель)
Временная динамика компонент, представленная на рис. 10.8, явно демонстрирует, что нейронные ответы в разных областях коры находят отражение в разных фазах негативных и позитивных колебаний потенциалов, регистрируемых с поверхности головы27. С определенной осторожностью эти позитивные и негативные отклонения могут соотноситься с периодами активации или торможения нейронов соответствующих нейронных сетей. Такой вывод относительно фаз торможения и возбуждения хорошо соотносится с данными импульсной активности нейронов мозга животных. В исследованиях на животных зачастую первая фаза активации нейрона сменяется фазой подавления, выражающейся в отсутствии спайков. За этими фазами возбуждения и торможения обычно следует вторая активация.
Более детально последовательность процессов торможения и активации может быть описана на примере активности зрительной системы на уровне 17-го поля Бродмана. Возбуждающие нейроны первичной зрительной коры получают входы с релейных ядер таламуса (латеральное коленчатое тело). Посредством местных связей возбуждение распространяется по локальной нейрональной сети. В свою очередь, распространение возбуждения активирует короткоаксоновые внутрикор-ковые нейроны. Фаза возбуждения очень быстра по причине малого времени ге-
76 Мы не знаем, почему электрофизиологические корреляты обработки зрительной информации различаются у детей и взрослых столь радикально. Однако из фМРТ-исследований известно, что нейроанатомическая организация мозга претерпевает весьма существенные изменения в периоде взросления (Lenroot, Giedd, 2006). Тем не менее данный вопрос требует дальнейших исследований.
27 Первая компонента генерировалась в затылочной коре и характеризовалась тремя временными фазами: 1) очень небольшая и кратковременная позитивная фаза с латентностью пика 116 мс; 2) сильно выраженная негативность с ЛП150 мс у взрослых и сильно выраженная позитивность у детей (7-12 лет) той же латентности; 3) сильно выраженная позитивная фаза с пиковой латентностью 260 мс. Вторые две компоненты генерировались билатерально в областях, входящих в состав вентрального пути обработки зрительной информации (в области средней височной извилины). Эти компоненты также демонстрировали три фазы: 1) позитивное отклонение с пиковой латентностью 130 мс; 2) негативность с ЛП 184 мс; 3) небольшое позитивное отклонение с ЛП 260 мс у взрослых и 360 мс у детей (7-12 лет). Четвертая компонента была локализована в области теменной коры (дорсальный путь обработки зрительной информации) и состояла из двух позитивных фаз, пиковая латентность которых составляла 148 мс и 330 мс с негативностью между ними (ЛП 220 мс).
218
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
прямая повторная
первичная сенсорная вторичная сенсорная
область область
Рисунок 10.8. Схема последовательности событий, происходящих в первичных и вторичных областях коры
А. Позитивные и негативные потенциалы, ассоциирующиеся с возбуждающими или тормозными постсинаптическими потенциалами. Внизу — нейрональные сети первичных (слева) и вторичных (справа) областей коры. Пустыми овалами обозначены возбуждающие нейроны. Тормозные нейроны обозначены овалами черного цвета. См. пояснения в тексте.
нерации возбуждающего постсинаптического потенциала. При активации тормозных нейронов подавляется активность близлежащих нейронов, при этом фаза торможения длится дольше (относительно фазы активации), по причине более медленной по времени генерации тормозных постсинаптических потенциалов28. Возникновение второй фазы активации может быть обусловлено как локальным восстановительным эффектом, который следует за фазой торможения29, так
и повторной активацией, распространяющейся за счет обратных связей нейрональных сетей коры (рис. 10.8). В любом слу-
чае данные скальповых ВП свидетельствуют о том, что последовательные этапы обработки информации на разных уровнях организации мозговых систем харак-
теризуются двойной (или большей) переработкой информации, являющейся необходимым условием раскрытия содержания сенсорного входа.
V. Информационные потоки слуховой информации
А. Системы «что» и «где»
Мозговая слуховая система приматов состоит из «центра» первичной слуховой коры30, проецирующей связи на окружающий «слуховой пояс», связанный с «парапоясными» областями. Каждая из этих областей обладает гистологическими особенностями строения, специфическими таламокортикальными и внутр икортикальными связями, уникальными физиологическими и функциональными характеристиками.
28 Вполне вероятно, что в младшем возрасте (менее 13 лет) возбуждение преобладает над торможением и мы не можем видеть негативные фазы компонентов ВП. По мере взросления первоначальное доминирование возбудительных процессов постепенно уменьшается, и на ВП все больше проявляются процессы торможения.
29 Механизм такого восстановительного эффекта может быть сопоставим с эффектом кальциевых спайков кортико-таламических нейронов. *
* 30 «Центр» слуховой коры мозга человека, или первичная слуховая кора, локализуется в первой поперечной извилине Хешли (41-м ПБ), которая получает входы от медиального коленчатого тела таламуса.
Часть 2. Вызванные потенциалы
219
Сравнительно недавно была сформирована концепция множественных и параллельных путей обработки информации в слуховой сенсорной модальности, принципиально аналогичная используемой при описании зрительной системы мозга (рис. 10.9). Электрофизиологические исследования приматов, как и томографические исследования мозга человека, указывают на существование различий переднего и заднего потоков обработки информации. В поток «что», связанный с обработкой вокальных характеристик звуковой стимуляции (предположительно ответственной за идентификацию слухового объекта), вовлекаются передние «поясные» (belt) и «парапоясные» (parabelt) области слуховой коры, которые в дальнейшем проектируются в передние височные и вентролатеральные фронтальные (экспериментально не показано) области. В поток «где», предназначенный для локализации источников звуков, вовлекаются задние «поясные» и «парапоясные» области слуховой коры, проецирующиеся в задние височные и дорсолатеральные фронтальные (экспериментально не показано) области. Как было показано, задние слуховые области реагируют на пространственные характеристики звуковых стимулов подобно реакциям нейронов теменной коры в ответ на пространственные параметры зрительной стимуляции, что подтверждает существование заднего височно-теменного потока, предназначенного для обработки пространственных характеристик слуховых стимулов. В 2002 году Заторре и Белин (Zatorre et al., 2002) представили доказательства возможного участия «дорсального» пути обработки информации в восприятии спектральной динамики звуков, отражающей их быстрые перестройки во времени31.
Рисунок 10.9. «Что» и «где» потоки обработки информации на уровне слуховой коры
Центральная, поясная и парапоясная слуховые области изображены разными цветами. Стрелками обозначены пути «что» (передняя стрелка) и «где» (задняя стрелка), на уровне которых осуществляется обработка слуховой информации.
31 Способность отвечать на быстрые изменения спектральной мощности является ключевой для локализации и ориентирования при восприятии звуков.
220 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Б. Тонотопическая организация слуховой коры
Тонотопия (упорядоченные карты избирательности звуковых частот) представлена на всех этапах обработки слуховой информации. Это особенно проявляется на уровне первичной слуховой коры при восприятии чистых тонов. Топографическая организация сохраняется и на уровне вторичной слуховой коры, в соответствии с которой посылаются входы на базальные ганглии (хвостатое ядро и скорлупу).
В. Обработка речевой информации
Активации, специфичные к восприятию речи, наблюдаются в левой передней верхневисочной доле. Данная область коры является мультимодальной и получает множественные проекции от зрительных, сенсомоторных и слуховых областей коры, что, по всей видимости, обеспечивает возможность понимания смысла произносимых слов и высказываний. По данным клинических исследований, пациенты с повреждениями серого вещества передних областей слуховой коры характеризуются прогрессивным ухудшением понимания отдельных слов, что свидетельствует о важной роли этой области в обеспечении семантического анализа речи32. В свою очередь, теменно-височный стык, являющийся связующим звеном между слуховой корой и теменной и фронтальной системами, является анатомически гетерогенной областью. Недавние исследования мозга человека показали, что в структуре данной области различаются как минимум 4 типа областей.
Как ранняя детская речь, произношение слов требует активности соответствующих структур мозга, обеспечивающих контроль лицевой экспрессии (выражения лица), артикуляционных движений и действий. Известно, что такие структуры локализованы в области нижней моторной и смежной с ней нижней префронтальной коры. При активном участии артикуляции генерируются звуки, которые кодируются мозговой системой слуха, включая области верхней височной доли. Наличие большого количества связей между слуховой и префронтальной корой обеспечивает формирование ассоциативных связей между нейронами, контролирующими специфические моторные речевые программы, и нейронами, стимулируемыми звуками собственной речи.
Г. Анализ ВП методом независимых компонент («HBI database»)
Метод разложения индивидуальных ВП на независимые компоненты, описанный выше для зрительной модальности, позволяет сопоставимым образом разделить этапы обработки слуховой информации. В качестве примера мы будем использовать слуховое тестовое задание, которое было сделано специально для создания HBI базы данных. Предъявлялись тоны разной частоты, одинаковой длительности — 100 мс. В некоторых пробах случайным образом время экспозиции стимула увеличивалось до 400 мс. В задачу испытуемого входило реагировать на такие длительные стимулы нажатием кнопки. ВП регистрировались в
32 В данном контексте термин «семантический» означает доступ к смысловому значению слова.
Рисунок 10.10. Независимые компоненты ВП, зарегистрированных на нерелевантные слуховые стимулы
Слева направо: 1) топография компонент и значения их вариабельности (под голограммами); 2) вертикально уложенные тонкие горизонтальные окрашенные полосы (цвет обозначает амплитуду), каждая из которых представляет изменение во времени амплитуды соответствующего компонента ВП отдельного испытуемого: представлены данные 600 здоровых испытуемых возрастом от 7 (расположены внизу) до 89 лет (расположены вверху); 3) временная шкала компонент со стрелками и числами, обозначающими соответствующую латентность первого позитивного пика; 4) локализация генераторов компонент, вычисленная с помощью метода томографии низкого разрешения (sLORETA). Шкалы представлены вблизи соответствующих изображений.
Часть 2, Вызванные потенциалы_________________________________________________________221
222 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
ответ на предъявление коротких тонов. На рис. 10.10 представлен результат анализа независимых компонент ВП для коротких стимулов (нерелевантных, те. не требующих реакции), зарегистрированных у группы здоровых добровольцев возрастом от 7 до 89 лет.
Как видно из рис. 10.10, выявляются две компоненты, генерируемые в области первичной слуховой коры правого и левого полушария. Пиковая латентность правой височной компоненты — 84 мс33. Обнаруженные височные компоненты являются многофазными, аналогично компонентам зрительных ВП. Третья компонента генерируется в области задней поясной извилины, а четвертая — в теменной коре. Примечательно, что эти компоненты характеризуются противоположным характером изменений в зависимости от возраста: с возрастом становится более выраженной компонента в поясной извилине, тогда как амплитуда теменной снижается.
Анализируя всю совокупность выявленных компонент, можно предполагать, что височные компоненты отражают работу системы «где», а остальные связаны с активностью системы «что» слуховой модальности.
VI. Соматосенсорная модальность
Все наше тело покрыто чувствительными рецепторами, которые позволяют нам чувствовать боль, прикосновения, контролировать висцеральные функции и т.д. Рецепторы располагаются у поверхности и в глубоких слоях кожи, мышцах, суставах и сухожилиях. Соматосенсорные рецепторы можно разделить на три большие группы: 1) ноцирецепторы, обеспечивающие болевую и температурную чувствительность; 2) тактильные рецепторы, отвечающие за перцепцию легких прикосновений и давления; 3) про-приорецепторы, позволяющие определить положение тела и его конечностей. Кроме рецепторов внешнего мира, существуют также рецепторы внутренних органов, или висцеральные рецепторы34.
А. Соматосенсорная и инсулярная кора
Внешние соматосенсорные рецепторы проецируются в соматосенсорную кору мозга (1—3-е ПБ) через заднее вентральное ядро таламуса. Рецепторы висцеральных органов посылают информацию в область инсулярной коры через вентромедиальное ядро таламуса. Инсулярная кора локализуется достаточно глубоко в мозге, и на данный момент мало известно о характере ее ритмической активности. Соматосенсорная кора располагается на поверхности коры мозга, генерирует так называемые мю-ритмы, которые регистрируются центральными электродами СЗ, С4, Cz (см. гл. 2).
33 Следует отметить, что первые ранние компоненты в зрительной модальности регистрировались с ЛП 116 мс, что примерно на 30 мс позже слухового компонента. Данный факт демонстрирует как разницу в реактивности таламических ядер в зрительной и слуховой системах, так и разность в сложности обработки информации в этих модальностях.
34 В соматосенсорной модальности выделяюттакже и подмодальносгь, а именно чувство вибрации. Например, когда мы трогаем вибрирующий предмет. Частота механической вибрации, которую мы можем воспринимать, варьирует от 5 до 50 Гц. Детекция вибрации как у человека, так и у приматов осуществляется быстро адаптирующимися кожными механорецепторами.
Часть 2. Вызванные потенциалы
223
VII. Детекция изменений
А. Функциональный смысл детекции изменений
Окружающий мир отражается в постоянной активности нейронов сенсорных систем мозга. Некоторые части мира остаются неизменными в течение достаточно длительных периодов времени, на которые мы чаще всего не обращаем внимания35. Но когда нас вдруг кто-нибудь трогает за рукав, или нарушается регулярность работы автомобильного двигателя, или на глаза попадается гриб в лесу, эти изменения могут быть сознательно замечены. По-видимому, мозг постоянно сравнивает сенсорную картину мира с его моделью, сформированной на основе предварительной стимуляции. Похожим образом мозг контролирует реализацию действий и, если они не соответствуют запланированным, корректирует поведение. В настоящей книге мы будем называть эти базовые операции операциями сравнения. Исторически в сенсорной модальности операция сравнения называется детекцией.
Два типа операций сравнения соответствуют двум разновидностям моделей окружающего мира: сенсорной и моторной. Модели формируются при участии сенсорной и исполнительной систем мозга. Несмотря на сильную взаимосвязь между ними, каждая из этих систем представляет собой отдельную нейронную сеть с определенным механизмом переработки информации, служащую для реализации определенной функции. Соответственно, в поведении человека детекция изменений является важной д ля достижения двух основных целей.
Во-первых, изменения в сенсорной картине мира привлекают внимание как сознательно (селективное внимание), так и бессознательно (автоматически). Например, неожиданное изменение в шуме работающего двигателя может быть замечено водителем и когда внимание сконцентрировано на дороге, и при намеренном прослушивании. В этой ситуации сенсорная модель формируется на базе сенсорной системы, а текущий стимул, не соответствующий данной модели, вызывает активацию соответствующей части сенсорной системы. Важную роль в формировании модели играет и предыдущий сенсорный опыт. Таким образом, цель данного типа детекции изменений обусловлена функцией сенсорной системы — выбор источника изменения сенсорной информации для ее более детальной обработки36.
Во-вторых, факт детекции изменения может означать, что реализованное действие не является корректным, а значит, должно быть изменено или подавляться в дальнейшем. Так, человек, собирающийся переходить улицу на зеленый свет, останавливается при резкой смене сигнала светофора на красный. В этой ситуации модель действия — переходить дорогу — формируется на уровне исполнительной системы. Изменение текущей поведенческой ситуации означает; что подготавливаемое действие является несоответствующим и должно быть подавлено. Таким образом, основной целью данного типа детекции изменений является подавление исполнительной системы для обеспечения отмены реализации планируемого действия.
35 В частности, мы не ощущаем давления давно носимой одежды, шум двигателя собственной машины и не замечаем минимальных изменений зрительной сцены, находясь в лесу, будучи погруженными в размышления.
36 В дальнейшем изменение может быть детектировано исполнительной системой, которая обеспечит наведение взгляда или перемещение тела (явно или нет) в направлении выбранного источника сенсорной информации. Однако не всякое изменение обязательно приводит к активации исполнительной системы.
224
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Б. Механизм детекции изменений (модель)
Наиболее изученными являются механизмы мониторинга изменений окружающей среды в слуховой модальности, с рассмотрения которой мы и начнем. Лидирующие позиции в исследовании этого вопроса занимают исследования Центра когнитивных исследований мозга Университета Хельсинки (Финляндия), созданного Ристо Наатаненом в 1980-х годах. Наатанен был первым исследователем, предположившим существование двух механизмов детекции изменений для слуховой модальности37.
Первый механизм отражается изменениями ранних компонент ВП и ассоциируется с обработкой информации на уровне первичной и, скорее всего, вторичной слуховой коры (рис 10.11). Работу первого механизма Ристо Наатанен связывает с N1 компонентой ВП. Предполагается, что в составе слуховой коры существует две разновидности нейронов — стабильные и рефрактерные. Стабильные нейроны не меняют свои ответы при повторных появлениях стимула. Активность рефрактерных нейронов снижается по мере того, как несколько раз предъявляется один и тот же стимул. Рефрактерность означает феномен снижения частоты ответов при повторяющейся стимуляции. Хотя синаптическая депрессия, как кажется, является основным клеточным механизмом затухания нейронных ответов на повторяющиеся стимулы38, сложные взаимодействия между нейронами, по-видимому, также играют некоторую роль (пока еще неопределенную) в реализации этого феномена.
Кроме существования двух типов нейронов, предполагается наличие и второго механизма, называемого детекцией изменений. Гипотетические нейроны, ответственные за обеспечение работы механизма, реагируют только на изменения стимулов. Если принять во внимание нелинейный характер активности нейронов39, то легко можно представить возможное устройство нейронной сети для детекции изменений (рис. 10.11 справа). Исходя из предложенной модели, ассоциативная кора (кортикальная область 3 на рис. 10.11) получает два входа: 1) из первичной сенсорной коры, ответственной за обработку физических свойств стимула и не изменяющей своей активности при повторении стимула; 2) из вторичной, активность которой снижается при повторении. Нейроны ассоциативной области посредством латерального торможения осуществляют сравнение информации, поступающей с этих двух входов. В результате некоторые нейроны этой области активируются только при появлении нового стимула4*’.
Аудиостимулы характеризуются не только физическими свойствами (частотой и интенсивностью), но и более интегративными свойствами, например, таким как регулярность. В частности, в слуховой модальности регулярность представлена во множестве звуковых сигналов нашей повседневной жизни: шуме работающего автомобильного двигателя, компьютерного венти-
37 Эта гипотеза представлена в книге «Внимание и функции мозга», впервые опубликованной в 1992 году в Lawrence Erlbaum Associates. Данная книга содержит понятный и детальный обзор всех литературных данных, доступных в то время, и заслуживает прочтения даже сейчас.
38 На синаптическом уровне явление депрессии связано со снижением величины генерируемых постсинаптических потенциалов в ответ на повторяющиеся стимулы.
39 В данном случае под нелинейностью подразумевается факт того, что возбуждающие нейроны в 3-й кортикальной области (рис. 10.1) разряжаются только в случае, когда активирующие влияния на эти нейроны превосходят тормозные и превышают некоторый порог.
* 40 Следует отметить, что представленная схема является одной из многих возможных. Структура и механизм работы реальных
нейронных сетей до сих пор не полностью понятны.
Часть 2. Вызванные потенциалы
225
Рисунок 10.11. Схема работы механизма детекции изменений
А. Нейрональные ответы. Б. Связи между нейронами трех предполагаемых областей коры. Черным цветом обозначены возбуждающие нейроны и связи, а серым — тормозные нейроны и связи. В соответствии с представленной моделью предполагается, что операции сравнения осуществляются сетью нейронов, состоящей из трех кортикальных областей. Первая из этих областей получает входы непосредственно от таламуса, и ее активность является наиболее устойчивой, т.е. стабильна при повторении стимулов. Далее проекции от этой области следуют на возбудительные нейроны второй области, активность которой более пластична, и тормозные нейроны третьей области, где осуществляется детекция изменений (т.е. область детекторов изменений). В свою очередь, нейроны третьей области, которые находятся под воздействием локальных тормозных нейронов, получают входы от возбудительных нейронов второй области. Таким образом, происходит сравнение, т.е. вычисление разности двух входов — пластичного и более ригидного. Если предъявляемый стимул является новым, то возбуждение превышает торможение, что приводит к разрядке нейронов. В обратном случае, когда стимулы повторяются, наблюдается преобладание торможения над возбуждением, и нейроны не реагируют на такие стимулы. Обозначения: станд. — стандартный стимул.
лятора или монотонной речи лектора... Как известно из нейрофизиологии, разные аспекты стимула обрабатываются нейронными элементами, локализованными в различных областях сенсорных систем. Регулярность как отдельное свойство стимула, по всей видимости, кодируется обособленной областью слуховой коры. Более того, весьма вероятно, что это свойство оценивается с помощью специального мозгового механизма. В действительности в отличие от физических характеристик регулярность как интегративное свойство формируется на основе достаточно длительного отрезка времени (десятков секунд). Однако механизм этой интеграции неизвестен. Несмотря на это, ЭЭГ-корреляты реакции мозга на изменение регулярности за последние 30 лет хорошо исследованы. Для слуховой модальности этим коррелятом является негативность рассогласования (HP).
В. HP в oddball-парадигме
Негативность рассогласования — электрический ответ на различаемое изменение регулярности слуховой стимуляции. Обычно этот ответ регистрируется на ВП с латентностью пика 150—200 мс с момента начала изменений, даже при отвлечении внимания. Феномен HP впервые был продемонстрирован для слуховой модальности, а его наличие в зрительной модальности является спорным вопросом41.
41 Причина этого заключается в природе операции сравнения, которая основана на сравнении со следом памяти. След в краткосрочной сенсорной памяти формируется в результате процесса реверберации и синаптической депрессии (или потенциации). Известно, что эхоическая память достаточно длительна — до 10 с, тогда как иконическая крайне коротка — до нескольких сотен миллисекунд. Таким образом, регулярность может быть сформирована в рамках традиционной исследовательской oddball парадигмы в слуховой, но не зрительной модальности. Следует подчеркнуть, что HP не регистрируется, если межстимульный интервал длится больше чем 10 с, что соответствует представлениям о временных характеристиках краткосрочной сенсорной памяти.
226 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Изучение HP в слуховой модальности традиционно проводится с использованием так называемой oddball-парадигмы. В упрощенном варианте тестовое задание выглядит следующим образом: в ряду повторяющихся стандартных стимулов (например, тон 1000 Гц длительностью 100 мс) редко предъявляются девиантные (например, тон 1100 Гц длительностью 100 мс)42. Более детальное описание приведено в главе «Методы» части 2.
Феномен негативности рассогласования был открыт в 1978 году в классической работе Наатанена, Джилларда (Gaillard) и Мантисало (Mantysalo). В соответствии с общепринятой интерпретацией, HP генерируется в результате детекции рассогласования между девиантным стимулом и следом памяти, сформированным повторно предъявляемыми стандартными слуховыми стимулами43. Усредненные ВП, зарегистрированные в рамках oddball-парадигмы при исследовании здоровых испытуемых в возрасте от 15 до 89 лет, представлены на рис. 10.12 (слева). По сравнению со стандартными стимулами предъявление девиантных вызывало увеличение амплитуды негативного отклонения потенциала в области лобно-центральных отведений.
С использованием метода независимых компонент было произведено разложение полученных ВП и выделены три независимых компоненты (рис. 10.12 справа). Первая компонента генерировалась в области первичной слуховой коры с латентностью пика 116 мс. Фактически эта компонента была одинакова и для стандартного, и для девиантного стимулов. Вторая компонента, генерировавшаяся в ассоциативных областях слуховой коры с латентностью пика 132 мс, характеризовалась вдвое большей амплитудой для девиантного стимула по сравнению со стандартным. Наконец, третья компонента была обнаружена в области передней поясной извилины. Латентность его пика составляла 236 мс. Компонента при девиантном стимуле (относительно стандартного) характеризовалась дополнительной позитивностью.
Г. Внутримозговые корреляты HP
В исследовании, проведенном в нашей лаборатории совместно с финским коллективом, возглавляемым Р. Наатаненом (Kropotov et al., 2000), было показано существование по крайней мере трех отдельных мозговых механизмов генерации HP. На рис. 10.13 представлены локальные потенциалы, зарегистрированные с электродов, имплантированных в слуховую кору в области 41-, 42- и 22-го ПБ. Как видно из рис. 10.13, стабильная активность демонстрируется только для первичной слуховой коры (ПБ 41) и не изменяется при повторно предъявляющихся стимулах. Следовательно, можно считать, что нейроны этой области обеспечивают кодирование физических характеристик стимулов (например, частоту сигнала). Вторичная слуховая кора (ПБ 42) снижает активность при повторении одинаковых стимулов. Вполне вероятно, что определенный вклад в наблюдаемый эффект может вносить синаптическая депрессия. По всей видимости, операция сравнения происходит на уровне ассоциативной области слу-
42 Во избежание эффекта рефрактерносги в генерации HP обычно используется небольшая (но тем не менее различаемая) разница между стандартными и девиантными стимулами.
43 Для обзора см. книгу Ристо Наатанена «Внимание и функции мозга», 1992. Hillsdale, NJ: Erlbaum.
Часть 2. Вызванные потенциалы
227
а ВП g Компоненты
Рисунок 10.12. Разложение негативности рассогласования на компоненты
Л. Усредненные по группе здоровых добровольцев ВП. Б. Топография, временная динамика и локализация генераторов трех независимых компонент, составляющих полученные ВП, вычисленная с помощью метода томографии низкого разрешения (sbORETA).
ховой коры (ПБ 22): если стимул не соответствует следу памяти, сформированному предварительной однотипной стимуляцией, то генерируется сильно выраженная негативная компонента, за которой следует позитивная44. Следует оговориться, что слуховая кора не является единственной областью, влияющей на генерацию HP. Так, кроме височной области, были найдены несколько источников генерации в префронтальной коре45.
Д. Детекция изменений в задаче по дискриминации двух стимулов
Негативность рассогласования представляется как частный случай более общей операции, ассоциирующийся с детекцией изменения регулярности звуковой стимуляции. Однако регулярность как свойство сенсорной стимуляции наиболее выражена именно в слуховой модальности46.
В общем смысле операция сравнения осуществляется с любой характеристикой сенсорного стимула и соответствующим следом памяти. Мы называем эту операцию детекцией изменения. Если задачей исследования не яв-
44 Важно уточнить, что этот позитивный компонент наблюдается только в oddball парадигме и отсутствует в ситуации, когда редким стимул предъявляется один.
4<> Генераторы HP, локализуемые в лобных долях, активируются после височных, спустя небольшой промежуток времени и играми важную роль в процессах инициации бессознательного переключения внимания на изменение звука, детектируемого в височном коре. Предполагается, что процесс детекции изменения происходит именно в слуховой коре (отражается субкомпонентом HP в нисочной области). Это является триггером активации фронтальных областей коры (генерируется фронтальный субкомпонент HP и |’3<1 компонент, следующий за ним), которые, вероятно, обеспечивают переключение внимания и сознательное восприятие изменения стимула.
Вероятно, именно поэтому вопрос существования HP в зрительной модальности вызывает активные споры.
228
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
ODDBALL
— стандартный юм Гц
— девиантный 1зоо Гц
Отдельное предъявление регулярных и редких стимулов
— стандартным юм Гц — девиантный 1зм Гц
Стабильные ответы
Рефрактерные ответы
Детекция изменений регулярности
Б
Рисунок 10.13. Три механизма операции сравнения (регистрация активности нейронов с долгосрочных имплантированных электродов в слуховой модальности)
Внутримозговые локальные полевые потенциалы регистрировались у пациентов, страдающих эпилепсией, которым по клиническим показаниям, с целью диагностики и лечения, в различные области височной доли имплантировались долгосрочные электроды (области Бродмана 41,42 и 22). В одном исследовании активность нейронов регистрировалась в момент чтения книги пациентом, при параллельном предъявлении последовательностей тонов. А. Тоны предъявлялись в рамках oddball-парадигмы: стандартные тоны 1000 Гц были перемешаны с девиантными 1300 Гц. В другом исследовании (Б) предъявлялись только стандартные и девиантные стимулы с соответствующими длинными и короткими межстимульными интервалами (Kropotov etal., 2000). Обозначения: HP — негативность рассогласования; ТС — область слуховой коры.
ляется изучение процессов, связанных с неожиданными изменениями регулярности стимулов, то нет необходимости использовать задание со стимулами, повторяющимися много раз. Следовательно, детекция изменений может быть экспериментально исследована не только в рамках oddball-парадигмы. Так, нами было разработано тестовое задание по дискриминации двух стимулов для изучения процессов детекции изменений свойств зрительной стимуляции. В этом тестовом задании стимулы предъявлялись парами, в которых второй стимул мог в некотором смысле совпадать или не совпадать с первым, соответственно задачей испытуемого было по-разному реагировать на пары проб этих двух категорий.
В исследовании обезьян (Romo et al., 2000) изучались процессы различения поверхностей при регистрации импульсной активности нейронов (ИАН), расположенных в соматосенсорных областях S1 и S2, вентральной префронтальной и медиальной премоторной областях коры. Для этого на кончике одного из пальцев размещался механический стимулятор, производивший короткие колебания. Каждая проба состояла из стимуляций с разными частотами fl и f2, разделенными коротким интервалом времени. Обезьяна должна была нажимать определенные кнопки в зависимости от того, была ли частота стимуляции f2 выше или ниже, чем fl. Последовательность психологических операций представлялась следующим образом: 1) кодирование частоты первой вибрации fl; 2) удержание следа в сенсорной памяти; 3) кодирование частоты второй вибрации f2; 4) сравнение результата кодирования со следом памяти П; 5) передача результата операции сравнения моторным нейронам.
Часть 2, Вызванные потенциалы
229
Полученные в этом исследовании результаты свидетельствовали о том, что операция сравнения обеспечивалась нейронами вторичной (а не первичной) соматосенсорной коры. То есть «вычислялась» разница между двумя типами вибраций, которая отражалась в частоте разрядов этих нейронов в ответ на второй из пары стимулов пробы47. Время, необходимое для реализации этой операции, составляло 200 мс. Доказательств участия нейронов первичной сенсомоторной коры (S1) в обеспечении операции сравнения выявить не удалось, а профиль их ответов выглядел как функция зависимости от частоты второго стимула (f2).
Е. Модальная специфичность
Начиная с 1990-х годов операции сравнения являются предметом исследований нашей лаборатории. Нами была разработана так называемая двухстимульная GO/NOGO-парадигма. Исследования проводились как в слуховой, так и в зрительной модальности. В качестве слуховых стимулов использовались высокие (В) и низкие тона (Н) в следующих сочетаниях: ВВ, ВН, НВ, НН. В качестве зрительных стимулов использовались цифры 6 и 9 в сочетаниях: 99, 96, 69, 66. Задачей испытуемого являлось выделять пары стимулов и нажимать кнопку, если второй стимул в паре (высокий тон или цифра 9) соответствовал первому (высокий тон или 9 соответственно). Мы полагали, что после инструкции и тренировочной сессии у испытуемого формируется поведенческая модель соответствия пары стимулов выбранной категории (например, два изображения цифры 9) с определенным действием (нажатие кнопки). Считалось, что модель состоит из двух частей: 1) сенсорная модель — шаблон категории релевантных стимулов; 2) моторная модель — шаблон действия (нажатие кнопки), которое должно быть исполнено как можно быстрее и точнее.
Выполнение тестового задания состояло из ряда психологических операций. Предъявление первого стимула индуцировало: 1) кодирование релевантных характеристик стимула; 2) сравнение характеристик с сенсорным шаблоном; 3) в случае совпадения — обновление сенсорной подготовительной установки (preparatory set) и инициация моторной подготовительной установки; 4) в случае рассогласования — подавление формирования моторной подготовительной установки. Психологические операции, вызываемые предъявлением второго стимула пары (GO или NOGO): 1) кодирование релевантных свойств стимула; 2) сравнение эти свойств с характеристиками предыдущего (первого) стимула48; 3) в случае совпадения — инициация выбора действия и установочной реакции; 4) при рассогласовании — подавление процессов автоматического выбора действия и обновление установочной реакции; 5) сравнение результатов NOGO-пробы (когда действие не реализуется) с шаблоном действия (когда следует нажимать кнопку) и коррекция дальнейшего поведения.
Для упрощения мы рассмотрим ВП только при предъявлении вторых стимулов проб. С учетом целей настоящей главы будут анализироваться только первые этапы
В данном случае определение частоты механической вибрации плоскости кодировалось ИАН S1 области: чем выше частота нибрации, тем выше частота импульсной активности. Вероятно, данный тип кодирования является простым способом репрезентации частотных свойств стимула.
Эти две операции сопоставимы с соответствующими операциями, индуцируемыми предъявлением первого стимула.
230
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Рисунок 10.14. Модальная специфичность детекции изменений
В верхней части расположены графики скальповых ВП для проб GO (тонкая линия) и NOGO (толстая линия), а также разностных ВП NOGO-GO (пунктирная линия), которые были зарегистрированы в F7- и Т5-отведениях. Слева представлены ВП в зрительной модальности, а справа — в слуховой. В нижней части рисунка представлены топограммы разностных ВП. Обозначения: выс., низк. — тоны высокой и низкой частоты.
Часть 2. Вызванные потенциалы
231
обработки сенсорной информации. Ранними отличиями между GO и NOGO являются: 1) модальная специфичность; 2) разный компонентный состав (пространственно перекрывающиеся, но разделенные по времени), отражающий соответственно физические и смысловые изменения стимулов; 3) отличие от компонент, связанных с моторной реакцией.
На рис. 10.14 показано, что ранние компоненты разностных ВП для GO- и NOGO-состояний модально специфичны. Для зрительных стимулов максимальная позитивность регистрировалась в области Т5-отведения, тогда как в слуховой модальности максимальные значения компоненты наблюдались в F7. Следует отметить и большую скорость обработки слуховой информации: пиковая латентность разностного ВП в слуховой модальности — 200 мс, а в зрительной — 240 мс.
100мс
HL - целевой стимул
НН -целевой стимул ----физическое изменение
семантическое
изменение
1-й стимул 2-й стимул
------ LH+LL --------NOGO HL + HH ------------ GO
Средняя височная кора
Рисунок 10.15. Процесс детекции изменений включает физический и семантический этапы
Запись, сделанная с имплантированных электродов, расположенных в средней височной доле (вверху) у пациента, страдающего эпилепсией. Электрическая активность, регистрировавшаяся в течение двух разных дней, представлена в верхней и средней области рисунка. Пациент выполнял двухстимульный слуховой тест, в котором с равной вероятностью в случайном порядке предъявлялись тоны высокой (Н) и низкой частоты (L). В течение первого исследовательского дня задачей пациента являлось нажимать кнопку в случае предъявления пары стимулов высокой частоты (НН). На второй день исследования нужно было нажимать кнопку при парном предъявлении стимулов высокого и низкого тонов (HL).
Ж. Детекция физических и смысловых изменений
Рис. 10.15 иллюстрирует тот факт, что ранние компоненты разностных ВП для GO- и NOGO-состояний действительно слагаются из двух составляющих. Исследования проводились на больных эпилепсией, которым были вживлены электроды в целях диагностики и терапии. Исследование осуществлялось в двухстимульной парадигме в разные дни: в один из дней требовалось нажимать кнопку только в пробах с парой слуховых стимулов ВВ (GO-условие), в другой при предъявлении ВН-пары (GO-условие). В результате NOGO-условие в обоих случаях характеризовалось позитивной компонентой, регистрируемой с ЛП около 200 мс. В то же время ранняя негативная компонента с ЛП 100 мс генерировалась в ответ на изменение физических свойств ожидаемого стимула. Следует также отметить, что ранняя
Компонента 2
Рисунок 10.16. Компоненты ВП, связанные с детекцией изменения, отличаются от компонент подавления моторного ответа
Представлены результаты анализа разностного ВП (GO-NOGO), произведенного методом независимых компонент. Изображены данные для более чем 800 здоровых испытуемых возрастом от 7 до 89 лет. Слева направо: 1) топография компонент и значения их вариабельности (под топограммами); 2) вертикально уложенные тонкие горизонтальные окрашенные полосы (цвет обозначает амплитуду), каждая из которых представляет изменение во времени амплитуды соответствующей компоненты ВП отдельного испытуемого: представлены данные приблизительно 800 здоровых испытуемых возрастом от 7 (расположены внизу) до 89 лет (расположены вверху); 3) временная шкала компонент со стрелками; 4) локализация генераторов компонент, вычисленная с помощью метода томографии низкого разрешения (sLORETA). Шкалы представлены вблизи соответствующих изображений.
Часть 2, Вызванные потенциалы
233
негативная компонента регистрировалась и в ситуации, когда первый стимул предъявляемой пары не соответствовал ожидаемому (т.е. Н-стимул на первом месте в паре вместо ожидаемого В-стимула).
3. Детекция изменений и подавление моторной реакции
Для демонстрации различий в топографии компонент, связанных с детекцией изменений и подавления моторного ответа, мы рассмотрим результаты анализа зрительных ВП в GO/NOGO-задании49, выполненном с помощью метода независимых компонент (рис. 10.16). Как видно из рис. 10.16, первая из наиболее выраженных компонент генерируется в левой височной области и, вероятно, отражает детекцию как физических, так и смысловых изменений стимулов50. Вторая компонента генерируется в области левой премоторной коры51 и, по всей видимости, характеризует процессы подавления выполнения подготовленного действия.
Сравнительно недавно группой ученых из Университета Хоккайдо в Японии (Kimura et al., 2006) в исследовании, проводимом в двухстимульной парадигме (SI—S2) в зрительной модальности, были зарегистрированы ВП, связанные с детекцией изменений. Как и в наших работах, в этом исследовании было показано, что изменение стимулов сопровождалось генерацией позитивного компонента с ЛП 100—200 мс в задних областях мозга. Варьирование различных характеристик зрительных стимулов (детекция изменения цвета и направления движения) характеризовалось специфическими топографическими и временными паттернами распределения компонент.
VIII. 1йпы сенсорных систем
А. U-образные кривые реактивности систем
Общая активность сенсорной системы, как и любой нейронной сети, зависит от потока входящей информации и обычно описывается сигмоидальной функцией (рис. 10.17). В то же время реакция системы на предъявление одинаковых стимулов подчиняется инвертированному U-закону (см. «Введение»). Вкратце данный закон гласит, что реакция системы на стимул мала в тех ситуациях, когда ее общая активность или слишком мала (пороговый эффект), или излишне высока (эффект насыщения).
Б. Увеличение и снижение активности сенсорных систем
Как будет вести себя сенсорная система при увеличении интенсивности стимуляции? Напрашивающийся ответ — увеличением своей активности (рис. 10.17). Однако характер этого увеличения будет зависеть от текущего функционального состояния системы. Если общий уровень активности и
49 Данное тестовое задание было специально разработано для нормативной базы данных «НВ1 Database». Вместо простых цифр предъявлялись изображения животных и растений. Использование этих стимулов было обусловлено соображением экологического соответствия, поскольку цифры, используемые первоначально, были слишком скучны.
50 Похожий компонент генерировался в области правой височной доли. Данный компонент не показан на рис. 10.16.
51 Испытуемые инструктировались нажимать кнопку правым (контралатеральным) пальцем.
234
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Выход
Рисунок 10.17. Два типа сенсорных систем
Зависимость результирующей активности сенсорной системы от характеристик входа может быть представлена сигмоидальной функцией. При возрастании интенсивности сенсорного входа увеличивается и результирующая активность сенсорной системы. Однако это увеличение неоднородно и зависит от изначального состояния сенсорной системы. Система, активность которой увеличивается медленнее (быстрее) по сравнению с увеличением интенсивности входа, называется редуцирующей (усиливающей) сенсорной системой.
приток входящей сенсорной информации невелики (точка в нижней части кривой), то относительные изменения ответа системы будут больше относительных изменений сенсорного входа. Такие системы будут относиться к усиливающим (augmenting) сенсорным системам (рис. 10.17.). В обратном случае, когда уровень общей активности системы высок, относительные изменения ответа будут меньше относительных изменений интенсивности стимуляции. Этот тип сенсорных систем можно охарактеризовать как редуцирующие сенсорные системы.
Например, в исследованиях на животных было обнаружено, что серотонинергические нейроны ствола мозга иннервируют слуховую кору. Такая иннервация, в свою очередь, приводит к зависимости общего уровня активности от уровня продукции серотонина. Хорошим индикатором функционального состояния слуховой системы является комплекс компонент ВП N1/P2. Так, если уровень серотонина и соответствующего притока входящей сенсорной информации увеличивается, то зависимость слуховых ВП от громкости снижается (на кривой рис. 10.17 изменение состояния системы отображается перемещением от левой точки на кривой направо). В исследованиях Джилланта (Gallinat et al., 2000) это свойство сенсорной
Часть 2. Вызванные потенциалы
235
системы и N1 /Р2 компоненты использовалось в качестве предиктора мощного ответа на ингибиторы обратного захвата серотонина при депрессии.
В. Компонента Р2 слуховых ВП в усиливающих (augmenting) и редуцирующих (reducing) системах
Одним из способов определения типа сенсорной системы является предъявление слуховых стимулов с увеличивающейся интенсивностью. Оценка изменения амплитуды компоненты Р2 относительно увеличения громкости стимуляции с использованием графика на рис. 10.17 может служить индикатором типа сенсорной системы испытуемого.
DG Диагностическая ценность сенсорных ВП
А. Негативность рассогласования
Одной из наиболее изученных компонент ПСС является негативность рассогласования. За последние 30 лет феномен HP интенсивно исследовался как с практической, так и с теоретической точки зрения. Сравнительно недавно HP стала использоваться для оценки фонологических и слуховых дисфункций при дислексии. В частности, в одном из таких исследований (Schulte-Korne et al., 1998) у двух групп подростков (здоровых и с диагнозом дислексии) проводился сравнительный анализ HP в ответ на речевые и неречевые стимулы. В качестве речевых стимулов использовались слоги (слог «да» — в качестве стандартного, «ба» — в качестве девиантного). Неречевыми стимулами были тоны (виды гармонической волны). Было показано, что амплитуда HP в ответ на тоны не отличалась между группами испытуемых. Однако при предъявлении слогов отмечалось относительное снижение амплитуды HP при дислексии по сравнению с HP здоровых испытуемых. Авторы интерпретировали полученные данные как отражение дисфункций в работе специфических фонологических систем, а не как генеральное нарушение процессов обработки слуховой информации. При этом когда дети, неспособные читать, тренировались с помощью зрительнослуховых упражнений, отмечалось увеличение амплитуды HP до нормальных значений.
Исследования шизофрении являются одним из интересных направлений клинических исследований с использованием HP. В частности, исследовалось влияние шизофрении на два типа генераторов HP, локализующихся во фронтальной и височной коре. Оказалось, что аффективное влияние заболевания больше выражено именно для фронтальных генераторов HP, чем для височных.
Исследования влияния процессов старения на амплитуду HP свидетельствуют о ее снижении с возрастом. Следовательно, как было показано в работах с использованием слуховых стимулов разной частоты, старение, по всей видимости, снижает объем слуховой памяти (не влияя на качество слу-
236
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Рисунок 10.18. Компонента сравнения и негативность рассогласования
Негативность рассогласования (HP) рассчитывалась как разностная волна, получаемая при сравнении ВП при девиантном и стандартном тоне, предъявляемых в рамках oddball-парадигмы. Аналогично компонента сравнения рассчитывалась как разница между ВП для GO- и NOGO-стимулов, регистрировавшихся в двухстимульном GO/NOGO-тесте. Представлены усредненные по группе здоровых испытуемых (от 14 до 80 лет) ВП. Данные были взяты из нормативной HBI базы данных.
хового восприятия в целом). Пациенты с диагнозом «болезнь Альцгеймера» при предъявлении стимулов с большим межстимульным интервалом (3 с) характеризуются сильным снижением амплитуды HP. HP используется и при прогнозах выхода из состояния комы. Наличие HP-подобной негативности в лобных областях при широком диапазоне изменений частоты стимуляции является предвестником возвращения сознательного состояния пациента.
Б. Компонента сравнения
Даже короткий обзор литературных данных по практическому применению негативности рассогласования показывает, что с помощью HP можно определить психиатрические и неврологические отклонения от нормы. Однако величина эффекта в терминах амплитудных значений ВП достаточно мала, что ограничивает практическое применение HP в клинике. Незначительная вели
Часть 2. Вызванные потенциалы
237
чина эффекта частично может быть объяснена с позиций природы HP, средний уровень амплитуды которой 1 мкВ, что в 30—50 раз меньше фоновой ЭЭГ-активности.
С учетом этого более мощным диагностическим инструментом является компонента сравнения в зрительной GO/NOGO-парадигме. По крайней мере, амплитуда такой компоненты сравнения в 4 раза больше амплитуды HP (рис. 10.18). Более того, использование пространственной фильтрации с помощью метода независимых компонент позволяет дополнительно увеличить соотношение сигнала к шуму и обеспечить более эффективное применение в оценке индивидуальных ВП. Конечно, необходимы дальнейшие исследования в этом направлении, однако уже сейчас имеющиеся данные свидетельствуют о том, что данный метод является более мощным в сравнении с более старыми подходами к определению биологических маркеров (эндофенотипов) некоторых психических и неврологических расстройств (например, таких как СНВГ, см. ч. 3).
X. Заключение
Мы оцениваем изменения во внутреннем и внешнем мире при помощи рецепторов разного типа. Существует несколько сенсорных модальностей, позволяющих воспринимать образы, звуки, движения тела и чувствовать боль, прикосновения, вкус, запах. Наша осведомленность о внешних или внутренних объектах не сводится к восприятию объектов как таковых, а заключается в обработке импульсной активности выходных нейронов рецепторных органов. Области мозга, нейроны которых отвечают на стимуляцию рецепторов конкретного типа, обычно рассматриваются как элементы соответствующих сенсорных систем (зрительной, слуховой системы и т.д.). В любой сенсорной модальности существует иерархическое подразделение на первичную, вторичную и ассоциативную кору. Ассоциативные области сенсорной коры отвечают за обработку наиболее сложных характеристик стимулов. Нейроны, кодирующие конкретные свойства сенсорного мира, формируют топографические организованные вычислительные карты. Например, нейроны, избирательная активность которых обеспечивает специфическую способность человека — восприятие лиц, локализуются в области височной коры и характеризуются специфическими компонентами скальпового сенсорного вызванного потенциала. Каждый класс объектов окружающего мира и каждый класс пространственных соотношений, как оказалось, обладает отдельной формой памяти. Категории памяти выступают в роли модели окружающей среды (Anticipating schemata). ВП, регистрируемые при активации этих моделей, могут быть разложены на несколько составляющих компонент. Мощным методом такого разложения ВП является недавно открытый метод независимых компонент. Выявляемые с помощью этого метода сенсорные компоненты ха-
238 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия растеризуются разной временной динамикой и пиковой латентностью, варьирующей от 110 мс в зрительной и от 80 мс в слуховой модальности. Разные компоненты генерируются в разных областях коры, начиная с первичных сенсорных, и распространяются вплоть до ассоциативных зон, включая переднюю поясную извилину. Психологические операции первичной зрительной (слуховой) обработки, сравнения (подразделяемые на детекцию физических и смысловых изменений) и вовлечения характеризуются разными компонентами. В зрительной модальности компоненты, отражающие операции сравнения и вовлечения, генерируются структурами вентрального и дорсального путей обработки зрительной информации. Операции сравнения в слуховой модальности, на иерархически низшем уровне, отображаются генерацией негативности рассогласования. HP отображает автоматический процесс оценки изменений, заключающийся в детекции рассогласования девиантного стимула со следом памяти, сформированным предшествующей повторяющейся слуховой стимуляцией. HP обнаружена в исследованиях, проводимых в рамках слуховой oddball -парадигмы, и используется в диагностических целях при разного рода психических и неврологических расстройствах.
Часть 2. Вызванные потенциалы
239
Глава 11. Система внимания
Мой опыт — это то, на что я согласен обратить внимание.
Вильям Джеймс, 1890
I. Психология
А. Внимание как операция выбора
Представьте, что вы находитесь в комнате с механическими часами с закрытыми глазами. В тот момент, когда вы только что закрыли глаза, вы не слышите, как идут часы, но спустя некоторое время вы обращаете на них внимание. Внимание к звукам в некотором роде выглядит как «увеличение громкости» громкоговорителя: тиканье часов становится как бы громче. Переключение внимания на другой источник звука, например шепот вашего собеседника, сидящего в углу комнаты, способствует «выключению» звука часов. Именно так и работает наше внимание! Как видно из этого примера, внимание усиливает релевантную сенсорную информацию и подавляет нерелевантную. Эти операции усиления/подавле-ния обозначаются термином «селективные операции» (операции выбора). Аналогично внимание как психологический процесс может быть определено как селективные операции в сенсорной модальности, целью которых является более тщательная (аккуратная) обработка релевантной сенсорной информации.
В начале XX века в мировой науке доминировала теория условных рефлексов Ивана Петровича Павлова и теория оперантного обучения52 Бурху-са Скиннера (Burhhus Skinner). В конце 1940-х и начале 1960-х в психологии было сформировано представление о различной природе глобальных состояний поведения, таких как возбуждение (активации) и сон. В исследованиях Джузеппе Моруззи (Giuseppi Moruzzi) и Хорасе Магуна (Horace Magoun) было показано, что стимуляция ретикулярной формации ствола мозга кошки вызывает пробуждение ото сна, тогда как стимуляция других, наоборот, индуцировала сон53. В литературу был введен термин «пробуждение», обозначающий физиологическое состояние реактивности субъекта, и открыты его ЭЭГ-корреляты. В конце 1960-х новая научная парадигма, названная информаци-онной обработкой, основанная на компьютерной метафоре, заменила бихе
52 Эти исследовательские подходы лежат в основе бихевиоризма. Основой бихевиоризма является утверждение, что все действия, выпол-। <яемые организмом, могут быть объективно описаны без учета внутренних психических процессов или таких гипотетических конструкций, как разум, сознание и тд. Бихевиористская концепция противоречит психоаналитическому и гештальт-подходу в психологии.
53 G. Moruzzi и Н. W. Magoun в 1949 году опубликовали свою книгу «Ретикулярная формация ствола мозга и активация ЭЭГ».
240
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
виоризм54. Внимание стало одним из основных предметов этого направления исследований в психологии.
Б. Сенсорные и моторные селективные операции
Селективные операции в более широком смысле не являются специфической характеристикой процесса обработки сенсорной информации, но в узком смысле представлены и на уровне моторных систем. Репертуар всевозможных моторных действий человека огромен. Мы можем танцевать, играть на музыкальных инструментах, говорить, читать и писать (некоторые из нас на нескольких языках)... Число элементарных действий, составляющих каждую из этих больших категорий действий, может быть также огромным. Просто представьте себе количество известных вам слов55. Таким образом, в любой интервал времени доступно большое количество потенциальных действий. Целью нашего поведения является выбор действий, в том числе и тех, которые подходят наилучшим образом для текущего момента. В то же же время селективные операции не ограничены. Операции выбора выполняются и на когнитивном уровне — на уровне мыслей. Как говорил Вильям Джеймс, «думать — значит выбирать»56.
В. Подготовительная установка
В попытке объединения разных психологических сущностей в рамках единой концепции в классическом труде Эдварда Эвартса (Edvard Evarts), Ешиказу Шинода (Yoshikazu Shinoda) и Стивена Вайза (Steven Wise) «Neurophysiological approaches to higher brain functions», опубликованном в 1984 году, было введено понятие подготовительной установки. Под подготовительной установкой понимали состояние готовности к появлению определенного стимула или совершению движения. С учетом современных концепций можно говорить, что установочные реакции в сенсорной области связаны с вниманием, а в моторной или когнитивной — с исполнительными функциями57.
Поведенческий паттерн в любой момент времени может быть разделен на две составляющие: исполнение когнитивно-моторного действия и его сенсорное восприятие58. Естественно, сенсорные и моторные компоненты поведения взаимосвязаны. В действительности почти любое произвольное действие ассоциируется с восприятием того, как оно выполнено (восприятие напряжения мышц,
54 Информационная обработка — психологический подход в психологии, который рассматривает познавательную функцию как вычислительную по своей природе. При этом разум рассматривается как программное обеспечение, а мозг — как компьютер. Информационная обработка может быть последовательной или параллельной, централизованной или распределенной. В середине 1980-х параллельно распределенный механизм обработки информации в нейронных сетях стал основной объяснительной парадигмой в когнитивной психологии.
55 Словарный запас Вильяма Шекспира составлял около 30 000 слов!
56 В книге "Принципы психологии», опубликованной первым американским психологом Вильямом Джеймсом в 1890 году, содержались главы, посвященные привычкам, вниманию, восприятию, памяти, ассоциациям, рассудку, инстинктам, эмоциям и воображению.
57 Более детально исполнительные функции будут рассмотрены в следующей главе.
58 Например, моторная составляющая речи проявляется в виде комплекса сочетанных двигательных актов языка, губ и гортани. В то же время сенсорная составляющая проявляется как восприятие производимых звуков и соматосенсорных сигналов, сопровождающих движения при речи. Из исследований пациентов с нарушениями ткани мозга известно, что представительство моторной составляющей локализуется в зоне Брока, а сенсорной — в зоне Вернике. Несмотря на реципрокную взаимосвязанность этих областей мозга, специфические нарушения речи наблюдаются только при повреждении одной из них и называются афазией Брока или Вернике, в зависимости от местоположения.
Часть 2, Вызванные потенциалы
241
позиции конечностей и т.д.) и как происходит взаимодействие с окружающим миром во время его реализации (восприятие изменения освещенности, акустических волн в окружающем воздухе и т.д.). И наоборот, почти любой перцептивный акт (оценка цвета, положения, конкретного объекта) связан с движением субъекта, его головы или глаз относительно внешнего мира (позиционирование взгляда в определенной точке видимого пространства как при слежении за летящим объектом, так и при рассматривании сложной фигуры)59.
С психологической точки зрения внимание, моторная подготовительная установка и селекция мыслей могут быть рассмотрены как похожие сущности, связанные с предварительной настройкой активности нейронных сетей. Несмотря на это, в условиях эксперимента такие элементы поведения исследуются отдельно. Исследования показали, что исполнительные и перцептивные функции реализуются разными нейронными механизмами. В обеспечении процессов выбора действий ключевую роль играют базальные ганглии в составе таламокортикальной системы. Селекция источников сенсорной информации обеспечивается за счет прямых и обратных связей разных кортикальных представительств сенсорных стимулов. Эти представительства имеют реципрокные связи с подкорковыми структурами (таламус, бугорки четверохолмия), что позволяет осуществлять операции по переключению внимания60.
Г. Обработка множества объектов
В повседневной жизни окружающий нас мир состоит из разнообразного множества загромождающих друг друга объектов разной формы, цвета, а также особенностей движения, локализованных в разных частях пространства. Наш опыт подсказывает нам, что мы не обрабатываем все окружающие объекты одновременно. В качестве демонстрации попытайтесь определить положение вертикальной серой полоски на рис. 11.1 (в верхней части слева). Как видно, на это требуется определенное время. Фокус вашего внимания последовательно перебирает несколько положений, пока не найдет нужный объект.
Здесь вполне уместна метафора «луч прожектора», когда мы рассматриваем процесс поиска цели. Это выглядит так, словно возможности нашей зрительной системы при обработке информации множества объектов в любой текущий момент времени ограниченны. На рис. 11.1 видно, как расположенные слева полоски разной ориентации и цвета конкурируют за внимание.
Д. Операции вовлечения, отвлечения и переключения
Исходя из приведенного выше примера, внимание является динамическим процессом. В соответствии с мнением Михаэля Познера, мирового эксперта в психологии внимания, можно выделить три типа операций в этом динамическом процессе (рис 11.1 внизу). Во-первых, это операция вовлечения — усиление нейронного представительства источника информации, являющегося объектом
59 При прогулке по лесу (движение) нам необходимо следить за тем, куда поставить ногу (перцепция). Смотря телевизор (перцепция), мы постоянно перемещаем взгляд (движения головы и глаз).
60 Переключение внимания может происходить неявно, без заметных движений, что, однако, обеспечивается теми же областями. которые вовлекаются в реализацию явных действий.
242
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
вовлечение высвобождение
переключение
В
Рисунок 11.1. Операции внимания
А. Изображения, иллюстрирующие два разных способа привлечения внимания. В соответствии с инструкцией внимание должно быть сконцентрировано на вертикальных полосках серого цвета. Поскольку возможности зрительной системы в обработке информации ограничены, то при одновременном предъявлении множества стимулов они будут конкурировать между собой. Очевидно, что процесс конкуренции сложен, если взглянуть на левое изображение (последовательный поиск цели), и относительно прост, если перевести взгляд на правое изображение (параллельный режим поиска цели). Следует отметить, что в первом случае нахождение цели обеспечивается в процессе поиска, контролируемого нисходящими влияниями от высших областей мозга, а во втором цель находится в результате управляемых стимулом восходящих процессов обработки информации. Б. Серыми стрелками обозначены перемещения внимания. Перемещения внимания могут быть явными и, соответственно, измеренными, например, при регистрации движений глаз, а именно — позиции центральной ямки глаза (фовеа) при рассматривании испытуемым изображения в поисках цели. Техника регистрации движений глаз во время рассматривания изображений впервые была использована отечественным ученым Александром Ярбусом. Перемещение внимания может быть также скрытным, т.е. осуществляться без видимых движений глаз. В. Три типа операций внимания, впервые описанные Майклом Познером (Posner et al., 1988): вовлечение, высвобождение и переключение.
внимания61. Второй элемент внимания — операция отвлечения — заключается в подавлении источника информации, на который внимание не направлено62. Третья операция — это операция перемещения или движения как промежуточное состояние при обработке двух разных источников информации. Упомянутые три типа реакций легко различимы на примерах, приведенных на рис. 11.1 (в середине), представляющих наложение визуальной сцены и траекторий движений глаз во время ее просмотра. Как можно заметить, во время поиска цели последовательные фиксации взгляда располагаются на разных полосках.
Е. Нисходящие и восходящие процессы
Важной предпосылкой существования внимания является наличие множества источников информации (например, несколько зрительных образов в зрительной модальности), которые конкурируют между собой63. Ситуация борьбы разных стимулов за ресурсы может быть разрешена двумя способами: нисходящими или восходящими факторами влияния. Пример нисходящего фактора представлен в верхней левой части рисунка 11.1. Инструкция смотреть только на вертикальные серые полоски инициирует про-
61 Важная особенность внимания заключается в том, что операция вовлечения может длиться достаточно долгое время, позволяя удерживать внимание в течение длительного периода. Хорошей иллюстрацией этой способности может быть прослушивание му-
зыки, решение математической задачи или игра в шахматы. Зачастую решение конкретной проблемы требует времени (и иногда достаточно долгого).
63 Особым случаем операции отвлечения является феномен подавления возвращения», проявляющийся как малая вероятность возвращения к объекту, на котором уже было сконцентрировано внимание. Например, при просматривании изображения на рис. 11.1. (в середине) внимание редко переключается на область, уже бывшую в фокусе внимания.
63 Это выглядит так, словно окружающий нас мир слишком сложен — внешняя среда, содержащая избыточное количество информации, слишком велика, чтобы единовременно воспринять ее целиком.
Часть 2. Вызванные потенциалы
243
цессы высшего порядка, которые управляют саккадическими движениями глаз, определяющими перемещение внимание от одного стимула к другому, пока не будет найден искомый объект. Другой пример, приведенный на рис. 11.1 вверху справа, иллюстрирует действие восходящих факторов. Важным условием выраженности восходящих процессов является высокая степень отличимости целевого стимула по отношения к остальным (например, форма, цвет, специфическая ориентация). На рис. 11.1 (вверху справа) вертикальная серая полоска легко выделяется на фоне других стимулов и, соответственно, быстро и без усилий детектируется, обеспечивая ее высокую конкурентоспособность в борьбе за внимание. Этот девиантный стимул словно появляется на изображении. Таким образом, восприятие зрительных сцен происходит при взаимодействии нисходящих и восходящих влияний. Это взаимодействие составляет сущность внимания.
II. Анатомия
А. Сенсорные системы
Внимание работает с сенсорной информацией и, следовательно, вовлекает элементы сенсорной системы (рис. 11.2). Помимо нисходящих и восходящих влияний на уровне иерархически организованных сенсорных областей мозга, обработка сенсорной информации контролируется подкорковыми мозговыми структурами — таламусом и верхними бугорками четверохолмия. Контроль информационных процессов в первичных и ассоциативных сенсорных областях коры осуществляется за счет их реципрокных связей с ядром подушки таламуса и ядрами ретикулярной формации, в соответствующей ее части. В обеспечение произвольных и непроизвольных перемещений взгляда вовлекаются верхние бугорки четверохолмия. Детальная обработка выбранных элементов окружающего трехмерного пространства контролируется ядрами подушки за счет реципрокных связей с сенсорными зонами коры.
Б. Исполнительные системы
Функция внимания подразумевает реализацию операций выбора в сенсорной модальности. Однако сенсорное событие, включающее модуляцию внимания, не может быть отделено от действия. В частности, при переключении внимания с одной части визуальной сцены на другую должны быть выполнены также и действия, связанные с ориентацией глаз и расширением зрачка. Области, входящие в состав сенсорных систем, локализующиеся в задней части коры головного мозга, имеют реципрокные внутрикортикальные связи с передними областями исполнительных систем, вовлекаемых в процессы выбора действий, необходимых для реализации детальной обработки релевантной зрительной информации. Как уже упоминалось выше, различные механизмы выбора действий обеспечиваются фронтально распределенной исполнительной системой мозга (см. гл. 12).
244
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Pill
Рисунок 11.2. Мозговые пути обеспечения функции внимания
Стрелками обозначены нисходящие и восходящие взаимодействия между задними и передними областями коры. Эти взаимодействия поддерживаются проекциями, берущими начало от ядра подушки и проецирующимися в соответствующие области коры.
III. Модуляция процессов обработки сенсорной информации
А. Взаимное подавление в исследованиях животных
Большинство наших представлений о нейронных коррелятах внимания было сформировано на основе исследований обезьян64. Эти исследо
64 Вернон Маунткастл (Vernon Mountcastle) в 1970-х годах впервые провел экспериментальные исследования внимания при регистрации импульсной активности нейронов дорсального пути обработки сенсорной информации. В частности, было показано, что и нейроны 7-й области теменной коры, и нейроны подушки демонстрировали увеличение ИАН в ситуации, когда обезьяны концентрировали внимание на стимулах.
Часть 2. Вызванные потенциалы
245
вания чаще всего проводились в зрительной модальности65. Схематический результат типичного исследования обезьян представлен на рис. 11.3 (для более подробного ознакомления см. Reynolds et al., 1999). Импульсная активность нейронов вентрального пути регистрировалась в ответ на предъявление зрительного стимула (например, полоска определенной ориентации) в рецептивном поле нейрона. В случае предъявления одного стимула, когда внимание было сфокусировано за пределами исследуемого рецептивного поля, регистрируемые ответы состояли из двух частей: ранний и поздний ответ. При одновременном предъявлении второго стимула в пределах этого же рецептивного поля наблюдалось подавление позднего ответа. Концентрация внимания обезьяны на первом из двух стимулов при игнорировании второго приводила к общему увеличение активности в ответ на такое сочетанное предъявление стимулов. Таким образом, полученные данные свидетельствуют о том, что одновременно предъявляемые одинаковые стимулы в пределах одного рецептивного поля, обрабатываются не независимо, а взаимодействуют друг с другом в режиме взаимного подавления. Как видно, внимание тормозит подавляющий эффект стимула, находящегося вне фокуса внимания, тем самым усиливая в ответ активность стимула, на который оно направлено. Коротко говоря, выявленные данные позволили считать, что внимание разрешает проблему противоборства среди множества стимулов, уравновешивая взаимное торможение между разными источниками информации. Это позволяет усилить обработку информации с тех источников, на которых сконцентрировано внимание, и параллельно ослабить влияние других66.
Б. Вовлечение подкорковых структур
Обработка сенсорной информации и модулирующее действие внимания на информационные потоки обеспечивается вовлечением двух подкорковых образований — ядер подушки таламуса и верхних бугорков. Верхние бугорки четверохолмия вовлекаются в генерацию саккадических движений глаз при произвольном поиске релевантных источников сенсорной информации. Их активность контролируется базальными ганглиями через окулярно-моторный проводящий путь. В ряде последних исследований приводится доказательство участия верхних бугорков четверохолмия не только в обеспечении целенаправленного поведения и
65 Как описано ранее, зрительная кора обезьян содержит более 20 различимых областей, организованных в два функционально специализированных маршрута: вентральный и дорсальный пути обработки информации. Вентральный путь важен для идентификации объектов, а дорсальный ответственен за оценку пространственных отношений между объектами, а также направление движения в сторону объектов. Нейроны вентрального пути демонстрируют избирательность ответов для атрибутов стимулов, важных для зрительного восприятия (форма, цвет, текстура). Нейроны дорсального пути избирательно реагируют на пространственное положение, скорость, направление движения объекта, находящегося в фокусе внимания. №итывая эти экспериментальные факты, вполне оправданно ожидать, что модулирующий эффект внимания к пространственным свойствам стимула будет проявляться на уровне структур дорсального пути. Аналогично внимание к визуальным характеристикам (цвет или форма) должно модулировать ответы нейронов в области вентрального пути.
66 Конкуренция между стимулами может быть объяснена с позиций механизма латерального торможения. Модель, которая может объяснить вышеописанные результаты, представлена в нижней части рис. 113. Необходимо отметить, что механизм латерального торможения играет важную роль не только в обеспечении внимания, но и в обработке специфических характеристик стимула (см. «Обработка информации сетями нейронов», гл. 8).
246
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Ст1
Один стимул. Внимание вне рецептивного поля
Два стимула. Внимание вне рецептивного поля
Два стимула. Внимание на первый стимул
8
Рисунок 11.3. Внимание разрешает ситуацию взаимного подавления двух конкурирующих стимулов
А. Рецептивные поля нейрона зрительной системы (обозначены квадратом) в трех разных ситуациях: слева—внутри рецептивного поля предъявлен только один зрительный стимул (горизонтальная полоска), а внимание сфокусировано вне рецептивного (обозначено серым цветом); посередине—в пределах рецептивного поля предъявлен дополнительный стимул (горизонтальная полоска), а внимание также направлено вне рецептивного поля; справа—внимание сфокусировано на горизонтальной полоске. Б. Типы реакцией нейронов на быстрое предъявление стимулов в трех разных ситуациях. Обращает на себя вниманието, что конкурирующие за внимание стимулы снижают позднюю компоненту ответа нейрона, которая восстанавливается при фокусировании внимания на одном из стимулов (в данном случае - горизонтальной полосе). В. Схематическое изображение сети нейронов, ответственной за обеспечение наблюдаемого эффекта. Пустыми кругами обозначены возбуждающие нейроны, а черными —тормозные нейроны. Слезет отметить, что внимание активируеттормозный нейрон, соответствующий первому стимулу, тем самым обеспечивая усиление детектирующих свойств нейронов, а также подавляя смежные нерелевантные нейроны. Обозначения: Ст. 1 — стимул 1; Ст. 2—стимул 2.
саккад, но и в обеспечении скрытого внимания, осуществляемого при отсутствии наблюдаемых движений глаз.
Ядра подушки таламуса, благодаря реципрокным связям со структурами дорсального и вентрального путей обработки зрительной информации, а также другими сенсорными областями, играют важную роль в обеспечении внимания. За счет связей с областями префронтальной коры координируется работа исполнительных и сенсорных систем. При предъявлении стимула, на котором сконцентрировано внимание, обычно регистрируется увеличение активности нейронов подушки.
Часть 2, Вызванные потенциалы
247
В. Негативные компоненты ВП, связанные с вниманием
Одной из первых парадигм исследования внимания с помощью метода вызванных потенциалов являлся тест дихотического прослушивания, разработанный Броадбентом в 1954 году67. При одновременном предъявлении в правое и левое ухо стандартных и девиантных тонов (в рамках oddball-парадигмы) внимание испытуемого направлено на один из слуховых каналов. Сравнение ВП, зарегистрированных при предъявлении иррелевантных стимулов игнорируемого и целевого канала, характеризуется разностной компонентой — «разностной негативностью». Схематическое изображение полу-
чаемых результатов в такого рода исследованиях представлено на рис. 11.4 (слева). Более детальную информацию можно найти в книге Р. Наатанена «Внимание и функции мозга».
Результаты исследований, проводимых в другой парадигме, проиллюстрированы на рис. 11.4 (справа). Задачей испытуемых, которым в случайном порядке предъявлялись изображения шахматной доски (с красными и голубыми клетками), было попеременно обращать внимание на клетки
тест дихотического прослушивания (внимание на правое или левое уко)
А
Внимание на клетки определенного цвета (красные или синие)
Рисунок 11.4. Типы негативностей, связанные с функцией внимания (схематическое представление)
А. Процессная негативность в слуховой модальности; топограмма разностного ВП. полученного в результате сравнения ВП, зарегистрированных при стимуляции игнорируемого и прослушиваемого уха в условиях дихотического предъявления слуховых стимулов. Б. Селективная негативность в зрительной модальности; топограмма разностного ВП, полученного в результате сравнения ВП, зарегистрированных в ответ на предъявление изображения релевантного и игнорируемого цвета. Быстро и в случайном порядке в центре поля зрения предъявлялось изображение шахматной доски с клетками красного или синего цвета. В отдельных исследовательских сессиях внимание концентрировалось либо на синих, либо на красных клетках доски.
Разностные ВП формировались при вычитании ВП на игнорируемый стимул из ВП на стимул релевантного цвета. В состав компонент, связанных вниманием, входит селективная негативность (см. Hillyard and Anllo-Vento, 1988).
одного или другого цвета. Разностный потенциал (при сравнении ВП стимула, на котором концентрировалось внимание, и ВП игнорируемого стимула) характеризовался негативной компонентой — «негативностью выбора». «Негативность выбора» считается ВП-показателем активности системы внимания на уровне вентрального пути обработки зрительной информации68.
67 В тесте дихотического прослушивания разная слуховая информация одновременно подается на разные уши. Испытуемые инструктируются концентрировать внимание только на одном из слуховых каналов. В эксперименте, проведенном Черри в 1953 г., было показано, что испытуемые могли воспроизвести лишь малую часть вербальной информации, подаваемой в игнорируемое ухо, за исключением базовых характеристик, таких как, например, пол человека, произносящего слова.
68 Разностные ВП характеризовались не только негативными компонентами. В зависимости от специфических условий тестовых заданий регистрировались также и позитивные компоненты. Некоторые исследователи поддаются искушению напрямую связывать негативные компоненты таких разностных ВП (в зрительной модальности) с тормозными постсинаптическими потенциалами нейронов средних слоев коры, а позитивные компоненты, наоборот, ассоциируют с возбуждающими постсинаптическими потенциалами.
248
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Г. Теменнофронтальная система в ПЭТ и фМРТ-исследованиях
Несмотря на то что настоящая глава посвящена контролю процессов обработки информации, осуществляемой мозговыми структурами задней части мозга, нельзя не упомянуть о взаимодействиях этих областей с фронтальными зонами мозга, традиционно связываемыми с рабочей памятью. В действительности понятия рабочей памяти и внимания тесно связаны: для того чтобы удерживать элемент в памяти, требуется внимание, и наоборот, внимание к определенным стимулам требует удержания в памяти ключевой информации о них. Кроме того, данные ПЭТ и фМРТ-исследований свидетельствуют об общности нейронных сетей, обеспечивающих эти операции. Традиционной находкой в разнообразных исследованиях со зрительно-пространственными заданиями, требующими затраты ресурсов внимания и рабочей памяти, являются активации в области теменных и лобных областей коры.
IV. Нейрофизиология
А. Сенсорный дефицит и повреждения правой теменной коры
Как уже было сказано в предыдущей главе, нейроны теменной коры входят в состав дорсального пути обработки зрительной информации (системы «где»), ответственного за кодирование пространственного расположения объектов. Активность нейронов теменной коры ощутимо модулируется в задачах на пространственное внимание, когда внимание испытуемых концентрируется на разных участках окружающего пространства. Эти знания дают нам основания полагать, что повреждения теменной коры могут приводить к нарушениям пространственного внимания. Это подтверждается клиническими данными, в соответствии с которыми унилатеральное повреждение теменной коры приводит к сенсорному дефициту для объектов в контралатеральной части зрительного поля (относительно стороны повреждения). В более тяжелых случаях пациенты полностью игнорируют происходящее в определенной части зрительного поля, контралатеральной относительно повреждения. Например, такие больные могут читать только одну (правую или левую) часть книги, наносить макияж только на одну из половин лица или есть только с одной стороны тарелки69. В менее тяжелых случаях сенсорный дефицит выражен меньше. В частности, дефицит может проявляться только в случаях предъявления конкурентных стимулов: как, например, в случае синдрома ослабления зрительного восприятия (visual extinction)70. Необходимо отметить, что пространственно-зрительный дефицит
69 Сенсорный дефицит чаще всего наблюдается при повреждениях теменной коры правого полушария (по сравнению с левосторонними повреждениями), что является свидетельством ключевой роли правой теменной коры в процессах управления вниманием. На основе этой полушарной асимметрии было выдвинуто предположение о том, что правая теменная кора вовлекается в мозговое обеспечение процессов внимания для обеих сторон зрительной сцены. Левая же теменная кора ответственна за внимание к стимулам в контралатеральной (правой) части зрительного поля (Mesulam, 1981). 8 соответствии с этим мнением, при повреждении левой теменной коры, интактное правое полушарие принимает на себя функцию поврежденной области. При этом повреждение в правой теменной коре будет приводить к сенсорному дефициту в левой стороне зрительного поля.
70При визуальном ослаблении пациенты могут управлять вниманием при взаимодействии с одним объектом в полуполе зрения, контралатеральном стороне повреждения; однако при одновременном предъявлении двух стимулов, один из которых находится в интактном зрительном полуполе, пациенты детектируют только «интактный» стимул и не замечают в «поврежденной» стороне зрительного пространства.
Часть 2. Вызванные потенциалы
249
может быть вызван не только унилатеральным повреждением теменной коры, но и повреждениями в области лобной коры, передней поясной извилины, базальных ганглиев и ядер подушки таламуса (см. Kastner and Ungerleider, 2000).
Б. Синдром Балинта
Полная утрата чувства пространства и способности выбора пространственных отношений между объектами наблюдается при билатеральном повреждении теменных долей. Поведенческие нарушения, вызываемые такими повреждениями, получили название синдрома Балинта. Пациенты с синдромом Балинта утрачивают способность воспринимать пространственную информацию (за пределами своего собственного тела) и с функциональной точки зрения слепы, за исключением сохранности перцепции одного зрительного объекта зрительной сцены. Они не могут ни определить положение видимого объекта, ни сказать, куда он движется (в их сторону или в другом направлении). Таким образом, полностью теряется пространственное восприятие71.
V. Нейрональные сети
А. Рекуррентная деполяризация апикальных дендритов
Повторяющаяся циклическая активность (как внутрикортикальная, так и таламокортикальная), по всей видимости, является важным условием обеспечения удержания внимания на определенном участке пространства (пространственное внимание) или объекте (непространственное внимание). Известно несколько моделей, имитирующих работу механизма внимания. Однако имеющихся эмпирических доказательств явно недостаточно для выбора наиболее адекватной модели. Вместе с тем один из элементов нейронной активности системы внимания наиболее, а именно поддерживаемая деполяризация апикальных дендритов пирамидных нейронов коры при различных установочных реакциях, часто подтверждается экспериментальными данными и входит в состав большинства моделей. Ключевую роль в обеспечении такой деполяризации играют возвратные связи, берущие начало от глубоких слоев областей коры высшего порядка с верхними слоями иерархически низших областей (рис. 11.5, на котором связи обозначены толстой стрелкой). Возвратные связи, активирующие по своей природе, вызывают деполяризацию апикальных слоев коры. По всей видимости, эта апикальная активность проявляется на ВП в виде медленной негативности при подготовительной установке72. Апикальная деполяризация модулирует активность пирамидных клеток и снижает их порог активации. Пирамидные клетки средних слоев коры могут работать как детекторы совпа
71 Существуют аргументы в пользу того, что синдром Балинта — тип двойного сенсорного дефицита. Так, пациенты с сенсорным дефицитом могут не замечать как левую часть комнаты, так и левую сторону цветка. Однако при синдроме Балинта не замечаются обе части комнаты, но может целиком восприниматься единичный объект. Фактически такие бальные не видят ничего, кроме отдельных объектов. Таким образом, соотношение между двойным сенсорным дефицитом и синдромом Балинта не столь прямое, как может показаться на первый взгляд.
72 Эти разновидности активности обычно объединяются общим термином — «условное негативное отклонение» (УНО). УНО проявляет зависимость от модальности установочной реакции и регистрируется вблизи соответствующих сенсорных областей коры.
250
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
дения, активируясь в случае деполяризации как апикальных, так и базальных дендритов73.
Б. Внимание и реакция активации
Из нашего опыта известно, что процессы сенсорного восприятия и распознавания объектов зависят от уровня общей активации. В состоянии бодрствования самому низкому уровню активации соответствует состояние дремоты. Противоположное экстремальное состояние — гипербдителъность (hypervigilance). В со
Рисунок 11.5. Схематическое представление сети нейронов, обеспечивающих функцию внимания
Слева направо — сети нейронов первичных и вторичных сенсорных областей. Контроль процессов внимания осуществляется за счет возвратного проводящего пути (обозначенных жирными линиями черного цвета) и деполяризации апикальных дендритов пирамидальных клеток (тонкие линии). Процессы деполяризации апикальных дендритов проявляются в виде негативности на регистрируемой со скальпа электрической активности.
стоянии дремоты нам едва ли удается обратить на что-нибудь свое внимание, потому как пороги активации сенсорных нейронов настолько высоки, что даже появление значимого, релевантного стимула не вызывает их активации. Следовательно, работоспособность системы внимания крайне мала в таком состоянии. В состоянии гипервозбудимости наше внимание отвлекается различными нерелевантными стимулами, так как пороги срабатывания сенсорных нейронов крайне низки. В силу этого и в данном случае работоспособность мала. Таким образом, если попытаться опреде
лить способность сенсорной системы дискриминировать конкретные стимулы на фоне шума, приняв за «Р» работоспособность и за «А» общий уровень ак-
тивации системы, то в результате получится так называемая инвертированная U-образная взаимосвязь.
Понятие возбудимости отражает фундаментальное свойство поведения и отчасти ассоциируется со способностью сенсорной системы к обработке информации. Складывается впечатление, что эффективные пороги активации сенсорных нейронов отражают способность мозга обрабатывать сенсорные стимулы (рис. 11.6). Установка этих порогов при общем уровне активации отлична от аналогичного механизма при подготовительной установке. В случае последней пороги выставляются весьма специфично, а действие такой настройки ограничено небольшим набором релевантных нейронов (относящихся к конкретным стимулам). Напротив, изменение общего уровня возбудимости приводит к общему
73 Однако не следует исключать и иные возможности модуляции активности с помощью механизма внимания. Один из механизмов модуляции связан с активностью тормозных нейронов, входящих в состав тех же путей возвратных связей. Тормозные нейроны локализуются в средних слоях коры мозга и демонстрируют внутрикортикальную ингибирующую активность, формируя рецептивные поля пирамидных клеток и настраивая их на ожидаемые стимулы. Тормозные нейроны с помощью механизма латерального торможения оказывают угнетающее действие на конкурирующие области коры, обеспечивая подавление процессов обработки информации, поступающей через нерелевантные сенсорные каналы. Тормозные постсинаптические потенциалы обеспечивают появление негативного потенциала, регистрируемого с поверхности коры и сопоставимого по своим характеристикам с потенциалом, генерируемым при апикальной деполяризации.
Часть 2. Вызванные потенциалы
251
Рисунок 11.6. Пюбальный эффект уровня бодрствования и локальный эффект внимания
А. Пороги активации сенсорных нейронов: при высоком уровне бодрствования наблюдается неспецифическое снижение порогов как для релевантных, так и для нерелевантных стимулов; внимание же избирательно снижает пороги активации для релевантного стимула. Б. Слишком высокий или слишком низкий уровень бодрствования одинаково приводят к уменьшению соотношения сигнала к шуму.
«неспецифическому» изменению порогов. Эти изменения вызываются отдельной «неспецифической» системой.
В. Тонические и фазические реакции голубого пятна ствола мозга
Ключевым звеном системы неспецифической возбудимости являются ядра голубого пятна ствола мозга74. Общий уровень активности нейронов голубого пятна строго коррелирует с изменением возбудимости в диапазоне состояний от глубокого сна до гипервозбудимости. Нейроны голубого пятна находятся в состоянии покоя в период сна, а именно фазы быстрых движений глаз (стадия парадоксального сна, или REM-стадия), и демонстрируют прогрессивную активацию при пробуждении. Фактически молчание нейронов голубого пятна является одним из физиологических параметров, которые используются для различения бодрствования и фазы парадоксального сна75.
Нейроны голубого пятна показывают две разновидности активности: тоническую, или фоновую, активность, которая медленно изменяется вслед за медленными изменениями состояний мозга, и фазическую, которая отражает быстрый ответ системы на краткосрочную сенсорную стимуляцию. Считается, что фазическая компонента активности нейронов выполняет функцию временного усилителя (или фильтра), который интенсифицирует обработку наиболее значимых стимулов.
Ниже представлены свойства нейрона голубого пятна, доказывающие это мнение. Во-первых, латентность нейронных реакций, возникающих в ответ на
74 Свое название эти скопления нейронов получили благодаря цвету пигментов. Голубое пятно ствола мозга человека содержит небольшое количество нейронов (по 16 000 в каждом полушарии).
75 Следует напомнить, что фаза REM, или парадоксальная фаза сна, характеризуется активациями на уровне коры и соответствующими изменениями паттернов ЭЭГ-активности, включая осцилляции тета-ритма срединной лобной топографии.
252 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия стимул, весьма мала — около 100 мс. Во-вторых, только целевые, т.е. поведенчески значимые стимулы, вызывают активацию нейронов. В силу того что скорость проведения импульсов по немиеленизированным волокнам мала, активность нейронов голубого пятна может достигать коры в пределах 60—70 мс. Теоретически эти ответы могут усиливать поздние этапы76 обработки информации на уровне сенсорных систем.
Г. Норэпинефрин как модулятор внимания
Нейроны голубого пятна являются единственным источником норэпинефрина77 в мозге. Норэпинефринергические нейроны немиелинизирова-ны и, следовательно, обладают медленной проводимостью. Норэпинефрин может высвобождаться экстрасинаптически, производя нелокальный эффект. Эти свойства отражают тот факт, что эффект норэпинефрина на нейроны-мишени носит модуляторный характер. Как известно, модулирующий эффект регулятора зависит от рецепторов синапса. На уровне коры мозга существует два вида адренорецепторов — al и a2. При их активации генерируется возбуждающий постсинаптический потенциал, что приводит к снижению порога активации нейронов-мишеней78. Аксоны клеток голубого пятна иннервируют кору мозга неравномерно. Наибольшая плотность терминалей наблюдается в теменной, моторной и премоторной коре (рис. 11.7). Подкорковые структуры, принимающие участие в обеспечении внимания (подушка таламуса и верхние бугорки четверохолмия), также получают значительную часть входов от голубого пятна. Роль норэпинефрина в модуляции внимания объясняет существующий интерес к исследованию с помощью фармакологических подходов к изучению влияния препаратов на функции норэпинефрина. Один из недавно разработанных препаратов, атомоксетин, является блокатором обратного захвата норэпинефрина, что позволяет уменьшить симптоматику расстройства дефицита внимания79.
VI. Поздние позитивные компоненты ВП
Известно, что в интервале от 250 до 400 мс в ВП регистрируются позитивные компоненты, обозначающиеся как «Р300» или «РЗ». РЗ генерируется при предъявлении значимых стимулов. Одна из компонент, принадлежащих к этой
76 Происходящие позже 200 мс с момента появления стимулы.
77 Норэпинефрин также называется норадреналином. Рецепторы этого модулятора называются адренорецепторами.
78 На уровне голубого пятна а2-адренорецепторы являются доминатными, выполняющими роль ауторецепторов. Их активация связывается с подавлением отрицательной обратной связи саморегуляции норэпинефрина.
79 Атомоксетин является блокатором обратного захвата норэпинефрина. Он производится компанией Eh Lilly and Company, продается в форме гидрохлоридной соли атомоксетина и носит название Strattera®. Изначально планировалось, что Strattera будет новым антидепрессантом. Однако клинические испытания не подтвердили успешность такого применения. Поскольку считается, что норэпинефрин играет важную роль в регуляции процессов внимания, были успешно проведены тестирования, позволившие утвердить данный препарат в качестве средства лечения нарушения внимания с гиперактивностью (Attention Deficit Hyperactivity Disorder (ADHD)). Это оказался первый препарат-нестимулянт, используемый при лечении ADHD. Его преимуществами перед стимулянтами являются отсутствие привыкания у пациентов, а также быстрое наступление терапевтического эффекта (в течение 24 часов) у детей и взрослых.
Часть 2. Вызванные потенциалы 253
группе, РЗЬ, регистрируется в ответ на стимулы-мишени, т.е. стимулы, после которых следует моторное или когнитивное действие, например нажатие кнопки или подсчет количества стимулов-мишеней80. Более ранняя компонента, называемый РЗа, увеличивается в амплитуде в ответ на неожиданное и заметное изменение в сенсорной стимуляции и отражает ориентировочную реакцию на подобное изменение стимула81 (рис. 11.7).
А. РЗЬ-компонента
Первое экспериментальное доказательство того, что компонента РЗЬ может отражать активность голубого пятна, было получено в работе Пинеды в 1989 году. В дополнение к этому существуют данные об общих характеристиках фазического ответа голубого пятна и РЗ-компоненты: 1) обе реакции наблюдаются после предъявления стимулов-мишеней в рамках oddball-парадигмы; 2) обе реакции зависят от поведенческой значимости и внимания, привлекаемого к появляющимся стимулам; 3) наблюдается более выраженная активность в ситуации корректного ответа на стимул-мишень по сравнению с ошибочными ответами или пропусками в задачах на детекцию стимулов. С учетом вышесказанного была выдвинута гипотеза о том, что РЗЬ-компонента отражает фазическое увеличение притока входящей активности в кору, индуцируемое нейронами голубого пятна посредством продукции норэпинефрина (см. Jones and Cohen, 2005).
Проводимые нами в 1990-х годах исследования по изучению импульсной активности, регистрируемой с имплантированных электродов, выявили удивительный факт: РЗЬ-подобные компоненты могут быть обнаружены во множестве корковых и подкорковых структур (рис. 11.8)82. Паттерн мультиклеточной активности сильно зависел от локализации регистрирующего электрода. Так, области височной коры характеризовались активностью, сопоставимой с негативностью рассогласования, пиковая латентность которой варьировала в пределах 80— 120 мс и не проявляла зависимости от внимания83. Компоненты, похожие на РЗЬ-компоненту, регистрировавшиеся в базальных ганглиях, ядрах таламуса и зонах лобно-затылочной коры, напротив, наблюдались только в активной модификации тестового задания, когда внимание испытуемого было сконцентрировано на детекции девиантных стимулов (требующих нажатия кнопки). Эти компоненты мультиклеточной активности по своим временным характеристикам
80 Обычно РЗб регистрируется в oddball-парадигме: например, тоны разной частоты (300 и 1000 Гц) предъявляются случайным образом с интервалом в 500 мс и вероятностью 0,1 и 0,9 соответственно. Задача испытуемого заключается в нажатии кнопки при появлении редкого стимула.
81 Типичными экспериментальными условиями регистрации РЗа-компоненты является предъявление трех типов стимулов: стандартных, девиантных и редко появляющихся новых стимулов (например, собачий лай или телефонный звонок). Именно предъявление новых стимулов вызывает ориентировочную реакцию, переключение внимания и приводит к генерации РЗа-компоненты.
82 Эти исследования проводились в рамках трехстимульной парадигмы, с предъявлением в случайном порядке следующих типов стимулов: стандартных (тон 1000 Гц, предъявлялся в 80 % проб), девиантных (тон 1300 Гц, в 10 % проб) и новых (случайная смесь различных тонов). В пассивном варианте тестового задания испытуемые читали книгу. В активной разновидности требовалось нажимать кнопку мыши при появлении девиантного стимула. При регистрации скальповой ЭЭГ как в активной, так и пассивной модификации тестового задания предъявление девиантных стимулов вызывало генерацию негативности рассогласования с максимальной амплитудой в Fz. Кроме этого, активное задание дополнительно характеризовалось РЗЬ-компонентой теменной топографии, предшество-вавшимся компонентом Н2.
83 Подробное описание HP можно найти в ч.2 гл. 1, в которой речь шла об операциях сравнения на уровне сенсорных систем мозга.
254
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
РЗа-ответ на целевой стимул РЗЬ ответ на новый стимул
Б
Рисунок 11.7. Взаимосвязь компонент ВП РЗа и РЗЬ и адренергической иннервации
А. Голубое пятно (locus coereleus — LC) является источником норэпинефрина (НЭ) в коре. Аксоны нейронов голубого пятна обширно иннервируют кору головного мозга (не показано на рисунке). Наиболее плотно иннервируются теменные области, а также моторная и премоторная кора (обозначено скоплениями рассеянных точек серого цвета). Б. РЗа-компонента, генерируемая в тестовых условиях oddball-парадигмы стремя слуховыми стимулами: в типичном исследовании ВП в последовательности стандартных стимулов (тоны) редко предъявляются девиантные (целевые стимулы), перемежающиеся с редкими новыми отвлекающими стимулами (например, собачий лай). Испытуемые при предъявлении целевого стимула должны нажимать кнопку. Компонента РЗа генерируется при предъявлении отвлекающих новых стимулов. Целевые стимулы вызывают появление РЗЬ-компонент. Компоненты РЗа и РЗЬ характеризуются разной топографией и локализациями генераторов, вычисленной с помощью метода томографии низкого разрешения (sLORETA).
совпадали со скальповой РЗЬ-компонентой. Однако по сравнению с присущей РЗЬ позитивностью регистрируемая активность была как положительной, так и отрицательной полярности. Более того, результаты исследования одного пациента показали, что характер РЗЬ-подобной активности существенно варьировал в зависимости от локализации электрода. По сути этот факт является свидетельством в пользу наличия нескольких разновидностей подобной активности, генерируемой в ответ на стимул-мишень, наблюдаемой в интервале 200—400 мс от момента его предъявления. Можно заключить, что скальповая РЗЬ-компонента
Часть 2. Вызванные потенциалы
255
в основном распространена над теменными областями коры. Однако при регистрации активности с имплантированных электродов картина меняется — выявляется широко распределенная мозговая система, объединяющая теменную, фронтальную кору, базальные ганглии и таламус.
В соответствии с литературными данными как в области клинических, так и теоретических исследований, компонента РЗЬ является наиболее изученной. Этому есть ряд причин. Во-первых, сама oddball-парадигма проста для применения практически в любых условиях исследования неврологических и психических больных. Во-вторых, РЗЬ — относительно большая компонента, которая легко выделяется на разностных ВП при сравнении ВП на девиантные стимулы мишени и игнорируемые стандартные стимулы. В-третьих, по всей видимости, РЗЬ характеризуется диагностической значимостью, убедительным доказательством которой являются значительные отклонения в величине этой компоненты при целом ряде дисфункций исполнительной системы, таких как шизофрения или синдром нарушения внимания с гиперактивностью.
Существует несколько предположений относительно функционального значения РЗЬ-компоненты. Наиболее популярная концепция, предложенная Дон-чиным в 1981 году, гласит, что РЗЬ отражает обновление рабочей памяти. То есть появление любого целевого стимула вызывает действие, после которого мозг обновляет содержимое рабочей памяти. В своей статье Дончин назвал этот процесс «обновление контекста»84. Концепция объясняла процессы на уровне психологии, но не предлагала конкретных нейрофизиологических или клеточных механизмов и в результате была раскритикована85.
Б. РЗа-компонента
Представьте, что вы управляете автомобилем, слушая музыку и наслаждаясь пейзажем за окном. Но неожиданное небольшое изменение в такте работы двигателя привлекает ваше внимание. Вы задаетесь вопросом: «Все ли в порядке?» Вы останавливаетесь, выходите из машины, открываете капот, смотрите и прислушиваетесь к двигателю. Данный пример демонстрирует то, что мозг создает сенсорную модель окружающего мира, удерживаемую в сенсорной системе. Когда происходит неожиданное изменение внешней среды, система обнаруживает это новое событие, что приводит к переключению внимания для его более тщательного изучения. Данный механизм позволяет людям (и животным) адаптироваться к постоянно меняющейся окружающей среде посредством так называемого ориентировочного ответа. Для того чтобы выделить бессознательную природу этого механизма, Иван Петрович Павлов называл его «ориентировочным рефлексом».
84 Однако, к сожалению, последующие исследования не подтвердили эту концепцию, которая была раскритикована в работе Верлегера в 1988 году (см. также ответ на критические замечания Donchin and Coles, 1988). До сих пор концепция «обновления контекста» остается заманчивой, но недоказанной гипотезой.
85 Основываясь на опыте наших исследований, можно говорить о существовании нескольких РЗЬ-подобных компонентов электрической активности, характеризующихся разной топографией и временными паттернами. Вероятно, эти компоненты ассоциированы с разными видами циклической активности и, следовательно, разным функциональным значением. Мы, как правило, определяем теменную РЗЬ-компоненту как компоненту «вовлечения». «Компонента вовлечения- рассматривается как результат активации групп нейронов, расположенных в теменных и лобных областях коры при реализации явных или скрытых действий. По всей видимости, в основе наблюдаемых феноменов лежат процессы преобразования сенсорной модели, которая отражается этой теменно-височнолобной циклической активностью, в действия.
256
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
стандартный девиантный
пассивный активный
стандартный девиантный
Рисунок 113. Разнообразие РЗЬ-подрбных компонент, регистрируемых при помощи имплантированных электродов
Вверху — скальповые ВП, зарегистрированные с Fz-отведения, усредненные по группе из 20 пациентов, которым по клиническим показаниям были имплантированы долгосрочные электроды. Амплитудная шкала — 1 мкВ. Ниже, сразу после верхнего ряда графиков ВП, представлены индивидуальные ВП, зарегистрированные с имплантированных электродов (амплитудная шкала—10 мкВ). ВП регистрировались во время выполнения акустического тестового задания в рамках oddball-парадигмы: стандартные стимулы (тоны длительностью 100 мс и частотой 1000 Гц) предъявлялись совместно с редкими и случайными девиантными стимулами (длительностью 100 мс и частотой 1300 Гц). Активный вариант теста (В) предполагал реакцию пациента в виде нажатия кнопки при предъявлении девиантного стимула. Пассивный вариант (Д) — чтение книги и игнорирование слуховой стимуляции. Графики разностных ВП в пассивной и активной модификации тестового задания представлены в среднем столбце (Б). Обозначения: ст.—стимул; дев.—девиантный; станд.—стандартный.
Часть 2. Вызванные потенциалы
257
Существенный прорыв в понимании природы ориентировочного ответа был достигнут благодаря изучению скальповых ВП человека. Впервые компонента ВП, связанная с ориентировочной реакцией, а именно РЗООа или РЗа, была описана в работе Саттона и др. в 1965 году. Несмотря на то что РЗа наиболее интенсивно исследовалась у людей, аналогичные компоненты регистрируются и у животных: макак, беличьих обезьян (саймири), кошек, собак, кроликов, дельфинов. Таким образом, РЗа может отражать мозговые процессы, свойственные млекопитающим.
В типичном для регистрации РЗа исследовании испытуемые выполняют тестовое задание по детекции стимула-мишени (тона), но при этом иногда среди нерелевантных стимулов случайным образом предъявляется новый неожиданный стимул (например, лай собаки). В отличие от стандартных стимулов и стимулов-мишеней появление нового стимула вызывает генерацию потенциала с пиковой латентностью около 200—300 мс (с момента предъявления начала стимула) и максимумом в центральных и лобных отведениях86 (рис. 11.7).
Следует отметить, что, как показывает sLORETA, нейрональные генераторы РЗа не локализуются исключительно на уровне премоторных областей. В нашем исследовании и работе, проведенной Эриком Халгреном (Eric Halgren) и др., РЗа-подобная активность встречается в различных корковых и подкорковых структурах: префронтальной, теменной, латеральной и медиальной височной коре, базальных ганглиях и таламусе (рис. 11.9). Очевидно, эта гетерогенная нейронная сеть объединяет несколько систем, выполняющих различные функции. Некоторые из этих структур, как, например, вторичные и ассоциативные области слуховой коры, ответственны за детекцию новизны посредством выявления характеристик стимуляции, которые весьма отличаются от фона. Другие (например, гиппокамп) вовлекаются в обеспечение кодирования контекстуально новых событий и их запоминание. И наконец, передняя поясная извилина и префронтальная кора играют важную роль в перераспределении внимания для более детальной обработки отклонений от фоновой стимуляции.
Наравне с фактом множественной генерации РЗа на сегодняшний день известно, что по крайней мере два нейромедиатора влияют на амплитуду РЗа: норадреналин (нейротрансмиттер, распространенный на уровне теменной и моторно-премоторных областей коры) и ацетилхолин, играющий важную роль в регуляции активности гиппокампа (связанной с процессами памяти).
В. Диагностическая ценность РЗа и РЗб-компонент
Как считается, РЗа и РЗЬ компоненты отражают активность системы внимания, что обусловливает их широкое применение в диагностике расстройств, в которых предполагается нарушение функции внимания. Боль-
86 Функциональное значение компонента РЗа и когнитивные процессы, связанные с ним, достаточно активно исследовались. В результате этих исследований стало возможным выявить четыре важных особенности РЗа. Во-первых, РЗа свойственно привыкание к последовательно предъявляемым новым стимулам, что выражается в снижении ее амплитуды в ситуации, когда эти стимулы становятся более ожидаемыми. Во-вторых, РЗа модально неспецифична — сопоставимые компоненты регистрируются при новых стимулах в зрительной, слуховой и соматосенсорной модальности. В-третьих, несмотря на то что обычно РЗа генерируется в ответ на сложные звуки, похожие потенциалы могут быть зарегистрированы и при предъявлении простых стимулов, отличающихся от текущего контекста. В-четвертых, РЗа регистрируется и в ответ на новый стимул в игнорируемом канале, что свидетельствует о выявлении нового стимула относительно фоновой стимуляции.
258
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Стандартный Девиантный Новый Дев.- Станд. Нов.- Станд.
Рисунок 11.9. Разнообразие РЗа-подобных компонент, регистрируемых при помощи имплантированных электродов Вверху — скальповые ВП, зарегистрированные с Fz-отведения, усредненные по группе из 20 пациентов, которым по клиническим показаниям были имплантированы долгосрочные электроды. Амплитудная шкала — 4 мкВ. Ниже, сразу после верхнего ряда графиков ВП, представлены индивидуальные ВП, зарегистрированные с имплантированных электродов (амплитудная шкала — 20 мкВ). ВП регистрировались во время выполнения акустического тестового задания в рамках трехстимульной oddball-парадигмы: стандартные стимулы (тоны длительностью 100 мс и частотой 1000 Гц) предъявлялись совместно с редкими девиантными и новыми стимулами. Во время предъявления пациенты читали книгу. Графики разностных ВП, полученные при сравнении ВП на стандартные и девиантные, а также стандартные и новые стимулы представлены в двух правых столбцах. Обозначения: HP — негативность рассогласования; дев.—девиантный; станд. — стандартный; нов. — новый.
Часть 2. Вызванные потенциалы
259
шинство исследований синдрома нарушения внимания с гиперактивностью свидетельствует о снижении амплитуды РЗЬ-компоненты у пациентов по сравнению с нормой.
Однако по сравнению с РЗЬ до сих пор не существует единого мнения о диагностической эффективности РЗа-компоненты. В некоторых исследованиях демонстрируется отсутствие значимой разницы между нормальными значениями РЗа и при СНВГ (например, см. Jonkman et al., 2000). Тем не менее существуют экспериментальные данные, согласно которым патологические изменения в генерации РЗа-компоненты (относительно нормы) могут отражать повышенную отвлекаемость детей с СНВГ (Gumenyuk et al., 2004).
VII. Заключение
С физиологической точки зрения внимание ассоциируется с усилением обработки релевантной сенсорной информации и подавлением нерелевантной. Данные регистрации импульсной активности нейронов у животных указывают на то, что внимание решает проблему конкурирования'стимулов за ресурсы с помощью механизма взаимного подавления репрезентаций множества одновременно представленных стимулов. В зрительной модальности пространственное внимание и внимание, относящееся непосредственно к объекту, реализуются на уровне дорсального и вентрального путей обработки информации соответственно. Нарушения пространственного внимания, называемые сенсорным дефицитом, ассоциируются с повреждениями в области правой теменной коры, структуры, входящей в состав дорсального зрительного информационного процессинга. Исследования ВП свидетельствуют о существовании ранних и поздних этапов обработки информации, которые по-разному модулируются процессами внимания. Ранние этапы характеризуются так называемыми негативностями выбора, а поздние — позитивными компонентами ВП: РЗа и РЗЬ. Механизм влияния внимания на генерацию ранних и поздних компонент ВП связывают с тонической или фазической активностью нейронов голубого пятна ствола мозга. Нейроны голубого пятна продуцируют регулирующий нейромедиатор норэпинефрин и обширно иннервируют кору головного мозга. На ранних этапах обработки информации тоническая активность этих нейронов играет важную |юль в регуляции активности областей коры, влияя на генерацию ранних компонент ВП и внося существенный вклад в генерацию процессной негативности, регистрируемой на ВП. Атомоксетин, недавно открытый препарат, блокирует обратный захват норэпинефрина, снижая симптоматику нарушений внимания при СНВГ. Нейроны, продуцирующие норэпинефрин, характеризуются также сильно выраженными связями с теменной и премоторной областями коры, что обусловливает регуляцию поздних этапов процессов обработки информации. ('читается, что фазическая модуляция активности коры этими нейронами отражается в компонентах РЗа и РЗЬ, которые широко используются как при исследовании здоровых испытуемых, так и в диагностических целях.
260
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Глава 12.
Исполнительные системы
Дело не в дороге, которую мы выбираем; то, что внутри нас, заставляет нас выбирать дорогу О’Генри. Дороги, которые мы выбираем
I. Психология
А. Необходимость исполнительного контроля
Термин «исполнительный контроль» используется для обозначения контроля моторной функции и когнитивных действий, направленных на достижение определенных целей. Долгое время в нейропсихологии с исполнительным контролем связывали функцию лобных долей мозга. Как было предложено, необходимость в механизме контроля исполнения деятельности возникала в нестереотипных ситуациях, требующих вмешательства контролирующих систем: при выборе определенного действия из множества возможных, при подавлении несоответствующего действия, при удержании в рабочей памяти планируемого действия и его ожидаемого результата.
Исполнительный контроль также требуется и для оптимизации поведения. Принятие решения в пользу определенного действия происходит с учетом ожидаемого результата. Рассогласование между прогнозируемым и действительным результатом действия используется для оптимизации и коррекции поведения. Так, если не последовало ожидаемого вознаграждения, возникающая ошибка ожидания обусловливает изменение ранее сформированного паттерна поведения.
Б. Типы исполнительных операций
В соответствии с современными представлениями при рассмотрении процессов исполнительного контроля выделяют несколько подкомпонентов. В одной из наиболее распространенных классификаций (Smith and Jonides, 1999) исполнительные процессы подразделяются: 1) на внимание и подавление; 2) управление задачами; 3) планирование; 4) мониторинг; 5) кодирование. Вместе с тем количество и точные функции выделяемых подкомпонент до сих пор являются предметом дискуссий. Последние исследования были сконцентрированы на изучении тех подкомпонент, которые относительно хорошо определяются как эмпирически, так и теоретически.
Часть 2. Вызванные потенциалы 261
В рамках настоящей главы мы различаем следующие операции над действиями: 1) селективные операции — вовлечение в действия и отвлечение от них; 2) рабочая память; 3) операции мониторинга (табл. 12.1). Эти операции хорошо определяются на психологическом уровне. Предполагается, что данные операции обеспечиваются разными нейронными механизмами и отражаются в различных компонентах ВП, вызванных действиями.
Таблица 12.1. Классификация исполнительных операций
Основные операции Исполнительная функция
Операция вовлечения Активация нейронов фронтально-теменной коры, ответственных за формирование репрезентации планируемого действия, для обеспечения возможности реализации данного действия
Операция отвлечения Подавление активности нейронов фронтальной коры, ответственных за формирование репрезентации планируемого действия, для воздержания от исполнения действия
Рабочая память Временное хранение информации о планируемых действиях (от нескольких минут до нескольких часов) для активного использования ее при операциях вовлечения и отвлечения
Операция мониторинга Сравнение результата реализованного действия с ожидаемым. При наличии рассогласования инициируется новое действие для его устранения
В. Связь с селекцией действий
На уровне нейронных сетей исполнительные функции определяются как операции, выполняемые над репрезентациями действий, которые хранятся в коре головного мозга. В действительности эти репрезентации — память о действиях и планы, связанные с ними. Фронтально-теменная нейронная сеть является местом локализации репрезентаций действий, активация которых обеспечивает доступ к хранящейся информации об этих действиях. Исполнительные функции рассматриваются как вычислительные процессы или алгоритмы обработки информации, которые вызывают активацию определенного действия в заданном интервале времени и сложившихся обстоятельствах или подавляют репрезентацию действия, ненужного в данной ситуации.
Как и любая сложная функция, исполнительные функции реализуются сложной системой мозга, состоящей из корковых и подкорковых структур, связанных между собой. В состав кортикальных структур входят теменные и фронтальные области. Работа нейронов этих областей регулируется параллельной циклической активностью, охватывающей структуры коры, имеющие связи с соответствующими частями базальных ганглиев, проецирующие свои связи на соответствующие ядра таламуса, которые, в свою очередь, реципрокно связаны с корой.
II. Базальные ганглии как «темный подвал» мозга
А. Анатомия
Согласно мнению, высказанному Киннеаром-Вильсоном (Kinnear-Wilson) в 1920-х годах, ясность в работе базальных ганглиев подобна темному подвалу. И так
262
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Прямой путь
Рисунок 12.1. Циклическая активность на уровне базальных ганглиев
А. Анатомическая локализация таламуса и базальных ганглиев на коронарном срезе. В состав базальных ганглиев входят стриатум и бледный шар. Базальные ганглии и таламус расположены в непосредственной близости друг от друга. Связи менщу корой, базальными ганглиями и таламусом обозначены стрелками черного (возбуждающие) и серого (тормозные) цвета. Б. Схематическое изображение прямого проводящего пути базальных ганглиев. В. Непрямые и гиперпрямые проводящие пути базальных ганглиев.
было большую часть прошлого века. Только сравнительно недавно удалось продвинуться в понимании механизмов вовлечения базальных ганглиев в обеспечение реализации моторных, когнитивных и аффективных актов. В соответствии с классическим определением в состав базальных ганглиев входят пять ядер: хвостатое ядро, скорлупа, бледный шар (подразделяемый на внутреннюю и внешнюю части), субталамическое ядро и черная субстанция87 (рис. 12.1). Базальные ганглии являются самыми большими подкорковыми образованиями переднего мозга. Входными ядрами базальных ганглиев являются хвостатое ядро, скорлупа (вместе формируют стриатум) и прилежащее ядро. Эти ядра получают входы практиче
87 Представление о связях базальных ганглиев расширилось в 1980 году, когда были открыты прилежащее ядро (также называемое вентральным стриатумом) и вентральный бледный шар.
Часть 2. Вызванные потенциалы
263
ски со всех областей коры, за исключением первичных. Выходные ядра базальных ганглиев, которые посылают результаты пространственного рекартирования и временной обработки к ассоциативным ядрам таламус, локализуются во внутренней части бледного шара. Ассоциативные таламические ядра также получают дополнительные входы от префронтальной коры, и затем реципрокно проецируют связи обратно в кору. Таким образом, базальные ганглии получают информацию фактически от всех областей коры обрабатывают ее и передают результаты обработки обратно в кору через таламические ядра88.
Каждый из базальных ганглиев имеет важное клиническое значение. В частности, дегенерация нейронов стриатума приводит к болезни Нантингтона и соответствующим гиперкинетическим расстройствам. Ослабление продукции дофамина, возникающее в результате потери дофаминергических нейронов черной субстанции, вызывает паркинсонизм89. Переизбыток О2-рецепторов дофамина является одним из признаков шизофрении. Синдром гиперактивности внимания характеризуется повышенным количеством DAT-рецепторов — рецепторов, ответственных за обратный захват дофамина в стриатуме. Бледный шар является структурой-мишенью при стереотаксических операциях и стимуляциях, применяемых в лечении парксинсонизма. Большое значение при лечении болезни Паркинсона имеют и субталамические ядра, являющиеся ключевыми структурами в процессах контроля функционирования выходных ядер базальных ганглиев (внутренняя часть бледного шара), электрическая стимуляция которых используется при лечении заболевания.
Б. Прямые проводящие пути
Проводящие пути базальных ганглиев подразделяются на два вида: прямые и непрямые. Эти пути выполняют разные функции. Прямой проводящий путь (рис. 12.1 вверху справа) обеспечивает фокусированное подавление активности нейронов выходных ядер базальных ганглиев. Внутренняя часть бледного шара, являющаяся выходной структурой базальных ганглиев, оказывает тормозные контролирующие влияния на таламус (рис. 12.1 вверху справа), моторные ядра ствола мозга и верхние бугорки четверохолмия (на рис. 12.1 не показаны). В отличие от нейронов стриатума фоновая импульсная активность нейронов бледного шара очень интенсивна, что лежит в основе угнетающего действия на нейроны таламуса90. Подавление этих тормозных влияний (называемых растормаживанием) выполняет воротную функцию, т.е. позволяет нейронам таламуса активироваться в ответ на внешнюю стимуляцию.
Следовательно, прямой проводящий путь обеспечивает положительную обратную связь с префронтальной корой. И действительно, увеличение активно
88 В дополнение к проекциям обратной связи к фронтальной коре базальные ганглии посылают результаты своей обработки к ядрам ствола мозга, обеспечивающим контроль моторных функций. В частности, базальные ганглии имеют проекции на верхние бугорки четверохолмия, которые участвуют в обеспечении контроля осевой ориентации головы и тела, а также саккадических движений глаз.
89 Компактная часть черной субстанции содержит дофаминергические нейроны, которые на уровне стриатума регулируют процесс обработки информации путем установки порогов срабатывания нейронов.
90 Спонтанная активность нейронов бледного шара человека составляет др 200 спайков в секунду, тогда как аналогичная активность нейронов стриатума достаточна низка—до 20 спайков в секунду. Эта разница в скорости разрядов нейронов иногда используется для точного определения области расположения бледного шара, когда при стереотаксических операциях необходимо произвести разрушение в этой области. Такие операции похожи на палидотомию, но являются более локальными.
264 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
сти префронтальной коры приводит к увеличению активности нейронов стриатума и подавлению нейронов бледного шара, что вызывает растормаживание нейронов таламуса и в конечном итоге способствует еще большей активации префронтальной коры. На уровне таламуса операция растормаживания может быть сравнима со снятием «тормоза», «удерживающего» активность нейронов таламуса. Таким образом, архитектура прямого проводящего пути базальных ганглиев предполагает, что его основная функция — управление активностью группы нейронов целевой системы через механизм растормаживания.
Нажатие кнопки после третьего стимула
Пробамтмормруемая
после первого стимула
Проба кг яюрмруевхая после второго стимула
Нейрон “о первого
Ь
1ст 2 ст Зет
Рисунок 12.2. Два типа активности нейронов на уровне петли проводящего базально-таламокортикального пути
А. Пример импульсной активности нейронов при обработке сенсорной информации. Б. Пример нейронной активности при подготовке к моторной реакции. На графиках по горизонтальной оси отмечено время, по вертикальной — частота разрядов нейронов (в относительных величинах). Регистрация импульсной активности при помощи имплантированных электродов производилась у пациента, страдающего болезнью Паркинсона. Каждая проба тестового задания состояла из трех стимулов, представленных в нижней части рисунка: 1-й стимул определял, будет ли пациент ждать следующий стимул пробы или игнорировать ее целиком; 2-й стимул определял, нужно или нет нажимать кнопку, 3-й стимул служил триггерным сигналом для запуска моторной программы ответа (см. Kropotov et al., 1997).
В. Регистрация активности глубоких структур мозга у пациентов
В нашей лаборатории91 на протяжении почти 20 лет изучалась импульсная активность нейронов и локальные потенциалы поля у пациентов, страдающих болезнью Паркинсона, эпилепсией и навязчивыми состояниями (обсессивно-компульсивное расстройство, ОКР). Все исследования проводились в диагностических и терапевтических целях, после стереотаксической имплантации электродов в структуры мишени. Стереотаксические операции проводились только в тех случаях, когда традиционные и более щадящие методы были малоэффективны в лечении. Пациенты с вживленными электродами принимали участие в различных исследованиях, построенных таким образом, чтобы изучать исполнительные функции.
При регистрации импульсной активности в базальных ганглиях, вентральном таламусе и премоторной коре мы столкнулись с большим количеством подготовительных компонент, т.е. компонент, относящихся к подготовке реализации действия или восприятия стимула. Такие вызванные нейронные ответы наблюдались только на поведенчески значимых стимулах (рис. 12.2).
Другой удивительной находкой был разный характер импульсной активности таламуса при предъявлении сигнальных GO и NOGO-сгимулов. Так, GO-стимулы обычно вы-
91 В 1970-1990 годах лаборатория принадлежала Институту экспериментальной медицины РАМН. Наталья Петровна Бехтерева, выдающийся советский и российский нейрофизиолог, была инициатором развития этого направления исследований, в котором использовался комплексный полиметодический подход, заключающийся в комбинировании разных методов регистрации мозговой активности: импульсной активности мозга, локальных потенциалов поля, сверхмедленной электрической активности, а также анализа результатов влияния терапевтических электростимуляций на поведение человека.
Нейрон второго
Часть 2, Вызванные потенциалы
265
Рисунок 12.3. Локальный потенциал поля, отражающий реакцию нейронов на уровне вентрального таламуса
А. Интракраниальные ВП. Б. Ответы таламических нейронов на GO-, NOGO- и игнорируемые стимулы трехстимульной oddball-парадигмы (описанной на рис. 12.2). (А): по оси Y — усредненные потенциалы, которые были зарегистрированы с электродов, имплантированных в вентральную область таламуса в ответ на GO*- (черная линия), NOGO- (серая линия) и игнорируемые (Ignore, внизу) стимулы. (Б): по оси Y — усредненная импульсная активность нейронов, зарегистрированная с тех же электродов. По оси X — время (неопубликованные данные из архивов лаборатории).
зывали активацию нейронов таламуса с латентностью около 200 мс, тоща как NOGO-стимулы, напротив, подавляли их активность (рис. 12.3). Эти реакции нейронов часто сопровождались соответственно позитивными (GO) и негативными (NOGO) компонентами локальных потенциалов, регистрируемых с одних и тех же электродов.
На основании ранее полученных данных нами была сформулирована теория программирования действий (Kropotov, 1987)92. В соответствии с данной теорией ключевую роль в выборе действий играет система таламокортикальных связей банальных ганглиев. Схематическая иллюстрация теории представлена на рис. 12.4.
Г. Модель выбора действий
Давайте представим ситуацию, когда из всего доступного репертуара действий нам необходимо выбрать одно, как, например, при правом или левом повороте на перекрестке. Пространственно распределенная активность, соответствующая кортикаль
92 Расширенный обзор основных результатов нашей лаборатории по изучению импульсной активности представлен в ciaibe, опубликованной в 1999 году (Kropotov, Etlinger, 1999). Сьюзан Этлингер в то время работала в Венском универ-си iere. В 1984-1986 гг. в нашей лаборатории она проводила исследования по изучению сверхмедленной активности нейронов, регистрируемой с имплантированных электродов у неврологических больных.
266 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
ному представительству каждого из действий, схематично представлена на рис. 12.4 (справа вверху). Данные активации существенно перекрываются на уровне премоторной и моторной коры (на рис. 12.4 горизонтальная ось обозначает пространственное расстояние). Полученные нами данные свидетельствуют о том, что каждая из распределенных нейронных сетей, соответствующая разным действиям, проецируется в разные области стриатума93. На представленном рисунке две программы действий перекрываются на уровне коры, но по-разному проецируются в стриатум. Таким образом, стриатум сам по себе может быть представлен как «карта» действий.
Выходные нейроны стриатума по характеру воздействия тормозные. Они проецируются на нейроны бледного шара и одновременно посылают коллатерали на соседние нейроны стриатума, выполняя функцию латерального торможения94. Механизм латерального торможения позволяет стриатуму выполнять специфическую фильтрующую функцию по принципу «победитель получает все». Фильтрация позволяет выбрать наиболее активированные репрезентации потенциальных действий и одновременно блокировать менее активированные (т.е. менее значимые в контексте данной ситуации).
Д. Дофамин как модулятор активности базальных ганглиев
Дофамин, продуцируемый нейронами компактной части черной субстанции, модулирует активационные пороги эфферентных нейронов стриатума. Нейроны черной субстанции проецируются на проксимальные части синаптических шипиков95, что обеспечивает положение, позволяющее существенно модулировать поток активаций от нейронов коры, которые имеют окончания на удаленных частях тех же синаптических шипиков (рис. 12.4, вставка). Известно, что дофаминовые рецепторы нейронов прямого проводящего пути базальных ганглиев — рецепторы В2-типа. Связывание дофамина с этими рецепторами вызывает активацию — медленную деполяризацию нейронов стриатума. Эффекторные нейроны, в свою очередь, реагируют только на активность, превосходящую порог срабатывания. Событие, превышающее этот порог, наблюдается в случаях, когда нейроны стриатума селективно деполяризуются нейронами коры (тогда как активность соседних нейронов подавлена) и глобально деполяризуется дофаминергическими проекциями из черной субстанции96. Таким образом, наличие подавляющих латеральных связей стриатума формирует основу функции выбора действий. Кроме того, дофамин, в свою очередь, благоприятствует операциям по выбору действий, снижая пороги актива-
93 Это свойство было названо сегрегацией действий и фактически означает, что репрезентация разных действий по-разному картируется разными частями стриатума.
94 Исходя из теории нейронных сетей (см. «Методы» ч. 2) известно, что латеральное торможение вместе с нелинейными свойствами активности нейронов позволяет осуществить выбор наиболее активированного входа в соответствии с принципом -победитель получает все». Как показывают наши исследования, нейроны стриатума демонстрируют низкий уровень активности, когда испытуемый находится в состоянии бодрствования. Одной из причин низкого уровня спонтанной активности являются сильная прямая связь и латеральное торможение, возникающие благодаря тормозному влиянию нейронов самого стриатума.
95 Шипик — небольшая выпуклость мембраны дендрита, соединенная с последней, еще меньшей по размеру шейкой шипика.
96 Следует напомнить, что дофаминергические нейроны черной субстанции активируются в ответ на поведенчески значимые стимулы, как, например, вознаграждение, таким образом способствуя выбору только релевантных кданной ситуации программ. Тонические и фазические свойства дофаминергической передачи по-разному изменяются при различных заболеваниях мозга, например, таких как болезнь Паркинсона, шизофрения или СНВГ.
Часть 2. Вызванные потенциалы
267
Рисунок 12.4. Процессы выбора действия на уровне нейрональной системы таламокортикальных связей базальных ганглиев
А. Схематическое отображение нейронов коры, стриатума, бледного шара и таламуса, а также их связей. Возбуждающие связи обозначены «+», тормозные обозначены «-». В овальной вставке изображен дендрит нейрона стриатума с дистальным синапсом от глугаматергического нейрона коры и синапсом от дофаминергического нейрона черной субстанции, расположенным проксимально. Б. Схематическое представление пространственных паттернов активаций на разных уровнях системы, объединяющей базальные ганглии и их таламокортикальные связи. На уровне коры мозга — перекрывающиеся отображения двух активированных действий. На уровне стриатума — две активированные программы действий картированы в разных частях стриатума. В силу сильной выраженности эффекта латерального торможения выбирается только одна из программ действий в соответствии с принципом «победитель получает все». На уровне бледного шара — спонтанная активность нейронов, которая соотносится с выбранной программой действия, ингибируется. На уровне таламуса соответствующие нейроны растормаживаются, что лежит в основе воротной функции таламуса, и при участии таламокортикального пути происходит интенсификация выбранной программы действия. Ось X — пространство. Порог — порог активации разрядов нейронов (см. Kropotov, Etlinger, 1999). Обозначения: SN — Substantia Nigra.
ции нейронов стриатума. В частности, при паркинсонизме отмечается наличие слишком высоких порогов активации, обусловленных дефицитом дофамина, что существенно затрудняет инициацию действий. При шизофрении, напротив, пороги срабатывания ненормально низки в силу высокой концентрации рецепторов дофамина в стриатуме, что приводит к одновременной активации нескольких программ и дезинтеграции сознания^_______________
97 Название «шизофрения» происходит от греческого слова охФфрёуеш, означающего «расщепленный мозг». Шизофрения является психическим расстройством, которое характеризуется нарушениями восприятия или выражения реальности и значимой социальной или трудовой дисфункцией. Люди, страдающие шизофренией, обычно характеризуются дезорганизацией мышления, галлюцинациями и бредом.
268 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Е. Растормаживание нейронов таламуса
Нейроны стриатума, входящие в состав системы выбора программ, проецируют свои связи в соответствующие области комплекса «бледный шар/черная субстанция» и подавляют их активность. Нейроны бледного шара и черной субстанции по сравнению с нейронами стриатума характеризуются высокочастотной спонтанной активностью. Следовательно, выбранное действие вызывает активацию нейронов стриатума и подавление высокочастотной активности нейронов бледного шара.
Нейроны бледного шара, в свою очередь, имеют связи с таламусом, за счет которых подавляют активность последнего. Так как нейронам бледного шара свойственна спонтанная активность, то таламус находится под постоянным тормозным влиянием. При ингибировании нейронов бледного шара уровень их импульсной активности снижается, что приводит к растормаживанию нейронов таламуса и активации таламокортикальных проводящих путей. Таким образом, таламус выполняет функцию «ворот», открывая доступ к коре для выбранного действия.
Используя известную «прожекторную» метафору Фрэнсиса Крика, мы можем сказать, что таламическая функция аналогична лучу прожектора, который выделяет и интенсифицирует выбранную программу на уровне коры.
Почему природа создала столь сложный механизм селекции действий? В соответствии с разработанной нами теорией (Kropotov, 1987) данный механизм обусловлен невозможностью одновременной активации (выделения) одного действия и торможения других исключительно на уровне коры. Дело в том, что все широко распределенные внутренние связи коры по своей природе активирующие, а кортикальное торможение может происходить только локально98. Таким образом, мозг использует дополнительные механизмы, позволяющие выбирать действия на подкорковом уровне, а лишь потом усиливать и выделять действия на уровне коры.
Ж. Непрямые и гиперпрямые проводящие пути
В результате базально-таламокортикальных взаимодействий не только инициируются выбранные программы, но и подавляются нерелевантные. Торможение неподходящих действий осуществляется через непрямые проводящие пути, распространение активаций по которым приводит к противоположным эффектам в сравнении с прямыми путями (рис. 12.1).
Суть нейронного механизма непрямого пути заключается в обеспечении фокусированных активаций на уровне выходных структур базальных ганглиев, оказывающих тормозные воздействия на возбудительные корковые и подкорковые структуры. Так, активация внутренней части бледного шара может приводить к подавлению подготовленной программы действий прямым способом, посредством торможения активности моторных ядер ствола мозга и верхних бугорков четверохолмия, или косвенным, посредством подавления нейронов таламуса, имеющих проекции на премоторные области
98 Следует упомянуть, что тормозные нейроны коры имеют слишком короткие аксоны, обеспечивающие только локальное торможение.
Часть 2. Вызванные потенциалы
269
коры. Известно также, что дофамин может модулировать распространение активаций по непрямым проводящим путям. Однако по сравнению с влиянием на прямые пути действие дофамина приводит к противоположным эффектам. Имеется в виду, что непрямые пути берут начало от нейронов стриатума, которые обладают D1-рецепторами (модуляция прямого пути осуществляется через О2-рецепторы). Связывание дофамина с D1-рецептором приводит к гиперполяризации мембраны, что и обусловливает торможение передачи. Таким образом, дофамин препятствует проведению информации на уровне непрямых проводящих путей (в то же время активируя передачу в прямом проводящем пути).
Гиперпрямой проводящий путь обеспечивает возбудительный эффект, оказываемый корой на бледный шар, в обход стриатума и характеризуется более короткой по времени проводимостью по сравнению с непрямыми путями. По всей видимости, функцией этого проводящего пути является подавление всех нерелевантных действий в момент выбора релевантного, осуществляемого с помощью проводящего пути.
3. Выходное влияние на ствол мозга
Нарушения в работе базальных ганглиев проявляются как неспособность не только инициировать и терминировать произвольные движения, но и прекращать непроизвольные. Кроме того, такие расстройства характеризуются ненормальными скоростью и количеством движений и мышечным тонусом. По всей видимости, базальные ганглии вносят вклад в поддержание мышечного тонуса и выполняют функцию «ворот» для моторной активности, что реализуется через влияние на моторную систему ствола мозга — базовую моторную систему, ответственную за поддержание тонуса мышц, поддерживающих положение тела, и локомоторную функцию.
И. Параллельные цепи
Известно, что практически любая область коры (за исключением первичных сенсорных зон) имеет проекции на стриатум. Наиболее обильными стриарными проекциями обладает префронтальная кора. Грубо говоря, эти проекции можно назвать топографически организованными. Однако они то-пографичны только при первом рассмотрении, поскольку более детальный анализ указывает на мозаичную форму проекций, позволяя любому нейрону префронтальной коры проецироваться на пространственно разные области стриатума. В то же время функционально связанные области коры (за счет внутренних кортикальных связей) проецируются в одну и ту же зону стриатума. Иными словами, на уровне стриатума представлена функциональная карта коры, а не ее топографические проекции. В соответствии с такой функциональной картой функционально связанные нейроны проецируются на конкретные области стриатума". *
99 Данные, полученные в возглавляемой мной лаборатории, подтверждают это. В действительности, как показывают наши исследования с регистрацией импульсной активности нейронов у пациентов, нейроны, ответственные за сенсорную и моторную функцию, на уровне стриатума пространственно разделены.
270 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
В соответствии с точкой зрения о разделении функций на уровне стриатума выделяют несколько параллельно работающих функциональных путей регуляции активности коры. Каждый из этих путей представлен пространственно разными областями на уровне коры, стриатума, бледного шара и таламуса. На основании своих анатомических и физиологических исследований Александер и Делонг в 1980-х годах выделили несколько типов параллельных проводящих путей: моторный, пространственный, зрительный и аффективный (рис. 12.5). Каждый из этих функциональных путей занимает специфическую часть стриатума и получает множественные, частично перекрывающиеся входы от нескольких анатомически связанных областей, участвующих в обеспечении одинаковых функций. Принимая во внимание названия этих путей, можно заключить, что они выполняют различные функции, а именно: выполнение и планирование действий (моторные), организация пространственной и объектной рабочей памяти (пространственные и зрительные), обеспечение мотиваций и эмоций (аффективные)100.
Моторный проводящий путь через ядро подушки осуществляет регуляцию активности моторной, премоторной и добавочной моторной коры. Также данный проводящий путь получает входы и от соматосенсорной коры. Эти цепи участвуют в инициации и подавлении моторных действий, а также в их подготовке. Некоторые специалисты отдельно рассматривают моторный и окуломоторный проводящий пути. С помощью окуломоторного пути обеспечивается регуляция активности области фронтальной коры, ответственной за движения глаз (фронтальное глазодвигательное поле), получающей также входы от задней части теменной коры (аналогичного теменного центра). Данный проводящий путь участвует в обеспечении ориентации тела и положения глаз в направлении выбранного источника сенсорной информации.
Удержание в памяти информации, касающейся пространственных стимулов, реализуется на уровне пространственного проводящего пути, объединяющего области дорсолатеральной префронтальной и задней теменной коры.
На уровне зрительного пути модулируется активность в области вентролатеральной префронтальной и латеральной височной коры, которые участвуют в реализации процессов регуляции рабочей памяти для объектов (в особенности зрительных).
В регуляцию настроения и эмоций вовлекаются структуры, входящие в состав аффективного проводящего пути: передняя поясная извилина, прилежащее ядро, орбитофронтальная кора и лимбические структуры аллокортекса (гипоталамус и энториальная кора).
Базальные ганглии, помимо проекций в вентральное, дорсомедиальное и переднее вентральное ядра таламуса, имеют связи с интраламинарными ядрами и ядрами срединной линии. Дополнительно эти ядра имеют сильные связи с холинергическими нейронами ретикулярной формации ствола мозга и об-
100 Как уже отмечалось выше, процессы, выполняемые базальными ганглиями, включают планирование действий, инициацию и подавление действий, а также хранение результатов действий в рабочей памяти. Все эти операции вместе взятые составляют основу исполнительных функций мозга.
Часть 2. Вызванные потенциалы
271
Рисунок 12.5. Параллельные проводящие пути нейрональной системы таламокортикальных связей базальных ганглиев
А—Г: моторная, пространственная, зрительная и аффективная петли связи коры и стриатума, которые участвуют в обеспечении выполнения и планирования моторных действий (моторная), организации пространственной и объектной рабочей памяти (пространственная, зрительная) и поддержании мотиваций и эмоций (аффективная). Обозначения: SMA — добавочная моторная область; РМС — премоторная кора; SSC — соматосенсорная кора; DLPFC — дорсолатеральная префронтальная кора; РРС — задняя теменная кора; VLPFC — вентралатеральная префронтальная кора; IT — нижняя височная кора; ST — верхняя височная кора; OFC — орбитофронтальная кора; CG — передняя поясная кора; НС — гиппокамп; BLA—базолатеральная амигдала; VP — вентральный паллидум; Gpi — внутренний сегмент бледного шара; SNpr — ретикулярная часть черной субстанции; VL — вентролатеральный таламус; VA — передний вентральный таламус; MD — медиодорсальный таламус; STN — субталамическое ядро; GPe — внешний сегмент бледного шара. Разных типы циклической активности включают разные части вышеперечисленных ядер (см. Alexander et al., 1986).
272
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
ратные проекции как в область базальных ганглиев, так и во многие области коры. Высокочастотная стимуляция рассматриваемых ядер вызывает десинхронизацию скалытовой ЭЭГ, сопровождающуюся реакцией возбуждения. К противоположным результатам приводит низкочастотная стимуляция — наблюдается постепенное нарастание медленной и веретенообразной волновой активности, ассоциирующейся с ослаблением внимания, сонливостью или сном.
К. ЭЭГ-корреляты нарушений базальных ганглиев
Известно, что функция базальных ганглиев тесно связана с движениями. С теоретической точки зрения ритмическая активность на уровне парал
лельных таламокортикальных проводящих систем активирует соответству-
ющие области коры, что сопровождается десинхронизацией на скалытовой ЭЭГ. Нарушения же, затрагивающие структуры, входящие в состав этих проводящих систем, должны приводить к синхронизации медленных осцилляций, регистрируемых непосредственно вблизи коры. Данные теоретические предположения подтверждаются экспериментальными данными.
В действительности было показано, что повреждение хвостатого ядра и нейронов скорлупы обезьян приводит к увеличению регу-
Рисунок 12.6. Спектральный анализ ЭЭГ при паркинсонизме
А. Спектральная мощность ЭЭГ с пиком для тета-ритма. Топограмма (Б) и локализация генераторов активности (В), вычисленная с помощью метода томографии низкого разрешения (sLORETA), относятся к тета-диапазону ЭЭГ (6 Гц), зарегистрированной у пациента на ранней стадии болезни Паркинсона.
лярной высокоамплитудной волновой активности частотой от 5 до 10 Гц. Сопоставимые эффекты наблюдаются и у пациентов, страдающих паркинсонизмом, вне зависимости от возраста или факта наличия деменции (рис. 12.6). Выраженность медленной активности зависит от степени моторной дисфункции в целом и акинезии в частности. Отмечается также снижение вызванной десинхронизации в ответ на произвольные движения пациентов с паркинсонизмом. При этом известно, что употребление L-DOPA может снимать эти патологические прояв-
ления, приводя ЭЭГ к нормальным значениям.
Часть 2. Вызванные потенциалы
273
III. Префронтальная кора и исполнительный контроль
А. Анатомия
Лобные доли составляют примерно треть от всей коры больших полушарий мозга человека. Граница с теменной корой проходит по центральной борозде, а с височной — по сильвиевой борозде. Фронтальная кора не является унитарной структурой и подразделяется на первичную моторную кору (ПБ 4), электрические стимуляции которой могут приводить к мышечным сокращениям или простым движениям, премоторную и добавочную моторную область (ПБ 6), заднюю часть поясной извилины и префронтальную кору. Нейроны префронтальной коры реципрокно связаны дорсомедиальным ядром таламуса. Премоторная кора получает входы и проецируется обратно в латеральные таламические ядра. Поясная извилина имеет реципрокные связи с передним ядром таламуса.
Несмотря на то что вполне ясным представляется факт связи префронтальной коры с высшими когнитивными функциями, до сих пор остается непонятным, каким образом эти функции реализуются. Нейронная архитектура префронтальной коры, пожалуй, самая сложная по сравнению с другими областями коры, что позволяет рассматривать ее как субстрат высших когнитивных функций, получивших большее развитие у человека по сравнению с животными101.
Префронтальная кора в составе нескольких проводящих путей получает множественные входы от мультимодальных сенсорных областей коры и лимбических структур мозга, проецирующиеся в разные зоны префронтальной коры. На рис. 12.7 схематично иллюстрирован информационный поток на уровне вентрального зрительного пути, достигающего лобной доли в области вентролатеральной префронтальной коры. В отличие от топографической организации проекций между областями задней части коры проекции префронтальной коры, строго говоря, не являются топографичными. Напротив, одна и та же область сетчатки имеет несколько представительств на поверхности префронтальной коры. Известны анатомические свидетельства, подтверждающие существование высокоупорядоченного паттерна терминалей аксонов, берущих начало в задних ассоциативных областях коры и оканчивающихся в префронтальной коре, формирующих чередующиеся, переплетенные полосы102.
Б. Сложность организации нейронных сетей
Первые исследователи в области сравнительной нейрофизиологии были впечатлены схожестью организации коры мозга млекопитающих. К несчастью, «схожесть» многими была понята как «одинаковость», и это привело к тому, что на протяжении второй половины прошлого столетия сложи
101 Префронтальная кора приматов определяется как кора, расположенная кпереди от центральной борозды, и может быть весьма грубо разделена на три основные составляющие: латеральную, медиальную и орбитальную части. В свою очередь, латеральная префронтальная кора может быть подразделена на дорсолатеральную и вентролатеральную части. Медиальная префронтальная кора делится на переднюю поясную извилину и медиальную фронтальную кору. А в орбитофронтальной коре выделяют медиальную, вентральную, латеральную и фронтополярную части.
102 До сих пор не существует определенного ответа на вопрос, для чего на уровне префронтальной коры нужны множественно по-нюряющиеся репрезентации стимулов или моторных (поведенческих) целей. Впрочем, теоретические исследования предоставляют ряд возможных объяснений. Так, известно, что мозг может достаточно гибко и вариабельно реагировать на идентичные стимулы в зависимости от контекста их появления. Отсутствие четкого соответствия между сенсорными стимулами и моторными ответами (по типу •один в один») делает множественное повторение соответствующих репрезентаций фактически неизбежным.
274 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
лось мнение об однообразном строении коры мозга. Считалось, что в целом кора (за исключением первичной зрительной коры) состоит из однородных и повторяющихся базовых элементов. В соответствии с этой догмой региональные отличия в обеспечении зрения, слуха и соматосенсорной чувствительности относили исключительно к разному составу входов и выходов.
Первые доказательства против эквипотенциальности были получены в исследованиях сенсорных систем. В период с 1980 по 1990 гг. было показано, что зрительная кора обезьян содержит около 30 областей. Исследования последних 10 лет свидетельствуют о том, что входящие и исходящие связи не являются единственной характеристикой специфичности процессов обработки информации зрительными областями коры. Особенности информационной обработки определяются также структурой связи нервных клеток (плотностью шипиков, паттерном ветвления дендритов и аксонов). Более того, внутренняя кортикальная структура различна для разных областей. Например, дендритное дерево третьего слоя пирамидных клеток в области ТЕ мозга макак содержит в среднем в 11 раз больше шипиков по сравнению с нейронами V1 зоны зрительной коры. Аналогичная картина характерна и для нейронов фронтальных областей: пирамидальные клетки префронтальной коры у человека и макаки в целом характеризуются наиболее разветвленными дендритами и большим количеством шипиков в сравнении с нейронами затылочной, теменной и височной долей (рис. 12.7).
В. Репрезентации сложных действий
Исходя из того что префронтальная кора получает множественные проекции от моторных областей коры, можно предполагать ее участие в хранении сложных репрезентаций действий. Считается, что дорсолатеральная префронтальная кора развивалась из моторных областей. Следовательно, вполне обоснованно считать, что функции более «новых» префронтальных моторных областей будут связаны с филогенетически более старыми структурами моторной системы. Недавно состоявшееся открытие зеркальных нейронов, активирующихся в ответ на реализацию как собственных действий, так и наблюдаемых действий других субъектов, поддерживает такую точку зрения.
Следует обратить внимание на то, что префронтальная кора ассоциируется не с действиями как таковыми, а с исполнительными функциями, т.е. контролем исполнения действий. При этом сложность поведения человека отождествляется со сложностью действий. Современные представления о степени функциональной дифференциации подобластей префронтальной коры, т.е. о том, насколько разные когнитивные функции могут картироваться разными областями, остаются противоречивыми. С одной стороны, в ситуациях, требующих когнитивного контроля, наблюдаются одновременные активации в дорсолатеральной, вентролатеральной и медиальной префронтальной коре. А с другой, рискуя увлечься неофренологией, некоторые исследователи склонны рас-
Часть 2. Вызванные потенциалы
275
Рисунок 12.7. Повышение «сложности» от зрительной до префронтальной коры
Представленные нейроны зрительной (поля VI, V2, V4,), височной и префронтальной коры мозга обезьян иллюстрируют различия в размере и паттернах ветвления базальных дендритов. Относительное количество дендритных шипиков III слоя пирамидальных клеток каждой из областей отображено вертикальной полоской со шкалой в правом верхнем углу (см. Elston, 2003). Нейроны зрительной коры демонстрируют только активацию в ответ на стимул, тогда как нейроны префронтальной коры характеризуются высоким уровнем спонтанной активности. ЭЭГ же человека характеризуется наличием выраженной альфа-активности в затылочной области и ее отсутствием в области префронтальной коры.
сматривать разные подразделения префронтальной коры в качестве ключевых структур обеспечения разнообразных когнитивных функций, которые взаимодействуют между собой с целью оптимизации деятельности (например, при выполнении конкретных поведенческих задач)103.
Г. Гиперфронтальность
Разная структура нейронных связей обусловливает специфичность импульсной активности нейронов. Исследования обезьян, проводимые Фастером и коллегами, позволили выявить различия свойств активности нейронов, локализующихся в VI и нижневисочных областях. Импульсная активность нейронов зрительной области носит фазический характер, тогда как клетки височной коры характеризуются тонической активностью (рис. 12.7)104. Такой устойчивый характер импульсной активности ассоциируется с процессами рабочей памяти и проявляется как гиперфронтальность — свойство, отражающее повышенную активированность фронтальной коры в сравнении с остальными кортикальными структурами. Этот эффект был открыт в работах шведского ученого Дэвида Ингвара в 1970-х годах.
103 Даже если мы и признаем некоторую степень функциональной специализации в пределах префронтальной коры, все же угверждение о том, что любая ее область обеспечивает только одну функцию, не может считаться верным. Например, существуют доказательства вовлечения вентролатеральной префронтальной коры в подавление ответов, в переключение с одной задачи на другую. в процессы процедурной памяти и извлечение информации из эпизодической памяти.
104 Первое электрофизиологическое доказательство вовлечения префронтальной коры в обеспечение исполнительных функций было получено английским ученым Грэем Уортером в 1964 г. Он исследовал условное негативное отклонение, медленный негативный компонент, регистрирующийся с передних областей головы при подготовке к восприятию стимула или совершению движения.
276 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Д. Специфичность ЭЭГ
При рассмотрении ЭЭГ можно наблюдать разницу в активности между передними и задними отделами мозга (рис. 12.7). В частности, имеются в виду спектральные характеристики ЭЭГ. В состоянии покоя альфа-ритм регистрируется только в центрально-задних отведениях. В норме альфа-ритм не представлен в лобных областях. Ранние (до 200 мс) компоненты ВП, характеризующие зрительные процессы, обнаруживаются в затылочных и теменных отведениях, а поздние позитивные компоненты, ассоциирующиеся с операциями вовлечения и мониторинга действий, регистрируются во фронтальных и теменных отведениях.
IV. Операции вовлечения и отвлечения
А. РЗЬ как компонента, отражающая операции вовлечения
Компонента РЗЬ, или, точнее говоря, РЗЬ-компоненты105, подробно рассматривались в предыдущей главе, посвященной описанию компонент ВП, характеризующих работу системы внимания. Несмотря на то что наибольшая плотность генераторов РЗЬ локализуется в области теменной коры, что и позволяет связывать данную компоненту с системой внимания, в генерацию этой волны вовлечена нейронная сеть, объединяющая фронтальные, височные, теменные области коры и базальные ганглии (таламокортикальными взаимодействиями).
Б. Сенсорное сравнение
В нашей повседневной жизни мы всегда стараемся «смотреть вперед»106 и заняты подготовкой к реализации предстоящих действий. Однако контекст окружающей действительности быстро меняется, что иногда требует от нас подавления ранее планируемого действия. Подавление подготовленных действий обеспечивается в результате сложных мозговых процессов, одним из элементов которых является активность латеральной префронтальной коры. Как уже было сказано выше, префронтальная кора получает информацию от сенсорных систем (зрительной, слуховой и соматосенсорной), на основе которой и принимает решение в пользу действия (GO) или подавления (NOGO). Далее возможна следующая спекуляция: для осуществления выбора действия мозгу в первую очередь необходимо выполнить сравнение текущей сенсорной ситуации с сенсорной моделью и выявить в случае наличия рассогласование. Операции сравнения сенсорных сигналов, как нам уже известно из предыдущей части книги, протекают на уровне сенсорных систем, располагающихся в задней части мозга. Продолжая рассуждения, логично предположить, что результаты таких операций сравнений передаются в префронтальную кору и активируют мозговые процессы, ответственные за подавление подготовленных действий.
105 Следует упомянуть, что РЗЬ в действительности является семейством компонент, генерируемых в ответ на стимулы-мишени, характеризующиеся разным распределением (от теменных до премоторных зон).
106 Как писал великий русский поэт А.С. Пушкин, «Сердце будущим живет...»
Часть 2. Вызванные потенциалы
277
Примечательно, что данные предположения получают экспериментальное подтверждение. Так, на рис. 12.8 продемонстрирован результат анализа ВП с применением метода независимых компонент, полученных в исследовании, проводимом в рамках GO/NOGO-парадигмы. Как видно из рис. 12.8, NOGO- и GO-стимулы вызывают генерацию разных независимых компонент ВП, генерируемых в височной и премоторной коре. На уровне височной коры NOGO-стимул характеризуется дополнительной компонентой (в сравнении с GO-пробой). Однако в области левой премоторной коры предъявление этих стимулов приводит к прямо противоположным реакциям. Разный характер временной динамики компонент свидетельствует о наличии функциональных отличий между ними. Вполне вероятно, что первая компонента, характеризующаяся позитивностью в височной области, может отражать операцию сравнения сенсорных сигналов — детекцию изменения (рис. 10.16).
В. Подавление моторной активности
Можно также предположить, что вторая компонента (рис. 12.8) связана с моторным подавлением. Если сравнить эту компоненту с паттерном вызванной активности таламуса (рис. 12.3), то обнаруживается удивительное сходство между скальповым ВП и локальным потенциалом на таламическом уровне. Факт этого сходства, а также локализация генераторов компоненты является подтверждением наших предположений107. Следует оговориться, что компонента, связанная с моторным подавлением во фронтальных отведениях, характеризуется негативными значениями амплитуды.
Нейронные механизмы подавления моторного ответа, как было показано выше, вовлекают процессы, происходящие в базальных ганглиях. Особенностью организации связей коры мозга является наличие длинных возбуждающих внутрикортикальных связей и отсутствие таковых для тормозных влияний, что обусловливает исключительно локальный характер процессов торможения, инициируемых структурами коры. Это означает, что сама кора не в состоянии полностью подавить программу действий. Наиболее вероятный механизм подавления подготовленных действий подразумевает вовлечение таламокортикальных взаимодействий, контролируемых базальными ганглиями посредством непрямых проводящих путей, с важным звеном системы, локализующимся в премоторной коре108.
Г. Подавление действия
Одним из примечательных компонент ВП, получаемых в результате анализа независимых компонент ВП в рамках парадигмы GO/NOGO, является третья NOGO-компонента (рис. 12.8). Пространственное распределение, временная динамика данной компоненты, а также результат прямого сравнения с ВП
107 Ранние участки компоненты, ассоциированной с начальными этапами подавления моторной активности, с возрастом увеличиваются в латентности.
108 Я вспоминаю пациента, страдавшего паркинсонизмом и проходившего лечение в нашей клинике, который испытывал трудности при пересечении улицы по регулируемому светофором пешеходному переходу. Он говорил: «Я знаю, что я должен остановиться на красный свет (процессы принятия решений не были нарушены), но попросту не могу перестать идти» (следовательно, было нарушено подавление действий, которое отчасти осуществляется с помощью системы связей вида: базальные ганглии — таламус — кора).
278
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
при GO-стимулах существенно отличаются от всех остальных компонент. Кроме того, NOGO-компонента является симметричной, характеризуется единственной выраженной пиковой латентностью в 340 мс, а ее генераторы широко распределены на уровне премоторной и моторной коры. Важно, что этот эффект
Моторное подавление
Рисунок 12.8. Независимые компоненты ВП, связанных с операциями сенсорного сравнения и моторного подавления
Метод независимых компонент применялся при анализе ВП, зарегистрированных в ответ на GO- и NOGO-стимулы, предъявлявшиеся в рамках двухстимульного GO/NOGO-теста. ВП-данные здоровых испытуемых возрастом от 1 до 89 лет были взяты из нормативной HBI базы данных. А. Компонента сравнения. Б. Компонента моторного подавления. Вертикально уложенные тонкие горизонтальные окрашенные полосы (цветом обозначается амплитуда), каждая из которых представляет изменение во времени амплитуды соответствующей компоненты ВП отдельного испытуемого. Представлены GO- и NOGO-условия. Топограммы соответствующих компонент, локализация генераторов компонент, вычисленная с помощью метода томографии низкого разрешения (sLORETA), а также временная динамика ВП представлены в виде отдельных графиков для GO- и NOGO-сти мулов.
наблюдается только в случае предъявления NOGO-стимулов. Также существует зависимость показателей компоненты от возраста: с возрастом отмечается снижение амплитуды и увеличение латентности. При этом амплитуда NOGO-компоненты негативна во фронтальных областях. Для различения асимметричной компоненты, связанной с моторным подавлением, мы назвали данную компоненту «компонентой подавления действия». Ситуация такова, что нам не известно, почему мозг генерирует эту компоненту после продукции специфической компоненты подавления моторной реакции. Мы не знаем, генерируется ли данная компонента посредством ганглионарноталамокортикальных взаимодействий или за счет активности самой коры. Но то, что нам действительно понятно, — что в терминах амплитуды компонента подавления действий в три раза превосходит
Часть 2. Вызванные потенциалы
279
А Б
Рисунок 12.9. Отражение операций сравнения и подавления действий в активности, зарегистрированной при помощи имплантированных электродов
Элетроды были имплантированы пациенту по терапевтическим показаниям (для проведения лечения и диагностики). Пациент выполнял двухстимульный GO/NOGO-тест. Электроды были локализованы в ассоциативной слуховой коре (А) и дорсолатеральной префронтальной коре (Б). Регистрировались ВП для GO, NOGO и GO/NOGO (разность) условий. Внизу рисунка проиллюстрированы области имплантации электродов.
компоненту моторного подавления и, следовательно, вполне вероятно, обладает лучшей диагностической ценностью (в сравнении с компонентой моторного подавления).
Д. Регистрация активности с имплантированных электродов
Проводя лечение различных больных с помощью имплантированных электродов, мы могли регистрировать внутримозговые слуховые ВП в условиях двухстимульной исследовательской GO/NOGO-парадигмы. Электроды имплантировались в разные корковые и подкорковые структуры исходя из диагностических и терапевтических целей. Пациентам, страдающим паркинсонизмом, электроды имплантировались в область таламуса и базальных ганглиев. Другие области-мишени применялись при лечении эпилепсии — медиальная (включая гиппокамп) и латеральная височная кора. При обсессивно-компульсивном расстройстве электроды локализовались в области передней поясной извилины и смежных областях префронтальной коры.
280
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
На рис. 12.9 представлены ВП, зарегистрированные с двух электродов, располагавшихся в височной доле и префронтальной коре. Пациенты выполняли тестовое задание — модифицированную версию двухстимульного теста GO/NOGO-парадигмы. Как видно из рис. 12.9, предъявление NOGO-стимула на уровне височной коры характеризовалось негативно-позитивной компонентой с соответствующими пиковыми латентностями 100 и 200 мс. В префронтальной коре наблюдался сопоставимый паттерн активности в ответ на NOGO стимул, но с большими значениями ЛП, а именно на 150 мс позже своего височного аналога. Следует также отметить и наличие ранней компоненты с латентным периодом 70 мс, зарегистрированного с височного электрода. Впрочем, данная ранняя компонента была одинакова для GO- и NOGO-стимулов и, по всей видимости, отражает ранние этапы сенсорной обработки стимулов. ВП префронтальных отведений, напротив, характеризовались только поздними компонентами и только при предъявлении NOGO-стимулов1 °9. Если проводить сравнение внутримозговых и скальповых ВП, то
можно говорить о том, что в конкретном рассматриваемом случае ВП, зарегистрированные с височной коры, отражают операцию сравнения. В то же время локальный потенциал префронтальной коры ассоциируется с операциями подавления действий.
В изучении процессов мозга человека, связанных с подавлением действий, кроме GO/NOGO-парадигмы также применяется так называемая задача стоп-сигнала. Аналогично GO/NOGO-парадигме в задачах со стоп-сигналами выполнение текущей деятельности должно быть прекращено по предъявлении соответствующего сигнального стимула. Полученные за последние время экспериментальные данные (в рамках обоих исследовательских подходов) свидетельствуют в пользу того, что ключевая роль в подавлении действий принад лежит 109 Здесь я хотел бы напомнить, что внутримозговые ВП (называемые также локальными потенциалами поля) являются весьма локальными и могут существенно отличаться один от другого даже при дистанции между ними в несколько миллиметров.
в 260 мс 420 мс
Рисунок 12.10. Волна N200, отражающая процесс моторного подавления Представлены усредненные по группе испытуемых ВП (испытуемые от 7 до 89 лет), рассчитанные на основе ВП-данных из нормативной HBI базы данных. А. ВП для GO- и NOGO-стимулов двухстимульного GO/NOGO-теста в Fz-отведении. Б. Разностный потенциал (NOGO-GO). В. Топографии разностных ВП, вычисленных для двух событий — операции сравнения и подавления.
Часть 2. Вызванные потенциалы
281
вентролатеральной префронтальной коре"0. В онтогенезе данная область развивается одной из последних и именно поэтому неполное развитие этой части префронтальной коры может рассматриваться как возможная причина синдрома импульсивности, по крайней мере у некоторых подгрупп детей с СНВГ. Так, в работах Адама Арона1" из Кембриджского университета (Англия) была показана корреляция выраженности повреждения правой вентролатеральной префронтальной коры (но никакой другой области) и показателей процессов подавления действий (время реакции при предъявлении стоп-сигналов): при более обширных повреждениях наблюдалось большее замедление времени реакции.
Е. N200 как компонента моторного подавления
Исследования нейронных механизмов подавления моторной деятельности берут начало в 70-х годах прошлого столетия. Наиболее важные открытия были сделаны японскими исследователями Гемба и Сасаки. Ими были выявлены специфические механизмы моторного подавления на уровне моторной и премоторной коры. Так, в исследованиях на обезьянах было показано, что возбуждение нейронов центральной извилины (principle sulcus) на фоне регулярных ответов может приводить к снижению активности премоторной и моторной коры и даже задержке или полному подавлению ответа. При этом прямая электрическая стимуляция нейронов этой области вызывает подавление ответа на GO-стимул. В исследованиях человека нейронный механизм процесса подавления изучался с помощью метода ВП (скальповые). Во множестве таких исследований регистрировалась негативная компонента N200, распределенная в лобных областях и достигающая максимальных значений спустя 200—260 мс после предъявления NOGO-стимула. В сравнении с GO-стимулом N200 при NOGO характеризуется большей амплитудой и считается индикатором процессов подавления в парадигме GO/NOGO. Сопоставимая М2-компонента наблюдается и при предъявлении стоп-сигналов. Однако следует отметить, что во всех классических работах ^-компонента получается в результате вычисления разностной волны между ВП для GO- и NOGO-стимулов.
Ниже мы собираемся показать, что классическая ^-компонента также наблюдается и при анализе полученных нами данных в виде фронтально распределенной негативности, регистрируемой в ответ на NOGO-стимул. На рис. 12.10 представлен ВП, усредненный по той же группе пациентов (исследовавшийся в рамках GO/NOGO-парадигмы), что и ВП, иллюстрированный на рис. 12.8 и 12.11. Как видим, NOGO-стимул характеризуется выраженным негативным отклонением с ЛП 260 мс, который может считаться аналогом классической N2-компоненты. Негативная компонента сменяется позитивной, также характеризующейся фронтальной топографией. Если провести сравнение ВП на рис. 12.10 и трех независимых компонент, полученных при анализе ВП методом независимых компонент (рис. 12.8 и 12.11), то можно заметить одну особенность: негативная компонента традиционного ВП в действительности является суммой
110 Вентролатеральная префронтальная кора мозга человека, иногда называемая нижней префронтальной корой, объединяет 44-е, 45-е и 47/2 поля Бродмана.
111 Для подробного ознакомления см. Aron et al., 2004.
282
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Подавление действия
А
Рисунок 12.11. Независимые компоненты ВП, связанные с подавлением действия
Метод независимых компонент применялся при анализе ВП, зарегистрированных в ответ на GO- и NOGO-стимулы, предъявлявшихся в рамках двухстимульного GO/ NOGO-теста. ВП-данные здоровых испытуемых возрастом от 7 до 89 лет были взяты из нормативной HBI базы данных. А. Топография и локализация генераторов компоненты подавления, вычисленная с помощью метода томографии низкого разрешения (sUORETA). Вертикально уложенные тонкие горизонтальные окрашенные полосы (цветом обозначается амплитуда), каждая из которых представляет изменение во времени амплитуды соответствующей компоненты индивидуального ВП. Б. Временная динамика ВП представлена в правой нижней части рисунка.
этих трех разных независимых компонент112.
Выявленная взаимосвязь компонент, получаемых разными методами, приводит по крайней мере к двум логичным заключениям. Во-первых, это говорит о преимуществе метода независимых компонент в дискриминации отдельных психологических операций над традиционными методами анализа ВП — построением разностных ВП: метод независимых компонент позволяет разбить ВП на статистически независимые компоненты, каждая из которых обладает специфическим пространственно-
временным паттерном и функциональным смыслом. Во-вторых, выявляемые независимые компоненты являются лучшим биологическим маркером изучаемых процессов по сравнению с традиционными подходами анализа ВП. Это, возможно, поможет эффективнее проводить диагностику различных мозговых дисфункций, ассоциируемых с нарушениями таких операций, как сравнение,
моторное подавление и подавление действий.
V. Операции мониторинга
А. Р400 как компонента мониторинга в GO/NOGO-парадигме
Набор компонент, связанных с NOGO-стимулом, не ограничивается только упомянутыми выше, отражающими активность височной (сенсорные операции сравнения), моторно-премоторной (операции подавления) коры. В действительности, как показывает анализ ВП с применением томографии низкого разрешения (sLORETA), наибольшие по амплитуде NOGO-компоненты генерируются в области медиальной префронтальной коры и поясной извилины
112 Следует отметить, что только компоненты моторного подавления и подавления действий ассоциируются с негативностью, наиболее выраженной в Fz. В то же время негативные компоненты, отражающие операции сравнения, наиболее представлены в Cz.
Часть 2. Вызванные потенциалы 283
(рис. 12.12). Это симметричные позитивные компоненты, локализующиеся центрально, а именно в области Cz- и Fz-отведений. При этом известно, что латентность этих компонент существенно меняется с возрастом — от 370 мс в среднем возрасте до 420 мс в раннем и 460 в пожилом возрасте.
Б. Функция передней поясной извилины
Компонента ВП, отражающая операции мониторинга, генерируется в передней поясной извилине. И сейчас самый подходящий момент для того, чтобы детально описать анатомию и физиологию передней поясной извилины (ППИ). ППИ не является гомогенной структурой. Она состоит из нескольких морфо
логически разных полей, имеющих различные «входные» и «выходные связи». Прежде всего следует отметить наличие моторных областей (рис. 12.13), полу
чающих входы от первичной моторной коры, премоторной и добавочной моторных областей. Как известно, эти области ответственны за хранение точных образов планируемых действий. Моторная область поясной извилины также принимает участие в инициации новых действий. Далее ППИ в своей вентральной части получает входы преимущественно от аффективной мозговой систе-
мы (лимбической) либо с помощью прямых путей (например, от миндалины), либо опосредованно (через переднее ядро таламуса). Благодаря такой организации связей вентральная часть ППИ называется также лимбической. И наконец, дорсальная часть передней поясной извилины обладает обширными реципрокными связями с латеральной, передней и медиальной областями префронтальной коры — областями, как это считается, вовлекаемыми в обеспечение когнитивных функций. Именно поэтому дорсальная часть ППИ носит название когнитивной.
Существует надежное доказательство в пользу правомерности такой
Операция
Рисунок 12.12. Независимые компоненты ВП, связанные с операцией мониторинга
Метод независимых компонент применялся при анализе ВП, зарегистрированных в ответ на GO- и NOGO-стимулы, предъявлявшиеся в рамках двухстимульного GO/NOGO-теста. ВП-данные здоровых испытуемых возрастом от 7 до 89 лет были взяты из нормативной HBI базы данных. А. Топография и локализация генераторов компонент, вычисленная с помощью метода томографии низкого разрешения (sLORETA). Вертикально уложенные тонкие горизонтальные окрашенные полосы (цветом обозначается амплитуда), каждая из которых представляет изменение во времени амплитуды соответствующей компоненты ВП отдельного испытуемого (для GO- и NOGO-стимулов). Б. Временная динамика ВП представлена в правой нижней части рисунка. Чтобы получить информацию о компоненте ВП, регистрируемой на поверхности скальпа, необходимо вычислить ее топографию и временную динамику, что в результате позволит выявить позитивную компоненту в области Cz- и Fz-отведений.
284
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
эмоциональная когнитивная
Рисунок 12.13. Отделы и связи передней поясной извилины
Передняя поясная извилина реципрокно связана практически со всеми областями префронтальной коры. Передняя поясная извилина может быть подразделена на три части (обозначены цветом) — эмоциональную, когнитивную и моторную.
анатомически обусловленной дифференциации передней поясной извилины. Данные многочисленных ПЭТ и фМРТ-исследований свидетельствуют об активациях дорсальной ППИ в условиях когнитивных тестовых заданий. Аналогично аффективные состояния, определяемые эмоциями и мотивациями, характеризуются активациями вентральной ППИ. Например, если испытуемые представляют ситуации с разным эмоциональным контекстом (например, злость или печаль), наблюдаются активации в вентральной части ППИ113. С другой стороны, дорсальная ППИ обнаруживается активируемой при совершении ошибок, допускаемых испытуемыми при выполнении классического теста Струпа.
В. Акинетический мутизм
Результаты нейрохирургических стимуляций вентральной части передней поясной извилины мозга пациентов могут приводить к проявлению эмоций страха или удовольствия, в зависимости от места воздействия. Напротив, такое же воздействие на дорсальную ППИ вызывает чувство ожидания движения. Пациенты с повреждениями в области медиальной фронтальной коры, затрагивающими ППИ, характеризуются дефицитом спонтанной инициации движений и речи, а также неспособностью подавления стереотипных моторных программ, запускаемых извне114. Дамазио и Ван Хойсен наблюдали нескольких пациентов, у кото
113 Когда, например, пациентам с навязчивыми состояниями, фобиями или посттравматическим стрессом предъявляются стимулы, являющиеся триггерами симптомов, активируется именно вентральная часть ППИ.
114 Акинетический мутизм, возникающий в результате билатерального повреждения медиальной коры и поясной извилины, является экстремальным случаем такой симптоматики. Акинетический мутизм — это клиническое состояние, при котором пациенты проявляют склонность к неспособности как говорить (мутизм), так и двигаться (акинезия). Унилатеральные повреждения приводят к более слабой форме мутизма: спонтанная речь скудна и, даже после восстановления, монотонна.
Часть 2, Вызванные потенциалы
285
рых в результате инсульта значительно повреждалась область передней поясной извилины. Сразу после инсульта такие пациенты лежали на кровати, очень мало разговаривали или двигались. Исследователи так описывали одну из пациенток с левосторонним повреждением передней поясной извилины спустя месяц после инсульта: «Пациентка заметно восстановилась. Она достаточно хорошо помнила острый период болезни и могла весьма точно описать свое состояние в тот момент. В частности, она отрицала наличие негативных эмоций или страдания в тот период болезни, когда была практически неспособна разговаривать. По ее словам, она ничего не говорила лишь потому, что ей нечего было сказать. Ее разум был пуст. Она, несомненно, могла следовать за нитью наших разговоров даже на ранних стадиях восстановления после инсульта, но не желала отвечать на наши вопросы»115. В недавнем исследовании Когена и др. исследовались 18 пациентов с некупируемой болью, для снятия которой применялась операция по небольшому лизису ткани передней поясной извилины (около 5 мм в диаметре). По прошествии не менее года с момента операции оценивались поведенческие данные этих пациентов и контрольной группы больных с хронической болью, но не подвергавшихся хирургическому вмешательству. Сравнение двух групп пациентов свидетельствовало в пользу того, что оперированные больные освободились от мучивших их хронических болей. Они сообщали, что хотя боль все еще присутствовала, но более не беспокоила столь сильно, как до операции116.
Г. Концепция мониторинга
Какова же функциональная роль когнитивной части передней поясной извилины? Все имеющиеся на сегодняшний день исследования говорят о том, что дорсальная часть ППИ ответственна за обеспечение функции мониторинга. Дело в том, что гибкая адаптация поведения человека и животных требует постоянного сравнения и оценки текущих действий и их результатов. Способность к такому мониторингу и сравнению результатов текущих действий с внутренне формируемыми целями и стандартами является критичной для оптимизации процессов принятия решения. Именно эта способность и носит название мониторинга действий, функции, выполняемой дорсальной частью передней поясной извилины.
В течение последних нескольких лет концепция мониторинга действий стала весьма влиятельной в когнитивной науке. Следует отличать процессы мониторинга действий и функцию контроля, осуществляемую вниманием. Этот вид контроля реализуется за счет нисходящего и ограниченного в ресурсах когнитивного механизма, модулирующего процессы обработки сенсорной информации. В то же время мониторинг действий подразумевает когнитивный механизм оценки качества исполнительного контроля, который активирует исполнительные системы в
115 Я вспоминаю наши попытки зарегистрировать ВП у пациентки с билатеральным повреждением передней поясной извилины. Она отвечала на вопросы и, казалось, полностью осознавала, что находится в больнице и почему, но не могла выполнить ни одного тестового задания. Она просто теряла всякий интерес к происходящему и чувствовала сонливость (засыпала).
116 Прооперированные пациенты характеризовались сниженным уровнем спонтанного поведения. Они произносили меньше фраз в течение опроса по сравнению с контрольной группой пациентов. В письменной версии интервью такие пациенты писали сравнительно короткими предложениями. Когда их просили создавать изображения предметов из разных элементов (известная игра для детей «Тинкертой»), пациенты создавали меньшее их число, и они отличались простотой конструкции. Таким образом, результатом небольшого хирургического повреждения ППИ явилось уменьшение спонтанности поведения, проявляющееся в разных его элемен-iax (речь, письмо, конструирование чего-либо и тщ.).
286 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
случае рассогласования между наличествующим результатом действия и ожидаемым.
Для того чтобы регистрируемая компонента ВП могла рассматриваться как показатель процессов мониторинга действий, необходимо выполнение по крайней мере трех условий. Во-первых, латентность компоненты должна быть достаточно большой, т.е. должна регистрироваться после нейронного ответа, связанного с инициацией действия, поскольку необходимо сравнить активацию исполняемого действия с планируемым. Во-вторых, компонента должна генерироваться в области, на уровне которой сходится информация об исполняемом и планируемом действии, для осуществления операции сравнения этих сигналов. И наконец, компонента должна наблюдаться в условиях конфликта, когда необходимо воздержаться от реализации планируемого действия в силу несоответствия текущей ситуации с ожидаемой или рассогласования результата выполненного действия и планируемого. Таким образом, примером подобной компоненты может являться N400, компонента ВП, регистрирующаяся в ответ на NOGO-стимулы и генерируемая передней поясной извилиной. N400 вполне может называться компонентой мониторинга, поскольку удовлетворяет всем перечисленным выше требованиям.
Д. Негативность, связанная с ошибкой
Компонента мониторинга также может быть зарегистрирована в особых условиях трудного и длительного тестового задания, при выполнении которого в некоторых пробах испытуемый непроизвольно совершает ошибки117. Совершение ошибки сопровождается генерацией компоненты, называемой негативностью, связанной с ошибкой (НСО), за которой регистрируется позитивная компонента. НСО наблюдается сразу после реализации некорректного ответа, примерно через 100 мс, и характеризуется центрально-фронтальной топографией (иногда эта компонента называется негативностью ошибки). Моделирование дипольного источника НСО (проводимое во многих исследованиях) указывает на его расположение вблизи ППИ. В подтверждение исследований по моделированию диполей следует упомянуть и данные фМРТ-исследований, свидетельствующих о повышенной активности дорсальной части ППИ в ошибочных пробах по сравнению с корректными.
Компоненты ВП, связанные с процессами мониторинга ошибок, также могут наблюдаться и в исследованиях, проводимых в рамках двухстимульной GO/ NOGO-парадигмы. Для выделения этих компонент нужно провести сравне-
117 В действительности исследования на людях повторяют экспериментальные работы, проводимые на животных. В 1979 г. Ники и Ватанабе в исследовании на обезьянах регистрировали изменения в активности одиночных нейронов передней поясной коры при совершении животными ошибок. Развитие этого направления исследований позволило выявить, что помимо нейронов ППИ, активирующихся в моменты времени, когда обезьяна осознавала совершение ошибки, некоторые нейроны избирательно реагировали на уменьшение подкрепления в тех ситуациях, когда требовалось изменение паттерна поведения для оптимизации ожидания вознаграждения. Причем, когда активность ППИ временно подавлялась с помощью инъекций агонистов гамма-аминомасляной кислоты (Shima and Tanji, 1998), нейроны обезьян переставали демонстрировать чувствительность к уменьшению вознаграждения. Эти исследования явились элегантным доказательством вовлечения передней поясной извилины в обеспечение мониторинга выполнения и адаптации деятельности, осуществляемого по типу управления поведением для оптимизации вознаграждения.
Часть 2. Вызванные потенциалы
287
Рисунок 12.14. Негативность, связанная с ошибкой
Метод независимых компонент применялся при анализе ВП, зарегистрированных у более чем 200 испытуемых (из нормативной HBI базы данных). ВП регистрировались при корректных и ошибочных ответах математического тестового задания. На рисунке представлены три наиболее крупные независимые компоненты (сверху вниз). Слева направо — топограммы, вертикально уложенные тонкие горизонтальные окрашенные полосы, обозначающие изменение амплитуды компонент во времени и локализация генераторов компонент, вычисленная с помощью метода томографии низкого разрешения (sLORETA). Под каждым рядом изображений располагаются наложенные графики ВП для корректных (толстая линия) и ошибочных (тонкая линия) проб, а также их разностные волны.
ние в двух условиях: при некорректном нажатии в NOGO-пробах (совершение ошибки) и корректном в GO-пробах. Пример разложения ВП на компоненты при таком сравнении представлен на рис. 12.14 (данные взяты из HBI базы данных). В представленном случае испытуемые выполняли математическое тестовое задание. Задание было достаточно сложным, что обусловливало совершение существенного количества некорректных ответов. Для каждого испытуемого
288 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
высчитывался ВП для корректных нажатий в GO-пробах118 и некорректных ответов в NOGO-пробах119. Нажатие кнопки испытуемым являлось синхронизирующим событием при построении ВП. Отдельно высчитывалась разностная компонента ВП (корректный ВП — некорректный ВП). Как видно из рис. 12.14, НСО в действительности состоит из двух независимых компонент, одна из которых локализована в дорсальной (когнитивной) части ППИ, а другая расположена несколько кзади от нее (в моторной части ППИ). В дополнение к упомянутым компонентам, генерируемым сразу после некорректного моторного ответа, НСО содержит также компоненту, возникающую до ответа, локализующуюся в левой сенсомоторной области коры120.
Говоря об использовании НСО в диагностических целях в клинике, следует отметить как минимум два ограничения. Во-первых, ошибка является очень субъективным событием и зависит от мотивации испытуемого, состояния внимания и возможностей сенсорной системы. В том числе и потому, что количество совершаемых ошибок в одном и том же тесте заметно варьирует от испытуемого к испытуемому. Следовательно, количество «ошибочных» усредняемых проб не является стандартным, что существенно влияет на индивидуальные параметры НСО. Во-вторых, для накопления достаточного количества проб с некорректными ответами, для надежного измерения НСО нужно использовать весьма сложные тестовые задания. Однако не все испытуемые (например, дети или пациенты, страдающие паркинсонизмом) способны выполнять сложные задания.
VI. Рабочая память
А. Активные манипуляции со следом памяти
Одним из удивительных свойств нейронов префронтальной коры является их способность к сохранению частоты разрядов нейронов на протяжении длительного интервала времени, включающего и происходящее событие, и длительное время после его окончания. Это может отражать способность префронтальной коры участвовать в поддержании репрезентаций стимулов и действий достаточно долгое время, позволяя субъекту инициировать поведение, направленное на достижение долгосрочных целей. Данная функция префронтальной коры отражена в концепции рабочей памяти. В соответствии с данной концепцией под рабочей памятью понимают психологические процессы, позволяющие удерживать как текущие планы действий, так и сенсорную информацию, необходимую для их реализации. Представление о рабочей памяти также подразумевает активные манипуляции с временно сохраненной информацией, что является центральным аспектом когнитивных действий, например таких, как язык, планирование, принятие решений и т.д.
118 Следует уточнить, что в этом типе проб второй стимул пробы (число) равнялся сумме двух чисел, предъявляемых в качестве первого стимула, а испытуемые корректно нажимали кнопку.
119 В этих пробах второй стимул не равнялся сумме чисел, предъявляемых в качестве первого стимула, а испытуемые совершали некорректное нажатие.
120 Локализация активности в сенсомоторной коре левого полушария соответствует тому факту, что большинство испытуемых нажимали на кнопку мыши пальцами правой руки.
Часть 2. Вызванные потенциалы
289
Восходящие связи
Дорсаль ная
Брока
Префронтальная кора
Нисходящие связи
Теменная
Вернике Теменно-височная ________ кора
Рисунок 12.15. Три системы рабочей памяти
Справа — система вербального повторения, которая является наиболее эффективной и преобладающей разновидность рабочей памяти человека. Осуществляется этот вид рабочей пямяти преимущественно премоторной (зона Брока) и теменной (зона Вернике) областями левого полушария. Посередине — рабочая память зрительных объектов (т.е. форма, цвет). Слева — пространственно-зрительная рабочая память (положение в пространстве).
Б. Анатомия реципрокных проводящий путей
С точки зрения нейрофизиологии процессы рабочей памяти реализуются за счет множества реципрокных связей внутри коры. Эти реципрокные связи являются основой реверберации нейронной активности, сохраняющейся после окончания действия стимула или после совершения действия. В состав реципрокных связей входят корковые пути прямой и обратной связи, а также петли обратной связи в составе базально-таламокортикальных путей. Несмотря на то что общим свойством префронтальной коры является способность к удержанию информации на протяжении длительного времени, типы этой информации и, следовательно, нейронные сети подразделяются на три модальности: зрительно-пространственную, зрительно-образную и цветовую, а также вербальную. Такая дифференциация получила отражение в популярной модели рабочей памяти, предложенной Аланом Бадделеем в 1974 году и дополненной в 2003-м121.
121 Концепция рабочей памяти была представлена в 1974 году Бадделеем и Хитчем и создавалась как альтернатива концепции краткосрочной памяти, разработанной Хеббом в 1949 г. Главное отличие этих концепций заключается в том, что рабочая память подразумевает не только временное хранение информации, но и активные манипуляции с ней на протяжении длительных интервалов времени. По Бадделею (2003), теоретическая концепция рабочей памяти предполагает существование системы с ограниченными ресурсами, которая временно удерживает и сохраняет информацию, что обеспечивает поддержку процессов мышления. Эта система выполняет функцию интерфейса между восприятием, долгосрочной памятью и деятельностью.
290 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
В. Три системы рабочей памяти
В соответствии с моделью нейронная сеть, ответственная за поддержание модально специфических следов рабочей памяти, состоит из реципрокно связанных префронтальных и ассоциативных височно-теменных областей коры (рис. 12.15). Такая сеть обеспечивает удержание планов действий, формируемых на основе синтеза сенсорной информации, доставляемой в префронтальную кору через восходящие проводящие пути. Также на уровне этих нейронных сетей посредством нисходящих влияний происходит интенсификация процессов обработки специфических аспектов сенсорной информации, требующихся для выбора действия.
Г. Условно негативное отклонение (УНО) как коррелят рабочей памяти
Рабочая память является медленным процессом, который может длиться от нескольких секунд до нескольких минут. Следовательно, оправданно ожидать, что эти процессы могут отражаться медленными компонентами ВП. Такие медленные компоненты обнаруживаются в условиях двухстимульной задачи, когда первый из двух следующих друг за другом стимулов является сигнальным, а второй — императивным. И именно в промежутке между этими стимулами и регистрируется медленная компонента, о которой идет речь (рис. 12.16). В нашем совместном с Ристо Наатаненом (Университет Хельсинки) исследовании мы регистрировали ЭЭГ (в частотном диапазоне от 0 до 70 Гц) во время выполнения здоровыми добровольцами зрительного и слухового варианта тестового задания в парадигме GO/NOGO. В соответствии с данной исследовательской парадигмой испытуемые инструктировались при предъявлении определенной пары стимулов нажимать кнопку и воздерживаться от нажатия в случае других сочетаний стимулов.
На рис. 12.16 представлены ВП (построенные на основе записи ЭЭГ в частотном диапазоне начиная от 0 Гц)122, характеризующие те пробы, в которых испытуемые находились в состоянии ожидания следующего стимула, когда первый (сигнальный) стимул пробы сигнализировал о том, что моторный ответ не требуется. В зрительной модальности наибольшая амплитуда негативных отклонений была обнаружена в задних височных областях левого полушария. В слуховой модальности аналогичный эффект наблюдался в среднезадней височной области. Эти УНО были связаны с ожиданием сенсорного события. Однако пробы, в которых требовалась моторная реакция, вне зависимости от модальности теста характеризовались максимальным в центральных областях УНО. Следовательно, можно предполагать, что формирование планов поведения происходит на уровне префронтальной коры. Высшие центры (префронтальная кора) через фронтально-височные связи контролируют иерархически низшие области (височная кора) путем деполяризации дистальных частей дендритов пирамидальных нейронов и за счет этого подготавливают их к быстрому ответу на ожидаемый побуждающий стимул123.
123 Данные медленные изменения ВП были названы Грэем Уолтером условным негативным отклонением.
123 Как нами было отмечено выше, в состав проводящих путей подготовительной активности входят базально-таламокортикальные связи. Однако одних ВП-данных недостаточно для того, чтобы дифференцировать вклад этих подкорковых связей в эффект работы кортикальных систем проводящих путей (префронтально-височных, префронтально-моторных).
Часть 2. Вызванные потенциалы
291
Внимание
Подготовка моторной реакции
Рисунок 12.16. Условно негативное отклонение, связанное с подготовкой к восприятию стимула или подготовкой действия
11|юдставлены усредненные ВП по группе здоровых испытуемых (из 10 человек), которые выполняли двухстимуль-ныи GO/NOGO-тест отдельно в зрительной и слуховой модальности. В исследовании использовались 4 комбинации нар из двух видов стимулов (GO и NOGO), а от испытуемого требовалось нажимать кнопку при предъявлении пары GO-стимулов. В зрительной модальности в качестве GO-стимула использовалась цифра 9, a NOGO — цифра 6. В слуховой модальности высокочастотный тон (1300 Гц) использовался как GO-стимул, а низкочастотный тон (1000 Гц) мнпялся NOGO-стимулом. На графиках ВП черными стрелками (1 и 2) обозначены моменты ожидания появления стимулов. Стрелка под номером 3 обозначает период, во время которого подготавливается действие. Следует обратить внимание на то. что ожидание появления стимула характеризуется негативностью в областях, соответствующих мо-дал|,нос1и сжмуляции. Подготовка к моторному ответу характеризуется негативностью, генерируемой в ценграль-..... областях коры — вблизи прецентральной извилины.
VII. Дофамин как медиатор исполнительных систем
А. Корковое и подкорковое распределение
Основным медиатором исполнительной системы является дофамин. Он продуцируется нейронами коры (префронтальная кора), а также подкорковыми структурами, входящими в состав исполнительной системы (например, стриатум). Распределение рецепторов к дофамину (ДА) на \ ровне префронтальной коры и стриатума показано на рис. 12.17. Наибо-к г ионн ую дофаминергическую иннервацию получает передняя поясная и пт lima
292
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Б. Дофаминергические системы
Сложность организации дофаминергической системы во многом обу-
словлена гетерогенностью исполнительной системы (рис. 12.17). Большинство дофаминсодержащих нейронов развиваются из одного типа
Рисунок 12.17. Распределение дофаминовых рецепторов в мозгу
Черная субстанцая (SN) волокнами, входящими в состав дофаминергического проводящего пути, иннервирует стриатум (Striatum). Дофаминергический путь от вентральной области покрышки (VT) иннервирует области префронтальной коры (PFC), при этом наибольшее количество входов получают передние отделы поясной извилины (ACC).
эмбриональных клеток мезенцефально-диэнцефального стыка. Эти нейроны могут быть условно подразделены на два ядра с разными проекциями, которые и дают начало различным дофаминергическим системам мозга. Первая носит название нигростриар-ной дофаминергической системы, которая происходит из компактной области черной субстанции и проецируется в область стриатума. Другая система ассоциируется с вен-
тральным тегментумом и известна как мезокорти-кальная дофаминергическая система. Нейроны медиальной области вен-
трального тегментума посылают обширные проекции в область медиальной префронтальной коры и поясной извилины124. Оба типа нейронов получают обратную связь от областей, которые они иннервируют, таким образом формируя циклы реверберации соответствующих (префронтальной и стриарной) систем.
В. D1- и 02-рецепторы дофаминергических систем
Описанные выше дофаминергические системы различаются и по типу рецепторов, наиболее характерных для каждой из них. Так, префронтальная кора характеризуется обилием рецепторов D1-семейства (D1 и D5). На уровне стриатума превалируют рецепторы О2-семейства125 (D2, D3 и D4; рис. 12.18).
124 Часть нейронов вентрального тегментума проецируется в область прилежащего ядра, септума. миндалины, гиппокампа и пе-риренальной коры. Эта система получила название мезолимбической дофаминергической системы и, по всей видимости, принимает участие в процессах консолидации памяти.
125 Исследования процессов связывания лигандов с рецепторами показывают, что наиболее плотная концентрация лигандов, специфических к Dl-рецепторам ДА, наблюдается во внешних (слои I—Illa) и глубоких слоях коры (V и VI), а не в средних кортикальных слоях (Illb-IV). Экспериментальные данные свидетельствуют о большей концентрации D1-рецепторов по сравнению с 05-рецепторами на уровне коры мозга человека и приматов.
PFC
Рисунок 12.18. Циркуляция дофамина в нейронах и дофаминергические проводящие пути
А. Схематическое изображение дофаминергический путей: 1) берущих свое начало в черной субстанции (SN) и иннервирующих стриатум с 02-рецепторами к дофамину; 2) от вентральной области покрышки (VT) до префронтальной коры (PFC) с Dl-рецепторами. Б. Динамика дофамина в нейроне — появление значимого стимула вызывает высвобождение молекул дофамина из везикул, которые диффундируют в синаптическую щель и связываются с D-рецепторами постсинаптической терминали. «Вымывание» дофамина из синаптической щели осуществляется за счет механизма обратного захвата дофамина DAT-рецепторами пресинаптической терминали. В. В норме любой поведенчески значимый стимул вызывает формирование следа, время существования которого определяется временем присутствия дофамина в синаптической щели; при чрезмерной активности D-транспортеров, обеспечивающих слишком быстрый обратный захват дофамина, время удержания следа события сокращается; при недостатке D-рецепторов амплитуда следа уменьшается. Обозначения: DA — дофамин.
асть 2. Вызванные потенциалы___________________________________________________________293
294 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Г. Функции дофаминергических систем
Считается, что две дофаминергические системы (кортикальные и стриарные) выполняют разные функции. Нигростриарные дофаминергические проводящие пути устанавливают пороги активации нейронов стриатума и в соответствии с теорией, предложенной нами, вовлекаются в процессы селекции действий и рабочей памяти. Дофаминергическая система среднего мозга модулирует мембранный потенциал нейронов префронтальной коры и передней поясной извилины и вовлекается в поддержание ряда функций, обеспечиваемых этими областями коры. Так, если говорить о ППИ, то это функция мониторинга действий. Функция префронтальной коры основывается на одном из основных качеств нейронов этой области, которое заключается в удержании следа действия или стимула в рабочей памяти126. В действительности уже около 20 лет известно, что дофамин оказывает самое критическое действие на процессы рабочей памяти, основной вклад в обеспечение которых вносит префронтальная кора. Исследования обезьян и крыс показывают, что слишком слабая или, наоборот, сильная стимуляция дофаминергических рецепторов может приводить к нарушениям пространственной памяти127. Клеточной основой хранения информации в рабочей памяти являются нейроны префронтальной коры, устойчивая активность которых обеспечивается оптимальным режимом активации D1-рецепторов.
Здесь следует упомянуть, что в отличие от глутамата (основного активирующего медиатора мозга) и ГАМК (основного тормозного медиатора), характеризующихся высокой скоростью передачи и короткой длительностью (несколько миллисекунд) действие дофамина в синаптической щели может длиться секунды или даже минуты, что определяется механизмом обратного захвата, обеспечиваемого специализированными рецепторами-транспортерами дофамина пресинаптической терминали (рис. 12.18). Причем чем больше таких рецепторов расположено на пресинаптической мембране, тем быстрее осуществляется захват дофамина и, следовательно, сокращается время активации, вызываемой значимым стимулом128.
В отличие от DAT-рецепторов, определяющих динамические свойства действия дофамина, концентрация рецепторов D1 - и Э2-семейства модулирует ам
126 Здесь необходимо упомянуть что другие функции передней поясной извилины заключаются в обеспечении эмоций и мотиваций. И в этом контексте дофамин ассоциируется с системой вознаграждения. В общих словах вознаграждение (или подкрепление) является состоянием мозга, при котором наличествующий результат действия соответствует желаемому или ожидаемому. Из исследований на животных нам известно, что стимулы подкрепления (как, например, пища, вода), стимуляция латерального гипоталамуса, а также злоупотребление некоторыми наркотическими веществами перестают выполнял, роль подкрепления, если животному дать антагонист дофамина. Известно также, что дофаминовые нейроны среднего мозга начинают генерировать спайки непосредственно в ответ на подкрепление, а также при предъявлении стимулов, предсказывающих его появление. Функция подкрепления заключается в выделении наиболее эффективной программы текущего поведения. Таким образом, высвобождение дофамина в стриатуме устанавливает наиболее низкий порог срабатывания для программ, которые с большей вероятностью приведут к получению подкрепления.
127 Обратившись к «Введению» данной части книги, слезет вспомнить о существовании инвертированной U-образной функции, описывающей отношение между входом и выходом нейронных систем. Аналогичная ей деятельность рабочей памяти зависит от степени активации дофаминергических рецепторов Dl-типа, локализующихся в префронтальной коре: максимальная работоспособность памяти наблюдается при средних уровнях стимуляции Dl-рецепторов, но ограничения в ее работе появляются при снижении или увеличении стимуляции.
128 Было обнаружено, что подгруппа пациентов с СНВГ характеризуется повышенной концентрацией DAT-рецепторов в структурах стриатума. Это в конечном счете может приводить к укорачиванию «жизни» следа памяти значимого стимула (рис. 12.18, средний график справа). В свою очередь, это может приводить кследующему: 1) ослаблению рабочей памяти; 2) невозможности концентрироваться на конкретном действии длительное время (невнимание); 3) быстрому переключению между разными моторными действиями (гиперактивность); 4) быстрому переключению между визуальными объектами и звуками (отвлекаемость); 5) устойчивому изменению процессов мышления (импульсивность).
Часть 2, Вызванные потенциалы
295
плитуду эффекта дофамина. Так, например, снижение количества рецепторов к ДА на постсинаптической терминали обусловливает низкий уровень возбудимости нейронов стриатума и коры (или высокий порог активации). Наоборот, повышенная концентрация этих рецепторов на постсинаптической мембране (как, например, при шизофрении, когда отмечается избыточное содержание П2-рецепторов в стриатуме) приводит к снижению порогов активации нейронов стриатума.
VIII. Заключение
Под термином «исполнительные функции» понимают функцию контроля и координации моторных и когнитивных действий, направленных на достижение специфических целей деятельности. Считается, что механизмы исполнительного контроля необходимы в нестереотипных (нерутинных) ситуациях, требующих вмешательства контролирующих систем. Примерами таких ситуаций являются выбор адекватного действия из множества возможных, подавление несоответствующих действий, а также удержание в рабочей памяти как плана действий, так и представлений об ожидаемом результате. Исполнительный контроль необходим также и для оптимизации поведения. Исполнительные функции обеспечиваются сложной мозговой системой, состоящей из различных кортикальных и подкорковых структур, взаимосвязанных между собой. В состав кортикальных структур входит префронтальная кора, связанная с соответствующими областя-ми теменно-височной коры. Нейронная активность этих областей регулируется параллельной циклической активностью, которая охватывает соответствующие области базальных ганглиев. Базальные ганглии проецируют свои связи в соответствующие ядра таламуса, которые, в свою очередь, имеют реципрокные связи с корой. Фактически замыкается петля обратной связи, охватывающей кору, ба-KuibHbie ганглии и таламус. Стриатум является ключевым звеном этой системы и может рассматриваться как когнитивная карта репрезентаций действий, распределенных по соответствующим областям коры. Стриатум обеспечивает вы-бор действий с помощью механизма латерального торможения, оказывающего < вое !и!ияние на длительные дистанции. Фронтальные области коры характери-iviotch сложной архитектурой связей нейронов, что получает отражение в повышенном уровне их активности по сравнению с задними областями мозга. Со-вмссгпо с базальными ганглиями области префронтальной коры вовлекаются в «икч'нечение исполнительных функций, заключающихся в операциях вовлечения, отвлечения, моторных операциях и рабочей памяти. Эти операции отра-♦ ли иен соответствующими компонентами ВП, которые могут генерироваться н м ювпях, модулируемых разными исследовательскими парадигмами, Напри-H. р 1лкими как oddball, GO/NOGO, а также тестовыми заданиями с привле-|« uni-м рабочей памяти. Несмотря на известное перекрывание во времени и п|нм ip.inciBe компонент ВП, связанных с исполнительными функциями, не-i.i in in разработанный метод анализа независимых компонент является мощ-
296
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
ним средством, позволяющим проводить их разделение. Использование нормативной HBI базы данных при регистрации ВП в рамках двухстимульной GO/ NOGO-парадигмы позволило нам выделить ряд компонент, связанных с исполнительными функциями: компоненты подавления моторной реакции и действия характеризовались фронтальной негативностью с латентным периодом 200 мс (известная М2-компонента подавления), компонента вовлечения — позитивностью в теменных областях и латентным периодом 300 мс (известная РЗЬ-компонента) и, наконец, компоненте, связанной с операциями мониторинга, соответствовала фронтально-центральная негативность с латентным периодом 400 мс. Использование метода томографии низкого разрешения (sLORETA) позволило выявить локализацию генераторов выделенных компонент ВП. Так, N2-KOMnoHeHTa моторного подавления генерировалась в ипсилатеральной премоторной коре, а источник генерации компоненты мониторинга действий Р400 локализовался в области передней поясной извилины. Передняя поясная извилина также вовлекается в обеспечение генерации негативности, связанной с ошибкой — компонент, который регистрируется при навязанных тестовыми условиями ошибочных моторных ответах в условиях GO/NOGO-парадигмы. Главным медиатором исполнительной системы является дофамин. Продукция дофамина осуществляется на уровне нейронов префронтальной коры, а его действие опосредуется специфическими рецепторами D1 -типа. Дофамин также обнаруживается и в базальных ганглиях, где его действие связано с рецепторами Э2-типа. Препараты, влияющие на разные этапы дофаминергической передачи, применяются для компенсации дисфункции исполнительной системы при шизофрении и СНВГ.
Часть 2. Вызванные потенциалы
297
Глава 13.
Аффективная система
Все счастливые семьи похожи друг на друга; каждая несчастливая семья несчастлива по-своему.
Лев Толстой. Анна Каренина
I. Психология
А. Эмоции и рассуждения
Благодаря эмоциям в жизни человека существует измерение, отличное от рассудочной деятельности. Природа создала аффективную систему мозга для обеспечения выживаемости в окружающем мире посредством формирования измерения, не связанного с рассудком. Когда мыслительной деятельности недостает для принятия решения, мозг полагается на аффективную систему. Позвольте привести простой пример: мы редко руководствуемся разумными доводами при выборе мужа или жены, когда болеем за любимую команду или когда играем с любимой собакой. Иногда при совершении важных шагов в нашей жизни (образование семьи, развод, увольнение с работы и т.д.) нами движут эмоции. Другим примером работы аффективной системы мозга является реализация фундаментальных поведенческих актов, например, таких как поиск пищи, воды или полового партнера.
При определенных событиях аффективная система продуцирует врожденные ответы, называемые эмоциональными реакциями. Эти реакции возникают, когда мозг детектирует эмоционально значимые стимулы129. «Привычки ра зума» — это стереотипные реакции мозга, которые позволяют формировать ассоциации определенных эмоций (и чувства, их сопровождающие) и соответствующих аспектов нашего жизненного опыта. Эволюционно мозг предрасположен отвечать на эмоционально значимые стимулы специфическим репертуаром действий. Несмотря на существование эволюционно детерминированных эмоционально значимых стимулов, по мере развития нашего опыта некоторые из событий и стимулов окружающего нас мира приобретают эмоциональную окраску130.
1" Эта концепция была разработана Антонио Дамазио. Научно-популярное описание его работы представлено в книге под на-1к<1ни(!м «The feeling of what happens», вышедшей в 1999 году.
1 *’ Таким образом, эмоция является сложным набором химических и нейронных ответов на определенные внешние и внутренние < оСн.пия. которые формируются в виде различных паттернов и отражают определенные поведенческие реакции (включая, например, ныражение лица). Реакции нашего тела (в частности, лицевая экспрессия) могут восприниматься другими людьми, а мимика отража-»’| их соответствующие реакции, что обеспечивается специфическими мозговыми системами так называемых зеркальных нейронов. Именно эмоции делают из нас людей. Все искусство (музыка, живопись, литература) пронизано эмоциями.
298 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Б. Наказание и вознаграждение
Принято разделять эмоционально значимые стимулы на два основных класса — стимулы наказания и вознаграждения (подкрепления). Два класса эмоциональных стимулов характеризуются двумя разными типами аффективных состояний: избеганием и приближением. Далее аффективные реакции, связанные с этими аффективными состояниями, могут быть подразделены на отдельные эмоции, такие как страх, тревога, радость... Один из вариантов классификации эмоционально значимых стимулов и реакций представлен в табл. 13.1. Антонио Дамазио, специалист мирового уровня в области изучения мозговых механизмов эмоций, выделяет три основных типа эмоций: 1) фоновые эмоции; 2) первичные эмоции; 3) социальные эмоции.
Таблица 13.1. Классификация аффективных состояний и эмоций
Событие (внешнее и/или внутреннее) Аффективное состояние Аффективная реакция (эмоция)
Наказание Избегание Страх Тревога Злость Отвращение Печаль Презрение Смущение Вина Стыд
Вознаграждение Приближение Радость Удовольствие Восторг Гордость Половая любовь Материнская любовь
К фоновым эмоциям относят разочарование и восторг. Правда, их достаточно сложно отличить от аффективных состояний. Первичными или основными эмоциями являются страх, злость, отвращение, печаль, удивление и радость. Основные эмоции хорошо определяемы и у людей, принадлежащих к разным культурам, и у животных разных видов. Большинство из того, что нам известно об эмоциях, было добыто в исследованиях первичных эмоций. При этом больше всего исследовалась эмоция страха. Под социальными эмоциями понимают сострадание (симпатия), смущение, стыд, чувство вины, гордость, зависть, ревность, благодарность, восхищение, возмущение и презрение.
В. Потребности и мотивации
Говоря об аффективной системе, с одной стороны, следует проводить разделение потребности и мотивации, а с другой — различать эмоции и чувства. Потребности и мотивации формируют аффективные состояния, которые необходимо отличать от эмоциональных ответов. Лимбическая система мозга (а значит, и эмоции) впервые появилась у млекопитающих, несмотря на то что, например, птицы обладают некоторыми атрибутами эмоций. Лимбическая система практически не присуща рептилиям, амфибиям и другим низшим животным131.
131 Пол Маклин обычно говорил, что очень сложно представить более одинокое и более беззмоциональное существо, чем крокодил.
Часть 2. Вызванные потенциалы
299
Наиболее яркими примерами потребностей и мотиваций являются голод, жажда, любопытство и влечение к противоположному полу Поведение регулируется мотивациями с помощью ряда механизмов. Известно, что главную роль в мотивационном поведении играет гипоталамус. Нейроны гипоталамуса характеризуются высокой чувствительностью к целому ряду биохимических веществ, при этом их импульсная активность проявляет зависимость от концентрации основных веществ, растворенных в крови и цереброспинальной жидкости. В свою очередь, нейроны гипоталамуса регулируют секреторную активность гипофиза, а также действуют на другие части аффективной системы, обеспечивая запуск определенных типов регулируемого поведения, как, например, пищевое или питьевое поведение. Поведение человека управляется не только простыми мотивациями, такими как питание или питье, но и более сложными, а подчас и весьма абстрактными потребностями.
II. Анатомия
А. Лимбическая система
Благодаря существованию аффективной системы мозга, обеспечивающей существование эмоций и мотиваций, в нашем восприятии и действиях возникает дополнительное измерение, новая характеристика. Это новое измерение перестраивает наше поведение, стимулируя поиск положительных эмоций и избегание негативных. С информационной точки зрения аффективная система функционирует таким образом, чтобы усилить след решающего для выживания события, ассоциируя его с определенным эмоциональным значением. Как можно заметить, описываемая нами система с функциональной точки зрения определяется как аффективная132. Однако исторически первое понятие, близкое к представлению об аффективной системе, возникло в виде идеи о существовании лимбической системы133. В основу этого представления легли особенности анатомии, на основании которых под лимбической системой понимался набор структур, окружающих (те. образующих круг) мозолистое тело. Термин «лимбическая система» приобрел большую популярность в психологии и психиатрии и служит в качестве объяснительной концепции при описании нейронных сетей, вовлекаемых в генерацию эмоций и аффективных состояний134.
Б. Круг Папеца
В 1937 году была предпринята первая попытка научного подхода в описании аффективной системы. Джеймс Папец предположил, что лимбическая доля работает совместно с гипоталамусом, который в те времена считался ключевой
137 Впрочем, так же, как и другие системы, которые описываются в данной книге.
133 Латинское слово limbus означает «граница». Впервые термин «лимбическая доля» (le grand lobe limbique) был введен в литературу Полем Брока в 1878 году для определения извилин коры, формирующих круг вокруг мозолистого тела. В соответствии с его концепцией лимбическая доля включает поясную извилину и гиппокамп с прилегающими к нему парагиппокампальной и энтори-альной корой.
134 В то же время нейрофизиология сконцентрирована на изучении всего разнообразия функций анатомических структур, окружающих мозолистое тело. С нейрофизиологической точки зрения эмоция является лишь одной из многих функций, присущих лимбической системе. В частности, к функциям лимбической системы относяттакже и память, и исполнительные функции.
300
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
структурой, вовлекаемой в выражение эмоций. Он особенно выделял дорсальную часть лимбической доли Брока, называемую тогда цингуломом135. На самом деле гипоталамус136 с переключением в передних ядрах таламуса посредством мамиллярно-таламического тракта посылает сигналы в поясную кору. Оттуда дальше информация перенаправляется в гиппокамп, который, в свою очередь, в составе пучка аксонов, называемого сводом, посылает свои проекции в гипоталамус. Таким образом, образуется циркуляция активности: гипоталамус и поясная кора за счет реципрокных связей позволяют эмоциям, запускаемым при участии гипоталамуса, достигать сознания (уровень поясной коры). С другой стороны, такая организация связей внутри круга Папеца способствует влиянию высших когнитивных функций на процессы, связанные с генерацией эмоций. Эта система реципрокных связей между гипоталамусом и поясной корой получила название круга Папеца (рис. 13.1). Позже, в 1949 году, Пол Маклин в нескольких работах расширил концепцию круга Папеца, дополнительно включив несколько анатомически связанных структур, а именно миндалину, септальную область, прилежащее ядро и орбитофронталъную кору.
В. Корковые и подкорковые элементы
Современные анатомические и физиологические исследования показывают, что с функциональной точки зрения лимбическая система не может рассматриваться как единая сущность. Это справедливо прежде всего потому, что лимбическая система может быть подразделена на несколько подсистем, функциональная роль которых связана не только с эмоциями и мотивациями, но также
Рисунок 13.1. Круг Папеца
А. Схема связей между анатомическими структурами, входящими в круг Папеца. Б. Изображение мозга человека со структурами мозга, входящими в состав круга Папеца, окрашенными серым цветом. Обозначения: VLPF — вентролатеральная префронтальная кора; DLPF — дорсолатеральная префротнальная кора; М — моторные области коры; SS — соматосенсорная кора; PC — теменная кора; VI, V2, V4 — области зрительной коры; IT — нижняя височная кора; ЕС — энториальная кора; Hip — гиппокамп; МВ — мамиллярные тела гипоталамуса; А — переднее ядро таламуса; VL — вентролатеральное ядро таламуса; DM — дорсомедиальное ядро таламуса.
135 Эта часть коры получила название поясной коры за счет того, что образует пояс, или ожерелье, или воротник вокруг мозолистого тела. Поясная кора (извилина), в свою очередь, делится на переднюю и заднюю части.
136 В действительности обособленные области гипоталамуса называются мамиллярными телами.
Часть 2. Вызванные потенциалы
301
с эпизодической памятью и исполнительными функциями. Во-первых, в дополнение к предыдущим концепциям новые исследования указывают на то, что гиппокамп обладает связями с ассоциативными сенсорными областями коры, а его билатеральное повреждение приводит к ретроградной или антероградной амнезии. Во-вторых, многократно было показано, что передняя поясная извилина вовлекается в обеспечение внимания и процессы коррекции ошибок — суть элементов испол-
НИТеЛЬНЫХ функций137. Рисунок 13.2. Анатомические структуры аффективной системы
R ыягтпсттттрй kaki О6означения: — орбитофронтальная кора; Ат — амигдала; Ас — при-
D Настоящей книге МЫ лежащее ядро (вентральный стриатум). Структуры коры мозга: АСС — пе-
рассматриваем аффективную редняя поясная кора; SS/lnsular — соматосенсорная и инсулярная кора.
Остальные обозначения, как на рис. 13.1.
систему как комплекс свя-
занных между собой корковых и подкорковых структур, образующих функциональную структуру, которая позволяет субъекту генерировать эмоции и чувствовать эмоциональные и мотивационные реакции на стимулы внешней и внутренней среды организма (рис 13.2)138. Области коры, входящие в состав аффективной системы: орбитофронтальная кора, передняя поясная извилина, инсулярная и соматосенсорная кора. Набор подкорковых структур аффективной системы включает миндалину, передний гипоталамус и вентральный стриатум (прилежащее ядро).
Обращает на себя внимание тот факт, что большинство структур аффективной системы располагается глубоко в мозге. Такая «внутренняя» локализация аффективной системы существенно затрудняет регистрацию ЭЭГ-коррелят ее функциональной активности с поверхности.
III. Физиология
А. Орбитофронтальная кора и картирование вознаграждения и наказания
Орбитофронтальная кора (ОФК) расположена в вентральной части лобной доли, включая 11-е и 47-е поля Бродмана (рис. 13.1). Иногда в состав ОФК включают и 10-е ПБ, которое, впрочем, чаще рассматривается как отдельная
1*' Разнообразие функций, атрибутируемых лимбической системой, ставит под вопрос использование самого термина «лимбиче-< кии», поскольку, на наш взгляд вполне справедливо рассматривать память, исполнительные и аффективные системы как отдельные < vни<осги. В данной книге мы руководствуемся представлениями о функциональной организации мозга и рассматриваем эти системы к. »< [кнныс. Здесь необходимо напомнить читателю, что любая анатомическая структура зачастую ответственна не за одну, а за несколько '|>у>1кции, тогда как конкретная функция реализуется при участии сложно организованной системы множества структур мозга.
1" Отметим, что круг Папеца является самой главной составляющей аффективной системы.
302 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
структура — передняя лобная кора. Важной характеристикой ОФ К является ее выраженная межсубъектная вариабельность. ОФК получает полимодаль-ные входы фактически от всех сенсорных систем: зрительная, слуховая и соматосенсорная системы снабжают ОФК информацией о внешнем мире, о текущем статусе нашего тела, состоянии глубоких структур и висцеральных органов. ОФК за счет фундаментальных филогенетически древних примитивных модальностей, таких как хемочувствительность, принимает участие в восприятии запаха и вкуса, сигнализирующих о притягательности или опасности пищи или питья. Миндалина посылает орбитофронтальной коре информацию, связанную с эмоциями и преимущественно ассоциирующуюся с эмоцией страха. На уровне передней поясной извилины обрабатывается высшая когнитивная и эмоциональная информация, в особенности в тех случаях, когда детектируется ее рассогласование с моделью ожидаемого. ОФК получает входы и от других структур аффективной системы, например, таких как гипоталамус. Кроме того, ОФК, как и любая область коры, саморегулируется за счет системы базально-таламокортикальных связей, с прилегающим ядром в качестве ведущего центра на уровне стриатума139.
Одним из наиболее важных симптомов избирательного повреждения орбитофронтальной коры является эффект отсутствия эмоций140. Браян Колб и Иан Вишоу в своей книге «Ап introduction to brain and behavior» («Введение в науку о мозге и поведении») описали случай пациентки, которая была подвергнута фронтальной лейкотомии. Как пишут авторы, первая особенность, которая обращала на себя внимание, заключалась в отсутствии эмоций и каких-либо признаков эмоциональной лицевой экспрессии. Пациентка вполне осознавала свой дефицит эмоций, отмечая, что более не испытывает никаких чувств в отношении людей и предметов, ощущает пустоту подобно зомби. Пациенты с повреждениями ОФК характеризуются нарушениями в идентификации социальных сигналов, таких как эмоциональное выражение лица и интонация речи. В дополнение к этому отмечаются глобальные изменения личности, проблемы с самообладанием, социальное несоответствие и безответственность, а также трудности в принятии решений в контексте повседневной жизни.
Метаанализ фМРТ и ПЭТ-исследований141 позволяет прийти к выводу о том, что на уровне орбитофронтальной коры могут картироваться репрезентации вознаграждений и наказаний в виде пространственного распределения паттерна активаций коры (рис. 13.3). Медиальные области ОФК принимают участие в обеспечении процессов, связанных с действием вознаграждений. Близкое расположение и обильные связи с передней поясной извилиной являются отраже-
139 Напомним, что эти базально-таламокортикальные связи известны под названием лимбических, функция которых заключается в регуляции эмоций и мотиваций.
140 Классическим примером разрушения ОФК (включающим также повреждение в области передней поясной извилины) является известный случай Финеаса Кэйджа, у которого в результате несчастного случая — прохождения металла сквозь голову — оказались повреждены лобные доли. После этого случая Кэйдж стал другим человеком, его эмоциональные процессы и процессы, связанные с принятием решений, кардинально изменились.
141 Крингелбах и Рол в своей обзорной работе, посвященной описанию функций орбитофронтальной коры, провели обширный метаанализ томографических исследований, позволивший сделать вывод о том, что разные подразделения ОФК выполняют разные функции.
Часть 2. Вызванные потенциалы
303
нием важной роли медиальной ОФК в организации поведения, направленного на достижение вознаграждения (подкрепления). Латеральная же часть ОФК вовлекается в обеспечение процессов, связанных с отрицательными подкрепляющими стимулами (наказание). Близкое расположение по отношению к латеральной префронтальной коре, а также сильные связи с этой областью свидетельствуют о возможной роли латеральной ОФК в подавлении поведения, ассоциирующегося с наказанием. В соответствии с этой точкой зрения пациенты с повреждениями ОФК не способны правильно ассоциировать стимулы положительного (вознаграждения) и отрицательного (наказание) подкрепления с определенными действиями. Именно поэтому у таких больных наблюдаются патологические изменения про-
Рисунок 13.3. Схематическое изображение процессов положительного и отрицательного подкрепления на уровне орбитофронтальной коры
Анализ литературы, посвященной исследованию функций орбитофронтальной коры (Kringelbach and Rolls, 2004), показал, что медиальные области коры принимают участие в обработке стимулов положительного подкрепления, а латеральные — в обработке стимулов отрицательного подкрепления (наказание). Наряду с этим было показано, что области, связанные с обработкой сложных и абстрактных подкрепляющих стимулов (как, например, денежный выигрыш или проигрыш), характеризуются выраженной «передней» локализацией относительно областей, обеспечивающих обработку более простых стимулов (как, например, вкус). Обозначения: АСС — передняя поясная извилина; LPFC — латеральная префронтальная кора.
цессов принятия адекватных решений в пользу выбора действий, приводящих к вознаграждению, и избегания действий, вызывающих наказание.
Б. Позитивный аффект, негативный аффект и мониторинг
Кажется весьма правдоподобным, что вознаграждение и, соответственно, положительные эмоциональные реакции на него формируют одно из измерений аффективной системы. Другое измерение, следовательно, формируется наказанием и связанными с ним эмоционально негативными реакциями (рис. 13.4). По всей видимости, эти два измерения и формируют темперамент142. В более широком смысле темперамент можно рассматривать как отражение не только эмоциональных, мотивационных наклонностей, но и мониторинга аффективных состояний. Тогда в дополнение к перечисленному выше появляется другое и змерение аффективной системы (рис. 13.4) — мониторинг эмоций, мотиваций и потребностей143. Обращает на себя внимание, что это третье измерение в действительности является атрибутом исполнительной системы, подробно описанной несколько выше. Как было продемонстрировано в предыдущей главе, основной вклад в поддержание операции мониторинга вносит сложная мозговая система, критическим элементом которой является передняя поясная кора. * 141
142 Не следует путать темперамент и личность. Темперамент в большей степени является атрибутом аффективной системы. «Лич-н(и:ть» же является более всеобъемлющим термином, объединяющим комбинацию атрибутов всех мозговых систем, включающих аффективную, исполнительную, сенсорную системы, а также внимание и память.
141 Эти три измерения по смыслу очень близки к трем основным измерениям темперамента — негативная эмоциональность, по-1И1ивная эмоциональность и сдерживание, которые были представлены в обзоре нейробиологических основ темперамента, опубли-koimhhom Сарой Вайтл и ее коллегами из Австралийского университета Мельбурна (Whittle at al., 2005).
304 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Дорсальная часть передней поясной извилины вовлекается в обеспечение мониторинга когнитивных действий, связанных с целенаправленным поведением и мышлением. По аналогии с идеей мониторинга при рассмотрении функций дорсальной части передней поясной извилины можно предполагать, что функция ее вентральной части связана с мониторингом эмоций.
Индивидуумам с «положительной» эмоциональностью свойственно испытывать широкий диапазон положительных эмоций, например, таких как ра
Рисунок 13.4. Три независимые характеристики аффективной системы В трехмерном пространстве схематично представлены три характеристики (позитивный аффект, негативный аффект и мониторинг). Нейтральное состояние или эмоциональная реакция соответствует началу координат. В данном пространстве координат могут быть схематично представлены: депрессия, тревожность и аддикция (отмеченные серым цветом).
дость, счастье, восторг и гордость. Благодаря связям орбитофронтальной коры, которые обеспечивают распространение активаций, связанных с положительным подкреплением, с передней поясной извилиной, ответственной за мониторинг поведения, субъекты с «положительной» эмоциональностью вовлекаются в активное и положительное социальное и семейное поведение. При этом некоторые типы депрессивных состояний могут быть связаны с низким уровнем эмоциональности этого типа.
Напротив, люди, которым свойственна негативная эмоциональность, чаще
испытывают широкий спектр негативных эмоций: страх, тревогу, печаль или вину. Посредством соответствующих связей (относящихся к наказанию) орбитофронтальной коры с латеральной фронтальной корой, вовлекаемой в подавление поведенческих паттернов, обеспечивается склонность таких людей к избеганию социальных взаимодействий, подавлению новых и неизвестных стремлений. Примером высоких показателей негативной эмоциональности может являться тревожность в виде психического расстройства.
Индивидуумы, у которых сильно выражено измерение аффективной системы, связанное с мониторингом, могут эффективно контролировать свои эмоции и аффективные импульсы. Такие люди способны ждать (и достаточно долгое время) вознаграждения или удовлетворения потребности. Основными чертами субъектов с высокими показателями этой размерности аффективной системы являются высокая трудоспособность, упорство, надежность и ответственность. Напротив, низкие показатели соответствуют импульсивности и агрессивности поведения. Такие люди не способны длительное время ожидать положительного
Часть 2. Вызванные потенциалы
305
подкрепления и живут настоящим моментом. Наличие патологических склонностей как разновидности психического расстройства также является отражением низких показателей этой размерности аффективной системы. Считается общепринятым, что типологические особенности темперамента детерминированы генетически, выявляются в раннем возрасте144 и сохраняются на протяжении всей жизни, претерпевая умеренные изменения, обусловленные действием окружающей среды.
Важной функциональной характеристикой орбитальной коры является способность к саморегуляции посредством системы базально-таламокортикальных связей, наличие которой свойственно любой из фронтальных областей коры. ОФК топографически картируется на уровне вентрального стриатума, а именно прилежащего ядра. Организация нейронных связей стриатума способствует особому режиму его работы по типу латерального торможения, что лежит в основе принципа функционирования, известного как «победитель получает все». В силу механизма латерального торможения активация определенной репрезентации (скажем, вознаграждения) на уровне орбитофронтальной коры приводит к подавлению противоположной репрезентации (скажем, одного из наказаний). Фактически такое представление об организации системы позволяет подходить к рассмотрению склонностей к определенным эмоциональным состояниям (депрессия или мания) с позиций дисбаланса процессов активации и подавления на уровне базально-таламокортикальной нейронной сети.
В. Асимметрия в картировании эмоций
Другой важной особенностью функционирования орбитофронтальной коры является асимметричность мозгового представительства эмоций. Вопреки многочисленным литературным данным (МРТ, фМРТ, ПЭТ-исследования), указывающим на асимметрию в мозговом обеспечении эмоций145, нам не удаюсь обнаружить действительно убедительного доказательства в пользу асимметричного распределения репрезентаций позитивных и негативных эмоций на уровне орбитофронтальной коры. Однако существует большое количество доказательств такой асимметрии относительно латеральной префронтальной коры. Идея о латерализации аффективной системы впервые появилась при рассмотрении клинических проявлений повреждений левой префронтальной коры, которые зачастую проявляются в виде депрессий. При этом аналогичные повреждения префронтальной коры правого полушария не приводят к таким последствиям146.
В 1993 году Ричардом Дэвидсоном, ученым из Университета Висконсина, была выдвинута гипотеза о том, что левая префронтальная кора проявляет наи
144 Это справедливо, пожалуй, за исключением процессов мониторинга, которые окончательно развиваются несколько позже, hi и/и» исполнительная система мозга формируется в достаточной степени.
,4‘' Имеется в виду мозговая асимметрия мозга человека (и животных) в терминах структуры, функции и поведения. Считается, <•<> 1.нерализация отражает действие эволюционных, наследственных, онтогенетических, индивидуальных и патологических факто-t и >п /Цтя более подробного рассмотрения мы может рекомендовать обзорную работу Артура В. Тога и Поля М. Томсона (2003).
'* *• Так, например, в исследовании Морриса и др. (1996) у пациентов с поврежденной префронтальной корой или базальными
• пиями левого полушария с большой частотой диагностировалась депрессия (9 из 12; 75 %) по сравнению с пациентами, у кото-PI.K оказывались поврежденными либо другие области коры (1 из 12; 8%), либо те же отделы, но в правом полушарии (5 из 17; 29 %), г о(Х)2).
306 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
большую активность при положительных эмоциях. И напротив, правая префронтальная кора более активна при эмоциях избегания (те. негативных). Проще говоря, предполагалось, что левая префронтальная кора отвечает преимущественно на эмоционально положительные события, а правая, соответственно, на эмоционально негативные.
Учитывая тот факт, что мощность альфа-ритма ЭЭГ обратно коррелировала с уровнем активации соответствующей области мозга, Дэвидсон с коллегами утверждали, что асимметрия альфа-ритма, наблюдаемая в лобных отведениях, отражает относительную разницу в активности между правым и левым полушариями. Исследования Дэвидсона демонстрировали стабильный характер асимметрии фронтального альфа-ритма. На основании этих данных было выдвинуто предположение о том, что проведенные измерения отражают характерологические особенности в реагировании на эмоционально положительные (приближение) и эмоционально отрицательные (избегание) стимулы. В этих исследованиях ЭЭГ регистрировалась в условиях просмотра видеоклипов, вызывающих либо положительные (радость, смех), либо отрицательные эмоции (страх, отвращение). В результате было обнаружено, что фронтальная ЭЭГ-асимметрия является стабильной характеристикой испытуемых и неплохо предсказывала как тип эмоциональной реакции, так и выраженность эмоций. Вместе с тем асимметрия фоновой ЭЭГ не проявляла такой зависимости от текущего настроения147.
Г. Миндалина
Миндалина — это относительно небольшое ядро, которое располагается внутри передненижней части медиальной височной коры148. Правомерно рассматривать миндалину как интрефейс между сенсорным миром149 и эмоциями. От полимодальных областей височной и теменной коры, опосредованно через гиппокамп, миндалина получает сенсорную информацию, извлекает хранящиеся на уровне миндалины следы памяти и через дорсомедиальное ядро таламуса пересылает их дальше, в префронтальную кору.
При регистрации импульсной активности непосредственно с миндалины наибольшее количество нейронов активируется в ответ на предъявление отталкивающих стимулов. Гораздо меньше нейронов активируется в ответ на привлекательные стимулы. Таким образом, миндалина может быть описана как структура, ответственная за детекцию угрозы или потенциального наказания и тем самым за генерацию негативных эмоций страха или тревоги. Иными словами миндалина, по всей видимости, определяет «негативное измерение» аффектив
147 Поскольку мощность альфа-ритма инвертированно коррелирует с метаболической активностью соответствующей области мозга, Дэвидсон с коллегами предложили использовать показатель мощности альфа-ритма для оценки относительной разницы в активности между полушариями мозга. Данная теория инициировала появление протокола обратной связи, называемого протоколом асимметрии альфа-ритма, который применяется в лечении депрессии (см. чЗ).
148 Представление о миндалине как о ядре является некоторым упрощением. Слово «миндалина- представляется наиболее точным обозначением в терминах описания структуры и функции этой части аффективной системы (Heimer, Van Hoesen, 2006). Миндалина состоит из центрального и медиального амигдалоидного ядер и их продолжения в субпаллидарную область. Миндалины являются стратегической системой, получающей полимодальный вход и координирующей активность многих структур лимбической системы.
149 Большей частью миндалина получает информацию от слуховой и зрительной систем.
Часть 2, Вызванные потенциалы
307
ной системы. Примечательно, что пациенты с повреждениями в области миндалины не испытывают ни страха, ни тревоги150.
Д. Медиальное и переднее ядра таламуса
Известно, что эмоциональная реактивность может существенно изменяться вследствие стимуляции или разрушения медиодорсального и переднего ядер таламуса. Важно отметить, что сами ядра не участвуют в процессах генерации эмоций, а выполняют транзитную функцию при распространении соответствующей информации до префронтальной коры. При этом переднее ядро таламуса перенаправляет информацию, приходящую от мамиллярных тел гипоталамуса, в область передней поясной извилины. А через медиодорсальное ядро осуществляется связь между миндалиной и префронтальной корой. Следует отметить одну важную особенность переднего таламуса — отсутствие скопления нейронов, окружающих остальной таламус, а именно ретикулярных ядер. По всей видимости, эта анатомическая характеристика играет определенную важную роль в работе аффективной системы мозга. В действительности отсутствие ретикулярных ядер в передней части таламуса проявляется в виде невозможности ингибирования активности по типу обратной связи, как раз осуществляемому тормозными нейронами этих ядер. Отсутствие этих нейронов приводит к невозможности блокирования распространения информации, связанной с эмоциями, до префронтальной коры. Фактически что бы ни происходило в окружающем нас мире, эмоциональное значение этих событий обязательно доходит до коры151!
Е. Гипоталамус
Гипоталамус представляет собой небольшое, располагающееся достаточно глубоко ядро (меньше 1 % объема мозга) без четко выделенного направления расположения нейронов. Вследствие такого строения с поверхности гипоталамуса невозможно зарегистрировать локальный потенциал, надежно отражающий активность определенной популяции нейронов. Гипоталамус (по крайней мере некоторые из его частей) принимает участие в регуляции многих основных программ, например, таких как температура тела, потребление пищи и воды, а также половое поведение. Известно, что повреждение ядер гипоталамуса может приводить к существенным изменениям некоторых вегетативных функций и некоторых видов так называемого мотивированного поведения: температурной регуляции, сексуальности, агрессии, голода или жажды.
Важную роль играет гипоталамус и в обеспечении эмоций. Как известно, многие области мозга вносят вклад в поддержание не одной, а нескольких функций. Это справедливо и в отношении гипоталамуса. И действительно, латеральные части гипоталамуса могут вовлекаться в обеспечение таких эмоций, как удо
150 В своей книге «The feeling of what happens» Антонио Дамазио описывает пациентку с билатеральным повреждением минда-1ин Он пишет: .. .способность запоминать новые факты не была нарушена у С... Но, с другой стороны, история ее социальных взаимодействий была удивительна. Проще говоря, С. воспринимала людей и события с изначально позитивных позиций». У этой пациентки. р<1нно так же, как и у других пациентов с аналогичными повреждениями, словно утратилась память о страхе и тревоге, по крайней мере для слуховых и зрительных стимулов.
,,>1 В частности, мать может пробудиться от сна, даже на глубоких его стадиях, если услышит короткий крик своего ребенка.
308
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
вольствие и ярость. В свою очередь, активность в медиальной области гипоталамуса ассоциируется с отвращением, неприятием и реакцией неконтролируемого и громкого смеха. Однако гипоталамус в большей степени вовлекается в обеспечение выражения эмоций (т.е. симптоматических проявлений), чем в генерацию аффективных состояний.
Ж. Вентральная область передней поясной извилины.
Как уже говорилось в предыдущей главе, передняя поясная извилина может быть подразделена на две части: вентральную (аффективную) и дорсальную (исполнительную). В частности, стимуляция вентральной ППИ может приводить к возникновению эмоций страха или удовольствия. Стимуляция дорсальной ППИ способствует появлению чувства ожидания движения. Таким образом, вентральная часть ППИ принимает участие как в обеспечении эмоциональных реакций на боль, так и в регуляции агрессивного поведения152. ППИ имеет сильно выраженные связи с миндалиной, которая, по всей видимости, доставляет информацию, связанную с эмоцией страха.
3. Функциональное картирование эмоций
С целью изучения нейрофизиологических основ эмоций и чувств Антонио Дамизио на базе Университетского медицинского колледжа Айовы в 2000 г. провел ряд ПЭТ-исследований. Во время ПЭТ-сканирования испытуемые вспоминали события из собственной жизни, связанные с разными эмоциями: печалью, радостью, злостью и страхом. Результаты этих исследований показали, что в обеспечение всех видов эмоций вовлекались структуры мозга, входящие в состав аффективной системы, а именно: инсулярная кора, вторичная соматосенсорная кора, поясная извилина, ядра покрышки ствола мозга и гипоталамус153. Эти паттерны активации позволяют судить о существовании разных «профилей восприятия» внутреннего состояния организма. Впрочем, именно на этом и основано логическое обоснование того, что разные эмоции переживаются по-разному.
Следует обратить внимание на то, что временное разрешение метода ПЭТ не позволяет анализировать паттерны временной динамики разных этапов эмоциональных реакций, чувств и процессов мониторинга эмоций. Для этих целей наиболее подходящими являются методы ЭЭГ и ВП. Однако известно, что мозговые структуры аффективной системы располагаются довольно-таки глубоко и, следовательно, могут генерировать только слабые потенциалы у поверхности скальпа. Кроме того, ядрам, входящим в состав аффективной системы, не свойственна какая-либо преобладающая ориентация клеток (аналогично ориентации пирамидных клеток коры), что обусловливает генерацию достаточно малых потенциалов в непосредственной близости от этих ядер.
152 Дикие животные, подвергнутые оперативному удалению поясной извилины (цингулотомии), становятся полностью прирученными и неопасными.
153 Так, индуцированная печаль характеризовалась билатеральными, но не симметричными активациями инсулярной коры и смешанным паттерном активации и деактивации поясной извилины (передние области активировались, а задние деактивировались). Эмоция радости индуцировала активацию задней части поясной извилины правого полушария и подавление активности передней ее трети, а также активацию левой инсулярной коры и правой соматосенсорной коры.
Часть 2. Вызванные потенциалы
309
И. Медиафронтальный тета-ритм ЭЭГ и эмоции
У некоторых испытуемых с достаточной степенью надежности активность передней поясной извилины может быть зарегистрирована с поверхности скальпа. Большой набор ЭЭГ-данных свидетельствует в пользу того, что основной вклад в генерацию тета-ритма, характеризующегося фронтальномедиальной топографией, вносит активность передней поясной извилины154. Тета-ритм проявляется в виде всплесков ритмической активности частотой 5—8 Гц, максимальная амплитуда которых регистрируется в области Fz. Ритм обычно регистрируется в условиях, когда испытуемый выполняет тестовое задание, требующее концентрации внимания, а его амплитуда увеличивается по мере увеличения нагрузки. Если испытуемый неспокоен или встревожен, то тета-ритм может снижаться или даже пропадать. Однако на фоне действия анксиолитических препаратов наблюдается восстановление ритма. Поэтому считается, что медиафронтальный тета-ритм отражает снятие тревожного состояния. Тем самым передняя поясная извилина вовлекается в регуляцию эмоционального состояния от неспокойной тревожности до сфокусированного расслабления. Это также согласуется с известным эффектом снижения тревожности при концентрации на какой-либо когнитивной проблеме155.
IV. Этапы реакций аффективной системы
Схематическое изображение этапов обработки эмоционально значимых стимулов представлено на рис. 13.5. Распознавание эмоциональных выражений лиц и движений тела обеспечивается на уровне височной и теменной коры. Миндалина, вентральный стриатум (прилежащее ядро) и орбитофронтальная кора участвуют в формировании двухмерного аффективного пространства с негативным (наказание) и позитивным (вознаграждение) аффектом в качестве измерений аффективной системы. Моторные области коры обеспечивают реализацию эмоционального ответа (а именно эмоциональную мимику и движения тела). Соматосенсорная кора получает информацию о реализующихся эмоциональных реакциях от висцеральных органов, а также от лица и тела. А передняя поясная извилина ответственна за сравнение ожидаемого действия и реального, а также за генерацию сигналов о результате операций сравнения, которые через моторную кору модифицируют поведение по типу избегания наказания или достижения вознаграждения.
Сами этапы могут быть классифицированы следующим образом: репрезентативный этап (восприятие), эмоциональная реакция, этап чувств и этап мониторинга (коррекция). Ниже подробно рассмотрен каждый из этапов.
154 Следует напомнить, что, как показали японские исследователи, испытуемые с фронтально-медиальным тета-ритмом встречаются в 11-43 % случаев, в зависимости от возраста.
155 Этот факт может быть объяснен близким расположением аффективной и исполнительной областей передней поясной извилины. По-видимому, активация, например, исполнительной области за счет механизма латерального торможения базальных ганглиев подавляет активность в аффективной области ППИ, и наоборот.
310
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Чувство
в г
Рисунок 13.5. Этапы обработки информации в аффективной системе См. объяснения в тексте.
А. Восприятие
Импульсная активность, запускаемая стимулом, практически одновременно достигает первичных сенсорных областей коры и миндалины156. Это ранний этап простого восприятия стимула.
Б. Эмоциональная реакция
Зрительная информация после обработки на уровне теменной и височной коры157 достигает моторной коры. Также информация обрабатывается
156 Близкие связи между восходящей активирующей системой и миндалиной позволяют мозгу быстро реагировать на отрицательные подкрепляющие стимулы, что необходимо для выживания в некоторых опасных ситуациях.
157 Например, фузиформная кора височных долей играет важную роль в распознавании лиц и обработке эмоциональной составляющей выражения лица.
Часть 2, Вызванные потенциалы
311
на уровне гиппокампа, откуда на ранних этапах сенсорной обработки стимула перенаправляется в миндалину, орбитофронтальную кору и вентральный стриатум. На данном этапе происходит «декомпозиция» стимула на два пространственных паттерна, соответствующих двухмерному аффективному пространству: позитивный и негативный аффект. При этом в ответ на такие «разложенные» стимулы могут наблюдаться моторные реакции (частично автоматические).
В. Чувство
На этой стадии информация достигает соматосенсорной коры и инсулы, где происходит картирование внутренних (висцеральных) и внешних (тело и лицо) атрибутов эмоциональных реакций в виде разных репрезентаций, соответствующих различным чувствам, например чувству радости или веселья158.
Г. Этап мониторинга
И наконец, на уровне передней поясной извилины происходит сравнение реальной и ожидаемой эмоциональной реакции. В зависимости от результата такой операции сравнения передняя поясная кора корректирует поведение (через моторную кору) таким образом, чтобы устранить выявленное рассогласование или оставаться в нынешнем состоянии дальше.
V. Серотонин как основной медиатор аффективной системы
Известно, что ядра ствола мозга модулируют процессы обработки информации, протекающие на уровне разных систем мозга. Говоря об аффективной системе мозга, следует отметить выраженное преобладание медиатора серотонина (5-гидрокситриптамин) в структурах, входящих в состав этой системы. Кроме того, традиционными мишенями при терапевтическом (фармакологическом) лечении расстройств аффективной системы являются как раз серотонинергические нейроны и рецепторы серотонина.
А. Функционирование и нарушения в работе серотонинергической системы
Серотонинергические проекции берут начало от дорсального и медиального ядер шва ствола мозга. В результате образуются две разные серотонинергические системы, отличающиеся по своим электрофизиологическим характеристикам, топографической организации, морфологии, а также чувствительности к нейротоксинам и, возможно, психоактивным агентам (рис. 16.6). Система тонких аксонов берет начало от серотонинергических клетчатых тел дорсального ядра шва, волокна которого обильно ветвятся в иннерви-
158 Так. при простом изменении выражения лица (скажем, при появлении улыбки) мы можем почувствовать эмоцию радости. В связи с этим мне вспоминается история об одном тибетском монахе, который лечил депрессивные состояния похожим способом. Однако радость может проявляться не только на уровне лицевой экспрессии, но и изменять частоту сердечных сокращений, скорость дыхания и других висцеральных процессов, которым соответствуют активации в разных областях коры, включая, в частности, инсулярную кору.
312 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Ядра шва
Рисунок 13.6. Двойная иннервация переднего мозга серотонинергической системой
Схема создана на основе данных, описанных в работе Hensler (2006).
руемых областях. Преимущественно иннервируются префронтальная кора, прилежащее ядро, амигдала и вентральный гиппокамп. Система толстых аксонов происходит от серотонинергических клетчатых тел медиального ядра шва, волокна которого иннервируют преимущественно гипоталамус и дорсальный гиппокамп159. На основе представлений о серотонинергической модуляции работы аффективной системы сформулирована моноаминная гипотеза депрессии. И действительно, если антидепрессанты вызывают увеличение концентрации серотонина в синапсе, то, например, ре
зерпин, являясь препаратом, разрушающим катехоламины, может вызывать похожие на депрессию синдромы.
VI. Заключение
С точки зрения психологии аффективная система добавляет дополнительное измерение (эмоции и мотивации) в наше восприятие и наши действия. Благодаря этому в общем виде регуляция нашего поведения сводится к поиску положительных эмоций и избеганию отрицательных. С информационной точки зрения аффективная система призвана усиливать и укреплять следы крайне важных для выживания событий, ассоциируя эти следы с определенным эмоциональным значением. Анатомическая организация аффективной системы, представляющая собой группу тесно взаимосвязанных корковых и подкорковых структур, обеспечивает возможность генерировать и чувствовать эмоции в ответ на сенсорную стимуляцию. Физиологически некоторые из этих структур (например,
159 Существует несколько типов серотониновых рецепторов, например: рецепторы 5-ГТ1А, 5-ГТ2А. 5-ПГ1А встречается только в ядре шва и выполняют ауторецепторную функцию, регулируя (подавляя) активность серотонинергических нейронов. Рецепторы 5-ГТ2А обнаруживаются в большинстве структур аффективной системы. Действие серотонина во внеклеточном пространстве ограничивается белками — переносчиками серотонина (SERT).
Часть 2, Вызванные потенциалы
313
орбитофронтальная кора и миндалина) приводят в соответствие события внешнего и внутреннего мира и эмоциональное значение этих событий: подкрепление (вознаграждение), наказание (отрицательное подкрепление), мотивации и потребности. Другие структуры аффективной системы являются ключевыми в обеспечении выражения эмоциональных реакций (лицевая экспрессия) в ответ на эмоционально значимые стимулы. Выделяются и структуры, ответственные за переживание эмоциональных реакций и состояний (инсулярная кора). В 1993 году Ричардом Дэвидсоном из Университета Висконсина была предложена гипотеза, в соответствии с которой лобные области коры левого полушария наиболее активны в ситуации эмоций, связанных с достижением (т.е. положительных эмоций). И напротив, предполагалась большая активность лобных областей правого полушария в случае эмоций, связанных с избеганием (отрицательные эмоции). Проще говоря, идея заключалась в том, что левое полушарие преимущественно реагирует на эмоционально положительные события, а правое, соответственно, — на эмоционально отрицательные. В силу того что мощность ЭЭГ в альфа-диапазоне инвертировано коррелирует с уровнем активации соответствующей области мозга, регистрируемая в лобных областях асимметрия альфа-ритма, по крайней мере для некоторых групп пациентов, может служить показателем депрессии или тревожности. Процессы обработки информации на уровне аффективной системы модулируются серотонинергическими системами, берущими начало от ядер шва ствола мозга. Некоторые из антидепрессантов вызывают увеличение концентрации серотонина в синапсе.
314
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Глава 14.
Системы памяти
I. Психология
А. Виды памяти
Лишь некоторые люди помнят о том, что древние греки помимо разных видов искусств изобрели искусство памяти, обычно называемое мнемотехникой. Во времена, когда еще не была изобретена техника печати, такой вид искусства был жизненно необходим. Несмотря на то что в современном мире мы полагаемся на информацию, хранимую в книгах, компьютерах и интернете, важность нашей собственной памяти сложно переоценить.
Главным достижением последних лет в исследованиях мозговых процессов научения и памяти является понимание того, что существует несколько видов памяти, обеспечиваемых разными системами мозга. Одним из показателей, который может быть использован в выделении видов памяти, является временная динамика следа памяти (время хранения). Соответственно, память можно подразделить на ультракороткую (длительностью около сотен миллисекунд), краткосрочную (длительностью порядка нескольких секунд) и долгосрочную (может сохраняться всю жизнь). Долгосрочная память, в свою очередь, может также подразделяться на две большие категории в зависимости от типа сохраняемой информации и мозговых механизмов ее обеспечения: имплицитную и эксплицитную память (табл. 14.1).
Декларативность памяти подразумевает осознание извлекаемой информации. В случае имплицитной памяти, напротив, процессы памяти протекают преимущественно бессознательно. Процедурная память, будучи имплицитной, обеспечивает научение и вспоминание моторных и когнитивных навыков. Различие между эксплицитным и имплицитным обучением может быть продемонстрировано на примере функции речи160.
Процессы, связанные с запоминанием любой информации, обеспечиваются целым набором мозговых систем. Однако в большинстве случаев существует конкретная мозговая система, играющая важную роль в обеспечении определенных форм
““Память играет важную роль в формировании и поддержании лексикона (словарного запаса) и грамматики. Декларативным видом памяти на примере речевой функции человека является запас специфических знаний, связанных со значениями слов (лексикон), в обеспечение которого вовлекаются мозговые структуры теменно-височно-затылочного стыка. Данный вид декларативной памяти подразумевает хранение и использование знаний о фактах или событиях, которые выражаются в словесной форме. Повреждения мозга в области теменно-височно-затылочного стыка могут приводить к существенному нарушению памяти лексикона, синдрому, известному как афазия Вернике. Грамматика, под которой понимаются процессы составления лексических элементов в сложные предложения (управляемые соответствующими правилами), обеспечивается разными мозговыми системами, включающими лобные и теменные области коры, базальные ганглии и структуры мозжечка. Грамматика является процедурным видом памяти. Нарушения речи, связанные с грамматикой, называются афазией Брока, которая может наблюдаться при повреждении левой фронтальной коры и подкорковых структур.
Часть 2. Вызванные потенциалы
315
Таблица 14.1. Виды памяти
Категория Подкатегория Содержание следа памяти Структуры мозга, ответственные за формирование и хранение
Декларативная (эксплицитная) Эпизодическая События личной жизни Гиппокампсенсорные области всех модальностей — префронтальная кора
Декларативная (эксплицитная) Семантическая Основные факты без явной связи с событиями личной жизни Гиппокампсенсорные области всех модальностей — префронтальная кора
Недекларативная (имплицитная) Процедурная Действия, например, такие как речь или вождение автомобиля Префронтальная кора — базальные ганглии — система таламических связей
Недекларативная (имплицитная) Инструментальные и условные рефлексы Ассоциативная память при ответах на условный стимул, с одной стороны, и безусловные ответы в ответ на стимулы положительного и отрицательного подкрепления (вознаграждение и наказание), с другой Сенсорные области всех модальностей, мозжечок, миндалина
Недекларативная (имплицитная) Прайминг* Активация в памяти частей определенных репрезентаций или ассоциаций непосредственно перед осуществлением действия или выполнением задания Вероятно, краткосрочные модификации синаптической связи нейронов, обеспечивающих активацию репрезентации стимула
Недекларативная (имплицитная) Габитуация Неассоциативный след памяти, формирующийся в результате снижения количества ответов при повторяющемся появлении стимулов Краткосрочная синаптическая депрессия
Примечание: * — прайминг является экспериментальной техникой, при которой предъявление стимула-праймера вызывает сенситизацию нейронной репрезентации этого стимула для дальнейшей презентации такого же или похожего стимула. Например, представим ситуацию, при которой испытуемому перед началом тестового задания предъявляется набор слов, среди которых есть слово «память». Позже на просьбу продолжить набор букв «пам» так, чтобы получилось слово, испытуемый скажет. «Память». То есть предварительный набор слов «праймировал» ответ испытуемого.
научения и памяти, которые могут существенно нарушаться при патологических изменениях в работе этой системы. Многие читатели могут вспомнить пессимистические выводы, сделанные Лешли (1950) по результатам серии исследований, посвященных изучению памяти, которые были опубликованы в известной статье под названием «В поисках энграммы» («In Search of the Engram»). При всем уважении к исследовательскому гению Лешли следует отметить, что существование разных видов памяти с разными нейронными субстратами не признавалось ни во времена Лешли, ни после появления современных аналитических средств анализа данных.
II. Декларативная память
А. Анатомия
Классический случай сильной амнезии, наблюдаемый у пациента Г.М. (одного из пациентов161 с билатеральным удалением медиальной височной коры), свидетельствует о важной роли этой области в обеспечении консолидации деклара-
161 Пациенту Г.М. в целях лечения устойчивой формы эпилепсии была сделана операция по удалению правой и левой медиальной нисочной коры, включая гиппокамп. В результате операции пациент больше не мог формировать новую декларативную память даже на высокоэначимые события.
316
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
тивной памяти. Медиальная височная кора характеризуется иерархической организацией (рис. 14.1): гиппокамп обладает мощными связями с энториальной корой, которая имеет сильные связи как с периренальной, так и парагиппокампальной корой; в свою очередь, эти области посылают свои проекции в область теменной и височной коры. Кроме того, гиппокамп связан с мамиллярными телами гипоталамуса и передним таламическим ядром. Известно, что повреждение
этих двух структур может приводить к амнестическому синдрому, известному как корсаковский синдром162. Исследования по оценке активности этих неболь-
ших близко расположенных и глубоко локализованных структур, проводимые с использованием томографии высокого разрешения (фМРТ), продемонстри-
ровали факт их активации во время выполнения тестовых заданий на память.
Аналогичные эксперименты на животных не только воспроизводят данные, полученные при исследовании людей, но и расширяют представление о мозговом обеспечении памяти: в частности, показано, что медиальная височная кора не является единственной гомогенной структурой, вовлекаемой в обеспечение процессов формирования памяти. Иными словами, процессы, связанные с памя-
Рисунок 14.1. Компоненты системы эпизодической памяти ТЬЮ, реализуются В
Обозначения, как на рис. 13.1 и 13.2. ’
результате сложного взаимодействия множества различных подструктур самой медиальной височной коры (рис 14.1), каждая из которых вносит свой индивидуальный вклад в формирование памяти.
В соответствии с современными представлениями медиальная височная кора рассматривается как сложная система, ответственная за реализацию ряда функций, связанных с памятью, включающих кодирование, консолидацию и извлечение новых следов памяти. Важно отметить, что хранение запоминаемой информации осуществляется на уровне височной, теменной и лобной областей
162 Сергей Сергеевич Корсаков, известный русский клиницист, впервые в 1887 году описавший вид амнезии (как ретроградную, так и антероградную) которая была связана с трофической (тиаминовой) недостаточностью при алкоголизме.
Часть 2. Вызванные потенциалы
317
коры, тогда как ее извлечение реализуется при активном вовлечении гиппокампа, который всякий раз «обращается» к хранилищу информации. В конечном итоге сами по себе следы памяти становятся независимыми от структур медиальной височной коры и находятся под влиянием структур неокортекса. В частности, считается, что декларативная память на зрительные объекты реализуется на уровне вентрального пути обработки зрительной информации163.
Похожие объекты, например чье-то лицо или определенная сцена, могут спонтанно запустить извлечение следа памяти. Однако в большинстве своем процессы вспоминания начинаются с целенаправленной попытки вспомнить. По всей видимости, активный процесс извлечения запоминаемой информации связан с вовлечением областей префронтальной коры. И действительно, с одной стороны, префронтальная кора получает от задних областей коры мозга в высокой степени обработанную сенсорную информацию, а также обеспечивает хранение планов действий и реализацию исполнительных функций (рабочая память, выбор, инициация и подавление действий). А с другой — префронтальная кора за счет нисходящих связей инициирует активацию запомненной информации, связанной с конкретным действием. Нейрофизиологические данные фактически подтверждают правдоподобность этой точки зрения: у пациентов с повреждениями в области левой префронтальной коры значительно нарушаются процессы произвольного вспоминания недавно запомненной информации164.
Б. Операции кодирования и извлечения
Память как единое целое проявляется в виде трех основных этапов: кодирования, хранения и извлечения. Хранение, в свою очередь, может быть долгосрочным и краткосрочным. Механизмы долгосрочного хранения исследуются клеточной биологией, изучающей структурные изменения синапсов нейронов. Краткосрочные механизмы хранения (или рабочей памяти) подробно рассматривались в гл. 11. В рамках данной главы мы рассмотрим операции кодирования и извлечения.
Вполне логично предположить, что в целях минимизации усилий и ресурсов следы памяти кодируются и извлекаются при участии тех же областей, которые вовлекаются в обработку предъявляемых сенсорных стимулов. Первые доказательства участия сенсорных областей в обеспечении процессов извлечения из памяти были представлены в 1940—1950-х годах в работах Вилдера Пенфилда из Неврологического института Монреальского университета (W Penfield & Р. Perot, 1963). Во время открытых операций на мозге, проводимых для лечения >11 иле пени, Пенфилд проводил электрические стимуляции областей затылоч
1ЬЗ Ключевыми областями в обеспечении этого вида декларативной памяти являются области нижней и латеральной височной коры, на уровне которых осуществляется формирование репрезентаций зрительных объектов. Формируемые репрезентации ассо-। (иируются с распознаванием, идентификацией объектов и долгосрочным хранением знаний, связанных сними. Таким образом, венчальная система обработки зрительной информации, будучи в функциональном плане тесно связанной с памятью, обеспечивает Формирование долгосрочных следов памяти (декларативной), а также их сравнение с сохраненной ранее информацией.
1М Более того, такие пациенты не используют стратегию организованного поиска элементов памяти. То есть если здоровые испытуемые склонны последовательно группировать конкретные предъявляемые слова пробы (например, «ложка» и «тарелка»), то пациенты с поврежденной левой фронтальной корой пользуются этим в меньшей степени, по существу вспоминая слова в случайном порядке. По всей видимости, по причине сложности усилий при таких нарушениях в извлечении информации из памяти некоторые пациенты начинают фантазировать. При этом поведенческий паттерн конфабуляций этого типа существенно отличается от паттерна, обычно наблюдаемого при потере памяти.
318
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
ной и височной коры. Во время электрического воздействия испытуемые находились в сознании. Оказалось, что такие стимуляции иногда приводили к появлению воспоминаний165, причем их модальность зависела от сенсорной специализации области.
Уже выполненные позже, в 1990-х годах, ПЭТ и фМРТ-исследования подтвердили эти выводы. Проведение этих исследований позволило получить прямое доказательство, что паттерны активаций метаболических процессов на разных этапах выполнения тестов на память, а именно при кодировании и извлечении информации, значительно перекрывались между собой. Так, в исследовании Вилера и др. (Wheeler et al., 2000) испытуемые запоминали зрительно предъявляемые слова в паре со звуком или изображением. Для выполнения задания необходимо было вспомнить, существовала ли смысловая связь (ассоциация) между предъявляемыми словами и сопутствующим стимулом. Такая конструкция тестового задания способствовала извлечению четких, модально-специфических следов памяти. В итоге процессы кодирования изображений ассоциировались с увеличением активности в области зрительной коры (от шпорной до веретенообразной извилины). Соответственно, кодирование слуховых стимулов вызывало активации в височной коре (от извилины Гешли до средней височной извилины). Извлечение информации об изображениях снова сопровождалось активацией в области веретенообразной извилины, а вспоминание звуков характеризовалось билатеральной активацией верхней височной извилины вблизи вторичных областей слуховой коры.
В свою очередь, операции извлечения могут быть разделены на два типа — операции сходства и воспоминания166. В одной из обзорных работ Майкл Ругг и Эндрю Йонелинас (2003) представили результат сравнительного анализа данных томографических, ВП и нейрофизиологических исследований, в соответствии с которым два разных типа операций извлечения обеспечиваются разными мозговыми механизмами.
В. Нейронные модели
Описание одной из наиболее популярных моделей работы гиппокампа, в соответствии с которой на уровне гиппокампа происходит кодирование и хранение информации о событии (эпизоде) (McNaughton, 1989), можно найти в гл. 14 (рис. 14.5). Вкратце, сенсорная компонента события кодируется височной и теменной корой, а компонента, связанная с действием, кодируется в лобных долях. Без гиппокампа следы памяти, относящиеся к конкретным событиям, могут храниться лишь непродолжительное время за счет реверберации периодической активности в нейронных сетях этих областей.
165 В этих исследованиях по электрической стимуляции верхняя и средняя височная кора считались ответственными за обеспечение слуховой памяти («... я слышу пение.. да, точно, это «Белое Рождество»»), а задние области височной коры и зрительная кора ассоциировались со зрительной памятью («... я видел кого-то приближающегося по направлению ко мне, словно он собирался ударить меня»).
166 Для понимания того, на чем основано такое разделение, достаточно вспомнить не самую комфортную ситуацию, когда мы распознаем кого-то как знакомого, но не можем извлечь из памяти ни имени, ни где могли ранее видеть этого человека. Наличие такого опыта является свидетельством разделения процесса вспоминания на две составляющие: 1) контекстуальное чувство сходства (знакомства); 2) извлечение детальной информации о событиях прошлого.
Часть 2. Вызванные потенциалы
319
Электрическая природа временного следа памяти обусловливает его высокую чувствительность к различным помехам (интерференции), что и лежит в основе его недолговременности. Именно на уровне гиппокампа происходит конвергенция компонент (в виде образа пространственно распределенной активности — репрезентации) в единый активационный паттерн. Образуемый таким образом след памяти характеризуется химически обусловленной долгосрочной потенциацией, индуцируемой на фоне всплесков тета-ритма. В силу этих изменений такой след может храниться достаточно долгое время. Таким образом, гиппокамп выполняет функцию временного хранения запоминаемой информации. Сохраненные следы памяти в дальнейшем могут извлекаться как сознательно при целенаправленном вспоминании, так и бессознательно, например, во время сна. Процессы извлечения обусловливают упорядочивание соответствующих следов памяти, в результате которого происходит формирование долгосрочных нейронных связей — то есть долгосрочной памяти.
III. Ацетилхолин как основной медиатор системы обеспечения декларативной памяти
А. Ядро перегородки как элемент восходящей холинергической системы мозга
Как было выяснено выше, ключевым элементом мозговой системы эпизодической памяти является гиппокамп. Гиппокамп генерирует уникальные ритмы, называемые гиппокампальными тета-ритмами. Процесс генерации этих ритмов контролируется системами мозга, основным нейротрансмиттером которых является ацетилхолин (АХ). Ацетилхолин продуцируется в ядрах перегородки и транспортируется в гиппокамп посредством септо-гиппокампального проводящего пути (рис. 14.2). От интенсивности транспорта АХ зависит амплитуда осцилляций тета-активности. Следует подчеркнуть, что ядра перегородки являются продолжением холинергической системы ствола мозга и обильно иннервируются из него.
Остальные области коры (за исключением гиппокампа), включая префронтальную, медиальную височную и инсулярную кору, получают входы от другого ядра ацетилхолинергической системы — базального ядра Мейнерта. Базальное ядро преимущественно связано с лимбической корой и, следовательно, может рассматриваться как интерфейс между лимбической системой и корой больших полушарий мозга. Эфферентные связи этого ядра также направляются в область ретикулярного ядра таламуса, что обеспечивает регуляцию информационных потоков от таламуса к коре.
Известны два вида рецепторов к АХ: никотиновые и мускариновые рецепторы167. Мускариновые рецепторы широко распространены по мозгу. Мускариновые рецепторы Ml-типа являются активирующими, а М2-типа соответственно тормозными. Н икотиновые рецепторы локализуются преимущественно на уровне ствола мозга.
167 Они получили свое название благодаря чувствительности к соответствующим алкалоидам.
320 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Б. Долгосрочная потенциация и гиппокампальный тета-ритм
Теоретически в соответствии со схемой, представленной на рис. 14.2, ацетил
холин может модулировать процессы эпизодической памяти с помощью двух механизмов: 1) увеличение амплитуды тега-ритма в гиппокампе и, соответственно,
усиление долгосрочной потенциации; 2) через активацию базального ядра, усиление активности многих областей коры в ответ на поведенчески значимый стимул168. Множество литературных данных свидетельствует о правомерности этой схемы.
Рисунок 14.2. Холинергические проводящие пути системы эпизодической памяти
Базальное ядро Мейнерта (BN) и септальное ядро (Septum) являются продолжением холинегрических нейронов восходящей ретикулярной формации ствола мозга. Холинергические нейроны перегородки проецируются в гиппокамп и соответствующие области коры. В то же время холинергический проводящий путь, берущий начало от базальных отделов переднею мозга, более диффузно проецируется в префронтальные области коры и ретикулярное ядро таламуса (Rt).
Так, фармакологические исследования показывают, что воздействия на холинергическую систему здоровых испытуемых могут приводить к существенным изменениям декларативной памяти. Например, ингибиторы ацетилхолинэстеразы168 169 улучшают декларативную память170. Аналогично введение скополамина, антихолинерги-ческого агента, вызывает нарушения процессов обучения при запоминании лиц (т.е. формирование ассоциаций «лицо — имя») и сопровождается снижением уровня насыщения крови кислородом в области вентролатеральной префронтальной и нижней височной коры, а также в гиппокампе. При введении этого препарата наблюдается снижение амплитуды РЗа-компоненты 171, регистрируемого в ответ на предъявление редкого девиантного стимула. Существование зависимости между
168 Базальное ядро в основном получает входы от лимбической системы, что формирует интерфейс между лимбической системой и корой.
169 Отметим, что ацетилхолинэстераза обеспечивает обратный захват молекул ацетилхолина из синаптической щели и тем самым продлевает активирующий эффект действий АХ на систему памяти.
170 Систематическое введение антагонистов мускариновых рецепторов АХ. напротив, может приводить к амнезии. Данный эффект наблюдался в исследованиях человека, приматов и грызунов. Известно, что блокировка АХ-передачи на уровне фронтальной и височной коры может приводить к дефициту памяти как при зрительном распознавании, так и при обучении, когда происходит формирование ассоциативных связей между объектом и положительным подкреплением. Кроме того, в случае иммунотоксического поражения базального ядра может наблюдаться снижение уровня продукции АХ на уровне фронтальных и височных отделов коры, степень которого коррелирует с сопутствующим нарушением процессов обучения.
171 В гл. 11, посвященной описанию мозговой системы внимания, упоминалась структурная гетерогенность, присущая нейронным структурам, ответственным за генерацию РЗа-компоненты ВП. Мозговая система внимания сильно взаимосвязана с системой памяти. По сути внимание усиливает ответ на выбранный источник информации и таким образом создает условия для кодирования и запоминания следа памяти. Известно также и то, что норадреналин, будучи основным медиатором мозговой системы внимания, может вызвать увеличение амплитуды РЗ-компоненты.
Часть2. Вызванныепотенциалы
321
продукцией АХ и амплитудой РЗа-компоненты демонстрирует факт контроля холинергической системой мозга усиления нейронного ответа на новые или контекстуально девиантные стимулы.
IV. ВП-показатели эпизодической памяти
А. Процессы вспоминания и эффект «старое — новое»
Существует несколько парадигм исследования нейробиологических основ эпизодической памяти. Принципиальная схема исследований универсальна — стимулы предъявляются на предварительной стадии запоминания, а непосредственно тестирование происходит во время стадии вспоминания. В исследованиях здоровых испытуемых применялись различные техники регистрации мозговой активности: фМРТ, ПЭТ и ЭЭГ. Гораздо реже применялся метод регистрации локальных потенциалов с имплантированных в структуры мозга электродов. Имплантация электродов осуществлялась исключительно в лечебно-диагностических целях.
В рамках исследовательской парадигмы «старое — новое» на этапе вспоминания показатели активности мозга регистрировались для двух видов предъявляемых стимулов. Первый тип стимула считался «старым», т.е. это стимулы, заученные испытуемым на предварительной стадии исследования. Второй тип стимула обозначался «новым», т.е. в рамках исследования такие стимулы предъявлялись впервые. Обычно наблюдаемое различие между ВП для «нового» и «старого» стимулов называется эффектом «старого — нового»172, а иногда — эффектом декларативной памяти. Получаемый в результате разностный ВП характеризуется выраженной височнотеменной топографией преимущественно в левом полушарии и отражает вовлечение сенсорных систем в обеспечение процессов эпизодической памяти.
Б. Эффект «запоминания — забывания» на стадии кодирования информации
В исследовании активности мозга при процессах запоминания и забывания регистрация осуществляется на этапе кодирования стимулов, которые необходимо запомнить. В частности, параметры ВП и ЭЭГ регистрируются во время кодирования стимулов (а не на стадии вспоминания), которые впоследствии вспоминаются или забываются. Эффект наблюдаемых различий этих параметров называется эффектом «запоминания — забывания». Известно лишь несколько исследований по регистрации импульсной активности с имплантированных электродов, проводимых в рамках данной парадигмы. Результаты таких исследований схематично представлены на рис. 14.3. Как видно из приведенных графиков, успешное запоминание стимулов (относительно забывания) проявляется: 1) как усиление
172 Данный эффект никогда не наблюдался на ВП, зарегистрированных в ответ на «старый» стимул, неверно определяемый испытуемым как «новый» (промахи), или «новый» стимул, ошибочно определяемый как «старый» («ложная тревога»). Значит, традиционно наблюдаемый на ВП эффект декларативной памяти в действительности не вызван ни повторением стимулов, ни процессами, связанными с принятием решения и ответа на стимул.
322
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Рисунок 14.3. Эффект «запоминания — забывания»
ЭЭГ-активность, зарегистрированная в ответ на стимулы, которые впоследствии будут запомнены (А) или забыты (Б). Вверху — тета-ритм скальповой ЭЭГ, зарегистриванной у здорового испытуемого. В середине — сочетанная активность в гамма-диапазоне ЭЭГ менаду обонятельной корой и гиппокампом, зарегистрированная с имплантированных электродов у пациента, страдающего эпилепсией. На рисунке представлено схематическое изображение результатов, полученных в работах Klimesch, 1999 (вверху), Fell et al., 2001 (в середине) и Fernandez et al., 1999 (внизу).
ответов на уровне гиппокампа; 2) увеличение когерентности между обонятельной корой (обладающей полимодальными связями с гиппокампом) и гиппокампом в гамма-диапазоне; 3) увеличение амплитуды тета-ритма173, регистрируемого со скальпа.
V. Система процедурной памяти
А. Сенсорная память и память, связанная с действием
Мозговая система процедурной памяти174 вовлекается в обеспечение процессов обучения новым комбинированным действиям и контроля реализации сформированных (приобретенных) сенсомоторных и когнитивных действий. Можно привести несколько примеров сенсомоторных действий: управление автомобилем, игра в теннис, игра на пианино, письмо или печать, артикуляция слов, пение... Такие действия также называются навыками, привычками, стереотипами (рутинными действиями)175. Чаще всего действия составляются из приобретенных ранее более элементарных действий. Таким образом, одной из функциональных задач процедурной памяти является координация и запоминание сложной последовательности простых действий. Как уже неоднократно упоминалось выше, мозг состоит их сильно взаимосвязанных, но
173 Функциональное значение тета-осцилляций ЭЭГ-активности описано в гл. 4.
174 Здесь термин «процедурная память» используется для обозначения только имплицитной, недекларативной мозговой системы памяти (см. табл. 14.1.).
175 Я вспоминаю моего хорошего друга, известного актера, который имел привычку говорить, что все актеры, когда играют, используют шаблоны и стереотипы. А хороший актер от плохого отличается лишь количеством этих шаблонов, выученных за время обучения.
Часть 2. Вызванные потенциалы 323
отдельных образований: сенсорных и связанных с действиями исполнительных систем176. Проще говоря, эпизодическая память связана с сенсорными системами мозга, а процедурная система ассоциируется с системами, имеющими отношение к действиям.
Б. Анатомия процедурной памяти
Области мозга, связанные с обеспечением действий, в первую очередь оперируют действиями. В гл. 12 уже говорилось о том, что стриатум, получая входы от областей коры переднего мозга, формирует «карту» действий. Овладение новым действием требует перестройки старой карты действий. Механизм такой перестройки и составляет основу процедурной памяти. Таким образом, процедурная память находится в сильной зависимости от работы базальных ганглиев и связанных с ними мозговых структур.
В отличие от эпизодической памяти системе процедурной памяти не требуется наличие отдельных систем кодирования и консолидации события. Сама исполнительная система располагает необходимыми элементами памяти. Энграмма процедурной памяти может быть представлена в виде медленно изменяющихся синаптических связей исполнительной системы (рис. 14.4). Представленная система состоит из петель положительных и отрицательных обратных связей фронтально-базально-таламокортикальной нейрональной сети. Таким образом, помимо исполнительных функций базальные ганглии вовлекаются: в обеспечение сенсомоторного сопряжения; организацию последовательных действий, подкрепленных обучением (в том числе с использованием эмоционально значимых стимулов в качестве подкрепления); в моторное планирование, особенно если оно характеризуется четкой временной структурой последовательности действий; в реализацию множества моторных программ177. В дополнение к рассмотренным мозговым структурам в обеспечении работы исполнительной системы участвует и мозжечок (не представлено на рис. 14.4)’78.
В. Базальные ганглии и речевая функция
Как было показано в гл. 12, базальные ганглии получают входы от фронтальных, теменных и височных областей коры, которые участвуют в параллельной обработке информации, осуществляемой на уровне разных систем проводящих путей. Эти параллельные пути обработки информации реализуют одни и те же базовые функции — выбор, инициация и подавление действий. В частности, зона Брока, как и любой отдел коры, сходным образом посылает проекции в область стриатума. Эго
' ” Следует напомнить, что области, преимущественно связанные с сенсорной информацией (сенсорные системы, системы вни-«м>и«м и. частично. аффективные системы), сохраняют информацию в виде различных фрагментов, элементов следа памяти. Важным именном системы сенсорной памяти является гиппокамп. Используя метафору «фрагмент памяти», можно говорить о том, что гип-««ммм1 обеспечивает ассоциативную связь фрагмента и сенсорного события (эпизода). Сенсорная система «удерживает» фрагмент «чмаии ми протяжении определенного промежутка времени, пока на уровне фронтально-височно-теменной системы не завершится «••дем жяяплидвции следа памяти. За д остаточно короткое время, необходимое для кодирования фрагмента памяти на уровне гип-«ьмммшльмоА системы, успевает произойти несколько осцилляций тета-ритма.
* ЙЛ» аишминается один из лечившихся у нас пациентов (в Институте мозга человека РАН), (предающих паркисонизмом, у которого «ижтное нарушение процесса обучения при выполнении простой GO/NOGO-задани. Он совершенно не мог запомнить про-♦•W жъиость действий: нажимал, кнопку при предъявлении СОсгимулов и воздерживаться от нажатия при ЮбОстимулах.
* * * Миме энмо мозжечок дополнительно вовлекается в обеспечение координации сложных бессознательных и воображаемых мммммае рук а темже мысленных вращений. Мозжечок важен для расчета времени и, по всей видимости, для координации после-««•«мпгмй действий, Подобно базальным ганглиям, мозжечок через таламус посылает свои проекции в область фронтальной
I Ipifim мвщдея область мозжечка четко проецируется на определенную область фронтальной коры.
Рисунок 14.4. Проводящие пути системы процедурной памяти
Базальные ганглии: 1) широко «картируют» действие на всем протяжении фронтально-теменной коры; 2) проецируют связи в область таламуса посредством прямых и непрямых путей (А); 3) через таламус проецируют связи обратно в область фронтальной доли (Б).
324____Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Часть 2, Вызванные потенциалы
325
указывает на то, что базальные ганглии участвуют в обеспечении всех функций, свойственных зоне Брока, включая процедурное обучение и грамматические правила179. Кажется вполне логичным, что повреждения в зоне Брока проводят к известным трудностям, называемым атрамматизмом. Таким пациентам сложно инициировать речь. Речь не беглая, затрудненная и с паузами. При этом происходит обеднение словарного запаса, а при составлении предложений используются разобщенные слова, которые с большим трудом собираются в подходящее предложение180. Более того, сопоставимые речевые нарушения могут наблюдаться и при повреждениях или электростимуляциях любой из структур, входящих в состав системы базально-таламических связей. В частности, в исследованиях Джорджа Ойд-жманна, ученого из Вашингтонского университета, сообщалось о похожих нарушениях в продукции речи при стимуляции доминантного вентролатерального таламуса. Вместе с тем стимуляция недоминантного вентролатерального таламуса не приводила к таким эффектам. Таким образом, характер речевых нарушений варьирует в зависимости от локализации стимулируемой o6nacTH(Johnson, Ojemann, 2000)181.
Г. Процесс постепенного запоминания
В отличие от быстрого кодирования информации на уровне декларативной памяти процессы обучения в рамках мозговой системы процедурной памяти протекают постепенно и требуют наличия множества ассоциативных связей между контекстом и осваиваемым действием. Вспомните, сколько времени заняло у вас обучение вождению автомобиля или игре в теннис, и сравните с теми несколькими секундами, которые потребовались на то, чтобы навсегда запомнить эпизод дорожного происшествия. И аналогично извлечение информации из эпизодической памяти связано с сознательным вспоминанием, которое в зависимости от типа сенсорной информации, занимает несколько сотен миллисекунд. А извлечение информации из процедурной памяти происходит быстрее и может осуществляться автоматически, без контроля со стороны сознания.
Д. ВП-корреляты процессов извлечения информации из процедурной памяти
ВП при извлечении информации из процедурной памяти в сравнении с процессами сознательного вспоминания представлены на рис. 14.5. Как видно из рис. 14.5, Р2-компонента ВП, регистрируемая в ответ на GO-стимул зрительного GO/NOGO-теста, отражает процесс вспоминания, реализуемый на уровне процедурной памяти. А результат сознательного вспоминания отображается РЗЬ-компонентой, связанной с операцией вовлечения (см. гл. 12). Таким образом, процессы процедурной памяти по крайней мере на 100 мс быстрее процессов эпи-юдической памяти, контролируемых сознанием.
*'* Ребенок постигает грамматическое строение речи матери постепенно и без специальных усилий. Однако, стал-миваясь с необходимостью изучить новый язык, нам приходится прилагать огромные усилия для формирования нового товарного запаса. При этом наибольшие трудности возникают в применении грамматических правил в беглой речи.
1-0 Например, так пациент с афазией Брока пытается объяснить, как он попал в госпиталь: «Да... ээ... Понедельник... ээ... Дэн и Питер Г... (его собственное имя), и Дэд.... ээ... госпиталь... и ээээ... Среда... Среда, 9 часов... и ох... Четверг... десять часов, rtOHiopa... два... доктора... и ээээ... зубы... да» (Goodlas, Geschwind, 1976).
Рвений профессор Владимир Смирнов (с которым автор работал в течение 15 лет) написал книгу «Стереотаксическая невроло-< и» (Smimov, 1976), в которой объединил все свои наблюдения за проявлениями электрических стимуляций подкорковых структур мозга, ।Ч1мк1димых у пациентов, которым по клиническим показателям в лечебных и диагностических целях имплантировались электроды.
326
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Рисунок 14.5. Процесс вспоминания из процедурной памяти протекает быстрее, чем сознательно контролируемое вспоминание стимула
Сравнение двух независимых компонент, вычисленных при анализе ВП, зарегистрированных в ответ на GO-стимул двухстимульного GO/NOGO-теста: (А, Б) — компонента, связанная с воспоминанием из процедурной памяти; (В, Г) -то же для сознательного воспоминания GO-события. Перед исследованием испытуемые выполняли тренировочный тест из 20 проб. Во время самого исследования тест выполнялся с несколькими ошибками. Первая компонента генерировалась в области премоторной коры фронтальной доли, а вторая (РЗЬ) — в области теменной коры.
VI. Основные медиаторы мозговой системы процедурной памяти
Теоретически можно выделить два основных медиатора, регулирующих работу системы процедурной памяти. Во-первых, это дофамин, транспортируемый в стриатум из черной субстанции. Во-вторых, это ацетилхолин, продуцируемый холинергическими клетками самого стриатума (рис. 14.6). Эти два медиатора модулируют информационные потоки на уровне стриатума. Дофамин изменяет пороги возбуди
Часть 2. Вызванные потенциалы
327
мости нейронов стриатума, а АХ активирует или деактивирует близлежащие эфферентные нейроны182.
Известные на сегодняшний день литературные данные подтверждают эту точку зрения. Свидетельством в пользу важной роли дофамина в реализации процессов процедурной памяти может служить ухудшение качества выполнения тестовых задач на процедурное обучение, которое наблюдается при паркинсонизме,
Рисунок 14.6. Медиаторные проводящие пути системы процедурной памяти Проекционные нейроны стриатума (ключевой элемент системы процедурной памяти отмечен черным цветом) получают входы от дофаминергических нейронов черной субстанции и от холинергических нейронов, локализованных в самом стриатуме.
характеризующемся истощением дофамина на уровне стриатума. Исследования на мы
шах показывают (Kitabatake et al., 2003), что селективная ампутация (повреждение) холинергических нейронов стриатума нарушает Процессы процедурного обучения в условиях управляемого звуком Т-образного лабиринта (звук указывает на наличие пищи в правом или левом рукаве лабиринта), а значит, это может являться доказательством участия АХ в обеспечении процедурной
памяти.
VII. Заключение
Существует несколько видов памяти, в обеспечение которых вовлекаются разные системы мозга. Один из принципов классификации памяти основан на представлениях о временной динамике процессов памяти — ультракороткая (сотни миллисекунд), краткосрочная (несколько секунд) и долгосрочная (вплоть до всей жизни). В свою очередь, в зависимости от механизмов обеспечения и типа сохраняемой информации долгосрочная память может быть подразделена на две большие категории. Это эксплицитная, или декларативная, память (в которой выделяют эпизодический и семантический подтип) и имплицитная, или процедурная, память (на уровне которой выделяются разновидно
Дофамин уже рассматривался нами в качестве основного медиатора исполнительной системы мозга, а ацетилхолин — как основной медиатор системы эпизодической памяти.
328
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
сти в вице процедурной памяти, инструментального и условного рефлексов). В состав структур системы обеспечения декларативной памяти входит гиппокамп, а также связанные с ним мамиллярные тела гипоталамуса и переднее ядро таламуса. ПЭТ и фМРТ-исследования свидетельствуют о совместной активации этих областей мозга при кодировании и извлечении информации из эпизодической памяти. Процессы кодирования сопровождаются генерацией гиппокампального тета-ритма, на фоне которого происходит долгосрочная потенциация и сохранение фрагментов следов памяти. ВП-корреляты процессов эпизодической памяти проявляются в виде двух основных эффектов: «старое — новое» при кодировании информации; «вспоминание — забывание» при извлечении информации. Мощность осцилляций тета-ритма регулируется медиатором ацетилхолином. Ацетилхолин продуцируется на уровне ядер перегородки и по септо-гиппокампальному пути транспортируется в гиппокамп. Процедурная память ассоциируется с научением новым видам моторных и когнитивных действий и обеспечивается базальными ганглиями (и связанными с ними структурами). Извлечение информации из процедурной памяти осуществляется гораздо быстрее, чем из эпизодической памяти. Процессы, относящиеся к извлечению информации из разных видов памяти, отражаются разными компонентами ВП, генерируемыми в ответ на GO-стимулы в условиях GO/NOGO-теста. Работа мозговой системы процедурной памяти регулируется двумя основными медиаторами: дофамином, транспортируемым в стриатум из черной субстанции, и ацетилхолином, который продуцируется собственными холинергическими клетками стриатума.
Часть 2. Вызванные потенциалы
329
Глава 15.
Методы: нейронные сети и вызванные потенциалы
I. Информационные процессы нейронных сетей
А. Аналитический подход
Одним из способов изучения информационных процессов мозга является аналитический подход, основная идея которого заключается в разложении сложных систем на более элементарные составляющие183. Успешным примером применения аналитического подхода являются исследования сенсорных систем мозга. В соответствии с ними восприятие рассматривается как отдельная операция, отличающаяся от движения. Более того, восприятие представляется как последовательность вычислительных операций, осуществляемых нейронами, разных иерархических уровней процесса обработки сенсорной информации.
Основы аналитического подхода были заложены в 50-х годах прошлого века. Несколько лабораторий стали использовать метод регистрации импульсной активности отдельных нейронов и изучать характер их ответов на стимуляцию разных модальностей. Большую известность приобрели исследования Хьюбелом и Визелем184 зрительной модальности восприятия. Их подход явился развитием концепции рецептивных полей, разработанной Стивом Куффлером также в 1950-х. Куффлер описал организацию рецептивных полей ганглиозных клеток сетчатки в форме ON-центра и OFF-периферии, а также ввел понятие латерального торможения. Хьюбел и Визель в своих работах показали, что рецептивные поля в первичной зрительной коре (VI) имеют более сложную организацию с протяженными ON- и OFF-областями, что позволяет нейронам селективно отвечать как на ориентацию предъявляемых полосок, так и на пространственные частоты двухмерных решеток.
После того как были описаны приблизительные свойства нейронов, исследователями завладела идея изучения того, как взаимодействие между отдель-
По смыслу аналитической подход противоположен синтетическому, в рамках которого изучается появление новых свойств (ложной системы, возникающих в результате коллективного поведения ее элементов.
Торстен Нильс Визель и Дэвид X. Хьюбел в 1981 году были удостоены Нобелевской премии по физиологии и медицине за открытие процессов обработки информации на уровне зрительной системы. В одном из экспериментов они регистрировали импульсную активность нейронов мозга кошки во время предъявления темных и светлых зрительных паттернов, появлявшихся перед кошкой на экране. Было обнаружено, что некоторые из нейронов коры демонстрируют быструю спайковую активность в ответ на предъявление линий, расположенных под определенным углом. Регистрируемые ответы весьма сильно отличались от ответов нейронов сетчатки и коленчатого тела таламуса. Хьюбел и Визель назвали эти реагирующие на линии нейроны простыми. Другие же нейроны, которые были названы исследователями сложными, лучше активировались при предъявлении движущихся в определенном направлении линий или решеток, позиционированных под определенным углом. Далее было показано, что различные виды нейронов рассеяны по зрительной коре не в случайном порядке, а образуют колонки и блоки (например, колонки ориентации или гпаэодрминантности).
330
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
(й
Б
Рисунок 15.1. Пространственная фильтрация в нейронных сетях с латеральным торможением
А. Структура нейронной сети с латеральным торможением. Сферы схематически обозначают нейроны. Каждый нейрон в положении х получает вход 1(х) и оказывает тормозное влияние на соседние нейроны. Б. Амплитудно-частотная характеристика ответа нейрональной сети на входящий сигнал синусоидальной формы (sin сох). По оси Y — F^oJ/F^JA — соотношение входа и выхода, со — пространственная частота входа. Чем шире латеральное торможение, тем более четкие
ними нейронами мозга может приводить к появлению новых свойств, например памяти, распознавания образов и даже сознания. Основоположниками этого нового направления исследований стали Маккалок и Пите, которые разработали новые методы регистрации активности нейронов. Это направление получило название исследований «нейронных сетей». В период с 1950 по 1980 гг. появились различные подходы к симуляции работающих нейронных сетей. Эти подходы впоследствии были объединены в общую концепцию нейронных сетей185.
Б. Сети с латеральным торможением
Латеральное торможение — это принцип работы нейронной сети, когда при активации определенного нейрона (группы нейронов) подавляется активность соседних (рис. 15.1 вверху). Такой вид нейронных сетей был впервые обнаружен Кеффером Хартлайном и коллегами из Рокфеллеровского университета при исследовании строения глаза и зрительной системы мечехвоста (Limulus polyphemus).
Обозначим вход, который получает нейрон,
амплитудно-частотныехарактеристики. в виде где х _ это координата определенного нейрона. В свою очередь, нейрон через связи с соседними нейронами
оказывает на них тормозное воздействие, распределение которого может быть описано весовой функцией W(y — х). W называется функцией связанности и отображает изменение силы связей между нейронами в зависимости от расстояния. Обычно распределение тормозных связей моделируется в соответствии с распределением Гаусса (нормальное распределение, см. формулу ниже) и имеет два параметра: амплитуду А и радиус действия При больших значениях А распределение изображается в виде высокого пика, при больших значениях кривая распределения имеет более широкий вид.
W(y-х) = А ехр(- (у-х)2/2а).
В. Пространственная фильтрация в нейронных сетях
Если получаемый вход клетка линейно переводит в выход (конечно, такая ситуация является явным приближением и правомочна тогда, когда потенциал мембраны удерживается у порога активации), то выходная функция Fo(x) ней
185 Симуляция нейронных сетей является быстро развивающимся направлением современной нейронауки. Данный подход преследует две цели: 1) создание общей теории обработки информации мозгом; 2) применение теоретических открытий в компьютерной науке (нейрокомпьютеры) и искусственном зрении, в сенсорной кибернетике.
Часть 2. Вызванные потенциалы
331
рона, занимающего положение х, может быть описана следующим образом:
f0(x) =k[fi(x) - fw(x -y)fjy)dyj.
Знак «—» перед интегралом означает, что клетки получают тормозные входы от соседних нейронов, Fo — выходящий сигнал, к—коэффициент усиления входящего и выходящего сигналов.
В терминах пространственной частоты (с использованием теоремы свертки) выше приведенное равенство будет иметь вид:
F (<») = к [Ffr)) - W((o)Fe((o)],
где F(w) — это преобразование Фурье функции/^, или:
Fo (ш) = kF.(m)/(l + kW(a>)).
Другими словами, такая нейронная сеть, благодаря механизму латерального торможения, работает как частотный фильтр — подавляя низкие частоты и оставляя высокие.
Преобразование Фурье гауссовской функции будет иметь вид:
W(w) = у!2л(г Ехр( -4л2ш27а2).
Таким образом, если латеральное торможение достаточно локально, то G будет небольшим, а фильтрация неэффективна, поскольку будут подавляться и низкие, и высокие частоты. Однако если латеральное торможение глобально, то подавляются только низкие частоты (рис. 15.1 внизу).
По всей видимости, такой тип нейронных сетей представлен на уровне стриатума, который характеризуется дальним действием своих тормозных связей. Значит, правомочно говорить о том, что стриатум, который, будучи структурой базальных ганглиев186, получает афференты от разных областей мозга, выполняет функцию фильтра. Режим работы этого фильтра модифицируется при обучении с вовлечением процедурной памяти и таким образом может настраиваться для обеспечения запоминания новых действий.
Г. Усиление высоких пространственных частот на уровне зрительной системы
Одним из свойств нейронных сетей с латеральным торможением является усиление наиболее выделяющейся информации, что делает ее еще более заметной. В терминах частотности усиление важной информации представляет собой подавление пространствен-t юго сигнала малой частоты. На уровне зрительной системы низкие частоты сигнала наи-
Стриатум получает входы от коры больших полушарий мозга. Если на уровне коры репрезентации разных действий могут 1Н'()екрываться, то на уровне стриатума этого не происходит (экспериментальные свидетельства этого можно найти в гл. 12 ч.2).
332
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Рисунок 15.2. Пример латерального торможения (полоса Маха) в зрительной системе
А. Изображение Маха — яркость меняется следующим образом: по внешнему кругу цвет белый, центр круга черный, а яркость линейно меняется в области между ними. Однако внешний круг мы воспринимаем как более яркий, а центр как более темный. Б. Кривая восприятия, получаемая при применении пространственного фильтра латерального торможения к обработке изображения.
менее значимы, поскольку содержат информацию об общем уровне яркости зрительной сцены. В свою очередь, высокочастотные пространственные сигналы кодируют небольшие детали изображения, важные для распознавания сложных объектов. Особенности функционирования нейронных сетей с латеральным торможением могут объяснить некоторые зрительные иллюзии (рис. 15.2).
Более реалистичная схема нейронной сети, состоящей из двух слоев нейронов, представлена на рис. 15.3. Помимо латерального торможения, для данной сети характерно наличие латерального возбуждения. Ситуация, при которой радиус тормозных влияний шире,
можно описать следующими уравнениями.
Выходная функция fo(x) нейрона локализации х нейронной сети, представленной на рис. 15.3, будет определяться следующим образом:
f0(x) = к[ fwjx -y)f(y)dy - Ц/х -y)f0(y)dy].
В частотном представлении получаем:
F» =kFi((0)+kWa(tO)Fo((O)-kWbi(tO)Fo((D).
Тогда соотношение между входом и выходом будет иметь вид:
Fo(a>) = kF.(o>)/(l + k(W.n(to) - WJv)).
Зависимость функции входа/выхода от входящего сигнала представлена на рис. 15.3 (справа). Видно, что нейронные сети с комбинацией механизма латерального торможения и латерального возбуждения малого радиуса работают как частотные фильтры, которые усиливают сигнал только в относительно узкой полосе частот. На протяжении всего проводящего пути зрительной системы, а в особенности на уровне сетчатки и латерально
Рисунок 153. Сеть нейронов с латеральным торможением и латеральным возбуждением
Взаимодействие менщу торможением и возбуждении приводит к появлению амгититуцно-пространственнь1х характеристик, изображенных на графике справа
го коленчатого тела, т.е. на ранних стадиях обработки информации, механизмы латерального торможения позволяют усиливать наиболее выделяющиеся аспекты сенсорных образов.
Д. Канонические кортикальные цепи
Широко распространенной моделью нейрона является модель Ходжкина — Хаксли187. Однако до сих пор не суще
187 Моделью нейрона Ходжкина — Хаксли являются нелинейные дифференциальные уравнения, описывающие инициацию и распространение потенциалов действия в нейроне. Алан Ллойд Ходжкин и Эндрю Хаксли за свою работу, выполненную в 1952 году, в 1963 году были удостоены Нобелевской премии в области физиологии и медицины.
Часть 2, Вызванные потенциалы
333
ствует общепринятой модели строения коры. Но большинство нейрофизиологов все же сходятся во мнении, что циклическая активность коры организуется в виде цепей связей тормозных и активирующих проводящих путей. В результате такой организации появляется ряд важных свойств, которые были описаны в первой модели подобного рода, разработанной в 1989 году Родни Дугласом и Кевином Мартином (Douglas et al., 1989, Douglas, Martin, 1991) и получившей название модели канонической кортикальной цепи (рис. 15.4).
В соответствии с этой моделью четыре популяции нейронов взаимодействуют между собой. Первой популяцией являются тормозные ГАМКергические нейроны. Остальные популяции нейронов возбуждающие: звездчатые нейроны IV слоя коры, пирамидальные нейроны верхнего слоя коры (слои II, III), а также пирамидальные клетки глубоких слоев (V и VI). В рамках предложенной модели процессы торможения и возбуждения не рассматриваются как независимые, а, напротив, предполагается их взаимодействие, что и является предпосылкой для возникновения уникальных свойств коры. Синхронная активация нейронов коры неизбежно приводит к запуску последовательности возбуждающих и тормозных событий. Времен-11ая д инамика этих событий зависит от локализации нейронов (слоя коры) и, по всей видимости, отображается положительными и отрицательными компонентами ВП.
Слой 2, 3.4
Слой 5,6
Таламус
Рисунок 15.4. Канонические кортикальные нейронные сети
Блок-диаграмма сети, моделирующая внутриклеточные ответы кортикальных нейронов на стимуляцию таламических афферентов. Взаимодействие происходит между тремя популяциями нейронов: одна популяция состоит из тормозных нейронов (обозначены закрашенным кругом), а две другие представлены возбуждающими нейронами, представляющими поверхностный (Р2 + 3) и глубокий (Р5 + 6) слои пирамидальных нейронов. На представленной схеме звездчатые нейроны IV слоя коры объединены с пирамидальными нейронами внешнего слоя. Нейроны каждой из популяций получают возбуждающие входы от таламуса. Соотношение толстых и тонких линий указывает на то, что большее активирующее влияние таламус оказывает на поверхностный слой пирамидальных клеток. Тормозные входы активируют ГАМК-А- и ГАМК-В-рецепторы пирамидальных нейронов. Толстые линии, соединящие нейроны на уровне Р5 + 6, слоя указывают на то, что тормозной вход к более глубоким слоям популяции пирамидальных клеток относительно больший по сравнению с поверхностым пирамидальным слоем. Однако увеличенное тормозное влияние обеспечивается только повышенной активностью ГАМК-А-рецепторов. Воздействие, оказываемое при активации ГАМК-В-рецепторов, одинаково для всех слоев клеток (см. Douglas, Martin, 1991).
Е. Торможение как следствие высокочастотной активности
Возбуждающие связи канонических кортикальных цепей доминируют над тормозными и характеризуются наличием петель положительной обратной связи. За счет реципрокного характера связей на уровне локальных цепей создаются условия для реверберации активности. В действительности чем больше нейронов активируется, тем больше постсинаптических потенциалов посылается на соседние клетки, т.е. увеличивается их деполяризация, и, следовательно, увеличивается количество разрядов. Таким образом, возбуждающие связи обеспечивают лавинообразный, экспоненциальный рост нейронной активности.
Однако тормозные связи позволяют подавить лавинообразное распространение активности. Тормозные нейроны вызывают гиперполяризацию соседних
334 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
и снижают их активность. Так, в период действия тормозного постсинаптического потенциала нейроны не генерируют спайки, но при его угасании возобновляют свою активность и т.д. Временная последовательность смены возбуждения и торможения продуцирует осцилляции определенной частоты, которая определяется длительностью тормозных постсинаптических потенциалов188. Исследования по регистрации внутриклеточной активности в сочетании с ио-нофорезом агонистов и антагонистов ГАМК (Douglas et al., 1989, Douglas, Martin, 1991) показали, что процессы внутрикортикального торможения опосредуются ГАМК-рецепторами А и В типа. Компонента тормозной активности, связанная с работой ГАМК А-рецепторов, наблюдается в ранней фазе импульсного ответа — гиперполяризации длительностью около 50 мс, которая сменяет фазу возбуждения. Вероятно, что реакция гиперполяризации этого типа отражается осцилляциями с частотой около 20 Гц, что соответствует бета-ритму ЭЭГ. Поздняя фаза ответа характеризуется гиперполяризацией длительностью около 200— 300 мс, которая связана с деятельностью ГАМК-рецепторов типа В.
Ж. Синаптическая депрессия как основа низкочастотных осцилляций
Другим механизмом подавления лавинообразного распространения активации является синаптическая депрессия. По сравнению с тормозными постсинаптическими потенциалами синаптическая депрессия — медленный процесс, длительность которого достигает нескольких сотен миллисекунд (400—500 мс). Следовательно, теоретически этот механизм может лежать в основе генерации осцилляций в частотном диапазоне дельта-ритма. В1990-х годах мы проводили исследования по моделированию поведения реалистичной нейронной сети с такой организацией внутренних связей, которая позволяла имитировать синаптическую депрессию. Компьютерные симуляции показывали, что в определенных условиях сеть начинала генерировать осцилляции с частотой, определяемой константой синаптической депрессии189. На рис. 15.5 представлены результаты этих исследований: видно, что при повторяющейся стимуляции сеть, состоящая из клеток, взаимосвязанных посредством синапсов, находящихся в состоянии депрессии, может продуцировать осцилляции. При этом частота осцилляций будет определяться константой синаптической депрессии.
3. Канонический кортикальный модуль
В 1980-х годах в нашей лаборатории была разработана математическая модель так называемого канонического кортикального модуля (Kropotov, Ponomarev, 1993). В основу базовой идеи модели легло предположение о том, что кора состоит из небольших модулей190. Каждый модуль занимает область, приблизительно соответ-
188 Как уже было продемонстрировано в гл. 3 ч.1, взаимодействие процессов торможения и возбуждения на уровне канонических кортикальных цепей лежит в основе генерации бета-ритма ЭЭГ.
ш Однако остается непонятным, встречаются ли такие условия в реальных нейронных сетях. Мы можем лишь спекулятивно предполагать, что в случае разрушения связей между областями коры и таламусом (например, вызванного опухолью) может возникнуть ситуация, при которой соответствующие области начнут генерировать ритмическую активность в частотном диапазоне дельта-ритма.
190 Организация модулей может зависеть от расположения области коры и, следовательно, может быть разной, например, в первичной зрительной коре и нижней височной области. Я был вдохновлен результатами работы, проводимой в то время Вадимом Глезером в Институте физиологии РАН им. И.П. Павлова (Санкт-Петербург). В своих исследованиях он продемонстрировал существование противоположных подполей рецептивных полей сложных и простых клеток зрительной коры кошки. Он предположил, что эти клетки обеспечивают преобразование Габора зрительных образов, приходящих от сетчатки.
Часть 2. Вызванные потенциалы
335
А Б
Рисунок 15.5. Осциляторная активность нейронных сетей, связанная с синаптической депрессией
Нейронная сеть с синаптической депрессией была симулирована на компьютере. Сеть состояла из 50 нейронов с сильно выраженными латеральными связями. А. Графики, отображающие изменения мембранного потециала всех 50 нейронов. Следует отметить, что возрастания мембранного потенциала происходили практически синхронно. Б. (рафик усредненной частоты разрядов нейронов. Обращает на себя внимание и периодический характер спайков.
336 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
ствующую квадрату размером 500 х 500 микрон. Предполагалось, что модуль демонстрирует весь набор операций, которые реализуются на уровне зрительной коры. Это операции кодирования всех возможных ориентаций и всех возможных пространственных частот, обеспечиваемые соответствующими областями зрительной коры. Предложенная модель основана на следующих принципах организации коры:
7. Принцип оппонентности клеток. В соответствии с этим принципом в состав нейронных цепей входят оппонентные нейроны, кодирующие комплементарные (оппонентные) характеристики входящей информации. Примером оп-понентных нейронов могут считаться отдельные ON- и OFF-каналы ретино-геникулярно-стриатной системы. Эти нейроны избирательно активируются в ответ на противоположные свойства зрительного образа: яркость кодируется ON-клетками191, а темнота — OFF-клетками.
2. Принцип совокупности. В соответствии с этим принципом клетки с одинаковыми характеристиками объединяются. Примером этого являются соматосенсорная, слуховая и зрительная кора. На уровне стриарной коры пластины глазодоминантности имеют форму параллельных полосок, а пластины пространственной частоты структурированы в форме спиц в колесе.
3. Принцип канонической кортикальной цепи. Одна и та же область коры (например, первичная зрительная кора) характеризуется одинаковой структурой микросвязей (цепей нейронов). В большинстве кортикальных областей некоторые из компонент и типов связей доминируют над остальными. Они складываются в так называемую каноническую кортикальную цепь (рис. 15.4).
4. Модульный принцип. Данный принцип подразумевает разделение кортикальных областей на модули с одинаковой внутренней организацией. В 1974 году Хьюбел и Визель первыми представили концепцию гиперколонок, которые являются функциональными единицами зрительной коры192.
Принимая во внимание представленные выше принципы, мы сформулировали представление о базовой архитектуре канонического кортикального модуля первичной зрительной коры. Можно заметить, что данные принципы не предлагают однозначного и единственного решения проблемы кортикальной организации. Вполне вероятно, что разные отделы зрительной коры (ПБ 17—19) и организованы по-разному, что позволяет им лучше выполнять свои специфические функции. Одно из возможных решений представлено на рис. 15.6.
Позвольте перейти к описанию основных операций, выполняемых каноническим кортикальным модулем (рис. 15.6). Пусть сигнал, поступающий на сетчатку, описывается функцией s(x, у). В соответствии с принципом оппонентности клеток на уровне таламуса приходящая от сетчатки информация кодируется на уровне двух каналов оппонентных клеток. Для упрощения дальнейших вычислений, введем новую переменную: S(x, у) = s(x, у) - saverage, где saverage— это средний уровень освещенности светового потока, попадающего на сетчатку. Функция S(x, у) отобража-
191 Яркость (темнота) определяется как локальное увеличение (уменьшение) освещенности по отношению к ее общему уровню.
192 Гиперколонка выполняет все возможные операции с информацией, приходящей из соответствующего участка зрительного поля. Гиперколонки повторяются в пределах зрительной коры.
Часть 2. Вызванные потенциалы
337
ет отклонение активности локального входа (локального участка сетчатки) s от среднего уровня фоновой активности Saverage. Тогда канал ON-клеток кодирует S(x, у), если S(x, у) > 0. При локальном снижении S(x, у), активируются OFF-клетки, т.е. происходит кодирование у>| = ~$(х> У), если S(x, у) < О193.
Далее предположим, что на таламическом уровне ON- и OFF-клетки распределены одинаково и расположены в разных слоях.
R| | Сетчатка
LGB I---------
т <>n Таламус
LGBoff
тормозные клетки простые клетки
Тормозные клетки простые клетки
сложные клетки
Рисунок 15.6. Канонический кортикальный модуль
А. Слои канонического кортикального модуля. С — сетчатка, LGB — латеральное коленчатое тело таламуса, Inl...ln4 — слои тормозных нейронов коры, S1...S4 — слои простых клеток коры, С — слой сложных клеток. Б. Распределение ON- и OFF-клеток в тормозном слове канонического кортикального модуля в виде паттерна «спиц в колесе». Белый цвет паттерна отображает расположение нейронов, получающих входы по ON-каналу. Черным цветом отображается положение нейронов, получающих входы по OFF-каналу. При появлении светового пятна активируются ON-клетки. OFF-клетки отвечают активацией на предъявление темных пятен изображения.
Огранизация тормозных ON и
OFF клеток в виде паттерна «спиц в колесе»
Тогда, в соответствии с принципом совокупности эфференты ON- и OFF-
клеток конвергируются на уровне тормозного слоя коры, формируя специфические мозаики. Теоретически возможно существование многих разных типов мозаик проекций ON- и OFF-клеток. Из всех вероятных пространственных паттернов мы опишем паттерн «спиц в колесе» (рис. 15.6, справа)194.
В соответствии с принципом канонической кортикальной цепи возбуждающие клетки кортикального слоя простых клеток получают прямые
позитивные входы от таламических нейронов и тормозные входы от тормозных клеток коры. Мы создали имитационную модель, состоящую из
четырех слоев тормозных нейронов, активность которых смещена относительно друг друга на величину п/2. Плотность распределения тормозных нейронов каждого их четырех слоев In. (I = 1, ..., 4) можно представить следующим образом:
П1„Ыо. = 1 -sin(©v +Jti/2) ni„l,wfr=1+sin(“'»' + 7n/2)
(1)
где ш — циклическая частота, определяющая число секторов мозаики; ц/ — полярный угол тормозного нейрона на уровне слоя In.
193 Известно, что мозг одновременно использует оба канала ON и OFF-типа вместо одного, поскольку разная информация распространяется по нервной системе за счет импульсной активности нейронов, значения которой всегда позитивны.
194 Спустя несколько лет после нашей публикации, посвященной модульной организации коры по типу «спиц в колесе», с помощью недавно разработанного метода оптической визуализации в первичной зрительной коре обезьян были выявлены несколько типов областей одинаковой ориентации. Полученные карты высокого разрешения позволили обнаружить преобладающий тип модульной организации, характеризующийся радиальным расположением, при котором формируется подобная «спицам в колесе» структура вокруг центральной точки (Bartfeld, Grinvald, 1992). Таким образом, наши теоретические предположения получили экспериментальное подтверждение.
338
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Каждый модуль содержит четыре слоя простых кортикальных нейронов (Sp 1=1,4) и один слой сложных нейронов (С). На уровне слоя сложных клеток интегрируются входы соответствующих простых клеток слоев простых клеток. Слой сложных клеток считается эфферентным слоем канонического кортикального модуля.
И. Фильтрация Габора в каноническом кортикальном модуле
Мы не будем приводить здесь все произведенные математические преобразования. Математически образованные читатели без труда смогут вывести конечную формулу информации, поступающей от сетчатки. Использование этой формулы позволяет определить пространственную частоту и ориентационные характеристики сложных нейронов, расположенных в разных частях модуля. В частности, эти характеристики представлены на рис. 15.7, из которого видно, что канонический кортикальный модуль выполняет частотно-пространственное разложение входящего сигнала. При этом разные частоты и ориентации отображаются в разных, отделенных пространственно областях модуля.
К. Кодирование текстуры в каноническом кортикальном модуле
Модель канонического кортикального модуля может применяться в различных практических приложениях компьютерного распознавания образов. Одно
Рисунок 15.7. Пространственная частота и селективность ориентации сложных клеток канонического кортикального модуля
А. Пространственно-частотные характеристики сложного нейрона — ответ нейронов с разными удалениями от центра модуля по радиусу на синусоидальную решетку с разными пространственными частотами со. Б. Ориентационные характеристики сложного нейрона — ответ нейрона на разные ориентацию (ц/) входа (т.е. части предъявляемой решетки).
Часть 2. Вызванные потенциалы
339
из вероятных свойств модели заключается в использовании алгоритма локального разложения Габора при обработке текстур входящего сигнала195. В
серии проводимых нами компьютерных симуляций для тестирования разработанной модели в качестве входящих сигналов были использованы разные текстуры. При этом анализировались ответы сложных нейронов (рис. 15.8): разным текстурам (значительно
Предъявляемая текстура
Результирующая активация сложных Б клеток
Рисунок 15.8. Кодирование текстуры в каноническом кортикальном модуле
А. Предъявляемая на входе текстура. Б. Результирующая активность слоя сложных клеток канонического кортикального модуля в ответ на соответствующую текстуру.
перекрывающимся на входе) соответствовали активации разных непере-крывающихся областей канонического кортикального модуля.
Из сказанного выше можно заключить, что нейронные сети со сложными
паттернами процессов торможения и возбуждения могут выполнять сложную пространственную фильтрацию входящих сигналов. Кроме того, если на входе разные элементы информации входящего сигнала могут накладываться друг на
друга, то на выходе будут наблюдаться неперекрывающиеся паттерны активности. На уровне зрительной коры локальное разложение Габора зрительных образов выполняется небольшими модулями (размером 500 х 500 микрон).
Л. Иерархическая организация
Информационные потоки коры головного мозга иерархически организованы. В частности, при рассматривании поведенчески значимого зрительного объекта сенсорная информация распространяется по иерархически организованной кортикальной системе — от первичных сенсорных (низших) областей к высшим областям. Латентности нейронных ответов на разных уровнях иерархии зрительной системы экспериментально оценивались в исследованиях на обезьянах. В частности, в исследовании Метью Смоленски и др. из Университета Юты (Schmolesky et al., 1998), проводимого на большом количестве приматов, были выявлены латентности ответов зрительных областей коры. Так, было продемонстрировано отличие латентностей ответов структур, входящих в состав двух основных путей обработки информации зрительной системой. Нейроны дорсального пути, берущего начало от магноцеллю-лярных слоев коленчатого тела таламуса, преимущественно вовлекаются в обеспечение восприятия пространственных отношений и движения196 и характеризуются меньшими значениями времени реакции. Нейроны вентрального пути, берущего начало от парвоцеллюлярных слоев коленчатого тела таламуса, преимущественно кодируют цвета объекта и обеспечивают распознание. Ответы этих нейронов характеризуются большей латентностью по сравнению с нейронами дорсального пути.
195 Под текстурой понимается набор элементов поверхности, заполняющих ее в более или менее периодической форме. Таким образом, сама по себе текстура может быть представлена как наложение небольших элементов разных пространственных частот и ориентаций.
196 Информация по дорсальному пути перенаправляется в зрительные области лобной и теменной коры для обеспечения навигации и манипуляций со зрительными объектами.
340
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
М. Прямые и обратные связи
На уровне иерархически организованных дорсального и вентрального путей помимо прямых синаптических связей, которые обеспечивают распространение информации в восходящем направлении, существуют и обратные, обеспечивающие возвращение информации, т.е. распространение в нисходящем направлении. Прямые и обратные связи берут начало и терминируются в разных слоях коры (рис. 15.9).
Прямые связи между низшими и высшими областями коры образуются аксонами пирамидных клеток. Преимущественно проекции оканчиваются на IV слое коры иерархически высшей области (аналогично таламическим проекциям). Основными мишенями проекций прямых связей являются шиловидные и
звездчатые нейроны, которые, в свою очередь, иннервируют базальные области
дендритного древа пирамидных нейронов на уровне II и III слоя коры.
Аксоны пирамидных нейронов V и VI слоев коры иерархически высшей обла-
сти дают начало обратным связям. В основном эти проекции оканчиваются в I и
Рисунок 15.9. Прямые и обратные связи коры
Слева и справа — низшие и высшие области коры соответвенно. Числа обозначают соответствующие слои коры. Прямые связи обозначены тонкими линиями. Обратные связи обозначены толстыми линиями. Пояснения см. в тексте.
VI слоях иерархически низших областей коры и подкорковых структурах (базальные ганглии). Отавными мишенями обратных связей являются апикальные дендриты (располагаются в I слое коры) нейронов II, III и V197 кортикальных слоев.
Таким образом, связь разных областей коры преимущественно носит реципрокный характер благодаря прямым и обратным связям. Такой тип связи (согласно Edelman, 1987198) также принято называть возвратным паттерном. Несмотря на то что о существо-
вании обратных связей было известно еще на ранних этапах развития нейрофизиологии, их функциональный смысл до сих пор окончательно не ясен. Извест-
но множество спекуляций относительно роли обратных связей. В соответствии с
одним из предположений нисходящие связи важны для осуществления процессов внимания и рабочей памяти. Другое мнение указывает на особую роль обратных связей в системе зрительных проводящих путей, а именно в обеспечении усиления контраста и выделения объекта из фона.
“7 Следеет напомнить, что медленные негативные потенциалы, регистрирующиеся со скальпа в условиях ожидания сенсорного стимула или подготовленного события, как это уже было сказано в m. 1 ч. 1, генерируются как раз за счет обратных связей. Нейроны обратной связи обеспечивают деполяризацию апикальных дендритов и снижение амплитуды электрического тока в верхних слоях коры.
“в Джералд Морис Эдельман (лауреат Нобелевской премии по физиологии и медицине в 1972 году за работы в области иммунологии) предложил теорию сознания, названную им «Нейронный дарвинизмом, или Теория отбора групп нейронов» (Edelman, 1987, 1993). Базовой концепцией его теории явилось представление о «возвратной сигнализации». По мнению Эдельмана, возвратная сигнализация является не только процессом коррекции ошибок, который основан на принципе обратной связи, но и механизмом создания новых групп нейронов, ответственных за реализацию действий.
Часть 2. Вызванные потенциалы 341
Н. ВП-проявления возвратных связей
Латентный период ответа нейронов зрительной коры обезьян варьирует от 76 до 110 мс, в зависимости от локализации. У человека аналогичные значения пиковой латентности компонент зрительных ВП больше. На рис. 15.10 представлены независимые компоненты ВП (и изображения плотности распределения их источников, полученные методом томографии малого разрешения — sLORETA), зарегистрированных у здоровых взрослых испытуемых (из HBI базы данных) в ответ на предъявление нерелевантных зрительных стимулов. На рис. 15.10 латентности пиков усредненных нейронных ответов зрительной коры обезьян на презентацию зрительного стимула приведены в овалах. Хотя сравниваемые результаты были получены на разных группах (люди и приматы) и с применением разных методик (регистрация импульсной активности отдельных нейронов у обезьян и использование метода независимых компонент199 у человека), с дидактической точки зрения такое сравнение представляется полезным. Во-первых, можно заметить, что независимая компонента ВП является суммой негативных и позитивных флуктуаций, отражающих последовательную генерацию возбуждающих и тормозных постсинаптических потенциалов коры. Полярность компонент при этом зависит от трех факторов: 1) тип постсинаптического потенциала (возбуждающий или тормозной), доминирующего в данный момент времени; 2) локализация; 3) ориентация активной области коры, генерирующей определенные компоненты; 4) слой коры, в котором генерируются постсинаптические потенциалы. Несмотря на то что эти факторы тяжело поддаются оценке, существует соблазн рассматривать позитивные компоненты зрительных ВП как отражение возбуждающих постсинаптических потенциалов и аналогично негативные компоненты ассоциировать с тормозными постсинаптическими потенциалами. Во-вторых, по ВП сложно судить о латентности появления компонент, поскольку мы можем оперировать только пиковыми латентностями, которые могут быть больше по сравнению с усредненной латентностью реакций импульсной активности. И наконец, самое важное — негативные компоненты ВП действительно отражают различия в скоростях информационного потока вентрального и дорсального пути (дорсальный путь «быстрее» вентрального).
II. Нейротрансмиттеры и нейромодуляторы
А. Быстрые трансмиттеры
Как уже было сказано в данной части книги, основу процессов обработки информации мозгом составляет преобразование импульсной активности пре-синаптического нейрона в более медленный мембранный потенциал200 постсинаптического нейрона. В свою очередь, информационные процессы могут быть подразделены на два класса операций нейронных сетей: информационный поток и информационная модуляция201. Реализация этих операций обеспечивается
199 Детальное описание можно найти в гл. «Методы» ч. 2.
200 Постсинаптические тормозные и возбуждающие потенциалы.
201 Под информационным потоком понимается быстрый процесс преобразования пресинаптической активности в постсинап-гическую. Этот процесс модулируется медленными изменениями мембранного потенциала, которые в данной книге объединяются тормином «модуляторный процесс» или, проще, «информационная модуляция».
342
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
188 мс
Рисунок 15.10. Латентности иерархически организованного процесса обработки информации зрительной корой В середине — схематическое изображение дорсального и вентрального путей обработки информации в зрительной системе со средними значениями латентностей нейрональных ответов на предъявление зрительных стимулов (в мс). Усредненные данные о латентности ответов получены в исследованиях на обезьянах, проводимых Мэтью Смоленски (Schmolesky et al., 1998). Значения латентностей расположены в овалах рядом с соответствующими таламическими или кортикальными структурами. Для сравнения представлены также локализация генераторов компонент, вычисленная с помощью метода томографии низкого разрешения (sUDRETA), и графики временной динамики независимых компонент. Независимые компоненты рассчитывались на массиве из 576 ВП-данных, зарегистрированных у испытуемых при предъявлении второго стимула Р-Р пробы двухстимульной GO/NOGO-парадигмы. Пиковые латентности основных флуктуаций амплитуды ЭЭГ — под соответствующими компонентами.
с помощью нейромедиаторов двух разных классов: медиаторов быстрого и медленного действия. Они выполняют разную функциональную роль в обеспечении процессов обработки информации.
Информационные потоки поддерживаются быстрыми медиаторами (рис. 15.11). Благодаря тому что эти медиаторы могут за несколько миллисекунд связываться с соответствующими рецепторами мембраны постсинаптических нейронов и передавать информацию, они называются нейротрансмиттерами. Обычно такие рецепторы являются лигандуправляемыми каналами и очень быстро открываются в ответ на пресинаптический сигнал. Так, спустя всего несколько миллисекунд после прихода сигнала на постсинаптическую тер
Часть 2. Вызванные потенциалы
343
миналь нейротрансмиттер, связываясь с рецептором постсинаптической мембраны, регулирует ионный поток (открывая или закрывая канал). К классу быстродействующих медиаторов принадлежат глутамат и гамма-аминомасляная кислота202.
Б. Медленные модуляторы
Медиаторы, ответственные за информационную модуляцию, действуют медленнее по сравнению с нейротрансмитеррами (рис. 15.11). Для того чтобы изменить мембранный потенциал постсинаптического нейрона, модуляторам требуется время порядка сотен миллисекунд или нескольких секунд. Функция этих медиаторов заключается в модуляции (медленном изменении) действия быстродействующих медиаторов. Действие нейромодуляторов по сравнению с нейротрансмиттерами может длиться долгое время (но при этом оно слабое), поскольку запускаемые ими каскадные реакции могут протекать в течение нескольких дней. К классу нейромодуляторов принадлежат: моноамины (норэпинефрин, дофамин и серотонин), ацетилхолин и нейропептиды203.
В. Модуляторные системы
В мозге существует четыре модуляторные системы: холинергическая, норадренергическая, серотонинергическая и дофаминергическая. Модуляторные си-
стемы характеризуются общими признаками: 1) берут начало от относительно небольших отделов ствола мозга и базальных отделов переднего мозга; 2) проецируются на ретикулярное ядро таламуса; 3) обильно иннервируют кору, базальные ганглии, мозжечок204 и центры ствола мозга205.
Холинергические и мо-ноаминергические нейроны мозга формируют глобальные сети. Хотя такие ней-
роны происходят из достаточно локальных областей
мозга, они занимают отделы
Рисунок 15.11. Нейротрансмиттеры и нейромедиаторы
А. Схематически представлено действие двух типов нейромедиаторов. В обоих случаях высвобождение медиатора в синаптическую щель вызывается спайном, приходящим на пресинагпическую терминаль. Как нейротрансмиттер, так и нейромодулятор действуют на соответствующие каналы мембраны, что приводит к их открытию. При этом действие нейромедиатора по времени более длительно по сравнению с нейротрансми-терром. Б. График временной динамики постсинаптических потенциалов, индуцируемых нейротрансмиттером и нейромедиатором. Обозначения: ПСП — постсинаптический потенциал.
203 Глутамат является общим возбуждающим медиатором мозга. В результате его действия открываются постсинаптические Na+ каналы, что, естественно, приводит к деполяризации. Как считается, ГАМК является тормозным нейротрансмиттером, который активирует СГ каналы и гиперполяризует мембрану. Эти два медиатора участвуют в передаче сенсорной информации о внешней и внутренней среда организма иерархически организованным сенсорным областям мозга. Они также ответственны за реализацию действий в ответ на постоянно меняющиеся условия окружающей среды.
303 Проводя аналогию с компьютерами и упрощая представление, можно говорить, что трансмиттеры выполняют функцию процессора, а модуляторы предоставляют необходимую для его работы энергию.
304 Однако иннервация не единообразна и неодинаково распределена. Для каждого медиатора существует несколько областей мозга, в иннервацию которых медиатор вносит больший вклад по сравнению с остальными.
305 Эти центры участвуют в поддержании основных функций—дыхания, сердечных сокращений и тд.
344 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия спинного мозга, ствола мозга и базальных отделов конечного мозга (рис. 15.12) и иннервируют кору и подкорковые структуры, моделируя информационные процессы. Нейроны работают как единая сеть, генерируя широко распространяющиеся паттерны активности совместно в фазах сна и бодрствования, настроения (радость и печаль), когнитивных состояний внимания и медитации.
Холинергические нейроны сконцентрированно локализуются на уровне ядер среднего мозга, ядер базальных отделов переднего мозга и базальных ганглиев. Из септальной области холинергические нейроны проецируются в гиппокамп. Нейроны базальных отделов переднего мозга посылают проекции в ретикулярное ядро таламуса, орбитофронтальную и другие области коры. Считается, что холинергическая система играет ключевую роль в регуляции общей возбудимости и формировании памяти (активируя гиппокампальную систему). Ацетилхолин рассматривается как основной регулятор мощности гиппокампального тета-ритма, при этом предполагается, что причиной болезни Альцгеймера может являться гибель нейронов, продуцирующих ацетилхолин. Именно этим обусловлено использование препаратов, увеличивающих уровень продукции ацетилхолина мозгом (ингибиторы ацетилхолинэстеразы) в лечении нарушений памяти у пациентов пожилого возраста (при болезни Альцгеймера).
Большинство норадренергических нейронов206 локализуется на уровне ствола мозга в области голубого пятна. Нейроны голубого пятна проецируются в лобные области коры, через бета1-рецепторы, регулируя настроение. Проекции, для которых характерно преобладание альфа2-рецепторов, участвуют в модуляции процессов, связанных с вниманием и рабочей памятью. Норадренергические нейроны также имеют связи с лимбической корой (регуляция эмоций), мозжечком (регуляция моторных действий), кардиоваскулярными центрами среднего мозга (регуляция кровяного давления). Считается, что депрессия может быть обусловлена снижением продукции норадреналина и, наоборот, ее увеличение (относительно нормы) может приводить к мании.
Серотонинергические нейроны в основном локализуются в ядрах шва ствола мозга и иннервируют лобные отделы коры и структуры лимбической системы, обеспечивая регуляцию настроения и эмоций (тревожность и эмоции при боли). Серотонинергические нейроны ядер шва, иннервируя базальные ганглии, участвуют в модуляции движений, а также навязчивых состояний и действий. Проецируясь в гипоталамус, нейроны, продуцирующие серотонин, регулируют аппетит и пищевое поведение. Дополнительно, благодаря связям с центрами сна на уровне ствола мозга, модулируются состояния сна и бодрствования.
В дофаминергической системе мозга выделяют мезолимбический, мезокорти-кальный и нигростриарный проводящие пути207. Дофаминергические нейроны черной субстанции в составе нигростриарного пути проецируются в базальные ганглии, которые играют определяющую роль в процессах выбора действий. Мезолимбический дофаминергический проводящий путь соединяет вентральную область покрышки ствола мозга с прилежащим ядром, которое, входя в состав ба-
206 Норадреналин также называют норэпинефрином.
207 Существует также тубероинфундибулярный дофаминергический путь, который берет свое начало в гипоталамусе и оканчивается в передних отделах гипофиза, регулирующих секрецию пролактина.
Часть 2. Вызванные потенциалы
345
Рисунок 15.12. Холинергические и моноаминергические нейроны мозга
Следует обратить внимание на то, что кластеры нейронов организованы на всем протяжении от спинного мозга до базальных отделов конечного мозга. Проекции от этих сравнительно немногочисленных нейронных образований иннервируют весь мозг и все периферические мышцы, органы и железы. Подробное описание см. в тексте.
зальных ганглиев, принимает участие в обеспечении работы лимбической системы и регулирует эмоциональное поведение — ощущение удовольствия, сильное чувство эйфории при употреблении наркотиков. Проекции мезокортикального дофаминергического пути, берущие начало от вентральной области покрышки ствола мозга, достигают дорсолатеральной префронтальной коры и поясной извилины (лимбической коры). Результатом фармакологических исследований, 1 гроводившихся в течение 30 лет, явилось создание препаратов, способных изменять концентрацию дофамина в мозге человека. Было показано, что препараты, приводящие к увеличению концентрации дофамина, вызывают появление положительных психотических симптомов. Тогда как действие препаратов, снижающих концентрацию дофамина, проявляется в снижении выраженности этих симптомов. В частности, употребление веществ, характеризующихся стимулирующим действием (как, например, амфетамин или кокаин, увеличивающие уровень продукции дофамина), может вызвать параноидальный психоз, сравнимый с шизофренией. С другой стороны, все известные антипсихотические вещества
346 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
(т.е. редуцирующие психотические симптомы) блокируют рецепторы дофамина. В частности, блокируются В2-рецепторы дофамина. Именно на этих представлениях и основана принимаемая многими специалистами теория психозов, известная как дофаминовая теория шизофрении.
III. Методы анализа вызванных потенциалов
А. Техника усреднения
Подавляющее большинство исследовательских парадигм, разработанных д ля изучения реакций мозга в ответ на стимулы или действия, подразумевают получение так называемых вызванных потенциалов, сигналов в форме волны, выделяемых из фоновой ЭЭГ посредством процедуры усреднения. Существует соблазн считать, что усреднение позволяет извлекать из фоновой ЭЭГ постоянный сигнал, связанный с мозговыми процессами обработки информации208. Предполагается также, что фоновая ЭЭГ является физиологическим параметром, отражающим процессы модуляции работы мозга. На рис. 15.13 представлены варианты записей ЭЭГ, сделанных в условиях последовательного предъявления двух видов стимулов в рамках GO/NOGO-парадигмы. Испытуемый был случайно отобран из нормативной базы. Отведение Fz выбрано не случайно, поскольку в лобных отведениях высокоамплитудный альфа-ритм фактически не наблюдается. Из рис. 15.13 видно, что участки ЭЭГ практически не отличаются. Для выявления тонкой разницы между ними необходимо произвести усреднение большого количества таких фрагментов209. Операция усреднения подразумевает суммирование паттернов ЭЭГ, синхронизированных с предъявлением стимула. Поскольку пробы не синхронизированы по времени с осцилляциями ЭЭГ и предъявляются случайно и независимо от фазы текущей ЭЭГ-активности, негативные и позитивные флуктуации в интервал времени, предшествующий появлению стимула, взаимно подавляются. В результате предстимульный интервал может приблизительно считаться прямой со средним значением потенциала, равным нулю.
Б. Количество проб
После усреднения фрагментов ЭЭГ, соответствующих пробам одной категории, получается ВП, представляющий волну, состоящую из нескольких пиков положительной и отрицательной полярности (рис. 15.13). Амплитуда пиков может варьировать в пределах от 1 до 15—20 мкВ в зависимости от задачи, испытуемого, его возраста и монтажа электродов. Следует напомнить, что средняя амплитуда ЭЭГ, зарегистрированной с электродов, располагающихся над задними областями коры, относительно ушных референтных электродов, у детей может
208 Однако теоретически, если сигнал меняется по мере выполнения тестового задания (например, привыкание или научение), то усредненный ВП может не соответствовать реальным потенциалам отдельных проб. Именно по этой причине исследователи до записи ЭЭГ предлагают испытуемым выполнить короткое тренировочное тестовое задание с целью удостовериться в правильном понимании инструкций теста, способности выполнить тест как можно быстрее и устранить ориентировочную реакцию.
209 Это именно та ситуация, когда визуальный анализ результатов не помогает. Для получения данных необходима вычислительная процедура (усреднение), при которой сигнал выделяется из спонтанной фоновой ЭЭГ-активности.
Часть 2. Вызванные потенциалы
347
быть около 70 мкВ, а у взрослых — около 50 мкВ. На рис. 15.13 представлены фрагменты ЭЭГ и ВП здоровых женщин в возрасте 32 лет. Обращает на себя внимание тот факт, что амплитуда спонтанных осцилляций в Fz-отведении превосходит среднюю амплитуду компонент ВП.
Если флуктуации ЭЭГ до предъявления стимула считаются шумом210 211, то усредненные ВП на интервале записи после начала предъявления стимула представляются сигналом. Интуитивно ясно, что уровень соотношения «сигнал — шум» зависит от количества усредняемых фрагментов ЭЭГ: чем больше проб усредняется, тем выше соотношение «сигнал — шум». Количество усредняемых проб, необходимое для достижения оптимального соотношения сигнала к шуму, зависит от амплитуды компонент ВП (сигнала) и амплитуды осцилляций фоновой ЭЭР11. Более того, считается, что усреднение не оказывает влияния на сигнал (s), а отношение сигнала к шуму (s/n) увеличивается с возраста-
нием числа (N) усредняемых проб (рис. 15.14). Теоретически можно доказать, что при определенных условиях отношение (s/n) увеличивается в л/N раз, где N — количество проб. Например, представьте эксперимент, в котором измеряется амплитуда компоненты РЗ, при этом реальная амплитуда компоненты составляет 20 мкВ. Если учесть, что амплитуда шума ЭЭГ-фрагмента одной пробы
ют, полученный в результате 5 усреднения 100 проб
Рисунок 15.13. Техника усреднения, используемая при вычислении ВП
Представлены фрагменты ЭЭГ, зарегистрированные при так называемых игнорируемых пробах двухстимульного GO/NOGO-теста (предъявления пары изображений типа «растение — растение», которое испытуемый должен игнорировать). А. Временная шкала пробы, обозначенной серым прямоугольником. Б. ВП, полученный в результате усреденния 100 проб. Вызванный потенциал, зарегистрированный в Fz-отведении, с высоким временным разрешением отображает паттерн нейрональной активности, вызванной предъявлением зрительного стимула. Для построения ВП из нормативной HBI базы данных были отобраны фрагменты ЭЭГ здоровых 32-летних женщин.
составляет 50 мкВ, нетрудно представить, что соотношение сигнала к шуму будет 20/50 или 0,4. Полученное значение соотношения s/n, следовательно, говорит о том, что шум превышает сигнал. Далее, если усреднить уже 5 проб, то соотношение изменится и будет составлять 2,2 (что не вполне удовлетворительно). Однако, усредняя 100 проб, мы в 10 раз увеличиваем сигнал относительно шума. Такая зависимость количества усредняемых проб и соотношения «сигнал — шум»
представляется весьма необнадеживающей, поскольку достижение существенного усиления сигнала относительно шума требует очень большого количества
210 В идеале амплитуда предстимульного интервала должна быть равна нулю, что может быть достигнуто только при усреднении бесконечного количества проб. На практике потенциал ВП до стимула не равен нулю, а степень такого отклонения от нулевого значения является показателем уровня шума.
211 Опыт — лучший помощник в выборе количества усредняемых проб. Нормативная база HBI Database создавалась исходя из эмпирически выведенного правила, в соответствии с которым при планировании исследования достаточно установить, что будет предъявляться не менее 100 проб для стимула каждой из категорий (за исключением исследований с регистрацией негативности рассогласования).
348 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
проб для усреднения. Например, чтобы достичь разумного соотношения s/n для регистрации негативности рассогласования, амплитуда которой колеблется в пределах 1—4 мкВ, необходимо предъявить около 2000 стандартных и 200 девиантных стимулов.
Соотношение «сигнал — шум» зависит по крайней мере от трех факторов: 1) вариабельность фоновой спонтанной ЭЭГ активности (альфа-, бета-, тета-осцилляции); 2) изменения самого сигнала, обусловленные привыканием, флуктуациями уровня возбуждения и т.д.; 3) артефакты, не связанные с ЭЭГ-активностью (движения глаз, мышечная активность и т.д.). Для минимизации влияния первых двух факторов соответствующим образом подходят к организации тестового задания. Для борьбы с третьим фактором применяют методы коррекции артефактов (см. гл. «Методы», ч. 1).
Рисунок 15.14. При построении ВП соотношение «сигнал — шум» от количества усредненных проб как квадратный корень из числа проб
А. Топография компоненты ВП, зарегистрированного спустя 160 мс с момента предъявления стимула. Б. ВП, полученные при усреднении 5,10,20 и 100 проб. ЭЭГ регистрировалась с Fz-отведения на тех же здоровых испытуемых, что на рис. 15.13.
В. Независимые компоненты ВП одной пробы
Считается общепринятым, что усредненные ВП могут быть представлены в виде набора отдельных компонент, генерируемых разными источниками коры. В случае если отношение сигнала к шуму недостаточно велико, теоретически существует возможность разложения фрагмента ЭЭГ одной пробы на отдельные компоненты. В нормативной HBI базе данных такая возможность предоставляется специальной вычислительной процедурой, в основе которой реализован алгоритм анализа независимых компонент. На рис. 15.15 приведен результат применения этого алгоритма к анализу нескольких фрагментов ЭЭГ последовательно предъявляемых проб с разными стимулами GO/NOGO-теста. Для анализа был отобран тот же испытуемый, что и на рис. 15.13.
Как видно из рис. 15.13, на усредненном ВП выделяется индивидуальная P1/N2-компонента, которая может был» разложена на две отдельные компоненты, одна из которых генерируется в левой, а другая — в правой нижней височной доле (пра
Часть 2. Вызванные потенциалы
349
вая часть рис. 15.15, на которой изображен результат применения томографии низкого разрешения). Следует упомянуть, что на обычном ВП максимальная амплитуда Р1/М2-компоненты составляет всего 2,3 мкВ (в Fz), однако применение метода независимых компонент позволило разложить ВП на две отдельные компоненты. Более того, эти две компоненты также могут выделяться и при анализе единичных проб (рис. 15.15. в центре)212.
При использовании вышеуказанного метода выполняется определенная последовательность действий, описываемых ниже. Сначала с помощью метода независимых компонент (у одного испытуемого) анализируются фрагменты ЭЭГ в единичных пробах. Далее, используя топографии выявленных независимых компонент, с помощью метода томографии низкого разрешения (sLORETA) вычисляется локализация генераторов компонент.
Возможен и другой подход, при котором в первую очередь выполняется анализ независимых компонент для записи ЭЭГ, включающей весь временной интервал выполнения тестового задания (не синхронизированного с предъявлением стимула). В результате анализа выявляются компоненты, связанные с разными типами ЭЭГ-осцилляций (те. фронтальный срединный тета-ритм, затылочный и теменной альфа-ритм, мю-ритм). Далее, используя пространственные фильтры, сформированные на основе соответствующих топографических особенностей, выявляются компоненты ЭЭГ-активности. И наконец, для выявленных ритмов выполняется построение ВП213.
Г. Звон (ringing) альфа-ритма
Пример применения второго из упомянутых выше подходов проиллюстрирован на рис. 15.16. В данном случае ЭЭГ регистрировалась в ситуации выполнения двухстимульного GO/NOGO-теста. С использованием метода независимых компонент были выявлены компоненты ЭЭГ. Следует оговориться, что эти выявляемые компоненты отличаются от тех, которые вычисляются для отдельных фрагментов ЭЭГ, синхронизированных с предъявлением стимула и соответствующих отдельным пробам. Топография и локализация генератора выявленных компонент (методом топографии низкого разрешения) свидетельствуют о теменном источнике этой компоненты. Далее проводилась фильтрация фоновой ЭЭГ с учетом выявленной компоненты и высчитывались ВП. Полученные ВП представлены на рис. 15.16, где показано, что в предстимульном интервале усредненные флуктуации ЭЭГ весьма малы. С предъявлением зрительного стимула картина меняется — за счет вызванной стимулом регуляции фазы флуктуаций увеличивается ам-। титула ВП — эффект, получивший название «звон альфа-ритма». Похожий подход был предложен группой исследователей, возглавляемой Скоттом Макейгом из Калифорнийского университета Сан-Диего, США (Jung et al., 2001).
717 Сравнительно недавно группой исследователей, возглавляемой Скоттом Макейгом из Калифорнийского университета (Сан-Диего). был предложен похожий метод анализа (Tsai et al., 2006). В этом исследовании был представлен статистический подход для < щновременной оценки как временного паттерна независимых компонент ЭЭГ, так и их пространственного распределения по коре. В iwiMKax предложенного подхода для изображения кортикальных источников характерных проявлений ЭЭГ-активности использовался метод пространственно-временного анализа, основанный на теореме Байеса. Данный подход позволяет исследователям учитывать пприорное знание о пространственном расположении и пространственно-временной независимости разных источников ЭЭГ. Метод получил название метода электромагнитного пространственно-временного анализа независимых компонент (EMSICA).
711 Данный метод был разработан той же группой из Калифорнийского университета (Сан-Диего, США).
Рисунок 15.15. Разложение фрагмента ЭЭГ одной пробы на независимые компоненты
Метод независимых компонент применялся к анализу ЭЭГ, регистрировавшейся у здоровых испытуемых при выполнении двухстимульного GO/NOGO-теста, ВП которых пред1 ставлены на рис. 15.13. В верхней и нижней части рисунка представлены две независимые компоненты, соответствующие Р1/М2-компоненте индивидуальных усредненных ВП. Слева — топографии и графики временной динамики компонент. В середине — вертикально уложенные тонкие горизонтальные окрашенные полосы, отображающие изменение во времени амплитуды компоненты отдельных проб. Всего представлено 400 проб. Справа — локализация генераторов компонент, вычисленная с помощью метода томографии низкого разрешения (sLORETA).
350____Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Часть 2. Вызванные потенциалы
351
Д. Анализ усредненных по группе ВП с помощью метода независимых ком
понент
Индивидуальный ВП отдельного испытуемого состоит из пиков положительной и отрицательной полярности. Усредненное значение амплитуды пика является отражением совокупности отдельных компонент. Предполагается, что эти компоненты генерируются в разных областях мозга и ассоциируются с определенными психологическими операциями (факторами). Следует, однако, отметить, что компоненты вовсе не обязательно связаны с позитивными и негативными пиками ВП214.
Идея разложения ВП на отдельные компоненты эквивалентна идее редукции многомерных ВП до меньшего количества составляющих. Предполагается, что выявляемые потенциалы отображают различные психологические операции, реализуемые специализированными мозговыми структурами со специфической
временной динамикой активности.
Первая попытка разложения ВП на компоненты была предпринята в 1970-х годах215. Для этого использовался метод главных компонент. Как уже было сказано в гл. «Методы» ч. 1 книги, метод главных компонент вы-
Рисунок 15.16. Проявление звона (ringing) альфа-ритма на ВП
ЭЭГ регистрировалась у здорового испытуемого, характеризующегося альфа-ритмом ЭЭГ высокой амплитуды в теменной области. Испытуемый выполнял двухстимульный зрительный GO/NOGO-тест. Анализировались только пробы игнорируемого типа (т.е. «растение — растение»). Представлены ответы только на первые стимулы проб. Компоненты ЭЭГ (ритмы) выявлялись с помощью метода независимых компонент. А. Топография наиболее мощных компонент. Б. Локализация генератора теменной компоненты альфа-ритма (третьего из представленных), вычисленная с помощью метода томографии низкого разрешения (sLORETA). На основе топографии этой компоненты был получен пространственный фильтр, с помощью которого осуществлялась фильтрация фоновой ЭЭГ. В. ВП, полученный в результате стандартной процедуры усреднения. В предсти-мульном интервале ритмическая активность выражена слабо. Однако после предъявления стимула меняется фаза ритмической активности, что приводит к существенному увеличению амплитуды ВП.
являет только ортогональные компоненты216. Однако физиологически значимые компоненты не обязательно являются ортогональными. В 1980-х годах мы в нашей лаборатории довольно-таки безу-
спешно пытались использовать этот метод для анализа импульсной нейронной активности. Основную сложность составляла физиологическая интерпретация
выделяемых компонент.
214 Напомним, что в данной книге позитивные флуктуации ЭЭГ отображаются как отклонение потенциала вверх по оси ординат, а негативные флуктуации, соответственно, вниз.
215 Я помню, как в нашей лаборатории в исследованиях по регистрации импульсной активности нейронов мы применяли метод главных компонент для выявления физиологически значимых факторов. Мы использовали тогда французский компьютер IN-110, и для того, чтобы выполнить все расчеты, мне требовалась вся ночь. В те дни было обычным делом использовать ночное время для вычислений на компьютере.
216 В двухмерном или трехмерном пространстве ортогональными называют векторы, располагающиеся перпендикулярно относительно друг друга (формируя угол 90°).
352
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
И только недавно был разработан метод, позволяющий успешно разрешить проблему разложения на компоненты. Решение этой задачи стало возможным с появлением метода независимых компонент (ICA)217, краткое описание которого приведено в гл. 8.
Метод независимых компонент применяется в анализе усредненных по группе ВП с учетом следующих допущений: 1) линейный характер суммации электрических кортикальных токов, регистрируемых скальповыми электродами и связанных с разными психологическими операциями; 2) стабильное во времени пространственное распределение генераторов компонент; 3) временная независимость разных операций и, следовательно, их генераторов, активность которых регистрируется скальповыми электродами; 4) распределение потенциалов не является нормальным, т.е. не соответствует распределению Гаусса. Конечно, эти предположения сложно проверить экспериментально, и только на практике можно оценить корректность метода в целом. Тем более что опыт нашей работы не подтвердил необходимости выполнения четвертого условия (В.А. Пономарев). И до настоящего времени применение метода независимых компонент не только соответствовало всему набору экспериментальных данных, накопленному в предыдущих исследованиях, но и способствовало возникновению новых идей в представлении о принципах информационных мозговых процессов, исследуемых в лабораторных условиях. Кроме того, благодаря этому методу появились новые возможности в клинической диагностике с использованием регистрации электрической активности мозга.
Пример применения метода независимых компонент на массиве ВП, зарегистрированных у более 800 здоровых человек, представлен на рис 15.17. Независимые компоненты ВП приведены в столбце, сверху вниз, в зависимости от латентности пика максимальной амплитуды. Как видно из рис. 15.17, только лишь несколько компонент, расположенных сверху, могут быть достаточно хорошо аппроксимированы эквивалентным единичным диполем (параметр RRE — относительная остаточная энергия — меньше 0,1 )218. Выявлены следующие компоненты: 1) компонента, связанная с обработкой зрительной информации, которая генерируется в области затылочной коры (компонента 2 на рис. 15.17); 2) компоненты, связанные с подавлением подготовленного моторного действия (компоненты 1 и 11). Остальные компоненты, по всей видимости, генерируются пространственно распределенной системой генераторов и не могут быть описаны единичными диполями.
На рис. 15.18 вместо дипольных источников представлены паттерны, полученные с помощью томографии низкого пространственного разрешения
217 В обоих методах главных и независимых компонент используется корреляционная структура набора ВП-данных (так называемые ковариационные матрицы). Таким образом, предполагается, что извлечение компонент, осуществляемое этими методами, основано на функциональных взаимоотношениях между ними. Следует оговориться, что любой метод, основанный на корреляции, имеет свои ограничения. Так, когда два отдельных когнитивных процесса коварьируют, то, несмотря на то что они протекают в разных областях мозга и отражают разные операции, они могут быть определены как единый компонент. Это, несомненно, является ограничением. Например, если детекция изменения в физической и смысловой модальности коррелирует, то метод независимых компонент скорее выявит суперпозицию компонент, чем обнаружит две отдельные компоненты с разной временной структурой и топографией. Другое ограничение метода независимых компонент проявляется в тех случаях, когда латентность одной и той же компоненты существенно варьирует между разными условиями: в такой ситуации метод независимых компонент будет рассматривать эту единую компоненту как множество разных компонент.
218 Принято считать, что остаточная относительная энергия (Residual Relative Energy — RRE) в таких случаях должна быть меньше 0,1, т.е. меньше 10 % (RRE < 0,1).
Часть 2. Вызванные потенциалы
353
(sLORETA). Пиковые латентности компонент отмечены вертикальными линиями, а числа обозначают значения Л П соответствующих компонент. По рис. 15.18 несложно проследить поток распространения информации по зрительной коре в условиях NOGO-стимула, который включает несколько этапов. Первый этап: активация зрительной коры в районе первичной зрительной области с латентным периодом 116 мс. Данная компонента отображает повторные возбуждения затылочной коры. Второй этап: спустя 144 мс с момента предъявления стимула активируется затылочно-височная область, входящая в состав вентрального проводящего пути зрительной информации. Третья компонента регистрируется непосредственно сразу после детекции изменения семантического значения стимула и ассоциируется с активацией левой премоторной области коры. Данная компонента отражает активацию нейронов, ответственных за подавление подготовленного движения. Четвертый этап: с латентным периодом 276 мс наблюдается активация в медиальной области теменной коры, которая соответствует традиционной РЗЬ-компоненте, отражающей операцию обновления содержимого рабочей памяти (в соответствии с Дончиным). Пятая компонента генерируется в медиальной префронтальной коре с латентностью 336 мс и ассоциируется с подавлением действия в целом. Данная компонента явно отличается от третьей компоненты, связанной с подавлением движения. И наконец, на уровне передней поясной извилины происходит конвергенция всей информации, касающейся текущего поведения, где рассогласование между ожидаемым действием (нажатие кнопки — ответ) и наличествующим (воздержание от нажатия кнопки — игнорирование) приводит к активации нейронов.
IV. Вызванные потенциалы в фармакологических исследованиях
Во введении к настоящей книге мы коротко рассмотрели так называемый фармакологический ЭЭГ-подход. Основная идея подхода заключается в использовании количественных показателей ЭЭГ для оценки изменений мозговой активности, вызываемой приемом фармацевтических препаратов. На протяжении последних 50 лет в многочисленных исследованиях было показано, что разные классы фармацевтических агентов по-разному влияют на пространственно-временные характеристики фоновой ЭЭГ. Вместе с тем индивидуальные профили чувствительности ЭЭГ к разным препаратам могут существенно перекрываться, что значительно снижает «размер эффекта» и автоматически делает невозможным применение такого подхода в клинической диагностике и лечении. Это является одним из множества препятствий, ограничивающих применение фармакологического ЭЭГ-подхода в клинической практике. Однако определенный оптимизм внушает использование метода независимых компонент в анализе ВП, который может наиболее эффективно оценивать функционирование мозговых систем и, следовательно, характери-
354
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Рисунок 15.17. Метод независимых компонент в анализе усредненных по группе ВП
Независимые компоненты извлекались из массива состоящих из 800 индивидуальных ВП, полученных в условии NOGO, на второй стимул пробы (т.е. пара стимулов «животное — растение») двухстимульного GO/ NOGO-теста. Компоненты последовательно представлены в соответствии с латентностями максимальных пиков. В середине — вертикально уложенные тонкие горизонтальные окрашенные полосы, отображающие изменение во времени амплитуды соответствующих компонент индивидуальных ВП и их временнную динамику. Справа — аппроксимация эквивалентным диполем представленных компонент. Качество аппроксимации отображено в показателе остаточной относительной энергии. Амплитудные шкалы расположены рядом с соответствующими изображениями.
Часть 2. Вызванные потенциалы
355
Рисунок 15.18. Локализация генераторов независимых компонент, вычисляемая с помощью метода томографии низкого разрешения (sLORETA)
Представлены независимые компоненты (те же, что и на рис. 15.17), а также локализации их генераторов, полученные томографией малого разрешения. Названия областей коры, для которых фиксировалась максимальная плотность компонент ВП, представлены рядом с паттернами локализации источников компонент. Следует обратить внимание на поток распространения зрительной информации на протяжении от затылочной области коры до передней поясной извилины.
юваться большим размером эффекта. Развитие этого нового, появившегося сравнительного недавно направления фармакологических ЭЭГ-исследований требует дальнейших исследований и экспериментальных проверок. Ниже мы представим методологию, которая может быть использована в рамках данного подхода.
Предлагаемый нами подход состоит из нескольких этапов. В первую очередь у репрезентативной группы здоровых испытуемых (норма) в условиях определенного тестового задания производится регистрация ВП. На втором папе с помощью метода независимых компонент осуществляется разложение на отдельные компоненты усредненных по группе испытуемых ВП. Выявляемые компоненты, генерируемые в разных областях коры, характеризуются разной динамикой временной активности и являются независимыми друг от друга. На третьем этапе пациенты выполняют тестовое задание то же самое тестовое задание, что и в контрольной группе, до и некоторое время спустя после фармакологического воздействия. На четвертом этапе с помощью пространственной фильтрации, основанной на топографии компонент, выявленных при анализе ВП испытуемых контрольной группы, производится
356
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Рисунок 15.19. ВП в фармакологических исследованиях
А. Компонента новизны, или М1/Р2-компонента (тонкая линия), полученная при построении ВП ребенка (мальчика) с СНВГ, до и спустя 1 час после принятия риталина в сравнении с независимой компонентой, вычисленной из массива ВП-данных группы здоровых испытуемых (толстая линия). А. Разностная волна (пунктирная линия), полученная в результате сравнения компоненты новизны пациента и компоненты, полученной на группе здоровых испытуемых. Б. Топограммы отношения спектральной мощности тета- и бета-ритмов, вычисленные у группы нормальных испытуемых и у пациента до и после принятия риталина.
разложение на компоненты ВП, зарегистрированных у пациентов. На следующем этапе до лечения у пациентов выявляются компоненты, отклоняющиеся от нормы. И наконец, оценивается влияние лечения на компоненты, которые до фармакологического воздействия отклонялись от нормы.
Результат применения предлагаемого нами подхода показан на рис. 15.19, где представлены ВП ребенка (мальчика) с СНВГ. До лечения у пациента наблюдалось избирательное отклонение от нормы в амплитуде 1Ч1/Р2-компоненты слухового ВП. Данная компонента регистрировалась на ВП, получаемых в ответ на второй стимул предъявляемой пары (растение, человек + новый звуковой стимул) и, как предполагается, отражала эффект новизны — ответ мозга на неожиданный новый стимул. Как видно из рис. 15.19, спустя час после приемы риталина Ы/М1/Р2-компонента этого пациента почти достигла нормальных значений.
V. Поведенческие парадигмы
А. Классификация парадигм
Ниже мы собираемся представить короткие описания некоторых тестовых задач, которые применяются в исследованиях ВП. Все задачи подразделяются на две категории. Конечно, перечень тестовых задач не является полным (табл. 15.1), но позволяет сформировать представление о попытках, предпринимавшихся с целью анализа этапов процессов переработки информации, протекающих на уровне разных систем мозга.
Ниже приведено более детальное описание парадигм.
Часть 2. Вызванные потенциалы
357
Б. Сенсорные системы и система внимания
I. Oddball-парадигма. В рамках данной парадигмы последовательно и случайно предъявляются стимулы двух типов. Вероятности предъявления стандартных (St) и девиантных (Dev) стимулов существенно отличаются: стандартные стимулы предъявляются намного чаще. Обычно стандартные стимулы предъявляются в 90 % случаев, а девиантные, соответственно, составляют 10 % от общего количества стимулов. Девиантные стимулы «нетипичны» («oddball»), поэтому исследовательская парадигма получила название oddball-парадигмы. Стимулы воспринимаются в достаточно монотонном режиме за счет коротких межстимульных интервалов. При такой организации предъявления получается, что время от времени стандартные стимулы «заменяются» девиантными и прерывается монотонность стимуляции. Обычно отличие между стандартными и девиантными стимулами относительно невелико и затрагивает одну из характеристик. Например, в слуховой модальности стандартными стимулами могут быть тоны частотой 1000 Гц и длительностью 100 мс. Девиантными же стимулами могут быть тоны той же длительности, но частотой в 1100 Гц. Стандартным стимулом в зрительной модальности может быть цифра 6, а девиантным — цифра 9. Межстимульный интервал обычно составляет около 1 с, и в одних исследованиях стабилен, а в других может варьировать.
Oddball-парадигма принадлежит к разновидности «активных» парадигм, т.е. от испытуемого требуется реагировать на предъявление девиантного стимула. Это может быть нажатие кнопки мыши или счет (в уме) количества девиантных стимулов. В слуховой модальности, однако, обычно применяется пассивная модификация теста, когда на фоне предъявления испытуемые инструктируются выполнять другую задачу, как, например, просмотр видеоклипов или чтение книги. Такая модификация тестового задания является наиболее благоприятной для регистрации негативности рассогласования — небольшая компонента ВП, которая представляет собой дополнительную негативную компоненту (относительно ВП при стандартном стимуле); регистрируемую в области Fz.
2. Oddball-парадигма с новыми стимулами. Данная парадигма является успешно применяемой разновидностью oddball парадигмы, по сравнению с которой в тестовую задачу вводится дополнительный вид стимулов, а именно новые стимулы (Nov). В силу того что новые стимулы являются достаточно уникальными (например, звон ключей, лай собаки и т.д.), они предъявляются с целью переключения внимания испытуемого. Существует также активная и пассивная модификации тестовых шданий. В пассивной модификации выполняется другая задача, а в активной испытуемые реагируют определенным действием на девиантный стимул. В рамках активной модификации парадигмы обычно регистри-
358 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Таблица 15.1. Тестовые задания для исследования информационных процессов мозга
мае система match mismatch Л □ □ _ test delay probe test delay probe Сначала предъявляется тестовый стимул (образец), после чего следует пауза и предъявляется стимул-проба. Задачей испытуемого является сравнить стимул-пробу с тестовым стимулом и решить, соответствуют ли они друг другу Парадигма отсроченного сопоставления с образцом Во время паузы между двумя стимулами регистрируется условно негативное отклонение
different same Предъявляется серия стимулов. От испытуемого требуется определить, соответствует ли текущий стимул предыдущему, предъявлявшемуся некоторое (N) количество проб до него N-обратная задача ВП-корреляты рабочей памяти
SSHSS sssss HHHHH HHSHH П П П П Задача испытуемого заключается в наиболее быстром и корректном реагировании на центрально расположенные стимулы (в данном случае буквы) в последовательности из 5 стимулов. Например, при (Н) нужно нажимать кнопку левым указательным пальцем руки, а при (S) соответственно правым Парадигма детекции конфликта (фланговый тест Эриксона, тест Струпа здесь не показан) Негативность, связанная с ошибкой, генерируемая в некорректных пробах
NOGO GO NOGO GO И пип В случайном порядке и с равной вероятностью предъявляются два типа стимулов (GO и NOGO). Длительность межстимульного интервала достаточна для того, чтобы испытуемый успел подготовить нужный ответ GO/NOGO-парадигма (двухстимульный GO/NOGO-тест является вариантом этой парадигмы) N2 NOGO-компонента генерируется в ответ на NOGO-стимулы. Р400 компонента мониторинга также генерируется при NOGO-стимулах
Аффективная система О п Сначала через наушники предъявляются стимулы, связанные с прошлым испытуемого и провоцирующие возникновение определенных эмоций. После чего предъявляются изображения лиц, выражающие разные эмоции (радость, печаль или эмоционально нейтральные) Парадигма провокации настроения Разность ВП при эмоциях радости и печали
Эпизодическая память Old new new ВПП За день до непосредственной регистрации ВП, испытуемому предъявляется список слов для запоминания. Во время тестирования, испытуемому предъявляются «старые» (запомненные накануне) и «новые» стимулы. Парадигма старое — новое Эффект старого — нового
Часть 2. Вызванные потенциалы_______________________________________________________359
360
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
руется две РЗ-компоненты: РЗа-компонента, генерируемая в ответ на появление нового стимула, и РЗЬ-компонента, наблюдаемая при предъявлении целевого стимула (т.е. девиантного стимула, требующего определенных действий).
3. Парадигма пространственного селективного внимания. Общая методология парадигмы исследования селективного внимания заключается в одновременном предъявлении двух разных потоков стимуляции, характеризующихся разной пространственной локализацией. При этом стимулы двух потоков отличаются между собой одним из своих базовых свойств219. Внимание испытуемого избирательно направляется на один из источников стимуляции. Достаточно простым вариантом тестового задания в рамках парадигмы пространственного селективного внимания в слуховой модальности является тест дихотическо-го прослушивания. Тест дихотического прослушивания был создан в 1950-х годах для исследования известного «эффекта вечеринки». Одним из вариантов этого теста является одновременное предъявление в правое и левое ухо речевых стимулов (речь, разговор) с концентрацией внимания на одном из слуховых каналов. В ВП-исследованиях в каждое ухо независимо посылаются две разные последовательности стандартных и девиантных звуковых стимулов220. Тест дихотического прослушивания является активным, т.е. испытуемый при предъявлении девиантного стимула в одном из слуховых каналов (на котором сконцентрировано внимание) выполняет определенное действие (например, нажатие кнопки). На протяжении исследования в соответствующих отдельных исследовательских сессиях внимание фокусируется то на правом, то на левом ухе. Для каждого уха рассчитывается разностный ВП, получаемый при вычислении разности ВП на стандартные стимулы двух типов: когда внимание сконцентрировано на данном канале и когда нет. Выявляемый разностный ВП ассоциируется с процессами селективного внимания для разных пространственных локализаций221. В слуховой модальности такой разностный потенциал является отрицательным по полярности и поэтому называется «процессной негативностью».
Аналогично тесту с дихотическим прослушиванием достаточно несложно создать зрительный аналог такого тестового задания. Для этого необходимо предъявлять зрительные стимулы в двух разных областях зрительного поля (в правой и левой части) и инструктировать испытуемого концентрировать внимание на одной из них в каждой отдельной сессии.
4. Парадигма пространственной сигнализации. Данная парадигма была создана с целью изучения процессов селективного пространственно
219 Потоки стимуляции могут различаться не только по признаку пространственной локализации, но и по относительно простым характеристикам, например цвету.
230 Дискриминация потоков информации упрощается за счет различия звуковых стимулов по ряду параметров (громкость, длительность и частота).
221 Пространственное внимание является одним из многих видов селективного внимания.
Часть 2. Вызванные потенциалы
361
го внимания. Суть ее заключается в том, что перед целевыми стимулами предъявляются сигнальные. Существует два вида сигнальных стимулов: одни стимулы корректно указывают место в зрительном поле (пространстве), в котором появляется целевой стимул, а другие нет. Наблюдаемая разница во времени реакции ответов на целевой стимул, предваряемый валидным и невалидным сигнальным стимулом (так называемый эффект валидности), как считается, отражает процессы пространственного внимания, управляемые сигнальными стимулами. Примером тестовой задачи такого вида является тест Познера: испытуемые концентрируются на центральной точке зрительного поля и должны детектировать быстро возникающие на периферии зрительные стимулы. Эти целевые стимулы предъявляются после направляющего внимание сигнального стимула (к центру или периферии зрительного поля) в течение нескольких сотен миллисекунд. Время реакции в пробах с корректными сигнальными стимулами короче по сравнению с пробами, в которых сигнальный стимул неверно указывает место появления целевого стимула. Выявляемая разница во времени реакции между пробами двух видов ассоциируется с процессами пространственного внимания к определенной указываемой области пространства222.
5. Тестовые задания на непространственное селективное внимание. Не-нространственное селективное внимание может быть исследовано в условиях последовательного предъявления стимулов, отличающихся по одному из параметров (например, цвет, направление движения и т.д.). В зрительной модальности целевые стимулы (т.е. стимулы, на которые стоит обращать внимание) могут быть синего цвета, а стимулы желтого цвета должны игнорироваться. Зарегистрировав ВП на целевые и игнорируемые стимулы, высчитывают разностный потенциал между двумя типами ВП, который характеризует эффект селективного внимания к стимулу определенного цвета. Такие компоненты разностных ВП традиционно называют негативностью селекции.
В. Исполнительные функции
/. Парадигма отсроченного сравнения с образцом. Для исследования процессов рабочей памяти, связанных с операциями кодирования, удержания и воспроизведения информации, часто применяется парадигма отсроченного сравнения. Первым предъявляется тестовый стимул, после которого следует пауза и появляется стимул-проба, а испытуемый должен решить (и должным образом отреагировать), соответствует пробный стимул тестовому или нет.
2. N-обратная задача. В условиях данного тестового задания испытуемый должен определить наличие или отсутствие соответствия между стимулом-
737 Парадигма с пространственными сигнальными стимулами позволяет исследователю оценить эффект внимания с помощью пычисления разности во времени реакции. Однако применение данной парадигмы в исследованиях ВП сталкивается с некоторыми 1РУДНОСТЯМИ. Это обусловлено тем, что ВП проб с валидными и невалидными сигнальными стимулами характеризуются наличием моюрных компонент, которые отличаются между собой при верно и неверно указанных целевых стимулах. Фактически эти моторные компоненты накладываются на компоненты, связанные с эффектом внимания, тем самым маскируя его.
362
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
пробой и стимулом, предъявлявшимся ранее, некоторое количество стимулов (N) до него. Таким образом, появляется возможность манипулировать нагрузкой на память: нагрузка увеличивается с увеличением N. В этих условиях испытуемый сталкивается с двойной задачей: кодировать стимул-пробу и сравнить его с предыдущим стимулом, который предъявлялся на N стимулов раньше.
3. Парадигма пространственной сигнализации. Тестовые задания в рамках данной парадигмы построены таким образом, чтобы исследовать процессы, связанные с операциями мониторинга на уровне исполнительной системы мозга. Считается, что ключевой областью мозговой системы, ответственной за процессы детекции конфликта, является передняя поясная извилина. Множество тестовых задач с конфликтными условиями характеризуются активациями в этой области мозга. Известный тест Струпа является примером такой задачи. Этот тест был разработан в 1935 году аспирантом Дж.Р. Струпом и стал одним из наиболее популярных исследовательских тестов в когнитивной психологии. В данном тесте перед испытуемым ставится задача как можно быстрее называть цвет последовательно предъявляемых слов. В случае, когда цвет чернил слова соответствует его значению, поставленная задача выполняется достаточно легко. Однако, если слово (скажем, зеленый) написано, например, чернилами красного цвета, то репрезентации значения слова и цвета будут конфликтовать между собой. Поведенчески такой конфликт будет выражаться в увеличении времени реакции и количества совершаемых ошибок.
Другим вариантом тестового задания в рамках парадигмы мониторинга конфликта является фланговый тест Эриксона, который широко применяется в ВП-исследованиях «негативности, связанной с ошибкой». Тест Эриксона был разработан в 1979 году (Eriksen, Eriksen, 1979) и выглядит следующим образом: вслед за предупреждающим стимулом следует предъявление одной из четырех последовательностей букв (ННННН, HHSHH, SSHSS или SSSSS). Испытуемый должен как можно быстрее и точнее реагировать на центрально расположенную букву, например нажимать на кнопку указательным пальцем левой руки в случае буквы Н и правой руки при S.
4. Парадигма GO/NOGO. Базовой идеей настоящей парадигмы является создание условий для исследования одной из операций исполнительной системы мозга — подавления действия. Таким образом, в рамках GO/ NOGO-парадигмы предполагается, что испытуемый готовится совершить действие (GO) в каждой пробе, но в некоторых из них должен подавить подготовленное действие (NOGO). Следует отметить, что в oddball-парадигме при одних стимулах необходимо действовать, а при других воздерживаться от реакции, но низкая вероятность появления девиантных стимулов и короткие межстимульные интервалы не позволяют подготовить действие к каждому стимулу. Таким образом, стандартные стимулы в oddball-парадигме формируют фон, а девиантные стимулы (GO) появ
Часть 2. Вызванные потенциалы
363
ляются достаточно редко. По сравнению с условиями oddball-парадигмы GO/NOGO-парадигма создает условия, характеризующиеся большими межстимульными интервалами и высокой вероятностью предъявления GO-стимулов.
В простом варианте GO/NOGO-теста в случайном порядке последовательно предъявляются стимулы двух видов (например, красного и зеленого цвета). Межстимульный интервал составляет 2 секунды, а вероятности появления каждого из стимулов одинаковы. В задачу испытуемого входит нажимать кнопку мыши в случае предъявления одного из стимулов, называемых GO-стимулом (это может быть, например, стимул зеленого цвета), и воздерживаться от нажатия при стимуле красного цвета (N OGO-стимул).
Существуют и другие модификации тестовых заданий в рамках GO/ NOGO-парадигмы. В частности, в «С-Х» варианте GO/NOGO-теста испытуемым предъявляются последовательности букв. В 20 % случаев появляется буква «С», за которой в 50 % случаев следует буква «X», а в остальных — другие буквы (R, V, Т и т.д.). В случае «С-Х» последовательности (NOGO-стимулы) испытуемые должны подавлять ответы. При остальных комбинациях требуется нажатие кнопки (C-R, C-V и т.д., т.е. GO-стимулы).
Другая версия GO/NOGO-парадигмы (так называемый стоп-сигнал тест) подразумевает предъявление проб, начинающихся или с буквы А, или с В, в каждой из которой испытуемый выполняет тест на время реакции двойного выбора, нажимая соответствующую кнопку при определенном стимуле, с которого начинается проба. В 25 % проб после стимула А или В через определенный интервал времени, варьирующий от 200 до 400 мс (интервал стоп-сигнала), предъявляется стоп-сигнал (буква S), информирующий испытуемого о необходимости воздержаться от нажатия кнопки. Известно, что здоровым испытуемым проще подавить действие, если межстимульный интервал колеблется в пределах 200—400 мс. При увеличении этого интервала до 400—600 мс подавление действия существенно усложняется, что приводит к значительному снижению вероятности воздержания от реакции. Для того чтобы графически измерить контроль подавления действий, достаточно по оси ординат отобразить вероятность подавления, а по оси абсцисс — временной интервал появления стоп-сигнала.
Двухстимульный GO/NOGO также является подтипом основной GO/NOGO-парадигмы. Суть теста заключается в таком предъявлении пар стимулов, которое позволило бы испытуемому непроизвольно быть готовым к реализации действия после первого стимула в последовательности из двух стимулов и как можно быстрее реагировать в ответ на предъявление второго. Существует два варианта данного исследовательского теста. В одном случае первый стимул пробы выполняет функцию обычного предупреждающего стимула. В другой модификации по первому стимулу для испытуемого становится ясно, следует ре
364
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
агировать на второй стимул или нет. Второй вариант двухстимульного теста в GO/NOGO-парадигме был специально разработан для наших исследований и использовался при создании нормативной HBI базы данных (см. гл. 12).
Г. Аффективная система
Тестовые задания, создаваемые для исследования аффективной системы мозга, обычно формируют условия для возникновения двух основных типов эмоций: радости и грусти. Для индукции этих эмоций используются две категории эмоционально значимых стимулов — предъявления изображений соответствующих эмоций и ситуаций. Это могут быть изображения грустных и радостных лиц людей, эмоционально значимых сцен человеческой жизни. Следует принять во внимание тот факт, что для генерации и выражения эмоций необходимо некоторое время. Следовательно, предполагается, что межстимульный интервал должен быть достаточно большим. Более того, эмоции крайне субъективны: одни и те же изображения у одних испытуемых могут вызывать положительные эмоции, а у других — отрицательные. Поэтому стимулы должны подбираться с учетом индивидуальных особенностей. Парадигма провокации настроения удовлетворяет всем указанным выше требованиям.
Парадигма провокации настроения. Тестовая задача, сформированная в рамках данной парадигмы, базируется на двух допущениях223: 1) все люди разные, и если у одних определенные ситуации могут приводить к возникновению положительных эмоций, то у других они же могут вовсе не вызывать эмоций или даже приводить к негативным переживаниям; 2) восприятие как эмоциональной экспрессии других людей, так и различных событий жизни сильно зависит от текущего фонового эмоционального состояния субъекта. И действительно, из собственного опыта мы знаем, что, будучи в плохом расположении духа, все происходящее воспринимаем в серых тонах. Наоборот, в приподнятом настроении все проблемы «кажутся столь далекими». Более того, переход от одного настроения к другому требует определенного времени. В тестовых условиях испытуемым предлагается пройти опрос, касающийся их собственной жизни. В частности, просят вспомнить по пять жизненных ситуаций (историй) каждого эмоционального типа (позитивную, негативную, нейтральную), сопровождавшихся либо соответствующими эмоциональными переживаниями радости, грусти, либо их отсутствием. Далее формируют так называемое напоминание длительностью около 8 секунд, которое предъявляется перед презентацией изображения лица с определенной эмоцией. Напоминание может выглядеть следующим образом: «С помощью предъявляемого изображения грустного лица вспомните свои
223 Тест разработан группой исследователей из Медицинской школы Питсбурского университета и Лондонского института психиатрии (Keedwell et al., 2005).
Часть 2. Вызванные потенциалы
365
переживания, когда на похоронах вашего отца плакала мама». Изображения лиц с выражением разных эмоций (например, 100% радостное, грустное или нейтральное) выбираются из стандартизованных специализированных наборов (Ekman and Friesen, 1976). Испытуемые участвуют в двух 6-минутных сессиях, в рамках которых предъявляется по десять 36-секундных блоков эмоционально значимых и нейтральных стимулов. Последовательность блоков в каждой сессии, как и самих сессий, обычно рандомизирована и уравновешена по всем испытуемым.
Д. Эпизодическая память
Парадигма старого — нового. За день до исследования испытуемым предъявляется список слов, которые необходимо запомнить. Предполагается, что в течение ночи они консолидируются в эпизодической памяти. В день исследования испытуемым предъявляются слова из предварительно показанного списка (т.е. «старые») и слова, не вошедшие в него («новые»). ВП, зарегистрированные в ответ на «старые» и «новые» слова, вычитаются друг из друга, получаемый разностный потенциал характеризует так называемый эффект старого — нового.
366
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Глава 16. Практика
I. Введение
Настоящая книга снабжена обучающей программой. Некоторые методы спектрального анализа спонтанной ЭЭГ были представлены в предыдущих главах книги. Основная идея спектрального анализа заключается в разложении ЭЭГ на простые компоненты — синусоидальные волны. Как было показано в ч. 1, осцилляции разных частотных диапазонов отражают разные режимы саморегуляции коры. Для разложения фоновой ЭЭГ на простые компоненты используется быстрое преобразование Фурье, вейвлет-преобразование и метод независимых компонент. В настоящей главе мы представим также основные методы анализа вызванных потенциалов, которые представляют несколько иной способ наблюдения за активностью мозга по сравнению с анализом длительной ЭЭГ. Если ВП отображают этапы информационных процессов, протекающих на уровне нейронных сетей, то фоновая ЭЭГ отражает модуляцию потоков информации. Несмотря на столь качественное различие, основные идеи анализа ВП и ЭЭГ-данных сопоставимы. Анализ ВП заключается в разложении на простые составляющие элементы — независимые компоненты, каждая из которых отражает специфические операции процессов обработки информации мозгом.
Методы вычисления и обработки вызванных потенциалов могут быть подразделены на следующие этапы: 1) организация психологических задач; 2) предварительная обработка ЭЭГ, как, например, установка монтажа электродов, коррекция и устранение артефактов; 3) применение метода независимых компонент к набору единичных проб одного испытуемого224; 4) усреднение фрагментов ЭЭГ-проб разных категорий индивидуально для каждого испытуемого225; 5) размещение получаемых ВП заданной группы испытуемых или пациентов во встроенную базу данных для обеспечения возможности их дальнейшего анализа: усреднения ВП по группе выделенных исследований или анализа независимых компонент для усредненных ВП226; 6) получение ВП, усредненных по группе испытуемых; 7) выявление общих для всей группы испытуемых независимых компонент, характеризующих заданные условия исследовательского те
224 Хотя данный вид анализа ВП реализован в нашем основном программном обеспечении, пока не вполне понятно, каким образом его применение может быть полезным на практике. Мы собираемся опустить рассмотрение этого метода анализа в последующем изложении и в описании обучающих программ.
225 Например, получение ВП для одного испытуемого при усреднении фрагментов ЭЭГ со всех отведений отдельно для проб со стимулами типа GO в рамках GO/NOGO-теста.
226 Например, размещение ВП, полученных в GO/NOGO-тесте для всех пациентов отдельно от группы пациентов невнимательного подтипа» с СНВГ.
Часть 2. Вызванные потенциалы 367
ста, а также формирование пространственного фильтра для каждой компоненты с учетом топографии компонент; 8) сравнение индивидуальных ВП или компонент ВП с нормативными данными, выявленными при анализе репрезентативной выборки здоровых испытуемых того же возраста и пола; 9) составление заключений, т.е. представление результатов обработки данных в краткой и доступной форме, с основными выводами и рекомендациями к лечению.
Все перечисленные этапы обработки вызванных потенциалов продемонстрированы на рис. 16.1. Они представлены также и в нормативной HBI базе данных. Во множестве известных коммерческих ЭЭГ-систем (Neuroscan, Nicolet и т.д.) есть некоторые из этих методов анализа ВП, а остальные могут быть также реализованы в среде Matlab (приложение EEG-lab). Следует подчеркнуть, что в рамках настоящей книги мы не собираемся полностью рассматривать всю систему обработки данных. А цель обучающего программного обеспечения — дать возможность пользователю освоить основные методы обработки вызванных потенциалов.
Создание файлов в \ 1нпр и wav формат е
Задание порядка у следования стимулов в пробе
Задание порядка проб V
Создание тестов
.п...п..пл......ал..
проба I £1 S2 проба 3
Установка монтажа
Коррекция артефактов
Усреднение проб
Размещение ВИ в базу дязпзых : РА ] )!»«.. | РМжх 1 : Sw
"оНы Vcpt sutowt 'i "
IMJM Subject 2 satoisss I
Усреднение ВП по группе испытуемых
Применение метода независимых компонент в анализе фрагме нте 331' отдельных проб
У среднение фрагментов у' ЭЭГ проб определенной категории
Получение разностных \' воли путем вычнтятшя ВП одной катез ории нз ВП друг ой
Создание базы данных ВП, классифицированных в соответствии с возрастом, иолом, зилом заболевания и тл»
Получение усредезпп.гх по з руине испытуемых ВП путем усреднения индивидуальных ВП из базы данных
Применение метода независимых компонент в анализе набора ВП зв базы данных
Вышслезше пространственных фильтров с учетом топографии независимых компонент
Применение пространственных фильтров к анализу зшдивзздуяльззых ВП
Сравнение зшдзтздуяльньзх ВП е нормапшиой базой данных /
Сравнешзе ко.хязонези
ИНДНКЗ|ДУЯЛЬНЫХ ВП с нормативной базой ДШПП.ТХ
Рисунок 16.1. Этапы анализа вызванных потенциалов
Ряды сверху вниз: 1) формирование психологических заданий; 2) предварительная обработка ЭЭГ; 3) использование метода независимых компонент в анализе отдельных проб одного испытуемого; 4) усреднение фрагментов ЭЭГ отдельных проб; 5) размещение файлов с ВП-данными каждого испытуемого в базу данных; 6) усреднение ВП по группе испытуемых; 7) применение метода независимых компонент в анализе групповых ВП; 8) сравнение индивидуальных Bl 1 или компонент ВП с нормативными данными. Элементы анализа, которые представлены в программном обеспечении EdEEG, отмечены галочками.
368 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
II. Формирование тестового задания
В состав обучающего программного обеспечения входит приложение Psytask222, которое позволяет исследователю создавать психологические те-
стовые задания и осуществлять их запуск с одновременной регистрацией ЭЭГ. Psytask работает синхронно с программным обеспечением по регистрации ЭЭГ и обеспечивает подачу синхронизирующих меток во время ее записи. Для этого предъявляющий {Подчиненный) и регистрирующий (Основ-
Рисунок 16.2. Окно приложения Psytask
Сверху вниз, кнопки: 1>Начатъ новый тест» — запускает психологические тестовые задания из списка созданных заданий; 2) «Просмотреть базу данных» — открывает базу данных с результатами выполнения психологических заданий; 3) •Пона зать список тестов» — отображает список стандартных заданий, поддерживающихся программой; 4) •Переключиться в режим подчиненного* — переводит предъявляющий компьютер в режим подчиненного; 5) •Установить базу данных* — используется для создания базы данных поведенческих параметров (количество ошибок, время реакции); 6) •Изменить параметры синхронизации* — применяется для изменения параметров COM-порта, связывающего предъявляющий и основной компьютер; 7) •Выйти» — закрывает программу.
ной) компьютеры соединяются с помощью СОМ-портов. Программа Psytask может быть также использована для измерения времени реакции, пропусков и совершения ошибок при выполнении психологических тестов.
После установки программы Psytask™ появится следующее диалоговое окно (рис. 16.2). Кнопка Начать новый тест используется для запуска психологического тестового задания из списка созданных заданий. Кнопка Просмотреть базу данных открывает встроенную базу данных результатов тестирования (время реакции, пропуски и т.д.). Кнопка Показать список тестов используется для запуска встроенного редактора протоколов психологических тестов. Эта же кнопка используется и при создании новых тестовых заданий. Кнопка Переключиться в режим подчиненного позволяет переводить предъявляющий компьютер в режим подчиненного, который во время записи ЭЭГ получает синхронизирующие команды от регистрирующего компьютера. Для этого оба компьютера соединяются через COM-порт. Кнопка Установить базу данных используется для изменения имени (пути) базы данных с целью ее открытия или создания новой. Кнопка Изменить параметры синхронизации ис-
пользуется для изменения параметров COM-порта, используемых для синхронизации. Выход из программы производится нажатием кнопки Выйти.
Ядром программы Psytask является протокол предъявления тестового задания. Протокол определяет стимулы, которые будут предъявляться в задании, время их
предъявления и длительность, последовательность проб, а также способ реагирования испытуемых на стимулы пробы. Вся эта информация, за исключением стимулов, хранящихся в отдельных файлах, содержится в файлах-протоколах пред ъявления стимулов с расширением .PRO229. Файлы протоколов и соответствующие фай-
227 Эта программа была написана Валерием Пономаревым — старшим научным сотрудником нашей лаборатории в Институте мозга человека РАН (Санкт-Петербург)» Данное программное обеспечение распространяется свободно.
228 Программа устанавливается с учебного диска (запустите программу Setup и следуйте инструкциям).
229 Файлы с расширением .PRO являются простыми текстовыми файлами (ASCII), которые могут читаться и редактироваться в любом текстовом редакторе.
Часть 2. Вызванные потенциалы
369
лы стимулов хранятся в отдельной папке. Зрительные стимулы хранятся в файлах формата .BMP или .JPEG. Звуковые стимулы сохраняются в файлах формата WAV При создании нового тестового задания задается его имя (например, GONOGO или какое-либо другое), а также имя папки (например, GONOGO). Папка, содержащая все соответствующие данному тесту файлы, создается в корневой папке программы Psytask (рис. 16.3). Туг же содержится файл PSYTASK. CFG, содержащий список всех имеющихся тестовых заданий. Протокол тестового задания автоматически генерируется при нажатии Новый в окне, вызываемом командой Показать список тестов, и состоит из следующих закладок Свойства теста (имя теста, имя папки, разрешение экрана), Список стимулов, Список проб, Список предъявления проб и команд, Обработка ответной реакции™.
Для создания списка стимулов (Список стимулов)
Psytask папка
GONOGO папка
Рисунок 163. Размещение файлов протокола двухстимульного GO/NOGO тестового задания
В папке приложения Psytask содержатся соответствующие папки с описанием всех тестовых заданий. Папка GONOGO представлена в качестве примера. Файл GONOGO.PRO содержит информацию о всех параметрах задания — списке стимулов, времени предъявления стимулов в пробе. Звуковые и зрительные стимулы (изображения) представлены отдельными файлами.
сначала нужно приготовить все необходимые звуковые файлы и файлы изображений. Далее эти файлы стимулов должны быть помещены в папку с тестовым заданием с помощью кнопки Добавить из файла в закладке Список стимулов.
После определения списка стимулов следует задать временную структуру проб, используя закладку Список проб. Типично проба включает предъявление одного или нескольких стиму-
лов. Список стимулов в пробе описывается именами стимулов, временем включения стимула от начала пробы и экспозициями. Параметры проб представлены на рис. 16.4.
В меню Список предъявления проб и команд необходимо определить: 1) порядок предъявления проб, паузы между ними (т.е. временные интервалы между пробами); 2) метку конкретной категории проб
В качетстве примера представлены параметры двухстимульного задания. Параметры задаются пользователем с помощью приложения Psytask. Обозначения: Ст. 1 — стимул 1, Ст. 2 — стимул 2.
(например, 1 для GO-категории и 2 для NOGO-категории). В меню Обработка ответной реакции необходимо определить следующие параметры: 1) устройство, регистрирующее ответы испытуемого — это может быть микропереключатель (ком-
730 В некоторых случаях необходимо перенести созданное тестовое задание на другой предъявляющий компьютер. Для этого в окне Слисок тестов следует нажать кнопку Добавить из и найти протокол, который вам нужен.
370 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
пьютерная мышь) с правой и левой кнопкой, а также некоторые из кнопок клавиатуры предъявляющего компьютера; 2) временной интервал для корректных ответов (например, от 200 до 1000 мс); 3) способ реагирования (например, нажатие левой кнопки или его отсутствие).
В качестве примера на рис. 16.5 представлен протокол двухстимульного GO/ NOGO-теста, использовавшийся при создании нормативной HBI базы данных. Для более детального ознакомления с программой Psytask используйте руководство пользователя, которое расположено в папке DOC корневого каталога программы.
III. Программа EdEEG
Шаг 1. Открытие ЭЭГ-файла
Установите программное обеспечение EdEEG в соответствии с процедурой, описанной в гл. «Практика» ч. 1. Запустите EdEEG.exe файл. В окне главного меню вы увидите меню: Файл, Правка, Вид, Формат, Запись, Анализ, Настройка и др. Каждое меню имеет свои собственные команды. В дополнение к окну основного меню изображены также меньшие по размеру окна — Окно топограмм и Окно диполей. Для просмотра 19-канальной записи ЭЭГ во время выполнения двухстимульного GO/NOGO-теста в меню Файл выберите команду Открыть файл.
Шаг 2. Просмотр ЭЭГ
Окно ЭЭГ используется для просмотра записей ЭЭГ и содержит следующие разделы: Панель названия электродов в левой части окна, Строка текущего состояния, расположенная внизу, а также Панель фильтров в верхней части окна. Непосредственно над кривыми записи ЭЭГ в виде прямоугольников с числами, отображающими номер пробы и категорию стимула, располагаются отметки проб (на рис. 16.6, например, в верхнем левом прямоугольнике Т23[ 1 ] обозначает, что предъявлена 23-я проба категории 1 — проба с GO-стимулом). Левая часть прямоугольника соответствует началу пробы. В нижней части окна ЭЭГ представлены отметки нажатий кнопки. Левая часть метки нажатия кнопки соответствует времени начала ответа испытуемого.
Шаг 3. Предварительная обработка ЭЭГ — установка монтажа
Зарегистрированная ЭЭГ по умолчанию сохраняется на диске в монтаже с «объединенными ушами» в качестве референтов и полной полосой пропускания усилителей ЭЭГ. Для просмотра и анализа ЭЭГ предусмотрена возможность выбора другого монтажа электродов. Для этого в меню Вид нужно выбрать команду Выбор монтажа. После чего появится окно Задание монтажа. При создании нормативной базы данных HBI Database мы использовали три разных варианта монтажа: монтаж с «объединенными ушами», средний референт и взвешенный средний референт (по Лемосу). В качестве примера на рис. 16.6 был выбран взвешенный средний референт.
Часть 2. Вызванные потенциалы
371
Шаг 4. Предварительная обработка записи ЭЭГ — коррекция артефактов движения глаз
В ч. 1 мы описали три метода коррекции артефактов: 1) ручное удаление; 2) автоматическое вырезание; 3) автоматическая коррекция артефактов посредством пространственной фильтрации. Более детальное описание представлено в гл. 9.
Шаг 5. Расчет вызванных потенциалов посредством усреднения проб
Индивидуальные ВП получаются в результате усреднения фрагментов ЭЭГ, синхронизированных с началом проб. На рис. 16.7 проиллюстрирован ВП, по-
строенный при усреднении 92 проб для одного из группы здоровых испытуе-
мых. ВП вычислялись в Cz-отведении для двух условий с GO- и NOGO-стимулами (изображения животных) в GO/NOGO-парадигме. Необходимо отметить, что до момента предъявления второго стимула пробы ВП для двух состояний фактически идентичны, поскольку первый стимул в GO- и NOGO-пробах один и тот же — изображение животного. Сопоставимость ВП на временном отрезке от первого стимула пробы до начала предъявления второго является так называемым показателем теста/ ретеста. Этот показатель может быть использован для оценки степени надежности ВП, рассчитываемых для определенного испытуемого.
Для осуществления процедуры усреднения нужно в меню Анализ выбрать команду Вызванные потенциалы. В результате появится окно с параметрами ВП (рис. 16.8).
TaskName "VCPT”
StnnuliLisr
Image SI STTM1JPG
Image S2 STIM2.JPG
Image S3 «ТШЗЛв
Image BkGr BkGr.JPG
Image Task Task.JPG
Sound S42 STEM3.WAV
Sound S63 ST1M4.WAV
EudStimuli
Trial T1 3000
SI 300 100
SI 1400 100
EndTiial
Trial T2 3000
S2 300 100
S2 1400 100
EndTiial
PsyTest 640x4S0\Tme(olor
LeitTop
Showpicture Task
SetBaekgrouliil BkGr
Wait 3000
T02 200 4
T2 200 1
T23 200 3
EmlTest
Список всех стимулов: Изображений,. Звуков, Фона и Изображения с описание теста
Список всех проб: Имя, Длительность первого стимула пробы, время предъявления от начала пробы, длительность.
Второй стилфл пробы, время предъявления от от начала пробы, длительность
Отображение порядка предъявления изображения с описанием теста, установки фона, последовательности проб, интервала между ними и количеством категорий проб
Respons еРгое easing
"Go" 2 100 1000 VKRIGHT Press None None 1
"Nog»’* 2 100 1000 VK_RIGHT Skip None None 2
"Ignore Г’ 2 100 1000 VKJRIGHT Slap None None 3
"Ignore!" 2 100 1000 VK RIGHT Skin None None 4
Endprocessing Реакции на пробы которые следует рассматривать как корректные Название категории пробы, временное окно для ответа, действия для трех кнопок, количество категорий проб
Рисунок 16.5. Пример протокола двухстимульного GO/NOGO тестового задания
На рисунке изображен файл из рис. 16.3. Файл включает описание стимулов, проб и последовательности проб тестового задания.
372
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
lOOjtV/cm FbIAvW Fb2AxW f7AvW
FzAAv
F4-AAM
F8AAM
13A.W
C3-A-.W
CMvW
CMvW
T4AvW
T5AvW
PSAsAV
PiAvW
PU«W
TSAvW
OlAvW
02A\AV
Burton
Фрагмент исходной ЭЭГ при выполнении GO/NOGO теста
Рисунок 16.6. Коррекция движений глаз
Пример 19-канальной ЭЭГ, зарегистрированной при выполнении двухстимульного GO/NOGO тестового задания: до (вверху) и после (внизу) процедуры коррекции артефактов движения глаз.
В верхней части подокна располагается список групп проб {Группы проб). Для двухстимульного GO/NOGO-теста (рис. 16.8) будут высчитываться ВП для следующих категорий (групп) проб: GO-пробы (АД), обозначенные цифрой 1; NOGO-пробы (АР), обозначенные цифрой 2; пробы, в которых предъявление первого стимула пробы сигнализирует об отмене реакции на второй в зрительной модальности (РР), обозначенные цифрой 3, и аналогичные пробы с дополни
Часть 2. Вызванные потенциалы
373
тельным предъявлением «нового» (неожиданного) стимула в слуховой модальности (PH), маркированные цифрой 4. Пробы 1-й и 2-й категории объединены в группу, обозначенную «+» как группа проб с подготовкой действия. Пробы 3-й и 4-й категории объединены в группу, в которой подготовка к действию отменяется после предъявления первого стимула («—»). Во время операции усреднения программа рассчитывает также количество корректных и некорректных ответов (пропуски и ошибки), а также удаляет из обработки пробы с артефактами (см. секцию Удаление артефактов). Дополнительно будут рассчитываться и
Рисунок 16.7. ВП в GO" и NOGO-пробах
Пример ВП, усредненных в условиях проб GO (тонкая линия) и NOGO (толстая линия) при выполнении двухстимульного тестового задания GO/NOGO. ВП рассчитывались для отведения Cz. Следует обратить внимание на то, что паттерны ВП для GO- и NOGO-условий выглядят одинаковыми. Это обусловлено тем, что первый стимул двух проб одинаков — изображение животного.
разностные вызванные потенциалы для выявления компонент, связанных с разными психологическими операциями231. Пары сравниваемых ВП перечислены в подокне Разности групп проб. Также с помощью команды Ъыбрать можно выбирать любые сочетания сравниваемых ВП из списка доступных ВП.
Содержание других полей касается коррекции артефактов (Удаление артефактов), параметров синхронизации (Синхронизация проб) и обработки ответов испытуемых (Обработка ответной реакции). Реализованная в данной программе процедура устранения артефактов основана на сравнении текущей амплитуды сигнала ЭЭГ с предустановленным порогом. Мы рекомендуем устанавливать значение порога, равное 100 мкВ. Обратите внимание на то, что для удаления артефактов следует выбирать только каналы записи ЭЭГ. В противном случае все пробы с ответами испытуемых могут быть выброшены из анализа ВП. у
Обычно рекомендуется выбирать синхронизацию по первому стимулу пробы. Другие установки параметров синхронизации могут применяться в тех случаях, когда проба состоит из двух и более стимулов с разными межстимульными интервалами и существует необходимость синхронизировать ВП со вторым или третьим стимулом пробы. ВП могут быть также синхронизированы и с нажатием кнопки испытуемым, т.е. с инициацией ответов на предъявляемые стимулы.
Также в отдельном подокне определяются параметры обработки ответов испытуемых, которые включают: 1) интервал времени, в пределах которо-го ответ испытуемого считается корректным (Интервал)', 2) группы проб,
331 Ранее (5-10 лет назад) вычисление разностных волн было единственным способом выявления компонент ВП. Специальная организация тестовых заданий предполагала наличие в одной из групп проб конкретной психологической операции, а в других нет, а процедура вычитания ВП соответствующих групп проб позволяла выявлять электрофизиологические корреляты этой психологической операции. Например, предполагалось, что операция подавления моторного ответа наличествует в NOGO-пробах и отсутствует в GO-пробах и в результате вычитания ВП для GO-проб из ВП при NOGO позволяет обнаружить показатель процессов подавления. Однако, как выяснилось, разные компоненты перекрываются во времени и пространстве, а простая процедура вычитания вызванных потенциалов может привадить к неверному представлению о распределении компонент. В действительности для GO-условия характерно наличие компонент вовлечения и инициации моторного ответа, которые перекрываются во време'ни и пространстве с компонентой подавления. Таким образом, операция вычитания не является универсальной, но может быть полезной при анализе ряда простых операций, происходящих в ходе исследуемой деятельности.
374 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
^2-1. «6-5
Choose
------Artifact processes------< LevetpOO* * W
Channel: jor^EEG 3
Thresholds fa charnels j
i—— Synchronization--------------1 r
Type: [stimulus jJ
Stimulus.:«i H
• Button channel |NoH«faefd
----Subject responce procetsins) Oetned
Define|
Compression: [off 3
Time interval before (m*t |o* P Calculate statistical s^ptficance 0>.
Time interval after (ms) fo~
P Use as default
100X
load [ Save | Load horn database| | OK ~| Cancel [
Рисунок 16.8. Окно построения вызванных потенциалов
Пользователь определяет названия категорий проб. Например, для пробы GO выбрана категория (метка) «1», а разница между GO- и NOGO-пробами обозначается как разница категорий «2 - 1». Метки задаются в протоколе приложения Psytask. Дополнительно пользователь устанавливает время корректного ответа и способ синхронизации, т.е. каким образом будет запускаться усреднение ВП — синхронно с предъявлением стимула или ответом испытуемого.
при предъявлении которых ожидается ответ; 3) стимулы, после которых должен реализовываться ответ; 4) категория ответа, т.е. нажатие правой или левой кнопки. Обращаем внимание на то, что в нашей версии оборудования существует много устройств регистрации времени реакции, таким образом, использованное устройство должно быть определено как канал для правой или левой кнопки (Канал кнопки для ответа 1 (2)).
Определив все параметры вычисления ВП, вы можете сохранить их в отдельном файле (срасширением .par) и при необходимости загрузить снова. Параметры для вычисления ВП для всех тестов нормативной HBI базы данных хранятся в отдельной папке (Parameters).
При использовании нормативной HBI базы данных нет необходимости всякий
раз выставлять параметры обработки ВП: они могут быть загружены из базы данных простым нажатием кнопки Загрузить из базы данных (рис. 16.8 внизу).
Шаг 6. Сравнение с нормативными данными
Теперь, когда мы получили индивидуальный вызванный потенциал пациента, нам может понадобиться выяснить, насколько он отличается от
S Event Related Potentials Patient 1 (13708/20071
g EEG Spectra
Eyes Closed
Eyes Opened
g Event Related Potentials
Visual CPI
S-r Event Related Potentials Components
00SEI
Load Database
Рисунок 16.9. Окно сравнения индивидуальных ВП с нормативной базой данных
Данное окно открывается при выборе команды Сравнение результатов меню анализа данных Анализ после нажатия кнопки Загрузить базу данных.
нормативных ВП, зарегистрированных в группе здоровых испытуемых того же возраста. Существует два варианта такого сравнения: 1) сравнение индивидуального ВП с ВП, усредненным по группе здоровых испытуемых; 2) сравнение индивидуальных компонент ВП с компонентами ВП, полученными методом анализа независимых компонент и усредненных по группе испы-
Часть 2. Вызванные потенциалы
375
--- --------— ----------Active Groups—-—----
0 1: jl-pNoGO ni jj О 2: [aPNoGO[2JjJ ° * | а-p N^O Й™*] О 4: [None
None а-s GO [D] a-pNoGOfOl РФ (DI
Ю)
а-p NoGO - a a GO ph • p-p (D| — PI
Сг-AvW
C3-AvW
P3-Avw
PfrAvW
рфП1 p-Mi]
-П1
а-p NoGO • a-a GO
-Г: p-h-p-рП)
П1
a-a GO [2]
T5A a-pNoGO [2]
РФ И
p-hRi
а-p NoGO - a-a GO p-h-p-p(2) biC-—
T4-AvW
------ a*p NoGO (1) ----а-p NoGO (2] ----- а-p NoGO [D)
Рисунок 16.10. Окно сравнения индивидуальных ВП с ВП нормативной базы
Окно появляется после нажатия кнопки ОК окна Озавнение ВП. Сравнение может быть сделано, если для испытуемого, базы данных и разностного ВП выбрать категории стимулов проб, для которых рассчитывались ВП (GO, NOGO). Обозначения: [1] — ВП испытуемого; [2] — ВП нормативной базы; [D] — разностный ВП, получаемый при вычитании ВП нормативной базы из индивидуального ВП.
туемых. В обоих случаях необходимо в меню анализа данных (Анализ) выбрать команду сравнения результатов (Сравнение результатов). Далее при нажатии кнопки Загрузить базу данных появится окно с содержанием базы данных (рис. 16.9).
Как видно из рис. 16.9, существует две опции сравнения ВП, зарегистрированных в условиях тестового задания GO/NOGO. Для сравнения вызванных потенциалов следует выбрать Event Related Potentials/Visual С РТ, а для сравнения компонент ВП, выявленных у пациентов с помощью пространственной фильтрации232, — опцию Event Related Potentials Components/Visual CPT.
Позвольте начать с простого сравнения ВП. На рис. 16.10 представлена часть окна, которое’появляется после нажатия кнопки ОКь окне сравнения результатов (рис. 16.9). В верхней части рис. 16.10 расположены на-
233 Пространственные фильтры формируются с учетом топографии компонентов, выявляемых в результате использования методе независимых компонент при анализе индивидуальных ВП большой контрольной группы здоровых испытуемых. Слезет отметить, что топография компонент может различаться между разными возрастными группами. Сравнение ВП, накопленных в нормативной базе HBI Database, свидетельствует о том, что группы детей и пожилых испытуемых характеризуются сопоставимыми по топографии независимыми компонентами. При этом структура временной динамики некоторых из этих топографически похожих компонент может существенно меняться с возрастом, а других—оставаться неизменной. Таким образом, оказывается, что пространственное распределение компоненты ВП, связанной со специфической психологической операцией, является весьма стабильной характеристикой по сравнению с временной динамикой, которая может зависеть от возраста.
376
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Active Groups
® 1 [pZNoGO m 2 ° 2 |P2NcG0 Й 3° 3 |P2NoGO (О) 3 ° 4; Rone
T3A
P1N1 vO[D] vcomTL [D] N1P2 aC |D] РЗЬ P [D] P3 supF |D] P4 mon CC |D] P1N1v0m vcomTL [1]
N1P2aC 11] P3bP(1) P3supF[1) P4 mon ССП] P1N1 vO(2] vcomFL [2] N1P2aC [2] РЗЬ P [2|
02-AvW
□1-AvW
P2NoGD [1]
P2NoGO|2] ............. P2NoG0|D]
Рисунок 16.11. Окно сравнения независимых компонент индивидуальных ВП с компонентами нормативной базы
После нажатия кнопки ОН в окне Сравнение компонент ВП появится окно сравнения. Для проведения сравнения нужно выбрать компоненты ВП (P3bG0, P3N0G0) для испытуемого, нормативной базы и расчета разности копмонент ВП. Обозначения: [1] — независимая компонента индивидуального ВП; [2] — соответствующие данные нормативной базы; [D] — разница между независимой компонентой ВП испытуемого и данными нормативной базы.
звания категорий ВП. Нажимая 3, можно выбирать различные категории стимулов, для которых рассчитывались ВП («а-а GO», «а-p N0G0» и т.д.). Символ в квадратных скобках справа от названия ВП означает следующее: [1] — индивидуальный ВП; [2] — ВП из нормативной базы данных; [D] — разностный потенциал, получаемый при вычитании [2] из [1].
Кроме сравнения ВП, обучающая программа (EdEEG) позволяет читателю сравнить независимые компоненты ВП, выявленные у отдельных испытуемых, с компонентами ВП нормативной базы группы испытуемых того же возраста. На рис. 16.11 представлена часть окна, появляющаяся в данном виде анализа ВП. В верхней части рис. 16.12 расположены названия компонент ВП. Нажимая 3, можно выбирать различные независимые компоненты ВП, зарегистрированных в GO/NOGO-тесте: например, P3GO, P3NOGO, P2NOGO и др. Символы в квадратных скобках справа от имени категории означают: [1] — независимая компонента индивидуального ВП; [2] — соответствующие нормативные данные; [D] — разница между независимой компонентой индивидуального ВП и данными нормативной базы.
Часть 2. Вызванные потенциалы
377
Рисунок 16.12. Сравнение независимых компонент индивидуального ВП и данных нормативной базы
Слева — независимые компоненты ВП пациента (тонкая линия) и ВП, усредненного по группе здоровых испытуемых одинакого с пациентом возраста. Справа — топография компонент сравнения (P2NOGO) и мониторинга (P3NOGO). В середине — отклонения от нормальных значений. Вертикальными столбцами, расположенными под графиком разностной волны, отмечены временные интервалы, на которых уровень значимости девиации достигает р < 0,001.
Пример сравнения индивидуальных независимых компонент с нормативными данными представлен на рис. 16.12.
В левой части рис. 16.12 изображены независимые компоненты одного пациента (тонкая линия графика) и компоненты усредненного по группе ВП (здоровые испытуемые такого же возраста). В правой части представлены топограм-мы двух компонент сравнения (P2NOGO) и мониторинга (P3NOGO). Разница между соответствующими компонентами ВП пациента и компонентами ВП нормативной базы данных приведены в средней части рисунка. Обращает на себя внимание сильное отклонение от нормальных значений компоненты ВП, связанной с операцией сравнения. При этом компонента ВП, ассоциирующаяся с операцией мониторинга, не отличается от нормы. В средней части рис. 16.12 под графиком ВП столбцами черного цвета обозначен уровень статистической значимости отклонения компоненты индивидуального ВП от нормативных значений. Столбец наименьшего размера соответствует уровню значимости р < 0,05, а максимального размера — р < 0,001.
IV. Упражнения
Обучающее программное обеспечение EdEEG снабжено записями ЭЭГ, зарегистрированными у здорового испытуемого и пациента (оба в возрасте 13—14 лет) в условиях выполнения двухстимульного GO/NOGO исследовательского теста.
Мы предлагаем читателю потренироваться в обработке этих двух файлов с ЭЭГ и ответить на следующие вопросы:
/. Какой из файлов относится к ЭЭГздорового испытуемого и пациента?
378
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
2. Какая из независимых компонент соответствует нормативным данным, а какая нет?
3. Какой тип дисфункции мозга виден на полученных ВПпациента?
4. С каким типом заболевания (синдром нарушения внимания с гиперактивностью, навязчивое состояние или дислексия) может быть связана наблюдаемая дисфункция?
Для ответа на поставленные вопросы выполните все шаги обработки и анализа ВП, представленные выше. Краткое описание необходимых действий:
1. Запустите программу EdEEG.exe.
2. В меню Файл выберите команду Открыть файл.
3. Откройте файлы S3 VCPT и S4_VCP, которые находятся в подпапке Data папки EdEEG (при обычной установке располагается на диске С). Далее анализируйте каждый файл по отдельности.
4. Просмотрите запись ЭЭГ, используя кнопки панели управления ввода ( ◄ ► ►►). Функциональное значение кнопок следующее: Остановка, Бы-
страя перемотка назад, Медленная перемотка назад, Медленное воспроизведение записи, Быстрое воспроизведение записи. Вы также можете просматривать запись ЭЭГ, используя полосу прокрутки, расположенную в нижней части, под окном ЭЭГ
5. Выберите команду Выбрать монтаж из меню Вид. В списке Имя монтажа выберите DataBase Version 1 montage. Это имя монтажа — последнее в списке монтажей. Нажав диалоговое окно Референты в окне Параметры монтажа, убедитесь в том, что выбран монтаж Взвешенный средний по Lemos. После этого нажмите Принять. Запись ЭЭГ появится в конфигурации выбранного монтажа.
6. Самостоятельно просмотрите запись ЭЭГ на предмет артефактов вертикальных и горизонтальных движений глаз.
7. Выделите всю запись ЭЭГ целиком. Это можно сделать, кликнув левой кнопкой мыши на начале записи и после этого — на окончании записи. Нажатия должны осуществляться на шкале времени, расположенной сверху записи ЭЭГ. После нажатия правой кнопки мыши шкала времени должна приобрести желтый цвет.
8. Выберите команду Коррекция артефактов из меню Анализ. На голограммах в правой части появившегося окна красным кругом обозначена первая голограмма (сверху вниз), отображающая наиболее выраженную компоненту, обычно соответствующую артефакту вертикального движения глаз. Убедитесь в том, что это действительно так, — нажмите кнопку ICA в списке Метод. Нажмите Принять. Программа отфильтрует этот тип артефактов из всей записи ЭЭГ. Внимательно просмотрите результат фильтрации и убедитесь в том, что артефакты скорректированы.
9. В меню Анализ выберите команду Отметить артефакты. Из базы данных загрузите параметры удаления артефактов, выбрав команду Загрузить из базы данных. После нажатия кнопки Принять программа отметит все артефакты, соответствующие заданным критериям.
10. В меню Анализ выберите команду Вызванные потенциалы. Из базы данных загрузите параметры расчета ВП, выбрав команду Загрузить из базы данных.
Часть 2. Вызванные потенциалы
379
Программа рассчитает ВП для условий VCPT233 теста отдельно для каждого электрода и представит в форме 19 графиков ВП, расположенных с учетом схемы локализации отведений на скальпе.
11. В меню Анализ выберите команду Сравнение результатов. Появится окно сравнения результатов Сравнение результатов обработки. Для загрузки ВП из нормативной базы нажмите Загрузить базу данных. В левой части появившегося окна будет расположено название индивидуального ВП, а в правой его части будут представлены две опции: Event Related Potentials (ВП) и Event related Components computed (выявленные компоненты ВП) для VCPT-теста.
12. Сначала сравните ВП с данными нормативной базы. Слева выберите из списка Event Related Potentials исследуемые вызванные потенциалы, а справа нажмите Event Related Potentials/Visual VCPT. Нажмите Принять.
13. В окне вызванных потенциалов (см. рис. 16.10) появится разностный ВП (индивидуальный ВП — нормативный ВП) с четырьмя категориями стимулов (a-a GO, а-p NOGO, р-р and p-h). Статистическая значимость отклонения от нормативных значений отображается под графиком ВП вертикальными столбцами черного цвета. Для рассмотрения ВП и разностных ВП на графике ВП одного из состояний (например, GO) сделайте следующее: а) нажмите 3 в первом списке Группы и выберите условие a-a GO[ 1] — ВП пациента в GO-условии; б) далее во втором списке Группы нажмите 3 и выберите условие a-a GO[2] — ВП усредненные по группе здоровых испытуемых того же возраста из нормативной базы в GO-условии; в) выберите разностный ВП — условие a-a GO[D], нажав 3 в третьем окне списка Группы.
14. Далее сравните независимые компоненты индивидуального ВП с нормативной базой. В меню Анализ выберите команду Сравнение результатов. Снова появится окно Сравнение результатов обработки. Загрузите нормативные ВП из базы данных, нажав Загрузить базу данных. В левой части окна будут расположены названия ВП испытуемых, а в правой — две опции выбора: Event Related Potentials и Event Related Components.
15. Выберите вторую опцию. Слева выберите из списка Event Related Potentials исследуемые вызванные потенциалы, а справа нажмите Event Related Potentials. Components/Visual VCPT. Нажмите Принять.
16. В окне вызванных потенциалов (рис. 16.12) появятся разности независимых компонент ВП (компонента индивидуального ВП — компонента нормативного ВП) для четырех независимых компонент (Pl, P3GO, P2NOGO и P3NOGO). Уровень статистической значимости отклонения от нормативных значений отображается под графиком ВП вертикальными столбцами черного цвета. Для рассмотрения компонент ВП и их разностей на графике ВП одного из компонент (например, P3GO) сделайте следующее: а) нажмите 3 в первом списке Группы и выберите компоненту P3GO[7/; б) далее во втором списке Группы нажмите 3 и выберите компоненту P3GO[2]\ в) выберите разность компонент — P3GO[D], нажав 3 в третьем списке Группы.
233 VCPT — просто другое название двухстимульного GO/NOGO-теста.
ЧАСТЬ 3.
Расстройства систем мозга
Введение
Как было показано во 2-й части, мозг может быть разделен на системы. Эти системы состоят из отдельных кортикальных и субкортикальных анатомических структур и играют различные функциональные роли в обработке сенсорной информации и организации действий. В предыдущей части мы рассматривали следующие системы: сенсорную, систему внимания, систему управления, эмоциональную систему, систему памяти. Мы предполагаем, что отдельные психические расстройства связаны с дисфункциями отдельных систем мозга.
Однако психические расстройства принято классифицировать на основе поведения, а не на нейрофизиологической базе. Два главных диагностических руководства: DSM-IV и ICD-101 основаны только на поведенческой симптоматике. Несмотря на успешные исследования и появление нескольких объективных диагностических параметров2 для определенных заболеваний, до сих пор нет общепринятых надежных биологических маркеров или эндофенотипов для отдельных расстройств. Эта часть книги посвящена следующим тесно взаимосвязанным задачам: 1) анализу дисфункций вышеупомянутых систем при ряде психических и неврологических расстройств; 2) изучению взаимосвязи дисфункций мозговых систем и маркеров количественной ЭЭГ и когнитивных ВП, описанных в 1-й и 2-й частях этой книги; 3) рекомендациям по выбору эффективной нейротерапии для коррекции дисфункций мозговых систем.
Из 2-й части мы узнали, что работа отдельных мозговых систем контролируется конкретными нейромедиаторами, синтезируемыми мозгом, следовательно, ее можно корректировать с помощью лекарственных3 воздействий на эти медиаторы. Эти вмешательства заключаются в увеличении или снижении эффекта нейромедиаторов: 1) путем введения в мозг биохимических предшественников медиаторов (как, например, L-dopa — предшественника дофамина); 2) блоки-
1 The Diagnostic and Statistical Manual of Mental Disorders (DSM) — Руководство по диагностике и статистике психических расстройств — содержит перечень различных категорий психических и психологических расстройств и диагностические критерии для
них согласно Американской психиатрической ассоциации. Оно используется повсеместно клиницистами и исследователями. Это руководство пережило пять изданий с 1952 года. Последнее пересмотренное и исправленное издание, DSM-IV, было опубликовано в 1994 г. В настоящее время подготавливается DSM-V, оно ориентировочно будет издано в 2011 г. Раздел психических расстройств содержится и в International Statistical Classification of Diseases and Related Health Problems (ICD) — Международной статистической классификации болезней и проблем, связанных со здоровьем — другом общепринятом руководстве, использующем те же диагностические нормы.
3 Например, индекс невнимательности (отношение мощности «тета/бета») по данным спектров ЭЭГ в диагностике нарушений внимания.
3 Фармакология (от греческого yapfiaxov (фармакон) — лекарство, яд и Муо; (логос) — слово) изучает воздействие различных биохимических субстанций на функции органов живого организма. Если эти вещества имеют лечебные свойства, они называются лекарствами. Эта область включает в себя создание лекарств, изучение их свойств и клинических эффектов. Нейро- и психофармакология (изучение эффектов лечения на поведение и работу нервной системы) рассматриваются как подразделы фармакологии.
Часть 3. Расстройства систем мозга
381
рованием постсинаптических рецепторов (как в случае с антипсихотическими средствами, имеющими сродство с дофаминовыми рецепторами); 3) блокированием механизмов обратного захвата медиатора (например, Риталин подавляет механизмы обратного захвата дофамина) или изменением клеточных механизмов нейротрансмиссии другими путями4.
Следует отметить, что до 1950-х предпринимались попытки использования некоторых нефармакологических вмешательств, таких как инвазивные и неинвазивные воздействия электрическим током, а также анатомические деструкции в процессе нейрохирургических, в том числе стереотаксических операций. От большинства этих вмешательств впоследствии отказались в связи с революционными открытиями в психофармакологии. Однако применение достижений психофармакологии на протяжении 50 лет сопровождалось неудовлетворенностью и вызывало споры. Отчасти полемика была связана с неудачными попытками поиска генотипов, ответственных за механизмы развития определенных заболеваний5. Недавно в опытах по молекулярной генетике было установлено, что наследование большей части психических расстройств не подчиняется простым правилам Менделя и что возникновение конкретного расстройства не может быть приписано отдельному гену. Эти наблюдения способствовали выдвижению частью ученых концепции эндофенотипов как биологических маркеров заболеваний, немолекулярных по строению, но более тесно связанных с генотипами по сравнению с классификацией расстройств, основанной на поведенческих симптомах. Некоторые параметры количественной ЭЭГ и компоненты когнитивных ВП предложены в качестве кандидатов в эндофенотипы, по меньшей мере для некоторых заболеваний мозга.
Недавнее начало возрождения нелекарственных методов лечения можно расценить как обратный эффект после чрезмерного использования фармакологических препаратов в коррекции психических заболеваний. Нелекарственные методы включают ЭЭГ-БОС и методы электромагнитной стимуляции, такие как транскраниальная микрополяризация (ТКМП), транскраниальная магнитная стимуляция (ТМС), глубинная стимуляция мозга (ГСМ) и электроконвульсивная терапия (ЭКТ). Они объединены под общим названием «нейротерапия».
В этой части книги мы приведем наиболее распространенные психические расстройства в сопоставлении с данными количественной ЭЭГ и когнитивных ВП в качестве показателей работы мозговых систем, предположительно вовлеченных в патогенез этих заболеваний. Перечень этих заболеваний представлен в табл. III. 1. Мозговые системы, связанные с данными дисфункциями, выделены серым цветом. Таблица упрощает суть вопроса, не отражая комплексного взаи
4 Столетиями лекарства создавались на основе растений, но современная западная медицина использует очищенные биоактивные соединения, а не все вещество растения, как это делалось ранее. Настоящий прорыв в психофармакологии произошел в 1950-х при появлении первого ряда психотропных веществ и новых методологических подходов в фармакологии. Появилась возможность печения таких гетерогенных заболеваний, как мания, шизофрения, депрессия, биполярное расстройство.
5 Другая сторона полемики касалась практических вопросов. Прежде всего фармакотерапия помогает не всем: некоторые пациенты резистентны к лекарственным средствам. Более того, при подборе действенного лечения (особенно — при депрессии) используется метод проб и ошибок. Поскольку достижение клинического эффекта при приеме некоторых лекарств требует нескольких недель, то при наличии большого числа потенциально эффективных при данном заболевании препаратов методика проб и ошибок может занимать месяцы, что крайне дорого как в финансовом отношении, так и с точки зрения человеческих ресурсов. Помимо этого, большая часть лекарств имеет нежелательные побочные эффекты.
382 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
модействия между системами6 и гетерогенности отдельных заболеваний. Тем не менее концепция, приведенная в табл. III. 1, может служить исходным пунктом для представления на рассмотрение д анных и идей относительно использования параметров количественной ЭЭГ и компонент когнитивных ВП в качестве маркеров психических расстройств и нейротерапевтических подходов к коррекции этих расстройств.
Из этой книги мы узнали, что работа мозговых систем поддерживается комплексом нейронных сетей, включающих кортикальные и субкортикальные анатомические структуры. Рассмотрим теперь некоторую обобщенную нейронную Таблица 111.1. Взаимосвязь между психическими/неврологическими заболеваниями и расстройствами мозговых систем
Заболевание/система Чувствительная Эмоциональная Память Управление
Дислексия
Сенсорный дефицит
Депрессия
Тревожность
Болезнь Альцгеймера
Болезнь Паркинсона
Шизофрения
Обсессивно-компульсивное расстройство —
СНВГ
Аддиктивные расстройства
сеть7. Такая сеть характеризуется двумя параметрами: 1) уровнем активации; 2) реакцией на единичное повышение уровня входного сигнала. Взаимосвязь между реактивностью системы и уровнем ее активации описывается перевернутой U-образной кривой (рис. III. 1).
Мы предполагаем, что ре-
Риг/унок PIII.1. Зависимость ответа (амплитуды компоненты ВП) от уровня активации гипотетической нейронной сети
активность нейронной сети находит приближенное выра-
жение в независимых компонентах когнитивных ВП, генерируемых сетью в соответствующих психологических условиях8. Можно видеть, что максимум
6 При мозговой дисфункции взаимодействие мозговых систем ведет к тому, что, например, дисфункция системы управления может способствовать развитию дисфункции эмоциональной системы, и наоборот. Эго взаимодействие вносит сложности в поведенческий паттерн заболевания каждого конкретного пациента.
7 Например, нейронные сети, связанные с операциями контроля, включают в себя несколько зон лобной коры, причем передняя цингулярная кора является их ключевым элементом.
6 Например, ответ системы контроля оценивается путем измерения амплитуды компоненты мониторинга Р400 в двухстимульном GO/NOGO-тесте.
Часть 3. Расстройства систем мозга 383
амплитудных значений ВП соответствует оптимальному уровню активации нейронной сети. Из 1-й части мы знаем, что превышение оптимального уровня активации коры связано с чрезмерным увеличением в фоновой ЭЭГ бета-активности, генерируемой соответствующими областями коры, в то время как снижение уровня активации сочетается с избыточной альфа-активностью9. Обратите внимание на то, что в обоих случаях отмечается уменьшение амплитуд компонент когнитивных ВП. Это связано со снижением реактивности системы при ее смещении с точки максимальной эффективности10. Следовательно, чрезмерное увеличение как бета-, так и альфа-активности, сопровождающееся снижением амплитудных значений когнитивных ВП, отражает снижение эффективности работы системы. Эти теоретические соображения показывают необходимость одновременной оценки компонент ВП и характеристик фоновой ЭЭГ.
Пюссарий
Аддикция — непреодолимое, не поддающееся контролю употребление наркотических средств, невзирая на серьезные негативные последствия.
СНВГ— синдром нарушения внимания с гиперактивностью — наиболее распространенное нервно-психическое расстройство детского возраста, встречающееся, согласно скромным подсчетам, у 3—8 % детей в популяции и характеризующееся не соответствующими возрасту уровнями невнимательности, гиперактивности и импульсивности, обусловливающими низкую успеваемость в школе и впоследствии оказывающими негативное влияние на интеллектуальную и профессиональную деятельность и социальные взаимоотношения.
Болезнь Альцгеймера — дегенеративное заболевание центральной нервной системы, характеризующееся прогрессирующим расстройством памяти в начальной стадии процесса, переходящим в глубокое общее слабоумие в конечной стадии. По-видимому, решающим фактором в возникновении первой стадии процесса является утрата холинергических клеток базальными отделами переднего мозга.
Антипсихотические средства — фармакопрепараты, используемые в лечении психоза.
Ангедония — психическое состояние, характеризующееся общим отсутствием интереса к жизни, утратой способности наслаждаться и радоваться.
Когнитивно-поведенческая терапия — разновидность психотерапии, основанная на понимании проблем пациента и направленная на укрепление у него чувства собственного достоинства. Когнитивно-поведенческая терапия занима
* Вспомните взаимосвязь между метаболической активностью коры и уровнем активности альфа- и бета-диапазонов в ЭЭГ, генерируемой в соответствующих корковых зонах (рис. 111.1). Согласно этой взаимосвязи, чрезмерное (по сравнению с нормой) увеличение бета-активности соответствует повышению метаболической активности, в то время как увеличение альфа-активности проис-одит при снижении локальной метаболической активности.
ш Например, снижение амплитуды компонента мониторинга может быть связано как с чрезмерной активацией нейронной сети мониторинга, так и со снижением уровня ее активности.
384 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
ется непосредственным анализом проблем и выбором конкретных целей и решений, так что пациент может видеть прогресс в ходе лечения.
Компульсии — устойчивые навязчивые действия, повторяющиеся в ритуалисти-ческой стереотипной последовательности.
Постоянный ток — электрический ток, сила и направление которого не изменяются с течением времени. При воздействии на ткань мозга постоянный ток поляризует ее, т.е. изменяет мембранный потенциал клеток.
Депрессия (большая депрессия) — расстройство эмоциональной системы, проявляющееся сниженным, или депрессивным, настроением, ангедонией, постоянным чувством усталости.
Руководство по диагностике и статистике психических расстройств (DSM) — содержит перечень различных категорий психических и психологических расстройств и диагностические критерии для них, разработанные Американской психиатрической ассоциацией. Это руководство пережило пять изданий с 1952 г. и используется повсеместно клиницистами и исследователями. Последнее пересмотренное и исправленное издание, DSM-FV, опубликовано в 1994 г. В странах Европы чаще используется International Statistical Classification of Diseases and Related Health Problems (ICD) — Международная статистическая классификация болезней и проблем, связанных со здоровьем.
Электроконвульсивная терапия (ЭКТ), также известная как электрошок, используется в психиатрии. Во время прохождения мощного электрического разряда через головной мозг у пациента возникает судорога.
Величина эффекта — показатель некоторого эффекта, вычисляемый как отношение разности средних величин параметра в экспериментальной и контрольной группе к стандартному отклонению в контрольной группе или к среднему стандартному отклонению обеих групп. Величина эффекта — стандартизованный показатель, позволяющий сравнивать эффекты, обнаруженные в разных исследованиях и с использованием разных параметров.
Эндофенотип — биологический маркер заболевания.
Моноаминоксидаза — фермент внешней мембраны митохондрий, катализирует реакцию гидролиза биогенных аминов, таких как катехоламины и серотонин.
Заболеваемость — показатель распространенности заболевания в популяции.
ЭЭГ-биоуправление — техника саморегуляции с использованием биологической обратной связи по ЭЭГ (ЭЭГ-БОС). Параметры ЭЭГ (например, мощность ЭЭГ в определенном частотном диапазоне) в процессе биоуправления регистрируются со скальпа и предъявляются субъекту в виде сигналов зрительной, слуховой или тактильной модальности. Задача состоит в произвольном изменении этих параметров в заданном (ведущем к более эффективной работе мозга) направлении.
Обсессии — это беспрестанно и настойчиво повторяющиеся нежелательные мысли.
Часть 3. Расстройства систем мозга
385
Обсессивно-компульсивное расстройство (ОКР) — хроническое заболевание, характеризующееся повторяющимися навязчивыми идеями и компульсиями, вызывающее стресс и значительно нарушающее жизнь человека.
Оппозиционно-девиантное поведение проявляется в неоправданно рискованных поступках, нарушении законов, отказе следованию инструкциям и указаниям.
Психоз — психическое расстройство, сопровождающееся нарушениями мыслительных операций — маниями (ложными убеждениями) и расстройствами восприятия — галлюцинациями. Слуховые галлюцинации, обычно переживаемые как внутренние голоса, наиболее типичны для шизофрении.
Психофармакология — раздел фармакологии, в частности изучающий влияние различных биохимических субстанций на поведение и работу нервной системы.
Психохирургия — избирательное хирургическое удаление или разрушение элементов ткани мозга в целях коррекции эмоциональных или когнитивных нарушений, обусловленных психическими заболеваниями.
Шизофрения — психическое расстройство, характеризующееся: 1) нарушениями в работе мозговой системы управления в виде расстройства мышления и речи, утраты способности планировать, инициировать и регулировать целенаправленное поведение, а также 2) изменением состояния сенсорной системы в форме галлюцинаций и 3) связанной с этими нарушениями дисфункцией эмоциональной системы в виде притупления эмоций.
Трудности в обучении связаны с затруднениями в освоении речи, чтения и математики.
Транскраниальная микрополяризация (ТКМП) — новый метод в нейротерапии, использующий воздействие постоянного тока на мозг с помощью расположенных на голове электродов. Метод получил свое название в 1960—1970-х гг. в связи с поляризующим влиянием постоянного тока на ткани мозга.
Синдром Туретта характеризуется тиками — неконтролируемыми движениями в виде морганий, подергиваний мимических мышц или вокальных звуков.
Трициклические препараты — подавляют обратный захват биогенных аминов, продлевая период их нахождения в синаптической щели в активной форме.
386
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Глава 17.
Синдром нарушения внимания с гиперактивностью
I. Клинические симптомы
А. Процессы управления и компоненты когнитивных вызванных потенциалов
Рассмотрим расстройства системы управления. Основные операции системы управления — это рабочая память и внимание, процессы вовлечения в действие, контроля и подавления действий. Компоненты когнитивных ВП соответствуют этим процессам. В табл. 17.1 перечислены тесты, в которых генерируются эти компоненты. Вспомните, что компонента ВП отражает ответ определенной системы мозга (например, нейронной сети управления) на адекватный стимул в соответствующей поведенческой парадигме. Видно, что ни один отдельно выбранный тест не может использоваться в качестве «золотого стандарта» для оценки работы системы управления. Однако некоторые варианты двухстимульного GO/NOGO-теста можно рассматривать как приближение к «золотому стандарту».
Таблица 17.1. Компоненты когнитивных вызванных потенциалов как показатели процессов в системе управления
Операции управления Вовлечение Рабочая память/ внимание Подавление ответа Контроль
Компонента когнитивных вызванных потенциалов и психологический тест РЗЬ на значимые стимулы в тестах Oddball и GO/NOGO Компонента сравнения Р2 в GO/NOGO-тесте и в тестах с отсроченным ответом N2 на стоп-стимул в стоп-сигнал тестах или компонента подавления действия на NOGO-стимул в GO/NOGO-тестах Компонента мониторинга P4N0G0 в GO/ NOGO-тестах, негативностью, связанной с ошибкой (НСО), в тестах на непрерывное выполнение
Симптомы СНВГ согласно DSM-IVи ICD-10. Вообразите, что вы находитесь в быстро движущемся калейдоскопе, где окружающие предметы постоянно перемещаются и ваше внимание переключается с одного образа, звука или впечатления на другой. Представьте себе также, что вы легко отвлекаетесь на несущественные звуки и явления, ощущая скуку и беспомощность при выполнении данного вам задания. Это то, что испытывают дети с СНВГ11.
Синдром нарушения внимания с гиперактивностью (СНВГ) — одно из самых распространенных расстройств в детской психиатрии12. В прошлом для его обо
u Литература изобилует описаниями подобных характеров. В1845 г. немецкий доктор Г. Гофман дал портрет мальчика с СНВГ в •Истории непоседы Филиппа». Другой пример ребенка с СНВГ описан известным русским писателем Н. Носовым в его прекрасной книге «Приключения Незнайки и его друзей». Незнайка противопоставлен Знайке, которого можно назвать нормальным, здоровым ребенком. Интересно, что среди российских детей Незнайка пользуется большей популярностью, чем Знайка.
“ По осторожным оценкам, он встречается у 3-7,5 % детей повсеместно.
Часть 3. Расстройства систем мозга
387
значения использовались другие названия: синдром нарушения внимания, СНВ, гиперактивность, гиперкинез, гиперкинетический синдром, минимальная мозговая дисфункция и минимальное мозговое поражение13. В DSM-FV СНВГразделен на три типа: с преобладанием невнимательности, с преобладанием гиперактивности и импульсивности и смешанный тип. Эта классификация базируется на трех группах симптомов: невнимательности, гиперактивности и импульсивности14. Основания для клинического использования этих типов недостаточны и спорны. До настоящего времени не решен вопрос, должен ли тип с преобладанием невнимательности рассматриваться самостоятельно или в составе СНВГ
Несмотря на то что СНВГ — одно из самых распространенных расстройств, подходы к его диагностике довольно противоречивы. Эти противоречия состоят, например, в разнице между американскими критериями для СНВГ, по данным DSM-IV, и европейскими диагностическими критериями для гиперкинетического расстройства (ГКР, по определению ICD-10). Обе классификации включают детей с не соответствующими возрасту уровнями невнимательности, гиперактивности и импульсивности, обусловливающими низкую успеваемость в школе и впоследствии оказывающими негативное влияние на интеллектуальную и профессиональную деятельность и социальные взаимоотношения. Но критерии ICD-10 строже и требуют большей выраженности симптоматики для постановки диагноза по сравнению с критериями DSM-IV15. По данным клинических работ, преобладание этого расстройства у мальчиков по сравнению с девочками выражено меньше, чем по результатам социальных исследований. Это может быть связано с тем, что у женщин СНВГ оказывает менее разрушительное влияние на жизнь, к тому же мужчины больше подвержены несчастным случаям, например черепно-мозговым травмам.
Несмотря на то что и DSM-IV и ICD-10 содержат хорошо структурированные, основанные на четких критериях диагнозы для СНВГ и ГКР, они имеют ряд недостатков. Диагностические критерии не позволяют разграничить симптомы СНВГ и проявления невнимательности, гиперактивности и импульсивности у здоровых детей, обусловленные особенностями созревания. Клиницисты получают данные для диагностики из многих источников (например, от родителей и учителей), но ни одно из руководств не дает указаний на то, как интегрировать эту информацию. Недостатки диагностической системы СНВГ вызвали критическое отношение к ней, как к субъективной, и инициировали поиск объективных маркеров эндофенотипов этого расстройства16.
13 Описание СНВГ в DSM-IV можно найти по адресу http://en.wikipedia.org/wiki/DSM-IV_Codes.
14 Невнимательным детям трудно сосредоточиться на одном задании дольше нескольких минут, они испытывают сложности с концентрацией внимания, организацией и выполнением дел. Гиперактивные дети все время в движении, они не могу сидеть спокойно. пытаются все потрогать и говорят без умолку, могут испытывать беспокойство, хватаясь за несколько дел сразу и перескакивая с одного на другое. Импульсивные личности не способны сдерживать свои непосредственные реакции, не могут думать перед тем, как лгйствовать, им трудно дожидаться получения желаемого, например своей очереди в играх.
13 Поэтому при использовании DSM-IV, включающей «невнимательный» тип, СНВГ выявляется чаще, чем ГКР — при использовании ICD. В связи с этим иногда создается ложное впечатление, что в США СНВГ распространен в большей степени, чем в странах, f дл диагностика основывается на критериях ICD. Однако при использовании одних и тех же критериев частота встречаемости СНВГ (мжнаковв повсеместно.
|а Впервые понятие «эндофенотип» какоснованный на нейронауке маркер СНВГ предложили К. Кастелланос и Р. Таннок в 2002 г. Они представили три эндофенотипа, которые могли бы лежать в основе СНВГ: 1) специфическая аномалия нейронных сетей, свя-жнных с механизмами подкрепления; 2) временной дефицит в обработке информации; 3) нарушения рабочей памяти (Castellanos, Unnock, 2002).
388 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
II. Генетические и внешние факторы
А. Комплексное генетическое расстройство
СНВГ прослеживается в поколениях, и приблизительно у половины родителей с СНВГ рождаются дети с этим расстройством. Согласно исследованиям у близнецов, гены играют важную роль в передаче СНВГ по наследству, и наследуемость СНВГ оценивается как 0,76. В то же время, по данным генетических исследований, СНВГ, как и большинство психических расстройств, не следует законам наследования Менделя и должен расцениваться как комплексное генетическое заболевание17.
Б. Внешние факторы риска
Хотя предположения о причинной связи между возникновением СНВГ и некоторыми видами пищи или пищевых добавок активно обсуждались средствами массовой информации, современная наука не подтвердила основных посылок этих теорий. Однако установлено, что воздействие свинца способствует возникновению этого расстройства.
Установлено, что осложненное протекание беременности и родов повышает риск развития СНВГ. Базальные ганглии — один из отделов мозга с наиболее активным метаболизмом и поэтому уязвимый для гипоксии — как правило, вовлечены в процесс при СНВГ. Многими исследованиями подтверждено, что недоношенность — фактор риска в возникновении СНВГ. Выявлено, что употребление матерью алкоголя во время беременности ведет к поведенческим и когнитивным нарушениям и проблемам в обучении у ребенка, которые могут рассматриваться как проявления СНВГ. Подверженность плода воздействию никотина в критические стадии развития при курении во время беременности также может вызвать поражение мозга. К другим факторам внешней среды можно отнести эмоциональную депривацию, неблагоприятную обстановку в семье, черепно-мозговые травмы и кровоизлияния в мозг, особенно в случае поражения базальных ганглиев.
В. Коморбидность
Многочисленные данные клинических и эпидемиологических исследований демонстрируют подверженность детей с СНВГ риску развития других психических расстройств, включая наркотическую зависимость. Данные некоторых работ позволяют предположить, что СНВГ ведет к риску формирования личностных расстройств. Во все времена СНВГ ассоциировался с функциональными нарушениями, приводящими к проблемам в школе и во взаимоотношениях со сверстниками, конфликтам в семье, неудачам в профессиональной деятельности, травмам, антисоциальному поведению, нарушениям дорожных правил и авариям. Продолженные наблюдения показывают, что проявления СНВГ сохраняются при взрослении. С возрастом симптомы сглаживаются, но даже в случае, если они недостаточно выражены
17 У детей с тяжелыми формами СНВГ выявлена аномалия гена транспорта дофамина (DAT1), локализованного в 5-й хромосоме. Известно, что психостимуляторы, эффективные при СНВГ, блокируют перенос дофамина. Этот факт позволил предположить, что гены рецепторов дофамина D4 и D5 и гены, имеющие отношение к реализации}» свойств, имеют отношение к СНВГ.
Часть 3. Расстройства систем мозга
389
для постановки диагноза, они нередко свидетельствуют о значительных клинических нарушениях.
СНВГ часто сочетается с другими расстройствами: специфическими трудностями в обучении, оппозиционно-девиантным поведением, синдромом Туретта, тревожностью и депрессией (рис. 17.1). Оппозиционнодевиантное поведение и синдром Туретта — проявления дисфунк-
Рисунок 17.1. Заболевания, коморбидные с СНВГ
ции системы управления. Тревожность и депрессия обусловлены нарушениями в эмоциональной системе, а трудности в обучении (например, дислексия) связа-
ны с вовлечением сенсорной системы.
III. Офушурные и физиологические корреляты
А. ПЭТ и МРТ
Является ли СНВГ крайним вариантом нормы, декомпенсирующимся под влиянием неблагоприятных внешних факторов, или же это заболевание, связанное с дисфункцией определенных мозговых систем? Методы нейронауки помогают ответить на этот вопрос. Однако, как и для любого другого психического расстройства, начальная фаза исследования нейробиологии СНВГ основывается на чисто описательных методах. Нейроанатомические и нейрофизиологические параметры мозга пациентов с СНВГ сравниваются с группой контроля здоровых лиц, при этом наибольшее внимание, как правило, вызывают базальные ганглии и префронтальные отделы коры. В ходе сравнения обнаружены следующие факты:
1. При МРТ-исследовании обнаружено уменьшение размеров хвостатого ядра, бледного шара и префронтальных отделов коры. В частности, группа ученых из Национального института здоровья (NIH) в Бетезде, США (Catellanos et al., 2003), изучала MPT-изображения мозга монозиготных близнецов, дискор-дантных по СНВГ. У близнецов с проявлениями этого расстройства отмечено значительное уменьшение объема хвостатого ядра18. Эти данные подтверждают вовлечение стриатума — структуры, вводящей в состав базальных ганглиев, в патофизиологию СНВГ. Стоит упомянуть и другую работу, выполненную объединенной группой исследователей из NHI и Медицинского института Джона Хопкинса в Балтиморе. Анализировались МРТ детей без проявлений СНВГ в анамнезе спустя три месяца после закрытой черепно-мозговой травмы. У детей с последующим развитием вторичного (индуцированного травмой) СНВГ были
“ Например, у одного из близнецов с симптомами СНВГ неожиданно была обнаружена аномалия хвостатого ядра, скорлупы и Фмялвишцего белого вещества слева.
390 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
выявлены более выраженные повреждения скорлупы справа по сравнению с теми, у кого вторичный СНВГ не возник19.
2. Согласно данным фМРТ- и ПЭТ-исследований, у детей с СНВГ при сравнении с нормой обнаружено снижение уровня потребления глюкозы в лобной коре и базальных ганглиях, в особенности во время выполнения заданий.
Хотя величина эффекта сравнений данных МРТ и ПЭТ была качественно (статистически) значимой, она была небольшой в количественном выражении: разница между группами пациентов с СНВГ и здоровых детей составляла около 6 % (этот параметр называется величиной эффекта), что сопоставимо с величиной стандартного отклонения в нормативной группе. Два десятилетия исследований в области психиатрической генетики, проводимых примерно в то же время, также не принесли заметных успехов. Неудовлетворенность результатами этих работ способствовала пробуждению интереса к изучению количественных индексов предрасположенности к заболеваниям — эндофенотипов — по данным ЭЭГ и когнитивных ВП. Предполагается, что эндофенотипы определяют риск формирования СНВГ подобно тому, как наличие спайковой активности в ЭЭГ говорит о риске возникновения эпилепсии. Эндофенотипы должны поддаваться количественному определению и предсказывать вероятность развития заболевания.
Б. Количественная ЭЭГ
Ниже мы даем обзор литературы и наших собственных работ в рамках теории, приведенной во 2-й части. Мы предполагаем, что при СНВГ основная дисфункция касается системы управления. Система управления характеризуется двумя параметрами: уровнем общей активации и амплитудой ответа, связанной с четырьмя различными процессами: рабочей памятью, выбором действий, подавлением действий и контролем действий. Позвольте начать с описания спонтанной электрической активности в ЭЭГ, которую можно рассматривать в качестве показателя общей активации системы управления и, следовательно, фона для процессов управления.
Как было показано в 1-й части этой книги, ЭЭГ, зарегистрированная при открытых и закрытых глазах, является хорошим индикатором метаболической активности коры. Локальное увеличение медленной активности и уменьшение бета-активности говорит о снижении уровня метаболизма в соответствующей области коры. Эти отклонения чаще всего встречаются при СНВГ.
В. Отношение «тета/бета» как показатель невнимательности
В большей части работ по изучению количественной ЭЭГ сообщается об увеличении мощности медленноволновой активности при СНВГ по сравнению с нор-
19 Базальные ганглии — по-видимому, не единственная структура, вовлеченная в патогенез СНВГ. В исследовании, проведенном недавно в Медицинском колледже Стэнфордского университета (Tamm et al., 2004), сравнивались данные фМРТ в процессе выполнения GO/NOGO-теста у группы подростков с СНВГ и у их здоровых сверстников. У пациентов с СНВГ были обнаружены явные отклонения в уровне активации мозга при предъявлении NOGO-проб, включающие гипоактивацию передних/средних отделов цингулярной коры, распространяющуюся на дополнительную моторную эону.
Часть 3. Расстройства систем мозга
391
Рисунок 17.2. "Голограммы индекса невнимательности
Данные взяты из нормативной HBI базы данных. Использован монтаж с локальным средневзвешенным референтом (Lemos). Индекс рассчитан как отношение мощностей тета/бета, где тета — 4-8 Гц, бета —13-21 Гц.
мой20. В связи с тем, что показатели абсолютной мощности спектров сильно варьируют в зависимости от нескольких факторов, не связанных с активностью мозга (например, толщина костей черепа), был предложен к использованию относительный параметр: отношение мощности диапазонов «тета/бета». Этот параметр был назван индексом невнимательности труппой участниковмежуниверситетских исследований в США (Monastia et al., 1999)21. Наш опыт анализа количественной ЭЭГ у более чем 1000 детей с проявлениями СНВГ показывает, что индекс невнимательности является надежным электроэнцефалографическим критерием по меньшей мере у части детей с этим диагнозом. Следует отметить, что картирование лого индекса у здоровых лиц выявило значимое смещение максимума этого показателя из центрально-теменных отделов у 7—8-летних детей в лобно-центральные отделы у взрослых (рис. 17.2). Это наблюдение показывает, что для более надежного разграничения СНВГ и нормы этот показатель должен оцениваться в разных точках расположения электродов в зависимости от возраста.
Касательно параметра когерентности Р. Барри и его коллеги из университета Воллонгонга (Австралия) на основе собственного опыта и анализа литературы пришли к выводу о том, что в настоящее время не имеется достаточных данных
« Одна из самых ранних работ по выявлению отклонений в ЭЭГ у детей с минимальной мозговой дисфункцией (синоним СНВГ в IU30- 1950-х годах) относится к 1938 г. (Jasper et al, 1938). Подобные исследования проводились позднее, в 1950-х и 1960-х годах при использовании визуального анализа чернильной записи ЭЭГ. Наиболее частой качественной находкой было повышение активности дельта- и тета-диапазонов. В 1970-х годах и позднее это явление нашло подтверждение при количественном анализе ЭЭГ.
п Так называемый индекс невнимательности определялся как отношение мощности тета-ритма (4-8 Гц) к мощности бета-ритма (13-21 Щ). Использовался монополярный монтаж с активным электродом в отведении Cz и объединенным ушным электродом в нгостм референта. У 6-7-летних детей с преимущественно невнимательным и смешанным типом СНВГ отношение *тета/бета» в 1 рма превышало норму. Чувствительность этого индекса (процент детей с диагнозом СНВГ с превышающими норму значениями него покаэвтеля) составляла 86 %, а его специфичность (процент детей без признаков СНВГ с нормальными значениями этого от-чмямия) — 98 % (Monastra et al., 1999).
392 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
для оценки надежности показателя когерентности в диагностике СНВГ (Валу etal.,2003)22.
IV. Корреляты когнитивных вызванных потенциалов
Существует большое число эмпирических работ, посвященных изучению когнитивных ВП при СНВГ. Как и исследования показателей количественной ЭЭГ, они до недавнего времени носили чисто описательный характер. Основные задачи состояли: 1) в подборе поведенческой парадигмы, которая могла бы отражать процессы, предположительно нарушенные при СНВГ; 2) в выборе компоненты ВП, рассчитанного как разностная волна между двумя условиями теста23; 3) в измерении этой компоненты в двух группах (СНВГ по сравнению с нормой) или при разных типах СНВГ («невнимательный» или комбинированный тип по сравнению с нормой); 4) в оценке величины эффекта, т.е. относительных различий между средними значениями величин, рассчитанных для каждой из групп.
Начало исследованиям когнитивных ВП при СНВГ положила в 1970-х работа Саттерфилда и его группы (Satterfield et al., 1972). Огромное число работ, посвященных когнитивным ВП при СНВГ, породило многообразие результатов, порой противоречивых. Полный обзор статей по этой теме недавно был опубликован в Clinical Neurophysiology Journal (Barry et al., 2003). В этой книге мы представляем только исследования с устойчивыми результатами. Эти работы сгруппированы в соответствии с психологическими процессами, предположительно нарушенными при СНВГ.
А. Селективное внимание
Селективное внимание исследуется в тестах с пространственной сигнализацией. Вспомните, что в этой парадигме значимым стимулам предшествуют условные стимулы, достоверно или ложно указывающие на локализацию последующего значимого стимула. Предполагается, что эффект достоверности, измеряемый как разница в скорости и точности ответа на значимые стимулы между достоверными и недостоверными пробами, отражает процесс концентрации внимания на определении местоположения стимула. Однако результаты исследований, использующих эту парадигму у детей с СНВ/СНВГ, противоречивы. Недавно Хуанг-Поллок и Нигг (2003) провели метаанализ экспериментальных находок и не обнаружили достоверного дефицита зрительнопространственной ориентации ни при СНВГ в целом и ни при одном из типов СНВГ в отдельности.
22 С точки зрения статистики при оценке значений когерентности присутствует систематическая ошибка. Эта ошибка зависит от числа эпох, используемых для расчета параметра. Строго говоря, для статистически надежной оценки параметра когерентности и возможности его сравнения с базой данных необходимо наличие равного числа ЭЭГ-эпох в нормативной базе данных и в сравниваемой с нею индивидуальной ЭЭГ.
23 Эта волна, например, может быть получена как разность когнитивных ВП на стандартные и девиантные стимулы в Oddball-парадигме, на значимые и незначимые стимулы в тесте на селективное внимание или как разность NOGO- и GO-компонент в GO/ NOGO-парадигме.
Часть 3. Расстройства систем мозга 393
Другая парадигма, используемая при изучении селективного внимания, — дихотический слуховой тест или его зрительный аналог. Недавно при пространственно-временном, с высокой степенью разрешения картировании когнитивных ВП в зрительной парадигме теста на селективное внимание в группе детей с СНВГ были выявлены отклонения (Jonkman et al., 2004)24. У этих пациентов отмечалось уменьшение так называемой лобной негативности выбора — компоненты ВП, отражающей процессы селекции сенсорной информации. Однако в предыдущем исследовании, использовавшем процессную негативность в качестве показателя селективного внимания, не было выявлено различий между пациентами с разными типами СНВГ и здоровыми испытуемыми (Rothenberger et al., 2000). Разногласия в работах по изучению механизмов селективного внимания при СНВГ могут объясняться гетерогенностью популяций СНВГ. Мы можем только предположить, что подгруппа детей с повреждением механизмов селективного внимания сравнительно невелика.
Б. Рабочая память
Как мы узнали из 2-й части этой книги, рабочая память — это операция
управления, обеспечиваемая сложной системой, и один из ее элементов —
базальные ганглии. При очень грубом упрощении (которое мы допускаем лишь из дидактических соображений) рабочая память может рассматриваться как активный след, характеризующийся определенной амплитудой и степенью угасания (рис. 17.3). Чем выше амплитуда следа и чем дольше он сохраняется, тем лучше рабочая память. Мы предполагаем, что при СНВГ начало
Рисунок 17.3. Гипотетическое угасание следов рабочей памяти в норме и в популяции СНВГ
следа рабочей памяти не отличается от нормы, однако его угасание происходит гораздо быстрее, чем у здоровых лиц (рис. 17.3)25.
Существует несколько параметров ВП, связанных с рабочей памятью.
•>го: 1) условное негативное отклонение (contingent negative variation, CNV)
м В этом исследовании участникам предъявлялись изображения красных и желтых прямоугольников в случайной последовательности. Инструкция заключалась в наблюдении за прямоугольниками только одного цвета при случайном изменении их положения и игнори-|юмнии прямоугольников другого цвета. Чуть больше чем через 100 мсудетей из группы контроля возникала модуляция потенциала лоб-мых отделов мозга, сохранявшаяся в течение нескольких сотен миллисекунд—феномен, названный лобной негативностью выбора.
л Существует несколько фактов, подтверждающих это предположение. Один из них — увеличение числа рецепторов в стриатума выявленное в подгруппе детей с СНВГ. Повышенная плотность этих рецепторов ведет к ускоренному вымыванию дофамина из (миалгической щели. Быстрое снижение уровня дофамина в стриатуме может привести к дефициту рабочей памяти, обеспечиваемой лотлями обратной связи, включающими базальные ганглии, таламус и кору.
394 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Рисунок 17.4. Избирательное нарушение компоненты сравнения Р2 у ребенка с СНВГ
Слева — сопоставление четырех независимых компонент индивидуального когнитивного ВП с нормой; сверху вниз: Р2-сравнения, Nl/Pl-слуховой, РЗЬ- и Р4-мониторинга. Тонкая линия — индивидуальная компонента ребенка сСНВГ, толстая линия — компонента, усредненная для группы здоровых сверстников. Справа — голограммы компонент пациента и нормативной группы.
в двустимульном тесте как показатель активации мозга во время ожидания26; 2) негативность рассогласования (mismatch negativity, MMN) в oddball-тесте как индекс следа памяти о регулярности повторяющегося стимула; 3) компонента сравнения Р2 в GO/NOGO-тесте как показатель отличий текущего стимула от следа, хранящегося в рабочей памяти.
В нескольких работах оценивалась амплитуда «условного негативного отклонения у детей с СНВГ», и результаты этих работ противо
26 Напомним, что условное негативное отклонение может быть разделено на две компоненты: одна связана с подготовкой к восприятию стимула, а другая — с подготовкой к выполнению движения. В исследованиях СНВГ эти компоненты не различаются, что приводит к противоречиям в результатах этих работ.
Часть 3. Расстройства систем мозга
395
речивы: в одних выявлено снижение амплитуды «условное негативное отклонение у пациентов с СНВГ» по сравнению с контрольной группой, в то время как в других не обнаружено различий. Исследования негативности рассогласования при СНВГ, по последним литературным данным, также содержат разногласия. Хотя в некоторых из них сообщается о снижении амплитуды «негативности рассогласования» в группе пациентов с СНВГ по сравнению со здоровыми лицами, в других не найдено этих различий. Следует, однако, иметь в виду, что негативность рассогласования может рассматриваться только как косвенный критерий рабочей памяти или, точнее говоря, ее автоматической составляющей27.
Другой косвенный показатель рабочей памяти — компонента сравнения, возникающая в GO/NOGO-тесте. Эта компонента наблюдается в ответ на второй стимул в условии NOGO, когда предъявляемый стимул не совпадает с ожидаемым по параметрам сенсорной физической или семантической модальности. В нашей работе принимали участие 150 детей с диагнозом СНВГ. Пространственные фильтры для компоненты сравнения Р2 были получены при обработке 500 когнитивных ВП (для GO- и NOGO-условий) у группы здоровых детей. Затем пространственной фильтрации подвергались ВП детей с СНВГ и результирующая компонента сравнения Р2 каждого пациента сопоставлялась с нормативной базой данных для соответствующей возрастной группы. Результаты такого сравнения для одного из пациентов представлены на рис. 17.4. Показаны четыре независимые компоненты: компонента сравнения Р2, компонента N1/P2, вызванная слуховыми новыми стимулами, РЗЬ- и Р4-компоненты мониторинга. Обратите внимание на то, что у этого пациента отмечается снижение амплитуды только компоненты сравнения Р2. Это избирательное снижение амплитуды компоненты сравнения может служить показателем уменьшения следа стимула в рабочей памяти. Действительно, сокращение следа памяти может вести к менее эффективному выявлению отличий и, следовательно, к уменьшению компоненты сравнения Р228.
В. Операция вовлечения
С нейрофизиологической точки зрения операция вовлечения связана с активацией корковых и подкорковых структур, вовлеченных в выполнение выбранного действия. С психологической, функциональной точки зрения операция вовлечения обусловлена объединением всех мозговых ресурсов для выполнения данного действия. Коррелят этой операции — компонента РЗЬ. Oddball-парадигма — простейший и чаще всего используемый поведенческий тест для его получения. В этом тесте компонента РЗЬ определяется как разностная волна между ВП на значимые (т.е. со
Негативность рассогласования возникает при сравнении предъявляемого стимула со следом памяти, не связана с вниманием и. предположительно, формируется автоматически.
м У 36 пациентов (25 % всей группы) обнаружено избирательное уменьшение компоненты сравнения Р2. Пока не выявлены отличия в поведении у этой подгруппы детей с СНВГ по сравнению с нормой.
396 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
провождающиеся действием) и стандартные (т.е. не требующие последующего ответа) стимулы. Не будет преувеличением сказать, что большинство клинических исследований когнитивных ВП при СНВГ использует oddball-парадигму29.
В большинстве работ сообщается о снижении амплитуды компоненты РЗЬ в популяции СНВГ по сравнению со здоровыми сверстниками (см., например, одну из наших ранних работ, Kropotova et al., 1995). Из дедуктивных соображений стоит упомянуть работу, сопоставляющую компоненты РЗЬ и РЗ при оценке различий между пациентами с СНВГ и здоровыми детьми (Jonkman et al., 2000). В этом исследовании трехстимульный зрительный oddball-тест (предъявление стандартных, девиантных и «новых» (неожиданных) стимулов) был основой для двух зрительных тестов меняющейся степени сложности. Компоненты РЗЬ и РЗа рассчитывались как разностные волны. Было показано, что амплитуда компоненты РЗЬ была ниже в группе СНВГ; при сложных заданиях компонента РЗЬ увеличивалась в контрольной группе, чего не происходило в группе СНВГ. Напротив, амплитуда компоненты РЗа при выполнении сложных заданий уменьшалась в обеих группах и не различалась между группами. Метилфенидат увеличивал компоненту РЗЬ, но не влиял на компоненту РЗа. Был сделан вывод о том, что у детей с СНВГ нет дефицита объема внимания, связанного, по-видимому, с компонентой РЗа30.
Г. Подавление ответа
С нейрофизиологической точки зрения подавление ответа вовлекает отдельный участок в петле обратной связи: вентральная префронтальная кора справа — базальные ганглии — таламус. Вентральная префронтальная кора получает информацию из систем, связанных с восприятием и определяющих рассогласование между ожидаемым и реальным сенсорными стимулами. Например, она получает входные сигналы из зрительных областей вентрального зрительного пути, а также из передней цингулярной коры — области, вовлеченной в сопоставление выполняемых и подготовленных действий. В обоих случаях вентральная префронтальная кора активируется при необходимости прервать или подавить осуществляющийся поведенческий паттерн. Операция подавления также вовлекает соответствующую петлю: базальные ганглии — таламус — кора. С тех пор как в 1970-х японские исследователи Гемба и Сасаки обнаружили электрофизиологические корреляты операции подавления у обезьян и человека, считается, что этот процесс
29 Следует отметить, что слуховая oddball-парадигма используется чаще, чем зрительная. Единственное преимущество использования слуховой модальности этой парадигмы состоит в меньшем количестве глазных даижений во время теста. Однако применение методов пространственной фильтрации при коррекции артефактов позволяет не думать об артефактах движения глаз в ЭЭГ. Более того, компоненты когнитивных ВП при использовании зрительной модальности (включая компоненту РЗЬ) по величине превосходят соответствующие компоненты при использовании слуховой модальности, возможно, в связи с тем, что в обработку зрительной информации вовлечены более обширные зоны коры по сравнению с теми, что участвуют в обработке слуховой информации. Таким образом, зрительный oddball-тест следует считать предпочтительным для изучения компоненты РЗЬ.
30 Согласно нашей теории, эти результаты показывают, что операция переключения, те. способность перемещать внимание на новые стимулы, не страдает при СНВГ, в то время как операция вовлечения, те. способность к объединению мозговых ресурсов для выполнения конкретного действия, явно ослаблена.
Часть 3. Расстройства систем мозга
397
отражает компоненту N2, зарегистрированную в лобных отделах в процессе выполнения GO/NOGO-теста31.
По мнению многих авторов, включая Р. Баркли, лидера в области изучения СНВГ, в основе этого расстройства лежит нарушение операции подавления ответа. Однако попытки проверить эту гипотезу в эксперименте привели к противоречивым результатам. Недавно международная группа исследователей из Геттингенского университета в Германии и университета г. Цюрих (Banaschewski et al., 2004) сообщила об отсутствии различий в параметрах компоненты N2, зарегистрированной в варианте GO/NOGO-парадигмы — тесте А-Х (см. описание теста в методах 2-й части) между группой пациентов с СНВГ и контрольной группой. Напротив, исследование когнитивных ВП, выполненное в Техасском университете в другой разновидности GO/NOGO-парадигмы — тесте «Стоп-сигнал» — выявило заметное снижение амплитуды компоненты N2 в группе СНВГ по сравнению со здоровыми испытуемыми. В ответ на все «стоп-сигналы» у участников контрольной группы возникала выраженная негативная волна с латентностью 200 мс (N200) над областью нижней лобной коры справа. У детей с СНВГ эта волна была заметно редуцирована. Амплитуда волны N200 значимо коррелировала со способностью подавить Подготовленный ответ. Согласно этой работе, у детей с СНВГ, по-видимому, имеется аномалия компоненты обработки информации, необходимой для подавления действий.
Противоречие между вышеупомянутыми работами показывает важность выбора адекватной парадигмы для получения надежных величин эффекта. В целом то обстоятельство, что некоторые.тесты не выявляют различий между группами пациентов (например, с СНВГ) и здоровых испытуемых (возможно, чересчур легкие при выполнении, или компонента, генерируемая в данной психологической операции, слишком мала по величине), в то время как другие тесты выявляют статистически значимые отличия от нормы (возможно, эти тесты более сложные или генерирующие более надежные компоненты с большими амплитудными значениями) — это обычное явление при изучении когнитивных ВП32.
Д. Операция мониторинга
Операция мониторинга — это процесс управления, необходимый, например, для коррекции ошибок. С нейрофизиологической точки зрения операция мониторинга, по-видимому, основывается на общих нейрональных механизмах сравнения ожидаемого и реального событий. Раз-
11Следует подчеркнуть, что применение метода независимых компонент при анализе когнитивных ВП нормативной базы данных Института мозга человека позволило выявить несколько компонент, предположительно связанных с подавлением подготовленного действия (подробнее см. 3-ю главу 2-й части).
“ Вспомните, что некоторые неполадки в работе автомобиля можно выявить только при движении на высокой скорости. Эта ме-тпфора подчеркивает важность выбора подходящего теста для получения надежной величины эффекта при сравнении группы пациентов с группой контроля. В целом тест должен быть достаточно сложным, и компоненты, генерируемые при предъявлении стимулов, долины быть достаточной величины, чтобы обеспечить надежную воспроизводимость результатов.
398 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
норм» пациент
Рисунок 17.5. Избирательное нарушение компоненты мониторинга Р400 у ребенка с СНВГ
Слева — сопоставление четырех независимых компонент индивидуального когнитивного ВП с нормой; сверху вниз: Р2-сравнения. Nl/Pl-слуховой, РЗЬ- и Р4-мониторинга. Тонкая линия — индивидуальная компонента ребенка с СНВГ, толстая линия — компонента, усредненная для группы здоровых сверстников. Справа — голограммы компонент пациента и нормативной группы.
ница состоит лишь в том, что в случае операции мониторинга с фактическим поведенческим ответом сравнивается не сенсорный след (как для компоненты детекции различий Р2), а ожидаемое действие. Если выполняемое или подавляемое действие не соответствует ожиданиям, результат сравнения инициирует изменение поведенческого паттерна с целью коррекции этого расхождения. Этот процесс непосредственно можно оценить с помощью негативности, связанной с ошибкой, и последующей по
Часть 3. Расстройства систем мозга 399
зитивности (Ре) — компонент ВП, генерируемых мозгом сразу же после совершения ошибки33.
Другой параметр, связанный с процессом контроля, — поздняя компонента Р400, выявляющаяся в ответ на NOGO-стимулы и генерируемый передней цингулярной корой. В нашей работе принимали участие 150 детей с диагнозом СНВГ. Пространственные фильтры для компоненты сравнения Р4 были получены при обработке 500 когнитивных ВП (для GO- и NOGO-условий) у группы здоровых детей. Затем пространственной фильтрации подвергались ВП детей с СНВГ, и результирующая компонента сравнения Р4 каждого пациента сравнивалась с нормативной базой данных для соответствующей возрастной группы. Результаты такого сравнения для одного из пациентов представлены на рис. 17.5. Показаны четыре независимые компоненты: компонента сравнения Р2, компонента N1/P2, вызванная слуховыми новыми стимулами, РЗЬ- и Р4-компоненты мониторинга. Обратите внимание на то, что у этого пациента отмечается снижение амплитуды только компоненты мониторинга Р4 и что эта компонента имеет то же пространственное распределение, что и негативность, связанная с ошибкой, и, согласно данным LORETA, генерируется передней цингулярной корой.
V. Дофаминовая гипотеза СНВГ
Дофамин — один из основных медиаторов системы управления. Как мы показали в главе 12, дофамин регулирует уровень возбудимости нейронов стриатума и, следовательно, играет решающую роль в селекции действий. Дофаминовая гипотеза СНВГ очень популярна и получает все больше научных подтверждений. Она основывается на веских экспериментальных доказательствах, например на том обстоятельстве, что симптомы СНВГ хорошо поддаются лечению психостимуляторами, подавляющими механизм обратного захвата дофамина, а также на том факте, что у детей с тяжелыми проявлениями СНВГ обнаружены аномалии генов, ответственных за регуляцию работы дофаминергической системы.
А. Повышение уровня DAT
Для непосредственной проверки дофаминовой гипотезы СНВГ было проведено несколько исследований с использованием фотонной эмиссионной компьютерной томографии (SPECT) и позитронной эмиссионной томографии у детей, подростков и взрослых. На основе данных SPECT и использования различных радиолигандов три независимые группы сообщили о повышении плотности DAT в скорлупе и хвостатом
u Недавно ученые из университета Гента в Белыми (Wiersema et aL, 2005) применили GO/NOGO-тест с 25 % NOGO-проб для изучения отличий когнитивных ВП при СНВГ от нормы. Авторы показали, что дети с СНВГ делали в два раза больше ошибок по сравнению <» здоровыми сверстниками и не могли регулировать скорость ответа после совершения ошибки. Изучение потенциалов, связанных с ошибками, выявило, что негативность, связанная с ошибкой, не отличалась в обеих группах, но у детей с СНВГ отмечалось уменьшение связанной с ошибкой позитивности (Ре). На основе этих находок авторы делают вывод о том, что процессы контроля ошибок на ранних налах. связанные с детекцией ошибок, у детей с СНВГ не нарушены, но у них имеются отклонения в мониторинге ошибок на поздних налах, связанном с субъективной эмоциональной и сознательной оценкой ошибок.
400 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия ядре у детей и взрослых, однако эти результаты не были воспроизведены двумя другими независимыми группами, не выявившими нарушения связывания DAT. Несогласованность результатов может быть обусловлена гетерогенностью популяции СНВГ, где некоторые подтипы связаны с повышением уровня рецепторов обратного захвата дофамина, а у других нет зависимости симптомов от нарушений регуляции дофаминергической системы34.
Б. Переносчик норадреналина
Следует заметить, что в то время как система управления модулируется дофамином, сйстема внимания модулируется норадреналином (NE) — медиаторов!, отличающимся от дофамина и по распространению, и по функциональному значению. Подобно тому как уровень дофамина в синаптической щели контролируется переносчиком дофамина, уровень норадреналина контролируется переносчиком норадреналина (NET). Самая высокая степень плотности NET в коре обнаружена в премоторной зоне и в теменных отделах мозга35. В течение последних тридцати лет некоторые исследователи предполагали, что в патофизиологии СНВГ решающую роль играет дизрегуляция норадреналиновой системы. Изобретение препарата атомоксетин, эффективного при лечении СНВГ, по-видимому, подтверждает это предположение. Действительно, атомоксетин обладает 300-кратной селективностью к NET по сравнению с DAT и преимущественно воздействует на норадренергическую систему мозга.
VI. Лечение
А. Психостимуляторы
Большинство фармакопрепаратов, уменьшающих поведенческие нарушения при СНВГ, воздействуют на дофаминергическую или норадренергическую системы. В течение более чем 60 лет для краткосрочного подавления симптомов СНВГ в клинической практике успешно используются психостимуляторы36. Эффективность этих средств подтверждена многочисленными исследованиями с использованием случайных выборок и методов двойного слепого и плацебо-контроля. Метилфенидат — наиболее часто используемый психостимулятор. Он блокирует обратный захват дофамина (и отчасти — норадреналина) переносчиками DAT и NET. Амфетамины повышают устойчивость переносчиков дофамина и норадреналина. Уменьшение симптоматики начинается приблизительно через 30 мин после приема препарата внутрь. Внутривенное же введение метилфенида-
34 Тем, кто интересуется этой работой, мы рекомендуем обратиться к статье К.-Х. Краузе и его коллег из Университета Людаига-Максимилиана (Krause et al., 2000).
35 Вспомните: уровень норадреналина в коре значительно выше, чем в стриатуме.
36 Психостимуляторы—это препараты, повышающие психическую и двигательную активность. К этому классу относятся амфетамин, метамфетамин, кокаин и метилфенидат. Психостимуляторы часто воздействуют на механизмы регуляции уровня дофамина в мозге.
Часть 3. Расстройства систем мозга
401
та вызывает эйфорические переживания, сходные с ощущениями, индуцируемыми кокаином.
Несмотря на довольно отчетливую эффективность стимуляторов в лечении СНВГ, около 30 % детей с этим заболеванием не отвечают на эти препараты или не могут их принимать в связи с побочными эффектами. К тому же СНВГ часто сочетается с другими психическими расстройствами. В таких случаях применение стимуляторов ЦНС может усилить проявления сопутствующих заболеваний, например коморбидных тиков или эмоциональных расстройств. Стимуляторы оказывают побочные эффекты. Наиболее частыми из них являются головные боли, боли в животе, снижение аппетита, раздражительность, бессонница, гипертензия37.
Б. ЭЭГ-биоуп равнение
То обстоятельство, что 30 % пациентов с СНВГ не может использовать психостимуляторы, побудило исследователей к поиску альтернативных методов лечения. Метод ЭЭГ-биоуправления основан на данных многочисленных нейрофизиологических исследований и анализе количественной ЭЭГ у пациентов с СНВГ Один из ведущих ученых в этой области, Б. Стерман, в своем обзоре (Sterman, 1996) указал, что «изменения уровня бодрствования и контроль поведения, по-видимому, напрямую связаны со специфическими таламокортикальными механизмами генерации ритмов и эти изменения проявляются определенными ритмами ЭЭГ, регистрирующимися над конкретными областями мозга». Он предположил, что психопатология (например, СНВГ) может коррелировать с изменением этих ритмов и что ЭЭГ-БОС, направленная на их нормализацию, может обусловить стойкий положительный клинический эффект.
Отклонения параметров количественной ЭЭГ от нормы при СНВГ описаны в данной главе. Ориентировочно можно выделить четыре различных нида этих отклонений38: 1) повышение уровня медленной активности (дельта-, тета-диапазонов) в центральных или лобно-центральных отделах; 2) аномальное увеличение продолжительности (более 1 с) вспышек тета-ритма частотой 5,5—8 Гц, максимально выраженного в Fz (лобный тета-ритм средней линии), усугубляющееся при усложнении заданий; 3) чрезмерное увеличение уровня бета-активности (13—30 Гц) в лобных отделах; 4) повышение уровня альфа-активности в задних, центральных или лобных (реже) отведениях, например: аномальное увеличение мю-ритма в СЗ и С4 — паттерн «лицо обезьяны», напханный так Б. Стерманом39. Обратите внимание: в первых двух группах отношение мощности «тета/бета» увеличено, что не характерно для 3-й группы. Очевидно, что каждой из этих подгрупп свойственно специфичное нарушение ме-
*' Мнения о возможной связи задержки роста с приемом стимулянтов противоречивы.
Выводы сделаны на основе нашего опыта использования нормативной базы данных HBI Database для оценки нейрофизиоло-। жжских коррелятов СНВГ.
" Основываясь на данных литературы и собственном опыте, Б. Стерман (Sterman, 1996) выделил следующие отклонения в ЭЭГ у я»той с СНВГ: 1) локальное увеличение тета-активности частотой 4-8 Гц в префронтальной, лобной и сенсомоторной коре; 2) диффуз-•« ж увеличение тега- или медленной альфа-активности, более выраженное при выполнении тестов; 3) значительное увеличение мощности альфа-ритма в передних отделах коры; 4) выраженное уменьшение ритмичной активности частотой 12-20 Гц в сенсомоторной же, 5) гиперкогерентность ЭЭГ между лобными областями правого и левого полушарий и между лобными и височными отделами (много полушария, а также межполушарная асимметрия мощности ЭЭГ в задневисочных и теменных областях.
402 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия ханизмов регуляции работы мозга и, следовательно, для коррекции нарушений в каждом случае нужен особый протокол ЭЭГ-биоуправления. Необходимость в выборе протоколов на основе данных количественной ЭЭГ была осознана довольно давно, но лишь недавно это стало нормой.
Благодаря многочисленным исследованиям, проведенным за последнее десятилетие, теперь мы знаем о существовании разных ЭЭГ-подгипов при СНВГ и осознаем необходимость использования разных протоколов ЭЭГ-БОС, но в первые годы эпохи ЭЭГ-биоуправления выбор протоколов основывался на эмпирическом подходе, а не на знании об ЭЭГ-подгипах СНВГ. Обычно пациенты, отобранные для исследования, выполняли один определенный вид тренинга, получая положительное подкрепление (слуховой или зрительный сигнал) в случае изменения активности коры в заданном направлении (например, при уменьшении низкочастотной активности или повышении уровня быстрых частот). Для получения подкрепления пациенты должны были удерживать заданные изменения в течение 0,5 с.
Как упоминалось ранее, в основе ЭЭГ-биоуправления лежит гипотеза, согласно которой способность пациента «нормализовать» паттерн ЭЭГ в областях, отвечающих за внимание и контроль поведения, дает ему возможность улучшить концентрацию внимания и поведенческий контроль. Впервые продемонстрировать, что ЭЭГ-биоуправление может вызвать изменение активности коры, сопровождающееся заметным улучшением поведения/функций, удалось Стерману и его коллегам40.
Большинство известных нам протоколов биоуправления, используемых в лечении СНВГ, было разработано и протестировано в лабораториях США. В эти протоколы традиционно были включены частоты ЭЭГ выше 0,1 Гц. ЭЭГ в низкочастотных диапазонах (называемых медленными корковыми потенциалами — SCP) использовалась в работах группы немецких ученых из Университета г. Тюбенген. Тренинг (сдвиг постоянного потенциала в положительном или отрицательном направлении) проводился для оптимизации функций лобных долей у детей с СНВГ41.
В. Повышение мощности бета-акгивности/подавление тета-активности
Этот протокол связан с именем Б. Стермана. В 1960-х в экспериментах с кошками он выявил «сенсомоторный ритм» (СМР), генерируемый над роландической корой. Пиковая частота СМР у кошек составляла 12—14 Гц42. Б. Стерман и его сотрудники обнаружили, что путем тренинга можно обучить животных генерировать этот ритм произвольно. Позднее они применили этот тип условно-рефлекторной связи для лечения пациентов с некоторыми формами эпилепсии. В своем обзоре (2000) Стерман сообщает, что этот тип ЭЭГ-биоуправления был, в частности, эффективен у пациен-тов с судорожными расстройствами, резистентными к фармакотерапии. В
40 В этом обзоре Б. Стерман обобщил свой опыт изучения аномалий количественной ЭЭГ, разработки протоколов для ЭЭГ-биоуправления, а также использования фармакопрепаратов у детей с СНВГ (Sterman, 2000).
41А также для снижения частоты эпилептических приступов у больных эпилепсией.
42 У человека частота альфа-активности, соответствующей холостому режиму работы сенсомоторной коры, находится в пределах 9-13 Гц у детей и взрослых (согласно нормативной базе данных HBI Database).
Часть 3. Расстройства систем мозга
403
1970-х Д. Любар впервые использовал CMP-тренинг в лечении СНВГ. Эффективность этого вида тренинга в коррекции гиперактивности стимулировала заметный интерес к нему как к потенциальному средству в лечении СНВГ.
Позднее, в 1980-х годах, после получения данных о выраженном замедлении активности коры при СНВГ в лобных и центральных отделах, Любар и его коллеги добавили к протоколам ЭЭГ-биоуправления повышение мощности активности в более высоких частотных диапазонах (бета 16—20 Гц) при одновременном подавлении низкочастотной активности (тета 4—8 Гц)43. В дальнейшем этот протокол использовался в нескольких контрольных групповых исследованиях эффективности ЭЭГ-БОС в лечении СНВГ44. При этом виде тренинга задача пациентов заключается в увеличении выработки бета-1-активности (16—20 Гц) и подавлении тета-активности (4—8 Гц). Активность регистрировалась в отведении Cz относительно ушного референта (см. Monastra et al., 2005)45.
Г. Относительный бета-тренинг
Общим процессом для вышеупомянутых протоколов является повышение мощности активности в высокочастотных диапазонах и подавление — в низкочастотных. В верхней части рис. 17.6 представлено сравнение спектров мощности ЭЭГ у группы пациентов с СНВГ и контрольной группы. ЭЭГ зарегистрирована в области Fz-Cz. Схематично представлены спектры мощности пациентов с наиболее распространенным (избыток тета-активности) типом СНВГ. Видно, что для группы СНВГ характерно повышение мощности ЭЭГ в низкочастотном (тета) диапазоне и снижение (по сравнению с нормой) мощности в бета-диапазоне. В результате отмечается снижение относительной мощности бета-активности по сравнению с нормой, что показано в нижней части рис. 17.6, где представлены инвертированные карты относительной мощности бета-активности для группы пациентов с СНВГ и контрольной группы.
Д. Нормализация компонент ВП, отражающих работу системы управления
В нашей работе, выполненной недавно (Kropotov et al., 2005), при ЭЭГ-биоуправлении мы использовали относительный бета-тренинг46. ЭЭГ регистрировалась в биполярном монтаже СЗ-Fz в соответствии с международной системой 10—20. Обычно сеанс включал 20 минут относительного бета-тренинга. Спектры мощности рассчитывались для 1-секундной эпохи каждые 250 мс с помощью быстрого преобразования Фурье. Сигналом обратной связи служило предъявление на экране компьютера столбика голубого цвета на сером фоне. Высота столбика отражала динамику управляемого параметра. Задача пациента
° Обычно ЭЭГ регистрировалась с одного активного электрода относительно объединенного ушного референта, с частотой дискретизации не менее 128 Гц. Слуховая (тон) и зрительная (движение элементов паззлов, графики или мультипликация) обратная связь основывалась на успешности пациента в контроле мощности или амплитуды соответствующего ритма, или процентного отношения времени, в течение которого этот ритм не превышал заданных пороговых значений.
44 Хотя недавнее обнаружение подтипа пациентов с СНВГ, характеризующегося увеличением мощности бета-активности в лобным областях, пробудило интерес к созданию протоколов, направленных на подавление избыточной бета-активности, пока нет сообщений о контрольных групповых исследованиях этого типа ЭЭГ-биоуправления.
44 В некоторых вариантах этого протокола использовался биполярный монтаж, например FCz-PCz или Cz-Pz.
44 Параметром ЭЭГ-биоуправления служило отношение мощности ЭЭГ в бета-диапазоне (15-18 Гц) к суммарной мощности
«таль ной части ЭЭГ-спекгра, т.е. диапазонам 4-14 Гц и 19-30 Гц.
404 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Топограммы инвертированной мощности относительной бетя-активностн
СНВГ Контроль
Рисунок 17.6. Обоснование относительного бета-протокола
Схематично представлены спектры ЭЭГ группы контроля (жирная линия) и пациентов с СНВГ и соответствующие им карты отношения мощностей тета/бета.
состояла в удержании столбика выше порогового уровня, выбранного на основе 2-минутной предгре-нинговой регистрации ЭЭГ
В дополнение к простому сигналу зрительной обратной связи использовалась так называемая ви-деомода. В этом случае параметр ЭЭГ-биоуправления контролировал уровень шума, генерируемого специальным электронным прибором (Jammer), созданным в нашей лаборатории. Амплитуда шума достигала максимума при минимальных
значениях параметра биоуправления и постепенно снижалась до нулевых значений при приближении параметра к порогу. Шум смешивался с видеосигналом видеоплеера и поступал на экран телевизора. Таким образом, пациент контролировал качество изображения на экране с помощью электрической активности своего мозга: если параметр биоуправления превышал порог, изображение было четким, в противном случае оно искажалось шумом.
Обычно в течение первых 5—8 сеансов пациенты выполняли тренинг со столбиком — простым сигналом зрительной обратной связи — для получения представления о процедуре47. Выбор порога зависел от уровня тренируемой активности в ЭЭГ пациента, регистрируемой в течение 2 минут перед тренингом при отрытых глазах. Порог обычно устанавливался в пределах 0,03—0,05 и 0,05—0,1 соответственно для младшей и старшей воз-
47 Пациенту предоставлялись необходимые сведения о процедуре, объяснялась зависимость параметров сигнала обратной связи от состояния мозга, уровня внимания. Перед процедурой пациенту предлагали расслабиться, постараться уменьшить мышечное напряжение, стремиться поддерживать медленное спокойное диафрагмальное дыхание. Перед ним ставилась задача постараться осознать свое внутреннее состояние, ощущения в моменты, когда высота столбца превышает порог, запомнить это состояние, научиться воспроизводить его и удерживать в течение периода тренинга. Разные пациенты использовали различные стратегии достижения этой цели. Для инициации состояния сосредоточения им рекомендовалось сфокусировать внимание на какой-либо точке вблизи от монитора, но так, чтобы при этом видеть сигнал обратной связи. Сеансы проводились от даух до пяти раз в неделю в течение 5-8 недель. Число сеансов для каждого пациента зависело от нескольких факторов, таких как возраст, тип СНВГ, характер кривых тренинга, отчеты родителей о состоянии ребенка, и составляло от 15 до 22 (в среднем 17). Критериями для завершения курса были: 1) стабилизация результатов в процессе тренинга в течение последних 3-5 сеансов; 2) стабилизация клинических симптомов пациента согласно мнению родителей.
Часть 3. Расстройства систем мозга
405
растной групп. Динамика управляемого параметра (кривая тренинга) регистрировалась для каждого сеанса у каждого пациента. В верхней части рис. 17.7 представлена характерная кривая динамики параметра БОС в процессе одного сеанса у одного из испытуемых. Видно, что во время
тренировки относительная бета-активность существенно возрастает по сравнению с таковой во время* отдыха.
В нижней части рис. 17.7 показано сравнение средних значений относительной мощности бета-активности (усредненной у 22 пациента) в периоды тренинга и отдыха для 19 отве
Покои
-♦-Тренинг
Рисунок 17.7. Относительный бета-тренинг
А. Кривая тренинга во время отдельного сеанса у пациента с СНВГ (вертикальная ось — параметр ЭЭГ-биоуправления, горизонтальная ось — время в секундах). Б. Средние значения относительной мощности бета-активности в течение периодов тренинга и отдыха во время отдельных сеансов, рассчитанные для 19 электродов и усредненные для 22 пациентов.
дений. Записи были выполнены во время одного из сеансов в конце курса лечения. Регистрация ЭЭГ с 19 электродов во время сеанса — длительная процедура; поэтому для этого исследования мы случайно выбрали 22 пациентов. Видно статистически значимое отличие мощности ЭЭГ в бета-
диапазоне между периодами тренинга и отдыха во всех отведениях, с максимальной выраженностью — в лобных.
Следует отметить, что не все пациенты могли увеличивать относительную мощность бета-активности даже после 10—20 сеансов. Мы проводили оценку успешности тренинга, т.е. способности пациента повышать управляемый параметр во время периодов тренинга. Мы считали тренинг успешным, если пациент увеличивал этот параметр во время периодов тренинга не менее чем на 25 % по сравнению с периодами отдыха. Успешными пациентами мы считали тех, кто справился с этой задачей не менее чем в 60 % сеансов, и к их числу мы отнесли 71 пациента (82,5 %). Тех же, кто выполнил гренинг успешно менее чем в 60 % сеансов, мы отнесли к группе неуспешных, и их число составило 15 (17,5 %). Эту группу мы рассматривали как контрольную при последующем анализе данных.
Работу системы управления мы оценивали с помощью когнитивных ВП, зарегистрированных в двухстимульном слуховом GO/NOGO-тесте до лечения и после курса ЭЭГ-биоуправления. На рис. 17.8 представлены когнитивные ВП для NOGO-проб, зарегистрированные до и после курса
406 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
ЭЭГ-БОС. Видно увеличение положительной компоненты в лобных отведениях после 20 сеансов относительного бета-тренинга.
На рис. 17.9 показана разность усредненных значений когнитивных ВП и карты разностных кривых, рассчитанные путем вычитания ВП, полученных до тренинга из показателей ВП после 20 сеансов относительного бета-тренинга. Для сравнения приведены те же данные для группы неуспешных пациентов. Обратите внимание на статистически значимое увеличение амплитуды когнитивных ВП в ответ на NOGO-стимулы в группе успешных пациентов при отсутствии существенной динамики этого показателя в группе неуспешных пациентов. В нашей работе для объективной оценки параметров внимания мы использовали двухстимульный GO/NOGO-тест. Следует отметить, что относительный бета-тренинг не влияет на ранние (с латентностью 80—180 мс) компоненты ВП, но вызывает значительное увеличение поздних положительных компонент. Таким образом, наше исследование показывает, что относительный бета-тренинг не влияет на обработку слуховой информации в мозге человека, в то время как он значительно изменяет работу системы управления, что
-------перед тренингом ...... после тренинга
Рисунок 17.8. Увеличение поздних положительных компонент когнитивных ВП в ответ на NOGO-пробы после относительного бета-тренинга
Усредненные когнитивные ВП в ответ на NOGO-стимулы в двухстимульном слуховом GO/NOGO-тесте в группе успешных пациентов до и после 20 сеансов относительного бета-тренинга. Тонкая линия — когнитивные ВП до тренинга; жирная линия — когнитивные ВП, зарегистрированные после 20 сеансов тре-uuura Оаиигтплпаил u<a kmrwMrn/ et al
находит отражение в поздних компонентах ВП.
Теоретически наш протокол отличается от общепринятых протоколов, т.к. увеличение значений управляемого параметра в нашем исследовании достигалось при повышении мощности бета-активности и/ или при уменьшении мощности тета- а также альфа-активности. Но результаты нашей работы показывают, что относительный бета-тренинг так же эффективен, как и общепринятые протоколы. В самом деле, 80 % наших пациентов могли значительно повышать параметр ЭЭГ-
Часть 3. Расстройства систем мозга
407
биоуправления более чем в 60 % сеансов. Более того, согласно результатам опроса родителей с помощью шкалы SNAP-IV, после курса ЭЭГ-БОС значительно улучшилось поведение детей, снизились показатели невнимательности и импульсивности.
Е. Транскраниальная микропо-ляризация
В нашей лаборатории помимо ЭЭГ-БОС для коррекции симптомов СНВГ мы ис-
Успешные пациенты
Неуспешные пациенты
Рисунок 17.9. Увеличение поздних положительных ком* понент когнитивных ВП после курса ЭЭГ-биоуправления обнаружено только в группе успешных пациентов
Вверху — разностные кривые когнитивных ВП до и после
Рисунок 17.10. Нормализация показателей ЭЭГ-когерентности после курса ТКМП
А. Значения когерентности, усредненные для группы (N = 12) детей с СНВГ в возрасте 7-12 лет. Значения ЭЭГ-когерентности для каждого электрода со всеми остальными электродами усреднялись для каждого пациента, а затем — для всех 12 пациентов. Б. Карты усредненных значений ЭЭГ-когерентности до и после 7 сеансов ТКМП по сравнению с контрольной группой.
408 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
ризация впервые была применена для подавления патологических ритмичных нейронных разрядов в подкорковых структурах у пациентов с болезнью Паркинсона* 48. Несколько лет экспериментов на кошках и собаках в 1970-х подтвердили, что подача отрицательных постоянных потенциалов к электродам, имплантированным в глубокие структуры, подавляет импульсную активность нейронов вблизи электрода, в то время как применение анодного тока вызывает противоположный эффект49. После экспериментов на животных метод был введен в клиническую практику для лечения различных неврологических дисфункций у взрослых. Недавно мы стали использовать слабую анодную стимуляцию у детей с СНВГ.
В работах одного из моих коллег, С. Сараева50, проводимых в 1990-х, анод устанавливался в зоне F8, а катод — поблизости от Fp2. Сила тока в пределах 700—1000 цА подбиралась индивидуально, с тем чтобы пациент не ощущал воздействия тока. Сеансы ТКМП продолжительностью 20 минут проводились каждые 2—3 дня, курс состоял из 7 сеансов. В качестве показателя функционирования мозга использовалась когерентность ЭЭГ в альфа-диапазоне. Усредненная когерентность рассчитывалась для каждого электрода как отношение ее значения под данным электродом к среднему значению под всеми остальными электродами. На рис. 17.10 приведены результаты этого исследования. Видно, что 7 сеансов ТКМП нормализовали показатели когерентности. Также психологическая оценка внимания (с помощью опросника SNAP-IV и корректурной пробы) выявила улучшение показателей внимания в группе СНВГ. Невролог Л.С. Чутко продолжает использовать ТКМП в нашей лаборатории не только в лечении СНВГ, но и при задержках речевого и психического развития у детей, используя модификацию метода, предложенного С. Сараевым, и расположение электродов: F7 — сосцевидный отросток слева и F8 — сосцевидный отросток справа.
VII. Заключение
Синдром нарушения внимания с гиперактивностью (СНВГ) — одно из самых распространенных психических расстройств детского возраста. В DSM-IV СНВГ разделен на три типа: с преобладанием невнимательности, с преобладанием гиперактивности и импульсивности и смешанный тип. Эта классификация базируется на трех группах симптомов: невнима
48 Мы подавали электрический ток к электродам, имплантированным с лечебной и диагностической целью пацинтам с болезнью Паркинсона. Регистрация импульсной активности нейронов с этих электродов показала уменьшение степени спайковой активности нейронной популяции в случае, если электрод служил катодом, и увеличение спайковой активности, если электрод служил анодом. Профессор Бехтерева предложила использовать этот метод в диагностических целях.
48 Я хочу упомянуть имена И. Данилова, Г. Галдинова и Г. Вартаняна, внесших большой вклад в понимание механизма влияния транскраниальной микрополяризации на нейрофизиологию мозга.
90 К несчастью, С. Сараев погиб в автокатастрофе и не смог опубликовать результаты своих исследований. Здесь я привожу дан-
ные нашей неопубликованной статьи.
Часть 3. Расстройства систем мозга
409
тельности, гиперактивности и импульсивности. По данным генетических исследований, СНВГ не следует законам наследования Менделя и должен расцениваться как комплексное генетическое заболевание. Внешние факторы, такие как осложнения течения беременности и родов, повышают риск возникновения СНВГ. СНВГ часто сочетается с другими расстройствами: специфическими трудностями в обучении, оппозиционнодевиантным поведением, синдромом Туретта, тревожностью и депрессией. По данным MPT-исследований, при СНВГ отмечается уменьшение размеров хвостатого ядра, бледного шара и префронтальной коры, в то время как ПЭТ- и фМРТ-исследования показали снижение уровня метаболизма в этих структурах. В большинстве работ по изучению количественной ЭЭГ сообщается об увеличении мощности медленноволновой активности в популяции СНВГ по сравнению с нормой. Как следствие этого при СНВГ повышено отношение мощностей «тета/бета» (обозначаемое как индекс невнимательности). В нескольких работах оценивалась амплитуда условно негативного отклонения у детей с СНВГ, и их результаты не согласуются. Данные исследований негативности рассогласования при СНВГ также противоречивы. Показано избирательное уменьшение компоненты Р2, полученной в двухстимульном тесте GO/NOGO и связанной с детекцией отличий, у детей с СНВГ. Неоднократно подтверждалось, что компонента РЗЬ, как параметр процесса вовлечения в действие, редуцирован при СНВГ. Метилфенидат увеличивает амплитуду РЗЬ-компоненты, но не влияет на РЗа-компоненту. В некоторых исследованиях обнаружена редукция компоненты N2 в GO/NOGO-тесте, что можно интерпретировать как дисфункцию подавления движений в популяции СНВГ. Снижение амплитуды компоненты мониторинга Р400 найдено в одной из подгрупп детей с СНВГ. Дофаминовая гипотеза СНВГ основана на том факте, что симптомы СНВГ хорошо поддаются лечению психостимуляторами, блокирующими рецепторы обратного захвата дофамина. Большая часть фармакопрепаратов, уменьшающих поведенческую симптоматику СНВГ, воздействует на дофаминергическую или норадренергическую систему. В течение более чем 60 лет для краткосрочного подавления симптомов СНВГ в клинической практике успешно используются психостимуляторы. Однако то обстоятельство, что 30 % пациентов с СНВГ не могут использовать эти препараты, побудило исследователей к поиску альтернативных методов лечения. Это ЭЭГ-биоуправление и транскраниальная микрополяризация. Метод ЭЭГ-биоуправления основан на данных многочисленных нейрофизиологических исследований и анализе количественной ЭЭГ у пациентов с СНВГ. В ЭЭГ-биоуправлении при СНВГ используется несколько протоколов, например повышение мощности бета-активности/подавление тета-активности и относительный бета-тренинг. Показано, что они уменьшают симптомы СНВГ, а также нормализуют спектральные характеристики ЭЭГ и компоненты когнитивных ВП, связанные с работой системы управления.
410
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Глава 18. Шизофрения
I. Клинические симптомы
А. Вовлечение трех мозговых систем
Шизофрения51 — психическое расстройство, характеризующееся дисфункцией трех мозговых систем: 1) системы управления, что проявляется паранойей, иллюзиями52, расстройством мышления и речи; 2) системы восприятия в форме слуховых галлюцинаций; 3) эмоциональной системы в виде притупления эмоций. Вовлечение в патогенез этого заболевания различных мозговых систем дало повод для дискуссии: является ли шизофрения самостоятельным заболеванием или этот диагноз включает ряд отдельных синдромов. По этой причине Э. Блейлер53 при выборе названия этому расстройству употребил форму множественного числа. Шизофрения — разрушительное заболевание, охватывающее приблизительно 0,4—0,6 % населения. Его клиническая манифестация обычно приходится на поздний подростковый или ранний взрослый возраст. Большая часть больных шизофренией — безработные, бедные и бездомные люди, около 10 % из них оканчивает жизнь самоубийством. Таким образом, это заболевание ложится тяжелым грузом на плечи членов семьи пораженных этой болезнью.
А. Дисфункция системы управления
Несмотря на то что самое выраженное проявление шизофрении — психоз, сам по себе он не является решающим диагностическим критерием. Среди многих клинических симптомов этого расстройства основными являются нарушение внимания, планирования, способности к инициации и регулированию целенаправленного поведения в результате дисфункции системы управления. Нарушения мышления прослеживаются на протяжении всей жизни пациентов, включая детство и подростковый период. У здоровых родственников этих пациентов наблюдаются сходные, хотя и выраженные в более мягкой форме когнитивные нарушения, что явно указывает на влияние генетических факторов. До настоящего времени не найдено диагностических критериев шизофрении по данным лабораторных или психологических тестов и методов нейрокартиро
51 Термин «шизофрения» дословно в переводе с греческого означает «расщепление души».
52 Иллюзия означает заблуждение. В психиатрии этот симптом обычно связывается с психическими расстройствами. Несмотря на то что иллюзии встречаются и при других психических состояниях, при шизофрении они имеют особое диагностическое значение.
53 П.Э. Блейлер — швейцарский психиатр, введший термин «шизофрения» в 1908 г. Он также известен своим участием в лечении известного русского балетного танцора Вацлава Нижинского.
Часть 3. Расстройства систем мозга 411
вания. Диагноз шизофрении, как и любого другого психического расстройства, ставится только на основе клинической симптоматики54. В целях надежности диагностики эти симптомы объединяются в критериальные группы55.
II. Генетические и внешние факторы
А. Вовлечение множества генов
В возникновении шизофрении важную роль играют генетические факторы56. Выявлено несколько генов предрасположенности к этому заболеванию со слабой выраженностью фактора риска для каждого из них57. Более того, для большинства генов биологическая основа повышенного риска развития заболевания неясна. Эти факты указывают на то, что для развития клинической картины заболевания влияния отдельных генетических факторов недостаточно. Возможно, что риск развития шизофрении обусловлен комплексным взаимодействием нескольких генов. В одном из недавних исследований высказано предположение о том, что гены, связанные с этим риском, неспецифичны и могут также повышать вероятность возникновения других психических расстройств, например биполярного расстройства.
Б. Внешние факторы
Жизнь в городе—один из серьезных факторов риска развития шизофрении. Социальные бедствия, расовая дискриминация, семейное неблагополучие, безработица и неблагоприятные жилищные условия, предположительно, вносят вклад в возникновение заболевания. По этой причине риск формирования шизофрении повышен у эмигрантов первого и второго поколений, у лиц, перенесших травму или подвергавшихся насилию в детстве. Есть также данные в пользу увеличения вероятности развития заболевания в результате пренатальных инфекций58.
III. Структурные и физиологические корреляты
А. МРТ
Как отмечено ранее, основная дисфункция при шизофрении затрагивает систему управления. Это проявляется, в частности, в нарушении направленного внимания. Традиционно в изучении функций и дисфункций
54 В психиатрии термин «синдром» относится к сочетанию нескольких клинически выраженных признаков, особенностей (выявленных доктором) и симптомов (со слов пациента).
55 Первый диагностический критерий требует наличия двух или более следующих клинических симптомов: иллюзии, галлюцинации, дезорганизация мышления и речи, выраженное нарушение поведения или кататония и таких негативных симптомов, как притупление эмоций, обеднение речи и утрата способности инициировать и поддерживать целенаправленное поведение. Второй диагностический критерий основан на данных о социальной или профессиональной дисфункции, например ухудшении межличностных взаимоотношений, рабочих навыков, несоблюдении личной гигиены. Третий критерий — признаки нарушений должны присутствовать в течение не менее 6 месяцев. Четвертый критерий — должно быть выяснено, что клинические симптомы не относятся к другому расстройству, например наркотической зависимости.
58 Оценки наследуемости шизофрении варьируют в связи со сложностями в разграничении влияния генетики и внешней среды. По данным исследований с участием близнецов, наследуемость составляет 80 %.
57 Фактор риска — параметр, связанный с повышенным риском возникновения заболевания. Факторы риска взаимосвязаны, но эта связь не обязательно носит причинный характер.
58 Амфетамины, способствуя освобождению дофамина в мозге, усугубляют симптомы шизофрении. Злоупотребление другими стимуляторами, например, галлюциногенами, также провоцирует ее возникновение.
412 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
системы управления используется трехстимульная oddball-парадигма, включающая стандартный, девиантный и новый стимулы. Недавно группой ученых из Медицинского колледжа Университета Пенсильвании в Филадельфии изучались изменения BOLD-сигнала у пациентов с шизофренией в фМРТ-исследовании во время трехстимульного oddball-теста. Было обнаружено, в частности, что при предъявлении значимых стимулов у этих пациентов по сравнению со здоровыми испытуемыми снижена активация верхней височной и лобной извилин, цингулярной коры, таламуса и базальных ганглиев. Эти данные указывают на гипоактивацию системы управления при шизофрении.
Б. Количественная ЭЭГ
Данные о количественной ЭЭГ при шизофрении довольно противоречивы. В то время как в некоторых работах сообщается о повышении уровня бета-активности и редукции альфа-ритма, в других показано отсутствие отклонений и даже противоположные результаты. Эта несогласованность обусловлена несколькими взаимосвязанными факторами: 1) неоднородностью популяции пациентов с шизофренией; 2) недостаточным количеством пациентов; 3) слабой величиной эффекта. В некоторых исследованиях пациенты принимали препараты, влиявшие на ЭЭГ-паттерн.
Хотя в фоновой ЭЭГ не найдено достоверных показателей шизофрении, выявлены ЭЭГ-корреляты такого состояния, как психоз. Например, галлюцинации связаны с повышением мощности бета-активности над левой височной областью (Lee at al., 2006).
В. Негативность рассогласования
Предполагается, что негативность рассогласования — это автоматический ответ височной коры на редкие изменения повторяющегося звукового сигнала. Негативность рассогласования связана с операцией сравнения в коре сенсорных сигналов на входе со следом памяти. Самым устойчивым наблюдением в исследованиях когнитивных ВП при шизофрении является снижение амплитуды негативности рассогласования у этих пациентов по сравнению со здоровыми испытуемыми. В некоторых работах сообщается, что эта разница достигает 50 % по сравнению с нормой (McCarleyet al., 1997). Примечательно, что величина эффекта уменьшения негативности рассогласования у пациентов с шизофренией по сравнению с контрольной группой зависит от параметров oddball-парадигмы (в частности, от межстимульного интервала: с увеличением интервала растет эффект величины)59 *. Важно отметить, что различия в величине негативности рассогласования зависят не только от параметров теста, но и от выбранной группы пациентов. Например, во время первого эпизода заболевания снижение негативности рассогласования не выявляется (Umbricht et al., 2006) в противоположность случаям с недавним началом заболевания или хроническим его течением.
59 Полный обзор клинического использования негативности рассогласования при шизофрении и других психических и невроло-
гических расстройствах представлен в статье Ристо Наатанена (Naatanen, 2003).
Часть 3. Расстройства систем мозга 413
Г. Условно-негативное отклонение
Как мы узнали из 2-й части, рабочая память зависит от состояния дорсолатеральной префронтальной коры и связана с петлей обратной связи: базальные ганглии — таламус — кора. У пациентов с шизофренией снижена успешность выполнения тестов на исследование рабочей памяти и обнаружено уменьшение активации дорсолатеральной префронтальной коры (по данным ПЭТ- и фМРТ-исследований) при попытках выполнения этих тестов. Более того, степень снижения активации этого отдела коры в тестах рабочей памяти предсказывает тяжесть когнитивных нарушений при шизофрении. Гипоактивация дорсолатеральной префронтальной коры, по-видимому, специфична для шизофрении, т.к. при других психотических расстройствах и при большой депрессии уровень активации этого отдела коры в подобных тестах соответствует норме. Таким образом, дефицит рабочей памяти может являться основной особенностью шизофрении, и выявление дисфункции дорсолатеральной префронтальной коры важно для понимания патофизиологических механизмов шизофрении.
Рабочая память как подготовительная активность отражена в условном негативном отклонении — медленной компоненте, генерируемой в двухстимульном тесте. Как было показано в 1970-х группой немецких исследователей из Университета Тюбенгена (Rochstough et al., 1980), у пациентов с шизофренией отмечается уменьшение амплитуды условно-негативного отклонения и увеличение так называемого постимперативного негативного отклонения — другой медленной волны, следующей за императивным стимулом.
Д. Операция вовлечения
Операция вовлечения системы управления отражена в компоненте РЗЬ-когнитивных ВП, зарегистрированных в тестах, требующих активного выполнения действия60. При шизофрении снижение амплитуды РЗЬ многократно воспроизводилось со времени первой работы Рота и Каннона, выполненной в 1972 г., несколько лет спустя после обнаружения этой компоненты. Уменьшение РЗЬ-компоненты, по-видимому, отражающее снижение ресурсов, необходимых для выполнения действия, классифицируется как маркер предрасположенности к шизофрении61.
Недавно группой ученых из Института психиатрии в Лондоне был выполнен метаанализ статей, опубликованных с января 1994-го по август 2003-го (Elvira Bramon et al., 2004). Стандартизованная величина эффекта для амплитуды РЗЬ, определенная как разность между средними ее значениями в двух группах (больные шизофренией и группа контроля), деленная на среднее стандартное отклонение обеих групп, составила 0,8562.
В работе, выполненной недавно группой исследователей из Университета Альберта-Людвига во Фрайбурге, Германия (Olbrich et al., 2005), когнитивные ВП, зарегистрированные в GO/NOGO-тесте у больных * 63
” В изучении компоненты РЗЬ чаще всего используется oddbal-парадигма, однако оба типа стимулов в GO/NOGO-ларадигме вызывают сходные РЗЬ-компоненты.
81 Обзор исследований РЗЬ-компоненты см. в статье Ю. Форд (Font 1999).
63 Для латентности компоненты РЗЬ объединенная стандартизованная величина эффекта была меньше и составляла 0,57.
414
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
шизофренией и здоровых испытуемых, были подвергнуты анализу независимых компонент. РЗЬ — одна из компонент, взятых для анализа — была локализована в теменных отделах с несколько большей длительностью для GO-проб по сравнению с NOGO-пробами63 и был значительно редуцирован у пациентов с шизофренией по сравнению с группой контроля (рис. 18.1).
Е. Операция контроля
Операция мониторинга системы управления отражена в компоненте Р400 когнитивных ВП, полученной при предъявлении NOGO-стимулов в двухстимульной зрительной GO/NOGO-парадигме. Как мы показали во 2-й части, эта компонента выделена с помощью анализа независимых компонент. Картирование с помощью sLORETA по-
казывает, что эта компонента
Рисунок 18.1. Уменьшение амплитуды компоненты РЗЬ у пациентов с шизофренией
А. Карта распределения компоненты РЗЬ, построенная с помощью анализа независимых компонент когнитивных ВП в ответ на GO-стммулы в GO/NOGO-тесте у здоровых испытуемых (данные из нормативной HBI базы данных). Б. Компоненты вызванных потенциалов на GO-стимулы в группе пациентов с шизофренией и у здоровых лиц. Заимствовано из Olblich et al. (2005).
генерируется на широком протяжении цингулярной коры, включая передние и субкаллозальные отделы. Метод анализа независимых компонент позволил вышеупомянутой группе немецких ученых (Olbrich et al., 2005) выделить РЗ большей амплитуды, возникающий в ответ на NOGO-пробы и названный P3ng. Эта компонента имела лобно-центральное распределение и была подобна и по пространственным, и по временным характеристикам компоненте мониторинга Р400, выделенной в наших ис
следованиях с нормативной базой данных. В работе немецкой группы
было выявлено снижение амплитуды компонентьгРЗ^ у больных шизофренией.
В работе, выполненной недавно группой исследователей во главе с Ю. Форд из Университета Стэнфорда, Калифорния, фМРТ и когнитивные ВП регистрировались в простом GO/NOGO-тесте. Было обнаружено, что амплитуда компоненты РЗ в ответ на NOGO-пробы у больных
шизофренией снижена. Соответствующая активация метаболизма, по данным фМРТ, у больных шизофренией была ограничена передней цингулярной корой, в то время как у здоровых испытуемых активация охватывала также дорсальную латеральную префронтальную кору, нижнюю
теменную дольку справа и хвостатое ядро. 63
63 Компонента РЗЬ, выделенная в этом исследовании, была идентична компоненте, полученной с помощью независимого компонентного анализа когнитивных ВП нормативной базы данных HBI Database для G0 и NOGO-условий.
Часть 3. Расстройства систем мозга
415
IV. Дофаминовая гипотеза шизофрении
А. Избыточное количество дофаминовых рецепторов в стриатуме
Предполагается, что один из ключевых патофизиологических механизмов шизофренического психоза состоит в относительной гиперфункции подкорковой дофаминовой системы. Несмотря на то что данные исследований в этой области весьма противоречивы, дофаминовая гипотеза шизофрении пока остается самой популярной и продолжает получать веские эмпирические под тверждения.
Эмпирические данные, полученные к настоящему времени, можно обобщить следующим образом: во-первых, недавние исследования различных моделей психоза на животных, в частности сенситизация поведения животных с помощью амфетамина, показывают, что у этих животных отмечается сверхчувствительность поведения к дофамину и явное повышение числа дофаминовых П2-рецепторов в мозге. Во-вторых, у человека психостимуляторы, такие как амфетамин и кокаин, повышают уровень дофамина в мозге и при длительном применении вызывают психоз (называемый амфетаминовым или кокаиновым), практически неотличимый от позитивных симптомов при шизофрении. В-третьих, антипсихотические препараты блокируют 02-рецепторы дофамина и подавляют позитивные (психотические) симптомы.
ПЭТ-исследование, выполненное недавно группой финских ученых из университета г. Турку, представило дополнительное доказательство дофаминовой гипотезы шизофрении (Hirvonen et al., 2005). В исследовании моно- и гетерозиготных близнецов, больных шизофренией и здоровых, использовался углерод 11, меченный ракроптидом (вещество, имеющее сродство с D2-рецепторами). У здоровых монозиготных близнецов — пациентов с шизофренией было обнаружено увеличение плотности 02-рецепторов в хвостатом ядре по сравнению с дизиготными близнецами и контрольной группой. Повышение связывания 02-рецепторов в хвостатом ядре коррелировало со снижением эффективности выполнения когнитивных тестов. Авторы сделали вывод о том, что нарушения регуляции дофаминовых 02-рецепторов в хвостатом ядре указывает на генетическую предрасположенность к шизофрении.
Б. Модель нейронной сети
Согласно нашей теории селекции действий, в системе «базальные ганглии — таламус — кора» (см. главу 11) стриатум является картой программ действий, на которой представительства различных действий имеют свою отдельную локализацию. Выбор необходимого действия внутри стриатума осуществляется на основе механизма латерального торможения по принципу «победитель забирает все». Согласно этой модели, конкретная программа действий выбирается, если активация ее представительства превышает определенный порог (рис. 18.2). Порог активации нейронов стриатума регулируется дофамином64. Влияние дофа-
64 В данном контексте порог означает минимальное количество возбуждающих постсинаптических потенциалов, исходящих из активированных корковых представительств и запускающих нейронные разряды в стриатуме. Он отличается от классического определения порога нейрона (являющегося постоянным), поскольку в этом контексте нейронный вход разделяется на две части: быструю активацию со стороны корковых нейронов и длительную модулирующую активацию со стороны дофаминергических нейронов черной субстанции. Следовательно, повышение модулирующего входа может вести к снижению коркового входа, необходимого для активации нейронов стриатума.
Рисунок 18.2. Нарушение механизма селекции действий в стриатуме у пациентов с шизофренией
А. Параллельные петли «базальные ганглии — таламус — кора» с модулирующим дофаминергическим входом в стриатум из черной субстанции. Б. Два представительства потенциальных действий частично перекрываются в коре, но изолированы в стриатуме. Избыток активирующих модулирующих дофаминергических влияний со стороны черной субстанции в связи с повышенной плотностью постсинаптических дофаминовых рецепторов при шизофрении ведет к тому, что неуместное действие (изображенное в виде купала с меньшей амплитудой) выбирается вместе с нужным действием (дающим больший вход в стриатум) — состояние, называемое расщеплением сознания. DA — дофамин, D-рецепторы — постсинаптические рецепторы к DA в стриатуме. Антипсихотическое средство — схематическое изображение молекулы антипсихотического препарата, блокирующего D-рецепторы и уменьшающего симптоматику шизофрении. Схематическая зависимость уменьшения психотических симптомов от степени сродства антипсихотического средства с О2-рецепторам (способности блокировать 02-рецепторами).
416____Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротератти
Часть 3. Расстройства систем мозга 417
мина определяется плотностью дофаминовых рецепторов на постсинаптической мембране нейронов стриатума.
У здоровых лиц порог достаточно высок, так что из возможных действий выбираются необходимые на данный момент, в то время как остальные подавляются. Информация о выбранном действии поступает в сознание. При шизофрении плотность D-рецепторов повышена и, следовательно, порог снижен, что создает возможность одновременного выбора нескольких действий. Конкуренция выбранных действий ведет к «расщеплению сознания*65. Антипсихотические препараты повышают порог выбора действий, блокируя О2~рецепторы, что ведет к уменьшению проявлений психоза. Эффективность препарата в уменьшении психотических симптомов четко коррелирует со степенью его сродства с D-рецепторами, т.е. способностью их блокировать (рис. 18.2, внизу справа). Объем ресурсов мозга для выполнения выбранного действия отражен в РЗЬ-компоненте когнитивных ВП. При распределении ресурсов на выполнение конкурирующих действий амплитуда компоненты РЗЬ снижается. Этим объясняется уменьшение РЗЬ-компоненты у больных шизофренией по сравнению со здоровыми лицами.
V. Лечение
А. Антипсихотические средства
Все фармакологические препараты, используемые в настоящее время в лечении шизофрении, главным образом подавляют психотические симптомы, но не устраняют когнитивных нарушений. Препараты, купирующие психоз, называются антипсихотическими средствами и используются уже более 50 лет. Один из них, хлорпромазин, был открыт в 1950-х. Эго открытие позволило значительно сократить число пациентов, нуждающихся в стационарном психиатрическом лечении. Антипсихотическая терапия остается основным направлением в лечениц шизофрении, облегчающим позитивную симптоматику психоза. В большинстве случаев антипсихотический эффект достигается через 7—14 дней от начала приема I ipenapaia. Считается, что из двух классов антипсихотических средств, одинаково эффективных в лечении позитивных симптомов, атипичные антипсихотики вызывают меньше побочных эффектов со стороны экстрапирамвдной системы. Ти-11ичные антипсихотики имеют сродство с дофаминовыми рецепторами D2 и блокируют их. Редукция психотических симптомов строго коррелирует со степенью сродства этих препаратов с О2-рецепторами. Об этой корреляции впервые сообщили Philip Seeman и его коллеги в 1975 г. (Seeman et al., 1975; Seeman et al., 1976). Следует подчеркнуть, что, несмотря на клиническую значимость когнитивных нарушений, до сих пор не созданы средства для их коррекции.
* В некоторых ситуациях (например, при научной работе) полезен выбор нетривиальных (и, следовательно, менее активированных) действий. Я вспоминаю своего одноклассника из физико-математического лицея в С.-Петербурге (известного как школа № 239), (лличавшегося выдающимися успехами в математике. Позднее он окончил математический факультет С.-Петербургского государственного университета и стал известным математиком. Половину времени он проводит в психиатрической больнице для лечения ым-юфрении, а другую половину пишет статьи для престижных математических журналов.
418
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Б. Электроконвульсивная терапия (ЭКГ)
Электроконвульсивная терапия, известная также как электрошок, — спорный вид психиатрического лечения, при кагором используется мощный электрический разряд для стимуляции судорогу пациента. ЭКТ была изобретена в Италии в 1937 г и получила распространение в 1940-х и 1950-х годах. Считается, что ЭКТ выполняемая современными методами, не вызывает повреждений мозга. В настоящее время ЭКТ чаще всего используется в лечении тяжелых форм большой депрессии, не отвечающих на другие виды терапии, и в ряде случаев — при шизофрении. Отмечено (Fink and Sackeim, 1996), что ЭКТ эффективна на ранних стадиях лечения острого психоза, особенно у пациентов с первым эпизодом заболевания.
В. Психохирургия
Психохирургия — избирательное хирургическое удаление или разрушение элементов ткани мозга в целях коррекции эмоциональных или когнитивных нарушений, обусловленных психическими заболеваниями. Впервые этот вид лечения был предложен Э. Моницем в 1936 г В то время не существовало эффективных фармакологических средств для лечения психических расстройств. На пике энтузиазма психохирургия была рекомендована для лечения шизофрении, депрессии, криминального поведения и некоторых других психических заболеваний. В промежутке между 1936 г. и серединой 1950-х в США было выполнено около 50 000 таких вмешательств. Последствия этих операций связаны с многочисленными осложнениями, такими как нарушения личности и интеллекта, судорожные приступы, параличи и смертельные исходы. Несмотря на эти осложнения, операции помогли большинству пациентов и в 1949 г. Мониц был удостоен Нобелевской премии в медицине «за открытие терапевтического значения префронтальной лейкотомии при некоторых видах психоза». С появлением в 1954 г. антипсихотических средств значение психохирургии снизилось. Тем не менее в случаях резистентности к лекарственной терапии некоторым пациентам рекомендуется нейрохирургическое вмешательство в специализированных центрах.
В настоящее время психохирургия применяется в лечении болезни Паркинсона, эпилепсии и ОКР — заболеваниях мозга с известной (до некоторой степени) патофизиологией66. Однако биологические основы большинства психических расстройств остаются недостаточно изученными, и биологические маркеры психических симптомов остаются предметом исследований. Эти обстоятельства ограничивают использование психохирургии в лечении шизофрении. Между тем использование данных ПЭТ, фМРТ, количественной ЭЭГ и когнитивных ВП в качестве биологических маркеров психических заболеваний вместе с тенденцией к использованию менее инвазивных методов67 в современной
“ Эти пациенты могут рассматриваться в качестве кандидатов для стереотаксической нейрохирургии в Институте мозга человека в С.-Петербурге. По моему личному опыту работы с этими пациентами, операции для них были единственным выбором, т.к. они не отвечали на другие общепринятые виды лечения. После щадящего стереотаксического вмешательства у довольно многих из них отмечено заметное уменьшение симптомов и операция была расценена как успешная. Подробнее см. Anichkov et al., 2005.
67 Один из таких метод ов — стереотаксическая нейрохирургия. С помощью специального оборуд ования (называемого стереотаксическим аппаратом) можно достичь самых труднодоступных областей мозга, минуя вскрытие черепа на широком протяжении и не разрушая здоровые унастки мозга на пути к целевой эоне. Стереотаксические аппараты позволяют точно поместить голову пациента в геометрическую систему координат; так что доступ к каждой структуре мозга может быть д остигнут снаружи. С помощью нейрохирургических игл под местной анестезией делаются маленькие отверстия в черепе, и тонкий зонд помещается в целевую область мозга.
Часть 3. Расстройства систем мозга 419
нейрохирургии открывает новые горизонты в лечении бальных шизофренией, резистентных ко всем видам фармакотерапии.
Г. ЭЭГ-биоуправление
Было предпринято несколько попыток лечения шизофрении с помощью ЭЭГ-биоуправления. Но не только серьезность расстройства, а и тот факт, что спектры ЭЭГ при этом заболевании не содержат статистически значимых и клинически различимых отклонений от нормы, не позволяет считать этот вид лечения шизофрении научно обоснованным. Однако, по-видимому, при шизофрении имеет место дисрегуляция медленных корковых потенциалов. Действительно, у больных шизофренией обнаружено снижение амплитуды условнонегативного отклонения и повышение так называемого постимперативного негативного отклонения — другой медленной волны, следующей за императивным стимулом68.
На основе этих наблюдений был сделан вывод о том, что при шизофрении имеет место нарушение регуляции возбудимости коры (отражающееся в медленных корковых процессах), а не просто снижение ее возбудимости (что нашло бы выражение в ЭЭГ-спектрах). В работе, выполненной группой исследователей из Университета г. Тюбенген в Германии (Schneider et al., 1992), находящиеся на лечении пациенты с шизофренией сравнивались с группой контроля. Несмо-гря на то что пациенты были способны освоить технику саморегуляции медленных потенциалов (при этом им требовалось большее количество сеансов, чем здоровым испытуемым), они испытывали затруднения в использовании освоенных навыков в повседневных ситуациях.
Позднее другая группа исследователей из Имперского медицинского колледжа Чаринг-кросского госпиталя в Лондоне (Gruzelier et al.‘, 1999) использовала подобную технику, обучая пациентов с шизофренией смещению негативности из одного полушария в другое. Предполагалось, что асимметрия с преобладанием активации левого полушария связана с повышением ментальной активности и эмоциональными иллюзиями, в то время как преобладание активации в правом полушарии коррелирует с социальным синдромом отмены (уходом в себя) при шизофрении. Эти работы показали, что, несмотря на дисфункцию системы yi фавления, больные шизофренией могли использовать ЭЭГ-БОС в саморегуляции. Однако до сих пор нет сведений о том, что этот метод облегчает симптомы шизофрении.
VI. Заключение
Шизофрения — психическое расстройство, характеризующееся поражением системы управления в виде паранойи, расстройства мышления и речи, нарушением системы восприятия в форме слуховых галлюцинаций и дисфункцией >моциональной системы в виде притупления эмоций. Вовлечение в патогенез
* Вспомните, что в парадигме условно-негативного отклонения предъявляются пары стимулов: предупреждающий стимул, гото-мщии пациента к последующему действию, и императивный стимул, запускающий подготовленное действие.
420
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
этого заболевания различных мозговых систем дало повод для дискуссии: является ли шизофрения самостоятельным заболеванием или этот диагноз включает ряд отдельных синдромов. Несмотря на то что в возникновении шизофрении важную роль играют генетические факторы, заболевание развивается при сочетании генетической предрасположенности и внешних факторов риска. Предполагается, что взаимодействие этих факторов изменяет процесс развития нервной системы, предшествующий возникновению симптомов. Все фармакологические препараты, используемые в настоящее время в лечении шизофрении, главным образом подавляют психотические симптомы, но не устраняют когнитивных нарушений. ФМРТ-исследования с использованием BOLD-сигнала свидетельствуют о гипоактивации системы управления при шизофрении. Результаты исследований количественной ЭЭГ при шизофрении довольно противоречивы. В то время как в некоторых работах сообщается о повышении уровня бета-активности и редукции альфа-ритма, в других показано отсутствие отклонений и даже противоположные результаты. Наиболее устойчивые наблюдения при исследовании когнитивных ВП у больных шизофренией: снижение амплитуды негативности рассогласования, условно-негативного отклонения и редукция компоненты вовлечения РЗЬ. В некоторых работах сообщается о редукции поздней позитивной компоненты мониторинга в ответ на NOGO-пробы в GO/ NOGO-тесте, генерируемой в передней цингулярной коре. Предполагается, что один из ключевых патофизиологических механизмов шизофренического психоза заключается в относительной гиперфункции подкорковой дофаминовой системы. Данные ПЭТ-исследований, так же как и выявленная способность антипсихотических препаратов блокировать рецепторы дофамина, по-видимому, подтверждают эту гипотезу. Согласно нашей модели, шизофрения может быть обусловлена нарушением селекции действий в стриатуме. Появление антипсихотических средств в 1950-х позволило значительно сократить число электроконвульсивных вмешательств и полностью прекратить использование психохирургии в лечении шизофрении. Было предпринято несколько попыток лечения шизофрении с помощью ЭЭГ-биоуправления.
Часть 3. Расстройства систем мозга
421
Глава 19.
Аддиктивные расстройства
I. Описание поведения
А. Симптомы
Аддиктивное расстройство связано с непреодолимым, не поддающимся контролю злоупотреблением наркотическими средствами69, невзирая на серьезные негативные последствия. Поведение адциктивной личности сосредоточено на получении и использовании наркотических средств и восстановлении своего состояния после их употребления. Этот паттерн сохраняется, несмотря на расстройство здоровья, нарушение взаимоотношений и жизненные провалы. Компульсивное поведение может неверно расцениваться членами семьи как намеренное саморазрушение.
Характерная черта аддикции — высокий риск рецидивов употребления наркотиков. Вероятность рецидива сохраняется даже при длительной ремиссии после прекращения их приема и обусловлена двумя факторами: провоцирующими ситуациями и стрессами70. Провоцирующие ситуации могут быть внешними или внутренними. Например, риск рецидива повышается при встрече людей или посещении мест, связанных с прежним приемом наркотиков. Современные методы лечения остаются неудовлетворительными и помогают лишь некоторым пациентам.
Исходя из теоретических принципов, изложенных во 2-й части, ад-дикция может рассматриваться как следствие нарушения операции контроля, ведущего к неконтролируемому, компульсивному употреблению наркотических средств. В этом отношении аддикция похожа на обсессивно-компульсивное расстройство (глава 20). Действительно, клинические проявления и ОКР, и аддикции связаны'с неспособностью подавления повторяющихся навязчивых идей (обсессии и непреодолимое желание соответственно) и ритуалистическим поведением (компульсии или активный поиск/употребление наркотических средств соответственно).
Б. Наркотические вещества
Существует много типов наркотических веществ. Они перечислены ниже в табл. 19.1.____
м В свою очередь, поведенческий паттерн при злоупотреблении алкоголем или наркотиками определяется существенным влиянием на жизнь человека этих веществ, занимающих в ней центральное место.
Продолжаются дискуссии касательно того, воздействуют ли провоцирующие ситуации на подсознательном условнорефлекгор-иом уровне или они опосредуют сознательное побуждение. Под влиянием провоцирующих факторов рецидивы возникают даже у лиц, рпшительно отказавшихся от наркотиков, при этом они могут не давать себе отчета в происходящем.
422 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Таблица 19.1
наркотическое вещество Основной эффект на клеточном уровне
Опиаты (морфин, героин) Действуют как агонисты опиатных рецепторов
Психостимуляторы (кокаин, амфетамины) Подавляют переносчики дофамина
Барбитураты и бензодиазепины Повышают проводимость CI—каналов мембраны нейрона, действуют как агонисты GABA-рецепторов
Никотин Агонист ацетилхолиновых рецепторов
Алкоголь Агонист GABA-рецепторов
В. Толерантность, зависимость и абстиненция
Помимо зависимости наркотические вещества могут вызывать толерантность и абстиненцию71. Следует подчеркнуть, что толерантность, зависимость и абстиненция могут наблюдаться также при приеме других препаратов (например, средств, используемых в лечении астмы и гипертензии). Не все наркотики вызывают физическую зависимость: к их числу относятся кокаин и амфетамин. У психостимуляторов отсутствуют физические симптомы абстиненции, а эмоциональные симптомы отмены непостоянны или слабо выражены. Физическая зависимость и абстиненция клинически значимы, но не лежат в основе механизмов «физиологической» зависимости (аддикции) — непреодолимой тяги к наркотикам с последующими рецидивами.
II. Структурные и физиологические корреляты
А. ПЭТ и МРТ
В ряде работ по изучению реакций на условные провоцирующие факторы с помощью позитронно-эмиссионной томографии и функциональной магнитно-резонансной томографии выявлена активация нескольких областей мозга, таких как префронтальная кора, прилежащее ядро в стриатуме и миндалина в лимбической системе72. Группа исследователей из Медицинского колледжа университета Эмори использовала ПЭТ для локализации нарушений синаптической активности, связанной с непреодолимым влечением к наркотикам, вызванным провоцирующими факторами (Kilts et al., 2001). Образы, взятые из личных воспоминаний, использовались в качестве индивидуальных факторов, провоцировавших состояние сильного влечения к кокаину и два контрольных состояния (гнев и нейтральное воспоминание). По сравнению с контрольным состоянием, связанным с воспоминанием нейтрального образа, состояние, индуцированное образом, вызывавшим влечение к наркотику, коррелировало с двусторонней активацией миндалин, островковой доли слева, передней цингулярной извилины, а также субкаллозальной извилины и прилежащего ядра справа.
71 Толерантность — это снижение эффекта наркотика при неизменной дозе или необходимость в повышении его дозы для поддержания стабильного эффекта. Некоторые наркотики вызывают сенситизацию (усиление) собственного эффекта. Зависимость в узком смысле проявляется в адаптации мозга к чрезмерной стимуляции наркотиками. После прекращения их приема это адаптированное состояние может приводить к появлению симптомов отмены на когнитивном, эмоциональном или физическом уровне.
72 Заметьте, что зоны активации определялись по разности ответа на факторы, провоцировавшие влечение к наркотику, и на нейтральные факторы.
Часть 3. Расстройства систем мозга 423
Б. Повышение уровня дофамина в прилежащем ядре
Недавние исследования показывают, что наркотические вещества непосредственно (например, кокаин) или косвенно (например, опиаты) повышают уровень синаптического дофамина в прилежащем ядре, например, блокируя DA-рецепгоры (рис. 19.1, справа вверху). Более того, повышение уровня дофамина в стриатуме положительно коррелирует с уровнем «эйфории», вызванным метилфенидатом (или кокаином)73 (рис. 19.1, справа внизу). Вспомните, что дофамин играет важную роль в мотивационных процессах, обеспечивающих «награду» и «подкрепление». Таким образом, по-видимому, он играет решающую роль в генерации мощного положительного подкрепления и инициации аддикции.
В современной литературе также высказываются предположения о том, что помимо мозговой системы позитивного подкрепления при адцикциях дисфункции также подвержены два отдела лобной коры (передняя цингулярная и орбитофронтальная кора), играющие решающую роль в контроле поведения, связанного с положительным подкреплением. Это те самые области, которые вовлечены при компульсивных состояниях, характеризующихся нарушением ингибиторного контроля над неадекватным поведением, как при обсессивно-компульсивном расстройстве (см. главу 20). В работе С. Формана (Forman et al., 2004) из университета Питтсбурга в исследовании пациентов с опиатной зависимостью и здоровых лиц из группы контроля использовалась фМРТ в процессе выполнения Go/NoGo-теста. По сравнению с нормой лица с опиатной зависимостью значительно хуже выполнили тест и у них был ослаблен сигнал ошибки в передней цингулярной коре.
III. Этапы аддиктивного процесса
Обширный эмпирический и теоретический материал по нейрофизиологии аддиктивных расстройств позволяет нам предположить, что аддикции — это сложный психологический процесс, включающий несколько стадий, каждая из которых характеризуется собственным нейрональным механизмом.
А. Этап ожидания
На начальном этапе мотивация к первому употреблению наркотика определяется естественным желанием улучшить настроение или испытать новые ощущения. Действительно, в большинстве случаев первый прием наркотика вызывает желаемый эффект — положительные субъективные переживания — удовольствие, приподнятость. Нейронные механизмы данного этапа — это стандартные механизмы мотивированного поведения.
Б. Этап консолидации
На втором этапе (запоминание опыта использования наркотика) субъективный опыт приема наркотика закрепляется как эмоционально значимое собы-
п См. недавний обзор (Р. Kalivas and N. Volkow, 2005).
Орбитофронтальная кора
Контрольное состояние
Состояние «подъема» после приема наркотика
Рисунок 19.1. Влияние кокаина на дофамин в добавочном ядре
А. Лимбический круг «базальные ганглии — таламус — кора» с модулирующим дофаминергическим входом в прилежащее ядро из черной субстанции. Б. Кокаин блокирует переносчики дофамина (DAT), что ведет к повышению его концентрации в прилежащем ядре. Уровень концентрации дофамина четко коррелируете субъективными ощущениями подъема у кокаиновых наркоманов. DA — дофамин, D-рецепторы (D-receptors) — посгсинаптические рецепторы к DA в стриатуме.
424 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Часть 3. Расстройства систем мозга
425
тие — награда. Как при любом процессе обучения, на этом этапе происходит объединение мысленного представления объекта и действий, связанных с употреблением наркотика, с вознаграждением. В результате этого ассоциативного обучения в дальнейшем любой объект или действие, связанное с употреблением наркотика, становятся провоцирующим сигналом к получению удовольствия. Нейрональные механизмы данного этапа — это стандартные механизмы обучения — сложные процессы, вероятно, включающие процедурную и эпизодическую память. Что отличает обучение при аддикции от нормального обучения, так это чрезвычайно выраженный эмоциональный контекст (высокий уровень удовольствия, эйфории) в основе первого приема наркотика. Но, по-видимому, самый важный фактор — прямое влияние наркотика на синаптическую трансмиссию, связанное с увеличением выделения дофамина в синаптическую щель. Как нам известно из нейронауки (см. главу 14), орбитофронтальная кора получает мощные проекции из мультимодальных областей коры и из лимбической системы. В коре хранятся образы вознаграждений как эмоционально значимых событий. Мы также знаем из нейронауки о том, что дофамин — это медиатор, закрепляющий след памяти, формируемый в соответствующей области коры. Таким образом, на этапе консолидации употребление наркотиков ведет к интенсивному запоминанию новых положительных подкреплений, связанных с их приемом, в орбитофронтальной коре74.
В. Этап привыкания/сенситизации
На третьем этапе начинается привыкание к эффекту наркотика (удовольствию или подавлению дисфории). При употреблении таких наркотических средств, как алкоголь, никотин или героин, удовольствие может существенно уменьшаться с течением времени в связи с осложнениями со стороны здоровья. Героиновые наркоманы часто, несмотря на очень плохое самочувствие и понимание того, что наркотики разрушают их жизнь, все же желают их принимать. Аддиктивные личности нередко объясняют продолжение употребления наркотических средств стремлением вновь испытать приподнятость и удовольствие, зачастую — безуспешно. Возникает вопрос: почему же они продолжают принимать наркотические вещества, если те уже больше не вызывают удовольствия? Для ответа на этот вопрос Робинсон и Берридж в 1993 г предложили теорию «стимул — сенситизация» (или просто потребность — удовольствие). Согласно этой теории, наркотическое вещество на второй стадии (консолидации) стимулирует определенный уровень активности, связанной с «потребностью» и «удовольствием» в различных системах мозга. После нескольких приемов толерантность к наркотику возрастает, а удовольствие снижается — наступает привыкание. В то же время «потребность», уже связанная не с самим наркотиком, а с удовольствием, связанным с позитивными ощущениями, вызванными наркотиком, возрастает.
'* В нейронную сеть, вовлеченную во второй этап аддикции, входят лимбическая система, включая миндалевидное ядро как структуру, ответственную за эмоциональный компонент, вентральный стриатум, включая прилежащее ядро как образование, участвующее в запоминании поведенческого паттерна, и орбитофронтальная кора как область представительства всего события в виде значительного эмоционального вознаграждения.
426
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Рисунок 19.2. Нейронные сети и стадии наркотической зависи!
См. объяснение в тексте.
Часть 3. Расстройства систем мозга 427
Г. Модель нейронной сети
Согласно теории работы мозга, приведенной в этой книге (часть 2), нейронные механизмы разных этапов аддикции выглядят следующим образом (рис. 19.2).
На этапе ожидания большая часть префронтального отдела коры оказывает активирующее влияние на моторную кору, инициируя новое и, возможно, ведущее к удовольствию действие.
На этапе консолидации орбитофронтальная и передняя цингулярная кора получают мощный и эмоционально значимый поток информации из лимбической системы. Образ вознаграждения в орбитофронтальной коре в норме консолидируется путем усиления синаптических связей в системе «орбитофронтальная кора —► прилегающее ядро —► бледный шар —> переднее ядро таламуса». В нормальных условиях этот дофаминергический круг является основным субстратом эффекта вознаграждения и положительного подкрепления в ответ на естественные стимулы, связанные с выживанием, такие как пища и возможность размножения. Под прямым и косвенным воздействием наркотических веществ освобождение дофамина значительно увеличивается. Высокая концентрация дофамина на уровне дендритов нейронов прилегающего ядра ведет к повышению активности нейронов стриатума и «запечатлению» образа вознаграждения и поведенческого паттерна, связанного с ним. Таким образом, стимуляция наркотическими веществами нейронных кругов, связанных с вознаграждением, по силе, длительности и надежности превышает почти все естественные стимулы, усиленно закрепляя ответы на стимулы, ассоциированные с наркотиками. После сохранения воспоминания о вознаграждении в лимбических кругах любой провоцирующий фактор (внутренний, как, например, субъективное воспоминание, или внешний, связанный с наркотиком), активирует эти круги75.
На этапе привыкания/сенситизации активность орбитофронтальной коры, вызванная воспоминанием о вознаграждении (например, как было замечательно во время первого приема наркотика...) сравнивается с образом, соответствующим реальной ситуации (например, жизнь серая и скучная) в передней цингулярной коре. Результат сравнения, отражающий это расхождение, активирует петлю «базальные ганглии—таламус — кора», связанную с передней цингулярной корой. В свою очередь, активация передней цингулярной коры запускает поведение, направленное на поиск наркотика, активируя премоторную и моторную зоны коры.
IV. Лечение
А. Стереотаксическая передняя цингулотомия при героиновой зависимости
По прогнозу, вытекающему из описанной выше модели, на последней стадии аддикция связана с высоким уровнем активации нейронов передней цингуляр-
* Основная нейронная сеть, участвующая в консолидации, включает лимбическую систему с миндалиной в качестве важнейшего элемента генерации эмоций, прилежащее ядро как основной элемент запоминания поведенческого паттерна, связанного с приемом наркотика, и орбитофронтальную кору гак гарту сохраненных образов вознаграждений. Действия, повышающие уровень д офамина в этих «кругах вознаграждение, в мозге имеют тенденцию к повторению вне зависимости оттого, связаны они с приемом наркотиков или выживанием.
428 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
ной коры. Эта гиперактивация обусловлена постоянным расхождением между реальностью76 и устойчивым следом памяти о поведении, связанном с приемом наркотика, и направляет поведение наркомана на поиск наркотика. Гиперактивное состояние передней цингулярной коры сходно с таковым при обсессивно-компульсивном расстройстве.
В Институте мозга человека в С.-Петербурге для облегчения компульсивного поведения у героиновых наркоманов была предложена стереотаксическая операция и к настоящему времени успешно прооперировано более 300 наркозависимых пациентов. На рис. 19.3 представлены результаты исследований группы наркоманов, полученные в нашей лаборатории. Регистрация когнитивных ВП со скальпа при выполнении двухстимульного зрительного GO/NOGO-теста проводилась до (тонкая линия) и после (жирная линия) стереотаксической операции передней цингулотомииу 13 пациентов, зависимых от героина77. Было обнаружено селективное подавление компоненты мониторинга Р400 после операции78. Эти данные указывают на то, что эта компонента является хорошим индикатором функционирования передней цингулярной коры и может быть использована для оценки результатов стереотак
Рисунок 19.3. Когнитивные ВП при аддиктивных расстройствах
Усредненные когнитивные ВП, зарегистрированные в двухстимульном GO/NOGO-тесте у группы пациентов с героиновой зависимостью до (жирная линия) и после (тонкая линия) стереотаксической деструкции участка цингулярной коры. Обращает внимание, что после операции избирательно уменьшается компонента мониторинга Р4 в NOGO-условии.
сических операций. Результаты операций также свидетельствуют о том, что разрушение нейронных кругов, вовлеченных в компульсивное поведение, столь выраженное при аддиктивных расстройствах, является мощным инструментом в лечении этих расстройств.
Б. ЭЭГ-биоуправление
Самый известный протокол ЭЭГ-биоуправления при аддикциях — это альфа-тета-тренинг. Этот метод состоит в одновременном измерении мощности альфа- (8—13 Гц) и тета-активности(4— 8 Гц) в затылочной области и предъявлении сигнала обратной связи79 при превышении заданного порога мощности для каждого из этих диапазонов80.
76 Информация о реальных событиях поступает в переднюю цингулярную кору из нескольких источников, включая кору островковой доли и сенсомоторной области, в то время как информация о высоко активированных программах, связанных с употреблением наркотиков, вероятно, исходит из орбитофронтальной коры.
77 Исследование было поддержано этическим комитетом Института мозга человека. Пациенты дали письменное согласие на проведение стереотаксической операции и на участие в исследовании.
78 Вспомните, что эта компонента генерируется в передней цингулярной коре.
79 Сигналами обратной связи для каждого диапазона были звуковые тоны.
80 Обратите внимание на то, что активность альфа-диапазона максимально выражена в задних отделах коры при закрытых тазах, в то время как уровень тета-активности повышается при глубоком расслаблении. Альфа-тета-тренинг проводился при закрытых глазах с использованием слуховых сигналов обратной связи.
Часть 3. Расстройства систем мозга 429
Испытуемый, находясь в состоянии расслабления, должен стараться как можно дальше удерживать сигнал обратной связи.
Альфа-тета-тренинг впервые был использован Грином и его сотрудниками в клинике Меннингера в 1975 г. Этот метод был основан на изучении Грином одноканальной записи ЭЭГ в процессе медитации у опытных медитаторов. При предъявлении испытуемому, находящемуся в расслабленном состоянии с закрытыми глазми, отдельных независимых звуковых сигналов обратной связи для альфа- и тета-диапазонов он погружался в состояние глубокой релаксации и транса. При углублении транса начальное повышение мощности в альфа-диапазоне сменялось повышением мощности тета-диапазона, на следующей стадии обычно происходило снижение мощности альфа-активности (тета-альфа-перекрест). В то время как ЭЭГ-паттерн соответствует 1-й стадии сна, испытуемые находятся в расслабленном и в то же время сфокусированном состоянии, субъективно сходным с гипнотическим трансом с утратой чувства времени. Эти эффекты неспецифичны для альфа-тета-ЭЭГ-биоуправления и могут быть достигнуты с помощью других техник биоуправления или медитации.
Следует заметить, что альфа-тета-тренинг при аддиктивных расстройствах не используется изолированно. Грин и его коллеги применяли эту технику для усиления эффекта психотерапии. Первые работы по альфа-тета-биоуправлению при аддикциях были направлены на увеличение эффективности психотерапевтического воздействия у бальных алкоголизмом, принимавших участие в 12-ступенчатой программе реабилитации на базе госпиталя Veterans Administration. В течение 6 недель ежедневно проводился 20-минутный альфа-тета-тренинг (в комплексе с биоуправлением по ЭМГ и контролю температуры), что вело к освобождению от травмирующих ассоциаций, увеличению восприимчивости и внушаемости. Пациенты коллективно обсуждали свои переживания и ощущения в процессе биоуправления несколько раз в неделю, что увеличивало воздействие психотерапии.
Пенистом и Кулкоски провели рандомизированное контрольное исследование с использованием альфа-тета-тренинга у бальных хроническим алкоголизмом, резистентных к обычному лечению (Peniston, Kulkosky, 1989). При сравнении с контрольной группой алкоголиков, получавших традиционное лечение, и группой не-алкоголиков, в группе больных алкоголизмом, прошедших курс биоуправления по ЭЭГ и температуре, наблюдалось увеличение процентного содержания альфа-и тета-ритмов в ЭЭГ и снижение показателей Шкалы депрессии по Беку. Данные катамнеза на протяжении 13 месяцев свидетельствовали об отсутствии рецидивов в группе алкоголиков, получивших полный курс альфа-тета-тренинга. Этот протокол известен под названием «Протокол Пенистона — Кулкоски».
V. Заключение
Аддиктивное расстройство связано с непреодолимым, не поддающимся контролю злоупотреблением наркотическими средствами, невзирая на серьезные
430 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
негативные последствия. Поведение аддиктивной личности сосредоточено на получении и приеме наркотических средств и восстановлении после их употребления. Аддикция — это сложный психологический процесс, включающий такие стадии, как ожидание, консолидация и привыкание/сенситизация, каждая из которых характеризуется собственным нейрональным механизмом. Недавние исследования показывают, что наркотические вещества непосредственно (например, кокаин) или косвенно (например, опиаты) повышают уровень дофамина в прилежащем ядре. Уровень концентрации дофамина четко коррелирует с ощущением эйфории у лиц, зависимых от кокаина. Таким образом, по-видимому, дофамин играет решающую роль в генерации мощного положительного подкрепления и инициации аддикции. На поздних этапах аддиктивного процесса возникает гиперактивация передней цингулярной коры, взаимосвязанной с прилегающим ядром. Передняя цингулярная кора играет решающую роль в контроле поведения, таким образом, ее гиперактивация ведет к формированию неконтролируемого компульсивного поведения, направленного на поиск наркотика. Согласно нашей теории, аддикция, по крайней мере у части пациентов, может рассматриваться как нарушение операции контроля. Этот вывод подтверждается нашими исследованиями компонент когнитивных ВП в GO/ NOGO-тесте у героиновых наркозависимых до и после стереотаксической цин-гулотомии, выполненной у этих пациентов с целью облегчения их навязчивых мыслей и компульсивных действий. У этих пациентов обнаружено селективное подавление компоненты мониторинга Р400 при разрушении передней цингулярной коры. Теоретически любой протокол биоуправления, направленный на релаксацию, может быть полезен при наркозависимости. Из них наиболее известен альфа-тета-тренинг в соответствии с протоколом Пенистона — Кулкоски. Он состоит в одновременном измерении уровня альфа- и тета-активности в затылочной области, при этом испытуемый, находясь в состоянии расслабления, должен стараться как можно дольше увеличивать сигнал обратной связи в обоих диапазонах.
Часть 3. Расстройства систем мозга
431
Глава 20.
Обсессивно-компульсивное расстройство
I. Описание поведения
А. Симптомы
Название «обсессивно-компульсивное расстройство» (ОКР) связано с симптомами этого заболевания: нарушением мышления (хода мыслей) и двигательного поведения (действий). Обсессии — это беспрестанно и настойчиво повторяющиеся нежелательные мысли. Компульсии — навязчивые действия, повторяющиеся в ритуалистической стереотипной последовательности. Обычно определенные компульсивные действия выполняются в ответ на определенные обсессии с целью нейтрализации связанной с ними тревожности. В то же время у некоторых пациентов отмечаются только обсессии, а другие страдают преимущественно от компульсий.
Люди с проявлениями ОКР обычно осознают, что обсессии и компульсии бессмысленны, но, несмотря на большие усилия, не могут их контролировать. Обычно обсессии и компульсии заключаются в многократной проверке завершенности определенных действий или правильности их выполнения в связи с постоянными сомнениями в этом. Другой тип ОКР проявляется в навязчивом мытье, наведении порядка... Обсессии и компульсии могут длиться часами. Например, при бесконечно повторяющихся проверках доминирует вопрос «Что, если?», мешая прекращению этого поведения. Обсессии и компульсии «достаточно серьезны, чтобы отнимать много времени и вызывать явный стресс или существенное нарушение здоровья» (Американская психиатрическая ассоциация, 1994)81. Непрекращающиеся сомнения и последующие проверки, сопровождающие каждое действие, — наиболее характерные признаки ОКР. В рамках модели мозга, описанной в этой книге, обсессии и следующие за ними компульсии можно рассматривать как нарушение контроля ответа. Но поскольку обсес-сии/компульсии сочетаются с тревожностью, ОКР может быть также связано с дисфункцией эмоциональной системы.
Установлено, что обсессивно-компульсивное расстройство затрагивает 1—3 % популяции вне зависимости от страны или культуры. Симптомы обычно появляются в детском или подростковом возрасте и связаны со значительными нарушениями в социальных взаимоотношениях или в сфере деятельности. По дан
81 Каждый из нас имеет привычки, манеры, и иногда нашим поведением движут предрассудки. Но в большинстве случаев это поведение не является разрушительным, оно является частью нашей индивидуальности, фоном мыслительной и двигательной активности. При обсессивно-компульсивном расстройстве паттерны этих привычек не поддаются контролю.
432 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
ным исследования, проведенного в 1998 г. Всемирной организацией здравоохранения, ОКР входит в список 10 лидирующих медицинских и психиатрических причин инвалидности в развитых странах. Даже при применении оптимальной медицинской или поведенческой терапии у 10 % пациентов с ОКР симптомы заболевания сохраняются.
II. Генетические факторы и коморбидностъ
А. Низкая наследуемость
Исследования на близнецах отчетливо указывают на то, что предрасположенность к ОКР может передаваться по наследству, однако у многих пациентов это заболевание не прослеживается в семьях. Только 10 % родителей детей с ОКР страдают этим расстройством.
Б. Коморбидные расстройства
Пациенты с обсессивно-компульсивным расстройством часто страдают от других (сопутствующих) заболеваний, включая депрессию, тревожность и синдром Туретта. У 60—70 % пациентов с ОКР симптомы большой депрессии превалируют на протяжении жизни, в то время как 90 % пациентов с синдромом Туретта страдают от ОКР.
III. Структурные и физиологические корреляты
А. ПЭТ, МРТ
Теоретически обсессивно-компульсивное расстройство может быть связано с нарушением различных операций в системе управления, в частности: 1) операции подавления, т.е. способности подавлять нежелательное действие; 2) операции селекции, т.е. способности к переключению на другое действие; 3) операции контроля, т.е. способности сравнивать выполняемую операцию с желаемой и завершать действие. Все эти операции опосредуются петлями связей в системе «лобная кора — базальные ганглии — таламус — кора» (рис. 20.1).
В подтверждение этих теоретических представлений в большей части работ по картированию мозга (ПЭТ, фМРТ) у пациентов с обсессивно-компульсивным расстройством была выявлена аномальная активация лобно-стриарной системы, включающей следующие анатомические структуры: 1) на уровне коры: орбитофронтальную, дорсолатеральную префронтальную и переднюю цингулярную кору; 2) на уровне хвостатого ядра: вентромедиальный стриатум (включая прилежащее ядро), бледный шар/ретикулярную часть черной субстанции; 3) на таламическом уровне: переднее и медиодорсалыюе ядра. В частности, повышенная активность устойчиво выявлялась в орбитофронтальной коре и хвостатом ядре и в состоянии покоя, и при провокации симптомов. Эта гиперактивность коррелировала с симптоматологией ОКР и исчезала под влиянием терапии.
Часть 3. Расстройства систем мозга
433
Б. Количественная ЭЭГ
В работе исследователей из Университета Неаполя в Италии (Paola Bucci et al., 2004) количественная ЭЭГ при обсессивно-компульсивном расстройстве использовалась для выявления признаков гиперактивации лобной коры. У пациентов с ОКР при сравнении с контрольной группой было обнаружено уменьшение мощности медленной альфа-активности преимущественно в лобных областях. Кроме того, наблюдалась выраженная негативная корреляция между мощностью медленной альфа-активности и временем
Рисунок 20.1. Нейронная модель ОКР
Входы из префронтальных областей коры (включая орбитопрефронтальную кору) значительно активируют переднюю цингулярную кору (ACC). АСС, в свою очередь, управляет работой двигательной коры или непосредственно через корково-корковые связи, или опосредованно через петлю «АСС -> NA -> GP ->А, где NA — прилежащее ядро. GP — соответствующая часть бледного шара, А — переднее ядро таламуса. Через эти связи к двигательной коре со стороны префронтальной коры поступает побуждение к выполнению компульсивных действий.
выполнения нейрофизиологического теста, направленного на исследова-
ние функций управления: с уменьшением мощности медленной альфа-
активности выполнение теста замедлялось. Эта взаимосвязь согласуется с позитивной корреляцией между мощностью бета-активности и этой характеристикой ОКР. Однако поскольку гиперактивация лобной коры была обнаружена не только у пациентов с ОКР, но также при некоторых фоби-
ях и при посттравматическом стрессовом расстройстве при провокации его симптомов, предположение о том, что она может быть связана с компонентой тревожности при ОКР в большей степени, чем его специфические нейрональные корреляты, оказалось спорным. Л. Шерлин и М. Конджедо из Novatech с помощью LORETA-картирования обнаружили избыточную плотность источников бета-активности в цингулярной коре у пациентов с ОКР при сравнении со здоровыми лицами (Sherlin, Congedo, 2005).
В. Компонента мониторинга когнитивных ВП
Как нам известно, передняя цингулярная кора вовлечена в операции контроля, отраженные в компоненте мониторинга Р400, возникающей в ответ на NOGO-пробы в двухстимульном GO/NOGO-тесте. Мы также знаем, что ги-псрактивация системы ведет к такой же редукции компонент ВП, как и ее ги-
434 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия поактивация82. Результаты работы, проведенной группой исследователей из Вюрцбургского университета в Германии (Herrmann et al., 2003), согласуются с этими теоретическими соображениями. В этом исследовании у пациентов с обсессивно-компульсивным расстройством и здоровых испытуемых проводилась многоканальная регистрация когнитивных ВП в процессе выполнения GO/NOGO-теста. Авторы показали уменьшение РЗ NOGO-компоненты у пациентов с ОКР по сравнению с контрольной 1руппой. Уменьшение компоненты РЗ NOGO проявлялось в уменьшении параметра антериоризации, рассчитанного как разница между локализациями позитивного центроида в GO- и NOGO-условиях. Этот параметр негативно коррелировал с симптомами Шкалы обсессий и компульсий Иеле-Брауна. В целом гиперактивация передней цингулярной коры может рассматриваться в качестве основы этого заболевания. Поскольку эта область коры входит и в систему управления, и в эмоциональную систему, в лечении ОКР, по-видимому, должны быть эффективны два базовых медиатора.
IV. Медиаторы
Имеются четкие доказательства того, что серотонинергическая система модулирует симптомы обсессивно-компульсивного расстройства. Сильнодействующие ингибиторы транспорта серотонина (ингибиторы обратного захвата серотонина) — единственный вид антидепрессантов, вызывающих некоторое облегчение симптомов у многих пациентов с ОКР. Интересен тот факт, что содержание и переносчика серотонина и некоторых подтипов серотониновых рецепторов, вовлеченных при ОКР, существенно увеличено в вентральном стриатуме, где они могут влиять на функционирование системы «базальные ганглии — таламус — кора». Теоретически другие нейротрансмиттеры в этой системе также могут оказывать влияние на восприимчивость к терапии ОКР и на ее течение и эффективность. Например, изучение дофаминергических механизмов в контрольных исследованиях показало, что нейролептики, неэффективные при монотерапии ОКР, оказывают позитивное влияние при подключении к терапии ингибиторами обратного захвата серотонина.
V. Лечение
А. Стереотаксическая передняя цингулотомия
В 1951 г. Фултон впервые предположил, что передний цингулум может быть целесообразной мишенью для психохирургического вмешательства. Впоследствии Баллантин (Ballantine et al., 1987) показал эффективность и безопасность цингулотомии у многих пациентов83. В течение последних 30 лет цингулотомия
Вспомните, что взаимосвязь между активацией системы и ее функциональными возможностями определяется U-образной кривой.
83 Хотя пациент с ОКР может испытывать уменьшение тревожности сразу после операции, обычно этот эффект проявляется с
A
Топограмм» отклонений
от нормы
Б
Спектр
Компонента Компонента
Норма
«LORETA
►5cm (XI (Y) *5 0 5 Wcm * ° ^Cm
LORETA для компоненты мониторинга
LORETA для бета-ритма фоновой ЭЭГ
Рисунок 20.2. Данные количественной ЭЭГ и когнитивных ВП у пациента с ОКР
Молодой пациент, страдающий ОКР, был подготовлен к стереотаксической передней цин гул отомни в нейрохирургической клинике Института мозга человека Российской академии наук. А Слева—фрагмент фоновой ЭЭГ с соответствующим спектром мощности, зарегистрированный при открытых глазах в точке Cz. Б. Результаты сравнения спектра мощности ЭЭГ с нормативной базой данных: голограмма отличий от нормы и разница спектров мощности. В. sLORETA-изображение сигнала фоновой ЭЭГ в бета-диапазоне. Г. Сопоставление компонент когнитивных ВП пациента (тонкая линия) с нормативными данными (жирная линия): компонента мониторинга Р400 слева и компонента сравнения Р2 справа. Д. sLORETA-изображение компоненты мониторинга Р400.
асть 3. Расстройства систем мозга________________________435
436 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
являлась операцией выбора в Северной Америке, и она до сих пор используется в лечении ОКР84.
Б. Данные количественной ЭЭГ и когнитивных ВП при ОКР
В лечении обсессивно-компульсивных расстройств в нейрохирургической клинике Института мозга человека в С.-Петербурге используется цингулотомия. Недавно мы стали использовать количественную ЭЭГ и когнитивные ВП для локализации источников гиперактивации коры у пациентов с ОКР, которым в связи с резистентностью ко всем традиционным видам лечения была рекомендована стереотаксическая операция. Данные одного из этих пациентов представлены на рис. 20.2. Видно выраженное (р < 0,0001) увеличение бета-акгивности в центральных медиальных отделах с максимумом в Cz, генерируемой, согласно sLORETA, в срединных отделах префронтальной коры и в передней цингулярной коре. Помимо этого, анализ независимых компонент когнитивных ВП показывает статистически значимое (р < 0,02) селективное отклонение от нормы компоненты мониторинга Р4 при отсутствии значимых отклонений со стороны других независимых компонент. Все эти находки согласованно указывают на гиперактивацию зоны коры, расположенной вокруг передней цингулярной коры. Обратите также внимание на то, что вовлеченный отдел перекрывается префронтальной областью, участвующей в подавлении действий.
В. ЭЭГ-б иоуправление
В обзоре литературы по использованию ЭЭГ-биоуправления в лечении тревожных расстройств (Мооге, 2000) упомянуто два опубликованных исследования по применению этого метода при обсессивно-компульсивном расстройстве. В обоих случаях использовался альфа-тренинг, причем безрезультатно. Оценивая эти работы, выполненные в середине 1970-х, с современных позиций, можно сказать, что в них использовался упрощенный подход, состоявший лишь в увеличении уровня альфа-активности. С тех пор было обнаружено, по данным литературы, по меньшей мере два типа паттернов ЭЭГ при ОКР. Имеется несколько сообщений об успешном лечении с длительным катамнезом трех случаев ОКР. В каждом из этих случаев протоколы ЭЭГ-биоуправления были составлены индивидуально, согласно нейрофизиологическим характеристикам каждого пациента по данным количественной ЭЭГ.
VI. Заключение
Название «обсессивно-компульсивное расстройство» связано с симптомами этого заболевания: нарушением мышления (хода мыслей) и двигательного поведения (действий). Обсессии — это беспрестанно и настой
84 А также при резистентных формах большого аффективного расстройства и хронической тревожности.
Часть 3. Расстройства систем мозга 437
чиво повторяющиеся нежелательные мысли, в то время как компульсии — это навязчивые действия, повторяющиеся в ритуалистической стереотипной последовательности. Люди с проявлениями ОКР обычно осознают, что обсессии и компульсии бессмысленны, но, несмотря на большие усилия, не могут их контролировать. В большей части работ по картированию мозга (ПЭТ, фМРТ) у пациентов с ОКР была выявлена аномальная активация лобно-стриарной системы, включая орбитофронтальную, дорсолатеральную префронтальную и переднюю цингулярную кору, прилежащее ядро и передние ядра таламуса. При изучении количественной ЭЭГ обнаружен паттерн гиперактивации лобной коры, в то время как исследование когнитивных ВП выявило редукцию NOGO-компоненты мониторинга при ОКР при сравнении со здоровыми лицами. Цингулотомия долгое время была операцией выбора при ОКР, и она до сих пор используется в лечении этого расстройства. ЭЭГ-БОС, направленная на увеличение альфа-активности, в середине 1970-х не принесла положительных результатов. В последнее время для повышения эффективности ЭЭГ биоуправления протоколы тренинга составляются индивидуально, в соответствии с нейрофизиологическими характеристиками пациента по данным его количественной ЭЭГ.
438 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Глава 21. Депрессия
I. Клиническая картина
А. История
Первое упоминание о депрессии под названием «меланхолия» (от греч. цгХхк; и коХсх; — черная желчь) встречается у Гиппократа 24 столетия назад. В конце XIX века Эмиль Крепелин создал новую нозологическую систему классификации психических расстройств, в которой был представлен термин «инволюционная меланхолия». Обращаясь к прошлому, можно видеть большие колебания в системах классификации — от описательных стратегий у Крепелина, до подходов, основанных на интерпретациях, как у Фрейда, рассматривавшего депрессию как проявление скрытого страха или утраты. В большинстве западных стран более популярны описательные методы Крепелина.
Б. Симптомы
Большая депрессия — это расстройство эмоциональной системы85, характеризующееся триадой симптомов: 1) сниженным, или депрессивным, настроением; 2) ангедонией (утратой интереса и удовольствия при любой деятельности); 3) низким уровнем энергии или усталостью. К другим симптомам относятся нарушение сна и психомоторной активности, пессимизм, чувство вины, низкая самооценка, суицидные настроения, нарушение аппетита и веса. Распространенность депрессии высока во всем мире, это расстройство возникает приблизительно у 7—18 % населения по меньшей мере один раз в жизни до 40 лет. Заболевание чаще встречается у женщин — соотношение «женщины/муж-чины» может достигать 5 : 2. Для депрессии характерно рецидивирующее течение; периоды развернутой симптоматики чередуются с периодами ремиссии86. Поскольку частым следствием этого заболевания является суицид, депрессия — одна из 10 основных причин смертности во всем мире87.
В. Разновидности депрессии
Согласно общепринятым представлениям, в основе симптоматики депрессии лежит не одна структура, и в зависимости от вовлеченных образований вы
85 К расстройствам эмоциональной системы также относятся дистимия (хроническая малая депрессия), биполярное расстройство (маниакально-депрессивное расстройство) и циклотимия (мягкая форма биполярного расстройства).
* “ Однако приблизительно у 20 % пациентов ремиссии отсутствуют.
87 Процент суицида зависит от возраста: в США это четвертая из лидирующих причин смертности среди лиц в возрасте 25-44 лет и третья — в возрастной группе от 15 до 24 лет. Заболеваемость большой депрессией растет, и начало возникновения симптомов приходится на более ранний возраст, чем прежде.
Часть 3. Расстройства систем мозга 439
деляют несколько типов депрессии. В исследованиях ПЭТ и фМРТ обнаружена гетерогенность структур мозга, проявляющих дисфункцию при этом заболевании88. Эта гетерогенность отражается в многообразии симптомов, изменчивости течения и высокоиндивидуальной реакции на лечение. Гетерогенность большой депрессии позволяет большинству экспертов рассматривать это расстройство не в качестве заболевания, а как синдром, предполагая, что в возникновении его симптомов участвуют различные нейронные круги.
Г. Наследственность
Депрессия — заболевание с высокой степенью наследования, при котором генетический риск составляет 40—50 %. Однако специфичные гены в основе этого расстройства пока не идентифицированы. Другие 60—50 % риска также недостаточно изучены. Предположительно, негенетическими факторами риска могут быть психические травмы в раннем возрасте, эмоциональный стресс, соматические заболевания и даже вирусные инфекции.
Д. Необходимость объективной системы диагностики
Ключевая проблема диагностики состоит в том, что существующая система классификации базируется только на субъективном описании симптомов. Такая детальная феноменология включает описание множества клинических подтипов, но при этом не определены биологические черты, которые бы отличали один подтип от другого. Разграничение депрессии и обыденного уныния базируется на устойчивости симптомов депрессии и обусловленной ею нетрудоспособности. Поскольку ни один из симптомов депрессии не отличается по качеству от переживаний, которые каждый из нас иногда испытывает в жизни, депрессия часто не распознается, ложно диагностируется, а иногда неоправданно гиперди-агностируется. Необходим метод объективной диагностики депрессии.
II. Структурные и физиологические корреляты
А. ПЭТ, МРТ
Первые серьезные работы по исследованию церебрального кровообращения и метаболизма глюкозы в покое с помощью ПЭТ выявили снижение префронтальной активности у пациентов с депрессией. Эта гипоактивация коррелировала со степенью тяжести депрессии и исчезала при выздоровлении. Эти находки впоследствии подтвердились фМРТ- и ЭЭГ-исследованиями, показавшими повышение мощности альфа-активности (считается, что мощность альфа-активности находится в обратной зависимости от нейрональной активности) в левой лобной области у пациентов с депрессией.
Клинические наблюдения показывают, что пациенты с депрессией фокусируют внимание в основном на печальных событиях (сниженное настроение) и
" Многочисленные исследования по нейровизуализации указывают на повреждение гиппокампа и префронтальной коры (вероятно, участвующих в возникновении когнитивных симптомов), миндалины (участвующей в происхождении эмоциональных симптомов) и гипоталамуса (ответственного за симптомы, связанные с нарушением аппетита, сна и циркадных ритмов).
440 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
не умеют радоваться счастливым событиям (ангедония). Один из этиологически значимых подходов к изучению эмоциональной системы состоит в изучении метаболических или электроэнцефалографических ответов мозга на эмоционально значимые (радостные и печальные) события. В работе исследователей из Медицинского колледжа Питтсбургского университета и Института психиатрии в Лондоне (Keedwell et al., 2005) эта парадигма (см. методы 3-й части) использовалась для сравнения ответов мозга на позитивно и негативно окрашенные стимулы89. Основная находка состояла в двойной диссоциации ответа вентромедиальной префронтальной коры на различные стимулы при депрессии по сравнению с группой контроля: относительное увеличение ответа этой области на радостные события по сравнению с печальными событиями у пациентов с депрессией и обратный паттерн — относительное уменьшение ответа на радостные события по сравнению с печальными событиями у здоровых лиц90.
Недавно у группы резистентных к терапии пациентов с депрессией помимо снижения активности префронтальной коры было обнаружено увеличение метаболической активности (по данным ПЭТ) части передней цингулярной извилины (Maybuig et al., 2005). У здоровых испытуемых в этой области возрастает кровоток при стимуляции печального настроения. Эта область также реагирует на терапию антидепрессантами.
Б. Асимметрия параметров количественной ЭЭГ
ЭЭГ-корреляты депрессии, по данным межполушарной асимметрии альфа-активности в лобных отделах, опираются на идею о латерализации эмоциональных реакций с тенденцией отрицательных эмоций к активации правого полушария, а положительных — в большей степени левого полушария91. Принимая во внимание то, что основная черта пациентов с депрессией — повышенная восприимчивость к негативным стимулам и избегание позитивных стимулов, асимметричное распределение коррелятов положительных и отрицательных эмоций в префронтальной коре может иметь важное значение в диагностике и лечении депрессии. Действительно, некоторые данные о префронтальной асимметрии (см. главу 13) позволяют предположить, что асимметрия ЭЭГ в лобных отделах может отражать предрасположенность, лежащую в основе индивидуальных различий реактивности на эмоционально значимые стимулы92. Идея об эмоциональной асимметричности мозга исходит из клинических наблюдений поражений левой лобной доли с последующим развитием депрессии.
Принимая во внимание относительную временную стабильность альфа-асимметрии (Tomarken et al., 1992) и ее возможную связь с пове
89 Они использовали зависимость сигнала от степени насыщения крови кислородом (blood-oxygenation-level-dependent, BOLD) по данным фМРТ в качестве показателя метаболической активности.
90 Параллельно со снижением метаболической активности главным образом в вентральном отделе префронтальной коры была обнаружена гиперакгивация (увеличение метаболической активности) в дорсальном отделе префронтальной коры, ассоциирующаяся с гиперактивностью миндалины и аномалиями гиппокампального круга.
91 Гипотеза об асимметричном представительстве положительных и отрицательных эмоций (и вероятно — поощрений и наказаний) подкреплена клиническими доказательствами. Поражение левой лобной области вследствие инсульта чаще сопровождается депрессией, чем правостороннее поражение.
92 Более специфичен факт, что у лиц с гипоактивацией левой лобной области, демонстрирующих в связи с этим повышенную реактивность при предъявлении негативных стимулов, может быть повышен риск переживания эпизодов депрессии.
Часть 3. Расстройства систем мозга 441
дением, направленным на избегание контактов с окружающим миром, можно ожидать наличия гипоактивации левой лобной доли у пациентов с депрессией и у лиц с повышенным риском формирования этого расстройства. Действительно, Хенрике и Дэвидсон (1991), исследуя асимметрию ЭЭГ в передних отделах полушарий93 у пациентов с симптомами депрессии и здоровых лиц, обнаружили увеличение мощности альфа-активности слева (F3 > F4) у первых. Некоторые исследователи показали, что индивидуальные различия в лобной асимметрии формируются на ранних этапах жизни и связаны с индивидуальными особенностями аспекта «приближения — избегания» в поведении детей94. Обобщая эти находки, можно предположить, что лобная асимметрия может быть биологическим маркером семейного и, возможно, генетического риска развития эмоциональных расстройств и может учитываться при проведении генетических исследований в психиатрии95 *.
Следует подчеркнуть, что значения фронтальной асимметрии очень невелики и в нормативной группе составляют в среднем около 1 % со стандартным отклонением приблизительно 5 %%. С этими данными согласуются результаты работы, выполненной в Питтсбургском университете (Vuga et al., 2006). Исследуя записи ЭЭГ у пациентов с депрессией и здоровых испытуемых с интервалом 1 —3 года, авторы показали, что асимметрия ЭЭГ в лобных отделах в покое отражает в среднем стабильные (межклассовая корреляция от 0,4 до 0,6) индивидуальные отличительные признаки у взрослых вне зависимости от пола и наличия депрессии в анамнезе. Обобщение этих данных показывает, что лобная асимметрия может быть связана только с одним (относительно небольшим) подтипом депрессии, что обусловливает несогласованность результатов и их зависимость от процедуры выбора пациентов.
В. Асимметрия когнитивных ВП
В недавних работах показано, что когнитивные ВП могут служить в качестве эндофенотипов для отличия пациентов с депрессией от здоровых лиц. Работа группы исследователей из Центра нейровизуализации в Лондоне (Sumich et al., 2006) выявила у здоровых испытуемых, в отличие от пациентов с субклинической депрессией, асимметрию (П > Л) амплитуды компоненты N200 когнитивных ВП, зарегистрированной слуховом oddball-тесте97.
93 Вспомните, что асимметрия спектров мощности ЭЭГ в лобных отделах обычно рассчитывается как ((Л - П)/((Л + П)/2)))х 100%, <де Л и П — квадратные корни значений мощности в гомологичных отведениях левого и правого полушарий (F3 и F4, F7 и F8).
м Например, в этих работах обнаружена гипоактивация левой лобной области у детей, чьи матери страдали депрессией, у поведенчески заторможенных детей.
* Однако в недавнем исследовании с участием 246 молодых женщин-близнецов, выполненном в Медицинском колледже при вашингтонском университете (Anokhin et al., 2005), был выявлен очень низкий (хотя и статистически значимый) параметр наследуемости лобной асимметрии, измеренной в точках F3/F4, позволивший предположить, что лишь 27 % наблюдавшихся различий могло быть обусловлено генетическими факторами. Напротив, показатель мощности альфа-ритма был высоконаследуемым и составлял М5-87 % во всех четырех лобных отведениях.
“ Вспомните также, что показатель лобной асимметрии предполагает расчет отношения параметров, в то время как стандартное отклонение отношения — это сумма стандартных отклонений обеих частей отношения. Это создает сложности в достижении надежности этого индекса у конкретного пациента.
•' Эти данные были получены в семи клиниках Brain-Resource Company (BRC), расположенных в Лондоне, Ниймегене, Сиднее, Аделаиде, Мельбурне, Нью-Йорке и Род-Айленде. Клиники BRC используют стандартный протокол и обнаруживают высокую межла-Рораторную согласованность результатов, не зависящую от пола и возраста (см. более детально www.brainresource.com).
442 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Г. Оценка количественной ЭЭГ и когнитивных ВП при депрессии
Пример использования нормативной базы данных HBI Database при исследовании пациента с депрессией показан на рис. 21.1. Наблюдается два типа асимметрии: первый касается спектров мощности ЭЭГ при открытых глазах, а второй — когнитивных ВП, зарегистрированных в двухстимульном GO/NOGO-тесте. Для сравнения представлены спектры мощности и когнитивные ВП, рассчитанные для симметричных лобных отведений (F7 и F8). Обратите внимание на то, что у этого пациента при открытых глазах амплитуда альфа-волн в лобном отведении левого полушария (F7) выше по сравнению с лобным отведением правого полушария (F8). По данным sLORETA генераторы этого аномального98 альфа-ритма находятся в вентролатеральной префронтальной коре и верхней височной коре левого полушария. Зная о том, что наличие альфа-ритма в определенной области коры связано с гипофункцией (бездействием) этой области, мы можем сделать вывод о гипоактивации вентролатеральной префронтальной коры левого полушария у этого пациента. Наряду с данными количественной ЭЭГ анализ компонент когнитивных ВП показывает асимметричную активацию лобных долей в ответ на GO- и NOGO-стимулы. На рис. 21.1 ввдно снижение амплитуды ВП в ответ на NOGO-стимулы99 по сравнению со здоровыми испытуемыми слева (F7) и отсутствие этого феномена справа (F8). Этот факт свидетельствует о гипофункции медиального и вентролатерального отделов префронтальной коры левого полушария и согласуется с альфа-асимметрией.
Д. Предсказание ответа на антидепрессанты поданным количественной ЭЭГ
Параметры количественной ЭЭГ и когнитивных ВП могут использоваться у пациентов с большой депрессией для предсказания реакции на антиде-рессанты. Это очень важное обстоятельство, поскольку 50—75 % эффективности антидепрессантов объясняется влиянием плацебо: у многих пациентов с депрессией состояние улучшается при приеме как фармакопрепаратов, так и плацебо. В работе, выполненной недавно в Лаборатории количественной ЭЭГ Медицинского колледжа UCLA, Лос Анджелес (Leuchter et al., 2002), при исследовании работы мозга у пациентов с депрессией, принимающих фармакопрепараты или плацебо, предпринималась попытка выявить различия в функционировании мозга между этими двумя группами с помощью количественной ЭЭГ Использовались как спектры мощности, так и корданс100, новый параметр, по-видимому, отражающий кровоснабжение лучше, чем спектры мощности ЭЭГ. Авторы показали раннее снижение корданса в префронтальных областях у пациентов, отвечавших на лечение, и ее увеличение — у тех, кто отвечал на плацебо. Судя по этим данным, корданс может использоваться не только в качестве параметра, помогающего разграничить пациентов, отвечающих на лечение, и тех, кто на него не реагирует, но и для выявления лиц, отвечающих на плацебо.
98 Обратите внимание на то, что мощность этого ритма значительно (р < 0,01) выше, чем у здоровых сверстников из группы контроля.
99 Представлено в качестве примера, в то время как ВП на GO-стимулы не показаны.
100 Корданс отражает отклонение от нормы и относительной, и абсолютной мощности, измеренной в особого рода локальном усредненном монтаже (более подробно см. методы 1-й части).
Рисунок 21.1. Спектры и асимметрия когнитивных ВП у пациента с депрессией
А. Фрагмент фоновой ЭЭГ в точках F7, F8 относительно локального средневзвешенного референта. Б. Сопоставление ЭЭГ-спектров под электродами F7 и F8 и отклонение от нормы в этих точках. Г. Когнитивные ВП в ответ на NOGO-пробы в двухстимульном зрительном GO/NOGO-тесте. Г. sLORETA-иллюстрации асимметрии альфа-активности и отклонения от нормативных значений когнитивных ВП э ответ на NOGO-пробы.
444 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
III. Нейрональная модель
А. Моноаминовая гипотеза депрессии
Моноаминовая гипотеза депрессии возникла на ранних этапах стремления понять механизмы этого расстройства. Согласно этой гипотезе, депрессия отражает дефицит или дисбаланс норадреналина (или серотонина) в мозге. Гипотеза основывалась на наблюдениях, показывавших, что некоторые антидепрессанты повышали синаптическую концентрацию норадреналина или серотонина, и что резерпин (препарат, истощающий запасы катехоламинов) мог вызывать симптомы, сходные с депрессией.
Лекарства, влияющие на катехоламины, начинают воздействовать на соответствующие нейротрансмиттерные системы не позднее чем через час после приема препарата, но позитивные изменения могут появиться не ранее чем через несколько недель после начала курса лечения! Одна из теорий, пытающихся объяснить это несоответствие, указывает на медленное развитие приспособительных изменений в работе синаптических ауторецепторов ствола мозга, приводящих в результате к их десенситизации101. Другая гипотеза утверждает, что депрессия обусловлена повреждением нейрональных кругов эмоциональной системы, а не дисфункцией медиаторов. Реорганизация этих патологически измененных кругов требует долговременной модификации, как и при вовлечении кругов, ответственных за процедуральную память.
Б. Нейронные круги при депрессивном настроении
По данным экспериментальной нейровизуализации, при депрессии, являющейся мультисимптомным расстройством, вовлечены и система управления, и эмоциональная система мозга102. При большой депрессии вовлечены такие элементы эмоциональной системы, как орбитофронтальная и медиальная кора, островковая доля, миндалина и структуры гипоталамо-гипофизарного комплекса. Элементы системы управления включают переднюю цингулярную кору и соответствующую петлю «базальные ганглии — таламус —- кора». Позвольте описать круги, вовлеченные при депрессивном настроении (рис. 21.2). Вспомните, что этот симптом служит наиболее распространенным проявлением депрессии и на его коррекцию чаще всего направлена фармакотерапия.
Большое число работ по нейровизуализации показывает, что грусть и депрессивное настроение связаны с аномальной активацией нейронов медиальной префронтальной коры, включая переднюю цингулярную и орбитофронтальную кору. Эти области коры получают входы из гиппокампа, миндалины и мамиллярных тел гипоталамуса через переднее ядро таламуса. Активность этих областей опосредована преимущественно серотонинер
101 Дело в том, что систематический прием лекарств влияет не только на целевые структуры эмоциональной системы, но и на ауторецепторы ядер ствола мозга, регулирующих, в свою очередь, продукцию медиаторов в целевых структурах. Таким образом, сетевой эффект в начале курса лечения может быть незначительным или даже нулевым. Считается, что отсроченный эффект антидепрессантов можно объяснить медленно развивающейся десенситизацией ауторецепторов ствола мозга.
102 Это хорошо согласуется с тем фактом, что симптомы аффективных расстройств помимо эмоциональных имеют также когнитивные, моторные и нейроэндокринные компоненты. Каждый из этих компонентов депрессии связан с собственными специфическими нейронными сетями.
Часть 3. Расстройства систем мозга
445
DLPF
VLPF
Ганглии
Рисунок 21.2. Нейронная модель депрессивного настроения
Выходы от миндалины (Ат), сосочковых тел гипоталамуса (МВ) и гиппокампа (Hip) через переднее ядро таламуса проецируются к островковой доле, орбитофронтальной коре (OF) и передней цингулярной коре (ACC). Печальные события, вероятно, картируются в подколенном отделе цингулярной извилины (представленном точечным паттерном), так что гипоактивация этой области приводит к депрессивному настроению.
Нейронная сеть, характеризующая депрессивное состояние
Базальные
гической иннервацией из ядра шва среднего мозга и частично — норадренергической иннервацией из голубого пятна. Антидепрессанты, уменьшающие проявления грусти и депрессивное настроение, влияют на эти медиаторные системы и нормализуют активность этих областей. Длительная глубинная стимуляция мозга103 субгенуального отдела передней цингулярной коры оказывает подобный эффект.
В отличие от депрессивного настроения, связанного с гиперактивностью медиального отдела лобной коры, дисфункция си
стемы управления при большой депрессии, вероятно, отражается в гипоактивации дорсолатеральной префронтальной коры. Эта гипоактивность может быть результатом взаимного подавления системы управления и эмоциональной системы. Нейрональные основы этого подавления могут обеспечиваться латеральными ингибиторными связями в стриатуме. Вспомните, что локализация отделов стриатума, получающих входы из корковых областей эмоциональной системы, отличается от локализации отделов, получа-\ ющих входы из системы управления104. Как нам известно, система управления контролирует широкий ряд когнитивных операций, включая рабочую память, внимание и социальную мотивацию. Таким образом, когнитивные симптомы депрессии связаны в основном с дофаминергической иннервацией системы управления и могут быть целью соответствующей медикаментозной терапии. Подавление системы управления происходит главным образом в доминантном полушарии и отражается альфа-асимметрией в фоновой ЭЭГ (больше альфа-активности в левом полушарии) и снижением компонент когнитивных ВП, связанных с системой управления.
103 Вспомните, что глубинная стимуляция мозга в высокочастотном диапазоне и при больших значениях тона подавляет активность в стимулируемой области.
104 Эго свойство отражено в существовании параллельных путей в базальных ганглиях и таламусе, в частности путях, возникающих в когнитивных и лимбических областях коры.
446 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
IV. Лечение
А. Когнитивная поведенческая терапия
Когнитивная поведенческая терапия — психологическое лечение, восходящее своими корнями к эпохе 3. Фрейда. В наши дни психотерапия обычно рекомендуется в качестве лечения мягких форм депрессии или как дополнение к лекарственной терапии при более тяжелых формах заболевания. Этот тип лечения базируется на идее о том, что депрессивные симптомы основаны на деструктивных убеждениях и мыслях, возникших на ранних этапах развития. Такие шаблоны активируются ситуациями или событиями, имеющими особое значение для индивида. Таким образом, когнитивная поведенческая терапия сфокусирована главным образом на выявлении негативных автоматических шаблонов и их подавлении. В нескольких работах продемонстрировано преимущество когнитивной поведенческой терапии перед другими психологическими техниками и плацебо105.
Б. ЭКГ и психохирургия
Используемые в настоящее время физические методы лечения депрессии описаны еще в 1937 г., начиная с введения электроконвульсивной терапии, которая, несмотря на беспокойство как общественности, так и профессионалов, остается наиболее эффективным средством лечения депрессии106. Основные возражения против использования ЭКТ связаны с провоцируемыми ею когнитивными нарушениями. Однако при недавнем метаанализе ЭКТ, выполненном в Великобритании и опубликованном в журнале Lancet в 2003 г., установлено, что повреждения после ЭКТ состоят преимущественно в кратковременной антероградной и ретроградной амнезии.
В 1940— 1950-х в лечении депрессии стала использоваться психохирургия, более точно — фронтальная лоботомия. Позднее, в 1960-х, были открыты первые лекарства, способные уменьшать симптомы большой депрессии. Изобретение антидепрессантов и изучение их фармакологии привело к коренным преобразованиям в этой области107. К тому же психохирургия, влекущая за собой массивные деструкции ткани мозга, была заменена современным стереотаксическим методом, позволяющим точно вводить электроды и являющимся инструментом для глубинной стимуляции мозга108. Группа исследователей из Университета г. Торонто, Канада (Maybeig et al., 2005), при лечении шести пациентов с тяжелой, резистентной к лечению депрессией109 использовала введение электродов в белое вещество латерально от субгенуального отдела передней цингулярной извилины. Авторы сообщают о поразительно стойкой ремиссии заболевания у
105 Результаты метаанализа эффективности когнитивной терапии у пациентов с депрессией представлены в статье Gloaguen etal., 1998.
106 Заметьте, что ЭКТ также применяется при лечении шизофрении с учетом наибольшей эффективности метода в случаях, если депрессия сочетается с психотическими симптомами, такими как мания и галлюцинации.
107 В настоящее время существуют десятки усовершенствованных средств, относящихся к четырем различным классам—трициклические препараты, селективные ингибиторы обратного захвата серотонина, ингибиторы МАО и смешанные антидепрессанты.
108 Глубинная стимуляция мозга, являясь в настоящее время экспериментальным методом лечения, представляет собой вмешательство, сходное с нейрохирургическим, но в отличие от него обратимое и позволяющее проводить плацебо-контроль для отдельных пациентов.
109 Это были пациенты, не отвечавшие на антидепрессанты, психотерапию и электроконвульсивную терапию.
Часть 3. Расстройства систем мозга
447
четырех из шести пациентов. ПЭТ-сканирование показало снижение исходно повышенного метаболизма в субгенуальной области у этих пациентов по сравнению со здоровыми испытуемыми.
В. Антидепрессанты
Антидепрессанты —- это гетерогенная группа препаратов, повышающих содержание моноаминов в синаптической щели. Понимание их фармакологии позволило сформулировать моноаминовую гипотезу депрессии и расширило возможности для создания новых лекарств, таких как селективные ингибиторы обратного захвата серотонина.
Эффективность многих известных антидепрессантов составляет около 60 %. На настоящий момент не существует возможности с уверенностью предсказать клинический эффект при применении отдельных препаратов. Так, если пациент не отвечает на одно средство, его переводят на другое, обычно относящееся к другому классу, и так до тех пор, пока не будут испытаны разные классы препаратов. Поскольку клинический эффект появляется через 6—8 недель, на подбор эффективного лекарства может уйти несколько месяцев. Этот процесс требует больших временных и финансовых затрат. Более того, он ведет к повышению вероятности суицида, поскольку частичный эффект может проявляться в увеличении психомоторной активности при сохранении депрессивных симптомов. И наконец, несмотря на метод проб и ошибок, используемый в лекарственном лечении депрессии, многие пациенты остаются резистентными ко всем известным препаратам.
Недостаточная эффективность фармакотерапии побудила медицинскую науку к поиску альтернативных методов лечения, таких как ЭЭГ-биоуправление, транскраниальная микрополяризация, глубинная стимуляция мозга, транскраниальная магнитная стимуляция. Все эти подходы (наряду с традиционными методами) нуждаются в надежной объективной оценке заболевания для диагностики и контроля процесса лечения.
В табл. 21.1 представлена большая часть из доступных в настоящее время методов лечения большой депрессии.
Г.ТМС
Транскраниальная магнитная стимуляция в качестве потенциального метода лечения депрессии впервые была применена в конце 1990-х. Однако начальный энтузиазм сменился более умеренной оценкой метода в наши дни. Метаанализ всех доступных данных, проведенный в 2006 г. (Ebmeieret al., 2006), указывает на тенденцию к существенному снижению величины эффекта с 1996 по 2004 г.
Д. ЭЭГ-биоуправление
ЭЭГ-биоуправление в лечении депрессии базируется главным образом на теории Дэвидсона об асимметричном распределении альфа-активности в латеральных лобных областях. На основе этой теории Розенфельд разработал протокол ЭЭГ-биоуправления для нормализации
448 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Таблица 21.1
Тип вмешательства Механизм действия
Трициклические антидепрессанты Подавление обратного захвата серотонина и норадреналина
Ингибиторы моноаминоксидазы (ИМАО) Подавление МАОА и связанное с ним подавление обратного захвата биогенных аминов
Литий Неизвестен
Атипичные антидепрессанты Неизвестен
Селективные ингибиторы обратного захвата серотонина (СИОЗС) Селективное подавление обратного захвата серотонина
Ингибиторы обратного захвата норадреналина (НИОЗС) Селективное подавление обратного захвата норадреналина
Ингибиторы обратного захвата серотонина и норадреналина (СНИОЗС) Подавление обратного захвата серотонина и норадреналина
Электроконвульсивная терапия Глобальная электрическая активация нейронных сетей с интенсивным высвобождением различных медиаторов
Транскраниальная магнитная стимуляция Последействие локальной электромагнитной активации нейронных сетей
Глубинная стимуляция мозга Постоянная стимуляция цингулярной коры в субгенуаль-ном отделе
Когнитивная поведенческая терапия Неизвестен
Внутриличностная терапия Неизвестен
аномальной асимметрии при депрессии. Так называемый ALAY-протокол (созданный для альфа-асимметрии) в качестве параметра обратной связи использует отношение (F4 — F3)/(F3 + F4). Предварительные результаты использования этого протокола в ряде случаев обнадеживают, но контрольные исследования пока не проводились. Например, Байер и его коллеги, изучив 1—5-летний катамнез пациентов, использовавших ALAY-протокол, установили, что изменения клинической картины после тренинга оставались стойкими и что фронтальная альфа-асимметрия не только изменилась в конце лечения, но и оставалась на обратном, физиологическом уровне в течение длительного периода наблюдения. К сожалению, исследований с Использованием плацебо-контроля и двойного слепого метода, как при доказательстве эффективности фармакологического лечения, не проводилось.
V. Заключение
Большая депрессия характеризуется триадой симптомов: 1) сниженным, или депрессивным, настроением; 2) ангедонией (утратой интереса и удовольствия при любой деятельности); 3) низким уровнем энергии или усталостью. Согласно общепринятым представлениям, в основе симптоматики депрессии лежит не одна структура, и в зависимости от вовлеченных отделов мозга выделяют несколько типов депрессии. Гетерогенность структур мозга, проявляющих дисфункцию, отражается в многообразии симптомов, изменчивости течения и высокоиндивидуальной реакции на лечение. ЭЭГ-корреляты депрессии, по данным межполушарной асимметрии альфа-активности в лобных отделах, опираются на идею о латерализации
Часть 3. Расстройства систем мозга
449
эмоциональных реакций с тенденцией отрицательных эмоций активировать правое полушарие, а положительных — в большей степени левое полушарие. Некоторые данные о фронтальной альфа-асимметрии позволяют предположить, что этот параметр может отражать предрасположенность к реактивности на эмоционально значимые стимулы. Во множестве работ по нейровизуализации депрессивное настроение связано с повышенной активацией нейронов медиальной префронтальной коры. В противоположность гиперактивности медиальной лобной коры дисфункция системы управления при большой депрессии отражается в гипоактивации дорсолатеральной префронтальной коры. Моноаминовая гипотеза предполагает, что депрессия отражает дисбаланс в регуляции медиаторов норадреналина и серотонина. С этой гипотезой согласуется тот факт, что некоторые антидепрессанты повышают синаптическую концентрацию норадреналина или серотонина. Электроконвульсивная терапия остается наиболее эффективным методом лечения депрессии. Эффективность антидепрессантов составляет не более 60 %, и это побудило медицинскую науку к поиску альтернативных методов лечения, таких как ЭЭГ-биоуправление, транскраниальная микрополяризация, глубинная стимуляция мозга, транскраниальная магнитная стимуляция. Так называемый ALAY-протокол был создан для нормализации аномальной альфа-асимметрии при депрессии.
450
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Глава 22.
Болезнь Альцгеймера
I. Описание поведения
А. Симптомы
С возрастом нервные клетки медленно умирают. Но в некоторых, к счастью, редких случаях этот естественный медленный процесс значительно ускоряется и ведет к дегенеративным изменениям. В 1906 г. немецкий доктор А. Альцгеймер опубликовал данные наблюдения 51-летней пациенткой с тяжелой деменцией. Он описал поведенческие симптомы и связал их с обнаруженными у этой пациентки патологическими изменениями в коре и лимбической системе. В настоящее время мы знаем о двух основных нейрональных нарушениях при болезни Альцгеймера: утрате холинергических клеток в базальном отделе переднего мозга и развитии сенильных бляшек в коре, локализованных преимущественно в области височных долей110.
II. Медиаторы
А. Связь с холинергическими и ГАМК-септально-гиппокампальными кругами
Как мы уже знаем, эпизодическая память связана с холинергическими и ГАМК-эргическими септально-гиппокампальными кругами. Эти круги, по-видимому, ответственны за запуск обработки информации в гиппокампе и связанных с ним структурах. Этот гипотетический запуск проявляется в кратковременных вспышках тета-ритма, генерируемого в гиппокампальных кругах. Нарушение генерации вспышек гиппокампального тета-ритма может быть следствием дисфункции перегородки в результате ее анатомической деструкции или воздействия антагонистов ацетилхолина. С гипотезой о роли запуска тета-акгивности в кодировании информации согласуется наличие связи между подавлением лимбического тета-ритма и нарушением эпизодической памяти.
Б. Холинергическая гипотеза болезни Альцгеймера
Идея о том, что некоторые симптомы болезни Альцгеймера обусловлены дефицитом нейромедиатора ацетилхолина в мозге, впервые возникла в 1976 г., когда Дэвис и Малонэ опубликовали свою статью в журнале Lancet. Последующие работы подтвердили их находки. Согласно холинергической гипотезе болезни
110 Бляшка состоит из расположенной в центре белковой субстанции — амилоида, окруженной дегенеративными клеточными фрагментами.
Часть 3. Расстройства систем мозга 451
Альцгеймера, некоторые когнитивные отклонения у пациентов с этим заболеванием обусловлены дефицитом холинергической нейротрансмиссии. Эта гипотеза стимулировала серьезные исследования в области экспериментальной фармакологии. Большинство препаратов, одобренных к использованию при болезни Альцгеймера Управлением по контролю над пищевыми продуктами и лекарственными средствами США (Food and Drug Administration, FDA) повышает уровень ацетилхолина путем подавления его распада.
III. Модель нейронной сети
А. Вспышки тета-волн в здоровом мозге
На рис. 22.1 изображены гипотетические нейронные сети, ответственные за консолидацию эпизодической памяти и, возможно, поврежденные при болезни Альцгеймера. Как нам известно из 2-й части, эпизодическая память формируется долговременной потенциацией в гиппокампальных кругах путем повторяющихся вспышек активности гиппокампальных нейронов. Кратковременная активация гиппокампальных нейронов во время привлекающих внимание событий отражается во вспышках тета-ритма в гиппокампе. Эта ритмическая активность передается в переднюю цингулярную кору по мамилло-таламическому тракту и видна в ЭЭГ в форме лобного тета-ритма средней линии. Гиппокампальный тета-ритм управляется септальным входом, так что амплитуда вспышек тета-ритма строго зависит от уровня активации холинергических нейронов в перегородке. Механизм генерации тета-ритма включает тормозные связи между перегородкой и гиппокампом.
Б. Увеличение спонтанной тета-активности в больном мозге
Дегенерация септальных нейронов при болезни Альцгеймера, вероятно, ведет к снижению септального тормозного контроля над гиппокампальными нейронами и, как следствие, к их растормаживанию111 *. Это растормаживание ведет к увеличению спонтанной нейронной активности в гиппокампе (рис. 22.1). Повышенная гиппокампальная активность, по-видимому, проявляется медленными осцилляциями, обнаруживаемыми в Fz 'b форме колебаний корковых потенциалов в тета-дельта, диапазонах. Эти колебания существенно отличаются от нормального гиппокампального тета-ритма, поскольку они не привязаны по времени к привлекающим внимание стимулам и не имеют отношения к консолидации эпизодической памяти. Более того, утрата тормозного контроля из септального региона может обусловить возникновение эпилептиформной активности в гиппокампе, а в некоторых случаях — эпилептических приступов1,2. В дополнение к этому дегенерация холинергических клеток в базальных ядрах (BN на рис. 22.1) при болезни Альцгеймера приводит к уменыпе-нию холинергического входа в префронтальную кору из восходящей активи
111 Экспериментальные доказательства и теоретические основы представлены в статье Л. Колома из Техасского университета в Ьраунсвилле.
IU Действительно, при болезни Альцгеймера 10-22 % пациентов страдает от приступов, обусловливающих необходимость антикон-аульсивной терапии (Mendez and Lim, 2003).
452 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
рующей системы, что, в свою очередь, может отражаться в повышении медленной активности и уменьшении бета-активности в префронтальных отделах.
IV. Структурные и физиологические корреляты
А. Количественная ЭЭГ
В работе, выполненной в Университете Тюбенгена А. Стевенсом и его коллегами (Stevens et al., 2001), регистрация 19-канальной ЭЭГ проводилась при закрытых и открытых глазах и при наблюдении за маятником113 у пациентов с мягкими формами болезни Альцгеймера и у их здоровых сверстников114. Значимый эффект различий двух групп по абсолютной мощности и когерентности был выявлен только для тета-диапазона и выражался в увеличении мощности и снижении когерентности у пациентов. Самым надежным параметром для отличия
Рисунок 22.1. Нейронная модель болезни Альцгеймера
А. Нейронная сеть, вовлеченная в генерацию гиппокампального тета-ритма. Б. Ответы в ЭЭГ (зарегистрированные в Fz) и гипотетическая импульсная активность нейронов в гиппокампе здорового испытуемого (вверху) и у пациента с болезнью Альцгеймера.
.....1-
Передне центральный тет.т
Болстнь Альцгеймера
Б
пациентов был параметр когерентности при открытых глазах.
В работе, выполненной недавно в лаборатории Р. Джона в Медицинском колледже Нью-Йоркского университета (Prichep et al., 2005), исследовались данные здоровых пожилых людей с субъективными жалобами на когнитивные расстройства с целью оценки возможности использования ЭЭГ для предсказания клинического ухудшения в течение последующих 7 лет115. Спектральные характеристики ЭЭГ у лиц с последующим нарастанием нарушений значимо (р < 0,0001) отличались от таковых улиц со стабильной клинической картиной увеличением абсолютных и относительных значений мощности в тета-диапазоне, замедлением основного ритма и изменениями межрегиональной ковариации.
ш Последний был создан с целью удержания внимания испытуемых и фиксации взора.
114 Групповые различия спектров мощности ЭЭГ и когерентности были максимальными в покое при открытых тазах, вкладывая почти 80 % в параметр дискриминации. Уже на ранних этапах болезни Альцгеймера изменения были топографически широко распространенными.
“в 44 здоровых пожилых испытуемых прошли основательное клиническое, нейрокогнитивное и ЭЭГ-обследобание. У всех отмечались только субъективные жалобы, но объективных доказательств когнитивного дефицита выявлено не было.
Часть 3. Расстройства систем мозга 453
Б. Когнитивные ВП
Компоненты когнитивных ВП также могут использоваться для разграничения пациентов с болезнью Альцгеймера и здоровых лиц. В работе, выполненной в Стэнфордском университете, возрастные и обусловленные деменцией изменения когнитивных ВП оценивались с помощью теста на название картинок (Mathalon et al., 2003). Авторы использовали синхронизированные с ответом ВП в качестве маркеров мониторинга ответа. Старение ассоциировалось с замедлением поведенческих ответов и снижением амплитуды негативности, связанной с ошибкой, в то время как для деменции было характерно уменьшение точности ответов и снижение амплитуды негативности по сравнению со здоровыми ровесниками.
В. Анализ когнитивных ВП методом главных компонент
В работе, недавно выполненной в Рочестерском университете в США (Chapman et al., 2006), была произведена попытка использования метода главных компонент для анализа когнитивных ВП с целью разграничения пациентов с болезнью Альцгеймера и здоровых лиц. Авторы использовали тест «Число — буква». Помимо известных компонент — РЗЬ, условного негативного отклонения и медленной волны (SW) — авторы сумели выделить другие компоненты, включая относительно ранние пики с латентностью 145 и 250 мс (компонента хранения в краткосрочной памяти). В дискриминантном анализе для создания функции, которая позволила бы успешно разделять индивидов с начальными этапами болезни Альцгеймера и здоровых лиц, использовались параметры когнитивных ВП в ответ на значимые и незначимые стимулы. Применение дискриминантной функции к половине данных показало, что приблизительно 90 % индивидов были корректно отнесены или к группе болезни Альцгеймера или к контрольной группе с чувствительностью 1,00.
V. Лечение
А. Ингибиторы ацетилхолинэстеразы
Основная идея современной фармакотерапии болезни Альцгеймера состоит в повышении уровня ацетилхолина в мозге, что обычно достигается путем подавления холинэстераз. Препараты, известные как ингибиторы ацетилхолинэстеразы (ИАХЭ), были одобрены Управлением по контролю над пищевыми продуктами и лекарственными средствами США в 1995 г. после того, как клинические испытания показали умеренное симптоматическое улучшение когнитивных, поведенческих и общих показателей116 в результате их применения.
Б. ЭЭГ-биоуп равнение
Поиск в PubMed с использованием комбинации ключевых слов «Болезнь Альцгеймера» и «ЭЭГ-биоуправление» не обнаружил ни одной статьи касательно использования ЭЭГ-БОС для коррекции симптомов болезни Альцгеймера.
1 '• В 2004 FDA одобрила применение мемантина, антагониста NMDA, для лечения симптомов деменции в среднетяжелых и тяжелы* случаях. В клинической практике мемантин может сочетаться с ИАХЭ, хотя и при использовании в отдельности, и в комбинации не оказывает влияния на патофизиологические механизмы деменции.
454 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
VI. Заключение
Болезнь Альцгеймера — это дегенеративное заболевание мозга, сопровождающееся прогрессирующим снижением памяти. Утрата холинергических клеток в базальном отделе переднего мозга, по-видимому, ответственна за первый этап развития заболевания. Согласно холинергической гипотезе болезни Альцгеймера, некоторые когнитивные отклонения у пациентов с этим заболеванием обусловлены дефицитом холинергической нейротрансмиссии. Большинство препаратов, одобренных к использованию при болезни Альцгеймера Управлением по контролю над пищевыми продуктами и лекарственными средствами США (Food and Drug Administration, FDA), повышают уровень ацетилхолина путем подавления его распада. Дегенерация септальных нейронов при болезни Альцгеймера, по-видимому, ведет к снижению тормозного контроля над гиппокампальными нейронами и, как следствие, к повышению фоновой активности в гиппокампе. Спонтанная гиппокампальная активность проявляется в лобных отведениях в виде тета-колебаний, отличающихся от лобного тета-ритма средней линии. ЭЭГ пациентов с болезнью Альцгеймера характеризуется увеличением абсолютных и относительных значений мощности в тета-диапазоне.
Часть 3. Расстройства систем мозга
455
Глава 23.
Методы нейротерапии
I. Плацебо
А. Плацебо как ожидание результатов
Впервые я столкнулся с этой проблемой в начале моей карьеры в1970-х. В то время мы работали с пациентами с болезнью Паркинсона, которым с диагностической и лечебной целью были имплантированы электроды117. Мой коллега (В.М. Смирнов) проводил пациентам глубокую электрическую стимуляцию мозга, в то время как я регистрировал у этих пациентов с этих же электродов импульсную активность нейронов и медленные электрические потенциалы. Однажды мы должны были докладывать о результатах нашей работы руководителю отдела, профессору Н.П. Бехтеревой. Поскольку мы работали с одним и тем же пациентом, во время демонстрации результатов нам нужно было переключать наш прибор (режимы регистрации и стимуляции). Сначала я продемонстрировал влияние двигательного акта на активность нейронной популяции, зарегистрированной одним из электродов. Затем мой коллега собрался показать влияние электростимуляции с того же электрода, и я должен был переключить прибор. Я отвлекся на вопрос и забыл это сделать. Коллега и пациент, не зная о моей ошибке, прекрасно продемонстрировали эффект: у пациента снизился тремор, когда В.М. Смирнов нажал на кнопку стимуляции. Дело было в том, что пациент ожидал воздействия, и, когда он увидел, как доктор нажимает на кнопку, произошел запуск выполнения ожиданий.
Б. Нейронная основа плацебо
На рис. 23.1 схематически представлен плацебо-эффект, наблюдавшийся в вышеупомянутом случае. Стимуляция проводилась в двухстимульном тесте с использованием предупреждающего и запускающего стимулов, следовавших с 2-се-кундными интервалами. Регистрация проводилась у пациента с болезнью Паркинсона с электродов, имплантированных с диагностической и терапевтической целью. Для регистрации импульсной активности мультинейронов использовались те же электроды, при подаче на которые высокочастотной электрической стимуляции у пациента подавлялся тремор. Можно видеть, что предъявление предупреждающего стимула изолированно увеличивает фоновую нейронную активность и повышает нейрональную реакцию на моторные действия. Глубинная стимуляция мозга с помощью этих электродов повышает фоновую активность и реакцию на действие, но не меняет подготовительную активность в промежутке
Автор работал над своей диссертацией по применению медленных процессов для оценки локальной метаболической активности у пациентов на базе клиники Института экспериментальной медицины в Ленинграде-
456
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
между предупреждающим и запускающим стимулами. Реакции, предшествующие глубокой стимуляции мозга, представлены в верхней части рис. 23.1, в то время как реакции, непосредственно сопровождающие ГСМ, показаны в средней части. «Сетевой» эффект стимуляции, представляющий собой разность между обеими реакциями, представлен внизу. Обратите внимание на то, что плацебо индуцирует подготовительную активность подобно тому, как ее вызывает ГСМ. Это так называемый плацебо-эффект, хотя ГСМ индуцирует более выраженный ответ на моторное действие. Сетевой эффект ГСМ проявляется как повышение фоновой активности и увеличение реактивности таламических нейронов.
Рисунок 23.1. Плацебо-эффект (модель)
А. Схема расположения имплантированных электродов у пациента с болезнью Паркинсона. Б. Гистограммы в постстимульном периоде (ось Y—усредненная величина разряда, ось X — время). Гистограммы в постстимульном периоде рассчитаны для двух случаев: 1) плацебо, т.е. в отутствие ГСМ (вверху); 2) ГСМ (середина). Сетевой эффект представлен в виде разницы двух гистограмм.
Это простое наблюдение повторилось 30 лет спустя в работе Бенедетти и его коллег в 2004 г. Авторы регистрировали импульсную активность нейронов субталамического ядра у пациентов с болезнью Паркинсона с помощью электродов, имплантированных для глубинной стимуляции мозга. Активность отдельных нейронов регистрировалась до и после приема плацебо с целью выявления связи изменений нейронной активности с клиническим плацебо-ответом. В качестве плацебо пациентам давали соленый раствор, говоря при этом, что это — лекарство от болезни Паркинсона. Эта процедура выполнялась в процессе операции после предоперационного фармакологического лечения апоморфином, мощным средством при болезни Паркинсона. У пациентов, отвечавших на плацебо, было обнаружено значимое уменьшение нейронных разрядов и редуцирование вспышек активности субталамических нейронов, в то время как у пациентов, не отвечавших на плацебо, этих изменений не было. Следует отметить
Часть 3. Расстройства систем мозга 457
выраженную корреляцию между субъективными отчетами пациентов, клинической неврологической оценкой и электрической активностью отдельных нейронов.
В. Необходимость в двойных слепых плацебо-контролируемых исследованиях
Эти наблюдения показывают, что любое лечение (включая ГСМ, фармакотерапию и ЭЭГ-биоуправление) сопровождается психологическими ожиданиями, которые могут влиять на терапевтический результат. Эффект ожидания называется эффектом плацебо (или ложным эффектом). Вспомните из главы 12, что во время подготовительных операций активируется вся широко распространенная система управления (включая дорсолатеральную префронтальную кору и базальные ганглии). Более того, нейрональные реакции во время подготовки к движению в некоторых случаях сходны с реакциями на сами двигательные акты118.
В контексте этой книги мы излагаем проблему плацебо-эффекта из двух соображений. Во-первых, плацебо-эффекты демонстрируют величайшую способность человека к саморегуляции. Эти эффекты имеют реальную нейрофизиологическую основу, и могут быть обнаружены с помощью ЭЭГ и других методов визуализации. Во-вторых, плацебо-эффект должен быть исключен при доказательстве эффективности любого нового терапевтического метода. Поэтому любой новый метод должен быть проверен в двойных слепых плацебо-контролируемых исследованиях. «Двойной слепой» означает, что ни испытуемый, ни экспериментатор не знают, является ли тестирование истинным или ложным. В таких исследованиях дается ложное (плацебо) лечение, но пациент вериг в то, что оно эффективно, и ожидает клинического улучшения. Плацебо-эффект — результат ложного лечения. Обзор на тему плацебо-эффекта представлен Ф. Бенедетги и его коллегами в Journal of Neuroscience в 2005 г. (Benedetti et al., 2005).
II. ЭЭГ-биоуправление
Эго все еще начальный период эпохи ЭЭГ-биоуправления. Каждый день мы находим новые пути понимания нарушений регуляции мозговой деятельности и новые способы более эффективной саморегуляции работы мозга... Мы в поиске. Так было и так будет...
Сью Отмер в своих заметках 8-го августа 2001 г.
А. История
Основные события истории ЭЭГ-биоуправления кратко представлены в табл. 23.1. Первая попытка представить большой обзор исследований в этой области была сделана в книге «Введение в количественную ЭЭГ и ЭЭГ-биоуправление», изданной Д. Эвансом и А Абарбанелем в 1999 г.
1“ Классический павловский рефлекс можно рассматривать как другой пример плацебо-эффекта. Действительно, после многократного связывания условного стимула с подкреплением собака реагирует на условный стимул, как на подкрепление.
458 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Физиология условных рефлексов создает объективные основы для ЭЭГ-биоуправления119. Эксперименты по условным рефлексам были выполнены И.П. Павловым в Институте экспериментальной медицины в С.-Петербурге. Институт был основан в 1890 г. принцем Ольденбургским — членом русской царской семьи120. Плодотворные идеи Павлова позднее продолжали развивать его ученики. В конце 1940-х П.С. Купалов, в прошлом студент Павлова, разработал методологию ситуационных условных рефлексов. На Западе этот метод был назван оперантным обусловливанием. В этом методе не внешние стимулы, а поведенческие реакции животных служили безусловными стимулами. Во времена Павлова обычно это были моторные реакции, например животное бежало в определенную секцию экспериментального помещения. В 1930 г американский математик Н. Винер в сотрудничестве с мексиканским психологом А Ро-зенблютом представил концепцию обратной связи применительно к биологическим системам. Эта концепция в дальнейшем развилась в науку, названную Винером «кибернетика» в его книге, опубликованной в 1948 г. Приблизительно в то же время русский ученый П.К. Анохин, ученик Павлова и Бехтерева, в 1935 г. разработал теорию функциональных систем. Ключевым элементом этой теории была нейрональная обратная связь — взаимодействие между так называемым «акцептором действия» и поведенческим приспособлением животного.
Следуя традициям Российской школы физиологической науки, Н.Н. Василевский, профессор Института экспериментальной медицины, в конце 1960-х— начале 1970-х гг. начал исследовать клеточные механизмы ЭЭГ-биоуправления. Н.В. Черниговская, работавшая в то время в отделе физиологии человека этого же института, стала использовать метод биологической обратной связи в лечении неврологических и психических заболеваний. Идея была простой: тренировать мозг или мышцы (как в случае церебрального паралича), используя для обратной связи физиологический параметр. Для биоуправления использовались паттерны ЭЭГ, электрическая активность мышц и медленные метаболические процессы. Приблизительно в то же время американский исследователь Д. Камия обнаружил, что испытуемые могли научиться произвольно контролировать уровень альфа-активности. Для достижения этой цели каждый раз, когда испытуемый генерировал альфа-ритм, исследователь сообщал ему об этом. Поэтому, хотя Н. Винер и П.К. Анохин сформулировали идею обратной связи в 1930—40-х годах, только в 1960-х годах впервые было показано, что данные ЭЭГ могут служить в качестве параметров обратной связи для саморегуляции работы мозга. Позднее Б. Стерман в своей работе с кошками представил ритм, связанный с сенсомоторной системой и поэтому названный «сенсомоторным ритмом». С
Мы предполагаем, что читатели знакомы с павловским методом условных рефлексов. Известный русский физиолог И.П. Павлов первым в начале XX столетия использовал этот метод в изучении физиологических механизмов психологических функций. По методу Павлова, некий условный стимул, изначально имеющий незначительное поведенческое значение для животного, во времени связывается с неким подкреплением или вознаграждением (по терминологии Павлова, безусловным стимулом). Условный стимул мог быть зрительным или слуховым. Безусловным стимулом мог быть кусочек мяса для собаки или небольшое количество сока для обезьяны. Подкрепление, имевшее жизненно важное значение для животного, вызывало существенные изменения в его поведении и физиологические реакции (например, слюноотделение). Теперь, если кормление собаки многократно сопровождалось звуком метронома, некоторое время спустя у нее будет отдаляться слюна в ответ на сам звук—так формируется условный рефлекс.
120 Наша лаборатория расположена всего лишь в 200 метрах от лаборатории Института экспериментальной медицины, где И.П. Павлов проводил исследования условных рефлексов. В музее его имени находится гармония (старинный музыкальный инструмент, похожий на орган), которую И.П. Павлов использовал в экспериментах для генерации условных стимулов.
Часть 3. Расстройства систем мозга 459
помощью оперантного обусловливания кошки обучались генерировать этот ритм с использованием пищевого подкрепления. Также было обнаружено, что тренинг предотвращал развитие экспериментальных судорожных приступов у этих животных. Вскоре после этого сенсомоторный тренинг был применен при эпилепсии. По данным этих работ, ЭЭГ-биоуправление значительно снизило судорожные проявления и нормализовало ЭЭГ.
Таблица 23.1. История ЭЭГ-биоуправления
Год Событие
Начало XX столетия И.П. Павлов разработал методику изучения условных рефлексов у собак
Конец 1940-х П.С. Купалов разработал методику ситуационных условных рефлексов у животных
1930-40-е Н. Винер и П.К. Анохин сформулировали идею обратной связи в саморегуляции живых организмов
1960-е Б. Стерман (США) проводил опыты по экспериментальному рефлексу на кошках, а результат использовал для лечения эпилепсии
1962 Д. Камия (США) показал, что испытуемые могут управлять альфа-ритмом в условиях обратной связи
1960-70-е В ИЭМе в Ленинграде Н.Н. Василевский и Н.В. Черниговская заложили основы современного метода биологической обратной связи в теории и на практике
1970 Немецкий ученый Н. Бирмбаум использовал медленные электрические процессы мозга для биологической обратной связи и успешно начал применять этот метод в лечении эпилепсии
1970-80-е Американский ученый Д. Любар доказал эффективность тренинга для лечения СНВГ
Середина 1980-х Применение нейрометрики (Р. Джон, США) для создания протоколов ЭЭГ-биоуправления
Настоящее время Начало использования данных LORETA и фМРТ для создания протоколов ЭЭГ-БОС. В 2006 International Journal for Neurotherapy отпраздновал 10-летний юбилей
В 1969 г. метод саморегуляции работы мозга с помощью ЭЭГ и других физиологических параметров получил название биологической обратной связи (biofeedback). ЭЭГ-биоуправление использовалось для лечения неврозов и эпилепсии, а также в антистрессовой реабилитации у ветеранов Вьетнамской войны в США121. В Германии в 1970-х Н. Бирмбаум и его коллеги стали использовать медленные электрические процессы мозга для биоуправления при эпилепсии и шизофрении.
Это был период эйфории, когда казалось, что биологическая обратная связь может стать панацеей от всех заболеваний мозга. Однако в 1974 г. в статье Лйнча и соавт. было показано, что испытуемые, научившиеся управлять альфа-ритмом при открытых глазах, не могли увеличить его в большей степени, чем просто закрывая глаза. Статья продемонстрировала пределы возможностей человека при ЭЭГ-биоуправлении, но эти ограничения были истолкованы слишком буквально и привели к снижению популярности этого метода122. Все же несколько энту
ш ЭЭГ-биоуправление при антистрессовой реабилитации включает тренинг тета-ритма ЭЭГ с целью погружения испытуемого в состояние измененного сознания.
122 По-видимому, было несколько причин плохой репутации ЭЭГ-биоуправления в те годы. Один из них — некорректное применение этого метода в клинической практике. Заметьте, что легко ошибиться в правильном использовании ЭЭГ-биоуправления по нескольким причинам: 1) в связи с недостаточным опытом; 2) из-за недостатка объективных критериев для выбора правильного протокола (вспомните, что в те годы количественная ЭЭГ редко использовалась перед ЭЭГ-биоуправлением); 3) в связи с недостат-мом солидной экспериментальной поддержки и обоснованности выбранных протоколов. Таким образом, переход от лабораторных экспериментов к клинической практике произошел слишком быстро. Явные ограничения ЭЭГ-биоуправления использовались оппонентами в критике этого метода. Следует также помнить, что оборудование для регистрации ЭЭГ в те дни было довольно дорогим, а сам метод — довольно чувствительным к артефактам и поэтому требовавшим больших затрат времени и усилий. Для клинициста было гораздо проще использовать в практике лекарственные средства, чем тратить время и деньги на этот сложный и довольно бездоказательный подход.
460 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
зиастов продолжали заниматься ЭЭГ-БОС. Важную роль сыграли исследования американского ученого Д. Любара, выполненные в 1970—80-х годах в Университете штата Теннесси. Он показал, что сеансы увеличения мощности бета-ритма с одновременным уменьшением мощности тета-ритма у пациентов с СНВГ значительно уменьшали у них проявления гиперактивности и улучшали концентрацию внимания. Однако в те годы в научных исследованиях использовался метод проб и ошибок и за ними не было никакой солидной теории.
В то же самое время (в начале 1970-х) в области ЭЭГ был разработан новый подход. Исследователем из Медицинского центра Университета Нью-Йорка Р. Джоном он был назван нейрометрикой123. Основная идея нейрометрики состояла в сравнении количественных параметров индивидуальной ЭЭГ с теми же параметрами, рассчитанными для нормативной группы. Эта идея была революционной, поскольку до тех пор стандартная оценка электроэнцефалограммы состояла только в визуальном анализе сигналов сырой ЭЭГ
В середине 1980-х годов произошло слияние двух подходов — ЭЭГ-биоуправления и нейрометрики, что сформировало новое направление, названное нейротерапией124 *. Американская фирма «Лексикор» первой организовала выпуск нейротерапевтических систем. В настоящее время этот подход активно развивается. Новые представления о генезе ритмов ЭЭГ позволили создать теоретическую базу этого подхода. Только в США существует несколько компаний, специализирующихся на анализе ЭЭГ и на разработке индивидуальных протоколов нейротерапии.
Б. Принцип«бульдозера» в ЭЭГ-биоуп равнении
ЭЭГ-БОС базируется на двух фактах: 1) состояние мозга (включая любую дисфункцию и дисрегуляцию) объективно отражается в параметрах ЭЭГ, зарегистрированной со скальпа; 2) мозг человека обладает способностью к запоминанию желательных (и поэтому поощряемых) состояний. При ЭЭГ-биоуправлении некоторые текущие параметры ЭЭГ, зарегистрированной со скальпа (например, мощность ЭЭГ в определенном частотном диапазоне, или отношение мощностей ЭЭГ разных частотных диапазонов) предъявляются индивиду в виде сигналов зрительной, слуховой или тактильной модальности с заданием произвольно изменять эти параметры в желательном (ведущем к более эффективной работе мозга) направлении. Локализация электродов и параметры ЭЭГ (называемые параметрами ЭЭГ-биоуправления) изменяются в зависимости от целей ЭЭГ-биоуправления. Вместе взятые локализация электродов и параметр ЭЭГ-БОС определяют так называемый протокол ЭЭГ-биоуправления. В лечебных целях большинство специалистов по ЭЭГ-БОС прямо или косвенно использует принцип «бульдозера». Согласно этому принципу, задача ЭЭГ-биоуправления состоит в нормализации патологически измененного паттерна ЭЭГ Так, при избыточной представленности какого-либо ЭЭГ-параметра у кон
123 Он использовал это название по аналогии с психометрикой — отраслью психологии, изучающей различия психологических параметров между пациентами и контрольной группой.
124 В данной книге мы используем концепцию нейротерапии в более широком смысле, включая в нее все другие техники, осно-
ванные на нейрофизиологии, такие какфМРТ-БОС, ТКМП, ТМС, ГСМ.
Часть 3. Расстройства систем мозга
461
кретного пациента в определенной области коры задача ЭЭГ-биоуправления состоит в снижении этого параметра, при недостаточности какой-либо другой характеристики ЭЭГ соответствующий ЭЭГ-параметр при тренинге повышается. Метод работает, как бульдозер, заполняя ямы и выравнивая ухабы.
В. Сравнение с базой данных
В настоящее время практика ЭЭГ-биоуправления в клинике предполагает несколько ступеней его применения. Первая ступень включает создание протокола ЭЭГ-биоуправления на основе параметров количественной ЭЭГ125. Как мы показали в этой книге, грандиозный объем эмпирических знаний по анализу ЭЭГ помогает выявить связь некоторых аномальных паттернов ЭЭГ с различными неврологическими и психическими расстройствами. Например, для части популяции СНВГ характерно увеличение отношения «тета/бета» в теменных-центральных-лобных (в зависимости от возраста) отведениях126.
Для анализа количественной ЭЭГ обычно требуется запись активности с по меньшей мере 19 электродов127. Спектральные характеристики ЭЭГ при открытых и закрытых глазах и во время некоторых психологических тестов (например, Oddball-тест или арифметический тест) статистически сопоставляются с нормативной базой данных. Это сопоставление дает объективную информацию об отличиях спонтанной активности ЭЭГ пациента от таковой у здоровых сверстников. Статистически значимое отклонение от нормы на кривых спектров или когерентности определяет параметры процедуры ЭЭГ-биоуправления, такие как локализация электродов (например, Fz, Cz), и параметры ЭЭГ-биоуправлеия (например, отношение «тета/бета»). Этот этап схематично изображен на рис. 23.2.
Г. Определение положения электродов
На рис. 23.2 вверху слева представлен фрагмент сырой ЭЭГ 13-летней девочки с симптомами СНВГ. Спектры целого фрагмента ЭЭГ при открытых глазах были сосчитаны (не показано) и сравнены с нормативными спектрами. Разность спектров (пациент-норма) вместе с Z-значениями представлены в средней части рис. 23.2. Обратите внимание на избыточную (р < 0,001) медленную активность под электродом Cz. Наряду с увеличением медленной активности отношение мощности «тета/бета» (индекс невнимательности), рассчитанное для Cz128, у этой пациентки в 2,5 раза выше, чем в группе здоровых сверстников. Этот факт указывает на локализацию одного электрода для протокола ЭЭГ-биоуправления, а именно в точке Cz. Положение второго электрода может быть определено на основе градиента спектрального параметра. Карта градиента отклонения от нормы в тета-диапазоне (6 Ш) показывает, что волна различий рез-
В течение последних нескольких лег некоторые лаборатории и специалисты по ЭЭГ-биоуправлению в дополнение к оценке количественной ЭЭГ стали использовать независимый компонентный анализ когнитивных ВП. Похоже, что этот под ход скоро станет золотым стандартом, поскольку он предоставляет совершенно новые данные касательно этапов обработки информации в мозге человека.
Отношение мощности «тета/бета» было обозначено как индекс невнимательности.
12719 — по-видимому, минимальное количество электродов, позволяющее пользователю получить 2-мерные карты и slDRETA изображения с приемлемым (для практических целей) пространственным разрешением.
““ Заметьте, что отношение ^тета/бета» рассчитывалось для локального среднего (согласно Лемосу) монтажа и поэтому несколько отличается от отношения по данным статьи Монастра с соавт. (Monastra et aL, 1999).
462 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Д s LORETA медленной активности
Рисунок 23.2. Первый шаг при создании протокола ЭЭГ-биоуправления — сравнение с базой данных
Шаг включает построение спектров фоновой ЭЭГ, сравнение спектров с базой данных и определение точек расположения электродов. А. Фоновая ЭЭГ 13-летней девочки с СНВГ с sUDRETA-изображением медленной активности внизу. Б. Разница ЭЭГ-спектров (пациент — нормативные данные) со значительным уровнем отклонения от нормы (внизу — разностная волна для Cz) и голограмма различий спектров. В. Отношение тета/бета у пациента и в группе здоровых сверстников.
шаг. 1
< равнение спектров < нормой, опреде ление поипщй электродов
ко снижается в Fz. Эго наблюдение указывает на локализацию второго электрода для протокола ЭЭГ-биоуправления129.
Д. Определение параметра ЭЭГ-биоуправления
После определения локализации электродов следует выбрать параметр ЭЭГ-биоуправления — спектральные характеристики ЭЭГ, которые пациент будет изменять при тренинге. Для этого нужно реконструировать паттерн сырой ЭЭГ пациента для уже установленного расположения электродов. Для сравнения с нормативной базой данных HBI Database мы выбираем фрагмент ЭЭГ пациента при открытых глазах. Такая процедура позволяет реконструировать, как будет выглядеть паттерн ЭЭГ у данного больного при данном биполярном монтаже. Результат ремонтажа представлен на рис. 23.3 внизу. Спектры фрагмента ЭЭГ при открытых глазах представлены справа. Опираясь на спектры, мы можем выбрать частотные диапазоны для расчета параметра ЭЭГ-биоуправления. Низко
139 Обратите внимание на то, что для процедуры ЭЭГ-биоуправления мы используем биполярный монтаж. По нашемуопыту, биполярный монтаж менее чувствителен к артефактам (например, движениям глаз). Однако у этого монтажа есть один недостаток—тренируемая область охватывает большие размеры коры (под обоими электродами) по сравнению с монополярным монтажом. Все же, если градиент отклоняющегося от нормы параметра довольно велик, биполярный монтаж предпочтительнее.
Часть 3. Расстройства систем мозга
463
частотный диапазон может составлять от 3 до 7 Гц, в то время как высокочастотный — от 13 до 21 Гц. Для нормализации спектров пациента (т.е. для приближения их к нормативным данным) необходимо увеличивать высокочастотную активность и подавлять низкочастотную. Процедура «нормализации» может проводиться с использованием лишь одного параметра: отношения мощностей ЭЭГ, рассчитанных для низкочастотного и высокочастотного диапазонов130.
Спектр в выбранном монтаже
Отклонение от нормы
Низкочастотный диапазон: J-“Hz Высокочастотный диапазон: 13-21 Hz
Параметры ЭЭГ бноуправления: отношение диапазонов мощностей ЭЭГ выскочастотного к низкочастотному Тренировка по увеличению данного отношения
Шаг!
Вычисление спектров в выбранном монтаже и определение параметров ЭЭГ-бноуправлештя
В
Рисунок 23.3. Первый шаг при создании протокола ЭЭГ-биоуправления — определение параметра ЭЭГ-биоуправления
А. Топограмма отклонений спектра от нормы. Б. Спектр, рассчитанный для монтажа Cz-Fz для всей эпохи ЭЭГ при открытых глазах по сравнению с нормативными данными. В. Фрагмент ЭЭГ пациента в монтаже Cz-Fz.
Е. Процедура тренинга
Второй этап — это сам тренинг, при котором обычно на скальпе размещаются два электрода, согласно локализации, установленной на первом этапе. Заземляющий электрод может быть установлен в любом месте, обычно — на одной из ушных мочек. В отличие от таких техник, как ТМС, ГСМ и ТКМП, мозг не подвергается воздействию тока. С помощью электродов лишь регистрируется текущая ЭЭГ, при этом компьютер постоянно рассчитывает параметр ЭЭГ и предъявляет его в определенном виде пациенту. В обычной жизни без помощи приборов для ЭЭГ-биоуправления мы не способны влиять на ЭЭГ в желательном направлении, поскольку человек, как и животные, не осознает паттерны своей ЭЭГ.
Существует много систем для ЭЭГ-биоуправления, разработанных разными производителями, такие как Lexicor, Brain Master, Thought Technology, Brain Inquiry. Важнейшими в любой процедуре ЭЭГ-биоуправления являются две характеристики, описывающие: 1) насколько надежно рассчитанный параметр
ш Некоторые системы ЭЭГ-биоуправления используют для тренинга два или больше параметров. Они основываются на способности человека одновременно следить за несколькими показателями. Однако наша практика показывает, что процедура ЭЭГ-БОС с одним параметром биоуправления работает по меньшей мере не хуже, чем процедура с несколькими параметрами.
464 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Высоко/ншко -частотное отношение
Шаг 2: расчет параметра ЭЭГ-биоуправления с помощью быстрого преобразования Фурье
250 мс бнны
Рисунок 23.4. Второй шаг — расчет в реальном времени параметра ЭЭГ-биоуправления с помощью быстрого преобразования Фурье
А. Фрагмент фоновой ЭЭГ в выбранном монтаже. Б. Спектры, вычисленные с помощью быстрого преобразования Фурье в 1-секундном скользящем окне со сдвигом каждые 250 мс. В. Параметр ЭЭГ-биоуправления рассчитан и представлен на экране (внизу справа).
ЭЭГ-биоуправления отражает текущее состояние мозга; 2) насколько часто обновляется параметр, предъявляемый испытуемому, те. насколько точно он следует изменениям состояния мозга.
Первая характеристика — это в действительности период интеграции текущих колебаний ЭЭГ, также называемый скользящим окном. Вторая характеристика — это интервалы времени, в течение которых интегрированный параметр предъявляется испытуемому. Из исследований когнитивных ВП мы знаем, что состояние мозга может изменяться с интервалом времени в 100 мс, в то время как индивид реагирует на стимул в течение 300 мс. Эти данные позволяют определить, с какой частотой рассчитанный параметр должен предъявляться испытуемому131. В современных компьютерах вычисление происходит очень быстро, и теоретически скорость предъявления параметра ЭЭГ-биоуправления может быть очень высокой. Однако для надежного измерения спектров ЭЭГ необходима по меньшей мере одна секунда записи, т.е. 10 циклов основного ритма, что определяет параметр интеграции ЭЭГ-биоуправления132.
Ж. Расчет параметров ЭЭГ-биоуправления
Различные производители предлагают разные способы вычисления спектральных характеристик параметра ЭЭГ-биоуправления. Эго следующие методы: 1) разложение с использованием быстрого преобразования Фурье (Fast Fourier Transformation — FFT) (рис. 23.4); 2) цифровая фильтрация с использова-
ш В примере на рис. 233 интервал «обновления» составляет 250 мс.
132 В примере на рис. 233 скользящее окно интеграции составляет 1 с. Заметьте, что 1 с — это в действительности интервал времени, достижимый в современном оборудовании для МРТ. Это позволяет МРТ-биоуправлению стать надежным инструментом коррекции мозговых дисфункций.
Часть 3. Расстройства систем мозга
465
ЭЭГ-сшнял при выбранном монтаже
FAw Ressxwe Fonctson inipufes response
50 100 150 200
Фильтр низких частот Фильтр высоких частот
4yW\A/^WW|!yvV\MA Шаг 2: расчет параметра ЭЭГ-биоуправления с помощью КИХ-фильтра
скользящее интегрирующее окно
высоко-низко
частотное отношение
J i I V'
250 мс бины
Рисунок 23.5. Второй шаг — расчет параметра ЭЭГ-биоуправления с помощью КИХ-фильтра
Сверху вниз—фоновая ЭЭГ, параметры КИХ-фильтра, фильтрованная ЭЭГ, фрагменты фильтрованной ЭЭГ, величины потенциалы которой возведены в квадрат для 1-секундного скользящего окна. Усреднение возведенных в квадрат потенциалов дает оценку мощности в заданном диапазоне. Параметр ЭЭГ-биоуправления определен как отношение мощностей ЭЭГ в двух частотных диапазонах.
нием фильтров с конечной переходной характеристикой (КИХ-фильтры) (рис. 23.5); 3) цифровая фильтрация с использованием фильтров с бесконечной переходной характеристикой (БИХ-фильтры). Любой из этих методов имеет свои преимущества и недостатки. Не вникая в технические детали и исходя из соображений теории и из опыта нашей лаборатории, предпочтительнее использовать БИХ-фильтры.
Таким образом, когда испытуемые получают обратную связь от своего мозга, т.е. могут видеть или слышать образ своего параметра, предполагается, что они учатся изменять этот параметр в желаемом направлении. Пациент обычно находится перед экраном компьютера. Изображение на дисплее может быть очень простым, например вертикальной полосой, или комплексным и многомерным, с использованием различных цветов и форм. В качестве сигнала БОС могут служить компьютерные игры, разработанные компаниями-производителями, или профессиональные компьютерные игры (например, Need-for-Speed), в которой
466
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
параметр игры (например, скорость автомобиля) контролируется с помощью ЭЭГ-биоуправления. В нашей практике мы используем технологию, включающую компьютер, видеоплейер (он может быть встроен в компьютер) и телевизор или компьютерный монитор. Пациенты смотрят любимые фильмы133, при этом параметр ЭЭГ-биоуправления, вычисляемый компьютером, контролирует качество видеоизображения: чем ближе к норме параметр, тем лучше изображение.
При любом из способов предъявления параметра ЭЭГ-биоуправления пациент концентрирует внимание на сигнале обратной связи и пытается запомнить связь между параметром и своим состоянием. Это непростая задача. Более того, она может быть решена разными способами. При такой неопределенности возникает вопрос: как помочь пациенту наиболее эффективно справиться с заданием. Некоторые специалисты по биоуправлению предпочитают не давать никаких инструкций пациентам, рекомендуя лишь «делать это». Другие же дают рекомендации в зависимости от типа процедуры: релаксации или активации. Процесс обучения при ЭЭГ-биоуправлении можно сравнить с техникой обучения езде на велосипеде. Обучение езде на велосипеде требует времени!
3. Кривая тренинга
Второй этап процедуры ЭЭГ-биоуправления предполагает оценку результатов тренинга после каждого сеанса. Дело в том, что не все пациенты могут научиться контролировать параметр ЭЭГ-биоуправления. Для этого существует несколько причин: 1) неправильный выбор параметра — ошибка инструктора; 2) неспособность связать изменения параметра с субъективным состоянием — ошибка пациента. Для оценки успешности пациента в контроле параметра компании-производители предлагают инструменты для расчета так называемых кривых тренинга. Эти кривые представляют собой сглаженную динамику параметра в процессе отдельного сеанса. Если параметр во время периода тренинга статистически отличается (р < 0,05) от параметра во время отдыха, сеанс можно рассматривать как успешный (рис. 23.6). Если в течение 5—7 сеансов не наблюдается никаких успехов, следует прервать тренинг и выбрать новый протокол.
И. Кривая обучения
Третий этап процедуры ЭЭГ-БОС включает оценку положительных изменений, вызванных сеансами ЭЭГ-биоуправления. Обычно для обучения изменению параметра биоуправления необходимо приблизительно пять сеансов, в то время как для «консолидации» приобретенного навыка требуется 20—35 дополнительных сеансов. Динамика параметра ЭЭГ отражается в так называемой кривой обучения, представляющей собой динамику абсолютных или относительных показателей тренинга для каждого сеанса. На рис. 23.6 показана техника вычисления относительного показателя успешности процедуры тренинга для отдельного сеанса. Этот показатель рассчитан как отношение усредненных величин параметра ЭЭГ-биоуправления во время периодов тренинга и отдыха.
133 Для лучшей мотивации к тренингу и повышения ело эффективности им предлагали приносить особой видеозаписи любимых фильмов.
Часть 3. Расстройства систем мозга
467
| Фон
Параметры
Обратной связи
Тренинг
Покой
А
Тренируемые параметры
ИВ Ml JN 409 ~ЯЮ ЫМ> 789 «И» <Ж> 1Й90 flWH’OnIBM I4W1ЯЮ
Время сессии
Кривая тренинга
Тренируемые параметры:
среднее значение во
i время грешшга /
среднее значение * в состоянии покоя
Кривая обучения
1.5
Шаг 3: определение качества тренинга
р 0,05
1.0 J-,----,-,-т--,--,---,---,-,--,---,--,---,--,--,--,--г
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
Число сессий Б
Рисунок 23.6. Третий шаг — определение качества сеанса нейротерапии
А. Кривая тренинга — сглаженная динамика параметра ЭЭГ во время отдельного сеанса. Успешность тренинга определяется как отношение средней величины параметра ЭЭГ в период тренинга к среднему значению этого параметра в период отдыха. Это отношение, рассчитанное для ряда сеансов и представленное в виде диаграммы, образует кривую обучения (Б).
Можно оценить уровень достоверности различий между этими величинами.
К. Методы вычисления параметра ЭЭГ-биоуправления
Следует подчеркнуть, что протоколы ЭЭГ-биоуправления базируются не только на FFT или цифровой фильтрации ЭЭГ. Могут быть предложены другие протоколы ЭЭГ-биоуправления. Один из возможных параметров — величина корреляции ЭЭГ в различных зонах коры, в частности когерентность или ко-модуляция (предложенная в нормативной базе данных Б. Стермана и Д. Кайзера).
Другой подход — использование электромагнитной томографии низкого разрешения (low resolution electromagnetic tomography — LORETA или sLORETA) для тренинга в определенных зонах коры. Этот подход основан на том факте, что ЭЭГ, зарегистрированная под отдельным электродом, отражает активность не толь
468 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
ко лежащей под ним коры, но и отдаленных корковых зон. Таким образом, если мы хотим проводить ЭЭГ-биоуправление в локальной области коры, нам нужно использовать томографические методы, позволяющие рассчитывать параметры корковой ЭЭГ на основе скальповой ЭЭГ, зарегистрированной с множества электродов. Локальная корковая ЭЭГ, извлеченная из многоканальной скальповой ЭЭГ, называется виртуальной ЭЭГ. Метод конструирования внутрикорковой ЭЭГ для тренинга в локальной области коры был впервые применен М. Конджедо в лаборатории Д. Любара (Congedo et al., 2004)134. Авторы назвали этот метод LORETA-биоуправление. Недавно в нашей лаборатории была разработана компьютерная программа, позволяющая вычислять плотность локальных источников тока в коре с помощью метода sLORETA Сам метод не дает систематической ошибки локализации и был недавно разработан Р. Паскуале-Марки (Rascual-Marqui, 2006). Все известные техники биоуправления перечислены в табл. 23.2.
Л. Типы протоколов ЭЭГ-биоуправления
Таблица 23.2. Техники ЭЭГ-биоуправления
Название Параметры ЭЭГ-биоуправления Примеры использования
Спектры ЭЭГ Мощность ЭЭГ в определенных частотных диапазонах, или отношение мощностей ЭЭГ в двух частотных диапазонах для одного канала ЭЭГ, или отношение мощностей ЭЭГ одного диапазона под разными электродами. Для извлечения этих параметров используются FFT, вэйвлет-преобразование или цифровая фильтрация Sterman, 1971; Lubar and Shouse, 1976
ЭЭГ-когерентность Параметр когерентности отражает корреляцию ЭЭГ в определенном частотном диапазоне между различными точками на скальпе Horvat, 2004
LORETA Виртуальная ЭЭГ, рассчитанная с помощью LORETA для интересующей области и соответствующие спектральные характеристики Congedo et al. 2004
sLORETA Плотность локальных источников тока в коре, рассчитанная с помощью метода sLORETA Ponomarev, Kropotov, 2007
С точки зрения физиологии протоколы ЭЭГ-биоуправления можно разделить на две большие категории: активирующие и релаксирующие. Это разделение основано на данных о корреляции между активностью ЭЭГ различных частотных диапазонов и метаболической активностью по результатам ПЭТ или фМРТ в соответствующих зонах коры135 136 (рис. 23.7). Активирующие протоколы заключаются в увеличении высоких (> 13 Гц) частот ЭЭГ, а релаксирующие — в увеличении мощности (8—12 Гц) альфа-диапазона и в некоторых случаях — частот ЭЭГ ниже 8 Пх. Хотя важность локализации электродов трудно переоценить, ниже из дидактических соображений мы рассматриваем протоколы вне связи с электродной локализацией.
134 Электромагнитная томография низкого разрешения является инструментом для вычисления платности генераторов тока, локализованных на поверхности коры, включая борозды и извилины. Решение представляет собой матрицу, которая при умножении
на данные многоканальной ЭЭГ дает в результате плотность кортикальных микродиполей. Когда определена внугрикорковая зона, становится возможным найти так называемую инвертированную матрицу. М. Конджедо и его коллеги использовали биоуправление, основанное на данных LORETA, для повышения величины (длины вектора) плотности источников низкочастотной бета-активности (16-20 Гц) и снижения платности источников (8-10 Гц) альфа-активности в области, приблизительно соответствующей когнитивной области передней цингулярной коры. Интересующий регион включал 38 векселей из общего числа 2394 векселей.
136 См., например, данные исследования (Cook et aL, 1998), в котором изучалась взаимосвязь между величиной локальной перфузии по данным ПЭТ и относительной мощностью ЭЭГ.
Часть 3. Расстройства систем мозга
469
1. Протоколы активации
Активирующий бета-тренинг — общепринятый метод, используемый для разнообразных целей. Протокол направлен на активацию определенной области коры. В нашей практике в процессе относительного бета-тренинга у детей с СНВГ мы иногда сталкиваемся с феноменом «ага», при котором инструктор вдруг обнаруживает конкретное ментальное состояние, связанное с генерацией высокоамплитудной бета-активности в лобных областях. Дети обычно
Корреляция с метаболической
Рисунок 23.7. Частотные диапазоны ЭЭГ в протоколах активации и релаксации
Схема показывает взаимосвязь между метаболической активностью и мощностью ЭЭГ в различных частотных диапазонах. В альфа-диапазоне (8-12 Гц) эта корреляция отрицательна, и поэтому альфа-тренинг является протоколом «релаксации». В то же время в бета-диапазоне корреляция позитивна, в связи с чем увеличение бета-активности представляет собой протокол «активации».
говорят: «Ага, теперь я знаю, что значит быть внимательным!» Бета-тренинг обычно выполняется в левом полушарии, поскольку в соответствии с теорией Дэвидсона (см. главу 13) активация левого полушария мозга, особенно лобной доли, связана с улучшением настроения и положительными эмоциями. Возможные негативные побочные эффекты могут включать возбуждение и раздражительность. Поэтому продолжительность сеанса может составлять лишь 10 минут, но все же обычно доходит до 20—30 минут. Часто бета-тренинг проводится в конце сеанса, чтобы вызвать у пациента состояние бодрости и энергичности136.
В другом типе активирующих протоколов в качестве параметра обратной связи используется когерентность. Когерентность — это индекс количественной ЭЭГ, отражающий корреляцию между показателями мощности ЭЭГ в определенном частотном диапазоне для двух отдельных регионов (см. методы 1-й части). Поданным наших исследований, у детей с СНВГ снижена когерентность в альфа-диапазоне в лобно-центральных областях136 137. СНВГ как расстройство системы управления,по-видимому, характеризуется нарушением координации в лобно-моторных зонах коры, что проявляется в снижении когерентности ЭЭГ в этих областях. То обстоятельство, что популяция СНВГ с проявлениями гипофронтальности характеризуется гипокогерентностью, может являться нейрофи-
136 Другой тип активирующих протоколов — это так называемые squash-протоколы, суть которых состоит в подавлении активности мозга в широком диапазоне. Предполагается, что при тренинге, направленном на подавление колебаний ЭЭГ (особенно высокоамплитудных), подкрепляется десинхронизированное поведение нейронов, связанное с активацией. Этот протокол может использоваться для любой области мозга, но некоторые специалисты придерживаются определенных зон, например лобных или центральных.
137 Один из особых видов альфа-активности в этих областях относится к мю-ритмам, отражающим холостой режим работы сенсомоторной области коры мозга человека.
470 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
энологической основой для протоколов «когерентности». Эти протоколы, по-видимому, связаны с активацией тренируемых зон.
2, Протоколы релаксации
Альфа-тренинг (8—12 Гц) — общепринятый метод, используемый для многих целей. Принимая во внимание то, что альфа-ритмы—это «холостые» ритмы мозга, мы можем заключить, что протоколы ЭЭГ-биоуправления, связанные с увеличением их мощности, это — релаксирующие протоколы, т.е. протоколы, направленные на деактивацию, торможение соответствующих зон коры. При релаксирующем тренинге часто предъявляются слуховые стимулы, поскольку зрительные стимулы активируют и десинхронизируют ЭЭГ в большей степени, чем слуховые.
3. Протокол Пенистона — Кулкоски
Протокол Пенистона — Кулкоски — это разновидность релаксирующего протокола, представляющая собой следующее. Сначала пациенты обучаются глубокой релаксации с помощью обратной связи по кожной температуре с использованием аутогенных приемов в течение не менее 5 сеансов. Во время последующих сеансов они занимаются ЭЭГ-биоуправлением с активным электродом в точке 01, при этом, находясь в состоянии релаксации с закрытыми глазах, они получают звуковые сигналы обратной связи. Для настроя на расслабление и погружение в себя используется стандартный текст. Когда уровень альфа-активности превышает установленный порог, предъявляется приятный тон, и при обучении произвольно вызывать этот тон пациент становится все более расслабленным. Когда амплитуда тета-волн (4—8 Гц) достигает достаточно высокого уровня, предъявляется другой тон, эффект релаксации усиливается, и пациент, согласно Пенистону, погружается в гипнагогическое состояние свободных ассоциаций и высокой внушаемости. Переход из состояния преобладания альфа-активности в состояние преобладания тета-активности называется перекрестом. Находясь некоторое время в тета-состоянии, пациенты могут быть чрезвычайно расслабленными и даже погрузиться в ступор. Впоследствии часто необходимо побеседовать с пациентом, чтобы убедиться в том, что он вышел из состояния повышенной сенситивности и погруженности в себя.
М. ЭЭГ-биоуправление и нейротерапия
В этой книге мы отделяем концепцию ЭЭГ-биоуправления от более общего понятия нейротерапии. Нейротерапия, помимо ЭЭГ-биоуправления, включает другие методы модуляции состояния мозга, основанные на электрофизиологии/Это транскраниальная микрополяризация, стимуляция переменным током, включающая глубинную стимуляцию мозга, транскраниальная магнитная стимуляция. ЭЭГ-БОС, в свою очередь, можно разделить на два подтипа: ЭЭГ-биоуправление и DC (или медленноволновая) БОС. ЭЭГ-биоуправление (как подтип ЭЭГ-БОС) — это метод коррекции или регулирования активности мозга с помощью электрофизиологической обратной связи. При такой обратной связи некоторые показатели ЭЭГ, например мощность ЭЭГ в конкретном частотном диапазоне, когерентность между корковыми областями или отношение «тета/бета», используются в качестве параметров БОС.
Часть 3. Расстройства систем мозга
471
Н. Восточные техники саморегуляции
Можно провести параллель между нейротерапией и восточными техниками саморегуляции. Древние восточные искусства саморегуляции, такие как йога и Ци-Гун138, в действительности базируются на способности человека сознательно влиять на внутренние психологические процессы. Однако для того чтобы овладеть этими искусствами, требуются большие затраты времени и энергии. Более того, часто необходимо непосредственное взаимодействие учителя и ученика. Говоря языком современной технологии ЭЭГ-биоуправления, в восточной технике саморегуляции и релаксации учитель (гуру) является своего рода агентом обратной связи. Процесс нейротерапии занимает существенно меньше времени и управляется обратной связью, исходящей непосредственно из мозга индивида. Более того, ЭЭГ-биоуправление не ограничивается протоколами релаксации.
О. Ложный эффект
Следует уделить особое внимание плацебо-эффекту нейротерапии (см. методы выше). Плацебо-эффект, как мы знаем из вышеизложенного, имеет определенные нейрональные механизмы, составляет до 30 % «чистого» эффекта фармакопрепаратов, и не имеет смысла отрицать его присутствие при ЭЭГ-биоуправлении. На ранних этапах становления нейротерапии плацебо-эффект неоднократно проверялся для двух протоколов — сенсомоторного тренинга и бета-тренинга. В настоящее время новые протоколы сравниваются не с плацебо-условием, а с ситуацией, в которой пациент принимает лекарство, эффективное при его заболевании. Это делается из этических соображений: было бы негуманно оставить пациента без лечения при наличии эффективных методов терапии.
П. Минимизация побочных эффектов
Как скальпель в хирургии, биоуправление может как лечить, так и наносить вред. Метод не может использоваться без знания о нарушениях работы мозга в каждом конкретном случае. Нейротерапия — это метод коррекции специфических симптомов, основанный на комбинации биоуправления и диагностической системы нейрометрики. Нейрометрика помогает установить электрофизиологические основы дисфункции мозга и вид протокола ЭЭГ-биоуправления139, показанного в конкретном случае.
138 Ци-Гун — китайская техника медитации. Ци означает -сила жизни-, а Гун — «работа», т.е. в переводе — работа ци. В отличие от западного здравоохранения Ци-Гун фокусируется на релаксации — ментальных упражнениях, а не на транировке мышц. Согласно китайской доктрине, Ци-Гун помогает снять -блоки-, мешающие циркуляции ци в теле человека, путем открытия -энергетических каналов-. По понятиям современной медицины, это не вполне ясное определение. Однако следует согласиться с тем, что мышцы, окружающие позвоночник, могут спазмироваться и находиться в этом состоянии долгое время. Этот спазм может приводить к разного рода проблемам — от головной боли до нарушений работы сердца. Следовательно, расслабление этих мышц будет благоприятным для пациента. Согласно дальнейшим рассуждениям, наше эмоциональное состояние тесно связано с соматическими ощущениями, иными словами, когда мы счастливы, мы улыбаемся, но когда мы улыбаемся — мы также можем получить ощущение счастья. Следовательно, тренинг эмоционального состояния также будет благоприятен для индивида.
ш Как и любой другой метод, биоуправление не может использоваться без предварительного диагностического обследования. Несмотря на то что сообщений о негативных последствиях применения биоуправления довольно немного, важно помнить, что изменение организации ЭЭГ-ритмов в неверном направлении может вести к дисрегуляции работы мозга и ухудшению клинической симптоматики.
472 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Поэтому нейротерапия может проводиться только сертифицированным специалистом, следующим базовым принципам нейротерапии на практике. Только в нескольких университетах проводятся курсы по количественной ЭЭГ и нейротерапии. Один из них находится в Техасе, США, другой — в Тронхейме, Норвегия140.
Одно из достоинств нейротерапии состоит в том, что побочные эффекты метода минимальны. Не секрет, что многие фармакопрепараты влияют не только на целевую нейротрансмиссию, но и на другие биохимические процессы в мозге и в организме в целом. Поэтому инструкция по применению любого лекарства включает длинный перечень побочных эффектов. По сравнению с фармакопрепаратами нейротерапия влияет на процессы мозга селективно. Более того, индивид активно участвует в процессе коррекции, сознательно или бессознательно избегая вредных эффектов с целью самосохранения. Поэтому о побочных воздействиях нейротерапии в литературе практически не упоминается, хотя это не значит, что они совсем не встречаются.
Р. Устойчивость эффекта
Другое преимущество нейротерапии состоит в устойчивости эффекта. Суть ней-ротерапевгической процедуры — это в действительности обучение. Эта процедура часто сравнивается с процессом приобретения специфического навыка, например умения ездил» на велосипеде. Когда человек впервые садится на велосипед, ему трудно держать равновесие, но после нескольких попыток он может с уверенностью держать руль; после довольно долгой тренировки некоторым даже удается ездить, не касаясь руля. Похожая ситуация складывается при нейротерапии: вначале индивиду сложно уловить связь между своими ощущениями и показателями биоэлектрической активности, предъявляемыми на экране. Но когда постепенно эта связь становится понятной, человека осеняет: «Ага, я знаю, как это делать!» Далее навык закрепляется, и ивдивцд обучается удерживать нужное состояние долгое время и не только в лабораторных условиях, но также дома и в школе. После 20—40 сеансов курс нейротерапии может быть завершен, но навык сохраняется на всю жизнь141.
С. Ограничения ЭЭГ-биоуправления
Означает ли вышесказанное, что у нейротерапии нет недостатков? Конечно, нет. Один из факторов, ограничивающих использование нейротерапии, — это большая продолжительность сеансов, занимающая от 20 до 40 часов рабочего времени клинициста, и такой же длительности интенсивный тренинг у пациента142. Помимо этого, перед началом курса нейротерапии техник должен заре
140 Как упоминалось выше, автор в настоящее время является профессором Норвежского университета науки и технологии в Тронхейме и проводит там курсы количественной ЭЭГ и нейротерапии.
141Д. Любар смог проследить 10-летний катамнеэ более чем 50 своих пациентов, используя объективную оценку поведенческих параметров. Положительные результаты лечения сохранялись в течение всего периода наблюдения почти у всех пациентов, однако некоторые из них нуждались в дополнительных сеансах для освежения в памяти полученных навыков, и после одной или двух процедур навык возвращался. Многие люди заявляют, что нейротерапия изменила их жизнь. В нашей работе мы смогли проследить эффект нейротерапии в течение одного года. Все дети сохранили показатели внимательности на высоком уровне. В нескольких случаях ослабления эффекта в результате физической травмы или заболевания было достаточно одного-двух сеансов для восстановлен ия прежних результатов.
142 Чтобы уменьшить потери времени, связанные с поездками в клинику, современные центры нейротерапии предлагают использовать так называемые домашние тренажеры. При этом несколько первых сеансов нейротерапий проводится совместно со специалистом и после приобретения пациентом первичных навыков тренинг продолжается дома. Пациент может купить или взять напрокат прибор для нейротерапии, который можно использовать в комплексе с домашним компьютером или телевизором.
Часть 3. Расстройства систем мозга
473
гистрировать ЭЭГ пациента. Затем подготовленный специалист устанавливает тип дисрегуляции мозговой деятельности143 и разрабатывает индивидуальный протокол нейротерапии. Это занимает 3—5 рабочих часов квалифицированных специалистов. На данное время в мире создано довольно небольшое количество приборов для нейротерапии с соответствующим программным обеспечением144. Для большинства из этих систем дополнительно требуется ЭЭГ-оборудование для регистрации многоканальной ЭЭГ для диагностических целей145.
Другой важный фактор для успешности нейротерапии — это мотивация пациента. Нейротерапия, как любая другая процедура обучения, базируется на настрое и мотивации. Чем выше уровень мотивации, тем выше приобретенный навык. В некоторых нейротерапевтических центрах мотивация у детей стимулируется с помощью призов в форме денежных вознаграждений, игрушек или жетонов, которые впоследствии можно обменять на деньги или игрушки. Важную роль в успешности нейротерапии играет помощь семьи. Действительно, в процессе нейротерапии мозг постепенно изменяется, но для того чтобы эти небольшие изменения привели к сдвигам поведенческого паттерна, требуются дополнительные усилия. Например, предполагается, что параметр внимания улучшается под влиянием относительного бета-тренинга, но внимательность — не единственный параметр, определяющий поведение. Самого по себе хорошего внимания недостаточно, чтобы дети могли использовать этот новый навык во время уроков в школе или лучше адаптировать свое поведение к общению с родителями или сверстниками. Некоторые пациенты с СНВГ, по-видимому, не могут сами изменить сложившийся имидж «трудного» ребенка и нуждаются для этого в помощи родителей и учителей. Чтобы помочь ребенку преодолеть эти социальные трудности, родителям может потребоваться специальный тренинг.
Т. Медицинское и немедицинское применение
Биоуправление используется как в лечебных, так и в нелечебных целях. В настоящее время Управление по контролю над пищевыми продуктами и лекарственными средствами США допускает использование ЭЭГ-биоуправления только в целях релаксации. Использование ЭЭГ-БОС в лечебных целях, например при депрессии, эпилепсии, головных болях, аутизме, рассматривается как эксперимент и считается необоснованным. Однако применение протоколов ЭЭГ-биоуправления в немедицинских целях для общего улучшения концентрации внимания и повышения успешности при различного рода деятельности не встречает возражений146.
143 Этот специалист должен уметь анализировать не только клиническую ЭЭГ с целью выявления в ней спайковой и другой пароксизмальной активности, но также должен владеть анализом количественной ЭЭГ, когнитивных ВП и основами нейротерапии, чтобы быть в состоянии оценить данные пациента и дать лечащему врачу рекомендации по фармако- и нейротерапии. Лишь несколько университетов в мире обучают студентов этим предметам.
144 Эти приборы производятся Thought ТесЬпоЬф, Daymed, Brain Inquiry, Brain Master и некоторыми другими компаниями.
146 В нашем институте (Институт мозга человека Российской академии наук) разработана многофункциональная систе-мя для анализа количественной ЭЭГ, когнитивных ВП, вызванной десинхронизации и для проведения нейротерапии. Система протестирована в Центре нейротерапии института и выпускается Российской производственной компанией «ООО Мицар» (С.-Петербург).
14в Любая ЭЭГ-система, созданная для тренинга в образовательных, развлекательных целях или для релаксации, — это не медицинский инструмент. FDA и аналогичные организации в других странах разрешают свободный выпуск и распространение таких приборов, пока они не представляют особого риска для здоровья пользователей.
474 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Любое оборудование для ЭЭГ-биоуправления может быть представлено в двух версиях и адресовано медицинскому и немедицинскому сообществам. Если оно предназначено для клинических целей, к нему предъявляются специальные требования, и эти требования должны быть рассмотрены и разъяснены FDA в США и подобными организациями — в других странах. В немедицинских объединениях это оборудование рассматривается в качестве приборов для обучения и отдыха147.
Опубликованные на настоящий момент исследования относятся к таким мозговым дисфункциям, как СНВ/СНВГ, расстройства поведения, трудности в обучении, тревожность, депрессия, синдром хронической усталости, эпилепсия, аутистические расстройства. Нейротерапия также используется с целью облегчения релаксации при аддиктивных расстройствах, при реабилитации после инсульта и травм мозга, а также для улучшения когнитивной деятельности при ее возрастных нарушениях. При немедицинском использовании ЭЭГ-биоуправление направлено на улучшение концентрации внимания и эффективности выполнения различной деятельности, помощь при медитации и релаксации, повышение личной осознанности и умственного здоровья.
У. Типы ЭЭГ-биоуправления
Термин «ЭЭГ-биоуправление» предполагает использование параметров ЭЭГ, таких как показатели спектров и когерентности, плотность источников тока в коре по данным LORETA или sLORETA. Теоретически любой физиологический параметр мозга может быть предъявлен пациенту и использован для изменения мозговой активности в нужном направлении. Перечень физиологических параметров, используемых в ЭЭГ-биоуправлении, представлен ниже в табл. 23.3.
7. ЭЭГ-БОС, основанная на когнитивных ВП
Априори компоненты когнитивных ВП в качестве показателей обработки информации могут использоваться в интерфейсе «мозг — компьютер». Однако технические сложности, такие как низкое отношение сигнала к шуму и отсутствие нормативной базы данных для компонент когнитивных ВП, были препятствием к использованию в интерфейсе «мозг — компьютер». Недавно в нашей лаборатории была создана первая версия программы в интерфейсе «мозг — компьютер» на основе когнитивных ВП (Kropotov, Murashev, 2007). Исходный пункт этого подхода — разложение когнитивных ВП на независимые компоненты148, характеризующиеся определенной топографией149 и временной динамикой. Топография необходима для создания пространственных фильтров для извлечения компонент из индивидуальных ВП. Следующий шаг — сравнение амплитуды или латентности индивидуальных компонент с нормативной базой данных. Статистически значимое отклонение от нормы определенной компоненты ис-
147 Подобно тому, как скальпель продается и для хирургических целей, и для использования в научной лаборатории. В обоих случаях используется один и тот же инструмент, но с разными намерениями.
148 Заметьте, что анализ независимых компонент с надежностью может применяться только при большом числе индивидуальных когнитивных ВП, что означает, что нормативная база данных должна включать как минимум несколько сотен здоровых испытуемых.
148 Топография компоненты представлена в виде sLORETA-изображения, дающего 3-мерную локализацию области коры, ответственной за генерацию этой компоненты.
Часть 3. Расстройства систем мозга 475
Таблица 23.3. Параметры ЭЭГ-биоуправления
Название Параметр ЭЭГ-биоуправления Примеры использования
ЭЭГ-БОС, основанная на вызванной десинхронизации Относительные изменения мощности ЭЭГ в конкретном диапазоне в ответ на реальные или воображаемые действия Pfurtscheller, 1944
ЭЭГ-БОС, основанная на когнитивных ВП Когнитивные ВП группы здоровых лиц разлагаются на независимые компоненты, характеризующиеся определенной топографией и временной динамикой. Топография необходима для создания пространственных фильтров для извлечения компонент из индивидуальных ВП. Амплитуда или латентность индивидуальных компонент сравниваются с нормативными данными и в дальнейшем используются в качестве параметров обратной связи Kropotov, Murashev, 2007
Саморегуляция вызванных медленных потенциалов коры Негативные и позитивные сдвиги медленных потенциалов коры в ответ на предупреждающие стимулы Birmbauer et al., 1991
Саморегуляция на основе фМРТ (функциональной магнитно-резонансной томографии Уровень гемодинамики в мозге на основе данных BOLD-ответа (ответа, зависящего от степени насыщения крови кислородом) Weiskopf et al., 2005
Саморегуляция на основе гемоэнцефалографии Уровень локальной оксигенации мозга, измеряемый с помощью активной или пассивной инфракрасной спектрофотометрии Toomim and Carmen, 1999
пользуется для выявления нарушений обработки информации в соответствующей области коры по данным sLORETA. Далее компонента может быть извлечена из многоканальной сырой ЭЭГ в каждой отдельной пробе и предъявлена пациенту. В течение последующих нескольких лет я планирую провести экспериментальное исследование по использованию этого метода у детей с СНВГ
2, Саморегуляция медленных потенциалов коры
Как нам известно, поверхностно-негативные медленные сдвиги потенциалов возникают на уровне апикальных дендритов коры и отражают синхронную деполяризацию апикальных дендритов нейронов коры. Эти негативные сдвиги потенциалов связаны с повышением возбудимости корковых нейронов150. Бирбаумер и его коллеги (1991) первыми предположили, что у пациентов с резистентными к лечению формами эпилепсии нарушена способность регулировать уровень возбудимости коры в корково-таламических петлях обратной связи. Исследования на здоровых испытуемых продемонстрировали способность человека обучаться саморегуляции медленных потенциалов коры. Это привело к мысли, что пациенты с эпилепсией могут овладеть навыком саморегуляции активности коры. Согласно этой гипотезе был разработан метод ЭЭГ-биоуправления, в котором реальные изменения медленных потенциалов коры предъявляются пациентам с эпилепсией в форме движущегося на экране объекта. В действительности, используя этот метод, большинство пациентов с устойчивой к лекарствам эпилепсией смогли научиться контролировать медленные потенциалы коры, что привело к значительному и длительному снижению частоты приступов (Birbaumeret al., 1991).
Похожая идея той же группы получила развитие при создании так называемого прибора трансляции мыслей, разработанного с целью восстановления способности к общению у пациентов с последствиями тяжелых инсультов. Ра
ш Вспомните, что эти негативные сдвиги регистрируются во время ожидания стимула и при подготовив к действию.
476 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
бота прибора основана на саморегуляции медленных потенциалов коры, т.е. на произвольных негативных и позитивных сдвигах потенциалов. После достижения пациентом надежного контроля над своими медленными корковыми потенциалами ответы могут использоваться для выбора изображений на экране компьютера. Программа позволяет пациенту выбирать отдельные буквы при последовательном предъявлении наборов букв пяти уровней, составленных по дихотомическому принципу. Несколько полностью парализованных пациентов с диагнозом бокового амиотрофического склероза могли составлять достаточно длинные сообщения с помощью потенциалов своего мозга151.
3. Биоуправление на основе фМРТ
В недавней работе исследователей из Тюбенгена (Weiskopf et al., 2005) был применен новый метод биоуправления. Метод осуществляет обработку данных фМРТ и предъявление пациенту показателя гемодинамики мозга в течение 1,3 с. Использование этой техники дает возможность осуществления дифференциальной обратной связи и саморегуляции, что было показано на примере дополнительной моторной зоны и парагиппокампальной области. Методология позволяет изучать поведенческие эффекты и локальную саморегуляцию у здоровых и больных лиц.
4. Биоуправление на основе гемоэнцефалографии
Термин «гемоэнцефалография» впервые был введен Туминым в 1997 г. в описании системы для инфракрасной спектрофотометрии. Другой тип системы — пассивная инфракрасная гемоэнцефалография — эволюционировал из применения технологии инфракрасного излучения для биоуправления по периферической температуре (Carmen, 2002). В обеих системах источником данных является динамика кровотока. Система пассивной инфракрасной гемоэнцефалографии была специально разработана для вмешательства при мигрени. Концепция тренинга цереброваскулярной регуляции для контроля над мигренью базировалась на предположении, что обучение контролю цереброваскулярной реактивности (особенно чрезмерной вазодилатации) должно облегчить симптомы мигрени. К сожалению, обучение непосредственному сужению чрезмерно расширенных цереброваскулярных структур не работало. Большинство испытуемых с легкостью обучались повышать сигнал пассивной инфракрасной гемоэнцефалографии, но только некоторые могли его снижать. Поэтому процедура была модифицирована для тренинга на повышение, а не снижение параметров гемоэнцефалографии. Эффект биоуправления на основе гемоэнцефалографии был позитивным как для профилактики мигрени, так и при купировании приступов этого заболевания.
161В работе исследователей из Тюбенгена (Hinterberge et al., 2005) взаимосвязь между негативными и позитивными медленными потенциалами и изменения BOLD-сигнала фМРТ изучались у испытуемых, успешно обучившихся саморегуляции своих медленных потенциалов. Поданным фМРТ генерация негативности (повышенной возбудимости коры) сопровождалась широко распространенной активацией центральных, префронтальных и теменных областей коры, а также базальных ганглиев. Позитивность (сниженная возбудимость коры) связывалась с широко распространенной деактивацией в нескольких участках коры, а также некоторой активацией преимущественно лобных и теменных структур и, кроме того, островковой доли и скорлупы. С помощью регрессионного анализа было обнаружено, что корковая позитивность предсказывалась активацией бледного шара и скорлупы и деактивацией дополнительной моторной эоны и моторной коры, при этом дифференциация между корковой негативностью и позитивностью выявлялась главным образом в парагиппокампальном регионе. Согласно этим данным, предполагается, что негативные и позитивные сдвиги корковых потенциалов в ЭЭГ связаны с отчетливыми и распространенными различиями паттернов церебральной активации/деактивации.
Часть 3. Расстройства систем мозга 477
Ф. Интерфейс «мозг — компьютер»
Если процесс модуляции активности мозга направлен на контроль внешних приборов, то такой подход называется «интерфейс «мозг — компьютер». Иногда он также называется прямым нейральным интерфейсом или интерфейсом «аппарат — мозг»152. В этом определении под словом «мозг» подразумевается мозг или нервная система, а под словом «компьютер» (аппарат) — любой прибор для обработки и расчета данных: от простых схем до сложных компьютеров. Исследования на тему «интерфейс «мозг — компьютер» начались в 1970-х, но первый работающий экспериментальный имплант у человека появился только в середине 1990-х. Существующие сейчас импланты для человека разработаны для восстановления слуха, зрения и движений. Общепринятая идея, стоящая за этим исследованием, сходна с базовыми идеями ЭЭГ-биоуправления — это удивительная пластичность коры мозга. Как и при ЭЭГ-БОС, в интерфейсе «мозг — компьютер» может использоваться любой физиологический параметр. Из них наиболее изученными являются электрические потенциалы, записанные со скальпа. Например, Н. Бирбаумер из университета г. Тюбинген в Германии использовал электроэнцефалографические записи медленных потенциалов коры, чтобы дать возможность парализованным пациентам контролировать курсор компьютера. Д. Бейлис в университете Рочестера показала, что добровольцы, надев шлемы «виртуальной реальности», могли контролировать элементы виртуального мира с помощью компонент РЗОО когнитивных ВП. Г Пфуртшеллер из Технического университета Граза в Австрии использовал различные параметры ЭЭГ для управления протезами у пациентов с параплегиями или для прогулок по виртуальному городу у здоровых испытуемых (Leeb, Pfurtscheller, 2004).
III. дубинная стимуляция мозга
А. Психохирургия
Когда в середине 20-х годов XX столетия возникла психохирургия как метод хирургического лечения тяжелых психических расстройств, единственным доступным для нейрохирургов способом модуляции активности мозга была полная деструкция целевого участка нервной ткани. От Буркхарда к Фултону, Мо-ницу и Фриману накапливался опыт резекции ткани лобной доли для лечения психических расстройств. Однако неразборчивое использование грубых и обширных хирургических вмешательств часто вело к страшным последствиям, таким как полное изменение личности и нарушения интеллекта153. Появление эффективной фармакотерапии в 1960-х годах, казалось бы, явилось панацеей. Увы, это было не так. Множество пациентов по-прежнему страдает от психических
ш Вообще термин -интерфейс -мозг — компьютер» можно определить как прямой путь сообщения между мозгом человека или животного (или культурой клеток мозга) и внешним прибором. В односторонних интерфейсах «мозг — компьютер» компьютеры или принимают команды из мозга (например, двигать протезом), или посылают сигналы в мозг (например, послать зрительную информацию к нейронам коры у слепых). Двухсторонние интерфейсы «мозг — компьютер» могли бы позволить мозгу и внешнему прибору обмениваться информацией в обоих направлениях.
153 В середине 1950-х годов только в США было выполнено более 20 000 фронтальных лоботомий. Хотя некоторым пациентам операция помогла, многие из них пострадали. Поэтому во многих странах эта практика запрещена законом. Таким образом, рождение и последующая кончина первого этапа психохирургии в начале и середине XX столетия в дальнейшем обусловили более осторожный подход к вмешательствам в мозг с целью облегчения симптомов трудноизлечимых психических заболеваний.
478
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
расстройств без надежды на улучшение, кроме как с помощью хирургического вмешательства.
Б. Стереотаксическая нейрохирургия
Хотя развитие стереотаксиса во 2-й половине XX столетия позволило нейрохирургам усовершенствовать вмешательства, сделав их более точными, все же разрушение ткани мозга оставалось единственным способом изменения функций нервной системы. Стереотаксическое разрушение, хотя и является эффективным, рассчитано на оказание длительного воздействия и поэтому связано с риском развития длительных побочных эффектов. При введении электродов в мозг остается слишком малое пространство для ошибок, и если они случаются, то нет другого выхода, кроме как другое или еще большее разрушение154.
В. ГСМ как обратимая деструкция
Экспериментальный поиск методов обратимого выключения локальной активности мозга начался в 1960-х годах. В отделе нейрофизиологии человека Института экспериментальной медицины (где автор работал в 1970—1980-х) мы использовали методы стимуляции прямым током и высокочастотной стимуляции для временного выключения активности нейронов вокруг электродов, имплантированных для диагностических и терапевтических целей в мозг неврологических пациентов155. Однако лишь недавно, с развитием новых электронных технологий, глубинная стимуляция мозга была введена в рутинную клиническую практику и привела к коренным изменениям нейрохирургических методов (рис. 23.8). ГСМ сначала применялась в лечении двигательных расстройств, а сейчас используется при резистентных формах психических заболеваний.
По сравнению с нейрохирургическими процедурами глубинная стимуляция мозга имеет ряд преимуществ. Во-первых, эффекты, вызванные ГСМ, полностью обратимы. Во-вторых, параметры стимуляции можно подобрать индивидуально, в соответствии с изменением симптоматики и динамикой заболевания пациента. В-третьих, можно включать или выключать стимуляцию, не сообщая об этом пациенту, что открывает уникальную возможность для двойных слепых исследований. Наряду с достоинствами у ГСМ есть некоторые недостатки, в первую очередь связанные с необходимостью длительного нахождения импланта в мозге, что создает опасность инфицирования.
154 Пациент остается жить как с положительными, так и с отрицательными последствиями однажды выполненной операции. Тем не менее профили результатов и побочных эффектов при современных вмешательствах в целом благоприятны.
155 Хирургические вмешательства при неврологических и психических заболеваниях должны выполняться яо строгим соблюдением мер безопасности для обеспечения правильности их выполнения. В случае психического расстройства симптоматика пациента должна соответствовать критериям конкретного психического заболевания поданным DSM-IV или ICD-10 (например, ОКР или большой депрессии). Далее кандидат должен отвечать строго определенным критериям тяжести заболевания, хронического течения, инвалидности и резистентности к лечению. Например, перед тем как рассматривать возможность хирургического лечения, пациент должен безрезультатно пройти несколько курсов медикаментозного лечения, включающего разнообразные препараты в комбинации с соответствующей психотерапией.
Часть 3. Расстройства систем мозга
479
Источник переменного юкя
IV. "фанскраниальная магнитная стимуляция
А. Физические основы ТМС
Транскраниальная магнитная стимуляция впервые была применена А. Баркером в Шеффилдском университете в Великобритании в 1985 г. Затем ТМС стала использоваться в качестве неинвазивного, безопасного и безболезненного метода активации моторной коры человека и для оценки целостности центральных двигательных путей. Со времени возникновения ТСМ использование этого метода в клинической нейрофизиологии, неврологии и психиатрии приобрело широкое распространение, в основном — в исследовательских работах, но с растущей тенденцией его применения в клинической практике. Однако на настоящее время не собрано достаточно данных для выработки объективных показаний к систематическому применению ТМС в качестве терапевтического инструмента.
ТМС базируется на принципе электромагнитной индукции, открытом Фарадеем в 1838 г Если импульс электрического тока, проходящего через катушку, размещенную вокруг головы индивида, имеет достаточную величину и короткую продолжительность, он генерирует быстро меняющиеся магнитные импульсы, проникающие через скальп и кости черепа в мозг с не
значительным ослаблением. Эти магнитные импульсы, в свою очередь, вызывают вторичные ионные токи в мозге156 (рис. 23.8).
Б. Диагностическое и терапевтическое применение
Воздействие ТМС на разных уровнях двигательной системы может дать информацию о возбудимости моторной коры, функциональной целостности внутрикорковых нейронных структур, проводимости кортико-спинальных,
156 Способность ТМС к деполяризации нейронов зависит от-функции активации», которая обусловливает трансмембранные токи и может быть описана математически как пространственная производная электрического поля вдоль нерва. В зависимости от направления вызванного в мозге тока ТМС будет активировать пирамидные клетки косвенно (те. транссинаптически) или прямо, воздействуя на аксонные холмики и вызывая прямые волны. При ТМС у быстрых аксонов (> 75 м/с) отмечается более низкий порог к прямым волнам, а у медленных (< 55 м/с) — к непрямым.
ТМС
Рисунок 23.8. (дубинная стимуляция мозга (ГСМ) и транскраниальная магнитная стимуляция (ТМС)
А. ГСМ — переменный ток подается через имплантированные электроды с целью включения и выключения (в зависимости от частоты тока) нейронной активности поблизости от электрода. Б. ТМС — электрический импульс в катушке генерирует переменное магнитное поле, индуцирующее ток в ткани.
480 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия кортико-нуклеарных и каллозальных волокон, а также о функции корешков и периферических двигательных путей, идущих к мышцам157. Примеры находок в такого рода исследованиях могут помочь в локализации уровня поражения в пределах нервной системы, дифференциальной диагностике между преимущественно демиелинизирующими или аксональными поражениями моторных трактов или в предсказании функциональных двигательных нарушений вследствие травмы.
Для повышения эффективности магнитной стимуляции ТМС может применяться повторно, с частотой от 1 до более чем 20 импульсов в секунду. Эффект повторной ТМС может варьировать от торможения к облегчению в зависимости от параметров стимуляции. Низкочастотная ТМС может уменьшать возбудимость двигательной коры, в то время как стимуляция с частотой 20 Гц, по-ввдимому, ведет к временному повышению ее возбудимости. Эффект повторной низкочастотной ТМС продолжительный и выраженный. Этот факт открывает возможности для терапевтического применения повторной ТМС с целью «нормализации» патологически сниженного или повышенного уровня активности коры.
У.Ткэанскраниальная микрополяризация
А. История
Из основ физиологии мы знаем, что постоянный ток (DC)158 смещает мембранный потенциал нейрона в сторону или гипо-, или гиперполяризации в зависимости от направления тока. Хотя о влиянии электрического тока на живую ткань было известно еще столетия назад159, только в середине XX века начались систематические исследования этого феномена. Данные о том, что воздействие слабого постоянного тока может вызывать стойкие изменения возбудимости мозга, открыли новые возможности в коррекции неврологических нарушений. Однако даже теперь, когда накоплено достаточно данных по поляризации мозга в экспериментах на животных, очень мало известно о том, каким образом слабый постоянный ток, подаваемый через скальповые электроды, влияет на возбудимость мозга у человека. В настоящее время процедура называется транскраниальной микрополяризацией.
Слабые постоянные токи при ТКМП лишь незначительно изменяют мембранные потенциалы нейронов коры. Эти токи гораздо меньшей величины, чем
157 При воздействии ТМС соответствующей интенсивности на моторную кору моторные вызванные потенциалы (МВП) можно зарегистрировать с мышц контралатеральных конечностей. Двигательный порог представляет собой наименьшую интенсивность воздействующего на моторную кору одиночного импульса ТМС, необходимую для вызова МВП в целевой мышце. Считается, что двигательный порог отражает возбудимость мембраны кортико-спинальных нейронов и межнейронных проекций к этим нейронам в моторной коре, а также возбудимость двигательных нейронов в спинном мозге, нервно-мышечных соединениях и мышцах.
158 DC — поток электрических зарядов, не меняющий направления. Он может быть вызван при приложении двух полюсов электрической батареи к ткани мозга.
150 Идея использования электрического тока для лечения мозговых расстройств была настолько простой, что ее возникновение относится к 43-48 г. до нашей эры, когда Скрибониус Ларгус наблюдал, как при прикладывании живой рыбы Торпедо, вырабатывающей электрический ток высокой мощности, к голове пациента, страдающего головной болью, у него возник внезапный прекодящий ступор с последующим исчезновением боли. Нейрофизиологи относятся с особым уважением к электрической рыбе Торпедо, поскольку ее систематическое изучение, начатое Уолшем в 1773 гч послужило отправным пунктом электрофизиологии. Спустя несколько лег итальянские ученые Гальвани и Вольта приступили к исследованию влияния электричества на живые органы. Вскоре гальванические токи (DC) стали применяться в клинической медицине, а именно при расстройствах умственной деятельности. В1804 г. племянник Гальвани, Альдини, сообщил об успешном лечении пациентов, страдающих от меланхолии, с помощью гальванических токов, приложенных к голове.
Часть 3. Расстройства систем мозга 481
при так называемой электроконвульсивной терапии. ЭКТ была разработана в 1930-е годы итальянскими учеными Бини и Черлетги и в те времена казалась значительным прорывом в контроле над умственными расстройствами. ЭКТ вызывает заметное и устойчивое улучшение состояния пациентов. Этот метод коренным образом отличается от ТКМП. В то время как при ЭКТ мощный ток вызывает судорожную активность в мозге, ТКМП индуцирует токи гораздо меньшей величины, мягко модулирующие мозговые функции, изменяя спонтанную нейронную активность160, не вызывая судорожных приступов. Полученные на настоящее время сведения указывают на то, что в отличие от ЭКТ ТКМП не вызывает нарушений памяти или утраты сознания, и при этом пациенту не требуется принимать седативные средства или миорелаксанты. ТКМП — настолько мягкое воздействие, что пациенты субъективно не могут отличить ощущения, вызываемые ТКМП, от плацебо.
Б. Процедура
Во время сеанса ТКМП прямой электрический ток небольшой силы воздействует на мозг с помощью двух размещенных на скальпе электродов: один из них — «стимулирующий», расположенный над областью, которую предполагается стимулировать, а другой — «референт», расположенный над «немой» областью на голове или теле, например на мочке уха или плече (рис. 23.9). Два электрода, соединенные соответственно с положительным и отрицательным полюсами источника DC (например, батареей) и расположенные на скальпе, созда-
Рисунок 23.9. Транскраниальная микрополяризация (ТКМП)
Справа — схема ТКМП: два электрода закреплены на голове, электрический ток подается от прибора, питающегося от батареи. Обычно ток не превышает 1 мА, причем лишь небольшая его часть поступает к серому веществу. В слоях коры «анодный» ток деполяризует пирамидные клетки на уровне базальной мембраны (см. направление тока в пирамидном нейроне внутри коры).
“° Хотя поляризующие токи не могут вызвать возбуждение «молчащих» клеток, они несколько изменяют поляризацию нейронной мембраны, что в результате коллективного кумулятивного эффекта обеспечивает модуляцию спонтанных клеточных разрядов.
482
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
ют электрическое поле и, в соответствии с законом Ома, электрические токи на протяжении этого поля.
В. Нейрофизиологические механизмы поляризации мембраны
Лишь небольшая часть электрического тока проходит через кору. Нейроны коры ориентированы перпендикулярно к поверхности, так что апикальные и базальные дендриты располагаются в разных слоях. Электрическое поле и соответствующий ток под анодом вызывают дефицит положительных ионов в базальном отделе нейронной мембраны, что ведет к деполяризации этого отдела. Деполяризация активирует потенциалзависимые Са++- и Ка+-каналы, что приводит к повышению возбудимости нейрона. Вследствие повышения возбудимости возрастает фоновая активность деполяризованных нейронов. Сетевой эффект состоит в так называемой анодной активации нейронов. Напротив, ток под катодом приводит к избытку положительных ионов у внешней части базальной мембраны, что ведет к гиперполяризации этого участка. Гиперполяризация инактивирует Са++- и Na+- каналы. Возбудимость нейрона снижается, и частота фоновой активности уменьшается. Сетевой эффект проявляется в подавлении нейронной активности под катодом.
Несмотря на то что изменения потенциала базальной мембраны под действием постоянного тока величиной 1 мА слишком малы, чтобы вызвать спайковую активность, они запускают процессы усиления в нейронных сетях. Эти факторы усиления в конечном счете индуцируют выраженные физиологические эффекты. Нам известен по меньшей мере один фактор усиления в нервной системе — коллективный эффект, обусловленный возвратными возбуждающими связями в коре. В возвратных сетях небольшие изменения мембранного потенциала могут вести к повсеместной спайковой нейронной активности. С этой точки зрения ТКМП можно рассматривать как технику подпороговой стимуляции, модулирующей спонтанную кортикальную активность и таким образом вызывающую преходящие функциональные изменения в мозге человека. ТКМП вызывает минимальный дискомфорт без негативных ощущений. Одно из недавних исследований (Gandiga et al., 2006) показало, что плацебо (ситуация, в которой исследователь собирался включить прибор, но в действительности не сделал этого) неотличимо от ТКМП. Последнее обстоятельство означает, что ТКМП — прекрасный инструмент для двойных слепых клинических исследований с использованием плацебо-контроля.
В начале 1970-х автор сотрудничал с группой, проводившей исследования по ТКМП в Институте экспериментальной медицины Академии медицинских наук СССР. Эти работы проводились в отделе нейрофизиологии человека на пациентах с имплантированными в лечебных и диагностических целях электродами. Было замечено, что под воздействием анодного тока повышалась активность нейронных популяций, в то время как под влиянием катодного тока она снижалась. Постоянный ток под катодом использовался с целью диагностики для временного выключения нейронных сетей вокруг электрода перед принятием решения о дальнейшем разрушении данного участка мозга. В 1970—80-х
Часть 3. Расстройства систем мозга
483
эксперименты по поляризации ткани мозга в Институте экспериментальной медицины были продолжены на кошках и собаках. Эти эксперименты доказали, что анодные токи оказывают активирующее действие на нейроны, в то время как катодные тормозят их активность. В 1980-х метод поляризации стал использоваться в нескольких клиниках Ленинграда для лечения неврологических и психиатрических пациентов. К сожалению, несколько трагических событий (смерть двух ведущих ученых, развал Советского Союза и последующее прекращение выделения средств на фундаментальные исследования) помешали продвижению этих перспективных работ. Только в начале XXI века эти исследования продолжились. В настоящее время методы поляризации успешно используются в коррекции поведения при СНВГ и при лечении речевых задержек у детей. В течение последних 5 лет интерес к ТКМП возобновился и в западном мире. Возрождение метода было отчасти связано с поиском альтернативных (нефармакологических) методов стимуляции мозга161.
Г. Физиологические основы
Из нейрофизиологических исследований нам известно, что медленные изменения потенциалов, записанных со скальпа, отражают распространенные сдвиги мембранных потенциалов нейронов коры. Идея о том, что модуляция этих медленных потенциалов с помощью тока может влиять на физиологические процессы в мозге, была столь очевидной, что в 1960—1970-х годах в нескольких лабораториях это предположение было протестировано. Вопрос заключался в том, достаточно ли изменения потенциалов на уровне отдельных нейронов для модуляции нейронной возбудимости. Доказательство того, что подача прямого тока с помощью электродов, наложенных на скальп, может влиять на возбудимость мозга, было получено на пациентах с височной эпилепсией при записи разности потенциалов с имплантированных для предоперационной диагностики электродов в процессе поляризации прямым током через скальп с использованием силы тока до 1,5 мА. (Dymond et al., 1975). Под влиянием тока такой величины градиент потенциала в коре колебался от 6,4 до16,4 мВ/см. Таких изменений градиента потенциала вполне достаточно для модуляции спонтанных нейронных разрядов, как это было показано Терцуоло и Буллоком (1956) в экспериментах на препаратах из речного рака и омара.
Одно из прямых доказательств эффективности поляризации прямым током недавно было получено в экспериментах с использованием изменений мышечных подергиваний, вызванных ТМС, в качестве индикаторов возбудимости коры. В этих исследованиях анод (или катод) размещался над моторной корой. Эффект ТКМП тестировался с помощью регистрации мышечных потенциалов, вызванных импульсами транскраниальной магнитной стимуляции. На рис 23.10 видно, что изменения возбудимости моторной коры, вызванные 5-минутным воздействием ТКМП, сохраняются в течение только 5 минут. Дальнейшее исследование показало, что увели
Исследовательские группы, в настоящее время занимающиеся ТКМП, работают в Национальном институте исследования неврологических расстройств и апоплексии, NIH, в Бетезде (США), Гарвардском центре неинвазивной стимуляции мозга в Востоке (США), Институте неврологии, Университетском колледже в Лондоне (Англия), Университете здоровья в Японии, Миланском университете в Италии и в нескольких университетах Германии: Университете Геттингена, Университете Любека, Универси-1Я1« Киля, Университете Эберхарда-Карла в Тюбингене.
484 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
чение продолжительности ТКМП ведет к продлению (др часа) эффекта. Следует отметить, что изменения возбудимости коры имеют локальный характер. Например, ТКМП только моторной, но не затылочной или контралатеральной коры оказывала активирующее или тормозное влияние на моторную кору162.
Рисунок 23.10. Специфичность влияния ТКМП на возбудимость двигательной коры в зависимости от полярности
Ось Y — относительная амплитуда двигательного потенциала, вызванного ТМС после ТКМП двигательной коры в течение 5 минут. Ось X — время после завершения ТКМП. <1оловы> вверху справа схематически изображают процедуру ТКМП и ТМС. Заимствовано из Nitsche and Paulus (2001).
Д. Поведенческие эффекты
Исторически применение ТКМП началось на здоровых добровольцах. В одной из работ; выполненной в 1964 е (Uppold et al., 1964), прямой ток силой 0,5 мА подавался с помощью двух электродов, расположенных в области лба, и одного — на правом колене. В исследовании показано, что под влиянием анодных токов повышались уровень бодрствования, настроение и двигательная активность, в то время как катодная поляризация вызывала состояние покоя и апатии. Однако потребовалось почти 40 лет для того, чтобы ТКМП вновь стала инструментом для исследований. В работе, выполненной группой ученых из университета в Геттингене (Antal, Nitsche and Paulus, 2001), было обнаружено, что стимуляция постоянным катодным током величиной 1мА в затылочной области значительно снижала контрастную чувствительность во время и непосредственно после процедуры163. Следует заметить, что во всех последних работах проводилось сравнение эффектов ТКМП и плацебо.
ТКМП считается довольно безопасной процедурой. Ни в одном из исследований не сообщается о побочных эффектах.
Ницше и Паулюс (2001) обнаружили отсутствие изменений сывороточной концентрации нейронспецифйчной энолазы, чувствительного маркера нейронных поражений, после применения постоянного тока. Авторы считают безопасной процедуру ТКМП с использованием силы тока 1 мА в течение нескольких минут при площади электродов 3,5 см2 (Nitsche, Paulus, 2000,2001)l64.
162 В работе, выполненной группой ученых из Университета Геттингена (Antal et al, 2005), анализировалась активность ЭЭГ в зрительной коре здоровых испытуемых после недельного курса ТКМП. Исследование показало, что катодная стимуляция значимо уменьшает, а анодная — незначительно увеличивает нормированную мощность активности в бета- и гамма-диапазонах. Это наблюдение согласуется с предположением о том, что анодная стимуляция повышает метаболическую активность соответствующей области коры, в то время как катодная стимуляция оказывает на кору тормозное воздействие.
183 Метод идеально подходил для этих целей. Процедура ТКМП сопровождается лишь слабыми ои^щениями при резком включении тока, при постепенном же повышении его интенсивности она не вызывает никаких ощущений. В условиях плацебо обычно ток сначала включается (для имитации процедуры ТКМП) на несколько секунд а затем медленно выключается.
184 Наш собственный опыт показывает, что применение ТКМП с силой тока 0,3-0,7 мА в течение 20-40 минут при площади электрода 4 см2 может вызывать умеренно выраженные побочные эффекты в виде повышения возбудимости, нарушения сна и головных болей.
Часть 3. Расстройства систем мозга
485
Е. Применение в клинике
Параллельно с экспериментальными работами с участием здоровых испытуемых проводились исследования метода на пациентах. В одной из ранних работ было показано, что применение транскраниальной анодной микрополяризации вызывало улучшение настроения у пациентов с депрессией (Costain at al., 1964). В наших исследованиях, выполненных в Институте мозга человека, было показано, что ТКМП положительно влияет на симптоматику СНВГ.
Экспериментальные находки, обнаруженные группой исследователей из Геттингенского университета при применении ТКМП в области двигательной коры, стимулировали применение этого метода в реабилитации пациентов с последствиями инсультов. В недавней работе Хаммель и сотр. (2005) ТКМП успешно использовалась в лечении пациентов с последствиями инсультов, сопровождавшихся двигательными нарушениями, спустя 1—2 года от начала заболевания165. Позитивные сдвиги при выполнении движений, возникавшие в процессе ТКМП, сохранялись по меньшей мере в течение 25 минут после заверше-
Рисунок 23.11. ТКМП у пациента с сенсорным дефицитом
А. sLORETA-изображения ВП пациента, зарегистрированных в разное время и в различных условиях (до, после). Б. Когнитивные ВП в ответ на GO-стимулы у пациента с сенсорным дефицитом др (тонкая линия) и после (пунктирная линия) нескольких сеансов ТКМП в сравнении с когнитивными ВП, рассчитанными для группы здоровых лиц того же возраста (жирная линия). В. Расположение электродов. Белым цветом схематично изображена область поражения.
Прямой ток подавался через два электрода (площадью 2,5 см2), покрытых губкой, пропитанной солевым раствором. Анод помещался на скальп над проекцией представительства кисти в области первичной моторной коры пораженного полушария, а катод— не кожу над контралатеральным супраорбитальным регионом. В процессе сеанса ТКМП анодный ток подавался в течение 20 минут, в при сеансе плацебо—не дольше 30 с.
486 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
ния сеанса и возвращались к исходному уровню лишь через несколько дней.
В наших опытных исследованиях166 мы использовали ТКМП при лечении трех пациентов, перенесших инсульт с поражением правой теменной доли, сопровождавшийся сенсорным дефицитом. На рис. 23.11 представлены данные одного из пациентов. Как видно, компонента Р1 когнитивных ВП генерируется асимметрично в левой затылочной области с отклонениями от нормы в правой затылочной области. Компонента N1 также генерируется асимметрично, но ла-теральнее и спереди, в височной области и отсутствует справа. Третья, поздняя компонента с латентностью 300 мс была значительно увеличена справа167. В результате сеансов ТКМП значительно уменьшилась латентность компоненты N1 слева и произошло смещение генератора поздней позитивной компоненты. Что касается психологических характеристик, то после курса ТКМП пациенты значительно лучше выполняли тесты с оценкой эффекта литерализации168. Необходимы дальнейшие клинические исследования, но представленные данные свидетельствуют о возможном терапевтическом эффекте ТКМП и о пользе оценки компонент когнитивных ВП для контроля эффективности лечения.
ш Работа была выполнена в Центре реабилитации в Мунквол в городе "фонхейм (Норвегия). Нейрофизиологи Ян Бруно и К. Хестад проводили психологическое обследование и тестирование пациентов до, во время и после 20-минутного сеанса ТКМП.
107 Увеличение компоненты РЗ справа объясняется растормаживанием эон коры вокруг области поражения. Вспомните, что латеральное торможение — одно из свойств кортико-таламических путей, и при поражении определенной эоны коры тормозные влияния на нее снимаются, что растормаживает, в свою очередь, окружающие участки.
ш Один из пациентов получил водительские права, что для него было невозможно в течение 2 лет после инсульта.
Заключение
I. Общие принципы анализа ЭЭГ и нейротерапии
Ниже мы обобщаем основные принципы методологии оценки мозговых функций и дисфункций, представленных в этой книге. Эта методология была разработана в лаборатории, возглавляемой автором, в Институте мозга человека Российской академии наук и основана на фундаментальных и прикладных исследованиях, выполненных в лаборатории в течение последних 30 лет. Эта методология была испытана в Центре нейротерапии института при оценке и лечении тысяч пациентов с различными мозговыми дисфункциями, такими как СНВГ, задержки речевого развития, ОКР, аддиктивные расстройства, эпилепсия.
Принцип 1
Колебания ЭЭГ и когнитивные вызванные потенциалы связаны с различными и не зависящими друг от друга процессами в мозге: 1) колебания ЭЭГ отражают процессы саморегуляции в коре и связаны с модулированием потока информации в корковоподкорковых нейронных сетях; 2) когнитивные ВПсвязаны с потоком сигналов, вызванных внешним событием, и отражают реакции мозга нй них в частности, извлечение физических, семантических и эмоциональных параметров стимула, выбор действий и управление.
Эти два независимых процесса исследуются различными методами. Спонтанные колебания ЭЭГ обычно разделяются на частотные составляющие с помощью анализа Фурье, в то время как их динамика в процессе выполнения тестов оценивается с помощью волнового (вейвлет) анализа. Для вычисления ВП используется техника усреднения: спонтанные колебания, не связанные с ответом на стимул, взаимно погашаются. Следовательно, усредненные потенциалы не зависят от фоновой активности и отражают поток сигналов в нейронных сетях мозга.
Принцип 2
Колебания ЭЭГ и когнитивные ВП делятся на отдельные компоненты. Компоненты спектров связаны с различными ритмами ЭЭГ, каждый из которых отражает различные нейронные механизмы корковой регуляции. Компоненты когнитивных ВПсвязаны с различными психологическими операциями, такими как обработка первичной сенсорной информации, извлечение семантического значения сигналов и их пространственная локализация, кодирование эмоциональных реакций и ощу-
488 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
щений, определение физических и семантических отличий, выбор и подавление действий, контроль результатов выполнения или подавления действий.
Для разделения компонент активности ЭЭГ и когнитивных ВП могут использоваться разные методы. Один из современных методов — анализ независимых компонент.
Анализ независимых компонент может применяться как к данным сырой ЭЭГ, так и к «очищенным» спектрам мощности. В активности здорового мозга анализ независимых компонент позволяет выделить несколько типов «нормальных» ритмов ЭЭГ, таких как теменной и затылочный альфа-ритмы, мю-ритмы, бета-ритмы и лобный тета-ритм средней линии. Нормальные ритмы генерируются различными механизмами и отражают различные свойства модуляции нейронных сетей. Альфа-ритмы генерируются реципрокными связями в кортико-таламических сетях и отражают «холостой» режим работы сенсорных систем. Бета-ритмы обусловлены взаимодействием возбуждающих и тормозных связей в кортикальных сетях и выполняют функцию обновления (reset) для удаления следов предшествующей активации коры. Лобный тета-ритм средней линии генерируется септально-гиппокампальными кругами и связан с процессами кодирования эпизодической памяти. Любое повреждение нейронных кругов может вызвать снижение или повышение мощности нормального ритма или появление патологической активности.
Анализ независимых компонент можно использовать для единичных проб нативной ЭЭГ, а также при анализе однородных групп индивидуальных ВП пациентов или здоровых испытуемых. Извлеченные компоненты когнитивных ВП отражают этапы обработки сенсорной информации, операции сравнения и контроля, вовлечения в действие и его подавления, а также некоторые другие психологические процессы.
Принцип 3
В состав мозга входят основные системы: сенсорная система зрительной, слуховой, соматосенсорной модальностей, система внимания, эмоциональная система, системы управления и памяти. Модуляция потока информации в каждой из этих систем характеризуется специфичными ритмами, в то время как стадии обработки информации в этих системах отражены в специфичных компонентах когнитивных ВП.
В табл. 3.1,3.2 представлено краткое описание ритмов ЭЭГ и компонент когнитивных ВП, их локализации, механизмов генерации, функционального значения и связи с системами мозга.
Принцип 4
Использование различных тестов и модальностей позволяет исследовать функции практически всех областей мозга. Для каждого пациента выбор теста определяется на основе его жалоб и соответствующих нейропсихологических нарушений.
Этот принцип отражен на рис. 3.1. Мы выбрали только наибольшую по величине из компонент, извлеченных в ответ на зрительные стимулы в трех раз-
Заключение
489
Таблица 3.1. Нормальные ритмы мозга человека
Название Чаете та (ГЦ) Топография Механизм генерации Функция Система мозга
Затылочный альфа-ритм 8-12 01,02 Са++-спайки, следующие за гиперполяризацией таламокортикальных клеток латерального коленчатого тела Отражает «холостой» режим работы первичных зрительных областей. Первичная зрительная информация игнорируется, но кора в состоянии готовности к ее немедленной оценке Сенсорная система
Теменной альфа-ритм 8-12 Pz Са++-спайки, следующие за гиперполяризацией таламокортикальных клеток в подушечном ядре таламуса Отражает «холостой» режим работы вторичных зрительных областей Сенсорная система
Мю-ритм 9-13 СЗ, С4 Са++-спайки, следующие за гиперполяризацией таламокортикальных клеток в заднем вентральном ядре Отражает «холостой» режим работы соматосенсорной системы Сенсорная система
Бета-ритмы 13-30 Лобные или центр, отделы Взаимодействие между возбуждающими и тормозными связями внутри кортикальных сетей в состоянии деполяризации Связана с обновлением (reset) следовой активации коры Любая система
Лобный срединный тета-ритм 5-8 Fz Деполяризация в септальногиппокампальных кругах в ответ на привлекающие внимание стимулы Отражает перевод информации в эпизодическую память Система эпизодической памяти
личных тестах (двухстимульный GO/NOGO-тест, чтение и математический тест) из нормативной базы данных HBI. Примите во внимание, что почти вся поверхность коры покрыта генераторами компонент, расположенными над различными кортикальными областями — от затылочной до передней цингулярной коры. Заметьте также, что мозг — это нейронная сеть с мощными связякш, в которой нейроны разных областей коры функционируют одновременно при выполнении определенной операции. Изображение sLORETA, представленное на рис. 3.1, показывает только вершину айсберга широко распространенной активности коры.
Принцип 5
Каждая из систем мозга подчиняется закону перевернутой U-кривой. Закон гласит, что ответы системы максимальны, когда ее активность соответствует среднему уровню, и анормально снижаются, если уровень активации системы ниже или выше среднего.
Уровень активации системы можно оценить с помощью спектрального анализа спонтанной ЭЭГ, генерируемой кортикальным отделом системы. В целом
490
Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Таблица 3 2. Основные компоненты когнитивных ВП
Название Тест Латентность пика (мс) Топография Генератор Функциональное значение Система мозга
Негативность рассогласования Слуховой oddball-тест 140 Fz Височная или лобная кора Автоматическое сравнение регулярных акустических стимулов с сенсорным следом Сенсорная система
Р1 и N1 Любое предъявление стимула 100 Зависит от модальности стимула Области по соседству с первичными сенсорными зонами Обработка информации в сенсорных системах Сенсорная система
РЗЬ Активный oddball-тест любой модальности 300 Pz Широко распространенные круги: «базальные ганглии — таламус — кора» Активация передних и задних областей коры, необходимая для выполнения действия Система управления
Р2 сравнения Дифференциальные тесты 240-300 Т5,Т6зри-тельн., F7, F8 слуховой Ассоциативные зрительные и слуховые зоны Активация нейронов, выявляющих отличия физических или семантических характеристик повторяющихся стимулов Сенсорная система
Р400 NOGO GO/NOGO 400 Fz Передняя цин-гулярная кора и медиальная префронтальная кора Операция мониторинга, т.е. активация нейронов, сравнивающих выполненное действие с подготовленным Система управления
РЗа Трехстимульный oddball-тест 260-300 Fz Обширные зоны префронтальной коры Смещение внимания, ориентировочная реакция Система управления
избыточная, по сравнению с нормой, альфа-активность связана с гипоакгива-цией системы, а избыточная бета-активность — с ее гиперактивацией. Ответы системы отражены в амплитуде и латентности компонент, генерируемых кортикальными областями системы1.
Принцип 6
Нарушения мозговой деятельности можно классифицировать на основе их связи с расстройствами мозговых систем и диагностировать согласно с отклонениями от нормы соответствующих параметров спектров ЭЭГ и когнитивных ВП.
Каждая из существующих в настоящее время систем классификации, таких как DSM-IV или ICD-10, основана на описании поведения пациентов и поэтому субъективна. В качестве возможных объективных физиологических маркеров заболеваний — эндофенотипов — были предложены некоторые компоненты количественной ЭЭГ и когнитивных ВП. Несмотря на то что необходимы более систематические исследования, первые результаты вполне обнадеживающие (табл. 3.3).
1 Например, аномально высокая амплитуда бета-активности, генерируемой в медиальной префронтальной коре, указывает на чрезмерную активацию этого отдела системы управления, в то время как аномальное снижение компонента мониторинга РЗ свидетельствует об уменьшении реактивности этой системы в ситуациях, где необходим контроль действий.
Заключение
491
Кодирование первичных свойств
GONOGO-тест
Нерелевантный стимул пробы
Кодирование формы
GO/NOGO-тест
Второй нерелевантный стимул пробы
Кодирование слова Задача на чтение Первый стимул пробы
Кодирование цифры Математический тест Первый стимул пробы
Детекция изменений GO/NOGO-тест Разностный ВП между GO-и NOGO-условиями
Вовлечение в задание (РЗЬ)
GO/NOGO-тест
GO-стнмул
Подавление действия GO/NOGO^recT NOGO-стимул
Мониторинг действия GO/NOGO-тест NOGO-стимул
Рисунок 3.1. Независимые компоненты в ответ на зрительные стимулы
Максимальная амплитуда независимых компонент в ответ на зрительные стимулы в трех тестах, использованных в нормативной HBI базе данных: зрительном GO/NOGO-тесте, тесте с чтением и математическом тесте. Слева направо: 1) топография компонент; 2) кривые активации компонент; 3) sLORETA-изображения корковых генераторов компонент.
492 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
Таблица 3.3. Классификация заболеваний мозга и их эндофенотипов
Диагностическая категория DSM-IV Примеры расстройств Система мозга ЭЭГ-эндофенотип Эндофенотип по когнитивным ВП
Расстройства, обычно впервые диагностируемые в младенческом, детском и подростковом возрасте СНВГ, аутизм, расстройства обучения, расстройства поведения Система управления Повышение отношения тета/бета в лобно-центральных отделах Уменьшение компонента сравнения Р2 и компонента мониторинга Р4
Психотические расстройства Шизофрения Система управления Увеличение бета-активности в лобных отделах Уменьшение компонента РЗЬ
Эмоциональные расстройства Большая депрессия, биполярное расстройство Эмоциональная система Асимметрия альфа-активности в лобных отделах слева > справа Увеличение когнитивных ВП на негативные стимулы, асимметрия когнитивных ВП
Тревожные расстройства ОКР, генерализованное тревожное расстройство, посттравматическое стрессовое расстройство Система управления Увеличение бета-активности в центральных отделах Уменьшение компонента мониторинга Р4
Деменция, бредовые, амнестические и другие когнитивные расстройства Болезнь Альцгеймера Система эпизодической памяти Увеличение бета-активности в лобноцентральных отделах Уменьшение компонента РЗЬ и компонента мониторинга Р4
Аддиктивные расстройства Героиновая аддик-ция, алкоголизм Эмоциональная система и система управления Увеличение бета-активности в центральных отделах Уменьшение компонента мониторинга Р4
Принцип 7
ТКМП и ЭЭГ-биоуправление представляют собой основанные на электрофизиологии инструменты для активации или подавления кортикальных нейронных сетей.
При ТКМП используются слабые постоянные токи, деполяризующие (анодные токи) и гиперполяризующие (катодные токи) пирамидные клетки коры под стимулирующим электродом. В процессе ЭЭГ-биоуправления индивид активно вовлечен в изменение параметров ЭЭГ, зарегистрированной данным электродом. Наши исследования показали, что при комбинации этих двух методов можно достичь лучших результатов в активации или подавлении пораженной системы мозга.
II. Предметы дальнейших исследований
Методы количественной ЭЭГ и когнитивных ВП в оценке функций и дисфункций мозга, описанных в книге, представляют собой относительно новую, но быстро развивающуюся область фундаментальных и клинических исследований. Методы коррекции дисфункций мозга, основанные на электрофизиологии, такие как ТКМП и ЭЭГ-биоуправление, находятся пока на этапе становления. Они ограничены рамками лабораторий и рассматриваются в основном в качестве экспериментальных методов. Ниже мы представляем темы наиболее значимых, с нашей точки зрения, проектов в данной области, которые необходимо выполнить в ближайшем будущем.
Заключение
493
Тема 1. Создание стандартных парадигм для оценки систем памяти и эмоций
Методы оценки функционирования мозговых систем внимания и управления, представленные в книге2, могли бы послужить образцами в создании стандартизованных парадигм для оценки систем памяти и эмоций. Например, на основе стандартной GO/NOGO-парадигмы можно предложить новый тест для оценки эмоциональной системы мозга. В этом тесте изображения грустных, радостных и нейтральных выражений лиц могли бы предъявляться вместо изображений растений, животных и людей, используемых в двухстимульном GO/NOGO зрительном тесте базы данных HBI Database. Задача пациента состояла бы в распознавании эмоций по выражению лица и нажатии на кнопку при предъявлении изображений двух радостных лиц3. Различия в реакции ЭЭГ на предъявление радостных и грустных лиц, так же как различия между реакциями на радостные/грустные и нейтральные стимулы, могли бы служить показателями работы эмоциональной системы мозга. Эти параметры могли бы служить эндофенотипами аутизма, депрессии и других эмоциональных расстройств.
Тема 2. Регистрация независимых компонент когнитивных ВП в поведенческих тестах
Анализ независимых компонент, применяемый к ряду индивидуальных ВП многочисленной группы здоровых испытуемых, представляет собой мощный инструмент создания пространственных фильтров для извлечения компонент ВП, отражающих основные психологические процессы, происходящие в мозгу человека. Независимые компоненты, извлеченные в ответ на зрительные стимулы в трех тестах базы данных HBI Database (GO/NOGO, математический и чтение), представлены на рис. 3.1. Обратите внимание на то, что компоненты, полученные в условиях трех тестов, связаны с активацией различных областей коры. Например, нейроны, кодирующие информацию о первичных зрительных признаках, форме, арифметических свойствах и письменных словах, находятся в разных (практически не перекрывающихся) задних отделах коры, в то время как нейроны, отвечающие за психологические операции с участием действий (например, подавление и контроль действий), локализованы в лобноцентральных отделах мозга.
Подобный подход может быть использован к когнитивным ВП, рассчитанным для гомогенной группы испытуемых в других поведенческих парадигмах4. Программное обеспечение базы данных HBI Database предоставляет пользователю все вычислительные процедуры для выполнения этого методологического подхода, включая разработку протоколов, регистрацию и хранение записей ЭЭГ во встроенной базе данных, автоматическую
2 Это oddbal, трехстимульный и GO/NOGO-тесты, которые в настоящее время можно рассматривать как стандартные и хорошо изученные парадигмы.
3 Другим выбором могла бы стать стандартизация существующих парадигм, например создание тестов «старое/новое» или «по-ложительное/отрицательное» для изучения систем памяти и эмоций соответственно.
4 В электрофизиологических лабораториях всего мира отдается предпочтение собственным поведенческим парадигмам, разработанным на местах для определенных научных целей. Повсеместное использование одних и тех же поведенческих парадигм представляется невозможным, но обеспечение всех лабораторий единым инструментом для их применения в исследовании работы мозга вполне осуществимо.
494 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
коррекцию артефактов, усреднение ВП и подсчет числа ошибок в тестах, выполнение анализа независимых компонент над группой вызванных потенциалов, расчет LORETA и sLORETA-изображений извлеченных компонент.
Тема 3. Использование независимых компонент ВП в качестве эндофенотипов в диагностике различных заболеваний мозга
В этой книге мы обсуждали возможность использования параметров когнитивных ВП и количественной ЭЭГ в качестве маркеров различных психоневрологических расстройств, таких как СНВГ, ОКР, шизофрения, депрессия. Однако методология, представленная в книге, все еще находится на стадии становления. Через несколько лет после создания нормативной базы данных HBI Database нами были собраны и проанализированы спектры ЭЭГ и когнитивные независимые компоненты в различных группах пациентов. Эти данные продолжают накапливаться в различных научных и клинических центрах всего мира. Таким образом, этот проект начал осуществляться. Следует отметить, что основная его проблема состоит в поиске пациентов, не использующих лекарственную терапию, в частности, пациентов с депрессией, которые никогда не принимали антидепрессанты5. Заметьте также, что количество пациентов с каждым отдельным заболеванием составляет приблизительно 1000, т.е. сопоставимо с числом здоровых испытуемых. Таким образом, предполагается, что проект по этой теме объединит усилия различных центров в создании единой базы данных количественной ЭЭГ/когнитивных ВП при патологии, которая будет включать несколько тысяч пациентов различных категорий.
После создания базы данных при патологии следующей ступенью стал бы поиск эндофенотипов, специфичных для определенных заболеваний или групп заболеваний. Эндофенотипами не обязательно должны быть отдельные параметры количественной ЭЭГ или когнитивных ВП6. Вполне возможно, что разделение между различными видами расстройств можно произвести с помощью так называемых дискриминационных плоскостей в многомерном пространстве параметров. Каждая дискриминационная плоскость может представлять собой линейную комбинацию параметров с определенными весами. Первая попытка дифференцирования специфического расстройства от нормы была предпринята Р. Джоном в 1980-х7 и позднее продолжена Б. Тэтчером. Однако заметьте, что во всех этих случаях дифференциация проводилась только для отдельных расстройств, например определение отличий количественной ЭЭГ пациентов с черепно-мозговой травмой от количественной ЭЭГ здоровых индивидов в базе данных Тэтчера. Пока нет возможности объединить данные ЭЭГ пациентов с отдельным заболеванием.
5 Следует заметить, что при сборе данных в детской популяции имеется больше шансов найти пациентов с СНВГ, диелексией и трудностями в обучении, не принимающих лекарств.
8 Даже если мы будем использовать независимые компоненты и для ЭЭГ, и для когнитивных ВП.
7 К сожалению, использование параметров количественной ЭЭГ ограничивалось несколькими стандартными диапазонами при регистрации ЭЭГтолько при закрытых глазах.
Заключение
495
Тема 4. Использование эндофенотипов по данным когнитивных ВП в фармакотерапии (фармо-ВП)
Эта тема предполагает изучение влияния различных антипсихотических и нейротропных средств на независимые компоненты когнитивных ВП. Используя такой подход, мы получим ключи к правильному использованию различных фармацевтических средств при разных расстройствах, связанных с нарушениями отдельных компонент когнитивных ВП. Источники этой темы относятся к фармако-ЭЭГ, возникшей в 1950—1960-х. Однако большое разнообразие спектральных характеристик ЭЭГ у здоровых людей вносит некоторые ограничения в использование подхода, основанного на количественной ЭЭГ. Применение когнитивных ВП, и особенно компонент когнитивных ВП, открывает новые горизонты использования физиологических методов для контроля эффектов фармацевтических средств на мозг человека.
Тема 5. Использование двойных слепых плацебо-контролируемых исследований для демонстрации поведенческих и физиологических изменений, вызванных ТКМП и ЭЭГ-биоуп равнением
Хотя несколько предпринятых в ряде лабораторий попыток таких исследований дали положительные результаты, работы были выполнены на ограниченном числе пациентов. Более того, в большинстве исследований использовались одни и те же протоколы для одних и тех же групп пациентов вне зависимости от типа расстройства по данным количественной ЭЭГ и когнитивных ВП. Как показано в этой книге, протоколы для ЭЭГ-биоуправления и ТКМП должны основываться на анализе количественной ЭЭГ и когнитивных ВП и соответствовать паттернам специфических дисфункций. Следует подчеркнуть, что ТКМП и ЭЭГ-биоуправление представляют собой два разных подхода, направленных на достижение одной цели — активации или подавления соответствующей системы мозга. По-видимому, комбинация этих двух методов может способствовать достижению лучших клинических результатов. Действительно, комбинируя эти методы, мы используем: 1) способность индивида произвольно изменять состояние собственного мозга; 2) способность нейронов мозга к пластическим изменениям под влиянием электрических токов.
Литература
1. Гнездицкий В.В. Вызванные потенциалы мозга в клинической практике. — Таганрог: Изд-во ТРТУ, 1997. — 252 с.
2. Гречин В.Б., Кропотов Ю.Д. Медленные неэлектрические процессы головного мозга человека. — Л.: Наука, 1979. — 128 с.
3. Кропотов Ю.Д., Пономарев В.А. Нейрофизиология целенаправленной деятельности. —- Л.: Наука, 1993. — 130 с.
4. Кропотов Ю.Д. Мозговая организация восприятия и памяти: гипотеза программирования действий // Физиология человека. — 1989. — № 3. — С. 19.
5. Кропотов Ю.Д. Роль стриатума в селекции действий // Физиол. журн. — 1997.-№ 1.-С. 45-48.
6. Лурия АР. Основы нейропсихологии. — М.: Изд-во МГУ, 1973. — 374 с.
7. Переслени Л.И., Рожкова ЛА. Психофизиологические механизмы дефицита внимания у детей разного возраста с трудностями обучения // Физиология человека. — 1994. — № 4. — С. 5-9.
8. Смирнов В.М. Стереотаксическая неврология. — М.: Медицина, 1976. — 264 с.
9. Фарбер Д.А., Дубровинская Н.В. Функциональная организация развивающегося мозга (возрастные особенности и некоторые закономерности) // Физиология человека. — 1991. — № 5. — С. 17.
10. ШтаркМ.Б., СкокАБ. Применение электроэнцефалографического биофидбека в клинической практике: (Лит. обзор) // Биоуправление-3: Теория и практика. — Новосибирск, 1998. — С. 131-148.
11. Alexander G.E., DeLong M.R., Strick P.L. Parallel organization of functionally segregated circuits linking basal ganglia and cortex // Annu Rev. Neurosci. 1986.9: 357-381.
12. Allan K., Rugg M.D. An erent-related potential study of explicit memory on tests of word-stem cued recall and recognition memory // Neuropsychologia. 1997.35:387-397.
13. Anichkov AD, Polonsky J.Z., Nizkovolos V.B. Stereotactic Systems. «Nauka» («Science») Publishing House, Saint-Petersburg. 2005.142 p. (In Russian).
14. Anokhin AP., Heath AC., Myers E. Genetic and environmental influences on frontal EEG asymmetry: a twin study // Biol. Psychol. 2006. 71(3): 289-295. Epub 2005 Jul 28.
15. Antal A, Nitsche M.A, Paulus W. External modulation of visual perception in humans//Neuroreport. 2001.12:3553-3555.
16. Aron A.R., Robbins T.W., Poldrack R.A Inhibition and the right inferior frontal cortex//TRENDS in Cognitive Sciences. 2004.8(4): 170-176.
17. Asada H., Fukuda Y, Tsunoda S., Yamaguchi M., Tonoike M. Frontal midline theta rhythms refect alternative activation of prefrontal cortex and anterior cingulate cortex in humans // Neuroscience Letters. 1999.274:29-32.
18. Aston-Jones G., Cohen J.D. An Integrative Theory of Locus Coeruleus-Norepineph-rine Function: Adaptive Gain and Optimal Performance //Annu. Rev. Neurosci. 2005. 28:403-450.
Литература 497
19. Atwater F.H. The Hemi-Sync Process // The Monroe Institute. 1997. http://www. monroeinstitute.com/content.php?content_id=24
20. Baddeley A Working Memory: Looking Back and Looking Forward // Nat. Rev. Neurosci. 2003.4:929-939.
21. Baddeley AD., Hitch G.J. Working memory. — In: G.A Bower. Ed.: Recent advances in learning and motivation, vol. 8. Academic Press, New York. 1974.47-90.
22. Ballantine H.T. Jr, Bouckoms AJ., Thomas E.K., Giriunas I.E. Treatment of psychiatric illness by stereotactic cingulotomy// Biol. Psychiatry. 1987.22(7): 807-819.
23. Ballantine H.T., Giriunas I.E. Treatment of intractable psychiatric illness and chronic pain by stereotactic cingulotomy. —- In: Schmidek HH and Sweet WH (eds). Operative Neurosurgical Techniques. Grune& Stratton, New York. 1982. 1069-1075.
24. Banaschewski T, Brandeis D., Heinrich H, Albrecht B., Brunner E., Rothenberger A Questioning inhibitory control as the specific deficit of ADHD — evidence from brain electrical activity//J Neural Transm. 2004. Ill: 841-864.
25. Barry R.J., Clarke AR., Johnstone S.J. A review of electrophysiology in attention-deficit/hyperactivity disorder I. Qualitative and quantitative electroencephalography//Clinical Neurophysiology. 2003. 114:171-183.
26. Barry R.J., Clarke AR., Johnstone S.J. A review of electrophysiology in attention-deficit/hyperactivity disorder I. II. Event-related potentials // Clinical Neurophysiology. 2003.114: 184-198.
Tl. Bartfeld E., Grinvald A Relationships between Orientation-Preference Pinwheels, Cytochrome Oxidase Blobs, and Ocular-Dominance Columns in Primate Striate Cortex. Proceedings of the National Academy of Sciences of the United States of America. 1992.89(24): 11905-11909.
28. Bazhenov M., Timofeev I., Steriade M., Sejnowski TJ. Model of thalamocortical slow-wave sleep oscillations and transitions to activated states//J. Neurosci. 2002.22:8691-8704.
29. Beauducel A, Brocke B., Leue A Energetical bases of extraversion: effort, arousal, EEG, and performance // Int J. Psychophysiol. 2006.62(2): 212-223. Epub. 2006 Jan 19.
30. Benedetti F, Colloca L., Torre E., Lanotte M., Melcame A, Pesare M., Bergamas-co B., Lopiano L. Placebo-responsive Parkinson patients show decreased activity in single neurons of subthalamic nucleus // Nat. Neurosci. 2004.7:587-588.
31. Benedetti E, Mayberg H.S., Wager T.D., Stohler C.S., Zubieta J-K. Neurobiological Mechanisms of the Placebo Effect//J. Neurosci. 2005.25(45): 10390-10402.
32. Bland B.H., Oddie S.D. Theta band oscillation and synchrony in the hippocampal formation and associated structures: the case for its role in sensorimotor integration // Behavioural Brain Research. 2001.127:119-136.
33. Bramon E., Rabe-Hesketh S., Sham P., Murray R.M., Frangou S. Meta-analysis of the P300 and P50 waveforms in schizophrenia // Schizophrenia Research. 2004.70:315-329.
34. Broadbent D.E. The role of auditory localization in attention and memory span // J. Exp. Psychol. 1954.47(3): 191-196.
35. Bucci P., Mucci A, \blpe U., Merlotti E., Galderisi S., Maj M. Executive hypercontrol in obsessive-compulsive disorder electrophysiological and neuropsychological indices//Clinical Neurophysiology. 2004. 115:1340-1348.
36. Buzsdki G. Rhythms of the brain. Oxford University Press. 2006.448 p.
37. Buzsaki G. Two-stage model of memory formation: a role for noisy brain states // Neuroscience. 1989.31:551-570.
498 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
38. Calcagnotto М.Е., Paredes М.Е, Tihan Т, Barbara N.M., Baraban C.S. Dysfunction of Synaptic Inhibition in Epilepsy Associated with Focal Cortical Dysplasia//J. Neu-rosci. 2005.25(42): 9649-9657.
39. Cantero J.L., Atienza M., Stickgold R., Kahana M.J., Madsen J.R., Kocsis B. Sleep-Dependent Oscillations in the Human Hippocampus and Neocortex // J. Neurosci. 2003.23(34): 10897-10903.
40. Carmen J. A. Passive Infrared Hemoencephalography, 4 Years and 100 migraines later. Presented at 2002 Society for Neuronal Regulation conference, Scottsdale, AZ. 2002.
41. Castellanos F.X., Tannock R. Neuroscience of attention-deficit/hyperactivity disorder the search for endophenotypes // Nature Reviews. 2002.3:617-628.
42. Chapman RM., Nowlis G.H., McCrary J.W, Chapman JA, Sandoval TC., Guiltily M.D., Gardner M.N., Reilly LA Brain event-related potentials: Diagnosing eariy-stage Alzheimer’s disease//Neurobiology of Aging. 2007.28(2): 194-201. Epub 2006 Jan 20.
43. Colom L.V. Septal networks: relevance to theta rhythm, epilepsy and Alzheimer’s disease//Journal of Neurochemistry. 2006.96:609-623.
44. Congedo M., Lubar J.F., Joffe D. Low-Resolution Electromagnetic Tomography Neurofeedback // IEEE Transactions on Neural Systems and Rehabilitation Engineering. 2004.12(4): 387-397.
45. Cook I.A, O’Hara R., Uijtdehaage S.H., Mandelkem M., Leuchter AF. Assessing the accuracy of topographic EEG mapping for determining local brain function // Electroe ncephalogr Clin. Neurophysiol. 1998.107(6): 408-414.
46. Costain R., Redfeam J.W., Lippold O.C. A controlled trial of the therapeutic effects of polarization ofthe brain in depressiw illness //Br. J. Psychiatry. 1964.110:786-799.
47. Crawford H.J. Brain dynamics and hypnosis: attentional and disattentional processes//Int. J. Clin. Exp. Hypn. 1994.42(3): 204-232.
48. Damasio AR. The Feeling of What Happens. Body and Emotion in the Making of Consciousness // Harcourt Brace, New York, 1999.
49. Davies P, Maloney AJ.F. Selective loss of central cholinergic neurons in Alzheimer’s disease//Lancet. 1976.2(8000): 1403.
50. Donchin E. Surprise! Surprise // Psychophysiology. 1981.18(5): 493-513.
51. Donchin E., Coles M.G. Is the P300 component a manifestation of context updating// Behavioral Brain Science. 1988.11:357-374.
52. Douglas R.J., Martin KA A functional microcircuit for cat visual cortex // J. Physiol. 1991.440:735-769.
53. Douglas R. J., Martin KA, Whitteridge D. A canonical microcircuit for neocortex // Neural. Comp. 1989.1:480-488.
54. Dymond AM., Coger R.W., Serafetinides EA Intracerebral current levels in man during electrosleep therapy//Biol. Psychiatry. 1975.10(1): 101-104.
55. Ebmeier K.P, Donaghey C., Steele J.D. Recent developments and current controversies in depression//Lancet. 2006.367:153-167.
56. Edelman G.M. Neural Darwinism: The Theory of Neuronal Group Selection. Basic Books, New York. 1987.
57. Edelman G.M. Neural Darwinism: Selection and reentrant signaling in higher brain function//Neuron. 1993.10:115-125.
58. Ekman P, Friesen W.V. Pictures of Facial Affect. Palo Alto, CA Consulting Psychologists. 1976.
Литература 499
59. Elston G.N. Cortex, Cognition and the Cell: New Insights into the Pyramidal Neuron and Prefrontal Function// Cerebral Cortex. 2003.13:1124-1138.
60. Engel A.K., Singer W. Temporal binding and neuronal correlates of sensory awareness//TRENDS in Cognitive Sciences. 2001. 1(1): 16-25.
61. EigenogluT., DemiralpT., BayraktarogluZ., EigenM., Beydagi H., UresinY. Alpha rhythm of the EEG modulates visual detection performance in humans // Cognitive Brain Research. 2004.20:376-383.
62. Eriksen C.W., Eriksen BA. Target redundancy in visual search: do repetitions of the target within the display impair processing? // Percept Psychophys. 1979.26:195-205.
63. Evarts E.V., Shinoda Y, Wise S.P. Neurophysiological approaches to higher brain functions // N.Y, Wiley, 1984.
64. Fallgatter AJ., Strik WK. The NoGo-anteriorisation as a neurophysiological standard-index for cognitive response control//Int. J. Psychophysiol. 1999.32:115-120.
65. Fell J., Klaver P., Lehnertz K., Grunwald T, Schaller C., Eiger C.E., Fernandez G. Human memory formation is accompanied by rhinal-hippocampal coupling and decoupling//Nat. Neurosci. 2001.4:1259-1264.
66. Fernandez G., Effem A, GrunwaldT., PezerN., Lehnertz К, DUmpelmann M.,\hn Roost D., Eiger C.E. Real-time tracking of memory formation in the human rhinal cortex and hippocampus // Science. 1999.285:1582-1585.
67. Fink M., Sackeim H.A. Convulsive therapy in schizophrenia? // Schiz. Bull. 1996. 22:27-39.
68. Ford J.M. Schizophrenia: The broken P300 and beyond // Psychophysiology. 1999. 36:667-682.
69. Forman S.D., Dougherty G.G.; Casey B.J., Siegle G.J., Braver T.S., Barch D.M., Stenger VA, Wick-Hull C., Pisarov L.A, Lorensen E. Opiate Addicts Lack Error-Dependent Activation of Rostral Anterior Cingulate // Biol. Psychiatry. 2004.55:531-537.
70. Fox M.D., Raichle ME. Spontaneous fluctuations of brain activity observed with functional magnetic resonance imaging // Nature Reviews. Neuroscience. 2007. V. 8, 700-711.
71. Foxe J.J., Simpson G.V. Flow of activation from VI to frontal cortex in humans: A framework for defining «early» visual processing //Exp. Brain. Res. 2002.142:139-150.
72. Fulton J.E. Frontal lobotomy and affective behavior a neurophysiological analysis. WW Norton, New York, 1951.
73. Gallinat J., Bottlender R., Juckel G., Munke-Puchner A, Stotz G., Kuss H.J. The loudness dependency of the auditory evoked N l/P2-component as a predictor of the acute SSRI response in depression// Psychopharmacology. 2000,148(4): 401-411.
74. Gandiga P.C., Hummel F.C., Cohen L.G. Transcranial DC stimulation (tDCS): a tool for double-blind sham-controlled clinical studies in brain stimulation // Clin. Neurophysiol. 2006.117(4): 845-50. Epub 2006 Jan 19.
75. Gevins A, Smith M.E., McEvoy L., Yu D. High-resolution EEG Mapping of Cortical Activation Related to Wbrking Memory: Effects of Task Difficulty, Type of Processing, and Practice I I Cerebral. Cortex. 1997.7:374-385.
76. Gloaguen V., Cottraux J., Cucherat M., Blackbum I.M. A metaanalysis of the effects of cognitive therapy in depressed patients // J. Affect. Disord. 1998.49:59-72.
77. Gottesman I.I., Gould T.D. The endophenotype concept in psychiatry, etymology and strategic intentions //Am. J. Psychiatry. 2003.160(4), 636-645.
500 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
78. Gnizelier J., Hardman Е., Wild J., Zaman R. Learned control of slow potential interhemispheric asymmetiy in schizophrenia//Int. J. Psychophysiol. 1999.34(3): 341-348.
79. Guaranha M.S.B., Garzon E., Buchpiguel CA, Tazima S., Yacubian E.M.T., Sakamoto AC. Hyperventilation revisited: Physiological effects and efficacy on focal seizure activation in the era of video-EEG monitoring // Epilepsia. 2005.46(1):69-75.
80. Gumenyuk V., Korzyukov O., Escera С.» Hamalainen M., Huotilainen M., Hayrinen T., Oksanen H., NWdnen R., vonWfendt L., Alho K. Electrophysiological evidence of enhanced distractibility in ADHD children // Neurosci Lett. 2005. 374(3): 212-217. Epub 2004 Nov 24.
81. Hagemann D. Individual differences in anterior EEG asymmetry: methodological problems and solutions// Biological Psychology. 2004.67:157-182.
82. Haider B., Duque A, Hasenstaub AR., McCormick DA Neocortical Network Activity In Vivo Is Generated through a Dynamic Balance of Excitation and Inhibition The Journal of Neuroscience. 2006.26(17): 4535-4545.
83. Hari R., Salmelin R. Human cortical oscillations: a neuromagnetic view through the skull// Trends Neurosci. 1997.20(1): 44-49.
84. Hebb D.O. The Organization of Behavior. Wiley, New York, 1949.
85. Heimer L., Vhn Hoesen G.W The limbic lobe and its output channels: implications for emotional functions and adaptive behavior // Neuroscience and Biobehavioral Reviews. 2006. 30:126-147.
86. Henriques J.B., Davidson R.J. Left frontal hypoactivation in depression // Journal of Abnormal Psychology. 1991.100:535-545.
87. Hensler J.G. Serotonergic modulation of the limbic system // Neuroscience and Biobehavioral Reviews. 2006.30:203-214.
88. Herrmann M.J., Jacob C., Unterecker S., Fallgatter AJ. Reduced response-inhibition in obsessive-compulsive disorder measured with topographic evoked potential mapping I I Psychiatry Research. 2003.120:265-271.
89. Hillyard SA, Anllo-Vento L. Event-related brain potentials in the study of visual selective attention// Proc. Natl. Acad. Sci. 1998. USA 95:781-787.
90. Hirvonen J., van ErpTG.M, Huttunen J., Aalto S., NJgren K, Huttunen M., Lonnqvist J., Kaprio J., Hietala J., Cannon T.D. Increased Caudate Dopamine D2 Receptor Availability as a Genetic Marker for Schizophrenia//Arch Gen Psychiatry. 2005.62:271-278.
91. Huang-Pollock C.L, Nigg J. T. Searching for the attention deficit in attention deficit hyperactivity disorder the case of visuospatial orienting // Clin. Psychol. Rev. 2003. Nov; 23(6): 801-30.
92. Huerta P.T., Lisman J.E. Heightened synaptic plasticity of hippocampal CAI neurons during a cholinergically induced rhythmic state // Nature. 1993.364(6439): 723-725.
93. Hughes S.W., Cope D.W., Blethyn K.L., Crunelli V. Cellular mechanisms of the slow (< 1 Hz) oscillation in thalamocortical neurons invitro/l Neuron. 2002.33(6): 947-958.
94. Hughes S.W., Lorincz M., Cope D.W., Blethyn K.L., Kekesi KA, Parri H.R., Ju-hasz G., Crunelli V. Synchronized oscillations at alpha and theta frequencies in the lateral geniculate nucleus // Neuron. 2004.42(2): 253-268.
95. Itier RJ., Taylor M.J. N170 or Nl? Spatiotemporal Differences between Object and Face Processing Using ERPs // Cerebral Cortex. 2004. February, 14:132-142.
96. Inanaga K. Frontal midline theta rhythm and mental activity// Psychiatry Clin. Neurosci. 1998.52(6): 555-566.
Литература 501
97. Jasper Н., Solomon Р., Bradley С. Electroencephalographic analyses of behaviour problem children //Am. J. Psychiatry. 1938.95:641-658.
98. Jenike M.A., Baer L., Ballantine H.T., Martuza R.L., Tynes S., Giriunas I., But-tolph M.L., Cassem N.H. Cingulotomy for refractory obsessive compulsive disorder. A long term follow-up of 33 patients//Arch. General. Psychiatry. 1991.48:548-555.
99. Johnson M.D., Qjemann G.A. The role of the human thalamus in language and memory: evidence from electrophysiological studies I I Brain Cogn. 2000.42(2): 218-230.
100. Jonkman L.M., Kemner C., \ferbaten M.N., Van Engeland H., Camfferman G., Buitelaar J.K., Koelega H.S. Attentional capacity, a probe ERP study: differences between children with attention-deficit hyperactivity disorder and normal control children and effects of methylphenidate // Psychophysiology. 2000.37:334-346.
101. Jung Т-P., Makeig S., Westerfield M., Townsend J., Courchesne E., Sejnowski T.J. Analysis and Visualization of Single-Trial Event-Related Potentials // Human Brain Mapping. 2001.14:166-185.
102. Kalivas P.W, Volkow N.D. The Neural Basis of Addiction: A Pathology of Motivation and Choice//Am. J.Psychiatry. 2005.162(8): 1403-1413.
103. Kandel E., Schwartz J., Jessel T. Principles of Neural Science. 4th ed. McGraw-Hill, New York 2000. (Also: http://en.wikipedia.org/wiki/Principles_of_Neural_Science)
104. Kastner S., Ungerleider L.G. Mechanisms of Visual Attention in the Human Cortex//Annu Rev. Neurosci. 2000.23:315-341.
105. Keedwell P.A., Andrew C., Williams S.C.R., Brammer M.J., Phillips M.L. A Double Dissociation of Ventromedial Prefrontal Cortical Responses to Sad and Happy Stimuli in Depressed and Healthy Individuals// Biol. Psychiatry. 2005.58:495-503.
106. Kilts C.D., Schweitzer J.B., Quinn C.K., Gross R.E., Faber T.L., Muhammad F., Ely T.D., Hoffman J.M., Drexler K.P. Neural activity related to drug craving in cocaine addiction//Arch. Gen. Psychiatry. 2001.58(4): 334-341.
107. Kimura M., Katayama J., Murohashi H. An ERP study of visual change detection: Effects of magnitude of spatial frequency changes on the change-related posterior positivity /I International Journal of Psychophysiology. 2006.62:14-23.
108. Kitabatake Y, Hikida T, Watanabe D., Pastan I., Nakanishi S. (2003) Impairment of reward-related learning by cholinergic cell ablation in the striatum // Proc. Natl. Acad. Sci. USA. 2003.100(13): 7965-7970.
109. Klimesch W. EEG alpha and theta oscillations reflect cognitive and memory performance: a review and analysis // Brain Res. Rev. 1999.29:169-195.
110. KramisR., VinderwolfC.H., Bland B.H. Two types ofhippocampal rhythmical slow activity in both the rabbit and the rat: relations to behavior and effects of atropine, diethyl ether, urethane, and pentobarbital // Exp. Neurol. 1975.49(1 Pt 1): 58-85.
111. Krause K.-H., Dresel S.H., Krause J., Kung H.F., Tatsch K. Increased striatal dopamine transporter in adult patients with attention deficit hyperactivity disorder effects of methylphenidate as measured by single photon emission computed tomography//Neuroscience Letters. 2000.285:107-110.
112. Kringelbach M.L., Rolls E.T. The functional neuroanatomy erf the human orbito-frontal cortex: evidence from neuroimaging and neuropsychology // Prog. Neurobiol. 2004.72:341-372.
113. Kropotov J.D, Edinger S.C. Selection of actions in the basal ganglia-thalamocortical circuits: Review arxJ Model//International Journal of PSychophysiotogy 1999.31(3): 197-217.
502 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
114. Kropotov J.D., Alho К., NaatSnen R., Ponomarev VA, Kropotova O.V, Anichkov A.D., Nechaev VB. Human auditory-cortex mechanisms of preattentive sound discrimination //Neuroscience Letters. 2000.280(2): 87-90.
115. Ktonas P.Y. Automated spike and sharp wave (SSW) detection. In: Methods of analysis of brain electrical and magnetic signals. EEG handbook (revised series, ЛЫ. 1) AS.GevinsandARemorid (Eds) //Elsevier Science Publishers. 1987. B.V 211-241.
116. Lashley K.S. In Search of the Engram // Psychological Mechanisms in Animal Behavior. Society for Experimental Biology. N.Y. 1950.454-482.
117. Laukka S.J., Jarvilehto T., Alexandrov Yu.L, Lindqvist J. Frontal midline theta related to learning in a simulated driving task // Biol. Psychol. 1995. 40(3): 313-320.
118. Lee S.H., Wynn J.K., Green M.F., Kim H., Lee K.J., Nam M., P^rkJ.K., Chung Y.C. Quantitative EEG and low resolution electromagnetic tomography (LORETA) imaging of patients with persistent auditory hallucinations // Schizophrenia Research. 2006.83 (2-3): 111-119. Epub 2006 Mar 9.
119. Leuchter AF., Sebastian U., Uijtdehaageb H.J., Cook I. A., O’Harae R., Mandelk-emd M. Relationship between brain electrical activity and cortical perfusion in normal subjects Psychiatry Research: Neuroimaging Section. 1999.90.125-140.
120. Leuchter A.F., Cook I A, Witte EA, Morgan M., Abrams M. Changes in Brain Function of Depressed Subjects during Treatment with Placebo // Am. J. Psychiatry. 2002.159:122-129.
121. Lippold O.C., Redfeam J.W Mental changes resulting from the passage of small direct currents through the human brain // Br. J. Psychiatry. 1964.110:768-772.
122. Luck S.J. An introduction to the event-related potential technique. The МГГ Press, Cambridge, MA. 2005.374 p.
123. Marshal M.P., Molina B.S. Antisocial behaviors moderate the deviant peer pathway to substance use in children with ADHD//J. Clin. Child. Adolesc. Psychol. 2006.35(2): 216-226.
124. Marshall L., Molle M., Hallschmid M., Bom J. Transcranial direct current stimulation during sleep improves declarative memory // J. Neurosci. 2004. 24(44): 9985-9992. Erratum in: J Neurosci. 2005,25(2): 1 p. following 531.
125. Mathalon D.H., Bennett A., Askari N., Gray E.M., Rosenbloom M.J., Ford J.M. Response-monitoring dysfunction in aging and Alzheimer’s disease: an event-related potential study // Neurobiology of Aging. 2003.24:675-685.
126. Mayberg H.S., Lozano AM., \bon V, McNeely H.E., Seminowicz D., Hamani C., Schwalb J.M., Kennedy S.H. Deep brain stimulation for treatment-resistant depres-' sion// Neuron. 2005.45:651-660.
127. McCarley R.W., O’Donnell B.F., Niznikiewicz MA, Salisbury D.F., Potts G.F., Hirayasu Y., Nestor P.G., Shenton M.E. Update on electrophysiology in schizophrenia // International Review of Psychiatry. 1997.9:373-386.
128. McCormick DA, Pape H.C. Properties of a hyperpolarization-activated cation current and its role in rhythmic oscillation in thalamic relay neurons // Journal of Physiology. 1990.431:291-318.
129. McNaughton B.L. The neurobiology of spatial computation and learning. In: Stein D J. (ed.) Lectures on complexity: Santa Fe institute studies in the sciences of complexity. Redwood, CA Addison-Vfesley. 1989.389-437.
Литература 503
130. Mendez М., Um G. Seizures in elderly patients with dementia: epidemiology and management // Drugs. Aging. 2003.20:791-803.
131. Meyer-Lindenbeig A., Wfeinbeiger D.R. Intermediate phenotypes and genetic mechanisms of psychiatric disorders // Nat. Rev. Neurosci. 2006.7(10): 818-827.
132. Monastra V, Lubar J., Linden M., VanDeusen P., Green G., Wing W, Phillips A., Fenger T. Assessing attention deficit hyperactivity disorder via quantitative electroencephalography: an initial validation study//Neuropsychology. 1999.13:424-433.
133. Monastra V.J., Lynn S., Linden M., Lubar J.F., GruzeUer J., LaVaque T.J. Electroencephalographic Biofeedback in the Treatment of Attention-Deficit/Hyperactivity Disorder //Applied Psychophysiology and Biofeedback. 2005. 30(2): 95-114.
134. Moore N.C. A review of EEG biofeedback treatment of anxiety disorders // Clin. Electroencephalogr. 2000.31(1): 1-6.
135. Morris P.L., Robinson R.G., Raphael B., Hopwood M.J. Lesion location and poststroke depression //J. Neuropsychiatry Clin. Neurosci. 1996.8:399-403.
136. Naatanen R. Mismatch negativity: clinical research and possible applications // Int. J. Psychophysiol. 2003.48:179-188.
137. Niedermeyer E. Alpha rhythms as physiological and abnormal phenomena // Int. J. Psychophysiol. 1997.26(1-3): 31-49. Review.
138. Niki H., Watanabe M. Prefrontal and cingulate unit activity during timing behaviour in the macaque//Brain Res. 1979.171:213-224.
139. Nitsche M.A., Paulus W. Excitability changes induced in the human motor cortex by weak transcranial direct current stimulation // J. Physiol. 2000.527 Pt 3:633-639.
140. Nitsche M.A., Paulus W. Sustained excitability elevations induced by transcranial DC motor cortex stimulation in humans //Neurology. 2001.57(10): 1899-1901.
141. O’Keefe J., Recce M.L. Phase relationship between hippocampal place units and the EEG theta rhythm // Hippocampus. 1993.3(3): 317-330.
142. Olbrich H.M., MaesH., Valerius G., Langosch J.M., Feige B. Event-related potential correlates selectively reflect cognitive dysfunction in schizophrenics // J. Neural. Transm. 2005.112:283-295.
143. Penfield W., Perot P. The brain’s record of auditory and visual experience // Brain. 1963.86:595-696.
144. Peniston E.G., Kulkosky PJ. Alpha-theta brainwave training and beta-endorphin levels in alcoholics// Alcohol. Clin. Exp. Res. 1989.13(2): 271-279.
145. Pfurtscheller G., Lopes Da Silva F.H. Event-related EEG/MEG synchronization and desynchronization: basic principles// Clinical Neurophysiology. 1999.110:1842-1857.
146. Pineda J.A., Foote S.L., Neville H.J. Effects of locus coeruleus lesions on auditory, long latency, event-related potentials in monkey // J. Neurosci. 1989.9:81-93.
147. Pizzagalli D.A., Oakes TR., Davidson R.J. Coupling of theta activity and glucose metabolism in the human rostral anterior cingulate cortex: An EEG/PET study of normal and depressed subjects // Psychophysiology 2003.40:939-949.
148. Pliszka S.R., Liotti M., X\bldorff M.G. Inhibitory Control in Children with Atten-tion-Deficit/Hyperactivity Disorder Event-Related Potentials Identify the Processing Component and Timing of an Impaired Right-Frontal Response-Inhibition Mechanism// Biol. Psychiatry. 2000.48:238-246.
149. PogarellO., MulertC., HegerlU. Event Related Potentials and fMRI in Neuropsychopharmacology // Journal of Clinical EEG & Neuroscience. 2006. V. 37, № 2,99-107.
504 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
150. Porjesz В., Rangaswamy М., Kamarajan С., Jones КА., Padmanabhapillai А, Begleiter Н. The utility of neurophysiological markers in the study of alcoholism // Clinical Neurophysiology. 2005.116:993-1018.
151. Posner M.I., Petersen S.E., Fbx P.T. Raichle M.E. Localization of cognitive operations in the human brain//Science. 1988.240:1627-1631
152. Prichep L.S., John E.R., Ferris S.H., Rausch L, Fang Z., Cancro R., Torossian C., Reisberg B. Prediction of longitudinal cognitive decline in normal elderly with subjective complaints using electrophysiological imaging // Neurobiology of Aging. 2006.27(3): 471-481. Epub 2005 Oct 6.
153. Reynolds J.H., Chelazzi L., Desimone R. Competitive Mechanisms Subserve Attention in Macaque Areas V2 and V4// J. Neurosci. 1999.19(5): 1736-1753.
154. Robinson D.L., Petersen S.E. The pulvinar and visual salience. TINS. 1992. 15(4): 127-132.
155. Robinson T.E., Berridge K.C. The neural basis of drug craving: an incentive-sensitization theory of addiction//Brain Res. Rev. 1993.18(3): 247-291.
156. Romo R. Salinas S. Flutter Discrimination: Neural codes, perception, memory and decision making// Nature Reviews, Neuroscience. 2003. V. 4:203-218.
157. Roth W.T., Cannon E.H. Some features of the auditory evoked response in schizophrenics // Arch. Gen. Psychiatry. 1972.27:466-471.
158. Ropohl A, Sperling W., Elstner S., Tomand B., Reulbach U., Kaltenh, Komhuber J., Maihofner C. Cortical activity associated with auditory Hallucinations // Neuroreport. 2004. W15, № 3,523-526.
159. Rothenberger A, Banaschewski T, Heinrich H., Moll G.H, Schmidt M.H., van’t Klooster B. Comorbidity in ADHD-children: effects of coexisting conduct disorder or tic disorder on event-related brain potentials in an auditory selective-attention task// Eur. Arch. Psychiatry Clin. Neurosci. 2000.250:101-110.
160. Rugg M.D., Yonelinas AP. Human recognition memory: a cognitive neuroscience perspective // TRENDS in Cognitive Sciences. 2003.7(7): 313-319.
161. Satterfield J.H., Cantwell D.P., Lesser L.I., PodosinR.L. Physiological studies of the hyperkinetic child //Am. J. Psychiatry. 1972.128:102-108.
162. SchmoleskyM.T, WangY.C., Hanes D.P., Thompson K.G., LeutgebS., SchallJ.D., Leventhal A.G. Signal timing across the macaque visual system // J. Neurophysiol. 1998.79:3272-3278.
163. Schneider E, Heimann H., Mattes R., Lutzenberger W., Birbaumer N. Self-regulation of slow cortical potentials in psychiatric patients: depression // Biofeedback Self Regul. 1992.17:203-214.
164. Schneider F., Rockstroh B., Heimann H., Lutzenberger W., Mattes R., Elbert T, Birbaumer N., Bartels M. Self-regulation of slow cortical potentials in psychiatric patients: schizophrenia // Biofeedback Self Regul. 1992.17(4): 277-292.
165. Schulte-Kome G., Deimel W, BartlingJ., RemschmidtH. Auditory processing and dyslexia: evidence for a specific speech processing deficit//Neuroreport 1998.9(2): 337-340.
166. Seeman P., Chau-Wbng M., Tedesco J., W6ng К Brain receptors for antipsychotic drugs and dopamine: direct binding assays // Proc. Natl Acad. Set USA 1975.72(11): 4376-4380.
167. Seeman P., Lee T, Chau-Wong M., Wbng K. Antipsychotic drug doses and neuro-leptic/dopamine receptors//Nature. 1976.261 (5562): 717-719.
Литература
505
168. Sherlin L, Congedo M. Obsessive-compulsive dimension localized using low-resolution brain electromagnetic tomography (LORETA) // Neurosci. Lett. 2005. 387: 72-74.
169. Sherman S.M., Guilleiy R.W. Exploring the thalamus and its role in cortical function I IМГГ Press. 2006.484 p.
170. Shima K., Tanji J. Role for cingulate motor area cells in voluntaiy movement selection based on reward//Science. 1998.282:1335-1338.
171. Singh K.D., Barnes G.R., Hillebrand A, Forde E.M., Williams A.L. Task-Related Changes in Cortical Synchronization Are Spatially Coincident with the Hemodynamic Response //NeuroImage. 2002.16:103-114.
172. Smit D.J., Posthuma D., Boomsma D.I., Geus E.J. Heritability of background EEG across the power spectrum // Psychophysiology. 2205.42(6): 691-697.
173. Smith E.E., JonidesJ. Storage and executive processes in the frontal lobes//Science. 1999.283(5408): 1657-1661.
174. Sterman M.B. Physiological origins and functional correlates of EEG rhythmic activities: Implications for self-regulation//Biofeedback and Self-Regulation. 1996.21:3-33.
175. Stevens A., KircherT., Nickola M., Bartels M., Rosellen N., Wbrmstall H. Dynamic regulation of EEG power and coherence is lost early and globally in probable DAT // Journal of Neurochemistry. 2001.96:609-623.
176. Suetsugi M., Mizuki Y, Ushijima I., Kobayashi T., Tsuchiya К.» Aoki T, Watanabe Y. Appearance of Frontal Midline Theta Activity in Patients with Generalized Anxiety Disorder // Neuropsychobiology. 2000.41:108-112.
177. Sumich A.L., Kumari V, Heasman B.C., Gordon E., Brammer M. Abnormal asymmetry of N200 and P300 event-related potentials in subclinical depression // Journal of Affective Disorders. 2006.92(2-3): 171-83. Epub 2006 Mar 9.
178. Talairach J., Bancaud J., Geier S., Bordas-Ferrer M., Bonis A, Szikla G., Rusu M. The cingulate gyrus and human behaviour // Electroencephalography & Clinical Neurophysiology. 1973.34:45-52.
179. Tallon-Baudry C., Bertrand O. Oscillatory gamma activity in humans and its role in object representation//Trends. Cogn. Sci. 1999.3(4): 151-162.
180. Tallon-Baudry d, Bertrand O., Hertiaff M.-A, Isnard J., Fischer C. Attention Modulates Gamma-band Oscillations Differently in the Human Lateral Occipital Cortex and Fusiform Gyrus // Cerebral. Cortex. 2005.15:654-662.
181. Tamm L., Menon V., Ringel J., Reiss A.L. Event-Related fMRI Evidence of Frontotemporal Involvement in Aberrant Response Inhibition and Task Switching in At-tention-Deficit/Hyperactivity Disorder// J. Am. Acad. Child. Adolesc. Psychiatry 2004.43(11): 1430-1440.
182. Terzuolo C.A, Bullock Т.Н. Measurement of Imposed Voltage Gradient Adequate to Modulate Neuronal Firing // Proc. Natl. Acad. Sci. USA 1956.42(9): 687-694.
183. Toga AW., Thompson P.M. Mapping Brain Asymmetry // Nat. Rev. Neurosci. 2003.4:37-48.
184. Tomarken AJ., Davidson R.J., Wheeler R.E., Kinney L. Psychometric properties of resting anterior EEG asymmetry: temporal stability and internal consistency // Psychophysiology. 1992.29:576-592.
185. Toomim H., Carmen J. Hemoencephalography (HEG) // Biofeedback 1999. 27 (4): 10-14.
506 Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия
186. Tsai AC., Uou М., Jung Т.Р., Onton J A., Cheng P.E., Huang C.-С., Duann J.-R., Makeig S. Mapping single-trial EEG records on the cortical surface through a spatiotemporal modality//Neuroimage. 2006.32:195-207.
187. UK ECT Review Group. Efficacy and safety of electroconvulsive therapy in depressive disorders: a systematic review and metaanalysis // Lancet. 2003.361:799-808.
188. Umbricht D.S.G., Bates J A, Lieberman J A, Kane J.M, Javitt D.C. Electrophysiological Indices of Automatic and Controlled Auditory Information Processing in First-Episode, Recent-Onset and Chronic Schizophrenia // Biol. Psychiatry. 2006. Apr 15; 59(8): 762-772. Epub 2006 Feb 21.
189. van\feen V., Carter C.S. The anterior cingulate as a conflict monitor fMRI and ERP studies// Physiology & Behavior. 2002.77:477-482.
190. Vanhatalo S., Palva J.M., Holmes M.D., Miller J.W., Vbipio J., Kaila K. Infraslow oscillations modulate excitability and interictal epileptic activity in the human cortex during sleep Ц Proc. Natl. Acad. Sci. USA 2004. 101(14): 5053-5057. Epub 2004 Mar 24.
191. Xferleger R. Event related potentials and cognition: a critique of context updating hypothesis and alternative interpretation of P3 // Behavioral Brain Science. 1988. 11: 343-427.
192. Vuga M., Fox NA, Cohn J.F., George C.J., Levenstein R.M., Kovacs M. Longterm stability of frontal electroencephalographic asymmetry in adults with a history of depression and controls // Int. J. Psychophysiol. 2006.59:107-115.
193. Whiter G. Contingent negative variation: An electrical sign of sensorimotor association and expectancy in the human brain // Nature. 1964.203:380-384.
194. Weiskopf N., Mathiak K., Bock S.W., Schamowski E, Veit R., Grodd W., Goebel R., Birbaumer N. Principles of a Brain-Computer Interface (BCI) Based on Real-Time Functional Magnetic Resonance Imaging (fMRI) // IEEE Transactions on Biomedical Engineering. 2004.51(6): 966-970.
195. Wheeler M.E., Petersen S.E., Buckner R.L. Memory’s echo: vivid remembering reactivates sensory-specific cortex // Proc. Natl. Acad. Sci. USA 2000. 97: 11125-11129.
196. Wickelgren WA Chunking and consolidation: a theoretical synthesis of semantic networks, configuring in conditioning, S-R versus congenitive learning, normal forgetting, the amnesic syndrome, and the hippocampal arousal system//Psychol. Rev. 1979.86(1): 44-60.
197. Wiersema J.R., van der Meere J J., Roeyers H. ERP correlates of impaired error monitoring in children with ADHD // J. Neural. Transm. 2005. 112(10): 1417-30. Epub 2005 Feb 22.
198. Yarbus A Eye Movements and Vision // Plenum Press, 1967.
199. Yerkes R.M., Dodson J.D. The relation of strength of stimulus to rapidity of habitformation//Journal of Comparative Neurology and Psychology. 1908.18:459-482.
200. Zatorre RJ., Bouffard M., Ahad P., Belin P. Where is "where’ in the human auditory cortex? // Nat. Neurosci. 2002.5:905-909.