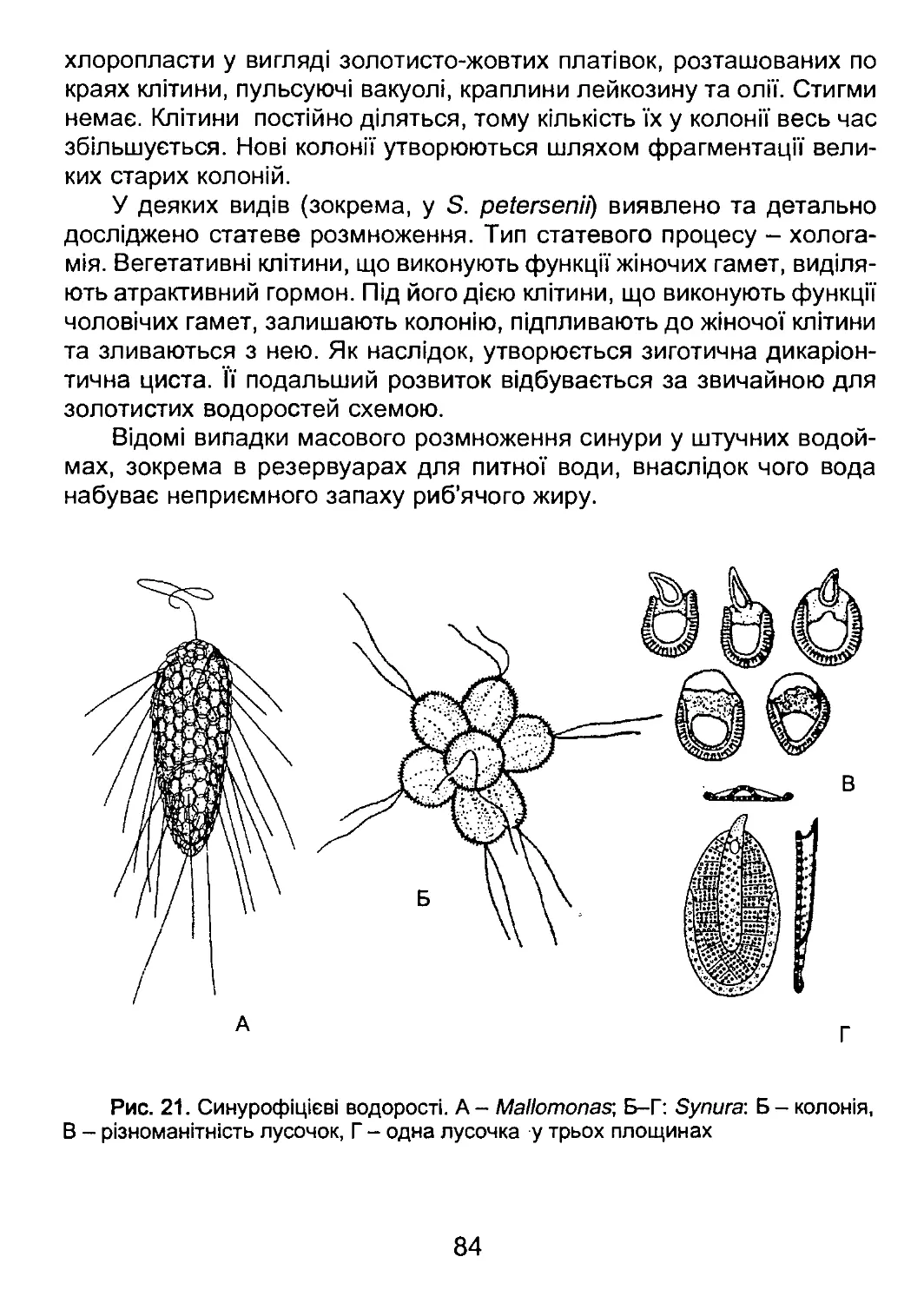
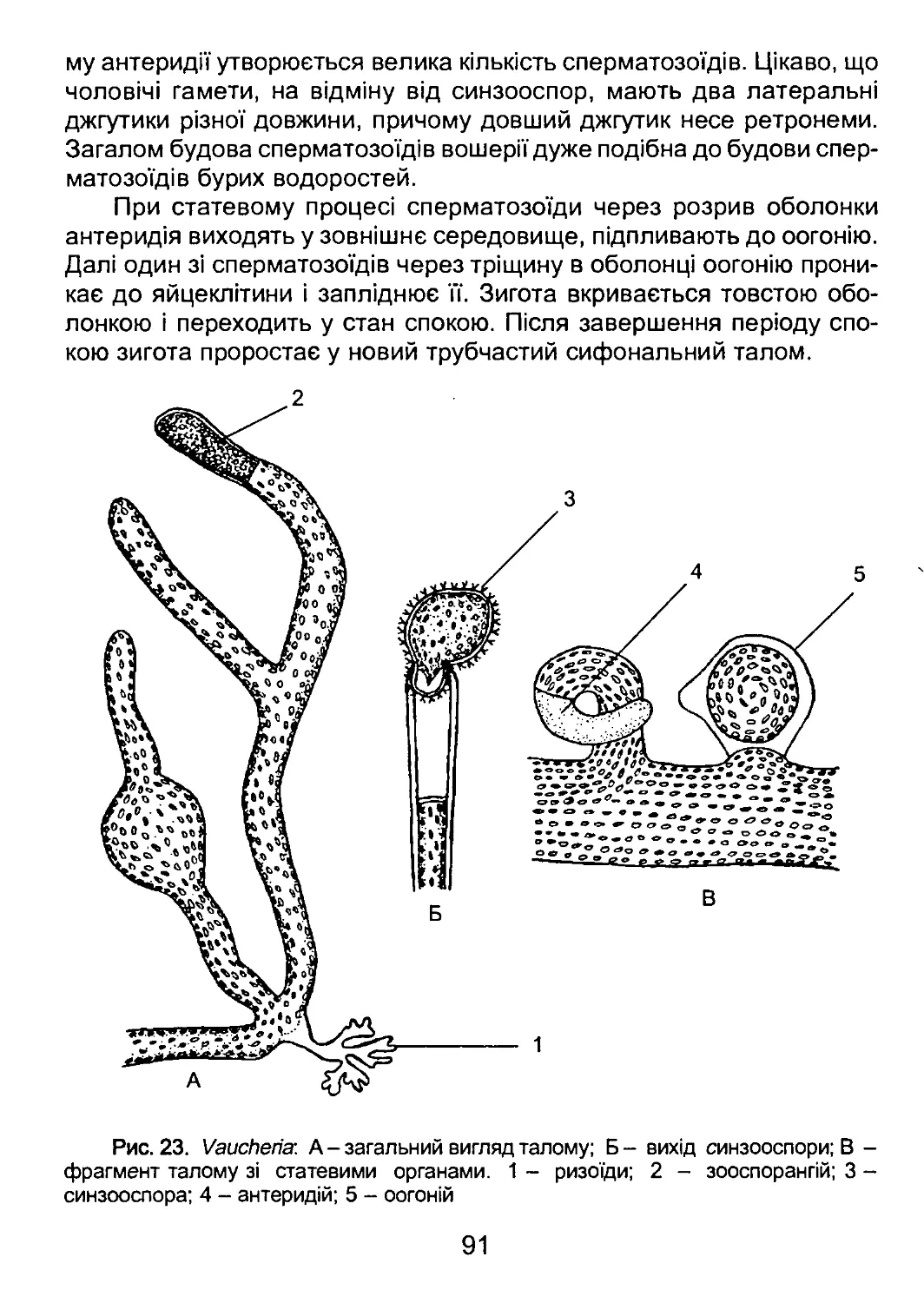
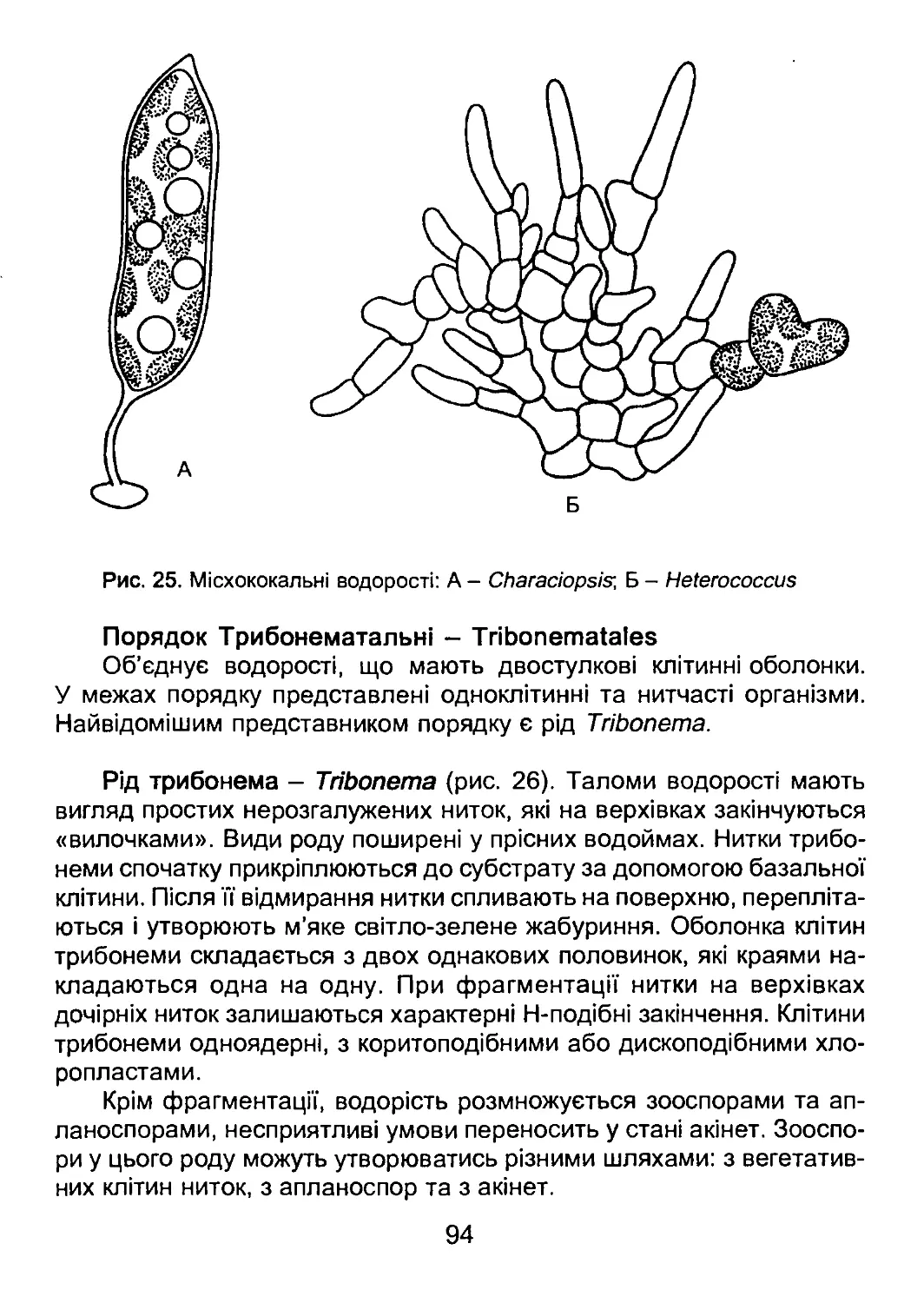
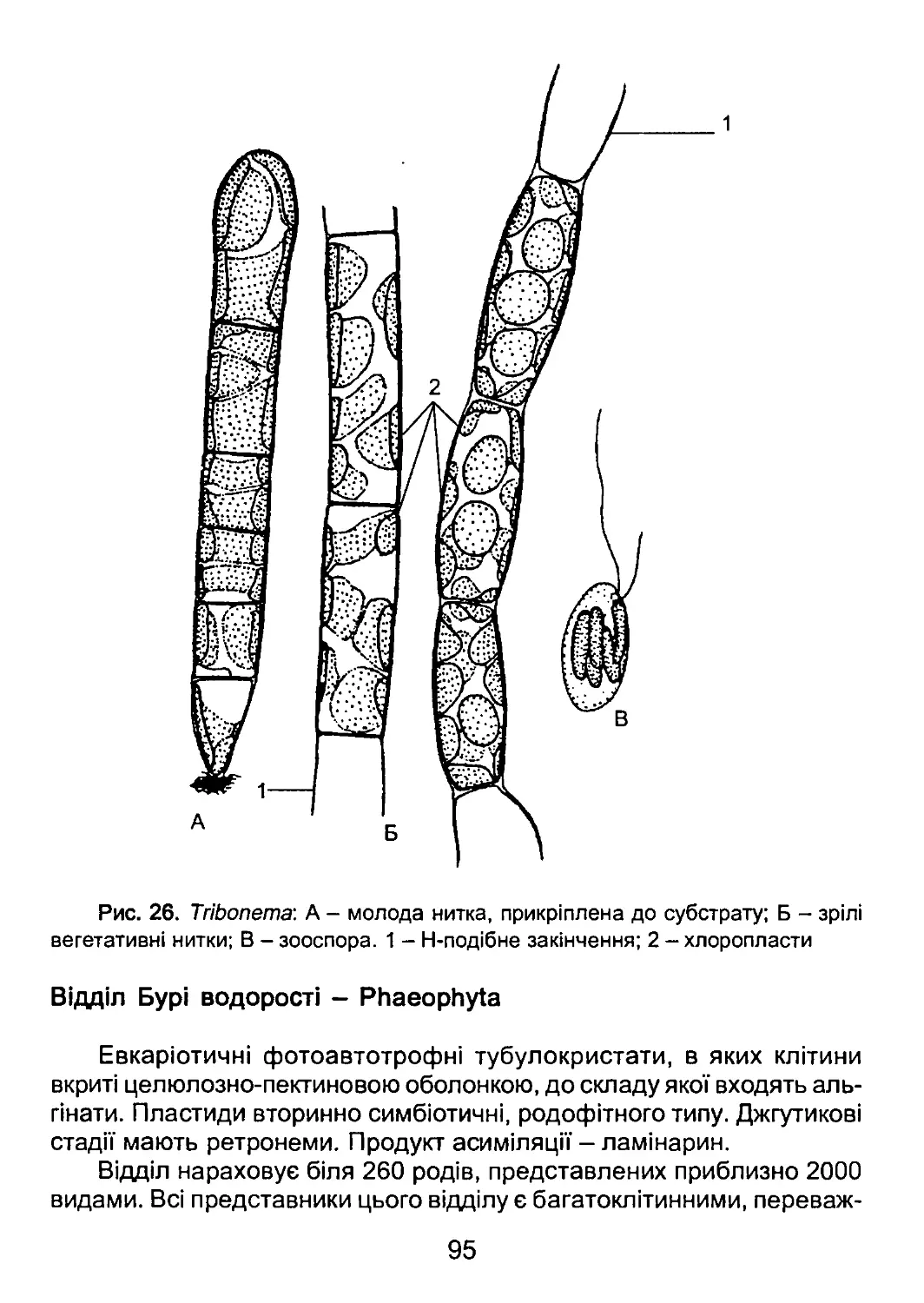
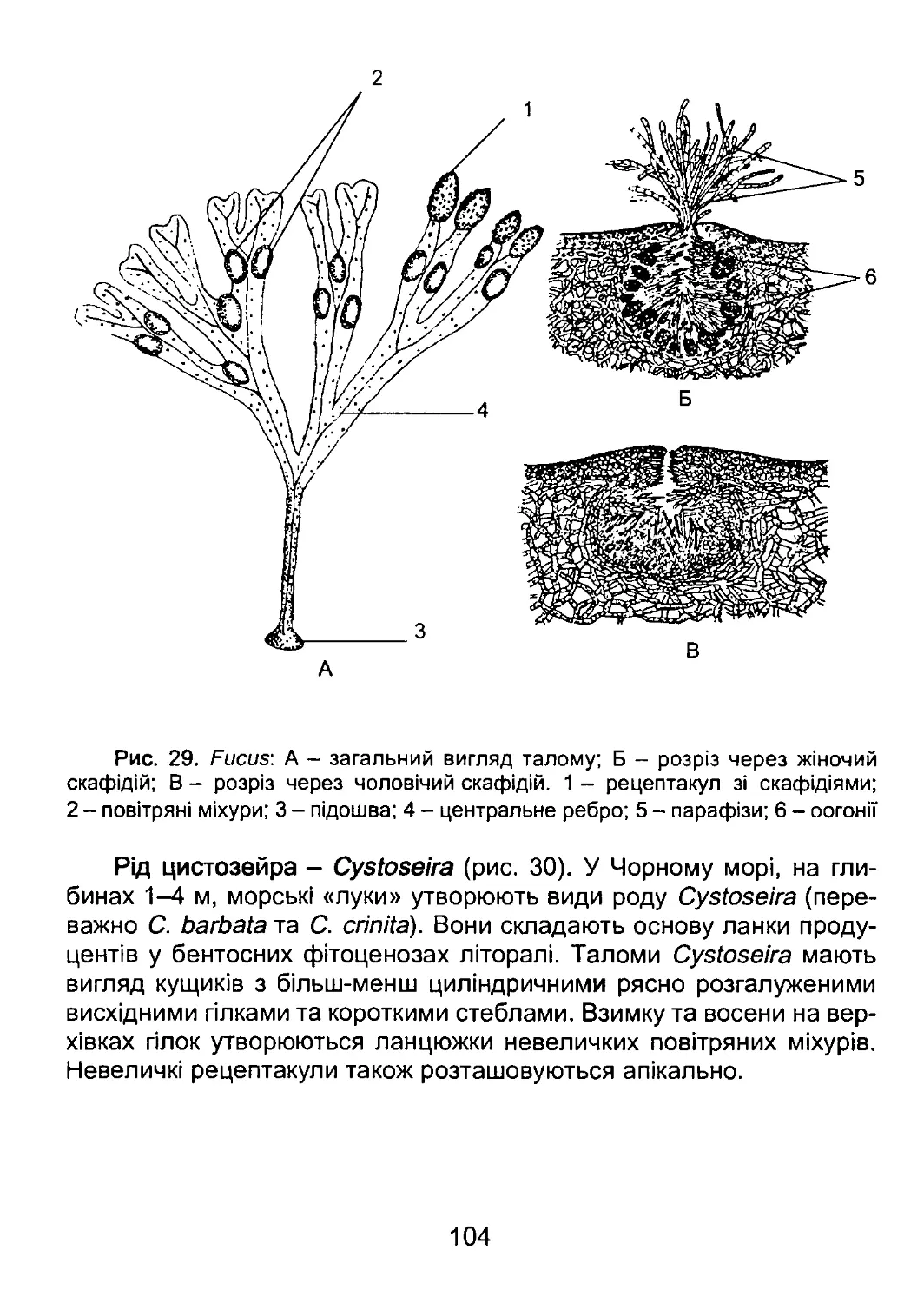
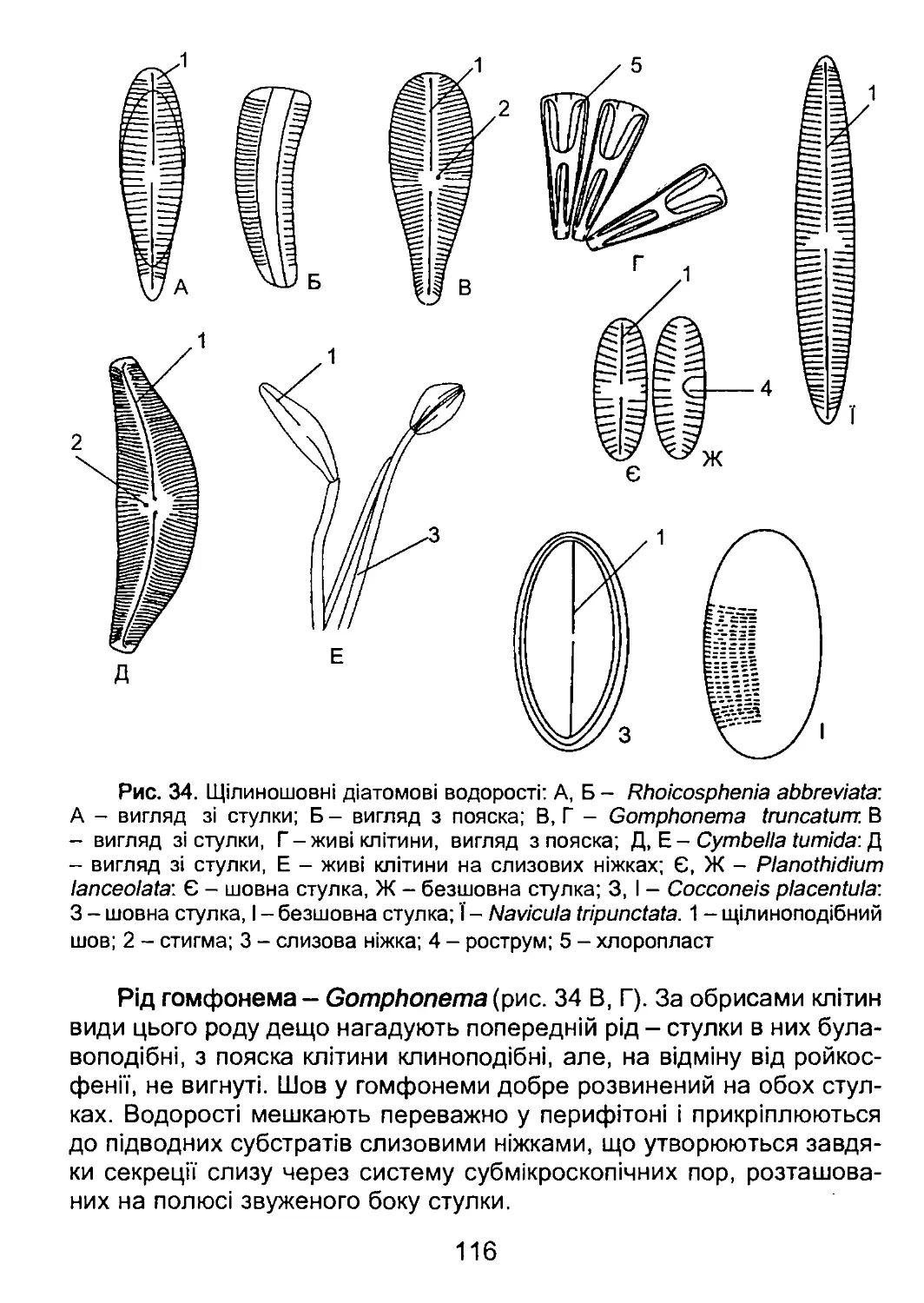
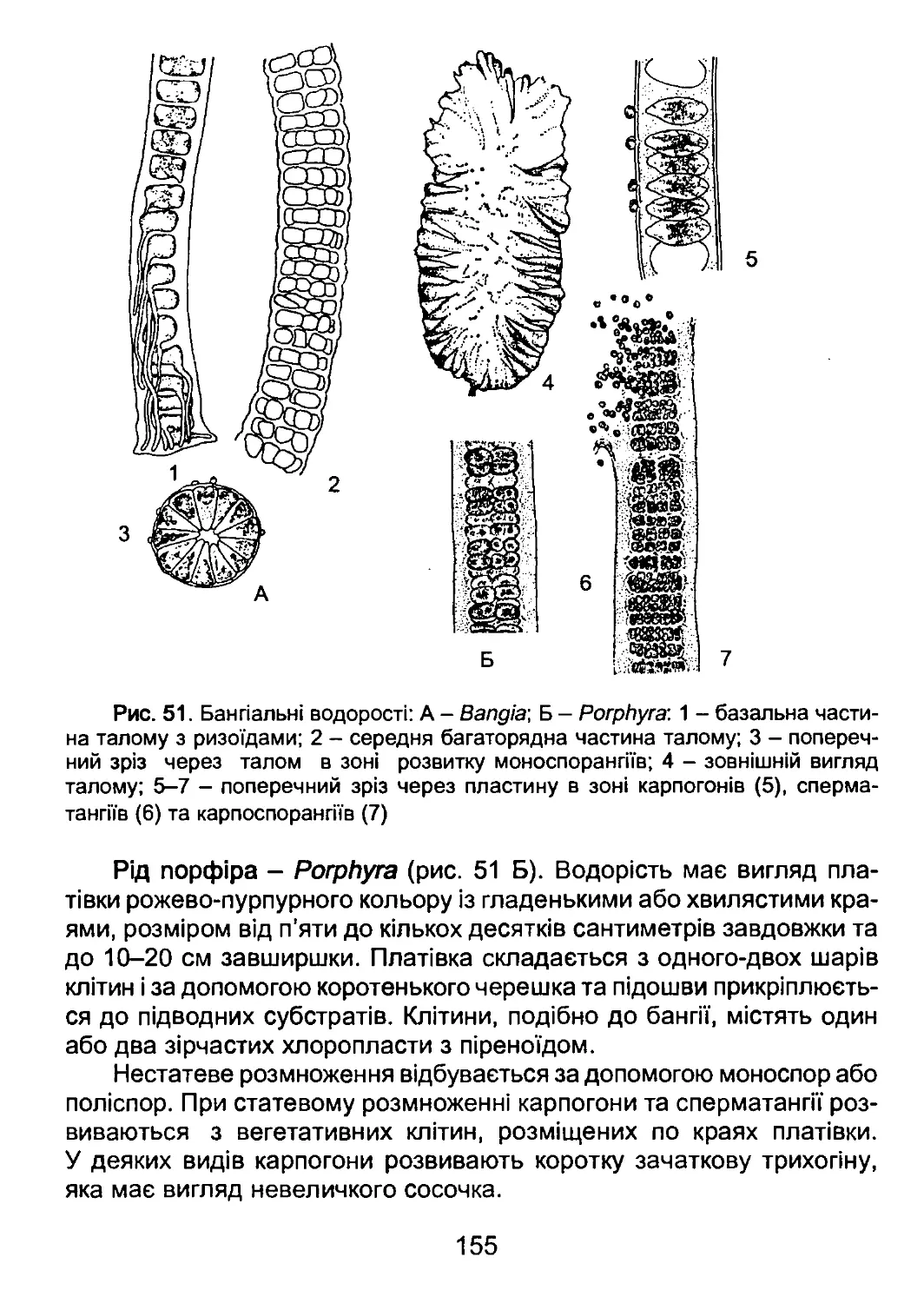
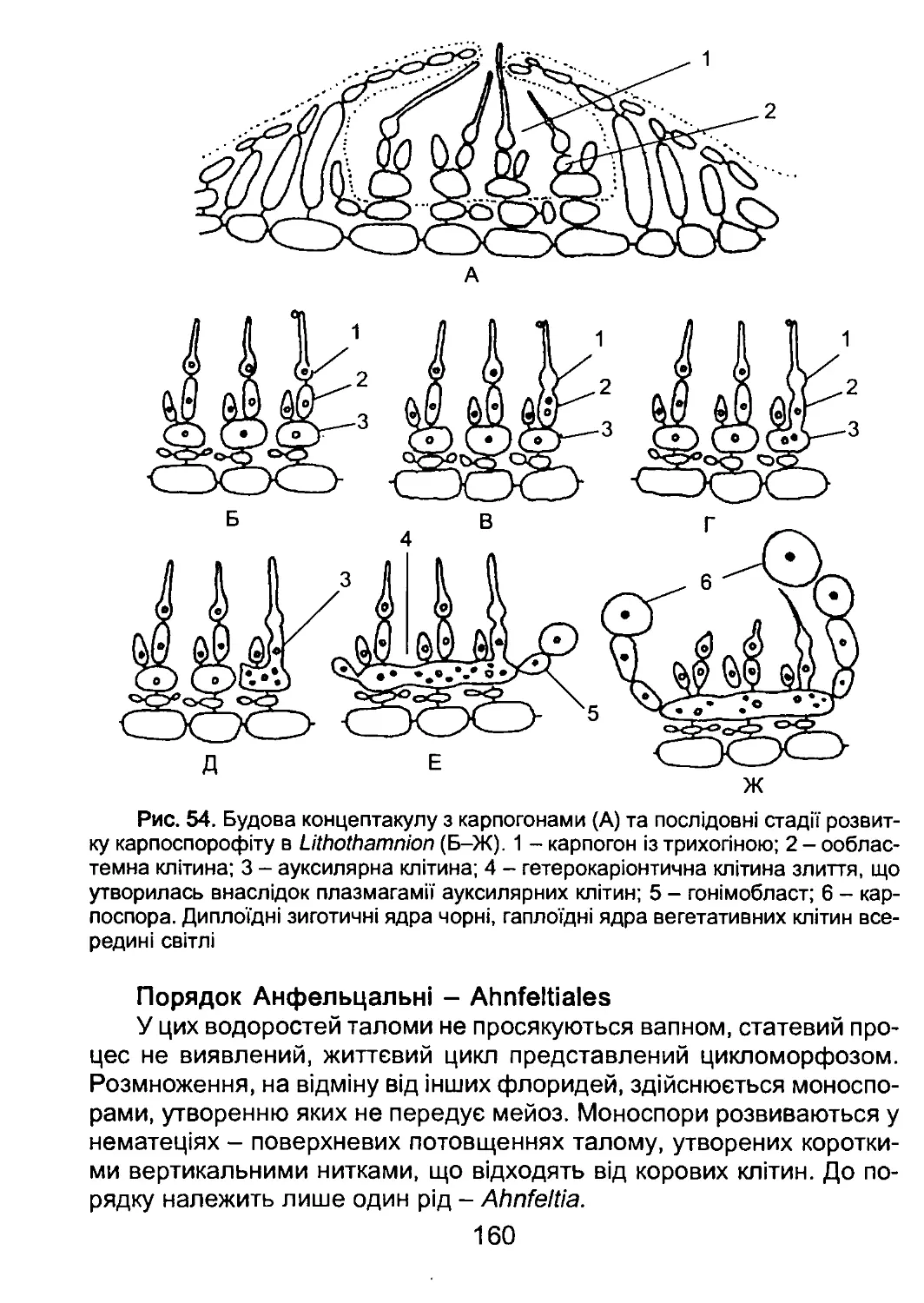
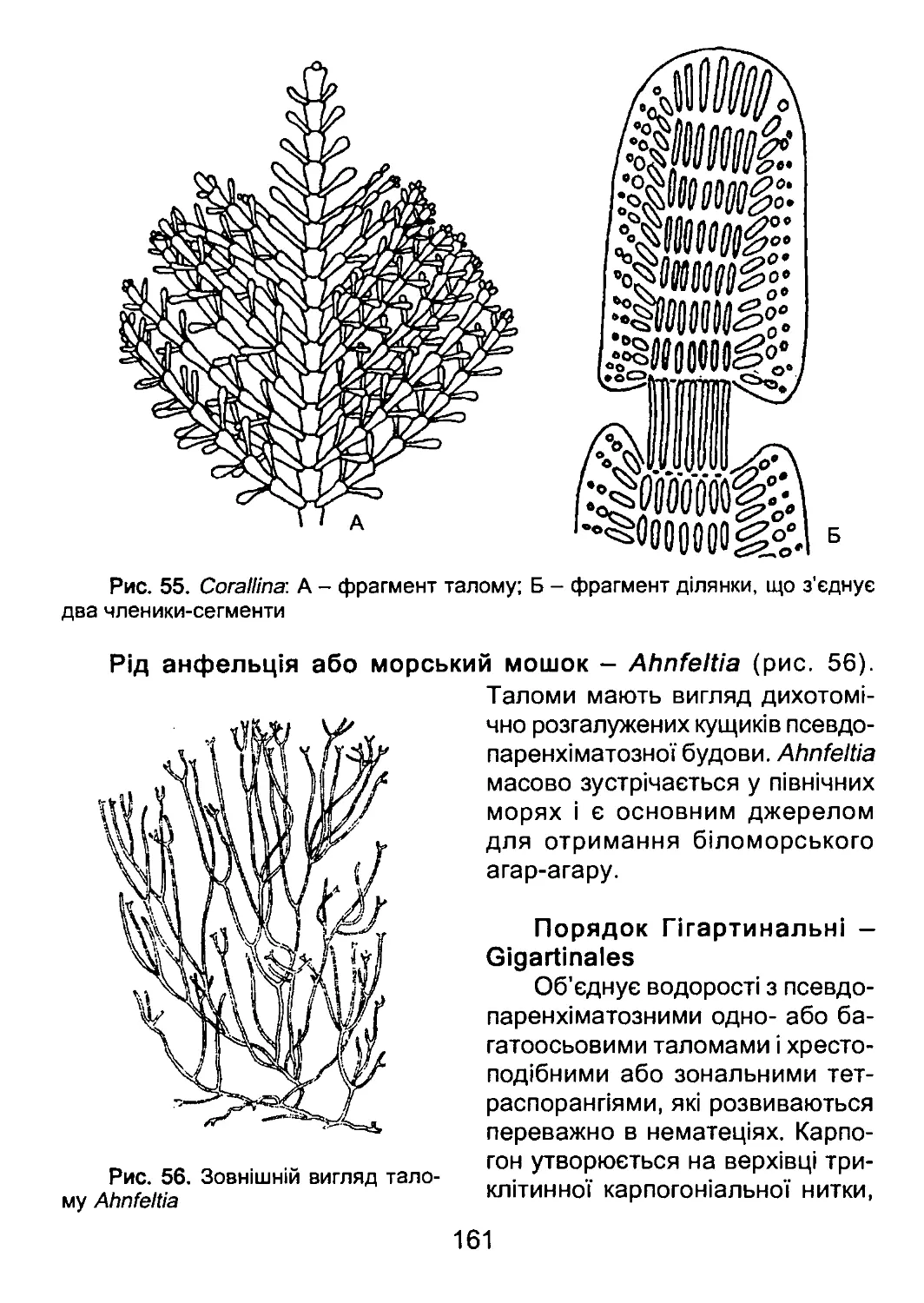
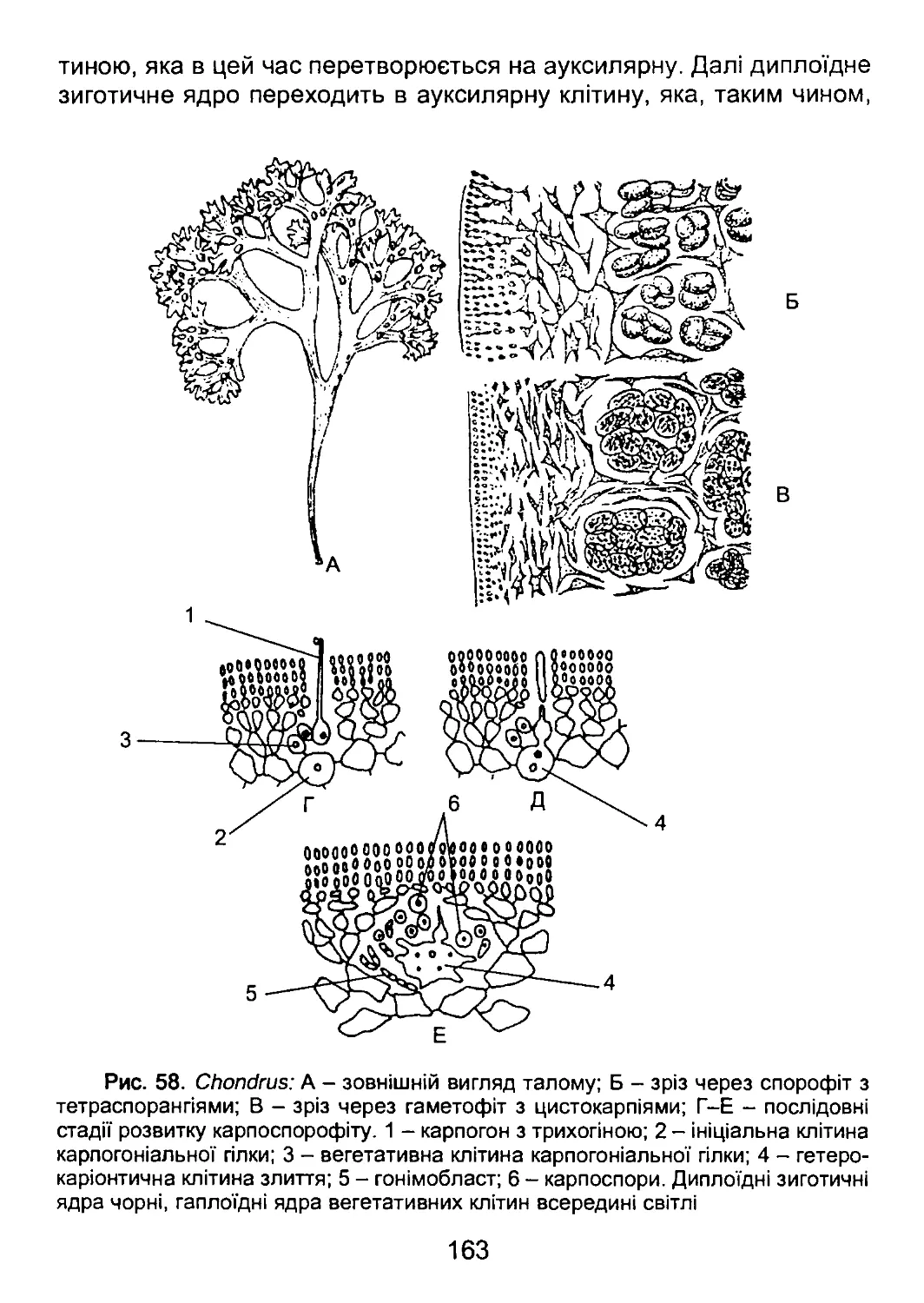
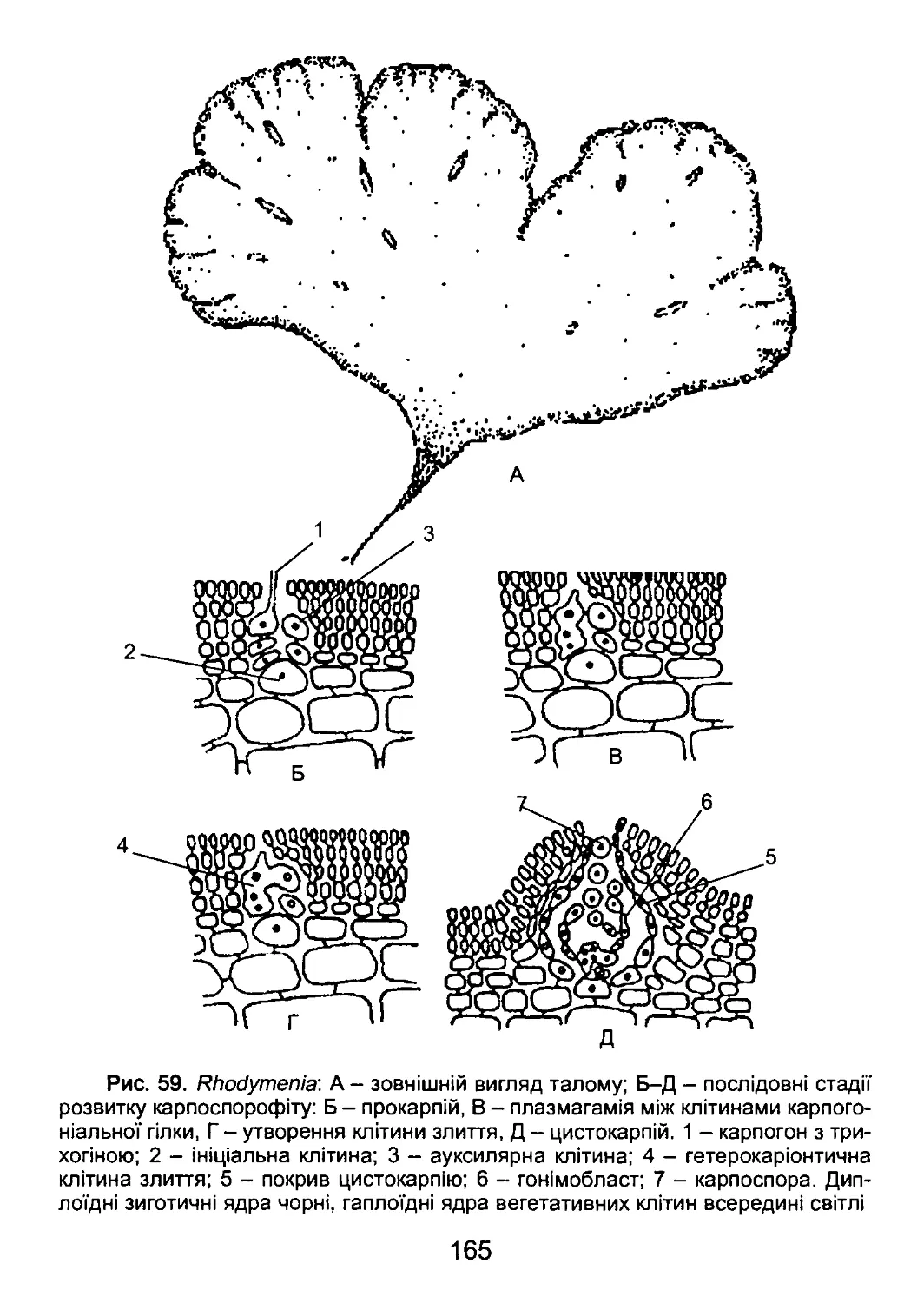
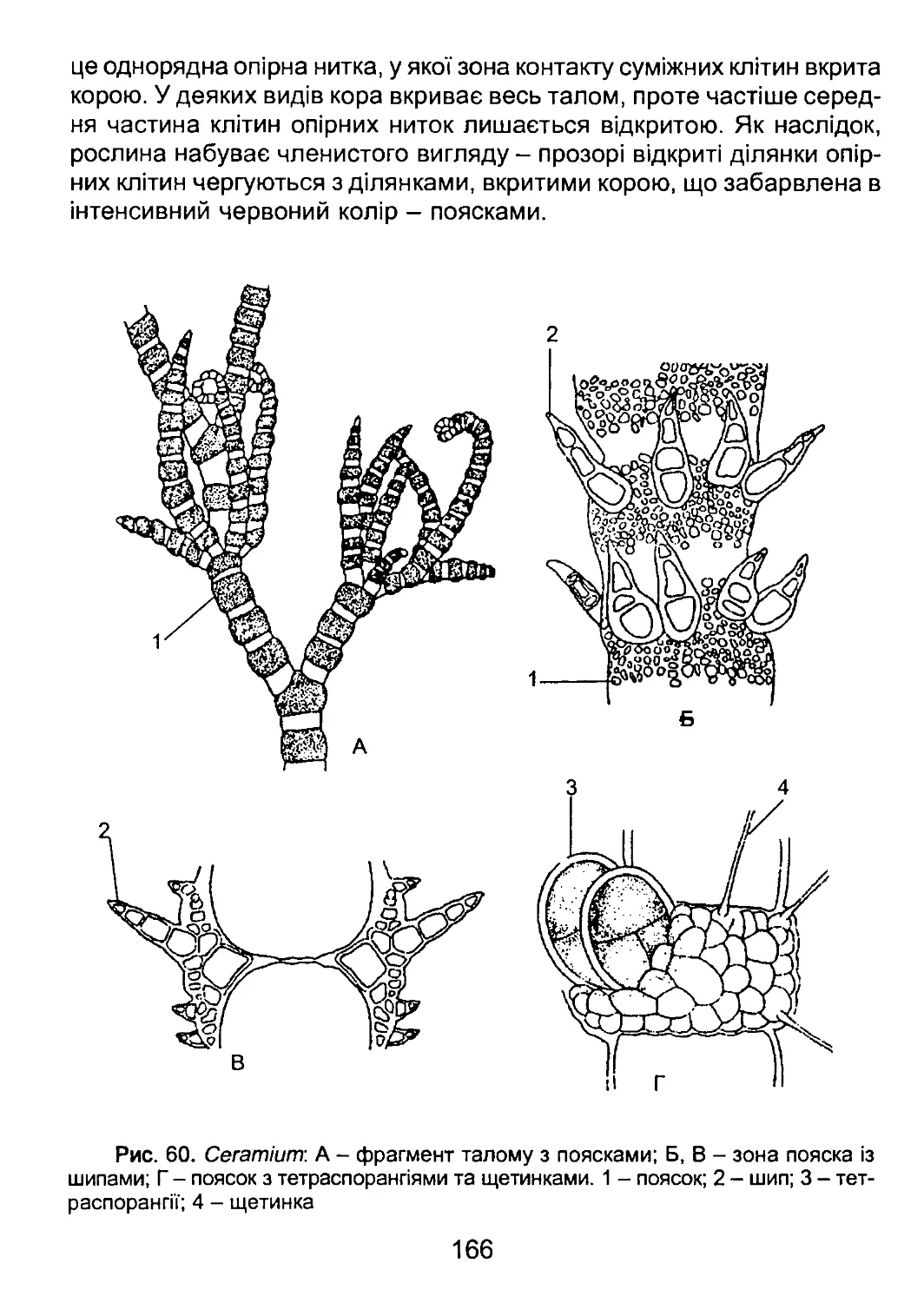
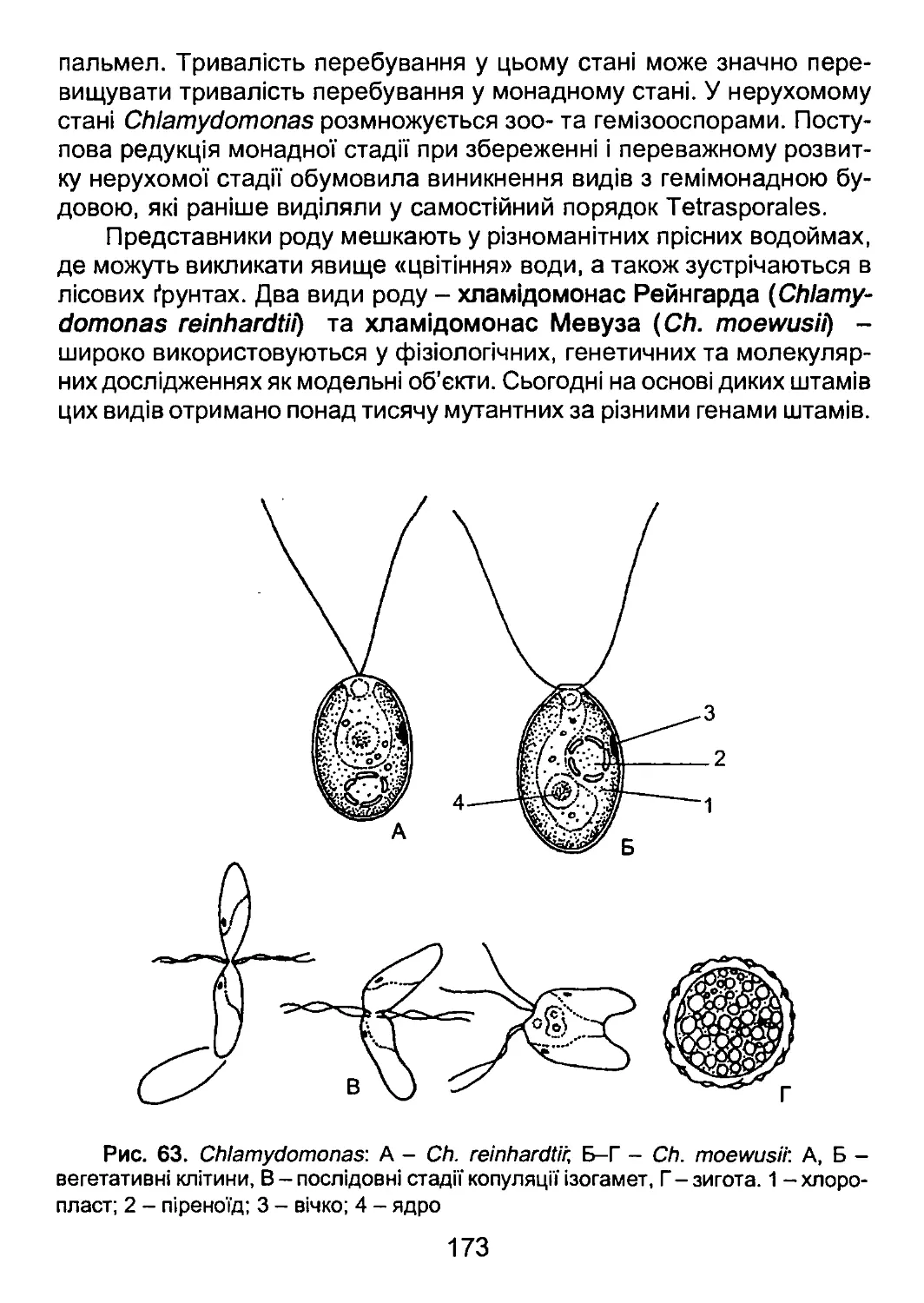
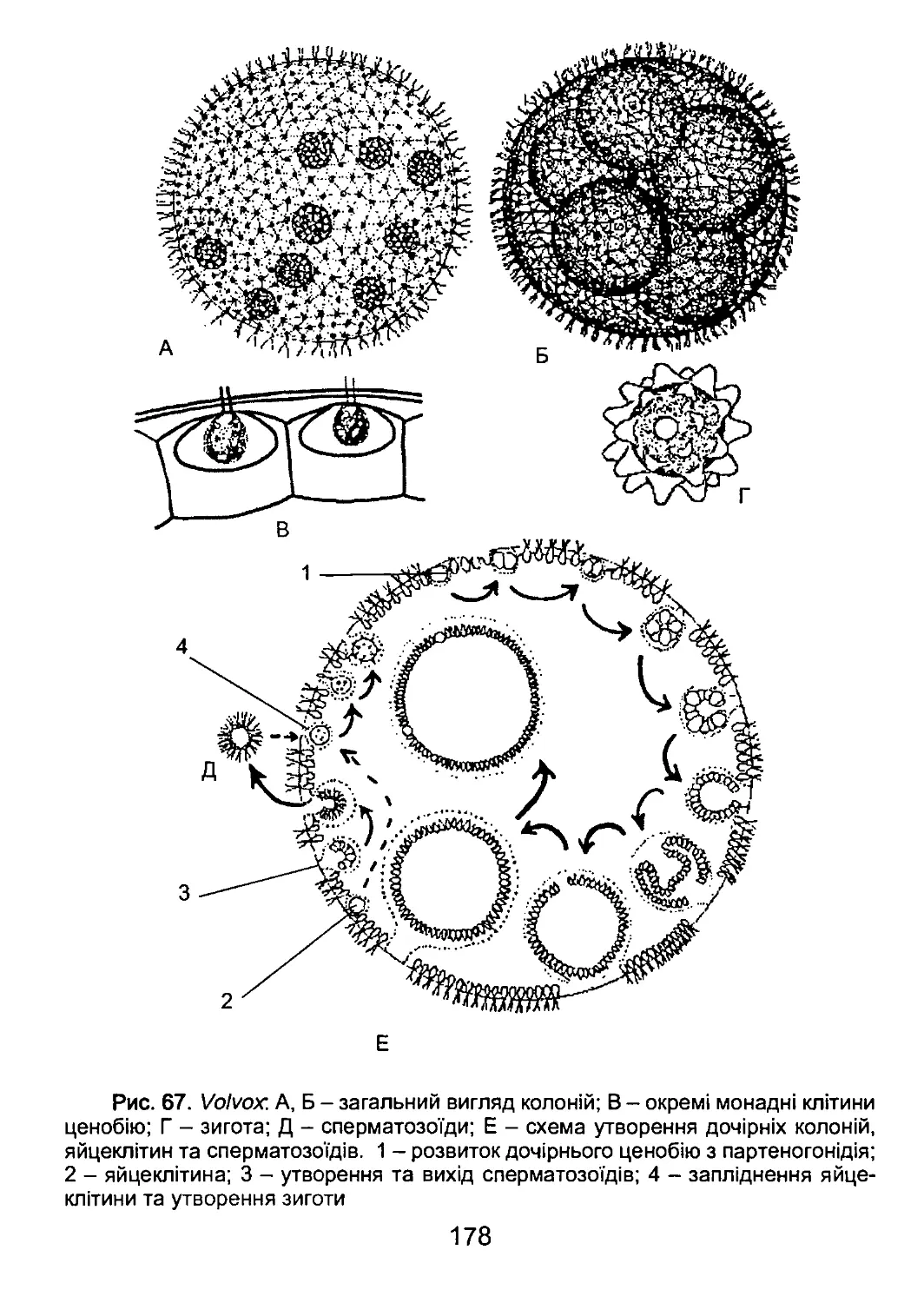
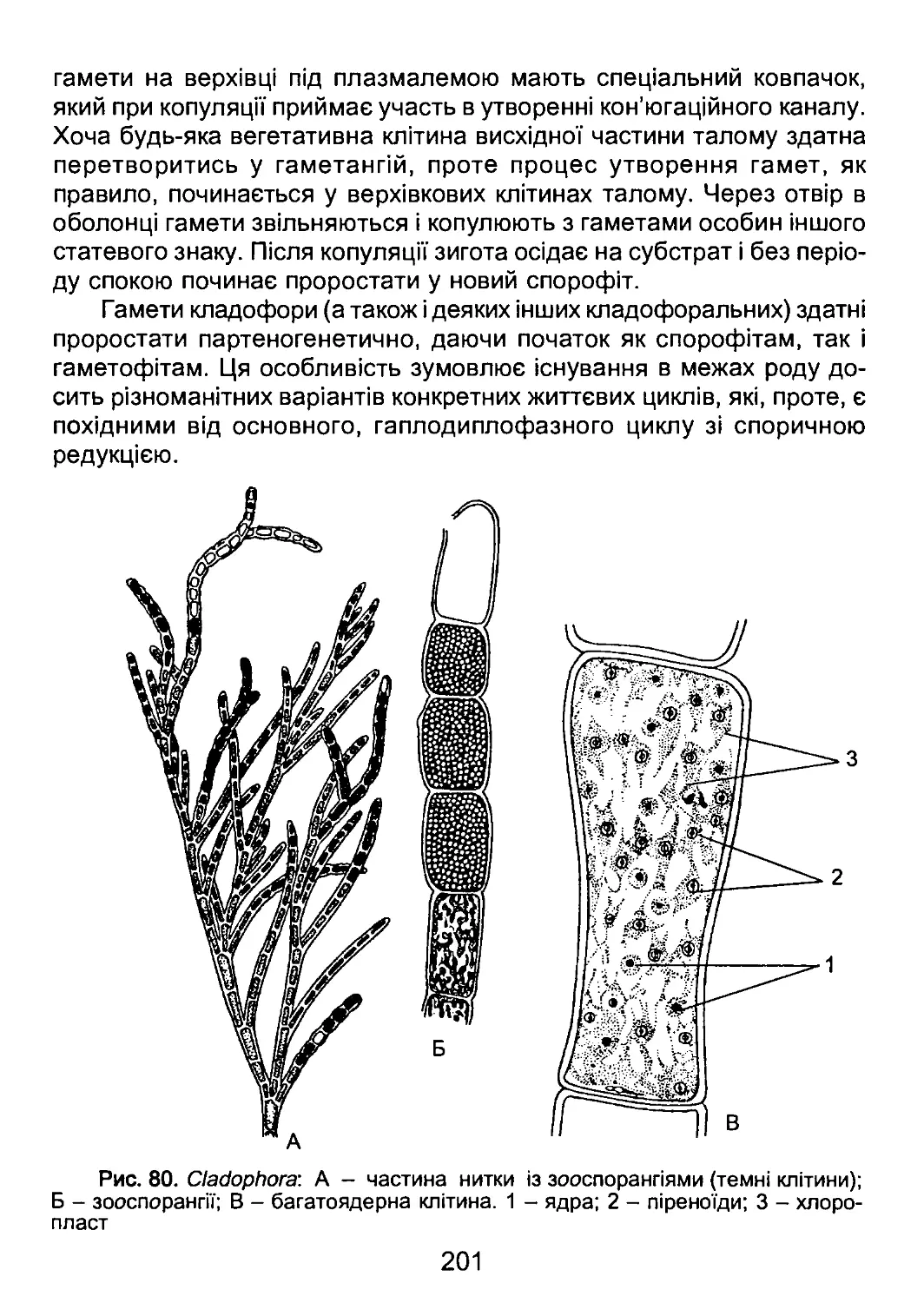
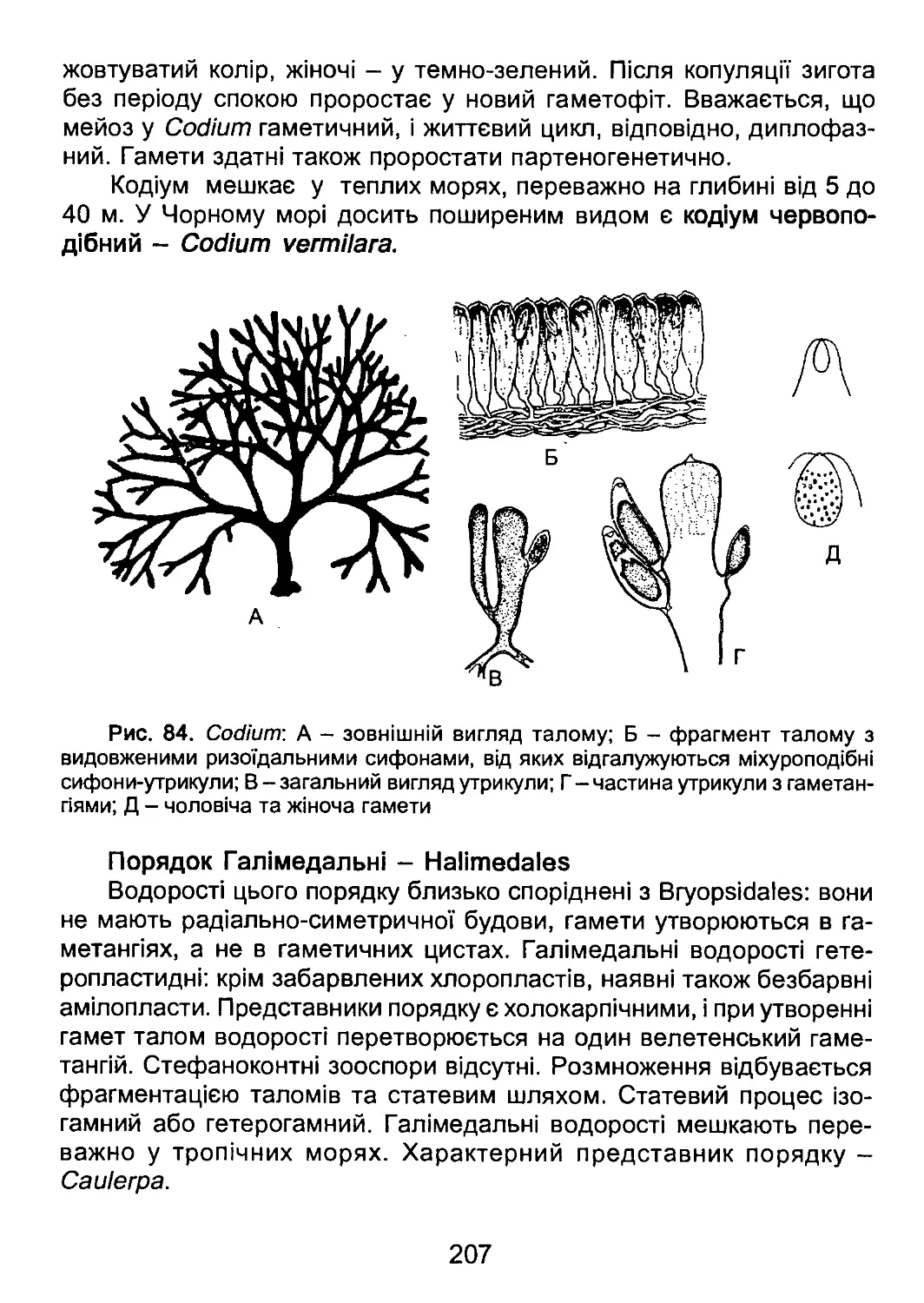
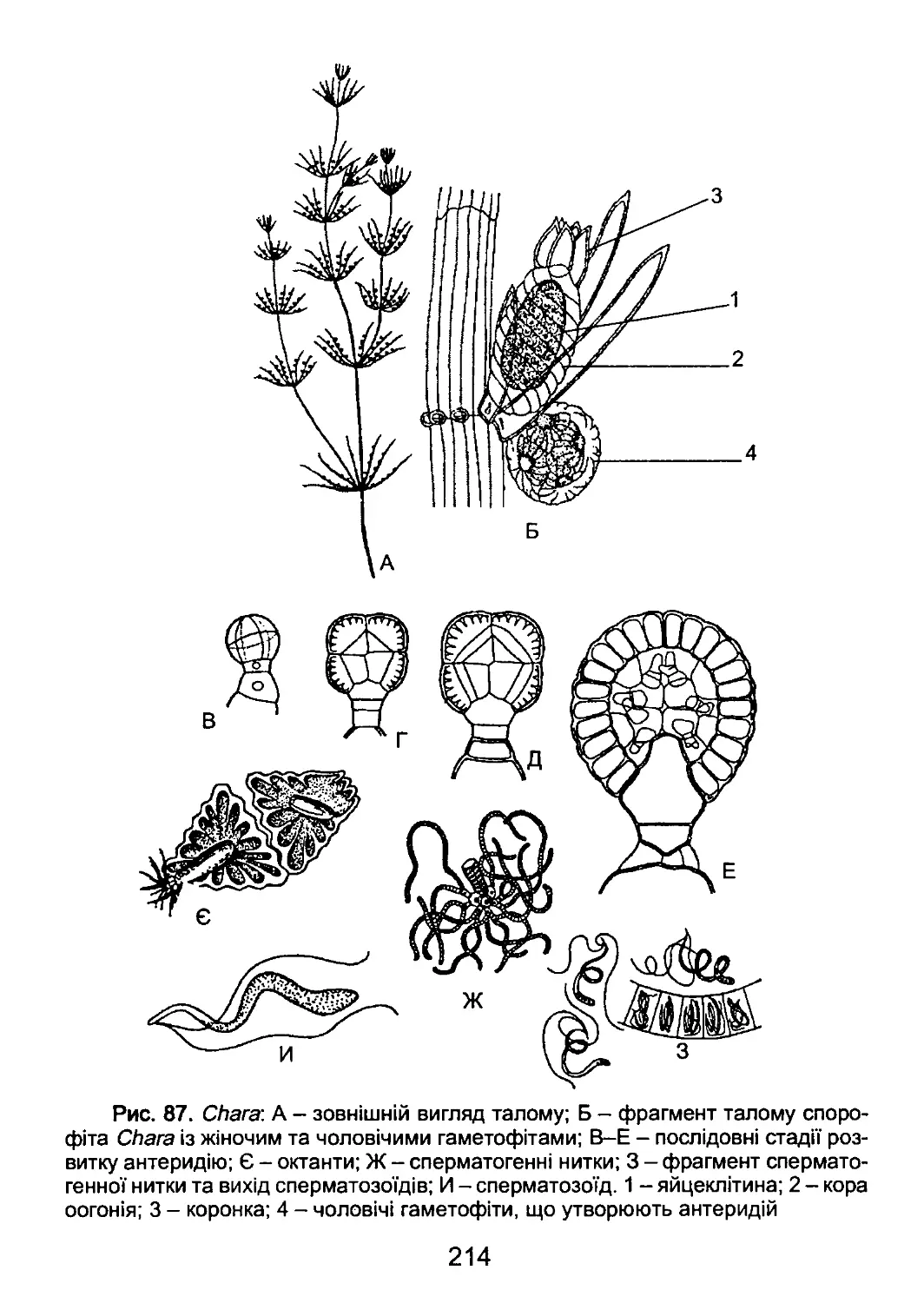
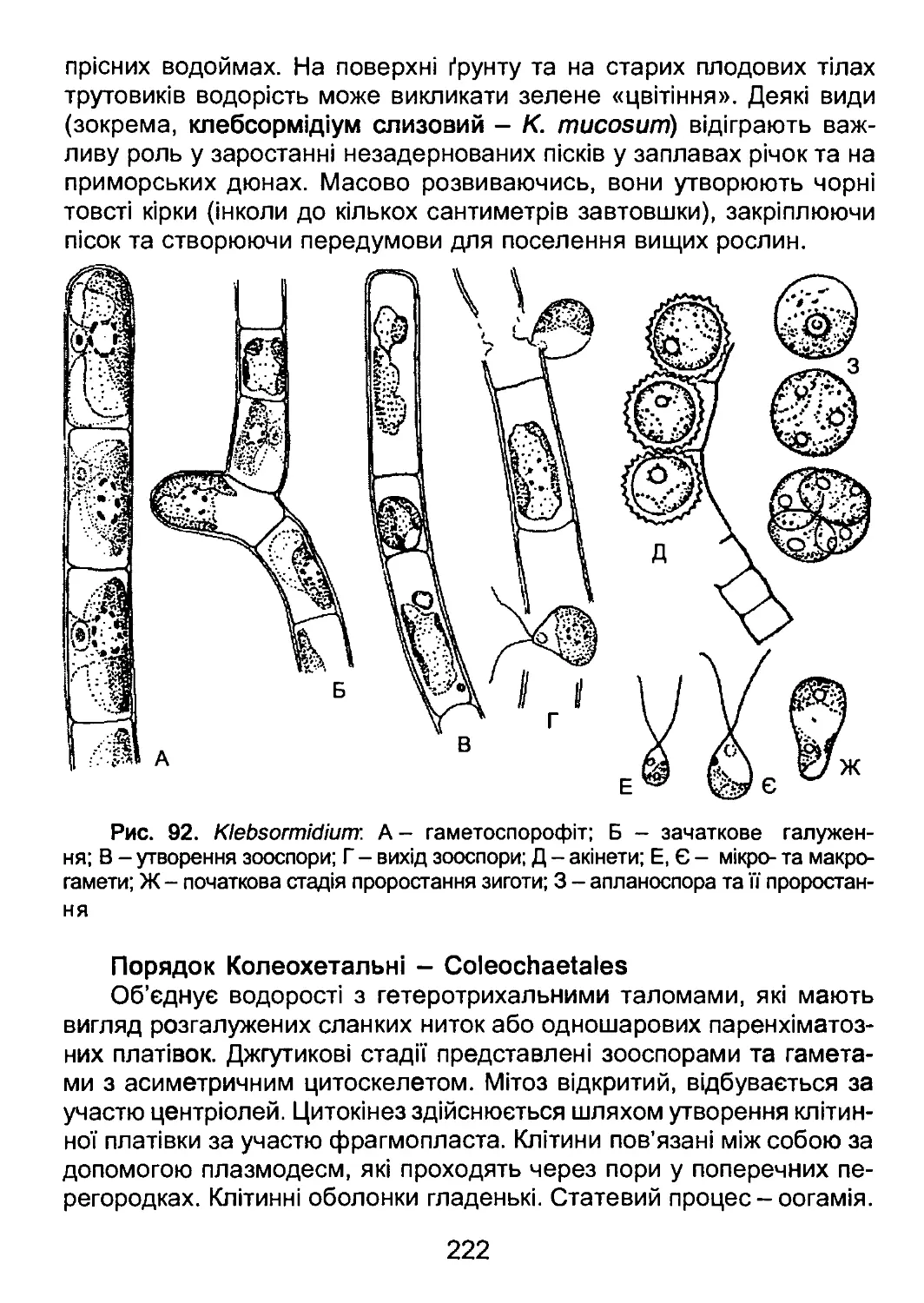
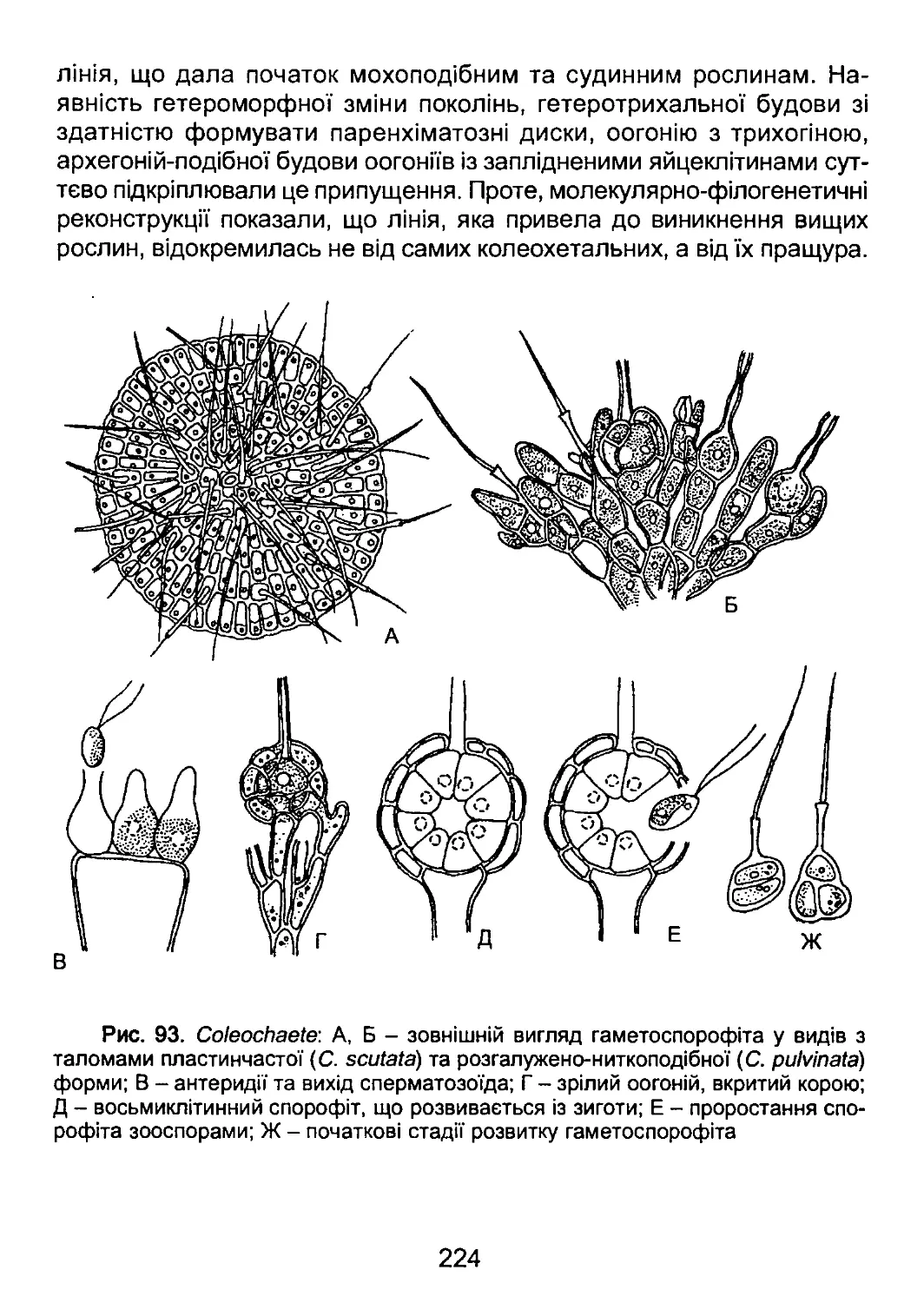
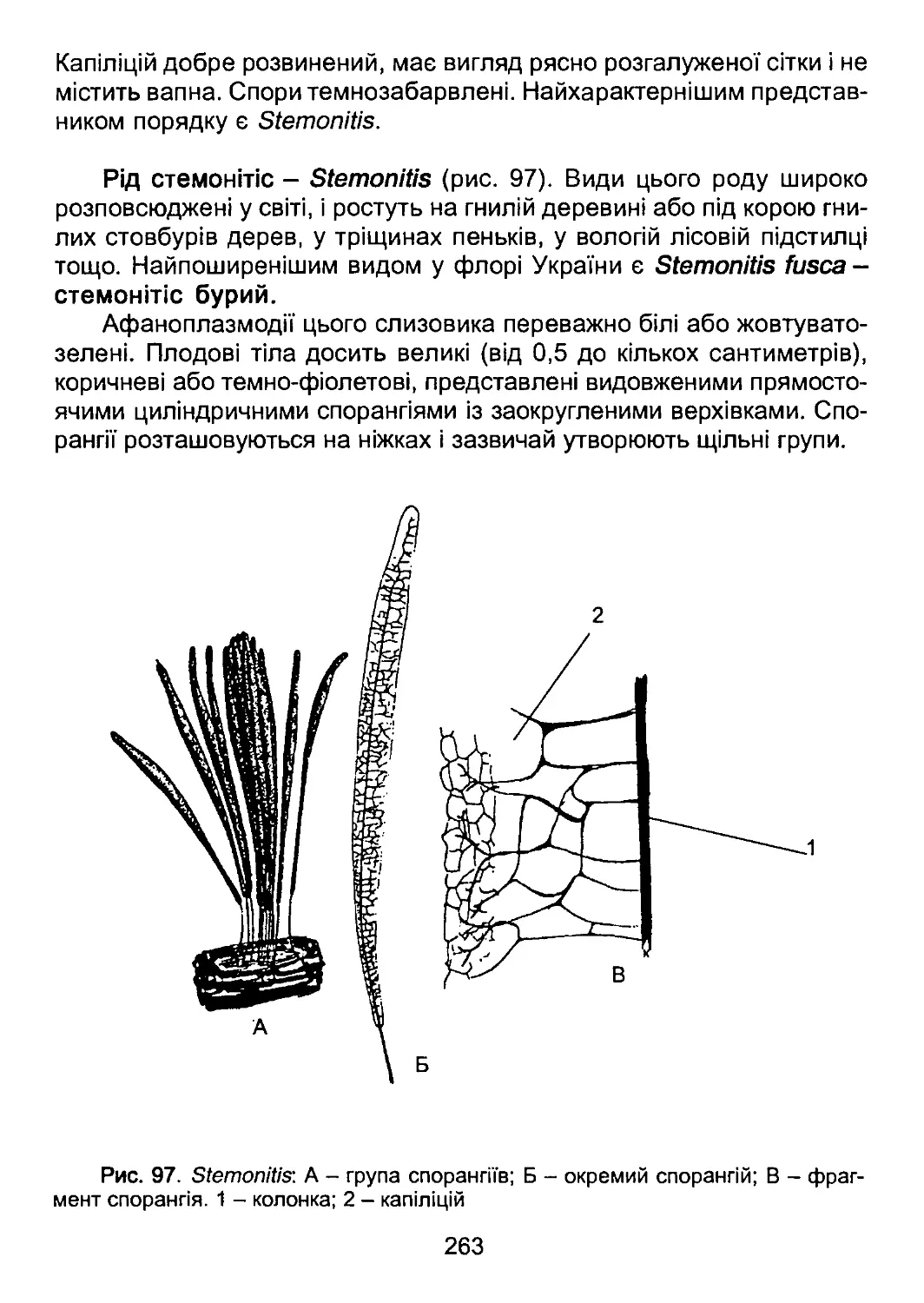
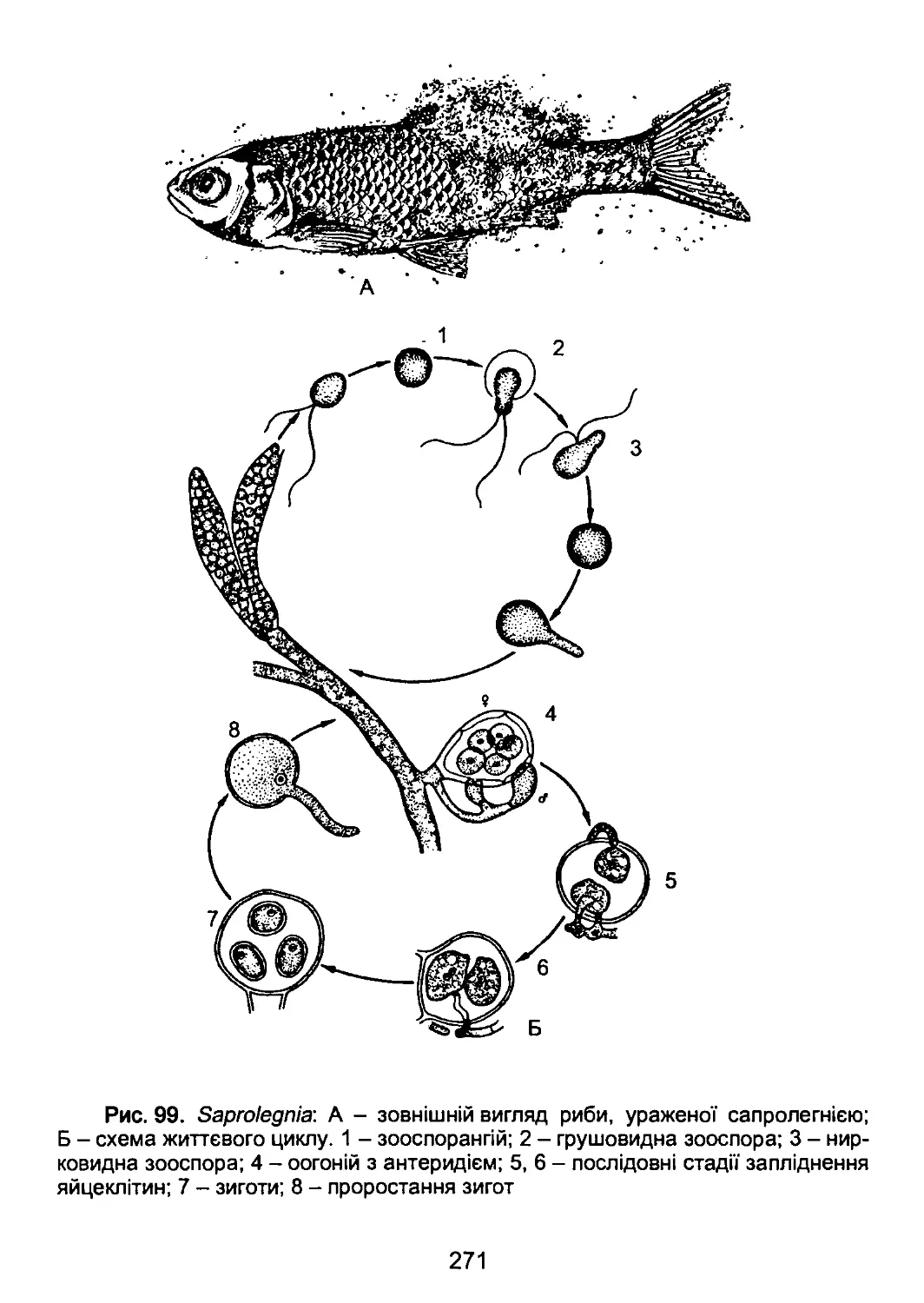
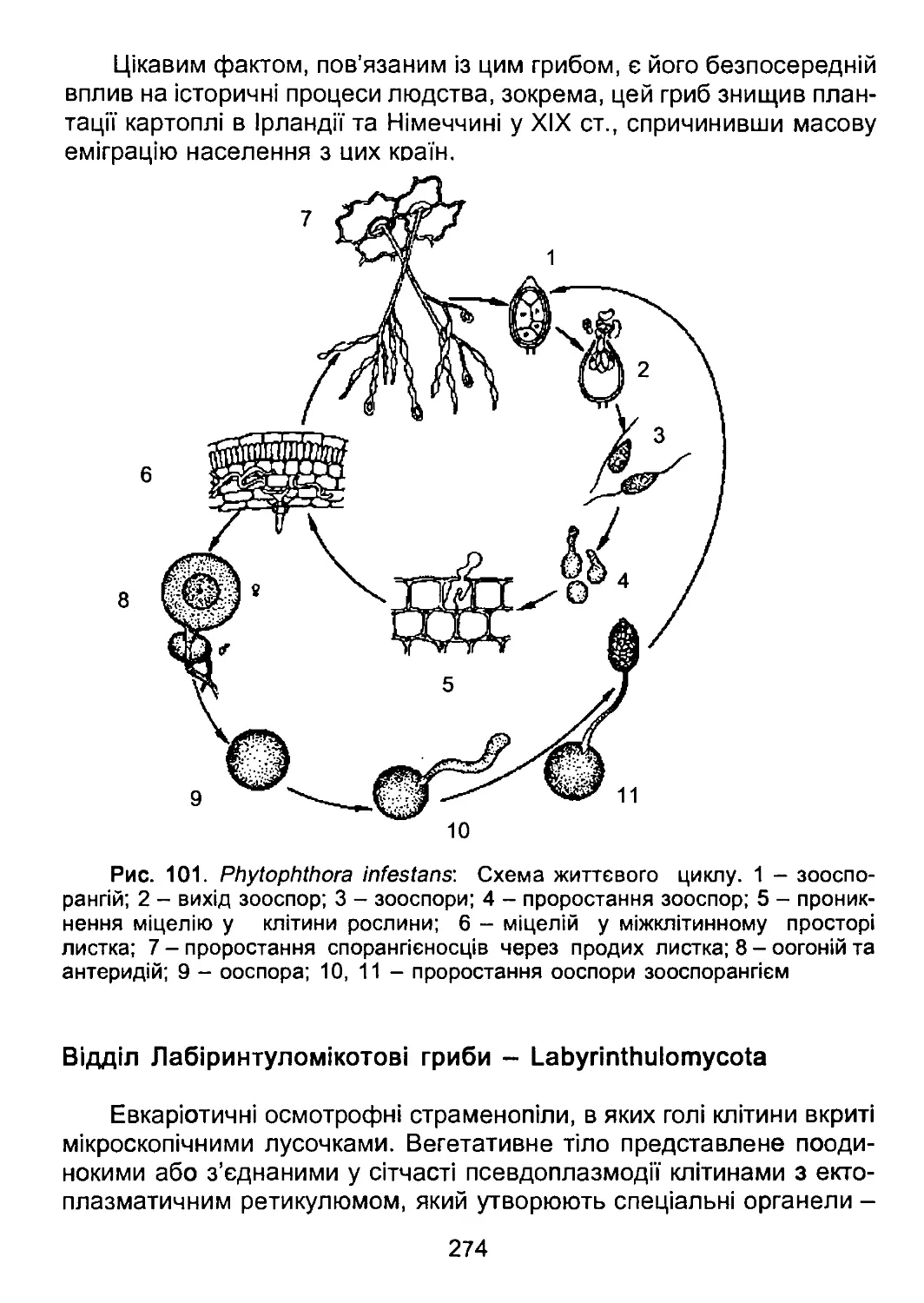
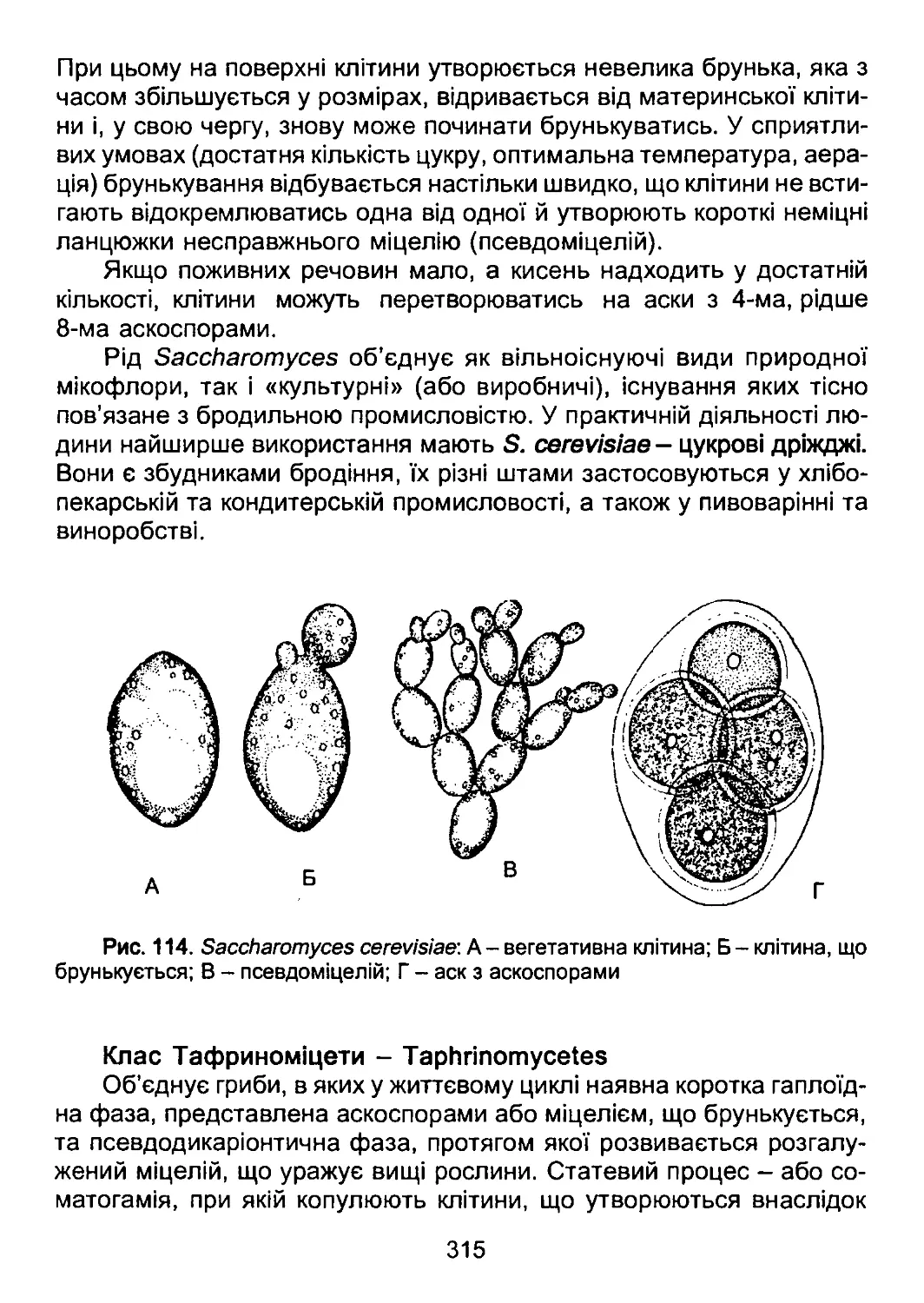
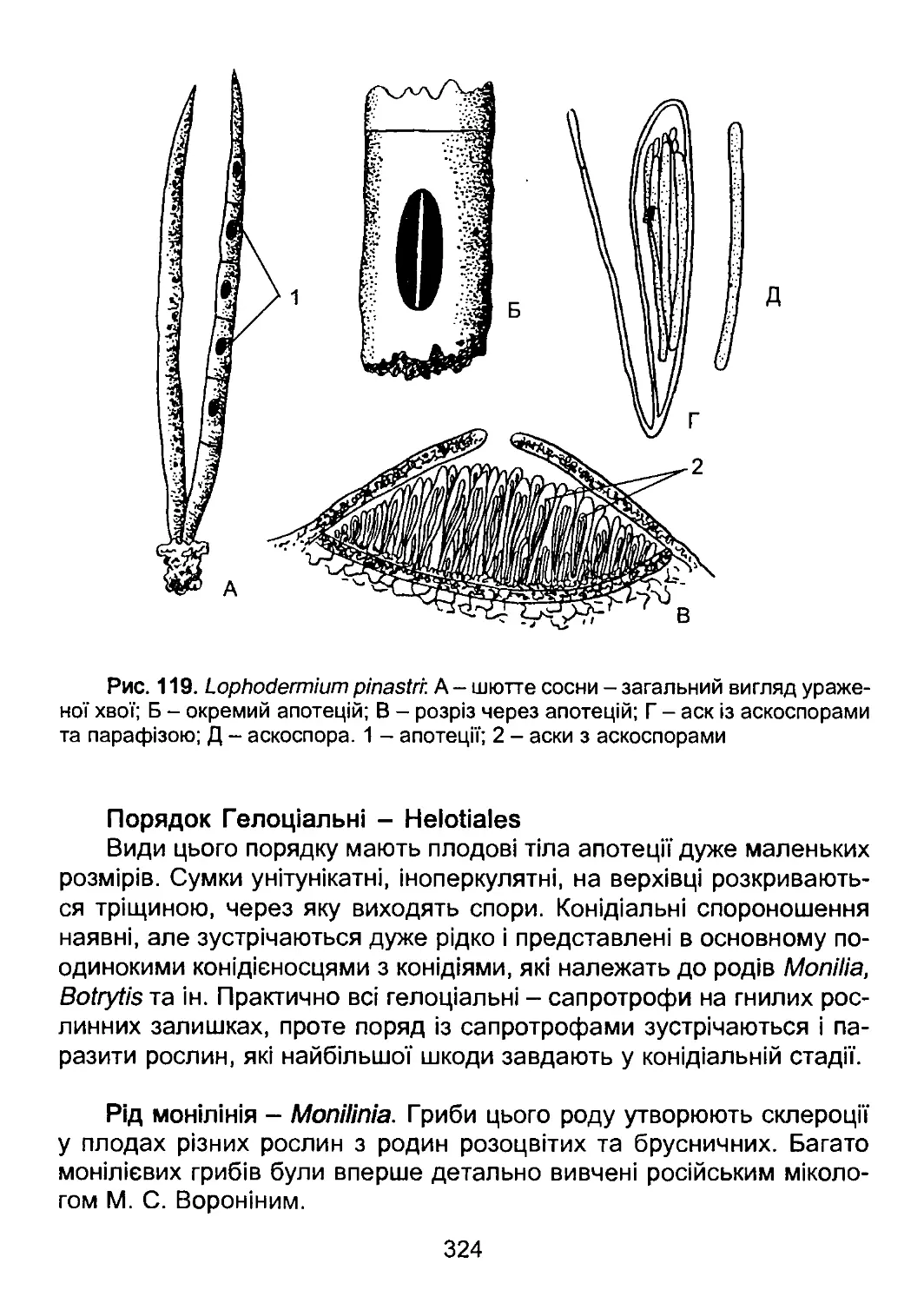
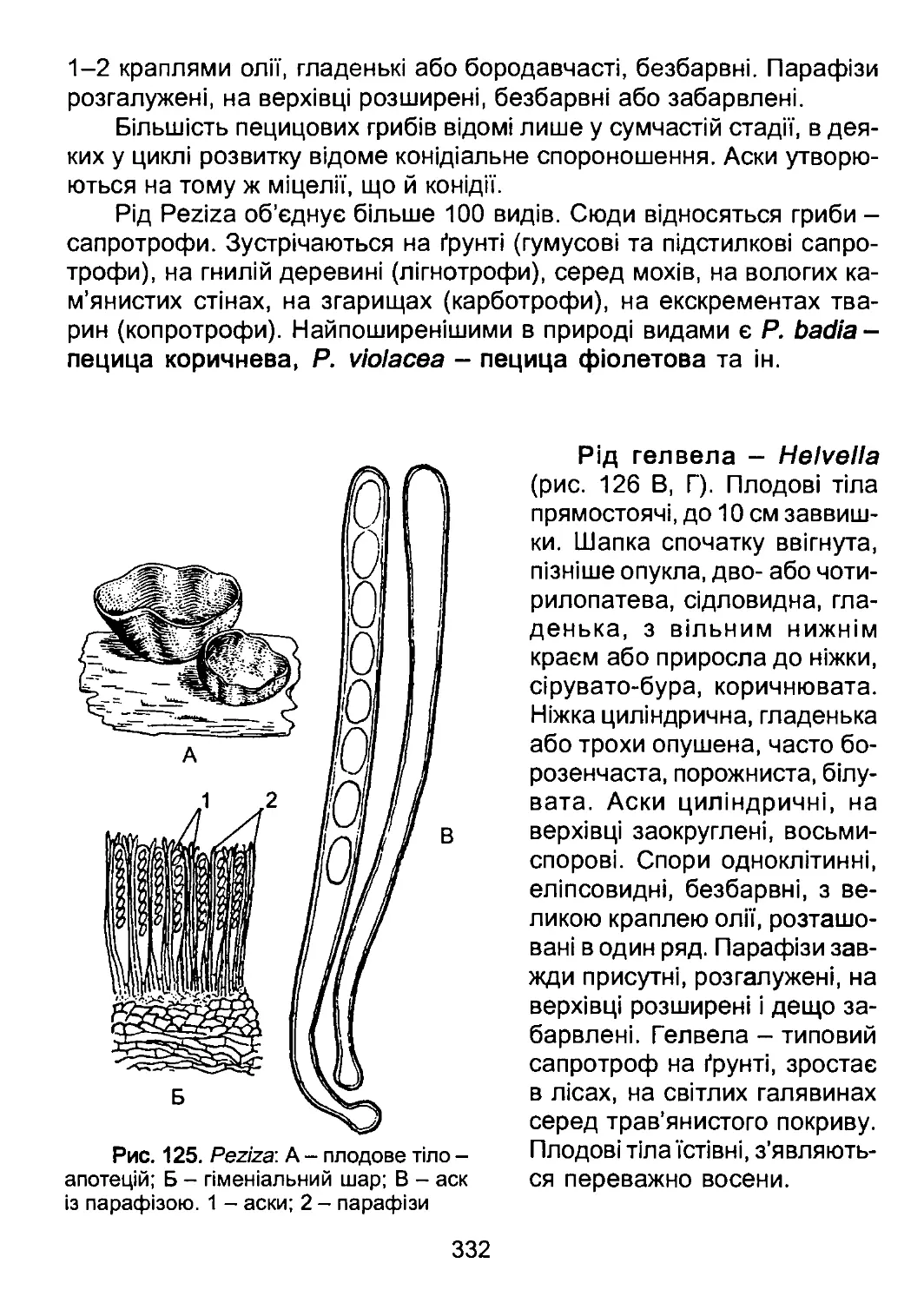
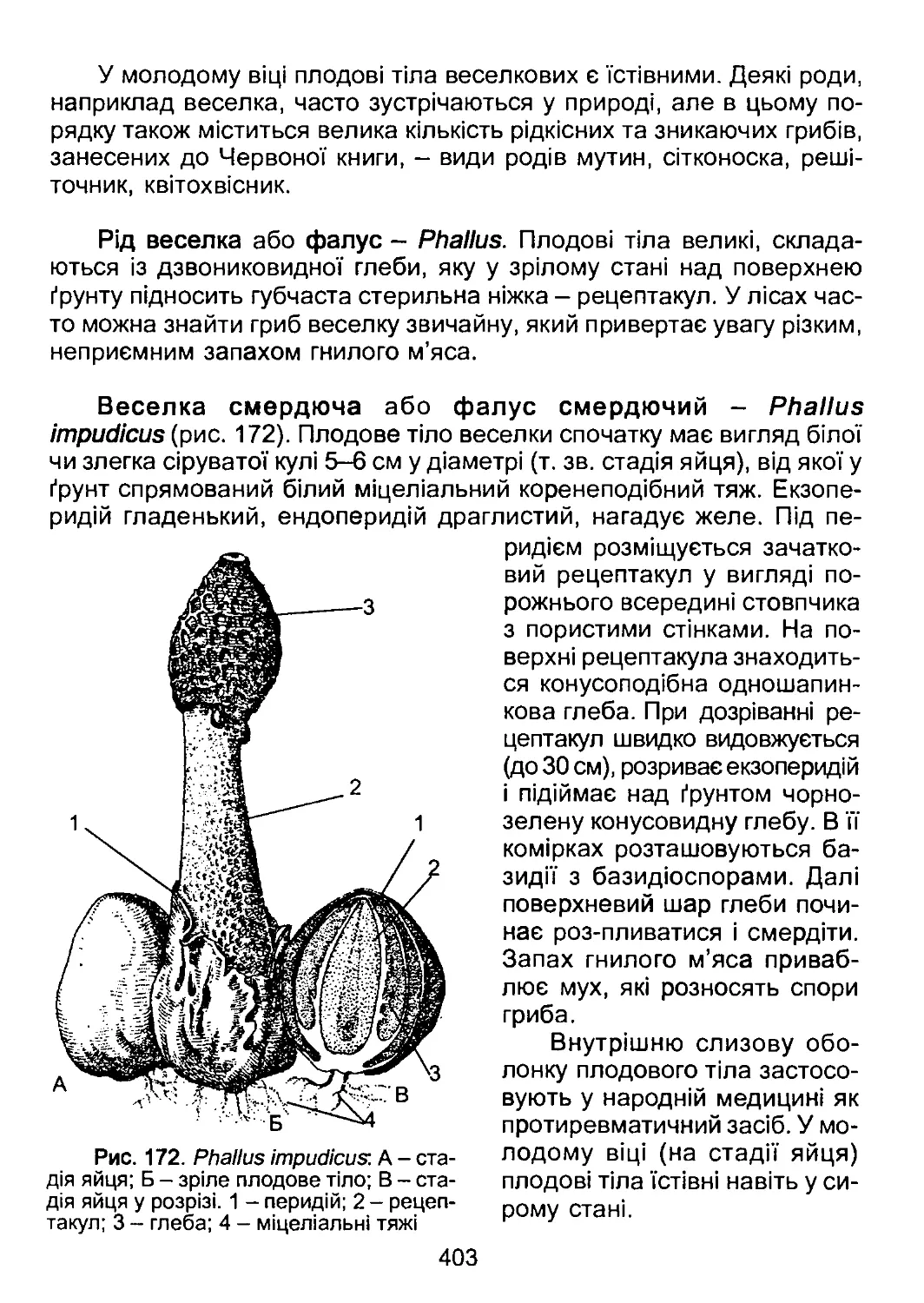
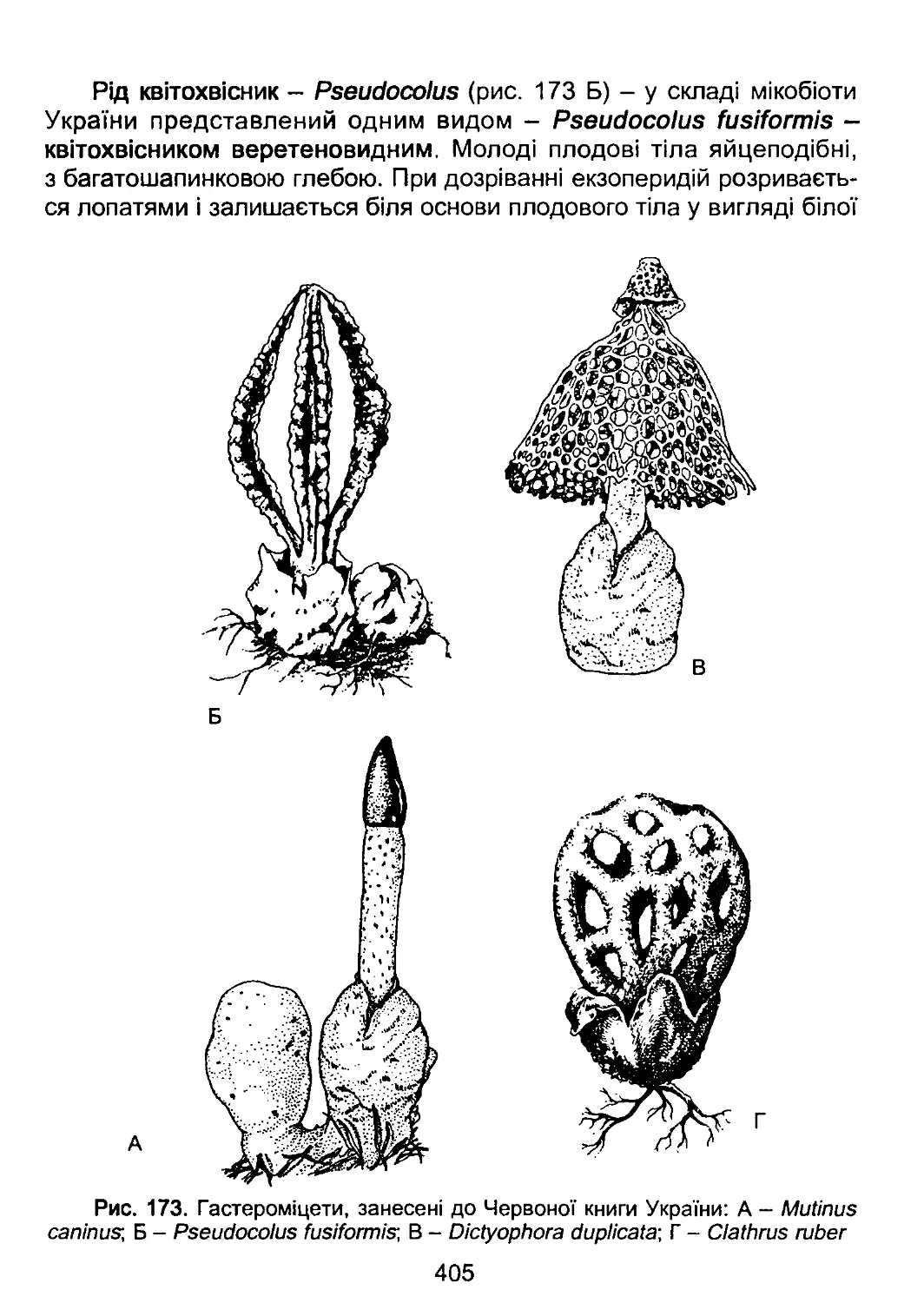
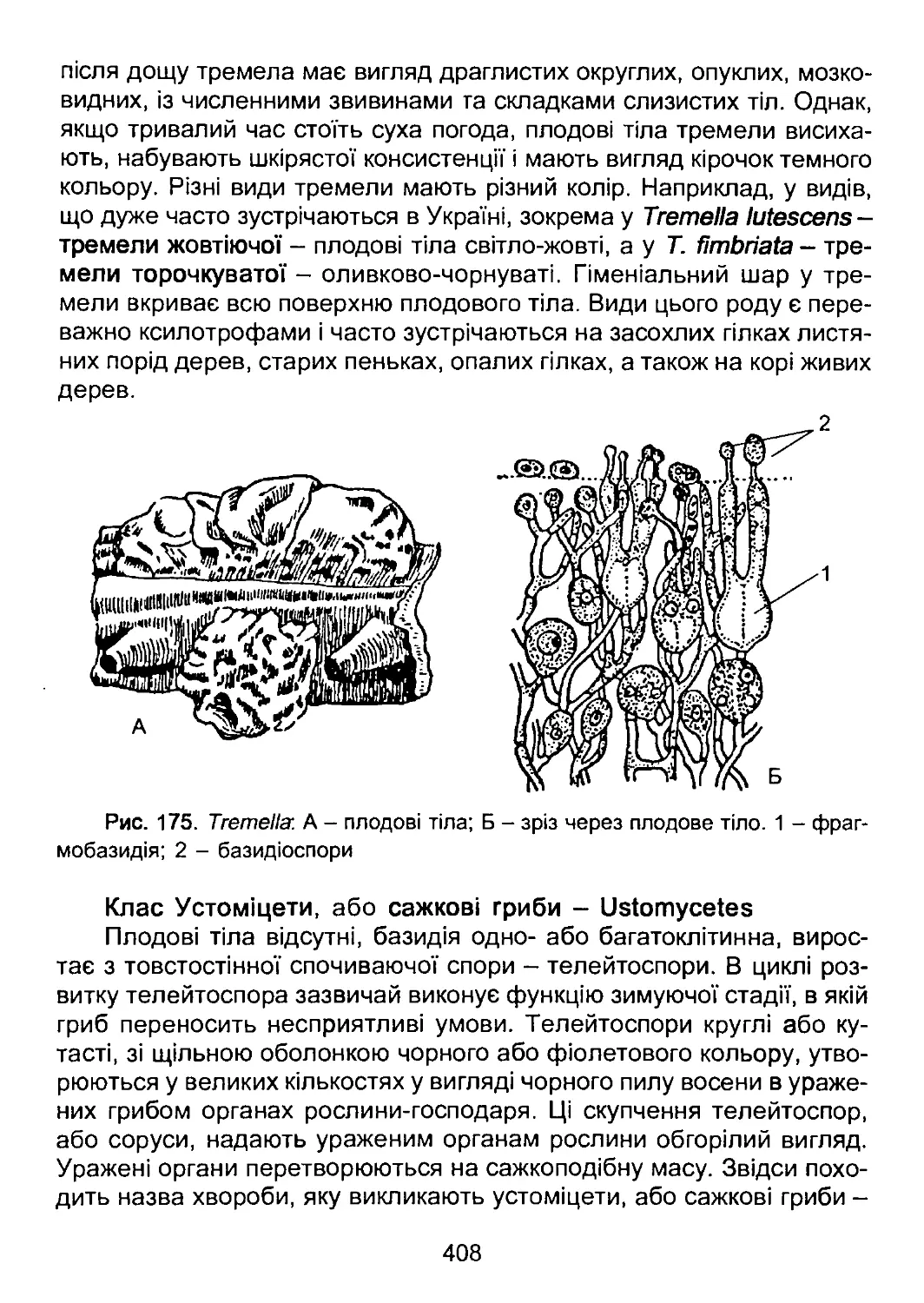
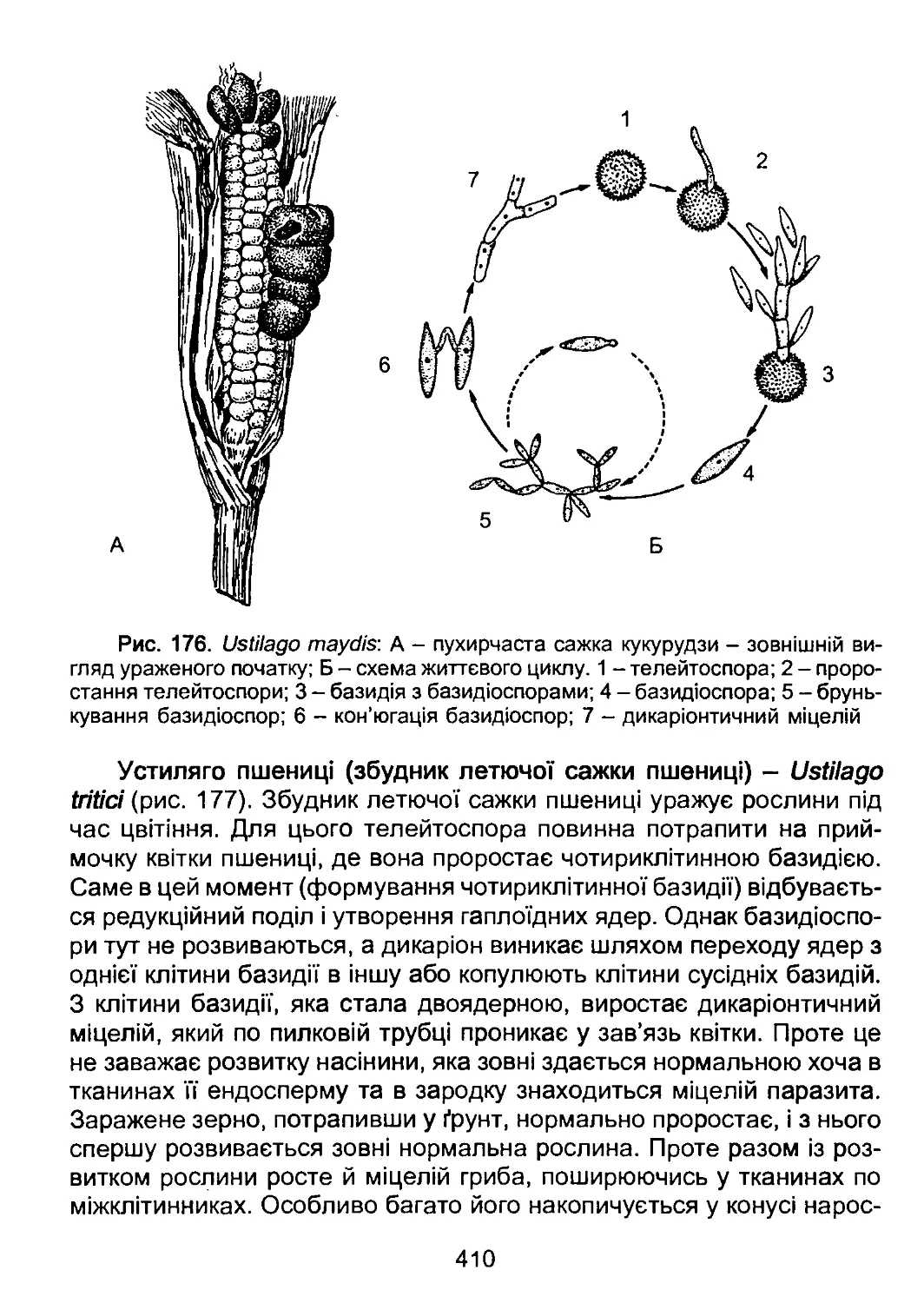
/


























































































































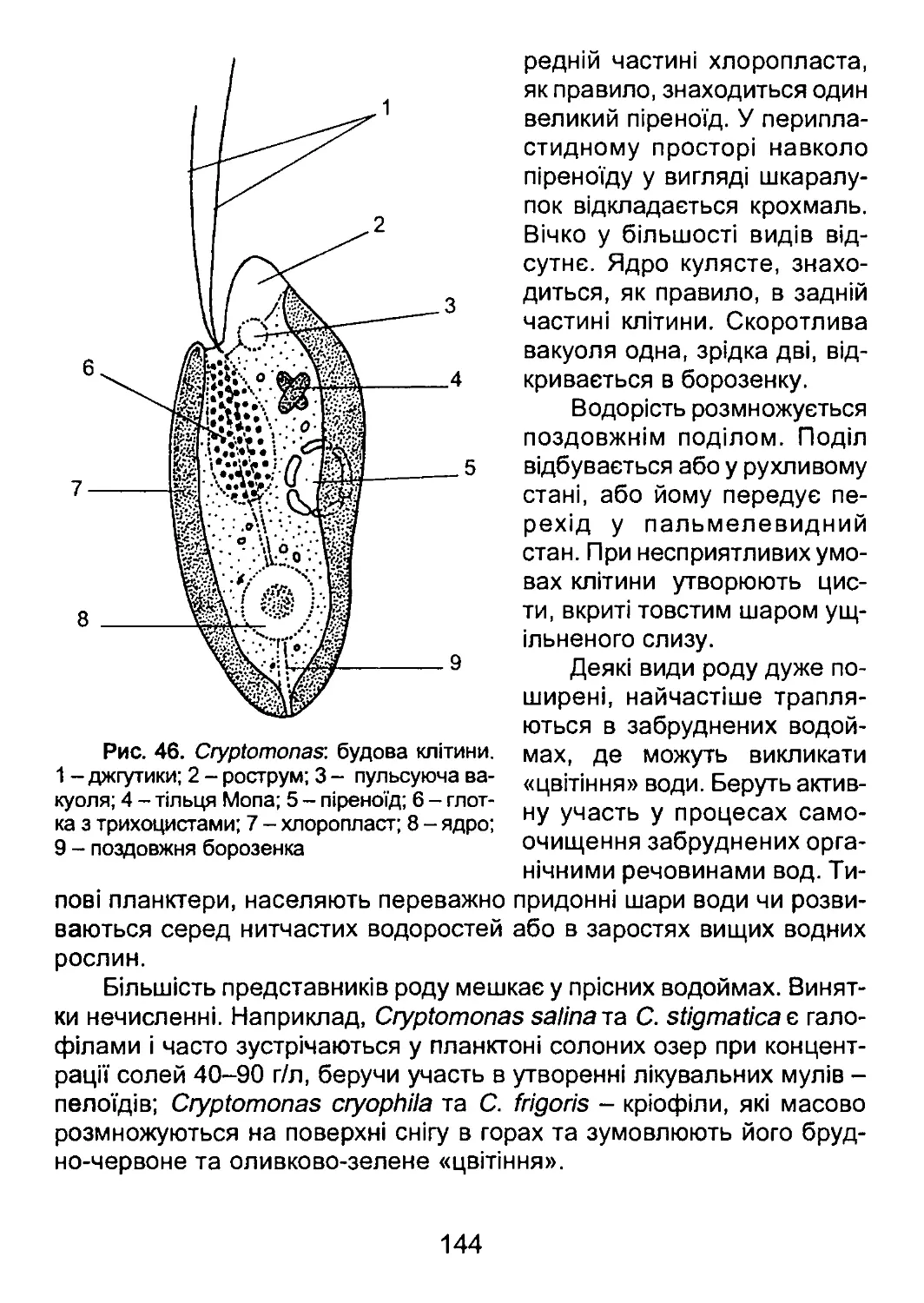



































































































































































































































































































Author: Костікоа І.Ю. Джаган В.В. Демченко Е.М. Бойко О.А. Бойко В.Р. Романенко П.О.
Tags: біологія
ISBN: 966-8458-67-2
Year: 2006




Text
Міністерство освіти і науки України
Київський національний університет
імені Тараса Шевченка
І. Ю. КОСТІКОВ, В. В. ДЖАГАН, Е. М. ДЕМЧЕНКО,
О. А. БОЙКО, В. Р. БОЙКО, П. О. РОМАНЕНКО
Ботаніка
Водорості та гриби
НАВЧАЛЬНИЙ ПОСІБНИК
2-ге видання, перероблене
За редакцією доктора біологічних наук,
професора і. Ю. Костікова
та кандидата біологічних наук В. В. Джаган
Рекомендовано Міністерством освіти і науки України
як навчальний посібник для студентів
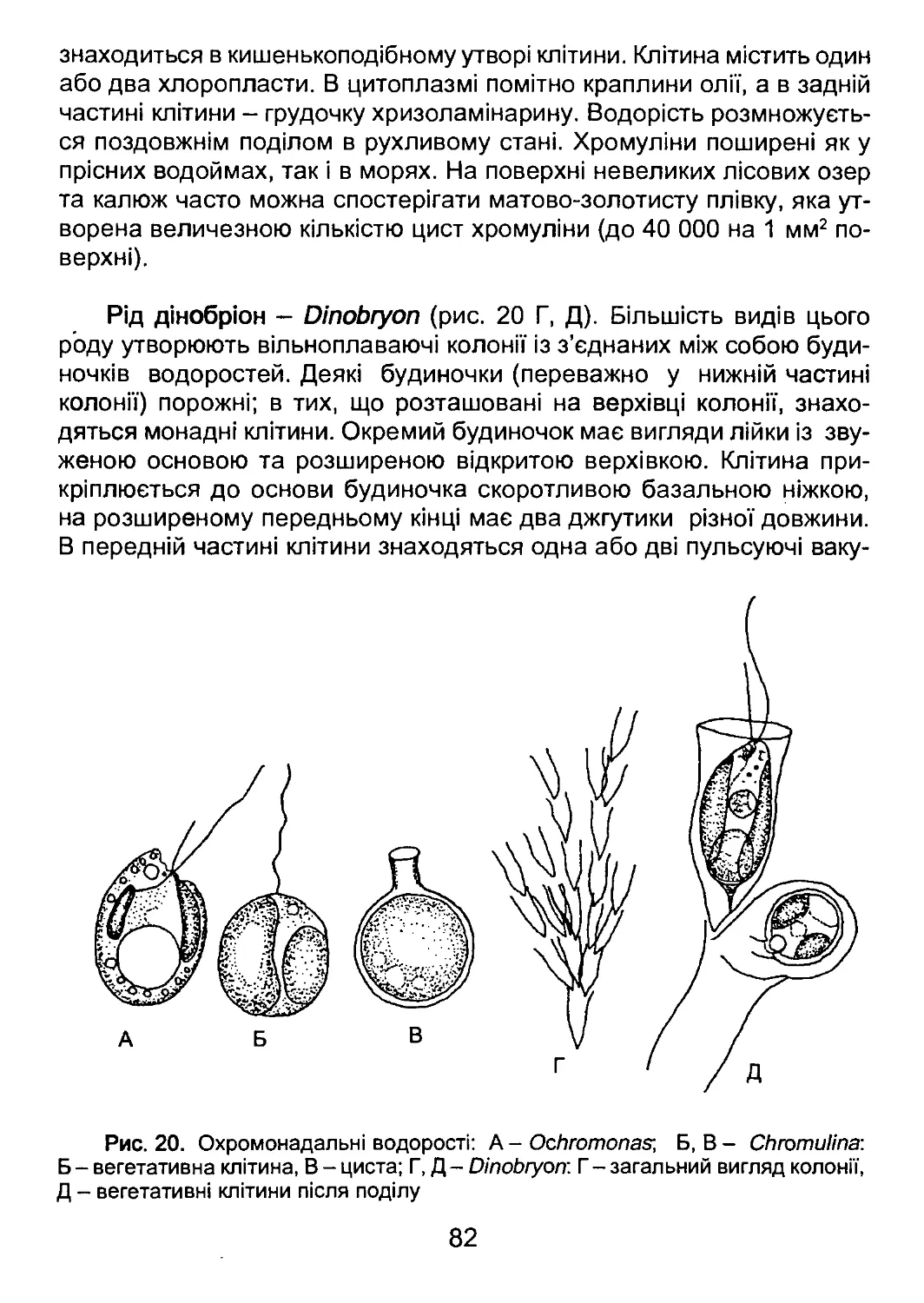
вищих навчальних закладів
Київ
2006
УДК 582.26/.28(075.8)
ББК 28.591 я73
Б86
Рекомендовано Міністерством освіти і науки України як навчальний посібник для
студентів вищих навчальних закладів
(лист № 14/18.2-70 від 17.01.2005 р.)
Рецензенти: П. М. Царанко - доктор біологічних наук, професор кафедри ботаніки НАУ,
зав. відділом фікології Інституту ботаніки ім. М. Г. Холодного НАНУ;
С. Я. Кондратюк - доктор біологічних наук, зав. відділом ліхенології і бріо-
логії Інституту ботаніки ім. М. Г. Холодного НАН України;
І. О. Дудка-доктор біологічних наук, професор, зав. відділом мікології Інсти-
туту ботаніки ім. М. Г. Холодного НАН України.
Костікоа І. Ю., Джаган В. В., Демченко Е. М., Бойко О. А., Бойко В. Р.,
Романенко П. О.
Б86 Ботаніка. Водорості та гриби: Навчальний посібник, 2-ге видання, переробл. - К.:
Арістей, 2006. - 476 с.
ІЗВИ 966-8458-67-2
Посібник містить обов’язковий мінімум програмного матеріалу у частині, що стосуєть-
ся об'єктної підготовки фахівця-біолога (лабораторний практикум, учбова практика) з нор-
мативного курсу "Ботаніка” освітньо-кваліфікаційного рівня “бакалавр” за розділами, що
охоплюють т. зв. “нижчі рослини”. У посібнику наводяться сучасні дані щодо місця водоро-
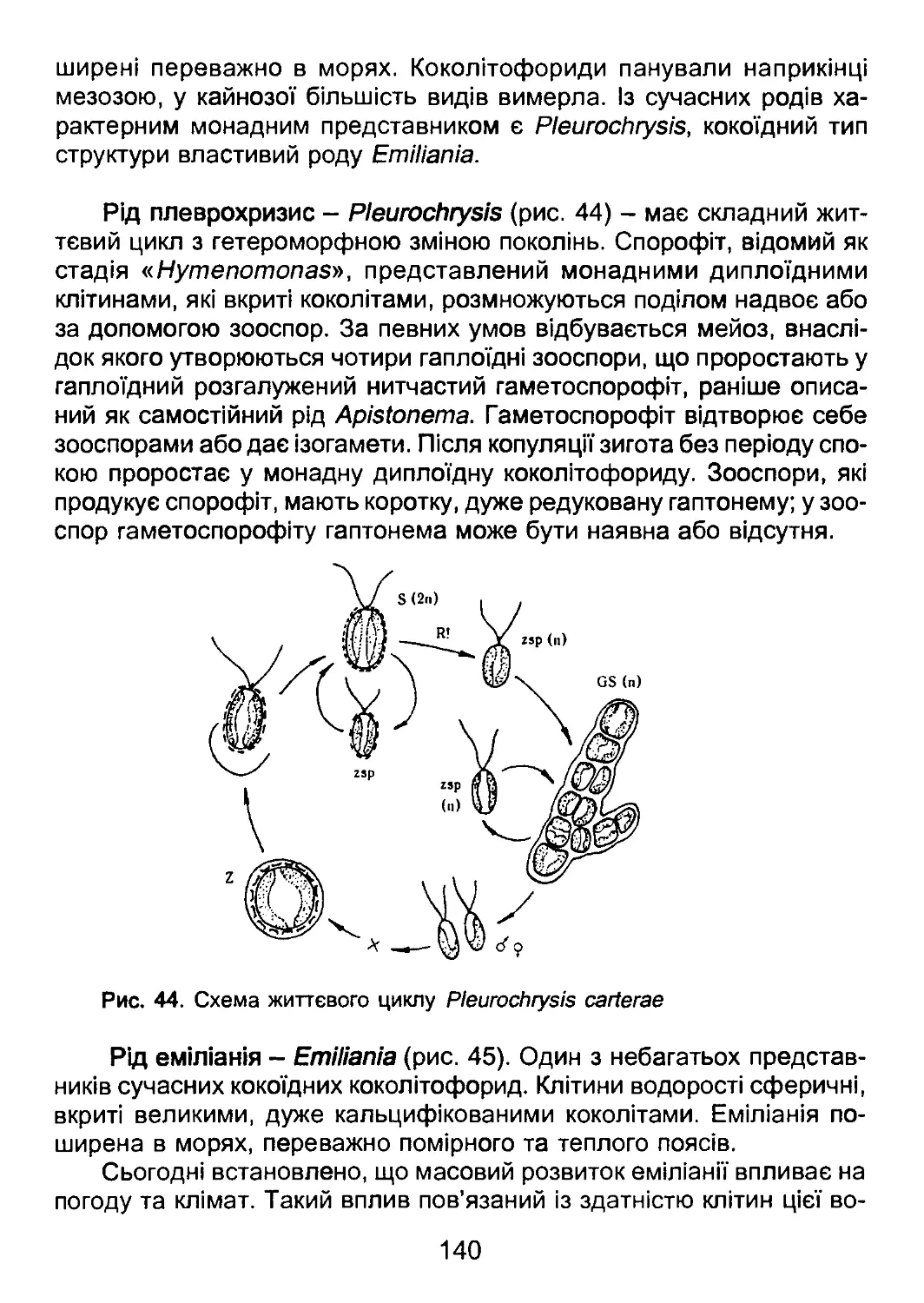
стей та грибів у системі органічного світу, розглядаються комплекси ознак відділів прока-
ріотичних та евкаріотичних водоростей, грибоподібних організмів (слизовиків та псевдо-
грибів) та справжніх грибів на морфолого-онтогенетичному, цитологічному та біохімічному
рівнях. Конспективно викладено принципи сучасної таксономії всіх відділів водоростей та
грибів на рівні класів та основних порядків. Детально розглянуто морфологію, розмножен-
ня, життєві цикли, поширення та практичне значення основних представників.
Для студентів та викладачів університетів та педагогічних вищих навчальних закладів,
що здійснюють підготовку за напрямом "Біологія".
УДК 582.26/.28(075.8)
ББК 28.591я73
© І. Ю. Костіков, В. В. Джаган,
Е. М. Демченко, О; А. Бойко,
В. Р. Бойко, П. О. Романенко, 2006
IЗВN 966-8458-67-2 © Арістей, 2006
Зміст
Передмова................................................................5
Вступ....................................................................6
Місце водоростей та грибів у системі органічного світу...................9
РОЗДІЛ 1. Водорості......................................................25
Основні систематичні ознаки водоростей різних відділів.............25
Біохімічні ознаки...........................................-25
Цитологічні ознаки..........................................29
Морфологічна різноманітність................................40
Розмноження водоростей......................................44
Життєві цикли водоростей....................................-48
Прокаріотичні водорості............................................52
Відділ Синьозелені водорості - Суалорітуїа.................. 52
Водорості дискокристати............................................64
Відділ Евгленофітові водорості - Еидіелоріїуіа..............64
Водорості тубулокристати...........................................72
Амебо-флагеляти.................................................72
Відділ Хлорарахніофітові водорості - Сіїїогагасітпіорітуіа..72
Страменопіли. Група відділів хромофітових водоростей............74
Відділ Рафідофітові водорості - Карітісіорітуїа..............74
Відділ Золотисті водорості - СІтгуворІтуїа...................78
Відділ Евстигматофітові водорості - Еивіідтаіорітуїа........35
Відділ Жовтозелені водорості - Халііторітуїа................ 87
Відділ Бурі водорості - Рііаеорітуіа........................95
Відділ Діатомові водорості - ВасіІІагіорІтуіа...............105
Відділ Диктіохофітові водорості - Оісіуосіторітуїа..........121
Альвеоляти......................................................124
Відділ Динофітові водорості - Оіпоріїуіа....................124
Водорості платикристати............................................136
Відділ Гаптофітові водорості - Наріорітуїа..................136
Відділ Криптофітові водорості - Сгуріорітуіа................141
Відділ Глаукоцистофітові водорості - (ЗІаисосувіорІтуїа.....145
Відділ Червоні водорості - КІтосіорІтуїа....................149
Відділ Зелені водорості - Сіїїогорітуїа.....................168
РОЗДІЛ 2. Гриби..........................................................225
Основні систематичні ознаки грибів різних відділів.................226
Біохімічні ознаки...........................................226
Фізіологічні ознаки, пов’язані з живленням..................227
Цитологічні ознаки..........................................228
Морфологія вегетативного тіла...............................234
З
Розмноження............................................... 238
Життєві цикли..............................................246
Еколого-трофічні групи грибів..............................248
Гриби дискокристати...............................................249
Відділ Акразіомікотові слизовики - Асгазіотусоіа...........249
Гриби тубулокристати..............................................253
Амебо-флагеляти...............................................253
Відділ Міксомікотові слизовики - Мухотусоіа................254
Страменопіли. Гпула відділів псевдогрибів.....................265
Відділ Оомікотові гриби - Оотусоіа.........................266
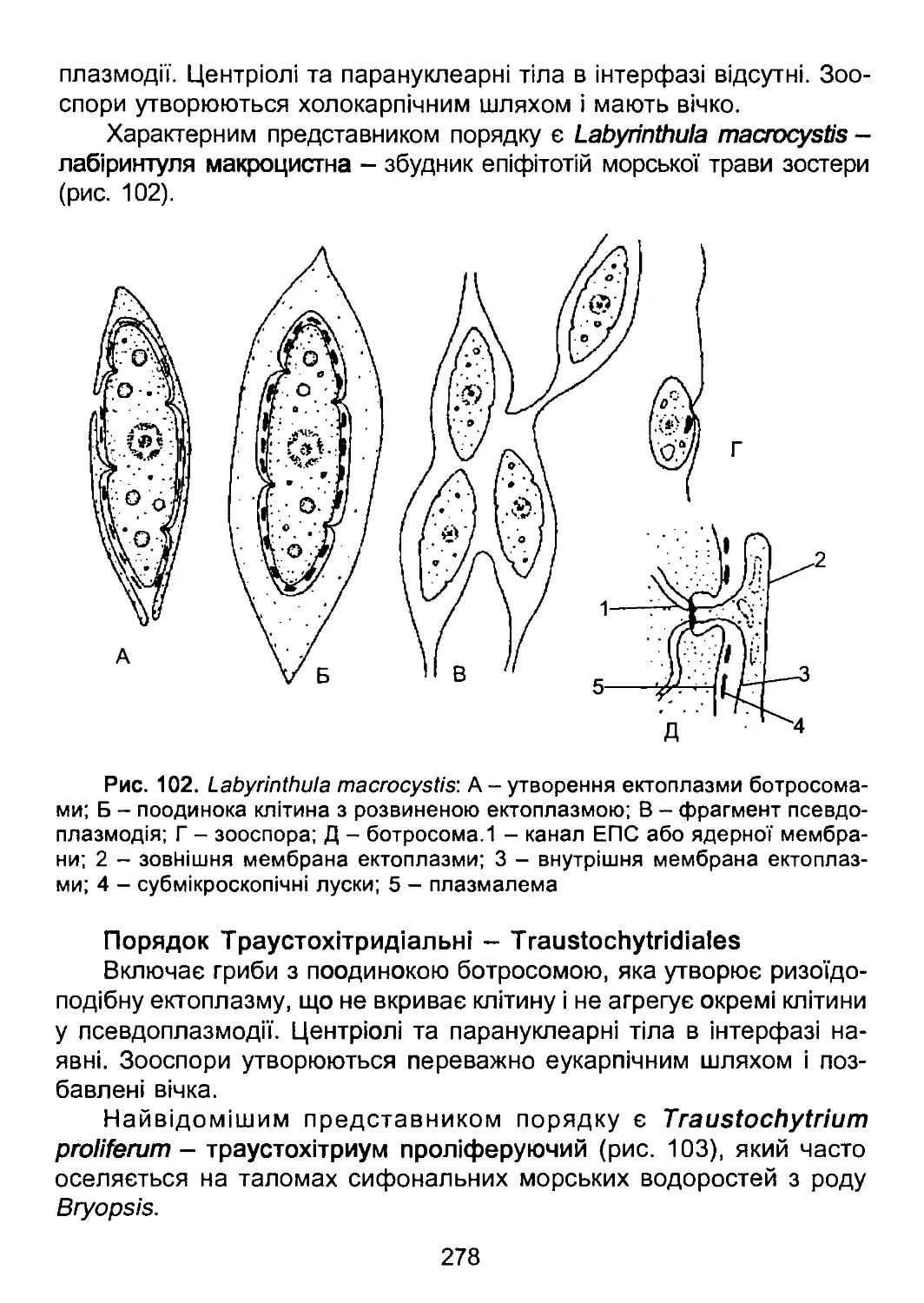
Відділ Лабіринтуломікотові гриби - ІаЬугіпФиіотусоІа.......274
Відділ Гіфохітриомікотові гриби - Нурітосіїуігіотусоіа.....279
Гриби платикристати...............................................282
Відділ Плазмодіофоромікотові слизовики -
РІазтосІіорІтоготусоіа.....................................283
Справжні гриби................................................288
Відділ Хітридіомікотові гриби - Сіїуігісііотусоіа..........289
Відділ Зигомікотові гриби - 2удотусо1а.....................299
Відділ Аскомікотові гриби - Азсотусоїа.....................305
Відділ Базидіомікотові - Вазісііотусоіа....................338
Анаморфні гриби або “Оеиіеготусеіез”..................... 416
Ліхенізовані гриби або лишайники.......................... 424
Покажчик латинських назв таксонів......................................444
Покажчик українських назв таксонів.....................................453
Покажчик термінів......................................................462
Рекомендована література...............................................471
4
ПЕРЕДМОВА
Підручники та навчальні посібники із систематики водоростей та
грибів для студентів університетів за напрямом підготовки "біологія”
в Україні та країнах СНД не видавались майже чверть сторіччя. За
цей час альгологія та мікологія пережили дві революції, пов'язані з
широким впровадженням у 70-80-х роках у практику систематичних
досліджень методів порівняльної цитології, та у 90-х роках - методів
молекулярної таксономії. Це докорінно змінило уявлення про систе-
му прокаріот та нижчих евкаріот, та про місце у цій системі водорос-
тей, грибоподібних організмів та грибів. У методичному забезпеченні
нормативного курсу ботаніка за розділами, що стосуються цих груп
організмів, утворився інформаційний вакуум.
Даний посібник спрямований на часткове розв’язання цієї про-
блеми, але не претендує на повноту викладу нормативних частин
альгології та мікології. Він має забезпечити лише необхідну інформа-
ційну підтримку процедурним формам навчальної роботи - лабора-
торному практикуму та учбовій практиці, і лише у тій частині, що сто-
сується об’єктної підготовки студента з основ альгології та мікології.
Крім того, посібник містить програмний матеріал, який на біологічно-
му факультеті Київського національного університету студенти ма-
ють опрацьовувати самостійно.
Автори щиро вдячні всім викладачам, науковцям і студентам, які
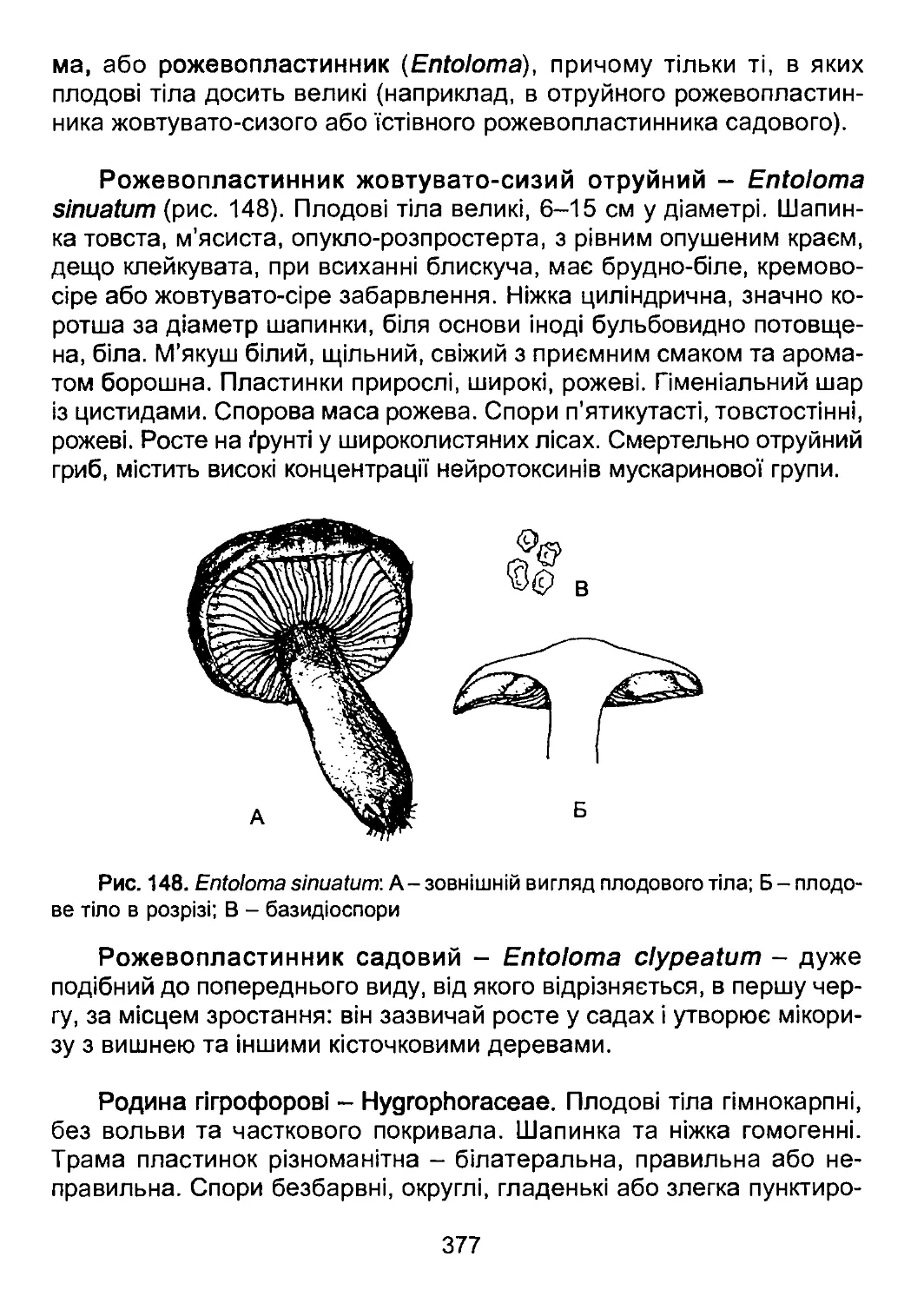
надали консультативну допомогу, висловили критичні зауваження та
побажання. Особлива подяка О. В. Рибчинському, А. А. Кривенді та
О. М. Іваненко за технічну допомогу при оформленні тексту рукопису
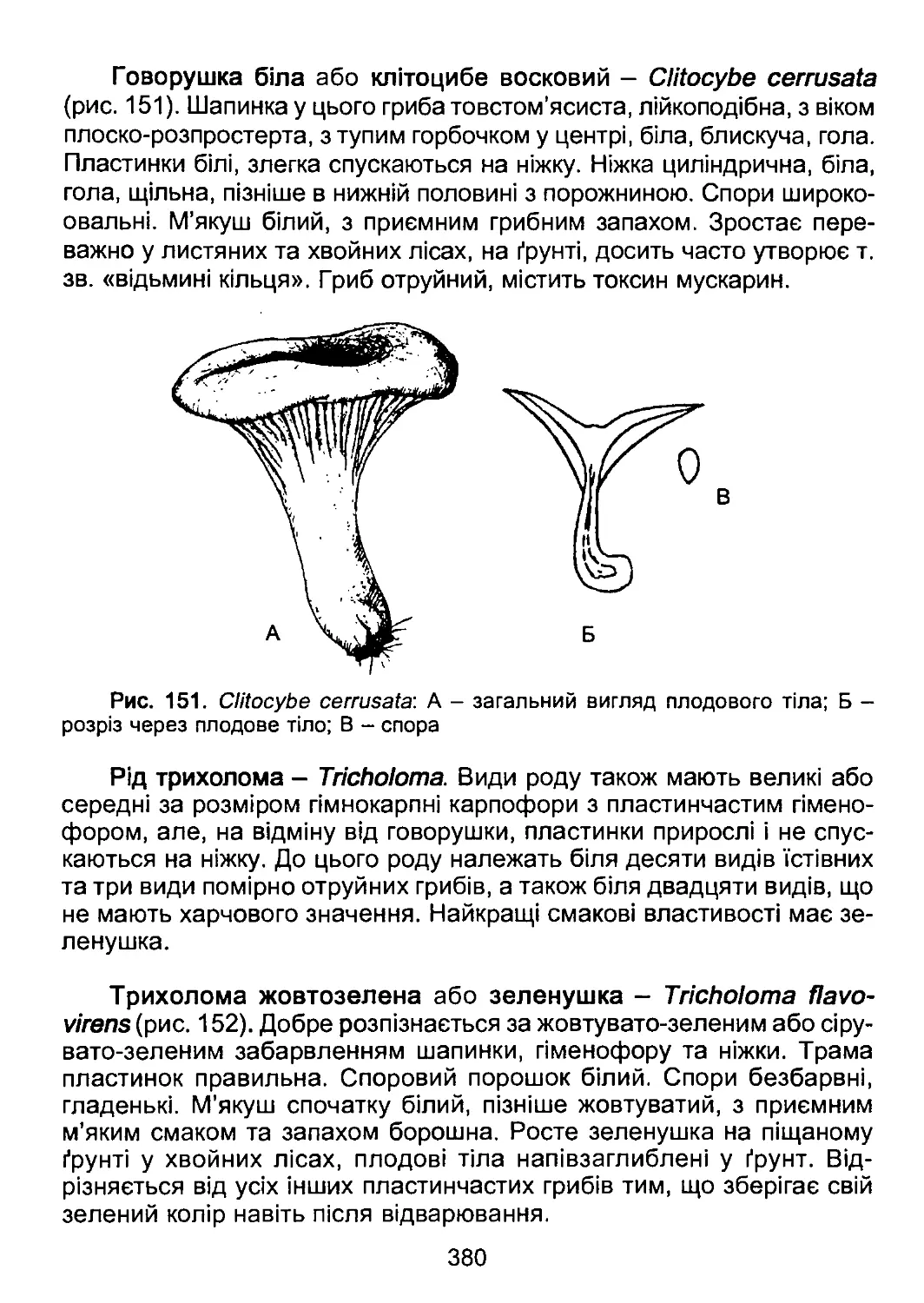
й ілюстрацій та С. Я. Кондратюку за цінні поради і зауваження сто-
совно розділу, присвяченому лишайникам.
5
ВСТУП
З античних часів, згідно з поглядами давньогрецького філософа
та натураліста Арістотеля (384-322 р. до н. е.), світ живих істот стали
розділяти на тваринний та рослинний. «Батьком» науки про рослин-
ний світ - ботаніки - прийнято вважати учня Арістотеля - Теофраста
(370-285 р. до н. е.), який написав дві перші фундаментальні ботанічні
праці - «Причини рослин» у шести книгах та дев’ятитомну працю «До-
слідження рослин». Теофраст також ввів у наукову літературу термін
«тусоз» - гриб, від якого далі пішла назва одного з розділів ботаніки
- мікології (від грецьк. тусоз - гриб та Іодоз - наука), тобто науки
про гриби.
Перші наукові описи макроскопічних водоростей були виконані
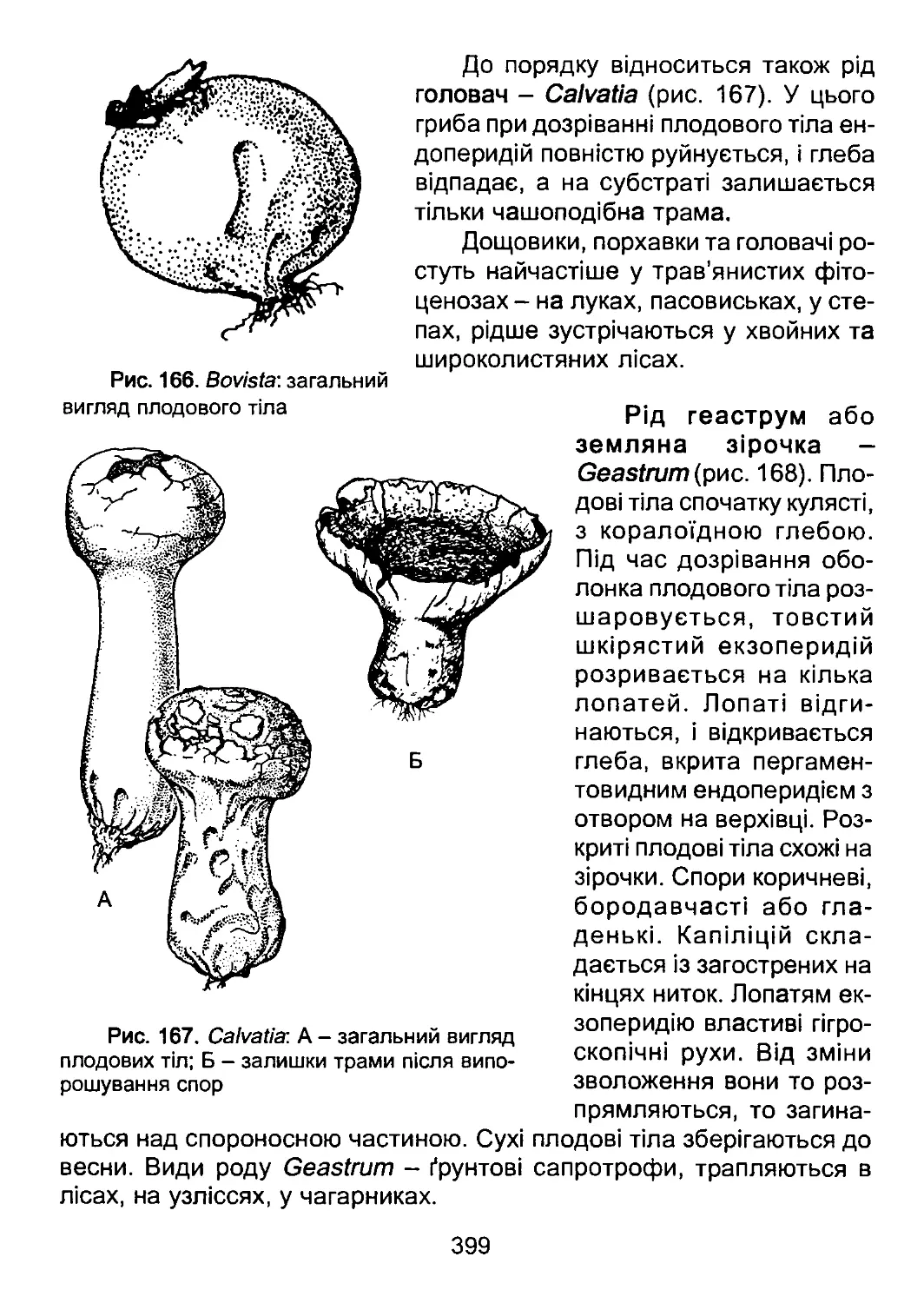
давньоримським натуралістом Плінієм Старшим (23-70 рр.) у фунда-
ментальній 37-томній праці «Природня історія». З цією роботою по-
в’язують появу у науковій літературі латинського терміну «аідае» -
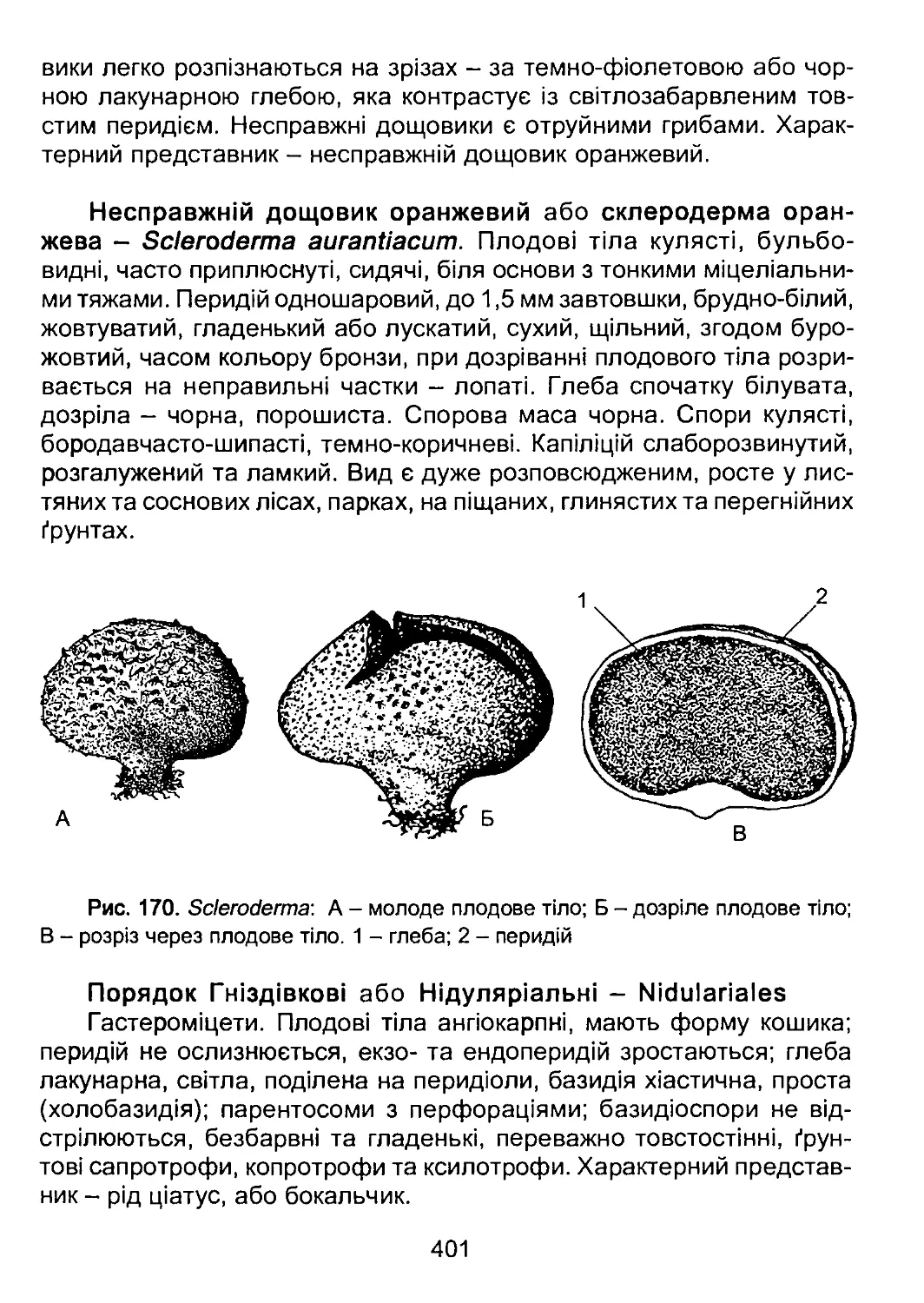
водорість, від якого пізніше виникла назва розділу ботаніки, що ви-
вчає водорості, - альгології.
У середині XIX ст. водорості та гриби об’єднують під назвою «нижчі
рослини». Всі інші рослини, починаючи від мохоподібних і закінчуючи
покритонасінними, відносять до «вищих рослин». Ознаками нижчих
рослин, на відміну від вищих, є відсутність провідної системи та ди-
ференціації тіла на листо-стеблові органи, а також одноклітинний план
будови жіночих статевих органів. Відповідно, до вищих включають ті
рослини, у яких є провідна система, тіло диференційоване на листо-
стеблові органи, а жіночі статеві органи багатоклітинні.
Слід відзначити, що поділ рослин на нижчі та вищі відображує
уявлення середини XIX ст. про місце водоростей та грибів у системі
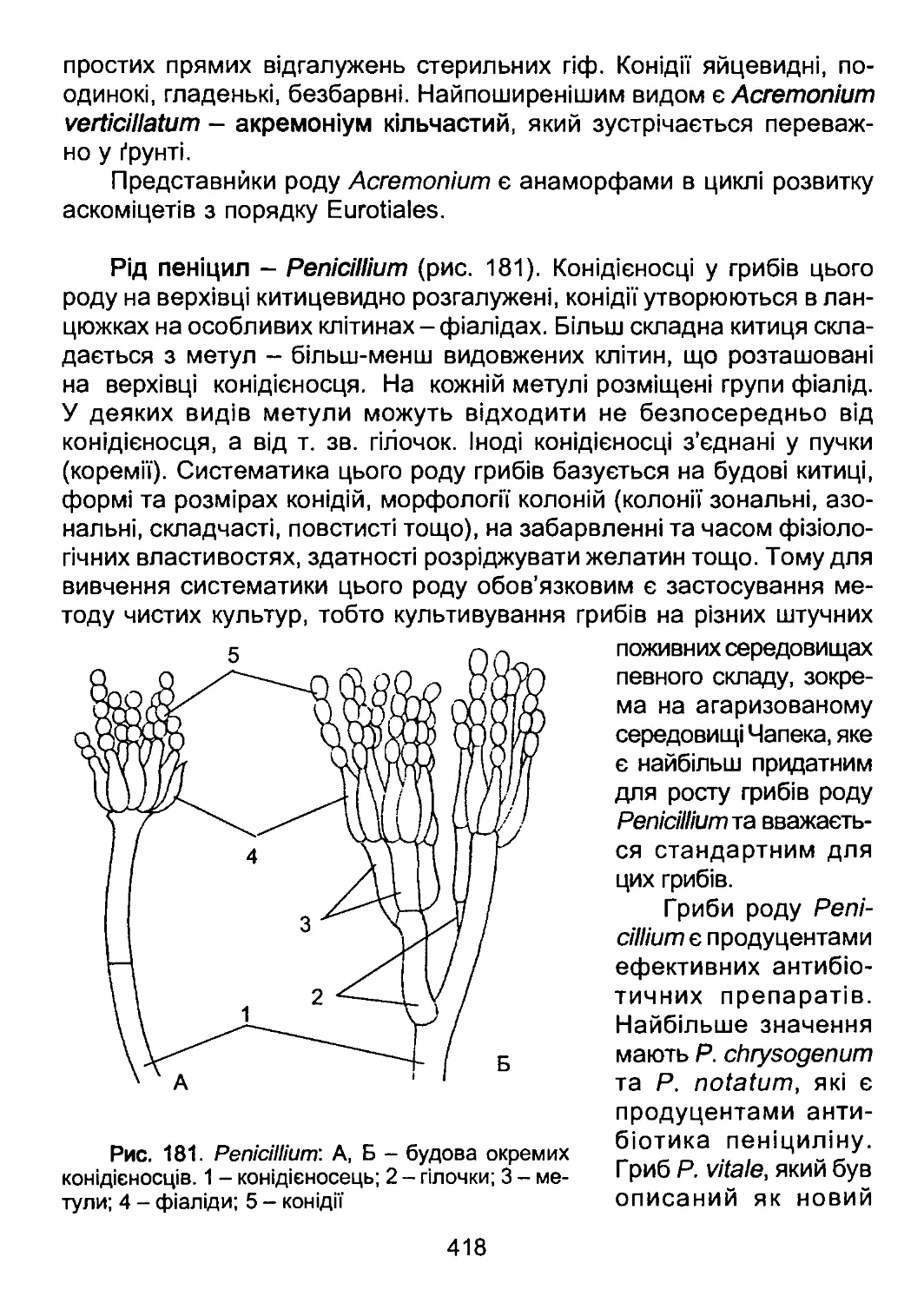
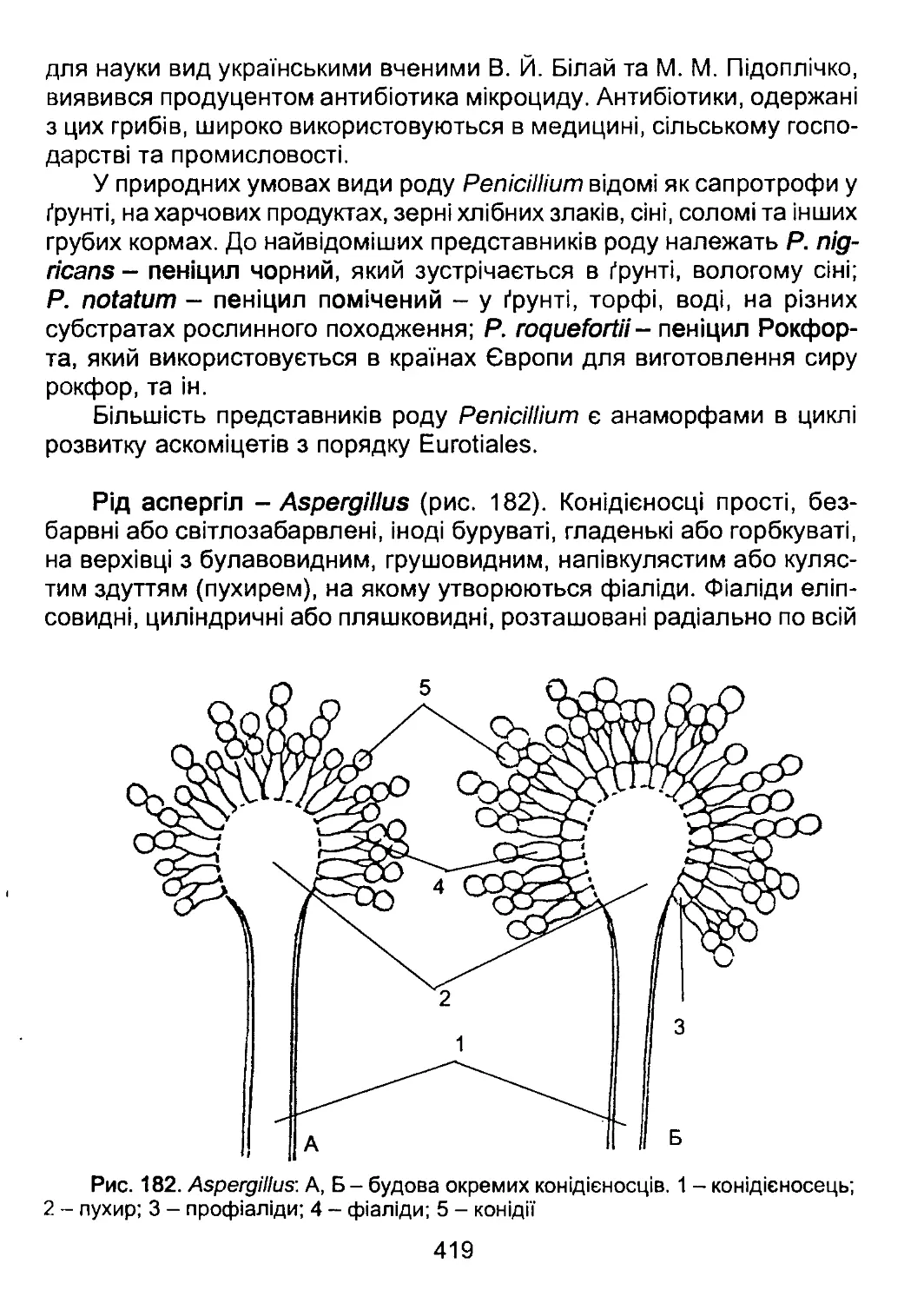
органічного світу. У XX та XXI ст. надзвичайний прогрес у філогенії
органічного світу позбавив поділ рослин на «нижчі» та «вищі» науко-
вого змісту і перетворив терміни «нижчі рослини» та «вищі рослини»
лише на історичні назви певних груп об’єктів.
За найобережнішими оцінками сьогодні на нашій планеті зареє-
стровано біля 1,8 млн. видів живих організмів. З них біля 450 тис.
видів - це рослини (з них біля 60 тис. видів представлені водоростя-
ми), біля 100 тис. - гриби, біля 1,2 млн. - тварини (з них біля 1 млн. -
комахи).
Це різноманіття виникло внаслідок тривалого процесу еволюції,
в результаті якого одні види давали початок іншим, деякі зникали.
Види, що утворилися від спільного предка, зберігають чимало рис
6
схожості. Чим віддаленішим є історичний зв'язок між видами, тим сут-
тєвішими є відміни між ними. Таким чином, всі види, що населяють
нашу планету, пов’язані між собою родинними зв’язками, тобто утво-
рюють природну систему. Дослідженням цієї системи і спробами
відтворити її, відобразити послідовність еволюційних подій на пла-
неті займається систематика.
Природна система має як наукове, так і прикладне значення. На-
укова цінність системи полягає в тому, що при її побудові необхідним
є синтез та узагальнення знань з усіх галузей біології - біохімії, біо-
фізики, генетики, молекулярної біології, цитології, екології. Таким чи-
ном, у системі в концентрованому вигляді представлені здобутки су-
часної біології в цілому. Прикладне значення системи полягає в її про-
гностичності. Знання ступеня спорідненості об’єктів дозволяє за ме-
тодом аналогій прогнозувати властивості інших об’єктів.
Систематика (синонім - таксономія) оперує двома основними по-
няттями: таксонами та таксономічними ознаками. Таксон - це група
дискретних об’єктів, пов’язаних між собою ознаками певного ступеня
схожості, завдяки чому таксони ієрархічно класифікуються і можуть
бути віднесені до певної таксономічної категорії. Таксономічна кате-
горія, на відміну від таксона, є поняттям логічним і характеризує ранг,
до якого може бути віднесений певний таксон на основі комплексу
встановлених таксономічних ознак. Основними таксономічними ка-
тегоріями в ботаніці є надцарство, царство, відділ, клас, порядок, ро-
дина, рід та вид. Крім основних таксономічних категорій, існують та-
кож допоміжні, наприклад, підвідділ, підклас, підпорядок, підродина.
Кожний таксон, у відповідності з тим, до якої таксономічної кате-
горії він належить, має власну унікальну назву. Порядок надання так-
сонам правильних та законних назв регламентується міжнародними
правилами, обов’язковими для будь-якого біолога - Міжнародними
кодексами номенклатури. Існує три таких кодекси - Міжнародний ко-
декс ботанічної номенклатури (МКБН), Міжнародний кодекс зоологіч-
ної номенклатури (МКЗН) та Міжнародний кодекс мікробіологічної но-
менклатури (МКМН). Назви таксонів водоростей, вищих рослин та
грибів регламентуються МКБН.
Згідно з МКБН, правильною науковою назвою таксона є латинсь-
ка назва. Причому для таксонів рангу від відділу до родини встанов-
люються спеціальні закінчення, які вказують, до якої таксономічної
категорії належить даний таксон. Спеціальні закінчення таксонів ран-
гу відділів та класів у водоростей, вищих рослин та грибів відмінні;
7
закінчення порядків та родин однакові (табл. 1). Родова назва пред-
ставляє собою іменник без спеціального закінчення, написаний з ве-
ликої літери. Видова назва є біномінальною, тобто складається з двох
слів, з яких перше є назвою роду, а друге - видовим епітетом. Прави-
ла написання назв царств та надцарств окремо в МКБН не обумов-
лені.
Таблиця 1
Спеціальні закінчення таксонів, що належать до основних
таксономічних категорій
Таксономічна категорія Водорості Вищі рослини Гриби Приклади таксонів
Відділ -рЬуІа -рЬуІа -гпусоїа СЬІогорІїуІа - зелені водорості МацпоІіорНуїа - покритонасінні Вазідіотусоїа - базидіомікотові гриби
Клас -рЬусеае -орзіба -тусеГез ТгсЬоихіорЬуссас - трсбуксієфіцієві МацпоІіор8І(1а - дводольні Вазібіотуссісз - базидіоміцетові
Порядок -аіез -аіез -аіез СЬІогсІІаІсз - хлореляльні Ра^аісз - букоцвітні Адагісаісз - агарикальні гриби
Родина -асеае -асеае -асеае СЬІогсІІассас - хлорелові Еа§ассас - букові Киззиіассас - сироїжкові
РІД іменник однини без спеціального закінчення СЬІогсІІа - хлорела Оисгсиз - дуб Киззиіа - сироїжка
Вид біномен з назви роду та видового епітета без спеціальних закінчень СЬІогсІІа уиІ§агІ5 - хлорела звичайна Оисгсиз гоЬиг - дуб черещатай Киззиіа стсііса - сироїжка блювотна
8
Місце водоростей та грибів у системі органічного світу
За фенотипічними системами, органічний світ поділяється на два
надцарства - Ргосагуоіа - прокаріоти, та Еисагуоіа - евкаріоти. До
прокаріот відносяться всі бактерії та один з відділів нижчих рослин -
синьозелені водорості. Всі інші організми - тварини, гриби та росли-
ни - належать до евкаріот.
Відміни між прокаріотами та евкаріотами на геномному рівні поля-
гають у тому, що прокаріотична клітина є системою, яка містить лише
один геном, зосереджений у нуклеоїді, тобто є моногеномною. Евка-
ріотична клітина є системою з кількома (двома, трьома або навіть
чотирма) неспорідненими геномами, тобто є полігеномною. Геноми
евкаріотичної клітини представлені: а) ядерним геномом, зосередже-
ним у ядрі і представленим ядерною ДНК; б) мітохондріальним гено-
мом, зосередженим у мітохондрії і представленим мітохондріальною
ДНК; в) пластидним (хлоропластним) геномом, що розташовується у
пластиді і представлений хлоропластною ДНК (генофором); г) гено-
мом нуклеоморфу, який виявлений лише у кількох відділах водорос-
тей у ядроподібній структурі, розташованій між оболонкою пластиди
та особливою клітинною системою - хлоропластною ендоплазма-
тичною сіткою. В нуклеоморфі виявлена власна, нуклеоморфна ДНК.
Двогеномні клітини, в яких представлено ядерний та мітохондріаль-
ний геноми, характерні для грибів та тварин; тригеномні - з ядерним,
мітохондріальним та пластидним геномами-для майже всіх евкаріо-
тичних рослин; чотиригеномні - з ядерним, мітохондріальним, плас-
тидним та нуклеоморфними геномами, виявлені у хлорарахніофіто-
вих та криптофітових водоростей, а також у деяких видів з відділу
ОіпорИуІа.
Клітини прокаріот та евкаріот на фенотипічному рівні схожі за на-
явністю ДНК та білок-синтезуючого апарату, представленого рибосо-
мами; за наявністю клітинних мембран, зокрема - зовнішньої мемб-
рани, яка називається плазмалемою; ферментних комплексів, що за-
безпечують процеси реплікації, транскрипції, трансляції та синтезу
АТФ. До складу клітин прокаріот та евкаріот входять білки, жири, вуг-
леводи, нуклеїнові кислоти, мінеральні речовини та вода.
Відміни у будові клітин прокаріот та евкаріот пов’язані, в першу
чергу, з тим, що ДНК прокаріот не має хромосомної організації і не
пов’язана з білками-гістонами; гени організовані у групи, які назива-
ють оперонами, ДНК нуклеоїду прикріплюється до плазмалеми, і про-
цес поділу нуклеоїду зумовлений не роботою веретена, а роботою
9
плазмалеми. Рибосоми у прокаріот менші та легші, ніж рибосоми ци-
топлазми евкаріот (умовна вага прокаріотичних рибосом становить
708, тоді як евкаріотичних - 808). Прокаріоти не здатні до фаго- та
піноцитозу, не мають морфологічно оформленого ядра, мітохондрій,
пластид, ендоплазматичної сітки, комплексу Гольджі, лізосом, перок-
сисом, а також органел, що побудовані з мікротрубочок - джгутиків,
базальних тіл джгутиків, клітинного центру з центріолями, джгутико-
вих коренів, веретена поділу. У прокаріот відсутні мітоз, мейоз, ста-
тевий процес, а обмін генетичною інформацією здійснюється пара-
сексуально - шляхом трансформацій та кон’югацій. Прокаріотичні
клітини за розмірами значно менші, ніж евкаріотичні - їх середній
діаметр становить біля 0,5-2 мкм, тоді як у евкаріот - 5-20 мкм.
За палеонтологічними даними, прокаріоти на нашій планеті ви-
никли близько 3,2 млрд років тому, тоді як евкаріоти є набагато мо-
лодшими -їх вік складає лише біля 1,6 млрд років. Сьогодні в межах
надцарства прокаріот відомо приблизно 40 тис. видів, тоді як у над-
царстві евкаріот - понад 1,7 млн видів.
У XIX ст. та першій половині XX ст. переважна більшість біологів
оминала питання походження евкаріот, вважаючи, що вони виникли
внаслідок дарвінівської еволюції, проте перехідні форми вимерли.
Гіпотеза, що описувала походження евкаріот за таким сценарієм, от-
римала назву автогенетичної.
Згідно з цією гіпотезою, евкаріотична клітина утворилася з прока-
ріотичної внаслідок розвитку плазмалемою системи інвагінацій, які
надалі замкнулись навколо ділянок плазмалеми з ферментами ди-
хального ланцюжка, фотосинтетичними пігментами, нуклеоїда, відпо-
відно утворивши мітохондрії, пластиди та ядро. Розвиток системи інва-
гінацій мембран призвів також до виникнення ендоплазматичної сітки,
комплексу Гольджі, лізосом та ін. Проте автогенетична гіпотеза не
змогла пояснити чимало фактів. Наприклад, чому мітохондрії та хло-
ропласти містять власну ДНК і розмножуються поділом, чому в міто-
хондріях та хлоропластах наявні легкі прокаріотичні рибосоми, чому
поровий апарат мітохондрій та пластид подібний до порового апара-
ту прокаріот і відрізняється від порового апарату плазмалеми евка-
ріот, як виник мітоз та з’явилися мікротрубочки та ін.
Наприкінці XIX ст. видатний ботанік та фізіолог рослин М. С. Цвєт
на основі спостережень поділу хлоропласту та ядерця у зеленої во-
дорості Зрігодуга висловив припущення, що клітини рослин, можли-
во, представляють собою симбіотичне утворення з клітини-господа-
ря та бактеріальних внутрішньоклітинних симбіонтів, які відомі як хло-
10
ропласти та ядерця. Це припущення у 20—40-х роках XX ст. було де-
іально розроблено, в першу чергу, К. С. Мережковським у так звану
ендосимбіотичну гіпотезу походження евкаріотичної клітини. Про-
те ідеї К. С. Мережковського не знайшли підтримки у широкого загалу
біологів. Гіпотезу ендосимбіотичного походження евкаріот називали
«вражаючим витвором людської фантазії», «луна-парком», «неприс-
тойною темою у колі освічених біологів». У 60-х роках гіпотеза ендо-
симбіотичного походження евкаріотичної клітини була заново сфор-
мульована американською дослідницею Л. Маргеліс.
Згідно з гіпотезою Мережковського-Маргеліс, евкаріотична кліти-
на виникла внаслідок кількох ендосимбіозів: гіпотетична прокаріотична
анаеробна клітина, здатна до фагоцитозу, захопила, проте не пере-
травила, а зберегла в цитоплазмі, аеробну гетеротрофну бактерію,
яка трансформувалась у мітохондрію. Далі клітина-господар, що місти-
ла мітохондрію, вступила в симбіоз із рухливою спірохетоподібною
гетеротрофною бактерією, яка дала початок джгутику. Після цього
внаслідок автогенетичного процесу, пов'язаного з утворенням глибо-
ких інвагінацій плазмалеми, навколо нуклеоїду клітини-господаря ут-
ворилось ядро. Далі базальні тіла джгутиків трансформувались у цен-
три організації веретена поділу, внаслідок чого виник спочатку мітоз,
а потім його модифікація - мейоз, разом зі статевим процесом. Ви-
никла перша гетеротрофна евкаріотична клітина. Нащадки цієї кліти-
ни, еволюція яких пішла шляхом вдосконалення фагоцитозу, дали
початок тваринам. Ті ж нащадки, які еволюціонували в напрямку вдос-
коналення осмотрофного живлення, дали початок грибам. Симбіоз
гетеротрофної евкаріотичної клітини із синьозеленою водорістю
призвів до появи рослин, оскільки симбіотична синьозелена водорість
надалі трансформувалась у пластиду.
Численні перевірки ендосимбіотичної гіпотези, виконані з широ-
ким використанням електронної мікроскопії, мікрохімічних та молеку-
лярно-генетичних методів, з одного боку, підтвердили гіпотезу ендо-
симбіотичного походження мітохондрій та пластид, з іншого-автоге-
не гичну гіпотезу походження ядра та більшості одномембранних
клітинних органел.
Таким чином, на початку XXI ст. теорія походження евкаріот сфор-
мувалась на основі синтезу обох гіпотез - як автогенетичної, так і
ондосимбіотичної - і отримала назву синтетичної гіпотези похо-
дження евкаріот.
Сьогодні вважають, що першим кроком на шляху виникнення ев-
кііріог було утворення у гіпотетичного прокаріотичного пращура чис-
11
ленних внутрішніх інвагінацій плазмалеми, які, з одного боку, замкну-
ли прокаріотичний нуклеоїд у двомембранну оболонку (тобто утвори-
ли морфологічно оформлене ядро), а з іншого - призвели до утво-
рення ендоплазматичної сітки та похідного від неї комплексу Гольджі,
а також травних вакуолей та їх похідних - лізосом (рис. 1).
Рис. 1. Виникнення евкаріот та їх дивергенція згідно із синтетичною гіпоте-
зою. Суцільними стрілками позначені напрямки еволюції клітини, штриховими
стрілками - ендосимбіози, штрих-пунктирною стрілкою - горизонтальний пере-
нос гена, що кодує тубулін, внаслідок чого виникли мікротрубочкові клітинні сис-
теми. І - автогенетичне утворення ядра та одномембранних евкаріотичних орга-
нел; II - виникнення мікротрубочкових систем, джгутикового апарату, мітозу, ме-
йозу, статевого процесу; III - ендосимбіоз, внаслідок якого виникли мітохондрії з
дископодібними кристами; IV - виникнення платикристат та тубулокристат;
V - ендосимбіоз, внаслідок якого виникла первинна пластида; VI - велика радіа-
ція евкаріот та виникнення в межах платикристат рослин, грибів та тварин, а в
межах тубулокристат - амебо-флагелят, альвеолят та страменопілів
На другому етапі, внаслідок горизонтального переносу гена, що
кодує білок тубулін від спірохетоподібних бактерій, примітивні евка-
ріоти набули здатності до синтезу тубулінових мікротрубочок1. В ре-
’ Сьогодні у біологів немає одностайної думки стосовно того, який комплекс органел
виник раніш - ядерний чи мікротрубочковий. Щодо цього питання існують дві альтерна-
тивні гіпотези - про первинність ядра і вторинність мікротрубочкової системи, і навпаки.
12
зультаті в евкаріот з'явився цитоскелет, джгутики з базальними тіла-
ми, веретено поділу, мітоз. Надалі базальні тіла джгутиків у частини
представників трансформувалися у клітинний центр, а порушення нор-
мального мітозу (зокрема, скорочення інтерфази) призвели до виник-
нення мейозу і пов’язаного з ним статевого процесу. Група первинно
безмітохондріальних евкаріот сьогодні розглядається як перше, най-
примітивніше царство евкаріот - Нуросізопсігіа (домітохондріальні, або
гіпохондріати). Всі гіпохондріати є одноклітинними гетеротрофами із
тваринною стратегією живлення.
На третьому етапі евкаріотична клітина утворила симбіотичний
комплекс з прокаріотичною клітиною, схожою із сучасними альфа-
протеобактеріями. Ця прокаріотична клітина надалі трансформува-
лась у мітохондрію. За результатами як цитологічних, так і молеку-
лярних досліджень, найдавнішими мітохондріальними евкаріотами
вважаються ті, які мають мітохондрії з особливими дископодібними
кристами. Група мітохондріальних евкаріот з дископодібними крис-
тами прийнята як таксон рангу царства - Оізсісгізіаіез (дискокриста-
ти). В основі царства дискокристат знаходяться первинно гетеротрофні
організми.
На четвертому етапі евкаріоти розділилися на дві великі групи.
Одна з цих груп має мітохондрії з трубчастими кристами і складає
царство ТиЬиІосгізіаіез (тубулокристати, трубчастокристні), друга -
мітохондрії з переважно пластинчастими кристами і приймається як
царство РІаіусгізіаіез (платикристати, платівчастокристні). Подібно до
дискокристат, в основі філ тубуло- та платикристат знаходяться пер-
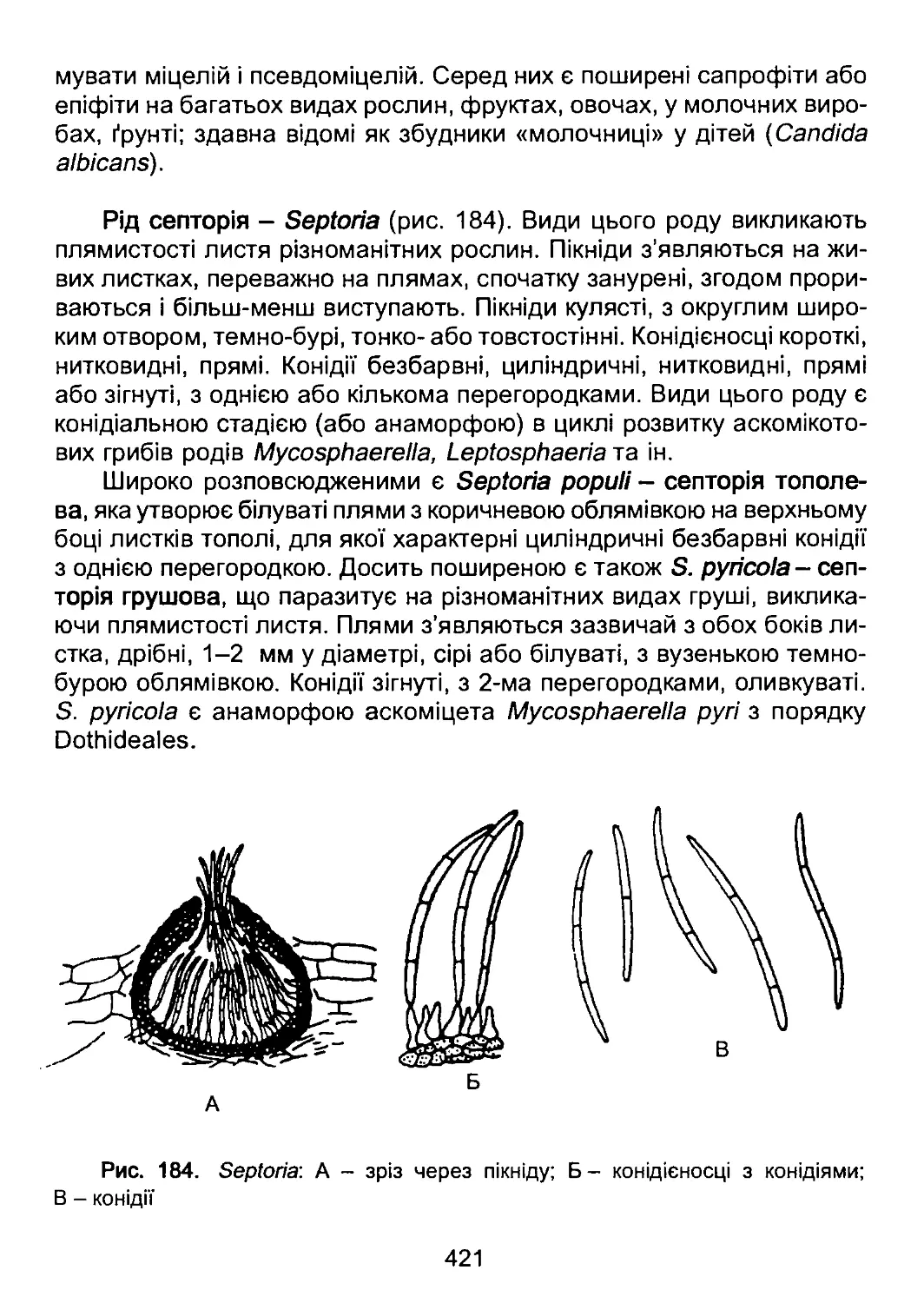
винно гетеротрофні організми з тваринною стратегією живлення.
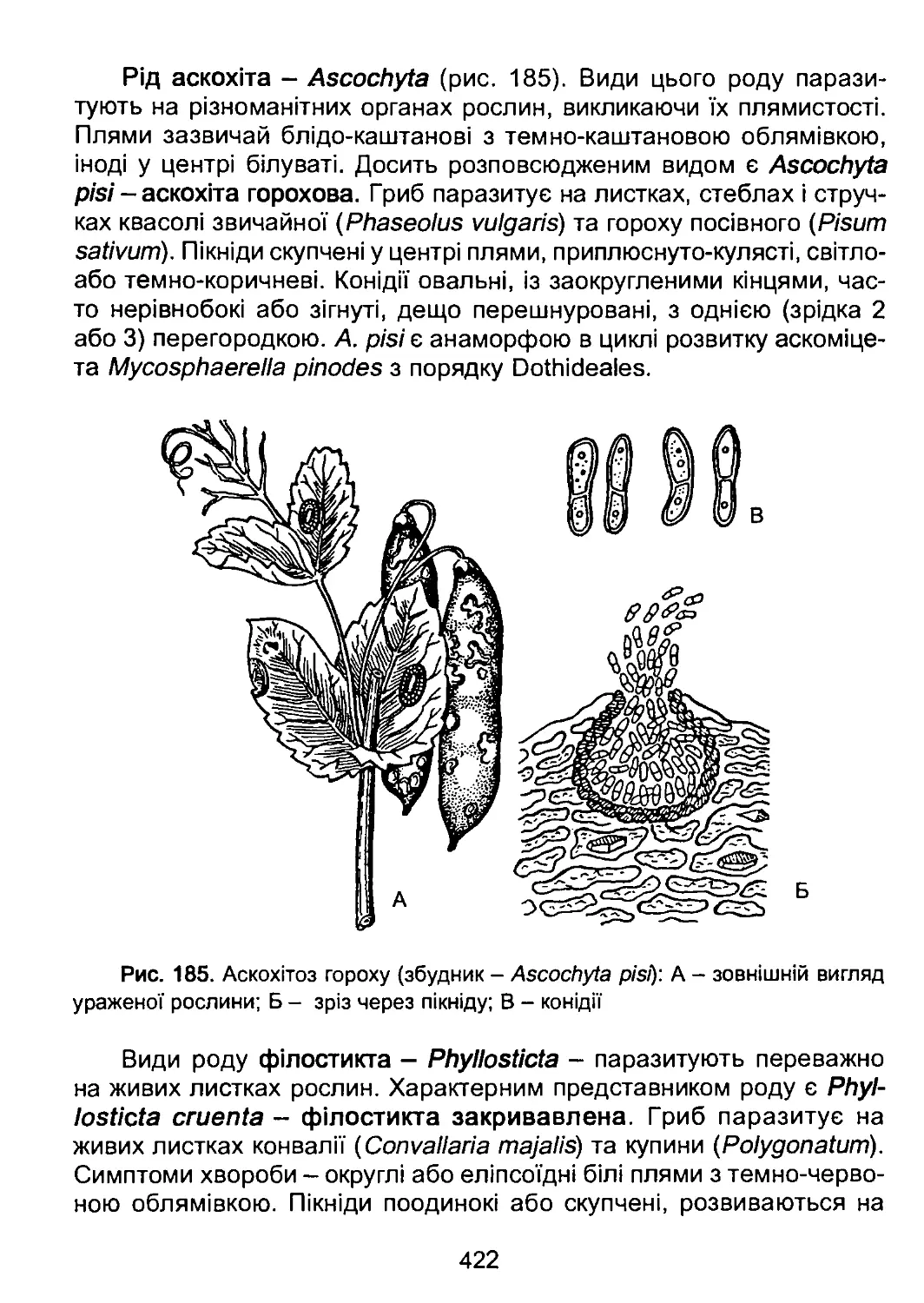
На п’ятому етапі в евкаріотичному світі з’явилися перші рослини.
За молекулярними та цитологічними даними, ця подія пов’язана із
симбіозом гетеротрофної евкаріоти - платикристати з фотоавтотроф-
і іим прокаріотом - синьозеленою водорістю. Внаслідок цього симбіозу
уїворилася пластида, оточена двома мембранами, яка отримала на-
шу первинно симбіотичної пластиди. Подальша дивергенція організмів
і первинно симбіотичними пластидами обумовила виникнення у ме-
жах філи платикристат групи фотоавтотрофних відділів, які склали
підцарство РІапіае - рослини. Дивергенція гетеротрофних платикри-
і.іііі за типами живлення зумовила виникнення, з одного боку, відділів
і осмотрофним живленням - підцарства Еипді (гриби), з іншого-так-
сонів із фаготрофним живленням, що склали підцарство Апігпаііа -
івприпи.
13
Велика радіація тубулокристат відбувалася майже паралельно з
платикристатами, проте була пов’язана не стільки з еволюцією за ти-
пами живлення, скільки з еволюцією клітинних покривів та джгутико-
вого апарату. В основі тубулокристат залишились амебоїдні організми,
які складають підцарство АтоеЬоГІадеІІаіез (амебо-флагеляти). Дві
більш розвинуті філи представляють, з одного боку, таксони зі спе-
цифічними, альвеольованими покривами - АІуеоІаіез (альвеоляти),
та з іншого - зі специфічними субмікроскопічними тричленними во-
лосками на поверхні клітин або джгутиків (т. зв. ретронемами) -
Зігагпепоріїез (страменопіли).
Первинні та вторинні ендосимбіози. Ендосимбіози, при яких в
евкаріотичну клітину-господаря включався прокаріотичний симбіонт,
що надалі трансформувався в клітинну органелу, називають первин-
ними. На основі молекулярно-таксономічних досліджень наприкінці
XX ст. було встановлено, що у світі евкаріот первинні симбіози відбу-
лися лише двічі. При першому ендосимбіозі прокаріотична бактерія,
подібна до альфа-протеобактерій, вступила у симбіоз із фаготроф-
ним джгутиконосцем із царства гіпохондріат і трансформувалась у
мітохондрію з трьома типами крист - дископодібними, трубчастими
та пластинчастими. Подальша морфологічна еволюція мітохондрії
відбувалася поступово, майже паралельно з клітиною-господарем.
Так, у тубулокристат дископодібні кристи були редуковані, а зберег-
лися трубчасті та інколи разом з ними - пластинчасті кристи. У пла-
тикристат були втрачені як дископодібні, так і трубчасті кристи, а збе-
реглися лише пластинчасті (за винятком відділів РІазгпосііорґіо-
гогпусоїа та Наріорґіуіа).
Другий первинний ендосимбіоз відбувся між прокаріотичним ок-
сигенним фотоавтотрофом - синьозеленою водорістю, та гетеро-
трофним організмом із царства платикристат. Як наслідок, виникла
перша евкаріотична клітина, здатна до оксигенного фотосинтезу -
так звана «протоводорість», тобто утворились евкаріотичні рослини.
У «протоводорості» пластида була вкрита двома мембранами, з яких
внутрішня являла собою мембрану прокаріотичного ендосимбіонту -
синьозеленої водорості, а зовнішня була похідною від плазмалеми
клітини-господаря і, ймовірно, представляла собою видозмінену мем-
брану травної вакуолі, в яку потрапив ендосимбіонт.
«Протоводорість» дала початок трьом паралельним гілкам рос-
лин з первинно симбіотичними пластидами - глаукоцистофітовим во-
доростям (СІаисосузіорґіуіа), червоним водоростям (ЯКосІоріїуіа) та
зеленим водоростям (СНІогорґіуІа). Всі ці три відділи зберігають плас-
14
тиди, що оточені лише двомембранною оболонкою. У глаукоцисто-
фітових водоростей є також цікава атавістична ознака - між зовніш-
ньою та внутрішньою мембранами пластиди розташовується шар му-
реїну - речовини, характерної для клітинних оболонок більшості
евбактерій, зокрема, синьозелених водоростей. Відділ глаукоцисто-
фітових є сліпою гілкою еволюції рослин.
Пластиди червоних водоростей - родопласти - також зберігають
деякі яскраві ознаки спорідненості із синьозеленими водоростями,
зокрема, особливі так звані фікобілінові пігменти. Пластиди зелених
водоростей - хлоропласти - ані муреїну, ані фікобілінових пігментів
не мають. Від зелених водоростей беруть початок вищі рослини, при-
чому всі вони зберігають первинно симбіотичні хлоропласти.
Червоні та зелені водорості неодноразово ставали ендосимбіон-
тами гетеротрофних евкаріот з різних царств - дискокристат, тубу-
локристат та платикристат. Симбіози, що відбувалися за сценарієм
«евкаріотичний господар + евкаріотичний ендосимбіонт», називають
вторинними.
Внаслідок вторинних ендосимбіозів виникла велика група відділів
водоростей із вторинно симбіотичними пластидами. Вторинно сим-
біотичні пластиди, на відміну від первинно симбіотичних, одягнені пе-
реважно чотирма мембранами, з яких внутрішня мембрана є залиш-
ком плазмалеми синьозеленої водорості (первинного ендосимбіон-
та), друга мембрана - видозміненою мембраною травної вакуолі гос-
подаря первинного ендосимбіонта (червоної або зеленої водорості),
який є водночас вторинним ендосимбіонтом, третя мембрана - влас-
не плазмалемою вторинного ендосимбіонта, четверта, зовнішня мем-
брана - плазмалемою господаря вторинного ендосимбіонта.
Цитоплазма, яка знаходиться у проміжку між другою та третьою
мембранами, так званий перипластидний простір, є редукованою ци-
юплазмою вторинного ендосимбіонта і в деяких випадках зберігає
Н05 рибосоми та редуковану ендоплазматичну сітку. Зовнішні (третю
ні четверту) мембрани вторинно симбіотичних пластид називають
хлоропластною ендоплазматичною сіткою.
Проте особливо цікавим є той факт, що у двох відділах водорос-
іой із вторинно симбіотичними пластидами - хлорарахніофітових
(СНІогагасйпіорНуІа) та криптофітових (Сгуріорґіуіа) - у перипластид-
іюму просторі зберігається навіть редуковане ядро вторинного ендо-
< имбіонта - нуклеоморф. Нуклеоморф оточений двомембранною
оболонкою і містить ДНК, яка за наявними у нуклеоморфі генами у
хпорарахніофітових споріднена з ядерною ДНК зелених водоростей,
її у криптофітових - з ядерною ДНК червоних водоростей.
15
У деяких водоростей, зокрема, у евгленофітових, вторинно сим-
біотичні пластиди вкриті не чотирма, а лише трьома мембранами.
Вважається, що у цих водоростей одна із зовнішніх мембран пласти-
ди була вторинно втрачена.
Відомі випадки, коли пластиди (як первинно, так і вторинно сим-
біотичні) редукувалися, і тоді виникали вторинно гетеротрофні таксо-
ни. Зокрема, доведена вторинна редукція пластид у тварин з типу
Арісогпріеха, пращурами яких були фотоавтотрофні динофітові во-
дорості (Оіпорґіуіа).
Є також дані, які дозволяють припустити, що оомікотові гриби
(Оогпусоіа) утворилися внаслідок вторинного переходу до гетеро-
трофного типу живлення від однієї з груп водоростей-тубулокристат.
Крім того, вторинно гетеротрофні таксони низьких та середніх
рангів (від видів до порядків і навіть класів) відомі майже в кожному
відділі водоростей, а також серед вищих рослин.
Поширення пластид через вторинні ендосимбіози обумовило ви-
никнення значної кількості відділів водоростей. Наприклад, якщо во-
дорості з первинно симбіотичними пластидами представлені лише
трьома відділами (Сіаисосузіорґіуіа, Пґюсіорґіуіа, Сґііогорґіуіа), то во-
дорості із вторинно симбіотичними пластидами належать до 12 відді-
лів. Серед них два відділи зберігають нуклеоморф (Сґііогагасґіпіорґіуіа,
Сгуріорґіуіа), 8 відділів мають чотиримембранні вторинно симбіотичні
пластиди без нуклеоморфу, що походять від червоних водоростей
(ПарКіборґіуІа, Сґігузорґіуіа, Еизіідгпаіоріїуіа, Хапіґюрґіуіа, Рґіаеорґіуіа,
ВасіІІагіоріїуіа, Оісіуосґюрґіуіа, Наріорґіуіа). Евгленофітові водорості
мають тримембранні пластиди, пращурами яких є зелені водорості.
У ОіпорЬуіа пластиди утворювались неодноразово, як внаслідок сим-
біозів із зеленими водоростями, так і внаслідок третинних симбіозів із
вторинно симбіотичними фотоавтотрофами.
Внаслідок розглянутих процесів та подій організми, відомі під тра-
диційною назвою «нижчі рослини», потрапили у найрізноманітніші
філи органічного світу. Сьогодні біологи виділяють до ЗО відділів ниж-
чих рослин, з яких ми наводимо 26. До нижчих рослин з філогенетич-
ної точки зору слід віднести також один тип тварин (Арісогпріеха),
який вторинно походить від водоростей внаслідок редукції пластиди,
проте зберігає пластидний геном.
Згідно зі схемою, що відображає напрямки еволюції клітин-госпо-
дарів (рис. 2), водорості та гриби належать до двох надцарств:
Ргосагуоіа (1 відділ) та Еисагуоіа (25 відділів).
16
Багатоклп ипні тварини
Деякі найпростіші
Тварини
Вищі рослини
22. Зелені водорості
21. Червоні водорості
20 Глаукоцистофггові
* водорості
19. Криптофітові водорості
18. Гаптофітові водорості
Рослини
26. Базидіомікотові
25. Аскомікотові
24. Змгомікотові
23. Хііридіомікотові
Справжні
гриби
Евкаріоти
6-12. ХромофІтові
водорост і
13-15. Псевдогрибн
Деякі найпростіші
Страменопіли
17. Плазмодіофоромікотові
слизовики
Амебо-флагеляти
Амебоїдні найпростіші
5'Хлорарахніофітові
водорості
4. Міксомікотові слизовики
Інфузорії
Апікомплекси
16. Дннофітові
водорості
Альвеоляти
Платикристати Тубулокристати
2. Акразієві слизовики Кіпетопластиди 3. Евгленофітові водорості
Дискокристати
Прокаріоти
І
Деякі найпростіші
Гіпохондріати
[ 1. Синьозелені водорості
Архебактерії Еубактерії
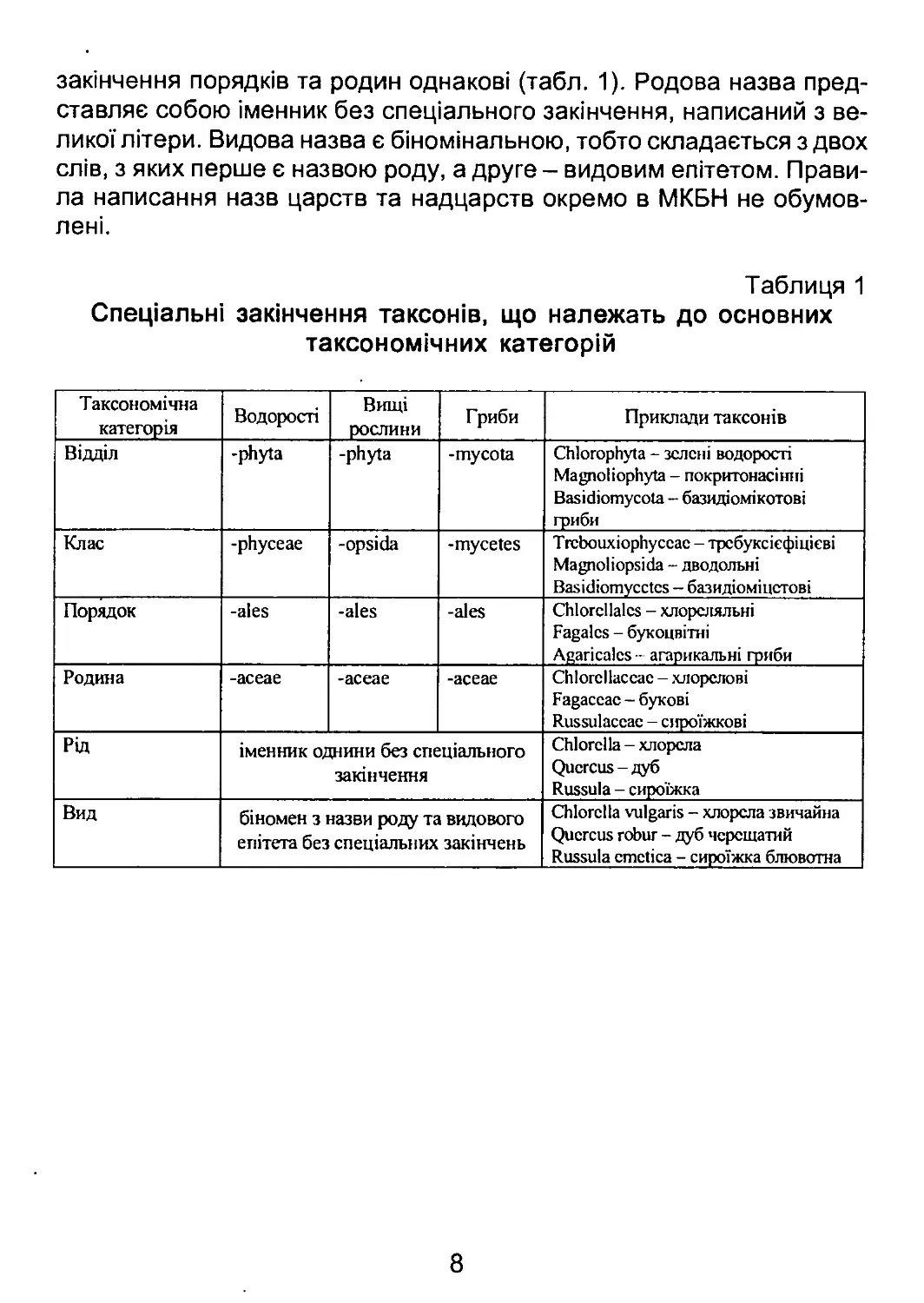
Рис. 2. Місце водоростей та грибів у системі органічного світу. Номерами
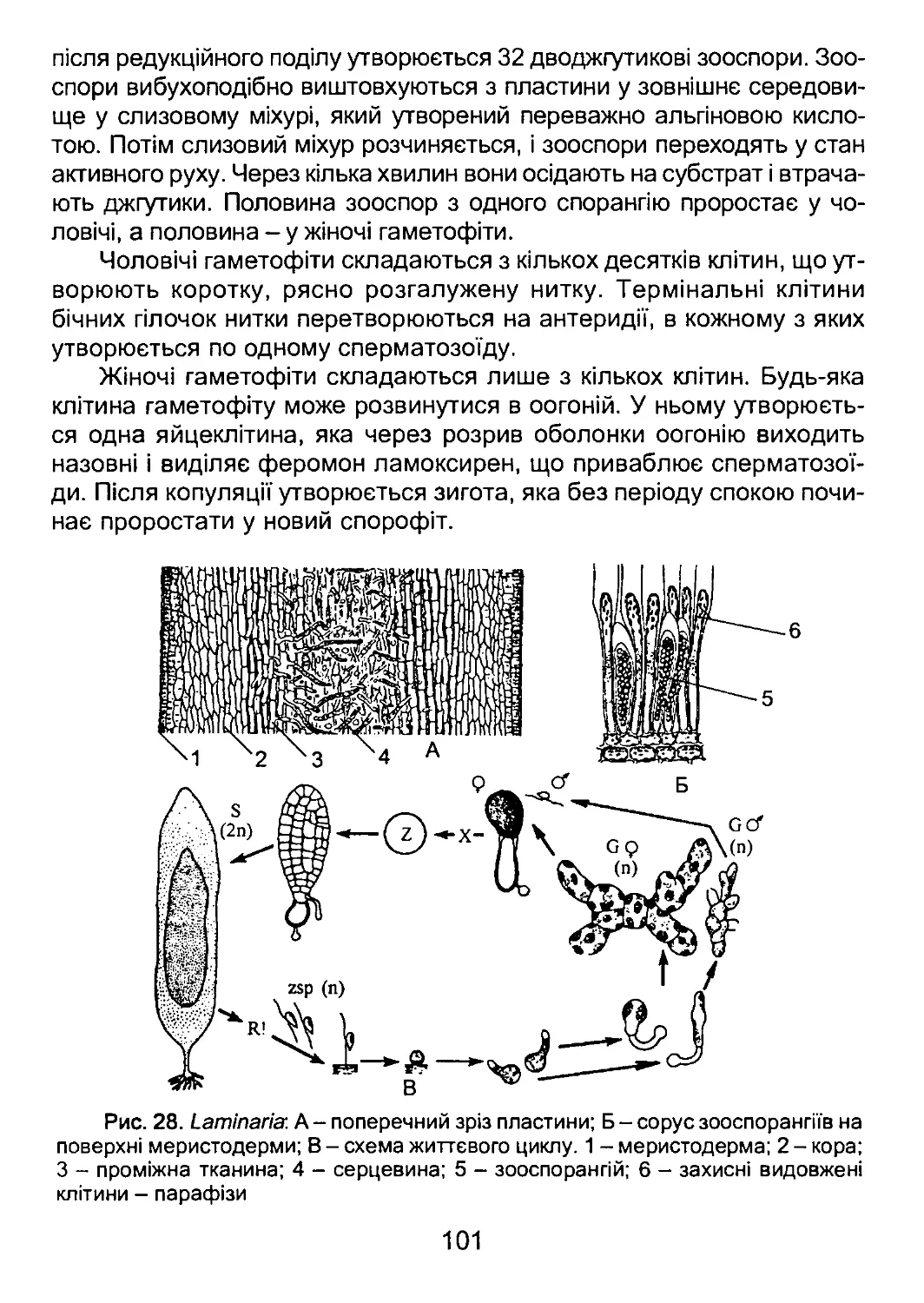
позначені відділи та групи відділів, які прийнято вважати нижчими рослинами
Серед нижчих рослин 16 відділів включають фотоавтотрофних
представників, тобто представляють водорості. З них один відділ -
Суапорґіуіа - належить до прокаріот, три відділи - СІаисосузіорґіуіа,
Кґюсіоріїуіа, Сґііогорґіуіа - до водоростей з первинно симбіотичними
пластидами, решту представляють 11 відділів із вторинно симбіотич-
ними пластидами.
Виключно гетеротрофні нижчі рослини представлені десятьма
відділами, які, за винятком Мухотусоіа, є облігатними гетеротроф-
ними осмотрофами. Представники міксомікотових слизовиків поєдну-
ють два типи живлення - осмотрофне та фаготрофне.
Відділи оомікотових, лабіринтуломікотових та гіфохітриомікото-
них утворюють філогенетично споріднену групу в межах підцарства
Страменопілів царства Тубулокристат. Вони лише віддалено спорід-
нені з іншими відділами грибів і, ймовірно, виникли від вторинно ге-
17
теротрофного пращура. Тому Оотусоіа, ІаЬугіпіґіиІотусоіа та НурКо-
сґіуігіотусоіа сьогодні називають псевдогрибами.
Чотири відділи - Сґіуігісііотусоіа, 2удотусоіа, Азсотусоіа та
Вазісііотусоіа - є філогенетично близькими первинними гетеротро-
фами - платикристатами, і утворюють групу справжніх грибів, або
підцарство Еипді.
Решту відділів грибів - Асгазіотусоіа, Мухотусоіа та РІазто-
біорґюготусоіа - традиційно називають слизовиками. На філогене-
тичних деревах кожний відділ слизовиків займає уособлене положен-
ня: Асгазіотусоіа - серед дискокристат, Мухотусоіа - серед нижчих
тубулокристат в групі амебо-флагелят, РІазтосііорКоготусоіа - у ба-
зальній частині дерева платикристат.
Нижче наведено систематичне положення відділів нижчих рос-
лин у системі органічного світу та стисла характеристика кожного з
відділів.
Надцарство Прокаріоти - Ргосагуоіа
Царство Еубактерії - ЕиЬасіегіа
1. Суапорґіуіа - синьозелені водорості.
Прокаріотичні фотоавтотрофні рослини.
Надцарство Евкаріоти - Еисагуоіа
Царство Дискокристати - Оізсісгізіаіез
Представники царства характеризуються наявністю різноманітних
крист, серед яких обов'язково представлені кристи дископодібної
форми. Поділ ядра представлений закритим ортомітозом.
2. Асгазіотусоіа - акразіомікотові слизовики.
Евкаріотичні первинно гетеротрофні твариноподібні дискокристати,
у яких клітини голі, а вегетативне тіло представлене амебоїдами, здат-
ними об’єднуватися у псевдоплазмодії.
3. Еидіепорґіуіа - евгленофітові водорості.
Евкаріотичні первинно гетеротрофні, фотоавтотрофні та вторинно
гетеротрофні дискокристати, що мають клітинний покрив, представ-
лений пелікулою. Фотоавтотрофні евгленофітові мають вторинно сим-
біотичні пластиди хлорофітного типу.
Царство Тубулокристати - ТиЬиІосгізіаіез
Представники царства характеризуються наявністю трубчастйх, пе-
решнурованих при основі мітохондріальних крист. Всі фотоавтотрофні
представники мають пластиди виключно вторинно симбіотичного типу.
18
Підцарство Амебо-флагеляти - АтоеЬоїіадеІІаіез
Об’єднує організми, в яких клітини голі, і в життєвому циклі представ-
лені лише амебоїдні або монадні стадії (причому амебоїдна стадія,
як правило, переважає). Джгутикові стадії позбавлені тричленних
мастигонем - ретронем.
4. Мухогпусоіа - слизовики.
Евкаріотичні первинно гетеротрофні твариноподібні тубулокристати,
в яких клітини голі, а вегетативне тіло представлене амебоїдом у ви-
гляді багатоядерного плазмодію.
5. Сґііогагасґіпіорґіуіа - хлорарахніофітові водорості.
Евкаріотичні фотоавтотрофні тубулокристати, в яких клітини голі, а
вегетативне тіло представлене амебоїдами, здатними об’єднуватися
у плазмодії. Пластиди вторинно симбіотичні, хлорофітного типу, з нук-
леоморфом.
Підцарство Страменопіли - Зігатепоріїез
Об’єднує організми з різноманітними типами покривів (оболонками,
панцирами, плазмалемою з додатковими захисними структурами).
Головна ознака - наявність у клітини (переважно - на поверхні джгу-
іиків) особливих тричленних мастигонем - ретронем, утворення яких
починається між мембранами ядерної оболонки або хлоропластної
ендоплазматичної сітки.
Група відділів хромофітових водоростей
Об'єднує всіх фотоавтотрофних страменопілів. Група монофілітична
за всіма геномами - ядерним, мітохондріальним, пластидним. Для
всіх відділів спільною ознакою є наявність вторинно симбіотичних
пластид родофітного типу. Представники всіх відділів хромофітових
водоростей мають хлорофіли а та с, чотиримембранні хлоропласти з
«поропластною ендоплазматичною сіткою; ретронеми утворюються
між мембранами як ядерної оболонки, так і хлоропластної ендоплаз-
ми іичної сітки. Відміни між відділами пов’язані, в першу чергу, з будо-
вою клітинних покривів, наявністю фукоксантину, продуктами асимі-
ляції, будовою фоторецепторного апарату, особливостями організації
циюскелета, а також із типами морфологічної структури тіла.
п Нлріїісіорґіуіа - рафідофітові водорості.
І вклріотичні фотоавтотрофні тубулокристати, у яких клітини голі (проте
н» лмебоїдні). Пластиди вторинно симбіотичні, родофітного типу.
19
Джгутикові стадії мають ретронеми. Продукт асиміляції - олія. Харак-
терні особливості - наявність глотки та супрануклеарного апарату,
представленого особливою видозміною комплексу Гольджі, а також
відсутність стигми та структурного зв’язку між ядерною оболонкою та
хлоропластною ендоплазматичною сіткою. Всі представники мають
виключно монадний тип структури тіла.
7. Сґігузорґіуіа - золотисті водорості.
Евкаріотичні фотоавтотрофні та вторинно гетеротрофні тубулокрис-
тати, в яких клітини голі (з кремнеземовими лусочками або без них)
або вкриті пектиновою оболонкою. Пластиди вторинно симбіотичні,
родофітного типу. Джгутикові стадії з ретронемами. Продукт асимі-
ляції - хризоламінарин. Характерна особливість - наявність у життє-
вому циклі стадії ендогенних кремнеземових цист.
8. Еизіідтаіорґіуіа - евстигматофітові водорості.
Евкаріотичні фотоавтотрофні тубулокристати, в яких клітини вкриті
пектиновою оболонкою. Пластиди вторинно симбіотичні, родофітно-
го типу. Джгутикові стадії з ретронемами. Продукт асиміляції - хризо-
ламінарин. Характерна особливість - наявність у монадних стадій
унікального фоторецепторного апарату, розташованого у цитоплазмі
біля основи джгутиків. Всі представники мають виключно кокоїдний
тип структури тіла.
9. Хапіґюрґіуіа - жовтозелені водорості.
Евкаріотичні фотоавтотрофні тубулокристати, в яких клітини вкриті
пектиновою або целюлозно-пектиновою оболонкою. Пластиди вто-
ринно симбіотичні, родофітного типу. Джгутикові стадії з ретронема-
ми. Продукт асиміляції - хризоламінарин. Характерна особливість -
відсутність жовтого ксантофілу фукоксантину.
10. Рґіаеорґіуіа - бурі водорості.
Евкаріотичні фотоавтотрофні тубулокристати, в яких клітини вкриті
целюлозно-пектиновою оболонкою, до складу якої входять альгіна-
ти. Пластиди вторинно симбіотичні, родофітного типу. Джгутикові стадії
з ретронемами. Продукт асиміляції - ламінарин. Характерна особ-
ливість - виключно багатоклітинна будова таломів.
11. ВасіІІагіорґіуІа - діатомові водорості.
Евкаріотичні фотоавтотрофні тубулокристати, в яких клітини вкриті
кремнеземовим панциром. Пластиди вторинно симбіотичні, родофіт-
20
ного типу. Монадні, як правило, стадії нерухомі, джгутики мають ретро-
неми. Продукт асиміляції-хризоламінарин. Всі діатомові водорості ма-
ють виключно кокоїдний тип структури тіла.
12. ПісІуосґюрИуіа - диктіохофітові водорості.
Евкаріотичні фотоавтотрофні та вторинно гетеротрофні тубулокрис-
іати, в яких клітини голі (частина представників має зовнішній крем-
неземовий скелет). Пластиди вторинно симбіотичні, родофітного типу.
Монадні стадії мають ретронеми. Продукт асиміляції - хризоламіна-
рин. Характерна особливість - асоційованість базальних тіл джгутиків
безпосередньо з ядерною мембраною, без участі джгутикових коренів.
Група відділів псевдогрибів
Об’єднує гетеротрофних страменопілів, які живляться осмотрофним
шляхом. Представники відділів, які належать до псевдогрибів, мають
джгутикові стадії, представлені зооспорами з двома або одним джгу-
іиком. На поверхні локомоторного джгутика розташовуються ретро-
номи, які утворюються між мембранами ядерної оболонки. Відділи
псевдогрибів відрізняються, в першу чергу, будовою джгутикового
апарату, типами вегетативного тіла, наявністю або відсутністю екто-
плазматичного ретикулюма, а також способами розмноження, в пер-
шу чергу - статевого. Псевдогриби, ймовірно, є вторинними гетерот-
рофами і походять від хромофітових водоростей, які втратили фото-
синтетичний апарат.
13. Оотусоїа - оомікотові гриби.
Евкаріотичні (ймовірно - вторинно гетеротрофні) осмотрофні стра-
монопіли, в яких клітини вкриті целюлозно-глюкановою оболонкою, а
вегетативне тіло представлене багатоядерним розгалуженим неклітин-
ним міцелієм. Монадні стадії дводжгутикові.
14. ІаЬугіпіИиІотусоІа - лабіринтуломікотові гриби.
Евкаріотичні (ймовірно - вторинно гетеротрофні) осмотрофні страме-
нопіли, в яких голі клітини вкриті мікроскопічними лусочками. Вегета-
іивне тіло представлене поодинокими або з’єднаними у псевдоплаз-
модії клітинами з ектоплазматичною сіткою, яку утворюють спеціальні
пріїїнели - ботросоми. Монадні стадії дводжгутикові.
Іб і ІурґюсКуІгіотусоіа - гіфохітриомікотові гриби.
І ШШ І ііотич ні (ймовірно - вторинно гетеротрофні) осмотрофні страме-
ні н Ні їй. в яких клітини у дорослому стані повністю або частково вкриті
21
целюлозно-хітиновою оболонкою, а вегетативне тіло представлене
поодинокими багатоядерними клітинами або ризоміцелієм. Монадні
стадії одноджгутикові.
Підцарство Альвеоляти - Аіуеоіаіез
Об’єднує організми, в яких клітини мають альвеольовані покриви, в
утворенні яких значну роль відіграють мікротрубочки. Для альвеолят
характерні різні варіанти ядерного апарату, які помітно відрізняють-
ся від типової схеми будови ядра евкаріот. Ретронеми у альвеолят
відсутні.
16. ОіпорИуіа - динофітові водорості.
Евкаріотичні первинно гетеротрофні, фотоавтотрофні та вторинно
гетеротрофні тубулокристати, в яких клітини вкриті альвеольованою
амфієсмою. Пластиди вторинно симбіотичні і досить різноманітні -
хлорофітного та родофітного типів. У примітивних представників ядро
типово евкаріотичне, в еволюційно розвинутих є особливим варіан-
том ядерного апарату - динокаріоном.
Додаток: тип Арісотріеха - апікомплекси.
Евкаріотичні вторинно гетеротрофні облігатні паразити, які втратили
мітохондрії, проте зберігають редуковані пластиди та пластидний
геном.
Царство Платикристати - РІаіусгізіаіез
Представники царства характеризуються наявністю пластинчастих
(інколи - також трубчастих) неперешнурованих при основі мітохонд-
ріальних крист.
17. РІазтосііорґюготусоіа - плазмодіофоромікотові слизовики.
Евкаріотичні первинно гетеротрофні твариноподібні платикристати,
вегетативне тіло яких представлене голим амебоїдом, а мітохондрії
мають крім пластинчастих ще й трубчасті кристи, проте неперешнуро-
вані при основі. Облігатні внутрішньоклітинні паразити вищих рослин.
Підцарство Рослини - РІапіае
18. НарІорИуіа - гаптофітові водорості.
Евкаріотичні фотоавтотрофні та вторинно гетеротрофні платикрис-
тати, що мають мітохондрії з трубчастими кристами (проте не пере-
шнурованими при основі), клітинний покрив - плазмалема, яка із зов-
нішнього боку вкрита субмікроскопічними органо-мінеральними лу-
22
сочками, а з внутрішнього підстелена оперезуючою цистерною ен-
доплазматичної сітки. Пластиди вторинно симбіотичні, родофітного
типу.
19. СгуріорИуіа - криптофітові водорості.
Евкаріотичні фотоавтотрофні та вторинно гетеротрофні платикрис-
тати, що мають клітинний покрив, представлений перипластом. Пла-
стиди вторинно симбіотичні, родофітного типу, з нуклеоморфом.
20. СІаисосузіорИуіа - глаукоцистофітові водорості.
Евкаріотичні фотоавтотрофні платикристати, що мають амфієсмопо-
дібний клітинний покрив. Пластиди первинно симбіотичні, глаукоцис-
тофітного типу (ціанели), між зовнішньою та внутрішньою мембрана-
ми хлоропластної оболонки зберігається шар муреїну. Продукт аси-
міляції - крохмаль, що відкладається у цитоплазмі.
21. КИосіорИуІа - червоні водорості.
Евкаріотичні фотоавтотрофні платикристати, клітини яких вкриті це-
люлозно-пектиновою оболонкою з фікоколоїдами. Пластиди первин-
но симбіотичні, родофітного типу (родопласти), з поодинокими тила-
коїдами. Продукт асиміляції-багрянковий крохмаль, що відкладаєть-
ся у цитоплазмі. Специфічна ознака - повна відсутність джгутикових
стадій.
22. СИІогорИуІа - зелені водорості.
Евкаріотичні фотоавтотрофні платикристати, вкриті плазмалемою або
клітинною оболонкою. Пластиди первинно симбіотичні, хлорофітно-
го типу (хлоропласти). Продукт асиміляції - крохмаль, що відкладаєть-
ся у хлоропласті.
Підцарство Гоиби - Еипді
І вкаріотичні первинно гетеротрофні платикристати, що живляться
осмотрофно, розмножуються за допомогою спор. Вегетативне тіло
і;і / або спори мають клітинні покриви, до складу яких входить хітин.
23. Сїіуігісііотусоіа - хітридіомікотові гриби.
І риби, в яких вегетативне тіло представлене амебоїдом, ризоміцелієм
цію нерозгалуженим неклітинним міцелієм. Специфічна ознака - на-
иііііісіь монадних репродуктивних клітин. Ці клітини мають один задній
іпиденький батогоподібний джгутик.
23
24. 2удотусо1а - зигомікотові гриби.
Гриби, в яких вегетативне тіло представлене розгалуженим неклітин-
ним або клітинним міцелієм. Вегетативні клітини гаплоїдні. Статевий
процес - зигогамія. Статеве спороношення - спорангій зі спорангіос-
порами. Монадні стадії повністю відсутні.
25. Азсотусоіа - аскомікотові гриби.
Гриби, в яких вегетативне тіло представлене розгалуженим клітин-
ним міцелієм (зрідка - міцелієм, що брунькується). Вегетативні кліти-
ни гаплоїдні або дикаріонтичні, причому гаплоїдна стадія за тривалі-
стю переважає. Статевий процес - гаметангіогамія. Статеве споро-
ношення - аск з аскоспорами. Монадні стадії повністю відсутні.
26. Вазібіотусоіа - базидіомікотові гриби.
Гриби, в яких вегетативне тіло представлене розгалуженим клітин-
ним міцелієм. Вегетативні клітини гаплоїдні або дикаріонтичні, причому
дикаріонтична стадія за тривалістю переважає. Статевий процес -
соматогамія. Статеве спороношення - базидія з базидіоспорами. Мо-
надні стадії повністю відсутні.
Розвиток комплексу пристосувань, пов'язаний із вдосконаленням
певної стратегії живлення - фотоавтотрофної або гетеротрофної -
обумовив існування у світі так званих «нижчих рослин» численних
проявів конвергентної морфологічної та біохімічної схожості між орга-
нізмами філогенетично віддалених груп. Це є однією з головних при-
чин, що зумовили подальше подання матеріалу із збереженням тра-
диційного поділу нижчих рослин на дві частини: водорості та гриби
(включаючи грибоподібні організми). В межах кожної з цих частин
збережено систематичний порядок викладення матеріалу.
24
РОЗДІЛ 1
ВОДОРОСТІ
Філогенетично гетерогенна група фотоавтотрофних організмів, у
яких відсутні провідна система та диференціація на листо-стеблові
органи, жіночі статеві органи, коли вони наявні, одноклітинні. На ци-
тологічному рівні відрізняються від грибів та грибоподібних організмів
наявністю фотосинтетичного апарату, який представлений тилакої-
дами з вбудованими в них молекулами хлорофілу, на біохімічному -
здатністю здійснювати оксигенний фотосинтез. Проте, в межах
більшості відділів водоростей відомі представники, які вторинно втра-
тили здатність до фотосинтезу, а серед евгленофітових та динофіто-
вих - навіть первинні гетеротрофи.
Водорості поширені в усіх типах біотопів - морях, континенталь-
них водоймах, ґрунтах, на надземних субстратах, на поверхні снігу та
льоду. У водних біотопах водорості входять до складу угрупувань
планктону, який мешкає у товщі води, донних угрупувань - бентосу,
угрупувань різноманітних обростань - перифітону, або угрупувань,
що розвиваються на поверхні води, - нейстону. Відомі також водо-
рості - симбіонти грибів, тварин та рослин, а також їх паразити (хоча
паразитизм для водоростей в цілому нетиповий).
Серед водоростей сьогодні описано біля 60 тис. видів (проте очі-
куване різноманіття за оцінками різних авторів знаходиться у межах
500 тис. -10 млн. видів). Водорості об’єднують у 16 відділів. Один з цих
відділів - СуапорНуіа - належить до прокаріот, решта - до евкаріот.
Основні систематичні ознаки водоростей різних відділів
Біохімічні ознаки
До комплексу біохімічних ознак на рівні відділів входять, в першу
чергу дві групи ознак: склад пігментів та склад запасних поживних
речовин (продуктів асиміляції).
Пігменти за хімічною природою, розчинністю та функціями по-
міняють на три основні групи - хлорофіли, фікобіліни та каротиноїди.
25
Таблиця 2
Основні пігменти водоростей різних відділів
1 і СуапорЬуіа Еи^ІепорЬуІа СЬІогагаскпіорЬу ВарИісіорЬуіа СЬгузорЬуіа ХапіЬорЬуіа Еизіі^таіоркуіа РЬаеорЬуіа ВасіІІагіорЬуІа ОісіуосЬорЬуіа ОіпорЬуіа НарЮрЬуіа СгуріорЬуіа С1 аисосузіорЬуіа КЬодорЬуіа СЬІогорЬуіа Вищі рослини
Хлорофіли
хлорофіл а ♦ * * * * * * * * ♦ ♦ * * * * * *
хлорофіл Ь ± * * + * *
хлорофіл с + * * ? * * * * * * ±
Фікобілінові п гменти
фікоціанін * * * *
алофікоціанін * * *
фікоеритрин * * * *
Каротиноїди
каротини
а-каротин ♦ * ♦ 4- 4- 4-
Р-каротин + 4- 4- 4- 4- 4- 4- 4- 4- 4- 4- 4- 4- 4- 4-
у-каротин * 4- 4-
є-каротин * * * *
ксантофіли лютеїнового ряду: * * 4- 4- 4- 4- * * 4- 4- * * * *
лютеїн 4- 4- 4- * *
зеаксантин * + + 4- + 4- 4- * * 4-
віолаксантин 4- 4- 4- * * ± * *
неоксантин * 4- 4- 4- 4- 4- 4- 4- * *
антераксантин 4- 4- 4- + 4- 4-
ксантофіли діатоксантино- вого ряду: + * * * * * * * *
діатоксантин ± * 4- * 4- * *
діадіноксантин 4- * 4- * 4- * 4- * *
діноксантин 4- ± 4-
фукоксантин * * * * * + *
ксантофіли специфічні:
схіненон * + + ± 4-
каитаксантин * 4- 4- ± 4-
осцилоксантин +
міксоксантофіл +
27
Хлорофіли мають зелене забарвлення, добре розчиняються у
полярних розчинниках, виконують функцію вловлення світла і є голов-
ною речовиною, що забезпечує світлову стадію фотосинтезу. В хіміч-
ному відношенні хлорофіли являють собою магнієвмісні тетрапіроли.
У водоростей відомі три типи хлорофілів: а, Ь, с (останній - з двома
формами - с, та с2).
Різні хлорофіли мають дещо відмінні спектри поглинання. Так,
хлорофіл а в ацетонових екстрактах найбільш інтенсивно поглинає
світло як у синій, так і в червоній областях спектра (довжина хвилі
у зоні максимуму поглинання становить 430 та 660 нм, відповідно). У
хлорофілу Ь максимум поглинання припадає на синю частину спектра
(435 нм), втричі менший пік спостерігається в червоній області (643
нм). Хлорофіл с також найінтенсивніше поглинає сині промені; до-
датковий максимум припадає на червону область.
У клітині може бути наявний або тільки хлорофіл а, або комбіна-
ція двох хлорофілів, з яких основний - лише хлорофіл а, додатковий -
Ь або с (табл. 2).
Друга група пігментів - фікобіліни - дещо нагадують хлорофіли,
оскільки теж являють собою тетрапіроли. Проте, на відміну від хло-
рофілів, фікобіліни є лінійними тетрапіролами, які не містять магнію,
а натомість з’єднуються ковалентними зв’язками з білками. Фікобілі-
ни належать до пігментів, що добре розчиняються у воді.
Виділяють три основні групи фікобілінів: червоний фікоеритрин
та сині фікоціанін і алофікоціанін.
Третя група пігментів - каротиноїди - мають жовте, червоне або
буре забарвлення і являють собою ізопреноїдні полієнові сполуки.
До каротиноїдів належать каротини та їх окислені похідні - ксанто-
філи.
У водоростей виявлено чотири типи каротинів - а, р, у, є. Кароти-
ни розчиняються у неполярних розчинниках. Найбільш поширеним є
р-каротин, який наявний майже в усіх відділах водоростей. Інші типи
каротинів зустрічаються лише у певних макротаксонах, через що склад
каротинів використовується як одна з додаткових ознак на рівні відділу.
Ксантофіли є водорозчинними пігментами, які поділяють на три
основні групи: а) ксантофіли лютеїнового ряду (лютеїн, віолаксантин,
неоксантин, зеаксантин, антераксантин); б) ксантофіли діатоксанти-
нового ряду (діатоксантин, діадіноксантин, діноксантин, фукоксантин);
в) специфічні, переважно мінорні, ксантофіли.
Склад та кількість хлорофілів, каротиноїдів та фікобілінів зумов-
люють забарвлення водоростевих клітин у певний колір.
26
Продовження таблиці 2
криптоксантин + + 4- + + 4- 4- 4- ±
астаксантин +
вошеріоксантин 4- * 4-
гетероксантин 4- *
алоксантин *
крококсантин
монадоксантин 4-
тараксантин 4-
сифонеїн +
сифоноксантин +
Примітка. Зірочкою (*) позначено переважаючі пігменти; знаком (+) - додаткові пігмен-
ти, представлені у відносно малих кількостях; знаком (±) - пігменти, що зустрічаються
лише в окремих представників.
Продукти асиміляції. Як кінцевий продукт фотосинтезу утворю-
ються вуглеводи, які накопичуються у клітинах як основні резервні
поживні речовини. У водоростей запасаються полісахариди двох ос-
новних груп: по-перше, крохмаль- та глікогенподібні полісахариди,
що являють собою а-1,4-глюкани (тваринний глікоген, рослинний крох-
маль, крохмаль синьозелених водоростей, багрянковий крохмаль);
по-друге, хризоламінарин-подібні р-1,3-глюкани, до яких належать
хризоламінарин, ламінарин та парамілон.
Продукти асиміляції у водоростей різних відділів різні (табл. 3) і
накопичуються або в цитоплазмі (у більшості відділів), або в пери-
пластидному просторі (криптофітові водорості), або в пластиді (зе-
лені водорості та вищі рослини).
Виявити певний продукт асиміляції найпростішим шляхом мож-
на за допомогою цитохімічних забарвлень. Зокрема, з розчином йоду
в йодистому калії крохмаль забарвлюється в темно-синій колір, крох-
маль синьозелених водоростей буріє, а багрянковий крохмаль чер-
воніє. Парамілон розпізнається за реакцією з їдким калієм або їдким
натрієм - під дією лугів нарамілон дуже набрякає. Ламінарин та хри-
золамінарин з йодом та лугами специфічних забарвлень або набря-
кання не дають.
Крім полісахаридів, клітини майже всіх водоростей при старінні
здатні накопичувати олію. В деяких відділах (зокрема, у синьозеле-
них водоростей) можуть запасатися також азот- та фосфоровмісні
сполуки: ціанофіцин (полімер амінокислот аргініну та аспарагіну) та
поліфосфатні тіла, відповідно.
28
Таблиця З
Основні продукти асиміляції водоростей різних відділів
СуапорЬуіа СЗ +-• Сі. о а о од □ ш СНІ огагасЬ піорЬу Іа КарЬідорЬуіа СЬгузорЬуіа ХапіИорЬуіа Еизіі^таіорЬуіа РЬаеорЬуіа ВасіІІагіорЬуіа ОісіуосЬорЬуіа ОіпорИуіа НарІорЬуІа СгуріорЬуіа СІаисосузіорЬуІа КИогіорИуІа СЬІогорЬуІа Вищі рослини
глікоген ±
крохмаль * * * * *
крохмаль синьозелених водоростей *
багрянковий крохмаль ♦
хризоламінарин ? * * * * * *
ламінарин *
парамілон ♦ * ±
Примітка. Зірочкою (*) позначено основні продукти асиміляції; знаком (±) - асиміля-
ти, що зустрічаються лише в окремих представників.
Цитологічні ознаки
До комплексу цитологічних ознак на рівні відділів належать чоти-
ри групи ознак: а) ознаки, що відображують спорідненість таксонів за
клітиною-господарем, тобто характеризують особливості нуклеоци-
топлазми (в першу чергу, покриви, ядерний апарат); б) система мікро-
трубочкових органел (в першу чергу, джгутиковий апарат та верете-
но поділу); в) фотосинтетичний апарат; г) мітохондріальний апарат.
Клітинні покриви. Клітини водоростей, подібно до інших
організмів, складаються з протопласта, який оточений фосфоліпід-
ною мембраною - плазмалемою. Клітини, що вкриті лише плазмале-
мою, називають голими. Проте частіше протопласт, крім плазмале-
ми, оточений ще й певним типом клітинного покриву. Клітинні покри-
ви водоростей можуть бути внутрішніми або зовнішніми.
Внутрішні покриви створюють каркас клітини, що розташовуєть-
ся під плазмалемою. Каркас може бути утворений стрічками або пла-
29
тівками білків, сплощеною цистерною ендоплазматичної сітки, спло-
щеними мембранними пухирцями. Деякі типи каркасних елементів
підстилаються також цитоскелетними мікротрубочками. Зовнішні по-
криви розташовуються виключно над плазмалемою, поза межами про-
топласту. Ці покриви за хімічним складом поділяють на органічні -
клітинні оболонки2, та мінеральні - панцири та скелети.
Внутрішні покриви характерні для вегетативних клітин евглено-
фітових, динофітових, криптофітових, гаптофітових та глаукоцисто-
фітових водоростей. У евгленофітових плазмалема з внутрішнього
боку підстелена білковими стрічками і називається пелікулою. Покрив
криптофітових-перипласт-утворений прямокутними білковими пла-
тівками, що розташовуються з внутрішнього боку плазмалеми і при-
кріплюються до неї за допомогою особливих білкових штифтів; у бага-
тьох видів криптофітових над внутрішніми платівками із зовнішнього
боку плазмалеми знаходяться також і зовнішні білкові платівки або
зовнішні субмікроскопічні лусочки.
У гаптофітових водоростей плазмалема з внутрішнього боку
підстилається велетенською сплощеною цистерною ендоплазматич-
ної сітки, а на поверхні плазмалеми відкладаються целюлозні лусоч-
ки або гранули, які у деяких видів просочуються вапном і перетворю-
ються на т. зв. коколіти.
У динофітових водоростей плазмалема підстелена сплощеними
мембранними пухирцями, всередині яких може розташовуватись тон-
ка органічна платівка. Цей тип покривів називають амфієсмою. Дуже
схожі з амфієсмою покриви глаукоцистофітових - тут плазмалема
також підстилається мембранними пухирцями, що містять всередині
платівку, але, на відміну від динофітових, під пухирцями розташову-
ються також цитоскелетні мікротрубочки. Покриви глаукоцистофіто-
вих називають амфієсмоподібною пелікулою.
Зовнішні покриви поділяють на оболонки, панцирі та зовнішні ске-
лети. Каркасну частину оболонок утворюють переважно полісахари-
ди, які полімеризуються у ферментних комплексах на поверхні плаз-
малеми. В оболонках усіх типів у значних кількостях виявляється пек-
тин, часто - целюлоза або геміцелюлоза, в окремих відділів -
спеціфічні каркасні речовини - муреїн, альгінова кислота та її солі,
фікоколоїди, спорополенін, лігнін, хітин.
2 Особливий тип органічних покривів - несправжня тека - наявний у деяких зелених
водоростей з класу празинофіцієвих. Морфологічно цей покрив нагадує оболонку, але
утворюється внаслідок злиття на поверхні плазмалеми субмікроскопічних органічних
лусочок.
ЗО
Пектини, альгінати та фікоколоїди виконують функцію клею, що
з’єднує інші речовини клітинних оболонок; крім того, пектини здатні
легко гідролізуватися і в такий спосіб формувати зовнішній слиз. Це-
люлоза та геміцелюлоза утворюють мікрофібрили, що виконують пе-
реважно каркасні функції. Хімічно стійкий муреїн, крім того, ще й за-
хищає протопласт від дії несприятливих хімічних чинників. Споропо-
ленін та лігнін зустрічаються як додаткові компоненти в оболонках
зелених водоростей та вищих рослин і підвищують механічну стійкість
оболонок. Хітин здатний утворювати різноманітні зовнішні вирости
клітинних оболонок - шипи, шипики, щетинки, роги.
У відповідності з хімічною природою головної каркасної речови-
ни оболонки поділяють на муреїнові (у синьозелених водоростей),
пектинові (у золотистих, евстигматофітових, жовтозелених водорос-
тей), целюлозні та целюлозно-пектинові (зелені та деякі динофітові
водорості), целюлозно-пектинові з альгінатами (бурі водорості), це-
люлозно-пектинові з фікоколоїдами (червоні водорості).
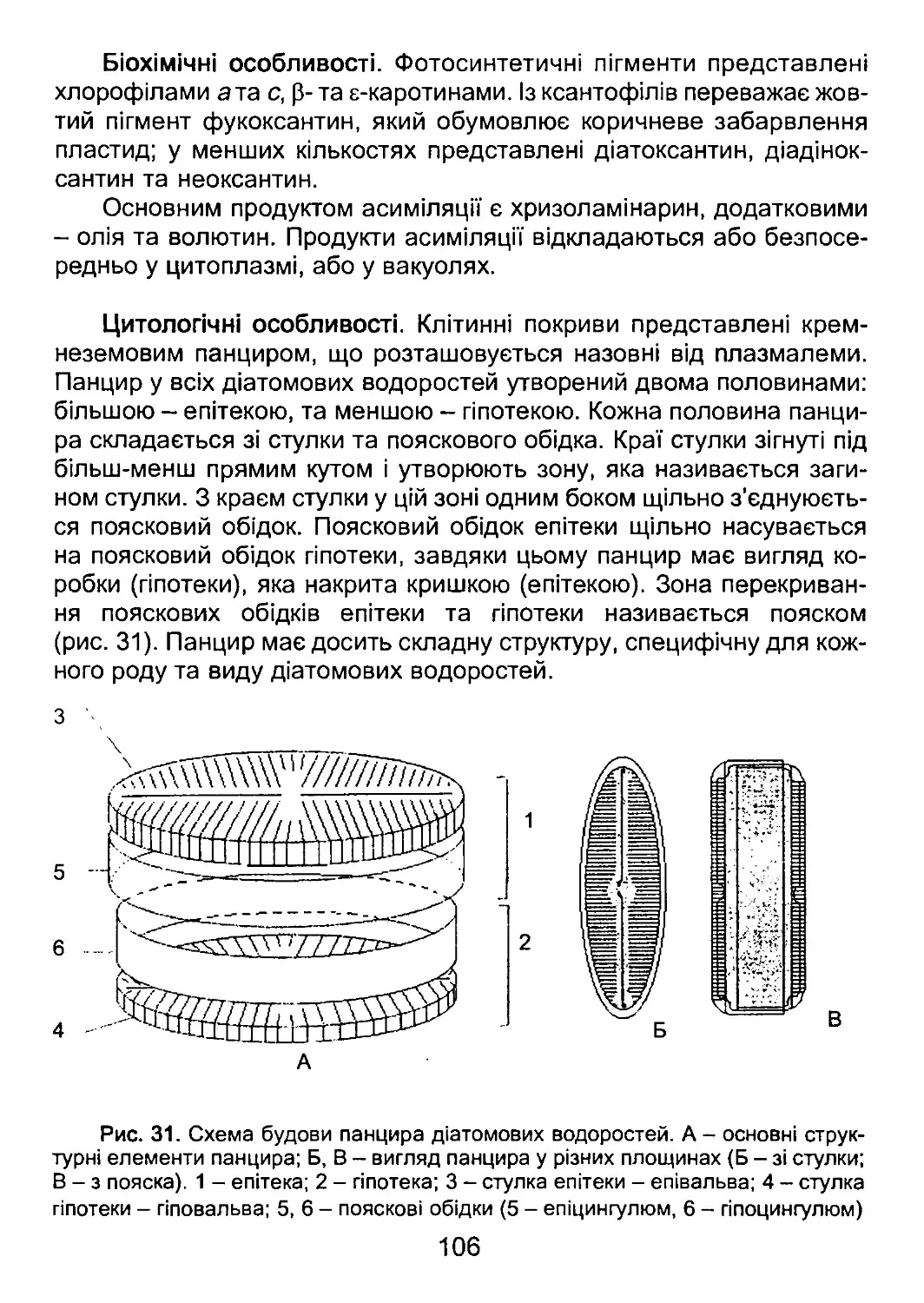
Мінеральні зовнішні покриви, основу яких складає кремнезем,
характерні для діатомових та диктіохофітових водоростей. У діато-
мових водоростей кремнезем утворює панцир у вигляді двостулкової
коробки. У диктіохофітових водоростей кремнезем формує систему
зовнішніх трубок, що з’єднуються між собою у вигляді кошика. Крем-
неземовий кошик вкритий шаром слизу і називається кремнеземо-
вим скелетом.
За ядерним апаратом водорості поділяються на дві групи: про-
каріотичні та евкаріотичні. Ядерний апарат прокаріотичних водорос-
тей представлений однією молекулою ДНК, що замкнена у кільце,
прикріплена до плазмалеми і не оточена ядерною оболонкою. Ця ДНК,
яку називають нуклеоїдом, не пов’язана з білками-гістонами і є ана-
логом однієї хромосоми.
Ядерний апарат евкаріотичних водоростей представлений ядром,
яке включає ядерну оболонку, хромосоми, ядерце. Головний компо-
нент ядра - це ядерна ДНК, яка має хромосомну організацію. Вона
пов’язана з білками-гістонами і утворює мікрофібрили хроматину.
Ядерний апарат класифікують за різними ознаками: за вмістом
гістонів, за наявністю морфологічно оформлених хромосом у інтер-
фазному ядрі, за здатністю ядерець до поділу, за особливостями ор-
ганізації зовнішньої мембрани ядерної оболонки, за поведінкою ядер-
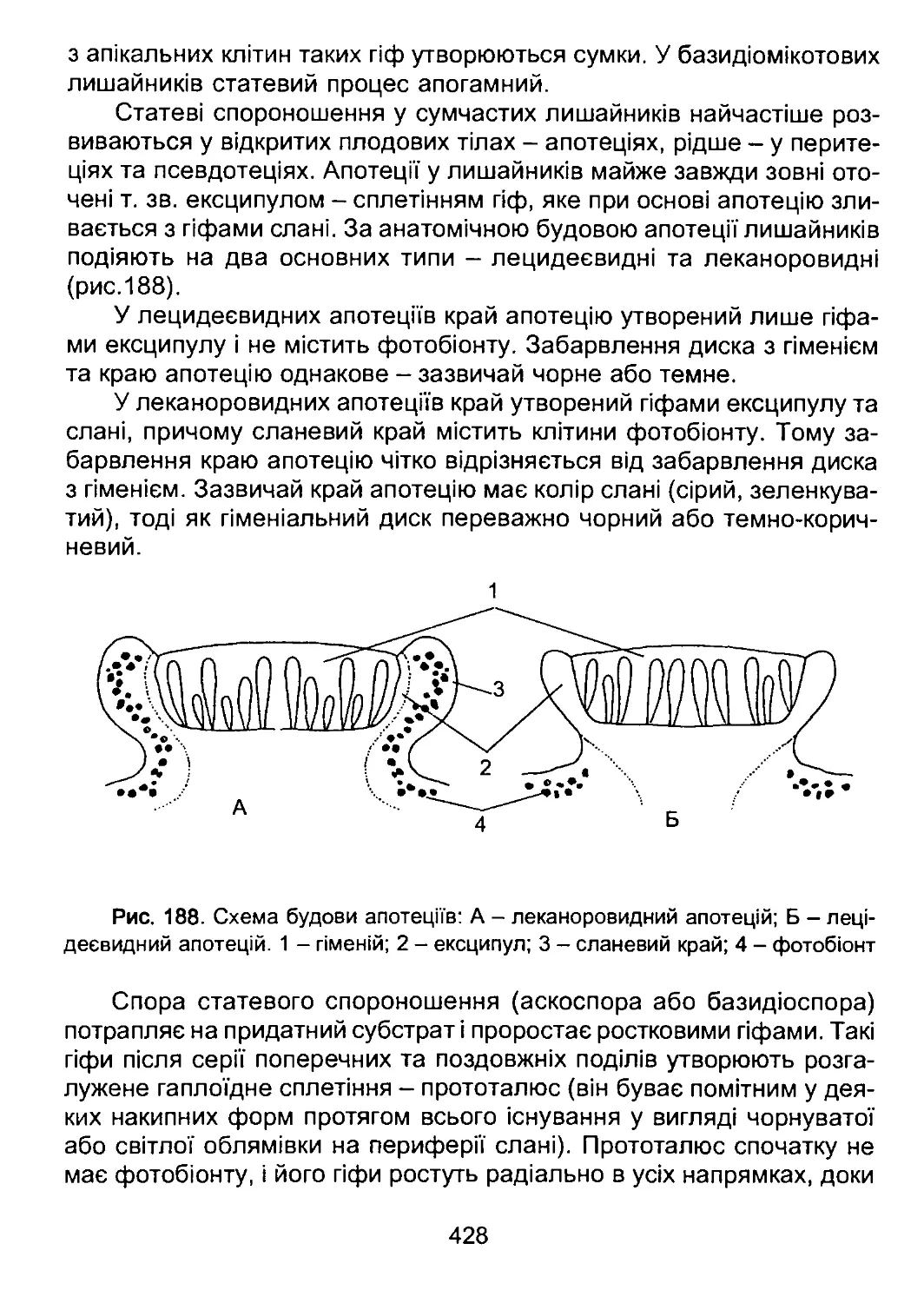
ної оболонки під час мітозу та ін.
31
За вмістом гістонів ядра водоростей поділяють на типово евка-
ріотичні та динокаріонтичні. В абсолютної більшості водоростей спів-
відношення «ядерна ДНК: гістони» становить приблизно 1:1. Проте
у більшості динофітових водоростей вміст гістонів низький, і спів-
відношення «ядерна ДНК: гістони» становить в середньому 1 : 0,04.
Ядра з таким низьким вмістом гістонів називають динокаріонами. Ціка-
во, що до 90-х років XX ст. в ядрах такого типу гістони виявити не
вдавалось, і цей тип ядра вважали перехідним між ядерним апара-
том прокаріот та справжнім евкаріотичним ядром, що відображено у
застарілій назві такого ядра - мезокаріотне.
За наявністю морфологічно оформлених інтерфазних хромосом
ядра поділяють на типово евкаріотичні (притаманні більшості евка-
ріот) та ті, в яких хромосоми перебувають у надспіралізованому стані
протягом всього клітинного циклу, включаючи інтерфазу. Останній тип
ядер характерний для відділів ЕидІепорЬуїа, Оіпорїіуіа та СгуріорЬуІа.
Ядерця зазвичай спостерігаються в інтерфазному ядрі і зника-
ють перед початком мітозу або мейозу. Проте у евгленофітових водо-
ростей ядерце перед початком поділу ядра ділиться, і дочірні ядерця
надалі стають центрами організації мікротрубочок веретена поділу.
Таке ядерце називають ендосомою. Здатність ядерця до поділу ви-
явлена також у частини зелених водоростей, що належать до поряд-
ку зигнематальних.
Ядерна оболонка. Зовнішня мембрана ядра в деяких відділах
(гаптофітові, червоні водорості) може утворювати випини у бік цитоп-
лазми, які морфологічно подібні до каналів ендоплазматичної сітки
— т. зв. перинуклеарний ендоплазматичний ретикулюм. Інколи кана-
ли перинуклеарного ретикулюму переходять у канали ендоплазма-
тичної сітки.
У більшості водоростей з вторинно симбіотичними пластидами
зовнішня мембрана ядра переходить у зовнішню, четверту мембрану
оболонки хлоропласта - хлоропластну ендоплазматичну сітку. В цьо-
му випадку ядро та пластида утворюють єдине структурне ціле.
За поведінкою ядерної оболонки під час мітозу останній поділя-
ють на відкритий, закритий та напіввідкритий. У більшості евкаріот
ядерна оболонка у профазі руйнується, і хромосоми під час поділу
розміщуються безпосередньо у цитоплазмі. Мітоз із таким типом по-
ведінки ядерної оболонки називають відкритим. Проте у частини во-
доростей оболонка при поділі ядра залишається інтактною. Тоді мітоз
називають закритим. У деяких водоростей оболонка при поділі ядра
32
руйнується лише на полюсах ядра, і через отвори в ядро проходять
мікротрубочки веретена поділу. Мітоз із частковим руйнуванням ядер-
ної оболонки називають напіввідкритим.
Інші нуклеоцитоплазматичні органели та структури. Ендо-
плазматична сітка (ендоплазматичний ретикулюм) у водоростей різних
відділів не має принципових відмінностей. Деякі особливі модифікації
цієї структури відмічені у гаптофітових, у яких периферичні канали
ендоплазматичної сітки утворюють велику сплощену цистерну, що
підстилає плазмалему і, таким чином, є одним із компонентів клітин-
ного покриву та особливої джгутикоподібної структури - гаптонеми.
Ендоплазматична сітка здатна переходити у канали перинуклеарно-
го ретикулюму, а також структурно поєднуватися з комплексом Гольджі.
У всіх евкаріотичних водоростей в клітинах наявний один або
кілька комплексів Гольджі, зазвичай розташованих поблизу ядра. У
рафідофітових водоростей комплекс Гольджі дуже великий, тісно при-
лягає до ядерної оболонки, може бути помітним в оптичний мікрос-
коп і отримав назву супрануклеарного апарату.
У водоростей, здатних до фаготрофного типу живлення, в цито-
плазмі можуть спостерігатися травні вакуолі. Травні вакуолі особливо
характерні для первинно гетеротрофних водоростей з відділів
ЕидІепорЬуіа та ОіпорЬуіа, а також для деяких хлорарахніофітових,
рафідофітових, золотистих та криптофітових водоростей, здатних до
міксотрофного або облігатно гетеротрофного живлення.
До одномембранних нуклеоцитоплазматичних органел належать
також лізосоми. Подібно до травних вакуолей, лізосоми містять ком-
плекс гідролітичних ферментів і беруть участь у процесах внутрішньо-
клітинного травлення.
Майже в усіх відділах водоростей виявлені представники, що
містять одномембранні пухирці з електронно-щільним вмістом, які от-
римали назву мікротільця, або пероксисоми. Встановлено, що перок-
сисоми містять фермент пероксидазу, який розкладає перекис вод-
ню на кисень та воду і, таким чином, відіграє важливу роль у світловій
фазі фотосинтезу.
У водоростей, вкритих клітинними оболонками, у цитоплазмі час-
то можна спостерігати вакуолі з клітинним соком. Ці вакуолі відмежо-
вуються від цитоплазми звичайною фосфоліпідною мембраною, яку
називають тонопласт. Вакуолі з клітинним соком містять воду, органічні
кислоти та амінокислоти, мінеральні речовини і відіграють важливу
роль у регуляції осмотичного тиску в клітині, а також акумулюють ряд
33
продуктів метаболізму клітини. Вакуолі з клітинним сс;ком особливо
характерні для багатоклітинних воДоростей з відділів‘бурих, черво-
них та зелених, а також для деяких одноклітинних, зокрема - діато-
мових водоростей.
У більшості прісноводних водоростей-джгутиконосців, а також у
деяких нерухомих представників, осморегуляторну функцію викону-
ють скоротливі (пульсуючі) вакуолі, які виводять назовні надлишкову
воду, що постійно надходить у клітину. Скоротливі вакуолі у більшості
водоростей парні, розташовуються на передньому кінці клітини, і ви-
плескують зайву воду безпосередньо в оточуюче середовище. В ев-
гленофітових та рафідофітових водоростей скоротливі вакуолі чис-
ленні, розміщуються на дні глотки; в міру заповнення водою дрібні
вакуолі зливаються в одну велику, що виплескує свій вміст у глотку.
Далі вода виводиться назовні через глотковий канал. У динофітових
водоростей функції скоротливих вакуолей виконують системи вели-
ких розгалужених інвагінацій плазмалеми, що отримали назву пузул.
Пузули не здатні до пульсації, відкриваються назовні вузьким кана-
лом. Вода, що надходить у пузули з клітини, виводиться через канал
назовні пасивно.
Система мікротрубочкових органел та структур. Головним
структурним елементом цієї системи є мікротрубочки, що побудовані
із субодиниць білку тубуліну. Мікротрубочки мають здатність подов-
жуватись за рахунок приєднання молекул альфа- та бета-тубуліну,
або розбиратися шляхом відщеплення цих мономерів. Як наслідок,
мікротрубочкові системи в клітині є утворами вельми динамічними.
Мікротрубочки є основою джгутикового апарату та його похідних -
центріолей, з мікротрубочок утворюються нитки веретена поділу, вони
також складають каркас цитоскелета.
Джгутиковий апарат. У водоростей, подібно до інших евкаріот,
джгутиковий апарат включає джгутики, їх базальні тіла та систему
джгутикових коренів. Джгутик утворений зовнішнім випином плазма-
леми, в якому розташовуються дев’ять периферичних пар мікротру-
бочок та дві поодинокі центральні мікротрубочки. Таке розміщення
мікротрубочок описується формулою «9+2», а сама мікротрубочкова
частина джгутика називається аксонемою. Будова джгутикової аксо-
неми за типом «9+2» вважається універсальною для всіх евкаріот.
Проте у водоростей відомі деякі модифікації цього плану будови. Зок-
рема, у чоловічих гамет діатомових водоростей відсутня центральна
пара мікротрубочок (формула «9+0»), а у багатьох динофітових во-
34
доростей периферична частина аксонеми містить не дуплети, а трип-
лети мікротрубочок.
Поверхня джгутика може бути гладенькою або нести на собі суб-
мікроскопічні волоски - мастигонеми, чи субмікроскопічні лусочки.
Якщо мастигонеми на джгутику розташовані по всій поверхні, то та-
кий джгутик називають пірчастим, якщо вони розташовуються одно-
бічно-гребінчастим. Мастигонеми у водоростей досить різноманітні,
та у загальному вигляді поділяються на прості, двочленні та тричленні.
Прості та двочленні мастигонеми синтезуються переважно на плаз-
малемі джгутика та/або в пухирцях комплексу Гольджі. Тричленні
мастигонеми - ретронеми - утворюються внаслідок багатоступінча-
того процесу синтезу та збірки окремих частин, в яких беруть участь
ядерна оболонка, хлоропластна ендоплазматична сітка, комплекс
Гольджі та плазмалема.
При основі джгутика знаходиться перехідна зона, за якою розта-
шовується його базальне тіло. В перехідній зоні центральні мікротру-
бочки зникають, і у представників різних відділів можуть з’являтися
додаткові структури - спіралі (у СЬгузоріїуіа, ЕизіідтаіорЬуіа,
ХапіЬоріїуІа, бісіуосіїоріїуіа), диски (Оіпорїіуіа, СгуріорЬуіа), перехідні
кільця (ОіпорЬуіа, Наріорїіуіа), зірчаста структура (СЬІогорЬуіа). Бу-
дова перехідної зони джгутика вважається важливою таксономічною
ознакою на рівні відділу.
Базальні тіла джгутиків утворені периферичними мікротрубочка-
ми аксонеми, де до кожного дуплету додається ще одна мікротрубоч-
ка (формула «3(9) + 0»). Відхилення від цього плану будови мають
лише базальні тіла чоловічих гамет діатомових водоростей, у яких
базальні тіла складаються не з дев’яти триплетів, а з дев’яти дуп-
летів мікротрубочок.
До базальних тіл приєднані джгутикові корені, за допомогою яких
джгутики закріплюються у клітин та які поєднують із джгутиковим апа-
ратом інші органели. До системи джгутикових коренів входять мікрот-
рубочкові та мікрофібрилярні корені. Мікрофібрилярні корені побудо-
вані переважно або з білка актину (такі корені під електронним мікрос-
копом виглядають гладенькими), або зі скоротливого білка центрину
(такі корені виглядають як поперечно-смугасті). Центринові попереч-
но-смугасті корені часто з’єднують базальні тіла джгутиків з ядром і в
цьому випадку називаються ризопластами.
Типи кореневих систем джгутиків у водоростей дуже різноманітні.
Відміни між різними типами систем полягають у кількості та співвідно-
шеннях мікротубулярних та мікрофібрилярних коренів, у кількості
35
мікротрубочок, що входять до складу мікротубулярних коренів, у наяв-
ності на джгутикових коренях додаткових структур (наприклад, т. зв.
багатошарової структури), у розташуванні коренів (зокрема, в тому, з
якими органелами асоціюється нижня частина кожного кореня).
У частини водоростей із джгутиковим апаратом пов’язані фото-
рецепторні системи. Зокрема, в евгленофітових, золотистих, евстиг-
матофітових, жовтозелених та бурих водоростей функції фоторецеп-
тора виконує потовщення при основі джгутика - парабазальне тіло,
до складу якого входять фоторецепторні флавін-подібні речовини.
У криптофітових та зелених водоростей функції фоторецептора ви-
конує ділянка однієї з мембран пластиди, у деяких динофітових та
гаптофітових - ущільнена ділянка цитоплазми, а у деяких динофіто-
вих - складна цитоплазматична вічкоподібна структура - оцелоїд.
З фоторецептором може бути скоординована стигма (вічко), що
виконує функції ширми, за допомогою якої клітина визначає напрям
падіння світла на фоторецептор. Стигма забарвлена у червоний колір
і може розташовуватися або в цитоплазмі (евгленофітові, евстигма-
тофітові, динофітові водорості), або у пластиді (золотисті, жовтозе-
лені, деякі динофітові, гаптофітові, криптофітові та зелені водорості).
У клітин, що мають джгутики, мікротрубочкові корені складають
основу внутрішнього скелета клітини - цитоскелета. До цитоскелета,
крім мікротрубочкових коренів та цитоплазматичних поодиноких мік-
ротрубочок, належать тонкі ^трічки актину (т. зв. мікрофіламенти) та
короткі з’єднувальні білкові стрічки (т. зв. проміжні філаменти). Ком-
плекс із мікротубулярних коренів, цитоплазматичних мікротрубочок,
актинових мікрофіламентів та проміжних філаментів утворює в сере-
дині клітини складну та динамічну внутрішню сітку, за допомогою якої
клітина з усіма органелами виступає як єдине структурне ціле.
Органелами, які походять від базальних тіл джгутиків, є центрі-
олі. Центріолі виявлені у представників відділів Сїігуворїіуіа, Хап-
іііорЬуіа, РЬаеорЬуіа та СЬІогоріїуІа. Центріолі виступають центрами
організації мікротрубочок веретена поділу. Будова центріолей повто-
рює план будови базальних тіл джгутиків - центріолі складаються з
дев’яти триплетів мікротрубочок, що розташовуються колом. Спорід-
неність центріолей з базальними тілами джгутиків підтверджується
тим, що у багатьох водоростей, які позбавлені центріолей, проте ма-
ють джгутики, саме базальні тіла виконують функцію центра органі-
зації мікротрубочок веретена поділу (наприклад, у ЕидіепорЬуіа,
РаріїісІорЬуїа, ЕизіідтаіорЬуіа, СІаисосузіорЬуІа, деяких Сїііогорїіуіа).
36
У ряду відділів водоростей центрами організації мікротрубочок
веретена можуть виступати відмінні від базальних тіл та центріолей
досить специфічні структури, у складі яких мікротрубочки взагалі не
виявлені: полярні платівки (діатомові водорості), полярні кільця (чер-
воні водорості), кіноплазматична сфера-ущільнена цитоплазма, що
оточена діктіосомами та ендоплазматичною сіткою (динофітові), ци-
стерни ендоплазматичної сітки та ризопласти (гаптофітові, крипто-
фітові, деякі зелені водорості).
Мітохондріальний апарат у евкаріотичних водоростей виглядає
досить одноманітним: мітохондрії відмежовуються від цитоплазми
двомембранною оболонкою; внутрішня мембрана утворює інвагінації
- кристи, на яких розташовуються ферменти та переносники елект-
ронів, що забезпечують дихання, в результаті якого відбувається син-
тез переважної кількості АТФ.
Мітохондрії мають власну мітохондріальну ДНК, яка організова-
на за принципом прокаріотичного нуклеоїду: вона замкнена в кільце,
не пов’язана з гістонами, гени організовані в оперони. Проте, на відміну
від геному прокаріот, мітохондріальний геном дуже редукований і
містить у середньому 15-80 тис. пар нуклеотидних основ замість 1,5-
4 млн. пар у прокаріот. Мітохондрії мають власний білок-синтезую-
чий апарат, представлений прокаріотичними 708 рибосомами. Под-
ібно до прокаріот, мітохондрії розмножуються поділом надвоє.
Різноманітність мітохондріального апарату проявляється, в пер-
шу чергу, за морфологією мітохондріальних крист. Так, у евгленофі-
тових у мітохондріях наявні три типи крист - пластинчасті, трубчасті
та дисковидні. У глаукоцистофітових, зелених, червоних та крипто-
фітових водоростей - лише пластинчасті кристи, в інших відділах у
мітохондріях переважають кристи трубчастої форми.
Фотосинтетичний апарат у евкаріотичних водоростей представ-
лений пластидами, що містять хлорофіл - хлоропластами. Хлоро-
пласти, подібно до мітохондрій, є ендосимбіотичними органелами з
власним хлоропластним геномом та білок-синтезуючим апаратом. Від
цитоплазми хлоропласт відмежовується дво-, три- або чотиримемб-
ранною оболонкою. Кількість мембран оболонки певним чином відоб-
ражає походження даної пластиди (первинно симбіотичне або вто-
ринно симбіотичне).
Первинно симбіотичні пластиди мають двомембранну оболонку.
У мембранах оболонки наявні субмікроскопічні пори прокаріотичного
37
типу, до складу яких входять особливі мембранні білки - порини. На
зовнішньому боці мембрани порини утворюють три канали, які на
внутрішньому боці зливаються в єдиний канал мембранної порино-
вої пори. Такі пори виявлені також в оболонці мітохондрій та в плаз-
малемі прокаріот.
Під хлоропластною оболонкою розташовуються сплощені інвагі-
нації внутрішньої мембрани - тилакоїди. В мембрани тилакоїдів вбу-
довані молекули хлорофілів, а також додаткових фотосинтетичних
пігментів: каротинів, ксантофілів, у деяких водоростей - також фі-
кобілінів. Через це тилакоїди вважаються елементарною структур-
ною фотосинтетичною одиницею пластиди. Тилакоїди можуть розмі-
щуватися поодиноко або об’єднуватись у групи. Такі групи тилакоїдів
називають ламелами. Якщо до складу ламели входить багато тила-
коїдів, розташованих один над одним, і такі стопки тилакоїдів пов’я-
зуються між собою за допомогою поодиноких тилакоїдів, то утворюєть-
ся грана. Тилакоїди можуть бути поодинокими (наприклад, у черво-
них водоростей), утворювати ламели з двох або трьох тилакоїдів (у
переважної більшості відділів) чи групуватися у грани (у частини зеле-
них водоростей). Тип організації тилакоїдів є однією з таксономічних
ознак на рівні відділу.
У хлоропластах багатьох водоростей можна спостерігати особ-
ливу структуру - піреноїд. Піреноїд - це білкове тіло, що утворене
ферментом рибульозо-дифосфат карбоксилазою (КІЛЗІ8СО). Робо-
та цього ферменту контролює початкові етапи темнової фази фото-
синтезу. Піреноїд може бути облямований крохмалем і тоді добре
помітним в оптичний мікроскоп. Проте у більшості водоростей піре-
ноїд голий, і для його спостереження необхідно проводити спеціальні
цитохімічні забарвлення або використовувати методи електронної
мікроскопії.
Подібно до мітохондрій, у хлоропласті є власний геном, представ-
лений хлоропластною ДНК, що організована за прокаріотичним ти-
пом, зокрема, гени групуються в оперони, ДНК не пов’язана з гістона-
ми. Хлоропластна ДНК - генофор - може бути розташована компакт-
но або представлена кількома фрагментами, розсіяними по всьому
матриксу пластиди. Хлоропластна ДНК більша від мітохондріальної,
проте менша від ДНК нуклеоїду прокаріот, її розмір коливається в
межах 100-300 тис. пар нуклеотидних основ. У матриксі хлоропласта
є власний білок-синтезуючий апарат, представлений прокаріотични-
ми 708 рибосомами.
Первинно симбіотичні пластиди наявні у трьох відділів водорос-
тей: Сіаисосузіоріїуіа, Кґіосіорґіуіа, СЬІогорЬуіа. Спільними ознаками
38
первинно симбіотичних пластид є наявність лише двох мембран у
складі оболонки хлоропласта. Проте у глаукоцистофітових водорос-
тей між зовнішньою та внутрішньою мембранами зберігається шар
муреїну, що являє собою залишок клітинної оболонки прокаріотичної
водорості-ендосимбіонта. В червоних та зелених водоростей муреї-
ну в оболонках пластид немає. Для відділу КЬосІорЬуІа характерні
пластиди з поодиноко розміщеними тилакоїдами, які не організовані
у ламели і мають на поверхні фікобілісоми, утворені фікобіліновими
пігментами. Хлоропласти зелених водоростей фікобілісом не мають,
тилакоїди зібрані в ламели, які у багатьох представників утворюють
грани, що морфологічно схожі з гранами вищих рослин.
Вторинно симбіотичні пластиди притаманні всім іншим відділам
евкаріотичних водоростей. Ці пластиди утворилися внаслідок ендо-
симбіозу евкаріотичної гетеротрофної клітини-господаря з евкаріо-
тичною водорістю з відділу КЬосіорґіуіа або СЬІогоріїуіа, що мала
первинно симбіотичну пластиду. Відповідно, вторинно симбіотичні
пластиди за походженням поділяють на вторинно симбіотичні ро-
допласти та вторинно симбіотичні хлоропласти.
Всі відомі на сьогодні вторинно симбіотичні родопласти оточені
чотирма мембранами, з яких дві внутрішні є мембранами оболонки
хлоропласту ендосимбіотичної червоної водорості. Третя мембрана
представляє собою видозмінену плазмалему ендосимбіонта, а чет-
верта - залишок мембрани травної вакуолі клітини-господаря, який
захопив червону водорість, проте не перетравив її. Простір між дру-
гою та третьою мембранами називають перипластидним. Він являє
собою залишок цитоплазми червоної водорості. У перипластидному
просторі часто виявляються евкаріотичні 808 рибосоми, у ньому в
представників деяких відділів можуть відкладатися продукти асимі-
ляції. Третю та четверту мембрани називають хлоропластною ендоп-
лазматичною сіткою. Зовнішня мембрана хлоропластної ендоплаз-
матичної сітки, як правило, переходить у зовнішню мембрану ядра.
У криптофітових водоростей в перипластидному просторі збері-
гається редуковане ядро червоної водорості - ендосимбіонта, яке
називається нуклеоморф. Нуклеоморф оточений двома мембрана-
ми, причому зовнішня мембрана має пори, морфологічно подібні до
ядерних пор. У нуклеоморф! виявлена ДНК, яка за молекулярними
даними представляє біля десяти генів, споріднених з аналогічними
ядерними генами РЬосІорїіуіа. Тилакоїди у криптофітових водорос-
тей зібрані парами (утворюють двотилакоїдні ламели).
39
В інших водоростей із вторинно симбіотичними родопластами -
всіх хромофітових водоростей (КарЬісІорЬуіа, СЬгузорЬуіа, Еизіід-
гпаіоріїуіа, Хапіііоріїуіа, РЬаеорЬуіа, ВасіІІагіоріїуіа, ОісіуосіїорЬуіа),
гаптофітових та деяких динофітових - нуклеоморф відсутній, наме-
ли утворені трійками тилакоїдів, фікобілінових пігментів немає.
Вторинно симбіотичні хлоропласти притаманні водоростям з
ЕидіепорЬуІа, СЬогагасЬпіорЬуіа та деяким представникам ОіпорЬуіа.
У хлорарахніофітових водоростей хлоропласт вкритий чотирма
мембранами, з яких дві зовнішні утворюють хлоропластну ендоплаз-
матичну сітку. В перипластидному просторі наявні евкаріотичні 808
рибосоми та нуклеоморф. Подібно до криптофітових, він відмежова-
ний від перипластидного простору двомембранною оболонкою і
містить ДНК, яка за молекулярними даними представляє дуже реду-
кований ядерний геном зеленої водорості. Тилакоїди групуються в
ламели по два або по три.
Два типи вторинно симбіотичних хлоропластів виявлено у дино-
фітових водоростей. Вторинно симбіотичні хлоропласти першого типу
нагадують пластиди хлорарахніофітових, оскільки теж вкриті чотир-
ма мембранами і в перипластидному просторі зберігають нуклеоморф.
Вторинно симбіотичні хлоропласти другого типу не мають нуклеомор-
фу і вкриті лише трьома мембранами. Тримембранні вторинно симбіо-
тичні хлоропласти без нуклеоморфу характерні також для всіх фото-
автотрофних евгленофітових водоростей.
Таким чином, пластиди у водоростей значно різноманітніші, ніж у
вищих рослин. Тип та особливості ультратонкої будови пластид є
важливими таксономічними ознаками на рівні відділу і характеризу-
ють походження цієї органели.
Морфологічна різноманітність
У надзвичайному ступені різноманітність водоростей проявляєть-
ся в їх зовнішньому вигляді. Розміри водоростей коливаються від
кількох десятих мікрометра до десятків метрів. Тіла водоростей мо-
жуть мати просту будову і бути представлені поодинокими клітинами
або бути складно розчленованими на тканини та органи. Частина
водоростей здатна до активного руху і цим нагадує тварин. Інші до
активного руху не здатні і ведуть прикріплений, суто рослинний спосіб
життя або ширяють у товщі водної маси, пасивно мігруючи за течія-
ми. Таломи водоростей можуть складатися з одноядерних клітин або
40
клітини містять велику кількість ядер і сягають таких розмірів, при яких
їх вже можна добре розрізнити неозброєним оком.
Різноманітність морфології водоростей відображає основні на-
прямки еволюції їх тіла, причому генеральний напрям пов’язаний із
вдосконаленням справжньої рослинної стратегії життя - збільшення
фотосинтезуючої поверхні, перехід до прикріпленого способу існу-
вання, збільшення розмірів для захисту від виїдання і т. п. Морфоло-
гічна еволюція за генеральним напрямом у схожих умовах середови-
ща обумовлювала конвергентний розвиток тіла у водоростей філоге-
нетично віддалених груп. Наприклад, магістральний напрям, пов'я-
заний зі збільшенням фотосинтезуючої поверхні та захистом від хи-
жаків, обумовив виникнення багатоклітинності як у прокаріотичних,
так і в евкаріотичних водоростей. Явище конвергентної еволюції тіла
водоростей отримало назву морфологічного паралелізму.
Певні плани будови водоростей, які відображають основні етапи
морфологічної еволюції, називають типами морфологічної структури
тіла, або скорочено - типами морфологічної структури. У водоростей
виділяють 8 типів морфологічних структур, з яких три представлені в
одноклітинних форм (монадний, гемімонадний, кокоїдний), три -
у багатоклітинних одноядерних водоростей (нитчастий, гетеротри-
хальний, тканинний), два - у багатоядерних, або т. зв. неклітинних
водоростей (сифональний та сифонокладальний). Поняття про типи
морфологічної структури застосовують лише стосовно евкаріотичних
водоростей.
Еволюція тіла в одноклітинних евкаріотичних фотоавтотрофів
відбувалась у напрямку втрати рухливості і переходу до нерухомого,
рослинного способу існування.
Найпростішим типом структури вважають монадний тип. Голов-
ною ознакою цього типу є наявність рухливих джгутиків. Монадні во-
дорості одноядерні і, як правило, мають також стигму та пульсуючі
вакуолі.
Водорості гемімонадного типу є нерухомими, проте зберігають
деякі ознаки, притаманні монадним організмам - або стигму, або пуль-
суючі вакуолі, або джгутики, що не здатні до активного руху - псевдо-
цилії.
У водоростей з кокоїдним типом структури повністю відсутні будь-
які ознаки монадної будови. Кокоїдні форми є одноклітинними, неру-
хомими, не мають стигми, джгутиків та пульсуючих вакуолей і, таким
чином, представляють одноклітинні організми із суто рослинною стра-
тегією життя.
41
Монадні, гемімонадні та кокоїдні водорості можуть бути представ-
лені поодинокими клітинами або можуть об’єднуватись у колонії. Най-
частіше колонії формуються за рахунок утримання водоростевих
клітин разом за допомогою спільного слизу. Кількість клітин у колонії
може змінюватись у процесі росту колонії або лишатися незмінною
протягом всього часу існування колонії, незалежно від її віку. Колонії,
в яких кількість клітин залишається постійною, а збільшення розмірів
відбувається лише завдяки росту кожної індивідуальної клітини, на-
зивають ценобіями.
У багатоклітинних водоростей морфологічна еволюція тіла була
пов’язана з поступовою диференціацією окремих частин талому за
функціями.
Так, у водоростей із нитчастим (трихальним) типом структури
талом зазвичай має вигляд нитки з послідовно розміщених одна за
одною одноядерних клітин. Клітини у нитчастих водоростей функціо-
нально та морфологічно однакові. Винятком можуть бути лише ба-
зальна та апікальна клітини: перша інколи видозмінюється у при-
кріплювальну клітину, що може утворювати підошву, друга - витягу-
ватись у волосок з частково редукованими хлоропластами. Нитки у
представників з нитчастим типом структури найчастіше нерозгалу-
жені (т. зв. прості), рідше можуть галузитися, проте всі гілки викону-
ють однакові функції і за морфологією між собою не відрізняються.
Ознакою гетеротрихального типу структури є наявність дифе-
ренціації ниток за функціями. У найпростішому варіанті нитки дифе-
ренціюються на сланкі, що виконують функції прикріплення до суб-
страту, та висхідні, клітини яких виконують функції фотосинтезу та
розмноження. Висхідні нитки, в свою чергу, можуть бути диференці-
йованими на опірні, в яких клітини мають товсті оболонки і редуко-
вані хлоропласти, та асиміляторні, що утворені переважно дрібними
клітинами з масивними хлоропластами. У деяких водоростей окремі
гілки асиміляторів видозмінюються у захисні корові нитки або утво-
рюють волоски чи паренхіматозні диски. Відомі випадки, коли кліти-
ни функціонально однакових ниток гетеротрихальних таломів зрос-
таються між собою, утворюючи несправжні тканини. Таломи з не-
справжніх тканин особливо характерні для багатьох червоних водо-
ростей.
Тканинна структура є наслідком процесів функціональної дифе-
ренціації груп клітин, що утворюються від спеціалізованих твірних
клітин. У водоростей твірними є переважно клітини кори або так зва-
ної меристодерми, в той час, як у вищих рослин - меристематичні
клітини. Водоростеві таломи тканинного типу структури можуть бути
42
простими або складними. Прості таломи утворені лише двома типа-
ми тканин - корою та серцевиною, складні таломи можуть містити до
чотирьох типів тканин - меристодерму, кору, проміжну тканину, сер-
цевину. Таломи з тканинним типом структури досить широко пред-
ставлені у відділі бурих водоростей, а також у деяких червоних водо-
ростей.
Таломи, що утворені багатоядерними клітинами, називають та-
ломами неклітинної будови. Термін «неклітинна будова» з’явився у
XIX ст., тоді, коли були відкриті перші багатоядерні водорості. В цей
час вже були сформульовані основні положення клітинної теорії, зокре-
ма, положення про одноядерність елементарної структурної одиниці
живого - клітини. Протиріччя цього теоретичного положення резуль-
татам фактичних спостережень стало причиною введення поняття
«неклітинна будова», хоча й у випадку неклітинної будови елемен-
тарною одиницею залишається клітина. З початку XX ст. і по теперішній
час термін «неклітинна будова» використовують стосовно багатоя-
дерних клітин. Відомо два типи неклітинних структур тіла - сифональ-
ний та сифонокладальний.
У водоростей сифонального типу структури весь талом являє
собою одну багатоядерну клітину, часто досить великих розмірів.
У багатьох водоростей сифональні таломи можуть мати досить склад-
ну диференціацію, зокрема, утворювати прикріплювальні структури,
що нагадують ризоїди та висхідні асиміляторні частини (наприклад, у
жовтозеленої водорості \/аисіїегіа, у зеленої водорості Вгуорзіз). Проте
такі частини не відокремлюються одна від одної поперечними пере-
городками, і, таким чином, складно диференційовані таломи все ж
таки являють собою лише одну велетенську багатоядерну клітину.
Таломи сифонокладального типу структури є багатоклітинни-
ми, а кожна клітина є багатоядерною. Сифонокладальні таломи ут-
ворюються внаслідок особливого типу поділу - сегрегативного, при
якому поділ ядра (каріокінез) випереджає поділ цитоплазми (цитокі-
нез). Часто сифонокладальні таломи мають складну функціональну
диференціацію, наприклад - на сланку та висхідну частини, на ризої-
ди та опірні багатоядерні нитки і т. п.
У систематиці водоростей для різних відділів типи морфологіч-
ної структури використовуються як важлива таксономічна ознака на
рівні родин та порядків, а в деяких випадках - навіть класів. Різні
типи морфологічних структур у різних філах виникали неодноразово,
причому в більшості відділів водоростей вихідним типом морфологіч-
ної структури був монад ний тип.
43
Розмноження водоростей
Розмноження у водоростей може відбуватися нестатевим або
статевим шляхом. При нестатевому розмноженні дочірні особини ус-
падковують геном, ідентичний до геному материнського організму, за
винятком випадків, коли мають місце генні або хромосомні мутації.
При статевому розмноженні геноми дочірніх організмів відрізняються
від батьківських, оскільки утворенню дочірнього покоління зазвичай
передують процеси обміну генетичною інформацією між різними осо-
бинами внаслідок кросинговеру, що відбувається після статевого про-
цесу.
Статевийипроцес та пов’язані з ним кросинговер і мейоз обумов-
люють рекомбінантну мінливість і сприяють зростанню рівня генетич-
ної гетерогенності популяцій. Рівень рекомбінантної мінливості може
зростати також внаслідок обміну частинами геномів при парасексу-
альних процесах (наприклад, під час кон’югації або трансформації у
прокаріот), проте парасексуальні процеси безпосередньо не пов'я-
зані з відтворенням дочірніх особин і тому до способів розмноження
не відносяться.
Нестатеве розмноження у водоростей поділяється на два ос-
новних типи: а) вегетативне, що здійснюється шляхом поділу вегета-
тивних клітин або фрагментами вегетативного тіла; б) розмноження
за допомогою спеціалізованих клітин - спор, яке інколи називають
справжнім нестатевим розмноженням або споруляцією.
Вегетативне розмноження. В одноклітинних водоростей веге-
тативне розмноження відбувається переважно поділом клітини на-
двоє. У багатоклітинних та колоніальних представників вегетативне
розмноження частіше пов’язане з фрагментацією таломів та колоній.
Фрагментацію можуть спричинювати механічні чинники (наприклад,
вітро-хвильові удари, течія) або фрагментація може відбуватися вна-
слідок відмирання частин таломів чи колоній. Наприклад, відмиран-
ня окремих інтеркалярних клітин у багатоклітинних трихомах синьо-
зелених водоростей часто обумовлює розпад трихому на репродук-
тивні фрагменти - гормогонії. У небагатьох водоростей вегетативне
розмноження може здійснюватися за допомогою спеціалізованих діля-
нок тіла - вивідкових бруньок (бура водорість ВрНасеІІагіа) або вивід-
кових бульбочок (зелені водорості з порядку Сїіагаїез). Особливим
варіантом вегетативного розмноження є розмноження за допомогою
44
акінет. Останні являють собою клітини, що утворюються із звичайних
вегетативних клітин при погіршенні умов існування. Акінети мають
потовщену клітинну оболонку, містять багато запасних поживних ре-
човин і, на відміну від нормальних вегетативних клітин, за несприят-
ливих умов здатні переходити у стан анабіозу або криптобіозу, і зав-
дяки цьому переживати несприятливі періоди. При покращенні умов
вегетації акінети проростають у нові вегетативні особини.
Нестатеве розмноження шляхом споруляцїї здійснюється спе-
ціалізованими клітинами - спорами, які утворюються в спорангіях.
Спорангії у водоростей одноклітинні, кількість спор в одному спорангії
може коливатися від однієї (наприклад, у зеленої нитчастої водорості
Оесіодопіит) до кількох десятків або навіть сотень (у багатьох бурих
водоростей).
У відповідності зі здатністю до активного руху спори поділяють на
рухливі із джгутиками - зооспори, нерухомі з клітинними покривами -
гемізооспори, апланоспори та автоспори, та малорухливі амебоїдні,
що позбавлені джгутиків - моноспори, біспори та тетраспори.
Зооспори завжди мають джгутики і часто - вічко та пульсуючі ва-
куолі. Зооспори можуть бути голими або вкритими клітинними покри-
вами.
Гемізооспори джгутиків не мають, проте зберігають ознаки монад-
ної організації - стигму, пульсуючі вакуолі, базальні тіла джгутиків.
Зазвичай гемізооспори спостерігаються в амфібіальних водоростей,
які у водному середовищі утворюють зооспори, проте у позаводному
припиняють процес формування джгутиків на стадії їх базальних тіл.
Апланоспори джгутиків також не мають, їх початкові етапи роз-
витку нагадують ранні стадії процесу утворення зооспор, зокрема, в
протопластах майбутніх апланоспор утворюються базальні тіла. Проте
на кінцевих етапах розвитку, незалежно від наявності у середовищі
води, базальні тіла або зникають, або мігрують вглиб клітини і пере-
творюються на центріолі. Інколи в апланоспор можна спостерігати
пульсуючі вакуолі, проте стигми у них не розвиваються. Зазвичай зрілі
апланоспори не повторюють морфології материнських клітин.
Автоспори у своєму розвитку не проходять навіть початкових
стадій зооспороутворення. В автоспор відсутні джгутики, їх базальні
тіла, стигми, пульсуючі вакуолі. Морфологічно автоспори подібні до
материнських клітин і відрізняються від них лише меншими розмірами.
Апланоспори та автоспори, крім функції розмноження, часто здатні
виконувати й функцію перенесення несприятливих умов існування.
45
Амебоїдні безджгутикові спори - моноспори, біспори, тетра-
спори- відомі у червоних та деяких бурих водоростей. Клітина такої
спори являє собою голий протопласт, одягнений лише плазмалемою
і здатний до повільного амебоїдного руху. Утворенню цих спор у пере-
важній більшості випадків передує мейоз, через що моно-, бі- та
тетраспори мають гаплоїдний набір хромосом. Назви цих спор відоб-
ражають те, в якій кількості утворюються спори у спорангії - по одній
(моноспори), по дві (біспори) чи по чотири (тетраспори).
За типом поділу ядра в клітині-спорангії (мітотичний чи мейотич-
ний) спори також часто поділяють на мітоспори та мейоспори.
Статеве розмноження відбувається внаслідок статевого проце-
су, який створює умови для кросинговеру, що, у свою чергу, забезпе-
чує певний рівень генетичної гетерогенності популяцій на основі ре-
комбінантної мінливості. Статеві процеси у водоростей поділяють на
дві основні групи - соматогамні та гаметогамні.
При соматогамних статевих процесах зливаються дві вегетативні
(інакше - соматичні) клітини. Якщо такі клітини представлені монад-
ними клітинами, позбавленими клітинних покривів, то цей тип сома-
тогамії називають хологамією.
Якщо копуляція відбувається між вегетативними клітинами, які
мають клітинні покриви, то цей тип соматогамії називають кон’юга-
цією. При кон’югації клітини, що мають копулювати, зближуються та
вкриваються спільним слизом. Далі кожна клітина утворює копуля-
ційний відросток в напрямку до іншої. Коли копуляційні відростки сти-
каються один з одним, оболонка між ними розчиняється й утворюєть-
ся кон’югаційний канал, по якому ядро з однієї клітини переходить в
іншу. Далі ядра зливаються і утворюється зигота.
При гаметогамії копуляція відбувається між спеціалізованими
клітинами статевого розмноження - гаметами, які утворюються в ста-
тевих органах - гаметангіях. У відповідності з морфологією гамет
виділяють три основі типи гаметогамії: ізогамію, гетерогамію та оога-
мію.
Ізогамією називають такий статевий процес, при якому копулю-
ють дві монадні гамети, які морфологічно виглядають однаковими.
Проте фізіологічно та на ультратонкому рівні гамети розрізняються:
одна з гамет, що при копуляції приймає ядро іншої гамети, вважаєть-
ся жіночою і позначається знаком «+». Друга гамета, ядро якої при
копуляції мігрує у цитоплазму жіночої гамети, вважається чоловічою
і позначається знаком «-».
46
При гетерогамії (синонім - анізогамії) копулюють дві монадні га-
мети, які відрізняються розмірами та ступенем рухливості. Чоловіча
гамета дрібна, рухається дуже швидко. Жіноча гамета має більші
розміри, ніж чоловіча, і її рухливість обмежена.
Оогамія - це статевий процес, при якому копуляція відбувається
між великою, нерухомою та позбавленою джгутиків жіночою гаметою
- яйцеклітиною - та дрібною чоловічою гаметою. Чоловічі гамети, що
мають джгутики і здатні до активного руху, називають сперматозої-
дами. Якщо ж чоловіча гамета позбавлена джгутиків і є нерухомою,
то її називають спермацієм. Гаметангії, в яких утворюються яйцеклі-
тини, називають оогоніями, а сперматозоїди чи спермації - антери-
діями. У водоростей оогонії та антеридії, на відміну від гаметангіїв
вищих рослин, є одноклітинними (за винятком деяких зелених водо-
ростей з класу харофіцієвих).
Крім основних типів гаметогамії (тобто ізогамії, гетерогамії та
оогамії), у водоростей відомо кілька додаткових специфічних типів
статевого процесу. Наприклад, у діатомових водоростей дуже поши-
реним типом статевого процесу є автогамія. При автогамії диплоїд-
не ядро клітини редукційно ділиться, далі з чотирьох дочірніх гаплої-
дних ядер два дегенерують, а два інших зливаються і дають початок
диплоїдному ядру зиготи. Таким чином, при автогамії гамети не утво-
рюються, а статевий процес відбувається без статевого партнера.
Інший специфічний варіант статевого процесу - атактогамія,
яка спостерігається у деяких зелених вольвокальних водоростей (на-
приклад, у роду СІїІогодопіит). Тут в гаметангіях представників навіть
однієї популяції утворюються монадні гамети різних розмірів і копу-
люють як морфологічно однакові, так і морфологічно відмінні гамети.
Причому гамета, що приймає ядро від іншої (тобто фізіологічно жіно-
ча гамета), може за розмірами дорівнювати чоловічій, бути більшою
від неї або меншою.
У водоростей, що мають ізогамні статеві процеси, розрізняють
види гомоталічні та гетероталічні. У ґомоталічних видів як «+», так і
«-» гамети утворюються на одному .таломі і здатні копулювати між
собою, тобто такі види є фізіологічно двостатевими. У гетероталіч-
них видів «+» та «-» гамети розвиваються на різних таломах, і тому
гетероталічні представники фізіологічно є роздільностатевими (од-
ностатевими).
Водорості, що мають статеві процеси гетеро- та оогамного типу,
поділяють також на однодомні та дводомні. В однодомних водорос-
тей антеридії та оогонії розміщуються на одному таломі, тобто такі
47
види є і фізіологічно, і морфологічно двостатевими. У дводомних видів
>кіночі та чоловічі статеві органи утворюються на різних таломах, тоб-
то має місце фізіологічна та морфологічна роздільностатевість (од-
ностатевість).
Після злиття цитоплазм вегетативних клітин чи гамет, що прий-
мали участь у статевому процесі (т. зв. плазмогамії) зазвичай відбу-
вається злиття ядер - каріогамія - і утворюється диплоїдне ядро зи-
готи. Проте з цього правила у водоростей відомі й виключення. На-
приклад, у багатьох золотистих водоростей після плазмогамії ядра
одразу не зливаються, і, як наслідок, утворюється клітина, що містить
два генетично відмінні ядра - дикаріон. Така дикаріонтична клітина
надалі перетворюється на дикаріонтичну зиготичну цисту, що пере-
ходить у стан спокою. Каріогамія і утворення власне зиготи відбу-
вається лише після закінчення періоду спокою.
Життєві цикли водоростей
Зміни життєвих форм, які відбуваються з індивідом протягом його
онтогенезу, складають життєвий цикл. Життєвий цикл охоплює всі
стадії розвитку індивіда між однаковими життєвими формами (на-
приклад, від зиготи до зиготи або від гамети до гамети, від спорофіта
до спорофіта).
Типи життєвих циклів. Звичайно розрізняють два основні типи
життєвих циклів: життєві цикли водоростей, позбавлених статевого
процесу, та життєві цикли водоростей, які мають статевий процес.
Водорості, які позбавлені статевого процесу, називають агамними, а
їх життєві цикли - цитоморфозом. У водоростей, що мають стате-
вий процес (т. зв. еугамні водорості), звичайно розрізняють три ос-
новні типи життєвого циклу - гаплофазний, диплофазний та гапло-
диплофазний.
Життєві цикли еугамних водоростей. У життєвих циклах водо-
ростей із статевим процесом обов’язково присутні принаймні три
стадії: вегетативна стадія, гамети (або вегетативні клітини, що вико-
нують їх функцію), зигота. Назву життєвим циклам дають, в першу
чергу, за наборами хромосом (плоїдністю) вегетативних стадій роз-
витку. Якщо вегетативна стадія має гаплоїдний набір хромосом, жит-
тєвий цикл називають гаплофазним, диплоїдний набір - диплофаз-
ним. Якщо ж у життєвому циклі чергуються вегетативні стадії як з гап-
лоїдним, так і з диплоїдним набором хромосом, то життєвий цикл
розглядається як гаплодиплофазний. Послідовність змін плоїдності
протягом життєвого циклу називають зміною ядерних фаз.
48
Під вегетативними стадіями (поколіннями) розуміють ті стадії,
протягом яких водорості активно вегетують та утворюють репродук-
тивні клітини. Під репродуктивними клітинами розуміють спеціалізо-
вані клітини як нестатевого, так і статевого розмноження - спори та
гамети. Залежно від типу розмноження (нестатеве або статеве) се-
ред вегетативних стадій розрізняють спорофіт - покоління, що утво-
рює спори, гаметофіт - покоління, що утворює гамети, та гаметоспо-
рофіт - покоління, яке здатне утворювати як гамети, так і спори.
У життєвому циклі може бути присутнє тільки одне покоління (на-
приклад, тільки гаметофіт або тільки гаметоспорофіт) або різні поко-
ління нерегулярно чи регулярно змінюються одне одним (наприклад,
спорофіт змінюється гаметофітом, а гаметофіт, у свою чергу, споро-
фітом). Залежно від цього розрізняють життєві цикли без зміни по-
колінь, з неправильним чергуванням поколінь та з правильним чергу-
ванням поколінь. Якщо при чергуванні поколінь різні покоління мор-
фологічно подібні (наприклад, гаметофіт зовні не відрізняється від
спорофіту), то таке чергування поколінь називають ізоморфним, у про-
тилежному випадку (наприклад, коли спорофіт макроскопічний, а га-
метофіт мікроскопічний) чергування поколінь називають гетеромо-
рфним.
Види, в яких одна й та ж сама особина здатна продукувати гаме-
ти обох статевих знаків, називають двостатевими (однодомними), а
ті, в яких чоловічі та жіночі гамети утворюються на різних особинах -
роздільностатевими (дводомними). Коли чоловіча та жіноча рослини
помітно відрізняються за зовнішнім виглядом, то таке явище назива-
ють статевим диморфізмом.
Плоїдність поколінь і, відповідно, тип життєвого циклу за чергу-
ванням ядерних фаз визначаються тим, на якій стадії життєвого цик-
лу відбувається мейотичний (редукційний) поділ. При мейозі кількість
хромосом зменшується вдвічі. Таким чином, всі стадії розвитку про-
тягом відрізка життєвого циклу від мейозу до статевого процесу бу-
дуть мати гаплоїдний набір хромосом, а всі стадії після статевого
процесу до мейозу - диплоїдний.
Типи мейозу та зміна ядерних фаз. Мейоз (редукційний поділ)
може відбуватися на одній із трьох стадій життєвого циклу: 1) на стадії
проростання зиготи - т. зв. зиготична редукція; 2) на стадії утворення
гамет (гаметична редукція); 3) при утворенні спор (спорична редук-
ція) (рис. 3). Крім того, відомі випадки, коли мейоз відбувається у веге-
тативних (соматичних) клітинах (соматична редукція).
49
(зигота)
Рис. 3. Можливі типи мейозу
(редукційного поділу) та умовні по-
значення основних стадій життєво-
го циклу.
При зиготичній редукції
вегетативне покоління завжди
буде гаплоїдним, відповідно,
життєвий цикл буде гаплофаз-
ним. Звичайно перед мейозом
зигота проходить період спо-
кою, протягом якого відбу-
вається підготовка до редук-
ційного поділу (рис. 4 А, Б)
Рис. 4. Типи життєвих циклів у водоростей за зміною ядерних фаз та чергу-
ванням поколінь: А, Б - гаплофазний цикл із зиготичною редукцією (А - без
чергування поколінь, Б - з чергуванням поколінь); В, Г - диплофазний цикл з
гаметичною редукцією (В - без чергування поколінь, Г- з чергуванням поколінь);
Д, Е - гаплодиплофазний цикл зі споричною редукцією (Д - з чергуванням двох
поколінь - спорофіту та гаметофіту, Е - з чергуванням трьох поколінь - гаметофіту,
спорофіту та карпоспорофіту). Позначення: п - гаплоїдний набір хромосом, 2п -
диплоїдний набір хромосом, К8 - карпоспорофіт, кзр - карпоспори, ПС - період
спокою. Інші позначення подані згідно з рис. З
50
При гаметичній редукції вегетативне покоління завжди буде дип-
лоїдним, відповідно, життєвий цикл - диплофазним. У цих випадках
зигота проростає, як правило, без періоду спокою. Таким чином, на-
явність періоду спокою дозволяє встановити тип мейозу і тим самим
визначити життєвий цикл (рис. 4 В, Г).
При споричній редукції диплоїдна зигота проростає без періоду
спокою у диплоїдний спорофіт. На спорофіті утворюються спорангії,
в яких відбувається мейоз і далі формуються гаплоїдні спори. Спори
проростають у гаплоїдний гаметофіт, що продукує гамети. Після ко-
пуляції гамет утворюється диплоїдна зигота і цикл замикається. Ос-
кільки при споричній редукції спорофіт є диплоїдним, а гаметофіт -
гаплоїдним, життєвий цикл буде гаплодиплофазним (рис. 4 Д, Е).
Таким чином, тип редукції визначає тип життєвого циклу за ядер-
ними фазами вегетативних поколінь: гаплофазний, диплофазний, гап-
лодиплофазний.
Різноманітність життєвих циклів. Різноманітність типів стате-
вого розмноження, редукційного поділу, чергувань поколінь обумов-
люють велику різноманітність життєвих циклів у водоростей.
Так, у деяких відділів водоростей (зокрема, Карїіісіорїіуіа,
Еизіідтаіорїіуіа, Оісіуосїіорїіуіа, СІаисосузіорїіуіа) статевого роз-
множення не виявлено, і життєвий цикл являє собою цикломорфоз,
хоча часом і досить складний.
Серед еугамних форм у кількох відділів життєвий цикл проходить
без чергування поколінь, з фіксованим типом редукції кількості хромо-
сом. Так, у еугамних Еидіепорїіуіа, Хапіїїорїіуіа, Сїігузорїіуіа редукція
зиготична і життєвий цикл гаплофазний. У ВасіІІагіорїіуІа життєвий
цикл завжди диплофазний, з гаметичною редукцією і особливою ста-
дією, що отримала назву ауксоспори.
У Оіпорїіуіа цікавою особливістю є те, що зигота навіть в одного
й того ж виду може проростати як з редукційним поділом, так і без
нього, тобто тип редукції жорстко не фіксований, на відміну від пред-
ставників інших відділів.
Складні цикли з чергуванням двох поколінь зустрічаються лише
у чотирьох відділах водоростей - Рїіаеорїіуіа, Наріорїіуіа, Сїііогорїіуіа
та примітивних КЬосіорйуІа. Цікаво, що всі ці відділи (за винятком
Наріорїіуіа), містять багато складних за морфологічною будовою ви-
дів. У розвинутих в еволюційному відношенні червоних водоростей із
класу Флоридей у життєвому циклі чергуються три покоління - спо-
рофіт, гаметофіт та карпоспорофіт (рис. 4 Е).
51
ГІрокаріотичні водорості
Серед прокаріот організми, здатні до оксигенного фотосинтезу,
представлені лише відділом синьозелених водоростей. Цей відділ,
разом із бактеріями-грацилікутами, фірмакутами та моллікутами, вхо-
дить до царства справжніх бактерій.
Відділ Синьозелені водорості - СуапорНуІа
Синьозелені водорості (Суапорїіуіа), або ціанобактерії, об’єдну-
ють всі прокаріотичні фотоавтотрофні рослини і нараховують при-
близно 2000 видів. Синьозелені водорості є однією з найдавніших
груп на планеті. Вважають, що Суапорїіуіа виникли біля 3,5-3,8 млрд.
років тому.
Біохімічні особливості. Синьозелені водорості забарвлені пе-
реважно у блакитно-зелений колір, в екстремальних умовах частіше
мають забарвлення з різними відтінками червоного кольору. Зелено-
го відтінку клітинам надає хлорофіл «а». Червоний або блакитний
колір обумовлений наявністю значної кількості фікобілінових пігментів
- фікоціаніну, алофікоціаніну (сині пігменти) та фікоеритрину (черво-
ний пігмент). Каротиноїди представлені лише р-каротином, ксанто-
філами лютеїнового циклу (лютеїном та зеаксантином) та специфіч-
ними ксантофілами синьозелених водоростей - переважно осцилок-
сантином, міксоксантином, афаніцином та афанізофілом.
До Суапорїіуіа належить також невелика, проте цікава група зе-
лених прокаріотичних водоростей, відкрита у 1976 р. - т. зв. прохло-
рофітові водорості. Ці водорості, крім хлорофілу «а», мають також
хлорофіл «Ь» або хлорофіл-подібний пігмент (дивініл-хлорофіл а-по-
дібний Мд-порфірин), інколи також а-каротин.
Майже у всіх Суапорїіуіа основним продуктом асиміляції є гліко-
геноподібний полісахарид-крохмаль синьозелених водоростей. Крім
вуглеводів, більшість синьозелених водоростей запасає також ціано-
фіцин та волютин.
Цитологічні особливості. Клітини Суапорїіуіа мають типову про-
каріотичну будову: вони позбавлені морфологічно оформленого ядра,
мітохондрій, хлоропластів, ендоплазматичної сітки, комплексу
Гольджі, лізосом. У Суапорїіуіа відсутні будь-які структури, побудо-
вані з мікротрубочок-джгутики, їх базальні тіла, мікротубулярні еле-
52
менти цитоскелета, центріолі, веретено поділу. Синьозелені водо-
рості не здатні до мітозу, мейозу та типового для евкаріот статевого
процесу.
Клітинні покриви у Суапорїіуіа - це двошарова клітинна оболон-
ка. Внутрішній шар утворений пептидогліканом - муреїном. Зовнішній
шар побудований переважно з пектинових речовин та мікрофібрил,
до складу яких входять скоротливі білки. Останні обумовлюють
здатність багатьох синьозелених водоростей до активного ковзаючо-
го або обертального руху. Назовні від клітинної оболонки у більшості
видів виділяються пектинові речовини, що утворюють колоніальний
слиз або слизові піхви.
Справжнього ядра немає, а його функції виконує нуклеоїд. ДНК
нуклеоїду не пов’язана з білками-гістонами, не має хромосомної орга-
нізації, хоча і розглядається як аналог однієї хромосоми. Звичайно
основна маса нуклеоїду зосереджується у центрі клітини, через що
цю зону цитоплазми називають нуклеоплазмою або центроплазмою.
Фотосинтетичний апарат представлений тилакоїдами, що є по-
хідними інвагінацій плазмалеми, але не відокремлюються від цито-
плазми двомембранною оболонкою. Таким чином, фотосинтетичний
апарат не організований у вигляді пластид. Тилакоїди розташовуються
поодиноко, у периферичному шарі цитоплазми. В оптичний мікрос-
коп зона розташування тилакоїдів виглядає інтесивно забарвленою,
через що її називають хроматоплазмою. На поверхні тилакоїдів є
особливі структури - фікобілісоми, що складаються з фікобілінових
пігментів.
До елементів фотосинтетичного апарату в Суапорїіуіа опосеред-
ковано відносять також поліедральні тіла, які є попередниками піре-
ноїду евкаріотичних водоростей.
Включення представлені дрібними гранулами крохмалю синьо-
зелених водоростей, великими ціанофіциновими та поліфосфатни-
ми гранулами. У деяких (переважно - планктонних) видів у цитолазмі
є газові вакуолі. При сприятливих умовах вони заповнюються газом і
зменшують питому вагу клітини. При несприятливих умовах газ ди-
фундує, газові вакуолі зменшуються в об’ємі та злипаються (колапсу-
ють), питома вага клітини збільшується, і водорість осідає на дно, де
«очікує» покращення умов для фотосинтезу.
Фізіологічні особливості. Суапорїіуіа мають принаймні одну
важливу фізіолого-біохімічну особливість, яку можна розглядати як
фізіологічний атавізм: це здатність фіксувати атмосферний азот.
53
При азотфіксації молекулярний азот відновлюється до сполук
амонію і в цій формі включається в основні шляхи клітинного метабо-
лізму. Процес азотфіксації каталізує нітрогеназний ферментний ком-
плекс, робота якого повністю інгібується молекулярним киснем. Тому
азотфіксація здійснюється лише в анаеробному середовищі. Проте,
серед багатоклітинних Суапорїіуіа є чимало видів, що фіксують ат-
мосферний азот також у присутності кисню. У цієї групи нітрогеназ-
ний комплекс локалізований в особливих клітинах - гетероцистах,
які мають кілька пристосувань, що запобігають проникненню вільно-
го кисню у клітину, і тим самим на фоні аеробного зовнішнього
середовища створюють у цитоплазмі клітини-гетероцисти анаеробні
умови.
Будова талому. Синьозелені водорості бувають як одно-, так і
багатоклітинними, поодинокими або в різноманітних колоніях, де ін-
дивіди найчастіше утримуються разом за допомогою колоніального
слизу.
Обов’язковим елементом тіла багатоклітинних водоростей є три-
хом - сукупність фізіологічно пов’язаних клітин. Зв’язок відбувається
за допомогою плазмодесм, що проходять через пори поперечних
перегородок клітин. Зовні трихому можуть знаходитись структуровані
слизові утвори - піхви. Трихом разом із піхвою називають ниткою. У
видів, позбавлених піхв, трихом і нитка є синонімічними поняттями.
Трихоми можуть бути нерозгалуженими та розгалуженими. Кліти-
ни нерозгалужених трихомів діляться лише в одній площині. Якщо
клітини здатні до поділу в кількох площинах, трихоми галузяться. Цей
тип галуження називають справжнім. Коли ж галузяться нитки, а самі
трихоми лишаються нерозгалуженими, то галуження вважають не-
справжнім.
У синьозелених водоростей розрізняють три основні типи клітин:
а) вегетативні клітини, що здійснюють фотосинтез та здатні до поділу;
б) гетероцисти-спеціалізовані клітини, що виконують функцію фіксації
атмосферного азоту в аеробних умовах; в) акінети - спочиваючі кліти-
ни, за допомогою яких водорості переносять несприятливі умови.
Вегетативні клітини властиві всім Суапорїіуіа, гетероцисти та акінети
-лише частині багатоклітинних представників. Трихоми, що склада-
ються лише з вегетативних клітин, називають гомоцитними, а ті, які
складаються з вегетативних клітин, гетероцистта акінет- гетероцит-
ними.
54
Поділ клітин та розмноження. В основі процесів розмноження
у всіх синьозелених водоростей лежить клітинний поділ. При
клітинному поділі спочатку плазмалема, а за нею і муреїновий шар
клітинної оболонки доцентрово вростають у протопласт, розділяючи
клітину на дві (зрідка - більше) рівні або нерівні частини. Клітинний
поділ у синьозелених водоростей може бути повним або неповним.
При повному поділі плазмалема та клітинна перегородка повністю
відокремлюють дочірні клітини одна від одної. При цьому пори та
плазмодесми не утворюються, і кожна дочірня клітина являє собою
фізіологічно самостійний індивід. Повний поділ властивий одноклітин-
ним Суапорїіуіа.
При неповному поділі поперечна перегородка не повністю розді-
ляє дочірні клітини, і вони лишаються з'єднаними плазмодесмами,
що проходять через пори в поперечній клітинній перегородці. Внас-
лідок такого поділу утворюються багатоклітинні ниткоподібні трихо-
ми. Короткі рухливі фрагменти трихомів, що складаються з 2-50 клітин,
називають гормогоніями, нерухомі - гормоцитами. Гормогонії та гор-
моцити є спеціалізованими репродуктивними структурами багатоклі-
тинних Суапорїіуіа.
Колоніальні одно- та багатоклітинні синьозелені водорості здатні
розмножуватися також шляхом фрагментації колоній.
Система відділу. Відділ включає лише один клас - Суапо-
рЬусеае, що за типом поділу клітин, типами трихомів та їх здатністю
до справжнього галуження поділяється на чотири порядки -
Сїігоососсаіез, ОзсіІІаіогіаІез, Мозіосаіез та Зіідопетаіаіез.
Порядок Хроококальні - Сїігоососсаіез
Включає всі одноклітинні синьозелені водорості. Поділ клітин
повний. Хроококальні водорості можуть бути представлені поодино-
кими клітинами або утворювати різноманітні колонії. Характерними
представниками порядку є роди Місгосузііз, Мегізторесііа та СІоео-
сарза.
Рід мікроцистіс - Місгосузііз (рис. 5) - мікроскопічна прісно-
водна водорість, що складається з великої кількості поодиноких клітин,
розташованих у слизових колоніях неправильної форми. Колоніаль-
ний слиз м’який, і тоді невиразний, або досить щільний, і в цьому ви-
падку добре помітний в оптичний мікроскоп навіть без спеціального
забарвлення. Кожна клітина має сферичну форму (за винятком тих,
55
що перебувають у стані поділу). Розмноження відбувається фрагмен-
тацією колоній та поділом клітин навпіл. Площини поділу клітин не
фіксовані - клітини можуть ділитися у будь-якому напрямку. Це зу-
мовлює неправильну та різноманітну форму колоній.
Найвідомішим представником роду є Місгосузііз аегидіпоза -
мікроцистіс синювато-зелений. Під мікроскопом клітини М. аеги-
діпоза здаються майже чорними через наявність в них великої
кількості газових вакуолей. Колонії у цього виду, від кулястих до нит-
коподібних та неправильних, можуть бути суцільними або містити
отвори, набуваючи в останньому випадку вигляду сітки.
М. аегидіпоза є небезпечним збудником «цвітіння» води у сто-
ячих та повільно текучих прісних водоймах, особливо у рівнинних
водосховищах. У процесі життєдіяльності водорість виділяє токсини
з групи гепатотоксинів (зокрема - мікроцистіни), що класифікуються
як фактор швидкої смерті. Розвитку «цвітіння» сприяють підвищена
температура води, високий вміст біогенних елементів (азоту, фосфо-
ру, калію), відсутність перемішування водної товщі.
фєю#..........ш
.‘50/ .•0’О‘®^ал®Ої‘*
'ОЬО'.о.^рОооі.
?о.о°-о р.°.о о9о«
о % о.ОрО<£ЯІ>Оі
••••ЯоХ>0<5о\..
'ОмПйв*’*
г .Ж
Рис. 5. Місгосузііз'. А -загальний вигляд колонії; Б - вегетативні клітини при
великому збільшенні. 1 - перфорації; 2 - колоніальний слиз; 3 - вегетативні
клітини; 4 - газові вакуолі
56
Рід мерисмопедія - Мегізторесііа (рис. 6 А). Ця колоніальна
водорість має клітини, що діляться почергово у двох площинах, при-
чому площина кожного наступного поділу проходить перпендикуляр-
но до площини попереднього. Клітини утримуються разом колоніаль-
ним слизом; як наслідок, колонії мають вигляд прямокутних одноша-
рових платівок, у яких клітини розміщуються правильними взаємно
перпендикулярними рядами. Мерисмопедія мешкає у планктоні
прісних водойм та приморських солоних озер.
Рід глеокапса - ОІоеосарза (рис. 6 Б). Клітини глеокапси більш-
менш сферичні, діляться у трьох взаємно перпендикулярних площи-
нах, здатні до швидкого багаторазового поділу й утворюють харак-
терні слизові колонії, що складаються із системи вкладених один в
одного слизових міхурів. Види роду зустрічаються в біотопах всіх типів
(морські та континентальні води, ґрунти), проте найчастіше оселя-
ються на зрошуваних бризками скелях, стінах водоспадів, вологому
камінні.
Рис. 6. Хроококальні водорості: А - Мегізторесііа-, Б - ОІоеосарза
57
Порядок Осциляторіальні - ОзсіІІаїогіаІез
Об’єднує багатоклітинні гомоцитні водорості, що мають нерозга-
лужені трихоми. Поділ клітин неповний і відбувається у площині, пер-
пендикулярній до поздовжньої осі трихому. Розмноження здійснюєть-
ся гормогоніями або гормоцитами. Типовим представником є рід
ОзсіІІаіогіа.
Рід осциляторія - ОзсіІІаіогіа (рис. 7). Водорість мікроскопічна,
має прямі або дещо викривлені чи злегка спіралевидні нерозгалу-
жені довгі трихоми однакової ширини. Інколи трихоми знаходяться у
слизових піхвах. Завдяки наявності в оболонці розміщених під кутом
скоротливих мікрофібрил, трихоми здатні до по-
вільного обертального (осциляторного) руху, від
якого походить назва роду. Кожний трихом скла-
дається з багатьох коротких циліндричних або дис-
ковидних клітин, однакових за формою та функці-
ями. Винятком є лише верхівкові клітини, які за фор-
мою можуть бути дещо відмінні, наприклад, мати
ковпачковидно-потовщену оболонку - каліптру.
Внутрішній вміст клітин гомогенний або зернистий,
іноді з газовими вакуолями. Зернисті включення -
ціанофіцинові гранули - розташовуються біля по-
перечних перегородок і добре помітні в оптичний
мікроскоп.
Ріст водорості відбувається завдяки неповно-
му поділу клітин, який проходить у площині, пер-
пендикулярній поздовжній осі трихому. Розмно-
ження здійснюється внаслідок розпаду трихомів на
малоклітинні фрагменти - гормогонії та гормоци-
ти. Гормогонії, подібно до трихомів, здатні до ак-
тивного ковзаючого руху. Гормоцити нерухомі.
Види роду мешкають у стоячих та повільноте-
кучих прісних континентальних водоймах, морях і
ґрунтах. При масовому розвитку ці водорості ут-
ворюють синьозелені плівки та шкуринки, які вільно
плавають або обростають різні субстрати. Деякі
морські види здатні викликати токсичні «цвітіння»
води.
Рис. 7. ОзсіІІаіогіа: загальний вигляд частини трихому. 1 - вегетативні кліти-
ни; 2 - ціанофіцинові зерна; 3 - відмерла клітина (некрида)
58
Рід артроспіра - Агіїїгозріга (рис. 8) - нагадує осциляторію, від
якої відрізняється трихомами, скрученими у вигляді правильної ши-
рокої спіралі. Два види цього роду, які зустрічаються у тропіках та
субтропіках, у прісних водоймах з високим вмістом карбонатів (А.
ріаіепзіз, А. тахіта), введені у промислову культуру і вирощуються у
багатьох країнах як харчові водорості, джерело біодобавок (зокрема
вітамінів та гормону тироксину Т4) та фармакологічна сировина для
виготовлення препаратів радіопротекторної дії.
Рис. 8. Трихом Агііїгозріга ріаіепзіз, яка є об’єктом промислового
культивування
Порядок Ностокальні - Мозїосаіез
Об’єднує багатоклітинні гетероцитні водорості з нерозгалужени-
ми трихомами. Як і у ОзсіІІаіогіаІез, поділ клітин є неповним і відбу-
вається у площині, перпендикулярній поздовжній осі трихому, а роз-
множення здійснюється гормогоніями або гормоцитами.
Ностокальні водорості поширені у морях, континентальних во-
доймах, у ґрунтах і, завдяки здатності до фіксації атмосферного азо-
ту, в аеробних умовах відіграють важливу роль у планетарному кру-
гообігу азоту. Деякі види ностокальних є небезпечними збудниками
«цвітіння» води.
Примітивніші ностокальні водорості (наприклад, СІоеоігісіїіа) ма-
ють асиметричні нитки, еволюційно розвинуті - симетричні (Аріїапі-
хотепоп, АпаЬаепа, №>зіос).
Рід глеотрихія - (ЗІоеоігісїііа (рис. 9). Водорість утворює ма-
сивні слизові колонії, що складаються з нерозгалужених асиметрич-
них ниток. При основі трихому знаходиться базальна гетероциста, за
нею (у зрілих індивідів) - видовжена акінета, далі - вегетативні кліти-
ни. Останні біля гетероцисти широкі та короткі, при наближенні до
верхівки звужуються та витягуються у довжину, і на самій верхівці
закінчуються довгим безбарвним волоском. СІоеоігісіїіа мешкає пе-
59
реважно у прісних стоячих водоймах. На початку розвитку колонії
прикріплені до підводних субстратів (переважно до стебел та листків
вищих водних рослин), згодом відриваються від них і вільно плава-
ють на поверхні води. У деяких країнах Азії місцеві жителі збирають
колонії глеотрихії та вживають у їжу.
Рис. 9. біоеоігісіїіа: А - зовнішній вигляд колонії; Б - розташування ниток у
колоніальному слизу; В - фрагмент колонії з групою ниток; Г - окрема нитка.
1 - гетероциста; 2 - пробка; 3 - акінета; 4 - слизова піхва; 5 - вегетативні клітини;
6 - волосок
Рід афанізоменон - Аріїапіготепоп (рис. 10). Трихоми Аріїа-
піхотепоп з’єднуються у шкуринки, що плавають у товщі води. Кож-
ний трихом дещо звужується до верхівок і закінчується безбарвними
витягнутими клітинами. Центральна частина трихому утворена веге-
тативними клітинами, між якими розташовуються поодинокі інтерка-
лярні гетероцисти та акінети. Клітини виповнені численними газови-
ми вакуолями. Всі види роду є небезпечними збудниками токсичного
«цвітіння» води. Так, у водосховищах Дніпровського каскаду приблиз-
но 20% випадків «цвітіння» зумовлює А. Поз-адиае- афанізоменон
цвітіння води. Окремі форми цього виду також спричинюють
«цвітіння» води в Азовському морі (починаючи із середини 90-х років
XX ст. це «цвітіння» спостерігається регулярно).
60
Рис. 10. Арїіапіготепоп Лоз-ариае: А - зовнішній вигляд колонії; Б - фраг-
мент центральної частини колонії з акінетами; В - центральна частина трихому;
Г - верхівка трихому з безбарвними видовженими термінальними клітинами.
1 - акінета; 2 - гетероциста
Рід анабена - АпаЬаепа (рис. 11). Це мікроскопічна багатоклі-
тинна водорість, що має поодинокі прямі або різноманітно зігнуті си-
метричні нерозгалужені трихоми однакової ширини. Трихоми оточені
м’якими, погано помітними слизовими піхвами, складаються з веге-
тативних клітин та розміщених між ними гетероцист і акінет. Вегета-
тивні клітини темні від газових вакуолей. Гетероцисти товстостінні,
безбарвні, з гомогенним вмістом. Кожна гетероциста має пробки, які
замикають пори в поперечних перегородках, і в оптичному мікроскопі
мають вигляд блискучих зерняток. Поодинокі або розміщені ланцюж-
ками акінети виникають із вегетативних клітин внаслідок потовщення
їх оболонок і накопичення запасних поживних речовин. Акінети збе-
рігають хлорофіл, їх вміст звичайно зернистий, що обумовлено у
більшості випадків наявністю гранул ціанофіцину.
61
Поділ клітин у анабени неповний. Розмноження здійснюється гор-
могоніями або гормоцитами.
Види роду АпаЬаепа широко розповсюджені в планктоні прісних і
солонуватих, стоячих або повільнотекучих водойм, нерідко розвива-
ються у ґрунтах та на їх поверхні. Деякі види роду влітку часто викли-
кають токсичне «цвітіння» води. До таких небезпечних збудників на-
лежать А. зсґіегетеііеуі - анабена Шереметьєвої, А. зрігоісіез -
анабена спіралевидна та А. ґіоз-адиае - анабена «цвітіння» води.
Рис. 11. АпаЬаепа: загальний вигляд нитки. 1 - вегетативні клітини з газови-
ми вакуолями; 2 - гетероциста; 3 - пробка; 4 - акінета
Рід носток - Иозіос (рис. 12) - здебільшого водорості з макро-
скопічними слизовими колоніями (розміри останніх у окремих видів
можуть сягати кількох сантиметрів). Колонії за консистенцією м’які
або тверді, за формою - різноманітні. У колоніальному слизу знахо-
дяться викривлені, переплетені та досить часто згорнуті у щільну
спіраль симетричні трихоми. Вегетативні клітини біля поперечних пе-
регородок зазвичай дуже перетягнуті. Між вегетативними клітинами
розміщуються гетероцисти. Акінети за розміром та формою лише
трохи відрізняються від вегетативних клітин і часто з’єднані в лан-
цюжки. Водорості розмножуються гормогоніями, акінетами, бруньку-
ванням та фрагментацією колоній.
Представники роду досить широко розповсюджені в прісних во-
доймах і на ґрунті. Серед них відомі азотфіксуючі та їстівні види. Най-
звичайнішими видами роду є N. Ііпскіа — носток Пінка, N. ргипіїогте
— носток сливовидний та N. соттипе — носток звичайний.
62
Рис. 12. N05(00: А - зріз через колонію; Б - загальний вигляд колоній на
субстраті. 1 - нитки; 2 - перидерм; 3 - колоніальний слиз; 4 - гетероциста;
5 - вегетативні клітини
Порядок Стигонематальні - Зіідопетаїаіез
Об’єднує гетероцитні водорості з розгалуженими трихомами.
Представники порядку, що морфологічно не відрізняються від су-
часних, відомі з кембрію (570-500 млн. років). Клітини стигонема-
тальних здатні до поділу в кількох площинах, внаслідок чого трихоми
утворюють гілки, а іноді - навіть паренхіматозні структури, що на-
гадують справжні тканини. Мешкають стигонематальні переважно у
тропічних та субтропічних регіонах. На території України з цього по-
63
рядку досить часто зустрічається лише рід стигонема - Зіідопета
(рис. 13).
Рис. 13. Зіідопета іпіегтесііа: зовнішній вигляд зрілої розгалуженої нитки
Водорості дискокристати
Представники царства дискокристат (Оізсісгізіаіез) мають евка-
ріотичну будову і характеризуються наявністю різноманітних крист,
серед яких обов’язково є кристи дископодібної форми. Поділ ядра -
закритий ортомітоз. Водорості в межах цього царства представлені
одним відділом - ЕидІепорНуіа.
Відділ Евгленофітові водорості - Еидіепоріїуіа
До евгленофітових водоростей належать евкаріотичні первинно
гетеротрофні, фотоавтотрофні та вторинно гетеротрофні дискокрис-
тати, що мають клітинний покрив, представлений пелікулою. Фотоав-
тотрофні евгленофітові мають вторинно симбіотичні пластиди хло-
рофітного типу.
Відділ нараховує біля 1000 видів мікроскопічних водоростей, по-
ширених переважно у прісних континентальних водоймах. Окремі
представники ведуть паразитичний спосіб життя. За винятком роду
64
Соїасіит, всі евгленофітові є монадними одноклітинними організма-
ми. За палеоальгологічними даними, найдавніші викопні рештки
Еидіепорїіуіа датуються віком біля 65 млн. років (кайнозой, третин-
ний період).
Біохімічні особливості. Гетеротрофні евгленофітові водорості
безбарвні. Для забарвлених представників характерна наявність хло-
рофілів а- та Ь-, 0- та у-каротинів. Основні ксантофіли - неоксантин
та діадіноксантин. Оскільки хлорофіли не маскуються ксантофілами,
хлоропласти фотоавтотрофних евгленофітів забарвлені у зелений
колір.
Основний продукт асиміляції - парамілон. Він накопичується зав-
жди поза межами хлоропласту у вигляді дрібних гранул або великих,
добре помітних парамілій. Парамілон може відкладатись також на
виступаючих за межі хлоропласту поверхнях піреноїдів. Додатковим
асимілятом у ЕидІепорНуіа є олія.
Цитологічні особливості. Клітини евгленофітових водоростей
поодинокі, на передньому кінці мають глотку, що складається з кана-
па та резервуара. Біля стінок резервуара розташовуються кілька пуль-
суючих вакуолей, які поступово заповнюються водою та зливаються
в одну велику вакуолю, яка виплескує свій вміст у резервуар. З дна
глотки підіймаються два джгутики.
У частини гетеротрофних евгленофітових водоростей у цито-
плазмі в районі глотки є органели для захоплення їжі - паличкоподі-
бна органела, або сифон. У цьому випадку часточки їжі, що потрап-
ляють у глотку, захоплюються і перетравлюються у травних вакуо-
лях. Неперетравлені рештки виштовхуються назовні на задньому кінці
клітини.
Клітинні покриви представлені пелікулою. Вона утворена плаз-
малемою, під якою розташовуються поздовжні або спірально вигнуті
білкові стрічки. У зоні стикання сусідніх стрічок утворюються виступи,
які зумовлюють характерну поздовжню або спіральну посмугованість
поверхні клітин. Пелікула звичайно еластична, тому клітини здатні в
процесі руху змінювати свою форму, наприклад, витягуватись або
стискатись. Клітини, які не зберігають у процесі руху постійну форму,
називають метаболічними. У видів з неметаболічними клітинами пе-
лікула має жорстку конструкцію.
У деяких евгленофітових водоростей монадні клітини розміщу-
ються всередині будиночків, які утворені слизовими тяжами, просякну-
тими солями марганцю та заліза.
65
Ядро досить велике і добре помітне без спеціальних забарвлень.
Хромосоми завжди знаходяться у конденсованому стані. В центрі ядра
розташовується велике ядерце, здатне до самостійного поділу - ен-
досома. Ядерна оболонка при поділі ядра лишається інтактною,
справжнє веретено не утворюється, центріолі відсутні, а їх функції
виконують базальні тіла джгутиків та ендосома.
Пластиди евгленофітових водоростей вкриті тримембранною
оболонкою і представляють вторинно симбіотичні хлоропласти. Ти-
лакоїди групуються переважно по три. Інколи в хлоропластах спосте-
рігаються голі напівзанурені піреноїди.
Мітохондрії. Звичайно клітина евгленофітових водоростей має
одну, досить велику розгалужену мітохондрію - т. зв. мітохондріон.
Будова мітохондріону евгленофітів унікальна серед водоростей, ос-
кільки внутрішня мембрана утворює кристи специфічної дископодіб-
ної форми.
Джгутиковий апарат. Клітини мають переважно два джгутики не-
рівної довжини, що підіймаються із дна глотки, при цьому один із них
дуже короткий і не виходить за межі глотки; його верхній кінець
з’єднується з аксонемою довшого джгутика. Такі джгутики називають
роздвоєними при основі. Інколи вони за довжиною майже однакові.
У деяких евгленофітових один із джгутиків редукований, і від нього
залишається тільки базальне тіло.
Аксонеми джгутиків вкриті мастигонемами двох типів: короткими,
розташованими по спіралі, та довгими, розміщеними гребінчасто. При
основі одного із джгутиків (рідше - обох) знаходиться потовщення -
парафлагелярне тіло, що виконує фоторецепторні функції. Біля па-
рафлагелярного тіла у багатьох Еидіепорґіуіа розміщується стигма.
Вона виконує функцію ширми, яка регулює кількість світла, що по-
трапляє на фоторецептор - парафлагелярне тіло.
У деяких видів джгутики відсутні, хоча зберігаються їх базальні
тіла. Такі клітини рухаються червоподібно, за рахунок метаболічних
вигинів клітини.
Розмноження та життєві цикли. Розмноження евгленофітових
водоростей відбувається шляхом поділу клітини навпіл у рухливому
або нерухомому стані. У трьох видів (Зсуіотопаз зиЬШіз, Ріїасиз
ругит, Еидіепа зр.) описаний хологамний та автогамний статевий
процес із зиготичною редукцією. Проте, в цілому, статеве розмно-
ження для Еидіепоріїуіа не характерне.
66
Життєві цикли евгленофітових водоростей досить прості, зде-
більшого вони представлені цикломорфозом. Найчастіше цикломор-
фоз евгленофітів полягає у зміні монадного та пальмелевидного
станів. Пальмелевидний стан спостерігається, зазвичай, у несприят-
ливих умовах.
Особливості живлення. Живлення різних представників Еид-
Іепорїіуїа може відбуватись за рахунок фотосинтезу та осмотрофно-
го або голозойного споживання органічних речовин. Фотоавтотрофні
види здатні рости на мінеральних поживних середовищах, проте най-
кращий ріст спостерігається при додаванні у середовище органічних
сполук. У темряві, на органічних середовищах, клітини деяких видів
можуть втрачати хлорофіл, накопичувати феофітин, зберігаючи при
цьому здатність до росту та розмноження. В знебарвлених клітинах
замість хлоропластів спостерігаються лейкопласти. При перенесенні
таких культур на світло, клітини відновлюють хлорофіл, і лейкопласти
перетворюються на хлоропласти. Знебарвлені культури можна отри-
мати і в умовах освітлення - дією антибіотиків, екстремальних тем-
ператур, ультрафіолетовим опроміненням.
Паразитичні безбарвні водорості з порядку Еидіепотогрїіаіез при
перенесенні на штучні мінеральні середовища на світлі синтезують
хлорофіл і переходять на фотоавтотрофне живлення.
Вільноіснуючі безбарвні Еидіепорїіуїа є облігатними гетеротро-
фами. Серед цієї групи частина видів живиться лише осмотрофно і
за морфологією нагадує знебарвлених фотоавтотрофних представ-
ників. Друга частина видів живиться осмотрофно та голозойно, не
має морфологічних «двійників» серед забарвлених форм, у багатьох
є паличкоподібна органела або сифон.
Система відділу. Відділ включає один клас - Еидіепорїіусеае,
та три порядки - Еидіепаїез, Регапетаіаіез, Еидіепотогрґіаіез. В ос-
нову поділу на порядки покладено тип живлення та ступінь спорідне-
ності безбарвних форм із забарвленими.
Порядок Перанематальні - Регапетаіаіез
Включає лише безбарвні види, які живляться осмотрофно та го-
лозойно і позбавлені забарвлених аналогів; багато представників
мають пристосування для активного захоплення їжі - паличкоподіб-
ну органелу або сифон. До порядку входять види як з одним нероз-
67
двоєним при основі джгутиком, так і з двома гетеродинамічними
джгутиками. Типовий представник порядку - рід Регапета.
2-паличкоподібна органела; 3-джгу-
тики; 4 - ядро з ендосомою; 5 - вклю-
чення парамілону
Рід перанема - Регапета (рис. 14). Водорість безбарвна, жи-
виться осмотрофно та голозойно, має дуже метаболічні клітини дещо
дорзовентральної будови. На передньому кінці клітини розташовується
глотка, канал якої відкривається субапікально, на черевному боці. З
дна глотки піднімаються два джгутики, кожний з яких розташовується
у власній короткій борозенці. Один джгутик направлений вперед і є
рушійним, другий загинається у напрямку заднього кінця клітини і
виконує функції керма. До глотки прилягає паличкоподібна органела,
яка обумовлює здатність водорості до голозойного живлення і утво-
Перанему можна знайти у во-
доймах, забруднених органічною
речовиною, наприклад, у відстій-
никах очисних споруд, в полісап-
робній зоні озер, а також у прибе-
режному мулі річок та ставків.
Порядок Евгленальні -
Еидіепаіез
Об’єднує всіх забарвлених
представників та безбарвні види,
які вторинно втратили хлороплас-
ти. Останні живляться осмотроф-
но і за морфологією нагадують за-
барвлені форми. Основними рода-
ми є Еидіепа, Тгасїіеіотопаз,
Ріїасиз.
Рід евглена - Еидіепа
(рис. 15). Об’єднує вільноіснуючі
прісноводні водорості з монадним
типом структури. Клітини евглени
витягнуті у довжину, в різному сту-
пені метаболічні, мають найрізно-
манітнішу форму: веретеновидну,
циліндричну, еліпсоїдну, стрічко-
видну та ін. На передньому кінці
68
клітини знаходиться глотковий канал, який переходить у великий ре-
зервуар. На черевному боці резервуара розташовується кілька ско-
ротливих вакуолей. У місці переходу резервуара у глотковий канал,
на спинному боці, знаходиться жовто-гаряче вічко. З дна глотки
підіймаються два джгутики, з яких довгий виходить назовні, короткий
у каналі зростається з довгим і не виходить за межі клітини.
На пелікулі у багатьох видів добре помітна спіральна посмуго-
ваність, зрідка - поздовжні штрихи або ряди виступаючих горбків.
Під пелікулою інколи можна розрізнити слизисті тільця, які у вигляді
пухирців або вузьких веретенець розташовуються рядами або роз-
сіяно.
Хлоропласти різноманітної форми та розміру, від декількох ве-
ликих платівок до багаточисленних дрібних дисків, зазвичай мають
піреноіди. Парамілон в клітинах
Рис. 15. Еидіепа: загальний
вигляд клітини. 1 - глотка; 2 - віч-
ко; 3 - хлоропласти з піреноїдами;
4 - ядро з ендосомою; 5 - джгутик
відкладається у вигляді або числен-
них дрібних зерняток, або неба-
гатьох великих кільцевидних па-
рамілій. Ядро велике, помітне
без спеціального забарвлення,
з великою ендосомою посере-
дині.
Розмноження у видів роду
Еидіепа відбувається по-
здовжнім поділом у рухливому
стані, джгутик при цьому ски-
дається. При несприятливих
умовах клітини округлюються,
втрачають джгутик і, виділяючи
слиз, переходять у пальмелепо-
дібний стан. У цьому стані во-
дорість може продовжувати роз-
множуватись поділом надвоє.
Життєвий цикл - цикломорфоз.
Рід об’єднує понад 150 ви-
дів водоростей, які існують пе-
реважно у невеликих прісних
(часто гумінізованих) водоймах,
зрідка - на вологому ґрунті або
у спріснілих ділянках морів. Де-
69
які види евглен за сприятливих умов можуть викликати нетоксичні
зелені або червоні «цвітіння» води.
На території України найпоширенішими видами цього роду є Е.
уігісііз - евглена зелена, Е. асиз - евглена гольчата та Е. зрі-
годуга - евглена спірогірна.
Рід трахеломонас - ТгасРеІотопаз (рис. 16) - включає вільно-
плаваючі одноклітинні організми з метаболічними клітинами, які знахо-
дяться в міцних будиночах, що утворені слизовими тяжами, просяк-
нутими мінеральними солями. На верхівці вони мають отвір, через
який назовні виходить один джгутик. Будиночки частини видів навко-
ло джгутикового отвору утворюють виріст - комірець. їх стінки зви-
чайно бурого кольору, гладенькі або орнаментовані порами, ямка-
Рис. 16. Тгасїіеіотопаз: загальний вигляд
клітини. 1 - джгутик; 2 - комірець; 3 - будино-
чок; 4 - шипи; 5 - ядро; 6 - ендосома; 7 - мо-
нада; 8 - хлоропласти; 9 - зерна парамілону
ми, малими чи велики-
ми виростами - сосоч-
ками, гранулами, шипа-
ми, зморшками. Задній
кінець будиночка заок-
руглений або звужений,
іноді з довгим та вузьким
порожнистим відрост-
ком. Деякі види утворю-
ють навколо будиночка
додаткову слизову об-
гортку.
У цитоплазмі кліти-
ни міститься від двох до
багатьох хлоропластів з
піреноїдами або без них,
червоне вічко, дрібні або
великі парамілонові
зерна. Деякі види вто-
ринно втратили хлоро-
філ і є безбарвними ге-
теротрофами, але збе-
рігають лейкопласт.
Розмноження водо-
ростей відбувається по-
здовжнім поділом мона-
ди всередині будиночка.
70
Далі, одна з дочірніх клітин виходить назовні крізь джгутиковий отвір і
будує собі новий будиночок. Друга клітина отримує у спадок мате-
ринський будиночок. У молодих індивідуумів будиночки безбарвні.
Рід належить до найрозповсюдженіших на території України. Його
представники зустрічаються переважно у невеликих водоймах зі сто-
ячою водою або слабкою течією, в ефемерних водоймах, болотах,
озерах, ставках та ін. Часто трахеломонас масово розмножується, і
тоді вода набуває бурого кольору з різними відтінками.
Найчастіше зустрічаються Т. уоіуосіпз — трахеломонас воль-
воксовий, Т. іпіегтесііа - трахеломонас проміжний, Т. ґіізрісіа -
трахеломонас дрібнощетинистий та Т. агтаіа - трахеломонас
озброєний.
Рід факус- РІіасиз (рис. 17) - має забарвлені та сильно стисну-
ті з боків клітини, які зазвичай на задньому кінці закінчуються безбар-
вним прямим або зігнутим відростком. Клітини мають щільну пеліку-
лу з косими або поздовжніми смугами, переважно численні диско-
видні пристінні хлоропласти без піреноїдів, одну-дві великі парамілїї
і багато маленьких парамілонових зерняток, розсіяних по цитоплазмі.
Водорості мешкають переважно в невеликих водоймах, забруднених
органічними речовинами.
Рис. 17. Рїіасиз: 1 - глотка; 2 - хло-
ропласти; 3- хвостовий відросток; 4 - па-
рамілія
Порядок Евгленаморфа-
льні - Еидіепагпогріїаіез
Порядок включає види,
що паразитують на тваринах
і мешкають у кишечниках олі-
гохет, нематод, копепод, ам-
фібій, на зябрах риб. У тілі
тварини-господаря ці водо-
рості безбарвні і позбавлені
джгутиків. При вилученні з
тіла тварини-господаря та пе-
ренесенні на світло, клітини
евгленаморфальних водоро-
стей відрощують джгутики,
синтезують хлорофіл та набу-
вають зеленого забарвлення.
Типовим представником цьо-
го порядку є рід евгленамор-
фа - Еидіепатогрїіа.
71
Водорості тубулокристати
Представники царства тубулокристат (ТиЬиІосгізіаїез) характе-
ризуються наявністю трубчастих, перешнурованих при основі міто-
хондріальних крист. Всі фотоавтотрофні представники мають плас-
тиди виключно вторинно симбіотичного типу. В межах цього царства
водорості розподілені між трьома підцарствами - Амебо-флагелята-
ми, Страменопілами та Альвеолятами.
Амебо-флагеляти
Підцарство амебо-флагеляти (АтоеЬоЛадеІІаіез) об’єднує орга-
нізми, в яких клітини голі, і в життєвому циклі представлені лише аме-
боїдні або монадні стадії (причому амебоїдна стадія, як правило, пе-
реважає). Джгутикові стадії позбавлені тричленних мастигонем - рет-
ронем. У цьому підцарстві до водоростей належить відділ СМога-
гасіїпіорНуіа.
Відділ Хлорарахніофітові водорості - СЬІогагасЬпіорЬуІа
Евкаріотичні фотоавтотрофні тубулокристати, в яких клітини голі,
а вегетативне тіло представлене амебоїдами, здатними об’єднува-
тися у плазмодії. Пластиди вторинно симбіотичні, хлорофітного типу,
з нуклеоморфом. Відділ включає чотири роди із шістьма видами.
Біохімічні особливості. Хлоропласти хлорарахніофітових за-
барвлені у зелений колір і містять хлорофіли а та Ь, які не маскують-
ся додатковими пігментами. Продукт асиміляції - парамілон.
Цитологічні особливості. Клітини хлорарахніофітових амебоїдні,
утворюють велику кількість тонких розгалужених ризоподій, за допо-
могою яких клітини з’єднуються у сітчасті колонії, які інколи назива-
ють плазмодієм. Кількість клітин у колонії коливається від кількох до
150. Кожна клітина колонії здатна до активного руху за допомогою
псевдоподій.
Покриви представлені лише плазмалемою. Під плазмалемою
знаходяться дифузно розміщені трихоцисти.
Клітини хлорарахніофітових одноядерні. Ядро має евкаріотичну
будову, хромосомну організацію, ядерна ДНК зв’язана з гістонами.
У клітині міститься кілька пристінних дископодібних хлоропластів,
вкритих чотиримембранною оболонкою. Дві внутрішні мембрани є
72
власними мембранами хлоропласту, дві зовнішні - утворюють хлоро-
пластний ендоплазматичний ретикулюм. Між власними хлоропласт-
ними мембранами та мембранами хлоропластного ендоплазматично-
го ретикулюму є перипластидний простір, у якому знаходиться унікальна
ДНК-вмістна структура - нуклеоморф, відмежований від перипластид-
ного простору власного двомембранною оболонкою.
Тилакоїди в хлоропласті зібрані переважно по два або по три.
У стромі хлоропласта також міститься великий, виступаючий за його
межі піреноїд. Навколо піреноїду, але завжди за межами пластиди,
накопичується парамілон.
Мітохондріальний апарат представлений розгалуженим мітохонд-
ріоном з трубчастими кристами.
Джгутикові стадії у хлорарахніофітів - це зооспори з одним су-
бапікальним, спірально закрученим навколо клітини джгутиком, спря-
мованим назад. Аксонема джгутика несе прості, дуже тонкі пірчасто
розміщені мастигонеми. Стигма завжди відсутня.
Розмноження та життєвий цикл. Розмноження відбувається
поділом клітин надвоє або за допомогою зооспор. За несприятливих
умов амебоїдні клітини можуть набувати сферичної форми, вкрива-
тися оболонкою і переходити у кокоїдний стан.
У СНІогагасНпіорНуїа спостерігалося статеве розмноження, про-
те його особливості, місце проходження мейозу та зміна ядерних фаз
лишаються нез’ясованими.
Система відділу. Всі відомі хлорарахніофітові належать до кла-
су СНІогагасНпіорНусеае, порядку СНІогагасНпіаІез. Типовим представ-
ником є рід хлорарахніон - СІїІогагасіїпіоп (рис. 18), що мешкає у
морському мулі.
Рис. 18. Сїііогагасіїпіоп: А - колонія; Б - окрема амебоїдна клітина; В -
зооспора; Г - кокоїдна стадія
73
Страменопіли. Група відділів хромофітових
водоростей
Підцарство страменопілів (Зігатепоріїез) включає організми з
різноманітними типами покривів (оболонками, панцирами, плазма-
лемою з додатковими захисними структурами). Головна ознака - на-
явність у монадних клітин (переважно - на поверхні джгутиків) особ-
ливих тричленних мастигонем - ретронем, утворення яких починаєть-
ся або між мембранами ядерної оболонки, або між мембранами хло-
ропластної ендоплазматичної сітки.
Водорості в межах цього підцарства утворюють компактну моно-
філітичну групу відділів, відомих під загальною назвою хромофітових
водоростей. Ця група включає сім відділів-КарИИорНуїа, СНгузорНуїа,
ЕизіідтаіорНуіа, ХапіііорНуіа, РНаеорбуіа, ВасіІІагіорНуіа та Оісіуо-
сіїорНуіа.
Об’єднує всіх фотоавтотрофних страменопілів. Група моно-
філітична за всіма геномами - ядерним, мітохондріальним, пластид-
ним. Для всіх відділів спільною ознакою є наявність вторинно симбіо-
тичних пластид родофітного типу. Представники всіх відділів хромо-
фітових водоростей мають хлорофіли а та с, чотиримембранні хло-
ропласти, в яких дві зовнішні мембрани утворюють хлоропластну
ендоплазматичну сітку, ретронеми утворюються як між мембранами
ядерної оболонки, так і між мембранами хлоропластної ендоплазма-
тичної сітки. Відміни між відділами пов'язані, в першу чергу, з будо-
вою клітинних покривів, наявністю фукоксантину, продуктами асимі-
ляції, будовою фоторецепторного апарату, особливостями організації
цитоскелета, а також із типами морфологічної структури тіла.
Відділ Рафідофітові водорості - КарЬідорЬуІа
Евкаріотичні фотоавтотрофні тубулокристати, в яких клітини голі
(проте не амебоїдні). Пластиди вторинно симбіотичні, родофітного
типу. Джгутикові стадії мають ретронеми. Продукт асиміляції - олія.
Характерні особливості - наявність глотки та супрануклеарного апа-
рату, представленого особливою видозміною комплексу Гольджі, а
також відсутність стигми та структурного зв’язку між ядерною обо-
лонкою та хлоропластною ендоплазматичною сіткою.
У цьому відділі сьогодні відомо близько 50 видів з 10 родів, усі
представники є виключно одноклітинними монадними водоростями,
які мешкають у прісних континентальних водоймах та у морях.
74
Рафідофітові належать до небезпечних водоростей, оскільки у них
виявлено три групи токсинів - нейротоксини, гемолітичні та гемаглю-
тинуючі токсини. Навіть при малій кількості клітин (3-10 тис. кл./ мл)
рафідофітові порушують координацію руху у риб, спричинюють їх ано-
мальну поведінку і протягом 50-80 хвилин призводять до їх загибелі.
Біохімічні особливості. Рафідофітовим притаманні хлорофіли
а та с, р-каротин, ксантофіли лютеїнового ряду - віолаксантин, лю-
теїн. Крім того, у прісноводних рафідофітових присутні діадиноксан-
тин, диноксантин, гетероксантин, вошеріоксантин, а у морських форм
- фукоксантин. Через це хлоропласти у прісноводних видів забарв-
лені звичайно у зелений або жовтозелений колір, а у морських-у
жовтий або жовто-коричневий.
Продуктом асиміляції є олія, яка накопичується в цитоплазмі.
В окремих випадках дрібні поодинокі краплини олії виявляються та-
кож у стромі хлоропласта.
Цитологічні особливості. Клітини рафідофітових досить великі
(30-100 мкм), звичайно дорзовентральної будови, з трикутною глот-
кою, двома гетероконтними та гетероморфними джгутиками, вузь-
кою поздовжньою борозенкою, що розташовується на вентральному
боці клітини.
Клітинні покриви представлені плазмалемою, під якою можуть
знаходитися слизові тільця та/або трихоцисти.
Ядро одне, велике, з одним або кількома ядерцями. Над ядер-
ною оболонкою у вигляді широкого ковпачка розташовується т. зв.
супрануклеарний апарат - система з кількох комплексів Гольджі, які
щільно притиснуті до ядерної оболонки. Мітоз закритий, веретено є
внутрішньоядерним. Функції центріолей виконують базальні тіла джгу-
тиків.
Хлоропласти дрібні, численні, розташовані на периферії клітини.
Кожний хлоропласт вкритий чотиримембранною оболонкою. Між дво-
ма зовнішніми та двома внутрішніми мембранами хлоропласта є пе-
рипластидний простір з перипластидним ретикулюмом. Тилакоїди
групуються по три у кілька ламел. Хлоропластна ДНК - генофор -
замкнена у кільце і знаходиться поблизу хлоропластної оболонки.
Мітохондрії розгалужені, утворюють мітохондріон. Мітохондріальні
профілі на зрізах мають трубчасті кристи.
На передньому кінці клітини або трохи збоку розташовуються два
джгутики нерівної довжини. Довший джгутик пірчастий, локомоторний,
75
на кінці спірально закручений, вкритий ретронемами. Короткий джгу-
тик гладенький, звичайно виконує функцію керма і лежить у поздовжній
черевній борозенці. Парабазальні потовщення на джгутиках та вічко
у рафідофітових не виявлені. Перехідна зона джгутика не має
спіралеподібної структури.
Вакуолярний апарат представлений однією великою та кількома
дрібними скоротливими вакуолями, що виштовхують свій вміст у глотку.
Типи морфологічних структур тіла. Всі рафідофітові водорості
мають монадний тип структури тіла. Клітини злегка метаболічні, пе-
реважно дорзовентральні або стиснуті з боків, з поздовжньою боро-
зенкою, від якої у безбарвних форм можуть відходити ризоподії.
Розмноження та життєві цикли. Розмноження відбувається по-
ділом клітини у поздовжньому напрямку в рухливому стані. Зооспори
та статеве розмноження у РаргіісіорНуіа не відомі. Життєвий цикл про-
стий, представлений цикломорфозом.
Особливості живлення. Переважна більшість рафідофітових -
фотоавтотрофи. Проте деякі представники відділу позбавлені хло-
ропластів і є облігатними гетеротрофами. Живлення гетеротрофних
видів відбувається осмотрофним шляхом. Зустрічаються також види,
здатні до голозойного живлення. Вони захоплюють їжу (бактерії, час-
тки детриту, інші водорості) ризоподіями; травлення відбувається у
травних вакуолях.
Система відділу. Всі КарЬісіорГіуіа належать до одного класу -
Раргіісіоріїусеае. Прісноводні рафідофітові складають порядок
Каргіісііаіез (=УасиоІагіаІез), морські - СгіаПопеІІаІез. Представники
СбайопеІІаІез у флорі України поки що не виявлені.
Порядок Рафідіальні - Каріїісііаіез
Найпоширенішими родами порядку є Сопіозіотит та \/асиоІагіа.
Рід гоніостомум - Сопіозіотит (рис. 19) - є одним з найбільш
характерних родів рафідофітових водоростей. Клітини гоніостомуму
мають досить великі розміри - до 100 мкм - і тому є зручними об'єк-
тами для мікроскопічних досліджень.
Водорість монадна, представлена поодинокими клітинами. Кліти-
ни сплощені, дещо метаболічні, мають обернено-яйцеподібну фор-
76
му, вкриті лише плазмалемою, під якою розташовуються паличко-
подібні еджективні органели -трихоцисти. На передньому кінці клітини
знаходиться велика трикутна глотка, що відкривається назовні вивід-
ним каналом. Біля каналу починається довга поздовжня борозенка.
З невеличкої заглибини на початку борозенки виходять два джгутики
нерівної довжини. Довший джгутик направлений вперед і є локомо-
торним, коротший розташовується у борозенці і виконує функцію
керма. Електронно-мікроскопічні дослідження показали, що довгий
джгутик несе на поверхні пірчасто-розташовані тричленні волоски -
ретронеми, тобто має типову страменопільну будову. Вакуолярний
апарат складається з численних дрібних пульсуючих вакуолей, що
наповнюють водою одну велику вакуолю. Остання при скороченні
виплескує свій вміст у глотку.
Під плазмалемою розташовується велика кількість дрібних при-
стінних хлоропластів, забарвлених у жовтозелений колір. Під хлороп-
ластами, ближче до переднього кінця клітини, добре помітно велике
ядро з ядерцем. Між ядром та дном глотки можна спостерігати неве-
личку лінзоподібну структуру - супрануклеарний апарат. Електрон-
но-мікроскопічні дослідження показали, що супрануклеарний апарат
- це два великих комплекси Гольжді, майже притиснуті до поверхні
ядерної оболонки.
Розмножується гоніостомум поділом клітин навпіл у рухливому
стані. Цитокінезу передує мітотичний поділ ядра, який відбувається у
напрямку, перпендикулярному до поздовжньої осі клітини. Цікаво, що
центріолі у Оопіозіотит, як і в інших рафідофітових водоростей,
відсутні, а центрами організації веретена поділу є базальні тіла джгу-
тиків. Мітоз закритий і на живому матеріалі можна спостерігати всі
стадії каріокінезу в динаміці.
Після закінчення поділу ядра на передньому кінці клітини утво-
рюється борозна, яка швидко розділяє материнську клітину на дві
дочірні. Статевий процес, цисти та пальмели у Оопіозіотит не вияв-
лені. Таким чином, життєвий цикл представлений найпростішим вар-
іантом цикломорфозу.
На прикладі гоніостомуму цікаво спостерігати рух клітини та ро-
боту еджективних структур - трихоцист. За допомогою переднього
джгутика клітина рухається повільно, злегка обертаючись навколо
поздовжньої осі. Але при контакті з будь-якою перепоною відстрілює
у її напрямку з кількох трихоцист пружні слизові нитки. Це викликає
реактивне стрибкоподібне переміщення гоніостомуму в напрямку від
перепони.
77
Рис. 19. бопіозіотипг. 1 - глот-
ка; 2 - вакуоля; 3 - супрануклеарний
апарат; 4 - ядро; 5 - хлоропласти;
6 - трихоцисти
Види роду Сопіозіотит є ти-
повими мешканцями сфагнових
боліт.
Рід вакуоля рія - \/асио!агіа
- досить подібний до гоніостому-
ма. Проте у вакуолярії замість три-
хоцист під плазмалемою знахо-
дяться дрібні сферичні слизові
тільця, які при подразненні виді-
ляють слиз. Вакуолярія здатна пе-
реходити у пальмелевидний стан,
а при дуже несприятливих умовах
утворювати цисти, що вкриті тов-
стою шаруватою слизовою обо-
лонкою. Таким чином, цикломор-
фоз у вакуолярії є складнішим, ніж
у гоніостомума, і включає три ста-
ни - вегетативний монадний, па-
льмелевидний та цисти.
Відділ Золотисті водорості -
СЬгузорЬуіа
Евкаріотичні фотоавтотрофні
та вторинно гетеротрофні тубу-
локристати, в яких клітини голі
(з кремнеземовими лусочками
або без них) або вкриті пектино-
вою оболонкою. Пластиди вторин-
но симбіотичні, родофітного типу.
Джгутикові стадії мають ретроне-
ми. Продукт асиміляції - хризола-
мінарин. Характерна особливість
- наявність у життєвому циклі стадії ендогенних кремнеземових цист.
СИгузорНуІа вважається відносно молодим відділом хромофіто-
вих водоростей. Найдавніші викопні рештки датуються віком біля
240 млн. років (тріасовий період мезозою). Сучасну флору СбгузорЬуІа
представляють близько 1000 видів, які виникли у палеогені та нео-
гені. Золотисті водорості мешкають переважно у прісних водоймах і
є, в основному, монадними організмами.
78
Біохімічні особливості. Для СНгузорНуїа характерні хлорофіли
а та с, р-каротин, дві групи ксантофілів: по-перше, ксантофіли діаток-
сантинового ряду (фукоксантин, діатоксантин, діадиноксантин, динок-
сантин), по-друге, лютеїнового ряду (лютеїн, неоксантин, зеаксантин,
антераксантин, віолаксантин). Зелені хлорофіли та жовті ксантофіли
(особливо - фукоксантин) надають хлоропластам СбгузорНуіа яск-
раво-золотистого відтінку.
Основним продуктом асиміляції є хризоламінарин, додатковими -
олія та волютин. Хризоламінарин накопичується у спеціальних одно-
мембранних вакуолях, що розташовуються на задньому кінці клітини.
Цитологічні особливості. Клітини у СНгузорНуіа або голі (у мо-
надних та амебоїдних представників), або вкриті пектиновою оболон-
кою (у гемімонадних, кокоїдних та нитчастих форм). Голі клітини мо-
жуть бути одягнені лише плазмалемою або мати на поверхні крем-
неземові лусочки. Деякі голі клітини знаходяться всередині буди-
ночка з органічних речовин. Зрідка під плазмалемою розміщені
еджективні структури - дискоболоцисти. У багатьох колоніальних
водоростей під плазмалемою є дрібні сферичні слизові тільця. При
подразненні вони «вистрілюють» назовні, утворюючи навколо кліти-
ни слизову капсулу.
Ядро евкаріотичне, вкрите двомембранною оболонкою, зовніш-
ня мембрана якої безпосередньо переходить у зовнішню мембрану
хлоропластної ендоплазматичної сітки. Мітоз відкритий. Справжні
центріолі відсутні. Центрами організації веретена поділу є попереч-
но-смугасті джгутикові корені - ризопласти.
Пластиди у СИгузорИуіа звичайно поодинокі, розташовують-
ся на периферії клітини, вкриті чотирма мембранами. Дві внутрішні
утворені мембранами власне хлоропласта, а дві зовнішні є мем-
бранами хлоропластної ендоплазматичної сітки. Між другою та
третьою мембранами знаходиться перипластидний простір. Зов-
нішня мембрана хлоропластної ендоплазматичної сітки відіграє
суттєву роль у процесах формування ретронем та поверхневих
кремнеземових лусочок. Тилакоїди у стромі зібрані по три, на пе-
риферії хлоропласта розташовується оперезуюча тритилакоїдна
ламела. Інколи у стромі хлоропласта містяться також голі піре-
ноїди та стигма.
Мітохондріальний апарат представлений розгалуженим мітохонд-
ріоном із трубчастими кристами.
79
Монадні клітини мають два джгутики нерівної довжини. Довгий
джгутик локомоторний, вкритий численними ретронемами, короткий
- гладенький або вкритий простими мастигонемами. У багатьох Сбгу-
зорбуіа короткий джгутик редукований, і від нього лишається тільки
базальне тіло. Перехідна зона кожного джгутика містить спіралеподіб-
ну структуру. При основі базального тіла короткого джгутика знахо-
диться потовщення - парабазальне тіло, яке виконує функцію фото-
рецептора і положення якого скоординоване з положенням стигми.
Джгутикові корені золотистих водоростей складаються з одного
поперечно-смугастого кореня - ризопласта - та чотирьох мікротубу-
лярних коренів. Ризопласт зв’язує базальні тіла з ядром і побудова-
ний зі скоротливого білка центрину. Біля основи джгутиків також роз-
ташовується одна-дві скоротливі вакуолі.
Типи морфологічних структур тіла. Серед СНгузорНуїа пере-
важають одноклітинні водорості з монадною та амебоїдною будовою,
рідше зустрічаються гемімонадні, кокоїдні та нитчасті представники.
Одноклітинні водорості нерідко утворюють колонії (в тому числі це-
нобії), багатоклітинні мають вигляд розгалужених або нерозгалуже-
них ниток чи паренхіматозних платівок.
Розмноження та життєві цикли. Розмножуються СНгузорНуіа
переважно нестатевим та зрідка - статевим шляхами. Нестатеве
розмноження відбувається поділом клітини надвоє у рухливому стані
та зооспорами. Крім того, багатоядерні амебоїдні форми (т. зв. плаз-
модіальні водорості) можуть розпадатися на багато одноядерних
амебоїдів, кожний з яких у процесі росту та за рахунок мітозів утво-
рює новий плазмодій. Багатоклітинні представники здатні до фраг-
ментації та нестатевого розмноження за допомогою зооспор. Деякі
гемімонадні, кокоїдні та нитчасті СНгузорНуїа розмножуються також
за допомогою апланоспор.
Статевий процес зустрічається зрідка і представлений переваж-
но хологамією, причому клітини під час копуляції з’єднуються задні-
ми кінцями. Спостерігалися також автогамний, ізо- та гетерогамний
статеві процеси. Еугамні золотисті водорості мають гаплофазний
життєвий цикл із зиготичною редукцією та з особливою стадією дво-
ядерних зиготичних цист.
Характерною особливістю хризофітових є здатність клітин при
несприятливих умовах утворювати спочиваючі ендогенні кремнезе-
80
мові цисти - статоспори. Вони мають сферичну або еліпсоїдну фор-
му, їх поверхня може бути інкрустована щетинками, шипиками, реб-
рами. На верхівці цисти знаходиться великий отвір, закритий полі-
сахаридною пробкою, вона звичайно не має кремнезему. При
проростанні цисти пробка руйнується, протопласт через отвір вихо-
дить назовні і розвивається у нову вегетативну клітину.
Система відділу. За наявністю на поверхні клітин кремнеземо-
вих лусочок відділ поділяють на два класи - СЬгузорЬусеае та
ЗупигорЬусеае.
Клас Хризофіцієві - СЬгузорЬусеае
Об'єднує одноклітинні та багатоклітинні водорості з усіма харак-
терними для відділу типами морфологічних структур - монадним
(включаючи амебоїдний та плазмодіальний варіанти), гемімонадним,
кокоїдним та нитчастим. Ознакою класу є відсутність здатності утво-
рювати кремнеземові лусочки на поверхні плазмалеми. Клас об’єднує
переважно прісноводні водорості. У відповідності до морфологічної
будови хризофіцієві поділяються на чотири порядки, серед яких про-
відним є порядок ОсЬготопасіаІез.
Порядок Охромонадальні - ОсЬготопасіаІез
До порядку входять одноклітинні, колоніальні та ценобіальні види,
які мають типову монадну будову. Поширені вони переважно в план-
ктоні чистих прісних водойм. Провідними родами порядку є Осіїго-
топаз, Сґіготиііпа, ОіпоЬгуоп.
Рід охромонас - ОсЬготопаз (рис. 20 А) - характеризується
наявністю голих, злегка метаболічних поодиноких монадних клітин з
двома нерівними джгутиками та одним жовтим хлоропластом.
Осіїготопаз найчастіше зустрічається взимку, у планктоні водойм, у
сфагнових болотах. Охромонас вважають найбільш примітивною зо-
лотистою водорістю. Цікаво, що деякі види роду здатні утворювати
короткі псевдоподії і таким чином живитися не тільки фотоавтотрофно,
але й голозойно.
Рід хромуліна - СЬготиІіпа (рис. 20 Б, В). Водорість одноклітин-
на. Клітини вкриті плазмалемою, округлі або веретеновидні, здатні
змінювати форму й інколи утворювати псевдоподії. В оптичному
мікроскопі видно лише один джгутик. Другий джгутик дуже короткий,
81
знаходиться в кишенькоподібному утворі клітини. Клітина містить один
або два хлоропласти. В цитоплазмі помітно краплини олії, а в задній
частині клітини - грудочку хризоламінарину. Водорість розмножуєть-
ся поздовжнім поділом в рухливому стані. Хромуліни поширені як у
прісних водоймах, так і в морях. На поверхні невеликих лісових озер
та калюж часто можна спостерігати матово-золотисту плівку, яка ут-
ворена величезною кількістю цист хромуліни (до 40 000 на 1 мм2 по-
верхні).
Рід дінобріон — ОіпоЬгуоп (рис. 20 Г, Д). Більшість видів цього
роду утворюють вільноплаваючі колонії із з’єднаних між собою буди-
ночків водоростей. Деякі будиночки (переважно у нижній частині
колонії) порожні; в тих, що розташовані на верхівці колонії, знахо-
дяться монадні клітини. Окремий будиночок має вигляди лійки із зву-
женою основою та розширеною відкритою верхівкою. Клітина при-
кріплюється до основи будиночка скоротливою базальною ніжкою,
на розширеному передньому кінці має два джгутики різної довжини.
В передній частині клітини знаходяться одна або дві пульсуючі ваку-
Рис. 20. Охромонадальні водорості: А - ОсЬготопаз; Б, В - СЬготиІіпа:
Б - вегетативна клітина, В - циста; Г, Д - ОіпоЬгуоп: Г - загальний вигляд колонії,
Д - вегетативні клітини після поділу
82
олі. Продукт асиміляції - хризоламінарин - відкладається в задній
частині клітини у вигляді однієї великої або декількох дрібних
грудочок.
Органічні речовини будиночка-целюлозні мікрофібрили з доміш-
ками амінокислот - синтезуються у комплексі Гольджі, далі транс-
портуються на поверхню плазмалеми і там полімеризуються. При
розмноженні протопласт материнської клітини ділиться, утворюючи
дві голі дочірні клітини. Вони виходять з будиночка, осідають на його
край біля отвору і далі будують власні будиночки. Внаслідок цього
звичайно утворюються деревоподібні дихотомічно розгалужені ко-
лонії.
Види роду є типовими мешканцями планктону прісних водойм
(найчастіше - озер) і зустрічаються протягом всього року.
Клас Синурофіцієві - ЗупигорЬусеае
Об’єднує одноклітинні, переважно монадні водорості, плазмале-
ма яких вкрита спірально розташованими кремнеземовими лусочка-
ми. Звичайно ці лусочки добре помітні в оптичний мікроскоп. Кожна
складається з платівки та з’єднаної з нею щетинки. Платівка перфо-
рована дрібними порами, має потовщений край та інколи - дрібні
шипики і бородавки. Щетинка приєднана до нижнього краю платівки і
має вигляд порожнистої трубки.
Клас включає один порядок - Вупигаїез. Найвідомішими пред-
ставниками класу є роди МаІІотопаз та Зупига.
Рід маломонас - МаІІотопаз (рис. 21 А) - характеризується по-
одинокими вільноплаваючими клітинами, які мають лише один джгу-
тик. На поверхні плазмалеми розташовуються численні черепично
розміщені кремнеземові лусочки з довгими щетинками. Цисти також
мають шар лусочок, звичайно з довгими шипами. Види цього роду
можна часто зустріти у лісових озерах з високим вмістом гумінових
речовин, особливо у холодну пору року. МаІІотопаз є найбагатшим у
видовому відношенні родом класу.
Рід синура - Зупига (рис. 21 Б - Г). Водорості цього роду утво-
рюють сферичні колонії без слизової обгортки, які містять багато
клітин, з’єднаних задніми кінцями. Кожна клітина має два джгутики
неоднакової довжини. Плазмалема вкрита типовими для 8упи-
горЬусеае кремнеземовими лусочками, кожна з яких складається з
платівки та короткої щетинки. У протопласті знаходиться ядро, два
83
хлоропласти у вигляді золотисто-жовтих платівок, розташованих по
краях клітини, пульсуючі вакуолі, краплини лейкозину та олії. Стигми
немає. Клітини постійно діляться, тому кількість їх у колонії весь час
збільшується. Нові колонії утворюються шляхом фрагментації вели-
ких старих колоній.
У деяких видів (зокрема, у 8. реіегзепіі) виявлено та детально
досліджено статеве розмноження. Тип статевого процесу - холога-
мія. Вегетативні клітини, що виконують функції жіночих гамет, виділя-
ють атрактивний гормон. Під його дією клітини, що виконують функції
чоловічих гамет, залишають колонію, підпливають до жіночої клітини
та зливаються з нею. Як наслідок, утворюється зиготична дикаріон-
тична циста. Її подальший розвиток відбувається за звичайною для
золотистих водоростей схемою.
Відомі випадки масового розмноження синури у штучних водой-
мах, зокрема в резервуарах для питної води, внаслідок чого вода
набуває неприємного запаху риб’ячого жиру.
Рис. 21. Синурофіцієві водорості. А - МаІІотопаз; Б-Г: Зупига: Б - колонія,
В - різноманітність лусочок, Г - одна лусочка у трьох площинах
84
Відділ Евстигматофітові водорості - ЕизІідтаіорЬуІа
Евкаріотичні фотоавтотрофні тубулокристати, у яких клітини
вкриті пектиновою оболонкою. Пластиди вторинно симбіотичні, родо-
фітного типу. Джгутикові стадії мають ретронеми. Продукт асиміляції
- хризоламінарин. Характерна особливість - наявність у монадних
стадій унікального фоторецепторного апарату, який розташований
у цитоплазмі поблизу основи джгутиків.
Відділ нараховує біля ЗО видів одноклітинних та колоніальних,
виключно кокоїдних організмів, поширених переважно у фунтах та
морському планктоні.
Біохімічні особливості. Евстигматофітові характеризуються на-
явністю хлорофілу а, 0-каротину, ксантофілів лютеїнового ряду, се-
ред яких переважає віолаксантин; інші ксантофіли цієї групи - лю-
теїн, неоксантин, зеаксантин, антераксантин - зустрічаються у не-
значних кількостях. Зі специфічних ксантофілів у евстигматофітових
виявлено вошеріоксантин. Зелений хлорофіл та жовтий віолаксан-
тин надають хлоропластам евстигматофітових водоростей жовтува-
то-зеленого забарвлення.
Продукти асиміляції евстигматофітових - хризоламінарин та олія,
а також полісахарид нез’ясованої природи, що відкладається у ци-
топлазмі навколо піреноїду та у вакуолях.
Цитологічні особливості. Клітинні покриви представлені пекти-
новою оболонкою. Оболонка може бути гладенькою або орнаменто-
ваною і нести потовщення, шипики, бородавки.
Ядерний апарат подібний до золотистих водоростей: ядро евка-
ріотичне, вкрите двомембранною оболонкою. У вегетативних клітин
зовнішня мембрана ядра безпосередньо переходить у зовнішню мем-
брану хлоропластної ендоплазматичної сітки. Мітоз, на відміну від
золотистих водоростей, закритий-ядерна оболонка лишається інтакт-
ною. Справжні центріолі відсутні, у монадних клітин їх функцію вико-
нують базальні тіла джгутиків.
Хлоропласти звичайно поодинокі, переважно розсічені або лопа-
теві, розташовуються на периферії клітини, вкриті чотиримембран-
ними оболонками. Дві зовнішні мембрани утворюють хлоропластну
ендоплазматичну сітку. Між другою та третьою мембранами є пери-
пластидний простір.
Тилакоїди у стромі хлоропласта зібрані по три, оперезуюча ла-
мела відсутня. У вегетативних клітинах звичайно є піреноїд. Він ви-
85
ступаючий, кутастий, непронизаний тилакоїдами. За межами хлоро-
пласту, навколо піреноїду, відкладаються тонкі платівки асиміляту не-
з’ясованої природи.
Мітохондрії мають трубчасті кристи.
Джгутикові стадії представлені зооспорами з двома джгутиками
неоднакової довжини - довгим пірчастим локомоторним, що несе два
ряди численних ретронем, та коротким гладеньким. Досить часто ко-
роткий джгутик редукований і від нього лишається тільки базальне
тіло. Перехідна зона джгутиків містить спіралеподібну структуру.
При основі базального тіла пірчастого джгутика знаходиться пара-
базальне тіло. Його положення скоординоване з положенням стигми.
Остання лежить поза межами хлоропласту і складається з різних за
розмірами пігментованих ліпідних глобул, що не оточені плазматич-
ною мембраною. Найбільша глобула огортає з одного боку параба-
зальне тіло, за нею розташовуються дрібніші. У потовщенні параба-
зального тіла виявлено електронно-щільну платівкоподібну структуру,
яка і є справжнім фоторецептором. Спеціальні дослідження показа-
ли, що стигма у евстигматофітових виконує не функцію ширми, як це
спостерігається у більшості водоростей, а навпаки, фокусує світлові
промені на платівкоподібну структуру парабазального тіла. Саме за
унікальною будовою та функцією стигми відділ отримав свою назву-
евстигматофітові.
Типи морфологічної структури тіла. Для всіх відомих евстиг-
матофітових водоростей характерний тільки кокоїдний тип структу-
ри. Більшість видів має поодинокі клітини, деякі представники утво-
рюють колонії.
Розмноження та життєві цикли. Розмножуються евстигматофі-
тові тільки за допомогою зооспор та апланоспор. Статеве розмно-
ження не виявлено. Життєві цикли представлені цикломорфозами.
Стадії типу цист або акінет у ЕизіідтаіорЬуіа не відомі.
Система відділу. Відділ включає один клас - ЕизіідтаіорЬусеае,
та один порядок - Еизіідтаіаіез. Характерним представником відділу
є рід Еизіідтаіоз.
Рід евстигматос - Еизіідтаіоз (рис. 22) - широко представле-
ний у різних типах ґрунтів всіх регіонів, за винятком тропічних та суб-
тропічних. Найпоширеніший вид - Еизіідтаіоз тадпиз - легко мож-
на виділити у культуру з лісових, лучних або степових ґрунтів. Вегета-
86
тивні клітини сферичні, поодинокі, мають великий дво- або трилопа-
тевий хлоропласт, звичайно з добре помітним кутастим піреноїдом,
який раніше називали білковим кристалом. Розмноження у грунті та
в культурах на агаризованому середовищі відбувається аплано-
спорами, які по 2-4 утворюються у материнській клітині. У водних
культурах водорість здатна утворювати одноджгутикові зооспори з
досить великою, добре помітною червоною стигмою, що знаходить-
ся біля основи джгутика і не пов’язана з хлоропластом.
Рис. 22. Евстигматофітові водорості: А-схема будови стигми у зооспори;
Б - зооспора та вегетативні клітини Еизіідтаїоз тадпиз
Відділ Жовтозелені водорості - ХапІЬорЬуіа
Евкаріотичні фотоавтотрофні тубулокристати, у яких клітини вкриті
пектиновою або целюлозно-пектиновою оболонкою. Пластиди вторин-
но симбіотичні, родофітного типу. Джгутикові стадії мають ретронеми.
Продукт асиміляції - хризоламінарин. Характерна особливість -
відсутність жовтого ксантофілу фукоксантину.
Відділ нараховує близько 600 видів одноклітинних, багатоклітин-
них та неклітинних сифональних водоростей, що мешкають у прісних
континентальних водоймах, ґрунтах та в аерофітоні. Достовірні ви-
копні рештки відомі з тріасових відкладів мезозою (біля 240 млн. років
тому), ймовірні - з кам'яновугільного періоду (300-340 млн. років
тому), тобто виникнення жовтозелених водоростей у часі приблизно
87
співпадає з часом появи перших рештків голонасінних рослин.
Більшість жовтозелених водоростей - організми мікроскопічні, поши-
рені виключно у прісних водоймах та ґрунтах.
Біохімічні особливості. Жовтозелені водорості мають хлоро-
філи а та с, 0-каротин. Переважаючими каротиноїдами є ксантофіли
діатоксантинового ряду - діатоксантин та діадіноксантин. У значно
менших кількостях у жовтозелених водоростей виявлені специфічні
ксантофіли - гетероксантин та вошеріоксантин, а також ксантофіли
лютеїнового ряду, зокрема неоксантин. Хлоропласти жовтозелених
водоростей забарвлені у жовтувато-зелений колір без коричневих
відтінків.
Продукт асиміляції - хризоламінарин, що завжди відкладається
позапластидно, і тому хлоропласти жовтозелених водоростей при
оптичній мікроскопії виглядають гладенькими та шовковистими, без
ознак гранульованості. При старінні у клітинах може накопичуватися
олія.
Цитологічні особливості. Клітинні покриви у жовтозелених во-
доростей представлені пектиновою або целюлозно-пектиновою обо-
лонкою. Клітинна оболонка може бути гладенькою або орнаментова-
ною, суцільною або складатися з двох половинок, що насуваються
одна на одну. У видів роду \/аисіїегіа оболонка, крім пектину, містить
також мікрофібрили целюлози.
Особливістю ядра є те, що зовнішня мембрана ядерної оболонки
переходить у зовнішню мембрану хлоропластної ендоплазматичної
сітки. Мітоз закритий, центрами організації веретена є центріолі. За-
звичай, на полюсах ядра, що ділиться, біля центріолей розташовуєть-
ся комплекс Гольджі. У зооспор, після завершення періоду активного
руху, при першому поділі ядра функції центріолей виконують базальні
тіла джгутиків.
Кількість хлоропластів у клітинах жовтозелених водоростей ва-
ріює у широких межах - від одного-десяти у кокоїдних та нитчастих
форм до кількох тисяч у сифональних представників. Хлоропласти
переважно займають пристінне положення. Хлоропластна оболонка
має чотири мембрани, дві зовнішні мембрани утворюють хлороплас-
тну ендоплазматичну сітку, яка переходить у ядерну оболонку. Між
зовнішніми та внутрішніми мембранами хлоропласта є перипластид-
ний простір. Тилакоїди групуються по три у кілька ламел і зазвичай
мають оперезуючу ламелу. Інколи у пластиді є голий піреноїд, що не
88
пронизаний тилакоїдами. Хлоропластна ДНК - генофор - розташо-
вується компактно на полюсах пластиди.
У зооспор багатьох жовтозелених водоростей у хлоропласті на-
явна також стигма, яка розташовується поблизу парабазального тіла
короткого джгутика.
Мітохондрії поодинокі або численні, їх профілі на зрізах мають
трубчасті кристи.
Монадні стадії у жовтозелених водоростей представлені зооспо-
рами та гаметами, на передньому кінці клітини або трохи збоку ма-
ють два джгутики різної довжини. Довший джгутик пірчастий, вкритий
ретронемами. Короткий джгутик гладенький, звичайно виконує функ-
цію керма. При основі короткий джгутик має потовщення - параба-
зальне тіло, яке є фоторецептором. Стигма у ХапіЬорЬуіа розташо-
вується у хлоропласті і виконує функцію ширми, яка з одного боку
затіняє фоторецептор. Короткий джгутик може бути редукованим,
проте його базальне тіло зберігається. Перехідна зона джгутика
містить спіралеподібну структуру.
Відхилення від типового плану будови мають зооспори сифональ-
ної водорості \/аисІіегіа. Вони великі, багатоядерні; вся поверхня зоо-
спори рівномірно вкрита парами майже однакових за довжиною гла-
деньких джгутиків. Вважають, що такі сифональні багатоджгутикові
зооспори є наслідком злиття у спорангії багатьох молодих дводжгу-
тикових зооспор. Таку об’єднану зооспору називають синзооспорою.
Вакуолярний апарат представлений вакуолями з клітинним со-
ком, що відмежовуються від цитоплазми мембраною - тонопластом.
У монадних стадій - зооспор та гамет - біля базальних тіл джгутиків,
як правило, розташовуються дві скоротливі вакуолі.
Типи морфологічних структур тіла. Більшість жовтозелених
водоростей є кокоїдними, нитчастими, гетеротрихальними або сифо-
нальними організмами.
Розмноження та життєві цикли. Жовтозелені водорості, за ви-
нятком одного роду, розмножуються лише нестатевим шляхом: по-
ділом клітин надвоє, фрагментацією таломів або колоній, апланос-
порами, зооспорами або за допомогою акінет.
Статевий процес представлений лише оогамією і відомий тільки
у видів \/аисІіегіа. У цієї водорості життєвий цикл диплофазний, з га-
метичною редукцією і без зміни поколінь.
89
Система відділу. Відділ включає один клас - ХапіЬорЬусеае, та
чотири порядки - Х/аисЬегіаіез, Воігудіаіез, МізсЬососсаІез та
ТгіЬопетаіаІез. Поділ на порядки проводиться за типами морфоло-
гічної струкутри тіла, наявністю синзооспор та статевого процесу,
особливостями будови оболонок.
Порядок Вошеріальні - УаисЬегіаІез
Об’єднує види з сифональним типом структури, які мають нитко-
подібні таломи із необмеженим ростом та суцільні целюлозно-пекти-
нові оболонки. Нестатеве розмноження відбувається за допомогою
синзооспор. Всі представники порядку можуть розмножуватись ста-
тевим шляхом, статевий процес-оогамія. Сперматозоїди мають два
джутики майже рівної довжини, поверхня яких позбавлена ретронем.
Порядок містить лише один рід - \/аисІіегіа- до якого входить близь-
ко 60 видів. Види роду мешкають на вологому фунті та у солонуватих
і прісних водоймах.
Рід вошерія - \/аисНегіа (рис. 23). Таломи вошерії мають вигляд
довгих сланких розгалужених трубок, які позбавлені поперечних пе-
регородок і здатні до необмеженого росту. До ґрунту трубки прикріплю-
ються за допомогою коротких розгалужених ризоїдів. У центральній
частині трубок розташовується вакуоля з клітинним соком, між нею
та оболонкою - цитоплазма. Безпосередньо під клітинною оболон-
кою знаходиться велика кількість дрібних дископодібних хлоропластів,
між хлоропластами та вакуолею - численні дрібні ядра.
При нестатевому розмноженні деякі верхівкові ділянки талому
відокремлюються поперечними перегородками і перетворюються на
зооспорангії. В кожному зооспорангії утворюється лише одна гола
синзооспора, вкрита парами майже рівних джгутиків. Під базальними
тілами кожної пари джгутиків розташовується одне ядро. Після дозрі-
вання синзооспори оболонка спорангію розривається, синзооспора
звільняється; після деякого періоду активного руху вона осідає на
субстрат і проростає у новий трубчастий сифональний талом.
Статеві органи - антеридії та оогонії - утворюються на одному й
тому ж (в однодомних) або на різних (у дводомних) таломах як бічні
інтеркалярні вирости. Вони, як і зооспорангії, відокремлюються від
основної частини талому поперечною перегородкою. Після утворен-
ня перегородки у гаметангіях відбувається мейоз.
Оогонії вошерії яйцеподібні; в кожному оогонії утворюється по
одній яйцеклітині. Антеридії мають вигляд трубчастих гачків. У кожно-
90
му антеридії утворюється велика кількість сперматозоїдів. Цікаво, що
чоловічі гамети, на відміну від синзооспор, мають два латеральні
джгутики різної довжини, причому довший джгутик несе ретронеми.
Загалом будова сперматозоїдів вошерії дуже подібна до будови спер-
матозоїдів бурих водоростей.
При статевому процесі сперматозоїди через розрив оболонки
антеридія виходять у зовнішнє середовище, підпливають до оогонію.
Далі один зі сперматозоїдів через тріщину в оболонці оогонію прони-
кає до яйцеклітини і запліднює її. Зигота вкривається товстою обо-
лонкою і переходить у стан спокою. Після завершення періоду спо-
кою зигота проростає у новий трубчастий сифональний талом.
Рис. 23. УаисЬегіа-. А-загальний вигляд талому; Б- вихід синзооспори; В -
фрагмент талому зі статевими органами. 1 - ризоїди; 2 - зооспорангій; 3 -
синзоослора; 4 - антеридій; 5 - оогоній
91
Порядок Ботридіальні - Воігусііаіез
Об’єднує сифональні водорості з обмеженим ростом та суціль-
ною пектиновою оболонкою. Таломи ботридіальних можуть мати
різноманітну, але не ниткоподібну форму. Зооспори дводжгутикові,
гетероконтні, довший джгутик несе ретронеми. Інколи короткий джгу-
тик редукований. Характерними представниками порядку є роди
Воігудіит та Воігусііорзіз.
Рід ботридіум - Воігусііит (рис. 24 А). Види цього роду присто-
совані до життя на вологому ґрунті. Талом представлений однією
багатоядерною клітиною і має вигляд системи дихотомічно розгалу-
жених безбарвних ризоїдів, за допомогою яких водорість закріплюєть-
ся у ґрунті, та розширеної забарвленої надземної частини, діаметр
якої досягає 1-2 мм. У надземній частині під оболонкою знаходяться
численні дрібні дископодібні хлоропласти, під якими розташовується
багато ядер. При наявності крапельно-рідкої вологи водорість роз-
множується дводжгутиковими зооспорами. За посушливих умов про-
топласт стискається, втягується в ризоїдальну частину і розпадаєть-
ся на одно- або багатоядерні апланоспори. Апланоспори вкривають-
ся міцною, часто товстою шаруватою оболонкою і переходять у стан
криптобіозу. При настанні сприятливих умов одноядерні апланоспо-
ри проростають безпосередньо у нові таломи, а багатоядерні пере-
творюються на зооспорангії з дводжгутиковими зооспорами.
Б - Воігусііорзіз (вегетативна клітина, зооспорангій та вихід зооспор)
92
Рід ботридіопсис - Воігусііорзіз (рис. 24 Б). У представників
цього роду вегетативні клітини сферичні, можуть досягати 0,1 мм у
діаметрі. У периферичному шарі цитоплазми розташовується багато
дрібних пристінних дископодібних хлоропластів, під ними-до кількох
десятків дрібних ядер. Центр клітини зайнятий вакуолею з клітинним
соком. При наявності крапельно-рідкої вологи водорість розмножуєть-
ся за допомогою численних зооспор. При відсутності води замість
зооспор утворюються одноядерні апланоспори. Види цього роду меш-
кають переважно у ґрунтах лісових та тундрових фітоценозів, а також
у полярних та високогірних пустелях.
Порядок Місхококальні - Мізсїіососсаіез. Об’єднує одноядерні
водорості із суцільними пектиновими оболонками. Примітивніші місхо-
кокальні мають кокоїдну будову, складніші - гетеротрихальну. Най-
поширенішими родами порядку є СІїагасіорзіз та Неіегососсиз.
Рід харациопсис - Сіїагасіорзіз (рис. 25 А). Види роду населя-
ють переважно перифітонні угруповання прісних водойм. Клітини цих
водоростей полярні, прикріплюються до підводних субстратів за до-
помогою коротких чи довгих ніжок, що утворені клітинною оболон-
кою. Розмноження відбувається переважно за допомогою зооспор,
які по 2-4 утворюються в материнській клітині. Потрапивши на при-
датний субстрат, вони виробляють ніжку і перетворюються на веге-
тативну клітину.
Рід гетерокок - Неіегососсиз (рис. 25 Б). В порядку Мізсіїосос-
саіез існують види, що здатні розмножуватись не лише спорами, але
й шляхом вегетативного поділу клітин. Поява у частини місхококальних
водоростей вегетативного поділу обумовила виникнення в межах
порядку численної групи видів з гетеротрихальним тилом структури,
серед яких найпоширенішими є види роду Неіегососсиз. Представ-
ники цього роду мешкають виключно в наземних умовах: переважно
у ґрунтах, рідше - на корі дерев та вологих скелях, і є характерним
компонентом альгофлори гірських ґрунтів.
93
Рис. 25. Місхококальні водорості: А - СЬагасіорзіз; Б - Неіегососсиз
Порядок Трибонематальні - ТгіЬопетаіаІез
Об’єднує водорості, що мають двостулкові клітинні оболонки.
У межах порядку представлені одноклітинні та нитчасті організми.
Найвідомішим представником порядку є рід ТгіЬопета.
Рід трибонема - ТгіЬопета (рис. 26). Таломи водорості мають
вигляд простих нерозгалужених ниток, які на верхівках закінчуються
«вилочками». Види роду поширені у прісних водоймах. Нитки трибо-
неми спочатку прикріплюються до субстрату за допомогою базальної
клітини. Після її відмирання нитки спливають на поверхню, перепліта-
ються і утворюють м’яке світло-зелене жабуриння. Оболонка клітин
трибонеми складається з двох однакових половинок, які краями на-
кладаються одна на одну. При фрагментації нитки на верхівках
дочірніх ниток залишаються характерні Н-подібні закінчення. Клітини
трибонеми одноядерні, з коритоподібними або дископодібними хло-
ропластами.
Крім фрагментації, водорість розмножується зооспорами та ап-
ланоспорами, несприятливі умови переносить у стані акінет. Зооспо-
ри у цього роду можуть утворюватись різними шляхами: з вегетатив-
них клітин ниток, з апланоспор та з акінет.
94
Рис. 26. ТгіЬопета: А - молода нитка, прикріплена до субстрату; Б - зрілі
вегетативні нитки; В - зооспора. 1 - Н-подібне закінчення; 2 - хлоропласти
Відділ Бурі водорості - Рїіаеорїіуіа
Евкаріотичні фотоавтотрофні тубулокристати, в яких клітини
вкриті целюлозно-пектиновою оболонкою, до складу якої входять аль-
гінати. Пластиди вторинно симбіотичні, родофітного типу. Джгутикові
стадії мають ретронеми. Продукт асиміляції - ламінарин.
Відділ нараховує біля 260 родів, представлених приблизно 2000
видами. Всі представники цього відділу є багатоклітинними, переваж-
95
но макроскопічними водоростями і поширені в основному в морях.
Найдавніші достовірні викопні рештки бурих водоростей мають вік
біля 400 млн. років, а їх вірогідні рештки - приблизно 570 млн. років.
Біохімічні особливості. Бурі водорості мають хлорофіли а та с,
р- та є-каротини, та дві основні групи ксантофілів: по-перше, ксанто-
філи жовтої пігментної групи - фукоксантин, діатоксантин, діадіно-
ксантин, та, по-друге, ксантофіли лютеїнового ряду - віолаксантин,
зеаксантин, антераксантин, неоксантин. Фукоксантин та віолаксан-
тин накопичуються у великій кількості і маскують хлорофіли, нада-
ючи хлоропластам яскраво-жовтого або бурого забарвлення.
Основними продуктами асиміляції є полісахарид ламінарин та
олія, рідше - багатоатомні спирти (наприклад, манніт). Всі продукти
асиміляції відкладаються поза межами хлоропластів.
Цитологічні особливості. Клітинні покриви представлені дво-
шаровими оболонками. Зовнішній шар складається з пектинових ре-
човин та розчинних солей альгінової кислоти (переважно - альгінату
натрію). Внутрішній шар має фібрилярну структуру, і його основою є
мікрофібрили целюлози та нерозчинний альгінат кальцію.
Ядро типово евкаріотичне. Як і у переважної більшості водорос-
тей-страменопілів, зовнішня мембрана ядерної оболонки переходить
у хлоропластну ендоплазматичну сітку. Біля ядерної оболонки розта-
шовуються кілька комплексів Гольджі та дві центріолі. Останні при
каріокінезі стають центрами організації мікротрубочок веретена по-
ділу. Мітоз напівзакритий. При утворенні зооспор на заключних ста-
діях цитокінезу в деяких видів центріолі мігрують до поверхні дочірніх
клітин і перетворюються на базальні тіла джгутиків.
Хлоропласти бурих водоростей дрібні, численні, вкриті чотирма
мембранами, з яких дві зовнішні утворюють хлоропластну ендоплаз-
матичну сітку, що переходить у ядерну оболонку, а дві внутрішні є
власне оболонкою хлоропласта. Між зовнішніми та внутрішніми мем-
бранами існує перипластидний простір. Тилакоїди розміщуються пе-
реважно по три, на периферії знаходиться оперезуюча ламела. У ба-
гатьох бурих водоростей у хлоропласті є піреноїд. Хлоропластна ДНК
організована у кільцеподібно замкнений нуклеоїд, що розташовуєть-
ся під оперезуючою ламелою. У монадних клітин в стромі хлоропла-
ста розміщена також стигма. Її положення скоординоване із положен-
ням парабазального тіла, розташованого при основі гладенького джгу-
тика.
96
Мітохондріальний апарат представлений розгалуженим мітохон-
дріоном, внутрішня мембрана якого утворює трубчасті кристи.
Джгутикові стадії представлені зооспорами та гаметами, що ма-
ють два джгутики різної довжини, розташовані на черевному боці кліти-
ни. Довший джгутик пірчастий і вкритий ретронемами. Коротший джгу-
тик- гладенький. У сперматозоїдів бурих водоростей на передньому
кінці клітини є складчаста структура - хоботок, за допомогою якого
відбувається дихання та пошук яйцеклітини.
Типи морфологічних структур та будова талому. В межах
відділу представлено нитчастий, гетеротрихальний та справжній тка-
нинний типи структур тіла. Всі бурі водорості, за винятком деяких видів
роду Загдаззит, ведуть прикріплений спосіб життя. Органами при-
кріплення є ризоїди або дископодібні підошви.
Таломи нитчастих представників переважно утворені системою
однорядних розгалужених ниток. Гетеротрихальні водорості мають
вигляд досить товстих шнурів. Такі шнури мають або одноосьову, або
багатоосьову будову.
Різноманітна форма таломів характерна для водоростей з тка-
нинною будовою. У найпростішому випадку такий талом складається
лише з двох типів тканин - кори та серцевини. Кора утворена дрібни-
ми забарвленими клітинами, що виконують функцію фотосинтезу та
з яких утворюються органи розмноження. Серцевина складається з
великих, переважно безбарвних клітин, які виконують провідну та за-
пасаючу функції.
У складніших випадках талом утворений більшою кількістю тка-
нин: меристодермою, корою, проміжною тканиною та серцевиною.
Меристодерма - це поверхнева тканина, що виконує захисну та ре-
продуктивну функції. Клітини меристодерми звичайно дрібні і здатні
до активного поділу. Кора утворена більшими за розміром клітинами
з добре розвиненими хлоропластами. Основною функцією кори є
здійснення процесу фотосинтезу. У клітинах проміжної тканини нако-
пичуються продукти асиміляції. Самі клітини великі, звичайно з реду-
кованими хлоропластами. Серцевина утворена ситовидними трубка-
ми та трубчастими нитками.
Ріст таломів бурих водоростей відбувається кількома шляхами:
за рахунок поділу клітин середньої частини талому (інтеркалярний
ріст), групи верхівкових клітин (апікальної меристеми), лише однієї
великої верхівкової клітини або трихоталічно - шляхом поділу групи
клітин, що розташовуються субапікально - між верхівковими волос-
97
ками та основною частиною «простого» тканинного талому. Тип на-
ростання талому є однією із систематичних ознак на рівні порядків.
Розмноження та життєві цикли. Розмножуються бурі водорості
нестатевим та статевим шляхами. Нестатеве розмноження може
відбуватися фрагментацією таломів, вивідковими бруньками, зооспо-
рами та тетраспорами.
Нестатеве -розмноження за допомогою зооспор властиве пере-
важній більшості бурих водоростей. Зооспорангії бурих водоростей
можуть бути одногніздні або багатогніздні. Одногніздний спорангій яв-
ляє собою збільшену за розмірами клітину, вміст якої після серії поділів
ядра розпадається на велику кількість зооспор. Перший поділ ядра
звичайно редукційний, і тому зооспори, що утворюються в одно-
гніздних спорангіях, мають гаплоїдний набір хромосом. Багатогніздні
спорангії утворюються з великої клітини, в якій перед формуванням
спор протопласт розділяється тонкими клітинними перегородками на
багато одноядерних камер (гнізд). У кожному гнізді утворюється одна
зооспора.
Статевий процес - ізо- або оогамія, зрідка зустрічається гетерога-
мія. Гаметангії бурих водоростей багатогніздні (за винятком представ-
ників Еисаіез). У нитчастих форм гаметангії утворюються на особли-
вих бічних гілках, у гетеротрихальних - з клітин асиміляторних ниток,
у тканинних - з кори або меристодерми.
У бурих водоростей життєвий цикл гаплодиплофазний з чергуван-
ням поколінь (ізоморфним або гетероморфним) або диплофазний без
зміни поколінь.
При гаплодиплофазному циклі на диплоїдному спорофіті в одно-
гніздних спорангіях відбувається мейоз, і утворюються гаплоїдні зоо-
спори. Вони проростають у гаплоїдні гаметофіти. Після копуляції зиго-
та без періоду спокою розвивається у новий спорофіт. Якщо чергуван-
ня поколінь гетероморфне, то гаметофіти, як правило, мікроскопічні,
а спорофіти макроскопічні.
Представники з диплофазним життєвим циклом не мають зоо-
спор, мейоз відбувається при утворенні гамет, а статевий процес
переважно оогамний. Оогонії та антеридії утворюються на спеціаль-
них плодючих гілочках (рецептакулах) у напівзамкнутих порожнинах
- скафідіях. Далі сперматозоїди, що утворились в антеридіях чоло-
вічих скафідіїв, виходять назовні. В жіночих скафідіях оболонки
оогоніїв розриваються, яйцеклітини потрапляють у порожнину ска-
98
фідія і виділяють атрактанти, які приваблюють сперматозоїди. Після
запліднення зигота без періоду спокою проростає.
Система відділу. Відділ включає лише один клас - РІіаеорИусеае
-та вісім порядків3. Поділ на порядки проводиться за такими ознака-
ми, як наявність та будова піреноїду, особливості розмноження, на-
явність зооспор, тип статевого процесу, життєвий цикл. Нижче роз-
глядаються два порядки - і-атіпагіаіез та Еисаіез.
Порядок Ламінаріальні - 1_атіпагіаІез
Об’єднує водорості, спорофіти яких мають вигляд нерозгалуже-
них шнурів або великих пластин складної тканинної будови. Як пра-
вило, у серцевині є елементи провідної системи. Ріст таломів інтер-
калярний. Зміна поколінь гетероморфна, правильна, статевий про-
цес-оогамія. Гаметофіти мікроскопічні, звичайно у вигляді коротких
однорядних розгалужених ниток.
Ламінаріальні водорості є досить важливими у практичному відно-
шенні, оскільки саме до них належить переважна більшість бурих
водоростей, які використовуються людиною.
Найбільш відомий представник порядку - Ьатіпагіа, або морсь-
ка капуста.
Рід ламінарія або морська капуста - Ьатіпагіа (рис. 27, 28).
Дорослі спорофіти ламінарії сягають 3-4 м завдовжки і складаються
з пластини, «стебла» та розгалуженої системи ризоїдів, за допомо-
гою яких водорість прикріплюється до підводних кам’янистих суб-
стратів. На межі переходу пластини у «стебло» розташовуються ме-
ристемні клітини, за рахунок поділу яких відбувається інтеркалярний
ріст талому.
3 Систему відділу подано за: 1_.О.ОгиеЬІ, С.Мауез, І.Н.Тап, С.УУ.ЗаипсІегз. Моіесиїаг
апсі тогрГіоіодісаІ рііуіодепезіз оі кеір апсі аззосіаіесі Ьго\л/п аідае. Іп: Огідіп ої АІдае апсі
ІЬеіг РІазМз (есі.О.ВГіайасГіагуа). Зргіпдег-\/егі., \А/іеп - ¥огк, 1997: 221-236; А.Реіегз,
М.СІауїоп. Моіесиїаг апсі тогріїоіодісаі іпуезіідаїіоп оГЙігее Ьгоюп аідаі депега \л/ііН зіеііаіе
ріазіісіз: еуісіепсе Гог Зсуіойїатпаіез огсі. поу. (РГіаеорГіусеае). - РКусоІодіа (1998), 37:106-
113; Р.Коиззеаи, В. сіє Рєуієгз. Сігситзсгірііоп оГ Ніе огсіег Есіосаграіез (РКаеоріїусеае):
ЬІЬІІодгарГіісаі зупіііезіз апсі тоїесиїаг еуідепсе. - Сгуріодатіе, АІдоІ. (1999), 20 (1): 5-18;
Р.Роиззеаи, В. сіє Рєуієгз. Рііуіодепеііс геїаііопзііірз «ІІІііп іііе Рисаіез (РКаеоріїусеае) Ьазед
оп сотЬіпесІ рагііаі 330+130 гОІМАзедиепсе сіаіа.— Еиг.З.РКусоІ. (1999), 34: 53-64. Не роз-
глядається порядок Азсозеігаїез (див.: Жизнь растений. Т. 3. Водоросли, лишайники. - М.,
Просвещение, 1977), представники якого нечисленні і мешкають у морях Південної півкулі.
99
Меристема ламінарій активна у холодні пори року, і ріст споро-
фіту відбувається з січня по квітень-травень у помірних широтах, з
лютого по червень-серпень у високих широтах. Поділ клітин мери-
стеми індукується короткою тривалістю світлової фази доби. Це яви-
ще отримало назву фотоперіодичного відгуку. В напрямку до верхів-
ки талому інтеркалярна меристема відчленовує клітини, що надалі
диференціюються у тканини пластини, у протилежному напрямку -
клітини тканин «стебла». Наприкінці вегетаційного сезону пластина
звичайно відмирає і знову відростає у наступному році. «Стебло» та
ризоїди у ламінарії є багаторічними.
Нестатеве розмноження відбувається за допомогою зооспор. Зоо-
спорангії одногніздні, утворюються восени щільними групами (соруса-
ми) з клітин кори пластини спорофіту. Соруси забарвлені у темніший
колір, порівняно із стерильною частиною пластини. У кожному спорангії
Рис. 27. Деякі види роду іатіпагіа-. А - іатіпагіа зассЬагіпа-, Б - і., сіідіїаіа;
В - і.. ЬурегЬогеа
100
після редукційного поділу утворюється 32 дводжгутикові зооспори. Зоо-
спори вибухоподібно виштовхуються з пластини у зовнішнє середови-
ще у слизовому міхурі, який утворений переважно альгіновою кисло-
тою. Потім слизовий міхур розчиняється, і зооспори переходять у стан
активного руху. Через кілька хвилин вони осідають на субстрат і втрача-
ють джгутики. Половина зооспор з одного спорангію проростає у чо-
ловічі, а половина - у жіночі гаметофіти.
Чоловічі гаметофіти складаються з кількох десятків клітин, що ут-
ворюють коротку, рясно розгалужену нитку. Термінальні клітини
бічних гілочок нитки перетворюються на антеридії, в кожному з яких
утворюється по одному сперматозоїду.
Жіночі гаметофіти складаються лише з кількох клітин. Будь-яка
клітина гаметофіту може розвинутися в оогоній. У ньому утворюєть-
ся одна яйцеклітина, яка через розрив оболонки оогонію виходить
назовні і виділяє феромон ламоксирен, що приваблює сперматозої-
ди. Після копуляції утворюється зигота, яка без періоду спокою почи-
нає проростати у новий спорофіт.
Рис. 28. іатіпагісг. А - поперечний зріз пластини; Б - сорус зооспорангіїв на
поверхні меристодерми; В - схема життєвого циклу. 1 - меристодерма; 2 - кора;
З - проміжна тканина; 4 - серцевина; 5 - зооспорангій; 6 - захисні видовжені
клітини - парафізи
101
Ламінарія - відома їстівна водорість. До середини 70-х років
XX ст. її збирали за допомогою драг та спеціальних тралів. Наразі
більшість ламінарії, яка використовується людиною, є об’єктом аква-
культури і вирощується на спеціальних морських фермах.
Рід макроцистіс - Масгосузііз. Вздовж узбережжя Америки до-
мінантом багатьох морських фітоценозів є близький до ламінарії рід
Масгосузііз. Довжина таломів цієї водорості становить до 60 м. Довге
тонке стебло під кутом підіймається із дна до поверхні і несе подвійний
ряд довгих (до 1-2 м) «листових» пластин. При основі кожної пласти-
ни знаходиться повітряний міхур.
Порядок Фукусові - Еисаіев
Представники порядку мають складну тканинну будову, переваж-
но оогамний (як виняток - гетерогамний) статевий процес, диплофаз-
ний життєвий цикл без зміни поколінь. У фукальних статеві органи
утворюються в особливих порожнинах кори та проміжної тканини, що
отримали назву скафідіїв або концептакулів. Нестатевого розмножен-
ня у представників порядку немає. Фукальні водорості поширені як у
холодних, так і в теплих морях, і звичайно є домінантами фітоценозів
літоралі.
Характерні представники порядку - роди Рисиз, Загдаззит,
Сузіозеіга.
Рід фукус - Рисиз (рис. 29). У холодних північних морях (на-
приклад, Білому, Баренцевому, Охотському) дно, що оголюється під
час відпливів, часто вкрите суцільним килимом таломів Рисиз. Його
таломи великі (30-70 см заввишки), складаються з дископодібної при-
кріплювальної підошви, циліндричного стебельця та дихотомічно роз-
галужених пластин, що розширюються до верхівки. Стебельце у пла-
стинах переходить у центральне ребро. Фотосинтез здійснюють пе-
реважно клітини пластин. Стебельце та центральне ребро викону-
ють опірну та провідну функції. Поблизу верхівки у більшості видів
роду є добре помітні здуття, виповнені повітрям - повітряні міхури.
Завдяки їм водорість перебуває у воді у вертикальному положенні.
Ріст таломів відбувається внаслідок поділу апікальної клітини, що
відчленовує від себе клітини субапікальної меристеми. Останні на-
далі дають початок всім іншим тканинам.
Статеві органи розвиваються на особливих плодючих гілочках -
рецептакулах. Ці гілочки, порівняно зі стерильною частиною плас-
102
тин, потовщені та мають шерхату поверхню. Шерхатість обумовлена
наявністю в рецептакулах великих (до 1-3 мм у діаметрі) порожнин -
скафідіїв, в яких розвиваються жіночі та чоловічі статеві органи.
Скафідії виповнені слизом, іх внутрішня поверхня вкрита шаром
вистилаючих клітин, що дають початок коротким стерильним ниткам
та ініціальним репродуктивним клітинам, з яких утворюються анте-
ридії та оогонії. Скафідії у фукуса двостатеві або роздільностатеві.
Коли ініціальна репродуктивна клітина починає розвиватися у
статевий орган, її ядро мейотично ділиться. Далі гаплоїдні клітини
діляться мітотично і диференціюються або в оогонії, або в антеридії.
Оогонії еліпсоїдні, мають коротку одноклітинну ніжку та вісім яйце-
клітин. Антеридії утворюються з верхівкових клітин коротких, рясно
розгалужених антеридіальних ниток, що являють собою видозмінені
парафізи. У кожному антеридії утворюється 64 дводжгутикових спер-
матозоїди.
Перед заплідненням антеридії та оогонії через отвір скафідія зви-
чайно виштовхуються слизом у зовнішнє середовище. Далі оболонки
антеридіїв та оогоніїв розпливаються, звільняючи гамети. Яйцекліти-
ни виділяють феромон фукосератин, завдяки якому навколо кожної
яйцеклітини швидко концентрується велика кількість сперматозоїдів.
Після запліднення зигота без періоду спокою починає проростати у
новий талом.
Процес розвитку антеридіїв та оогоніїв у представників Еисаіез
різні автори інтерпретують по-різному. Так, за класичними погляда-
ми (Куііп, 1940; ЕгіїзсЬ, 1945), статеві органи у цих водоростей утво-
рюються внаслідок звичайної гаметичної редукції спеціалізованих
клітин талому, і тому життєвий цикл є диплофазним та відбувається
без зміни поколінь. На думку інших дослідників (ЗігазЬигдег, 1906;
уап бел Ноек, Мапп, ОаЬпз, 1995), ініціальні клітини, з яких утворю-
ються статеві органи, є гомологами одногніздних спорангіїв, а самі
оогонії та антеридіальні нитки - мікроскопічними гаплоїдними гаме-
тофітами. Тому життєвий цикл фукальних гаплодиплофазний, зі спо-
ричною редукцією та правильною гетероморфною зміною поколінь.
Цікаву гіпотезу висловив Ю. Е. Петров (1986). На його думку, весь
шар вистилаючих клітин скафідіїв є видозміненим диплоїдним гаме-
тофітом, і тому життєвий цикл слід розглядати як диплофазний, з га-
метичною редукцією та гетероморфним чергуванням поколінь4.
4 Розгляд аргументів «за» та «проти» кожної з цих гіпотез може бути темою окремого
семінарського заняття. Детальніше див.: Петров Ю. Е. Зволюция циклов развития у водо-
рослей (Комаровские чтения, XXXV). - Л., Наука, 1986. - 61 с.
103
2
Рис. 29. Рисиз: А - загальний вигляд талому; Б - розріз через жіночий
скафідій; В - розріз через чоловічий скафідій. 1 - рецептакул зі скафідіями;
2 - повітряні міхури; 3 - підошва; 4 - центральне ребро; 5 - парафізи; 6 - оогонії
Рід цистозейра - Сузіозеіга (рис. ЗО). У Чорному морі, на гли-
бинах 1-4 м, морські «луки» утворюють види роду Сузіозеіга (пере-
важно С. ЬагЬаіа та С. сгіпііа). Вони складають основу ланки проду-
центів у бентосних фітоценозах літоралі. Таломи Сузіозеіга мають
вигляд кущиків з більш-менш циліндричними рясно розгалуженими
висхідними гілками та короткими стеблами. Взимку та восени на вер-
хівках гілок утворюються ланцюжки невеличких повітряних міхурів.
Невеличкі рецептакули також розташовуються апікально.
104
Рис. ЗО. Сузіозеіга: А, Б - Сузіозеіга ЬагЬаіа: А - фрагмент талому; Б - вер-
хівка талому зі скафідіями; В - Сузіозеіга сгіпіїа (верхівка талому зі скафідіями)
Рід саргас - Загдаззит. У літоральній зоні Атлантичного та Ти-
хоокеанського узбереж домінуючою водорістю є Загдаззит. Окремі
види цього роду, що мешкають у відкритих акваторіях Атлантики (зок-
рема, у т. зв. Саргасовому морі), на відміну від інших бурих водорос-
тей, здатні розмножуватися у неприкріпленому стані. Таломи
Загдаззит кущисті і диференційовані на підошву, стебло та основні
гілки. Рецептакули та повітряні міхури у саргасу розвиваються на ко-
ротких гілочках, у пазухах основних гілок.
Відділ Діатомові водорості - ВасіІІагіоріїуіа
Евкаріотичні фотоавтотрофні тубулокристати, в яких клітини
вкриті кремнеземовим панциром. Пластиди вторинно симбіотичні, ро-
дофітного типу. Монадні стадії переважно нерухомі, джгутики мають
ретронеми. Продукт асиміляції - хризоламінарин.
Відділ налічує більше 20 тис. видів. Діатомові водорості виникли
порівняно недавно - біля 130 млн. років тому, у крейдяний період
фанерозою, майже одночасно з квітковими рослинами. У теперішній
час діатомові перебувають у стані біологічного прогресу і є доміну-
ючою групою у морських та прісноводних біотопах. Всі діатомові во-
дорості - це кокоїдні одноклітинні або колоніальні організми, розміри
яких коливаються в діапазоні 4-2000 мкм при середніх значеннях
20-50 мкм.
105
Біохімічні особливості. Фотосинтетичні пігменти представлені
хлорофілами а та с, р-та є-каротинами. Із ксантофілів переважає жов-
тий пігмент фукоксантин, який обумовлює коричневе забарвлення
пластид; у менших кількостях представлені діатоксантин, діадінок-
сантин та неоксантин.
Основним продуктом асиміляції є хризоламінарин, додатковими
- олія та волютин. Продукти асиміляції відкладаються або безпосе-
редньо у цитоплазмі, або у вакуолях.
Цитологічні особливості. Клітинні покриви представлені крем-
неземовим панциром, що розташовується назовні від плазмалеми.
Панцир у всіх діатомових водоростей утворений двома половинами:
більшою - епітекою, та меншою - гіпотекою. Кожна половина панци-
ра складається зі стулки та пояскового обідка. Краї стулки зігнуті під
більш-менш прямим кутом і утворюють зону, яка називається заги-
ном стулки. З краєм стулки у цій зоні одним боком щільно з’єднуюєть-
ся поясковий обідок. Поясковий обідок епітеки щільно насувається
на поясковий обідок гіпотеки, завдяки цьому панцир має вигляд ко-
робки (гіпотеки), яка накрита кришкою (епітекою). Зона перекриван-
ня пояскових обідків епітеки та гіпотеки називається пояском
(рис. 31). Панцир має досить складну структуру, специфічну для кож-
ного роду та виду діатомових водоростей.
з
А
Рис. 31. Схема будови панцира діатомових водоростей. А - основні струк-
турні елементи панцира; Б, В - вигляд панцира у різних площинах (Б - зі стулки;
В - з пояска). 1 - епітека; 2 - гіпотека; 3 - стулка епітеки - епівальва; 4 - стулка
гіпотеки - гіповальва; 5, 6 - пояскові обідки (5 - епіцингулюм, 6 - гіпоцингулюм)
106
В
Форма панцира у діатомових водоростей досить різноманітна і
характеризується, в першу чергу, типом симетрії стулки. Якщо через
стулку можна провести багато осей симетрії, таку стулку називають
радіально-симетричною, або актиноморфною. Коли ж через стулку
можна провести лише дві, одну або не можна провести жодної осі
симетрії, стулку називають зигоморфною. Зигоморфні стулки бува-
ють симетричними як у поздовжньому, так і в поперечному напрямку
(бісиметричні), симетричними лише по одній осі - або поздовжній,
або поперечній (моносиметричні стулки), дзеркально симетричними
(8-подібними) або асиметричними. На вигляд панцири діатомових
зазвичай (але не завжди) більш-менш прямокутні.
Стулка має гомогенні ділянки, які називають гіаліновими, систе-
му перфорацій, через яку відбувається взаємодія протопласта із
зовнішнім середовищем (ареоли, шви і т. п.), систему внутрішніх та
зовнішніх виростів та потовщень панцира (ребра, вирости і т. п.). Ве-
ликі гіалінові ділянки називаються полями. Якщо поле розташовуєть-
ся в центрі стулки, його називають центральним. Поле, яке прохо-
дить від одного полюса стулки до протилежного, називають осьовим.
Найпоширенішим типом перфорацій є ареоли. При оптичній
мікроскопії ареоли мають вигляд дрібних крапок. Якщо такі крапки
розташовуються щільними правильними рядами, то система крапок
набуває вигляду штриха. У значної кількості діатомових водоростей
стулка має систему потовщених ділянок, що надають її поверхні виг-
ляду пральної дошки. Такі потовщення називають ребрами. При виг-
ляді зі стулки ребра дещо нагадують штрихи, проте відмінність ребер
від перфорацій стає добре помітною при розгляді панцира з пояска.
Крім ребер, стулка може мати й інші потовщення, наприклад: труб-
часті вирости з опорами - фултопортули, двогубі вирости - римопор-
тули, короткі шипи та довгі щетинки і т. п.
У водоростей із зигоморфним панциром досить часто наявний
шов. У шві циркулює цитоплазма, і завдяки шву діатомові водорості
здатні до активного ковзаючого руху. Шви поділяють на щілиноподібні
та каналоподібні.
Щілиноподібний шов в оптичний мікроскоп має вигляд чіткої лінії,
яка починається на полюсах клітини, проходить по осьовому полю і
переривається в центральній частині стулки - у зоні центрального
поля. Ділянка шва від полюса до центрального поля називається
гілкою шва. У товщі стулки в зоні центрального поля кожна гілка шва
закінчується вертикальним каналом. Канали обох гілок, в свою чергу,
з’єднуються одним горизонтальним каналом. Вся ця система з двох
107
вертикальних та одного з’єднувального горизонтального каналу роз-
ташовується у потовщенні стулки, яке називається центральним вуз-
ликом. На полюсах зовнішня щілина шва розширюється, утворюючи
полярну щілину. Потовщення стулки, в якому розташовується поляр-
на щілина, називається полярним вузликом.
Каналоподібний шов має вигляд суцільної лінії, що, не перерива-
ючись, проходить від одного полюса клітини до іншого. Шов цього
типу складається з вузької зовнішньої щілини та трубчастого каналу,
який є аналогом внутрішньої щілини щілиноподібного шва. Трубчас-
тий канал відкривається у порожнину клітини добре помітними отво-
рами - фібулами.
Поясковий обідок кожної половини панцира може бути простим
або складним. Простий обідок складається лише з однієї замкненої у
кільце стрічки. Складні - зустрічаються у значно меншої кількості
видів. Вони утворені кількома послідовно вкладеними одна в одну
кільцеподібними стрічками. Кільце, яке безпосередньо прилягає до
стулки, називають стулковим кільцем, або вальвокопулою. Кільце, яке
з'єднується з вальвокопулою, та всі наступні послідовно приєднані
кільця пояскового обідка називають вставними обідками. Вставні обід-
ки половин панцира, за допомогою яких епітека з’єднується з гіпоте-
кою, називають з’єднувальними. У деяких діатомових водоростей по-
ясковий обідок може розростатися в порожнину клітини, утворюючи
розташовану паралельно до стулки неповну перегородку - септу.
До внутрішнього боку панцира щільно притиснута плазмалема.
Чимало видів діатомових водоростей здатні виділяти слиз. За
допомогою слизу водорості утворюють слизові трубки, різноманітні
слизові ніжки, з’єднуються у колонії. У водоростей, які мають шов,
слиз відіграє надзвичайну роль у процесі активного ковзаючого руху
клітини.
Ядро одне, розташовується в центрі клітини. Як і в інших водоро-
стей-страменопілів, воно структурно зв’язане з оболонкою хлоро-
пластів, оскільки зовнішня мембрана ядра переходить у зовнішню
мембрану хлоропластної ендоплазматичної сітки. Біля поверхні ядра,
як правило, розташовуються 1-2 комплекси Гольджі. Мітоз у діато-
мових водоростей відкритий, центріолі відсутні. Замість них функції
центру організації мікротрубочок виконують особливі полярні диски.
Фотосинтетичний апарат представлений вторинно симбіотични-
ми пластидами родофітного типу. Хлоропласти переважно займають
пристінне положення, їх кількість і форма специфічні для різних родів.
Хлоропластна оболонка має чотири мембрани, з яких дві зовнішні
108
утворюють хлоропластну ендоплазматичну сітку, що переходить у
ядерну оболонку. Між зовнішніми та внутрішніми мембранами хло-
ропласта є перипластидний простір.
Тилакоїди по три зібрані у ламели; безпосередньо під оболон-
кою хлоропласта розташовується оперезуюча ламела. Досить часто
в хлоропласті є голий піреноїд, який, на відміну від багатьох інших
водоростей-страменопілів, може бути пронизаний парами тилакоїдів.
На полюсах пластид компактно розташовується замкнена у кільце
хлоропластна ДНК.
Мітохондрій зазвичай кілька. Вони розміщуються у периферичній
частині клітин і мають лише трубчасті кристи.
Вакуолярний апарат включає 4 типи вакуолей: з клітинним со-
ком, волютином, хризоламінарином та олією.
Монадні стадії представлені виключно голими одноджгутикови-
ми сперматозоїдами і відомі лише у кількох родів діатомових водоро-
стей. Джгутик вкритий ретронемами, що утворюються в хлоропласт-
ному ендоплазматичному ретикулюмі та міжмембранному просторі
ядерної оболонки. Аксонема нетипова, оскільки представлена лише
дев’ятьма дублетами периферичних мікротрубочок, а дві центральні
мікротрубочки відсутні (формула 9+0). Є відміни і в будові базально-
го тіла - замість триплетів воно утворене дублетами мікротрубочок і
притиснуте до ядерної мембрани.
Типи морфологічних структур тіла. Діатомові водорості мають
виключно кокоїдний тип морфологічної структури тіла, хоча в деяких
випадках ниткоподібні колонії діатомей можуть бути помилково іден-
тифіковані як нитчасті форми.
Розмноження та життєві цикли. Представники відділу розмно-
жуються вегетативним поділом клітин надвоє та статевим шляхом.
Всі діатомеї мають єдиний тип життєвого циклу - диплофазний цикл
з гаметичною редукцією, без зміни поколінь та зі специфічною росто-
вою стадією - ауксоспорою.
Вегетативний поділ надвоє є основним типом розмноження і
притаманний всім видам діатомових. Перед поділом клітина набря-
кає. Внаслідок цього починається розсування епітеки та гіпотеки. Далі
діляться хлоропласти, відбувається мітоз. Після мітозу кожна дочір-
ня клітина отримує одну половину материнського панцира і далі бу-
дує другу половину. Материнська половина завжди стає епітекою до-
чірньої клітини.
109
Таким чином, внаслідок вегетативного поділу надвоє, клітина, що
отримала у спадок материнську епітеку, зберігає розміри материнсь-
кої клітини. У другої клітини половина панцира, яка у материнської
клітини була гіпотекою, стає епітекою. Оскільки кремнезем не здат-
ний розтягуватися, то в принципі розміри клітин у популяції мали б
швидко зменшуватись. Проте в дійсності такого не відбувається, ос-
кільки у діатомових водоростей існують певні механізми підтримання
сталих розмірів. Серед них до провідних належать механізми відтво-
рення оптимальних розмірів через стадію ауксоспори, переважно
внаслідок статевого процесу.
Ауксоспора (або спора, що росте) - це особлива клітина, яка здат-
на до росту. Клітина, яка перетворюється на ауксоспору, скидає пан-
цир і починає швидко збільшуватись у розмірах. Після закінчення про-
цесу росту протопласт ауксоспори утворює епітеку та гіпотеку, фор-
муючи т. зв. ініціальну клітину, розміри якої зазвичай наближуються
до максимально можливих для даного виду. Хоча ауксоспори можуть
утворюватись внаслідок різних процесів (наприклад, при досягненні
клітинами мінімально можливих розмірів або при пошкодженнях пан-
цира), проте найчастіше вони виникають після статевого процесу-із
зиготи, що проростає.
При статевому процесі дві клітини вкриваються спільним слизом,
протопласти збільшуються у розмірах і розсувають половинки пан-
цирів. Подальший розвиток у різних видів відбувається різними шля-
хами, зумовлюючи прояв певного типу статевого процесу. Проте
спільним в усіх випадках є те, що ядра кожної з клітин від одного до
кількох разів діляться, причому один з поділів обов’язково є редукцій-
ним.
У водоростей з актиноморфними стулками найчастіше зустрі-
чається оогамний статевий процес. У водоростей із зигоморфними
стулками найпоширенішими типами статевого процесу є ізо- та гете-
рогамія. Внаслідок ізогамії утворюються дві зиготи, а гетерогамії -
або дві, або одна.
Особливим способом статевого процесу в діатомей є автогамія.
У такому процесі бере участь лише одна клітина. Її ядро редукційно
ділиться, далі два гаплоїдних ядра дегенерують, а два інших злива-
ються, відновлюючи диплоїдне ядро і утворюючи зиготу, що перетво-
рюється на ауксоспору.
Стадії спокою представлені двостулковими кремнеземовими ци-
стами. Утворення цисти починається з поділу протопласта клітини.
Далі материнський панцир скидається, кожний дочірній протопласт
110
вкриваєтся двома опуклими стулками, при цьому пояскові обідки не
розвиваються. Стулка, яка утворилась першою, несе на поверхні
різноманітні потовщення (гребні, шипи, шипики). Друга стулка зали-
шається гладенькою.
Система відділу. Поділ на класи проводиться, в першу чергу, за
типом симетрії стулок (актиноморфні чи зигоморфні) та наявністю шва.
З цим комплексом ознак корелює чимало інших ознак, пов’язаних з
будовою протопласта та особливостями статевого процесу. Відділ
поділяють на три класи - СозсіпосІізсорИусеае, ЕгадіІагіорИусеае та
ВасіІІагіорИусеае.
Клас Косцинодискофіцієві або Центричні - Созсіпобізсо-
рНусеае (СепІгорНусеае)
Об’єднує водорості з актиноморфними стулками, які завжди поз-
бавлені шва. У більшості випадків стулки косцинодискофіцієвих круглі,
радіально-симетричні, через що представників класу часто назива-
ють центричними діатомеями. Лише у центричних діатомей виявле-
но оогамні статеві процеси та рухливі сперматозоїди. Представники
цього класу мешкають переважно у планктоні морів та прісних во-
дойм. У класі виділяють 22 порядки, серед яких у прісних водоймах
найпоширенішими є представники Меіозігаїез.
Порядок Мелозиральні - Меіовігаїев
Об’єднує водорості з актиноморфними стулками, в яких немає
виростів із підпорками - фултопортул, ареоли закриті кремнеземо-
вою платівкою з зовнішнього боку і переважно не утворюють чітких
правильних штрихів. Пластиди у мелозиральних численні, дрібні,
лопатеві, розміщуються у периферичному шарі цитоплазми. Типовий
представник порядку - Меіозіга.
Рід мелозіра - Меіозіга (рис. 32). Водорість надзвичайно харак-
терна для планктонних угруповань прісних водойм. Колонії мають
вигляд нерозгалужених ниток, які складаються з циліндричних клітин,
з’єднаних між собою за допомогою слизових валиків, шипів або зубців.
Панцири мелозіри мають високі загини стулки та складні пояскові
обідки, через що в препаратах клітини лежать переважно з пояска.
Стулки панцира завжди круглі, вкриті дрібними ареолами, які часто
утворюють радіальні ряди. На стулці розташовується кілька двогубих
виростів - римопортул, інколи - кільце дрібних шипів. Через римо-
111
портули виділяється слиз, за допомогою якого дочірні клітини після
поділу залишаються з’єднаними стулками, утворюючи пари клітин.
Ці пари, залишаючись разом, обумовлюють формування типових для
цієї водорості ниткоподібних колоній. На великому збільшенні в кліти-
нах добре помітні невеликі дисковидні або лопатеві хлоропласти, які
розміщуються біля бічних країв стулки.
У Меіозіга досить детально досліджено статевий процес, який
представлений оогамією. На початку статевого процесу в одній з
клітин відбувається мейоз. При цьому після першого поділу (мейоз 1)
одне з ядер дегенерує і не переходить у фазу мейозу 2. Друге, життє-
здатне ядро, ділиться, і знов одне з ядер дегенерує. При поділах у
фазах мейоз 1 та 2 цитокінез не відбувається. Таким чином, у жіночо-
му гаметангії утворюється одна яйцеклітина.
У клітині, що виконує функцію чоловічого гаметангію, протопласт
набрякає, обумовлюючи скидання панцира. Далі на поверхні прото-
пласта з’являються чотири джгутики, відбувається мейоз і утворюєть-
ся чотириядерна гола сперматогенна клітина, що зберігає всі чотири
1
А
джгутики. Далі одночасно від спер-
матогенної клітини відбрунькову-
ються чотири одноджгутикові одно-
ядерні сперматозоїди. Брунькуван-
ня відбувається таким чином, що в
гамети не потрапляють пластиди:
вони залишаються в центрі спер-
матогенної клітини, утворюючи т.
зв. залишкову клітину, що позбав-
лена ядра і незабаром руйнується.
Далі сперматозоїди запліднюють
яйцеклітину, яка перетворюється
на ауксоспору.
Види роду мелозіра широко
представлені в планктоні ставків,
озер, річок, заплавних водойм. Се-
ред представників роду є також і
літорально-бентосні організми, де-
які види здатні розвиватися на зро-
шуваних скелях.
Рис. 32. Меіозіга: А - загальний вигляд колонії
Б з пояска; Б - вигляд зі стулки. 1 - хлоропласти
112
Клас Фрагілярієфіцієві, або Безшовні - ЕгадіІагіорКусеае.
Об’єднує водорості із зигоморфними стулками, які позбавлені швів,
але мають осьове поле. На стулці фрагілярієфіцієвих, як правило,
розташовується один двогубий виріст - римопортула, який вважається
попередником шва. В межах цього класу, на відміну від Созсіпо-
сіізсоріїусеае, повністю відсутні джгутикові стадії. В класі виділяють
12 порядків. Типовим є порядок Егадіїагіаіез.
Порядок Фрагіляріальні - Егадіїагіаіез
Характерними ознаками порядку є наявність на полюсі стулки
однієї римопортули, розміщеної під кутом до поздовжньої осі; вставні
обідки відсутні або наявні, проте в останньому випадку позбавлені
септ. Клітини містять від двох великих до багатьох дрібних хлоро-
пластів, розміщених переважно в периферичному шарі цитоплазми,
під стулкою. Більшість представників цього порядку утворює колонії,
за формою яких роди цих водоростей можна розпізнавати навіть без
виготовлення постійних препаратів панцирів. До найпоширеніших у
континентальних водоймах родів належать Егадіїагіа, АзіегіопеІІа,
Оіаіота.
Рід фрагілярія - Егадіїагіа (рис. 33 А, Б). Клітини водорості зде-
більшого утворюють колонії у вигляді стрічок, в яких панцири з’єдну-
ються стулками. При вигляді з пояска окремий панцир має видовже-
но-прямокутну форму; зі стулки панцир видовжений, білатерально-
симетричний, на полюсах більш-менш дзьобоподібний. У постійних
препаратах, виготовлених з очищених від цитоплазми панцирів, на
стулці добре розрізняються осьове поле та чергово розміщені штри-
хи. Види роду є переважно мешканцями перифітонних та бентосних
угруповань прісних водойм.
Рід астеріонела - АзіегіопеІІа (рис. 33 В, Г). За будовою панци-
ра вельми подібна до фрагілярії прісноводна планктонна водорість
АзіегіопеІІа. її клітини з’єднані у зірчасті колонії за допомогою слизу
та дрібних шипиків, розташованих на полюсі стулки.
Рід діатома - Оіаіота (рис. 33 Д, Е). У живому стані представни-
ки цього роду розрізняються за наявністю зигзагоподібних колоній.
В них з'єднання клітин також здійснюється за допомогою слизу та
полярних шипиків на стулці. Крім того, яскравою ознакою є наявність
на внутрішньому боці стулки грубих поперечних ребер, які надають
113
клітині характерної поперечної посмугованості. Діатому часто можна
зустріти у планктонних та бентосних угрупованнях прісних водойм.
Рис. 33. Фрагілярієфіцієві діатомові водорості: А, Б - Ргадіїагіа уаисіїегіае.
А - окрема клітина, Б - колонія; В, Г - АзІегіопеІІа їогтоза: В - окрема клітина,
Г - фрагмент колонії; Д, Е - Оіаіота уиїдаге: Д - окрема клітина, Е - колонія
114
Клас Бацилярієфіцієві, або Шовні - ВасіІІагіорКусеае
До цього класу включені водорості із зигоморфними стулками, в
яких на одній або на обох стулках є шов. У водоростей з ВасіІІа-
гіоріїусеае джгутикові стадії повністю відсутні, статеві процеси пред-
ставлені ізо-, гетеро- та автогамією.
Згідно із сучасними поглядами на еволюцію діатомових водорос-
тей, шов утворився з римопортули, що поступово видовжувалася. Шов
є структурою, з якою пов’язана здатність бацилярієфіцієвих водорос-
тей до активного ковзаючого руху. Поява такої здатності розглядається
як крупний ароморфоз. Тому ознаки, пов’язані зі швом (зокрема, його
тип, кількість, положення на стулці, ступінь розвитку і т. д.), покладені
в основу поділу ВасіІІагіоріїусеае на порядки.
ВасіІІагіоріїусеае поділяють на десять порядків, з яких нижче роз-
глядаються п’ять -СутЬеІІаІез, АсЬпапІІіаІез, Иауісиїаіез, ВасіІІагіаіез
та ЗигігеІІаІез.
Порядок Цимбеляльні - СутЬеІІаіез
Об’єднує види, в яких на одній зі стулок є розвинений щілинопо-
дібний шов, за допомогою якого клітини можуть активно рухатись. На
протилежній стулці також є шов, який у деяких видів недорозвинений
(наприклад, РІіоісозрІїепіа), в інших - розвинений повністю
(Сотріїопета, СутЬеІІа). Стулки у цимбеляльних водоростей аси-
метричні принаймні щодо однієї з осей.
Види із СутЬеІІаіез населяють переважно бентос та перифітон
прісних водойм.
Рід ройкосфенія - РНоісозрЬепіа (рис. 34 А, Б). Водорість час-
то трапляється в обростаннях вищих водних рослин. Її клітини си-
метричні зі стулки лише щодо поздовжньої осі і мають булавоподібну
форму. З пояска панцир клиноподібний й вигнутий, прикріплюється
до субстрату довгою слизовою ніжкою. Цей слиз секретується через
відкриті пори, розташовані на звуженому кінці стулки. Шов повністю
розвинений лише на одній стулці, а на іншій - лише біля полюсів.
Хлоропласт у ройкосфенії один, чотирилопатевий; лопаті починають-
ся від притиснутого до пояска піреноїда.
115
.1
.1
5
Рис. 34. Щілиношовні діатомові водорості: А, Б - РЬоісозрЬепіа аЬЬтіаІа:
А - вигляд зі стулки; Б - вигляд з пояска; В, Г - ОотрЬопета Ігипсаіит В
- вигляд зі стулки, Г-живі клітини, вигляд з пояска; Д, Е - СутЬеІІа іитісіа'. Д
- вигляд зі стулки, Е - живі клітини на слизових ніжках; Є, Ж - РІапоІЇїідіит
Іапсеоіаіа'. Є - шовна стулка, Ж - безшовна стулка; З, І - Соссопеіз ріасепіиіа:
З - шовна стулка, І - безшовна стулка; ї - №мсиІа ігірипсіаіа. 1 - щілиноподібний
шов; 2 - стигма; 3 - слизова ніжка; 4 - рострум; 5 - хлоропласт
Рід гомфонема- (Зотрїюпета (рис. 34 В, Г). За обрисами клітин
види цього роду дещо нагадують попередній рід - стулки в них була-
воподібні, з пояска клітини клиноподібні, але, на відміну від ройкос-
фенії, не вигнуті. Шов у гомфонеми добре розвинений на обох стул-
ках. Водорості мешкають переважно у перифітоні і прикріплюються
до підводних субстратів слизовими ніжками, що утворюються завдя-
ки секреції слизу через систему субмікроскопічних пор, розташова-
них на полюсі звуженого боку стулки.
116
Рід цимбела - СутЬеІІа (рис. 34 Д, Е). Види роду СутЬеІІа
нагадують півмісяці, що знаходяться на верхівках розгалужених сли-
зових ніжок. Стулки цимбели симетричні щодо поперечної осі і тому
розглядаються як дорзовентральні. На вигнутому боці клітини розта-
шовується хлоропласт із піреноїдом. Майже посередині стулки про-
ходить шов, полярні щілини якого відігнуті на дорзальний бік. В оп-
тичному мікроскопі на постійних препаратах біля центрального вуз-
лика можна спостерігати одну-кілька крапок, що є особливим типом
перфорацій - стигмою. Представників цього роду можна знайти в будь-
якій прісній водоймі, переважно в обростаннях вищих водних рос-
лин.
Порядок Ахнантальні - АсКпапІНаІев
Найяскравішою особливістю представників порядку є наявність
щілиноподібного шва лише на одній зі стулок - або на епітеці, або на
гіпотеці. Розташування шва лише на одній стулці пов’язане з присто-
суванням цих водоростей до перифітонного способу життя. Клітини
ахнантальних водоростей поодинокі, розташовуються на поверхні
субстратів, притискаючись до них стулкою зі швом. До найпоширені-
ших представників порядку належать роди РІапоіЬісІіитта Соссопеіз.
Рід планотидіум - РІапоіїїідіит (рис. 34 Є, Ж). Види цього роду
масово розвиваються у перифітонних та бентосних угрупованнях
прісних водойм. Панцири мають бісиметричні стулки, з яких шовна
стулка увігнута, а безшовна - опукла. За рахунок секреції слизу по-
лярною щілиною клітини утворюють ніжку, якою під час вегетації при-
кріплюються до субстрату. Пластида у представників роду одна, роз-
ташовується вздовж одного з боків стулки. На постійних препаратах
планотидіум легко розпізнається за підковоподібною структурою - ро-
струмом - біля центрального поля безшовної стулки.
Рід коконеїс - Соссопеіз (рис. 34 З, І). У прісних водоймах цей
рід є найхарактернішим домінантом обростань вищих водних рослин
та зелених водоростей-макрофітів. Стулки коконеїсу широкоеліп-
соїдні, пояскові обідки низькі. Клітини Соссопеіз не утворюють слизо-
вих ніжок, а міцно притискаються до субстрату тією стулкою, на якій
розміщений шов. Кожна клітина містить один лопатевий пристінний
хлоропласт у вигляді літери «С», з одним або кількома дрібними ви-
довженими голими піреноїдами.
117
Порядок Навікуляльні - Мауісиїаіез
Водорості цього порядку мають розвинені щілиноподібні шви на
обох стулках. Самі стулки або бісиметричні, або 8-подібні (дзеркаль-
но симетричні). Представники №уісиІаІев поширені майже в усіх ти-
пах морських та прісноводних угруповань. Характерними приклада-
ми навікуляльних водоростей є роди Мауісиіа та Ріппиіагіа.
Рід навікула - Мауісиїа (рис. 34 і). Види одного з найчисленні-
ших родів порядку - Мауісиїа - мають досить просту будову: стулки
за обрисами ланцетоподібні і нагадують човник. По середині осьово-
го поля проходить прямий щілиноподібний шов. На поверхні стулок
розташовуються паралельні, радіальні або конвергентні штрихи, ут-
ворені правильними рядами ареол. Центральне поле відносно неве-
лике і не доходить до країв стулки. При вигляді з пояска клітини ви-
довжено-прямокутні. Пояскові обідки прості і не мають вставних
обідків. Хлоропластів два, вони пластинчасті, розташовуються вздовж
пояскових обідків на різних боках стулки і містять по одному паличко-
подібному піреноїду.
Рід піннулярія - Ріппиіагіа (рис. 35). Стулки видів роду Ріппиіагіа
мають форму, наближену до видовжено-еліпсоїдної. Шов у піннулярії
дещо хвилястий. Із внутрішнього боку стулки панцир має систему гру-
бих потовщень - ребер, які дещо нагадують поверхню пральної дош-
ки, і в оптичний мікроскоп можуть бути помилково прийняті за штри-
хи. Між ребрами панцир перфорований рядами дрібних ареол. При
вигляді з пояска панцир правильно-прямокутний, причому в цій по-
зиції добре помітно, що ребра є саме системою внутрішніх потов-
щень стулки, а не її перфорацій.
Хлоропластів два, стрічковидні, розташовуються вздовж поясків
з протилежних боків стулки і мають дуже порізані краї. У деяких видів
протилежні стрічки хлоропласта під гіповальвою з’єднуються неве-
ликим містком, завдяки чому хлоропласт набуває Н-подібної форми.
В центральній частині кожної стрічки у багатьох видів розташовується
по одному голому еліпсоїдному піреноїду. Оскільки панцири у видів
цього роду переважно великі, клітини не здатні утримуватись у товщі
води, через що мешкають або у бентосних, або у перифітонних угру-
пованнях прісних водойм.
118
Рис. 35. Ріппиіагіа: А - вигляд зі стулки; Б - вигляд з пояска. 1 - щілинопо-
дібний шов; 2 - полярний вузлик; 3 - центральний вузлик; 4 - ребра; 5 - осьове
поле; 6 - епітека; 7 - гіпотека
Порядок Бациляріальні - ВасіІІагіаІев
До порядку включено водорості, на стулках яких розташовується
по одному відносно прямому каналовидному шву. Шов бациляріаль-
них складається з вузької зовнішньої щілини, яка сполучається з ка-
налом, розміщеним у внутрішньому потовщенні стулки - кілі. Канал
відкривається у порожнину панцира великими отворами - кільовими
точками. Цей канал є гомологом внутрішньої щілини шва щілиношов-
них діатомей. Водорості з ВасіІІагіаіез поширені у водоймах всіх типів
та у багатьох ґрунтах. Характерним представником порядку є рід
Міхзсіїіа.
Рід ніцшия - Міхзсіїіа (рис. 36 А). Водорість має бісиметричні
або 3-подібні стулки, які переважно досить витягнуті у довжину. Ка-
наловидний шов зміщений до краю стулки, причому шов епітеки роз-
119
ташовується на протилежному боці від шва гіпотеки. Зазвичай види
цього роду мають дві пластинчасті цілісні або злегка розсічені плас-
тиди, розміщені по різні боки від поперечної площини клітини. За ви-
довим багатством рід Мігзсіїіа є одним з найчисленніших у відділі.
Види з вузькими та дрібними панцирами поширені переважно у планк-
тоні, а з великими та грубими - у бентосі та перифітоні континенталь-
них водойм та морів.
2
Рис. 36. Канало-
шовні діатомові водо-
рості: А - Шгзсіїіа раїеа-,
Б - ЗигігеІІа ЬгеЬіззопіі;
В - Сутаіоріеига зоїеа.
1 - кільові точки, 2 - осьо-
ве поле; 3 - крило
Порядок Суриреляльні - ЗигігеІІаІез
У представників цього порядку на кожній стулці розташовано по
два каналовидних шва. Вони розміщуються у зовнішніх складках біля
загину стулки, які називають крилами. Стулки суриреляльних мають
складні системи ребер, що підсилюють крила. Між ребрами на стулці
знаходяться системи ареол. Оскільки панцири представників поряд-
ку великі та важкі, ці водорості населяють переважно бентосні угру-
повання. Характерними представниками порядку є роди ЗигігеІІа та
Сутаіоріеига.
Рід сурірела - ЗигігеІІа (рис. 36 Б) має еліпсоїдні або яйцеподібні
стулки з добре розвиненими крилами, в яких знаходяться канало-
120
видні шви. На поверхні стулки розташовуються грубі поперечні реб-
ра, що закінчуються на крилах. Клітина має два великих пластинчас-
тих хлоропласти, які розташовуються на периферії клітини таким чи-
ном, що один з них притиснутий до епітеки, другий - до гіпотеки. Види
роду населяють як моря, так і прісні водойми.
Рід циматоплевра - Сутаіоріеига (рис. 36 В). Панцир цієї во-
дорості при вигляді зі стулки перетягнутий і дещо нагадує гітару. По-
верхня стулок хвиляста, а крила дуже вузькі. Хлоропласт водорості
один, складається із двох пластин, що притиснуті до епі- та гіпотеки,
відповідно, і на одному з полюсів з’єднані між собою тонким містком.
Види роду населяють прісні водойми, переважно з лужною реакцією
середовища.
Відділ Диктіохофітові водорості - ОісіуосНорНуіа
Евкаріотичні фотоавтотрофні та вторинно гетеротрофні тубуло-
кристати, у яких клітини голі, часто із зовнішнім кремнеземовим ске-
летом (силікофлагеляти). Пластиди вторинно симбіотичні, родофіт-
ного типу. Монадні стадії мають ретронеми. Продукт асиміляції - хри-
золамінарин. Характерна особливість - асоційованість базальних тіл
джгутиків безпосередньо з ядерною мембраною, без участі джгути-
кових коренів.
Відділ об’єднує біля 40 видів мікроскопічних одноклітинних водо-
ростей з монадною або амебоїдною будовою. Відома також значна
кількість викопних диктіохофітових. Найдавніші представники мають
вік біля 120 млн. років. Поширені диктіохофітові переважно у планк-
тоні морів та прісних водойм.
Біохімічні особливості. Представники відділу характеризують-
ся наявністю хлорофілів а та с, які маскуються додатковими пігмен-
тами, в першу чергу - фукоксантином. Через це хлоропласти диктіо-
хофітів забарвлені у жовтий або світло-коричневий колір. З інших ксан-
тофілів виявлено діатоксантин, діадіноксантин, лютеїн. З каротинів
знайдено лише 0-каротин. Запасними поживними речовинами є олія
та хризоламінарин.
Цитологічні особливості. Клітинні покриви представлені плаз-
малемою, над якою може знаходитися кремнеземовий скелет, пок-
ритий слизовою капсулою.
121
Диктіохофітові водорості одноядерні. Ядро евкаріотичного типу,
відмежоване від цитоплазми двомембранною оболонкою, з одним
або кількома ядерцями. З апікального боку до ядерної оболонки при-
кріплюються два базальних тіла джгутиків. До ядерної оболонки та-
кож безпосередньо прилягає комплекс Гольджі. Крім того, у диктіохо-
фітових наявні особливі цитоплазматичні мікротрубочкові структури,
які відходять від поверхні ядра і утворюють у цитоплазмі додаткові
опірні елементи, а у амебоїдних форм заходять у ризоподії і беруть
участь у їх русі.
На ультратонких зрізах мітохондріальні профілі мають трубчасті
кристи.
У здатних до фотосинтезу представників клітина звичайно містить
кілька пристінних хлоропластів. Кожний хлоропласт вкритий чотири-
мембранною оболонкою. Дві внутрішні мембрани є власними мем-
бранами хлоропласту, дві зовнішні мембрани утворюють хлороплас-
тний ендоплазматичний ретикулюм. Під хлоропластною оболонкою
розташовується оперезуючий тилакоїд, далі кілька тритилакоїдних
ламел.
Монадні стадії дводжгутикові. Джгутики гетероконтні, гетеро-
морфні та гетеродинамічні. Інколи короткий джгутик редукований, і
від нього зберігається лише базальне тіло. Довгий джгутик несе два
ряди ретронем. Унікальною особливістю джгутикового апарату є по-
вна відсутність мікротрубочкових джгутикових коренів та прикріплен-
ня базальних тіл джгутиків безпосередньо до ядерної мембрани.
Розмноження відбувається вегетативним поділом клітин надвоє
та зооспорами. Зооспори голі, позбавлені кремнеземового скелету і
мають два гетероконтні та гетеродинамічні джгутики, що виходять з
апікальної заглибини.
Система відділу. Відділ включає два класи - Оісіуосіюріїусеае
та РесііпеІІорІїусеае. У Оісіуосіїоріїусеае кремнеземовий скелет є, у
РесііпеІІорІїусеае він відсутній.
Клас Диктіохофіцієві або Силікофлагеляти - ОісіуосНо-
рНусеае
У межах класу представлено лише один порядок - Оісіуосіїаіез.
Типовим родом є Оісіуосїіа.
Рід диктіоха - Оісіуосіїа (рис. 37). Водорість одноджгутикова, з
протопластом, диференційованим на дві зони - периферичну та цен-
122
тральну перинуклеарну. У периферичній частині, назовні від плазма-
леми, розміщується кремнеземовий скелет. Він має вигляд кошика,
який утворений з’єднаними між собою порожнистими кремнеземови-
ми трубкам. Периферична зона складається з численних радіально
розміщених тяжів цитоплазми, які анастомозують між собою та роз-
ширюються на верхівці. У цих розширеннях розташовуються хлоро-
пласти та мітохондрії. У перинуклеарній зоні міститься ядро, комп-
лекси Гольджі, від центральної зони відходить джгутик, інколи -
псевдоподія, яку називають щупальцем.
Диктіоха надзвичайно широко розповсюджена у морях та океа-
нах високих широт. Однією з яскравих екологічних особливостей є її
холодолюбність. Температурний оптимум вегетації для представників
цього роду знаходиться у діапазоні 5-1І2°С.
Рис. 37. Оісіуосіїа: А - зовнішній вигляд клітини; Б - кремнеземовий скелет
(вигляд збоку). 1 - трубка кремнеземового скелету; 2 - слизова капсула; 3 - хло-
ропласти; 4 - джгутик
Клас Педінелофіцієві - РебіпеІІорНусеае
Об'єднує монадні одноджгутикові водорості, які позбавлені крем-
неземового скелета. Зазвичай на передньому кінці клітина здатна ут-
ворювати псевдоподії, а на задньому - стебельце, яке у частини видів
123
здатне скорочуватися. Протопласт не диференційований на перифе-
ричну та перинуклеарну зони. Типовим родом є педінела - РесІіпеІІа
(рис. 38), яка зустрічається у прісних водоймах, зокрема в озерах.
Альвеоляти
Рис. 38. РесІіпеІІа: зовнішній вигляд
клітин. 1 -джгутик; 2-хлоропласт; 3-псев-
доподії; 4 - скоротливе стебельце
Підцарство альвеолят
(АІуеоІаіез) об’єднує органі-
зми з альвеольованими клі-
тинними покривами, в утво-
ренні яких значну роль ві-
діграють мікротрубочки. Для
альвеолят характерні різні
варіанти ядерного апарату,
які помітно відрізняються від
типової схеми будови ядра
евкаріот. Ретронеми у аль-
веолят відсутні. До підцар-
ства належать динофітові
водорості, апікомплекси та
інфузорії.
Відділ Динофітові водо-
рості - ОіпорЬуіа
Евкаріотичні первинно
гетеротрофні, фотоавто-
трофні та вторинно гетеро-
трофні тубулокристати, в
яких клітини вкриті особли-
вим типом покриві в - ал ьве о-
льованою амфієсмою. Пластиди вторинно симбіотичні і дуже різнома-
нітні - хлорофітного та родофітного типів. У примітивних представників
ядро типово евкаріотичне, у еволюційно продвинутих представляє
особливий варіант ядерного апарату - динокаріон.
До відділу відносять біля 2000 видів сучасної флори. Ще близько
2000 видів відомі у викопному стані. Найдавніші рештки динофітових
водоростей представлені переважно цистами (т. зв. гістрихосфери-
ди), які знайдені у силурійських (435-460 млн. років тому) та докемб-
рійських (600 млн. років) осадових породах.
124
Біохімічні особливості. Для Оіпорґіуіа склад пігментів не є уні-
версальною систематичною ознакою. Приблизно 50% видів є об-
лігатними гетеротрофами і не мають фотосинтетичних пігментів. У
фотоавтотрофних представників сьогодні відомо принаймні чотири
варіанти комбінацій пігментів: перша вважається основною, інші -
додатковими.
Основна комбінація пігментів властива переважній більшості
фотоавторофних динофітових. Вона включає хлорофіли а та с,
р-каротин, ксантофіли перидинін, діадиноксантин та диноксантин.
Хлоропласти у представників з основною комбінацією забарвлені у
різні відтінки жовтого кольору.
Продуктами асиміляції є крохмаль та олія, рідше - хризоламіна-
рин, а у облігатно гетеротрофних видів - глікоген. Всі асиміляти відкла-
даються поза межами хлоропластів.
Цитологічні особливості. Клітинні покриви представлені амфі-
єсмою, що розташовується під плазмалемою. Амфієсма складаєть-
ся з шару сплощених мембранних пухирців, що щільно прилягають
один до одного. Кількість пухирців у різних представників різна - від
двох до кількох десятків. Пухирці заповнені або зернистим матрик-
сом, або всередині кожного пухирця відкладається полісахаридна
платівка. У першому випадку амфієсма еластична, клітини здатні до
слабких метаболічних змін форми. Якщо у пухирцях утворюються
платівки, то клітини мають сталу форму. Цей варіант амфієсми нази-
вають текою. Зрідка клітинні покриви представлені лише плазмале-
мою (наприклад, у ОіпатоеЬісІіит), і тоді клітини мають амебоїдну
будову, здатні утворювати псевдоподії. У нерухомих (гемімонадних,
кокоїдних та нитчастих) динофітових водоростей є справжня целю-
лозно-пектинова оболонка.
Клітини переважно дорзовентральні. Характерна особливість -
наявність двох борозенок - поперечної та поздовжньої. У зоні боро-
зенок амфієсма відсутня, і протопласт відмежовується від зовнішнього
середовища лище плазмалемою. Поздовжня борозенка знаходиться
на черевному боці, її пересікає поперечна борозенка. Остання роз-
діляє клітину на верхню частину - епікон, та нижню - гіпокон. З місця
перехрещення борозенок виходять два гетероконтні, гетероморфні
та гетеродинамічні джгутики. Один джгутик знаходиться у поперечній
борозенці і є рушійним, другий - у поздовжній і виконує функції
стерна.
Біля основи джгутиків розташовується вакуолярний апарат,
представлений непульсуючими вакуолями-пузулами. У зоні поздовж-
125
ньої борозни клітини деяких видів утворюють псевдоподії, за допомо-
гою яких активно захоплюють часточки їжі. Перетравлювання здо-
бичі відбувається у травних вакуолях.
Ядро у переважної більшості динофітових водоростей має суттєві
відміни від типово евкаріотичних ядер: хромосоми майже позбавлені
гістонів, протягом всього клітинного циклу знаходяться у надспіралі-
зованому стані та прикріплюються до ядерної оболонки за допомо-
гою центромерів.
Ядерна оболонка при поділі лишається інтактною, мітоз закри-
тий. На початку поділу в ядрі утворюються цитоплазматичні тунелі, в
які занурюються пучки мікротрубочок. Більшість мікротрубочок про-
ходить тунелі наскрізь, формуючи інтерзональне веретено.
Хлоропласт основного типу покритий трьома мембранами. Дві
внутрішні мембрани складають власне оболонку хлоропласта, зов-
нішня утворена мембраною, похідною від ендоплазматичної сітки. Ти-
лакоїди групуються по три, оперезуючі тилакоїди відсутні.
Мітохондрїі динофітових водоростей мають трубчасті кристи.
Монадні клітини динофітових несуть по два гетероконтні, гетеро-
морфні та гетеродинамічні джгутики. Вони виходять з невеличких
заглибин - т. зв. джгутикових каналів, що зазвичай розташовані на
черевному боці клітини, в місці перехрещення поздовжньої та попе-
речної борозенок. Один джгутик - поперечний - направлений право-
руч і знаходиться у поперечній борозенці, другий - поздовжній - орі-
єнтований у напрямку гіпокону і лежить у поздовжній борозенці.
Поперечний джгутик виконує локомоторну функцію. Він має ви-
гляд хвилястої стрічки і нагадує ундулюючу мембрану. На поверхні
джгутика розташовується один ряд простих мастигонем. При основі,
у джгутиковому каналі, поперечний джгутик має потовщення - пара-
флагелярне тіло, що є фоторецептором; з цією структурою звичайно
скоординовано положення стигми. Поздовжній джгутик виконує
функції стерна. Він дещо сплощений, з аксонемою, що відповідає фор-
мулі 9+2. На поверхні поздовжнього джгутика розташовані 1-3 ряди
простих мастигонем.
У деяких динофітових джгутики майже однакові, а поперечна бо-
розенка редукована до апікальної заглибини. В окремих видів по-
здовжній джгутик редукований, хоча зберігається його базальне тіло
та мікротубулярний корінь.
Особливості живлення. У динофітових водоростей широко пред-
ставлені два типи живлення - фотоавтотрофний та гетеротрофний.
126
Приблизно половина видів ОіпорЬуїа - це забарвлені форми, що ма-
ють хлоропласти та здатні до оксигенного фотосинтезу. Інші пред-
ставники відділу безбарвні і є облігатними гетеротрофами.
Гетеротрофні Оіпоріїуїа можуть споживати розчинені у воді орга-
нічні речовини осмотрофним шляхом, всмоктуючи їх всією поверх-
нею тіла. Проте основним способом споживання органічних речовин
є фаготрофне живлення.
Захоплення часточок їжі здійснюється за допомогою особливих
псевдоподій, що утворюються на черевному боці клітини у зоні по-
здовжньої борозенки.
Плани будови тіла. Переважна більшість видів є одноклітинни-
ми монадними організмами. Приблизно 10% видів мають амебоїдну
будову. Існує також невеличка група динофітових водоростей з гемі-
монадним, кокоїдним та нитчастим типами морфологічної структури
тіла.
Розмноження та життєві цикли. Розмножуються динофітові
водорості переважно нестатевим шляхом, приблизно у 50 видів ви-
явлено статевий процес.
Основний спосіб нестатевого розмноження - вегетативний поділ
клітин надвоє. У видів, що не мають панцира, дочірні клітини при поділі
отримують від материнської клітини амфієсму епікону або гіпокону і
потім добудовують половину, якої не вистачає. Значна частина видів,
що мають панцир, перед початком поділу скидають амфієсму, і тоді
дочірні клітини виробляють новий панцир. Досить часто нестатеве
розмноження відбувається за допомогою зооспор. Деякі види утво-
рюють також апланоспори.
Статевий процес відбувається у формі ізо- або гетерогамії, у де-
яких видів функції гамет виконують вегетативні клітини, і тоді стате-
вий процес в принципі нагадує хологамію. Гамети можуть бути голі
або вкриті міцною амфієсмою.
У Оіпоріїуіа наявні гаплофазні, примітивні гаплодиплофазні та
диплофазні життєві цикли, а редукція кількості хромосом може бути
зиготичною, споричною та гаметичною.
Система відділу. За екологічними особливостями та комплек-
сом цитологічних ознак (зокрема, за будовою ядра, його поділом, бу-
довою джгутикового апарату), відділ поділяється на п’ять класів -
Охуггііісіоріїусеае, ЗупсІіпіорЬусеае, Мосіііисіріїусеае, ВІазіосІІ-
пірйусеае, Оіпоріїусеае. Всі класи, за вийнятком ОіпорІїусеае, є мо-
нотипними і включають по одному порядку.
127
Клас Оксиридофіцієві - ОхуггІїісІорЬусеае
Представлений лише родом оксиріс (Охуггіїіз), який має по два
ізоконтні, ізоморфні та ізодинамічні джгутики. Ядро типово евкаріо-
тичне, в інтерфазі хроматин деспіралізований, а ядерна ДНК пов’я-
зана з гістонами. Веретено поділу внутрішньоядерне. Водорість без-
барвна, живиться осмотрофно, мешкає у морях та приморських ли-
манах.
Клас Синдиніофіцієві - ЗупсііпіорНусеае
Об’єднує високоспеціалізованих внутрішньоклітинних паразитів,
що мають амебоїдну будову та евкаріотичне ядро. У синдиніофіціє-
вих наявні деякі ознаки «типових» динофітових: так, у монадних стадій
є поперечний джгутик характерної «динофітової» будови, веретено
поділу утворюється позаядерно, у цитоплазматичних тунелях, що
проходять крізь ядро. Всі синдиніофіцієві - облігатні гетеротрофи.
Переважаючим типом живлення є фаготрофія. Синдиніофіцієві пара-
зитують у клітинах морських інфузорій, ракоподібних, риб та великих
(т. зв. «справжніх») динофітових водоростей.
Клас Ноктилюціфіцієві - МосііІисірЬусеае
Включає облігатно гетеротрофні вільноіснуючі морські водорості,
в яких вегетативні клітини мають типово евкаріотичне ядро, а гамети
- динокаріон. Клас є монотипним, типовий представник - ИосіПиса.
Рід ноктилюка, або ночесвітка - Мзсіііиса (рис. 39). Особливо-
стями ноктилюки є, по-перше, здатність до біолюмінісценції, по-дру-
ге - до утворення симбіозів з дрібними одноклітинними зеленими во-
доростями. Завдяки останньому деякі популяції ноктилюки фактично
переходять до автотрофного живлення.
У вільноіснуючому стані ця водорість живиться фаготрофно, за-
хоплюючи часточки їжі щупальцем, у якому є мікротрубочкові опірні
елементи. Останні дещо нагадують мікротубулярний кошик. Вегета-
тивні клітини великі (до 1 мм у діаметрі), вкриті амфієсмою, пухирці
якої заповнені рідиною і не містять платівок. Джгутик лише один, за
будовою відповідає типовому поздовжньому джгутику. Цитоплазма
містить велику вакуолю, пронизану цитоплазматичними тяжами. Ядро
у клітині одне,- з диплоїдним набором хромосом, що організовані за
евкаріотичною схемою: інтерфазний хроматин деспіралізований, ДНК
пов’язана з гістонами. Мітоз також евкаріотичного типу, проте здійс-
нюється за допомогою характерного для динофітових позаядерного
веретена.
128
Монадні стадії представлені гаметами, що утворюються у веге-
тативній клітині після редукційного поділу. Гамети, на відміну від ве-
гетативних клітин, мають динокаріотне ядро зі спіралізованими інтер-
фазними хромосомами, що не пов'язані з гістонами. Джгутиковий
апарат у гамет типової динофітової будови, поздовжній та попереч-
ний джгутики виходять з місця перехрещення поздовжньої та попе-
речної борозенок.
Статевий процес представлений ізогамією. При утворенні гамет
ядро вегетативної клітини редукційно ділиться. Після копуляції гамет
планозигота без періоду спокою швидко проростає у нову диплоїдну
вегетативну клітину. Таким чином, життєвий цикл ночесвітки дипло-
фазний, з гаметичною редукцією та без чергування поколінь.
ІЧосііІиса вважається космополітною водорістю, що зустрічаєть-
ся в морях помірного, субтропічного та тропічного поясів. Розвива-
ючись масово, вона викликає нічне свічення морів. Це явище обу-
мовлено наявністю у клітинах ночесвітки люмінофору люциферину.
Рис. 39. ЬІосНІиса: А - вегетативна клітина; Б - гамета
129
Клас Бластодиніфіцієві - Віазіосііпірґіусеае
Об’єднує водорості, що є внутрішньотканинними паразитами вод-
них тварин, переважно - з класу ракоподібних (Сгизіасеа). Монадні
клітини бластодиніфіцієвих мають справжній динокаріотний ядерний
апарат. У вегетативних клітин мітоз типово динофітовий. Переважній
більшості представників властиві справжні хлоропласти основного
типу. Лише один вид безбарвний і живиться виключно гетеротрофно.
Таким чином, майже всі види класу живляться міксотрофно.
Характерним представником є ВІазіодіпіит зріпиіозит. Водорість
паразитує у кутикулярному шарі травної системи веслоногих рачків
(ряд Сореросіа) і має особливий цикл розвитку зі стадіями клітин-тро-
фоцитів, гоноцитів, зооспор (спороцитів) та цист.
Клас Динофіцієві - ОіпорНусеае
Головна ознака класу - наявність динокаріону та типово дино-
фітового мітозу на всіх стадіях життєвого циклу. Переважна більшість
динофіцієвих - це монадні водорості з динофітовим планом будови
клітин - поздовжньою та поперечною борозенками, диференціацією
клітини на епікон та гіпокон, двома гетероморфними джгутиками,
здатністю до фаготрофного живлення, причому жертва захоплюєть-
ся псевдоподіями, що утворюють кошики з мікротрубочок. Еугам-
ним динофіцієвим водоростям властивий гаплофазний життєвий
цикл. Динофіцієві ведуть переважно вільний спосіб життя, рідше є
внутрішньотканинними або внутрішньоклітинними симбіонтами тва-
рин та водоростей. Клас поділяють на п'ять порядків-АтрИісІіпіаІез,
бопіаиіасаіез, ЗутЬіосііпіаІез, Регісііпіаіез та Ргогосепігаїез, серед яких
два останніх є провідними.
Порядок Амфідиніальні - АтрИісІіпіаІез
Об’єднує монадні водорості з м’якою амфієсмою, що не утворює
теки. Поперечна борозенка в амфідиніальних зміщена на верхівку
клітини. Характерний представник - рід амфідініум (Атріїкіїпіит)
- мешкає переважно у морському планктоні.
Порядок Гоніаулакальні - Сопіаиіасаіез
Включає динофіцієві водорості, що вкриті міцною текою та ма-
ють специфічний тип поділу клітин: дочірні клітини отримують поло-
вину амфієсми від материнської клітини і самостійно добудовують
лише частину, якої не вистачає. Через це часто утворюються лан-
цюжки клітин, особливо - у культурі. Додатковою ознакою порядку
130
вважають наявність довгої поздовжньої борозенки, що звичайно за-
ходить на епікон, сягаючи навіть апексу клітини. Життєві цикли гап-
лофазні або гаплодиплофазні, з тривалою стадією рухливої плано-
зиготи. Характерні представники порядку-роди (Зопіаиіахта Сегаїіит.
Рід гоніаулакс - Оопіаиіах - має більш-менш еліпсоїдні кліти-
ни, у яких тека не утворює великих виростів. Види роду дуже поши-
рені у планктоні морів. Деякі види здатні викликати токсичні «цвітіння»
води, відомі під назвою «червоні припливи».
Рід цераціум - Сегаііит (рис. 40) характеризується дуже витяг-
нутими в поздовжньому напрямку клітинами, які помітно дорзовент-
рально стиснуті. Верхівка клітини - апекс - витягнута в довгий відро-
сток. Нижня, антапікальна частина утворює два або три роговидних
придатки. Поперечна борозенка проходить через найширшу частину
Рис. 40. Сегаііит: 1 - епі-
вальва; 2 - гіповальва
клітини. Поздовжня борозенка, що почи-
нається від поперечної, йде донизу нав-
кіс. З місця перехрещення борозенок
виходять два джгутики - поперечний та
поздовжній. Протопласт вкритий міцною
текою. Розмноження відбувається в рух-
ливому стані шляхом поділу протоплас-
та разом з текою в косому напрямку.
Одна з дочірніх клітин отримує більшу
частину верхньої половини материнсь-
кої клітини і невелику частину нижньої, а
друга - більшу частину нижньої і неве-
лику частину верхньої. Відсутні частини
доростають і добудовують частини теки,
яких не вистачає. Наприкінці вегетацій-
ного сезону водорість утворює цисти і в
цьому стані переживає несприятливі
умови.
Еугамні види роду мають гетерогам-
ний статевий процес з особливим типом
копуляції гамет: мікрогамета потрапляє
у цитоплазму макрогамети через розта-
шований на черевному боці останньої
отвір. Платівка теки макрогамети в зоні
отвору перед копуляцією руйнується.
131
Тека мікрогамети поступово розчиняється в цитоплазмі макрогаме-
ти; через деякий час від чоловічої гамети у цитоплазмі лишається тільки
ядро. Пізніше воно зливається з ядром макрогамети. Таким чином,
гамети не зливаються (у звичайному розумінні), а жіноча гамета «по-
жирає» чоловічу.
Більшість видів цього роду мешкає у морях, і лише окремі пред-
ставники зустрічаються у планктоні прісних континентальних водойм.
Порядок Симбіодиніальні - ЗутЬіоФпіаІез
Специфічна група динофіцієвих, що є ендосимбіонтами корало-
утворюючих кишковопорожнинних. Характерним представником по-
рядку є рід ЗутЬіосІіпіит.
Вегетативні клітини симбіодініуму (ЗутЬіодіпіит) - зооксанте-
ли - позбавлені джгутиків, містять кілька жовтих або коричневих хло-
ропластів. Мешкають зооксантели у цитоплазмі клітин господаря і
забезпечують останнього низькомолекулярними метаболітами, що
утворюються у процесі фотосинтезу (гліцеролом, глюкозою, алані-
ном, гліколатом). Натомість водорість отримує від поліпа азот у формі
амонію або сечовини.
Звичайно зооксантели діляться у клітинах ендодерми господаря
і передаються молодим поліпам у складі яєць.
Порядок Перидиніальні - РегіФпіаІез
Об’єднує представників з різноманітними типами морфологічних
структур-монадною, амебоїдною, кокоїдною, нитчастою-та з різно-
манітними варіантами будови амфієсми. У перидиніальних амфієс-
ма материнської клітини не передається дочірнім - під час поділу
або після його закінчення амфієсма скидається, і дочірні клітини по-
вністю будують нову; в еугамних видів при статевому процесі відбу-
вається типова копуляція гамет, що з’єднуються черевними боками, і
утворюється планозигота, яка тривалий час (від кількох годин до
кількох тижнів) перебуває у рухливому стані. Найпоширенішими пред-
ставниками порядку є бутпосііпіит, Регісііпіит та Оіпоріїузіз.
Рід гімнодиніум - бутподіпіит (рис. 41 А). Водорість має най-
простіший план будови. Вона одноклітинна, монадна, при розгляді в
оптичний мікроскоп здається голою, хоча насправді вкрита м’якою
амфієсмою. Від близьких родів відрізняється тим, що поперечна бо-
розенка розташовується приблизно в екваторіальній зоні клітини.
Всі види роду здатні до гетеротрофного живлення, що здійснюєть-
ся осмотрофним та фаготрофним способами. В останньому випадку
132
водорість утворює псевдоподії з характерним кошиком мікротрубо-
чок. Невелика кількість видів є облігатними гетеротрофами. Проте
переважна більшість видів - міксотрофи, що мають хлоропласти, за-
барвлена у жовтуватий або коричнюватий колір. Деякі представники
містять хлоропласти інших типів. Так, у забарвленого в синій колір
гімнодиніуму синювато-зеленого (бутпосііпіит авгидіпозит)
хлоропласт утворився внаслідок ендосимбіозу з криптофітовою во-
дорістю. Гімнодиніум зелений - в. уігісіє (=ЬерісІо(1іпіит уігісіє) -
забарвлений у зелений колір і має хлоропласт, попередником якого
була зелена водорість.
Види роду населяють переважно морський планктон, частина
видів мешкає у прісних та солонуватих водоймах.
Рис. 41. Перидиніальні водорості: А - Сутпосііпіит; Б - Регісііпіит;
В - СіпорГіузіз. 1 - поперечна борозенка; 2 - поздовжня борозенка; 3 - хлоро-
пласт; 4 - ядро; 5 - краплина олії; 6 - вічко; 7 - платівки теки; 8 - комірець;
9 - криловидний виріст
133
Рід перидиніум - Регісііпіит (рис. 41 Б) - має добре розвине-
ну теку. Вона складається з двох частин - верхньої (епівальви) та
нижньої (гіповальви). Епівальва утворена зрослими апікальними, про-
міжними та приекваторіальними, гіповальва - приекваторіальними
та антапікальними платівками амфієсми. В зоні стикання платівок тека
утворює грубі виразні шви. Епівальва та гіповальва розділені попе-
речною борозенкою, що проходить в екваторіальній площині клітини.
Поздовжня борозенка розташовується на гіповальві і лише злегка
заходить на епівальву.
Переважна більшість видів роду має жовто-бурі хлоропласти ос-
новного типу. В деяких видів, наприклад, перидиніуму балтійсько-
го (Регісііпіит Ьаііісит), хлоропласти виникли внаслідок ендосимб-
іозу із золотистою водорістю. Незначна кількість видів живиться ви-
ключно гетеротрофно.
Види роду Регісііпіит мешкають в основному у морському планк-
тоні, і лише біля 15% видів зустрічаються у континентальних прісних
водоймах.
Рід динофізис - Оіпоргіузіз (рис. 41 В) - також об’єднує види з
міцною текою. Проте у цих водоростей поперечна борозенка зміще-
на майже на апекс, а клітина дуже стиснута з боків. Тека навколо
поперечної борозенки утворює лійкоподібний комірець, а по боках
поздовжньої - великі крилоподібні вирости. Види Оіпоріїузіз мешка-
ють у планктоні морів холодного, помірного та субтропічного поясів.
Порядок Пророцентральні - Ргогосепїгаїез
Характеризується наявністю двостулкової теки, поділом клітин
виключно у поздовжньому напрямку, передніми джгутиками майже
однакової довжини. Типовий рід порядку - Ргогосепігит.
Рід пророцентр - Ргогосепігит (рис. 42) - має монадні кліти-
ни з міцною текою, що складається з двох платівок, з’єднаних між
собою як шкаралупки горіха. На верхівці клітини знаходиться апі-
кальна заглибина. Її дно вкрите кількома дрібними платівками, дві з
яких пронизані джгутиковими порами. Із джгутикових пор виходять
два гетероморфні та майже ізоконтні джгутики, направлені вперед.
Проте ультратонка будова джгутиків та їх коренева система відповідає
типовому плану будови динофіцієвих водоростей. У цитоплазмі міс-
титься одне динокаріонтичне ядро та жовто-коричневі хлоропласти.
134
Поділ клітини відбувається у поздовжньому напрямку, причому
дочірні клітини, подібно до Сопіаиіасаіез, отримують половину теки
від материнської клітини, а половину, якої не вистачає, добудовують.
Рід об’єднує більше 100 видів морських планктонних водорос-
тей. Деякі з них викликають токсичне «цвітіння» у морях та приморсь-
ких лиманах. Зокрема, у Чорному морі небезпечним збудником
«цвітіння» є Ргогосепігит тісапз - пророцентр блискучий.
Рис. 42. Ргогосепігит: А - вигляд з широкого боку; Б - вигляд з вузького
боку. 1 - шип; 2 - джгутик; 3 - пузула; 4 - ядро; 5 - з’єднувальний шов
135
Водорості платикристати
Платикристати (Ріаіусгієїаіеє) є провідним царством у системі
евкаріот та в органічному світі в цілому. Представники царства ха-
рактеризуються наявністю пластинчастих (інколи-також трубчастих)
неперешнурованих при основі мітохондріальних крист. До царства
входять три підцарства - РІапіае, Апітаїіа та Гипді.
Підцарство рослин (Ріапіае) об’єднує платикристат, здатних до
оксигенного фотосинтезу, та похідні від них вторинно гетеротрофні
організми. В межах підцарства відділи НаріорЬуіа та СгуріорЬуіа ма-
ють вторинно симбіотичні пластиди. Іншим представникам підцар-
ства, до яких належать три відділи водоростей (ОІаисосузІорЬуІа,
РїіосіорЬуІа та СИІогорИуіа) та всі вищі рослини, притаманні пласти-
ди первинно симбіотичного типу.
Відділ Гаптофітові водорості - НаріорЬуіа
Евкаріотичні фотоавтотрофні та вторинно гетеротрофні плати-
кристати, що мають мітохондрії з трубчастими кристами (проте не
перетягнутими при основі), клітинний покрив, представлений плаз-
малемою, яка із зовнішнього боку вкрита субмікроскопічними органо-
мінеральними лусочками, а із внутрішнього підстелена оперезуючою
цистерною ендоплазматичної сітки. Пластиди вторинно симбіотичні,
родофітного типу. В багатьох представників є джгутикоподібна струк-
тура - гаптонема.
Відділ нараховує біля 500 видів з 80 родів. Переважна більшість
гаптофітових - це морські одноклітинні організми. Викопні гаптофі-
тові утворили основні відклади крейди.
Біохімічні ознаки. Хлоропласти гаптофітових звичайно забарв-
лені у жовтий або коричневий колір завдяки наявності хлорофілів а
та с, які маскуються жовтими ксантофілами: фукоксантином та його
дериватами. З каротинів виявлено лише р-каротин. Основний про-
дукт асиміляції - хризоламінарин (у павловальних - парамілон), що
відкладається поза межами пластид.
Цитологічні особливості. Клітини вкриті плазмалемою, на по-
верхні якої знаходиться один чи кілька шарів субмікроскопічних це-
люлозних лусочок або гранул, а з боку цитоплазми розташовується
одна велика оперезуюча цистерна ендоплазматичної сітки, що утво-
136
рює два додаткові внутрішні шари покривних мембран. У деяких видів
лусочки просякуються вапном, видозмінюючись у органомінеральні
покривні елементи, що отримали назву коколітів.
Ядро має евкаріотичну будову. Зовнішня ядерна мембрана без-
посередньо переходить у зовнішню мембрану хлоропласта. Зазви-
чай ядерце не спостерігається. Центріолі відсутні, а їх функцію вико-
нує система пухирців комплексу Гольджі або (у водоростей з порядку
Рауіоуаіез) електронно-щільні структури, асоційовані із джгутикови-
ми коренями. Мітоз відкритий, напівзакритий або закритий.
Пластиди представлені вторинно симбіотичними родопластами.
Вони вкриті чотирма мембранами, з яких дві зовнішні складають хло-
ропластну ендоплазматичну сітку, а дві внутрішні - власне хлоро-
пластну оболонку. Зовнішня мембрана безпосередньо продовжуєть-
ся у зовнішню мембрану ядра. Тилакоїди зібрані по три, оперезуючої
ламели немає. Досить часто наявний голий піреноїд.
Мітохондрії мають трубчасті кристи, які, на відміну від тубулокри-
стат і подібно до інших платикристат, не перешнуровані при основі.
Монадні стадії (вегетативні клітини, зооспори, гамети) мають два
джгутики рівної або нерівної довжини, що виходять із субапікальної
або латеральної заглибини. Джгутики гладенькі або вкриті простими
мастигонемами.
Крім джгутиків, у частини гаптофітових водоростей є унікальна
джгутикоподібна нерухома органела - гаптонема. Вона являє собою
виріст плазмалеми, в якому розташована одна цистерна ендоплаз-
матичної сітки, що обгортає 6-7 мікротрубочок.
Між базальними тілами джгутиків та ядром розташовується один
великий комплекс Гольджі, який, на відміну від водоростей-страме-
нопілів, не притиснутий до ядерної мембрани.
План будови тіла. Переважна більшість гаптофітових мають мо-
надну будову. Рідше зустрічаються кокоїдні форми. Гаметоспорофіти
деяких примнезіальних, ізохризидальних та коколітальних водоростей
мають нитчастий тип структури. За несприятливих умов більшість видів
Наріоріїуіа здатні втрачати джгутики та переходити у пальмелеподіб-
ний стан.
Розмноження та життєві цикли. Гаптофітові розмножуються
переважно вегетативним поділом клітин надвоє, зрідка - зооспора-
ми. У кількох видів описаний ізогамний статевий процес. У більшості
життєвий цикл являє собою цикломорфоз. У небагатьох еугамних видів
137
життєвий цикл гаплодиплофазний, з гетероморфною зміною поколінь
та споричною редукцією.
Система відділу. У межах відділу виділяють лише один клас -
НаріорЬусеае, який поділяють на чотири порядки - Ргутпезіаіез,
ІзосЬгузісіаІез, СоссоІИЬаіез, Рауіоуаіез. В основі поділу на порядки
лежить план будови клітин, особливості будови клітинних покривів та
наявність гаптонеми.
Порядок Примнезіальні - Ргутпезіаіез
Об’єднує одноклітинні та колоніальні монадні та амебоїдні рівно-
джгутикові водорості з довгою або короткою гаптонемою. Коколіти
відсутні, на поверхні плазмалеми є лише мікроскопічні целюлозні
лусочки. Представники порядку мешкають як у морському планктоні,
так і у прісних континентальних водоймах. Деякі примнезіальні водо-
рості здатні зумовлювати токсичне «цвітіння» води. Найпоширеніши-
ми є монадні водорості, в першу чергу - види родів Сіїгузосіїготиііпа
та Ргутпезіит.
Рід хризохромуліна - СНгузосИготиІіпа (рис. 43 А). Водорість
дрібна, одноклітинна, з двома майже рівними апікальними джгутика-
ми, довжина яких більш-менш дорівнює клітині, та довгою гаптоне-
мою, яка набагато довша за джгутики. У клітині при оптичній мікро-
скопії помітні два парієтальних жовтих або коричнюватих хлороплас-
ти, між ними - одне ядро, на задньому кінці - від однієї до кількох
гранул хризоламінарину.
На прикладі цього роду було відкрито токсичні гаптофітові водо-
рості. Зокрема, перше токсичне «цвітіння» води у морях, обумовлене
розвитком гаптофітових, зареєстроване наприкінці 80-х років XX ст. Воно
охопило узбережжя Норвегії та Швеції, тривало протягом майже двох
місяців і спричинило масову загибель лососевих риб на морських
фермах та у рибоводних господарствах. Збудником «цвітіння» вия-
вилася СІігузосІїготиІіпа роїуіеріз.
Рід примнезіум - Ргутпезіит (рис. 43 Б, В). Представники цьо-
го роду мають еліпсоїдні косо зрізані та дещо стиснуті клітини, на
передньому кінці яких розташовуються два довгих ізоконтних джгу-
тики та коротка пряма гаптонема. За допомогою гаптонеми водорість
здатна на деякий час прикріплюватися до субстрату. Клітини містять
по два пристінних бічних хлоропласти з піреноїдом або без нього.
138
В передній частині клітини розташовані дві пульсуючі вакуолі. Запас-
на поживна речовина - хризоламінарин. Він відкладається в задній
частині клітини у вигляді одного великого або багатьох дрібних зер-
няток. Розмноження відбувається шляхом поздовжнього поділу клітини
в рухливому стані.
Деякі види примнезіуму відіграють негативну роль як у природі,
так і в господарській діяльності людини. Один з видів роду - примне-
зіум дрібний (Ргутпезіит рагуит) - є збудником небезпечного ток-
сичного «цвітіння» солонуватих континентальних водойм та естуаріїв.
При масовому розвитку він виділяє у воду суміш протеофосфоліпідів,
що отримала назву іхтіотоксину. Іхтіотоксин після активації іонами
натрію, магнію або кальцію змінює проникність зябер і здатний спри-
чинювати масову загибель риб. Спалахи «цвітіння» Ргутпезіит за-
реєстровані у Болгарії, Ізраїлі, Данії, Нідерландах.
Рис. 43. Примнезіальні водорості: А - СЬгузосЬготиІіпа; Б, В - вигляд кліти-
ни Ргутпезіит в різних положеннях. 1 - гаптонема; 2 - вакуоля з хризоламіна-
рином; 3 - хлоропласт
Порядок Ізохризидальні - ІзосЬгузісІаІез
Об’єднує одноклітинні та колоніальні монадні та кокоїдні водорості,
що не мають гаптонеми і не утворюють коколітів. Монадні клітини ізох-
ризидальних мають однакові або злегка нерівні джгутики. Мешкають
переважно в морях. Характерним родом є хризотіла (СЬгузоІПа).
Порядок Коколітальні або Коколітофориди - Соссоїііііаіез
Об’єднує монадні або кокоїдні водорості, що позбавлені гаптоне-
ми, проте мають кальцифіковані поверхневі лусочки - коколіти. По-
139
ширені переважно в морях. Коколітофориди панували наприкінці
мезозою, у кайнозої більшість видів вимерла. Із сучасних родів ха-
рактерним монадним представником є РІеигосіїгузіз, кокоїдний тип
структури властивий роду Етіїіапіа.
Рід плеврохризис - РІеигосіїгузіз (рис. 44) - має складний жит-
тєвий цикл з гетероморфною зміною поколінь. Спорофіт, відомий як
стадія «Нутепотопаз», представлений монадними диплоїдними
клітинами, які вкриті коколітами, розмножуються поділом надвоє або
за допомогою зооспор. За певних умов відбувається мейоз, внаслі-
док якого утворюються чотири гаплоїдні зооспори, що проростають у
гаплоїдний розгалужений нитчастий гаметоспорофіт, раніше описа-
ний як самостійний рід Арізіопета. Гаметоспорофіт відтворює себе
зооспорами або дає ізогамети. Після копуляції зигота без періоду спо-
кою проростає у монадну диплоїдну коколітофориду. Зооспори, які
продукує спорофіт, мають коротку, дуже редуковану гаптонему; у зоо-
спор гаметоспорофіту гаптонема може бути наявна або відсутня.
Рис. 44. Схема життєвого циклу РІеигосіїгузіз сагіегае
Рід еміліанія - Етіїіапіа (рис. 45). Один з небагатьох представ-
ників сучасних кокоїдних коколітофорид. Клітини водорості сферичні,
вкриті великими, дуже кальцифікованими коколітами. Еміліанія по-
ширена в морях, переважно помірного та теплого поясів.
Сьогодні встановлено, що масовий розвиток еміліанії впливає на
погоду та клімат. Такий вплив пов’язаний із здатністю клітин цієї во-
140
дорості в процесі метаболізму виділяти в атмосферу леткий диме-
тилсульфід. Ця речовина після фотохімічної трансформації у верхніх
шарах атмосфери утворює в хмарах центри конденсації вологи, що
призводить до випадіння дощів.
Рис. 45. Клітина Етіїіапіа з коколіта-
ми
Порядок Павловальні
- Рауіоуаіез
Об’єднує монадні водо-
рості, в яких наявні два джгу-
тики різної довжини (обидва
- з простими мастигонема-
ми), стигма, парамілон,
особливі «шишкоподібні» ко-
коліти, редукована гаптоне-
ма. До порядку входить
лише один рід - павлова
(Рау/оуа), поширений у
планктоні морів та естуаріїв.
Відділ Криптофітові водорості - СгурІорЬуІа
Евкаріотичні фотоавтотрофні та вторинно гетеротрофні плати-
кристати, що мають клітинний покрив, представлений перипластом.
Пластиди вторинно симбіотичні, родофітного типу, з нуклеоморфом.
Відділ нараховує близько 200 видів одноклітинних мікроскопічних
монадних водоростей, поширених як у континентальних водоймах,
так і в морях.
Біохімічні особливості
Для фотоавтотрофних криптофітових характерні хлорофіли а та
с, а- та є-каротини, фікобілінові пігменти - фікоціанін та фікоеритрин,
ксантофіли лютеїнового ряду (лютеїн, віолаксантин, зеаксантин) та
специфічні ксантофіли (алоксантин, крококсантин, монадоксантин).
Кількісні співвідношення пігментів у різних родів варіюють у широко-
му діапазоні. Це обумовлює велику різноманітність забарвлення крип-
тофітових водоростей. Так, хлоропласти можуть бути червоними,
блакитними, жовтими, коричневими, зеленими. Вторинно гетеро-
трофні представники безбарвні.
Основний продукт асиміляції - крохмаль, який відкладається у
перипластидному просторі, часто - поблизу піреноїда. Додатковий
асимілят - олія.
141
Цитологічні особливості. Клітини криптофітових звичайно дор-
зовентральні, на передньому кінці косо зрізані, мають глибоку чи не-
велику глотку, два гетероконтні джгутики, що відходять від дорзаль-
ного боку глоткового отвору. При основі глотки у прісноводних видів
є одна-дві, рідше п’ять-вісім скоротливих вакуолей. На вентрально-
му боці розташовується вузька поздовжня борозенка.
Клітинні покриви представлені перипластом. Він складається з
плазмалеми, яка з внутрішнього боку несе прямокутні чи полігональні
білкові платівки, що прикріплюються до плазмалеми за допомогою
білкових шипів. На зовнішньому боці плазмалеми зазвичай є шар
зовнішніх білкових платівок, що за формою повторюють внутрішні.
Платівки утворюють правильні ряди, які надають клітинним покри-
вам криптофітових характерної поздовжньої посмугованості. Під пе-
рипластом знаходяться трихоцисти. Великі трихоцисти звичайно ви-
стилають глотку, дрібні - розсіяні під усією поверхнею клітини.
Ядро розташовується в задній частині клітини, має досить вели-
ке ядерце. Зовнішня ядерна мембрана безпосередньо переходить у
зовнішню мембрану хлоропласта. Центріолі відсутні, їх функцію ви-
конують базальні тіла джгутиків та ризопласти. Хромосоми в інтер-
фазі перебувають у конденсованому стані. Мітоз відкритий.
Зазвичай у клітині є один-два великі пристінні хлоропласти, вкриті
чотиримембранною оболонкою. Дві зовнішні мембрани утворюють
хлоропластну ендоплазматичну сітку. Зовнішня мембрана хлоропла-
стного ендоплазматичного ретикулюму безпосередньо переходить у
зовнішню мембрану ядра. Перипластидний простір між двома зовніш-
німи та двома внутрішніми мембранами хлоропласта містить крох-
маль та особливу органелу - нуклеоморф.
Від перипластидного простору нуклеоморф відмежований по-
двійною мембраною, яка, подібно до ядерної, має пори та містить
ДНК і РНК.
Тилакоїди у хлоропласті зібрані парами. На внутрішніх боках кож-
ної пари локалізовано фікобілінові пігменти, які, проте, не організо-
вані у фікобілісоми. Більшість криптофітових мають великий піреноїд
напівзануреного типу. В зоні перипластидного простору, що безпосе-
редньо прилягає до піреноїда, відкладається крохмаль. У деяких крип-
тофітових у хлоропласті є вічко, положення якого не скоординоване
із джгутиковим апаратом. Вторинні гетеротрофи безбарвні, але збе-
рігають редуковану пластиду - лейкопласт.
Мітохондріальний апарат представлений двома великими міто-
хондріями - дорзальною та вентральною. Обидві мають пластинчасті
кристи.
142
Монадні клітини дводжгутикові, джгутики злегка гетероконтні, спря-
мовані вперед. На довшому джгутику мастигонеми розташовуються
у два ряди, на коротшому вони однорядні. У перехідній зоні джгутики
мають по дві поперечні платівки.
Специфічними органелами криптофітових є тільця Мопа - великі,
добре помітні одномембранні структури, що знаходяться поблизу глот-
ки і містять переважно залишки трихоцист. Припускають, що тільця
Мопа є гомологами лізосом і їх функція полягає в перетравленні над-
лишкових трихоцист. Звичайно між глоткою та тільцями Мопа розта-
шовується один комплекс Гольджі.
Типи структури тіла. Майже всі криптофітові водорості мають
монадний тип структури тіла. Переважна більшість видів здатна втра-
чати джгутики, покриватися шаром слизу і переходити у пальмеле-
видний стан.
Розмноження та життєві цикли. Розмноження відбувається по-
ділом клітин надвоє у поздовжньому напрямку в рухливому стані. В од-
ного виду (Ргоіеотопаз зиісаіа) описаний гаплодиплофазний життє-
вий цикл з гетероморфною зміною поколінь. У Сіїгоотопаз асиіа
спостерігався хологамний статевий процес.
Система відділу. Всі криптофітові належать до одного класу -
СгуріорИусеае, та одного порядку - Сгуріогпопасіаіез. Характерними
представниками є роди Сгуріотопаз, Ріюсіотопаз та Сіїііотопаз.
Рід криптомонас - Сгуріотопаз (рис. 46). Найчисельніший рід
відділу, налічує більше 50 видів. Клітини криптомонасу латерально
стиснуті, дорзовентральні, більш-менш метаболічні, з опуклим дор-
зальним та плоским або злегка ввігнутим вентральним боками. Спе-
реду клітини косо зрізані, із заглибиною, на дорзальній стороні мають
випин - рострум. Перипласт поздовжньо посмугований або має ви-
гляд скошеної сіточки. По вентральному боку клітини від заднього кінця
до переднього проходить поздовжня борозенка, яка біля роструму у
вигляді сліпої трубки входить у середину клітини, утворюючи широку
глотку. Глотку вистилають трихоцисти.
Між отвором глоткового каналу та рострумом з клітини виходять
два нерівних, спрямованих вперед джгутики. Вздовж клітини розмі-
щуються один-два пристінних великих, часто лопатевих хлороплас-
ти, забарвлених переважно в оливковий або коричневий колір. В се-
143
Рис. 46. Сгуріотопаз: будова клітини.
1 - джгутики; 2 - рострум; 3 - пульсуюча ва-
куоля; 4 - тільця Мопа; 5 - піреноїд; 6 - глот-
ка з трихоцистами; 7 - хлоропласт; 8 - ядро;
9 - поздовжня борозенка
пові планктери, населяють переважно
ваються серед нитчастих водоростей
редніи частині хлоропласта,
як правило, знаходиться один
великий піреноїд. У перипла-
стидному просторі навколо
піреноїду у вигляді шкаралу-
пок відкладається крохмаль.
Вічко у більшості видів від-
сутнє. Ядро кулясте, знахо-
диться, як правило, в задній
частині клітини. Скоротлива
вакуоля одна, зрідка дві, від-
кривається в борозенку.
Водорість розмножується
поздовжнім поділом. Поділ
відбувається або у рухливому
стані, або йому передує пе-
рехід у пальмелевидний
стан. При несприятливих умо-
вах клітини утворюють цис-
ти, вкриті товстим шаром ущ-
ільненого слизу.
Деякі види роду дуже по-
ширені, найчастіше трапля-
ються в забруднених водой-
мах, де можуть викликати
«цвітіння» води. Беруть актив-
ну участь у процесах само-
очищення забруднених орга-
нічними речовинами вод. Ти-
придонні шари води чи розви-
або в заростях вищих водних
рослин.
Більшість представників роду мешкає у прісних водоймах. Винят-
ки нечисленні. Наприклад, Сгуріотопаз заііпа та С. зіідтаіісає гало-
філами і часто зустрічаються у планктоні солоних озер при концент-
рації солей 40-90 г/л, беручи участь в утворенні лікувальних мулів -
пелоїдів; Сгуріотопаз сгуоріїііа та С. ігідогіз - кріофіли, які масово
розмножуються на поверхні снігу в горах та зумовлюють його бруд-
но-червоне та оливково-зелене «цвітіння».
144
Рід родомонас - Кіїосіотопаз. Водорість дуже, схожа з крип-
томонасом, від якого відрізняється забарвленням хлоропластів та роз-
ташуванням трихоцист у глотці. Хлоропласт один, від іржасто-черво-
ного до темно-червоного кольору, з піреноїдом. Глотка звичайно з
двома поздовжніми рядами трихоцист, які вистилають лише задню її
половину. Трихоцисти можна добре роздивитись після забарвлення
клітини розчином Люголю.
Більшість видів роду є холодолюбними планктонними організма-
ми, поширеними як у прісних водоймах, так і в морях. Трапляються
переважно поодиноко, і є індикаторами чистих вод.
Рід хіломонас - Сіїііотопаз - за морфологією клітин схожий з
криптомонасом та родомонасом, від яких відрізняється облігатно ге-
теротрофним живленням. Хлоропласт відсутній, замість нього у клітині
є безбарвний лейкопласт, часто - з піреноїдом, навколо якого у пе-
рипластидному просторі зрідка може відкладатись крохмаль. Крім
того, в цитоплазмі є багато великих або дрібних крапель олії. Види
цього роду мешкають переважно в континентальних стоячих водой-
мах, забруднених органічними речовинами.
Відділ Глаукоцистофітові водорості - бІаисосузіорИуіа
Евкаріотичні фотоавтотрофні платикристати, що мають амфієс-
моподібний клітинний покрив. Пластиди первинно симбіотичні, глау-
коцистофітного типу (ціанели). Між зовнішньою та внутрішньою мем-
бранами оболонки пластиди зберігається шар муреїну. Продукт аси-
міляції- крохмаль, що відкладається у цитоплазмі.
Нараховує близько 10 видів вільноіснуючих мікроскопічних одно-
клітинних водоростей, поширених у прісних континентальних водой-
мах. Викопні рештки глаукоцистофітових поки що не знайдені.
Найяскравіша особливість відділу - це наявність унікального
фотосинтетичного апарату, представленого ціанелами - видозміне-
ними симбіотичними синьозеленими водоростями, які втратили
здатність до самостійного існування поза межами клітини-господаря.
Біохімічні особливості. Пігменти представлені хлорофілом а,
р-каротином, фікобілінами (фікоціаніном та алофікоціаніном), р-крип-
токсантином. Завдяки великій кількості синіх фікобілінів ціанели глау-
коцистофітових забарвлені у блакитно-зелений колір. Продукт асим-
іляції- крохмаль, який відкладається безпосередньо у цитоплазмі.
145
Цитологічні особливості. Клітини вкриті амфієсмоподібною пе-
лікулою: під плазмалемою розташовуються сплощені мембранні пу-
хирці - лакуни, всередині яких можуть знаходитись тонкі платівки.
Лакуни з боку цитоплазми підстилаються мікротрубочками. У деяких
представників (наприклад, у роді Суапоріусіїе) із зовнішнього боку
амфієсмоподібної пелікули є клітинна оболонка, утворена сотоподіб-
но розміщеними субодиницями, над якими розташовується кілька
шарів мікрофібрил поки що нез’ясованої природи.
Ядерний апарат евкаріотичний. Всередині ядра міститься вели-
ке ядерце. Цікавою особливістю ядра є наявність в ньому пучків мікро-
філаментів.
Мітохондрії глаукоцистофітових розгалужені, на зрізах вигляда-
ють як численні невеликі пухирці з пластинчастими кристами.
Фотосинтетичний апарат являє собою ціанели і не має аналогів
серед евкаріотичних водоростей. Кожна ціанела відмежована від
цитоплазми однією цитоплазматичною мембраною. Під цією мемб-
раною знаходиться тонкий електронно-щільний шар, утворений му-
реїном. Із внутрішнього боку цього шару розташовується ще одна мем-
брана. Всередині ціанели концентричними колами розміщуються по-
одинокі тилакоїди, на поверхні яких є численні фікобілісоми. У цент-
ральній частині ціанели знаходиться одне велике або багато дрібних
поліедральних тілець, утворених рибульозо-дифосфат-карбоксила-
зою.
Ціанели мають свою власну ДНК, яка замкнена в кільце та не
пов'язана з гістонами. Встановлено, що їх геном містить приблизно в
10 разів менше нуклеотидів, ніж геном синьозелених водоростей.
Монадні стадії глаукоцистофітових дводжгутикові, дорзовент-
ральні, можуть бути представлені як вегетативними клітинами, так і
зооспорами. Джгутики гетероконтні та гетеродинамічні, виходять із
субапікального заглиблення, яке є початком поздовжньої борозенки.
Коротший джгутик направлений вперед і є рушійним. Довший - ле-
жить у поздовжній борозенці і виконує функцію стерна. Кожний джгу-
тик має два ряди простих мастигонем. Стигма відсутня.
Плани будови тіла. Хоча сьогодні відомо лише чотири роди гла-
укоцистофітових, проте вони представляють два типи морфологіч-
них структур: монадний та гемімонадний.
Розмноження та життєві цикли. Монадні глаукоцистофітові во-
дорості розмножуються вегетативним поділом клітини надвоє в рух-
ливому стані. Поділу клітини передує поділ ціанел. Представники з
146
гемімонадною будовою можуть розмножуватись простим поділом
клітин надвоє та за допомогою зооспор. Статеве розмноження у гла-
укоцистофітових не виявлене. Життєвий цикл являє собою цикло-
морфоз.
Система відділу. Відділ включає один клас - СІаисосузіорИусеае,
і два порядки - Суапоріїогаїез та СІаисосузіаІез. Порядки розрізняють-
ся за типами морфологічної структури тіла представників.
Порядок Цианофоральні - СуапорЬогаІез
Об’єднує водорості з монадним типом морфологічної структури
тіла. Включає лише один рід - Суапоріїога.
Рід цианофора - Суапорїюга (рис. 47). Клітини цианофори по-
одинокі, з двома джгутиками трохи різної довжини: коротшим переднім
Рис. 47. Суапоріїога рагадоха:
1 - джгутики; 2 - пульсуюча вакуоля;
З - ціанела; 4 - карбоксисома; 5 - ядро;
6 - ядерце; 7 - гранули крохмалю
та довшим заднім. Останній
проходить у невеликій по-
здовжній борозенці. Біля ос-
нови джгутиків розташову-
ється одна пульсуюча вакуо-
ля. В цитоплазмі добре по-
мітні дві великі синьозелені
еліпсоїдні ціанели. В центрі
кожної ціанели є світла ділян-
ка, яка відповідає місцю роз-
ташування попередника
піреноїду - карбоксисомі.
Ближче до заднього кінця клі-
тини розміщується ядро. Про-
дукт асиміляції - крохмаль -
розташовується дифузно у
цитоплазмі у вигляді неве-
личких зерняток. Цианофора
розмножується вегетативним
поділом клітини надвоє у рух-
ливому стані. Види роду роз-
виваються переважно у
планктоні ефемерних во-
дойм, забруднених органіч-
ною речовиною.
147
Дослідження геному цианофори відіграли важливу роль у фор-
муванні сучасних уявлень про принципові відміни пластид від фото-
автотрофних ендосимбіонтів на геномному рівні. Внаслідок секвену-
вання ДНК ціанел Суапоріїога рагасіоха було встановлено, що розмір
геному ціанел майже співпадає з розміром геному хлоропластів інших
евкаріот і є в середньому в 10 разів меншим від геному СуапорИуіа.
Крім того, у геномі ціанел було відкрито дві ділянки т. зв. інвертова-
них послідовностей нуклеотидів. Ці ділянки являли собою гени, що
кодують хлоропластну рибосомальну РНК. Транскрипція цих фраг-
ментів відбувається у зворотніх напрямках (тобто інвертовано). Інвер-
товані послідовності, що кодують хлоропластну рибосомальну РНК,
було виявлено також у хлоропластній ДНК вищих рослин, проте не
знайдено у синьозелених водоростей. З цього часу наявність інвер-
тованих послідовностей та розмір геному в межах 70-200 тис. пар
нуклеотидних основ вважають ознакою пластиди, а відсутність таких
інвертованих послідовностей та більший розмір геному - ознаками
ендосимбіотичного фотоавтотрофного організму, навіть у тих випад-
ках, коли геном останнього частково редукований і ендосимбіонт не
може мешкати за межами клітини-господаря.
Порядок Глаукоцистальні - СІаисосузїаІез
Включає водорості з гемімонадним типом морфологічної струк-
тури тіла, в яких зовні від амфієсмоподібної пелікули є додаткові за-
хисні утворення - слизові капсули або оболонка. До порядку входить
три роди, з яких нижче розглядається типовий представник - СІаи-
сосузііз.
Рід глаукоцистіс - біаисосузііз (рис. 48). Об’єднує мікроскопічні
гемімонадні водорості, що утвоюють невеличкі слизові колонії. Кож-
на колонія складається з 2-4-8 клітин широкоеліпсоїдної форми, інко-
ли має вигляд системи вкладених один в одного слизових міхурів.
Клітини глаукоцистісу вкриті амфієсмоподібною пелікулою, над
якою знаходиться клітинна оболонка. Між цією оболонкою та пеліку-
лою на латеральному боці клітини є невеличка порожнина, в яку ви-
ходять два дуже редуковані джгутики. Біля їх базальних тіл розташову-
ється пульсуюча вакуоля. Поблизу джгутиків розміщується ядро. У
цитоплазмі знаходяться 8-16 синьозелених видовжених ціанел. Ціа-
нели з’єднуються між собою одним із полюсів, утворюючи один або
два зірчастих комплекси, по 4-8 ціанел у кожному. В цитоплазмі роз-
ташовуються також численні дрібні зернятка крохмалю.
148
Водорість розмножується поділом клітини надвоє, а також дво-
джгутиковими зооспорами, які морфологічно подібні до вегетативних
клітин цианофори.
Представники роду вважаються рідкісними водоростями, які меш-
кають у континентальних прісних водоймах різного типу. На території
України знайдено лише один вид цього роду - глаукоцистіс носто-
ковидний (О. позіосМпеагит).
Рис. 48. ОІаисосузііз позіосіїіпеагит: А - окрема вегетативна клітина;
Б - колонія. 1 - ціанели; 2 - ядро; 3 - клітинна оболонка; 4 - амфієсмоподібна
пелікула
Відділ Червоні водорості - КНодорЬуіа
Евкаріотичні фотоавтотрофні платикристати, клітини яких вкриті
целюлозно-пектиновою оболонкою з фікоколоїдами. Пластиди пер-
винно симбіотичні, родофітного типу (родопласти), з поодинокими
тилакоїдами. Продукт асиміляції - багрянковий крохмаль, що відкла-
дається у цитоплазмі. Специфічна ознака - повна відсутність джгути-
кових стадій.
Відділ нараховує біля 5000 видів переважно багатоклітинних мак-
роскопічних водоростей. Майже всі червоні водорості є мешканцями
бентосних та перифітонних морських угруповань, і, лише як виняток,
деякі види зустрічаються у прісних водоймах та у ґрунтах. Викопні
рештки ККосІорИуІа відомі з раннього палеозою (кембрій, 600 млн.
років тому).
149
Біохімічні особливості. Клітини забарвлені переважно у черво-
ний колір, значно рідше - у сірий або синьозелений (як правило, у
водоростей, що мешкають у прісних водоймах або у ґрунтах). Забар-
влення визначається, в першу чергу, наявністю хлорофілу а та фіко-
білінових пігментів - фікоціаніну, аллофікоціаніну та фікоеритрину.
Звичайно фікоеритрин переважає над двома іншими фікобілінами.
Каротиноїди представлені а- та р-каротинами, криптоксантином, та-
раксантином, зеаксантином та лютеїном.
Основний продукт асиміляції-багрянковий крохмаль. У примітив-
них бангієвих водоростей продуктом асиміляції є глікоген. Додатко-
вими запасними поживними речовинами можуть бути галактозид,
флоризид, багатоатомні спирти (маніт), у незначній кількості - олія.
Всі продукти асиміляції відкладаються поза межами хлоропластів.
Цитологічні особливості. Клітинні покриви представлені триша-
ровими пектиновими та целюлозно-пектиновими оболонками.
Зовнішній шар - кутикула - утворений згущеною полімеризованою
манозою; середній шар - фікоколоїдами (агар-агаром, агароїдом,
карагеніном) та пектиновими речовинами. Внутрішній шар складаєть-
ся з целюлози. Примітивні бангієві водорості целюлозного шару поз-
бавлені.
Зв’язок між клітинами відбувається за допомогою плазмадесм,
що проходять крізь первинні та вторинні пори. Первинні пори утво-
рюються внаслідок неповного змикання країв поперечної перегород-
ки після закінчення поділу клітини і закриті спеціальними пробками.
Тип пробки є важливою ознакою на рівні порядків.
Ядерний апарат типово евкаріотичний. Мітоз закритий, ядерна
оболонка у профазі не зникає, а перфорується. Центріолі відсутні, а
їх функцію виконують особливі білкові полярні кільця.
Пластиди червоних водоростей вкриті двомембранною оболон-
кою, тилакоїди поодинокі. На їх поверхні зосереджені фікобілісоми,
будова яких подібна до фікобілісом синьозелених водоростей.
Джгутикові стадії у червоних водоростей повністю відсутні.
Плани будови талому. В межах відділу представлено кокоїд-
ний, нитчастий та гетеротрихальний типи структур. Кокоїдні форми
нечисленні, зустрічаються серед найпримітивніших бангієфіцієвих
водоростей. Таломи водоростей з нитчастим типом структури можуть
мати ниткоподібну або пластинчасту форму, бути одно- або багато-
рядними. Найпоширеніші в межах відділу представники з гетеротри-
150
хальним типом морфологічної будови тіла. Ріст таких водоростей
відбувається переважно за рахунок поділу апікальної клітини. Бічні
нитки-асимілятори інколи щільно зростаються між собою, утворюючи
псевдопаренхіматозні структури. Залежно від кількості опірних ниток
гетеротрихальні таломи поділяють на одноосьові та багатоосьові
(фонтанні).
Розмноження та життєві цикли. Червоні водорості розмножу-
ються нестатевим та статевим шляхами. Нестатеве розмноження
відбувається вегетативно (поділом клітини надвоє, фрагментацією
багатоклітинних таломів та додатковими «пагонами», які можуть відро-
стати від сланких ниток або підошви) та за допомогою спор.
В останньому випадку розмноження здійснюється переважно
моноспорами та тетраспорами. Спори червоних водоростей позбав-
лені оболонки і здатні до амебоїдного руху. При утворенні спор у спо-
рангіях, як правило, відбувається редукційний поділ. Таким чином,
спори гаплоїдні. Моноспори утворюються поодиноко, з поверхневих
(корових) клітин, внаслідок відокремлення частини протопласта кліти-
ни-спорангія косою перегородкою. Тетраспори утворюються по чоти-
ри; залежно від напрямку площини поділу виділяють хрестоподібні,
зональні та тетраедричні тетраспорангії. При проростанні тетраспо-
ри утворюють гаметофіти.
Статевий процес у червоних водоростей виключно оогамний.
У чоловічих статевих органах - сперматангіях - утворюється по од-
ному безджгутиковому спермацію, у жіночих статевих органах - кар-
погонах - по одній яйцеклітині.
Карпогон, як правило, складається із розширеної частини - че-
ревця, та трубкоподібного виросту - трихогіни, який вловлює спер-
мації. У найпростішому випадку карпогони утворюються з будь-якої
клітини талому, у складніших випадках - з верхівкової клітини спеці-
альних карпогоніальних ниток. Клітину, від якої відгалужується карпо-
гоніальна нитка, називають несучою (ініціальною). У багатьох черво-
них водоростей із класу флоридей ця клітина надалі трансформується
у спеціальну ауксилярну клітину, з якою карпогон зливається після
заплідення.
При статевому процесі спермації виходять із сперматангіїв, па-
сивно переносяться током води і, у випадку потрапляння на карпо-
гон, запліднюють яйцеклітину. Одразу після запліднення зигота без
періоду спокою проростає в особливе покоління, присутнє лише у
151
червоних водоростей - карпоспорофіт. На карпоспорофіті утворю-
ються карпоспори, які дають початок новому спорофіту.
Утворення карпоспор може відбуватися трьома основними шля-
хами, які обумовлюють три основні плани будови карпоспорофіта.
У найпростішому випадку зигота одразу починає ділитися, утво-
рюючи карпоспори безпосередньо у карпогоні. В цьому варіанті кар-
поспорофіт дуже простий і представлений карпогоном з карпоспо-
рами.
При складнішому варіанті від карпогону відростають диплоїдні
нитки із зиготичними ядрами - гонімобласти. Далі клітини таких гоні-
мобластів перетворюються на карпоспори. Тут покоління карпоспо-
рофіта представляють карпогон, гонімобласти та карпоспори.
Третій варіант найскладніший: карпогон із заплідненою яйцеклі-
тиною зливається з ауксилярною клітиною і утворює гетерокаріон-
тичну (різноядерну) клітину злиття. Остання містить одне або кілька
власних гаплоїдних «гаметофітних» ядер та диплоїдне зиготичне
ядро, що потрапило у клітину злиття із заплідненого карпогону. Дип-
лоїдне ядро кілька разів мітотично ділиться, дочірні ядра мігрують у
периферичні випини клітини злиття. Кожний випин відокремлюється
перегородкою і проростає в гонімобласт, клітини якого перетворю-
ються на карпоспори. В цьому варіанті карпоспорофіт складається з
карпогону, клітини злиття, ауксилярних клітин, гонімобластів та кар-
поспор.
Таким чином, життєвий цикл у Кїіоборїіуіа переважно гаплодип-
лофазний, зі споричною редукцією і чергуванням трьох поколінь -
гаметофіта, карпоспорофіта та спорофіта.
Система відділу. За будовою карпогону, хлоропластів, способа-
ми нестатевого розмноження, продуктами асиміляції та наявністю
вторинних пор відділ поділяють на два класи - Вапдіорїіусеае та
ПогібеорКусеае.
Клас Бангієфіцієві - ВапдіорЬусеае
Об’єднує водорості, в яких карпогон позбавлений трихогіни, кар-
поспори утворюються безпосередньо із зиготи, хлоропласти зірчасті
або стрічкоподібні, як правило - з піреноїдом; продуктом асиміляції,
крім багрянкового крохмалю, може бути також глікоген; нестатеве
розмноження відбувається переважно моноспорами; первинні пори
або позбавлені пробок, або мають прості пробки першого чи другого
типу; вторинні пори відсутні.
152
За типами морфологічної будови та розмноженням клас поділя-
ють на 3 порядки - РогрЬугісііаІез, Сотрзородопаіез та Вапдіаіез.
Порядок Порфіридіальні - РогрЬугісііаІез
Об’єднує кокоїдні водорості. Типовий представник порядку -
Рогріїугісііит.
Рід порфіридіум - Рогріїугісііит (рис. 49) - утворює слизові ко-
лонії. Клітини сферичні, з великим центральним хлоропластом, в
центрі якого знаходиться піреноїд. Водорість мешкає переважно у
прісних водоймах та у ґрунті. Види цього роду (зокрема, Рогріїугісііит
ригригеит) інтенсивно досліджуються у багатьох лабораторіях, ос-
кільки вважаються перспективними фікотехнологічними об'єктами для
промислового культивування з метою отримання фікоколоїдів.
Рис. 49. Рогріїугісііит ригригеит: А - слизова колонія; Б - окремі клітини
Порядок Компсопогональні - Сотрзородопаіез
Включає водорості з гетеротрихальним типом структури. Роз-
множуються моноспорами, статевий процес невідомий. Компсопо-
гональні водорості мешкають переважно у прісних водоймах тропіч-
ного поясу. Типовий представник - Сотрзородоп.
Рід компсопогон - Сотрзородоп (рис. 50) завезений в Україну
разом із тропічними акваріумними рослинами. Водорість добре відо-
ма акваріумістам під назвою «синя борода». Компсопогон розвиваєть-
ся як епіфіт на вищих водних рослинах, переважно - на криптокорі-
нах. Таломи мають вигляд розгалужених кущиків, диференційованих
153
на систему сланких ниток, занурених у тканини рослини, і висхідних
одно- та багаторядних ниток. Клітини висхідної частини мають довгі
пристінні стрічкоподібні хлоропласти. Окремі клітини після поділу ко-
сою перегородкою перетворюються на моноспорангії.
Рис. 50. Сотр8ородоіт. А - зовнішній вигляд талому; Б - базальна частина
талому із ризоїдальними нитками, зануреними у тканини рослини криптокоріни;
В - висхідна частина нитки; Г - фрагмент талому (показані клітини, що містять
стрічкоподібні хлоропласти, у багаторядній висхідній частині спостерігаються
поодинокі моноспорангії, що мають трикутно-заокруглену форму)
Порядок Бангіальні - Вапдіаіез
До цього порядку входять водорості, які мають нитчасті, як пра-
вило, багаторядні нерозгалужені або пластинчасті таломи. Розмно-
жуються бангіальні моноспорами (у деяких видів - поліспорами) та
додатковими «пагонами». Статевий процес відомий, мейоз зиготич-
ний, відбувається одразу ж після запліднення яйцеклітини. Представ-
ники порядку живуть переважно у морях. Найпоширенішими є роди
Вапдіа та Рогріїуга.
Рід бангія - Вапдіа (рис. 51 А) - має багаторядний нитчастий
нерозгалужений талом. Розростання бангії на камінні тендітні та
слизькі і нагадують червону повсть. До субстрату водорість при-
кріплюється ризоїдальними відростками. В верхівковій частині тало-
му розвиваються моноспорангії, сперматангії та карпогони. У вегета-
тивних клітинах бангії добре помітно один або два великих централь-
них зірчастих хлоропласти з голим піреноїдом.
154
Рис. 51. Бангіальні водорості: А - Вапдіа; Б - Рогрґіуга: 1 - базальна части-
на талому з ризоїдами; 2 - середня багаторядна частина талому; 3 - попереч-
ний зріз через талом в зоні розвитку моноспорангіїв; 4 - зовнішній вигляд
талому; 5-7 - поперечний зріз через пластину в зоні карпогонів (5), сперма-
тангіїв (6) та карпоспорангіїв (7)
Рід порфіра - Рогрїіуга (рис. 51 Б). Водорість має вигляд пла-
тівки рожево-пурпурного кольору із гладенькими або хвилястими кра-
ями, розміром від п’яти до кількох десятків сантиметрів завдовжки та
до 10-20 см завширшки. Платівка складається з одного-двох шарів
клітин і за допомогою коротенького черешка та підошви прикріплюєть-
ся до підводних субстратів. Клітини, подібно до бангії, містять один
або два зірчастих хлоропласти з піреноїдом.
Нестатеве розмноження відбувається за допомогою моноспор або
поліспор. При статевому розмноженні карпогони та сперматангії роз-
виваються з вегетативних клітин, розміщених по краях платівки.
У деяких видів карпогони розвивають коротку зачаткову трихогіну,
яка має вигляд невеличкого сосочка.
155
Види цього роду поширені як у північних, так і в південних морях
(зокрема, в Чорному морі), населяючи літораль та сублітораль. Жи-
вуть у прикріпленому стані на камінні, скелях, таломах великих морсь-
ких водоростей. У Чорному морі порфіра вегетує з листопада по тра-
вень.
Порфіра є їстівною водорістю (це відображено у її народній назві
- червоний морський салат) і введена у промислову аквакультуру.
Зокрема, порфіру вирощують у Чорному морі на спеціальних морсь-
ких фермах. Червоний морський салат вважається делікатесом, смак
якого обумовлюють вільні амінокислоти та ізофлоризиди.
Клас Флоридеєфіцієві - ЕІогібеорЬусеае
Включає водорості, в яких карпогон має трихогіну, карпоспори
утворюються різними шляхами у різних порядках, хлоропласти чис-
ленні, дископодібні, без піреноїдів. Продукт асиміляції - багрянковий
крохмаль (глікоген не утворюється). Первинні пори закрити пробка-
ми третього-сьомого типів (у порядку Аїіпїеіііаіез пробки прості, пер-
шого типу), клітини пов’язані між собою за допомогою як первинних,
так і вторинних пор. До класу входять водорості виключно з гетерот-
рихальним типом будови, причому в багатьох випадках утворюються
псевдопаренхіматозні структури.
У межах класу за типами пробок первинних пор, наявністю та
особливостями ауксилярних клітин, наявністю карпогоніальних гілок,
типом чергування поколінь, морфологією тетраспор, екологічними
особливостями виділяють вісім порядків, з яких нижче розглядають-
ся шість - Иетаїіаіез, СогаІІіпаІез, АЬпїеШаїез, бідагііпаїез,
Кґюсіутепіаіез, Сегатіаіез.
Порядок Немаліальні - Метаїіаіез
Таломи у немаліальних переважно гетеротрихальні або псевдо-
паренхіматозні, як одно-, так і багатоосьові. Характерні ознаки по-
рядку - наявність хрестоподібних тетраспорангіїв, відсутність аукси-
лярних клітин; карпогони можуть утворюватись як на коротких карпо-
гоніальних гілочках, так і зі звичайних вегетативних клітин. Життєві
цикли гаплодиплофазні, зі споричною редукцією та зміною поколінь.
Характерний представник - Ваігасіюзрегтит.
Рід батрахоспермум - Ваігасїюзрегтит (рис. 52). Всі види
роду є прісноводними водоростями, що мешкають переважно у хо-
лодних гірських річках або великих озерах. Таломи цих водоростей
156
можуть мати різні відтінки як червоного, так і синьозеленого забарв-
лення.
У Ваігасіїозрегтит гаплоїдний макроскопічний гаметофіт розви-
вається на диплоїдному мікроскопічному спорофіті. Спорофіт у цієї
водорості представлений однорядними розгалуженими нитками, у
верхівкових клітинах яких може відбуватися мейоз. Після мейозу такі
клітини не перетворюються на тетраспорангії, а починають безпосе-
редньо проростати у гаметофіти. Таломи гаметофітів оточені товстим
шаром слизу, диференційовані на однорядну опірну нитку та кільча-
сто розміщені бічні гілочки-асимілятори. Зовні гаметофіти вигляда-
ють як тендітні невеличкі (3-8 см заввишки) синювато-зелені кущики,
що складаються з окремих пагонів. Останні схожі на нитки з наниза-
ним на них намистом.
Кожний пагін являє собою центральну опірну нитку з кільцем ни-
ток-асиміляторів. Опірна нитка складається з одного ряду великих
безбарвних клітин. Від клітин опірних ниток беруть початок кільця аси-
міляторів, що рясно розгалужені і складаються з коротких діжкоподі-
бних клітин із численними хлоропластами. Великі клітини опірних ни-
ток при основі талому вкриті нещільно розміщеними тонкими бага-
токлітинними коровими нитками, що відгалужуються від основи ни-
ток-асиміляторів.
Рис. 52. Ваігасіїозрегтит: А - диплоїдний спорофіт; на верхівці розташо-
вуються клітини-спорангії та тетради клітин, гомологічні тетраспорангіям, з яких
починається розвиток гаметофіту; Б - зовнішній вигляд гаметофіту; В - фраг-
мент гаметофіту зі сферичними карпоспорофітами. 1- цистокарпій з карпоспо-
рами
157
Сперматангії утворюються з апікальних клітин асиміляторних
ниток, карпогони - на коротеньких карпогоніальних гілочках, які у різних
видів розміщуються на різних частинах талому гаметофіту. Після зап-
ліднення зигота без періоду спокою ділиться і утворює щільну групу
малоклітинних гонімобластів, у яких верхівкові клітини перетворюються
на карпоспори. Структура, утворена гонімобластами з карпоспора-
ми, має сферичну форму (вона трохи нагадує плід малини), зовні
вкрита тонкою слизовою обгорткою і називається цистокарпієм. Кар-
поспори проростають в одноосьові розгалужені кущисті спорофіти.
Порядок Коралінальні - СогаІІіпаІез
Найяскравішою особливістю представників порядку є кальцифі-
кація клітинних покривів. Таломи багатоосьові, звичайно просякнуті
вапном і мають вигляд червоного каміння, невеликих кущиків або
нагадують корали. Чергування поколінь у представників порядку ізо-
морфне. Органи як нестатевого, так і статевого розмноження (тет-
распорангії, карпогони та сперматангії) розвиваються у спеціальних
заглибинах талому, що називаються концептакулами. Тетраспорангії
хрестоподібні або зональні. Карпоспорофіти складаються з багатьох
елементів - заплідненого карпогону, ообластемної клітини, ініціаль-
ної клітини, ауксилярної клітини, гетерокаріонтичної клітини злиття та
карпоспор.
Найпоширенішими родами порядку є иіііоіііатпіоп та СогаІІіпа.
Обидва роди зустрічаються у різних морях, зокрема в Чорному морі.
Рід літотамніон - иіїюіїїатпіоп (рис. 53). Водорість має звап-
няковілі таломи і нагадує більш-менш розгалужене червоне каміння.
В різних частинах талому розвиваються заглибини - концептакули, в
яких у спорофітів утворюються тетраспорангії, а у гаметофітів - орга-
ни статевого розмноження.
У жіночіх концептакулах (рис. 54) карпогон закладається на
верхівці двоклітинної карпогоніальної нитки. Клітина, розташована під
карпогоном, називається ообластемною. Карпогоніальна нитка, утво-
рена карпогоном та ообластемною клітиною, бере початок від несу-
чої (ініціальної) клітини, від якої також розвивається ще одна коротка,
проте стерильна нитка. Ініціальна клітина надалі перетворюється на
ауксилярну.
Після запліднення карпогон зливається з ообластемною кліти-
ною, яка, в свою чергу, зливається з ауксилярною клітиною. Через
158
Рис. 53. Зовнішній вигляд талому иіїюіЬатпіоп
ообластемну клітину диплоїдне ядро зиготи мігрує в ауксилярну кліти-
ну. Далі всі ауксилярні клітини одного концептакулу зливаються між
собою, і в концептакулі утворюється одна велика багатоядерна ге-
терокаріонтична клітина злиття, яка містить диплоїдні зиготичні та
гаплоїдні вегетативні ядра. Диплоїдні ядра мігрують у бічні випини
клітини злиття і відокремлюються від останньої клітинними пере-
городками. Далі такі диплоїдні клітини діляться, перетворюючись
на гонімобласти, з верхівкових клітин яких розваваються карпо-
спори. Зрілі карпоспори через отвір виходять з концептакулу на-
зовні та проростають у спорофіти.
Рід кораліна - СогаІІіпа (рис. 55) - має таломи у вигляді пірчас-
то та вильчасто розгалужених кущиків. Кожна гілка талому складається
з вапнякових сегментів-члеників, які утворені багаторядними опірни-
ми нитками та бічними нитками-асиміляторами. Ділянки між сусідні-
ми сегментами позбавлені асиміляторів, а клітини опірних ниток до-
сить видовжені. Органи розмноження та карпоспорофіти розвивають-
ся у концептакулах подібно до літотамнію.
Кораліну використовують як лікарську водорість, виготовляючи з
неї препарати для усунення печії.
159
1
Рис. 54. Будова концептакулу з карпогонами (А) та послідовні стадії розвит-
ку карпоспорофіту в ііїїюіїїатпіоп (Б-Ж). 1 - карпогон із трихогіною; 2 - ооблас-
темна клітина; 3 - ауксилярна клітина; 4 - гетерокаріонтична клітина злиття, що
утворилась внаслідок плазмагамії ауксилярних клітин; 5 - гонімобласт; 6 - кар-
поспора. Диплоїдні зиготичні ядра чорні, гаплоїдні ядра вегетативних клітин все-
редині світлі
Порядок Анфельцальні - АЬпГеКіаІез
У цих водоростей таломи не просякуються вапном, статевий про-
цес не виявлений, життєвий цикл представлений цикломорфозом.
Розмноження, на відміну від інших флоридей, здійснюється моноопе-
рами, утворенню яких не передує мейоз. Моноопери розвиваються у
нематеціях - поверхневих потовщеннях талому, утворених коротки-
ми вертикальними нитками, що відходять від корових клітин. До по-
рядку належить лише один рід - АІігіїеІііа.
160
Рис. 55. СогаІІіпгг. А - фрагмент талому; Б - фрагмент ділянки, що з’єднує
два членики-сегменти
Рід анфельція або морський мошок - Аіїп/еіііа (рис. 56).
Рис. 56. Зовнішній вигляд тало-
му Аіїпїеііїа
Таломи мають вигляд дихотомі-
чно розгалужених кущиків псевдо-
паренхіматозної будови. АЬп/еІііа
масово зустрічається у північних
морях і є основним джерелом
для отримання біломорського
агар-агару.
Порядок Гігартинальні -
бідаїТіпаІез
Об’єднує водорості з псевдо-
паренхіматозними одно- або ба-
гатоосьовими таломами і хресто-
подібними або зональними тет-
распорангіями, які розвиваються
переважно в нематеціях. Карпо-
гон утворюється на верхівці три-
клітинної карпогоніальної нитки,
161
яка відгалужується від ініціальної клітини. Після запліднення ініціаль-
на клітина перетворюється на ауксилярну.
Гігартинальні водорості цікаві, в першу чергу, тим, що до них на-
лежить більшість представників, з яких у промислових масштабах
отримують фікоколоїди, зокрема РіїуІІоріїога та Сїіопгігиз.
Рід філофора - РїіуІІорїіога (рис. 57). З видів цього роду в Ук-
раїні отримують агароїд, який називають «чорноморським агаром».
Уздовж північно-західного узбережжя Чорного моря, між Одесою та
Опаковим, знаходиться зона, де на глибині 5-60 м суцільні зарості
утворює РІіуІІорїіога пег/оза. Це найбільший у світі філофорний фіто-
ценоз, який називають філофорним полем Зернова.
Таломи РРуІІорґіога кущисті, представлені сланкими «пагонами»,
від яких підіймаються вертикальні стебельця. Верхня частина кожно-
го стебельця сплощена, розростається по краях і утворює пластину з
потовщеною середньою частиною - ребром. Пластина рясно галу-
зиться, часто по краях та на ребрі несе дочірні, молодші пластини.
Тетраспорангії та статеві органи розвиваються у нематеціях, розта-
шованих на поверхні пластин вздовж ребра.
Рис. 57. РїіуІІорїюга пегчоза - основ-
не джерело отримання агароїду в Україні
Рід хондрус або ірланд-
ський мох - СЬопсІгиз
(рис. 58). Ця водорість, що
мешкає у північній частині Ат-
лантичного океану та у мо-
рях Північного льодовитого
океану, є основним джере-
лом сировини для отриман-
ня карагеніну.
Таломи водорості спло-
щені, рясно дихотомічно роз-
галужені. При статевому роз-
множенні у таломі розви-
вається ініціальна клітина,
яка дає початок триклітинній
карпогоніальній нитці. Вер-
хівкова клітина нитки пере-
творюється на карпогон.
Після запліднення карпогон
з’єднується з ініціальною клі-
162
тиною, яка в цей час перетворюється на ауксилярну. Далі диплоїдне
зиготичне ядро переходить в ауксилярну клітину, яка, таким чином,
Рис. 58. СНопсІгиз: А - зовнішній вигляд талому; Б - зріз через спорофіт з
тетраспорангіями; В - зріз через гаметофіт з цистокарпіями; Г-Е - послідовні
стадії розвитку карпоспорофіту. 1 - карпогон з трихогіною; 2 - ініціальна клітина
карпогоніальної гілки; 3 - вегетативна клітина карпогоніальної гілки; 4 - гетеро-
каріонтична клітина злиття; 5 - гонімобласт; 6 - карпоспори. Диплоїдні зиготичні
ядра чорні, гаплоїдні ядра вегетативних клітин всередині світлі
163
перетворюється на клітину злиття. Клітина злиття гетерокаріонтична,
оскільки, крім набутого із заплідненого карпогону ядра, зберігає власне
гаплоїдне вегетативне ядро. Клітина злиття утворює гонімобласти з
диплоїдними ядрами, які надалі розпадаються на карпоспори. Кар-
поспори проростають у спорофіти, що утворюють гаплоїдні тетрас-
пори. З останніх знов виростають гаплоїдні гаметофіти.
Порядок Родіменіальні - КІіобутепіаІез
Представники порядку мають псевдопаренхіматозні багатоосьо-
ві таломи пластинчастої форми, переважно з порожниною всередині.
Тетраспорангії хрестоподібні та тетраедричні. Ініціальна клітина, з
якої починається розвиток жіночих статевих органів, дає початок двом
ниткам - триклітинній карпогоніальній та двоклітинній ауксилярній з
ауксилярною клітиною на верхівці. Система з ініціальної клітини, кар-
погоніальноїта ауксилярної ниток отримала назву прокарпія. Вегета-
тивні клітини, що знаходяться поруч із прокарпієм, під час розвитку
гонімобластів розростаються і утворюють асиміляційний та трофіч-
ний покрив цистокарпію. У процесі дозрівання карпоспор цистокарпії
збільшуються у розмірі і на таломах гаметофітів утворюють сферичні
здуття. Типовий представник порядку - Еіїїосіутепіа.
Рід родіменія - Кіюсіутепіа (рис. 59) - поширений у літоральній
зоні північних морів і належить до їстівних багрянок. Родименія за
формою талому дещо нагадує порфіру і має таку ж саму народну
назву - червоний морський салат. Водорість збирають як супутній
продукт при промислі морської капусти.
Порядок Цераміальні - Сегагпіаіез
Об’єднує водорості з ниткоподібними одноосьовими таломами,
що прикріплюються до субстрату підошвою, яку утворює базальна
клітина; сланкі нитки таломів редуковані. Спорофіти цераміальних
мають тетраедричні тетраспорангії, гаметофіти утворюють прокарпії,
карпоспорофіти - цистокарпії. Карпоспорофіт розвивається подібно
до родименіальних, за винятком того, що ауксилярні клітини форму-
ються лише після запліднення яйцеклітини. Характерні представни-
ки - Сегатіит та Роїузіріюпіа.
Рід цераміум - Сегатіит (рис. 60). Види роду мають вигляд
невеличких кущиків, утворених дихотомічно розгалуженими нитками
із загнутими всередину вильчастими верхівками. Талом цераміуму-
164
Рис. 59. Еіїїосіутепіа: А - зовнішній вигляд талому; Б-Д - послідовні стадії
розвитку карпоспорофіту: Б - прокарпій, В - плазмагамія між клітинами карпого-
ніальної гілки, Г - утворення клітини злиття, Д - цистокарпій. 1 - карпогон з три-
хогіною; 2 - ініціальна клітина; 3 - ауксилярна клітина; 4 - гетерокаріонтична
клітина злиття; 5 - покрив цистокарпію; 6 - гонімобласт; 7 - карпоспора. Дип-
лоїдні зиготичні ядра чорні, гаплоїдні ядра вегетативних клітин всередині світлі
165
це однорядна опірна нитка, у якої зона контакту суміжних клітин вкрита
корою. У деяких видів кора вкриває весь талом, проте частіше серед-
ня частина клітин опірних ниток лишається відкритою. Як наслідок,
рослина набуває членистого вигляду - прозорі відкриті ділянки опір-
них клітин чергуються з ділянками, вкритими корою, що забарвлена в
інтенсивний червоний колір - поясками.
Рис. 60. Сегатіит: А - фрагмент талому з поясками; Б, В - зона пояска із
шипами; Г - поясок з тетраспорангіями та щетинками. 1 - поясок; 2 - шип; 3 - тет-
распорангії; 4 - щетинка
166
Кора являє собою асиміляторні короткі нитки, що щільно притис-
каються до оболонок клітин опірної нитки. Деякі асиміляторні нитки
видозмінюються у щетинки або шипи. Сперматангії, карпогоніальні
нитки у гаметофітів та тетраспорангії у спорофітів розвиваються з
клітин кори.
Сегатіит зустрічається в усіх широтах, але масово розвиваєть-
ся переважно у теплих морях, зокрема в Чорному та Азовському.
Рід полісифонія - Роіузірїіопіа (рис. 61). Таломи цієї водорості
також кущисті і мають членисту будову, але іншого типу, ніж у
Сегатіит. Нитку утворюють два типи клітин - центральні, що викону-
ють опірну функцію, та перицентральні, які здійснюють фотосинтез і
утворюють репродуктивні органи: тетраспорангії у спорофітів, спер-
матангії та карпогоніальні гілки у гаметофітів.
Перицентральні клітини розташовуються правильними ярусами,
причому один ярус перицентральних клітин оточує однорядним
кільцем одну центральну клітину. Перицентральні та центральну кліти-
ни одного ярусу називають, відповідно, периферичними та централь-
ним сифонами.
Рис. 61. РоїувірНопіа: А - загальний вигляд талому; Б - фрагмент талому з
короткими бічними гілочками; В - поперечний зріз талому; Г - цистокарпій; Д - гілка
зі сперматангіями; Е - гілка з тетраспорангіями. 1 - перицентральні клітини (си-
фони); 2 - центральна клітина; 3 - сперматангій; 4 - тетраспорангії
167
Відділ Зелені водорості - СНІогорНуіа
Евкаріотичні фотоавтотрофні платикристати, клітини яких вкриті
плазмалемою або клітинною оболонкою. Пластиди первинно симбіо-
тичні, хлорофітного типу (хлоропласти). Продукт асиміляції - крох-
маль, що відкладається у хлоропласті.
Відділ є найчисленнішим серед водоростей. За даними різних
авторів, він нараховує 20-25 тис. видів. Найдавніші рештки відомі з
верхнього докембрію - протерозою (біля 1400 млн. років). Зелені во-
дорості поширені у прісних та гіпергалінних водоймах, у морях та оке-
анах, у наземних біотопах, на снігу та льоду.
Від зелених водоростей походять вищі рослини.
Біохімічні особливості. До складу фотосинтетичних пігментів
входять майже всі пігменти, властиві вищим рослинам: хлорофіли а
та Ь, всі відомі каротини (а, р,у та е-каротин), ксантофіли лютеїнового
ряду (лютеїн, зеаксантин, неоксантин, віолаксантин, антераксантин).
Специфічні ксантофіли, виявлені у сифонофіцієвих, ульвофіцієвих та
празинофіцієвих водоростей - це сифонеїн, сифоноксантин, лорок-
сантин, празиноксантин. Завдяки переважанню хлорофілів, більшість
видів СІїІогорІїуіа забарвлені у зелений колір. В екстремальних
умовах у деяких зелених водоростей «вмикається» механізм гіпер-
синтезу каротину, і тоді клітини набувають червоного забарвлення.
Основний продукт асиміляції-крохмаль, який завжди відкладаєть-
ся у хлоропласті, часто - навколо піреноїду. Додаткові асиміляти -
олія та лейкозин. У сифонофіцієвих водоростей основним асимілятом
може бути полісахарид інулін.
Цитологічні особливості. Клітини переважної більшості видів
вкриті целюлозно-пектиновою оболонкою. Зрідка зустрічаються пред-
ставники з голими клітинами. У водоростей з класу празинофіцієвих,
а також у монадних репродуктивних клітин деяких ульвофіцієвих та
харофіцієвих водоростей на поверхні плазмалеми відкладаються від
одного до кількох шарів субмікроскопічних органічних лусочок.
Ядерний апарат зелених водоростей має типову евкаріотичну
будову. Зовнішня мембрана ядерної оболонки ізольована і не має
структурного зв’язку з хлоропластами.
Мітоз закритий, напівзакритий або відкритий. У монадних клітин
центріолі звичайно відсутні, їх функцію виконують базальні тіла джгу-
168
тиків. У клітинах представників, позбавлених здатності до активного
руху, центріолі наявні (за винятком більшості харофіцієвих водорос-
тей).
Фотосинтетичний апарат представлений первинно симбіотич-
ними хлоропластами. Вони майже тотожні із хлоропластами вищих
рослин: вкриті двомебранною оболонкою, тилакоїди групуються у
ламели по 3-6 або утворюють грани, оперезуючі тилакоїди відсутні.
Звичайно у хлоропластах є один-кілька піреноїдів, переважно обля-
мованих крохмалем. У більшості монадних зелених водоростей у
хлоропласті знаходиться стигма. У місці розташування стигми обо-
лонка хлоропласта тісно прилягає до плазмалеми. Ця ділянка плаз-
малеми виконує функції фоторецептора, а стигма - функцію шир-
ми, яка регулює кількість світла, що падає на фоторецептор.
Мітохондріям притаманні пластинчасті кристи.
Джгутикові стадії. Переважна більшість монадних клітин зеле-
них водоростей має два ізоконтні та ізоморфні джгутики. Рідше зустрі-
чаються чотири- або багатоджгутикові (стефаноконтні) форми. Се-
ред примітивних СИІогорИуІа є види з одним, двома-трьома неод-
наковими, шістьома або вісьмома однаковими за довжиною джгутика-
ми. Поверхня джгутиків звичайно гладенька або вкрита органічними
субмікроскопічними лусочками. У деяких родів наявні прості трубчасті
мастигонеми. Перехідна зона джгутика має специфічну зірчасту струк-
туру (т. зв. «візкове колесо»), що виявлена тільки в монадних кліти-
нах зелених водоростей та вищих рослин. Система джгутикових ко-
ренів складається з мікротубулярних та мікрофібрилярних коренів.
Плани організації кореневої системи у різних класах відділу різні.
Вакуолярний апарат представлений справжніми вакуолями з
клітинним соком. У клітинах багатьох монадних та гемімонадних
прісноводних та наземних водоростей звичайно є дві, зрідка - кілька
скоротливих вакуолей. Деяким празинофіцієвим водоростям властиві
трихоцисти та слизові тільця.
Типи морфологічної структури тіла. В межах відділу представ-
лені майже всі типи морфологічної структури тіла - монадний, гемі-
монадний, кокоїдний, нитчастий (включаючи прості паренхіматозні ва-
ріанти), гетеротрихальний, сифональний та сифонокладальний. Як
одно-, так і багатоклітинні водорості можуть утворювати різноманітні
колонії, вести вільний або прикріплений спосіб життя.
169
Розмноження та життєві цикли. У зелених водоростей спос-
терігаються всі основні способи нестатевого розмноження і статево-
го процесу, відомі в евкаріотичних водоростей. В цілому, в межах
відділу спостерігається тенденція заміни рухливих репродуктивних
клітин нерухомими та редукція нестатевого розмноження спорами
за рахунок інтенсифікації вегетативного і статевого розмноження. В
окремих тупікових гілках еволюції (наприклад, хлорококальні або сце-
недесмальні) наявна тенденція вторинної втрати статевого процесу
та перехід до апогамії.
Зеленим водоростям властиві різні типи життєвих циклів. Близь-
ко чверті відомих видів є агамними або апогамними, із життєвими
циклами за типом цикломорфозу.
До еугамних належить більшість видів відділу, і їх життєві цикли
представлені гаплофазними, гаплодиплофазними та диплофазними
циклами із зиготичною, гаметичною, споричною і соматичною редук-
ціями, без зміни поколінь або з ізо- та гетероморфною змінами по-
колінь. Життєві цикли можуть бути однотипними у всіх представників
таксону рангу класу або порядку, а можуть відрізнятись навіть у різних
видів одного роду.
Система зелених водоростей. За сучасними системами, відділ
поділяють на шість класів: РгазіпорИусеае, Сйіогорїіусеае, ТгеЬои-
хіорИусеае, ІЛуорЬусеае, ЗірИопорИусеае та СіїагорИусеае. В основі
поділу відділу на класи покладено комплекс ознак, пов’язаних з особ-
ливостями будови клітинних покривів, типами кореневих систем у
монадних стадій, мітозу та цитокінезу, деякими біохімічними особли-
востями. З цими ознаками корелюють також екологічні особливості
представників різних класів.
Клас Празинофіцієві - РгазіпорНусеае
Об’єднує виключно одноклітинні, здебільшого морські монадні
водорості. Характерна ознака - наявність голих клітин, вкритих суб-
мікроскопічними органічними лусочками. Джгутики виходять із апікаль-
ної, рідше - латеральної заглибини. На відміну від інших класів, для
РгазіпорИусеае характерні різноманітні плани будови клітин, різні типи
кореневих систем джгутиків, мітозу та цитокінезу. Прикладами празі-
нофіцієвих є роди Ругатітопаз та Мезозіідта з порядку Руга-
тітопасіаіез.
Рід пірамімонас - Ругатітопаз (рис. 62 А) - має чотири, вісім
або навіть шістнадцять джгутиків, що починаються в апікальній за-
170
глибині. Клітина містить один великий парієтальний хлоропласт з ба-
зальним піреноїдом.
Рис. 62. Празинофіцієві водорості: А - Ругатітопаз; Б-В - Мезозіідта
(Б - латеральний вигляд, В - фронтальний вигляд). 1 - хлоропласт; 2 - піреноїд;
З - крохмальна обгортка піреноїда; 4 - вічко; 5 - пульсуючі вакуолі; 6 - джгутики;
7 - ядро
Рід мезостигма - Мезозіідта (рис. 62 Б, В) - має два злегка
гетероконтні джгутики, які виходять із заглибини на черевному боці
клітини. Біля заглибини розташовується вічко. У пристінному шарі ци-
топлазми розміщений хлоропласт з двома піреноїдами.
Клас Хлорофіцієві - СНІогорґіусеае
Об’єднує водорості з одноклітинною, багатоклітинною, зрідка -
неклітинною будовою, які мешкають переважно у прісних водоймах.
Клітини вкриті оболонками. У монадних клітин у складі кореневої си-
стеми джгутиків є чотири мікротрубочкових корені, прикріплених до
базальних тіл джгутиків хрестоподібно або джгутиковий апарат органі-
зований за стефаноконтним (багатоджгутиковим) типом. Базальні тіла
джгутиків зміщені одне відносно іншого за годинниковою стрілкою (орі-
єнтація 1-7) або не зміщені (орієнтація 12-6). Мітоз закритий. Цитокі-
нез у багатоклітинних форм відбувається за типом клітинної платів-
ки, що розвивається за участю фікопласту. Більша частина представ-
ників здатна до гіперсинтезу вторинних каротиноїдів і, як наслідок,
171
на певних стадіях життєвого циклу може змінювати забарвлення із
зеленого на червоне або жовтогаряче.
Система класу. Клас поділяють на вісім порядків відповідно до
типу морфологічної структури тіла, типу зооспор, орієнтації базаль-
них тіл джгутиків, наявності у клітинних оболонках шару кристалічно
впорядкованої целюлози, здатності до гіперсинтезу вторинних каро-
тиноїдів, особливостей поділу клітин тощо. Нижче розглядаються
шість основних порядків-Уоіуосаіез, СИІогососсаІез, Сїіаеіоріпогаїез,
Зсепесіезтаїез, Місгозрогаїез, Оесіодопіаіез.
Порядок Вольвокальні - Уоіуосаіез
Об’єднує одноклітинні та ценобіальні водорості з монадним ти-
пом морфологічної структури тіла. Шар кристалічно впорядкованої
целюлози в оболонці відсутній, і каркасною основою оболонки є гідро-
ксипроліни. Частина вольвокальних водоростей здатна до гіперсинте-
зу вторинних каротиноїдів. У монадних вегетативних клітин є ризоп-
ласт, побудований зі скоротливого білка центрину, мітоз відбуваєть-
ся за особливим, хламідомонадним типом (СИ-мітоз). При такому
мітозі веретено формується з участю базальних тіл, в анафазі дочірні
ядра розходяться, у пізній анафазі та ранній телофазі ядра дещо збли-
жуються за рахунок скорочення залишків інтерзонального веретена,
а цитокінез здійснюється кільцевою борозною.
Зооспори у вольвокальних вкриті оболонкою або голі, базальні
тіла джгутиків зміщені за годинниковою стрілкою (орієнтація 1-7).
Одноклітинні форми у порядку представляють роди СІїІату-
сіотопаз, ОипаІіеІІа та Наетаіососсиз, ценобіальні - бопіит,
Рапсіогіпа, Еисіогіпа, \/оіуох.
Рід хламідомонас, або хламідомонада, - СІїІатусіотопаз
(рис. 63) - є провідним у порядку і нараховує біля 500 видів. Водорість
дводжгутикова, вкрита гідроксипроліновою оболонкою, яка, зазвичай,
у місці виходу джгутиків утворює потовщення у вигляді носика. Кліти-
на має хлоропласт з піреноїдом та стигмою, одне ядро, яке при вели-
ких збільшеннях мікроскопа помітне навіть без спеціального забарв-
лення. Біля основи джгутиків розміщуються дві пульсуючі вакуолі. Роз-
множення відбувається за допомогою зооспор або статевим шляхом
(статевий процес - ізо-, гетеро- та оогамія).
При несприятливих умовах клітина втрачає джгутики і перехо-
дить до нерухомого способу життя. У деяких видів перехід до неру-
хомого стану супроводжується ослизненням оболонки і утворенням
172
пальмел. Тривалість перебування у цьому стані може значно пере-
вищувати тривалість перебування у монадному стані. У нерухомому
стані СІїІатусІотопаз розмножується зоо- та гемізооспорами. Посту-
пова редукція монадної стадії при збереженні і переважному розвит-
ку нерухомої стадії обумовила виникнення видів з гемімонадною бу-
довою, які раніше виділяли у самостійний порядок Теігазрогаїез.
Представники роду мешкають у різноманітних прісних водоймах,
де можуть викликати явище «цвітіння» води, а також зустрічаються в
лісових ґрунтах. Два види роду - хламідомонас Рейнгарда (СІїІату-
сіотопаз геіпіїагсіііі) та хламідомонас Мевуза (С/). тоешизіі) -
широко використовуються у фізіологічних, генетичних та молекуляр-
них дослідженнях як модельні об’єкти. Сьогодні на основі диких штамів
цих видів отримано понад тисячу мутантних за різними генами штамів.
Рис. 63. СМатусІотопаз: А - С/і. геіпЬагсІ(іг, Б-Г - СЛ. тоешизіі: А, Б -
вегетативні клітини, В - послідовні стадії копуляції ізогамет, Г - зигота. 1 - хлоро-
пласт; 2 - піреноїд; 3 - вічко; 4 - ядро
173
Рід дюналієла - ОипаїіеІІа (рис. 64). Представники цього роду
подібні до хламідомонад, але відрізняються відсутністю клітинної
оболонки. Крім того, у видів, що мешкають у солоних континенталь-
них водоймах та морях, відсутні пульсуючі вакуолі. Оскільки клітини
дюналієли вкриті лише плазмалемою, вони здатні до метаболічної
зміни форми. Розмноження водорості відбувається шляхом поздовж-
нього поділу клітини в рухомому стані. Для деяких видів роду відомий
статевий процес, який відбувається в результаті копуляції двох веге-
тативних клітин, тобто представлений хологамією. Зигота, яка утво-
рюється внаслідок статевого процесу, переходить у стан спокою. Після
його завершення відбувається мейоз і утворюються молоді гаплоїдні
вегетативні клітини.
Переважна більшість представників цього роду мешкає в дуже
солоних (гіпергалійних) водоймах, де може викликати червоне
«цвітіння» ропи. Майже з тридцяти виявлених на території України
видів найпоширенішою є дюналієла солоноводна (ОипаїіеІІа
заііпа), яка мешкає в гіпергалійних водоймах на півдні країни.
О. заііпа належить до провідних фікотехнологічних об’єктів: з біо-
маси цієї водорості отримують р-каротин та гліцерин. Технологія про-
мислового культивування дюналієли вперше розроблена саме в
Україні, професором Київського університету Н. П. Масюк. Сьогодні
ця технологія широко використовується у різних країнах світу.
Рис. 64. ОипаїіеІІа заііпа: А - вегетативна клітина; Б - поділ клітини; В -
циста; Г - зигота
Рід гематокок - Наетаіососсиз (рис. 65) - має клітини, в яких
протопласт компактно розташовується в центрі клітини, а до клітин-
ної оболонки підходять лише тонкі цитоплазматичні тяжі. Вегетативні
клітини гематококу дводжгутикові, одноядерні, з одним хлороплас-
том та кількома піреноїдами. У центральній частині протопласта за-
174
звичай швидко накопичуються краплини олії, забарвлені каротиної-
дами у жовтогарячий колір. При несприятливих умовах водорість втра-
чає джгутики і вегетує або в кокоїдному стані, або утворює червоні
акінети.
Види цього роду, зокрема гематокок дощовий (Наетаіососсиз
ріиуіаііз), подібно до дюналієли, здатні до гіперсинтезу каротиноїдів
і швидко набувають червоного забарвлення. Гематокок масово роз-
вивається на снігу, в калюжах з дощовою водою, у криницях, на по-
верхні ґрунту в пустелях, зумовлюючи «червоні цвітіння». Клітини,
підняті з ґрунту або води у повітря, випадають разом з осадами. Це
явище відоме в народі під назвою «криваві дощі» та «кривавий сніг».
Сьогодні гематокок інтенсивно досліджується у багатьох лаборато-
ріях світу як перспективний об’єкт для введення в промислову куль-
туру з метою отримання каротину.
Рід гоніум -
бопіит (рис. 66 А) - на-
лежить до ценобіальних
водоростей. Ценобії ма-
ють вигляд платівки, що,
залежно від виду, скла-
дається з 4-х або 16-ти
клітин. Клітини утриму-
ються разом за допомо-
гою спільної слизової об-
гортки - інволюкруму. В
окремої клітини ценобію
помітні два ізоконтні джгу-
тики, пристінний хлороп-
ласт із піреноїдом та
вічком, дві пульсуючі ва-
куолі, одне ядро.
Рис. 65. Вегетативна клітина Наета-
іососсиз ркл/іаііз. 1 - цитоплазматичні тяжі;
2 - зони накопичення каротиноїдів
При нестатевому розмноженні кожна клітина два-вісім разів
ділиться, і дочірні клітини ще всередині материнської оболонки скла-
даються у новий ценобій. Статевий процес ізогамний: з кожної веге-
тативної клітини утворюється 16 однакових гамет, що попарно ко-
пулюють. Зигота вкривається товстою гладенькою оболонкою і пе-
реходить у стан спокою. Після його завершення відбувається мей-
оз, утворюються чотири гаплоїдні клітини, які ще в зиготі складають-
ся у дочірній ценобій.
175
1
1
1
Рис. 66. Ценобіальні вольвокальні водорості: А - Сопіит ресіогаїе; Б -
Рапсіогіпа тогит; В - Еисіогіпа еіедапз. 1 - інволюкрум
Рід пандоріна - Рапсіогіпа (рис. 66 Б). Ценобіальна водорість з
овальними ценобіями, що складаються з 16-ти або 32-х клітин. Це-
нобії вкриті інволюкрумом, а клітини щільно прилягають одна до од-
ної, набуваючи гранчастої форми. Кожна окрема клітина має два джгу-
тики, пару пульсуючих вакуолей, пристінний хлоропласт з піреноїдом,
вічко та одне ядро, дещо нагадуючи, таким чином, клітину хламідо-
монади. При нестатевому розмноженні протопласт клітини кілька різів
ділиться, утворюючи 16 або 32 зооспори, які, не залишаючи мате-
ринської оболонки, складаються у новий ценобій.
Статевий процес - гетерогамія. В клітинах чоловічих ценобіїв
утворюються дводжгутикові сперматозоїди, в жіночих - малорухливі
яйцеклітини. Після запліднення зигота одягається товстою гладень-
кою оболонкою, накопичує велику кількість олії та каротиноїдів і пе-
реходить у стан спокою. Після його закінчення у зиготі відбувається
мейоз. З чотирьох гаплоїдних ядер три дегенерують, а одне пророс-
тає великою зооспорою, в якій через деякий час відбувається серія
мітозів і дочірні клітини складаються у новий ценобій.
Пандоріна дуже поширена у планктоні прісних водойм, особливо
ефемерних.
Рід евдоріна - Еисіогіпа (рис. 66 В). Ценобії водорості нагаду-
ють пандоріну, але, на відміну від останньої, клітини розміщені в пра-
вильному порядку, на значній відстані одна від одної, п'ятьма кільця-
ми. Переднє і заднє кільця мають по чотири клітини, три середні - по
вісім. Клітини ценобію кулясті, з тонкою оболонкою, без носика. Кож-
176
на окрема клітина має два джгутики, як правило, дві пульсуючі ваку-
олі, пристінний хлоропласт з одним або кількома піреноїдами, вічко
та одне ядро. При нестатевому розмноженні протопласти клітин це-
нобію починають одночасно ділитися, утворюючи дочірні ценобії по-
дібно до пандорини.
Статевий процес - оогамія, причому в одних ценобіях утворю-
ються лише яйцеклітини, а в інших - сперматозоїди. Запліднені яй-
цеклітини швидко вкриваються міцною гладенькою оболонкою, чер-
воніють через накопичення великої кількості олії та каротиноїдів і
переходять у стан спокою. При проростанні із зиготи виходить одна
гола зооспора, яка потім утворює перший ценобій, вкриваючись пе-
ред початком поділів ніжною слизовою оболонкою.
Види цього роду поширені в планктоні прісних водойм.
Рід вольвокс - Уоіуох (рис. 67). Водорість утворює ценобії ку-
лястої, еліпсоїдної або яйцевидної форми, розмір яких може сягати
2 мм. Ценобій складається з великої кількості дрібних дводжгутико-
вих клітин, що розміщуються периферично під тонким одношаровим
слизовим інволюкрумом. Кожна клітина вкрита власною оболонкою,
яка зростається з оболонками сусідніх клітин, набираючи полігональ-
ної (5-6-кутної) форми. Крім того, апікальний бік оболонки притис-
кається та щільно з’єднується з інволюкрумом. Як наслідок, безпосе-
редньо під інволюкрумом утворюється периферичний шар камер-обо-
лонок, в яких вільно лежать протопласти клітин, ніби підвішені на джгу-
тиках. Джгутики крізь канали в інволюкрумі виходять назовні. Простір
між протопластами та клітинними оболонками, а також вільна від
клітин порожнина ценобію виповнені водянистим слизом. Ці деталі
будови ценобію стають помітними при фарбуванні препаратів слаб-
ким розчином метиленового синього. У деяких видів вольвоксу про-
топласти сусідніх клітин з’єднані між собою плазмодесмами, що про-
ходять крізь латеральні пори клітинних оболонок.
Протопласт кожної окремої вегетативної клітини нагадує клітину
дюналієли: в ньому помітно пристінний хлоропласт з піреноїдом та
стигмою, дві апікальні пульсуючі вакуолі, одне ядро.
Нестатеве розмноження відбувається за допомогою спеціалізо-
ваних клітин нестатевого розмноження - партеногонідій. Партено-
гонідії більші за розміром, ніж вегетативні клітини і, на відміну від
останніх, здатні до вегетативного поділу. Після перших восьми
поділів з партеногонідію утворюється сферичний порожній у центрі
комплекс з 16-ти клітин. На оберненому до інволюкруму боці є отвір.
177
Е
Рис. 67. Уо/уох А, Б - загальний вигляд колоній; В - окремі монадні клітини
ценобію; Г - зигота; Д - сперматозоїди; Е - схема утворення дочірніх колоній,
яйцеклітин та сперматозоїдів. 1 - розвиток дочірнього ценобію з партеногонідія;
2 - яйцеклітина; 3 - утворення та вихід сперматозоїдів; 4 - запліднення яйце-
клітини та утворення зиготи
178
Клітини в такому комплексі полярні - вони містять пульсуючі вакуолі
та базальні тіла джгутиків, орієнтовані до центру майбутнього цено-
бію. Подальший поділ клітин супроводжується розвитком великої інва-
гінації шару клітин з боку, протилежному до інволюкруму, і вивертан-
ням молодого ценобію таким чином, що скоротливі вакуолі та базальні
тіла джгутиків орієнтуються назовні. Далі молодий дочірній ценобій
занурюється в порожнину материнського. Цікаво, що в молодому
ценобії вже можна розрізнити майбутні партеногонідії. Звільнення
дочірніх ценобіїв відбувається після руйнування материнського.
Статевий процес - оогамія. Окремі клітини ценобію, що за роз-
мірами нагадують партеногонідії, перетворюються на антеридії та
оогонії. В межах роду відомі однодомні та дводомні види. В антери-
діях утворюється від 32 до 64 дводжгутикових сперматозоїдів, які до
моменту звільнення щільно з’єднані боками і утворюють жовтувату
платівку. Після дозрівання платівка виштовхується назовні з антери-
дію слизом, і сперматозоїди роз’єднуються. В оогоніях утворюється
по одній великій яйцеклітині, яка не покидає ценобію. Оболонка оого-
нію в місці контакту з інволюкрумом ослизнюєтся, утворюючи отвір.
Один із сперматозоїдів через цей отвір проникає в оогоній і заплід-
нює яйцеклітину.
Після статевого процесу зигота одягається товстою оболонкою із
шипами і переходить у стан спокою. Після його завершення ядро зи-
готи редукційно ділиться. Три дочірні ядра дегенерують, життєздат-
ним лишається тільки одне. Як наслідок, зигота проростає однією зоо-
спорою, яка після серії поділів розвивається у новий ценобій.
Представники роду мешкають у планктоні озер, річок, боліт та
калюж. В Україні найпоширеніший вольвокс кулястий (V. дІоЬаіог),
який може викликати «цвітіння» води калюж.
Порядок Хлорококальні - Сїііогососсаіез
Об'єднує поодинокі або зібрані у тетради кокоїдні водорості. Шар
кристалічно впорядкованої целюлози в оболонці відсутній, і, подібно
до вольвокальних, каркасною основою оболонки є гідроксипроліни.
Представники порядку не здатні до гіперсинтезу вторинних кароти-
ноїдів і залишаються зеленими на всіх стадіях життєвого циклу. Мітоз
у хлорококальних хламідомонадного типу (СК-мітоз). Зооспори вкриті
оболонкою, базальні тіла джгутиків зміщені за годинниковою стрілкою
(орієнтація 1-7). Типовим представником порядку є СІїІогососсит.
Рід хлорокок - СІїІогососсит (рис. 68). Клітини хлорокока по-
одинокі, сферичні, мають одне ядро, великий пристінний хлоропласт
179
з піреноїдом, дві скоротливі вакуолі. Нестатеве розмноження відбу-
вається дводжгутиковими зооспорами, що вкриті оболонками, або
гемізооспорами. Зооспори за ультраструктурою (зокрема за будовою
оболонки) подібні до вегетативних клітин хламідомонад. При неспри-
ятливих умовах вегетативні клітини можуть покриватися товстою
оболонкою із шипами і перетворюватись на спочиваючі клітини -
гіпноспори. Статевий процес у більшості видів роду ізогамний, життє-
вий цикл гаплофазний із зиготичною редукцією (в одного з видів -
хлорококу диплобіонтного (СМогососсит сіірІоЬіопіісит) - жит-
тєвий цикл диплофазний з гаметичною редукцією).
Види роду належать до амфібіальних організмів - вони дуже ча-
сто зустрічаються в ґрунтах, переважно лісових фітоценозів, в зоні
заплеску прісних водойм та в ефемерних водоймах.
Рис. 68. Сіїїогососсит Ьурпозрогиггг. А - зооспора; Б - вегететивна кліти-
на; В - гіпноспора
Порядок Хетофоральні - Сїіаеіорїіогаїез
Об’єднує багатоклітинні водорості з нитчастим та гетеротрихаль-
ним типом морфологічної структури тіла. Каркас клітинної оболонки
хетофоральних утворений фібрилами кристалічно впорядкованої
целюлози. Таломи хетофоральних представляють фізіологічно цілісні
структури, оскільки клітини у нитках сполучаються між собою за до-
помогою плазмодесм. Останні проходять через первинні пори, які ут-
ворюються внаслідок уронемового типу мітозу.
При такому мітозі веретено формується з участю центріолей, в
анафазі дочірні ядра розходяться, центріолі розташовуються на дис-
тальних полюсах дочірніх ядер і із залишків веретена утворюється
180
фікопласт. У телофазі ядра помітно зближуються за рахунок скоро-
чення залишків інтерзонального веретена. У фікопласті скупчуються
пухирці комплексу Гольджі з матеріалом майбутньої перегородки.
Далі вони частково зливаються, утворюючи клітинну платівку, в якій
лишаються пори з плазмодесмами.
Монадні клітини - зооспори та гамети - голі. Зооспори мають
чотири джгутики, гамети дводжгутикові. Базальні тіла джгутиків у мо-
надних клітин зміщені за годинниковою стрілкою (орієнтація 1-7). Всі
хетофоральні водорості мешкають у перифітоні прісних континенталь-
них водойм.
У межах порядку простежується поступове ускладнення плану
будови тіла: від нитчастих нерозгалужених водоростей до гетеротри-
хальних представників, що складно диференційовані у морфологіч-
ному та функціональному відношеннях. Цей ряд добре простежуєть-
ся на прикладі родів СІгопета, Зіідеосіопіит, Огарагпаїсііа.
Рід уронема - игопета (рис. 69 А, Б) - має талом у вигляді про-
стої нерозгалуженної нитки, що складається з морфологічно та фун-
кціонально однакових клітин (за винятком базальної та апікальної).
Кожна клітина одноядерна, має пристінний хлоропласт у вигляді не-
замкненого кільця, в якому розташовується один, рідше-два-чотири
піреноїди. Базальна клітина утворює диск, за допомогою якого во-
дорість прикріплюється до підводних субстратів. Апікальна клітина
має звужену та загострену верхівку. Розмноження відбувається за
допомогою чотириджгутикових зооспор або апланоспорами.
Рис. 69. Хетофоральні водорості: А, Б - игопета (А - вегетативна нитка;
Б - вихід зооспор); В, Г - Зіідеосіопіит (В - фрагмент талому, Г - зооспора та її
проростання); Д - Огарагпакіїа
181
Рід стигеоклоніум - 8(ідеосІопіит (рис. 69 В, Г) має гетерот-
рихальні таломи, що складаються з розгалужених висхідних та слан-
ких ниток. Перші здійснюють функції фотосинтезу та розмноження,
за допомогою других водорість прикріплюється до субстрату. Висхідні
нитки на верхівці звужені і досить часто закінчуються одно- або бага-
токлітинними тонкими безбарвними волосками - хетами. Рясність
галуження та кількість хет залежить від концентрації азоту в середо-
вищі: якщо у воді міститься велика кількість азоту, галуження зустрі-
чається зрідка, хети майже відсутні (в цьому випадку водорість нага-
дує ІІгопета) і навпаки. Нестатеве розмноження відбувається чоти-
риджгутиковими зооспорами та апланоспорами, статевий процес -
ізогамія, здійснюється дводжгутиковими гаметами. Цікаво, що гаме-
ти здатні проростати партеногенетично і, таким чином, виконувати
також функції зооспор. Життєвий цикл гаплофазний, із зиготичною
редукцією і без зміни поколінь.
Рід драпарнальдія - Огарагпакііа (рис. 69 Д). Водорість макро-
скопічна, має вигляд тендітного кущика, вкритого м’яким прозорим
слизом, може досягати 10-20 см у висоту. Таломи Огарагпаїсііа ди-
ференційовані на три системи ниток - сланкі, висхідні опірні та аси-
мілятори. Сланки нитки виконують функції прикріплення до субстра-
ту, асимілятори - фотосинтезу та розмноження. Опірні нитки підно-
сять над субстратом всю масу асиміляторів.
Клітини опірних ниток мають товсті клітинні оболонки, редукова-
ний хлоропласт у вигляді поясковидної, по краю зазубреної стрічки,
іноді продірявленої або майже сітчастої, з численними піреноїдами.
Нитки-асимілятори короткі, рясно розгалужені, на верхівках часто
закінчуються волоском (хетою). Клітини ниток-асиміляторів містять
великий хлоропласт з кількома піреноїдами і здатні утворювати зоо-
спори та гамети. Клітини сланких ниток безбарвні.
Нестатеве розмноження здійснюється мікро- і макрозооспорами,
розвиток яких відбувається одночасно в усіх клітинах окремої нитки-
асимілятора або навіть одночасно в усіх асиміляторах. Крім того, кліти-
ни асиміляторів можуть утворювати апланоспори, які іноді залиша-
ються з’єднаними у вигляді рядів або купок. Апланоспори мають дуже
потовщену оболонку і червоне забарвлення. Статеве розмноження
відбувається шляхом ізогамії, так само, як у стигеоклоніуму.
Нечисленні представники цього роду трапляються в Україні в
різних водоймах здебільшого навесні. Найпоширенішим видом роду
є ОгарагпаІРІа діотегаіа - драпарнальдія грудкувата.
182
Порядок Сценедесмальні - Зсепесіезтаїез
Об’єднує одноклітинні, колоніальні та ценобіальні водорості з
кокоїдним типом морфологічної структури тіла. Оболонки містять шар
кристалічно впорядкованої целюлози. Представники порядку здатні
до гіперсинтезу вторинних каротиноїдів (особливо при нестачі азоту)
і на певних стадіях життєвого циклу можуть набувати червоного за-
барвлення. Мітоз циліндрокапсового типу: веретено формується за
участю центріолей, в анафазі дочірні ядра розходяться, центріолі
розташовуються на проксимальних полюсах дочірніх ядер. У тело-
фазі помітного зближення ядер не спостерігається, а цитокінез
здійснюється кільцевою борозною, що росте за рахунок злиття пу-
хирців ендоплазматичної сітки з інвагінацією плазмалеми.
Зооспори завжди голі, базальні тіла джгутиків не зміщені одне
відносно іншого (орієнтація 12-6). В межах порядку окреслені тен-
денції переходу від одноклітинних форм до ценобіальних, втрати ста-
тевого процесу та зооспор, пов'язані з пристосуванням сценедесмаль-
них до планктонного способу існування. Характерними представни-
ками порядку є роди Вгасіеасоссиз, Нусігосіісіуоп, Ресііазігит,
Всепесіезтиз.
Рід брактеакок - Вгасіеасоссиз (рис. 70 А). Дорослі клітини
поодинокі, сферичні, мають багато пристінних дископодібних хлоро-
пластів, позбавлені піреноїдів і містять кілька ядер. Розмноження
відбувається за допомогою одноядерних дводжгутикових голих зоо-
спор або апланоспорами. Зооспори та апланоспори досить часто не
одразу звільняються з оболонки материнської клітини, а починають
рости безпосередньо у спорангії. В цьому випадку вони з’єднуються
боками і звільняються зі спорангія у вигляді агрегатів, що нагадують
сферичні ценобії. Розпад таких агрегатів на поодинокі клітини відбу-
вається лише на початку спороутворення, коли кожна клітина стає
багатоядерною. Види роду є амфібіальними організмами, проте час-
тіше зустрічаються у ґрунтах, ніж у водоймах.
Рід гідродикціон або водяна сіточка - Нудгодісіуоп (рис. 70 Б,
В). Ценобії Нусігосіісіуоп макроскопічні, складаються з кількох тисяч
великих циліндричних багатоядерних клітин, з’єднаних на кожному
полюсі з двома-чотирма сусідніми клітинами. Внаслідок цього утво-
рюються п’яти-шестигранні комірки, що у сукупності надають цено-
бію вигляду сітки (звідси походить народна назва водорості - водяна
сіточка).
183
Рис. 70. Сценедесмальні водорості: А-Вгасіеасоссиз(вегетативна клітина,
вихід зооспор, зооспори та їх проростання, апланоспорангій, ценобій-подібний
агрегат молодих клітин); Б-В - Нусігосіісіуогг Б - комірка з молодих клітин; В - стара
клітина з дочірнім ценобієм; Г - ценобій Редіазігипт, Д - ценобій Зсепесіезтиз.
1 - дочірній ценобій; 2 - рогоподібний виріст клітинної оболонки; 3 - піреноїд;
4 - хлоропласт
Оболонка дорослих клітин целюлозна, з кутинізованим зовнішнім
шаром. Тонкий шар цитоплазми з хлоропластом і ядрами вистилає
внутрішню поверхню оболонки й оточує велику вакуолю з клітинним
соком. Хлоропласт неправильно-сітчастий, з численними дрібними
піреноїдами, розміщений безпосередньо під оболонкою. Під хлоро-
пластом, у глибшому шарі цитоплазми, розташовуються численні
дрібні ядра. При наявності великої кількості крохмалю у хлоропласті
будова останнього стає невиразною.
184
При нестатевому розмноженні протопласт кожної клітини водя-
ної сіточки розпадається на кілька тисяч голих дводжгутикових зоо-
спор, які, не виходячи за межі материнської клітини, з’єднуються між
собою і утворюють дочірній ценобій. Після ослизнення материнської
оболонки такий ценобій починає існувати як самостійний організм.
При статевому розмноженні протопласт клітини розпадається на
кілька тисяч голих дводжгутикових ізогамет. Через отвір в оболонці
материнської клітини гамети виходять назовні. Далі гамети різних
статевих знаків копулюють. Утворюється зигота, що переходить у стан
спокою. Після його закінчення відбувається мейоз і зигота проростає
чотирма великими гаплоїдними зооспорами. Вони здатні відпливати
від місця утворення на значну відстань і, таким чином, виконують фун-
кцію розселення. Після закінчення руху кожна зооспора проростає у
багатокутну багатоядерну клітину - поліедр, що являє собою спорофіт
водяної сіточки. Протопласт поліедра ділиться на велику кількість зоо-
спор, які, не виходячи за межі поліедра, складаються у новий ценобій.
Таким чином, життєвий цикл Нусігосіісіуоп гаплофазний, із зиготич-
ною редукцією та гетероморфною зміною поколінь - ценобіального
гаметоспорофіта та одноклітинного спорофіта-поліедра.
Єдиний у флорі України вид цього роду - Н. геіісиїаіит - гідро-
дикціон сітчастий - часто можна знайти влітку в невеличких калю-
жах, затоках річок у заростях вищих водних рослин тощо.
Рід педіаструм - Ресііазігит (рис. 70 Г). У видів роду ценобії
сплощені, мають вигляд зіріФ або коліщатка, що складається з 4-
128 клітин, з’єднаних боками. Кожна доросла клітина містить при-
стінний хлоропласт з одним-кількома піреноїдами та кілька ядер.
Нестатеве та статеве розмноження у Ресііазігит майже повністю ана-
логічне до водяної сіточки: ценобії утворюються із зооспор, статевий
процес - ізогамія, зигота проростає після періоду спокою, в життєво-
му циклі існує стадія поліедра. Види роду є надзвичайно характер-
ним компонентом планктонних угруповань прісних континентальних
водойм.
Рід сценедесмус - Зсепесіезтиз (рис. 70 Д). Ценобії у пред-
ставників цього роду мають вигляд платівок з 4 або 8, рідше 2 або 16
видовжених клітин, з’єднаних боками паралельно одна до одної. Це-
нобії інколи вкриті слизом, який можна побачити при забарвленні пре-
парату розчином туші. Клітини у ценобії розміщуються в один чи два
ряди або розташовуються альтернативно - по черзі вище і нижче від
умовної повздовжньої вісі ценобію. Оболонка клітин має різноманітні
185
«прикраси» у вигляді ребер, шипів, шипиків, щетинок, бородавок,
довгих рогів. Тип таких «прикрас» є таксономічною ознакою на рівні
видів. Часто довгі шипи та роги забарвлені гідроокисом заліза у бу-
рий колір. Цікаво, що залежно від хімічного складу води морфологія
«прикрас» може суттєво змінюватися (наприклад, замість рогів утво-
рюватись щетинки або бородавки). В окремій клітині можна розрізни-
ти пристінний хлоропласт з піреноїдом та одне центральне ядро.
Розмноження відбувається автоспорами, які ще у материнській
клітині з’єднуються у новий ценобій. Хоча Всепесіезтиз вважається
типовим прикладом автоспорових хлорофіцієвих водоростей, проте
сьогодні в культурі у кількох видів цього роду вдалось отримати та-
кож зооспори. Електронно-мікроскопічне дослідження цих зооспор по-
казало, що їх базальні тіла орієнтуються за типом 12-6, тобто так само,
як у Вгасіеасоссиз, Ресііазігит, Нусігосіісіуоп.
Рід об’єднує переважно прісноводні планктонні організми і пред-
ставлений значною кількістю видів. Найвідомішим серед них є 8. ри-
асігісаисіа - сценедесмус чотирихвостий.
Порядок Мікроспоральні - Місгозрогаіез
Об’єднує водорості з нитчастим типом структури. Оболонка містить
шар кристалічно впорядкованої целюлози і складається з двох Н-под-
ібних сегментів. Зооспори голі, дводжгутикові, злегка гетероконтні;
базальні тіла не зміщені й орієнтовані за схемою 12-6. Порядок пред-
ставлений лише одним родом Місгозрога, що зустрічається у прісних
континентальних водоймах.
Рід мікроспора - Місгозрога (рис. 71). Таломи мають вигляд
простих нерозгалужених ниток, що закінчуються «вилками», схожи-
ми на аналогічні структури жовтозеленої водорості ТгіЬопета. Плаз-
модесми між клітинами в нитках не виявлені. Кожна клітина містить
одне ядро, пристінний сітчастий хлоропласт, який у деяких видів здат-
ний розпадатися на дископодібні сегменти. Піреноїди відсутні.
Розмноження відбувається фрагментацією ниток та зрідка - зоо-
спорами. У кількох видів спостерігали ізогамний статевий процес,
проте його деталі лишаються недослідженими.
Порядок Едогоніальні - Оесіодопіаіез
Об’єднує нитчасті та гетеротрихальні водорості, які за цитокіне-
зом з участю клітинної платівки нагадують хетофоральні водорості,
проте суттєво відрізняються від них наявністю особливого типу міто-
186
зу (едогонієвий мітоз), клітинним поділом з утворенням ковпачків, сте-
фаноконтними монадними клітинами (зооспорами, андроспорами та
сперматозоїдами) та високорозвиненим оогамним статевим проце-
сом.
Едогоніальні водорості можуть бути однодомними або дводом-
ними. Життєві цикли в едогоніальних водоростей бувають двох типів:
гаплофазні без зміни поколінь та гаплофазні зі стадією мікроскопіч-
ного чоловічого гаметофіту - нанандрію. Види, які мають перший тип
життєвого циклу, називають макрандрієвими, другий - нанандріє-
вими.
До порядку входить близько 500 видів водоростей трьох родів, з
яких нижче розглядаються два - Оесіодопіит та ВиІЬосІїаеіе.
Рід едогоніум - Оесіодопіит (рис. 72, 73 А-Е). Таломи мають
вигляд простих нерозгалужених ниток, що складаються з циліндрич-
них або злегка головчастих клітин. Базальні клітини утворюють ри-
зоїдоподібні вирости, якими водорість прикріплюється до субстрату.
Верхівкова клітина тупа або загострена, зрідка витягнута в щетинку.
Клітинна оболонка гладенька або злегка хвиляста і на окремих
клітинах має характерні для даного порядку ковпачки. Утворення ков-
пачків є наслідком особливого мітозу та специфічного цитокінезу.
На початку поділу біля верхівки клітини з'являються складка з
диктіосом, які, зливаючись, утворюють увігнуте в цитоплазму кільце.
Рис. 71. Місгозрога: А - фрагмент нитки; Б - вихід зооспори
187
Далі ядро мітотично ділиться. Між дочірніми ядрами за способом
фікопласту починає формуватись клітинна перегородка, материнсь-
ка оболонка у зоні кільця розривається. Кільце починає швидко роз-
прямлятися, за рахунок чого відбувається ріст верхньої нової кліти-
ни. Залишки оболонки материнської клітини лишаються на дочірній у
вигляді ковпачка. Кількість ковпачків відповідає кількості поділів да-
ної клітини.
Рис. 72. Оедодопіит. А-Г - клітинний поділ; Д - вихід зооспори; Е - проро-
стання зооспори
При едогонієвому типі мітозу центріолі відсутні, центрами орга-
нізації веретена виступають елементи ендоплазматичного ретикулю-
му. В анафазі дочірні ядра розходяться, а із залишків веретена утво-
рюється фікопласт. У телофазі ядра помітно зближуються за рахунок
скорочення залишків інтерзонального веретена. У фікопласті скупчу-
ються пухирці, що відшнуровуються від ендоплазматичної сітки і
містять матеріал майбутньої перегородки. Пухирці частково злива-
ються, і утворюється клітинна платівка з первинними порам.
У протопласті вегетативних клітин розрізняється пристінний сітча-
стий хлоропласт з дрібними піреноїдами, облямованими крохмаль-
ними платівками, та одне ядро. Розмноження едогонію відбувається
нестатевим або статевим шляхом.
Зооспори едогоніуму стефаноконтні. При утворенні зооспор одна
з вегетативних клітин дещо здувається, клітинна оболонка розриваєть-
ся; протопласт клітини виходить назовні й незабаром виробляє фіб-
розне кільце з кількома десятками пар джгутиків, перетворюючись на
стефаноконтну зооспору. Експериментальним шляхом встановлено,
188
Рис. 73. Характерні представники Оесіодопіаіез: А-Е - Оесіодопіит: А - ве-
гетативна нитка; Б - нитка з антеридіями (макрандрієвий вид), В - вихід сперма-
тозоїда з антеридія; Г - сперматозоїд; Д - нитка з оогонієм; Е - нитка з оогонієм
та двома нанандріями (нанадрієвий вид); Є, Ж - ВиІЬосЬаеІе: фрагменти таломів
з хетами, оогоніями та нанандріями. 1 - ковпачки; 2 - антеридій; 3 - спермато-
зоїд; 4 - оогоній; 5 - нанадрій; 6 - хета
що на зооспори здатні перетворюватись будь-які вегетативні кліти-
ни, якщо отримати їх ізольовані протопласти.
Статеве розмноження у макрандрієвих та нанандрієвих видів
відбувається дещо різними шляхами.
У макрандрієвих видів як антеридії, так і оогонії розвиваються на
звичайних вегетативних нитках. В антеридіях формується по два спер-
матозоїди, в оогонії - одна яйцеклітина. Сперматозоїди виходять з
антеридія, підпливають до оогонія, через пору або тріщину в оболонці
проникають до яйцеклітини та запліднюють її. Зигота покривається
товстою оболонкою і переходить у стан спокою. Після його закінчен-
189
ня в зиготі відбувається редукційний поділ, з кожного гаплоїдного ядра
утвоюється зооспора, яка надалі проростає у нову нитку.
У нанандрієвих видів на нитках розвиваються оогонії та особливі
спорангії, в яких утворюється чоловіча зооспора -андроспора. Вона
підпливає до оогонія, осідає на нього або на сусідню клітину та про-
ростає у мікроскопічний чоловічій гаметофіт, який називається на-
надрієм і складається з двох-чотирьох клітин. Верхівкова клітина
нанандрію перетворюється на антеридій, в якому розвиваються два
сперматозоїди. Далі цикл розвитку відбувається так само, як і в мак-
рандрієвих видів.
Оесіодопіит нараховує приблизно 400 видів, які широко розпов-
сюджені у прісних водоймах. Як правило, розвиток цих водоростей
починається у перифітоні або бентосі, проте нитки, які досягли знач-
ної довжини, можуть відриватися від субстрату і продовжувати рости
у вільному стані, утворюючи м’яке неслизьке жабуриння на поверхні
води.
Рід бульбохете - ВиІЬосіїаеіе (рис. 73 Є, Ж) - таломи розгалу-
жені, гетеротрихальні, диференційовані на висхідні нитки та кінцеві
гілки. На останніх, як правило, утворюються бульбоподібно потов-
щені при основі волоски - хети. Поділ клітин, ядра, будова монадних
стадій, життєві цикли у цього роду такі ж, як в едогонію.
Рід нараховує біля 100 видів, які поширені переважно у бентос-
них угрупованнях озер.
Клас Требуксієфіцієві - ТгеЬоихіорїіусеае
Нараховує приблизно 250 видів кокоїдних, нитчастих та гетеро-
трихальних водоростей, що мешкають переважно у позаводних еко-
топах - у ґрунтах, в аерофітоні, входять до складу лишайників, рідше
зустрічаються як симбіонти прісноводних безхребетних тварин (най-
частіше - інфузорій та губок).
Клітини требуксієфіцієвих завжди вкриті целюлозно-пектиновою
оболонкою, яка у багатьох представників містить додатковий шар
спорополеніну. Монадні клітини завжди голі, дводжгутикові (зрідка
чотириджгутикові), представлені зооспорами, зрідка - гаметами. Ко-
ренева система джгутиків хрестоподібна. їх базальні тіла зміщені про-
ти годинникової стрілки (орієнтація 11-5). Мітоз напівзакритий (як ви-
няток - закритий). Цитокінез відбувається за участю фікопласта. Всі
представники не здатні до гіперсинтезу вторинних каротиноїдів і про-
тягом всього життя залишаються зеленими.
190
Система класу. За наявністю монадних клітин, наявністю та по-
ведінкою центріолей при мітозі, типами морфологічної структури тіла,
полярністю клітин, здатністю до статевого розмноження клас поділя-
ють на п’ять порядків, з яких нижче розглядаються два - ТгеЬоихіаіев
та СІїІогеІІаІев.
Порядок Требуксіальні - ТгеЬоихіаІез
Об’єднує одноклітинні та зібрані в сарциноїдні пакети або тетра-
ди водорості з кокоїдним або нитчастим типом морфологічної струк-
тури тіла. Центріолі в клітинах присутні протягом усього клітинного
циклу, але не беруть безпосередньої участі в утворенні веретена по-
ділу. Більшість видів здатні розмножуватися за допомогою зооспор; у
деяких видів зооспори недорозвинені, і зооспорогенез завершується
утворенням гемізооспор або апланоспор. Клітини мають лопатевий
пристінний або центральний хлоропласт без піреноїду або з погано
помітним піреноїдом, навколо якого крохмаль не відкладається. Тре-
буксіальні мешкають переважно в аерофітоні, часто є фікобіонтами
лишайників. Найпоширенішими представниками порядку є роди
ТгеЬоихіа та Оезтососсиз.
Рід требуксія - ТгеЬоихіа (рис. 74). Клітини требуксії поодинокі
або зібрані у тетради, діади або октади, чи багатоклітинні комплекси,
що складаються з тетрад, діад та октад. Окремі клітини сферичні, з
масивним центральним зірчастим хлоропластом, який має численні
короткі радіальні вирости і один голий піреноїд. Ядро одне, розташо-
ване латерально в невеликій заглибині хлоропласта.
Розмноження відбувається зооспорами, апланоспорами та авто-
спорами. Зооспори голі, мають два джгутики й утворюються у великій
кількості (від 16 до 64). Після закінчення періоду руху зооспори втрача-
ють джгутики і набувають сферичної форми. Апланоспори починають
розвиватися, як зооспори, по 16-64 у спорангії, але без джгутиків. Ав-
тоспори також не мають джгутиків, проте утворюються переважно по
чотири, рідше - по дві або вісім, і при проростанні з’єднуються у тетра-
ди, діади або октади.
Є відомості, що один із видів роду має ізогамний статевий про-
цес, але його деталі та особливості життєвого циклу залишаються
недослідженими.
Рід містить близько 50 видів, які є найтиповішими фікобіонтами
лишайників.
191
Рис. 74. ТгеЬоихіа сгепиіаіа: А - вегетативна клітина з центральним хлоро-
пластом та піреноїдом; Б - зооспорангій; В - зооспори
Рід десмокок - Оезтососсиз (рис. 75). Водорість утворює па-
кети з двох-чотирьох клітин та багатоклітинні більш-менш кубічні аг-
регати, що складаються з пакетів. Інколи по краях агрегатів утворю-
ються короткі нерозгалужені нитки. Клітинна оболонка тонка, проте
щільна і неослизнена. В окремій клітині розрізняється один великий
пристінний хлоропласт, який має погано помітний піреноїд, позбав-
лений крохмальної обгортки. Ядро одне, непомітне без спеціального
забарвлення.
Розмноження відбувається, в першу чергу, внаслідок послідов-
ного вегетативного поділу клітин у двох або трьох взаємно перпенди-
кулярних напрямах. Дочірні клітини лишаються щільно з’єднаними
одна з одною, внаслідок чого утворюються пакети. Якщо клітини кілька
разів підряд діляться в одному напрямку, то по краях агрегатів утво-
рюються короткі нитки.
Водорість здатна також утворювати апланоспори. При цьому одна
з вегетативних клітин збільшується у розмірах і перетворюється на
спорангій. На оболонці спорангія після забарвлення метиленовим
синім стають помітні невеличкі шипики. Протопласт спорангія бага-
торазово ділиться, утворюючи до 32-64 еліпсоїдних апланоспор. Після
звільнення зі спорангія апланоспори проростають у нові вегетативні
клітини і дають початок новим пакетам. Вважають, що апланоспори
десмокока є недорозвиненими зооспорами, подібно до апланоспор
требуксії.
Десмокок зустрічається повсюдно. Він розвивається на корі де-
рев, обумовлюючи її зелене аерофітне «цвітіння». Водорість досить
чутлива до атмосферного забруднення, через що у країнах Західної
192
Європи використовується як тестовий організм при здійсненні моні-
торингу стану повітряного басейну. Найпоширенішим та відомим ви-
дом роду є Оезтососсиз оііуасеиз — десмокок оливковий, більш
відомий в учбовій літературі за синонімічними назвами протокок
(Ргоіососсиз) та плеврокок (РІеигососсиз).
Рис. 75. Резтососсиз оііуасеиз: А - пакет; Б - агрегат з пакета та мало-
клітинної нитки
Порядок Хлореляльні - СІтІогеІІаІез
Включає кокоїдні та нитчасті водорості, які ніколи не утворюють
зооспор, а також не здатні до статевого розмноження. Центріолі
відсутні. Клітини мають парієтальний хлоропласт з добре помітним
піреноїдом або без нього. Типовий представник - СіїІогеІІа.
Рід хлорела - СІїІогеІІа (рис. 76). Водорість одноклітинна, ко-
лоній або агрегатів не утворює. Клітини хлорели дрібні, сферичні або
широкоеліпсоїдні, з гладенькою оболонкою. В оболонці багатьох видів
часто наявний шар спорополеніну, який надає їй механічної міцності
та хімічної стійкості. В клітині розрізняється один пристінний хлороп-
ласт, який має форму напівсфери, платівки або широкої стрічки і, як
правило, містить піреноїд, облямований невеличкими гранулами крох-
малю або кількома крохмальними платівками.
Розмноження відбувається тільки автоспорами, які утворюються
в материнській клітині по 2-8 і звільняються при розриві оболонки
спорангія. Для деяких видів описані спочиваючі клітини - акінети. Вони
утворюються зі звичайних вегетативних клітин внаслідок потовщен-
ня клітинної оболонки, можуть містити багато олії, проте ніколи не
накопичують великої кількості вторинних каротиноїдів і тому залиша-
ються зеленими.
Хлорела зустрічається у найрізноманітніших умовах - у прісних
водоймах, ґрунтах, аерофітоні, може бути симбіонтом найпростіших,
193
наприклад, інфузорій та губок. Симбіотичні хлорели в зоологічній літе-
ратурі відомі під назвою зоохлорели.
Хлорела є першою мікроскопічною водорістю, яка була введена
в масову промислову культуру і започаткувала новий напрямок біо-
технології - фікотехнологію. На жаль, хлорела виявилась не досить
вдалим об'єктом: хоча водорість відносно легко культивується, про-
те біомасу хлорели важко переробляти, оскільки клітинна оболонка
містить вельми стійкий до хімічних та механічних обробок шар спо-
рополеніну.
Рис. 76. СЬІогеІІа Уиїдагіз: А - вегетативна клітина; Б - автоспорангій;
В - вихід автоспор
Клас Ульвофіцієві - ІЛуорІтусеае
Об’єднує переважно морські бентосні та перифітонні водорості,
хоча в межах класу представлені також і прісноводні та аерофітні
форми. Майже всі ульвофіцієві водорості мають багатоклітинні тало-
ми. Переважаючі типи морфологічної структури тіла - нитчастий, ге-
теротрихальний та сифонокладальний.
Клітини ульвофіцієвих завжди вкриті целюлозно-пектиновою або
целюлозною оболонкою. Монадні клітини завжди голі, чотириджгути-
кові або дводжгутикові, представлені зооспорами та гаметами. Коре-
нева система джгутиків хрестоподібна, базальні тіла зміщені проти
годинникової стрілки (орієнтація 11-5). Мітоз закритий. Фікопласт
відсутній, цитокінез відбувається шляхом утворення кільцевої бороз-
ни (у трентеполіальних водоростей - за участю примітивного фраг-
мопласта).
До класу входить близько 80 родів, що об’єднують більш ніж 1000
видів.
194
Система класу. Ознаки, за якими клас поділяють на порядки,
пов’язані з типами морфологічної структури тіла, життєвими цикла-
ми, ультратонкою будовою зооспор та гамет, а також з особливостя-
ми поширення. До класу входить шість порядків, з яких нижче розгля-
даються чотири - ІЛоігісІтаІев, ІЛуаІез, СІасІорИогаІез, Тгепіероііііаіез.
Порядок Улотрихальні - ІЛоігісіїаіез
Об’єднує морські та прісноводні водорості з нитчастим та гете-
ротрихальним типом структури. Життєвий цикл гаплодиплофазний, з
гетероморфною зміною поколінь. Спорофіт представлений диплоїд-
ною одноклітинною рослиною, відомою під назвою кодіолюм
(Собіоіит). Клітинні оболонки містять велику кількість пектинових
речовин і здатні ослизнюватися. Монадні клітини голі, але на плазма-
лемі є шар субмікроскопічних органічних лусочок. Зооспори переважно
чотириджгутикові, гамети дводжгутикові. Типовий представник -
ІЛоіїїгіх.
Рід улотрикс - ІЛоіїніх (рис. 77). Більшу частину життя водорість
проводить у стані гаметоспорофіта, який має вигляд довгих нерозга-
лужених однорядних ниток. У кожної клітини нитки (за винятком ба-
зальної") є пристінний хлоропласт у вигляді незамкненого кільця, кілька
піреноїдів, одне ядро; центр клітини зайнятий вакуолею з клітинним
соком. Всі клітини здатні до поділу, що обумовлює постійне нарос-
тання талому в довжину, а також утворення репродуктивних клітин.
Базальна клітина, за допомогою якої водорість прикріплюється до
субстрату, відрізняється від звичайних вегетативних клітин нитки: вона
витягнута у короткий ризоїд, не здатна до поділу, має редукований
хлоропласт.
Вегетативне розмноження відбувається фрагментацією нитки.
При нестатевому розмноженні у будь-якій клітині утворюються 2-4 (8)
чотириджгутикові зооспори. Зооспороутворення розпочинається в
апікальній частині нитки і просувається до її основи. Зрілі зооспори
виходять назовні у слизовому міхурі через боковий отвір клітинної
стінки. Міхур швидко розчиняється. Після певного періоду руху зоо-
спора зупиняється, відкидає один за одним усі чотири джгутики, при-
кріплюється боком до субстрату і проростає в нову нитку.
Статевий процес ізогамний. Будь-яка клітина нитки здатна утво-
риш велику кількість (8-64) дводжгутикових ізогамет, які для копу-
ляції виходять у зовнішнє середовище. Нитки ІЛоіїїгіх є роздільноста-
іоними, оскільки, незважаючи на морфологічну схожість гамет, одні
195
нитки продукують лише фізіологічно жіночі гамети («плюс»-гамети),
інші - фізіологічно чоловічі («мінус»-гамети). Після копуляції зигота
деякий час зберігає джгутики і рухається, тобто перебуває у стані
планозиготи. Далі вона зупиняється, втягує джгутики, виробляє тов-
сту оболонку і переходить у стан спокою. Після його завершення зи-
гота проростає у булавоподібний одноклітинний спорофіт (стадія
Собіоіит): вона формує трубковидний випин, куди переходить ци-
топлазма та ядро; далі випин здувається, набуваючи грушоподібної
форми; потім диплоїдне ядро спорофіта редукційно ділиться і він ут-
ворює 4-8 чотириджгутикові зооспори. Останні через отвір у клітинній
оболонці виходять назовні у слизовому міхурі; слиз швидко розчи-
няється, і зооспори звільняються. Після деякого періоду активного
руху зооспори осідають на субстрат і проростають, даючи початок
новим нитчастим гаметоспорофітам.
Види роду поширені переважно у морях, де зустрічаються у суп-
раліторальних ваннах або в обростаннях літоральної зони. Кілька
видів досить часто зустрічаються у прісних водоймах - переважно в
холодних гірських річках та струмках, а також у весняний сезон в річках
та озерах помірного поясу.
Рис. 77. ІЛоНіпх: А - гаметоспорофіт; Б - утворення гамет; В - послідовні
стадії копуляції гамет; Г - зигота; Д - спорофіт, стадія СоФоіит; Е - утворення
зооспор спорофітом; Ж - зооспора спорофіта; 3 - утворення зооспор гаметоспо-
рофітом; І - зооспора гаметоспорофіта
196
Порядок Ульвальні - ІІІуаІев
Включає переважно морські водорості з гетеротрихальною або
примітивною тканинною будовою. Життєвий цикл гаплодиплофазний,
з ізоморфною зміною поколінь. Оболонки целюлозно-пектинові, здатні
до ослизнення. Монадні клітини ульвальних позбавлені субмікроско-
пічних лусочок. Зооспори переважно чотириджгутикові, гамети дво-
джгутикові. Характерні представники - О^а та Епіеготогріїа.
Рід ульва або зелений морський салат - ІЛуа (рис. 78). Спо-
рофіти та гаметофіти водорості морфологічно однакові і мають ви-
гляд двошарової платівки з гофрованими краями, довжина якої ста-
новить кілька сантиметрів. Кожна клітина містить одне ядро, при-
стінний пластинчастий хлоропласт з одним-кількома піреноїдами.
Таломи гаметофітів розвиваються із чотириджгутикових зооспор,
спорофітів - із щойно утворених зигот, які зберігають джгутики гамет,
тобто із планозигот. Розвиток пластин спорофітів та гаметофітів відбу-
вається із зооспор або планозигот однаковим способом: монадна
клітина прикріплюється до субстрату, втрачає джгутики і ділиться спо-
чатку в поперечній площині, утворюючи однорядну нерозгалужену
первинну нитку, яка нагадує талом улотриксу. Далі клітини нитки
діляться повздовжньо-радіально, утворюючи одношарову трубку з по-
рожниною в центрі. Стінки трубки змикаються, і крайові клітини про-
довжують ділитися, внаслідок чого утворюється двошарова платівка
зі складчастими краями.
Хоча таломи спорофітів та гаметофітів морфологічно однакові,
проте плоїдність їх різна, і тому зооспори і гамети ніколи не утворю-
ються на одному й тому ж таломі. Зооспори утворюються шляхом
послідовного поділу протопласта будь-якої клітини на чотири-вісім
клітин, причому перший поділ є редукційним, і, таким чином, споро-
фіти є диплоїдними, зооспори - гаплоїдними.
Статеве розмноження відбувається за допомогою дводжгутико-
вих гамет. Статевий процес, залежно від виду, може бути ізогамним
або гетерогамним. Для ульви характерна гетероталічність - гамето-
фіти відрізняються в статевому відношенні: одні продукують гамети
зі знаком «+», інші - зі знаком «-». Копуляція відбувається лише між
гаметами різних статевих знаків. Якщо статевий процес ізогамний,
«+» та «-» таломи розрізнити не вдається. У випадку гетерогамії та-
ломи різних статевих знаків розпізнаються навіть неозброєним оком,
за забарвленням плодючих ділянок. Так, у чоловічих таломів ці ділянки
жовтуваті, у жіночих - темно-зелені.
197
Зигота, що утворюється внаслідок копуляції гамет, містить дип-
лоїдне ядро і без періоду спокою одразу проростає в диплоїдний спо-
рофіт. Таким чином, в ульви спостерігається правильне чергування
ізоморфних поколінь: диплоїдного спорофіта та гаплоїдного гамето-
фіта.
Види роду ІЛуа мешкають в морях теплого та помірного поясів,
зокрема в Чорному морі. Водорість є їстівною, в деяких країнах навіть
введена в аквакультуру і вирощується на спеціалізованих морських
фермах.
Б
Рис. 78. 1//уа; А-зовнішній вигляд талому; Б - поперечний зріз талому; В
- зооспорангій; Г - зооспора; Д - гаметангій; Е - копуляція ізогамет; Є - плано-
зигота
Д
198
Рід ентероморфа або кишечниця - Епіеготогріїа (рис. 79).
У цього близького до ульви роду таломи видовжено-пластинчасті або
трубчасті. За типом життєвого циклу, будовою клітин та розмножен-
ням цей рід подібний до ІІІуа. Відміни полягають у морфогенезі тало-
му, що й відбивається на зовнішньому вигляді водорості.
Зокрема, в ентероморфи талом розвивається з зооспори або пла-
нозиготи через стадію однорядної первинної нитки, далі - одношаро-
вої трубки з порожниною в центрі. Як тільки стінки трубки у верхів-
ковій частині змикаються, морфологічні трансформації талому, за
винятком його лінійного збільшення у довжину, припиняються. Як
наслідок, молода водорість має вигляд кишки (звідси її народна на-
зва - кишечниця), а доросла у нижній частині представлена одноша-
ровою трубкою з порожниною, у верхній - двошаровою видовженою
платівкою.
Кишечниця поширена переважно в морях, де часто зустрічаєть-
ся разом з ульвою. Проте деякі види роду добре переносять значне
опріснення води, завдяки чому піднімаються у гирла річок і навіть
розвиваються у прісних континентальних водоймах.
Рис. 79. Епіеготогріїа: А - трубчастий талом; Б - пластинчастий талом;
В-Д - поперечні зрізи талому: В - зріз у зоні двошарової платівки, Г - зріз у
середній частині талому з великою порожниною, Д - зріз у базальній частині
199
Порядок Кладофоральні - СІаборІїогаІез
Об’єднує водорості із сифонокладальним типом морфологічної
структури тіла, хоча таломи у більшості морських та прісноводних
представників мають вигляд нерозгалужених ниток або розгалуже-
них кущиків. У деяких морських родів таломи представлені системою
розгалужених міхурів, або складаються з центральної осі та з’єдна-
них з нею бічних сегментів, або набувають псевдопаренхіматозного
вигляду.
Життєвий цикл гаплодиплофазний, з ізоморфною зміною поколінь.
Клітинні оболонки целюлозні, позбавлені пектинових речовин. Веге-
тативні клітини мають багато ядер, пристінний сітчастий хлоропласт
або велику кількість дрібних дископодібних хлоропластів, з’єднаних
між собою дуже тонкими тяжами; у хлоропластах розташовуються
дрібні піреноїди. Кожний піреноїд поділений на дві напівсфери, на-
вколо яких відкладається суцільна напівсферична платівка крохма-
лю. Мітози у різних ядрах однієї клітини відбуваються синхронно, проте
цитокінез та каріокінез між собою не скоординовані. Монадні клітини
позбавлені субмікроскопічних лусочок; зооспори переважно чотири-
джгутикові, гамети-дводжгутикові. У кладофоральних виявлено спе-
цифічний ксантофіл - сифоноксантин. Характерні представники -
СІасІорІїога та КІіігосІопіит.
Рід кладофора - СІасІорІїога (рис. 80) - є провідним родом по-
рядку. Таломи кладофори кущикоподібні, складаються з багатоядер-
них клітин, що утворюють системи розгалужених ризоїдальних та
висхідних ниток. Клітини вкриті товстими шаруватими оболонками,
до яких прикріплюються численні епіфітні організми. В цитоплазмі
окремої клітини міститься пристінний сітчастий хлоропласт з багать-
ма піреноїдами та численні ядра. В живому стані ядра не помітні у
світловий мікроскоп, тому, щоб їх роздивитися, рекомендується за-
барвити препарати ацетокарміном. Водорість розмножується фраг-
ментацією таломів, зооспорами, акінетами та статевим шляхом. Ста-
тевий процес - ізогамія.
При нестатевому розмноженні у верхівкових клітинах спорофіта
відбувається мейоз, після якого утворюється велика кількість чоти-
риджгутикових зооспор. Через отвір в оболонці спорангія зооспори
виходять у зовнішнє середовище і проростають у гаметофіт. Гамето-
фіти кладофори є роздільностатевими. Чоловічі та жіночі гамети мор-
фологічно не відрізняються між собою, проте різняться фізіологічно
та за ультратонкою будовою апікальної частини клітини. Так, жіночі
200
гамети на верхівці під плазмалемою мають спеціальний ковпачок,
який при копуляції приймає участь в утворенні кон'югаційного каналу.
Хоча будь-яка вегетативна клітина висхідної частини талому здатна
перетворитись у гаметангій, проте процес утворення гамет, як
правило, починається у верхівкових клітинах талому. Через отвір в
оболонці гамети звільняються і копулюють з гаметами особин іншого
статевого знаку. Після копуляції зигота осідає на субстрат і без періо-
ду спокою починає проростати у новий спорофіт.
Гамети кладофори (а також і деяких інших кладофоральних) здатні
проростати партеногенетично, даючи початок як спорофітам, так і
гаметофітам. Ця особливість зумовлює існування в межах роду до-
сить різноманітних варіантів конкретних життєвих циклів, які, проте, є
похідними від основного, гаплодиплофазного циклу зі споричною
редукцією.
Рис. 80. СІасіорїіога: А - частина нитки із зооспорангіями (темні клітини);
Б - зооспорангії; В - багатоядерна клітина. 1 - ядра; 2 - піреноїди; 3 - хлоро-
пласт
201
Найпоширенішим видом морів помірних та теплих широт є кладо-
фора розкидиста (СІасіоріїога уадаЬипсІа). У прісних водоймах
найчастіше трапляється кладофора клубочковидна (С. діотегаіа).
Морфологічно ці два види не відрізняються між собою, але перший має
гаплодиплофазний життєвий цикл зі зміною поколінь, тоді як у другого
статевий процес відсутній, а нестатеве розмноження відбувається дво-
джгутиковими зооспорами. Спеціальні дослідження показали, що зоо-
спори С. діотегаіа за походженням є гаметами, що втратили здатність
до копуляції, проте зберегли здатність до партеногенетичного розвитку.
Таким чином, вегетативні таломи найпоширенішого прісноводного виду
роду - С. діотегаіа - це гаплоїдні гаметофіти.
Рід ризоклоніум - КІіігосІопіит (рис. 81) - має вигляд довгих
нерозгалужених ниток. Кожна клітина містить незначну кількість ядер
- звичайно, не більш чотирьох. Молоді особини ведуть прикріплений
спосіб життя, утримуючись на субстраті за допомогою безбарвних
ризоїдів. Дорослі нитки звичайно вільноплаваючі, їх скупчення утво-
Рис. 81. ЕІМгосІопіит:
фрагмент нитки. 1 - хлоро-
пласт; 2 - піреноїд; 3 - ядра
рюють жорстке неслизьке жабуриння, в
яких ширина клітин становить 30-50 мкм,
а довжина окремої нитки - до кількох
метрів. Спорофіт ризоклоніуму продукує
дводжгутикові зооспори, гаметофіт-двод-
жгутикові анізогамети. У прісних водоймах
найпоширенішим видом є ризоклоніум
ієрогліфічний (/?. іїіегодіуріїісит), у мо-
рях - ризоклоніум прибережний (/?. гі-
рагіит).
Порядок Трентеполіальні - Тгеп-
іероііііаіез
Порядок об’єднує виключно наземні
гетеротрихальні водорості, таломи яких
мають вигляд розгалужених ниток або
одно- чи багатошарових паренхіматозних
платівок. У трентеполіальних відсутні піре-
ноїди, крохмаль синтезується у незначній
кількості і відкладається у вигляді дрібних
гранул у стромі хлоропласта. Цитокінез
відбувається за участю фрагмопласта, зу-
мовлюючи утворення клітинної платівки зі
справжніми порами та плазмодесмами.
Мітоз закритий.
202
Розмноження відбувається фрагментацією ниток, акінетами, чо-
тириджгутиковими зооспорами та статевим шляхом. Життєвий цикл
гаплодиплофазний, з ізоморфною зміною поколінь, причому зооспо-
ри утворює як гаметоспорофіт, так і спорофіт. Статевий процес ізо-
гамний, здійснюється дводжгутиковими гаметами, які також здатні
проростати партеногенетично.
Трентеполіальні викликають червоне цвітіння кори дерев, пара-
зитують на листках різноманітних деревних порід або є фікобіонтами
лишайників. Біохімічною особливістю є здатність накопичувати вели-
ку кількість вторинних каротиноїдів (переважно р-каротину) та по-
лігідроксиспиртів (поліолу та альдітолу). Типовий представник -
Тгепіероііііа.
Рід трентеполія - ТгепіероМа (рис. 82). Таломи Тгепіероііііа
мають вигляд коротких розгалужених сланких ниток, висхідні нитки
редуковані. У багатьох видів кінцеві клітини помітно збільшені за роз-
міром і являють собою або зооспорангії, або гаметангії. Після приго-
тування водних препаратів можна спостерігати, як протягом кількох
хвилин з кінцевих клітин виходять гамети або зооспори.
рангій; Д - зооспора
203
Види роду часто зустрічаються на корі листяних порід дерев, ви-
кликаючи цегляно-червоне аерофітне «цвітіння».
Клас Сифонофіцієві - Зірїіопорїіусеае
Об’єднує сифональні, переважно морські макроскопічні водорості.
Подібно до ІЛуорЬусеае, клас представляє лінію еволюції зелених
водоростей, у яких коренева система джгутиків хрестоподібна, ба-
зальні тіла зміщені проти годинникової стрілки (орієнтація 11-5), а
фікопласт відсутній. Зооспори часто стефаноконтні, джгутики розмі-
щуються по спіралі; гамети дводжгутикові. Мітоз закритий. Цитокінез
у вегетативних стадій відсутній, оскільки представники мають неклітин-
ну будову. В сифонофіцієвих виявлено специфічні ксантофіли - си-
фонеїн та сифоноксантин. Продуктом асиміляції замість крохмалю
може бути полімер фруктози - полісахарид інулін.
У деяких сифонофіцієвих талом має два типи пластид - забарв-
лені хлоропласти та безбарвні амілопласти. Амілопласти не беруть
безпосередньої участі у фотосинтезі, а виконують функції накопичення
запасних полісахаридів.
Розмноження здійснюється за допомогою дво-, чотириджгутико-
вих та стефаноконтних зооспор і статевим шляхом. При утворенні
репродуктивних клітин на спорангій або гаметангій може перетвори-
тися або весь талом, або лише його частина. Перший шлях розвитку
називають холокарпічним, другий - еукарпічним. При еукарпічному
шляху розвитку сифони, де починається утворення монадних клітин,
відмежовуються від основної частини талому пробками. У дазикла-
дальних гамети утворюються лише з особливих гаметичних цист (див.
нижче).
Система класу. Клас поділяють на чотири порядки - Вгуорзісіаіез,
Наїітесіаіез, Оісіїоіотозіріїопаїез та Оазусіасіаіез. В основу поділу
на порядки покладено тип симетрії талому, наявність стефаноконт-
них зооспор, особливості фотосинтетичного апарату (зокрема, на-
явність амілопластів), спосіб утворення гамет (за холо- чи еукарпіч-
ним типом, або через гаметичні цисти), тип статевого процесу. Ниж-
че розглядаються три порядки - Вгуорзісіаіез, Наїітесіаіез та Оазу-
сіабаїез.
Порядок Бриопсидальні - Вгуорзісіаіез
Об’єднує водорості, таломи яких не мають радіально-симетрич-
ної будови і являють собою одну велетенську клітину з багатьма (інколи
204
- до кількох тисяч) ядрами. Центріолі зазвичай відсутні. Фотосинте-
тичний апарат представлений лише хлоропластами, амілопластів
немає. Характерна особливість бриопсидальних - наявність у життє-
вому циклі стефаноконтних зооспор.
Бриопсидальні - це водорості, в яких гамети утворюються еукар-
пічним шляхом, тоді як зооспори - як еу-, так і холокарпічно. Розмно-
жуються бриопсидальні фрагментацією талому, апланоспорами, сте-
фаноконтними зооспорами та статевим шляхом. Статевий процес -
гетерогамія. Типовий представник порядку - Вгуорзіз.
Рід бриопсис - Вгуорзіз (рис. 83) - поширений у морях тропіч-
ного, субтропічного та помірного поясів. Гаметофіти цієї водорості
диференційовані на сланкі ризоїдальні та висхідні асиміляторні си-
фони. Висхідні сифони зазвичай пірчасто розгалужені і нагадують пта-
шине пір’я. При утворенні гамет деякі бічні сифони відокремлюються
перегородкою. Із сифонів, забарвлених в інтенсивно зелений колір,
утворюються жіночі гаметангії, зі світліших, жовтуватих сифонів - чо-
ловічі. Далі вміст гаметангіїв розпадається на численні дводжгути-
кові жіночі та чоловічі гамети, що різняться за розмірами. Через отвір
в оболонці гамети виходять у зовнішнє середовище, де попарно ко-
пулюють, утворюючи планозиготу.
Планозигота осідає на субстрат, втрачає джгутики і без періоду
спокою починає проростати у диплоїдний розгалужений ниткоподіб-
ний спорофіт, що отримав назву стадії ОегЬезіа. Розвиток стадії
ОегЬезіа відбувається 2-4 місяці. Ядро у 20-40 разів збільшується у
діаметрі, переходячи зі стадії диплоїдного зиготичного ядра у стадію
первинного ядра. Далі у первинному ядрі відбувається серія мітотич-
них поділів, дочірні ядра зменшуються у діаметрі до 5-10 мкм. Ос-
танній поділ ядра є редукційним. Після мейозу завершується форму-
вання вторинних ядер, і спорофіт холо- або еукарпічним шляхом дає
початок одноядерним стефаноконтним гаплоїдним зооспорам. Ці зоо-
спори проростають або у нові гаметофіти, які складаються із сифонів
пірчастої будови, або у нові спорофіти стадії ОегЬезіа.
Цікаво, що за певних умов планозигота може безпосередньо про-
ростати у гаметофіт, не проходячи стадію ОегЬезіа. Проте процеси,
що відбуваються з ядром при цьому шляху розвитку, залишаються
недослідженими.
Таким чином, Вгуорзіз має гаплодиплофазний життєвий цикл з
неправильним гетероморфним чергуванням поколінь та споричною
редукцією (принаймі, при типовому шляху розвитку зиготи).
205
Рис. 83. Стадії розвитку Вгуорзіз: А - зовнішній вигляд гаметофіта; Б -
верхівкова частина «гілки» гаметофіта; В - чоловічій гаметангій; Г - чоловіча
гамета; Д - жіночий гаметангій; Е - жіноча гамета; Є - зовнішній вигляд спороф-
іта (стадія ОегЬезіа); Ж - молодий одноядерний спорофіт з первинним ядром;
З - спорофіт з багатьма первинними ядрами; И - зооспорангій з протопласта-
ми майбутніх зооспор, що мають вторинні ядра; І - зооспора
Рід кодіум - Сосііит (рис. 84) - є прикладом водорості зі складно
диференційованим сифональним таломом. Центральна частина та-
лому складається зі щільно переплетених видовжених ризоїдальних
сифонів. На периферії від ризоїдальних сифонів відгалужуються пу-
хирчасті вирости - утрикули, що утворюють шар, подібний до кори.
Система ризоїдальних сифонів та сифонів-утрикул має вигляд тов-
стих (до 2 см у діаметрі), дихотомічно розгалужених шнурів, поверх-
ня яких завдяки утрикулам здається губчастою.
Сосііит розмножується лише статевим шляхом. Статевий про-
цес - гетерогамія. Дводжгутикові жіночі та чоловічі гамети форму-
ються у гаметангіях, що відокремлюються від утрикул перегородка-
ми, тобто водорість є еукарпічною. Чоловічі гаметангії забарвлені у
206
жовтуватий колір, жіночі - у темно-зелений. Після копуляції зигота
без періоду спокою проростає у новий гаметофіт. Вважається, що
мейоз у Сосііит гаметичний, і життєвий цикл, відповідно, диплофаз-
ний. Гамети здатні також проростати партеногенетично.
Кодіум мешкає у теплих морях, переважно на глибині від 5 до
40 м. У Чорному морі досить поширеним видом є кодіум червопо-
дібний - Сосііит уегтіїага.
Рис. 84. Сосііит: А - зовнішній вигляд талому; Б - фрагмент талому з
видовженими ризоїдальними сифонами, від яких відгалужуються міхуроподібні
сифони-утрикули; В - загальний вигляд утрикули; Г - частина утрикули з гаметан-
гіями; Д - чоловіча та жіноча гамети
Порядок Галімедальні - Наїітебаїез
Водорості цього порядку близько споріднені з Вгуорзісіаіез: вони
не мають радіально-симетричної будови, гамети утворюються в га-
метангіях, а не в гаметичних цистах. Галімедальні водорості гете-
ропластидні: крім забарвлених хлоропластів, наявні також безбарвні
амілопласти. Представники порядку є холокарпічними, і при утворенні
гамет талом водорості перетворюється на один велетенський гаме-
тангій. Стефаноконтні зооспори відсутні. Розмноження відбувається
фрагментацією таломів та статевим шляхом. Статевий процес ізо-
гамний або гетерогамний. Галімедальні водорості мешкають пере-
важно у тропічних морях. Характерний представник порядку -
Саиіегра.
207
Рід каулерпа - Саиіегра (рис. 85). Водорість макроскопічна,
складно розчленована і нагадує невеличку трав’янисту рослину. Та-
лом диференційований на систему сланких ризомів, ризоїди та
висхідні асиміляторні «пагони», що у різних видів мають різну форму.
Наприклад, у каулерпи проліферуючої (Саиіегра ргоііїега) «пагін»
має вигляд видовженої ланцетоподібної платівки до ЗО см завдовжки.
Окремий «пагін» складається з багатоосьового більш-менш цилінд-
ричного «стебельця» та звичайно сплощеної верхівки. Кожний «пагін»
утворений ниткоподібними висхідними сифонами, що несуть на собі
утрикули. Ниткоподібні сифони щільно переплетені і з’єднуються між
собою численними анастомозами. Всередині сифонів є система ка-
лозних балок, яка виконує функцію внутрішнього скелета. Ризоми утво-
рені товстими багатоосьовими ниткоподібними сифонами, від яких у
субстрат заглиблюються рясно розгалужені сифони-ризоїди.
Функцію фотосинтезу виконують утрикули висхідних сифонів. Вони
містять численні хлоропласти та амілопласти. Вдень пластиди зосе-
реджуються у периферичній частині утрикул, вночі вони мігрують у
глибину талому і концентруються у ниткоподібних сифонах.
Вегетативне розмноження відбувається при розриві ризомів. Тоді
пошкоджені сифони перекриваються пробками, а ділянка талому, що
відокремилася, здатна прикріпитися до субстрату і прорости у нову
рослину.
При статевому розмноженні протопласт всієї рослини розпадаєть-
ся на велику кількість дводжгутикових жіночих або чоловічих гамет.
В
А
Рис. 85. Саиіегра: А - зовнішній вигляд талому; Б - калозні опірні балки
(поздовжній та поперечний зрізи через ризом); В - чоловіча та жіноча гамети
208
Гамети виходять назовні і копулюють. Статевий процес гетерогам-
ний. Зигота без періоду спокою проростає в одноядерну сферичну
рослину - протосферу. У протосфері ядро мітотично ділиться, утво-
рюючи велику кількість первинних ядер. Далі протосфера утворює
численні випини, розвиває систему ризомів, ризоїдів і, нарешті, висхід-
них «пагонів». На цій стадії ядра вже вторинні. Вважають, що завер-
шення розвитку протосфери й утворення вторинних ядер відбуваєть-
ся внаслідок мейозу.
Порядок Дазикладальні - Оазусіабаїез
Таломи мають радіально-симетричну будову і являють собою вісь
із системою апікальних кільцевих сифонів. Клітинні оболонки склада-
ються переважно з маннану і просякнені карбонатами, найчастіше -
у формі арагоніту. Фотосинтетичний апарат гомопластидний, пред-
ставлений лише хлоропластами. Запасні полісахариди відкладають-
ся як у стромі хлоропласта, так і безпосередньо у цитоплазмі.
В онтогенезі дазикладальні проходять послідовно дво- або три-
річну стадію одноядерної рослини з первинним ядром та коротко-
часну стадію власне сифональної рослини з багатьма вторинними
ядрами. Статевий процес ізогамний, причому гамети утворюються
не у гаметангіях, а в особливих гаметичних цистах.
Рід ацетабулярія - АсеіаЬиІагіа (рис. 86). Водорість поширена
в теплих морях. Зокрема, в Середземному морі зустрічається ацета-
булярія середземноморська (Д. тесіііеггапеа). Талом ацетабулярії
має вигляд парасольки до 18 см заввишки, яка утворена вертикаль-
ною віссю та кільцем верхівкових сифонів. Водорість прикріплена до
субстрату лопатевим ризоїдом. Нерозгалужена вертикальна вісь, яку
часто називають пагінцем, спочатку утворює на верхівці стерильні
гілочки, де наприкінці циклу розвитку формується плодюче кільце.
Промені парасольки спершу сполучаються з порожниною пагінця.
До цієї стадії ацетабулярія не лише одноклітинна, а й одноядерна, на
відміну від більшості сифональних водоростей. Одне велетенське ядро
знаходяться в ризоїді. Лише після того як сформувалось плодюче
кільце, відбувається редукційний поділ ядра, дочірні ядра в ризоїді
продовжують ділитись мітотично. Тисячі новоутворених дрібних ядер
з током цитоплазми по пагінцю переходять у промені кільця.
Промені кільця відділяються від пагінця перегородками, а їх ба-
гатоядерний вміст розпадається на сферичні клітини - цисти, які по-
криваються оболонками. У цистах, звільнених після руйнування про-
менів кільця, утворюються дводжгутикові ізогамети. Цисти відкрива-
209
ються кришечкою. Гамети через отвір виходять назовні і попарно ко-
пулюють. Зигота розвивається у нову диплоїдну особину. Отже, мей-
оз в ацетабулярії відбувається безпосередньо перед утворенням га-
метангіїв (цист). Таким чином, життєвий цикл водорості диплофаз-
ний, з гаметичною редукцією, без чергування поколінь та специфіч-
ною стадією - цистою, яка виконує функцію гаметангія.
Рис. 86. Послідовні стадії циклу розвитку АсеіаЬиІагіа: А, Б-стерильні таломи
першого та другого років із первинним ядром у ризоїдальній частині; В - фертиль-
ний талом третього року з сегментами, що утворюють парасольку; Г - поздовжній
зріз через парасольку; Д - послідовні стадії утворення гаметичних цист; Е - зріла
гаметична циста; Є - вихід гамет із цисти; Ж - ізогамета; 3 - планозигота;
И - зигота; І - проростання зиготи; ї - молодий стерильний талом першого року
210
Протягом більшої частини життєвого циклу - до настання роз-
множення - ацетабулярія не лише одноклітинна, але й одноядерна,
причому ядро локалізоване в певному місці - ризоїді. Це зробило аце-
табулярію ідеальним об’єктом для дослідження взаємовідносин між
ядром і цитоплазмою в живій клітині. Цьому сприяють швидка реге-
нерація і надзвичайна живучість велетенського ядра ацетабулярії.
Ядро можна дістати з клітини, відмити розчином сахарози та імплан-
тувати в іншу клітину. Саме в результаті подібних маніпуляцій з аце-
табулярією вдалось з'ясувати функцію ядра в клітині та відкрити яви-
ще позаядерної спадковості.
Клас Харофіцієві - СЬагорЬусеае
Об’єднує одноклітинні та багатоклітинні прісноводні, солонува-
товодні та наземні водорості з кокоїдним, нитчастим та гетеротри-
хальним типами морфологічної структури тіла. Монадні клітини (коли
вони є) вкриті субмікроскопічними органічними лусочками і мають
асиметричний цитоскелет. Мітоз відкритий або напіввідкритий. Цито-
кінез відбувається за участю фрагмопласта. Для вегетативних клітин
характерна наявність комплексу пероксисома-ядро-хлоропласт-міто-
хондрія.
Система класу. За будовою талому, наявністю джгутикових
стадій, типом статевого процесу, особливостями організації клітин-
них оболонок, наявністю порового апарату, будовою жіночих стате-
вих органів клас за різними системами поділяють на 7-10 порядків, з
яких нижче розглядається п’ять, які представляють три різні лінії ха-
рофіцієвих водоростей. До першої входить порядок СКагаІез; до дру-
гої - так звані кон’югати - 2удпетаіаІЄ8 та Оезтісііаіез; до третьої -
КІеЬзогтісІіаІез та СоІеосКаеіаІез. За молекулярно-генетичними да-
ними, найдавнішим є порядок СКагаІез. Пізніше в межах СКагорКусеае
відокремилися дві гілки: одна дала початок кон’югатам (2удпетаіаіе8
та Оезтісііаіез), друга - порядкам КІеЬзогтісІіаІез та СоІеосКаеіаІез,
що мають спільного пращура з вищими рослинами.
Порядок Харальні - СЬагаїез
Об’єднує водорості з гетеротрихальним типом морфологічної
структури тіла, які мають макроскопічні членисто-кільцеві таломи,
джгутикові стадії, що представлені сперматозоїдами, оогамний ста-
тевий процес, багатоклітинні жіночі статеві органи. Харальні водо-
211
рості мешкають у бентосі озер та приморських солонуватоводних
лиманів. Характерними представниками порядку є СІїага та МіеІІа.
Рід хара - СІїага (рис. 87). Спорофіти хари мають вигляд розга-
лужених кущиків, що складаються з прямостоячих опірних ниток нео-
бмеженого росту-т. зв. «стебел», бічних кільцево розміщених асим-
іляторних ниток обмеженого росту- «листків», та ниток-ризоїдів. Кож-
на нитка («стебло», «листок», ризоїд) диференційована на клітини
вузлів та міжвузлів, що правильно чергуються одна з одною.
Вузли та міжвузля утворюються внаслідок поділу апікальної кліти-
ни. Вона ділиться у поперечній до осі нитки площині, по черзі відок-
ремлюючи від себе клітини двох типів - подвійно-опуклу та подвійно-
увігнуту. Подвійно-опукла клітина до подальшого поділу не здатна -
вона лише росте та розтягується, поступово диференціюючись у ве-
летенську багатоядерну клітину міжвузля. Подвійно-увігнута здатна
до подальших поділів. Ці поділи відбуваються у радіальних напрям-
ках у поздовжній до осі клітини площині. Дочірні клітини утворюють
вузол. Кожна його клітина, в свою чергу, здатна до поділу, який відбу-
вається так само, як і в апікальній клітині, внаслідок чого утворюють-
ся «листки» та бічні гілки - «стебла».
З клітин стеблових вузлів розвиваються стеблова кора та при-
листники, з листових вузлів - спорангії, що дають початок чолові-
чим або жіночим гаметофітам. На ризоїдальних вузлах утворюють-
ся вивідкові бруньки (бульбочки), що є органами вегетативного роз-
множення.
Клітини вкриті оболонками, в яких мікрофібрили целюлози син-
тезуються розетковими термінальними ферментними комплексами,
і зазвичай просякуються вапном. Центр клітини зайнятий великою
вакуолею з клітинним соком. Цитоплазма з органелами утворює тон-
кий периферичний шар. Вона диференційована на дві зони: стабіль-
ну та лабільну. Стабільна зона прилягає до плазмалеми, і в ній од-
ним шаром у правильних поздовжніх рядах розташовуються численні
дрібні дископодібні хлоропласти, що позбавлені піреноїдів. Під ста-
більною зоною знаходиться лабільна зона. Тут зосереджені численні
ядра, мітохондрії, елементи ендоплазматичної сітки та прикріплені
до них мікрофібрили міозину; весь цей комплекс потоком цитоплаз-
ми швидко переміщується вздовж осі клітини. Швидкість руху цито-
плазми становить 50-100 мкм/секунду. Стабільна та лабільна зони
розділені шаром паралельно розташованих пучків актину, що утво-
рюють направляючі «рейки», по яких ковзають мікрофібрили міозину
212
разом зі з'єднаними з ними органелами. Потоки цитоплазми проти-
лежних напрямків не змішуються між собою, і межа між ними має
вигляд безбарвної смуги.
Клітини міжвузлів багатоядерні. їх ядра мають характерну лопа-
теву форму, крім звичайних ядерних структур (хроматину, ядерець,
прорибосом), вони містять пучки мікротрубочок і діляться амітотич-
но. Клітини вузлів, навпаки, одноядерні, сферичні, їх поділ відбуваєть-
ся шляхом мітозу. Всі клітини спорофіта позбавлені центріолей. У кліти-
нах чоловічих гаметофітів, навпаки, центріолі є і беруть участь у форму-
ванні веретена поділу при утворенні сперматозоїдів. Мітоз відкритий.
Цитокінез відбувається за участю фрагмопласта, причому у попереч-
них перегородках між дочірніми клітинами утворюються первинні пори
з плазмодесмами.
Гаметофіти розвиваються зі спорангіальних клітин, що утворю-
ються у вузлах бічних гілок обмеженого росту. Жіночий гаметофіт скла-
дається з великої яйцеклітини та дрібних супутніх клітин. Супутні кліти-
ни з’єднують гаметофіт зі спеціалізованою клітиною міжвузля споро-
фіта, яку називають клітиною-ніжкою. Від вузла, розташованого під
клітиною-ніжкою відходять п’ять клітин кори оогонія. Вони по спіралі
щільно обплітають гаметофіт і на верхівках відокремлюють по одній
замикаючій клітині, які разом утворюють одноярусну п’ятиклітинну
коронку. Таким чином, дво- або чотириклітинний жіночий гаметофіт
захищається десятьма спеціалізованими клітинами спорофіта. Сис-
тема з гаметофіта, його корових клітин та коронки утворює оогоній.
Чоловічі гаметофіти - октанти - багатоклітинні, вони з’єднуються
по вісім, утворюючи сферичні антеридії. Октант складається із щитка,
головок та сперматогенних ниток. Зовнішні клітини октантів - щитки -
з’єднуються між собою боками, утворюючи замкнену сферу. Від щитка
у порожнину сфери відходить одна клітина-рукоятка з однією
первинною та шістьма вторинними клітинами-головками. Вторинні
головки несуть на собі по чотири двохсотклітинні сперматогенні нит-
ки, кожна клітина яких дає початок одному сперматозоїду. Спермато-
зоїди спірально скручені, мають два джгутики, асиметричну кореневу
систему і вкриті субмікроскопічними лусочками.
Статевий процес оогамний. Зрілі щитки антеридію відокремлю-
ються один від одного, оболонки клітин сперматогенних ниток ослиз-
нюються, і сперматозоїди потрапляють у воду. Далі вони підплива-
ють до оогонія, клітини коронки при цьому розсуваються, утворюючи
отвір, через який сперматозоїд потрапляє до яйцеклітини. Після ко-
213
Рис. 87. Сіїага: А - зовнішній вигляд талому; Б — фрагмент талому споро-
фіта Сіїага із жіночим та чоловічими гаметофітами; В—Е - послідовні стадії роз-
витку антеридію; Є - октанти; Ж - сперматогенні нитки; 3 - фрагмент спермато-
генної нитки та вихід сперматозоїдів; И - сперматозоїд. 1 - яйцеклітина; 2 - кора
оогонія; 3 - коронка; 4 - чоловічі гаметофіти, що утворюють антеридій
214
пуляції утворюється ребриста зигота - ооспора, яка переходить у стан
спокою. Після закінчення періоду спокою в зиготі відбувається мей-
оз, причому з чотирьох гаплоїдних ядер три дегенерують. Далі про-
ходить мітоз, одна з утворених клітин стає апікальною клітиною май-
бутнього стебла, друга - апікальною клітиною ризоїдів. Внаслідок
поділу та подальшої диференціації клітин утворюється сланка (ри-
зоїдальна) та висхідна частини нового талому. Таким чином, життє-
вий цикл харальних водоростей гаплофазний, із зиготичною редук-
цією, гетероморфною зміною поколінь та статевим диморфізмом.
Рід нітелла - МіеІІа (рис. 88) - зовні нагадує хару, але відрізняєть-
ся від останньої відсутністю стеблової кори та наявністю коронки, що
складається з десяти клітин, розміщених у два яруси - по п’ять клітин
у кожному.
2
Рис. 88. МіеІІа: А - спорофіт; Б - фрагмент талому з
гаметофітами; В - оогоній. 1 - яйцеклітина; 2 - кора оого-
нія; 3 - коронка; 4 - супутні клітини жіночого гаметофіта
215
Порядок Зигнематальні - 2удпета1аІе8
Об’єднує водорості з кокоїдним та нитчастим типами морфологічної
структури тіла, в яких джгутикові стадії та центріолі відсутні, статевий
процес - кон’югація, мітоз напіввідкритий. Клітинні оболонки гладенькі,
без порового апарату. Клітини не диференційовані на напівклітини.
Життєвий цикл гаплофазний, без чергування поколінь, із зиготич-
ною редукцією.
На роди зигнематальні поділяються, в першу чергу, за типом
морфологічної структури та формою хлоропласта. До найпоширені-
ших зигнематальних водоростей належать нитчасті водорості з родів
Зрігодуга, Худпета та Моидеоііа.
Рід спірогіра - Зрігодуга (рис. 89 А, Б). Нитки цієї водорості скла-
даються з витягнутих або коротких циліндричних клітин. Клітинна обо-
лонка целюлозна, зовні вкрита слизовим шаром, виразно помітним у
розчині туші, а також відчутним на дотик. Поперечні клітинні перего-
родки часто утворюють різноманітні зморшки.
У кожній клітині міститься від одного до кількох (здебільшого 1-4)
стрічковидних спіральних хлоропластів з піреноїдами. Краї хлоро-
пластів нерівні, інколи розсічені. У багатьох видів середина хлоро-
пластної стрічки з внутрішнього боку потовщена у вигляді т. зв. гребе-
ня. По гребеню на більш-менш однаковій відстані розміщені добре
помітні піреноїди з крохмальною обгорткою. Центр клітини займає
велика вакуоля з клітинним соком. Ядро велике, з добре помітним
ядерцем. У частини видів воно кулясте, в інших має лінзовидну фор-
му. Ядро «підвішене» в центрі вакуолі в цитоплазматичному мішечку,
від якого на різні боки до периферії клітини відходять цитоплазма-
тичні тяжі.
Наростання нитки відбувається внаслідок скоординованих поділів
ядра та цитоплазми. На початку мітозу ядерце не зникає, а ділиться
як самостійна органела, через що його інколи називають ендосомою.
Цитокінез у видів роду Зрігодуга відбувається внаслідок одночасного
росту кільцевої борозни та клітинної платівки. Остання утворюється
за участю фрагмопласта. Клітинна платівка збільшується у розмірах
внаслідок злиття пухирців з матеріалом майбутньої клітинної перего-
родки, що відшнуровуються від комплексу Гольджі. В платівці немає
пор, і тому дочірні клітини не з’єднуються між собою плазмодесма-
ми.
Вегетативне розмноження у спірогіри здійснюється фрагмента-
цією ниток внаслідок відмирання окремих інтеркалярних клітин. Ці
фрагменти, і навіть окремі клітини, можуть проростати у нові нитки.
216
Статевий процес - кон’югація. Розрізняють драбинчасту та бічну
кон’югацію. Більш поширена драбинчаста кон’югація, яка відбуваєть-
ся між клітинами двох ниток. У цьому випадку нитки розміщуються
паралельно одна до одної і об’єднуються спільним слизом. Далі від
клітин протилежних ниток назустріч одна одній починають рости ко-
пуляційні відростки, які стикаються своїми кінцями. Оболонки в зоні
контакту розчиняються, і утворюється кон’югаційний канал. По ньо-
му протопласт однієї клітини поступово проштовхується в іншу. Там
протопласти та ядра зливаються, і утворюється зигота. Оскільки по-
ведінка клітин при кон’югації відрізняється, то сприймаючу клітину на-
зивають жіночою, а ту, яка віддає протопласт - чоловічою.
Зигота виробляє товсту тришарову оболонку і переходить у стан
спокою. При проростанні диплоїдне ядро зиготи редукційно ділиться,
причому з чотирьох гаплоїдних ядер лише одне залишається життє-
здатним. Оболонка зиготи розривається, і протопласт проростає у
нову нитку.
Рис. 89. Нитчасті зигнематальні водорості: А, Б - Зрігодуга: А - вегетативна
нитка; Б - драбинчаста кон’югація; В - 7удпета\ Г - Моидеоііа
217
При бічній кон’югації популяційні відростки утворюються між су-
сідніми клітинами однієї нитки.
Види роду Зрігодуга поширені у стоячих або повільно текучих
водах: заводях, старицях річок, ставках, болотах, канавах, калюжах.
Часто спірогіра утворює великі маси яскраво-зеленого жабуриння,
яке слизьке на дотик.
Рід зигнема - Худпета (рис. 89 В). Види цього роду за зовнішнім
виглядом жабуриння нагадують спірогіру, але при мікроскопії відрізня-
ються будовою хлоропластів: в кожній клітині їх два, вони розміщені
в центрі, мають зірчасту форму. В кожному хлоропласті добре по-
мітний один великий піреноїд.
Зигнема розвивається в таких самих біотопах, як і спірогіра, час-
то разом з нею.
Рід мужоція - Моидеоііа (рис. 89 Г). Види цього роду також ут-
ворюють темно-зелене слизьке жабуриння і часто суцільно вкрива-
ють поверхню водойм. Від зігнеми та спірогіри мужоція відрізняється,
в першу чергу, наявністю в кожній клітині лише одного хлоропласта,
який має вигляд платівки з двома або кількома піреноїдами і розмі-
щується по осі клітини.
Таломи багатьох видів мужоції легко розпадаються на поодинокі
клітини. При цьому водорість стає схожою на кокоїдну водорість роду
Мезоіаепіит. Цікаво, що в обох родах хлоропласт здатний змінюва-
ти своє положення залежно від інтенсивності освітлення. Так, при
слабкому освітленні він повертається анфас, перпендикулярно до
падаючих променів; при інтенсивному - навпаки, у профіль, у пло-
щину, паралельну до джерела світла.
Порядок Десмідіальні - Оезтібіаіез
Включає кокоїдні водорості, в яких джгутикові стадії та центріолі
відсутні, статевий процес - кон’югація, мітоз напіввідкритий. Клітинні
оболонки переважно скульптуровані, обов’язково мають простий або
складний поровий апарат. У примітивніших десмідіальних клітини не
диференційовані на напівклітини, у більш розвинених клітина поділена
на дві напівклітини, що з’єднуються між собою перешийком, в якому
розташовується ядро. Подібно до зигнематальних, життєвий цикл гап-
лофазний, без чергування поколінь, із зиготичною редукцією.
Найпоширенішими та багатими у видовому відношенні представ-
никами порядку є СІозіегіит та Созтагіит.
218
Рід клостеріум - СІозіегіит (рис. 90). Водорість має пооди-
нокі, веретеноподібно зігнуті клітини. Клітинна оболонка гладенька
або злегка штрихувата і містить багато дрібних та дві великі пори.
Останні знаходяться на полюсах клітини і виконують функції локомо-
торних органів - через них клітина виділяє слиз. Виділення слизу або
спричинює повільний реактивний рух, або слиз утворює швидкорос-
тучі слизові ніжки, за допомогою яких клітина змінює своє положення
у просторі. При слабкому освітленні у клітин спостерігається пози-
тивний фототаксис, при інтенсивному - негативний. Хлоропласти на-
гадують осьові конуси з радіальними поздовжніми ребрами; по осі
хлоропласта розташовується кілька піреноїдів. На полюсах клітин ут-
ворюються вакуолі з кристалами гіпсу.
Водорість розмножується поділом клітин надвоє та внаслідок ста-
тевого процесу - кон’югації. Зигота, що утворюється внаслідок кон’ю-
гації, переходить у стан спокою, після завершення якого відбуваєть-
ся мейоз. Далі два з чотирьох гаплоїдних ядер дегенерують, і зигота
проростає двома проростками.
Рід налічує більше двохсот видів, серед яких близько 70 знай-
дені на території України. Види роду широко розповсюджені в різно-
манітних прісних водоймах, рідше зустрічаються на вологих скелях
та мохах.
Рис. 90. СІозіегіит А - вегетативна клітина; Б - виділення слизу термі-
нальними порами (препарат забарвлений розчином туші)
219
Рід космаріум - Созтагіит (рис. 91). До цього роду належать
кокоїдні мікроскопічні прісноводні водорості, що мають клітини з двох
симетричних напівклітин. Останні з’єднуються між собою широким
або вузьким перешийком. Напівклітини не розсічені на лопаті, хоча їх
форма може бути дуже різноманітною (наприклад, округлою, еліп-
соїдною, майже квадратною, кутастою тощо).
Клітинні оболонки зазвичай скульптуровані бородавками, шипа-
ми, шипиками, нерідко просякуються солями заліза. Клітини мають
складний поровий апарат, що складається з багатьох дрібних та двох-
чотирьох великих слизових пор, розташованих на полюсах клітин.
Слизові пори забезпечують рух клітини, механізм якого такий самий,
як і у СІозіегіит. Кожна напівклітина містить один або два хлоропла-
сти з піреноїдами, а в зоні перешийка - одне ядро.
Розмножується космаріум вегетативним поділом або внаслідок
кон’югації. При вегетативному поділі у перешийку за способом фраг-
мопласта утворюється клітинна перегородка. Кожна молода клітина
розростається у зоні перешийка, внаслідок чого половини переший-
ка перетворюються на нові напівклітини. Під оболонкою кожної моло-
дої напівклітини утворюється типова для десмідіальних водоростей
тришарова оболонка. Далі зовнішній шар оболонки нової напівкліти-
ни скидається, і клітини роз’єднуються.
Життєвий цикл водоростей роду Созтагіит гаплофазний, без
чергування поколінь, із зиготичною редукцією.
Рис. 91. Созтагіит'. А - клітина в оптичному розрізі; Б - вигляд з поверхні.
1 - хлоропласт; 2 - піреноїди; 3 - ядро
220
На території України водорості цього роду дуже поширені і різно-
манітні за видовим складом. Більшість видів належить до р-мезосап-
робів і мешкає у прісних водоймах різного типу (річках, озерах, став-
ках, калюжах). Проте найчастіше види космаріуму зустрічаються у
сфагнових болотах. Частіше за інші трапляються С. гепі/огте - кос-
маріум ниркоподібний, С. ругатісіаіит - космаріум пірамідаль-
ний, С. тагдагіііїегит - космаріум перламутровий та С. саеіаіит
- космаріум вирізаний.
Порядок Клебсормідіальні - КІеЬзогтібіаіез
Об’єднує водорості з нитчастим типом структури. Джгутикові стадії
представлені зооспорами та гаметами з асиметричним цитоскеле-
том. Мітоз відкритий, відбувається за участю центріолей. Цитокінез
здійснюється шляхом утворення кільцевої борозни, напрямок руху
пухирців з матеріалом поперечної клітинної перегородки визначається
залишками телофазного веретена. Пори з плазмодесмами не утво-
рюються. Клітинні оболонки гладенькі та суцільні. Статевий процес-
гетерогамія. До порядку належить біля 40 видів, більшість з яких пред-
ставляє рід КІеЬзогтісІіит.
Рід клебсормідіум - КІеЬзогтісІіит (рис. 92). Талом має ви-
гляд нерозгалуженої однорядної нитки, що зовні дуже подібна до
ІЛоіЬгіх (значна частина видів клебсормідію була спочатку описана
саме як види роду улотрикс, і лише спеціальні цитологічні та молеку-
лярні дослідження дозволили визначити їх справжнє систематичне
положення). Інколи у ниток можна спостерігати зачаткове галуження,
проте всі клітини нитки залишаються морфологічно та функціональ-
но однакові. Кожна клітина містить один пристінний хлоропласт з піре-
ноїдом, одну-дві великі вакуолі з клітинним соком, розміщені пере-
важно на полюсах клітини; в центральній частині є досить широкий
цитоплазматичний місток, в якому знаходиться ядро.
Розмножується КІеЬзогтісІіит фрагментацією ниток, акінетами,
зооспорами та статевим шляхом. При утворенні монадних клітин -
зооспор або гамет-одна материнська клітина дає початок лише одній
рухливій клітині.
Статевий процес гетерогамний. Гамети, крім того, здатні парте-
ногенетично проростати у нові гаметоспорофіти, тобто виконувати
функцію зооспор. Життєвий цикл гаплофазний, із зиготичною редук-
цією і без чергування поколінь.
Види КІеЬзогтісІіит надзвичайно широко поширені у ґрунтах
(особливо степових та лучних), в аерофітоні, зрідка зустрічаються у
221
прісних водоймах. На поверхні ґрунту та на старих плодових тілах
трутовиків водорість може викликати зелене «цвітіння». Деякі види
(зокрема, клебсормідіум слизовий - К. тисозит) відіграють важ-
ливу роль у заростанні незадернованих пісків у заплавах річок та на
приморських дюнах. Масово розвиваючись, вони утворюють чорні
товсті кірки (інколи до кількох сантиметрів завтовшки), закріплюючи
пісок та створюючи передумови для поселення вищих рослин.
Рис. 92. КІеЬзогтісііипг. к- гаметоспорофіт; Б - зачаткове галужен-
ня; В - утворення зооспори; Г - вихід зооспори; Д - акінети; Е, Є - мікро- та макро-
гамети; Ж - початкова стадія проростання зиготи; 3 - апланоспора та її проростан-
ня
Порядок Колеохетальні - СоїеосЬаеіаІев
Об’єднує водорості з гетеротрихальними таломами, які мають
вигляд розгалужених сланких ниток або одношарових паренхіматоз-
них платівок. Джгутикові стадії представлені зооспорами та гамета-
ми з асиметричним цитоскелетом. Мітоз відкритий, відбувається за
участю центріолей. Цитокінез здійснюється шляхом утворення клітин-
ної платівки за участю фрагмопласта. Клітини пов’язані між собою за
допомогою плазмодесм, які проходять через пори у поперечних пе-
регородках. Клітинні оболонки гладенькі. Статевий процес-оогамія.
222
Типовий представник порядку - Соїеосіїаеіе.
Рід колеохете - Соїеосіїаеіе (рис. 93). До цього роду належать
прісноводні, переважно епіфітні гетеротрихальні водорості, які ма-
ють макроскопічні таломи у вигляді розгалужених сланких ниток або
одношарових паренхіматозних платівок. Від окремих клітин таломів,
як правило, догори піднімаються волоски (хети). У кожної хети при
основі є комірець. Таломи водорості прикріплюються до субстрату
без допомоги ризоїдів.
У вегетативних клітинах, які мають ізодіаметричну або злегка
видовжену форму, розрізняється пристінний пластинчастий хлоро-
пласт з одним-двома піреноїдами та одне ядро. При тривалих спос-
тереженнях за живим матеріалом можна помітити, що протопласти
клітин під оболонкою здатні до обмеженого обертального руху.
Розмножуються колеохетальні водорості фрагментацією таломів,
голими дводжгутиковими зооспорами та статевим шляхом.
Статевий процес - оогамія. Яйцеклітина розвивається в оогонії,
який у деяких видів має довгу шийку - трихогіну. В одноклітинних
антеридіях утворюється по одному дводжгутиковому сперматозоїду.
Цікаво, що сперматозоїди, як і у КІеЬзогткііит, здатні проростати
партеногенетично.
Після запліднення зигота переходить у стан спокою, при цьому
від клітини з оогонієм та сусідніх клітин починають відростати гілоч-
ки, щільно притиснені до поверхні оогонія. Через це оогоній із заплід-
неною яйцеклітиною дещо нагадує архегоній примітивних вищих рос-
лин. Після закінчення періоду спокою у зиготі відбувається мейоз і
зигота проростає у гаплоїдний спорофіт. Спорофіт має вигляд диска
з 8-16 клиноподібних клітин. З кожної такої клітини утворюється по
одній дводжгутиковій зооспорі, яка проростає у новий гаплоїдний га-
метоспорофіт. Таким чином, життєвий цикл у Соїеосіїаеіе гаплофаз-
ний, із зиготичною редукцією та гетероморфним чергуванням поколінь.
На території України зустрічається шість видів цього роду, які меш-
кають переважно в обростаннях вищих водних рослин у прісних кон-
тинентальних водоймах різних типів. Найпоширенішими серед них є
С. зсиіаіа - колеохете щитовидна з таломами у формі паренхіма-
тозних дисків, та С. риМпаіа - колеохете подушковидна з нитко-
подібними розгалуженими таломами.
Дослідження видів роду колеохете відіграли важливу роль у фор-
муванні сучасних уявлень про походження вищих рослин: порядок
Соїеосіїаеіаіез після відкриття схожості будови джгутикових стадій
харофіцієвих водоростей та вищих спорових рослин розглядався як
223
лінія, що дала початок мохоподібним та судинним рослинам. На-
явність гетероморфної зміни поколінь, гетеротрихальної будови зі
здатністю формувати паренхіматозні диски, оогонію з трихогіною,
архегоній-подібної будови оогоніїв із заплідненими яйцеклітинами сут-
тєво підкріплювали це припущення. Проте, молекулярно-філогенетичні
реконструкції показали, що лінія, яка привела до виникнення вищих
рослин, відокремилась не від самих колеохетальних, а від їх пращура.
Рис. 93. Соїеосїіаеіе: А, Б - зовнішній вигляд гаметоспорофіта у видів з
таломами пластинчастої (С. зсиіа(а) та розгалужено-ниткоподібної (С. риМпа(а)
форми; В - антеридії та вихід сперматозоїда; Г - зрілий оогоній, вкритий корою;
Д - восьмиклітинний спорофіт, що розвивається із зиготи; Е - проростання спо-
рофіта зооспорами; Ж - початкові стадії розвитку гаметоспорофіта
224
РОЗДІЛ 2
ГРИБИ
У широкому розумінні гриби - це філогенетично гетерогенна гру-
па евкаріотичних безхлорофільних гетеротрофних організмів, які жив-
ляться переважно осмотрофно, здатні розмножуватися за допомо-
гою спор та мають, як правило, гіфальну будову вегетативного тіла.
Від водоростей гриби відрізняються, в першу чергу, відсутністю
фотосинтетичного апарату; від тварин - переважанням осмотроф-
ного типу живлення, розмноженням за допомогою спор та наявністю
клітинних оболонок. Проте такі відміни грибів від тварин не є універ-
сальними. Наприклад, до об’єктів, які за назвами визнаються Міжна-
родним кодексом ботанічної номенклатури (МКБН) грибами, належать
міксомікотові слизовики. Вони позбавлені клітинних оболонок і здатні
до фаготрофного живлення (хоча розмножуються за допомогою спор).
Крім того, серед грибів, що живляться осмотрофно, виділяються
відділи, що походять від гетеротрофних пращурів (т.зв. справжні гри-
би - СИуігісІіотусоіа, 2удотусоіа, Азсотусоіа, Вазісііотусоіа), та
відділи, що, ймовірно, походять від водоростей, які вторинно втрати-
ли пластиди. Останніх часто називають псевдогрибами, і включають
у цю групу відділи Оотусоїа, НурИосИуігіотусоіа та 1_аЬугіпіііиІотусоіа.
На філогенетичному дереві відділи грибів представлені серед
царств Дискокристат, Тубулокристат та Платикристат. Зокрема, до
дискокристат належить відділ акразіомікотових слизовиків
(Асгазіотусоіа). Серед тубулокристат відділ справжніх слизовиків
(Мухотусоіа) входить до підцарства Амебо-флагелят, а всі три відділи
псевдогрибів - оомікотові (Оотусоїа), гіфохітриомікотові (НурИосіїу-
ігіотусоіа) та лабірінтуломікотові гриби (ІаЬугіпіііиІотусоіа) - до
підцарства Страменопілів. Гриби з царства Платикристат є найчис-
ленішими. До них належить відділ плазмодіофоромікотових слизо-
виків (РІазтосІіорІїоготусоіа) та чотири відділи справжніх грибів -
хітридіомікотові (СИуігісІіотусоІа), зигомікотові (2удотусоіа), аскомі-
котові (Азсотусоіа) та базидіомікотові (Вазісііотусоіа) гриби.
Зазвичай, слизовики та псевдогриби об'єднують під загальною
назвою «грибоподібні організми». Справжні гриби розглядаються як
самостійне підцарство Еипді, яке разом з підцарствами РІапіае та
Апітаїіа складає царство Платикристат. Нижче, оперуючи терміном
«гриби», ця група приймається у широкому розумінні і включає відділи
слизовиків, псевдогрибів та справжніх грибів.
225
Гриби поширені в усіх типах біотопів - ґрунтах, морях, континен-
тальних водоймах, у повітрі тощо. Вони розвиваються на різноманіт-
них природних субстратах рослинного та тваринного походження, на
штучних матеріалах, створених людиною, та ін. Серед грибів відомі
сапротрофи, симбіонти рослин та тварин, а також паразити рослин,
грибів, комах, тварин та людини. Багато грибів є їстівними, проте відо-
мо також чимало їх отруйних видів.
Сьогодні описано приблизно 100 тис. видів грибів, які об’єднані у
10 відділів різних царств. Проте їх очікуване різноманіття, за оцінка-
ми різних авторів, становить від 300 тис. до 1,5 млн. видів.
Основні систематичні ознаки грибів різних відділів
Біохімічні ознаки
До комплексу провідних біохімічних ознак відділів грибів нале-
жать, в першу чергу, шляхи біосинтезу амінокислоти лізину, а також
способи синтезу триптофану, здатність до синтезу сидерамінів, склад
продуктів асиміляції.
Біосинтез лізину. Ця амінокислота належить до незамінних, ба-
гатоклітинними тваринами не синтезується. В евкаріотичному світі
відомо два основних шляхи біосинтезу лізину: через а-аміноадипіно-
ву кислоту (т. зв. ААА-шлях) та через диамінопімелінову кислоту (ДАП-
шлях).
Лізин за ААА-шляхом синтезується з ацетилкофермента А та а-ке-
тоглутарової кислоти. Вони утворюють а-аміноадипінову кислоту, що
надалі перетворюється на лізин. Цей шлях називають грибним шля-
хом біосинтезу, оскільки за ним лізин синтезується у справжніх грибів
(відділи СИуїгісІіотусоІа, 2удотусо1а, Азсотусоіа та Вазісііотусоіа),
хоча також притаманний евгленофітовим водоростям.
Синтез за ДАП-шляхом відбувається з аспартата та пірувата че-
рез диамінопімелінову кислоту. Цей шлях біосинтезу називають рос-
линним, оскільки саме за ним відбувається біосинтез лізину у всіх
евкаріотичних водоростей (крім евгленофітових) та вищих рослин, а
також у псевдогрибів із відділів Оотусоїа та НурИосіїуїгіотусоІа.
Для частини грибів лізин є незамінною амінокислотою, яку
організм сам не здатний синтезувати. Зокрема, це спостерігається у
псевдогрибів з І_аЬугіпіи!отусоІа.
Біосинтез триптофану. У грибів ця амінокислота утворюється
внаслідок роботи специфічних груп ферментів. Загалом у грибів відо-
мо п'ять таких груп ферментів. Перші чотири групи притаманні гри-
226
бам - платикристатам, п'ята група - оомікотовим грибам. Ферментні
комплекси першої групи виявлено у плазмодіофоромікотових, хітри-
діомікотових та аскомікотових грибів. Друга група - у шапинкових
базидіомікотових грибів. Третя група характерна для дріжджеподіб-
них аскомікотових, а четверта - для зигомікотових та базидіомікото-
вих грибів (крім шапинкових).
Утворення сидерамінів. Сидерамінами називають складні
органічні сполуки, які транспортують у клітину залізо. Сидераміни
синтезують всі справжні гриби, крім зигомікотових. Далі молекули цих
речовин виділяються у зовнішнє середовище, зв’язуються з іонами
заліза і знов поглинаються клітиною гриба. У псевдогрибів (зокрема
оомікотових) сидераміни не утворюються, і поглинання іонів заліза
здійснюється іншими способами.
Продукти асиміляції. Основними продуктами асиміляції майже
в усіх грибів є р-1,4-глюкани, представлені тваринним глікогеном. У ак-
разіомікотових слизовиків основним продуктом асиміляції є близький
до парамілону р-1,3-глюкан або р-1,6-глюкан. В оомікотових грибів
основний продукт асиміляції - це також р-1,3-глюкан, який, проте,
близький до ламінарину та хризоламінарину і має назву міколаміна-
рин. Найпоширенішим додатковим асимілятом виступає олія, яка за-
пасається у великих кількостях переважно у спорах та старіючих кліти-
нах.
Фізіологічні ознаки, пов’язані із живленням
Всі гриби є облігатно гетеротрофними організмами. Проте погли-
нання органічних речовин у них може здійснюватися двома шляхами
- осмотрофно та фаготрофно.
При осмотрофному живленні організм або його окремі клітини
(наприклад, клітини трофічних гіф - гаусторій) поглинають розчине-
ну органічну речовину шляхом абсорбції, без утворення травних ва-
куолей. Зазвичай абсорбуються низькомолекулярні органічні речо-
вини, які утворюються при розщепленні високомолекулярних сполук
різноманітними гідролітичними екзоферментами. Грибні екзофермен-
ти здатні розкладати до мономерів такі біополімери, як целюлозу,
лігнін, хітин, білки, нуклеїнові кислоти тощо.
При фаготрофному живленні органічна речовина поглинається у
вигляді твердих часток за допомогою псевдоподій і надалі перетрав-
люється або у травних вакуолях, або в лізосомах. Осмотрофний тип
227
живлення притаманний всім грибам, тоді як фаготрофний - лише
міксомікотовим слизовикам.
Цитологічні ознаки
До комплексу провідних цитологічних ознак, за якими гриби
відрізняються на рівні відділів, належать: тип клітинних покривів, особ-
ливості організації ядерного апарату, тип мітохондріального апара-
ту, наявність та будова джгутикових стадій.
Клітинні покриви. У справжніх грибів та грибоподібних організмів
клітини можуть бути: а) голими; б) вкритими ектоплазматичним рети-
кулюмом; в) мати клітинну оболонку.
Голі клітини, які вкриті лише плазмалемою, зазвичай здатні до
амебоїдного руху та фаготрофного живлення. Відсутність клітинних
покривів характерна для вегетативних тіл акразіомікотових, міксомі-
котових та плазмодіофоромікотових слизовиків, а також для частини
справжніх грибів з відділу хітридіомікотових. Голі клітини мають і зоо-
спори псевдогрибів та справжніх грибів.
У плазмодіофоромікотових слизовиків вегетативне тіло, представ-
лене багатоядерним плазмодієм, перед початком утворення спор вкри-
вається тонкою оболонкою нез’ясованої хімічної природи, розташова-
ною назовні плазмалеми. Цей покрив помітний лише при електронній
мікроскопії. Плазмодії плазмодіофоромікотових хоча і є голими, але
не здатні до фаготрофного живлення і не утворюють псевдоподій.
У лабіринтуломікотових псевдогрибів над плазмалемою розта-
шовується ектоплазматичний ретикулюм. Він являє собою додатко-
вий зовнішній одномембранний покрив, що утворює численні довгі
тонкі випини, які анастомозують між собою та з ектоплазматичним
ретикулюмом інших клітин, утворюючи складну сітчасту структуру.
Простір між мембраною ектоплазматичного ретикулюму та плазма-
лемою виповнений однорідним аморфним матриксом. Мембрани ек-
топлазматичного ретикулюму продукуються особливими одномемб-
ранними органелами-ботросомами (сагеногенетосомами). Крім того,
на поверхні клітин лабіринтуломікотових виявлені субмікроскопічні
лусочки, які утворюються у везикулах, похідних від комплексу Гольджі.
Найпоширенішим типом клітинних покривів, який притаманний
більшості псевдогрибів та справжніх грибів, є клітинна оболонка. У гри-
бів вона здійснює не лише захисну, але й ряд інших важливих функцій,
зокрема, є місцем локалізації гідролітичних екзоферментів, бере
228
участь в абсорбції поживних речовин із субстрату, в морфогенетич-
них та ростових процесах, надає форму клітинам грибних гіф та орга-
нам розмноження. Зовні клітинна оболонка може бути вкрита шаром
слизу, який утворює капсулу.
Основу клітинної оболонки складають мікрофібрилярні скелетні
компоненти-хітин або целюлоза. Над ними розташовується зовнішній
аморфний матрикс, утворений глюканами, хітозаном або маннаном.
З мікрофібрилярними та аморфними компонентами пов'язані різні
білки та ліпіди, а також пігменти (меланіни, хінони), розчинні цукри,
амінокислоти, різноманітні іони та солі.
В межах конкретних відділів грибів хімічний склад оболонки вва-
жається консервативною ознакою високої таксономічної ваги. Зокре-
ма, скелетним компонентом оболонок псевдогрибів є целюлоза, а
справжніх грибів - хітин. Структурні компоненти аморфного матрик-
су у різних відділах кожної з цих груп різні.
Загалом, за хімічним складом мікрофібрилярного та аморфного
компонентів, оболонки грибів поділяють на целюлозно-глюканові
(Оотусоїа), целюлозно-хітинові (НурИосіїуігіотусоіа), хітин-глюканові
(СИуігісІіотусоїа, Вазісііотусоїа), хітин-глюканові (інколи з мананами)
(Азсотусоіа) та хітин-хітозанові (2удотусоїа). Проте відомо також ряд
виключень з правила сталості типу клітинної оболонки в межах відділу.
Зокрема, у дріжджів, які належать до аскомікотових грибів, в оболон-
ках може бути відсутній хітин, а у представників порядку моноблефа-
ридальних з відділу хітридіомікотових виявлено целюлозу.
У слизовиків, вегетативні стадії яких позбавлені клітинних по-
кривів, оболонками вкриті спори. Зокрема, у спор слизовиків із
Мухотусоіа та представників порядку Асгазіаіез з Асгазіотусоіа ос-
нову оболонки складає целюлоза, а у слизовиків з РІазто-
сііоріїоготусоїа - хітин.
Ядерний апарат. Ядро грибів має типову евкаріотичну будову -
воно оточене двомембранною ядерною оболонкою, містить одне або
рідше - кілька ядерець; ядерна ДНК пов’язана з гістонами і має хро-
мосомну організацію.
Різноманітність ядерного апарату в грибів різних відділів прояв-
ляється, в першу чергу, в кількості ядер у клітині та в особливостях
міюзу.
Кількість ядер. Вегетативні тіла грибів можуть бути одно- та бага-
токлітинними або мати неклітинну будову. Клітини одно- та багатоклі-
тинних грибів зазвичай або одноядерні, або містять два генетично не-
229
однакових ядра, які називають дикаріоном. Дикаріони є асоціацією двох
ядер, що утворилися після злиття цитоплазм двох клітин без подаль-
шої каріогамії. Одноядерні та дикаріонтичні клітини характерні для ве-
гетативних стадій лабіринтуломікотових, аскомікотових та базидіомі-
котових грибів. Крім того, одноядерними є вегетативні амебоїдні кліти-
ни акразіомікотових слизовиків, а також спори більшості грибів.
Неклітинний план будови тіла, при якому в одній клітині містить-
ся багато ядер, відомий у ряді відділів слизовиків, псевдогрибів та
справжніх грибів. Багатоядерні клітини за ступенем генетичної неод-
норідності ядер поділяють на три групи: а) ті, що містять лише гене-
тично однакові ядра; б) ті, що містять багато пар дикаріонів, тобто є
полідикаріонтичними; в) ті, що містять багато генетично відмінних
ядер, тобто гетерокаріонтичні.
Багатоядерні клітини з генетично однаковими ядрами утворюють-
ся тоді, коли багатоядерність виникає внаслідок серії мітозів лише
одного вихідного ядра (наприклад, ядра спори). Такі вегетативні кліти-
ни мають оомікотові, гіфохітриомікотові, хітридіомікотові, зигомікотові
та деякі аскомікотові гриби.
Клітини з багатьма дикаріонами утворюються тоді, коли при ста-
тевому процесі після плазмагамії каріогамія одразу не відбувається,
а ядра щойно утвореного дикаріону починають синхронно мітотично
ділитися. Саме такий тип клітин мають вторинні плазмодії плазмодіо-
форомікотових слизовиків. Первинні плазмодії представників цього
відділу утворюються внаслідок серії послідовних мітозів ядра однієї
зооспори, і, таким чином, всі ядра в первинному плазмодії, на відміну
від вторинного, є генетично однаковими.
Гетерокаріонтичність характерна для плазмодіїв більшості міксо-
мікотових слизовиків, оскільки багатоядерність у цих організмів роз-
вивається не лише внаслідок багатьох мітозів, але й внаслідок злит-
тя із цитоплазмами інших (часто багатьох) одно- та багатоядерних
особин.
Різноманітність мітозу у грибів простежується, в першу чергу, за
наявністю центріолей, поведінкою ядерної оболонки та ядерця.
В акразіомікотових та плазмодіофоромікотових слизовиків, всіх
псевдогрибів, а також хітридіомікотових грибів мітоз відбувається за
участю центріолей. Цікаво, що представники цих відділів у життєво-
му циклі мають репродуктивні джгутикові стадії. У тих відділах
справжніх грибів, де монадні стадії повністю відсутні, а саме - у
2удотусоіа, Азсотусоїа та Вазісііотусоіа - відсутні також і центрі-
олі. У слизовиків із Мухотусоіа центріолі переважно є, але стають
230
центрами організації веретена поділу і беруть участь у мітозі лише
при поділі ядра одноядерних клітин - зооспор чи міксамеб; в багато-
ядерних плазмодіях мітоз відбувається без участі центріолей навіть
у тих випадках, коли вони наявні.
Більшість грибів під час мітозу зберігає ядерну оболонку. В акра-
зіомікотових, міксомікотових, оомікотових, зигомікотових та аскомі-
котових грибів вона залишається інтактною, і мітоз, таким чином, є
закритим. Крім того, на зовнішній ядерній мембрані у зигомікотових
грибів у метафазі з’являється специфічна дископодібна структура.
У плазмодіофоромікотових, лабіринтуломікотових, гіфохітриомікото-
вихта хітридіомікотових біля кожного полюса веретена в ядерній обо-
лонці утворюється по одній великій перфорації, тобто мітоз є на-
піввідкритим. Відкритий мітоз виявлений лише у міксомікотових сли-
зовиків (на стадії одноядерних міксамеб) та у деяких шапинкових ба-
зидіомікотових грибів.
Ядерце у грибів при мітозі демонструє три варіанти поведінки.
При першому варіанті воно зникає на початку мітозу і відновлюється
після його закінчення (Мухотусоіа, І_аЬугіпіиІотусоіа, Нуріїосіїуі-
гіотусоіа). При другому варіанті ядерце не зникає, і на певній стадії
ділиться перешнуровкою: у профазі - в акразіомікотових, у метафазі
- у плазмодіофоромікотових, на початку телофази - у оомікотових та
більшості видів зигомікотових грибів. У частини видів з Сґіуїгісііотусоїа,
Азсотусоіа та Вазібіотусоіа наявний третій варіант поведінки ядер-
ця: на початку мітозу воно виштовхується з ядра у цитоплазму, де і
зберігається до закінчення поділу.
Мітохондріальним апарат. Всі гриби мають мітохондрії типово-
го для евкаріот плану будови - вони оточені двомембранною обо-
лонкою, внутрішня мембрана утворює інвагінації - кристи. Матрикс
мітохондрій містить мітохондріальну ДНК, прокаріотичні рибосоми з
коефіцієнтом седиментації 708, комплекс різноманітних ферментів,
більшість з яких забезпечує процес дихання.
Різноманітність організації мітохондріального апарату, як і у во-
доростей, простежується, в першу чергу, за морфологією крист. Зок-
рема, акразіомікотові слизовики, подібно до евгленофітових водоро-
стей, мають кристи трьох типів - дископодібні, трубчасті та пластин-
часті. Міксомікотові слизовики та псевдогриби містять мітохондрії з
трубчастими та пластинчастими кристами або виключно з трубчасти-
ми. У плазмодіофоромікотових слизовиків та всіх справжніх грибів
(Сіїуігісііотусоіа, 2удотусоіа, Азсотусоіа, Вазібіотусоіа) кристи лише
пластинчасті.
231
Джгутикові стадії. Представники всіх відділів слизовиків, псев-
догрибів та одного з відділів справжніх грибів (СНуїгісііотусоіа) в жит-
тєвому циклі здатні утворювати джгутикові стадії, представлені зоо-
спорами або гаметами. У справжніх грибів з відділів 2удотусо1а,
Азсотусоїа та Вазібіотусоїа відсутні не лише джгутикові стадії, але
й центріолі - органели, похідні від базальних тіл джгутиків.
У найзагальнішому вигляді джгутикові стадії грибів поділяють на
три основні типи: а) з двома ізо- або гетероконтними передніми
гладенькими джгутиками (інколи один із джгутиків може бути реду-
кованим, але зберігається його базальне тіло); б) з двома гетеро-
контними передніми або бічними джгутиками, з яких довший вкри-
тий тричленними волосками - ретронемами (другий джгутик може
бути редукованим до базального тіла); в) з одним заднім батогопо-
дібним гладеньким джгутиком.
Джгутики першого типу характерні для всіх слизовиків. При цьо-
му акразіомікотові мають монадні клітини з двома ізоконтними джгу-
тиками, міксомікотові - з двома гетероконтними, плазмодіофоромі-
котові - з двома гетероконтними або одним джгутиком.
Монадні клітини другого, страменопільного плану будови, харак-
терні для псевдогрибів. Утворення ретронем, які вкривають один із
джгутиків, починається в міжмембранному просторі ядерної оболон-
ки. Далі від зовнішньої мембрани ядра у цитоплазму відшнуровують-
ся пухирці із зачатковими ретронемами. Після дозрівання везикули
рухаються до поверхні клітини, зливаються з плазмалемою біля ба-
зальних тіл джгутиків і ретронеми виводяться, таким чином, на плаз-
малему при основі аксонеми джгутика. Монадні клітини у різних
відділах псевдогрибів відрізняються, в першу чергу, за кількістю джгу-
тиків. У оомікотових та лабіринтуломікотових монадні стадії дводжгу-
тикові, довший джгутик вкритий ретронемами і направлений вперед,
коротший гладенький і орієнтований назад. Монадні клітини гіфохіт-
риомікотових мають лише один передній пірчастий джгутик, від дру-
гого залишається тільки базальне тіло, видозмінене в поодиноку цен-
тріолю.
Монадні стадії третього типу - з гладеньким заднім батогоподіб-
ним джгутиком - називають інколи опістоконтними. Ультратонка бу-
дова таких клітин дуже подібна до будови сперматозоїдів багатоклі-
тинних тварин. Вважають, що опістоконтні клітини виникли від дво-
джгутикових внаслідок редукції одного з джгутиків. Це підтверджуєть-
ся наявністю у клітині поодинокої центріолі, розташованої поблизу
базального тіла джгутика, яка, ймовірно, є видозміненим базальним
232
тілом другого джгутика. Опістоконтні джгутикові стадії у грибів пред-
ставлені лише у відділі СНуігісііотусоіа і розглядаються як один із
доказів походження справжніх грибів та багатоклітинних тварин від
спільного пращура.
Інші нуклеоцитоплазматичні органели та структури. Ендо-
плазматична сітка представлена в усіх грибів, але розвинена в різних
відділах неоднаково. Зокрема, у справжніх грибів із відділів аскоміко-
тових та базидіомікотових вона, крім каналів, часто утворює цистер-
ни, які можуть асоціюватись одна з одною, набуваючи деякої по-
дібності до комплексу Гольджі.
У деяких хітридіомікотових грибів ендоплазматична сітка утво-
рює систему сотоподібно розміщених цистерн, що прилягають до
одного з її каналів. Ця структура відома під назвою румпосоми, але
функція її не з’ясована. У зооспор представників цього відділу ядро
повністю або частково оточене каналами ендоплазматичної сітки, що
відмежовують ділянку цитоплазми, відому під назвою ядерний ков-
пачок. Ядерний ковпачок виповнений великою кількістю рибосом,
рибосомальною РНК та рибосомальними білками і, ймовірно, вико-
нує функцію ядерця, яке, проте, розташоване поза межами ядра. Си-
стема, яка нагадує ядерний ковпачок, виявлена також у зооспор гіфо-
хітриомікотових грибів.
Для базидіомікотових грибів досить характерною є видозміна
ендоплазматичної сітки, відома під назвою парентосоми. Вона являє
собою одномембранну структуру, яка закриває пору у поперечній
перегородці та, ймовірно, бере участь у транспортуванні речовин між
сусідніми клітинами. До аналогічних структур належать тільця Воро-
ніна - електронно-щільні структури, які закривають пору у попереч-
них перегородках деяких аскомікотових грибів.
Комплекс Гольджі розвинений не у всіх грибів. Зокрема, його не-
має у акразіомікотових слизовиків та справжніх грибів (хітридіоміко-
тових, зигомікотових, аскомікотових та базидіомікотових). В аскомі-
котових та базидіомікотових у клітинах замість комплексу Гольджі інко-
ли спостерігаються цистерни ендоплазматичної сітки, що утворюють
невеликі стопки. Проте такі комплекси цистерн обов’язково асоцію-
ються принаймі з одним із каналів ендоплазматичної сітки. Добре
розвинені типові комплекси Гольджі наявні у плазмодіофоромікото-
вих слизовиків та псевдогрибів (оомікотових, гіфохітриомікотових та
лабіринтуломікотових). Зазвичай у цих грибів комплекс Гольджі при-
тиснутий до поверхні ядерної мембрани.
До органел, які беруть участь у процесі росту апікальних клітин
гіф аскомікотових та базидіомікотових грибів, належать літичні вези-
233
кули та хітосоми. Літичні везикули - це дрібні численні одномемб-
ранні везикули, які містять гідролітичні ферменти, здатні розчиняти
хітинову оболонку в зоні росту, обумовлюючи тим самим можливість
апікального росту протопласта. Хітосоми - це великі везикули, ви-
повнені матеріалом майбутньої клітинної стінки, з якого утворюється
хітинова оболонка. Завдяки хітосомам протопласт, що росте, фор-
мує нові ділянки оболонки.
Специфічною органелою справжніх грибів є ломасоми - одно-
мембранні електронно-прозорі везикули, що розміщуються між плаз-
малемою та клітинною оболонкою. Функція ломасом поки що не вста-
новлена.
Мікротільця, що мають вигляд одномембранних везикул з елект-
ронно-щільним вмістом, виявлені у більшості відділів грибів. У хітри-
діомікотових вони асоційовані з великими ліпідними глобулами і ут-
ворюють специфічний везикулярно-ліпідний комплекс. У гіфохітрио-
мікотових мікротільця притиснуті до ядерної оболонки.
Клітини майже всіх грибів при старінні здатні утворювати велику
вакуолю із клітинним соком. Вона відмежовується від цитоплазми ва-
куолярною мембраною - тонопластом - і містить резервні поживні
речовини, а також токсичні продукти метаболізму. В молодих кліти-
нах цитоплазма заповнює весь простір, і в ній можна спостерігати
лише дрібні вакуолі. До найпоширенішого типу включень у грибів на-
лежать ліпосоми, які являють собою великі або дрібні краплини ліпідів.
Морфологія вегетативного тіла
Вегетативні тіла грибів можуть бути макро- або мікроскопічними і
мають один із двох основних варіантів будови: або амебоїдний, або
міцеліальний.
Амебоїдна будова. Амебоїдні таломи позбавлені твердих клітин-
них покривів, здатні до метаболічних змін форми, можуть утворюва-
ти справжні або несправжні псевдоподії. Амебоїдні таломи характерні
для всіх слизовиків, лабіринтуломікотових та гіфохітриомікотових, а
також для багатьох хітридіомікотових грибів. Залежно від кількості
ядер амебоїдні вегетативні тіла поділяють на міксамеби, міксофла-
геляти, псевдоплазмодії та плазмодії.
Міксамеби завжди мікроскопічні, одноядерні або зрідка дикаріон-
тичні, позбавлені джгутиків, здатні до активного руху за допомогою
псевдоподій. За відносними розмірами та кількістю псевдоподії мікса-
меб поділяють на видовжені вузькі і зазвичай численні філоподії та
234
короткі широкі поодинокі лобоподії. Філоподії здатні захоплювати
тверді частинки їжі, тобто, крім локомоторної, виконують ще й тро-
фічну функцію. Міксамеби з філоподіями притаманні міксомікотовим
слизовикам.
Міксамеби з поодинокою лобоподією, яка здійснює лише локо-
моторну функцію, характерні для акразіомікотових слизовиків. Лобо-
подія у міксамеб акразіомікотових розташовується на передньому кінці
клітини. Задня частина міксамеби утворює лопать, в якій розташову-
ються видільні вакуолі, і називається уроїдною зоною.
Міксофлагеляти також мікроскопічні, утворюють філоподії, але,
крім того, мають два гетероконтних джгутики і, таким чином, пред-
ставляють особливий варіант монадних клітин. Залежно від наявності
крапельно-рідкої води та концентрації іонів кальцію міксофлагеляти
здатні перетворюватись на міксамеби і навпаки. Серед грибів веге-
тативні клітини, представлені міксофлагелятами, зустрічаються лише
у відділі Мухотусоіа.
Псевдоплазмодії являють собою агрегати міксамеб, які зберіга-
ють свою індивідуальність. Утворюються псевдоплазмодії внаслідок
об’єднання у колонії поодиноких міксамеб, при цьому цитоплазми
останніх між собою не зливаються. Псевдоплазмодії відомі в акразіо-
мікотових слизовиків, лабіринтуломікотових псевдогрибів та міксомі-
котових слизовиків із класу диктіостеліоміцетів. Псевдоплазмодії кож-
ного з цих таксонів відрізняються за способом агрегації клітинних
індивідів, їх здатністю до руху взагалі та скоординованого руху зок-
рема.
Плазмодії мають неклітинну будову: вони схожі на міксамеби, але
містять багато ядер - від чотирьох до десятків та навіть сотень тисяч.
Розміри плазмодіїв коливаються в широких межах - від мікроскопіч-
них (наприклад, у плазмодіофоромікотових слизовиків) до макроско-
пічних (у багатьох міксомікотових). Плазмодії можуть бути нерухоми-
ми або здатними рухатися за допомогою псевдоподій. Відомо два
способи утворення плазмодіїв: по-перше, з одноядерних міксамеб або
репродуктивних клітин (зокрема, зооспор) внаслідок серії послідов-
них мітотичних поділів ядра; по-друге, внаслідок злиття між собою
кількох (інколи - багатьох) амебоїдних одно- або багатоядерних
індивідів. Плазмодії можуть також утворюватися внаслідок комбінації
обох цих шляхів.
Зі способом утворення плазмодію пов’язаний ступінь генетичної
однорідності його ядер, у відповідності з чим плазмодії поділяють на
генетично однорідні, полідикаріонтичні та гетеродикаріонтичні (див.
235
вище). За розмірами, формою та наявністю в плазмодіях закономірно-
го руху цитоплазми (т. зв. човникових рухів), плазмодії поділяють на
протоплазмодії, афаноплазмодії та фанероплазмодії.
Протоплазмодії мікроскопічні, зазвичай без певної форми або
неправильно сітчасті, без човникових рухів цитоплазми. Такий тип
плазмодіїв притаманний деяким міксомікотовим, плазмодіофоромі-
котовим слизовикам та частині хітридіомікотових грибів.
Афаноплазмодії макроскопічні, неправильно-віялоподібної фор-
ми, із цитоплазмою, що здійснює човникові рухи. Афаноплазмодії зовні
нагадують губчасту протоплазматичну масу. Характерні для частини
міксомікотових слизовиків.
Фанероплазмодії також макроскопічні. На відміну від афаноплаз-
модіїв вони мають досить щільну консистенцію і майже правильну
віялоподібну форму. У фанероплазмодіях є система розгалужених
жилок, по яких цитоплазма здійснює човникові рухи. Зовні фанеро-
плазмодії вкриті шаром ущільненого слизу. Як і афаноплазмодій, цей
тип вегетативного тіла зустрічається лише у представників Мухо-
тусоіа.
Особливим варіантом вегетативного тіла, який часто розгляда-
ють як перехідний між амебоїдним та міцеліальним планами будови,
є ризоміцелій. Ризоміцелій складається з багатоядерної, вкритої
клітинною оболонкою, центральної частини, від якої відходять голі
тонкі розгалужені ризоподії, що позбавлені ядер. Такий тип вегета-
тивного тіла відомий у представників відділів НурНосНуігіотусоіа та
СИуігісііотусоіа.
Міцеліальна будова. Вегетативне тіло переважної більшості
грибів являє собою систему вкритих клітинними оболонками ниток,
яка називається міцелій. Окрема нитка є елементарною одиницею
міцелію і називається гіфою. Зрідка міцелій складається лише з однієї
гіфи, тобто є нерозгалуженим (у деяких хітридіомікотових грибів).
Проте у більшості випадків міцелій галузиться і складається з вели-
кої кількості гіф.
Гіфи мають вигляд циліндричних трубок, діаметр яких коливається
в межах 2-150 мкм, проте найчастіше становить 5-10 мкм. Гіфа здатна
до необмеженого росту в довжину, причому цей ріст завжди апікаль-
ний і обумовлений, у першу чергу, роботою літичних пухирців та хіто-
сом або їх аналогів. За будовою гіфи поділяють на два типи - несеп-
товані та септовані.
Несептовані гіфи багатоядерні і позбавлені поперечних перего-
родок, тобто мають неклітинну будову. Міцелій, який утворений не-
236
септованими гіфами, називають неклітинним міцелієм. Такий тип міце-
лію притаманний майже всім оомікотовим грибам, частині хітридіо-
мікотових та більшості зигомікотових грибів.
Міцелій, який складається з гіф, що рівномірно поділені на кліти-
ни поперечними перегородками - септами, називається клітинним,
або септованим міцелієм. Клітини гіф септованого міцелію можуть
містити або лише одне ядро, або один дикаріон, або кілька ядер.
Септи, які розмежовують сусідні вегетативні клітини гіфи, майже зав-
жди мають поровий апарат, завдяки якому міцелій являє собою фізіо-
логічно цілісний багатоклітинний організм. Структура порового апа-
рату септ у грибів різних таксономічних груп різна. За будовою пор
септи поділяють на мікропорові, прості та доліпорові.
Мікропорові септи перфоровані багатьма дрібними порами. Сеп-
ти такого типу зустрічаються зрідка і відомі лише у деяких представ-
ників хітридіомікотових, зигомікотових та аскомікотових грибів.
Прості септи мають лише одну досить велику центральну пору.
Септа у напрямку до пори потоншується. Пора в простій септі може
бути відкрита (у сажкових грибів з Вазібіотусоіа) або прикрита спе-
ціальними структурами - дрібними вакуолями (наприклад, в іржастих
базидіомікотових грибів) чи тільцями Вороніна (у багатьох аскоміко-
тових).
Доліпорові септи також мають лише одну велику центральну
пору, проте, на відміну від простих септ, поперечна перегородка на-
вколо неї потовщена. Пора в доліпорових септах може бути відкри-
тою, закритою пробкою з аморфної електронно-щільної речовини або
прикрита пористим мембранним ковпачком - парентосомою. Доліпо-
рові септи з парентосомами характерні для більшості базидіомікото-
вих грибів з макроскопічними плодовими тілами. Доліпорові септи без
парентосом поширені переважно серед зигомікотових грибів із сеп-
тованим міцелієм.
У деяких грибів вегетативне тіло має вигляд поодиноких клітин,
що брунькуються. Якщо дочірні клітини після утворення септи не відок-
ремлюються від материнської, то утворюється ланцюжок фізіологіч-
но не пов’язаних між собою клітин, який називають псевдоміцелієм.
Такий тип вегетативного тіла властивий деяким аскоміцетам, зокре-
ма, він характерний для дріжджів.
237
Розмноження
Гриби розмножуються нестатевим та статевим шляхами. Спосо-
би розмноження грибів, в цілому, різноманітніші, ніж у водоростей.
Нестатеве розмноження, аналогічно до водоростей, поділяють
на два основних типи - вегетативне, яке здійснюється переважно за
рахунок різних варіантів фрагментації вегетативного тіла, та справжнє
нестатеве, яке відбувається за допомогою спеціалізованих репродук-
тивних клітин. Зазвичай в мікології під терміном «нестатеве розмно-
ження» розуміють лише справжнє нестатеве розмноження, не при-
єднуючи до нього вегетативне.
Вегетативне розмноження може відбуватися внаслідок поділу
одноклітинних індивідів (міксамеб та міксофлагелят), фрагментацією
плазмодіїв та міцелію. До способів вегетативного розмноження відно-
сять також розмноження шляхом утворення артроспор та хламідо-
спор. Артроспори виникають внаслідок фрагментації гіфи на окремі
короткі клітини. Хламідоспори утворюються подібно до артроспор,
але, на відміну від останніх, мають темнозабарвлені, переважно по-
товщені клітинні оболонки, і є аналогом акінет у водоростей. Брунь-
кування, яке супроводжується відокремленням дочірніх клітин, також
вважається одним із способів вегетативного розмноження.
Нестатеве розмноження здійснюється спеціалізованими кліти-
нами - спорами, які, на відміну від водоростей, можуть утворюватись
не лише ендогенно у спорангіях, але й екзогенно, відокремлюючись
від спеціалізованих гіф - конідієносців.
До спеціалізованих клітин нестатевого розмноження, що мають
ендогенне походження, належать спори міксомікотових слизовиків (за
винятком диктіостеліоміцетових), міксамеби, зооспори, а також спо-
рангіоспори зигомікотових грибів.
Спори міксомікотових нерухомі, вкриті целюлозною оболонкою і
утворюються у спорангіях або всередині плодових тіл. В еугамних
слизовиків утворенню спор передує редукційний поділ ядер; такі спо-
ри мають гаплоїдний набір хромосом. Залежно від наявності у сере-
довищі крапельно-рідкої води спора проростає поодинокою міксаме-
бою, міксофлагелятою або зооспорою.
Розмноження за допомогою міксамеб притаманне акразіомікото-
вим та міксомікотовим слизовикам. Міксамеби утворюються при про-
ростанні ендогенних (у міксомікотових) або екзогенних (у акразіомі-
котових) спор.
Зооспори відомі в усіх відділах слизовиків, псевдогрибів та у
справжніх грибів з відділу хітридіомікотових. Зооспори грибів завжди
238
позбавлені клітинної оболонки. їх ультратонка будова є однією з най-
вагоміших таксономічних ознак, яка використовується при поділі грибів
на відділи (див. вище). В різних відділах грибів зооспори утворюють-
ся в спорангіях різних типів. Зокрема, у слизовиків зооспори розвива-
ються із ендогенних (міксомікотові) або екзогенних (акразіомікотові
та плазмодіофоромікотові) нерухомих спор, причому з однієї спори
виходить лише одна зооспора (фактично нерухомі спори слизовиків
являють собою спорангій з однією зооспорою або міксамебою). Зоо-
спори лабіринтуломікотових утворюються із сорусів цист. Такі цисти
виникають всередині однієї клітини після серії мітозів, які завершу-
ються редукційним поділом. Одна циста дає початок одній зооспорі з
гаплоїдним набором хромосом.
У гіфохітриомікотових, оомікотових та хітридіомікотових грибів в
одному зооспорангії утворюється багато зооспор. Гриби, в яких на
зооспорангій перетворюється все вегетативне тіло, називають холо-
карпічними. Представників, у яких спорангієм стає лише частина ве-
гетативного тіла, належать до еукарпічних форм. Зокрема, всі гіфо-
хітриоміцети є організмами холокарпічними. У хітридіомікотових відомі
як холо-, так і еукарпічні представники.
Оомікотові належать до еукарпічних грибів. Причому спорангії
оомікотових здатні розвиватись або безпосередньо з верхівкової ча-
стини вегетативної гіфи, яка відокремлюється від решти міцелію сеп-
тою (переважно у тих видів, що мешкають у водному середовищі),
або відшнуровуватись від спеціалізованої гіфи - спорангіофора5 (у
більшості наземних представників). Спорангій, який розвивається на
спорангіофорі, є багатоядерним і, залежно від умов зволоження, про-
ростає або зооспорами, або амебоїдним протопластом, або неклітин-
ною гіфою. У водних оомікотових зооспори здатні змінювати свою
морфологію. Це явище отримало назву дипланетизму: зооспора, яка
виходить зі спорангія, має апікальні джгутики; після деякого періоду
активного руху вона інцистується; при проростанні зооспора вихо-
дить з оболонки цисти, але тепер її джгутики розташовуються лате-
рально.
Спорангіоспори також утворюються ендогенно, але, на відміну
від зооспор, міксамеб та спор слизовиків, позбавлені джгутиків, вкриті
міцною оболонкою і проростають у гіфи міцелію. Спорангіоспори
формуються всередині спорангіїв, які знаходяться на верхівках спе-
ціалізованих гіф - спорангієносців, які піднімають органи нестатево-
го спороношення над субстратом.
5 Спорангіофор та спорангій наземних оомікотових грибів у вітчизняній літературі ча-
сто називають конідієносцем із конідією, хоча ці репродуктивні утворення принципово
відрізняються від конідієносців та конідій справжніх грибів.
239
До спеціалізованих клітин нестатевого розмноження, що мають
екзогенне походження, належать спори плазмодіофоромікотових та
акразіомікотових слизовиків і конідії.
Спори плазмодіофоромікотових та акразіомікотових слизовиків
розвиваються внаслідок трансформації та фрагментації частин плаз-
модіїв та псевдоплазмодіїв без утворення спорангіїв. Подібно до спор
міксомікотових, вони проростають поодинокими міксамебами або
зооспорами.
Конідії. Розмноження конідіями є основним і найпоширенішим
способом нестатевої репродукції переважної більшості справжніх
грибів. Конідії - це нерухомі, вкриті оболонкою клітини, що утворю-
ються екзогенно, відокремлюючись від спеціалізованих гіф міцелію -
конідієносців. На відміну від артроспор та хламідоспор, які проходять
стадію диференціації лише після відокремлення від вегетативної гіфи,
диференціація конідій розпочинається ще на міцелії: верхівкова час-
тина конідієносця перетворюється на зачаток конідії, часто збільшуєть-
ся у розмірах, інколи синтезує додаткові шари оболонки і далі відок-
ремлюється від гіфи.
Конідії, які спочатку відокремлюються від конідієносця септою, а
далі дозрівають на ньому, називають алевроконідіями. Перед відок-
ремленням від материнської гіфи пори, які з’єднували зачаткову алев-
роконідію з протопластом конідієносця, закриваються пробками.
Конідії, які спочатку проходять стадію диференціації і лише потім
відокремлюються септою від конідієносця, називають бластоконі-
діями.
Конідієносці мають різноманітну форму - від поодиноких нероз-
галужених до складно багаторазово розгалужених. Конідії на конідіє-
носцях можуть бути поодинокими або утворювати довгі ланцюжки.
Відносно один одного конідієносці можуть розташовуватись різни-
ми способами. За характером розташування конідієносців виділяють
кілька типів конідіальних спороношень: поодинокі конідієносці, коремії,
спородохії, ложа та пікніди.
Поодинокі конідієносці щільних груп не утворюють. Якщо конідіє-
носці розвиваються щільною групою, склеюючись боками за допомо-
гою слизу і утворюючи колонку з головкою конідій на верхівці, то таке
спороношення називають коремієм. Суцільні шари конідієносців, які
розвиваються у вигляді подушечок на опуклій системі щільно пере-
плетених гіф, утворюють спородохії. Ложа нагадують спородохії, але
тут конідієносці розвиваються не на опуклому, а на плоскому спле-
тенні гіф. Якщо сплетення гіф, на якому розвиваються конідієносці,
240
глибоко увігнуте і нагадує горщик, занурений у субстрат або у струк-
туру з видозмінених вегетативних гіф (т. зв. строму), то таке спороно-
шення називають пікнідою.
Типи конідіальних спороношень є важливою ознакою в таксономії
справжніх грибів на рівнях, починаючи з класів і закінчуючи видами.
Проте особливого значення морфологія конідіальних спороношень
набуває у таксономії т. зв. мітоспорових грибів - тих аскомікотових та
базидіомікотових, у яких статеві спороношення не відомі, і місце у
системі поки що є нез'ясованим. У справжніх грибів вегетативну ста-
дію, яка утворює лише нестатеве (переважно - конідіальне) споро-
ношення, називають анаморфною, на відміну від вегетативної стадії,
яка бере участь у статевому процесі і утворює статеве спороношен-
ня - т. зв. телеоморфної. Разом анаморфна та телеоморфна стадії
складають повний цикл розвитку гриба - голоморфу.
Статеве розмноження відбувається внаслідок статевого проце-
су, який, як і в інших евкаріот, обумовлює підтримання певного рівня
рекомбінантної мінливості популяцій. Статевий процес серед грибів
не виявлений лише в акразіомікотових слизовиків, і достовірно не
описаний у лабіринтуломікотових (хоча ультраструктурні дані свідчать
про наявність в останньому відділі мейозу, який відбувається при ут-
воренні цист).
Як і у випадку нестатевого розмноження, статеві процеси у грибів
різноманітніші, ніж у водоростей, і представлені хологамією, різними
варіантами гаметогамії, гаметангіогамією та соматогамією.
Хологамія характерна для слизовиків з відділу Мухотусоіа і відбу-
вається внаслідок злиття вегетативних амебоїдів (переважно мікса-
меб).
Гаметогамія, як і у водоростей, представлена ізо-, гетеро- та оога-
мією. Ізогамія виявлена у міксомікотових та плазмодіофоромікото-
вих слизовиків, а також у гіфохитріомікотових псевдогрибів та
більшості еугамних хітридіомікотових. Гетерогамія в цілому у грибів
зустрічається зрідка і відома лише у деяких хітридіомікотових.
Оогамний статевий процес зустрічається у частини хітридіомі-
котових з порядку моноблефаридальних та в абсолютної більшості
оомікотових (останній відділ отримав назву саме за типом статевого
процесу). В цих відділах оогамія здійснюється по-різному. В хітридіо-
мікотових нерухома яйцеклітина запліднюється рухливими
сперматозоїдами, що мають джгутики, тобто представлена типовим
варіантом. У оомікотових нерухомі яйцеклітини запліднюються не
сперматозоїдами, а вмістом недиференційованих на клітини багато-
241
ядерних чоловічих гаметангіїв, які переливають свій вміст в яйцеклі-
тини через спеціальні вирости клітинної оболонки.
Гаметангіогамія являє собою процес злиття вмісту недиферен-
ційованих на гамети двох гаметангіїв. Варіанти статевого процесу цьо-
го типу відомі у зигомікотових (під назвою зигогамії) та у аскомікотових.
При зигогамії зливаються дві багатоядерні клітини, які розташо-
вуються на верхівках гіф неклітинного міцелію, від якого відокремлю-
ються септами. Морфологічно ці клітини виглядають однаково, але
їх ядра мають різні статеві знаки. Після плазмогамії ядра різних ста-
тевих знаків попарно зливаються, і утворюється зигота з багатьма
диплоїдними ядрами. Вона одягається багатошаровою щільною обо-
лонкою і перетворюється на зигоспору. Після періоду спокою у зиго-
спорі відбувається мейоз, і вона проростає гаплоїдною гіфою, на
верхівці якої формується спорангій із гаплоїдними спорангіоспорами.
Гаметангіогамія з утворенням дикаріонів притаманна аскоміко-
товим грибам. Тут також зливаються гаметанпї, вміст яких не дифе-
ренційований на гамети. Проте, на відміну від зигогамії, плазмогамія
не супроводжується каріогамією. Крім того, хоча гаметангії багато-
ядерні, в статевому процесі беруть участь лише по одному ядру з
кожного гаметангію, які утворюють дикаріон. З такої клітини злиття з
дикаріонтичним ядром, як правило, розвиваються гіфи дикаріонтич-
ного міцелію. Згодом ядра дикаріону зливаються, утворюючи диплої-
дне зиготичне ядро. Далі відбувається мейоз і розвивається статеве
спороношення, представлене сумкою (аском). В аску ендогенно роз-
виваються гаплоїдні аскоспори.
Соматогамія, при якій також утворюються дикаріони, характерна
для базидіомікотових грибів. Спеціалізовані статеві органи в цьому
випадку повністю втрачені, а їх функцію виконують соматичні клітини
вегетативного міцелію. Копуляція відбувається між двома вкритими
оболонками клітинами гаплоїдних гіф. При цьому плазмогамія не су-
проводжується каріогамією, натомість утворюються дикаріони. З та-
ких дикаріонтичних клітин розвивається основний тип міцелію бази-
діомікотових - дикаріонтичний септований міцелій. Наприкінці дика-
ріонтичної фази ядра дикаріонів зливаються, утворюється диплоїдне
зиготичне ядро, відбувається мейоз і розвивається статеве спороно-
шення - базидія із розташованими на її поверхні екзогенними гаплоїд-
ними базидіоспорами.
Серед грибів із соматогамним статевим процесом відомі як гомо-
талічні, так і гетероталічні форми. У гомоталічних грибів статевий
процес може відбуватися між клітинами одного й того ж міцелію.
242
У гетероталічних зливаються тільки клітини різних статевих знаків, які
утворюються на різних міцеліях. Гетероталізм може бути двох типів:
біполярний, при якому стать визначається однією парою алелей, та
тетраполярний, коли стать визначають дві пари алелей, локалізова-
них в різних хромосомах. Такі алельні пари комбінуються незалеж-
но, і, як наслідок, клітини, які беруть участь у копуляції, мають не два
(чоловіча або жіноча), а чотири статевих знаки.
Таким чином, відділи грибів добре відрізняються за типом стате-
вого процесу. Зокрема, серед слизовиків у акразіомікотових він
відсутній, у міксомікотових - хологамний або ізогамний, у плазмодіо-
форомікотових - ізогамний. Серед псевдогрибів статевий процес у
гіфохітриомікотових представлений ізогамією, а у оомікотових-оога-
мією, яка здійснюється без участі чоловічих гамет. У справжніх грибів
в межах відділу хітридіомікотових статевий процес являє собою різні
типи класичної гаметогамії - ізо-, гетеро- та оогамію; у зигомікотових
- гаметангіогамію за типом зигогамії, в аскомікотових - також гаме-
тангіогамію, але без утворення справжніх зигот. Для базидіомікото-
вих характерна соматогамія.
Статеві спороношення. Кінцевим продуктом статевого процесу
у грибів є утворення диплоїдних або гаплоїдних спор, які внаслідок
рекомбінації ДНК у диплоїдній фазі стають генетично відмінними від
батьківських геномів. Такі спори «тиражують» результати статевого
процесу. Структури, де утворюються ці спори, називають органами
статевого спороношення грибів, або статевими спороношеннями.
В різних відділах грибів статеві спороношення відмінні і завдяки цьо-
му широко використовуються в систематиці як одна з головних так-
сономічних ознак на рівні відділів.
Статеві спороношення відсутні в акразіомікотових слизовиків.
У міксомікотових статеве спороношення представлене спорокарпа-
ми зі спорами. Спорокарпи являють собою плодові тіла, в яких утво-
рюються спори. Статеве спороношення розвивається після статево-
го процесу не одразу, оскільки зигота без періоду спокою перетво-
рюється на міксамебу або міксофлагеляту з диплоїдним ядром і внас-
лідок серії мітозів або злиття з іншими амебоїдами дає початок гомо-
або гетерокаріонтичному плазмодію з диплоїдними ядрами. В таких
плазмодіях надалі відбувається мейоз, і плазмодії перетворюються
на плодові тіла з гаплоїдними спорами.
У плазмодіофоромікотових статеве спороношення - це зооспо-
рогенні цисти, що утворюються із вторинного плазмодія. Подібно до
243
міксомікотових, статеве спороношення після статевого процесу
розвивається не одразу: після злиття ізогамет утворюється дика-
ріонтична чотириджгутикова клітина злиття, яка надалі внаслідок
синхронних поділів ядер дикаріону розвивається в дикаріонтичний
вторинний плазмодій. З часом ядра дикаріонів зливаються, утво-
рюючи диплоїдне зиготичне ядро. Далі відбувається мейоз, і плаз-
модій повністю розпадається на одноядерні, вкриті клітинними обо-
лонками гаплоїдні цисти, які також називають спочиваючими спора-
ми. На відміну від міксомікотових, плодові тіла у плазмодіофороміко-
тових не утворюються.
В оомікотових зиготи (ооспори) після періоду спокою або безпо-
середньо перетворюються на зооспорангій з гаплоїдними зооспора-
ми, або проростають у спорангіофор з одним чи кількома зооспоран-
гіями на верхівці. Таким чином, статеве спороношення оомікотових
представлено зооспорангіями із зооспорами.
Статеве спороношення гіфохітриомікотових залишається оста-
точно не з’ясованим. Відомо, що після статевого процесу зигота
збільшується у розмірах, її диплоїдне ядро багаторазово ділиться, і
один з поділів є редукційним. Далі протопласт розпадається на одно-
ядерні ділянки, подальша доля яких незрозуміла - за даними одних
авторів, ці ділянки перетворюються на одноядерні спочиваючі спори,
з яких врешті-решт утворюються зооспори; за даними інших дослід-
ників, такі ділянки одразу перетворюються на зооспори. Таким чи-
ном, статеве спороношення гіфохітриомікотових являє собою спо-
рангій, але нез’ясованого типу.
Лабіринтуломікотові внаслідок статевого процесу утворюють ба-
гато диплоїдних цист. У кожній цисті відбувається мітоз, потім - ме-
йоз, і з цисти розвивається вісім зооспор з гаплоїдними ядрами. Та-
ким чином, статеве спороношення представлене диплоїдними зоо-
спорогенними цистами.
У хітридіомікотових відомо два варіанти поведінки після статево-
го процесу. При першому варіанті (хітридіальні гриби) після статево-
го процесу утворюються дикаріонтичні зимові цисти; в них ядра дика-
ріонів при проростанні зливаються, далі редукційно діляться, після
чого цибта перетворюється на зооспорангій з гаплоїдними зооспора-
ми. При другому варіанті (бластокладіальні та моноблефаридальні
гриби) ядра гамет зливаються одразу, без утворення дикаріонів; зи-
гота після періоду спокою проростає гіфою - спорофітом, на якій ут-
ворюються зооспорангії з диплоїдними зооспорами. Таким чином, ста-
теве спороношення у хітридіомікотових представлене або зооспоро-
244
генними цистами, або зооспорангіями із зооспорами, які утворюють-
ся на гіфах спорофіта.
Статеве спороношення зигомікотових являє собою спорангій зі
спорангіоспорами, який розвивається із зигоспори. Спорангіоспори
нерухомі, вкриті клітинною оболонкою і мають гаплоїдний набір хро-
мосом.
Аскомікотовим притаманне статеве спороношення, яке називаєть-
ся аском з аскоспорами. Після злиття гаметангіїв одна пара несест-
ринських ядер утворює дикаріон, з якого, як правило, розвивається
дикаріонтичний міцелій. У клітинах гіф такого міцелію відбувається
каріогамія, і після певних морфогенетичних процесів (наприклад, з
утворенням гачка, див. нижче), під час яких здійснюється редукцій-
ний поділ ядра, клітина перетворюється на ендогенний статевий спо-
рангій - аск (сумку), всередині якої розвиваються гаплоїдні аскоспори.
Аски можуть розвиватися безпосередньо на міцелії або розташову-
ватися на поверхні чи всередині спеціалізованих видозмін міцелію,
які називають плодовими тілами і поділяють на закриті (клейстотеції),
напіввідкриті (перитеції), відкриті (апотеції) чи несправжні (псевдо-
теції). Гриби, в яких плодові тіла макроскопічні, називають макроміце-
тами (на відміну від мікроскопічних грибів - мікроміцетів).
У базидіомікотових статеве спороношення представлене бази-
дією з базидіоспорами. Після злиття цитоплазм вегетативних клітин
міцелію (соматогамії), як і в аскоміцетів, каріогамія одразу не відбу-
вається, а утворюється дикаріон, з якого розвивається дикаріонтич-
ний міцелій. Згодом окремі клітини гіф такого міцелію починають
перетворюватись на органи статевого спороношення. Під час цього
процесу відбувається каріогамія, далі - мейоз, і, врешті-решт, з таких
клітин розвивається гаплоїдна базидія. У випини, які утворює обо-
лонка базидії, мігрують дочірні гаплоїдні ядра. Випини відділяються
від базидії септами і перетворюються на одноядерні, вкриті оболон-
кою гаплоїдні базидіоспори. Таким чином, базидіоспори, на відміну
від аскоспор, утворюються екзогенно. Базидії можуть розташовува-
тись безпосередньо на міцелії або на поверхні відкритих (гімнокарп-
них), напіввідкритих (геміангіокарпних) чи всередині закритих (ангіо-
карпних) плодових тіл. У багатьох базидіомікотових базидії розвива-
ються зі спочиваючих дикаріонтичних клітин - телейтоспор.
245
Життєві цикли
Подібно до водоростей, у грибів розрізняють два основних типи
життєвих циклів: цикли агамних грибів, які представлені цикломор-
фозом, та цикли еугамних грибів, які мають статевий процес.
Цикломорфоз є єдиним типом життєвого циклу лише в акразіомі-
котових грибів. Здатність до статевого розмноження не виявлена або
повністю втрачена у багатьох грибів, які за комплексом морфологіч-
них, цитологічних та молекулярно-генетичних ознак належать до ас-
комікотових та базидіомікотових грибів. Оскільки системи цих відділів
базуються, в першу чергу, на ознаках, пов’язаних з органами стате-
вого спороношення, для агамних представників не вдається визна-
чити чіткого місця у системі цих відділів. Такі гриби об'єднані у штуч-
ну групу грибів з нез’ясованим систематичним положенням, яку
сьогодні називають групою анаморфних або мітоспорових грибів, а
нещодавно їх розглядали в межах класу дейтероміцетів (Оеиіе-
готусеіез) або називали незавершеними грибами (Еипді Ітрегїесй).
Анаморфні гриби розмножуються виключно нестатевим шляхом, пе-
реважно за допомогою конідій. Таким чином, цикломорфоз є основ-
ним типом життєвого циклу в анаморфних грибів.
Хоча у більшості анаморфних або мітоспорових грибів статевий
процес відсутній, рівень рекомбінантної мінливості є досить високим.
Він обумовлений явищами гетерокаріозису (різноядерності) та пара-
сексуальним циклом. При гетерокаріозисі в різних клітинах одного
міцелію можуть знаходитися генетично неоднакові ядра, які через
анастомози переходять з однієї клітини міцелію в іншу. Хоча «мігру-
ючі» ядра зазвичай не зливаються з ядрами клітини-реципієнта, про-
те вони приносять нову генетичну інформацію і розширюють тим
самим адаптивні можливості гриба щодо умов зовнішнього сере-
довища.
В окремих випадках «мігруючі» ядра зливаються з ядрами кліти-
ни-реципієнта. Таке явище називають парасексуальним циклом. Воно
призводить до утворення гетерозиготних диплоїдів за рахунок злиття
ядер в гетерокаріонах. Наступним етапом є мітотичне розщеплення
цих диплоїдів з утворенням гаплоїдних або диплоїдних рекомбінантів.
В інших відділах, принаймі в окремих представників, виявлено
статеве розмноження, тобто існують життєві цикли зі зміною ядерних
фаз. Проте конкретні типи життєвих циклів не завжди визначені че-
рез брак експериментальних даних щодо окремих стадій онтогенезу.
Зокрема, така ситуація має місце для лабіринтуломікотових та гіфо-
246
хітриомікотових псевдогрибів, де спостереженнями охоплені не всі
стадії, незважаючи на відомості про навність статевого процесу.
Життєві цикли еугамних грибів, порівняно з водоростями, мо-
жуть бути ускладнені наявністю в онтогенезі вегетативної стадії, пред-
ставленої особинами з дикаріонтичними клітинами. В таких випадках
після статевого процесу утворюється не зигота, а клітина злиття з
дикаріонтичними ядрами. Зигота у класичному розумінні цього тер-
міну в онтогенезі може бути взагалі відсутня, хоча замість неї після
каріогамії обов’язково утворюються диплоїдні зиготичні ядра. Такі ядра
є гомологами зиготи, і при розрахунку схеми життєвого циклу саме їх
визначають як зиготу. Присутність у життєвому циклі грибів дикаріон-
тичних поколінь обов'язково відзначають у назві типу життєвого
циклу.
В межах конкретних відділів грибів життєві цикли за їх типом є
переважно досить одноманітними, що дозволяє використовувати цю
ознаку при характеристиці грибних таксонів найвищого рангу.
Зокрема, для міксомікотових характерні диплофазні життєві цик-
ли з гаметичною редукцією та без зміни поколінь або гаплодипло-
фазні цикли зі споричною редукцією та неправильним чергуванням
ізоморфних поколінь без дикаріонтичних фаз. У плазмодіофоромі-
котових життєві цикли гаплофазні, із зиготичною редукцією та чергу-
ванням одноядерного гаметофіта та дикаріонтичного спорофіта. Оомі-
котові гриби мають диплофазні життєві цикли з гаметичною редук-
цією та без зміни поколінь.
Для справжніх грибів у більшості випадків характерні життєві цикли
з наявністю дикаріонтичних поколінь, і лише у частини хітридіоміко-
тових та зигомікотових грибів такі покоління відсутні. Аскомікотові та
базидіомікотові мають переважно гаплофазні життєві цикли із зиго-
тичною редукцією та чергуванням міцеліїв з одноядерними та дика-
ріонтичними клітинами. У частини хітридіомікотових грибів життєвий
цикл гаплофазний із зиготичною редукцією та чергуванням одноядер-
ного плазмодія з літніми цистами та дикаріонтичного плазмодія із
зимовими цистами. В іншої частини представників цього відділу жит-
тєвий цикл диплофазний з гаметичною редукцією та неправильним
чергуванням поколінь і без стадій дикаріонів. Для зигомікотових ха-
рактерні гаплофазні життєві цикли із зиготичною редукцією та без зміни
поколінь.
Часто один і той же гриб у життєвому циклі може мати декілька
типів спороношення: нестатеве та статеве, які послідовно змінюють
одне одного. Таке послідовне чергування анаморф та телеоморф у
247
грибів отримало назву плеоморфізму. Явище плеоморфізму широко
представлене у грибів різних відділів, проте найбільшого розвитку воно
набуло в аскомікотових.
Еколого-трофічні групи грибів
Гриби зустрічаються у біотопах всіх типів. За середовищем існу-
вання гриби поділяють на наземні та водні (прісноводні та морські).
За трофічною приуроченістю до субстрату гриби поділяють на
еколого-трофічні групи. У найзагальнішому випадку гриби поділяють
на сапротрофів, паразитів та симбіотрофів. Сапротрофи отримують
поживні речовини з мертвої органічної речовини, оселяючись на різно-
манітних рослинних та тваринних залишках і розкладаючи її. Гриби-
паразити оселяються на інших організмах (рослинах, тваринах, інших
грибах) і споживають органічну речовину господаря. Симбіотрофи
отримують органічні речовини внаслідок симбіозу з іншими організ-
мами (переважно вищими рослинами та водоростями), не завдаючи
партнеру помітної шкоди, а навпаки, забезпечуючи його водою та міне-
ральними речовинами чи надаючи йому укриття.
Загально визнаними групами сапротрофів є, зокрема, гриби-кси-
лотрофи, які розвиваються на деревині, гумусові та підстилкові сап-
ротрофи, що зростають у ґрунті або у лісовій підстилці відповідно,
копротрофи, що утилізують екскременти тварин, та ін.
Серед паразитичних грибів основними групами вважаються фіто-
патогенні гриби, які паразитують на вищих рослинах, - фітотрофи;
альготрофи, які живляться за рахунок водоростей; зоотрофи, що роз-
виваються на різноманітних тваринах (в межах цієї групи окремо ви-
діляють ентомофільні гриби, що уражують комах); мікотрофи, що па-
разитують на інших грибах, а також гриби-паразити людини. За ха-
рактером зв’язку з господарем гриби поділяють на факультативних
та облігатних паразитів. Факультативні паразити для господаря є не-
безпечнішими від облігатних, оскільки, спричинивши загибель госпо-
даря, продовжують свій розвиток, споживаючи його органічну речо-
вину як сапротрофи. За топологією паразитичні гриби поділяють на
внутрішньоклітинних та внутрішньотканинних паразитів.
Найбільш відомими та численими групами грибів-симбіотрофів є
мікоризоутворюючі та ліхенізовані гриби. Перші розвиваються в ри-
зосфері вищих рослин, утворюючи зовнішню (екторофну), внутріш-
ню (ендотрофну) або змішану мікоризи. Ліхенізовані гриби живуть у
248
симбіозі з мікроскопічними водоростями і утворюють симбіотичні асо-
ціації, відомі під назвою лишайники.
Існує ряд специфічних екологічних груп грибів, які розвиваються
на різноманітних матеріалах, створених людиною: металах, пласт-
масах, полімерних плівках, тканинах, клею, гумових виробах, склі,
лакофарбових покриттях, а також на папері, книгах, рукописах, кар-
тинах тощо, зумовлюючи їх псування або біологічне пошкодження.
Гриби дискокристати
У межах дискокристат присутні три основні групи таксонів, серед
яких евгленофітові представляють рослинний світ, кінетопластиди -
тваринний, акразіомікотові - грибний. Віднесення акразіомікотових
до грибів є вельми дискусійним через їх високу морфологічну схожість
з амебами. До аргументів на користь грибної природи акразіомікото-
вих належать їх грибний (осмотрофний) тип живлення та розмножен-
ня за допомогою спор. За науковими назвами таксонів різного рівня,
які запропоновані для даних організмів, Асгазіотусоіа підпадає під
керівництво Міжнародного Кодексу Ботанічної Номенклатури, тобто
розгляд цієї групи серед грибів номенклатурно є законним.
Відділ Акразіомікотові слизовики - Асгазіотусоіа
Евкаріотичні первинно-гетеротрофні дискокристати, в яких клітини
голі, а вегетативне тіло представлене амебоїдами (міксамебами).
Відділ нараховує менше 20 видів мікроскопічних амебоїдних
організмів. На початку репродуктивної фази клітини цих слизовиків
утворюють псевдоплазмодії, які перетворюються на спороношення -
соруси або сорокарпи зі спор. Акразіомікотові є сапротрофами, що
мешкають на рослинних рештках, у ґрунті, на екскрементах травоїд-
них тварин та на плодових тілах грибів-макроміцетів.
Біохімічні особливості та живлення. В акразіомікотових сли-
зовиків основним продуктом асиміляції є схожий із парамілоном
Р-1,3-глюкан або специфічний полісахарид, представлений р-1,6-глю-
каном. Відомості щодо шляхів біосинтезу лізину та триптофану
відсутні.
Живлення в акразіомікотових відбувається осмотрофно, шляхом
поглинання розчиненої органічної речовини всією поверхнею клітин.
Фагоцитоз та травні вакуолі у цих слизовиків не виявлені.
249
Цитологічні особливості. Вегетативні амебоїдні клітини акра-
зіомікотових вкриті лише плазмалемою. Проте у монадних стадій по-
верхня клітини має тонку спіральну посмугованість і в оптичний мікрос-
коп нагадує пелікулу евгленофітових. При цьому схожість з клітина-
ми первинно гетеротрофних евгленофітових підсилюється наявністю
поздовжньої борозенки.
Міксамеби акразіомікотових здатні до активного руху за допомо-
гою однієї широкої передньої лобоподії. Вона прозора і не містить
органел чи включень. При електронній мікроскопії у псевдоподії ви-
явлено пучки цитоскелетних мікрофіламентів, побудованих з актину
(і, ймовірно, - центрину). З мікрофіламентами також асоційовані
мікротрубочки. Задній кінець клітини називають уроїдною зоною, ос-
кільки там розташовуються одна-дві видільні вакуолі з рідкими про-
дуктами метаболізму. Центральна частина клітини містить одне ядро,
зазвичай з одним ядерцем.
Ядерний апарат евкаріотичний. У процесі мітозу ядерна оболон-
ка залишається інтактною. Ядерце у профазі переважно ділиться пе-
решнуровкою. Веретено поділу внутрішнє, в його утворенні бере
участь пара центріолей.
Мітохондрії акразіомікотових мають щонайменше два типи крист
- дископодібні та трубчасті. Специфічною особливістю мітохондрі-
ального апарату є те, що кожний мітохондріальний профіль оточе-
ний шерхатою ендоплазматичною сіткою, яка утворює структуру,
схожу на додаткову мітохондріальну оболонку. Комплекс Гольджі в
акразіомікотових не виявлений.
Джгутикові стадії, представлені зооспорами, відомі у двох родах
- Асгазіз та Росіїепіа. Монадні клітини мають два передні гладенькі
джгутики неоднакової довжини. На зооспори здатні перетворюватись
міксамеби або ними за певних умов проростають спори сорусів чи
сорокарпів.
Вегетативне тіло акразіомікотових представлене міксамебами,
які перед початком спороношення агрегуються разом, утворюючи
нерухомі псевдоплазмодії. При агрегації міксамеби рухаються одна
до одної, зближуються і врешті-решт утворюють щільну масу у ви-
гляді щитка. У стані псевдоплазмодія клітини не зливаються між со-
бою, кожна зберігає свою індивідуальність. Об’єднавшись у псевдо-
плазмодій, міксамеби припиняють живитись. Перехід у стан псевдо-
250
плазмодія є зворотнім, проте механізми, що контролюють агрегацію
та дезінеграцію в акразіомікотових, залишаються нез’ясованими.
Всі представники акразіомікотових є мікроскопічними організмами.
Розмноження та життєвий цикл. Акразіомікотові розмножують-
ся поділом міксамеб надвоє та за допомогою спор, що екзогенно ут-
ворюються з псевдоплазмодіїв. Псевдоплазмодії розвиваються при
нестачі поживних речовин або води.
При утворенні спор пседоплазмодій підіймається над субстратом,
набуває напівсферичної форми. У найпростішому випадку (зокрема,
у роду Росїіепіа) на цій стадії міксамеби, які утворили псевдоплаз-
модій, округлюються, вкриваються целюлозними оболонками і пере-
творюються на спори. Скупчення спор при такому типі спорогенезу
називають сорусом.
У складніших випадках міксамеби переміщуються у напівсфе-
ричній масі від її основи до верхівки, через що спороношення набу-
ває вигляду сфери на стебельці, тобто диференціюється на споро-
носний відділ та ніжку. Таким тип спороношення в акразіомікотових
називають сорокарпом. Інколи міксамеби, що утворюють ніжку, виді-
ляють багато слизу (наприклад, у Сиііиііпорзіз). У роду Асгазіз мікса-
меби спороносного відділу після утворення головки продовжують
рухатись таким чином, що врешті-решт утворюють деревовидно роз-
галужений сорокарп.
Після завершення формування спороношення всі міксамеби
(включаючи клітини ніжки) перетворюються на спори, і сорокарпи, як
і соруси, розпорошуються. Спори поширюються вітром, водою, тва-
ринами. Потрапивши у сприятливе для живлення середовище, спо-
ри проростають поодинокими міксамебами. У деяких видів зі спор,
що потрапили у воду, після мітотичного поділу ядра виходить дві зоо-
спори.
Статевий процес в акразіомікотових не виявлений, і життєвий
цикл, таким чином, представлений цикломорфозом. Він включає стадії
міксамеби, псевдоплазмодія та спори. При несприятливих умовах
поодинокі міксамеби можуть вкриватись оболонкою і перетворюва-
тись на спочиваючі клітини - мікроцисти, минаючи стадію агрегації.
Мікроцисти є гомологами спор псевдоплазмодія.
Система відділу. Відділ включає один клас - Асгазіотусеїез, та
два порядки - Асгазіаіез та бийиііпаїез. Поділ на порядки проводять
за будовою оболонки спор та наявністю слизу в ніжці сорокарпу.
251
Порядок Акразіальні - Асгазіаіез
Об’єднує види, у яких оболонка спор одношарова, ніжка соро-
карпів позбавлена слизу і складається лише із щільно розташованих
міксамеб. Типовий представник порядку - рід Асгазіз.
Рід акразіс - Асгазіз (рис. 94). Характерним видом цього роду є
дуже поширений слизовик Асгазіз гозеа - акразіс рожевий. Він по-
всюдно мешкає у вологих умовах на гниючих рослинних залишках,
але через мікроскопічні розміри зазвичай залишається непоміченим.
За результатами спостережень у природі відомо, що спороношення
акразіс утворює вранці, після випадання роси. Вдень, коли вологи не
достатньо, слизовик перебуває у стані мікроцист.
Акразіс можна легко виявити культуральним методом, якщо у
вологій камері у чашці Петрі провести інкубацію залишків листя, гни-
лої деревини або опалої деревної кори.
Міксамеби акразіса на передньому кінці мають широку безбарв-
ну лобоподію, за допомогою якої рухаються. Решта цитоплазми за-
барвлена у рожевий колір. Задній кінець клітини відтягнутий у дещо
звужену лопать з кількома вакуолями, що утворюють уроїдну зону.
При виведенні з вакуолей їх вмісту на кінці лопаті з’являються короткі
тонкі волоскоподібні випини цитоплазми, що нагадують ризоподїї.
В центрі міксамеби можна інколи розрізнити досить велике світле ядро
з темнуватим ядерцем. При підсиханні міксамеби припиняють рух,
одягаються оболонкою і перетворюються на неправильні або більш-
менш сферичні мікроцисти. Розсіяне світло та зволоження субстра-
ту, на якому знаходяться мікроцисти, зумовлюють їх проростання
міксамебами. Вони починають активно рухатись та ділитись.
Міксамеби при агрегації об’єднуються в рожеві псевдоплазмодії
0,5-1 мм у діаметрі, з яких утворюється один або кілька сорокарпів.
Вони піднімаються над субстратом на ніжках-колонках, які склада-
ються з багатьох щільно з’єднаних між собою міксамеб. На верхівці
ніжки знаходяться прості або розгалужені ланцюжки спор, де кожна
спора являє собою вкриту оболонкою міксамебу. В місцях з’єднання
сусідніх спор на оболонках залишаються характерні круглі рубці -хілу-
ми. При підвищенні вологи спори проростають міксамебами.
Деякі спори, потрапивши у краплинки води, здатні проростати
парою зооспор з двома нерівними передніми джгутиками. Після дея-
кого періоду активного руху зооспори перетворюються на міксамеби.
252
2
Рис. 94. Асгазіз гозегг. міксамеба. 1-лобоподія; 2-ядро з ядерцем;
З - уроїдна зона з вакуолями
Порядок Гуттулінальні - биііиііпаїез
Об’єднує два роди, в яких оболонка спор двошарова (утворена
полісахаридами, але целюлоза серед них не виявлена). Ніжка соро-
карпів складається з нечисленних міксамеб, занурених у слиз. Ха-
рактерний представник - рід гуттулінопсис (Сиіїиііпорзіз) - є коп-
ротрофом, який мешкає на екскрементах тварин, а також у ґрунті.
Гриби тубулокристати
Мікологічні об’єкти в межах цього царства представлені, з одного
боку, міксомікотовими слизовиками, що належать до підцарства аме-
бо-флагелят і споріднені з евгліптовими амебами та хлорарахніофі-
товими водоростями, з іншого - трьома відділами псевдогрибів, які
входять до підцарства страменопілів і споріднені з групою відділів
хромофітових водоростей.
Амебо-флагеляти
Підцарство амебо-флагелят об’єднує переважно гетеротрофні
організми, за винятком відділу хлорарахніофітових водоростей.
У життєвому циклі представників цього підцарства обов’язково є аме-
боїдна вегетативна стадія. Система родинних зв'язків таксонів висо-
ких рангів у межах цієї групи поки що з’ясована недостатньо. Ряд так-
253
еонів (наприклад, евгліптові амеби, формаменіфери, радіолярії, со-
нячники-актинофрідії) одностайно визнаються тваринами і підпоряд-
ковані Міжнародному Кодексу Зоологічної Номенклатури. Інші так-
сони - наприклад, церкомонади та диктіостеліди - розглядаються
переважно у системі тваринного світу, хоча відомі системи, де ці
організми віднесені до ботанічних об'єктів. Міксомікотові слизовики
розглядаються як в зоологічних, так і в ботанічних системах, причому
експериментальні дослідження цих організмів традиційно здійснюють
ботаніки.
Відділ Міксомікотові слизовики - Мухотусоіа
Евкаріотичні первинно-гетеротрофні тубулокристати, вегетатив-
не тіло яких представлене багатоядерними плазмодіями, які розви-
ваються з міксамеб або зооспор. Статеве спороношення - спорокар-
пи з ендогенними спорами. Відділ нараховує біля 800 видів.
Слизовики були описані у системі грибів у 1833 р. під назвою
Мухотусеіез. Після з’ясування типу живлення і доведення здатності
до фаготрофїї видатний міколог А. Де Барі у 1887 р. запропонував
для слизовиків назву Мусеїогоа, тобто «грибо-тварини», підкреслю-
ючи їх подвійну природу. В 1969 р. видатним американським філоге-
нетиком Уіттеккером слизовики були виділені у відділ Мухотусоїа.
Біохімічні особливості та живлення. Живляться міксомікотові
осмотрофно та фаготрофно. В останньому випадку джерелом їжі є
бактерії, клітини грибів та найпростіші. Захоплення їжі здійснюється
псевдоподіями, а її перетравлення - у травних вакуолях. Вирощу-
вання міксомікотових на штучних поживних середовищах із глюко-
зою та без зовнішніх джерел амінокислот свідчить про здатність цих
організмів синтезувати лізин, хоча конкретний шлях його біосинтезу
залишається нез’ясованим. Продукт асиміляції - це справжній тва-
ринний глікоген.
Цитологічні особливості. Міксамеби та плазмодії міксомікото-
вих вкриті лише плазмалемою, через що здатні до метаболічних змін
форми та активного амебоїдного руху. Локомоторними структурами
є тонкі філоподії, у складі яких виявлено скоротливий цитоскелетний
білок актин. У деяких видів на поверхні клітин виявлені субмікрос-
копічні лусочки, що утворюються в пухирцях, похідних від комплексу
Гольджі. Спори міксомікотових вкриті клітинними оболонками, осно-
ву яких складає целюлоза.
254
Ядерний апарат типово евкаріотичний. Процес мітозу у міксамеб
та зооспор, з одного боку, і плазмодіїв, з іншого, - різний. Міксамеби
та зооспори одноядерні, мітоз відкритий, центрами організації вере-
тена є центріолі. На стадії багатоядерних плазмодіїв ядерна оболон-
ка або лишається інтактною, або на її полюсах утворюється по одній
великій перфорації. Центріолі в плазмодіях участі в утворенні вере-
тена не беруть, натомість в середині ядра, в місцях, що відповідають
полюсам веретена виявляються електронно-щільні структури - по-
лярні тіла, які стають центрами організації мікротрубочок. Таким чи-
ном, мітоз у плазмодіях або закритий, або напіввідкритий, а веретено
внутрішньоядерне. Ядерце у міксомікотових на початку мітозу зни-
кає і відновлюється після його завершення.
Мітохондрії у плазмодіях численні і мають трубчасті кристи. Ком-
плекс Гольджі у клітинах наявний.
Джгутикові стадії представлені зооспорами з двома гладенькими
нерівними джгутиками. Базальні тіла орієнтовані одне до одного під
майже прямим кутом. У перехідній зоні розташовується базальна
платівка або циліндричне тіло. Цитоскелетна система містить зазви-
чай п’ять мікротубулярних коренів. Один з них дуже широкий, він скла-
дається з великої кількості мікротрубочок, огортає базальні тіла і про-
стягається в напрямку до заднього кінця клітини. Один із джгутикових
коренів пов’язує базальні тіла з ядром, виконуючи тим самим функ-
цію ризопласта (проте такий корінь не виявлений у видів роду
Сегаііотуха). Залежно від умов зволоження, зооспори можуть втягу-
вати джгутики і перетворюватись на міксамеби, і навпаки, міксамеби
здатні виробляти джгутики і перетворюватись на зооспори.
Вегетативне тіло міксомікотових - одноядерні гаплоїдні мікса-
меби та багатоядерні гетерокаріонтинчі плазмодії з диплоїдними яд-
рами. Плазмодії розвиваються внаслідок багаторазових синхронних
поділів ядер, які починаються з поділу ядра однієї міксамеби та вна-
слідок злиття між собою міксамеб або зооспор.
Плазмодій на 75% складається з води, іншу частину вегетативно-
го тіла становлять органічні речовини та солі. Для деяких міксоміко-
тових характерна наявність вапна. Плазмодії також містять пігменти,
які надають їм різноманітного забарвлення - від жовтого та білого до
рожевого та червоного.
Плазмодії здатні до активного руху за допомогою псевдоподій і
володіють різними таксисами - позитивними гідро- та трофотаксиса-
ми, негативним фототаксисом. Тому плазмодії у вегетативному стані
255
мешкають у темних вологих місцях, багатих розчинною органічною
речовиною або об’єктами фаготрофного живлення. Розміри пла-
змодіїв коливаються від мікро- до макроскопічних (інколи - до 50 см у
діаметрі) і можуть бути представлені прото-, афано- або фанеро-
плазмодіями.
Розмноження та життєвий цикл. Міксамеби та зооспори здатні
розмножуватись вегетативним поділом надвоє. При цьому зооспори
перед поділом втягують джгутики і перетворюються на міксамеби.
Плазмодії, які являють собою спорофіти, утворюють гаплоїдні
спори, що розвиваються ендогенно в плодових тілах - спорокарпах.
Переходячи в стадію спороношення, плазмодії припиняють живитись,
їх позитивний гідротаксис та негативний фототаксис змінюються на
протилежні. Як наслідок, плазмодії виповзають на світлі сухі ділянки
і починають перетворюватись на спорокарпи.
Нижня частина плазмодію дає початок стерильній основі споро-
карпу - плівчастому або сітчастому гіпоталюсу (підшарку). На ньому
розвивається спороносна частина спорокарпу, яка вкривається хря-
щуватою оболонкою - перидієм. Він складається з ущільненого сли-
зу та мікрофібрил целюлози. Цитоплазма плазмодія на початку спо-
роутворення вакуолізується і розпадається на фрагменти з поодино-
кими диплоїдними ядрами. Вміст вакуолей частково кристалізується
та випадає в осад, утворюючи капіліцій - систему ниток, здатних до
гігроскопічних рухів. Кожне диплоїдне ядро редукційно ділиться, ут-
ворюючи чотири гаплоїдних ядра. Три з них дегенерують, а четверте
перетворюється на ядро спори. Цитоплазма навколо нього ущіль-
нюється, і розвивається целюлозна оболонка зі специфічною орна-
ментацію.
Спорокарпи поділяють на три основні типи: плазмодіокарпи, спо-
рангії та еталії. Плазмодіокарпи мають неправильну, розгалужену або
сітчасту форму і нагадують плазмодії, вкриті перидієм.
Спорангії характеризуються формою, сталою для певного виду, і
обов’язково мають більш-менш округлу спороносну головку. Спорангії
можуть бути сидячими або розташовуватись на ніжках-стебельцях.
Ніжка може утворюватись або зі спороносної частини плазмодія як
продовження перидію (субгіпоталічний спосіб), або зі стерильного гіпо-
талюса і заходити у спороносну частину спорангія, утворюючи колон-
ку (колумелу) - епігіпоталічний спосіб.
Якщо кілька спорангіїв з’єднуються між собою боками, зберіга-
ючи при цьому власні перегородки, то такі скупчення спорангіїв інко-
ли називають псевдоеталіями.
256
Справжні еталії являють собою групи спорангіїв, які зливаються у
велике сидяче, переважно сферичне або напівсферичне плодове тіло,
вкрите загальним покривом - кортексом. Залишки перидію окремих
спорангіїв, що злилися, зберігаються в еталіях у вигляді псевдокапіл-
іцію. Псевдокапіліцій, на відміну від капіліцію, не здатний до гігроско-
пічних рухів і, таким чином, не бере безпосередньої участі у розсію-
ванні спор, а лише надає еталіям додаткової механічної міцності.
Після руйнування перидію або кортексу спори висіваються зі спо-
рокарпів і проростають гаплоїдними міксамебами або зооспорами.
Через кілька годин вони виділяють назовні т. зв. фактор статевої зрілості
- речовину з гормональною дією, яка ініціює початок статевого про-
цесу. Міксамеби та зооспори різних статевих знаків попарно копулю-
ють, утворюючи диплоїдну зиготу. Її ядро незабаром починає міто-
тично ділитися, даючи початок новому плазмодію. Молоді плазмодії
здатні зливатись один з одним, утворюючи гетерокаріонтичні плаз-
модії.
В деяких випадках міксамеби можуть проростати у плазмодії апо-
міктично- без копуляції з іншими одноядерними індивідами. В таких
випадках внаслідок ендомітозу гаплоїдне ядро міксамеби диплоїди-
зується і далі ділиться звичайним мітотичним способом, утворюючи
диплоїдний плазмодій.
Таким чином, у життєвому циклі слизовиків чергується стадія гап-
лоїдних одноядерних (зрідка - багатоядерних) міксамеб чи похідних
від них зооспор, які при цьому виконують функцію гамет, та диплоїд-
них спорофітів - плазмодіїв. Тобто життєвий цикл міксомікотових є
диплогаплофазним, зі споричною редукцією, гетероморфною зміною
поколінь і без стадії дикаріонів.
При несприятливих умовах міксомікотові здатні утворювати спо-
чиваючі стадії. Зокрема, поодинокі міксамеби можуть вкриватись обо-
лонкою і перетворюватись на сферичні мікроцисти, що проростають
зооспорами. Плазмодії здатні розпадатися на окремі багатоядерні
фрагменти, які вкриваються оболонкою і перетворюються на макро-
цисти (сферули). Фанероплазмодії здатні цілком вкриватись оболон-
кою із галактозамінів і перетворюватись на спочиваючі інцистовані
плазмодії - склероції.
Екологічні особливості. Слизовики поширені на рослинних,
рідше на тваринних залишках, екскрементах тварин, базидіальних
грибах, мохах, лишайниках, на корі живих дерев та чагарників, на
травах, у ґрунті.
257
Система відділу. У відділі Мухотусоіа розглядаються 2 класи -
Ргоіозіеііотусеїез та Мухотусеїез. Класи відрізняються за перева-
жаючими типами плазмодіїв, кількістю спор, які утворює поодинокий
спорокарп, наявністю спеціалізованих структур для відокремлення
спор від спорокарпів (апофіз та газових пухирців). Крім того, до міксо-
мікотових умовно відносять також Оісіуозіеііотусеіез.
Клас Протостеліоміцети - Ргоіозіеііотусеіез
Об'єднує біля 40 видів слизовиків, у яких спорофіт представле-
ний мікроскопічним багатоядерним протоплазмодієм. Органами спо-
роношення є спорангії на ніжках, що утворюються зі спороносної ча-
стини плазмодія, тобто субгіпоталічним шляхом. Спорангії можуть бути
поодинокими або утворюватись у великій кількості на макроскопіч-
ному комірчастому підшарку. В одному спорангії розвивається лише
одна спора, яка може бути двоядерною або восьмиядерною. Відділен-
ня спори від ніжки в багатьох випадках відбувається завдяки спеціа-
лізованим структурам - апофізам або газовим пухирцям. Капіліцій у
протостеліоміцетів відсутній.
Клас включає два основні порядки. Перший порядок- протосте-
ліоміцетальні (Ргоіозіеііотусеїаіез) - об’єднує слизовиків, у яких
підшарок нерозвинений, і спорангії є поодинокими. Для другого - це-
раціоміксальні (Сегаїіотухаїез) - характерна наявність великих мак-
роскопічних розгалужених гіпоталюсів, на яких розвивається багато
поодиноких спорангіїв.
Найпоширенішим видом класу є Сегаііотуха іїиіісиїоза - це-
раціомікса фруктова (рис. 95). Цей слизовик зустрічається повсюд-
но на гнилій деревині та в лісовій підстилці. Протоплазмодій цераціо-
мікси утворює макроскопічній гіпоталюс, що має вигляд системи білих
розгалужених губчатих трубок, на яких на довгих ніжках знаходяться
спорангії з поодинокими спорами.
Клас Міксоміцети - Мухотусеіез
Об’єднує понад 700 видів слизовиків, у яких спорофіт представ-
лений мікроскопічними або макроскопічними прото-, афано- та фа-
нероплазмодіями. Органами спороношення є плазмодіокарпи, спо-
рангії та еталії, в яких утворюється багато спор. Спори одноядерні.
В межах класу за морфологією плазмодія, типом розвитку споро-
фору, планом будови капіліцію, наявністю вапна у структурах споро-
карпу виділяють до восьми порядків, з яких нижче розглядаються
чотири - Псеаіез, Тгісіїіаіез, Рііузагаїез та Зіетопііаіез.
258
Рис. 95. Сегаііотуха іїиіісиїоза: А - зовнішній вигляд спороносних гілочок;
Б - спорокарпи на верхівці спороносної гілочки; В - окремі спорокарпи.
Порядок Ліцеальні - Нсеаіез
Порядок об’єднує види, в яких спорофіт представлений прото-
або фанероплазмодієм. Спорокарпи різноманітні і являють собою
плазмодіокарпи, спорангії (сидячі або на ніжці, що розвивається суб-
гіпоталічно і не утворює колонки) або еталії. Капіліцій завжди відсутній,
проте у видів з еталіями часто наявний псевдокапіліцій. Спори утво-
рюються у великій кількості, є безбарвними або лише злегка забарв-
лені. Характерний представник порядку - Ьусодаїа.
Рід лікогала - іусодаїа (рис. 96). Слизовик мешкає у лісових
фітоценозах під поваленими деревами, пеньками та у підстилці. Спо-
роношення лікогали, представлені еталіями, часто можна зустріти на
пеньках, повалених стовбурах або навіть на грунті протягом усього
літа. Зазвичай плодові тіла слизовика напівкулясті, до 1,5 см у діа-
метрі, і переважно утворюють щільні групи, які розташовуються на
дуже тонкому, майже непомітному підшарку.
Молоді еталії добре помітні завдяки яскравому червонувато-ро-
жевому забарвленню, м’які на дотик. При найменшому ушкодженні з
плодових тіл виступає червонувата або рожева рідина. При дозрі-
ванні перидій еталію твердіє і набуває оливкового, коричневого або
майже чорного забарвлення. Далі перидій потоншується і після до-
зрівання спор розкривається на верхівці неправильною тріщиною, че-
рез яку висіваються спори. їх оболонки мають сітчастий візерунок,
який добре помітний на великих збільшеннях мікроскопа.
259
Еталії розвиваються з груп щільно розміщених спорангіїв, які час-
тково зливаються боками. Залишки перидію окремих спорангіїв утво-
рюють цілу систему розгалужених стрічок псевдокапіліцію, що про-
низує вміст еталію. Кожна стрічка одним кінцем прикріплюється до
кортексу, інший кінець залишається вільним.
Найпоширенішим видом роду є і-усодаїа ерісіепсігит - лікога-
ла деревна, відома під народною назвою “вовче вим'я”, зустрічаєть-
ся в Україні повсюдно у лісових фітоценозах різних типів.
Рис. 96. Ьусодаїа'. скупчення еталіїв.
Порядок Трихіальні - ТгісЬіаІез
Спорофіт представлений афано- або фанероплазмодієм, спо-
рокарпи - плазмодіокарпами, спорангіями (сидячими або на суб-
гіпоталічних ніжках) та еталіями. Найяскравішою ознакою порядку є
наявність.у плодових тілах розвиненого суцільного або трубчастого
розгалуженого капіліцію, нитки якого скульптуровані спіралями,
кільцями, рідше - шипами та шипиками. Інколи нитки капіліцію
представлені поодинокими нерозгалуженими елатерами. Спорангії
завжди позбавлені колонки. Спори трихіальних яскраво забарвлені,
переважно у жовтий або червоний колір.
Характерними представниками порядку є роди Агсугіа та Тгісіїіа.
Рід арцирія - Агсугіа - розрізняється за будовою спорангіїв та
капіліцію. Спорангії невеликі, до 2 мм заввишки, утворюються на ко-
роткій ніжці. Перидій тоненький, плівчастий і після дозрівання спор
260
руйнується, за винятком базальної частини спорангія. Тут залишки
перидію зберігаються у вигляді маленької чашечки. Капіліцій утворе-
ний багатьма дуже розгалуженими нитками, які інкрустовані шипами,
шипиками та напівкільцями. Типовим видом роду є Агсугіа сіеписіаіа
- арцирія гола, яка має яскраво-червоні спорангії.
Рід трихія - Тгісіїіа - також утворює спорангії на ніжках, а за-
лишки перидію зберігаються у вигляді чашечок. Проте, на відміну від
арцирії, капіліцій складається з багатьох нерозгалужених елатер, які
орнаментовані спіральними потовщеннями. На корі дерев часто мож-
на знайтй забарвлені у жовтий колір спорангії Тгісіїіа уагіа - трихії
варіюючої, висота яких становить біля 1 мм.
Порядок Фізаральні - Рііузагаїез
Найчисленніший порядок класу, який характеризується наявні-
стю макроскопічних фанероплазмодїїв, яскраво забарвлених у різні
відтінки червоного, жовтого, рожевого або коричневого кольорів.
Плодові тіла представлені плазмодіокарпами, еталіями та спорангія-
ми. Капіліцій майже нерозгалужений, гладенький або має вузлико-
подібні потовщення, утворені відкладами вапна. Перидій та ніжки
спорангіїв також містять вапно. У видів, що утворюють спорангії на
ніжках, часто наявна колонка, яка, проте, як і ніжка, утворюється суб-
гіпоталічним шляхом. Спори темнозабарвлені.
До порядку належать види, які зустрічаються зазвичай на гнилій
деревині, в лісовій підстилці або на піднятих над землею гілочках.
Найвідомішими представниками є роди РІіузагит, Риіідота Мисіїадо.
Рід фізарум - Ріїузагит - утворює макроскопічні, добре помітні
фанероплазмодії. Вони мають віялоподібну форму і складаються з
системи розгалужених жилок. Жилки товсті при основі, далі рівномір-
но, переважно дихотомічно галузяться і потоншуються. Кожна жилка
складається з периферичного шару гелеподібної, обмежено рухомої
цитоплазми та центральної зони з рідкої і дуже рухливої цитоплазми,
яка почергово пульсуюче рухається вперед та назад, обумовлюючи
явище, відоме під назвою човникових рухів цитоплазми. Рух у перед-
ньому напрямку дещо довший, ніж у зворотньому, і завдяки цьому на
передньому боці цитоплазми утворюються псевдоподії, за допомо-
гою яких плазмодій рухається.
Плодові тіла фізаруму - це переважно сферичні спорангії на ви-
довженій ніжці, рідше - плазмодіокарпи. Перидій та капіліцій споро-
261
ношення слизовика містять багато вапна, яке легко виявляється про-
стою реакцією на скипання зі слабким розчином соляної кислоти.
Одним з найпоширеніших видів є Ріїузагит уігісіє - фізарум зеле-
ний, що утворює великі скупчення із численних зеленкуватих спо-
рангіїв на видовжених ніжках. Перидій розтріскується на неправильні
фрагменти, капіліцій у місцях галуження має вузлики з аморфного
вапна.
Рід фуліго - Риіідо - є одним з найпоширеніших фізарових сли-
зовиків. Він належить до відомих їстівних представників міксомікото-
вих. Яскраво-жовті фанероплазмодії Риіідо зерііса - фуліго сеп-
тичного, які сягають 20 см у ширину та до 5 см завтовшки, на тери-
торії Українських Карпат місцеві жителі вживають у їжу. В смаженому
вигляді вегетативне тіло цього слизовика за смаком нагадує яєшню.
Плодові тіла слизовика представлені еталіями. Вони мають по-
душковидну форму, вкриті досить товстим щільним і ламким кортек-
сом, який може бути забарвлений у білуватий, сірий, жовтий, бурий і
навіть червонуватий колір. Кортекс просякнутий вапном, яке відкла-
дається на поверхні у вигляді добре помітних дрібних кристалів. Все-
редині еталій виповнений спорами, скупчення яких мають зеленува-
то-сірий або чорно-фіолетовий колір. Нитки капіліцію розгалужені,
безбарвні і в місцях галужень інкрустовані веретеновидно здутими
вапняковими вузликами. Еталії фуліго часто можна зустріти на пень-
ках, відмерлих рослинних залишках (листі, опалих гілках) або прямо
на землі влітку та на початку осені.
Рід муциляго - Мисіїадо - також утворює еталії. Вони забарв-
лені у білий, сірий або кремовий колір і утворюються після злиття
багатолопатевих спорангіїв, внаслідок чого набувають характерної
форми намету з багатьма поверхневими складками. В середині ета-
лію розташовуються темнуваті спори та капіліцій, утворений звапня-
ковілими платівками, які позбавлені вапнякових вузликів.
Муциляго часто можна зустріти в лісах, особливо хвойних. Пере-
ходячи у стан спороношення, слизовик часто виповзає на гілочки, які
щодо поверхні ґрунту розташовуються більш-менш вертикально.
Порядок Стемонітальні - Зіетопііаіез
Включає види, в яких спорофіти представлені афаноплазмодія-
ми, плодові тіла - переважно спорангіями на ніжках, що утворюють-
ся зі стерильного підшарка, тобто епігіпоталічним способом, і, як пра-
вило, продовжуються у колонку (рідше спорангії зливаються в еталії).
262
Капіліцій добре розвинений, має вигляд рясно розгалуженої сітки і не
містить вапна. Спори темнозабарвлені. Найхарактернішим представ-
ником порядку є Зіетопіііз.
Рід стемонітіс - Зіетопіііз (рис. 97). Види цього роду широко
розповсюджені у світі, і ростуть на гнилій деревині або під корою гни-
лих стовбурів дерев, у тріщинах пеньків, у вологій лісовій підстилці
тощо. Найпоширенішим видом у флорі України є Зіетопіііз їизса —
стемонітіс бурий.
Афаноплазмодії цього слизовика переважно білі або жовтувато-
зелені. Плодові тіла досить великі (від 0,5 до кількох сантиметрів),
коричневі або темно-фіолетові, представлені видовженими прямосто-
ячими циліндричними спорангіями із заокругленими верхівками. Спо-
рангії розташовуються на ніжках і зазвичай утворюють щільні групи.
Рис. 97. Зіетопіііз'. А - група спорангіїв; Б - окремий спорангій; В - фраг-
мент спорангія. 1 - колонка; 2 - капіліцій
263
Ніжка кожного спорангія має всередині порожнину і відходить від
стерильного тонкого плівчастого підшарка- гіпоталюсу. Тонка плівча-
ста оболонка спорофора - перидій - дуже рано зникає, тому спори у
вигляді буруватого пилу вільно розносяться вітром. Циліндрична ніжка
продовжується у спорогенну частину спорангія, утворюючи колонку
(колумелу), яка, поступово витончуючись, доходить до вершини пло-
дового тіла. Від колонки беруть початок темно-бурі розгалужені гла-
денькі нитки капіліцію, що при зміні вологості здатні здійснювати гігро-
скопічні рухи, сприяючи, таким чином, рівномірному висіванню спор.
Спори стемонітісу округлі і забарвлені у коричневий колір.
Потрапивши у придатні умови, спори стемонітісу проростають
міксамебами або дводжгутиковими зооспорами, які здатні до вегета-
тивного поділу. Якщо зустрічаються міксамеби або зооспори різних
статевих знаків, вони копулюють, утворюючи диплоїдну зиготу. Вна-
слідок багатьох мітотичних поділів ядра із зиготи розвивається диплої-
дний спорофіт, представлений афаноплазмодієм. Різні афаноплазмодії
здатні зливатися між собою, внаслідок чого утворюється гетерокар-
іонтичний плазмодій з різними диплоїдним ядрами. Після завер-
шення періоду росту, як правило, внаслідок зниження вологи та
зменшення кількості об’єктів живлення плазмодій переходить до
утворення плодових тіл. На цій стадії відбувається мейоз, і, врешті-
решт, утворюються спорангії з гаплоїдними спорами.
Додаток до відділу Мухотусоїа: клас Диктіостеліоміцети -
□ісіуозіеііотусеіез
Представників цього класу раніше розглядали серед акразіомі-
котових слизовиків. Проте, за сучасними молекулярними даними, цей
клас являє собою одну з гілок найпримітивніших тубулокристат з гру-
пи амебо-флагелят, яка віддалено споріднена з міксомікотовими сли-
зовиками та деякими філозними амебами.
Для диктіостеліомікотових характерними ознаками є наявність
рухливих псевдоплазмодіїв, які утворюються внаслідок агрегації одно-
ядерних міксамеб. Агрегація відбувається за допомогою хемотактич-
них реакцій, пов’язаних із секрецією окремими міксамебами циклічно-
го аденозинмонофосфату - акразину, та схожих з ним за характером
дії інших органічних сполук. Органами спороношення є сорокарпи на
ніжках. Спори утворюються екзогенно і мають целюлозну оболонку.
Джгутикові стадії та центріолі в життєвому циклі відсутні.
Клас представлений одним порядком Оісіуозіеііаіез, в межах яко-
го відомо біля 60-ти видів. Типовим представником порядку є Оісіуоз-
264
іеііит сіізсоісіеит - диктіостеліум дисковидний, який зустрічаєть-
ся на рослинних залишках. Цей вид добре розвивається в культурі на
агаризованих середовищах, через що його широко використовують у
різноманітних експериментах з дослідження механізмів таксисів, мор-
фогенезу, поведінки ядер при агрегації міксамеб та ін. (рис. 98).
Страменопіли. Група відділів псевдогрибів
Рис. 98. Оісіуозіеііит: 1 - псевдо-
плазмодій; 2— спорокарп; 3- спора;
4 - проростання спори; 5 - міксамеба;
6 - мікроцисти; 7 - проростання мікро-
цист
Страменопіли являють со-
бою досить чітко молекулярно
відокремлену гілку тубулокри-
стат. Організми, які належать
до страменопілів, мають яск-
раву фенотипічну ознаку: їх
монадні клітини на поверхні
джгутиків несуть мастигонеми
тричленної будови (ретроне-
ми), що утворюються у пери-
нуклеарному просторі - між
зовнішньою та внутрішньою
мембранами ядерної оболон-
ки. У фотоавтотрофних
організмів ретронеми можуть
утворюватись також у перип-
ластидному просторі хлороп-
ластної еноплазматичної сітки,
який є похідним від перинукле-
арного простору.
До страменопілів нале-
жать організми з рослинною,
тваринною та грибною страте-
гіями живлення, а саме група
відділів хромофітових водоро-
стей, група відділів псевдо-
грибів (зокрема оомікотові, лабіринтуломікотові та гіфохітриомікотові)
та деякі найпростіші (опаліни та бікосоециди).
Питання про походження псевдогрибів поки що залишається дис-
кусійним - за однією із гіпотез, псевдогриби є первинними гетеротро-
фами, що близько споріднені з найпростішими із групи страменопілів,
265
за іншою - вторинними гетеротрофами, що походять від хромофіто-
вих водоростей, які з різних причин втратили здатність до фотосин-
тезу.
Відділ Оомікотові гриби - Оотусоіа
Евкаріотичні осмотрофні страменопіли, у яких клітини вкриті целю-
лозно-глюкановою оболонкою, а вегетативне тіло представлене ба-
гатоядерним розгалуженим неклітинним міцелієм. Монадні стадії
дводжгутикові. Відділ нараховує понад 800 видів прісноводних, морсь-
ких та наземних організмів, які ведуть паразитичний або сапротроф-
ний спосіб життя.
Біохімічні особливості та живлення. За комплексом провідних
біохімічних ознак оомікотові більше подібні до рослин, ніж до справжніх
грибів. Зокрема, оомікотовим притаманний рослинний шлях біосин-
тезу лізину - через диамінопімелінову кислоту. У представників
відділу, так само, як і у хромофітових водоростей, не виявлено здат-
ності до синтезу сидерамінів, й іони заліза поглинаються рослинним
способом. На відміну від справжніх грибів, у оомікотових відсутні такі
багатоатомні спирти (поліоли), як манніт та арабіт. Основним
продуктом асиміляції оомікотових є близький до хризоламінарину
р-1,3-глюкан - міколамінарин.
До унікальних ознак оомікотових належить спосіб синтезу аміно-
кислоти триптофану, який відбувається за участю комплексу фер-
ментів т. зв. п’ятої групи. Цікавою особливістю оомікотових є
відсутність у них цитохрому С1, який характерний для інших грибів та
багатьох рослин. Відсутня також здатність до синтезу стероїдів. Гор-
моном, який індукує статевий процес у оомікотових, є антеридіол.
Живлення представників цього відділу відбувається виключно
осмотрофним шляхом.
Цитологічні особливості. Оомікотові гриби мають міцеліальну
будову. Гіфи оомікотових вкриті клітинною оболонкою, каркасною
основою яких є целюлоза, а поверхневий шар утворений глюканами.
Є також відомості про знахідки у деяких оомікотових хітину. Однією з
характерних особливостей оомікотових є наявність у цитоплазмі мем-
бранних везикулів із шаруватою периферичною та гомогенною елект-
ронно-щільною центральною частиною. Ці везикули містять глюкани
266
та фосфоглюкани і беруть участь у побудові клітинної стінки, особли-
во - у молодих зигот.
Ядерний апарат оомікотових евкаріотичний. Гіфи цих грибів не
поділені септами на окремі клітини і містять багато ядер. При утво-
ренні зооспор у перинуклеарному просторі починається збірка рет-
ронем, які надалі дозрівають у цитоплазмі у везикулах, похідних від
зовнішньої мембрани ядра, і виводяться на поверхню монади побли-
зу місця розташування джгутиків. До ядра щільно притиснутий один
або два комплекси Гольджі.
Мітоз закритий, здійснюється за участю центріолей. Ядерце у
профазі, метафазі та анафазі залишається інтактним, а на початку
телофази ділиться перетяжкою.
Мітохондрії поодинокі або численні і мають трубчасті кристи.
Джгутикові стадії представлені зооспорами з двома гетероконт-
ними гетероморфними та гетеродинамічними джгутиками. Один із
джгутиків спрямований вперед і є локомоторним. На його поверхні
розташовуються ретронеми. Другий джгутик гладенький, спрямова-
ний назад і виконує функцію керма.
Перехідна зона джгутиків містить кілька кілець і позбавлена спіра-
леподібної структури, характерної для багатьох хромофітових водо-
ростей. Цитоскелет, досліджений на прикладі видів роду РЬуіорЬіога,
складається з чотирьох мікротубулярних коренів. Два з них спрямо-
вані вперед і складаються з трьох та двох мікротрубочок, два інших-
назад і містять відповідно три та вісім мікротрубочок. Від базальних
тіл джгутиків також віялоподібно розходяться поодинокі мікротрубоч-
ки, частина з яких поєднує базальні тіла з ядерною оболонкою, вико-
нуючи таким чином функції ризопласта. Самі базальні тіла утриму-
ються разом поперечно-смугастим волокном, побудованим зі скорот-
ливого білка центрину.
Вегетативне тіло представлене багатоядерним розгалуженим
неклітинним міцелієм. Септами відділяються лише спорангії та гаме-
тангії. У представників, які ведуть паразитичний спосіб життя, окремі
гіфи міцелію видозмінюються у короткі трофічні гіфи - гаусторії. Вони
проникають у цитоплазму клітини-господаря і споживають органічну
речовину клітини, виконуючи функцію органа живлення.
Розмноження здійснюється нестатевим та статевим шляхами.
Нестатеве розмноження відбувається за допомогою зооспор,
що утворюються або у спорангіях, розташованих на верхівках веге-
267
тативних гіф міцелію, або в екзогенних спорангіях, які розвиваються
на спеціалізованих гіфах - спорангіофорах. Спорангіофори ооміко-
тових зі спорангіями в літературі часто називають конідієносцями з
конідіями, хоча ці структури принципово відрізняються від тих, які при-
таманні справжнім грибам.
Зооспорогенез в оомікотових еукарпічний - при утворенні зоо-
спор весь вміст спорангія витрачається на побудову репродуктивних
клітин. Зооспори оомікотових можуть бути двох типів - первинні та
вторинні. Первинні зооспори завжди розвиваються у багатоядерних
спорангіях, вони овальні і мають джгутики, розташовані апікально.
Такі зооспори або безпосередньо проростають у гіфу нового міце-
лію, або переходять у стан цисти, після чого проростають однією вто-
ринною зооспорою, яка має нирковидну форму, і джгутики у неї роз-
міщені латерально. При проростанні вторинні зооспори завжди да-
ють початок гіфам нового міцелію. Явище чергування у життєвому
циклі первинних та вторинних зооспор отримало назву дипланетиз-
му. Воно спостерігається переважно у водних оомікотових, наприк-
лад, у представників порядку Заргоіедпіаіез, у яких зооспорангії ут-
ворюються на верхівках вегетативних гіф міцелію.
У тих оомікотових, які паразитують на наземних рослинах, най-
частіше розвиваються зооспорангії на спорангієносцях. Такі зооспо-
рангії проростають зооспорами лише тоді, коли потрапляють у водне
середовище. Поза межами води, але при високій вологості повітря,
зооспорангій проростає голим багатоядерним протопластом, який за
рахунок амебоїдного руху потрапляє у міжклітинний простір майбут-
нього господаря, далі синтезує оболонку і починає розвиватися у гіфу
неклітинного міцелію. У посушливих умовах спорангій одразу проро-
стає неклітинною гіфою.
У деяких оомікотових існує додатковий спосіб нестатевого роз-
множення за допомогою хламідоспор. Хламідоспори оомікотових
(геми) - це неправильної форми фрагменти гіфи, які відокремлюють-
ся від талому і ростуть, утворюючи зародкові трубки. Такі трубки на-
далі проростають у гіфи.
Статевий процес - оогамія. Жіночі статеві органи - оогонії -
утворюються переважно з коротких бічних здутих гіф, що відокрем-
люються від міцелію септою. У водних оомікотових весь вміст оого-
нія використовується на побудову кількох яйцеклітин. У більшості на-
земних оомікотових в оогонії утворюється лише одна яйцеклітина,
причому частина цитоплазми оогонія зберігається навколо яйцеклі-
тини у вигляді т. зв. периплазми. Периплазма забезпечує зиготу, що
268
розвивається після запліднення, поживними речовинами та бере
участь у формуванні її оболонки.
Антеридії також утворюються як короткі бічні багатоядерні гіфи і
відокремлюються від основної частини міцелію септами. Вміст анте-
ридію не диференціюється на гамети. Антеридіальні гіфи ростуть у
напрямку оогонія. Далі антеридії утворюють короткі копуляційні відро-
стки, які контактують з оогонієм, проростають крізь оболонку жіночо-
го гаметангію до яйцеклітини і переливають у неї свій вміст. Чоловіче
та жіноче ядра зливаються, і утворюється зигота-ооспора. Вона вкри-
вається товстою оболонкою і переходить у стан спокою. Після завер-
шення періоду спокою ооспора проростає або новим міцелієм, або
спорангіофором із зооспорангіями.
Життєвий цикл в оомікотових диплофазний, без зміни поколінь.
Мейоз відбувається під час формування статевих органів, тобто є
гаметичним.
Екологічні особливості. Більшість представників відділу є па-
разитами прісноводних тварин та наземних вищих рослин. Значно
менша кількість видів належить до прісноводних та ґрунтових сап-
ротрофів. Чимало видів завдають значної шкоди промисловому риб-
ництву та сільському господарству. Зокрема, водні ооміцети спричи-
нюють небезпечні хвороби риб - сапролегніози. Наземні паразити
вищих рослин викликають захворювання, відомі під загальною на-
звою несправжньої борошнистої роси.
Система відділу. Відділ включає лише один клас - ооміцетові
(Оотусеіез), у межах якого виділяють вісім порядків. Провідними та
найважливішими у практичному відношенні серед них є три порядки
- Заргоіедпіаіез, Регопозрогаіез та Руііііаіез. Вони розрізняються, в
першу чергу, за особливостями нестатевого розмноження, будовою
оогоніїв, а також за екологічними особливостями їх представників.
Порядок Сапролегніальні - Заргоіедпіаіез
Міцелій добре розвинутий, багатоядерний, несептований. Неста-
теве розмноження дводжгутиковими зооспорами, часто двох типів,
які формуються в циліндричних видовжених зооспорангіях. Переваж-
но паразити водоростей та водних тварин, поширені у прісних водой-
мах, рідше в морях.
269
Рід сапролегнія - Заргоіедпіа (рис. 99). Міцелій сапролегнії не-
септований, складається із слабо розгалужених безбарвних гіф, які
вкриті тонкою оболонкою. Цитоплазма зерниста, з добре помітними
продовгуватими вакуолями. На кінцях гіф утворюються зооспорангії
-довгі булавовидні здуття, відокремлені клітинною перегородкою від
решти гіфи. Зооспори після дозрівання виходять через отвір на
верхівці зооспорангія. Після виходу зооспор від зооспорангія лишаєть-
ся порожній чохол, в який може вростати наступний зооспорангій від
цієї ж гіфи. Цей процес називається проліферацією, іноді він може
повторюватися 5-6 разів. Зооспори, що вийшли із спорангія, мають
грушовидну форму і два передні джгутики різної довжини та будови
(один гладенький, інший пірчастий). Після нетривалого періоду ак-
тивного руху ці зооспори зупиняються, вкриваються оболонкою і, вна-
слідок проростання, дають початок вторинним зооспорам нирковид-
ної форми (явище дипланетизму). Нирковидні зооспори прямують до
субстрату і проростають міцелієм.
Оогонії та антеридії утворюються на коротких бокових гіфах по-
близу субстрату. Перш за все, помітні кулясті оогонії, які у зрілому
стані виповнені темними сферичними яйцеклітинами. У сапролегнії в
одному оогонії утворюється до восьми яйцеклітин. Антеридії розта-
шовуються поряд з оогоніями. Окремий антеридій являє собою роз-
ширений кінець бічної гіфи і відокремлений від неї перегородкою. Він
прикладається до оогонія, дає кілька запліднюючих відростків, які
проходять через пори у товстій оболонці оогонія, і переливає в яй-
цеклітини частину свого вмісту з одним ядром. Зазвичай один оогоній
обростає кількома антеридіями. Після запліднення яйцеклітини утво-
рюється зигота, яка вкривається щільною двошаровою оболонкою
(такі зиготи часто називають ооспорами), і після деякого періоду спо-
кою проростає, утворюючи зооспорангій із зооспорами. В деяких оого-
ніях ооспори утворюються партеногенетично.
Представників цього роду завжди можна зустріти у вигляді вато-
подібного пуху на органічних залишках тваринного походження в річці
або ставку. Найчастіше вони розвиваються сапротрофно на мертвій
рибі, комахах, жабах, раках, але можуть і паразитувати на ікрі риби
та жаб, а також молоді риб, наносячи цим значних збитків рибним
господарствам. Найпоширенішим видом, що паразитує на рибах та
їх ікрі, є 5. рагазіііса - сапролегнія паразитична.
270
Рис. 99. Заргоіедпіа: А - зовнішній вигляд риби, ураженої сапролегнією;
Б - схема життєвого циклу. 1 - зооспорангій; 2 - грушовидна зооспора; 3 - нир-
ковидна зооспора; 4 - оогоній з антеридієм; 5, 6 - послідовні стадії запліднення
яйцеклітин; 7 - зиготи; 8 - проростання зигот
271
Порядок Пероноспоральні - Регопозрогаіез
Найбільший за чисельністю видів порядок у складі відділу
Оотусоіа. Об’єднує біля 250 видів. Нестатеве розмноження - спо-
рангіями (конідіями), які утворюються на спеціалізованих гіфах-спо-
рангієносцях (або конідієносцях).
Рід плазмопара - РІазторага (рис. 100). Види цього роду ви-
кликають захворювання вищих рослин - несправжню борошнисту
росу. Один із видів - Р. уііісоїа - плазмопара виноградова - зумов-
лює захворювання винограду, яке відоме також під назвою «мільдью».
Міцелій гриба міжклітинний, розпростертий, з дрібними присосками.
Конідієносці (спорангієносці) виходять пучками через продихи, ближ-
че до верхівки моноподіально розгалужені майже під прямим кутом.
Кінцеві розгалуження мають зубоподібні короткі гілочки. Зооспорангії
яйцеподібні, на верхівці без горбочка. Оогонії круглі, оболонка бага-
тошарова, безбарвна або жовтувата, ооспори - жовтуваті або корич-
неві із гладенькою або складчастою оболонкою. Ооспори пророста-
ють навесні ниткоподібною гіфою із здуттям на кінці - первинним
зооспорангієм. У ньому утворюється 8-10 дводжгутикових зооспор.
Зооспори, потрапляючи в краплину води на листку чи іншій тканині
рослини, прямують до продихів, втягують у себе джгутики, покрива-
ються тонкою оболонкою і проростають нитковидною гіфою. Гіфа про-
никає в тканини рослини через продихи і за допомогою гаусторіїв при-
смоктується до клітин листка. Ріст міцелію в рослині продовжується
тривалий час. Потім утворює спорангієносці із зооспорангіями (т. зв.
конідіями). За вегетаційний період гриб дає 7-11 поколінь нестате-
вих спороношень. Зооспорангії можуть проростати зооспорами, якщо
потрапляють у краплину води, або безпосередньо гіфою, якщо вони
не у водному середовищі. Ооспори утворюються всередині ураже-
них тканин і зимують в опалому листі.
Гриб уражує всі нездерев'янілі наземні органи винограду. Захво-
рювання проявляється навесні появою на молодому листі блідо-зе-
лених або жовтуватих плям, які з часом буріють. На нижньому боці
листків, на пошкоджених ділянках листа, з’являється рясний білува-
тий наліт-пучки спорангієносців. Основні заходи боротьби з мільдью
винограду-оприскування виноградників бордоською рідиною (суміш
мідного купоросу та гашеного вапна) та виведення стійких до захво-
рювання сортів винограду.
272
Рис. 100. РІазторага VІ^ісоIа: А- листок винограду, уражений мільдью;
Б - уражене гроно; В - ооспори в тканині листка; Г - проростання спорангіє-
носців через продих листка; Д - окремий спорангієносець із зооспорангіями;
Е - зооспора
Порядок Пітіальні - РуФіаІев
Порядок об’єднує види, які мають нестатеве спороношення у виг-
ляді зооспорангіїв, які, залежно від умов, проростають зооспорами
або гіфами. Спорангієносці розгалужені, добре відрізняються від веге-
тативних гіф міцелію.
Рід фітофтора - РІіуіорГііІюга (рис.101). Найпоширеніший вид
роду - Рй. ігіїезіапз - фітофтора інфекційна або фітофтора кар-
топлі - дуже небезпечний паразит картоплі та інших пасльонових,
зокрема, помідорів, може спричинити повну втрату врожаю. Захво-
рювання, яке викликає цей гриб, має назву фітофтороз. Збудник зах-
ворювання поширений скрізь, найбільш інтенсивно розвивається
у вологу погоду.
273
Цікавим фактом, пов’язаним із цим грибом, є його безпосередній
вплив на історичні процеси людства, зокрема, цей гриб знищив план-
тації картоплі в Ірландії та Німеччині у XIX ст., спричинивши масову
еміграцію населення з цих коаїн.
Ю
Рис. 101. РіїуіорМіїога іпїезіапз: Схема життєвого циклу. 1 - зооспо-
рангій; 2 - вихід зооспор; 3 - зооспори; 4 - проростання зооспор; 5 - проник-
нення міцелію у клітини рослини; 6 - міцелій у міжклітинному просторі
листка; 7 - проростання спорангієносців через продих листка; 8 - оогоній та
антеридій; 9 - ооспора; 10, 11 - проростання ооспори зооспорангієм
Відділ Лабіринтуломікотові гриби - ЬаЬугіпНіиІотусоіа
Евкаріотичні осмотрофні страменопіли, в яких голі клітини вкриті
мікроскопічними лусочками. Вегетативне тіло представлене пооди-
нокими або з’єднаними у сітчасті псевдоплазмодїі клітинами з екто-
плазматичним ретикулюмом, який утворюють спеціальні органели -
274
ботросоми. Монадні стадії дводжгутикові. Відділ включає біля 50-ти
видів псевдогрибів, поширених переважно у морських біотопах.
Біохімічні особливості та живлення. За метаболізмом лізину
лабіринтуломікотові подібні до тварин, оскільки самі не синтезують
лізин, а отримують його у складі білків у процесі живлення. Основний
продукт асиміляції - глікоген, додатковий - олія.
Органічні речовини поглинаються осмотрофно. Процеси абсорбції
здійснюються за участю ектоплазматичного ретикулюму.
Цитологічні особливості. Вегетативне тіло лабіринтуломіко-
тових представлено поодинокими клітинами, які здатні утворювати
надклітинні агрегати у вигляді псевдоплазмодіїв. Найяскравіша особ-
ливість лабіринтуломікотових - наявність ектоплазматичного рети-
кулюму, що розташовується назовні плазмалеми і утворюється
унікальними клітинними структурами - ботросомами (сагеногене-
тосомами, сагеногенами). Ботросоми мають вигляд невеликих
внутрішніх інвагінацій плазмалеми, на дні яких розташовується елект-
роннощільне тіло, до якого із внутрішнього боку клітини підходить
один-два мембранних канали, а із зовнішнього боку у вигляді спло-
щеної цистерни - мембрана ектоплазматичного ретикулюму. Такі ци-
стерни сусідніх ботросом зливаються одна з одною, утворюючи
навколо клітини додатковий зовнішній двомембранний покрив - екто-
плазматичну обгортку. В середині цієї обгортки знаходиться позбав-
лений органел матрикс ектоплазми. В ньому розміщуються пучки ско-
ротливих мікрофібрил, одним із компонентів яких є актин, завдяки
чому клітина здатна пересуватися всередині ектоплазматичної об-
гортки. Скорочення мікрофібрил є кальцій-залежним.
Зовнішні мембрани ектоплазматичного ретикулюму різних клітин,
контактуючи між собою, частково зливаються, утворюючи анастомо-
зи і об’єднуючи клітини у сітчастий псевдоплазмодій. У деяких лабі-
ринтуломікотових ектоплазматичний ретикулюм не охоплює всю кліти-
ну, а розташовується лише на одному її боці, трансформуюючись у
сітчасту ризоїдоподібну структуру. В цих випадках клітини залиша-
ються поодинокими і псевдоплазмодіїв не утворюють.
На поверхні плазмалеми, під внутрішньою мембраною ектоплаз-
матичного ретикулюму знаходяться субмікроскопічні лусочки, які
утворюються у комплексі Гольджі та секретуються на поверхню плаз-
малеми.
Ядерний апарат евкаріотичний, клітини у вегетативному стані
одноядерні. В інтерфазі в ядрі є одне ядерце та периферичні скуп-
275
чення рибонуклеопротеїду, до ядерної оболонки прилягає один-два
розвинених комплекси Гольджі, а в одноклітинних (траустохітридіаль-
них) представників - також пара центріолей та електронно-щільне
лопатеве парануклеарне тіло, утворене комплексом сплощених цис-
терн ендоплазматичної сітки. Як і в інших пседогрибів, при утворенні
джутикових стадій ядерна оболонка бере участь у формуванні рет-
ронем.
Мітоз напіввідкритий, здійснюється за участю центріолей. Останні
або присутні у клітині протягом всього клітинного циклу, або з’явля-
ються безпосередньо перед початком поділу ядра. Ядерце у профазі
зникає і відновлюється лише після завершення каріокінезу.
Мітохондріальні профілі у клітинах численні, кристи трубчасті.
Цікавими цитоплазматичними структурами лабіринтуломікотових є
мультивезикулярні тіла та кортикальні глобули. Мультивезикулярні
тіла являють собою одномембранні пухирці, всередині яких знахо-
диться кілька дрібніших мембранних пухирців. Кортикальні глобули
утворені ліпідами, які розташовуються під плазмалемою на периферії
клітини.
Монадні стадії представлені зооспорами з двома латеральними
гетероконтними гетероморфними та гетеродинамічними джгутиками.
Довший джгутик пірчастий, він направлений вперед і вкритий ретро-
немами. Коротший - гладенький і спрямований назад. У деяких видів
(зокрема І_аЬугіпіііиІа) при основі коротшого джгутика знаходиться
парабазальне потовщення, до якого прилягає вічко. Парабазальне
потовщення є фоторецептором, а вічко виконує функцію ширми. Ядра
зооспор мають гаплоїдний набір хромосом.
Вегетативне тіло може бути представлене двома варіантами.
Перший - поодинокими клітинами з однією ризоїдоподібною екто-
плазматичною цистерною (у траустохітридіальних). Ці вегетативні тіла
нагадують ризоміцелій, через що таких лабіринтуломікотових ще
наприкінці XX ст. розміщували серед оомікотових або навіть хітриді-
омікотових грибів. Другий варіант - це сітчастий псевдоплазмодій,
характерний для лабіринтуляльних псевдогрибів.
Розмноження відбувається нестатевим та статевим шляхами.
Найпростіший спосіб нестатевого розмноження - це вегетативний
поділ клітин надвоє. У складніших випадках лабіринтуломікотові ут-
ворюють зооспори. У представників з поодинокими клітинами зоо-
спорангії розвиваються переважно еукарпічно: ядро клітини багато-
276
разово мітотично ділиться, далі навколо більшості ядер уособлюють-
ся ділянки цитоплазми, які надалі трансформуються у голі нерухомі
спори. Решта частини цитоплазми відмежовується від спорангія плаз-
малемою. Після розриву покриву спорангія спори звільняються, швид-
ко виробляють джгутики і перетворюються на зооспори. Після періо-
ду активного руху зооспори осідають на субстрат (переважно - на
вищу водну рослину або макроскопічну водорість), втягують джгути-
ки і розвивають ризоїдоподібну ектоплазматичну сітку.
У видів із псевдоплазмодіями на спорангій перетворюється вся
вегетативна клітина, тобто розвиток холокарпічний. Після серії міто-
тичних поділів ядра весь вміст клітини розпадається на одноядерні
диплоїдні цисти. Після руйнування покривів материнської клітини
цисти звільняються, в них відбувається один мітоз та мейоз, і з кож-
ної цисти виходить вісім гаплоїдних зооспор.
Статевий процес, за деякими спостереженнями, нагадує ізога-
мію, при якій зооспори можуть виконувати функції гамет. Проте де-
талі цього процесу залишаються нез’ясованими.
Екологічні особливості. Лабіринтуломікотові мешкають пере-
важно на морських рослинах: макроскопічних водоростях, морській
траві зостері, а також на їх відмерлих рештках. Більшість лабіринту-
ломікотових є сапротрофами, які розкладають детрит та рослинні
залишки. Проте відомі випадки паразитизму, коли розвиток цих
організмів у естуаріях спричинював хворобу та загибель водоростей
із родів Вгуорізіз, 1_атіпагіа та епіфітотії морської квіткової рослини
Хозіега тагіпа. Є відомості про знахідки лабіринтуломікотових у
прісних водоймах та на вологому ґрунті, де ці гриби розвиваються на
таломах жовтозеленої водорості Уаисіїегіа.
Система відділу. Відділ включає один клас - лабіринтуломіце-
тові (І_аЬугіпіІіиІотусеіе8), та два порядки - лабіринтуляльні (1_аЬу-
гіпіїїиіаіез) та траустохітридіальні (ТгаизІосНуІгісІіаІез). Порядки
відрізняються за кількістю ботросом в однієї клітини, будовою екто-
плазматичної сітки (утворює вона псевдоплазмодій чи ризоїдоподібні
вирости), цитологічними особливостями (наявність в інтерфазі цент-
ріолей, парануклеарного тіла), способом утворення зооспор (еукар-
пічно без стадії цисти чи холокарпічно зі стадією цисти).
Порядок Лабіринтуляльні - ЬаЬугіпіЬиіаІез
Об’єднує організми, в яких ботросоми численні, ектоплазма пов-
ністю вкриває клітину і здатна агрегувати клітини у сітчасті псевдо-
277
плазмодії. Центріолі та парануклеарні тіла в інтерфазі відсутні. Зоо-
спори утворюються холокарпічним шляхом і мають вічко.
Характерним представником порядку є 1_аЬугіпіїіиІа тасгосузііз -
лабіринтуля макроцистна - збудник епіфітотій морської трави зостери
(рис. 102).
Рис. 102. 1.аЬугіпІЇюІа тасгосузііз: А - утворення ектоплазми ботросома-
ми; Б - поодинока клітина з розвиненою ектоплазмою; В - фрагмент псевдо-
плазмодія; Г - зооспора; Д - ботросома.1 - канал ЕПС або ядерної мембра-
ни; 2 - зовнішня мембрана ектоплазми; 3 - внутрішня мембрана ектоплаз-
ми; 4 - субмікроскопічні луски; 5 - плазмалема
Порядок Траустохітридіальні - ТгаивіосЬуігісііаІев
Включає гриби з поодинокою ботросомою, яка утворює ризоїдо-
подібну ектоплазму, що не вкриває клітину і не агрегує окремі клітини
у псевдоплазмодії. Центріолі та парануклеарні тіла в інтерфазі на-
явні. Зооспори утворюються переважно еукарпічним шляхом і поз-
бавлені вічка.
Найвідомішим представником порядку є Тгаизіосіїуігіит
ргоііїегит - траустохітриум проліферуючий (рис. 103), який часто
оселяється на таломах сифональних морських водоростей з роду
Вгуорзіз.
278
Рис. 103. Тгаизіосіїуігіитргоііїегит: А-талом; Б -зооспорангій із зооспора-
ми; В - вихід зооспор
Відділ Гіфохітриомікотові гриби - НурІїосЬуігіотусоіа
Евкаріотичні осмотрофні страменопіли, в яких клітини у дорослому
стані повністю або частково вкриті целюлозно-хітиновою оболонкою,
а вегетативне тіло представлене поодинокими багатоядерними кліти-
нами або ризоміцелієм. Монадні стадії одноджгутикові. Відділ нара-
ховує лише біля ЗО видів, які належать до п’яти родів. Більшість видів
є ґрунтовими сапротрофами (розвиваються на мертвих тілах безхре-
бетних тварин та грибів, на відмерлих рослинах) або паразитами на-
земних та водних організмів (оомікотових та аскомікотових грибів,
зелених та бурих водоростей, ракоподібних).
Біохімічні особливості та живлення. Представники відділу син-
тезують лізин рослинним шляхом - з аспартата та пірувата через
діамінопімелінову кислоту (ДАП-шлях). На відміну від оомікотових,
гіфохітриомікотові здатні до біосинтезу стероїдів. Споживання орга-
нічних речовин здійснюється виключно шляхом абсорбції. Фагоцитоз
та травні вакуолі, незважаючи на наявність у ряду представників ри-
зоподієподібних виростів ризоміцелію, відсутні. Як джерело вуглецю
гіфохітриомікотові здатні використовувати різноманітні прості цукри,
а також целюлозу та крохмаль. Азот ці організми засвоюють виключ-
но у вигляді органічних сполук, зокрема у складі пептону або аспара-
279
гіну. Продукт асиміляції - олія. Відомості про запасні полісахариди
відсутні.
Цитологічні особливості. У дорослому стані всі представники
відділу повністю або частково вкриті целюлозно-хітиновими оболон-
ками. На ранніх стадіях розвитку вегетативне тіло гіфохітриомікото-
вих може бути представлене голими протопластами.
Ядерний апарат евкаріотичний. Зооспори одноядерні, дорослі
вегетативні клітини містять багато ядер з одним ядерцем у центрі.
Особливістю інтерфазних ядер у зооспорах та дорослих вегетатив-
них клітинах є наявність навколо зовнішньої мембрани ядерної обо-
лонки щільного кільця рибосом, які утворюють т. зв. перинуклеарну
рибосомальну зону. До ядерної оболонки також притиснутий комп-
лекс Гольджі та одне або кілька мікротілець (припускають, що це пе-
роксисоми). На периферії рибосомальної зони розташовується кілька
мітохондріальних профілів, які мають трубчасті кристи.
Мітоз напіввідкритий, веретно утворюється за участю центріолей.
Ядерце у профазі зникає і відновлюється лише після закінчення карі-
окінезу.
Зооспори гіфохітриомікотових мають лише один апікальний джгу-
тик, вкритий двома рядами ретронем. Перехідна зона джгутика містить
термінальну платівку. Біля його базального тіла знаходиться одна
центріоля, яка є залишком базального тіла другого, редукованого
джгутика. Від базального тіла беруть початок три джгуткових корені,
два з яких спрямовані вперед і з’єднані із гребінчасто розташованою
системою мікротрубочок, третій - назад і проходить вздовж ядерної
оболонки. До базального тіла щільно притиснута верхівка ядра.
У цитоплазмі зооспор розміщені мультивезикулярні тіла, подібні до
тих, що є у лабіринтуломікотових псевдогрибів, а також велика крап-
лина олії та електронно-щільні дрібні везикули.
Вегетативне тіло гіфохітриомікотових у зрілому стані представ-
лене або поодинокою багатоядерною сферичною клітиною, або ри-
зоміцелієм. Останній може бути простим або складним. Простий
ризоміцелій складається із вкритої оболонкою багатоядерної цент-
ральної частини та голого розгалуженого ризоїда. У складного ризо-
міцелію від центральної багатоядерної частини відходить кілька
ризоїдів. На верхівці кожного з них може розвиватися нова здута ба-
гатоядерна частина, яка надалі утворює нові ризоїди. Як наслідок,
вегетативне тіло поступово набуває вигляду справжнього міцелію.
280
Розмноження нестатевим шляхом відбувається зооспорами, які
утворюються холокарпічно. Статевий процес - ізогамія. Гамети, що
копулюють, представлені двома голими протопластами, які виходять
з молодих вегетативних клітин. Статеве спороношення - багатоядерна
спочиваюча спора, яка згодом перетворюється на зооспорангій.
Життєвий цикл агамних видів та цикл нестатевого розмноження
еугамних представників включає стадії: а) зооспори; б) молодої кліти-
ни, вкритої оболонкою; в) дорослого талому; г) зооспорангія.
Наприклад, у видів роду анісольпідіум (Апізоірідіит), які роз-
виваються переважно на водоростях, зооспора осідає на придатний
субстрат, втрачає джгутик, вкривається целюлозно-хітиновою оболон-
кою, розчиняє оболонку господаря. Через такий отвір протопласт
молодої клітини перетікає у клітину господаря. Гриб починає живи-
тись та рости. Ядра при цьому мітотично діляться, талом стає бага-
тоядерним. Згодом ріст уповільнюється, протопласт вкривається обо-
лонкою і перетворюється на дорослу багатоядерну сферичну кліти-
ну. Вона утворює шийку, яка прориває назовні оболонку господаря.
Вміст клітини розпадається на багато зооспор. Після розриву шийки
спорангія зооспори виходять назовні і уражують нові водоростеві
клітини.
Якщо на одну клітину осідає дві зооспори, то їх протопласти після
проникнення у клітину можуть копулювати, утворюючи зиготу. Вона
одразу розвивається в багатоядерний зооспорангій і далі переходить
у стан спокою. Після його завершення за непрямими даними відбу-
вається мейоз і утворюються гаплоїдні зооспори.
Види роду ризидіоміцес (КІііхідіотусез) уражують оомікотові
та хітридіомікотові гриби. їх зооспори осідають на тіло господаря, втра-
чають джгутик та покриваються оболонкою, за винятком місця кон-
такту із субстратом. З цієї точки гриб утворює ризоїд простого ризо-
міцелію, що проникає у тіло господаря і споживає його органічні ре-
човини. Зовнішня частина паразита розростається, стає багатоядер-
ною і згодом перетворюється на зооспорангій.
Схожий цикл розвитку має Нуріїосіїуігіит репіііае - гіфохітри-
ум пенілли, який викликає масову загибель планктонного рачка
Репіііа ауігозігіз. Відміни полягають у тому, що під час росту гіфохіт-
риум утворює складний ризоміцелій, ріст якого і обумовлює смерть
господаря (рис. 104).
281
Рис. 104. Нуріїосіїуігіит: А - зооспора; Б - проростання зооспори; В - ризо-
міцелій
Система відділу включає лише один клас гіфохітридіоміцето-
вих (Нуріїосіїуігіотусеіез) та один порядок гіфохітридіальних
(НуріїосНуігісІіаІез).
Гриби платикристати
До грибів-платикристат належать п’ять відділів осмотрофних ге-
теротрофів - плазмодіофоромікотові, хітридіомікотові, зигомікотові,
аскомікотові та базидіомікотові. Серед цих відділів за комплексом
фенотипічних ознак, які корелюють з молекулярно-філогенетичними
даними, відокремлену позицію на філогенетичному дереві займають
плазмодіофоромікотові слизовики, які більш-менш рівновіддалені від
підцарства рослин, з одного боку, та підцарства справжніх грибів, з
іншого. У грибів-платикристат у клітинних оболонках вегетативних тіл
або спор каркасним елементом є хітин. Основний продукт асиміля-
ції - глікоген. Мітохондріальні кристи у плазмодіофоромікотових двох
типів - пластинчасті та трубчасті, у всіх справжніх грибів - лише пла-
стинчасті. Кристи при основі, незалежно від їх форми, не мають ба-
зальної перетяжки.
282
Відділ Плазмодіофоромікотові слизовики -
РІазтосііорНоготусоіа
Евкаріотичні первинно-гетеротрофні твариноподібні платикрис-
тати, вегетативне тіло яких представлене голим внутрішньоклітин-
ним плазмодієм. Джгутикові стадії - зооспори з двома гладенькими
нерівними джгутиками. Відділ об’єднує біля п’ятидесяти видів облігат-
них внутрішньоклітинних паразитів вищих рослин, водоростей та
грибів.
За результатами молекулярно-філогенетичних реконструкцій,
плазмодіофоромікотові розташовуються при основі дерева платик-
ристат. Найближчими родичами цього відділу є, з одного боку, гапто-
фітові водорості, з іншого - хітридіомікотові гриби. Цікавим є те, що,
подібно до гаптофітових, у багатьох плазмодіофоромікотових у міто-
хондріях наявні кристи двох типів - як пластинчасті, так і трубчасті.
Проте за наявними електронограмами трубчасті кристи при основі
не перетягнуті, тобто гомологічні типовим пластинчастим кристам
інших платикристат.
Біохімічні особливості та живлення. Відомості щодо ранніх
стадій метаболізму лізину у плазмодіофоромікотових відсутні. Біосин-
тез триптофану відбувається за допомогою ферментів першої групи,
аналогічно до хітридіомікотових та частини аскомікотових грибів.
Основний продукт асиміляції - глікоген, додатковий - олія.
Незважаючи на наявність плазмодіїв, живлення плазмодіофоромі-
котових відбувається виключно осмотрофно, абсорбційним шляхом.
Ці гриби не утворюють травних вакуолей, хоча при розростанні плаз-
модії здатні неповністю охоплювати своєю цитоплазмою протопласт
клітини-господаря.
Цитологічні особливості. Вегетативне тіло, яке представлене
внутрішньоклітинним плазмодієм, вкрите лише плазмалемою, тобто
є голим. Перед утворенням спор таке тіло оточується тонким шару-
ватим зовнішнім покривом, але його природа залишається нез’ясо-
ваною. Спори плазмодіофоромікотових вкриті хітиновою оболонкою.
Ядерний апарат евкаріотичний. У репродуктивних клітин (зооспор
та міксамеб) ядро одне, вегетативні стадії- первинні та вторинні плаз-
модії - багатоядерні, причому первинні плазмодії мають ядра гене-
тично однакові, а вторинні містять багато пар дикаріонів. У інтерфазі
ядра плазмодіїв мають одне ядерце; до поверхні ядерної мембрани
283
притиснута пара центріолей, які розташовуються на одній осі і май-
же стикаються базальними частинами. Кут між центріолями стано-
вить 180°. До протилежного боку ядерної оболонки прилягає комп-
лекс Гольджі.
Мітоз напіввідкритий, здійснюється за участю центріолей. Під час
мітозу в метафазі ядерце ділиться перешнуровкою, причому поділ
відбувається у площині, перпендикулярній до осі веретена. В ана-
фазі, коли хромосоми розходяться до полюсів веретена, як наслідок,
всередині ядра спостерігається структура, яка нагадує хрест (т. зв.
хрестоподібний мітоз плазмодіофоромікотових): на полюсах ядра біля
отворів ядерної оболонки розташовується хроматин, а латерально
до осі веретена - по одному ядерцю.
Мітохондріальні профілі численні і мають трубчасті та пластин-
часті кристи, які не перетягнуті при основі.
Джгутикові стадії представлені зооспорами з двома передніми
гладенькими джгутиками нерівної довжини. Базальні тіла джгутиків
орієнтовані одне до одного під кутом 40-60°. У зооспорах на стадії
підготовки до інфікування клітини рослини-господаря з’являється
специфічна паличкоподібна органела, яка надалі розвивається у
складний апарат проникнення в клітину-господаря - адгезорій.
Вегетативне тіло представлене внутрішньоклітинними багато-
ядерними первинними та дикаріонтичними вторинними плазмодія-
ми. Плазмодії не здатні до активного руху за допомогою псевдоподій,
хоча у молодому віці можуть переноситися в інші клітини господаря
через пори по плазмодесмах.
Розмноження відбувається нестатевим шляхом за допомогою
первинних зооспор, що утворюються з нерухомих спор. Статевий
процес - ізогамія. Гамети морфологічно подібні до первинних зоо-
спор і називаються вторинними зооспорами, завдяки їх здатності про-
ростати партеногенетично.
Життєвий цикл гаплофазний із зиготичною редукцією та чергу-
ванням первинних гаплоїдних та вторинних дикаріонтичних плаз-
модіїв.
У загальному вигляді життєвий цикл наступний: вторинний плаз-
модій холокарпічно розпадається на вкриті оболонками спори. Після
руйнування клітини-господаря спори звільняються, і за певних умов
окрема спора проростає дводжгутиковою первинною зооспорою.
284
Вона після періоду активного руху осідає на придатний субстрат, вкри-
вається оболонкою, розвиває адгезорій, за допомогою якого просвер-
длює оболонку клітини-господаря і переливає в неї свою цитоплазму.
Протопласт первинної зооспори росте, споживаючи органічні ре-
човини господаря; його ядро багаторазово мітотично ділиться, вна-
слідок чого розвивається первинний плазмодій. Після споживання
органічних речовин господаря первинний плазмодій вкривається
шаруватою оболонкою, часто утворює вивідні трубки, які проривають
покриви мертвої клітини-господаря і створюють канали для виводу
назовні майбутніх репродуктивних клітин. Після цього вміст первин-
ного плазмодію розпадається на одноядерні ізогамети (т. зв. вторинні
зооспори), які здатні також проростати партеногенетично.
Гамети виходять у зовнішнє середовище. Далі вони попарно ко-
пулюють, утворюючи дикаріонтичну клітину злиття. Така клітина збе-
рігає дві пари джгутиків, активно рухається, далі осідає на придатний
субстрат, розвиває адгезорій і переливає свій вміст у клітину госпо-
даря. Клітина злиття росте, її ядра синхронно діляться. Як наслідок,
розвивається вторинний дикаріонтичний плазмодій. При вичерпанні
поживних речовин клітини-господаря дикаріони вторинного плазмо-
дію зливаються, відбувається мейоз, і плазмодій розпадається на од-
ноядерні, вкриті оболонками спори. Після руйнування покривів мерт-
вої клітини-господаря спори звільняються і можуть проростати пер-
винними зооспорами.
Система відділу. Відділ включає лише один клас Плазмодіофо-
роміцетових (РІазтосІіорНоготусеіез) та один порядок - плазмодіо-
форальних (РІазтосІіорНогаІез). До найпоширеніших та найбільш
важливих у практичному відношенні належать роди РІазтосііорІїога,
Роїутуха та Зропдозрога.
Рід плазмодіофора - РІазтодіорІюга (рис. 105). Типовий пред-
ставник роду - РІазтосІіорІїога Ьгаззісае — плазмодіофора капус-
тяна, або збудник захворювання коренів капусти та інших хрестоцві-
тих, яке відоме під назвою “кіла”, - був описаний російським вченим
М. С. Вороніним у 1887 р. На коренях ураженої рослини спочатку з’яв-
ляються здуття, які розростаються до великих пухлин найрізноманіт-
нішої форми з нерівною поверхнею, пізніше корені рослини набува-
ють потворного вигляду. Паразит, перебуваючи в клітинах рослини,
призводить до збільшення їх розмірів (гіпертрофії) та інтенсивного
ділення (гіперплазії). Гриб може уражувати капусту в будь-якому віці,
285
тому такі деформовані рослини можна знаходити з ранньої весни се-
ред розсади і до пізньої осені, коли капусту зрізають та збирають.
При ураженні капусти в молодому віці рослина, як правило, гине, а
на більш пізніх стадіях розвитку качан або зовсім не розвивається,
або дуже затримується у розвитку. Захворювання знижує урожай ка-
пусти на 30-40% і більше.
Зооспори або міксамеби проникають у кореневий волосок росли-
ни, де відбувається їх злиття та утворення первинного плазмодію.
Пізніше плазмодій розпадається на кілька зооспорангіїв, в яких після
багаторазових поділів ядер утворюються гаплоїдні зооспори, що по-
трапляють у ґрунт. Вони копулюють, перетворюючись на дикаріон-
тичні амебоїди, які знов уражують клітини кореня капусти. Амебоїди
розростаються, зливаються і формують багатоядерні вторинні плаз-
модії, які наповнюють клітини хазяїна. Оточуючі клітини, що не містять
плазмодіїв паразита, активно діляться і розростаються. Потім у плаз-
модії ядра зливаються попарно, утворюючи нетривалу диплоїдну ста-
дію, далі діляться мейотично, і формуються спори. Ці спори звільня-
ються при згниванні кореня і в наступному році, при сприятливих умо-
Рис. 105. Ріазтосііоріїога Ьгаззісае: А- уражений корінь капусти; Б - схе-
ма життєвого циклу. 1 - спочиваюча спора; 2 - проростання спори; 3 - первинна
зооспора; 4 - міксамеба; 5 - гаплоїдний первинний плазмодій; 6 - літній спо-
рангій; 7 - вторинна зооспора (ізогамета); 8 - копуляція; 9 - плазмогамія;
10 - дикаріонтична клітина злиття; 11 - вторинний плазмодій; 12 - спочиваючі
спори у клітинах пухлин ураженого кореня
286
вах вологи та температури, проростають міксамебою або зооспорою
з двома джгутиками різної довжини. Цикл повторюється. Спори мо-
жуть зберігати життєздатність у ґрунті тривалий час (6-7 років).
Рід полімікса - Роїутуха. Найроширенішим видом є Роїутуха
Ьеіае - полімікса бурякова (рис. 106), яка завдає значної шкоди
бурякам, знижуючи їх урожайність у 10-20 разів, та призводить до
зменшення цукристості коренеплодів. Уражені рослини мають дефор-
мовані, недорозвинуті коренеплоди з великою кількістю дрібних ко-
рінчиків, що переплітаються між собою. Уражені поліміксою корене-
плоди буряка загнивають під час вегетації. Цікавим фактом, пов’яза-
ним з цим видом, є здатність Р. Ьеіае переносити вірус некротичного
пожовтіння жилок цукрового буряка, який викликає захворювання на
ризоманію.
Рис. 106. Роїутуха Ьеіае. плазмодії (А) та цистосоруси (Б) у клітинах росли-
ни-господаря
Рід спонгоспора - Зропдозрога. Серед представників цього
роду важливе практичне значення має Зропдозрога зиЬіеггапеа -
спонгоспора підземна, яка паразитує на коренях, бульбах і столо-
нах картоплі (Зоїапит іиЬегозит), викликаючи небезпечне захворю-
вання під назвою “порошиста парша бульб картоплі”. Паразит спри-
чиняє утворення на бульбах опуклих блідих або темнуватих пухлин
287
(пустул), які у міру підсихання зірчасто розтріскуються, оголюючи по-
рошисту масу спочиваючих спор, які щільно з’єднані у кулясті або
еліпсовидні клубочки (т. зв. спорокупки).
Серед інших представників порядку, які є облігатними ендопара-
зитами водоростей та грибів, слід згадати види роду вороніна (ІУо-
гопіпа), що паразитують на жовтозеленій водорості \/аисііегіага гри-
боподібних організмах з порядку Заргоіедпаііез, а також рід соро-
дискус (Зогосіізсиз), види якого паразитують на зеленій водорості
Сіїага, на оомікотових грибах та вищих рослинах.
Справжні гриби
Справжні гриби (підцарство Еипді) є філогенетично відособле-
ною великою групою евкаріотичних первинно гетеротрофних платик-
ристат, що живляться осмотрофно. Для справжніх грибів характер-
ний грибний шлях синтезу лізину - через р-аміноадипінову кислоту
(ААА-шлях), біосинтез триптофану за допомогою ферментів першої-
четвертої груп. Крім того, справжні гриби (за винятком зигомікотових)
здатні синтезувати спирт манітол (його наявність у крові або екстрак-
тах тваринних тканин є індикатором грибної інфекції). У справжніх
грибів відсутній комплекс Гольджі (його функції виконують різноманітні
видозміни ендоплазматичної сітки). В межах цієї філи чітко окреслю-
ються тенденції прогресуючого розвитку дикаріонтичної фази у жит-
тєвому циклі.
Справжні гриби мають чимало спільних рис зі справжніми твари-
нами (включаючи багатоклітинних - Меіагоа): утворення сечовини
як одного з кінцевих продуктів метаболізму, накопичення глікогену як
основного резервного полісахариду, наявність клітинних покривів,
каркасною основою яких є хітин. Монадні клітини справжніх грибів,
представлені у найпримітивнішому в даній групі відділі хітридіоміко-
тових, подібно до сперматозоїдів багатоклітинних тварин мають лише
один задній гладенький джгутик.
На молекулярних деревах справжні гриби утворюють компактну
монофілітичну групу. За системами різних авторів відділи, що нале-
жать до справжніх грибів, виділяються у таксон найвищого система-
тичного рангу - царство або підцарство.
Нижче справжні гриби (відділи СИуігісІіотусоіа, 2удотусоїа,
Азсотусоіа та Вазісііотусоїа) розглядаються як окреме підцарство
Еипді, яке є сестринським до підцарства тварин (Апітаїіа). Філа, яка
надалі розділяється на Еипді та Апітаїіа, споріднена також із підцар-
288
ством рослин (РІапіае), до якого належать фотоавтотрофні платик-
ристати.
Цікаво, що у молекулярну кладу, яку утворюють представники
підцарства Рипді, потрапляє також тип внутрішньоклітинних парази-
тичних тварин - мікроспоридії. Проте цей таксон досі не перебуває
під керуванням МКБН, тому нижче характеристка мікроспоридій не
наводиться.
Відділ Хітридіомікотові гриби - СКуігісІіотусоіа
Справжні гриби, специфічною ознакою яких є наявність монад-
них репродуктивних клітин з одним заднім гладеньким джгутиком.
Відділ нараховує біля тисячі видів. Включає переважно морських та
прісноводних (рідше - ґрунтових) паразитів водоростей, квіткових рос-
лин, безхребетних та грибів, а також сапротрофів, що розвиваються
на субстратах, багатих на хітин, целюлозу, кератин.
Біохімічні особливості та живлення. Хітридіомікотові синтезу-
ють лізин через р-аміноадипінову кислоту (ААА-шлях), триптофан -
виключно за допомогою ферментів першої групи. Іони заліза поглина-
ються за участю сидерамінів. Як джерело вуглецю використовують
широкий спект оліго- та полісахаридів (від глюкози та фруктози до це-
люлози та хітину). Азот здатні засвоювати у вигляді амінокислот, пеп-
тидів, мінеральних сполук (солей амонію, нітратів, деякі види - також і
нітритів).
Поглинання органічних сполук здійснюється виключно шляхом
абсорбції. Травні вакуолі у хітридіомікотових не утворюються.
Цитологічні особливості. Клітини хітридіомікотових принаймні
на заключних стадіях розвитку вкриті хітин-глюкановими клітинними
оболонками.
Ядерний апарат евкаріотичний. Кількість ядер у різних представ-
ників варіює від одного до багатьох. Ядра зазвичай мають по одному
ядерцю.
Мітоз напіввідкритий, відбувається за участю центріолей. При
мітотичному поділі ядерце у різних представників веде себе по-різно-
му. У більшості випадків ядерце у профазі зберігається і зникає у ме-
тафазі або анафазі. Рідше воно у профазі виштовхується з ядра у
цитоплазму і там наприкінці мітозу зникає. Відомі також поодинокі
289
представники, в яких ядерце одразу зникає у профазі і відновлюєть-
ся у дочірніх ядрах лише в ранній інтерфазі.
Мітохондрії хітридіомікотових мають пластинчасті кристи. Комп-
лекс Гольджі відсутній. До специфічних структур належать асоціації
ліпідних глобул з мікротільцями. У деяких видів виявлено цікаві сото-
подібні мембранні структури - румпосоми, проте їх функції залиша-
ються нез’ясованими.
Монадні стадії представлені голими зооспорами та гаметами, які
мають один гладенький задній джгутик. Крім того, в таких клітинах
зберігається видозмінене базальне тіло другого джгутика, яке нази-
вають нефункціонуючою центріолею. Перехідна зона джгутика містить
термінальну платівку, інколи також спіралеподібну структуру. Від
мікротрубочок верхівки базального тіла до плазмалеми в місці її джгу-
тикового випину відходять дев'ять радіальних підпорок. Така струк-
тура дещо нагадує опори базального тіла у гіфохітриомікотових, хоча
не є їх гомологом. Базальне тіло з’єднується з ядерною оболонкою
або кількома мікротубулярними коренями, або поперечно-смугастим
ризопластом, або безпосередньо контактує з поверхнею ядра.
Вегетативні тіла хітридіомікотових досить різноманітні і мо-
жуть бути представлені амебоїдом, ризоміцелієм або несептованим
міцелієм. Амебоїди мікроскопічні, містять багато гаплоїдних ядер і
наприкінці ростової фази покриваються хітин-глюкановою оболон-
кою, перетворюючись на зооспорангії.
Складнішим варіантом є ризоміцелій. Він утворений великою
одно- або багатоядерною, вкритою оболонкою клітиною, від якої відга-
лужуються тонкі, позбавлені ядер та клітинних оболонок трофічні
ризоїди, схожі на ризоподії. Зазвичай ризоїди заглиблюються в середи-
ну субстрату, звідки абсорбують поживні речовини.
Найскладнішим варіантом вегетативного тіла є міцелій, що скла-
дається з багатоядерних трубчастих гіф. Окремі ділянки таких гіф
можуть відмежовуватись перегородками і перетворюватись на зоо-
спорангії. Тоді такий міцелій здається септованим і злегка розгалу-
женим.
Розмноження. Нестатеве розмноження відбувається одноджгу-
тиковими зооспорами. Статевий процес - гаметогамія, яка представ-
лена різними варіантами (ізо-, гетеро- або оогамією), та досить про-
ста гаметангіогамія. Крім того, в окремих видів виявлено соматогам-
ний статевий процес, при якому копулюють ризоїди двох різних ризо-
290
міцеліїв. У зону контакту мігрують ядра. Там вони зливаються, утво-
рюючи диплоїдне зиготичне ядро.
Життєві цикли у хітридіомікотових досить різноманітні. За ядер-
ними фазами, наявністю дикаріонтичних стадій та наявністю чергу-
вання поколінь вони представлені п’ятьма варіантами:
1. Гаплофазні без дикаріонтичних стадій та без чергування по-
колінь (наприклад, у МопоЬІерІїагіз)-,
2. Гаплофазні без дикаріонтичних стадій та з гетероморфною
зміною поколінь (КІіігорІїусІіиггіу,
3. Гаплофазні з дикаріонтичною стадією зиготичної цисти, без
чергування поколінь (Роїуріїадиз)-,
4. Гаплофазні з дикаріонтичною стадією та гетероморфним чер-
гуванням поколінь: гаметоспорофітів, що утворюють зооспорангії (на-
приклад, літні цисти), та дикаріонтичних спорофітів, що перетворю-
ються на дикаріонтичні зиготичні цисти (зимові цисти) (типовий пред-
ставник - Зупсіїуігіиту,
5. Гаплодиплофазні без дикаріонтичних стадій та з ізоморфною
зміною поколінь (АІІотусез).
При гаплофазних циклах мейоз зиготичний, при гаплодиплофаз-
них- споричний. Зазвичай гамети морфологічно не відрізняються від
зооспор.
Система відділу. Відділ представлений класом Сіїуігісііотусеїез.
На основі ультраструктурних особливостей зооспор (взаємного по-
ложення базального тіла та нефункіонуючої центріолі, організації зони
навколо ядра, цитоскелетної системи, наявності румпосом, кількості,
розмірів та характеру асоціації ліпідних глобул з мікротільцями та міто-
хондріями), частково - будови вегетативного тіла, статевого проце-
су, клас поділяють на 4 порядки - Сґіуігісііаіез, ЗрігеІІотусеІаІез, ВІа-
зіосіасііаіез, МопоЬІеріїагісіаІез.
Порядок Хітридіальні - Сґіуігісііаіез
Нефункціонуюча центріоля розміщується паралельно до базаль-
ного тіла джгутика. Центр зооспори зайнятий комплексом компактно
розміщених та притиснутих одна до одної органел - ядра, мітохондрії,
великої ліпідної глобули, мікротіла та багатьох рибосом - і оточений
оболонкоподібною цистерною ендоплазматичної сітки. На цій цистерні
з боку ліпідної глобули розташовується притиснута до плазмалеми
291
румпосома. Базальне тіло пов’язане з румпосомою мікротубулярним
коренем.
До порядку входять гриби з вегетативним тілом у вигляді або аме-
боїду, або ризоміцелію.
Більшість видів паразитує на водоростях, вищих наземних
рослинах та інших грибах. Деякі види є водними та ґрунтовими сапро-
трофами і розвиваються на субстратах, що містять переважно целю-
лозу, хітин та кератин. До найпоширеніших родів належать Зупсіїу-
ігіит, Еіїїіхоріїус/іит та Роїуріїадиз.
Рід синхітріум - ЗупсНуіпит (рис. 107). Представники цього роду
- внутрішньоклітинні паразити вищих рослин. їх вегетативне тіло пред-
ставлене багатоядерним плазмодієм, який наприкінці фази росту вкри-
вається оболонкою. Ці гриби зумовлюють утворення галів у вигляді бо-
родавок, ґуль чи наростів, розмір та колір яких залежить від виду росли-
ни. Найважливішим у практичному відношенні видом є ЗупсНуіпит
епсІоЬіоІІсит — синхітріум ендобіотичний, або збудник раку картоплі.
Життєвий цикл цього гриба розпочинається зі стадії зооспори.
Вона осідає на клітину епідермісу бульби картоплі, вкривається влас-
ною оболонкою, розчиняє оболонку клітини-господаря і переливає в
неї свій вміст. Протопласт паразита починає рости, його ядро міто-
тично ділиться. Утворюється мікроскопічний багатоядерний плаз-
модій. Під час росту плазмодію клітина господаря збільшується у роз-
мірах - гіпертрофується. Далі плазмодій вкривається оболонкою і
перетворюється на літню цисту - заключну стадію розвитку вегета-
тивного тіла, гомологічну нерозгалуженому неклітинному міцелію.
Циста виділяє фітогормони, які стимулюють прискорений поділ су-
сідніх здорових клітин тканини господаря - гіперплазію. Клітини гос-
подаря, що інтенсивно діляться, поступово виштовхують цисту на
поверхню вегетативного тіла господаря, утворюючи характерну ро-
зетку навколо хворої клітини із цистою. Потім оболонка цисти розри-
вається, з отвору виходить міхуроподібний виріст. Протопласт цисти
переходить у цей виріст і розпадається на 5-7 багатоядерних діля-
нок, кожна з яких перетворюється на зооспорангій з кількома сотня-
ми зооспор. Таку групу спорангіїв називають сорусом. Оболонка со-
руса руйнується, далі розриваються оболонки зооспорангіїв, і назовні
виходять зооспори, що знов уражують здорові клітини.
Наприкінці вегетаційного сезону літня циста замість зооспор ут-
ворює ізогамети, морфологічно ідентичні до зооспор. Гамети копу-
люють, утворюючи дикаріонтичну рухливу клітину злиття - планози-
292
готу. Вона, як і зооспора, уражує здорову клітину господаря, розрос-
тається у дикаріонтичний плазмодій і далі перетворюється на вкриту
шаруватою кутастою оболонкою темно-жовту зимову цисту, що пе-
реходить у стан спокою. Навесні ядра дикаріонів зливаються, далі
відбувається мейоз, і циста перетворюється на зооспорангій. Розет-
ки та соруси при цьому не утворюються. Таким чином, у життєвому
циклі ЗупсЬуігіит чергуються гаплоїдний гаметоспорофіт, представ-
лений плазмодієм, що утворює літні цисти, та дикаріонтичний спо-
рофіт, який представлений плазмодієм, що утворює зимові цисти.
Синхітріум ендобіотичний є надзвичайно небезпечним патогеном.
Втрати врожаю від раку картоплі можуть сягати 40-60%. Основний захід
боротьби з цією хворобою - виведення стійких до ураження патогеном
сортів картоплі. Ефективним заходом профілактики захворювання є та-
кож правильне ведення сівозмін. У надзвичайних випадках вдаються до
протруювання ґрунту дезинфікуючими речовинами, зокрема розчином
формальдегіду.
Рис. 107. Зупсіїуігіит епдоЬіоіісиггг. А - рак картоплі - уражені бульби; Б-
зимові цисти в тканині ракової пухлини бульб картоплі; В — схема життєвого цик-
лу. 1 - зимова циста; 2 - проростання зимової цисти зооспорами; 3 - зооспори;
4-6 - ураження клітин епідермісу бульби картоплі; 7 - літня циста; 8 - пророс-
тання літньої цисти; 9 - утворення сорусу зооспорангіїв із літньої цисти; 10-
сорус зооспорангіїв; 11 - зооспори; 12 - копуляція зооспор, які виконують функ-
цію гамет; 13 - планзигота; 14 - ураження клітин епідермісу планзиготою; 15 -
зимова циста в тканині рослини
293
Рід ризофідіум - Кїіухорїіусііит - об'єднує гриби, що парази-
тують переважно на водоростях, найпростіших та пилку вищих рос-
лин. У межах роду відомі також види-сапротрофи, що мешкають на
різноманітних рослинних та тваринних залишках. Вегетативне тіло цих
грибів представлене ризоміцелієм. До найпоширеніших та детально ви-
вчених видів належить КІіухорІїусііит роїііпіз - ризофідіум пилко-
вий, який масово розвивається на пилку сосни, що потрапив у воду
(рис.108 А).
Зооспора гриба осідає на пилкове зерно, втягує джгутик, вкрива-
ється оболонкою і утворює голі ризоїдальні вирости, що проривають
покриви пилку і занурюються в його цитоплазму. Зовнішня, вкрита
оболонкою, частина колишньої зооспори росте, стає багатоядерною
і разом із ризоїдами утворює ризоміцелій. Далі зовнішня (екстра-
матрикальна) частина ризоміцелію перетворюється на зооспоран-
гій, вміст якого розпадається на зооспори. Через пору на верхівці
зооспорангія зооспори виходять назовні і уражують нові пилкові
зерна.
При статевому розмноженні, яке зазвичай пов’язане із зменшен-
ням на поверхні води кількості пилкових зерен, одна із зооспор (її
умовно вважають чоловічою) осідає на клітину пилку, вкривається
оболонкою і без поділу ядра розвиває ризоїди, проростаючи в одно-
ядерний чоловічий гаметофіт. Він виділяє феромони, що приваблю-
ють жіночу зооспору. Вона сідає на чоловічий гаметофіт, вкриваєть-
ся оболонкою і перетворюється на жіночій гаметофіт. У зоні контакту
між гаметофітами утворюється отвір, через який протопласт чоло-
вічої клітини перетікає в жіночу. Таким чином, гаметофіти підчас
статевого процесу ведуть себе, як гаметангії, вміст яких не диферен-
ціюється на гамети, тобто статевий процес представлений гаметан-
гіогамією. Ядра гаметангіїв зливаються, і зигота перетворюється на
товстостінну цисту, яка є гомологом зимової цисти синхітріуму. Після
періоду спокою, який може тривати рік і більше, ядро зиготи редук-
ційно ділиться, далі відбувається серія мітозів, і вміст цисти розпа-
дається на численні гаплоїдні зооспори. Через пору в оболонці цисти
вони виходять назовні і уражують новий пилок, що потрапляє у воду.
Таким чином, у життєвому циклі КІііхорІїусІіит наявна стадія гаплоїд-
ного спорофіта, представленого багатоядерним ризоміцелієм, та гап-
лоїдних одноядерних чоловічих і жіночих гаметофітів. Стадія дикарі-
онів відсутня, а статевий процес - гаметангіогамія.
294
Рис. 108. Хітридіальні гриби: А - КЬугоріїусііит роїііпіз на пилку сосни; Б - Д -
Роїуріїадиз. Б - вегетативне тіло гриба із захопленими цистами евглени; В - ста-
тевий процес; Г - зигота; Д - вихід зооспор із зооспорангія
Рід поліфагус- Роїуріїадиз (рис. 108 Б-Д). Найбільш відомим та
дослідженим видом цього роду є Роїуріїадиз еидіепае - поліфагус
евгленовий, який паразитує на евгленах. Вегетативне тіло гриба - ри-
зоміцелій з багаторазово розгалуженими ризоїдами.
Розвиток гриба починається зі стадії зооспори. Вона знаходить
скупчення інцистованих евглен, зупиняється, втягує джгутик, вкри-
вається оболонкою і розвиває багато ризоїдів ризоміцелію, що про-
никають всередину багатьох клітин господарів (інколи уражується
водночас до кількох десятків евглен). Згодом від центральної час-
тини ризоміцелію відбруньковується випин, в який переходить ядро.
Воно багаторазово ділиться, і випин перетворюється на великий
міхуроподібний зооспорангій. Зооспори надалі уражують нових ев-
глен.
Статеве розмноження починається при нестачі їжи. В цьому ви-
падку від двох різних ризоміцеліїв у напрямку один до одного почина-
ють утворюватись довгі копуляційні відростки. Відросток одного із
ризоміцеліїв на верхівці здутий, має шипасту оболонку, і в ньому зна-
ходиться ядро. Після контакту відростків між ними утворюється кон’-
югаційний канал, через який ядро з другого ризоміцелію переходить
у здуття із шипастою оболонкою. Здуття відмежовується від батьків-
ських ризоміцеліїв септами і перетворюється на дикаріонтичну цис-
ту. При проростанні цисти ядра зливаються, відбувається мейоз та
серія мітозів, і циста перетворюється на зооспорангій з багатьма зоо-
спорами. Таким чином, життєвий цикл гаплофазний, із зиготичною
295
редукцією, без чергування поколінь та зі стадією дикаріонтичної зиго-
тичної цисти; статевий процес - соматогамія.
Порядок Спіцеломіцетальні - ЗрігеІІотусеіаІез
Нефункціонуюча центріоля розміщується до базального тіла джгу-
тика під кутом біля 45°. Центральний комплекс органел, оточених ци-
стерною ендоплазматичної сітки, відсутній. Рибосоми та мікротільця
розміщені в цитоплазмі дифузно. Мітохондрїі розташовані біля ба-
зального тіла джгутика і пов'язані з ним багатьма поодинокими мікро-
трубочками. Ядро має довгий випин, який майже контактує з базаль-
ним тілом або переходить у поперечно-смугастий ризопласт. Ліпідних
глобул кілька, вони дрібні, розташовуються на передньому (проти-
лежному від джгутика) боці клітини. Румпосома відсутня. Вегета-
тивне тіло - плазмодій або ризоміцелій. Характерний представник
порядку - рід Оірісііит.
Рід ольпідіум - ОІрісііит. Види роду уражують переважно різно-
манітні органи наземних рослин. До найбільш небезпечних фітопа-
тогенних видів належить ОІрідіит Ьгаззісае - ольпідіум капустя-
ний, який паразитує на хрестоцвітих і викликає хворобу капусти,
відому під назвою «чорна ніжка капустяної розсади». На коренях,
кореневих волосках та кореневій шийці рослини-господаря з’являють-
ся бурі плями, тканини в цих місцях швидко загнивають, і рослина гине.
Вегетативне тіло цього гриба - плазмодій, який розташовується в
середині клітини рослини-господаря. Життєвий цикл досить схожий
із таким у синхітріуму, проте, на відміну від попереднього представни-
ка, види роду ольпідіум не утворюють соруси зооспорангіїв. Вміст плаз-
модію за кількістю ядер розпадається на окремі одноджгутикові фраг-
менти - майбутні зооспори. Після цього в оболонці плазмодію утво-
рюється вивідна протока, яка пробиває оболонку клітини-господаря, і
зооспори через неї виходять назовні. Зооспора деякий час плаває, після
чого прикріплюється до клітини кореня рослини, переливає в неї свій
вміст, і весь процес починається знову. В цьому випадку вся особина
(все вегетативне тіло) перетворює свій вміст на зооспорангій. Такий
шлях утворення зооспорангіїв називають холокарпічним або монокар-
пічним, а особини - відповідно холокарпічними або монокарпічними.
Порядок Бластокладіальні - ВІазіосІаФаІез
Нефункціонуюча центріоля розміщується під прямим кутом до ба-
зального тіла джгутика. Центральний комплекс органел включає ядро,
яке з одного боку асоційоване з базальним тілом, а з протилежного -
296
т. зв. ядерним ковпачком (рибосомальною зоною, оточеною ендо-
плазматичною сіткою і притиснутою до поверхні ядерної оболонки).
На латеральному боці клітини є складний комплекс, утворений при-
тиснутою до плазмалеми цистерною ендоплазматичної сітки, видов-
женим мікротільцем, яке асоціюється з кількома дрібними лінійно
розміщеними ліпідними глобулами; останні асоційовані з видовже-
ною мітохондрією, верхівка якої оперезує базальне тіло.
Вегетативне тіло - плазмодій, ризоміцелій або несептований мі-
целій. Статевий процес - ізо- та гетерогамія. Життєвий цикл - гаплоди-
плофазний.
Характерним представником порядку є рід алломіцес - АІІоту-
сез (рис. 109), вегетативне тіло якого має міцеліальну будову. Гриб
розвивається як водний або ґрунтовий сапротроф, оселяючись на
рослинних залишках та трупах тварин. Спорофіт гриба - це мало
розгалужений неклітинний міцелій з перетяжками та несправжніми
септами. Ядра спорофітів диплоїдні. На верхівках бічних відгалужень
після відокремлення їх від міцелію мікропоровими септами розвива-
ються тонкостінні або товстостінні зооспорангії, які також часто нази-
вають цистами. В кожному зооспорангії утворюється багато зооспор.
Зооспори з тонкостінних зооспорангіїв проростають у нові диплоїдні
спорофіти.
Рис. 109. АІІотусез: А-гаметофіт; Б- спорофіт. 1 - жіночий гаметангій,
2 - чоловічий гаметангій; 3 - зооспорангій; 4 - циста
297
У товстостінних зооспорангіях перед утворенням зооспор відбу-
вається мейоз, і, як наслідок, в них утворюються гаплоїдні зооспори.
Вони проростають у гаплоїдні гаметофіти, що морфологічно не
відрізняються від спорофітів. Проте на верхівках бічних відгалужень
замість зооспорангіїв утворюються або чоловічі, або жіночі гаметангії,
які відокремлюються від гіфи мікропоровими септами. Після виходу у
зовнішнє середовище гамети, що морфологічно не відрізняються від
зооспор, копулюють. Копуляція пов’язана з реакцією на статевий гор-
мон - сиренін. Далі ядра гамет зливаються, і зигота без періоду спо-
кою розвивається у новий диплоїдний спорофіт. Таким чином, життє-
вий цикл гаплодиплофазний зі споричною редукцією та ізоморфною
зміною поколінь.
Порядок Моноблефаридальні - МопоЬІерґіагісіаІез
Нефункціонуюча центріоля розміщується паралельно до базаль-
ного тіла джгутика. Центральний комплекс органел наявний, але пред-
ставлений лише ядром та рибосомальною зоною, які оточені цистер-
нами ендоплазматичної сітки. Ліпідних глобул кілька, вони дрібні,
розташовуються на передньому боці клітини і асоціюються з багать-
ма видовженими мікротільцями. Мітохондрії розташовані біля базаль-
ного тіла джгутика і пов’язані з ним багатьма поодинокими мікротру-
бочками. Ядро не має прямого зв’язку з базальним тілом. Румпосома
наявна, розташовується латерально і асоційована з плазмалемою
та одним із мікротілець.
Вегетативне тіло - нерозгалужений або мало розгалужений не-
клітинний міцелій. Статевий процес - оогамія.
Типовим представником є рід моноблефаріс - МопоЬІеріїагіз.
Гриб належить до прісноводних сапротрофів і розвивається на різно-
манітних рослинних та тваринних рештках, утворюючи на них білу-
ваті повсті.
Гаметоспорофіт гриба представлений мало розгалуженим неклі-
тинним міцелієм, диференційованим на довгі тонкі ризоїди та широкі
висхідні нерозгалужені гіфи. При нестатевому розмноженні верхівка
висхідної гіфи відмежовується від міцелію септою, видовжується і
перетворюється на зооспорангій. Зооспори після періоду активного
руху знаходять придатний субстрат, осідають на нього і проростають
у новий міцелій.
При статевому розмноженні верхівка висхідної гіфи відмежовує
мікропоровими септами дві клітини, з яких одна розвивається в оогоній
298
з однією яйцеклітиною, а друга - в антеридій з чотирма-вісьмома
сперматозоїдами. Після звільнення з антеридію сперматозоїди підпли-
вають до оогонія, осідають на нього, один з них проникає в оогоній і
запліднює яйцеклітину. Утворюється диплоїдна зигота, яка вкриваєть-
ся товстою оболонкою і переходить у стан спокою. Після завершення
періоду спокою ядро редукційно ділиться, оболонка зиготи тріскаєть-
ся і зигота проростає у новий міцелій. Таким чином, життєвий цикл
гаплофазний, із зиготичною редукцією, без чергування поколінь та
без дикаріонтичних стадій.
Відділ Зигомікотові гриби - 2удотусоіа
Справжні гриби, для яких характерні вегетативні тіла у вигляді
несептованого, рідше септованого міцелію. Ядра у вегетативних кліти-
нах гаплоїдні. Статевий процес - зигогамія, статеве спороношення -
спорангій зі спорангіоспорами. Монадні стадії повністю відсутні.
Відділ нараховує біля 1100 видів та майже 200 родів. Зигоміко-
тові - це переважно сапротрофи, що розвиваються на різноманітних
субстратах. Рідше паразитують на різних організмах або вступають
із ними у симбіоз. Частина зигомікотових представлена хижими гри-
бами.
Біохімічні особливості та живлення. За комплексом біохіміч-
них ознак зигомікотові є справжніми грибами. Зокрема, вони синтезу-
ють лізин грибним шляхом - через а-аміноадипінову кислоту. Синтез
триптофану відбувається за допомогою четвертої групи ферментів,
подібно до деяких базидіомікотових грибів. Кінцевим продуктом ме-
таболізму є сечовина, а запасною поживною речовиною - глікоген.
Живлення зигомікотових виключно абсорбційне, причому як дже-
рело вуглецю та азоту використовується широкий спектр органічних
сполук.
На відміну від інших відділів справжніх грибів, зигомікотові не
синтезують сидерамінів, а серед поліолів, які продукують види цього
відділу, маніт не виявляється взагалі або утворюється лише у неве-
ликих кількостях.
Гормоном, який індукує статевий процес - зигогамію, - у зигомі-
котових є триспорова кислота, яка не виявлена в інших справжніх
грибів. З іншого боку, у зигомікотових не знайдені мікоспорини - гор-
мони, що ініціюють утворення органів спороношення, хоча в інших
відділах справжніх грибів вони є.
299
Цитологічні особливості. Клітинні покриви зигомікотових най-
частіше представлені хітин-хітозановими оболонками, хоча в одного
з порядків (2оорадаІез) оболонки хітин-глюканові. Міцелій або не-
септований, або поділений септами на окремі клітини. В останньому
випадку септи можуть бути мікропоровими, хоча частіше вони доліпо-
рові (або відкриті, або з електронно-щільною пробкою, проте завжди
без парентосом).
Ядро евкаріотичне. Центріолі на всіх стадіях клітинного циклу
відсутні. Мітоз закритий. Його цікавою особливістю є поява у мета-
фазі на ядерній оболонці дископодібної структури. Ядерце зберігаєть-
ся майже на всіх стадіях каріокінезу і дезінтегрується лише у тело-
фазі.
Мітохондрії мають пластинчасті кристи. Комплекс Гольджі, як і у
всіх справжніх грибів, відсутній. У клітинах наявні літичні везикули та
хітосоми, які концентруються на верхівках вегетативних гіф і обумов-
люють їх апікальний ріст. Крім того, в цитоплазмі виявляються мікро-
тільця, а також є відомості щодо наявності ломасом.
Джгутикові стадії у зигомікотових повністю відсутні.
Вегетативне тіло найчастіше представлене розгалуженим бага-
тоядерним несептованим міцелієм. При старінні у такому міцелії
можуть утворюватися септи. У деяких зигомікотових міцелій септо-
ваний, причому клітини є одноядерними.
Гіфи міцелію можуть бути функціонально та морфологічно ди-
ференційованими, наприклад, на темнозабарвлені повітряні гіфи -
столони, на системи ризоїдів, на спорангієносці, ловчі гіфи хижих
грибів та ін.
Розмноження. Нестатеве розмноження здійснюється як вегета-
тивним шляхом внаслідок фрагметації міцелію, так і за допомогою
ендогенних спор, що утворюються у спорангіях - спорангіоспор.
Статевий процес - зигогамія - здійснюється шляхом копуляції
двох багатоядерних клітин, що утворюються на верхівках гіф. У резуль-
таті статевого процесу розвивається товстостінна зигота з багатьма
диплоїдними ядрами - зигоспора. Після періоду спокою і редукційно-
го поділу ядер вона проростає у спорангій з гаплоїдними спорангіо-
спорами.
Життєвий цикл гаплофазний, із зиготичною редукцією, без зміни
поколінь та без дикаріонтичних стадій.
300
Екологічні особливості. Зигомікотові гриби є сапротрофами
або паразитами рослин, тварин та грибів. Деякі з них утворюють міко-
ризу з трав'янистими рослинами. Серед зигомікотових грибів зустрі-
чаються ендо- та екзосимбіонти членистоногих, а також хижі гриби,
що вловлюють ґрунтових безхребетних (амеб, нематод, кліщів) за до-
помогою спеціальних ловчих гіф.
Система відділу. Відділ включає один клас - зигоміцети (2у-
дотусеїез). Крім того, до зигомікотових як групу з нез’ясованим місцем
у системі відносять т. зв. трихоміцетів, яких часто розглядають як
формальний клас Тгісіїотусеїез.
Клас Зигоміцети - 2удотусеІез
Об’єднує зигомікотові гриби, в яких наявний статевий процес зи-
гогамія. Цей клас різні автори поділяють на 7-10 порядків, серед яких
найпоширенішими є чотири - Мисогаїез, ЕпОіоторіїОіогаІез,
Епсіодопаїез та 2оорадаіез. В основу поділу на порядки покладено, в
першу чергу, тип нестатевого спороношення, будову повітряних гіф,
наявність чи відсутність септ та їх ультраструктуру, морфологію зи-
госпор, а також екологічні особливості.
Рис. 110. РЬІгориз'. А - споран-
гієносці на міцелії; Б - спорангіо-
спори. 1 - ризоїди; 2 - столони; 3 -
зрілий спорангій; 4-звільнення спо-
рангіоспор
Порядок Мукоральні - Ми-
согаїез
Представники порядку мають
ценоцитний міцелій. Органи не-
статевого розмноження - багато-
спорові або декількаспорові спо-
рангії (спорангіолі). Переважно
сапротрофи (зустрічаються у ґрун-
ті, на рослинних залишках), інко-
ли паразитують на мукорових та ін-
ших грибах, деякі викликають
мікози тварин та людини. Найпо-
ширенішим родом порядку є /?/)/-
хориз.
Рід ризопус - КМхориз (рис.
110). Деякі види цього роду спри-
чиняють гниття плодів, овочей та
хлібопродуктів при їх зберіганні.
301
Вони утворюють цвіль темно-сірого кольору. Спорангієносці ризопу-
са утворюються пучечком на видозмінених частинах міцелію - ета-
лонах, які розповсюджуються над поверхнею субстрату. В місцях, де
еталони стикаються із субстратом, розвиваються розетки спорангіє-
носців, які піднімаються над поверхньою субстрату, та розгалужені
ризоїди, які занурюються у субстрат. Зрілі спорангії мають чорний
колір від спорової маси, яка просвічується крізь оболонку спорангія.
В народі гриби цього роду відомі під збірною назвою «чорна цвіль».
Деякі представники роду Гіїїігориз мають високу ферментативну ак-
тивність або здатні синтезувати різноманітні органічні кислоти, завдя-
ки чому мають практичне застосування.
Спорідненими родами є мукор (Мисоґ) та пілоболус (РПоЬоІиз).
Представники роду мукор, на відміну від ризопуса, не утворюють ета-
лони, а їхні спорангієносці розвиваються на розгалуженому міцелії
вегетативного тіла (рис.111). Види роду пілоболус є копрофілами і
розвиваються на екскрементах травоїдних тварин. Спорагієносці цих
грибів здатні відстрілювати спорангії на значну відстань (рис. 112).
Порядок Ентомофторальні - Епііюторіїііюгаїез
Порядок об’єднує вузькоспеціалізовану групу облігатних паразитів
членистоногих. Для ентомофторальних характерне активне звільнен-
ня спор (конідій) нестатевого розмноження, що утворюються екзо-
генно. Типовим представником порядку є рід ЕпіоторМюга.
Рис. 111. Мисог. А - загальний вигляд міцелію з поодинокими спорангієно-
сцями; Б - група спорангіїв на міцелії; В - будова спорангія; Г - звільнення спо-
рангіоспор. 1 - спорангій; 2 - колонка; 3 - спорангієносець; 4 - спорангіоспо-
ри; 5 - залишки перидію спорангія (комірець)
302
Рис. 112. РІІоЬоІиз: А - група спорангіїв, орієнтованих на джерело світла;
Б - окремий спорангій на спорангієносці; В - відстрілювання спорангія. 1 - спо-
рангій; 2 - спорангієносець
Рід ентомофтора - ЕпіоторМІюга. Найбільш відомим видом
роду є Е. тизсае - ентомофтора мушина (рис. 113). Цей парази-
тичний гриб викликає хворобу кімнатних мух, яка має назву «осінньої
або сонної хвороби мух». Зараження комах відбувається конідіями,
Рис. 113. ЕпІЇїоторМіога тизсае: А - уражена муха в ореолі конідій;
Б - конідієносець із конідією; В - відстрілювання конідії
303
які, потрапивши на муху, проростають у септовану гіфу. В жировому
тілі мухи гіфа розпадається на багатоядерні клітини неправильної
форми - гіфенні тільця. Кількість цих тілець збільшується в результаті
їхнього поділу або брунькування, з потоком крові вони розносяться
по всьому тілу комахи. Через декілька днів після зараження комаха
гине, а на ній з'являються булавовидні конідієносці, які виходять че-
рез дихальні отвори та тонкі місця хітинового покриву. На конідієнос-
цях утворюються кульовидні конідії, оточені шаром слизу, які від-
стрілюються на 1-2 сантиметри, утворюючи борошнистий наліт біля
мухи, який добре помітний на віконному склі. Якщо конідія не потра-
пила на муху, то вона може проростати новим конідієносцем з коні-
дією, але меншого розміру, ніж попередня, яка також відстрілюється.
Так може відбуватися декілька разів.
Порядок Ендогональні - Епбодопаїез. Види порядку існують
як сапротрофи у ґрунті, у підстилці, на рослинних залишках або на
підземних частинах рослин. Міцелій більшості ендогональних зигомі-
цетів здатен утворювати ендотрофну мікоризу з корінням багатьох
вищих рослин, зокрема полуниць, яблунь, томатів, пшениці (всього
біля 80% трав’янистих рослин). При цьому міцелій гриба проникає в
клітини кореня рослини, утворюючи в них деревовидні розгалужені
гіфи - арбускули або округлі здуття - везикули. Такий тип мікоризи
називають везикулярно-арбускулярною, або ендомікоризою. Серед
представників порядку види роду гломус ((ЗІотиз) є облігатними сим-
біотрофами, які утворюють у корінні рослин арбускули та везикули.
Характерною ознакою порядку є утворення ними підземних плодо-
вих тіл або т. зв. спорокарпів. Вони розвиваються в результаті спле-
тення гіф несептованого, ценоцитного міцелію та являють собою дуже
щільні сферичні тільця жовтуватого кольору розміром від декількох
міліметрів до 2-3 см. Всередині спорокарпів знаходяться декілька
круглих багатоспорових спорангіїв без колонок або зиготи, або хла-
мідоспори. Найпоширенішим видом у складі порядку є Епсіодопе
Іасііїїиса, на прикладі якого вперше було простежено статевий про-
цес в ендогональних грибів.
Порядок Зоопагальні - 2оорадаіез
Види цього порядку мають дуже тонкий, слабо розвинутий міцелій.
Нестатеве розмноження відбувається за допомогою конідій, що утво-
рюються поодиноко або ланцюжками на верхівках конідієносців.
Конідії, на відміну від конідій ентомофторових грибів, активно не
304
відстрілюються. Статевий процес - типова зигогамія. Порядок об'-
єднує облігатних паразитів тварин, у тому числі амеб, ризопод та не-
матод, а також грибів.
Представники порядку використовуються в народному госпо-
дарстві як біологічний засіб боротьби з нематодами - шкідниками
сільськогосподарських культур. Типовим прикладом є Зіуіораде Ііасіга,
гіфи міцелію якого виділяють особливу клейку речовину, за допомо-
гою якої «хижак» захоплює нематоду. Потім гіфи безпосередньо міце-
лію гриба (або за допомогою гаусторій) проникають в середину тіла
нематоди і живляться за її рахунок, досить швидко вбиваючи «жерт-
ву» (нематоду).
Додаток до відділу 2удотусо1а: клас Трихоміцети - ТгісКо-
тусеіез.
До зигомікотових також традиційно включають групу організмів
нез’ясованого систематичного положення, які є облігатними екто- або
ендосимбіонтами, що мешкають у шлунку або на хітинових покривах
членистоногих, переважно комах - т. зв. трихоміцетів.
Характерна ознака трихоміцетів - будова вегетативного тіла. Воно
являє собою одноклітинний або багатоклітинний розгалужений міцелій
з мікропоровими септами, який, на відміну від зигоміцетів, при-
кріплюється до кутикули членистоногих особливою клітиною-підош-
вою. Органами нестатевого розмноження є односпорові спорангіолі
або багатоспорові спорангії. У грибів цього класу, за винятком пред-
ставників порядку НагреІІаІев, статевий процес не виявлений.
Хімічний склад оболонок досить різноманітний і використовуєть-
ся як систематична ознака на рівні порядку. Зокрема, в порядках
НагреІІаІев та АвеІІагіаІев оболонки хітин-глікозамінові, в Ессгіпаїев -
целюлозні, а в АтоеЬісІіаІев - галактанові та галактозамінові.
Відділ Аскомікотові гриби - Авсотусоіа
Справжні гриби, в яких вегетативне тіло представлене розгалу-
женим клітинним міцелієм (зрідка - міцелієм, що брунькується). Ве-
гетативні клітини з генетично однаковими гаплоїдними ядрами (го-
мокаріонтичні) або дикаріонтичні, причому гаплоїдна стадія за трива-
лістю, як правило, переважає. Статевий процес - гаметангіогамія, як
виняток - соматогамія. Статеве спороношення - аск з аскоспорами.
Монадні стадії повністю відсутні.
305
За видовим багатством аскомікотові є найчисельнішим відділом
грибів. До нього належить біля 50 тис. видів (включаючи 16 тис. видів,
що утворюють лишайники). Аскомікотові поширені на всіх континен-
тах та практично в усіх основних біотопах.
Біохімічні особливості та живлення. Біосинтез лізину відбу-
вається грибним шляхом (ААА-шлях). Триптофан в аскомікотових син-
тезується за допомогою ферментів або першої (у більшості представ-
ників), або третьої групи (у сахароміцетів). Іони заліза поглинаються
за допомогою сидерамінів. Поліоли (включаючи маніт) наявні у до-
сить-значних кількостях і є речовинами, у вигляді яких по гіфах транс-
портується вуглець.
Живлення виключно осмотрофне. Абсорбції органічних речовин
зазвичай передує їх частковий розклад за допомогою екзоферментів
та ферментних комплексів клітинних оболонок. Джерелом вуглецю є
різноманітні органічні сполуки, а у деяких анаморфних аскоміцетів
(зокрема в родах аспергіл, кладоспоріум) - навіть вуглекислий газ.
Азот поглинається у складі органічних сполук.
Гормонами, що здатні ініціювати спороношення, є мікоспорини.
Функції гормонів, що ініціюють статевий процес, у аскомікотових мо-
жуть виконувати ацетат та гліцин.
Важливими вторинними метаболітами аскомікотових можуть бути
антибіотики, мікотоксини, речовини психотропної дії, речовини-інгібіто-
ри розвитку вищих рослин, фітогормони.
Біотехнологічно важливою властивістю є здатність деяких аско-
мікотових (особливо-серед сахароміцетів) отримувати необхідну для
розвитку енергію внаслідок процесів бродіння (в першу чергу - спир-
тового).
Цитологічні ознаки. Клітини аскомікотових вкриті оболонками,
які утворені переважно хітином та глюканами, а у сахароміцетів -
мананом та глюканами. Оболонки аскомікотових двошарові: зовнішній
шар тонкий та електронно-щільний; внутрішній - товстий та елект-
ронно-прозорий. Септи між клітинами можуть бути простими (часто-
з тільцями Вороніна), мікропоровими або доліпоровими з простими
пробками.
Ядерний апарат евкаріотичний. Переважаючий у життєвому циклі
тип міцелію складається з гіф, утворених одноядерними клітинами.
Проте після статевого процесу з клітини злиття, як правило, розвива-
ються гіфи, що складаються з дикаріонтичних клітин. На окремих
306
фазах розвитку клітини інколи стають багатоядерними, проте в ціло-
му неклітинна будова для аскомікотових нетипова.
Мітоз закритий. Центріолі відсутні, а центрами організації мікро-
трубочок веретена є аморфні скупчення нуклеопротеїду. Ядерце заз-
вичай є; на початку мітозу воно, як правило, виштовхується в цито-
плазму, де і зберігається до завершення поділу ядра.
Мітохондрії мають пластинчасті кристи. Комплекс Гольджі відсутній,
проте у деяких аскомікотових (зокрема у лабульбеніальних грибів)
ендоплазматична сітка відшнуровує цистерни, що нагадують типові
диктіосоми. Аскомікотові мають розвинену систему хітосом та літич-
них везикулів, що концентруються на верхівках гіф і обумовлюють ап-
ікальний їх ріст. Під оболонкою наявні ломасоми, в цитоплазмі - чис-
ленні мікротільця, ліпідні глобули, вакуолі з клітинним соком, що відме-
жовуються тонопластом.
Джгутикові стадії повністю відсутні.
Вегетативне тіло найчастіше представлене розгалуженим сеп-
тованим міцелієм, рідше (особливо у сахароміцетів) - псевдоміцелієм.
Окремі гіфи міцелію можуть бути видозмінені, найчастіше - в ап-
ресорії та гаусторії. Апресорії є органами прикріплення міцелію до
субстрату, вони одноклітинні і нагадують присоски. Гаусторії - видо-
змінені гіфи, що проникають всередину клітини господаря і абсорбу-
ють з неї поживні речовини. До видозмінених гіф також належать пер-
форуючі гіфи - багатоклітинні утвори, що виконують одразу кілька
функцій - прикріплення до субстрату, проникнення в нього та погли-
нання поживних речовин. Перфоруючі гіфи характерні для багатьох
грибів - збудників дерматомікозів тварин і людини.
В аскомікотових спостерігається чимало варіантів видозмін не лише
гіф, але й міцелію. Серед останніх найпоширенішими є склероції, строми
та плодові тіла. Всі ці видозміни пов’язані з утворенням гіфами щільних
переплетень, які називають несправжньою тканиною - плектенхімою.
Склероції - це щільні переплетення міцелію, у стані яких гриб
переносить несприятливі умови. Зазвичай у склероціях спостерігаєть-
ся диференціація плектенхіми на кору та внутрішню плектенхіму. Гіфи
кори товстостінні, забарвлені меланіном у темний колір. Внутрішні
гіфи світлі, тонкостінні. У клітинах склероціїв накопичується велика
кількість запасних поживних речовин.
Строми також утворені щільним переплетенням гіф, але їх ос-
новна функція - це захист (а інколи - і піднесення над субстратом)
307
органів нестатевого або статевого спороношень. Тому анатомічна
будова стром складніша і включає захисний шар корових гіф, внутрішні
гіфи та органи спороношень - пікніди з конідіями або плодові тіла з
асками, які занурені у строми.
Плодові тіла - це також видозміни міцелію, утворені плектенхі-
мою. В аскомікотових розрізняють чотири типи плодових тіл: закрите
(клейстотецій), напівзакрите (перитецій), відкрите (апотецій) та не-
справжнє (псевдотецій). Плодові тіла аскомікотових переважно мікро-
скопічні (виняток становлять пецицальні гриби).
Справжні плодові тіла утвоюються з гаплоїдних гіф, з яких части-
на надалі розвивається в аскогони та антеридії (після статевого про-
цесу з клітин злиття утворюються аскогенні гіфи, що дають початок
аскам з аскоспорами), а решта - у захисні структури органів статево-
го спороношення - плектенхіму плодового тіла. Вона може повністю
обростати аски з аскоспорами (тоді утворюються замкнуті плодові
тіла - клейстотеції), обростати статеве спороношення лише частково
(в цьому випадку утворюються напівзакриті плодові тіла - перитеції,
що зазвичай мають вигляд глечика) або розростатися і підносити аски
з аскоспорами над субстратом (відкриті плодові тіла - апотеції).
Зовнішній шар плектенхіми плодового тіла видозмінюється у його
захисну оболонку - перидій. Із внутрішнього шару утворюються до-
даткові захисні структури (найчастіше - стерильні парафізи).
Несправжні плодові тіла - псевдотеції - на відміну від справжніх,
не мають перидію. Перед їх утворенням гаплоїдні гіфи міцелію роз-
ростаються і видозмінюються у строму. Далі в ній утворюються аско-
гони та антеридії, відбувається статевий процес, і утворюються аско-
генні гіфи та аски. Вони розсувають гіфи строми або частково їх руй-
нують, внаслідок чого утворюються порожнини - локули. Саме в цих
локулах і розміщуються аски.
Розмноження. У більшості аскомікотових у життєвому циклі спо-
стерігаються два типи розмноження - нестатеве та статеве, внаслі-
док яких розвиваються відповідно два типи спороношень - нестате-
ве (анаморфа) та статеве (телеоморфа).
Нестатеве розмноження відбувається вегетативним шляхом
(фрагментацією справжнього міцелію, брунькуванням, артроспора-
ми, хламідоспорами) або за допомогою різних типів конідій - алевро-
конідій та бластоконідій. З особливих варіантів бластоконідій в аско-
мікотових досить поширеними є пороспори (конідії, які виникають че-
рез пори в оболонці конідієносця) та фіалоспори (конідії, які утво-
308
рюються на верхівці особливих виростів конідієносця - фіалідах).
Конідії можуть утворюватись на поодиноких конідієносцях, в коремі-
ях та спородохіях, на ложах або в пікнідах.
Статеве розмноження відбувається аскоспорами, які утворю-
ються в асках (сумках) внаслідок статевого процесу.
Статевий процес у більшості представників відділу - це гаметан-
гіогамія, при якій зливаються протопласти двох недиференційова-
них на гамети гаметангіїв - аскогону та антеридію. Рідше плазмога-
мія відбувається між аскогоном та вегетативною клітиною міцелію,
оскільки антеридій не утворюється. У деяких аскомікотових статевий
процес - це соматогамія, при якій копулюють або звичайні вегета-
тивні гіфи, або аскоспори (інколи навіть не виходячи за межі аску).
В окремих групах аски утворюються партеногенетично.
Життєвий цикл та розвиток статевих спороношень. У життє-
вому циклі аскомікотових розрізняють три головні події, які інколи на-
зивають кардинальними: статевий процес, внаслідок якого відбуваєть-
ся злиття протопластів клітин статевих партнерів (плазмогамія). Після
нього або одразу, або після певного періоду існування у дика-
ріонтичному стані гаплоїдні ядра зливаються (каріогамія), утворюю-
чи диплоїдне зиготичне ядро. Воно одразу або після певного періоду
існування у диплоїдному стані редукційно ділиться (мейоз), після чого
починається утворення статевого спороношення - аску з гаплоїдни-
ми аскоспорами. Аскоспори або проростають гаплоїдним міцелієм,
на якому утворюються статеві органи, або одразу копулюють, розпо-
чинаючи новий життєвий цикл.
В аскомікотових життєві цикли різноманітні, але найпоширеніши-
ми та детально дослідженими є три типи: а) гаплодиплофазні без
утворення дикаріонтичних поколінь; б) гаплофазні з партеногенетич-
ним утворенням псевдодикаріонтичного покоління; в) гаплофазні з
чергуванням гаплоїдного та дикаріонтичного поколінь.
Перший тип характерний для сахароміцетів: дві гаплоїдні клітини
псевдоміцелію копулюють, їх ядра зливаються, утворюючи диплоїд-
ну зиготу. Вона брунькується, утворюючи диплоїдний псевдоміцелій.
Далі в одній з клітин відбувається мейоз, і клітина перетворюється на
молодий аск. У ньому цитоплазма розмежовується на одноядерні
ділянки, що покриваються оболонками і перетворюються на аскоспо-
ри. Після звільнення з аску вони діляться або брунькуються, розви-
ваючи новий гаплоїдний псевдоміцелій.
Другий тип досліджений на прикладі тафринальних грибів. Тут
гаплоїдні клітини, що утворюються внаслідок брунькування аскоспор,
309
після одного з мітозів стають двоядерними. Хоча вони нагадують ди-
каріон, проте містять два генетично однакових ядра, тобто є псевдо-
дикаріонтичними. Внаслідок поділу таких клітин утворюється розга-
лужений клітинний псевдодикаріонтичний міцелій, який у життєвому
циклі є поколінням, що переважає. В окремих клітинах такого міце-
лію згодом відбувається каріогамія, утворюється зиготичне дип-
лоїдне ядро, і клітина перетворюється на молоду сумку. Її ядро
редукційно ділиться, навколо гаплоїдних дочірніх ядер виникає
власна цитоплазма, розвиваються клітинні оболонки, і в клітині-
аску утворюються аскоспори.
Третій тип життєвого циклу серед аскомікотових є найпоширені-
шим. На гіфах гаплоїдного міцелію, який є переважаючою вегетатив-
ною стадією гриба, розвиваються багатоядерні антеридії та аскогони
з трихогінами (у грибів з порядку евроціальних трихогіна відсутня).
Ядра в гаметангіях гаплоїдні. Аскогон (або його трихогіна) контактує
з антеридієм, у місці контакту утворюється кон’югаційний канал, по
якому ядра з антеридію переходять в аскогон і утворюють дикаріон-
тичні пари. Із заплідненого аскогону розвиваються дикаріонтичні гіфи,
що дають початок аскогенним гіфам, верхівки яких згодом перетво-
рюються на аски з аскоспорами. Такий життєвий цикл притаманний
більшості аскомікотових, що мають плодові тіла.
В межах цієї групи у деяких грибів плазмогамія може відбуватися
між аскогоном та вегетативною клітиною міцелію, тобто антеридій не
утворюється, і статевий процес є проміжним між гаметангіогамією та
соматогамією (наприклад, у сордаріальних (Зогсіагіа іїтісоїа) та сфе-
ріальних (Меигозрога зііоріїііа) грибів). Крім того, досить поширеною
є справжня соматогамія, при якій не утворюється ані аскогон, ані ан-
теридій, і плазмогамія відбувається між двома клітинами вегетатив-
них гаплоїдних гіф (наприклад, у гелоціальних (Зсіегоііпіа зсіегоііогит)
та дотидеальних (СотосІаІЇїгиз) грибів).
Розвиток аску. Цикл розвитку телеоморф закінчується утворен-
ням асків з аскоспорами. Під час цього процесу зазвичай дикаріон-
тичні або псевдодикаріонтичні ядра аскогенних гіф зливаються (відбу-
вається каріогамія), утворюється зиготичне ядро, яке редукційно
ділиться, і далі в аску утворюються гаплоїдні аскоспори.
У більшості аскомікотових аск розвивається за способом гачка:
верхівкова дикаріонтична клітина аскогенної гіфи загинається (утво-
рює т. зв. гачок), причому ядра дикаріону розташовуються у зоні за-
гину. Далі вони одночасно мітотично діляться. Пара ядер різної статі
залишається в зоні загину гачка, третє ядро переходить у кінчик гач-
310
ка, а четверте мігрує до базальної частини клітини. Після цього гачок
розділяється двома поперечними перегородками на три клітини -
верхівкову одноядерну, середню дикаріонтичну, базальну одноядер-
ну. Далі верхівкова та базальна клітини гачка зливаються, ядро з вер-
хівкової клітини переходить у базальну, відновлюючи дикаріон. Така
клітина надалі може утворювати новий гачок. Середня дикаріонтич-
на клітина гачка розвивається в сумку: вона збільшується в розмірах,
ядра дикаріона зливаються, утворюючи зиготичне диплоїдне ядро.
Воно одразу редукційно ділиться. Далі зазвичай відбувається один
мітоз. Навколо кожного з ядер утворюється цитоплазма, яка вкри-
вається оболонкою. Як наслідок, в аску виникає вісім гаплоїдних ас-
коспор.
У тафриноміцетів розвиток аску простіший: тут ядра верхівкової
клітини псевдодикаріонтичного міцелію зливаються, диплоїдне ядро
один раз мітотично ділиться, клітина розділяється поперечною пере-
городкою на нижню диплоїдну клітину-ніжку та верхівкову клітину
майбутнього аску. В останній відбувається мейоз і утворюється чоти-
ри гаплоїдні аскоспори. Вони здатні брунькуватись як у середині аску,
так і після виходу з нього, обумовлюючи розвиток гаплоїдного псев-
доміцелію.
У сахароміцетів, які не мають у циклі розвитку дикаріонтичної
стадії, спосіб утворення аску є найпростішим - диплоїдна клітина, що
утворилась після статевого процесу, одразу або після кількох циклів
мітозів редукційно ділиться і перетворюється на аск із чотирма аско-
спорами.
Розвиток аскоспор. У молодому аску з ядрами майбутніх аско-
спор починається формування їх оболонок. Плазмалеми аскоспор
можуть утворюватись двома шляхами: з інвагінацій плазмалеми аску
та з інвагінацій мембранного навколоядерного мішка.
Перший тип характерний для сахароміцетів та тафриноміцетів.
Тут плазмалема аску утворює внутрішні інвагінації, які оточують кож-
не ядро та частину цитоплазми майбутньої аскоспори і далі відшну-
ровуються від плазмалеми сумки. Внутрішня мембрана відшнурова-
ної інвагінації стає плазмалемою аскоспори, простір між внутрішньою
та зовнішньою мембранами наповнюється полісахаридами, які полі-
меризуються, утворюючи оболонку аскоспори. Після цього зовнішня
мембрана інвагінації руйнується.
При розвитку за типом ядерного мішка навколо диплоїдного зи-
готичного ядра молодої сумки вздовж плазмалеми ендоплазматична
сітка утворює численні цистерни, що зливаються між собою, утво-
311
рюючи внутрішній відкритий двомембранний циліндр. Після мейозу
та утворення гаплоїдних ядер майбутніх аскоспор верхівки циліндра
замикаються, і всі гаплоїдні ядра майбутніх аскоспор виявляються
оточеними великим двомембранним навколоядерним мішком. Внут-
рішня мембрана мішка утворює інвагінації, які аналогічно до попе-
реднього типу оточують кожне ядро з частиною цитоплазми подвійною
мембраною. Внутрішня мембрана стає плазмалемою аскоспори, у
просторі між нею та зовнішньою мембраною утворюється оболонка
аскоспори.
Таким чином, аскоспори в аску розвиваються ендогенно, проте
плазмалеми перших можуть бути похідними або від плазмалеми аску,
або від ендоплазматичної сітки.
Типи асків. Залежно від будови аску, аскоспори з нього звільня-
ються або пасивно (внаслідок ослизнення або автолізу оболонки сум-
ки), або активно викидаються з аску.
Механізм активного звільнення пов’язаний з тим, що після завер-
шення утворення аскоспор в цитоплазмі аску розпочинається гідроліз
глікогену до простих цукрів. Як наслідок, аск за рахунок осмосу почи-
нає стрімко наповнюватись водою, тургорний тиск в ньому швидко
зростає, оболонка аску розривається, і аскоспори активно викида-
ються назовні, інколи на досить значну відстань.
Аски, з яких аскоспори звільняються пасивно, називають про-
тотунікатними, а ті, з яких аскоспори викидаються примусово, -
еутунікатними. Еутунікатні аски, залежно від будови їх оболонки,
поділяють на унітунікатні та бітунікатні.
Унітунікатними називають такі аски, які мають лише одну обо-
лонку. Вона може бути одно- або багатошаровою, проте в останньо-
му випадку шари зростаються між собою і не можуть зміщуватись
один відносно іншого.
На верхівках унітунікатних асків часто знаходяться додаткові спе-
ціальні структури, які обумовлюють лише апікальний розрив оболон-
ки сумки, і тим самим визначають напрям відстрілу аскоспор. Сумки,
які відкриваються на верхівці кришечкою, називають оперкулятними,
а ті, які розриваються щілинами - іноперкулятними. Особливий варі-
ант унітунікатного аску притаманний лишайниковим грибам-лекано-
роміцетам: тут на верхівці аску знаходиться спеціальне фібрилярне
кільце, яке при розриві оболонки аску розпрямляється, як пружина, і
витягує за собою назовні плазмалему аску разом з наявними у неї
аскоспорами (т. зв. леканоровий тип унітунікатної сумки).
Бітунікатні аски мають дві оболонки - зовнішню (екзоаск) та внут-
рішню (ендоаск). На верхівці сумки у просторі між цими оболонками
312
також знаходиться мікрофібрилярне кільце. При розриві оболонки
екзоаску фібрилярне кільце розпрямляється і частково підтягує до
отвору ендоаск з аскоспорами. Останній під впливом внутрішнього
тиску розтягується, далі розривається і знову скорочується, додаючи
додатковий імпульс аскоспорам, що викидаються.
Екологічні особливості. Хоча аскомікотові гриби представлені
в усіх еколого-трофічних групах та в усіх основних типах біотопів, проте
їх переважна більшість мешкає на суходолі, де веде сапротрофний
спосіб життя, або паразитує на різних організмах (найчастіше - на
вищих рослинах), або вступає у різноманітні симбіози (в першу чергу,
з водоростями, утворюючи лишайники).
Система відділу. За молекулярними даними, з якими корелю-
ють важливі фенотипні ознаки, в межах відділу виділяється шість
досить чітко відокремлених філ, які розглядаються як таксони рангу
класів4. До провідних фенотипних ознак цих класів належать: а) на-
явність справжніх плодових тіл; б) тип асків. Крім того, важливими
систематичними ознаками класів є специфічні особливості, пов’язані
зі здатністю утворювати симбіотичні комплекси з водоростями (ліхе-
нізація), хімічним складом клітинних оболонок, особливостями пове-
дінки аскоспор (зокрема їх здатністю до брунькування).
За цими ознаками, з якими корелює ряд інших ознак, виділяють-
ся наступні класи:
1. Сахароміцети - ЗассНаготусеїез. Плодові тіла відсутні. Аски
прототунікатні. Покриви аскоспор утворюються з плазмалеми аску.
Специфічна особливість - мананово-глюканові клітинні оболонки.
2. Тафриноміцети - ТарНгіпотусеіез. Плодові тіла відсутні. Аски
унітунікатні, проте двошарові. Покриви аскоспор утворюються з плаз-
малеми аску. Специфічна особливість - здатність аскоспор до брунь-
кування.
3. Сордаріоміцети, або аскоміцети - Зогсіагіотусеіез (Азсоту-
сеіез). Плодові тіла наявні (клейстотеції, перитеції, апотецїї). Аски
унітунікатні. Покриви аскоспор утворюються з мембран навколоядер-
ного мішка.
4. Леканороміцети - І_есапоготусе1ез. Плодові тіла наявні (апо-
теції, рідше перитеції). Аски унітунікатні, леканорового типу. Покри-
ви аскоспор утворюються з мембран навколоядерного мішка. Спе-
цифічна особливість-утворення симбіотичних комплексів із водоро-
стями - лишайників.
313
5. Локулоаскоміцети, або дотідеоміцети - Іосиїоазсотусеїез (Оо-
ІЬІсІеотусеІез). Плодові тіла наявні (переважно псевдотеції). Аски біту-
нікатні. Покриви аскоспор утворюються з мембран навколоядерного
мішка.
6. Евроціоміцети- Еигоііотусеїез. Плодові тіла наявні (переваж-
но клейстотеції). Аски прототунікатні. Покриви аскоспор утворюють-
ся з мембран навколоядерного мішка. Специфічна особливість -
відсутність у карпогону трихогіни.
Клас Сахароміцети - Зассіїаготусеїез
До цього класу включені гриби, які мають міцелій у вигляді по-
одиноких клітин, що розмножуються вегетативним поділом надвоє
та брунькуванням. Плодові тіла та дикаріонтична фаза в циклі роз-
витку відсутні.
На біохімічному рівні сахароміцети відрізняються від інших класів
наявністю третього комплексу ферментів синтезу триптофану, мана-
ново-глюкановими клітинними оболонками. Багато представників
класу здатні отримувати енергію шляхом зброджування простих
цукрів. Септи сахароміцетів мають мікропори. Статевий процес-со-
матогамія. Аски розвиваються найпростішим способом, без стадії
гачка та без утворення диплоїдних клітин-ніжок. Аски прототунікатні,
аскоспори звільняються пасивно, переважно внаслідок ослизнення
оболонки сумки. Покриви аскоспор розвиваються з інвагінацій плаз-
малеми аску. Анаморфи представлені псевдоміцелієм, що брунькуєть-
ся. Справжні конідіальні спороношення відсутні.
Типовим порядком є Бассіїагогпусеїаіез.
Порядок Сахароміцетальні - Зассітаготусеіаіез
Ознаки порядку повторюють ознаки класу. Сахароміцетальні гри-
би є переважно сапротрофами, що мешкають у ґрунті, на поверхні
плодів та на інших субстратах, багатих на прості цукри. Деякі сахаро-
міцети у стадії анаморфи здатні викликати хвороби людей (наприк-
лад, рід кандіда - Сапсіісіа), а також спричиняти біокорозію пластмас,
біопошкодження бітумних покриттів тощо. Значна кількість видів має
важливе практичне значення та використовується в різних галузях
промислової діяльності людини.
Рід сахароміцес або дріжджі - Зассіїаготусез (рис. 114).
Представники роду Зассіїаготусез не мають справжнього міцелію,
існують у вигляді поодиноких клітин, розмножуються брунькуванням.
314
При цьому на поверхні клітини утворюється невелика брунька, яка з
часом збільшується у розмірах, відривається від материнської кліти-
ни і, у свою чергу, знову може починати брунькуватись. У сприятли-
вих умовах (достатня кількість цукру, оптимальна температура, аера-
ція) брунькування відбувається настільки швидко, що клітини не всти-
гають відокремлюватись одна від одної й утворюють короткі неміцні
ланцюжки несправжнього міцелію (псевдоміцелій).
Якщо поживних речовин мало, а кисень надходить у достатній
кількості, клітини можуть перетворюватись на аски з 4-ма, рідше
8-ма аскоспорами.
Рід Зассіїаготусез об’єднує як вільноіснуючі види природної
мікофлори, так і «культурні» (або виробничі), існування яких тісно
пов’язане з бродильною промисловістю. У практичній діяльності лю-
дини найширше використання мають 8. сегеуізіае- цукрові дріжджі.
Вони є збудниками бродіння, їх різні штами застосовуються у хлібо-
пекарській та кондитерській промисловості, а також у пивоварінні та
виноробстві.
Рис. 114. Зассіїаготусез сегеуізіае: А - вегетативна клітина; Б - клітина, що
брунькується; В - псевдоміцелій; Г - аск з аскоспорами
Клас Тафриноміцети - Таріїгіпотусеіез
Об’єднує гриби, в яких у життєвому циклі наявна коротка гаплоїд-
на фаза, представлена аскоспорами або міцелієм, що брунькується,
та псевдодикаріонтична фаза, протягом якої розвивається розгалу-
жений міцелій, що уражує вищі рослини. Статевий процес - або со-
матогамія, при якій копулюють клітини, що утворюються внаслідок
315
брунькування аскоспор, або партеногамія, при якій дикаріонтизація
відбувається внаслідок мітотичного поділу гаплоїдного ядра клітини
міцелію, що брунькується. Плодові тіла відсутні, аски унітунікатні, проте
двошарові, й аскоспори звільняються активно. При утворенні аску гачок
не розвивається, але під сумкою часто зберігається диплоїдна кліти-
на-ніжка. Покриви аскоспор формуються з плазмалеми аску. Клас
представляє єдиний порядок ТарЬгіпаІез.
Порядок Тафринальні - ТарЬгіпаїев
Об’єднує виключно облігатних паразитів судинних рослин. Пере-
важаючою стадією в життєвому циклі є розгалужений міцелій з дво-
ядерними клітинами, який розвивається у міжклітинному просторі тка-
нин рослини-господаря. Аски з аскоспорами утворюються щільним
шаром на поверхні такого міцелію. Спочатку вони прикриті епідермі-
сом та кутикулою рослини-господаря, далі проривають ці покриви та
виступають через розриви назовні. Справжні конідіальні спороношен-
ня, як і у сахароміцетів, відсутні.
Більшість видів тафринальних грибів порушують гормональний
баланс рослини - вони або безпосередньо синтезують 0-індолілоц-
тову кислоту та цитокініни, або спричиняють гіперсинтез цих гормонів
самою рослиною. Як наслідок, уражені органи гіпертрофуються, на
них розвиваються різноманітні деформації або починається ріст спля-
чих бруньок, через що розвиваються симптоми хвороби, відомої в
народі під назвою «відьмині мітли».
Порядок включає лише один рід - Таріїгіпа, до якого входить біля
100 видів.
Рід тафрина - ТарМпа (рис. 115). До найвідоміших представ-
ників роду належить Таріїгіпа ргипі- тафрина сливова. Гриб є об-
лігатним паразитом рослин, який завдає великої шкоди плодівницт-
ву, уражуючи сливу, вишню, аличу, терен та ін. На сливі спричиняє
утворення здутих, неїстівних плодів.
Плодові тіла відсутні, сумки з аскоспорами утворюються щільним
шаром безпосередньо на міцелії, під кутикулою уражених органів
рослини. Гіфи сливової тафрини проникають через квітконіжки у за-
в’язь і при сприятливих умовах (підвищена волога, температура
17-19°С), через два-чотири тижні після зараження, викликають появу
деформованих плодів. Перші ознаки захворювання проявляються
через 12-15 днів після цвітіння. Заражені плоди вкриваються білува-
316
тим воскоподібним нальотом (сумчасте спороношення), в них роз-
ростається перикарпій, кісточка та зародок не утворюються, замість
них виникає велика порожнина.
Внаслідок брунькування аскоспор утворюються гаплоїдні кліти-
ни, які після одного з мітозів стають двоядерними, тобто містять два
генетично однакових ядра, і є псевдодикаріонтичними. Такі клітини
діляться і утворюють розгалужений клітинний псевдодикаріонтичний
міцелій. Такий тип міцелію є переважаючим у життєвому циклі таф-
рини сливової. Згодом в окремих (верхівкових) клітинах такого міце-
лію відбувається каріогамія, утворюється зиготичне диплоїдне ядро,
і клітина перетворюється на майбутню сумку. Її диплоїдне ядро один
раз мітотично ділиться, клітина розділяється поперечною перегород-
кою на нижню диплоїдну клітину-ніжку та верхівкову клітину майбут-
нього аску. В останній відбувається редукційний поділ і утворюється
чотири гаплоїдні аскоспори. Навколо гаплоїдних дочірніх ядер вини-
кає власна цитоплазма, розвиваються клітинні оболонки, і в клітині-
аску утворюються аскоспори. Вони здатні брунькуватись як у сере-
дині аску, так і після виходу з нього, обумовлюючи розвиток гаплоїд-
Рис. 115. ТарМпа ргипг. А - «кишеньки
женими плодами); Б - аски з аскоспорами
слив» (фрагмент гілки сливи з ура-
317
ного псевдоміцелію, який є короткочасним і згодом починає бруньку-
ватись.
Через подібність уражених плодів до роздутих кишень, хворобу,
яку викликає Т. ргипі, називають «кишеньки плодів», або «дуті пло-
ди». Наприкінці літа уражені плоди підсихають і опадають з дерев.
Вони непридатні в їжу, їх слід збирати і знищувати. Зимує гриб у тріщи-
нах кори, бруньках і тканинах уражених пагонів.
До інших поширених збудників хвороб рослин належать тафрина
деформуюча (Таріїгіпа с/еґогтапз), що паразитує на персиках і ви-
кликає деформацію їх листків (ця хвороба відома під назвою «куче-
рявість листків персика»), та тафрина грабова (Т. сагріпі), що уражує
широколистяні деревні породи, переважно - граб, і викликає на них
утворення «відьминих мітел».
Клас Сордаріоміцети або Аскоміцети - Зогбагіотусеіез (Аз-
сотусеіез)
Переважаючою стадією життєвого циклу є гаплоїдний розгалу-
жений септований міцелій, гіфи якого складаються з одноядерних
клітин. Септи між клітинами прості, їх центральні пори закриті тільця-
ми Вороніна. Статевий процес-гаметангіогамія, аскогони мають три-
хогіну. Аски унітунікатні, аскоспори звільняються активно. Сумки
утворюються з дикаріонтичних аскогенних гіф, переважно - за спосо-
бом гачка. Покриви аскоспор розвиваються з мембран навколоядер-
ного мішка аску. Аски у сордаріоміцетів розвиваються у плодових тілах
різних типів - клейстотеціях, перитеціях або апотеціях.
У межах класу анаморфи представників більшості порядків (за
винятком пецицальних) мають добре розвинені конідіальні спороно-
шення, які утворюються на гаплоїдному міцелії.
В екологічному відношенні сордаріоміцети дуже різноманітні -
серед них багато сапротрофів (як фунтових, так і ксилотрофів), па-
разитів вищих рослин та комах, є певна кількість мікоризоутворюючих
симбіотрофів, проте відсутні ліхеноутворюючі гриби.
Сордаріоміцетів поділяють на порядки, в першу чергу, за типами
плодових тіл, видозмінами міцелію (зокрема наявністю стром та їх
забарвленням), наявністю конідіальних спороношень та частково -
за екологічними особливостями. Провідними та найбільш важливи-
ми у практичному відношенні є сім порядків, які представляють три
філогенетичні лінії в межах цього класу. Ці лінії розглядають як підкласи
- Ьеоііотусеїісіае, Зогсіагіотусеіісіае та Регіготусеіісіае:
318
а) Ьеоііотусеіісіае - гриби з мікроскопічними плодовими тілами
клейстотеціями та апотеціями, в яких строми або відсутні, або мікро-
скопічні; аски іноперкулятні; конідіальні спороношення наявні; ана-
морфи є переважно облігатними паразитами рослин, тоді як теле-
оморфи - як паразити, так і сапротрофи. Цікавою особливістю є чут-
ливість цих грибів до сірки, на чому базується один з найпоширені-
ших засобів боротьби з ними - обкурювання сірчаним газом або об-
робка сірковмісними препаратами. Характерні порядки - ЕгузірЬаІез,
НЬуІізтаіаіез та Неіоііаіез;
б) Зогсіагіотусеїісіае - гриби з мікроскопічними плодовими тіла-
ми перитеціями, що розвиваються у стромах (часто макроскопічних);
аски іноперкулятні; конідіальні спороношення наявні; сапротрофи та
паразити рослин. Підвищена чутливість до сірковмісних препаратів у
цих грибів не зареєстрована. Провідні порядки - Нуросгеаіез та
Хуіагіаіез.
в) Регіготусеїісіае - гриби з макроскопічними плодовими тілами
апотеціями; аски оперкулятні; строми та конідіальні спороношення
відсутні; переважно сапротрофи та мікоризоутворюючі гриби. Вклю-
чають порядки Регігаїез та ТиЬегаїез.
Порядок Еризіфальні або Борошнисторосяні гриби - Егу-
зірНаіез
Порядок об’єднує виключно облігатних паразитів, які викликають
захворювання вищих рослин, відоме під назвою «борошниста роса».
У класі сордаріоміцетів це єдиний порядок, що має закриті плодові
тіла - клейстотеції. Вони мікроскопічні, утворюються на міцелії, що
розвиваються на поверхні листків вищих рослин. Сумки еризіфаль-
них унітунікатні, але їх стінка має два шари, що не здатні рухатись
один відносно іншого. Строми завжди відсутні. Конідіальні спроно-
шення наявні. Анаморфи представлені переважно видами родів
Оісііит та Рзеисіоісііит. Телеоморфи в межах порядку розглядають-
ся на прикладі родів Місгозрґіаега, ВІитегіа, РЬуІІасІіпіа.
Рід мікросфера - МісгозрЬаега. Найпоширенішим видом роду
є Місгозрґіаега аІрМоісІез - мікросфера дубова (рис. 116). На ли-
стках і стеблах молодих пагонів дуба утворюється характерний білий
борошнистий наліт з міцелію гриба та конідій. Живлення гриба відбу-
вається за допомогою гаусторіїв, які проникають у тканину рослин.
До поверхні листя та інших органів рослин грибниця прикріплюється
спеціальними присосками - апресоріями.
319
У циклі розвитку мікросфери є дві стадії: конідіальна і сумчаста.
Конідіальна з’являється на початку літа і може продовжуватися до
кінця серпня залежно від кліматичних умов регіону. Поширюється гриб
еліпсовидними конідіями, які утворюються поодиноко на верхівках
видовжених циліндричних конідієносців.
Сумчаста стадія розвивається в кінці періоду вегетації. На міцелії
гриба утворюються статеві органи. Чоловічий орган (антеридій) скла-
дається з двох клітин, жіночий (аскогон) - з однієї клітини без трихо-
гіни. Вміст верхньої клітини антеридія переливається через спеціаль-
ний отвір в аскогон, відбувається запліднення. Після статевого про-
цесу на поверхні листків утворюються чорні клейстотеції з 6-20-ма
асками всередині. В кожному аску утворюється 4-8 аскоспор. Клей-
стотеції мають 8-15 коротких, дихотомічно розгалужених придатків.
Розвиток клейстотеціїв та сумок - тривалий процес. Сумки дозріва-
ють тільки восени і до весни залишаються у клейстотеціях.
Гриб є дуже небезпечним, особливо для молодої порослі дуба.
Часто хвороба набуває характеру епіфітотій, призводячи до всихан-
ня дубових насаджень.
Рис. 116. Місгозрїіаега аїріїііоісіез'. А - листок дуба, уражений борошнис-
тою росою; Б - плодове тіло - клейстотецій; В - аски з аскоспорами; Г - коніді-
альне спороношення. 1 - придатки
320
Рід блюмерія - ВІитегіа (рис. 117 А). Серед нечисленних
паразитів на однодольних рослинах велику шкоду наносить збудник
борошнистої роси злаків - ВІитегіа дгатіпіз - блюмерія злакова.
Міцелій гриба спочатку павутинистий, білий, борошнистий від конідій,
згодом ущільнюється. Клейстотеції занурені в міцелій, мають численні,
короткі, світло-коричневі прості придатки. Аски від 9 до ЗО у клейсто-
теціях, циліндричні або яйцевидні чи еліпсоїдні, з ніжкою.
Рід філактинія - РїіуІІасііпіа (рис. 117 В). Представники цього
роду борошнисторосяних грибів мають клейстотеції з придатками
складної будови. Придатки двох типів: на верхівці клейстотецію у ви-
гляді тонких гілок, що рясно галузяться та виділяють слиз, а по еква-
тору клейстотецію - у вигляді шипів, які загострені на кінцях та надуті
біля основи у вигляді цибулини. Зазвичай придатки другого типу роз-
ташовані горизонтально. При зменшенні вологості частина оболонки
розширеної частини придатків зморщується, придатки опускаються
та впираються гострими кінцями в субстрат, піднімаючи клейстотецій
над поверхнею субстрату. Клейстотецій легко здувається вітром,
відривається від субстрату та переноситься на інше місце. При цьо-
му в повітрі клейстотецій перевертається верхівкою донизу і, потрап-
ляючи на будь-який придатний субстрат, приклеюється до нього за
допомогою слизу, який виділяють придатки першого типу. Види роду
РІіуІІасІіпіа паразитують на деревах та чагарниках, переважно на
представниках родини березових (Веіиіасеае).
Рис. 117. Клейстотеції борошнисторосяних грибів: А - ВІитегіа; Б -
РІіуІІасІіпіа. 1 - аски з аскоспорами; 2 - придатки; 3 - придатки першого типу;
4 - придатки другого типу
321
Порядок Ритизматальні - КЬуіізтаіаІез
Порядок об’єднує гриби з плодовими тілами - мікроскопічними
апотеціями, які бувають округлі або лінійні, спочатку закриті, а при
дозріванні розкриваються шляхом щілиновидного або лопатевидного
розриву верхньої частини оболонки. Аски утворюються за способом
гачка, розкриваються щілиною або кільцевидним розривом, проте
кришечка відсутня. Конідіальні спороношення наявні, представлені
переважно пікнідіальними родами Меіазтіа та Ьеріозігота. Розви-
ваються ритизматальні гриби як сапротрофи або паразити на різно-
манітних субстратах рослинного походження. Важливе практичне
значення мають представники родів Ріїїуіізта та Ьоріюсіегтіит.
Рід ритизма - Кіїуіізта. Найпоширенішим видом роду є
іізта асегіпит - ритизма кленова (рис. 118). Гриб паразитує на
листках різних видів клену, утворюючи округлі, смолисто-чорні, блис-
кучі плями - строми. Якщо плям настільки багато, що для гриба не
вистачає поживних речовин, то вони не стають смолисто-чорними, а
залишаються недорозвиненими, неправильно крапчастими або виг-
лядають як побурілі ділянки листкової пластинки. Влітку в цих стро-
мах розвивається конідіальне спороношення - анаморфна стадія
Меіазтіа асегіпа. Хворе листя передчасно всихає та опадає. На пля-
Рис. 118. Ріїуіізта асегіпит: А-чорна плямистість листя клену-зовнішній
вигляд ураження; Б - апотеції на стромі; В - аск із аскоспорами та парафізами.
1 - апотеції; 2 - строма; 3 - аск; 4 - аскоспора; 5 - парафізи
322
мах опалого листя, яке перезимувало, навесні утворюється сумчас-
те спороношення - лінійно-звивисті апотеції з восьмиспоровими бу-
лавовидними сумками. Апотеції розкриваються щілиною, краї апоте-
цію відгортаються назовні. Спори одноклітинні, безбарвні, голковидні.
Між сумками завжди присутні нитковидні, зігнуті на верхівці парафізи.
Гриб широко розповсюджений в парках та широколистяних лісах, є
збудником чорної плямистості листя клена. Найчастіше зустрічаєть-
ся в насадженнях, де не здійснюється прибирання опалого листя.
Інший вид цього роду - Ріїї. рипсіаіит - в анаморфній стадії, яка
має назву Меіазтіа рипсіаіит, є паразитом клена татарського (Асег
іаіагісит), на листі якого утворює округлі, невеличкі строми, що роз-
ташовані групами на жовтих плямах.
Рід лофодерміум - 1_орію(іегтіит. Серед нечисленних пара-
зитів голонасінних збудником дуже небезпечного захворювання хвої
різних видів сосен, яке відоме під назвою «шютте», є І-орІюсіегтіит
ріпазігі - лофодерміум сосновий (рис. 119). Хвороба вперше була
виявлена в Німеччині наприкінці XVIII ст., тому свою назву отримала
від німецького слова «зсьйеп», що означає «сипати». Гриб пошире-
ний у розсадниках та молодих соснових культурах. Дуже небезпеч-
ний особливо для молодих рослин сосни віком до 5 років, може при-
зводити до повного відмирання хвої. Зараження сосни відбувається
в середині літа, коли дозрівають апотеції та звільняються спори із
сумок. При проростанні спор міцелій гриба проникає у хвоїнки крізь
продихи. Восени на хвої можна спостерігати перші ознаки хвороби -
дрібні жовтуваті плями в місцях проникнення спор та пожовтіння
кінчиків хвої. В березні-квітні після першого потепління хвоя набуває
червоно-бурого забарвлення, і в цей період з’являються пікніди у виг-
ляді дрібних чорних крапок. Це анаморфна стадія гриба, яка пред-
ставлена видом іеріозігота ріпазігі. Проте роль конідій у зараженні
сосни не відома. Наприкінці весни хвоя опадає, протягом літа на опа-
лих хвоїнках у місцях колишніх пікнід утворюються апотеції. Вони еліп-
соїдні, лінійні, занурені у субстрат, чорні, при дозріванні з отвором у
вигляді щілини. Сумки в апотеціях багаточисленні, булавовидні. Кож-
на сумка містить 8 спор. Спори нитковидні, довжина їх дорівнює дов-
жині аска, безбарвні. Парафізи на кінцях загнуті. Характерною озна-
кою хвороби є також утворення на хвоїнках чорних поперечних смуг,
що відокремлюють апотеції один від одного. Апотеції дозрівають в
кінці червня, після чого відбувається звільнення спор та зараження
хвої. Хвороба «шютте» відома в усіх районах, де вирощують сосну.
323
Рис. 119. Ьоріюсіегтіит ріпазігг. А - шютте сосни - загальний вигляд ураже-
ної хвої; Б - окремий апотецій; В - розріз через апотецій; Г - аск із аскоспорами
та парафізою; Д - аскоспора. 1 - апотеції; 2 - аски з аскоспорами
Порядок Гелоціальні - Неіоііаіез
Види цього порядку мають плодові тіла апотеції дуже маленьких
розмірів. Сумки унітунікатні, іноперкулятні, на верхівці розкривають-
ся тріщиною, через яку виходять спори. Конідіальні спороношення
наявні, але зустрічаються дуже рідко і представлені в основному по-
одинокими конідієносцями з конідіями, які належать до родів Мопіііа,
Воігуііз та ін. Практично всі гелоціальні - сапротрофи на гнилих рос-
линних залишках, проте поряд із сапротрофами зустрічаються і па-
разити рослин, які найбільшої шкоди завдають у конідіальній стадії.
Рід монілінія - Мопіііпіа. Гриби цього роду утворюють скпероції
у плодах різних рослин з родин розоцвітих та брусничних. Багато
монілієвих грибів були вперше детально вивчені російським міколо-
гом М. С. Вороніним.
324
Цей рід включає один з найпоширеніших паразитичних видів -
Мопіііпіа /гисіідепа - монілінія плодова (рис. 120 А-В). В циклі
розвитку гриба завжди зустрічається конідіальна стадія типу монілія
(Мопіііа) - подушечки коротких конідієносців, на яких утворюються
довгі ланцюжки конідій. М. /гисіідепа викликає плодову гниль яблук і
груш. Гриб проникає в плоди через пошкодження. На уражених пло-
дах з’являються плями відмерлої тканини, від них концентричними
кільцями розвиваються жовтуваті подушечки конідіального спороно-
шення. Конідії поширюються вітром, дуже часто переносяться жуком
казаркою. Заражені плоди опадають з дерев і слугують джерелом
інфекції. Плоди, що залишаються на деревах, перетворюються у скле-
роції. Кулясті склероції чорного кольору утворюються під епідермі-
сом, внаслідок чого плоди мають чорне забарвлення. Склероції зи-
мують, а весною в них знову розвивається конідіальне спороношен-
ня. Апотеції у цього виду утворюються рідко.
Рід склеротинія - Зсіегоііпіа . Найбільш важливий вид цього
роду - Зсіегоііпіа зсіегоііогит - збудник білої гнилі різних сільсько-
господарських рослин (рис. 120 Г). Цей гриб уражує вегетативні орга-
ни рослин, а також овочі при зберіганні. Гриб розвивається на стеблах
і суцвіттях соняшника, на плодах кабачків, томатів, моркви, буряка і
качанах капусти при зберіганні. На поверхні уражених частин рослин
утворюється повстистий білий міцелій, на якому формуються склеро-
ції. Апотеції утворюються завжди зі склероціїв весною, поодинокі
Рис. 120. Гелоціальні гриби: А-В - Мопіііпіа /гисіідепа: А - зовнішній вигляд
ураженого яблука; Б - конідіальна стадія - Мопіііа; В - окремі конідієносці;
Г - Зсіегоііпіа зсіегоііогит - склероції гриба на моркві
325
або групами, коричнюваті, сіруваті або жовтуваті, на видовженій ци-
ліндричній ніжці. Аски циліндричні. Спори еліпсовидні або видовжені,
прямі або зігнуті, одноклітинні, безбарвні. Парафізи нитковидні, роз-
галужені або на верхівці розширені. Конідіальне спороношення у видів
цього роду відсутнє.
Порядок Гіпокреальні - Нуросгеаіез
Порядок об’єднує види, в яких плодові тіла - перитеції, переваж-
но світло забарвлені, м'якої консистенції, завжди занурені у строми
найрізноманітнішого забарвлення, які складаються виключно з гіф
гриба. Сумки еутунікатні. У складі порядку переважають паразити рос-
лин, зокрема збудники хвороб культурних рослин, паразити членис-
тоногих, паразити на плодових тілах базидіальних грибів тощо; зуст-
річаються також сапротрофи на рослинних залишках.
Рід клавіцепс - СІауісерз. Багато представників цього роду роз-
виваються на злаках. Найпоширенішим і важливим у господарсько-
му відношенні є СІауісерз ригригеа - клавіцепс пурпуровий, а хво-
роба, яку він викликає, має назву «ріжки злаків» (рис. 121). На ураже-
них цим збудником суцвіттях рослин добре помітні склероції, які ма-
ють вид ріжків чорно-фіолетового кольору. Вони являють собою зи-
муючу стадію гриба. Спочиваючі склероції містять біля 3-4% цукрів
(трегалози, глюкози та ін.), до 1% багатоатомних спиртів і велику
кількість ліпідів. Склероції зимують у ґрунті, куди вони потрапляють
при зборі урожаю із хворих культурних або дикоростучих злаків. Вес-
ною склероції проростають головчастими стромами (червонуваті,
мають голівку до 0,5 см у діаметрі, кулясті, горбкуваті). Довжина ос-
танніх залежить від рослини-живителя та умов розвитку і становить
від кількох міліметрів до 5 см. По периферії головок стром розвива-
ються плодові тіла - перитеції. Перитеції пляшковидні або ширококо-
нусовидні, виступають на поверхню строми верхівкою з округлим от-
вором. Аски вузько-булавовидно-циліндричні. Спори нитковидні.
Клавіцепс пурпуровий має досить складний життєвий цикл. Ас-
коспори уражують злаки в період цвітіння. Після звільнення з пери-
теціїв аскоспори поширюються вітром і потрапляють на рослини. Якщо
вони потрапили на квіткові луски, їх росткова трубка не здатна про-
никнути через них, і зараження не відбувається. У випадку, коли
квіткові луски відкриті, аскоспори потрапляють на приймочку маточки
або у нектар, проростають, і їх росткові трубки досягають зав’язі. Че-
рез декілька днів після ураження на рослинах розвивається коні-
326
діальна стадія гриба - сфацелія (Зріїасеїіа). У зав’язі утворюється
щільна маса міцелію, вкрита шаром конідієносців, на яких утворюється
велика кількість конідій, що занурені у краплини «медяної роси».
«Медяна роса» відіграє суттєву роль у поширенні конідій гриба. Вона
має неприємний запах і містить велику кількість цукрів. Конідії мо-
жуть поширюватися краплинами дощу, вітром та комахами. Склероції
розвиваються повільно. Спочатку вони жовтувато-коричневі, потім
мають сіро-фіолетове або чорно-фіолетове забарвлення. Повне їх
дозрівання відбувається при достиганні зерна. Розвиваючись на куль-
турних злаках, гриб значно знижує їх урожай. Проте практичне зна-
чення збудника «ріжків злаків» визначається не тільки зниженням уро-
жаю або погіршенням якості насіння, а також токсичною дією алка-
лоїдів, які містяться в його склероціях.
С. ригригеа - один із давно відомих токсичних грибів. У наш час
токсикози у людей, викликані алкалоїдами гриба, який потрапляє в
Рис. 121. СІауісерз ригригеа: А - «ріжки злаків» - уражений колос; Б - скле-
роцій із головчастими стромами; В - розріз через головчасту строму; Г - окремий
перитецій у стромі; Д - аск із аскоспорами; Е - конідіальна стадія - Зріїасеїіа.
1 - склероцій; 2 - перитеції; 3 - аски; 4 - конідії
327
зерно, а потім борошно та продукти з нього, спостерігаються дуже
рідко. Однак у минулому токсикоз був дуже поширений у Європі і в
періоди сильних спалахів забирав життя великої кількості людей. Кла-
віцепсотоксикоз, або ерготизм, обумовлений здатністю алкалоїдів
гриба викликати скорочення гладеньких м’язів, судин, а також діяти
на нервову систему. Відомі дві його форми: гангренозна («антонів
вогонь») і конвульсивна («злі корчі»).
Алкалоїди клавіцепса пурпурового широко застосовують у су-
часній медичній практиці для лікування серцево-судинних і нервових
захворювань. В офіційну медицину гриб був введений на початку XIX
сторіччя, тоді ж було розпочато вивчення алкалоїдів. Однак в аку-
шерстві склероції широко застосовували ще раніше.
На Україні цей вид широко розповсюджений, особливо у Карпа-
тах, на Поліссі, Правобережному та Лівобережному Лісостепу.
Рід епіхлое - Ерісіїїол (рис. 122). Найбільш відомим видом роду
є Ерісіїїол (уріїіпа - епіхлое рогозовидний. Характерним для цього
Рис. 122. Ерісіїїол: А - «чохловидна» хвороба злаків - зовнішній вигляд
ураженої рослини; Б - зовнішній вигляд строми; В - розріз через строму з пери-
теціями; Г - аск із аскоспорами
328
гриба є утворення на стеблах злаків широко розпростертих стром у
вигляді чохлика. На його поверхні навесні утворюються дрібні од-
ноклітинні конідії. У липні-серпні конідіальна строма ущільнюється,
потовщується, набуває оранжевого кольору; в ній розвиваються
багаточисельні перитеції. Міцелій паразита розповсюджується в
рослині дифузно, по міжклітинниках. Гриб є збудником чохловид-
ної хвороби багаторічних злаків, зокрема, найчастіше зустрічаєть-
ся на грястиці збірній (Оасіуііз діотегаіа) та на видах тонконогу (Роа).
Хвороба завдає значної шкоди при культивуванні трав на насіння.
Рід кордицепс - Согсіусерз (рис. 123). Представники цього роду -
паразити комах, павуків та інколи грибів.
Строма кордицепсу розвивається на муміфікованих комахах або
грибах, має циліндричну ніжку й округлу (циліндричну, булавовидну)
голівку, яка завжди м’ясиста, світло або яскраво забарвлена - пурпу-
рова, жовтогаряча, сіра, жовта, рідше червона. Перитеції занурені у
строму, еліпсовидні. Аски циліндричні. Спори нитчасті, з численними
Рис. 123. Согсіусерз:
строми на трупі комахи
поперечними перегородками.
Одним з найпоширеніших видів роду
є Согсіусерз тіїііагіз - кордицепс вій-
ськовий, який розвивається на личин-
ках та лялечках метеликів, що зимують
у ґрунті. Аскоспори цього гриба потрап-
ляють на зовнішні покриви лялечки, про-
ростають гіфальними трубками, гідролі-
зують хітинові покриви та занурюються
в тіло господаря. Гіфи гриба інтенсивно
розвиваються в тілі комахи, перетворю-
ючи його на тверду муміфіковану масу
гіф, т. зв. псевдосклероцій. Восени псев-
досклероцій проростає жовтогарячими
головчастими стромами.
Рід нектрія - Месігіа (рис. 124 А).
Види цього роду мають світлі перитеції
м’ясистої консистенції, які сидять група-
ми на поверхні стерильної подушки -
строми, утвореної виключно гіфами гри-
ба, або занурені в строми. Типовим ви-
дом роду є нектрія кіноварно-червона
329
(Месігіа сіппаЬагіпа), яка поширена на сухих гілках дерев. Проте у
стадії анаморфи, яку називають туберкулярією звичайною (ТиЬег-
сиіагіа уиїдагіз), гриб викликає небезпечні захворювання рослин та
призводить до їх загибелі.
Рис. 124. Строми аскоміцетів: А - Хуіагіа роІутогрЬа; Б - Нурохуіоїт, В -
Месігіа. 1 - перитеції
Інший важливий рід у складі порядку Нуросгеаіез - гіберела
(СіЬегеІІа). Він характеризується м’якими шкірястими перитеціями
чорного кольору, які утворюються безпосередньо на субстраті або на
стромах темно-синього кольору. Широко поширеним видом цього роду
є СіЬегеІІа Ацікигоі - гіберела фуйкуроє, або фуджикуроє, яка зу-
стрічається переважно в анаморфній стадії - Еизагіит топіііїогте.
Гриб є збудником дуже небезпечної хвороби рису та інших злаків,
яка отримала назву «бакане», або «хвороби дурних пагонів». Ця хво-
роба була виявлена на початку минулого століття на рисових планта-
ціях Японії, Китаю, Індії. Окремі стеблини рису виростали, видовжую-
чись до небаченої довжини, та відмирали, не даючи зерна. У 1926 р.
японський вчений Курозава вперше ізолював з ураженої рослини збуд-
ника хвороби та довів, що саме цей гриб є причиною такого ненор-
мального росту рису. Гриб своїми продуктами обміну впливає на ко-
реневу систему рослини, спричиняє обмежене відмирання тканин і
опадання зав’язі. При цьому міжвузля витягуються, листки стають тон-
шими та вужчими, уражені рослини стають хлоротичними і гинуть до
цвітіння. Такі симптоми зумовлюють виникнення ростових речовин -
гіберелінів (від назви аскової стадії гриба), які гриб синтезує в ураженій
рослині. Пбереліни за фізіологічною дією являють собою фітогормони,
які характеризуються високою фізіологічною активностю, здатністю
різко активізувати ріст рослин та навіть змінювати його характер.
330
Порядок Ксиляріальні - Хуіагіаіез
Перитеції темного забарвлення, шкірястої або вуглистої консис-
тенції, сферичні або глекоподібні, з вузьким отвором на верхівці, роз-
ташовані переважно у великих стромах. Сумки унітунікатні. Переважно
сапротрофи.
Рід ксилярія -Хуіагіа (рис. 124 Б). Характерною ознакою роду є
наявність добре розвинутих стром, що утворені виключно гіфами гри-
ба. Строми прямостоячі, булавовидні, темнозабарвлені, чорні, вуг-
листої консистенції, зазвичай утворюються по декілька. Перитеції
занурені у строму. Дуже поширений рід, зокрема в листяних лісах,
розвивається на гнилій деревині, переважно на старих стовбурах та
пеньках дерев.
Рід гіпоксилон - Нурохуіоп (рис. 124 В). У представників цього
роду строми червоні, бурі, коричневі, круглої та напівсферичної фор-
ми, поодинокі або зростаються, утворюючи суцільні кірки. Перитеції
занурені у строму і відкриті назовні. Отвори перитеціїв оточені вали-
ком плектенхіми і нагадують бугорки на поверхні строми. Всі види
цього роду є ксилотрофами.
Порядок Пецицальні - Рехіхаїез
Порядок характеризується наявністю плодових тіл - апотеціїв,
розміри яких можуть коливатись від 1 мм до 10 см. У деяких видів
плодові тіла великих розмірів, диференційовані на шапинку та ніжку,
інші мають підземні (гіпогейні) апотеції. Аски еутунікатні, оперкулятні -
на верхівці відкриваються кришечкою.
Анаморфи зустрічаються дуже рідко, представлені поодинокими
конідієносцями, які належать до родів Оесіосерїіаіит, Созіапііпеїіа
та ін. Строми відсутні. Представники порядку - переважно гумусові
сапротрофи.
Рід пецица - Рехіха (рис. 125). Для розвитку цих грибів потрібна
підвищена вологість, тому основна їх кількість розвивається весною.
Для цього роду характерні блюдцевидні або чашовидні апотеції діа-
метром 1-5 см, переважно бурого або коричневого кольору, розта-
шовані групами, сидячі, м’ясисті, спочатку кулясті, пізніше розкрива-
ються, нерідко з нерівним краєм. Аски циліндричні, оперкулятні,
8-спорові. Спори еліпсовидні, рідше веретеновидні, одноклітинні, з
331
1-2 краплями олії, гладенькі або бородавчасті, безбарвні. Парафізи
розгалужені, на верхівці розширені, безбарвні або забарвлені.
Більшість пецицових грибів відомі лише у сумчастій стадії, в дея-
ких у циклі розвитку відоме конідіальне спороношення. Аски утворю-
ються на тому ж міцелії, що й конідії.
Рід Ре/і/а об’єднує більше 100 видів. Сюди відносяться гриби -
сапротрофи. Зустрічаються на фунті (гумусові та підстилкові сапро-
трофи), на гнилій деревині (лігнотрофи), серед мохів, на вологих ка-
м’янистих стінах, на згарищах (карботрофи), на екскрементах тва-
рин (копротрофи). Найпоширенішими в природі видами є Р. Ьадіа-
пецица коричнева, Р. уіоіасеа - пецица фіолетова та ін.
Рис. 125. Регіга: А - плодове тіло -
апотецій; Б - гіменіальний шар; В - аск
із парафізою. 1 - аски; 2 - парафізи
Рід гелвела - НеІуеІІа
(рис. 126 В, Г). Плодові тіла
прямостоячі, до 10 см заввиш-
ки. Шапка спочатку ввігнута,
пізніше опукла, дво- або чоти-
рилопатева, сідловидна, гла-
денька, з вільним нижнім
краєм або приросла до ніжки,
сірувато-бура, коричнювата.
Ніжка циліндрична, гладенька
або трохи опушена, часто бо-
розенчаста, порожниста, білу-
вата. Аски циліндричні, на
верхівці заокруглені, восьми-
спорові. Спори одноклітинні,
еліпсовидні, безбарвні, з ве-
ликою краплею олії, розташо-
вані в один ряд. Парафізи зав-
жди присутні, розгалужені, на
верхівці розширені і дещо за-
барвлені. Гелвела - типовий
сапротроф на фунті, зростає
в лісах, на світлих галявинах
серед трав’янистого покриву.
Плодові тіла їстівні, з’являють-
ся переважно восени.
332
Рід зморшок - МогсІїеІІа (рис. 126 Б). Це весняні гриби, що ро-
стуть у хвойних та листяних лісах, на галявинах, у парках та садах.
Всі представники роду їстівні. Плодові тіла - апотеції, великі (6-12 см
заввишки), м’ясисто-восковидні, мають конусовидну або яйцевидну
шапинку, яка зростається з ніжкою. Шапинка має сітку складок, які
утворюють комірки. Поверхня цих комірок вистелена гіменієм. Колір
шапинки зморшок коричневих відтінків, каротиноїди відсутні. Ніжка
біла, складчаста або гладенька, порожниста. Аски циліндричні, 8-спо-
рові. Спори еліпсоїдні, закруглені, гладенькі, одноклітинні, безбарвні
або трохи забарвлені, розташовані в один ряд. Парафізи розгалу-
жені, на верхівці розширені й забарвлені.
Найбільш розповсюдженим видом є МогсІїеІІа езсиїепіа- змор-
шок їстівний, який має приємні смакові якості та використовується в
їжу гурманами.
Рід строчок - Оуготііга (рис. 126 А). Для цього роду характерні
прямостоячі, м’ясисті апотеції. Шапинка неправильно куляста або
яйцевидна, місцями зростається з ніжкою, мозковидна, з глибокими
звивистими складками, бура або темно-бура, рідше світла. Ніжка по-
товщена, порожниста. Аски циліндричні, 8-спорові. Спори еліпсоїдні
або веретеновидні, одноклітинні, з 1-2 краплинами олії, на кінцях з
бородавчастими потовщеннями, гладенькі, безбарвні або жовтуваті,
розташовані в один ряд. Парафізи розгалужені, на верхівці розши-
рені і трохи забарвлені.
Види цього роду - ґрунтові сапротрофи, зустрічаються переваж-
но весною. На відміну від зморшок, строчки є отруйними грибами зав-
дяки наявності в їхніх плодових тілах токсину гіромітрину. Цей ток-
син розчиняється у воді та швидко зникає, проте ні виварювання, ні
висушування не видаляє його остаточно. Тому строчок вважається
небезпечним отруйним грибом.
Рис. 126. їстівні та отруйні пецицальні гриби: А - Суготііга; Б - МогсІїеІІа',
В - НеІуеІІа еіазііса-, Г - Нєіуєііз сгізра
333
Порядок Туберальні - ТиЬегаІез
Апотеції вторинно замкнуті, макроскопічні, підземні. Симбіотро-
фи, утворюють ектотрофну мікоризу з деревними породами.
Рід трюфель - ТиЬег (рис. 127). Трюфелі утворюють підземні
(гіпогейні) плодові тіла, які в дозрілому віці замкнуті. Вони округлі,
бульбовидні, м’ясисті або роговидні, гладенькі або з бородавчастою
поверхнею, всередині із жилками, між якими знаходиться гіменіаль-
ний шар. Аски грушовидні, широко еліпсоїдні, рідше кулясті, сітчасті
або щетинисті, жовтувато-коричневі або коричневі, які містять 2, 4
або 8 спор. Аскоспори завжди одноклітинні, безбарвні або бурі, куле-
видні або еліпсоїдні. Звільнення спор відбувається пасивно.
На молодих стадіях розвитку гриба зачатки плодових тіл закла-
даються у вигляді відкритого блюдцевидного утворення, на увігнутій
і дещо складчастій поверхні якої формуються зачатки гіменіального
шару. В подальшому таке плодове тіло, яке знаходиться у ґрунті, не
може рости в ширину, на поверхні його гіменія розвиваються багато-
чисельні складки (зморшки), а потім воно замикається. Зморшки пе-
ретворюються у внутрішні, а щілини між ними - у зовнішні вени. Ос-
танні заповнюються пухкою тканиною з переплетених парафіз. Біля
основи парафіз пізніше утворюються сумки.
Трюфелі - гриби, що дуже цінуються гурманами багатьох країн. Серед
них найбільше практичне значення має ТиЬег теїапозрогит - чорний
французький трюфель. На Україні поширені два види - Т. Ьгитаїе - чор-
ний трюфель, Т. аезіїліт - літній трюфель. Всі трюфелі є облігатними
мікоризними грибами.
Рис. 127. ТиЬег аезііуит: А - загальний вигляд плодового тіла; Б - розріз
через плодове тіло; В - фрагмент плодового тіла з асками та аскоспорами
334
Клас Леканороміцети - Ьесапоготусеіез
Плодові тіла наявні (апотеції, рідше перитеції). Аски унітунікатні,
леканорового типу. Покриви аскоспор утворюються з мембран на-
вколоядерного мішка. Специфічна особливість - утворення симбіо-
тичних комплексів із водоростями - лишайників (див. ліхенізовані гри-
би, або лишайники).
Клас Локулоаскоміцети або Дотідеоміцети - Ьосиїоаз-
сотусеіез (ОоНіісІеотусеіез)
Плодові тіла наявні, представлені псевдотеціями. Аски бітунікатні.
Покриви аскоспор утворюються з мембран навколоядерного мішка.
Конідіальні спороношення розвинуті.
Порядок Плеоспоральні - РІеозрогаІез
Псевдотеції кулеподібні або грушоподібні, з однією або декілько-
ма локулами, занурені у тканину рослини-господаря. Сумки в локу-
лах або утворюють шар гіменію, або зібрані у пучки. Вони завжди
бітунікатні. Відомі як сапротрофи на різноманітних рослинних залиш-
ках, а також паразити рослин.
Рід вентурія - Уепіигіа. Об’єднує паразитів трав’янистих та де-
ревних рослин, які уражують листя та плоди. На живих листках та
плодах утворюється конідіальне спороношення, на відмерлих - сум-
часте. Найбільш розповсюдженим видом є Уепіигіа рігіпа — венту-
рія грушова. Гриб паразитує на груші і викликає захворювання, яке
відоме під назвою «парша». Паразитує гриб у конідіальній стадії, яка
має назву фузикладій грушовий (Ризісіасііит рігіпит). Уражує гриб
листя, плоди, пагони. На листках утворюються плями невизначеної
форми, чорнувато-оливкові, на них з’являються дернинки конідіально-
го спороношення. Конідієносці прості, оливкові. Конідії утворюються
на їх верхівці, поодинокі, обернено-булавовидні, видовжено-грушо-
видні, одноклітинні, згодом двоклітинні, оливкові або майже безбарвні.
На плодах спочатку утворюються дрібні чорно-оливкові плями. Зго-
дом вони розростаються, зливаються. Тканина плода під плямою кор-
ковіє. Уражені плоди розвиваються ненормально, викривляються,
розтріскуються, втрачають смакові якості й товарну цінність.
Сумчасте спороношення розвивається в сапротрофних умовах
на опалих уражених листках, що перезимували. Плодові тіла -
псевдотеції - виступають назовні верхівкою, оточеною бурими ще-
335
тинками. Сумки 8-спорові, оточені парафізами. Спори оливкові,
видовжені, з однією перегородкою. Верхня клітина спори часто дов-
ша і ширша.
Інший вид цього роду - V. іпаедиаііз - вентурія яблунева - зу-
мовлює паршу яблунь (рис. 128).
Рис. 128. \/епіигіа іпаедиаііз: А - уражений листок яблуні; Б - уражений
плід яблуні; В - конідіальне спороношення (стадія Ризісіасііит сіепсігііісіт); Г -
псевдотецій
Клас Евроціоміцети - Еигоііотусеіез
Плодові тіла наявні (переважно клейстотеції). Аски прототунікатні.
Покриви аскоспор утворюються з мембран навколоядерного мішка.
Специфічна особливість - відсутність у карпогону трихогіни. Тенден-
ція до втрати телеоморф і переважного розвитку анаморф.
Порядок Евроціальні - Еигоііаіез
Характеризується наявністю плодових тіл - клейстотеціїв. Сумки
прототунікатні, розташовані у плодових тілах безладно. Аскоспори
звільняються пасивно, після руйнування оболонки сумки.
Клейстотеції з безладно розташованими сумками утворюються
безпосередньо на міцелії. В життєвому циклі більшості евроцієвих
грибів суттєву роль відіграє безстатеве розмноження за допомогою
конідій. Конідіальні стадії належать до родів АзрегдіІІиз, РепісіІІіит,
Асгетопіит (див. анаморфні гриби, або Оеиіеготусеїез).
336
Типовим представником порядку є рід евроцій (Еигоііит), що зу-
стрічається в природі у ґрунті, на рослинних залишках, на продуктах,
що зберігаються, викликаючи їх цвіль.
Інший рід з цього порядку - елафоміцес (ЕІаріїотусез) - харак-
теризується наявністю макроскопічних клейстотеціїв, які занурені у
ґрунт. Типовим представником роду є Е. дгапиіаіиз - елафоміцес
зернистий, або трюфель оленячий (рис. 129). Гриб є рідкісним,
знайти його в природі досить важко, оскільки його макроскопічні пло-
дові тіла, вкриті товстим щільним перидієм, розвиваються глибоко у
ґрунті. Клейстотеції кулясті, жовто-коричневі, 1-5 см, з дрібно боро-
давчастою поверхнею. Міцелій елафоміцеса характерного жовтого
кольору, обплітає коріння дерев. Зустрічається гриб переважно во-
сени, у хвойних лісах, особливо на піщаних ґрунтах. Він утворює міко-
ризу із хвойними та деякими листяними деревами. Знайти цей гриб у
природі можна за допомогою іншого аскоміцета - Согсіусерз оріїіо-
діоззоісіез (з порядку Нуросгеаіез), який є облігатним паразитом і ут-
ворює свої булавовидні строми, що підіймаються над землею, на клей-
стотеціях елафоміцеса.
Рис. 129. ЕІарІїотусеє. А- клейстотецій серед коренів дерева; Б-зовнішній
вигляд клейстотецію; В - розріз через клейстотецій
337
Відділ Базидіомікотові гриби - Вазісііотусоіа
Гриби, в яких вегетативне тіло представлене розгалуженим клітин-
ним міцелієм. Вегетативні клітини гаплоїдні або дикаріонтичні, при-
чому дикаріонтична стадія за тривалістю переважає. Статевий про-
цес - соматогамія. Статеве спороношення - базидія з базидіоспора-
ми. Монадні стадії повністю відсутні.
Відділ нараховує близько ЗО тис. видів і є другим за кількістю видів
після аскомікотових. Еволюція базидіомікотових відбувалася пара-
лельно до аскомікотових.
У складі відділу представлені майже всі екологічні групи грибів, у
тому числі сапротрофи на різноманітних субстратах, паразити вищих
рослин, мікоризоутворювачі, дереворуйнуючі гриби тощо. Зустріча-
ються також копротрофи та мікотрофи. Серед базидіальних значна
кількість видів належить до їстівних або отруйних грибів. Деякі пред-
ставники мають цінні лікарські властивості.
Біохімічні особливості та живлення. Базидіомікотові синте-
зують лізин за грибним шляхом (ААА-шлях). Синтез триптофану
відбувається по-різному - у шапинкових агарикоїдних грибів він син-
тезується за допомогою другого комплексу ферментів, у всіх інших -
за допомогою четвертого комплексу. Іони заліза поглинаються за
допомогою сидерамінів. Поліоли (включаючи маніт) наявні у досить
значних кількостях.
Споживання органічних речовин відбувається виключно шляхом
абсорбції. Багатьом базидіомікотовим притаманні потужні екзофер-
ментні системи, за допомогою яких поза межами клітин відбувається
гідроліз целюлози та лігніну (целюлази, пероксидази, оксидази), білків
(різноманітні пептидази), жирів (ліпази). Джерелом вуглецю та азоту
є різноманітні органічні сполуки.
Метаболізм стероїдів регулюється мікоспоринами, які у багатьох
базидіомікотових здатні ініціювати статевий процес.
Грибам цього відділу притаманний широкий спектр вторинних
метаболітів, до яких належать антибіотики, мікотоксини, інгібітори
росту вищих рослин, фітогормони та ін.
Цитологічні ознаки. Клітини базидіомікотових вкриті оболонка-
ми, основу яких складають хітин та глюкани. Оболонки, на відміну від
аскомікотових, багатошарові. Септи між клітинами прості або доліпо-
рові з відкритою порою (в устоміцетів), прості з порою, закритою
дрібними вакуолями (теліоміцети), або доліпорові з парентосомою
338
(базидіоміцетові). Парентосома може бути суцільною і не мати пер-
форацій, бути перфорованою або складатись із численних фраг-
ментів-пухирців (ця ознака використовується при поділі базидіоміце-
гових на порядки).
Ядерний апарат евкаріотичний. Міцелій, що розвивається з ба-
зидіоспор, має клітини переважно з одним ядром та існує лише ко-
роткий час. Переважаюча стадія у циклі розвитку - це дикаріонтич-
ний септований міцелій, де пари ядер утворюються при статевому
процесі внаслідок плазмогамії, що одразу не супроводжується каріо-
гамією.
Мітоз відкритий (у шапинкових грибів) або закритий (у всіх інших),
центріолі відсутні, а центрами організації мікротрубочок, подібно до
аскомікотових, є полярні скупчення аморфного нуклеопротеїду. Ядер-
це на початку мітозу переважно дезінтегрується, рідше виштовхуєть-
ся у цитоплазму, де зберігається до початку або завершення тело-
фази.
Мітохондрії з пластинчастими кристами. Комплекс Гольджі від-
сутній. Одномембранні органели подібні до аскомікотових - це літичні
пухирці, хітосоми, ломасоми, мікротільця, вакуоля з клітинним соком.
Найпоширенішими типами включень є краплини олії та глобули гліко-
гену.
Джгутикові стадії повністю відсутні.
Вегетативне тіло представлене короткоіснуючим гаплоїдним та
довгоіснуючим дикаріонтичним міцелієм. Міцелій, що розвивається
із спор статевого спороношення - базидіоспор, - гаплоїдний і часто
утворений клітинами, що здатні до брунькування, внаслідок чого може
мати габітус псевдоміцелію. Переважаючим у циклі розвитку типом
вегетативного тіла є дикаріонтичний міцелій, що розвивається після
статевого процесу. Такий міцелій, як правило, рясно розгалужений,
багатоклітинний та септований.
У більшості базидіомікотових дикаріонтичний міцелій має пряж-
ки - особливі дугоподібні клітини, що розташовані біля поперечної
перегородки гіфи та з’єднують її дві клітини. Пряжка гомологічна
гачку аскогенної гіфи і виконує функцію відновлення двоядерності
клітини. Наявність пряжок на міцелії є стабільною ознакою для груп
різного рангу - від порядку до виду (наприклад, пряжки відсутні у
представників порядків Огебіпаїез, та Яиззиіаіез, багатьох видів
роду Тгісїюіота).
Найбільш відомою видозміною міцелію у базидіомкотових є пло-
дові тіла (див. нижче). На відміну від аскомікотових, вони утворюють-
339
ся виключно гіфами дикаріонтичного міцелію. До інших видозмін на-
лежать ризоморфи - тяжі зі щільно переплетених гіф, за допомогою
яких міцелій здатний швидко переміщуватись у просторі. Для деяких
базидіомікотових характерне утворення склероціїв, за допомогою яких
гриб зберігає життєдіяльність у несприятливих для нього умовах.
Розмноження. Основним способом розмноження базидіомікото-
вих є розмноження за допомогою спор статевого спороношення -
базидіоспор. У меншому ступені представлене вегетативне роз-
множення - частинами міцелію та поодинокими спеціалізованими
клітинами, що утворюються внаслідок фрагментації гіф - телей-
тоспорами, уредоспорами або спочиваючими акінетоподібними
товстостінними клітинами - хламідоспорами. Значно рідше зустрі-
чається нестатеве розмноження за допомогою конідій.
Статеве розмноження. На відміну від аскоміцетів, у базидіаль-
них грибів відсутні диференційовані статеві органи. Статевий процес-
соматогамія. Вона відбувається або шляхом злиття двох вегетативних
клітин гаплоїдного міцелію, або двох базидіоспор, або продуктів їх
брунькування. У гомоталічних видів відбувається злиття гіф одного й
того ж міцелію. У гетероталічних, до яких відноситься більшість бази-
діомікотових грибів, копулюють клітини гіф, що беруть початок від спор
протилежних знаків: «+» та «-». Після плазмогамії ядра об’єднують-
ся в дикаріони, які згодом синхронно діляться, утворюючи дикаріон-
тичний міцелій. Такий міцелій може існувати довгий час і є переважа-
ючою стадією життєвого циклу базидіомікотових. Із дикаріонтичних
клітин розвивається статеве спороношення - базидія з базидіоспо-
рами.
Життєвий цикл та розвиток статевих спороношень. У най-
більш загальному вигляді життєвий цикл базидіомікотових включає
наступні етапи: з гаплоїдної базидіоспори розвивається гаплоїдний
міцелій. Його клітини копулюють, при цьому відбувається плазмога-
мія, що не супроводжується каріогамією, і утворюється дикаріонтич-
на клітина злиття. З неї розвивається дикаріонтичний міцелій, що є
основним вегетативним поколінням базидіомікотових. Окремі дика-
ріонтичні клітини розвиваються у базидію: в них ядра дикаріону зли-
ваються (каріогамія), відбувається мейоз, і на базидії утворюються
гаплоїдні базидіоспори. Таким чином, життєвий цикл гаплофазний, із
зиготичною редукцією та гетероморфною зміною поколінь - гаплоїд-
ного та дикаріонтичного міцеліїв, причому дикаріонтична фаза є пе-
реважаючою.
340
У деяких грибів (наприклад, іржастих) дикаріонтична фаза може
утворювати додаткові нестатеві спороношення - ецидіоспори (вес-
няні спори), уредоспори (літні спори), телейтоспори (зимові спори) -
або розвивати конідіальні спороношення.
Розвиток базидій. Відомо два основні варіанти розвитку базидій:
з поодиноких спочиваючих клітин - телейтоспор - та з верхівкових
клітин гіф дикаріонтичного міцелію (переважно за способом пряжки).
При розвитку базидії з телейтоспор її дикаріонтичні ядра злива-
ються, з телейтоспори виростає диплоїдна базидія, далі в ній відбу-
вається мейоз, і оболонка утворює випини - стеригми, в кожний з
яких заходить одне гаплоїдне ядро з частиною цитоплазми. Стериг-
ма відділяється від базидії септою, розростається на верхівці і пере-
творюється на одноядерну гаплоїдну базидіоспору.
При розвитку з дикаріонтичних гіф міцелію на базидії перетворю-
ються верхівкові дикаріонтичні клітини таких гіф: від оболонки верхів-
кової клітини гіфи відходить невеликий бічний виріст - пряжка, яка
загинається донизу. В цей же час відбувається поділ ядер дикаріону,
і в клітині утворюються чотири ядра. Два різностатевих ядра пряму-
ють до верхівки гіфи, третє пересувається до основи клітини, а чет-
верте заходить у виріст пряжки. Одночасно з поділом ядер відбуваєть-
ся поділ самої клітини, утворюються дві перегородки - одна відділяє
пряжку, друга ділить клітину навпіл. Таким чином, утворюється три
клітини: верхня, яка містить два різностатевих ядра, нижня клітина з
одним ядром та клітина-виріст, або пряжка, яка також містить одне
ядро протилежного статевого знаку. Згодом виріст пряжки доростає
до стінки нижньої клітини, і через отвір ядро мігрує в цю клітину, віднов-
люючи її двоядерність або дикаріонтичний стан.
З верхньої клітини з двома різностатевими ядрами утворюється
базидія. Ця клітина поступово розростається, в ній відбувається
каріогамія і утворюється диплоїдне ядро. Воно майже одразу ре-
дукційно ділиться, утворюючи чотири гаплоїдних ядра. Оболонка
майбутньої базидії розвиває випини - стеригми. В кожну стеригму
переходить частина цитоплазми базидії та одне ядро: Поступово
розростаючись, верхівки стеригм здуваються і перетворюються на
базидіоспори.
Таким чином, в обох варіантах базидіоспори розвиваються екзо-
генно, на відміну від аскоспор, що утворюються ендогенно.
Базидії можуть бути поодинокими (якщо вони розвиваються з те-
лейтоспор) або утворюватися на гіфах розвиненого дикаріонтичного
міцелію, складаючи основу гіменію.
341
Гіменій у базидіомікотових - це шар базидій з базидіоспорами та
стерильних захисних елементів. Стерильні елементи, що утворюються
в гіменіальному шарі, представлені, в першу чергу, парафізами (вони
відокремлюють базидії одну від одної та запобігають злипанню бази-
діоспор) і цистидами (захищають гіменіальний шар відтиску зверху).
Зрідка гіменій розвивається на вегетативному міцелії (зокрема в ек-
зобазидіальних грибів), проте у більшості випадків він формується
на поверхні або всередині плодових тіл. Наявність плодових тіл є
головною ознакою класу базидіоміцетів (ВазИіотусеІез).
Типи базидій. За морфологічною будовою базидії поділяють на
одноклітинні - холобазидії, та на базидії із септами - фрагмобазидії.
Септи у фрагмобазидії можуть бути поздовжніми або поперечними.
Фрагмобазидії з поздовжніми перегородками зазвичай поділені сеп-
тами на чотири клітини (така будова характерна для тремеляльних
грибів). Варіанти з поперечними септами найрізноманітніші: в деяких
порядках базидія має лише одну септу і поділена на дві клітини -
верхню епібазидію та нижню гіпобазидію. В аурікуляріальних, бага-
тьох сажкових та іржастих грибів кількість поперечних септ дорівнює
трьом, тобто базидія складається з чотирьох клітин, розміщених одна
над одною.
За характером розташування веретена поділу під час мейозу
(фаза мітоз І) та рівнями розташування ядер після другого поділу (фаза
мітоз II) базидії поділяють на хіастичні та стихічні.
В хіастичних базидіях під час мейозу диплоїдне ядро розташо-
вується на верхівці базидії, веретено орієнтується перпендикулярно
до її поздовжньої осі. Як наслідок, після другого поділу (фаза мітоз II)
всі чотири гаплоїдних ядра розташовуються на верхівці базидії при-
близно на одному рівні.
У стихічних базидіях під час мейозу ядро знаходиться приблизно
у центрі базидії, а веретено орієнтується паралельно до її поздовж-
ньої осі. Як наслідок, після другого поділу дочірні гаплоїдні ядра роз-
міщуються у базидії на різних рівнях.
Типи базидіоспор. Хоча всі базидіоспори утворюються екзогенно
і мають одне гаплоїдне ядро, вони є досить різноманітними за ба-
гатьма морфологічними ознаками (зокрема за формою, кольором,
розмірами, поверхнею оболонки, за наявністю проросткової пори
та ін.). До провідних ознак базидіоспор, що використовуються при
поділі базидіомікотових на таксони високого та середнього рангів (зок-
рема на порядки та родини), належить їх здатність активно відокрем-
люватися від базидії та забарвлюватися певними барвниками, в пер-
шу чергу, тими, що містять йод.
342
Базидіоспори, що активно відстрілюються від базидій, називають
балістоспорами. Такий відстріл пов’язаний з утворенням у стеригмах
вакуолі, де відбувається гідроліз глікогену до простих цукрів і, як на-
слідок, розвивається високий тургорний тиск.
За здатністю до забарвлення розчином Люголя або реактивом
Мельцера базидіоспори поділяють на амілоїдні, неамілоїдні та дек-
стриноїдні. В амілоїдних спорах наявний розчинний крохмаль, який з
йодом дає брудно-фіолетове забарвлення. Декстриноїдні спори при
забарвленні реактивом Мельцера набувають винно-червоного або
коричневого кольору.
Система відділу. За наявністю плодових тіл, типами базидій,
місцем їх утворення, типами септ, здатністю базидіоспор до бруньку-
вання, наявністю у циклі розвитку стадії псевдоміцелію, відділ по-
діляють на три класи: Вазісііотусеїез, Теііотусеіез та Іізіотусеіез.
Клас Базидіоміцети - Вазібіотусеіез
Провідний клас, до якого входить переважна більшість представ-
ників відділу. Головна ознака - наявність плодових тіл (саме їх у по-
буті зазвичай називають грибами). Базидії одноклітинні (холобазидії),
рідше септовані (фрагмобазидії), розвиваються у гіменії. Базидіоспо-
ри не здатні до брунькування. Стадія псевдоміцелію в цілому не ха-
рактерна, оскільки базидіоспори дають початок розвиненому септо-
ваному міцелію (хоча ця фаза є короткочасною і швидко переходить
у дикаріонтичну). Клітинам базидіоміцетів притаманні доліпорові септи
з парентосомами різних типів (суцільними, перфорованими, фрагмен-
тованими на системи пухирців).
Плодові тіла базидіоміцетів - карпофори - завжди диференцій-
овані на стерильну та фертильну частини. Стерильну частину плодо-
вого тіла - траму - утворюють дикаріонтичні вегетативні гіфи. Фер-
тильна частина утворена гіменієм. Відкрита поверхня плодового тіла,
на якій розташовується гіменій, називається гіменофором. Якщо
гіменій розвивається всередині плодового тіла, то гіменофор не ут-
ворюється, а плектенхіму з гіменієм називають спороносною «ткани-
ною» - глебою. Базидіомікотові гриби, в яких плодові тіла закриті і
замість гіменофора мають глебу, називають гастероміцетами, а ті, в
яких плодові тіла мають гіменофор, - гіменоміцетами.
Плодове тіло закладається на дикаріонтичному міцелії у вигляді
зачатків - примордіїв - і далі розвивається за одним із трьох спо-
собів: гімнокарпним, геміангіокарпним або ангіокарпним.
343
При гімнокарпному типі розвитку гіменіальний шар із самого по-
чатку закладається на поверхні примордіїв і до дозрівання спор зали-
шається відкритим. При такому способі розвитку плодове тіло поз-
бавлене будь-яких покривал, і називається відкритим, або гімнокарп-
ним. Прикладом є плодові тіла білого гриба, лисички, сироїжки.
При геміангіокарпному способі розвитку гіменій закладається на
бічній поверхні примордію, але зовні захищається шкірястими спле-
теннями гіф - покривалами. Якщо покривало охоплює весь примордій,
його називають загальним, або вольвою. Покривало, яке закриває
лише гіменій, є частковим. У процесі росту плодового тіла під покри-
валами диференціюється гіменофор, далі вони розриваються. Залиш-
ки загального покривала часто залишаються на верхньому боці пло-
дового тіла у вигляді пластівців та при його основі у вигляді вольви
(або піхви). Часткове покривало після розриву, як правило, зберігаєть-
ся під гіменофором у вигляді кільця на ніжці (наприклад, у мухомо-
ра). Інколи гіменофор прикритий лише тонким павутинистим спле-
тенням гіф, які називають кортиною (такі плодові тіла характерні для
грибів-павутинників).
При ангіокарпному розвитку гіменій закладається в середині при-
мордіїв. Гіменофор при цьому не утворюється, а замість нього розви-
вається глеба. Такі плодові тіла називають закритими - ангіокарпни-
ми. Звільнення базидіоспор відбувається лише після їх повного дозрі-
вання та подальшого руйнування зовнішніх оболонок плодового тіла.
Як один із варіантів розвитку гіменію інколи виділяють також псев-
доангіокарпний спосіб, при якому гіменофор спочатку завжди відкри-
тий, а згодом стає закритим внаслідок загинання країв шапинки.
Плодове тіло не лише захищає гіменій від пошкоджень, але й
підіймає його над субстратом, сприяючи тим самим розповсюджен-
ню базидіоспор. Ступінь прояву останньої функції простежується за
формою плодового тіла. Найпростішим морфологічним типом пло-
дового тіла є розпростерте (ресупінатне, кортиціоїдне). Воно має ви-
гляд щільної плівки, на поверхні якої розташовується гіменій. Якщо
така шкірка відгинається від бічної поверхні субстрату або набуває
вигляду копитоподібного утвору, прикріпленого до субстрату боком,
то карпофор називають консолевидним. Ще вище підносяться над
субстратом клаваріоїдні плодові тіла, що мають вигляд паличок або
розгалужених кущиків з гіменофором у верхній частині. У шапинко-
вих грибів гіменофор розташовується на нижньому боці шапинки, яку
підіймає над субстратом стерильна ніжка (т. зв. агарикоїдний карпо-
фор). Плодові тіла у вигляді закритих сидячих або піднесених на ніжці
344
кулястих утворів із глебою всередині називають гастероїдними та се-
котоїдними відповідно. Ресупінатні, консолевидні, клаваріоїдні та ага-
рикоїдні плодові тіла притаманні лише гіменоміцетам, а гастероїдні
та секотоїдні - гастероміцетам.
За консистенцією плодові тіла поділяють на здерев’янілі, кор-
ковидні, м’ясисті та желеподібні. За тривалістю існування - на ба-
гаторічні та однорічні. Серед однорічних виділяють ті, що існують
протягом короткого часу (від кількох годин до 10-15 діб) - їх часто
називають загниваючими та ті, термін існування яких зазвичай
відповідає більшій частині одного вегетаційного сезону, а їхня заги-
бель не супроводжується загниванням.
Типи гіменофору. Поверхня плодових тіл, яка у гіменоміцетів несе
гіменій - гіменофор - різноманітна за формою і використовується як
одна із систематичних ознак при поділі їх на порядки. Прийнято вва-
жати, що еволюція гіменофору в напрямку збільшення його поверхні
певним чином пов’язана з напрямками еволюції класу. За загально-
вживаними класифікаціями, до основних морфологічних типів гіме-
нофору належать: гладенький, складчастий, шипастий, дедалієвид-
ний, трубчастий та пластинчастий.
Типи глеби. У гастероміцетів розташування у глебі гіменію є ос-
новою для виділення певних морфологічних типів, аналогічних до
типів гіменофору. Глебу поділяють на однорідну (базидії розміщуються
рівномірно і не утворюють правильного гіменію), лакунарну (у глебі
без певного порядку розвиваються порожнини - лакуни, на поверхні
яких безладно розташовуються базидії), коралоїдну (центральна ча-
стина глеби стерильна і суцільна, до периферії від неї радіально відхо-
дять щілиноподібні лакуни, на поверхні яких розташовується правиль-
ний одношаровий гіменій), багатошапинкову (має кілька правильно
розміщених ізольованих лакун, вкритих одношаровим гіменієм), од-
ношапинкову (має лише одну кільцевидну конусоподібну лакуну і ди-
ференційована на плодоносну ніжку - рецептакул-та шапинкоподібну
частину, на поверхні якої розвивається гіменій).
Система класу. За сучасними молекулярними даними, в межах
класу виділяється вісім основних ліній та 14 великих відокремлених
молекулярних клад, які розглядають як порядки.
Фенотипічна відокремленість порядків простежується, в першу
чергу, за наступними ознаками: тип базидії (холо- чи фрагмобазидія;
хіастична чи стихічна), наявність перфорацій у парентосомі, тип пло-
дового тіла (ангіокарпне чи ні), його консистенція (загниваюче чи ні),
345
тип гіменофору, особливості базидіоспор, еколого-трофічна приуро-
ченість, а також деякі додаткові специфічні особливості (наявність
телефорової кислоти, щетинок у гіменії, сфероцист та ін.).
Система фенотипічних ознак, яка відображує родинні зв’язки між
порядками, сьогодні зрозуміла недостатньо і лише обмежено узго-
джується із класичними морфологічними системами. На молекуляр-
но-філогенетичних деревах порядки групуються парадоксально: са-
мостійними гілками розташовуються Роїурогаїез та ТКеІерКогаІез.
Досить компактну групу утворюють Адагісаіез, Різіиііпаїез, І_усо-
регбаїез та Місіиіагіаіез. Близько спорідненими є Воіеіаіез та 8сІе-
гобегтаіаіез. Уособлену позицію займає Риззиіаіез. В одному клас-
тері розташовуються такі морфологічно несхожі порядки, як Ну-
тепосКаеіаІез, СапіКагеІІаІез та РКаІІаіез. Найвіддаленішими від
інших базидіоміцетів є Аигісиїагіаіез та ТгетеІІаІез.
Нижче для зручності викладання матеріалу порядки згруповані за
традиційними морфологічними групами відповідно до типу базидії -
холо- чи фрагмобазидія (з цією ознакою добре корелює консистен-
ція плодового тіла - драглиста вона чи ні). Холобазидіоміцети розді-
лені на гіменоміцети та гастероміцети. Гіменоміцети поділені на тих,
що мають незагниваючі плодові тіла (більшість з них традиційно на-
зивають трутовиками) -т. зв. афілофороїдні гіменоміцети, та гриби із
загниваючими плодовими тілами (традиційна назва - шапинкові гри-
би), або агарикоїдні гіменоміцети.
1. Афілофороїдні гіменоміцети (Роїурогаїез, ТКеІерКогаІез,
Еізкіїіпаїез, Нутепосіпаеіаіез, СапіКагеІІаІез);
2. Агарикоїдні гіменоміцети (Воіеіаіез, Адагісаіез, Киззиіаіез);
3. Гастероміцети (І_усорегсіаіез, Зсіегосіегтаіаіез, Місіиіагіаіез,
РІіаІІаІез).
4. Фрагмобазидіоміцети з драглистими плодовими тілами (Аигі-
сиїагіаіез, ТгетеІІаІез).
Слід пам’ятати, що такий поділ не відповідає системі родинних
зв’язків між порядками, а відображає переважно ступінь їх морфоло-
гічної конвергентної схожості.
Афілофороїдні гіменоміцети
Афілофороїдні гіменоміцети представляють групу грибів, які
відіграють надзвичайно важливу роль не лише у природі, але й у прак-
тичній діяльності людини. Велике практичне значення афілофороїд-
них гіменоміцетів обумовлене, в першу чергу, здатністю багатьох їх
346
представників спричиняти руйнування деревини. Звідси походить і їх
традиційна назва - дереворуйнуючі гриби.
Дереворуйнуючі гриби розкладають здерев’янілі стінки клітинних
оболонок дерев, руйнують клітковину, целюлозу, лігнін, пектини та
інші компоненти оболонки. Внаслідок цього відбувається деструкція
деревини та втрата її якості. Крім того, в результаті руйнування клітин-
ної оболонки змінюються хімічний склад, фізичні та технологічні вла-
стивості деревини.
За фізіологічною ознакою дереворуйнуючі гриби поділяють на дві
основні великі групи: целюлозо- та лігнінрозкладаючі. Гриби першої
групи спричиняють т. зв. буру (або червону) деструктивну гниль де-
ревини: за допомогою ферменту целюлази вони розкладають целюло-
зу клітинних стінок, внаслідок чого деревина стає бурою від лігніно-
вих речовин та дуже крихкою, її об’єм значно зменшується. Найчасті-
ше бура гниль поширюється від периферії стовбура до центру. До
целюлозорозкладаючих грибів належать види родів Сопіоріїога, 8ег-
риіа, Тгатеіез, іепііпиз та ін. Найбільш небезпечними збудниками
бурої гнилі є справжній домовий гриб (Зегриіа Іасгутапз), трутовик
сірчано-жовтий (1_аеіірогиз зиірііигеиз), губка березова (Ріріорогиз Ье-
іиііпиз), губка дубова (Оаесіаіеа диегсіпа), печіночниця звичайна
(Різіиііпа Ьераііса).
Лігнінрозкладаючі гриби руйнують лігнін та частково целюлозу
деревини. В цьому випадку деревина не темніє, в ній іноді з’являють-
ся світлі смуги або комірці, які заповнені білою целюлозою, що не
розклалася. Деструкція деревини здійснюється за допомогою фер-
ментів целюлаз та оксидаз. Така деревина зазвичай розпадається
на волокна та пластини. Цей тип розкладу називають білою (інколи -
строкатою) гниллю. Типовими представниками цієї групи грибів є тру-
товик справжній (Ротез /отепіагіиз), трутовик лускатий (Роїурогиз
здиатозиз), лензитес березовий (іепхііез Ьеіиііпа), трутовик не-
справжній (РіїеІІІпиз ідпіагіиз) та ін.
У той же час деякі афілофороїдні гіменоміцети з м’ясистими кар-
пофорами є цінними їстівними грибами (наприклад, гриб-баран, тру-
товик розгалужений, печіночниця, більшість рогатикових грибів та ін.).
Окремі види таких трутовиків наразі стали рідкісними і занесені до
Червоної книги України.
Серед афілофороїдних гіменоміцетів відомо чимало лікарських
грибів (наприклад, трутовик лаковий, чага, лентин та ін.). Деякі з них
(наприклад, шиітаке та трутовик лаковий) вже введені у промислову
культуру як джерело сировини для фармакології.
347
Проте у філогенетичному відношенні група афілофороїдних гіме-
номіцетів є збірною. Нижче наводяться п’ять основних порядків, се-
ред яких деякі (наприклад, фістулінальні), є ближчими до агарикоїд-
них гіменоміцетів, ніж до інших афілофороїдних.
Порядок Поліпоральні - Роїурогаїез
Плодові тіла гімнокарпні, незагниваючі (однорічні або багаторічні,
здерев’янілі, корковидні або м’ясисті незагниваючі, з гіменофором
різних типів); щетинки в гіменофорі відсутні; базидія хіастична, про-
ста (холобазидія); парентосоми з перфораціями; спори гладенькі,
неамілоїдні, циліндричні. Ксилотрофи та факультативні паразити.
Значна кількість видів є дереворуйнівними грибами, які викликають
різні типи гнилей деревини.
Поліпоральні гриби, за результатами молекулярно-філогенетич-
них реконструкцій, поділяються на три основні групи родів. З цим по-
ділом на фенотипічному рівні корелюють ознаки, пов’язані, по-пер-
ше, з анатомічною будовою плодових тіл (зокрема наявністю сполуч-
них гіф), по-друге, з консистенцією карпофорів (м’ясисті вони чи ні). З
цими ознаками корелюють також інші особливості, зокрема, тип схре-
щування (біполярне чи тетраполярне), типи гнилей, які викликають
певні трутовики, наявність специфічних вторинних метаболітів (утому
числі таких, що мають лікувальні властивості). В межах окремих груп
роди розрізняються, в першу чергу, за типом гіменофора, консистен-
цією та часом існування плодових тіл.
Нижче основні роди поліпоральних згруповані наступним чином:
група 1: гриби з ресупінатними плодовими тілами (т. зв. домові
гриби);
група 2: гриби з м’ясистими плодовими тілами без сполучних гіф
(роди 2рагаззіз та 1_аеіірогиз)\
група 3: гриби з корковидними або здерев’янілими плодовими тіла-
ми (одно- чи багаторічними) без сполучних гіф (роди Ріріорогиз,
Тгатеіез, Оаесіаіеа, Ротііорзіз);
група 4: гриби з м’ясистими, корковидними або здерев’янілими
плодовими тілами (одно- чи багаторічними) та зі сполучними гіфами
(роди Роїурогиз, іепгііез, іепііпиз, Ігрех).
група 5: гриби зі здерев’янілими багаторічними плодовими тіла-
ми та зі сполучними гіфами (роди Ротез та ваподегта)
Група 1. Включає гриби, які мають ресупінатні плодові тіла і спри-
чинюють буру деструктивну гниль ділової деревини (т. зв. домові гри-
348
би). До групи входить найнебезпечніший дереворуйнуючий домо-
вий гриб - Зегриіа Іасгутапз (рис. 130).
Плодові тіла гриба широко розпростерті, до 50 см у діаметрі, м’я-
систо-губчасті, до 1-4 см завтовшки, прирослі до субстрату. Гімено-
фор переважно сітчасто-комірчастий, вохряно-жовтий, яскраво- або
темно-іржавий. Розростаючись по субстрату, домовий гриб виділяє
на поверхні плодового тіла, наче сльози, краплинки прозорої рідини.
Спори гладенькі, вохряно-жовті або іржаві.
Гриб оселяється на дерев’яних деталях підвалів та нижніх поверхів
будинків. За допомогою ферменту целюлази він споживає целюлозу,
зумовлюючи цим буру деструктивну гниль. Деревина зменшується в
об’ємі і швидко руйнується. За допомогою видозмінених та щільно пе-
реплетених гіф, що мають вигляд довгих шнурів, міцелій поширюється
на нові ділянки, часом долаючи значні відстані (до 1,5 м).
Незважаючи на відносну простоту плодового тіла, належність
домового гриба до порядку поліпоральних є спірною. Зокрема, ре-
зультати молекулярно-генетичних досліджень дозволяють припусти-
ти, що домовий гриб є більш спорідненим з агарикоїдними гіменомі-
цетами з порядку Воіеїаіез, ніж з іншими поліпоральними грибами.
Рис. 130. Зегриіа: зовнішній вигляд міцелію гриба на деревині
349
Група 2. Об’єднує гриби з м’ясистими однорічними плодовими
тілами без сполучних гіф. Викликають буру гниль деревини. Більшість
представників є їстівними. Характерні роди - Зрагаззізта 1_аеіірогиз.
Рід летипор - іаеіірогиз. Плодові тіла однорічні. Шапинки си-
дячі, водянисто-м’ясисті, жовтогарячі або сірчано-жовті, зібрані у ве-
ликі групи, які розташовуються у вигляді черепиці на стовбурах де-
рев. Гіменофор трубчастий, трубочки короткі, сірчано-жовті. Спори
безбарвні. Найвідомішим представником роду є іаеіірогиз
зиІрПигеиз - летипор сірчано-жовтий, або трутовик сірчано-жов-
тий (рис. 131). Гриб є паразитом багатьох листяних дерев, найчаст-
іше розвивається на стовбурах дуба, тополі, верби, акації білої, рідше
уражує хвойні дерева. Спричиняє центральну буру деструктивну
гниль. Після відмирання дерева продовжує свій розвиток як сап-
ротроф. Плодові тіла в молодому віці їстівні.
Рід спарассіс - Зрагаз-
Рис. 131. іаеіірогиз зиіріїигеиз:
зовнішній вигляд плодового тіла
5/5. Об’єднує представників з
однорічними плодовими тіла-
ми кораловидної форми. Кар-
пофори великі, зазвичай від
основи сильно розгалужені,
м’ясисті, білуваті, жовті або
кремові. Гілочки широкі, сплю-
щені, з кучерявими краями.
Спори безбарвні, гладенькі,
неамілоїдні, споровий відби-
ток кремового кольору. Кси-
лотрофи, зростають, як пара-
зити, на коренях дерев.
Характерним представни-
ком роду є Зрагаззіз сгізра -
спарассіс кучерявий, або
гриб-баран. Зустрічається
переважно у хвойних лісах на
коренях або біля основи стов-
бурів сосни, ялини, ялиці. Ви-
кликає червону гниль. Гриб
їстівний, з дуже приємним
смаком. Занесений до Черво-
ної книги України.
350
Група 3. Включає гриби з корковидними або здерев’янілими пло-
довими тілами (одно- чи багаторічними), позбавленими сполучних гіф.
Викликають переважно буру гниль деревини. Завдяки коркоподібній
консистенції карпофорів майже всі представники раніше використо-
вувались для виготовлення труту, звідки походить назва трутовики.
Прикладами є роди Ріріорогиз, Тгатеіез, Оаесіаіеа, Ротііорзіз.
Рід піптопор - Ріріорогиз. Плодові тіла однорічні, подушковидні
або округлі, сидячі, м’ясисто-коркові. Шапинки зовні вкриті жовтою
або буруватою шкіркою. Тканина біла або жовтувата. Пори округлі.
Спори циліндричні. Досить поширеним є РіріорогизЬеіиііпиз-піпто-
пор березовий, або губка березова (рис. 132). Гриб має м’ясисті,
м’ясисто-коркові, приплюснуто-копитовидні, подушковидно-опуклі,
сидячі або із зачатковою ніжкою шапинки. Верхня поверхня шапинки
гладенька, з тонкою шкірочкою, жовтувато-коричнева або жовтува-
то-сіра. Тканина біла, м’якокоркова. Гіменофор трубчастий, трубочки
спочатку білі, потім жовтіють, розташовані в один ряд, у старих грибів
іноді відділяються від тканини плодового тіла. Спори циліндричні,
зігнуті, безбарвні. Губка березова - факультативний паразит. Зрос-
тає у листяних та мішаних лісах, на мертвих та живих стовбурах і
гілках берези. Спричиняє жовто-буру гниль, яка швидко поширюєть-
ся від периферії стовбура до центру.
Рис. 132. Ріріорогиз Ьеіиііпиз: загаль-
ний вигляд плодових тіл
Рід траметес -
Тгатеіез. Плодові тіла
однорічні. Шапинки си-
дячі, округлі, коркові,
поодинокі або розмі-
щені черепичасто. По-
верхня шапинок опу-
шена, замшева або
бархатиста, білого, кре-
мового або сірого коль-
орів. Трама має силь-
ний анісовий запах.
Гіменофор трубчастий.
Пори округлі. Спори
безбарвні. Значно по-
ширений на живих сто-
вбурах сосни Тгатеіез
351
ріпі - траметес сосновий, що спричиняє інтенсивну білу гниль де-
ревини.
Рід дедалеа - Оаесіаіеа. Плодові тіла багаторічні, коркові, ве-
ликі, сидячі, білі, жовтуваті, з віком коричнево-бурі. Найпоширенішим
видом роду є Оаесіаіеа диегсіпа - дедалея дубова, або губка дубо-
ва (рис. 133). Трапляється гриб переважно на пеньках дуба та на
обробленій деревині. Плодові тіла плоскі, у вигляді напівокруглих або
копитовидних, розпростерто-відігнутих або розпростертих шапинок,
прикріплених боком, поодинокі або часто зібрані черепичастими гру-
пами. Шапинки голі або дещо опушені, шорсткі, зверху горбкуваті,
кольору свіжої деревини, з нечіткими концентричними зонами щоріч-
ного наростання. Гіменофор лабіринтовидний (у вигляді товстостін-
них звивистих ходів, що проникають у плодове тіло на глибину
3-5 см). Гіменій поновлюється щороку в тих самих ходах, стінки яких
наростають знизу. Спори еліпсоїдні, біля основи косо загострені, без-
барвні. Тканина плодового тіла пружна, корковидна, світло-коричню-
вата або сірувата, при розрізанні ножем скрипить (що зумовило на-
родну назву гриба - скрипуха). Гриб викликає буру деструктивну гниль
деревини.
Рис. 133. Оаесіаіеа диегсіпа'. А - загальний вигляд плодового тіла; Б - лабі-
ринтовидний гіменофор
Рід фомітопсіс - Ротііорзіз. Плодові тіла багаторічні, копито-
видні, подушковидні, розпростерто-відігнуті, вкриті дещо смолистою
кіркою.Тканина світлих відтінків, корково-дерев’янистої консистенції.
Гіменофор трубчастий, без щетинок. Типовим видом є Р. ріпісоіа.
Фомітопсіс сосновий, або трутовик облямований -
Ротііорзіз ріпісоіа. Плодові тіла подушковидні, пізніше копито-
подібні, сидячі, переважно дерев’янистої консистенції. Поверхня пло-
дового тіла нерівна, борозенчасто-зональна, світло-жовта, рудувата,
352
кіноварно-червона. Тканина корково-дерев’яниста, біла або кольору
деревини. Р. ріпісоїа - факультативний паразит, зустрічається на
відмерлих та живих стовбурах багатьох хвойних та листяних порід.
Швидко й інтенсивно руйнує деревину, спричинюючи буру гниль.
Група 4. Представлена грибами з м’ясистими, корковидними або
здерев’янілими плодовими тілами (одно- чи багаторічними), які ма-
ють т. зв. сполучні гіфи. Спричиняють виключно білу гниль. У карпо-
форах накопичуються специфічні вторинні метаболіти, через що до
цієї групи потрапляє чимало лікарських грибів. Найвідоміші представ-
ники належать до родів Роїурогиз, іепхііез, іепііпиз, Ігрех.
Рід поліпор - Роїурогиз. Плодові тіла однорічні, у вигляді ша-
пинки з ніжкою, поодинокі або зростаються по кілька екземплярів.
Шапинки округлі, увігнуто-розпростерті, віялоподібні. Ніжка зачатко-
ва або добре сформована, бічна, ексцентрична або центральна. За
консистенцією плодові тіла шкірясто-м’ясисті, коркові, часто лускаті,
щетинисто-волосисті, рідше голі. Гіменофор трубчастий. Спори без-
барвні, гладенькі. Досить часто зустрічається Роїурогиз зриатозиз —
поліпор лускатий, або трутовик лускатий (рис. 134) - зокрема на
живих та мертвих стовбурах широколистяних порід. Шапинки округлі,
увігнуто-віялоподібні, з прямою або зігнутою, товстою бічною або екс-
центричною ніжкою, донизу чорно-бархатистою. Зовні шапинки жов-
туваті, з коричневими
або темно-бурими, кон-
центрично розташова-
ними лусочками; з тон-
ким, злегка хвилястим,
іноді підгорнутим краєм.
Тканина спочатку б.-м.
м’ясиста, м’яка, потім
жорстко-м’ясиста, у су-
хому стані крихка, біла.
Гіменофор трубчастий,
трубочки одного кольо-
ру з тканиною, спуска-
ються на ніжку, при ви-
сиханні буріють або ста-
ють коричневі. Спори
безбарвні, видовжено-
еліпсовидні. Трутовик
Рис. 134. Роїурогиз здиатозиз: загаль-
ний вигляд плодового тіла
353
лускатий є небезпечним руйнівником деревини, спричиняє інтенсив-
ну білу центральну гниль, деревина розпадається на волокна і плас-
тини. Молоді плодові тіла трутовика лускатого можна вживати в їжу.
Рід лензитес - Ьепгііез. Плодові тіла однорічні, сидячі. Шапин-
ки зовні густоволосисті або повстисті, зональні, різнозабарвлені. Гіме-
нофор пластинчастий. Спори безбарвні, циліндричні. Типовим видом
роду є іепгііез Ьеіиііпа - лензитес березовий (рис. 135). Плодові
тіла сидячі, черепичасті, іноді зростаються по довжині. Шапинки
напівокруглі, половинчасті, прикріплені по всій довжині або цент-
ральною дисковидною основою до субстрату. Зовні шапинки по-
встисто-волосисті, звичайно невиразно зональні, брудно-білі, сірі
або буруваті. Тканина плодового тіла шкіряста, при розриві вато-
подібна, волокниста, потім жорстка або трохи пружна, біла або
палева. Пластинки розходяться радіально, трохи розгалужені. Спо-
ри безбарвні, циліндричні, дещо зігнуті. Росте гриб на пеньках, пова-
лених стовбурах і гілках листяних порід, особливо берези, осики,
вільхи. Дуже поширений. Спричиняє білу гниль деревини.
Рис. 135. іепгііез Ьеіиііпа'. А-загаль-
ний вигляд плодового тіла; Б - загальний
вигляд гіменофора
Рід лентин - Ьепііпиз
(рис. 136). Плодові тіла од-
норічні, жорстко-м’ясисті,
коркуваті, при підсиханні
значно твердіють. Гімено-
фор пластинчастий, плас-
тинки мають зубчастий край.
Шапинка та ніжка гомогенні,
пружні. Шапинки опукло-,
плоско- або увігнуто-розпро-
стерті, лійковидні, із закруг-
леним краєм. Ніжка ексцен-
трична, бічна, луската або з
кільцевидно розміщеними
пластівцями. Пластинки спу-
скаються на ніжку або, при
відсутності ніжки, віялоподі-
бно розміщені, край пласти-
нок зазубрений. Трама пла-
стинок неправильна. М’я-
куш жорсткий, пізніше у
більшості видів коркуватий.
354
У деяких видів роду є часткове покривало. У молодому стані їстівні.
Цистиди відсутні. Спорова маса біла. Спори гладенькі, видовжено-
еліпсовидні. Ксилотрофи. Найпоширенішим є І_епііпиз іідгіпиз - лен-
тин тигровий, що зростає щільними групами на старих пеньках та
стовбурах листяних дерев, особливо тополі та верби. Шапинка лійко-
видна, світла, вкрита численними темно-коричневими лусками. Ніжка
центральна або ексцентрична, пружна, з кільцем, що швидко зникає,
з дрібними темними лусками. Цей гриб є їстівним лише у дуже моло-
дому стані.
Інший представник роду - есіосіез- відомий у світі під назвою
«сіі-таке», або «шиітаке». Він є цінним їстівним грибом, що має та-
кож численні й унікальні лікарські властивості. Антиканцерогенні, то-
нізуючі та імуностимулюючі властивості сіі-таке відомі у традиційній
азіатській фітотерапії вже протягом багатьох сотень років. Протягом
кількох останніх десятиріч цей гриб став об’єктом детальних дослі-
джень і в сучасній медицині. Крім вищеназваної протиракової та іму-
ностимулюючої дії, препарати із сіі-таке ефективно застосовуються з
метою попередження і лікування вірусних та бактеріальних захворю-
вань, нормалізації процесу обміну речовин та рівня холестерину в
крові. Вживання в їжу плодових тіл сіі-таке сприяє покращенню іму-
нітету, виведенню з організму іонів важких металів та шлаків. Висока
вартість та популярність сіі-таке в країнах Європи та Америки також
пояснюються високими смаковими якостями, високим вмістом віта-
міну О та лікувально-профілактичними властивостями, які широко ви-
світлюються у східній літературі. Сіі-таке є одними з найбільш перс-
пективних для вирощування їстивним грибом з лікувальними власти-
востями. За обсягом виробництва сіі-таке займає на сьогодні друге
місце у світі після печериці двоспорової. Більше ніж 2000 років сіі-
таке культивують в країнах Південно-Східної Азії, особливо в Японії.
Зараз Японія належить до найбільших виробників у світі. Широко
культивують сіі-таке і в інших країнах південно-східної Азії. Ці гриби
високо цінуються в Японії та Китаї завдяки приємному смаку та запа-
ху, як у сирому, так і сушеному вигляді. Плодові тіла гриба, зібрані у
природі та вирощені на обрубках деревини різних видів, мало відрізня-
ються між собою за смаком чи запахом. Високі смакові якості і фар-
макологічні властивості сіі-таке та ряду продуктів, отриманих з нього,
зумовлюють зростання попиту на цей гриб на світовому рівні.
Шапинка сіі-таке, залежно від штаму й інтенсивності освітлення
під час формування плодового тіла, світло- або темно-коричнева, в
центрі дещо темніша. Поверхня шапинки частково вкрита дрібними
355
лусочками з волосками. На початку утворення шапинка куляста із за-
горнутим всередину краєм. Плодове тіло часто з ексцентричною
ніжкою, пізніше з притиснутою шапинкою, діаметр якої досягає 5-
20 см залежно від штаму, ступеня зрілості та умов вирощування. Пла-
стинки білого кольору і змінюють свій колір у перезрілих плодових тіл
на жовтуватий. Ніжка білувата, з ніжними лусочками. М’якуш плодо-
вих тіл білий, пружної консистенції, причому ніжка жорсткіша. Гриб
має специфічний смак і запах, що часто нагадує редьку. Сіі-таке ха-
рактеризується довгим строком зберігання у свіжому вигляді і, по-
рівняно з іншими їстівними грибами, значним вмістом сухої речовини
у плодових тілах.
Рід ірпекс- Ігрех. Плодові тіла
розпростерто-відігнуті або у виг-
ляді сидячих, іноді з ніжковидно
звуженою основою, черепичасто
розташованих шкірястих шапок.
Гіменофор шипастий, зубчастий,
комірчасто-лабіринтовидний, бі-
лий, вохряний. Цистиди цилінд-
ричні, булавовидні, здебільшого
інкрустовані по всій поверхні. Спо-
ри безбарвні, видовжено-еліп-
соїдні, гладенькі, неамілоїдні. Ти-
повим видом є Ігрех Іасіеиз —
ірпекс молочно-білий. Гриб має
шкірясті, розпростерто-відігнуті,
зрідка у вигляді напівокруглих ша-
пок плодові тіла. Поверхня опуше-
Рис. 136. іепііпиз-. загальний на- часто3 концентричними боро-
вигляд плодових Тіл зенками, біла, з часом жовтувата.
Гіменофор спочатку комірчастий,
потім у вигляді довгих, плоских шипів, зубців, які розташовані пра-
вильними послідовними рядами, білий, згодом бруднувато-вохряний.
Цистиди на верхівці часто інкрустовані. Спори безбарвні, еліпсовидні,
до основи звужені. На відмерлих, інколи на живих стовбурах та гілках
листяних дерев. Гриб є незначним руйнівником деревини.
Група 5. Об’єднує гриби зі здерев’янілими багаторічними плодо-
вими тілами, що мають сполучні гіфи. Спричиняють різні варіанти білої
гнилі. Подібно до попередньої групи, містить чимало лікарських видів.
Основні роди - Ротез та Саподегта).
356
Рід фомес - Ротез. Плодові тіла зазвичай великих розмірів,
багаторічні, копитовидні. Тканина трутовидна, корковидна, рудувата
або іржаво-бура. Гіменофор трубчастий, трубочки розташовані ша-
рами одного кольору з тканиною або дещо світліші. Пори округлі.
Спори безбарвні, гладенькі, видовжено-еліпсовидні.
Надзвичайно поширеним є Ротез /отепіагіиз - трутовик
справжній (рис. 137). Міцелій гриба розвивається в середині дерев-
ного субстрату, а на поверхні ураженого дерева формуються бага-
торічні плодові тіла. Оселяючись на живому дереві, трутовик справ-
жній паразитує на ньому, спричиняє задуху деревини або побуріння,
підпар з білими смугами, мармуровидну змішану стовбурну гниль.
Хвороба, розвиваючись, призводить до загибелі дерева. Деструкція
деревини здійснюється за допомогою целюлази та оксидаз.
Після загибелі дерева трутовик справжній продовжує розвивати-
ся вже на мертвій деревині, живлячись при цьому сапротрофно. Пло-
дові тіла трутовика справжнього здерев’янілі, копитовидної форми,
однією стороною прикріплюються до субстрату. Здерев’яніла стериль-
на частина плодового тіла - трама, до якої кріпиться гіменофор. Гіме-
нофор трубчастий, розташований на нижньому боці плодового тіла.
Щороку, починаючи з весни, минулорічний гіменофор починає вкри-
ватися шаром нових переплетених між собою гіф, який з часом ущі-
льнюється. В цьому ущільненому шарі нових гіф формується трубча-
стий гіменофор. Завдяки такому наростанню, на поверхні плодового
тіла формуються концентричні зони, за якими, певною мірою, можна
судити про вік плодового тіла.
Рис. 137. Ротез /отепіагіиз'. А - загальний вигляд плодового тіла; Б - по-
здовжній розріз через плодове тіло; В - поперечний розріз через гіменофор.
1 - базидія з базидіоспорами
357
Рід ганодерма - Сапосіегта. Плодові тіла одно- або бага-
торічні, приплюснуто-плоскі або копитовидні, сидячі або з добре сфор-
мованою ніжкою. Шапка і ніжка зовні вкриті специфічною кіркою, час-
то яскраво забарвленою, блискучою, ніби полакованою. Тканина пло-
дового тіла корковидна, часом дерев’яниста, бурих відтінків. Пори
дрібні, округлі. Спори яйцевидно-еліптичні, з подвійною оболонкою:
зовнішня - безбарвна, гладенька; внутрішня - бура, бородавчаста
або сітчаста. Деякі види роду мають цінні лікарські властивості, на-
приклад ганодерма блискуча або трутовик лакований.
Трутовик лакований - Сапосіегта Іисісіит (рис. 138). Плодові
тіла у вигляді плескатої, нирковидної або віялоподібної шапинки, яка
кріпиться до субстрату за допомогою добре помітної, досить довгої
ніжки, що може бути боковою, іноді вертикальною, рідко централь-
ною або ексцентричною. Шапинка і ніжка вкриті червоною або темно-
бурою, блискучою, ніби лакованою, кіркою. Край шапинки білуватий,
жовтуватий, звичайно гострий, іноді хвилястий і трохи загнутий вниз.
Поверхня трубчастого шару білувата, кремова, потім коричнева, при
натискуванні і висиханні буріє. Спори жовто-бурі, яйцевидні або май-
же еліпсоїдні, ніби зрізані, з безбарвним еліпсоїдним виразним при-
датком, бородавчасті. Трутовик лакований є сапротрофом, розви-
вається на пеньках переважно листяних дерев. Зустрічається у при-
роді поодиноко, зрідка, тому відноситься до категорії рідкісних грибів.
Спричиняє повільну білу гниль деревини.
Рис. 138. Сапосіегта Іисісіит: загальний
вигляд плодового тіла
З давніх-давен відомі
лікарські властивості С.
Іисісіит, або гриба
«рейши», як його тради-
ційно називають у східній
медицині. Сьогодні він є
одним з основних об’єктів
досліджень медичної
мікології. Його застосову-
ють для профілактики та
боротьби з онкологічними
захворюваннями завдяки
високому вмісту в ньому
Ь-О-глюканів, а також при
серцево-судинних пато-
логіях. Гриб має високі
358
антидіабетичні та гепатопротекторні властивості. Крім того, «рейши»
активізує імунну систему, має значний антистресовий та антиокси-
дантний вплив за рахунок вмісту в ньому органічного германію. Відо-
мо також, що С. Іисісіит є ефективним радіопротектором. Не випад-
ково цей гриб називають панацеєю від усіх хвороб.
Порядок Телефоральні - ТНеІерНогаІез
Плодові тіла гімнокарпні, однорічні (переважно коркоподібної кон-
систенції або м’які, з гладеньким або шипастим гіменофором); ще-
тинки у гіменофорі відсутні; базидія хіастична, проста (холобазидія);
парентосоми з перфораціями; спори темнозабарвлені, орнаменто-
вані, в обрисах кутасті. Утворюють ектотрофну мікоризу. Специфічна
особливість - наявність телефорової кислоти, яка має характерний
неприємний запах.
Рід телефора - Тїіеіеріюга. Плодові тіла шкірясті, розпростерті,
булавовидні, віялоподібні, іноді розгалужені, часом лійковидні, з роз-
січеними краями, сидячі або на добре помітній ніжці; переважно ко-
ричневі, із фіолетовим відтінком, рідше білі. Гіменіальний шар не-
рівногорбкуватий, без цистид. Спори коричнюваті, щетинисті. Харак-
терним представником роду є телефора наземна.
Телефора наземна - Пеіеріюга іеггезігіз (рис. 139). Плодові
тіла з ніжкою, м’які, шкірясті, часто розпростерті та розташовуються
розетками або піднімаються по субстрату, з відігнутими, черепичас-
тими, лопатевими, розірваними або суцільними білуватими краями.
Плодові тіла темно-коричневі, волосисті. Гіменіальний шар гладень-
кий або нерівногорбкуватий. Спори округлі або кутасті, щетинисті,
коричневі. Зростає гриб на
ґрунті, серед трав’янистих
рослин по всій території Ук-
раїни. На відміну від телефо-
ри наземної, інший вид цього
роду - Т. раїтаіа - телефо-
ра пальчаста - має дуже роз-
галужені, приплюснуті, корич-
нево-шоколадні плодові тіла,
які характеризуються досить
неприємним запахом, що збе-
рігається навіть при висиханні
гриба.
Рис. 139. Тіїеіеріїога іеггезігіз: за-
гальний вигляд плодових тіл
359
Рід гіднелюм - НусіпеІІит. Плодові тіла щільні й товстом’ясисті,
у вигляді шапинки з ніжкою. Шапинка лійковидна, плоско- або опук-
ло-розпростерта, не диференційована від ніжки. Гіменофор шипас-
тий, досить низько спускається на ніжку. Шипи коричневі. Спори жов-
туваті, коричнюваті, кутасто-кулясті, грубобородавчасті або шипуваті.
Характерними представниками роду є Н. гопаіит - гіднелюм зо-
нальний — та Н. зиауеоіепз - гіднелюм запашний.
Рід саркодон - Загсосіоп. Плодові тіла щільном’ясисті, у вигляді
шапинки із центральною або ексцентричною ніжкою. Шапинка опук-
ло- або увігнуто-розпростерта, здебільшого з опуклим краєм, гладень-
ка, луската. Гіменофор шипастий, дещо спускається на ніжку. Шипи
світло- або темнозабарвлені. Спори безбарвні, коричнюваті, буруваті,
кутасто-кулясті, грубобородавчасті або шипуваті. Найбільш відомим
є Загсосіоп ітЬгісаіиз - саркодон черепичастий, або їжовик лус-
катий, який має опукло- або увігнуто-розпростерті світло-коричневі
шапинки, з великими, особливо в центрі шапинки, бурими черепича-
сто розташованими лусочками. Це пізньоосінній гриб, який є їстівним.
Порядок Гіменохетальні - Нутепосіїаеіаіез
Плодові тіла гімнокарпні, однорічні та багаторічні, здерев’янілі або
шкірясті, незагниваючі, ресупінатні, відігнуті або копитовидні; гіменофор
гладенький, складчастий або трубчастий. Характерна особливість - на-
явність у гіменофорі щетинок. Парентосоми без перфорацій; плодові
тіла від розчину КОН -темніють; спори безбарвні або темнозабарв-
лені, гладенькі, зрідка амілоїдні. Види порядку викликають білу гниль
деревини.
Рід гіменохете — Нутепосіїаеіе. Плодові тіла розпростерті, роз-
простерто-відігнуті або у вигляді бічних шапинок, тонкі, твердо-
плівчасті, шкірясті. Гіменіальний шар має довгі бурі товстостінні гла-
денькі щетинки, загострені на кінці. Спори гладенькі, безбарвні. Дуже
розповсюдженим є Нутепосіїаеіе гиЬідіпоза - гіменохете іржаво-
червона. Гриб має шкірясто-коркуваті, розпростерті плодові тіла з
відігнутими краями або має вигляд черепичасто розташованих бічних
шапинок. Поверхня шапинок темно-коричнева, майже чорна, з кон-
центричними смугами. Гіменій іржаво-коричневий, з товстостінними
загостреними щетинками, що помітно виступають над базидіями.
Росте гриб переважно на пеньках листяних дерев.
360
Рід фелін - РІіеІІіпиз. Плодові тіла багаторічні, сидячі, копито-
видні, розпростерто-відігнуті або розпростерті, корковидної або де-
рев’янистої консистенції. Гіменофор трубчастий, шаруватий, щетин-
ки присутні. Пори дрібні. Спори безбарвні або злегка забарвлені, май-
же кулясті до еліпсовидних, гладенькі.
Дещо подібні до трутовика справжнього плодові тіла має РІіеІІіпиз
ідпіагіиз - трутовик несправжній (рис. 140). Плодові тіла цього гри-
ба багаторічні, копитоподібні, сидячі. Шапинки зовні у молодих грибів
рудуваті, із сіруватим нальотом, потім - концентрично борозенчасті,
гладенькі, із слабо блискучою кіркою, яка з часом робиться сірувато-
чорною, чорно-бурою, нерідко з глибокими тріщинами. Край шапинки
притуплений, від іржаво-коричневого до сірого кольору. Тканина пло-
дового тіла дерев’яниста, дуже тверда, рудувато- або каштаново-бура.
Щетинки яйцевидно-шиловидні, коричнево-бурі або каштанові, із зду-
тою основою.
Трутовик несправжній є небезпечним паразитом багатьох листя-
них дерев, найчастіше уражує березу, вербу, вільху, осику та деякі
інші породи. Спричиняє центральну білу або злегка жовтувату гниль
з характерними чорними лініями.
Рис. 140. РііеІІіпиз ідпіагіиз: А - загальний вигляд плодового тіла; Б - розріз
через гіменіальний шар. 1 - щетинка; 2 - базидія з базидіоспорами
Порядок Кантареляльні - СапШагеІІаІез
Плодові тіла гімнокарпні, однорічні, м’ясисті, але незагниваючі;
гіменофор гладенький, складчастий (переважно у вигляді пластинок
з анастомозами) або шипастий; базидія стихічна, проста (холобази-
дія); парентосоми з перфораціями; спори забарвлені або безбарвні,
не амілоїдні, гладенькі або шипасті. Ектомікоризні гриби.
361
Роди кантареляльних грибів добре розрізняються за формою
карпофорів та типами гіменофору. Так, Катагіа та СІауагіа, які в Ук-
раїні часто називають також «грибною локшиною», мають розгалу-
жені клаваріоїдні плодові тіла з гладеньким гіменофором. У видів роду
СІауиІіпа карпофори перехідні між клаваріоїдними та агарикоїдними,
гіменофор гладенький. Для родів СгаіегеІІиз та СапіїїагеІІиз харак-
терні агарикоїдні карпофори із складчастим гіменофором. Рід Нудпит
також має агарикоїдні плодові тіла, але з гіменофором шипастого типу.
Цікаво, що за молекулярними даними деякі кантареляльні гриби (зок-
рема з групи «грибної локшини») близькі до агарикоїдних гіменомі-
цетів з порядку Адагісаіез.
Рід рамарія - Ратагіа (рис. 141 А). Плодові тіла видів цього роду
м’ясисті, дуже розгалужені - кущиковидні, деревовидні, переважно
великих розмірів, здебільшого яскраво забарвлені. Гіменофор на всій
поверхні плодового тіла гладенький. Ніжка б.-м. диференційована,
товста, дещо зморшкувата. Тканина плодового тіла волокниста, жор-
стка та ламка. Спори видовжені, бородавчасті або шипасті, жовтих
відтінків, забарвлюються солями заліза і часто містять одну велику
Рис. 141. Рогатикові гриби: А - Патагіа; Б - СІа-
уагіа
краплину олії. Рос-
туть у лісах, на ґрунті
або гнилій деревині.
Серед видів роду Ка-
тагіа відомі їстів-
ні гриби, наприклад
Я Ьоігуїіз - рамарія
гроновидна.
Рід клаварія -
СІауагіа (рис. 141 Б).
Плодові тіла видів
клаварії прості, рідше
радіально розгалу-
жені, поодинокі, час-
тіше досить тісними
групами, часто м’я-
систі, крихкі, різ-
номанітних кольорів
- від сірих до роже-
вих та фіолетових,
362
рідше жовті. Ніжка здебільшого нечітко відмежована від спороутво-
рюючої частини плодового тіла. Гіменофор гладенький. Спори без-
барвні, гладенькі, злегка шорсткі, часто із численними краплями. Ро-
стуть переважно на ґрунті, рідше на залишках деревини.
Рід клавуліна - СІауиІіпа. Плодові тіла деревовидно, кущико-
видно, пальцевидно розгалужені, інколи прості, циліндричні або
видовжено-булавовидні, забарвлені переважно у світлі кольори.
Тканина плодового тіла щільна, крихкувата, білувата. Базидії од-
ноклітинні, циліндричні, з 1-2 спорами, які розташовані на дуже
вигнутих стеригмах. Псля відділення спор базидії зазвичай стають
дво- або багатоклітинні, через утворення в них поперечних перего-
родок. Спори безбарвні, але при тривалому зберіганні гербарних
зразків стають вохряними, кулясті, гладенькі, з великою краплею. Види
роду є їстівними грибами низької якості.
Рід кантарел або лисичка - СапіїїагеІІиз. До цінних їстівних
грибів належить СапііїагеІІиз сіЬапиз - лисичка звичайна (рис. 142 А).
Яскраво-жовті плодові тіла цього гриба можна зустріти у хвойних та
мішаних лісах з червня по жовтень. Плодові тіла пружном’ясисті, ніжка
дуже щільна, гладенька, поступово потовщується і переходить у ша-
пинку. Шапинка щільном'ясиста, спочатку опукла, із загорнутим краєм,
згодом лійковидна. Гіменофор складчастий, складочки рідкі, вузькі,
низько спускаються по ніжці, з чисельними анастомозами. М’якуш
щільний, неламкий, має приємний аромат та гострий смак.
Лисичка - один з найвідоміших їстівних грибів, який використову-
ють у свіжому або консервованому вигляді. Цікаво, що м'якуш цих
грибів майже ніколи не буває поточений хробаками або іншими без-
хребетними.
Плодові тіла лисички багаті на вітаміни РР та Вг мікроелементи,
у тому числі цинк та мідь.
Рід кратерел або чорна лисичка - СгаіегеІІиз. Плодові тіла
тонком’ясисті. Шапинка лійковидна, сіра, коричнева або чорна. Ніжка
з порожниною, здебільшого відкритою в центрі шапинки. Гіменофор
гладенький або зморшкувато-горбкуватий, кольору шапинки. Спори
безбарвні, гладенькі. Серед видів роду також є їстівні гриби, наприк-
лад СгаіегеІІиз сіпегеиз - кратерел попелясто-сірий (рис. 142 Б),
який має темно-сіру шапинку, котра при підсиханні світлішає. В центрі
лійка шапинки переходить у відкриту порожнину ніжки. Поверхня
363
шапинки волокнисто-кошлата. Гіменофор складчастий (з пластинко-
видними анастомозованими складочками), низько спускається по
ніжці, сірий. М’якуш із приємним фруктовим запахом. Спори гладенькі,
безбарвні. Рідкісний гриб, зростає у лісах, на ґрунті.
Рис. 142. Кантареляльні гриби: А - СапіІіагеІІиз', Б - СгаіегеІІиз
Рід гіднум - Нусіпит. Плодові тіла у вигляді шапинки з ніжкою,
м’ясисті, при висиханні крихкі. Тканина плодового тіла біла, згодом
кремова або жовтувата. Гіменофор шипуватий, спускається на ніжку.
Шипи білі, кремові або рудуваті. Спори безбарвні, в масі жовтуваті,
Рис.143. Нусіпипт. загальний вигляд
плодового тіла
гладенькі. Найпоширенішим
є Нусіпит герапсіит -
гіднум виїмчастий, або їжо-
вик жовтий (рис. 143).
Росте цей гриб на фун-
ті у хвойних, листяних та
мішаних лісах. Він має жов-
туватий м’якуш, який у моло-
дих плодових тіл солодкий, у
старих - пекуче-гіркий. У мо-
лодому віці плодове тіло їс-
тівне.
У складі групи афілофо-
роїдних гіменоміцетів деякі
родини або навіть порядки
не мають чітко визначеного
364
місця в системі базидіальних грибів. Дані молекулярно-біологічних
досліджень останніх років дещо суперечать класичній картині розпо-
ділу певних таксонів у межах відділу Вазібіотусоіа. Ці таксони з оста-
точно нез’ясованим положенням потребують подальшого поглибле-
ного вивчення. До таких таксономічних одиниць належить порядок
фістулінальних грибів.
Порядок Фістулінальні - Еівіиііпаїез
Плодові тіла гімнокарпні, однорічні, гіменофор трубчастий і, на
відміну від інших афілофороїдних гіменоміцетів, легко відділяється
від трами. Трубочки вільні, між собою не зростаються. Характерний
представник - печіночниця звичайна.
Печіночниця звичайна - Еізіиііпа ііераііса. Плодові тіла од-
норічні, у вигляді бічної, сидячої або з короткою ніжкою шапинки, на-
півкруглі, язиковидні. Шапинки соковиті, м’ясисті, потім сухуваті,
щільні, поодинокі або зростаються основами по 2-5 екземплярів,
криваво-червоні або м’ясо-червоні, з віком темно-бурі. Тканина
плодового тіла соковита (з червонуватим соком), з радіально розта-
шованими світлішими прожилками, кислувата на смак. Спори світло-
жовто-рожеві, гладенькі, яйцевидні, біля основи гостро звужені. Гіме-
нофор трубчастий. Трубочки на нижньому боці шапинки вільні (не
з’єднані між собою). Зростає печіночниця на живих стовбурах дерев,
переважно біля основи старих дубів та каштанів, спричиняє буру гниль
деревини, відому під назвою «бурий дуб». У природі зустрічається
рідко. В молодому віці гриб їстівний.
Не виключено, що фістулінальні гриби можуть бути перехідною
ланкою між афілофороїдними та агарикоїдними гіменоміцетами. Зок-
рема, за даними молекулярно-генетичних досліджень, фістулінальні
гриби дуже близькі до болетальних. Цікаво, що така схожість підтвер-
джується і морфологічними ознаками, зокрема за анатомічною будо-
вою гіменофору, трубочки якого легко відділяються від стерильної
частини карпофору.
Агарикоїдні гіменоміцети
Шапинкові гриби як фактор ризику
Агарикоїдних гіменоміцетів із загниваючими карпофорами -т. зв.
шапинкові гриби - у повсякденному житті називають грибами і части-
ну видів цієї групи традиційно використовують як цінний харчовий
продукт. З іншого боку, саме ця група містить переважну більшість
365
представників, які належать до небезпечних грибів, здатних спричи-
няти різноманітні отруєння. Грибні отруєння бувають первинними та
вторинними. Первинні пов’язані із вживанням у їжу тих видів, що
містять токсини. Вторинні - із вживанням у їжу їстівних видів, в яких,
проте, або розпочалися процеси розпаду білків, або вторинно нако-
пичилися токсичні речовини - отрутохімікати, важкі метали, радіоак-
тивні речовини тощо.
Отруйні гіменоміцети та їх токсини. Сьогодні відомо біля 90
видів агарикоїдних гіменоміцетів, які здатні викликати первинні от-
руєння різної тяжкості. Тяжкість отруєння залежить від двох основ-
них факторів: від типу токсину, який містить даний гриб, та кількості
отрути, яка потрапила в організм. Загалом при грибних отруєннях
простежується наступна тенденція: отруєння є тим тяжчим, чим дов-
ший латентний період, тобто часовий проміжок між споживанням
грибів та проявом перших симптомів отруєння.
За характером дії на людину та тривалістю латентного періоду
отруйні грибні умовно поділяють на чотири групи: а) гриби з локаль-
ною збуджуючою дією і коротким (до 0,5 год.) латентним періодом;
б) гриби із чітко окресленим впливом на нервові центри та середнім
латентним періодом (0,5-8 год.); в) гриби з чітко окресленою плаз-
молітичною дією та тривалим латентним періодом (8-72 год.); д) гри-
би з гемолітичною дією та дуже довгим латентним періодом (від
кількох тижнів до кількох років).
Гоиби з локальною збуджуючою дією містять різноманітні слаб-
ко токсичні речовини, які майже одразу після вживання спричинюють
нудоту, блювання, спітніння, пронос, головний та шлунковий біль.
Зазвичай хвороба швидко проходить без будь-яких негативних
наслідків. Прикладами грибів цієї групи є Воіеіиз заіапаз (сатанинсь-
кий гриб), Воіеіиз егуіїїгориз (синяк зернистоногий), Киззиіа етеііса
(сироїжка блювотна), Ьасіагіиз Ьеіуиз (хрящ-молочник звичайний),
СІііосуЬе пеЬиіагіз (говорушка сіра), І_ерізіа писіа (лепіста фіоле-
това).
Окремі групи грибів з локальною збуджуючою дією становлять
гриби, що містять коприн - токсин, який спричиняє отруєння при вжи-
ванні з алкоголем, та гриби, що містять галюциногенні речовини пси-
лоцин та псилоцибін.
Коприн виявлений у Соргіпиз аігатепіагіиз (гнойовик чорниль-
ний), С. тісасеиз (гнойовик мерехтливий), СІііосуЬе сіауірез (гово-
рушка булавиднонога), Воіеіиз Іигісіиз (дубовик оливково-бурий). При
вживанні з алкоголем коприн, що міститься у плодових тілах, розчи-
366
няється, всмоктується і блокує роботу ферменту алкогольдегідроге-
нази. Як наслідок, спирт, що надійшов в організм, окислюється не
повністю, в тканинах накопичуються альдегіди, які й обумовлюють
типову симптоматичну картину отруєння - шкіра червоніє і вкриваєть-
ся синюшними плямами, прискорюється серцебиття, починається
пронос та блювота. Через кілька годин функції організму відновлю-
ються. Але при повторному вживанні алкоголю симптоми отруєння
повторюються знов.
Галюциногенні речовини псилоцин та псилоцибін, що є фосфат-
ними ефірами похідних індолу, містять види родів РзііосуЬе (псило-
цибе) та Рапаеоіиз (панеол). Отруєння цими грибами при надходженні
в організм незначних доз токсинів (1-3 мг) викликає появу зорових та
слухових галюцинацій, відчуття сп'яніння. У більших дозах (10-20 мг)
виникає ілюзорний стан, втрата відчуття часу, відтворюються давно
забуті події. Проте у продуцентів псилоцибіну та псилоцину у плодо-
вих тілах наявні також алкалоїди беоцистин та норбеоцистин, які спри-
чинюють гіперсинтез серотоніну в корі головного мозку, що призво-
дить до розвитку патологічних процесів у мозку і спричинює психічні
захворювання.
Гоиби із чітко окресленим впливом на нервові центри містять
токсини, в яких летальна доза (Ю100) для людини становить від 0,1 г
і вище. Це, переважно, низькомолекулярні органічні сполуки - муска-
рин, мусцимол, буфотенін, іботенова кислота. Найчастіше симптоми
отруєння наступають через 0,5-2 год. і пов’язані з порушенням функцій
нейромедіаторів у нервових синапсах. Симптомами отруєння є по-
ява холодного поту, уповільнення серцебиття, зниження тиску, зву-
ження зіниць, задуха, різкий біль у шлунку. Антидотом при отруєнні
мускарин-подібними токсинами є атропін. Найвищий вміст мускари-
ну виявлений у ІпосуЬе раіоиіііагсііі (плютка Патуйяра), Атапііа
рапіїїегіпа (мухомор пантерний), СІіїосуЬе деаІЬаіа (говорушка білу-
вата) - 40-300 мг/кг плодових тіл (для людини летальна доза муска-
рину (Ю100) становить біля 500 мг). Споживання цих грибів навіть у
незначній кількості спричинює тяжкі отруєння, а у значній - отруєння
з летальними наслідками.
На один-два порядки менший вміст мускарин-подібних токсинів у
плодових тілах Атапііа тизсагіа (мухомор червоний) та Мусепа рига
(міцена чиста). Ці гриби спричинюють отруєння, які, проте, не при-
зводять до смерті людини.
У грибів із плазмолітичною дією виявлено дві групи токсинів:
а) циклічні поліпептиди - аматоксини та фалотоксини; б) ореланін-
подібні токсини (ореланін, гржималін, кортинарин).
367
Аматоксини (аманітин, аманін) та фалотоксини (фалоїдин) є
надзвичайно небезпечною групою токсинів, летальна доза яких для
людини складає 20-30 мг. Найвищий вміст цих токсинів мають пло-
дові тіла Атапіїа рііаііоісіез (бліда поганка), А. Уігоза(мухомор смер-
дючий), А. уегпа (мухомор білий). Ці три види належать до найне-
безпечніших грибів, споживання яких навіть у малих дозах майже
завжди призводить до смерті. Значно менша кількість цих токсинів
у карпофорах Нуріїоіота /азсісиїаге (несправжній опеньок сірчано-
жовтий), Н. зиЬІаіегіііит (несправжній опеньок цегляно-червоний),
Ьеріоіа Ьеіуеоіа (лепіота отруйна), Ьгиппео-іпсагпаіа (лепіота ко-
ричнево-червонувата). Отруєння цими грибами проходить важко, але
у багатьох випадках при своєчасній медичній допомозі людину вдаєть-
ся врятувати.
Симптоми отруєння ама- та фаллотоксинами з’являються лише
через кілька годин після надходження цих токсинів в організм (інколи
навіть через 2-4 доби). Протягом латентного періоду ніяких ознак от-
руєння не спостерігається, але в цей час токсини порушують регуля-
цію роботи окремих органів, ініціюють набряк та жирове переродження
тканин печінки, нирок, серця. Після закінчення латентного періоду
з’являються перші ознаки отруєння - зневоднення організму, си-
нюшність, блювота, пронос. Далі наступає параліч нервів, що регу-
люють роботу кровоносних судин. Стан хворого різко погіршується, і
майже завжди наступає смерть.
Механізми дії аматоксинів та фалоїдинів дещо відмінні. Зокрема,
фалоїдин діє швидше, і є менш токсичним порівнянно з аманітином.
Фалоїдин порушує функції натрій-калієвого насоса і зумовлює відтік
іонів калію, а також вихід ферментів лізосом у цитоплазму. Крім того,
фалоїдин полімеризує молекули актину, порушуючи тим самим функції
цитоскелета.
Аматоксини діють повільніше, проте є більш токсичними. Ці ток-
сини зв’язуються з РНК-полімеразою, яка забезпечує синтез т-РНК.
Як наслідок, порушується регуляція біосинтезу білків.
Ореланін-подібні токсини (ореланін, гржималін, кортинарин), що
виявлені у видів роду Согііпагіиз (павутинник) - Согііпагіиз огеїіапиз
(павутинник оранжево-червоний отруйний), С. зресіоззітиз (павутин-
ник найкрасивіший), подібно до аманітину та фалоїдину, є високомо-
лекулярними поліпептидами, дія яких схожа з наведеними вище ток-
синами. Проте при отруєнні павутинниками латентний період дуже
довгий - від 3 до 14 діб (найчастіше - 8-11 діб). Зафіксовано, що в
середньому 15% випадків отруєння павутинниками закінчуються
летально.
368
Гоиби з гемолітичною дією та дуже довгим латентним періодом
(від кількох тижнів до кількох років) представляє РахіІІиз іпуоіиіиз (сви-
нуха тонка). Свинуха містить антиген, що викликає в організмі утво-
рення антитіл, які поступово накопичуються у плазмі крові та руйну-
ють еритроцити. При регулярному споживанні свинух розвивається
жовтуха, анемія, зрідка хвороба закінчується смертю пацієнтів.
Слід відмітити, що простих тестів для виявлення отруйних грибів
не існує, і єдиним підґрунттям для профілактики отруєнь токсинами
гіменоміцетів є вміння ідентифікувати конкретні отруйні та їстівні види.
Найменшу кількість небезпечних видів містять представники поряд-
ку болетальних, найбільшу - агарикальних (у першу чергу, з родин
мухоморових, павутинникових та строфарієвих).
Порядок Болетальні - Воіеіаіеа
Плодові тіла гімнокарпні, однорічні, загниваючі. Гіменофор труб-
частий або пластинчастий з анастомозами. Щетинки в гіменофорі
відсутні. Базидії хіастичні, прості (холобазидії). Парентосоми з пер-
фораціями. Спори темнозабарвлені, гладенькі або з дрібними бо-
родавками. Утворюють ектотрофну мікоризу. Включає три основні
родини, що відрізняються, в першу чергу, за типом гіменофору та
наявністю несправжнього покривала.
Більшість видів порядку є їстівними. До отруйних грибів з локаль-
ною збуджуючою дією належать деякі види роду болет (сатанинсь-
кий гриб, синяк, дубовик оливково-бурий), а до небезпечних грибів із
гемолітичною дією - свинуха тонка. Представники з нейро- та гепато-
токсинами на території Європи не відомі.
Родина болетові - Воіеіасеае. Об’єднує представників з труб-
частим гіменофором. До найважливіших родів належать болет, баб-
ка, маслюк, моховик, тилопіл.
Рід болет - Воіеіиз. Має плодові тіла з м’ясистими шапинками
та товстими ніжками. Шапинка гладенька, суха, у вологу погоду лише
злегка клейкувата, її шкірка від м’якуша легко не відділяється. Ніжка
переважно з ніжним сітчастим візерунком, не луската. Гіменофор
дорослих карпофорів світло-жовтий, жовтий або червоний. Спорова
маса оливкового кольору. М’якуш білий, на зламі не змінює кольору
або жовтіє, синіє чи червоніє.
Всі представники цього роду належать до їстівних грибів. Проте
слід пам’ятати, що види, в яких м’якуш карпофорів на зламі змінює
369
колір, у сирому вигляді містять токсини локальної збуджуючої дії (на-
приклад, сатанинський гриб, синяк, дубовик оливково-бурий). Ці ток-
сини руйнуються при будь-якій термічній обробці. Види, в яких м’я-
куш на зламі не змінює кольору, можна вживати в їжу навіть у сирому
стані (наприклад, білий гриб).
Білий гриб або боровик - Воіеіиз есіиііз (рис. 144 А) - є од-
ним з найвідоміших та найпопулярніших їстівних грибів. Плодові тіла
великі, товстом’ясисті. Шапинка 6-20 (ЗО) см у діаметрі, у молодих
грибів напівсферична, у дорослих-опукло-розпростерта, коричнева з
різними відтінками, гладенька або злегка зморшкувата, гола та суха, в
сиру погоду трохи клейка. Гіменофор легко відділяється від м’якоті ша-
пинки. У молодих грибів він білий або світло-кремовий, згодом жовтіє.
Спорова маса оливкова, спори світло-коричневі. Ніжка світла (брудно-
біла або сірувата), у верхній частині з тонким білим сітчастим візерун-
ком. М’якуш щільний, білий, на зламі не змінює кольору, має приємний
запах і смак.
Росте білий гриб у листяних та хвойних лісах, утворюючи мікори-
зу з дубом, буком, грабом, березою, ліщиною, сосною, ялиною. Білий
гриб має лікарські властивості, містить алкалоїд герцинін, який влас-
тивий також столовим бурякам. Плодові тіла білого гриба (в першу
чергу - у вигляді свіжих грибних салатів) рекомендують вживати при
стенокардії, туберкульозі, зниженні загального тонусу, а також для
профілактики онкологічних захворювань.
Боровик зернистоногий або синяк - Воіеіиз егуіГігориз
(рис. 144 Б). За формою плодового тіла подібний до білого гриба,
проте добре відрізняється від нього забарвленням шапинки, ніжки та
гіменофору, а також зміною кольору м’якуша на зламі. Шапинка у си-
няка темно-коричнева. Гіменофор у молодих карпофорів жовто-чер-
вонуватий, у дорослих - темно-червоний або пурпуровий, при дотику
синіє. Спорова маса оливкова. Ніжка зерниста, жовта, з яскраво-чер-
воним візерунком у вигляді сітки або дрібних цяточок, при дотику синіє.
М’якуш дуже щільний, жовтий, на зламі швидко синіє, з приємним
запахом і добрий на смак.
Росте синяк у хвойних та листяних лісах і утворює мікоризу пере-
важно із сосною та буком. У сирому стані гриб отруйний, проте після
термічної обробки його вживають у їжу.
Сатанинський гриб - Воіеіиз заіапаз (рис. 144 В) - вельми
схожий із синяком. Відміни полягають, у першу чергу, в забарвленні
370
шапинки - у сатанинського гриба вона біла або світло-сіра. Дещо
відмінна також зміна кольору м’якуша на зламі - він спочатку злегка
червоніє, потім синіє. Ніжка має добре помітний червоний сітчастий
візерунок. Росте сатанинський гриб у листяних, рідше мішаних лісах.
У сирому вигляді плодові тіла містять токсини локальної збуджуючої
дії, які, проте, повністю руйнуються при відварюванні.
Польський гриб - Воіеіиз Ьасііиз (рис. 144 Г). Від інших видів
роду відрізняється однотонною темно-каштановою гладенькою ша-
пинкою, щільним м’якушем та коричнюватою ніжкою. Гіменофор жов-
туватих відтінків. Ніжка та гіменофор при натисканні забарвлюється
в синювато-зелений колір. М’якуш світло-жовтий, на зламі синіє, з
приємним запахом та смаком. Росте на піщаному ґрунті у хвойних та
мішаних лісах, де утворює мікоризу із сосною.
Рід бабка або лекцин - 1_ессіпит - об’єднує гриби, в яких
шапинка гладенька, шкірка не знімається. Ніжка циліндрична, волок-
ниста та луската. Гіменофор не приростає до ніжки, сірий, жовтува-
тий або світло-коричневий, з дуже довгими трубочками. Спорова маса
оливкового кольору. М’якуш білий, на зламі спочатку злегка рожевіє,
потім темніє або стає чорним. Всі види цього роду вважаються гарни-
ми їстівними грибами, проте, як і у всіх болетових грибів з темніючим
на зламі м’якушем, споживання сирих плодових тіл може викликати
шлункові розлади. Найвідомішими представниками роду є підосич-
ник та підберезовик.
Рис. 144. Плодові тіла болетових грибів: А - Воіеіиз есіиііз', Б - Воіеіиз
сгуііїгориз; В - Воіеіиз заіапаз; Г - Воіеіиз Ьасііиз', Д - Ьессіпит зсаЬгит
371
Підосичник або бабка червона - іессіпит аигапііасит -
легко впізнати за шапинками, які забарвлені у червоний або жовтога-
рячий колір. Поверхня шапинки гладенька. Ніжка біла, вкрита темни-
ми волокнистими лусками. М’якуш з приємним запахом та смаком,
на зламі спочатку стає рожево-оливковим, потім темніє. Росте підо-
сичник на ґрунті в листяних та мішаних лісах, переважно під осика-
ми, з якими утворює мікоризу.
Підберезовик або бабка темна - Ьессіпит зсаЬгит (рис. 144 Д)
- відрізняється від підосичника коричневими шапинками без черво-
них відтінків, а також тим, що утворює мікоризу переважно з березою.
Рід моховик або ксероком - Хегосотиз. Для видів цього роду
характерні плодові тіла середнього розміру. Шапинки у моховиків гла-
денькі, злегка повстисті або розтріскані. Ніжки зазвичай циліндричні
або звужені донизу, гладенькі, без сітчастих візерунків або волокнис-
тих лусок. Гіменофор забарвлений у жовті відтінки, його трубочки
злегка спускаються на ніжку, і при натисканні синіють. Спорова маса
оливково-бура. М’якуш жовтий, на зламі синіє. Всі види роду нале-
жать до їстівних грибів переважно низької якості.
Найпоширенішим видом є Хегосотиз сіїгузепіегоп - моховик
тріщинуватий. Він добре розпізнається за дуже розтрісканою поверх-
нею шапинки, великими порами гіменофору, що при дотику синіють,
та почервонінням м’якуша у місцях ушкодження личинками.
Рід маслюк - ЗиіІІиз. Має клейку шапинку, в якої шкірка легко
знімається. Ніжка гладенька, інколи з кільцем. Спорова маса оливко-
во-жовта. М’якуш жовтий, солодкий, на зламі або не змінює забарв-
лення, або злегка синіє. Характерний представник - маслюк звичай-
ний.
Маслюк звичайний - ЗиіІІиз Іиіеиз (рис. 145). Шапинка опукло-
розпростерта, жовто-коричнево-бура, у вологу погоду вкрита слизом,
клейка. Краї шапинки з’єднані з ніжкою білою щільною плівкою, яка з
віком розривається, утворюючи навколо ніжки кільце. Ніжка централь-
на, щільна, гладенька, жовто-коричнева. М’якуш білий, на зламі не
змінює кольору і має приємний смак та запах. Трубчастий шар ша-
пинки жовтий, при дотику не змінює кольору. Споровий порошок жов-
то-вохряний. Спори темно-жовті, гладенькі. Росте маслюк переваж-
но на піщаному ґрунті у соснових лісах і утворює мікоризу із сосною.
Належить до цінних їстівних грибів.
372
Рис. 145. ЗиіІІиз Іиіеиє. А - молоде плодове тіло; Б - зріле плодове тіло.
1 - несправжнє покривало; 2 - залишки несправжнього покривала у вигляді
кільця на ніжці
Рід тилопіл - Туїоріїиз. За зовнішнім виглядом та забарвлен-
ням шапинки дуже схожий на білий гриб. Проте відрізняється від ньо-
го, в першу чергу, рожевуватим кольором гіменофору. Рисунок на
ніжці коричневий і виразніший, ніж у білого гриба. Спорова маса ро-
жева. М’якуш білий, на зламі злегка рожевіє. В Україні зустрічаються
два види цього роду. Найпоширенішим є Туїоріїиз ГеІІеиз - жовч-
ний гриб, або гірчак. М’якуш у карпофорів цього виду дуже гіркий,
через що гриб є неїстівним. Гірчак росте у мішаних та хвойних лісах і
утворює мікоризу із сосною, дубом, грабом. Інший вид цього роду-
Т. аіиіагіиз - тилопіл солодкий - має солодкий м’якуш і є їстівним.
Тилопіл солодкий росте на лісових галявинах, на полонинах у Карпа-
тах. Цей вид є рідкісним і занесений до Червоної книги України.
Родина свинухові або паксилові - Рахіііасеае. Об’єднує
болетальні гриби із пластинчастим гіменофором, причому пластинки
низько спускаються на ніжку й анастомозують одна з одною (особли-
во біля ніжки), інколи з’єднуючись виразними перегородками. В ос-
танньому випадку на нижніх ділянках гіменофору наявні видовжені
пори, що розглядаються як видозмінені трубки трубчастого гімено-
фору. Гіменофор завжди відкритий, несправжні покривала відсутні,
поверхня шапинки суха і забарвлена в темно-жовтий або коричне-
вий колір. Трама пластинок білатеральна. Провідний рід - свинуха.
373
Рід свинуха або корбан - РахіІІиз - об'єднує неїстівні та от-
руйні гриби. Вживання сирих плодових тіл свинухи викликає швидке
отруєння, спричинене токсинами з локальною збуджуючою дією. Після
відварювання та видалення відвару плодові тіла вважаються умовно
їстівними. Проте принаймні у найпоширенішого виду - свинухи тон-
кої - карпофори навіть після відварювання проявляють гемолітичну
дію і, таким чином, зберігають отруйні властивості.
Свинуха тонка - РахіІІиз іпуоіиіиз (рис. 146) - має увігнуту лійко-
подібну темно-жовту або коричневу гладеньку та злегка клейкувату
шапинку, часто досить великих розмірів (до 15-20 см у діаметрі),
краї якої підігнуті донизу. Пластинки гіменофору також темно-жовті,
товсті, з чисельними анастомозами, спускаються на ніжку, яка дещо
світліша від шапинки. М’якуш буруватий, на зламі не змінюється, має
приємний смак та слабкий запах гнилої деревини. Спорова маса ко-
ричнева. Спори охристо-бурі, гладенькі. Росте свинуха тонка на ґрунті
у хвойних та листяних лісах.
Довгий час гриб вважався умовно їстівним, але зараз його от-
руйність доведено. Тривале вживання в їжу свинухи тонкої призво-
дить до тяжких отруєнь, особливо у людей із хворими нирками.
Рис. 146. РахіІІиз іпуоіиіиз: А — загальний вигляд плодового тіла; Б - розріз
через плодове тіло; В - спори
374
Родина мокрухові або гомфідієві - БотрНісІіасеае. Вклю-
чає болетальні гриби з пластинчастим гіменофором (анастамози між
пластинками виражені нечітко), пластинки якого спускаються на
ніжку. В молодих карпофорах гіменофор закритий плівчастим не-
справжнім покривалом, яке утворено клейкими гіфами шкірки шапин-
ки. Шкірка легко відділяється від шапинки і також клейка. Трама пла-
стинок білатеральна. Всі представники родини є їстівними. Провідний
рід - мокруха.
Рід мокруха - СотрМсііиз (рис. 147). У видів цього роду пло-
дові тіла м’ясисті, досить великі. Шапинки клейкі і на поверхні мають
тонкий шар слизу. Краї шапинки з’єднуються з ніжкою слизисто-па-
вутинистим несправжнім покривалом, від якого пізніше на ніжці зали-
шається слизьке невиразне кільце. Пластинки гіменофору товсті, вос-
кової консистенції, рідкі, спочатку світлі, згодом темні (переважно сірі
або коричневі). Спорова маса майже чорна. Спори темно-бурі, гла-
денькі. Всі види мокрух утворюють мікоризу із хвойними деревами.
Добрим їстівним грибом є Оотрїіісііиз діиііпозиз - мокруха клейка.
Порядок Агарикальні - Адагісаіез
Плодові тіла гімнокарпні або геміангіокарпні, однорічні, загнива-
ючі; гіменофор пластинчастий, без сфероцист; щетинки в гіменофорі
відсутні; базидія хіастична, проста (холобазидія); парентосоми з пер-
фораціями; спори різноманітні,
Рис. 147. Оотріїісііиз: А - зріле
плодове тіло; Б - молоде плодове
тіло; В - спори
але без сітчастої орнаментації. Ґрун-
тові сапротрофи, ксилотрофи або
мікоризні гриби. Триптофан синте-
зується за допомогою другої гру-
пи ферментів (на відміну від інших
порядків).
За ступенем однорідності гіф
у ніжці та шапинці агарикальні гри-
би поділяють на тих, у яких шапка
та ніжка гомогенні (утворені одно-
типними скелетними гіфами, які з
ніжки переходять у шапинку), та
тих, у яких вони гетерогенні. Заз-
вичай гомогенність та гетероген-
ність добре розрізняються навіть
неозброєним оком на поздовжніх
зрізах карпофорів.
375
Загальне покривало в агарикальних може бути відсутнім або
наявним. В останньому випадку загальні покривала поділяють
на: а) стійкі - якщо його залишки зберігаються тривалий час, утворю-
ючи при основі ніжки мішковидний залишок - вольву (піхву), а на
шапинці - пластівці або лусочки; б) нестійкі - якщо його залишки швид-
ко зникають у дорослих карпофорів.
Часткове покривало, коли воно наявне, складається з більш-менш
щільної плектенхіми, і його залишки, як правило, зберігаються у ви-
гляді кільця на ніжці. У деяких агарикальних часткове покривало тон-
ке і павутинчасте, утворене легко зникаючим плетивом - кортиною.
Різні роди та родини агарикальних мають різну анатомічну бу-
дову стерильної тканини, що знаходиться у пластинці між поверх-
невими шарами її гіменія. Цю тканину називають трамою пласти-
нок. Розрізняють наступні основні типи трам: білатеральна - гіфи
трами розміщені більш-менш симетрично в напрямку від центру
пластинки вперед до периферії; інверсна - гіфи трами розміщені
більш-менш симетрично в напрямку від центру пластинки назад до
периферії; правильна - паралельно вздовж пластинки; неправиль-
на - гіфи трами розміщені без певного порядку; ділянки правильної
та неправавильної трами в межах однієї пластинки розглядають як
траму мішану.
В агарикальних виділяють чотири основних типи забарвлення ба-
зидіоспор: а) безбарвні або світло забарвлені; б) рожевуваті; в) тем-
но-жовті та коричневі; г) темно-фіолетові та чорні. За формою бази-
діоспори поділяють на багатокутні та заокруглені (в останніх також
виділяють додаткові категорії - циліндричні, еліпсоїдні, субсферичні
та сферичні). За наявністю інкрустацій оболонок виділяють гладенькі
та бородавчасті. Із забарвленням базидіоспор зазвичай корелює за-
барвлення пластинок та спорової маси.
Наведені вище ознаки є провідними при поділі агарикальних на
родини. Система порядку розглядається на прикладі 10 родин, до
яких входить переважна більшість їстівних та отруйних агарикальних
грибів.
Родина ентоломові або рожевопластинникові - Епіо-
Іотаіасеае. Плодові тіла гімнокарпні, без вольви та часткового по-
кривала. Шапинка та ніжка гомогенні. Трама пластинок правильна.
Спори з рожевим відтінком, багатокутні, з товстою оболонкою. Спо-
рова маса також рожева. Містить як їстівні, так і отруйні гриби. Прак-
тичне значення у межах цієї родини мають лише види роду ентоло-
376
ма, або рожевопластинник (Епіоіота), причому тільки ті, в яких
плодові тіла досить великі (наприклад, в отруйного рожевопластин-
ника жовтувато-сизого або їстівного рожевопластинника садового).
Рожевопластинник жовтувато-сизий отруйний - Епіоіота
зіпиаіит (рис. 148). Плодові тіла великі, 6-15 см у діаметрі. Шапин-
ка товста, м’ясиста, опукло-розпростерта, з рівним опушеним краєм,
дещо клейкувата, при всиханні блискуча, має брудно-біле, кремово-
сіре або жовтувато-сіре забарвлення. Ніжка циліндрична, значно ко-
ротша за діаметр шапинки, біля основи іноді бульбовидно потовще-
на, біла. М’якуш білий, щільний, свіжий з приємним смаком та арома-
том борошна. Пластинки прирослі, широкі, рожеві. Гіменіальний шар
із цистидами. Спорова маса рожева. Спори п’ятикутасті, товстостінні,
рожеві. Росте на ґрунті у широколистяних лісах. Смертельно отруйний
гриб, містить високі концентрації нейротоксинів мускаринової групи.
Рис. 148. Епіоіота зіпиаіит'. А- зовнішній вигляд плодового тіла; Б - плодо-
ве тіло в розрізі; В - базидіоспори
Рожевопластинник садовий - Епіоіота сіуреаіит - дуже
подібний до попереднього виду, від якого відрізняється, в першу чер-
гу, за місцем зростання: він зазвичай росте у садах і утворює мікори-
зу з вишнею та іншими кісточковими деревами.
Родина гігрофорові - Нудгорїіогасеае. Плодові тіла гімнокарпні,
без вольви та часткового покривала. Шапинка та ніжка гомогенні.
Трама пластинок різноманітна - білатеральна, правильна або не-
правильна. Спори безбарвні, округлі, гладенькі або злегка пунктиро-
377
вані. Спорова маса біла. Характерні особливості - наявність великих
базидій (їх довжина більше ніж у десять разів перевищує довжину
спор), товстих пластинок та яскравого забарвлення карпофорів. От-
руйних видів у межах родини немає, хоча смакові якості цих грибів
низькі. Провідний рід родини - гігрофор (Нудгорїюгиз).
Родина трихоломові - Тгісїіоіотаїасеае. Плодові тіла гімно-
карпні або геміангіокарпні, без вольви та переважно (але не завжди)
без часткового покривала. Шапинка та ніжка гомогенні. Трама плас-
тинок правильна або неправильна. Спори безбарвні або світлозабарв-
лені, округлі, гладенькі або бородавчасті, але не кутасті. Спорова маса
біла або блідозабарвлена. Від близької родини гігрофорових
відрізняється, в першу чергу, нормальними розмірами базидій. До
родини входить чимало їстівних та деякі отруйні види. Багато пред-
ставників через незначні розміри практичного значення не мають.
Найпоширенішими та найвідомішими родами є міцена, маразміус, го-
ворушка, трихолома, опеньок.
Рід міцена - Мусепа- об’єднує гриби з дуже дрібними гімнокар-
пними карпофорами. Пластинки у видів роду прирослі, шапинки пе-
реважно дзвоникоподібні або розпростерті, проте з горбочком у центрі.
Ніжка тонка, переважно капіляроподібна. Майже всі види міцен є
підстилковими сапротрофами.
Рис. 149. Мусепа Ьает-
аіорода: А-загальний вигляд пло-
дового тіла; Б - спори
Міцена чиста - Мусепа рига -
є одним з найбільших видів роду за
розміром карпофорів. Діаметр її ша-
пинки може сягати 7 см. Шапинка за-
барвлена у бузково-рожевий колір, гла-
денька, з рубчастим краєм. Ніжка од-
ного кольору із шапинкою, напівпро-
зора, гладенька. М’якуш тонкий, з не-
приємним запахом та смаком гнилої
картоплі. Пластинки широкі, злегка ро-
жевуваті. Росте гриб на ґрунті в листя-
них та хвойних лісах. Гриб є отруйним,
містить мускарин-подібний токсин.
У деяких видів плодові тіла на
зламі виділяють червоний молочний
сік (наприклад, у М. їіаетаіоросіа -
міцени кривавоногої, яка росте на
гнилій деревині (рис. 149).
378
Рід маразміус - Магазтіиз - також має дрібні гімнокарпні
плодові тіла. їх особливістю є здатність переходити у стан анабіозу
при відсутності вологи і поновлювати ріст при настанні сприятливих
умов. Шапинка у видів роду опукла або дзвоникоподібна, часто руб-
часта. Ніжка тонка, капіляровидна, з порожниною. Пластинки приро-
стають до ніжки. Найбільш відомим видом є опеньок лучний.
Опеньок лучний - Магазтіиз огеадез (рис. 150). Шапинка може
сягати 5-8 см у діаметрі, спочатку опукла, згодом плоска, з бугорком,
гладенька, зі слабо рубчастим краєм, забарвлена у світлі кремові
відтінки. Ніжка щільна, одного кольору із шапинкою. М’якуш білий, на
зламі кольору не змінює, з приємним запахом та смаком. Пластинки
прирослі до ніжки, рідкі, білі або світло-кремові. Споровий порошок
білий. Спори гладенькі, безбарвні. Дуже поширений вид в Україні. Він
зростає поодиноко або великими групами переважно на відкритих тра-
в'янистих місцях, зокрема на пасовищах, на лісових галявинах, на лу-
ках, у парках. Часто утворює т. зв. «відьмині кільця». їстівний гриб ви-
соких смакових якостей. Містить ряд цінних лікарських речовин.
Рис. 150. Магазтіиз огеасіез:
зовнішній вигляд плодових тіл
Деякі види роду характери-
зуються різким часничним запа-
хом (наприклад, Магазтіиз
аіііасеиз - часничник вели-
кий). Такі гриби, незважаючи на
їх дрібні розміри, також нале-
жать до їстівних і використову-
ються як ароматична приправа
до страв.
Рід говорушка - СІИосуЬе.
Види цього роду мають великі
або середні за розміром гімно-
карпні плодові тіла з пластинча-
стим гіменофором, що спус-
кається на ніжку. Серед видів із
великими плодовими тілами є як
їстівні, так і отруйні. Найбільш
небезпечні види роду мають
карпофори, забарвлені у білий
колір (наприклад, говорушка
біла).
379
Говорушка біла або клітоцибе восковий - СІііосуЬе сеггизаіа
(рис. 151). Шапинка у цього гриба товстом’ясиста, лійкоподібна, з віком
плоско-розпростерта, з тупим горбочком у центрі, біла, блискуча, гола.
Пластинки білі, злегка спускаються на ніжку. Ніжка циліндрична, біла,
гола, щільна, пізніше в нижній половині з порожниною. Спори широко-
овальні. М’якуш білий, з приємним грибним запахом. Зростає пере-
важно у листяних та хвойних лісах, на ґрунті, досить часто утворює т.
зв. «відьмині кільця». Гриб отруйний, містить токсин мускарин.
Рис. 151. СІііосуЬе сеггизаіа: А - загальний вигляд плодового тіла; Б -
розріз через плодове тіло; В - спора
Рід трихолома - Тгісіюіота. Види роду також мають великі або
середні за розміром гімнокарпні карпофори з пластинчастим гімено-
фором, але, на відміну від говорушки, пластинки прирослі і не спус-
каються на ніжку. До цього роду належать біля десяти видів їстівних
та три види помірно отруйних грибів, а також біля двадцяти видів, що
не мають харчового значення. Найкращі смакові властивості має зе-
ленушка.
Трихолома жовтозелена або зеленушка - Тгісіюіота Лауо-
уігвпз (рис. 152). Добре розпізнається за жовтувато-зеленим або сіру-
вато-зеленим забарвленням шапинки, гіменофору та ніжки. Трама
пластинок правильна. Споровий порошок білий. Спори безбарвні,
гладенькі. М’якуш спочатку білий, пізніше жовтуватий, з приємним
м’яким смаком та запахом борошна. Росте зеленушка на піщаному
фунті у хвойних лісах, плодові тіла напівзаглиблені у ґрунт. Від-
різняється від усіх інших пластинчастих грибів тим, що зберігає свій
зелений колір навіть після відварювання.
380
Рис. 152. ТгісЬоІота ї^о-
VІгеп8. А - загальний вигляд пло-
дового тіла; Б - спори
Рис. 153. АгтіІІагіа теїіеа:
група плодових тіл на деревині
Рід опеньок - АгтіІІагіа -
один з небагатьох родів трихоломо-
вих грибів, що має часткове покри-
вало, залишки якого на зрілих кап-
рофорах зберігаються у вигляді
кільця на ніжці, тобто плодове тіло є
геміангіокарпним. Більшість видів є
сапротрофами, які розвиваються на
відмерлій деревині, або факультатив-
ними паразитами. Деякі види нале-
жать до добре відомих їстівних грибів,
зокрема опеньок осінній.
Опеньок осінній - АгтіІІагіа
теііеа(рис. 153). Плодові тіла серед-
нього розміру, рідше великі, з част-
ковим покривалом. Шапинка напівку-
ляста, згодом плоско-розпростерта,
іноді з горбочком у центрі, з тонким
краєм, рудувато-коричнювата, тем-
нолуската. Шкірка легко знімається.
Пластинки густі, тонкі, білуваті, при-
рослі, іноді злегка спускаються на
ніжку. Трама правильна. Спорова ма-
са кремова. Спори безбарвні, еліп-
соїдні, гладенькі, неамілоїдні. Ніжка
центральна, циліндрична або злегка
потовщена донизу, щільна, волокни-
ста, пластівчастолуската, з часом
майже гола, кольору шапки або
світліша, біля основи жовтувата, гу-
стоволокниста або повстиста. Кільце
на ніжці білувате, з віком здебільшо-
го зникає. Довгі чорні ризоморфи роз-
виваються під корою уражених де-
рев. М’якуш білий, з приємним гриб-
ним запахом. їстівний гриб, зростає
переважно великими групами у лис-
тяних та хвойних лісах. Паразитує на
багатьох видах дерев та чагарників,
належить до небезпечних дереворуй-
нуючих грибів.
381
Родина гливові, або плевротові, - Ріеигоіасеае - за гімно-
карпним розвитком плодових тіл, гомогенними шапинкою та ніжкою,
трамою, спорами та споровою масою повторює ознаки трихоломо-
вих. Проте ніжка у гливових або ексцентрична, або взагалі відсутня, і
тоді плодові тіла консолевидні. Гриби цієї родини є ксилотрофами,
розвиваються на мертвій деревині або на стовбурах та гілках ослаб-
лених дерев. Сапротрофний спосіб життя, не пов’язаний з необхідн-
істю утворювати мікоризу, та високі смакові якості карпофорів окре-
мих видів обумовили вибір деяких видів гливових у якості об’єктів про-
мислового грибівництва. Серед них найбільш популярним видом є
плеврот черепичастий, або глива.
Глива або плеврот черепичний - РІеигоіиз озігеаіиз (рис.
154). Карпофори гімнокарпні, сидячі, ніжка відсутня. Шапинка аси-
метрична, мушле-, вухо-, віялоподібна, широковоронковидна, гла-
денька, темнозабарвлена, сірувата, сіро-бура, фіолетово-коричнева,
із сизим відтінком. М’якуш білий, спочатку м’який, пізніше жорсткува-
тий, з приємним запахом та смаком. Пластинки часті, білі, внизу інко-
ли з анастомозами. Споровий порошок білий. Глива - типовий кси-
лотроф, зростає на деревині листяних та хвойних порід, часто утво-
рює зростання до ЗО та більше карпофорів, нагадуючи черепицю,
їстівний гриб з гарними смаковими якостями. Вирощується у промис-
лових масштабах у багатьох країнах світу, в тому числі в Україні.
Рис. 154. РІеигоіиз озігеаіиз: загаль-
ний вигляд плодових тіл
Існує декілька методів виро-
щування гливи. Найпоширеніши-
ми є два: екстенсивний та інтен-
сивний.
Екстенсивний спосіб виро-
щування. Передбачає вирощу-
вання грибів у природних умо-
вах. Його недоліком є сезонність
збору плодових тіл і залежність
величини врожаю від кліматич-
них умов. Технологія вирощуван-
ня грибів екстенсивним спосо-
бом не вимагає великих затрат.
При цьому використовують
відходи лісозаготівельної про-
мисловості (низькоякісна дере-
вина, пеньки, стружка, кора тощо).
382
Глива звичайна може рости на стовбурах багатьох листяних де-
рев, однак найкращими для неї є різні види тополі, верби, граба, бука
й дуба. На листяних породах з м’якою деревиною (тополя, верба,
граб) міцелій гливи розростається швидше, але врожайність її ниж-
ча, ніж на деревах з більш твердою деревиною (бук, дуб), на яких
грибниця розвивається повільніше.
Для культивування гливи найкраще використовувати свіжо зру-
бану деревину, яка містить достатню кількість вологи для розвитку
міцелію гриба. При використанні давно зрубаної деревини її слід вимо-
чувати у воді протягом тижня. Розрізати стовбур на бруски (обрубки)
треба в день інокуляції або напередодні. Оптимальний їх діаметр -
30-40 см, висота брусків повинна становити 30-35 см. Після цього
бруски інокулюють посівним міцелієм. У природних умовах глива зви-
чайна плодоносить наприкінці вересня - у жовтні. Тому обрубки, іно-
кульовані міцелієм, переносять у кінці серпня для плодоношення на
лісові галявини, де достатньо вологи і немає прямих сонячних про-
менів. Бруски висаджують у ґрунт, прикопуючи на 10-12 см.
Екстенсивним способом гливу звичайну можна культивувати і на
пеньках. Для інокуляції використовують пеньки листяних порід тільки
в рік рубки, бажано зимово-весняної. Найкращий діаметр їх 40-70 см.
Посівний міцелій наносять у травні-червні на заболонну частину по-
верхні пенька, попередньо зрізавши з нього диск товщиною 3-5 см.
Після посіву диск прибивають двома цвяхами. Норма витрати міце-
лію - 70-100 г на один пеньок.
Інтенсивний спосіб вирощування. Передбачає культивування
плодових тіл гливи у спеціальних приміщеннях, де є можливість регу-
лювання умов мікроклімату. Перевагою цього способу над екстен-
сивним є те, що процес вирощування проводиться круглорічно; уро-
жайність більш висока і стабільна; використовується більша кількість
субстратів з ряду целюлозо- і лігніновмісних відходів сільського гос-
подарства і промисловості у зв’язку з наявністю в технологічному про-
цесі фази їх теплової обробки; більш короткий виробничий цикл, рівний
8-10 тижням; можливість застосування механізації і автоматизації тех-
нологічних процесів.
Традиційним субстратом для вирощування плодових тіл гливи
звичайної є солома злакових культур: пшениці, жита, ячменю, вівса,
проса. Широко використовуються також подрібнені стрижні і качани
кукурудзи, рисова солома, відходи бавовнопереробної промисловості.
Підготовка субстрату передбачає його часткову стерилізацію за-
мочуванням, ступінчастою термічною обробкою або ферментацією.
383
Далі оброблений субстрат набивають у мішки, касети та ін. з
одночасним внесенням посівного міцелію в кількості 3-5% від маси
субстрату. Субстрат після сівби необхідно оберігати від висихання,
найкраще для цього підходить перфорована поліетиленова плівка.
Найчастіше для вирощування гливи звичайної використовують про-
зорі поліетиленові мішки на 15-30 кг субстрату.
Після інокуляції (посіву) мішки чи касети розміщують у камерах
для росту міцелію. Температура повітря в них становить 22-24°С,
вологість-60-65%, вентиляція -1-2 об’єми за годину. Оскільки світло
сповільнює ріст грибниці гливи, приміщення повинно бути темним.
У випадку використання мішків їх складають один на інший по чотири
в ряд, а касети встановлюють попарно, залишаючи проходи 1,5-2 м.
Залежно від виду субстрату, що використовується, його обростання
міцелієм триває 10-20 діб.
Родина печерицеві або агарикові - Адагісасеае. Плодові тіла
геміангіокарпні, завжди без вольви. Гіменофор до моменту дозріван-
ня базидіоспор закритий частковим покривалом, залишки якого збері-
гаються у вигляді кільця на ніжці. Шапинка та ніжка гетерогенні.
Трама пластинок правильна або неправильна. Спори - від майже
безбарвних до коричневих, гладенькі, округлі. Спорова маса-від без-
барвної до темно-коричневої, проте без фіолетових відтінків. Родина
об’єднує як ґрунтових сапротрофів, так і мікоризоутворюючі гриби.
Найвідомішими родами є печериця та макролепіота.
Рід печериця - Адагісиз. Різні види печериць добре відрізня-
ються від інших агарикових грибів кольором пластинок (у молодих
карпофорів - рожевуваті, у дорослих - темно-коричневі), темно-ко-
ричневим споровим порошком, темними спорами та збереженням
залишків часткового покривала у вигляді прирослого до ніжки кільця.
Ростуть печериці на багатому органічними речовинами лісовому
та лучному перегної. Найчастіше карпофори цих грибів зустрічають-
ся на галявинах, уздовж доріг, у лісових полезахисних смугах, пар-
ках, садах, на пасовищах. Більшість видів печериць не утворюють
мікоризи, що робить їх зручним об’єктом промислового грибівництва.
Зокрема, у промислову культуру більш ніж у 70-ти країнах світу вве-
дено печерицю двоспорову. Деякі види (наприклад, печериця зви-
чайна) належать до цінних їстівних грибів, які поки що у промисло-
вому масштабі не культивуються і збираються лише у природних
місцезростаннях. Відомі також отруйні представники (наприклад,
печериця рудіюча).
384
Печериця двоспорова - Адагісиз Ьізрогиз (рис. 155) - має
карпофори з центральною ніжкою та шапинкою. Спочатку шапинка
напівкуляста, згодом опукло або плоско розпростерта, брудно-біла
або світло-коричнювата, гола або з малопомітними притиснутими лу-
сочками, блискуча, шовковиста. Пластинки вільні, широкі, густі, у мо-
лодого гриба блідо-рожеві, згодом темнішають і набувають темно-
бурого, майже чорного кольору. Трама пластинок спочатку правиль-
на, згодом неправильна. Ніжка гладенька, циліндрична, щільна або з
порожниною та із залишками часткового покривала. М’якуш білий, на
зламі злегка червоніє, з приємним запахом та смаком. Базидії одно-
клітинні, прості, булавоподібні і, на відміну від інших видів роду, не-
суть лише по дві базидіоспори.
Печериця двоспорова культивується вже близько 300 років. Ос-
новним середовищем її культивування є різні компости, які готують з
тих матеріалів, що задовольняють потреби нормального росту та
розвитку міцелію печериці. Компост повинен містити компоненти
органічних речовин загального та білкового азоту, фосфору, магнію
та ін. На перших етапах культивування печериці у штучних екосисте-
мах часто використовували компост із 15-процентним вмістом кінсько-
го гною (такий компост називається натуральним). Він може мати ви-
сокий або низький вміст соломи і, відповідно до цього, бути світлим
або темним. Для кращої ферментації співвідношення гною та соломи
повинно становити 4:1. Для певної структури компосту та регуляції
рН до нього додають гіпс (26-30 кг/т гною).
Крім натурального компосту, нині у спеціалізованих господар-
ствах, де вирощують печериці, готують «напівсинтетичні» та «синте-
тичні» компости із гною або курячого посліду з домішкою соломи.
Рис. 155. Адагісиз Ьізрогиз: А-В - зовнішній вигляд плодового тіла;
Г - розріз через пластинку; Д - фрагмент трами та гіменію з двоспоровими бази-
діями. 1 - цистиди; 2 - базидії
385
Весь процес включає такі послідовні етапи: ферментація вихідних ком-
понентів; формування буртів для ферментації печеричного компосту
(декілька фаз); інокуляція компосту міцелієм (посівним матеріалом -
грибницею); нанесення покривного ґрунту (найчастіше торфового);
збір грибів під час трьох-шести «хвиль» утворення плодових тіл про-
тягом 50-65 днів при вологості приміщень біля 85-90% та темпера-
турі 15-16°С.
Печериця степова - Адагісиз сатрезігіз, яка масово зростає
на полях та луках і часто утворює «відьмині кільця», за смаковими
якостями та зовнішнім виглядом карпофорів (за винятком білого за-
барвлення шапинки) подібна до печериці двоспорової. Найсуттєвіші
відміни полягають у наявності чотириспорових базидій.
Печериця рудіюча - Адагісиз хапіїїодегтиз - представляє
отруйні види роду. Вона за забарвленням подібна до їстівної пече-
риці степової. Проте її шапинка при дотику жовтіє, м’якуш також жов-
туватий і має неприємний запах карболової кислоти. Карпофори цього
гриба містять мускарин-подібні токсини, що можуть спричинювати
тяжкі отруєння, проте без летальних наслідків.
Рід макролепіота - Масгоіеріоіа. Карпофори мають великі роз-
міри - до 40-50 см заввишки, із шапинками, діаметр яких може сяга-
ти 50 см. Пластинки завжди білі, не змінюють забарвлення навіть при
старінні. Споровий порошок також білий. Характерною ознакою є збе-
реження залишків часткового покривала у вигляді кільця, що вільно
рухається по ніжці. Ніжка при основі бульбовидно потовщена. Всі види
роду є маловідомими гарними їстівними грибами. Вони зустрічаються
переважно на відкритих місцях - галявинах, узбіччях доріг, у садках,
розріджених акацієвих насадженнях. Один із видів - М. риеііагіз -
макролепіота дівоча - занесена до Червоної книги України. Найпо-
ширенішим видом є макролепіота строката.
Макролепіота строката або парасолька велика - Масгоіеріоіа
ргосега (рис. 156). Карпофори геміангіокарпні, досить великих
розмірів. У молодих грибів шапинка яйцевидна, конусовидна, у до-
рослих - розпростерта, парасолеподібна, з тупим горбочком, з кон-
центричними зонами з темних прилеглих лусок. У центрі коричнева,
гола, до краю світліша, повстисто-волокниста. Пластинки вільні, ши-
386
рокі, густі, білі. Трама пластинок правильна. Ніжка циліндрична, біля
основи з бульбою, спочатку коричнева, згодом сірувата, пізніше кор-
кувата, з порожниною та із широким подвійним нестійким кільцем,
яке швидко опадає. М’якуш білий, з віком дуже щільний, у ніжці корку-
ватий. Страви, виготовлені із шапинок, обсмажених у яйці, вважають-
ся делікатесом.
Рис. 156. Масгоіеріоіа рго-
сега: загальний вигляд плодово-
го тіла
Родина мухоморові або аман-
ітові - Атапііасеае. Плодові тіла ге-
міангіокарпні, із загальним покрива-
лом, залишки якого зберігаються при
основі ніжки у вигляді стійкої вольви
(піхви), інколи також на поверхні ша-
пинки у вигляді білих пластівців. Част-
кове покривало є, рідше відсутнє. Ша-
пинка та ніжка гетерогенні. Трама пла-
стинок білатеральна. Спори безбарвні,
гладенькі, переважно з однією вели-
кою краплиною зеленкуватої олії. Спо-
рова маса світла. Провідним є рід ама-
ніта, або мухомор.
Рід аманіта або мухомор -
Атапііа. Серед усіх грибів цей рід вва-
жається найнебезпечнішим, оскільки
містить найбільшу кількість отруйних
та смертельно отруйних видів. Харак-
терними ознаками роду є наявність
загального та часткового покривал, білого (як виняток-лимонно-жов-
того) гіменофору з вільними, не прирослими до ніжки пластинками.
Ніжка при основі бульбовидно потовщена. Трама білатеральна.
Всі види роду утворюють ектотрофну мікоризу з багатьма дерев-
ними породами.
Три види роду - бліда поганка, білий мухомор та мухомор смер-
дючий - містять токсини з групи циклічних поліпептидів з плазмолі-
тичною дією і належать до найнебезпечніших смертельно отруйних
грибів мікофлори України. Чимало інших видів (наприклад, мухомор
пантерний) належать до високоотруйних грибів. Деякі (зокрема мухо-
мор червоний) є помірно отруйними грибами. Відомі також і їстівні
представники (зокрема мухомор цезарів).
387
Бліда поганка або мухомор зелений - Атапііа рНаІІоідез
(рис. 157). Карпофори геміангіокарпні. Шапинка зеленувата, оливку-
вата, кольору бронзи, до центру темніша, гола. Пластинки завжди
білі, густі, тонкі. Трама пластинок білатеральна. Ніжка до основи по-
ступово потовщується, біля основи з бульбовидним розширенням,
білувата, з порожниною. На ніжці, у верхній частині, завжди є широ-
ке, зверху гладеньке, жовтувате, із внутрішнього боку рубчасте біле
кільце - залишок часткового покривала. Біля основи ніжки у вигляді
білої піхви зберігається залишок загального покривала. Він вільний,
широкий, з нерівним лопатевим краєм. Споровий порошок білий.
Спори гладенькі, безбарвні, широкоеліпсоїдні. М’якуш білий, солод-
кий, з неприємним запахом. Росте бліда поганка на ґрунті у листя-
них, переважно дубових, букових з ліщиною та мішаних лісах, зазви-
чай групами.
Бліда поганка є смертельно отруйним грибом, який містить плаз-
молітичні токсини - аматоксини (аманітин, аманін) та фалотоксини
(фалоїдин), що представляють надзвичайно небезпечну групу ток-
синів. Для людини летальна доза цього гриба складає 2-3 г плодово-
го тіла.
Блідою поганкою найчастіше отруюються, помилково приймаючи
її за їстівну зеленушку, хоча зеленушка від блідої поганки візуально
добре відрізняється відсутністю кільця та вольви, а також жовтувато-
Рис. 157. Атапііа рііаііоісіеа: А-В - стадії розвитку плодового тіла; Г - розріз
через пластинку. 1 - загальне покривало; 2 - часткове покривало; 3 - залишки
загального покривала; 4 - залишки часткового покривала; 5 — білатеральна трама
388
зеленим кольором пластинок. Проте обидва гриби часто ростуть по-
руч, особливо на піщаних ґрунтах. У цих умовах більша частина ніжки
блідої поганки може залишатись зануреною у ґрунт, включаючи част-
кове покривало. Тому при зборі трапляються випадки, коли грибни-
ки, навіть досвідчені, зрізаючи лише шапинку, не помічають ознак
геміангіокарпного плодового тіла. З випадками отруєнь блідою по-
ганкою в деяких регіонах пов’язана традиція збирати гриби, не зріза-
ючи, а викручуючи їх із ґрунту разом з основою ніжки. При такому
способі ймовірність помилкового збору гриба з вольвою та частко-
вим покривалом різко зменшується.
Мухомор білий - Атапііа уегпа - відрізняється від блідої
поганки білим забарвленням шапинки та ніжки, що знайшло відобра-
ження у його назві. Росте в листяних лісах. Токсичні властивості
цього виду ідентичні до блідої поганки.
Мухомор смердючий - Атапііа уігоза. Росте на ґрунті у листя-
них та хвойних лісах. За зовнішнім виглядом схожий на бліду поган-
ку, а за забарвленням - на мухомор білий. Від останнього виду мухо-
мор смердючий відрізняється текстурою поверхні ніжки, яка є волок-
нистою; волокна можуть відстовбурчуватись, набуваючи вигляду
численних лусок. Характерна особливість гриба - неприємний запах
та смак м’якуша, через що випадки отруєнь ним нечисленні. Проте за
складом та концентрацією токсинів мухомор смердючий майже іден-
тичний до блідої поганки та білого мухомора.
Мухомор червоний - Атапііа тизсагіа (рис. 158 Б) - має ша-
пинки, забарвлені у різні відтінки цегляно-червоного кольору. На їх
поверхні у вигляді білих пластівців зберігаються залишки загального
покривала. Ніжка при основі бульбовидно потовщена, з прирослою
піхвою, яка має вигляд концентричних бородавчасто-лускатих зон.
Мухомор червоний росте у листяних та хвойних лісах. Плодові тіла
містять нейротоксин мускарин.
Мухомор цезарів - Атапііа саезагеа (рис. 158 А). Цінний
їстівний гриб, який занесено до Червоної книги України. Шапинка оран-
жево-жовта або оранжево-червона, залишки загального покривала
дуже швидко зникають. Пластинки, ніжка та кільце жовтого кольору.
Піхва біла, вільна або напіввільна, шкіряста. Гриб поширений у бага-
тьох країнах світу, зростає в листяних світлих лісах. В Україні зустрі-
чається лише на Закарпатті та в Криму.
389
Рис. 158. Плодові тіла аманітових грибів: А - Атапііа саезагеа; Б - Атапііа
тизсагіа
Родина плютейні - Ріиіеасеае. Плодові тіла геміангіокарпні, із
загальним покривалом, яке дуже швидко зникає (у видів роду плю-
тей - РІиіеиз) або зберігається у вигляді піхви біля основи ніжки (рід
вольварієла - УоІуагіеІІа). Часткове покривало відсутнє. Шапинка
та ніжка гетерогенні. Трама пластинок інверсна, а самі пластинки ма-
ють рожевуватий відтінок. Спори також рожевуваті, гладенькі. Споро-
ва маса світло-рожева. До родини належать виключно види-ксилот-
рофи, що зазвичай розвиваються на відмерлій деревині. Отруйні пред-
ставники відсутні. Найбільш відомим видом є РІиіеиз сегуіпиз - плю-
тей оленячий, для якого характерні великі плодові тіла з коричне-
вою більш-менш розпростертою шапинкою та білуватою щільною
волокнистою ніжкою.
Родина павутинникові, або кортинарієві - Согііпагіасеае. Пло-
дові тіла геміангіокарпні, загальне покривало переважно відсутнє. Ча-
сткове покривало представлене кортиною, яка, як правило, швидко
зникає. Шапинка та ніжка гетерогенні. Трама пластинок правильна.
Спори світлі або темні, коричневих відтінків, гладенькі, бородавчасті
або кутасті. Спорова маса від жовтої до темно-коричневої. Частина
видів родини є їстівними, але більшість належить або до неїстівних,
390
або до отруйних. Найбільш небезпечні види належать до родів паву-
тинних та плютка, які відрізняються між собою за морфологією спор
та частково - за текстурою поверхні шапинки.
Рід павутинних або кортинаріус - Согііпагіиз. Хоча карпо-
фори павутинників досить різноманітні, проте шапинки у цих грибів
зазвичай з невеликим тупим горбочком у центрі, пластинки прирослі,
в молодому віці закриті кортиною. Гіменіальний шар, як правило, з
численними цистидами. Від близького роду плютка відрізняється бо-
родавчастими спорами, а також більш-менш гладенькою, не волок-
нистою текстурою поверхні шапинки. Деякі види павутинників (на-
приклад, Согііпагіиз уіоіасеиз - павутинних ліловий) їстівні, але
є й небезпечні отруйні представники, серед яких до найвідоміших
належить павутинник оранжево-червоний.
Павутинних оранжево-червоний - Согііпагіиз огеііапиз. Гриб
має досить великі карпофори, в яких шапинка забарвлена в рудий
колір. Жовта кортина прикриває рідкі оранжеві прирослі до ніжки пла-
стинки, які.при старінні набувають коричнювато-червоного відтінку.
Ніжка забарвлена у золотисто-жовтий колір і при основі переходить у
коренеподібний виріст. Павутинник оранжево-червоний містить не-
безпечний токсин спазмолітичної дії - ореланін - і належить до смер-
тельно отруйних грибів.
Рід іноцибе або плютка - ІпосуЬе - за наявністю корти-
ни, горбочка у центрі шапинки, коричневими тонами забарвлення пла-
стинок, різноманітними та численними цистидами дещо нагадує па-
вутинник. Проте спори у плюток гладенькі, а поверхня шапинки ра-
діально розтріскана та радіально волокниста. Більшість видів роду
отруйні, оскільки містять високі концентрації мускарин-подібних ней-
ротоксинів. Найбільш небезпечною є плютка Патуйяра.
Плютка Патуйяра - ІпосуЬе раіоиіііагдіі (рис. 159). Карпофори
у цієї плютки щільном’ясисті. Шапинки тупоконусовидні, згодом роз-
простерті, з горбочком у центрі, спочатку білі або кремові, пізніше
нерівномірно забарвлені у різні червоно-коричневі відтінки. Поверхні
шапинок радіально тріщинуваті. Пластинки червонуваті, згодом ко-
ричнюваті. Трама пластинок правильна, з великими цистидами. Спо-
ри гладенькі, нерівнобоко-еліпсоїдні. Ніжка циліндрична, кольору ша-
пинки, щільна. М’якуш білий, на зламі ніжки може злегка червоніти, з
приємним фруктовим запахом.
391
Росте у листяних лісах, на узліссях, галявинах, у парках, найчас-
тіше під дубом та липою, з якими утворює мікоризу. Отруйний гриб.
Родина гнойови-
кові або копринові -
Соргіпасеае. Плодові
тіла геміангіокарпні, із
загальним покривалом,
яке дуже швидко зникає.
Часткове покривало є
або відсутнє. Шапинка та
ніжка гетерогенні. Трама
правильна. Спори тем-
но-коричневі, гладенькі
або бородавчасті. Спо-
рова маса чорна. Харак-
Рис. 159. ІпосуЬе раіоиіііагсіїг. зовнішній терна особливість пред-
вигляд плодових тіл ставників родини -
здатність пластинок та
плодових тіл до автолізису після дозрівання спор, внаслідок чого зрілі
карпофори швидко розпливаються. Провідними родами є гнойовик
та псатирела.
Рід гнойовик або копринус - Соргіпиз. Види цього роду до-
сить легко розпізнаються за характерною формою карпофорів - ша-
пинки переважно дзвоникоподібні, ніжки тонкі, з порожниною всере-
дині. Пластинки швидко набувають темно-фіолетового забарвлення,
і у зрілих плодових тілах розпливаються внаслідок процесів автолізи-
су. Поселяються на гної, рідше на ґрунті, збагаченому органічною
речовиною, завдяки чому отримали свою назву. Більшість видів роду
через дрібні розміри карпофорів харчової цінності не мають. Винят-
ком є гнойовик чубатий та гнойовик чорнильний.
Гнойовик чубатий - Соргіпиз сотаіиз (рис. 160 А). Плодові
тіла досить великі, вкриті загальним покривалом, яке швидко зникає
при дозріванні гриба. Пластинки у молодому віці прикриті частковим
покривалом. Шапинка видовжено-еліпсоїдна, дзвониковидна, біла,
спочатку гладенька, з часом волокнисто-луската. Пластинки темно-
фіолетові, при дозріванні плодового тіла розпливаються. Трама пла-
стинок правильна, без цистид. Ніжка циліндрична, біла, біля основи
потовщена. М'якуш світлий. Спорова маса чорна. Спори яйцевидні.
392
Росте гнойовик чубатий у лісонасадженнях, на відкритих місцях, на
ґрунті. Молоді плодові тіла гриба їстівні, проте при вживанні з алкого-
лем спричинюють отруєння, оскільки містять токсин локальної збуд-
жуючої дії - коприн.
Гнойовик чорнильний - С. аігатепіагіиз (рис. 160 Б). Карпо-
фори, як і у попереднього представника, досить великі. Але шапинка
не овальна, а дзвоникоподібна, не має великих лусок, більш-менш
гладенька і лише в центрі несе дрібні лусочки. Пластинки білуваті, з
часом чорні, при дозріванні гриба розпливаються. Трама пластинок
правильна, без цистид. Ніжка з вузьким, зникаючим кільцем, розта-
шованим у нижній її частині. М’якуш білуватий, у молодих плодових
тіл з приємним грибним запахом. Спорова маса буро-коричнева. Спо-
ри широкоовальні, коричневі. Гриб вважається їстівним, проте при
вживанні з алкоголем спричиняє отруєння, оскільки подібно до гно-
йовика чубатого містить коприн. Росте гриб на угноєному ґрунті, час-
то на узбіччі доріг.
Рис. 160.
аігатепіагіиз
Гнойовикові гриби: А - Соргіпиз сотаіиз; Б - Соргіпиз
Рід псатирела - РзаіНугеІІа (рис. 161). Види роду за зовнішнім
виглядом карпофорів подібні до дрібних гнойовиків, але в їх пластин-
ках швидкий автолізис не відбувається, і, як наслідок, гіменофори не
393
розпливаються до моменту загнивання плодового тіла. Цистиди зав-
жди присутні і особливо численні на вістрі пластинок (т. зв. хейлоци-
стиди).
Зростають псатирели у лісах, садах, часто на відкритих місцях,
на угноєному ґрунті, зрідка на пеньках і належать до мало відомих
їстівних грибів низької якості. Найпоширенішим видом є РзаііїугеІІа
сапсіоііеапа - псатирела Кандолля, що росте на ґрунті в лісах та
парках, переважно на гнилій деревині, часто великими групами.
Рис. 161. РзаОїугеІІа: зовнішній
вигляд плодових тіл
Рис. 162. Нурїюіота /азсісиїаге:
зовнішній вигляд плодових тіл
Родина строфарієві -
ЗігорЬагіасеае. Плодові тіла ге-
міангіокарпні, із загальним покри-
валом, що зазвичай швидко зникає.
Часткове покривало переважно є,
рідше відсутнє. Шапинка та ніжка
гомогенні. Трама пластинок пра-
вильна. Спори темні, мають забар-
влення з фіолетовими відтінками,
гладенькі. Пластинки також темні
з фіолетовими або брудно-зелени-
ми тонами. Спорова маса фіоле-
това, до чорної. Більшість видів є
неїстівними або отруйними. До
найбільш небезпечних строфаріє-
вих грибів належить несправжній
опеньок сірчано-жовтий.
Несправжній опеньок сір-
чано-жовтий, або гіфолома гру-
пова - Нуріїоіота /азсісиїаге
(рис. 162). Карпофори геміангіо-
карпні. Шапинка тонком’ясиста, на-
півсферична, згодом опукло- або
плоско-розпростерта, часто з гор-
бочком, сірчано-жовта, в центрі
темніша, гола, по краю зазвичай із
залишками покривала. Пластинки
вузькі, густі, сірчано-жовті, з віком
зеленкувато-коричневі. Трама пла-
стинок правильна. Ніжка цилінд-
рична, сірчано-жовта, до основи
394
коричнювата, темноволокниста, з порожниною. М’якуш сірчано-жов-
тий, гіркий, з неприємним запахом. Спори еліпсоїдні, гладенькі.
Росте опеньок несправжній у листяних та хвойних лісах, на ста-
рих пеньках, щільними групами. Гриб отруйний, містить плазмолітичні
токсини з групи ама- та фалотоксинів і, таким чином, належить до
дуже небезпечних грибів. Проте вміст токсинів нижчий, ніж у блідої
поганки, білого та смердючого мухоморів, через що випадки смер-
тельних отруєнь несправжнім опеньком відносно нечисленні.
Порядок Сироїжкові або Русуляльні - Киззиіаіез
Плодові тіла гімнокарпні, однорічні, загниваючі; гіменофор плас-
тинчастий, зі сфероцистами; щетинки в гіменофорі відсутні; базидія
хіастична, проста (холобазидія); парентосоми з перфораціями; спо-
ри амілоїдні, із сітчастою орнаментацією. Мікоризні гриби.
Характерною особливістю грибів цього порядку є наявність у пло-
дових тілах великих сферичних клітин - сфероцист. Завдяки сферо-
цистам карпофори сироїжкових ламкі та хрусткі. Специфічною є та-
кож сітчаста орнаментація базидіоспор, завдяки чому вони вигляда-
ють гребінчасто-бородавчастими.
Переважна більшість сироїжкових-облігатні мікоризні гриби, що
утворюють ектотрофну або ендо-ектотрофну мікоризу із хвойними або
листяними деревами. Серед русуляльних грибів є цінні їстівні, умовно
їстівні та неїстівні види. Порядок
включає лише два роди -
сироїжка та хрящ-молочник, які,
проте, багаті у видовому відно-
шенні. Роди добре розрізняють-
ся, в першу чергу, за наявністю
молочного соку та за характе-
ром приростання пластинок
(прирослі чи спускаються на
ніжку).
Рід сироїжка - Пиззиіа
(рис. 163). Види роду Риззиіа
позбавлені молочного соку і
мають пластинки, що прироста-
ють до ніжки. Трама неправиль-
Рис. 163. Киззиіа: А - зовнішній на- СПОР„И із ^частою орнамен-
вигляд плодового тіла; Б — спори тацією. Рід нараховує понад сто
395
видів. До їстівних грибів належить сироїжка луската, до неїстівних, які
у сирому вигляді здатні спричинювати шлункові розлади, - сироїжка
блювотна.
Сироїжка луската, товстуха - Пиззиіа уігезсепз - має великі
хрусткі плодові тіла. Шапинка зелена, луската. Пластинки білі або
кремові, приростають до ніжки. Спори мають сітчастий орнамент та
короткі шипики. Ніжка товста, біла. М’якуш також білий, із приємним
запахом та смаком, без молочного соку. Росте у листяних та мішаних
лісах.
Сироїжка блювотна - Пиззиіа етеііса. Карпофори середнього
розміру. Шапинка рожева або червона, гладенька. Пластинки білі, з
віком кремові, приростають до ніжки і в середній частині мають виїм-
ку. Спори з тонким сітчастим орнаментом та невеликими шипиками.
Ніжка біла, місцями червонувата, у дорослих плодових тіл з порож-
нинами. М’якуш дуже гіркий та пекучий на смак. Карпофори, крім гірких
речовин, містять невелику кількість мускарин-подібних токсинів, че-
рез що у сирому вигляді є отруйними. Після відварювання токсини
інактивуються, але пекучий гіркий смак зберігається. Росте у листя-
них та соснових лісах.
Рід хрящ-молочник - іасіагіиз (рис. 164). Відрізняється від си-
роїжки наявністю молочного соку, який виступає на зламі карпофорів.
Молочний сік містять товстостінні молочні гіфи - латицифери. Білий
Рис. 164. Ьасіагіиз: А, Б - зовнішній вигляд плодових тіл; В - спора
396
молочний сік у багатьох видів забарвлюється на повітрі. Інша харак-
терна відміна хрящ-молочників від сироїжок пов’язана з тим, що пла-
стинки у перших спускаються на ніжку. Рід за видовим багатством
наближується до сироїжки. Серед хрящ-молочників немає отруйних
видів, хоча через пекуче-гіркий молочний сік більшість видів роду
вважаються лише умовно їстівними. Кілька видів хрящ-молочників, у
яких молочний сік солодкий або пряний, проте не гіркий (наприклад,
рижик), вважаються делікатесними грибами, які вживають у їжу у си-
рому вигляді. Інші види (наприклад, чорний груздь, гірчак) потребу-
ють видалення гіркоти, наприклад, вимочуванням, відварюванням або
солінням.
Рижик або хрящ-молочник смачний - іасіагіиз сіеіісіозиз -
має шапинки, забарвлені у рудувато-оранжевий колір, жовто-
оранжеві пластинки, що при натисканні зеленіють, оранжеві ніжки та
оранжевий солодкий молочний сік з пікантним гоструватим присма-
ком. Росте у хвойних лісах і утворює мікоризу із сосною та ялиною.
Вважається одним з найцінніших їстівних грибів.
Чорний груздь або хрящ-молочник оливково-чорний - Ьа-
сіагіиз іигріз. Карпофори цього гриба оливково-коричневі, з концен-
тричними зонами. Молочний сік білий, пекучий. Утворює мікоризу з
березою. їстівний і використовується переважно на засолювання.
Гірчак або хрящ-молочник гіркий - Ьасіагіиз гиїиз - дуже
часто зустрічається у хвойних та мішаних лісах, де утворює мікоризу
із сосною. Карпофори гриба червонувато-коричневі, із шапинкою, яка
має горбочок у центрі. Молочний сік білий, пекуче-їдкий. Гриб є
їстівним, проте стає придатним до вживання у їжу після тривалих про-
цедур видалення молочного соку, при яких значно втрачає свою хар-
чову цінність.
Гастероміцети
Порядок Дощовикові або Лікопердальні - Ьусорегдаїев
Гастероміцети; плодові тіла ангіокарпні; перидій не ослизнюється,
складається із зовнішнього шару - екзоперидію - та розташованого
під ним ендоперидію; екзо- та ендоперидій не зростаються; глеба
коралоїдна або однорідна, світла, базидія хіастична, проста (холо-
базидія); парентосоми з перфораціями; базидіоспори не відстрілю-
397
ються, переважно темнозабарвлені, гладенькі або орнаментовані, по-
ширюються вітром. Ґрунтові сапротрофи та мікоризні гриби.
Рід лікопердон або дощовик - Ьусорегсіоп (рис. 165). Пло-
дові тіла дощовика грушовидні, із звуженою несправжньою ніжкою.
Якщо плодове тіло розрізати, то можна побачити, що під зовнішнім
покривом - перидієм - розташовується коралоїдна глеба - споро-
носна тканина, і трама - стерильна тканина, яка розміщується в нижній
частині плодового тіла і формує несправжню ніжку. На розрізі помітно,
що перидій складається з двох шарів - зовнішнього (екзоперидія) та
внутрішнього (ендоперидія). При дозріванні плодового тіла екзопе-
ридій розтріскується та осипається. Дозріла глеба складається з ни-
ток ка-піліцію, серед яких розміщуються спори. Після дозрівання пло-
дового тіла ендоперидій розривається на верхівці і плодове тіло по-
рошить, розсипаючи темну спорову масу.
Плодові тіла їстівні в молодому віці. Також білий м’якуш дощо-
виків (стерильний і гігроскопічний) можна використовувати для зупинки
кровотечі.
Рис. 165. Ьусорегсіоп: А - загальний вигляд плодових тіл; Б - розріз через
плодове тіло; В - ділянка капіліцію зі спорою; Г - спора. 1 - трама; 2 - глеба;
З - перидій
Схожу будову, але без ніжки, мають плодові тіла грибів з роду
порхавка - Воуізіа (рис. 166).
398
Рис. 166. Воуізіа: загальний
До порядку відноситься також рід
головач - Саіуаііа (рис. 167). У цього
гриба при дозріванні плодового тіла ен-
доперидій повністю руйнується, і глеба
відпадає, а на субстраті залишається
тільки чашоподібна трама.
Дощовики, порхавки та головачі ро-
стуть найчастіше у трав'янистих фіто-
ценозах - на луках, пасовиськах, у сте-
пах, рідше зустрічаються у хвойних та
широколистяних лісах.
вигляд плодового тіла
Рис. 167. Саіуаііа: А - загальний вигляд
плодових тіл; Б - залишки трами після випо-
рошування спор
Рід геаструм або
земляна зірочка -
(Зеазігит (рис. 168). Пло-
дові тіла спочатку кулясті,
з коралоїдною глебою.
Під час дозрівання обо-
лонка плодового тіла роз-
шаровується, товстий
шкірястий екзоперидій
розривається на кілька
лопатей. Лопаті відги-
наються, і відкривається
глеба, вкрита пергамен-
товидним ендоперидієм з
отвором на верхівці. Роз-
криті плодові тіла схожі на
зірочки. Спори коричневі,
бородавчасті або гла-
денькі. Капіліцій скла-
дається із загострених на
кінцях ниток. Лопатям ек-
зоперидію властиві гігро-
скопічні рухи. Від зміни
зволоження вони то роз-
прямляються, то загина-
ються над спороносною частиною. Сухі плодові тіла зберігаються до
весни. Види роду Сеазігит - ґрунтові сапротрофи, трапляються в
лісах, на узліссях, у чагарниках.
399
Рис. 168. Сеазігит: загальний
вигляд плодового тіла
Спори бородавчасті, щетинисті а
Рід тулостома -
Тиіозіота (рис. 169). Молоді
плодові тіла кулясті, вкриті ендо-
та екзоперидієм. Глеба рівномі-
рна, і у нижній частині залишаєть-
ся стерильною. При дозріванні
екзоперидій розкривається, сте-
рильна частина плодового тіла
видовжується у ніжку, яка підно-
сить над субстратом кулясту гле-
бу, вкриту тонким перетинчастим
ендоперидієм з отвором на вер-
хівці. Ніжка циліндрична, до 6 см
завдовжки. Капіліцій та спорова
маса вохряні або коричнюваті,
гладенькі; висипаються з плодо-
вого тіла, як з маківки, через отвір, від коливань плодового тіла на
ніжці і від поштовхів. Види цього роду є ґрунтовими сапротрофами.
Рис. 169. Тиіозіота:
загальний вигляд плодо-
вого тіла
Більшість з них росте на сухих сонячних
місцях: у степах, напівпустелях та пустелях.
Порядок Несправжньодощовикові
або Склеродерматальні - Зсіегосіег-
таіаіез
Гастероміцети. Плодові тіла ангіокарпні;
перидій не ослизнюється, екзо- та ендопе-
ридій зростаються; глеба лакунарна, темно-
фіолетова або чорна, базидія хіастична,
проста (холобазидія); парентосоми з перфо-
раціями; базидіоспори не відстрілюються,
темнозабарвлені, гладенькі або орнаменто-
вані, поширюються ґрунтовими тваринами.
Ґрунтові сапротрофи та мікоризні гриби.
В Україні поширений лише один рід - не-
справжній дощовик.
Рід несправжній дощовик або
склеродерма - Зсіегосіегта (рис. 170) -
інколи помилково вважається трюфелем, ос-
кільки розвиток плодового тіла починається
під землею. Проте зрілі несправжні дощо-
400
вики легко розпізнаються на зрізах - за темно-фіолетовою або чор-
ною лакунарною глебою, яка контрастує із світлозабарвленим тов-
стим перидієм. Несправжні дощовики є отруйними грибами. Харак-
терний представник - несправжній дощовик оранжевий.
Несправжній дощовик оранжевий або склеродерма оран-
жева - Зсіегосіегта аигапііасит. Плодові тіла кулясті, бульбо-
видні, часто приплюснуті, сидячі, біля основи з тонкими міцеліальни-
ми тяжами. Перидій одношаровий, до 1,5 мм завтовшки, брудно-білий,
жовтуватий, гладенький або лускатий, сухий, щільний, згодом буро-
жовтий, часом кольору бронзи, при дозріванні плодового тіла розри-
вається на неправильні частки - лопаті. Глеба спочатку білувата,
дозріла - чорна, порошиста. Спорова маса чорна. Спори кулясті,
бородавчасто-шипасті, темно-коричневі. Капіліцій слаборозвинутий,
розгалужений та ламкий. Вид є дуже розповсюдженим, росте у лис-
тяних та соснових лісах, парках, на піщаних, глинястих та перегнійних
ґрунтах.
Рис. 170. Зсіегосіегта: А - молоде плодове тіло; Б - дозріле плодове тіло;
В - розріз через плодове тіло. 1 - глеба; 2 - перидій
Порядок Гніздівкові або Нідуляріальні - NібиіагіаІез
Гастероміцети. Плодові тіла ангіокарпні, мають форму кошика;
перидій не ослизнюється, екзо- та ендоперидій зростаються; глеба
лакунарна, світла, поділена на перидіоли, базидія хіастична, проста
(холобазидія); парентосоми з перфораціями; базидіоспори не від-
стрілюються, безбарвні та гладенькі, переважно товстостінні, ґрун-
тові сапротрофи, копротрофи та ксилотрофи. Характерний представ-
ник - рід ціатус, або бокальчик.
401
Рід ціатус або бокальчик - Суаііїиз (рис. 171). Плодові тіла
нагадують мініатюрний кошик до 1,5 см заввишки і 0,5-1 см завшир-
шки, прикритий хустиною, під якою щільно укладено 10-15 білих
дрібних кульок. «Кошик» утворений перидієм, хустина - тонким спле-
тенням стерильних гіф, похідних від екзоперидія, - епіфрагмою. Куль-
ки (перидіоли) - це фрагменти глеби, вкриті додатковою оболонкою,
що походить від ендоперидія. Перидіоли прикріплюються до
внутрішніх стінок «бокальчика» пуповиноподібним шнуром - фуніку-
люсом. На розрізі перидіоли видно щілиновидну камеру із гладеньки-
ми яйцевидними базидіоспорами, які випадають, коли руйнується
оболонка перидіоли. Спори розповсюджуються оригінально: від ударів
дощових крапель по «бокальчику» перидіоли разом із фунікулюсом
відриваються від ендоперидія і вилітають із кошика. Далі фунікулюс
прилипає до навколишніх предметів (гілочок, стеблин і т. п.), набря-
кає і видовжується. Гідроліз глікогену водночас із поглинанням води
гіфами фунікулюсу призводить до того, що в них розвивається знач-
ний тургорний тиск. Як наслідок, фунікулюс відстрілює від себе пери-
діолу. Від удару об ґрунт або інший більш-менш твердий субстрат
оболонка перидіоли розривається, і базидіоспори розсипаються на-
вколо неї.
Залишки порожніх кошиків часто зберігаються 1-2 роки. Гриб ро-
сте в лісах, садах, на рештках деревини, пеньках, сухій траві, зазви-
чай щільними групами.
Рис. 171. СуаІЇїиз: зовнішній вигляд
плодових тіл
Порядок Веселкові або
Фаляльні - РГіаііаІез
Гастероміцети. Плодові
тіла ангіокарпні, зі складною
диференціацією; ендоперидій
ослизнюється, екзоперидій
розривається; глеба багато-
шапинкова або одношапинко-
ва, темна або яскравозабар-
влена, часто з різким непри-
ємним запахом; базидія хіас-
тична, проста (холобазидія);
парентосоми з перфораціями;
базидіоспори не відстрілю-
ються, поширюються комаха-
ми. Ґрунтові сапротрофи та
мікоризні гриби.
402
У молодому віці плодові тіла веселкових є їстівними. Деякі роди,
наприклад веселка, часто зустрічаються у природі, але в цьому по-
рядку також міститься велика кількість рідкісних та зникаючих грибів,
занесених до Червоної книги, - види родів мутин, сітконоска, реші-
точник, квітохвісник.
Рід веселка або фалус - РіїаІІиз. Плодові тіла великі, склада-
ються із дзвониковидної глеби, яку у зрілому стані над поверхнею
ґрунту підносить губчаста стерильна ніжка - рецептакул. У лісах час-
то можна знайти гриб веселку звичайну, який привертає увагу різким,
неприємним запахом гнилого м’яса.
Веселка смердюча або фалус смердючий - РЬаІІиз
ітрисіісиз (рис. 172). Плодове тіло веселки спочатку має вигляд білої
чи злегка сіруватої кулі 5-6 см у діаметрі (т. зв. стадія яйця), від якої у
ґрунт спрямований білий міцеліальний коренеподібний тяж. Екзопе-
ридій гладенький, ендоперидій драглистий, нагадує желе. Під пе-
Рис. 172. РііаІІиз ітрисіісиз: А - ста-
дія яйця; Б - зріле плодове тіло; В - ста-
дія яйця у розрізі. 1 - перидій; 2 - рецеп-
такул; 3 - глеба; 4 - міцеліальні тяжі
ридієм розміщується зачатко-
вий рецептакул у вигляді по-
рожнього всередині стовпчика
з пористими стінками. На по-
верхні рецептакула знаходить-
ся конусоподібна одношапин-
кова глеба. При дозріванні ре-
цептакул швидко видовжується
(до ЗО см), розриває екзоперидій
і підіймає над ґрунтом чорно-
зелену конусовидну глебу. В її
комірках розташовуються ба-
зидії з базидіоспорами. Далі
поверхневий шар глеби почи-
нає роз-пливатися і смердіти.
Запах гнилого м’яса приваб-
лює мух, які розносять спори
гриба.
Внутрішню слизову обо-
лонку плодового тіла застосо-
вують у народній медицині як
протиревматичний засіб. У мо-
лодому віці (на стадії яйця)
плодові тіла їстівні навіть у си-
рому стані.
403
Рід мутин - Миііпиз (рис. 173 А) - дещо нагадує веселку, але
має дрібніші плодові тіла, і глеба не набуває вигляду шапинки. Вона
розташовується на забарвленій у жовто-гарячий колір верхівці ре-
цептакулу у вигляді зеленувато-оливкового слизистого шару. Глеба
після ослизнення не тхне гнилим м’ясом, а має запах свіжого борош-
на. Гриб зустрічається на ґрунті, у вологих листяних лісах Закарпаття
й Лісостепу та на Південному узбережжі Криму. В Україні рід пред-
ставлено одним видом - мутином собачим (Миііпиз сапіпиз), який
занесений до Червоної книги України.
Рід сітконоска або диктіофора - Оісіуоріїога (рис. 173 В) -
включає лише один вид - Оісіуоріюга сіиріісаіа — сітконоска
подвоєна. В Україні відоме лише одне місцезнаходження цього гри-
ба - на ґрунті у мішаних насадженнях інтродукованих рослин у
Нікітському ботанічному саду в Криму. Плодові тіла сітконоски нага-
дують веселку, за винятком того, що з під глеби над рецептакулом
звисає міцеліальна жовтувато-біла сітка - індузій. Він вночі здатний
світитися, оскільки містить люциферин-подібні речовини. У зрілому
стані гриб має різкий, неприємний запах, який, як і у веселки, приваб-
лює деяких комах. Диктіофора подвоєна занесена до Червоної книги
України під категорією І - «зникаючі види».
Рід решіточник або клатрус - СІаіїїгиз (рис. 173 Г) - у міко-
флорі України також представлений лише одним видом, а саме
СІаІЇїгиз гиЬег - клатрусом червоним. Плодові тіла існують на по-
верхні ґрунту лише кілька годин.
Молоді плодові тіла кулясті або яйцевидні, 5-10 см заввишки та
5 см завширшки, вкриті білуватим або жовтуватим перидієм, який
розривається на верхівці неправильними лопатями. Перидій з віком
швидко зникає, оголюючи товстий драглистий середній шар. Зріле
плодове тіло має рецептакул у вигляді решіткоподібного червоного
сітчастого купола, на внутрішньому боці якого розташовується ба-
гатошапинкова глеба. Вона має неприємний різкий запах, який при-
ваблює комах. Спори гладенькі, безбарвні.
Ґрунтовий сапротроф. Дуже рідкісний гриб. Його плодові тіла з’яв-
ляються нерегулярно, часто з інтервалом у кілька років. Трапляється
виключно на узбережжі Чорного моря, зокрема в Україні - у садах,
парках та листяних лісах південного берега Криму в районі Ялти. Клат-
рус червоний занесено до Червоної книги України під категорією
«рідкісні види».
404
Рід квітохвісник - Рзеисіосоїиз (рис. 173 Б) - у складі мікобіоти
України представлений одним видом - Рзеисіосоїиз Гизіїогтіз -
квітохвісником веретеновидним. Молоді плодові тіла яйцеподібні,
з багатошапинковою глебою. При дозріванні екзоперидій розриваєть-
ся лопатями і залишається біля основи плодового тіла у вигляді білої
Б
Рис. 173. Гастероміцети, занесені до Червоної книги України: А - Миііпиз
сапіпиз, Б - Рзеисіосоїиз їизіїогтіз; В - ОісІуорЬога диріісаіа; Г - СІаІіігиз гиЬег
405
піхви, ендоперидій ослизнюється. Нижня частина рецептакулу утво-
рює коротку ніздрювату ніжку, яка не виходить за межі піхви, верхня
його частина поздовжньо розщеплена на три-вісім червоних лопа-
тей. На внутрішній поверхні лопатей розташовується ослизнена тем-
но-оливкова глеба. Після повного ослизнення глеби лопаті рецепта-
кулу на верхівці роз’єднуються, через що карпофори набувають виг-
ляду великої червоної квітки, що розпустилася (звідси й походить на-
родна назва гриба). Плодові тіла з'являються періодично, з інтерва-
лом у кілька років.
Гумусовий сапротроф. Зростає лише на південному узбережжі
Криму (зокрема на території Нікітського ботанічного саду). Квітохвіс-
ник веретеновидний занесений до Червоної книги України.
Фрагмобазидіоміцети з драглистими плодовими тілами
Порядок Аурікуляріальні - Аигісиїагіаіез
Гіменоміцети. Плодові тіла гімнокарпні, драглистої консистенції,
мають переважно форму вушної раковини; базидія стихічна, поділе-
на на клітини трьома поперечними перегородками (фрагмобазидія).
Базидіоспори проростають гачковидно зігнутими конідіями, які зго-
дом розвиваються у міцелій (т. зв. репетитивне проростання базиді-
оспор). Септи доліпорові, з неперфорованою парентесомою. Типо-
вий рід порядку — аурикулярія.
Рід аурикулярія - Аигісиїагіа - характеризується наявністю ву-
хоподібних, забарвлених у коричневий колір плодових тіл драглистої
консистенції, які прикріплюються боком до деревних субстратів - по-
валених стовбурів, гнилих пеньків, засохлих гілок тощо. Гіменіаль-
ний шар гладенький або злегка складчастий. Найвідомішим представ-
ником роду є іудине вухо - цей гриб належить до їстівних і навіть
промислово культивується у деяких країнах Азії.
Іудине вухо або аурикулярія Іуди - Аигісиїагіа аигісиіа-іисіае
(рис. 174). Плодові тіла здебільшого вухоподібні, черепашковидні або
у вигляді чашечки, прикріплені звуженою основою до субстрату,
1-8 см у діаметрі, драглистої консистенції, зовні сірувато-коричневі,
з короткими волосками. У суху погоду карпофори іудиного вуха вси-
хають і набувають шкірястого вигляду, проте при наявності вологи
швидко відновлють нормальну драглисту консистенцію. Гіменіальний
406
шар складчастий. Базидії циліндричні, розділені на чотири клітини
трьома поперечними перегородками.
Зустрічається на деревних залишках - відмерлих гілках, стовбу-
рах та пеньках листяних порід дерев з ранньої весни до пізньої осені.
В Україні цей гриб поширений здебільшого в тих регіонах, де є ліси -
Карпати, Прикарпаття, Полісся, Лісостеп, Крим.
Гриб широко використовується в їжу в традиційній китайській та
корейській кухні, а сьогодні його можна зустріти на продуктових при-
лавках України. Є відомості, що іудине вухо спеціально вирощували
в Китаї та Кореї ще у VI столітті нашої ери.
Рис. 174. Аигісиїагіа аигісиІа-]идае: А - плодове тіло; Б - зріз через плодо-
ве тіло: 1 - фрагмобазидія; 2 - базидіоспори
Порядок Тремеляльні - ТгетеІІаІез
Гіменоміцети; плодові тіла гімнокарпні, драглистої консистенції;
базидія хіастична, поділена на чотири клітини двома поздовжніми
перегородками, розташованими хрестоподібно, тобто являє собою
один з варіантів фрагмобазидії. Септи доліпорові, парентосоми скла-
даються з окремих фрагментів-пухирців. Базидіоспори проростають
конідіями (репетитивне проростання). Провідний рід порядку - тре-
мела.
Рід тремела - ТгетеІІа (рис. 175). Плодові тіла, залежно від во-
логості оточуючого середовища, можуть виглядати по-різному. Так,
407
після дощу тремела має вигляд драглистих округлих, опуклих, мозко-
видних, із численними звивинами та складками слизистих тіл. Однак,
якщо тривалий час стоїть суха погода, плодові тіла тремели висиха-
ють, набувають шкірястої консистенції і мають вигляд кірочок темного
кольору. Різні види тремели мають різний колір. Наприклад, у видів,
що дуже часто зустрічаються в Україні, зокрема у ТгетеІІа Іиіезсепз —
тремели жовтіючої - плодові тіла світло-жовті, а у Т. іїтЬгіаіа - тре-
мели торочкуватої - оливково-чорнуваті. Гіменіальний шар у тре-
мели вкриває всю поверхню плодового тіла. Види цього роду є пере-
важно ксилотрофами і часто зустрічаються на засохлих гілках листя-
них порід дерев, старих пеньках, опалих гілках, а також на корі живих
дерев.
Рис. 175. ТгетеІІа. А - плодові тіла; Б - зріз через плодове тіло. 1 - фраг-
мобазидія; 2 - базидіоспори
Клас Устоміцети, або сажкові гриби - изіотусеіез
Плодові тіла відсутні, базидія одно- або багатоклітинна, вирос-
тає з товстостінної спочиваючої спори - телейтоспори. В циклі роз-
витку телейтоспора зазвичай виконує функцію зимуючої стадії, в якій
гриб переносить несприятливі умови. Телейтоспори круглі або ку-
тасті, зі щільною оболонкою чорного або фіолетового кольору, утво-
рюються у великих кількостях у вигляді чорного пилу восени в ураже-
них грибом органах рослини-господаря. Ці скупчення телейтоспор,
або соруси, надають ураженим органам рослини обгорілий вигляд.
Уражені органи перетворюються на сажкоподібну масу. Звідси похо-
дить назва хвороби, яку викликають устоміцети, або сажкові гриби -
408
сажка. Базидіоспори здатні брунькуватися або попарно копулювати,
утворюючи дикаріонтичний міцелій.
Пори у септах гіф міцелію прості, шаруваті, рівномірної товщини,
проте тільця Вороніна відсутні.
Сажкові гриби є збудниками небезпечних захворювань більшості
культурних рослин, переважно злаків. У межах класу, за способом
проростання телейтоспор, виділяють два порядки - ІІзІІІадіпаїез та
ТІІІеІіаІез, представники яких - виключно облігатні паразити вищих
рослин.
Порядок Устилягінальні - ІІзііІадіпаІез
Телейтоспори проростають чотириклітинною базидією (або про-
міцелієм), по боках якої розташовані базидіоспори. Найбільш відо-
мими збудниками захворювань злаків є види роду устілаго (ІІзііІадо),
зокрема СІзіііадо таусііз - збудник пухирчастої сажки кукурудзи,
СІзіііадо ігііісі- збудник летючої сажки пшениці, СІзіііадо писіа - збуд-
ник летючої сажки ячменю та ін.
Устиляго маїсовий (збудник пухирчастої сажки кукурудзи) -
СІзіііадо таусііз (рис. 176). Збудник пухирчастої сажки кукурудзи може
уражувати різні органи рослини: міжвузля стебел, листки, жіночі та
чоловічі квітки. Уражуються як молоді, так і дорослі рослини, проте у
дорослих — лише молоді тканини, що знаходяться у процесі росту.
В телейтоспорах збудника пухирчастої сажки кукурудзи — СІ. таусііз
- відбувається каріогамія, після чого утворюється проросткова гіфа,
в ній проходить мейоз, і гіфа перетворюється в чотириклітинний про-
міцелій, який відповідає базидії. По боках кожної клітини базидії роз-
вивається тонкостінна базидіоспора, яка починає брунькуватись ще
на базидії, утворюючи ланцюжки клітин, що поступово відпадають. Ці
клітини переносяться потоками повітря на різні органи кукурудзи, де
вони копулюють, маючи різні статеві знаки («+» та «-»). Дикаріон-
тичний міцелій, що виникає після копуляції спор, заражує рослину.
Потрапивши у тканини, міцелій розростається, зумовлюючи місцеві
ураження у вигляді пухлин чи галів, заповнених телейтоспорами па-
разита. Патологічний ріст пухлин стимулюють утворені грибом рос-
тові речовини. Всередині пухлин міцелій рясно розростається, руй-
нує тканини пухлини і пізніше розпадається на окремі телейтоспори.
На початку утворення пухлини маса спор прикрита епідермісом, який
потім розривається, і спори розносяться потоками повітря. Телейто-
спори здатні одразу проростати, викликаючи нові зараження рослин.
409
Рис. 176. изіііадо таусііз: А - пухирчаста сажка кукурудзи - зовнішній ви-
гляд ураженого початку; Б - схема життєвого циклу. 1 -телейтоспора; 2 - проро-
стання телейтоспори; 3 - базидія з базидіоспорами; 4 - базидіоспора; 5 - брунь-
кування базидіоспор; 6 - кон’югація базидіоспор; 7 - дикаріонтичний міцелій
Устиляго пшениці (збудник летючої сажки пшениці) - изіііадо
ігііісі (рис. 177). Збудник летючої сажки пшениці уражує рослини під
час цвітіння. Для цього телейтоспора повинна потрапити на прий-
мочку квітки пшениці, де вона проростає чотириклітинною базидією.
Саме в цей момент (формування чотириклітинної базидії) відбуваєть-
ся редукційний поділ і утворення гаплоїдних ядер. Однак базидіоспо-
ри тут не розвиваються, а дикаріон виникає шляхом переходу ядер з
однієї клітини базидії в іншу або копулюють клітини сусідніх базидій.
З клітини базидії, яка стала двоядерною, виростає дикаріонтичний
міцелій, який по пилковій трубці проникає у зав’язь квітки. Проте це
не заважає розвитку насінини, яка зовні здається нормальною хоча в
тканинах її ендосперму та в зародку знаходиться міцелій паразита.
Заражене зерно, потрапивши у ґрунт, нормально проростає, і з нього
спершу розвивається зовні нормальна рослина. Проте разом із роз-
витком рослини росте й міцелій гриба, поширюючись у тканинах по
міжклітинниках. Особливо багато його накопичується у конусі нарос-
410
тання. Потім міцелій потрапляє у колос, що закладається, рясно там
розростається, руйнує зерна і колоски та, зрештою, розпадається на
окремі телейтоспори. У колоска, що виходить із листкової піхви, збе-
рігається лише стержень. Решта його частин перетворюється на
сажкоподібну масу телейтоспор. Останні здатні до проростання без
періоду спокою, розносяться вітром на квітучі в цей час колоски, за-
ражуючи зав’язь так, як зазначено вище.
Рис. 177. СІзіПадо Ігіїісг. А - колос пшениці, уражений летючою сажкою;
Б - схема життєвого циклу. 1 - розсіювання і перенесення телейтоспор на квіту-
ючі рослини; 2 - спори на приймочці квітки; 3 - проростки гриба проникають у
зародок насінини; 4 - уражене зерно (зовні не відрізняється від здорового); 5 -
зріз через уражену зернину (в оболонці видно міцелій гриба); 6 - проростання
грибниці
Порядок Тілеціальні - ТіІІеііаІез
Телейтоспори проростають одноклітинною базидією-проміцелієм,
на верхівці якої розташовані базидіоспори. Характерним представ-
ником порядку є ТіІІеііа сагіез — збудник захворювання, відомого під
назвою «тверда сажка пшениці».
411
Тілеція пшениці (збудник твердої сажки пшениці) - ТіІІеііа
сагіез (рис. 178). Збудник небезпечного захворювання пшениці. При
цьому захворюванні в колосках уражених рослин замість зернівок
утворюється щільно складена маса телейтоспор, які під час обмоло-
ту зерна висипаються, потрапляють на здорове насіння і зимують на
ньому. Навесні телейтоспори проростають базидіями, де ядро ділить-
ся редукційно, а гаплоїдні ядра переходять до видовжених базидіос-
пор різних знаків («+» та «-»). Базидіоспори або безпосередньо ко-
пулюють між собою завдяки утворенню копуляційних містків, або
спершу брунькуються, а потім копуляція відбувається між клітинами,
що брунькуються. В обох випадках після копуляції розвивається ди-
каріонтичний міцелій. Гриб прориває тканини молодих проростків.
Потім міцелій проникає у конус наростання, просувається вгору, про-
низуючи вузли та міжвузля. На цій стадії ураження ніяких видимих
Рис. 178. ТіІІеііа сагіез: А - колос пшениці, уражений твердою сажкою;
Б - схема життєвого циклу. 1 - колоски пшениці, уражені сажкою; 2 - зерна, ви-
повнені телейтоспорами (сажкові мішечки); 3 - розпилення телейтоспор із роз-
давленого сажкового мішечка; 4 - телейтоспори на зерні пшениці; 5 - пророс-
тання телейтоспор на зерні; 6 - проникнення гіф гриба у проростки пшениці
412
ознак ураження рослина не виявляє. На момент формування колос-
ка в уражених рослинах замість зернівок розвивається міцелій, ак-
тивно використовуючи поживні речовини рослини. Згодом міцелій
розпадається на окремі товстостінні телейтоспори. Візуально уражені
колоски відрізняються тим, що не схиляються до землі під вагою зріло-
го зерна, до того ж мають дещо «неохайний» пом’ятий вигляд.
Клас Теліоміцети - Теїіотусеіез
Плодові тіла відсутні, базидія багатоклітинна (фрагмобазидія),
утворюється з телейтоспори. Базидіопори завжди проростають гіфою.
Септи прості, шаруваті, біля пори дещо звужені.
Порядок Урединальні або іржасті гриби - Іігесііпаїез
Пукцинія злакова або збудник стеблової, лінійної або чор-
ної іржі злаків - Риссіпіа дгатіпіз (рис. 179). Гриб паразитує на
барбарисі та багатьох хлібних, кормових чи дикоростучих злаках. Види
іржастих грибів, що розвивають стадії спороношення на різних рос-
линах-господарях, називають дводомними. Повний цикл розвитку
пукцинії злакової включає п’ять стадій спороношень, кожна з яких ви-
конує певну функцію.
Розвиток гриба починається ранньою весною проростанням зи-
муючих на стерні або соломі двоклітинних товстостінних дикаріон-
тичних спор - телейтоспор. Проростанню телейтоспори передує ка-
ріогамія, після якої відразу відбувається редукційний поділ. Кожна
клітина телейтоспори проростає чотириклітинною фрагмобазидією і
утворює базидіальне спороношення. Кожна базидія утворює чотири
одноядерні гаплоїдні базидіоспори різних статевих знаків («+» та «-»).
Для подальшого розвитку базидіоспори повинні потрапити на
листок проміжного господаря - барбарису. Гіфа, що виростає з бази-
діоспори, проникає у міжклітинний простір листка і дає початок гап-
лоїдному одноядерному міцелію. Згодом на верхньому боці листка
міцелій формує пікнідіальне спороношення - пікніди з пікноспорами.
Пікніда має вигляд глечика і виповнена одноядерними гаплоїдними
пікноспорами. Пікноспори звільняються через отвір пікніди в цукристій
пахучій рідині, яка приваблює комах. Вони на своєму тілі переносять
пікноспори різних статевих знаків. Нових уражень пікноспори не ви-
кликають, а забезпечують відновлення дикаріонтичної стадії: з пікно-
спор різних статевих знаків розвиваються гаплоїдні гіфи «+» та «-»
міцелію, що знову проникають у листок барбарису. При контакті клітин
413
гіф різних статевих знаків їх цитоплазми зливаються і утворюються
дикаріонтичні клітини, з яких розвивається дикаріонтичний міцелій.
На нижньому боці листка барбарису цей міцелій формує наступ-
не спороношення - ецидіальне. Воно має вигляд стовпчикоподібних
ецидіїв, виповнених одноклітинними дикаріонтичними ецидіоспора-
ми. Для подальшого розвитку гриба ецидіоспори повинні потрапити
на листок або стебло злаку, тобто уразити основного господаря.
Ецидіоспори проростають дикаріонтичними гіфами, які через про-
дихи проникають у тканини рослини і розвивають дикаріонтичний
міцелій з гаусторіями. Цей міцелій утворює наступне спороношення -
пустули з уредоспорами. Пустули утворюються під епідермісом, у неве-
личких порожнинах, наповнених одноклітинними тонкостінними уре-
доспорами цегляно-червоного кольору, і мають вигляд лінійних
іржастих плям, звідки й походить назва хвороби - іржа. Згодом епі-
дерміс над пустулами розривається, і уредоспори звільняються. Вони
розносяться вітром, потрапляють на злаки, уражуючи нові рослини, і
знов утворюють уредоспороношення. Протягом одного сезону пере-
зараження може відбуватися багаторазово. Таким чином, за допомо-
гою уредоспор гриб поширюється і спричинює масові ураження та
захворювання рослин епіфітотії.
Гриб, поглинаючи поживні речовини господаря, уповільнює ріст
рослини і поступово її виснажує. Врешті-решт, на таких рослинах у
пустулах замість уредоспор починають утворюватися темно-бурі дво-
клітинні товстостінні телейтоспори на ніжках, формуючи останнє спо-
роношення - телейтоспороношення. У стані телейтоспор на відмер-
лих рослинних рештках господаря гриб зимує. Навесні у телейтоспо-
рах знову відбувається каріогамія, мейоз, і розвиваються нові базидії
з гаплоїдними базидіоспорами.
Тамим чином, цикл розвитку збудника стеблової іржі включає п’ять
типів спороношень: а) базидіальне, призначене для ураження про-
міжного господаря; б) пікнідіальне, яке забезпечує відновлення дика-
ріонтичної фази; в) ецидіальне, що забезпечує перше ураження ос-
новного господаря; г) уредоспороношення, за допомогою якого гриб
розселюється і здатний викликати епіфітотії; д) телейтоспороношен-
ня, призначене для перенесення несприятливих умов і подальшого
утворення базидій.
Ці спороношення мають певні позначення: 0 - пікнідіальне; І - еци-
діальне; II - уредоспороношення; III - телейтоспороношення; IV - ба-
зидіальне. Цикл, в якому представлені всі типи спороношень, у іржа-
стих грибів називають повним.
414
Специфічним заходом боротьби з Риссіпіа дгатіпіз є знищення
проміжного господаря - барбарису. Інші заходи боротьби передба-
чають, з одного боку, знищення рослинних решток, де зимують те-
лейтоспори (глибока оранка, спалювання), а з іншого - виведення
стійких до захворювання сортів рослин.
Рис. 179. Риссіпіа дгатіпіз. А - зовнішній вигляд уражених листків барба-
рису; Б - поперечний зріз через листок барбарису; В - уредопустули на стеблі та
листках злаку; Г - зріз через уредопустулу; Д - уредоспори; Е - телейтопустули на
стеблі та листках злаку; Є - зріз через телейтопустулу; Ж - проростання телей-
тоспори. 1 - пікніди (спермогонії); 2 - ецидій з ецидіоспорами; 3 - базидія з
базидіоспорами
На розрив-траві дрібноквітковій (Ітраііепз рагуіїїога) повсюдно
зустрічається інший вид іржастих грибів - пукцинія Комарова
(Риссіпіа котагоуії). Цей гриб всі спороношення утворює на одному
господарі і за цією ознакою вважається однодомним. Ецидії щільни-
ми групами розвиваються на стеблах і черешках. Пустули з уредо-
спорами зазвичай утворюються на нижньому боці листків. Оскільки
уредоспори забарвлені у жовтий колір, уредопустули не руді, а ко-
ричневі. Клітини уредоспор майже кулясті, шипуваті. Пустули з те-
лейтоспорами мають вигляд дрібних жовтих плям із каштаново-ко-
ричневими ділянками у центрі і розташовуються по колу на нижньо-
415
му боці листків. Телейтоспори еліпсоїдні, двоклітинні, на верхівці з без-
барвним сосочком, майже не перешнуровані, гладенькі, каштаново-
коричневі. Ніжка телейтоспори безбарвна, коротка та ламка.
Інший представник роду Риссіпіа, який паразитує на копитняку
європейському (Азагит аигораеит) - пукцинія копитнякова (Р.
азагіпа) - має скорочений цикл розвитку. Цей гриб утворює лише
телейто- та базидіальне спороношення. Пустули з телейтоспорами
розвиваються на обох боках листка копитняка у вигляді дрібних, зібра-
них щільними групами округлих коричневих плям. Телейтоспори двок-
літинні, більш-менш еліпсоїдні або коротковеретеновидні, із сосоч-
ковидною звуженою верхівкою та більш-менш заокругленою основою,
яка переходить у безбарвну ламку ніжку. Обидві клітини телейтоспори
майже однакові, з гладенькою світло-жовто-коричневою оболонкою.
Рід фрагмідіум - Ріїгадтісіїит (рис. 180) - об’єднує однодомні
види з повним циклом розвитку, які паразитують на представниках
родини розових (Возасеае), зокрема на трояндах, шипшині, ожині,
перстачі, суниці тощо. Ецидіальна стадія розвивається переважно на
стеблах. Ецидії руді, при дозріванні розривають покривні тканини рос-
лини і рясно порошать золотисто-оранжевими ецидіоспорами. Ця
Рис. 180. Ріїгадтіс/іит: зовнішній
вигляд телейтоспор різних видів
стадія найнебезпечніша і спричи-
нює відмирання пагонів, що розта-
шовуються вище від місць утворен-
ня спороношень. Далі на листках ут-
ворюються уредопустули, в яких
уредоспори оточені парафізами.
Потім на нижньому боці листків роз-
вивається телейтоспороношення.
Телейтоспори видовжені, багато-
клітинні, складаються з 8-13 клітин,
розміщених стовпчиком одна над
одною, з безбарвним сосочком на
верхівці. Оболонка телейтоспор
темно-бура, бородавчаста.
Анаморфні гриби або «Оеи-
іеготусеіез»
Під цією назвою, тобто ана-
морфні гриби (або незавершені гри-
416
би відділу □еиіеготусоіа, або група Еипді Ітрегїесіі, або мітос-
порові гриби) розглядають традиційну групу дейтероміцетів або кон-
ідіальних грибів, яка налічує понад 15 тис. назв. Група об’єднує гриби
з багатоклітинним міцелієм, які розмножуються виключно нестате-
вим шляхом за допомогою спор - конідій. Конідії утворюються на відга-
луженнях міцелію - конідієносцях - або шляхом розчленування гіф
на окремі клітини-артроспори. Вищі форми спороношення, які пов’я-
зані зі статевим процесом, відсутні. Більшість представників анамор-
фних грибів є стадією розвитку (анаморфою) у життєвому циклі аско-
міцетів, рідше-базидіоміцетів.
Анаморфні гриби широко представлені у природі і мають велике
значення в практичній діяльності людини. Вони мешкають як сап-
ротрофи у ґрунті та на рослинних залишках, беручи участь в проце-
сах розкладу органічної речовини в ґрунті. Значна кількість їх видів
паразитує на рослинах, зумовлюючи різноманітні плямистості (не-
крози) листя, в'янення та всихання гілок, гниття плодів та ін. Ряд
видів анаморфних грибів є збудниками алергій у людини та тварин
або викликають досить небезпечні захворювання, утому числі різно-
манітні мікотоксикози та мікози. Деякі представники цієї групи грибів
синтезують антибіотики або є антагоністами фітопатогенних мікро-
організмів та використовуються як біологічні агенти боротьби зі шкідни-
ками.
Класифікація цієї групи грибів є штучною і базується, головним
чином, на морфології конідіального спороношення, зокрема спосо-
бах утворення та біології органів нестатевого розмноження.
У деяких видів анаморфних грибів конідієносці поодинокі, утво-
рюються безпосередньо на гіфах добре розвиненого, септованого
міцелію. Інколи конідієносці можуть об’єднуватися у групи - коремії.
Такий тип конідіального спороношення властивий родам Асгетопіит,
АзрегдіІІиз, РепісіІІіит, Сегсозрога та ін.
У більш розвинених форм конідії виникають на конідієносцях, які
утворюють суцільний шар на стінках закритих кулястих споровмісти-
лищ - пікнідах. Характерними представниками з пікнідіальним типом
конідіального спороношення є види родів Азсосіїуіа, Зеріогіа,
РііуІІозіісіа та ін.
Рід акремоніум - Асгетопіит. Види цього роду є типовими сап-
ротрофами, які розвиваються у ґрунті, на різноманітних залишках
рослинного походження. Стерильні гіфи міцелію повзучі, розгалужені,
з перегородками, утворюють білі дернинки. Конідієносці у вигляді
417
простих прямих відгалужень стерильних гіф. Конідії яйцевидні, по-
одинокі, гладенькі, безбарвні. Найпоширенішим видом є Асгетопіит
уегіісіїїаіит — акремоніум кільчастий, який зустрічається переваж-
но у ґрунті.
Представники роду Асгетопіит є анаморфами в циклі розвитку
аскоміцетів з порядку Еигоііаіез.
Рід пеніцил - РепісіІІіит (рис. 181). Конідієносці у грибів цього
роду на верхівці китицевидно розгалужені, конідії утворюються в лан-
цюжках на особливих клітинах-фіалідах. Більш складна китиця скла-
дається з метул - більш-менш видовжених клітин, що розташовані
на верхівці конідієносця. На кожній метулі розміщені групи фіалід.
У деяких видів метули можуть відходити не безпосередньо від
конідієносця, а від т. зв. гілочок. Іноді конідієносці з’єднані у пучки
(коремії). Систематика цього роду грибів базується на будові китиці,
формі та розмірах конідій, морфології колоній (колонії зональні, азо-
нальні, складчасті, повстисті тощо), на забарвленні та часом фізіоло-
гічних властивостях, здатності розріджувати желатин тощо. Тому для
вивчення систематики цього роду обов’язковим є застосування ме-
тоду чистих культур, тобто культивування грибів на різних штучних
Рис. 181. РепісіІІіит: А, Б - будова окремих
конідієносців. 1 - конідієносець; 2 - гілочки; 3 - ме-
тули; 4 - фіаліди; 5 - конідії
поживних середовищах
певного складу, зокре-
ма на агаризованому
середовищі Чапека, яке
є найбільш придатним
для росту грибів роду
РепісіІІіит та вважаєть-
ся стандартним для
цих грибів.
Гриби роду Репі-
сіїїіит є продуцентами
ефективних антибіо-
тичних препаратів.
Найбільше значення
мають Р. сіігузодепит
та Р. поіаіит, які є
продуцентами анти-
біотика пеніциліну.
Гриб Р. уіїаіе, який був
описаний як новий
418
для науки вид українськими вченими В. Й. Білай та М. М. Підоплічко,
виявився продуцентом антибіотика мікроциду. Антибіотики, одержані
з цих грибів, широко використовуються в медицині, сільському госпо-
дарстві та промисловості.
У природних умовах види роду РепісіІІіит відомі як сапротрофи у
фунті, на харчових продуктах, зерні хлібних злаків, сіні, соломі та інших
грубих кормах. До найвідоміших представників роду належать Р. під-
гісапз — пеніцил чорний, який зустрічається в фунті, вологому сіні;
Р. поіаіит - пеніцил помічений - у ґрунті, торфі, воді, на різних
субстратах рослинного походження; Р. годиеїогііі- пеніцил Рокфор-
та, який використовується в країнах Європи для виготовлення сиру
рокфор, та ін.
Більшість представників роду РепісіІІіит є анаморфами в циклі
розвитку аскоміцетів з порядку Еигоііаіез.
Рід аспергіл - АзрегдіІІиз (рис. 182). Конідієносці прості, без-
барвні або світлозабарвлені, іноді буруваті, гладенькі або горбкуваті,
на верхівці з булавовидним, грушовидним, напівкулястим або куляс-
тим здуттям (пухирем), на якому утворюються фіаліди. Фіаліди еліп-
совидні, циліндричні або пляшковидні, розташовані радіально по всій
419
Рис. 183. СапМа: А - псевдоміцелій;
Б - конідії, що брунькуються
поверхні здуття або тільки на верхній його частині, прості або дво-
ярусні. У деяких видів аспергілів фіаліди утворюються не на самому
пухирі, а на спеціальних клітинах - профіалідах. Конідії кулясті, еліп-
совидні, овальні або бобовидні, гладенькі, горбкуваті або шипуваті,
безбарвні або світлозабарвлені, утворюють на верхівці фіалід лан-
цюжки, які потім складаються у радіальну голівку або склеєні у ви-
гляді колонки.
Систематика в межах роду базується на комплексі ознак, зокре-
ма на морфології конідій та конідієносців, а також на забарвленні ко-
лоній і спороношень. Стандартним середовищем для культивування
і визначення видів роду АзрегдіІІиз є агаризоване або рідке середо-
вище Чапека.
Представники роду є сапротрофами на різноманітних субстра-
тах, сільськогосподарських продуктах, у ґрунті тощо; іноді виклика-
ють захворювання людини і тварин. Види роду АзрегдіІІиз широко
використовуються у промисло-
вості для одержання різних кис-
лот, зокрема лимонної, ітаконо-
вої та ферментів.
Найбільш розповсюджени-
ми видами є А. огугае - ас-
пергіл рисовий, А. Лауиз - ас-
пергіл жовтий, А. уєгзісоіог -
аспергіл різнобарвний, А. ге-
репз - аспергіл повзучий, А.
їитідаіиз - аспергіл димчас-
тий та ін. Два останні види є па-
тогенними для людини та тварин,
зокрема А. герепз відомий як
збудник вушних мікозів людини,
а А. /итідаіиз -т. зв. аспергиль-
озів.
Більшість представників
роду АзрегдіІІиз є анаморфами
в циклі розвитку аскоміцетів з
порядку Еигоііаіез.
Рід СапМа - кандіда, ас-
порогенні дріжджеподібні
гриби (рис. 183) - здатні фор-
420
мувати міцелій і псевдоміцелій. Серед них є поширені сапрофіти або
епіфіти на багатьох видах рослин, фруктах, овочах, у молочних виро-
бах, ґрунті; здавна відомі як збудники «молочниці» у дітей (Сапгіігіа
аІЬісапз).
Рід септорія - Зеріогіа (рис. 184). Види цього роду викликають
плямистості листя різноманітних рослин. Пікніди з’являються на жи-
вих листках, переважно на плямах, спочатку занурені, згодом прори-
ваються і більш-менш виступають. Пікніди кулясті, з округлим широ-
ким отвором, темно-бурі, тонко- або товстостінні. Конідієносці короткі,
нитковидні, прямі. Конідії безбарвні, циліндричні, нитковидні, прямі
або зігнуті, з однією або кількома перегородками. Види цього роду є
конідіальною стадією (або анаморфою) в циклі розвитку аскомікото-
вих грибів родів МусозріїаегеІІа, іеріозргіаегіа та ін.
Широко розповсюдженими є Зеріогіа рориіі — септорія тополе-
ва, яка утворює білуваті плями з коричневою облямівкою на верхньому
боці листків тополі, для якої характерні циліндричні безбарвні конідії
з однією перегородкою. Досить поширеною є також 3. ругісоїа - сеп-
торія грушова, що паразитує на різноманітних видах груші, виклика-
ючи плямистості листя. Плями з’являються зазвичай з обох боків ли-
стка, дрібні, 1-2 мм у діаметрі, сірі або білуваті, з вузенькою темно-
бурою облямівкою. Конідії зігнуті, з 2-ма перегородками, оливкуваті.
8. ругісоїа є анаморфою аскоміцета МусозріїаегеІІа ругі з порядку
Ооіііісіеаіез.
А
Рис. 184. Зеріогіа: А - зріз через пікніду; Б - конідієносці з конідіями;
В - конідії
421
Рід аскохіта - Азсосіїуіа (рис. 185). Види цього роду парази-
тують на різноманітних органах рослин, викликаючи їх плямистості.
Плями зазвичай блідо-каштанові з темно-каштановою облямівкою,
іноді у центрі білуваті. Досить розповсюдженим видом є Азсосіїуіа
різі - аскохіта горохова. Гриб паразитує на листках, стеблах і струч-
ках квасолі звичайної (Рііазеоіиз уиїдагіз) та гороху посівного (Різит
заііуит). Пікніди скупчені у центрі плями, приплюснуто-кулясті, світло-
або темно-коричневі. Конідії овальні, із заокругленими кінцями, час-
то нерівнобокі або зігнуті, дещо перешнуровані, з однією (зрідка 2
або 3) перегородкою. А. різі є анаморфою в циклі розвитку аскоміце-
та МусозріїаегеІІа ріпосіез з порядку Ооіііібеаіез.
Рис. 185. Аскохітоз гороху (збудник - Азсосіїуіа різі): А - зовнішній вигляд
ураженої рослини; Б - зріз через пікніду; В - конідії
Види роду філостикта - РііуІІозіісіа - паразитують переважно
на живих листках рослин. Характерним представником роду є Рііуі-
Іозіісіа сгиепіа - філостикта закривавлена. Гриб паразитує на
живих листках конвалії (СопуаІІагіа та]аііз) та купини (Роіудопаіит).
Симптоми хвороби - округлі або еліпсоїдні білі плями з темно-черво-
ною облямівкою. Пікніди поодинокі або скупчені, розвиваються на
422
плямах, а в пікнідах утворюються конідії, які занурені у слиз. Конідії
одноклітинні, гладенькі, безбарвні. Хвороба, яку викликає гриб, має
назву філостиктоз. Слід зазначити, що Р. сгиепіа є незавершеною
стадією - анаморфою - в циклі розвитку аскоміцета биідпагсііа
геіісиїаіа з порядку ОоіНісІеаІез.
Рід церкоспора - Сегсозрога - паразитує на широкому колі
рослин-господарів. Основні симптоми хвороби-бруднувато-сірі, бу-
руваті або бурувато-жовті розсіяні плями з обох боків листків. Най-
розповсюдженішим є гриб Сегрозрога Ьеіісоїа - церкоспора бу-
рякова (рис. 186), який паразитує на листках видів буряка (Веіа).
При висиханні плями у центрі стають майже білі, оточені червону-
вато-бурою облямівкою. Конідієносці прямі, світло-коричневі, розта-
шовані пучками з обох боків листка. Конідії обернено булавовидні,
майже голковидні, до 360 мкм завдовжки, з багатьма перегородками,
безбарвні.
С. Ьеіісоїа є небезпечним збудником церкоспорозу листків буря-
ка, який значно знижує врожай коренеплодів та їх цукристість.
Рис. 186. Церкоспороз буряка (збудник - Сегсозрога Ьеіісоїа): А - зовнішній
вигляд ураженого листка; Б - конідієносці; В - конідії
423
Ліхенізовані гриби або лишайники
Ліхенізованими грибами, або лишайниками називають філогене-
тично гетерогенну групу грибів-симбіотрофів, для яких характерним
є утворення симбіотичних комплексів із водоростями (переважно зе-
леними або синьозеленими) на основі мутуалістичного типу відно-
син. Лишайники характеризуються цілим комплексом морфологічних,
анатомічних та біохімічних особливостей, які обумовлені саме їх сим-
біотрофним способом існування.
Гриб, який утворює слань лишайника, називають мікобіонтом,
водоростевий компонент лишайника - фотобіонтом. Фотобіонт по-
стачає грибу органічні речовини, які утворюються у водоростевій кліти-
ни внаслідок фотосинтезу. Мікобіонт, у свою чергу, забезпечує во-
дорість елементами мінерального живлення та захищає її від впливу
несприятливих умов середовища. Паритетні відносини між міко- та
фотобіонтом не є абсолютно сталими - за умов нестачі поживних
речовин мікобіонт часто паразитує на фотобіонті і може навіть по-
вністю знищувати останнього.
Поняттю «лишайник» можна дати таке визначення: лишайник-
це асоціація одного або двох (рідше більше) мікобіонтів та одного
або двох (рідше більше) фотобіонтів, яка характеризується комплек-
сом морфологічних, анатомічних та біохімічних особливостей.
Сьогодні відомо понад 20 тис. видів ліхенізованих грибів, пере-
важна більшість яких належить до аскомікотових (98% видів). Біля
100 видів входять до відділу базидіомікотових. Понад 300 видів пред-
ставлені анаморфними стадіями, тобто є мітоспоровими грибами.
Фотобіонти представлені приблизно 300 видами водоростей, се-
ред яких понад 200 видів належать до зелених водоростей з класу
ТгеЬоихіорїіусеае (переважно види родів ТгеЬоихіа та Рзеисіо-
соссотуха) та ІЛуорїіусеае (в першу чергу, Тгепіероііііа). Приблизно
чверть видів фотобіонтів є синьозеленими водоростями, серед яких
переважають види роду І\Іозіос.
Морфологічні типи слані лишайників
Слань (або талом) лишайника на 90% складається з гіф мікобіон-
ту. За зовнішнім виглядом та характером прикріплення до субстрату
слані лишайників поділяють на лепрозні (порошкоплодні), накипні,
листуваті та кущисті.
Лепрозна слань нагадує сірувату або зеленкувату цвіль, яка роз-
вивається на поверхні ґрунту або при основі стовбурів дерев. Утво-
424
рена лепрозна слань рихлими скупченнями гіф, які нещільно при-
кріплені до субстрату і легко від нього відокремлюються.
Накипна слань має вигляд кірки або нальоту, щільно прирослого
до субстрату. Зазвичай більшість гіф накипної слані розвивається на
поверхні субстрату, менша частина занурена у субстрат. В окремих
випадках слань може бути повністю занурена у субстрат - на поверхні
залишаються лише плодові тіла, а іноді й плодові тіла розвиваються
всередині субстрату. Накипні лишайники не відокремлюються від суб-
страту і гербаризуються разом із ним.
Листувата слань за формою нагадує платівки з лопатевими чи
розсіченими краями, які у багатьох точках прикріплені до поверхні
субстрату. Майже всі гіфи гриба розвиваються на поверхні субстра-
ту, за винятком тих, що виконують функцію прикріплення. Тому лис-
тувата слань при гербаризації майже цілком і без пошкоджень може
бути відокремлена від субстрату.
Кущиста слань має вигляд прямостоячих чи звисаючих розгалу-
жених або нерозгалужених кущиків. Для неї характерна наявність
лише однієї точки прикріплення.
Забарвлення слані зумовлене наявністю різноманітних хімічних
речовин - як водоростевих пігментів, так і забарвлених вторинних
метаболітів грибів. Через це слань може бути забарвлена різномані-
тно: у сіруваті, сизі, зеленкуваті, коричневі, буро-чорні, жовті, вохряні,
помаранчеві, червоні кольори, і лише як виняток слань буває біла.
Анатомічна будова слані
У більшості випадків основу слані лишайника складає мікобіонт і
він же виконує формоутворюючу функцію. Водоростевий партнер
одного й того ж виду лишайника може бути представлений різними
видами, хоча часто дуже близькими. Лише у групі слизистих лишай-
ників основну масу слані формує симбіотична водорість.
Слань лишайників за типами гіф та їх роташуванням може мати
просту або складну будову. У випадку простої будови слань на попе-
речному перерізі складається з однакових гіф, тобто утворена про-
стою плектенхімою (рис.187).
У складніших випадках сплетення гіф утворюють несправжні тка-
нини (плектенхіми) різних типів. Зокрема, на верхній та нижній по-
верхнях слані розташовуються покривні плектенхіми - верхній та
нижній корові шари відповідно. Середня частина слані утворена пух-
ким плетивом гіф - серцевиною. Між гіфами серцевини розташову-
ються клітини водоростей.
425
Прикріплюються лишайники до субстрату різноманітними спо-
собами. У примітивно організованих форм слань приростає до суб-
страту гіфами серцевинного шару. У більш високоорганізованих ли-
шайників вона прикріплюється пучками гіф - ризинами. Для части-
ни видів характерне прикріплення масивним темнозабарвленим
виростом слані - гомфом. За характером розміщення фотобіонту
слані лишайників поділяють на гомеомерні та гетеромерні.
У гомеомерній слані фотобіонт розміщується дифузно по всій плек-
тенхімі і не має певної зони розташування. У деяких випадках вона
може мати коровий шар. Гомеомерна слань характерна для тих ли-
шайників, у яких фотобіонтом є синьозелені водорості, особливо сли-
зоутворюючі види роду Мозіос.
У гетеромерній слані фотобіонт формує окрему, як правило, добре
виражену зону фотобіонту, зазвичай безпосередньо під верхньою ко-
ровою плектенхімою, утворюючи фотосинтезуючу зону слані. В гете-
ромерній слані фотобіонтом найчастіше є одноклітинні зелені водо-
рості.
Рис. 187. Анатомічна будова слані лишайників: А- проста гомеомерна слань;
Б - гомеомерна слань із коровим шаром; В - гетеромерна слань без нижнього
корового шару; Г - гетеромерна слань із верхнім та нижнім коровими шарами
426
Розмноження лишайників
Ліхенізовані гриби розмножуються переважно вегетативним та
статевим способами, причому при вегетативному розмноженні час-
то утворюються структури, що забезпечують розселення мікобіонта
разом з його фотобіонтом.
Вегетативне розмноження здійснюється фрагментацією слані,
соредіями та ізидіями.
Соредії - це структура з однієї-кількох клітин водорості, оточених
гіфами серцевинної плектенхіми. Останні щільно обростають кілька
водоростевих клітин фотосинтезуючого шару. Внаслідок росту гіф
соредії проривають корову плектенхіму і виходять назовні у вигляді
порошковидного нальоту. Таким чином, власних покривів соредії не
мають. Щільні скупчення соредіїв називають соралями.
Ізидії нагадують соредії - вони також утворені клітинами фотобі-
онту та гіфами мікобіонту. Закладаються ізидії у вигляді горбиків або
паличок на поверхні слані. Центр таких утворень зайнятий гіфами
серцевинної плектенхіми, далі розташовується шар водоростей. Зовні
ізидій вкритий гіфами корового шару плектенхіми. Одним із пошире-
них варіантів ізидій є побули - лусочковидні ізидії, розташовані по
краях слані або плодових тіл ліхенізованих грибів.
Вітер, удари дощових крапель або інші фактори відокремлюють
соредії чи ізидії від слані і сприяють їх потраплянню на новий суб-
страт. За сприятливих умов соредії та ізидії розвиваються у нову
слань.
Нестатеве розмноження відбувається за допомогою пікноконідій.
Вони утворюються в замкнених вмістилищах, розташованих під ко-
ровою плектенхімою - пікнідах. Зовні пікніди нагадують невеличкі
горбочки, розташовані на поверхні слані. Коли пікніда виповнюється
пікноконідіями, то останні проривають корову плектенхіму і виходять
назовні. Проте питання про можливість утворення нової слані лишай-
ників з пікноконідій залишається відкритим.
Статеве розмноження лишайників відбувається за допомогою
аскоспор (у сумчастих лишайників, до яких належить більшість видів)
або базидіоспор (у базидіомікотових лишайників).
Статевий процес у сумчастих лишайників має чимало відмін від
класичної схеми. Зокрема, гачки не утворюються, запліднення кар-
погону не відбувається, натомість сестринські ядра карпогону групу-
ються у дикаріонтичні пари. Далі від карпогону відростають дика-
ріонтичні аскогенні гіфи, в яких відбувається каріогамія та мейоз. Потім
427
з апікальних клітин таких гіф утворюються сумки. У базидіомікотових
лишайників статевий процес апогамний.
Статеві спороношення у сумчастих лишайників найчастіше роз-
виваються у відкритих плодових тілах - апотеціях, рідше - у перите-
ціях та псевдотеціях. Апотеції у лишайників майже завжди зовні ото-
чені т. зв. ексципулом - сплетінням гіф, яке при основі апотецію зли-
вається з гіфами слані. За анатомічною будовою апотеції лишайників
подіяють на два основних типи - лецидеєвидні та леканоровидні
(рис.188).
У лецидеєвидних апотеціїв край апотецію утворений лише гіфа-
ми ексципулу і не містить фотобіонту. Забарвлення диска з гіменієм
та краю апотецію однакове - зазвичай чорне або темне.
У леканоровидних апотеціїв край утворений гіфами ексципулу та
слані, причому сланевий край містить клітини фотобіонту. Тому за-
барвлення краю апотецію чітко відрізняється від забарвлення диска
з гіменієм. Зазвичай край апотецію має колір слані (сірий, зеленкува-
тий), тоді як гіменіальний диск переважно чорний або темно-корич-
невий.
Рис. 188. Схема будови апотеціїв: А - леканоровидний апотецій; Б - леці-
деєвидний апотецій. 1 - гіменій; 2 - ексципул; 3 - сланевий край; 4 - фотобіонт
Спора статевого спороношення (аскоспора або базидіоспора)
потрапляє на придатний субстраті проростає ростковими гіфами. Такі
гіфи після серії поперечних та поздовжніх поділів утворюють розга-
лужене гаплоїдне сплетіння - прототалюс (він буває помітним у дея-
ких накипних форм протягом всього існування у вигляді чорнуватої
або світлої облямівки на периферії слані). Прототалюс спочатку не
має фотобіонту, і його гіфи ростуть радіально в усіх напрямках, доки
428
не зустрінуть клітину необхідного виду водорості. Тоді гіфи оточують
останню і починають формувати слань. Якщо ж гіфи прототалюса не
знайдуть необхідної водорості, то, зазвичай, гриб гине.
Екологічні групи лишайників
Лишайники можна знайти на різноманітних субстратах - на ґрунті,
стовбурах дерев, камінні тощо. Відповідно до субстратної причетності
лишайники поділяють на епігейні, епіфітні та епілітні відповідно.
Зазвичай слань лишайника розвивається на поверхні субстрату
(в епігейних, епіфітних та епілітних видів), проте у деяких видів вона
повністю занурена у субстрат, і на поверхні розвиваються лише пло-
дові тіла або й останні залишаються всередині субстрату. В таких
випадках серед лишайників виділяють також ендофлеодні та ендолітні
види.
Практичне значення лишайників
Основні аспекти практичного використання лишайників пов’язані
з їх здатністю синтезувати специфічні лишайникові речовини та ви-
сокою чутливістю до антропогенного забруднення середовища.
Лишайникові речовини утворюються внаслідок взаємодії вторин-
них метаболітів мікобіонту та фотобіонту. Ці речовини часто (особли-
во у старій літературі) називають «лишайниковими кислотами», сьо-
годні їх відомо близько 700 і вони мають важливе значення для сис-
тематики лишайників. Лишайникові речовини не виявлені в інших
організмів, у тому числі й у вільно існуючих водоростей та грибів. Для
багатьох лишайникових речовин встановлені цінні властивості, зок-
рема антибіотичні, бактерицидні, онкостатичні. Проте широке вико-
ристання лишайникових речовин обмежується труднощами отриман-
ня біомаси лишайників у промислових масштабах. Наприклад, цінний
антибіотик уснеїн, який було відкрито ще в середині XX століття, можна
отримати лише з видів роду СІзпеа. Проте всі представники цього
роду ростуть надзвичайно повільно і, крім того, дуже чутливі до ат-
мосферного забруднення.
Чимало видів ліхенізованих грибів реагують на забруднення се-
редовища. Зокрема, висока чутливість лишайників до забруднення
атмосферного повітря була відмічена вченими понад 200 років тому.
З того часу почались інтенсивні дослідження лишайників як біоінди-
каторів, і сформувався цілий напрямок практичного використання ли-
шайників - ліхеноіндикація. Сьогодні при дослідженні стану повітря-
ного басейну розробляються ліхеноіндикаційні шкали, карти, створе-
429
но декілька ліхенологічних методик оцінки стану атмосферного по-
вітря.
В деяких випадках лишайники використовують для визначення
віку гірських порід чи кам'яних споруд. Ця галузь використання нази-
вається ліхенометрією. Відомо, що тривалість життя різних лишай-
ників різна: деякі лишайники живуть в межах від кількох десятків до
кількох сотень років, але є й такі, що існують усього кілька місяців.
Проте відомі епілітні представники, що можуть розвиватися на кам’я-
нистому субстраті по кілька тисяч років. Саме останню групу лишай-
ників використовують при ліхенометрії.
Основні таксони ліхенізованих грибів та їх місце у системі
Абсолюта більшість ліхенізованих грибів належить до відділу
Азсотусоіа, де розташовується переважно у класах Зогбагіотусеїез
та Іесапоготусеіез. Серед сордаріоміцетів розглядається один по-
рядок - Агіїїопіаіез, серед леканороміцетів - чотири - Теїозсїіізіаіез,
І_есапогаІез, Озігораіез, Реііідегаїез. Крім того, до Іесапоготусеіез
як додатки включено два порядки з недостатньо з'ясованими родин-
ними зв’язками - Рїііусіібаїез та Саіісіаіез.
Лишайники - сордаріоміцети
Порядок Артоніальні - АгіНопіаІез
Слань накипна. Плодові тіла є особливим варіантом апотеціїв -
вони відкриті, неправильної форми, зі слаборозвиненим ексципулом.
Аски товстостінні, мають зовнішній та внутрішній шари, які перед ви-
ходом аскоспор повністю роз’єднуються, і з цього моменту аск стає
схожим на бітунікатний. На відміну від бітунікатних сумок, у асків ар-
тоніальних між зовнішнім та внутрішнім шарами не розвинена систе-
ма пружних мікрофібрил, яка висуває назовні ендоаск. Сумки мають
куполовидний апарат для відкривання асків. Загалом будова асків у
артоніальних вельми схожа із такою у еризіфальних грибів. Аскоспори
здуті, булавовидні до сферичних. У порядку представлені епіфітні,
ендофітні та епілітні лишайники. Характерний рід - Оредгаріїа.
Рід опеграфа - Оредгаріїа (рис. 189). Має накипну просту по-
верхневу або занурену слань без корового шару, що прикріплена до
субстрату гіфами серцевинної плектенхіми. Апотеції лецидеєвидні,
430
чорні, притиснуті до слані, іноді напівзанурені, овальні до широкове-
ретеновидних, рідше круглі, з вузьким щілиновидним або більш-менш
розширеним диском (ліреловидний апотецій). Ексципул чорний. Сум-
ки булавовидні або видовжені, тонкостінні, але на вершині часто по-
товщені, з 8 яйцевидними або видовженими, поперечно-багатоклі-
тинними незабарвленими спорами. Види роду поширені на гладенькій
корі листяних дерев.
Лишайники - лекано-
роміцети
Лишайники з класу лекано-
роміцетів характеризуються
особливим типом аску, який
називають леканоровидний. В
оптичний мікроскоп такий аск
може виглядати або як унітуні-
катний, або як бітунікатний, про-
те за ультратонкою будовою він
є багатошаровим. При розриві
аску в одних порядках шари
роз’єднуються, в інших - зали-
шаються зрослими. Аск має апікальний апарат для розривання, пред-
ставлений найчастіше різноманітними амілоїдними апікальними
структурами. Нижче в межах класу леканороміцетів розглядаються
представники п’яти порядків, серед яких один (Саіісіаіез) наводиться
як додаток, оскільки його родинні зв’язки та коректне місце у системі
сумчастих грибів залишаються нез’ясованими. Відміни між порядка-
ми пов'язані з типом та розташуванням апотеціїв (зокрема, лекано-
ровидні вони чи лецидеєвидні, занурені у слань чи ні), будовою обо-
лонок аску, морфологією парафіз та аскоспор, анатомічною будо-
вою слані.
Порядок 1. Телосхістальні - ТеїозсЬізіаІез
Слань накипна, листувата або кущиста, з розвиненою з обох боків
коровою плектенхімою. Плодові тіла - виступаючі леканоровидні яс-
кравозабарвлені (жовті, жовтогарячі та червоні) апотеції. Аски лека-.
норовидні, з оболонкою з двох зрослих шарів, які при звільненні аско-
спор на верхівці розділяються. Апікальний апарат аску амілоїдний і
характерний лише для представників даного порядку. Парафізи
прості, розширені на верхівці. Аскоспори зазвичай двоклітинні, біпо-
431
лярні, з товстою септою, безбарвні. Фотобіонтами є переважно зелені
водорості з роду ТгеЬоихіа. Характерний представник - рід Хапіїїогіа.
Рід ксанторія - Хапіїїогіа. Слань листувата, дорзовентральна,
вкрита коровим шаром з обох боків, прикріплюється до субстрату
ризоїдами. Апотеції розташовані на поверхні лопатей, рідше по їх
краях, сидячі. Сумки з 8-ма спорами. Спори біполярні, безбарвні. На
території України одним з найпоширеніших лишайників є Хапіїїогіа
рагіеііпа - ксанторія настінна, або золотянка настінна (рис. 190).
Слань золотянки настінної має вигляд правильних округлих розеток
жовтогарячого кольору, складається з великих, широких лопатей. Краї
лопатей виїмчасто-порізані. Апотеції леканорового типу, нагадують
дрібні жовті або жовтогарячі тарілочки, зосереджені в центральній
частині слані. Поверхня слані від КОН миттєво забарвлюється у вин-
но-червоний колір. Фотобіонтами золотянки є різні види роду
ТгеЬоихіа.
Золотянка настінна повсюдно зустрічається на корі дерев, але
може зростати й на обробленій деревині (особливо часто на парка-
нах) та стінах будівель. Поширена по всій земній кулі.
Рис. 190. Хапіїїогіа рагіеііпа: зовнішній
вигляд слані з апотеціями
Порядок 2. Лекано-
ральні - Ьесапогаїез
Слань різноманітна - на-
кипна, листувата, кущиста,
майже завжди з розвиненою
коровою плектенхімою. Пло-
дові тіла - виступаючі лекано-
ровидні або лецидеєвидні
апотеції, переважно забарв-
лені у темний колір. Аски ле-
каноровидні, з товстою бага-
тошаровою оболонкою, шари
якої між собою зростаються і
частково розділяються на
верхівці перед виходом аскоспор. Апікальний апарат аску амілоїд-
ний. Парафізи розгалужені, на верхівках розширені. Аскоспори пере-
важно одноклітинні без септ або двокпітинні з однією товстою сеп-
тою, безбарвні або темно-коричневі. Фотобіонтами є різноманітні роди
зелених водоростей, переважно з класу ТгеЬоихіорїіусеае.
432
Найчисельніший порядок серед грибів - леканороміцетів. Нижче
леканоральні лишайники згруповані за морфологічними типами слані
та субстратною причетністю у чотири групи: накипні (І_есісІеа,
1_есапога, СапсІеІагіеІІа), листуваті (РІіузсіа, Рагтеїіа), кущисті епігейні
(СІасіопіа, Сеігагіа) та кущисті епіфітні (Ратаїіпа, Еуегпіа, Апаріусіїіа,
ІІзпеа).
Накипні леканоральні
Рід лецідея - Ьесісіеа (рис. 191). Має накипну слань у вигляді
цілісної або потрісканої кірки, з рівною, бородавчастою, зернистою
або порошистою поверхнею, рідше у вигляді тонкого борошнистого
нальоту. Апотеції лецідеєвого типу округлі або від взаємного тиску
кутасті, чорні, твердої консистенції. Сумки з 8-ма спорами (дуже рідко
16-ма). Спори прості, безбарвні, здебільшого еліптичні, рідше кулясті
чи яйцевидні. Більшість видів є епілітними ліхенізованими грибами.
Рис. 191. Ьесідеа: А - загальний вигляд накипної слані з апотеціями;
Б - розріз через апотецій
Рід леканора - і_есапога (рис. 192). Слань однорідна, накипна,
гладенька, зерниста або бородавчаста, іноді у вигляді окремих гор-
биків або лусочок, часто малопомітна, прикріплена до субстрату. Зро-
стає на різноманітних субстратах. Апотеції сидячі, з плоским або ви-
пуклим, рідко увігнутим диском, зазвичай оточені сланевим краєм
(леканоровий тип). Сумки з 8-ма спорами, рідко 16-32-ма. Спори Од-
ноклітинні, безбарвні, здебільшого еліптичні, важко піддаються ви-
значенню. Дуже поширеним видом, що часто зустрічається на корі
листяних дерев, є Ьесапога аііоріїапа - леканора різноманітна.
433
Рис. 192. Ьесапога: А - загальний вигляд накипної слані з апотеціями;
Б - розріз через апотецій
Рід канделарієла - СапсІеІапеІІа. Лишайник має накипну гете-
ромерну жовтувату слань, яка щільно прикріплюється до субстрату.
Поверхня слані зерниста або бородавчаста. Верхній коровий шар
слаборозвинений. Апотеції леканорового типу, округлі або кутасті,
сидячі. Спори по 8-32 у сумках, безбарвні, одно- або двокпітинні, ви-
довжені. Одним з найпоширеніших видів є СапсІеІапеІІа аигеїіа— кан-
делярієла золотистенька. Цей вид часто зустрічається на вапнякових
субстратах та на бетонних спорудах. Слань чорнувато-жовтого кольо-
ру, часто так зливається із субстратом, що стає майже непомітна.
Апотеції 0,2-1,5 мм, жовтого кольору. Канделярієла золотистенька
досить стійка до забруднення оточуючого середовища і тому, на
відміну від багатьох інших лишайників, є типовим представником ліхе-
нофлори великих міст, де розвивається переважно на бетонних
стовпах.
Листуваті леканоральні
Рід фісція, або стрільчатка - РІіузсіа (рис. 193). Слань листу-
вата, у вигляді правильних розеток, рідше - листуватих кірок, що
щільно приростають до субстрату за допомогою чисельних ризоїдів.
Якщо розглядати слань зверху, ризоїди помітні у вигляді дрібних війок
по краю лопатей. Поверхня слані зазвичай щільна, рівна, гладенька,
без зморшок та нерівностей, сіра або коричнева, часто з білуватим
або сизим нальотом. Апотеції леканорового типу. Спори двокпітинні
коричневі або сіро-коричневі, з нерівномірно потовщеною оболон-
кою, еліптичні або бобовидні.
434
Одним із найпоширеніших видів є Рііузсіа зіеііагіз (фісція зірча-
ста), яка має слань у вигляді щільно прирослих до субстрату розеток.
Зверху розетки сірі, а знизу - світлі, з густим шаром сірих ризоїдів.
Лопаті слані вузькі, витягнуті. Зверху слань від КОН жовтіє. Апотеції
численні, з темно-коричневим диском. Часто зустрічається на корі
листяних, рідше - хвойних порід, а також на камінні.
Рис. 193. Рїіузсіа: зовнішній вигляд
слані
Рід пармелія -
Рагтеїіа. Слань листувата,
розрізано-лопатева, у виг-
ляді великих розеток, при-
кріплена до субстрату ризи-
нами, рідше вільна. Лопаті
різноманітні: вузькі або ши-
рокі, сильно або слабо роз-
галужені, плоскі або випуклі,
щільно зімкнені або розді-
лені. Верхня сторона від
білувато-сіруватої та жовту-
ватої до коричневої та чор-
ної, матова або блискуча;
нижня - від білуватої або
світло-коричневої до чорної.
Ризини добре розвинені, прості або розгалужені. Часто розвивають-
ся соредії та ізидії різної форми. Апотеції леканорового типу, сидячі
або на ніжках. Спори одноклітинні, безбарвні, від еліптичних до куля-
стих, по 8 у сумці. Часто види роду розвиваються як епіфіти на корі
дерев, рідше - на ґрунті, скелях. В Україні найпоширенішим видом
роду є пармелія борозенчаста (Р.зиісаіа). Характерною ознакою
цього виду є розтріскані вузькими щілинами сітчасті зморшки верхнь-
ого корового шару (боріздчасті соралі), в яких виступають соредії.
Кущисті епігейні леканоральні
Рід кладонія - СІадопіа. Види цього роду мають первинну та
вторинну слань. Первинна слань накипна або луската. У частини видів
вона зберігається протягом всього існування лишайника, в інших -
швидко зникає. В останньому випадку зберігається лише вторинна
слань (подеції), що розвилася на первинній. Подеції всередині по-
рожнисті, мають різноманітну форму: паличковидні, сцифовидні (у ви-
435
гляді бокальчика) або кущисті з розгалуженими округлими в перерізі
гілками. Сцифовидні подеції часто проліферують-зцентруабозкраю
дорослої сцифи розвиваються дочірні. На подеціях утворюються чер-
воні, біло- або темно-коричневі плодові тіла, які представляють особ-
ливий варіант лецидеєвидних апотеціїв - т. зв. біаторові. Спори од-
ноклітинні, дуже рідко дво- до чотириклітинних, безбарвні, яйцевидні
або видовжені, по 8 у сумці.
Численні види роду кладонія поширені майже по всій земній кулі.
Деякі види у північних районах Євразії складають основну частину
раціону оленів і відомі під загальною назвою оленячого моху або яге-
ля. Найпоширенішими видами в Україні є кладонія бахромчаста, кла-
донія лісова та кладонія оленяча.
Кладонія бахромчаста - СІадопіа /ітЬгіаіа (рис. 194). Цей ли-
шайник має мішаний тип слані — листувата та кущиста. Первинна
слань листувата, довго зберігається у вигляді дрібних лусочок 2-
10 мм завдовжки, що нерівномірно виїмчасто посічені, зверху мають
сірувато-зелений, а знизу - білий колір. Подеції кущисті, складають-
ся з правильних простих сірувато-зеленкуватих сциф 1—4 см заввиш-
ки. Останні вкриті тонким, проте густим нальотом соредіїв. Апотеції
коричневі, розміщені по краю сциф. Від КОН слань не змінює свій
колір. Лишайник часто зустрічається на гнилих пеньках, повалених
стовбурах, рідше - на ґрунті (піщаному, гумусному, торф’янистому),
в усіх типах лісів.
Рис. 194. СІадопіа (ітЬгіаіа: будо-
ва слані. 1 - первинна листувата слань;
2 - вторинна слань - подеції (сцифи)
У кладонії лісової -
СІадопіа зуіуаііса - дуже швид-
ко розвивається вторинна слань,
і тому в неї первинну слань спос-
терігати не вдається. Подеції кла-
донії лісової сірувато- або жовту-
вато-зелені, до 10 см заввишки,
дуже розгалужені і в місцях галу-
ження часто з отворами, від КОН
злегка жовтіють або зовсім не
змінюють колір. Кінцеві гілочки
чисельні, односторонньо відігнуті
донизу, з коричнюватими кінчика-
ми. Апотеції коричневі. Характер-
на особливість цього виду - слань
пекучо-гірка на смак. Лишайник
436
часто розвивається на піщаних ґрунтах, торф’яниках, гнилих пеньках
як у лісі, так і на відкритих місцях.
Кладонія оленяча - СІадопіа гапдіїегіпа - також має первинну
слань, що швидко зникає. Подеції сіруваті або сірувато-білуваті, до
20 см заввишки, дуже розгалужені, особливо у верхній частині. Кінцеві
гілочки нахилені в один бік, і їх верхівки зазвичай темно-коричневі.
Утворює щільні дернинки. Від КОН слань жовтіє. Апотеції дуже дрібні,
коричневі. Розвивається на піщаних ґрунтах, торф’яниках, у світлих
соснових лісах. Кладонія оленяча має найбільшу за розмірами слань
серед тих видів роду, які мають деревовидно розгалужені подеції.
Рід цетрарія - Сеігагіа. Лишайники з гетеромерною листуватою
(у більшості видів) або кущистою дорзовентральною сланню. Апо-
теції леканорового типу. Спори одноклітинні, прямі, еліптичні до ку-
лястих, безбарвні, по 6-8 у сумці. Види цього роду ростуть переваж-
но на ґрунті та на продуктах вивітрювання гірських порід. До найпо-
ширеніших на території України видів належать цетрарія ісландська
та цетрарія сніжна.
Цетрарія ісландська або ісландський мох - Сеігагіа ізіапсііса
(рис. 195). Слань має вигляд білувато- або зеленкувато-коричневих
кущиків, що складаються з плоских, коритовидно згорнутих лопатей
до 10 см заввишки та 0,5—4 см завширшки. Знизу лопаті червонуваті.
Рис. 195. Сеігагіа ізіапсііса:
загальний вигляд кущистої слані
з війками по краях лопатей
їх краї зазвичай з війками. Від КОН
слань жовтіє. Зрідка на ній розви-
ваються соредії та ізидії. Апотеції
також зустрічаються зрідка - вони
розташовуються на краях або на
верхівках розширених лопатей, і
забарвлені так само, як слань. Ро-
сте на ґрунті в соснових лісах, на
болотах серед мохів. В Ісландії та
інших північних країнах цетрарію
ісландську після спеціальної об-
робки використовують як домішку
до борошна при випіканні хліба. У
народній медицині відвари ісланд-
ського моху використовують як сти-
мулюючий засіб для підвищення
загального тонусу організму. Вхо-
дить у раціон північних оленів.
437
Кущисті епіфітні леканоральні
Рід рамаліна - Катаїіпа (рис. 196). Для видів цього роду харак-
терна кущиста прямостояча або звисаюча, досить жорстка слань, при-
кріплена до субстрату гомфом. Апотеції леканорового типу, які у дея-
ких видів трапляються дуже рідко. Спори двокпітинні, безбарвні, ви-
довжено-еліптичні, по 8 у сумці. Ростуть на корі дерев, ґрунті, камінні.
Речовини, що екстрагують з видів роду рамаліна, використовують у
парфумерній промисловості.
Найпоширенішим видом цього роду є рамаліна борошниста
(/?. ґгахіпеа), яка має дуже жорстку слань у вигляді прямостоячих
слабо розгалужених кущиків до 4-15 см заввишки. Стрічковидні ло-
паті до 3 см завширшки, однакові сірувато-зеленкуваті з обох боків,
складчасто-зморшкуваті, без соредіїв. Апотеції сидять на коротких
ніжках. Диски апотеціїв 3-5 мм діаметром, рідше до 15 мм. Зустрі-
чається на корі листяних дерев у гарно освітлених місцях.
Рис. 196. Ратаїіпа: загальний
вигляд слані з подеціями на верхів-
ках
Рід евернія - Еуетіа. Одні
з найбільш розповсюджених на
земній кулі лишайників. Мають
м’яку кущисту дорзовентральну
слань, яка прикріплюється до суб-
страту гомфом або (дуже рідко)
ризинами. Апотеції леканорово-
го типу, трапляються рідко. Спо-
ри одноклітинні, еліптичні,
безбарвні, по 8 у сумках. Зустрі-
чаються на корі дерев, оголеній
деревині, ґрунті. На території Ук-
раїни заготовляють евернію
сливову (дубовий мох) - Е.
ргипазігі (рис. 197), екстракти з
якої використовують у парфу-
мерній промисловості. Для цьо-
го лишайника характерна кущис-
та, прямостояча або звисаюча,
щільно прикріплена до субстра-
438
ту гомфом м’яка слань, до 10 см довжиною, з плоскими або трохи
жолобчастими неширокими дихотомічно розгалуженими лопатями.
Зверху слань зеленувата, а знизу білувата.
Рис. 197. Еуегпіа ргипазігї. за-
гальний вигляд слані
Рис. 198. ІІзпеа: загальний
вигляд слані з апотеціями на вер-
хівках
Рід анаптіхія - АпаріусМа.
Слань кущиста (у деяких видів ли-
стувата), у вигляді розеток або зви-
саючих дуже розгалуджених ку-
щиків, які прикріплюються до суб-
страту ризоїдами. Лопаті часто по
краю з вієчками. Апотеції лекано-
рового типу. Спори двокпітинні,
темні. Ростуть на корі дерев та
камінні. Види роду досить стійкі до
забруднення навколишнього сере-
довища.
Рід уснея або бородач -
ІІзпеа (рис. 198). Слань лишай-
ників кущиста, складається з однієї
або частіше з багатьох прямостоя-
чих чи звисаючих гілочок, що більш-
менш дихотомічно розгалужені. На
поперечному зрізі слані добре по-
мітне щільне сплетення гіф, що
утворюють так звану хондроїдну
плектенхіму, яка виконує механічні
функції. Апотеції леканорового
типу. Росте у хвойних та мішаних
рівнинних і гірських лісах. Деякі
види, зокрема бородач шорсткий
(ІІзпеа Ііігіа), продукують антибіо-
тичні речовини - уснеїнову кисло-
ту. Види роду дуже чутливі до ат-
мосферного забруднення.
Порядок 3. Остропальні -
Озігораіез
Слань переважно накипна, без
корової плектенхіми. Плодові тіла
439
глибоко занурені у слань лецидеєвидні апотеції, з твердим ексципу-
лом, який часто просякується вапном. Аски леканоровидні, багатоша-
рові, шари зрослі. Апікальний апарат аску неамілоїдний. Парафізи
прості, часто ослизнюються. Аскоспори видовжені, переважно нитко-
подібні, багатоклітинні, з поздовжніми та поперечними септами (т. зв.
муральні). Фотобіонтами є зелені водорості з роду Тгепіероііііа
(ІЛуорїіусеае).
Рід графіс- вгаріїіз- як й інші остропальні лишайники, має на-
кипну дуже просту в анатомічному відношенні слань, зі слабо або
зовсім нерозвиненим коровим шаром. Апотеції занурені або сидячі,
витягнені у вигляді рисочок. Найпоширенішим видом роду є Сгаріїіз
зсгіріа — графіс письменний (рис. 199), який зустрічається по всій
земній кулі на корі дерев та кущів у широколистяних, рідше хвойних
лісах. Слань графісу письменного дуже тонка, має вигляд поодино-
ких або таких, що зливаються між собою, плям сірого, сизого або
оливково-сірого кольору. По краю слань оточена чорним вінчиком
прототалюса. Апотеції чисельні, видовжені, прямі або звивисті, зо-
середжені переважно у центральній частині слані, один відносно
іншого розміщені більш-менш паралельно або утворюються зірчасті
чи неправильної форми групи і нагадують арабські ієрогліфи. Молоді
апотеції занурені у слань, старі - напівзанурені, до 3 мм завдовжки
та 0,3-0,4 мм завширшки, можуть бути простими або розгалужени-
ми. Спори циліндричні, звичайно загострені на одному кінці 7-10
(15)-клітинні, безбарвні. Від КОН слань не змінює свій колір. Фотобі-
онтом у цього виду є ульвофіцієві водорості з роду Тгепіероііііа.
Порядок 4. Пельтигеральні - РеІІідегаІеа
Слань листувата, з коровою плектенхімою, розвиненою або лише
Рис. 199. бгарїііз зсгі-
ріа: зовнішній вигляд слані
з апотеціями
на верхньому боці, або на обох сторонах
слані. Плодові тіла - глибоко занурені у
слань лецидеєвидні апотеції, які на почат-
кових стадіях розвитку прикриті покривною
плектенхімою. Аски леканоровидні, мають
оболонку з двох незрослих між собою
шарів і нагадують бітунікатні (проте поз-
бавлені системи пружних мікрофібрил, яка
висуває ендоаск назовні). Апікальний апа-
рат аску амілоїдний. Парафізи прості. Ас-
коспори безбарвні або коричневі, видов-
440
жені, з кількома поперечними септами. Фотобіонтами є переважно
зелені водорості з роду Рзеисіососсотуха (ТгеЬоихіорНусеае) або си-
ньозелені водорості (найчастіше види роду №>8іос).
Рід пелтігера - Реііідега (рис. 200). Слань листувата, горизон-
тально розпростерта, загнута угору по краях, знизу помітна сітка жи-
лок, яка у багатьох видів зливається у суцільну повстяну масу і має
чисельні пучки ризин. Нижня кора ніколи не розвивається. Апотеції
сидячі, великі, забарвлені в коричневий колір і розташовуються на
верхньому боці звужених лопатей. На молодих стадіях диск має по-
крив, який з віком розривається і зберігається лише на бережках апо-
теціїв, утворюючи несправжній край. Сумки із 6-8 спорами. Спори
безбарвні до буруватих, веретеновидні до голковидних. На території
України повсюдно зустрічається пельтигера собача.
Пелтігера собача - Реііідега сапіпа. Слань у цього епігейного
лишайника листувата, крупна, лопаті до 11 см завдовжки і до 3,5 см
завширшки. Верхня сторона слані попелясто- або коричнювато-сіра,
з тонким повстистим нальотом; нижня — світла, рожевувато-біла, до
центру темнішає, з густою мережею вузьких світло-коричневих жи-
лок та з білими довгими (до 1 см) ризинами. Апотеції розвиваються
зрідка і розташовуються на піднятих догори лопатях; вони є опукли-
ми, з червоно-коричневим диском. Спори восьмиклітинні. Лишайник
росте в затінених вологих місцях, переважно на ґрунті, рідше на по-
рослих мохом скелях та пеньках.
Рис. 200. Реііідега'. загальний вигляд
слані
Рід лобарія - ЬоЬагіа -
на території України пред-
ставлений лише рідкісними та
зникаючими видами. Мають
великолистувату слань, вкри-
ту з нижнього боку низеньким
пушком. У Гірському Криму та
в Українських Карпатах мож-
на знайти вид ЬоЬагіа
риїтопагіа - лобарія леге-
нева (рис. 201). Лишайник
має листувату слань до 10-
30 см завдовжки та 5-20 см
завширшки, що зростається із
субстратом лише одним кра-
441
єм. Лопаті слані 1-5 см завширшки, по краю вирізблені. Зверху слань
світло-коричнева, сітчасто-складчаста, з глибокими западинами, що
розділені ребрами. Цим западинам із нижнього боку слані відповіда-
ють білуваті голі опуклості, які розділені ділянками з коротким густим
опушенням. Уздовж ребер на верхньому боці слані розміщені великі
соралі. Апотеції зустрічаються зрідка і розміщуються вздовж країв ло-
патей. Спори дво-, чотириклітинні, веретеновидні, безбарвні. Зу-
стрічається на стовбурах листяних дерев, рідше на порослих мохом
скелях. Використовується у парфумерній промисловості.
Додаток. Порядок Каліціальні - Саіісіаіез
Слань соредіозна. Апотеції (коли наявні) лецидеєвидні. Аски з
тонкою оболонкою, близькі до протоунікатних, аскоспори звільняються
пасивно. Парафізи відсутні. Залишки оболонок асків разом з аско-
спорами на поверхні апотеціїв утворюють порошистий наліт - ма-
цедій. Прикладом теломорфних каліціальних грибів є рід каліцій.
Рід каліцій - Саіісіит. Характеризується соредіозною, дуже сла-
бо розвиненою сланню - рихлим сплетенням гіф, між якими розта-
шовуються клітини фотобіонту. Апотеції сидячі або піднімаються від
окремих гіф вертикально догори на довгих ніжках (до 2 мм). Сумки
циліндричні, з 8-ма спорами. Спори двокпітинні. Оболонки асків швид-
ко руйнуються, через що апотеції зверху вкриті скупченнями аскос-
пор та залишками стінок сумок - мацедієм. Каліцій зустрічається на
вологій корі дерев та на оголеній деревині і нагадує сіро-зелений ви-
сип. Види роду надзвичайно чутливі до змін екосистем і є надійними
індикаторами пралісових масивів.
А Б
Рис. 201. 1_оЬагіа риїтопагіа: А — верхній бік слані; Б — нижній бік слані
442
Додаток. Штучна група лишайників, які не мають органів
розмноження (ЬісНепез Ітрегїесіі)
Це збірна група ліхенізованих грибів, які постійно залишаються
стерильними, плодові тіла у них не утворюються або досі не знай-
дені, а філогенетичні зв’язки залишаються нез’ясованими. Для окре-
мих лишайників з цієї групи за вторинними ознаками (цитолого-мор-
фологічними, біохімічними) вдалося знайти двійників серед
теліоморфних грибів і з’ясувати їх систематичне положення. Водно-
час для ряду лишайників не вдається встановити не лише система-
тичне положення на рівні роду або родини, а навіть на рівні відділів
Азсотусоіа чи Вазісііотусоіа.
Слань у вигляді порошистої поволоки вкриває субстрат. Апотеції
невідомі.
Рід лепрарія - 1_ергагіа. Слань виглядає майже ідентично до
каліцію - вона соредіозна, порошковидна, але не утворює плодових
тіл і являє собою анаморфну стадію. Сірувато-зелені нальоти леп-
рарії зустрічаються повсюдно у широколистяних лісах на оголеному
вологому ґрунті та при основі стовбурів дерев.
Покажчик латинських назв таксонів
АсеіаЬиІагіа, 209
АсеіаЬиІагіа тесіііеггапеа, 209
АсЬпапіЬаІез, 117
Асгазіаіез, 252
Асгазіотусеіез, 251
Асгазіотусоіа, 18, 249
Асгазіз, 252
Асгазіз гозеа, 252
Асгетопіит, 336, 417
Асгетопіит уегіісіїїаіит, 417
Адагісасеае, 384
Адагісаіез, 375
Адагісиз, 384
Адагісиз Ьізрогиз, 385
Адагісиз сатрезігіз, 386
Адагісиз хапіЬосІегтиз, 386
АЬпТеїііа, 161
АЬпТеІІіаІез, 160
АІІотусез, 297
АІуеоіаіез, 14, 22, 124
Атапііа, 387
Атапііа саезагеа, 389
Атапііа тизсагіа, 367, 389
Атапііа рапіЬегіпа, 367
Атапііа рЬаІІоісІез, 368, 388
Атапііа уета, 368, 389
Атапііа уігоза, 368, 389
Атапііасеае, 387
АтоеЬісііаІез, 305
АтоеЬоПадеііаіез, 14, 19, 72
АтрЬісІіпіаІез, 130
АтрЬісІіпіит, 130
АпаЬаепа, 61
АпаЬаепа Поз-адиае, 62
АпаЬаепа зсгіегетеііеуі, 62
АпаЬаепа зрігоісіез, 62
АпаріусЬіа, 439
Апітаііа, 13
Апізоірісііит, 281
АрЬапіготепоп, 60
Аргіапіготепоп Поз-адиае, 60
Арісотріеха, 22
Арізіопета, 140
Агсугіа, 260
Агсугіа сіеписіаіа, 261
АгтіІІагіа, 381
АгтіІІагіа теііеа, 381
АгіЬопіаіез, 430
АгіЬгозріга, 59
АзсосЬуіа, 422
АзсосЬуіа різі, 422
Азсотусеіез, 318
Азсотусоіа, 24, 305
АзеІІагіаіез, 305
Азрегдіііиз, 336, 419
Азрегдіііиз Лауиз, 420
Азрегдіііиз Титідаіиз, 420
Азрегдіііиз огу/ае, 420
Азрегдіііиз герепз, 420
Азрегдіііиз уєгзісоіог, 420
АзіегіопеІІа, 113
Аигісиїагіа, 406
Аигісиїагіа аигісиїа-іисіае, 406
Аигісиїагіаіез, 406
ВасіІІагіаІез, 119
ВасіІІагіорЬусеае, 113
ВасіІІагіорЬуіа, 20, 105
Вапдіа, 154
Вапдіаіез, 154
ВапдіорЬусеае, 152
Вазісііотусеіез, 343
Вазісііотусоіа, 24, 338-
Ваігасгіозрегтит, 157
Веіа, 423
Віазіосіасііаіез, 296
ВІазіосІіпірЬусеае, 130
ВІазіосІіпіит зріпиіозит, 130
ВІитегіа, 321
ВІитегіа дгатіпіз, 321
Воіеіасеае, 369
Воіеіаіез, 369
Воіеіиз, 369
444
Воіеіиз Ьасііиз, 371 СЬІатусІотопаз тоем/изіі, 173
Воіеіиз есіиііз, 370 СЬІатусІотопаз геіпЬагсНіі, 173
Воіеіиз егуіЬгориз, 370, 371 СЬІогагасЬпіаІез, 73
Воіеіиз Іигісіиз, 366 СЬІогагасЬпіоп, 73
Воіеіиз заіапаз, 366, 370 СЬІогагасЬпіорЬусеае, 73
Воігусііаіез, 93 СЬІогагасЬпіорЬуІа, 19, 72
Воігусііорзіз, 93 СЬІогеІІа, 193
Воігусііит, 92 СЬІогеІІаІез, 193
Воігуііз, 324 СЬІогососсаІез, 179
Вгасіеасоссиз, 183 СЬІогососсит, 179
Вгуорізіз, 277 СЬІогососсит сІірІоЬіопіісит, 180
Вгуорзісіаіез, 204 СЬІогорЬусеае, 171
Вгуорзіз, 205 СЬІогорЬуІа, 23, 168
ВиІЬосЬаеіе, 189 СЬопсігиз, 162
Саіісіаіез, 442 СЬготиІіпа, 81
Саіісіит, 442 СЬгоососсаІез, 55
Саіуаііа, 399 СЬгоотопаз асиїа, 143
СапсІеІагіеІІа, 434 СЬгузосЬготиІіпа, 138
СапсІеІагіеІІа аигеїіа, 434 СЬгузосЬготиІіпа роїуіеріз, 138
Сапсіісіа, 314, 420 СЬгузорЬусеае, 81
Сапсіісіа аІЬісапз, 421 СЬгузорЬуІа, 23, 78
СапіЬагеІІаІез, 361 СЬгузоііІа, 139
СапіЬагеІІиз, 363 СЬуігісІіаІез, 291
СапіЬагеІІиз сіЬагіиз, 363 СЬуІгісІіотусеіез, 291
Саиіегра, 208 СЬуІгісІіотусоіа, 23, 289
Саиіегра ргоіііега, 208 СІасІопіа, 435
СепігорЬусеае, 111 СІасІопіа ЛтЬгіаіа, 436
Сегатіаіез, 164 СІасІопіа гапдИегіпа, 437
Сегатіит, 164 СІасІопіа зуіуаііса, 436
Сегаїіотуха (гиіісиїоза, 258 СІасІорЬога, 200
Сегаїіотухаїез, 258 СІасІорЬога діотегаїа, 202
Сегаііит, 131 СІасІорЬога уадаЬипсІа, 202
Сегаіотуха, 255 СІасІорЬогаїез, 200
Сегсозрога, 423 СІаІЬгиз, 404
Сегрозрога Ьеіісоїа, 423 СІаіЬгиз гиЬег, 404
Сеігагіа, 437 СІауагіа, 362
Сеігагіа ізіапсііса, 437 Сіауісерз, 326
СЬаеіорЬогаІез, 180 СІауісерз ригригеа, 326
СЬага, 212, 288 СІауиІіпа, 363
СЬагасіорзіз, 93 СІіІосуЬе, 379
СЬагаІез, 211 СІіІосуЬе сеггизаіа, 380
СЬагорЬусеае, 211 СІіІосуЬе сіауірез, 366
СЬіІотопаз, 145 СІіІосуЬе сІеаІЬаіа, 367
СЬІатусІотопаз, 172 СІіІосуЬе пеЬиІагіз, 366
445
Сіозіегіит, 219
Соссоїіігіаіез, 139
Соссопеіз, 117
Сосііоіит, 195
Сосііит, 206
Собіит уегтіїага, 207
Соїасіит, 65
Соїеосгіаеіаіез, 222
Соїеосгіаеіе, 223
СоїеосЬаеІе риіуіпаіа, 223
Соїеосгіаеіе зсиіаіа, 223
Сотосіаігігиз, 310
Сотрзородоп, 153
Сотрзородопаїез, 153
СопіорЬога, 347
СопуаІІагіа та]аііз, 422
Соргіпасеае, 392
Соргіпиз, 392
Соргіпиз аігатепіагіиз, 366, 393
Соргіпиз сотаїиз, 392
Соргіпиз тісасеиз, 366
Согаіііпа, 158
СогаІІіпаІез, 158
Согсіусерз, 329
Согсіусерз тіїііагіз, 329
Согсіусерз орґііодіоззоісіез, 337
Согііпагіасеае, 390
Согііпагіиз, 368, 391
Согііпагіиз огеїіапиз, 368, 390
Согііпагіиз зресіоззітиз, 368
Согііпагіиз уіоіасеиз, 391
Созсіпосіізсоргіусеае, 111
Созтагіит, 220
Созтагіит саеіаіит, 221
Созтагіит тагдагііИегит, 221
Созтагіит ругатісіаіит, 221
Созтагіит гепііогте, 221
СозІапііпеІІа, 331
СгаіегеІІиз, 363
СгаіегеІІиз сіпегеиз, 363
Сгуріотопасіаіез, 143
Сгуріотопаз, 143
Сгуріотопаз сгуорЬііа, 144
Сгуріотопаз (гідогіз, 144
Сгуріотопаз заііпа, 144
Сгуріотопаз зіідтаііса, 144
Сгуріорґіусеае, 143
Сгуріоргіуіа, 23, 141
СуапорЬога, 147
СуапорЬога рагасіоха, 148
Суапоргіогаїез, 147
СуапорЬусеае, 55
Суапоргіуіа, 18
Суапоріусгіе, 146
Суаігіиз, 402
Сутаіоріеига, 121
СутЬеІІа, 117
СутЬеІІаІез, 115
Сузіозеіга, 104
□аесіаіеа, 348, 352
□аесіаіеа диегсіпа, 347, 352
□азусіасіаіез, 209
□егЬезіа, 205
□езтісііаіез, 218
□езтососсиз, 192
□езтососсиз оііуасеиз, 193
□еиіеготусеїез, 246, 416
□еиіеготусоїа, 417
□іаіота, 113
□ісіуосгіа, 122
□ісІуосгіорЬусеае, 122
□ісіуосгіоргіуіа, 21
□ісіуоргіога, 404
□ісіуоргіога сіиріісаіа, 404
□ісіуозіеііаіез, 265
□ісіуозіеііотусеіез, 264
□ісіуозіеііит сіізсоісіеит, 264
□іпогігуоп, 82
□іпоргіусеае, 130
□іпоргіузіз, 134
□іпоргіуіа, 22, 124
□ізсісгізіаіез, 13, 18, 64
□оігіісіеотусеїез, 335
□гарагпаїсііа, 182
□гарагпаїсііа діотегаіа, 182
□ипаїіеііа, 174
□ипаїіеііа заііпа, 174
Ессгіпаїез, 305
446
ЕІарЬотусез, 337
ЕІарЬотусез дгапиіаіиз, 337
Етіїіапіа, 140
Епсіодопаїез, 304
Епсіодопе ІасІІЛиса, 304
ЕпІеготогрЬа, 199
ЕпіЬоІотаіасеае, 376
ЕпіЬоторЬІЬогаІез, 302
ЕпІЬоторЬіога тизсае, 303
Епіоіота сіуреаіит, 377
Епіоіота зіпиаіит, 377
ЕпіоторЬІЬога, 303
ЕрісЬІое, 328
ЕрісЬІое іурЬіпа, 328
ЕгузірЬаІез, 319
ЕиЬасіегіа, 18
Еисагуоіа, 9, 18
Еисіогіпа, 176
Еидіепа, 68
Еидіепа асиз, 70
Еидіепа зрігодуга, 70
Еидіепа уігісііз, 70
Еидіепаіез, 68
ЕидІепатогрЬа, 71
ЕидІепатогрЬаІез, 71
ЕидІепорЬусеае, 68
ЕидІепорЬуІа, 18, 64
Еигоііаіез, 336
Еигоііотусеіез, 336
Еигоііит, 337
Еизіідтаіаіез, 86
ЕизіідтаїорЬусеае, 86
ЕизіідтаїорЬуІа, 20, 85
Еизіідтаїоз, 86
Еизіідтаіоз тадпиз, 86
Еуегпіа, 438
Еуегпіа ргипазігі, 438
Еізіиііпа Ьераііса, 347, 365
Еізіиііпаїез, 365
ЕІогісІеорЬусеае, 156
Еотез, 348, 357
Еотез (отепіагіиз, 347, 357
Еотііорзіз, 348, 352
Еотіїорзіз ріпісоїа, 352
Егадіїагіа, 113
Егадіїагіаіез, 113
Еисаіез, 102
Еисиз, 102
Еиіідо, 262
Еиіідо зерііса, 262
Еипді, 13, 23, 288
Еипді Ітрегїесіі, 246, 417
Еизагіит топіІИогте, 330
Еизісіасііит рігіпит, 335
бапосіегта, 348, 358
бапосіегта Іисісіит, 358
беазігит, 399
біЬегеІІа, 330
біЬегеІІа Ги)ікигоі, 330
бідагііпаїез, 161
біаисосузіаіез, 148
біаисосузііз, 148
біаисосузііз позіосЬіпеагит, 149
бІаисосузІорЬусеае, 147
бІаисосузіорЬуІа, 23, 145
біоеосарза, 57
бІоеоігісЬіа, 59
біотиз, 304
ботрЬісІіасеае, 375
ботрЬісііиз, 375
ботрЬісііиз діиііпозиз, 375
ботрЬопета, 116
бопіаиіасаіез, 130
бопіаиіах, 131
бопіозіотит, 76
бопіит, 175
бгарЬіз, 440
бгарЬіз зсгіріа, 440
биідпагдіа геїісиїаіа, 423
биііиііпаїез, 253
биііиііпорзіз, 253
бутпосііпіит, 132
бутпосііпіит аегидіпозит, 133
бутпосііпіит уігісіе, 133
буготііга, 333
Наетаіососсиз, 174
Наетаіососсиз ріиуіаііз, 175
Наїітесіаіез, 207
447
НаріорЬуіа, 22, 136
Нагреііаіез, 305
Неіоііаіез, 324
НеІуеІІа, 332
Неіегососсиз, 93
НусІпеІІит, 360
НусІпеІІит зиауеоіепз, 360
НусІпеІІит гопаіит, 360
Нусіпит, 364
Нусіпит герапсіит, 364
Нусігосіісіуоп, 183
Нусігосіісіуоп геіісиїаіит, 185
НудгорЬогасеае, 377
НудгорЬогиз, 378
НутепосЬаеіаІез, 360
НутепосЬаеіе, 360
Нутепосіїаеіе гиЬідіпоза, 360
Нутепотопаз, 140
НурЬосіїуігісііаІез, 282
НурЬосіїуігіотусеіез, 282
НурЬосЬуігіотусоіа, 21, 279
НурЬосЬуігіит репіііае, 281
НурЬоІота Тазсісиїаге, 368, 394
НурЬоІота зиЬІаіегіііит, 368
Нуросіїопсігіа, 13
Нуросгеаіез, 326
Нурохуіоп, 331
ІпосуЬе, 391
ІпосуЬе раіоиіііагсііі, 367, 391
Ігрех, 348, 356
Ігрех Іасіеиз, 356
ІзосЬгузісІаІез, 139
КІеЬзогтісІіаІез, 221
КІеЬзогтісІіит, 221
КІеЬзогтісІіит тисозит, 222
ЬаЬугіпіИиІа тасгосузііз, 278
ЬаЬугіпіЬиІаІез, 277
ЬаЬугіпіЬиІотусеіез, 277
ЬаЬугіпіііиІотусоіа, 21, 274
Іасіагіиз, 396
Ьасіагіиз сіеіісіозиз, 397
Ьасіагіиз Ьеіуиз, 366
і-асіагіиз гиіиз, 397
Іасіагіиз іигріз, 397
Ьаеіірогиз, 350
Ьаеіірогиз зиіргіигеиз, 347, 350
Іатіпагіа, 99, “211
Ьатіпагіаіез, 99
Ьесапога, 433
Ьесапога аііоргіапа, 433
Ьесапогаїез, 432
Ьесапоготусеіез, 335
Ьессіпит, 371
Ьессіпит аигапііасит, 372
Ьессіпит зсаЬгит, 372
Ьесісіеа, 433
Ьепііпиз, 347, 348, 354
Ьепііпиз есіосіез, 355
Ьепііпиз іідгіпиз, 355
І.ЄП2ІІЄЗ, 348, 354
Ьепгііез Ьеіиііпа, 347, 354
Ьеоііотусеіісіае, 319
Ьерісіосііпіит уігісіє, 133
Ьеріоіа Ьгиппео-іпсагпаіа, 368
Ьеріоіа гіеіуеоіа, 368
Серізіа писіа, 366
Ьергагіа, 443
І_еріО8рі)аегіа, 421
(.еріозігота, 322
Ьеріозігота ріпазігі, 323
і-ісеаіез, 259
Ьііііоігіатпіоп, 158
ЬоЬагіа, 441
і-оЬагіа риїтопагіа, 441
Ьосиїоазсотусеіез, 335
Соріїосіегтіит, 323
І-оріюсІегтіит ріпазігі, 323
і-усодаїа, 259
І-усодаїа ерісіепсігит, 260
і-усорегсіаіез, 397
і-усорегсіоп, 398
Масгосузііз, 102
Масгоіеріоіа, 386
Масгоіеріоіа ргосега, 386
Масгоіеріоіа риеііагіз, 386
МаІІотопаз, 83
Магазтіиз, 379
Магазтіиз аіііасеиз, 379
448
Магазтіиз огеасіез, 379 МііеІІа, 215
Меіазтіа, 322 Мігзсіїіа, 119
Меіазтіа асегіпа, 322 Мосіііиса, 128
Меіазтіа рипсіаіит, 323 Мосіііисіріїусеае, 128
Меіозіга, 111 Мозіос, 62, 424
Меіозігаїез, 111 Мозіос соттипе, 62
Мегізторесііа, 57 Мозіос Ііпскіа, 62
Мезозіідта, 171 N03(00 ргипііогте, 62
Мезоіаепіит, 218 N03(003168, 59
Місгосузііз, 55 Осіїготопасіаіез, 81
Місгосузііз аегидіпоза, 56 Осіїготопаз, 81
Місгозріїаега, 319 Оесіосеріїаіит, 331
Місгозріїаега аіргіііоісіез, 319 Оесіодопіаіез, 186
Місгозрога, 186 Оесіодопіит, 187
Місгозрогаіез, 186 Оісііит, 319
Мізсітососсаіез, 93 ОІрісІіит, 296
Мопіііа, 324 ОІрісІіит Ьгаззісае, 296
Мопіііпіа, 324 Оотусеіез, 269
Мопіііпіа Тгисіідепа, 325 Оотусоіа, 21, 266
МопоЬІергіагісІаІез, 298 Оредгаріїа, 430
МопоЬІеріїагіз, 298 'ОзсіІІаіогіа, 58
МогсІїеІІа, 333 ОзсіІІаіогіаІез, 58
МогсІїеІІа езсиїепіа, 333 Озігораіез, 439
Моидеоііа, 218 Охуггйісіоріїусеае, 128
Мисіїадо, 262 Охуггіііз, 128
Мисог, 302 Рапаеоіиз, 367
Мисогаїез, 301 Рапсіогіпа, 176
Миііпиз, 404 Рагтеїіа, 435
Миііпиз сапіпиз, 404 Рагтеїіа зиісаіа, 435
Мусепа, 378 Рауіоуа, 141
Мусепа гіаетаіоросіа, 378 Рауіоуаіез, 141
Мусепа рига, 367, 378 РахіІІасеае, 373
МусозріїаегеІІа, 421 РахіІІиз, 374
МусозрІїаегеІІа ріпосіез, 422 РахіІІиз іпуоіиіиз, 369, 374
МусозріїаегеІІа ругі, 421 Ресііазігит, 185
Мухотусеіез, 258 РесІіпеІІа, 124
Мухотусоіа, 19, 254 РесІіпеІІорІїусеае, 123
№уісиіа, 118 Реііідега, 441
№уісиіаіез, 118 Реііідега сапіпа, 441
Несігіа, 329 Реііідегаїез, 440
Месігіа сіппаЬагіпа, 330 РепісіІІіит, 336, 418
Метаїіаіез, 156 РепісіІІіит сіігузодепит, 418
Меигозрога зііоріїііа, 310 РепісіІІіит підгісапз, 419
Кіісіиіагіаіез, 401 РепісіІІіит поіаіит, 419
449
РепісіІІіит годиеіогііі, 419
РепісіІІіит Уііаіе, 418
Репіііа ауігозігіз, 281
Регапета, 68
Регапетаіаіез, 67
Регісііпіаіез, 132
Регісііпіит, 134
Регісііпіит Ьаііісит, 134
Регопозрогаїез, 272
Регіга, 331
Регіга Ьасііа, 332
Регіга уіоіасеа, 332
Регігаїез, 331
Регіготусеіісіае, 319
РЬасиз, 71
РЬасиз ругит, 66
РЬаеорЬусеае, 99
РЬаеорЬуіа, 20, 95
РЬаІІаІез, 402
РЬаІІиз, 403
РЬаІІиз ітридісиз, 403
РЬазеоІиз уиїдагіз, 422
РЬеІІІпиз, 361
РЬеІІІпиз ідпіагіиз, 347, 361
РЬгадтісІіит, 416
РЬуІІасііпіа, 321
РЬуІІорЬога, 162
РЬуІІозіісіа, 422
РЬуІІозіісіа сгиепіа, 422
РЬузагаїез, 261
РЬузагит, 261
РЬузагит уігісіє, 262
РЬузсіа, 434
РЬузсіа зіеііагіз, 434
РЬуіорЬіЬога, 273
РЬуіорЬіЬога іпіезіапз, 273
РіІоЬоІиз, 302
Ріппиіагіа, 118
Ріріорогиз, 348, 351
Ріріорогиз Ьеіиііпиз, 347, 351
Різит заііуит, 422
РІапоіЬідіит, 117
РІапіае, 13, 22
РІазтосІіорЬога, 285
РІазтосІіорЬога Ьгаззісае, 285
РІазтосііорЬогаїез, 285
РІазтосііорЬоготусеіез, 285
РІазтосііорЬоготусоіа, 22, 283
РІазторага, 272
РІазторага уііісоїа, 272
РІаіусгізіаіез, 13, 22, 136
РІеозрогаІез, 335
РІеигосЬгузіз, 140
РІеигососсиз, 193
РІеигоіасеае, 382
РІеигоіиз озігеаіиз, 382
РІиіеасеае, 390
РІиіеиз, 390
РІиіеиз сеп/іпиз, 390
РосЬепіа, 251
Роїудопаіит, 422
Роїутуха, 287
Роїутуха Ьеіае, 287
РоїурЬадиз, 295
РоїурЬадиз еидіепае, 295
Роїурогаїез, 348
Роїурогиз, 348, 353
Роїурогиз здиатозиз, 347, 353
РоїузірЬопіа, 167
РогрЬуга, 155
РогрЬугісІіаІез, 153
РогрЬугісІіит, 153
РогрЬугісІіит ригригеит, 153
РгазіпорЬусеае, 170
Ргосагуоіа, 9, 16
Ргогосепігаїез, 134
Ргогосепігит, 134
Ргогосепігит тісапз, 135
Ргоіеотопаз зиісаіа, 143
Ргоіососсиз, 193
Ргоіозіеііотусеіаіез, 258
Ргоіозіеііотусеіез, 258
Ргутпезіаіез, 138
Ргутпезіит, 138
Ргутпезіит рап/ит, 138
РзаіііугеІІа, 393
РзаіЬугеІІа сапсіоііеапа, 394
Рзеисіососсотуха, 424
450
Рзеидосоїиз, 405
Рзеидосоїиз іизііогтіз, 405
Рзеидоідіит, 319
РзіїосуЬе, 367
Риссіпіа азагіпа, 415
Риссіпіа дгатіпіз, 413
Риссіпіа котагоуіі, 415
Ругатітопадаїез, 170
Ругатітопаз, 170
РуіИіаІез, 273
Ратаїіпа, 438
Катаїіпа Ггахіпеа, 438
Катагіа, 362
Катагіа Ьоігуііз, 362
КарИідіаІез, 76
Каріїідоріїусеае, 76
КарИідорИуіа, 19, 74
КИІгідіотусез, 281
Кііігосіопіит, 202
КИігосІопіит ИіегодІурИісит, 202
КИігосІопіит гірагіит, 202
КИігориз, 301
КИодотопаз, 145
КИодоріїуіа, 23, 149
КИодутепіа, 164
КИодутепіаІез, 164
КИоісозріїепіа, 115
КИуіізта, 322
КИуіізта асегіпит, 322
КИуіізта рипсіаіит, 323
КИуіізтаіаІез, 322
КИугорИудіит, 294
КИугорИудіит роїііпіз, 294
Киззиїа, 395
Киззиіа етеііса, 366, 396
Киззиіа уігезсепз, 396
Киззиіаіез, 395
ЗассИаготусез, 314
ЗассИаготусез сегеуізіае, 315
ЗассИаготусеіаІез, 314
ЗассИаготусеіез, 314
Заргоіедпіа, 270
Заргоіедпіа рагазіііса, 270
Заргоіедпіаіез, 269
Загсодоп, 360
Загсодоп ітЬгісаіиз, 360
Загдаззит, 105
Зсепедезтаїез, 183
Зсепедезтиз, 185
Зсепедезтиз диадгісаида, 186
Зсіегодегта, 400
Зсіегодегта аигапііасит, 401
Зсіегодегтаіаіез, 400
Зсіегоііпіа, 325
Зсіегоііпіа зсіегоііогит, 310, 325
Зсуіотопаз зиЬііііз, 66
Зеріогіа, 421
Зеріогіа рориіі, 421
Зеріогіа ругісоіа, 421
Зегриіа, 347
Зегриіа Іасгутапз, 347, 349
ЗірйопорИусеае, 204
Зогдагіа Гітісоіа, 310
Зогдагіотусеіез, 318
Зогдагіотусеіідае, 319
Зогодізсиз, 288
Зрагаззіз, 348, 350
Зрагаззіз сгізра, 350
ЗрИасеііа, 327
Зрігодуга, 216
ЗрігеІІотусеіаІез, 296
Зропдозрога, 287
Зропдозрога зиЬіеггапеа, 287
Зіетопііаіез, 262
Зіетопіііз, 263
Зіетопіііз іизса, 263
Зіідеосіопіит, 182
Зіідопета, 64
Зіідопетаіаіез, 63
Зігатепоріїез, 14, 19, 74
ЗігорИагіасеае, 394
Зіуіораде Иадга, 308
Зиіііиз, 372
Зиіііиз Іиіеиз, 372
Зигігеііа, 120
ЗигігеІІаіез, 120
ЗутЬіодіпіаіез, 132
ЗутЬіодіпіит, 132
451
ЗупсИуігіит, 292 ТгісЬоІота, 380
ЗупсИуігіит епдоЬіоіісит, 292 ТгісИоІота Лауо-уігепз, 380
8упдіпіорИусеае, 128 ТгісИоІотаіасеае, 378
Зупига, 83 ТгісЬотусеіез, 305
Зупигаїез, 83 ТиЬег, 334
ЗупигорИусеае, 83 ТиЬег аезїіуит, 334
ТарИгіпа, 316 ТиЬег Ьгитаїе, 334
ТарИгіпа сагріпі, 318 ТиЬег теїапозрогит, 334
ТарИгіпа ЬеГогтапз, 318 ТиЬегаІез, 334
ТарИгіпа ргипі, 316 ТиЬегсиїагіа уиїдагіз, 330
ТарИгіпаІез, 316 ТиЬиІосгізіаІез, 13,18, 72
ТарИгіпотусеіез, 315 Тиіозіота, 400
Теїіотусеїез, 413 Туїоріїиз, 373
Теїозсйізіаіез, 431 Туїорііиз аіиіагіиз, 373
Теігазрогаіез, 173 Туїоріїиз ТеІІеиз, 373
ТИеІерИога, 359 ІЛоііігіх, 195
ТИеІерИога раїтаїа, 359 ІЛоІгісИаІез, 195
ТИеІерЬога іеггезігіз, 359 ІЛуа, 197
ТЬеІерИогаІез, 359 иіуаіез, 197
ТіІІеііа сагіез, 411 ІЛуорИусеае, 194
ТіІІеІіаІез, 411 ІІгедіпаІез, 413
ТгасЬеІотопаз, 70 ІІгопета, 181
ТгасЬеІотопаз агтаїа, 71 изпеа, 429, 439
ТгасИеІотопаз Иізрісіа, 71 □зпеа ЬіПа, 439
ТгасЬеІотопаз іпіегтедіа, 71 изіііадіпаїез, 409
ТгасИеІотопаз уоіуосіпа, 71 изіііадо таусііз, 409
Тгатеїез, 347, 348, 351 изіііадо Ігіїісі, 410
Тгатеіез ріпі, 351 изіотусеіез, 408
ТгаизІосИуігісііаІез, 278 Уасиоіагіа, 78
ТгаизіосИуігіит ргоІИегит, 278 УаисИегіа, 90, 277, 288
ТгеЬоихіа, 191,424, 432 УаисИегіаІез, 90
ТгеЬоихіаІез, 191 \/епіигіа, 335
ТгеЬоихіорЬусеае, 189 Уепіигіа іпаедиаіез, 336
ТгетеІІа, 407 Х/епіигіа рігіпа, 335
ТгетеІІа ТітЬгіаіа, 408 УоІуагіеІІа, 390
ТгетеІІа Іиіезсепз, 408 Уоіуосаіез, 172
ТгетеІІаІез, 407 Уоіуох, 177
ТгепІероИІіа, 202, 424 Уоіуох дІоЬаІог, 179
ТгепіероИІІаІез, 202 Могопіпа, 288
ТгіЬопета, 94 ХапІИорИуіа, 20, 87
ТгіЬопетаїаІез, 94 ХапІИогіа, 432
ТгісИіа, 261 ХапІИогіа рагіеііпа, 432
ТгісИіа уагіа, 261 Хегосотиз, 372
ТгісИіаІез, 260 Хегосотиз сИгузепІегоп, 372
452
Хуіагіа, 331
Хуіагіаіез, 331
2оорадаіе8, 304
7озІега тагіпа, 277
2удпета, 218
2удпетаІаІе8, 216
2удотусеІез, 301
2удотусоІа, 24, 299
Покажчик українських назв таксонів
агарикальні, 375
агарикові, 384
акразіальні, 252
акразіомікотові слизовики, 18, 249
акразіс, 252
акразіс рожевий, 252
акремоніум, 417
акремоніум мутовчастий, 417
алломіцес, 297
альвеоляти, 12, 22, 124
альфа-протеобактерії, 13
аманіта, 387
аманітові, 387
амебо-флагеляти, 19, 72, 253
амфідиніальні, 130
амфідініум, 130
анабена, 61
анабена спіралевидна, 62
анабена цвітіння води, 62
анабена Шереметьєвої, 62
анаморфні гриби, 416
анаптіхія, 439
анісольпідіум, 281
анфельцальні, 160
анфельція, 161
апікомплекси, 22
артоніальні, 430
артроспіра, 59
арцирія, 260
арцирія гола, 261
аскомікотоаі гриби, 24, 305
аскоміцети, 318
аскохіта, 422
аскохіта горохова, 422
аспергильоз, 419
аспергіл, 420
аспергіл димчастий, 420
аспергіл жовтий, 420
аспергіл повзучий, 420
аспергіл рисовий, 420
аспергіл різнобарвний, 420
астеріонела, 113
аурикулярія, 406
аурикулярія Іуди, 406
аурікуляріальні, 406
афанізомен цвітіння води, 60
афанізоменон, 60
ахнантальні, 117
ацетабулярія, 209
ацетабулярія середземноморська, 209
бабка, 371
бабка темна, 371
бабка червона, 372
базидіомікотові гриби, 24, 338
базидіоміцети, 343
бакане, 336
бангіальні, 154
бангієфіцієві, 152
бангія, 154
батрахоспермум, 156
бациляріальні, 119
бацилярієфіцієві, 115
білий гриб, 370
бластодиніфіцієві, 130
бластокладіальні, 296
бліда поганка, 388
блюмерія, 321
блюмерія злакова, 321
бокальчик, 402
болет, 369
болетальні, 369
болетові, 369
453
боровик, 370
боровик зернистоногий, 370
бородач, 439
бородач шорсткий, 439
борошнисторосяні гриби, 319
ботридіальні, 92
ботридіопсис, 93
ботридіум, 92
брактеакок, 183
бриопсидальні, 204
бриопсис, 205
бульбохете, 189
бурі водорості, 20, 95
вакуолярія, 78
вентурія, 335
вентурія грушова, 336
вентурія яблунева, 336
веселка, 403
веселка смердюча, 403
веселкові, 402
водяна сіточка, 183
вольварієла, 390
вольвокальні, 172
вольвокс, 177
вольвокс кулястий, 179
вороніна, 288
вошеріальні, 90
вошерія, 90
галімедальні, 207
ганодерма, 358
гаптофітові водорості, 22, 136
гастероміцети, 345, 397
геаструм, 399
гелвела, 322
гелоціальні, 324
гематокок, 174
гематокок дощовий, 175
гетерокок, 93
гіберела, 330
гіберела фуйкуроє, 330
гігартинальні, 161
гігрофор, 378
гігрофорові, 377
гіднелюм, 360
гіднелюм запашний, 360
гіднелюм зональний, 360
гіднум, 364
гіднум виїмчастий, 364
гідродикціон, 183
гідродикціон сітчастий, 185
гіменоміцети, 343
гіменохетальні, 360
гіменохете, 360
гіменохете іржаво-червона, 360
гімнодиніум, 132
гімнодиніум синювато-зелений, 133
гімнодініум зелений, 133
гімнокарпне плодове тіло, 245, 343
гіпокреальні, 326
гіпоксилон, 331
гірчак, 373, 397
гіфолома групова, 394
гіфохітридіальних, 282
гіфохітридіоміцетових, 282
гіфохітриомікотові гриби, 21, 279
гіфохітриум пенілли, 281
глаукоцистальні, 148
глаукоцистіс, 148
глаукоцистофітові водорості, 23, 145
глеокапса, 57
глеотрихія, 59
глива, 382
глибові, 382
гломус, 304
гніздівкові, 401
гнойовик, 392
гнойовик чорнильний, 393
гнойовик чубатий, 392
гнойовикові, 392
говорушка, 379
говорушка біла, 380
головач, 399
гомфідієві, 375
гомфонема, 116
гоніаулакальні, 130
гоніаулакс, 131
гоніостомум, 76
гоніум, 175
454
графіс, 440
графіс письменний, 440
гриб-баран, 350
гриби, 21, 225
грибоподібні організми, 225
губка березова, 351
губка дубова, 352
гуттулінальні, 253
гуттулінопсис, 253
дазикладальні, 209
дедалеа, 352
дедалея дубова, 352
дейтероміцети, 246
десмідіальні, 218
десмокок, 192
десмокок оливковий, 193
диктіостеліоміцети, 264
диктіостеліум дисковидний, 264
диктіофора, 404
диктіоха, 122
диктіохофітові водорості, 21, 121
диктіохофіцієві, 122
динофізис, 134
динофітові водорості, 22, 124
динофіцієві, 130
дискокристати, 13, 18, 64, 249
діатома, 113
діатомові водорості, 20, 105
дінобріон, 82
домітохондріальні евкаріоти, 13
домовий гриб, 349
дотідеоміцети, 335
дощовик, 398
дощовикові, 397
драпарнальдія, 182
драпарнальдія грудкувата, 182
дріжджі, 314
дубовий мох, 438
дюналієла, 174
евглена, 68
евглена гольчата, 70
евглена зелена, 70
евглена спірогірна, 70
евгленальні, 68
евгленаморфа, 71
евгленаморфальні, 71
евгленофітові водорості, 18, 64
евдоріна, 176
евернія, 438
евернія сливова, 438
евкаріоти, 9, 18
евроціальні, 336
евроцій, 337
евроціоміцети, 336
евстигматос, 86
евстигматофітові водорості, 20, 85
едогоніальні, 186
едогоніум, 187
елафоміцес, 337
елафоміцес зернистий, 337
еміліанія, 140
ендогональні, 304
ентероморфа, 199
ентоломові, 376
ентомофтора, 303
ентомофтора мушина, 303
ентомофторальні, 302
епіхлое, 328
епіхлое рогозовидний, 328
еризіфальні, 319
еубактерїї, 18
жовто-зелені водорості, 20, 87
жовчний гриб, 373
зелений морський салат, 197
зелені водорості, 23, 168
зеленушка, 380
земляна зірочка, 399
зигнема, 218
зигнематальні, 216
зигомікотові гриби, 24, 299
зигоміцети, 301
зморшок, 333
зморшок їстівний, 333
золотисті водорості, 20, 78
золотянка настінна, 432
зоопагальні, 304
зоохлорела, 194
ізохризидальні, 139
455
іноцибе, 391
іржасті гриби, 413
ірландський мох, 162
ірпекс, 356
ірпекс молочно-білий, 356
ісландський мох, 437
іудине вухо, 406
їжовик жовтий, 364
їжовик лускатий, 360
каліціальні, 442
каліцій, 442
канделарієла, 434
канделярія золотистенька, 434
кандіда, 314, 420
кантарел, 363
кантареляльні, 361
каулерпа, 208
каулерпа проліферуюча, 208
квітохвісник, 405
квітохвісник веретеновидний, 405
кишечниця, 199
клаварія, 362
клавіцепс, 326
клавіцепс пурпуровий, 326
клавуліна, 363
кладонія, 435
кладонія бахромчаста, 436
кладонія лісова, 436
кладонія оленяча, 437
кладофора, 200
кладофора клубочковидна, 202
кладофора розкидиста, 202
кладофоральні, 200
клатрус, 404
клатрус червоний, 404
клебсормідіальні, 221
клебсормідіум, 221
клебсормідіум слизовий, 221
клітоцибе восковий, 380
клостеріум, 219
кодіум, 206
кодіум червеподібний, 207
коколітальні, 139
коколітофориди, 139
коконеїс, 117
колеохетальні, 222
колеохете, 223
колеохете подушковидна, 223
колеохете щитовидна, 223
компсопогон, 152
компсопогональні, 153
кон’югати, 211
копринові, 392
копринус, 392
кораліна, 159
коралінальні, 158
корбан, 374
кордицепс, 329
кордицепс військовий, 329
кортинарієві, 390
кортинаріус, 391
космаріум, 220
космаріум вирізаний, 220
космаріум ниркоподібний, 220
космаріум перламутровий, 220
космаріум пірамідальний, 220
косцинодискофіцієві, 111
кратерел, 363
кратерел попелясто-сірий, 363
криптомонас, 143
криптофітові водорості, 23, 141
ксанторія, 432
ксанторія настінна, 432
ксероком, 372
ксиляріальні, 331
ксилярія, 331
лабіринтуломікотові гриби, 21, 274
лабіринтуломіцетові, 277
лабіринтуля макроцистна, 277
лабіринтуляльні, 272
ламінаріальні, 99
ламінарія, 99
леканора, 433
леканора різноманітна, 433
леканоральні, 432
леканороміцети, 335, 431
лекцин, 371
лензитес, 354
лензитес березовий, 354
456
лентин, 355
лентин тигровий, 355
лепрарія, 443
летипор, 350
летипор сірчано-жовтий, 350
лецідея, 433
лисичка, 363
лисичка звичайна, 363
лишайник, 249, 424
лікогала, 259
лікогала деревна, 260
лікопердальні, 397
лікопердон, 398
літній трюфель, 334
літотамніон, 153
ліцеальні, 259
лобарія, 441
лобарія легенева, 441
локулоаскоміцети, 335
лофодерміум, 323
лофодерміум сосновий, 323
макролепіота, 386
макролепіота дівоча, 386
макролепіота строката, 386
макроцистіс, 102
маломонас, 83
маразміус, 379
маслюк, 372
маслюк звичайний, 372
мезостигма, 171
мелозиральні, 111
мелозіра, 111
мерисмопедія, 57
мікроспора, 186
мікроспоральні, 186
мікросфера, 319
мікросфера дубова, 319
мікроцистіс, 56
мікроцистіс синювато-зелений, 56
міксомікотові слизовики, 254
міксоміцети, 258
міксофлагеляти, 234
місхококальні, 93
мітоспорові гриби, 246, 417
міцена, 378
міцена чиста, 378
міцени кривавоногої, 378
мокруха, 375
мокруха клейка, 375
мокрухові, 375
монілінія, 324
монілінія плодова, 325
монілія, 324
моноблефаридальні, 298
моноблефаріс, 298
морська капуста, 99
морський мошок, 161
моховик, 372
моховик тріщинуватий, 372
мужоція, 218
мукор, 302
мукоральні, 301
мутин, 404
мутин собачий, 404
мухомор, 387
мухомор білий, 389
мухомор зелений, 388
мухомор смердючий, 389
мухомор цезарів, 389
мухомор червоний, 389
мухоморові, 387
муциляго, 262
навікула, 118
навікуляльні, 118
незавершені гриби, 416
нектрія, 329
нектрія кіноварно-червона, 329
немаліальні, 156
несправжній дощовик оранжевий, 401
несправжній опеньок сірчано-жовтий, 394
несправжньодощовикові, 400
нідуляріальні, 401
нітелла, 215
ніцшия, 119
ноктилюка, 128
ноктилюціфіцієві, 128
носток, 62
носток звичайний, 62
457
носток Пінка, 62
носток сливовидний, 62
ностокальні, 59
ночесвітка, 128
оксиридофіцієві, 128
оксиріс, 128
оленячий мох, 436
ольпідіум, 296
ольпідіум капустяний, 296
оомікотові гриби, 21, 266
ооміцетові, 269
опеграфа, 430
опеньок, 381
опеньок лучний, 379
опеньок осінній, 381
остропальні, 439
осциляторіальні, 39
осциляторія, 58
охромонадальні, 81
охромонас, 81
павлова, 141
павловальні, 141
павутинник, 391
павутинник ліловий, 391
павутинник оранжево-червоний, 391
павутинникові, 390
паксилові, 373
паличкоподібна органела, 65
пандоріна, 176
парасолька велика, 386
пармелія, 435
пармелія боріздчата, 435
педіаструм, 185
педінела, 124
педінелофіцієві, 123
пелтігера, 441
пельтигера собача, 441
пельтигеральні, 440
пеніцил, 418
пеніцил помічений, 418
пеніцил Рокфорта, 419
пеніцил чорний, 419
перанема, 68
перанематальні, 67
перидиніальні, 132
перидиніум, 134
перидиніум балтійський, 134
перидій, 256, 308
пероноспоральні, 272
пецица, 331
пецица коричнева, 332
пецица фіолетова, 332
пецицальні, 331
печерицеві, 384
печериця, 384
печериця двоспорова, 385
печериця звичайна, 386
печериця рудіюча, 386
печіночниця звичайна, 365
підберезовик, 372
підосичник, 372
пілоболус, 302
пінулярія, 118
піптопор, 351
піптопор березовий, 351
пірамімонас, 170
пітіальні, 273
плазмодіофора, 285
плазмодіофора капустяна, 285
плазмодіофоральні, 285
плазмодіофоромікотові
слизовики, 22, 283
плазмодіофороміцетові, 285
плазмопара, 272
плазмопара виноградова, 272
планотидіум, 117
плеврокок, 193
плевротові, 282
плеврохризис, 140
плеоспоральні, 335
плютей, 390
плютей оленячий, 390
плютейні, 390
плютка, 391
плютка Патуйяра, 391
повітряні міхури, 102
полімікса, 287
полімікса бурякова, 287
458
поліпор, 353
поліпор лускатий, 353
поліпоральні, 348
полісифонія, 167
поліфагус, 295
поліфагус евгленовий, 295
польський гриб, 371
порфіра, 155
порфіридіальні, 153
порфіридіум, 153
празинофіцієві, 17
примнезіальні, 138
примнезіум, 138
прокаріоти, 9, 18
пророцентр, 134
пророцентр блискучий, 135
пророцентральні, 134
протокок, 193
протостеліоміцетальні, 258
протостеліоміцети, 258
прохлорофітові водорості, 52
псатирела, 393
псатирела Кандолля, 394
псевдогриби, 21, 225
пукцинія злакова, 413
пукцинія Комарова, 415
пукцинія копитнякова, 415
рамаліна, 438
рамаліна борошниста, 438
рамарія, 362
рамарія гроновидна, 362
рафідіальні, 76
рафідофітові водорості, 19, 76
решіточник, 404
рижик, 397
ризидіоміцес, 281
ризоклоніум, 202
ризоклоніум ієрогліфічний, 202
ризоклоніум прибережний, 202
ризопус, 301
ризофідіум, 294
ризофідіум пилковий, 294
ритизма, 322
ритизма кленова, 323
ритизматальні, 322
родіменіальні, 164
родіменія, 164
родомонас, 145
рожевопластинник жовтувато-сизий
отруйний, 377
рожевопластинник садовий, 377
рожевопластинникові, 376
ройкосфенія, 115
руссуляльні, 395
сажкові гриби, 409
сапролегніальні, 270
сапролегнія, 269
сапролегнія паразитична, 270
саргас, 105
саркодон, 360
саркодон черепичастий, 360
сатанинський гриб, 370
сахароміцес, 314
сахароміцетальні, 314
сахароміцети, 314
свинуха 374
свинуха тонка 374
свинухові 373
септорія, 421
септорія грушова, 421
септорія тополева, 421
сидераміни, 225, 226
силікофлагеляти, 122
симбіодиніальні, 132
симбіодініум, 132
симбіотрофи, 248
синдиніофіцієві, 128
синура, 83
синурофіцієві, 83
синхітріум, 292
синхітріум ендобіотичний, 292
синьо-зелені водорості, 18
синяк, 370
сироїжка, 395
сироїжка блювотна, 396
сироїжка луската, 396
сироїжкові, 395
сифонофіцієві, 204
459
цимбела, 117
цимбеляльні, 115
цистозейра, 104
ціанобактерії, 52
ціатус, 402
цукрові дріжджі, 315
часничник великий, 379
червоний морський салат, 156
червоні водорості, 23, 149
чорна лисичка, 363
чорна ніжка капустяної розсади, 296
чорна цвіль, 302
чорний груздь, 397
чорний трюфель, 334
чорний французький трюфель, 334
шиітаке, 355
шовні діатомеї, 115
ягель, 436
Покажчик термінів
а-1,4-глюкан, 27
(3-1,3-глюкан, 27, 227
р-1,6-глюкан, 227
р-індолілоцтова кислота, 316
ААА-шлях біосинтезу лізину, 227
автогамія, 47, 110
автогенетична гіпотеза походження
евкаріот, 10
автоспора, 45
агар-агар, 150
агарикоїдний карпофор, 344
агарикоїдні гіменоміцети, 365
агароїд, 150
адгезорій, 284
азотфіксація, 54
аквакультура, 102,156,198
акінета, 45, 54
акразин, 264
аксонема, 34
актин, 35
алевроконідія, 240, 308
алофікоціанін, 28
альгінова кислота, 31, 96
альгінати, 96
альготрофи, 248
альдітол, 113
аманін, 268
аманітин, 268
аматоксини, 268
амебоїд, 72
амебоїдний план будови, 234
амілопласти, 205
амфієсма, ЗО, 124
амфієсмоподібна пелікула, ЗО, 146
анаморфа, 241
ангіокарпне плодове тіло, 245, 343
анізогамія, 47
антераксантин, 26
антеридій, 47
антеридіол, 266
антибіотики, 417
апланоспора, 45
апотецій, 245, 308
апотецій леканоровидний, 428
апотецій лецидеєвидний, 428
апофізи, 258
апресорій, 307, 319
арбускули, 304
ареоли, 107
артроспори, 238
архегоній, 223
аск, 242, 245, 308
аск бітунікатний, 312
аск еутунікатний, 312
аск іноперкулятний, 312
аск леканорового типу, 312
аск оперкулятний, 312
аск прототунікатний, 312
аск унітунікатний, 312
аскоспора, 245, 311
атактогамія, 47
атропін, 367
462
ауксилярна клітина, 152
ауксоспора, 110
афаноплазмодій, 236
афілофороїдні гіменоміцети, 346
багатоосьовий талом, 151
багрянковий крохмаль, 27
базальне тіло, 34, 232
базидіоспора, 241, 245, 340, 342
базидіоспора амілоїдна, 342
базидіоспора декстриноїдна, 342
базидія, 245, 340, 342
базидія стихічна, 342
базидія хіастична, 342
балістоспора, 343
безшовні діатомеї, 113
бентос, 25
біла гниль, 325
білковий кристал, 87
бластоконідія, 240, 308
бордоська рідина, 272
ботросоми, 228, 275
бродіння, 306
бульбочки, 212
буфотенін, 367
бура деструктивна гниль, 350
вакуолі, 34
вакуолі скоротливі, 34
вальвокопула, 108
вегетативне розмноження, 44
везикулярно-ліпідний комплекс, 234
везикулярно-арбускулярна мікориза, 304
вивідкова брунька, 44, 212
вивідкова бульбочка, 44
видільні вакуолі, 250
вольва, 344
віолаксантин, 26
відьмині мітли,.316
вошеріоксантин, 88
вставні обідки, 106
вузли, 212
газові вакуолі, 53
гаметангій, 46
гаметангіогамія, 242, 309
гамети, 46
гаметична редукція, 49, 247
гаметичні цисти, 204
гаметогамія, 47, 241
гаметоспорофіт, 49
гаметофіт, 49
гаплодиплофазний цикл, 48, 247
гаплофазний цикл, 48, 247
гаптонема, 33, 136
гастероїдний карпофор, 345
гаусторія, 267, 307
гемаглютинуючі токсини, 75
геми, 268
геміангіокарпне плодове тіло,
245, 343
гемізооспора, 45
гемімонадний тип морфологічної
структури, 41
геміцелюлоза, ЗО
гемолітичні токсини, 75
генофор, 9, 38
гетерогамія, 46, 241
гетерокаріозис, 246
гетерокаріонтичність, 230
гетероксантин, 88
гетеромерна слань, 426
гетероморфне чергування
поколінь, 49
гетероталічність, 47, 242
гетеротрихальний тип
морфологічної структури, 42
гетероциста, 54
гетероцитний трихом, 54
гібереліни, 330
гідроксипроліни, 172
гідротаксис, 255
гіменій, 342
гіменофор, 343, 344
гіперплазія, 286
гіпертрофія, 286
гіпобазидія, 342
гіпокон, 125
гіпоталюс, 256
гіпотека, 106
гіромітрин, 333
463
сіі-таке, 355
сітконоска, 404
сітконоска подвоєна, 404
склеродерма, 400
склеродерма оранжева, 401
склеродерматальні, 400
склеротинія, 325
слизовики, 19, 225
сордаріоміцети, 318
сородискус, 288
спарассіс, 350
спарассіс курчавий, 350
спірогіра, 216
спіцеломіцетальні, 296
спонгоспора, 287
спонгоспора підземна, 287
справжні гриби, 225, 288
стемонітальні, 262
стемонітіс, 263
стемонітіс бурий, 263
стигеоклоніум, 182
стигонема, 64
стигонематальні, 63
стрільчатка, 434
строфарієві, 294
строчок, 333
суриреляльні, 120
сурірела, 120
сфацелія, 327
сценедесмальні, 183
сценедесмус, 185
сценедесмус чотирихвостий, 186
тафрина, 316
тафрина грабова, 318
тафрина деформуюча, 318
тафрина сливова, 316
тафринальні, 316
тафриноміцети, 315
телефора, 359
телефора наземна, 359
телефора пальчаста, 359
телефоральні, 359
теліоміцети, 413
телосхістальні, 431
тилопіл, 373
тилопіл солодкий, 373
тілеціальні, 411
тілеція пшениці, 411
товстуха, 396
траметес, 351
траметес сосновий, 351
траустохітридіальні, 278
траустохітриум проліферуючий, 278
трахеломонас, 70
трахеломонас вольвоксовий, 71
трахеломонас дрібнощетинистий, 71
трахеломонас озброєний, 71
трахеломонас проміжний, 71
требуксіальні, 191
требуксієфіцієві, 189
требуксія, 191
тремела, 407
тремела жовтіюча, 408
тремела торочкувата, 408
тремеляльні, 407
трентеполіальні, 202
трентеполія, 203
трибонема, 94
трибонематальні, 94
трихіальні, 260
трихія, 261
трихія варіююча, 261
трихолома, 380
трихолома жовто-зелена, 380
трихоломові, 378
трихоміцети, 305
трутовик лакований, 358
трутовик лускатий, 353
трутовик несправжній, 361
трутовик облямований, 352
трутовик сірчано-жовтий, 350
трутовик справжній, 357
трюфель, 334
трюфель оленячий, 337
туберальні, 334
туберкулярія звичайна, 330
тубулокристати, 13, 17, 72, 253
тулостома, 400
460
улотрикс, 195
улотрихальні, 195
ульва, 197
ульвальні, 197
ульвофіцієві, 194
урединальні, 413
уронема, 181
уснея, 439
устилягінальні, 409
устиляго маїсовий, 409
устиляго пшениці, 410
устоміцети, 408
факус, 71
фалус, 403
фалус смердючий, 403
фаляльні, 402
фелін, 361
фізаральні, 261
фізарум, 261
фізарум зелений, 262
філактинія, 321
філостикта, 422
філостикта закривавлена, 422
філофора, 162
фістулінальні, 365
фісція, 434
фісція зірчаста, 434
фітофтора, 273
фітофтора інфекційна, 273
флоридеєфіцієві, 156
фомес, 357
фомітопсіс, 352
фомітопсіс сосновий, 352
фрагіляріальні, 113
фрагілярієфіцієві, 113
фрагілярія, 113
фрагмідіум, 416
фрагмобазидіоміцети, 406
фуджикуроє, 330
фузикладій грушовий, 335
фукус, 102
фукусові, 102
фуліго, 262
фуліго септичний, 262
хара, 212
харальні, 211
харациопсис, 93
харофіцієві, 211
хетофоральні, 180
хіломонас, 145
хілум, 252
хітридіальні, 291
хітридіомікотові гриби, 23, 289
хламідомонада, 172
хламідомонас, 172
хламідомонас Мевуза, 173
хламідомонас Рейнгарда, 173
хлорарахніон, 73
хлорарахніофітові водорості, 19, 72
хлорела, 193
хлореляльні, 193
хлорокок, 179
хлорокок диплобіонтний, 180
хлорококальні, 179
хлорофіцієві, 171
хондрус, 162
хризофіцієві, 81
хризохромуліна, 138
хромофітові водорості, 19
хромуліна, 81
хроококальні, 55
хрящ-молочник, 396
хрящ-молочник гіркий, 397
хрящ-молочник оливково-чорний, 397
хрящ-молочник смачний, 397
центричні діатомеї, 111
цераміальні, 164
цераміум, 164
цераціомікса фруктова, 258
цераціоміксальні, 258
цераціум, 131
церкоспора, 423
церкоспора бурякова, 423
цетрарія, 437
цетрарія ісландська, 437
цианофора, 147
цианофоральні, 147
циматоплевра, 121
461
гістрихосфериди, 124
гіфа, 236
гіфенні тільця, 304
гіфи несептовані, 236
гіфи придатки, 321
глеба, 343, 345
глікоген, 27, 225
глюкани, 229
гниль деревини, 347
голоморфа, 241
гомеомерна слань, 426
гомоталічність, 47, 242
гомоцитний трихом,54
гомф, 426
гонімобласти, 152
гормогоній, 44, 58
гормоцит, 55
грана, 38
гржималін, 367
ДАП-шлях біосинтезу лізину, 226
дводомність, 47
дереворуйнуючі гриби, 347
джгутик гребінчастий, 35
джгутик пірчастий, 35
джгутики, 34
джгутики гетероконтні, 232
джгутики ізоконтні, 232
джгутиковий апарат, 34
джгутикові корені, 35
дикаріон, 42, 230
динокаріон, 48, 124
дипланетизм, 239, 270
диплофазний цикл, 48, 247
дискоболоцисти, 79
діадіноксантин, 26
діатоксантин, 26
діноксантин, 26
екзоаск, 312
екзоперидій, 397
еколого-трофічні групи грибів, 248
ексципул, 428
ектоплазма, 275
ектоплазматичний ретикулюм, 228
ендоаск, 312
ендолітні лишайники, 429
ендомікориза, 304
ендоперидій, 397
ендоплазматична сітка, 33, 233
ендосимбіоз вторинний, 14
ендосимбіоз первинний, 14
ендосимбіотична гіпотеза походження
евкаріот, 11
ендосома, 32, 66, 216
ендофлеодні лишайники, 429
епібазидія, 342
епігейні лишайники, 429
епікон, 125
епілітні лишайники, 429
епітека, 106
епіфітні лишайники, 429
епіфітотія, 414
епіфрагма, 402
ерготизм, 328
еталій, 256
еукарпічність, 204, 239
ецидіальне спороношення, 414
ецидіоспора, 414
життєвий цикл, 48
ецицій, 414
загин стулки, 106
закритий ортомітоз, 64
зеаксантин, 26
зигогамія, 242, 300
зигоспора, 242, 300
зиготична редукція, 49, 247
зиготичні цисти, 80
зірчаста структура перехідної зони
джгутика, 169
зміна ядерних фаз, 48
зооспора, 45, 238
зоотрофи, 248
іботенова кислота, 367
ізидії, 427
ізогамія, 46, 242
ізоморфне чергування поколінь, 49
інволюкрум, 175
індузій, 404
інтеркалярний ріст, 99
464
інулін, 168, 204
іхтіотоксин, 139
каліптра, 58
капіліцій, 256
каріогамія, 48, 309
каріокінез, 43
каротини, 26
каротиноїди, 26
карпогон, 152
карпоспори, 152
карпоспорофіт, 51, 152
карпофор, 343
каррагенін, 150
кишеньки плодів, 318
кіла, 285
кільцева борозна, 194
клаваріоїдний карпофор, 345
клавіцепсотоксикоз, 328
клейстотецій, 245, 308
клітина злиття, 152, 244
клітинна оболонка, ЗО, 228
клітинні покриви водоростей, 29
клітинні покриви грибів, 228
кокоїдний тип морфологічної
структури, 41
коколіти, ЗО, 137, 138
колумела, 256
комплекс Гольджі, 33, 233
кон’югація, 46
кон’югація бічна, 217
кон’югація драбинчаста, 217
конідієносець, 240
конідія, 240
консолевидний карпофор, 345
концептакули, 158
коприн, 306
копротрофи, 248
коремії, 240, 417
коронка, каральні, 213
кортекс, 257
кортикальні глобули, 276
кортина, 344
кортинарин, 367
кортиціоїдний карпофор, 343
кремнеземова циста, 78, 110
кремнеземовий панцир, 106
кремнеземовий скелет, 31,121
кремнеземові лусочки, 79
кристи дисковидні, 37
кристи пластинчасті, 37
кристи трубчасті, 37
крохмаль, 27
крохмаль синьо-зелених
водоростей, 27
ксантофіли, 26
ксантофіли діатоксантинового ряду, 26
ксантофіли лютеїновго ряду, 26
ксилотрофи, 248
кутикула, 150
кущиста слань, 424
кучерявість листків персика, 318
ламели, 38
ламінарин, 27, 96
лейкозин, 84
лейкопласт, 67
лепрозна слань, 424
летюча сажка пшениці, 409
листувата слань, 424
лишайникові кислоти, 429
лишайникові речовини, 429
лігнін, 31
лігнінрозкладаючі гриби, 347
лізин, 226
лізосоми, 33
лікарські гриби, 347
лінійна іржа злаків, 413
ліпосоми, 234
літичні везикули, 234
ліхенізовані гриби, 248, 424
ліхеноіндикація, 429
ліхенометрія, 430
лобоподії, 235
побули, 427
ложе, 240
ломасоми, 234
лороксантин, 162
лютеїн, 26
люциферин, 129, 404
465
макроміцети, 245
маннан, 229
манніт, 96
мастигонеми, 35
мацедій, 442
мейоспора, 46
меристодерма, 42, 97
метаболія, 65
метули, 418
міжвузля, 212
мікобіонт, 424
міколамінарин, 227, 266
мікориза, 248
мікоспорини, 299, 306, 338
мікотрофи, 248
мікроміцети, 245
мікротільця, 33, 234
мікротрубочки, 34
мікроцид, 419
мікроциста, 251
мікроцистін, 56
міксамеба, 234, 239
мільдью, 272
міозин, 212
мітоз, 32, 231
мітоз відкритий, 32, 231
мітоз закритий, 32, 231
мітоз напіввідкритий, 33, 231
мітоспора, 46
мітохондріальний апарат, 37
мітохондріальний геном, 9
мітохондрії, 231
мітохондріон, 66
міцеліальний план будови, 234
міцелій, 237
міцелій клітинний, 237
міцелій неклітинний, 237
міцелій псевдодикаріонтичний, 310
міцелій септований, 237
монадний тип морфологічної
структури, 41
моноспора, 45, 151
морфологічний паралелізм, 41
мультивезикулярні тіла, 276
муреїн, 31, 146
мускарин, 367
мусцимол, 367
навколоядерний мішок, 311
накипна слань, 424
нанандрій, 187
нейротоксини, 75
нейстон, 25
неклітинна будова, 41
неоксантин, 26
несправжні тканини, 41
несправжня борошниста роса, 272
нестатеве розмноження, 44, 238
нитка асиміляторна, 42
нитка висхідна, 42
нитка проста, 42
нитка сланка, 42
нитчастий тип морфологічної
структури, 42
нітрогеназний ферментний
комплекс, 54
нуклеоїд, 31
нуклеоморф, 9, 15, 72, 39, 141
нуклеоплазма, 53
оболонка хітин-глюканова, 229
оболонка хітин-хітозанова, 229
оболонка целюлозно-глюканова, 229
оболонка целюлозно-хітинова, 229
оболонки з альгінатами, 31
оболонки з фікоколоїдами, 31
оболонки муреїнові, 31
оболонки пектинові, 31
одноосьовий талом, 151
болонки целюлозно-пектинові, 31
однодомність, 47
оболонки целюлозні, 31
октант, 213
огамія, 46, 242
оогоній, 47
ооспора, 215, 244
оперезуюча ламела, 79
оперон, 9
опістоконтні клітини, 232
ореланін, 367
466
ореланін-подібні токсини, 367
осіння хвороба мух, 303
осмотрофне живлення, 227
отруєння вторинні, 366
отруєння первинні, 366
отруйні гіменоміцети, 366
панцир, 31
парабазальне тіло, 88
паразити, 248
парамілія, 65
парамілон, 27, 227
парануклеарне тіло, 276
парасексуальний цикл, 246
парафізи, 308, 342
парафлагелярне тіло, 66
парентосома, 233, 237, 338
партеногонідії, 177
парша, 335
пектин, 31
пелікула, ЗО
пелоїди, 144
пеніцилін, 418
пептидоглікан, 53
перехідна зона джгутика, 35
перидіоли, 401
перинуклеарна рибосомальна
зона, 280
перинуклеарний ендоплазматичний
ретикулюм, 32
периплазма, 268
перипласт, ЗО, 141
перипластидний простір, 15, 39
перитецій, 245, 308
перифітон, 25
пероксисоми, 33
перфоруючі гіфи, 307
пігменти водоростей, 26, 27, 28
пігменти грибів, 229
пікніда, 241, 413, 417
пікнідіальне спороношення, 414
пікноконідія, 427
пікноспора, 413
піреноїд, 38
плазмалема, 29
плазмогамія, 48, 309
плазмодій, 72, 235
плазмодіокарп, 256
планктон, 25
планозигота, 196
пластиди вторинно симбіотичні, 39
пластиди первинно симбіотичні, 15, 37
пластидний геном, 9
платикристати, 13, 22, 136, 282
плектенхіма, 308
плеоморфізм, 247, 248
плодові тіла аскомікотових, 245, 308
плодові тіла базидіомікотових, 245, 343
плоїдність, 48
подецій, 435
подецій сцифовидний, 436
покривало загальне, 344
покривало часткове, 344
поле, 107
полідикаріонтичність, 230
поліедр, 165
поліедральні тіла, 53
поліол, 203
поліфосфатні гранули, 53
поліфосфатні тіла, 27
полярний вузлик, 108
порини, 38
пороспори, 308
порошиста парша, 287
полярні тіла, 255
порошкоплодні лишайники, 424
поясковий обідок, 106
поясок, 106
празиноксантин, 168
продукти асиміляції, 27, 29, 424
проліферація, 270
проміжна тканина, 97
протоплазмодій, 236
протосфера, 209
прототалюс, 428
пряжки, 339
псевдоеталій, 256
псевдокапіліцій, 257
псевдоміцелій, 237
467
псевдоплазмодії, 235
псевдоподії, 72, 235
псевдосклероцій, 329
псевдотецій, 245, 308
псевдоцилії, 41
псилоцибін, 367
псилоцин, 367
пузула, 34,125
пухирчаста сажка кукурудзи, 409
пустула, 288, 414
рак картоплі, 292
ребра, 107
ресупінатний карпофор, 343
редукційний поділ, 49
ретронема, 35, 72, 232, 265
рецептакул, 98, 345, 403
рибосоми,9
ризини, 426
ризом, 208
ризоманія, 287
ризоміцелій, 236
ризоморфи, 340
ризопласт, 35, 79
ризоподії, 72
римопортула, 111, 113
ріжки злаків, 326
родопласт, 39
рослини, 22
рострум, 143
румпосоми, 233, 292
сагеногенетосома, 228
сагеногени, 228
сапролегніоз, 269
сапротрофи, 248
сегрегативний поділ, 43
секотоїдний карпофор, 345
септа доліпорова, 237, 300, 338
септа проста, 237, 338
септа мікропорова, 237, 300
септи грибів, 237
септи, діатомові водорості, 108
синзооспора, 89
синтетична гіпотеза походження
евкаріот, 11
сиренін, 298
ситовидні трубки, 97
сифон, 67
сифональний тип морфологічної
структури, 43
сифонеїн, 168
сифони (зелені водорості), 204
сифони (червоні водорості), 168
сифонокладальний тип морфологічної
структури, 43
сифоноксантин, 168
скафідій, 98
скелет, ЗО
склероцій, 257, 307
слань, 424
слиз, 31
слизові піхви, 54
слизові тільця, 75
соматична редукція, 49
соматогамія, 46, 242, 309
сораль, 427
соредії, 427
сорокарп, 251
сорус, 100, 251, 292
сперматозоїд, 47
спермацій, 47
спіралеподібна структура, 86
спорангієносець, 240
спорангії багатогніздні, 98
спорангії одногніздні, 98
спорангій, 45, 256
спорангіоспори, 239
спорангіофор, 239
спори, 45
спорична редукція, 49, 247
спородохії, 240
спорокарп, 243, 304
спорополенін, 31
спорофіт, 49
споруляція, 45
статеве розмноження, 46, 241
статевий диморфізм, 49
статоспори, 81
стеригми, 341
468
стефаноконтність, 187, 169
столони, 300
страменопіли, 14, 19, 74, 265
строма, 307
сумка, 242
стулка, 106
супрануклеарний апарат, 33, 74, 75
сфероцисти, 395
таксон, 7
таксономічна категорія, 7
талом, 42
тверда сажка пшениці, 411
тека, 125
телейтоспора, 245, 340, 413
телейтоспороношення, 414
телеоморфа, 240
телефорова кислота, 359
тетраспора, 45, 151
тилакоїди, 38
типи трами, 376
тільця Вороніна, 233
тільця Мопа, 143
тканинний тип морфологічної
структури, 42
тонопласт, 33, 234
травні вакуолі, 33
трама, 343
триптофан, 226
триспорова кислота, 299
трихогіна, 151, 223
трихом, 54
трихоталічний ріст, 97
трихоцисти, 72, 75, 142
трофотаксис, 265
тубулін, 34
ундулююча мембрана, 126
уредоспора, 340, 414
уредоспороношення, 414
уроїдна зона, 235, 250
уснеїн, 429
уснеїнова кислота, 439
утрикули, 208
фаготрофне живлення, 227
фалоїдин, 368
фалотоксини, 368
фанероплазмодій, 236
фіаліди, 309
фіалоспори, 308
фібули, 108
фікобіліни, 26
фікобілісоми, 39, 53
фікоеритрин, 26
фікоколоїди, 31, 150
фікопласт, 171
фікотехнологія, 194
фікоціанін, 28
філоподїї, 234, 235
фітопатогенні гриби, 248
фітотрофи, 248
фотобіонт, 191, 424
фотосинтетичний апарат, 37
фототаксис, 256
фрагмопласт, 194, 202, 211
фрагмобазидія, 342
фукоксантин, 26
фултопортула,107
фунікулюс, 402
хети, 182, 223
хітин, 31, 229
хітозан, 229
хітосоми, 234
хламідоспора, 240, 340
хлоропласт, 15
хлоропластна ендоплазматична
сітка, 15, 32, 39
хлорофіли, 26
холобазидія, 342
хологамія, 46, 241
холокарпічність, 204, 239
хрестоподібний мітоз, 284
хризоламінарин, 27
хроматоплазма, 53
целюлоза, 31, 229
целюлозорозкладаючі гриби, 347
ценобій, 42
центральний вузлик, 108
центрин, 35, 80, 172
центріолі, 36, 230
469
центроплазма, 53
цикломорфоз, 48, 246
циста зооспорогенна, 243
цистиди, 342
цистокарпій, 158
цитокінез, 43
цитокініни, 316
цитоскелет, 36
ціанели, 146
ціанофіцин, 52
ціанофіцинові гранули, 53, 58
чергування поколінь, 48, 247
членисто-кільцевий талом, 211
човниковий рух цитоплазми, 236
чохловидна хвороби злаків, 329
шов каналоподібний, 107
шов щілиноподібний, 107
штрихи, 107
шютте, 323
ядерна оболонка, 32, 230
ядерний апарат, 31, 229
ядерний геном, 9
ядерний ковпачок, 233, 297
ядерце, 32, 230
ядро мезокаріотне, 32
яйцеклітина, 47
Рекомендована література
Основна:
Вилай В. И. Основні общей микологии. - К.: Наук, думка, 1980. - 392 с.
Вассер С. П., Кондратьева Н. В., Масюк Н. П. и др. Водоросли. Справочник. - К.:
Наук, думка, 1989. - 606 с.
Великанов Л. Л., Успенская Г. Д. Некоторьіе вопросьі зкологии грибов // Итоги
науки и техники. Серия ботаника. -Т. 4. - М., 1980. - С. 49-105.
Все о грибах / Горленко М. В., Гарибова Л. В., Сидорова И. И. и др. - М.: Лесная
промьішленность, 1986. - 280 с.
Гарибова Л. В., Сидорова И. И. Грибні. Знциклопедия грибов России. - М.: изд-во
АВЕ, 1997.-352 с.
Глущенко В. И. и др. Слизевики: Учеб. пособ. / В. И. Глущенко, Д. В. Леонтьев,
А. Ю. Акулов. - Харьков: ХНУ, 2002. - 135 с.
Дудка И. А., Вассер С. П. Грибні: справочник миколога и грибника. - К.: Наук,
думка, 1987. - 535 с.
Еленкин А. А. Понятия “лишайник” и “лишайниковьій симбиоз” // Новости систе-
матики низших растений. - 1975. -Т. 12. - С. 3-81.
Жизнь растений. Т. 2. Грибні / Под ред. М. В. Горленко. - М.: Просвещение, 1976. -
479 с.
Жизнь растений. Т. 3. Водоросли, лишайники / Под ред. М. М. Голлербаха. - М.:
Просвещение, 1977. -488 с.
Зеров Д. К. Очерк филогении бессосудистьіх растений. - К.: Наук, думка, 1972. -
315 с.
Каратьігин И. В. Козволюция грибов и растений. - СПб.: Гидрометеоиздат, 1993. -
118с.
Каратьігин И. В. Макросистематика грибов на современном зтапе // Ботан. журн. -
2000. - Т. 85, № 6. - С. 19-34.
Костіков І. Ю. та ін. Водорості грунтів України (історія та методи дослідження,
система, конспект флори). - К.: Фітосоціоцентр, 2001. - 300 с.
Курс низших растений / Под ред. М. В. Горленко. - М.: Вьісшая школа, 1981.-
520 с.
Кусакин О. Г., Дроздов А. Л. Филема органического мира. Насть 2: Ргокагуоіа,
Еикагуоіа: МісгозрогоЬіопІез, АгсЬетопасіоЬіопіез, ЕидіепоЬіопіез, МухоЬіопіез,
КЬосіоЬіопІез, АІУеоіаІез, Неіегокопіез. - СПб.: Наука, 1997. - 381 с.
Масюк Н. П., Костіков І. Ю. Водорості в системі органічного світу. - К.: Академпе-
ріодика, 2002. - 178 с.
Методьі зкспериментальной микологии / Под ред. В. И. Вилай. - К.: Наук, думка,
1982.-582 с.
Мюллер 3., Леффлер В. Микология. - М.: Мир, 1995. - 343 с.
Новое в систематике и номенклатуре грибов / Под. ред. Ю. Т. Дьякова,
Ю. В. Сергеева. - М.: Национальная академия микологии; Медицина для всех,
2003. - 496 с.
471
Окснер А. Н. Определитель лишайников СССР. Вьіп. 2. Морфология, системати-
ка и географическое распределение. - Ленинград: Наука, 1974. - 284 с.
Окснер А. М. Флора лишайників України в двох томах. Том 1. - Київ: Вид-во
АН УРСР, 1956.-494 с.
Саут Р., Уиттик А. Основні альгологии. - М.: Мир, 1990. - 595 с.
Седова Т. В. Основні цитологии водорослей. - Л.: Наука, 1977. - 172 с.
Топачевский А. В., Масюк Н. П. Пресноводнніе водоросли Украинской ССР. - К.:
Вища школа, 1984. - 334 с.
Хоуксворт Д. Л. Общее количество грибов и их значение в функционировании
зкосистем, сохранение и значение для человека // Микология и фитопатология.
- 1992. - Том 26, № 2. - С. 152-166.
Шапиро И. А. Загадки растения-сфинкса. Лишайники и зкологический монито-
ринг. - Ленинград: Гидрометеоиздат, 1991. - 80 с.
АІехороііІоз С. 1, МІГП8 С. \Л/., ВІаскмеІІ М. Іпігосіисіогу МусоІоду (4ііі есі.) - Иєуу
Уогк. С8А, 1996.-868 р.
Оопаїсі 3. 8. Вагг. Еуоіііііоп апсі кіпдсіотз ої огдапізтз їгот Іііе регзресііуе ої а
тусоїодізі // МусоІодіа. - 1992. - УоІ. 84, № 1. - Р. 1-11.
Науукзууогїїі О. І., Кігк Р. М., Зийоп В. С., Редіег О. N. Аіпзууогііі & ВізЬу’з Оісііопагу
оНіїе Рипді. 81Н есі.- Кеуу-Зиггеу: СМІ. - 1995. - 616 р.
Наїе М. Тіїе Ьіоіоду оі іііе ІісЬепз. - Іопсіоп: Есіхл/агсІ Агпоісі, 1974. - 181 р.
Кігк Р. М., Саппоп Р. Р., ОауісІ 3. С. апсі Зіаірегз 3. А. АіпзчуогіЬ апсі ВізЬу’з Оісііопагу
ої ІЬе Рипді. 91Н есі. - САВ ІпіегпаііопаІ, \Л/аІІіпдк>гсі, ІІК. - 2001. - 655 р.
ИазН Т. Н. ІісНеп Ьіоіоду. - СатЬгісіде: ІіпІУегзііу Ргезз, 2001. - 303 р.
Уап сіеп Ноек С., Мапп О. С., Заііпз Н. М. АІдае. Ап іпігосіисііоп іо рНусоіоду. -
СатЬгісіде: Ііпіуегзіїу Ргезз, 1995. - 627 с.
Додаткова:
Вилай В. Й. Биологически активнніе вещества микроскопических грибов и их при-
менение. - К.: Наук, думка, 1965. - 267 с.
Василнева Л. Н. Пиреномицетні и локулоаскомицетні // Низшие растения, грибні
и мохообразнніе Далннего Востока России / Под ред. 3. М. Азбукиной. - СПб.:
Наука, 1998.-Т. 4.-419 с.
Визначник грибів України. Т. 1-5. - К.: Наук, думка, 1967-1979.
Визначник прісноводних водоростей Українснкої РСР. - Т. 1-11,- К.: Наук, дум-
ка, 1938-1989.
Гарибова Л. В. Обзор и анализ современнніх систем грибов. - Петрозаводск:
Изд-во Карелнского НУ, 1999. - 134 с.
Голлербах М. М., Штина 3. А. Почвеннніе водоросли. - Л.: Наука, 1969. - 228 с.
ГромовБ. В. Микроорганизмні - паразитні водорослей. - Л.: изд-во Ленингр.
ун-та, 1976. - 144 с.
Дудка И. А. Место грибов в системе органического мира и современнніе концеп-
ции их классификации // Ботаника и микология на пути в третне тнісячелетие. -
К.: Ин-т ботаники НАНУ, 1996. — С. 295-308.
Заика В. Е. Симбиоз воднніх животнніх с водорослями. - К.: Наук, думка, 1991. -
241 с.
472
Кондратьева Н. В. Морфогенез и основньїе пути зволюции гормогониевьіх водо-
рослей. - К.: Наук, думка, 1975. - 302 с.
Кондратьева Н. В. Прокариотические зеленьїе водоросли - РгосЬІогорЬуіа ієууіп
(обзор литературньїх данньїх) // Альгология. - 1991. -Т. 1, № 3. - С. 87-101.
Масюк Н. П. Зволюционньїе аспектьі морфологии зукариотических водорослей. -
К.: Наук, думка, 1993.-232 с.
Морозов А. И. Лекарственньїе грибьі. - М.: АСТ; Донецк: Сталкер, 2003. - 207 с.
Новожилов Ю. К. Определитель грибов России. Отдел Мухотусоїа. Вьіп. 1. Класе
Мухотусеїез. - СПб.: Наука, 1993. - 288 с.
Определитель пресноводньїх водорослей СССР / Под ред. М. М. Голлербаха. -
М.-Л.: Наука.-Т. 1-13.-1951-1980.
ГІаламарь-Мордвинцева Г. М. Десмидиевьіе водоросли Украинской ССР. - К.:
Наук, думка, 1982. - 238 с.
Рейвн П., Зверт Р., Айкхорн С. Современная ботаника (в 2-х томах). - М.: Мир,
1990.
Сьітник К. М„ Вассер С. П. Современньїе представлення о биологическом разно-
образии // Альгология. - 1992. - Том 2, № 3. - С. 3-17.
Токсинообразующие микроскопические грибьі и вьізьіваемьіе ими заболевания
человека и животньїх / В. И. Вилай, Н. М. Пидопличко. - К.: Наук, думка, 1970. -
291 с.
Червона книга України. Рослинний світ / Під ред. Ю. Р. Шеляг-Сосонка. - К.: Вид-
во “Укр. енциклопедія” ім. М. П. Бажана, 1996. - 608 с.
Царенко П. М. Краткий определитель хлорококковьіх водорослей Украинской ССР. -
К/. Наук, думка, 1990.-207 с.
Штина 3. А., Голлербах М. М. Зкология почвенньїх водорослей. - М.: Наука, 1976. -
144 с.
Сіїгізіепзеп М. Ауієуу оИііпдаї ееоіоду // МусоІодіа. -1989. - УоІ. 81, №1.-
Р. 1-19.
Сигіііе М. 3., Маікіпзоп 8. С. Тіїе Рипді. - Ьопсіоп: Асасіетіс Ргезз, 1994. -482 р.
Рипді оШкгаіпе: Аргеіітіпагу Сіїескіізі / Т. V. Апбгіапоуа, І. О. Оисіка, V. Р. Науоуа,
V. Р. Неіиїа, В. Іпд еі аі. / ЕФ Ьу О. \А/. Міпіег & І. О. Оисіка. - САВ Іпіегпаііопаї &
М. 6. КЬоІосІпу Іпзіііиіе ої Воіапу, 1996. - 361 р.
8«апп Е. С., ТауІогЗ. \А/. Рііуіодепеііс регзресііуез оп Ьазісііотусеіе зузіетаїісз:
єуісієпсє (гот іііе 188 гРИА депе. // СапаФ 3. Воі.-1995. УоІ. 73, ЗиррІ. 1. -
Р. 862-868.
473
Навчальне видання
Костіков Ігор Юрійович
Джаган Вероніка Володимирівна
Демченко Едуард Миколайович
Бойко Ольга Анатоліївна
Бойко Вікторія Ростиславівна
Романенко Петро Олександрович
БОТАНІКА
Водорості та гриби
Навчальний посібник
За редакцією доктора біологічних наук, професора І. Ю. Костікова
та кандидата біологічних наук В. В. Джаган
Літературний редактор - П. Коробчук
Технічний редактор - Т. Демчук
Комп’ютерна верстка - О. Мельник
Коректори - Н. Лелікова, О. Сидорук
Підписано до друку 08.09.2005.
Папір офсетний. Формат 60x84/16. Друк офсетний. Гарнітура АгіаІ.
Ум. друк. арк. 27,67. Обл.-вид. арк. 28,84.
Зам. № 1455.
Видавництво “Арістей”
Свідоцтво про внесення суб’єкта видавничої справи до
Державного реєстру видавців, виготівників і розповсюджувачів
видавничої продукції ДК № 1066 від 27.09.2002 р.
02105 м. Київ, вул. Тампере, 13 Б
т/ф (+38 044) 451-44-66 (багатоканальний)
агі8Іеу@орІіта.сот.иа (комерційний відділ)
агі8іеу1@орііта.сот.иа (видавничий відділ)
ілллдл/.агізіеу.кіеу.иа