



















































































































































































































































































































































































































































































































































Author: Андреев М.П. Ахти Т. Гагарина Л.В. Гимельбрант Д.Е. Войцехович А.А. Давыдов Е.А.
Tags: биология флора лишайники семейство лишайников
ISBN: 978-5-87317-935-0
Year: 2014




ФЛОРА ЛИШАИНИКОВ
РОССИИ
биология, экология,
разнообразие, распространение
и методы изучения лишайников
РОССИЙСКАЯ АКАДЕМИЯ НАУК
БОТАНИЧЕСКИЙ ИНСТИТУТ им. В. Л. КОМАРОВА
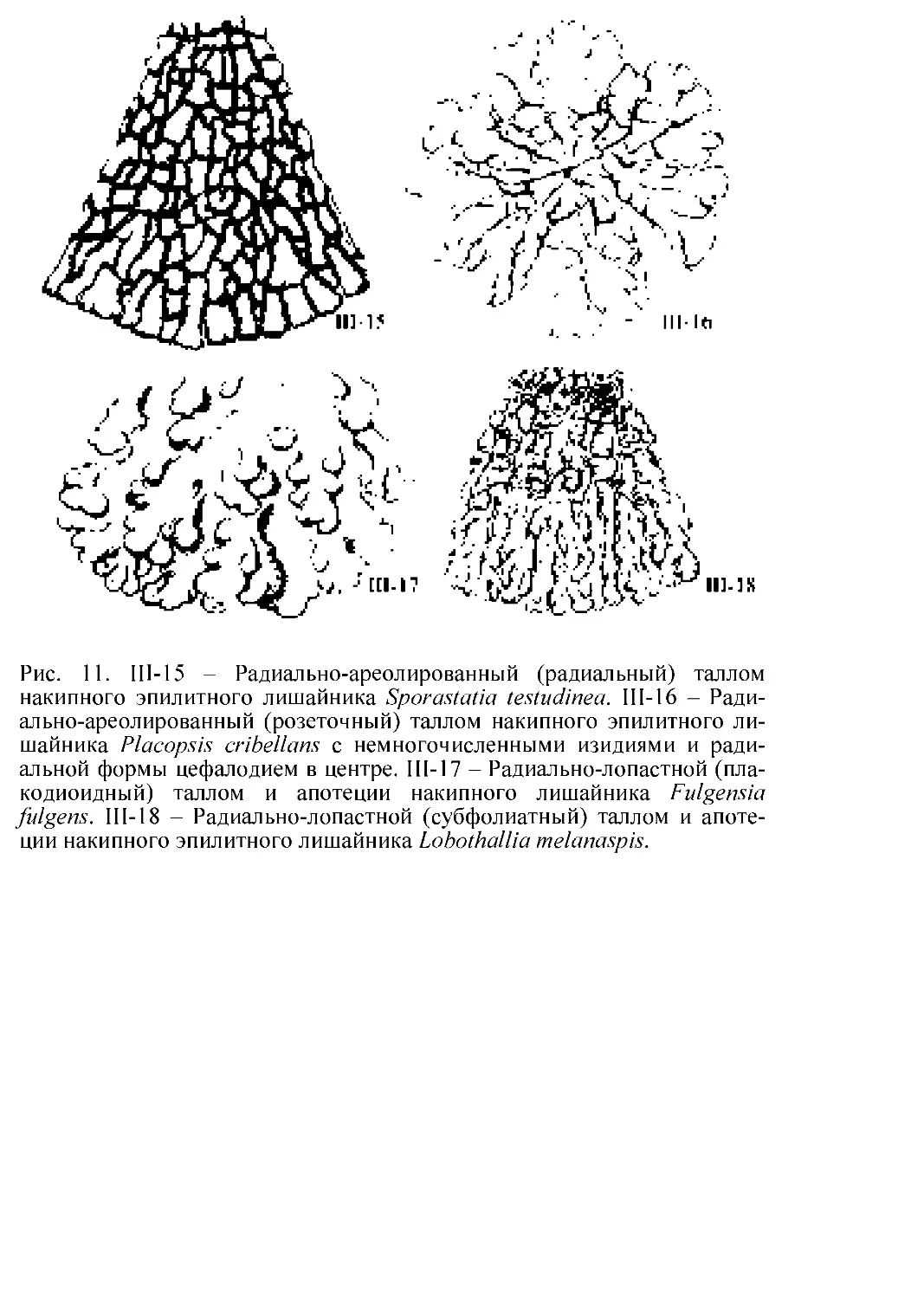
Флора лишайников России
Биология, экология, разнообразие, распространение и методы изучения
ISBN 978-5-87317-935-0
Товарищество науч. изд. КМК
2014
Андреев М.П., Ахти Т., Войцехович А.А., Гагарина Л.В.,
Гимельбрант Д.Е., Давыдов Е.А., Конорева Л.А., Кузнецова
Е.С., Макрый Т.В., Надеина О.В., Рандлане Т., Сааг А.,
Степанчикова И.С., Урбанавичюс Т.П.
ПРЕДИСЛОВИЕ
Многотомное издание «Флора лишайников России» является первой
фундаментальной сводкой по лихенофлоре России. Это издание включает
сведения по таксономическому разнообразию флоры лишайников России
и сопредельных территорий на семейственном, родовом и видовом уров¬
нях. Приводятся также сведения по морфологии таксонов, их экологии и
географическому распространению на территории России и за ее преде¬
лами.
Создание этого издания было инициировано проф. Н. С. Голубко¬
вой, а решение о ее написании было принято на Международном совеща¬
нии, посвященном 120-летию со дня рождения Всеволода Павловича Са-
вича «Флора лишайников России: состояние и перспективы исследова¬
ний», проходившем в Ботаническом институте в Санкт-Петербурге 24-27
октября 2006 г.
Справочный том «Флоры лишайников России» является универ¬
сальным справочным пособием для изучения лишайников. Книга содер¬
жит материалы по истории изучения лишайников России и данные по ос¬
новным лихенологическим гербариям России и мира, где хранятся кол¬
лекции лишайников, собранные на территории России. В книге дается
описание анатомического строения и морфологии талломов и репродук¬
тивных структур лишайников и характеристики их жизненных форм. Са¬
мостоятельные разделы посвящены водорослям-фотобионтам лишайни¬
ков, особенностям вторичного метаболизма и хемосистематики, молеку¬
лярно-генетическим методам их изучения, разнообразию и особенностям
распространения лишайников на территории России, их экологии, мето¬
дам сбора и определения лишайников и рекомендациям по хранению ли-
хенологических коллекций, дана информация по номенклатуре и типифи-
кации лишайников. В работе приведена современная иерархическая си¬
стематическая классификация таксонов лихенофлоры России, вниз, до
уровня родов и, впервые, ключи для определения 397 родов лишайников
флоры России, включающих 3540 известных на территории страны видов,
а также ключи для определения стерильных лишайников и для определе¬
ния водорослей-фотобионтов. В приложениях даются схема деления тер¬
ритории Российской Федерации на регионы, принятое в настоящем изда¬
нии, терминологический словарь, стандартные сокращения фамилий ав¬
торов при таксонах и указатель латинских названий. Для каждой главы
приводится исчерпывающий список литературы. Издание иллюстрирова¬
но 87 оригинальными рисунками, фотографиями и картами. Рисунки пре¬
имущественно иллюстрируют особенности морфологии и анатомии тал¬
ломов и репродуктивных структур лишайников.
В основу настоящей обработки, кроме оригинальных данных, поло¬
жена критическая ревизия современных и исторических литературных
данных не только отечественных, но и зарубежных.
Издание рассчитано на студентов, аспирантов и специалистов био¬
логов, ботаников и микологов, а также любителей, занимающихся изуче¬
нием флоры лишайников России. Настоящий выпуск должен стать осно¬
вой для работы над последующими систематическими выпусками много¬
томной «Флоры лишайников России».
В составлении выпуска приняли участие сотрудники Ботанического
института им В. Л. Комарова РАН и Санкт-Петербургского государствен¬
ного университета М. П. Андреев, Л. В. Гагарина, Д. Е. Гимельбрант, Л.
А. Конорева, Е. С. Кузнецова, И. С. Степанчикова, Центрального Сибир¬
ского ботанического сада СО РАН Т. В. Макрый, Алтайского государ¬
ственного университета Е. А. Давыдов, Института проблем промышлен¬
ной экологии Севера КНЦ РАН Г. П. Урбанавичюс, Института ботаники
им. Н. Г. Холодного НАН Украины А. А. Войцехович и О. В. Надеина,
Тартуского университета (Эстония) Т. Рандлане и А. Сааг и Хельсинского
университета (Финляндия) Т. Ахти.
Оригинальные рисунки выполнены Е. С. Кузнецовой, рисунки водо-
рослей-фотобионтов - А. А. Войцехович.
Книга подготовлена и опубликована при финансовой поддержке
Российского фонда фундаментальных исследований, гранты № 08-04-
00569, 11-04-00901, 11-04-10017-К, 12-04-10076-К и 13-04-00000.
Глава I.
Три века русской лихенологии.
История изучения лишайников России.
Андреев М. П., Ахти Т.
Русское государство, Россия, Российская империя, Советский Союз
и Российская Федерация в разные периоды своего существования объеди¬
няли разные территории, которые в различной степени были включены в
сферу русской культуры. Период наиболее интенсивного развития ли¬
хенологической науки в России пришелся на годы существования Совет¬
ского Союза, когда государственных и национальных границ внутри
страны практически не было, а культурные различия, особенно в научной
сфере, были минимальны. Поэтому крайне сложно отделить русскую ли¬
хенологию от общесоветской, поскольку ученые, работавшие на террито¬
рии собственно России в ее нынешних границах и их коллеги в союзных
республиках часто работали в одних и тех же институтах и гербариях, со¬
бирали материал по всей территории Союза и публиковались, часто вме¬
сте, в одних и тех же журналах. Кроме того, почти все советские ученые-
лихенологи - это, работавшие на всем пространстве огромной страны,
выходцы из трех наиболее авторитетных лихенологических школ - ле¬
нинградской, киевской и тартуской.
В настоящей публикации мы постараемся ограничиться, насколько
это возможно, описанием состояния лихенологической науки России в ее
современных границах, но обращаясь, тем не менее, в тех случаях, когда
это необходимо, к важнейшим событиям и явлениям, происходившим и в
независимых ныне государствах. Вне предмета нашего исследования ока¬
зываются лишь Польша и Финляндия, поскольку они получили независи¬
мость и оказались вне российского и советского социумов на самой ран¬
ней стадии развития лихенологии в России, вследствие чего лихенологи-
ческая наука там в дальнейшем развивалась самостоятельно. Кроме того
финская лихенологическая наука - сама по себе, это такое огромное и са¬
мобытное явление, со своей собственной долгой и богатой историей, вы¬
дающимися учеными мирового уровня и грандиозными достижениями,
что описание ее должно быть предметом самостоятельного исследования.
Что касается Прибалтийских государств - Эстонии, Латвии и Лит¬
вы, то, несомненно являясь частью западноевропейской цивилизации,
равно, и в области науки, они, после краткого 20-летия независимости, в
послевоенный период почти на 50 лет стали частью Советского Союза и
лихенологическая наука в Прибалтике, как и в других союзных республи¬
ках развивалась в рамках общего культурного и научного пространства. В
силу сказанного, в настоящем очерке мы не можем не упоминать о рабо¬
тах и достижениях прибалтийских ученых-лихенологов, равно как и об
исследованиях ученых из других Союзных республик бывшего СССР.
Русская лихенология в XVIII веке.
Русская ботаника, как наука возникла в начале XVIII столетия. До
этого систематического изучения ботанических объектов на территории
России не было. Указом от 11 февраля 1714 года Пётр I повелел учредить
на Вороньем острове, лежащем близ Санкт-Петербурга Аптекарский ого¬
род. Этим было положено начало будущему Императорскому Ботаниче¬
скому саду - нынешнему Ботаническому институту им. В J1. Комарова
Российской Академии наук, и самому существованию ботанической
науки в России.
Первым среди русских ботаников следует назвать академика Иоган¬
на Христиана Буксбаума (1694-1730). Он прибыл в Россию в 1721 г. для
работы в Медицинской коллегии и оказался в числе тех лиц, которые по¬
могли Петру Великому основать Академию наук, был академиком и пер¬
вым в ней ботаником. В 1724 г. в составе посольства X. X. Румянцева И.
С. Буксбаум был отправлен в Турцию с заданием «делать тщательные
разыскания в трех царствах природы», в особенности, по части лекар¬
ственных растений. Кроме Малой Азии, через которую следовала миссия,
Буксбаум посетил Грецию и Армению, а на обратном пути в Петербург -
Баку, Дербент и Астрахань, собирая при этом ботанические материалы и,
среди прочего - лишайники. После возвращения в 1726 г. в Петербург он
обрабатывал в Академии наук свои богатые ботанические коллекции, но
болезнь и ранняя смерть в 1730 г. помешала ему закончить свое произве¬
дение «Plantarum minus cognitarum Centuria I-V, complectens plantas circa
Byzantium in Oriente observatas» (Buxbaum, 1728, 1728, 1729, 1733,1740),
огромный для своего времени труд, в котором было описано и изображе¬
но несколько сот растений, не только обнаруженных им в Турции и на
Кавказе, но и в России. После смерти И. К. Буксбаума окончание его тру¬
да было издано И. Гмелином. Готовилось и второе издание, для которого
было приготовлено несколько сот уже раскрашенных рисунков, храня¬
щихся сейчас в Архиве Академии наук в Санкт-Петербурге. В двух пер¬
вых центуриях своего труда Буксбаум приводит соответственно 4 и 8 ви¬
дов лишайников, сопровождая описания отлично выполненными рисун¬
ками, на которых некоторые виды легко узнаваемы (рис. 1). Кроме описа¬
ний лишайников в работе приводятся сведения об их распространении.
Организация первого гербария в Кунсткамере и создание Ботаниче¬
ских садов сыграли важную роль в развитии ботаники в России, но среди
ботанических исследований, проводившихся Академией наук в XVIII ве¬
ке, первостепенное значение, бесспорно, имели знаменитые «академиче¬
ские» экспедиции, маршрутами которых были покрыты значительные
пространства России. Благодаря героическим усилиям их участников, до¬
стоянием мировой науки становились сведения о географии, геологии,
животном и растительном мире гигантской Terra incognta, какой остава¬
лась до этого территория Российской империи.
Рис. 1. Рисунки лишайников, подготовленные ко второму изданию книги И. X. Бук¬
сбаума «Plantarum minus cognitarum Centuria 11, complectens plantas circa Byzantium in
Oriente observatas» (1728). a. Lichen coralloides aculeatus, nigricans (=Cetraria
aculeata); 6. Lichen coralloides aculeatus, nigricans, parvus, repens (=Cetraria aculeata f.
?); e. накипной лишайник на камне; г. Lichen terrestris cinerei Rai (=Cetraria islandica).
Первое такое путешествие по Сибири, Даурии и Китаю совершил в
1720-1727 гг. врач из Данцига Даниель Готлиб Мессершмидт (1685-1735).
15 ноября 1718 г. Петр I издал указ о направлении Мессершмидта в Си¬
бирь «для сбора всяких трав, цветов, корений, семян и прочих аптекар¬
ских вещей». По собственной инициативе путешественник, кроме изуче¬
ния животного и растительного мира, включил в план своих работ иссле¬
дования в области географии, истории, археологии и этнографии. По его
совету Григорием Акинфиевичем Демидовым в Соликамске был заложен
первый в России научно организованный ботанический сад, а в 1720 г.
Мессершмидтом был составлен первый в стране научный каталог расте¬
ний, встреченных им по пути. В своих исключительно подробных, ча¬
стично изданных дневниках Мессершмидт приводит и лишайники. (Mes-
serschmidt, 1962, 1966).
Грандиозным научным мероприятием XVIII столетия стала Великая
Северная или Вторая Камчатская экспедиция под руководством капитана-
командора Витуса Беринга, продолжавшаяся в течение 11 лет с 1733 по
1743 г. Число ее участников доходило до 2000 человек. Это были морские
офицеры, учёные, художники, переводчики, административные и техни¬
ческие работники. Из натуралистов в экспедиции участвовали — И. Г.
Гмелин, Г. В. Стеллер и С. П. Крашенинников. Силами различных отря¬
дов были составлены первые карты и описания арктического побережья
России от Архангельска до Колымы, Охотского моря и полуострова Кам¬
чатки. Были описаны природа, народы и история Сибири. Совершены
плавания к берегам Японии и Америки, обследованы Курильские и Але¬
утские острова. Блестящие молодые ученые, участники экспедиции, впер¬
вые изучали флору отдаленных регионов империи и собрали там среди
прочего первые коллекции лишайников.
Наиболее обширны и продуктивны были флористические работы
Иоганна Георга Гмелина (1709-1755), который с 1728 г., еще девятнадца¬
тилетним юношей работал в Петербургской Академии наук, а уже в 1731
г. стал академиком. Ученый-энциклопедист И. Гмелин, путешествовал по
России с 1734 по 1742 г., собирая преимущественно ботанические кол¬
лекции. Он обследовал Урал, Западную и Восточную Сибирь, Алтай и
Якутию и за почти десять лет странствий, до возвращения в Петербург,
покрыл расстояние около 34 тыс. км.
Плодом этой поездки стали опубликованные дневники (Gmelin,
1753) и основной труд И. Г. Гмелина - четырехтомная «Flora sibirica, sive
Historia plantarum sibiricae» (Гмелин, 1747, 1749, 1768, 1769), содержащая
описания 1178 видов растений, из них около 500 новых для науки, с при¬
ложением 300 рисунков. Линней в 1744 г. писал, что Гмелин «открыл
столько растений, сколько другие ботаники все вместе». К сожалению,
пятый том его «Флоры», посвященный криптогамным растениям, так и не
был опубликован, его рукопись хранится в Архиве Академии наук в Пе¬
тербурге. Черновик написан рукой Гмелина, а сама рукопись переписана
или написана Степаном Крашенинниковым на основе материалов Гмели¬
на уже после его смерти. Рукопись тома из 130 страниц включает описа¬
ния 312 видов растений, среди которых 35 видов лишайников, названия
которых автор приводит в старой еще долиннеевской номенклатуре. В
Архиве, среди других иллюстраций к пятому тому «Флоры», выполнен¬
ных художником Люрсениусом, хранятся цветные рисунки двух видов
лишайников - Cladoniapyxidata и Peltigera dactylina. (рис. 2).
Другой участник экспедиции - Георг Вильгельм Стеллер (правиль¬
нее - Штеллер) (1709-1746), адъюнкт кафедры натуральной истории -
широко образованный и талантливый ученый. В 1738 г. он был направлен
во Вторую Камчатскую экспедицию помощником И. Г. Гмелина и до 1740
г. самостоятельно работал в Сибири, а потом до 1743 г. - на Камчатке.
Вместе с В. Берингом он участвовал в плавании к берегам Аляски и во
время трагической зимовки на острове Беринга умело руководил отрядом
после смерти капитана-командора. Георг Стеллер стал первым ученым
естествоиспытателем, ступившим на западное побережье Северной Аме¬
рики. В его неопубликованных работах, в частности, в книге «Flora
Irkutiensis», приводится 37 видов лишайников, встреченных им в районе
Байкала и Ангары, даны сведения об их экологии и распространении.
Гербарий, собранный Г. Стеллером, его рукописи и дневники поступили в
Академию наук и после его смерти были частично опубликованы (Steller,
1793) и использованы С. Крашенинниковым и П. С. Палласом.
Степан Петрович Крашенинников (1711-1755) был студентом Сла-
вяно-греко-латинской академии в Москве, когда в 1732 г. его включили в
состав Второй Камчатской экспедиции, также в качестве помощника И.
Гмелина. С 1737 по 1741 г. Крашенинников уже самостоятельно работал
на Камчатке, результатом чего явилась его книга «Описание Земли Кам¬
чатки», изданная в 1755 г и переведенная затем на многие языки. В этом
произведении имеется и описание растительности Камчатки. По возвра¬
щении в Петербург С. П. Крашенинников вскоре был избран академиком
и работал в университетском (академическом) Ботаническом саду на Ва¬
сильевском острове, которым заведовал в 1746-1749 гг. Последние годы
своей жизни он посвятил изучению флоры Петербургской губернии, со¬
брал свыше 500 видов растений и оставил рукопись «De plantis Ingricis»,
вышедшую в свет уже после его смерти (Крашенинников, 1761).
Рис. 2. Портрет
Иоганна Георга
Гмелина и рисунки
лишайников худож¬
ника Люрсениуса,
выполненные для
пятого неизданного
тома «Flora sibrica».
а. Cladonia pyxidata;
б. Pel tiger a dactyli-
na.
В годы царствования императрицы Екатерины Великой был осу¬
ществлен грандиозный проект Российской Академии наук — проведение
многолетних комплексных обследований обширных районов Российской
империи. С 1768 по 1774 г. несколько отрядов академических экспедиций
работали в разных уголках империи: отряд И. И. Лепехина (1740-1802),
оренбургские отряды П. С. Палласа (1741 -1811), И. П. Фалька (1732-1774)
и И. Г. Георги (1729-1802) и астраханские отряды С. Г. Гмелина (младше¬
го) (1774) и И. А. Гильденштедта. Ученые работали на русском Севере - в
Архангельске и на Кольском полуострове, обследовали европейскую и
азиатскую части России, Поволжье, Урал, Юг России, Кавказ и Закавка¬
зье. Ценность научных открытий, сделанных в ходе академических экспе¬
диций, была огромна. Было собрано огромное количество разнообразных
материалов: отчетов, рапортов, путевых записок, исторических реликвий,
J* . f'i ' . i
образцов минералов, животных и растений, в том числе и коллекции ли¬
шайников.
В те же годы исследованием флоры окрестностей столицы занимал¬
ся профессор ботаники Григорий Федорович Соболевский (1741-1807),
автор первой Петербургской флоры, составленной научно и написанной
на русском языке, оставивший после своей смерти гербарные коллекции,
насчитывавшие более 6500 видов и более 2000 семян. В его труде «Flora
Petropolitana» (Sobolewskj, 1799) приводится, среди прочего, 29 видов ли¬
шайников, а во втором издании - «Санктпетербургская флора» (Соболев¬
ский, 1803), опубликованном на русском языке - уже даны описания 72
видов и многочисленные комментарии к ним, в частности, об использова¬
нии лишайников в медицине, кулинарии и хозяйстве.
Практика академических экспедиций продолжались и в более позд¬
нее время - в конце XVIII и в начале XIX века.
Натуралисты были непременными участниками и морских походов.
Например, участник Северо-Восточной астрономической и географиче¬
ской экспедиции капитана И. И. Биллингса судовой врач Карл Мерк в
1791-1792 гг. впервые собрал лишайники в нижнем течении Колымы.
К сожалению, большая часть наиболее ценных коллекций лишайни¬
ков XVIII века не сохранилась. Часть материалов, например, коллекция Д.
Г. Мессершмидта погибли при пожаре в Кунсткамере в 1747 г., а коллек¬
ции лишайников, хранившихся в гербарии Ботанического института им.
В. J1. Комарова, бесследно пропали в годы Великой Отечественной вой¬
ны, при эвакуации в Казань. Возможно, некоторые образцы были вывезе¬
ны П. С. Палласом в Германию.
В тот же период в Москве работали немецкие ботаники X. Стефан,
Ф. Фишер и Г. Гофман. В 1798г. граф А. К. Разумовский основал в своем
подмосковном имении Горенки ботанический сад, для разработки кол¬
лекций которого был приглашен ряд ботаников из-за границы и в том
числе Ф. Фишер, ставший впоследствии первым директором ботаниче¬
ского сада в Петербурге.
В 1804 г. в Москву приехал немецкий ботаник и лихенологГеорг
Франц Гофман (1760-1826), профессор Эрлангенского и Гёттингенского
университетов, чьи лекции слушали И. В. Гёте и А. Гумбольдт. В 1792-
1803 годах он возглавлял в Гёттингене университетский ботанический
сад. Гофману принадлежит ряд выдающихся трудов по систематике, в
частности, первая германская флора - Deutschlands Flora (Hoffman, 1794)
и монография о лишайниках - Enumeratio lichenum (Hoffman, 1784).
В Москве он обосновался уже будучи известным ботаником, в осо¬
бенности благодаря его работам по лишайникам, где стал руководить ка¬
федрой ботаники в Московском университете, а также Ботаническим са¬
дом, став первым его директором. Работая в Москве, Гофман собрал
большой гербарий, вошедший в гербарий Московского университета, и
описал флору университетского Ботанического сада.
Русская лихенология в XIX веке.
В середине и в конце XIX века огромный размах приобрели иссле¬
дования Туркестана и Центральной Азии. В эти годы в Императорский
Ботанический сад стали поступать обширные коллекции растений, со¬
бранные П. П. Семеновым в Тянь-Шане, И. Т. Борщевым в Арало-
Каспийской низменности, Н. Г. Потаниным у оз. Зайсан, коллекции Н. М.
Пржевальского, X. X. Певцова, X. X. Роборовского, X. X. Козлова из ра¬
нее недоступных районов Центральной Азии. Всего за 30 лет Русское гео¬
графическое общество организовало в Центральную Азию 12 крупных
экспедиций, ботанические коллекции которых представляли собой ис¬
ключительную ценность по их новизне и множеству типового материала
для новых видов и родов растений и стали основой для многочисленных
флор, книг и статей. В числе прочих ботанических материалов в Петер¬
бург поступали и коллекции лишайников. К сожалению, из-за отсутствия
специалистов лихенологов, лишь часть материалов была обработана и
нашла свое отражение в публикациях. Некоторые коллекции отправля¬
лись для изучения в уже существовавшие тогда признанные европейские
лихенологические центры, преимущественно в Швецию и Финляндию,
известным специалистам Теодору Фризу, Вели Нюландеру и Эдуарду
Вайнио.
Исследованиями были охвачены и необъятные пространства Сибири
и Крайнего Севера. В 1837 г. состоялось организованное Ботаниче-
скимСадом путешествие А. Г. Шренка в тундры европейского Севера и к
Полярноу Уралу. В отчете, в частности, приводятся и 33 вида лишайников
(Schrenk, 1854)/
Первые сведения о лишайниках северо-восточной Азии появилась в
книге А. Ф. Миддендорфа «Путешествие на Крайний Север и в Восточ¬
ную Сибирь в 1843-44 гг. » (Middendorff,l 867). По определениям В.
Нюландера для побережья Охотского моря он приводил 47 видов лишай¬
ников, для полуострова Таймыр 27 видов.
Особенно ценные ботанические коллекции, и прежде всего - ли¬
шайников, были собраны шведским ботаником Э. Альмквистом (1844-
1923) в русской Арктике и на побережье Чукотки зимой 1878-1879 гг.
(Almquist, 1879, 1883, 1887). Э. Альмквист был участником Шведской по¬
лярной экспедиции А. Е. Норденшельда, впервые прошедшей на судне
«Вега» Северо-восточным проходом вдоль арктического побережья Рос¬
сии. Он собирал материал в нескольких пунктах арктического побережья,
но наиболее полную коллекцию лишайников собрал в районе Колючин-
ской губы на Чукотке, на мысе Инретлен у становища Питлекай. Обшир¬
ные сборы были сделаны также и на восточном побережье Чукотки - в
Кониамской бухте близ пос. Провидения и в заливе Св. Лаврентия. Сотни
образцов, собранных Э. Альмквистом, и обработанных выдающимися
финскими и шведскими лихенологами В. Нюландером, Э. Вайнио и Г.
Мальме послужили основой для многочисленных таксономических и
флористических публикаций. После их изучения были составлены списки
лишайников Берингийской Чукотки и описаны многие новые таксоны
(Malme, 1932; Nylander, 1884, 1885, 1888; Vainio, 1909).
Сборы Э. Альмквиста в настоящее время хранятся в Стокгольме в
гербарии Шведского королевского музея естественной истории, а наибо¬
лее важные дублеты и типовые образцы - в гербариях В. Нюландера в
Хельсинки и Э. Вайнио в Турку. Во время путешествия на «Веге» Аль-
мквист собрал также богатые коллекции лишайников на о-ве Беринга в
Командорском архипелаге и в Порт-Кларенсе на американском побережье
Берингова пролива (Nylander, 1885, 1888), положив тем самым начало
изучению флоры лишайников Аляски.
С наступлением XX века интенсивность ботанических исследований
в России еще более возросла, а исследования распространились на Юг
России, Среднюю Азию, Кавказ и Крым, а на востоке - на нынешний Ка¬
захстан, Западную и Восточную Сибирь и Дальний Восток. Это объясня¬
лось началом функционирования Переселенческого управления, а также
уже накопленным опытом и сложившимися традициями ботанических
исследований.
Русская и советская лихенология в XX веке.
Начало изучения лишайников в Императорском Ботаническом Саду
Петра Великого и новый этап развития русской лихенологической науки
связаны с именем А. А. Еленкина (1873-1942). (Рис. 3)
Александр Александрович Еленкин поступил на службу в Импера¬
торский ботанический сад в качестве консерватора в 1897 году. В то вре¬
мя научная деятельность в Саду почти не велась, а научный персонал не
справлялся с разборкой колоссальных коллекций, привезенных со всех
концов России и из других стран. Лабораторного оборудования не было, и
научная деятельность ограничивалась только хранением старых коллек¬
ций и некритической инсерацией новых.
Образцы споровых растений для определения тогда посылали за
границу, так как своих специалистов в Саду в тот период еще не было. В
Архиве Ботанического института хранится письмо шведского ботаника
Теодора Фриза 1887 года с благодарностью за присылку для определения
большой коллекции лишайников, собранных Н. Г. Потаниным и Н. М.
Пржевальским в Центральной Азии и сыном директора Сада А. Э. Реге¬
лем в горных областях Туркестана, где тот служил врачом.
Споровая часть гербария, помещавшаяся в одном помещении с гер¬
барием сосудистых растений, с 1895 по 1898 год находилась в ведении Г.
А. Надсона, занимавшегося изучением водорослей. Обработанные и
необработанные коллекции споровых растений складывались на чердаке
здания гербария, поскольку площадей для хранения тоже не хватало.
Рис. 3. «Отец русской лихено¬
логии» Александр Алексан¬
дрович Еленкин (1873-1942).
Фото 1905-10 гг.
В 1900 году Гербарий низших споровых растений был выделен в
самостоятельное подразделение. Одновременно была основана Централь¬
ная Фитопатологическая Станция, которую до 1906 года возглавлял А. А.
Ячевский, а после его ухода и до 1913 года-А. А. Еленкин.
В 1901 году, после приобретения микроскопа, в гербарии началась
настоящая научная деятельность и интенсивность изучения поступающего
материала существенно возросла. С самого начала самостоятельного су¬
ществования гербария работа была направлена к возможно более плано¬
мерному и систематическому изучению споровых растений на всей тер¬
ритории России. Приоритетными А. А. Еленкин считал задачи изучения
споровых растений Европейской, особенно Средней России. Однако мно¬
гочисленные экспедиции, прежде всего, Переселенческого Управления,
также доставляли для обработки в Гербарий богатейшие коллекции ли¬
шайников с дальневосточных и южных окраин страны, собранные такими
известными учеными как Ф. Е. Фишер, А. Г. Шренк, Г. С. Карелин, Г. Н.
Потанин, А. Е. Регель, В. J1. Комаров, Б. А. Федченко и многими другими
ботаниками и путешественниками.
В те годы в Арктику отправлялись многочисленные русские поляр¬
ные экспедиции, поставлявшие материалы с северных окраин России,
например, сборы лишайников, сделанные на Новосибирских островах
Бялыницким-Бируля, участником экспедиции барона Толя (Elenkin, 1909;
Еленкин, 1909), коллекции, собранные И. В. Палибиным во время плава¬
ния ледокола «Ермак» в Северном Ледовитом океане (Еленкин, Савич,
1912). Поступили и были обработаны коллекции И. Шульги, участника
экспедиции С. А. Бутурлина на нижнюю Колыму, сборы Н. Сокольникова
с Анадыря и Г. А. Борисова с Восточной Чукотки (Савич, 1911, 1915),
лишайники, собранные Р. Н. Ниманом в Лапландии и Новой Земле, Р. Р.
Поле на крайнем севере Европейской России, И. И. Тржемеским в поляр¬
ной Сибири, Б. Н. Городковым в Тобольской губ. (Савич, 1911, 1912,
1916а, 19166) и другие.
Сам А. А. Еленкин, начиная с 1899 года, также неоднократно со¬
вершал поездки в разные области России с целью сбора и изучения ли¬
шайников, мхов, грибов, пресноводных и морских водорослей. В 1899 г.
он посетил Кавказ, в 1900 г. - Крым, в 1902 г. вместе с В. Л. Комаровым
работал в Саянских горах - в Иркутской губ. и северной Монголии, с 1903
по 1911 гг. собирал материал в Средней России, в 1905-1906 гг - на Мур-
мане, в 1912 г. - на Черноморском побережье Кавказа, в 1898-1922 гг. со¬
вершил многочисленные поездки в окрестности Петербурга, пополняя
растущую коллекцию лишайников Ботанического Сада.
Кроме того Гербарий пополнялся коллекциями, поступавшими для
обработки от коллекторов-любителей, в результате обмена с зарубежны¬
ми научными центрами и приобретёнными у частных лиц. К определению
лишайников А. А. Еленкин привлекал таких крупных зарубежных ученых,
как Т. Фриз, В. Нюландер, Е. Альмквист и других. Появляются первые
публикации (Еленкин, 1901, 1904).
В 1901-1904 гг. А. А. Еленкин три выпуска (всего 150 номеров) эк¬
сикат «Lichenes Florae Rossiae et regionum confinium orientalium». Вскоре
вышли в свет первые фундаментальные работы А. А. Еленкина по флоре
и систематике лишайников «Флора лишайников средней России» в четы¬
рех частях (Еленкин, 1911). Это была первая региональная лишайниковая
флора России.
В 1906 году А. А. Еленкин принял на себя заведование Фитопатоло¬
гической станцией и перевел Споровый гербарий в соседнее с ней поме¬
щение. Это дало сотрудникам возможность пользоваться хорошо обору¬
дованной лабораторией и привело к интенсификации анатомических и
физиологических исследований лишайников. Расширение лабораторных
работ сразу же сказалось на продуктивности, а у А. А. Еленкина появи¬
лось несколько учеников - студентов естественного отделения Петер¬
бургского университета и других институтов: В. П. Савич, А. Н. Данилов,
Jl. Г. Раменский, Б. Ф. Кашменский, И. Е. Вереитинов, А. А. Юницкий, Г.
К. Крейер, Л. И. Оль, Л. И. Любицкая, Э. Леман, В. А. Петров, И. А. Беке¬
тов. Их трудами и усилиями многочисленных коллекторов - А. С. Ботки¬
на, Н. А. Буша, А. А. Бялыницкого-Бирули, Н. А. Димо, В. А. Дубянского,
Б. А. Келлера, В. Л. Комарова, С. И. Коржинского, П. Н. Крылова, Н. И.
Кузнецова, В. И. Липского, Р. Ф. Нимана, И. В. Палибина, Р. Р. Поле, Н.
И. Прохорова, Н. Сокольникова, В. Н. Сукачева, П. С. Сюзева, Г. И. Тан-
фильева, Б. О. Федченко, И. И. Шираевского, В. И. Роборовского, А. М.
Фетисова и многих других, было положено начало изучению флоры ли¬
шайников России. Только в предреволюционные годы они совершили
многочисленные экспедиции в разные уголки России: В. П. Савич - в Пе¬
тербургскую губернию и Эстляндию (1907), на Камчатку(1908-1909), в
Уссурийский край (1909), в Минскую губернию (1907-1910), в Новгород¬
скую (1910), В Сувалкскую и Люблинскую губернии в Польше (1914), В
Терскую обл. на Кавказе (1915), В Архангельскую (1917) и Петроград¬
скую губернии (1908-1922); Л. И. Савич-Любицкая - в Минскую губер¬
нию (1909-1913) и в Ново-Николаевск (1912); А. Н. Данилов - в За¬
каспийскую обл. Туркестана (1914); И. А. Вереитинов - в Гродненскую
(1904), Петроградскую (1905) и Псковскую (1907) губернии и в Крым
(1908); Э. Леман. - в г. Вознесенск Херсонской губ. (1905); В. Ф. Кашмен¬
ский - в Курскую и Харьковскую губернии (1904-1905); А. А. Юницкий -
в Костромскую губернию (1908); Г. К. Крейер - в Могилевскую (1908 -
1913) и Петроградскую губернии (1914) и в Крым (1915); В. А. Петров и
И. А. Бекетов - в Петроградскую губернию (1912 и 1918). Их трудами и
усилиями многих коллекторов было положено начало изучению флоры
лишайников России.
Но кроме работ по флоре, систематике и морфологии лишайников
А. А. Еленкин, совместно с А. Н. Даниловым, проводили на базе Споро¬
вого гербария исследования по анатомии, физиологии и биологии споро¬
вых растений, в частности занимались проблемой симбиоза и изучением
гонидий лишайников.
В 1900 году Отдел низших (споровых) растений был организован и
в Ботаническом музее Академии наук. Им руководил Вольдемар Генрихо¬
вич (Владимир Андреевич) Траншель (1868-1942). Сам Ботанический му¬
зей был учрежден в 1823 году по ходатайству академика К. А. Триниуса.
До этого, на протяжении XVIII столетия ботанические коллекции Акаде¬
мии наук, вместе со всеми прочими, принадлежавшими Кабинету нату¬
ральной истории, хранились в старом здании петровской Кунсткамеры, по
свидетельству академика И. И. Георги 1790 года - «в нетронутых связках,
в ожидании рассмотрения». Вскоре многие из этих связок из-за недостат¬
ка места были вынесены на чердак, где и находились «в безобразном со¬
стоянии». Академик Триниус собрал остатки коллекций и перенес их в
сухое, отапливаемое помещение. Позднее в 1835 году Музей перебрался
из Кунсткамеры в новое здание Академии наук, где он и находился до
1931 года. В 1855 г. директором Ботанического Музея стал Ф. И. Рупрехт,
который, кроме прочего, занимался изучением лишайников и других низ¬
ших растений.
При В. А. Траншеле музей пополнился ценными собраниями, обра¬
батывать которые приглашались А. А. Еленкин, В. П. Савич и другие спе¬
циалисты. Из лихенологов, работавших в Отделе низших растений Бота¬
нического музея, можно назвать К. А. Рассадину, отдельные исследова¬
ния, связанные с лишайниками, выполнял Н. Н. Воронихин.
Но помимо русских исследователей в России работали и иностран¬
ные ученые. Участник шведский экспедиции в устье Енисея, М. Бреннер
(Brenner, Marten Magnus Widar (1843-1930), собрал чрезвычайно богатый
материал, на основе которого, прежде всего А. Г. Магнуссоном, были
описаны многие виды лишайников и который до сих пор еще остается до
конца необработанным. Известный финский ботаник и будущий прези¬
дент Финляндии А. К. Каяндер, путешествовал по Западной Сибири и
Якутии. Для устья Лены, в частности, он приводит 9 видов лишайников
(Cajander, 1903). Его материалы изучали выдающиеся финские лихеноло-
ги Э. Вайнио и В. Нюландер. Сами они в те годы обследовали территорию
Карелии и находившиеся тогда в пределах Финляндии Южную Карелию,
Карельский перешеек и острова Финского залива. Австро-венгерский
ученый X. Лойка (Hugo Lojka (1844-1887), путешествовал по Кавказу. Ма¬
териалы его также были изучены Э. Вайнио (ссылка). На Дальнем Восто¬
ке - на Сахалине и Курильских о-вах, принадлежавших тогда Японии, ра¬
ботал французский исследователь аббат Фурье (Abbe Faurie = Faurie,
Urbain Jean (1847-1915)?), известный своими обширными коллекциями из
Восточной Азии - Кореи, Японии и Формозы (Тайваня). Его материалы,
обработанные французским лихенологом А. Гю (Hue, Auguste-Marie
(1840-1917), хранятся в Париже, а многие дублеты - в гербарии Ботаниче¬
ского института в Петербурге. На их основе описаны многие новые виды.
Позднее на Южном Сахалине и Курильских островах работали японские
исследователи. Лишайники германской провинции Восточная Пруссия и
окрестностей Кёнигсберга (ныне Калининград и Калининградская область
РФ), продолжая исследования О. Олерта (1816-1875) (Ohlert, 1863, 1870,
1871), изучал немецкий лихенолог Г. Леттау (1878-1951) (Lettau, 1912,
1919).
Следует также упомянуть, работавшего несколько позже, шведского
ботаника Эрика Гультена, участника Шведской камчатской экспедиции
1920-1922 гг. Его сборы лишайников из южной части Камчатки были об¬
работаны Э. Дюрье (Du Rietz, 1929). Специалистам хорошо известны пуб¬
ликации выдающегося шведского ученого А. Г. Магнуссона, обрабаты¬
вавшего коллекции лишайнков с Севера Сибири (Магнуссон, 1927; Mag-
nusson, 1936).
На островах Баренцева моря в Арктике работали норвежские экспе¬
диции, по результатам которых норвежский ученый Бент Люнге (Lynge,
Bernt Arne (1884-1942) опубликовал ряд фундаментальных работ (Lynge,
1928, 1931), в которых для Новой Земли приводится 413 видов лишайни¬
ков, а для Земли Франца Иосифа - 64 вида.
Кроме Петербурга и Гельсинфорса в Финляндии, лихенологические
исследования в России в начале века проводились только в Казани (Шель,
1883). В Казанском университете в эти годы работал выдающийся рус¬
ский ученый Константин Сергеевич Мережковский (1855-1921). Мереж¬
ковский изучал лишайники Поволжья, Крыма, Эстляндии. В Ботаниче¬
ском музее Казанского университета он собрал обширный гербарий ли¬
шайников, описал много новых видов, издавал эксикаты и готовил к изда¬
нию Флору лишайников России, которая, к сожалению, так и не была
опубликована. Мережковский известен также своими теоретическими
трудами, он является основоположником Теории симбиогенеза.
В 1913 году А. А. Еленкину удалось выделить Споровый гербарий в
отдельное учреждение со своим собственным бюджетом - был создан
Институт Споровых Растений. (Рис. 4).
Рис. 4. Сотрудники Института споровых растений в 1914 г. Сидят: А. Н. Данилов, А.
А. Еленкин и неизвестный. Стоят: И. А. Бекетов и В. П. Савич, уходящий на фронт
Первой мировой войны, в форме прапорщика.
На оборудование Института была выделена солидная сумма, давав¬
шая возможность устроить образцовую лабораторию и приобрести за гра¬
ницей ряд ценных коллекций. Увеличилось число консерваторов. Научная
деятельность еще более расширилась. В 1916 году коллекция лишайников
насчитывала уже 4800 образцов (2500 видов), которые хранились в 178
пачках и еще в 388 пачках размещались разобранные, но неопределенные
коллекции. В 1919 году было отремонтировано старое здание Гербария и
Библиотеки, расположенное по Песочной (улица Профессора Попова)
улице, куда и был переведен Институт. Здесь гербарий лишайников нахо¬
дится и по сию пору.
Долгие и тяжелые годы революции и войны сказались на деятельно¬
сти Института, тем не менее, научная работа не прекращалась, а с окон¬
чанием Гражданской войны начался постепенный процесс восстановле¬
ния и роста. К 1921 году гербарий Института Споровых Растений состоял
уже из 4 отделов и насчитывал около 100 тыс. образцов и 18 тыс. видов. В
первые годы Советской власти обрабатывались в основном материалы по
местной Петроградской флоре и изучались коллекции, доставляемые се¬
верными экспедициями. Возникла необходимость подготовки общедо¬
ступных руководств и определителей по споровым растениям местной
флоры.
В 1922 г. под редакцией А. А. Еленкина начинает издаваться сбор¬
ник «Новости систематики Института Споровых Растений Главного бота¬
нического сада», позднее переименованный в «Новости систематики низ¬
ших растений» и существующий и поныне.
В 1930 году Главный ботанический сад был преобразован в Ботани¬
ческий институт и передан в ведение Академии наук СССР. В нем был
образован единый Отдел споровых растений, куда влились Институт Спо¬
ровых Растений, Отдел фитопатологии и Отдел гидробиологии Ботаниче¬
ского сада, а также Отдел низших (споровых) растений Ботанического
музея Академии наук. Полное слияние двух отделов с переводом персо¬
нала и всего имущества на территорию Ботанического Сада произошло в
1931 году. Споровый гербарий Музея был влит в Отдел Споровых расте¬
ний, а из его персонала в Отдел перешли В. А. Траншель, К. А. Рассадина,
Н. Н. Воронихин и J1. И. Савич-Любицкая.
Заведующим Отделом с 1932 года становится В. П. Савич, который
возглавлял Отдел споровых растений и Лабораторию лихенологии и
бриологии до 1963 года. (Рис. 5).
Всеволод Павлович Савич (1885-1972) начал работать в Споровом
гербарии в 1906 г., еще будучи студентом Петербургского университета,
сначала «по вольному найму», как практикант, а в 1907 г. был зачислен в
штат. Тогда же совместно с Л. Г. Раменским и под руководством В. Л.
Комарова он провел свое первое экспедиционное исследование расти¬
тельности Ямбургского и Гдовского уездов Петербургской губернии, а в
1908 г. прочитал свой первый доклад в Обществе естествоиспытателей на
тему «Из жизни лишайников в западной части Петербургской губернии и
прилегающей части Эстляндской». В 1908-1909 гг. вместе с В. J1. Комаро¬
вым Савич принял участие в экспедиции Ф. П. Рябушинского на Камчат¬
ку. В гербарий поступили богатые лихенологические коллекции, на осно¬
ве которых В. П. Савич опубликовал статьи, посвященные различным
таксономическим группам лишайников.
Рис. 5. Заведующий От¬
делом споровых расте¬
ний Ботанического ин¬
ститута АН СССР В. П.
Савич. Ок. 1932 г.
В дальнейшем В. П. Савич был участником и организатором многих
экспедиций: Олонецкой (1920-1924), Белорусской (1923-1924), Астрахан¬
ской (1926), Мурманской (1927) и др. В 1930 г. в составе Арктической
экспедиции на ледокольном пароходе «Г. Седов», руководимой О. Ю.
Шмидтом, он изучал лишайники на островах Новой Земли, Земли Франца
Иосифа и Северной Земли.
По результатам обработки собственных сборов, а также коллекций
лишайников, собранных другими исследователями, В. П. Савич опубли¬
ковал многочисленные статьи, в которых приводятся обширные сведения
о видовом составе лишайников различных районов России. Он был в чис¬
ле первых лихенологов, которые начали изучать лишайниковые сообще¬
ства. В. П. Савич занимался также монографическими исследованиями,
опубликовав «Конспект к флоре лишайников сем. Umbilicariaceae СССР»
(Савич, 1950), одну из первых отечественных лихенологических моно¬
графических работ.
Работа с гербарием занимала особое место в деятельности В. П. Са-
вича. В частности, с 1924 г. им было издано 22 декады эксикат «Licheno-
theca Rossica», тиражом от 70 до 100 экземпляров.
Всего В. П. Савичем было опубликовано 180 научных работ. Благо¬
даря его публикациям, а также работам других специалистов в области
споровых растений в первой половине ХХ-го столетия был заложен проч¬
ный фундамент для развития флористических, систематических, морфо¬
логических, экологических и биохимических исследований в области
низших растений в нашей стране (Голубкова, 2003).
В 30-50-е годы XX века задачи развития оленеводства в районах
Крайнего Севера потребовали детального изучения растительного покро¬
ва арктических районов страны. Изучая кормовую базу оленя, многочис¬
ленные ботанические экспедиции работали в разных секторах Арктики. К
этой работе были привлечены значительные научные силы и, в частности,
такие известные ученые, как Б. Н. Городков, собиравший лишайники в
Сибири, Якутии, на Чукотке и на о-ве Врангеля (Городков, 1939, 1943,
1958). Его образцы и материалы других исследователей изучали А. Н.
Окснер, К. А. Рассадина и В. П. Савич. Для Залива Креста на Восточной
Чукотке А. Н. Окснер опубликовал 17 видов лишайников (Окснер, 1929).
К. А. Рассадина по сборам В. Б. Сочавы 1929 г. привела для Анадырского
края 71 вид (Рассадина, 1934). В 1950 году Рассадина выпустила первое в
нашей стране монографическое исследование, посвященное роду Cetraria
(Рассадина, 1950).
В 50-е годы в Ботаническом институте на базе Спорового отдела
был создан самостоятельный Отдел лихенологии и бриологии, позже пре¬
образованный в лабораторию. Сначала его возглавлял В. П. Савич, а с
1963 года - выдающийся бриолог Иван Иванович Абрамов. Споровый
гербарий превратился в четыре самостоятельных гербария — лишайни¬
ков, мохообразных, грибов и водорослей. В эти годы в лаборатории тру¬
дились такие исследователи, как К. А. Рассадина, Е. К. Штакенберг и др.
Лаборатория и огромный гербарий были центрами притяжения для спе¬
циалистов изо всех уголков Советского Союза. Здесь, в частности, часто
работал выдающийся украинский лихенолог Альфред Николаевич Окснер
и многочисленные другие ученые. Под руководством В. П. Савича в лабо¬
ратории проходили подготовку молодые специалисты из республик Сою¬
за ССР и из-за рубежа. В те годы велся активный обмен гербарными кол¬
лекциями с зарубежными гербариями и обширная переписка.
В 1982 году лабораторию возглавила Н. С. Голубкова (Рис. 6), кото¬
рая руководила ею до 2003 года. Нина Сергеевна Голубкова (1932-2009),
выдающийся русский и советский лихенолог, выпускница Ленинградско¬
го университета и ученица В. П. Савича, работала в Ботаническом инсти¬
туте с 1955 г. Ее первые работы были посвящены изучению флоры ли¬
шайников Московской области. Позже она изучала флоры лишайников
Памира, Монголии и Антарктики, как систематик занималась такими
сложными и интересными группами лишайников, как сем. Acarosporaceae
и Umbilicariaceae, роды Usnea, Mycobilimbia, Heterodermia, Bacidia и
Bacidina и др.
Рис. 6. Глава ленинградской ли¬
хенологической школы Нина Сер¬
геевна Голубкова (1932-2009).
Автор более сотни научных публикаций, в том числе таких книг, как
«Конспект флоры лишайников Монгольской Народной Республики»
(1981), «Анализ флоры лишайников Монголии» (1983) и «Лишайники се¬
мейства Acarosporaceae в СССР» (1988), она долгие годы была признан¬
ным руководителем петербургской лихенологической школы, воспитате¬
лем целой плеяды учеников, которые работают не только в России, но в
других странах. Проф. Голубкова руководила и была движущей силой ра¬
бот по изданию «Определителя лишайников СССР» и инициатором нача¬
ла работ над «Флорой лишайников России». Ее заслуги перед наукой бы¬
ли отмечены Международной ассоциацией лихенологов, Н. С. Голубкова
была награждена высшей наградой ассоциации - медалью Э. Ахариуса.
Все годы своего существования Лаборатория лихенологии и брио¬
логии Ботанического института и гербарий лишайников были не только
основным центром лихенологической науки в России и СССР, но и глав¬
ной базой подготовки кадров для других научных учреждений нашей
страны, особенно для Академий Наук союзных республик, а также неко¬
торых зарубежных стран. Так, в 50-60-х годах под руководством В. П.
Савича в аспирантуре института учились и успешно защитили диссерта¬
ции Ц. Н. Инашвили (Грузия), А. В. Питеранс (Латвия), А. В. Домбров¬
ская (Полярно-альпийский ботанический сад-институт), К. А. Рябкова
(Свердловский педагогический институт), Вэй Цзян-чунь (Китай), кото¬
рые впоследствии зарекомендовали себя как высоко квалифицированные
специалисты в области лихенологии и стали создателями собственных
региональных научных школ и направлений. Академик Вэй Цзян-чунь,
например, долгие годы руководит лихенологической лабораторией в Пе¬
кине и стал основателем большой китайской лихенологической школы.
Независимо от старейшего ботанического учреждения страны, но в
тесном сотрудничестве с ним развивалась украинская лихенологическая
школа в Киеве. Ее основатель, Альфред Николаевич Окснер (1898-1973) -
это яркое явление в отечественной лихенологии (Рис. 7).
А. Н. Окснер учился в Киевском университете, который окончил в
1924 г. Тогда же была опубликована его первая лихенологическая работа
«Материалы к флоре лишайников Белоруссии» (Окснер, 1924), а уже с
1931 г. он был заведующим Отделом лихенологии в Институте ботаники
АН Украины. Во время Великой Отечественной войны А. Н. Окснер эва¬
куировался в Киров и в 1942 г. в Уфе, где находился тогда эвакуирован¬
ный институт, в тяжелейших условиях военного времени завершил и
успешно защитил докторскую диссертацию «Анализ и история проис¬
хождения лихенофлоры Арктики». После возвращения в 1944 г. Институ¬
та ботаники в Киев А. Н. Окснер возглавил в нем Отдел споровых расте¬
ний, с которым была неразрывно связаны вся его дальнейшая жизнь и
научная работа.
Деятельность А. Н. Окснера многогранна и плодотворна. Он был ав¬
тором первого пособия для определения лишайников Украины «Визнач-
ник лишайниюв УРСР» (Окснер, 1937). Итогом же исследований лихено¬
флоры Украины и флористических работ А. Н. Окснера является его фун¬
даментальная, но, к сожалению, не завершенная работа «Флора лишай-
ниюв Укра'ши» (1956, 1968, 1993). Этот труд, демонстрирующий высо¬
чайший уровень работы флориста, долгие годы оставался примером и
стандартом для других исследователей. Кроме подробного обсуждения
систематики таксонов во «Флоре» впервые дана географическая оценка и
выявлены ареалы всех приведенных видов, к каждому таксону приводятся
многочисленные таксономические и экологические примечания.
Рис. 7. Основатель украинской лихено¬
логической школы Альфред Николаевич
Окснер (1898-1973).
Но известность ученого А. Н. Окснер получил еще до написания
«Флоры лишайников Украины», за свой ощутимый вклад в изучении ли¬
шайников таких регионов Советского Союза, как Урал, Арктика, Средняя
Азия и Дальний Восток. По результатам обработки собранных им самим
интереснейших материалов, и присланных ему коллекций, А. Н. Оксне-
ром были опубликованы многочисленные работы (Окснер, 1929, 1934,
1939а, 19396, 1940а, 19406, 1940в, 1945). В 1939 году по заказу Института
полярного земледелия он подготовил рукопись «Определителя лишайни¬
ков Крайнего Севера», которая, к сожалению, так и не была опубликова¬
на. Эта работа содержала 1037 видов и не потеряла актуальности и в
настоящее время. В 1935 году была подготовлена «Лихенофлора Урала»,
включавшая 350 видов, которая также осталась неопубликованной.
Из разных регионов Советского Союза А. Н. Окснером описано
около 150 новых для науки таксонов лишайников и предложено свыше
230 новых номенклатурных комбинаций (Kondratyuk, 1998). А. Н. Окснер
одним из первых начал изучать лишайниковые группировки, опубликовав
еще в начале своей научной деятельности работу «До вивчення флори
обр1сник1в каменястих виход1в УкраУни» (Окснер, 1927).
С именем А. Н. Окснера связан историко-географический подход к
анализу региональных лихенофлор. Он впервые разработал принципы ис¬
торической лихеногеографии и систему эколого-географических элемен¬
тов, которая широко применяется в работах отечественных ботаников до
настоящего времени.
А. Н. Окснер стоял у истоков такого фундаментального издания, как
«Определитель лишайников СССР». Он был автором его 2-го выпуска
«Общая часть. Морфология, систематика и географическое распростране¬
ние» (Окснер, 1974) и нескольких таксономических обработок, например,
по роду Aspicilia, сем. Dermatocarpaceae, Endocarpaceae, Normandinaceae
и др. опубликованных в первых томах. Второй же том, написанный как
введение к многотомному изданию, по своей сути является первым в
нашей стране базовым курсом лихенологии, не имеющим аналогов по
полноте приводимых данных и глубине их анализа, в котором обобщены
и критически осмыслены важнейшие сведения о лишайниках.
А. Н. Окснер по праву считается основателем украинской лихеноло¬
гической школы, оставившей глубокий след в отечественной науке. Со¬
трудники и ученики А. Н. Окснера были авторами первых томов «Опре¬
делителя лишайников СССР». М. Ф. Макаревич и Е. Г. Копачевская под¬
готовили целый ряд разделов по сем. Ьесапогасеае для первого выпуска
Определителя, а М. Ф. Макаревич написала раздел по сем. Pertusariaceae.
Для третьего тома О. Б. Блюм написал разделы по стиктовым и гиалекто-
вым лишайникам, Е. Г. Рооме подготовила разделы по калициевым,
цифелиевым и сферофоровым. Украинскими лихенологами - самим А. Н.
Окснером, М. Ф. Макаревич и Е. Г. Копачевской полностью подготовлен
четвертый том Определителя, вышедший из печати в 1977 г., а позднее -
уже С. Я. Кондратюком и А. Е. Ходосовцевым почти полностью написан
девятый выпуск, вышедший в 2004 г.
Силы и энергия, потраченные А. Н. Окснером на воспитание и под¬
готовку научных кадров, не были потрачены зря. Он многое сделал для
развития лихенологической науки в республиках бывшего СССР. Воспи¬
танники школы Окснера, его ученики и сотрудники, которым он помогал
ценным советом, составляют гордость не только украинской науки, но и
плодотворно работают во многих уголках России и других республик
бывшего Союза. В Новосибирске - это Н. В. Седельникова, в Таджики¬
стане - И. Кудратов, на Украине дело Окснера продолжили О. Б. Блюм, И.
J1. Навроцкая, Е. Г. Копачевская С. Я. Кондратюк, А. Е. Ходосовцев и М.
Ф. Макревич, занимавшаяся изучением лишайников Украинских Карпат
(Макаревич, 1963), и другие, уже более молодые последователи.
Необходимо также упомянуть основателя белорусской лихенологи¬
ческой школы - Михаила Петровича Томина (1883-1967). М. П. Томин
окончил, работал в Воронеже, где концентрировался на изучении лишай¬
ников сухих областей. Приехал в Белоруссию и начал работу по деталь¬
ному выявлению белорусской лихенофлоры. Опубликовал статьи и Опре¬
делители, которые в течение многих лет были настольными книгами со¬
ветских лихенологов (Томин, 1956). После его смерти дело его продолжи¬
ла Горбач. В настоящее время в Минске, Гродно, Гомеле работают
наследники Томина - В. В. Голубков, Яцына и др.
Наиболее притягательными регионами для отечественной лихеноло¬
гии всегда были Арктика, Урал, Сибирь и Дальний Восток страны, как в
силу своей малой изученности, так и из-за богатства и разнообразия ли¬
шайниковой флоры. В 60-е годы сотрудниками Лаборатории лихенологии
и бриологии Ботанического института была организована большая экспе¬
диция для сбора и изучения лишайников Урала - последняя экспедиция, в
которой участвовал В. П. Савич. К сожалению, только небольшая часть
обширных материалов этой экспедиции была обработана и опубликована,
хотя многие уральские сборы оказались важным дополнением при подго¬
товке очередных выпусков Определителя лишайников СССР.
В 60-е и 70-е годы в восточных районах страны складывается не¬
сколько лихенологических центров. На Урале - это Свердловск (Екате¬
ринбург), где в Свердловском педагогическом институте работала К. А.
Рябкова (Рябкова, 1982а, б) составившая первый список лишайников Ура¬
ла (Рябкова,1991) и, вместе с И. И. Макаровой, первый список лишайни¬
ков Полярного и Приполярного Урала (Рябкова, Макарова, 1991). Позднее
в Екатеринбурге в Институте экологии растений и животных УрО АН
СССР работали М. А. Магомедова, занимавшаяся изучением экологии
эпилитных лишайников Северного Урала (Магомедова, 1979) и влияния
на них человека.
В Новосибирске под руководством ученицы А. Н. Окснера Н. В. Се-
дельниковой сформировалось ядро лихенологов, центром притяжения
научных интересов которых стали горные районы Южной Сибири - Ал¬
тай, Саяны, Кузнецкий кряж и другие и, кроме того - тундровые террито¬
рии Западной Сибири. Трудами Н. В. Седельниковой и ее сотрудников,
таких как Т. В. Макрый создан гербарий лишайников Центрального Си¬
бирского ботанического сада и проводится изучение лишайников обшир¬
ных территорий Сибири (Седельникова, 1985, 1990, 1993, 1996а, 19966,
2001; Макрый, 1990). Среди ученых этой группы следует назвать и дру¬
гих.
В Якутии в Институт биологических проблем криолитозоны АН
СССР выдающийся советский ботаник В. Н. Андреев, еще в 30-е годы за¬
нимавшийся изучением кормовой базы оленеводства в тундрах Европей¬
ской части СССР и на Ямале (Андреев, 1932, 1933), собрал группу иссле¬
дователей, начавших планомерное изучение флоры и растительности
Якутии. Результатом этой работы стали многочисленные публикации, ка¬
савшиеся в той или иной мере лишайников, прежде всего арктических
районов республики (Рыкова, 1978; Егорова и др., 1991; Порядина 2001).
Флору Северо-востока - Колымы и Чукотки, в 60-е годы изучала со¬
трудница Институте биологических проблем Севера в Магадане М. А.
Локинская (1970). Позднее в Магадане в СВКНИИ ДВО РАН изучением
лишайников занимался А. А. Галанин (Галанин, 2002).
Изучение флоры лишайников Камчатки началось еще в самом нача¬
ле XX века, когда В. П. Савич вместе с В. Л. Комаровым участвовал в
экспедиции Ф. П. Рябушинского. В 80-е годы сотрудник Биолого¬
почвенного института ДВО АН СССР во Владивостоке А. Г. Микулин
опубликовал первый определитель лишайников Камчатки, насчитывав¬
ший 250 видов (Микулин, 1990). Сейчас исследование флоры лишайников
Камчатки проводят сотрудники Лаборатории лихенологии и бриологии Д.
Е. Гимельбрант, Е. С. Кузнецова и И. С. Степанчикова (Davydov,
Himelbrant, Stepanchikova, 2011; Гимельбрант, Степанчикова, Кузнецова,
2009, 2012; Гимельбрант, Степанчикова, 2008 и др. ). В результате их ра¬
бот число известных науке видов камчатских лишайников достигло 460.
Изучением флоры лишайников Дальнего Востока занимались во
Владивостоке сотрудники Биолого-почвенного института ДВО АН СССР
Л. А. Княжева и Тихоокеанского института географии ДВО РАН И. Ф.
Скирина, в Лазовском заповеднике, а позже в Сахалинском ботаническом
саду ДВО РАН в Южно-Сахалинске - С. И. Чабаненко (Княжева, Семено¬
ва, 1979; Княжева, Скирина, 1982; Чабаненко, 1990; Чабаненко, Скирина,
Княжева, 2002). Существенный вклад в изучение флоры лишайников
Дальнего Востока внесли украинские (работы А. Н. Окснера) и эстонские
исследователи (X. X. Трасс и его коллеги). В 60-е годы X. X. Трассом бы¬
ла организована целая серия экспедиций на Байкал и на Дальний Восток,
результатом которых были публикации и гербарные материалы, хранящи¬
еся в Тартуском университете (Эстония).
В Союзных республиках в те же годы сложились собственные ли-
хенологические центры, работавшие, как правило, в контакте с Ботаниче¬
ским институтом в Ленинграде и с другими старейшими научными учре¬
ждениями страны. Тем более, что лихенологи Ботанического института
начали проводить масштабные работы в Средней и Центральной Азии. Н.
С. Голубкова занялась изучением лишайников Памира (Голубкова, 1972,
1973, 1975), а позже в течение многих лет работала над разработкой мон¬
гольской флоры, составив и опубликовав ее Конспект и проведя всесто¬
ронний анализ. Флору лишайников Киргизии изучали Л. А. Бредкина
(Бредкина, Макрова, 2005) и ученица Н. С. Голубковой - Н. Э. Байбулато-
ва (публикации).
В Алма-Ате в Институте ботаники АН Казахской ССР работала Е.
И. Андреева, издавшая первую сводку по лишайникам Казахстана (Ан¬
дреева, 1972). Лишайниками Казахстанского Алтая занималась аспирант¬
ка М. П. Андреева - Зафия Сеитовна Мырзакулова (Гёз) (Мырзакулова
1988; Андреев, Мырзакулова, 1992).
Таджикская лихенология была представлена сотрудником Институ¬
та ботаники Таджикской АН в Душанбе, учеником А. Н. Окснера -
Имомназаром Кудратовым. Его кандидатская диссертация была посвяще¬
на лишайникам Зеравшанского хребта (Кудратов 1968), позднее он зани¬
мался изучением флоры лишайников Таджикистана (Кудратов 1990; Ан¬
дреев, Кудратов, 1992).
В столице Туркмении - Ашхабаде работала X. X. Акрамова, изу¬
чавшая лишайники гор Копет-Дага (Акрамова, 1969).
В республиках Закавказья также сформировались свои собственные
лихенологические центры. В Азербайджане в Баку работал выдающийся
ученый Шабан Омарович Бархалов (1938-1999), составивший первый
сводный список лишайников Кавказа (Бархалов, 2000), в Кировабаде
(Гянджа) - В. С. Новрузов, изучавший лишайники Лагонахского нагорья
(Новрузов, 1972).
В Армении флору лишайников окрестностей озера Севан исследо¬
вала аспирантка Н. С. Голубковой Армене Абрамян (Абрамян, 1988).
В Тбилиси в Грузии работали такие видные лихенологи, как Цими
Инашвили. Им принадлежит целый ряд публикаций по лихенофлоре Гру¬
зии (Инашвили 1978), а Ц. Н. Инашвили написала несколько системати¬
ческих разделов для Определителя лишайников СССР, в частности сем.
Collemataceae, Pannariaceae и Heppiaceae.
Из прибалтийских республик, наибольшее развитие лихенологиче¬
ская наука получила в Эстонии. Здесь сложилось два научных центра:
тартуский - в Тартуском университете, возглавляемый Хансом Хартмуто-
вичем Трассом и таллинский - в Таллинском ботаническом саду, под ру¬
ководством Юри Мартина. Сотрудники обоих учреждений, кроме реше¬
ния вопросов, связанных с изучением лишайников республики, работали
и на территории Российской Федерации, в частности - на Урале, в Сибири
и на Дальнем Востоке, изучая флору лишайников России. Например, Тай-
ми X. Пийн (Таллин,) изучала лишайники полуострова Таймыр (Piin,
1977, 1979а, 19796; Пийн, Трасс, 1971), Ю. Л. Мартин работал на Поляр¬
ном Урале (Мартин, 1970), X. X. Трасс написал основополагающую рабо¬
ту по лихенофлоре Эстонии (Трасс, 1970) и важнейший раздел по сем.
Cladoniaceae для Определителя лишайников СССР.
Латвийская лихенологическая наука была представлена учеником В.
П. Савича - Альфонсом В. Питерансом, кроме изучения лихенофлоры
Латвии (Питеранс, 1982), внесшим существенный вклад в создание Опре¬
делителя лишайников СССР, написав для первых выпусков издания ряд
разделов, например, по сем. Diploschistaceae, Coenogoniaceae,
Lichinaceae, Pyrenopsidaceae и др.
Становление литовской лихенологической школы началось не¬
сколько позже, после того, как в Вильнюсе начала работать ученица Н. С.
Голубковой - Юрга Мотеюнайте.
После распада СССР судьба лихенологических школ в бывших рес¬
публиках Союза сложилась по-разному. На Украине, в Белоруссии и в
Прибалтике они получили дальнейшее развитие и сейчас занимают за¬
метное место в мировой лихенологической науке, а в республиках Сред¬
ней Азии и Закавказья, к сожалению, исчезли или почти перестали суще¬
ствовать.
Наиболее притягательными регионами для отечественной лихеноло¬
гии всегда были Арктика, Урал, Сибирь, Дальний Восток страны и Сред¬
няя Азия, как в силу своей малой изученности, так и из-за богатства и
разнообразия лишайниковой флоры. В 60-е годы сотрудниками Лаборато¬
рии лихенологии и бриологии Ботанического института была организова¬
на большая экспедиция для сбора и изучения лишайников Урала - по¬
следняя экспедиция, в которой участвовал В. П. Савич. Уральские сборы
оказались важным дополнением при подготовке очередных выпусков
Определителя лишайников СССР.
В 70-е годы многочисленные экспедиции работали в высокогорьях
Средней Азии, результатом чего стали публикации по флоре лишайников
Памира (Голубкова, 1972, 1973, 1975) и Таджикистана (Андреев, Кудра¬
тов, 1992), киргизского Тянь-Шаня (Бредкина, Макарова, 2005) и Казах¬
стана (Андреев, Мырзакулова, 1992).
В 60-80-е годы одним из основных объектов изучения ленинград¬
ских лихенологов становится Крайний Север. На Кольском полуострове в
Кировске в Полярно-альпийском ботаническом саду начинает работать А.
В. Домбровская. Она занималась изучением флоры лишайников Кольско¬
го полуострова (Домбровская, 1962) и рода Stereocaulon (Домбровская,
1990). А. В. Домбровская смогла создать в Кировске богатый гербарий и
собрать сильную группу сотрудников - И. В. Антонову и Т. А. Дудореву,
которая позже продолжила свои исследования в Саратовской области
(Дудорева, Гимельбрант, 2009, 2010).
После образования в Ботаническом институте Полярной экспедиции
специалисты лихенологи смогли непосредственно участвовать в полевых
выездах в районы Крайнего Севера. В эти годы в центральной и восточ¬
ной частях Чукотского полуострова работала И. И. Макарова (Макарова,
1979, 1983, 1986, 1987; Макарова, Катенин, 1983, 1987, 1992), внесшая
наиболее весомый вклад в изучение лихенофлоры Чукотки, она также об¬
рабатывала материалы из арктической Якутии и с Новосибирских остро¬
вов (Макарова, 1989; Макарова, Перфильева, 1984, 1985, 1989; Макарова,
Перфильева, Николин, 1988), лихенофлору Западной Чукотки и Ямала
изучал М. П. Андреев (Андреев, 1980, 1981, 1984а, 19846), лишайники
острова Врангеля - А. А. Добрыш (Добрыш, 1988, 1995), флору Северной
Земли и Полярного Урала исследовал Ю. В. Котлов (Котлов, 1993, 1994;
Kotlov, 1994), на Таймыре работали А. Н. Титов и М. П. Журбенко (Жур¬
бенко, 1992, 2000).
В 70-80-е годы, в связи с интенсивным освоением отдаленных обла¬
стей страны и районов Крайнего Севера, развернувшимся транспортным и
промышленным строительством, начались работы по изучению влияния
промышленного загрязнения и других типов антропогенного воздействия
на лишайниковую флору и растительность (Магомедова, 1984).
90-е годы прошлого века, после открытия внешних границ страны,
ознаменовались масштабными проектами, выполнявшимися в кооперации
с иностранными исследователями. В рамках проекта «Панарктическая
флора», был подготовлен первый список лишайников Русской Арктики,
объединивший все известные к тому времени данные о видовом составе
лишайников арктических и тундровых районов России и включавший
1102 вида (Andreev, Kotlov, Makarova, 1996). В эти же годы началось сов¬
местное с американскими исследователями изучение лишайников Берин-
гийской Чукотки и Аляски, причем выявленная в результате проделанной
работы лихенофлора региона, насчитывала 626 видов, оказавшись наибо¬
лее богатой в Арктике. В конце 90-х годов состоялись две совместные
российско-китайские экспедиции в горы китайского Тянь-Шаня
(Синьцзянь-Уйгурский автономный район КНР) и в Восточный Тибет
(пров. Сычуань, КНР) (Titov, 2003).
В те годы были организованы две совместные с зарубежными кол¬
легами экспедиции в ранее закрытые для иностранных ученых районы
России. В 1991 году - Советско-шведская экспедиция в Приморский край.
Со шведской стороны в ней участвовали такие известные ученые, как
академик Рольф Сантессон, профессора Роланд Моберг, Лейф Тибелл и
Ингвар Чернефельт. Работы проводились на территории Лазовского запо¬
ведника и позволили пополнить гербарные коллекции шведских и рос¬
сийских гебариев. В 1992 году состоялась Российско-финская экспедиция
в Карелию, участниками которой стали сотрудники Финского музея есте¬
ственной истории (Хельсинки) проф. Теуво Ахти, Орво Витикайнен и
Микко Куусинен.
Основными направлениями научных работ в области лихенологии в
России в конце XX столетия и сейчас являются фундаментальные иссле¬
дования, связанные с изучением флоры, систематики, экологии, физиоло¬
гии и биохимии лишайников, региональные флористические исследова¬
ния, которые проводились и проводятся как на территории России, так и
за рубежом и монографическая разработка отдельных таксономических
групп лишайников, предусматривающая изучение их морфологических,
эколого-географических и биохимических особенностей с целью решения
вопросов систематики, филогении и эволюции. Результатом этих исследо¬
ваний должно быть составление определителей, сводок, монографий.
В 1950 г. была опубликована первая отечественная монография по
одной из систематических групп лишайников - роду Cetraria (Рассадина,
1950), увидела свет работа В. П. Савича по сем. Umbilicariaceae (Савич,
1950), позднее появились монография Н. С. Голубковой по семейству
Acarosporaceae (Голубкова, 1988) и монография по микокалициевым гри¬
бам и лишайникам Голарктики (Титов, 2006), проводились исследования
лихенофильных грибов (Журбенко, 2010; Zhurbenko et al., 2012), законче¬
на монографическая работа по гиалектовым лишайникам России, (Гага¬
рина, 2009, 2012).
Большой интернациональный коллектив лихенологов всей страны в
течение многих лет работал над многотомным изданием «Определитель
лишайников СССР» (1971, 1974, 1975, 1977, 1978) и его продолжением
«Определитель лишайников России» (2006, 2008). Издание является пер¬
вой фундаментальной сводкой по флоре лишайников СССР и России и
содержит описания 1050 видов, 270 родов и 77 семейств, а также данные
об экологии и географическом распространении видов на территории Рос¬
сии и за ее пределами. Издание завершено последним 10-м томом в 2008
году.
Второе направление научных работ - региональные флористические
исследования, которые проводились и проводятся как на территории Рос¬
сии, например, в Ленинградской (Stepanchikovaet al., 2009, 2010, 2011;
Степанчикова, Гимельбрант, Кузнецова, 2011; Kuznetsova et al., 2012) и в
Новгородской областях (Катаева, 2002, 2009), в Мурманской области (Ко-
норева, 2010; Урбанавичюс, Урбанавичене, Мелехин, 2012 и др. ), в Каре¬
лии (Фадеева и др., 1997), в Средней России, Центрально-черноземной
области и в Поволжье (Конорева и др., 2010; Кузнецова, Сказкина, 2010;
Гудовичева, Гимельбрант, 2012; Muchnik, Konoreva, 2012), в Пермском
крае и на Урале (Котлов, 2003; Шаяхметова, Селиванов, Урбанавичене,
2009; Урбанавичене, 2011), в Калмыкии и на Сев. Кавказе (Очирова, 2005;
Урбанавичюс, Урбанавичене, Отте, 2012), на Алтае и в Зап. Сибири (Ка¬
таева и др., 2005), в Вост. Сибири, Якутии и на Дальнем Востоке (Бредки¬
на и др., 1991; Урбанавичене, Урбанавичюс, 1998; 1999; Вершинина и др.,
2012), так и за рубежом, например, в Антарктике (Голубкова, Савич, Си¬
монов, 1968; Голубкова, 1969; Голубкова, Симонов, 1972; Андреев, 1988,
1990, Андреев, Курбатова, 2009), США, Монголии (Голубкова, 1981,
1983), Израиле (Temina, Andreev, Barinova, Nevo, 2009), Китае (Titov,
2003), Корее (Joshi, Andreev, Hur, 2012), Югославии (Savic, Tibell,
Andreev, 2006), а также на территориях бывших республик Союза ССР: в
Литве (Motiejunaite et al., 2012), Абхазии (Гагарина, 20096). При финансо¬
вой поддержке зарубежных коллег были изучены и опубликованы резуль¬
таты исследований флоры лишайников Восточной части Ленинградской
области (Kuznetsova, Ahti, Himelbrant, 2007) и Мурманской области (Ur-
banavichus, Ahti, Urbanavichene, 2008) и др.
Флористическая работа в рамках совместных проектов сочеталась с
экологическими исследованиями. Так, в частности, был выполнен сов¬
местный российско-шведский проект по изучению биологически ценных
лесов Северо-Запада Европейской части России (Конечная и др., 2009)
Большое внимание уделялось изучению флоры лишайников запо¬
ведных и охраняемых территорий. Исследования проводились во много¬
численных ООПТ (Особо охраняемых природных территориях) на всем
пространстве России. В качестве одного из промежуточных результатов
этой работы можно назвать сводный список лишайников ООПТ России
(Урбанавичус, Урбанавичене, 1998)
В 2011 году наконец увидел свет первый «Список лишайников Рос¬
сии», в значительной мере, подготовленный Г. П. Урбанавичусом на базе
лаборатории лихенологии и бриологии и на материалах Гербария лишай¬
ников Ботанического института - наиболее полная сводка, обобщающая
все виды лишайников, отмеченные на территории страны. В списке при¬
веден 3331 вид и 476 родов лишайников, даны основные синонимы и в
краткой форме - регионы, где были отмечены те или иные таксоны (Спи¬
сок лишайников России, 2010).
Третье направление лихенологических исследований — физиолого¬
биохимическое изучение лишайников. Впервые изучение биохимических
свойств лишайников в нашей стране было начато в Ботаническом инсти¬
туте в 50-х годах XX века сотрудником Лаборатории лихенологии и брио¬
логии Е. Н. Моисеевой. Особое внимание в своих исследованиях она уде¬
ляла изучению состава и активности ферментов у лишайников, произрас¬
тающих в разных экологических условиях и на разных субстратах, а так¬
же исследованию особенностей лишайниковых кислот, их физико¬
химических свойств. Эти работы нашли и практический выход. Так, на
основе усниновой кислоты был получен лекарственный препарат
«БИНАН», выпускавшийся под названием «натрия уснинат» (Моисеева,
1957). С 1967 г. в лаборатории начали работать Е. А. Вайнштейн, А. П.
Равинская, И. А. Шапиро, которые, продолжая изучение биохимических
свойств лишайников, стали разрабатывать новые проблемы как теорети¬
ческого, так и прикладного плана. Эти исследования были связаны с изу¬
чением особенностей метаболизма лишайников с целью познания биоло¬
гической роли лишайниковых веществ и с разработкой теоретических ос¬
нов их использования в систематике и биологии симбиотических орга¬
низмов. Вскоре были получены интересные данные о сезонной динамике
содержания в талломах лишайников биохимических соединений и влия¬
нии экологических факторов на их аккумуляцию, о транспорте метаболи¬
тов между симбионтами и возможной регуляционной роли лишайниковых
веществ в отношениях между симбионтами, а также о влиянии лишайни¬
ковых веществ на фотосинтез фотобионта и дыхание обоих компонентов
лишайникового таллома.
Разрабатывались также вопросы, связанные с изучением роли ли¬
шайниковых веществ во взаимодействии лишайников с другими растени¬
ями и микроорганизмами. Было проведено изучение целого комплекса
ферментов азотного метаболизма, получены интересные данные в области
изучения азотного обмена лишайников и роли лишайниковых симбионтов
в усвоении азота.
В целях экологического мониторинга изучалось воздействие антро¬
погенных факторов на физиологические характеристики лишайников.
Проводилось исследование физиолого-биохимических механизмов ответа
на стрессовые воздействия у лишайников разных систематических групп.
Проводилось хемотаксономическое изучение отдельных таксономических
групп лишайников, данные этих исследований использовались система¬
тиками при работе над составлением «Определителя лишайников России»
(Шапиро, 1980). В лаборатории была создана единственная в стране эта¬
лонная коллекция лишайниковых веществ, которая широко использова¬
лась в хемотаксономических и физиологических исследованиях лишайни¬
ков. К сожалению, в последствии в Лаборатории лихенологии и бриоло¬
гии Ботанического института это направление исследований не получило
дальнейшего развития и в настоящее время проводятся работы только в
области прикладной хемотаксономии, в частности тонкослойной хрома¬
тографии, служащей целям идентификации лишайников.
Публикацией Определителя лишайников России завершился оче¬
редной этап в истории отечественной лихенологии и начался новый.
В новом тысячелетии лихенологи России рассчитывают сосредото¬
чить свои усилия в работе по проекту «Флора лишайников России», нача¬
ло которому было положено в 2008 г. В настоящее время в рамках проек¬
та ведется работа по созданию первого тома, посвященного пармелиевым
лишайникам. По-прежнему, большое внимание уделяется вопросам си¬
стематики лишайников и региональным флористическим исследованиям.
Наибольший интерес вызывают такие систематические группы как - пар-
мелиевые, бацидиевые, цианобионтные, гиалектовые, лецидеоидные ли¬
шайники, лепрарии, микареи и лихенофильные грибы. Экспедиционные и
флористические работы, в основном при поддержке Российского фонда
фундаментальных исследований, проводятся в Северо-западном регионе,
в Средней России, на Урале, Кавказе, Алтае, в Восточной Сибири и на
Камчатке, за пределами России - на Шпицбергене и в Антарктике. Ак¬
тивно изучается флора лишайников заповедных территорий, лихенологии
участвуют в создании региональных и общероссийской Красных Книг,
сводных флористических списков и чек-листов.
В добавление к существовавшим ранее, в России появилось доволь¬
но большое количество региональных центров, где работают и уже из¬
вестные и молодые ученые и существуют солидные гербарии лишайников
(см. Гл. II).
Для успешного выполнения всех задач, стоящих перед русской ли¬
хенологией в третьем тысячелетии, необходимо привлечь в науку моло¬
дые кадры будущих исследователей и лабораторных технических работ¬
ников, повысить качество их подготовки и радикально расширять коопе¬
рацию с зарубежными научными центрами, максимально интегрировав
отечественную науку в международное научное сообщество. Это позво¬
лило бы, с одной стороны заметно поднять уровень отечественных специ¬
алистов, с другой - привлечь иностранных ученых для решения общих
задач, но на нашем материале и на территории России. Для развития ис¬
следований в области систематики лишайников и решения вопросов их
происхождения и эволюции необходимо углублять исследования физио-
лого-биохимических особенностей лишайников и расширить методиче¬
ские основы систематики, в частности, использовать современные методы
молекулярной геносистематики и новые хроматографические методики.
Очевидно, что в гербарной работе необходим переход на качественно но¬
вый стиль, предусматривающий компьютеризацию и дигитализацию кол¬
лекций и обеспечение свободного доступа к гербарным базам данных че¬
рез Интернет, а в лабораторной работе - оснащение исследователей со¬
временным оборудованием и оптикой. Выполнение все вышеперечислен¬
ных условий в сочетании с опытом отечественных исследователей, богат¬
ством имеющихся в коллекциях материалов, размерами и разнообразием
природных условий России и, соответственно, богатством и разнообрази¬
ем ее лихенофлоры, позволит русской лихенологии достойно показать
себя в третьем тысячелетии.
Литература
Андреев В. Н. Кормовая база Ямальского оленеводства // Советское
оленеводство. 1933. Вып. 1. С. 99-164
Андреев В. Н. Типы тундр запада Большой Земли // Тр. Бот. муз. АН
СССР. Л.: Изд. АН СССР. 1932. Вып. 25. С. 121-268.
Андреев М. П. Лишайники родов Cladonia и Cladina Анюйского
нагорья // Ботан. журн. 1981. Т. 66, № 1. С. 31 -41.
Андреев М. П. Лишайники в растительном покрове горы Эломбал,
Чукотский автономный округ // Ботан. журн. 1980. Т. 65, № 4. С. 563-570.
Андреев М. П. Лишайники оазиса Бангера (Восточная Антарктида)
// Новости систематики низших растений. 1990. Т. 27. С. 85-93.
Андреев М. П. Лишайники полуострова Файлдс, остров Кинг
Джордж, Антарктика // Новости систематики низших растений. 1988. Т.
25. С. 111-118.
Андреев М. П. Лишайники полуострова Ямал // Новости системати¬
ки низших растений. 1984а. Т. 21. С. 127-136.
Андреев М. П. Систематический состав лихенофлоры Анюйского
нагорья // Новости систематики низших растений. 19846. Т. 21. С. 136-
140.
Андреев М. П., Кудратов И. Lecidea (Lecanorales) и близкие таксоны
лишайников флоры северо-западного и центрального Таджикистана //
Ботан. журн. 1992. Т. 77, № 1. С. 101-111.
Андреев М. П., Мырзакулова 3. С. Виды семейств Lecideaceae и
Lecanoraceae (Lichenes) Южного Алтая (Восточный Казахстан) // Ботан.
журн. 1992. Т. 77, № 3. С. 114-118.
Андреев М. П., Курбатова J1. Е. Новые данные о мхах и лишайниках
тихоокеанского сектора Антарктиды // Новости систематики низших рас¬
тений. 2009. Т. 42. С. 142-152.
Бредкина J1. И., Добрыш А. А., Макарова И. И., Титов А. Н. К флоре
лишайников острова Кунашир (Курильские острова) // Новости система¬
тики низших растений. 1991. Т. 28. С. 90-94.
Бредкина J1. И., Макарова И. И. Аннотированный список лишайни¬
ков Центрального Тянь-Шаня (Киргизия) // Новости систематики низших
растений. 2005. Том 39. СПб. С. 179-195.
Вершинина С. Э., Гимельбрант Д. Е., Кузнецова Е. С., Габышева Л.
М., Габышев Э. М. Первые сведения о лихенофлоре государственного за¬
поведника «Олёкминский» (Республика Саха-Якутия) // Вестник ТвГУ.
Серия «Биология и экология». 2012. Вып. 25. № 3. С. 138-149.
Гагарина J1. В. Гиалектовые лишайники: история изучения и совре¬
менное состояние изученности // Новости систематики низших растений.
2009а. Т. 42. С. 153-168.
Гагарина J1. В. Лишайники порядка Gyalectales Henssen ex D.
Hawksw. & О. Eriksson внетропической Евразии. Автореферат диссерта¬
ции на соискание ученой степени кандидата биологических наук. Санкт-
Петербург, 2012. 23 с.
Гагарина Л. В. Первые сведения о лишайниках порядка Gyalectales
(s. 1. ) в Абхазии // Вестник ТвГУ. Серия «Биология и экология». 20096. №
34. С. 161-167.
Галанин А. А. Лихенометрия: современное состояние и направления
развития метода (аналитический обзор). Магадан, СВКНИИ ДВО РАН,
2002. 74 с.
Гимельбрант Д. Е., Степанчикова И. С. Новые данные о пармелие-
вых лишайниках и калициоидных грибах Камчатки // Новости системати¬
ки низших растений. 2008 (2009). Т. 42. С. 169-172.
Гимельбрант Д. Е., Степанчикова И. С., Кузнецова Е. С. Лишайники
некоторых кустарников и кустарничков полуострова Камчатки // Новости
систематики низших растений. 2009. Т. 43. С. 150-171.
Гимельбрант Д. Е., Степанчикова И. С., Кузнецова Е. С. Лишайники
// Растительный покров вулканических плато Центральной Камчатки /
Ред. В. Ю. Нешатаева. М., Товарищество научных изданий КМК, 2012. С.
126-171.
Голубкова Н. С. Анализ флоры лишайников Монголии. Л.: Наука.
1983.248 с.
Голубкова Н. С. Всеволод Павлович Савич (1885-1972) - ученый и
организатор научных исследований в области криптогамной ботаники //
Ботан. журн. 2003. Т. 88, № 10. С. 142-150.
Голубкова Н. С. Конспект флоры лишайников Монгольской Народ¬
ной Республики. Л.: Наука. 1981. 200 с.
Голубкова Н. С. Лихенологические исследования в Ботаническом
институте им. В. Л. Комарова РАН в XX столетии // Микология и крипто¬
гамная ботаника в России: традиции и современность. Тр. междунар.
конф., посвящ. 100-лет. организ. иссл. по мик. и крипт, бот. в БИН РАН.
СПб, 2000, с. 20-32.
Голубкова Н. С. Лишайники Антарктиды // Атлас Антарктиды. Л.:
Гидрометеоиздат. 1969. Т. 2. С. 488-492.
Голубкова Н. С. Лишайники Восточного Памира // Новости систе¬
матики низших растений. 1975. Т. 12. С. 250-263.
Голубкова Н. С. Лишайники семейства Acarosporaceae Zahlbr. в
СССР. Л.: Наука. 1988. 127 с.
Голубкова Н. С. Новые для лихенофлоры СССР лишайники Восточ¬
ного Памира // Новости систематики низших растений. 1972. Т. 9. С. 214—
233.
Голубкова Н. С. Первый систематический список лишайников Во¬
сточного Памира // Новости систематики низших растений. 1973. Т. 10. С.
206-223.
Голубкова Н. С., Савич В. П., Симонов И. М. Лишайники западной
части Земли Эндерби // Тр. Советской антарктической экспедиции. 1968.
Т. 38. С. 247-253.
Голубкова Н. С., Симонов И. М. Лишайники оазиса Ширмахера //
Тр. Советской антарктической экспедиции. 1972. Т. 60. С. 317-327.
Городков Б. Н. Ботанико-географический очерк Чукотского побере¬
жья // Уч. зап. Лен. гос. пед. ин-та, каф. физ. геогр. 1939. Вып. 21. С. 99-
175.
Городков Б. Н. Полярные пустыни о. Врангеля // Ботан. журн. 1943.
Т. 28. №4. С. 127-143.
Городков Б. Н. Почвенно-растительный покров острова Врангеля //
Растительность Крайнего севера СССР и ее освоение. М. -Л.: Изд. АН
СССР, 1958. Вып. З.С. 5-58.
Гудовичева А. В., Гимельбрант Д. Е. Дополнение к лихенофлоре
Севера Среднерусской возвышенности // Вестник ТвГУ. Серия «Биология
и экология». 2012. Вып. 25. № 3. С. 150-164.
Давыдов Е. А. Аннотированный список лишайников западной части
Алтая (Россия) // Новости систематики низших растений. 2002. Т. 35. С.
140-161.
Добрыш А. А. Аннотированный список лишайников острова Вран¬
геля // Новости систематики низших растений. 1995. Т. 30. С. 52-60.
Добрыш А. А. Лишайники южного побережья острова Врангеля
(бухта Сомнительная) // Новости систематики низших растений. 1988. Т.
25. С. 124-126.
Дудорева Т. А., Гимельбрант Д. Е. Лишайники Арзянского бора
(Саратовская область) // Вестник ТвГУ. Серия «Биология и экология».
2010. Вып. 17. № 16. С. 136-141.
Дудорева Т. А., Гимельбрант Д. Е. Предварительный список лишай¬
ников окрестностей г. Хвалынска (Саратовская область) // Вестник ТвГУ.
Серия «Биология и экология». 2009. Вып. 16. № 36. С. 144-148.
Егорова А. А., Васильева И. И., Степанова Н. А., Фесько Н. Н. Фло¬
ра тундровой зоны Якутии. Якутск, ЯНЦ СО АН СССР, 1991. С. 1-186.
Еленкин А. А. А. А. Фишер фон-Вальдгейм. Известия Главного Бо¬
танического Сада. 1921. С. 48-54.
Еленкин А. А. Из итогов 25-летней деятельности Института Споро¬
вых Растений Главного Ботанического Сада. Петроград, 1923. С. 1-28.
Еленкин А. А. Лишайники полярного побережья Сибири // Научные
результаты русской полярной экспедиции барона Э. В. Толля. Отд. Д, бо¬
таники /Зап. Акад. Наук, сер. 8. СПб, 1909. Т. 26, вып. 1. С. 1-53.
Еленкин А. А. Лишайники русской флоры и пограничных восточ¬
ных областей. В кн.: Тр. Имп. Бот. Сада. СПб., 1904. Т. 24. Вып. 1. С. 118-
X.
Еленкин А. А. Материалы для лишайниковой флоры России // Тр.
Имп. Бот. Сада. СПб., 1901. Т. 19. С. 155-182.
Еленкин А. А. О деятельности Спорового Гербария за 14 лет (с 1899
г. по 1913 г. ) и о ближайших задачах деятельности "Института Споровых
Растений" - нового отдельного учреждения Императорского Ботаниче¬
ского Сада Петра Великого. Известия Императорского Ботанического Са¬
да Петра Великого. Т. 14. Вып. 1-2. 1914. С. 1-20.
Еленкин А. А. О текущей деятельности Института Споровых Расте¬
ний Главного Ботанического Сада. Известия Главного Ботанического Са¬
да. 1921. С. 55-58.
Еленкин А. А. Флора лишайников Средней России. Части 1 и 2.
СПб, 190Х; части 3 и 4. Юрьев. 1911. С. 361 -682.
Еленкин А. А., Савич В. П. Лишайники, собранные И. В. Палиби-
ным в плавание ледокола «Ермак» в Северном Ледовитом океане в 1901 г.
//Тр. Имп. Бот. Сада. СПб, 1912. Т. 32. С. 69-100.
Журбенко М. П. Лишайники и лихенофильные грибы Путоранского
заповедника // Флора и фауна заповедников. 2000. Вып. 89. 52 с.
Императорский С. -Петербургский Ботанический сад за 200 лет его
существования (1713-1913). Юбилейное издание, ч. 1.СП6., 1913.
Исторический очерк за последнее 25-летие Сада (1873-1898).
История Академии Наук СССР. Т. 2. (1803-1917). М. -JL, Наука,
1964. 772 с.
Катаева (Катенина) О. А. Аннотированный список видов лишайни¬
ков Новгородской области // Новости систематики низших растений.
2002. Т. 36. С. 114-143.
Катаева О. А. Лишайники и лихенизированные грибы // Кадастр
флоры Новгородской области / Ред. Юрова Э. А., Крупкина Л. И., Конеч¬
ная Г. Ю. Новгород, Изд. «ЛЕМА», 2009. С. 247—254.
Катаева О. А., Макарова И. И., Таран Г. С., Тюрин В. Н. Лишайники
поймы р. Обь в окрестностях г. Сургута (Тюменская область, Западная
Сибирь) // Новости систематики низших растений. 2005. Т. 38. С. 186—
199.
Княжева Л. А., Семенова Е. А. К лихенофлоре Уссурийского запо¬
ведника. Сем. Lecanoraceae II Изучение флоры и растительности Дальнего
Востока. Владивосток, ДВГУ, 1979. С. 83-90.
Княжева Л. А., Скирина И. Ф. Лишайники // Растительный и живот¬
ный мир Сихотэ-Алинского заповедника. М., Наука, 1982. С. 44-47.
Конечная Г. Ю., Курбатова Л. Е., Потемкин А. Д., Гимельбрант Д.
Е., Кузнецова Е. С., Змитрович И. В., Коткова В. М., Малышева В. Ф.,
Морозова О. В., Попов Е. С., Яковлев Е. Б., Andersson L., Кияшко П. В.,
Skujiene G. Выявление и обследование биологически ценных лесов на Се-
веро-Западе Европейской части России. Т. 2. Пособие по определению
видов, используемых при обследовании на уровне выделов / Отв. ред. Л.
Андерссон, Н. М. Алексеева, Е. С. Кузнецова. СПб, 2009. 258 с.
Конорева Л. А. Новые находки редких и охраняемых видов лишай¬
ников из тундровой зоны Мурманской области // Вестн. ТвГУ. Сер. Био¬
логия и экология. 2010. Вып. 20. С. 115-121.
Конорева Л. А., Мучник Е. Э., Урбанавичене И. Н., Урбанавичюс Г.
П. Виды лишайников, предлагаемые к включению в Красную книгу Кур¬
ской области // Исследования по Красной книге Курской области. Вып. 2.
Курск, 2010. С. 69-85.
Котлов Ю. В. Лишайники, собранные на птичьей колонии в северо-
западной части острова Большевик (Северная Земля) // Ботан. журн. 1993.
Т. 78. № 8. С. 34-36.
Котлов Ю. В. Новые для Полярного Урала виды лишайников с по¬
бережья Карского моря // Ботан. журн. 1994. Т. 79. № 7. С. 122-124.
Котлов Ю. В. Новые для Урала виды лишайников из Свердловской
области // Ботан. журн. 2003. Т. 88, № 4. С. 147-148.
Кузнецова Е. С., Сказина М. А. К изучению лихенофлоры Костром¬
ской области // Новости систематики низших растений. 2010. Т. 44. С.
200-209.
Локинская М. А. Наиболее распространенные виды лишайников на
северо-востоке СССР // Водоросли и грибы Сибири и Дальнего Востока.
Новосибирск: Наука, Сиб. отд., 1970. Вып. 1 (3). С. 233-246.
Магнуссон А. Г. Новые лишайники севера СССР // Изв. Главн. бот.
сада СССР. Л., 1927. Т. 26. Вып. 4. С. 359-370.
Магомедова М. А. Лишайниковый покров высокогорий Северного
Урала // Растительные сообщества Урала и их антропогенная деградация.
Свердловск, 1984 С. 91-101.
Магомедова М. А. Сукцессии сообществ литофильных лишайников
в высокогорьях Северного Урала // Экология. 1979. № 3. С. 29-38.
Макаревич М. Ф. Анал1з л1хенофлори УкраУнських Карпат. КиУв,
1963.263 с.
Макарова И. И. К флоре лишайников бухты Провидения (Чукотка) //
Новости систематики низших растений. 1986. Т. 23. С. 179-181.
Макарова И. И. К флоре лишайников низовьев реки Лены // Новости
систематики низших растений. 1989. Т. 26. С. 118-124.
Макарова И. И. Лишайники островов Айон и Большой Раутан
(Чаунская губа) // Новости систематики низших растений. 1987. Т. 24. С.
159-163.
Макарова И. И. Лишайники юго-востока Чукотского полуострова
(бухта Пенкигней) // Новости систематики низших растений. 1983. Т. 20.
С. 146-150.
Макарова И. И. Характеристика конкретных лихенофлор запада Чу¬
котского полуострова // Ботан. журн. 1979. Т. 64. № 3. С. 351-361.
Макарова И. П., Катенин А. Е. Лишайники в горах юго-востока Чу¬
котского полуострова // Ботан. журн. 1983. Т. 68. № 11. С. 1477-1487.
Макарова И. П., Катенин А. Е. Лишайники равнинного юго-
западного побережья Чукотского полуострова // Ботан. журн. 1987. Т. 72.
№ 4. С. 455-463.
Макарова И. П., Катенин А. Е. Лишайники средней части хребта
Искатень на западе Чукотского полуострова // Ботан. журн. 1992. Т. 77. №
1.С. 45-57.
Макарова И. П., Перфильева В. И. Дополнение к лихенофлоре севе-
ро-запада Якутии // Новости систематики низших растений. 1985. Т. 22. С.
180-184.
Макарова И. П., Перфильева В. И. К флоре лишайников северо-
запада Якутии // Новости систематики низших растений. 1984. Т. 21. С.
150-160.
Макарова И. П., Перфильева В. И. К флоре лишайников Усть-
Ленского заповедника // Водоросли, лишайники, грибы и мохообразные в
заповедниках РСФСР. М., 1989. С. 53-59.
Макарова И. П., Перфильева В. П., Николин Е. Г. К флоре лишайни¬
ков Новосибирских островов // Новости систематики низших растений.
1988. Т. 25. С. 127-133.
Макрый Т. В. Лишайники Байкальского хребта. Новосибирск, 1990.
199 с.
Мартин Ю. Л. Заметки о лихенофлоре и основных синузиях лишай¬
ников стационара «Харп» // Споровые растения Урала. Тр. ин-та экол.
раст. жив. УФ АН СССР. Свердловск, 1970. Вып. 70. С. 134-139.
Микулин А. Г. Определитель лишайников полуострова Камчатка.
Владивосток, 1990. 128с.
Мучник Е. Э. Конспект лишайников степных и остепненных место¬
обитаний Центрального Черноземья // Новости систематики низших рас¬
тений. 2002. Т. 35. С. 183-195.
Мырзакулова 3. С. Лишайники Южного Алтая // Тр. II молодежи,
конф. ботаников Ленинграда. Л., 1988. Ч. 3. С. 109-113. Деп. в ВИНИТИ
14. 07. 88, № 5684-В 88.
Новрузов X. X.,
Окснер А. Н. Визначник лишайниюв УРСР / АН УРСР. 1н-т бо¬
ташки. Кшв, 1937. 341 с.
Окснер А. Н. Де-що з флори обрюниюв Чукотського швострова //
BicH. Кшв. ботан. саду. 1929. Вип. 9. С. 87-91.
Окснер А. Н. До вивчення флори обр!сник1в каменястих виход1в
УкраУни // BicH. Кшв. ботан. саду. 1927. Вип. 5/6. С. 23-82.
Окснер А. Н. Лишайники басейну piK Iнд!г1рк1 Яни, Лени та П1вден-
ного Прибайкалля. I // Журн. 1н-ту боташки АН УРСР. 1939а. № 23 (31).
С. 117-139.
Окснер А. Н. Лишайники басейну р!к 1щцпрки, Яни, Лени та
П1вденного Прибайкалля. IV // Бот. журн. АН УРСР. 1940в. Т. 1, № 3-4. С.
31-57.
Окснер А. Н. Лишайники басейну piK Лени, Яни, Инд1прки та
П1вденного Прибайкалля. II // Бот. журн. АН УРСР. 1940а. Т. 1, № 1. С.
77-100.
Окснер А. Н. Лишайники басейну piK Лени, Яни, ИндИрки та
П1вденного Прибайкалля. III // Бот. журн. АН УРСР. 19406. Т. 1, № 2. С.
313-324.
Окснер А. Н. Матер1али для лихенофлоры Урала та прилеглих обла¬
стей // Бот. журн. АН УРСР. 1941-1945. Т. 2, № 3-4. С. 217-247.
Окснер А. Н. Материалы до лихенофлори Середньой A3iV // Журн.
1н-ту боташки АН УРСР. 19396. №20 (28). С. 111-136.
Окснер А. Н. Материалы к флоре лишайников Белоруссии: (Предва¬
рит. сообщ.) // BicH. Кшв. Ботан. саду. 1924. Вип. 1. С. 27-36.
Окснер А. Н. Определитель лишайников СССР. Вып. 2. Общая
часть. Морфология, систематика и географическое распространение. Л.,
Наука, 1974. 284 с.
Окснер А. Н. Рел1ктов1 обр1сники з Далекого Сходу // BicH. Кшв. бо¬
тан. саду. 1934. Вип. 17. С. 37-40.
Окснер А. Н. Флора лишайниюв УкраУни. В 2-х т. Т. 2 (Вип. 1) / АН
УРСР. 1н-т боташки. Кшв, Наук. Думка, 1968. 500 с.
Окснер А. Н. Флора лишайниюв УкраУни. В 2-х т. Т. 2 (Вип. 2) / АН
УРСР. 1н-т боташки. Кшв, Наук. Думка, 1993
Окснер А. Н. Флора лишайниюв УкраУни: В 2-х т. Т. 1 / АН УРСР.
1н-т боташки. Кшв, 1956. 495 с.
Определитель лишайников России / Под ред. Н. С. Голубковой.
СПб., 1996. Вып. 6. 203 с.; 1998. Вып. 7. 166 с.; 2003. Вып. 8. 277 с.; 2004.
Вып. 9. 339 с.; 20ХХ. Вып. 10. 450 с.
Определитель лишайников СССР / Под ред. Н. С. Голубковой. Л.,
Наука 1971. Вып. 1.203 с.; 1975. Вып. 3. 166 с.; 1977. Вып. 4. 277 с.; 1978.
Вып. 5. XXX с.
Очирова Н. Н. Материалы к изучению лихенофлоры Республики
Калмыкия // Новости систематики низших растений. 2005. Т. 38. С. 278-
285.
Пийн Т. X. Напочвенные лишайники мыса Челюскин // Арктические
тундры и полярные пустыни Таймыра. Л., Наука, 1979а. С. 61-73.
Пийн Т. X. Напочвенные лишайники окрестностей бухты Марии
Прончищевой (сверо-восточный Таймыр) // Арктические тундры и поляр¬
ные пустыни Таймыра. Л., Наука, 19796. С. 140-143.
Пийн Т. X., Трасс X. X. Напочвенные лишайники окрестностей Та-
реи (Западный Таймыр) // Биогеоценозы таймырской тундры и их продук¬
тивность. Л., Наука, 1971. С. 151-160.
Питеранс А. В. Лишайники Латвии. Рига, Зинатие, 1982. 352 с.
Порядина Л. Н. Лишайники заказника «Сунтар-Хаята» (Якутия) //
Новости систематики низших растений. 2001. Т. 34. С. 167-176.
Пыстина Т. Н. Систематический список лишайников равнинной ча¬
сти республики Коми (подзоны южной и средней тайги) // Новости систе¬
матики низших растений. 2001. Т. 34. С. 176-185.
Рассадина К. А. Лишайники, собранные В. Б. Сочавой в 1929 г. в
Анадырском крае // Тр. Бот. ин-та АН СССР, сер. 2, Споровые растения.
1934. Вып. 2. С. 321-338.
Рыкова Ю. В. Определитель листоватых лишайников Якутии.
Якутск, 1978. 112 с.
Рябкова К. А. Виды семейства Parmeliaceae на Урале // Новости си¬
стематики низших растений. 1982 Т. 19. С. 149-154.
Рябкова К. А. Лишайники Урала (список видов). Свердловск,
Свердловский гос. пед. инст. 1991. Деп. ВИНИТИ № 2504-В91. 30 с.
Рябкова К. А. Семейства Baeomycetaceae, Cladoniaceae, Stereocau-
laceae на Урале // Новости систематики низших растений. 1982 Т. 19. С.
154-164.
Рябкова К. А., Макарова И. И. Лишайники Приполярного и Поляр¬
ного Урала. Свердловск, Свердловский гос. пед. инст, 1991. Деп.
ВИНИТИ №2504-В91. 15 с.
Савич В. П. 1950. Конспект к флоре лишайников сем. Umbilicaria-
сеае СССР
Савич В. П. Лишайники собранные в Анадырском округе в 1903-07
гг. Н. Сокольниковым // Изв. Имп. Бот. сада. СПб., 1911. Т. 11. Вып. 3. С.
82-90.
Савич В. П. Лишайники, собранные И. И. Тржемеским в полярной
Сибири // Изв. Имп. Бот. Сада Петра Великого. Петроград, 1916а. Т. 16.
Вып. 1. С. 133-135.
Савич В. П. Лишайники, собранные Р. Р. Поле на крайнем севере
Европейской России //Тр. Имп. Бот. Сада. СПб, 1912. Т. 32. С. 15-67.
Савич В. П. Отдел споровых растений // От Аптекарского огорода
до Ботанического института. Очерки по истории Ботанического института
Академии наук СССР. М. -Л., Издательство АН, 1957. С. 115-133.
Савич В. П. Список лишайников Тобольской губ., собранных Б. Н.
Городковым в 1914 году // Изв. Имп. Бот. Сада Петра Великого.
Петроград, 19166. Т. 16. Вып. 1. С. 101-111.
Савич В. П. Список лишайников, собранных Р. Н. Ниманом в Ла¬
пландии и Новой Земле в 1903 и 1908-1909 гг. // Тр. Студенч. Научн.
Кружков Физ.-Мат. Фак. СПб. Унив. СПб, 1911. Вып. 3. С. 37-56.
Савич В. П. Чукотские лишайники из коллекции Г. А. Борисова //
Изв. Имп. Бот. Сада Петра Великого. СПб., 1915. Т. 15, вып. 5-6. С. 542-
548.
Седельникова Н. В. Лихенофлора нагорья Сангилен. Новосибирск,
Наука, 1985. 180 с.
Седельникова Н. В. Лишайники Алтая и Кузнецкого нагорья. Ново¬
сибирск, Наука, 1990. 175 с.
Седельникова Н. В. Лишайники Западного и Восточного Саяна. Изд.
СО РАН, Новосибирск, 2001. 190 с.
Седельникова Н. В. Лишайники Салаира // Флора Салаирского кря¬
жа. Новосибирск, ЦСБС, 1993. С. 32-78.
Седельникова Н. В. Особенности лихенофлоры Западного Саяна //
Ботан. журн. 1996а. Т. 81. № 3. С. 75-83.
Седельникова Н. В. Систематический список лишайников Восточ¬
ного Саяна // Новости систематики низших растений. 19966. Т. 31. С. 144-
151.
Соболевский Г. Санктпетербургская флора. Часть 2. 1803. С. XX.
Список лихенофлоры России. [Сост. Г. П. Урбанавичюс] Спб., 2010.
194 с.
Степанчикова И. С., Гимельбрант Д. Е., Кузнецова Е. С. Лишайники
// Природа Сестрорецкой низины и вопросы ее охраны / Ред. Е. А. Волко¬
ва, Г. А. Исаченко, В. Н. Храмцов. СПб, 2011. С. 155-166.
Титов А. Н. Микокалициевые грибы Голарктики. М., Товарищество
научных изданий КМК, 2006. 296 с.
Титов А. Н. Профессор А. А. Еленкин - основатель лихенологиче¬
ской школы России // Фундаментальные и прикладные проблемы ботани¬
ки в начале XXI века: Материалы Всероссийской конференции, 22-27
сентября. Ч. 2. Альгология, Микология, Лихенология, Бриология. Петро¬
заводск, 2008. С. 246-249.
Томин М. П. Определитель корковых лишайников европейской ча¬
сти СССР (кроме Крайнего Севера и Крыма). Минск, Изд-во АН БССР,
1956. 533с.
Трасс X. X. Элементы и развитие лихенофлоры Эстонии // Тр. по
ботанике. Тарту, 1970. Вып. 9. С. 5-225.
Урбанавичене И. Н. Первые сведения о лишайниках Национального
парка «Зюраткуль» (Челябинская область) // Новости систематики низших
растений. Т. 45. 2011. С. 223-236.
Урбанавичене И. Н., Урбанавичюс Г. П. К флоре лишайников
хребта Хамар-Дабан // Новости систематики низших растений. 1999. Т.
33. С. 161-171.
Урбанавичене И. Н., Урбанавичюс Г. П. Лишайники Байкальского
заповедника // Флора и фауна заповедников. М., 1998. Вып. 68. 53 с.
Урбанавичюс Г. П., Урбанавичене И. Н., Мелехин А. В. Лихенофло-
ра Лапландского государственного природного биосферного заповедника
(аннотированный список). Апатиты, Изд. КНЦ РАН. 2012. 148 с.
Урбанавичюс Г. П., Урбанавичене И. Н., Отте Ф. Лишайники //
Красная книга Адыгеи. Майкоп, 2012. С. 100-130.
Фадеева М. А., Голубкова Н. С., Витикайнен О., Ахти Т. Предвари¬
тельный список лишайников Карелии и обитающих на них грибов. Петро¬
заводск, 1997. 100 с.
Херманссон Я., Пыстина Т. Н., Кудрявцева Д. И. Предварительный
список лишайников Республики Коми. Сыктывкар, 1998. 134 с.
Чабаненко С. И. Лишайники // Флора, мико- и лихенобиота Лазов¬
ского заповедника (Приморский край). Владивосток, ДВО АН СССР,
1990. С. 167-191.
Чабаненко С. И., Скирина И. Ф., Княжева Л. А. Список лишайников
Приморского края и обитающих на них грибов. Южно-Сахалинск, Саха¬
линский ботанический сад, СахНЦ, ДВО РАН, 2002. 89 с.
Шапиро И. А. Обзор современного состояния хемосистематики ли¬
шайников // Новости систематики низших растений. 1980. Т. 17. С. 152—
167.
Шаяхметова 3. М., Селиванов А. Е., Урбанавичене И. Н. Лишайники
предгорных средне- и южно-таежных лесов Пермского края // Вестн.
ТвГУ. Сер. биология и экология. 2009. Вып. 12, 6. С. 139-153.
Шель Ю. Материалы для ботанической географии Уфимской и
Оренбургской губернии (Споровые растения) // Труды общества есте¬
ствоиспытателей при Казанском университете. 1883. Вып. 12. С. 1-93.
Шустов М. В. Лишайники Приволжской возвышенности // Новости
систематики низших растений. 2002. Т. 36. С. 185-203.
Almquist Е. Die Lichenenvegetation der Kiisten des Beringsmeeres //
Nordenskiold A. E. / Die wissenschaftliche Ergebnisse der Vega-Expedition
(Vega-expeditionis vetenskapliga jakttagelser). Stockholm, 1887. Vol. Bd 4. P.
509-542.
Almquist E. Lichenologische Beobachtungen an der Nordkiiste Sibiriens
// Nordenskiold A. E. / Die wissenschaftliche Ergebnisse der Vega-Expedition.
Leipzig, 1883. Bd 1. P. 50-74.
Almquist E. Lichenologiska iakttagelser pe Sibiriens nordkust // Otver-
sigt af Kongl. Vetenskaps-Akademiens Forhandlingar. 1879. № 9. P. 29-59.
Andreev M. Halecania santessonii, a new lichenicolous lichen from Rus¬
sia // Lichenologist. 2010. Vol. 42, № 2. P. 1-4.
Andreev M. P., Kotlov Yu. V., Makarova I. I. Checklist of lichens and li¬
chenicolous fungi of the Russian Arctic // The Bryologist. 1996. Vol. 99, № 2.
P. 137-169.
Buxbaum J. C. Plantarum minus cognitarum Centuria I-V, complectens
plantas circa Byzantium in Oriente observatas. Petropoli ex Typographia Aca-
demiae, 1728, 1728, 1729, 1733, 1740. P. XX. P. XX. P. XX.
Cajander A. K. Beitrage zur Kenntniss der Vegetation der Alluvionen des
nordlichen Eurasiens. I. Die Alluvionen des unteren Lena-Thales // Acta Soc.
Sci. Fenn. Helsingfors, 1903. Vol. 32. Iss. 1. P. 1-182.
Davydov E. A., Himelbrant D. E., Stepanchikova I. S. Contribution to the
study of Umbilicariaceae (lichenized Ascomycota) in Russia. II. Kamchatka
Peninsula // Herzogia. 2011. Bd. 24. Heft 2. P. 229-241.
Elenkin A. A. Species novae lichenum in Sibiria arctica a d. A. A. Biru-
la-Bialynizki collectae (expeditio baronis Toll) // Annal Mycol. 1906. Vol. 4.
№ 1. P. 36-38.
Gmelin J. G. Flora sibirica, sive Historia plantarum sibiricae Гмелин,
1747, 1749, 1768, 1769
Gmelin J. G. Reise durch Sibirien, von dem Jahr 1733 bis 1743, 4 vols.
Goettingen, 1751-52.
Joshi Y., Andreev М., Hur J. -S. Caloplaca lacinulata rediscovered for
the lichen flora of South Korea // Feddes Repertorium. 2012. Vol. 122. № 5-6.
P. 421-423.
Kondratyuk S. Prof. Dr. A. M. Oxner (22. 02. 1898-20. 11. 1973). On
occasion of 100-years anniversary of the birthday. Kiev: Phytosociocentre,
1998.64 p.
Kotlov Yu. V. Lichens from two bird colonies in the West Siberian Arc¬
tic // Graphis Scripta. 1994. № 6. P. 57-59.
Kuznetsova E., Ahti Т., Himelbrant D. Lichens and allied fungi of the
Eastern Leningrad Region //Norrlinia. 2007. Vol. 16. P. 1-62.
Kuznetsova E. S., Motiejunaite J., Stepanchikova I. S., Himelbrant D. E.,
Czarnota P. New records of lichens and allied fungi from the Leningrad Re¬
gion, Russia. Ill // Folia Cryptogamica Estonica. 2012. Fasc. 49. P. 31-37.
Lettau G. Beitrage zur Lichenenflora von Ost- und WestpreuBen // Fest-
schriften des PreuBischen Botanischen Vereins zu Konigsberg. 1912. Bd 53. S.
17-91.
Lettau G. Nachtrage zur Lichenenflora von Ost- und WestpreuBen //
Schriften der Koniglichen Physikalischokonomischen Gesellschaft zu Konigs¬
berg. 1919. Bd 60. S. 5-21.
Lynge B. Lichens collected on the Norvegian scientific expedition to
Franz Josef Land 1930 // Skrifter Svalbard og Ishavet. 1931. № 38. P. 1-31.
Lynge B. Lichens from Novaya Zemlya (excl. Acarospora and Lecanora)
II Rep. Sci. Results Norweg. Exped. Novaya Zemlya 1921. Oslo (Kristiania),
1928. Iss. 43. P. 1-299.
Magnusson A. H. Neue Flechten aus dem Jenisei-Gebiet // Svensk Bota-
nisk Tidskr. 1936. Vol. 30. № 3. P. 247-263.
Malme G. Lichenes orae Sibiriae Borealis inde ab insula Minin usque ad
promantorium Ryrkajpia in Expeditione Vegae lecti // Arkiv for Bot. 1932.
Vol. 25A. №2. P. 1-42.
Messerschmidt D. G. Forschungsreise durch Sibirien 1720-1727. Tage-
buchaufzeichnungen. T. 1. Januar 1721-1722. Berlin, 1962; T. 2. Januar 1723 -
Mai 1724. Berlin, 1966
Middendorff A. Th. Reise in den aussersten Norden und Osten Sibiriens
wahrend der Jahre 1843 und 1844 mit Allerhochster Genehmigung auf Veran-
staltung der keiserl. Akademie der Wissenschaften zu St. -Petersburg. Bd 4.
Ubersicht der Natur Nord- und Ost-Sibiriens. Theil 2. L. 1. Anhang 6. Zu den
Gewachsen Sibiriens. Lichenes Middendorffiani. Enumeravit prof. Dr. W.
Nylander. St. -Petersburg, 1867. Vol. 4, 2. Iss. 1, 6. P. 15-18.
Motiejunaite J., Berglund Т., Czarnota P., Himelbrant D., Hognabba F.,
Konoreva L. A., Korchikov E. S., Kubiak D., Kukwa М., Kuznetsova E., Lep-
pik E., Lohmus P., Prigodina Lukosiene I., Pykala J., Stoncius D., Stepan¬
chikova I., Suija A., Thell A., Tsurykau A., Westberg M. Lichens, lichenico¬
lous and allied fungi found in Asveja Regional park (Lithuania) // Botanica Li¬
thuania. 2012. In press.
Muchnik E. E., Konoreva L. A. The lichen biota of protected territories
in Ryazan region (Central Russia) // Lichen protection - Protected lichen spe¬
cies / Ed. L. Lipnicki. Sonar Literacki, Gorzow Wlkp. 2012. P. 213-221.
Nylander W. Enumeratio Lichenum Freti Behringii // Bulletin de la So-
ciete Linneanne de Normandie, 4 ser. 1888. Vol. 1. P. 1-91.
Nylander W. Lichenes novi e Freto Behringii. I. Lichenes novi e Konya-
mbay // Flora. 1884. Vol. 67. P. 211-223.
Nylander W. Lichenes novi e Freto Behringii. II. Lichenes novi e Beh-
ring-insula et e Lawrence-insula // Flora. 1885. Vol. 68. P. 439-446.
Ohlert A. Gruppierung der Lichenen der Provinz PreuBen nach Standort
und Substrat // Lichenologische Aphorismen II. Schriften der Naturforschenden
Gesellschaft zu Danzig. 1871. Bd 2. H. 3-4. S. 3-37.
Ohlert A. Verzeichnis PreuBischer Flechten // Schriften der Koniglichen
Physikalisch-okonomischen Gesellschaft zu Konigsberg. 1863. Bd 4. S. 6-34.
Ohlert A. Zusammenstellung der Lichenen der Provinz PreuBen // Schrif¬
ten der Koniglichen Physikalisch-okonomischen Gesellschaft zu Konigsberg.
1870. Bd 11. S. 1-51.
Piin T. A list of terricolous lichens of "Forest Island" of Ary Mas // Folia
Criptogamica Estonica. 1977. № 8. P. 3-8.
Savic S., Tibell L., Andreev M. New and interesting lichens from Serbia
// Mycologia Balcanica. 2006. Vol. 3. P. 99-106.
Schrenk A. G. Reise nach dem Nordosten des europaischen Russlands,
durch die Tundren der Samojeden, zum Arktischen Uralgebirge, auf Aller-
hochsten Befehl fur den keis. botanischen Garten zu St. -Petersburg im Jahre
1837 ausgefuhrt. 2 Theil. Wissenschaftliche Beilagen. Dorpat, 1854. 665 p.
Sobolewskj G. Flora Petropolitana. Petropoli, Typis Collegii Imperialis
Medici, 1799. XX p.
Steller. G. W. Reise von Kamtschatka nach Amerika mit dem comman-
der-kapitan Bering, Georg Wilhelm Steller, St. Petersburg, 1793.
Stepanchikova I. S., Himelbrant D. E., Kukwa М., Kuznetsova E. S. New
records of lichens and allied fungi from the Leningrad Region, Russia II // Folia
Cryptogamica Estonica. 2011. Fasc. 48. P. 85-94.
Stepanchikova I. S., Kukwa М., Kuznetsova E. S., Motiejunaite J.,
Himelbrant D. E. New records of lichens and allied fungi from the Leningrad
Region, Russia // Folia Cryptogamica Estonica. 2010. Fasc. 47. P. 77-84.
Stepanchikova I. S., Kuznetsova E. S., Himelbrant D. E. New records of
lichens and allied fungi from the Eastern Leningrad Region // Folia Cryptogam¬
ica Estonica. 2009. Fasc. 46. P. 75-78.
Temina М., Andreev M. P., Barinova S., Nevo E. The diversity and ecol¬
ogy of epiphytic lichens in «Evolution Canyon» II, Lower Nahal Keziv, Upper
Western Galilee, Israel // Turk. J. Bot. 2009. Vol. 33. P. 263-275.
Urbanavichus G. P., Ahti Т., Urbanavichene I. N. Catalogue of lichens
and allied fungi of Murmansk Region, Russia//Norrlinia. 2008. № 17. 80 p.
Vainio E. Lichenes in viciniis hibernae expeditionis Vegae prope pagum
Pittlekai in Sibiria septentrionali a D-re E. Almquist collecti // Ark. for Bot.
1909. Vol. 8. №4. 175 p.
Zhurbenko M. P. Lichens from the coast of Eniseiskii Gulf, W. Taimyr,
Siberia. Graphis Scripta. 1992. Vol. 4. № 1. P. 1-4.
Zhurbenko M. P., Himelbrant D. E., Kuznetsova E. S., Stepanchikova I.
S. Lichenicolous fungi from Kamchatka Peninsula, Russia // The Bryologist.
2012. Vol. 115, № 2. P. 295-312.
Глава II.
Основные лнхенологическне гербарии России и важнейшие зарубеж¬
ные коллекции российских лишайников. Коллекционные фонды.
Андреев М. П., Ахти Т.
Гербарий является основным инструментом ботанического исследо¬
вания и основным источником информации для систематической и фло¬
ристической работы. В гербариях России и ряда других стран хранятся
многочисленные образцы, являющиеся материальным документирован¬
ным подтверждением существования того или иного таксона на опреде¬
ленной территории или эталоном таксона, в тех случаях, когда речь идет о
типовых образцах. Эти бесценные материалы являются основой для со¬
здания Флоры лишайников России.
В России в настоящее время существует около 50 более или менее
крупных гербариев или коллекций лишайников, которые могут быть ис¬
пользованы в качестве основы для этой работы. В них, по грубой оценке,
хранится ок. 675 тыс. образцов лишайников, а с учетом не инсерирован-
ного, не разобранного и не до конца изученного материала, не поступив¬
шего в основные фонды - до 1 млн. образцов. Кроме того, более или ме¬
нее крупные коллекции лишайников, собранных на территории России,
хранятся, по крайней мере, в 29 зарубежных гербариях. Объем этого ма¬
териала можно оценить приблизительно в 50 тыс. образцов. Наиболее
крупные, интересные и важные для исследователей коллекции россий¬
ских лишайников находятся в гербариях Финляндии Гербарий им. Д. П.
Сырейщикова при МГУ им. М. В. Ломоносова. Гербарий лишайников,
Эстонии, Швеции, Литвы, в Белоруссии и на Украине.
Имеющиеся в нашем распоряжении сведения, собранные в резуль¬
тате опроса кураторов, сотрудников или владельцев большинства россий¬
ских и многих зарубежных собраний, позволяют оценить объем имеюще¬
гося в распоряжении исследователей материала, в какой-то мере система¬
тизировать и предоставить научному сообществу. Полностью результаты
опроса можно увидеть на сайте Ботанического института им. В. Л. Кома¬
рова по адресу www.binran.ru, а краткий предварительный анализ и
наиболее важную информацию - ниже.
Все российские собрания лишайников по объёму имеющегося мате¬
риала можно условно разделить на три группы: крупные, средние и не¬
большие коллекции. К крупным гербариям мы отнесли те, где хранится от
30 до 10 тыс. образцов и представлено, как правило, более 1 тыс. видов, к
средним - от 1 до 10 тыс. образцов и до 1 тыс. видов. Небольшие коллек¬
ции, как правило, содержат до 1000 образцов и до 500 видов лишайников.
Такое разделение достаточно условное и отражает лишь общую тенден¬
цию. Кроме того, ценность коллекции и гербария не всегда напрямую свя¬
зана с объемом хранящегося материала, а зависит и от других факторов -
разнообразия представленных в коллекции систематических групп, гео¬
графического охвата территории, наличию типовых образцов и эксикат,
состояния материала и доступности его для исследователей и многого
другого.
Из российских лихенологических гербариев, к первой группе -
крупных, можно отнести 15 коллекций основные данные о которых при¬
водятся ниже. Во второй группе - средних гербариев оказалось 13 кол¬
лекций и в третьей - 10. Данные по объему имеющегося материала в
остальных собраниях лишайников у нас отсутствуют. Вне списка остался,
не имеющий себе равных в России и по объему представленного материа¬
ла и по его разнообразию - Гербарий лишайников Ботанического инсти¬
тута им. В. Л. Комарова РАН (LE), основные фонды которого содержат
ок. 430 тыс. образцов, а не разобранные коллекции - не меньше (более
700 коробок). Самым старым в стране является собрание лишайников
Московского университета, основанное в 1767 г. А всего до начала XX
столетия в Российской империи гербарии лишайников существовали еще
лишь в Императорском ботаническом саду и в Ботаническом музее Ака¬
демии наук в Петербурге (слились позднее в Гербарий Ботанического ин¬
ститута им. В. Л. Комарова РАН), в Казанском, Гельсинфорском (Хель¬
синском), Виленском (Вильнюсском) и Юрьевском (Тартуском) и, веро¬
ятно, еще в Харьковском и Варшавском университетах. До Великой оте¬
чественной войны (1941 г.) в Российской федерации было всего пять гер¬
бариев лишайников, за 50 лет, прошедших после войны и до распада Со¬
ветского Союза (1991 г.) появилось еще 16, и уже после его распада, за
прошедшие двадцатилетие возникли еще 20 гербариев, причем, наиболее
«урожайными» были 1997 и 2003 годы, когда появились по 4 новых со¬
брания.
Если проанализировать географию расположения лихенологических
коллекций на территории России, то можно заметить, что наибольшая их
концентрация наблюдается в Москве, Петербурге и в крупных научных
центрах страны. В Петербурге, кроме основного Гербария лишайников
Ботанического института, существует большой и довольно старый Герба¬
рий лишайников Петербургского университета и сравнительно небольшие
и молодые частные коллекции. В Москве, кроме гербария Московского
университета, есть еще четыре гербария. На Северо-западе, кроме Петер¬
бурга имеется еще 8 собраний лишайников, в Средней и Южной России -
тоже 8, в Поволжье - 6, на Северном Кавказе - 3, на Урале - 6, в Сибири -
9, на Дальнем Востоке - 6, причем три из них во Владивостоке. В Евро¬
пейской части России сконцентрировано около 80% всего гербарного
фонда.
Ниже приводится список наиболее важных гербариев лишайников,
причем, для гербариев первой группы дана более развернутая характери¬
стика. К сожалению, статистические данные о некоторых коллекциях у
нас отсутствуют.
Крупнейшим в России и одним из старейших является Гербарий
лишайников Ботанического института РАН (LE) в Петербурге (Рис. 8).
Гербарий основан выдающимся русским ученым Александром Алексан¬
дровичем Еленкиным в 1898 г., хотя истоки формирования этого фонда
восходят первой половине XVIII-ro столетия. В 1931 г. в гербарий вли¬
лись обширные коллекции лишайников Ботанического музея Академии
Наук, переданные туда из «Кунсткамеры» Петра Великого, где хранились
материалы многочисленных экспедиций Академии Наук. Они содержали
образцы, собранные известными членами Российской Академии — Г.
Стеллером, П. С. Палласом, И. X. Буксбаумом, И. Георги, И. Г. Гмелином,
С. П. Крашенинниковым, Ф. Рупрехтом, М. С. Ворониным и др. Заметная
роль в становлении и развитии гербария принадлежит таким ученым как
К. А. Рассадина, В. П. Савич, который возглавлял Отдел споровых расте¬
ний и Лабораторию лихенологии и бриологии до 1963 г., и Н. С. Голубко¬
вой, руководившей лабораторией с 1982 по 2003 год.
Со дня основания гербария его фонды постоянно пополнялись об¬
разцами лишайников, собранными как на территории России, так и за ру¬
бежом, а также за счет обмена с отечественными и иностранными герба¬
риями. В настоящее время в гербарии лишайников насчитывается около
430 тысяч образцов. Основную часть коллекции составляют образцы ли¬
шайников из России и союзных республик, но имеются также богатые
гербарные материалы со всех континентов. Огромную ценность состав¬
ляют типовые материалы таксонов (ок. 500 типовых образцов, включая
типы видов, описанных А. А. Еленкиным, В. П. Савичем, К. С. Мереж¬
ковским, А. Н. Окснером, М. П. Томиным, К. А. Рассадиной, Н. С. Голуб¬
ковой, А. Н. Титовым и другими исследователями, в основном с террито¬
рии Советского Союза) и образцы многих серий эксикат, изданных как
отечественными, так и крупнейшими зарубежными лихенологами. В по¬
следние десятилетия, благодаря активной экспедиционной деятельности
сотрудников, гербарий значительно пополнился коллекциями, собранны¬
ми в Арктике, Сибири, на Дальнем Востоке и Северо-западе и в других
районах России, а также за рубежом — в США, Монголии, Китае, Ан¬
тарктике и других местах.
Фонды гербария открыты для работы и широко используются при
проведении фундаментальных исследований по систематике, морфологии
и географии лишайников, как российскими так и зарубежными специали¬
стами. Имеется постоянно пополняемая база данных, начитывающая 4500
записей. В БД внесены образцы типового гербария и небольшая часть ос¬
новного, преимущественно, новые поступления.
Гербарий лишайников Гербария им. Д. П. Сырейщикова при
Московском государственном университете им. М. В. Ломоносова
(MW).
Основан 1767 г. Г.-Ф. Гофманом. Находится на кафедре микологии
и альгологии биологического факультета МГУ. Ок. 10 тыс. образцов и ок.
1000 видов. Куратор и контактное лицо Т. Ю. Толпышева. Основные кол¬
лекторы: М. И. Назаров, И. П. Петров, В. Б. Куваев, В. Писаржевский, Ю.
Б. Королев, Н. А. Комарницкий, В. С. Говорухин, О. Д. Минаева, Т. Ю.
Толпышева. Представленные территории: средняя полоса европейской
части России, Архангельская обл., Кольский п-ов, Поволжье, Крым, Кав¬
каз, Сибирь, Дальний Восток, Чукотка, Украина, Киргизия.
Рис. 8. Здание Института Споровых Растений (быв. Главное здание Ботанического
Сада, где размещался Гербарий и библиотека. 1913 г. Празднование 200-летия Импе¬
раторского Ботанического Сада.
Образцы хранятся в пакетах, приклеенных на листы бумаги, в пач¬
ках в гербарных шкафах. Отдельно, на Каф. геоботаники хранится герба¬
рий Г.-Ф. Гофмана и X. X. Эрхарда. Имеются микрофиши этих гербариев.
Отдельно и в том виде, как они были изданы, хранятся эксикаты А. А.
Еленкина, К. С. Мережковского, X. X. Rabenhorst’a (полные серии), Ph.
Нерр’а (нет 13-15 томов). Помимо изданных А. А. Еленкиным эксикат,
гербарий располагает дубликатами образцов лишайников из гербария Бо¬
танического института им. В. Л. Комарова (LE), собранных и/или опреде¬
ленных А. А. Еленкиным и использованных им для составления «Флоры
лишайников Средней России». Большую роль в формировании гербария
лишайников сыграл крупнейший лихенолог России В. П. Савич. Благода¬
ря ему, гербарий имеет не только эксикаты, изданные В. П. Савичем -
«Lichenotheca Rossica», но и образцы лишайников, которые присылали
ему многие зарубежные лихенологи: А. Цальбрукнер, А. Г. Магнуссон и
др. Кроме того, гербарий располагает зарубежными эксикатами, преиму¬
щественно из Европы. Виды рода Stereocaulon проверены А. В. Домбров¬
ской, Umbilicaria - Вей Цзянь-чунем, Caloplaca (эпилиты) - I. Nordin.
Много неинсерированного материала. Состояние удовлетворительное.
Доступен для посещения в зимнее время.
Гербарий лишайников Полярно-альпийского ботанического са¬
да-института им. Н. А. Аврорина КНЦ РАН (KPABG).
Около 30 тыс. образцов, более 1000 видов. Основан А. В. Домбров¬
ской в 70-х годах 20 века. Куратор - А. В. Мелехин. Основные коллекто¬
ры: Домбровская А. В., Дудорева Т. А., Урбанавичюс Г. П., Урбанавичене
И. Н., Антонова И. М., Петрова О. В., Гудовичева А. В., Мелехин А. В.,
Конорева Л. А. Представленные территории: Мурманская область, архи¬
пелаг Шпицберген, Карелия, Алтай, Камчатка, Курилы, Сахалин, Кавказ,
Крым, Урал, Сибирь, Центральная Россия, Финляндия, Норвегия. Образ¬
цы хранятся в пакетах, вложенных в гербарные папки, в пачках в гербар-
ных шкафах. Гербарий располагает дубликатами образцов из гербария
Ботанического института (LE), гербария Ботанического Музея Универси¬
тета Хельсинки (Н). Коллекция лишайников рода Stereocaulon, легла в
основу монографии по этому роду для всего бывшего СССР (Домбров¬
ская, 1996). Коллекция рода Cladonia, собранная и обработанная, в основ¬
ном, Т. А. Дудоревой, так же охватывает все биоразнообразие этого рода
в пределах России. Гербарий доступен для посещения в любое время года
по предварительной договоренности. Данные об образцах гербария ли¬
шайников ПАБСИ (этикетки в структурированной форме) общедоступны
в информационной ситеме ISL (http://phpmybotan.ru/allsystem2). На
10.01.2013 информационная система содержала записи о 8481 гербарном
образце KPABG и 915-ти видах этого гербария.
Гербарий лишайников Тихоокеанского института географии
(Нет акронима). Основан в 1980 г. И. Ф. Скириной. Ок. 30 тыс. образцов и
ок. 1000 видов. Основные коллекторы: И. Ф. Скирина, Л. А. Княжева, И.
М. Родникова. Представленные территории: Приморский и Хабаровский
края, Амурская обл., Еврейская АО, Камчатский край, Сахалинская обл.
Небольшая историческая коллекция с п-ова Муравьева-Амурского (1953
г.). Образцы хранятся в гербарных шкафах в пакетах, упакованных в ко¬
робки. Имеется небольшое количество образцов, определенных или про¬
веренных специалистами - монографами (М. П. Томин, М. Ф. Макаревич,
Т. Randlane, R. Moberg, I. Karnefelt). Более 70 коробок неинсерированного
гербария. Имеется база данных части обычных образцов. Доступен для
посещения и работы.
Гербарий лишайников Сахалинского ботанического сада
(SAKH). Основан в 1992 г. С. И. Чабаненко. Ок. 30 тыс. обр. Основные
коллекторы А. А. Таран, С. И. Чабаненко. Представленные территории:
Дальний Восток, Урал, Зап. Сибирь, Байкал, Алтай, Зап. Европа, Крым,
Кавказ (Абхазия), Япония (Хоккайдо), Юго-Вост. Китай, Сев. Америка.
Образы хранятся в пакетах, сложенных в коробки и в гербарных шкафах.
Часть материала разложена по родам, большая часть - по местам сбора.
Около 50 коробок неинсерированного гербария. Доступен для посещения
и работы.
Гербарий лишайников при Лаборатории радиоэкологического
мониторинга в регионах АЭС и биоиндикации Института проблем
экологии и эволюции им. А. Н. Северцова РАН (ИПЭЭ РАН), Москва
(нет акронима). Основан Л. Г. Бязровым в 1965 г., первоначально в Лабо¬
ратории биогеоценологии Ботанического института им. В. Л. Комарова
АН СССР. Позднее научное подразделение было переведено в Институт
эволюционной морфологии и экологии животных им. А. Н. Северцова АН
СССР (ныне ИПЭЭ РАН). Ок. 30 тыс. образцов и ок. 500 (??) видов. Кон¬
тактное лицо Л. Г. Бязров. Основной коллектор: Л. Г. Бязров. Представ¬
ленные территории: б. ч. Монголия (Хангайское нагорье), Московская и
Смоленская обл., Республика Мари Эл. Образцы хранятся в пакетах, как в
гербарных шкафах, так и гербарных коробках, а также на стеллажах в
пачках. Поскольку гербарий создавался в ходе изучения роли лишайников
в различных биогеоценозах, то с позиций систематиков качество собран¬
ного материала разнородное, ибо довольно много невыразительных мел¬
ких образцов. Ряд видов трудно найти, поскольку на одном образце могут
быть представители несколько видов. Некоторые массовые виды пред¬
ставлены большим числом образцов. Примерно 2/3 образцов не иденти¬
фицированы, но все снабжены этикетками. Часть представителей рода
Stereocaulon из Монголии определены М. Lamb, сем. Umbilicariaceae из
Монголии - Вей Цзянь-чунем, Состояние разное, поскольку на часть ма¬
териала испорчена. Доступен для посещения по согласованию с Л. Г.
Бязровым.
Гербарий лишайников Алтайского государственного универси¬
тета (ALTB). Основан в 1994 г. Е. А. Давыдовым. Ок. 20 тыс. обр. и ок.
500 видов. Основные коллекторы: Е. А. Давыдов, Е. Ю. Скачко. Пред¬
ставленные регионы: Алтай (Россия, Монголия, Китай, Казахстан), Ал¬
тайский край. Значительная часть материала не инсерирована. Доступен
для посещения и работы.
Гербарий лишайников Ботанического Музея Казанского уни¬
верситета (KAZ). Основан в 1804 г. К. Ф. Фуксом. Ок. 20 тыс. обр. Ос¬
новные коллекторы: К. С. Мережковский, X. X. Хлебников, Л. Н. Василь¬
ева. Представленные регионы: Татарстан, Сев. Урал, Мурманская обл.,
Калмыкия, Крым, Эстония, Австрия, Франция, Италия, Греция. Первые
коллекции в 1804 создал Карл Федорович Фукс, занимавший в 1805-1818
г.г. должность профессора ботаники на Кафедре естественной истории и
ботаники. В 1835-1836 гг. заведующий кафедрой ботаники Петр Яковле¬
вич Корнух-Троцкий приобрел несколько крупных гербариев, сохранив¬
шихся и поныне. Типовой гербарий не выделен. Систематический герба¬
рий К. С. Мережковского выделен в отдельные коробки по родам, образ¬
цы хранятся в плоских коробках, каждый отдельный образец вида зани¬
мает отдельную плоскую коробку внутри «родовой». Имеются прислан¬
ные и приобретенные в начале XX в. коллекции лишайников из многих
районов Европы, а также обширные сборы (К. С. Мережковского) эпи¬
литных лишайников с Крымского п-ова и большая коллекция эксикатов
начала XX века. Доступен для посещения и работы.
Коллекция «Лишайники Байкальской Сибири» в Гербарии им.
М. Г. Попова Центрального сибирского ботанического сада (NSB).
Основана в 1977 г. Т. В. Макрый. Ок. 20 тыс. обр. и ок. 1000 видов. Ос¬
новные коллекторы: Т. В. Макрый, В. М. Буркова, С. Э. Воронюк, И. И.
Стецура. Представленные регионы: Байкальская Сибирь: Становое наго¬
рье - хребты Кодар, Удокан, Северо-Муйский, Южно-Муйский, Верхне-
Ангарский, Баргузинский; Прибайкалье - хребты Байкальский, Примор¬
ский, Олхинское плато, Хамар-Дабан, Голец Давыдов; Зап. Забайкалье -
Баргузинская долина, бассейн р. Джиды, хребты Мал. Хамар-Дабан, Джи-
динский, Малханский, Хэнтэй-Чикойское нагорье; Юго-Вост. Забайкалье
- Торейская котловина, массив Адун-Челон, долина р. Онон, хребты При¬
аргунский, Нерчинский, г. Алханай, окр. Нерчинского Завода, и др.; Во¬
сточный Саян - Тункинские Гольцы, Тункинская долина, Присаянье.
Дальний Восток: Приамурье - хр. Тукурингра, Еврейская авт. обл. (запо¬
ведник «Бастак»); Италия: Юго-Вост. Альпы. Основой коллекции стали
сборы лишайников разных коллекторов (в основном необработанные),
вывезенные в 1977 г. из СИФИБР (Сибирский институт физиологии и
биологии растений) СО РАН (Иркутск). Образцы хранятся в пакетах.
Имеются типовые образцы и образцы редких видов, известных в России
лишь из Байкальской Сибири, а также коллекция цианобионтных лишай¬
ников. Большая часть образцов определена и требует гербарной обработ¬
ки и инсерации. Часть коллекции (отдельные группы) доступна для изу¬
чения.
Лихенологический гербарий Института степи УрО РАН (ORIS).
Основан в 1999 г. О. С. Вондраковой (Меркуловой). Ок. 13.5 тыс. образ¬
цов и ок. 500 видов. Основные коллекторы: О. С. Вондракова (Меркуло¬
ва), В. Г. Кулаков, О. Г. Калмыкова. Представленные территории: Орен¬
бургская, Волгоградская, Астраханская обл., юг Челябинской обл., Кал¬
мыкия, Казахстан. Гербарий специализирован на представлении видового
разнообразия лишайников степной зоны Евразии. Значительная часть
коллекционного фонда сформирована образцами, собранными в особо
охраняемых природных территориях указанных регионов. В гербарий пе¬
реданы на временное хранение части коллекций В. П. Савича, К. С. Ме¬
режковского, В. А. Дубянского, М. Д. Спиридонова, И. М. Крашенинни¬
кова и др. В состав фонда также входят современные коллекции В. Г. Ку¬
лакова (Волгоград), Н. Н. Очировой (Калмыкия), Ю. В. Пережогина (Ка¬
захстан) и неопределённые сборы различных авторов. Образцы хранятся в
пакетах, находящихся в коробках (120 коробок), размещённых в шкафах.
Отдельные систематические группы - роды Acarospora, Aspicilia, Са-
loplaca, Xanthoparmelia и др. определены и проверены специалистами.
Большая часть фонда внесена в базу данных. Доступен для работы.
Коллекция лишайников Гербария Института биологии Коми
НЦ УрО РАН (SYKO). Основан в 1994 г. Т. Н. Пыстиной и Я. Херманс-
соном. Ок. 12тыс. образцов и ок. 800 видов. Основные коллекторы: Т. Н.
Пыстина, Я. Херманссон, А. А. Кустышева, С. Н. Плюснин, О. В. Лаври¬
ненко. Представленные территории: Республика Коми, Ненецкий АО, Ар¬
хангельская область. Разрозненные и незначительные сборы из Киров¬
ской и Мурманской областей, Краснодарского края, Украины, Швейца¬
рии, Финляндии и Швеции. Образцы хранятся в пакетах, размещенных на
листах бумаги в пачках в гербарных шкафах. Расположение видов в ос¬
новном фонде в алфавитном порядке, этикетки двуязычные (на англий¬
ском и русском языках). Часть старых образцов (около 2 тыс.) определена
бриологом И. Д. Кильдюшевским и сотрудниками Ботанического инсти¬
тута им. В. Л. Комарова лихенологами К. А. Рассадиной, В. П. Савичем и
Е. К. Штукенберг. Имеется ок. 9 тыс. неинсерированых образцов. Досту¬
пен для посещений и работы.
Гербарий лишайников Ботанического сада-института ДВО
РАН (VBGI). Основан в 2002 г. А. В. Галаниным. Ок. 10 тыс. образцов.
Основные коллекторы: А. В. Галанин, И. А. Галанина, Л. С. Яковченко.
Представленные территории: Приморский край (заповедники «Кедровая
падь», Лазовский и Сихотэ-Алинский, зап. побережье оз. Ханка, нац. парк
«Удыгейская Легенда», памятник природы «Борисовское плато»), Хаба¬
ровский край (Буреинский заповедник), Амурская область, Забайкальский
край (Сохондинский заповедник, заказник «Горная степь», нац. парк «Ал-
ханай», Агинский Бурятский АО), Бурятия, Камчатка, Сахалин, Якутия,
Монголия. Образы хранятся в пакетах, в специальных коробках и гербар¬
ных шкафах. Имеется небольшое количество образцов, определенных или
проверенных специалистами - монографами. Типовой гербарий не выде¬
лен. Много неинсерированного материала. Разложенные в шкафы образ¬
цы доступны.
Коллекция лишайников Гербария Псковского государственного
педагогического университета им. С. М. Кирова (PSK). Основан в
1960-е годы. Ок. 10 тыс. образцов и ок. 200 видов. Основные коллекторы:
Г. В. Недоспасова, Н. В. Недоспасова, Н. Б. Истомина, О. В. Лихачева.
Представленные территории: Псковская обл., включая особо охраняемые
природные территории (Полистовский государственный заповедник,
национальный парк «Себежский», музей-заповедник «Изборск»), Твер¬
ская область (Центрально-лесной государственный природный биосфер¬
ный заповедник). Большинство сборов датируются периодом с 90-х гг.
XX века по настоящее время. Образы хранятся в пакетах, приклеенных на
листы бумаги, а также в коробках в гербарных шкафах. Некоторые образ¬
цы проверены специалистами - монографами Ботанического института
им. В. Л. Комарова РАН и Московского государственного университета
им. М. В. Ломоносова. Имеется 20 коробок неинсерированного гербария.
Создается база данных гербарных образцов. Доступен для посещения и
работы.
Гербарий лишайников Кафедры ботаники Санкт-
Петербургского государственного университета (LECB). Основан в
1924 г. К. А. Рассадиной. Ок. 10 тыс. образцов. Основные коллекторы: Н.
М. Алексеева, А. А. Еленкин, А. А. Заварзин, Д. Е. Гимельбрант, Е. С.
Кузнецова, К. А. Рассадина, И. С. Степанчикова. Представленные терри¬
тории: Ленинградская обл., Камчатский край, Краснодарский край, Каре¬
лия, Мурманская обл., Волгоградская обл. и др. регионы, преимуществен¬
но в пределах РФ. Образы хранятся в пакетах, вложенных в папки, в пач¬
ках, в 2 гербарных шкафах и во многочисленных коробках. Типовой ма¬
териал отсутствует. В гербарии находится полное собрание изданных
Rabenhorst'oM эксикат лишайников Европы, а также часть коллекции эк¬
сикат, выпускавшихся отделом Споровых Растений Ботанического инсти¬
тута им. В. Л. Комарова РАН (Lichenotheca Rossica, Fasc. I - XVI). Значи¬
тельная часть гербария неинсерирована. Гербарная база данных отсут¬
ствует. Частично доступен для посещения и работы.
Гербарий Петрозаводского государственного университета
(PZV). Ок. 10 тыс. образцов. Основан в 1940 г. М. Л. Раменской. Основ¬
ные коллекторы: К. Алешкина, С. Хоровец, А. В. Сонина, В. Н. Тарасова,
В. И. Андросова (Степанова). Представленные территории: Карелия,
Мурманская и Волгоградская обл., Коми. Образцы из бывшей Финской
учительской школы в Сортавале. Образы хранятся в пакетах, приклеен¬
ных на листы бумаги, в пачках в гербарных шкафах. Имеются база дан¬
ных (электронная, карточная, общий список видов). Часть образцов, про¬
верена специалистами. Часть гербария в стадии инсерации. Доступен для
посещения и работы.
Ниже перечислены по регионам наиболее важные лишайниковые
коллекции России:
Санкт-Петербург.
Ботанический институт им. В. Л. Комарова РАН: Лаб. лихенологии
и бриологии (LE); Лаб. растительности Крайнего Севера (LE). Коллекция
О. В. Лавриненко.
Санкт-Петербургский государственный университет, Биолого¬
почвенный факультет, Кафедра ботаники (LECB).
Москва.
Московский государственный университет им. М. В. Ломоносова,
Биологический факультет, Кафедра микологии и альгологии. Гербарий
им. Д. П. Сырейщикова. (MW).
Институт проблем экологии и эволюции им. А. Н. Северцова РАН
(ИПЭЭ РАН), Лаборатории радиоэкологического мониторинга в регионах
АЭС и биоиндикации. Коллекция Л. Г. Бязрова.
Институт лесоведения РАН, Московская обл., Одинцовский р-н, с.
Успенское, Коллекция Е. Э. Мучник.
Институт глобального климата и экологии Росгидромета и РАН.
Коллекция А. В. Пчелкина.
Северо-запад России.
Полярно-альпийский ботанический сад-институт им. Н. А. Аврори¬
на, Кировск. Мурманская обл. (KPABG).
Институт леса Карельского НЦ РАН, Петрозаводск (PTZ).
Петрозаводский государственный университет, Кафедра ботаники и
физиологии растений, Петрозаводск (PZV).
Заповедник «Кивач», Карелия.
Институт биологии Коми научного центра УрО РАН, Отдел флоры
и растительности Севера, Сыктывкар (SYKO).
Дарвинский государственный природный биосферный заповедник,
Борок, Вологодская обл.
Псковский государственный педагогический университет им. С. М.
Кирова, Кафедра ботаники и экологии растений, Псков (PSK).
Средняя и Южная Россия.
Российский государственный университет им. Иммануила Канта,
Факультет биоэкологии, Кафедра ботаники и экологии растений, Кали¬
нинград (KLGU).
Тверской государственный университет, Ботанический сад ТвГУ,
Тверь (TVBG).
Институт биологии внутренних вод им. И. Д. Папанина РАН, пос.
Борок, Ярославская обл. (IBIW).
Воронежский государственный университет им. Ленинского Ком¬
сомола, Кафедра биологии и экологии растений, Воронеж (VOR).
Заповедник «Галичья Гора», Воронежского государственного уни¬
верситета, пос. Донское, Липецкая обл (VU).
Рязанский государственный университет им. С. А. Есенина, Кафедра
ботаники и агробиологии. Рязань (RSU).
Окский государственный природный биосферный заповедник, пос.
Лакаш, Рязанская обл.
Ростовский универитет. Ростов на Дону.
Повольжье:
Казанский (Поволжский) Федеральный Университет, Биологиче¬
ский факультет, Ботанический Музей, Казань (KAZ).
Марийский государственный университет, Йошкар-Ола.
Государственный природный заповедник «Большая Кокшага»,
Йошкар-Ола.
Коллекция Т. А. Дудоревой, с. Любимово, Саратовская обл.
Волгоградский государственный педагогический университет, Ка¬
федра ботаники и методики преподавания биологии Волгоград (VOLG).
Астраханский университет.
Сев. Кавказ:
Кубанский государственный аграрный университет, Краснодар
(KBAI).
Кабардино-Балкарский государственный университет, Нальчик.
Горный ботанический сад, Махачкала.
Урал:
Пермский государственный педагогический университет, Кафедра
ботаники, Пермь (PSU).
Институт экологии растений и животных УрО РАН, Екатеринбург.
(SVER).
Уральский федеральный университет, Екатеринбург.
Институт биологии Уфимского НЦ РАН.
СГПА, Стерлитамак.
Институт степи Уральского отделения РАН, Лаб. биогеографии и
мониторинга биоразнообразия, Оренбург (ORIS).
Сибирь:
Коллекция Т. В. Макрый «Лишайники Байкальской Сибири» в Гер¬
барии им. М. Г. Попова при Центральном сибирском ботаническом саде
СО РАН, Новосибирск (NSB).
Центральный сибирский ботанический сад СО РАН, Новосибирск
(NSB).
Алтайский государственный университет, Южно-Сибирский бота¬
нический сад, Барнаул (ALTB).
Институт леса им. В. Н. Сукачева СО РАН, Красноярск.
Саяно-Шушенский государственный природный биосферный запо¬
ведник, с. Шушенское, Красноярский край.
Байкальский государственный природный биосферный заповедник,
Иркутская обл. Урбанавичене И. Н.
Государственный природный биосферный заповедник «Баргузин-
ский», Лаб. биологического мониторинга, Улан-Удэ.
Музей природы Бурятии, Улан-Удэ.
Институт общей и экспериментальной биологии СО РАН, Улан-Удэ
(UUH).
Дальний Восток:
Магадан, Институт биологических проблем Севера или СВКНИИ?
Галанин А. А.
Институт биологических проблем криолитозоны, Якутск (SASY).
Якутский университет им. Аммосова.
Сахалинский ботанический сад ДВО РАН, Южно-Сахалинск
(SAKH).
Тихоокеанский институт географии ДВО РАН, Центр ландшафтно¬
экологических исследований, Владивосток (Нет акронима).
Ботанический институт Ботанического сада-института ДВО РАН,
Лаб. экспериментальной фитоценологии, Владивосток.
Биолого-почвенный институт ДВО РАН, Отдел ботаники, Владиво¬
сток (VLA).
По всей вероятности существуют и другие небольшие коллекции
лишайников, которые не попали в наш обзор. Для пополнения имеющейся
базы данных, просим сообщать нам сведения о заслуживающих внимания
научных коллекциях лишайников России.
Подавляющее большинство гербариев созданы при институтах си¬
стемы Российской Академии наук (22) или находятся в ведении универси¬
тетов (21). Имеются немногочисленные частные коллекции (3), коллекции
при музеях и ботанических садах (4) и заповедниках (7).
В Ботанических институтах бывших республик Союза ССР также
имеются гербарные коллекции лишайников, частью собранные и на тер¬
ритории России. Ниже приводится список и краткие сведения о таких, из¬
вестных нам коллекциях.
Украина.
Киев. Гербарий лишайников Института ботаники им. Н. Г. Холод¬
ного НАН Украины (KW). Всего более 50 тыс. образцов (второй по объе¬
му материала гербарий на территории бывшего СССР), из России - около
11 тыс. образцов: Калининградская, Ленинградская, Мурманская обл.,
Средняя Россия, Арктика, Урал, Западная и Восточная Сибирь, Алтай,
Забайкалье, Якутия, Приморский край. Много образцов, изученных А. Н.
Окснером и типовых образцов А. Н. Окснера.
Херсон. Херсонский государственный университет
Харьков.
Львов.
Белоруссия.
Минск. Коллекция лишайников Лаборатории флоры и систематики
растений Института экспериментальной ботаники НАН Беларуси (MSK).
Всего 20 тыс. образцов, из России: Карелия, Ленинградская и Астрахан¬
ская обл., п-ов Ямал, Кавказ и др.).
Гродно. Коллекция лишайников Кафедры ботаники Факультета
биологии и экологии Гродненского государственного университета им. Я.
Купалы. Всего около 1500 образцов.
Гомель. Научный гербарий Белорусского Полесья Кафедры ботани¬
ки и физиологии растений Гомельского государственного университета.
Около 1500 образцов из России: Карелия, Мурманская, Ленинградская и
Самарская обл., Красноярский край.
Эстония.
Тарту. Гербарий лишайников Ботанического и микологического му¬
зея Музея естественной истории Тартуского университета (TU). Всего 80
тыс. образцов, около 10-15 тыс. образцов из России: Кольский п-ов, Ка¬
релия, Ленинградская обл., Прибалтика, Урал, Ямал, Сибирь (в основном
- Прибайкалье), Дальний Восток (Камчатка, Приморский и Хабаровский
края, Чукотка, Сахалин), Казахстан.
Таллинн. Гербарий Таллиннского ботанического сада (TALL). Все¬
го 40 тыс. образцов, из России: Кольский п-ов, Полярный Урал, Урал, о-в
Большой Бегичев, о-в Диксон Таймыр, Чукотка.
Латвия.
Рига. University of Riga.
Рига. Botanical Institute.
Литва.
Вильнюс. Гербарий Вильнюсского университета. (WI). Всего 6 тыс.
образцов, около 300 обр. из России: Хибины, Коми, Урал, Алтай, Саяны.
ВИЛЬНЮС. Коллекция лишайников Гербария Института ботаники
Исследовательского центра природы (BILAS). Всего 9 тыс. образцов.
Армения.
Ереван. Ботанический институт.
Г рузия.
Тбилиси. Ботанический институт.
Сухуми (Абхазия). Сухумский ботанический сад.
Азербайджан.
Баку. Институт ботаники.
Гянджа. Педагогический институт.
Казахстан.
Алма-Ата. Институт ботаники.
Туркмения.
Ашхабад. Институт ботаники.
Узбекистан.
Ташкент. Институт ботаники.
Киргизия.
Бишкек. Институт ботаники.
Таджикистан.
Душанбе. Институт ботаники.
Более полная информация об этих и других коллекциях доступна на
сайте www.binran.ru.
Что касается иностранных гербариев, то наиболее важными с точки
зрения материалов, собранных на территории России, являются следую¬
щие:
Финляндия.
Хельсинки. Гербарий лишайников Ботанического сада и Ботаниче¬
ского музея Финского музея естественной истории Хельсинского универ¬
ситета (Н). Всего 450 тыс. образцов, около 20 тыс. образцов из России:
Карелия, Ленинградская, Мурманская, Архангельская, Вологодская, Ка¬
лининградская, Тюменская, Иркутская обл., Красноярский и Хабаровский
края, Татарстан, Алтай, Якутия, Чукотка. Много типов.
Турку. Университет Турку (TUR). Около 10 тыс. образцов из Каре¬
лии, Мурманской, Ленинградской и Тюменской обл., Ханты-Мансийской
АО, Красноярского края, Хакассии и Чукотки. Много типов.
Оулу. Гербарий университета Оулу (OULU). Около 3 тыс. образцов
из России: Карелия, Мурманская, Ленинградская и Архангельская обл.,
Ненецкий АО, Алтай, Якутия.
Куопио. Гербарий Музея естественной истории Куопио (KUO).
Около 1000 образцов из России: Карелия, Ленинградская и Иркутская
обл.
Швеция.
Упсала. Гербарий Шведского музея эволюции Упсальского универ¬
ситета (UPS). Около 10 тыс. образцов из Карелии, Ленинградской обл.,
Республики Коми, с Новой Земли, Таймыра, Тюменской обл., Краснояр¬
ского, Приморского и Камчатского краев. Много типов.
Стокгольм. Королевский музей естественной истории (S). Около 10
тыс. образцов из Русской Арктики, Карелии, Мурманской обл., Краснояр¬
ского края, Таймыра, Чукотки и Камчатки. Много типов.
Лунд. Университет Лунда (LD). Более 1000 образцов из Русской
Арктики, Чукотки, Красноярского и Приморского края.
Норвегия.
Осло. Университет (О). Более 1000 образцов с Новой Земли, Земли
Франца-Иосифа и из Якутии. Много типов.
Дания.
Копенгаген. Ботанический музей (С).
Германия.
Киль. Гербарий Кристиана Дольника в Институте охраны природы
и ресурсов Кильского университета. Всего 6 тыс. образцов, 1500 образцов
из России: Калининградская обл.
Мюнхен. Государственное ботаническое собрание Мюнхен (М). Из
России: Калининградская обл., Кавказ.
Австрия.
Грац. Университет Граца (GZU). Около 1000? Образцов из Хаба¬
ровского края (Гербарий Toby Spribille).
Вена. (W). Забайкалье.
Швейцария.
Женева (G). Из России: Иркутская обл., Бурятия.
Чехия.
Прага. (PR).
США.
Боулдер, Колорадо. (COLO). Из России материалы американской
экспедиции 80-х годов под руководством В. А. Вебера - Алтай и Иркут¬
ская обл.
Вашингтон. (US). Гербарий С. Wright’a из Хабаровского края.
МЭДИСОН, Висконсин (WIS). Из России: Мурманская обл., Карелия,
Алтай, Иркутская обл.
Япония.
Токио. National History Museum Tokyo (TNS): Около 1000? Образ¬
цов с Сахалина и Курильских островов.
Глава III.
Таллом и репродуктивные структуры лишайников
Гимельбрант Д. Е., Кузнецова Е. С.
Лишайники традиционно рассматривали и рассматривают как от¬
дельный объект исследований, изучаемый самостоятельным научным
направлением - лихенологией. Эта традиция сохраняется на протяжении
более чем двух столетий, начиная с основополагающих работ Эрика Аха-
риуса (Acharius, 1798, 1803, 1810, 1814) несмотря на то, что лишайники
уже длительное время относят к царству грибов (Fungi) в качестве лихе-
низированных его представителей.
В настоящее время известно примерно 17.5-20 тыс. видов лишайни¬
ков. Лишайники - полифилитическая и, следовательно, несистематиче¬
ская группа, в которую входят представители различных филогенетически
обособленных групп грибов. Известны полностью или практически пол¬
ностью лихенизированные роды, семейства и целые порядки грибов, то¬
гда как некоторые другие грибные порядки, семейства и даже роды лихе-
низированы лишь отчасти. Наиболее обоснованным подходом можно
считать отнесение лишайников к отдельной несистематической биологи¬
ческой группе грибов, выделяемой на основании трофических и физиоло-
го-биохимических особенностей. Существуют значительные сложности в
разграничении лишайников и нелихенизированных грибов, связанные с
полибионтной природой лишайниковой ассоциации, различной степенью
ее стабильности и лихенизации.
Лишайник - обычно двух или трехбионтная система, в состав ко¬
торой обязательно входит мицелиальный микобионт, а также экстрацел-
люлярно расположенный по отношению к микобионту одноклеточный
или нитчатый фотобионт (водоросль и/или цианобактерия). Однако фак¬
тически взаимоотношения в лишайниковом талломе могут быть более
сложными и полибионтными, например, в случае участия в ассоциации
одного или более видов лихенофильных паразитических, парасимбиоти-
ческих или сапротрофных грибов.
Сложности в установлении отчетливой границы между лихенизиро-
ванными и нелихенизированными грибами не позволяют оценить точное
количественное разнообразие лишайников и ясно указать, какие группы
относятся к компетенции лихенологов, а какие должны изучать микологи.
В этом вопросе как лихенологи, так и микологи во многом следуют в рус¬
ле сложившихся традиций. Степень лихенизации может существенно ва¬
рьировать: таллом лишайника может включать как несколько клеток фо-
тобионта, ассоциированных с рыхло рассеянным в субстрате мицелием
(некоторые калициоидные лишайники), так и хорошо выраженный аль¬
гальный слой в сложно организованном и ясно стратифицированном тал¬
ломе (например, сем. Parmeliaceae и Peltigeraceae). В ряде случаев водо¬
росли не полностью интегрированы в таллом лишайника (Botryolepraria
lesdainii). Кроме того, некоторые грибы лишь факультативно вступают в
симбиотическую ассоциацию с фотобионтом и могут существовать в ка¬
честве свободноживущих сапротрофов (например, ряд видов рода
Leptorhaphis). Помимо этого, существуют и облигатно нелихенизирован-
ные сапротрофные грибы (например, Agyrium, Sarea, некоторые виды ро¬
да Мусорогит), обитающие в тех же микроместообитаниях и на тех же
субстратах, что и лишайники, в связи с чем лихенологи традиционно счи¬
тают их объектом своего изучения. При этом степень родства таких нели-
хенизированных грибов с таксонами лишайников может быть различной.
К числу связанных с лишайниками грибов относят также лихенофильные
грибы.
Лихенофильные грибы, не будучи сами лихенизированными, по¬
селяются на талломах лишайников как паразиты (виды, паразитирующие
на микобионте), парасимбионты или комменсалы (виды, умеренно пара¬
зитирующие на фотобионте совместно с основным микобионтом талло¬
ма), либо сапротрофы (виды, использующие отмершие части таллома ли¬
шайника или экскретируемые микобионтом, либо фотобионтом продук¬
ты). К настоящему времени известно более 1640 видов лихенофильных
грибов. Отдельно стоит упомянуть грибы, способные поселяться на по¬
верхности или внутри талломов лишайников, но не связанные трофически
с микобионтом или фотобионтом (например, эндофитные грибы). Неко¬
торые грибы, паразитирующие или обитающие в качестве сапротрофов на
лишайниках, встречаются также и на более широком круге хозяев -
например, на колониях свободноживущих водорослей аэрофитона и на
эпифитных мхах (виды рода Athelia), другие пока известны только как
лихенофильные (например, виды родов Abrothallus, Lichenostigma). В со¬
став некоторых родов грибов (например, Tremella) входят лихенофильные
и микофильные представители.
Большое значение для понимания взаимоотношений между объек¬
тами изучения лихенологов играют лихенофильные лишайники. Их ми-
кобионт в течение всей жизни или, по крайней мере, части ее связан с
собственным фотобионтом и в этот период не связан с фотобионтом хозя¬
ина или, возможно, выступает в качестве парасимбионта по отношению к
нему. Фотобионт лихенофильного лишайника располагается в его соб¬
ственном талломе или погружен в таллом хозяина. Лихенофильные ли¬
шайники используют таллом хозяина не только в качестве субстрата, но и
паразитируют на его микобионте. Внешне это, как правило, проявляется в
заметных патологических изменениях таллома лишайника-хозяина. Ми-
кобионты некоторых лихенофильных лишайников лишь часть жизни свя¬
заны с собственным фотобионтом и в это время имеют совой лихенизиро-
ванный таллом. Так, целый ряд представителей рода Rhizocarpon начина¬
ет развиваться на талломах других накипных лишайников и лишь позднее
формирует собственный таллом с фотобионтом, уже не связанный с тал¬
ломом хозяина. На первых стадиях своего развития микобионты таких
видов являются паразитами или парасимбионтами на талломах хозяев.
Даже в пределах одного лихенофильного рода могут быть представлены
виды с различными стратегиями развития. Так Epilichen scabrosus форми¬
рует собственный поверхностный лихенизированный таллом на талломах
видов рода Baeomyces и позднее может существовать независимо от тал¬
лома хозяина. В то же время Е. stellatus имеет нелихенизированный в те¬
чение всей жизни таллом, погруженный в таллом и апотеции Lecidea
tessellata, и является, таким образом, лихенофильным грибом. Интерес¬
ным и редким примером взаимоотношений близкородственных таксонов
у лихенизированных грибов является комменсализм (адельфопаразитизм)
лишайника Cladonia luteoalba на различных видах из той же секции
Cocciferae рода Cladonia. Подобное явление можно отметить и в пределах
некоторых других групп лишайников (например, у рода Caloplaca).
Поскольку лишайники не являются филогенетически единой систе¬
матической группой, а представляют собой группу биолого-трофическую,
важное значение для понимания объема группы играет характер взаимо¬
отношений между бионтами. Таллом лишайника - полибионтная (2-5 и
более бионтов) симбиотическая ассоциация, в которой отношения между
бионтами строятся на основе устойчивого, сбалансированного (умеренно¬
го), биохимически контролируемого паразитизма микобионта на фото-
бионте. Такую форму отношений между бионтами часто интерпретируют
как мутуализм (Ainsworth..., 2008).
Таллом лишайника не может быть воспринят как единый организм
или особь с позиций молекулярно-генетических и в этом отношении ско¬
рее представляет собой сообщество организмов или экосистему. В то же
время таллом лишайника - стабильная во времени и неограниченно само-
воспроизводящаяся облигатная и физиологически тесная ассоциация фи¬
логенетически различных организмов, имеющая специфические черты
морфологии, анатомии, физиологии и вторичного метаболизма. В любых
сообществах лишайники устойчиво занимают характерные для них ме¬
стообитания и, наряду с водорослями, мохообразными и грибами, функ¬
ционируют как целостные организмы - элементы трофических цепей. По
этой причине в ряде исследований (флористические, экологические, мно¬
гие экофизиологические и биоиндикационные) лишайники изучают теми
же методами, что и многие растительные организмы, не акцентируя вни¬
мание на их комплексной полибионтной природе.
Таллом
Таллом лишайника представляет собой мицелиальное сплетение
(набор состоящих из грибных гиф плектенхим), в которое включены
клетки фотобионта. Такой таллом может быть с полным основанием ин¬
терпретирован как грибная строма. В результате интеграции клеток фото-
бионтов в талломы систематически различных грибов-микобионтов в хо¬
де длительной эволюции сформировался набор разнообразных и уникаль¬
ных жизненных форм лишайников, в совокупности не имеющих аналогов
в других группах живых организмов. Аналоги отдельных жизненных
форм лишайников, тем не менее, можно встретить в неродственных им
систематических группах организмов (талломы некоторых морских водо¬
рослей, клавариоидные плодовые тела грибов из отдела Basidiomycota,
постройки коралловых полипов).
В подавляющем большинстве случаев таллом лишайника по своей
морфологии и анатомии не имеет ничего общего с колониями выделен¬
ных в чистые культуры микобионта или фотобионта, а также свободно-
живущего фотобионта. В культуре изолированный микобионт либо не
удается выращивать, либо он образует медленно растущие аморфные сте¬
рильные колонии, а развитие свойственного лишайнику таллома начина¬
ется лишь при условии контакта с фотобионтом. В противовес этому тал¬
лом многих лишайников представляет собой достаточно сложно органи¬
зованную стратифицированную структуру.
Микобионт. Вступать в симбиотические отношения с фотобионтом
и формировать талломы лишайников могут представители сумчатых (отд.
Ascomycota, несколько более 40% его представителей), базидиальных
(отд. Basidiomycota, незначительное число представителей нескольких
родов) и анаморфных или митоспоровых, несовершенных грибов
(anamorphic fungi, mitosporic fungi, Fungi Imperfecti или отд.
Deuteromycota).
Как правило, микобионт доминирует в талломе лишайника: на его
долю в среднем приходится 80-90% биомассы таллома, микобионт пре¬
имущественно определяет общий габитус и детали строения таллома.
Лишь у некоторых лишайников фотобионт в талломах преобладает над
микобионтом по биомассе и во многом определяет жизненную форму
(например, у цианобионтных видов рода Ephebe и хлоробионтных -
Racodium). Однако, даже у таких высокоорганизованных лишайников, как
представители порядка Peltigerales, фотобионт также способен влиять на
морфогенез таллома и определять некоторые особенности его морфоло¬
гии и окраски. Наглядным примером этого могут служить талломы неко¬
торых видов рода Peltigera, сформированные на базе одного микобионта,
но разных фотобионтов. Такие талломы отличаются не только по окраске,
но и по некоторым особенностям морфологии. Иногда лопасти, содержа¬
щие в альгальном слое клетки разных фотобионтов (зеленой водоросли и
цианобактерии), входят в состав одного таллома, формируя так называе¬
мую фотосимбиодему.
Лихенизация является облигатной для большинства лишайниковых
грибов, вне ассоциации с фотобионтом микобионт в природе не встреча¬
ется или существует крайне непродолжительное время до момента есте¬
ственного ресинтеза таллома. Поскольку изолированный из талломов ли¬
шайников микобионт является симбиотрофом и растет исключительно
медленно, представляется маловероятным, чтобы он был способен вы¬
держивать конкуренцию с другими организмами и длительное время раз¬
виваться в свободноживущем состоянии.
В противоположность фотобионту, систематика грибного компо¬
нента лишайников разработана значительно лучше. Следует особо отме¬
тить, что систематика лишайников есть систематика их микобионтов и
научное латинское (как и тривиальное) название таксона лишайника от¬
носится только к микобионту, тогда как фотобионт в любом случае имеет
собственное латинское название.
Морфология таллома
До сих пор не существует единой общепринятой классификации
жизненных форм (биоморф) лишайников. Это связано с объективными
трудностями формализации всего внешнего (включая значительную внут¬
ривидовую адаптивную изменчивость) разнообразия талломов этих орга¬
низмов полифилитического происхождения и полибионтной природы.
Тем не менее, очевидно, что выделение условных типов жизненных форм
необходимо производить на анатомо-морфологической основе с учетом
морфогенетических особенностей и комплекса экологических адаптаций,
а не базироваться только на характеристиках внешнего облика (габитуса).
Жизненные формы (биоморфы) лишайников
Согласно классической и наиболее простой схеме жизненных форм
лишайников, широко используемой как в лихенологии, так и в других об¬
ластях биологии (геоботаника, экология, систематика и др.), их талломы
можно условно разделить на три основных морфологических типа -
накипные, листоватые и кустистые.
Накипные талломы представляют собой распростертые по поверх¬
ности субстрата (плагиотропные) и повторяющие его фактуру (реже по¬
груженные в субстрат) корочки различной формы, размеров и толщины.
Они характеризуются плотным контактом большей части своей нижней
поверхности с субстратом - прикрепляются к нему гифами сердцевины и
не образуют каких-либо специальных органов прикрепления, поэтому ме¬
ханически не могут быть отделены от субстрата без существенных по¬
вреждений. Нижний коровой слой у накипных лишайников не развит или
развит только на периферических участках таллома. Верхний коровой
слой хорошо развит или отсутствует, альгальный слой и сердцевина диф¬
ференцированы или дифференциация не выражена. Таким образом,
накипные талломы относятся к талломам дорсивентрального плана строе¬
ния. Прирост накипных талломов осуществляется по краю (перифериче¬
ски), в некоторых случаях может присутствовать вставочный (интерка-
лярный) рост центральных элементов таллома.
Листоватые талломы могут быть ориентированы преимущественно
вдоль поверхности субстрата (плагиотропно), но могут расти перпендику¬
лярно к его поверхности (ортотропный характер расположения) или даже
комбинировать в себе элементы плагиотропного и ортотропного роста
(плагио-ортотропный рост). Таллом листоватого лишайника состоит из
одной (монофильный) или многих (полифильный) лопастей, каждая из
которых представляет более или менее сложно организованную в про¬
странстве пластину. Вне зависимости от степени плотности контакта с
субстратом, листоватые талломы в течение всей жизни или на какой-то
стадии развития имеют те или иные органы прикрепления к нему и, в
большинстве случаев, могут быть отделены от субстрата без существен¬
ных механических нарушений. Нижний коровой слой у листоватых ли¬
шайников хорошо развит или отсутствует, однако вне зависимости от это¬
го наблюдаются характерные видимые отличия верхней и нижней по¬
верхностей таллома, проявляющиеся в окраске, фактуре поверхности,
присутствии и расположении поверхностных структур (таких, как органы
прикрепления, псевдоцифеллы, структуры размножения). Различия в ана¬
томическом строении верхней и нижней поверхностей также выражены.
Верхний коровой слой, как правило, хорошо развит, альгальный слой и
сердцевина дифференцированы или дифференциация отсутствует, кроме
верхнего альгального слоя иногда может присутствовать нижний. Могут
различаться и тонкие детали строения верхнего и нижнего корового слоя
(например, характер стратификации, особенности ориентации гиф). В лю¬
бом случае, можно говорить о наличии у талломов листоватых лишайни¬
ков дорсивентрального плана строения. Прирост листоватых талломов
осуществляется периферически, в некоторых случаях может присутство¬
вать интеркалярный рост центральных участков таллома (например, у
представителей сем. Umbilicariaceae).
Кустистые талломы могут быть ориентированы по отношению к по¬
верхности плагиотропно, ортотропно или плагио-ортотропно. Таллом ку¬
стистого лишайника состоит из неразветвленных, слабо или обильно раз¬
ветвленных более или менее цилиндрических или уплощенно-
цилиндрических элементов, в поперечном сечении имеющих радиальное
строение. Элементы обильно разветвленных кустистых талломов принято
называть ветвями. Вне зависимости от ориентации по отношению к суб¬
страту, кустистые талломы в течение всей жизни или на какой-то стадии
развития имеют органы прикрепления к нему и, в большинстве случаев,
могут быть отделены от субстрата без существенных механических нару¬
шений. У кустистых лишайников отсутствуют какие-либо явные морфо¬
логические или анатомические признаки, позволяющие говорить о нали¬
чии верхней и нижней поверхностей таллома или его ветвей (дорсивен-
тральное строение не выражено). Прирост кустистых талломов осуществ¬
ляется преимущественно апикально.
Однако такая упрощенная схема слишком слабо отражает суще¬
ствующее разнообразие морфологических типов талломов лишайников. В
ней не находит адекватного отображения как целый ряд промежуточных
форм, которые трудно отнести к одному из перечисленных типов, так и
разнообразие форм в пределах накипного, листоватого и кустистого ти¬
пов.
Еще более упрощенным является деление лишайников на так назы¬
ваемые макролишайники и микролишайники. По фактически сложившей¬
ся традиции к макролишайникам относят лишайники с листоватым и ку¬
стистым (иногда также с чешуйчатым) типами талломов, к микролишай¬
никам - с накипным. Однако такая интерпретация терминов не может
быть признана корректной, так как она основана не на размерных харак¬
теристиках, а на морфологических, что противоречит самому содержанию
терминов. Совершенно очевидно, что очень многие накипные лишайники
характеризуются макроскопическими размерами, а основные детали их
морфологии можно наблюдать без использования микроскопической тех¬
ники (невооруженным глазом). Вероятно, более корректным следует счи¬
тать такое понимание термина «микролишайники», при котором к по¬
следним относят только виды, все морфологические характеристики ко¬
торых видны только при использовании не менее чем 10х оптического
увеличения (Messuti, 1993). В таком случае к макролишайникам следует
относить не только большинство лишайников с кустистыми и листоваты¬
ми талломами, но и значительную часть накипных лишайников. Однако
при такой интерпретации использование терминов макролишайники и
микролишайники для описания жизненных форм представляется некор¬
ректным и бессмысленным.
Хотя в лихенологии и не существует единого общепринятого взгля¬
да на систему жизненных форм лишайников, можно выделить два основ¬
ных подхода, используемых наиболее часто. Один из них практически не
подразумевает иерархического соподчинения выделяемых жизненных
форм (см., например: Poelt, 1958; Определитель.., 1974; Clauzade, Roux,
1985; Brodo et al., 2001; Lichen..., 2002; Budel, Scheidegger, 2008; The
lichen..., 2009). Для традиционного в отечественной лихенологии подхода
характерна многоуровневая иерархическая система жизненных форм,
включающая разные их ранги (Голубкова, 1983; Голубкова, Бязров, 1989;
Бязров, 2009).
В таблице 1 сделана попытка представить оба варианта системы в
сравнительном плане, проиллюстрировав их изображениями жизненных
форм.
Таблица 1. Сопоставление наиболее распространенных систем жизненных форм лишайников.
Тривиальная
система
Неиерархическая система
Иерархическая система
Н. С. Голубковой
Рисунок
ПРИКРЕПЛЕННЫЕ ФОРМЫ
НАКИПНЫЕ
ЭНДОГЕННЫЕ (отдел)
ПЛАГИОТРОПНЫЕ (тип)
Эндосубстратные (эндогенные, криптоталлические)
НАКИПНЫЕ (класс)
Эндолитные (группа)
Рис. 9, III-1
Эндофлеодные (группа)
ЭПИГЕННЫЕ (отдел)
Эписубстратные (эпигенные)
ПЛАГИОТРОПНЫЕ (тип)
НАКИПНЫЕ (класс)
Однообразно-накипные
(группа)
Аталлические
Аталлические
Рис. 9, 111-2
Лепрозные (leprose)
Лепрозные
Рис. 9, I1I-3
Гониоцистные (гониоцистальные)
нет аналога
Рис. 9, 111-4
Зернисто-бородавчатые
Зер н исто-бородав чаты е
Рис. 9, III-5
Пленчатые (film-like)
нет аналога
Рис. 9, III-6
Плотнокорковые (однообразно-накипные)
Плотнокорковые
Рис. 9, I1I-7
Трещиноватые
Рис. 10, III-8
Т рещиновато-ареол ированные
Ареолированные
Рис. 10, III-9
Ареолированные (включая бородавчато-
ареолированные)
Рис. 10, III-10
Чешуйчатые (группа)
Булавовидно-ареолированные (булавовидно¬
чешуйчатые, вздуто-бородавчатые, тониниеобразные,
Тониниеобразные
Рис. 10, III-11
bullate)
Чешуйчато-ареолированные (пельтатные - peltate)
Однообразно-чешуйчатые
Рис. 10, 111-12
Чешуйчатые (squamulose)
Рис. 10, III-13
Чешуйчато-лопастные
Чешуйчато-лопастные
Рис. 10, III-14
Диморфные (группа)
Радиально-ареолированные (радиальные, розеточные,
эффигуратные - effigurate)
Радиальные
Рис. 11, III-15
Розеточные
Рис. 11, III-16
Радиально-лопастные плакодиоидные (placodioid)
Лопастные
Рис. 11, III-17
Радиально-лопастные субфолиатные (subfoliose)
Субфолиатные
Рис. 11, 111-18
ЛИСТОВАТЫЕ
Умбиликатные
УМБИЛИКАТНЫЕ (класс)
Умбиликатно-накипные
(группа)
Рис. 12, III-19
У мбил икатно-листоватые
(группа)
Рис. 12, III-20
ЛИСТОВАТЫЕ (класс)
Широколопастные (тип Peltigera)
Широколопастные
ризоидальные (группа)
Рис. 12,111-21
Узколопастные (тип Parmelia)
Рассеченнолопастные
ризоидальные (группа)
Рис. 12, II1-22
Узколопастные (тип Hypogymnia)
Вздутолопастные
неризоидальные (группа)
Рис. 12, III-23
Узколопастные повисающие (тип Pseudevernia)
Листовато-кустистые
повисающие (группа)
Рис. 12, 111-24
Узколопастные субфрутикозные (subfruticose)
ОРТОТРОПНЫЕ (тип)
КУСТИСТЫЕ (класс)
Кустистые прямостоячие
(группа)
Плосколопастные (ортотроп-
ные узколопастные листоватые
виды, например Flavocetraria
nivalis)
Рис. 12, III-25
КУСТИСТЫЕ
Карликово-кустистые (микрокустистые)
НАКИПНЫЕ КАРЛИКОВО¬
КУСТИСТЫЕ (класс)
Рис. 13, 111-26
Филаментозные, биссоидные (byssoid)
Рис. 13, III-27
Пульвинатные (pulvinate)
Рис. 13, III-28
КУСТИСТЫЕ (класс)
Кустистые прямостоячие
(группа)
Прямостоячие
Радиально-угловато-лопастные
Рис. 13, III-29
Радиально-лопастные
Рис. 13, III-30
Кустистые повисающие
(группа)
Повисающие
Плосколопастные
Рис. 14, Ш-31
Радиально-угловато-лопастные
Рис. 14, 111-32
Радиально-лопастные
Рис. 14, III-33
Кустистые стелющиеся
(группа)
Стелющиеся (распростертые)
Радиально-угловатые
Рис. 14, Ш-34
ПЛАГИО-ОРТОТРОПНЫЕ
(тип)
«Кладонииформные»
БОРОДАВЧАТО- ИЛИ
ЧЕШУЙЧАТО-КУСТИСТЫЕ
(класс)
Шиловидные
Рис.
15, III-35
Палочковидные
Шило- или сцифовидные
(группа)
Рис.
15, Ш-36
Сцифовидные
Рис.
15, III-37
Кустисто-разветвленные
Кустисто-разветвлен ны е
(группа)
Рис.
15, 111-38
Неприкрепленные (Свободноживущие кочующие) формы
Кочующие, странствующие или вагрантные (в т.ч.
СВОБОДНОЖИВУЩИЕ
эрратические)
(отдел)
НАКИПНЫЕ
НАКИПНЫЕ (класс)
Эгагропильные (фрутикулозные, щетинистые)
Эгагропильные (группа)
Рис. 16, 111-39
Пластинчатые
нет аналога
Рис. 16, 111-40
ЛИСТОВАТЫЕ
ЛИСТОВАТЫЕ (класс)
Узколопастные
Рассеченнолопастные (группа)
Рис. 16, Ш-41
КУСТИСТЫЕ (класс)
нет аналога
Листоватые плосколопастные
(группа)
Рис. 16, 111-42
КУСТИСТЫЕ
Кустистые
Радиально-угловатолопастные
(группа)
Рис. 16, 111-43
Рисунки, иллюстрирующие наиболее распространенные
жизненные формы лишайников (см. Таб. 1).
*
&
*
ini
си-:
-jC-e ^ ;■
не-1.|
"I A
J* I
• уч.
311--^
с о
о О
С.-
111
Рис. 9. III-l - Накипной эндосубстратный (эндолитный) лишайник
Bagliettoa baldensis (на поверхности субстрата видны только перитеции).
III-2 - Немногочисленные элементы таллома и апотеции накипного эпи¬
литного лишайника Lecidea diducens. 111-3 Лепрозные талломы накип¬
ных лишайников из рода Lepraria: а - с периферическими лопастями, б
- без периферических лопастей. III-4 - Гониоцистный таллом и апоте¬
ции накипного лишайника Micarea prasina. Ш-5 - Зернисто¬
бородавчатый таллом и апотеции накипного эпифитного лишайника
Bacidia rubella. III-6 - Пленчатый таллом и апотеции накипного лишай¬
ника Absconditella lignicola. III-7 - Плотнокорковый (однообразно¬
накипной) таллом и апотеции накипного эпифитного лишайника Buellia
disciformis.
Рис. 10. III-8 - Трещиноватый таллом и апотеции накипного лишайника
Bacidina inundata. III-9 - Трещиновато-ареолированный таллом и по¬
груженные апотеции накипного эпилитного лишайника Aspicilia cinerea.
III-10 - Ареолированный таллом накипного эпилитного лишайника Rhi-
zocarpon alpicola. Показаны светлые ареолы на темном гипоталломе. III-
11 - Булавовидно-ареолированный таллом накипного лишайника из ро¬
да Toninia. III-12 - Чешуйчато-ареолированный таллом и погруженные
апотеции накипного эпилитного лишайника Acarospora fuscata. 111-13 -
Чешуйчатый таллом накипного лишайника Нуросепотусе scalaris. III-14
- Чешуйчато-лопастной таллом и погруженные апотеции накипного
эпилитного лишайника Acarospora sinopica.
II] 1<
Рис. 11. II1-15 Радиально-ареолированный (радиальный) таллом
накипного эпилитного лишайника Sporastatia testudinea. 111-16 - Ради-
ально-ареолированный (розеточный) таллом накипного эпилитного ли¬
шайника Placopsis cribellans с немногочисленными изидиями и ради¬
альной формы цефалодием в центре. 111-17 - Радиально-лопастной (пла-
кодиоидный) таллом и апотеции накипного лишайника Fulgensia
fulgens. 111-18 - Радиально-лопастной (субфолиатный) таллом и апоте¬
ции накипного эпилитного лишайника Lobothallia melanaspis.
к
111.ЗПЛ
-Г, . -
, /*■!'■ ff
v*.W
|[|r?|
v
Шн;
Q-
K?5,^""s
л> I
v-:.
ч V
:Л '■
С/4 '■
J
111-21
■.,
. )
[II-??
Рис. 12. 111-19 - Умбиликатный (умбиликатно-накипной) таллом и апо¬
теции листоватого эпилитного лишайника Rhizoplaca chrysoleuca. 111-20
- Умбиликатный (умбиликатно-листоватый) монофильный таллом и
апотеции листоватого лишайника Umbilicaria proboscidea: а - вид свер¬
ху, б - вид сбоку. 111-21 - Широколопастной таллом и апотеции листова¬
того лишайника Lobaria pulmonaria. 111-22 - Узколопастной (рассечен-
нолопастной ризоидальный) таллом и апотеции листоватого эпилитного
лишайника Melanelia commixta. III-23 - Узколопастной (вздутолопаст¬
ной неризоидальный) таллом листоватого эпифитного лишайника Ну-
pogymnia fragillima. 111-24 - Узколопастной повисающий таллом листо¬
ватого лишайника Pseudevernia furfuracea. 111-25 - Узколопастной суб-
фрутикозный таллом листоватого эпигейного лишайника Flavocetraria
nivalis.
Рис. 13. 111-26 - Карликово-кустистый таллом и апотеции эпилитного
лишайника Lichina confinis. 111-27 Филаментозный таллом эпилитного
кустистого лишайника Cystocoleus ebenus. 111-28 - Пульвинатный таллом
эпилитного кустистого лишайника Aspicilia transbaicalica. 111-29 - Пря¬
мостоячий радиально-угловато-лопастной таллом кустистого эпигейно¬
го лишайника Evernia perfragilis. 111-30 Прямостоячий радиально¬
лопастной таллом кустистого лишайника Thamnolia vermicularis.
Рис. 14. 111-31 - Повисающий плосколопастной таллом и апотеции эпи¬
фитного лишайника Ramalina fraxinea. 111-32 - Повисающий радиально-
угловато-лопастной таллом кустистого эпифитного лишайника Evernia
divaricata. 111-33 - Повисающий радиально-лопастной таллом кустисто¬
го эпифитного лишайника из рода Usnea. 111-34 - Стелющийся таллом
эпилитного кустистого лишайника Pseudephebe minuscula.
Рис. 15. III-35 - Шиловидный чешуйчато-кустистый таллом эпигейного
лишайника Cladonia cornuta. 111-36 - Палочковидный чешуйчато¬
кустистый таллом эпигейного лишайника Cladonia cariosa. 111-37 -
Сцифовидный чешуйчато-кустистый таллом эпигейного лишайника
Cladonia fimbriata. III-38 - Кустисто-разветвленный таллом эпигейного
лишайника Cladonia arbuscula.
Рис. 16. III-39 - Эгагропильные талломы неприкрепленных накипных
эпигейных лишайников: а - Circinaria esculenta, б - С. fruticulosa. 111-40
Пластинчатые талломы неприкрепленного накипного эпигейного ли¬
шайника Circinaria emiliae. III-41 - Узколопастной (рассеченнолопаст-
ной) таллом неприкрепленного листоватого эпигейного лишайника Хап-
thoparmelia camtschadalis: а - вид сверху, б - вид снизу. III-42 - Плоско¬
лопастной таллом неприкрепленного листоватого эпигейного лишайни¬
ка Allocetraria stracheyi. III-43 - Радиально-угловато-лопастной таллом
неприкрепленного кустистого эпигейного лишайника Cetraria steppae.
Накипные лишайники
Эндогенные лишайники (например, ряд видов рода
Verrucaria) имеют полностью погруженную в субстрат вегета¬
тивную часть таллома (клетки фотобионта также погружены в
субстрат). На поверхности субстрата в большей или меньшей
степени заметны только генеративные структуры таллома.
Аталлические лишайники (некоторые виды рода
Lecidea) характеризуются хотя и поверхностным, но очень
слабо развитым и слабо стратифицированным или нестрати-
фицированным лихенизированным талломом, состоящим
лишь из немногих отдельных и плохо заметных бугорков. Ча¬
сто такие элементы таллома очень рассеянны и преимуще¬
ственно приурочены к апотециям. Название «аталлические»
представляется не вполне корректным (слабо развитый эпи-
генный таллом присутствует), но является вполне традицион¬
ным (см., например: Определитель..., 1974; Голубкова, 1983).
Лепрозные лишайники (виды рода Lepraria) образуют
таллом, состоящий из покрывающего субстрат нестратифици-
рованного рыхлого слоя более или менее плотных, зернистых,
округлых, мелких элементов, лишенных сформированного ко¬
рового слоя и состоящих из переплетенных гиф микобионта
клеток фотобионта (обычно такие элементы называют сореди-
ями). В некоторых случаях нижняя (лежащая на субстрате)
часть таллома лепрозных лишайников представлена более или
менее развитой сердцевиной, не содержащей клеток фотобио¬
нта. Коровой слой полностью отсутствует на всех стадиях раз¬
вития таллома.
Гониоцистные лишайники (некоторые виды рода
Micarea) образуют более плотный нестратифицированный
таллом, состоящий из плотных, зернистых, округлых, мелких
элементов - гониоцист, покрытых тонкой параплектенхимной
оболочкой из клеток микобионта, которая окружает одну
клетку или сформированную из нее плотную группу клеток
фотобионта.
Зернисто-бородавчатые лишайники (некоторые виды ро¬
да Bacidia) образуют однородные талломы, состоящие из
плотных зернистых или бородавковидных элементов, покры¬
тых коровым слоем. Хотя элементы таких талломов могут
быть стратифицированы на срезе, в целом таллом производит
впечатление нестратифицированного.
Пленчатые лишайники (Absconditella lignicola) имеют
таллом, в увлажненном состоянии приобретающий вид более
или менее тонкой нестратифицированной слизистой пленки, в
сухом состоянии напоминающий тонкую, порой слабо замет¬
ную корку на поверхности субстрата.
Плотнокорковые лишайники (некоторые виды рода
Buellia) образуют хорошо развитый, обычно стратифициро¬
ванный таллом, морфологически не расчлененный на отдель¬
ные элементы.
Трещиноватые лишайники (некоторые виды родов
Bacidina или Pertusaria) имеют таллом, пересеченный глубо¬
кими трещинами, не расчленяющими его, однако, на отдель¬
ные обособленные элементы. Борта таких трещин не покрыты
коровым слоем.
Трещиновато-ареолированные лишайники (некоторые
виды рода Aspicilia) формируют таллом, разбитый глубокими
трещинами на отдельные обособленные элементы - уплощен¬
ные или выпуклые ареолы, сидящие на общем гипоталломе.
Борта таких трещин, как и в предыдущем случае, не покрыты
коровым слоем.
Ареолированные лишайники (виды рода Rhizocarpon)
характеризуются таллом, состоящим из отдельных обособлен¬
ных элементов - уплощенных или выпуклых ареол, сидящих
на общем гипоталломе. Борта таких ареол покрыты коровым
слоем.
Булавовидно-ареолированные лишайники (виды рода
Toninia) имеют таллом, представленный сидящими на общем
гипоталломе более или менее вытянутыми элементами с за¬
уженным основанием и расширенной апикальной частью.
Борта таких булавовидных элементов также покрыты коровым
слоем.
Чешуйчато-ареолированные лишайники (некоторые ви¬
ды рода Acarospora) образуют таллом, представленный со¬
бранными вместе одинаковыми дорсивентральными чешуйка¬
ми, имеющими вогнутую до уплощенной или слабо выпуклой
верхнюю поверхность и прикрепленными к субстрату сужаю¬
щимся основанием. Края и периферическая часть нижней по¬
верхности таких чешуек обычно покрыты коровым слоем.
Чешуйчатые лишайники (виды рода Нуросепотусе) ха¬
рактеризуются талломом из одинаковых дорсивентральных
чешуек, прикрепленных к субстрату одной из сторон. Нижняя
поверхность чешуек обычно лишена корового слоя.
Чешуйчато-лопастные лишайники (некоторые виды рода
Acarospora) формируют таллом, центральная часть которого
носит характер чешуйчато-ареолированной, тогда как перифе¬
рическая представлена вытянутыми лопастями. Края и нижняя
поверхность чешуек и лопастей покрыты коровым слоем
только по внешнему контуру, поэтому такие лопасти не отде¬
лить от субстрата без повреждений.
Радиально-ареолированные (радиальные) лишайники
(некоторые виды рода Sporastatia) имеют таллом, в центре бо¬
родавчатый или ареолированный, по периферии ясно радиаль¬
ного строения. Периферические радиальные элементы также
состоят из бородавчатых элементов или ареол и могут напо¬
минать лопасти (настоящие периферические лопасти не фор¬
мируются).
Розетковидные радиально-ареолированные (розеточные)
лишайники (виды рода Placopsis) имеют таллом, в центре бо¬
родавчатый или ареолированный, по периферии ясно радиаль¬
ного строения благодаря развитию лопастей. Нижняя поверх¬
ность лопастей плотно связана субстратом и только по внеш¬
нему контуру покрыта коровым слоем, поэтому такие лопасти
невозможно отделить без повреждений.
Радиально-лопастные плакодиоидные лишайники (неко¬
торые виды рода Fulgensia) образуют таллом, в центре чешуй¬
чатый или состоящий из мелких лопастинок, по периферии
представленный хорошо выраженными настоящими лопастя¬
ми, снизу покрытыми коровым слоем или уплотненными
гифами сердцевины. Такие лопасти достаточно легко отделя¬
ются от субстрата.
Радиально-лопастные субфолиатные лишайники
(ХХХХ) характеризуются талломом, мелколопастным в цен¬
тральной части, по периферии с хорошо развитыми лопастями,
снизу покрытыми коровым слоем. Хотя практически весь та¬
кой таллом легко отделяется от субстрата и должен быть отне¬
сен к числу листоватых, его лопасти не несут настоящих ризин
и прикрепляются к субстрату отдельными гаптерами или ри¬
зоидами.
Листоватые лишайники
Умбиликатные (умбиликатно-накипные [тип Rhizoplaca\
и умбиликатно-листоватые [тип Umbilicaria]) лишайники
формируют моно- или полифильный таллом, прикрепленный к
субстрату в центральной части гомфом (реже несколькими
гомфами).
Широколопастные (широколопастные ризоидальные,
тип Peltigera) лишайники отличаются крупным листоватым
талломом, широкие лопасти которого слабо или совершенно
не разветвлены и прикреплены к субстрату хорошо развитыми
ризинами.
Узколопастные (рассеченнолопастные ризоидальные,
тип Parmelia) лишайники имеют таллом, состоящий из узких
хорошо разветвленных лопастей, прикрепленных к субстрату
развитыми ризинами.
Узколопастные (вздутолопастные неризоидальные, тип
Hypogymnia) лишайники образуют таллом из узких хорошо
разветвленных лопастей (обычно имеющих внутри полость),
прикрепленных к субстрату гаптерами.
Узколопастные повисающие (тип Pseudevernia) лишай¬
ники образуют таллом из узких хорошо разветвленных и сви¬
сающих вдоль субстрата дорсивентральных вторичных лопа¬
стей, обычно первоначально формирующихся на более широ¬
ких первичных лопастях, распростертых по субстрату и при¬
крепленных к нему ризинами. В ряде случаев первичные лопа¬
сти быстро исчезают или не формируются.
Узколопастные субфрутикозные (тип Cetraria islandica)
лишайники развивают ортотропный таллом, состоящий из уз¬
ких хорошо разветвленных дорсивентральных лопастей, пер¬
воначально прикрепленных к субстрату (грунту) ризинами,
позднее утрачивающих органы прикрепления и удерживаю¬
щихся на грунте за счет горизонтальных связей с другими
участниками дерновины.
Кустистые лишайники
Карликово-кустистые лишайники (тип Lichina) образуют
отдельные пучки, мелкие кустики или подушковидные талло¬
мы, полностью состоящие из тонких более или менее гладких
веточек (распростертая накипная часть таллома не формирует¬
ся).
Филаментозные лишайники (тип Cystocoleus) имеют
таллом, состоящий из еще более тонких, нитевидных или во¬
лосовидных элементов, часто состоящих из нитей фотобионта,
окруженных чехлом из гиф микобионта (распростертая накип¬
ная часть таллома также не формируется). Отделение этой
жизненной формы от предыдущей представляется очень
условным и, вероятно, излишним.
Пульвинатные лишайники (Aspicilia transbaicalica) ха¬
рактеризуются выпуклым подушковидным талломом, состав¬
ленным из плотно собранных вместе мелких вытянутых эле¬
ментов-веточек, произрастающих на общем распростертом
накипном талломе, более заметном по периферии.
Кустистые прямостоячие лишайники имеют крупный,
хорошо развитый таллом, состоящий из угловато-округлых
(радиально-угловато-лопастные, Evernia perfragilis) или более
или менее округлых (радиально-лопастные, Thamnolia
vermicularis) ветвей, растущих перпендикулярно (ортотропно)
по отношению к субстрату.
Кустистые повисающие лишайники формируют круп¬
ный, хорошо развитый таллом, состоящий из более или менее
уплощенных (плосколопастные, Ramalina fraxinea), угловато¬
округлых (радиально-угловато-лопастные, Evernia divaricata)
или округлых (радиально-лопастные, виды рода Usnea) ветвей.
Такой таллом обычно начинает расти перпендикулярно (орто¬
тропно) по отношению к субстрату, но в дальнейшем приобре¬
тает форму свисающего вдоль субстрата кустика.
Кустистые стелющиеся лишайники образуют крупный,
хорошо развитый таллом, состоящий из более или менее
округлых или угловато-округлых (радиально-угловатые, виды
рода Pseudephebe) ветвей и довольно быстро приобретающий
форму распростертого по субстрату кустика.
Кустистые плагио-ортотропные лишайники представле¬
ны распростертым по субстрату накипным (обычно чешуйча¬
тым или бородавчатым), часто со временем исчезающим пер¬
вичным талломом (первичным слоевищем), и обычно хорошо
развитым разветвленным или неразветвленным растущим
перпендикулярно к субстрату кустистым вторичным талломом
(вторичным слоевищем), существующим длительное время.
Вторичный таллом представлен подециями (развиваются из
нижней бесплодной части примордиев апотециев или пикнид
после их закладки на первичном талломе: виды рода Cladonia)
или псевдоподециями (развиваются как выросты первичного
таллома вне прямой зависимости от процесса развития апоте¬
циев или пикнид, которые обычно закладываются позднее на
псевдоподециях: виды рода Stereocaulon). Подеции могут
иметь внутри полость, псевдоподеции никогда не формируют
полость. По форме вторичного таллома выделяют шиловид¬
ные (подеции неразветвленные с шиловидно заостренными
окончаниями: Cladonia cornuta), палочковидные (подеции не-
или слаборазветвленные с притупленными окончаниями: С.
cariosa), сцифовидные (подеции не- или слаборазветвленные с
воронковидными расширениями на концах: С. fimbriata) и ку¬
стисто-разветвленные (подеции более или менее разветвлен¬
ные: С. uncialis) плагио-ортотропные лишайники.
Неприкрепленные лишайники
Эгагропильные лишайники (накипные) начинают расти
как эпилиты на мелких камешках и в процессе развития фор¬
мируют шаровидной или овальной формы талломы, поверх¬
ность которых представлена бородавчатыми, чешуйчато-
ареолированными, чешуйчатыми, сосочковидными или корот¬
кокустистыми коралловидно разветвленными элементами
('Circinaria esculenta и другие неприкрепленные кочующие ви¬
ды рода).
Пластинчатые лишайники (накипные) образуют таллом,
внешне напоминающий различным образом свернутые пла¬
стинки неправильной формы, верхняя поверхность которых
покрыта плотным коровым слоем, а нижняя большей частью
лишена его (Circinaria emiliae).
Листоватые узколопастные лишайники (рассеченноло-
пастные; Xanthoparmelia camtschadalis) имеют таллом, типич¬
ный для листоватых узколопастных прикрепленных видов
(тип Parmelia), однако края их лопастей и сами лопасти в су¬
хом состоянии более или менее подвернуты вниз, что придает
таллому округлую форму, наиболее адаптивную для переме¬
щения по открытым пространствам. Во влажном состоянии
лопасти таллома расправляются и лишайник приобретает бо¬
лее уплощенный вид. Ризины на нижней поверхности лопа¬
стей обычно редки и развиты слабо.
Листоватые плосколопастные лишайники формируют
разветвленный таллом, состоящий из более или менее плос¬
ких, направленных в разные стороны лопастей, края которых,
как и сами лопасти, не подвернуты вниз даже в сухом состоя¬
нии (Allocetraria stracheyi). На нижней поверхности лопастей
местами присутствуют короткие редкие ризины.
Кустистые (радиально-угловато-лопастные) лишайники
имеют хорошо разветвленный таллом, образованный углова-
то-округлыми ветвями, направленными в разные стороны, что
придает ему форму рыхлого неправильно-шаровидного кусти¬
ка (Cetraria steppae).
Реже используют и некоторые другие термины для
наименования жизненных форм лишайников. Ниже приведем
те из них, употребление которых представляется недостаточно
удачным или избыточным:
Зонированные - выделяют для однообразно-накипных
лишайников на основе образования по периферии их таллома
молодых внешне отличающихся (преимущественно по цвету и
структуре корового слоя) зон нарастания, чаще всего со вре¬
менем приобретающих сходство с более старыми участками
таллома (Окснер, 1974). Такая дифференциация таллома носит
временный возрастной характер, часто связана с ростовыми
процессами гипоталлома или с переходом центральной более
старой части таллома в фертильное состояние. Она встречает¬
ся не только у однообразно-накипных лишайников, но и у не¬
которых лепрозных, трещиновато-ареолированных и ареоли-
рованных лишайников. Периферическое нарастание талломов
с морфологически выраженной возрастной дифференцировкой
характерно и для листоватых лишайников, а также для кусти¬
стых (растущие апикальные участки ветвей). Термин следует
применять для описания формы таллома (см. ниже);
Хлопьевидные или флокулозные (floccose) - централь¬
ная часть таллома теряет кору и дезинтегрируется на элементы
вплоть до полного отслоения таких элементов от субстрата.
Такая организация поверхности таллома является возрастной
стадией его развития, а не жизненной формой;
Слизистые (gelatinous) - термин иногда используют для
описания сборной группы жизненных форм цианобионтных
лишайников с гомеомерными талломами, способными погло¬
щать значительные количества воды и принимать желеобраз¬
ную консистенцию. Представляется, однако, более логичным
не выделять их в отдельную морфологически весьма гетеро¬
генную группу, а относить такие виды и роды в соответству¬
ющие группы жизненных форм вместе с остальными лишай¬
никами. Так, широко распространены слизистые лишайники с
широко- (некоторые виды рода Collema) и узколопастными
(некоторые виды рода Leptogium), карликово-кустистыми
(Lichina), чешуйчато-лопастными (.Placynthium flabellosum) и
ареолированными (Psorotichia schaereri) талломами.
Характерной деталью строения таллома для многих
накипных и части листоватых лишайников является гипотал¬
лом (hypothallus, подслоевище) - нелихенизированная распро¬
стертая по субстрату нижняя часть таллома, на которой в
дальнейшем развивается лихенизированная часть. У целого
ряда видов такой гипоталлом хорошо заметен по периферии
таллома (Рис. 05-044) и/или между его элементами (например
- ареолами), отличается по структуре и окраске от лихенизи-
рованной части и сохраняется на протяжении всей жизни (ви¬
ды рода Pannaria). У других он развит слабо или вовсе неза¬
метен. На плотных субстратах (особенно на силикатных поро¬
дах) периферическая часть гипоталлома может приобретать
радиально-древовидную структуру. Онтогенетически гипотал¬
лом является результатом разрастания гиф недифференциро¬
ванного и не содержащего клеток фотобионта проталлюса
(prothallus), формируемого в ранний период жизни таллома
для ускоренного захвата участка субстрата.
Рис. 17. Участок таллома накипного лишайника Phlyctis argena с хоро¬
шо заметным гипоталлом по периферии.
Форма таллома
Важной характеристикой накипного и распростертого по
субстрату листоватого лишайника является форма его талло¬
ма. Иногда гипоталлом и/или собственно лихенизированная
часть таллома постепенно утончаются к периферии или по¬
гружены в субстрат, что приводит к отсутствию хорошо види¬
мого и четко выраженного края (таллом неопределенной фор¬
мы). Неопределенная форма таллома может образоваться и в
результате развития системы связанных гипоталломом рассе¬
янных элементов (рассеянный таллом, Buellia ectolechioid.es).
Если же край хорошо выраженный и четкий, различают тал¬
ломы неправильной (виды рода Lepraria) и правильной формы
(некоторые виды рода Aspicilia), дендроидные (некоторые ви¬
ды рода Lecidea), радиальные (некоторые виды рода
Caloplaca), зонированные (некоторые виды рода Pertusaria,
см. выше). Иногда для описания формы накипных талломов с
дезинтегрирующейся и отслаивающейся центральной частью
используют термин флокулозные или хлопьевидные
{Caloplaca cirrochroa). Форма таллома часто связана с услови¬
ями местообитания (например, с особенностями субстрата и
наличием конкуренции за него со стороны других лишайни¬
ков), а также с ростовыми процессами, поэтому в некоторых
систематических группах она может меняться в течение жизни
лишайника и постоянным признаком не является.
Характер ветвления талломов кустистых и листоватых
лишайников
Ветвление таллома может носить иррегулярный (не под¬
чиняющийся какой-либо повторяющейся закономерности) и
регулярный (подчиняющийся какой-либо закономерности) ха¬
рактер. Регулярное ветвление по взаимному расположению
ветвей или лопастей может быть очередным, супротивным,
мутовчатым, а по числу ветвей, образуемых в точке ветвления,
дихотомическим, трихотомическим, тетрахотомическим или
политомическим. При этом в случае образования из точки
ветвления равнозначных ветвей или лопастей (более или ме¬
нее сходных по размерным характеристикам) ветвление назы¬
вают изотомическим, если же ветви неравнозначны - анизо-
томическим или симподиальным. В случае анизотомического
ветвления нарастание кустистого таллома идет с формирова¬
нием явно выраженного основного стволика, что не наблюда¬
ется при изотомическом ветвлении. Такое нарастание у мно¬
гих видов рода Usnea приводит к формированию обильно раз¬
ветвленного таллома, в пределах которого можно выделить
ветви первого, второго, а иногда и третьего порядка, и наибо¬
лее тонкие, далее неделимые ветви - фибриллы, располагаю¬
щиеся на более крупных ветвях. Фибриллы можно рассматри¬
вать как структуры вегетативного размножения.
Для листоватых лишайников возможны монофильный и
полифильный варианты организации таллома (оба реализова¬
ны, например, в пределах рода Umbilicaria).
Поверхностные структуры таллома.
Структуры прикрепления к субстрату и морфологически
близкие к ним образования.
Характер прикрепления таллома лишайника к субстрату
зависит от его жизненной формы, особенностей субстрата,
условий местообитания, а иногда и стратегии размножения.
Большинство жизненных форм накипных лишайников
(плотнокорковые, ареолированные, радиально-
ареолированные и им подобные) прикреплено к субстрату
гифами сердцевины по всей площади таллома (Рис. 18) или
только в его центральной части (плакодиоидная жизненная
форма; Рис. 19), либо краями элементов таллома (чешуйчатые;
Рис. 20). Закрепление осуществляется за счет плотного кон¬
такта гиф сердцевины со всеми неровностями поверхности
субстрата и за счет более или менее глубокого проникновения
гиф во все его поры и микроскопические трещины. Неровно¬
сти поверхности и менее плотный внешний слой субстрата
благоприятствуют успешному и более быстрому заселению и
росту лишайников. Отделить накипной лишайник от субстрата
без значительного механического повреждения таллома не¬
возможно, поэтому такие лишайники необходимо собирать
вместе с субстратом.
Рис. 19. Прикрепление плакодиоидного накипного лишайника к суб¬
страту гифами сердцевины.
Рис. 18. Прикрепление однооб¬
разно-накипного лишайника к
субстрату гифами сердцевины.
Y
с.
Рис. 20. Прикрепление
чешуйчатого накипного
лишайника к субстрату
гифами сердцевины.
Г
Некоторые жизненные формы накипных лишайников не
имеют развитой сердцевины и не образуют каких-либо специ¬
альных гиф для контакта с субстратом. Лишайники с нестра-
тифицированными лепрозными (Рис. 21), гониоцистными,
зернистыми и пленчатыми талломами просто покрывают уча¬
сток субстрата мелкими гранулами или слизью, что обеспечи¬
вает достаточно прочное закрепление на нем.
Рис. 21. Лепрозный таллом, покрывающий участок субстрата.
Функцию прикрепления накипных и даже некоторых ли¬
стоватых лишайников (например, из семейства Pannariaceae)
может выполнять плотный или рыхлый гипоталлом, гифы ко¬
торого непосредственно контактируют с поверхностью суб¬
страта и частично проникают в него (Рис. 22).
Степень связи с субстратом и структуры прикрепления у
талломов листоватых и кустистых лишайников могут варьиро¬
вать более значительно. Далее приведены наиболее распро¬
страненные структуры прикрепления, формирующиеся на
нижней поверхности талломов.
Рис. 22. Прикрепление листоватого лишайника к субстрату посредством
гипоталлома.
Ризины - макроскопические пучки плотно переплетен¬
ных и/или склеенных гиф, формирующиеся как выросты ниж¬
него корового слоя (у представителей сем. Parmeliaceae) или
сердцевины (у представителей сем. Peltigeraceae) и проника¬
ющие в рыхлый субстрат или цепляющиеся за неровности
плотного субстрата. В субстрате ризины могут разветвляться
на отдельные гифы или их группы. Ризины могут быть про¬
стые или разветвленные, в том числе в форме кисти или ерши¬
ка (fibrillose, squarrose) (Рис. 23). Иногда как отдельные струк¬
туры рассматривают ризиноидные тяжи, отличающиеся от ри-
зин менее плотной агрегацией гиф и более глубоким проник¬
новением в субстрат (Рис. 24).
., 44 и
Рис. 23. Ризины листоватых лишайников семейства Peltigeraceae.
V*— , -г-г-ч .-=^г Рис. 24. Ризиноидные
-яжи.
Ризоиды (ризогифы) - отдельные одноклеточные или
многоклеточные гифы, формирующиеся из клеток нижнего
корового слоя, проникающие в субстрат и обеспечивающие
закрепление на нем таллома лишайника (Рис. 25). Могут фор¬
мировать густую сеть на нижней поверхности таллома.
Рис. 25. Прикрепление
лишайника с гетеромер-
ным строением таллома
к субстрату посредством
ризоидов.
Гайтеры - короткие и неразветвленные выросты или
просто участки нижней коры таллома, прикрепляющиеся к
субстрату наподобие присосок или прикрепительных дисков
(Рис. 26). Структуры, подобные гаптерам, могут образовы¬
ваться на концах ресничек и на других частях таллома, в слу¬
чае их контакта с субстратом.
Рис. 26. При¬
крепление тал¬
лома лишайника
к субстрату по¬
средством гап-
тер.
Гомф (umbilicus) - короткая и очень плотная структура
прикрепления, расположенная в центральной части нижней
поверхности талломов эпилитных умбиликатных лишайников
и состоящая из гиф сердцевины и корового слоя (Рис. 27).
Рис. 27. Прикрепле¬
ние таллома листо¬
ватого лишайника
из рода Umbilicaria
к субстрату посред¬
ством гомфа.
Псевдогомф - структура прикрепления, более характер¬
ная для кустистых лишайников и отличающаяся от гомфа тем,
что в прикреплении к субстрату участвуют только гифы серд¬
цевины (Рис. 28).
Рис. 28. Псевдогомф ку¬
стистого лишайника из
рода Usnea.
Морфологически близкими к ризинам структурами, не
выполняющими, однако, функции прикрепления к субстрату,
являются реснички - плотные тяжи гиф, формирующиеся по
краям лопастей (Рис. 29) или апотециев. Реснички характерны,
например, для ряда представителей семейств Parmeliaceae и
Physciaceae.
Рис. 29. Реснички на краях лопастей листоватого лишайника, прикреп¬
ляющегося к субстрату ризинами.
У некоторых умбиликатных лишайников на нижней по¬
верхности лопастей могут формироваться ризиноморфы (Рис.
30) - структуры, сходные по внешнему виду и строению с ри¬
зинами.
Рис. 30. Ризиноморфы на нижней поверхности таллома лишайника из
рода Umbilicaria.
Ризиноморфы могут быть простыми или разветвленны¬
ми, иногда с более или менее уплощенными участками. Дру¬
гие характерные образования умбиликатных лишайников,
также не выполняющие функции прикрепления к субстрату -
пластинчатые трабекулы разнообразной, обычно неправиль¬
ной формы, у некоторых видов покрывающие снизу все лопа¬
сти таллома.
Прочие поверхностные структуры
На поверхности талломов кустистых, листоватых и
накипных лишайников формируется целый ряд структур раз¬
личных функций и строения. К их числу относятся филлокла¬
дии (Рис. 31), представляющие собой выросты подециев (виды
рода Cladonia) или псевдоподециев (<Stereocaulon) кустистых
лишайников. Филлокладии покрыты коровым слоем (у видов
рода Cladonia нижний коровой слой отсутствует), под кото¬
рым лежат альгальный слой и сердцевина, однако в состав
филлокладиев, в отличие от веточек таллома, не входят эле¬
менты стереома. Филлокладии играют важную ассимиляцион¬
ную роль (так у многих видов рода Stereocaulon весь фотосин¬
тез осуществляется в филлокладиях), а также выполняют
функцию пропагул вегетативного размножения. Форма фил¬
локладиев очень разнообразна - от уплощенных чешуйчатых
до пальцевидных и коралловидных. Филлокладиоидные ве¬
точки у некоторых видов рода Stereocaulon хотя и напоминают
филлокладии и покрыты корой, но содержат в центральной
части ответвления стереома основных ветвей и, таким обра¬
зом, являются боковыми ветвями таллома.
Поверхность таллома может нести такие образования
неясного функционального назначения, как папиллы - мел¬
кие сосочковидные выросты на ветвях первого порядка у мно¬
гих видов рода Usnea (Рис. 32), формируемые только за счет
Рис. 31. Филло¬
кладии предста¬
вителей родов
Cladonia (а) и
Stereocaulon (б).
корового слоя, или пустулы - небольшие пузыревидные взду¬
тия на верхней поверхности талломов некоторых видов рода
Lasallia (Рис. 33), которым на нижней поверхности соответ¬
ствуют углубления. Другим примером неровностей таллома
могут являться фовеолы - неправильной формы вогнутые
участки коры на поверхности крупных ветвей у некоторых ви¬
дов рода Usnea (Рис. 34).
Рис. 32. Папиллы (а) и фибриллы (б) на ветвях первого порядка у видов
рода Usnea (внешний вид и срез).
Рис. 33. Пустулы
на верхней по¬
верхности таллома
видов из рода
Lasallia.
Рис. 34. Фовеолы на поверхности ветвей первого порядка у некоторых
видов рода Usnea.
В различных систематических группах циано-
хлоробионтных лишайников на поверхности (верхней или
нижней) или в толще талломов можно встретить характерные
образования - цефалодии (Рис. 35). Цефалодии содержат
группы клеток или нити цианобионта, окруженные более или
менее плотным слоем, сформированным гифами микобионта.
Поверхностные цефалодии имеют бородавковидную, чешуй¬
ковидную и даже коралловидную форму, погруженные же мо¬
гут быть заметны только по изменению окраски корового слоя
и формы поверхности (Nephroma arcticum) или на срезе через
таллом. Основная функция цефалодиев в трехбионтном тал¬
ломе - фиксация цианобионтом атмосферного азота, дополни¬
тельные - участие в процессе фотосинтеза и, возможно, веге¬
тативное размножение. Известен пример независимого разви¬
тия и даже более широкого географического распространения
свободноживущих карликово-кустистых цефалодиев Lobaria
amplissima (в таком состоянии их часто упоминают под соб¬
ственным названием Dendriscocaulon umhausense), которые
содержат в качестве фотобионта трихомы имеющей ложное
ветвление цианобактерии Scytonema.
Рис. 35. Цефалодии: а - Nephroma arcticum, б - Peltigera aphthosa, в -
Stereocaulon paschale.
Жилки (вены) - разветвленные структуры, развивающи¬
еся на лишенной коры нижней поверхности лопастей листова¬
тых лишайников и являющиеся уплотненными тяжами гиф
сердцевины (многие виды рода Peltigera, Рис. 36). Жилки мо¬
гут быть пигментированными, рельефными, контрастно выде¬
ляющимися на более светлом фоне нижней поверхности, или
светлыми, плоскими и слабо выраженными.
Рис. 36. Жилки (а), межжилковые (интерстициальные) пространства (б)
и ризины (в) на нижней поверхности лопастей листоватых лишайников
из рода Peltigera.
К поверхностным структурам относятся также цифеллы
и псевдоцифеллы, являющиеся структурами, облегчающими
газо- и водообмен таллома с окружающей средой. Цифеллы
встречаются на нижней поверхности лопастей лишайников из
рода Sticta и на поверхности ветвей видов рода Oropogon. Они
представляют собой обычно четко очерченные и хорошо за¬
метные углубления кратеровидной округлой или овальной
формы, полностью или полупогруженные в таллом (Рис. 37) и
выстланные рыхлым параплектенхимным ложнокоровым сло¬
ем (псевдокортексом), образованным из клеток сердцевины.
Рис. 37. Цифеллы на нижней поверхности лопастей у видов рода Sticta
(срез и внешний вид).
Псевдоцифеллы (Рис. 38) распространены гораздо шире
и характерны для целого ряд родов лишайников. Это округ¬
лые, вытянутые или сложной формы участки верхней или
нижней поверхности таллома, лишенные корового слоя или
какой-либо иной выстилки. Псевдоцифеллы могут быть по¬
груженными, плоскими или слегка выпуклыми и часто пред¬
ставляют собой мелкие более или менее хорошо или плохо
отграниченные структуры. Они выстланы или заполнены пуч¬
ками гиф сердцевины, выходящими на поверхность через от¬
верстия в коровом слое. Формироваться псевдоцифеллы могут
на поверхности таллома накипных лишайников, на поверхно¬
сти ветвей кустистых или на верхней или нижней поверхно¬
стях лопастей листоватых лишайников, а иногда даже на тал-
ломном крае апотециев.
* v’*V‘eV?3* в
*
V".
/
t,
V LiH
v>
"' J 1 ( j/ >7
>.' ■'■ j Щ V.4v‘, 'У >
'A* "■• M Lf^/^V,
77
Рис. 38. Псев¬
доцифеллы: срез
(а) и внешний вид
у представителей
рода Bryoria (б) и
Bryocaulon diver¬
ge ns (в).
Анатомия таллома
Основной структурный элемент, из которого построены
как вегетативная, так и генеративная части таллома лишайни¬
ка - гифы гриба. Их сплетения, как и у нелихенизированных
грибов, не образуют настоящих тканей (паренхим), поэтому в
отношении функционально и анатомически различных слоев
мицелиальных сплетений, входящих в состав таллома, исполь¬
зуют термин плектенхима (ложная ткань).
В зависимости от формы слагающих клеток выделяют
два типа плектенхим, из которых составлены все анатомиче¬
ски выраженные слои талломов лишайников. В состав парап¬
лектенхимы (псевдопаренхимы) входят гифы, составленные
более или менее изодиаметрическими округлыми или углова¬
то-округлыми клетками, поэтому на продольном и поперечном
срезах отдельные гифы плохо различимы, сложно определить
и направление их заложения (Рис. 39).
Рис. 39. Параплектенхима.
Прозоплектенхима (прозенхима) состоит из гиф, образо¬
ванных из вытянутых (анизодиаметрических) клеток, поэтому
в этом случае проще различить отдельные гифы и определить
их ориентацию в пространстве (Рис. 40). По отношению друг к
другу и к поверхности таллома выделяют несколько типов
ориентации гиф прозоплектенхимы. При антиклинальной (па¬
лисадной) ориентации гифы располагаются параллельно друг
другу, но перпендикулярно поверхности таллома. При пери-
клинальной ориентации гифы также располагаются парал¬
лельно друг другу и в то же время параллельно поверхности
таллома. В случае иррегулярно ориентированных (интрикат-
ных) гиф какой-либо закономерности в их укладке относи¬
тельно друг друга и поверхности таллома обнаружить невоз¬
можно, прозоплектенхима состоит из перепутанных и направ¬
ленных в разные стороны элементов.
Как в случае пара-, так и прозоплектенхимы гифы могут
быть уложены очень плотно или крайне рыхло. Обычно гифы
погружены в более или менее развитый межклеточный мат¬
рикс, однако в некоторых случаях такой матрикс практически
не развивается и промежутки между гифами заполнены возду¬
хом или впитываемыми из окружающей среды водными рас¬
творами (например, плектенхимы сердцевины некоторых
представителей сем. Parmeliaceae). Клеточные стенки гиф мо¬
гут быть тонкими или значительно утолщенными, просветы
клеток - широкими или узкими.
Рис. 40. Прозоплектенхима с антиклинальной (а), периклинальной (б) и
иррегулярной (в) ориентацией гиф.
Плектенхиму, гифы которой погружены в развитый
плотный желатинозный или хрящеватый матрикс и составле¬
ны из толстостенных клеток, часто называют склероплектен-
химой. Однако поскольку форма клеток, слагающих гифы та¬
кой плектенхимы, может быть как изодиаметрической (парап¬
лектенхима), так и вытянутой (прозоплектенхима), то в соот¬
ветствующих случаях предпочтительно использовать термины
склеропараплектенхима (собственно склероплектенхима в по¬
нимании Yoshimura, Shimada, 1980) и склеропрозоплектенхи-
ма (Рис. 41). Склеротизация является модификацией пара- или
прозоплектенхимы, обычно встречающейся в структурах, от¬
вечающих за придание таллому прочности и/или жесткости
(Рис. 416).
•Г"\
С: X:
Рис. 41. Склеропараплектенхима (а) и склеропрозоплектенхима (б).
Стратификация таллома лишайника.
Талломы лишайников могут быть достаточно четко
стратифицированы на выраженные слои или же наоборот
практически лишены какой-либо стратификации. В талломах
гомеомерных лишайников клетки фотобионта более или ме¬
нее равномерно распределены между гифами гриба по всему
объему (Рис. 42). Как правило, у них отсутствуют выраженные
слои, однако представители рода Leptogium формируют тон¬
кий, сложенный одним слоем гиф коровый слой (верхний и
нижний или только верхний). Гомеомерный тип организации
таллома характерен преимущественно для цианобионтных
лишайников (именно некоторых из них часто называют слизи¬
стыми или желатинозными) и может быть встречен у предста¬
вителей различных жизненных форм.
Рис. 42. Срез гомеомерного таллома лишайника с верхним и нижним
коровыми слоями: а - коровый слой, б - клетки фотобионта, в - гифы.
В талломах гетеромерных лишайников (Рис. 43) обычно
можно выделить коровый слой (или слои - верхний и ниж¬
ний), альгальный (водорослевый) слой и сердцевину. У неко¬
торых лишайников развит только верхний коровый слой, а
нижний не формируется (род Peltigera).
Рис. 43. Срез гетеромерного таллома лишайника с верхним и нижним
коровыми слоями: а - коровый слой, б - альгальный слой, в - сердцеви¬
на.
Коровый слой (cortex) является поверхностной плектен¬
химой таллома и может быть представлен пара- или прозо-
плектенхимой с пери- или антиклинальной ориентацией гиф.
В некоторых случаях верхний коровый слой состоит из двух
слоев, один из которых представлен пара-, а другой - прозо-
плектенхимой (например, у многих представителей сем.
Parmeliaceae). Обычно гифы корового слоя расположены
очень плотно, что способствует защите таллома от механиче¬
ских повреждений и препятствует быстрой потере талломом
воды в результате ее испарения. Плотная укладка гиф позво¬
ляет именно коровому слою у ряда родов кустистых лишайни¬
ков (например, Alectoria или Bryoria) брать на себя роль един¬
ственной плектенхимы, обеспечивающей механическую проч¬
ность таллома.
Важную роль в удержании влаги в талломе играют раз¬
личные модификации или производные корового слоя, форми¬
рующиеся на его поверхности. Так желатинизация кортикаль¬
ных гиф приводит к формированию глянцевой, блестящей по¬
верхности, отражающей часть светопотока и, таким образом,
снижающей ее нагрев. В то же время поверхность таллома
может быть матовой, неровной, мелкозернистой и даже ска-
брозной (грубозернисто- или почти чешуйчато-
ареолированной) из-за формирования многочисленных очень
мелких выростов корового слоя. У некоторых лишайников
клетки корового слоя синтезируют кнаружи гомогенный поли¬
сахаридный слой - эпикортекс. У видов рода Xanthoparmelia
он пронизан порами диаметром до 25 мкм, тогда как у пред¬
ставителей рода Parmelia s. str. такие поры отсутствуют. При
отмирании гиф поверхностной части корового слоя у некото¬
рых лишайников формируется матово-блестящий эпинекраль-
ный слой, состоящий из остатков желатинизированных корти¬
кальных клеток (например, у видов рода Miriquidica). Другим
неклеточным поверхностным производным корового слоя яв¬
ляется светлый налет (pruina), образуемый из остатков
эпинекрального слоя, кристаллов оксалатов, карбонатов и ли¬
шайниковых веществ. К числу клеточных производных коро¬
вого слоя относятся одноклеточные или многоклеточные,
обычно неразветвленные, хрупкие и бесцветные тонкие волос¬
ки - гифы (например, на верхней поверхности концов лопа¬
стей Melanelixia subargentifera). На верхней или, чаще, на
нижней поверхности талломов лишайников может формиро¬
ваться плотный слой беспорядочно переплетенных, ветвящих¬
ся, мягких гиф, называемый войлоком (tomentum). Такой слой
не только снижает нагрев поверхности таллома, но и умень¬
шает скорость испарения воды с его поверхности, задерживает
в себе некоторое количество влаги.
У многих лишайников верхний (а часто - и нижний) ко¬
ровый слой окрашен благодаря синтезу в его клетках разнооб¬
разных по цвету кортикальных лишайниковых веществ-
пигментов, отлагающихся на поверхности клеточных стенок
(см. главу VI). Пигментация корового слоя может играть важ¬
ную роль в защите фотосистемы фотобионта от переэкспони-
рования на солнце. На окраску поверхности таллома может
влиять и окраска клеток фотобионта, располагающихся ниже в
альгальном слое. Неравномерность окраски поверхности мо¬
жет быть вызвана различными причинами. Так пестрая окрас¬
ка талломов некоторых лишайников (например, ряда видов
рода Physcia) обусловлена неравномерным распределением
фотобионта («островками») в альгальном слое, расположен¬
ном под более светлым коровым. Участки корового слоя, под
которыми располагаются группы клеток фотобионта, имеют
несколько более темную окраску, тогда как участки, под кото¬
рыми фотобионт отсутствует, светлее и носят название маку-
лы (Рис. 44). Иногда в таких участках коровой слой может
быть более тонким, но не несет следов дезинтеграции. Другой
причиной пестроты окраски поверхности может быть точеч¬
ное обесцвечивание гиф корового слоя.
Рис. 44. Макулы у Physcia dubia. Внешний вид и срез гетеромерного
таллома с неравномерным распределением фотобионта в альгальном
слое.
Альгальный (водорослевый) слой обычно располагается
под коровым и включает клетки фотобионта (хлоробионта или
цианобионта) и иррегулярно расположенные гифы микобион¬
та. Именно в этом слое в клетках фотобионта происходит про¬
цесс фотосинтеза, здесь же осуществляется трофический кон¬
такт между бионтами. В гетеромерных талломах этот слой до¬
статочно тонок и у некоторых видов может быть разбит на от¬
дельные островки-фрагменты. У некоторых лишайников
аридных местообитаний альгальный слой может быть инвер¬
тирован и располагаться непосредственно над нижним коро¬
вым, что является дополнительной защитой фотобионта от из¬
быточного освещения.
Сердцевина (medulla) состоит из гиф микобионта, распо¬
лагается под альгальным слоем и снизу обычно прикрыта
нижним коровым слоем (виды рода Parmelia), реже может
быть лишена его (первичный таллом многих видов рода
Cladonia). В последнем случае гифы сердцевины могут фор¬
мировать жилки и ризины (см. часть «Поверхностные струк¬
туры таллома»). Функцией сердцевины является не только
накопление запасных питательных веществ в клетках гиф ми¬
кобионта. Она активно участвует в газо- и водообмене талло¬
ма, поскольку может накапливать значительные запасы воды и
атмосферного воздуха в межгифальном пространстве. Синте¬
зируемые в сердцевине многих видов медуллярные лишайни¬
ковые вещества откладываются на поверхности гиф и, обладая
гидрофобными свойствами, препятствуют полной гидратации
таллома и уходу из сердцевины атмосферного воздуха. Это
обеспечивает в талломе нормальное поддержание всех основ¬
ных физиологических процессов.
У большинства гетеромерных лишайников сердцевина
более или менее рыхлая, у видов рода Hypogymnia и некото¬
рых Ramalina в ней даже формируются значительные по объ¬
ему полости. В то же время, именно гифы сердцевины у неко¬
торых групп лишайников способны формировать склеротизи-
рованные механические плектенхимы, придающие таллому
значительную упругость, прочность на разрыв и жесткость -
так называемый стереом. Пространственная организация и
расположение механических плектенхим могут быть различ¬
ными (Рис. 45). Так у видов родов Usnea в центральной части
сердцевины формируется плотный, но эластичный и упругий
даже в сухом состоянии стереом (центральный тяж или осевой
цилиндр). В то же время у видов рода Cladonia стереом хотя и
занимает такое же центральное положение в талломе, но имеет
форму цилиндра (внутри несет полость), что придает таллому
в целом большую жесткость, но, вместе с тем, и хрупкость. У
большинства видов рода Stereocaulon псевдоподеций, по сути,
представляет собой стереом, так как лишен покрывающих его
снаружи альгального и корового слоев (они присутствуют
только в филлокладиях, расположенных на стереоме). У пред¬
ставителей рода Letharia стереом носит рассеянный характер и
представлен набором отдельных анастомозирующих тяжей,
располагающихся в сердцевине и на уровне альгального слоя.
Рис. 45. Пространственная организация стереома у видов родов Usnea
(a), Letharia (б) и Stereocaulon (в).
Репродуктивные структуры
Вегетативное размножение характерно для значительной
части видов лишайников и является единственным способом
их размножения как целостных, единых организмов. При веге¬
тативном размножении происходит увеличение числа генети¬
чески идентичных особей, поскольку соматические мутации
гаплоидного мицелия, вероятно, довольно редки. Этот способ
в простейшем случае основан на механической и часто слу¬
чайной фрагментации таллома, либо осуществляется посред¬
ством специализированных вегетативных пропагул - не¬
больших по размерам, легко отделяющихся от таллома и до¬
статочно легко перемещаемых в пространстве структур веге¬
тативного размножения, объединяющих в себе клетки водо¬
росли и гифы гриба. Хотя вегетативное размножение гаранти¬
рует репродукцию полибионтных талломов, практика показы¬
вает, что естественный их ресинтез на основе развивающегося
из грибной споры проталлюса и находящихся на субстрате
свободноживущих колоний фотобионта является не менее ре¬
зультативной, хотя и более зависимой от случайных факторов
стратегией. Хорошо известно, что многие лишайники никогда
не образуют каких-либо специализированных структур вегета¬
тивного размножения. В то же время немало видов лишайни¬
ков успешно использует обе стратегии размножения - вегета¬
тивную и половую.
Структуры вегетативного размножения лишайника
К числу более или менее специализированных типов ве¬
гетативных пропагул относятся соредии, бластидии, изидии,
спинулы, филлидии, шизидии, кортикальные чешуйки, гонио¬
цисты и гормоцисты (Рис. 46).
ip
« с
P.'S
Рис. 46. Специализированные вегетативные пропагулы лишайников: а -
соредии, б - бластидии, в - изидии, г - спинулы, д - филлидии, е - ши¬
зидии, ж кортикальные чешуйки, з гониоцисты, и гормоцисты.
Соредии представляют собой микроскопические или
субмикроскопические (от 20 до 100-200 мкм в диаметре), бо¬
лее или менее плотные или рыхлые, зернистые, округлые
структуры, лишенные сформированного корового слоя и со¬
стоящие из переплетенных гиф микобионта и клеток фотобио¬
нта. Соредии на гетеромерных талломах образуются в аль¬
гальном слое до или после дезинтеграции корового слоя, но¬
сящей, как правило, направленный характер. Часто соредии
условно подразделяют на более мелкие мучнистые (диаметром
от 25 до 40-50 мкм) и более грубые зернистые или грануляр¬
ные (диаметром от 45-55 до 80-100 мкм). Агрегаты из не¬
скольких соредий называют консоредиями (80-400 мкм в диа¬
метре). Окраска соредий может быть различной в зависимости
от состава лишайниковых веществ, продуцируемых микобио¬
нтом. Иногда при формировании соредий образуются лишай¬
никовые вещества, отсутствующие в остальных частях талло¬
ма. Интересно отметить, что цианобионтные лишайники с го-
меомерным талломом никогда не формируют соредии, хотя
способны формировать изидии или гормоцисты.
Талломы некоторых лишайников, не формирующих ко¬
рового слоя, могут быть представлены практически исключи¬
тельно самопродуцирующейся соредиозной массой (многие
виды родов Chrysothrix, Lepraria). В других случаях заклады¬
вающийся коровой слой полностью дезинтегрируется в про¬
цессе роста таллома или таллом полностью распадается на
гранулы (некоторые виды рода Lecanora, Lecidea), также при¬
обретая характер лепрозного. У видов, большую часть жизни
имеющих развитый коровой слой, соредии развиваются ло¬
кально, формируя на талломе ограниченные кортексом участ¬
ки различной формы - соралии. По характеру расположения
на талломе выделяют поверхностные (ламинальные), краевые
(маргинальные) и концевые (терминальные) соралии (Рис. 47),
по взаимному расположению - сгруппированные или рассеян¬
ные (диффузные), по форме - плоские, выпуклые, вогнутые,
кратеровидные (с сильно вогнутой погруженной поверхно¬
стью, ограниченные хорошо выраженным приподнятым краем
из гиф корового слоя), головчатые, губовидные, шлемовид¬
ные, вытянутые щелевидные (фиссуральные) (Рис. 48) и неко¬
торые другие. Необычный вариант развития соралей на концах
коротких палочковидных выростов коры - туберкул - наблю¬
дается у некоторых видов рода Usnea.
_-гт—
уу К С4 -г
L>—и/-'1 " '■11
Рис. 47. Расположение соралей: а - поверхностное (ламинальное), б -
краевое (маргинальное), в концевое (терминальное).
Развитие молодого лишайника из соредий происходит
через раннюю стадию роста достаточно рыхлой и недиффе¬
ренцированной плектенхимы, что позволяет формировать бу¬
дущий таллом из нескольких пропагул, иногда сформировав¬
шихся на разных талломах и генетически неидентичных. Это
создает условия для образования «химерных» талломов, в том
числе на основе пропагул различных хеморас и даже видов в
современном понимании (например, у представителей рода
Lepraria). Эту возможность необходимо учитывать при интер¬
претации результатов хемотаксономических и молекулярно¬
генетических исследований.
Целый ряд пропагул вегетативного размножения иногда
относят к так называемым соредияподобным структурам, имея
ввиду как механизм их образования из внутренних слоев тал¬
лома или путем отпочковывания от им подобных, так и отсут¬
ствие покрывающего такие пропагулы настоящего корового
слоя. Так гониоцисты несколько напоминают соредии, но
развиваются на основе одиночных клеток фотобионта, плотно
окруженных тонким параплектенхимным чехлом гиф (некото¬
рые виды родов Micarea, Lichenomphalia). Они образуют четко
ограниченные скопления (гониоцистангии) или формируют
целые гониоцистные талломы путем отпочковывания друг от
друга.
Гормоцисты формируются только на талломах некото¬
рых цианобионтных гомеомерных лишайников. Они пред¬
ставляют собой одиночные участки нитей цианобионта - гор-
могонии, окруженные тонким чехлом гиф гриба. Скопления
гормоцист формируют желатинозные кластеры - гормоци-
стангии (например, у карликово-кустистых видов рода
Lempholemma).
Рис. 48. Форма соралей: а - плоские, б - выпуклые, в - вогнутые, г -
кратеровидные, д - головчатые, е - губовидные, ж - шлемовидные, з -
щелевидные (фиссуральные).
Изидии формируются как мелкие поверхностные выро¬
сты таллома или в результате вторичного роста корового слоя,
покрывающего лишенные коры вегетативные пропагулы
(например, соредии). В хорошо развитых изидиях можно раз¬
личить коровой и альгальный слои, сердцевину, т.е. наблюда¬
ется стратификация, подобная таковой в талломе. Изидии
очень разнообразны по форме и могут быть от бородавковид¬
ных, шпателевидных и палочковидных до сложно разветвлен¬
ных коралловидных. Их размеры колеблются от сотых до де¬
сятых долей миллиметра в диаметре до 2-3 мм в длину. Бу¬
дучи более крупными пропагулами, изидии имеют меньший
радиус распространения по сравнению с соредиями. Как и со¬
ралии, изидии могут иметь различное взаимное расположение
на талломе (сгруппированные или диффузно расположенные
изидии). В связи с тем, что отделение изидий от таллома
обычно требует некоторого механического воздействия,
настоящие изидии достаточно тонки или имеют зауженное ос¬
нование. В некоторых случаях к изидиям не вполне корректно
относят структуры, функционально не имеющие отношения к
вегетативному размножению. Так у Melanohalea exasperata
изидиями часто называют низкие прочно сидящие на талломе
бородавковидные выросты с широким основанием, в которые
обычно погружены пикниды лишайника. Похожие на изидии
короткие палочковидные структуры вегетативного размноже¬
ния, развивающиеся одиночно или группами в соралиях ряда
видов рода Usnea, называют изидиоморфами и к настоящим
изидиям также не относят.
К числу близких к изидиям структур относятся филли¬
дии (лобули) - мелкие чешуйковидные лопастинки, образую¬
щиеся на талломах или на талломном крае апотециев некото¬
рых листоватых и кустистых лишайников как поверхностные
выросты. У листоватых лишайников филлидии обычно напо¬
минают лопасти таллома и имеют такую же стратификацию и
дорсивентральный план строения. Так у Peltigera praetextata
они лишены нижнего корового слоя и часто образуются по
краям трещин таллома и раневых поверхностей. У кустистых
лишайников (род Cladonia) подобные по строению структуры
называют филлокладиями.
Шизидии имеют иной механизм образования и форми¬
руются не как выросты поверхности, а путем локального от¬
слаивания от таллома небольших участков корового и связан¬
ного с ним альгального слоя (встречаются, например, по краям
лопастей Peltigera elisabethae, а также на поверхности подеци-
ев Cladonia scabriuscula).
Вероятно, промежуточными структурами между филли-
диями и шизидиями являются кортикальные чешуйки, обра¬
зующиеся, например, внутри сциф Cladonia pyxidata. Они яв¬
ляются, по сути, прикрепленными к таллому большей частью
нижней поверхности уплощенно-округлыми элементами че¬
шуйчато-ареолированного корового слоя и, в случае механи¬
ческого воздействия, могут легко отделяться от поверхности.
Бластидии (кортикальные гранулы) - округлые вегета¬
тивные пропагулы мелких размеров (70-300 мкм в диаметре),
содержащие клетки фотобионта и гифы микобионта и покры¬
тые более или менее сформированным коровым слоем. Фор¬
мирование бластидий происходит путем отпочковывания от
краев лопастей таллома или от ранее образовавшихся бласти¬
дий (например, у ряда видов рода Caloplaca). Эти структуры
занимают промежуточное положение между настоящими со-
редиями и изидиями.
Спинулы по характеру развития не являются изидиями
и представляют собой короткие и неразветвленные боковые
веточки с перетяжкой при основании, формирующиеся на тал¬
ломах видов некоторых родов кустистых лишайников (напри¬
мер, Bryoria). Однако, как и изидии, при механическом воз¬
действии они сравнительно легко отламываются от таллома и
переносятся на небольшие расстояния.
Фибриллы - наиболее тонкие и короткие, далее неде¬
лимые веточки, располагающиеся на более крупных ветвях
талломов видов рода Usnea. Анатомическое строение фибрилл
практически не отличается от строения более крупных ветвей,
однако центральная часть их сердцевины - стереом - не свя¬
зана со стереомом основной части таллома. На этом основа¬
нии фибриллы можно рассматривать в качестве своеобразных
структур вегетативного размножения. По характеру развития
фибриллы наиболее близки к спинулам. После обламывания
фибрилл на остающихся рубцах могут продуцироваться соре¬
дии.
Для многих видов лишайников единственно возможный
способ вегетативного размножения - это неспециализирован¬
ная и часто случайная фрагментация таллома. Размножение
неспециализированными частями таллома наиболее характер¬
но и результативно в первую очередь для многих кустистых
напочвенных (виды родов Cladonia и Stereocaulon) и эпифит-
ных повисающих лишайников (виды рода Bryoria). В то же
время у некоторых эпифитных лишайников имеются образо¬
вания, облегчающие такую фрагментацию - поперечные бо¬
розды, перетяжки на основных и боковых ветвях (Usnea
articulata), специфические кольцевые разрывы коры и аль¬
гального слоя, обнажающие рыхлую сердцевину (Evernia
divaricata) (Рис. 49).
Рис. 49. Перетяжки на ветвях
Usnea articulata (а) и кольце¬
вые разрывы коры и альгально¬
го слоя у Evernia divaricata,
обнажающие рыхлую сердце-
вину (б).
Структуры полового размножения микобионта
Половое размножение является весьма обычным и ши¬
роко распространенным элементом жизненного цикла различ¬
ных систематических групп лихенизированных сумчатых и
базидиальных грибов (микобионт). Хлоробионт в лихенизиро-
ванном состоянии не реализует своих потенциальных способ¬
ностей к половому размножению, для цианобионта половое
размножение не характерно как в лихенизированном, так и в
нелихенизированном состоянии. Для обозначения половой
стадии жизненного цикла нелихенизированных грибов широко
используют термин телеоморфа в противоположность ана-
морфной стадии (стадия, не несущая структур полового раз¬
множения, но способная нести структуры бесполого размно¬
жения). Использование этих терминов возможно и в случае
лишайников, однако следует помнить, что их талломы часто
несут структуры полового и конидиального спороношения од¬
новременно, а конидии далеко не всегда выполняют функцию
спор бесполого размножения. В этой связи необходимо учесть,
что лихенизированные сумчатые грибы могут иметь различ¬
ные варианты полового процесса - гаметангиогамию, сперма-
циогамию (функцию спермациев выполняют конидии) или
соматогамию (последняя более характерна для базидиальных
грибов). Половой процесс часто может происходить путем
слияния структур, образующихся на одном талломе (автога¬
мия). В случае лишайников не использовали и не используют
отдельные латинские названия для обозначения телеоморф и
анаморф, как это было широко принято для нелихенизирован-
ных грибов.
Для некоторых таксонов лишайников, филогенетически
связанных с отделом Ascomycota, половое размножение неиз¬
вестно. Многие из них обычно формируют те или иные струк¬
туры вегетативного размножения. Такие таксоны принято от¬
носить к числу стерильных. В то же время часть видов лишай¬
ников, для которых известны структуры полового размноже¬
ния, образует их редко или крайне редко и обычно встречается
только со структурами вегетативного размножения. Относить
такие виды к числу стерильных некорректно, как и виды,
единственным известным способом размножения которых яв¬
ляется бесполое (конидиальное) спороношение микобионта.
Половое размножение микобионта лишайников следует
за проходящей на гаплоидном (в случае сумчатых грибов) или
дикариотическом (в случае базидиальных грибов) мицелии
кариогамией, являющейся завершающей частью полового
процесса. Обычно половое размножение сопровождается об¬
разованием плодовых тел микобионта - мицелиальных спле¬
тений, несущих формирующиеся в результате мейоза в сумках
(сумчатые грибы) или на базидиях (базидиальные грибы) спо¬
ры полового размножения - эндогенные аско- или экзогенные
базидиоспоры соответственно. После попадания зрелых аско-
или базидиоспор микобионта в благоприятные для их прорас¬
тания условия развитие лишайникового таллома возможно,
как правило, только через стадию естественного ресинтеза с
участием подходящего свободноживущего фотобионта (фото¬
бионтов).
В случае сумчатых грибов для обозначения несущего
сумки мицелиального сплетения используют термины плодо¬
вое тело, аскома или аскокарп. Для обозначения несущего ба-
зидии мицелиального сплетения базидиальных грибов упо¬
требляют термины плодовое тело, базидиома или базидио-
карп. Плодовые тела сумчатых грибов образованы гаплоид¬
ным мицелием и дикариотическими аскогенными гифами, ба¬
зидиальных грибов - дикариотическим мицелием. Формиро¬
вание плодовых тел сумчатых грибов происходит по одному
из двух возможных путей: асколокулярному или аскогимени-
альному. Более редкий у лишайников асколокулярный тип
развития подразумевает первоочередное формирование из ве¬
гетативных гиф таллома примордиев строматических образо¬
ваний - псевдотециев, в полостях которых несколько позднее
происходит закладка генеративных аскогенных гиф, формиро¬
вание сумок, а также элементов хаматеция. При реализации
более обычного для сумчатых лишайников аскогимениального
пути строматические образования не формируются, развитие
примордия плодового тела инициируется закладкой аскоген¬
ных гиф, а сумки и элементы хаматеция образуют гимений. В
итоге развиваются перитеции или апотеции.
Особый путь развития плодовых тел без образования
примордиев известен у видов родов Lichinella и Gonohymenia.
Аскогониальные гифы закладываются непосредственно среди
вегетативных гиф таллома и формируют вокруг себя мицели-
альные сплетения, называемые таллинокарпами. В случае за¬
кладки аскогониальных гиф в пикнидах (у видов родов
Ephebe, Thyrea) формирующиеся плодовые тела напоминают
апотеции и называются пикноаскокарпами.
Зрелые плодовые тела большинства лихенизированных
сумчатых грибов - апотеции и перитеции - имеют общий план
строения, мало отличающийся от такового у нелихенизиро-
ванных грибов. Однако важной особенностью плодовых тел
многих лишайников является их способность к многолетнему
функционированию. У некоторых лишайников, в то же время,
плодовые тела эфемерны (например, у видов рода
Absconditella).
Апотеции представляют собой открытые в зрелости
плодовые тела чашевидной или блюдцевидной формы и со¬
стоят из генеративной части и защитных покровов. Экспони¬
рованную поверхность генеративной части называют диском
апотеция.
Лецидеиновые (лецидеевые) апотеции полностью лише¬
ны клеток фотобионта (Рис. 50). Генеративная часть представ¬
лена гимением (тецием), составленным сумками и элементами
хаматеция, погруженными в более или менее развитый жела¬
тинозный матрикс. Тонкий поверхностный слой гимения, со¬
стоящий из апикальных частей элементов хаматеция и защи¬
щающий сверху обычно более короткие сумки, называется
эпигимением или эпитецием. В эпитеции, преимущественно в
его верхней части, могут содержаться окрашенные или бес¬
цветные гранулы, кристаллы или капли масла, формирующие
так называемую эпипсамму. Окрашенными могут быть и апи¬
кальные части элементов хаматеция или желатинозный мат¬
рикс. Окраска этих компонентов хаматеция часто определяет
цвет диска апотеция. Ниже лежит собственно гимений - тол¬
стый слой, в котором располагаются сумки и основная по
длине часть элементов хаматеция. Окраска этого слоя может
определяться цветом гранул (если они есть) или желатинозно¬
го матрикса; здесь также могут присутствовать капли масла.
Тонкую нижнюю часть гимения называют субгимением. В ней
располагаются гифы, формирующие элементы лежащей выше
основной части гимения. Субгимений также может быть
окрашен. Защитные покровы лецидеинового апотеция пред¬
ставлены производными паратеция: расположенным под ги-
мением стерильным гипотецием, а также его периферической
частью - эксципулом (собственным краем апотеция). Гипоте-
ций, как и эксципул, может быть окрашен. В ряде случаев
окраска эксципула неоднородна - его периферическая часть
окрашена темнее или светлее внутренней. У некоторых ли¬
шайников (род Cladonia) разрастающееся основание гипоте-
ция приводит к формированию хорошо развитых подециев,
воспринимаемых как кустистая часть таллома.
Рис. 50. Схема строения лецидеинового (лецидеевого) апотеция: а ги¬
мений (теций), б - сумки, в - элементы хаматеция (парафизы), г - эпи-
гимений (эпитеций), д -эпипсамма, е - субгимений, ж - гипотеций, з -
эксципул.
Иногда из лецидеиновых выделяют и отдельно рассмат¬
ривают биаториновые (биаторовые, псевдолеканоровые, псев-
долеканориновые) апотеции, характеризующиеся светлой
окраской, более мягкой во влажном состоянии консистенцией.
Диск таких апотециев быстро становится выпуклым, а соб¬
ственный край постепенно исчезает. Однако все эти признаки
в той или иной комбинации могут быть характерны как для
типично лецидеиновых, так и для леканориновых апотециев, а
у некоторых таксонов меняются с возрастом апотеция. Как и
все лецидеиновые, биаториновые апотеции лишены несущего
водорослевый слой талломного края.
Леканориновые (леканоровые, эулеканоровые) апотеции
(Рис. 51) отличаются слабым развитием эксципула (собствен¬
ного края) и наличием хорошо развитого талломного края
(амфитеция), лежащего снаружи от эксципула. Талломный
край, как и формирующий его таллом, составлен коровым и
водорослевым слоями, а также сердцевиной. В амфитеции мо¬
гут содержаться кристаллы различной формы, размеров и ха¬
рактера расположения. У большинства лишайников с лекано-
риновым апотецием эксципул можно наблюдать только на
срезе, тогда как у немногих групп (например, виды рода
Diploschistes) видны при внешнем осмотре как амфитеций, так
и эксципул. Апотеции с хорошо развитыми и заметными как
эксципулом, так и талломным краем иногда называют зеори-
новыми (зеороидными). Если эксципул покрыт снаружи толь¬
ко коровым слоем талломного края, такой апотеций иногда
называют суперлецидеиновым (талломный край и гипотеций
не содержат клеток фотобионта). В случае если фотобионт
присутствует, но только в гипотеции, апотеций называют ми-
колеканориновым. Погруженные в таллом леканориновые
апотеции иногда именуют криптолеканориновыми (аспици-
лиоидными).
Рис. 51. Схема строения леканоринового (леканорового) апотеция: а
эксципул, б - амфитеций.
Существуют различные морфолого-анатомические мо¬
дификации апотециев лишайников (Рис. 52). Так у предста¬
вителей родов Graphis и Opegrapha апотеции напоминают по
строению лецидеиновые, но имеют узкий щелевидный диск,
линейную вытянутую форму и часто даже ветвятся (гистеро-
теции - от нелихенизированного рода Hysterium, лирелловид-
ные апотеции). У многих видов родов Arthonia и Micarea
округлые апотеции лишены как развитого амфитеция, так и
эксципула, присутствуют только гимений и гипотеций. В про¬
тивоположность этому у некоторых видов рода Pertusaria ам¬
фитеций столь развит, что скрывает большую часть зрелого
гимения, оставляя открытым лишь точковидный диск. У
большинства видов родов Calicium и Chaenotheca апотеции
располагаются на более или менее длинных ножках, представ¬
ляющих собой гипертрофированную нижнюю часть гипоте-
ция. Для многих представителей семейства Umbilicariaceae
характерен диск апотеция, имеющий сложную лабиринтооб-
разно- или концентрически-ребристую поверхность (гирозный
диск).
Рис. 52. Некоторые наиболее распространенные модификации апотеци¬
ев у лишайников (внешний вид и срезы): а-б - лирелловидные апотеции
видов рода Opegrapha с темным эксципулом, в - лишенные амфитеция и
эксципула апотеции видов рода Arthonia, г-д - погруженные в таллом-
ные бородавочки апотеции видов рода Pertusaria, е - апотеций с гироз-
ным диском видов рода Umbilicaria, ж- апотеций на ножке (с мазедием)
видов рода Calicium.
Перитеции имеют достаточно сходный план строения с
апотециями лецидеинового типа, однако являются полуза¬
мкнутыми плодовыми телами округлой или кувшиновидной
формы (Рис. 53). Их гимений окружен пигментированным или
бесцветным защитным покровом - эксципулом и открывается
во внешнюю среду апикально расположенной остиолой (усть¬
ицем). Под оболочкой эксципула над гимением находится по¬
лость перитеция. Особенностью строения перитециев у мно¬
гих видов пиренокарпных лишайников является наличие ин-
волюкреллюма (покрывальца) - пигментированной структуры,
лежащей над эксципулом и выполняющей защитную функ¬
цию. Дополнительную защитную роль может играть таллом, в
который более или менее значительно погружены перитеции
многих видов.
Рис. 53. Схема строения и внешний вид перитециев: а - эксципул, б -
остиола (устьице), в - инволюкреллюм, г - перифизы, д - сумки, е -
субгимений.
Плодовые тела базидиальных лишайников сходны по
строению с таковыми у представителей наиболее близких
нелихенизированных систематических групп (Рис. 54). Они
также несут гимений, в котором, однако, формируются бази-
дии с экзогенными базидиоспорами. В таких сезонных плодо¬
вых телах агарикоидного (род Lichenomphalia) или клаварио-
идного (род Multiclavula) плана строения фотобионт от¬
сутствует (он содержится в однообразно-накипном или че¬
шуйчатом талломе), тогда как у некоторых лихенизированных
афиллофороидных грибов (род Dictyonema, в России не изве¬
стен) он локализован в отдельном слое непосредственно в
многолетнем плодовом теле.
Рис. 54. Плодовые тела базидиальных лишайников: а - агарикоидное
(.Lichenomphalia hudsoniana); б - клавариоидное (род
Multiclavula).
Под хаматецием понимают совокупность стерильных
элементов гимения (интераскальных филаментов), а также
гифы, выступающие в полость плодового тела (Рис. 55). У
многих систематических групп лишайников интераскальные
филаменты формируют матрикс гимения, защищающий раз¬
вивающиеся сумки и поддерживающий их в вертикальном по¬
ложении. Хаматеций может быть представлен различными
типами, отличающимися по характеру развития и форме со¬
ставляющих его элементов. При асколокулярном пути разви¬
тия плодового тела в псевдотеции еще до или в процессе фор¬
мирования сумок может формироваться интераскальная псев¬
допаренхима - масса клеток, плотно заполняющих полость
аскома. Такая масса остается недифференцированной на фи¬
ламенты в течение всего времени существования плодового
тела, а сумки развиваются между ее клетками, уплотняя ее.
Более часто формируются парафизоиды, являющиеся произ¬
водным интераскальной псевдопаренхимы и представляющие
собой сеть из ветвящихся и анастомозирующих филаментов, в
которой развиваются сумки (напоминают псевдопарафизы).
Рис. 55. Хаматеций в плодовых телах лишайников: а - настоящие пара¬
физы, б - перифизы, в - перифизоиды, г - парафизоиды (также выглядят
псевдопарафизы), д интераскальная псевдопаренхима.
При аскогимениальном пути развития в плодовом теле
после закладки сумок могут формироваться настоящие пара¬
физы (филаменты, вырастающие из субгимения по направле¬
нию к поверхности гимения) или псевдопарафизы (ветвящиеся
и анастомозирующие филаменты, растущие из купола еще за¬
мкнутого молодого плодового тела в направлении субгимения
и врастающие сверху в гимений; позднее теряют связь с купо¬
лом аскома). Также выделяют перифизы - короткие филамен¬
ты, формирующиеся в остиоле перитеция или пикниды и рас¬
тущие вверх, к выходу из остиолы (могут формироваться в до¬
полнение к парафизам, псевдопарафизам или перифизоидам).
В противоположность перифизам растущие с купола плодово¬
го тела перифизоиды направлены вниз. Деление на клетки,
форма, характер ветвления и окраска элементов хаматеция
сильно варьируют и могут являться важными систематиче¬
скими признаками.
Сумка (аск, ascus) является результатом полового про¬
цесса у сумчатых грибов, развивается из концевой клетки ас-
когона и служит вместилищем эндогенно формирующихся и
созревающих аскоспор. Строение сумок лихенизированных
грибов довольно сложное, весьма разнообразное и во многом
определяется механизмом высвобождения зрелых спор. Со¬
гласно строению клеточной стенки и механизму высвобожде¬
ния спор у лишайников выделяют несколько основных типов
сумок (Рис. 56):
— сумки прототуникатного типа имеют недифференци¬
рованную на слои клеточную стенку и, как следствие, апи¬
кальный аппарат у сумок отсутствует. Высвобождение спор
происходит путем разрыва клеточной стенки. Такой тип сумок
характерен, например, для видов рода Heppia, а также
Calicium, Chaenotheca. В последних двух случаях разрушение
сумок происходит до полного созревания спор и их масса ока¬
зывается открыто экспонированной в чашевидных защитных
покровах апотеция на ножке, формируя так называемый мазе-
дий (Рис. 52 ж). Споры при этом дозревают за счет своих
внутренних резервов;
— сумки битуникатного типа имеют клеточную стенку,
состоящую из двух отличных по составу и функциям слоев.
При разрыве наружного слоя (экзоаска) внутренний слой (эн-
доаск) моментально вытягивается вверх и образует канал, до¬
стигающий поверхности гимения. При этом зрелые споры вы¬
брасываются по каналу из сумки благодаря ее внутриклеточ¬
ному давлению (виды родов Arthonia, Peltigera). Такой тип
сумок также называют фисситуникатным, так как при вскры¬
тии сумки эндо- и экзоаск ведут себя независимо;
— сумки унитуникатного типа также имеют двухслой¬
ную клеточную стенку, однако оба слоя при вскрытии сумки
вытягиваются согласованно и ведут себя как один (такой ме¬
ханизм вскрытия - Teloschistes-тип - противоположен фисси-
туникатному и встречается, например, у видов родов
Baeomyces и Pertusaria) или реже не вытягиваются (эверсион-
ный механизм вскрытия, характерен для видов рода
Placynthiella);
i
Рис. 56. Основные типы сумок у лишайников: а - прототуникатный, б -
унитуникатный, в - леканоровый, г - битуникатный.
— сумки леканорового типа более всего напоминают
унитуникатные, однако после разрыва экзоаска эндоаск не от¬
рывается от него, но заметно вытягивается в короткую трубку
(хемифисситуникатный тип сумок), формируя рострум (канал
для выведения спор).
Сумки леканорового типа наиболее распространены у
лишайников и свойственны большинству представителей по¬
рядка Lecanorales. Именно для этого типа сумок характерно
развитие апикального аппарата или толуса (apical dome, tholus)
- сложно организованной структуры, являющейся апикальным
утолщением эндоаска (Рис. 57). Аскопласт (протопласт сумки)
может вдаваться апикально в собственно толус центральным
выпячиванием - окулярной камерой (ocular chamber), которая
может еще более проникать в толус посредством аксиального
канала (axial canal). Центральная часть толуса над окулярной
камерой носит название аксиального тела (axial body, axial
mass, apical cushion) и может быть окружена характерным
уплотнением - полисахаридной трубкой. Особенности строе¬
ния апикального аппарата и реакция его элементов с люголем
являются важными систематическими признаками уровня ро¬
дов, семейств и даже порядков.
Рис. 57. Верхняя часть сумки
леканорового типа: а - апи¬
кальный аппарат (толус), б -
аскопласт (протопласт сумки),
в - окулярная камера, г - акси¬
альный канал, д - аксиальное
тело.
В настоящее время известно большое число типов стро¬
ения апикального аппарата и реакции его компонентов с лю¬
голем. Такие типы носят названия согласно названиям родов,
для которых они характерны (например Bacidia-тип, Lecanora-
тип и другие, Рис. 58). Изображения значительного числа та¬
ких типов можно найти в литературе (см. например Hafellner,
1984; The Lichen..., 1992; Malcolm, Galloway, 1997).
Рис. 58. Некоторые типы строения апикального аппарата сумок лекано¬
рового типа: а - Biatora-тип, б - Bacidia-тип, в - Catillaria-тип, г - Fus-
cidea-тип, д - Lecanora-тип, е - Lecidea-тип, ж - Porpidia-тип, з - Rhizo-
carpon-тт, и - Rimularia-тип, к - Peltigera-тип, л -Xanthoria-тип.
В сумках лихенизированных сумчатых грибов в резуль¬
тате мейоза эндогенно формируется 8 (наиболее распростра¬
ненный вариант) или, реже, иное количество (от одной споры
до нескольких сотен) аскоспор. Зрелые аскоспоры микобион¬
та крайне разнообразны по форме, размерам (длина варьирует
от менее чем 2 мкм у некоторых видов рода Acarospora до бо¬
лее чем 300 мкм у некоторых Pertusaria) и количеству состав¬
ляющих их клеток (Рис. 59). В большинстве случаев аскоспо¬
ры лишайников не окрашены (например, у видов родов
Lecanora, Lecidea и у большинства представителей семейства
Parmeliaceae), однако представители целого ряда таксонов
имеют меланизированные споры (например, виды родов
Calicium и Cyphelium, многие виды рода Rhizocarpon).
Спора в процессе созревания в сумке окружена двумя
клеточными мембранами, в просвете между которыми форми¬
руется ее клеточная стенка. У зрелой споры клеточная стенка
состоит из трех или четырех основных слоев, которые в ряде
случаев можно разглядеть в световой микроскоп. Самым
внутренним, лежащим кнаружи от клеточной мембраны, явля¬
ется эндоспорий, развитый в аскоспорах представителей ряда
систематических групп лишайников (например, у видов рода
Buellia). Внешним по отношению к нему является эписпорий
(собственно стенка споры). Над эписпорием сильно меланизи-
рованных аскоспор бывает заметен тонкий промежуточный
слой, несущий пигмент. Поверхностный слой - периспорий -
развит у ряда таксонов и бывает хорошо дифференцирован на
плотную внутреннюю часть (ее гетерогенность определяет ор¬
наментацию поверхности спор) и аморфную слизистую внеш¬
нюю (способна при сильном развитии формировать заметную
в световой микроскоп слизистую оболочку или гало).
Рис. 59. Разнообразие строения зрелых аскоспор микобионтов лишайни¬
ков: а - Strangospora, б - Schaereria, в - Micarea, г - Japewia, д -
Lecania, е - Leptorhaphis, ж - Caloplaca, з - Rinodina, и - Calicium, к -
Pertusaria, л - Arthonia, м - Collema, н-п - Gyalecta, р - Porina, с - Scoli-
ciosporum, т - Bacidia, у - Arthotelium, ф - Polyblastia, х - Graphis, ц -
Phlyctis, ч - Diploschistes, ш - Belonia.
Перегородки между клетками аскоспор могут формиро¬
ваться как без участия слоев клеточной стенки, так и при уча¬
стии экзоспория и/или эндоспория. Обычно в перегородке
присутствует центральная пора, видимая только в электрон¬
ный микроскоп. У представителей семейств Physciaceae и
Teloschistaceae в перегородке есть центральный канал, кото¬
рый можно наблюдать в световой микроскоп. Просвет клетки
между перегородками и клеточной стенкой (полость клетки)
иногда называют люминой. Многие лишайники имеют про¬
стые (несептированные, без перегородок) аскоспоры, однако
многоклеточные споры также очень распространены во мно¬
гих систематических группах. Среди многоклеточных разли¬
чают поперечно многоклеточные (с одной или более попереч¬
ными перегородками) и муральные (кроме поперечных при¬
сутствует несколько продольных перегородок) споры. Под
субмуральными понимают споры, способные кроме попереч¬
ных перегородок формировать 1-2 продольные.
Конидиальные спороношения микобионта
Микобионт более чем половины всех известных видов
лихенизированных грибов формирует структуры, которые от¬
носят к конидиальным спороношениям. В конидиальных спо-
роношениях в результате митоза продуцируются конидии -
экзогенно формирующиеся неподвижные специализированные
споры с двухслойной клеточной стенкой, функции которых
могут заметно различаться в разных группах лишайников. Ве¬
роятно, наиболее распространенной является ситуация, когда
конидии являются пропагулами бесполого размножения мико¬
бионта. При этом формирование молодого таллома происхо¬
дит в результате естественного ресинтеза при контакте про¬
растающей конидии с подходящим фотобионтом, как и в слу¬
чае аскоспор. Однако у целого ряда таксонов (например, род
Cladonia) отмечено участие конидий в половом процессе в ка¬
честве «мужских» гамет - спермациев. В ходе полового про¬
цесса в форме спермациогамии спермаций копулирует с «жен¬
ским» органом - аскогоном через гифоподобный вырост аско-
гона - трихогину, в которую переходит ядро спермация.
Конидии формируются на специализированных кони-
диогенных клетках в результате специфического митотическо¬
го деления - конидиогенеза. У лишайников отмечен только
конидиогенез бластического типа, при котором будущая кони¬
дия начинает расти и заметно увеличивает свой объем еще до
отделения перегородкой от конидиогенной клетки. При талли-
ческом типе конидиогенеза, не известном у лишайников, бу¬
дущие конидии отделяются перегородками до начала увеличе¬
ния их объема.
При голобластическом онтогенезе формирование кле¬
точной стенки конидии идет за счет внешнего и внутреннего
слоев клеточной стенки конидиогенной клетки, тогда как при
энтеробластическом онтогенезе формирование клеточной
стенки конидии идет только за счет внутреннего слоя клеточ¬
ной стенки конидиогенной клетки с достройкой внешнего или
вовсе без участия клеточной стенки конидиогенной клетки
(достраиваются внутренний и внешний слои). В случае энтер-
областического развития каждая следующая конидия может
отделяться перегородкой от конидиогенной клетки несколько
дальше, чем располагалась перегородка ранее отделившейся
конидии (Рис. 60), в результате чего можно наблюдать коль¬
цеобразные телескопические рубцы - аннеляции - по числу
ранее образовавшихся конидий (конидиогенез аннелидного
типа) или такие рубцы незаметны, поскольку все конидии от¬
деляются от конидиогенной клетки в том же месте, где и самая
первая из них (конидиогенез фиалидного типа).
Г\
о
о
О
г\
> i
ь(
* \
А
f-\
о
О
Г\
п.
S L
А
А
Г\
а
51
...п
f\ Г"'
Рис. 60. Развитие конидий (конидиогенез) у лишайников: а - энтеробла-
стическое фиалидного типа, б - энтеробластическое аннелидного типа, в
- холобластическое.
Конидиогенные клетки у лишайникового микобионта,
как и у других грибов, обычно располагаются на специальных
разветвленных или неразветвленных гифах - конидиеносцах
(конидиофорах). Одиночные конидиеносцы редко удается об¬
наружить на талломе лишайника (гифомицетный тип, некото¬
рые виды рода Chaenotheca), чаще же они объединены в более
или менее сложные структуры - конидиомы. У лишайников
встречаются следующие типы конидиом (Рис. 61):
Пикниды (пикнидии) являются наиболее обычным ти¬
пом конидиом у лишайников. Они напоминают по форме
округлые или кувшиновидные перитеции и, также как и пери¬
теции, открываются наружу одной или несколькими остиола-
ми (устьицами). Внутри пикниды имеют одну или несколько
камер, в которых расположены направленные к центру или к
остиоле конидиеносцы. По положению относительно поверх¬
ности таллома пикниды могут быть полностью или в разной
степени погруженными, сидячими или даже приподнятыми
посредством ножек;
Рис. 61. Основные типы конидиом у лишайников: а - пикниды, б - спо-
родохий, в - синнемы, г - гифофоры, д - кампилидии.
Спородохии (ложа) - открытые подушковидно распро¬
стертые конидиомы, состоящие из массы коротких конидие-
носцев. Среди лишайников известны только у Micarea adnata.
Синнемы (коремии) - кистевидные приподнятые кони¬
диомы, состоящие из плотной массы длинных, иногда срос¬
шихся между собой гиф, формирующих ножку, на вершине
которой открыто располагаются конидиеносцы и конидии
(Dictyocatenulata alba). Иногда конидии могут быть обнаруже¬
ны и по бокам ножки;
Гифофоры (иногда рассматривают в качестве частного
случая или синонима синнем) - конидиомы на ножках или си¬
дячие, на или в которых формируются пучки диагиф. Форма
гифофоров может существенно различаться: наиболее обычны
кистевидные и щитковидные на ножках, а также сидячие или
прижатые к таллому чешуйчатые гифофоры, реже встречают¬
ся сферические или цилиндрические (тлазидии). Гифофоры
характерны для некоторых преимущественно тропических и
субтропических эпифильных лишайников (представители се¬
мейства Gomphillaceae, например виды рода Gyalectidium);
Кампилидии - шлемовидные полуоткрытые конидиомы,
внутри которых расположены конидиеносцы, прикрытые
сверху лопастью. Такие конидиомы характерны для нерод¬
ственных друг другу эпифильных тропических и субтропиче¬
ских лишайников (например, некоторых представителей родов
Calopadia, Byssoloma, Sporopodium). Иногда кампилидии мо¬
гут развиваться из пикнид (отдельные представители родов
Arthonia, Badimia), гифофоров (Ferraroa) и даже апотециев
(Badimia). Иногда содержимое кампилидий может быть ассо¬
циировано с клетками фотобионта, что, вероятно, может бла¬
гоприятствовать расселению вида.
Совершенно особняком в ряду конидиальных спороно-
шений лишайников стоят таллоконидии (таллоспоры) (Рис.
62). Они образуются на нижней поверхности талломов умби-
ликатно-листоватых лишайников (род Umbilicaria) или на
проталлюсе некоторых накипных (виды рода Protoparmelia) и
представляют собой гранулярные структуры, состоящие из
одной или нескольких сильно меланизированных толстостен¬
ных клеток микобионта. Их формирование предположительно
связано с деятельностью конидиогенных клеток и происходит
за счет выходящих на поверхность таллома слабоспециализи¬
рованных гиф.
Конидии большинства лишайников мелкие, одноклеточ¬
ные и одноядерные, бесцветные, округлые или вытянутой
формы, с гладкой поверхностью стенки, однако они могут
быть многоклеточными, окрашенными или сложной разветв¬
ленной формы. Иногда на одном талломе формируется 2 или
даже 3 типа конидий одновременно. Так у Micarea denigrata
(Fr.) Hedl. в различных типах пикнид формируются изогнутые
2-4-клеточные нитевидные макроконидии размерами 12-24 х
1 мкм, цилиндрические или яйцевидные одноклеточные мезо-
конидии 2.5-5 х 1-2 мкм и веретеновидные или бутылковид¬
ные одноклеточные микроконидии 4.5-7.5 х 0.5-1 мкм. В по¬
добных случаях макроконидии, вероятно, выполняют функ¬
цию спор бесполого размножения, тогда как микроконидии -
спермациев. Интересным случаем является распространение
диагиф в качестве пропагул бесполого размножения (у неко¬
торых эпифильных представителей семейства Gomphillaceae).
Диагифами называют более или менее разветвленные кониди¬
еносцы с цепочками конидий (Рис. 63), расположенные на или
внутри гифофоров апикально или субапикально. Диагифы
свисают пучками из-под щитковидных апикальных частей
гифофоров на ножках, иногда располагаются в сферических
или цилиндрических, чешуйчатых или прижатых к таллому
гифофорах без ножек. В некоторых случаях диагифы форми¬
руются открыто прямо на поверхности таллома. Нити диагиф
могут быть тонкими, без перетяжек, однако более распростра¬
нены монилиформные диагифы, клетки которых отделены
друг от друга перетяжками. Клетки диагиф могут быть ассо¬
циированы с клетками фотобионта, что способствует более
успешному расселению вида.
Рис. 63. Диагифа.
В ряде таксонов лишайников, образующих пикниды,
важным систематическим признаком являются особенности
строения конидиеносцев. Выделяют 8 основных типов кони-
диеносцев (Рис. 64) на основе расположения и формы кони-
диогенных клеток, их терминальной или интеркалярной лока¬
лизации на конидиеносцах, характера ветвления конидиенос¬
цев:
'Wm
•J ?
'■У//?*'
if
4Wy-r‘'i ifYf ;,.-f
' V L v" L'
VIII
Рис. 64. Основные типы конидиеносцев: I VIII.
I. Конидиогенные клетки бутылковидной формы, фор¬
мируются прямо на параплектенхимной выстилке внутренней
поверхности пикниды (конидиеносцы не формируются) и сла¬
бо отличимы от ее клеток. Этот тип представлен, например, у
видов родов Microcalicium и Mycocalicium;
II. Конидиогенные клетки вытянутые палочковидные,
формируются также практически прямо на плектенхимной вы¬
стилке пикниды (настоящие конидиеносцы не формируются),
но хорошо отличимы от ее клеток. Отмечен, например, у ви¬
дов родов Microcalicium и Peltigera;
III. Конидиогенные клетки вытянутые палочковидные,
формируются по нескольку на несептированной разветвлен¬
ной апикальной клетке короткого конидиеносца. Конидиоген¬
ные клетки и клетки конидиеносца хорошо отличимы от кле¬
ток выстилки пикниды. Такие конидиеносцы встречаются,
например, у видов родов Dirina и Roccella;
IV. Конидиогенные клетки располагаются терминально,
конидии формируются апикально. Конидиеносцы обильно
разветвленные, септированные, с явно просматривающейся
основной осью и боковыми ветвями. Встречается у видов из
родов Cladonia и Ramalina;
V. Конидиогенные клетки располагаются терминально,
однако небольшое число интеркалярных клеток также встре¬
чается, поэтому большинство конидий формируются апикаль-
но, но некоторые - латерально. Конидиеносцы также септиро-
ванные, но менее правильно ветвятся. Известен у видов родов
Cetraria и Cyphelium;
VI. Конидиеносцы септированные, разветвленные и ана-
стомозирующие. Конидиогенные клетки располагаются тер¬
минально, однако большее их число имеет интеркалярное по¬
ложение. Встречается у представителей семейств
Parmeliaceae, Physciaceae и Umbilicariaceae;
VII. Конидиеносцы образуют внутри пикниды плотную и
сильно разветвленную сеть, включающую ряды интеркалярно
расположенных конидиогенных клеток. Большинство кони¬
дий, таким образом, формируется латерально. Отмечен,
например, у представителей семейств Lobariaceae и
Physciaceae;
VIII. В зрелой пикниде отдельные конидиеносцы не про¬
сматриваются, ее полость заполнена параплектенхимой, со¬
держащей отдельные мелкие локулы. Локулы выстланы кони-
диогенными клетками, причем выделить интеркалярные и
терминальные среди них невозможно. Встречается у предста¬
вителей родов Dermatocarpon и Xanthoria.
Пикниды лишайников очень разнообразны, однако на
основании их анатомии, онтогенеза и характера конидиенос¬
цев (типы конидиеносцев) традиционно принято выделять 5
основных типов по названию родов, у представителей которых
они обнаружены:
Lecanactis-тип. Однокамерная более или менее погру¬
женная в таллом конидиома чашевидной формы, открываю¬
щаяся широким отверстием и неприкрытая сверху краями, что
отличает ее от настоящей пикниды. Тип конидиеносцев II. Из¬
вестен, например, в пределах родов Arthonia и Lecanactis;
Roccella-тип. Однокамерная пикнида яйцевидной фор¬
мы, открывающаяся узкой и хорошо оформленной остиолой.
Полость камеры заполнена конидиеносцами III типа и ните¬
видными конидиями. Встречается у представителей родов
Dirina и Roccella;
Lobaria-тип. Однокамерная пикнида шаровидной формы,
открывающаяся узкой и хорошо оформленной остиолой. По¬
лость камеры заполнена конидиеносцами VII типа и мелкими
конидиями. Встречается у представителей родов Lobaria и
Nephroma;
Umbilicaria-тип. Первично однокамерная, но способная с
возрастом становиться многокамерной (из-за разрастания ко¬
нидиеносцев) пикнида сливовидной или яйцевидной формы,
открывающаяся узкой и хорошо оформленной остиолой. Типы
конидиеносцев V или VI. Известен у ряда представителей се¬
мейства Parmeliaceae;
Xanthoria-тип. Первично многокамерная пикнида, каме¬
ры которой закладываются еще в раннем онтогенезе. Камеры
открываются в общую полость пикниды по мере созревания,
остиола общая. Тип конидиеносцев VIII. Встречается у пред¬
ставителей родов Dermatocarpon и Xanthoria.
Литература
Бязров Л. Г. Эпифитные лишайники г. Москвы: совре¬
менная динамика видового разнообразия. М.: КМК, 2009. 146
с.
Голубкова Н. С. Анализ флоры лишайников Монголии.
Л.: Наука, 1983. 248 с.
Голубкова Н. С., Бязров Л. Г. Жизненные формы лишай¬
ников и лихеносинузии // Ботанический журнал. 1989. Т. 74, №
6. С. 794-805.
Жизнь растений. Т. 3. Водоросли. Лишайники / Под. ред.
М. М. Голлербаха. М.: Просвещение, 1977. 487 с.
Каратыгин И. В. Коэволюция грибов и растений // Труды
Ботанического института им. В. Л. Комарова. Вып. 9. СПб.:
Гидрометеоиздат, 1993. С. 1-119.
Кутафьева Н. П. Морфология грибов. Новосибирск: Си¬
бирское университетское издательство, 2003. 215 с.
Мир растений. Т. 2. Грибы / Под. ред. М. В. Горленко.
М.: Просвещение, 1991. 475 с.
Определитель лишайников СССР / Под ред. И. И. Абра¬
мова. Л.: Наука, 1974. Вып. 2. 284 с.
Aguirre-Hudson В. A taxonomic study of the species re¬
ferred to the ascomycete genus Leptorhaphis // Bull. Brit. Mus.
Nat. Hist. Botany. 1991. Vol. 21. P. 85-192.
Ahmadjan V. The lichen symbiosis. New York: Jhon Wiley
& Sons, 1993. 250 p.
Ainsworth & Bisby’s Dictionary of the Fungi / Ed. by P. M.
Kirk, P. F. Cannon, D. W. Minter, J. A. Stalpers CAB Internation¬
al, 2008 (10th edition). 771 p.
Brodo I. М., Duran-Sharnoff S., Sharnoff S. Lichens of
North America. New Haven & London: Yale University Press,
2001. 795 p.
BUdel B., Scheidegger C. Thallus morphology and anatomy
// Lichen Biology / Ed. by Т. H. Nash III. Cambridge: Cambridge
University Press, 2008 (2th edition). P. 40-68.
Cannon P. F., Kirk P. M. Fungal Families of the World. CAB
International, 2007. 455 p.
Clauzade G., Roux C. Likenoj de Okcidenta Eflropo. Ilustrita
Determinlibro // Bull. Soc. Bot. du Centre-Ouest, Nouvelle Serie.
1985. Numero Special 7. 892 p.
CRC Handbook of Lichenology. Vol. 1-3 // Ed. by M. Ga-
lun. Boca Raton: CRC Press, 1988.
Flechten: Geschichte, Biologie, Systematik, Okologie, Na-
turschutz und kulturelle Bedeutung // Ed. by H. Scholler. Frankfurt
am Main: Kramer, 1997. 247 p.
Galloway D. J. Flora of New Zealand lichens: Lichens, in¬
cluding lichen-forming and lichenicolous fungi. Vol. 1-2. Lincoln,
2007. 2262 p.
Goward T. The lichens of British Columbia, illustrated keys.
Part 2. Fruticose species. // Special Report Series 9. Victoria, 1999.
319 p.
Goward Т., McCune B., Meidinger D. The lichens of British
Columbia, illustrated keys. Part 1. Foliose and Squamulose species
// Special Report Series 8. Victoria, 1994. 181 p.
Hafellner J. Studien in Richtung einer natUrlicheren Gleide-
rung der Sammelfamilien Lecanoraceae und Lecideaceae // Beih.
Nowa Hedwigia. 1984. Bd. 79. S. 241-371.
Hale М. E. The Biology of Lichens. London: Edward Ar¬
nold, 1983 (3th edition). 190 p.
Hawksworth D. L. The variety of fungal-algae symbioses,
their evolutionary significance and the nature of lichens // Botan.
Journ. Linn. Soc. 1988. Vol. 96. P. 3-20.
Hawksworth D. L., Hill D. J. The Lichen-Forming Fungi.
Glasgow & London: Blackie, 1984. 158 p.
Honegger R. Lichen-Forming Fungi and Their Photobionts //
Plant Relationships. The Mycota V. / Ed. by H. Deising. Berlin,
Heidelberg: Springer-Verlag, 2009 (2th edition). P. 308-333.
Horizons in Lichenology // Botan. Journ. Linn. Soc. 1988.
Vol. 96. / Ed. by D. H. Dalby, D. L. Hawksworth, S. L. Jury. Lon¬
don: Linnean Society of London and Academic Press, 1988. P. 1-
100.
Kershaw K. A. Physiological ecology of lichens. Cambridge:
Cambridge University Press, 1985. 293 p.
Lichen Biology / Ed. by Т. H. Nash III. Cambridge: Cam¬
bridge University Press, 1996 (1th edition). 303 p.
Lichen Biology / Ed. by Т. H. Nash III. Cambridge: Cam¬
bridge University Press, 2008 (2th edition). 486 p.
Lichen Flora of the Greater Sonoran Desert Region. Vol. 1-
3. Arizona: Arizona State University, 2002-2007.
Malcolm W. М., Galloway D. J. New Zealand Lichens:
Checklist, Key, and Glossary. Wellington: Museum of New Zea¬
land Те Papa Tongarewa, 1997. 192 p.
Nordic Lichen Flora. Vol. 1-4. Uddevalla, 1999-2011.
Orange A., James P. W., White F. J. Microchemical methods
for the identification of lichens. British Lichen Society, 2001. 101
P-
Poelt J. Die lobaten Arten der Flechtengattung Lecanora
Ach. sensu. ampl. in der Holarktis // Mitt. Bot. StSamml. Mun-
chen. 1958. Bd. 2. S. 411-589.
Protocols in lichenology. Culturing, Biochemistry, Ecophy-
siology and Use in Biomonitoring / Ed. by Kranner I., Beckett R.
P., Varma A. K. Berlin: Springer-Verlag, 2002. 580 p.
Rambold G., Triebel D. The inter-lecanoralean associations
// Bibl. Lich. 1992. Vol. 48. P. 1-201.
Rikkinen J. What’s behind the pretty colors? A study on the
photobiology of lichens // Bryobrothera. 1995. Vol. 4. P. 1-239.
Stenroos S., Ahti Т., Lohtander K., Myllys L. Lichen flora of
Finland // Norrlinia. 2011. Vol. 21. 534 p.
The Lichen Flora of Great Britain and Ireland / Ed. by Purvis
O. W., Coppins B. J., Hawksworth D. L., James P. W., Moore D.
М.. London, 1992. 493 p.
The Lichens of Great Britain and Ireland / Ed. by Smith C.
W., Aptroot A., Coppins B. J., Fletcher A., Gilbert O. L., James P.
W., Wolseley P. A. London, 2009. 1046 p.
Yoshimura I., Shimada R. Fine structures of lichen plecten-
chymas viewed with the scanning electron microscope // Bulletin
of Kochi Gakuen Junior College, 1980. Vol. 11. P. 13-28.
Глава IV.
Водоросли-фотобионты лишайников
Войцехович А. А., Надеина О. В.
Разнообразие фотобионтов
Несмотря на стремительное развитие лихенологии, как
науки в целом, водоросли, входящие в состав лишайников,
остаются недостаточно исследованными. В настоящее время
точное определение большинства фотобионтов требует обяза¬
тельного выделения фотобионтов в культуру, морфологиче¬
ских исследований культурального материала, сравнений с
аутентичными штаммами, а также подтверждения результатов
идентификации молекулярно-филогенетическими методами. В
то же время, в определителях лишайников (например, The
Lichen Flora of Great Britain..., 1992; Окснер, 1993) и водорос¬
лей (например, Мошкова, 1979; Андреева, 1998; Ettl, Gartner,
1995) для большинства фотобионтов приведены описания и
иллюстрации, составленные на основании исследований при¬
родного или гербарного материала. В связи с этим, даже в уз¬
коспециализированных обзорных работах, посвящённых ис¬
следованиям фотобионтов (Tschermak-Woess, 1988; Ahmadjian,
1993; Friedl, Budel, 2008), сведения о разнообразии последних
и корреляции их с микобионтом остаются весьма неполными,
а по некоторым позициям - противоречивыми и неточными.
Широкое использование молекулярно-филогенетических ме¬
тодов способствовало заметному прогрессу в исследованиях
фотобионтов лишайников и привело к накоплению значитель¬
ного количества новых данных, которые нуждаются в анализе
и обобщении.
В последних обобщающих работах (Войцехович и др.,
2011 а, б)1 приводится 148 видов лишайниковых фотобионтов
из 50 родов. Однако, в связи с интенсивным развитием
направления исследования фотобионтов, уже сегодня эту циф¬
ру можно считать несколько устаревшей. Так, согласно нашим
подсчётам, число водорослей, известных в качестве фотобион¬
та лишайников составляет 155 видов из 55 родов, которые от¬
носятся к 5 отделам: Chlorophyta - 114 видов (73.6 % от обще¬
1 В работах проанализировано 210 литературных источников, в которых
приводятся точные данные о составе фотобионтов 670 видов лишайни¬
ков, что составляет около 3 % мирового разнообразия лишайникообра¬
зующих грибов. Данные о составе фотобионтов из определителей ли¬
шайников сознательно учтены не были из-за их неточности, а порой и
сомнительности.
го разнообразия фотобионтов), Cyanophyta (= Cyanoprokaryota)
- 35 (22.6 %), Streptophyta - 3 (1.9 %), Xanthophyta - 2 (1.3 %),
Phaeophyta - 1 (0.6 %).
Большинство зелёных фотобионтов относятся к классу
Trebouxiophyceae, родам Asterochloris Tscherm.-Woess (рис.
65а), Соссотуха Schmidle (рис. 656), Dictyochloropsis Geitler
em. Tscherm.-Woess, Diplosphaera Bial., Elliptochloris (рис. 65в)
Tscherm.-Woess, Gloeocystis Nageli, Myrmecia Printz (рис. 65r),
Stichococcus Nageli и, конечно, Trebouxia Puym. (рис. 65д). Ре¬
же встречаются представители класса Ulvophyceae - Blidingia
Kylin, Cephaleuros Kun. ex Fries, Dilabifdum Tscherm.-Woess,
Phycopeltis Mill., Printzina Thompson et Wujek и Trentepohlia C.
F. P. Martius (рис. 65e). В отдельных случаях в составе лишай¬
ников встречаются водоросли класса Chlorophyceae -
Chlamydomonas Ehrenberg, Chlorosarcinopsis Herndon,
Neocystis F.Hindak, Radiococcus Schmidle, Trochiscia KUtz. От¬
дел Streptophyta представлен всего двумя родами Interfdum
R. Chodat и Mesotaenium Nageli, обнаруженными в лишайни¬
ках в качестве дополнительных фотобионтов.
Около четверти разнообразия фотобионтов приходится
на Cyanoprokaryota, среди которых представлены хроокок-
кальные и ностокальные виды, способные к фиксации атмо¬
сферного азота. К наиболее распространённым цианобионтам
относятся Calothrix С. Agardh ex Bornet et Flahault,
Chroococcidiopsis Geitler, Gloeocapsa KUtz., Nostoc Vaucher ex
Bornet et Flahault, Scytonema C.Agardh ex Bornet et Flahault,
Stigonema C. Agardh ex Bornet et Flahault.
В единичных случаях как фотобионты приводятся жел-
тозелёные водоросли Heterococcus caespitosus Visch. и
Xanthonema montanum (Visch.) Silva (Tschermak, 1941; Zeitler,
1954), а также бурая водоросль Petroderma maculiforme
(Wollng.) Kuck (Мое, 1997). Все три указанных вида были
найдены в слоевищах лишайников семейства Verrucariaceae.
Следует отметить, что некоторые виды родов
Asterochloris, Соссотуха, Trebouxia и, вероятно,
Dictyochloropsis, пока известны исключительно как компонен¬
ты лишайников и в свободноживущем состоянии не найдены.
Такие виды водорослей относятся к облигатным фотобионтам.
На сегодняшний день известны лишь их единичные находки в
свободноживущем состоянии (Garty, 1990; Mikhailyuk et al.,
2003; KocTiKOB и др., 2001; Дар1енко и др., 2004; Войцехович,
2008 и др.), однако их достоверность весьма сомнительна из-за
определённых методических сложностей. Все другие водорос¬
ли, выступающие партнёрами лишайниковой ассоциации, яв¬
ляются также свободноживущими наземными видами и отно¬
сятся к факультативным фотобионтам (Войцехович и др., 2011
а).
Рис. 65. Микрофотографии наиболее распространённых зелёных водо-
рослей-фотобионтов: а - Asterochris excentrica, б - Соссотуха sp., в -
Elliptochloris biloobata, г - Myrmecia biatorellae, д - Trebouxia crenulata,
e - Trentepohlia sp.
Однако, вопрос облигатности/факультативности среди
фотобионтов не так прост, как кажется на первый взгляд. Су-
ществует достаточно примеров водорослей, первоначально
описанных из лишайника, и найденных позднее в свободно¬
живущем состоянии. Классическими примерами такой ситуа¬
ции являются Myrmecia biatorellae, описанная из лишайника
Biatorella simplex (Taylor) Branth & Rostr. как Myrmecia
pyriformis Tscherm.-Woess (Tschermak-Woess, Plessl, 1948);
Elliptochloris bilobata - описанный из Catolechia wahlenbergii
(Ach.) Flot. (Tschermak-Woess, 1980), а также Diplosphaera
chodatii Bial. em. Vischer - из Lecanora tartarea (L.) Ach.
(Vischer, 1960). Позднее эти виды были были приведены в ка¬
честве широко распространённых наземных водорослей (Ан¬
дреева, 1998; Кост1ков и др., 2001; Mikhailyuk et al., 2003;
Войцехович и др., 2009).
За последние несколько десятилетий в систематике во¬
дорослей произошли значительные изменения, были проведе¬
ны многочисленные номенклатурные комбинации, описаны
нове виды и роды. В связи с этими изменениями лихенологам,
не специализирующимся в исследованиях фотобионтов, тяже¬
ло распознавать фотобионты, которые в разное время приво¬
дились под несколькими разными названиями. Так, например,
под хорошо известными лихенологам, хоть и весьма устарев¬
шими названиями Protococcus dermatocarponis miniati Zeitler и
Hyalococcus dermatocarponis Waren., скрываются, соответ¬
ственно, Desmococcus olivaceus и Diplosphaera chodatii, а под
загадочным названием «микареоидный фотобионт» подразу¬
меваются виды рода Elliptochloris. Водоросль, известная ранее
под названием Myrmecia reticulata, сегодня относится к роду
Dictyochloropsis (D. reticulata). Однако, наиболее сложной, на
наш взгляд, является ситуация с представителями рода
Trebouxia sensu lato. В наиболее ранних работах мы находим
их под названием Cystococcus humicola, в более поздних -
Trebouxia и Pseudotrebouxia. Впрочем, совсем недавно, на ос¬
новании результатов исследования жизненных циклов, мор¬
фологических и молекулярно-филогенетических признаков,
часть видов данного рода была переведена в род Asterochloris
(Skaloud, Peksa, 2010). В лихенизированном состоянии заме¬
тить отличия между представителями этих двух родов практи¬
чески невозможно, т.к. различия проявляются только после
нескольких недель культивирования. Таким образом, в насто¬
ящее время в лихенологии назрела острая необходимость с
большей внимательностью подходить к вопросу определения
фотобионтов и, по возможности, принимать во внимание из¬
менения, произошедшие в систематике и таксономии водорос¬
лей за последние декады.
К сожалению, ключи для определения фотобионтов,
приведённые в определителях лишайников (например, Окснер,
1974), являются достаточно запутанными, в большинстве сво¬
ём содержат устаревшие или непроверенные данные, что по¬
рою приводит к невозможности определения фотобионта. В
некоторых случаях для одного лишайника приводятся не¬
сколько совершенно разных родов фотобионта, которые при
тщательной проверке оказываются всего лишь синонимами.
Потому, для облегчения определения фотобионтов, мы пред¬
лагаем использовать графические ключи для определения ци¬
анопрокариотических (Рис. 66) и эукариотических (Рис. 67)
фотобионтов. В таблицы включены схематические изображе¬
ния наиболее распространённых водорослей-фотобионтов с их
новыми и старыми названиями, а также примеры лишайников,
в состав которых входят указанные фотобионты.
Корреляция между таксономической принадлежностью
мико- и фотобионта
Долгое время считалось, что каждому виду лишайника
соответствует отдельный вид фотобионта, однако позже было
выяснено, что разнообразие водорослей-фотобионтов значи¬
тельно ниже разнообразия лишайникообразующих грибов, по¬
скольку, один и тот же вид фотобионта может встречаться в
разных лишайниках (Tschermak-Woess, 1988). Более того, со¬
став фотобионтов одного и того же вида лишайникообразую¬
щего гриба может быть непостоянным или включать несколь¬
ко видов или даже родов фотобионтов одновременно (Geitler,
1955; Voytsekhovich et al., 2011; Войцехович, 2012). За послед¬
ние несколько лет наметилась общая тенденция в изучении
коэволюционных взаимоотношений лишайникообразующих
грибов и их фотобионтов с помощью молекулярно¬
филогенетических методов. На сегодняшний день в этом от¬
ношении довольно хорошо изучены семейства Physciaceae
(Bhattacharya et al., 1996; Dahlkild et al, 2001; Helms, 2003),
Cladoniaceae (Piercey-Normore, DePriest, 2001), Teloschistaceae
(Beck, 2002; Honegger et al., 2004; Nyati, 2006), Graphidaceae
(Nakano, 1988), Verrucariaceae (ThUs et al., 2011), а также роды
Letharia (Th.Fr.) Zahlbr. (Kroken, Taylor, 2000), Lecanora Ach.
(Blaha et al., 2006), Umbilicaria Hoffm. (Romeike et al., 2002).
Отдельные работы посвящены исследованию разнообразия
трентеполиальных фотобионтов определённых видов лишай-
ников, принадлежащих разным родам и семействам (Meier,
Chapman, 1983; Rands, Davis, 1993; Nelsen et al., 2011).2
ЦианфпрокариОтичССкиС фотобКОНТЫ
t,
v 5
Ё Sf
Щр
Ф
m
I
Ия LuaicHr
NuSUM':
Gtococopsa
ftyciia
< HHDHH4
щ [ "h [ViiK ft: С к] i й [H i i я-
Наа.чн с
я cTk i in;
f WJriru. i j-ftlrrfcitun.
S.-aixi rid (ОтДигм-ииЛзл
Stphroma. t'лчмгц.
Гг'{^гг,7. lT4-JTVJr4iH/<''T
Лагяш. A'umj.'Ili.
1, SifftpiO, ^LVdn-Jj,
{■‘cinla. PiwvtKbiii.
FysnrujHJi. ,'7_1 П1Д1
.ViffWlh/rt*
Arihopyrf*\ii
h
II
'iLb
%
M
4:|
■^--1 ■* '
4^-
ITftiuHat
Seyfonvmtt
Stiganema
CalutHrix
( И НОННЧ
-I.H-cbaLhri* -
Наа.чн с
k. c^k-i ah;
f чнччн-иrpiu, licpftiu
iJlETrtWlfJHd, i'Wrii/d,
ftmwrw, Piotopsn,
Stcnocmiton
i.fctda1. Ffacopus,
Xpi{iV\(M4T. ■5(rrrc\iM‘iw
i VpiiTnUrrjriiT. iirxirlUi.
X.riJtlrtd. Il0SOi,y|DlllkJl
Гул flAlJ b'iW.
Рис 66. Графический ключ для определения прокариотических фото¬
бионтов лишайников.
2 Более подробные данные об основных семействах лишайников и их
фотобионтах представлены в работе Войцехович и др., 2011 а, б.
)} КЯ|>1101 H'llTKIIl' 1
№ i uGiiuii f ы
S н
?• ■
\ 1
©
0
о
t ■ -J
■ 4
и
ш Е
£ ^
4$
VF . ■ ■-/
*4*-
(§>
7T.ruiK«Ji'
ВДтлгпн
JJ 1C fh.- iflruwip ч гл
f.tl/pt^K Marti
С
тН Silu.inridS-
- Ч7 |-»*n 11'
-F4H irnid1
R"* 1И«
ll^rTLUO. (
ifdMAm f 1Ы.
'liwAffM
l! Л Н^Та'Л 11Ш
Г ptfH
h
ч
9
%
€>■ 3
If
ад
iPaiajaw
^i|fFT4*ik/«ynT
FmKtMMCctts
i'rjL I'ft.V-at iTbil
Ггеп/гроЛЛа
1 ЫМУЧ
- 1 Г Г !"■>* LU ■*
il.h.n'.Un-
-1 ЬгййЪрЬът
ILi* |уь
а. г«ш
•! ^fHWrUIV
Mrm mtm
№inihrih
С
<ьЫ ■ Гь^А |> L|*
ГЛЪНЪЛ. l"i*fcV|'IU
Jhnu. I'irjiiii.
tJprftipb*. Лл ггРл.
V
я
1 3
я 4
И
w ■
<Й
Оэ
(£0
/Jt\0
'B
ILnu«»-r
DiptvspHucra
/'^глпРО'-НЧ'-НТ
( <ЧЧ'1т|ДО
CAkwifa
■ |1‘рЬч.'Чччп-*
■Гг^'Ч¥Ч1 п ■
- Гьгвйс-:Н1*ггЯл-
IL'i* 1 IIP
a. i‘*i‘ 1ы
i r ГМ
.W^rud
Рис. 67. Графический ключ для определения эукариотических фотобио¬
нтов лишайников.
Центральным классом, включающим наибольшее число
видов лишайников, является Lecanoromycetes (Ainsworth and
Bisbey's Dictionary of fungi..., 2008). Значительная часть его
видов, слагающих семейства Thelotremataceae (Ostropales),
Pertusariaceae (Pertusariales), Lecanoraceae и Parmeliaceae
(Lecanorales), Physciaceae, Teloschistaceae и Rhizocarpaceae
(Teloschistales), Candelariaceae (Candelariales), а также
Umbilicariaceae (Umbilicariales), ассоциирована с Trebouxia.
Часть видов, относящихся к семействам Cladoniaceae и
Stereocaulaceae (Lecanorales) - с Asterochloris. В целом,
Trebouxia и Asterochloris являются наиболее распространён¬
ными фотобионтами лишайников, однако для некоторых клас¬
сов лишайников они всё же не являются типичными. Так, ви¬
ды класса Arthoniomycetes вступают в симбиоз с трентеполи-
альными водорослями, Lichinomycetes - с цианопрокариотами.
Виды базидиальных лишайников ассоциируются с цианопро¬
кариотами (Scytonema) и зелёными водорослями (Соссотуха
Schmidle, Gloeocystis Nageli, Mesotaenium Nageli и
Pseudococcomyxa Korschikov). Наиболее разнообразен состав
фотобионтов семейства Verrucariaceae (Eurotiomycetes), среди
которых отмечены зелёные (.Diplosphaera, Dilabifilum
Tscherm.-Woess, Myrmecia Printz, Stichococcus и др.), бурые
(.Petroderma Kuck.) и желтозелёные (Xanthonema P.С. Silva,
Heterococcus Chodat) водоросли. Представители семейств
Coenogoniaceae, Graphidaceae, Gyalectaceae и Porinaceae
(Ostropales) формируют ассоциации преимущественно с трен-
теполиальными водорослями. Однако, есть и семейства, у
представителей которых состав фотобионтов очень разнообра¬
зен и часто различные образцы одного и того же вида лишай¬
ника, содержат разные фотобионты. К таким семействам от¬
носятся Acarosporaceae (Acarosporales), Agyriaceae (Agyriales),
Pilocarpaceae, Ramalinaceae и Stereocaulaceae (Lecanorales), a
также Lecideaceae (Lecanoromycetidae families incertae sedis).
Интересен в этом отношении порядок Peltigerales, содержащий
лишайники, ассоциированные в основном с цианобионтом. В
семействах Lobariaceae, Nephromataceae и Peltigeraceae циа-
нобионт дополняется зелёным фотобионтом, роль которого
выполняют виды родов Dictyochloropsis (Lobariaceae) или
Соссотуха и Pseudococcomyxa (Nephromataceae и
Peltigeraceae). Таким образом, лишайники этих семейств со¬
держат 2 фотобионта, один из которых образует фотобионт-
ный слой лишайника, а другой находится в специализирован¬
ных структурах - цефалодиях. У представителей рода Lobaria
зелёная водоросль формирует фотобионтный слой, а циано-
бионт находится в цефалодиях. У некоторых представителей
рода Peltigera Willd., наоборот, цианобионт, находится в слое¬
вище, а зелёный фотобионт - в цефалодиях. Однако, в литера¬
туре отмечалось, что в результате вегетативного размножения
возможно формирование лишайника с зелёным фотобионтом в
слоевище и с цианобионтом в цефалодиях (Ahmadjian, 1993).
Такие варианты одного и того же вида, называемые фотосим-
биодемами, имеют различный цвет слоевища, но представля¬
ют собой один и тот же вид лишайника.
Таким образом, микобионт, как правило, проявляет из¬
бирательность по отношению к фотобионту, потому опреде¬
лённую корреляцию между мико- и фотобионтом можно найти
уже на уровне классов, порядков и семейств лишайников. В то
же время, уровни избирательности микобионта у разных групп
лишайников могут быть разными. В работе А. Бэка с соавто¬
рами (Beck et al., 2002) представлено 5 уровней избирательно¬
сти микобионта лишайников. К I (очень высокому) и II (высо¬
кому) уровням отнесены лишайникообразующие грибы, фор¬
мирующие ассоциацию с одним штаммом или с одним видом
водоросли-фотобионта. В качестве примеров лишайников с
такими уровнями избирательности можно привести Xanthoria
parietina (L.) Th. Fr., микобионт которого формирует слоевище
с Trebouxia arboricola Puym. или с видами подгруппы «Г. аг-
boricola» (Nyati, 2006); изученные виды рода Pertusaria DC.,
ассоциирующиеся с Т. potteri Ahmadjian ex Gartner (Ahmadjian,
1993) и Umbilicaria - с T. simplex Tscherm.-Woess (Beck, 2002;
Romeike et al., 2002). К лишайникообразующим грибам III
(среднего) уровня избирательности относятся те, которые
формируют постоянные ассоциации с разными видами одного
рода водоросли-фотобионта. Этот уровень характерен для ви¬
дов Cladonia P. Browne (ассоциированы с Asterochloris
(Ahmadjian, 1993; Piercey-Normore, DePriest, 2001)), Megalo-
spora Meyen (c Dictyochloropsis (Tschermak-Woess, 1984)), Col-
lema F. H. Wigg. (c Nostoc (Degelius, 1954)) и др. Этот уровень
избирательности, очевидно, также характерен для цефалоди-
альных лишайников, например, для Peltigera aphtosa (L.)
Willd., который формирует ассоциацию с видами Соссотуха
(Jaag, 1933) и Nostoc (O'Brien et al., 2005).
IV (низкий) и V (очень низкий) уровни характерны для
лишайников, избирающих фотобионты в пределах одного се¬
мейства или порядка (IV) или групп высшего таксономическо¬
го уровня (V). Примерами низкой избирательности могут слу¬
жить Stereocaulon ramulosum Rauschel (ассоциируется с вида¬
ми родов Gloeocapsa KUtz., Nostoc, Scytonema, Stigonema
(Lamb, 1951), или виды Coenogoniaceae, Graphidaceae,
Rocellaceae (формируют слоевище с представителями
Trentepohliaceae (Santesson, 1952; Uyenko, 1965; Meier,
Chapman, 1983). Очень низкий уровень избирательности фото¬
бионта характерен для видов Verrucaria Schrad., которые ассо¬
циируются с Dilabifilum prostratum (Ettl, Gartner, 1995),
Diplosphaera chodatii (Geitler, 1960), Heterococcus caespitosus
(Tschermak, 1941; Zeitler, 1954; Sanders, 2004), Petroderma
maculiforme (Мое, 1997; Sanders, 2004). Кроме того, известны
лишайники, например базидиолишайник Multiclavula mucida
(Pers.) R. H. Petersen, которые могут содержать несколько фо¬
тобионтов одновременно - виды Mesotaenium, Соссотуха,
Gloeocystis (Geitler, 1955).
Наивысшим уровнем избирательности к фотобионту об¬
ладают преимущественно лишайникообразующие грибы, ас¬
социированные с облигатными фотобионтами. Само явление
избирательности микобионта легко объясняется различными
физиолого-биохимическими и экологическими особенностями
водорослей, которые значительно отличаются у цианопрока¬
риотических и эукариотических представителей (Войцехович
и др., 2011 б), потому многие лишайникообразующие грибы
проявляют избирательность, в первую очередь, в рамках при¬
надлежности фотобионтов к определённой таксономической
группе водорослей. Высокая степень избирательности фото¬
бионта, вероятно, указывает на высокую специализацию сим¬
биотических отношений у этих лишайников и является след¬
ствием достаточно продолжительной коэволюции обоих ком¬
понентов лишайниковой ассоциации.
Лишайники с разными фотобионтами, в определённой
степени различаются своими эколого-географическими осо¬
бенностями, а также, отчасти, морфолого-анатомическим
строением слоевища. Так, лишайники с цианобионтом, перио¬
дически нуждающиеся в капельно-жидкой воде (Honegger,
1991, 2006) произрастают во влажных местообитаниях, где
скапливается вода - уступы, выемки скал, почва и т.д. (Lange
et al., 1998). Значительная часть лишайников с цианобионтом
характеризуется гомеомерным типом слоевища, в котором во¬
доросли не образуют оформленного слоя, а беспорядочно раз¬
бросаны по слоевищу и окутаны гифами. У таких гомеомер-
ных слоевищ коровой слой либо совсем отсутствует, либо вы¬
ражен слабо. Однако у цианолишайников встречается и другой
тип слоевища - гетеромерный, как, например, у Peltigera. Ви¬
ды этого рода имеют сложно устроенное слоевище, с разви¬
тыми коровыми слоями и оформленным слоем фотобионта
(Vitikainen, 1994). Учитывая, что цианопрокариоты и зелёные
водоросли имеют различную стратегию получения воды
(Lange et al., 1998), вероятно, эта группа лишайников проблему
водного баланса «решила» вполне успешно, используя в раз¬
ных условиях тот фотобионт, который наиболее подходит для
успешного существования в тех или иных условиях окружаю¬
щей среды. Несмотря на то, что в классических работах по ли¬
хенологии представители Peltigerales, наряду с остальными
цианолишайниками, считаются одними из наиболее древних,
есть мнение, что данная группа является достаточно молодой
и прогрессивной (Goward, 2009). Очевидно, именно благодаря
сложной организации слоевища, объединяющего в себе двух
фотобионтов с принципиально разными свойствами и страте¬
гиями, представители Peltigerales смогли приспособиться к
условиям меняющейся среды обитания.
Достаточно простая организация слоевища также харак¬
терна для многих лишайников с трентеполиальным фотобион¬
том. Так, у некоторых родов (Coenogonium Ehrenb., Racodium
Fr.) формируется лишь футляр гиф, оплетающих нити водо¬
рослей (Honegger, 2008). Лишайники с трентеполиальными
фотобионтами наиболее разнообразны в местах с влажным
тропическим климатом, где также находится центр разнообра¬
зия свободноживущих представителей Trentepohliales (Rindi et
al., 2009). Эти лишайники колонизируют кору и листья тропи¬
ческих растений, часто являются эндофитами и эндопаразита¬
ми высших растений, а также образуют разрастания на скалах
и фасадах зданий (Lakastos et al., 2006).
Многие лишайникообразующие грибы, вступающие в
симбиоз с зелёными водорослями (за исключением Trebouxia
и Asterochloris) или водорослями других групп, характеризу¬
ющиеся низкой избирательностью к фотобионту, также обра¬
зуют неоформленное слоевище. Так, слоевище большинства
базидиальных лишайников представляет собой мицелий,
оплетающий клетки водорослей (Oberwinkler, 1984). Гетеро-
мерное слоевище в наиболее простых своих вариантах может
быть характерным для некоторых лишайников с трентеполи¬
альным (Graphis Adans., Dirina Fr., Roccella DC.) или зелёным
фотобионтом. Однако, наиболее сложно устроенное гетеро-
мерное слоевище характерно для лишайников, содержащих
Trebouxia и Asterochloris. Исключение составляют лишь неко¬
торые цефалодиальные лишайники. При гетеромерном типе
слоевища слой фотобионта четко очерчен, имеются в разной
степени развитые коровые слои, а также сердцевинная часть и,
часто, специальные органы прикрепления к субстрату. Уро¬
вень симбиотических отношений у таких лишайников также
следует считать высокоорганизованным, по сложности близ¬
ким к эктомикоризе высших растений и базидиальных грибов
(Honegger, 1991). Такие симбиотические отношения являются
следствием длительного совместного проживания водоросли и
гриба (Honegger, 1991). Лишайники с Trebouxia, как правило,
колонизируют поверхности скал, хотя ряд видов растёт на ко¬
ре деревьев, а реже - на почве. Эти лишайники распростране¬
ны очень широко и колонизируют субстраты не только уме¬
ренной зоны, но также аридных и холодных регионов
(Galloway, 2008).
Представители Cladoniaceae также характеризуются до¬
статочно высокой степенью избирательности микобионта, ас¬
социированного с облигатным фотобионтом Asterochloris. Эти
лишайники образуют сложно устроенные (от чешуйчатого до
кустистого) слоевища и распространены, как правило, на поч¬
ве, куртинах мхов или у основания стволов деревьев.
Таким образом, состав фотобионтов лишайниковой ас¬
социации, как правило, не является случайным, а тесно корре¬
лирует с определёнными группами лишайникообразующих
грибов. Лишайники различных полифилетических линий про¬
являют разную избирательность к фотобионту, а симбиотиче¬
ские отношения в ассоциации находятся на разных ступенях
развития. Наиболее высоких ступеней специализации и спе¬
цифичности фотобионта, очевидно, достигли лишайники с
Trebouxia и Asterochloris.
Источники фотобионтов. Основные, дополнительные и
«потенциальные» фотобионты
Вопросы о том, каково происхождение фотобионтов, что
является источником фотобионта и существует ли какой-либо
обмен фотобионтами среди популяций лишайников, были
впервые подняты более 20 лет назад (например, Friedl, 1987;
Ott, 1987; Ihda et al., 1993; Helms, 2003), но до сих пор остают¬
ся нераскрытыми в полном объёме. Как было показано выше,
для большинства лишайников характерна определённая кос-
пецифичность компонентов, что обусловливает присутствие в
слоевище лишь одного определённого вида фотобионта. Меж¬
ду тем, как уже упоминалось ранее, известны случаи, когда в
одном и том же слоевище представлено несколько фотобион¬
тов одновременно, например, некоторые виды семейств
Parmeliaceae (Bhattacharya et al., 1996) и Physciaceae (Helms,
2003) могут содержать в своём слоевище несколько разных
видов рода Trebouxia. Существуют также сведения о некото¬
рой нестабильности состава фотобионтов у одного и того же
вида лишайника. Чтобы объяснить этот и другие случаи, необ¬
ходимо рассмотреть основные пути получения фотобионта
микобионтом.
Так, существует 2 основных пути получения фотобионта
- от материнского слоевища лишайника и из окружающей
среды (Окснер, 1937). Первый случай реализуется в процессе
вегетативного размножения лишайников, при формировании
изидий и соредий, представленных в виде грибных гиф, опле¬
тающих несколько клеток фотобионта. Такой способ получе¬
ния фотобионта является наиболее простым и продуктивным
для многих видов лишайников. Однако вегетативное размно¬
жение встречается далеко не у всех лишайников и служит для
распространения лишайника на близкие расстояния, с целью
колонизации уже освоенной территории (Ott, 1987).
Второй путь получения фотобионта - из окружающей
среды, осуществляется при половом и бесполом размножении
лишайников, когда спора, попав в благоприятную среду, про¬
растает в ростовую трубку. Разрастаясь, ростовая трубка раз¬
ветвляется и формирует гифы первичного слоевища. Если
вблизи формирования первичного слоевища нет подходящих
водорослей, оно отмирает. Отыскав соответствующую водо¬
росль в окружающем пространстве, гифы первичного слоеви¬
ща её обволакивают и начинают процесс релихенизации - т.е.
формирование слоевища лишайника. Существует определён¬
ная закономерность: виды, которые продуцируют соредии или
изидии, редко имеют плодовые тела и, наоборот, виды с пло¬
доношением, обычно редко формируют соредии либо изидии
(Окснер, 1937; Poelt, 1970, 1972). Таким образом, в реализации
этих двух путей размножения, а значит и получения фотобио¬
нта, существует некоторая специализация среди лишайников.
Для лишайников, ассоциированных с факультативным
фотобионтом, оба пути получения фотобионта являются эф¬
фективными. Для таких лишайникообразующих грибов источ¬
ником фотобионтов могут выступать популяции свободножи¬
вущих водорослей (Войцехович, 2012). Так, например, жиз¬
ненный цикл тропического лишайника Strigula sp. проходит
стадию, на которой он колонизирует свободноживущую водо¬
росль рода Cephaleuros Kunze ex Е. М. Fries и формирует с ней
своё слоевище (Chapman, Waters, 2001). Во многих случаях
нелихенизированные водоросли встречаются в тех же биото¬
пах, что и лишайники, в состав которых они входят
(Ahmadjian, 1962; Tschermak-Woess, 1976; Войцехович, 2008;
Войцехович, 2012). Более того, бывают случаи, когда морфо¬
логия свободноживущих водорослей и лишайников, в состав
которых они входят, настолько сходна, что различить эти объ¬
екты в полевых условиях не просто (Рис. 68). Для многих ли¬
шайников совместное произрастание с потенциальными фото¬
бионтами в одних и тех же биотопах связано с необходимо¬
стью их поиска в окружающей среде, а также совпадением
экологических оптимумов лихенизированных и свободножи¬
вущих популяций одного и того же вида фотобионта. Таким
образом, биологические особенности фотобионта в какой-то
степени определяют распространение и экологические пред¬
почтения всей лишайниковой ассоциации, что особенно ярко
проявляется у лишайников с факультативным фотобионтом.
Рис. 68. Морфологическое подобие макроскопических колоний свобод-
ноживущего цианопрокариота Nostoc commune (а) и лишайника, в со¬
став которого он входит Collema sp.(6).
У лишайников, ассоциированных с облигатным фото¬
бионтом, например, с Trebouxia - также реализуются два пути
получения фотобионта, однако с некоторыми особенностями.
Поскольку фотобионт таких лишайников очень редко встреча¬
ется в свободноживущем состоянии в природе (или не встре¬
чается вообще), то получить его из окружающей среды непро¬
сто. Наиболее простым путём получения фотобионта для пло¬
доносящих лишайников с Trebouxia является получение его из
слоевища другого лишайника - т.е. лишайниковый парази¬
тизм, широко распространённый среди лихенизированных
грибов (Ott, 1987, Ott et al., 1995; Friedl, 1987; Timdal, 1991;
Nyati, 2006; Poelt, 1958, 1962). Спора лишайника, попадающая
на субстрат, где нет представителей Trebouxia, прорастает и
формирует подслоевище. В этом состоянии лишайник может
находиться долгое время, иногда до нескольких лет, как это
происходит у некоторых видов Aspicilia A. Massal. (Окснер,
1937), может даже временно быть сапрофитом, как это описа¬
но для Xanthoria parietina (Ott, 1987), пребывая в ожидании
подходящего источника фотобионта - чужой соредии или изи¬
дии. В другом случае, если спора попадает на слоевище друго¬
го лишайника, ассоциированного с Trebouxia, разрастаясь,
гифы молодого микобионта внедряются в слоевище лишайни-
ка-хозяина и используют часть клеток его фотобионта для
формирования собственного слоевища.
Вероятность того, что спора попадёт на слоевище ли¬
шайника с совместимым фотобионтом невелика, потому в ли¬
тературе приводится значительное количество фактов непо¬
стоянства фотобионтного состава одного и того же вида ли¬
шайника. Так, например, из 32-х исследованных образцов ли¬
шайника Xanthoria parietina были выделены 5 видов фотобио¬
нтов, болшая часть которых 3 вида - это группа видов морфо¬
логически и генетически близких к Trebouxia arboricola и два
других вида - представители рода Asterochloris (Ahmadjian,
1993; Archibald, 1975; Nyati, 2006). Виды Xanthoria parietina и
X. polycarpa (Hoffm.) Rieber, формирующие слоевища с
Trebouxia arboricola и T. decolorans, на начальных стадиях
своего развития используют фотобионт лишайника Physcia
(Trebouxia impressa), на котором они поселяются (Nyati, 2006).
Caloplaca coronata (Krempelh. ex Korber) J. Steiner паразитиру¬
ет на слоевищах лишайников Aspicilia (фотобионт -
Trebouxia), Catapyrenium Flot. (Myrmecia), Rinodina (Ach.) Gray
(Trebouxia) и Verrucaria (Dilabifilum, Diplosphaera, Myrmecia и
др.); Caloplaca tetraspora (Nyl.) H. Olivier выявлена как пара¬
зит лишайника Cladoniapyxidata (L.) Hoffm. (Ходосовцев и др,
2004). Diploschistes muscorum (Scop.) R. Sant, на ранних стади¬
ях своего развития поселяется на эпигейном лишайнике
Cladonia sp., колонизирует его фотобионт, относящийся к роду
Asterochloris, а затем формирует слоевище с Trebouxia
showmanii (Friedl, 1987). Таким образом, довольствуясь «слу¬
чайным» фотобионтом, молодой микобионт паразитирует на
взрослом лишайнике-хозяине. Так может продолжаться до тех
пор, пока вблизи его слоевища не появится источник подхо¬
дящего фотобионта (будь-то слоевище другого лишайника,
изидии или соредии). Следовательно, один и тот же лишайник
на различных стадиях своего развития может характеризо¬
ваться разным уровнем избирательности фотобионта, потому в
его слоевище в процессе онтогенеза могут сменяться несколь¬
ко разных видов фотобионтов.
В пользу гипотезы о получении фотобионта лишайника¬
ми с Trebouxia в результате паразитического образа жизни
свидетельствует широкое распространение паразитизма имен¬
но среди леканоромицетовых лишайников (Ott, 1987), непо¬
стоянство их фотобионтного состава (Nyati, 2006; Helms, 2003
и др.), случаи развития нескольких фотобионтов в одном слое¬
вище (Friedl, 1987; Friedl et al., 1988), а также находки парази¬
тирующих лишайников, имеющих тот же вид фотобионта, что
и их хозяева (Gorbushina et al., 2006). По нашим подсчётам,
сделанным на основании анализа различных литературных
источников (Timdal, 1991; Ott, 1987; Poelt, 1958, 1962;
Hafellner, Poelt, 1980 и др.), на данный момент известно 34 ро¬
да лишайникообразующих грибов, которые в своём развитии
имеют паразитическую стадию и поселяются на слоевищах
других лишайников с целью получения фотобионта. Среди
этих паразитических лишайников преобладают представители
класса Lecanoromycetes и его порядков Candelariales
(Candelariella Mull. Arg.), Teloschistales (Caloplaca Th. Fr.,
Fulgensia A. Massal., Xanthoria (Fr.) Th. Fr., Rhizocarpon
Ramond ex DC., Rinodina, Rinodinella H. Mayrhofer et Poelt),
Lecanorales (Parmelia Ach., Protoparmelia M. Choisy, Physcia
(Schreb.) Michx., Psora Hoffm.) и Pertusariales (Schaereria
Korb., Ochrolechia A. Massal.), ассоциированные с фотобион¬
том Trebouxia.
Интересной в плане иллюстрации поиска облигатного
фотобионта является работа 3. Отт (Ott, 1987), посвящённая
исследованию жизненного цикла Xanthoria parietina. Автор
описывает и экспериментально доказывает несколько путей
релихенизации лишайника с совместимым фотобионтом. Про¬
росшая спора лишайника образует сеть гиф. При встрече сов¬
местимого фотобионта рода Trebouxia (из чужой изидии или
соредии) развивается нормальное слоевище. Если такой слу¬
чай не представился, гриб вступает в ассоциацию с любой
другой найденной водорослью, образуя зачаточное слоевище -
ареолированную корку. В частности автором детально проил¬
люстрирована такая ассоциация с широко распространёнными
аэрофитными водорослями рода Pleurococcus Meneghini (=
Apatococcus Brand em. Geitler, Desmococcus Brand em. Vischer).
В таком состоянии микобионт может существовать до тех пор,
пока не встретит подходящий фотобионт из чужой изидии или
соредии. Наконец, третий путь - колонизация слоевища друго¬
го лишайника (в данном случае, представителя рода Physcia),
несущего необходимый фотобионт. Так, при прорастании спо¬
ры X. parietina на слоевище Physcia, молодое слоевище
Xanthoria формируется с клетками фотобионта атакуемого
лишайника (лишайника-хозяина), образуя химерное слоевище.
Описаны интересные случаи паразитизма, при которых на
слоевище Physcia образовывались апотеции Xanthoria или же
в апотециях, сформированных Physcia происходило замеще¬
ние спор лишайника-хозяина на споры Xanthoria и, наконец,
при формировании соредий наблюдалось обволакивание кле¬
ток фотобионта гифами Xanthoria и распространение таким
образом соредий паразитирующего лишайника, а не лишайни-
ка-хозяина. Таким образом, жизненный цикл X. parietina пока¬
зывает, что лишайники с облигатным фотобионтом проявляют
низкую избирательность к фотобионту на ранних этапах раз¬
вития. Однако, в дальнейшем получают совместимый фото¬
бионт из вегетативных органов размножения других или слое¬
вищ других лишайников.
В отличие от лишайников с факультативным фотобион¬
том, лишайники с облигатным фотобионтом не следуют в
своём распространении за свободноживущей популяцией во¬
доросли. Более того, эколого-географическое распространение
ближайших родственников облигатных фотобионтов - пред¬
ставителей класса Trebouxiophyceae, не всегда совпадает с
распространением лишайников, обладающих этим фотобион¬
том (Войцехович и др., 2011а, б) За счёт симбиоза с лишайни¬
кообразующим грибом виды рода Trebouxia, вероятно, расши¬
ряют свои эколого-географические возможности. Выделение
двух групп лишайников - ассоциированных с облигатным или
факультативным фотобионтом, основывается не только на
различии их фотобионтного состава, но и на принципиально
разных жизненных стратегиях. В ассоциациях с факультатив¬
ным фотобионтом взаимоотношения водоросли и гриба не яв¬
ляются взаимозависимыми (Tschermak-Woess, 1988; Stenroos
et al., 2006), водоросль может существовать самостоятельно,
тогда как, поведение микобионта может выходить за рамки
симбиотических отношений, в некоторых случаях приводя к
гибели фотобионта (Chapman, 1976). Такие примеры свиде¬
тельствуют о невысокой специализации обоих компонентов
ассоциации, а также об относительной примитивности данной
группы лишайникообразующих грибов, что подтверждается
молекулярно-филогенетическими исследованиями (Hibbett et
al., 2007). Лишайниковые ассоциации с факультативным фо¬
тобионтом преимущественно заселяют поверхность почвы
('Collema, Peltigera, Placynthium (Ach.) Gray и др.), а также ко¬
ру и листья деревьев (Coenogonium, Graphis, Opegrapha Ach.,
Strigula Fr.n др.).
Лишайники с облигатным фотобионтом (например,
Trebouxia), считаются наиболее прогрессивным среди лишай¬
никообразующих грибов (Hibbett et al., 2007; Hofstetter et al.,
2007). Симбиотические отношения их компонентов являются
не только высокоспециализированными, но и взаимозависи¬
мыми. Основной жизненной стратегией таких лишайников при
бесполом и половом размножении, является паразитизм на
других лишайниках, обусловленный высокой конкуренцией за
субстрат и обладание фотобионтом. Основными местами про¬
израстания требуксиоидных лишайников являются камени¬
стый субстрат и кора деревьев, где они образуют обширные
пёстрые многовидовые разрастания, в которых чётко просле¬
живаются активные конкурентные взаимоотношения между
представителями нескольких видов.
Вышесказанное относится к основному фотобионту ли¬
шайников, т.е. водоросли определённого рода (вида), которая
является характерной для того или иного лишайника. Именно
с основным фотобионтом лишайники формируют постоянное
слоевище и, как правило, именно с его клетками формируются
их вегетативные органы размножения. Однако, кроме основ¬
ного фотобионта, в слоевище лишайников могут присутство¬
вать и другие водоросли, являющиеся дополнительными фо¬
тобионтами. Среди таких дополнительных фотобионтов,
прежде всего, хотелось бы отметить водоросли, присутствую¬
щие в цефалодиях. Чаще всего это цианопрокариоты - Nostoc,
Stigonema, и значительно реже зелёные водоросли -
Соссотуха (Vitikainen, 1994; Lohtander et al., 2003). Состав во¬
дорослей в цефалодиях непостоянен, потому, наряду с харак¬
терными для этих структур видами, в них могут присутство¬
вать и другие водоросли, например, Stichococcus или
Diplosphaera. Существует предположение, что образование
цефалодиев обусловлено внедрением в слоевище лишайника
свободноживущих лихеноэпифитных водорослей, которые,
раздражая грибной компонент, вызывают местные разраста¬
ния ткани (Окснер, 1937, 1974).
Недавние исследования некоторых видов Micarea и
Placynthiella (Voytsekhovich et al., 2011), показали, что допол¬
нительные фотобионты могут также находиться непосред¬
ственно в слоевище лишайника (Рис. 695а). Из 22 исследован¬
ных образцов указанных лишайников, 17 содержали дополни¬
тельные фотобионты, причём видовой состав последних зави¬
сел как от эпилихенофитного окружения исследуемого образ¬
ца, так и от фотобионтого состава лишайников, произрастав¬
ших в непосредственной близости с местом сбора образцов.
Так, к примеру, в составе одного из образцов лишайника
Placynthiella uliginosa (Schrad.) Coppins et P. James кроме ос¬
новного фотобионта (Radiococcus signiensis), были обнаруже¬
ны Asterochloris cf. excentrica (P. A. Archibald) Skaloud et
Peksa, Elliptochloris subsphaerica (Reisigl) Ettl et G. Gartner,
Interfdum massjukiae Mikhilyuk et al. и Trebouxia cf. incrustata
Ahmadjian ex G. Gartner. В данном случае Elliptochloris и
Interfilum (Рис. 696) (являющиеся факультативными фотобио¬
нтами), очевидно, попали в слоевище из свободноживущего
состояния, тогда как Asterochloris и Trebouxia (облигатные фо¬
тобионты), были «заимствованы» у близрастущих видов
Cladonia и Xanthoparmelia соответственно. Авторы предпола¬
гают, что свободноживущие популяции наземных водорослей,
находящиеся в непосредственной близости от слоевища ли¬
шайника, а также органы вегетативного размножения других
лишайников, которые содержат подходящие водоросли, могут
быть колонизированы микобионтом и включены непосред¬
ственно в слоевище лишайника. В данном случае гриб исполь¬
зует свободноживущие водоросли в качестве дополнительного
источника питания. В целом же, лабильность некоторых видов
лишайников в выборе и включении фотобионта в слоевище
играет адаптивную роль в освоении новых местообитаний
(напр., Scheidegger & Werth, 2009).
Рис. 69. Схематическое изображение срезов лишайников: а) основной и
дополнительные фотобионты Placynthiella uliginosa, масштаб 40 мкм; б)
дополнительный фотобионт Interfilum sp. в слоевище Micarea, масштаб
10 мкм.
Особый интерес представляют так называемые «гимени-
альные» и «эпитециальные» водоросли, обнаруженные в пери¬
тециях, в слизи между сумками и эпитециальном слое апоте-
циев некоторых лишайников. Данные водоросли характерны
для лишайников с многоклеточными, муральными, спорами и
являются неотъемлемым компонентом слоевища многих ви¬
дов. Считается, что их клетки распространяются вместе со
спорами и при их прорастании гифы микобионта уже имеют
необходимые водоросли для формирования слоевища (Окснер,
1974). В то же время, состав гимениальных и эпитециальных
водорослей неизвестен, есть лишь данные, что по размеру и
цвету, они существенно отличаются от тех, что расположены в
фотобионтном слое. Чаще всего гимениальные водоросли
представлены родами Diplosphaera или Stichococcus
(Ahmadjian, 1969; Ahmadjian, Heikkila, 1970). Таким образом,
на сегодняшний день неясно, что представляют собой эти во¬
доросли - клетки из фотобионтного слоя или другие виды во¬
дорослей, и какова их роль в лишайниковой ассоциации.
Наконец, слоевище лишайников может выступать суб¬
стратом для свободноживущих водорослей и зачастую на по¬
верхности содержит богатую лихеноэпифитную альгофлору,
по своему видовому составу близкую к альгофлоре, развива¬
ющейся на близлежащих субстратах (Михайлюк и др. 2003;
Войцехович и др., 2009; Войцехович, 2012). В складках и тре¬
щинах слоевища свободноживущие водоросли находят защиту
от черезмерной инсоляции и сухости. Сравнение видового со¬
става водорослей-эпифитов литофильных лишайников показа¬
ло, что количество видов и их обилие зависит от строения сло¬
евища, т. е. степени его трещиноватости и гигроскопичности
(Mikhailyuk, Kondratyuk, 2005). Вероятно, лихеноэпифитные
водоросли отчасти являются источником цефалодиальных,
эпитециальных и гимениальных водорослей, а также «потен¬
циальными» фотобионтами лишайников с факультативным
фотобионтом. Однако роль их в лишайниковой ассоциации до
сих пор окончательно не выяснена.
Изоляция и культивирование фотобионтов
Фотобионты лишайников являются сложным объектом
для исследований вследствие многих проблем, возникающих
при введении их в культуру, а также очень медленного роста
отдельных представителей. Жизнеспособность фотобионтов
варьирует в зависимости от типа лишайникового слоевища: в
гомеомерных (лепрозного типа) слоевищах она ниже, чем в
гетеромерных. Так, жизнеспособность фотобионта в слоевище
Lepraria sp., хранящегося при комнатной температуре, сохра¬
няется на протяжении 6 месяцев. В то же время, водоросли
лишайников рода Xanthoparmelia (Vain.) Hale могут сохра¬
няться до нескольких лет (Peksa, Skaloud, 2008).
Существует несколько методов изоляции фотобионтов в
культуру, которые хорошо описаны в литературе (Bubrick,
1989; Ahmadjian, 1993; Backor et al., 1998; Friedl, BUdel, 2008).
Для всех методов общими являются следующие этапы: разру¬
шение слоевища лишайника (путём центрифугирования, гомо¬
генизации и т.п.), извлечение клеток фотобионта и инокуляция
их на питательную среду. На сегодняшний день широко ис¬
пользуемым является метод «микропипетки» (Ahmadjian,
1993), позволяющий минимизировать контаминацию выделя¬
емого в культуру бионта. С помощью пипетки Пастера (с се¬
чением 50-75 мкм в диам.) клетки фотобионта переносятся в
каплю стерильной воды на предметное стекло, где из них по¬
лучают разбавленную суспензию. Затем отдельные клетки фо¬
тобионта переносятся пипеткой на питательную среду. Этот
метод довольно сложен, однако является наиболее успешным
в получении чистых культур. Он также может быть использо¬
ван для изоляции аскоспор, конидий, соредий и т.д.
Для культивирования цианобионтов используют среду
Болда (ВВМ - Bold’s Basal Medium) (Deason, Bold, 1960) или
BG-11 (Waterbury, Stanier, 1978: цит. no BUdel, Henssen, 1983),
для зелёных фотобионтов - модифицированные питательные
среды на основе ВВМ: с утроенным содержанием азота - 3N
ВВМ, а также с добавлением пептона и глюкозы - Trebouxia-
Organic-Medium (ТОМ) (Ahmadjian, 1967). Оптимальные усло¬
вия культивирования фотобионтов подбирают в зависимости
от местообитания лишайника, из которого они изолированы.
Так, оптимальная температура культивирования фотобионтов
альпийских, арктических и антарктических лишайников со¬
ставляет около +10°С, тогда как для фотобионтов лишайников
умеренной зоны температурный оптимум лежит в пределах
+18—21 °С. Культуры водорослей-фотобионтов, как правило, не
нуждаются в интенсивном освещении. Кроме того, многие зе¬
лёные фотобионты способны к росту на органических средах
при полном отсутствии света (Ahmadjian, 1993). Колонии циа¬
нобионтов, а также большинства зелёных водорослей-
фотобионтов (Asterochloris, Chlorella Beijerinck,
Pseudococcomyxa, Myrmecia и др.) появляются в течение 2-3
недель после посева, тогда как Trebouxia требует для своего
развития от 6 недель и более.
Выделенные в культуру фотобионты существенно изме¬
няют свой метаболизм. Так, клетки Соссотуха sp. (изолиро¬
ванной из Peltigera sp.), при росте на жидкой среде вскоре по¬
сле изоляции резко уменьшали синтез и выделение в среду ри-
битола. Вместо этого, синтезировалась сахароза и намного
больше нерастворимых в этаноле веществ, чем в лихенизиро-
ванном состоянии (Green, Smith, 1974; цит. по: Honegger,
1991).
Морфология большинства водорослей-фотобионтов в
культуре, по сравнению с лихенизированным состоянием,
имеет значительные отличия. В большей степени это касается
нитчатых водорослей, которые под влиянием микобионта рас¬
падаются на отдельные клетки и часто сильно деформируются
(Geitler, 1933), как это показано на примере водоросли рода
Trentepohlia в лихенизированном состоянии (Рис. 70). Слизи¬
стые обвёртки, характерные для свободноживущих цианопро-
кариот, таких как, например Gloeocapsa, в слоевищах лишай¬
ников едва развиты или совсем не развиваются. Хлоропласт
лихенизированных водорослей, как правило, несколько изме¬
нён, он более компактен и менее структурирован, а соотноше¬
ние объёма хлоропласта и пиреноида почти вдвое больше, чем
у свободноживущих водорослей (Skaloud, Peksa, 2008). В ли¬
хенизированном состоянии у большинства представителей
класса Trebouxiophyceae (Dictyochloropsis, Myrmecia,
Trebouxia) отсутствуют жгутиковые стадии в жизненном цикле
и размножение водорослей происходит за счёт автоспорообра¬
зования. Исключение может составлять Leptosira thrombii
Tscherm.-Woess, которая способна продуцировать зооспоры
даже в лихенизированном состоянии при условии длительных
ливневых дождей (Schiman, 1961). Известно, что представите¬
ли класса Ulvophyceae (Cephaleuros, Phycopeltis и Trentepohlia)
также способны продуцировать зооспоры и гаметы в слоевище
лишайника (Santesson, 1952; Meier, Chapman, 1983; Rands,
Davis, 1993). В свободноживущем состоянии бурая водоросль-
фотобионт Petroderma maculiforme формирует небольшой
дисковидный таллом с радиально расходящимися нитями
(Wilce et al., 1970: цит. по Sanders, 2004). В лихенизированном
состоянии нити водоросли растут в направлении сверху вниз,
внедряясь между гифами гриба. Наименее подвержены изме¬
нениям в лихенизированном состоянии виды рода Stichococcus
(Raths, 1938), имеющие простую морфологию и жизненный
цикл.
После изоляции фотобионтов и последующих несколь¬
ких недель культивирования клетки некоторых видов родов
Asterochloris, Trebouxia и Myrmecia, утрачивают характерную
для них в лихенизированном состоянии грушевидную форму и
утолщение оболочки, а их хлоропласт приобретает складча¬
тую структуру (Geitler, 1962; 1963). Определённые виды часто
образуют подвижные зооспоры и гаметы в культуре, в отличие
от лихенизированного состояния. Большинство морфологиче¬
ских признаков, такие как размеры вегетативных клеток, тип
хлоропласта, наличие или отсутствие стигмы у зооспоры,
форма и размер зооспорангиев или гаметангиев, могут наблю¬
даться только в условиях культуры.
Рис. 70. Результат изменения морфологических особенностей факульта¬
тивного фотобионта Trentepohlia sp. в процессе лихенизации.
Структура хлоропластов многих зелёных фотобионтов
сильно варьирует в зависимости от возраста и состояния куль¬
туры (Skaloud, Peksa, 2008). Так, авторы выделили несколько
стадий развития хлоропласта в ходе жизненного цикла пред¬
ставителей рода Asterochloris: осевой хлоропласт с удлинён¬
ными Т-образными лопастями; массивный зубчатый по краю
хлоропласт со многими простыми лопастями; осевой хлоро¬
пласт с уплощёнными лопастями, которые заканчиваются
плоскими периферическими краями различной формы. Для
видов рода Trebouxia было выделено 5 основных типов хлоро¬
пласта и пиреноида, каждый из которых соответствует опре¬
делённой группе видов (Friedl, 1989).
Многочисленные исследования ультраструктуры как зе¬
лёных фотобионтов, так и цианобионтов (Peveling, 1973;
Archibald, 1975; Hildreth, Ahmadjian, 1981; Honegger, 1984;
Konig, Peveling, 1980, 1984; Brunner, Honegger, 1985; Gartner,
1985; Chapman, Henk, 1986; Friedl, 1989; Matthews et al. 1989;
Ascaso et al., 1995; Van den Hoek et al., 1995), существенных
особенностей, связанных с лихенизацией, не обнаружили, т.е.
особенности ультраструктуры фотобионтов соответствуют
особенностям тех групп водорослей, к которым они принад¬
лежат. Кроме того, отмечено отсутствие существенных отли¬
чий ультраструктуры лихенизированных грибов по сравнению
с нелихенизированными (Honegger, 2008).
Наличие аксеничных культур фотобионтов значительно
расширило возможности исследователей и позволило осуще¬
ствить ряд опытов по релихенизации (синтезу лишайников в
лабораторных условиях), а также по изучению избирательно¬
сти микобионта в отношении фотобионта (Ahmadjian, 1967;
Ahmadjian, Heikkila, 1970; Ahmadjian, 1993). Для такого синте¬
за необходимы следующие условия: аксеничные культуры ми-
ко- и фотобионта; длительный период инкубации; отсутствие
органических добавок или остатков органических веществ в
среде или субстрате; высокая постоянная влажность; низкая
интенсивность света - 10-30 fimol • m"2 • s"1 PPFD (Плотность
фотосинтетического фотонного потока); специфическая тем¬
пература инкубации. Например, синтез антарктических ли¬
шайников проходит успешнее при температуре + 4° С, а синтез
лишайников умеренной зоны - при температуре + 15-20° С
(Ahmadjian, 1993). На сегодняшний день релихенизировано
более 30 видов лишайников, среди них представители родов
Caloplaca Th. Fr., Cladonia P. Browne, Lecanora Ach., Peltigera
Willd., Stereocaulon Hoffm., Usnea Dill, ex Adans, Xanthoria
(Fr.) Th. Fr. и др. (Ahmadjian, 1993; Schaper, Ott, 2003 и др.).
Таким образом, лихенизация приводит к изменениям
морфологии и жизненных циклов водорослей по сравнению с
их характеристиками в свободноживущем состоянии, тогда
как на особенности ультраструктуры лихенизированное состо¬
яние особого влияния не оказывает. Наличие чистых культур
фотобионтов является обязательным условием при исследова¬
нии их состава, таксономии, генетических особенностей и ко-
специфичности с лишайникообразующим грибом.
В заключение подчеркнем несколько основных направ¬
лений изучения фотобионтов лишайников, исходя из их зна¬
чимости:
1) для идентификации лишайников важную роль играет
умение определять фотобионты в лихенизированном состоя¬
нии до уровня рода;
2) выделение фотобионтов в культуру и исследования их
морфолого-анатомических особенностей позволяет сделать
определение более точным;
3) для исследования лишайниковых популяций и охраны
редких видов лишайников: важную роль играет изучение кос-
пецифичности с лишайникообразующим партнером, степень
облигатности/факультативности и циркуляция фотобионтов в
лишайниковой ассоциации;
4) для установления филогеографии лишайников, в част¬
ности для экстраполяции постгляциального освоения место¬
обитаний и адаптивных стратегий изучается генетическая и
видовая вариабельность, а также степень облигатности фото-
бионтного состава коспецифических лишайниковых ассоциа¬
ций.
Литература
Андреева В. М. Почвенные и аэрофильные зелёные во¬
доросли (Chlorophyta: Tetrasporales, Chlorococcales,
Chlorosarcinales). СПб.: Наука, 1998. 351 с.
Войцехович А. А. Фотобионты и водоросли-эпифиты ли-
тофильных лишайников Берегового хребта Карадагского при¬
родного заповедника (Крым, Украина) // Актуальш проблеми
боташки та еколопУ. Зб1рка наукових праць. Вип.2. Кшв:
Ф1тосоцюцентр, 2008. С. 46-51.
Войцехович А. О. Водоростевий компонент лишайниюв
Карадазького природного запов1дника: автореф. дис. на здо-
буття наук, ступеня канд. бюл. наук: спец. 03.00.05 боташка.
Кшв, 2012. 23 с.
Войцехович А. О., Михайлюк Т. I., Дар1енко Т. М. Водо-
рост1 наземних м!сцезростань хребта Карагач (Карадазький
природний запов1дник (УкраУна)) // Зб1рник наук, праць, приев.
95-р1ччю КарадазькоУ наук. станщУ та 30-р1ччю Карадазького
прир. заповщника НАНУ шд ред. А. В. ГаевськоУ, А. Л. Моро-
зовоУ. Севастополь: Экос1-Пдроф1зика, 2009. С. 50-60.
Дар1енко Т. М., Войцехович А. О., Кондратюк С. Я. Фо-
тобюнти деяких представниюв родини Teloschistaceae II Укр.
ботан. журн. 2004. Т. 61, № 2. С. 49-59.
Войцехович А. А., Михайлюк Т. П., Дариенко Т. М. Фо¬
тобионты лишайников. 1: разнообразие, экологические осо¬
бенности, взаимоотношения и пути совместной эволюции с
микобионтом // Альгология. 201 la. Т. 21, № 1. С. 3-26.
Войцехович А. А., Михайлюк Т. П., Дариенко Т. М. Фо¬
тобионты лишайников. 2: происхождение и корреляция с ми¬
кобионтом // Альгология. 20116. Т. 21, № 2. С. 151-177.
Кост1ков I. Ю., Романенко П. О., Демченко Е. М.,
Дар1енко Т. М., Михайлюк Т. I., Рибчинський О. В., Солонен-
ко А. М. Водорост’1 грушчв УкраУни (icTopiH та методи до¬
ел щження, система, конспект флори). Кшв: Ф'1тосоц’юцентр,
2001. 300 с.
Михайлюк Т. П., Дариенко Т. М., Демченко Э.
Н. Водоросли гранитных обнажений регионального ланд¬
шафтного парка “Гранитно-степное Побужье” (Николаевская
обл., Украина) // Новости систематики низших растений. Т. 37.
2003. С. 53-71.
Мошкова Н. О. Визначник пр1сноводних водоростей
УкраТнськоТ РСР. VI. Улотриксов1 й кладофоров1 водоростг К.:
Наук.думка, 1979. 500 с.
Окснер А. М. Визначник лишайниюв УРСР. К.: Вид-во
Акад. наук УРСР, 1937. 341 с.
Окснер А. Н. Определитель лишайников СССР. Вып. 2:
Морфология, систематика и географическое распространение.
Л.: Наука, 1974. 284 с.
Окснер А. М. Флора лишайниюв Укра'ши. Т. 2., вип. 2.
К.: Наук. Думка, 1993. 544 с.
Ходосовцев А. Е., Кондратюк С. Я., Макарова И.
И., Окснер А. Н. Определитель лишайников России. 9. Fuscid-
eaceae, Teloschistaceae. СПб.: Наука. 2004. 339 с.
Ahmadjian V. Lichens // Physiology and Biochemistry of
Algae / R. A. Lewin [ed.]. - New York-London: Academic Press,
1962. P. 817-822.
Ahmadjian V. A guide to the algae occuring as lichen sym¬
bionts: Isolation, culture, cultural physiology, and identification //
Phycologia. 1967. Vol. 6. P. 127-160.
Ahmadjian V. Lichen Synthesis // Osrerr. Bot. Z. 1969. Vol.
116. P. 306-311.
Ahmadjian V. The Lichen Symbiosis. New York: John
Wiley and Sons, Inc., 1993. 250 p.
Ahmadjian V., Heikkila H. The culture and synthesis of En-
docarpon pusillum and Staurothele clopima II Lichenologist. 1970.
Vol. 4. P. 259-267.
Ainsworth and Bisby's Dictionary of the Fungi. 10th edition
/ Eds. P. M. Kirk et al. Trowbridge: Cromwell Press, 2008. 771 p.
Archibald P. A. Trebouxia de Puymaly (Chlorophyceae,
Chlorococcales) and Pseudotrebouxia gen. nov. (Chlorophyceae,
Chlorococcales)//Phycologia. 1975. Vol. 14. P. 125-137.
Ascaso C., Valladares F., De los Rios A. New ultrastructurel
aspects of pyrenoidsof the lichen photobiont Trebouxia (Mi-
crothamniales, Chlorophyta) // J. Phycol. 1995. №. 31. P. 114—119.
Backor М., Hudak J., Ziegler W., Backorova M. Methods for
isolation and cultivation of the eukaryotic lichen photobionts look¬
ing for a general method. A review. // Thaiszia J. Bot., Kosice.
1998. № 8. P. 1-6.
Beck A. Selektivitat der Symbionten schwermetalltoleranter
Flechten. Ludwig-Maximilians-Universitat, Munchen. PhD thesis.
Fischer GmbH, 2002. 196 s.
Beck A., Kasaiicky Т., Rambold G. Myco-photobiontal se¬
lection in a Mediterranean cryptogam community with Fulgensia
fulgida // New Phytol. 2002. Vol. 153. P. 317-326.
Bhattacharya D., Friedl Т., Damberger S. Nuclear-encoded
rDNA group I introns: origin and phylogenetic relationships of in¬
sertion site lineages in the green algae // Mol. Biol. Evol. 1996.
Vol. 13. P. 978-989.
Blaha J., Baloch E., Grube M. High photobiont diversity as¬
sociated with the euryoecious lichen-forming ascomycete Lecano¬
ra rupicola II Biol. Journ. Lin. Soc. 2006. Vol. 88, №2. P. 283-
293.
Brunner U., Honegger R. Chemical and ultrastuctural studies
on the distribution of sporopolleninlike biopolymers in six genera
of lichen phycobionts // Can. J. Bot. 1985. 63. P. 2221-2230.
Bubrick P. Methods for cultivating lichens and isolated bi-
onts // CRC Handbook of Lichenology / M. Galun, (ed.). III., Boca
Raton, Fla.: CRC Press, 1989. P. 127-138.
Budel B., Henssen A. Chroococcidiopsis (Cyanophyceae), a
phycobiont in the lichen family Lichinaceae // Phycologia. 1983.
Vol. 22, № 4. P. 367-375.
Chapman R. L. Ultrastructural investigation on the fo-
liicolous pyrenocarpous lichen Strigula elegans (Fee) Mull. Arg. //
Phycologia. 1976. Vol. 67. P. 191-196.
Chapman R. L., Henk М. C. Phragmoplasts in cytokinesis of
Cephaleuros parasiticus (Chlorophyta) vegetative cells // J. Phy-
col. 1986. №22. P. 83-88.
Chapman R. L. Waters D. A. Lichenization of the Tren-
tepohliales complex algae and odd relationships // Symbiosis:
Mechanisms and Model Systems / J. Seckbach [ed.]. Kluwer Aca¬
demic Press, 2001. P. 361-371.
Dahlkild A., Kallersjo М., Lohtander K. et al. Photobiont di¬
versity in the Physciaceae (Lecanorales) // Bryologist. 2001. Vol.
104, №4. P. 527-536.
Deason T. R., Bold H. C. Phycological Studies I. Explorato¬
ry Studies of Texas Soil Algae. University of Texas, 1960. 70 p.
Degelius G. The lichen genus Collema in Europe // Symb.
Bot. Upsal. 1954. Vol. 13, № 2. P. 1-499.
Ettl H., Gartner G. Syllabus der Boden-, Luft-, und Flechten-
algen. Stuttgart, Jena, New York: Gustav Fischer, 1995. 710 S.
Friedl T. Thallus development and phycobionts of the para¬
sitic lichen Diploschistes muscorum II Lichenologist. 1987. Vol.
19. P. 183-191.
Friedl T. Comparative ultrastructure of pyrenoids in Tre¬
bouxia (Microthamniales, Chlorophyta) // PI. Syst. Evol. 1989.
164. P. 145-159.
Friedl Т., Budel B. Photobionts // The Lichen Symbiosis /
Nash III T. (ed.). Cambridge University Press, 2008. P. 9-26.
Friedl Т., Gartner G. Trebouxia (Pleurastrales, Chlorophyta)
as a phycobiont in the lichen genus Diploschistes II Arch. Pro-
tistenk. 1988. Vol. 135. P. 147-158.
Galloway D. J. Lichen biogeography // The Lichen Symbio¬
sis /Nash III T. (ed.). Cambridge University Press, 2008. P. 315-
335.
Gartner G. Die Gattung Trebouxia Puymaly (Chlorellales,
Chlorophyceae) // Arch. Hydrobiol. Suppl. Algol. Studies. 1985.
71, №4. S. 495-548.
Garty J. Re-establishment of lichens on chalk rocks after a
forest fire // Fire in Ecosystem Dynamics. 1990. P. 77-84.
Geitler L. Beitrage zur Kenntnis der Flechtensymbiose I—III
// Arch. Protistenk. 1933. 80. S. 378-409.
Geitler L. Clavaria mucida eine extratropische Basidioliche-
ne // Biologisches Zentralblatt Band. 1955. Vol. 74. S. 145-159.
Geitler L. Schizophyceen II. Umgearbeitete Aflage // Encyc¬
lopedia of Plant Anatomy / W. Zimmermann, P. Ozenda [eds.].
Berlin: Borntraeger, 1960. P. 98-112.
Geitler L. Uber die Flechtenalge Myrmecia biatorellae II Os-
terr. Bot. Z. 1962. № 109. S. 41-47.
Geitler L. Uber Haustorien bei Flechten und uber Myrmecia
biatorellae in Psora globifera II Osterr. Bot. Z. 1963. № 110. S.
270-276.
Gorbushina A. A., Beck A., Schulte A. Microcolonial rock
inhabiting fungi and lichen photobionts: evidence for mutualistic
interactions // Mycol. Res. 2006. Vol. 109. P. 1288-1296.
Goward T. Twelve Readings on the Lichen Thallus. VII.
Species// Evansia. 2009. Vol. 26, №4. P. 153-162.
Hafellner J., Poelt J. Der «Flechtenparasit» Buellia pulver-
ulenta eine bleibend interne parasitische Flechte // Phyton [Austri-
a]. 1980. Vol. 20. S. 129-133.
Helms G. Taxonomy and symbiosis in associations of
Physciaceae and Trebouxia. PhD thesis. University of Gottingen,
Germany, 2003. 155 p.
Hibbett D. S., Binder М., Bischoff J. F. et al. A higher-level
phylogenetic classification of the Fungi // Mycological Research.
2007. Vol. 111. P. 509-547.
Hildreth К. С., Ahmadjian V. A study Trebouxia and
Pseudotrebouxia isolates from different lichens // Lichenologist.
1981. 13. P. 65-86.
Hofstetter V., Miadlikowska J., Kauff F. et al. Phylogenetic
comparison of protein-coding versus ribosomal RNA-coding se¬
quence data: A case study of the Lecanoromycetes (Ascomycota) //
Molecular Phyl. Evol. 2007. Vol. 44. P. 412-426.
Honegger R. Cytological aspects of the mycobiont-
phycobiont relationship in lichens // Lichenologist. 1984. Vol. 16,
№2. P. 111-127.
Honegger R. Functional aspects of the lichen symbiosis //
Annu. Rev. Plant Mol. Biol. 1991. Vol. 42. P. 553-578.
Honegger R. Water relations in lichens // Fungi in the Envi¬
ronment / Gadd G. М., Watkinson S. C., Dyer P. (eds.). Cambridge
Univ. Press, 2006. P. 185-200.
Honegger R. Mycobionts // The Lichen Symbiosis / Nash III
T. (ed.). Cambridge University Press, 2008. P. 27-39.
Honegger R., Zippier U., Gansner H. et al. Mating systems in
the genus Xanthoria (lichen-forming ascomycetes) // Mycological
Research. 2004. Vol. 108. P. 480-488.
Ihda Т., Nakano Т., Yoshimura I. et al. Phycobionts isolated
from Japanese species of Anzia (Lichenes) // Arch. Protistenk.
1993. Vol. 143. P. 163-172.
Jaag О. Соссотуха Schmidle. Monographic einer Algengat-
tung. // Beitr. Kryptogamenfl. Schweiz. Bot. Ges. 1933. Vol. 42.
132 S.
Konig J., Peveling E. Vorkommen von Sporopollenin in der
Zellwand des Phycobionten Trebouxia II Z. Pflanzenphysiol. 1980.
Vol. 98. S. 459-464.
Konig J., Peveling E. Cell walls of the phycobionts Tre¬
bouxia and Pseudotrebouxia: constituents and their location // Li¬
chenologist. 1984. Vol. 16. P. 129-144.
Kroken S., Taylor J. W. Phylogenetic species, reproductive
mode, and specificity of the green alga Trebouxia forming lichens
with genus Letharia II Bryologist. 2000. Vol. 103, №4. P. 645-
660.
Lakastos М., Rascher U., Budel B. Functional characteristics
of corticolous lichens in the understory of a tropical lowland rain
forest//New Phytologist. 2006. Vol. 172, № 4. P. 679-695.
Lamb I. M. On the morphology, phylogeny, and taxonomy of
the lichen genus Stereocaulon II Can. J. Bot. 1951. Vol. 29. P.
522-536.
Lange O. L., Belnap J., Reichenberger H. Photosynthesis of
the cyanobacterial soil-crust lichen Collema tenax from arid lands
in southern Utah, USA: role of water content on light and tempera¬
ture responses of C02 exchange // Functional Ecology. 1998. Vol.
12. P. 195-202.
Lohtander K., Oksanen I., Rikkinen J. Genetic diversity of
green algal and cyanobacterial photobionts in Nephroma (Peltiger¬
ales) // Lichenologist. 2003. Vol. 35. P. 325-339.
Meier J. L., Chapman R. L. Ultrastructure of the lichen Coe-
nogonium interplexum Nyl. // Amer. J. Bot. 1983. Vol. 70. P. 400-
407.
Matthews S., Tucker S., Chapman R. Ultrastructural features
of mycobionts and trentepohliaceous phycobionts in selected sub¬
tropical crustose lichens // Bot. Gaz. 1989. Vol. 150, № 4. P. 417-
438.
Mikhailyuk Т., Kondratyuk S. Epiphytic algae of some li¬
chen species // XVII Intern. Bot. Congr. Vienna, Austria 17-23 Ju¬
ly 2005: abstract. 2005. P. 445.
Mikhailyuk Т. I., Demchenko E. М., Kondratyuk S. Ya. Al¬
gae of granite outcrops from the left bank of the river Pivdennyi
Bug (Ukraine) // Biologia. Bratislava. 2003. Vol. 58, № 4. P. 589-
601.
Meier J. L., Chapman R. L. Ultrastructure of the lichen Coe-
nogonium interplexum Nyl. // Amer. J. Bot. 1983. Vol. 70. P. 400-
407.
Мое R. Verrucaria traversiae sp. nov., a marine lichen with
a brown algal photobiont // Bulletin of the California Lichen Socie¬
ty. 1997. №4. P. 7-11.
Nakano T. Phycobionts of some Japanese species of the
Graphidaceae II Lichenologist. 1988. Vol. 20, № 4. P. 353-360.
Nelsen M. P., Plata E. R., Andrew C. J. et al. Phylogenetic
diversity of Trentepohlialean algae associated with lichen-forming
fungi // J. Phycol. 2011. Vol. 47. P. 282-290.
Nyati Sh. Photobiont diversity in Teloschistaceae (Lecano¬
romycetes) / Erlangung der naturwissenscheftlichen Doktorwiirde.
Univ. Zurich. 2006. 130 p.
Oberwinkler F. Fungus-alga interactions in basidiolichens //
Nova Hedwigia. 1984. Bd 79. P. 739-774.
O’Brien H. E., Miadlikowska J., Lutzoni F. Assessing host
specialization in the symbiotic cyanobacteria associated with four
closely related species of the lichenfungus Peltigera II
Eur. J. Phycol. 2005. Vol. 40. P. 363-378.
Ott S. Reproductive strategies in lichens // Progress and
problems in lichenology in the eighties / E. Peveling [ed.]. Biblio-
theca Lichenologica. 1987. Vol. 25. P. 81-93.
Ott S., Meier Т., Jahns H. M. Development, regeneration,
and parasitic interactions between the lichens Fulgensia bracteata
and Toninia caeruleonigricans II Canad. J. Bot. 1995. Vol. 73. P.
595-602.
Peksa O., Skaloud P. Changes in chloroplast structure in li-
chenized algae // Symbiosis. 2008. Vol. 46. P. 1-8.
Peveling E. Fine structure // The Lichens / Ahmadjian V.,
Hale M. A., (Eds.). New York, London: Academic Press, 1973. P.
147-182.
Piercey-Normore М., De Priest P. T. Algal switching among
lichen symbioses // Amer. J. Bot. 2001. Vol. 88, № 8. P. 1490—
1498.
Poelt J. Uber parasitische Flechten. II // Planta. 1958. Vol.
51. S. 288-307.
Poelt J. Parasitische Flechten III // Osterreich. Bot. Zeitschr.
1962. Vol. 109. S. 521-528.
Poelt J. Das Konzept der Artenpaare bei der Flechten //
Deutsch. Bot. Ges. Neue Folge. 1970. Vol. 4. S. 187-198.
Poelt J. Die taxonomische Behandlung von Artenpaaren bei
den Flechten // Bot. Notis. 1972. Vol. 25, № 1. S. 77- 81.
Rands D. G., Davis J. S. Observations on lichenized and
free-living Physolinum (Chlorophyta, Trentepohliaceae) // J. Phy¬
col. 1993. Vol. 29. P. 819-825.
Raths H. Experimentelle Untersuchungen mit Flechtengoni-
dien aus der Familie der Caliciaceen II Ber. der Schweiz. Bot. Ges.
1938. Bd 48. S. 13-30.
Rindi F., Allali H. A., Lam D. W., Lopez-Bautista M. An
overview of the biodiversity and biogeography of terrestrial green
algae // Biodiversity Hotspots / V. Rescingo et al. (eds.). Nova Sci¬
ence Publ., Inc. 2009. P. 1-25.
Romeike J., Friedl Т., Helms G. et al. Genetic diversity of
algal and fungal partners in four species of Umbilicaria along a
transect of the Antarctic Peninsula // Mol. Biol. Evol. 2002.
Vol. 19. P. 1209-1217.
Sanders W. B. Bacteria, algae, and phycobionts: maintaining
useful concepts and terminology // Lichenologist. 2004. Vol. 36, №
5. P. 269-275.
Santesson R. Foliicolous lichens I. A revision of the taxono¬
my of the obligately foliicolous, lichenized fungi // Symb. bot. up-
sal. 1952. Vol. 12, № l.P. 1-590.
Schaper Т., Ott S. Photobiont selectivity and interspecific in¬
teractions in lichen communities. I. Culture experiments with the
mycobiont fulgensia bracteata // Plant Biology. 2003. № 5. P. 1-10.
Scheidegger С., Werth S.. Conservation strategies for li¬
chens: insights from population biology // Fungal Biology Re¬
views. 2009. №23. P. 55-66.
Schiman H. Uber die Entwickungmoglichkeiten von Lepto-
sira thrombii Tschermak-Woess als Algenkomponente in der
Flechte Thrombium epigaeum // Ost. bot. Z. 1961. Bd 108. S. 1-4.
Skaloud P., Peksa O. Comparative study of chloroplast mor¬
phology and ontogeny in Asterochloris (Trebouxiophyceae, Chlo¬
rophyta) // Biologia. 2008. Vol. 63, №5. P. 1-8.
Skaloud P., Peksa O. Evolutionary inferences based on ITS
rDNA and actin sequences reveal extensive diversity of the com¬
mon lichen alga Asterochloris (Trebouxiophyceae, Chlorophyta) //
Mol. Phyl. and Evol. 2010. Vol. 54. P. 36-46.
Stenroos S., Hognabba F., Myllys L., Hyvonen J., Thell A.
High selectivity in symbiotic associations of lichenized ascomy-
cetes and cyanobacteria // Cladistics. 2006. Vol. 22. P. 230-238.
The Lichen Flora of Great Britain and Ireland / Purvis O. W.,
Coppins B. J., Hawksworth D. L. et al. (eds.). London: Nat. Hist.
Mus. Publ. 1992. 710 p.
Timdal E. A monograph of the genus Toninia (Lecideaceae,
Ascomycetes) // Opera Bot. 1991. Vol. 110. P. 1-137.
Tschermak E. Untersuchungen uber die Beziehungen von
Pilz und Alge im Flechtenthallus // Ost. bot. Z. 1941. Bd 90. S.
233-307.
Tschermak-Woess E. Algal taxonomy and the taxonomy of
lichens: the phycobiont of Verrucaria adriatica II Lichenology:
Progress and Problems / Brown D. H., Hawksworth D. L., and Bai¬
ley R. H., (eds.). London: Academic Press. 1976. P. 79-88.
Tschermak-Woess E. Elliptochloris bilobata, gen. et spec,
nov., der Phycobiont von Catolechia wahlenbergii II Plant Syst.
Evol. 1980. Vol. 136. P. 63-72
Tschermak-Woess E. Uber die weite Verbeitung lichenisier-
ter Sippen von Dictyochloropsis und die systematische Stellung
von Myrmecia reticulata (Chlorophyta) // Plant. Syst. Evol. 1984.
Vol. 147. S. 299-307.
Tschermak-Woess E. The algal partner // CRC Handbook of
Lichenology / M. Galun, ed. Boca Raton, Fla.: CRC Press, 1988. P.
39-92.
Tschermak-Woess E., Plessl A. Uber zweierlei Typen der
sukzedanen Teilung und ein auffallendes Teilungsverhalten des
Chromatophors bei einer neuen Protococcale Myrmecia pyriformis
II Osterr. Bot. Z. 1948. Bd 95. S. 103-109.
Thus H., Muggia L., Perez-Ortega S., et al. Revisiting photo¬
biont diversity in the lichen family Verrucariaceae (Ascomycota)
// Eur. J. Phycol. 2011. Vol. 46, № 4. P. 399-415.
Uyenko F. R. Studies of some lichenized Trentepohlia asso¬
ciated in lichen thalli with Coenogonium II Trans. Am. microsc.
Soc. 1965. Vol. 84. P. 1-14.
Van den Hoek C., Mann D. G., Jahns H. M. Algae: an intro¬
duction to phycology. United Kingdom, Cambridge: Univ. Press,
1995.628 p.
Vischer B. Reproduction und systematische Stellung einiger
Rinden- und Bodenalgen // Schweizerische Zeitschrift fur Hydro-
logie. 1960. Bd 22. S. 329-349.
Vitikainen O. Taxonomic revision of Peltigera (lichenized
Ascomycotina) in Europe // Acta Bot. Fennica. 1994. 152. P. 1-96.
Voytsekhovich A., Dymytrova L., Nadyeina O. Photobiont
composition of some members of the genera Micarea and Placyn¬
thiella (Lecanoromycetes, lichenized Ascomycota) from Ukraine //
Folia Cryptogamica Estonica. 2011. Vol. 48. P. 135-148.
Zeitler I. Untersuchungen uber die Morphologie, Entwick-
lungsgeschichte und Systematik von Flechtengonidien // Osterr.
Bot. Z. 1954. Bd 101. S. 453-483.
Глава V.
Особенности вторичного метаболизма и хемосистематика
лишайников
Рандлане Т., Сааг А.
Лишайниковые вещества
Органические вещества, встречающиеся в лишайниках,
по своей природе подразделяются на две основные группы:
первичные лишайниковые вещества и вторичные метаболиты.
Первичные вещества - это белки, аминокислоты, поли¬
сахариды, витамины и пр., которые, будучи синтезированы
или фотобионтом или грибом, находятся внутри их клеток,
часто они растворимы в воде. Подобные соединения встреча¬
ются также у свободноживущих грибов, водорослей и высших
растений.
Вторичные лишайниковые вещества синтезируются ми¬
кобионтом, хотя необходимый для их синтеза углерод, гриб¬
ной компонент симбиоза получает от производящего их фото¬
бионта.
Вторичные соединения находятся в талломе лишайника
экстрацеллулярно, накапливаясь на поверхности грибных гиф.
Обычно они нерастворимы в воде. Большинство этих соеди¬
нений специфично для лишайников и лишь немногие встре¬
чаются у других грибов или высших растений (Elix, Stocker-
Worgotter, 2008).
Экспериментальные иследования последнего времени
показали, что микобионты разных хемотипов, выращенные
апосимбиотически, т. е. без присутствия водорослевого ком¬
понента, способны продуцировать те же самые или сходные
композиции химические веществ, что и родительский таллом,
однако это в высшей степени зависит от условий культивиро¬
вания. Число известных в настоящее время вторичных лишай¬
никовых веществ, превышает 800 (Stocker-Worgotter et al.,
2004).
Разнообразие вторичных метаболитов лишайниках до¬
статочно велико и в части путей их биосинтеза и по химиче¬
ской структуре. Лишайниковые вещества подразделяются на
основные группы, в соответствии с возможным путем их био¬
синтеза (Shibata, 1965; Culberson et al., 1977; Elix, Stocker-
Worgotter, 2008). Существует три основных пути биосинтеза
лишайниковых веществ: (а) ацетил-полималоновый путь; (б)
путь мевалоновой кислоты и (в) путь шикимовой кислоты
(рис. 71).
ФОТОБИОНТ
I
Углеводы
__ ГРИБ
Ампнсжнслоты
ГМ нт ад q-ф ос фв тми й
ПуП. ШИМИИНОЙ М4СЛ01
Ацетил-С оА
I
Путь нналано-
■он нм илоты
№пОИ№П'С4А
Ац9т№т-пОлиы||п(и<|тн ы)
путь
ДериВЭТЫ ПуЛЬйИНйЙОЙ КИСЛОТЫ
терфеыилхиноны
стероиды зрометич&ские высшие эпифэтические
ТВрП^НОг1ДЬ| ПОЛИкфТИДы НЛСПОТы
j СЦГСИНОЛЬНЫЙ ТИП | | А-ОрСННОЛЫЧ ый тип]
пар>-двпс¥1д
трцдтсцд
м«1#Д4ПСЦа
ДОПСмДОч
ir днбанлофуран
гшра-депснд
&в«1И ЛС***й 1ф**р
Ы#ТД-ДМ(ЦД
допсиДОН
усы*шо*#-п ш?Д0Т*
Рис. 71. Упрощенная схема возможных путей образования основных групп лишайниковых веществ (по Андрееву, 2003).
Подавляющее большинство вторичных лишайниковых
веществ, таких как алифатические кислоты и ароматические
вещества образуются по ацетил-полималоновому типу. Ди- и
тритерпены, также как и стероиды образуются по типу мева-
лоновой кислоты. И только небольшое число веществ - пуль-
виновая кислота и ее производные синтезируются по пути ши-
кимовой кислоты.
Наиболее многочисленный класс лишайниковых ве¬
ществ (более 300), характерных только для лишайников, со¬
ставляют ароматические продукты ацетил-полималонового
происхождения, которые образуются путем объединением
двух или трех фенольных колец орсинольного или |3-
орсинольного типа посредством эфирных и сложноэфирных
связей или связи углерод-углерод. Это депсиды, депсидоны,
дибензофураны и группа усниновой кислоты.
Эти вещества играют важную роль в систематике ли¬
шайников. Другие ароматические вещества, образование ко¬
торых идет по тому же пути (антрахиноны и ксантоны, всего
около 100 веществ) часто идентичны или аналогичны продук¬
там нелихенизированных грибов или высших растений (Elix,
Stocker-Worgotter, 2008). Представители большинства классов
вторичных лишайниковых веществ и их химическая структура
представлены в Табл. 2.
Вторичные метаболиты встречаются в разных частях
лишайниковых талломов, в основном в виде водонераствори¬
мых кристаллов на наружной поверхности грибных гиф, в ко¬
ре и в сердцевине. Один и тот же вид может содержать не¬
сколько веществ и в коре и в сердцевине. Лишайниковые ве¬
щества могут находиться также в плодовых телах, например в
гимении или в эксципуле. Концентрация их варьирует внутри
таллома и зависит от возраста тканей, но обычно колеблется
от 1 до 5% от сухого веса таллома (Lawrey, 2009).
Роль лишайниковых веществ в лишайниковых талломах.
Химическая структура лишайниковых веществ вариа¬
бельна, столь же многообразна и биологическая роль этих со¬
единений. Некоторые из них считаются отходами метаболиз¬
ма, в то время как другие определенно имеют биологическое
значение. Признаются следующие адаптивные функции вто¬
ричных метаболитов лишайников (Huneck, Yoshimura, 1996;
Lawrey, 2009):
Таблица 2.
Некоторые представители основных классов вторичных лишайниковых веществ (по Elix, Stocker-Worgotter, 2008) и их хими¬
ческие формулы.
1.2.1. Моноциклические
Орцинол
с7н8о2
фенольные соединения
си,
Гематоммовая кислота
С9Н8О5
1.2.2. Двойные и тройные производные простых фенольных колец
1.2.2.3. Депсоны
Пикролихеновая кислота
С25Н30О7
1.2.2.4. Дибензофураны и
усниновые кислоты
Паннаровая кислота
C,6H|2Ov
Стрепсилин
Усниновая кислота
CisHmOs
С|8Н|бС>7
1.2.5. Нафтахиноны
Родокладоновая кислота
CisHioOg
1.2.6. Ксантоны
Лихексантон
С16Н14О5
Тиофановая кислота
СиНбС^СЬ
т
2. Путь мевалоновой кислоты
2.1. Ди- и тритерпены
Урсоловая кислота
С30Н48О3
■сн,.
2.2. Стероиды
Эргостерол
С28Н44О
3. Путь шикимовой кислоты
3.1. Терфенилхиноны
Телофоровая кислота
CigHgOg
ОН
о (
_л/гмон
н°^5' J'0
но
3.2. Пульвиноая кислота и
Пульвиновая кислота
С18Н12О5
г-\
производные
6
Ризокарповая кислота
c28h23no6
(а) Регуляторы внутреннего водного баланса. Боль¬
шинство лишайниковых веществ обладают гидрофобными
свойствами и практически нерастворимы в воде. Их присут¬
ствие на поверхности гиф сердцевины может служить цели
поддержания воздушных «карманов», необходимых для газо¬
обмена клеток фотобионта. Лишайниковые вещества препят¬
ствуют насыщению сердцевины водой, обеспечивая тем са¬
мым непрерывный газообмен с атмосферой (Armaleo et al.,
2008).
(б) Регуляторы метаболизма фотобионта. Хотя ли¬
шайниковые вещества не участвуют в первичном метаболиз¬
ме, известно, что они могут подавлять рост и метаболическую
активность водоросли, регулируя деятельность ферментов,
участвующих в первичном метаболизме. Например, усниновая
кислота, одно из наиболее широко распространенных лишай¬
никовых веществ, встречающихся в коровом слое, подавляет и
фотосинтез и дыхание фотобионта Trebouxia, в то время, как
аккумуляция в коровом слое другого лишайникового вещества
- атранорина, часто связана с уменьшением плотности водо¬
рослевых клеток в старых частях лишайникового таллома. Та¬
ким образом, синтез и аккумуляция некоторых фенольных со¬
единений создает основу для механизма контроля грибом хода
клеточного обмена в водорослевом слое (Honegger, 1987).
(в) Минерализация основных (первичных) элементов.
Некоторые лишайниковые вещества способны связывать в
химические соединения основные минеральные элементы, ко¬
торые существенны для снабжения лишайников минералами
из субстрата. Например, для лишайников, растущих на меде¬
содержащих субстатах было отмечено связывание меди нор-
стиктовой и псоромовой кислотами (Purvis et al., 1987, 1990).
(г) Защита от света. Вещества, образующиеся в верхнем
коровом слое, обеспечивают защиту нижележащего фотобио¬
нта от избыточного освещения. Наиболее обычные вещества
корового слоя, это депсиды - атранорин и хлоратранорин, ус-
ниновые кислоты, антрахиноны, ксантоны и производные
пульвиновой кислоты (Elix, Stocker-Worgotter, 2008). Все эти
вещества являются пигментами, способными фильтровать, т.е.
селективно абсорбировать солнечную радиацию, достигаю¬
щую чувствительных клеток фотобионта, находящихся ниже.
Известно, что степень концентрации лишайниковых веществ в
коровом слое зависит от интенсивности светового воздей¬
ствия. Кроме основного прямого эффекта - ослабления излу¬
чения видимой части спектра, эти вещества выполняют и до¬
бавочную функцию, защищая таллом от чрезмерного ультра¬
фиолетового излучения (Fahselt, 1994; Lawrey, 1984, 2009).
(д) Аллелопатический фактор. Лишайники, как прави¬
ло, очень медленно растущие организмы, и чтобы иметь шанс
на выживание в соревновании с грибами, мхами и сосудисты¬
ми растениями, они должны как-то воздействовать на рост
этих организмов. Ранее отмечалось подавление лишайниками
роста бриофитов, сосудистых растений и других лишайников.
Лишайниковые вещества известны также как ингибиторы ро¬
ста микоризных грибов, оказывающие таким образом влияние
на растения конкуренты. Подавление бриофитов и сосудистых
растений достигается ингибированием прорастания и развития
спор и проростков (Lawrey, 1984).
(е) Защита от патогенных микроорганизмов и поеда-
телей (консументов). Некоторые фенольные соединения за¬
щищают лишайники от поедания насекомыми, улитками и
другими беспозвоночными, другие вещества обладают анти¬
биотическими свойствами, которые способствуют защите ли¬
шайника от различных микроорганизмов (Lawrey, 1984, 2009).
В качестве эксперимента (Gauslaa, 2005), у 17 видов лишайни¬
ков одна половина таллома была обработана ацетоном, чтобы
удалить фенольные соединения, а другая оставлена нетрону¬
той, после чего лишайники были предложены улиткам. Во
всех случаях, части обработанные ацетоном и свободные от
фенолов, были более привлекательными для улиток, чем
необработанный таллом. Этот эксперимент показывает, что
лишайники достаточно хорошо защищены химически. Анти¬
биотический эффект лишайниковых веществ против патоген¬
ных бактерий и микроскопических грибов был проверен еще в
1950-60-х годах, на основе чего в разных странах Европы бы¬
ли созданы и выпущены на рынок медицинские препараты,
содержащие лишайниковые вещества (например «Evosin» в
Германии, «Usno» в Финляндии и «Натрия уснинат» или «Би-
нан» в СССР и др.).
(ж) Реакция на стресс. Лишайники, как долгоживущие
и медленно растущие организмы, способные выживать при
экстремальных условиях окружающей среды, являются субъ¬
ектами многочисленных биотических и абиотических стрес¬
сов. Было показано, что растения в условиях стресса способны
синтезировать так называемые метаболиты стресса, которые
помогают им справиться со стрессом. Вероятно некоторые из
вторичных метаболитов лишайников действуют также как ме¬
таболиты стресса (Lange, 1992; Huneck, Yoshimura, 1996; Law¬
rey, 2009). Например, отмечалось, что концентрация физода-
ловой (physodalic) кислоты существенно возрастает в талломе
Hypogymnia physodes перенесенной из незагрязненного района
в район загрязненный тяжелыми металлами, что указывает на
возможную роль этого вещества в противодействии послед¬
ствиям загрязнения (Bialoriska, Dayan, 2005). Фумарпрото-
цетраровая кислота, вероятно, обеспечивает защиту от метал¬
лов в устойчивом к загрязнению лишайнике Lecanora conizae-
oides (Hauck, Huneck, 2007).
Большинство авторов признают, что вторичные лишай¬
никовые вещества, по всей вероятности, приспособлены к раз¬
нообразным воздействиям, причем одно и то же вещество мо¬
жет играть при этом разные роли (Lawrey, 2009).
Хемотаксономия лишайников (включая краткую историю
метода).
Вульпиновая кислота была первым вторичным метабо¬
литом, выделенным из лишайников еще в 1831 году (Huneck,
Yoshimura, 1996). Первые цветные реакции на лишайниковом
талломе для таксономических целей были проведены финским
лихенологом В. Нюландером, использовавшим для этих целей
раствор йода (7), гидроокись калия (К) и гипохлорит кальция
(С) в 1860 г. (Vitikainen, 2001). Первые серьезные исследова¬
ния лишайниковых веществ проводились в Германии Ф. В.
Цопфом (Zopf) и Р. Гессе (R. Hesse). В фундаментальной рабо¬
те Ф. В. Цопфа (Zopf, 1907) описано более 150 лишайниковых
веществ. Структура многих лишайниковых веществ была вы¬
явлена в течение первой половины 20-го века японскими хи¬
миками Ю. Асахина (Y. Asahina) и С. Шибата (S. Shibata), ко¬
торые предложили также b метод идентификации лишайнико¬
вых веществ по микрокристаллам и описали предполагаемые
пути синтеза этих веществ (Asahina, Shibata, 1954).
Важной отправной точкой в химии лишайников стала
публикация книги Никиты Калберсон (Chicita Culberson) с со¬
авторами «Chemical and botanical guide to lichen products» и
двух дополнений к Hefi(Culberson, 1969, 1970; Culberson et al.,
1977). Эта трилогия объединила данные о более чем 400, из¬
вестных к 1976 г., веществах и их присутствии у видов лишай¬
ников. Эта же группа исследователей предложила стандарти¬
зованный метод для определения лишайниковых веществ -
метод тонкослойной хроматографии (thin layer chromatography
- TLC) - наиболее доступную и точную процедуру для рутин¬
ной идентификации лишайниковых метаболитов, широко ис¬
пользуемую до настоящего времени. В последнее время по¬
явились новые дополнительные данные о лишайниковых ве¬
ществах и более развитая техника для их идентификации.
Большинство морфологически обособленных видов об¬
ладают устойчивым набором лишайниковых веществ, незави¬
симо от района произрастания, субстрата и экологических
условий. Это позволяет использовать присутствие или отсут¬
ствие определенных лишайниковых кислот в качестве таксо¬
номического признака в систематике лишайников. У морфо¬
логически одинаковых видов могут наблюдаться три наиболее
распространенных модели изменений химического состава: (а)
замещение основных веществ, (б) хемосиндромные вариации,
и (в) дополнительные вещества (Elix, 1996).
При замещении типичных веществ, у разных хемотипов
(химических расе), обладающих одинаковыми морфологиче¬
скими признаками, происходит замещение одного метаболита
другим. В случаях, когда два метаболита биосинтетически
близки (т. е. один может синтезироваться из другого в резуль¬
тате одной биосинтетической реакции), хемотипы рассматри¬
ваются в качестве представителей одного вида. В тех же слу¬
чаях, когда вещества биосинтетически далеки и биохимиче¬
ские различия подкрепляются географической изоляцией, хе¬
мотипы следует считать разными видами. Классический при¬
мер этого рода - Pseudevernia furfuracea, вид, в котором были
выявлены три хеморасы: раса, распространенная в северной
Европе, содержащая оливеторовую кислоту, раса из южной
Европы и северной Африки, содержащая физодовую кислоту,
и содержащая леканоровую кислоту раса из северной Амери¬
ки. Поскольку оливеторовая и физодовая кислоты биохимиче¬
ски близки (одна является биологическим производным дру¬
гой) и европейские расы частично перекрывают ареалы рас¬
пространения друг друга, не смотря на то, что у них не выяв¬
лена корреляция с экологией местообитаний, эти расы рас¬
сматриваются как хеморасы одного вида. Североамериканский
таксон, напротив, считается самостоятельным видом - Р. соп-
socians, поскольку его основной продукт леканоровая кислота,
биосинтетически далека от двух других веществ и распростра¬
нение таксона обособленное (Elix, 1996).
Хемосиндромные вариации [хемосиндромная изменчи¬
вость] (Culberson, Culberson, 1976) демонстрируют более
сложные ситуации, когда хемотипы различаются содержанием
в сердцевине постоянных для определенного хемотипа основ¬
ных веществ (одного или двух), в то время как состав допол¬
нительных веществ может каким-то образом варьировать. Ос¬
новные вещества одного хемотипа часто выступают в качестве
дополнительных в другом, и при этом большая часть этих ве¬
ществ являются биогенетически близко родственными. Соот¬
ветственно, хемосиндромы относятся к группе биосинтетиче¬
ски родственных метаболитов, которые в разных хемотипах
представлены в различных комбинациях и концентрациях
(Таб. 3). Среди лихенологов нет согласия в таксономической
трактовке хемотипов в случаях хемосиндромных вариаций.
Например, хемотипы соредиозных лишайников рода Cetrelia
разными авторами рассматривались как один, три или четыре
самостоятельных вида (Randlane, Saag, 1991; Obermayer,
Mayrhofer, 2007). В тех случаях, когда репродуктивная изоля¬
ция вида приводит к его экологической дифференциации, хе¬
мотипы, с хемосиндромными вариациями могут также иногда
рассматриваться в качестве сестринских виды, (Culberson,
1986).
Таблица 3. Химические вариации рода Cetreliopsis (Randlane, Saag,
2003). Присутствие лишайниковых веществ в сердцевине таллома: М -
основное вещество, m— (minor) дополнительное вещество, + - следы
вещества.
Хемотипы
Виды
Лишайниковые вещества, содержа¬
щиеся в сердцевине таллома
(3-орсинольные
депсидоны без
лактонового
кольца
(3-орсинольные деп¬
сидоны с лактоно¬
вым кольцом
Протоцетраровая к-та
Фумарпротоцетраровая к-та
Суккинпротоцетраровая к=та
Физодаловая к-та
Салациновая к=та
9а-0-метил-салациновая к-та
Кваезитиковая к-та
Конкваезитиковая к-та
Норстиктовая к-та
I
С. asahinae
М
m
m
II
С. рариае
+
М
+
+
+
m
+
С. rhytidocarpa
+
М
+
+
+
m
+
III
С.
endoxanthoides
+
m
+
М
m
С. laeteflava
+
m
+
М
m
С. thailandica
+
m
+
+
М
m
IV
С. hypotrachyna
М
m
Наличие дополнительных веществ может усложнить ин¬
терпретацию химических признаков. В таксоне у некоторых
образцов дополнительные вещества иногда присутствуют вме¬
сте с постоянным основным веществам, но могут отсутство¬
вать у остальных. Наличие этих или отсутствие этих веществ
не коррелирует с какими-нибудь морфологическими призна¬
ками или с географическим распространением и, следователь¬
но, не имеет таксономического значения. Тем не менее, преж¬
де чем судить о химии таксона необходимо определить, явля¬
ется вещество постоянным или дополнительным.
Лишь немногие исследования касались вопросов фило¬
генетического сходства разных хемотипов, обладающих сход¬
ной морфологией. Сравнение двух хемотипов вида Ramalina
farinacea, проведенное на основе их ITS-последовательностей,
с другим видом Ramalina показало, что наиболее обычный хе¬
мотип R. farinacea (содержащий протоцетраровую кислоту)
образует монофилетическую группу вместе с очень редким
хемотипом (содержащим циклографин/виренсовую кислоту
(cyclographin/virensic acid), и, следовательно, их объединение в
одном виде оправдано (Stocker-Worgotter et al., 2004).
Основные методики определения лишайниковых веществ.
Определение и изучение вторичных лишайниковых ве¬
ществ может быть независимым субъектом научного исследо¬
вания, но в большинстве случаев эти исследования проводятся
лишь для определения лишайниковых таксонов.
Существует много различных методик, используемых в
химии лишайников: (а) цветные реакции; (б) флюоресценция в
УФ-излучении; (в) микрокристаллизация; (г) стандартная тон¬
кослойная хроматография (TLC); (д) тонкослойная хромато¬
графия высокой точности (HPTLC); (е) жидкостная хромато¬
графия высокой точности (HPLC); (ж) специфические химиче¬
ские методы определения структуры вновь выявленных ли¬
шайниковых продуктов, например, масс-спектрометрия, ядер-
ная магнитно-резонансная спектроскопия, рентгеноструктур¬
ный анализ и др. Ниже детально описаны наиболее удобные и
широко используемые методы. Описания других методов
имеются в литературе (Culberson et al., 1987; Huneck,
Yoshimura, 1996; Orange et al., 2001).
1. Цветные реакции («spot»-test).
Метод цветных реакций - универсальный, быстрый и
наиболее простой метод для определения присутствия некото¬
рых лишайниковых веществ в талломе. Наиболее часто для
этого метода используются следующие реагенты:
(а) 10% раствор едкого кали (гидроокись калия, КОН,
сокр. К). Для его приготовления необходимо 10 г едкого кали
растворить в 100 мл дистиллированной воды. Раствор посте¬
пенно абсорбирует углекислый газ из воздуха и становится
неэффективным, и его следует менять, когда он становится
мутным. Каустическая сода (гидроокись натрия, NaOH) может
быть использована в качестве замены (Orange et al. 2001). С
некоторыми депсидами, депсидонами и хромонами (о-
гидроксил-альдегидами) К вызывает желтую или красную ре¬
акции. Это наиболее удобный реагент для различения между
веществами группы пульвиновой кислоты и желтоватыми
пигментами антрохинонового типа. Хиноны от желтого до
оранжевого цвета под действием реагента становятся ярко-
красными, в то время как производные пульвиновой кислоты
реакции не дают.
(б) Раствор гипохлорита кальция или натрия [Са(С10)2,
сокр. С]. Обычный домашний отбеливатель, без каких либо
добавок, является лучшим источником этого реагента. Раствор
быстро портится и егоследует менять, как только он потеряет
отчетливый запах хлора. Использование темных бутылочек и
содержание в холодильнике продлевает период использования
реагента. С дает розовую, оранжевую или красную реакцию с
некоторыми депсидами и ксантонами (имеющими в метапо¬
ложении две свободные гидроксильные группы) и зеленую
реакцию с определенными дибензофуранами (например,
стрепсилин). Реакции как правило продолжаютсялишь не¬
сколько секунд и быстро проходят. Иногда используется реак¬
ция КС, то есть на образец капают раствором К, а затем быст¬
ро добавляют в каплю раствор С.
(в) Парафенилендиамин [СбН4(ТМН2)2, сокр. PD или Р].
Используется в виде кристаллов, 2-3 кристалла следует разве¬
сти на часовом стекле в нескольких каплях этилового спирта.
Раствор быстро испаряется, но может быть восстановлен до¬
бавлением новой порции алкоголя. Для того чтобы раствор
меньше испарялся, кристаллы можно развести в небольшом
шприце, после чего использовать шприц, как пипетку. При
этом капля раствора будет очень небольшого размера. Аль¬
тернативой может служить более стабильный раствор PD,
именуемый «Раствором Штайнера», которым можно пользо¬
ваться постоянно в течение нескольких месяцев. Для его при¬
готовления необходимо 1 г парафенилендиамина и 10 г суль¬
фита натрия (Na2SC>3), а также ок. 5 мл моющего средства
(например, средство для мытья посуды) развести в 100 мл ди¬
стиллированной воды. PD токсичен и работать с ним следует с
большой осторожностью, избегая контакта с кожей и попада¬
ния на одежду! Небольшие количества реагента приводят к
образования несмываемых коричневых пятен, поэтому фраг¬
менты таллома, тестированные раствором PD, следует удалять
из гербарного образца. PD вызывает желтую или красной ре¬
акцию с ароматическими веществами, содержащими альде¬
гидную (СНО) группу. Полное проявление окраски может за¬
нять несколько минут.
(г) Иод (IKI, сокр. I или J). Йодный раствор реагирует не
со вторичными метаболитами лишайников, а скорее с полиса¬
харидами крахмального типа, имеющимися в талломе и в пло¬
довых телах. Настоящая амилоидная реакция характеризуется
интенсивным синим окрашиванием, вследствие наличия изо-
лихенана в талломе. Ткани аскомы (гимений, сумки или спо¬
ры) также часто окрашиваются йодом. Для тестирования этих
тканей часто используется раствор Люголя (0.5-1 г йода и 1.5-
3 г калия в 100 мл дистиллированной воды (Orange et al. 2001).
При изучении апикального аппарата сумок рекомендуется
предварительно обработать препарат 10% К: капнуть каплю К
под покровное стекло и вытягивая фильтровальной бумагой
воду с другой стороны стекла, после этого таким же образом
промыть срез чистой водой перед воздействием йода. Это дей¬
ствие обозначается как К/1.
(д) 50% азотная кислота (HNO3, сокр. N), смесь концен-
трированой азотной кислоты и дистиллированной воды 1:1.
Этот реактив очень опасен для кожи, глаз и одежды! N с раз¬
ными пигментами, содержащимися в апотециях, дает красную
реакцию разных оттенков, либо зеленую реакцию с другими
пигментами в эпитеции. Поэтому он применяется при микро¬
скопическом изучении срезов аскомы.
Тест на цветные реакции можно проводить и на коровом
слое и на сердцевине (после удаления кусочка корового слоя
бритвенным лезвием) таллома лишайника. Реагенты наносятся
на сухой лишайник в виде маленькой капли, после чего все
изменения цвета аккуратно фиксируются. Реакцию следует
проводить под лупой (х10), а еще лучше - наблюдая в стерео¬
микроскоп. Присутствие вторичных метаболитов можно опре¬
делить, сравнивая результаты теста с данными таблицы из¬
вестных цветных реакций (Таб. 07-003).
Для удобства работы и увеличения срока действия реак¬
тивов их можно помещать в шприцы для инъекций (объемом
2-5 мл), предварительно обрезав острый конец иглы.
2. Ультрафиолетовая флюоресценция.
Облучение некоторых лишайниковых веществ длинно¬
волновым ультрафиолетом с длиной волны 366 нм вызывает
флюоресценцию разной интенсивности и цвета (Таб. 4). В не¬
которых случаях флюоресценция сразу видна на лишайнико¬
вом талломе, а у некоторых видов заметна только на хромато¬
графических пластинках. Флюоресценцию лучше всего
наблюдать в специальном закрытом боксе. Пигменты корово¬
го слоя могут маскировать флюоресценцию, поэтому для
наблюдения флюоресценции веществ сердцевины, чтобы при¬
открыть ее, необходимо удалять верхний коровой слой талло¬
ма.
Таблица 4. Цветные реакции наиболее распространенных лишайнико¬
вых веществ с К, С, КС и Р и их флюоресценция в ультрафиолетовом
свете с длинной волны 366 нм (Randlane, Saag, 2004). Для пигментов
цвет вещества добавлен в скобках.
Лишайниковое
вещество
Реагенты
UV
К
С
КС
Р
Алекториаловая
(alectorialic) к-та
желт.
красн.
красн.
темно¬
желт.
желт.
Алектороновая
(alectoronic) к-та
-
-
красн.
-
голуб.-
бел.
Атранорин
(atranorin)
светло-
желт.
-
-
светло-
желт.
-/бел.
Барбатовая
(barbatic) к-та
-
/оранж
/оранж
-
белова
т.
Барбатоловая
(barbatolic) к-та
желт.
-
-
желт.
-
Вульпиновая
(vulpinic) к-та
(желт.)
-
-
-
-
-
Г ирофоровая
(gyrophoric) к-та
-
розов./
красн
красн.
-
сер./бе
л.
Диварикатовая
(divaricatic) к-та
-
-
-
-
бел.
Констиктовая
(constictic) к-та
желт.
-
-
оранж.
-
Леканоровая
(lecanoric) к-та
-
красн.
красн.
-
-
Лихексантон
(lichexanthone)
-
-
-
-
желт.
Лобаровая (lobaric)
к-та
-
-
красн./
фиол.
-
бел.
Норстиктовая
(norstictic) к-та
желт. ->■
темно-
красн.
-
красн.
желт./к
расн.
-
Оливеторовая
(olivetoric) к-та
-
красн.
красн.
-
бел.
Паннарин (pannarin)
-
-
-
оранж.
-
Паннаровая
(pannaric) к-та
-
оливк.
темн.
зелен.
-
-
Париетин (parietin)
(желт., оранж.)
красн.
-
-
-
-
Перлатоловая
(perlatolic) к-та
-
-
-
-
бел.
Пикролихеновая
(picrolichenic) к-та
-
-
фиолет
-
белова
т.
Протоцетраровая
(protocetraric) к-та
-
-
розов.
оранж.
/красн.
-
Псоромовая
(psoromic) к-та
-
-
-
ярко-
желт.
-
Салациновая
(salazinic) к-та
красн./
желт.
-
-
оранж.
-
Сордидон
(sordidone) (желт.)
-
желт.
желт.
-
-
Стиктовая (stictic) к-
та
желт.
-
-
оранж.
-
Тамноловая
(thamnolic) к-та
темно¬
желт.
-
-
оранж.
-
Тиофановая
(thiophanic) к-та
-
оранж.
оранж.
-
-
Усниновая (usnic) к-
та (светл. желт.)
-
-
оранже
ват.
-
темн.
Физодаловая
(physodalic) к-та
желт,-
коричн
-
-
красн.
-
Физодовая
(physodic) к-та
-
-
розов.
-
белова
т.
Фу м арпрото цетраро
вая
(fumarprotocetraric)
к-та
коричн
еват.
-
-
красн.
-
Эверновая (evernic)
к-та
-
-
-
-
бел.
3. Микрокристаллизация.
Методика определения лишайниковых веществ по мик¬
рокристаллам была в основном разработана в 1930-е годы
японским лихенологом Ю. Азахиной (Y. Asahina). Лишайни¬
ковые вещества в этой методике определяются по характеру
(форме и цвету) кристаллов, формирующихся лишайниковыми
веществами в разных реагентах. Облучение некоторых ли¬
шайниковых веществ длинноволновым ультрафиолетом с
длиной волны 366 нм вызывает флюоресценцию разной ин¬
тенсивности и цвета (Таб. 07-003). В некоторых случаях флю¬
оресценция сразу видна на лишайниковом талломе, а у неко¬
торых видов заметна только на хроматографических пластин¬
ках. Флюоресценцию лучше всего наблюдать в специальном
закрытом боксе. Пигменты корового слоя могут маскировать
флюоресценцию, поэтому для наблюдения флюоресценции
веществ сердцевины, чтобы приоткрыть ее, необходимо уда¬
лять верхний коровой слой таллома.
Фотографии характерных кристаллов около 200 вторич¬
ных лишайниковых веществ опубликованы (Huneck,
Yoshimura, 1996) и доступны для сравнения с целью определе¬
ния лишайниковых веществ. После утверждения тонкослой¬
ной хроматографии в качестве рутинного метода определения
лишайниковых веществ, метод микрокристаллизация перестал
использоваться широко, но по прежнему удобен для различе¬
ния некоторых пар веществ, которые плохо разделяются хро¬
матографически, напримет, таких как гирофоровая и лекано¬
ровая кислоты или барбатовая и диффрактовая (Orange et al.,
2001). Метод практичен в тех случаях, когда требуется разде¬
лить образцы относящиеся к небольшому числу видов с уже
известной химией. Прекрасный пример реализации этого
представлен в «Ключе для определения группы Cladonia
chlorophaea в Европе с использованием микрокристалличе¬
ских реакций» (Orange, 1992).
4. Тонкослойная хроматография (TLC).
Тонкослойная хроматография - это сравнительно про¬
стой и недорогой метод изучения лишайниковых метаболитов,
хотя он требует наличия обычного лабораторного оборудова¬
ния, включая вытяжной шкаф и ультрафиолетовую лампу. Он
был разработан в 1960-70-х годах группой исследователей под
руководством Никиты Калберсон, в качестве стандартного ме¬
тода для определения лишайниковых веществ (Culberson,
Kristinsson, 1970; Culberson, 1972b; Culberson, Amman, 1979;
Culberson et al., 1981) и превратился в настоящее время в
обычную для большинства лихенологов процедуру. Ацетоно¬
вый экстракт, полученный из лишайниковых образцов капают
на стеклянную или алюминиевую пластинку покрытую сили¬
кагелем. Пластинка помещается в герметичную кювету так,
чтобы основание пластинки было неглубоко погружено в
определенную смесь органических растворителей. Для рутин¬
ной работы используется четыре системы растворителей (А,
В, С и G) и еще около десяти дополнительных - для специаль¬
ных целей. Лишайниковые вещества, имеющиеся в образце,
разделяются одно от другого во время движения растворителя
в тонком слое силикагеля, продвигаясь с раствором на разное
расстояние от точки старта, и позднее проявляются в виде пя¬
тен при опрыскивании пластинки серной кислотой после ко¬
роткого нагревания («обугливания»). Полученные пятна пред¬
варительно определяются по их цвету и относительному по¬
ложению на пластинке, а окончательная идентификация про¬
изводится по расположению пятен от исследуемых образцов
по отношению к пятнам контрольных образцов с известным
химическим составом (Orange et al., 2001). Таблицы, содер¬
жащие характеристики хроматографических данных лишай¬
никовых веществ (например, Rf - относительное расстояние
пятна от стартовой линии в разных растворителях, цвет пятна
до и после сушки и при ультрафиолетовом свете, рекомендуе¬
мые контрольные виды и др.) доступны в многочисленных по¬
собиях и позволяют определять вещества по пятнам, получен¬
ным на хромотографических пластинках (Orange et al. 2001;
Vainshtein et al. 1990). Для более точного определения веществ
мы рекомендуем сравнение четырех цифровых фотографий
одной и той же пластинки (Рис. 07-002) - (а) в коротковолно¬
вом (254 нм) ультрафиолетовом свете до сушки; (б) в длинно¬
волновом (366 нм) УФ свете до сушки; (в) в дневном свете по¬
сле сушки; (г) в длинноволновом УФ после сушки. Использо¬
вание цифровых фотографий при ультрафиолетовом облуче¬
нии увеличивает чувствительность метода и позволяет проще
производить обработку химической информации (Saag, 2008).
5. Тонкослойная хроматография высокой точности
(HPTLC).
Тонкослойная хроматография высокой точности - это
метод, основанный на тех же принципах, что и TLC. Он был
предложен в 1990-х годах группой лихенологов из Лундского
университета в Швеции (Arup et al. 1993). Преимущества этого
метода перед TLC выражается в более высоком разрешении
пятен, большей чувствительности, более высокой скорости
анализа и в меньшем расходе растворителей. Более высокое
разрешение пятен достигается использованием пластинок с
тонкозернистым силикагелем.
чем пластинки для TLC, размеры (обычно 10 х 10 см) и
должны высушиваться при 50°С. Для нанесения капель ис¬
пользуются капиллярные трубочки объемом 1 цл и оборудо¬
вание, позволяющее точно наносить капли. Пластинки прояв¬
ляются в горизонтальном положении внутри специальной ма¬
ленькой камеры, с использованием лишь нескольких милли¬
литров растворителя. Но поскольку горизонтальные камеры
для HPTLC довольно дороги, метод этот не столь широко ис¬
пользуется, как TLC. Более того, одна из стандартных систем
растворителей (В), в целях сохранения здоровья исследовате¬
лей, для использования ее в HPTLC была немного изменена.
Это привело к различиям хроматографических данных ве¬
ществ, от результатов, полученных стандартным TLC мето¬
дом.
6. Жидкостная хроматография высокой точности
(HPLC).
Жидкостная хроматография высокой точности - это ме¬
тод для анализа веществ, которые не могут быть разделены
газовой хроматографией, поскольку они или не достаточно
летучие или слишком нестабильны при высоких температурах
(Huneck, Yoshimura 1996). Для этого метода необходимо иметь
специальное хроматографическое оборудование, включая
насос, дроссель (restrictor), колонку и датчик. Впервые метод
HPLC для определения лишайниковых веществ был предло¬
жен Никитой Калберсон (Culberson С. F., 1972а), а наиболее
подробно использование этого метода описано в работе Г.
Файге (Feige et al., 1993), в которой представлены данные для
определения более 330 лишайниковых веществ. Для иденти¬
фикации веществ используется индекс удерживания, рассчи¬
танный по элюционному времени (времени вымывания из ад¬
сорбента) для соответствующего пика, в сравнении со стан¬
дартами. Не смотря на то, что TLC вне всякого сомнения бу¬
дет оставаться простейшим и недорогим методом для рутин¬
ной идентификации вторичных лишайниковых метаболитов,
HPLC - это удобное дополнение, особенно для количествен¬
ного измерения содержания веществ в смесях, в тех случаях,
когда разделение их методом TLC затруднительно.
Рис. 72. TLC-хроматограммы видов Lepraria (1-10) в растворителе А
(Saag, 2008). А - в УФ 254нм до нагревания; В - в УФ 366 нм до нагре¬
вания; С - в видимом свете после нагревания; D - в УФ 366 нм после
нагревания. Вещества: а - атранорин; al - алекториаловая к-та; b - бео-
мицесовая к-та; f - фумарпротоцетраровая к-та; п - норстиктовая к-та;
pnm -6-метилэфир паннаровой кислоты; ро - порфириловая к-та; гп -
рангиформовая к-та; го - роцелловая к-та; sq - скваматовая к-та; st -
стиктовая к-та; t - тамноловая к-та.
Литература
Андреев М. П. Эпилитные лецидеоидные лишайники
России (сем. Lecanoraceae): Дис. ... док. биол. наук. Санкт-
Петербург, 2004. 331 с.
Вайнштейн Е. А., Равинская А. П., Шапиро И. А. Спра¬
вочное пособие по хемотаксономии лишайников. Ботаниче¬
ский институт им. В. Л. Комарова АН СССР. Л., 1990. 152 с.
Armaleo D., Zhang Y., Cheung S. Light might regulate di¬
vergently depside and depsidone accumulation in the lichen Par-
motrema hypotropum by affecting thallus temperature and water
potential // Mycologia. 2008. Vol. 100, № 4. P. 565-576.
Arup U., Ekman S., Lindblom L., Mattsson J.-E. High per¬
formance thin layer chromatography (HPTLC), an improved tech¬
nique for screening lichen substances // Lichenologist. 1993. Vol.
25. P. 61-71.
Asahina Y., Shibata S. Chemistry of Lichen Substances. Jap¬
anese Society for the Promotion of Science, Tokyo. 1954. 240 pp.
Bialonska D., Dayan F. E. Chemistry of the lichen Hy-
pogymnia physodes transplanted to an industrial region // Journal
of Chemical Ecology. 2005. Vol. 31, № 12. P. 2975-2991.
Culberson C. F. Chemical and Botanical Guide to Lichen
Products. University of North Carolina Press, Chapel Hill. 1969.
628 pp.
Culberson C. F. Supplement to "Chemical and Botanical
Guide to Lichen Products" // Bryologist. 1970. Vol. 73. P. 177—
377.
Culberson C. F. High-speed liquid chromatography of lichen
extracts // Bryologist. 1972a. Vol. 75. P. 54-62.
Culberson C. F. Improved conditions and new data for the
identification of lichen products by a standardized thin-layer chro¬
matographic method // Journal of Chromatography. 1972b. Vol.
72. P. 113-125.
Culberson C. F., Ammann K. Standardmethode zur Dunn-
schichtchromatographie von Flechtensubstanzen // Herzogia. 1979.
Vol. 5. P. 1-24.
Culberson C. F., Culberson W. L., Gowan S., Johnson A.
New depsides from lichens: microchemical methodologies applied
to the study of new natural products discovered in herbarium spec¬
imens // American Journal of Botany. 1987. Vol. 74. P. 403-414.
Culberson C. F., Culberson W. L., Johnson A. Second Sup¬
plement to "Chemical and Botanical Guide to Lichen Products".
American Bryological and Lichenologial Society, Missouri Botan¬
ical Garden, St. Louis. 1977. 400 pp.
Culberson С. F., Culberson W. L., Johnson A. A standarized
TLC analysis of P-orcinol depsidones // Bryologist. 1981. Vol. 84.
P. 16-29.
Culberson C. F., Kristinsson H. A standardized method for
the identification of lichen products // Journal of Chromatography.
1970. Vol. 46. P. 85-93.
Culberson W. L. Chemistry and sibling speciation in the li-
chen-forming fungi: ecological and biological considerations //
Bryologist. 1986. Vol. 89. P. 123-131.
Culberson W. L., Culberson C. F. Chemosyndromic varia¬
tion in lichens // Systematic Botany. 1977. № 1. P. 325-339.
Elix J. A. Biochemistry and secondary metabolites / Nash 111
Т. H. Lichen biology. Cambridge University Press. 1996. P. 154—
180.
Elix J. A., Stocker-Worgotter E. Biochemistry and secondary
metabolites / Nash III, Т. H. Lichen biology. 2nd Ed. Cambridge
University Press. 2008. P. 104-133.
Fahselt D. Secondary biochemistry of lichens // Symbiosis.
1994. Vol. 16. №2. P. 117-165.
Feige G. B., Lumbsch H. Т., Huneck S., Elix J. A. Identifica¬
tion of lichen substances by a standardized high-performance liq¬
uid chromatographic method // Journal of Chromatography. 1993.
№646. P. 417-427.
Gauslaa Y. Lichen palatability depends on investments in
herbivore defence // Oecologia. 2005. No 143. P. 94-105.
Hauck М., Huneck S. Lichen substances affect metal absorp¬
tion in Hypogymnia physodes II Journal of Chemical Ecology.
2007. Vol. 33. P. 219-223.
Honegger R. Questions about pattern formation in the algal
layer of lichens with stratified (heteromerous) thalli / Peveling E.
(ed.): Progress and Problems in Lichenology in the Eighties. Bibli-
otheca Lichenologica. 1987. Vol. 25. P. 59-71.
Huneck S., Yoshimura I. Identification of lichen substances.
Springer-Verlag. Berlin, Heidelberg, New York. 1996. 493 pp.
Lange O. L. Pflanzenleben unter Stress. Flechten als Pioniere
der Vegetation an Extremstandorten der Erde. Rostra Universitatis
Wirceburgensis, Wurzburg. 1992. 59 pp.
Lawrey J. D. Biology of lichenized fungi. Praeger Publishers,
New York. 1984. 407 pp.
Lawrey J. D. Chemical defense in lichen symbioses / White
J. F., Torres M. S. (eds). Defensive Mutualism in Microbial Sym¬
biosis. Mycology. 2009. Vol. 27. P. 167-181.
Obermayer W., Mayrhofer H. Hunting for Cetrelia chicitae
(lichenized Ascomycetes) in the Eastern European Alps // Phyton
(Horn, Austria). 2007. Vol. 47. P. 231-290.
Orange A. A key to the Cladonia chlorophaea group in
Europe, using microcrystal tests // British Lichen Society Bulletin.
1992. № 70, Summer. P. 36-42.
Orange A., James P. W., White F. J. Microchemical methods
for the identification of lichens. British Lichen Society. 2001. 101
pp.
Purvis O. W., Elix J. A., Broomhead J. A., Jones G. C. The
occurrence of copper-norstictic acid in lichens from cuprificerous
substrata// Lichenologist. 1987. Vol. 19. P. 193-203.
Purvis O. W., Elix J. A., Gaul K. L. The occurrence of cop-
per-psoromic acid in lichens from cuprificerous substrata. Lichen¬
ologist. 1990. Vol. 22. P. 345-354.
Randlane Т., Saag A. Chemical and morphological variation
in the genus Cetrelia in the Soviet Union // Lichenologist .1991.
Vol. 23. P. 113-126.
Randlane Т., Saag A. Taxonomic notes on some cetrarioid
lichens // Mycotaxon. 2003. Vol. 87. P. 479-487.
Randlane Т., Saag A. (eds) Eesti pisisamblikud (Estonian
microlichens; in Estonian). Tartu Ulikooli Kirjastus, Tartu. 2004.
581 pp.
Saag L. Taxonomic and ecologic problems in the genus
Lepraria (,Stereocaulaceae, lichenised Ascomycota) //
Dissertationes Biologica Universitatis Tartuensis. 2008. № 146 P.
1-167.
Shibata S. Biogenetical and chemotaxonomical aspects of li¬
chen substances // Beitrage zur Biochemie und Physiologie von
Naturstoffen. Festschrift Kurt Mothes zum 65. Geburstag. VEB
Gustav Fischer Verlag, Jena. 1965. P. 451-465.
Stocker-Worgotter E., Elix J. A., Grube M. Secondary chem¬
istry of lichen-forming fungi: chemosyndromic variation and
DNA-analyses of cultures and chemotypes on the Ramalina farina¬
cea complex // Bryologist. 2004. Vol. 107. № 2. P. 152-162.
Vitikainen O. William Nylander (1822-1899) and lichen
chemotaxonomy // Bryologist. 2001. Vol. 104. P. 263-267.
Zopf W. Die Flechtenstoffe in chemischer, botanischer,
pharmakologischer und technischer Beziehung. Gustav Fischer
Verlag, Jena. 1907. 449 pp.
Глава VI.
Разнообразие и распространение лишайников России
Урбанавичюс Г. П., Урбанавичене И. Н.
Уровень богатства лихенофлоры России.
В целом, в мире на начало 2006 г. описано около 19 тыс.
видов лишайников, включая порядка 1 600 видов лихенофиль¬
ных грибов (Feurer, Hawksworth, 2007), при этом Т. Feurer и D.
L. Hawksworth полагали, что эту цифру можно увеличить до 20
тыс. видов за счет недоучтенных таксонов. В тоже время по
нашим подсчетам только за 6 лет (2006-2011 гг.) было описа¬
но еще свыше 1700 видов лишайников и лихенофильных гри¬
бов, т.е., в среднем, за год описывалось от 200 до 400 видов
(http://www.mycobank.org). Американские микологи J. Р.
Schmit и G. М. Mueller дают оценку возможного разнообразия
лихенофильных грибов в 3 тыс. видов при 1 600 известных
(Schmit, Mueller, 2007) - т. е. неописанными, по их мнению,
остаются чуть ли не половина видов (по нашим оценкам, ис¬
ходя из соотношения числа видов лишайников и лихенофиль¬
ных грибов, число последних следует ожидать на уровне
3.5-4.5 тыс. видов). Поэтому на 2011 г. число известных в ми¬
ре лишайников можно оценить примерно в 22 тыс. видов.
Сколько всего видов лишайников обитает на Земле? R.
Lucking с коллегами (Lucking et al., 2009), оценивает все ожи¬
даемое разнообразие лишайников в 28 тыс. видов, причем, ос¬
новное разнообразие неописанных видов ожидается в тропи¬
ческих регионах Земли. С учетом скорости появления и коли¬
чества новых описаний, глобальное разнообразие можно ожи¬
дать приблизительно в 30-32 тыс. видов, т. е. еще неописан¬
ными остаются порядка от 8-10 тыс. видов. Это означает, что
до 30 % таксонов лишайников остаются неописанными. Как
много видов еще может быть описано, продемонстрировано в
коллективной работе, объединившей свыше 100 авторов, спе¬
циально собравшихся и описавших в одной статье 100 новых
видов лишайников (Lumbsch et al., 2011).
В России на 2012 г., по нашим подсчетам, известно око¬
ло 3540 видов лишайников и систематически близких нелихе-
низированных сапротрофных и лихенофильных грибов. Много
это или мало и сколько всего видов можно ожидать для флоры
России? По одной из оценок, для территории России, при
уровне изученности лихенофлоры, сопоставимом со Швецией,
потенциально может насчитываться 4600-4650 видов (Урба¬
навичюс, 2011). Но это число можно считать лишь миниму¬
мом, т. к., например, для Северной Америки (севернее Мекси¬
ки) указывается более 5200 видов (Esslinger, 2011), а для Ев¬
ропы (без территории России) в настоящее время выявлено
около 7300 видов (http://www.checklists.de/). Можно ли сопо¬
ставить разнообразие природных условий Европы (несомнен¬
но, очень значительное на пространстве от Шпицбергена до
Средиземноморья) с разнообразием природных условий Рос¬
сии? И влияет ли то, что территория России в несколько раз
больше Европы? В зависимости от ответа может варьировать
и гипотетическая оценка таксономического разнообразия ли¬
хенофлоры России.
Многочисленные лихенофлористические данные (Таб. 5)
свидетельствуют, что не наблюдается явной зависимости ви¬
дового богатства от размеров территории. Видовое богатство
небольшой территории определяет разнообразие субстратов,
доступных для обитания лишайников и широкая амплитуда
микро- и мезоклиматических условий на конкретной площади.
То есть значительная часть лихенофлоры может быть выявле¬
на на основе даже на небольшого фрагмента территории. Так,
если на 1 кв. км площади будет максимально представлено все
разнообразие возможных местообитаний, приемлемых для
лишайников, то на этом участке может быть выявлено до 90 %
видового состава лихенофлоры территории в 100 или даже в
1000 раз большей по площади. Об этом свидетельствуют мно¬
гочисленные примеры, например, когда на 1 кв. км площади
было обнаружено до 50 % видов, выявленных во всей Антарк¬
тике (Smith, 2007), или почти 29 % состава лихенофлоры Фен-
носкандии (около 800 из почти 2800 видов) обнаружено на
площади примерно 200 кв. км, составляющей менее 0.02 %
территории Фенноскандии (Elvebakk, Bjerke, 2006; Alstrup et
al., 2008).
Таблица 5. Глобальное и региональное разнообразие лихенофлор
(значения округлены до десятков).
Регион
Число
видов
Площадь,
кв. км
Источник
Норд Пойнт на о-ве
-130
1
Smith, 2007
Сигни (Южные Орк¬
нейские о-ва, Ан¬
тарктика)
Израиль
-240
20800
Galun, Mukhtar,
1996
Антарктика
-250
*
Ovstedal, Smith,
2001
Сирия
-400
185200
John et al., 2004
Кипр
-430
9300
http://www.chec
klists.de/
Резерват
«Муньеллос»
(Испания)
-500
55
Barreno, Perez-
Ortega, 2005
Шпицберген
-740
62000
Ovstedal et al.,
2009
Национальный парк
«Иль Рояль» (США)
-610
2313
Bennett,
Wetmore, 2005
Литва
-640
65300
Motiejunaite
(личн. сообщ.,
2010)
Окрестности
Скиботн (Сев.
Норвегия)
-700
-200
Elvebakk,
Bjerke, 2006
Исландия
-830
103000
Kristinsson,
Heidmarsson,
2006
Крым
-900
27000
Ходосовцев,
2004
Провинция Рио
Гранде до Сул (Бра¬
зилия)
-900
282000
Spielmann,
2006
Провинция Лигурия
(Италии)
-930
5421
http://dbiodbs.ii
niv.trieste.it/
Дания
-950
44500
Sochting, Al-
strup, 2008
Тайвань
-1000
36000
http://www.chec
klists.de/
Монголия
-1000
1564000
Бязров, 2007
Канарские о-ва
-1100
7450
http://www.chec
klists.de/
Эстония
-1130
45000
Randlane et al.,
2011
Провинция Трентино
Альто Адидже (Ита¬
лии)
-1400
13607
http://dbiodbs.ii
niv.trieste.it/
Земля Баден-
Вюртенберг
(Германия)
-1500
35700
http://www.chec
klists.de/
Коста Рика
1500
51100
http://www.chec
klists.de/
Калифорния
-1510
410000
Tucker, Ryan,
2006
Япония
-1600
372800
Harada et al.,
2004
Финляндия
-1840
338000
Jaaskelainen et
al., 2010
Австрия
-2200
84000
Hafellner, Turk,
2001
Италия
-2350
301000
http://dbiodbs.u
niv.trieste.it/
Германия
-2360
357000
Wirth et al.,
2011
Швеция
-2410
450000
Thor et al., 2010
Бразилия
-2650
8547400
http://www.chec
klists.de/
Фенноскандия
-2860
1173300
Santesson et al.,
2004
Испания
-3000
506000
Llimona,
Hladun, 2001
Франция
-3440
549200
Roux, 2012
Россия
-3540
17075000
Список
лихенофлоры
..., 2010, с
дополн. (2012)
Австралия
-3600
7682300
McCarthy, 2012
США и Канада
-5200
19480000
Esslinger, 2011
Земля
-22000
149000000
Feuerer,
Hawksworth,
2007; данные
http://www.myc
obank.org
Примечание: * - площадь Антарктики (с островами), свободная ото льда, со¬
ставляет около 320-330 тыс. кв. км.
Специально подобранные примеры для разных природ¬
ных зон в пределах России (Таб. 6) с ранжированием исследо¬
ванных территорий по числу выявленных видов при макси¬
мальной степени изученности показывают следующее. На
участке, занимающем менее 0.001% от площади региона, мо¬
жет быть выявлено до 1/4 (заповедник «Галичья гора» в Цен¬
тральном Черноземье) или 1/7 (урочище Сейднотлаг по отно¬
шению к Мурманской обл.) видового состава лихенофлоры.
На площади, составляющей 0.01-0.1% от площади региона
выявляется от 25 до 60% лихенофлоры: остров Долгий по от¬
ношению к Ненецкому АО, локальная флора «Сейд» (в JIa-
пландском заповеднике) по отношению ко всему Лапландско¬
му заповеднику, заповедник «Кивач» - Республика Карелия,
Дальневосточный морской (или Лазовский заповедник) -
Приморский край. В горных районах с наиболее богатой ли-
хенофлорой может быть выявлено до 80-90% видового соста¬
ва лихенофлоры на площади всего 1-2 % от территории
огромного региона (Байкальский заповедник - Республика Бу¬
рятия, Печеро-Илычский заповедник - Республика Коми).
Необходимо отметить, что и на равнинных территориях с от¬
носительно бедной лихенофлорой на площади 0.1-0.2 % выяв¬
ляется до 70-75 % видового состава лихенофлоры (Волжско-
Камский заповедник - Республика Татарстан, заповедник «Бе-
логорье» - Белгородская обл.). Как можно увидеть из данных
таблицы 6, какая-либо зональная зависимость изменения
удельного видового богатства лишайников от размеров пло¬
щади исследования не выражена.
Таблица 6. Разнообразие лихенофлоры регионов России.
Регион
Число
видов
Площадь,
кв. км
Источник
о. Долгий (Баренцево
260
100
Урбанавичюс
море)
и др., 2008
Ненецкий АО
460
176700
Лавриненко и
др., 2006
Новая Земля
510
82600
Андреев и
др., 1996
Урочище Сейднотлаг
-170
-2
Урбанавичюс
(Лапландский
и др., 2012
заповедник)
Локальная флора
-300
-50
Урбанавичюс
«Сейд» (Лапландский
и др., 2012
заповедник)
Лапландский
-610
2780
Урбанавичюс
заповедник
и др., 2012
Мурманская обл.
-1200
145000
Urbanavichus
et al., 2008 с
дополнения¬
ми
Заповедник «Кивач»
326
109
Херманссон и
др, 2002
Онежская Карелия
656
-24000
Фадеева и
др., 2007
Респ. Карелия
1240
172000
Фадеева и
др., 2007
Нижне-Свирский
315
416
Kuznetsova et
заповедник
al., 2007
Восток Ленинградской
520
32600
Kuznetsova et
обл.
al., 2007
Ленинградская обл.
-900
85000
Компил.
данные
Печеро-Илычский
872
7210
Херманссон и
заповедник
др., 2006
Респ. Коми
-980
416000
Пыстина,
2008
Керженский
207
470
Урбанавичене
заповедник
5
Урбанавичюс,
2001
Нижегородская обл.
-400
77000
Шарапова М.
(личн.
сообщ.)
Волжско-Камский
244
100
Урбанавичюс,
заповедник
Урбанавичене
,2005
Респ. Татарстан
350
68000
Компил.
данные
Заповедник «Галичья
93
2
Мучник, 2000
Г ора»
Заповедник
180
21
Конорева Л.
«Белогорье»
(личн.
сообщ.)
Белгородская обл.
240
27100
Конорева Л.
(личн.
сообщ.)
Центральное
-400
168000
Мучник, 2003
Черноземье
Богдинско-
75
185
Компил.
Баскунчакский
данные
заповедник
Волгоградская обл.
220
114000
Веденеев,
2001
Кавказский
820
2800
Собств.
заповедник
данные
Краснодарский край +
-1100
83600
Компил.
Респ. Адыгея
данные
Байкальский
720
1650
Собств.
заповедник
данные
Респ. Бурятия
-950
351000
Компил.
данные
Дальневосточный
205
13
Скирина,
морской заповедник
2004
Лазовский заповедник
474
1210
Чабаненко,
2004; UPS**
Приморский край
-810
166000
Чабаненко и
др., 2002;
UPS
Примечание: * - Компилированные данные, в основе которых положена ревизия
многих источников на 2009 г.; ** - UPS - гербарий университета г. Упсала,
Швеция.
При примерно равных площадях и близкой степени изу¬
ченности хорошо выражена зависимость изменения видового
богатства от характера рельефа сравниваемых территорий.
Например, на равнинной территории юга Коми, площадью
примерно 170 тыс. кв. км, отмечено около 430 видов (Пысти-
на, 2003), а на гораздо меньшей по площади горной террито¬
рии Печеро-Илычского заповедника (ок. 7.2 тыс. кв. км) за¬
фиксировано ок. 870 видов (Херманссон и др., 2006). Вся рав¬
нинная территория Средней России, площадью примерно 1500
тыс. кв. км) насчитывает ок. 830 видов, а горный Южный Урал
(площадь примерно 250 тыс. кв. км) - около 840 видов.
Важнейшие таксономические группы лихенофлоры Рос¬
сии.
По относительному богатству (т. е. доле видов от состава
всей лихенофлоры) 15 наиболее полиморфных семейств ли¬
хенофлоры России следующие: Parmeliaceae (8.77 %),
Lecanoraceae (6.93 %), Physciaceae (6.76 %), Verrucariaceae
(6.39%), Teloschistaceae (5.85%), Ramalinaceae (4.81%),
Lecideaceae (4.38 %), Cladoniaceae (3.42 %), Megasporaceae
(3.08%), Acarosporaceae (2.32%), Stereocaulaceae (2.18%),
Rhizocarpaceae (2.15%), Lichinaceae (2.09%), Pertusariaceae
(2.04 %), Collemataceae (1.95 %). Следует заметить, что суще¬
ственные изменения в систематической принадлежности ро¬
дов могут привести к изменению ранга семейств и это необхо¬
димо учитывать при изучении систематического разнообразия
и, особенно, при сравнении лихенофлор разных регионов.
Наибольшим родовым разнообразием в лихенофлоре
России выделяется семейство Parmeliaceae (включающее
представителей 56 родов), которое является крупнейшим в
мировой лихенофлоре. Далее по убыванию располагаются се¬
мейства: Verrucariaceae - 31 род, Lichinaceae - 25 родов,
Physciaceae - 19 родов, Lecanoraceae - 18 родов, Roccellaceae
и Lecideaceae (включая Porpidiaceae) - 13 родов, Ramalinaceae
(включающее большинство представителей родов, ранее отно¬
сившихся к семейству Bacidiaceae) - 12 родов. Крупнейшие
роды лихенофлоры России, включающие более 50 видов, сле¬
дующие: Lecanora и Caloplaca - более 160 видов, Cladonia -
более 110 видов, Lecidea и Aspicilia - более 90, Verrucaria,
Pertusaria и Rhizocarpon - более 70, Rinodina, Arthonia и
Acarospora - более 60 видов.
Естественно ожидать некоторой географической диффе¬
ренциации в зависимости от физико-географических или при¬
родно-климатических условий того или иного региона, как по
разнообразию, так и по составу семейств и родов. Наглядно
продемонстрировать это можно на примере спектров ведущих
по числу видов семейств основных регионов.
Спектры ведущих семейств лихенофлоры в Арктической России
Европейский сектор
Сибирский сектор
Дальневосточный
сектор
Parmeliaceae
Parmeliaceae
Parmeliaceae
Cladoniaceae
Lecanoraceae
Lecanoraceae
Lecanoraceae
Physciaceae
Cladoniaceae
Verrucariaceae
Cladoniaceae
Verrucariaceae
Physciaceae
Lecideaceae
Lecideaceae
Lecideaceae
Verrucariaceae
Physciaceae
Teloschistaceae
Megasporaceae
Ramalinaceae
Rhizocarpaceae
Teloschistaceae
Rhizocarpaceae
Megasporaceae
Rhizocarpaceae
Teloschistaceae
Ramalinaceae
Ramalinaceae
Stereocaulaceae
Примечание: жирным шрифтом выделены семейства, входящие в 10
ведущих в спектре лихенофлоры России в целом, простым - входящие в
число 15 ведущих, курсивом - не входящие в число 15 ведущих.
Спектры ведущих семейств лихенофлоры в Европейской России
(без арктической части)
Север
Центр
Юг
Lecanoraceae
Parmeliaceae
Parmeliaceae
Parmeliaceae
Ramalinaceae
Lecanoraceae
Verrucariaceae
Cladoniaceae
Teloschistaceae
Lecideaceae
Physciaceae
Physciaceae
Ramalinaceae
Lecanoraceae
Cladoniaceae
Physciaceae
Teloschistaceae
Ramalinaceae
Cladoniaceae
Verrucariaceae
Verrucariaceae
Teloschistaceae
Lecideaceae
Megasporaceae
Rhizocarpaceae
Collemataceae
Acarosporaceae
Stereocaulaceae
Arthoniaceae
Arthoniaceae
Спектры ведущих семейств лихенофлоры Урала и Кавказа
Северный Урал
Южный Урал
Российский Кавказ
Parmeliaceae
Parmeliaceae
Parmeliaceae
Lecanoraceae
Physciaceae
Physciaceae
Cladoniaceae
Lecanoraceae
Teloschistaceae
Physciaceae
Teloschistaceae
Ramalinaceae
Ramalinaceae
Cladoniaceae
Lecanoraceae
Lecideaceae
Ramalinaceae
Verrucariaceae
Verrucariaceae
Verrucariaceae
Cladoniaceae
Teloschistaceae
Lecideaceae
Lecideaceae
Stereocaulaceae
Collemataceae
Collemataceae
Pertusariaceae
Peltigeraceae
Pertusariaceae
Спектры ведущих семейств лихенофлоры Сибири
Западная
Восточная
Южная
Parmeliaceae
Parmeliaceae
Parmeliaceae
Cladoniaceae
Lecanoraceae
Physciaceae
Ramalinaceae
Physciaceae
Lecanoraceae
Physciaceae
Cladoniaceae
Teloschistaceae
Lecanoraceae
Lecideaceae
Verrucariaceae
Teloschistaceae
Ramalinaceae
Ramalinaceae
Peltigeraceae
Teloschistaceae
Cladoniaceae
Lecideaceae
Verrucariaceae
Lecideaceae
Stereocaulaceae
Stereocaulaceae
Acarosporaceae
Caliciaceae
Rhizocarpaceae
Megasporaceae
Спектры ведущих семейств лихенофлоры Дальнего Востока России
Север
Юг
Parmeliaceae
Parmeliaceae
Cladoniaceae
Physciaceae
Lecanoraceae
Cladoniaceae
Lecideaceae
Ramalinaceae
Physciaceae
Lecanoraceae
Ramalinaceae
Teloschistaceae
Teloschistaceae
Pertusariaceae
Stereocaulaceae
Stereocaulaceae
Rhizocarpaceae
Mycocaliciaceae
Pertusariaceae
Collemataceae
Высоким разнообразием и высокими рангами в спектрах
ведущих семейств в большинстве регионов России, независи¬
мо от природных зон и характера рельефа, отличаются семей¬
ства Parmeliaceae, Lecanoraceae, Cladoniaceae и Physciaceae.
Следом за ними (редко поднимаясь выше) располагаются
Teloschistaceae, Verrucariaceae, Ramalinaceae и Lecideaceae
(несколько более сильно варьирующие от региона к региону,
как по видовому богатству, так и по рангу). В тоже время, се¬
мейства, отличающиеся высоким видовым разнообразием (по
сравнению с другими регионами), могут и не находиться в
числе первых. Например, в лихенофлоре Севера Европейской
России и Южной Сибири сем. Cladoniaceae уступает высокие
позиции другим семействам благодаря более значительному
их видовому разнообразию. Интересно, что сем.
Verrucariaceae и Lecideaceae, как и другие, представленные
преимущественно накипными эпилитными видами, более за¬
метную роль играют в лихенофлорах лучше изученных регио¬
нов, а также изобилующих выходами горных пород с много¬
численными каменистыми местообитаниями, что хорошо вид¬
но на примере Европейской России, где на Севере преоблада¬
ют именно эти два семейства. Значение семейств
Megasporaceae и Acarosporaceae в лихенофлорах зависит от
наличия каменистых субстратов, поскольку большинство
представителей этих семейств является облигатными эпили¬
тами.
Высокий ранг сем. Teloschistaceae и Megasporaceae
(включая род Aspicilia, ранее относившийся к семейству
Hymeneliaceae) в региональных флорах часто считается свиде¬
тельством близости (даже флорогенетической) с лихенофло-
рами Древнесредиземноморского подцарства Голарктики. Эти
семейства имеют высокий ранг в Средиземноморье, как, впро¬
чем, и в других южных и/или континентальных областях Зем¬
ли (сем. Teloschistaceae часто занимает1-3 места). В тоже вре¬
мя, сем. Teloschistaceae находится на втором по числу видов
месте, например, в арктической лихенофлоре архипелага
Шпицберген (0vstedal et al., 2009), опережая сем. Cladoniaceae
и Parmeliaceae, и уступая лишь сем. Lecanoraceae, занимаю¬
щему первое место. Это, безусловно, никак не свидетельствует
о каких-либо связях лихенофлоры Шпицбергена с лихенофло-
рами Древнесредиземноморского подцарства. Дело в том, что
на Шпицбергене семейство представлено существенно иным
набором видов, характерных именно для арктических и высо¬
когорных ландшафтов. Кроме того, необходимо иметь в виду
то, что высокое разнообразие семейства Teloschistaceae свой¬
ственно регионам с преобладанием кальцийсодержащих каме¬
нистых субстратов.
То же касается и семейства Megasporaceae. В спектрах
региональных лихенофлор России, в частности, в лихенофлоре
Юга Европейской России и в лихенофлоре Сибирской Аркти¬
ки, сем. Megasporaceae занимает относительно высокое место.
Если в первом случае связь с лихенофлорами Древнесреди¬
земноморского подцарства еще можно предположить, то во
вторым - заподозрить близкую связь лихенофлоры Арктиче¬
ской Сибири с лихенофлорами Средиземноморья невозможно.
Лихенофлора Юга Европейской России не отличается высо¬
ким разнообразием и не характеризуется большим числом ви¬
дов рода Aspicilia, хотя набор видов этого рода сходен с видо¬
вым составом рода в лихенофлорах Древнесредиземноморско¬
го подцарства Голарктики. В Арктической Сибири сложился
самостоятельный центр разнообразия рода Aspicilia со «свои¬
ми» арктическими (или, скорее, горно-арктическими) видами,
широко распространёнными также в горах Южной Сибири,
этот род намного богаче видами.
Не может служить основанием для утверждения о близ¬
ких связях исследуемой лихенофлоры с лихенофлорами
Древнесредиземноморского подцарства Голарктики и высокое
положение в спектре ведущих сем. Collemataceae и
Pertusariaceae. Высокий ранг этих семейств и их высокое так¬
сономическое разнообразие на Юге Дальнего Востока России
связаны, например, с тем, что регион находится вблизи во-
сточноазиатско-австралийской области, характеризующейся
высоким разнообразием пертузариевых и коллемовых. Следу¬
ет сделать вывод - о связях и сходстве лихенофлор можно су¬
дить не по высоким рангам семейств, а путем анализа видово¬
го (или родового) состава семейств, а еще надежнее - видово¬
го состава родов.
Относительные характеристики регионального разнооб¬
разия лихенофлоры России, представляемые в виде спектров
ведущих семейств, не отражают разнообразия того или иного
семейства или рода в регионе. В качестве примера, для каждо¬
го региона можно рассмотреть долю некоторых основных се¬
мейств и родов, от полного числа известных в России. Макси¬
мальное разнообразие видов семейства Parmeliaceae (Рис. 73)
отмечено на Дальнем Востоке. В одной только южной части
Дальнего Востока здесь встречается почти 70% видового со¬
става пармелиевых России, что вполне закономерно, посколь¬
ку Юг Дальнего Востока расположен ближе всего к Юго-
Восточной Азии, являющейся одним из центров разнообразия
пармелиевых в мировой лихенофлоре. Высокая доля пармели¬
евых (свыше 50% видового состава семейства) наблюдается
также в горах Южной Сибири и Кавказа. В равнинных лесо¬
степных, степных и пустынных районах Юга Европейской
России, а также в таежных и лесостепных районах Западной
Сибири разнообразие пармелиевых минимально. Невелико,
менее 30 % от состава пармелиевых России, видовое разнооб¬
разие и в арктических районах.
j V- "Va
■>гг
Рис. 73. Доля видов семейства Parmeliaceae в разных регионах России.
Особенности динамики видового разнообразия пармели¬
евых на уровне рода показано на примере рода Usnea (Рис.
74). Максимальное разнообразие наблюдается на Кавказе,
Юге Дальнего Востока, на Севере и в Центре Европейской
России. В целом же отмечается увеличение видового и родо¬
вого разнообразия пармелиевых в южных горных регионах
России по сравнению с равнинными и арктическими.
Наибольшее разнообразие семейства Verrucariaceae
(включая нелихенизированные виды) отмечается на Севере
Европейской России, в Южной Сибири и на Кавказе (Рис. 75),
хотя, возможно, это объясняется и сравнительно более высо¬
кой степенью изученности лихенофлоры в этих регионах.
Рис. 74. Доля видов рода Usnea в разных регионах России.
ъ
Рис. 75. Доля видов семейства Verrucariaceae в разных регионах России.
Из числа ведущих в лихенофлоре России, можно отме¬
тить сем. Physciaceae, почти 2/3 видового состава которого,
отмечено в горах Южной Сибири, и сем. Lecideaceae -2/3 ви¬
дов которого известно на Севере Европейской России.
Наибольшим разнообразием на Севере Европейской России
характеризуется семейство Lecanoraceae - здесь встречается
свыше 60% видового состава семейства лихенофлоры всей
России. Более 50% видов семейств Ramalinaceae и
Teloschistaceae отмечены на Северном Кавказе, около 50% ви¬
дов семейства Megasporaceae - в лихенофлоре Южной Сиби¬
ри. Из числа ведущих семейств максимального разнообразия в
одном регионе достигают семейства Acarosporaceae, 85% ви¬
дового состава которого встречается в Южной Сибири, и
Cladoniaceae - почти 82% видов которого, сосредоточены на
Юге Дальнего Востока.
Из числа многовидовых семейств, не входящих в 10 ве¬
дущих, можно выделить отличающиеся максимальным разно¬
образием для лихенофлоры Севера Европейской России сем.
Pilocarpaceae (включающие здесь почти 75% видового состава
семейства), Trapeliaceae (73%), Rhizocarpaceae (68%),
Arthoniaceae (62%), Stereocaulaceae (57%), Lichinaceae (51%).
Для Южной Сибири - это сем. Umbilicariaceae (почти 93%),
Collemataceae (72%), Pannariaceae (52%), Catillariaceae (52%).
Для Юга Дальнего Востока, кроме сем. Parmeliaceae и
Cladoniaceae, наивысшим разнообразием отличаются семей¬
ства Lobariaceae (89%), Mycocaliciaceae (71%) и
Pertusariaceae (60%). Остальные регионы - европейские и си¬
бирские, Север Дальнего Востока, все арктические и оба
уральских региона, не имеют ни одного семейства (из числа
многовидовых) с наибольшим разнообразием.
Не менее значительная дифференциация видового раз¬
нообразия наблюдается на уровне родов. В качестве примеров
контрастного географического распределения разнообразия
видов можно привести два рода: род Acarospora - преимуще¬
ственно южного и горно-степного распространения (Рис. 76) и
род Micarea - преимущественно северного и таежного (Рис.
77). Хотя, следует учитывать, что характер распределения
обилия видов последнего рода в значительной мере находится
в зависимости от степени изученности данной группы в реги¬
оне.
При анализе флоры существенное значение имеет учет
отрицательно специфичных семейств, т. е. семейств, отсут¬
ствующих в том или ином регионе. Естественно, что больше
всего таких семейств будет наблюдаться в регионах с бедной
лихенофлорой, таких, например, как Юг Европейской России
и Западная Сибирь. Важно при этом выявить связь таких от¬
рицательно специфичных семейств с географическим факто¬
ром. Наиболее яркий пример - отсутствие в лихенофлоре всех
арктических секторов России таких семейств, как
Pyrenulaceae, Monoblastiaceae, Coenogoniaceae, Phlyctidaceae,
Peltulaceae, Stictidaceae, Strigulaceae и др. (всего более 2 де¬
сятков). При этом всего одно семейство Mastodiaceae с одним
родом и видом Mastodia tessellata является положительно спе¬
цифичным для лихенофлоры Российской Арктики (сам вид
Mastodia tessellata не является сугубо арктическим).
. • &
-•■/v
V
Рис. 76. Доля видов рода Acarospora в разных регионах России.
■tr'
Рис. 77. Доля видов рода Micarea в разных регионах России.
Другой пример: в лихенофлоре одного из наиболее вы¬
сокоспецифичных регионов России - Юга Дальнего Востока -
известно лишь одно положительно специфичное семейство
Megalosporaceae и более двух десятков отрицательно специ¬
фичных, в том числе таких, как Peltulaceae, Stictidaceae,
Gomphillaceae, Biatorellaceae и др.
Приведенные примеры показывают географические раз¬
личия видового разнообразия родов и семейств. Причины та¬
кой дифференциации главным образом связаны с экологиче¬
скими факторами и для части таксонов определяется климати¬
ческими условиями региона, в то время как для других - суб¬
стратными.
Экологические основы региональных и зональных разли¬
чий лихенофлор России.
Положение территории России в схеме биогеографиче-
ского деления Земли и биоклиматические и экологические
условия разных регионов России оказывают определяющее
влияние на распространение и на биологическое разнообразие
лишайников.
Территория России охватывает практически весь спектр
природно-климатических условий внетропической зоны от
высокоарктических пустынь на севере (архипелаги Земли
Франца Иосифа и Северной Земли) до субтропических вечно¬
зеленых лесов на черноморском побережье и горно-аридных
ландшафтов на востоке Кавказа, горных черневых и высоко¬
горных гольцовых и горно-степных ландшафтов Южной Си¬
бири, богатейших хвойно-широколиственных горных лесов
(дериватов юго-восточноазиатской флоры) юга Дальнего Во¬
стока. Эти обстоятельства позволяют формироваться весьма
разнообразной флоре лишайников, существенно отличающей¬
ся по таксономическому составу от региона к региону. В тоже
время территория России почти полностью лежит в пределах
Циркумбореальной области Голарктики. Это накладывает на
таксономический состав лихенофлоры отпечаток относитель¬
ной однородности. Исключением являются Прикаспийская
низменность, относящаяся к Ирано-Туранской области, Юг
Дальнего Востока и небольшие участки в Забайкалье, относя¬
щиеся к Восточно-Азиатской области.
Климатические условия на значительной части террито¬
рии России - арктические или бореальные. Настоящий тепло¬
умеренный климат со средней положительной температурой в
зимние месяцы встречается только на Кавказе, на крайне
ограниченных территориях. Даже на самом юге Приморья в
зимний период средняя температура воздуха обычно ниже —8—
10 С). Более или менее однообразные на значительном протя¬
жении условия Восточно-европейской равнины и Западной
Сибири, при небольшом разнообразии субстратов и однооб¬
разных лесорастительных условиях, обуславливают невысокое
разнообразие локальных и региональных лихенофлор. Однако
на территориях с горным рельефом, особенно в районах с раз¬
витой системой высотно-климатических поясов и при макси¬
мальном разнообразии субстратных и климатических условий,
формируется особенно богатая лихенофлора, достигающая
наибольшего разнообразия в горах регионах на юге страны -
на Кавказе, в Южной Сибири и на юге Дальнего Востока.
Очевидно, что распространение лишайников в значи¬
тельной степени зависит от экологических условий, следова¬
тельно формирование региональных лихенофлор в первую
очередь зависит от климатических условий, наличия и доступ¬
ности определенныхсубстратов. При этом климатические
условия в какой-то мере могут определять тип или облик ли¬
хенофлоры, а доступность субстратов - богатство и разнооб¬
разие видового состава.
Поскольку лишайники - это преимущественно пойкило-
гигрические организмы, жизненные циклы и распространение
которых зависят от доступности влаги, то и распределение
таксонов с разной потребностью во влаге отражает характери¬
стики влажности воздуха на той или иной территории. Напри¬
мер, влаголюбивые виды лишайников, неспособные перено¬
сить периоды с низкой влажностью воздуха (например, виды
океанических местообитаний или дождевых лесов), не извест¬
ны или встречаются крайне редко там, где даже кратковре¬
менно, на один-два месяца в году устанавливается засушливый
период. Наоборот, аридные виды лишайников, приспособлен¬
ные к обитанию в условиях низкой влажности воздуха и по¬
вышенной температуры, не выживают или находятся в угне¬
тенном состоянии в избыточно влажных районах с прохлад¬
ным климатом. Естественно, что ярко выраженные гумидный
или аридный типы климата на обширной, хотя в биоклимати-
ческом аспекте более или менее однородной территории Рос¬
сии встречаются не столь часто.
Для большей части территории России характерен про¬
межуточный или индифферентный тип лихенофлоры, сложен¬
ной видами с широкой амплитудой в отношении факторов
влажности и температуры. Для регионов с хорошо развитым
горным рельефом более характерны варианты, несущие в себе
разные лихенофлористические комплексы, обусловленные
проявлением различных сочетаний мезо- или микроклимати¬
ческих условий. Например, на склонах разной экспозиции
обычно представлены и гумидный, и аридный типы лихено¬
флор.
Влажность воздуха, являясь одним из основных факто¬
ров, определяющих распределение видов лишайников, глав¬
ным образом зависит от трех факторов - близости территории
к океаническим побережьям континентов, рельефа земной по¬
верхности и господствующего направления переноса воздуш¬
ных масс. Океанический и, отчасти, субокеанический климат
характеризуется незначительными амплитудами годового хода
температур и влажности воздуха и в Северном полушарии, в
силу особенностей движения атмосферных фронтов и конфи¬
гурации континентов, характерен для северо-западных их
окраин. В Европе океанический тип климата наиболее резко
выражен на северо-западных побережьях Ирландии, Велико¬
британии и Норвегии, а в Северной Америке - на юге Аляски
и в Британской Колумбии. В пределах России субокеаниче¬
ский характер климата проявляется на Северо-западе России -
в Мурманской области, Карелии, Ленинградской, Калинин¬
градской и отчасти Псковской областях. На Дальнем Востоке
- на Камчатке, островах Курильской гряды, Сахалине и в
Приморье, отчасти на побережье Охотского моря в Хабаров¬
ском крае, ярко выражен один из вариантов океанического -
муссонный тип климата. В этих регионах наблюдается макси¬
мальное разнообразие некоторых океанических или субокеа-
нических родов и семейств лишайников, например, рода Usnea
или семейства Lobariaceae.
С продвижением вглубь континентов влияние океанов
ослабевает и климат становится все более континентальным,
т. е. с более высокими значениями амплитуды колебаний тем¬
пературы и влажности воздуха от сезона к сезону, а в глубине
континентов, на значительном удалении от побережий, стано¬
вится все более засушливым. В этих районах наиболее часто и
обильно встречаются виды, традиционно называемые арид¬
ными (степными, ксероконтинентальными), например, пред¬
ставители семейства Acarosporaceae и, в особенности, рода
Acarospora (Рис. 76).
Проявление фактора влажности часто носит меридио¬
нальный или субмеридиональный характер, также называемый
секторальным, с выраженным градиентом «океаничность -
континентальность», который обуславливает степень конти¬
нентальное™ климата. Следовательно, на глобальном уровне
разнообразие лишайников также имеет субмеридиональный
характер распределения.
Повышение влажности воздуха наблюдается при встрече
атмосферных потоков с горными поднятиями. На наветренных
склонах возвышенностей и горных массивов создаются кли¬
матические условия с более высокой влажностью воздуха,
большим количеством осадков, меньшей амплитудой колеба¬
ний климатических показателей, по сравнению с прилегаю¬
щими равнинными территориями и подветренными склонами.
Т. е. условия наветренных склонов будут в некоторой степени
приближаться к субокеаническим, а подветренных - к более
континентальным. В соответствии с характером распределе¬
ния влажности воздуха ведут себя и лишайники. Гумидный
характер лихенофлоры хорошо выражен на наветренных за¬
падных и северо-западных склонах хребтов Уральской горной
страны, Алтая, Саян, горных систем Прибайкалья и Забайка¬
лья. Тогда как на восточных, южных и юго-восточных склонах
хребтов и в межгорных котловинах часто более ярко выражен
аридный характер лихенофлоры. Особенно ярко проявляется
дифференциация между гумидным и аридным типами ли¬
хенофлор в более южных регионах, что связано с термическим
фактором. Например, на Западном Кавказе - на черноморском
побережье, в долинах рек Шахе, Сочи, Хоста, Кудепста,
Мзымта, обильны виды, ареалы которых в основном находят¬
ся в приокеанических областях теплоумеренных и тропиче¬
ских зон Северного и Южного полушария, например, Degelia
atlantica, D. plumbea, Fuscopannaria ignobilis, F. leucosticta,
Parmeliella parvula, Parmelinopsis afrorevoluta, Pyrenula
macrospora и др. Здесь же встречается около двух десятков
эпифильных видов лишайников, произрастающих на листьях
вечнозеленых растений (например, Echinoplaca epiphylla,
Gyalectidium caucasicum, Phyllogyalidea phyllophila, Porina
hoehneliana, P. rubentior, Vezdaea dawsoniae и др.). Большин¬
ство из них является теплолюбивыми гигрофитами и широко
распространены в тропических регионах Земли. В то время
как на Восточном Кавказе, в горах Дагестана сконцентрирова¬
ны виды родов Anaptychia, Апета, Caloplaca, Fulgensia,
Neocatapyrenium, Rinodina, Squamarina, Thyrea, Toninia и др.,
обычные в резко континентальных субтропических регионах
Центральной Азии, Малоазиатского и Ирано-Туранского
нагорьев.
В Южной Сибири, на наветренных склонах Кузнецкого
Алатау, Западного и Восточного Саяна, Хамар-Дабана и др.
горных систем, несмотря на глубоко внутриконтинентальное
положение, в условиях очень влажного климата, с наиболь¬
шим в континентальной Азии уровнем выпадения дождевых
осадков, весьма многочисленны виды, считающиеся в Запад¬
ной Европе и Северной Америке океаническими или субокеа-
ническими (например, представители родов Cetrelia, Collema,
Heterodermia, Leptogium, Lobaria, Usnea и др.). Напротив, в
межгорных котловинах и на подветренных склонах, в услови¬
ях засушливого континентального климата, наибольшего раз¬
нообразия достигают виды родов Acarospora, Aspicilia, Toninia
и пр.
Кроме влагораспределяющей функции, горный рельеф
обуславливает и высотный градиент изменения температуры,
позволяющий формироваться нескольким высотным расти¬
тельно-климатическим поясам, в определенной степени анало¬
гичным зональным. Формирование местных растительно¬
климатических условий под воздействием особенностей рель¬
ефа оказывает существенное влияние на состав локальных ли¬
хенофлор, обуславливая, таким образом, орографический ха¬
рактер распределения лишайников. Но поскольку рельеф зем¬
ной поверхности не имеет какой-либо зональной обусловлен¬
ности, поэтому и распределение лишайников и их разнообра¬
зие, в том случае, когда это связано с горным характером ре¬
льефа, не может быть зональным. Поскольку большая часть
территории, например, Азиатской России, является горной, то
и выявлять какие-либо зональные особенности в распределе¬
нии лишайников весьма затруднительно. Для этого необходи¬
мы обширные исследования на меридиональном градиенте.
В расселении лишайников в зависимости от характера
распределения влажности воздуха, наряду с субмеридиональ-
ными, наблюдаются и некоторые зональные закономерности.
Несмотря на то, что основным зональным фактором является
термический, непосредственно на расселение лишайников он
оказывает незначительное влияние. В пределах России встре¬
чается совсем мало термофильных видов. Влияние температу¬
ры проявляется, главным образом, опосредованно - через рас¬
пределение доступных для поселения лишайников субстратов,
связанных в первую очередь с древесной растительностью, т.
е. этот фактор правильнее называть биотическим. Лесная рас¬
тительность, от лесотундры и северной тайги до широколист¬
венных лесов и лесостепи, оказывает существенное влияние на
зональное распределение лишайников в пределах России.
Большая группа лишайников, обитающих на стволах и ветвях
деревьев, насчитывающая несколько сотен видов из многих
десятков родов, практически не выходит за пределы лесной
зоны, ни в Арктику, ни в степную зону. Это одно из наиболее
ярких проявлений зональности в характере распределения ли¬
шайников на территории России.
Зональный характер распространения многих видов ли¬
шайников является основанием для выделения географиче¬
ских элементов по зональному принципу, наиболее часто ис¬
пользуемому в отечественной лихенофлористике в географи¬
ческом анализе (Окснер, 1974 и др.; Макаревич, 1963, 1968;
Трасс, 1970; Голубкова, 1983; Седельникова, 1990; Макрый,
1990; Урбанавичюс, 2001; и др.). Не останавливаясь на осве¬
щении проблем и подходов выделения географических эле¬
ментов, отсылаем читателя к указанным источникам.
При сходных климатических условиях распространение
и разнообразие лишайников обусловлено преимущественно
субстратом обитания. Присутствие или отсутствие необходи¬
мого субстрата часто обуславливает характер расселения вида
в пределах занимаемого им ареала - сплошной или фрагмен¬
тарный, а разнообразие и широта спектра доступных субстра¬
тов в определенной местности или регионе определяют богат¬
ство и разнообразие лихенофлоры. Если вид поселяется на
субстрате, который присутствует повсеместно на всем протя¬
жении ареала, то ареал будет более или менее сплошным. Та¬
кое распространение свойственно многим видам, поселяю¬
щимся на деревьях или на почве. Виды, обитающие на каме¬
нистом субстрате, чаще имеют фрагментарный ареал, т. к. ка¬
менистые породы не образуют на земной сплошного покры¬
тия. Например, обширные заболоченные равнинные террито¬
рии Западной Сибири и степи юга Европейской России очень
бедны каменистыми субстратами или вовсе лишены их на
больших пространствах. В то же время любая горная террито¬
рия отличается обилием каменистых обнажений и характери¬
зуется широкой амплитудой микроместообитаний, что также
связано с орографическим фактором. Кроме того, для лишай¬
ников принципиальное значение имеет состав горных пород,
поскольку, например, облигатные кальцефильные виды не мо¬
гут произрастать на кислых горных породах, а ацидофильные
виды - на известняках. Широта распространения видов связа¬
на также и с экологией лишайников. Благодаря высокой эко¬
логической пластичности лишайников они смогли расселиться
от пустынь (полярных, бореальных, высокогорных или суб¬
тропических) до тропических вечнозеленых лесов.
Для стенотопных и приуроченных к определенному суб¬
страту и местообитанию видов лишайников распространение
часто ограничивается присутствием приемлемого (необходи¬
мого) субстрата или подходящих биотопов. Тогда как эври-
топные виды, способные поселяться на нескольких разных
типах субстрата, гораздо менее ограничены в распространении
и часто имеют более широкие ареалы. Однако доля эвритоп-
ных видов в лихенофлоре России не велика, составляя не бо¬
лее 30%. Поэтому широко и повсеместно распространенных
видов не так уж и много. В тоже время, нужно иметь в виду,
что свойство эвритопности/эврисубстратности для некоторых
лишайников может играть существенную роль в сохранении
популяций далеко за пределами климатических оптимумов
основных частей ареалов. Благодаря этому свойству в конти¬
нентальных условиях Южной Сибири - практически в центре
азиатского континента, в тысячах километрах от океанических
побережий и областей с благоприятными климатическими
условиями, выживают лишайники, обладающие пантропиче¬
ским ареалом, например, относящиеся к роду Coccocarpia (С.
erythroxyli, С. palmicola) и многим другие. Можно добавить то,
что небольшие размеры позволяют лишайникам заселять мик-
роэкотопы с оптимальными гидротермическими условиями
при весьма отличных макроэкологических условиях. Поэтому,
если для сохранения жизнеспособности любого вида высших
растений необходимы достаточно большие размеры популя¬
ции и занимаемые ею площади, то для выживания лишайни¬
ков, например, в экстремальных климатических условиях, в
которых высшие растения жить не способны, достаточно не¬
большой экологически приемлемой ниши, зачастую сравни¬
мой с размерами таллома лишайника (Урбанавичюс, 2002).
Характер распространения лишайников на территории
России. Эндемичные и эвритопные роды.
Сколько видов лишайников обладают практически
сплошным распространением в России? Лишь около 70 видов
присутствуют во всех основных регионах России - от Коль¬
ского полуострова и Кавказа до Чукотки и Приморья (Список
.., 2010). Большая часть из них - это макролишайники, в том
числе кустистые Bryoria fuseescens, Cladonia amaurocraea, С.
arbuscula, С. cariosa, С. cervicornis, С. chlorophaea, С.
coniocraea, С. cornuta, С. deform is, C. fimbriata, C. furcata, C.
gracilis, C. macilenta, C. phyllophora, C. pocillum, C. pyxidata, C.
rangiferina, C. squamosa, C. stellaris, C. subulata, C. verticillata,
Evernia mesomorpha, Ramalina pollinaria, Xanthoria candelaria
и листоватые Cetraria sepincola, Hypogymnia bitter i, H.
physodes, Leptogium tenuissimum, Melanohalea exasperatula, M.
olivacea, Parmelia saxatilis, P. sulcata, Parmeliopsis ambigua, P.
hyperopta, Peltigera aphthosa, P. canina, P. didactyla, P.
malacea, P. polydactylon, P. rufescens, Phaeophyscia sciastra,
Physcia aipolia, Ph. caesia. Ph. diibia. Ph. stellaris, Vulpicida
pinastri. Почти треть из перечисленных, представлена видами
всего одного рода - Cladonia и значительную долю составля¬
ют виды рода Peltigera - т.е . представители преимущественно
эпигейных лишайников.
Среди микролишайников всего около двух десятков ви¬
дов являются общими для всех регионов России - Acarospora
fuscata, Amandinea punctata, Aspicilia cinerea, Biatora vernalis,
Buellia disciformis, Caloplaca cerina, C. ferruginea, Candelariella
aurella, C. vitellina, C. xanthostigma, Diploschistes muscorum,
Lecanora hagenii, L. pulicaris, L. symmicta, L. varia, Lecidella
elaeochroma, L. euphorea, Rinodina exigua и 7?. sophodes. Несо¬
мненно, что при дальнейшем изучении лихенофлоры России
доля широко распространенных видов микролишайников, как,
впрочем, и макролишайников, будет существенно увеличи¬
ваться. Надо отметить, что среди широко распространенных,
подавляющее большинство видов является напочвенными и
эпифитными лишайниками, причем среди последних явно
преобладают не облигатные виды.
С другой стороны, из более чем 3 тыс. видов, известных
в Азиатской России (включая Российский Кавказ), порядка
1190 видов не обнаружены в Европейской России - т. е. их
можно считать специфичными для лихенофлоры Азиатской
России, и менее 500 видов (из примерно 2350 известных в Ев¬
ропейской России) не выявлены ни в одном из азиатских реги¬
онов. В сумме это составляет немногим менее 1700 видов или
46-47% от всей лихенофлоры России. Число видов, встреча¬
ющихся и в Европейской и в Азиатской России, составляет
около 1850. Если взглянуть на состав жизненных форм специ¬
фичных (европейских или азиатских) видов лишайников, то
окажется, что среди азиатских специфичных лишайников око¬
ло 720 видов относятся к микролишайникам и примерно 470
видов - к макролишайникам (60% и 40% соответственно, от
состава азиатских специфичных), а из европейских специфич¬
ных - микролишайников около 400 видов и макролишайников
менее 100 видов (80% и 20% соответственно). То есть основ¬
ное отличие российской азиатской и российской европейской
лихенофлор приходится на микролишайники, что, наверное,
закономерно, т. к. эта группа лишайников изучена хуже. Об¬
щее число специфичных лишайников каждого из основных
регионов России достигает порядка 1260 видов, что составляет
более 1/3 лихенофлоры. Наибольшим своеобразием, оценива¬
емым количеством специфичных видов, обладают лихенофло¬
ры Севера Европейской России, Юга Дальнего Востока, Рос¬
сийского Кавказа и Южной Сибири. Лихенофлора Российской
Арктики в целом также является весьма своеобразной.
По сравнению с небольшим числом широко распростра¬
ненных в большинстве регионов России видов, заметно боль¬
ше видов, имеющих обширные ареалы в пределах определен¬
ных природно-климатических зон (и/или заходящих в сосед¬
ние зоны), т . е. приуроченных к зональным природно-
климатическим поясам, а в горных регионах к их высотным
аналогам.
В пределах Российской Арктики около 35% видов, а из
макролишайников - даже более 50% из почти 1460 известных
там видов, встречаются на всем ее протяжении - от европей¬
ского до дальневосточного секторов. Более 97% из этих широ¬
ко распространенных видов имеют широкие ареалы, выходя¬
щие далеко за пределы Арктики, по крайней мере, в границах
России. Большинство этих видов встречаются в высокогорьях
Восточной Сибири, Севера Дальнего Востока и, отчасти, Се¬
верного Урала, но многие из них обычны также и в горах Юж¬
ной Сибири и на Кавказе, вполне вероятно они могут оказать¬
ся обычными и в высокогорьях Юга Дальнего Востока, пока
еще недостаточно изученных.
Настоящих арктических лишайников, распространенных
исключительно или хотя бы преимущественно в Арктике
очень мало. Лишь немногим более 100 видов можно считать
эндемичными для Арктики, пока они не выявлены за предела¬
ми арктической флористической области. Уровень эндемизма,
ранее во многих регионах Арктики считавшийся высоким, по
современным данным оказывается существенно более низким.
Так, для Новой Земли эндемичными можно считать всего око¬
ло 4-4.5% известных там лишайников, но никак не 12%, как
считалось в недавнем прошлом (Окснер, 1974; Пчелкин, 1991).
Эндемичными можно считать такие лишайники, как Arthonia
atomaria, Aspicilia ochrofusca, A. syernaiae, A. vaginans, Belonia
arctica, Buellia minutissima, Caloplaca ursina, Carbonea
invadens и др. - всего около двух десятков видов. В дальнево¬
сточном секторе Арктики доля эндемичных видов составляет
всего около 4%, ранее полагалась - 31% (!). В отличие от ев¬
ропейского сектора Арктики, эндемичными видами здесь яв¬
ляются представители иных таксономических групп лишайни¬
ков - Arthopyrenia humicola, Bacidia alborussula, В. allinita, В.
graminum, В. indigens, В. subabbrevians, Buellia septentrionalis,
Caloplaca geogena, Lecidea cupreorufa, L. delicatula, L.
lygotropa, L. speirococca и др., всего около 3-3.5 десятков ви¬
дов. При этом велика вероятность того, что многие виды, счи¬
тающиеся пока эндемичными в результате критических реви¬
зий таксономических групп могут оказаться широко распро¬
страненными.
Казалось бы, особые суровые арктические условия, на
самом деле играют очень незначительную роль в формирова¬
нии лихенофлор Арктики. На это указывают, например, такие
факты, как полное отсутствие в лихенофлоре архипелага Зем¬
ля Франца Иосифа, насчитывающей почти 140 видов, соб¬
ственно арктических видов. Известен лишь единственный ли¬
шайник, пока не отмеченный в других секторах Арктики и в
других регионах России: «Aspicilia» leprosescens - на самом
деле, нитрофильный супралиторальный вид, широко распро¬
страненный вдоль побережий Северной и Западной Европы и
Северной Америки. Это же подтверждают и последние данные
по лихенофлоре архипелага Шпицберген (0vstedal et al., 2009),
которые показывают всего 12 эндемичных видов (около 1.6%
от всей лихенофлоры). Из 742 лишайников, встречающихся на
Шпицбергене, 93% видов известны в Фенноскандии, а боль¬
шинство видов отмечены либо в Гренландии, либо в Арктиче¬
ской Сибири. Постоянно происходящее снижение доли энде¬
мичных видов в локальных лихенофлорах вполне закономер¬
но. Оно происходит по мере выявления этих видов в иных сек¬
торах Арктики и за ее пределами, либо в результате сведения
их в синонимы ранее описанных видов в ходе таксономиче¬
ских ревизий. Понятно, что после проведения критических
монографических исследований таксономических групп со¬
кращение числа малоизвестных и узко распространенных ви¬
дов будет происходить и далее. Это относится не только к арк¬
тическим лишайникам, но и в равной степени к лишайникам
всех других природных зон.
Лишайники лесной зоны России, расположенной южнее,
но к северу от Кавказа, гор Южной Сибири и Юга Дальнего
Востока, имеют не такое широкое распространение, как арк¬
тические. Из примерно 2200 лишайников, известных для тер¬
ритории, простирающейся от Калининграда до Камчатки,
лишь третья часть видов - около 30%, являются общими для
всех регионов лесной зоны, а среди макролишайников - до
33%. Большая, по сравнению с арктической лихенофлорой,
дифференциация видового состава лихенофлоры лесной зоны
России объясняется тем, что в отличие от Арктики, здесь нет
такой однородности природно-климатических условий. Субо-
кеанические климатические условия крайнего запада и северо-
запада России, соответственно, с более богатой и разнообраз¬
ной лихенофлорой, сменяются на востоке Восточно¬
европейской равнины умеренно континентальными, и далее -
очень разнообразными условиями Уральской горной страны.
Заболоченные равнины Западной Сибири отличаются менее
разнообразной, по сравнению с европейскими и тем более
горными территориями Восточной Сибири и Севера Дальнего
Востока лихенофлорой, хотя, возможно, пока недостаточно
изученной. При этом, регионы в отдельности обладают невы¬
сокой специфичностью флор - доля специфичных видов ко¬
леблется от 1.5% до 3%. Исключением является Северо-Запад
Европейской России, обладающий «благодаря» Балтийскому
щиту и высокой степени изученности, богатой и разнообраз¬
ной лихенофлорой. Однообразие лесных флор объясняется
сравнительной однородностью и однообразием условий на
всем протяжении полосы бореальных лесов. В тоже время,
необходимо учитывать, что горные территории на значитель¬
ной части Сибири остаются слабоизученными, а многие райо¬
ны - настоящими «белыми пятнами».
Граница распространения лишайников между немораль¬
ными широколиственными и бореальными (таежными) хвой¬
ными и мелколиственными лесами выражена менее отчетливо.
Это скорее всего связано с тем, что зона широколиственных
лесов в пределах России представлена незначительно, к тому
же почти полностью сведена вырубками. Кроме того, восточ¬
ноевропейские широколиственные леса расположены в конти¬
нентальных условиях, на крайнем пределе распространения
формаций. Характерный пример - Lobaria pulmonaria, тради¬
ционно считающийся неморальным видом, обычен в западно¬
европейских широколиственных лесах, в то время как в Рос¬
сии в широколиственных лесах этот вид почти не встречается,
но весьма обычен на обширных таежных пространствах
вплоть до лесотундры (Рис. 78). За Полярным кругом, в север¬
ных таежных лесах Мурманской области известно около де¬
сятка его местонахождений, но он так и не найден в зональных
широколиственных лесах Калужской, Тульской, Рязанской,
Брянской, Орловской и Курской областей. Хотя сейчас ло-
бария легочная нередко уже встречается в темнохвойно¬
широколиственных лесах Среднего Поволжья, например, в
Республике Марий Эл. В широколиственных и смешанных
темнохвойно-широколиственных лесах этот вид можно обна¬
ружить лишь на западных склонах Южного Урала.
Рис. 78. Ареал Lobaria pulmonaria на территории России.
Невысокая дифференциация между лихенофлорами се¬
верных и южных лесов в пределах равнинной части Европей¬
ской России (исключая Урал) связана с тем, что лихенофлора
южнолесной полосы практически не имеет своей специфики
видового состава. Например, из приблизительно 400 видов ли¬
стоватых лишайников, известных в лесной полосе Европей¬
ской России (исключая Урал), в широколиственных и сме¬
шанных лесах встречаются менее 30 видов, не известных в
таежной зоне. То есть более южные широколиственные и
смешанные леса равнинной части Европейской России отли¬
чаются крайне низкой специфичностью лихенофлоры.
Совсем иная картина наблюдается в самых южных реги¬
онах России - на Кавказе, в Южной Сибири и на Юге Дальне¬
го Востока. Из более чем 2500 видов, известных по совокуп¬
ности из трех этих регионов (это больше, чем во всех осталь¬
ных внеарктических регионах, вместе взятых), общими явля¬
ются около 22% видов. На фоне остальных регионов России,
за исключением, пожалуй, только Севера Европейской России,
лихенофлора каждого из этих регионов обладает достаточно
высокой специфичностью - по 200 и более специфичных ви¬
дов известно в лихенофлоре Южной Сибири (12% от общего
числа видов лихенофлоры Южной Сибири), на Кавказе (15%)
и на Юге Дальнего Востока (19%). Еще более возрастает доля
специфичных видов лихенофлор этих трех регионов друг от¬
носительно друга - 35%, 28% и 26%, соответственно, для каж-
дого региона, что, вероятно, объясняется тем, что эти регионы
наиболее удалены друг от друга. В то же время, лихенофлора
каждого из этих регионов обладает очень высоким разнообра¬
зием и ярко выраженным своеобразием. В частности, на Кав¬
казе, после наивысших значений в Южной Сибири и на Юге
Дальнего Востока, отмечены абсолютные и относительные
максимумы, видового разнообразия таких полиморфных се¬
мейств, как Teloschistaceae и Ramalinaceae (абсолютный мак¬
симум среди всех регионов России), Collemataceae,
Parmeliaceae, Pertusariaceae, Physciaceae, Pannariaceae,
Graphidaceae и др.
В южных регионах России проходят северные границы
распространения лишайников, основные части ареалов кото¬
рых расположены гораздо южнее. Большинство специфичных
видов Юга Дальнего Востока - это лишайники, ареалы кото¬
рых расположены в более южных областях Южной и Восточ¬
ной Азии. На юге Сибири нередко проходят северные границы
распространения видов, известных из Центральной Азии, цен¬
тральных и южных регионов Китая, Тибета, Гималаев и др. На
Северном Кавказе на северном пределе своего распростране¬
ния находится большинство специфичных видов лишайников,
основные ареалы которых зачастую охватывают тропические
области Земли (кроме эпифилльных видов, известных с Запад¬
ного Кавказа). На Восточном Кавказе встречается немало ви¬
дов, ближайшие местонахождения которых находятся в среди¬
земноморских регионах, Западной, Юго-Западной и Цен¬
тральной Азии. Можно утверждать, что «южные» лихенофло¬
ры Кавказа, Южной Сибири и Юга Дальнего Востока являют¬
ся более самостоятельными в своем развитии, чем располо¬
женные севернее бореальные и арктические. Все это позволяет
предположить, что историческое развитие лихенофлоры Кав¬
каза, Южной Сибири и юга Дальнего Востока шло более неза¬
висимыми путями (Урбанавичюс, 2011).
Литература
Андреев М. П., Котлов Ю. В., Макарова И. И. Биологи¬
ческое разнообразие лишайников Русской Арктики (таксоно¬
мический состав и предварительный анализ) // Новости систе¬
матики низших растений. СПб., 1996. Т. 31. С. 82-94.
Бязров JI. Г. Видовой состав лихенобиоты Монголии.
Версия 5. 2007. -
http://www.sevin.ru/laboratories/biazrov_mong.html
Веденеев А. М. Флора лишайников Волгоградской обла¬
сти: Автореф. дис. ... канд. биол. наук. СПб., 2001. 27 с.
Голубкова Н. С. Анализ флоры лишайников Монголии.
Л., 1983. 248 с.
Лавриненко О. В., Урбанавичюс Г. П., Плюснин С. Н.,
Урбанавичене И. Н., Журбенко М. П. Лихенофлора Ненецкого
автономного округа // Флора лишайников России: состояние и
перспективы исследований. Тр. междунар. совещ., посвящ.
120-летию со дня рождения В. П. Савича. СПб., 2006. С. 135—
140.
Макаревич М. Ф. Анал1з л1хенофлори УкраУнських Кар¬
пат. Киев, 1963. 263 с.
Макаревич М. Ф. Зональный географический элемент
как основа географического анализа // Материалы III закавказ¬
ской конференции по споровым растениям. Тбилиси, 1968. С.
215-221.
Макрый Т. В. Лишайники Байкальского хребта. Новоси¬
бирск, 1990. 200 с.
Мучник Е. Э. Лихенологические исследования в запо¬
веднике «Галичья гора» // Биоразнообразие и экологические
особенности природы Русской лесостепи / Сб. науч. статей,
посвящ. 75-летию гос. заповедника «Галичья гора». Воронеж,
2000. С. 50-57.
Мучник Е. Э. Лихенофлора Центрального Черноземья:
таксономический и эколого-географический анализы, вопросы
охраны: Автореф. дис. ... д-ра биол. наук. Воронеж, 2003. 40 с.
Пчелкин А. В. Использование принципа сопряженности
флоры сосудистых растений и лишайников для флористиче¬
ского районирования // Проблемы экологического мониторин¬
га и моделирования экосистем. Л.: Гидрометеоиздат, 1991. Т.
13. С. 176-188.
Пыстина Т. Н. Лишайники таежных лесов европейского
Северо-Востока (подзоны южной и средней тайги). Екатерин¬
бург, 2003. 240 с.
Пыстина Т. Н. Биота лишайников Республики Коми: со¬
временное состояние и перспективы дальнейших исследова¬
ний // Фундаментальные и прикладные проблемы ботаники в
начале XXI века: Матер. XII Делегатского съезда РБО и Все-
росс. конфер. Часть 2. Петрозаводск, 2008. С. 221-224.
Седельникова Н. В. Лишайники Алтая и Кузнецкого
нагорья. Конспект флоры. Новосибирск, 1990. 175 с.
Скирина И. Ф. Лишайники островов и прибрежных
участков. Лихеноиндикация загрязнения воздуха // Дальнево¬
сточный морской биосферный заповедник. Исследования. Т. 1.
Владивосток, 2004. С. 568-571.
Список лихенофлоры России. [Сост. Г. П. Урбанавичюс]
Спб, 2010. 194 с.
Трасс X. X. Элементы и развитие лихенофлоры Эстонии
// Учен. зап. Тарт. унив. Вып. 268. / Тр. по бот. 1970. Т. 9. С. 5-
233.
Урбанавичене И. Н., Урбанавичюс Г. П. Лишайники
Керженского заповедника // Природные условия Керженского
заповедника и некоторые аспекты охраны природы Нижего¬
родской области / Тр. Гос. природ, заповедника "Керженский".
Том 1. Нижний Новгород, 2001. С. 149-171.
Урбанавичюс Г. П. Как определить географический эле¬
мент лишайников // Труды Первой Русской лихенологической
школы. Петрозаводск, 2001. С. 223-238.
Урбанавичюс Г. П. Лихеноиндикация современных и па-
леобиоклиматических условий Южного Прибайкалья // Изве¬
стия РАН. Сер. геогр. 2002. № 1. С. 81-90.
Урбанавичюс Г. П. Исследования разнообразия лишай¬
ников России (успехи, достижения, тенденции и надежды) //
Флора лишайников России: состояние и перспективы исследо¬
ваний. Тр. междунар. совещ., посвящ. 120-летию со дня рож¬
дения В. П. Савича. СПб., 2006. С. 264-271.
Урбанавичюс Г. П. Особенности разнообразия лихено¬
флоры России // Известия РАН. Сер. геогр. 2011. № 1. С. 66-
78.
Урбанавичюс Г. П., Лавриненко О. В., Урбанавичене И.
Н. Лишайники острова Долгий и близлежащих островов юго-
востока Баренцева моря // Бот. журн. 2009. Т. 94. № 5. С. 32-
51.
Урбанавичюс Г. П., Урбанавичене И. Н. Аннотирован¬
ный список лишайников и близких к ним грибов Волжско-
Камского заповедника // Тр. Волжско-Камского гос. природ,
заповедника. Вып. 6. Казань, 2005. С. 160-187.
Урбанавичюс Г. П., Урбанавичене И. Н., Мелехин А. В.
Лихенофлора Лапландского государственного природного
биосферного заповедника (аннотированный список). (В печа¬
ти).
Фадеева М. А., Голубкова Н. С., Витикайнен О., Ахти Т.
Конспект лишайников и лихенофильных грибов Республики
Карелия / Под ред. Г. П. Урбанавичюса. Петрозаводск, 2007.
194 с.
Херманссон Я., Тарасова В. Н., Степанова В. И., Сонина
А. В. Лишайники заповедника «Кивач» // Флора и фауна запо¬
ведников. Вып. 101 / Под ред. Г. П. Урбанавичюса. М., 2002.
35 с.
Херманссон Я., Пыстина Т. Н., Ове-Ларссон Б., Журбен¬
ко М. П. Лишайники и лихенофильные грибы Печоро-
Илычского заповедника // Флора и фауна заповедников. Вып.
109 / Под ред. Г. П. Урбанавичюса. М., 2006. 79 с.
Ходосовцев А. Е. Лишайники каменистых обнажений
Крымского полуострова: Автореф. дис. ... д-ра биол. наук. Ки¬
ев, 2004. 36 с.
Чабаненко С. И. Лишайники Лазовского государствен¬
ного природного заповедника // Урбанавичюс Г. П., Урба¬
навичене И. Н. Лишайники заповедников России / Современ¬
ное состояние биологического разнообразия на заповедных
территориях России. Вып. 3. Лишайники и мохообразные /
Под ред. О. М. Афониной, Н. С. Голубковой. М., 2004. С. 31-
215.
Чабаненко С. П., Скирина И. Ф., Княжева Л. А. Список
лишайников Приморского края и обитающих на них грибов.
Южно-Сахалинск, 2002. 89 с.
Alstrup V., Grube М., Motiejunaite J., Nordin A., Zhurbenko
M. Lichenicolous fungi from the Skibotn area, Troms, Norway //
Graphis Scripta. 2008. Vol. 20. P. 1-8
Barreno E., Perez-Ortega S. The UNESCO-MAB Reserve of
Muniellos (Spain, Asturias), an example of high lichen diversity in
Europe and the success of conservations strategies // FI. Medit.
2005. Vol. 15. P. 453-460.
Bennett J. P., Wetmore С. M. Lichens of the U. S. National
Parks // Bryologist. 2005. Vol. 108. № 4. P. 544-553.
Elvebakk A., Bjerke J. W. The Skibotn area in North Nor¬
way - An example of very high lichen species richness far to the
north: A supplement with an annotated list of species // Mycotax-
on. 2006. Vol. 96. P. 141-146.
Esslinger T. L. A cumulative checklist for the lichen-
forming, lichenicolous and allied fungi of the continental United
States and Canada. North Dakota State University:
http://www.ndsu.edu/pubweb/~esslinge/chcklst/chcklst7.htm (First
Posted 1 December 1997, Most Recent Version (#17) 16 May
2011), Fargo, North Dakota.
Feuerer Т., Hawksworth D. L. Biodiversity of lichens, in¬
cluding a world-wide analysis of checklist data based on Takhta-
jan’s floristic regions // Biodivers. Conserv. 2007. Vol. 16. P. 85-
98.
Galun М., Mukhtar A. Checklist of the lichens of Israel //
Bocconea. 1996. Vol. 6. P. 149-171.
Hafellner J., Turk R. Die lichenisierten Pilze Osterreichs -
eine Checkliste der bisher nachgewiesenen Arten mit Verbrei-
tungsangaben // Stapfia. 2001. Vol. 76. P. 3-167.
Harada H., Okamoto Т., Yoshimura 1. A checklist of lichens
and lichen-allies of Japan // Lichenology. 2004. Vol. 2. № 2. P.
47-165.
Jaaskelainen K., Pykala J., Rama H. et al. Jakalat • Lichens //
Suomen lajien uhanalaisuus. Ppunainen Kirja. 2010. P. 278-310.
John V., Seaward M. R. D., Sipman H. J. М., Zedda L. Li¬
chens and lichenicolous fungi of Syria, including a first checklist //
Herzogia. 2004. Vol. 17. P. 157-177.
Kristinsson H., Heidmarsson S. Checklist of lichens in Ice¬
land. Listinn var sidast uppfaerdur 21 mai 2006. -
http://www.floraislands.is/fletlist.htm
Kuznetsova E., Ahti Т., Himelbrant D. Lichens and allied
fungi of the Eastern Leningrad Region // Norrlinia. 2007. Vol. 16.
P. 1-62.
Llimona X., Hladun N. L. Checklist of the lichens and li¬
chenicolous fungi of the Iberian Peninsula and Balearic Islands //
Bocconea. 2001. Vol. 14. P. 1-581.
Lumbsch H. Т., Ahti Т., Altermann S. et al. One hundred
new species of lichenized fungi: a signature of undiscovered global
diversity// Phytotaxa. 2011. Vol. 18. P. 1-127.
Lucking R., Rivas Plata E., Chaves J. L., Umana L., Sipman
H. J. M. How many tropical lichens are there ... really? // Biblio-
theca Lichenologica. 2009. N 100. P. 399-418.
McCarthy P. M. Checklist of the Lichens of Australia and its
Island Territories. Australian Biological Resources Study, Canber¬
ra. Version 10 March 2012
http://www.anbg.gov.au/abrs/lichenlist/introduction.html.
Ovstedal D. O., Smith R. I. L. Lichens of Antarctica and
South Georgia: a guide to their identification and ecology. Cam¬
bridge: Cambridge University Press, 2001. 411 p.
Ovstedal D. O., Tonsberg Т., Elvebakk A. The Lichen Flora
of Svalbard // Sommerfeltia. 2009. Vol. 33. 393 p.
Randlane Т., Saag A., Suija A. Lichenized, lichenicolous and
allied fungi of Estonia. Ver. November 1, 2011 -
http://esamba.bo.bg.ut.ee/checklist/est/home.php
Roux C. Liste des lichens et champignons lichenicoles de
France // Bulletin de la Societe linneenne de Provence. 2012. Nu-
тёго special 16. P. 1-220.
Santesson R., Moberg R., Nordin A., Tonsberg Т., Vitikai-
nen O. Lichen-forming and lichenicolous fungi of Fennoscandia.
Uppsala, 2004. 359 p.
Scholz P. Katalog der Flechten und Flechtenbewohnenden
Pilze Deutschlands // Schriftenreihe Vegetationsk. 2000. Hf. 31. S.
1-298.
Schmit J. P., Mueller G. M. An estimate of the lower limit of
global fungal diversity // Biodivers. Conserv. 2007. Vol. 16. P. 99-
111.
Smith L. R. 1. Half a continent in a square kilometer: the ex¬
ceptional lichen diversity of a small Antarctic island // Biblioth.
Lichenol. 2007. Vol. 95. P. 387-403.
Sochting U., Alstrup V. Danish lichen checklist. Version 2.
Copenhagen, 2008. 46 p.
Spielmann A. A. Checklist of lichens and lichenicolous fungi
of Rio Grande do Sul (Brazil) // Caderno de Pesquisa. Ser. Bio.
Santa Cruz do Sul, 2006. Vol. 18. № 2. P. 7-125.
Thor G., Arup U., Arvidsson L. et al. Lavar - Lichens //
Rodlistade arter i sverige. 2010. P. 285-300.
Tucker S. C., Ryan B. D. Revised catalog of lichens, li¬
chenicolous, and allied fungi in California // Constancea. 2006.
Vol. 84. P. 1-275.
Urbanavichus G. P., Ahti Т., Urbanavichene 1. N. Checklist
of lichens and allied fungi of Murmansk Region, Russia // Norrlin-
ia. 2008. Vol. 17. P. 1-80.
Wirth V., Hauck М., von Brackel W. et al. Checklist of
lichens and lichenicolous fungi in Germany. Ver. 2: 19 January
2011 - http://wwwuser.gwdg.de/~mhauck/
Глава VII.
Экология лишайников
Макрый Т. В.
Экология лишайников - это раздел лихенологии, изуча¬
ющий взаимоотношения лишайников с окружающей средой:
влияние экологических факторов (факторов окружающей сре¬
ды) на лишайники и влияние лишайников на экосистемы (био¬
геоценозы), в которых они обитают. Изучение влияния от¬
дельных экологических факторов на лишайники представляет
большой интерес, однако чрезвычайно трудоемко. Лучше все¬
го изучены особенности распределения разных видов лишай¬
ников по субстратам и экотопам (когда многие факторы дей¬
ствуют совместно), поскольку подобные исследования прово¬
дятся, как правило, параллельно с флористическим и таксоно¬
мическим изучением лишайников.
Экологические факторы
Основными экологическими факторами, влияющими на
развитие лишайников и особенности их распределения по
природным зонам, поясам гор, растительным сообществам, а
внутри биогеоценозов по микроэкотопам, являются влаж¬
ность, тепло, освещенность и субстрат. Причем, если пер¬
вые три фактора среды определяются в первую очередь соляр¬
ной зоной, климатическими особенностями региона (океани¬
ческий, континентальный, муссонный или др. климат) и рель¬
ефом местности (перераспределение осадков, неодинаковая
инсоляция склонов разных румбов и др.), то последний фактор
зависит, как от первых трех, влияющих на характер расти¬
тельного покрова, так и от строения земной коры, определяю¬
щего разнообразие и состав горных пород в регионе.
Особое влияние на распределение лишайников и разви¬
тие их талломов оказывают стихийные (например, лесные по¬
жары) и антропогенные факторы - атмосферное загрязнение,
рубки лесов, горные выработки и карьеры, распашка степей,
рекреационная нагрузка и т. д. Подробно вопросы антропоген¬
ного влияния и особенно атмосферного загрязнения на лишай¬
ники освещены, в частности, Л. Г. Бязровым (2002, 2009).
Влажность. Лишайники - пойкилогидридные организмы,
не имеющие никаких специальных приспособлений для регу¬
лирования водного обмена и удержания воды в талломе. Вод¬
ный режим их напрямую зависит от содержания воды в окру¬
жающей среде, в основном - от влажности воздуха. Поглоще¬
ние воды происходит всей поверхностью таллома, при этом
влагоемкость может составлять от 50-250% к сухому весу
таллома (у лепрозных лишайников), 300-500% (у накипных и
листоватых) и до 650-1700% (у слизистых лишайников)
(Barkman, 1958). Высыхание, как и намокание талломов, мо¬
жет происходить очень быстро - в течение нескольких минут.
Основными источниками воды для лишайников служат
атмосферные осадки (дождь, роса, туман), водяные пары воз¬
духа, а также субстрат (например, почва), который бывает
сильно или слабо увлажненным. В регионах, характеризую¬
щихся недостатком атмосферных осадков и сухостью субстра¬
та, преобладают аэрогидридные виды, способные хорошо впи¬
тывать водяной пар прямо из воздуха (например, лишайники
пустынь).
В талломах лишайников есть гифы с хорошо смачиваю¬
щимися стенками, способные к быстрому набуханию, погло¬
щению и удержанию воды, и гифы, стенки которых инкрусти¬
рованы нерастворимыми лишайниковыми веществами, благо¬
даря чему они не смачиваются и способны сохранять запас и
обеспечивать беспрепятственное проникновение воздуха к во¬
дорослям, что необходимо для нормальной жизнедеятельности
лишайника (Окснер, 1974).
Все процессы метаболизма лишайников (фотосинтез,
дыхание и т. п.) могут происходить лишь в условиях достаточ¬
ного увлажнения таллома, поэтому способность лишайников к
фотосинтезу напрямую зависит от влажности воздуха. Опти¬
мальная влажность таллома, при которой фотосинтез достига¬
ет наивысшей интенсивности, колеблется у разных видов от 30
до 90% к сухому весу (Шапиро, 1991). В сухом состоянии
происходит полное прекращение всех процессов жизнедея¬
тельности.
По отношению к влажности лишайники делятся на че¬
тыре основных группы: ксерофильные, мезофильные, гигро¬
фильные и гидрофильные. Эти группы соответствуют основ¬
ным типам местообитаний: ксерофильные лишайники обита¬
ют в сухих условиях (пустыни, сухие степи, саванны), мезо¬
фильные - в условиях умеренного увлажнения (например, леса
умеренного пояса), гигрофильные - в условиях повышенного
увлажнения (влажные леса в условиях постоянной высокой
влажности воздуха), гидрофильные - в воде (ручьи, озера).
Выделение групп основано главным образом на наблюдениях
того, к каким экотопам приурочены виды, что не всегда может
отражать истинные физиологические потребности лишайни¬
ков в воде, а указывать лишь насколько те или иные виды мо¬
гут переносить высушивание (полное и длительное, либо
кратковременное, периодическое). В конкретных биогеоцено¬
зах разнообразие лишайников по отношению к влажности
очень велико. Даже под пологом одного леса могут обитать
виды, с различными потребностями в увлажнении - мезо-
ксерофиты, ксеро-мезофиты, мезофиты, гигро-мезофиты, ме-
зо-гигрофиты, при этом все они занимают соответствующие
микрониши, различающиеся по условиям увлажнения.
Многие лишайники в весенний период (а в горах и в
летний), во время интенсивного таяния снегов, могут долгое
время находиться затопленными водой и не погибать, но это
не означает, что данные виды по своей природе - гидрофиль¬
ны. Среди них бывают и довольно ксерофильные лишайники
(например, Aspicilia transbaicalica). Нормальная жизнедея¬
тельность всех этих видов происходит в воздушной среде. К
типичным водным (гидрофильным) лишайникам относится
лишь небольшое число видов из родов Verrucaria, Staurothele,
Dermatocarpon, Hymenelia, Ionaspis и некоторых других.
Наиболее приспособлены к высушиванию цианобионт-
ные, преимущественно слизистые лишайники, содержащие в
качестве фотобионта цианобактерии (синезеленые водоросли)
из родов Gleocapsa, Chroococcus и др. Обезвоживание для та¬
ких лишайников является нормой, что обусловлено свойства¬
ми самих цианобактерий. Способность цианобактерий перено¬
сить засуху определяется физико-биологическими свойствами
их протоплазмы - большой вязкостью и устойчивостью к
плазмолизу, относительно высоким содержанием связанной
воды в клетке и большой сосущей силой, позволяющей ис¬
пользовать гигроскопическую влагу воздуха.
Температура. Тепловой режим того или иного региона
обусловлен количеством получаемой солнечной радиации.
Отношение лишайников к тепловому режиму проявляется че¬
рез их поясно-зональное распределение. Более холодолюби¬
вые или холодовыносливые виды распространены в арктиче¬
ских и антарктических широтах, а также высоко в горах (в
альпийском и гольцовом поясах), умеренно холодолюбивые
растут в таежной зоне, умеренно теплолюбивые - в зоне ши¬
роколиственных лесов, а наиболее теплолюбивые - в субтро¬
пиках и тропиках (в пустынях, саваннах и т. д.).
Прямое воздействие температуры на лишайники опреде¬
лить довольно сложно. У многих лишайников развитие и рост
таллома происходит в течение короткого периода (или двух
периодов) в году с оптимальными для дыхания и фотосинтеза
показателями температуры, освещенности и влажности возду¬
ха. Большую же часть времени лишайники пребывают в со¬
стоянии покоя - анабиоза, перенося неблагоприятные, в том
числе - и некомфортные температурные для них условия.
Нормальные процессы жизнедеятельности - дыхание и
фотосинтез, происходят у лишайников при невысоких темпе¬
ратурах. У видов умеренных широт пик метаболической ак¬
тивности приходится на весенний период. Это объясняется
тем, что условия, наиболее благоприятные для фотосинтеза у
лишайников, складываются в конце зимы - начале весны
(Smith, 1961). В это время лишайники постоянно увлажнены и
не затенены листвой деревьев и наземной растительностью. В
это же время происходит наиболее активный синтез и накоп¬
ление лишайниковых веществ (Равинская, 1991).
Многие виды не только арктических и антарктических,
но и умеренных широт фотосинтезируют при отрицательных
температурах. У лишайников умеренных широт фотосинтез
происходит в основном при температурах от -10°С до +20°С
(оптимальные значения - от +4 до +8°С). За пределами этих
температур наблюдается лишь незначительная метаболиче¬
ская активность. Арктические и высокогорные виды способны
фотосинтезировать и при более низких температурах.
В обезвоженном состоянии лишайники могут сохранять
жизнеспособность при воздействии на них высоких положи¬
тельных (в эксперименте - до +100°С) и низких отрицатель¬
ных (в эксперименте - до -196°С) температур, после чего в
увлажненном состоянии они восстанавливают свою жизнедея¬
тельность - дыхание и фотосинтез (Kappen, Lange, 1972;
Lange, 1992).
Многие виды лишайников, считающиеся экстремофила-
ми из-за своей способности противостоять экстремальным
температурам, радиации, дегидратации и другим факторам
внешней среды, рассматриваются в качестве наиболее удоб¬
ных кандидатов для изучения влияния факторов космического
пространства на живые организмы.
Лишайники обычны в полярных и альпийских регионах
и других районах со сложными климатическими условиями,
сходными с условиями космоса (экстремально низкие темпе¬
ратуры с длительным периодом ниже точки замерзания, высо¬
кий уровень УФ радиации в летний период и длительный тем¬
ный период, очень низкая относительная влажность воздуха),
где способны формировать доминантные типы растительно¬
сти. В эксперименте они показали способность восстанавли¬
вать метаболическую активность даже после 10 лет дегидра¬
тации, замораживания, отсутствия света и погружения в жид¬
кий азот. Обладание подобными качествами делает их пре¬
красным субъектами для астробиологических экспериментов.
Широкое распространение на планете, способность к выжива¬
нию и симбиотическая природа даже позволяет рассматривать
их в качестве потенциальных межпланетарных пропагул.
Европейское космическое агентство (ESA) в 2005 и 2007
годах провело в установках BIOPAN-5 и BIOPAN-6 на россий¬
ском спутнике Фотон М2 изучение воздействие на лишайники
солнечного ультрафиолетового излучения.
Два вида лишайников Rhizocarpon geographicum и Xant¬
horia elegans и сообщество эндолитных грибов, водорослей и
бактерий были экспонированы в открытом космосе в течение
16 дней. Образцы были отобраны в высокогорных районах
Иберийского полуострова и в области Сухие Долины в Ан¬
тарктиде. Эксперимент был построен так, чтобы отделить воз¬
действие жесткого УФ излучения от других космических фак¬
торов, таких как вакуум, резкий перепад температур и невесо¬
мость.
Уже через 48 часов после полета все образцы, включая
те, которые были экспонированы полностью (без защитных
фильтров), показали уровень фотосинтетической активности,
сравнимый с дополетным. Позднейший ультраструктурный
анализ подтвердил целостность грибных и водорослевых кле¬
ток, хотя мембраны их были существенно повреждены.
Если бы лишайники не обладали способностью быстро
отдавать воду, то при замораживании их талломы разрушались
бы ледяными кристаллами, а чрезмерное нагревание губи¬
тельно действовало бы на белки и клеточные структуры.
По потребностям к теплу лишайники обычно делят на
термофильные и термофобные, но при детальном анализе их
можно разделить на более мелкие группы: микротермные, ме-
зотермные, макротермные (и мегатермные). Однако, когда мы
говорим о макро- и мегатермных лишайниках, это не означает,
что для поддержания жизнедеятельности им необходимы вы¬
сокие температуры. Скорее, они могут выдерживать высокие и
очень высокие температуры в течение длительного времени,
сохраняя жизнеспособность.
В природе, в одном экотопе можно встретить растущие
рядом виды, различающиеся своими потребностями к теплу.
Например, эпифиты, обитающие в одном лесу, но на разных
деревьях, находятся в неодинаковых температурных условиях,
что зависит и от экспозиции стволов (северные и южные по¬
верхности получают разное количество тепла), и от особенно¬
стей коры (у разных видов деревьев кора по-разному нагрева¬
ется). То же самое можно сказать о скалах, находящихся на
склонах разных экспозиций и образованных разными горными
породами (например, известняки - более теплые, а граниты -
холодные и характеризуются резкими колебаниями темпера¬
туры).
Свет. Потребности лишайников в освещенности часто
сочетаются с потребностями их в тепле. Недостаток тепла мо¬
жет компенсироваться лучистой энергией, получаемой лишай¬
никами на экспонированных (освещенных) поверхностях. Од¬
нако при равных условиях температуры и влажности степень
освещенности может являться основным фактором регулиру¬
ющим фотосинтез - при недостатке света фотосинтез прекра¬
тится или его интенсивность будет минимальной. Поэтому
следует различать освещенность прямыми солнечными луча¬
ми, непосредственно вызывающую нагревание самого таллома
лишайника, и освещенность рассеянными лучами, при кото¬
рой температура лишайника определяется температурой
окружающего воздуха и субстрата.
Интересно, что многие теплолюбивые тропические и
субтропические лишайники встречаются и в более северных
по отношению к их основному ареалу широтах, где обитают,
например, в межгорных котловинах, на экспонированных по¬
верхностях южных склонов, компенсируя тем самым недоста¬
ток тепла. В подобных условиях эти виды могут считаться
светолюбивыми, тогда как в центрах своих ареалов - в тропи¬
ках или субтропиках, они могут характеризоваться как тене¬
любивые или теневыносливые. С другой стороны, в одних и
тех же широтах один и тот же вид в одних районах может
встречаться под пологом леса, как тенелюбивый лишайник, в
других - на экспонированных скалах, как светолюбивый. При
этом окраска талломов, обусловленная содержанием лишай¬
никовых веществ, накапливающихся в коровом слое, будет
сильно варьировать - в освещенных условиях она будет более
яркой.
В чистом виде определить влияние освещенности на
распределение лишайников довольно сложно, поскольку свет
не всегда влияет на распределение лишайников прямо, а чаще
опосредованно, через другие факторы. Например, видовое
разнообразие лишайников на освещенных и затененных по¬
верхностях скал, как правило, сильно различается. На осве¬
щенных поверхностях отдача лишайниками воды под дей¬
ствием солнечных лучей происходит значительно быстрее,
чем на затененных, поэтому обычно здесь развиваются виды,
которые либо более требовательны к свету и теплу, либо ме¬
нее чувствительны к высыханию таллома.
По отношению к свету лишайники, как и другие расте¬
ния, подразделяют на фотофильные (светолюбивые) и фото-
фобные, либо на гелиофильные (солнцелюбивые) и гелиофоб-
ные или ангелиофильные (то есть, боящиеся, нелюбящие
солнца).
Субстрат. По приуроченности к разным типам субстра¬
тов лишайники делятся на несколько больших субстратных
или эколого-субстратных групп: эпилыты - обитающие на ка¬
менистом субстрате, эпигеиды - на почве и грунте, эпифиты
(стволовые) - на стволах и ветвях деревьев и кустарников,
эпифиллы - развивающиеся на хвое и листьях вечнозеленых
растений, эпибриофиты - на мхах, эпилихенофиты - обитаю¬
щие на лишайниках, эпиксилы - на истлевшей и гниющей дре¬
весине, эпифитореликвиты - на сухих и гниющих раститель¬
ных остатках. Все эти группы внутри неоднородны и при бо¬
лее детальном рассмотрении подразделяются на более мелкие.
Среди эпилитов выделяется большая группа собственно
эпилитных видов с хорошо развитым талломом и группа эндо-
литных видов, которые живут внутри горной породы (в основ¬
ном известняков и доломитов), не формируя на поверхности
видимого таллома, а образуя на субстрате лишь слабо окра¬
шенные иногда лепрозные пятна и плодовые тела.
Группа эпигеидов также неоднородна. В тундрах и лесах
напочвенный покров, как правило, образуют кустистые и ли¬
стоватые виды, обитающие на грунте (дресве), скелетных гру¬
бых почвах или на мхах, в степях и пустынях эпигеиды пред¬
ставлены накипными и чешуйчатыми видами, растущими
непосредственно на суглинистых почвах. Первые было пред¬
ложено называть «субэпигеидами», вторые - «собственно эпи-
геидами» (Макрый, 1990).
Стволовые эпифиты подразделяются на эпифлеодные -
обитающие непосредственно на коре деревьев и эндофлеод-
ные - развивающиеся внутри коры лиственных (широколист¬
венных) деревьев и кустарников, а на поверхности образую¬
щие лишь плодовые тела (многие пиренокарпные виды). Кро¬
ме того, само дерево (форофит) представляет собой сложную
экосистему с условиями для обитающих там лишайников раз¬
личными - в основании ствола, в подкроновой части и в кроне
(Klement, 1955; Barkman, 1958; Бязров, 2002 и др.). Многие
виды с длинным повисающим волосовидным талломом
(Usnea, Bryoria) обычно обитают лишь на ветвях кроны.
Среди эпифитореликвитов выделяется группа видов,
обитающих на гниющих растительных остатках (в основном
мхов), а также группа видов, растущих на сухих истлевших
остатках мхов и сосудистых растений (например, на сухой се-
лагинелле).
Эпиксилы можно разделить на виды, поселяющиеся на
сухой истлевшей древесине, в основном на сушинах (предста¬
вители родов Calicium, Chaenotheca), и виды, обитающие на
сырой, гнилой древесине колодника (.Icmadophila, большая
группа видов рода Cladonia, обычных также на почвах с бога¬
той подстилкой и разлагающейся древесиной). Эти группы
могут рассматриваться как последовательные сукцессионные
стадии заселения меняющего свои свойства субстрата.
На камнях и скалах лишайники часто поселяются не на
обнаженной каменистой поверхности, а на тонких прослойках
мелкозема, или на так называемом «детрите» (измельченные
частицы грунта, песок, гумус и растительные остатки). Эти
лишайники нельзя считать эпилитами. Как правило, это сбор¬
ная группа видов, среди которых могут быть и напочвенные
лишайники - эпигеиды и субэпигеиды (в том числе ксило-
фильные), и эпифиты, и собственно эпилиты (некоторые ли¬
стоватые виды), и эпифитореликвиты.
Многие виды лишайников иногда могут выступать как
эврисубстратные в тех случаях, когда кроме типичных для
них субстратов они заселяют и другие, несвойственные им или
свойственные в меньшей степени. Например, вид Parmelia sul¬
cata, будучи типичным эпифитом, во многих районах встреча¬
ется и на камнях, и на древесине, реже - на растительных
остатках и почве. Некоторые виды, например, Lecanora
hagenii, не являются строго специфичными к тому или иному
типу субстрата. Возможно, это результат слабой изученности
видов, и при более детальных исследованиях на протяжении
всего ареала можно определить наиболее типичные для них
субстраты.
Некоторые широко распространенные виды легко посе¬
ляются на неприродных антропогенных субстратах, таких как
железные конструкции, бетон, кирпич, черепица. Характер¬
ным субстратом, особенно в Арктике, являются кости круп¬
ных млекопитающих, которые заселяются широко распро¬
страненными в регионе видами лишайников.
Отнесение того или иного вида лишайника к определен¬
ной эколого-субстратной группе не всегда бесспорно. Так, не¬
которые виды, в континентальных районах умеренных широт
обитающие на каменистом субстрате, в центрах своих ареалов
в субокеанических субтропических и тропических районах
являются эпифитами. Такие виды было предложено называть
«вторичными эпилитами» (Макрый, 1990). Как правило, это
широколопастные листоватые лишайники, которые к типич¬
ным эпилитам никакого отношения не имеют.
Узкоспециализированные виды и смена субстрата.
Эпифитные лишайники эволюционно связаны с древесными
породами, на которых они растут и, соответственно, с опреде¬
ленными типами лесов, образуемых этими породами. У них
наблюдается высокая степень специализации к разным типам
коры, что наиболее отчетливо проявляется у накипных видов,
многие из которых приурочены к деревьям с одним типом ко¬
ры или даже иногда к одному виду форофита. Среди листова¬
тых лишайников таких видов очень мало. Как правило, если
экологические условия леса благоприятны для данного вида
листоватого лишайника, то он может поселяться на несколь¬
ких или многих видах древесных растений, хотя в других
условиях предпочитает лишь один определенный вид форофи¬
та. Специализация обнаруживается на уровне семейств и ро¬
дов. Так, представители семейств Physciaceae и Teloschista¬
ceae поселяются преимущественно на лиственных деревьях, в
Сибири, в основном, - на тополях, а большинство видов Usnea
и Bryoria обитает на хвойных деревьях, особенно на темно¬
хвойных (пихта, ель и другие). Первостепенную роль в такой
избирательности играют физические и химические свойства
коры.
Кора может быть гладкой, шероховатой, лущащейся,
трещиноватой, сильно растресканной, отслаивающейся,
опробковевшей или с другими характеристиками. Виды ли¬
шайников, которые могут поселяться на гладкой коре, не
встречаются на сильно трещиноватой. Даже на одном форо-
фите виды, развивающиеся на гладкой молодой коре (в том
числе многие эндофлеодные лишайники), не могут поселяться
на старой растресканной (на которой обитают только эпиф-
леодные виды).
По степени кислотности кора подразделяется на кислую,
субнейтральную и нейтральную (Barkman, 1958). Кора хвой¬
ных деревьев характеризуется в основном кислой реакцией
(рН=3.4^1.5), а кора лиственных деревьев - нейтральной или
субнейтральной (рН=4.7-7.9). В то же время, у одного и того
же вида форофита физические свойства коры и ее кислотность
могут меняться с возрастом. Так у молодого дуба кора слабо¬
трещиноватая и субнейтральная, а у старого - сильно растрес-
канная, отслаивающаяся и более кислая. Этим объясняется
последовательная смена видового состава лишайниковых си-
нузий.
У эпилитных лишайников специализация проявляется в
первую очередь по отношению к химическому составу горных
пород, которые подразделяются на кислые, основные и карбо¬
натные, а кроме того - по отношению к типу горных пород,
которые при одинаковом химическом составе могут иметь со¬
вершенно различные физические свойства, например, кислые
породы могут быть - кристаллическими изверженными (гра¬
нит), метаморфизованными (гнейс) и осадочными (кремни¬
стый песчаник).
Самая многочисленная группа эпилитных лишайников -
это виды, обитающие на кислых породах. Большую группу
составляют кальцефильные лишайники - облигатные кальце-
филы, которые встречаются исключительно на карбонатных
породах. Но и внутри этой группы выделяются более мелкие
группы видов: обитающих либо на известняках, либо на доло¬
митах, либо на мраморах (твердая кристаллическая порода),
либо на осадочно-метаморфизованных кальцийсодержащих (в
том числе вторично карбонатизированных) породах.
Четкая специализация по отношению к субстрату
наблюдается, как правило, у накипных лишайников, большин¬
ство из которых не может менять субстрат на протяжении все¬
го своего ареала, и которые следует считать эволюционно
наиболее специализированной группой по сравнению с листо¬
ватыми и кустистыми лишайниками. Последние, напротив,
довольно часто меняют субстрат в несвойственных им экото-
пах - например, переходят со стволов деревьев на камни и
скалы, могут переходить с почвы на деревья.
Один и тот же вид лишайника даже в одном регионе, в
различающихся по климатическим особенностям районах или
в разных типах леса, может вести себя по отношению к суб¬
страту неодинаково. Например, виды рода кладония, являю¬
щиеся напочвенными в сухих сосняках, во влажных темно¬
хвойных лесах переходят на стволы деревьев и колодник, а
некоторые листоватые виды, во влажных лесах обитающие на
деревьях в качестве эпифитов, в сухих лесах могут встречаться
на камнях и скалах.
Многие лишайники, имеющие дизъюнктивные ареалы с
широкими разрывами, на разных континентах приурочены к
резко различным субстратам (Brodo, 1973). Лишайники эпи¬
фиты субокеанических районов умеренно-теплых, субтропи¬
ческих и тропических областей, в континентальных районах и
в областях с более холодным климатом переходят на скалы,
поселяясь на замшелой или обнаженной поверхности (напри¬
мер, Almborn, 1948; Barkman, 1958; Jorgensen, 1978). Такие
виды при изменении климата - например, общем похолодании
или усилении континентальное™ - могут сокращать свои аре¬
алы, исчезая в естественных для них экотопах, но сохраниться
в качестве реликтов на новых для них субстратах. Ярким при¬
мером этого служат виды Coccocarpia palmicola и С. erythroxy-
li, которые в тропиках обитают на листьях вечнозеленых рас¬
тений как эпифиллы и на коре деревьев как эпифиты
(Arvidsson, 1982). В умеренных широтах свойственные им в
центре массовости субстраты отсутствуют, и в горах Южной
Сибири эти виды встречаются как реликты исключительно на
камнях и скалах.
Адаптивные приспособления к экстремальным усло¬
виям среды. В пустынях, приполярных областях и высокого¬
рьях лишайники находятся в экстремальных условиях суще¬
ствования. Эпилитные лишайники высокогорий изначально
должны быть приспособлены к повышенной солнечной ради¬
ации, значительным температурным колебаниям и периодиче¬
скому недостатку влаги. Часто их приспособительные свой¬
ства являются многофункциональными (Макрый и др., 2000).
Одним из приспособлений лишайников к повышенному
уровню солнечной радиации является окраска их талломов.
Темную окраску (виды родов Umbilicaria, Melanelia, Cetraria и
другие) им придают меланины - высокомолекулярные соеди¬
нения, пигменты, которые выступают универсальными радио¬
протекторами, защищающими талломы от жесткого излуче¬
ния, и, в то же время, - фотоловушками, преобразующими лу¬
чистую энергию в тепловую.
В лишайниках кроме того содержится большой набор
низкомолекулярных метаболитов, отлагающихся в основном
на оболочках гиф верхнего корового слоя и уменьшающих его
светопроницаемость, предохраняя тем самым фотобионт от
чрезмерного освещения (Равинская, 1991). Среди них есть
окрашенные вещества, влияющие на цвет таллома, например -
ризокарповая кислота, содержащаяся в ярко-желтых талломах
фотофильных видов родов Rhizocarpon, Pleopsidium, Acarospo¬
ra, усниновая кислота, придающая лишайникам желто-
зеленый цвет (Alectoria ochroleuca, Asahinea chrysantha, Fla-
vopunctelia soredica, виды рода Cladonia и другие), неокра¬
шенный флюоресцирующий атранорин, известный у предста¬
вителей разных таксономических групп, и другие. Доказано,
что в условиях большей освещенности содержание этих ве¬
ществ значительно увеличивается. Известно также, что усни-
новая кислота способна усиливать процессы фотосинтеза у
симбиотических водорослей (Равинская, 1991).
Некоторые лишайники (представители родов Lecidea,
Miriquidica, Immersaria) образуют на поверхности коры тол¬
стый эпинекральный слой, придающий таллому стеклянный
или лаковый блеск. Этот слой резко увеличивает отражающую
способность поверхности таллома, что служит дополнитель¬
ной защитой от солнечной радиации и от физического (абра¬
зивного) воздействия песка и кристалликов льда, переносимых
ветром. Защитный эпинекральный слой в виде толстого, часто
зернисто растресканного налета, есть и у многих светлоокра¬
шенных лишайников, обитающих в экстремально-сухих усло¬
виях пустынь.
Приспособленность лишайников к засушливым услови¬
ям определяется в первую очередь способностью их погло¬
щать водяной пар из воздуха, но не только. Считалось, что в
клетках лишайников вода содержится и в свободном и в свя¬
занном состоянии. Однако для некоторых видов, обитающих в
условиях холодных пустынь Восточного Памира, было уста¬
новлено присутствие третьего состояния - подвижной изоли¬
рованной воды, способствующей регулированию водного ре¬
жима, а именно ксероустойчивости (Николаев и др., 1975;
Жданова, Василевская, 1982).
Адаптивной структурой, позволяющей многим циано-
бионтным лишайникам обитать в аридных условиях, является
обильная слизь, покрывающая клетки цианобактерии. Эта
слизь состоит из гидрофильных коллоидных полисахаридов,
способных быстро поглощать и удерживать большое количе¬
ство влаги.
Защитным приспособлением цианобактерий от избыточ¬
ной инсоляции является темная окраска слизистых чехлов, а
также дополнительные пигменты клетки. Именно цианобакте¬
рии наиболее засухоустойчивы, наиболее устойчивы к высо¬
ким и низким температурам, повышенной инсоляции и радиа¬
ционному излучению, что позволяет лишайникам, содержа¬
щим цианобактерии, выживать на голых каменистых субстра¬
тах в экстремальных аридных условиях.
Экологическая обусловленность жизненных форм
Эволюция лишайников в разных экологических услови¬
ях шла как путем дифференциации таксонов, так и выработкой
в разных семействах и родах общих морфологических типов
(конвергенция, параллелизм). Связь морфологического типа
лишаиников с окружающей средой проявляется очень ярко.
Формирование разных морфологических типов с характерным
строением таллома (жизненной формой) происходило в про¬
цессе приспособления (специализации) их к определенным
типам субстратов и экотопов.
При анализе лихенофлор обычно ограничиваются кон¬
статацией соотношения таких крупных морфотипов лишайни¬
ков, как накипные, листоватые и кустистые. Однако эти кате¬
гории не могут адекватно отражать все многообразие габитуса
лишайников и форм их приспособления к среде обитания (Го¬
лубкова, Бязров, 1989). Например, среди листоватых лишай¬
ников выделяются две формы: широколопастные мезо-
гигрофильные виды (Lobaria, Platismatia, Cetrelia и другие) и
рассеченнолопастные (узколопастные) мезофильные и мезо-
ксерофильные виды (такие, как Parmelia, Physcia).
Накипные эпифитные и эпилитные лишайники в боль¬
шинстве случаев морфологически четко различаются: первые
характеризуются цельным талломом, вторые - ареолирован-
ным, что также является результатом приспособления тех и
других к различным физическим свойствам субстрата - орга¬
нического (коры) или каменистого. Каменистый субстрат при
увлажнении не изменяется в размерах, тогда как таллом ли¬
шайника сильно разбухает. При нагревании на солнце горные
породы расширяются, а таллом лишайника от жары и сухости,
наоборот, сжимается. Таким образом, ареолированная струк¬
тура талломов эпилитных лишайников позволяет им избегать
опасных напряжений и разрывов (Окснер, 1974).
Семейства и роды являются полностью, или в подавля¬
ющем большинстве своих представителей, узкоспециализиро¬
ванными и имеют определенную жизненную форму. Так,
например, представители родов Psora, Endocarpon, Еп-
dopyrenium - эпигеиды с чешуйчатым талломом, виды родов
Bryoria и Usnea - эпифиты с кустистым повисающим (волосо¬
видным) талломом, виды Umbilicaria, Lasallia, Dermatocarpon
- эпилиты с умбиликатнолистоватым талломом, виды Rhi-
zoplaca, Peltula - эпилиты с умбиликатнонакипным (чешуйча¬
тым) талломом.
Таким образом, жизненная форма лишайника четко кор¬
релирует с его субстратной приуроченностью и той ролью,
которую лишайник играет в формировании растительного по¬
крова, т. е. слагаемых им синузий. Эколого-морфологическая
классификация жизненных форм лишайников, основанная на
одинаковом габитусе видов, исторически сформировавшемся
как приспособление к определенным условиям среды разрабо¬
тана Н. С. Голубковой и Л. Г. Бязровым (1989) на примере ли¬
хенофлоры Монголии.
Экология и особенности распространения лишайников
Климатические и микроклиматические условия.
Субстраты. Экологическая микрониша. Экотипы. Пред¬
ставление о лишайниках как об организмах с очень широкой
экологической амплитудой и исключительными адаптивными
способностями несколько преувеличено. Такое представление
возникает из-за недооценки экологической специфики микро¬
ниш, в которых обитают лишайники, когда на микронишу ав¬
томатически переносятся все те условия, что ее окружают.
Экологическую амплитуду того или иного вида лишай¬
ника часто понимают через его широкий ареал, а то, что во
всех частях ареала он приурочен либо к более или менее сход¬
ным экотопам (например, к одинаковым поясам гор, к лесам
одного типа, выходам карбонатных горных пород, или влаж¬
ным силикатным скалам), либо к резко различным экотопам,
но с равнодействующим комплексом экологических факторов
- это не принимается во внимание.
Действительно, в сравнении с высшими сосудистыми
растениями экологическая амплитуда лишайников несколько
шире из-за высокой их толерантности к неблагоприятным
факторам среды (! но в состоянии анабиоза). Это не противо¬
речит тому, что нормальная жизнедеятельность каждого вида
лишайника связана с более или менее ограниченной экологи¬
ческой средой. Если для сосудистых растений эта среда опре¬
деляется формацией, типом растительности, природной зоной,
то для лишайников, в первую очередь - микроэкотопом и суб¬
стратом. Зональная приуроченность видов проявляется через
наиболее характерный для данной зоны экотоп. На фоне ши¬
рокого распространения большинства видов, охватывающего
несколько природных зон, каждый вид лишайника имеет бо¬
лее или менее четкий оптимум в одной зоне, где обнаруживает
наибольшую встречаемость и обилие и заселяет типичные для
этой зоны экотопы и субстраты.
Иногда в разных климатических зонах (например, в тро¬
пиках и умеренных областях), резко различающихся по соляр-
ности, тепловому режиму, количеству выпадающих осадков и
влажности воздуха, обитают одни и те же виды лишайников.
Можно предполагать, что период активного фотосинтеза и ро¬
ста у одного и того же вида в разных регионах происходит в
разное время года, когда показатели влажности воздуха, тем¬
пературы и освещенности имеют примерно одинаковые значе¬
ния, тогда как в остальное время лишайник находится в состо¬
янии покоя. Но возможно также, что лишайники в разных ре¬
гионах обитают в одинаковых или сходных микроклиматиче¬
ских условиях, которые обусловлены особенностями микро¬
ниши.
Кроме того, в разных частях ареала один и тот же вид
может быть представлен популяциями, относящимися к раз¬
ным экотипам, потребности которых в тех или иных экологи¬
ческих факторах (влажность, тепло, свет) могут резко разли¬
чаться и которые могут по-разному реагировать на одинако¬
вые их значения. Особенно это касается видов с широкими
дизъюнктивными ареалами. Различия между экотипами воз¬
никли в результате длительного отбора и имеют наследствен¬
ную природу. Возникновение экотипов - это первый шаг к по¬
явлению новых физиологических и химических рас, приводя¬
щих впоследствии к возникновению подвидов и видов (Нин-
бург, 2005).
Поскольку характер и свойства субстрата для многих
лишайников являются первостепенными условиями их суще¬
ствования, то особенности распространения субстратов долж¬
ны обусловливать и особенности распространения лишайни¬
ков.
Распространение некоторых типов субстратов не зависит
от каких-либо климатических и ботанико-географических за¬
кономерностей. К таким субстратам относятся горные породы,
распространение которых связано с особенностями геологиче¬
ского строения материков. Многие лишайники, приуроченные
к силикатным или карбонатным горным породам, широко
распространены во всей Голарктике, как в средних поясах гор,
так и на равнине - везде, где имеются выходы соответствую¬
щих горных пород или обломочный материал, оставленный
покровным оледенением (морены).
Однако большинство субстратов имеет вполне опреде¬
ленные зональные (поясно-зональные) закономерности рас¬
пространения, связанные с климатическими условиями.
Например, эпифилльные лишайники, поселяющиеся на вечно¬
зеленых представителях тропической и субтропической денд¬
рофлоры, ограничены в своем распространении зоной обита¬
ния этих древесных растений и их конкретными ареалами.
Большинство эпифитных видов рода Usnea связаны с хвойны¬
ми и особенно с горными темнохвойными лесами и распро¬
странено в пределах ареалов этих лесов.
Многие чешуйчатые лишайники родов Psora, Toninia,
Endocarpon и некоторых других поселяются лишь на почвах с
высоким содержанием карбонатов кальция. Такие почвы, как
зональное явление, характерны для степной и пустынной зон и
обусловлены особенностями их водного режима - непромыв¬
ного и выпотного типов. В то же время экстразонально карбо¬
натные почвы могут развиваться и на близко залегающих кар¬
бонатных породах в горах, в таежной зоне и в Арктике, что
позволяет представителям указанных родов встречаться и в
этих регионах.
Таким образом, можно считать, что чем менее экологи¬
чески специализирован вид, тем шире его ареал и зона обита¬
ния, тогда как у узкоспециализированных видов зона распро¬
странения и ареал ограничены ареалом распространения спе¬
цифичного для них субстрата или экотопа.
Лишайники и растительный покров
Лишайники - слабые в конкурентном отношении орга¬
низмы. В формировании растительного покрова они в основ¬
ном играют второстепенную роль, формируя напочвенный по¬
кров и внеярусньге синузии, и лишь в тундрах могут выступать
основными ценозообразователями. В то же время лишайники
являются пионерами заселения обнаженного каменистого суб¬
страта и участвуют в образовании несомкнутых раститель¬
ных группировок на скалах и каменных россыпях.
В растительных сообществах лишайники встречаются на
тех же субстратах и в тех же экотопах, что и мохообразные. У
двух этих групп организмов одна общая экологическая ниша,
которую они делят в разных соотношениях в зависимости от
таких экологических показателей, как влажность и освещен¬
ность. Так, в напочвенном покрове лесов, в более влажных
условиях преобладают мохообразные - до полного выпадения
лишайников (ельники долгомошники, кедровые и пихтовые
зеленомошные леса), а в сухих условиях, наоборот, развивает¬
ся лишайниковый покров (боры беломошники на сухих песча¬
ных почвах). В гипновых болотах наблюдается полное доми¬
нирование мхов, а в сухих степях и пустынях - преобладание
лишайников. Стратегию мохообразных и лишайников при за¬
селении ими различных экотопов и субстратов можно охарак¬
теризовать как комплементарность (т. е. они дополняют друг
друга). При этом лишайники по своей природе в целом более
светолюбивы и сухолюбивы, а мохообразные - влаголюбивы и
тенелюбивы. Например, в любой лесной экосистеме законо¬
мерности распределения мохообразных и лишайников на
стволах деревьев одинаковы - в нижней более влажной и зате¬
ненной части ствола преобладают мохообразные, а в верхней
более сухой и освещенной - лишайники. Различия, обуслов¬
ленные климатическими и фитоценотическими причинами,
проявляются лишь в том, на какой высоте от уровня почвы
проявляется доминирование той или иной группы. В условиях
сухого резко континентального климата Южного Забайкалья
мохообразные поселяются на основаниях деревьев, а лишай¬
ники - в комлевой части до высоты 40-60 (редко более) см над
грунтом, тогда как в условиях влажного муссонного климата
Приморья мохообразные заселяют стволы деревьев до высоты
3-7 (и более) м, а лишайники - до высоты 10-15 (и более) м
над землей.
Роль лишайников в экосистемах
Лишайники существенно влияют на функционирование
наземных экосистем. Процессы разрушения горных пород и
первичное почвообразование, гумусонакопление, поддержание
водного и теплового балансов в лесах и тундрах за счет боль¬
шой влагоемкости и низкой теплопроводности напочвенного
лишайникового покрова, предохранение почвы от водной и
ветровой эрозии в результате формирования тонкой водорос¬
лево-лишайниковой корочки в степях и пустынях или мощно¬
го мохово-лишайникового покрова в лесах и тундрах, все это -
невидимая работа лишайников и других споровых организмов.
Защитные функции лишайников в экосистемах проявля¬
ются по-разному. При распашке степей, нарушении поверх¬
ностного почвенного слоя в пустынях водорослево¬
лишайниковая корочка уничтожается, и почвы легко разру¬
шаются и выветриваются.
В сибирских лесах при сплошных рубках лишайниковый
покров высыхает и в течение нескольких лет полностью де¬
градирует, многолетняя (вечная) мерзлота оттаивает, уровень
грунтовых вод под вырубками резко понижается, образуются
термокарсты, а на месте хвойных лишайниковых и мохово¬
лишайниковых лесов происходит возобновление березово¬
осиновых травяных лесов. Лишайники навсегда вытесняются
из указанных экосистем более конкурентоспособными, обра¬
зующими дернину травянистыми растениями и лишайниковый
покров на этих территориях более не восстанавливается. На
смену темнохвойным породам приходят светлохвойные или
мелколиственные, вследствие чего площади лишайниковых
(кладониевых) лесов катастрофически сокращаются.
Процессы почвообразования также зависят от лишайни¬
кового покрова. При первичном почвообразовании ризины и
гифы лишайников, проникающие в поверхностные слои гор¬
ной породы, в условиях переменной влажности сильно изме¬
няют свой объем, способствуя образованию трещин в породах
и их разрушению. Кроме того, растворимые лишайниковые
кислоты образуют комплексные соединения с катионами, вхо¬
дящими в состав горных пород. Лишайниковый покров влияет
также на развитие почвенной микрофлоры и, как было недав¬
но установлено (Заварзина, Заварзин, 2006), на образование
гумуса.
Наиболее заметно влияние лишайников на гидрологиче¬
ский режим экосистем. Помимо того, что они препятствуют
испарению воды и оттаиванию грунтов, что особенно сильно
проявляется в северных и арктических широтах, лишайники
способны также перехватывать значительную часть атмо¬
сферных осадков, которые в результате не попадают в почву.
Это касается как эпифитных лишайников в лесах, так и напоч¬
венных лишайников в лесах, степях, пустынях. Исследования,
проведенные в Калифорнии, показали, что в дубовых лесах
7.5% осадков перехватывается эпифитными лишайниками
(Nash, 1996).
Экология и эволюция
Экологические факторы являются универсальным меха¬
низмом эволюции лишайников, обеспечивающим отбор видов.
Под влиянием экологических факторов в прошлые геологиче¬
ские эпохи шла эволюция жизненных форм лишайников, про¬
исходило становление видов и формирование их ареалов.
В последующие эпохи, в складывающейся новой эколо¬
гической обстановке у лишайников шел процесс отыскания
подходящих экотопов и субстратов. Этому способствовали
небольшие размеры лишайников и их способность к вегета¬
тивному размножению. Уникальными адаптациями лишайни¬
ков к переживанию неблагоприятных условий меняющейся
среды и освоению новых экотопов объясняются их широкие
ареалы (равные ареалам родов и даже семейств цветковых
растений). Этим же можно объяснить и большое количество
реликтов среди лишайников, на порядок превышающее коли¬
чество реликтов того же геологического возраста среди цвет¬
ковых растений.
Изучение экологии лишайников - приуроченности их к
определенным растительным сообществам, экотопам и суб¬
стратам, неразрывно связано с решением проблем становления
флоры лишайников любого региона. Экологический анализ
позволяет выявлять однородные элементы - эколого¬
субстратные группы и экологические комплексы видов, шед¬
шие по какому-то общему пути или различными путям эволю¬
ции, адаптируясь к новым экологическим условиям и возмож¬
ной смене субстрата и экотопа. Он также с большей уверенно¬
стью дает возможность судить о возможных путях эволюции
отдельных эколого-флористических комплексов в геологиче¬
ском прошлом и в настоящее время.
Литература
Бязров Л. Г. Эпифитные лишайники г. Москвы: совре¬
менная динамика видового разнообразия. М.: Товарищество
научных изданий КМК, 2009. 146 с.
Бязров Л. Г. Лишайники в экологическом мониторинге.
М.: Научный мир, 2002. 336 с.
Голубкова Н. С., Бязров Л. Г. Жизненный формы ли¬
шайников и лихеносинузии // Ботан. журн. 1989. Т. 63. № 8. С.
1145-1154.
Жданова Н. Н., Василевская А. И. Экстремальная эколо¬
гия грибов в природе и эксперименте. Киев: Наукова думка,
1982. 168 с.
Заварзина А. Г., Заварзин А. А. Лакказная и тирозиназ-
ная активности у лишайников // Микробиология. 2006. Т. 75.
№5. С. 630-641.
Макрый Т. В. Лишайники Байкальского хребта. Новоси¬
бирск: Наука, Сиб. отд-ние, 1990. 200 с.
Макрый Т. В., Лиштва А. В., Скирина И. Ф. Особенности
строения, физиологии и химии эпилитных лишайников в экс¬
тремальных условиях высокогорий // Микология и криптогам¬
ная ботаника в России: традиции и современность / Труды
международ. конф., посвящ. 100-лению организации исслед.
по микологии и криптогамной ботанике в Ботаническом ин-те
им. В. Л. Комарова РАН (Санкт-Петербург, 24-28 апреля, 2000
г.). СПб: Изд-во Санкт-Петербургской государственной хими¬
ко-фармацевтической академии, 2000. С. 349-351.
Николаев Г. М., Маторин Д. Н., Аксенов С. И. Состояние
воды и функционирование первичных реакций фотосинтеза у
лишайника Placolecanora melanophtalma из холодных пустынь
Восточного Памира // Науч. докл. высш. школы, биол. науки.
1977. № 1.С. 106-106.
Нинбург Е. А. Введение в общую экологию (подходы и
методы). М.: Товарищество научных изданий КМК, 2005. 138
с.
Окснер А. Н. Морфология, систематика и географиче¬
ское распространение // Определитель лишайников СССР.
Вып. 2. Л.: Наука, 1974. 284 с.
Равинская А. П. Действие экологических факторов на
содержание усниновой кислоты и атранорина у лишайников //
Актуальные проблемы экспериментальной лихенологии в
СССР. Л., 1991. Вып. 1. С. 93-98.
Шапиро И. А. Загадки растения-сфинкса. Лишайники и
экологический мониторинг. Л.: Гидрометеоиздат, 1991. 80 с.
Almborn О. Distribution and ecology of some south Scandi¬
navian lichens // Bot. Notiser. Suppl. 1-2. Lund, 1948. P. 1-354.
Arvidsson L. A monograph of the lichen genus Coccocarpia
II Opera Bot. 1982. № 67. P. 1-96.
Smith D. C. The physiology of Peltigera polydactyla (Neck.)
Hoffm. // Lichenologist 1962. Vol. 1. P. 209-226.
Barkman J. J. Phytosociology and ecology of cryptogamic
epiphytes. Assen: van Gorcum, 1958. 628 p.
Brodo I. M. Substrate ecology // V. Ahmadjian, М. E. Hale
(eds.). The lichens. New York, London: Academic Press, 1973. P.
401-441.
Jorgensen P. M. The lichen family Pannariaceae in Europe //
Opera Bot. 1978. № 45. - 123 p.
Kappen L., Lange O. L. Die Kaltresistenz einiger
Makrolichenen//Flora. 1972. Bd 161. S. 1-29.
Klement O. Prodromus der mitteleuropaischen Flechtenge-
sellschaften // Feddes Repert. Beih. 135 (Beitrage zur Vegetations-
kunde). 1955. S. 5-194.
Lange O. L. Pflanzenleben unter Stress: Flechten als Pioniere
der Vegetation an Extremstandorten der Erde. Rostra Universitatis
Wirceburgensis, 1992. 59 s.
Nash Т. H. Lichen biology. Cambridge: University Press,
1996. 303 p.
Глава VIII.
Сбор и определение лишайников, хранение лихенологиче¬
ских коллекций
Степанчикова И. С., Гагарина Л. В.
Лишайники, в отличие от многих высших растений,
можно собирать практически в любое время года, хотя зимой
сбору напочвенных и некоторых других видов может препят¬
ствовать снежный покров. Большинство лишайников, в случае
благоприятных климатических условий, способны вегетиро¬
вать круглый год, у многих видов существуют долгоживущие
структуры размножения (например, плодовые тела, соралии).
Поэтому, как правило, время года существенно не влияет на
встречаемость видов и не сказывается на качестве образцов.
Однако нужно иметь в виду, что и среди лишайников есть ор¬
ганизмы с короткоживущими плодовыми телами и даже тал¬
ломами. Для таких видов может быть характерна сезонность в
размножении, например, для базидиальных лишайников из
родов Lichenomphalia, Multiclavula, а также, вероятно, для не¬
которых сумчатых лишайников - Absconditella sphagnorum,
Cryptodiscus gloeocapsa. Выраженная сезонность характерна
для жизненного цикла и некоторых лихенофильных грибов
(Lawrey, Diederich, 2003). Поэтому для наиболее полного вы¬
явления локальных лихенофлор полезно совершать полевые
выезды в разные периоды года.
Поиск и сбор лишайников в природе - достаточно слож¬
ная задача, и качество коллекций во многом зависит от опыта
исследователя. Многие внешне похожие виды различимы
только на уровне микроморфологических, микроскопических
или даже только химических признаков - в таких случаях
важно собрать достаточно материала для последующей иден¬
тификации. В то же время, многие виды специалист способен
легко распознать в поле с использованием лупы, поэтому при
флористических исследованиях зачастую вполне достаточно
ограничиться регистрацией в полевом дневнике данных о
нахождении таких видов. Важно помнить, что указанию любо¬
го вида с исследуемой территории должен соответствовать
хотя бы один гербарный образец. Одна из причин, по которым
необходима гербарная документация данных о нахождении
даже легко определимых и широко распространенных массо¬
вых видов, заключается в том, что представления о различиях
между таксонами и их объемах непостоянны. С получением
новых сведений о той или иной систематической группе, мо¬
жет значительно измениться понимание даже очень широко
известных и легко определимых видов (см., например:
Argiiello, del Prado et al., 2007; Crespo, Molina et al., 2002;
Crespo, Perez-Ortega, 2009; Grube, Kroken, 2000). Кроме того,
при работе с такими сложно идентифицируемыми организма¬
ми как лишайники, неизбежны ошибки определения. Крите¬
рий проверяемости исходных данных обязательно должен
быть соблюден при любых филогенетических и популяцион¬
ных исследованиях. Флористические данные также имеют
научную ценность только в том случае, если возможна ревизия
сопровождающего их гербарного материала.
Лишайники - это живые организмы, в большинстве сво¬
ем очень медленно растущие. Среди них немало сокращаю¬
щихся в численности и редких видов, заслуживающих статуса
охраняемых, некоторые уже внесены в Красные книги. При
сборе образцов лишайников нужно следить за тем, чтобы не
уничтожать местные популяции видов полностью, значитель¬
ная часть экземпляров всегда должна оставаться в природе. Не
следует собирать больше биологического материала, чем
необходимо для работы.
Методы полевой работы.
Работа с лишайниками как в поле, так и в лаборатории
занимает много сил и времени - поэтому в первую очередь
следует четко сформулировать задачи и продумать методику
исследования. Существуют различные подходы к тому, как
следует собирать лишайники, а стратегии поиска могут варьи¬
ровать в зависимости от поставленных целей.
Наиболее простой и распространенный метод лихено-
флористических и систематических исследований в поле -
маршрутный, когда исследователь учитывает и собирает ли¬
шайники на протяжении всего маршрута (пешего, велосипед¬
ного, конного). Ценность метода заключается в том, что он
дает возможность относительно быстро и равномерно обсле¬
довать большие территории. В то же время, маршрутный ме¬
тод не лишен определенных недостатков. Многие накипные
лишайники очень плохо заметны в поле, для многих видов ха¬
рактерна строгая приуроченность к определенным, часто
весьма специфическим, микроместообитаниям. Чтобы не про¬
пустить такие виды по ходу маршрута, коллектор должен
иметь не только большой опыт полевой лихенологической ра¬
боты, но также хорошо представлять себе лихенофлору изуча¬
емого региона. Существенно и то, что возможности статиста-
ческой обработки данных, полученных маршрутным методом,
весьма ограничены.
Другой подход, позволяющий получить сведения о ли¬
хенофлоре региона, предполагает полевой учет и сбор лишай¬
ников на пробных площадях (метод пробных площадей).
Пробные площади закладывают в пределах однородных
участков фитоценоза, при этом каждой такой площади должно
соответствовать хотя бы краткое геоботаническое описание.
Для выполнения различных задач используют и различные
схемы расположения пробных площадей (см., например,
Бязров, 2002; Tibell, 1992). При эколого-флористических ис¬
следованиях пробные площади следует выбирать так, чтобы
максимально охватить все разнообразие имеющихся фитоце¬
нозов и ландшафтных элементов. В таких случаях выбор
участков осуществляется, как правило, в ходе рекогносциро¬
вочных маршрутов. Размер пробных площадей варьирует в
зависимости от сложности состава и пространственной струк¬
туры фитоценоза. Чаще всего он составляет не менее 400 кв. м
(20x20 или 10^40 м) в лесных фитоценозах и не менее 100 кв.
м (10x10 м) в других сообществах - тундровых, стланиковых,
болотных, луговых и им подобных (Юнатов, 1964; Раменский,
1971; Сукачев, 1972).
На каждой пробной площади проводят детальную ин¬
вентаризацию, фиксируя видовой состав лишайников на всех
субстратах. В зависимости от конкретных задач исследования,
можно учитывать различные дополнительные параметры -
обилие отдельных видов лишайников, их встречаемость и
пространственную ориентацию на субстрате или в пределах
пробной площади, высоту и угол наклона стволов деревьев
(при изучении эпифитных сообществ) и другие характеристи¬
ки. Полное лихенофлористическое описание пробных площа¬
дей подразумевает детальное изучение видового состава ли¬
шайников и максимальное выявление микроместообитаний,
которые они могут занимать. Этот метод позволяет получать
данные, подходящие для обработки различными статистиче¬
скими методами, после чего можно делать обоснованные вы¬
воды, например, о встречаемости и обилии видов лишайников
на исследуемой территории, об их субстратной и фитоценоти-
ческой приуроченности, о видовой насыщенности различных
типов сообществ. Работа маршрутным методом не дает таких
статистически надежных и сравнимых данных, так как не ос¬
нована на учете лишайников в пределах каких-либо стандар¬
тизованных контуров. В то же время, очевидным недостатком
исследования лихенофлоры методом пробных площадей явля¬
ется высокая трудоемкость, большие затраты времени и, соот¬
ветственно, значительно более низкая скорость работы. Одно
детальное описание может занимать от 1-2 часов до полного
светового дня в зависимости от сложности пространственной
структуры фитоценоза. Объем собранного материала оказыва¬
ется часто очень большим и требует продолжительной лабора¬
торной обработки. При изучении лихенофлоры обширных
участков (более 100 кв. км), характеризующихся значительной
фитоценотической и ландшафтной пестротой, пробные пло¬
щади физически невозможно закладывать в таких количе¬
ствах, чтобы равномерно и репрезентативно обследовать всю
территорию.
Одним из возможных решений при изучении больших
территорий является сочетание маршрутного метода с деталь¬
но-маршрутным - последний предполагает заложение проб¬
ных площадей в ходе маршрутного обследования в наиболее
типичных или интересных фитоценозах с учетом их ланд¬
шафтного положения. При этом, однако, следует помнить, что
данные, полученные на пробных площадях и в ходе маршрут¬
ных наблюдений и сборов в большинстве случаев невозможно
использовать совместно при статистическом анализе результа¬
тов исследования.
Для изучения изменений в лихенофлоре, отражающих
влияние какого-либо очевидного или предполагаемого факто¬
ра, используют метод трансект. При этом пробные площади
или точки сбора материала располагают на местности вдоль
линии (трансекты), на которой можно наблюдать градиент
воздействия фактора. Чаще всего метод трансект используют в
различного рода лихеноиндикационных исследованиях (см.,
например, Bishop, Cook, 1975; Kivisto, Kuusinen, 2000; Бязров,
2008).
В некоторых случаях перед исследователем стоит задача
изучения не всей лихенофлоры территории, а только какой-то
ее части (например, определенной систематической или эко-
лого-субстратной группы). Например, при изучении эпифит-
ных сообществ в пределах пробной площади обследуют не¬
большие площадки количественного учета, закладываемые на
стволах деревьев (см., например, Asta et al., 2002; Скирина и
др., 2010). Данные, собранные таким образом, оказываются
удобными для сравнительного анализа с использованием ма¬
тематических методов. При изучении какой-либо таксономи¬
ческой группы лишайников необходимо в первую очередь
учитывать эколого-субстратные и эколого-ценотические осо¬
бенности ее представителей. Так, если представители изучае¬
мой группы предпочитают ненарушенные старовозрастные
леса (во многих регионах мира это представители родов
Cyphelium, Nephroma, Sclerophora, Sphinctrina, Sticta и другие),
то исследователь при планировании полевых работ должен
обращать особое внимание на охраняемые природные терри¬
тории и труднодоступные районы, где вероятность сохранения
старовозрастных лесов наиболее высока. Особенности лишай¬
ников как комплексных полибионтных организмов следует
учитывать при сборе материала для популяционных и популя¬
ционно-генетических исследований (подробнее см. Werth,
2011).
Сбор материала и необходимые инструменты. Хранение
образцов.
Стандартный набор инструментов для сбора лишайников
включает лупу кратностью Т или 10х (реже используют лупы
большей кратности), нож, зубило (предпочтительно с кожухом
для защиты рук) и молоток. Лупа позволяет обнаружить на
поверхности субстрата мелкие накипные виды, которые не
всегда видны невооруженным глазом, и выбрать наиболее раз¬
витые экземпляры. В лупу видны основные макроморфологи-
ческие признаки лишайников (например, характер, форма и
цвет талломов и плодовых тел, изидий, соралей, псев-
доцифелл, цефалодий, пикнид, органов прикрепления). При
помощи ножа можно срезать лишайники с коры деревьев и
древесины, собрать напочвенные виды. Для эффективной и
безопасной работы нож должен быть острым и прочным,
складной нож обязательно должен иметь надежный фиксатор
лезвия. Некоторые исследователи при работе с твердой корой
и древесиной используют стамеску, а для срезания ветвей -
садовый секатор (Brodo et al., 2001; Hinds, Hinds, 2007; Smith
et al., 2009). Собирать лишайники с живых деревьев следует
осторожно, не срезая толстого слоя корки, чтобы не повредить
дерево. Для сбора лишайников с каменистого субстрата ис¬
пользуют зубило и молоток, а также защитные очки. Суще¬
ствуют различные варианты инструментов для обработки кам¬
ня, которые в отдельных случаях также могут оказаться по¬
лезными - например, зубило с победитовым наконечником
более удобно для сбора лишайников с гранитных валунов. При
отборе образцов эпилитных лишайников в полустационарных
условиях возможно использование механизированной геоло¬
гической техники (Huhtinen, 2002).
При сборе лишайников необходимо выбирать наиболее
зрелые экземпляры с хорошо развитыми структурами размно¬
жения (апотеции, соралии) и систематически значимыми
структурами вегетативной части таллома (псевдоцифеллы,
цефалодии). Образец обязательно должен включать органы
прикрепления, поэтому в большинстве случаев лишайники
приходится собирать вместе с фрагментами субстрата. Со¬
бранные экземпляры сразу упаковывают в бумажные конвер¬
ты (предпочтительно, из плотной бумаги, не сильно изменяю¬
щей форму при намокании - например, крафт-бумага) или в
мешочки из ткани. При сборе лишайников с почвы следует
учитывать, что почва при высыхании рассыпается, и поэтому
лучше дополнительно заворачивать образец в мягкую бумагу
(салфетку) или укладывать в спичечный коробок или другую
мелкую тару (например, пластмассовые чашки Петри). Если в
дальнейшем необходимо извлекать из образцов ДНК, то их
предпочтительнее помещать в небольшие герметично закры¬
вающиеся полиэтиленовые пакеты. Собирать такой материал
лучше в сухую и прохладную погоду, так как при этом умень¬
шается вероятность его загрязнения бактериями и плесневыми
грибами. После предварительного просушивания образцов в
пакеты можно добавить силикагель. Очевидно, что при сборе
образцов лишайников для молекулярных исследований необ¬
ходимо избегать примесей других видов живых организмов,
особенно - лишайников и лихенофильных грибов.
В полевых условиях лишайники, собранные в одной точ¬
ке и с одного субстрата, можно складывать в один бумажный
полевой конверт, который изготавливают из листа формата А4
(или, по необходимости, любого другого). Наиболее распро¬
страненные варианты изготовления конвертов представлены
на Рис. 79, однако можно использовать и готовые пакеты про¬
мышленного изготовления. Полевой конверт является времен¬
ным и, как правило, для гербаризации образцов его в даль¬
нейшем не используют. Для каждого образца на конверте
необходимо указать этикеточные данные: место сбора (по
возможности, привязку на местности, географические коорди¬
наты и высоту над уровнем моря), характер растительного со¬
общества, субстрат, а также дату сбора и фамилию коллекто¬
ра. Важно дать подробную характеристику субстрата: если со¬
бирают с древесного субстрата, то обязательно указывают по¬
роду дерева и, если возможно, его возраст, если с каменистого
- название горной породы или, по крайней мере, ее отношение
к группе карбонатных или силикатных пород. При работе в
полевых условиях часто бывает более удобно пользоваться
системой кратких обозначений (например, номерами точек
сбора или пробных площадей, сокращенными названиями
субстратов). В ходе полевых работ все этикеточные данные
для каждого такого обозначения (номера) должны быть отра¬
жены в полевом дневнике или записаны на один из вариантов
мобильных электронных носителей информации, а координа¬
ты точки сбора следует фиксировать с помощью GPS-
навигатора.
L
Hypogymnia physodes а-гои
Сйн"г-П*Гербург. Tlfupfoqj.
я-f л СТ&нш*й Уникрсиггт пср«.
рОвдЭ. но Pirijj sylvrslnf
Рис. 79. Образец изготовления и этикетирования полевого конверта.
Сушка образцов лишайников может быть проведена на
открытом воздухе вне прямого действия солнечных лучей,
осадков и ветра, или в сухих закрытых помещениях с исполь¬
зованием печки или электроприборов. Чтобы не испортить об¬
разцы и соблюсти правила техники безопасности, не рекомен¬
дуется сушить материал на открытом огне и в условиях высо¬
ких температур (руке не должно быть горячо!). Слишком
быстрое высушивание талломов лишайников при высокой
температуре приводит к искажению их формы и цвета, может
привести к разрушению ДНК и лишайниковых веществ. В то
же время, долго хранящиеся во влажном состоянии образцы
становятся удобным субстратом для развития грибов и беспо¬
звоночных животных различных систематических групп, что
приводит к порче или полной утрате материала. По этой же
причине хранение свежих образцов лишайников в полиэтиле¬
новой или пластиковой упаковке более нескольких часов не¬
желательно.
В гербарные сетки конверты с лишайниками, как прави¬
ло, не закладывают, так как это приводит к изменению внеш¬
него облика многих видов. Высушенные образцы, если воз¬
можно, помещают в морозильную камеру (желательная тем¬
пература - около -20°С) на несколько дней. Такая практика
позволяет избежать попадания живых беспозвоночных живот-
ных-вредителей гербария в основные фонды вместе со свежим
материалом.
Гербарий лишайников следует по возможности хранить
при невысокой температуре и стабильно низкой влажности
воздуха. Хранение материалов для молекулярно-генетического
анализа осуществляется в морозильной камере (-20°С) где об¬
разцы могут храниться до 1-2 лет. В то же время, в ряде слу¬
чаев выделение ДНК микобионта возможно в течение пяти лет
после сбора образцов даже при обычных условиях хранения в
основных гербарных фондах. Специальной антисептической
обработки гербарий лишайников не требует (Скирина и др.,
2010). Учитывая, что при идентификации многих видов ли¬
шайников существенную роль играет состав вторичных мета¬
болитов, а определительными признаками являются цветные
реакции таллома, следует крайне аккуратно относиться к лю¬
бым вариантам химической обработки материала.
Определение лишайников.
Лишайники определять достаточно сложно, что связано
как со все еще ощущающимся недостатком литературы по ря¬
ду таксонов, так и со спецификой объекта. При определении
лишайников необходимо обращать внимание на весь комплекс
признаков (морфологических, анатомических, химических).
Исследуя свежесобранный материал, следует иметь в виду, что
некоторые микроскопические признаки изменяются в течение
первых дней после сбора - например, наличие масляных ка¬
пель в гимении некоторых представителей родов Lecidella и
Verrucaria, размеры и форма перегородок в спорах видов рода
Caloplaca. Характеристики, данные в описаниях видов и в
определителях, как правило, соответствуют сухому материалу
(Smith et al., 2009), если иное не оговорено отдельно.
Начинают определение лишайников с использования
стереоскопического микроскопа (так называемого «бинокуля-
ра») кратностью 4-100х (оптимальное рабочее увеличение на
большинстве моделей составляет примерно 16х). Такое увели¬
чение дает возможность определить тип плодовых тел, харак¬
теристики поверхности таллома, особенности органов при¬
крепления к субстрату и структур вегетативного размножения.
Для того, чтобы изучить микроанатомические признаки объ¬
екта (особенности строения, форму и размеры сумок, парафиз,
аскоспор, конидий, соредий), используют светлопольный мик¬
роскоп проходящего света с основным набором объективов
(10х, 40х, 60х, 100х). Процедура подготовки материала для
микроскопического изучения зависит от особенностей объекта
и может несколько различаться. Для определения некоторых
видов, систематическое положение которых исследователю
уже известно, часто достаточно лишь определить характер и
размеры спор. В других случаях для идентификации вида тре¬
буется знать комплекс признаков микроскопического строения
плодового тела или таллома, для чего необходимо изготовить
тонкий срез. Срезы толщиной около 25 мкм можно сделать от
руки лезвием безопасной бритвы под стереомикроскопом. Пе¬
ред тем, как резать плодовое тело или таллом, его нужно смо¬
чить водой и подождать около минуты. Если на препарате сре¬
за плодового тела анатомические признаки видны плохо, то в
препарат можно добавить 10% раствор КОН. При этом следу¬
ет иметь в виду, что в растворе КОН размеры клеток, особен¬
но с тонкими стенками, иногда существенно изменяются, так
же как и окраска некоторых структур плодового тела. Измере¬
ния удобнее проводить на спорах, лежащих вне сумок, так как
в сумках они могут быть деформированы и плохо видны -
причем измерять желательно зрелые споры. Для изучения ха¬
рактеристик эксципула, гипотеция и других структур плодово¬
го тела срез должен быть тонким, аккуратным и проходить
через его центр. При изготовлении препаратов с использова¬
нием замораживающего микротома рекомендуемая толщина
среза - 10-15 мкм (Smith et al., 2009). В тех случаях, когда ха¬
рактер скульптуры поверхности спор является важным таксо¬
номическим признаком, необходимо использовать сканирую¬
щий электронный микроскоп.
При изготовлении полупостоянных препаратов нередко
в качестве среды используют лактофенол хлопковый синий
(lactophenol cotton-blue), который окрашивает срез и делает
его детали более контрастными. При этом срез помещают в
каплю лактофенола хлопкового синего, затем накрывают по¬
кровным стеклом, слегка придавливают и нагревают до тем¬
пературы, немного не достигающей температуры кипения сре¬
ды. После охлаждения препарат можно герметизировать,
нанеся по периметру покровного стекла лак для ногтей или
другое подходящее вещество (Smith et al., 2009). Использова¬
ние лактофенола хлопкового синего особенно удобно при ра¬
боте с микроскопическими признаками и размерными харак¬
теристиками структур плодовых тел: коэффициент преломле¬
ния лактофенола близок к показателю преломления стекла,
что обеспечивает наименьшее оптическое искажение объекта.
С 60-х годов XX века систематизация и изучение разно¬
образия лишайников немыслимы без исследования состава их
лишайниковых веществ - гетерогенной группы ацетонраство-
римых вторичных метаболитов, ранее носивших название
«лишайниковые кислоты».
В последние десятилетия особое внимание в системати¬
ке и при изучении разнообразия лишайников уделяют молеку¬
лярно-генетическим методам исследований. Для систематиза¬
ции таксонов разного ранга наиболее удобным подходом явля¬
ется сравнительный анализ различных фрагментов генома.
Чаще всего анализируют консервативные нуклеотидные по¬
следовательности, кодирующие рибосомальные РНК (боль¬
шую и малую субъединицы «ядерных», цитоплазматических
рибосом - nuSSU, nuLSU и малую субъединицу митохондри¬
альных - mtSSU) (Mueller et al., 2004). Более подробная ин¬
формация о разнообразных методах лабораторных исследова¬
ний в ботанике, микологии и лихенологии содержится в ряде
работ (Юрченко, Синявская, 2007; Куцев, 2009; Zoller et al.,
1999; Orange et al., 2001; Kranner et al., 2002 и др.).
Регистрация данных об образце.
В ходе идентификации образцов лишайников исследова¬
тель получает разнообразную и часто значительную по объему
информацию об изучаемом материале. Все важнейшие резуль¬
таты изучения морфологических (например, характер псев-
доцифелл, цефалодиев, размеры плодовых тел и конидиаль-
ных спороношений), анатомических (толщина и структура ко¬
рового слоя таллома, окраска и высота различных частей ги¬
мения и эксципула апотеция, форма, окраска и размеры спор),
хемотаксономических (результаты проведенных цветных хи¬
мических реакций с талломом и плодовыми телами, данные
хроматографических исследований любого характера) и моле-
кулярно-генетических (результаты идентификации и номера
образца в базах данных генетических банков) признаков
должны быть записаны на отдельный листок бумаги и вложе¬
ны, с указанием авторства и даты, в гербарный конверт. В этот
же конверт в некоторых случаях вкладывают плотно закрытые
микропробирки с участками талломов, которые использовали
для хроматографических исследований (с целью обеспечения
повторимости результата и из соображений бережного отно¬
шения к ценному материалу).
При работе с определительными ключами (лучше - с не¬
сколькими!) необходимо помнить, что в них использованы
лишь ведущие признаки, тогда как идентификация образцов
проводится с учетом всего их комплекса. Это предполагает
внимательное изучение не только ключей, но и описаний ви¬
дов, наиболее подходящих к исследуемому образцу. Любые
несоответствия признаков образца описаниям вида должны
быть отмечены особо. Наибольший вес имеет комплекс ана-
томо-морфологических и хемотаксономических признаков,
используемых во всех современных определителях. Признаки
эколого-ценотические и эколого-субстратные являются важ¬
ными лишь для некоторых стенотопных видов (реже - таксо¬
нов более высокого ранга, например, сем. Umbilicariaceae) и
могут иметь значительную региональную специфику (см. гла¬
ву VI). Особенности географического распространения могут
быть с уверенностью использованы в качестве диагностиче¬
ских только для хорошо различимых и изученных видов, тогда
как для многих накипных лишайников и лихенофильных гри¬
бов надежными не являются. При работе с теми или иными
таксонами необходимо использовать как современные, так и
более ранние монографические обработки, а также литератур¬
ные данные из других регионов мира. Наиболее надежным
подтверждением предположений исследователя является зна¬
комство с хранящимися в гербариях типовыми материалами и
эксикатами, выпущенными известными специалистами в об¬
ласти таксономии или крупными лихенологическими центра¬
ми. При этом, однако, следует помнить, что какое-либо меха¬
ническое или химическое воздействие на такие образцы допу¬
стимо только при серьезных исследованиях и в минимально
необходимых объемах, а повреждающее изучение скудных
типовых материалов («олиготипов») недопустимо или его про¬
водят лишь в исключительных случаях (Huhtinen, 1994).
После того, как образец определен, его кладут в гербар-
ный конверт вместе со всеми результатами его изучения, а да¬
лее сдают на хранение в гербарий. При изготовлении гербар¬
ных конвертов можно воспользоваться той же схемой, что при
изготовлении простейшего варианта полевых конвертов (Рис.
80).
Для защиты хрупких образцов желательно использовать
также мягкие салфетки, небольшие коробочки и подложки из
картона. Всегда лучше, если один гербарный конверт содер¬
жит только один вид лишайника, однако в случае мелких
накипных (и особенно эпилитных) видов такое разделение
обеспечить сложно. Желательно, чтобы сопровождающие ви¬
ды также были идентифицированы, так как эта информация
может иметь важное значение для флористических и ценоло¬
гических исследований. На конверты наклеивают гербарные
этикетки, содержащие информацию о родовом и видовом
названии лишайника, его местонахождении, местообитании и
субстрате, дате сбора и коллекторах, а также о том, кем и ко¬
гда образец был определен. Гербарные этикетки в последнее
время принято печатать на принтере. Не рекомендуем исполь¬
зовать для этого струйные принтеры, так как их чернила не
только обладают низкой устойчивостью к действию влаги, но
и могут обесцвечиваться со временем даже при минимальном
доступе света.
Правила работы в гербарии.
Гербарием принято называть как непосредственно кол¬
лекцию подготовленных определенным образом образцов, так
и учреждение или помещение, где эта коллекция находится и
где ведется работа с ней. Главной задачей гербария является
хранение материалов, документирующих научные исследова¬
ния в области ботаники, лихенологии или микологии; герба¬
рий обеспечивает соблюдение одного из основных научных
принципов - верифицируемое™ (проверяемости) данных.
Нижнего предела размеров гербария не существует - любая
коллекция имеет ценность, если она должным образом оформ¬
лена и доступна для научного использования. Закрытые для
доступа специалистов коллекции имеют ограниченную науч¬
ную ценность, хотя и имеют шанс увеличить ее в последствии.
Гербарный образец, собранный в качестве документации
для одной работы, может быть в дальнейшем использован для
других исследований другими авторами. Например, при фло¬
ристических работах весьма желательно изучить не только
свои материалы, относящиеся к исследуемому району, но и
все доступные образцы предшественников и современных
специалистов. Это дает возможность не только «проверить
точность определения» ранее собранных и опубликованных
(или неопубликованных) образцов, но и выявить и идентифи¬
цировать значительную долю материала, собранного ранее, но
неидентифицированного и неопубликованного. В таком мате¬
риале часто содержится заметная доля видов редких или ранее
не известных как для района исследования, так и для более
обширных территорий. Так, в ходе ревизии коллекций с тер¬
ритории Северо-запада России, хранящихся в гербариях уни¬
верситетов городов Хельсинки и Турку (Финляндия), было
выявлено около сотни видов, которые присутствовали в гер¬
барных конвертах только в качестве дополнительных к основ¬
ным видам и ранее не были идентифицированы (данные авто¬
ров).
Гербарий является фондом накопления первичных науч¬
ных данных. На основе ревизии гербарного материала можно
получить обширные сведения о географическом распростра¬
нении и экологической приуроченности видов, о меж- и внут¬
ривидовой изменчивости организмов, о флоре регионов и мно¬
гом другом; гербарные коллекции являются основной базой
для исследований в области эволюции и систематики расте¬
ний, грибов и лишайников. Любой исследователь, допущен¬
ный к работе с гербарными фондами, имеет право использо¬
вать в своих исследованиях и публикациях любую информа¬
цию, полученную им в ходе ревизии гербария. При этом, од¬
нако, необходимо придерживаться правил научной этики, ка¬
сающихся уважения авторских прав других исследователей,
правил хранения гербарных материалов и работы с ними, по¬
рядка фиксации результатов исследований, цитирования гер¬
барных образцов и коллекций.
В разных гербарных коллекциях хранение образцов мо¬
жет иметь свои особенности. Гербарные конверты можно хра¬
нить в гербарных папках (Рис. 81) или размещать в ящиках
или коробках. Гербарные папки или коробки хранят в специ¬
альных гербарных шкафах или в компакторах, где материал
уложен в какой-либо (чаще алфавитной, иногда в системати¬
ческой) последовательности. Во многих гербариях мира каж¬
дому конверту присвоен уникальный гербарный номер, а дан¬
ные образца внесены в специальную базу. В некоторых герба¬
риях такие базы данных доступны через Интернет. Например,
базы данных гербариев Упсальского университета, Швеция
(http://www-hotel2.uu.se:8888/cgi-
bin/wwwdrive.fytotek/beginner) или Шведского королевского
музея естественной истории в Стокгольме
(http://www.nrm.se/kbo/saml/lichen.html.en). В перспективе
каждому образцу в такой базе может соответствовать фото¬
графия или сканированное изображение как самого образца,
так и его оригинальной гербарной этикетки. В настоящее вре¬
мя в ряде гербариев кроме этикеток каждому образцу присва¬
ивается оригинальный бар-код, позволяющий упростить и ча¬
стично автоматизировать работу с базами данных и техниче¬
ское обслуживание коллекций, в частности, контролировать
перемещение и обмен гербарными образцами. Внесение об¬
разца в уже существующий гербарий называют инсерацией.
Она включает не только помещение образца в соответствую¬
щую ему папку, но и предшествующее этому присвоение об¬
разцу уникального гербарного номера и внесение всей извест¬
ной информации о нем в гербарную базу данных.
Рис. 81. Схема гербарной папки.
При работе с гербарием следует соблюдать общие
принципы работы с биологическими коллекциями. Прежде
всего, следует бережно относиться к гербарному материалу, не
деформировать конверты и не ронять образцы - талломы су¬
хих лишайников крайне хрупкие. Категорически запрещается
нарушать порядок расположения гербарных папок или коро¬
бок - перемещать материал в шкафах можно только по согла¬
сованию с куратором гербария. Отбирать из гербарной кол¬
лекции дублеты для какой-либо другой коллекции (дублет
гербарного образца - это часть материала из того же конверта)
или выносить за пределы гербария какие-либо материалы
можно только с разрешения куратора гербария.
При исследовании признаков, требующих изготовления
препаратов, фрагментации образцов для хроматографических
или молекулярно-генетических исследований, важно помнить,
что каждый образец уникален и невосстановим в случае утра¬
ты, поэтому ни в коем случае нельзя расходовать весь матери¬
ал образца, в конверте всегда должен оставаться репрезента¬
тивный (определимый) материал. При работе с типовым мате¬
риалом требуется особая аккуратность. При проведении лю¬
бых цветных реакций с гербарным образцом следует обяза¬
тельно удалять из конверта участки таллома, загрязненные ре¬
активами, а лучше всего проводить химические тесты на пред¬
варительно отделенных от основного материала фрагментах.
Важнейшие признаки, выявленные в ходе работы с гербарным
образцом (как и в случае определения собственных сборов, см.
выше), необходимо фиксировать на листке бумаги (аннота¬
ции), который следует вложить в конверт, указав свое автор¬
ство и дату. На гербарный конверт, с которым работал иссле¬
дователь, должен быть наклеен листок бумаги («детерминант-
ка» или «notula criticae», см. варианты на Рис. 82), на котором
необходимо указать: название таксона согласно мнению ис¬
следователя и в соответствии с современной номенклатурой;
фамилию и инициалы исследователя; дату работы с образцом.
На «детерминантке» должно быть отмечено, переопределен ли
этот образец исследователем («det.», «determ.» - determinare),
или он подтверждает предшествующее определение
(«confirm.» - confirmare; использование аббревиатуры «conf.»
в данном случае нежелательно, так как она имеет несколько
значений). Если специалист только видел образец и не прово¬
дил его внимательного изучения или не смог прийти к какому-
либо выводу - можно использовать обозначение «vidi» (не
следует писать «vidi», если Вы обнаружили пустой конверт!).
Документация данных важна как для дальнейших исследова¬
ний, так и для понимания, какой именно материал исследова¬
тель использовал при работе. Помимо перечисленных выше,
на гербарных конвертах можно встретить некоторые другие
сокращения (см. главу «Номенклатура и типификация»).
Рис. 82. Образец де¬
терминации. (
Гербарный фонд постоянно пополняется как за счет но¬
вых сборов, так и за счет материалов (как правило, дублетных
образцов), присылаемых другими гербариями. Важно пом¬
нить, что порядок инсерации образцов в каждом гербарии
свой, поэтому при передаче гербарию на хранение материалов
следует выяснить у куратора гербария, какие дополнительные
условия должны быть соблюдены (например, при инсерации
образцов в гербарий Ботанического института им. В. Л. Кома¬
рова РАН в г. Санкт-Петербурге или университета г. Упсала
необходимо внести этикеточные данные в электронную базу
данных).
В каждом гербарии, помимо общепринятых принципов,
существуют свои установившиеся правила, с которыми необ¬
ходимо ознакомиться перед заранее согласованным визитом.
Например, во многих гербариях после работы с образцами
специалисту следует самостоятельно вернуть их на место, од¬
нако в некоторых случаях этим занимаются только сотрудники
гербария. В некоторых гербариях категорически запрещено
брать крупные фрагменты образца для каких-либо лаборатор¬
ных исследований, тогда как в других это разрешено при
условии наличия достаточного количества материала. Обычно
детерминантки к конвертам приклеивают сами исследователи,
но иногда этим вправе заниматься только персонал гербария.
Кроме своих основных задач, гербарий несет функцию
сохранения на этикетках и листочках-аннотациях образцов
почерков коллекторов и других специалистов, работавших с
материалом, а также интересных исторических, топографиче¬
ских и иных сведений, то есть является архивом, ценным еще
и с точки зрения истории науки. Соответственно, важно со¬
держать гербарий так, чтобы сохранить наиболее полно всю
(!) информацию с этикеток, аннотаций, гербарных конвертов,
детерминанток. При замене старого конверта или гербарной
этикетки обязательно нужно вкладывать в новый конверт как
старую этикетку, так и все фрагменты упаковки, несущие на
себе какие-либо записи об образце. Важно помнить, что все
аннотации, имеющие отношение к образцу, должны иметь
четкое авторство, дату и храниться в неизменном виде беско¬
нечно долгое время. Менять их содержание или уничтожать не
вправе даже сам автор.
К работе с гербарием может быть допущен любой чело¬
век - однако куратор гербария имеет право не допустить в
гербарий исследователя, не соблюдающего основные правила
работы с гербарными фондами.
Гербарные коллекции могут быть зарегистрированы в
Международной системе Index Herbariorum
(http://sweetgum.nybg.org/ih/). После регистрации гербарию
присваивается акроним (его краткая аббревиатура). Акроним
уникален для каждого гербария. Например, акроним LE соот¬
ветствует гербарию, хранящемуся в Ботаническом институте
им. В. Л. Комарова. Акронимы основных лихенологических
гербариев России и мира приведены в главе II.
Литература
Бязров Л. Г. Лишайники в экологическом мониторинге.
М.: Научный Мир, 2002. 336 с.
Бязров Л. Г. Современная динамика видового состава
эпифитной лихенобиоты г. Москвы // Современная микология
в России. Тезисы докладов второго съезда микологов России.
Том 2. М.: Национальная академия микологии, 2008. С. 523-
524.
Куцев М. Г. Фрагментарный анализ ДНК растений:
RAPD, DAF, ISSR. Барнаул: ARTHKA, 2009. 164 с.
Раменский Л. Г. Избранные работы. Проблемы и методы
изучения растительного покрова. Л.: Наука, 1971. 334 с.
Скирина И. Ф., Коженкова С. И., Родникова И. М. Эпи-
фитные лишайники Приморского края и использование их в
экологическом мониторинге. Владивосток: Дальнаука, 2010.
134 с.
Сукачев В. Н. Избранные труды. Т. 1. Основы лесной
типологии и биогеоценологии. Л.: Наука, 1972. 419 с.
Юнатов А. А. Типы и содержание геоботанических ис¬
следований. Выбор пробных площадей и заложение экологи¬
ческих профилей // Полевая геоботаника. Т. 3. / Ред. А. А.
Корчагин, Е. М. Лавренко. М.-Л.: Наука, 1964. С. 9-36.
Юрченко Е. О., Синявская М. Г. Основы молекулярного
маркирования грибной ДНК. Практическое руководство.
Минск: Право и экономика, 2007. 101 с.
Arguello A., del Prado R., Cubas P., Crespo A. Parmelina
quercina (Parmeliaceae, Lecanorales) includes four phylogenet-
ically supported morphospecies // Biological Journal of the Lin-
naean Society. 2007. Vol. 91(3). P. 455-467.
Asta J., Erhardt W., Ferretti M, Fornasier F., Kirschbaum U.,
Nimis P. L., Purvis O. W., Pirintsos S., Scheidegger C., van Ha-
luwyn C., Wirth V. Mapping Lichen Diversity as an Indicator of
Environmental Quality // Monitoring with Lichens - Monitoring
Lichens / Eds. P. L. Nimis, C. Scheidegger, P. A. Wolseley. Dor¬
drecht, Boston, London: Kluwer Academic, 2002. P. 273-279.
Bishop J. A., Cook L. М., Muggleton J., Seaward M. R. D.
Moths, Lichens and Air Pollution Along a Transect from Manches¬
ter to North Wales//Journal of Applied Ecology. 1975. Vol. 12(1).
P. 83-98.
Brodo I. М., Sharnoff S. D., Sharnoff S. Lichens of North
America. New Haven and London: Yale University Press, 2001.
795 p.
Crespo A., Molina М. C., Blanco O., Schroeter B., Sancho
L. G., Hawksworth D. L. DNA ITS and P-tubulin gene sequence
analyses reveal two monophyletic groups within the cosmopolitan
lichen Parmelia saxatilis // Mycological Research. 2002. Vol.
106(7). P. 788-795.
Crespo A., Perez-Ortega S. Cryptic species and species pairs
in lichens: A discussion on the relationship between molecular
phylogenies and morphological characters // Anales del Jardfn Bo-
tanico de Madrid. 2009. Vol. 66, S 1. P. 71-81.
Frizzell D. L. Terminology of types // The American Mid¬
land Naturalist. 1933. № 6. P. 637-668.
Grube М., Kroken S. Molecular approaches and the concept
of species and species complexes in lichenized fungi // Mycologi¬
cal Research. 2000. Vol. 104(11). P. 1284-1294.
Hinds J. W., Hinds P. L. The Macrolichens of New England.
New York: New York Botanical Garden Press, 2007. 608 p.
Huhtinen S. An effective technique for collecting epilithic
crustose lichens // Memoranda Societatis pro Fauna et Flora Fenni¬
ca. 2002. Vol. 77. P. 1-3.
Huhtinen S. Anyone for olygotypes? // Taxon. 1994. Vol.
43(8). P. 441-443.
Kivisto L., Kuusinen M. Edge effects on the epiphytic lichen
flora of Picea abies in middle boreal Finland // The Lichenologist.
2000. Vol. 32(4). P. 387-398.
Lawrey, J. D., Diederich P. Lichenicolous fungi: interac¬
tions, evolution and biodiversity // The Bryologist. 2003. Vol. 106,
№ 1. P. 80-120.
Orange A., James P. W., White F. J. Microchemical methods
for the identification of lichens. London: British Lichen Society,
2001. 101 p.
Protocols in lichenology: culturing, biochemistry, ecophysi-
ology and use in biomonitoring / Kranner I., Beckett R., Varma A.
(eds.). Berlin: Springer, 2002. 580 p.
Santesson R., Moberg R., Nordin A., Tonsberg Т., Vi-
tikainen O. Lichen-forming and lichenicolous fungi of Fen-
noscandia. Uppsala: Museum of Evolution, Uppsala University,
2004. 359 p.
The lichens of Great Britain and Ireland / Smith C. W., Ap-
troot A., Coppins B. J., Fletcher A., Gilbert O. L., James P. W.,
Wolseley P. A. (eds). The British Lichen Society, London, 2009.
1046 p.
Tibell L. Crustose lichens as indicators of forest continuity in
boreal coniferous forests // Nordic Journal of Botany. 1992. Vol.
12(4). P. 427-450.
Werth S. Optimal sample sizes and allelic diversity in studies
of the genetic variability of mycobiont and photobiont populations
// The Lichenologist. 2011. Vol. 43, № 1. P. 73-81.
Zoller S., Scheidegger Ch., Sperisen Ch. PCR primers for the
amplification of mitochondrial small subunit ribosomal DNA of
lichen-forming Ascomycetes // Lichenologist. 1999. Vol. 31(5). P.
511-516.
Глава IX.
Номенклатура и типификация лишайников
Гагарина Л. В., Степанчикова И. С.
Признак не определяет род, но род
признак...
Признак существует не для того, что¬
бы учредить род, а для того, чтобы
его познать.
К. Линней (Linnaeus, 1736)
До появления бинарной номенклатуры, единого, призна¬
ваемого всеми принципа построения названий биологических
видов не существовало. Название вида, как правило, состояло
из нескольких (иногда более десятка) слов и отражало харак¬
терные морфологические признаки, то есть, по сути, являлось
его кратким описанием. Зачастую такие названия были неточ¬
ными и расплывчатыми. Так, в работе Иоганна Кристиана
ByKc6ayMa«Observationes circa Quasdam Plantas Ingricas»
(Buxbaum, 1728) упомянут вид «Lichen arboreus membranaceus,
nigricans Buxb.» Название правильно пишется так, у вас был
другой падеж! («лишайник древесный пленчатый, черный»).
По названию организма и прилагаемой к нему иллюстрации
можно заключить, что речь идет о цианобионтном лишайнике,
скорее всего, из рода Collema. Это новый вид, описанный Бук-
сбаумом. Он есть в Historia Muscorum Dilenius’a, 1741, на стр.
54. В большинстве случаев в долиннеевских работах невоз¬
можно установить видовую принадлежность лишайника. Надо
смотреть по Historia Muscorum и Catalogus Plantarum J. J.
Dilenius’a 1741 и 1719, Theatr. Botanici Caspar’a Bauhin’a, 1623,
lnstitutiones Rei Herbariae J. P. Tournefort’a 1700 и по другим
до- и послелиннеевским каталогам, там эти лишайники часто
есть и можно установить видовую принадлежность, поскольку
есть четкая привязка долиннеевских и линнеевских названий!
Линней использовал эти книги и «раскладывал по своим по¬
лочкам» уже описанные виды. Можно даже найти соответ¬
ствие с видами, упоминаемыми у Плиния и Теофраста.
Карл Линней (Linne, Linnaeus) существенное упростил и
стандартизировал биологической номенклатуры (Nicolson,
1991). Попытки сделать это предпринимались и ранее. Андреа
Чезальпино (ссылка) первым начал разрабатывать единые
принципы для выделения систематических групп, а непосред¬
ственными предшественниками Линнея можно считать Джона
Рэя, Августа Бахмана (Ривинуса) и Жозефа Питтона де Турне-
фора (ссылки) (см. Куприянов, 2005). К. Линней осуществил
идею Ривиниуса, утверждая, что «наименование - второе ос¬
нование ботаники». Он тщательно проработал принципы си¬
стематики, в значительной степени формализовав их. Номен¬
клатура К. Линнея называется бинарной (двойной), в соответ¬
ствии с ней название вида строится из родового и видового
эпитетов. Основы номенклатуры Линнея изложены в его рабо¬
тах, в частности: «Критика ботаники» («Critica botanica», 1737
г.), «Философия ботаники» («Philosophia botanica», 1751 г.),
«Виды растений» («Species plantarum», 1753 г.). (сделать нор¬
мальные ссылки)
Работы К. Линнея были очень популярны и получили
широкое распространение (см. Куприянов, 2005). За несколько
десятилетий бинарная номенклатура вытеснила другие спосо¬
бы обозначения таксонов и была закреплена в виде Кодексов.
Первый формальный перечень основных номенклатурных
принципов в биологии (зоологии) - «Кодекс Стрикленда» -
появился в 30-е гг. XIX века и был назван в честь его разра¬
ботчика зоолога Хью Эдвина Стрикленда (Strickland, 1837; см.
Rules..., [2012]). Первый ботанический кодекс - «Законы бота¬
нической номенклатуры» Альфонса Декандоля - был опубли¬
кован в 1867 г. (ссылка).
Кодекс, определяющий принципы номенклатуры в бота¬
нике и микологии, до недавнего времени имел название
«Международный кодекс ботанической номенклатуры» - да¬
лее МКБН (McNeill et al., 2006; Международный..., 2009). На
Мельбурнском ботаническом конгрессе 2011 г. было принято
решение о переименовании кодекса, и в настоящий момент его
полное название - «Международный кодекс номенклатуры
водорослей, грибов и растений» («International Code of
Nomenclature for algae, fungi, and plants»), (ссылка).
Современная номенклатура базируется на работах К.
Линнея. Правила и алгоритмы действий в таксономии подроб¬
но проработаны и опубликованы в современном МКБН. В со¬
ответствии с существующими правилами, научные названия
таксонов разных рангов у лишайников необходимо приводить
в соответствии с версией МКБН, принятой на последнем по
времени Международном ботаническом конгрессе (проходит
раз в пять лет). Фамилии авторов названий таксонов цитируют
в соответствии с электронным источником Authors of Fungal
Names
(http://www.indexfungorum.Org/Names/AuthorsOfFungalNames.a
sp), который основан на работах Wright и Lois (1949),
Stevenson (1960), Viegas (1961), Barnhart (1965), Laundon
(1979).
На практике ситуация с обозначением таксонов в бота¬
нике, в частности в лихенологии, достаточно сложна, несмот¬
ря на жесткую формализацию таксономического процесса.
Лаконичная система таксонов ограничивает возможности от¬
ражения предполагаемых родственных связей между объекта¬
ми и их разнообразия в природе. Формально придерживаясь
принципов бинарной номенклатуры, исследователи на протя¬
жении почти двух столетий описывали внутри видов лишай¬
ников разнообразные подвиды, разновидности, формы. Зача¬
стую название объекта состояло не только из родового и видо¬
вого эпитетов, но включало в себя еще несколько эпитетов,
соответствующих названиям различных таксонов внутривидо¬
вого ранга (см. Rasanen, 1939; Савич, 1914; Vainio, 1878, 1921;
Nylander, 1877 и др.). Так, в работе Э. А. Вайнио «Lichenes in
viciniis Viburgi observati» (Vainio, 1878) для окрестностей Вы¬
борга приведен лишайник Lecidea pelidna Ach. var. compacta
(Korb.) f. asserculorum Korb., а В. П. Савич в 1914 г. описал с
Камчатки новую форму Gyrophora hyperborea Ach. f.
sublaevigata Savicz (Савич, 1914). До середины XX века под¬
виды или таксоны любого иного внутривидового ранга неред¬
ко обозначали буквами греческого алфавита а, Р, у, 5 - напри¬
мер, Cladonia squamosa (Scop.) Hoffm. у multibrachiata Florke f.
phyllopoda Vain.
В настоящее время формы лишайников, отличные от уже
известного типового материала, чаще описывают как новые
виды. Хотя таксоны в ранге подвида выделяют реже, тем не
менее, они не полностью утратили свое значение. Статус мно¬
гих морфо- и хемотипов, популяций, экологических форм ли¬
шайников в таксономическом отношении в настоящее время
неясен. Границы между биологическими видами и критерии
вида в ботанике, и особенно в микологии, в том числе в ли¬
хенологии были и остаются предметом дискуссий; нередко
сомнению подвергается сама идея монофилетического проис¬
хождения таксонов. Вопрос о существовании четко очерчен¬
ных биологических видов остается открытым.
Номенклатура имеет во многом формальный характер.
Тем не менее, она необходима как инструмент для работы с
объектами живой природы и как универсальный язык общения
специалистов.
Наиболее часто употребляемые понятия и термины
(см. также Международный кодекс ботанической номенклату¬
ры)
Бинарная номенклатура - система, согласно которой для
обозначения видов употребляются бинарные (двойные) назва¬
ния. Названия видов состоят из родового и видового эпитетов.
Базионим - первое название данного вида, впоследствии
замененное другим с использованием той же основы или эпи¬
тета вследствие изменения ранга таксона или его систематиче¬
ского положения.
Дата обнародования - дата опубликования работы, где
приведен тот или иной таксон, или дата опубликования перво¬
го названия таксона.
Действительное обнародование - обнародование, со¬
провождаемое описанием или диагнозом таксона, или ссылкой
на таковые, и удовлетворяющее другим правилам кодекса
(МКБН, статьи 35-45).
Диагноз - изложение того, что, по мнению автора, отли¬
чает данный таксон от другого или других (МКБН, статья
32.2), описание таксона.
Долиннеевские - названия таксонов или работы, опубли¬
кованные до 1 мая 1753 года - принятой даты публикации ра¬
боты К. Линнея «Species plantarum». Таким образом, в номен¬
клатурном отношении ранние публикации самого Линнея рас¬
сматриваются как долинневские.
Излишнее название (superfluous name) - название, давае¬
мое таксону, для которого уже имеется действительно обнаро¬
дованное другое название. Излишние названия незаконны и
должны отвергаться. Например, для современного названия
Coenogonium luteum (Dicks.) Kalb. & Lucking базионимом яв¬
ляется Lichen luteus Dicks. Однако в 1810 г. Э. Ахариусом (Е.
Acharius) было предложено название Lecidea melizea Ach. По¬
следнее название основано на том же образце, что и название
Lichen luteus, но опубликовано позже, чем видовое название,
опубликованное Д. Диксоном (J. Dickson). В данном случае,
Lecidea melizea Ach. является излишним названием.
Название голое (nomen nudum) - название, обнародован¬
ное без описания или диагноза и без ссылки на то и другое.
Например, так было опубликовано название Wilmsia latens
(Нерр in Arnold) J. Lahm (синоним Pachyphiale fagicola (Hepp
in Arnold) Zwackh).
Описание - перечисление признаков определенного так¬
сона.
Протолог - все элементы, связанные с описанием таксо¬
на при его первом действительном обнародовании, т. е. диа¬
гноз, иллюстрации, описание, ссылки, географические данные,
цитирование типа, комментарии.
Синоним - название, рассматриваемое как относящиеся
к тому же таксону, что и принятое название. Синонимы быва¬
ют номенклатурные и таксономические. Номенклатурные си¬
нонимы (или гомотипные) - это разные названия, основанные
на одном и том же типе. Таксономические синонимы (или ге-
теротипные) - это разные названия, основанные на разных ти¬
пах, рассматриваемых в качестве принадлежащих одному и
тому же таксону (МКБН, статья 14).
В лихенологической (как и ботанической) литературе
часто используют предлоги «et», «in» и «ех». Предлог «et»
обозначает «и» и применяется в том случае, когда имя таксона
опубликовано совместно двумя авторами. Вместо слова «et»
можно использовать знак «&». Однако не следует использо¬
вать оба символа одновременно. Например, Gyalecta croatica
[синоним Gyalecta derivata (Nyl.) H. Olivier] опубликована
совместно Шулером (Schuler) и Цальбрукнером (Zahlbruckner),
то есть Gyalecta croatica Schuler et Zahlbr., Ost. Bot. Zeitsch.
55(1): 5. 1905.
Цитирование названия Gyalecta friesii Flot. ex Korb. по¬
казывает, что Кёрбер (Korber) является автором действитель¬
ного обнародования названия Gyalecta friesii, которое было
указано Ю. Флотовым (J. Flotow) в письме (может быть указа¬
но на гербарной этикетке или обнародовано в противоречие
правилам МКБН). Автором таксона считается второй автор, т.
е. G. W. Korber. При цитировании фамилию автора, стоящую
перед «ех», можно опустить: Gyalecta friesii Korb. (МКБН, ста¬
тья 46).
Цитирование названия Bacidia fagicola Нерр in Arnold
[синоним Pachyphiale fagicola (Hepp in Arnold) Zwackh] пока¬
зывает, что И. Хепп (J. Нерр) является автором действительно
обнародованного названия в работе, изданной Ф. Арнольдом
(F. Arnold). В данном случае автором обнародованного таксо¬
на считается J. Нерр, то есть первый автор. При сокращенном
цитировании «in Arnold» можно опустить: Bacidia fagicola
Нерр. Однако следует помнить, что при таком сокращенном
цитировании будет труднее найти литературный источник, в
котором таксон был опубликован.
В тех случаях, когда название обнародовано совместно
более чем двумя авторами, вместо указания второго и после¬
дующих авторов можно использовать сокращение «et al.» (et
alii, aliorum).
Аббревиатуры и условные обозначения, используемые при
работе с гербарием.
При описании лишайников, работе с гербарным матери¬
алом и в публикациях наиболее часто встречаются также сле¬
дующие аббревиатуры и условные обозначения (см. также
Кирпичников М. Э., Забинкова, 1977):
! - на гербарных этикетках - «подтверждаю», знак того,
что специалист согласен с предшествующим определением.
Наиболее часто встречается в старых коллекциях [в частности,
этим обозначением нередко пользовался Э. А. Вайнио (Е. А.
Vainio)]. При цитировании типового материала в систематиче¬
ских публикациях (напр., Davydov et al., 2011; Kukwa, 2011),
знаком «!» принято отмечать образцы, которые видел автор
работы.
Aff. (affinis) - родственный (виду, роду, иному таксону)
или похожий на него.
Agg. (aggregatio) - совокупность. Обычно используют
для обозначения совокупности близкородственных таксонов.
Cf., cfr. (confer, conferatur) - сравните, подлежит срав¬
нению; используется также в значении сравнимый с..., похо¬
жий на... (Международный..., 2009).
Coll. (colligere) - собиратель.
Comb. nov. (combinatio nova) - новая номенклатурная
комбинация.
Conf. - данная аббревиатура имеет разные значения в
латинском языке (Кирпичников М. Э., Забинкова, 1977):
- Conf. / Conff. - то же, что cf. (в единственном числе /
во множественном числе);
- Conf. (conformis) - похож по форме;
- Conf. (confirmatio) - подтверждение. Данное сокра¬
щение не является общепринятым, однако в лихенологических
коллекциях и литературе достаточно широко используется как
указание на то, что специалист согласен с предыдущим опре¬
делением. Более корректным в данном случае является сокра¬
щение confirm.
Design, (designare) - указывать, отмечать (обращать
внимание).
Det., determ, (determinare, determinavit) - определять,
определен (кем-то).
Exs. (exsiccati) - эксикаты, изданные гербарные образ¬
цы.
F. / f. (forma) - форма.
ibid. (ibidem) - то же (место, страница, коллектор...).
In lit., in litt. (in litteris) - в письме.
Incl. (incluso) - включая, в том числе.
Ined. (ineditus) - не описанный, не опубликованный.
Leg. (legi, legit) - собрал.
N. п., пот. nud. (nomen nudum) - название без описа¬
ния, считается недействительным.
Nom. invalidum (nomen invalidum) - недействительное
название.
N. v. (non vidi) - не видел.
Sensu lat., s. 1. (sensu la to) - в широком смысле, в более
широком смысле.
Sp. п., sp. nov., spec, nova (species nova) - новый вид.
S. str. (sensu stricto) - в точном (узком) смысле.
V., var. (varietas) - разновидность.
Vid. (vidi) - видел.
Типификация
Типификация, т. е. обозначение или выделение типа -
эталона, соответствующего определенному названию таксона,
необходима для стабильности в применении названий живых
организмов.
В лихенологии (как и в ботанике, микологии) типом
названия семейства является род, типом названия рода являет¬
ся вид, а типом названия вида является высушенный гербар¬
ный образец (в некоторых случаях - рисунок). Тот образец,
который указал и описал автор названия таксона, опубликовав
описание, является голотипом (holotypus). Голотипу соответ¬
ствует только один образец. Голотип не может быть отверг¬
нут. Если сам автор позднее определяет какой-либо другой
элемент в качестве типа, то такой более поздний выбор не
имеет особого статуса.
В том случае, когда собранный автором описания мате¬
риал присутствует в виде более чем одного гербарного образ¬
ца, для всех образцов (кроме собственно голотипа) использу¬
ют термин изотип (isotypus). Изотип - это каждый дубликат
(часть одного сбора, произведенного коллектором единовре¬
менно) голотипа, однако само описание таксона основано
именно на голотипе. Как правило, изотипы рассылают в раз¬
ные гербарии мира для обеспечения большей гарантии со¬
хранности типового материала таксона. До 1 января 1958 года
авторы описаний новых видов не обязаны были (и часто не
делали этого) указывать, на каком именно типовом материале
основано описание (голотип, изотипы). Если в оригинальной
публикации (протологе) указан только один образец, то это
голотип.
Часто, однако, приходится сталкиваться с ситуацией, ко¬
гда автор использовал при описании вида несколько образцов,
собранных из одного места в одно и то же время. Такие образ¬
цы называют синтипами. Синтип (syntypus) - это каждый из
двух или большего числа образцов, цитированных автором
при описании таксона, когда голотип не был указан, или каж¬
дый из двух или большего числа образцов, одновременно ука¬
занных как типы.
Если голотип утерян, уничтожен или просто не обозна¬
чен автором при публикации, должен быть предложен лекто-
тип. Лектотип - это экземпляр, выбранный из имеющегося
оригинального материала автора для того, чтобы служить но¬
менклатурным типом до тех пор, пока голотип не обнаружен.
Важно, что ни сам автор, ни другие авторы не могут выбрать
голотип после обнародования названия таксона, этот выбран¬
ный тип будет считаться лектотипом. Если существует один
или более изотипов или синтипов, то в качестве лектотипа
должен быть выбран именно один из них.
Если нет ни изотипов, ни синтипов, ни какого-либо пер¬
воначального материала, на котором основано описание авто¬
ра, то может быть выбран неотип. Неотип - это экземпляр,
выбранный на основании авторского описания таксона, чтобы
служить его номенклатурным типом до тех пор, пока отсут¬
ствует какой-либо материал, на котором было основано автор¬
ское название таксона. Неотип может быть выделен только в
том случае, когда полагают, что весь первоначальный матери¬
ал испорчен или утерян. Неотип может быть выбран из экси-
катов (см. ниже), хранящихся в наиболее крупных гербариях
мира. Предпочтение всегда отдается лектотипу перед неоти¬
пом. Неотип может быть отвергнут любым более поздним ав¬
тором, если он после тщательного исследования установит,
что данное название неприменимо к таксону.
Иногда помимо образцов, собранных одновременно с
голотипом, автор цитирует в той же публикации другие мате¬
риалы, просмотренные им и относящиеся к тому же виду. Па¬
ратип - образец, цитированный автором в протологе помимо
голотипа и изотипа (изотипов). В большинстве случаев, когда
голотип не указан, не будет и паратипов, так как все цитируе¬
мые образцы будут синтипами.
Эпитипом (epitypus) называют образец или изображе¬
ние, используемые для обеспечения интерпретируемости типа,
если имеющийся типовой материал не соответствует в точно¬
сти описанию таксона.
Существует множество и других названий типового ма¬
териала, бывших в употреблении до выхода первого МКБН.
Так, топотипом (topotypus) считают образец, собранный в
том же месте (locus classicus), где был собран типовой матери¬
ал, но позднее. Типотипом (typotypus) является образец, на ос¬
новании которого сделано изображение, являющееся типом.
Весь типовой материал (голотипы, изотипы, паратипы,
лектотипы, синтипы, неотипы и другие типовые образцы)
должен бережно храниться. Типовой материал хранят либо в
основном гербарии, как, например, в лихенологическом герба¬
рии Ботанического музея университета Хельсинки (Н) в Фин¬
ляндии, либо в отдельном типовом гербарии, как в гербарии
лаборатории лихенологии и бриологии Ботанического инсти¬
тута им. В. JI. Комарова Российской Академии Наук (LE-L)
или в гербарии Ботанической секции Музея эволюции универ¬
ситета Упсалы (UPS) в Швеции.
Для работы систематика также очень важны эксикаты.
Эксикаты - это гербарные образцы одного вида, собранные
одновременно в одном и том же месте в определенном коли¬
честве экземпляров. Эксикаты к типовому материалу не отно¬
сятся, предназначены для рассылки в ведущие гербарии мира
и сопровождаются публикацией всех данных этикеток образ¬
цов в многотиражном научном издании. Без такой публикации
официально изданными эксикаты не считаются. Например, А.
Vezda в 1960-1991 гг. отдельными брошюрами издавал серию
Lichenes selecti exsiccati. Добавить сюда эксикаты Еленкина и
Савича и ссылки на них в список литературы!
Литература
Кирпичников М. Э., Забинкова Н. Н. Русско-латинский
словарь для ботаников. Л.: «Наука», 1977. 856 с.
Куприянов А. В. Развитие принципов систематики в
XVII-XVIII вв. Автореф. дис. канд. биол. наук. М., 2005. 26 с.
Международный кодекс ботанической номенклатуры
(Венский кодекс). М.-СПб.: Товарищество научных изданий
КМК, 2009. 282 с.
Савич В. П. Новые виды и формы лишайников Камчатки
// Известия Императорского Ботанического Сада Петра Вели¬
кого. 1914. Т. 14. С. 111-128.
Authors of Fungal Names. Версия 2. 1992. [Электронный
ресурс]. Код доступа:
http://www.indexfungorum.org/Names/AuthorsOfFungalNames.as
Р
Barnhart J. Н. Biographical Notes upon Botanists. 1965.
Vol. 1. 563 p.; Vol. 2. 259 p.; Vol. 3. 545 p.
Buxbaum I. Ch. Observationes circa Quasdam Plantas
Ingricas // Commentarii Academiae Scientiarum Imperialis
Petropolitanae. Tomus III. Petropoli, Typis Academiae: 1728. P.
270-273.
Davydov E. A., Himelbrant D. E., Stepanchikova I. S. Con¬
tribution to the study of Umbilicariaceae (lichenized Ascomycota)
in Russia. II. Kamchatka Peninsula // Herzogia. 2011. Bd 25. P.
229-241.
Kukwa M. The lichen genus Ochrolechia in Europe.
Gdansk: Fundacja Rozwoju Uniwersytetu Gdanskiego, 2011. 309
P-
Laundon J. R. Deceased lichenologists: their abbreviations
and herbaria. Lichenologist. 1979. Vol. 11. P. 1-26.
Linnaeus K. Fundamenta botanica, quae majorum operum
prodromi instar theoriam scientiae botanices per breves
aphorismos tradunt (1736). Kessinger Publishing, 2010. 226 p.
McNeill J., Barrie F. R., Burdet H. М., Demoulin V.,
Hawksworth D. L., Marhold K., Nicolson D. H., Prado J., Silva P.
C., Skog J. E., Wiersema J. H. & Turland N. J. (eds.). International
Code of Botanical Nomenclature (Vienna Code) adopted by the
Seventeenth International Botanical Congress Vienna, Austria, Ju¬
ly 2005. Regnum Vegetabile 146, A. R. G. Gantner Verlag KG,
Ruggell, Liechtenstein. [I]-XVIII. 2006. 568 p.
Nicolson D. H. A History of Botanical Nomenclature // An¬
nals of the Missouri Botanical Garden. 1991. Vol. 78, No. 1. P. 33-
56.
Nylander W. Addenda nova ad Lichenographiam
Europaeam. Continuatio octava et vicesima // Flora. 1977. Vol. 60.
P. 457-463 [in: William Nylander's Collected Lichenological Pa¬
pers (reprint J. Cramer, 1990). II. P. 113-1144].
Rasanen V. Die Flechtenflora der nordlichen Kustengegend
am Laatokka-See // Annales Botanici Societatis Zoologicae-
Botanicae Fennicae Vanamo. 1939. Vol. 12. 240 p.
Rules for Zoological Nomenclature by Hugh E. Strickland
[Правила зоологической номенклатуры, сочиненные Хью
Эдвином Стриклендом]. Оцифровка, перевод на русский язык
и предисловие А. В. Куприянова -
http://tinea.narod.ru/library/methodus/stricklandl 837/ (Дата об¬
ращения 18 февраля 2012).
Stevenson L. A. List of authors of plant parasite names; rec¬
ommended abbreviations // Agricultural Handbook, U.S. Forest
Service. 1960.№ 165. P. 517-531.
Strickland, Hugh E. Rules for Zoological Nomenclature //
The Magazine of Natural History and Journal of Zoology, Botany,
Mineralogy, Geology, and Meteorology. 1837. Vol. 1. New Series.
P. 173-176.
Vainio E. A. Lichenes in viciniis Viburgi observati // Medde-
landen af Societas Pro Fauna et Flora Fennica. 1878. Vol. 2. P. 35-
72.
Vainio E. A. Lichenographia Fennica II. Baeomyceae et
Lecidiales // Acta Societatis Pro Fauna et Flora Fennica. 1922. Vol.
53. 343 p.
Viegas A. P. Indice de Fungos da Ameica do Sul. Campinas,
Argentina; Instituto Agronomico. 1961. 921 p.
Wright J. E., Lois R. J. Lista da singlas de autores empleades
en micologiay fitopatologia// Lilloa. 1949. Vol. 21. P. 225-269.
Глава X.
Молекулярно-генетические методы в систематике
лишайников
Давыдов Е. А.
Биологическая систематика - раздел биологии, нацелен¬
ный на разработку принципов классификации живых организ¬
мов и практическое приложение этих принципов к построе¬
нию системы. Систематика включает в себя таксономию (вы¬
деление и описание таксонов), номенклатуру (принципы при¬
менения названий таксонов), филогению (эволюционные от¬
ношения между таксонами) и классификацию (иерархию так¬
сонов). Систематика - древнейшая биологическая дисциплина,
составляющая классический фундамент современной биоло¬
гии. В контексте эволюционной теории систематика занимает¬
ся идентификацией и прояснением филогенетических взаимо¬
отношений разных организмов, как современных, так и вы¬
мерших. Единицы классификации - таксоны выделяются и
сравниваются на основании изучения их признаков. Эволюци¬
онная теория утверждает, что сходство среди индивидуумов
или видов на основе гомологичных признаков указывает на их
общее происхождение. Однако бывает очень трудно отличать
гомологичные признаки от аналогичных, выделять признаки
наиболее важные для целей систематики, определять причину
отсутствия отдельных признаков или их непрерывность. Си¬
стематика стала наукой, практически зависящей от опыта и
знания отдельных экспертов - монографов. Технический уро¬
вень биологических методов при выявлении определенных
признаков сходства или отличия все время развивается. Сего¬
дня, наряду с морфологическими, экологическими, анатомиче¬
скими, кариологическими, биохимическими, иммунологиче¬
скими признаками все большую роль играют молекулярные
данные, полученные при изучении ДНК.
Для этого имеются теоретические основания. Организм
представляет собой единство фенотипа и генотипа. В ходе
эволюции генетическая изменчивость накапливается в геноти¬
пе, после чего морфологические и/или физиологические изме¬
нения принимают то или иное направление под воздействием
внешних условий. Систематика до сих пор базировалась в ос¬
новном на фенотипических признаках, проявление которых
подвержено влиянию со стороны окружающей среды, в то
время как молекулярно-генетические данные позволяют ре¬
конструировать эволюционный процесс на первичных - гено¬
типических признаках. Сама природа ДНК позволяет исполь¬
зовать ее как «документ» эволюционной истории. Поскольку
геномы эволюционируют через постепенное накопление мута¬
ций, количество отличий последовательностей нуклеотидов
между парой геномов разных организмов должно указать, как
давно эти два генома отделились от общего предка. Два гено¬
ма, которые разделились в недавнем прошлом, должны иметь
меньшие отличий, чем два генома, чей общий предок очень
давний. Поэтому, сравнивая геномы друг с другом, можно по¬
лучить сведения об эволюционных взаимоотношениях между
ними. Эволюция ДНК очень сложна, а изменения касаются
множества отдельных нуклеотидов, поэтому выявление гомо¬
логии часто возможно только с помощью методов компьютер¬
ной обработки данных. Адекватный анализ данных молеку¬
лярных методов становится задачей информационных техно¬
логий - биоинформатики. Геносистематика оперирует призна¬
ками, полностью независимыми от тех, что используются в
других эволюционных построениях и позволяет анализировать
гомологичные признаки там, где классические методы пока не
дали определенного результата. В эволюционной интерпрета¬
ции сродство понимается как родство по происхождению, а
признаки и сходства значимы как отражение генеалогических
(филогенетических) связей. Родство, таким образом, интер¬
претируется как преимущественно генотипическое сходство,
тем самым подчеркивается приоритет молекулярно¬
генетических данных. Молекулярные методы значительно
расширяют фактологическую базу систематики и являются
важнейшим критерием установления родственных связей, но,
как и любой инструмент, требуют осторожного и правильного
применения.
Для правильного получения и использования данных
молекулярных методов решающим является знание объекта
исследования, поскольку, только хорошо зная объект, можно
правильно поставить задачу и решить ее. Нужно всегда пом¬
нить, что молекулярные методы не заменяют классические
методы систематики, а только дополняют их!
Лишайник - обычно двух или трехбионтная система, в
состав которой обязательно входит несколько организмов:
мицелиальный микобионт, а также фотобионт (водоросль
и/или цианобактерия). В систематике с понятием «лишайник»
отождествляется грибной компонент этой системы - лихени-
зированный гриб, поэтому с таксономической точки зрения
лишайники рассматриваются в системе грибов, на изучении
которых в настоящем разделе делается акцент. Тем не менее,
молекулярно-генетические исследования водорослей и циа¬
нобактерий в талломах лишайников представляют несомнен¬
ный интерес как с точки зрения эволюции этих групп, так и
эволюции лишайникового симбиоза в целом. Для изучения
фотобионтов используются те же молекулярно-генетические
методы, которые затем интерпретируются в соответствии с
целями исследования.
Обзор раннего этапа развития молекулярных методов в
изучении лишайников был сделан DePriest (2004). Первое ис¬
следование с использование ДНК-методов было проведено в
СССР О. Б. Блюмом и Г. П. Кашеваровым (1986), которые ис¬
пользовали ДНК-гибридизацию для изучения обоснованности
выделения рода Lasallia, а затем опробовали этот метод и на
других объектах. В 1991 году Armaleo & Clerc изучали при
помощи гибридизации и ПЦР две пары фотосимбиодем из ро¬
дов Sticta и Рseudocyphellaria и доказали, что морфологически
значительно отличающиеся талломы образует один вид ли¬
шайникового гриба в симбиозе с зеленой водорослью либо
цианобактерией. Niu & Wei (1993) впервые использовали по¬
следовательности ITS2 для сравнения двух видов рода
Lasallia. Начиная с 1995 года, число работ, выполненных с
помощью молекулярно-генетических методов, стремительно
возрастает. Большинство из них посвящены филогенетиче¬
скому анализу отдельных таксонов разного ранга. Одной из
сложных задач геносистематики является выбор маркеров,
наиболее точно отражающих филогенез. В ранних работах в
качестве филогенетических маркеров использовались, в ос¬
новном, последовательности ядерной рибосомальной ДНК. На
уровне семейств и порядков использовали консервативную
последовательность SSU рДНК и консервативную часть LSU
рДНК, а на видовом и внутривидовом уровне - вариабельный
ITS фрагмент, иногда дополняемый соседними участками SSU
или LSU рДНК. В последующем набор маркеров пополнился
рибосомальной ДНК митохондрий и генами, кодирующими
такие белки как Р-тубулин, гистоны, хитин-синтетазу, РНК-
полимеразу и др., а также анонимными маркерами. В то же
время алгоритмы филогенетического анализа усложнились,
создано и оптимизировано для анализа последовательностей
ДНК программное обеспечение. Технологии секвенирования
стали точнее и дешевле, что привело к накоплению огромного
массива данных, и созданию сервисов для оперирования ими
(генетические банки). Поскольку эволюция разных генов и
некодирующих последовательностей идет неравномерно, кла-
дограммы, построенные на базе разных маркеров, также отли¬
чаются. Для получения более надежных данных было предло¬
жено сравнивать результаты, полученные при использовании
различных маркеров, а также использовать несколько марке¬
ров одновременно. Развитие технологий в настоящее время
вплотную подошло к возможности быстрого секвенирования
всего генома. Это дает новый импульс развитию информаци¬
онных технологий, так как массивы данных, которыми необ¬
ходимо оперировать исчисляются миллионами пар оснований
на один образец. Информация о геноме даст новые инструмен¬
ты исследователям, в том числе и систематикам для решения
стоящих перед ними вопросов классифицирования биологиче¬
ского разнообразия.
Основные вопросы систематики, решаемые при помощи
молекулярных методов
Описание новых таксонов с привлечением молекулярно¬
генетических данных стали гораздо убедительнее. Классиче¬
ский морфолого-эколого-географический метод даёт гипотезы,
которые могут быть проверены независимыми ДНК-методами.
Генетические отличия между особями, популяциями, потенци¬
альными видами, родами могут быть количественно выраже¬
ны, либо показаны на филогенетическом древе. Эти данные
могут подтвердить или опровергнуть гипотезу. Таким образом,
обоснован статус сотен видов лишайников, описанных в по¬
следнее время, и такая методология становится общепризнан¬
ной. Этому способствует значительный массив данных,
накопленных в генетических банках, который возможно при¬
влекать к исследованиям. В настоящее время описать таксон в
престижном издании, не подтвердив его молекулярными дан¬
ными, становится все сложнее. Подобные исследования про¬
водятся в отношении парных видов - сходных по морфологии,
но отличающихся способом размножения пар, фотосимбиодем
- морфотипов, образованных при участии разных фотобион¬
тов, хеморас, которые могут быть отдельными видами, а могут
быть выражением биохимической изменчивости в пределах
одного вида. Молекулярными методами возможно выявление
викарных видов, близких морфологически, но отличающихся
географически, изучение генетического единства дизъюнктив¬
ных видов, выявление «скрытых» (cryptic) видов. Молекуляр¬
ные методы дают инструмент тестирования гипотез, связан¬
ных с описанными выше случаями, когда фенотипические ме¬
тоды исчерпали свои возможности.
Молекулярный анализ дал возможность критически
взвесить и переосмыслить применимость тех или иных
морфологических и биохимических признаков в система¬
тике лишайников на разных иерархических уровнях. Его
результаты подтверждают взгляд, что универсальных феноти¬
пических признаков - «единого основания деления» не суще¬
ствует. Ключевые характеристики появлялись и многократно
изменялись независимо в разных группах аскомицетов в ходе
эволюции, применимость каждого признака для целей систе¬
матики необходимо в каждом случае обосновывать. Идея того,
что систематическое значение конкретных признаков обычно
не выходит за рамки отдельной группы и не может механиче¬
ски переноситься на другую группу, не нова. Однако в течение
долгого времени, классификация аскомицетов базировалась на
ограниченном числе признаков плодовых тел, которые ис¬
пользовались не только для описания таксонов, но и опреде¬
ляли филогенетические отношения между ними. С широким
вовлечением молекулярных методов систематика грибов
очень существенно изменилась. При этом, как правило,
наблюдается корреляция выделенных таксонов с морфологи¬
ческими признаками, однако не теми, которые использовались
ранее.
Происхождение лишайников. Начиная с ранних моле-
кулярно-филогенетических работ, появление лишайников или
процесс лихенизации в разных группах грибов вообще и среди
аскомицетов в частности становится доказанным фактом,
кроме того обсуждается и обратный процесс - происхождение
некоторых групп грибов в результате процесса делихенизации.
Напрямую не относится к систематике лишайников, но
является важным в контексте их биологической природы во¬
прос коэволюции микобионта и фотобионта, соотношение ге¬
нетического разнообразия участников симбиоза, селектив¬
ность гриба в выборе фотобионта. Эти вопросы также реша¬
ются преимущественно молекулярно-генетическими метода¬
ми.
Геном грибов и лишайников
В клетках грибов ДНК находится в ядрах (геном) и ми¬
тохондриях (хондром), в клетках водорослей, кроме того в
пластидах (пластом), а в клетках цианобактерий - в цитоплаз¬
ме. Размер генома измеряется миллионами пар оснований (Ьр),
а хондрома и пластома - первыми сотнями тысяч Ьр. Размер
генома грибов занимает промежуточное положение между ге¬
номом бактерий и высших эукариот. Наименьший геном среди
грибов имеет аскомицет Aeshbya gossypii - 9,7 Mbp, геном ве¬
шенки, Pleurotus ostreatus составляет 21 Mbp, а плютея олень¬
его, Pluteus cervinus - 90 Mbp (Дьяков и др. 2005). Размер ге¬
нома лихенизированного гриба Xanthoria parietina -31,9 Mbp;
Cladonia grayi - 28,6 Mb, что сравнимо со средним размером
геномов Peziziomycotina, а одноклеточной водоросли
Asterochloris sp. - 106,7 Mb, что находится на нижнем уровне
размера геномов у изученных представителей
Trebouxiophyceae (Armaleo, May, 2009). К настоящему времени
расшифрованы последовательности нуклеотидов геномов не¬
скольких десятков грибов (Galagan et al. 2005), близка к за¬
вершению работа по секвенированию генома Xanthoria
parietina.
Большой размер эукариотических геномов обусловлен
значительной долей некодирущих участков, повторяющихся
последовательностей ДНК. Поэтому выявление генов в геноме
и их клонирование требуют весьма чувствительных методов,
один из которых - полимеразная цепная реакция (ПЦР).
Выделение и клонирование ДНК
Для выделения ДНК из микобионтов лишайников берут
плодовые тела, часть таллома, а в ряде случаев культуру гри¬
ба, выращенную на питательной среде. Важнейшим фактором,
влияющим на успешность выделения и анализа ДНК, является
возраст образца, а также условия его высушивания и хранения.
Если материал собирается специально для ДНК исследований
рекомендуют коллектировать сухие талломы лишайников, в
которых ДНК естественным образом «законсервирована», ли¬
бо быстро высушивать их с применением силикагеля или про¬
сто на воздухе, но без нагревания. Для анализа всего генома
необходим свежий материал возрастом, как правило, не более
года, а для секвенирования отдельных фрагментов подходит
материал возрастом, в среднем, до 3-5 лет, хотя отдельные
случаи удачного секвенирования известны для гербарных об¬
разцов возрастом до нескольких десятков лет. Замораживание
высушенных образцов значительно увеличивает срок их при¬
годности для ДНК методов. При выборе участка таллома или
плодовых тел обращают внимание на отсутствие видимых по¬
вреждений, связанных с присутствием других грибов, ДНК
которых может загрязнять целевой продукт. По этой же при¬
чине стараются по возможности избавиться от ризиноморф, а
также фотобионтов.
Практически все молекулярно-генетические методы тре¬
буют выделение чистой и как можно менее поврежденной
ДНК.
Большинство современных методов выделения ДНК из
тканей грибов и лишайников состоят из следующих этапов:
1. механическое разрушение гиф;
2. лизис клеточных мембран;
3. очистка лизата от ингибиторов ферментативных ре¬
акций (белков, полисахаридов, полифенолов и т.д.).
Разрушение гиф осуществляется механически перетира¬
нием с оксидом кремния (или оксидом алюминия) или с ис¬
пользованием жидкого азота.
Лизис клеточных мембран происходит в лизис-буфере:
под воздействием поверхностно-активных веществ или хо-
атропных солей происходит разрушение липидного бислоя. В
состав лизис-буфера могут входить ферменты протеазы, раз¬
рушающие протеины или РНКаза для удаления РНК.
После разрушения клеточных стенок и лизиса мембран
образуется мультикомпонентная смесь, содержащая, в том
числе, ДНК.
После стадии лизиса возможны 3 основных принципи¬
альных подхода для очистки целевой ДНК:
1. Очистка раствора с помощью методов органической
экстракции (с помощью фенола, хлороформа) с последующим
осаждением ДНК спиртами и растворением в воде и ТЕ-
буфере (Doyle & Doyle, 1987). Метод был адаптирован для
лишайников (Armaleo, Clerc, 1995; Grube et al. 1995).
2. Дифференциальная сорбция ДНК на твердом носите¬
ле (чаще всего силикагели с повышенным отрицательным за¬
рядом или модифицированной поверхностью). Сорбент с ло¬
кализованной на его поверхности ДНК промывается органиче¬
скими растворителями, затем ДНК смывается водой или ТЕ-
буфером. Эта технология реализована в наборах реагентов
многих производителей, например NucleoSpin -
www.macherey-nagel.com; DNeasy - www.qiagen.com; PureLink
- www.invitrogen.com/plgenomic
3. Дифференциальная сорбция примесей, селективное
осаждение ДНК, с последующей промывкой органическими
растворителями и растворением в воде или ТЕ-буфере. Техно¬
логия реализована в наборах реагентов DiamondDNA, АВТ,
11с. www.diamond-dna.ru
Каждый из этих подходов имеет свои достоинства и не¬
достатки и выбирается исследователем исходя из объекта, за¬
дач и бюджета научного проекта.
Для оценки чистоты выделенной ДНК чаще всего ис¬
пользуют спектрофотомерию. Нуклеиновые кислоты имеют
характерный спектр поглощения в ультрафиолетовом излуче¬
нии с вершиной около 260 нм, в то время как белки около 280
нм. Оценку качества экстрагированной ДНК проводят на ос¬
новании измерения ДНК в области белкового и нуклеинового
спектров поглощения, т.е. при 280 и 260 нм, соответственно.
Чистая ДНК имеет соотношение оптической плотности рас¬
твора 260 нм/280 нм между 1.8 - 2.0.
Этот же принцип делает возможным расчет концентра¬
ции ДНК и используется в приборах ДНК-флюориметрах;
OD26o= 1 соответствует концентрации 50 мкг /мл.
2.1. Полимеразная цепная реакция.
Полимеразная цепная реакция - ПЦР (PCR - polimerase
chain reaction) - метод ферментного синтеза ДНК вне организ¬
мов (in vitro). С момента его открытия в 1983 году, этот метод
используется практически во всех молекулярно¬
биологических лабораториях. Исходным материалом для ПЦР
является раствор ДНК. Из одного фрагмента ДНК посред¬
ством ПЦР за несколько часов можно получить много милли¬
онов копий. Таким образом, ПЦР является быстрым, чувстви¬
тельным и избирательным методом. Он основывается на сле¬
дующем принципе. Относительно короткие (приблизительно
20 нуклеотидов) одноцепочечные олигонуклеотиды (т. н.
праймеры или затравки) находят комплементарные последо¬
вательности даже в весьма сложных геномах и гибридизируют
с ними (гибридизация фрагментов ДНК часто также называют
отжиг от англ. annealing). Для гибридизации фрагмент ДНК в
растворе также должен быть одноцепочечным. ДНК-
полимераза использует свободную ОН-группу на З'-конце
праймера в качестве стартовой и синтезирует вторую цепь
ДНК. Двойная цепь ДНК путем нагревания до 94°С опять раз¬
деляется (денатурация), что является условием для присоеди¬
нения праймеров. Поскольку один цикл дает две двойных цепи
ДНК, необходимо провести как минимум 25-30 циклов дена¬
турации и синтеза, чтобы получить достаточное количество
необходимого фрагмента ДНК, растущее в геометрической
пронрессии. Для синтеза используют ДНК-полимеразу, выде¬
ленную из термофильной бактерии Thermus aquaticus (Taq-
полимераза), которая стабильна даже при многократном
нагревании до 94°С. Это позволило проводить ПЦР in vitro в
так называемых «термоциклерах». Типичный цикл ПЦР со¬
стоит из 3 повторяющихся 25-40 раз шагов.
1. Денатурация при 94°С 30-60 секунд
2. Отжиг праймеров; в зависимости от длины и состава
праймеров при 37°С - 65°С (чем длиннее праймер, тем выше
температура связывания) - 30 - 60 секунд.
3. Синтез (репликация) ДНК при 72°С (оптимальная
температура для Taq-полимеразы) 1 -2 минуты.
Только при наличии подходящих двух праймеров воз¬
можна амплификация одного фрагмента определенной длины
и его амплификация.
ПЦР используется для решения самого широкого спек¬
тра вопросов, например для клонирования определенного
фрагмента ДНК; замены определенного нуклеотида (целевой
мутагенез); ДНК-баркодинг; определение количества повторов
генов, выделения новых генов и т. д. (Erlich, 1989).
2.2. Разделение фрагментов ДНК, имеющих разную
длину
ДНК имеет слабый отрицательный заряд и в электриче¬
ском поле движется к аноду. Эта особенность используется
для разделения ДНК фрагментов различной длины в полиак¬
риламидном или агарозном геле методом электрофореза (Рис.
Рис. 83. Разделение фрагментов ДНК, имеющих разную длину: а - за¬
полнение карманов в агарозном геле раствором ДНК с пробным краси¬
телем; б - разделение фрагментов ДНК в агарозном геле при электро¬
форезе (по: Фризен, 2007).
Разрешающая способность полиакриламидных гелей
выше, но их приготовление дороже и более трудоемко. Фраг¬
менты ДНК от 100 до 2000 нуклеотидов хорошо разделяются в
агарозном геле. Обычно используются концентрации от 0,6%
до 2%, при этом длинные фрагменты лучше разделяются в ге¬
лях с низкой концентрацией агарозы, а короткие - с повышен¬
83).
а -
L
ной. В кармашки геля помещается проба с ДНК (рис. 83),
предварительно смешанная с загрузочным буфером, содержа¬
щим утяжеляющий пробу глицерин и краситель бромфенол
голубой. Краситель делает пробу видимой и передвигается как
и ДНК к аноду со скоростью примерно равной фрагменту ДНК
из 100 нуклеотидов. Гель помещается в электрофорезный ап¬
парат, заливается на 1-2 мм специальным буфером. По окон¬
чанию электрофореза ДНК окрашивается этидиум бромидом
или другими красителями, которые светятся в ультрафиолето¬
вом излучении, что позволяет фотографировать результат
электрофореза. Длину фрагмента ДНК определяют, сравнивая
со стандартом - смесью фрагментов ДНК известного размера.
3. Основные молекулярные методы
Молекулярные методы, используемые в систематике
можно разделить на две группы: методы, базирующиеся на
анонимном полиморфизме ДНК и методы сравнения последо¬
вательности ДНК определенных фрагментов (метод секвени¬
рования участков ДНК). Для анализа родственных отношений
между близкородственными таксонами или внутривидового
разнообразия, часто более подходят методы, базирующиеся на
анонимном полиморфизме ДНК, поскольку дают «портрет»
всего генома, для анализа родственных отношений более вы¬
соких таксономических категорий (вид, род, семейство), неза¬
меним метод анализа последовательностей ДНК определен¬
ных генов или фрагментов ДНК (секвенирование). В любом
случае, выбор молекулярного метода зависит от объекта ис¬
следования и поставленной научной задачи. Ниже приводятся
краткие описания основ молекулярных методов, нашедших
применение в систематике лишайников. Для более глубокого
ознакомления и методами и конкретными методики необхо¬
димо обращаться к специальной литературе, которая предло¬
жена в конце раздела.
3.1. Молекулярные методы, базирующиеся на ано¬
нимном полиморфизме ДНК.
Методы позволяют проводить скрининг генома и полу¬
чать сравнимые данные о его полиморфизме без расшифровки
нуклеотидных последовательностей отдельных фрагментов. К
таким методам относятся следующие: RFLP, RAPD, AFLP,
DAF, SCAR, CAPS и STR. Методы используют широкое раз¬
нообразие ДНК-маркеров и сильно различаются по воспроиз¬
водимости, трудоемкости, стоимости и природе полиморфиз¬
ма, который они выявляют. Эти различия сильно влияют на
возможности их применения для различных исследований.
Они могут базироваться на гибридизации (например, RFLP),
или основаны на полимерной цепной реакции (PCR) (напри¬
мер, RAPD, DAF и AFLP); они могут выявлять уникальные
участки ДНК (например, SCAR, CAPS) или участки ДНК,
встречающиеся многократно в геноме (STR или микросател¬
литы). Выявленные признаки могут быть проанализированы в
матрицах на базе принципа присутствия/отсутствия, т.е. фене-
тическими методами анализа.
3.1.1. RAPD (random amplified polymorphic DNA): Слу¬
чайно амплифицированный полиморфизм ДНК.
RAPD метод появился в 1990 году и служит для изуче¬
ния генетического родства между близкородственными таксо¬
нами (Williams et al., 1990; Welsh, McClelland, 1990).
Для проведения RAPD используют короткие праймеры
обычно длиной 10 нуклеотидов со случайной последователь¬
ностью, желательно содержащие 50-70% ГЦ нуклеотидов. При
помощи ПЦР можно довольно быстро и недорого получить
большое количество фрагментов ДНК различной длины (при¬
знаков), результаты анализа которых можно использовать для
решения целого ряда исследований: выяснения родственных
отношений внутри и между видами, определения гибридов,
разрешения филогеографических вопросов и других.
На первом этапе выясняют, какие праймеры дают удоб¬
ное количество фрагментов ДНК на объекте исследования,
которые хорошо разделяются электрофорезом. Затем проводят
амплификацию всех образцов ДНК, с выбранными праймера¬
ми, разделяют амплифицированные фрагменты с помощью
электрофореза и документируют (см. 2.2). На основе присут¬
ствия или отсутствия фрагментов одинаковой длины составля¬
ется матрица для фенетического анализа. Чтобы увеличить
число признаков в матрице и получить более надежный ре¬
зультата обычно используется несколько праймеров. Эмпири¬
чески показано, что для достоверного анализа количество при¬
знаков в матрице должно быть не менее чем в три раза больше
чем исследуемых таксонов.
Ограничения метода: поскольку весьма трудно опреде¬
лить гомологические фрагменты, полученные в ходе разных
ПЦР, рекомендуется сравнение фрагментов, полученных с по¬
мощью одного праймера, из одной ПЦР и одного элекрофо-
резного геля (учитывая, что в самой широкой электрофорез-
ной камере помещается гребенка для 50 карманов, то и ана¬
лизировать можно не более 48 образцов).
Рис. 84. Результат электрофореза фрагментов ДНК, полученных при
использовании праймера gagccaaccg для видов рода Lasallia.
Составление матрицы RAPD-признаков. Приклады¬
ванием линейки для каждого образца отмечаются наличие или
отсутствие фрагментов ДНК одинаковой длины на фотогра¬
фии электрофорезного геля (подразумевается, что фрагменты,
имеющие одинаковое количество нуклеотидов, гомологичны),
наличие отмечается цифрой 1, отсутствие цифрой 0. Получен¬
ную матрицу анализируют с помощью программ для фенети-
ческого анализа (например: NTSYSpc, GelQuest, Treecon и
другие).
DAF (DNA amplification fingerprint):
Это модификация RAPD метода, при которой использу¬
ются праймеры длиной всего в 7 или 8 нуклеотидов. Они про¬
дуцируют достаточно сложный комплекс амплифицированных
фрагментов, поэтому амплификационный продукт разделяется
в полиакриламидном геле и окрашивается серебром. Этот ме¬
тод гораздо более чувствителен чем RAPD и позволяет полу¬
чить полиморфные фрагменты там, где нормальные RAPD
праймеры не дают никакого полиморфизма (Caetano-Annoles,
1994).
3.1.2. AFLP (Amplified Fragment Length Polymorphism) -
Амплифицированный полиморфизм длины фрагментов.
Метод похож на RAPD, но в отличие от него, праймеры
(затравки) состоят из длинной фиксированной части (около 15
пар нуклеотидов) и короткой случайной (2-4 нуклеотида).
Фиксированная часть дает праймеру стабильность (и следова¬
тельно повторяемость), а случайная часть позволяет выявить
много полиморфных фрагментов ДНК (два случайных нуклео¬
тида дают больший полиморфизм, чем три или четыре слу¬
чайных нуклеотида). AFLP технология основывается на вое-
производстве геномных фрагментов ДНК, предварительно
разрезанных с помощью рестриктаз при использовании ПЦР.
ДНК разрезается рестриктазой, и затем при помощи ДНК-
лигазы к разрезанным концам фрагментов ДНК присоединя¬
ются адаптеры, нуклеотидная последовательность которых
совпадает с фиксированной частью праймера. Случайные нук¬
леотиды (2-4 нуклеотида) прикрепляются к фиксированной
части праймера со стороны 3’ конца. Только те разрезанные
фрагменты, имеющие эти случайные 2-4 последовательности
перед присоединенным адаптером, будут амплифицированы в
ПЦР-ной реакции. Полиморфизм выявляется как присут¬
ствие/отсутствие. Матрицы признаков анализируются, как и в
случае RAPD, фенетическими методами.
AFLP признаки (фрагменты) часто являются сцепленны¬
ми группами повторной ДНК центромерной и теломерной об¬
ласти хромосом, но встречаются и в других областях хромо¬
сом. Метод достаточно сложный и дорогой, но при наличии
соответствующего оборудования и компьютерного обеспече¬
ния можно достаточно быстро получить большое количество
признаков для выявления родства между видами и для фило¬
географического анализа.
Подробный AFLP протокол можно получить на веб¬
странице: http : //b i owe b. u s u. ed и/wo 1 f/afl pprotoco 1. htm
3.1.3. SCAR (Sequence-Characterized Amplified Region) -
Амплифицированный регион с известной последовательно¬
стью.
ДНК фрагменты, амплифицированные в ПЦР с помощью
специфических праймеров, синтезированные по известной
нуклеотидной последовательности определенного фрагмента
ДНК. Например нуклеотидная последовательность, установ¬
ленная на основе клонирования определенного RAPD фраг¬
мента, представляющего интерес для анализа или нуклеотид¬
ная последовательность, взятая из базы данных (GenBank).
Синтезируются достаточно длинные праймеры (от 18 до 25
нуклеотидов), поэтому SCAR не сталкивается с проблемой
низкой воспроизводимости.
SCAR может использоваться для быстрого скрининга
большого количества образцов/таксонов, для определения об¬
разцов, имеющих уникальную последовательность ДНК, на
базе которой синтезированы праймеры.
3.1.4. CAPS (Cleaved Amplified Polymorphic Sequences) -
Расщепленный полиморфизм амплифицированных последова¬
тельностей; этот метод также часто называют PCR RFLP
ДНК фрагменты, амплифицированные в ходе ПЦР с по¬
мощью специфических праймеров, синтезированных по из¬
вестной нуклеотидной последовательности определенного
фрагмента ДНК. Эти праймеры, как и в случае SCAR прайме¬
ров, амплифицируют определенный фрагмент ДНК, но в дан¬
ном случае встречающийся у всех исследуемых таксонах, при
этом чаще всего амплифицируются различные спейсеры
(участки ДНК между генами), как наиболее полиморфные
участки ДНК. Они часто включают различные дупликации
(включения) или делеции (исключения). Полиморфные при¬
знаки выявляются при гидролизе амплифицированных фраг¬
ментов ДНК с помощью различных коротких ферментов - ре-
стриктаз. Полиморфизм проявляется в длине расщепленных
фрагментов. На основе присутствия или отсутствия фрагмен¬
тов одинаковой длины составляется матрица для фенетическо-
го анализа. Метод не требует секвенирования и с его помощью
можно достаточно быстро получить необходимое количество
признаков для фенетического анализа.
3.1.5. Микросателлиты
STMS (SequenceTagged Microsatellite Sites) или STR
(Short Tandem Repeats)
Микросателлиты представляют собой фрагменты ДНК с
повторяющимися «мотивами» - идентичными короткими по¬
следовательностями (тандемами) из нескольких нуклеотидов
(обычно 1-6 пар нуклеотидов), которые встречаются в геномах
почти всех изученных к настоящему времени таксонов. В свя¬
зи с тем что последовательности ДНК фланкирующих участ¬
ков, окружающих микросателлитные локусы, обычно консер¬
вативны, они идентичны у индивидуумов, относящихся к од¬
ному виду или близким видам. Поэтому существует возмож¬
ность определения конкретных микросателлитных локусов по
их фланкирующим участкам. Для связывания с фланкирую¬
щим участком специально разрабатываются праймеры для ам¬
плификации микросателлитного локуса с помощью ПЦР
В отличие от фланкирующих участков, сами микроса¬
теллитные повторы имеют более высокие темпы мутирования
(Dieringer, Schlotterer, 2003), что приводит, главным образом, к
изменению количества повторов, и следовательно, длины всей
последовательности повторов. Применение гель-
электрофореза высокого разрешения позволяет производить
быстрое генотипирование многих индивидуумов по большому
количеству локусов путем определения их генетической
«идентификационной метки».
Метод очень стабильный, амплифицирует уникальный
локус и выявляет высокополиморфные регионы в геноме. В
данном случае выбор специфической пары праймеров рас¬
сматривается как важный этап изоляции микросателлитных
маркеров, что требует довольно больших вложений времени и
средств в первоначальной стадии, но когда он адаптирован для
определенного вида, с его помощью можно быстро и недорого
выявить широкий полиморфизм внутри этого вида и его бли¬
жайших родственников.
Полиморфизм проявляется в длине амплифицированного
продукта, который зависит от количества повторов микроса¬
теллитов внутри уникального фрагмента. Метод прекрасно
подходит для картографирования генов, популяционного
скрининга и т.д.
3.2. Методы изучения последовательностей фрагментов
ДНК
Сравнение и анализ выравненных последовательностей
ДНК отдельных фрагментов генома и хондрома лихенизиро¬
ванных грибов становиться одним из важнейших молекуляр¬
ных методов при изучении родственных отношений в надви-
довой систематике. Для очень многих групп лишайников уже
расшифрованы многие гены или некодирующие последова¬
тельности как геномной, так и митохондриальной ДНК.
Методы расшифровки нуклеотидной последовательно¬
сти нуклеиновых кислот в отечественной литературе принято
называть методами секвенирования. Изучение последователь¬
ностей фрагментов ДНК включает нескольких этапов:
1. Выделение ДНК.
2. Амплификация заданного фрагмента ДНК всех вклю¬
ченных в анализ образцов.
3. Очистка амплификата.
4. Секвенирование.
5. Выравнивание всех полученных последовательностей
ДНК.
6. Филогенетический анализ и интерпретация результа¬
тов.
3.2.1. Амплификация
Выбор ДНК-маркеров и подбор праймеров
Выбор филогенетических маркеров зависит от постав¬
ленной задачи. Маркеры должны быть достаточно вариабель¬
ны для выбранного таксономического уровня (внутривидовой,
вид, семейство, класс), но не слишком вариабельны, чтобы
последовательность могли быть выровнены для соблюдения
принципа гомологии.
Правильно подобранные для конкретного фрагмента
ДНК праймеры играют важнейшую роль для успешной ам¬
плификации. Праймеры в паре должны быть сходными по оп¬
тимальной температуре отжига, которая зависит от длины и
состава нуклеотидов. Существуют специальные программы
расчета оптимальной температуры отжига, для ее определения
можно также поставить эксперимент на градиентном термо¬
циклере. При помощи подбора праймеров можно исключить
амплификацию ДНК сопутствующих организмов (фотобионта,
парасимбионтов), либо получать из одной пробы (часть тал¬
лома) амплификаты ДНК и микобионта и фотобионта. Разра¬
ботаны праймеры, специфичные для грибов, водорослей, а
также, например, для аскомицетов, базидиомицетов и отдель¬
ных таксономических групп лишайников (Gargas, Taylor, 1992;
Liu et al. 1999). Можно создавать праймеры самостоятельно,
исходя из знаний о консервативных последовательностях в
изучаемой группе таксонов.
Очистка амплификата
Секвенирование продуктов амплификации может быть
заблокировано наличием в растворе олигонуклеотидов и азо¬
тистых оснований. Для их удаления проводят очистку ампли¬
фиката, как правило, при помощи коммерческих наборов реа¬
гентов, большинство из которых работают на основе гель-
силикатных фильтров, встроенных в специальные пробирки-
колонки. Для более тщательной очистки амплификат разгоня¬
ют на 0,8% агарозном геле, вырезают целевой фрагмент из ге¬
ля и выделяют из него ДНК. Этот прием используют также в
случае если амплифицировались нескольких отличающихся по
длине копий фрагмента (например, гены ядерной ДНК от раз¬
личных родителей, с интроном и без него или гены парасим¬
бионтов). В ряде случаев секвенировать эти фрагменты воз¬
можно только после клонирования ПЦР амплификатов в плаз¬
мидах, что приводит к значительному удорожанию и замедле¬
нию процесса получения ДНК последовательностей.
3.2.2. Секвенирование амплифицированного фраг¬
мента
В настоящее время существует уже довольно большое
количество методов секвенирования отдельных фрагментов
ДНК, большинство из которых подробно описаны в книге Че-
мериса с соавт. (1999). Подробное описание процесса секве¬
нирования и принцип работы автоматических секвенаторов
можно прочитать также на веб-странице:
http://molbiol.ru/protocol/13_03.html Существует достаточное
количество лабораторий? в которых можно заказать секвени¬
рование. Как правило, для этого нужно отправить очищенный
ПЦР-продукт и необходимые для секвенирования праймеры и
получить последовательность нуклеотидов с кривыми, отра¬
жающими качество их распознавания (рис. 85). Рекомендуется
секвенирование обеих цепей двойной спирали ДНК, с прямым
и обратным праймерами. Результаты сопоставляются и, в слу¬
чае неуверенного распознавания нуклеотида на одной цепи,
можно верифицировать данные по другой. Даже незначитель¬
ное количество ошибок может повлиять на результат исследо¬
вания.
I I
Рис. 85. Фрагмент сиквенса с кривыми и автоматически расшифрован¬
ными нуклеотидами.
3.2.3. Выравнивание
При анализе нуклеотидных последовательностей в каче¬
стве отдельных признаков выступают отдельные нуклеотид¬
ные позиции. А поскольку принцип филогенетического анали¬
за предполагает сравнение гомологичных признаков, то необ¬
ходимо установить позиционную гомологию нуклеотидов в
разных последовательностях. К этому и сводится следующий
этап обработки данных после того, как определены последова¬
тельности гомологичных генов, - процедура выравнивания.
Процесс выравнивания подразумевает выстраивание по¬
следовательностей наиболее близких к исследуемому объекту
видов друг под другом с таким расчетом, чтобы они совпадали
как можно более полно, тогда одни консервативные участки
будут располагаться под другими. Зачастую имеется несколь¬
ко вариантов выравнивания последовательностей, а их субъек¬
тивный выбор отражается на результате. Более вариабельные
участки необходимо выравнивать до тех пор, пока идентичные
нуклеотиды по вертикали не будут наиболее максимально со¬
ответствовать друг другу. При этом возможно включение про¬
пусков, поскольку в некоторых последовательностях могут
быть как делеции, так и вставки. Существуют способы стати¬
стической оценки качества выравнивания пары последова¬
тельностей, например коэффициент сходства Нидлемана -
Вунша (Needleman & Wunsch, 1970) или логарифмическая си¬
стема (Gu & Li, 1995). Участки створа последовательностей,
которые не могут быть достоверно выровнены, чаще всего ис¬
ключаются из анализа, чтобы не привносить систематических
ошибок. В настоящее время, этот этап работы компьютеризи¬
рован, тем не менее, компьютерную версию выровненных по¬
следовательностей необходимо корректировать вручную. Ши¬
роко распространенная программа для выравнивания многих
последовательностей Clustal (Higgins & al. 1992) основана на
итерационном процессе построения модельных деревьев и
расчета генетических дистанций. Программу можно свободно
загрузить с веб-страницы: http://www.ebi.ac.uk/Tools/clustalw/.
Достаточно эффективен смешанный метод: сначала компью¬
терная, затем ручная корректировка. Некоторые исследовате¬
ли предпочитают с самого начала выравнивание вручную.
3.2.4. Генетические дистанции
Для отражения числа замещений в последовательностях
(сиквенсах) рассчитываются генетические дистанции. Про¬
стейшим показателем служит p-дистанция, которая показыва¬
ет отношение количества отличающихся нуклеотидов к длине
последовательности при попарном сравнении, при этом про¬
белы или неуверенно распознанные нуклеотиды не учитыва¬
ются. Для всех моделей замещений нуклеотидов в ДНК суще¬
ствуют выведенные формулы расчета генетических дистанций
(см. например Swofford & al. 1996: 456), наиболее простыми из
них являются Дюкес-Кантор и Кимура-2 дистанции.
4. Методы филогенетического анализа
Существуют две главные группы методов изучения фи¬
логенетических взаимоотношений: фенетические и кладисти-
ческие методы.
Филогенетические деревья
Систематика описывает взаимоотношения между таксо¬
нами и призвана помочь нам понять историю эволюции изуча¬
емых организмов. Но история не является чем-то, что мы мо¬
жем увидеть, она произошла один раз и оставила только кос¬
венные свидетельства фактических событий. В филогенетике
наиболее удобный путь визуального представления эволюци¬
онных взаимоотношений среди групп организмов осуществля¬
ется посредством графиков, которые называются филогенети¬
ческими деревьями (Рис. 86).
ветвь
— Usnea florid а
кЛ ЗЛЯ
Parmelta sulcata
Umbilicana deusla
* *
длина BCTBtt
Рис. 86. Пример филогенетического дерева. Узел - представляет таксо¬
номическую единицу. Он может быть или существующей группой или
предком; Ветвь - определяет взаимоотношение между таксонами в тер¬
минах потомков и предков; Топология - правило по которому разветв¬
ляется дерево; Длина ветви - представляет число изменений, которые
произошли между таксонами. Корень - общий предок всех рассматри¬
ваемых организмов; Масштаб расстояния масштаб, который отражает
число отличий между организмами или последовательностями генома;
Клада - группа двух или больше таксонов или последовательностей
ДНК, которая включает как своего общего предка, так и всех его потом¬
ков. Оперативная таксономическая единица (ОТЕ, OTU) - уровень дета¬
лизации, выбранный исследователем для данной работы, например ин¬
дивидуумы, популяции, виды, роды, семейства.
4.1. Фенетические методы
Первые математические методы построения филограмм
на основе преимущественно фенотипических данных, возник¬
ли еще в начале 60-х годов вместе с первыми вычислительны¬
ми программами. Все они были основаны на принципах нуме-
рической таксономии (Fitch, Margoliash, 1967; Wagner, 1969; и
др.). Следовательно, в данном случае родство определялось по
критерию "общего сходства" - чем выше значение этого пара¬
метра, тем ближе таксоны друг к другу. В основе современных
корень
дистанционно-матричных методов филогенетического анализа
лежат те же принципы с их модификациями, которые техниче¬
ски менее громоздки. С их помощью за короткое время можно
анализировать довольно массивные базы данных. Филограм¬
ма, полученная такими методами, не отражает эволюционного
процесса, а только демонстрирует конечную степень дивер¬
генции таксонов. Фенетика, или числовая таксономия, исполь¬
зует разные методики для определения наибольшего сходства
рассматриваемых видов. Нет никаких ограничений по числу
или типу данных, которые могут использоваться, хотя все
данные должны быть сначала преобразованы в числовые зна¬
чения, без любого «нормирования». Для создания филограммы
выровненные последовательности попарно сравниваются друг
с другом и для каждой пары структурные отличия трансфор¬
мируются в особый числовой коэффициент, являющийся ме¬
рой их сходства или различия. Создается числовая матрица
данных, которая затем преобразуется в графическую модель
эволюционного дерева на основе принципов кластерного ана¬
лиза. Пары наименее различающихся последовательностей
объединяются в кластеры первого порядка, наименее разли¬
чающиеся кластеры первого порядка - в кластеры второго по¬
рядка, и т.д. В результате получают таксономические класте¬
ры или фенограммы, которые не обязательно отображают ге¬
нетическое сходство или эволюционную связь. В этом заклю¬
чается принцип действия так называемых "дистанционно -
матричных" методов (UPGMA; NJ).
4.1.1. UPGMA (Unweighted Pair Group Method with
Arithmetic Mean) Sokal & Mitchener 1958
UPGMA является простейшим методом оценки филоге¬
нетических взаимоотношений на основе генетических дистан¬
ций. Шаг за шагом, похожие виды суммируются к новым опе¬
рационным таксономическим единицам. UPGMA предполага¬
ет постоянную эволюционную изменчивость (молекулярные
часы).
4.1.2. Метод связывания ближайших соседей (neigh¬
bour - joining - NJ)
Самый распространенный метод связывания ближайших
соседей (neighbour - joining или NJ) (Saitou, Nei, 1987). Метод
NJ основан на алгоритмической аппроксимации дерева мини¬
мальной эволюции. Из полностью неразрешенного дерева
(звезда-дерево), шаг за шагом суммируются пары оперативных
единиц до тех пор, пока не будет построено дихотомическое
дерево. При каждом конструкционом шаге используется
принцип минимальной эволюции. Те пары оперативных еди¬
ниц, у которых длина ветви наименьшая, связываются вместе.
Длина ветвей определяется с помощью Fitch-Margoliash мето¬
да определения генетической дистанции (число замещений на
нуклеотид) (Fitch & Margoliash, 1967).
На этом теоретические возможности NJ анализа не за¬
канчиваются. Кимура (Kimura, 1980) предложил метод, кото¬
рый учитывает химические особенности мутационного про¬
цесса. А именно, неравную вероятность транзиций (замен типа
пурин <-> пурин или пиримидин пиримидин) и трансверсий
(замен типа пурин пиримидин).
Существует довольно большое количество компьютер¬
ных программ для фенетического анализа. Наиболее часто ис¬
пользуемыми являются программы:
NTSYSpc, Numerical Taxonomy System, Version 2.2
ttp://www.exetersoftware.com/cat/ntsyspc/ntsyspc.html
GelQuest, Molecular Analysis Software for AFLP, RAPD, t-
RFLP, Microsatellites and Minisatellite Analysis
http://www.sequentix.de/software_gelquest.php
Treecon:
http://bioinformatics.psb.ugent.be/software_details.php?id=3
4.2. Кладистические методы
Основное предположение кладистики заключается в том,
что члены группы имеют общую эволюционную историю. По¬
тому они более близко относятся друг к другу, чем к другим
группам организмов. Связанные группы определяются по
наличию набора уникальных особенностей (апоморфий), ко¬
торые отсутствовали в отдаленных предках, но которые харак¬
терны для большинства или всех организмов в пределах груп¬
пы. Полученные характеристики, относящиеся к членам груп¬
пы, называются синапоморфиями. Потому, в отличие от фене-
тических, кладистические группы не зависят от того сходны
ли организмы по физическим чертам, а зависят от их эволю¬
ционных взаимоотношений. Действительно, в кладистических
анализах у двух организмов могут быть общими многочислен¬
ные характеристики, но они будут членами разных групп.
Кладистический анализ использует ряд предположений.
Например, считается, что виды появляются только в результа¬
те дивергенции - раздвоения предкового вида. В случае ги¬
бридизации или горизонтального переноса генетической ин¬
формации виды считаются исчезнувшими, а такие явления —
редкими или отсутствующими. Кроме того, кладистические
группы должны иметь следующие характеристики: все виды в
группе должны разделять общего предка и все виды, получен¬
ные от общего предка, должны войти в таксон. Соблюдение
этих требований отражается в терминологии, которая исполь¬
зуются для ссылки на состав выделяемых групп:
Монофилетическая группа (или клада), в которой все
виды разделяют общего предка и все виды, которые происхо¬
дят от этого общего предка, включаются в группу.
Парафилетическая группа, в которой все виды разде¬
ляют общего предка, но не все виды, которые происходят от
этого общего предка, включаются в группу.
Полифилетическая группа, в которой виды, которые
не разделяют непосредственного общего предка, складывают¬
ся в одну группу, исключая виды, которые бы связали их.
Плезиоморфия означает примитивный или изначальный
статус признака у определенного таксона при сравнении с его
предками.
Апоморфия означает производный, или специализиро¬
ванный признак.
Для выяснения эволюционного родства необходим ана¬
лиз состояния конкретных признаков, которые изменялись в
процессе эволюции. Обычно предполагают, что эволюцион¬
ный путь признака должен быть таким, который требует
наименьшего числа его преобразований. В этом и заключается
принцип "максимальной экономии", следуя которому, воз¬
можно выстроить схемы родственных отношений (Hennig,
1966). Это универсальный кладистический метод, позволяю¬
щий анализировать любые сравнительные базы данных -
морфологические, поведенческие, физиологические, молеку¬
лярные и т.д.
За последние десятилетия были разработаны компью¬
терные программы, использующие этот принцип для анализа
филогенетических связей на основе первичных структур нук¬
леиновых кислот (PAUP, Swofford, 1993, 2001). Этот подход
оказался очень информативным при реконструкции филоге¬
нии, поскольку позволил напрямую проследить приобретение
новых признаков родственными группами. Таким образом, мы
можем обнаружить явные плезиоморфии и апоморфии в эво¬
люционной истории уже на молекулярном уровне.
4.2.1. Метод максимальной экономии (maximum
parsimony, или МР) технически гораздо более трудоемок и за¬
нимает больше времени, чем все дистанционно-матричные
приемы. Это объясняется тем, что весь массив выровненных
последовательностей анализируется целиком.
Суть метода состоит в последовательном анализе всех
возможных филограмм, состоящих из всех возможных комби¬
наций представленных элементов (т.е. последовательностей).
Анализ проводится по каждому признаку (т.е. по каждой ко¬
лонке выравнивания). Цель его - найти среди всего множества
такую филограмму, для которой необходимое число измене¬
ний признака в процессе предполагаемой эволюции будет ми¬
нимальным. Этим и объясняется название метода: "макси¬
мально экономным" будет такое дерево, для которого в сумме
число изменений всех признаков будет наименьшим.
Использование МР анализа позволяет не только полу¬
чить вероятную картину действительного эволюционного про¬
цесса, но и обнаружить набор "максимально экономных" при¬
знаков для каждого узла ветвления филограммы, который за¬
писывается программой в специальном файле. Эта информа¬
ция незаменима при поиске участков молекулы, которые с
большей вероятностью содержат следы апоморфных приобре¬
тений того или иного таксона в процессе его естественной ис¬
тории.
В настоящее время, созданы специальные пакеты про¬
грамм, анализирующие различные типы данных с использова¬
нием этого метода (Williams, Fitch, 1990; Swofford, 1993, 2001;
Felsenstein, 1993).
Статистические подтверждения: Bootstrap и Jackknife
Bootstrap и Jackknife являются статистическими метода¬
ми проверки качества деревьев. В качестве статистической
единицы выборки принимается каждая колонка в реальном
выравнивании (из генома нескольких организмов). Bootstrap и
Jackknife являются различными вариантами взятия выборок,
чтобы затем составить их в новое виртуальное выравнивание.
Каждое полученное таким образом новое выравнивание ис¬
пользуется для построения новой филограммы. При много¬
кратном повторении этого процесса можно получить для каж¬
дой ветви (клады) процентное значение, которое показывает
долю филограмм, где клада проявлялась в качестве монофиле-
тической единицы. Это процентное значение отражает ста¬
бильность клад.
В бутстреп анализе из реального выравнивания случайно
выделяются колонки из которых составляется новый створ,
такой же длины, как и оригинальное выравнивание, в сравне¬
нии с которым некоторые колонки отсутствуют, а другие при¬
сутствуют многократно. В методе Джекнайф (Jackknife) слу¬
чайное исключение определенной части колонок в оригиналь¬
ном выравнивании, составляется новое более короткое вырав¬
нивание.
4.2.2. Метод максимального правдоподобия (maximum
likelyhood - ML) в математической статистике - это метод
оценивания неизвестного параметра путём максимизации
функции правдоподобия. Оценка максимального правдоподо¬
бия - это такая оценка, которая максимизирует функцию
правдоподобия при фиксированной реализации выборки. Так¬
же как и метод максимальной экономии, он основан на ис¬
пользовании дискретных признаков для построения филогене¬
тических деревьев, (Huelsenbeck, Grandall, 1997).
Наиболее популярной программой для анализа методом
максимальной экономии и методом максимального правдопо¬
добия является PAUP.
PAUP*: Phylogenetic Analysis Using Parsimony (and Other
Methods) 4.0 Beta http://www.sinauer.com/detail.php?id=8060
Метод Байесовой оценки филогении - Оценка степени
правдоподобия филогенетической реконструкции относитель¬
но априори заданной модели "эволюции" признаков на основе
байесовых вероятностей.
Предлагаются три программы использующие байесовые
вероятности:
MrBayes: Программа для Байесовой оценки филогении
(Redelings & Suchard, 2005).
http://sourceforge.net/projects/mrbayes/
BEAST: (Bayesian Evolutionary Analysis Sampling Trees) -
Байесов эволюционный анализ базы филогенетических дере¬
вьев
http://evolve.zoo.ox.ac.uk/beast/primateTutorial.html
BEST (Bayesian Estimation of Species Trees) Байесовые
вероятности филогенетических деревьев
http://www.stat.osu.edu/~dkp/BEST/
Информацию о большинстве филогенетических про¬
грамм можно получить на веб-странице
http://evolution.genetics.washington.edu/phylip/software.html
Стратегии поиска оптимальной топологии дерева -
эвристический анализ (Heuristics search).
Начиная от анализа более 20 последовательностей ста¬
новится технически невозможным за реалистичное время по¬
строить все возможные топологии деревьев, полученные с
применением определенного критерия (например максималь¬
ной экономии, максимального правдоподобия, минимальной
эволюции и др.), найти лучшую топологию. Поэтому необхо¬
димо применить процедуры эвристического анализа, которые
позволяют находить оптимальные деревья, не прибегая к пол¬
ному анализу. То, что найденные топологии глобально опти¬
мальны не всегда однозначно, но если при частом повторении
процедуры с использованием случайных критериев много¬
кратно встречаются деревья с одинаковой топологией, то ве¬
роятность, что эти деревья и в целом оптимальны, довольно
высока. Важными аспектами эвристического анализа является
построение стартового дерева и алгоритм трансформации де¬
ревьев.
5. Филогенетические сети
Представление данных в виде филогенетических деревь¬
ев не всегда достаточно для отражения процесса эволюцион¬
ной истории таксонов. Такие процессы как гибридизация, го¬
ризонтальный перенос генов, и другие, объединяемые в поня¬
тие «сетчатая эволюция», требуют построения более сложных,
чем дихотомические и политомические дендрограммы струк¬
тур. В настоящее время разработана методология построения
различных сетчатых графов (Huson et al. 2010). В лихенологии
наиболее активно используются сети гаплотипов (haplotype
network).
а
• 1
/■
t
Рис. 87. Сеть гаплотипов микобионта (а) и фотобионта (б).
6. Интерпретация результатов филогенетических по¬
строений
Анализируя молекулярно-филогенетическое древо все¬
гда необходимо помнить, что это не ответ на вопрос об эво¬
люции таксонов, а гипотеза эволюции генотипической инфор¬
мации, основанная на многих допущениях. «Принцип эконо¬
мии» a priori не является принципом, обязательно присущим
ходу эволюции, а является математической моделью. Даже
сложные алгоритмы, использующие критерий максимального
правдоподобия, используют теоретические модели, к которым
эволюционный процесс содержательно не сводится. Молеку¬
лярные данные объективно выделяют эволюционные группы,
в том смысле, что объединение происходит без предваритель¬
ных допущений о важности конкретного признака. В то же
время молекулярные методы дают гипотезы, проверка кото¬
рых требует скрупулёзного изучения морфологических, био¬
химических и онтогенетических признаков. В этом отношении
морфологические, биохиимические и молекулярно¬
генетические данные дополняют, а не подменяют друг друга,
поскольку молекулярно-филогенетическое древо должно вос¬
приниматься не как финальный результат филогенетической
реконструкции, а как ценный материал для интерпретаций,
биологический контекст которых формируется при участии
филогенетического мышления.
7. ДНК-баркодинг
ДНК-баркодинг - это экспресс-метод идентификации
видов, на основе полученного сиквенса определенного фраг¬
мента ДНК. При развитии ДНК-технологии, этот метод стано¬
вится ценным рабочим инструментом как систематика, так
флориста и геоботаника. Он позволяет быстро и точно опреде¬
лить видовую принадлежность образца, либо его наиболее
близкое родство. Он подходит как для идентификации, так и
для выявления потенциально новых видов. Метод дает уни¬
кальную возможность экспресс-определения сложных групп,
особенно стерильных лишайников.
Создание платформы для ДНК-баркодинга включает два
этапа: 1) создание базы данных - библиотеки ДНК-баркодов;
2) сравнение сиквенса неизвестного образца с библиотекой
ДНК-баркодов для идентификации. Первый этап достаточно
трудоемкий, поскольку для его осуществления необходимо
изучить таксономическое разнообразие и секвенировать фраг¬
менты, которые могли бы стать эталонными. Неверные опре¬
деления образцов, из которых выделяются такие эталонные
фрагменты, повлечет за собой ошибки в определении. Ведутся
попытки создания как национальных программ баркодинга,
так и глобальная программа в рамках проекта «Дерево жизни».
Для разных групп организмов подбираются адекватные
задаче генетические маркеры. Они должны быть универсаль¬
ными, легко амплифицируемыми, достаточно вариабельными.
Для лишайников в качестве такого маркера предложен ITS
фрагмент рДНК. Это многокопийная некодирующая последо¬
вательность, что делает ее легко амплифицируемой, даже из
небольшого или поврежденного материала. В то же время в
клетке могут присутствовать неодинаковые копии, а высокая
вариабельность затрудняет выравнивание.
В настоящее время ДНК-баркодинг как сервис недоста¬
точно разработан. Его функции выполняет инструмент поиска
ближайшего сиквенса в генетическом банке - BLAST (Basic
Local Alignment Search Tool), который позволяет отправить
сиквенс неизвестного образца и получить наиболее близкие
идентифицированные последовательности из Генбанка.
Литература.
Антонов А. С. Основы геносистематики высших расте¬
ний. - Москва, 2000. - 136 с.
Антонов А. С. Геносистематика растений. - Москва,
2006.-293 с.
Блюм О. Б., Кашеваров Г. П. Гомологи ДНК как доказа¬
тельство правомерности выделения рода Lasallia Merat в се¬
мействе Umbilicariaceae (Lichenes) // Докл. АН УССР, серия Б.
1986. № 12.-С. 61-64.
Куцев М.Г. Фрагментарный анализ ДНК растений:
RAPD, DAF, ISSR. - Барнаул, 2009. - 164 с.
Павлинов И. Я. Филогенетическое мышление в совре¬
менной биологии // Журнал общей биологии. 2007. Т.68. № 1.
С. 19-34.
Павлинов И. Я., Любарский Г.Ю. Биологиче¬
ская систематика. Эволюция идей. - Москва, 2011. - 632 с.
Фризен Н. Молекулярные методы, используемые в си¬
стематике растений. - Барнаул, 2007. - 64 с.
Чемерис А. В., Ахунов Э. Д., Вахитов В. А. Секвениро¬
вание ДНК. М.: Наука. 1999.
Armaleo D., Clerc P. A rapid and inexpensive method for
the purification of DNAfrom lichens and their symbionts // Li¬
chenologist, 1995. 27: 207-213.
Armaleo, D., Clerc P. Lichen chimeras: DNA analysis sug¬
gests that one fungus forms two morphotypes // Experimental My¬
cology, 1991. 15: 1-10.
Begerow D., Nilsson H., Unterseher М., Maier W. Current
state and perspectives of fungal DNA barcoding and rapid identifi¬
cation procedures // Applied Microbiology and Biotechnology
2010. 87: 99-108.
Blattner F. R. Direct amplification of the entire ITS region
from poorly preserved plant material using recombinant PCR //
Biotechniques, 1999. 27:1180-1186.
Caetano-Annoles G., Gresshoff P. M. DNA amplification
fingerprinting using arbitrary mini-hairpin oligonucleotide primers
// Biotechnology, 1994. Vol. 6. Pp.619-623.
DePriest P. T. Early molecular investigations of lichen-
forming symbionts: 1986-2001 //Annual Review of Microbiology,
2004.58:273-301.
Doyle J. J., Doyle J. L. A rapid DNA isolation procedure for
small quantities of fresh leaf tissue // Phytochemical Bulletin,
1987. 19: 11-15.
Felsenstein J. PHYLIP - Phylogeny Inference Package, ver¬
sion 3.5. University of Washington, Seattle, 1993.
Fitch W., Margoliash E. Constructing Phylogenetic Trees //
Science, 1967. 155: 279 - 284
Feuerer, T. Hawksworth, D. L. Biodiversity of lichens, in¬
cluding a world-wide analysis of checklist data based on Takhta-
jan's floristic regions // Biodiversity and Conservation 2007. 16(1):
85-98.
Gargas A., Taylor J.W. Polymerase chain reaction (PCR)
primers for amplifying and sequencing nuclear 18S rDNA from
lichenized fungi // Mycologia, 1992. 84 (5), 589-592.
Gherbawy Y, Voigt K, Molecular identification of fungi. -
Springer Verlag, Berlin, Heidelberg, New York, 2010. 501 pp.
Grube М., DePriest P.T., Gargas A., Hafellner J. DNA isola¬
tion from lichen ascomata// Mycol. Res. 1995. 99:1321-24
Hennig W. Phylogenetic Systematics. Chicago, Univ. of Illi¬
nois Press, 1966
Jukes Т.Н., Cantor C.R. Evolution of protein molecules. In:
"Mammalian protein metabolism". Acad. Press, NY - London,
1969.21 - 132.
Hixson J. E., Brown W. M. A Comparison of the Small Ri-
bosomal RNA Genes from the Mitochondrial DNA of the Great
Apes and Humans: Sequence, Structure, Evolution, and Phyloge¬
netic Implications// Mol. Biol. Evol. 1986. 3: I- 18.
Hogeweg P., Hesper B. The Alignment of Sets of Sequences
and the Construction of Phyletic Trees: An Integrated Method // J.
Mol. Evol. 1984. 20: 175-186.
Huson D. H., Rupp R., Scornavacca C. (eds.) Phylogenetic
Networks: concepts, algorithms and applications. - Cambridge
University Press, 2009. xii+362 pp.
Kelly L.J., Hollingsworth P.M., Coppins B.J., Ellis C.J., Har-
rold P., Tosh J., Yahr R., DNA barccoding of lichenized fungi
demonstrates high identification success in a floristic context //
New Phytologist, 2011. 191 (1 ):288-300.
Kimura M. A simple method for estimating evolutionary
rates of base substitution through comparative studies of nucleo¬
tides sequences // Journal of Molecular Evolution. 1980. 16:111-
120
Lemey P., Salemi М., Vandamme A.-M. The Phylogenetic
Handbook: A Practical Approach to Phylogenetic Analysis and
Hypothesis Testing. - Cambridge University Press, 2009. 723 pp.
Liu, Y. J., Whelen S. & Hall B. D. Phylogenetic relation¬
ships among ascomycetes: evidence from an RNA polymerase II
subunit// Mol. Biol. Evol. 1999. 16(12): 1799-1808.
Molecular Systematics / Hillis D. М., Moritz C., Mable В. K.
(eds.), Second Edition. - Sinauer Associates, Inc. Massachusetts,
1996-655 p.
Moore P. D. Molecular biogeography. Progress report. In.:
Progress in Physical Geography 2001. 24: 545-551.
Printzen C. Lichen Systematics: The Role of Morphological
and Molecular Data to Reconstruct Phylogenetic Relationships //
Progress in Botany, 2010: 71: 233-278.
Redelings B. D. & Suchard M. A. Joint Bayesian Estimation
of Alignment and Phylogeny In.: Systematic Biology, 2005, 54(3):
401-418,
Saitou N., Nei M. The neighbour-joining method: a new
method for reconstructing phylogenetic tree // Mol. Biol. Evol.
1987.4:406-425.
Sanger F., Coulson A.R. A rapid method for determining se¬
quences in DNA by primed syntesis with DNA polymerase // J.
Mol. Biol. 1975. 94: 444-448.
Sanger F., Niclein S., Coulson A.R. DNA sequencing with
chain-terminating inhibitors // Proc. Natl. Acad. Sci. USA, 1977.
74: 5463-5467.
Sokal R. R. & Mitchener C. D. A statistical method for eval¬
uating systematic relationships //. Univ. Kansas. Bull. 1958. 38:
1409-1438.
Swofford, D. L., P. J. Waddell, J. P. Huelsenbeck, P. G. Fos¬
ter, P. O. Lewis, and J. S. Rogers. Bias in phylogenetic estimation
and its relevance to the choice between parsimony and likelihood
methods // Syst. Biol. 2001. 50: 525-539.
Swofford, D. L. PAUP* Phylogenetic Analysis Using Parsi¬
mony (*and Other Methods). Version 4. Sinauer Associates, Sun¬
derland, Massachusetts. 2001.
Swofford D.L. PAUP: Phylogenetic Analysis Using Parsi¬
mony, ver. 3,1. Computer program distributed by the Illinois Histo¬
ry Survey, Champaign, Illinois. 1993
Taberlet P., Gielly L., Pautou G., Bouvet J. Universal pri¬
mers for amplification of three non-coding regions of chloroplast
DNA// PI. Mol. Biol. 1991. 17: 1105-1109.
van de Peer Y., de Wachter R. TREECON for Windows: a
software package for the construction and drawing of evolutionary
trees for the Microsoft; Windows environment // Comput. Appi.
Biosci. 1994. 10: 569 - 570
Vos P., Hogers R., Bleeker М., Reijans М., van de Lee Т.,
Hornes М., Frijters A., Pot J., Peleman J., Kuiper М., Zabeau M.
AFLP: a new technique for DNA fingerprinting. Nucleic Acids
Research, 1995. 23: 4407-4414.
Wagner W.H. The construction of classification. In: Sibley
CG (Eds) Systematic Biology, Washington DC: Publication 1962,
National Academy of Sciences, 1963. 67 - 90.
White T.J., Bruns Т., Lee S., Taylor J. Amplification and di¬
rect sequencing of fungal ribosomal RNA genes for phylogenetics.
In: Innis MA, Gelfand DH, Shinsky J.J., White T.J. (eds.) PCR
Protocols: A Guide to Methods and Applications, 315-322. Aca¬
demic Press, San Diego. 1990.
Welsh J., McClelland M. Fingerprinting genomes using PCR
with arbitrary primers. Nucleic Acids Research, 1990. 18: 7213-
7218.
Wiley E. O. Phylogenetics. The theory and practice of phy¬
logenetic systematics. -N.Y., Chichester, Brisbane, Toronto, 1981.
439 p.
Williams, J. G., Kubelik, A. R., Livak, K. J., Rafalski, J. A.,
and Tingey, S. V. DNA polymorphisms amplified by arbitrary pri¬
mers are useful as genetic markers. Nucleic Acids. 1990.
Глава XI.
Систематическая классификация таксонов лихенофлоры
России
Урбанавичус Г. П.
Поскольку лишайники не являются самостоятельной
эволюционной группой организмов, а представляют собой
экологические группы грибов, то и их классификация не мо¬
жет быть отделена от классификации нелихенизированных
грибов. В основе современной систематической классифика¬
ции грибов лежит филогенетический подход, отражающий
эволюционные связи (родственные отношения), выявление
которых стало возможным благодаря достижениям молеку¬
лярной филогении. При этом молекулярные методы дают воз¬
можность оценить полезность и важность тех или иных мор¬
фологических или химических признаков в систематике ли¬
шайников. Но в тоже время всегда следует быть осторожным
при интерпретации результатов молекулярных данных, осо¬
бенно полученных 15-20 лет назад, когда методы были недо¬
статочно совершенны, если они находятся в противоречии с
морфологическими, анатомическими или хемотаксономиче-
скими характеристиками. С ранними представлениями о клас¬
сификации лишайников и их эволюции можно познакомиться,
например, в работах А. Н. Окснера (1974), С. Printzen (2010).
Представленная здесь классификация таксонов лихено¬
флоры России основана на последнем издании Dictionary of the
Fungi (2008) с учетом последующих систематических измене¬
ний и включая часть пропущенных в Словаре новшеств, начи¬
ная с 2007 г. (отражены в концевых сносках). Несомненно, что
и в будущем будут публиковаться все новые и новые результа¬
ты исследований, в той или иной степени способствующих
дальнейшему совершенствованию и изменению систематиче¬
ской классификации.
В лихенофлоре России отдел Ascomycota с подотделом
Pezizomycotina включает 7 классов - Arthoniomycetes,
Dothideomycetes, Eurotiomycetes, Lecanoromycetes,
Leotiomycetes, Lichinomycetes и Sordariomycetes, отдел
Basidiomycota включает подотдел Agaricomyctina с классами
Agaricomycetes и Tremellomycetes и подотдел Pucciniomycotina
с классами Agaricostilbomycetes и Cystobasidiomycetes. Оче¬
видно, что в иерархической структуре таксоны должна распо¬
лагаться относительно друг друга следуя филогенетической
близости, но так как далеко не для всех таксономических
групп имеются достоверные филогенетические данные, в
настоящем разделе принят алфавитный порядок расположе¬
ния.
Условные обозначения:
+ - нелихенизированные сапротрофные грибы;
(+) - род включает наряду с лихенизированными также
нелихенизированные виды;
* - нелихенизированные лихенофильные грибы;
(*) - род включает наряду с лихенизированными также
нелихенизированные лихенофильные виды.
Вопросительный знак (?) указывает, что положение так¬
сона в данной таксономической категории не является оконча¬
тельно определенным.
Отдел Ascomycota Caval.-Sm., Biol. Rev. 73: 247 (1998).
Подотдел Pezizomycotina О. E. Erikss. & Winka, Myconet 1: 9
(1997).
Класс Arthoniomycetes О. E. Erikss. & Winka, Myconet 1: 4
(1997).
Порядок Arthoniales Henssen ex D. Hawksw. & О. E. Erikss.,
Syst. Ascom. 5: 177 (1986).
Семейство Arthoniaceae Rchb. (1841).
(*)Arthonia Ach. (1806).
Arthothelium A. Massal. (1852).
*Briancoppinsia Diederich, Ertz, Lawrey & van den Boom
(2011)3.
Семейство Chrysothricaceae Zahlbr. (1905).
Chrysothrix Mont. (1852).
?Семейство Melaspileaceae Walt. Watson (1929).
(+)MeIaspilea Nyl. (1857).
Семейство Roccellaceae Chevall. (1826).
Bactrospora A. Massal. (1852).
Cresponea Egea & Torrente p 993).
Dendrographa Darb. (1895) .
3 Diederich P., Lawrey J. D., Sikaroodi М., van den Boom P. P.
G., Ertz. D. Briancoppinsia, a new coelomycetous genus of Arthonia¬
ceae (Arthoniales) for the lichenicolous Phoma cytospora, with a key
to this and similar taxa//Fungal Diversity. 2012. vol. 52. N l.P. 1-
12.
4 Ertz D., Tehler A. The phylogeny of Arthoniales (Pezizomy¬
cotina) inferred from nucLSU and RPB2 sequences // Fungal Diversi¬
ty. 2010. Vol. 49. N l.P. 47-71.
Dirina Fr. (1825).
Enterographa Fee (1824).
Lecanactis Korb. (1855).
Lecanographa Egea & Torrente (1994).
(*)Opegrapha Ach. (1809).
*Phacographa Hafellner (2009)5.
*Plectocarpon Fee (1825).
Roccella DC. (1805).
Schismatomma Flot. & Korb. ex A. Massal. (1852).
Zwackhia Korb. (1855)6.
Arthoniales, рода неопределенного положения
Alyxoria Gray (1821)7.
*Arthophacopsis Hafellner (1998).
Класс Dothideomycetes О. E. Erikss. & Winka, Myconet 1: 5
(1997).
Подкласс Dothideomycetidae P. M. Kirk, P. F. Cannon, J. C. Da¬
vid & Stalpers ex Schoch et al., Mycologia 98: 1047 (2007)
["2006"].
Порядок Capnodiales Woron., Annls Mycol. 23: 177 (1925).
Семейство Davidiellaceae C. L. Schoch, Spatafora, Crous &
Shoemaker (2007).
*Cladosporium Link (1816)8,9.
Семейство Mycosphaerellaceae Lindau (1897).
*Sphaerellothecium Zopf (1897).
*Stigmidium Trevis. (1860).
Семейство Teratosphaeriaceae Crous & U. Braun (2007).
Cystocoleus Thwaites (1849)10.
5 Hafellner, J. Phacothecium resurrected and the new genus
Phacographa (Arthoniales) proposed // Biblioth. Lichenol. 2009. Vol.
100. P. 85-121.
6 Ertz D., Tehler A. The phylogeny of Arthoniales (Pezizomy-
cotina) inferred from nucLSU and RPB2 sequences // Fungal Diversi¬
ty. 2010. Vol. 49. N l.P. 47-71.
7 Ertz D., Tehler A. The phylogeny of Arthoniales (Pezizomy-
cotina) inferred from nucLSU and RPB2 sequences // Fungal Diversi¬
ty. 2010. Vol. 49. N l.P. 47-71.
8 Crous P. W., Schoch C. L., Hyde K. D., et al. Phylogenetic
lineages in the Capnodiales // Stud. Mycol. 2009. Vol. 64. P. 17-47.
9 Schoch C. L., Crous P. W., Groenewald J. Z., et al. A class-
wide phylogenetic assessment of Dothideomycetes // Stud. Mycol.
2009. Vol. 64. P. 1-15.
*Xanthoriicola D. Hawksw. (1973)11.
Capnodiales, род неопределенного положения
Racodium Pers.: Fr. (1829)12.
Порядок Dothideales Lindau in Engler & Prantl (eds.), Nat.
Pflanzenfam. 1(1): 373 (1897).
Семейство Dothioraceae Theiss. & P. Syd. (1918).
*Plowrightia Sacc. (1883).
Подкласс Pleosporomycetidae C. L. Schoch, Spatafora, Crous &
Shoemaker, Mycologia98: 1049 (2007) ["2006"].
Порядок Pleosporales Luttr. ex М. E. Barr, Prodr. Class Locu-
loasc.: 67 (1987).
?Семейство Arthopyreniaceae Walt. Watson (1929)1314.
Arthopyrenia A. Massal. (1852).
Семейство Cucurbitariaceae G. Winter (1885).
?*Pyrenochaeta DeNot. (1849)15,16.
Семейство Pleomassariaceae М. E. Barr (1979).
?+Peridiothelia D. Hawksw. (1985).
10 Ruibal C., Gueidan C., Selbmann L., et al. Phylogeny of
rock-inhabiting fungi related to Dothideomycetes // Stud. Mycol.
2009. Vol. 64. P. 123-133.
11 Ruibal C., Millanes A. М., Hawksworth D. L. Molecular
phylogenetic studies on the lichenicolous Xanthoriicola physciae re¬
veal Antarctic rock-inhabiting fungi and Piedraia species among clos¬
est relatives in the Teratosphaeriaceae // IMA Fungus. Vol. 2. N l.P.
97-103.
12 Ruibal C., Gueidan C., Selbmann L., et al. Phylogeny of
rock-inhabiting fungi related to Dothideomycetes // Stud. Mycol.
2009. Vol. 64. P. 123-133.
13 Zhang Y., Crous P. W., Schoch C. L., Hyde K. D. Pleospora¬
les // Fungal Diversity. 2011. DOI 10.1007/s 13225-011-0117-x
14 Nelsen M. P., Lucking R., Grube М., et al. Unravelling the
phylogenetic relationships of lichenised fungi in Dothideomyceta//
Stud. Mycol. 2009. Vol. 64. P. 135-144.
15 Gruyter J. D., Woudenberg J. H. C., Aveskamp М. М., et al.
Systematic reappraisal of species in Phoma section Paraphoma, Pyr-
enochaeta and Pleurophoma II Mycologia. 2010. Vol. 102. P. 1066—
1081.
16 Hyde K. D., McKenzie E. H. С., KoKo T. W. Towards in¬
corporating anamorphic fungi in a natural classification - checklist
and notes for 2010//Mycosphere. 2011. Vol. 2. N l.P. 1-88.
Семейство Pleosporaceae Nitschke (1869).
*Pleospora Rabenh. ex Ces. & De Not. (1863).
Dothideomycetes, порядок неопределенного положения
Порядок Trypetheliales Lucking, Aptroot & Sipman (2008)l7.
Семейство Trypetheliaceae Zenker (1827).
?+Julella Fabre (1879)18.
?+Mycomicrothelia Keissl. (1936)19.
Dothideomycetes, семейства неопределенного положения
Семейство Dacampiaceae Korb. (1855).
*Clypeococcum D. Hawksw. (1977).
(*)Dacampia A. Massal. (1853).
Eopyrenula R. C. Harris (1973).
*Polycoccum Saut. ex Korb. (1865).
*Pyrenidium Nyl. (1865).
*Weddellomyces D. Hawksw. (1986).
Семейство Lichenotheliaceae Henssen (1986).
*Lichenostigma Hafellner (1983).
+(*)Lichenothelia D. Hawksw. (1981).
Семейство Monoblastiaceae Walt. Watson (1929)20.
Acrocordia A. Massal. (1854).
Anisomeridium (Mull. Arg.) M. Choisy (1928).
Семейство Mycoporaceae Zahlbr. (1903).
+Mycoporum Flot. ex Nyl. (1855).
Семейство Naetrocymbaceae Hohn. ex R. C. Harris (1995).
(+)Leptorhaphis Korb. (1855).
+Naetrocymbe Korb. (1865).
?+TomaselIia A. Massal. (1856).
17 Aptroot A., Lucking R., Sipman H. J. М., et al. Pyrenocar-
pous Lichens with Bitunicate Asci: A First Assessment of the Lichen
Biodiversity Inventory in Costa Rica // Biblioth. Lichenol. 2008. N
97. 162 pp.
18 Nelsen M. P., Lucking R., Mbatchou J. S. et al. New insights
into relationships of lichen-forming Dothideomycetes // Fungal Di¬
versity. 2011. Vol. 51. P. 155-162.
19 Nelsen M. P., Lucking R., Grube М., et al. Unravelling the
phylogenetic relationships of lichenised fungi in Dothideomyceta//
Stud. Mycol. 2009. Vol. 64. P. 135-144.
20 Nelsen M. P., Lucking R., Grube М., et al. Unravelling the
phylogenetic relationships of lichenised fungi in Dothideomyceta //
Stud. Mycol. 2009. Vol. 64. P. 135-144.
Семейство Pseudoperisporiaceae Toro (1926).
?*Epibryon Dobbeler (1978).
*Neocoleroa Petr. (1934).
Семейство Strigulaceae Zahlbr. (1898)21.
Strigula Fr. (1823).
Семейство Trichothyriaceae Theiss. (1914)22.
*LichenopeltelIa Hohn. (1919).
Dothideomycetes, роды неопределенного положения
*Buelliella Fink (1935).
*Cercidospora Korb. (1865).
+Cyrtidula Minks (1876).
?*Didymocyrtis Vain. (1921).
?*Echinothecium Zopf (1898).
?*Endococcus Nyl. (1855).
?*Homostegia Fuckel (1870).
*Karschia Korb. (1865).
*Lichenoconium Petr. & Syd. (1927)23.
+Mycoglaena Hohn. (1909).
*Myxophora Dobbeler & Poelt (1978).
*Rosellinula R. Sant. (1986).
Класс Eurotiomycetes О. E. Erikss. & Winka, Myconet 1: 6
(1997).
Подкласс Chaetothyriomycetidae Doweld, Prosyllabus:
LXXVIII (2001).
Порядок Chaetothyriales М. E. Barr, Mycotaxon 29: 502 (1987).
Семейство Chaetothyriaceae Hansf. ex М. E. Barr (1979).
*Capronia Sacc. (1883)2 .
21 Nelsen M. P., Lucking R., Grube М., et al. Unravelling the
phylogenetic relationships of lichenised fungi in Dothideomyceta//
Stud. Mycol. 2009. Vol. 64. P. 135-144.
22 Wu Hai X., Schoch C. L., Boonmee S., Bahkali A. H.,
Chomnunti P., Hyde K. D. A reappraisal of Microthyriaceae // Fungal
Diversity. 2011. Vol. 51. P. 189-248.
23 Lawrey J. D., Diederich P., Nelsen M. P., et al. The obligate-
ly lichenicolous genus Lichenoconium represents a novel lineage in
the Dothideomycetes // Fungal Biol. 2011. Vol. 115. N 2. P. 176-187.
24 Untereiner, W. A., C. Gueidan, M.-J. Orr & P. Diederich The
phylogenetic position of the lichenicolous ascomycete Capronia pel-
tigerae II Fungal Diversity. 2011. Vol. 49. N 1. P. 225-233.
Порядок Pyrenulales Fink ex D. Hawksw. & О. E. Erikss., Syst.
Ascom. 5: 182 (1986).
Семейство Pyrenulaceae Rabenh. (1870).
Anthracothecium Hampe ex A. Massal. (1860).
(+)Pyrenula Ach. (1814).
Порядок Verrucariales Mattick ex D. Hawksw. & О. E. Erikss.,
Syst. Ascom. 5: 183 (1986).
Семейство Adelococcaceae Triebel (1993).
*Adelococcus Theiss. & Syd. (1918).
*Pseudopyrenidium Nav.-Ros., Zhurb. & Cl. Roux (2010)25.
*Sagediopsis (Sacc.) Vain. (1921).
Семейство Verrucariaceae Zenker (1827).
(=Mastodiaceae Zahlbr.)26
Agonimia Zahlbr. (1909).
Atla S. Savic & Tibell (2008)27.
Bagliettoa A. Massal. (1853).
Catapyrenium Flot. (1850).
Clavascidium Breuss (1996).
Dermatocarpon Eschw. (1824).
Endocarpon Hedw. (1798).
*Halospora (Zschacke) Tomas. & Cif. (1952)28.
Henrica B. de Lesd. (1921).
Heteroplacidium Breuss (1996).
Hydropunctaria Keller, Gueidan & Thus (2009)29.
25 Navarro-Rosines P., Zhurbenko M. P., Roux C. Pseu-
dopyrenidium nova genero por inkludi Weddellomyces tartaricola
(nelikeniginta fungo likenloga, Ascomycota) // Bull. Soc. Linn. Pro¬
vence. 2010. T. 61. P. 129-140.
26 Perez-Ortega S., de los Rios A., Crespo A., Sancho L. Sym¬
biotic lifestyle and phylogenetic relationships of the bionts of Masto¬
dia tessellata (Ascomycota, Incertae Sedis) // Amer. J. Bot. 2010.
Vol. 97. N 5. P. 738-752.
27 Savic S., Tibell L. Atla, a new genus in the Verrucariaceae
(Verrucariales) // Lichenologist. 2008. Vol. 40. N 4. P. 269-282.
28 Hafellner J. Halospora resurrected and segregated from Mer-
ismatium II Biblioth. Lichenol. 2011. N 106. P. 75-93.
29 Gueidan C., Savic S., Thus H., et al. Generic classification of
the Verrucariaceae (Ascomycota) based on molecular and morpho¬
logical evidence: recent progress and remaining challenges // Taxon.
2009. Vol. 58. N l.P. 184-208.
Lauderlindsaya J. С. David & D. Hawksw. (1989).
Leucocarpia \^zda (1969).
Mastodia Hook. f. & Harv. (1847).
*Merismatium Zopf (1898).
*MueIlerella Hepp (1862).
Neocatapyrenium H. Harada (1993).
Parabagliettoa Gueidan & Cl. Roux (2009)30.
?*Phaeospora Hepp ex Stein (1879).
Placidiopsis Beltr. (1858).
Placidium A. Massal. (1855).
Placocarpus Trevis. (1860).
Placopyrenium Breuss (1987).
Polyblastia A. Massal. (1852).
Psoroglaena Mull. Arg. (1891).
Sporodictyon A. Massal. (1852).
Staurothele Norman (1853).
Thelidium A. Massal. (1855).
Verrucaria Schrad. (1794).
Verrucula J. Steiner (1896).
Verruculopsis Gueidan, Nav.-Ros. & Cl. Roux (2007).
Wahlenbergiella Gueidan & Thus (2009)31.
Verrucariales, род неопределенного положения
Botryolepraria Canals, Hern.-Mar., Gomez-Bolea & Llimona
(1997)32.
Подкласс Mycocaliciomycetidae Tibell, Mycol. Res. 111(5): 528
(2007).
Порядок Mycocaliciales Tibell & Wedin, Mycologia 92: 579
(2000).
Семейство Mycocaliciaceae A. F. W. Schmidt (1970).
30 Gueidan C., Savic S., Thus H., et al. Generic classification of
the Verrucariaceae (Ascomycota) based on molecular and morpho¬
logical evidence: recent progress and remaining challenges // Taxon.
2009. Vol. 58. N l.P. 184-208.
31 Gueidan C., Savic S., Thus H., et al. Generic classification of
the Verrucariaceae (Ascomycota) based on molecular and morpho¬
logical evidence: recent progress and remaining challenges // Taxon.
2009. Vol. 58. N l.P. 184-208.
32 Kukwa М., Perez-Ortega S. A second species of Botry¬
olepraria from the Neotropics and the phylogenetic placement of the
genus within Ascomycota // Mycol. Progress. 2010. Vol. 9. N 3. P.
345-351.
Asterophoma D. Hawksw. (1981).
Chaenothecopsis Vain. (1927).
Mycocalicium Vain. (1890).
Phaeocalicium A. F. W. Schmidt (1970).
Stenocybe Nyl. ex Korb. (1855).
Семейство Sphinctrinaceae M. Choisy (1950).
Pyrgidium Nyl. (1867).
*Sphinctrina Fr. (1825).
Класс Lecanoromycetes О. E. Erikss. & Winka, Myconet 1: 7
(1997).
Подкласс Acarosporomycetidae Reeb, Lutzoni & Cl. Roux, Mol.
Phylogen. Evol. 32: 1053 (2004).
Порядок Acarosporales Reeb, Lutzoni & Cl. Roux, Mycol. Res.
111(5): 528 (2007).
Семейство Acarosporaceae Zahlbr. (1906).
Acarospora A. Massal. (1852).
Caeruleum K. Knudsen & L. Arcadia (2012)33.
Glypholecia Nyl. (1853).
Myriospora Nageli ex Uloth (1861) [=Silobia M. Westb. & Wedin
(2010), Trimmatothelopsis Zschacke (1934)]34’35’36.
Pleopsidium Korb. (1855).
Polysporina Vezda (1978).
Sarcogyne Flot. (1851).
Подкласс Lecanoromycetidae P. M. Kirk, P. F. Cannon, J. C.
David & Stalpers ex Mi^dl., Lutzoni & Lumbsch, Mycol. Res.
111(5): 529 (2007).
Порядок Caliciales Bessey, Univ. Stud. Nebraska 7: 299 (1907)37.
33 Arcadia L., Knudsen K. The name Myriospora is available
for the Acarospora smaragdula group // Opuscula Philolichenum.
2012. Vol. 11. P. 19-25.
34 Roux C., Navarro-Rosines P. Trimmatothelopsis (Acarospo¬
raceae, Ascomycota 1 ichenisati), le nom legitime de Silobia // Bull.
Soc. Linn. Provence. 2011. T. 62. P. 167-187.
35 Westberg М., Crewe A. Т., Purvis O. W., Wedin M. Silobia,
a new genus for the Acarospora smaragdula complex (Ascomycota,
Acarosporales) and a revision of the group in Sweden // Lichenolo¬
gist. 2011. Vol. 43. N l.P. 7-25.
36 Arcadia L., Knudsen K. The name Myriospora is available
for the Acarospora smaragdula group // Opuscula Philolichenum.
2012. Vol. 11. P. 19-25.
38
Семейство Caliciaceae Chevall. (1826) .
Calicium Pers. (1794).
(*)Cyphelium Ach. (1815).
Thelomma A. Massal. (1860).
Tholurna Norman (1861).
Семейство Physciaceae Zahlbr. (1898) 39.
Amandinea M. Choisy ex Scheid. & H. Mayrhofer (1993).
Anaptychia Korb. (1848).
Buellia De Not. (1846).
Dimelaena Norman (1853).
Diploicia A. Massal. (1852).
Diplotomma A. Massal. (1852).
Dirinaria (Tuck.) Clem. (1909).
Heterodermia Trevis. (1868).
Hyperphyscia Mull. Arg. (1894).
37 Gaya E., Hognabba F., Holguin A., et al. Implementing a cu¬
mulative supermatrix approach for a comprehensive phylogenetic
study of the Teloschistales (Pezizomycotina, Ascomycota) // Mol.
Phylogenet. Evol. 2012. doi:10.1016/j.ympev.2012.01.012
38 Несмотря на последние данные о необходимости вклю¬
чения родов семейства Caliciaceae в семейство Physciaceae
(Mi^dlikowska et al., 2006; и др.), здесь предлагается сохранить
самостоятельным статус этого семейства до получения более
обоснованных результатов. Аналогичной точки зрения поддер¬
живаются также в лаборатории лихенологии и филогенетики
Francois Lutzoni (Duke University, North Carolina, США), изучаю¬
щей порядок Teloschistales: «The delimitation of the Caliciaceae
and Physciaceae remains unclear to this day» (ULR
http://www.lutzonilab.net/teloschistales/index.html - 15 XII 2011),
Patrick McCarthy (Australian Biological Resources Study, Canberra,
Australia: ULR
http://www.anbg.gov.au/abrs/lichenlist/classification.html - 15 XII
201 П.
В тоже время, согласно Е. Gaya et al. (2012) к семейству
Caliciaceae отходят роды, ранее входившие в состав семейств
Buelliaceae Zahlbr. и Pyxineaceae Stizenb., образующие здесь сов¬
местно с родами Thelomma и Tholurna подсемейство Buellioideae.
39 Gaya Е., Hognabba F., Holguin A., et al. Implementing a cu¬
mulative supermatrix approach for a comprehensive phylogenetic
study of the Teloschistales (Pezizomycotina, Ascomycota) // Mol.
Phylogenet. Evol. 2012. doi:10.1016/j.ympev.2012.01.012
Phaeophyscia Moberg (1977).
Phaeorrhiza H. Mayrhofer & Poelt (1979).
Physcia (Schreb.) Michx. (1803).
Physciella Essl. (1986).
Physconia Poelt (1965).
Pyxine Fr. (1825).
Rinodina (Ach.) Gray (1821).
Rinodinella H. Mayrhofer & Poelt (1978).
(*)Tetramelas Norman (1853).
Tornabea 0sth. (1980).
Порядок Lecanorales Nannf., Nova Acta Regiae Soc. Sci. Upsal.
ser. 4 8(2): 68 (1932).
Семейство Biatorellaceae M. Choisy ex Hafellner & Casares
(1992).
Biatorella De Not. (1846).
Семейство Cladoniaceae Zenker (1827).
Cladonia P. Browne (1756).
Pilophorus Th. Fr. (1857).
Pycnothelia (Ach.) Dufoiir (1821).
Семейство Dactylosporaceae Bellem. & Hafellner (1982).
Dactylospora Korb. (1855).
Семейство Ectolechiaceae Zahlbr. (1905).
Lopadium Korb. (1855).
Семейство Gypsoplacaceae Timdal (1990).
Gypsoplaca Timdal (1990).
Семейство Haematommataceae Hafellner (1984).
Haematomma A. Massal. (1852).
Семейство Helocarpaceae Hafellner (1984).
Helocarpon Th. Fr. (1860).
Семейство Lecanoraceae Korb. (1855).
Arctopeltis Poelt (1983).
Bryodina Hafellner (2001).
Bryonora Poelt (1983).
Carbonea (Hertel) Hertel (1983).
?Claurouxia D. Hawksw. (1988)40.
Clauzadeana Cl. Roux (1984).
Frutidella Kalb (1994)4 .
40 Eriksson О. E. (ed.) Outline of Ascomycota - 2006 // My¬
conet. 2006. Vol. 12. P. 1-82.
41 Schmull М., Mi^dlikowska J., Pelzer M. et al. Phylogenetic
affiliations of members of the heterogeneous lichen-forming fungi of
Japewia Tonsberg (1990)42.
Lecanora Ach. in Luyken (1809). (=Protoparmeliopsis M.
Choisy).
Lecidella Korb. (1855).
Miriquidica Hertel & Ram bold (1987).
Myrionora R. C. Harris (1988).
Psorinia Gotth. Schneid. (1980).
?Pycnora Hafellner (2001).
Pyrrhospora Korb. (1855).
Ramboldia Kantvilas & Elix (1994).
Rhizoplaca Zopf (1905).
Семейство Megalariaceae Hafellner (1984).
Megalaria Hafellner (1984).
Семейство Mycoblastaceae Hafellner (1984).
Mycoblastus Norman (1853).
Семейство Parmeliaceae Zenker (1827).
Alectoria Ach. in Luyken (1809).
Allantoparmelia (Vain.) Essl. (1978).
Allocetraria Kurok. & M. J. Lai (1991).
Anzia Stizenb. (1861).
Arctocetraria Karnefelt & A. Thell (1993).
Arctoparmelia Hale (1986).
Asahinea W. L. Culb. & C. F. Culb. (1965).
Brodoa Goward (1987).
Bryocaulon Karnefelt (1986).
Bryoria Brodo & D. Hawksw. (1977).
Cetraria Ach. (1803).
Cetrariella Karnefelt & A. Thell (1993).
Cetrelia W. L. Culb. & C. F. Culb. (1968).
Cetreliopsis M. J. Lai (1980).
Cornicularia (Schreb.) Hoffm. (1792).
Dactylina Nyl. (1860).
Evernia Ach. in Luyken (1809).
Everniastrum Hale ex Sipman (1986).
Flavocetraria Karnefelt & A. Thell (1994).
Flavoparmelia Hale (1986).
Flavopunctelia (Krog) Hale (1984).
the genus Lecidea sensu Zahlbruckner (Lecanoromycetes, Ascomyco¬
ta)// Mycologia. 2011. Vol. 103. N 5. P. 983-1003.
42 Schmull М., Mi^dlikowska J., Pelzer M. et al. Phylogenetic
affiliations of members of the heterogeneous lichen-forming fungi of
the genus Lecidea sensu Zahlbruckner (Lecanoromycetes, Ascomyco¬
ta) // Mycologia. 2011. Vol. 103. N 5. P. 983-1003.
Gowardia Halonen, Myllys, Velmala & Hyvarinen (2009)43.
Hypogymnia (Nyl.) Nyl. (1896).
Hypotrachyna (Vain.) Hale (1974).
Imshaugia S. L. F. Mey. (1985).
Letharia (Th. Fr.) Zahlbr. (1892).
Lethariella (Motyka) Krog (1976).
Masonhalea Karnefelt (1977).
Melanelia Essl. (1978).
Melanelixia O. Blanco, A. Crespo, Divakar, Essl., D. Hawksw. &
Lumbsch (2004).
Melanohalea O. Blanco, A. Crespo, Divakar, Essl., D. Hawksw. &
Lumbsch (2004).
Menegazzia A. Massal. (1854).
Myelochroa (Asahina) Elix & Hale (1987).
Neofuscelia Essl. (1978).
Nephromopsis Mull. Arg. (1891).
*Nesolechia A. Massal. (1856).
Nipponoparmelia (Kurok.) К. H. Moon, Y. Ohmura & Kashiw. ex
A. Crespo & al. (2010)44.
Oropogon Th. Fr. (1861).
Parmelia Ach. (1803).
Parmelina Hale (1974).
Parmelinopsis Elix & Hale (1987).
Parmeliopsis (Nyl.) Nyl. (1866).
Parmotrema A. Massal. (1860).
*Phacopsis Tul. (1852).
Platismatia W. L. Culb. & C. F. Culb. (1968).
Pleurosticta Petr. (1931).
Protoparmelia M. Choisy (1929).
Pseudephebe M. Choisy (1930).
Pseudevernia Zopf (1903).
Punctelia Krog (1982).
Remototrachyna Divakar & A. Crespo (2010)45.
43 Halonen P., Myllys L., Velmala S., Hyvarinen H. Gowardia
(Parmeliaceae) - a new alectorioid lichen genus with two species //
Bryologist. 2009. Vol. 112. N l.P. 138-146.
44 Crespo A., Kauff F., Divakar P. K., et al. Phylogenetic gener¬
ic classification of parmelioid lichens (Parmeliaceae, Ascomycota)
based on molecular, morphological and chemical evidence // Taxon.
2010. Vol. 59. P. 1735-1753.
45 Divakar P. K., Lumbsch H. Т., Ferencova Z., et al. Remoto¬
trachyna, a newly recognized tropical lineage of lichens in the Hypo-
Tuckermannopsis Gyeln. (1933).
Usnea Dill, ex Adans. (1763).
Usnocetraria M. J. Lai & J. C. Wei (2007)46.
Vulpicida J.-E. Mattsson & M. J. Lai (1993).
Xanthoparmelia (Vain.) Hale (1974).
Семейство Pilocarpaceae Zahlbr. (1905).
Byssoloma Trevis. (1853).
Fellhanera Vezda (1986).
Fellhaneropsis Serus. & Coppins (1996).
Leimonis R. C. Harris (2010)47,48.
Micarea Fr. (1825).
Psilolechia A. Massal. (1860).
*Scutula Tul. (1852).
Семейство Psoraceae Zahlbr. (1898).
Protoblastenia (Zahlbr.) J. Steiner (1911).
Protomicarea Hafellner (2001).
Psora Hoffm. (1896).
Psorula Gotth. Schneid. (1980).
Семейство Ramalinaceae C. Agardh (1821).
Adelolecia Hertel & Hafellner (1984).
Arthrosporum A. Massal. (1853).
Bacidia De Not. (1846).
Bacidina Vezda (1991).
Biatora Ach. in Luyken (1809).
Catinaria Vain. (1922).
Cliostomum Fr. (1825).
Lecania A. Massal. (1853).
Ramalina Ach. in Luyken (1809).
Schadonia Korb. (1859).
(*)Toninia A. Massal. (1852).
trachyna clade (.Parmeliaceae, Ascomycota), originated in the Indian
subcontinent // Amer. J. Bot. 2010. Vol. 97. N 4. P. 579-590.
46 Lai M. J., Qian Z. G., Xu L. Synopsis of the cetrarioid lichen
genera and species (Parmeliaceae, lichenized Ascomycotina) in Chi¬
na//J. Natl. Taiwan M us. 2007. Vol. 60. N 1. P. 45-61.
47 Harris R. C. Four novel lichen taxa in the lichen biota of
eastern North America // Opuscula Philolichenum. 2009. N 6. P. 149—
156.
48 Schmull М., Mi^dlikowska J., Pelzer M. et al. Phylogenetic
affiliations of members of the heterogeneous lichen-forming fungi of
the genus Lecidea sensu Zahlbruckner (Lecanoromycetes, Ascomyco¬
ta) // Mycologia. 2011. Vol. 103. N 5. P. 983-1003.
Waynea Moberg (1990).
Семейство Scoliciosporaceae Hafellner (1984).
Scoliciosporum A. Massal. (1852).
Семейство Sphaerophoraceae Fr. (1831).
Bunodophoron A. Massal. (1861).
Sphaerophorus Pers. (1794).
Семейство Stereocaulaceae Chevall. (1826).
Hertelidea Printzen & Kantvilas (2004).
Lepraria Ach. (1803).
Squamarina Poelt (1958).
Stereocaulon Hoffm. (1796).
Семейство Tephromelataceae Hafellner (1984).
Calvitimela Hafellner (2001)49.
Tephromela M. Choisy (1929).
Violella T. Sprib. (2011) °.
Lecanorales, рода неопределенного положения
Myochroidea Printzen, T. Sprib. & Tonsberg (2008)51.
Puttea S. Stenroos & Huhtinen (2009)52.
Порядок Lecideales Vain., Acta Soc. Fauna Flora Fennica 53(1):
13 (1922)53.
Семейство Lecideaceae Chevall. (1826). (=Porpidiaceae Hertel
& Hafellner)54,5
49 Spribille Т., Goffinet B., Klug B., et al. Molecular support for
the recognition of the Mycoblastus fucatus group as the new genus
Violella (Tephromelataceae, Lecanorales) // Lichenologist. 2011.
Vol. 43. N5. P. 445-466.
50 Spribilie Т., Goffinet B., Klug B., et al. Molecular support
for the recognition of the Mycoblastus fucatus group as the new genus
Violella (Tephromelataceae, Lecanorales) // Lichenologist. 2011.
Vol. 43. N 5. P. 445-466.
51 Printzen C., Spribi lie Т., Tonsberg T. Myochroidea, a new
genus of corticolous, crustose lichens to accommodate the Lecidea
leprosula group // Lichenologist. 2008. Vol. 40. N 3. P. 195-207.
52 Stenroos S., Huhtinen S., Lesonen A., Palice Z., Printzen C.
Puttea, gen. nov., erected for the enigmatic lichen Lecidea mar-
garitella H Bryologist. 2009. Vol. 112. N 3. P. 544-557.
53 Schmull М., Mi^dlikowska J., Pelzer M. et al. Phylogenetic
affiliations of members of the heterogeneous lichen-forming fungi of
the genus Lecidea sensu Zahlbruckner (Lecanoromycetes, Ascomyco¬
ta) // Mycologia. 2011. Vol. 103. N 5. P. 983-1003.
Amygdalaria Norman (1852).
Bellemerea Hafellner & Cl. Roux (1984).
Cecidonia Triebel & Ram bold (1988).
Clauzadea Hafellner & Bellem. (1984).
Farnoldia Hertel (1983).
Immersaria Rambold & Pietschm. (1989).
Koerberiella Stein (1879).
Lecidea Ach. (1803).
Lecidoma Gotth. Schneid. & Hertel (1981).
Mycobilimbia Rehm (1890).
Porpidia Korb. (1855).
Romjularia Timdal (2007)56.
?Steinia Korb. (1873).
Порядок Peltigerales Walt. Watson, New Phytologist 28: 9
(1929).
Подпорядок Collematineae Mi^dl. & Lutzoni, Am. J. Bot. 91:
459 (2004).
Семейство Coccocarpiaceae (Mont. ex Mull. Stuttg.) Henssen
(1986).
Coccocarpia Pers. (1827).
Spilonema Bornet (1856).
Семейство Collemataceae Zenker (1827).
Collema F. H. Wigg. (1780).
Leptogium (Ach.) Gray (1821).
Семейство Pannariaceae Tuck. (1872).
Degelia Arv. & D. J. Galloway (1981).
Fuscopannaria P. M. Jorg. (1994).
Leciophysma Th. Fr. (186 5)57.
54 Lumbsch H. Т., Huhndorf S. M. (ed.) Notes on ascomycete
systematics. Nos. 4408 - 4750 // Myconet. 2007. Vol. 13. P. 59-99.
55 Schmull М., Mi^dlikowska J., Pelzer M. et al. Phylogenetic
affiliations of members of the heterogeneous lichen-forming fungi of
the genus Lecidea sensu Zahlbruckner (Lecanoromycetes, Ascomyco¬
ta)//Mycologia. 2011. Vol. 103. N5. P. 983-1003.
56 Timdal E. Romjularia / In: Т. H. Nash III, C. Gries, F. Bun-
gartz (eds.) Lichen Flora of the Greater Sonoran Desert Region. Vol.
3. Arizona State University, Tempe, 2007. P. 287-289.
57 Wedin М., Wiklund E., Jorgensen P. М., Ekman S. Slippery
when wet: phylogeny and character evolution in the gelatinous cya-
nobacterial lichens (.Peltigerales, Ascomycetes) // Mol. Phylogenet.
Evol. 2009. Vol. 53. P. 862-871.
Leptogidium Nyl. (18 7 3)58.
Moelleropsis Gyeln. (1939).
Pannaria Delise ex Bory (1828).
Parmeliella Mull. Arg. (1862).
Protopannaria (Gyeln.) P. M. Jerg. & S. Ekman (2000).
Psoroma Ach. ex Michx. (1803).
Santessoniella Henssen (1997).
Семейство Placynthiaceae A. E. Dahl (1950).
Koerberia A. Massal. (1854).
Placynthium (Ach.) Gray (1821).
Vestergrenopsis Gyeln. (1940).
Семейство Vahliellaceae Wedin, P. M. Jorg. & S. Ekman
(2010)59.
Vahliella P. M. Jorg. (2008)6 .
Подпорядок Peltigerineae sensu Mi^dlikowska & Lutzoni (2004).
Семейство Lobariaceae Chevall. (1826).
Dendriscocaulon Nyl. (1888).
Lobaria (Schreb.) Hoffm. (1796).
Pseudocyphellaria Vain. (1890).
Sticta (Schreb.) Ach. (1803).
Семейство Massalongiaceae Wedin, P. M. Jorg. & E. Wiklund
(2007)61.
Leptochidium M. Choisy (1952).
Massalongia Korb. (1855).
Polychidium (Ach.) Gray (1821).
Семейство Nephromataceae Wetmore ex J. C. David & D.
Hawksw. (1991).
Nephroma Ach. in Luyken (1809).
58 Muggia L., Nelson P., Wheeler T. et al. Convergent evolution
of a symbiotic duet: The case of the lichen genus Polychidium (Pelti¬
gerales, Ascomycota) // Amer. J. Bot. 2011. Vol. 98. N 10. P. 1647-
1656.
59 Wedin М., Jorgensen P. М., Ekman S. Vahliellaceae, a new
family of cyanobacterial lichens (Peltigerales, Ascomycetes) // Li¬
chenologist. 2011. Vol. 43. N 1. P. 67-72.
60 Jorgensen P. M. Vahliella, a new lichen genus // Lichenolo¬
gist. 2008. Vol. 40. N 3. P. 221-225.
61 Wedin М., Jorgensen P. М., Wiklund E. Massalongiaceae
fam. nov., an overlooked monophyletic group among the cyanobacte¬
rial lichens (Peltigerales, Lecanoromycetes, Ascomycota) // Lichen¬
ologist. 2007. Vol. 39. N 1. P. 61-67.
Семейство Peltigeraceae Dumort. (1822).
Peltigera Willd. (1787).
Solorina Ach. (1808).
Порядок Rhizocarpales Mi^dl. et al., Mycologia 98(6): 1097
(2007) [“2006”].
?Семейство Catillariaceae Hafellner (1984).
Catillaria A. Massal. (1852).
Halecania M. Mayrhofer (1987).
Placolecis Trevis. (1860).
Sporastatia A. Massal. (1854).
Семейство Rhizocarpaceae M. Choisy ex Hafellner (1984).
Catolechia Flot. (1850).
Epilichen Clem. (1909).
Rhizocarpon Ramond ex DC. (1805).
Порядок Teloschistales D. Hawksw. & О. E. Erikss., Syst.
Ascom. 5: 183 (1986).
Семейство Brigantiaeaceae Hafellner & Bellem. (1982) .
Brigantiaea Trevis. (1853).
Семейство Megalosporaceae \^zda ex Hafellner & Bellem.
(1982).
Megalospora Meyen (1843).
Семейство Teloschistaceae Zahlbr. (1898).
Caloplaca Th. Fr. (1861).
Cephalophysis (Hertel) H. Kilias (1985).
Fulgensia A. Massal. & De Not. (1853).
Oxneria S. Y. Kondr. & Karnefelt (2003).
Rusavskia S. Y. Kondr. & Karnefelt (2003).
Seirophora Poelt (1983).
Teloschistes Norman (1853).
Xanthomendoza S. Y. Kondr. & Karnefelt (1997).
Xanthoria (Fr.) Th. Fr. (1860).
Подкласс Ostropomycetidae Reeb, Lutzoni & Cl. Roux, Mol.
Phylogen. Evol. 32: 1055 (2004).
Порядок Baeomycetales Lumbsch, Huhndorf & Lutzoni, Mycol.
Res. 111(5): 529 (2007).
Семейство Anamylopsoraceae Lumbsch & Lunke (1995)63.
62 Gaya E., Hognabba F., Holguin A., et al. Implementing a cu¬
mulative supermatrix approach for a comprehensive phylogenetic
study of the Teloschistales (Pezizomycotina, Ascomycota) // Mol.
Phylogenet. Evol. 2012. doi: 10.1016/j.ympev.2012.01.012
Anamylopsora Timdal (1991).
Семейство Baeomycetaceae Dumort. (1829).
Ainoa Lumbsch & I. Schmitt (2001)64.
Baeomyces Pers. (1794).
Порядок Ostropales Nannf., Nova Acta Regiae Soc. Sci. Upsal.,
ser. 4 8(2): 68 (1932).
Семейство Coenogoniaceae (Fr.) Stizenb. (1862).
Dimerella Trevis. (1880).
Семейство Graphidaceae Dumort. (1822).
[=Gomphillaceae Walt. Watson ex Hafellner, Solorinellaceae
Vezda & Poelt, Thelotremataceae (Nyl.) Stizenb.]65’66
Echinoplaca Fee (1824).
Diploschistes Norman (1853).
Graphis Adans. (1763).
Gyalectidium Mull. Arg. (1881).
Gyalidea Lettau ex Vezda (1966).
Gyalideopsis Vezda (1972).
Leptotrema Mont. & Bosch (1855).
Phaeographis Mull. Arg. (1882).
63 Lumbsch H. Т., Schmitt I., Lucking R., et al. The phylogenet¬
ic placement of Ostropales within Lecanoromycetes (Ascomycota)
revisited // Mycol. Res. 2007. Vol. 111. P. 257-267.
64 Lumbsch H. Т., Schmitt I., Lucking R., et al. The phylogenet¬
ic placement of Ostropales within Lecanoromycetes (Ascomycota)
revisited // Mycol. Res. 2007. Vol. 111. P. 257-267.
65 Mangold A., Martin M. P., Lucking R., Lumbsch H. T. Mo¬
lecular phylogeny suggests synonymy of Thelotremataceae within
Graphidaceae (Ascomycota: Ostropales) // Taxon. 2008. Vol. 57. N
2. P. 476-486.
66 Plata E. R., Lucking R., Lumbsch H. T. A new classification
for the family Graphidaceae (Ascomycota: Lecanoromycetes: Ostro¬
pales) // Fungal Diversity. 2012. Vol. 52. P. 107-121. Согласно но¬
вой классификации семейства, перечисленные рода отнесены к
двум подсемействам Gomphilloideae (Walt. Watson ex Hafellner)
Rivas Plata, Lucking & Lumbsch (Echinoplaca, Gyalectidium, Gya¬
lidea, Gyalideopsis, Phyllogyalidea) и Graphidoideae Rivas Plata,
Lucking & Lumbsch, при этом в последнем выделены трибы
Graphideae Rivas Plata, Lucking & Lumbsch (Graphis, Phaeogra¬
phis), Ocellularieae Rivas Plata, Lucking & Lumbsch (Leptotrema) и
Thelotremateae Rivas Plata, Lucking & Lumbsch (Diploschistes,
Thelotrema .
Phyllogyalidea Lucking & Aptroot (2008)67.
Thelotrema Ach. (1803).
Семейство Gyalectaceae Stizenb. (1862)68.
?BeIonia Korb. (1856).
Gyalecta Ach. (1808).
?Pachyphiale Lonnr. (1858).
Семейство Odontotremataceae D. Hawksw. & Sherwood (1982).
*(+)Odontotrema Nyl. (1858).
*ThamnogalIa D. Hawksw. (1980).
Семейство Phlyctidaceae Poelt & Vezda ex J. C. David & D.
Hawksw. (1991).
Phlyctis (Wallr.) Flot. (1850).
Семейство Porinaceae Rchb. (1828).
Porina Ach. (1809).
Семейство Sagiolechiaceae Baloch, Lucking, Lumbsch & Wedin
(2010)6.
Rhexophiale Th. Fr. (1860).
Sagiolechia A. Massal. (1854).
Семейство Stictidaceae Fr. (1849).
(+)Absconditella Vezda (1965).
Cryptodiscus Corda (1838). (=Bryophagus Nitschke ex Arnold)70.
*Nanostictis M. S. Christ. (1954).
?Petractis Fr. (1845).
+Stictis Pers. (1799).
?Thelopsis Nyl. (1855).
Ostropales, род неопределенного положения
?Xyloschistes Vain. (1883).
67 Lucking R. Foliicolous Lichenized Fungi // Flora Neotropica
Monograph. 2008. Vol. 103. 866 pp.
68 Предлагается сохранить самостоятельным статус родов
Belonia и Pachyphiale, невзирая на филогенетическое родство
этих родов с родом Gyalecta (Baloch Е. et al., 2010). Неубедитель¬
ными выглядят попытки объединить в один род виды с плодовы¬
ми телами апотециями и перитециями.
69 Baloch Е., Lucking R., Lumbsch Н. Т., Wedin М. Major
clades and phylogenetic relationships between lichenized and non-
lichenized lineages in Ostropales (Ascomycota: Lecanoromycetes) //
Taxon. 2010. Vol. 59. N 5. P. 1483-1494.
70 Baloch E., Gilenstam G., Wedin M. Phylogeny and classifi¬
cation of Cryptodiscus, with taxonomic synopsis of the Swedish spe¬
cies // Fungal Diversity. 2009. Vol. 38. P. 51-68.
Порядок Pertusariales М. Choisy ex D. Hawksw. & О. E. Erikss.,
Syst. Ascom. 5: 181 (1986).
(=Agyriciles Clem. & Shear)71
Семейство Agyriaceae Corda (1838)72.
+Agyrium Fr. (1822).
Семейство Icmadophilaceae Triebel (1993).
Dibaeis Clem. (1909).
Icmadophila Trevis. (1853).
Siphula Fr. (1831).
Thamnolia Ach. ex Schaer. (1850).
Семейство Megasporaceae Lumbsch in Lumbsch, Feige & K.
Schmitz (1994).
Aspicilia A. Massal. (1852).
Circinaria Link. (1809)73.
Lobothallia (Clauzade & Cl. Roux) Hafellner (1991).
Megaspora (Clauzade & Cl. Roux) Hafellner & V. Wirth (1987).
Sagedia Ach. (1809)74.
Семейство Ochrolechiaceae R. C. Harris ex Lumbsch & I.
Schmitt (2006).
Ochrolechia A. Massal. (1852).
Varicellaria Nyl. (1858).
Семейство Pertusariaceae Korb. ex Korb. (1855).
Pertusaria DC. (1805).
Pertusariales, род неопределенного положения
Aspilidea Hafellner (2001) 5.
71 Hodkinson B. P., Lendemer J. C. The orders of Ostropomy¬
cetidae (Lecanoromycetes, Ascomycota): Recognition of Sarrame-
anales and Trapeliales with a request to retain Pertusariales over
Agyriales // Phytologia. 2011. Vol. 93. N 3. P. 407-412.
72 Hodkinson B. P., Lendemer J. C. The orders of Ostropomy¬
cetidae (Lecanoromycetes, Ascomycota): Recognition of Sarrame-
anales and Trapeliales with a request to retain Pertusariales over
Agyriales// Phytologia. 2011. Vol. 93. N 3. P. 407-412.
73 Nordin A., Savic S., Tibell L. Phylogeny and taxonomy of
Aspicilia and Megasporaceae II Mycologia. 2010. Vol. 102. N 6. P.
1339-1349.
74 Nordin A., Savic S., Tibell L. Phylogeny and taxonomy of
Aspicilia and Megasporaceae II Mycologia. 2010. Vol. 102. N 6. P.
1339-1349.
75 Nordin A., Savic S., Tibell L. Phylogeny and taxonomy of
Aspicilia and Megasporaceae II Mycologia. 2010. Vol. 102. N 6. P.
1339-1349.
Порядок Sarrameanales Hodkinson & Lendemer, Phytologia
93(3): 410 (201 1)76.
Семейство Sarrameanaceae Hafellner (1984).
Loxospora A. Massal. (1852).
Порядок Trapeliales Hodkinson & Lendemer, Phytologia 93(3):
410 (201 1)77.
Семейство Trapeliaceae M. Choisy ex Hertel (19 70)78,79.
Lithographa Nyl. (1857).
Placopsis (Nyl.) Linds. (1867).
Placynthiella Elenkin (1909).
Rimularia Nyl. (1868).
?+Sarea Fr. (1825)80.
Trapelia M. Choisy (1929).
Trapeliopsis Hertel & Gotth. Schneid. (1980).
Xylographa (Fr.) Fr. (1836).
Ostropomycetidae, семейства неопределенного положения
Семейство Arctomiaceae Th. Fr. (1860).
Arctomia Th. Fr. (1860).
Gregorella Lumbsch (2005).
76 Hodkinson B. P., Lendemer J. C. The orders of Ostropomy¬
cetidae (Lecanoromycetes, Ascomycota): Recognition of Sarrame¬
anales and Trapeliales with a request to retain Pertusariales over
Agyriales // Phytologia. 2011. Vol. 93. N 3. P. 407-412.
77 Hodkinson B. P., Lendemer J. C. The orders of Ostropomy¬
cetidae (Lecanoromycetes, Ascomycota): Recognition of Sarrame¬
anales and Trapeliales with a request to retain Pertusariales over
Agyriales // Phytologia. 2011. Vol. 93. N 3. P. 407-412.
78 Lumbsch H. Т., Schmitt I., Mangold A., Wedin M. Ascus
types are phylogenetically misleading in Trapeliaceae and Agyriaceae
(Ostropomycetidae, Ascomycota) // Mycol. Res. 2007. Vol. 111. N 9.
P. 1133-1141.
79 Hodkinson B. P., Lendemer J. C. The orders of Ostropomy¬
cetidae (Lecanoromycetes, Ascomycota): Recognition of Sarrame¬
anales and Trapeliales with a request to retain Pertusariales over
Agyriales // Phytologia. 2011. Vol. 93. N 3. P. 407-412.
80 Hodkinson B. P., Lendemer J. C. The orders of Ostropomy¬
cetidae (Lecanoromycetes, Ascomycota): Recognition of Sarrame¬
anales and Trapeliales with a request to retain Pertusariales over
Agyriales // Phytologia. 2011. Vol. 93. N 3. P. 407-412.
Семейство Arthrorhaphidaceae Poelt & Hafellner (1976).
Arthrorhaphis Th. Fr. (1860).
Семейство Hymeneliaceae Korb. (1855).
Eiglera Hafellner (1984).
Hymenelia Kremp. (1852).
Ionaspis Th. Fr. (1871).
Tremolecia M. Choisy (1953).
Семейство Protothelenellaceae Vezda, H. Mayrhofer & Poelt
(1985).
Protothelenella Rasanen (1943).
Thrombium Wallr. (1831 )81.
Семейство Schaereriaceae M. Choisy ex Hafellner (1984).
Schaereria Korb. (1855).
Семейство Thelenellaceae О. E. Erikss. ex H. Mayrhofer
(1987)82.
Thelenella Nyl. (1855).
Ostropomycetidae, рода неопределенного положения
Anzina Scheid. (19 82)83,8 .
Strangospora Korb. (I860)85.
Lecanoromycetes, порядки неопределенного положения
Порядок Candelariales Mi^dl., Lutzoni & Lumbsch, Mycol. Res.
1 11(5): 530 (2007).
Семейство Candelariaceae Hakul. (1954).
Candelaria A. Massal. (1852).
81 Schmitt I., Mueller G., Lumbsch H. T. Ascoma morphology
is homoplaseous and phylogenetically misleading in some pyrenocar-
pous lichens // Mycologia. 2005. Vol. 97. N 2. P. 362-374.
82 Schmitt I., Mueller G., Lumbsch H. T. Ascoma morphology
is homoplaseous and phylogenetically misleading in some pyrenocar-
pous lichens // Mycologia. 2005. Vol. 97. N 2. P. 362-374.
83 Lumbsch H. Т., Schmitt I., Mangold A., Wedin M. Ascus
types are phylogenetically misleading in Trapeliaceae and Agyriaceae
(Ostropomycetidae, Ascomycota) // Mycol. Res. 2007. Vol. 111. N 9.
P. 1133-1141.
84 Lumbsch H. Т., Schmitt I., Lucking R., et al. The phylogenet¬
ic placement of Ostropales within Lecanoromycetes (Ascomycota)
revisited // Mycol. Res. 2007. Vol. 111. P. 257-267.
85 Schmull М., Mi^dlikowska J., Pelzer M. et al. Phylogenetic
affiliations of members of the heterogeneous lichen-forming fungi of
the genus Lecidea sensu Zahlbruckner (Lecanoromycetes, Ascomyco¬
ta)//Mycologia. 2011. Vol. 103. N 5. P. 983-1003.
Candelariella Mull. Arg. (1894).
Порядок Umbilicariales J. C. Wei & Q. M. Zhou, Mycosystema
26(1): 44 (2007).
Семейство Elixiaceae Lumbsch (1997).
Elixia Lumbsch (1997).
Семейство Fuscideaceae Hafellner (1984).
Fuscidea V. Wirth & Vezda (1972).
?*Lettauia D. Hawksw. & R. Sant. (1990).
?Maronea A. Massal. (1856).
?Orphniospora Korb. (1874).
Семейство Ophioparmaceae R. W. Rogers & Hafellner (1988).
Boreoplaca Timdal (1994).
Hypocenomyce M. Choisy (1951).
Ophioparma Norman (1852).
Семейство Ropalosporaceae Hafellner (1984)86.
Ropalospora A. Massal. (1860).
Семейство Umbilicariaceae Chevall. (1826).
Lasallia Merat (1821).
Umbilicaria Hoffm. (1789).
Lecanoromycetes, рода неопределенного положения
Biatoridium J. Lahm ex Korb. (1860).
Bilimbia De Not. (1846).
?*Corticifraga D. Hawksw. & R. Sant. (1990).
Leprocaulon Nyl. (1878).
Maronella M. Steiner (19 5 9)87.
Piccolia A. Massal. (18 5 6)88.
?Stenhammarella Hertel (1967).
Класс Leotiomycetes О. E. Erikss. & Winka, Myconet 1(1): 7
(1997).
Порядок Helotiales Nannf. ex Korf & Lizon, Mycotaxon 75: 501
(2000).
(=Leotiales Carpenter).
86 Bylin A., Arnerup J., Hogberg N., Thor G. A phylogenetic
study of Fuscideaceae using mtSSU rDNA // Biblioth. Lichenol.
2007. N 96. P. 49-60.
87 Hafellner J. A revision of Maronella laricina and Piccolia
ochrophora II Symb. Bot. Upsal. 2004. Vol. 34. N 1. P. 87-96.
88 Hafellner J. A revision of Maronella laricina and Piccolia
ochrophora II Symb. Bot. Upsal. 2004. Vol. 34. N 1. P. 87-96.
Семейство Helotiaceae Rehm (1886).
*Unguiculariopsis Rehm (1909).
Семейство Hyaloscyphaceae Nannf. (1932).
*Pezizella Fuckel (1870).
*Polydesmia Boud. (1885).
?*Protounguicularia Raitv. & R. Galan (1986).
*Spirographa Zahlbr. (1903)89.
Helotiales, роды неопределенного положения90
*Echinodiscus Etayo & Diederich (2000).
*Geltingia Alstrup & D. Hawksw. (1990).
*Llimoniella Hafellner & Nav.-Ros. (1993).
*Phaeopyxis Rambold & Triebel (1990).
*Rhymbocarpus Zopf (1896).
*Skyttea Sherwood, D. Hawksw. & Coppins (1981).
*Skyttella D. Hawksw. & R. Sant. (1988).
Leotiomycetes, семейства неопределенного положения
Семейство Thelocarpaceae Zukal (1893)91.
Athelium Nyl. (1886)92.
Sarcosagium A. Massal. (1856).
Thelocarpon Nyl. (1853).
Семейство Vezdaeaceae Poelt & Vezda ex J. C. David & D.
Hawksw. (1991 )93.
Vezdaea Tscherm.-Woess & Poelt (1976).
89 Lawrey J. D., Diederich P. Lichenicolous fungi - worldwide
checklist, including isolated cultures and sequences available. 2011.
URL: http://www.lichenicolous.net [7/24/2011].
90 Lawrey J. D., Diederich P. Lichenicolous fungi - worldwide
checklist, including isolated cultures and sequences available. 2011.
URL: http://www.lichenicolous.net [7/24/2011].
91 Lumbsch H. Т., Zimmermann D. G., Schmitt I. Phylogenetic
position of ephemeral lichens in Thelocarpaceae and Vezdaeaceae
(Ascomycota) // Biblioth. Lichenol. 2009. N 100. P. 389-398.
92 Khodosovtsev A., Naumovich A., Vondrakova O., Vondrak
J. Athelium imperceptum Nyl. (Thelocarpaceae, Ascomycota), a little
known ephemeral lichen of biological soil crusts, new to Ukraine //
Chornomors'k. bot. z. 2010. Vol. 6. P. 385-389.
93 Lumbsch H. Т., Zimmermann D. G., Schmitt I. Phylogenetic
position of ephemeral lichens in Thelocarpaceae and Vezdaeaceae
(Ascomycota) // Biblioth. Lichenol. 2009. N 100. P. 389-398.
Класс Lichinomycetes Reeb, Lutzoni & Cl. Roux., Mol. Phy-
logen. Evol. 32: 1055 (2004).
Порядок Lichinales Henssen & Budel ex D. Hawksw. & О. E.
Erikss., Syst. Ascom. 5: 138 (1986).
Семейство Lichinaceae Nyl. (1854).
Anema Nyl. ex Forssell (1885).
Cryptothele Th. Fr. (1866).
Ephebe Fr. (1825).
Epiphloea Trevis. (1880).
Euopsis Nyl. (1875).
Gonohymenia J. Steiner (1902).
Heppia Nageli ex A. Massal. (1854).
Lemmopsis (Vain.) Zahlbr. (1906).
Lempholemma Korb. (1855).
Lichina C. Agardh (1817).
Lichinella Nyl. (1873).
Lichinodium Nyl. (1875).
Peccania A. Massal. ex Arnold (1858).
Phylliscum Nyl. (1855).
Porocyphus Korb. (1855).
Psorotichia A. Massal. (1855).
Pterygiopsis Vain. (1890).
Pyrenocarpon Trevis. (1855).
Pyrenopsis Nyl. (1858).
Synalissa Fr. (1825).
Thallinocarpon A. E. Dahl (1950)94.
Thelignya A. Massal. (1855).
Thermutis Fr. (1825).
Thyrea A. Massal. (1856).
Zahlbrucknerella Herre (1912).
Семейство Peltulaceae Budel (1986).
Peltula Nyl. (1853).
Класс Sordariomycetes О. E. Erikss. & Winka, Myconet 1:10
(1997).
Подкласс Hypocreomycetidae О. E. Erikss. & Winka, Myconet
1: 6(1997).
Порядок Coronophorales Nannf., Nova Acta Regiae Soc. Sci.
Upsal, ser.4 8(2): 54(1932).
Семейство Nitschkiaceae (Fitzp.) Nannf. (1932).
94 Jergensen P. M. Thallinocarpon 11 Nordic Lichen Flora. Vol.
3. Cyanolichens. Nordic Lichen Society, Uddevalla. P. 72-73.
*Acanthonitschkea Speg. (1908).
*Lasiosphaeriopsis D. Hawksw. & Sivan. (1980)95.
*Rhagadostoma Korb. (1865).
Порядок Hypocreales Lindau in Engler & Prantl (eds), Nat.
Pflanzenfam. 1(1): 343 (1897).
Семейство Bionectriaceae Samuels & Rossman (1999).
*Nectriopsis Maire (1911).
*Paranectria Sacc. (1878).
*Pronectria Clem. (1931).
*Trichonectria Kirschst. (1907).
Семейство Nectriaceae Tul. & C. Tul. (1865).
*Xenonectriella Weese (1919).
Семейство Niessliaceae Kirschst. (1939).
*NiessIia Auersw. (1869).
Hypocreales, род неопределенного положения
*Roselliniella Vain. (1921)96.
Подкласс Sordariomycetidae О. E. Erikss. & Winka, Myconet 1:
10 (1997).
Порядок Sordariales Chadef. ex D. Hawksw. & О. E. Erikss,
Syst. Ascom. 5: 182 (1986).
Sordariales, рода неопределенного положения
*Reconditella Matzer & Hafellner (1990).
*RoseIliniopsis Matzer & Hafellner (1990).
Sordariomycetes, порядки неопределенного положения
Порядок Phyllachorales М. Е. Barr, Mycologia 75: 10 (1983).
Семейство Phyllachoraceae Theiss. & H. Syd. (1915).
*Lichenochora Hafellner (1989).
Порядок Trichosphaeriales М. E. Barr, Mycologia 75: 1 1 (1983).
Семейство Trichosphaeriaceae G. Winter (1885).
Cresporhaphis М. B. Aguirre (1991).
*Trichosphaeria Fuckel (1870).
95 Eriksson О. E. (ed.) Outline of Ascomycota - 2006 // My¬
conet. 2006. Vol. 12. P. 1-82.
96 Hawksworth D. L, Millanes A. M, Wedin M. Roselliniella
revealed as an overlooked genus of Hypocreales, with the description
of a second species on parmelioid lichens // Persoonia. 2010. Vol. 24.
P. 12-17.
Sordariomycetes, род неопределенного положения
*Neolamya Theiss. & Syd. (1918).
Ascomycota, семейства неопределенного положения97
Семейство Coniocybaceae Rchb. (1837).
Chaenotheca (Th. Fr.) Th. Fr. (1860).
Sclerophora Chevall. (1826).
Семейство Epigloeaceae Zahlbr. (1903).
(+*)Epigloea Zukal (1889).
Семейство Microcaliciaceae Tibell (1984).
+Microcalicium Vain. (1927).
Семейство Xanthopyreniaceae Zahlbr. (1926).
Collemopsidium Nyl. (1881).
*Didymellopsis (Sacc.) Clem. & Shear (1931).
Frigidopyrenia Grube (2005).
*Zwackhiomyces Grube & Hafellner (1990).
Ascomycota, рода неопределенного положения
*Abrothallus DeNot. (1845).
Normandina Nyl. (1855).
98
Anamorphic Ascomycota
*Bachmanniomyces D. Hawksw. (1981).
Cheiromycina B. Sutton (1986).
*Cornutispora Piroz. (1973).
Dictyocatenulata Finley & E. F. Morris (1967).
*EpicIadonia D. Hawksw. (1981).
*EverniicoIa D. Hawksw. (1982).
*Graphium Corda (1837).
*Hansfordiellopsis Deighton (1960).
*Hobsoniopsis D. Hawksw. (2001).
*Illosporiopsis D. Hawksw. (2001).
*Illosporium Mart. (1817).
*Intralichen D. Hawksw. & M. S. Cole (2002).
*Karsteniomyces D. Hawksw. (1980).
*Lambinonia Serus. & Diederich (2005).
*LibertielIa Speg. & Roum. (1880).
97 Eriksson О. E. (ed.) Outline of Ascomycota - 2006 // My¬
conet. 2006. Vol. 12. P. 1-82.
98 Lawrey J. D, Diederich P. Lichenicolous fungi - worldwide
checklist, including isolated cultures and sequences available. 2011.
URL: http://www.lichenicolous.net [7/24/2011].
*Lichenodiplis Dyko & D. Hawksw. (1979).
*Lichenopuccinia D. Hawksw. & Hafellner (1984).
*Lichenosticta Zopf. (1898).
*Minutoexcipula V. Atienza & D. Hawksw. (1994).
*Monodictys S. Hughes (1958).
*Nigropuncta D. Hawksw. (1981).
*Phaeoseptoria Speg. (1908).
*Phaeosporobolus D. Hawksw. & Hafellner (1986).
*Phoma Sacc. (1880).
*PhylIosticta Pers. (1818).
*Pseudoseptoria Speg. (1910).
*Refractohilum D. Hawksw. (1977).
*Sclerococcum Fr. (1819).
*Taeniolella S. Hughes (1958).
*VouauxieIla Petr. & Syd. (1927).
*Vouauxiomyces Dyko & D. Hawksw. (1979).
Отдел Basidiomycota R. T. Moore, Bot. Mar. 23: 371 (1980).
Подотдел Agaricomycotina Doweld, Prosyllabus: LXXVII
(2001).
Класс Agaricomycetes Doweld, Prosyllabus: LXXVII (2001).
Подкласс Agaricomycetidae Parmasto, Windahlia 16: 16 (1986).
Порядок Agaricales Underw, Moulds, Mildews Mushrooms: 97
(1899).
Семейство Hygrophoraceae Lotsy (1907).
*Arrhenia Fr. (1849)".
Lichenomphalia Redhead, Lutzoni, Moncalvo & Vilgalyis (2002).
Порядок Atheliales Julich, Biblthca Mycol. 85: 343 (1981).
Семейство Atheliaceae Julich (1982).
+(*)Athelia Pers. (1822).
Agaricomycetes, порядки неопределенного положения
Порядок Cantharellales Gaum, Vergl. Morph. Pilze: 495 (1926).
Семейство Clavulinaceae Donk (1970).
Multiclavula R. H. Petersen (1967).
99 Lawrey J. D, Lucking R, Sipman H. J. M, et al. High con¬
centration of basidiolichens in a single family of agaricoid mush¬
rooms (Basidiomycota: Agaricales: Hygrophoraceae) // Mycol. Res.
2009. Vol. 113. N 10. P. 1154-1171.
Порядок Corticiales К. Н. Larss, Mycol. Res. 111(5): 540
(2007).
Семейство Corticiaceae Herter (1910).
*Marchandiobasidiuin Diederich & Schultheis (2003).
*Marchandiomyces Diederich & D. Hawksw. (1990).
Класс Tremellomycetes Doweld, Prosyllabus: LXXVII (2001).
Порядок Filobasidiales Julich, Biblioth. Mycol. 85: 324 (1981).
Семейство Syzygosporaceae Julich (1982) 10°.
*Syzygospora G. W. Martin (1937).
Порядок Tremellales Fr, Syst. Mycol. 1: 2 (1821).
Семейство Tremellaceae Fr. (1821).
*Biatoropsis Rasanen (1934).
*Tremella Pers. (1794).
Подотдел Pucciniomycotina R. Bauer, Begerow, J. P. Samp, M.
Weiss & Oberw, Mycol. Progr. 5(1): 45 (2006).
Класс Agaricostilbomycetes R. Bauer, Begerow, J. P. Samp, M.
Weiss & Oberw, Mycol. Progr. 5(1): 45 (2006).
Порядок Agaricostilbales Oberw. & R. Bauer, Sydowia 41: 240
(1989).
Семейство Chionosphaeraceae Oberw. & Bandoni (1982).
*Chionosphaera D. E. Cox (1976).
Класс Cystobasidiomycetes R. Bauer, Begerow, J. P. Samp, M.
Weiss & Oberw, Mycol. Progr. 5(1): 46 (2006).
Порядок Cystobasidiales R. Bauer, Begerow, J. P. Samp, M.
Weiss & Oberw, Mycol. Progr. 5(1): 46 (2006).
Семейство Cystobasidiaceae Gaum. (1926).
*Cystobasidium (Lagerh.) Neuhoff (1924).
100 Millanes A. M, Diederich P, Ekman S, Wedin M. Phylog¬
eny and character evolution in the jelly fungi (Tremellomycetes, Ba¬
sidiomycota, Fungi) // Mol. Phylogenet. Evol. 2011. Vol. 61. N l.P.
12-28.
Глава XII.
Ключи для определения родов лишайников флоры России.
Урбанавичюс Г. П, Макрый Т. В, Кузнецова Е. С, Гагарина
J1. В, Гимеьбрант Д. Е, Степанчикова И. С, Конорева J1. А,
Андреев М. П.
В настоящее время для территории России известно 3540
видов и 394 рода лишайников. Первым этапом идентификации
любого таксона является определение его родовой принад¬
лежности. Довольно часто это вызывает значительные трудно¬
сти, особенно для редких таксономических групп и в тех слу¬
чаях, когда отсутствуют монографические обработки по дан¬
ной группе. До настоящего времени для русской флоры не су¬
ществовало общего родового ключа, охватывающего все роды
лишайников. Исследователям лихенофлоры России приходи¬
лось пользоваться родовыми ключами, написанными для дру¬
гих, часто территориально удаленных областей Земли, что по¬
рой бывает неудобно, и часто в таких определительных клю¬
чах отсутствуют таксоны и таксономические группы русской
флоры. Либо для определения использовались ключи, состав¬
ленные для отдельных семейств, порядков и др. таксономиче¬
ских групп, разбросанные по многочисленным публикациям.
Наиболее удобными для работы являются ключи, опублико¬
ванные в выпусках Определителя лишайников СССР и России,
но и они уже частью устарели или не охватывают всего объе¬
ма известных на территории России родов лишайников.
Настоящий ключ является первой попыткой обобщить
все известные к началу 2013 г. данные и составить пособие
для определения родов лишайников, известных для террито¬
рии России. Для удобства работы с ключем, все известные в
России роды лишайников объединены в 15 морфологических
групп, более или менее различаемых уже на начальном этапе
работы с материалом. Для выхода на нужную группу, состав¬
лен Синопсис - краткий вспомогательный ключ. Стерильные
лишайники, определение которых вызывает наибольшие
трудности из-за отсутствия четких диагностических трудно¬
стей, вынесены в самостоятельный раздел. Деление лишайни¬
ков на морфологические группы довольно условно и некото¬
рые роды могут оказаться в двух или нескольких группах, но
это лишь облегчает работу исследователя, позволяя использо¬
вать для определения таксонов больше признаков и получить
более объемную картину. Составные части родового ключа
подготовлены разными исследователями и могут нести следы
разного представления специалиста о значимости тех или
иных признаков для идентификации того или иного таксона.
Это надо учитывать при работе с ключем и критически подхо¬
дить к имеющимся в нем данным, проверяя полученные ре¬
зультаты, если это возможно, по другим руководствам, моно¬
графиям и статьям, и обязательно сопоставляя вывод с макси¬
мально полным диагнозом таксона.
Морфологические группы родов лишайников флоры Рос¬
сии.
Ключ 1. Кустистые (микрокустистые, нитевидные - фи-
ламентозные, трубчатые, бородовидно-повисающие и др.) ли¬
шайники. а) Фотобионт - зеленые водоросли, б) фотобионт -
цианобактерии.
Ключ 2. Листоватые (лопастные) лишайники, а) Фото¬
бионт - зеленые водоросли, б) фотобионт - цианобактерии.
Ключ 3. Лишайники с плакоидным диморфным талло¬
мом - накипным в центре накипной и лопастным на перифе¬
рии, но всегда приросшим к субстрату гифами сердцевины.
Ключ 4. Чешуйчатые лишайники, а) Фотобионт - зеле¬
ные водоросли, б) фотобионт - цианобактерии.
Ключ 5. Накипные лишайники с плодовыми телами - не
перитециями и не апотециями, либо плодовые тела отсутству¬
ют.
Ключ 6. Накипные лишайники с плодовыми телами -
перитециями.
Ключ 7. Накипные лишайники с плодовыми телами -
апотециями и фотобионтом - цианобактериями.
Ключ 8. Накипные лишайники с апотециями на ножках
и фотобионтом - зелеными водорослями.
Ключ 9. Накипные лишайники с вытянутыми или не¬
правильной формы апотециями и фотобионтом - зелеными
водорослями.
Ключ 10. Накипные лишайники с круглыми апотециями
и фотобионтом - Trentepohlia.
Ключ 11. Нелихенизированные грибы (без фотобионта)
с круглыми апотециями.
Ключ 12. Накипные лишайники с круглыми апотециями,
фотобионтом - зелеными хлорококковыми водорослями и
окрашенными спорами. Андреев
Ключ 13. Накипные лишайники с круглыми апотециями,
фотобионтом - зелеными хлорококковыми водорослями и
многочисленными (более 8 в сумке) бесцветными спорами.
Ключ 14. Накипные лишайники с круглыми апотециями,
фотобионтом - зелеными хлорококковыми водорослями и
бесцветными 2-клеточными, поперечно-многоклеточными или
муральными спорами (не более 8 в сумке).
Ключ 15. Накипные лишайники с круглыми апотециями,
фотобионтом - зелеными хлорококковыми водорослями и
бесцветными одноклеточными спорами (не более 8 в сумке).
Андреев
Ключ 16. Стерильные лишайники.
Синопсис (вспомогательный ключ для определения групп
родов).
1. Таллом кустистый (микрокустистый, нитевидный - фила-
ментозный) 2.
— Таллом не кустистый 3.
2. Фотобионт - зеленые водоросли Ключ 1а.
— Фотобионт - сине-зеленые водоросли Ключ 16.
3. Таллом листоватый (лопастной) 4.
— Таллом без лопастей, плакоидный, чешуйчатый или накип¬
ной 5.
4. Фотобионт - зеленые водоросли Ключ 2а.
— Фотобионт - сине-зеленые водоросли Ключ 26.
5. Таллом плакоидный (диморфный: в центре накипной, на
периферии - лопастной, но всегда приросший к субстра¬
ту гифами сердцевины) или чешуйчатый 6.
— Таллом накипной (вкл. лепрозный, эндосубстратный, без
фотобионта и др.) 8.
6. Таллом плакоидный Ключ 3.
— Таллом чешуйчатый 7.
7. Фотобионт - зеленые водоросли Ключ 4а.
— Фотобионт - сине-зеленые водоросли Ключ 46.
8. Плодовые тела - апотеции или перитеции 9.
— Плодовые тела - не апотеции и не перитеции или отсут¬
ствуют Ключ 5.
9. Плодовые тела - перитеции Ключ 6.
— Плодовые тела апотеции 10.
10. Фотобионт - сине-зеленые водоросли Ключ 7.
— Фотобионт - зеленые водоросли 11.
11. Апотеции не округлые - бороздчатые или на ножках .... 12.
— Апотеции округлые, сидячие или погруженные 13.
12. Апотеции стеблевидные (на ножках) Ключ 8.
— Апотеции бороздчатые Ключ 9.
13. Фотобионт - Trentepohlia или отсутствует 14.
— Фотобионт - зеленые хлорококковые водоросли 15.
14. Фотобионт - Trentepohlia Ключ 10.
— Фотобионт отсутствует Ключ 11.
15. Споры окрашенные Ключ 12.
— Споры бесцветные 16.
16. Спор больше чем по 8 в сумке Ключ 13.
— Спор не более чем по 8 в сумке 17.
17. Споры 2-клеточные, многоклеточные, муральные
Ключ 14.
— Споры одноклеточные Ключ 15.
Родовые ключи.
Ключ 1а.
Кустистые (микрокустистые, нитевидные - филаментоз-
ные, трубчатые, бородовидно-повисающие и др.) лишайники.
Фотобионт - зеленые водоросли. Автор: Урбанавичус Г. П.
1. Фотобионт сине-зеленые водоросли Ключ 16.
— Фотобионт зеленые водоросли 2.
2. Таллом желто-оранжевого, оранжевого цвета, К+ пурпурно-
фиол 3.
— Таллом иного цвета, если желтого, то от К не окрашивается
в пурпурно-фиол. цвет 6.
3. Лопасти с полостью внутри, более или менее округлые в
сечении (заметно на поперечном срезе)
Rusavskia subfruticulosa.
— Лопасти плоские, без внутренней полости 4.
4. Коровой слой параплектенхимный, без ризин, ресничек, ци-
лий и волосков Xanthoria candelaria.
— Коровой слой прозоплектенхимный, с волосками, реснич¬
ками, цилиями или резинами 5.
5. Реснички и ризины отсутствуют, гифы корового слоя тол¬
стые, 3-7 мкм, с толстыми стенками, 1-2 мкм, сильно
склеенные после обработки К, перегородка в споре уз¬
кая, 2-4 мкм Seirophora.
— Реснички и резины присутствуют, гифы корового слоя тон¬
кие, 1.5-3 мкм, с тонкими стенками, 0.5 мкм, рыхлые и
расходящиеся после обработки К, перегородка в споре
широкая, 4-10 мкм Teloschistes.
6. Таллом от ярко желтого до желтовато-коричневого, желто-
вато-зеленого или бледно-зеленоватого цвета 7.
— Таллом иной окраски 21.
7. Таллом с полостью внутри, бледно-зеленоватый, желтова¬
тый или желтовато-коричневый 8.
— Таллом без внутренней полости, окраска различная 10.
8. Таллом представлен довольно толстыми, от 2-3 до 10(—15)
мм толщ, пальцевидными выростами, обычно несколько
уплощенными, всегда без соредий и горизонтальных че-
шуй Dactylina.
— Таллом из более узких с гладкой поверхностью и сильно
разветвленных веточек, либо наряду с вертикальными
ветвями (подециями) развиты горизонтальные чешуйки в
основании таллома и на ветвях, соредиозный или без со¬
редий 9.
9. Таллом с горизонтальными чешуйками (в основании или на
подециях), ветви обычно округые в поперечном сечении,
апотеции (биаториновые) и пикнидии темноокрашенные
на концах ветвей или подециев, соредии если развивают¬
ся, то покрывают подеции более или менее равномерно
Cladonia.
— Таллом без чешуек, ветви округлые или уплощенные, апо¬
теции леканориновые, светлоокрашенные, боковые или
конечные, соредии сгруппированы в соралии или при¬
урочены к шлемовидным расширениям концов лопастей
Ramalina.
10(7). Таллом из уплощенных лопастей 11.
— Таллом из нитевидных или округлых в сечении вет¬
вей/лопастей, или ветви/лопасти неровные в сечении, уг¬
ловато-округлые 15.
11. Сердцевина ярко желтая, содержит вульпиновую кислоту .
Vulpicida.
— Сердцевина не ярко желтая, беловатая, бледно-желтоватая,
без вульпиновой кислоты 12.
12. Таллом зеленовато-серый, без псевдоцифелл, соредиозный,
более или менее мягкий, содержит эверновую кислоту,
на стволах и ветвях деревьев в лесной зоне, преимуще¬
ственно в широколиственных и хвойно¬
широколиственных лесах Evernia prunastri.
— Таллом с иным набором признаков 13.
13. Таллом в виде длинных, до 20-50 см длин, неравномерно
разветвленных лентовидных лопастей с сильно неровной
поверхностью, на почве, скалах в арктических и высоко¬
горных районах, редко на основании стволов деревьев в
субарктических и субальпийских условиях, содержит
алектороновую и скваматовую кислоты
Alectoria sarmentosa subsp. vexillifera.
— Таллом с иными признаками, обычно меньших размеров,
если лопасти длинные, то произрастает на деревьях в ле¬
сах умеренной зоны, с другим набором лишайниковых
веществ 14.
14. Лопасти более или менее желобчато свернутые, произрас¬
тают на почве обычно в тундровых или альпийских
условиях, северо-таежных лесах, апотеции темно-
коричневые, споры простые Flavocetraria.
— Обычно произрастает на деревьях, редко на скалах, если на
почве, то тогда лопасти не желобчатые, апотеции свет¬
лые, споры 2-клеточные Ramalina.
15(10). Ветви неровные в сечении, угловато-округлые 16.
— Ветви нитевидные и/или округлые в сечении 19.
16. Сердцевина без просветов и полости, местами компактная,
концы лопастей заостренные, обычно на стволах и вет¬
вях деревьев, часть видов на почве 17.
— Сердцевина паутинистая, часто с просветами, кончики ло¬
пастей тупые, округлые, исключительно на почве в арк¬
тических и высокогорных районах 18.
17. Таллом ярко зеленовато-желтый (преобладает желтая
окраска), содержит вульпиновую кислоту
Letharia vulpina.
— Таллом светло желтовато-зеленый (преобладает зеленова¬
тая окраска), содержит усниновую и диварикатовую кис¬
лоты, без вульпиновой кислоты Evernia.
18. Таллом бледно желтовато-зеленоватый, без коричневого
оттенка, содержит протолихестериновую кислоту
Allocetraria madreporiformis.
— Таллом желтовато-коричневатый, с сизым налетом, не со¬
держит протолихестериновой кислоты Dactylina ramulosa.
19. Ветви с центральным тяжем, преимущественно без псев-
доцифелл (если псевдоцифеллы образуются, то мелкие,
слабо выраженные, в виде пятен), зрелые споры бес¬
цветные, таллом без алектороновой кислоты Usnea.
— Ветви без центрального тяжа, с псевдоцифеллами или без
них 20.
20. Псевдоцифеллы всегда развитые, хорошо заметные, вы¬
пуклые, апотеции коричневые, споры одноклеточные,
зрелые коричневые, таллом содержит алектороновую
кислоту в качестве главного вещества, иногда сквамато-
вую, барбатовую и тамноловую кислоты Alectoria.
— Псевдоцифеллы вогнутые, плоские или отсутствуют, апо¬
теции светлые, споры всегда бесцветные, двуклеточные,
состав лишайниковых веществ различный, но без выше¬
названных кислот Ramalina.
21(6). Таллом беловато-серый, розоватый, зеленовато-серый
или с серым оттенком 22.
— Таллом темноокрашенный, серо-коричневый, зеленовато¬
коричневый, коричневый, темно-коричневый, до черного
44.
22. Лопасти плоские, дорсивентральные 23.
— Лопасти округлые или угловато-округлые в сечении, но не
плоские 30.
23. Нижняя поверхность без корового слоя 24.
— Нижняя поверхность с коровым слоем 26.
24. Таллом образован псевдоподециями - более или менее
твердыми округлыми стволиками в основании, с вверх
расширяющими листовидными филлокладиями, прочно
прикрепляется к каменистому субстрату, обычно с цефа-
лодиями Stereocaulon.
— Таллом иного строения 25.
25. Таллом при действии К желтеет (атранорин) Heterodermia.
— Таллом при действии К не изменяется в окраске (атранорин
отсутствует) Anaptychia.
26. Таллом мягкий, слабо дорсивентральный, нижняя сторона
лопастей сероватая или беловатая, содержит эверновую
кислоту, без ризин Evernia prunastri.
— Таллом жесткий, ясно дорсивентральный (хотя бы у осно¬
вания), без эверновой кислоты 27.
27. Нижняя сторона лопастей светлая, беловатая, сероватая,
ветви заметно уплощены дорсивентрально у основания,
ближе к концам вальковато-округлые, на концах с апо¬
тециями, приуроченными к нижней стороне, с мазедием,
споры округлые, 1-клеточные, с изидеоподобными вы¬
ростами коротких веточек Bunodophoron.
— Таллом с иными характеристиками, нижняя сторона лопа¬
стей темно окрашенная, коричневая или фиолетовая 28.
28. Лопасти с краевыми цилиями и редкими короткими ризи¬
нами на нижней стороне лопастей
Everniastrum cirrhatum.
— Лопасти без краевых цилий и ризин 29.
29. Лопасти выпуклые, с завернутыми вниз краями, изидии
поверхностные; таллом прикрепляется к субстрату при
помощи псевдогомфа Pseudevernia furfuracea.
— Лопасти сильно желобчато вогнутые, изидии краевые; тал¬
лом прикрепляется к субстрату редкими черными ризи¬
нами, расположенными в базальной части
Platismatia herrei.
30(22). Таллом с полостью внутри 31.
— Таллом без внутренней полости 35.
32. Таллом простой, образован из неразветвленных или слабо
разветвленных, шиловидно заостренных, прямых или
изогнутых в разные стороны веточек мелово-белого цве¬
та, без соредий и чешуек, апотеции и пикнидии не разви¬
ваются Thamnolia.
— Развивается первичный таллом в виде корочки или чешуек
и вторичный в виде подеций 33.
33. Таллом из высоких, до 10-15 см выс, подециев, густо или
слабо разветвленных, без корового слоя, либо слабораз-
ветвленных, с коровым слоем и чешуйками, с соредиями
или без, прорастающих из первичного таллома, состоя¬
щего из горизонтальных чешуй Cladonia.
— Таллом их невысоких, до 0.5-1.5 см выс, пальцевидных
подециев, произрастающих из первичного таллома в ви¬
де корочки, без соредиев и чешуек 34.
34. Подеции низкие, до 1.5-2 мм выс, продольно сморщен¬
ные, не разветвленные, густо разрастающиеся в форме
полусферических подушечек, на концах подециев апоте¬
ции с черным мазедием, на тонких веточках сосны или
ели в открытых местообитаниях
Tholurna.
— Подеции до 0.5-1.5 см выс, гладкие, иногда слабо разветв¬
ленные, на кислых торфянистых или песчаных почвах,
редко на старых пнях или корнях деревьев, апотеции
темно-коричневые, без мазедия, развиваются редко
Pycnothelia.
35(30). Произрастают на стволах и ветвях деревьях, редко на
замшелых скалах 36.
— Произрастают на почве, замшелых скалах или каменистом
субстрате 38.
36. Таллом более или менее жесткий, не повисающий, распро¬
стертый, поникающий или торчащий, покрыт мелкими
волосками (войлочком), часто с блюдцевидными апоте¬
циями, с коричневым диском, споры коричневые, 2-
клеточные Tornabea scutellifera.
— Таллом обычно повисающий, мягкий, с соралями или без
них, без волосков, споры простые, бесцветные 37.
37. Таллом с центральным тяжем/цилиндром, кора часто про¬
дольно морщинистая, сетчато-складчатая, сердцевина
местами с желтым пигментом (содержит вульпиновую
кислоту) Lethariella togashii.
— Таллом без центрального тяжа, кора гладкая, сердцевина
без желтого пигмента (не содержит вульпиновой кисло¬
ты) Bryoria.
38. Кроме подециев или псевдоподециев иногда развивается
первичный накипной или чешуйчатый таллом; псевдо-
подеции с филлокладиями, подеции с ареалированным
или бугорчатым коровым слоем, всегда с цефалодиями
39.
— Таллом из ровных, гладких или с продольными морщинка¬
ми веточек, без каких-либо филлокладийных выростов и
цефалодий 41.
39. Апотеции и цефалодии всегда отсутствуют, псевдоподеции
очень мелкие и нежные, более или менее лепрозные,
первичный таллом напоминает таллом видов рода
Lepraria Leprocaulon.
— Апотеции развиваются часто, цефалодии имеются почти
всегда, подеции и псевдоподеции крупные, если мелкие,
то не такие нежные и не лепрозные, первичный таллом
накипной или чешуйчатый, не лепрозный 40.
40. Подеции простые или наверху вильчато или зонтиковидно
разветвленные, со сплошным, иногда прерывистым, аре-
олированным или бугорчатым коровым слоем, без фил-
локладий, апотеции шаровидные, полушаровидные или
вытянутые Pilophorus.
— Псевдоподеции сильно разветвленные, редко слабо ветвя¬
щиеся или простые, голые или покрыты войлочком, ред¬
ко со сплошным коровым слоем, всегда с филлокладия¬
ми или филлокладийными веточками, апотеции не ша¬
ровидные Stereocaulon.
41. Таллом с соредиями, от С краснеет, прикрепляется к суб¬
страту площадочкой прикрепления (базальной пластин¬
кой) с охристо-желтыми гифам, фотобионт трентеполиа,
произрастает на прибрежных скалах на Черноморском
побережье Кавказа Roccella phycopsis.
— Таллом без соредий, от С не краснеет, фотобионт не трен¬
теполиа, произрастает высоко в горах или в Арктике 42.
42. Таллом с выступающими над поверхностью ветвей свет¬
лыми, вытянутыми псевдоцифеллами, концы веточек
нитевидно вытянутые и немного завернутые, кора и
сердцевина от КС краснеют
Gowardia nigricans (светлые формы).
— Таллом без псевдоцифелл, концы веточек нитевидно не вы¬
тянутые, кора и сердцевина от КС не краснеют 43.
43. Таллом меловато-белый, матовый, не или слабо разветв¬
ленный, поверхность ветвей с продольными морщинка¬
ми, развитые апотеции не известны Siphula ceratites.
— Таллом сильно коралловидно разветвленный, поверхность
ветвей обычно блестящая, гладкая, на концах веточек
развиваются шаровидные апотеции, при созревании рас¬
крывающиеся с черным мазедием Sphaerophorus.
44(21). Таллом из тонких, черных, горизонтально распростер¬
тых нитевидных веточек, менее 20 мкм толщ, образую¬
щих рыхлую или более или менее плотную дернинку,
фотобионт трентеполиоидный, обитают на каменистом
субстрате в затененных условиях под нависающими ска¬
лами 45.
— Таллом из более толстых ветвей, свыше 100 мкм толщ, фо¬
тобионт не трентеполиоидный, местообитания различ¬
ные 46.
45. Гифы мелко-волнистые, неравномерно окутывают нити
водоросли, поверхность веточек неровная, узловатая
Cystocoleus.
— Гифа ровные, параллельно ориентированы, веточки гладкие
Racodium.
46. Лопасти округлые, угловато-округлые в сечении 47.
— Лопасти плоские и/или дорсивентральные 63.
47. Таллом с внутренней полостью Cladonia.
— Таллом без внутренней полости 48.
48. Таллом эпилитный, начинает развиваться от накипного,
затем карликово-кустистый, веточки до 10(—15) мм выс,
булавовидные, простые или 1 —3(— 10)-разветвленые, тес¬
но собранные в подушечки, на концах веточек псев¬
доцифеллы или апотеции, покрытые беловатым налетом
Aspicilia transbaicalica.
— Таллом с иными характеристиками 49.
49. Таллом свободноживущий, не прикрепляется, на почве в
аридных регионах 50.
— Таллом не свободноживущий, либо крпко прикрепляется к
субстрату, либо основания ветвей постепенно отмираю¬
щие 52.
50. Таллом в виде плотных шаровидных образований, 1—2(—
3.5) см шир, из коротких, толстых веточек, расходящих¬
ся во всех направлениях от общего центра, на концах
обычно утолщенные, с белыми точковидными углубле¬
ниями - псевдоцифеллами Aspicilia fruticulosa.
— Таллом не образует плотных шаровидных комочков, тонкие
раздельные веточки хорошо выражены, на концах утон¬
чающиеся 51.
51. Веточки широкие, до 2-4 мм диам, в основании сплюсну¬
тые, иногда ямчатые, псевдоцифеллы вытянутые, обра¬
зуются редко Cetraria steppae.
— Веточки узкие, 0.5-1 (-1.3) мм диам, округлые, не ямчатые,
псевдоцифеллы обычно правильно округлые, всегда хо¬
рошо заметные, белые, резко контрастирующие с тем¬
ным оливково-коричневым, буровато-серым цветом тал¬
лома Circinaria hispida.
52. Таллом из горизонтально распростертых, достаточно плот¬
но прирастающих к каменистому субстрату подушечек,
ветви многократно дихотомически разветвленные, сте¬
лющиеся, прикрепляются к субстрату многочисленными
гаптерами Pseudephebe
[у P. minuscula лопасти в центре уплощаются].
— Таллом с иными характеристиками 53.
53. Таллом сильно коралловидно разветвленный, обычно
жесткий, поверхность ветвей блестящая, гладкая, без
псевдоцифелл, на концах веточек развиваются шаровид¬
ные апотеции, при созревании раскрывающиеся с чер¬
ным мазедием Sphaerophorus.
— Таллом с иными характеристиками 54.
54. Таллом с центральным тяжем/цилиндром, светло-
коричневый с желтоватым оттенком или серый, лопасти
вальковатые, угловато-цилиндрические, местами сплюс¬
нутые, кора часто продольно морщинистая, сетчато¬
складчатая, без псевдоцифелл, сердцевина местами с
желтым пигментом (содержит вульпиновую кислоту)
Lethariella togashii.
— Таллом без центрального тяжа, лопасти с гладкой корой,
без вульпиновой кислоты (кроме Bryoria fremontii, сора-
ли и апотеции которой содержат вульпиновую кислоту)
55.
55. Псевдоцифеллы выпуклые, приподнятые над поверхно¬
стью таллома 56.
— Псевдоцифеллы погруженные, плоские или отсутствуют 57.
56. Таллом красно-коричневый, Р-, содержит оливеторовую
кислоту Bryocaulon divergens.
— Таллом черный, черно-коричневый, иногда светло корич¬
невато-серый, темно-серый, Р+ желт, содержит алекто-
риаловую кислоту Gowardia.
57. На почве, замшелых скалах 58.
— На стволах и ветвях деревьев, редко на замшелых скалах ...
60.
58. Веточки без псевдоцифелл, часто с тонким, паутинистым
войлочком, и длинными простыми или слабо разветлен-
ными фибриллами Anaptychia101.
— Веточки обычно с псевдоцифеллами, гладике, без войлочка
и фибрилл 59.
59. Таллом обычно невысокий, до 2-4 см выс, веточки тол¬
стые, 0.5-2 мм толщ, в основании и в местах ветвления
немного сплюснутые; псевдоцифеллы белые, ямчатые,
глубоко погруженные, или плоские, но расположенные
ниже уровня поверхности таллома; таллом не содержит
фумарпротоцетраровой кислоты Cetraria.
— Ветви таллома более 4-5 см длин, тонкие, 0.2-0.8 мм
толщ, постоянно округлые в поперечном сечении; псев¬
доцифеллы темные, обычно плоские, на поверхности
таллома; таллом содержит фумарпротоцетраровую кис¬
лоту Bryoria.
60. Сердцевина полая, споры коричневые, муральные, соредии
и изидии отсутствуют, апотеции обычны, редкий во¬
сточноазиатский вид, известен пока только с Южных
Курильских островов Oropogon asiaticus.
— Сердцевина заполненная 61.
61. Ветви таллома в сечении часто неправильно-округлые, уг¬
ловатые, иногда почти желобчатые, покрыты мелкими
волосками (войлочком), с блистидиями или без них, без
соредий и изидий, часто с блюдцевидными апотециями,
с коричневым диском, по краю с гиалиновыми волоска¬
ми, споры коричневые, 2-клеточные
Tornabea scutellifera.
— Ветви нитевидные, голые, обычно правильно округлые в
сечении, апотеции образуются редко, споры простые,
бесцветные, таллом часто с соредиями или изидиями
62.
62. Кора состоит из одного слоя, псевдоцифеллы (если при¬
сутствуют) вытянутые; таллом не содержит оливеторо-
вой кислоты, широко распространенные лесные виды ....
Bryoria.
— Кора двухслойная, внешний слой тонкий, параплектенхим¬
ный, внутренний слой толстый, прозоплектенхимный,
псевдоцифеллы точковидные; таллом содержит оливето-
101 Редкие формы, встречающиеся в горах или тундрах. Веточки из дорсивен¬
тральных часто превращаются в округые, с плотно сомкнувшимися снизу края¬
ми узких лопастей.
ровую кислоту, редкий восточноазиатско-
западносевероамериканский вид, известный в России с
юга Дальнего Востока Bryocaulon pseudosatoanum.
63(46). Таллом формирует компактный кустовидный пучок, до
2 см выс, плотно прирастающий к каменистому суб¬
страту широкой площадкой, плоские лопасти без диффе¬
ренциации верхней и нижней сторон (билатеральные)
Cornicularia normoerica.
— Таллом с иными характеристиками 64.
64. Таллом из тонких лентовидных ясно дорсивентральных
лопастей, нижняя сторона частично без коры, обычно не
прикрепляется к субстрату
Masonhalea richardsonii.
— Таллом с иными характеристиками 65.
65. Конидии почти бутылковидные; таллом содержит гирофо-
ровую, хиасциновую или алектороновую кислоты
Cetrariella.
— Конидии палочковидные, веретеновидные до удлиненно¬
лимоновидных; таллом содержит иные кислоты 66.
66. Верхний и нижний коровые слои обычно двухслойные, 20-
130 мкм толщ, таллом содержит среди прочих фумар-
протоцетраровую и норстиктовую кислоты
Cetraria.
— Верхний и нижний коровые слои однослойные, 20-50 мкм
толщ, таллом не содержит фумарпротоцетраровую и
норстиктовую кислоты 67.
67. Произрастает на почве, содержит норрангиформовую кис¬
лоту Arctocetraria.
— Произрастает преимущественно на ветвях деревьев или ку¬
старников, кустарничков, не содержит норрангиформо-
вой кислоты
Tuckermannopsis. / Masonhalea inermis.
Ключ 16.
Кустистые (микрокустистые, нитевидные - филаментоз-
ные, трубчатые, бородовидно-повисающие и др.) лишайники.
Фотобионт-цианобактерии. Автор Макрый Т. В.
Талломы нитевидные, мелко- и микрокустистые (веточ¬
ки тонкие, длина значительно превышает толщину), карлико-
во-кустистые (веточки толстые, длина в 1.2-3(5) раз превыша¬
ет толщину, иногда с лопастинками, ширина которых превы¬
шает длину), субкустистые (веточки или лобули вырастают из
чешуек), псевдокустистые (многочисленные вертикальные ве¬
точки или лобули вырастают из горизонтальных лопастей, ко¬
торые обычно не видны или отмирают); образуют кустики,
коврики, подушечки, розетки с радиально ориентированными
веточками или беспорядочно распластаны по субстрату.
1. Таллом нитевидный, мелко- или микрокустистый, субкусти-
стый; веточки тонкие, длина значительно превышает
толщину 2.
— Таллом карликово-кустистый, карликовый суб- и псевдоку-
стистый; веточки, лобули, лопастинки толстые и корот¬
кие, длина в 1.2-3(5) раза превышает толщину или не
превышает ее 13.
2. Таллом нежелатинозный, отчетливо гетеромерный с парап-
лектенхимным коровым слоем и плотным центральным
тяжем, серый, коричневый, 1-2 см выс, фотобионт Nos¬
toc Dendriscocaulon (Lobariaceae).
— Таллом другой, более или менее желатинозный 3.
3. Морские (обитающие на литорали), образующие коврики,
мелко-кустистые лишайники; апотеции шаровидные пе¬
ритециевидные терминальные (на концах веточек); фо¬
тобионт Calothrix Lichina (Lichinaceae).
— Не морские лишайники 4.
4. Фотобионт Nostoc в длинных или коротких цепочках, тал¬
лом с параплектенхимным коровым слоем 5.
— Фотобионт Scytonema, Stigonema или хроококковый 8.
5. Таллом с коровым слоем, нитевидный, слабо разветвлен¬
ный, распластанный, с бороздчатыми веточками; апоте¬
ции леканоровые, редкие; споры муральные; на извест¬
няковых скалах Leptogium (Collemataceae).
— Признаки другие 6.
6. Таллом без корового слоя, образующий коврики или рас¬
пластанные кустики, с цилиндрическими, разветвленны¬
ми, прижатыми, перепутанными лопастями; плодовые
тела - пикноаскокарпы, редкие, ламинальные, шаровид¬
ные до полупогруженных; споры одноклеточные
Lempholemma (Lichinaceae).
— Таллом с параплектенхимным коровым слоем 7.
7. Таллом мелко-кустистый (до 1 см выс.), образующий по¬
душковидные дернинки, с цилиндрическими, местами
плоскими или ямчатыми, дихотомически-
разветвленными (на концах часто дланевидными) веточ¬
ками; апотеции с собственным краем, споры 2-
клеточные; на замшелых камнях, скалах, почве
Polychidium (Massalongiaceae).
— Таллом субкустистый, с лопастинками до 0.2 мм толщ, 1
мм дл.; апотеции биаторовые с исчезающим краем; спо¬
ры поперечно-многоклеточные с 7-10 перегородками; на
мхах, почве, аркто-альпийский Arctomia (Arctomiaceae).
8. Фотобионт хроококовый; таллом микрокустистый, из пря¬
мостоячих коралловидных веточек 20-150 мкм толщ, до
3 мм выс, образующий отдельные кустики или ареоли-
рованную корочку до 2 см диам.; плодовые тела - талли-
нокарпы на концах веточек, шаровидные, с открытым
диском; эпитеций с отдельными клетками фотобионта;
споры одноклеточные по 16-32 в сумке
Lichinella s. str. (Lichinaceae).
— Фотобионт Scytonema или Stigonema 9.
9. Фотобионт Stigonema (с настоящим ветвлением) 10.
— Фотобионт Scytonema (с ложным ветвлением), сем. Lichina¬
ceae 11.
10. Таллом с ризоидами, микрокустистый (из вертикальных
или поникающих нитевидных разветвленных веточек, до
60 мкм толщ, 6 мм выс.), образующий дернинки; апоте¬
ции биаторовые, черно-коричневые
Spilonema (Coccocarpiaceae).
— Таллом без ризоидов, мелко-кустистый распростертый (из
сильно разветвленных нитевидных веточек, 20-200 мкм
в диам.), образующий коврики или розетки, 2-30 мм в
диам.; плодоношения - пикноаскокарпы, черно¬
коричневые, с собственным краем и точковидным до
плоского диском Ephebe (Lichinaceae).
11. Нити таллома очень тонкие (до 12 мкм толщ.), простые или
слабо разветвленные, с недифференцированными гифа¬
ми (окружающими нить цианобактерии, пронизывая ее
слизистое влагалище), образуют буроватые войлочные
подушечки; плодовые тела - пикноаскокарпы, краснова-
то-коричневатые, с собственным краем Thermutis.
— Нити таллома толще 12.
12. Эпилит; таллом розетковидный, до 0.5-2 см в диам, из ра¬
диально простирающихся нитей (80-100 мкм диам. у ос¬
нования, 10-20 мкм на концах), с центральным тяжем из
гиф с круглыми клетками, без ризин; плодовые тела -
пикноаскокарпы, красновато-коричневатые, со слоевищ¬
ным краем Zahlbrucknerella.
— Эпифит (на веточках ели), на мхах и лишайниках (на видах
Parmelia); нити таллома до 2 мм дл, полупрозрачные,
состоящие из нитей цианобактерии, окруженных гифами
гриба с круглыми клетками, образуют кустики и розетки;
апотеции желатинозные, без эксципула Lichinodium.
13. Фотобионт Nostoc 14.
— Фотобионт хроококковый; споры одноклеточные, сем. Li¬
chinaceae 17.
14. Таллом с коровым слоем; лопасти вальковатые, приподни¬
мающиеся, коричневые, глянцевые, сильно морщини¬
стые, образующие отдельные пучки, розетки или поду¬
шечки; апотеции леканоровые, очень редкие; споры му¬
ральные; на карбонатной почве, скалах, стенках
Leptogium (Collemataceae).
— Таллом без корового слоя 15.
15. Апотеции с собственным краем, обычны; споры однокле¬
точные; таллом с вертикальными лобулями ок. 1 мм дл,
1.5 мм шир.; на карбонатной почве и мхах, аркто-
альпийский Leciophysma (Collemataceae).
— Признаки другие 16.
16. Апотеции леканоровые, часто отсутствуют; споры (2)4-
клеточные или субмуральные; таллом из кластеров вер¬
тикальных (суб)цилиндрических, иногда складчатых ло¬
пастей, отходящих от первичных горизонтальных лопа¬
стей, обычно невидимых или разрушенных, либо не¬
определенной формы с неровными веточками; на карбо¬
натной почве, известняках и доломитах
Collema (Collemataceae).
— Плодовые тела - пикноаскокарпы, редкие, шаровидные на
концах веточек, с более или менее открытым диском;
споры одноклеточные; таллом черный или оливково¬
коричневый, с вальковатыми прижатыми или приподни¬
мающимися лопастями, образующий подушечки; на кар¬
бонатных скалах Lempholemma (Lichinaceae).
17. Плодовые тела - таллинокарпы, погруженные, на концах
лопастей; диск открытый, черный, эпигимений с клетка¬
ми фотобионта; гимений I + сине-зеленеет, затем корич-
нево-краснеет; сумки с 16-32 спорами; таллом с рыхлым
центральным тяжем и отходящими вееровидно гифами,
сетчато-переплетенными у поверхности; на карбонатных
скалах в ксеротических условиях
Gonohymenia s.str. (Lichinella s. 1.)
— Плодовые тела - апотеции 18.
18. Апотеции на концах лопастей, с открытым черным диском,
обычны; гимений с красно-коричневыми участками в
верхней части, / + синеет; сумки с 8 спорами; конидии
игловидные, нитевидные или изогнутые; таллом черный,
часто с налетом, с центральным тяжем из плотно пере¬
плетенных гиф и отходящими от него периклинально
гифами, сетчато-переплетенными на периферии; на поч¬
ве и в расщелинах карбонатных скал Peccania.
— Апотеции погруженные в расширенные шаровидные концы
лопастей, с толстым слоевищным краем и точковидным
или расширенным темно-красно-коричневым диском;
гимений /-; сумки с 8-32 спорами; конидии эллипсоид¬
ные или палочковидные; таллом темно-красно-
коричневый, из сетчато-переплетенных гиф; преимуще¬
ственно на карбонатных скалах Synalissa.
Ключ 2а.
Листоватые (лопастные) лишайники. Фотобионт - зеле¬
ные водоросли. Автор: Урбанавичус Г. П.
1. Фотобионт сине-зеленые водоросли Ключ 26.
— Фотобионт зеленые водоросли 2.
2. Таллом прикрепляется к субстрату при помощи гомфа 3.
— Таллом прикрепляется при помощи других структур, либо
свободноживущий 9.
3. Плодовые тела перитеции, погруженные в таллом
Dermatocarpon.
— Плодовые тела апотеции или отсутствуют 4.
4. Таллом темноокрашенный, темно-серый, коричневый, чер¬
новатый, редко беловатый (эпинекральный слой), без
желтовато-зеленоватой окраски 5.
— Таллом желтовато-зеленоватой, желтовато-буроватой
окраски 6.
5. Верхняя поверхность таллома с пустулами, более или менее
гладкая Lasallia.
— Верхняя поверхность таллома без пустул, от гладкой до не¬
ровной, иногда складчатой Umbilicaria.
6. Апотеции погруженные или слабо выступащие над поверх¬
ностью таллома 7.
— Апотеции сидячие, с зауженным основанием 8.
7. Апотеции сложные, состоят из многочисленных мелких
апотециев, каждый мелкий апотеций обособлен самосто¬
ятельным эксципулом, поверхность таллома разбита
глубокими трещинами на множество неправильной фор¬
мы ареол или чешуек, с густым голубовато-беловатым
налетом, споры по 50-100 в сумке Glypholecia.
— Апотеции не сложные, с черно-коричневым диском, покры¬
тым налетом, поверхность таллома с извилистыми ради-
альным складками, желтовато-оливковая, буровато¬
желтоватая, без налета, споры по 3-4(-8) в сумке
Aspicilia oxneriana.
8. Таллом желтовато-коричневый, С+ оранж. (содержит ксан-
тоны), обычно полностью покрыт апотециями
Arctopeltis.
— Таллом зеленовато-желтый, коричневатый, беловатый, С—
или С+ (не содержит ксантоны), апотеции обычно не
полностью покрывают таллом Rhizoplaca.
9(2). Таллом гомеомерный, желатинозный, полупрозрачный,
водоросль Prasiola преобладает на микобионтом, плодо¬
вое тело - перитеции, поселяется в приморских место¬
обитаниях, в местах, забразгиваемых водой и посещае¬
мых птицами (галофил, нитрофил)
Mastodia tessellata.
— Таллом с иными характеристиками 10.
10. Таллом ярко-желтый или оранжевый 11.
— Таллом иной окраски 16.
11. Таллом от К окрашивается в пурпурный цвет 12.
— Таллом от К не окрашивается в пурпурный цвет 15.
12. Таллом нижней поверхностью плотно прирастает к суб¬
страту Fulgensia.
— Таллом не всей нижней поверхностью прирастает к суб¬
страту, с гаптерами или ризинами 13.
13. Лопасти округло-вальковатые или уплощенные, с внутрен¬
ней полостью Rusavskia.
— Лопасти плоские, без полости внутри 14.
14. Без ризин на нижней поверхности лопастей, с гаптерами,
конидии широкоэллипсоидные Xanthoria.
— С ризинами или без на нижней стороне лопастей, без гап-
тер, конидии узкоэллипсоидные или палочковидные
(сложные роды, их разделение не общепризнано)
Oxneria / Xanthomendoza.
15. Таллом мелкий, до 1 см шир, лопасти узкие, 0.1-0.6 мм
шир, не содержит вульпиновую кислоту Candelaria.
— Таллом обычно свыше 1 см шир, лопасти широкие, 1 —3(—5)
мм шир, содержит вульпиновую кислоту Vulpicida.
16(10). В окраске таллома преобладают бледно-желтоватые
цвета, беловато-желтоватые, желтовато-зеленоватые, зе-
леновато-желтоватые, коричневато-желтоватые 17.
— Таллом без желтоватых оттенков 31.
17. Нижняя поверхность таллома голая, без ризин, волосков и
войлочка 18.
— Таллом с ризинами, волосками или войлочком
19.
18. Лопасти широкие, до 2-3 см шир, неровно-ямчатые, ниж¬
няя поверхность темно-коричневая, блестящая
Asahinea chrysantha.
— Лопасти узкие, лентовидные, узко-рассеченные, либо че¬
шуйчатые, часто с вертикальными выростами - подеци-
ями, нижняя поверхность без корового слоя, обычно бе¬
ловатая Cladonia.
19. Таллом с цефалодиями, содержащими сине-зеленый фото¬
бионт Nostoc, нижняя поверхность с черным войлочком
в центральной части Nephroma arcticum.
— Нижняя поверхность без войлочка, таллом не содержит це-
фалодий 20.
20. Таллом в центральной части чешуйчатый, плотно прикреп¬
ляющийся к субстрату при помощи ризогиф или ризои-
дальных тяжей, по краю отчетливо лопастной, нижняя
поверхность без корового слоя Squamarina.
— Таллом не чешуйчатый в центральной части, с настоящими
ризинами, с нижним коровые слоем 21.
21. Лопасти с псевдоцифеллами 22.
— Лопасти без псевдоцифелл 23.
22. Псевдоцифеллы на верхней и нижней поверхности лопа¬
стей Cetreliopsis asahinae.
— Псевдоцифеллы и апотеции на верхней поверхности лопа¬
стей, таллом с соредиями
Flavopunctelia102.
— Псевдоцифеллы на нижней поверхности лопастей, апоте¬
ции развиваются на краю нижней стороны лопастей,
таллом без соредий (кроме N. laureri) Nephromopsis.
23. Ризины густо дихотомически разветвленные
Hypotrachyna sinuosa.
— Ризины простые или слабо разветвленные 24.
24. Нижняя поверхность с очень мелкими бугорочками
(«шершавая»), со слабым или сильным беловатым или
сизым налетом Arctoparmelia.
— Нижняя поверхность без таких бугорочков (не «шерша¬
вая»), без налета 25.
25. Лопасти завернутые в трубочку, таллом свободно лежащий
на земле Xanthoparmelia.
— Лопасти плоские, узкие, менее 1-1.5 мм шир, с соредиями,
плотно прижаты к субстрату 26.
— Лопасти плоские, более 1 мм шир 27.
26. На коре и древесине, сердцевина К-, Р-
102
У Flavopunctelia soredica псевдоцифеллы часто плохо развиты.
Parmeliopsis ambigua.
— На каменистом субстрате, сердцевина K+ желт, Р+ оранж.
(в России не известен, но может быть обнаружен в Бал¬
тийском регионе) Xanthoparmelia mougeotii.
27. Таллом с соредиями 28.
— Таллом без соредий 30.
28. Лопасти 5-15 мм шир, соредии поверхностные, верхний
коровый слой прозоплектенхимный Flavoparmelia.
— Лопасти 1-4 мм шир, соредии краевые, верхний и нижний
коровый слой параплектенхимный 29.
29. Лопасти плоские, с беловато-сизоватым налетом и с крае¬
выми лобулями, на голых и замшелых скалах
“Cetraria” annae.
— Лопасти вогнутые, желобчатые, без лобулей и без налета,
на стволах деревьев
Usnocetraria oakesiana.
30. Пикнидии краевые, конидии 12-14 мкм дл, клеточные
стенки содержат Cetraria-тип лихенана
Allocetraria stracheyi.
— Пикнидии поверхностные, конидии 5-8 мкм дл, клеточные
стенки содержат Xanthoparmelia-тип лихенана
Xanthoparmelia.
31(16). В окраске таллома преобладают беловато-серый, се¬
рый, темно-серый, зеленовато-серый цвета 32.
— Таллом коричневый, оливково-коричневый, зеленовато¬
коричневый, серовато— или черновато-коричневый
71.
32. Таллом в виде широкой, торчащей лопастевид-
ной(вееровидной) пластины, прочно прикрепляется к
субстрату узкой площадкой, верхняя и нижняя поверх¬
ность сильно ребристые, нижняя с белыми удлиненными
псевдоцифеллами Ramalina asahinana.
— Таллом с переходом от накипного к листоватому, 1-3 см
шир, 0.5-1.2 мм толщ, твердый, морщинистый, светло¬
буроватый, рыжевато-серый, нижняя поверхность более
темная, морщинистая, с многочисленными толстыми
корнеподобными разветвленными выростами, апотеции
погруженные в таллом, споры коричневые, муральные,
на силикатных горных породах, юг Дальнего Востока ....
Leptotrema lithophila.
— Таллом с иными характеристиками 33.
33. Лопасти с ресничками (цилиями, фибриллами) по краю 34.
— Лопасти без ресничек по краю 41.
34. Нижняя поверхность лопастей темноокрашенная, черная,
кориневато-черноватая 35.
— Нижняя поверхность лопастей светлая, беловатая, светло¬
коричневатая, иногда с желтовато-оранжеватыми пятна¬
ми или более темная в центральной части 40.
35. Таллом кустовидный, из узких, сильно переплетенных ло¬
пастей, лопасти правильно дихотомически разветвлен¬
ные, часто желобчато завернутые вниз, по краю с длин¬
ными, хорошо заметными, черными цилиями и ризинами
Everniastrum.
— Таллом не кустовидный, обычно прирастает к субстрату
нижней поверхностью, более или менее розетковидный,
ризины фомируются не по краю лопастей 36.
36. Лопасти широкие, обычно около 1 см шир. или шире, края
лопастей восходящие, нижняя сторона лопастей с широ¬
кой голой зоной, без ризин по краю Parmotrema.
— Лопасти узкие, до 2-6 мм шир, редко шире, края прижаты
или распростерты по субстрату, ризины развиты до са¬
мого края лопастей 37.
37. Сердцевина с желтым пигментом Myelochroa.
— Сердцевина без желтого пигмента 38.
38. Таллом маленький, до 1 см шир, не образует розеток, ло¬
пасти менее 1 мм шир.
Myelochroa sayanensis.
— Таллом обычно свыше 2 см шир, более или менее розетко¬
видный, лопасти более 2 мм шир 39.
39. Лопасти по краю округлые, округло-городчатые, цилии
приурочены к пазухам между лопастей, ризины простые,
верхняя поверхность таллома часто пятнистая (с маку-
лами), без соредий Parmelina.
— Лопасти по краю не округлые, концы усеченные («срезан¬
ные»), цилии обычны также на концах лопастей, ризины
преимущественно простые или дихотомически разветв¬
ленные, верхняя поверхность таллома не пятнистая (без
макул), с соредиями
Parmelinopsis afrorevoluta (+Hypotrachyna revoluta).
40. Верхний коровой слой образован гифами с узкими вытяну¬
тыми клетками, преимущественно параллельно (пери-
клинально) ориентированных к поверхности (прозо-
плектенхимный), нижний коровой слой часто отсутству¬
ет Heterodermia.
— Верхний коровой слой образован гифами с округлыми или
изодиаметрическими клетками (параплектенхимный), с
нижним коровым слоем Physcia.
41(33). Нижняя поверхность без корового слоя 42.
— Нижняя поверхность с коровым слоем 44.
42. Нижняя поверхность представляет собой губчатый слой
рыхло и сильно переплетенных черных гиф, верхняя по¬
верхность гладкая, матовая или блестящая, апотеции ле¬
канориновые, споры бесцветные, простые, виды исклю¬
чительно юга Дальнего Востока Anzia.
— Таллом более или менее протно прирастает всей нижней
поверхностью к субстрату, лишь по периферии отчетли¬
во лопастной, нижняя поверхность без черного губчатого
слоя, верхняя поверхность матовая, всегда с налетом,
апотеции лецидеиновые, споры коричневые, с перего¬
родками 43.
43. Лопасти на концах утолщающиеся, расширяющиеся, глу¬
боко рассечены на доли, поверхность лопастей с густым
мучнистым, беловато-голубоватым налетом, апотеции
встречаются редко, с соредиями Diploicia canescens.
— Лопасти на концах не расширяющиеся, не рассечены на до¬
ли, поверхность лопастей с сильно гранулярным нале¬
том, обитают на богатой кальцием почве в аридных ре¬
гионах Buellia.
44(41). Таллом без ризин 45.
— Таллом с ризинами 50.
— Таллом с густым войлочком (волосками) на нижней по¬
верхности Lobaria amplissima.
45. Лопасти более или менее округлые в сечении 46.
— Лопасти плоские, ясно дорсивентральные 48.
46. Лопасти с плотной сердцевиной Brodoa.
— Лопасти полые внутри 47.
47. Верхняя поверхность лопастей без перфораций
Hypogymnia.
— Верхняя поверхность лопастей с перфорациями
Menegazzia.
48. Таллом плотно прирастает к субстрату участками нижнего
корового слоя, нижняя поверхность черная, лопасти на
большом протяжении слитые (почти сросшиеся в одно¬
родную бородавчато-ареолированную корочку), отделе¬
ны друг от друга лишь на периферии таллома, с белыми
головчатыми соралями Dirinaria applanata.
— Таллом с иными характеристиками 49.
49. Таллом кустовидный, обычно лопасти узкие, вытянутые,
редко лопасти широкие, нижняя поверхность лопастей
более или менее вогнутая, желобчатая, фиолетовая, бе¬
ловатая близ концов, обычно эпифит
Pseudevernia furfuracea.
— Таллом листоватый, лопасти широкие, по краю восходя¬
щие, нижняя поверхность лопастей черная, обитает на
почве и скалах в горах Asahinea sholanderi.
50(44). Нижняя поверхность лопастей светлая, беловатая,
светло-буроватая, у редких видов чернеющая, верхняя
поверхность без псевдоцифелл, лопасти обычно узкие,
до 5 мм шир 51.
— Нижняя поверхность лопастей черная, темно-коричневая,
черно-коричневая, верхняя поверхность с псевдоцифел-
лами или без них, лопасти разной ширины 57.
51. Таллом К-, не содержит атранорин 52.
— Таллом К+ желт, с атранорином, лопасти без гиалиновых
волосков (шипиков) на концах 55.
52. Верхний коровой слой прозоплектенхимный, лопасти с ко¬
роткими, до 50(—70) мкм длин, гиалиновыми волосками
(шипиками) на концах, споры Physconia-типа
Anaptychia.
— Верхний коровой слой параплектенхимный, без таких гиа¬
линовых волосков, споры не Physconia-типа 54.
54. Нижний коровой слой параплектенхимный, апотеции часто
на основании с «юбочкой» ризин Phaeophyscia.
— Ннижний коровой слой прозоплектенхимный, апотеции без
ризин на основании Physciella.
55. Верхний коровой слой прозоплектенхимный Heterodermia.
— Верхний коровой слой параплектенхимный 56.
56. С настоящими цилиндрическими изидями, сердцевина Р+
желт, переходящий в оранж.-красн, содержит гирофо-
ровую(лобаровую?) кислоту Imshaugia aleurites.
— Настоящие цилиндрические изидии не образуются, сердце-
вина Р-, не содержит гирофоровую кислоту Physcia.
57(50). Лопасти с псевдоцифеллами на верхней поверхности 58.
— Лопасти без псевдоцифелл 63.
58. Лопасти ширкие, около 1 см шир. и шире, по краю восхо¬
дящие, не распростертые 59.
— Лопасти менее 1 см шир, обычно до 5 мм шир, более или
менее плоские, распростертые, прижаты к субстрату или
нет 60.
59. Верхняя поверхность более или менее ровная, псев¬
доцифеллы отчетливые, таллом не содержит каперато-
вой кислоты Cetrelia.
— Верхняя поверхность неровная, сетчато-ямчатая или склад¬
чатая, псевдоцифеллы иногда слабо выражены, таллом
содержит каператовую кислоту Platismatia.
60. Псевдоцифеллы обычно линейные или неправильной фор¬
мы, но не точковидные 61.
— Псевдоцифеллы точковидные 62.
61. Верхний коровой слой прозоплектенхимный, сердцевина
белая, споры простые, бесцветные Parmelia.
— Верхний коровой слой параплектенхимный, сердцевина от
светло-желтой до ярко-желтой, желто-оранжевой, споры
2-клеточные, коричневые Pyxine.
62. Псевдоцифеллы на поверхности лопастей, таллом не со¬
держит салациновой кислоты Punctelia.
— Псевдоцифеллы по краю лопастей, таллом содержит сала-
циновую кислоту Nipponoparmelia.
63(57). Ризины интенсивно дихотомически разветвленные 64.
— Ризины простые, если густо разветвлены, то ветвление не
дихотомическое 65.
64. Лопасти узкие, вытянутые, концы усеченные, ветвление
дихотомическое или субдихотомическое, гимений 40-69
мкм выс, гипотеций двухслойный, плектенхима верхне¬
го корового слоя эксципула образована тонкостенными
клетками Hypotrachyna s. str.
— Лопасти широкие, концы округлые, ветвление нерегуляр¬
ное, гимений 50-100 мкм выс, гипотеций трехслойный,
стенки клеток плектенхимы верхнего корового слоя экс¬
ципула очень толстые Remototrachyna koyaensis.
— Виды с иными характеристиками Hypotrachyna s. lat.
65. Лопасти более 5 мм шир 66.
— Лопасти менее 5 мм шир 67.
66. Лопасти по краю округлые, более или менее плоские, рас¬
простертые по субстрату, содержит протопраесоредио-
зовую и праесоредиозовую кислоты
Parmotrema praesorediosum.
— Лопасти по краю рассеченные, восходящие, с иным соста¬
вом лишайниковых веществ Platismatia.
67. Верхний коровой слой параплектенхимный, споры 2-
клеточные, коричневые 68.
— Верхний коровой слой прозоплектенхимный, споры прсо-
тые, бесцветные 69.
68. Верхняя поверхность лопастей с густым беловатым нале¬
том (ближе к кончикам лопастей крапчатым), зрелые
апотеции лецидеиновые, без ризин по нижнему краю,
нижний коровой слой прозоплектенхимный Pyxine.
— Верхняя поверхность обычно без налета, апотеции лекано¬
риновые, часто с «юбочкой» ризин по нижнему краю,
нижний коровой слой параплектенхимный
Phaeophyscia.
69. Таллом с изидиями, лопасти сильно вогнутые, желобчатые,
до 3-5 мм шир, изидии краевые, обычно коралловид¬
ные, таллом рыхло прикрепляется к субстрату централь¬
ной частью, содержит каператовую кислоту, дальнево¬
сточный вид Platismatia herrei.
— Таллом без изидий, с соредиями или без, лопасти плоские,
распростертые, более или менее плотно прикрепляются
к субстрату 70.
70. Сердцевина от К и Р не изменяется, содержит диварикато-
вую кислоту Parmeliopsis.
— Сердцевина от К желтеет с переходом в красно-коричневый
цвет, от Р желтеет, не содержит диварикатовую кислоту
(цилии образуются редко) Myelochroa sibirica.
71(31). Свободноживущие виды, обитают на почве в безлес¬
ных ландшафтах, в тундровых или пустынно-степных
регионах 72.
— Несвободноживущие, всегда прикрепляются к субстрату,
местообитания различные 73.
72. Таллом из тонких лентовидных, ясно дорсивентральных
лопастей, лопасти желобчатые с вниз завернутыми края¬
ми, дихотамически разветвляющиеся, верхняя поверх¬
ность темно— или светло-коричневая, нижняя сторона
частично без коры, местами с налетом
Masonhalea richardsonii.
— Таллом в виде листоватой пластинки, неправильно изогну¬
той, иногда до почти трубчато свернутой, с сильно вол¬
нистой поверхностью, с утолщенным, иногда бугристым
краем, завернутым книзу, верхняя поверхность жетова-
то-серая, буроватая, грязно-желто-оранжевая, гладкая
или морщинистая, покрытая сетью мельчайщих трещин,
обитает на каменистых или солонцеватых почвах в пу¬
стынных и каменистых степях Circinaria emiliae.
73. Лопасти узкие, обычно до 5 мм шир, редко шире, без вой-
лочка на нижней поверхности 74.
— Лопасти широкие, обычно свыше 0.5 см - до 1 см шир, но
часто 2-3 см и шире, большинство видов с войлочком на
нижней поверхности 95.
74. Таллом без ризин 75.
— Таллом с ризинами 79.
75. Лопасти в поперечном сечении округлые, не плоские ... 76.
— Лопасти плоские, ясно дорсивентральные 77.
76. Лопасти внутри полые Hypogymnia.
— Лопасти внутри без полости (обычно серый, но иногда ко-
ричневато-черный) Brodoa.
77. Таллом светлый, серовато-зеленовато-коричневатый, но
чаще бледно-зеленовато-желтоватый, в центре чешуйча-
то-накипной, лопасти выражены лишь по краю
Squamarina.
— Таллом темный, коричневый, темно-серо-коричневый,
оливково-коричневый, лопасти узкие или не выражены .
78.
78. Виды тундровых или высокогорных местообитаний, верх¬
ний коровой слой прозоплектенхимный, сердцевина КС+
розов.-красн, содержат алекториаловую, алектороно-
вую, барбатоловую или оливеторовую кислоты
Allantoparmelia.
— Виды лесных или южных горных регионов, верхний коро¬
вой слой параплектенхимный, сердцевина КС-, иногда
содержит оранжевый пигмент (скирин),тогда К+ пур-
пурн, не содержит лишайниковых веществ
Hyperphyscia.
79(74). Нижняя поверхность светлая, беловатая, буроватая,
светло-коричневатая, на экспонированных участках тем¬
но-бурая 80.
— Нижняя поверхность обычно черная, черно-коричневая,
темно-коричневая (по краю лопастей может быть светло-
коричневая до буроватой, иногда беловатая, но быстро
темнеет до черного или черно-коричневого в централь¬
ной части таллома) 83.
80. Верхний и нижний коровой слой прозоплектенхимный,
споры Physconia-типа Anaptychia.
— Верхний коровой слой пара— или склероплектенхимный,
споры Physconia-типа или иные 81.
81. Таллом с густым белым налетом, споры споры Physconia-
типа Physconia.
— Таллом без налета, верхний коровой слой параплектенхим¬
ный, споры не Physconia-типа 82.
82. Нижний коровой слой параплектенхимный, апотеции часто
на основании с «юбочкой» ризин Phaeophyscia.
— Нижний коровой слой прозоплектенхимный, апотеции без
ризин на основании Physciella.
83. Таллом обычно серый, темно-серый, черновато-серый,
иногда коричневый, коричневато-серый или серо¬
коричневый, с хорошо заметными беловатыми линей¬
ными псевдоцифеллами (образуют на поверхности лопа¬
стей сеть, или четко линейные по краю лопастей), кора
содержит атранорин, К+ желт
Parmelia (P. omphalodes s. lat, P. skultii)
— Таллом всегда коричневый, черно-коричневый, серо¬
коричневый, зелено-коричневый, без таких псев¬
доцифелл 84.
84. Ризины густо-разветвленные, ершистые 85.
— Ризины простые или слабо-разветвленные, вильчатые ... 86.
85. Верхняя поверхность обычно с густым белым налетом,
нижний коровой слой прозоплектенхимный .. Physconia.
— Верхняя поверхность без налета, нижний коровой слой па¬
раплектенхимный Phaeophyscia.
86. Таллом плотно прирастает к субстрату, ризины очень ред¬
кие, часто почти не развиты, нижняя поверхность по
краю светло-буроватая, темнеющая к центру
Hyperphyscia.
— Таллом всегда с хорошо развитыми ризинами, нижняя по¬
верхность темно-коричневая, черно-коричневая, черная
87.
87. Таллом с густым беловатым или голубоватым налетом,
сердцевина часто с желтым пигментом, кора обычно от
К желтеет, сердцевина с С, КС, Р не реагирует, нижний
коровой слой прозоплектенхимный Pyxine.
— Таллом с иными характеристиками 88.
88. Кора и сердцевина не реагируют с N, К, С, КС, Р, у редких
видов сердцевина с оранжевым пигментом (скирин) от К
окрашивается в фиолетово-пурпурный цвет, верхняя по¬
верхность обычно матовая, псевдоцифеллы отсутствуют,
пикнидии погруженные, споры 2-клеточные, коричневые
Phaeophyscia.
— Кора или сердцевина реагируют с какими-либо реактивами,
верхняя поверхность обычно блестящая, редко не бле¬
стящая, тогда с налетом, с псевдоцифеллами или без,
пикнидии погруженные или поверхностные по краю ло¬
пастей, споры простые, бесцветные 89.
89. Верхний коровой слой от N окрашивается 90.
— Верхний коровой слой от N не окрашивается 91.
90. Верхний коровой слой от N фиолетовый, сердцевина
Р+желт.-оранж, /С+красн, лопасти вогнутые, с припод¬
нимающимися краями, часто с налетом
Pleurosticta.
— Верхний коровой слой от N сине-зеленый, сердцевина Р-,
К-, лопасти плоские, с прижатыми к субстрату краями,
без налета Neofuscelia.
91. Лопасти с длинными цилиндрическими пикнидиями по
краю, вогнутые, нижняя поверхность светло-коричневая,
сердцевина - все реакции негативные
Cetrariella commixta103.
— Лопасти без таких пикнидий, обычно плоские и вогнутые 92.
92. Сердцевина С+ красн 93.
— Сердцевина С- или редко С+ слабо желт 94.
93. Обычно эпифиты, лопасти часто с тонкими гиалиновыми
волосками, часто с изидиями, без псевдоцифелл, содер¬
жат леканоровую кислоту Melanelixia.
— Эпилит, без изидий, содержит гирофоровую кислоту
Melanelia tominii.
94. Обычно эпифиты, лесные виды, псевдоцифеллы бледные,
слабо выражены, часто на вершинках бородавочек или
изидий Melanohalea104.
— Обычно эпилиты, преимущественно высокогорно¬
арктические виды, псевдоцифеллы на поверхности или
по краю лопастей Melanelia105.
95(73). Нижняя поверхность с цифеллами или псевдоцифелла-
ми 94.
— Нижняя поверхность без цифелл и псевдоцифелл 95.
94. Нижняя поверхность с псевдоцифеллами
Pseudocyphellaria aurata.
— Нижняя поверхность с цифеллами Sticta.
95. Нижняя поверхность без корового слоя, обычно с ризина¬
ми и венами 96.
— Нижняя поверхность с коровым слоем, без вен 97.
96. Апотеции на концах лопастей, цефалодии поверхностные
мелкочешуйчатые, либо на нижней поверхности - на
темных, резко-выступающих венах, споры 4-клеточные,
узко-веретеновидные или игловидные Peltigera.
— Апотеции в центре таллома, урновдные, глубко погружен¬
ные, либо плоские, но тогда с ярко-оранжевой нижней
поверхностью, цефалодии внутренние или внешние, не
сидячие на поверхности таллома, споры 2-клеточные,
коричневые, широкэллипсоидные Solorina.
1(Ь Если сердцевина К+ желт., нижняя поверхность черно-коричневая, то это -
Melanelia hepatizon.
104 Melanohalea infumata нередко встречается в качестве эпилита, особенно в
горах и в тундрах.
105 У Melanelia agnata С+ слабо желт. У Melanelia panniformis псевдоцифеллы
отсутствуют или слабо выражены.
97. Нижняя поверхность голая, без волосков и войлочка, с хо¬
рошо выраженными простыми ризинами и широкой зо¬
ной без ризин по краю лопасти Pleurosticta.
— Нижняя поверхность томентозная - с волосками (войлоч¬
ком) 98.
98. Верхняя поверхность сетчато-ямчатая (лакунозная) или
ровная, нижняя - полностью томентозная до самого края
либо с голыми участками на выпуклостях (соответству¬
ющих ямкам верхней стороны), апотеции поверхностные
Lobaria.
— Верхняя поверхность гладкая, ровная, часто с налетом и
очень мелкими волосками (войлочком), нижняя - с шро-
кой, светлой голой зоной вдоль края, к центру темно-
коричневая, апотеции на концах нижней стороны лопа¬
стей Nephroma expallidum.
Ключ 26.
Листоватые лишайники. Фотобионт - цианобактерии.
Автор: Макрый Т. В.
Талломы крупно- и мелколистоватые до чешуйчато¬
листоватых, моно- и полифильные, листовато-кустистые, ум¬
биликатные и субумбиликатные, узколопастные и чешуйчато¬
лопастные розеточные с удлиненными краевыми лопастями.
1. Таллом нежелатинозный (не разбухающий во влажном со¬
стоянии), гетеромерный (с выраженными верхним коро¬
вым и сердцевинным слоями), реже гомеомерный парап¬
лектенхимный; различно, чаще светло окрашенный
2.
— Таллом желатинозный (более или менее разбухающий во
влажном состоянии), гомеомерный, черный, буровато-,
красновато- или зеленовато-черный, редко серый, иногда
с налетом 16.
2. Таллом мелколистоватый, узколопастной или чешуйчато¬
лопастной, лопасти (0.3) 1-3(5) мм шир 3.
— Таллом крупнолистоватый, лопасти более 5 мм шир 10.
3. Таллом гомеомерный параплектенхимный, чешуйчато¬
лопастной или узколопастной розетковидный (лопасти
0.3-1.5 мм шир.); фотобионт Scytonema- типа, сем. Р1а-
cynthiaceae 4.
— Таллом гетеромерный; фотобионт Nostoc 7.
4. Подслоевище не развито; лопасти узкие длинные, менее 0.5
мм шир.; слоевище в центре не отмирающее; апотеции
со слоевищным краем 5.
— Подслоевище обычно развито, сине-черное; лопасти более
0.5 мм шир.; слоевище в центре часто отмирающее; апо¬
теции без слоевищного края, коричнево-черные; споры
(1 —)2—6-клеточные Placynthium.
5. Эпифит; апотеции с красно-коричневым диском, споры од¬
ноклеточные, зрелые 2-клеточные по 8 в сумке
Koerberia.
— Эпилит 6.
6. Апотеции с коричнево-черным диском; споры одноклеточ¬
ные по 8-16 в сумке; на влажных, заливаемых силикат¬
ных скалах Vestergrenopsis.
— Апотеции с красно-коричневым диском; споры 2-
клеточные по 8 в сумке; на карбонатных породах в сухих
местообитаниях Placynthium.
7. Нижняя поверхность из плотно переплетенных продольных
гиф, без подслоевища; таллом мелколистоватый, обра¬
зующий дернинки; апотеции с собственным краем,
встречаются редко, споры 2-клеточные
Massalongia (Massalongiaceae).
— Нижняя поверхность без корового слоя, из рыхло перепле¬
тенных гиф с отходящими ризогифами, часто с сине-
черым подслоевищем; таллом плакоидный, мелколисто¬
ватый или чешуйчато-лопастной розетковидный; апоте¬
ции леканоровые или биаторовые, споры одноклеточ¬
ные, сем. Pannariaceae 8.
8. Верхняя кора склеренхимная (клетки с утолщенными стен¬
ками); подслоевище развито слабо; таллом Р—
Fuscopannaria.
— Верхняя кора параплектенхимная 9.
9. Апатеции леканоровые; таллом Р+ оранжевеет; подслоеви¬
ще хорошо развитое, войлочное, выступающее по краям
лопастей Pannaria.
— Апотеции биаторовые; таллом Р— ; подслоевище заметное .
Parmeliella .
10. Фотобионт Scytonema; апотеции биаторовые; нижняя по¬
верхность с коровым слоем и густыми черно-синими ри¬
зинами, образующими войлок; верхняя поверхность
свинцово-серая с концентрической волнистостью
Coccocarpia (Coccocarpiaceae).
— Фотобионт Nostoc 11.
11. Апотеции биаторовые; таллом монофильный, плакоидный,
по краям рассеченно-лопастной, голубовато-серый, с ра¬
диальной бороздчатостью и концентрической волни¬
стость на концах лопастей, снизу без корового слоя, с
ризогифами, образующими хорошо развитое сине¬
черное подслоевище Degelia (Pannariaceae).
— Признаки другие 12.
12. Апотеции леканоровые, на нижней стороне таллома - на
заворачивающихся кверху концах лопастей; нижняя по¬
верхность с коровым слоем, гладкая или опущенная
Nephroma (Nephromataceae).
— Апотеции на верхней стороне таллома 13.
13. Апотеции, на концах лопастей, часто отгибающихся квер¬
ху, на начальных стадиях закрытые оболочкой, впослед¬
ствии разрывающейся; нижняя поверхность без корового
слоя, плотно или рыхло паутинистая, с ризинами, и бо¬
лее или менее выраженными жилками
Peltigera (Peltigeraceae).
— Апотеции леканоровые, на поверхности лопастей или по
краям; нижняя поверхность с коровым слоем, более или
менее ворсисто-войлочная, сем. Lobariaceae 14.
14. Нижняя поверхность без цифелл или псевдоцифелл
Lobaria.
— Нижняя поверхность с цифеллами или псевдоцифеллами ...
15.
15. Нижняя поверхность с цифеллами Sticta.
— Нижняя поверхность с псевдоцифеллами
Pseudocyphellaria.
16. Фотобионт Nostoc; таллом из рыхло переплетенных гиф, с
корой или без нее 17.
— Фотобионт хроококковый (Chroococcidiopsis, Gleocapsa);
таллом лопастной или листовато-кустистый, умбиликат¬
ный, субумбиликатный, образующий подушечки, чер¬
ный, иногда с бело-серым налетом, сем. Lichinaceae
20.
17. Таллом мелколистоватый, с многорядной верхней и ниж¬
ней корой и рыхлой сердцевиной, серо-черный с белыми
стекловидными волосками по краям лопастей; фото¬
бионт в коротких цепочках; апотеции с собственным
краем, редкие, споры 2-клеточные; на влажных замше¬
лых скалах Leptochidium (Massalongiaceae).
— Таллом от мелко- до крупнолистоватого, широколопастно¬
го, гомеомерный, из рыхло переплетенных гиф 18.
18. Плодовые тела - пикноаскокарпы с точковидным или рас¬
ширенным диском; споры одноклеточные; таллом плен¬
чатый или узколопастной, сильно разбухающий, без ко¬
рового слоя Lempholemma (Lichinaceae).
— Плодовые тела - леканоровые апотеции; споры 2-
клеточные, поперечно-многоклеточные или муральные,
сем. Collemataceae 19.
19. Таллом с настоящим параплектенхимным (1-2-рядным)
коровым слоем, иногда частично развитым, или полно¬
стью параплектенхимное, иногда с опушенной нижней
поверхностью Leptogium.
— Таллом без настоящей коры, но может развиваться псевдо-
кортекс (типичный или примитивный Collema.
20. Таллом лопастной, с воздушными полостями внутри; пло¬
довые тела - перитециевидные апотеции, погруженные в
таллом, с точковидным диском; гимений без парафиз,
фотобионт Gleocapsa с красными оболочками Phylliscum.
— Таллом без воздушных полостей, лопастной или листовато¬
кустистый умбиликатный; плодовые тела - таллинокар-
пы или пикноаскокарпы; споры одноклеточные 21.
21. Плодовые тела - таллинокарпы, имеющие вид «ложных
апотециев», с выделяющимся слоевищным краем и сет¬
чато-щелистым диском, с точечными отверстиями, ве¬
дущими к парциальным гимениям (гимениальным каме¬
рам), разделенным широкими клиновидными участками
стерильного таллома; гимений / + сине-зеленеет, затем
коричнево-краснеет; сумки с 16-32 спорами; на карбо¬
натных и силикатных скалах Thallinocarpon.
— Плодовые тела - пикноаскокарпы, погруженные или вы¬
ступающие, с заметным слоевищным краем и точковид¬
ным диском, расположенные по краям лопастей и на по¬
верхности; сумки с 8 спорами; таллом с бело-серым
налетом; на карбонатных скалах Thyrea.
Ключ 3.
Лишайники с плакоидным диморфным талломом -
накипным в центре накипной и лопастным на периферии, но
всегда приросшим к субстрату гифами сердцевины. Автор:
Кузнецова Е. С.
1. Таллом оранжевый, желтый или яркий зелено-желтый 2.
— Таллом иного цвета 8.
2. Таллом К+ интенсивно красный (быстрая реакция) 3.
— Таллом К+ розовый до красноватого (постепенная реакция)
или К- 4.
3. Споры 2-клеточные, полярилокулярные, с утолщенной кле¬
точной стенкой, таллом без шизидий, преимущественно
на камнях Caloplaca.
— Споры 1-2(4)-клеточные, не полярилокулярные, клеточная
стенка не утолщена, таллом часто с шизидиями, пре¬
имущественно на мхах и почве Fulgensia.
4. Спор в сумках 8-32 5.
— Спор в сумках около 100 7.
5. Споры бесцветные, простые, редко 2-клеточные
Candelariella.
— Споры коричневые, 1-2(4)-клеточные 6.
6. Апотеции со слоевищным краем, таллом плотно приросший
к субстрату, без ризин Dimelaena oreina.
— Апотеции без слоевищного края, таллом с пучками ризин ..
Catolechia wahlenbergii.
7. Апотеции более или менее погруженные, диск апотециев
коричневый или черный, высота гимения, как правило,
превышает 100 мкм, апикальный аппарат/-
Acarospora.
— Апотеции сидячие или полупогруженные, диск апотециев
желтый, высота гимения не превышает 100 мкм, апи¬
кальный аппарат 1+ синий Pleopsidium.
8. Фотобионт - цианобактерии (нитчатые) Placynthium.
— Фотобионт - зеленая водоросль, в центральной части тал¬
лома могут присутствовать цефалодии 9.
9. Таллом с цефалодиями Placopsis.
— Таллом без цефалодиев 10.
10. Таллом без апотециев 11.
— Таллом с апотециями 15.
11. Таллом С- 12.
— Таллом С+ розовый или красный 14.
12. Таллом с густым налетом, таллом Р- Diploicia.
— Таллом без налета или налет развивается лишь по краям
лопастей, таллом Р+ желтый или оранжевый 13.
13. Таллом белый, беловато-серый или коричневато-серый,
шизидии не формируются Dirinaria.
— Таллом серо-зеленый, иногда коричневатый (ярко-зеленый
когда влажный), с шизидиями Baeomyces.
14. Таллом с четкими сильно рассеченными радиальными ло¬
пастями Placopsis.
— Таллом состоит из сливающихся ареол или компактных
чешуек, лишь по краю располагаются обособленные
ареолы или короткие лопасти, придающие таллому ра¬
диальный облик Trapelia, Trapeliopsis.
15. В сумках около 100 спор 16.
— В сумках 8-16 спор 17.
16. Апотеции со слоевищным краем (срез!) или погружены в
таллом, гипоталлом отсутствует, апикальный аппарат
сумок не окрашивается / Acarospora.
— Апотеции без слоевищного края (срез!), выражен гипотал¬
лом, апикальный аппарат сумок дает синее окрашивание
с / Sporastatia
17. Споры бесцветные, 2-клеточные, редко 4-клеточные, поля-
рилокулярные, с утолщенной клеточной стенкой
Caloplaca.
— Споры иные, если клеточная стенка утолщенная, то споры
коричневые 18.
18. Споры коричневые 19.
— Споры простые, бесцветные 22.
19. Таллом без налета, К- 20.
— Таллом сероватый, голубовато-серый до белого, без налета
или с хорошо выраженным налетом, К+ желтый 21.
20. Таллом светлый желто-зеленый или желтовато-белый,
плотно приросший к субстрату, без ризин Dimelaena.
— Таллом коричневато-серый до темно-коричневого, с ред¬
кими светлыми ризинами на концах лопастей
Hyperphyscia.
21. Таллом, как правило, с хорошо выраженным налетом, тал¬
лом Р- Diploicia.
— Таллом без налета или с фрагментарным налетом, таллом
Р+ желтый Dirinaria.
22. Апотеции красно-коричневые на коротких ножках, таллом
К+ желтый цвет Baeomyces.
— Апотеции сидячие или погруженные в таллом 23.
23. Таллом и соралии С+ розовый или красный 24.
— Таллом и соралии С- 25.
24. Споры 15-25 мкм Trapelia.
— Споры 7-14 мкм Trapeliopsis.
25. Парафизы с пигментированными кончиками, на карбонат¬
ных горных породах Placolecis ораса.
— Парафизы без пигментированных кончиков, на различных
субстратах 26.
26. Споры обычно длиннее 14 мкм, апотеции погруженные с
невыраженным краем, сумки I- или слабо синий, без вы¬
раженного апикального аппарата Lobothalia.
— Споры обычно короче 14 мкм, апотеции обычно сидячие с
выраженным краем, сумки с 1+ синий апикальным аппа¬
ратом Lecanora (вкл. Protoparmeliopsis).
Ключ 4а.
Чешуйчатые лишайники. Фотобионт - зеленые водорос¬
ли. Автор: Урбанавичус Г. П.
1. Фотобионт сине-зеленые водоросли Ключ 46.
— Фотобионт зеленые водоросли 2.
2. Чешуйки правильно округлые, 2-3(-5) мм диам, зрелые
округло-дольчатые, светло зелено-серые, во влажном со¬
стоянии темно-зеленые, с белым краем, фотобионт Сос¬
сотуха, плодовое тело базидаильный гриб, обитает на
почве, моховых дернинках Lichenomphalia.
— Чешуйки и плодовые тела иные 3.
3. Чешуйки округлые, ушковидные, 0.5—1.5(—2) мм диам, го-
лубовато-серые, серовато-зеленоватые, зеленовато¬
серые, с выступающим краем, часто с соредиями, по¬
верхность с концентрическими кругами, фотобионт Nan-
nochloris normandinae Normandina.
— Чешуйки жесткие, толстые, неровные, бугорчатые, выпк-
лые, с волнистыми краями, сросшиеся в довольно круп¬
ные комочки, 1 —3(—4) см шир, неправильной формы,
оливкового, буровато— или светло-серого, инога ржаво¬
красного цвета, с налетом или без, апотеции формиру¬
ются редко, свободно лежащие на почве, в аридных ре¬
гионах Aspicilia esculenta.
— Чешуйки иной формы и цвета с другими фотобионтом
4.
4. Обитают на древесном субстрате 5.
— Обитают на почве или каменистом субстрате 13.
5. Плодовые тела перитеции Agonimia.
— Плодовые тела апотеции 6.
6. Таллом желто-оранжевый 7.
— Таллом не оранжевый 8.
7. Чешуйки таллома плотно прирастают всей нижней поверх-
нстью, без ризин Caloplaca.
— Чешуйки таллома прикрепляются к субстрату одним краем,
черепитчато расположенные, с соредиями по краю, с ри¬
зинами на нижней стороне Xanthomendoza.
8. Чешуйки плотно приросшие к субстрату нижней поверхно¬
стью 9.
— Чешуйки восходящие по краю 10.
9. Апотеции леканориновые, таллом преимущественно серый,
серовато-беловатый до серовато-зеленого, от С и КС не
изменяется в окраске Rinodina.
— Апотеции не известны, таллом с желтовато-серым или
светло желтовато-коричневым оттенком, от С и КС
краснеет Руспога.
10. Чешуйки свыше 1.5-2 мм длин
Cladonia (молодые неразвитые особи).
— Чешуйки не превышают 1.5 мм длин 11.
11. Чешуйки с гиалиновыми волосками на поверхности
Waynea hirsuta.
— Чешуйки без таких волосков 12.
12. Верхний коровой слой параплектенхимный, без
эпинекрального слоя, таллом и сердцевина не реагируют
на К, С, КС, Р, сумки Bacidia-типа, эпитеций от К и N
окрашивается в пурпурный цвет
Waynea stoechadiana.
— Верхняя кора состоит из двух слоев, включая толстый
эпинекральный слой, таллом и сердцевина реагируют на
воздействие К, С, КС или Р, сумки не Bacidia-типа, эпи¬
теций не окрашивается в пурпурный цвет при действии
К и N Hypocenomyce.
13(4). Плодовые тела перитеции 14.
— Плодовые тела апотеции апотеции 20.
14. Споры муральные, коричневые или светлоокрашенные 15.
— Споры простые или 2-клеточные, бесцветные 16.
15. Гимениальные водоросли отсутствуют Agonimia.
— Гимениальные водоросли присутствуют Endocarpon.
16. Споры 2-клеточные Placidiopsis.
— Споры простые 17.
17. Верхний коровой слой тонкий, 5-40 мкм толщ, Cinerea-
типа, плохо отграничен от водорослевого слоя
Catapyrenium.
— Верхний коровой слой толстый, 20-100 мкм толщ, ясно
отграничен от водорослевого слоя 18.
18. С толстыми черными ризоидальными тяжами
Neocatapyrenium.
— Без таких тяжей 19.
19. Чешуйки мелкие, 0.5-1.5 мм шир, сумки булавовидные,
споры в сумках расположены в 2 ряда, обитают исклю¬
чительно на камнях, плотно прикрепляются гомфообраз-
ным выростом (относится только к видам, известным в
России) Heteroplacidium.
— Чешуйки крупные 2—10(—15) мм шир, сумки цилиндриче¬
ские, споры в сумках расположены в 1 ряд, обитают на
почве, прослойке почвы поверх камней, прикрепляются
с помощью ризогиф, иногда дополнительно ризинами ....
Placidium.
20(13). Апотеции красновато-коричневые, округлые, без вы¬
раженного края (ровно переходят в поверхность талло¬
ма), постепенно разрастаются по всей поверхности тал¬
лома, которая превращается в сплошной апотеций; ксе-
рофильный лишайник обитает в горах Азиатской России
Gypsoplaca.
— Лишайники с иными характеристиками 21.
21. Апотеции на ножке Baeomyces / Cladonia.
— Апотеции сидячие, полупогруженные или погруженные 22.
22. Сумки многоспоровые, >50 спор 23.
— Сумки с 2-8 спорами 25.
23. Апотеции сложные, состоят из многочисленных мелких
апотециев, каждый мелкий апотеций обособлен самосто¬
ятельным эксципулом Glypholecia.
— Апотеции не сложные 24.
24. Водорослевый слой непрерывный Acarospora.
— Водорослевый слой разрывается сердцевинными гифами
Trimmatothelopsis.
25. Апотеции с водорослью в крае 26.
— Апотеции без водоросли в крае 34.
— Апотеции кубковидные, глубоко погружены в округлые
чешуйки, узкой, 1-5 мм шир, полоской окружающие
апотеции, споры 2-клеточные, коричневые
Solorina spongoisa.
26. Споры 1-клеточные 27.
— Споры 2-4-клеточные 31.
27. Таллом прикрепляется к субстрату гомфом 28.
— Таллом без гомфа 29.
28. Таллом желтовато-коричневый, С+ оранж. (содержит ксан-
тоны), обычно полностью покрыт апотециями Arctopeltis.
— Таллом зеленовато-желтый, коричневатый, беловатый, С—
или С+ (не содержит ксантоны), апотеции обычно не
полностью покрывают таллом Rhizoplaca.
29. С внешними цефалодиями, содержащими Nostoc
Psoroma.
— Без цефалодий 30.
30. Верхняя поверхность чешуек коричневая, оливковая, чер¬
неющая, с оранжевым оттенком в центре, зеленоватая
при увлажнении, апотеции с лобулями по краю
Fuscopannaria.
— Верхняя поверхность желтовато-зеленая, беловатая, часто с
густым белым налетом, апотеции без лобулей
Squamarina.
31. Споры бесцветные
— Споры коричневые
32. Споры биполярные
32.
Phaeorrhiza.
Caloplaca.
— Споры не биполярные
33. Таллом желтый
— Таллом не желтый
Fulgensia.
Lecania.
33.
34(25). Таллом ярко-желтый, желто-зеленый или желтоватый .
35. Таллом крупный, обычно свыше 2 см диам, споры 2-
клеточные, коричневые, произрастает преимущественно
— Таллом мелкий, обычно мены 2 см диам, споры бесцвет¬
ные, многклеточные, произрастает преимущественно на
почве поверх лишайников, редко свободноживущий
Arthrorhaphis.
36. Споры коричневые, обычно паратизирует на коричнево-
окрашенных эпилитных видах семейства Parmeliaceae,
редко свободноживущий Buellia badia.
— Споры бесцветные, свободноживущие, если паразитируют,
то на лишайниках с сине-зеленым фотобионтом 37.
37. Обитает на почве 38.
— Обитает на каменистом субстрате 46.
38. Споры одноклеточные, эллипсоидные 39.
— Споры 2— или многоклеточные, узкоэллипсоидные или
подошвовидные 43.
39. Чешуйки сильно выпуклые, часто булавовидные, вздутые,
с углублением в центре Toninia.
— Чешуйки более или менее плоские 40.
40. Апотеции широкие (соизмеримые с размерами чешуек),
часто неправильной формы, довольно тонкие, распро¬
стертые по поверхности таллома или немного погружен¬
ные Lecidoma.
— Апотеции не уплощенные, не распростертые на талломе,
обычно с зауженным основанием 41.
41. Таллом серовато-белый, соредиозный Trapeliopsis.
— Таллом не серовато-белый, без соредиев 42.
42. Апотеции без выраженного края, эпитеций содержит ан-
трахиноны (главным образом париетин), гипотеций с
кристаллами оксалата кальция, пикнидии погружены в
верхнюю поверхность чешуек Psora.
— Молодые апотеции с хорошо выраженным краем, эпитеций
без антрахинонов, гипотеций без кристаллов оксалата
Таллом не желтой окраски
35.
36.
на скальной поверхности
Catolechia.
кальция, пикнидии сидячие, на краю или на нижней сто¬
роне чешуек близ края Romjularia.
43. Споры неравной величины, с перетяжкой по перегородке ..
Arthonia glebosa.
— Споры без перетяжки на перегородке, ровные, обычно уз¬
коэллипсоидные 44.
44. Чешуйки обычно сильно выпуклые, если не выпуклые, то
без темных центров, часто являются парасимбионтами
лишайников с синее-зеленым фотобионтом или циа¬
нобактерий Toninia.
— Чешуйки более или менее плоские, слабо выпуклые или
вогнутые, иногда с более темной центральной частью и
светлыми краями 45.
45. Таллом К-, споры 3-4-клеточные Bilimbia lobulata.
— Таллом К+ желт, споры 2-клеточные Stereocaulon.
46(37). Таллом от С краснеет, розовеет 47.
— Таллом с С не реагирует 51.
47. С цефалодиями Placopsis.
— Без цефалодий 48.
48. Таллом серовато-белый, соредиозный (в стерильном со¬
стоянии роды не различимы) 49.
— Таллом коричневый, желтовато-, зеленовато-, красновато-
коричневый, без соредиев 50.
49. Споры 15-25 мкм Trapelia.
— Споры 7-14 мкм Trapeliopsis.
50. Чешуйки плотно приросшие к субстрату, по периферии
таллома обычно крупнее, отчетливо вытянутые лопаст¬
ные, снизу черные, апотеции более или менее плоские, с
постоянным краем, сгруппированные в центре таллома ..
Boreoplaca.
— Чешуйки с восходящими краями, обычно черепитчато
налегающие друг на друга, или разрозненные, на пери¬
ферии не образуются вытянутые лопастевидные, снизу
беловатые, светло-коричневатые, апотеции без края,
сильно выпуклые, не группируются в центре таллома
Psora.
51. Споры 1-клеточные 52.
— Споры 2- или многоклеточные 57.
52. Произрастают на эпилитных цианобионтных лишайниках .
53.
— Не связаны с цианобионтными лишайниками 54.
53. Всегда произрастает на Spilonema revertens, поверх некар¬
бонатных горных пород, чешуйки мелкие, до 2(-3) мм
диам, тонкие, с темным до черного краем, верхний ко-
ровой слой содержит остатки клеток водорослей, эпите¬
ций и эксципул с темно-зеленым пигментом, гипотеций
темно красно-коричневый Psorula.
— Часто произрастает на представителях сем. Collemataceae и
Lichinaceae поверх карбонатных горных пород, чешуйки
более толстые, крупные, до 5 мм диам, край одноцвет¬
ный или светлее верхней поверхности, эпитеций и экс¬
ципул без зеленого пигмента, гипотеций светло¬
коричневатый Romjularia.
54. Гимений не амилоидный, сумки Trapelia-типа, содержит
алекториаловую кислоту Anamylopsora.
— Гимений амилоидный, сумки иного типа, без алекториало-
вой кислоты 55.
55. Чешуйки прикрепляются к субстрату с помощью коровых
ризин, ризогифы отсутствуют, сумки Lecanora-типа
Psorinia.
— Чешуйки пркрепляются всей нижней поверхностью, или
одним краем, ризогифы часто развиты, сумки Porpidia-
типа 56.
56. Апотеции без выраженного края, эпитеций содержит ан-
трахиноны (главным образом париетин), гипотеций с
кристаллами оксалата кальция, пикнидии погружены в
верхнюю поверхность чешуек Psora.
— Молодые апотеции с хорошо выраженным краем, эпитеций
без антрахинонов, гипотеций без кристаллов оксалата
кальция, пикнидии сидячие, на краю или на нижней сто¬
роне чешуек близ края Romjularia.
57(51). Чешуйки более или менее плоские, беловатые, серова¬
тые, обычно с темным центром, приросшие центральной
частью нижней поверхности или одним краем, тогда
восходящие, соредиозные, таллом не бывает розетко¬
видный, верхняя поверхность К+, Р+ Stereocaulon.
— Чешуйки обычно вздутые, булавовидные, редко уплощен¬
ные, не белые, но часто покрыты белым налетом, часто
розетковидные, верхняя поверхность от К и Р не изменя¬
ется Toninia.
Ключ 46.
Чешуйчатые лишайники. Фотобионт - цианобактерии.
Автор: Макрый Т. В.
Талломы крупно- и мелкочешуйчатые до чешуйчато¬
листоватых, пельтатные и полифильные умбиликатные (с
гомфом), чешуйчато-ареальчатые, плакоидные с радиально
ориентированными чешуйками или краевыми лопастинками;
талломы в виде одиночных или сближенных чешуек, рыхлых
дернинок или плотно сросшихся корочек из сомкнутых или
черепитчато-налегающих чешуек, или розетковидные.
1. Таллом нежелатинозный (не разбухающий во влажном со¬
стоянии), редко субжелатинозный, светло или темно се¬
рый, оливково-зеленый, оливково-коричневый, но не
черный, иногда с белым или серым налетом 2.
— Таллом желатинозный (разбухающий во влажном состоя¬
нии), черный, буровато-, красновато- или оливково¬
черный, редко серый, иногда с налетом 11.
2. Споры поперечно-многоклеточные с 4-10 перегородками;
таллом чешуйчато-накипной, из скопления чешуек с уз¬
ловатой поверхностью и иногда беспорядочно отходя¬
щими тонкими разветвленными субцилиндрическими
лопастинками, или ясно розетковидный с короткими
краевыми лопастинками; на мхах и почве, аркто-
альпийский Arctomia (Arctomiaceae).
— Споры одноклеточные или с 1-5 поперечными перегород¬
ками; признаки другие 3.
3. Таллом гетеромерный без нижнего корового слоя или го¬
меомерный параплектенхимный, чешуйчатый или че¬
шу йчато-лопастной, часто розетковидный; чешуйки по
краям кренулированные, дланевидно-рассеченные или
узко-лопастные; апотеции сидячие 4.
— Таллом гомеомерный или гетеромерный, из сетчато-
переплетенных гиф с толстым, тонким или прерывистым
коровым слоем, пельтатный или чешуйчато-
ареольчатый, реже розетковидный; чешуйки с ровными
краями, округлые или выемчато-округло-лопастные;
апотеции погруженные, с точковидным или чашевид¬
ным, редко плоским диском; споры одноклеточные .. 10.
4. Таллом гомеомерный параплектенхимный, оливковый до
темно-бурого, часто с сине-черным подслоевищем; фо¬
тобионт Scytonema-^wjxwhm в кластерах; апотеции биа¬
торовые, буро-черные, споры 2-6-клеточные; на залива¬
емых силикатных или сухих карбонатных скалах, редко
на почве Placynthium (Placynthiaceae).
— Таллом гетеромерный без нижнего корового слоя; фото¬
бионт - Nostoc, в кластерах; апотеции леканоровые и би-
торовые, не буро-черные; споры 1-2-клеточные 5.
5. Таллом чешуйчато-листоватый до чешуйчатого, коричне¬
вый; нижняя поверхность из плотно переплетенных про¬
дольных гиф; апотеции с собственным краем, редкие;
споры 2-клеточные; на замшелых силикатных скалах
Massalongia (Massalongiaceae).
— Таллом чешуйчатый до чешуйчато-лопастного, голубовато-
или желтовато-серый, до коричневого; нижняя поверх¬
ность из рыхло переплетенных гиф с ризогифами, часто
с подслоевищем; апотеции леканоровые или биаторовые;
красно-коричневые, реже коричнево-черные; споры од¬
ноклеточные, сем. Pannariaceae 6.
6. Таллом Р+ оранжевеет; верхняя кора параплектенхимная,
апотеции леканоровые, красно-коричневые или буро¬
черные; гимений 1+ синеет вокруг сумок, сумки без ами¬
лоидной структуры; подслоевище хорошо развитое,
сине-черное; на стволах деревьев и замшелых скалах
теплолюбивый, на скалах аркто-альпийский
Pannaria.
— Таллом Р- 7.
7. Сумки без амилоидной апикальной структуры; апотеции
всегда присутствуют, леканоровые, красно-коричневые;
гимений 1+ темно синеет; споры с бородавчатым экзо-
спорием; таллом из мелких срастающихся чешуек; под¬
слоевище незаметное; на почве, аркто-альпийский
Protopannaria.
— Сумки с амилоидной апикальной структурой; апотеции
обычны или отсутствуют; споры с гладкой оболочкой;
талом в виде плотных корочек или рыхлых дерновинок
из черепитчато-налегающих или прижатых чешуек до
чешуйчато-лопастного розетковидного; на скалах, почве,
стволах деревьев 8.
8. Сумки с амилоидной апикальной структурой в виде покры-
вальца; апотеции обычны, леканоровые или с исчезаю¬
щим слоевищным краем, от светло-красно-коричневых
до буро-черных; гимений 1+ сине-зеленеет, затем корич-
нево-краснеет; верхняя кора склеренхимная; подслоеви¬
ще сине-черное, более или менее развитое
Vahliella (? не Pannariaceae).
— Сумки с кольцевой амилоидной апикальной структурой;
апотеции леканоровые или биаторовые, часто отсут¬
ствуют 9.
9. Верхняя кора параплектенхимная; апотеции преимуще¬
ственно биаторовые, красно-коричневые; гимений 1+ си¬
неет; подслоевище заметное, сине-черное
Parmeliella.
— Верхняя кора склеренхимная (клетки с утолщенными стен¬
ками); апотеции коричневые, леканоровые или с исчеза¬
ющим слоевищным краем; гимений 1+ сине-зеленеет за¬
тем коричнево-краснеет; подслоевище слабо развито или
отсутствует Fuscopannaria.
10. Чешуйки округлые до слегка выемчато-округло-
лопастных, прирастающие всей нижней поверхностью;
таллом с цельным или прерывистым, верхним и/или
нижним коровым слоем; фотобионт Scytonema-Tvma. в
кластерах; апотеции с расширенным, обычно чашевид¬
ным диском, по 1-3 на чешуйке, сумки с 8 спорами; на
почве, часто карбонатной, в сухих и аридных условиях
Heppia (Heppiaceae).
— Чешуйки пельтатные или полифильные с гомфом, ареоль-
чатые до выпуклых и столбчатых, реже блюдцевидные,
одиночные или сближенные, до черепитчато-
налегающих; таллом с более или менее развитым коро¬
вым слоем; фотобионт хроококковый (Chroococcidiopsis,
Myxosarcina); апотеции точковидные до широкооткры¬
тых (иногда со слоевищным краем), от 1-2 до многочис¬
ленных; сумки многоспоровые; на скалах и почве, в су¬
хих и аридных условиях Peltula (Peltulaceae).
11. Фотобионт Nostoc в цепочках 12.
— Фотобионт хроококковый; споры одноклеточные, сем. Li¬
chinaceae 15.
12. Споры одноклеточные; плодовые тела - погруженные пик¬
ноаскокарпы с точковидным реже открытым диском,
редкие; таллом без корового слоя, черный, в виде умби¬
ликатных или черепитчато налегающих чешуек обычно с
горизонтально или вертикально отходящими валькова-
тыми лопастинками; на карбонатных (редко силикат¬
ных), периодически увлажняемых, освещенных скалах ..
Lempholemma (Lichinaceae).
— Споры поперечно 2-4-клеточные или муральные; плодовые
тела - апотеции, редкие или обычные; таллом с коровым
слоем или без него 13.
13. Таллом с многорядной верхней и нижней корой и рыхлой
сердцевиной, чешуйчато-листоватый, серо-черный с бе¬
лыми стекловидными волосками по краям лопастей; фо¬
тобионт в коротких цепочках; апотеции с собственным
краем, редкие, споры 2-клеточные; на влажных замше¬
лых скалах Leptochidium (Massalongiaceae).
— Таллом без многорядной коры, чешуйчато-листоватый,
мелкочешуйчатый, чешуйчато-изидиозный, чешуйчато-
накипной до плакоидного, образующего пятна до 1.5 см
диам.; фотобионт в длинных цепочках; апотеции лекано¬
ровые, сем. Collemataceae 14.
14. Таллом с коровым слоем или насквозь параплектенхим¬
ный; споры муральные; на карбонатных скалах и почве,
стволах деревьев и колоднике Leptogium.
— Таллом без корового слоя или с псевдокортексом; споры 2-
4-клеточные или муральные; на стволах деревьев, кар¬
бонатных скалах Collema.
15. Таллом мелкий, чешуйчато-умбиликатный или чешуйчато-
ареольчатый; чешуйки плоские или выпуклые, иногда с
гранулезной поверхностью 16.
— Таллом более крупный, пельтатно-умбиликатный или обра¬
зующий подушечки с вертикально направленными, часто
складчатыми лопастями 22.
16. Плодовые тела - таллинокарпы, с открытым диском; эпи-
гимений с клетками фотобионта; гимений 1+ сине-
зеленеет, затем коричнево-краснеет; сумки с 16-32 спо¬
рами; на карбонатных скалах, почве, в аридных и ксеро-
тических условиях
Gonohymenia s. str. (Lichinella s. 1.).
— Признаки другие 17.
17. Плодовые тела - погруженные перитециевидные апотеции
или пикноаскокарпы, открывающиеся узкой порой; ги¬
мений без парафиз; сумки с 8-24 спорами; фотобионт
Gloeocapsa с красно-коричневыми влагалищами; таллом
темно-красно-коричневый; на влажных силикатных ска¬
лах 18.
— Плодовые тела - леканоровые апотеции и апотециевидные
пикноаскокарпы, зрелые с более или менее расширен¬
ным диском; сумки с (4) 8 спорами 19.
18. Таллом с воздушными полостями, чешуйчато-умбиликат¬
ный; апотеции без перифиз
Phylliscum.
— Таллом без воздушных полостей, чешуйчато-ареольчатый;
апотеции с хорошо развитым эксципулом, с перифизами
в верхней части Cryptothele.
19. Диск апотеция глянцевый красно-коричневый; сумки с
толстыми стенками; таллом темно-красновато-
коричневый, из сетчато-переплетенных гиф; иногда
наряду с цианобактерией содержит зеленую водоросль;
на мокрых силикатных скалах, грунте, мхах, арктко-
альпийский Euopsis.
— Диск не глянцевый; таллом черный, оливково-коричневый
или коричнево-черный, иногда с налетом; на карбонат¬
ных и кальцийсодержащих, реже силикатных породах, в
более или менее сухих, теплых местообитаниях 20.
20. Диск черный; гимений с красно-коричневыми участками в
верхней части, 1+ синеет; сумки с 8 спорами; апотеции
на ножке или сидячие; конидии игловидные, нитевидные
или серповидно изогнутые; таллом и диск апотеция ча¬
сто с налетом; на почве и в расщелинах карбонатных
скал Peccania.
— Диск не черный; гимений без красных и коричневых участ¬
ков; конидии палочковидные или эллипсоидные 21.
21. Плодовые тела - пикноаскокарпы, редкие, вначале погру¬
женные, закрытые, зрелые с открытым красно¬
коричневым диском, тонким эксципулом и толстым
(200-250 мкм) слоевищным краем; парафизы двух ти¬
пов: неразветвленные длинноклеточные и разветвленные
в виде нитей жемчуга; таллом из сетчато-переплетенных
гиф с группами клеток цианобактерии, придающими ему
параплектенхимный вид Anema.
— Плодовые тела - апотеции, обычны, вначале погруженные,
зрелые сидячие или полупогруженные, с коричневым
диском, тонким незамкнутым внизу эксципулом и посто¬
янным слоевищным краем; парафизы другие; таллом
мелкоклеточно-параплектенхимный Psorotichia.
22. Плодовые тела - таллинокарпы, имеющие вид «ложных
апотециев», с выделяющимся слоевищным краем и сет¬
чато-щелистым диском, с точечными отверстиями, ве¬
дущими к парциальным гимениям (гимениальным каме¬
рам), разделенным широкими клиновидными участками
стерильного таллома; гимений I + сине-зеленеет, затем
коричнево-краснеет; сумки с 16-32 спорами; таллом с
рыхлым центральным тяжем и отходящими вееровидно
гифами, сетчато-переплетенными у поверхности; на кар¬
бонатных и силикатных скалах Thallinocarpon.
— Плодовые тела - пикноаскокарпы, погруженные или вы¬
ступающие, с заметным слоевищным краем и точковид¬
ным диском, расположенные по краям лопастей и на по¬
верхности; сумки с 8 спорами; таллом с налетом; на кар¬
бонатных скалах Thyrea.
Ключ 6.
Накипные лишайники с плодовыми телами - перитеция¬
ми или похожими на перитеции. Автор: Урбанавичус Г. П.
1. Фотобионт сине-зеленые водоросли 2.
— Фотобионт зеленый - требоуксоидный или трентеполиоид¬
ный 7.
2. Споры простые 3.
— Споры 2-клеточные 6.
3. Фотобионт Gloeocapsa, К + пурп 4.
— Фотобионт Calothrix, К- 5.
4. Без парафиз Cryptothele.
— С парафизами Pyrenopsis.
5. Таллом во влажном состоянии красновато-коричневый, ги¬
мений/+голуб, споры 10-18 х 7-12 мкм
Porocyphus coccodes.
— Таллом во влажном состоянии темно-зеленоватого оттенка,
гимений /-, споры мельче 8-10 х 6-7 мкм
Thelignya lignyota.
6. Таллом погруженный или отсутствует, на известняке (каме¬
нистом субстрате), раковинах морских моллюсков
Collemopsidium.
— Таллом чешуйчатый, на почве Frigidopyrenia.
7. Перитеции светлые (до коричневых), обычно не погружен¬
ные в субстрат или таллом, либо погруженные, но тогда
не настоящие перитеции 8.
— Перитеции/плодовые тела черные (редко темно-
коричневые), либо светлые и погружены в таллом ... 13.
8. Сумки многоспоровые Thelocarpon.
— Сумки с 1-8 спорами 9.
9. Плодовые тела не настоящие перитеции (апотеции почти с
сомкнутым эксципулом, погруженные в талломные бо-
родавочки); экспонированная часть апотеция темно-
окрашенная Pertusaria.
— Плодовые тела типичные перитеции 10.
10. Фотобионт не трентеполиоидный 11.
— Фотобионт трентеполиоидный 12.
11. Перитеции 80-120 мкм диам, споры 2-4-клеточные
Psoroglaena.
— Перитеции 140-220 мкм диам, споры слабомуральные
Leucocarpia.
12. Споры с 20-ю и более клетками, свыше 50 мкм длин.
Belonia.
— Споры 4-10-клеточные, менее 40 мкм длин Porina.
13. Сумки многоспоровые Thelopsis106.
— Сумки с 1-8 спорами 14.
14. Молодые плодовые тела перитециевидные, зрелые - глу¬
боко урновидные апотеции, погруженные в талломные
бородавочки Thelotrema
— Плодовые тела типичные перитеции 15.
15. Споры одноклеточные 16.
— Споры с перегородками 26.
16. Гифы (парафизы) в межеумочном пространстве присут¬
ствуют Thrombium.
— Отсутствуют 17.
17. Перитеции полностью глубоко погружены в субстрат, в
котором при отмирании их образуются хорошо выра¬
женные ямки Bagliettoa / Verrucaria.
— Перитеции сидячие или погружены в субстрат неглубоко,
не оставляют глубокие ямки после отмирания, либо по-
гружены/полупогружены в таллом 18.
18. Споры узковеретеновидные до игловидных, обычно эфи-
титы 19.
— Споры широкоэллипсоидные до округлых, обычно эпили¬
ты, редко на другом субстрате 20.
19. Сумки с одной стенкой, покрывальце не развито
Cresporhaphis.
— Сумки с двумя стенками, с покрывальцем .... Leptorhaphis.
20. Таллом почти чешуйчатый, ареолы часто сужены в «нож¬
ку», по краю лопастевидные 21.
— Таллом без таких ареол 22.
21. Верхняя и боковая поверхность чешуек коричневая, без
налета Heteroplacidium.
— Верхняя поверхность чешуек обычно серая с серовато¬
беловатым налетом, боковая поверхность черная
Placopyrenium.
22. Таллом толстый, до 2 мм толщ, с мощным сердцевинным
слоем, часто плакоидный по краю Placocarpus.
— Таллом обычно тоньше 23.
23. Обитает в воде, таллом без выраженного корового слоя, не
дифференированного от сердцевины, при намокании
ослизняется 24.
106 Видимость многоспоровых сумок создается также у видов ро¬
да Bcictrosporci (на самом деле в сумках по 8 очень длинных
многклеточных спор, которые часто разламываются на множе¬
ство коротких фрагментов).
— Обычно обитает в иных условиях, таллом с иными характе¬
ристиками 25.
24. Покрывальце с неровной поверхностью, споры обычно
свыше 12 мкм длин Hydropunctaria.
— Покрывальце с ровной поверхностью, споры обычно менее
12 мкм длин Wahlenbergiella.
25. Обычно паразитирует на других лишайниках
Verrucula / Verruculopsis
(иногда Heteroplacidium / Placopyrenium).
— Обычно не паразитирует на других лишайниках
Parabagliettoa / Verrucaria.
26(15). Зрелые споры коричневые 27.
— Зрелые споры бесцветные 41.
27. Фотобионт трентеполиоидный или отсутствует 28.
— Фотобионт не трентеполиоидный 36.
28. Споры муральные 29.
— Споры от 2-х до поперечно-многоклеточных 30.
29. Споры крупные, 40-60 х 15-21 мкм Anthracothecium.
— Споры мелкие, 20-40 х 8-12 мкм Mycoporum.
30. Споры от 4-х до многоклеточных, фотобионт присутствует,
редко отсутствует 31.
— Споры 2-4-клеточные, фотобионт обычно отсутствует
32.
31. Клетки спор линзовидные, ромбовидные, таллом часто с
псевдоцифеллами Pyrenula.
— Клетки спор не линзовидные и не ромбовидные, таллом без
псевдоцифелл Eopyrenula leucoplaca.
32. Перитеции сгруппированы под общим покрывалом, стро-
матические Tomasellia gelatinosa.
— Перитеции не в строме 33.
33. Содержимое перитеция от/синеет Peridiothelia.
— Содержимое перитеция от / не синеет 34.
34. Эпилит Lichenothelia.
— Эпифит 35.
35. Стенки перитеция от К+ оливково-зеленые или оливково¬
коричневые (смотреть в препарате), споры до 25 мкм
длин, 2-клеточные Mycomicrothelia.
— Нет такой реакции на стенках перитеция с К, споры круп¬
нее, 27-40 мкм длин, 2-4-клеточные
Mycoporum antecellens.
36(27). Гимениальные водоросли присутствуют ... Staurothele.
— Гимениальные водоросли отсутствую 37.
38. Парафизы присутствуют 39.
— Парафизы отсутствую 40.
39. Перитеции разрозненные, эксципул светлый в основании,
перифизы отсутствуют, перифизоиды присутствуют или
отсутствуют Thelenella.
— Перитеции строматические, эксципул черно-коричневый
целиком, перифизы присутствуют
Dacampia hookeri107
40. Покрывальце отсутствует, эксципул 3-х слойный, клетки
корового слоя часто с мелкими папиллами Agonimia.
— Покрывальце обычно развито, редко без него, эксципул 1-
или 2-слойный, клетки корового слая без папилл
Polyblastia s. 1.
=Atla / Henrica / Polyblastia / Sporodictyon.
41(26). Фотобионт не трентеполиоидный, редко отсутствует
42.
— Фотобионт трентеполиоидный, либо отсутствует 51.
42. Споры муральные 43.
— Споры от 2-х до поперечно-многоклеточных (иногда встре¬
чаются споры с единичной продольной перегородкой) ...
48.
43. Гимениальные водоросли присутствуют Staurothele.
— Гимениальные водоросли отсутствую 44.
44. Покрывальце отсутствует Agonimia.
— Покрывальце развито 45.
45. Парфизоиды отсутствуют 46.
— Парафизоиды присутствуют 47.
46. С цефалодиями, содержащими Nostoc, споры крупные, 50-
90 мкм длин Sporodictyon terrestris.
— Без цефалодий, споры мельче Polyblastia.
47. Сумки и содержимое гимения амилоидные Protothelenella.
— Сумки и содержимое гимения не амилоидные ... Thelenella.
48. Сумки узкоцилиндрические, до 15 мкм шир, споры в сум¬
ках расположены обычно в один ряд Acrocordia.
— Сумки широкоовальные, булавовидные, либо споры в сум¬
ках расположены иным образом 49.
49. Споры 2-х, редко 4-5-клеточные Thelidium.
— Споры обычно имеют свыше 4 перегородок 50.
50. Обитает на почве, таллом с соредиями, споры узкоэллип¬
соидные Lauderlindsaya.
— Обычно обитает на камнях (виды с бесцветными спорами с
преимущественно поперечными и 1-2 продольными пе-
107 Остальные виды рода представлены нелихенизирован-
ными лихенофильными грибами.
регородками), таллом всегда без соредий, споры широ¬
коэллипсоидные Polyblastia.
51(41). Споры узкие, обычно до 4-6 мкм шир, с 15—40 попе¬
речными перегородками 52.
— Споры с меньшим числом перегородок, если с 10-15 пере¬
городками, то споры шире, либо споры муральные
53.
52. Плодовое тело - перитециоидный апотеций, в зрелом со¬
стоянии полусферические, споры в сумках распадаются
на короткие фрагменты Bactrospora.
— Плодовое тело - типичный перитеций, споры не распада¬
ются на фрагменты Belonia.
53. Споры муральные 54.
— Споры только с поперечными перегородками (среди них
редко встречаются с единичными продольными) 58.
54. Стенки перитеция ярко-зеленые, N+ красн Mycoglaena.
— Стенки перитеция не зеленые, N- 55.
55. Парафизы обычно простые, редко слабо разветвленные,
постоянно с хорошо развитым лихенизированным тал¬
ломом Strigula.
— Парафизы сильно разветвленные, нелихенизированные ви¬
ды 56.
56. Перитеции сливаются, строматические Mycoporum elabens.
— Перитеции не сливаются, без стромы 57.
57. Споры с периспорием Julella.
— Споры без периспория Cyrtidula.
58. Перитеции сгруппированы под общим покрывалом, стро¬
матические Tomasellia.
— Перитеции одиночные 59.
59. Споры обычно ровные, прямые или слегка изогнутые, па¬
лочковидные, узкоэллипсоидные, с клетками равной
ширины, без перетяжек 60.
— Споры обычно неровные, булавовидные, каплевидные,
клетки часто неравной ширины и размеров, с перетяж¬
ками 63.
60. Сумки с одной стенкой, унитуникатные 61.
— Сумки с двумя стенками, битуникатные 62.
61. Покрывальце имеется, содержимое перитеция /-, споры с
хорошо выраженными перегородками, от 4-х до много¬
клеточных Porina.
— Покрывальце отсутствует, содержимое перитеция 1+ голу¬
бовато-зеленый, споры 1-клеточные, иногда неясно 2-4-
клеточные, факультативно лихенезированный
Cresporhaphis.
62. Споры обычно прямые, палочковидные, узкоэллипсоид¬
ные, преимущественно шире 3-4 мкм, если уже, то дли¬
на не превышает 15 мкм Strigula.
— Споры изогнутые, спирально завернутые, игловидные или
узковеретеновидные, в основном уже 4 мкм, длина пре¬
вышает 15 мкм, факультативно лихенезированный
Leptorhaphis.
63. Стенки перитеция ярко-зеленые, N+ красн
Mycoglaena myricae.
— Стенки перитеция не зеленые, N— 64.
64. Стенки перитеция имеют клеточное строение, без черных
клеток Anisomeridium.
— Стенки перитеция образованы гифами с черными клетками
(разделение родов сложное, не всеми признаваемое,
многие представители этой группы - нелихенизирован¬
ные или факультативно лихенизированные)
.. Arthopyrenia / Naetrocymbe / Mycoporum antecellens.
Ключ 7.
Накипные лишайники с плодовыми телами - апотециями
и фотобионтом - цианобактериями. Автор: Макрый Т. В.
Талломы аморфные, ареолированно-накипные, зернисто¬
накипные, чешуйчато-накипные, чешуйчато-ареольчатые
(ложно-ареолированные), плакоидные (с радиальными лопа¬
стинками по краю); рессеянные, образующие бесформенные
пятна или округлые розетки и кольца, в тех случаях, когда
таллом в центре отмирает.
1. Таллом эндолитный; фотобионт Scytonema; апотеции по¬
груженные в субстрат, сначала закрытые, зрелые - гиа-
лектовидные с растресканным слоевищным краем; спо¬
ры 4-клеточные (иногда с одной продольной перегород¬
кой); на известняках Petractis (?).
— Таллом эпилитный; признаки другие 2.
2. Фотобионт хроококковый (Gloeocapsa, Chroococcidiopsis,
Myxosarcina и др.); плодовые тела - пикноаскокарпы ли¬
бо перитециевидные или леканоровые апотеции; споры
одноклеточные (часто с плазматическими перемычками)
3.
— Фотобионт нитчатый (Nostoc, Scytonema, Callothrix и др.) в
цепочках или кластерах; плодовые тела - леканоровые
или биаторовые апотеции; споры различные 14.
3. Таллом нежелатинозный, коричневый, с коровым слоем,
ареолированно-накипной, розетковидный, с округлыми
ареолами, иногда с удлиненными радиальными лопа¬
стинками по краям, апотеции леканоровые с узким,
позднее расширенным диском; сумки многоспоровые ....
Peltula (Peltulaceae).
— Таллом обычно желатинозный, черный, красновато-, олив-
ково- или буро-черный или с беловато-серым налетом,
без радиальных лопастинок сем. Lichinaceae 4.
4. Плодовые тела - таллинокарпы, с открытым диском; эпиги-
мений с клетками фотобионта; гимений I + сине-
зеленеет, затем коричнево-краснеет; сумки с 16-32 спо¬
рами; таллом зернисто-накипной; на карбонатных ска¬
лах, почве, в аридных и ксеротических условиях
Gonohymenia s. str. (Lichinella s. 1.).
— Признаки другие 5.
5. Таллом с воздушными полостями, чешуйчато-накипной (-
ареольчатый); плодовые тела - перитециевидные апоте¬
ции с точковидным вдавленным диском; гимений без
парафиз; фотобионт - Gleocapsa с красно-коричневыми
влагалищами Phylliscum.
— Таллом без воздушных полостей; плодовые тела - сидячие
или погруженные апотеции или пикноаскокарпы, в том
числе перитециевидные 6.
6. Эксципул расширенный в верхней части до 100 мкм; апоте¬
ции с отчетливым собственным краем, имеют вид рыбь¬
их глаз; гимений I- ; на влажных, заливаемых силикат¬
ных скалах Pyrenocarpon.
— Эксципул не расширенный; плодовые тела с узким соб¬
ственным краем или без него, слоевищный край более
или менее хорошо развитый 7.
7. Апотеции с глянцевым красно-коричневым диском; таллом
красновато-коричневый, наряду с цианобактерией ино¬
гда содержит зеленую водоросль Trebouxia; таллом из
сетчато переплетенных гиф, вокруг Trebouxia - парап¬
лектенхимный; стенки сумок толстые; на мокрых сили¬
катных скалах, артко-альпийский Euopsis.
— Диск апотециев не глянцевый; никогда не содержит зеле¬
ную водоросль 8.
8. Таллом плотный, параплектенхимный, с вертикальными ря¬
дами (кластерами) мелкоклеточной цианобактерии,
окруженными коричневыми влагалищами; апотеции
редкие; гимений со стерильными участками, 1+ синеет;
слоевищный край ложный, отграничен от таллома тре¬
щинкой; на карбонатных скалах Metamelaena.
— Строение таллома другое, апотеции без стерильных участ¬
ков 9.
9. Гифы короткоклеточные, веерообразно направленные, что
придает таллому вид параплектенхимного; фотобионт
мелкоклеточный, 3-6 мкм в диам, в кластерах с корич¬
неватыми влагалищами; апотеции с отчетливым слое¬
вищным краем; гимений 1+ синеет; на заливаемых ска¬
лах Pterygiopsis.
— Гифы другие, не направлены веерообразно; таллом парап¬
лектенхимный или выглядит таким 10.
10. Плодовые тела - погруженные перитециевидные апотеции
или пикноаскокарпы, с точковидным в виде поры дис¬
ком, с перифизами или без них; на мокрых, периодиче¬
ски заливаемых силикатных скалах 11.
— Плодовые тела - леканоровые апотеции или апотециевид-
ные пикноаскокарпы; диск расширенный или узкий, но
не поровидный 12.
11. Апотеции всегда закрытые, с перифизами в верхней части;
эксципул хорошо развит; гимений без парафиз (иногда с
разветвленными, анастомозирующими парафизоидами);
сумки с 8 (редко 16-24) спорами; конидии игловидные;
фотобионт с красно-коричневыми влагалищами
Cryptothele.
— Апотеции с закрытым или немного открывающимся дис¬
ком, без перифиз; эксципул плохо развит; гимений с па-
рафизами или без них; сумки с 8 реже 16-64 спорами;
конидии эллипсоидные или палочковидные; фотобионт с
коричневыми или красно-коричневыми влагалищами,
сборный род Pyrenopsis (часть видов).
12. Фотобионт с красно-коричневыми влагалищами; таллом
красновато-коричневый; апотеции с вогнутым, позднее
плоским коричневым или черноватым диском; на зали¬
ваемых гранитных и карбонатных скалах; или фото¬
бионт с коричневатыми влагалищами, тогда на почве и
мхах в тундре Pyrenopsis (часть видов).
— Фотобионт с коричневатыми влагалищами, в кластерах или
одиночных клетках; таллом оливково-коричневый или
коричнево-черный (иногда с серым налетом); в более
или менее сухих местообитаниях 13.
13. Плодовые тела - пикноаскокарпы, редкие, сначала закры¬
тые, зрелые - с открытым красно-коричневым диском,
тонким эксципулом и толстым (200-250 мкм) слоевищ¬
ным краем; парафизы двух типов: неразветвленные
длинноклеточные и разветвленные в виде нитей жемчу-
га; таллом из сетчато-переплетенных гиф с группами
клеток цианобактерии, придающими ему параплектен¬
химный вид; на карбонатных породах Апеша.
— Плодовые тела - апотеции, обычные, зрелые от полупогру-
женных до сидячих, с плоским коричневым диском, тон¬
ким не замкнутым внизу эксципулом и постоянным сло¬
евищным краем; парафизы другие; таллом мелкоклеточ-
но-параплектенхимный; на карбонатных и силикатных
породах Psorotichia.
14. Таллом желатинозный, преимущественно без корового
слоя; фотобионт Callothrix, Nostoc или ностоковидный в
цепочках или кластерах; споры одноклеточные 15.
— Таллом нежелатинозный, с более или менее выраженными
коровым и сердцевинным слоями или полностью парап¬
лектенхимный; фотобионт Nostoc или Scytonema-Bwj\\\h\v\
в кластерах; споры одноклеточные, многоклеточные,
муральные 20.
15. Фотобионт Callothrix; лишайники, обитающие на мокрых и
влажных силикатных скалах, по берегам ручьев и озер
сем. Lichinaceae 16.
— Фотобионт Nostoc (или ностоковидный) в кластерах или
цепочках 17.
16. Таллом из рыхло переплетенных круглоклеточных гиф,
окружающих цианобактерию, слабо лихенизированный;
Callothrix в трихомах, клетки 4-10 мкм диам.; плодовые
тела - апотеции, маленькие, погруженные, 0.2-0.5 мм
диам.; эксципул чашевидный с точковидным диском на
дне; гимений I- Thelignia.
— Таллом из плотно переплетенных гиф; ареолы гранулезные
или изидиозные с фонтановидной (в разрезе) парап-
лектенхимной структурой до 500 мкм шир, Callothrix в
разорванных цепочках, клетки 6-9 х 3-6 мкм; плодовые
тела - пикноаскокарпы с исчезающим слоевищным кра¬
ем; гимений 1+ синеет (не всегда); парафизы двух типов:
анастомозирующие первичные, утолщенные наверху
вторичные Porocyphus.
17. Апотеции леканоровые, часто слабо развитым, с отчетли¬
вым эксципулом; таллом ареолированно- или зернистый
накипной, без корового слоя, вокруг апотеция парап¬
лектенхимный; на карбонатных скалах, глинистой почве
Lemmopsis (Lichinaceae).
— Апотеции биаторовые, зрелые выпуклые; таллом зернисто-
или чешуйчато-накипной с зернистой поверхностью 18.
18. Апотеции черно-коричневые, редкие; таллом без корового
слоя, оливково-черный; Nostoc в цепочках; на почве,
растительных остатках, мхах, растущих на карбонатном
грунте, аркто-альпийский
Leciophysma (Collemataceae).
— Апотеции оранжево- или красно-коричневые, обычны; тал¬
лом с коровым слоем, Nostoc в кластерах; таллом зерни¬
сто-накипной 19.
19. Таллом оливково-бурый, состоящий из гониоцист, окру¬
женных коровым слоем; апотеции красно-коричневые с
быстро исчезающим собственным краем; гимений /-;
споры с тонкой оболочкой; на свеженарушенной почве .
Gregorella (Arctomiaceae).
— Таллом коричневый, морщинистый, из рыхло переплетен¬
ной сети гиф с однослойной корой; апотеции оранжево¬
коричневые; споры со слабобородавчатым экзоспорием;
на мхах и растительных остатках, аркто-альпийский
Santessoniella (Pannariaceae).
20. Апотеции буро-черные, биаторовые; споры 2-6-клеточные;
таллом параплектенхимный, накипной или чешуйчато¬
накипной, часто с сине-черным подслоевищем; фото¬
бионт - Scytonema-типа; на сухих карбонатных и влаж¬
ных силикатных скалах, редко на карбонатной почве
сем. Placynthiaceae 21.
— Апотеции красно-коричневые, леканоровые или биаторо¬
вые; споры одноклеточные, поперечно-многоклеточные
или муральные; фотобионт-Nostoc; на почве 22.
21. Таллом накипной, комковато-трещиноватый, аморфный,
серый, с налетом из оксалата кальция, без подслоевища;
на карбонатных породах Collolechia.
— Таллом чешуйчато-накипной, розетковидный, часто с сине¬
черным подслоевищем Placynthium.
22. Споры муральные; таллом полностью параплектенхимный,
ареолированный, ареолы зернисто-чешуйчатые; апоте¬
ции леканоровые, редкие, с плоским или вогнутым ко¬
ричневым диском; фотобионт - Nostoc (или Scytonema);
на обнаженной, обычно карбонатной почве, в более или
менее сухих местообитаниях
Epiphloea (Heppiaceae).
— Споры не муральные; таллом с параплектенхимным или
примитивным коровым слоем, мелко-чешуйчатый или
зернистый; апотеции от светлых розово-коричневых до
темно-красно-коричневых, обычны; фотобионт - Nostoc;
в более или менее влажных местообитаниях 23.
23. Споры поперечно-многоклеточные с 4-10 перегородками;
апотеции биаторовые темно-красно-коричневые; таллом
коричневый, чешуйчато-накипной или розетковидный с
краевыми лопастинками; на почве, мхах, отмершей коре
деревьев Arctomia (Arctomiaceae).
— Споры одноклеточные; апотеции розовато-, или оранжево¬
коричневые; таллом голубовато или коричнево-серый,
сем. Pannariaceae 24.
24. Апотеции леканоровые, гимений 1+ темно синеет; споры с
бородавчатым экзоспорием, 25-30 х 9-12 мкм (19-25 х
8-10 мкм без экзоспория); таллом чешуйчато-накипной,
из дланевидных чешуек; на гумусной почве, мхах, во
влажных местообитаниях, аркто-альпийский
Protopannaria.
— Апотеции биаторовые, часто с ложным слоевищным краем
из зерен таллома; гимений 1+ сине-зеленеет, затем крас-
но-коричневеет; споры 10-15(20) х 5-8 мкм с гладкой
оболочной; таллом зернисто-накипной, из кластеров
Nostoc, оплетенных гифами, с примитивной корой; на
обнаженной глинистой почве Moelleropsis.
Ключ 8.
Накипные лишайники с апотециями на ножках и фото¬
бионтом - зелеными водорослями. Автор: Кузнецова Е. С.:
1. В апотециях формируется мазедий 2.
— Мазедий не формируется 9.
2. Споры одноклеточные, округлые или эллипсоидные 3.
— Споры двухклеточные, многоклеточные или субмуральные,
эллипсоидные 5.
3. Мазедий светлый, розоватый или желтоватый, оболочка
спор бесцветная (гиалиновая) Sclerophora.
— Мазедий коричневый или черный, оболчка спор пигменти¬
рованная, коричневая до почти черной 4.
4. Мазедий черный, головки апотециев яйцевидные с сильно
суженными краями эксципула, длина ножек сопоставима
с шириной головок Sphinctrina.
— Мазедий коричневый, головки апотециев иной формы, дли¬
на ножек в два и более раза превышает ширину головок
Chaenotheca.
5. Головки апотециев субсферической, булавовидной или об¬
ратнояйцевидной формы, с сильно суженными краями
эксципула 6.
— Головки апотециев иной формы 7.
6. Собственный лихенизированный таллом не образуется, па¬
разиты или парасимбионты на талломах накипных ли¬
шайников (преимущественно из родов Lecanora и Per¬
tusaria), мазедий не выходит за пределы апотециев, спо¬
ры покрыты желатинизированной оболочкой
Sphinctrina.
— Таллом соредиозный, фотобионт Trentepohlia, мазедий воз¬
вышается над апотециями, споры без желатинизирован¬
ной оболочки Pyrgidium.
7. Мазедий коричневый Chaenotheca.
— Мазедий черный или зеленый 8.
8. Мазедий зеленый или грязно зеленый, сумки широкоэллип¬
соидные, формирующиеся цепочками, собственный ли¬
хенизированный таллом не формируется, паразиты на
колониях водорослей или талломах накипных лишайни¬
ков Microcalicium.
— Мазедий черный, сумки цилиндрические или булавовид¬
ные, располагаются в гимении, таллом лихенизирован¬
ный Calicium.
9. Собственный лихенизированный таллом не формируется,
обитают в качестве сапротрофов на коре и древесине или
паразитируют на колониях водорослей или талломах
лишайников 10.
— Имеется лихенизированный таллом, обитают преимуще¬
ственно на почве, валеже, камнях и растительных остат¬
ках 11.
10. Длина сумок 20-70 мкм
Chaenothecopsis, Mycocalicium.
— Длина сумок более 70 мкм Phaeocalicium, Stenocybe.
11. Апотеции языковидные, на коротких ножках
Icmadophila.
— Апотеции иной формы 12.
12. Апотеции формируются на концах полых, вздутых, про¬
стых или слабо разветвленных ножек
Pycnothelia papillaria.
— Ножки апотециев иные 13.
13. Диаметр апотециев не превышает 0.5 мм, парафизы ветвя¬
щиеся, часто обвивающие сумки Vezdaea.
— Диаметр апотециев более 1 мм, парафизы простые, лишь
изредка разветвляющиеся, никогда не обвивающие сум¬
ки 14.
14. Апотеции розовые Dibaeis.
— Апотеции коричневые до красновато-коричневых
Baeomyces.
Ключ 9.
Накипные лишайники с вытянутыми или неправильной
формы апотециями и фотобионтом - зелеными водорослями.
Автор: Кузнецова Е. С.
1. Апотеции без собственного края, или собственный край
недоразвитый, развивающийся лишь по бокам апотециев
2.
— Апотеции с хорошо развитым собственным краем 3.
2. Споры 1-7 клеточные, яйцевидные или продолговато яйце¬
видные Arthonia.
— Апотеции субмуральные до муральных, эллипсодно-
яйцевидные Arthotelium.
3. Споры простые 4.
— Споры многоклеточные или муральные 6.
4. Споры мелкие 4-6 х 2-3 мкм Elixia flexella.
— Споры крупнее 5.
5. Внешняя поверхность апотециев черная, на скалах
Lithographa.
— Внешняя поверхность апотециев светло-коричневая, на
древесине, изредка на коре Xylographa.
6. Споры 2-клеточные Melaspilea.
— Споры многоклеточные или муральные 7.
7. Споры при действии I окрашиваются в фиолетовый цвет,
люмина клеток линзовидная, округлая или овальная
8.
— Споры при действии I не окрашиваются, люмина клеток
прямоугольная или квадратная 9.
8. Зрелые споры серо-коричневые, апотеции плоские с неза¬
метным, едва приподнимающимся краем и широким
диском Phaeographis.
— Зрелые споры бесцветные, с хорошо выраженным, часто
продольно бороздчатым краем и узким диском
Graphis.
9. Споры коричневые Opegrapha.
— Споры бесцветные 10.
10. Собственный край хорошо развитый, коричневый до чер¬
ного (срез!) 11.
— Собственный край плохо развит или отсутствует 13.
11. Собственный край состоит из рыхло переплетенных темно-
коричневых гиф, более светлых по периферии собствен¬
ного края Lecanactis.
— Собственный край состоит из плотно переплетенных, чер¬
ных гиф, не светлеющих к краю 12
12. Диск апотеция с выраженным беловатым, сероватым или
желтоватым налетом, апикальный аппарат сумок gramu-
losa-типа Lecanographa.
— Диск апотеция без налета, апикальный аппарат сумок vulga-
ta-типа Opegrapha.
13. Гипотеций темный, красно-коричневый, споры 4-
клеточные Schizmatomma.
— Гипотеций бесцветный или светло-коричневый, споры 4-
17-клеточные Enterographa.
Ключ 10.
Накипные лишайники с круглыми апотециями и фото¬
бионтом - Trentepohlia. Автор: Гагарина Л. В.
1. Сумки содержат более 8 спор Pachyphiale.
— Сумки содержат менее 8 спор 2.
2. Апотеции на коротких или длинных ножках, с хорошо раз¬
витым мазедием 3.
— Апотеции сидячие, мазедий отсутствует 5.
3. Аскоспоры коричневые 4.
— Аскоспорые не коричневые, бледные, шаровидной формы
Sclerophora.
4. Аскоспоры 2-х клеточные, таллом поверхностный, сореди¬
озный, коричневый или коричнево-черный
Pyrgidium.
— Аскоспоры 1-клеточные, реже 2-5 клеточные, таллом по¬
верхностный или погруженный, не соредиозный, серый,
зеленый, желтый или коричневый Chaenotheca.
5. Эксципул состоит из клеток угловатой формы, перифизы
присутствуют Ramonia.
— Клетки эксципула иной формы, перифизы отсутствуют
6.
6. Апотеции погруженные, похожи на перитеции 7.
— Апотеции погруженные или сидячие, не похожи на перите¬
ции 8.
7. Апротеции погружены в бородавки таллома, диск черный,
слоевищный и собственный крй хорошо выражены, сум¬
ки содержат от 1 до 8 спор, споры с поперечными пере¬
городками или муральные, без периспория
Thelotrema.
— Апотеции погружены в субстрат, диск не черный, слоевищ¬
ный и собственный край плохо различимы, слоевищный
край обычно плохо развит, сумки всегда с 8 спорами,
споры только с поперечными перегородками, с периспо¬
рием Petractis.
8. Аскоспоры 1-клеточные, апотеции аспициллевидные, с по¬
груженным диском и тонким слоевищным краем
Ionaspis и Hymenelia.
— Аскоспоры 2-клеточные или многоклеточные, апотеции
дисковидные или точковидные, часто без слоевищного
края 9.
9. Споры 2-клеточные Coenogonium.
— Споры многоклеточные 10.
10. Собственный край апотеция не развит или плохо выражен
11.
— Собственный край апотеция хорошо развит 13.
11. Гипотеций отсутствует или не заметен Arthonia.
— Гипотеций хорошо выражен, темно-оливковой или черной
окраски 12.
12. Апикальный аппарат сумок Opegrapha-типа, споры 4-
клеточные, клетки спор одинаковые по размерам Dirina.
— Апикальный аппарат сумок не Opegrapha-типа, споры бо¬
лее чем 4-клеточные, клетки спор не одинакового разме¬
ра Schismatomma.
13. Апотеции лиреллевидной или широковеретеновидной
формы, таллом часто соредиозный Opegrapha.
— Апотеции округлые, таллом не соредиозный 14.
14. Аскоспоры нитевидные, с многочисленными перегородка¬
ми, сумки из-за фрагментации спор выглядят как мно¬
госпоровые Bactrospora.
— Аскоспоры иной формы, сумки с 8 спорами, не распадаю¬
щимися на части 15.
15. Диск апотеция без налета 16.
— Диск апотеция покрыт налетом 18.
16. Слоевищный край цельный, редко радиально потрескав¬
шийся, диск не черный (искл. Gyalecta nigricans), эсци-
пул и гипотеций светлые, споры поперечно 4-клеточные,
многоклеточные или муральные Gyalecta.
— Слоевищный край глубоко звездчато потрескавшийся, диск
черно-коричневый или черный, эксципул в наружной ча¬
сти черно-бурый, внутри - желто-коричневый или бес¬
цветный, споры 4-клеточные 17.
17. Аскоспоры с тонким периспорием, апотеции сидячие
Rhexophiale rhexoblephara.
— Аскоспоры без периспория, апотеции погруженные или си¬
дячие Sagiolechia protuberans.
18. Гимений /+, приобретает грязно-желтую или оливково¬
зеленую окраску Lecanographa.
— Г имений /-, или /+, приобретает синюю окраску 19.
19. Гипотеций темно-бурый Lecanactis.
— Гипотеций бесцветный или в нижней части слабо бурый
Cresponea.
Ключ 11.
Нелихенизированные грибы (без фотобионта) с круглы¬
ми апотециями. Автор: Кузнецова Е. С.
1. Апотеции на ножках 2.
— Апотеции сидячие 5.
2. В апотециях формируется мазедий 3.
— Мазедий не формируется 4.
3. Головки апотециев субсферической, булавовидной или об¬
ратнояйцевидной формы, с сильно суженными краями
эксципула, мазедий черный, споры гладкие или с боро¬
давчатым орнаментом Sphinctrina.
— Головки апотециев иной формы, мазедий зеленый или
грязно-зеленый, споры с орнаментом из спирально ори¬
ентированных ребер Microcalicium.
4. Длина сумок 20-70 мкм
Chaenothecopsis, Mycocalicium.
— Длина сумок более 70 мкм Phaeocalicium, Stenocybe.
5. На смоле, апотеции розовые или темно-коричневые
Sarea.
— На других субстратах 6.
6. Молодые апотеции, погруженные в субстрат и открываю¬
щиеся порой, со временем прорывающиеся 7.
— Апотеции иные 8.
7. Диск апотеция в сухом состоянии отделяется от края, по
краю формируется густой белый налет Stictis.
— Диск апотеция в сухом состоянии от края не отделяется, по
краю белый налет не формируется Odontotrema.
8. Споры простые Agyrium.
— Споры 2-клеточные, многоклеточные или муральные 9.
9. Споры муральные или субмуральные Arthotelium.
— Споры 2-клеточные или многоклеточные 10.
10. Сумки широко-булавовидные или округлые Arthonia.
— Сумки вытянуто-булавовидные Melaspilea
Ключ 12.
Накипные лишайники с круглыми апотециями, фото¬
бионтом - зелеными хлорококковым и водорослями и окра¬
шенными спорами. Авторы: Конорева Л. А, Андреев М. П.
1. Споры темноокрашенные 2.
— Споры бесцветные 28.
2. Споры простые, до поперечно-многоклеточных 3.
— Споры муральные, иногда субмуральные 19.
3. При освобождении спор из сумок на поверхности апотециев
формируется порошистая масса - мазедий 4.
— Мазедий не формируется 7.
4. Апотеции на ножках 5.
— Апотеции сидячие или погруженные 6.
5. Мазедий коричневый Chaenotheca.
— Мазедий черный Calicium.
6. Таллом с черными, гранулярными изидиями, седцевина 1+
(синеет) Thelomma.
— Таллом без изидий, седцевина I- Cyphelium.
7. Паразитирует на талломах видов рода Baeomyces Epilichen.
— Свободноживущие или паразитирующие на других видах
лишайников 8.
8. Таллом плакодиоидный 9.
— Таллом накипной или мелкочешуйчатый 10.
9. Краевые лопасти хорошо развиты, до 1.2 мм в диам, зеле-
новато-желтые, на камнях Dimelaena.
— Краевые лопасти мельче, иначе окрашены, на различных
субстратах Buellia epigaea.
10. Слоевищный край апотеция содержит водоросли 11.
— Слоевищный край апотеция без водорослей, в эксципуле и
гипотеции фотобионт отсутствует 14.
11. Стенки спор тонкие, до 0.5 мкм толщиной Rinodinella.
— Стенки спор значительно толще, равномерно или неравно¬
мерно утолщенные 12.
12. Конидии нитевидные, до 30 мкм длиной Amandinea.
— Конидии палочковидные 13.
13. Зрелые споры от 4-клеточных до слабомуральных
Diplotomma.
— Зрелые споры 2-клеточные, редко 3-4-клеточные или сла¬
бомуральные Rinodina.
14. Споры с отчетливой слизистой оболочкой (гало)
Rhizocarpon.
— Споры без слизистой оболочки 15.
15. Апотеции лирелловидные, до широковеретеновидных, ре¬
же округлые, диск щелевидный Opegrapha.
— Апотеции другого строения 16.
16. Споры одноклеточные, редко 2-клеточные, стенки спор
тонкие Orphniospora.
— Споры от 2-клеточных до многоклеточных, стенки спор
утолщенные 17.
17. Конидии нитевидные, до 30 мкм длиной Amandinea.
— Конидии палочковидные 18.
18 Buellia.
— Tetramelas.
19. При освобождении спор из сумок на поверхности апотеци¬
ев формируется порошащая масса - мазедий
Cyphelium.
— Мазедий не формируется 20.
20. Споры с отчетливой слизистой оболочкой (гало)
Rhizocarpon.
— Споры без слизистой оболочки 21.
21. Сумки с одной спорой 22.
— Сумки 2-8-споровые 23.
22. Апотеции сидячие Lopadium.
— Апотеции полупогруженные или погруженные Xyloschistes.
23. Сумки 2-4 споровые 24.
— Сумки 8-споровые 26.
24. Таллом Р+ оранжевый, К+ сначала желтеет, потом красне¬
ет, ХС+красный, С- (норстиктовая кислота) Phlyctis.
— Реакции таллома другие 25.
25. Апотеции перитециевидные, кувшинообразные, на почве,
камнях, мхах
Diploschistes.
— Апотеции округлые, полупогруженные, на древесине
Xyloschistes.
26. Верхушки парафиз утолщенные 27.
— Верхушки парафиз не утолщены Diploschistes.
27. Эксципул плохо развит Diplotomma.
— Экципул хорошо развит, внешняя часть оранжево¬
коричневая Leptotrema.
28. Споры одноклеточные, до поперечно-многоклеточных 29.
— Споры муральные, до субмуральных 31.
29. Споры с отчетливой слизистой оболочкой (гало)
Rhizocarpon.
— Споры без слизистой оболочки 30.
30. Споры одноклеточные, редко 2-клеточные
Orphniospora.
— Споры имеют 4 или более клеток Opegrapha.
31. Споры с отчетливой слизистой оболочкой (гало)
Rhizocarpon.
— Споры без слизистой оболочки 32.
32. Апотеции лирелловидные, до широковеретеновидных,
звездчатые, удлиненные 33.
— Апотеции округлые, сидячие или перитециевидные, полу¬
погруженные 34.
33. Апотеции лирелловидные, до широковеретеновидных,
эксципул присутствует Opegrapha.
— Апотеции звездчатые, удлиненные или неправильной фор¬
мы, эксципул отсутствует Arthothelium.
34. Апотеции сидячие 35.
— Апотеции полупогруженные или погруженные 36.
35. Сумки с 1 спорой Lopadium.
— Сумки с 8 спорами Opegrapha.
36. Сумки с 2 (4) спорами Phlyctis.
— Сумки с 8 спорами Leptotrema.
Ключ 13.
Накипные лишайники с круглыми апотециями, фото¬
бионтом - зелеными хлорококковыми водорослями и много¬
численными (более 8 в сумке) бесцветными спорами. Автор:
Кузнецова Е. С.
1. Таллом оранжевый, желтый или яркий зелено-желтый 2.
— Таллом иного цвета 4.
2. Таллом К+ интенсивно красный (быстрая реакция)
Caloplaca.
— Таллом К- 3.
3. Спор в сумках не более 32 Candelariella.
— Спор в сумках более 100 Pleopsidium.
4. Спор в сумках не более 32 5.
— Спор в сумках более 32, до 100 и более 11
5. Споры простые 6.
— Споры состоят из двух и более клеток 8.
6. Апотеции со слоевищным краем Lecanora.
— Апотеции без слоевищного края 7.
7. Сумки бутылковидные или цилиндрические, апотеции часто
с желтым налетом Thelocarpon.
— Сумки продолговатые до булавовидно-цилиндрических,
апотеции черные или коричневые без желтого налета
Steinia geophana.
8. Споры полярилокулярные Caloplaca.
— Споры иные 9.
9. Споры веретеновидные или игловидные, изогнутые
Scoliciosporum.
— Споры эллипсоидные 10.
10. Слоевищный край присутствует, по крайней мере, у моло¬
дых апотециев Lecania.
— Слоевищный край отсутствует Catinaria.
11. Сумки бутылковидные, до цилиндрических, апотеции ча¬
сто с желтым налетом Thelocarpon.
— Сумки цилиндрические, булавовидные или грушевидные,
апотеции без желтого налета 12.
12. На коре, древесине или на эпифитных мхах 13.
— На почве и растительных остатках 16.
— На камнях 18.
13. Споры круглые 14.
— Споры эллипсоидные 15.
14. Апотеции светлые желтоватые или коричневатые, апи¬
кальный аппарат сумок 1+ синий Biatoridium.
— Апотеции красные, коричневые или черные, апикальный
аппарат сумок I- Strangospora.
15. Апотеции без слоевищного края, охристые или оранжево¬
красные с густым оранжевым налетом Piccola.
— Апотеции с хорошо развитым серым слоевищным краем и
черным диском Магопеа.
16. Собственный край очень тонкий или не развит
Biatorella.
— Собственный край хорошо развит 17.
17. Апотеции черные, на ранних стадиях развития напомина¬
ющие перитеции, позднее раскрываются и становятся
гирозными Polysporina.
— Апотеции темные красно-коричневые, не напоминающие
перитеции, взрослые не гирозные
Sarcosagium campestre.
18. Апотеции без слоевищного края (срез!) 19.
— Апотеции со слоевищным краем (срез!) 21.
19. Развит гипоталлом, слоевище в виде хорошо развитой аре-
олированной корочки Sporastatia.
— Гипоталлом отсутствует, слоевище иное 20.
20. Парафизы разветвленные, анастамозирующие, апотеции
гирозные Polysporina.
— Парафизы простые или лишь изредка ветвящиеся, не ана¬
стамозирующие, апотеции не гирозные Sarcogyne.
21. Апотеции более 0.3 мм в диаметре, таллом покрыт коро¬
вым слоем, апикальный аппарат сумок I-
Acarospora, Silobia (=Myriospora).
— Апотеции мелкие 0.2-0.4 мм в диаметре, таллом лишен ко¬
рового слоя, апикальный аппарат сумок 1+ синий
Myriospora.
Ключ 14.
Накипные лишайники с круглыми (сидячими) апотеция¬
ми, фотобионтом - зелеными хлорококковыми водорослями и
бесцветными многоклеточными (2-клеточными, поперечно¬
многоклеточными или муральными спорами (не более 8 в
сумке). Автор: Гагарина Л. В.
1. На поверхности листьев 2.
— На других субстратах 7.
2. Аскоспоры муральные, по 1-2 аскоспоре в сумке 3.
— Аскоспоры только с поперечными перегородками 4.
3. Эксципул сильно желатинозный, образован сетью анасто-
мизирующих гиф, внешняя часть эксципула «распро¬
странена» по таллому, фотобионт хлорококковый
Echinoplaca.
— Эксципул слабо развит, прозоплектенхимный, внешним
краем плотно срастается с талломом, фотобионт всегда
Trebouxia Gyalectidium.
4. Апикальный аппарат сумок Byssoloma-типа, перегородки у
спор всегда четкие 5.
— Апикальный аппарат сумок Bacidia-типа, перегородки спор
часто не четкие Bacidina.
5. Гипотеций не окрашен, конидии сливовидной формы, апо¬
теции часто с белым налетом Fellhanera.
— Гипотеций красно-коричневый, конидии колбовидной, па¬
лочковидной, эллипсоидной или нитевидной формы,
апотеции без налета 6.
6. Эксципул паутинистый, состоит из сложно переплетенных
толстостенных гиф, конидии всегда одного вида, про¬
стые, колбовидные, палочковидные или эллипсоидные,
суженные посередине Byssoloma.
— Эксципул тонкий, без плотно переплетенных толстостен¬
ных гиф, конидии двух типов: простые, длинные, ните¬
видные или палочковидные Felhaneropsis.
7. Аскоспоры одноклеточные или только с поперечными пере¬
городками 8.
— Аскоспоры муральные 10.
8. Аскоспоры одноклеточные 9.
— Аскоспоры только с поперечными перегородками 19.
9. Верхняя кора отсутствует или очень слабо развита, апоте¬
ции без налета, слоевищный край отсутствует, сумки
Catillaria-, Bacidia- или Biatora-типа Catillaria.
— Верхняя кора обычно развита, кристаллизирована или по¬
крыта эпинекральным слоем, апотеции часто с налетом,
сумки Bacidia или Biatora-типа Lecania (cyrtella).
10. Диск апотециев светлый, беловатый, светло-желтый или
коричнево-серый 11.
— Диск апотециев темно-бурый, красно-коричневый, черный
13.
11. Поверхность таллома ровная, мелкобородавчатая, ареоли-
рованная, обычно инкрустирована кристаллами оксалата
кальция, апотеции Calenia-типа Gyalectidium.
— Таллом бородавчатый, цельный или трещиноватый, апоте¬
ции иного типа 12.
12. Таллом бородавчатый, сумки 1-2 споровые, с толусом,
KJI+, приобретают синюю окраску Brigantiaea.
— Таллом цельный или трещиноватый, сумки 4-8 споровые,
слегка утолщены в апикальной части, К/I- Gyalidea.
13. Аскоспоры с тонким или толстым периспорием
Gyalideopsis.
— Аскоспоры без периспория 14.
14. Таллом с хорошо развитым подслоевищем 15.
— Таллом без хорошо развитого подслоевища 16.
15. Таллом К+, краснеет, подслоевище белое Phlyctis.
— Таллом К—, подслоевище черное, белое или коричнево¬
серое Rhizocarpon.
16. Сумки содержат 1 спору 17.
— В сумках содержится от 2 до 8 спор 18.
17. Аскоспоры 20-39 мкм длиной, на обработанной древесине
и веточках сосны лишенных коры Xyloshistes.
— Асксоспоры 40-135 мкм длиной, на мхах, коре или расти¬
тельных остатках Lopadium.
18. Апотеции без четко выраженного края, сумки содержат по
8 спор Arthothelium.
— Апотеции с четко выраженным черным или коричнево¬
черным краем, сумки содержат 2-8 спор Schadonia.
19. Апотеции на коротких ножках Ichmadophila.
— Апотеции сидячие или погруженные 20.
20. Таллом накипной, плакоидный, чешуйчатый, оранжево¬
желтой, реже серой или черной окраски, К+, становится
пурпурным, эпитеций с многочисленными кристаллами,
К+, становится пурпурным Caloplaca.
— Таллом накипной, К-, эпитеций без кристаллов, К- 21.
21. Подслоевище выражено, белое, темно-коричневое, корич¬
нево-серое или черное 22.
— Подслоевище не заметно 26.
22. В сумках содержится 1-8 спор 23.
— В сумках содержится 8-30 и более спор 25.
23. Таллом образует гониоцисты, апикальный аппарат сумок
Bacidia-типа Bacidina.
— Таллом не образует гониоцисты, апикальный аппарат сумок
иного типа 24.
24. Гипотеций коричневый, апикальный аппарат сумок Rhizo-
сагроп-типа, аскоспоры с периспорием Rhizocarpon.
— Гипотеций бесцветный, сумки булавовидные, не утолщен¬
ные в апикальной части, аскоспоры без периспория
Cliostomium.
25. Аскоспоры овальные или веретеновидные. Сумки не окру¬
жены амилоидным желатинозным матриксом. Апикаль¬
ный аппарат сумок Byssoloma типа. Конидии сливовид¬
ные Fellhanera.
— Аскоспоры игловидной формы, сумки окружены амилоид¬
ным желатинозным матриксом, апикальный аппарат су¬
мок состоит из наружного амилоидного слоя, внутренне¬
го амилоидного слоя и в меньшей степени амилоидного
слоя между ними, конидии палочковидные
Ropalospora.
26. Таллом лимонно-желтый или интенсивно желто-зеленый ..
27.
— Таллом иной окраски 28.
27. Апотеции черные, без налета, гимений с многочисленными
каплями масла Arthrorhaphis.
— Апотеции зелено-желтые, бледно-оранжевые или коричне¬
вые, часто с налетом, гимений без капель масла
Chrysothrix.
28. Апикальные части парафиз темно-коричневые . Halecania.
— Апикальные части парафиз бесцветные 29.
29. Аскоспоры с бородавковидным эписпорием Bilimbia.
— Аскоспоры без бородавковидного эписпория 30.
30. Апотеции располагаются в плодовых бородавочках по 1-5
31.
— Апотеции сидячие или погруженные, не располагаются в
плодовых бородавочках 32.
31. Сумки содержат 1 крупную спору, споры состоят из 2 кле¬
ток Varicellaria.
— Сумки содержат 8 спор, споры состоят из 2-4 клеток
Anzina.
32. Парафизы с темноокрашенными апексами 33.
— Парафизы с неокрашенными апексами 35.
33. Споры бобовидные, 4- клеточные, сумки Bacidia-типа
Arthrosporum.
— Сумки Catillaria-, Bacidia- или Biatora-типа. Споры эллип¬
соидные или широко эллипсоидные 34.
34. Сумки Catillaria-ти па. Аскоспоры с желатинозным обод¬
ком Catinaria.
— Сумки Catillaria-, Bacidia- или Biatora-типа, аскоспоры
без желатинозного ободка Catillaria.
35. Аскоспоры с 39-49 поперечными перегородками
Stictis.
— Аскоспоры с 38 и меньшим числом поперечных перегоро¬
док 36.
36. Апотеции со слоевищным краем 37.
— Апотеции без слоевищного края 40.
37. Конидии сливовидной формы Fellhanera.
— Конидии палочковидной, нитевидной или изогнуто¬
нитевидной формы 38.
38. Парафизы простые, не утолщенные в апикальной части, не
анастомозирующие, таллом образует гониоцисты
Bacidina.
— Парафизы простые или разветвленные, не утолщенные или
утолщенные в апикальной части, анастомозирующие или
нет, таллом не образует гониоцист 39.
39. Апикальный аппарат сумок Lecanora-типа
Haematomma.
— Сумки не имеют апикального аппарата Ophioparma.
40. Апотеции с сильно вогнутым диском, при увлажнении тал¬
лом ослизняется 41.
— Апотеции с выпуклым, плоским и вогнутым диском, тал¬
лом не ослизняется при увлажнении 42.
41. Гимений K/I+, синеет, сумки слегка утолщены в апикаль¬
ной части Cryptodiscus.
— Г имений K/I-, сумки с толстой апикальной зоной
Absconditella.
42. Клетки спор не одинакового размера, часто апикальная
клетка крупнее остальных, апикальный аппарат сумок
Arthonia-ти па, сумки грушевидной, обратнояйцевидной
или округлой формы Athonia.
— Клетки спор более или менее одинакового размера, апи¬
кальный аппарат сумок иного типа или отсутствует, сум¬
ки обычно булавовидной или широко булавовидной
формы 43.
43. Гимений с многочисленными каплями масла
Megalospora.
— Капли масла в гимении отсутствуют 44.
44. Настоящий эксципул рыхлый, состоит из переплетенных
тонкостенных гиф - «биссоидный» эксципул Byssoloma.
— Настоящий эксципул имеет иное строение 45.
45. Конидии не известны Scoliciosporum.
— Конидии имеются 46.
46. Эксципул состоит из радиальных гиф, часто с утолщенной
оболочкой Bacidia.
— Эксципул имеет иное строение 47.
47. Апотеции лиреллевидные или широковретеновидные, экс¬
ципул черный Opegrapha.
— Апотеции округлые, эксципул иной окраски 48.
48. Конидии извитые Lecania.
— Конидии палочковидной или эллипсоидной формы 49.
49. Аскоспоры с периспорием 50.
— Аскоспоры без периспория 51.
50. Парафизы разветвленные и анастомозирующие, аскоспоры
с 15-20 поперечными перегородками, часто с гифофора-
ми Gyalideopsis.
— Парафизы преимущественно простые, аскоспоры с 1-3 по¬
перечными перегородками, гифофоры отсутствуют
Gyalidea.
51. Аскоспоры 2-клеточные Megalaria.
— Аскоспоры 4-8-клеточные Loxospora.
Ключ 15.
Накипные лишайники с круглыми апотециями, фото¬
бионтом - зелеными хлорококковыми водорослями и бесцвет¬
ными одноклеточными спорами (не более 8 в сумке). Авторы:
Конорева Л. А, Андреев М. П.
1. Таллом бывает кустистым, листоватым, плакодиоидным или
чешуйчатым 2.
— Таллом накипной 6.
2. Таллом кустистый или листоватый 3.
— Таллом плакодиоидный или чешуйчатый 5.
3. Таллом листоватый Aspicilia.
— Таллом кустистый 4
4. Таллом диморфный, состоит из накипного первичного слое¬
вища и псевдоподециев Pilophorus.
— Таллом выглядит иначе Aspicilia.
5. Таллом плакодиоидный 7.
— Таллом чешуйчатый 11
6. Слоевищный край апотеция содержит водоросли (фото¬
бионт в эксципуле и гипотеции) 19.
— Слоевищный край апотеция без водорослей (в эксципуле и
гипотеции фотобионт отсутствует) 35.
7. Таллом желтоокрашенный, апотеции желтые, споры 1-2 кл,
К- Candelariella.
— Таллом иначе окрашен, иногда может иметь желтый отте¬
нок, но апотеции не одного цвета с талломом 8.
8. Таллом t/F+оранжевый, Р+ оранжевый, Х+желтый, С-, апо¬
теции на ножках Baeomyces placophyllus.
— Реакции таллома другие, апотеции сидячие 9.
9. Апотеции леканорового типа 10.
— Апотеции лецидеевого или биаторового типа 79.
10. Таллом с цефалодиями или без них, коровый слой от С
краснеет, апикальный аппарат сумок Trapelia - типа
Placopsis.
— Цефалодии отсутствуют, апикальный аппарат сумок
Lecanora-или Bacidia- типа, кора таллома С+ или С-
Lecanora.
11. Апотеции на ножках Baeomyces.
— Апотеции сидячие или погруженные 12.
12. Апотеции Aspicilia-типа Schaereria.
— Апотеции не Aspicilia-типа. 13.
13. Содержит алекториаловую кислоту и ее производные, ко¬
ровый слой и сердцевина таллома от К и Р желтеют, от С
и КС краснеют, UV- Руспога.
— Алекториаловая кислота отсутствует, реакции таллома и
сердцевины другие 14.
14. Цефалодии присутствуют Psoroma.
— Цефалодии отсутствуют 15.
15. Сумки Trapelia-типа 79.
— Сумки другого типа 16.
16. Слоевищный край присутствует Protoparmelia.
— Слоевищный край отстутствует 17.
17. Вехушки парафиз сильно утолщены, с ясными темными
слизистыми «шапочками» Toninia.
— Верхушки парафиз утолщенные или простые, окрашенные
или бесцветные, но без темных слизистых «шапочек» 18.
18. Сумки Lecanora-типа Psorinia.
— Сумки Porpidia-типа Lecidoma.
19. Споры с периспорием 20.
— Споры без периспория, стенки спор утолщенные или нет ...
21.
20. Таллом мелкочешуйчатый, до почти гранулярного, с цефа-
лодиями Psoroma.
— Таллом трещиновато-ареолированный, обычно с изидиями,
без цефалодий Koerberiella.
21. Таллом жетоокрашенный, апотеции желтые, споры 1-2 кл,
К- Candelariella.
— Таллом иначе окрашен, иногда может иметь желтый отте¬
нок, но апотеции не одного цвета с талломом 22.
22. Слоевищный край апотециев присутствует, хорошо заме¬
тен 23.
— Слоевищный край апотециев отсутствует или плохо развит,
почти не заметен 29.
23. Сумки Pertusaria-типа 24.
— Сумки другого типа 25.
24. Апотеции сидячие, в талломе присутствует гирофоровая
кислота Ochrolechia.
— Апотеции погружены в таллом, гирофоровая кислота отсут¬
ствует Pertusaria.
25. Споры крупные, более 30 мкм в длину Megaspora.
— Споры мельче 26.
26. Сумки Lecanora-типа 27.
— Сумки Bacidia-типа 28.
27. Таллом обычно коричневоокрашенный, атранорин отсут¬
ствует Protoparmelia.
— Таллом более светло окрашен, атранорин часто присутству¬
ет Lecanora.
28. Собственный край плохо развит, парафизы покрыты жела¬
тинозной оболочкой, набухающей в воде .... Tephromela.
— Собственный край плохо развит или присутствует, парафи¬
зы не набухают в воде Lecanora.
29. Апотеции погруженные, до полупогруженных,
Aspicilia-типа 30.
— Апотеции сидячие, до выпуклых, не Aspicilia-типа 32.
30. Апикальный купол сумок синеет в /, внутренний слой су¬
мок также синеет в I и формирует утолщение в апикаль¬
ной части Eiglera.
— Апикальный купол в I не изменяется, окрашивается только
тонкий внутренний слой сумки 31.
31. Гимений гимиамилоидный, апикальный купол сумок I не
изменяется, окрашивается только тонкий внутренний
слой сумки Aspicilia.
— Гимений эуамилоидный, латеральная часть толуса сумок
окрашивается от/в синий цвет Aspilidea.
32. На мхах Вгуопога.
— На других субстратах 33.
33. Слоевищный край присутствует 34.
— Слоевищный край отсутствует Stenhammarella.
34. Апотеции Aspicilia-типа Clauzadeana.
— Апотеции не Aspicilia-типа Lecanora.
35. Цефалодии присутствуют 36.
— Цефалодии отсутствуют 39.
36. Слоевищный край апотециев хорошо развит Placopsis.
— Слоевищный край отсутствует или плохо развит 37.
37. Апотеции погружены в ареолы слоевища Amygdalaria
— Апотеции сидячие или на ножках 38.
38. Апотеции сидячие, фотобионт Micarea-типа, состоит из
мелких, 4-7 мкм в диаметре, почти округлых клеток
Micarea.
— Апотеции сидячие или на ножках, фотобионт не
Micarea- типа Pilophorus.
39. Апотеции на ножках 40.
— Апотеции сидячие или погруженные в таллом 41.
40. Апотеции коричневые Baeomyces.
— Апотеции имеют розоватый оттенок Dibaeis.
41. Апотеции с хорошо развитым собственным краем 42.
— Собственный край апотециев отсутствует или почти не за¬
метен 91.
42. Апотеции светлоокрашенные, до темно-коричневых, но не
черные 43.
— Апотеции черные 57.
43. Апотеции Aspicilia-типа, погруженные или полупогружен¬
ные 73.
— Апотеции не Aspicilia-типа, сидячие 44.
44. Сумки Trapelia-типа 45.
— Сумки другого типа 48.
45. Апотеции 1-3.5 мм в диаметре, собственный край темно-
коричневый, до черного, парафизы легко разделяются ...
Ainoa.
— Апотеции мельче, собственный край более светлый, пара¬
физы плотно соединенные 46.
46. Таллом трещиновато-ареолированный, ареолированный ....
Trapelia.
— Таллом зернистый, с соредиями или без них 47.
47. Таллом зернистый, без соредий или очень редко с сореди¬
ями, парафизы в верхней части утолщенные, темно-
коричневые Placynthiella.
— Таллом зернистый, с соредиями или без них, парафизы как
правило, не утолщенные, без темных пигментов 79.
48. Растет на листьях печеночника Ptilidium pulcherrimum
Puttea
— Растет на других субстратах 49.
49. Фотобионт Micarea-типа, состоит из мелких, 4-7 мкм в
диаметре, почти округлых клеток Micarea.
— Фотобионт не Micarea-типа 50.
50. Сумки Biatora-типа. 51.
— Сумки другого типа 53.
51. Апотеции лецидиевые Adelolecia.
— Апотеции биаторовые 52.
52. Гимений без хорошо развитого эпигимения, вторичные ме¬
таболиты обычно присутствуют Biatora.
— Гимений с хорошо развитым эпигимением, вторичные ме¬
таболиты, определяемые с помощью TLC, отсутствуют .
Mycobilimbia.
53. Сумки Lecanora-типа 54.
— Сумки Fuscidea-иш Micarea-типа 56.
54. Эпитеций с коричневыми гранулами, от К становится пур¬
пурным Pyrrhospora.
— Эпитеций другой 55.
55. Эксципул окрашенный, темно-зеленый, до черного, или
коричневый Lecidella.
— Эксципул состоит из неокрашенных гиф Ramboldia.
56. Сумки Fuscidea-типа Fuscidea.
— Сумки Micarea-типа Hertelidea.
57. На мхах родов Andreaea, Grimmia, Rhacowiitrium
Frutidella.
— на других субстратах 58.
58. Апикальный аппарат сумок Lecanora-типа 59.
— Апикальный аппарат сумок другого строения 66.
59. Таллом зернисто-бородавчатый, до почти чешуйчатого,
/^красный, споры 1—4 клеточные
Bryodina rhypariza.
— Строение таллома, хим. реакции и споры иные 60.
60. Эксципул темный, до угольно-черного, гимений сильно
желатинизированный, в верхней части синий, сине-
зеленый, до изумрудно-зеленого, в нижней - светлый
Carbonea.
— Окраска внутренних частей апотеция иная 61.
61. Внутренние части апотеция окрашиваются в К 62.
— Внутренние части апотеция в К uq изменяются 64.
62. Эпитеций с коричневыми гранулами, от К становится пур¬
пурным Pyrrhospora.
— Эпитеций без коричневых гранул, с К не реагирует, или
окраска иная 63.
63. Содержит алекториаловую кислоту и ее производные, ко¬
ровый слой и сердцевина таллома от К и Р желтеют, от С
и КС краснеют, UV- Руспога.
— Алекториаловая кислота отсутствует, реакции таллома и
сердцевины различны Lecidella.
64. Содержит мириквидиковую кислоту Miriquidica.
— Мириквидиковая кислота отсутствует 65.
65. Эксципул темно-зеленый, голубовато-черный или корич¬
невый Lecidella.
— Эксципул состоит из неокрашенных гиф Ramboldia.
66. Апикальный аппарат сумок Porpidia-типа 67.
— Апикальный аппарат сумок другого строения 70.
67. Апотеции погруженные Immersaria.
— Апотеции сидячие или погруженные 68.
68. Собственный край с темным, до черного, ободом, внутрен¬
няя часть светлая, главным образом на кремнийсодер¬
жащих субстратах, очень редко на субстратах с кальцием
Porpidia.
— Собственный край от темно-коричневого до черного, растут
на кальцийсодержащих субстратах 69.
69. Эпитеций N- Clauzadea.
— Эпитеций N+ краснеет Farnoldia.
70. Споры среднего размера или крупные, 25-100 ><15-40 мкм,
одноклеточные, с сильно утолщенными стенками
Mycoblastus.
— Споры мельче, 1-или многоклеточные, строение стенок
различное 71.
71. Апотеции Aspicilia-типа, погруженные или полупогружен¬
ные 72.
— апотеции сидячие, до сильновыпуклых, не Aspicilia-типа 74.
72. Сумки Schaereria-типа, парафизы растворяются в К
Schaereria.
— Сумки не Schaereria-типа, парафизы растворяются или не
растворяются в К 73.
73. Эпитеций N- Ionaspis.
— Эпитеций N+ краснеет Hymenelia.
74. Вехушки парафиз сильно утолщены, с ясными темными
слизистыми «шапочками» Toninia.
— Верхушки парафиз утолщенные или простые, окрашенные
или бесцветные, но без темных слизистых «шапочек» 75.
75. Сумки Trapelia-типа 76.
— Сумки другого типа 80.
76. Апотеции 1-3.5 мм в диаметре, собственный край темно-
коричневый, до черного, парафизы легко разделяются ...
Ainoa.
— Апотеции мельче, собственный край более светлый, пара¬
физы плотно соединенные 77.
77. Таллом трещиновато-ареолированный, ареолированный ....
Trapelia.
— Таллом зернистый, с соредиями или без них 78.
78. Таллом зернистый, без соредий или очень редко с сореди¬
ями, парафизы в верхней части утолщенные, темно-
коричневые Placynthiella.
— Таллом зернистый, с соредиями или без них, парафизы как
правило, не утолщенные, без темных пигментов 79.
79. Споры 7-14 мкм длиной Trapeliopsis.
— Споры 15-25 мкм длиной Trapelia.
80. Сумки Biatora-типа 81.
— Сумки другого типа 82.
81. Апотеции лецидеевые, споры 1-2 клеточные
Adelolecia.
— Апотеции биаторовые, споры от 1 до многоклеточных
Mycobilimbia.
82. Фотобионт Micarea-типа, состоит из мелких, 4-7 мкм в
диаметре, почти округлых клеток Micarea.
— Фотобионт не Micarea-типа 83.
83. Сумки Teloschistes-тппа 84.
— Сумки другого типа 85.
84. Споры мелкие, 6-11 х 4-6.5 мкм, бесцветные, одноклеточ¬
ные, без утолщенной оболочки Cephalophysis.
— Споры крупнее, большей частью коричневые, до темно-
оливково-коричневых, редко бесцветные, простые или с
плохо заметной септой, с утолщенной оболочкой
Orphniospora.
85. Сумки Fuse idea-типа. Fuscidea.
— Сумки другого типа 86.
86. Сумки Rimularia-типа, споры вначале бесцветные, при со¬
зревании становятся коричневыми Rimularia.
— Сумки другого типа, споры бесцветные 87.
87. Сумки Micarea-типа Hertelidea.
— Сумки другого типа 88.
88. Сумки Candelariella-типа Claurouxia.
— Сумки другого типа 89.
89. Таллом ржавчинно-красный, апотеции погруженные в тал¬
лом Tremolecia atrata.
— Таллом иначе окрашенный, может иметь красноватую
окраску, но не ржавчинно-красную, апотеции сидячие ...
90.
90. Сумки Lecidea-ти па Lecidea.
— Сумки Byssoloma-типа Leimonis erratica.
91. Споры с периспорием 92.
— Споры без периспория 94.
92. Апотеции погруженные, до полупогруженных 93.
— Апотеции сидячие Japewia.
93. Кора таллома содержит коричневые пигменты, покрыта
эпинекральным слоем, сердцевина /+синеет Immersaria.
— Кора таллома не содержит коричневых пигментов,
эпинекральный слой отсутствует, сердцевина
7+фиолетовый Bellemerea.
94. Слоевищный край присутствует Placopsis.
— Слоевищный край отсутствует 95.
95. Апотеции оранжевые, до оранжево-красных, /^красный ...
Protoblastenia.
— Апотеции иначе окрашены, если присутствуют желто¬
оранжевые пигменты, то К- 96.
96. Вехушки парафиз сильно утолщены, с ясными темными
слизистыми «шапочками» Toninia.
— Верхушки парафиз утолщенные или простые, окрашенные
или бесцветные, но без темных слизистых «шапочек» ....
97.
97. Споры среднего размера или крупные, 25-100 х 15-40
мкм, одноклеточные, с сильно утолщенными стенками ..
Mycoblastus.
— Споры мельче, или многоклеточные, стенки не утолщены
98.
98. Сумки Lecanora-типа 99.
— Сумки различных типов 104.
99. Внутренние части апотеция окрашиваются в К 100.
— Внутренние части апотеция в К не изменяются 101
100. Эпитеций с коричневыми гранулами, от К становится
пурпурным Pyrrhospora.
— Эксципул от К становится фиолетовым Руспога.
101. Таллом зернисто-бородавчатый, до почти чешуйчатого,
Л’+красный, споры 1—4 клеточные
Bryodina rhypariza.
— Строение таллома, хим. реакции и споры иные 102.
102. Фотобионт хлорококкоидный Calvitimela.
— Фотобионт Trebouxia-типа, 103.
103. Содержит мириквидиковую кислоту Miriquidica.
— Мириквидиковая кислота отсутствует Ramboldia.
104. Споры 1-клеточные, каплевидные, 4-7 х 1-2 мкм
Psilolechia.
— Споры 1- или многоклеточные, если каплевидные, то более
крупные 105.
105. Растет на листьях печеночника Ptilidium pulcherrimum
Puttea.
— Растет на других субстратах 106.
106. Таллом гониоцистный 107.
— Таллом другого строения 109.
107. Апотеции часто бесформенные, настоящий эксципул и
гипотеций отсутствуют Vezdaea.
— Апотеции более-менее правильной формы, эксципул и ги¬
потеций присутствуют 108.
108. Фотобионт Micarea-типа, состоит из мелких, 4-7 мкм в
диаметре, почти округлых клеток Micarea.
— Фотобионт не Micarea-типа Myochroidea.
109. Сумки Micarea-типа 110.
— Сумки другого строения 112.
110. Фотобионт Micarea-типа, состоит из мелких, 4-7 мкм в
диаметре, почти округлых клеток Micarea.
— Фотобионт не Micarea-типа 111.
111. Фотобионт Trebouxia-типа Myochroidea.
— Фотобионт хлорококкоидный Hertelidea.
112. Гипотеций красновато-коричневый, Л’+оранжевый
Protomicarea.
— Гипотеций иначе окрашен, если коричневый, то с К не реа¬
гирует 113.
114. Сумки Fuscidea-типа Fuscidea.
— Сумки другого строения 115.
115. Таллом состоит из сливающихся, почти чешуйчатых,
смыкающихся краями ареол Lecidoma.
— Таллом не ареолированный Leimonis erratica.
Стерильные лишайники.
Ключ 16.
Ключ для определения стерильных лишайников. Авто¬
ры: Гимельбрант Д. Е, Степанчикова И. С.
1. Таллом с ясно выраженными краевыми лопастями до 2 мм
шириной 2.
— Краевые лопасти отсутствуют или редкие, неясные, до 0.5
мм шириной 5.
2. Таллом желтовато-серый, беловато-желтый, желтовато¬
зеленый, с ясным коричневым до серовато-черного ги¬
поталломом и четкой белой сердцевиной
Lepraria.
— Таллом без желтоватого оттенка, голубовато-серый, гипо¬
таллом и сердцевина ясные или неясные 3.
3. Соредии с длинными (до 100 мкм) выступающими гифами;
таллом с более или менее толстой, хорошо отграничен¬
ной белой сердцевиной Crocynia.
— Соредии без выступающих гиф или гифы короткие (до 10—
40 мкм); сердцевина не выражена или встречается ме¬
стами, тонкая, плохо отграниченная 4.
4. Таллом содержит леканоровую кислоту и зеорин
Botryolepraria.
— Леканоровая кислота и зеорин в талломе отсутствуют
Lecanora.
5(1). Таллом состоит из массы микрокустистых пучков гиф,
гроздевидно несущих на концах клетки фотобионта,
рыхлый, ватообразный; содержит лезданин; в сырых и
сильно затененных местообитаниях на карбонатных по¬
родах Botryolepraria lesdainii.
— Таллом состоит из массы грубых или тонкозернистых соре¬
дий, лежащих более или менее плотно; лезданин отсут¬
ствует; в различных местообитаниях на разнообразных
субстратах 6.
6. Соредии с длинными (до 100—120 мкм) выступающими
гифами 7.
— Соредии без выступающих гиф или гифы короткие, неза¬
метные (до 10—40 мкм) 10.
7. В UV заметна голубовато-белая флюоресценция таллома
(содержит диварикатовую кислоту); таллом толстый, с
хорошо отграниченной белой сердцевиной
Lepraria.
— В UV голубовато-белая флюоресценция таллома не наблю¬
дается (не содержит диварикатовой кислоты); таллом
толстый с хорошо отграниченной сердцевиной или тон¬
кий без явно выраженной сердцевины 8.
8. Сердцевина не выражена, таллом содержит нефростерано-
вую кислоту Lepraria rigidula.
— Сердцевина более или менее хорошо отграниченная, белая,
нефростерановая кислота в талломе отсутствует 9.
9. Таллом грубозернистый, зерна диаметром до 300 мкм,
окраска таллома беловатая до светло-серой; содержит
тамноловую и рокцелловую кислоты; вид распространен
в теплых районах Центральной и Южной Европы
Lepraria nylanderiana.
— Таллом мучнистый, соредии диаметром до 50 мкм, консо-
редии диаметром преимущественно до 100 мкм, окраска
таллома зеленоватая, с серым или голубовато-серым от¬
тенком; содержит стиктовую, констиктовую кислоты и
зеорин; широко распространенный вид
Lecanora lobificans.
10(6). Сердцевина белая, хорошо отграниченная 11.
— Сердцевина не выражена или неясная, слабо отграниченная
13.
11. Талломы мелкие, с редкими соредиями, белые, содержат
алекториаловую кислоту Ochrolechia.
— Талломы крупные, густо соредиозные, зеленоватых, желто¬
ватых, голубоватых тонов, алекториаловая кислота от¬
сутствует 12.
12. Таллом от С краснеет, в UV заметна голубовато-белая
флюоресценция, содержит диварикатовую, нордиварика-
товую кислоты и зеорин; цвет таллома зеленый, зелено-
вато-серый, голубоватый Lepraria crassissima.
— Таллом от С не изменяется, в UV голубовато-белая флюо¬
ресценция не наблюдается, содержит другие лишайни¬
ковые вещества; таллом серовато-желтый, зеленовато-
желтый или беловатый Lepraria vouauxii.
13(10). В UV заметна голубовато-белая флюоресценция талло¬
ма, содержит диварикатовую кислоту ... Lepraria incana.
— В UV голубовато-белая флюоресценция таллома не наблю¬
дается, диварикатовая кислота отсутствует 14.
14. Таллом белый, беловато-серый до темно-серого, иногда с
голубоватым оттенком; преимущественно виды с грубо¬
зернистым талломом 15.
— Таллом зеленоватых, зеленовато-желтоватых, желтоватых
тонов, реже с голубоватым оттенком; преимущественно
виды с тонкозернистым талломом 22.
15. Таллом грубозернистый, от С и КС ясно розовеет или
краснеет (содержит алекториаловую или леканоровую
кислоты) 16.
— Таллом грубо- или тонкозернистый, от С и КС не изменяет¬
ся или слабо желтеет (алекториаловая или леканоровая
кислоты отсутствуют) 17.
16. Гипоталлом заметный, войлочный, голубовато-серый до
темно-коричневого; таллом содержит леканоровую кис¬
лоту; азиатский тропический океанический вид, обита-
ющий в затененных сырых местообитаниях вне прямого
действия осадков Lecanora.
— Гипоталлом незаметный; таллом содержит алекториаловую
кислоту; широко распространенный вид, приуроченный
к открытым местообитаниям Lepraria neglecta.
17(15). Таллом грубо зернистый, по краю с зернами диаметром
до 200—400 мкм 18.
— Зерна таллома более мелкие, диаметром до 120(160) мкм ...
19.
18. Таллом содержит ангардиановую кислоту; обитает в от¬
крытых местах Lepraria caesioalba.
— Ангардиановая кислота отсутствует; обитает в затененных
местах Lepraria nivalis.
19(17). Таллом содержит порфириловую кислоту 20.
— Порфириловая кислота отсутствует 21.
20. Таллом содержит ангардиановую кислоту; широко распро¬
страненный вид, обитает в открытых местах
Lepraria alpina.
— Ангардиановая кислота отсутствует; приморский, возмож¬
но океанический вид, обитает в затененных местах
Pertusaria.
21(19). Соредии мучнистые, диаметром до 45 мкм (консоредии
— до 100 мкм); таллом от Р становится оранжевым, со¬
держит зеорин, стиктовую кислоту и ее производные
Mycoblastus.
— Соредии более грубые, диаметром до 75 мкм (консоредии
— до 160 мкм); таллом от Р не изменяется, содержит
рангиформовую кислоту Lecanora borealis.
22(14). Таллом от С и КС ясно розовеет или краснеет (содер¬
жит алекториаловую кислоту) Normandina.
— Таллом от С и КС не изменяется или слабо желтеет (алек¬
ториаловая кислота отсутствует) 23.
23. Таллом содержит стиктовую кислоту и ее производные
Micarea.
— Стиктовая кислота и ее производные отсутствуют 24.
24. Таллом содержит порфириловую кислоту
Lepraria alpina.
— Порфириловая кислота отсутствует 25.
25. Таллом содержит оксипаннаровую кислоту и другие ди-
бензофураны Bacidia.
— Оксипаннаровая кислота и другие дибензофураны отсут¬
ствуют 26.
26. Таллом содержит тамноловую кислоту Biatora.
— Тамноловая кислота отсутствует 27.
27. Таллом содержит обтузатовую кислоту, атранорин отсут¬
ствует Ainoa.
— Обтузатовая кислота отсутствует, атранорин обычно при¬
сутствует 28.
28. Жирные кислоты представлены только тонсбергиановой
кислотой Biatora toensbergiana.
— Таллом содержит другой набор жирных кислот
Lepraria jackii s. lat.
Литература
Андреев В. Н. Кормовая база Ямальского оленеводства //
Советское оленеводство. 1933. Вып. 1. С. 99-164
Андреев В. Н. Типы тундр запада Большой Земли // Тр.
Бот. муз. АН СССР. Л.: Изд. АН СССР. 1932. Вып. 25. С. 121—
268.
Андреев М. П. Лишайники родов Cladonia и Cladina
Анюйского нагорья // Ботан. журн. 1981. Т. 66, № 1. С. 31-41.
Андреев М. П. Лишайники в растительном покрове горы
Эломбал, Чукотский автономный округ// Ботан. журн. 1980. Т.
65, № 4. С. 563-570.
Андреев М. П. Лишайники оазиса Бангера (Восточная
Антарктида) // Новости систематики низших растений. 1990.
Т. 27. С. 85-93.
Андреев М. П. Лишайники полуострова Файлдс, остров
Кинг Джордж, Антарктика // Новости систематики низших
растений. 1988. Т. 25. С. 111-118.
Андреев М. П. Лишайники полуострова Ямал // Новости
систематики низших растений. 1984а. Т. 21. С. 127-136.
Андреев М. П. Систематический состав лихенофлоры
Анюйского нагорья // Новости систематики низших растений.
19846. Т. 21. С. 136-140.
Андреев М. П, Кудратов И. Lecidea (Lecanorales) и близ¬
кие таксоны лишайников флоры северо-западного и
центрального Таджикистана // Ботан. журн. 1992. Т. 77, № 1.
С. 101-111.
Андреев М. П, Мырзакулова 3. С. Виды семейств Le¬
cideaceae и Lecanoraceae (Lichenes) Южного Алтая (Восточ¬
ный Казахстан) // Ботан. журн. 1992. Т. 77, № 3. С. 114-118.
Андреев М. П, Котлов Ю. В, Макарова И. И. Биологи¬
ческое разнообразие лишайников Русской Арктики (таксоно¬
мический состав и предварительный анализ) // Новости систе¬
матики низших растений. СПб, 1996. Т. 31. С. 82-94.
Андреев М. П, Курбатова Л. Е. Новые данные о мхах и
лишайниках тихоокеанского сектора Антарктиды // Новости
систематики низших растений. 2009. Т. 42. С. 142-152.
Андреева В. М. Почвенные и аэрофильные зелёные во¬
доросли (Chlorophyta: Tetrasporales, Chlorococcales, Chlorosar-
cinales). СПб.: Наука, 1998. 351 с.
Бредкина Л. И, Добрыш А. А, Макарова И. И, Титов А.
Н. К флоре лишайников острова Кунашир (Курильские остро¬
ва) // Новости систематики низших растений. 1991. Т. 28. С.
90-94.
Бредкина Л. И, Макарова И. И. Аннотированный список
лишайников Центрального Тянь-Шаня (Киргизия) // Новости
систематики низших растений. 2005. Том 39. СПб. С. 179-195.
Бязров Л. Г. Видовой состав лихенобиоты Монголии.
Версия 5. 2007. -
http://www.sevin.ru/laboratories/biazrov_mong.html
Бязров Л. Г. Лишайники в экологическом мониторинге.
М.: Научный мир, 2002. 336 с.
Бязров Л. Г. Современная динамика видового состава
эпифитной лихенобиоты г. Москвы // Современная микология
в России. Тезисы докладов второго съезда микологов России.
Том 2. М.: Национальная академия микологии, 2008. С. 523-
524.
Бязров Л. Г. Эпифитные лишайники г. Москвы: совре¬
менная динамика видового разнообразия. М.: Товарищество
научных изданий КМК, 2009. 146 с.
Вайнштейн Е. А, Равинская А. П, Шапиро И. А. Спра¬
вочное пособие по хемотаксономии лишайников. Ботаниче¬
ский институт им. В. Л. Комарова АН СССР. Л, 1990. 152 с.
Веденеев А. М. Флора лишайников Волгоградской обла¬
сти: Автореф. дис. ... канд. биол. наук. СПб, 2001. 27 с.
Вершинина С. Э, Гимельбрант Д. Е, Кузнецова Е. С,
Габышева Л. М, Габышев Э. М. Первые сведения о лихено¬
флоре государственного заповедника «Олёкминский» (Рес¬
публика Саха-Якутия) // Вестник ТвГУ. Серия «Биология и
экология». 2012. Вып. 25. № 3. С. 138-149.
Войцехович А. А. Фотобионты и водоросли-эпифиты ли-
тофильных лишайников Берегового хребта Карадагского при¬
родного заповедника (Крым, Украина) // Актуальш проблеми
боташки та екологп. 36ipKa наукових праць. Вип.2. КиУв:
Ф1тосоцюцентр, 2008. С. 46-51.
Войцехович А. О. Водоростевий компонент лишайниюв
Карадазького природного заповщника: автореф. дис. на здо-
буття наук, ступеня канд. бюл. наук: спец. 03.00.05 боташка.
Кшв, 2012.23 с.
Войцехович А. О, Михайлюк Т. I, Дар1енко Т. М. Водо-
pocTi наземних м!сцезростань хребта Карагач (Карадазький
природний заповщник (У краУна)) // Зб1рник наук, праць, приев.
95-р1ччю КарадазькоУ наук. стан^У та 30-р1ччю Карадазького
прир. заповщника НАНУ шд ред. А. В. ГаевськоУ, А. Л. Моро-
зовоУ. Севастополь: ЭкоЫ-Пдроф1зика, 2009. С. 50-60.
Войцехович А. А, Михайлюк Т. И, Дариенко Т. М. Фо¬
тобионты лишайников. 1: разнообразие, экологические осо¬
бенности, взаимоотношения и пути совместной эволюции с
микобионтом // Альгология. 2011 а. Т. 21, № 1. С. 3-26.
Войцехович А. А, Михайлюк Т. И, Дариенко Т. М. Фо¬
тобионты лишайников. 2: происхождение и корреляция с ми¬
кобионтом // Альгология. 20116. Т. 21, № 2. С. 151-177.
Гагарина Л. В. Гиалектовые лишайники: история изуче¬
ния и современное состояние изученности // Новости система¬
тики низших растений. 2009а. Т. 42. С. 153-168.
Гагарина Л. В. Лишайники порядка Gyalectales Henssen
ex D. Hawksw. & О. Eriksson внетропической Евразии. Авто¬
реферат диссертации на соискание ученой степени кандидата
биологических наук. Санкт-Петербург, 2012. 23 с.
Гагарина Л. В. Первые сведения о лишайниках порядка
Gyalectales (s. 1. ) в Абхазии // Вестник ТвГУ. Серия «Биология
и экология». 20096. № 34. С. 161-167.
Галанин А. А. Лихенометрия: современное состояние и
направления развития метода (аналитический обзор). Магадан,
СВКНИИ ДВО РАН, 2002. 74 с.
Гимельбрант Д. Е, Степанчикова И. С. Новые данные о
пармелиевых лишайниках и калициоидных грибах Камчатки //
Новости систематики низших растений. 2008 (2009). Т. 42. С.
169-172.
Гимельбрант Д. Е, Степанчикова И. С, Кузнецова Е. С.
Лишайники некоторых кустарников и кустарничков полуост¬
рова Камчатки // Новости систематики низших растений. 2009.
Т. 43. С. 150-171.
Гимельбрант Д. Е, Степанчикова И. С, Кузнецова Е. С.
Лишайники // Растительный покров вулканических плато Цен¬
тральной Камчатки / Ред. В. Ю. Нешатаева. М, Товарищество
научных изданий КМК, 2012. С. 126-171.
Голубкова Н. С. Анализ флоры лишайников Монголии.
Л.: Наука, 1983. 248 с.
Голубкова Н. С. Всеволод Павлович Савич (1885—1972) —
ученый и организатор научных исследований в области крип¬
тогамной ботаники // Ботан. журн. 2003. Т. 88, № 10. С. 142—
150.
Голубкова Н. С. Конспект флоры лишайников Монголь¬
ской Народной Республики. Л.: Наука. 1981. 200 с.
Голубкова Н. С. Лихенологические исследования в Бо¬
таническом институте им. В. Л. Комарова РАН в XX столетии
// Микология и криптогамная ботаника в России: традиции и
современность. Тр. междунар. конф, посвящ. 100-лет. орга-
низ. иссл. по мик. и крипт, бот. в БИН РАН. СПб, 2000, с. 20-
32.
Голубкова Н. С. Лишайники Антарктиды // Атлас Ан¬
тарктиды. Л.: Гидрометеоиздат. 1969. Т. 2. С. 488-492.
Голубкова Н. С. Лишайники Восточного Памира // Но¬
вости систематики низших растений. 1975. Т. 12. С. 250-263.
Голубкова Н. С. Лишайники семейства Acarosporaceae
Zahlbr. в СССР. Л.: Наука. 1988. 127 с.
Голубкова Н. С. Новые для лихенофлоры СССР лишай¬
ники Восточного Памира // Новости систематики низших рас¬
тений. 1972. Т. 9. С. 214-233.
Голубкова Н. С. Первый систематический список ли¬
шайников Восточного Памира // Новости систематики низших
растений. 1973. Т. 10. С. 206-223.
Голубкова Н. С, Бязров Л. Г. Жизненные формы лишай¬
ников и лихеносинузии // Ботанический журнал. 1989. Т. 74, №
6. С. 794-805.
Голубкова Н. С, Савич В. П, Симонов И. М. Лишайники
западной части Земли Эндерби // Тр. Советской антарктиче¬
ской экспедиции. 1968. Т. 38. С. 247-253.
Голубкова Н. С, Симонов И. М. Лишайники оазиса
Ширмахера // Тр. Советской антарктической экспедиции.
1972. Т. 60. С. 317-327.
Городков Б. Н. Ботанико-географический очерк Чукот¬
ского побережья // Уч. зап. Лен. гос. пед. ин-та, каф. физ.
геогр. 1939. Вып. 21. С. 99-175.
Городков Б. Н. Полярные пустыни о. Врангеля // Ботан.
журн. 1943. Т. 28. № 4. С. 127-143.
Городков Б. Н. Почвенно-растительный покров острова
Врангеля // Растительность Крайнего севера СССР и ее освое¬
ние. М. -Л.: Изд. АН СССР, 1958. Вып. 3. С. 5-58.
Гудовичева А. В, Гимельбрант Д. Е. Дополнение к ли¬
хенофлоре Севера Среднерусской возвышенности // Вестник
ТвГУ. Серия «Биология и экология». 2012. Вып. 25. № 3. С.
150-164.
Давыдов Е. А. Аннотированный список лишайников за¬
падной части Алтая (Россия) // Новости систематики низших
растений. 2002. Т. 35. С. 140-161.
Дар1енко Т. М, Войцехович А. О, Кондратюк С. Я. Фо-
тобюнти деяких представниюв родини Teloschistaceae // Укр.
ботан. журн. 2004. Т. 61, № 2. С. 49-59.
Добрыш А. А. Аннотированный список лишайников
острова Врангеля // Новости систематики низших растений.
1995. Т. 30. С. 52-60.
Добрыш А. А. Лишайники южного побережья острова
Врангеля (бухта Сомнительная) // Новости систематики низ¬
ших растений. 1988. Т. 25. С. 124-126.
Дудорева Т. А, Гимельбрант Д. Е. Лишайники Арзян-
ского бора (Саратовская область) // Вестник ТвГУ. Серия
«Биология и экология». 2010. Вып. 17. №16. С. 136-141.
Дудорева Т. А, Гимельбрант Д. Е. Предварительный
список лишайников окрестностей г. Хвалынска (Саратовская
область) // Вестник ТвГУ. Серия «Биология и экология». 2009.
Вып. 16. №36. С. 144-148.
Егорова А. А, Васильева И. И, Степанова Н. А, Фесько
Н. Н. Флора тундровой зоны Якутии. Якутск, ЯНЦ СО АН
СССР, 1991. С. 1-186.
Еленкин А. А. А. А. Фишер фон-Вальдгейм. Известия
Главного Ботанического Сада. 1921. С. 48-54.
Еленкин А. А. Из итогов 25-летней деятельности Инсти¬
тута Споровых Растений Главного Ботанического Сада. Пет¬
роград, 1923. С. 1-28.
Еленкин А. А. Лишайники полярного побережья Сибири
// Научные результаты русской полярной экспедиции барона
Э. В. Толля. Отд. Д, ботаники / Зап. Акад. Наук, сер. 8. СПб,
1909. Т. 26, вып. 1.С. 1-53.
Еленкин А. А. Лишайники русской флоры и погранич¬
ных восточных областей. В кн.: Тр. Имп. Бот. Сада. СПб,
1904. Т. 24. Вып. 1.С. 118-Х.
Еленкин А. А. Материалы для лишайниковой флоры
России//Тр. Имп. Бот. Сада. СПб, 1901. Т. 19. С. 155-182.
Еленкин А. А. О деятельности Спорового Гербария за 14
лет (с 1899 г. по 1913 г.) и о ближайших задачах деятельности
"Института Споровых Растений" - нового отдельного учре¬
ждения Императорского Ботанического Сада Петра Великого.
Известия Императорского Ботанического Сада Петра Велико¬
го. Т. 14. Вып. 1-2. 1914. С. 1-20.
Еленкин А. А. О текущей деятельности Института Спо¬
ровых Растений Главного Ботанического Сада. Известия
Главного Ботанического Сада. 1921. С. 55-58.
Еленкин А. А. Флора лишайников Средней России. Ча¬
сти 1 и 2. СПб, 190Х; части 3 и 4. Юрьев. 1911. С. 361-682.
Еленкин А. А, Савич В. П. Лишайники, собранные И. В.
Палибиным в плавание ледокола «Ермак» в Северном Ледови¬
том океане в 1901 г. // Тр. Имп. Бот. Сада. СПб, 1912. Т. 32. С.
69-100.
Жданова Н. Н, Василевская А. И. Экстремальная эколо¬
гия грибов в природе и эксперименте. Киев: Наукова думка,
1982. 168 с.
Жизнь растений. Т. 3. Водоросли. Лишайники / Под. ред.
М. М. Голлербаха. М.: Просвещение, 1977. 487 с.
Журбенко М. П. Лишайники и лихенофильные грибы
Путоранского заповедника // Флора и фауна заповедников.
2000. Вып. 89. 52 с.
Заварзина А. Г, Заварзин А. А. Лакказная и тирозиназ-
ная активности у лишайников // Микробиология. 2006. Т. 75.
№5. С. 630-641.
Императорский С. -Петербургский Ботанический сад за
200 лет его существования (1713-1913). Юбилейное издание,
ч. 1.СП6, 1913.
История Академии Наук СССР. Т. 2. (1803-1917). М. -Л,
Наука, 1964. 772 с.
Караваев М. Н. Конспект флоры Якутии. М, Л, Изда¬
тельство АН СССР, 1958. 189 с.
Каратыгин И. В. Коэволюция грибов и растений // Труды
Ботанического института им. В. Л. Комарова. Вып. 9. СПб.:
Гидрометеоиздат, 1993. С. 1-119.
Катаева (Катенина) О. А. Аннотированный список видов
лишайников Новгородской области // Новости систематики
низших растений. 2002. Т. 36. С. 114-143.
Катаева О. А. Лишайники и лихенизированные грибы //
Кадастр флоры Новгородской области / Ред. Юрова Э. А,
Крупкина Л. И, Конечная Г. Ю. Новгород, Изд. «ЛЕМА»,
2009. С. 247—254.
Катаева О. А, Макарова И. И, Таран Г. С, Тюрин В. Н.
Лишайники поймы р. Обь в окрестностях г. Сургута (Тюмен¬
ская область, Западная Сибирь) // Новости систематики низ¬
ших растений. 2005. Т. 38. С. 186-199.
Кирпичников М. Э, Забинкова Н. Н. Русско-латинский
словарь для ботаников. Л.: «Наука», 1977. 856 с.
Княжева Л. А, Семенова Е. А. К лихенофлоре Уссурий¬
ского заповедника. Сем. Lecanoraceae // Изучение флоры и
растительности Дальнего Востока. Владивосток, ДВГУ, 1979.
С. 83-90.
Княжева Л. А, Скирина И. Ф. Лишайники // Раститель¬
ный и животный мир Сихотэ-Алинского заповедника. М,
Наука, 1982. С. 44^17.
Конечная Г. Ю, Курбатова Л. Е, Потемкин А. Д, Гиме-
льбрант Д. Е, Кузнецова Е. С, Змитрович И. В, Коткова В.
М, Малышева В. Ф, Морозова О. В, Попов Е. С, Яковлев Е.
Б, Andersson L, Кияшко П. В, Skujiene G. Выявление и об¬
следование биологически ценных лесов на Северо-Западе Ев¬
ропейской части России. Т. 2. Пособие по определению видов,
используемых при обследовании на уровне выделов / Отв. ред.
Л. Андерссон, Н. М. Алексеева, Е. С. Кузнецова. СПб, 2009.
258 с.
Конорева Л. А. Новые находки редких и охраняемых ви¬
дов лишайников из тундровой зоны Мурманской области //
Вестн. ТвГУ. Сер. Биология и экология. 2010. Вып. 20. С. 115-
121.
Конорева Л. А, Мучник Е. Э, Урбанавичене И. Н, Ур¬
банавичюс Г. П. Виды лишайников, предлагаемые к включе¬
нию в Красную книгу Курской области // Исследования по
Красной книге Курской области. Вып. 2. Курск, 2010. С. 69-
85.
KoctIkob I. Ю, Романенко П. О, Демченко Е. М,
Дар1енко Т. М, Михайлюк Т. I, Рибчинський О. В, Солонен-
ко А. М. Водорост1 грунт1в УкраУни (icTopiH та методи до-
слщження, система, конспект флори). КиУв: Ф1тосоцюцентр,
2001. 300 с.
Котлов Ю. В. Лишайники, собранные на птичьей коло¬
нии в северо-западной части острова Большевик (Северная
Земля) // Ботан. журн. 1993. Т. 78. № 8. С. 34-36.
Котлов Ю. В. Новые для Полярного Урала виды лишай¬
ников с побережья Карского моря // Ботан. журн. 1994. Т. 79.
№7. С. 122-124.
Котлов Ю. В. Новые для Урала виды лишайников из
Свердловской области // Ботан. журн. 2003. Т. 88, № 4. С. 147—
148.
Кузнецова Е. С, Сказина М. А. К изучению лихенофло¬
ры Костромской области // Новости систематики низших рас¬
тений. 2010. Т. 44. С. 200-209.
Куприянов А. В. Развитие принципов систематики в
XVII-XVIII вв. Автореф. дис. канд. биол. наук. М, 2005. 26 с.
Кутафьева Н. П. Морфология грибов. Новосибирск: Си¬
бирское университетское издательство, 2003. 215 с.
Куцев М. Г. Фрагментарный анализ ДНК растений:
RAPD, DAF, ISSR. Барнаул: ARTOKA, 2009. 164 с.
Лавриненко О. В, Урбанавичюс Г. П, Плюснин С. Н,
Урбанавичене И. Н, Журбенко М. П. Лихенофлора Ненецкого
автономного округа // Флора лишайников России: состояние и
перспективы исследований. Тр. междунар. совещ, посвящ.
120-летию со дня рождения В. П. Савича. СПб, 2006. С. 135—
140.
Локинская М. А. Наиболее распространенные виды ли¬
шайников на северо-востоке СССР // Водоросли и грибы Си¬
бири и Дальнего Востока. Новосибирск: Наука, Сиб. отд.,
1970. Вып. 1 (3). С. 233-246.
Магнуссон А. Г. Новые лишайники севера СССР // Изв.
Главн. бот. сада СССР. Л, 1927. Т. 26. Вып. 4. С. 359-370.
Магомедова М. А. Лишайниковый покров высокогорий
Северного Урала // Растительные сообщества Урала и их ан¬
тропогенная деградация. Свердловск, 1984 С. 91-101.
Магомедова М. А. Сукцессии сообществ литофильных
лишайников в высокогорьях Северного Урала // Экология.
1979. №3. С. 29-38.
Макаревич М. Ф. Анал1з л1хенофлори УкраУнських Кар¬
пат. Киев, 1963. 263 с.
Макаревич М. Ф. Зональный географический элемент
как основа географического анализа // Материалы III закавказ¬
ской конференции по споровым растениям. Тбилиси, 1968. С.
215-221.
Макарова И. И. К флоре лишайников бухты Провидения
(Чукотка) // Новости систематики низших растений. 1986. Т.
23. С. 179-181.
Макарова И. И. К флоре лишайников низовьев реки Ле¬
ны // Новости систематики низших растений. 1989. Т. 26. С.
118-124.
Макарова И. И. Лишайники островов Айон и Большой
Раутан (Чаунская губа) // Новости систематики низших расте¬
ний. 1987. Т. 24. С. 159-163.
Макарова И. И. Лишайники юго-востока Чукотского по¬
луострова (бухта Пенкигней) // Новости систематики низших
растений. 1983. Т. 20. С. 146-150.
Макарова И. И. Характеристика конкретных лихенофлор
запада Чукотского полуострова // Ботан. журн. 1979. Т. 64. №
3. С. 351-361.
Макарова И. И, Катенин А. Е. Лишайники в горах юго-
востока Чукотского полуострова // Ботан. журн. 1983. Т. 68. №
11. С. 1477-1487.
Макарова И. И, Катенин А. Е. Лишайники равнинного
юго-западного побережья Чукотского полуострова // Ботан.
журн. 1987. Т. 72. № 4. С. 455^-63.
Макарова И. И, Катенин А. Е. Лишайники средней ча¬
сти хребта Искатень на западе Чукотского полуострова // Бо¬
тан. журн. 1992. Т. 77. № 1. С. 45-57.
Макарова И. И, Перфильева В. И. Дополнение к ли¬
хенофлоре северо-запада Якутии // Новости систематики низ¬
ших растений. 1985. Т. 22. С. 180-184.
Макарова И. И, Перфильева В. И. К флоре лишайников
северо-запада Якутии // Новости систематики низших расте¬
ний. 1984. Т. 21. С. 150-160.
Макарова И. И, Перфильева В. И. К флоре лишайников
Усть-Ленского заповедника // Водоросли, лишайники, грибы и
мохообразные в заповедниках РСФСР. М, 1989. С. 53-59.
Макарова И. И, Перфильева В. И, Николин Е. Г. К фло¬
ре лишайников Новосибирских островов // Новости система¬
тики низших растений. 1988. Т. 25. С. 127-133.
Макрый Т. В. Лишайники Байкальского хребта. Новоси¬
бирск: Наука, Сиб. отд-ние, 1990. 200 с.
Макрый Т. В, Лиштва А. В, Скирина И. Ф. Особенности
строения, физиологии и химии эпилитных лишайников в экс¬
тремальных условиях высокогорий // Микология и криптогам¬
ная ботаника в России: традиции и современность / Труды
международ. конф, посвящ. 100-лению организации исслед.
по микологии и криптогамной ботанике в Ботаническом ин-те
им. В. Л. Комарова РАН (Санкт-Петербург, 24-28 апреля, 2000
г.). СПб: Изд-во Санкт-Петербургской государственной хими¬
ко-фармацевтической академии, 2000. С. 349-351.
Мартин Ю. Л. Заметки о лихенофлоре и основных си-
нузиях лишайников стационара «Харп» // Споровые растения
Урала. Тр. ин-та экол. раст. жив. УФ АН СССР. Свердловск,
1970. Вып. 70. С. 134-139.
Международный кодекс ботанической номенклатуры
(Венский кодекс). М.-СПб.: Товарищество научных изданий
КМК, 2009. 282 с.
Микулин А. Г. Определитель лишайников полуострова
Камчатка. Владивосток, 1990. 128с.
Мир растений. Т. 2. Грибы / Под. ред. М. В. Горленко.
М.: Просвещение, 1991. 475 с.
Михайлюк Т. И, Дариенко Т. М, Демченко Э.
Н. Водоросли гранитных обнажений регионального ланд¬
шафтного парка “Гранитно-степное Побужье” (Николаевская
обл., Украина) // Новости систематики низших растений. Т. 37.
2003. С. 53-71.
Мошкова Н. О. Визначник пр1сноводних водоростей
УкраУнськоУ РСР. VI. Улотриксов1 й кладофоров1 водорости К.:
Наук.думка, 1979. 500 с.
Мучник Е. Э. Конспект лишайников степных и
остепненных местообитаний Центрального Черноземья // Но¬
вости систематики низших растений. 2002. Т. 35. С. 183-195.
Мучник Е. Э. Лихенологические исследования в запо¬
веднике «Галичья гора» // Биоразнообразие и экологические
особенности природы Русской лесостепи / Сб. науч. статей,
посвящ. 75-летию гос. заповедника «Галичья гора». Воронеж,
2000. С. 50-57.
Мучник Е. Э. Лихенофлора Центрального Черноземья:
таксономический и эколого-географический анализы, вопросы
охраны: Автореф. дис. ... д-ра биол. наук. Воронеж, 2003. 40 с.
Мырзакулова 3. С. Лишайники Южного Алтая // Тр. II
молодежи, конф. ботаников Ленинграда. Л, 1988. Ч. 3. С. 109-
113. Деп. в ВИНИТИ 14. 07. 88, № 5684-В 88.
Николаев Г. М, Маторин Д. Н, Аксенов С. И. Состояние
воды и функционирование первичных реакций фотосинтеза у
лишайника Placolecanora melanophtalma из холодных пустынь
Восточного Памира // Науч. докл. высш. школы, биол. науки.
1977. № 1.С. 106-106.
Нинбург Е. А. Введение в общую экологию (подходы и
методы). М.: Товарищество научных изданий КМК, 2005. 138
с.
Окснер А. М. Флора лишайниюв УкраУни. Т. 2, вип. 2.
К.: Наук. Думка, 1993. 544 с.
Окснер А. Н. Визначник лишайниюв УРСР / АН УРСР.
1н-т боташки. КиУв, 1937. 341 с.
Окснер А. Н. Де-що з флори обр^ниюв Чукотського
швострова // BicH. КиУв. ботан. саду. 1929. Вип. 9. С. 87-91.
Окснер А. Н. До вивчення флори обрюниюв каменястих
виход1в УкраУни // BicH. КиУв. ботан. саду. 1927. Вип. 5/6. С.
23-82.
Окснер А. Н. Лишайники басейну piK 1нд1прю Яни, Лени
та веденного Прибайкалля. I // Журн. 1н-ту боташки АН
УРСР. 1939а. № 23 (31). С. 117-139.
Окснер А. Н. Лишайники басейну piK 1нд1прки, Яни, Ле¬
ни та веденного Прибайкалля. IV // Бот. журн. АН УРСР.
1940в. Т. 1,№ 3-4. С. 31-57.
Окснер А. Н. Лишайники басейну piK Лени, Яни,
Инд*1прки та веденного Прибайкалля. II // Бот. журн. АН
УРСР. 1940а. Т. 1, № 1. С. 77-100.
Окснер А. Н. Лишайники басейну piK Лени, Яни,
Индтрки та веденного Прибайкалля. III // Бот. журн. АН
УРСР. 19406. Т. 1, № 2. С. 313-324.
Окснер А. Н. Матер1али для лихенофлоры Урала та при-
леглих областей // Бот. журн. АН УРСР. 1941-1945. Т. 2, №
3-4. С. 217-247.
Окснер А. Н. Материалы до лихенофлори Середньой
Азн // Журн. 1н-ту боташки АН УРСР. 19396. № 20 (28). С.
111-136.
Окснер А. Н. Материалы к флоре лишайников Белорус¬
сии: (Предварит, сообщ.) // B'ich. КиУв. Ботан. саду. 1924. Вип.
1.С. 27-36.
Окснер А. Н. Определитель лишайников СССР. Вып. 2.
Морфология, систематика и географическое распространение.
Л, 1974. 284 с.
Окснер А. Н. Определитель лишайников СССР. Вып. 2.
Общая часть. Морфология, систематика и географическое
распространение. Л, Наука, 1974. 284 с.
Окснер А. Н. Рел1ктов1 обрюники з Далекого Сходу //
BicH. КиУв. ботан. саду. 1934. Вип. 17. С. 37-40.
Окснер А. Н. Флора лишайнимв УкраУни. В 2-х т. Т. 2
(Вип. 1) / АН УРСР. 1н-т боташки. КиУв, Наук. Думка, 1968.
500 с.
Окснер А. Н. Флора лишайниюв УкраУни. В 2-х т. Т. 2
(Вип. 2) / АН УРСР. 1н-т боташки. КиУв, Наук. Думка, 1993
Окснер А. Н. Флора лишайниюв УкраУни: В 2-х т. Т. 1 /
АН УРСР. 1н-т боташки. КиУв, 1956. 495 с.
Окснер А. М. Визначник лишайниюв УРСР. К.: Вид-во
Акад. наук УРСР, 1937. 341 с.
Окснер А. Н. Определитель лишайников СССР. Вып. 2:
Морфология, систематика и географическое распространение.
Л.: Наука, 1974. 284 с.
Определитель лишайников России / Под ред. Н. С. Го¬
лубковой. СПб, 1996. Вып. 6. 203 с.; 1998. Вып. 7. 166 с.;
2003. Вып. 8. 277 с.; 2004. Вып. 9. 339 с.; 20ХХ. Вып. 10. XXX
с.
Определитель лишайников СССР / Под ред. Н. С. Голуб¬
ковой. Л, Наука 1971. Вып. 1. 203 с.; 1975. Вып. 3. 166 с.;
1977. Вып. 4. 277 с.; 1978. Вып. 5. XXX с.
Очирова Н. Н. Материалы к изучению лихенофлоры
Республики Калмыкия // Новости систематики низших расте¬
ний. 2005. Т. 38. С. 278-285.
Пийн Т. X. Напочвенные лишайники мыса Челюскин //
Арктические тундры и полярные пустыни Таймыра. Л, Наука,
1979а. С. 61-73.
Пийн Т. X. Напочвенные лишайники окрестностей бух¬
ты Марии Прончищевой (сверо-восточный Таймыр) // Аркти¬
ческие тундры и полярные пустыни Таймыра. Л, Наука,
19796. С. 140-143.
Пийн Т. X, Трасс X. X. Напочвенные лишайники
окрестностей Тареи (Западный Таймыр) // Биогеоценозы тай¬
мырской тундры и их продуктивность. Л, Наука, 1971. С. 151 —
160.
Питеранс А. В. Лишайники Латвии. Рига, Зинатие, 1982.
352 с.
Порядина Л. Н. Лишайники заказника «Сунтар-Хаята»
(Якутия) // Новости систематики низших растений. 2001. Т. 34.
С. 167-176.
Пчелкин А. В. Использование принципа сопряженности
флоры сосудистых растений и лишайников для флористиче¬
ского районирования // Проблемы экологического мониторин¬
га и моделирования экосистем. Л.: Гидрометеоиздат, 1991. Т.
13. С. 176-188.
Пыстина Т. Н. Биота лишайников Республики Коми: со¬
временное состояние и перспективы дальнейших исследова¬
ний // Фундаментальные и прикладные проблемы ботаники в
начале XXI века: Матер. XII Делегатского съезда РБО и Все-
росс. конфер. Часть 2. Петрозаводск, 2008. С. 221-224.
Пыстина Т. Н. Лишайники таежных лесов европейского
Северо-Востока (подзоны южной и средней тайги). Екатерин¬
бург, 2003. 240 с.
Пыстина Т. Н. Систематический список лишайников
равнинной части республики Коми (подзоны южной и средней
тайги) // Новости систематики низших растений. 2001. Т. 34.
С. 176-185.
Равинская А. П. Действие экологических факторов на
содержание усниновой кислоты и атранорина у лишайников //
Актуальные проблемы экспериментальной лихенологии в
СССР. Л, 1991. Вып. 1.С. 93-98.
Разнообразие растительного мира Якутии. Новосибирск,
Издательство СО РАН, 2005. 328 с.
Раменский Л. Г. Избранные работы. Проблемы и методы
изучения растительного покрова. Л.: Наука, 1971. 334 с.
Рассадина К. А. Лишайники, собранные В. Б. Сочавой в
1929 г. в Анадырском крае // Тр. Бот. ин-та АН СССР, сер. 2,
Споровые растения. 1934. Вып. 2. С. 321-338.
Рыкова Ю. В. Определитель листоватых лишайников
Якутии. Якутск, 1978. 112 с.
Рябкова К. А. Виды семейства Parmeliaceae на Урале //
Новости систематики низших растений. 1982 Т. 19. С. 149—
154.
Рябкова К. А. Лишайники Урала (список видов). Сверд¬
ловск, Свердловский гос. пед. инст. 1991. Деп. ВИНИТИ №
2504-В91. 30 с.
Рябкова К. А. Семейства Baeomycetaceae, Cladoniaceae,
Stereocaulaceae на Урале // Новости систематики низших рас¬
тений. 1982 Т. 19. С. 154-164.
Рябкова К. А, Макарова И. И. Лишайники Приполярно¬
го и Полярного Урала. Свердловск, Свердловский гос. пед.
инст, 1991. Деп. ВИНИТИ № 2504-В91.15с.
Савич В. П. Лишайники собранные в Анадырском окру¬
ге в 1903-07 гг. Н. Сокольниковым // Изв. Имп. Бот. сада.
СПб, 1911. Т. 11. Вып. 3. С. 82-90.
Савич В. П. Лишайники, собранные И. И. Тржемеским в
полярной Сибири // Изв. Имп. Бот. Сада Петра Великого. Пет¬
роград, 1916а. Т. 16. Вып. 1. С. 133-135.
Савич В. П. Лишайники, собранные Р. Р. Поле на край¬
нем севере Европейской России // Тр. Имп. Бот. Сада. СПб,
1912. Т. 32. С. 15-67.
Савич В. П. Новые виды и формы лишайников Камчатки
// Известия Императорского Ботанического Сада Петра Вели¬
кого. 1914. Т. 14. С. 111-128.
Савич В. П. Отдел споровых растений // От Аптекарско¬
го огорода до Ботанического института. Очерки по истории
Ботанического института Академии наук СССР. М. -Л, Изда¬
тельство АН, 1957. С. 115-133.
Савич В. П. Список лишайников Тобольской губ, со¬
бранных Б. Н. Городковым в 1914 году // Изв. Имп. Бот. Сада
Петра Великого. Петроград, 19166. Т. 16. Вып. 1. С. 101-111.
Савич В. П. Список лишайников, собранных Р. Н. Нима-
ном в Лапландии и Новой Земле в 1903 и 1908—1909 гг. // Тр.
Студенч. Научн. Кружков Физ.-Мат. Фак. СПб. Унив. СПб,
1911. Вып. 3. С. 37-56.
Савич В. П. Чукотские лишайники из коллекции Г. А.
Борисова // Изв. Имп. Бот. Сада Петра Великого. СПб, 1915.
Т. 15, вып. 5-6. С. 542-548.
Седельникова Н. В. Лихенофлора нагорья Сангилен. Но¬
восибирск, Наука, 1985. 180 с.
Седельникова Н. В. Лишайники Алтая и Кузнецкого
нагорья. Новосибирск, Наука, 1990. 175 с.
Седельникова Н. В. Лишайники Западного и Восточного
Саяна. Изд. СО РАН, Новосибирск, 2001. 190 с.
Седельникова Н. В. Лишайники Салаира // Флора
Салаирского кряжа. Новосибирск, ЦСБС, 1993. С. 32-78.
Седельникова Н. В. Особенности лихенофлоры Западно¬
го Саяна // Ботан. журн. 1996а. Т. 81. № 3. С. 75-83.
Седельникова Н. В. Систематический список лишайни¬
ков Восточного Саяна // Новости систематики низших расте¬
ний. 19966. Т. 31. С. 144-151.
Скирина И. Ф. Лишайники островов и прибрежных
участков. Лихеноиндикация загрязнения воздуха // Дальнево¬
сточный морской биосферный заповедник. Исследования. Т. 1.
Владивосток, 2004. С. 568-571.
Скирина И. Ф, Коженкова С. И, Родникова И. М. Эпи-
фитные лишайники Приморского края и использование их в
экологическом мониторинге. Владивосток: Дальнаука, 2010.
134 с.
Соболевский Г. Санктпетербургская флора. Часть 2.
1803. С. XX.
Список лихенофлоры России. [Сост. Г. П. Урбанавичюс]
Спб, Издательство Наука, 2010. 194 с.
Степанчикова И. С, Гимельбрант Д. Е, Кузнецова Е. С.
Лишайники // Природа Сестрорецкой низины и вопросы ее
охраны / Ред. Е. А. Волкова, Г. А. Исаченко, В. Н. Храмцов.
СПб, 2011. С. 155-166.
Сукачев В. Н. Избранные труды. Т. 1. Основы лесной
типологии и биогеоценологии. Л.: Наука, 1972. 419 с.
Титов А. Н. Микокалициевые грибы Голарктики. М, То¬
варищество научных изданий КМК, 2006. 296 с.
Титов А. Н. Профессор А. А. Еленкин - основатель ли¬
хенологической школы России // Фундаментальные и при¬
кладные проблемы ботаники в начале XXI века: Материалы
Всероссийской конференции, 22-27 сентября. Ч. 2. Альголо¬
гия, Микология, Лихенология, Бриология. Петрозаводск, 2008.
С. 246-249.
Томин М. П. Определитель корковых лишайников евро¬
пейской части СССР (кроме Крайнего Севера и Крыма).
Минск, Изд-во АН БССР, 1956. 533с.
Трасс X. X. Элементы и развитие лихенофлоры Эстонии
// Учен. зап. Тарт. унив. Вып. 268. / Тр. по бот. 1970. Т. 9. С. 5-
233.
Урбанавичене И. Н. Первые сведения о лишайниках
Национального парка «Зюраткуль» (Челябинская область) //
Новости систематики низших растений. Т. 45. 2011. С. 223-
236.
Урбанавичене И. Н, Урбанавичюс Г. П. К флоре лишай¬
ников хребта Хамар-Дабан // Новости систематики низших
растений. 1999. Т. 33. С. 161-171.
Урбанавичене И. Н, Урбанавичюс Г. П. Лишайники
Байкальского заповедника // Флора и фауна заповедников. М,
1998. Вып. 68. 53 с.
Урбанавичене И. Н, Урбанавичюс Г. П. Лишайники
Керженского заповедника // Природные условия Керженского
заповедника и некоторые аспекты охраны природы Нижего¬
родской области / Тр. Гос. природ, заповедника "Керженский".
Том 1. Нижний Новгород, 2001. С. 149-171.
Урбанавичюс Г. П. Исследования разнообразия лишай¬
ников России (успехи, достижения, тенденции и надежды) //
Флора лишайников России: состояние и перспективы исследо¬
ваний. Тр. междунар. совещ, посвящ. 120-летию со дня рож¬
дения В. П. Савича. СПб, 2006. С. 264-271.
Урбанавичюс Г. П. Как определить географический эле¬
мент лишайников // Труды Первой Русской лихенологической
школы. Петрозаводск, 2001. С. 223-238.
Урбанавичюс Г. П. Лихеноиндикация современных и па-
леобиоклиматических условий Южного Прибайкалья // Изве¬
стия РАН. Сер. геогр. 2002. № 1. С. 81-90.
Урбанавичюс Г. П. Особенности разнообразия лихено¬
флоры России // Известия РАН. Сер. геогр. 2011. № 1. С. 66-
78.
Урбанавичюс Г. П, Лавриненко О. В, Урбанавичене И.
Н. Лишайники острова Долгий и близлежащих островов юго-
востока Баренцева моря // Бот. журн. 2009. Т. 94. № 5. С. 32-
51.
Урбанавичюс Г. П, Урбанавичене И. Н. Аннотирован¬
ный список лишайников и близких к ним грибов Волжско-
Камского заповедника // Тр. Волжско-Камского гос. природ,
заповедника. Вып. 6. Казань, 2005. С. 160-187.
Урбанавичюс Г. П, Урбанавичене И. Н, Мелехин А. В.
Лихенофлора Лапландского государственного природного
биосферного заповедника (аннотированный список). Апатиты,
Изд. КНЦ РАН. 2012. 148 с.
Урбанавичюс Г. П, Урбанавичене И. Н, Отте Ф. Ли¬
шайники // Красная книга Адыгеи. Майкоп, 2012. С. 100-130.
Фадеева М. А, Голубкова Н. С, Витикайнен О, Ахти Т.
Конспект лишайников и лихенофильных грибов Республики
Карелия / Под ред. Г. П. Урбанавичюса. Петрозаводск, 2007.
194 с.
Фадеева М. А, Голубкова Н. С, Витикайнен О, Ахти Т.
Предварительный список лишайников Карелии и обитающих
на них грибов. Петрозаводск, 1997. 100 с.
Херманссон Я, Пыстина Т. Н, Кудрявцева Д. И. Пред¬
варительный список лишайников Республики Коми. Сыктыв¬
кар, 1998. 134 с.
Херманссон Я, Пыстина Т. Н, Ове-Ларссон Б, Журбен¬
ко М. П. Лишайники и лихенофильные грибы Печоро-
Илычского заповедника // Флора и фауна заповедников. Вып.
109 / Под ред. Г. П. Урбанавичюса. М, 2006. 79 с.
Херманссон Я, Тарасова В. Н, Степанова В. И, Сонина
А. В. Лишайники заповедника «Кивач» // Флора и фауна запо¬
ведников. Вып. 101 / Под ред. Г. П. Урбанавичюса. М, 2002.
35 с.
Ходосовцев А. Е, Кондратюк С. Я, Макарова И.
И, Окснер А. Н. Определитель лишайников России. 9.
Fuscideaceae, Teloschistaceae. СПб.: Наука. 2004. 339 с.
Ходосовцев А. Е. Лишайники каменистых обнажений
Крымского полуострова: Автореф. дис. ... д-ра биол. наук. Ки¬
ев, 2004. 36 с.
Чабаненко С. И. Лишайники // Флора, мико- и лихено-
биота Лазовского заповедника (Приморский край). Владиво¬
сток, ДВО АН СССР, 1990. С. 167-191.
Чабаненко С. И. Лишайники Лазовского государствен¬
ного природного заповедника // Урбанавичюс Г. П, Урба¬
навичене И. Н. Лишайники заповедников России / Современ¬
ное состояние биологического разнообразия на заповедных
территориях России. Вып. 3. Лишайники и мохообразные /
Под ред. О. М. Афониной, Н. С. Голубковой. М, 2004. С. 31-
215.
Чабаненко С. И, Скирина И. Ф, Княжева Л. А. Список
лишайников Приморского края и обитающих на них грибов.
Южно-Сахалинск, Сахалинский ботанический сад, СахНЦ,
ДВО РАН, 2002. 89 с.
Шапиро И. А. Загадки растения-сфинкса. Лишайники и
экологический мониторинг. Л.: Гидрометеоиздат, 1991. 80 с.
Шапиро И. А. Обзор современного состояния хемоси¬
стематики лишайников // Новости систематики низших расте¬
ний. 1980. Т. 17. С. 152-167.
Шаяхметова 3. М, Селиванов А. Е, Урбанавичене И. Н.
Лишайники предгорных средне- и южно-таежных лесов Перм¬
ского края // Вестн. ТвГУ. Сер. биология и экология. 2009.
Вып. 12, 6. С. 139-153.
Шель Ю. Материалы для ботанической географии
Уфимской и Оренбургской губернии (Споровые растения) //
Труды общества естествоиспытателей при Казанском универ¬
ситете. 1883. Вып. 12. С. 1-93.
Шустов М. В. Лишайники Приволжской возвышенности
// Новости систематики низших растений. 2002. Т. 36. С. 185-
203.
Юнатов А. А. Типы и содержание геоботанических ис¬
следований. Выбор пробных площадей и заложение экологи¬
ческих профилей // Полевая геоботаника. Т. 3. / Ред. А. А.
Корчагин, Е. М. Лавренко. М.-Л.: Наука, 1964. С. 9-36.
Юрцев Б. А, Толмачев А. И, Ребристая О. В. Флористи¬
ческое ограничение и разделение Арктики // Арктическая
флористическая область. Л, Наука, 1978. С. 9-104.
Юрченко Е. О, Синявская М. Г. Основы молекулярного
маркирования грибной ДНК. Практическое руководство.
Минск: Право и экономика, 2007. 101 с.
Aguirre-Hudson В. A taxonomic study of the species re¬
ferred to the ascomycete genus Leptorhaphis // Bull. Brit. Mus.
Nat. Hist. Botany. 1991. Vol. 21. P. 85-192.
Ahmadjian V. A guide to the algae occuring as lichen sym¬
bionts: Isolation, culture, cultural physiology, and identification //
Phycologia. 1967. Vol. 6. P. 127-160.
Ahmadjian V. Lichen Synthesis // Osrerr. Bot. Z. 1969. Vol.
116. P. 306-311.
Ahmadjian V. Lichens // Physiology and Biochemistry of
Algae / R. A. Lewin [ed.]. - New York-London: Academic Press,
1962. P. 817-822.
Ahmadjian V. The Lichen Symbiosis. New York: John
Wiley and Sons, Inc, 1993. 250 p.
Ahmadjian V, Heikkila H. The culture and synthesis of En-
docarpon pusillum and Staurothele clopima // Lichenologist. 1970.
Vol. 4. P. 259-267.
Ahti T, Jorgensen P.-M, Kristinsson H, Moberg R, Socht-
ing U, Thor G. (eds.) Nordic Lichen Flora. Vol. 2. Physciaceae.
Uddevalla. 2002. 116 p.
Ainsworth and Bisby's Dictionary of the Fungi. 10th edition
/ Eds. P. M. Kirk et al. Trowbridge: Cromwell Press, 2008. 771 p.
Almborn O. Distribution and ecology of some south Scandi¬
navian lichens // Bot. Notiser. Suppl. 1-2. Lund, 1948. P. 1-354.
Almquist E. Die Lichenenvegetation der Kusten des Be-
ringsmeeres // Nordenskiold A. E. / Die wissenschaftliche Ergeb¬
nisse der Vega-Expedition (Vega-expeditionis vetenskapliga jakt-
tagelser). Stockholm, 1887. Vol. Bd 4. P. 509-542.
Almquist E. Lichenologische Beobachtungen an der Nord-
kuste Sibiriens // Nordenskiold A. E. / Die wissenschaftliche Er¬
gebnisse der Vega-Expedition. Leipzig, 1883. Bd l.P. 50-74.
Almquist E. Lichenologiska iakttagelser pe Sibiriens nord-
kust // Otversigt af Kongl. Vetenskaps-Akademiens Forhandlingar.
1879. №9. P. 29-59.
Alstrup V, Grube M, Motiejunaite J, Nordin A, Zhurbenko
M. Lichenicolous fungi from the Skibotn area, Troms, Norway //
Graphis Scripta. 2008. Vol. 20. P. 1-8
Andreev M. Halecania santessonii, a new lichenicolous li¬
chen from Russia // Lichenologist. 2010. Vol. 42, № 2. P. 1-4.
Andreev М. P, Kotlov Yu. V, Makarova I. I. Checklist of
lichens and lichenicolous fungi of the Russian Arctic // The Bryol¬
ogist. 1996. Vol. 99, № 2. P. 137-169.
Aptroot A, Lucking R, Sipman H. J. M, et al. Pyrenocar-
pous Lichens with Bitunicate Asci: A First Assessment of the Li¬
chen Biodiversity Inventory in Costa Rica // Biblioth. Lichenol.
2008. N 97. 162 pp.
Arcadia L, Knudsen K. The name Myriospora is available
for the Acarospora smaragdula group // Opuscula Philolichenum.
2012. Vol. 11. P. 19-25.
Archibald P. A. Trebouxia de Puymaly (Chlorophyceae,
Chlorococcales) and Pseudotrebouxia gen. nov. (Chlorophyceae,
Chlorococcales)//Phycologia. 1975. Vol. 14. P. 125-137.
Arguello A, del Prado R, Cubas P, Crespo A. Parmelina
quercina (Parmeliaceae, Lecanorales) includes four phylogenetical¬
ly supported morphospecies // Biological Journal of the Linnaean
Society. 2007. Vol. 91(3). P. 455-467.
Armaleo D, Zhang Y, Cheung S. Light might regulate di¬
vergently depside and depsidone accumulation in the lichen Par-
motrema hypotropum by affecting thallus temperature and water
potential // Mycologia. 2008. Vol. 100, № 4. P. 565-576.
Arup U, Ekman S, Lindblom L, Mattsson J.-E. High per¬
formance thin layer chromatography (HPTLC), an improved tech¬
nique for screening lichen substances // Lichenologist. 1993. Vol.
25. P. 61-71.
Arvidsson L. A monograph of the lichen genus Coccocarpia
// Opera Bot. 1982. № 67. P. 1-96.
Asahina Y, Shibata S. Chemistry of Lichen Substances. Jap¬
anese Society for the Promotion of Science, Tokyo. 1954. 240 pp.
Ascaso C, Valladares F, De los Rios A. New ultrastructure 1
aspects of pyrenoidsof the lichen photobiont Trebouxia (Mi-
crothamniales, Chlorophyta) Hi. Phycol. 1995. №. 31. P. 114—119.
Asta J, Erhardt W, Ferretti M, Fornasier F, Kirschbaum U,
Nimis P. L, Purvis O. W, Pirintsos S, Scheidegger C, van Ha-
luwyn C, Wirth V. Mapping Lichen Diversity as an Indicator of
Environmental Quality // Monitoring with Lichens - Monitoring
Lichens / Eds. P. L. Nimis, C. Scheidegger, P. A. Wolseley. Dor¬
drecht, Boston, London: Kluwer Academic, 2002. P. 273-279.
Authors of Fungal Names. Версия 2. 1992. [Электронный
ресурс]. Код доступа:
http://www.indexfungorum.org/Names/AuthorsOfFungalNames.as
Р
Backor М, Hudak J, Ziegler W, Backorova M. Methods for
isolation and cultivation of the eukaryotic lichen photobionts look¬
ing for a general method. A review. // Thaiszia J. Bot, Kosice.
1998. №8. P. 1-6.
Baloch E, Gilenstam G, Wedin M. Phylogeny and classifi¬
cation of Cryptodiscus, with taxonomic synopsis of the Swedish
species// Fungal Diversity. 2009. Vol. 38. P. 51-68.
Baloch E, Lucking R, Lumbsch H. T, Wedin M. Major
clades and phylogenetic relationships between lichenized and non-
lichenized lineages in Ostropales (Ascomycota: Lecanoromycetes)
//Taxon. 2010. Vol. 59. N 5. P. 1483-1494.
Barkman J. J. Phytosociology and ecology of cryptogamic
epiphytes. Assen: van Gorcum, 1958. 628 p.
Barnhart J. H. Biographical Notes upon Botanists. 1965.
Vol. 1. 563 p.; Vol. 2. 259 p.; Vol. 3. 545 p.
Barreno E, Perez-Ortega S. The UNESCO-MAB Reserve of
Muniellos (Spain, Asturias), an example of high lichen diversity in
Europe and the success of conservations strategies // FI. Medit.
2005. Vol. 15. P. 453-460.
Beck A. Selektivitat der Symbionten schwermetalltoleranter
Flechten. Ludwig-Maximilians-Universitat, Munchen. PhD thesis.
Fischer GmbH, 2002. 196 s.
Beck A, Kasalicky T, Rambold G. Myco-photobiontal se¬
lection in a Mediterranean cryptogam community with Fulgensia
fulgida // New Phytol. 2002. Vol. 153. P. 317-326.
Bennett J. P, Wetmore С. M. Lichens of the U. S. National
Parks // Bryologist. 2005. Vol. 108. № 4. P. 544-553.
Bhattacharya D, Friedl T, Damberger S. Nuclear-encoded
rDNA group I introns: origin and phylogenetic relationships of in¬
sertion site lineages in the green algae // Mol. Biol. Evol. 1996.
Vol. 13. P. 978-989.
Bialonska D, Dayan F. E. Chemistry of the lichen Hy-
pogymnia physodes transplanted to an industrial region // Journal
of Chemical Ecology. 2005. Vol. 31, № 12. P. 2975-2991.
Bishop J. A, Cook L. M, Muggleton J, Seaward M. R. D.
Moths, Lichens and Air Pollution Along a Transect from Manches¬
ter to North Wales//Journal of Applied Ecology. 1975. Vol. 12(1).
P. 83-98.
Blaha J, Baloch E, Grube M. High photobiont diversity as¬
sociated with the euryoecious lichen-forming ascomycete Lecanora
rupicola // Biol. Journ. Lin. Soc. 2006. Vol. 88, №2. P. 283-293.
Brodo I. M. Substrate ecology // V. Ahmadjian, М. E. Hale
(eds.). The lichens. New York, London: Academic Press, 1973. P.
401-441.
Brodo I. M, Duran-Sharnoff S, Sharnoff S. Lichens of
North America. New Haven & London: Yale University Press,
2001. 795 p.
Brunner U, Honegger R. Chemical and ultrastuctural studies
on the distribution of sporopolleninlike biopolymers in six genera
of lichen phycobionts // Can. J. Bot. 1985. 63. P. 2221-2230.
Bubrick P. Methods for cultivating lichens and isolated bi-
onts // CRC Handbook of Lichenology / M. Galun, (ed.). Ill, Boca
Raton, Fla.: CRC Press, 1989. P. 127-138.
Budel B, Henssen A. Chroococcidiopsis (Cyanophyceae), a
phycobiont in the lichen family Lichinaceae // Phycologia. 1983.
Vol. 22, № 4. P. 367-375.
Budel B, Scheidegger C. Thallus morphology and anatomy
// Lichen Biology / Ed. by Т. H. Nash III. Cambridge: Cambridge
University Press, 2008 (2th edition). P. 40-68.
Buxbaum I. Ch. Observationes circa Quasdam Plantas Ingri-
cas // Commentarii Academiae Scientiarum Imperialis Petropoli-
tanae. Tomus III. Petropoli, Typis Academiae: 1728. P. 270-273.
Buxbaum J. C. Plantarum minus cognitarum Centuria I-V,
complectens plantas circa Byzantium in Oriente observatas.
Petropoli ex Typographia Academiae, 1728, 1728, 1729, 1733,
1740. P. XX. P. XX. P. XX.
Bylin A, Arnerup J, Hogberg N, Thor G. A phylogenetic
study of Fuscideaceae using mtSSU rDNA // Biblioth. Lichenol.
2007. N 96. P. 49-60.
Cajander A. K. Beitrage zur Kenntniss der Vegetation der
Alluvionen des nordlichen Eurasiens. I. Die Alluvionen des unte-
ren Lena-Thales // Acta Soc. Sci. Fenn. Helsingfors, 1903. Vol. 32.
Iss. 1. P. 1-182.
Cannon P. F, Kirk P. M. Fungal Families of the World. CAB
International, 2007. 455 p.
Chapman R. L. Ultrastructural investigation on the fo¬
liicolous pyrenocarpous lichen Strigula elegans (Fee) Mull. Arg. //
Phycologia. 1976. Vol. 67. P. 191-196.
Chapman R. L. Waters D. A. Lichenization of the Tren-
tepohliales complex algae and odd relationships // Symbiosis:
Mechanisms and Model Systems / J. Seckbach [ed.]. Kluwer Aca¬
demic Press, 2001. P. 361-371.
Chapman R. L, Henk М. C. Phragmoplasts in cytokinesis of
Cephaleuros parasiticus (Chlorophyta) vegetative cells // J. Phycol.
1986. №22. P. 83-88.
Clauzade G, Roux C. Likenoj de Okcidenta Ейгоро. Ilustrita
Determinlibro // Bull. Soc. Bot. du Centre-Ouest, Nouvelle Serie.
1985. Numero Special 7. 892 p.
CRC Handbook of Lichenology. Vol. 1-3 // Ed. by M. Ga-
lun. Boca Raton: CRC Press, 1988.
Crespo A, Kauff F, Divakar P. K, et al. Phylogenetic ge¬
neric classification of parmelioid lichens (Parmeliaceae, Ascomy¬
cota) based on molecular, morphological and chemical evidence //
Taxon. 2010. Vol. 59. P. 1735-1753.
Crespo A, Molina М. C, Blanco O, Schroeter B, Sancho
L. G, Hawksworth D. L. DNA ITS and P-tubulin gene sequence
analyses reveal two monophyletic groups within the cosmopolitan
lichen Parmelia saxatilis // Mycological Research. 2002. Vol.
106(7). P. 788-795.
Crespo A, Perez-Ortega S. Cryptic species and species pairs
in lichens: A discussion on the relationship between molecular
phylogenies and morphological characters // Anales del Jardfn Bo-
tanico de Madrid. 2009. Vol. 66, S 1. P. 71-81.
Crous P. W, Schoch C. L, Hyde K. D, et al. Phylogenetic
lineages in the Capnodiales // Stud. Mycol. 2009. Vol. 64. P. 17-
47.
Culberson C. F. Chemical and Botanical Guide to Lichen
Products. University of North Carolina Press, Chapel Hill, 1969.
628 pp.
Culberson C. F. High-speed liquid chromatography of lichen
extracts // Bryologist. 1972a. Vol. 75. P. 54-62.
Culberson C. F. Improved conditions and new data for the
identification of lichen products by a standardized thin-layer chro¬
matographic method // Journal of Chromatography. 1972b. Vol.
72. P. 113-125.
Culberson C. F. Supplement to "Chemical and Botanical
Guide to Lichen Products" // Bryologist. 1970. Vol. 73. P. 177—
377.
Culberson C. F, Ammann K. Standardmethode zur Diinn-
schichtchromatographie von Flechtensubstanzen // Herzogia. 1979.
Vol. 5. P. 1-24.
Culberson C. F, Culberson W. L, Gowan S, Johnson A.
New depsides from lichens: microchemical methodologies applied
to the study of new natural products discovered in herbarium spec¬
imens // American Journal of Botany. 1987. Vol. 74. P. 403-414.
Culberson С. F, Culberson W. L, Johnson A. A standarized
TLC analysis of P-orcinol depsidones // Bryologist. 1981. Vol. 84.
P. 16-29.
Culberson C. F, Culberson W. L, Johnson A. Second Sup¬
plement to "Chemical and Botanical Guide to Lichen Products".
American Bryological and Lichenologial Society, Missouri Botan¬
ical Garden, St. Louis. 1977. 400 pp.
Culberson C. F, Kristinsson H. A standardized method for
the identification of lichen products // Journal of Chromatography.
1970. Vol. 46. P. 85-93.
Culberson W. L. Chemistry and sibling speciation in the li¬
chen-forming fungi: ecological and biological considerations //
Bryologist. 1986. Vol. 89. P. 123-131.
Culberson W. L, Culberson C. F. Chemosyndromic varia¬
tion in lichens // Systematic Botany. 1977. № 1. P. 325-339.
Dahlkild A, Kallersjo M, Lohtander K. et al. Photobiont di¬
versity in the Physciaceae (Lecanorales) // Bryologist. 2001. Vol.
104, №4. P. 527-536.
Davydov E. A, Himelbrant D. E, Stepanchikova I. S. Con¬
tribution to the study of Umbilicariaceae (lichenized Ascomycota)
in Russia. II. Kamchatka Peninsula // Herzogia. 2011. Bd. 24. Heft
2. P. 229-241.
Deason T. R, Bold H. C. Phycological Studies I. Explorato¬
ry Studies of Texas Soil Algae. University of Texas, 1960. 70 p.
Degelius G. The lichen genus Collema in Europe // Symb.
Bot. Upsal. 1954. Vol. 13, № 2. P. 1-499.
Dictionary of the Fungi / Ed. by Kirk P. M. et al. 10th ed.
Wallingford: CABI Publishing, 2008. 771 p.
Diederich P, Lawrey J. D, Sikaroodi M, van den Boom P.
P. G, Ertz. D. Briancoppinsia, a new coelomycetous genus of
Arthoniaceae (Arthoniales) for the lichenicolous Phoma cytospora,
with a key to this and similar taxa // Fungal Diversity. 2012. Vol.
52. N l.P. 1-12.
Divakar P. K, Lumbsch H. T, Ferencova Z, et al. Remoto-
trachyna, a newly recognized tropical lineage of lichens in the Hy¬
potrachyna clade (Parmeliaceae, Ascomycota), originated in the
Indian subcontinent // Amer. J. Bot. 2010. Vol. 97. N 4. P. 579-
590.
Elenkin A. A. Species novae lichenum in Sibiria arctica a d.
A. A. Birula-Bialynizki collectae (expeditio baronis Toll) // Annal
Mycol. 1906. Vol. 4. № 1. P. 36-38.
Elix J. A. Biochemistry and secondary metabolites / Nash III
Т. H. Lichen biology. Cambridge University Press. 1996. P. 154—
180.
Elix J. A, Stocker-Worgotter E. Biochemistry and secondary
metabolites / Nash III, Т. H. Lichen biology. 2nd Ed. Cambridge
University Press. 2008. P. 104-133.
Elvebakk A, Bjerke J. W. The Skibotn area in North Nor¬
way - An example of very high lichen species richness far to the
north: A supplement with an annotated list of species // Mycotax-
on. 2006. Vol. 96. P. 141-146.
Eriksson О. E. (ed.) Outline of Ascomycota - 2006 // My¬
conet. 2006. Vol. 12. P. 1-82.
Ertz D, Tehler A. The phylogeny of Arthoniales (Pezizomy¬
cotina) inferred from nucLSU and RPB2 sequences // Fungal Di¬
versity. 2010. Vol. 49. N 1. P. 47-71.
Esslinger T. L. A cumulative checklist for the lichen-
forming, lichenicolous and allied fungi of the continental United
States and Canada. North Dakota State University:
http://www.ndsu.edu/pubweb/~esslinge/chcklst/chcklst7.htm (First
Posted 1 December 1997, Most Recent Version (#17) 16 May
2011), Fargo, North Dakota.
Ettl H, Gartner G. Syllabus der Boden-, Luft-, und Flechten-
algen. Stuttgart, Jena, New York: Gustav Fischer, 1995. 710 S.
Fahselt D. Secondary biochemistry of lichens // Symbiosis.
1994. Vol. 16. №2. P. 117-165.
Feige G. B, Lumbsch H. T, Huneck S, Elix J. A. Identifica¬
tion of lichen substances by a standardized high-performance liq¬
uid chromatographic method // Journal of Chromatography. 1993.
№646. P. 417-427.
Feuerer T, Hawksworth D. L. Biodiversity of lichens, in¬
cluding a world-wide analysis of checklist data based on Takhta-
jan’s floristic regions // Biodivers. Conserv. 2007. Vol. 16. P. 85-
98.
Flechten: Geschichte, Biologie, Systematik, Okologie, Na-
turschutz und kulturelle Bedeutung // Ed. by H. Scholler. Frankfurt
am Main: Kramer, 1997. 247 p.
Friedl T. Comparative ultrastructure of pyrenoids in Tre¬
bouxia (Microthamniales, Chlorophyta) // PI. Syst. Evol. 1989.
164. P. 145-159.
Friedl T. Thallus development and phycobionts of the para¬
sitic lichen Diploschistes muscorum // Lichenologist. 1987. Vol.
19. P. 183-191.
Friedl T, Budel B. Photobionts // The Lichen Symbiosis /
Nash III T. (ed.). Cambridge University Press, 2008. P. 9-26.
Friedl T, Gartner G. Trebouxia (Pleurastrales, Chlorophyta)
as a phycobiont in the lichen genus Diploschistes // Arch. Pro-
tistenk. 1988. Vol. 135. P. 147-158.
Frizzell D. L. Terminology of types // The American Mid¬
land Naturalist. 1933. № 6. P. 637-668.
Galloway D. J. Flora of New Zealand lichens: Lichens, in¬
cluding lichen-forming and lichenicolous fungi. Vol. 1-2. Lincoln,
2007. 2262 p.
Galloway D. J. Lichen biogeography // The Lichen Symbio¬
sis / Nash III T. (ed.). Cambridge University Press, 2008. P. 315-
335.
Galun M, Mukhtar A. Checklist of the lichens of Israel //
Bocconea. 1996. Vol. 6. P. 149-171.
Gartner G. Die Gattung Trebouxia Puymaly (Chlorellales,
Chlorophyceae) // Arch. Hydrobiol. Suppl. Algol. Studies. 1985.
71, №4. S. 495-548.
Garty J. Re-establishment of lichens on chalk rocks after a
forest fire // Fire in Ecosystem Dynamics. 1990. P. 77-84.
Gauslaa Y. Lichen palatability depends on investments in
herbivore defence // Oecologia. 2005. No 143. P. 94-105.
Gaya E, Hognabba F, Holguin A, et al. Implementing a
cumulative supermatrix approach for a comprehensive phylogenet¬
ic study of the Teloschistales (Pezizomycotina, Ascomycota) //
Mol. Phylogenet. Evol. 2012. doi:10.1016/j.ympev.2012.01.012
Gaya E, Hognabba F, Holguin A, et al. Implementing a
cumulative supermatrix approach for a comprehensive phylogenet¬
ic study of the Teloschistales (Pezizomycotina, Ascomycota) //
Mol. Phylogenet. Evol. 2012. doi:10.1016/j.ympev.2012.01.012
Geitler L. Beitrage zur Kenntnis der Flechtensymbiose I—III
//Arch. Protistenk. 1933. 80. S. 378-409.
Geitler L. Clavaria mucida eine extratropische Basidioliche-
ne // Biologisches Zentralblatt Band. 1955. Vol. 74. S. 145-159.
Geitler L. Schizophyceen II. Umgearbeitete Aflage // Encyc¬
lopedia of Plant Anatomy / W. Zimmermann, P. Ozenda [eds.].
Berlin: Borntraeger, 1960. P. 98-112.
Geitler L. Uber die Flechtenalge Myrmecia biatorellae // Os-
terr. Bot. Z. 1962. № 109. S. 41^17.
Geitler L. Uber Haustorien bei Flechten und uber Myrmecia
biatorellae in Psora globifera // Osterr. Bot. Z. 1963. № 110. S.
270-276.
Gmelin J. G. Flora sibirica, sive Historia plantarum sibiricae
Гмелин, 1747, 1749, 1768, 1769
Gmelin J. G. Reise durch Sibirien, von dem Jahr 1733 bis
1743, 4 vols. Goettingen, 1751-52.
Gorbushina A. A, Beck A, Schulte A. Microcolonial rock
inhabiting fungi and lichen photobionts: evidence for mutualistic
interactions // Mycol. Res. 2006. Vol. 109. P. 1288-1296.
Goward T. The lichens of British Columbia, illustrated keys.
Part 2. Fruticose species. // Special Report Series 9. Victoria, 1999.
319 p.
Goward T. Twelve Readings on the Lichen Thallus. VII.
Species // Evansia. 2009. Vol. 26, №4. P. 153-162.
Goward T, McCune B, Meidinger D. The lichens of British
Columbia, illustrated keys. Part 1. Foliose and Squamulose species
// Special Report Series 8. Victoria, 1994. 181 p.
Grube M, et al. Unravelling the phylogenetic relationships
of lichenised fungi in Dothideomyceta // Stud. Mycol. 2009. Vol.
64. P. 135-144.
Grube M, Kroken S. Molecular approaches and the concept
of species and species complexes in lichenized fungi // Mycologi-
cal Research. 2000. Vol. 104(11). P. 1284-1294.
Gruyter J. D, Woudenberg J. H. C, Aveskamp М. M, et al.
Systematic reappraisal of species in Phoma section Paraphoma,
Pyrenochaeta and Pleurophoma // Mycologia. 2010. Vol. 102. P.
1066-1081.
Gueidan C, Savic S, Thus H, et al. Generic classification of
the Verrucariaceae (Ascomycota) based on molecular and morpho¬
logical evidence: recent progress and remaining challenges // Tax¬
on. 2009. Vol. 58. N 1. P. 184-208.
Hafellner J. A revision of Maronella laricina and Piccolia
ochrophora // Symb. Bot. Upsal. 2004. Vol. 34. N 1. P. 87-96.
Hafellner J. Halospora resurrected and segregated from Mer-
ismatium // Biblioth. Lichenol. 2011. N 106. P. 75-93.
Hafellner J. Studien in Richtung einer naturlicheren Gleide-
rung der Sammelfamilien Lecanoraceae und Lecideaceae // Beih.
Nowa Hedwigia. 1984. Bd. 79. S. 241 -371.
Hafellner J, Poelt J. Der «Flechtenparasit» Buellia pulver-
ulenta eine bleibend interne parasitische Flechte // Phyton [Austri-
a]. 1980. Vol. 20. S. 129-133.
Hafellner J, Turk R. Die lichenisierten Pilze Osterreichs -
eine Checkliste der bisher nachgewiesenen Arten mit Verbrei-
tungsangaben // Stapfia. 2001. Vol. 76. P. 3-167.
Hafellner J. Phacothecium resurrected and the new genus
Phacographa (Arthoniales) proposed // Biblioth. Lichenol. 2009.
Vol. 100. P. 85-121.
Hale М. E. The Biology of Lichens. London: Edward Ar¬
nold, 1983 (3th edition). 190 p.
Halonen P, Myllys L, Velmala S, Hyvarinen H. Gowardia
(Parmeliaceae) - a new alectorioid lichen genus with two species //
Bryologist. 2009. Vol. 112.N l.P. 138-146.
Harada H, Okamoto T, Yoshimura I. A checklist of lichens
and lichen-allies of Japan // Lichenology. 2004. Vol. 2. № 2. P.
47-165.
Harris R. C. Four novel lichen taxa in the lichen biota of
eastern North America // Opuscula Philolichenum. 2009. N 6. P.
149-156.
Hauck M, Huneck S. Lichen substances affect metal absorp¬
tion in Hypogymnia physodes // Journal of Chemical Ecology.
2007. Vol. 33. P. 219-223.
Hawksworth D. L. The variety of fungal-algae symbioses,
their evolutionary significance and the nature of lichens // Botan.
Journ. Linn. Soc. 1988. Vol. 96. P. 3-20.
Hawksworth D. L, Hill D. J. The Lichen-Forming Fungi.
Glasgow & London: Blackie, 1984. 158 p.
Hawksworth D. L, Millanes A. M, Wedin M. Roselliniella
revealed as an overlooked genus of Hypocreales, with the descrip¬
tion of a second species on parmelioid lichens // Persoonia. 2010.
Vol. 24. P. 12-17.
Helms G. Taxonomy and symbiosis in associations of
Physciaceae and Trebouxia. PhD thesis. University of Gottingen,
Germany, 2003. 155 p.
Hibbett D. S, Binder M, Bischoff J. F. et al. A higher-level
phylogenetic classification of the Fungi // Mycological Research.
2007. Vol. 11 l.P. 509-547.
Hildreth К. C, Ahmadjian V. A study Trebouxia and
Pseudotrebouxia isolates from different lichens // Lichenologist.
1981. 13. P. 65-86.
Hinds J. W, Hinds P. L. The Macrolichens of New England.
New York: New York Botanical Garden Press, 2007. 608 p.
Hodkinson В. P, Lendemer J. C. The orders of Ostropomy¬
cetidae (Lecanoromycetes, Ascomycota): Recognition of Sarrame¬
anales and Trapeliales with a request to retain Pertusariales over
Agyriales // Phytologia. 2011. Vol. 93. N 3. P. 407-412.
Hoffman, 1784. Enumeratio lichenum.
Hoffman, 1794. Deutschlands Flora.
Hofstetter V, Miadlikowska J, Kauff F. et al. Phylogenetic
comparison of protein-coding versus ribosomal RNA-coding se¬
quence data: A case study of the Lecanoromycetes (Ascomycota) //
Molecular Phyl. Evol. 2007. Vol. 44. P. 412^126.
Honegger R. Cytological aspects of the mycobiont-
phycobiont relationship in lichens // Lichenologist. 1984. Vol. 16,
№2. P. 111-127.
Honegger R. Functional aspects of the lichen symbiosis //
Annu. Rev. Plant Mol. Biol. 1991. Vol. 42. P. 553-578.
Honegger R. Lichen-Forming Fungi and Their Photobionts //
Plant Relationships. The Mycota V. / Ed. by H. Deising. Berlin,
Heidelberg: Springer-Verlag, 2009 (2th edition). P. 308-333.
Honegger R. Mycobionts // The Lichen Symbiosis / Nash III
T. (ed.). Cambridge University Press, 2008. P. 27-39.
Honegger R. Questions about pattern formation in the algal
layer of lichens with stratified (heteromerous) thalli / Peveling E.
(ed.): Progress and Problems in Lichenology in the Eighties. Bibli-
otheca Lichenologica. 1987. Vol. 25. P. 59-71.
Honegger R. Water relations in lichens // Fungi in the Envi¬
ronment / Gadd G. M, Watkinson S. C, Dyer P. (eds.). Cambridge
Univ. Press, 2006. P. 185-200.
Honegger R, Zippier U, Gansner H. et al. Mating systems in
the genus Xanthoria (lichen-forming ascomycetes) // Mycological
Research. 2004. Vol. 108. P. 480-488.
Horizons in Lichenology // Botan. Journ. Linn. Soc. 1988.
Vol. 96. / Ed. by D. H. Dalby, D. L. Hawksworth, S. L. Jury. Lon¬
don: Linnean Society of London and Academic Press, 1988. P. 1-
100.
Huhtinen S. An effective technique for collecting epilithic
crustose lichens // Memoranda Societatis pro Fauna et Flora Fenni¬
ca. 2002. Vol. 77. P. 1-3.
Huhtinen S. Anyone for olygotypes? // Taxon. 1994. Vol.
43(8). P. 441-443.
Huneck S, Yoshimura I. Identification of lichen substances.
Springer-Verlag. Berlin, Heidelberg, New York. 1996. 493 pp.
Hyde K. D, McKenzie E. H. С, KoKo T. W. Towards in¬
corporating anamorphic fungi in a natural classification - checklist
and notes for 2010 // Mycosphere. 2011. Vol. 2. N l.P. 1-88.
Ihda T, Nakano T, Yoshimura I. et al. Phycobionts isolated
from Japanese species of Anzia (Lichenes) // Arch. Protistenk.
1993. Vol. 143. P. 163-172.
Jaag О. Соссотуха Schmidle. Monographic einer Algengat-
tung. // Beitr. Kryptogamenfl. Schweiz. Bot. Ges. 1933. Vol. 42.
132 S.
Jaaskelainen K, Pykala J, Rama H. et al. Jakalat • Lichens //
Suomen lajien uhanalaisuus. Ppunainen Kirja. 2010. P. 278-310.
John V, Seaward M. R. D, Sipman H. J. M, Zedda L. Li¬
chens and lichenicolous fungi of Syria, including a first checklist //
Herzogia. 2004. Vol. 17. P. 157-177.
Jorgensen P. M. Thallinocarpon //Nordic Lichen Flora. Vol.
3. Cyanolichens. Nordic Lichen Society, Uddevalla. P. 72-73.
Jorgensen P. M. The lichen family Pannariaceae in Europe //
Opera Bot. 1978. № 45. - 123 p.
Jorgensen P. M. Vahliella, a new lichen genus // Lichenolo¬
gist. 2008. Vol. 40. N 3. P. 221-225.
Joshi Y, Andreev M, Hur J. -S. Caloplaca lacinulata redis¬
covered for the lichen flora of South Korea // Feddes Repertorium.
2012. Vol. 122. № 5-6. P. 421-423.
Kappen L, Lange O. L. Die Kaltresistenz einiger Makroli-
chenen//Flora. 1972. Bd 161. S. 1-29.
Kershaw K. A. Physiological ecology of lichens. Cambridge:
Cambridge University Press, 1985. 293 p.
Khodosovtsev A, Naumovich A, Vondrakova O, Vondrak
J. Athelium imperceptum Nyl. (Thelocarpaceae, Ascomycota), a
little known ephemeral lichen of biological soil crusts, new to
Ukraine // Chornomors'k. bot. z. 2010. Vol. 6. P. 385-389.
Kivisto L, Kuusinen M. Edge effects on the epiphytic lichen
flora of Picea abies in middle boreal Finland // The Lichenologist.
2000. Vol. 32(4). P. 387-398.
Klement O. Prodromus der mitteleuropaischen Flechtenge-
sellschaften // Feddes Repert. Beih. 135 (Beitrage zur Vegetations-
kunde). 1955. S. 5-194.
Kondratyuk S. Prof. Dr. A. M. Oxner (22. 02. 1898-20. 11.
1973). On occasion of 100-years anniversary of the birthday. Kiev:
Phytosociocentre, 1998. 64 p.
Konig J, Peveling E. Cell walls of the phycobionts Tre¬
bouxia and Pseudotrebouxia: constituents and their location // Li¬
chenologist. 1984. Vol. 16. P. 129-144.
Konig J, Peveling E. Vorkommen von Sporopollenin in der
Zellwand des Phycobionten Trebouxia // Z. Pflanzenphysiol. 1980.
Vol. 98. S. 459-464.
Kotlov Yu. V. Lichens from two bird colonies in the West
Siberian Arctic // Graphis Scripta. 1994. № 6. P. 57-59.
Kristinsson H, Heidmarsson S. Checklist of lichens in Ice¬
land. Listinn var sfdast uppfaerdur 21 maf 2006. -
http://www.floraislands.is/fletlist.htm
Kroken S, Taylor J. W. Phylogenetic species, reproductive
mode, and specificity of the green alga Trebouxia forming lichens
with genus Letharia // Bryologist. 2000. Vol. 103, №4. P. 645-660.
Kukwa M. The lichen genus Ochrolechia in Europe. Gdansk:
Fundacja Rozwoju Uniwersytetu Gdanskiego, 2011. 309 p.
Kukwa M, Perez-Ortega S. A second species of Botry¬
olepraria from the Neotropics and the phylogenetic placement of
the genus within Ascomycota // Mycol. Progress. 2010. Vol. 9. N
3. P. 345-351.
Kuznetsova E. S, Motiejunaite J, Stepanchikova I. S,
Himelbrant D. E, Czarnota P. New records of lichens and allied
fungi from the Leningrad Region, Russia. Ill // Folia Cryptogamica
Estonica. 2012. Fasc. 49. P. 31-37.
Kuznetsova E, Ahti T, Himelbrant D. Lichens and allied
fungi of the Eastern Leningrad Region // Norrlinia. 2007. Vol. 16.
P. 1-62.
Lai M. J, Qian Z. G, Xu L. Synopsis of the cetrarioid lichen
genera and species (Parmeliaceae, lichenized Ascomycotina) in
China // J. Natl. Taiwan Mus. 2007. Vol. 60. N 1. P. 45-61.
Lakastos M, Rascher U, Budel B. Functional characteristics
of corticolous lichens in the understory of a tropical lowland rain
forest // New Phytologist. 2006. Vol. 172, № 4. P. 679-695.
Lamb I. M. On the morphology, phylogeny, and taxonomy of
the lichen genus Stereocaulon // Can. J. Bot. 1951. Vol. 29. P.
522-536.
Lange O. L. Pflanzenleben unter Stress. Flechten als Pioniere
der Vegetation an Extremstandorten der Erde. Rostra Universitatis
Wirceburgensis. Wurzburg. 1992. 59 pp.
Lange O. L, Belnap J, Reichenberger H. Photosynthesis of
the cyanobacterial soil-crust lichen Collema tenax from arid lands
in southern Utah, USA: role of water content on light and tempera¬
ture responses of C02 exchange // Functional Ecology. 1998. Vol.
12. P. 195-202.
Laundon J. R. Deceased lichenologists: their abbreviations
and herbaria. Lichenologist. 1979. Vol. 11. P. 1-26.
Lawrey J. D. Biology of lichenized fungi. Praeger Publishers.
New York. 1984. 407 pp.
Lawrey J. D. Chemical defense in lichen symbioses / White
J. F, Torres M. S. (eds). Defensive Mutualism in Microbial Sym¬
biosis. Mycology. 2009. Vol. 27. P. 167-181.
Lawrey J. D, Diederich P. Lichenicolous fungi - worldwide
checklist, including isolated cultures and sequences available.
2011. URL: http://www.lichenicolous.net [7/24/2011].
Lawrey J. D, Diederich P, Nelsen М. P, et al. The obligate-
ly lichenicolous genus Lichenoconium represents a novel lineage
in the Dothideomycetes // Fungal Biol. 2011. Vol. 115. N 2.
P. 176-187.
Lawrey J. D, Lucking R, Sipman H. J. M, et al. High con¬
centration of basidiolichens in a single family of agaricoid mush¬
rooms (Basidiomycota: Agaricales: Hygrophoraceae) // Mycol.
Res. 2009. Vol. 113. N 10. P. 1154-1171.
Lawrey, J. D, Diederich P. Lichenicolous fungi: interac¬
tions, evolution and biodiversity // The Bryologist. 2003. Vol. 106,
№ 1. P. 80-120.
Lettau G. Beitrage zur Lichenenflora von Ost- und West-
preuBen // Festschriften des PreuBischen Botanischen Vereins zu
Konigsberg. 1912. Bd 53. S. 17-91.
Lettau G. Nachtrage zur Lichenenflora von Ost- und West-
preuBen // Schriften der Koniglichen Physikalischokonomischen
Gesellschaft zu Konigsberg. 1919. Bd 60. S. 5-21.
Lichen Biology / Ed. by Т. H. Nash III. Cambridge: Cam¬
bridge University Press, 1996 (1th edition). 303 p.
Lichen Biology / Ed. by Т. H. Nash III. Cambridge: Cam¬
bridge University Press, 2008 (2th edition). 486 p.
Lichen Flora of the Greater Sonoran Desert Region. Vol. 1-
3. Arizona: Arizona State University, 2002-2007.
Linnaeus K. Fundamenta botanica, quae majorum operum
prodromi instar theoriam scientiae botanices per breves aphoris-
mos tradunt (1736). Kessinger Publishing, 2010. 226 p.
Llimona X, Hladun N. L. Checklist of the lichens and li¬
chenicolous fungi of the Iberian Peninsula and Balearic Islands //
Bocconea. 2001. Vol. 14. P. 1-581.
Lohtander K, Oksanen I, Rikkinen J. Genetic diversity of
green algal and cyanobacterial photobionts in Nephroma (Peltiger¬
ales) // Lichenologist. 2003. Vol. 35. P. 325-339.
Lucking R. Foliicolous Lichenized Fungi // Flora Neotropica
Monograph. 2008. Vol. 103. 866 pp.
Lucking R, Rivas Plata E, Chaves J. L, Umana L, Sipman
H. J. M. How many tropical lichens are there ... really? // Biblio-
theca Lichenologica. 2009. N 100. P. 399-418.
Lumbsch H. T, Ahti T, Altermann S. et al. One hundred
new species of lichenized fungi: a signature of undiscovered global
diversity// Phytotaxa. 2011. Vol. 18. P. 1-127.
Lumbsch H. T, Huhndorf S. M. (ed.) Notes on ascomycete
systematics. Nos. 4408 - 4750 // Myconet. 2007. Vol. 13. P. 59-
99.
Lumbsch H. T, Schmitt I, Lucking R, et al. The phyloge¬
netic placement of Ostropales within Lecanoromycetes (Ascomy¬
cota) revisited // Mycol. Res. 2007. Vol. 111. P. 257-267.
Lumbsch H. T, Schmitt I, Mangold A, Wedin M. Ascus
types are phylogenetically misleading in Trapeliaceae and Agyr-
iaceae (Ostropomycetidae, Ascomycota) // Mycol. Res. 2007. Vol.
111.N 9. P. 1133-1141.
Lumbsch H. T, Zimmermann D. G, Schmitt I. Phylogenetic
position of ephemeral lichens in Thelocarpaceae and Vezdaeaceae
(Ascomycota) // Biblioth. Lichenol. 2009. N 100. P. 389-398.
Lynge В. Lichens collected on the Norvegian scientific ex¬
pedition to Franz Josef Land 1930 // Skrifter Svalbard og Ishavet.
1931. №38. P. 1-31.
Lynge B. Lichens from Novaya Zemlya (excl. Acarospora
and Lecanora) // Rep. Sci. Results Norweg. Exped. Novaya Zemlya
1921. Oslo (Kristiania), 1928. Iss. 43. P. 1-299.
Magnusson A. H. Neue Flechten aus dem Jenisei-Gebiet //
Svensk Botanisk Tidskr. 1936. Vol. 30. № 3. P. 247-263.
Malcolm W. M, Galloway D. J. New Zealand Lichens:
Checklist, Key, and Glossary. Wellington: Museum of New Zea¬
land Те Papa Tongarewa, 1997. 192 p.
Malme G. Lichenes orae Sibiriae Borealis inde ab insula
Minin usque ad promantorium Ryrkajpia in Expeditione Vegae
lecti // Arkiv for Bot. 1932. Vol. 25A. № 2. P. 1-42.
Mangold A, Martin М. P, Lucking R, Lumbsch H. T. Mo¬
lecular phylogeny suggests synonymy of Thelotremataceae within
Graphidaceae (Ascomycota: Ostropales) // Taxon. 2008. Vol. 57.
N 2. P. 476-486.
Matthews S, Tucker S, Chapman R. Ultrastructural features
of mycobionts and trentepohliaceous phycobionts in selected sub¬
tropical crustose lichens // Bot. Gaz. 1989. Vol. 150, № 4. P. 417-
438.
McCarthy P. M. Checklist of the Lichens of Australia and its
Island Territories. Australian Biological Resources Study, Canber¬
ra. Version 10 March 2012
http://www.anbg.gov.au/abrs/lichenlist/introduction.html.
McNeill J, Barrie F. R, Burdet H. M, Demoulin V,
Hawksworth D. L, Marhold K, Nicolson D. H, Prado J, Silva P.
C, Skog J. E, Wiersema J. H. & Turland N. J. (eds.). International
Code of Botanical Nomenclature (Vienna Code) adopted by the
Seventeenth International Botanical Congress Vienna, Austria, Ju¬
ly 2005. Regnum Vegetabile 146, A. R. G. Gantner Verlag KG,
Ruggell, Liechtenstein. [I]-XVIII. 2006. 568 p.
Meier J. L, Chapman R. L. Ultrastructure of the lichen Coe-
nogonium interplexum Nyl. // Amer. J. Bot. 1983. Vol. 70. P. 400-
407.
Messerschmidt D. G. Forschungsreise durch Sibirien 1720-
1727. Tagebuchaufzeichnungen. T. 1. Januar 1721-1722. Berlin,
1962; T. 2. Januar 1723 - Mai 1724. Berlin, 1966
Messuti, 1993. Это было где-то в Бюлл. Брит. лих.
общества...
Middendorff A. Th. Reise in den aussersten Norden und Os-
ten Sibiriens wahrend der Jahre 1843 und 1844 mit Allerhochster
Genehmigung auf Veranstaltung der keiserl. Akademie der Wis-
senschaften zu St. -Petersburg. Bd 4. Ubersicht der Natur Nord-
und Ost-Sibiriens. Theil 2. L. 1. Anhang 6. Zu den Gewachsen Si¬
biriens. Lichenes Middendorffiani. Enumeravit prof. Dr. W.
Nylander. St. -Petersburg, 1867. Vol. 4, 2. Iss. 1, 6. P. 15-18.
Mikhailyuk Т. I, Demchenko E. M, Kondratyuk S. Ya. Al¬
gae of granite outcrops from the left bank of the river Pivdennyi
Bug (Ukraine) // Biologia. Bratislava. 2003. Vol. 58, № 4. P. 589—
601.
Mikhailyuk T, Kondratyuk S. Epiphytic algae of some li¬
chen species // XVII Intern. Bot. Congr. Vienna, Austria 17-23 Ju¬
ly 2005: abstract. 2005. P. 445.
Millanes A. M, Diederich P, Ekman S, Wedin M. Phyloge¬
ny and character evolution in the jelly fungi (Tremellomycetes,
Basidiomycota, Fungi) // Mol. Phylogenet. Evol. 2011. Vol. 61. N
1. P. 12-28.
Мое R. Verrucaria traversiae sp. nov, a marine lichen with a
brown algal photobiont // Bulletin of the California Lichen Society.
1997. №4. P. 7-11.
Motiejunaite J, Berglund T, Czarnota P, Himelbrant D,
Hognabba F, Konoreva L. A, Korchikov E. S, Kubiak D, Kukwa
M, Kuznetsova E, Leppik E, Lohmus P, Prigodina Lukosiene I,
Pykala J, Stoncius D, Stepanchikova I, Suija A, Thell A, Tsu-
rykau A, Westberg M. Lichens, lichenicolous and allied fungi
found in Asveja Regional park (Lithuania) // Botanica Lithuanica.
2012. In press.
Muchnik E. E, Konoreva L. A. The lichen biota of protected
territories in Ryazan region (Central Russia) // Lichen protection -
Protected lichen species / Ed. L. Lipnicki. Sonar Literacki, Gorzow
Wlkp. 2012. P. 213-221.
Muggia L, Nelson P, Wheeler T. et al. Convergent evolu¬
tion of a symbiotic duet: The case of the lichen genus Polychidium
(Peltigerales, Ascomycota) // Amer. J. Bot. 2011. Vol. 98. N 10. P.
1647-1656.
Nakano T. Phycobionts of some Japanese species of the
Graphidaceae // Lichenologist. 1988. Vol. 20, № 4. P. 353-360.
Nash Т. H. Lichen biology. Cambridge: University Press,
1996. 303 p.
Navarro-Rosines P, Zhurbenko М. P, Roux C. Pseu-
dopyrenidium nova genero por inkludi Weddellomyces tartaricola
(nelikeniginta fungo likenloga, Ascomycota) // Bull. Soc. Linn.
Provence. 2010. T. 61. P. 129-140.
Nelsen М. P, Lucking R, Grube M, et al. Unravelling the
phylogenetic relationships of lichenised fungi in Dothideomyceta//
Stud. Mycol. 2009. Vol. 64. P. 135-144.
Nelsen М. P, Lucking R, Mbatchou J. S. et al. New insights
into relationships of lichen-forming Dothideomycetes // Fungal
Diversity. 2011. Vol. 51. P. 155-162.
Nelsen М. P, Plata E. R, Andrew C. J. et al. Phylogenetic
diversity of Trentepohlialean algae associated with lichen-forming
fungi // J. Phycol. 2011. Vol. 47. P. 282-290.
Nicolson D. H. A History of Botanical Nomenclature // An¬
nals of the Missouri Botanical Garden. 1991. Vol. 78, No. 1. P. 33-
56.
Nordic Lichen Flora. Vol. 1-4. Uddevalla, 1999-2011.
Nordin A, Savic S, Tibell L. Phylogeny and taxonomy of
Aspicilia and Megasporaceae // Mycologia. 2010. Vol. 102. N 6. P.
1339-1349.
Nyati Sh. Photobiont diversity in Teloschistaceae (Lecano¬
romycetes) / Erlangung der naturwissenscheftlichen Doktorwurde.
Univ. Zurich. 2006. 130 p.
Nylander W. Addenda nova ad Lichenographiam Euro-
paeam. Continuatio octava et vicesima // Flora. 1977. Vol. 60. P.
457-463 [in: William Nylander's Collected Lichenological Papers
(reprint J. Cramer, 1990). II. P. 113-1144].
Nylander W. Enumeratio Lichenum Freti Behringii // Bulle¬
tin de la Societe Linneanne de Normandie, 4 ser. 1888. Vol. 1. P.
1-91.
Nylander W. Lichenes novi e Freto Behringii. I. Lichenes
novi e Konyambay // Flora. 1884. Vol. 67. P. 211-223.
Nylander W. Lichenes novi e Freto Behringii. II. Lichenes
novi e Behring-insula et e Lawrence-insula // Flora. 1885. Vol. 68.
P. 439-446.
O’Brien H. E, Miadlikowska J, Lutzoni F. Assessing host
specialization in the symbiotic cyanobacteria associated with four
closely related species of the lichenfungus Peltigera //
Eur. J. Phycol. 2005. Vol. 40. P. 363-378.
Obermayer W, Mayrhofer H. Hunting for Cetrelia chicitae
(lichenized Ascomycetes) in the Eastern European Alps // Phyton
(Horn, Austria). 2007. Vol. 47. P. 231-290.
Oberwinkler F. Fungus-alga interactions in basidiolichens //
Nova Hedwigia. 1984. Bd 79. P. 739-774.
Ohlert A. Gruppierung der Lichenen der Provinz PreuBen
nach Standort und Substrat // Lichenologische Aphorismen II.
Schriften der Naturforschenden Gesellschaft zu Danzig. 1871. Bd
2. H. 3-4. S. 3-37.
Ohlert A. Verzeichnis PreuBischer Flechten // Schriften der
Koniglichen Physikalisch-okonomischen Gesellschaft zu Konigs¬
berg. 1863. Bd 4. S. 6-34.
Ohlert A. Zusammenstellung der Lichenen der Provinz Preu¬
Ben // Schriften der Koniglichen Physikalisch-okonomischen Ge¬
sellschaft zu Konigsberg. 1870. Bd 11. S. 1-51.
Orange A. A key to the Cladonia chlorophaea group in Eu¬
rope, using microcrystal tests // British Lichen Society Bulletin.
1992. № 70, Summer. P. 36—42.
Orange A, James P. W, White F. J. Microchemical methods
for the identification of lichens. British Lichen Society, 2001. 101
P-
Ott S. Reproductive strategies in lichens // Progress and
problems in lichenology in the eighties / E. Peveling [ed.]. Biblio-
theca Lichenologica. 1987. Vol. 25. P. 81-93.
Ott S, Meier T, Jahns H. M. Development, regeneration,
and parasitic interactions between the lichens Fulgensia bracteata
and Toninia caeruleonigricans // Canad. J. Bot. 1995. Vol. 73. P.
595-602.
Ovstedal D. O, Smith R. I. L. Lichens of Antarctica and
South Georgia: a guide to their identification and ecology. Cam¬
bridge: Cambridge University Press, 2001. 411 p.
Ovstedal D. O, Tonsberg T, Elvebakk A. The Lichen Flora
of Svalbard // Sommerfeltia. 2009. Vol. 33. 393 p.
Peksa O, Skaloud P. Changes in chloroplast structure in li¬
chenized algae // Symbiosis. 2008. Vol. 46. P. 1-8.
Perez-Ortega S, de los Rios A, Crespo A, Sancho L. Sym¬
biotic lifestyle and phylogenetic relationships of the bionts of Mas¬
todia tessellata (Ascomycota, Incertae Sedis) // Amer. J. Bot. 2010.
Vol. 97. N 5. P. 738-752.
Peveling E. Fine structure // The Lichens / Ahmadjian V,
Hale M. A, (Eds.). New York, London: Academic Press, 1973. P.
147-182.
Piercey-Normore M, De Priest P. T. Algal switching among
lichen symbioses // Amer. J. Bot. 2001. Vol. 88, № 8. P. 1490—
1498.
Piin T. A list of terricolous lichens of "Forest Island" of Ary
Mas // Folia Criptogamica Estonica. 1977. № 8. P. 3-8.
Plata E. R, Lucking R, Lumbsch H. T. A new classification
for the family Graphidaceae (Ascomycota: Lecanoromycetes: Os¬
tropales) // Fungal Diversity. 2012. Vol. 52. P. 107-121.
Poelt J. Das Konzept der Artenpaare bei der Flechten //
Deutsch. Bot. Ges. Neue Folge. 1970. Vol. 4. S. 187-198.
Poelt J. Die lobaten Arten der Flechtengattung Lecanora
Ach. sensu. ampl. in der Holarktis // Mitt. Bot. StSamml. Mun-
chen. 1958. Bd. 2. S. 411-589.
Poelt J. Die taxonomische Behandlung von Artenpaaren bei
den Flechten // Bot. Notis. 1972. Vol. 25, № 1. S. 77- 81.
Poelt J. Parasitische Flechten III // Osterreich. Bot. Zeitschr.
1962. Vol. 109. S. 521-528.
Poelt J. Uber parasitische Flechten. II // Planta. 1958. Vol.
51. S. 288-307.
Printzen C. Lichen Systematics: The Role of Morphological
and Molecular Data to Reconstruct Phylogenetic Relationships //
Progress in Botany. 2010. Vol. 71. P. 233-278.
Printzen C, Spribille T, Tonsberg T. Myochroidea, a new
genus of corticolous, crustose lichens to accommodate the Lecidea
leprosula group // Lichenologist. 2008. Vol. 40. N 3. P. 195-207.
Protocols in lichenology. Culturing, Biochemistry, Ecophys-
iology and Use in Biomonitoring / Ed. by Kranner I, Beckett R. P,
Varma A. K. Berlin: Springer-Verlag, 2002. 580 p.
Protocols in lichenology: culturing, biochemistry, ecophysi-
ology and use in biomonitoring / Kranner I, Beckett R, Varma A.
(eds.). Berlin: Springer, 2002. 580 p.
Purvis O. W, Elix J. A, Broomhead J. A, Jones G. C. The
occurrence of copper-norstictic acid in lichens from cuprificerous
substrata//Lichenologist. 1987. Vol. 19. P. 193-203.
Purvis O. W, Elix J. A, Gaul K. L. The occurrence of cop-
per-psoromic acid in lichens from cuprificerous substrata. Lichen¬
ologist. 1990. Vol. 22. P. 345-354.
Rambold G, Triebel D. The inter-lecanoralean associations
// Bibl. Lich. 1992. Vol. 48. P. 1-201.
Randlane T, Saag A. (eds) Eesti pisisamblikud (Estonian
microlichens; in Estonian). Tartu Ulikooli Kirjastus, Tartu. 2004.
581 pp.
Randlane T, Saag A. Chemical and morphological variation
in the genus Cetrelia in the Soviet Union // Lichenologist .1991.
Vol. 23. P. 113-126.
Randlane T, Saag A. Taxonomic notes on some cetrarioid
lichens // Mycotaxon. 2003. Vol. 87. P. 479-487.
Randlane T, Saag A, Suija A. Lichenized, lichenicolous and
allied fungi of Estonia. Ver. November 1, 2011 -
http://esamba.bo.bg.ut.ee/checklist/est/home.php
Rands D. G, Davis J. S. Observations on lichenized and
free-living Physolinum (Chlorophyta, Trentepohliaceae) // J. Phy¬
col. 1993. Vol. 29. P. 819-825.
Rasanen V. Die Flechtenflora der nordlichen Kustengegend
am Laatokka-See // Annales Botanici Societatis Zoologicae-
Botanicae Fennicae Vanamo. 1939. Vol. 12. 240 p.
Raths H. Experimentelle Untersuchungen mit Flechtengoni-
dien aus der Familie der Caliciaceen // Ber. der Schweiz. Bot. Ges.
1938. Bd 48. S. 13-30.
Rikkinen J. What’s behind the pretty colors? A study on the
photobiology of lichens // Bryobrothera. 1995. Vol. 4. P. 1-239.
Rindi F, Allali H. A, Lam D. W, Lopez-Bautista M. An
overview of the biodiversity and biogeography of terrestrial green
algae // Biodiversity Hotspots / V. Rescingo et al. (eds.). Nova Sci¬
ence Publ, Inc. 2009. P. 1-25.
Romeike J, Friedl T, Helms G. et al. Genetic diversity of
algal and fungal partners in four species of Umbilicaria along a
transect of the Antarctic Peninsula // Mol. Biol. Evol. 2002.
Vol. 19. P. 1209-1217.
Roux C. Liste des lichens et champignons lichenicoles de
France // Bulletin de la Societe linneenne de Provence. 2012. Nu-
mero special 16. P. 1-220.
Roux C, Navarro-Rosines P. Trimmatothelopsis (Acarospo¬
raceae, Ascomycota lichenisati), le nom legitime de Silobia // Bull.
Soc. Linn. Provence. 2011. T. 62. P. 167-187.
Ruibal C, Gueidan C, Selbmann L, et al. Phylogeny of
rock-inhabiting fungi related to Dothideomycetes // Stud. Mycol.
2009. Vol. 64. P. 123-133.
Ruibal C, Millanes A. M, Hawksworth D. L. Molecular
phylogenetic studies on the lichenicolous Xanthoriicola physciae
reveal Antarctic rock-inhabiting fungi and Piedraia species among
closest relatives in the Teratosphaeriaceae // IMA Fungus. Vol. 2.
N 1. P. 97-103.
Rules for Zoological Nomenclature by Hugh E. Strickland
[Правила зоологической номенклатуры, сочиненные Хью
Эдвином Стриклендом]. Оцифровка, перевод на русский язык
и предисловие А. В. Куприянова -
http://tinea.narod.ru/library/methodus/stricklandl 837/ (Дата об¬
ращения 18 февраля 2012).
Saag L. Taxonomic and ecologic problems in the genus
Lepraria (Stereocaulaceae, lichenised Ascomycota) // Disserta-
tiones Biologica Universitatis Tartuensis. 2008. № 146 P. 1-167.
Sanders W. B. Bacteria, algae, and phycobionts: maintaining
useful concepts and terminology // Lichenologist. 2004. Vol. 36, №
5. P. 269-275.
Santesson R. Foliicolous lichens I. A revision of the taxono¬
my of the obligately foliicolous, lichenized fungi // Symb. bot. up¬
sal. 1952. Vol. 12,№ 1. P. 1-590.
Santesson R, Moberg R, Nordin A, Tonsberg T, Vi-
tikainen O. Lichen-forming and lichenicolous fungi of Fen-
noscandia. Uppsala, 2004. 359 p.
Santesson R, Moberg R, Nordin A, Tonsberg T, Vi-
tikainen O. Lichen-forming and lichenicolous fungi of Fen-
noscandia. Uppsala: Museum of Evolution, Uppsala University,
2004. 359 p.
Savic S, Tibell L. Atla, a new genus in the Verrucariaceae
(Verrucariales) // Lichenologist. 2008. Vol. 40. N 4. P. 269-282.
Savic S, Tibell L, Andreev M. New and interesting lichens
from Serbia// Mycologia Balcanica. 2006. Vol. 3. P. 99-106.
Schaper T, Ott S. Photobiont selectivity and interspecific in¬
teractions in lichen communities. I. Culture experiments with the
mycobiont fulgensia bracteata // Plant Biology. 2003. № 5. P. 1-10.
Scheidegger C, Werth S.. Conservation strategies for li¬
chens: insights from population biology // Fungal Biology Re¬
views. 2009. № 23. P. 55-66.
Schiman H. Uber die Entwickungmoglichkeiten von Lepto-
sira thrombii Tschermak-Woess als Algenkomponente in der
Flechte Thrombium epigaeum // Ost. bot. Z. 1961. Bd 108. S. 1^4-.
Schmit J. P, Mueller G. M. An estimate of the lower limit of
global fungal diversity // Biodivers. Conserv. 2007. Vol. 16. P. 99-
111.
Schmitt I, Mueller G, Lumbsch H. T. Ascoma morphology
is homoplaseous and phylogenetically misleading in some pyre-
nocarpous lichens // Mycologia. 2005. Vol. 97. N 2. P. 362-374.
Schmull M, Mi^dlikowska J, Pelzer M. et al. Phylogenetic
affiliations of members of the heterogeneous lichen-forming fungi
of the genus Lecidea sensu Zahlbruckner (Lecanoromycetes, As¬
comycota) // Mycologia. 2011. Vol. 103. N 5. P. 983-1003.
Schoch C. L, Crous P. W, Groenewald J. Z, et al. A class-
wide phylogenetic assessment of Dothideomycetes // Stud. Mycol.
Vol. 64. P. 1-15.
Scholz P. Katalog der Flechten und Flechtenbewohnenden
Pilze Deutschlands // Schriftenreihe Vegetationsk. 2000. Hf. 31. S.
1-298.
Schrenk A. G. Reise nach dem Nordosten des europaischen
Russlands, durch die Tundren der Samojeden, zum Arktischen
Uralgebirge, auf Allerhochsten Befehl fur den keis. botanischen
Garten zu St. -Petersburg im Jahre 1837 ausgefUhrt. 2 Theil. Wis¬
senschaftliche Beilagen. Dorpat, 1854. 665 p.
Shibata S. Biogenetical and chemotaxonomical aspects of li¬
chen substances // Beitrage zur Biochemie und Physiologie von
Naturstoffen. Festschrift Kurt Mothes zum 65. Geburstag. VEB
Gustav Fischer Verlag. Jena. 1965. P. 451-465.
Skaloud P, Peksa O. Comparative study of chloroplast mor¬
phology and ontogeny in Asterochloris (Trebouxiophyceae, Chlo¬
rophyta) // Biologia. 2008. Vol. 63, №5. P. 1-8.
Skaloud P, Peksa O. Evolutionary inferences based on ITS
rDNA and actin sequences reveal extensive diversity of the com¬
mon lichen alga Asterochloris (Trebouxiophyceae, Chlorophyta) //
Mol. Phyl. and Evol. 2010. Vol. 54. P. 36^16.
Smith D. C. The physiology of Peltigera polydactyla (Neck.)
Hoffm. // Lichenologist 1962. Vol. 1. P. 209-226.
Smith L. R. I. Half a continent in a square kilometer: the ex¬
ceptional lichen diversity of a small Antarctic island // Biblioth.
Lichenol. 2007. Vol. 95. P. 387-403.
Sobolewskj G. Flora Petropolitana. Petropoli, Typis Collegii
Imperialis Medici, 1799. XX p.
Sochting U, Alstrup V. Danish lichen checklist. Version 2.
Copenhagen, 2008. 46 p.
Spielmann A. A. Checklist of lichens and lichenicolous fungi
of Rio Grande do Sul (Brazil) // Caderno de Pesquisa. Ser. Bio.
Santa Cruz do Sul, 2006. Vol. 18. № 2. P. 7-125.
Spribille T, Goffinet B, Klug B, et al. Molecular support
for the recognition of the Mycoblastus fucatus group as the new
genus Violella (Tephromelataceae, Lecanorales) // Lichenologist.
2011. Vol. 43. N 5. P. 445^166.
Steller. G. W. Reise von Kamtschatka nach Amerika mit
dem commander-kapitan Bering, Georg Wilhelm Steller, St. Pe¬
tersburg, 1793.
Stenroos S, Ahti T, Lohtander K, Myllys L. Lichen flora of
Finland // Norrlinia. 2011. Vol. 21. 534 p.
Stenroos S, Hognabba F, Myllys L, Hyvonen J, Thell A.
High selectivity in symbiotic associations of lichenized ascomy-
cetes and cyanobacteria // Cladistics. 2006. Vol. 22. P. 230-238.
Stenroos S, Huhtinen S, Lesonen A, Palice Z, Printzen C.
Puttea, gen. nov, erected for the enigmatic lichen Lecidea mar-
garitella // Bryologist. 2009. Vol. 112. N 3. P. 544-557.
Stepanchikova I. S, Himelbrant D. E, Kukwa M, Kuz¬
netsova E. S. New records of lichens and allied fungi from the Len¬
ingrad Region, Russia II // Folia Cryptogamica Estonica. 2011.
Fasc. 48. P. 85-94.
Stepanchikova I. S, Kukwa M, Kuznetsova E. S,
Motiejunaite J, Himelbrant D. E. New records of lichens and al¬
lied fungi from the Leningrad Region, Russia // Folia Cryptogami¬
ca Estonica. 2010. Fasc. 47. P. 77-84.
Stepanchikova I. S, Kuznetsova E. S, Himelbrant D. E.
New records of lichens and allied fungi from the Eastern Leningrad
Region // Folia Cryptogamica Estonica. 2009. Fasc. 46. P. 75-78.
Stevenson L. A. List of authors of plant parasite names; rec¬
ommended abbreviations // Agricultural Handbook, U.S. Forest
Service. 1960. № 165. P. 517-531.
Stocker-Worgotter E, Elix J. A, Grube M. Secondary chem¬
istry of lichen-forming fungi: chemosyndromic variation and
DNA-analyses of cultures and chemotypes on the Ramalina farina¬
cea complex // Bryologist. 2004. Vol. 107. № 2. P. 152-162.
Strickland, Hugh E. Rules for Zoological Nomenclature //
The Magazine of Natural History and Journal of Zoology, Botany,
Mineralogy, Geology, and Meteorology. 1837. Vol. 1. New Series.
P. 173-176.
Temina M, Andreev М. P, Barinova S, Nevo E. The diver¬
sity and ecology of epiphytic lichens in «Evolution Canyon» II,
Lower Nahal Keziv, Upper Western Galilee, Israel // Turk. J. Bot.
2009. Vol. 33. P. 263-275.
The Lichen Flora of Great Britain and Ireland / Purvis O. W,
Coppins B. J, Hawksworth D. L. et al. (eds.). London: Nat. Hist.
Mus. Publ. 1992.710 р.
The lichens of Great Britain and Ireland / Smith C. W, Ap¬
troot A, Coppins B. J, Fletcher A, Gilbert O. L, James P. W,
Wolseley P. A. (eds). The British Lichen Society, London, 2009.
1046 p.
Thor G, Arup U, Arvidsson L. et al. Lavar - Lichens //
Rodlistade arter i sverige. 2010. P. 285-300.
Thus H, Muggia L, Perez-Ortega S, et al. Revisiting photo¬
biont diversity in the lichen family Verrucariaceae (Ascomycota) //
Eur. J. Phycol. 2011. Vol. 46, № 4. P. 399-415.
Tibell L. Crustose lichens as indicators of forest continuity in
boreal coniferous forests // Nordic Journal of Botany. 1992. Vol.
12(4). P. 427-450.
Timdal E. A monograph of the genus Toninia (Lecideaceae,
Ascomycetes) // Opera Bot. 1991. Vol. 110. P. 1-137.
Timdal E. Romjularia / In: Т. H. Nash III, C. Gries, F. Bun-
gartz (eds.) Lichen Flora of the Greater Sonoran Desert Region.
Vol. 3. Arizona State University, Tempe, 2007. P. 287-289.
Tschermak E. Untersuchungen uber die Beziehungen von
Pilz und Alge im Flechtenthallus // Ost. bot. Z. 1941. Bd 90. S.
233-307.
Tschermak-Woess E. Algal taxonomy and the taxonomy of
lichens: the phycobiont of Verrucaria adriatica // Lichenology:
Progress and Problems / Brown D. H, Hawksworth D. L, and Bai¬
ley R. H, (eds.). London: Academic Press. 1976. P. 79-88.
Tschermak-Woess E. Elliptochloris bilobata, gen. et spec,
nov, der Phycobiont von Catolechia wahlenbergii // Plant Syst.
Evol. 1980. Vol. 136. P. 63-72
Tschermak-Woess E. The algal partner // CRC Handbook of
Lichenology / M. Galun, ed. Boca Raton, Fla.: CRC Press, 1988. P.
39-92.
Tschermak-Woess E. Uber die weite Verbeitung lichenisier-
ter Sippen von Dictyochloropsis und die systematische Stellung
von Myrmecia reticulata (Chlorophyta) // Plant. Syst. Evol. 1984.
Vol. 147. S. 299-307.
Tschermak-Woess E, Plessl A. Uber zweierlei Typen der
sukzedanen Teilung und ein auffallendes Teilungsverhalten des
Chromatophors bei einer neuen Protococcale Myrmecia pyriformis
// Osterr. Bot. Z. 1948. Bd 95. S. 103-109.
Tucker S. C, Ryan B. D. Revised catalog of lichens, li¬
chenicolous, and allied fungi in California // Constancea. 2006.
Vol. 84. P. 1-275.
Untereiner, W. A, C. Gueidan, M.-J. Orr & P. Diederich The
phylogenetic position of the lichenicolous ascomycete Capronia
peltigerae // Fungal Diversity. 2011. Vol. 49. N 1. P. 225-233.
Urbanavichus G. P, Ahti T, Urbanavichene I. N. Catalogue
of lichens and allied fungi of Murmansk Region, Russia // Norrlin-
ia. 2008. № 17. 80 p.
Uyenko F. R. Studies of some lichenized Trentepohlia asso¬
ciated in lichen thalli with Coenogonium // Trans. Am. microsc.
Soc. 1965. Vol. 84. P. 1-14.
Vainio E. A. Lichenes in viciniis Viburgi observati // Medde-
landen af Societas Pro Fauna et Flora Fennica. 1878. Vol. 2. P. 35-
72.
Vainio E. A. Lichenographia Fennica II. Baeomyceae et Le-
cidiales // Acta Societatis Pro Fauna et Flora Fennica. 1922. Vol.
53. 343 p.
Vainio E. Lichenes in viciniis hibernae expeditionis Vegae
prope pagum Pittlekai in Sibiria septentrionali a D-re E. Almquist
collecti // Ark. for Bot. 1909. Vol. 8. № 4. 175 p.
Van den Hoek C, Mann D. G, Jahns H. M. Algae: an intro¬
duction to phycology. United Kingdom, Cambridge: Univ. Press,
1995.628 p.
Viegas A. P. Indice de Fungos da Ameica do Sul. Campinas,
Argentina; Instituto Agronomico. 1961. 921 p.
Vischer В. Reproduction und systematische Stellung einiger
Rinden- und Bodenalgen // Schweizerische Zeitschrift fur Hydro-
logie. 1960. Bd 22. S. 329-349.
Vitikainen O. Taxonomic revision of Peltigera (lichenized
Ascomycotina) in Europe // Acta Bot. Fennica. 1994. 152. P. 1-96.
Vitikainen O. William Nylander (1822-1899) and lichen
chemotaxonomy// Bryologist. 2001. Vol. 104. P. 263-267.
Voytsekhovich A, Dymytrova L, Nadyeina O. Photobiont
composition of some members of the genera Micarea and Placyn¬
thiella (Lecanoromycetes, lichenized Ascomycota) from Ukraine //
Folia Cryptogamica Estonica. 2011. Vol. 48. P. 135-148.
Wedin M, Jorgensen P. M, Ekman S. Vahliellaceae, a new
family of cyanobacterial lichens (Peltigerales, Ascomycetes) // Li¬
chenologist. 2011. Vol. 43. N 1. P. 67-72.
Wedin M, Jorgensen P. M, Wiklund E. Massalongiaceae
fam. nov, an overlooked monophyletic group among the cyano¬
bacterial lichens (Peltigerales, Lecanoromycetes, Ascomycota) //
Lichenologist. 2007. Vol. 39. N 1. P. 61-67.
Wedin M, Wiklund E, Jorgensen P. M, Ekman S. Slippery
when wet: phylogeny and character evolution in the gelatinous cy¬
anobacterial lichens (Peltigerales, Ascomycetes) // Mol. Phyloge¬
net. Evol. 2009. Vol. 53. P. 862-871.
Werth S. Optimal sample sizes and allelic diversity in studies
of the genetic variability of mycobiont and photobiont populations
// The Lichenologist. 2011. Vol. 43, № 1. P. 73-81.
Westberg M, Crewe A. T, Purvis O. W, Wedin M. Silobia,
a new genus for the Acarospora smaragdula complex (Ascomyco¬
ta, Acarosporales) and a revision of the group in Sweden // Lichen¬
ologist. 2011. Vol. 43. N 1. P. 7-25.
Wirth V, Hauck M, von Brackel W. et al. Checklist of li¬
chens and lichenicolous fungi in Germany. Ver. 2: 19 January 2011
- http://wwwuser.gwdg.de/~mhauck/
Wright J. E, Lois R. J. Lista da singlas de autores empleades
en micologia у fitopatologia // Lilloa. 1949. Vol. 21. P. 225-269.
Wu Hai X, Schoch C. L, Boonmee S, Bahkali A. H,
Chomnunti P, Hyde K. D. A reappraisal of Microthyriaceae //
Fungal Diversity. 2011. Vol. 51. P. 189-248.
Yoshimura I, Shimada R. Fine structures of lichen plecten-
chymas viewed with the scanning electron microscope // Bulletin
of Kochi Gakuen Junior College, 1980. Vol. 11. P. 13-28.
Zeitler I. Untersuchungen uber die Morphologie, Entwick-
lungsgeschichte und Systematik von Flechtengonidien // Osterr.
Bot. Z. 1954. Bd 101. S. 453-483.
Zhang Y, Crous P. W, Schoch C. L, Hyde K. D. Pleospora-
les // Fungal Diversity. 2011. DOI 10.1007/s 13225-011-0117-x
Zhurbenko M. P. Lichens from the coast of Eniseiskii Gulf,
W. Taimyr, Siberia. Graphis Scripta. 1992. Vol. 4. № 1. P. 1-4.
Zhurbenko М. P, Himelbrant D. E, Kuznetsova E. S, Ste¬
panchikova I. S. Lichenicolous fungi from Kamchatka Peninsula,
Russia // The Bryologist. 2012. Vol. 115, № 2. P. 295-312.
Zoller S, Scheidegger Ch, Sperisen Ch. PCR primers for the
amplification of mitochondrial small subunit ribosomal DNA of
lichen-forming Ascomycetes // Lichenologist. 1999. Vol. 31(5). P.
511-516.
Приложения
Приложение I.
Деление территории Российской Федерации на регионы,
принятое в настоящем издании
Андреев М. П.
В настоящей работе выделение основных регионов Рос¬
сии (Рис. 87) соответствует в основном схеме, принятой в
«Списке лихенофлоры России» (2010). Принципиальным от¬
личием является выделение Арктики в отдельную область.
Рис. 87. Территориальное деление Российской Федерации.
Территория Российской Федерации в широтном отноше¬
нии подразделяется на арктическую область и внеарктические
территории. Граница Арктики принимается по Б. А. Юрцеву
(Юрцев, Толмачев, Ребристая, 1978), т.е. примерно совпадает
с южной границей безлесных территорий и соответствует
принятой в работе М. П. Андреева с коллегами (Andreev & al,
1996). Арктическая область подразделяется на европейскую
(без Кольского п-ова, но включая горы Полярного Урала и
арктические архипелаги - Земля Франца Иосифа и Новая Зем¬
ля), западносибирскую (от Ямала почти до Енисейского зали¬
ва), восточносибирскую (п-ов Таймыр и архипелаг Северная
Земля), якутскую (тундровые территории и Новосибирские о-
ва) и чукотскую (тундры севера и востока Чукотского авто¬
номного округа, включая о-в Врангеля).
Территория России, расположенная вне пределов Аркти¬
ки, разделена на три сектора - Европейский, Сибирский и
Дальневосточный, а в их пределах выделяются следующие 13
регионов: Север Европейской части России (для удобства -
Северная Россия), Средняя Россия, Южная Россия, Север¬
ный Кавказ, Северный Урал, Южный Урал, Север Запад¬
ной Сибири, Юг Западной Сибири, Средняя Сибирь, Юж¬
ная Сибирь, Якутия, Север Дальнего Востока и Юг Даль¬
него Востока.
Урал (без Полярного Урала и других арктических райо¬
нов), разделенный на Северный и Южный примерно по 58° с.
ш, отнесен к Европейской России. Западная и восточная гра¬
ницы Урала проходят по предгорьям Уральской горной стра¬
ны. Расположенные здесь административные области, вклю¬
чают как горные уральские, так и равнинные европейские или,
соответственно, сибирские территории. Граница Европейской
России, таким образом, проводится не по географической гра¬
нице Европы и Азии, а по восточным (сибирским) предгорьям
Уральского хребта. В пределах Европейской России рассмат¬
ривается и Северный Кавказ, хотя формально это территория
является частью Азии.
На обширной территории Сибири выделяются: равнин¬
ный таежный Север Западной Сибири, равнинный степной и
лесостепной Юг Западной Сибири, Средняя Сибирь (Сред¬
несибирское плоскогорье), горная Южная Сибирь, включа¬
ющая Алтай, Саяны и Забайкалье) и Якутия, внеарктические
районы которой, из-за размеров и разнообразия природных
условий, подразделяются для удобства на четыре более мел¬
ких региона: западный, восточный, центральный и южный.
Границы регионов соответствуют флористическим районам,
предложенным М. Н. Караваевым (1958), и уточненным позд¬
нее (Разнообразие растительного мира Якутии, 2005), за ис¬
ключением Верхне-Ленского и Алданского, объединенных
нами в Южный, и Яно-Индигирского и Колымского, объеди¬
ненных в Восточный.
Дальневосточный сектор подразделяется на Север
Дальнего Востока, включающий Магаданскую обл., горно¬
лесную южную часть Чукотского АО, Камчатский край и се¬
верную часть Хабаровского края, и Юг Дальнего Востока в
составе южной части Хабаровского края, Сахалинской обла¬
сти с Курильскими островами, Амурской области, Еврейской
АО и Приморского края.
Таким образом, территория Российской Федерации под¬
разделяется на 18 крупных регионов, присутствие видов в ко¬
торых в работе указывается отдельно для каждой из админи¬
стративных областей. Для удобства, авторы стремились мак¬
симально приблизить предлагаемую в работе схему террито¬
риального деления к схеме административных единиц Россий¬
ской Федерации, хотя в ряде случаев это оказалось невозмож¬
ным, поскольку физико-географические границы рассматри¬
вались, как более значимые.
Регионы Российской Федерации и принятые обозначения
Регионы Российской Федерации в работе обозначены
сокращениями их русского названия, приводимыми ниже. Ла¬
тинские обозначения областей предлагаются для возможных
иноязычных публикаций.
I. Арктика. - АРКТ. - ARC.
1. Арктика европейская - Аркт. (евр.) - Ае. Ненецкий
АО (кроме лесной крайней юго-западной части), включая ар¬
хипелаги Земля Франца-Иосифа и Новая Земля, полуостров
Канин и острова Колгуев и Вайгач), горная уральская часть
территории Ямало-Ненецкого АО, крайний северо-восток Рес¬
публики Коми.
2. Арктика западно-сибирская - Аркт. (зап.-сиб.) - Aws.
Равнинная тундровая внеуральская часть Ямало-Ненецкого
АО (п-ов Ямал, Тазовский и Гыданский (до границы Ямало-
Ненецкого АО).
3. Арктика таймырская - Аркт. (тайм.) - At. Тундровая
часть Таймырского АО Красноярского края, включая архипе¬
лаг Северная Земля.
4. Арктика якутская - Аркт. (як.) - Ауа. Арктические
районы Якутии (Анабарский, Аллаиховский и Нижнеколым¬
ский, тундровые части Булунского и Усть-Янского админи¬
стративных районов).
5. Арктика чукотская - Аркт. (чук.) - Ach. Чукотский
АО (тундровая часть).
II. Европейская Россия. - ЕР. - ER.
6. Европейский Север - Евр. Север - Еп. Мурманская
обл., Карелия, Архангельская обл., лесная юго-западная часть
Ненецкого АО, Республика Коми (кроме крайнего северо-
востока и западных склонов Уральского хребта), Ленинград¬
ская и Вологодская обл., северо-западная часть Кировской
обл. к сев. примерно от 60° с. ш.
7. Средняя Россия - Ср. Россия - Еш. Калининградская,
Псковская, Новгородская, Тверская, Ярославская, Костром¬
ская, Кировская обл. (кроме северо-западной части), Пермский
край (внеуральская часть), Смоленская, Московская, Влади¬
мирская, Ивановская, Нижегородская обл., Республика Марий-
Эл, Чувашия, Татарстан, Удмуртия, Брянская, Калужская, Ор¬
ловская, Тульская, Рязанская обл., Мордовия, Пензенская,
Ульяновская обл., Самарская обл. (северная часть, включая
Жигули), Башкирия (западная внеуральская часть), Курская,
Липецкая, Тамбовская, Белгородская обл. (кроме крайней юго-
восточной части), Воронежская обл. (кроме южной части).
8. Южная Россия - Юж. Россия - Es. Белгородская
(юго-восточная часть), Воронежская (южная часть), Саратов¬
ская, Самарская (южная часть) обл., Оренбургская (западная
внеуральская часть), Ростовская, Волгоградская, Астраханская
обл., Калмыкия.
9. Северный Кавказ - Сев. Кавказ - NC. Краснодарский
и Ставропольский края, Адыгея, Карачаево-Черкесия, Кабар-
дино-Балкария, Сев. Осетия, Ингушетия, Чеченская республи¬
ка, Дагестан.
10. Северный Урал - Сев. Урал - Un. Горные районы
Республики Коми, Пермского края (северная часть), Ямало-
Ненецкого и Ханты-Мансийского АО и Свердловской обл.
(северная часть).
11. Южный Урал - Юж. Урал - Us. Горные районы
Пермского края (южная часть), Свердловской обл. (южная
часть), Башкирии, Челябинской и Оренбургской обл.
III. Сибирь. - СИБ. - SIB.
12. Север Западной Сибири - Зап. Сиб. (сев.) - Swn.
Южная равнинная и лесная часть Ямало-Ненецкого АО, Хан-
ты-Мансийский АО (равнинная часть), восточная (внеураль¬
ская) часть Свердловской обл., северная лесная часть Тюмен¬
ской обл., Томская обл. (кроме самой юго-восточной части,
представляющей северные отроги Кузнецкого Алатау).
13. Юг Западной Сибири - Зап. Сиб. (юж.) - Sws. Степ¬
ные и лесостепные восточные зауральские районы Оренбург¬
ской и Челябинской обл., Курганская обл., южная часть Тю¬
менской обл., Омская обл., Новосибирская обл. (кроме горной
крайней восточной части), Алтайский край (кроме горных
юго-восточной и восточной частей).
14. Средняя Сибирь - Ср. Сиб. - Sm. Южная лесная
часть Таймырского АО, север Красноярского края (примерно
до 58° с. ш. и широтного течения р. Ангары), Эвенкийский
АО, северная часть Иркутской обл. (к северу примерно от 59°
с. ш.).
15. Южная Сибирь - Юж. Сиб. - Ss. Горная юго-
восточная и восточная часть Алтайского края, крайний восток
Новосибирской обл. (Салаирский кряж), юго-восточная часть
Томской обл. (северные отроги Кузнецкого Алатау), Кемеров-
ская обл. Республика Алтай, Хакасия, Тува, Бурятия, Красно¬
ярский край (южная часть - примерно южнее Ангары), Иркут¬
ская обл. (кроме крайнего севера области - т. е. южнее при¬
мерно 59° с. ш.), Забайкальский край.
16. Якутия - Як. - Sya. Внеарктическая часть, делится на
Западную, Центральную, Восточную и Южную:
Западная - внеарктическая часть Якутии к западу от р.
Лены и к северу примерно от р. Вилюй или от 64° с. ш. (почти
целиком Оленекский административный район и западные ча¬
сти Булунского и Жиганского районов).
Центральная - территории, расположенные к западу и
северу от рек Алдан и Лена и к югу примерно от 64° с. ш. (ад¬
министративные районы Горный, Сунтарский, Вилюйский,
Мирнинский, Вехневилюйский, Нюрбинский и многочислен¬
ные небольшие районы, расположенные вблизи Якутска).
Восточная - внеарктическая Якутия к востоку от рек
Лены и Алдана, в границах Яно-Индигирского и Колымского
флористических районов (Эвено-Бытантайский НО, админи¬
стративные районы Верхоянский, Абыйский, Среднеколым¬
ский, Верхнеколымский, Момский, Оймяконский, Томпон-
ский, части Усть-Янского, Кобяйского, Жиганского и Булун¬
ского).
Южная - горные области южной Якутии, расположен¬
ные южнее рек Алдана и верхней Лены, в верховьях рек Амги,
Олекмы, Витима и южных притоков Вилюя (административ¬
ные районы Усть-Майский, Алданский, Олекминский, Лен¬
ский и часть Мирнинского).
IV. Дальний Восток. - ДВ. - FE.
17. Север Дальнего Востока - Сев. Д. в. - Fn. Внеаркти¬
ческая часть Дальнего востока от границы леса на севере при¬
мерно до северной части бассейна р. Уда на юге (южная часть
Чукотского АО, Магаданская обл. Камчатский край, Хабаров¬
ский край (северная часть, включая Шантарские о-ва).
18. Юг Дальнего Востока - Юг Д. в. - Fs. Амурская
обл, южная часть Хабаровского края (примерно от р. Уда и
исключая Шантарские о-ва); Еврейская АО, Сахалинская обл,
включая Курильские о-ва, Приморский край.
Литература.
Andreev М. Р, Kotlov Yu. V, Makarova I. I. Checklist of
lichens and lichenicolous fungi of the Russian Arctic // The Bryol¬
ogist. 1996. Vol. 99. No 2. P. 137-169.
Караваев М. Н. Конспект флоры Якутии. М, Л, Изда¬
тельство АН СССР, 1958. 189 с.
Разнообразие растительного мира Якутии. Новосибирск,
Издательство СО РАН, 2005. 328 с.
Список лихенофлоры России. [Сост. Г. П. Урбанавичюс]
Спб, Издательство Наука, 2010. 194 с.
Юрцев Б. А, Толмачев А. И, Ребристая О. В. Флористи¬
ческое ограничение и разделение Арктики // Арктическая
флористическая область. Л, Наука, 1978. С. 9-104.
Приложение II.
Терминологический словарь
Автогамия - половой процесс, при котором копулируют
структуры, образующиеся на одном талломе.
Агарикоидные плодовые тела - плодовые тела агарикоидных
базидиальных грибов и лишайников, разделенные на
шляпку и ножку и характеризующиеся пластинчатым
гименофором.
Адельфопаразитизм - паразитизм на близкородственных ви¬
дах.
Акроним гербария - уникальный латинский акроним (краткая
аббревиатура) названия гербария, получаемый после ре¬
гистрации в Международной системе Index Herbariorum.
Аксиальное тело - центральная часть толуса сумки леканоро¬
вого типа, расположенная над окулярной камерой.
Аксиальный канал - вдающееся в толус сумки леканорового
типа продолжение окулярной камеры.
Альгальный (водорослевый) слой - слой таллома гетеромер-
ного строения, обычно располагающийся под коровым и
состоящий из клеток фотобионта и иррегулярно распо¬
ложенных гиф микобионта.
Амилоидный - окрашивающийся в голубой цвет от действия
раствора йода.
Амфитеций - см. Талломный край апотеция.
Анаморфа - бесполая стадия жизненного цикла грибов, не
несущая структур полового размножения, но способная
нести структуры бесполого размножения.
Анастомозирующие гифы - гифы, срастающиеся друг с дру¬
гом во многих точках и формирующие сеть (обычно
элементы хаматеция).
Аннелидный тип конидиогенеза - энтеробластический кони¬
диогенез, при котором каждая следующая конидия отде¬
ляется перегородкой от конидиогенной клетки несколько
дальше, чем располагалась перегородка ранее отделив¬
шейся конидии. При таком характере деления образуют¬
ся кольцеобразные телескопически расположенные руб¬
цы - аннеляции.
Антиклинальная (палисадная) ориентация гиф - гифы
плектенхимы располагаются параллельно друг другу и
перпендикулярно поверхности таллома.
Апикальный аппарат сумки (толус) - сложно организованная
структура, являющаяся утолщением эндоаска в верхней
части сумки леканорового типа и служащая для ее
вскрытия после созревания спор.
Апотеций - открытое в зрелости плодовое тело (аскома) ча¬
шевидной или блюдцевидной формы.
Ареолы - отдельные обособленные лихенизированные эле¬
менты мозаики таллома некоторых накипных лишайни¬
ков, расположенные на общем нелихенизированном ги¬
поталломе.
Аск (ascus) - см. Сумка.
Аскогенная гифа - дикариотическая гифа, клетки которой по¬
сле кариогамии в результате мейоза продуцируют сумки.
Аскогон - «женский» половой орган сумчатых грибов (недиф¬
ференцированный на гаметы гаметангий).
Аскокарп - см. Аскома.
Аскологимениальный тип развития - тип развития плодо¬
вых тел сумчатых грибов, при котором первоначально
происходит закладка аскогенных гиф, вокруг которых
позднее формируются примордии аском (будущих пери¬
тециев или апотециев).
Асколокулярный тип развития - тип развития плодовых тел
сумчатых грибов, при котором первоначально из вегета¬
тивных гиф таллома формируются примордии аском
(будущих строматических образований - псевдотециев).
В их полостях позднее происходит закладка генератив¬
ных аскогенных гиф, формирование сумок и элементов
хаматеция.
Аскома (аскокарп) - любая структура полового размножения
сумчатых грибов, содержащая сумки.
Аскопласт - протопласт сумки, в котором развиваются аско¬
споры.
Аскоспоры - споры полового размножения сумчатых грибов,
образующиеся эндогенно в сумках в результате мейоза.
Аспицилиоидный апотеций - см. Криптолеканориновый
апотеций.
Базидиокарп - см. Базидиома.
Базидиома (базидиокарп) - любая структура полового раз¬
множения базидиальных грибов, содержащая базидии.
Базидиоспоры - споры полового размножения базидиальных
грибов, образующиеся экзогенно на стеригмах (выро¬
стах) базидий в результате мейоза.
Базидия - клетка, на которой в результате мейоза экзогенно
(на специальных выростах - стеригмах) формируются
споры полового размножения базидиальных грибов - ба¬
зидиоспоры.
Биаториновый (биаторовый, псевдолеканоровый, псевдоле-
канориновый) апотеций - модификация лецидеинового
апотеция, характеризующаяся светлой окраской и более
мягкой во влажном состоянии консистенцией. Диск та¬
ких апотециев быстро становится выпуклым, а соб¬
ственный край может постепенно исчезать.
Биаторовый апотеций - см. Биаториновый апотеций.
Битуникатного (фисситуникатного) типа сумки - сумки,
клеточная стенка которых состоит из двух отличных по
составу и функциям слоев (эндоаск и экзоаск), которые
при вскрытии сумки ведут себя независимо: при разрыве
экзоаска эндоаск вытягивается вверх и образует канал,
достигающий поверхности гимения. Зрелые споры вы¬
брасываются по каналу из сумки благодаря ее внутри¬
клеточному давлению.
Бластидии (кортикальные гранулы) - мелкие округлые веге¬
тативные пропагулы, содержащие клетки фотобионта и
гифы микобионта и покрытые более или менее сформи¬
рованным коровым слоем. Формируются путем отпочко¬
вывания от краев лопастей таллома или от ранее образо¬
вавшихся бластидий. Занимают промежуточное положе¬
ние между настоящими соредиями и изидиями.
Бластический тип конидиогенеза - конидиогенез, при кото¬
ром будущая конидия начинает расти и заметно увели¬
чивает свой объем до отделения перегородкой от кони¬
диогенной клетки.
Вегетативные пропагулы - любые небольшие по размерам,
легко отделяющиеся от таллома и достаточно легко пе¬
ремещающиеся в пространстве структуры вегетативного
размножения, в случае лишайника объединяющие в себе
клетки водоросли и гифы гриба.
Вены - см. Жилки.
Водорослевый слой - см Альгальный слой.
Войлок (tomentum) - плотный слой беспорядочно переплетен¬
ных, ветвящихся, мягких гиф, формирующийся на по¬
верхности талломов некоторых лишайников.
Волоски - одноклеточные или многоклеточные, обычно не-
разветвленные, бесцветные тонкие одиночные гифы,
обычно развивающиеся на верхней поверхности концов
лопастей некоторых листоватых лишайников и являю¬
щиеся производными клеток корового слоя.
Вторичный таллом (подеции или псевдоподеции) - восхо¬
дящая кустистая часть таллома кустистых плагио-
ортотропных лишайников, которая формируется на рас¬
простертой по субстрату бородавчато- или чешуйчато¬
накипной части.
Гало - сильно развитая у некоторых таксонов аморфная сли¬
зистая внешняя часть периспория аскоспоры.
Галонатные аскоспоры - аскоспоры с сильно развитой сли¬
зистой внешней частью периспория, хорошо заметной в
оптический микроскоп.
Гаметангиогамия - у сумчатых грибов половой процесс, в
ходе которого копулируют «мужской» и «женский» по¬
ловые органы - недифференцированные на гаметы гаме-
тангии.
Гаптеры - короткие и неразветвленные выросты или просто
участки нижней коры таллома, прикрепляющиеся к суб¬
страту наподобие присосок или прикрепительных дис¬
ков.
Гетеромерный таллом - таллом лишайника, в котором клет¬
ки фотобионта более или менее сконцентрированы в от¬
дельном слое, в результате чего возможно выделить аль¬
гальный слой и сердцевину.
Гиалиновый - не окрашенный, прозрачный.
Гимений (теций) - генеративная часть апотеция или перите¬
ция, состоящая из совокупности сумок и элементов ха¬
матеция, погруженных в более или менее развитый же¬
латинозный матрикс.
Гипоталлом - нелихенизированная распростертая по суб¬
страту нижняя часть таллома некоторых накипных и ли¬
стоватых лишайников, на которой в дальнейшем разви¬
вается лихенизированная часть.
Гипотеций - расположенная под гимением стерильная часть
плодового тела, по периферии переходящая в эксципул.
Гирозный диск - диск апотеция, имеющий сложную лабирин¬
тообразно- или концентрически-ребристую поверхность.
Гистеротеций (лирелловидный апотеций) - линейно вытяну¬
тый лецидеиновый апотеций с узким щелевидным дис¬
ком.
Гифомицетный тип конидиального спороношения - пред¬
ставленный одиночными конидиеносцами, не объеди¬
ненными в какие-либо сложные структуры.
Гифофор - конидиома на ножке или сидячая, на или внутри
которой формируются пучки диагиф (иногда рассматри¬
вают в качестве частного случая или синонима син¬
немы).
Гомеомерный таллом - таллом лишайника, в котором клетки
фотобионта более или менее равномерно распределены
между гифами гриба по всему его объему, в результате
чего выделить отдельный альгальный слой и сердцевину
затруднительно.
Гомф - короткая и очень плотная структура прикрепления к
субстрату, расположенная в центральной части нижней
поверхности талломов эпилитных умбиликатных ли¬
шайников и состоящая из гиф сердцевины и корового
слоя.
Гониоцистангии - ограниченные коровым слоем и лишенные
его участки таллома, в пределах которых развивается
масса гониоцист, сформированная путем отпочковыва¬
ния их друг от друга.
Гониоцисты - микроскопические вегетативные пропагулы, а
также элементы таллома некоторых накипных лишайни¬
ков, покрытые тонкой параплектенхимной оболочкой из
клеток микобионта, которая окружает одну клетку или
сформировавшуюся из нее группу плотно агрегирован¬
ных клеток фотобионта.
Гормоцистангии - ограниченные участки таллома некоторых
цианобионтных гомеомерных лишайников, в пределах
которых формируются желатинозные кластеры - скоп¬
ления гормоцист.
Гормоцисты - вегетативные пропагулы некоторых циано¬
бионтных лишайников с гомеомерным талломом, по¬
крытые тонкой параплектенхимной оболочкой из клеток
микобионта, которая окружает одиночные участки нитей
цианобионта - гормогонии.
Детально-маршрутный метод полевых исследований - ме¬
тод, при котором исследователь, пользуясь схемой
маршрутного метода обследования территории, закла¬
дывает стандартизованные пробные площади в наиболее
типичных или интересных фитоценозах с учетом их
ландшафтного положения. При этом работа на пробных
площадях также проходит с использованием стандарти¬
зованной схемы действий, как и в случае метода проб¬
ных площадей.
Детерминантка (notula criticae) - небольшой листок бумаги,
на котором исследователь гербарного образца указывает,
согласно своему мнению, название содержащегося в
конверте таксона в соответствии с современной номен¬
клатурой, свои фамилию и инициалы, дату работы с об¬
разцом. Детерминантка должна быть наклеена на гер¬
барный конверт или вложена в него (в соответствии с
правилами данного гербария).
Диагифы - разветвленные повисающие конидиеносцы с це¬
почками конидий, расположенные апикально или суб-
апикально на или внутри гифофоров, реже непосред¬
ственно на талломе. Диагифы могут выполнять функцию
пропагул бесполого размножения.
Диск апотеция - экспонированная поверхность генеративной
части апотеция - гимения.
Дублет гербарного образца - часть материала, полученная
путем разделения исходного гербарного образца и
оформленная в соответствии со всеми требованиями,
предъявляемыми к оформлению гербарного материала.
Жилки - разветвленные структуры, развивающиеся на лишен¬
ной коры нижней поверхности лопастей листоватых ли¬
шайников и являющиеся уплотненными тяжами гиф
сердцевины (виды рода Peltigera).
Зеориновый (зеороидный) апотеций - апотеций с хорошо
развитыми и заметными как эксципулом, так и таллом-
ным краем. Является разновидностью леканоринового
апотеция.
Зеороидный апотеций - см. Зеориновый апотеций.
Изидии - мелкие поверхностные выросты таллома, в хорошо
развитых экземплярах которых можно различить коро¬
вой, альгальный слои и сердцевину. Другим вариантом
образования изидий является обрастание корой лишен¬
ных ее соредий. Один из наиболее распространенных
типов вегетативных пропагул.
Изидиоморфы - похожие на изидии короткие палочковидные
структуры вегетативного размножения, развивающиеся
одиночно или группами в соралиях ряда видов рода Us¬
nea.
Инволюкреллюм - пигментированная структура, лежащая над
эксципулом перитеция и выполняющая защитную функ¬
цию.
Инсерация - формализованный процесс внесения гербарного
образца в уже существующий гербарий. Включает не
только помещение образца в соответствующую ему пап¬
ку или коробку, но и предшествующее этому присвоение
образцу уникального гербарного номера, а также внесе¬
ние всей известной информации о нем в гербарную базу
данных.
Интераскальная псевдопаренхима - недифференцированная
на филаменты масса клеток, плотно заполняющих по¬
лость плодового тела.
Интераскальные филаменты - см. Хаматеций.
Иррегулярная ориентация гиф - какая-либо закономерность
в укладке гиф относительно друг друга и поверхности
таллома отсутствует, плектенхима состоит из перепутан¬
ных и направленных в разные стороны элементов.
Иррегулярное ветвление - ветвление, характер которого не
подчиняется какой-либо определенной закономерности.
Камеры - см. Локулы.
Кампилидии - шлемовидная конидиома полузамкнутого типа,
внутри которой расположены конидиеносцы, прикрытые
сверху лопастью.
Карбонизированный - окрашенный в черный цвет.
Кариогамия - слияние ядер копулирующих структур, следу¬
ющее за плазмогамией и являющееся завершающей ста¬
дией полового процесса.
Клавариоидные плодовые тела - булавовидные плодовые
тела клавариоидных базидиальных грибов и лишайни¬
ков, не разделенные на шляпку и ножку и характеризу¬
ющиеся гладким гименофором.
Компактор - специализированное крупногабаритное обору¬
дование для хранения гербарных папок или коробок, за¬
меняющее гербарные шкафы.
Конидиеносец (конидиофор) - неразветвленная или ветвящая¬
ся гифа, несущая специализированные конидиогенные
клетки, продуцирующие конидии.
Конидии - споры бесполого размножения грибов, образующи¬
еся митотически путем отпочковывания от конидиоген¬
ной клетки конидиеносца.
Конидиогенез - специфический вариант митотического деле¬
ния, в результате которого на конидиогенных клетках
конидиеносцев образуются споры бесполого деления -
конидии.
Конидиогенные клетки - специализированные клетки ветвей
конидиеносцев, митотически отпочковывающие споры
бесполого размножения грибов - конидии.
Конидиомы - более или менее сложные структуры бесполого
спороношения сумчатых и базидиальных грибов, объ¬
единяющие множество конидиеносцев, продуцирующих
конидии.
Конидиофор - см. Конидиеносец.
Консоредии - агрегаты из нескольких соредий.
Коровый слой - поверхностный слой таллома (верхний или
нижний), составленный плотно уложенными гифами ми¬
кобионта.
Кортикальные гранулы - см. Бластидии.
Кортикальные чешуйки - уплощенно-округлые элементы
чешуйчато-ареолированного корового слоя некоторых
кустистых лишайников, прикрепленные к таллому
большей частью нижней поверхности и способные вы¬
полнять функцию вегетативных пропагул.
Криптолеканориновый (аспицилиоидный) апотеций - лека-
нориновый апотеций лишайника, погруженный в таллом.
Кустистый таллом - состоит из более или менее цилиндри¬
ческих или уплощенно-цилиндрических ветвей, в попе¬
речном сечении имеющих радиальное строение и ли¬
шенных анатомо-морфологической дифференцировки на
верхнюю и нижнюю поверхности.
Лактофенол хлопковый синий (lactophenol cotton-blue) - сре-
да-краситель, используемая для изготовления и окраски
полупостоянных препаратов, а также контрастирования
их деталей.
Леканориновый (леканоровый, эулеканоровый) апотеций -
апотеций лишайника, характеризующийся слабым раз¬
витием эксципула и наличием хорошо развитого таллом¬
ного края, лежащего снаружи от эксципула. Талломный
край, подобно формирующему его таллому, содержит
коровой, альгальный слои и сердцевину.
Леканорового типа сумки - сумки, клеточная стенка которых
состоит из двух слоев (эндоаск и экзоаск), которые при
вскрытии сумки ведут себя частично согласованно: эн¬
доаск не отрывается от экзоаска, но заметно вытягивает¬
ся в трубку, формируя рострум (канал для выведения
спор). Для сумок этого типа характерен развитый апи¬
кальный аппарат.
Леканоровый апотеций - см. Леканориновый апотеций.
Лецидеевый апотеций - см. Лецидеиновый апотеций.
Лецидеиновый (лецидеевый) апотеций - апотеций лишайни¬
ка, все части которого полностью лишены клеток фото¬
бионта.
Лирелловидный апотеций (лирелла) - см. Гистеротеций.
Листоватый таллом - состоит из одной (монофильный) или
многих (полифильный) лопастей, каждая из которых
представляет пластину, характеризуется морфолого¬
анатомической дифференцировкой на верхнюю и ниж¬
нюю поверхности и формирует на нижней поверхности
органы прикрепления к субстрату.
Лихенизированные грибы - см. Лишайники.
Лихенофильные грибы - эколого-трофическая несистемати¬
ческая полифилитическая группа нелихенизированных
грибов, поселяющихся и в течение всей своей жизни
развивающихся на талломах лишайников в качестве па¬
разитов, парасимбионтов или комменсалов, либо сапро¬
трофов.
Лихенофильные лишайники - эколого-трофическая несисте¬
матическая полифилитическая группа лишайников (ли¬
хенизированных грибов), поселяющихся и в течение
всей или части своей жизни развивающихся на талломах
лишайников в качестве паразитов или парасимбионтов.
Лишайники (лихенизированные грибы) - эколого¬
трофическая несистематическая полифилитическая
группа грибов. Таллом лишайника - двух или
трехбионтная система, в состав которой обязательно
входит мицелиальный микобионт, а также экстрацеллю-
лярно расположенный по отношению к микобионту од¬
ноклеточный или нитчатый фотобионт - водоросль
и/или цианобактерия (мицелиальное сплетение с вклю¬
чением клеток фотобионта). Фактически взаимоотноше¬
ния в лишайниковом талломе могут носить более слож¬
ный полибионтный характер. В типичном случае они
представлены умеренным сбалансированным биохими¬
чески регулируемым паразитизмом микобионта на од¬
ном или двух фотобионтах.
Лишайниковые вещества - гетерогенная группа ацетонрас-
творимых вторичных метаболитов лишайников, форми¬
руемых микобионтом. Ранее носила название «лишайни¬
ковые кислоты», в настоящее время не употребляемое.
Лишайниковые кислоты - см. Лишайниковые вещества.
Лобули - см. Филлидии.
Ложе - см. Спородохий.
Ложная ткань - см. Плектенхима.
Локулы (камеры) - полости различного происхождения, раз¬
вивающиеся внутри плодовых тел и конидиом.
Люмина - просвет или полость клетки, окруженная клеточной
стенкой и вмещающая протопласт. Чаще употребляется
в отношении аскоспор.
Мазедий - масса спор, свободно лежащая и дозревающая в
чашевидных защитных покровах апотециев калициоид-
ных лишайников (унитуникатный тип сумок). Образует¬
ся в результате раннего (за долго до полного созревания
спор) разрушения всех структур гимения.
Макроконидии - более крупные конидии, выполняющие
функцию спор бесполого размножения (в случае форми¬
рования на талломах одного вида более чем одного типа
конидий).
Макролишайники - традиционно некорректно используемый
термин, призванный объединить в одну группу лишай¬
ники с кустистым и листоватым типами талломов.
Макулы - светлые элементы мозаичной окраски поверхности
некоторых лишайников, соответствующие участкам ко¬
рового слоя, под которыми отсутствует фотобионт.
Между ними располагаются более темные участки по¬
верхности, под которыми находятся группы клеток фо¬
тобионта.
Маршрутный метод полевых исследований - метод, при ко¬
тором исследователь учитывает и собирает материал на
протяжении всего маршрута, не стремясь стандартизо¬
вать выбор мест сборов и характер работ на них.
Мезоконидии - конидии средних размеров (в случае форми¬
рования на талломах одного вида более чем двух типов
конидий).
Меланизированный - темноокрашенный в результате содер¬
жания коричневого, черно-коричневого или оливково¬
коричневого пигмента.
Микобионт - гетеротрофный грибной мицелиальный компо¬
нент лишайникового таллома, представленный видами
из отделов Ascomycota или Basidiomycota.
Миколеканориновый апотеций - апотеций лишайника, в ко¬
тором фотобионт присутствует только в гипотеции.
Микроконидии - более мелкие конидии, выполняющие функ¬
цию спермациев (в случае формирования на талломах
одного вида более чем одного типа конидий).
Микролишайники - традиционно некорректно используемый
термин, призванный объединить в одну группу лишай¬
ники с накипным типом таллома. Корректным следует
считать понимание термина, при котором к этой группе
относят только виды, морфологические характеристики
которых видны только при использовании не менее чем
10х оптического увеличения.
Микрохимические тесты - см. Цветные реакции.
Монофильный таллом - таллом листоватого лишайника, со¬
стоящий из одной нерассеченной на лопасти более или
менее цельной пластинки.
Морфотип - нетаксономическая совокупность особей вида,
отличающихся от других особей того же вида характер¬
ным набором морфологических признаков.
Муральные споры - многоклеточные споры, у которых кроме
поперечных присутствуют несколько (не менее трех)
продольных перегородок.
Мутовчатое ветвление - ветвление, в результате которого
боковые ветви формируются группами по три и более и
располагаются на основной оси в узлах (результат моно-
подиального нарастания таллома).
Накипной таллом - представляет собой распростертую по
поверхности субстрата корочку (реже полностью погру¬
жен в субстрат) и характеризуется плотным контактом
большей части нижней поверхности с ним за счет гиф
сердцевины.
Налет (pruina) - неклеточный поверхностный слой светлой
окраски, являющийся производным корового слоя и
формируемый из остатков эпинекрального слоя, синте¬
зируемых талломом кристаллов оксалатов, карбонатов и
лишайниковых веществ.
Окулярная камера - апикальное выпячивание аскопласта, ко¬
торое может вдаваться в толус сумки леканорового типа.
Олиготип - скудно представленный типовой материал.
Осевой цилиндр - см. Стереом.
Остиола (устьице) - чаще всего апикально расположенное
отверстие, которым открывается во внешнюю среду по¬
лость перитеция или пикниды.
Очерёдное ветвление - ветвление, в результате которого бо¬
ковые ветви или лопасти формируются одиночно (т. е. не
сгруппированы в узлы) и располагаются на основной оси
последовательно (результат моноподиального нараста¬
ния таллома).
Палисадная ориентация гиф - см. Антиклинальная ориен¬
тация гиф.
Папиллы - мелкие сосочковидные выросты на ветвях первого
порядка у видов рода Usnea, формируемые за счет коро¬
вого слоя.
Параплектенхима (псевдопаренхима) - плектенхима, образо¬
ванная гифами, составленными более или менее изодиа-
метрическими округлыми или угловато-округлыми клет¬
ками.
Парасимбиоз - вторичный, дополнительный вариант симбио¬
тических взаимоотношений.
Парафизоиды - производные интераскальной псевдопарен¬
химы элементы хаматеция, представляющие собой сеть
из ветвящихся и анастомозирующих филаментов, в ко¬
торой развиваются сумки (внешне напоминают псевдо¬
парафизы).
Парафизы - элементы хаматеция, представленные филамен-
тами, вырастающими из субгимения по направлению к
поверхности гимения.
Первичный таллом - распростертая по субстрату бородавча-
то- или чешуйчато-накипная часть таллома кустистых
плагио-ортотропных лишайников, на которой формиру¬
ется восходящая кустистая часть (подеции или псевдо¬
подеции).
Периклинальная ориентация гиф - гифы плектенхимы рас¬
полагаются параллельно друг другу и параллельно по¬
верхности таллома.
Периспорий - наиболее поверхностный слой клеточной стен¬
ки аскоспор, развитый у ряда таксонов. Бывает диффе¬
ренцирован на плотную внутреннюю часть, определяю¬
щую орнаментацию поверхности спор, и аморфную сли¬
зистую внешнюю, способную при сильном развитии
формировать гало.
Перитеций - полузамкнутое в зрелости плодовое тело округ¬
лой или кувшиновидной формы.
Перифизоиды - элементы хаматеция, представленные корот¬
кими филаментами, растущими с купола плодового тела
и направленными вниз.
Перифизы - элементы хаматеция, представленные короткими
филаментами, формирующимися в остиоле перитеция
или пикниды и растущими вверх, к выходу из остиолы.
Пикнида (пикнидия) - конидиома полузамкнутого типа,
напоминающая по форме округлый или кувшиновидный
перитеций и открывающаяся наружу одной или несколь¬
кими остиолами (устьицами). Внутри пикниды имеют
одну или несколько камер, в которых расположены ко¬
нидиеносцы.
Пикнидия - см. Пикнида.
Пикноаскокарп - напоминающее апотеций плодовое тело ви¬
дов родов Ephebe и Thyrea, формирующееся без образо¬
вания примордия путем закладки аскогониальных гиф в
пикниде.
Плазмогамия - слияние протопластов и клеточных мембран
копулирующих структур, происходящее перед кариога¬
мией и являющееся начальной стадией полового процес¬
са.
Плектенхима (ложная ткань) - образованное из гиф гриба и
формирующее некоторый слой или структуру мицели-
альное сплетение, более или менее напоминающее
ткань, но, по характеру развития, тканью не являющееся.
Плодовые тела лишайников - мицелиальные сплетения, не¬
сущие споры полового размножения микобионта, фор¬
мирующиеся в результате мейоза в сумках у сумчатых
грибов (аскоспоры) или на базидиях у базидиальных
грибов (базидиоспоры).
Подеции - элементы вторичного таллома, развивающиеся у
видов рода Cladonia из нижней бесплодной части при¬
мордиев апотециев или пикнид после их закладки на
первичном талломе.
Полифильный таллом - таллом листоватого лишайника, со¬
стоящий из более чем одной пластинки или пластинка
таллома рассечена на некоторое количество лопасти.
Полярилокулярные аскоспоры - двухклеточные аскоспоры с
утолщенной перегородкой, пронизанной тонким цен¬
тральным каналом.
Поперечно многоклеточные споры - споры с одной или бо¬
лее поперечными перегородками.
Пробных площадей метод полевых исследований - метод,
при котором исследователь пользуется на местности
стандартизованной в соответствии с поставленной зада¬
чей схемой выбора обследуемых пробных площадей, в
пределах которых учитывает и собирает материал также
с использованием стандартизованной схемы действий.
Прозенхима - см. Прозоплектенхима.
Прозоплектенхима (прозенхима) - плектенхима, образован¬
ная гифами, составленными из вытянутых (анизодиамет-
рических) клеток.
Проталлюс - недифференцированный и не содержащий кле¬
ток фотобионта таллом, формируемый микобионтом на
ранних стадиях развития из споры и существующий до
стадии лихенизации.
Прототуникатного типа сумки - сумки, клеточная стенка
которых недифференцирована на слои, поэтому апи¬
кальный аппарат у таких сумок отсутствует. Высвобож¬
дение спор происходит путем разрыва клеточной стенки.
Псевдогомф - структура прикрепления к субстрату, более ха¬
рактерная для кустистых лишайников и отличающаяся
от гомфа тем, что контакте с субстратом участвуют
только гифы сердцевины.
Псевдоифеллы - слабо или четко очерченные участки различ¬
ной формы на верхней или нижней поверхности талло¬
ма, лишенные корового слоя или какой-либо иной спе¬
циализированной выстилки и, в отличие от цифелл, вы¬
стланные или заполненные рыхло расположенными пуч¬
ками гиф сердцевины. Являются структурами, облегча¬
ющими газо- и водообмен таллома с окружающей сре¬
дой.
Псевдолеканориновый апотеций - см. Биаториновый апо¬
теций.
Псевдолеканоровый апотеций - см. Биаториновый апоте¬
ций.
Псевдопарафизы - элементы хаматеция, представленные вет¬
вящимися и анастомозирующими филаментами, начи¬
нающими расти из купола еще замкнутого молодого
плодового тела в направлении субгимения и врастаю¬
щими сверху в гимений. К моменту созревания плодово¬
го тела теряют связь с его куполом.
Псевдопаренхима - см. Параплектенхима.
Псевдоподеции - элементы вторичного таллома, развивающи¬
еся у видов рода Stereocaulon как выросты первичного
таллома вне прямой зависимости от процесса закладки и
развития апотециев или пикнид.
Пустулы - небольшие пузыревидные вздутия на верхней по¬
верхности талломов некоторых видов рода Lasallia, ко¬
торым на нижней поверхности соответствуют углубле¬
ния
Регулярное ветвление - ветвление, характер которого подчи¬
няется какой-либо определенной закономерности.
Ресинтез таллома - естественный или организованный в ла¬
бораторных условиях (искусственный) процесс форми¬
рования таллома лишайника из клеток фото- и микобио¬
нта.
Реснички - плотные тяжи гиф, формирующиеся по краям ло¬
пастей или апотециев и обычно не выполняющие функ¬
цию прикрепления к субстрату.
Ризиноидные тяжи - макроскопические пучки рыхло агре¬
гированных гиф, формирующиеся как выросты нижнего
корового слоя или сердцевины и выполняющие функцию
закрепления таллома на субстрате. Проникают в суб¬
страт глубже, чем ризины.
Ризиноморфы - структуры нижней поверхности лопастей ум¬
биликатных лишайников, сходные по внешнему виду и
строению с ризинами, но не выполняющие функцию за¬
крепления таллома на субстрате.
Ризины - макроскопические пучки плотно переплетенных
и/или склеенных гиф, формирующиеся как выросты
нижнего корового слоя или сердцевины и выполняющие
функцию закрепления таллома на субстрате.
Ризогифы - см. Ризоиды.
Ризоиды - отдельные одноклеточные или многоклеточные
гифы, формирующиеся из клеток нижнего корового
слоя, проникающие в субстрат и выполняющие функцию
закрепления таллома на нем.
Сердцевина (medulla) - рыхлый или плотный слой таллома
гетеромерного строения, состоящий из гиф микобионта,
обычно располагающийся под альгальным слоем или, у
представителей некоторых родов (все виды рода Pelti¬
gera, некоторые виды рода Cladonia), более или менее
открыто экспонированный.
Симбиоз - совместное тесно взаимосвязанное сосуществова¬
ние двух или более организмов (без уточнения характера
взаимоотношения между ними!).
Синнема (коремия) - кистевидная приподнятая конидиома,
состоящая из плотной массы длинных, иногда сросших¬
ся между собой гиф, формирующих ножку, на вершине,
а иногда и по бокам которой открыто располагаются ко¬
нидиеносцы и конидии.
Скаброзная поверхность таллома - поверхность, фактура
которой носит грубозернистый или почти чешуйчато-
ареолированный характер ("шагреневая").
Склеропараплектенхима - склероплектенхима, гифы кото¬
рой состоят из изодиаметрических клеток.
Склероплектенхима - плектенхима, гифы которой погруже¬
ны в развитый желатинозный или хрящеватый матрикс и
составлены из толстостенных клеток.
Склеропрозоплектенхима - склероплектенхима, гифы кото¬
рой состоят из вытянутых анизодиаметрических клеток.
Слизистые лишайники - полифилитическая группа циано-
бионтных лишайников с гомеомерными талломами, спо¬
собными поглощать значительные количества воды и
принимать желеобразную консистенцию.
Собственный край апотеция - см. Эксципул.
Соматогамия - половой процесс, при котором копулируют
неспециализированные вегетативные гифы (более харак¬
терен для базидиальных грибов).
Соралии - ограниченные коровым слоем и лишенные его
участки таллома, в пределах которых на базе альгально¬
го слоя развивается масса соредий.
Соредии - микроскопические или субмикроскопические, зер¬
нистые, округлые вегетативные пропагулы, лишенные
сформированного корового слоя и состоящие из пере¬
плетенных гиф микобионта и клеток фотобионта. На ге¬
теромерных талломах соредии образуются в альгальном
слое до или после направленной дезинтеграции корового
слоя.
Спермации - одноклеточные или многоклеточные структуры
сумчатых грибов, участвующие в половом процессе в
качестве аналогов «мужских» гамет, но формирующиеся
по механизму образования конидий в типичных кони-
диальных спороношениях.
Спермациогамия - у сумчатых грибов половой процесс, в хо¬
де которого спермаций копулирует с «женским» поло¬
вым органом - аскогоном (недифференцированный на
гаметы гаметангий) через его гифоподобный вырост -
трихогину.
Спинулы - короткие неразветвленные боковые веточки с пе¬
ретяжкой при основании, формирующиеся на талломах
некоторых кустистых лишайников. При механическом
воздействии могут отламываться от таллома и выпол¬
нять функции вегетативных пропагул.
Спородохий (ложе) - открытая подушковидная распростертая
на талломе конидиома, состоящая из плотной массы ко¬
ротких агрегированных конидиеносцев.
Стереом (центральный тяж, осевой цилиндр) - часть сердце¬
вины, представленная склеротизированной плектенхи¬
мой и придающая таллому значительную упругость,
прочность и жесткость.
Стерильные лишайники - условное обозначение для филоге¬
нетически гетерогенной группы таксонов лишайников, у
которых не известно половое и бесполое размножение.
Строма - плотное мицелиальное сплетение, в которое могут
быть погружены аскомы и/или конидиомы грибов. Тал¬
лом лишайника можно рассматривать как лихенизиро-
ванную строму.
Субгимений - тонкая нижняя часть гимения, в которой распо¬
лагаются гифы, формирующие элементы лежащего вы¬
ше гимения.
Субмуральные споры - многоклеточные споры, у которых
кроме поперечных присутствуют одна или две продоль¬
ных перегородок.
Сумка (аск, ascus) - клетка, в которой в результате мейоза эн¬
догенно формируются споры полового размножения
сумчатых грибов - аскоспоры.
Су перле циде иное й апотеций - апотеций, эксципул которого
покрыт снаружи только коровым слоем талломного края,
при этом талломный край и гипотеций не содержат кле¬
ток фотобионта.
Супротивное ветвление - ветвление, в результате которого
боковые ветви или лопасти формируются попарно и рас¬
полагаются на основной оси в узлах супротивно друг
другу (результат моноподиального нарастания таллома).
Таллинокарп - плодовое тело видов родов Lichinella и Gono-
hymenia, формирующееся без образования примордия
путем развития мицелиального сплетения вокруг аского-
ниальных гиф, расположенных среди вегетативных гиф
таллома.
Таллический тип конидиогенеза - конидиогенез, при кото¬
ром будущая конидия отделяются перегородкой от ко¬
нидиогенной клетки до начала увеличения своего объе¬
ма.
Таллоконидии (таллоспоры) - специфические споры бесполо¬
го размножения, образующиеся на нижней поверхности
талломов умбиликатно-листоватых или на проталлюсе
некоторых накипных лишайников. Представляют собой
гранулярные структуры, состоящие из одной или не¬
скольких сильно меланизированных толстостенных кле¬
ток микобионта, и отчленяющиеся от концов гиф, выхо¬
дящих из таллома перпендикулярно его поверхности.
Таллом - вегетативное тело споровых организмов, не диффе¬
ренцированное на органы и ткани.
Талломный край апотеция (амфитеций) - периферическая
защитная часть леканоринового апотеция, лежащая сна¬
ружи от эксципула и снизу переходящая в таллом.
Сформирован талломом, поэтому составлен коровым,
альгальным слоями, и сердцевиной.
Таллоспоры - см. Таллоконидии.
Телеоморфа - половая стадия жизненного цикла грибов, не
несущая структур бесполого размножения, но способная
нести структуры полового размножения.
Теций - см. Гимений.
Тлазидий - гифофор сферической или цилиндрической фор¬
мы.
Толус - см. Апикальный аппарат сумки.
Трансектный метод полевых исследований - метод, при ко¬
тором пробные площади или точки сбора материала рас¬
полагают на местности вдоль условной линии (трансек-
ты), на которой можно наблюдать градиент воздействия
какого-либо очевидного или предполагаемого фактора
или вызванных им изменений в лихенофлоре. Работа на
пробных площадях проходит с использованием стандар¬
тизованной схемы действий.
Трихогина - гифоподобный вырост аскогона, через который
происходит копуляция со спермацием.
Туберкулы - короткие палочковидные выросты коры, на кон¬
цах которых у некоторых видов рода Usnea развиваются
соралии.
Унитуникатного типа сумки - сумки, клеточная стенка ко¬
торых состоит из двух слоев (эндоаск и экзоаск), кото¬
рые при вскрытии сумки ведут себя согласованно как
один слой, вытягиваясь по направлению к поверхности
гимения. Зрелые споры выбрасываются по каналу из
сумки благодаря ее внутриклеточному давлению.
Устьице - см. Остиола.
Фиалидный тип конидиогенеза - энтеробластический кони¬
диогенез, при котором каждая следующая конидия отде¬
ляется перегородкой от конидиогенной клетки в том же
месте, где и самая первая из них. При таком характере
деления кольцеобразные телескопически расположен¬
ные рубцы на от конидиогенной клетке отсутствуют.
Фибриллы - наиболее тонкие и короткие веточки, располага¬
ющиеся на более крупных ветвях талломов видов рода
Usnea. По строению сходны с более крупными ветвями,
однако их центральная часть - стереом - не связана со
стереомом основной части таллома. Могут выполнять
функцию структур вегетативного размножения.
Филлидии (лобули) - мелкие дорсивентрального плана строе¬
ния чешуйковидные выросты, образующиеся на поверх¬
ности талломов или на талломном крае апотециев неко¬
торых листоватых и кустистых лишайников. Как и изи¬
дии, являются вегетативными пропагулами и обладают
внутренней стратификацией, соответствующей страти¬
фикации самого таллома.
Филлокладии - выросты подециев или псевдоподециев кусти¬
стых лишайников, покрытые (хотя бы частично) коро¬
вым слоем, под которым лежат альгальный слой и серд¬
цевина, но в состав которых не входят элементы сте¬
реома. Филлокладии могут выполнять функцию пропа¬
гул вегетативного размножения.
Филлокладиоидные веточки (у некоторых видов рода Stere¬
ocaulon) - напоминают филлокладии, но являются боко¬
выми ветвями таллома, так как содержат в центральной
части ответвления стереома основных ветвей.
Фисситуникатного типа сумки - см. Битуникатного типа
сумки.
Фовеолы - неправильной формы вогнутые участки коры на
поверхности крупных ветвей некоторых видов рода Us¬
nea.
Фотобионт - автотрофный фотосинтезирующий однокле¬
точный или нитчатый компонент лишайникового талло¬
ма, обычно представленный зеленой водорослью и/или
цианобактерией.
Фотосимбиодема - единый морфологически химерный тал¬
лом вида рода Peltigera, в состав одних лопастей которо¬
го в качестве фотобионта входит зеленая водоросль, а
других - цианобактерия.
Фрагментация таллома - простейший способ вегетативного
размножения, при котором увеличение числа особей
происходит за счет разрастания неспециализированных
обломков таллома, образующихся в результате его меха¬
нического повреждения.
Хаматеций - совокупность стерильных элементов гимения
(интераскальных филаментов), а также гифы, выступа¬
ющие в полость плодового тела.
Хемотип - нетаксономическая совокупность особей вида, от¬
личающихся от других особей того же вида характерным
набором вторичных метаболитов.
Хлоробионтные лишайники - лишайники, в талломы кото¬
рых в качестве фотобионта входит только зеленая водо¬
росль.
Хлороцианобионтные лишайники - лишайники, в талломы
которых в качестве фотобионтов входят как зеленая во¬
доросль, так и цианобактерия одновременно.
Холобластический онтогенез конидий - онтогенез, при ко¬
тором формирование клеточной стенки конидий идет за
счет внешнего и внутреннего слоев клеточной стенки
конидиогенной клетки.
Цветные реакции (микрохимические тесты) - набор про¬
стейших цветных химических тестов, производимый с
различными вегетативными или генеративными струк¬
турами (или их частями) таллома лишайника посред¬
ством стандартного для лихенологических исследований
набора реактивов и призванный выявить качественное
содержание в них тех или иных групп лишайниковых
веществ.
Центральный тяж - см. Стереом.
Цефалодии - характерные образования на поверхности или в
толще талломов циано-хлоробионтных лишайников, со¬
держащие группы клеток или нити цианобионта, окру¬
женные более или менее плотным слоем, сформирован¬
ным гифами микобионта.
Цианобионтные лишайники - лишайники, в талломы кото¬
рых в качестве фотобионта входит только цианобакте¬
рия.
Цифеллы - четко очерченные углубления округлой или
овальной кратеровидной формы, полностью или полупо¬
груженные в таллом и выстланные рыхлым парап¬
лектенхимным слоем (псевдокортексом), образованным
из клеток сердцевины. Наиболее характерны для нижней
поверхности лопастей видов рода Sticta. Являются
структурами, облегчающими газо- и водообмен таллома
с окружающей средой.
Шизидии - вегетативные пропагулы, формирующиеся путем
локального отслаивания от таллома небольших участков
корового и связанного с ним альгального слоя.
Эксципул (собственный край апотеция) - периферическая за¬
щитная часть апотеция, снизу переходящая в гипотеций.
Эндогенные лишайники - лишайники, вегетативная часть
талломов которых полностью погружена в субстрат.
Эндоспорий - внутренний слой клеточной стенки аскоспоры,
лежащий непосредственно кнаружи от плазмалеммы.
Эндофитные грибы - эколого-трофическая несистематиче¬
ская полифилитическая группа нелихенизированных
грибов, поселяющихся внутри талломов лишайников, но
не связанных трофически с мико- или фотобионтами.
Энтеробластический онтогенез конидий - онтогенез, при
котором формирование клеточной стенки конидий идет
только счет внутреннего слоя клеточной стенки кони¬
диогенной клетки или совсем без участия ее клеточной
стенки. Недостающий слой или слои в этом случае до¬
страиваются самостоятельно.
Эпигенные лишайники - лишайники, вегетативная часть тал¬
ломов которых располагается преимущественно над по¬
верхностью субстрата.
Эпигимений (эпитеций) - тонкий поверхностный слой гиме¬
ния, состоящий из апикальных частей элементов хамате¬
ция и защищающий сверху обычно более короткие сум¬
ки.
Эпикортекс - тонкий поверхностный гомогенный полисаха¬
ридный слой, который синтезируют клетки коры некото¬
рых лишайников.
Эпинекральный слой - тонкий матово-блестящий поверх¬
ностный слой, формируемый у некоторых лишайников в
результате отмирании гиф поверхностной части корово¬
го слоя и состоящий из остатков желатинизированных
кортикальных клеток.
Эпипсамма - совокупность окрашенных или бесцветных гра¬
нул, кристаллов или капель масла, содержащихся в эпи-
теции и часто занимающих его верхнюю часть.
Эписпорий - слой клеточной стенки аскоспоры, лежащий
непосредственно кнаружи от внутреннего слоя - эндо-
спория.
Эпитеций - см. Эпигимений.
Эулеканоровый апотеций - см. Леканориновый апотеций.
Приложение III.
Принятые сокращения фамилий авторов при таксонах
Ach. — Acharius Е.
C. Agardh — Agardh С. А.
Almb. — Almborn О.
Almq. — Almquist S. О. I.
Anzi — Anzi M.
Aptroot — Aptroot A.
Arnold — Arnold F. Ch. G.
Arup — Arup U.
Asahina — Asahina Y.
D. D. Awasthi — Awasthi D. D.
C. Bab. — Babington C.
Bagl. — Baglietto F.
G. E. Baker — Baker G. E.
Barkh. — Бархалов LLI. O.
Baruffo — Baruffo M.
Bausch — Bausch W.
Bel. — Belanger C. P.
Bellem. — Bellemere A.
Beltr. — Beltramini de Casati F.
H. E. Bigelow — Bigelow How¬
ard E.
Bitter — Bitter F. A. G.
Boistel — Boistel A. B.
Bong. — Bongard A. (G.) H. von
Van den Boom — Boom Pieter
P.G. van den
Bornet — Bornet J.-B. Ё.
Borrer— Borrer W. J.
Bory — Bory J. B. G. G. M.
Bowler — Bowler P.
Beltr. — Beltramini de Casati F.
Botnen — Botnen A.
M. Brand — Brand M.
Brandt —
Branth — Branth J. S.
Bredkina — Бредкина Jl. И.
Brenner — Brenner М. M. W.
Bresinsky — Bresinsky A.
Britzelm. — Britzelmayr M.
Brodo — Brodo I. M.
Brusse — Brusse F. A.
Bull. — Bulliard J. B. F.
G. Buschardt — Burchard G.
Bystrek — Bystrek J.
Canals — Canals A.
Carestia — Carestia A.
Chaub. — Chaubard L. A.
Chaves — Chaves J. L.
B. Chen — Chen Jian Bin
Chevall. — Chevallier F. F.
Chodat — Chodat R.
M. Choisy — Choisy M. G. B.
St. Clair — Clair St.
Clauzade — Clauzade F. J. G. A.
Clem. — Clements F. E.
A. Crespo — Crespo (de las
Casas) A.
Coppins — Coppins B. J.
Corda — Corda A. K. J.
Cretz. — Cretzoiu P.
Cromb. — Crombie J. М. M.
C. F. Culb.— Culb C. F.
W. L. Culb. — Culb W. L.
Dalla Torre — Dalla Torre K. W.
von
J. C. David — J. C. David
DC. — Candolle A. P. de
Degel. — Degelius G. B. F.
Delise — Delise D. F.
Desf. — Desfontaines R. L.
Desm. — Desmazieres J. В. H. J.
J. P. Dey — Dey J. P.
Dicks. — Dickson J. J.
Diederich — Diederich P.
D. Dietr. — Dietrich D. N. F.
Dobbeler— Dobbeler P.
C. W. Dodge — Dodge C. W.
Dombr. — Домбровская A. B.
Du Rietz — Du Rietz G. E.
Duby — Duby J. E.
Dufour — Dufour J.-M. L.
Ehrh. — Ehrhart J. F.
Eitner — Eitner E.
Elenkin — Еленкин A. A.
Elix — Elix L. A.
Erichsen — Erichsen Ch. F. E.
Essl. — Esslinger T. L.
J. Favre — Favre J.
Fee — Fee A. L. A.
Fink — Fink B.
Flagey — Flagey C.
Florke — Floeke H. G.
Flot. — Flotow J. Ch. G. U. G. G.
A. E. F.
Follmann — Follmann G.
Fr. — Fries E. M.
Frey — Frey E.
Th. Fr. — Fries Th. M.
Fiirnr. — Fiirnrohr A. E.
Giralt — Giralt M.
J. F. Gmel. — Gmelin J. F.
N. S. Golubk. — Голубкова H.
C.
Gomez-Bolea — Gomez-Bolea A.
Graewe — Graewe P. H. F.
Gray — Gray S. F.
Grube — Grube M.
Grummann — Grummann V. J.
Gyeln. — Gyelnik V. K.
Hafellner — Hafellner J.
Hale — Hale М. E.
Hampe — Hampe G. E. L
Harm. — Harmand J. H. A. J.
R. C. Harris —Harris R. C.
Hav. — Havaas J. J.
D. Hawksw. — Hawksworth D. L.
Hazsl. — Hazslinszky von Hazs-
lin F. A.
Hedw. — Hedwig J.
Heibel — Heibel E.
R. Heim — Heim R.
Hellb. — Hellbom P. J.
D. M. Hend. — Henderson D. M.
Hepp — Hepp J. A. P.
Hem.-Mar. — Hernandez-Marine
M.
Herre Herre A. W. С. T.
Hertel — Hertel H.
Hillmann — Hillmann J.
Hochst. — Hochstetter C. F. F.
Hoffm. — Hoffmann G. F.
Hook. — Hooker W. J.
Hook f.— J. D. Hook
Horw. — Horwood A. R.
Houmeau — Houmeau J.-W.
G. R. Hu — HuG. R.
Huds. — Hudson W.
Hue — Hue A.-M.
Hulting— Hulting J.
Humb. — Humboldt F. W. H. A.
von
Imshaug — Imshaug H. A.
Inasch. — Inaschvili T. N.
P. James — James P. W.
Jatta — Jatta A.
H. S. Jenn. — Jennings H. S.
M. Joshi — Joshi M.
Kalb Kalb K.
Kantvilas — Kantvilas G.
Kashiw. — Kashiwadani H.
Keissl. — Keissl. K.
Kernst. — Kernstock E.
H. Kilias — Kilias H.
Klem. — Klement O.
Korb. — Korber G. W.
Kremp. — Krempelhuber A. von
Krog — Krog H.
Kukwa — Kukwa M.
Kulakov — Кулаков В. Г.
Kullh. — Kullhem H. A.
Kummerl. — Kummerling H.
Kurok. — Kurokawa S.
Kuyper — Kuyper T. W.
L. — Linnaeus C.
J. Lahm — Lahm J. G. Fr.-X.
Lam. — Lamarck J. B.
I. M. Lamb — Lamb I. M.
Lambinon — Lambinon J.
Lamy — Lamy P. М. E.
M. Lange — Lange M.
J. R. Laundon — Laundon J. R.
Leight. — Leighton W. A.
de Lesd. — Bouly de Lesdain M.
Lettau — Lettau G.
Leuckert — Leuckert Ch.
Lilj. — Liljeblad S.
Linds. — Lindsey W. L.
Link — Link J. H. F.
Llimona — Llimona X.
Lohtander — Lohtander K.
Lucking — Lucking R.
Lumbsch — Lumbsch H. Th.
Lunke — Lunke T.
Lutzoni — Lutzoni F. M.
Lynge — Lynge B. A.
Maas Geest. — Maas Geesteranus
R. A.
H. Magn. — Magnusson A. H.
Malme — Malme G. O. A.
A. Massal. — Massalongo A. B.
Tat. Matsumoto — Matsumoto
Tatsuo
H. Mayrhofer — Mayrhofer H.
Mereschk. — Мережковский К.
С.
G. Merr. — Merrill G. K.
Meyer — Meyer S. L. F.
Michx. — Michaux A.
Mig. — Migula E. Fr. A. W.
Moberg — Moberg R.
Moncalvo — Moncalvo J.-M.
Mong. — Monguillon E. L. H.
Mont. — Montagne J. P. F. C.
К. H. Moon — Moon К. H.
B. J. Moore — Moore B. J.
М. M. Moser — Moser Meinhard
(M.)
Motyka — Motyka J.
Mudd — Mudd W. A.
Muhr — Muhr L.-E.
Mull. Arg. — Muller Argoviensis
J.
Nadv. — Nadvorm'k J.
Nageli — Nageli C. W.
Nannf —Nannfeldt J. A. F.
Neck. — Necker N. M. J. de
Nimis — Nimis P. L.
Noble — Noble M.
Norman — Norman J. M.
Norrl. — Norrlin J. P.
A. Nyl. — Nylander A. E.
Nyl. — Nylander W.
Ohlert — Ohlert O. L. A.
H. Olivier — Olivier H. J. F.
Orange — Orange A.
0sth. — Osthagen H.
T. Otn. — Отнюкова Т. H.
Overeem — Overeem C. van
Ovstedal — Ovstedal D. O.
Peck — Peck С. H.
Pers. — Persoon С. H.
R. H. Petersen — Petersen R. H.
Pietschm. — Pietschmann M.
Poelt — Poelt J.
Poetsch — Poetsch I. S.
Pouzar — Pouzar Z.
Printzen — Printzen Ch.
Puntillo — Puntillo D.
Purvis — Purvis W.
Quel. — Quelet L.
Rabenh. — Rabenhorst G. L.
Rambold — Rambold G.
Ramond — Ramond de Carbon-
niere L. F. E.
Rasanen — Rasanen V. J. P. B.
Rass. — Рассадина К. A.
Redhead — Redhead S. A.
Rehm — Rehm H. S. L. Fr. F.
Reinke — Reinke J.
Retz. — Retzius A. J.
Ropin — Ropin K.
Rostr. — Rostrup E.
Cl. Roux — Roux Cl.
Rundel — Rundel P. W.
Runemark — Runemark H.
Rupr. — Ruprecht F. J.
Rutstr. — Rutstrom С. B.
D. Sacc. — Saccardo D.
Sacc. — Saccardo P. A.
Samp. — Sampaio G. A. da S. F.
Sandst. — Sandstede H.
Santha — Santha L.
R. Sant. — Santesson R.
Sarnth. — Sarnthein L. von
M. Sato —Sato M.
Savicz — Савич В. П.
Schaer. — Schaerer L. E.
T. Schauer — Schauer T.
Scheid. — Scheidegger C.
Schleich. — Schleicher J. C.
I. Schmitt — I. Schmitt
Gott. Schneid. — Schneider Gott-
hard
Schreb. — Schreber J. C. D. von
R. Schubert — Schubert R.
A. J. Schwab —Schwab A. J.
Schwein. — Schweinitz L. D. von
Scop. — Scopoli J. A.
Serus. — Serusiaux E.
Servit — Servit M.
Shear — Shear C. L.
Sheard — Sheard J. W.
Kr. P. Singh — Singh Krishna Pal
S. R. Singh — Singh Shri Ram
Sipman — Sipman H. J. M.
Skorepa — Skorepa A. C.
Slavikova — Slavikova E.
A. L. Sm. — Smith A. L.
Sm. — Smith J. E.
Sommerf. — Sommerfelt S. Ch.
Sparrius — Sparrius L. B.
Spier — Spier (J.) L.
Spreng. — Sprengel C. P. J.
M. Steiner — Steiner M.
Stangl — Stangl J.
Stein — Stein B.
J. Steiner — Steiner J.
Stenh. — Stenhammar Ch.
Stirt. — Stirton J.
Stizenb. — Stizenberg E.
Suza — Suza J.
Sw. — Swartz O. (P.)
Swinscow — Swinscow T. D. V.
Syd. — Sydow H.
Szatala — Szatala O.
Taylor — Taylor T.
J. W. Thomson — Thomson J. W.
G. Thor —Thor G.
Thorold — Thorold C. A.
Timdal — Timdal E.
Tomin — Томин М. П.
Tonsberg — Tonsberg T.
Torss. — Torssell G.
Trass — Trass H.
Tretiach — Tretiach M.
Trevis. — Trevisan de Saint-Leon
V. B. A.
Triebel — Triebel D.
Tscherm.-Woess — Tschermak-
Woess E.
Tuck. — Tuckerman E.
Turner — Turner D.
Umana — Umana L.
I. Urban. — Урбанавичене И. H.
G. Urban. — Урбанавичус Г. П.
Vain. — Vainio E. A.
Vezda — Vezda A.
Vilgalys — Vilgalys R.
Vivant — Vivant J.
Vouaux — Vouaux L.
Wahlenb. — Wahlenberg G.
Wallr. — Wallroth K. F. W.
Walt. Watson — Watson W.
W. R. Watson — Watson W. R.
Weber — Weber G. H.
Wedd. — Weddell H. A.
J. C. Wei — Wei J. C.
Werner — Werner R.-G.
Westr. — Westring J. P.
Wetmore — Wetmore C.
F. H. Wigg. — Wiggers F. H.
Willey — Willey H.
V. Wirth — Wirth V.
Wolseley — Wolseley P. A.
Woron. — Воронихин H. H.
Wulfen — Wulfen F. X.
Yasuda — Yasuda A.
Zahlbr. — Zahlbruckner A.
Zhurbenko — Журбенко М. П
Zopf—Zopf W.
Приложение IV.
Zukal — Zukal H.
Ziirn — Ziirn L.
Индекс латинских названий
Abies Mill.
Abrothallus De Not
— pulverulentus Anzi
Acarospora A. Massal.
Agaricus L.
— alpinus Britzelm.
— umbelliferus L. : Fr.
AGYRIACEAE Corda
Agyrium Fr.
— rufum (Pers.) Fr.
Ainoa Lumbsch & I. Schmitt
Alectoria Ach.
— thrausta Ach.
Amandinea M. Choisy ex Scheid. &
H. Mayrhofer
— cacuminum (Th. Fr.) H. Mayrho¬
fer & Sheard
— coniops (Wahlenb.) M. Choisy ex
Scheid. & H. Mayrhofer
— lecideina (H. Mayrhofer & Poelt)
Scheid. & H. Mayrhofer
— polyspora (Willey) E. Lay & P.
May
— punctata (Hoffm.) Coppins &
Scheid.
Amylora Rambold
Anamylopsora Timdal
— pulcherrima (Vain.) Timdal
ANAMYLOPSORACEAE Lumbsch
& Lunke
Anaptychia Korb.
— albicans Kurok.
— angustiloba (Mull. Arg.) Kurok.
— aquila (Ach.) A. Massal.
— bryorum Poelt
— casarettiana A. Massal.
— chilensis Kurok.
var. austroafricana Kurok.
— ciliaris (L.) Korb.
f. angusta A. Massal.
subsp. mamillata (Taylor) D.
Hawksw. & P. James
— — var. crinalis (Schleich. ex
Schaer.) Rabenh.
— — var. melanosticta (Ach.)
Boistel
var. solenaria (Duby) Frey
— crinalis (Schleich. ex Schaer.)
Vezda
— dendritica (Pers.) Vain.
var. dissecta (Kurok.) Kurok.
var.japonica M. Sato
var. propagulifera Vain.
— desertorum (Rupr.) Poelt
— detons a (Fr.) Jatta
— diademata (Taylor) Kurok.
— elbursiana (Szatala) Poelt
— esorediata (Vain.) Du Rietz &
Lynge
— ethiopica Swinscow & Krog
—fragilissima Kurok.
—fusca (Huds.) Vain.
var. stippaea (Ach.) Lynge
— granulifera (Ach.) A. Massal.
— hypocaesia Yasuda
— hypochraea Vain.
— hypoleuca (Ach.) A. Massal.
var. microphylla Kurok.
— intricata (Defs.) A. Massal.
— isidiata Tomin
— isidiophora (Nyl.) Vain.
— isidiza Kurok.
—japonica (M. Sato) Kurok.
— kaspica Gyeln.
— leucomela (L.) A. Massal.
— lutescens Kurok.
— mamillata (Taylor) D. Hawksw.
— melanosticta (Ach.) Trass
— mereschkowskii (Tomin)
Kulakov
— microphylla ( Kurok.) Kurok.
— obscurata (Nyl.) Vain.
— palmatula auct.
— palmulata (Michx.) Vain.
var. caucasica Vain.
var. isidiata Zahlbr.
— podocarpa (Bel.) A. Massal.
— pseudospeciosa Kurok.
— roemeri Poelt
— runcinata (With.) J. R. Laundon
— setifera (Mereschk.) Rasanen
— solenaria sensu Savicz
— speciosa (Wulfen) A. Massal.
var. angustiloba Mull. Arg.
— spinulosa Kurok.
— stippaea (Ach.) Nadv.
— subascendens Asahina
— subheterochroa Kurok.
var. dissecta Kurok.
— ulotrichoides (Vain.) Vain.
Anzina Scheid.
APHANOPSIDACEAE Printzen &
Rambold
Aphanopsis Nyl. ex Syd.
— coenosa (Nyl.) Coppins & P.
James
— lutigena J. Lahm ex Syd.
— terrigena (Ach.) Nyl.
Argopsis Th. Fr.
Arthonia Ach.
— subtilis (Vezda) Vezda
ARTHRORHAPHIDACEAE Poelt
& Hafellner
Arthrorhaphis Th. Fr.
— alpina (Schaer.) R. Sant.
— anzi ana (Lynge) Poelt
— citrinella (Ach.) Poelt
— fuscireagens (Vain.) Poelt
— grisea Th. Fr.
— vacillans Th. Fr. & Almq. ex Th.
Fr.
Arthrospora Th. Fr.
— chlorococca H. Olivier
Ascomycota
Aspicilia A. Massal.
— badio-atra Hepp ex Kremp.
— bockii (Fr.) Boistel
BACIDIACEAE Walt. Watson
Bacidia De Not.
— alpina (Schaer.) Vain.
— anziana Lynge
— asserciilorum Th. Fr.
— bouteillei (Desm.) Arnold
— bouteillei (Desm.) Hulting
— buxi Vezda & Vivant
— chlorococca (Graewe ex Stenh.)
Lettau
— chlorococca Sandst.
— citrinella (Ach.) Branth & Rostr.
— corticola (Anzi) Dalla Torre &
Sarnth.
—flavovirescens (Dicks.) Anzi
— flavovirescens (Dicks.) Anzi var.
alpina (Schaer.) R. Sant.
— flavovirescens (Dicks.) Anzi var.
grisea (Th. Fr.) Vain.
—fuscireagens (Vain.) Vain.
— gorgonea Vezda & Poelt
— holophaea Anzi
— interspersula (Nyl.) Zahlbr.
— kuopioensis (Vain.) Vain.
— myriocarpa Erichsen
— myrtillicola Erichsen
— ophiospora (Hellb.) Th. Fr.
— perpusilla (J. Lahm) Th. Fr.
— podicellata Hellb.
— pruinosa P. James
— sarothamni Vain.
— schadeana Erichsen
— subtilis Vezda
— turgida (Korb.) Hellb.
— umbrina (Ach.) Bausch var. com-
pacta (Korb.) Th. Fr.
— umbrina (Ach.) Bausch var. ma¬
rina Degel.
— umbrina (Ach.) Bausch var. pso-
tina (Fr.) Th. Fr. (“psorot-
ind’)
— umbrina (Ach.) Bausch var. tur¬
gida (Korb.) Th. Fr.
— umbrina (Ach.) Bausch var. um¬
brina
— vacillans (Th. Fr. & Almq. ex Th.
Fr.) Rostr.
— vezdae Coppins & P. James
Baeomyces Pers.: Fr.
— roseus Pers.
— rubiformis Ach.
— rufus (Huds.) Rebent.
Biatora Fr.
— anthracophila (Nyl.) Hafellner
— borealis Hepp
— boreella (Nyl.) Hellb.
— bot ry os a Fr.
— bouteillei (Desm.) A. Massal.
— coniasis A. Massal. in Atti
— cyclospora Hepp ex Korb.
— demissa (Rutstr.) Fr.
—friesii (Ach.) Tuck.
— geophana (Nyl.) Th. Fr.
— himalayana C. Bab.
— humigena (Taylor) Walt. Watson
— hypnophila Graewe ex Stenh.
var. chlorococca Graewe
var. chlorococca Stenh.
— incrustans Arnold
— os treat a (Hoffm.) Fr.
var. cladonioides Fr.
— quernea Fr.
— rufonigra Tuck.
— rupestris Fr.
var. alpina Arnold
var. terricola Anzi
— siebenhaariana Korb.
— subduplex
— tabescens Korb.
— terricola (Anzi) Th. Fr.
— vernalis
Biatorella De Not.
— deplanata Almq.
— geophana (Nyl.) Rehm
— rubidula (Nyl.) Zahlbr.
Biatorina
— hohenbiihelii Poetsch
Bilimbia De Not.
— bouteillei (Desm.) Hulting
— caradocensis (Leight. ex Nyl.) A.
L. Sm.
— chlorococca Graewe
— sabuletorum (Schreb.) Arnold
Blastenia A. Massal.
— siebenhaariana (Korb.) Lettau
— terricola (Anzi) Lindau
Borrera Spreng.
— boryi Fee
— crinalis Schleich. ex Schaer.
— dendritica Pers.
Botryolepraria (Hue) Canals, Hern.-
Mar., Gomez-Bolea &
Llimona
— lesdainii (Hue) Canals, Hern.-
Mar., Gomez-Bolea &
Llimona
BRIGANTIAEACEAE Hafellner &
Bellem.
Brigantiaea Trevis.
— ferruginea (Mull. Arg.) Kashiw.
& Kurok.
— fuscolutea (Dicks.) R. Sant.
— leucoxantha (Spreng.) R. Sant. &
Haffelner
— nipponica (M. Sato) Haffelner
— praetermissa Hafellner & St. Clair
— purpurata (Zahlbr.) Hafellner &
Bellem.
Buellia De Not.
— aethalea (Ach.) Th. Fr.
— aethaleoides (Nyl.) H. Olivier
— alboatra (Hoffm.) Th. Fr.
var. zabotica (Korb.) Th. Fr.
var. venusta (Korb.) Th. Fr.
— alboatra (Hoffm.) Branth &
Rostr.
— ambigua (Ach.) Malme
— arborea Coppins & Tonsberg
— arnoldii Servit
— asterella Poelt & Sulzer
— atrocinerea (Anzi) Zahlbr.
— badia (Fr.) A. Massal.
— baltica Erichsen
— bayrhofferi (Schaer.) H. Olivier
— betulina (Zw.) Th. Fr.
— bryophyla Korb.
— candidula Arnold
— canescens (Dicks.) De Not.
— chionea (Th. Fr.) Sheard
— chloroleuca Korb.
— chlorophaea (Hepp ex Leight.)
Lettau
— concinna Th. Fr.
— con i ops (Wahlenb.) Th. Fr.
— contermina Arnold
— convex a Th. Fr.
— crystallifera (Vain) Hav.
— disciformis (Fr.) Mudd
— f. muscorum (Korb.) Lynge
var. insignis (Nageli) Flag.
f. muscorum (Korb.) Vain.
var. nodulosa Lynge
— dubia (Vain.) Malme
— ectolechioides (Vain.) Erichsen
— elegans Poelt
— elenkinii Tomin
— epigaea (Pers.) Tuck.
var. intermedia (Schrad.) Anzi
— epipolia (Ach.) Mong.
— erubescens Arnold
—fimbriata (Tuck.) Scheard
—frisiaca Erichsen
— geophila (Florke) Lynge
— gevrensis Th. Fr.
— gotlandica J. Steiner
— griseovirens (Turn. & Borr. ex
Sm.) Almb.
— hypopodioides (Nyl.) Arnold
— immersa Lynge
— impressula (Leight.) A. L. Sm.
— injucunda (Vain.) H. Magn.
— insignis (Nageli) Th. Fr.
var. muscorum Korb.
— italica var. lactea A. Massal.
— jugorum (Arnold) Arnold
— lactea (A. Massal.) Korb.
var. atrocinerea Anzi
— leptocline (Flot.) A. Massal.
— lutosa (A. Massal.) Anzi
— malmei Lynge
— margaritacea Lynge
— maritima (A. Massal.) Bagl.
— microplaca (Vain.) Erichsen
— minutissima Lynge
— miriquidica Scheid.
— modica (Nyl.) Mig.
— mongolica H. Magn.
— myriocarpa (DC.) De Not.
P polyspora Willey
— nigritula (Nyl.) Mudd
— nitida Eitner
— nivalis (Bagl. & Carest.) Hertel
— nodulosa (Lynge) H. Magn.
— notabilis Lynge
— ocellata (Flot.) Korb.
var. tenella Mull. Arg.
— olivaceofusca (Anzi) Zahlbr.
— papillata (Sommerf.) Tuck.
var. albocincta Th. Fr.
var. nodulosa (Lynge) Degel.
— parasema (Ach.) De Not.
var. microcarpa Korb.
var. muscorum (Korb.) Th. Fr.
var. rugulosa Korb.
var. saprophila (Ach.) Korb.
subsp. dives Th. Fr.
— pharcidia (Ach.) Malme
— polyspora (Willey) Vain.
— praecavenda (Nyl. ex Cromb.)
Arnold
— pulverulenta (Anzi) Jatta
— punctata (Hoffm.) A. Massal.
var. polyspora (Willey) Tuck.
— punctiformis (Hoffm.) A. Massal.
— ryssolea (Leight.) A. L. Sm.
— scalpturata Vain.
— schaereri De Not
— schisticola H. Magn.
— septentrionalis Vain.
— sororia Th. Fr.
— sororioides Erichsen
— spuria (Schaer.) Anzi
— stellulata (Taylor) Mudd
— stigmatea Korb.
— subatra Erichsen
— subcanescens Werner
— subconcinna (Vain.) Zahlbr.
— subdisciformis (Leight.) Vain.
f. corticola (Nyl.) Sandst.
— subdispersa Mig.
— subviridescens (Nyl.) Vain.
— tesserata Korb.
— trifracta J. Steiner
— triphragmia (Nyl.) Arnold
subsp. thionella Norman
— triphragmioides Anzi
— uberior Anzi
— venusta (Korb.) Lettau
— verruculosa auct. non (Sm.)
Mudd
— verruculosa (Sm.) Mudd
\m.jugorum Arnold
— vilis Th. Fr.
— zabotica (Korb.) Rasanen
— zahlbruckneri auct. non J. Steiner
— zahlbruckneri}. Steiner
Byssocaulon Mont.
Byssoloma Trevis.
— leucoblepharum (Nyl.) Vain.
— marginatum (Arnold) Serus.
— rotuliforme (Mull. Arg.) R. Sant.
in Thorold
— subdiscordans (Nyl.) P. James
Byssus L.
— in сапа L.
Calenia Mull. Arg.
— caucasica (Elenkin & Woron.)
Vezda
Caloplaca Th. Fr.
— demissa (Korb.) Arup & Grube
Calvitimela Hafellner
— armeniaca (DC.) Hafellner
— testaceoatra (Vain.) Hafellner
Candelariella Mull. Arg.
— vitellina (Hoffm.) Mull. Arg.
Carbonea (Hertel) Hertel
— intrusa (Th. Fr.) Rambold &
Triebel
— vorticosa (Florke) Hertel
Catillaria A. Massal.
— bouteillei (Desm.) Zahlbr.
— epiphylla Fink
— intrusa (Th. Fr.) Th. Fr.
— littorella (Nyl.) Zahlbr.
— oospora Vain.
— schroderi Zahlbr.
— synothea (Ach.) Beltr.
CatolechiaFlot.
— lactea A. Massal.
— maritima A. Massal.
Chlorophyta
CHRYSOTHRICHACEAE Zahlbr.
Chrysopsora (Vain.) M. Choisy
— testacea (Hoffm.) M. Choisy
Chrysothrix Mont.
— candelaris (L.) J. R. Laundon
— chlorina (Ach.) J. R. Laundon
— chrysophthalma (P. James) P.
James & J. R. Laundon
— flavovirens Tonsberg
Circinaria Link
— berteriana Fee
Clavaria Fr.
— mucida Pers.
— vernalis Schwein.
CLAVARIACEAE Chevall.
Clavulinopsis Overeem
— corynoides (Peck) Comer
— septentrionalis Comer
— vernalis (Schwein.) Corner
Coccobotrys Chodat
Collema Weber ex F. H. Wigg.
— coenosum Ach.
Conida A. Massal.
— intrusa (Th. Fr.) Sacc. & D. Sacc.
Coppinsia Lumbsch & Heibel
Coriscum Vain.
— viride (Ach.) Vain.
Crocynia auct.
— alpina de Lesd.
— arctica Lynge
— caesioalba de Lesd.
— cupressicola Hue
—fmkii de Lesd.
— lanuginosa Hue
— lesdainii Hue
— murorum de Lesd.
— neglecta (Nyl.) Hue
— rigidula de Lesd.
— subaeruginosa Rasanen
— vouauxii Hue
Dactylospora Korb. emend Hafellner
— pulverulenta (Anzi) Arnold
Dimelaena Norman
— oreina (Ach.) Norman
Diploicia A. Massal.
— cacuminum A. Massal.
— canescens (Dicks.) A. Massal.
— epigaea (Pers.) A. Massal.
— — var. intermedia (Schrad.)
Korb.
— subcanescens (Werner) Hafellner
& Poelt
Diploschistella Vain.
— lithophila (G. Thor & Vezda)
Lucking, Serus. & Vezda
— urceolata Vain.
Diplotomma Flot.
— alboatrum (Hoffm.) Flot.
— — var. epipolium (Ach.) A.
Massal.
— ambiguum (Ach.) Flagey
— betulinum (Hepp) Arnold
— chlorophaeum (Hepp ex Leight.)
Szatala
— dispersum (Kremp.) Arnold
— epipolium (Ach.) Arnold
— geophillum (Florke) S. R. Singh
& D. Awasthi
— lutosum A. Massal.
— nivale (Bagl. & Carest.) Hafellner
— pharcidium (Ach.) M. Choisy
— pulverulentum (Anzi) Crespo &
D. Hawksw.
— triphragmoides (Anzi) Szatala
— venustum Korb.
Dirinaria (Tuck.) Clem.
— applanata (Fee) D. D. Awasthi
— picta (Sw.) Clem. & Shear
Echinoplaca Fee
— epiphylla Fee
ECTOLECHIACEAE Zahlbr.
Endocarpon Hedw.
— crenatum Taylor
Epigloea Zukal
Fellhanera Vezda
— bouteillei (Desm.) Vezda
— buxi (Vezda & Vivant) Vezda
— christiansenii Serus. & Vezda
— fuscatula (Mull. Arg.) Vezda
— gyrophorica Serus., Coppins,
Diederich & Scheid.
— margaritella (Hulting) Hafellner
— myrtillicola (Erichsen) Hafellner
— ochracea Sparrius & Aptroot
— subtilis (Vezda) Diederich &
Serus.
— vezdae (Coppins & P. James) V.
Wirth
— viridiosorediata Aptroot, Brand &
Spier
Fellhaneropsis Serus. & Coppins
— myrtillicola (Erichsen) Serus. &
Coppins
— australiana Lucking
— kurokawana G. Thor, Lucking &
Tat. Matsumoto
— vezdae (Coppins & P. James)
Serus. & Coppins
Fistulariella Bowler & Rundel
— minuscula (Nyl.) Bowler & Run¬
del
Fraxinus L.
Frutidella Kalb
— caesioatra (Schaer.) Kalb
Fuscidea V. Wirth & Vezda
— praeruptorum (Du Rietz & H.
Magn. in Du Rietz) V. Wirth
& Vezda
GOMPHILLACEAE W. R. Watson
ex Hafellner
Gyalecta Ach.
— aethalea Ach.
— piceicola (Nyl.) Arnold
Gyalectidium Mull. Arg.
— caucasicum (Elenkin & Woron.)
Vezda
— colchicum Vezda
— filicinum Mull. Arg.
—filicinum auct. fl. cauc. non Mull.
Arg.
— puntilloi Serus.
— setiferum Vezda & Serus.
Gyalidea Lettau
— psammoica (Nyl.) Lettau
— piceicola (Nyl.) Vezda
Gyalideopsis Vezda
— alnicola Noble & Vezda
— anastomosans P. James & Vezda
— athalloides (Nyl.) Vezda
— calabrica Puntillo & Vezda
— helvetica van den Boom & Vezda
— muscicola P. James & Vezda
var. muscicola P. James &
Vezda
var. alba Vezda & Tonsberg
— peruviana G. Merr. ex Vezda
— piceicola (Nyl.) Vezda & Poelt
— scotica P. James
— tuerkii Vezda
Gypsoplaca Timdal
— macrophylla (Zahlbr.) Timdal
GYPSOPLACACEAE Timdal
Haematomma A. Massal.
— caesium Copins & P. James
Hafellia Kalb, H. Mayrhofer &
Scheid.
— arnoldii (Servit) Hafellner &
Tuck.
— disciformis (Fr.) Marbach & H.
Mayrhofer
Hafellnera Houmeau & Cl. Roux
— parasemella (Nyl.) Houmeau &
Cl. Roux
Heterothecium Mont.
— pezizoideum Stizenb.
var. disciforme Flot.
Hertelidea Printzen & Kantvilas
— botryosa (Fr.) Printzen & Kantvi¬
las
Heterodermia Trevis.
— albicans (Pers.) Swinscow &
Krog
— angustiloba (Mull. Arg.) D. D.
Awasthi
— boryi (Fee) Kr. P. & S. R. Singh
— casarettiana (A. Massal.) Trevis.
— chilensis (Kurok.) Swinscow &
Krog
— corallophora (Taylor) Skorepa
— dendritica (Pers.) Poelt
— — var. propagulifara (Vain.)
Poelt
— diademata (Taylor) D. D. Awas¬
thi
— dissecta (Kurok.) D. D. Awasthi
— fragilissima (Kurok.) Wei
— granulifera (Ach.) W. L. Culb.
— hypocaesia (Yasuda) D. D. Awas¬
thi
— hypochraea (Vain.) Swinscow &
Krog
— hypoleuca (Ach.) Trevis.
var. divergens Trass
— intermedia Trass
— isidiophora (Nyl.) D. D. Awasthi
var. coralligera Trass
— japonica (M. Sato) Swinscow &
Krog
— leucomela (L.) Poelt
var. boryi (Fee) Kr. P. & S. R.
Singh.
— leucomelaena (L.) Vain.
— lutescens (Kurok.) Follmann
— microphylla (Kurok.) Skorepa
— neoleucomelana Kurok.
— obscurata (Nyl.) Trevis.
— podocarpa (Bel.) D. D. Awasthi
— propagulifera (Vain.) J. P. Dey
— pseudospeciosa (Kurok.) W. L.
Culb.
— speciosa (Wulfen) Trevis.
— spinulosa (Kurok.) J. C. Wei
— subascendens (Asahina) Trass
Hygrophorus Fr.
— hudsonianus H. S. Jenn.
Hyperphyscia Mull. Arg.
— adglutinata (Florke) H.
Mayrhofer & Poelt
— crocata Kashiw.
— pandani (H. Magn.) Moberg
— granulata (Poelt) Moberg
— isidiata Moberg
— minor (Fee) Kalb
— minor (Fee) D. D. Awasthi
— pruinosa Moberg
— syncolla (Tuck, ex Nyl.) Kalb
Hypocenomyce M. Choisy
— anthracophila (Nyl.) P. James &
Gott. Schneid.
— australis Timdal
— caradocensis (Leight. ex Nyl.) P.
James & Gott. Schneid.
— castaneocinerea (Rasanen)
Timdal
— foveata Timdal
— friesii (Ach.) P. James & Gott.
Schneid.
— isidiosa Elix
— leucococca R. Sant.
— oligospora Timdal
— praestabilis (Nyl.) Timdal
— scalaris (Ach. ex Lilj.) M. Choisy
— sierrae Timdal
— sorophora (Vain.) P. James &
Poelt
— xanthococca (Sommerf.) P.
James & Gotth. Schneid.
Immersaria Rambold & Pietschm.
— athroocarpa (Ach.) Rambold &
Pietschm.
Jamesiella Lucking, Serus. & Vezda
— anastomosans (P. James &
Vezda) Lucking, Serus. &
Vezda
— chaverriae Chaves, Umana &
Lucking
— perlucida (Vezda & Hafellner)
Lucking, Serus. & Vezda
— scotica (P. James) Lucking,
Serus. & Vezda
Karschia Korb. emend Hafellner
— puiveruienta (Anzi) Korb.
Lambiella Hertel
— psephota (Tuck.) Hertel
Lecanora Ach.
— adglutinata Florke
— atrocinerella Nyl.
— atropallidula Nyl.
— badioatra (Kremp.) Zahlbr.
— bicincta Ramond
— bockii (Fr.) Rabenh.
— bouteillei (Desm.) Harm.
— calva Nyl.
— contracta (Th. Fr.) Zahlbr.
— corrugatula (Arnold) Nyl.
— demissa Korb.
— expallens Ach.
— intercedens H. Magn.
— irrubata Nyl.
— nephaea Sommerf.
var. isidiosa Erichsen
— os treat a (Hoffm.) Rabenh.
— pharcidia Ach.
— rubiginans Nyl.
— rupicola (L.) Zahlbr.
LECANORACEAE Korb.
Lecidea Ach. emend Hertel.
— acutula Nyl.
— aethaleoides Nyl.
— affmis Schaer.
— albilabra auct.
— alboatra (Hoffm.) Chevall.
var. epipolia (Ach.) Schaer.
— alpestris Th. Fr.
— alpinus Fr.
— ambigua Ach.
— amphilecta Stirt.
— anthracophila Nyl.
— aphanoides Nyl.
— artyta Ach.
— atroalba Fr.
— atrocinerea (Schaer.) Vain.
— atrorufa (Dicks.) Ach.
— badia Fr.
— badioatra (Kremp.) Arnold
— borealis (Korb.) Anzi
— boreella Nyl.
— botryosa (Fr.) Th. Fr.
— bouteillei (Desm.) Nyl.
— calcarea Leight.
— caradocensis Leight. ex Nyl.
— chlorococca Stizenb.
— cinereorufa Schaer.
— cinnabarina Sommerf.
— cladonioides Th. Fr.
— conglomerascens Nyl.
— conglomerata Ach.
— coniops Wahlenb.
— conioptiza Nyl.
— contrusa Vain.
— corrugatula Arnold
— corticola (Ach.) Ach.
— cyclospora (Korb.) Mull. Arg.
— decipiens (Hedw.) Ach.
— demissa (Rutstr.) Ach.
— disciformis (Fr.) Nyl.
var. enteroleucoides Nyl.
— dovrensis Nyl.
var. limosa Nyl.
— elabens Fr.
— elenkinii (Rass.) Cretz.
— epiiodiza Nyl.
— euphoroides Nyl.
— exigua Chaub.
subsp. parasemella (Nyl.) Nyl.
—ferruginea Vain.
—flavovirescens (Dicks.) Borrer
var. alpina Schaer.
— freyi H. Magn.
—friesii Ach.
—fucata Stirt.
—furvella Nyl. ex Mudd
—fuscireagens Vain.
—fuscocinerea auct.
—fuscocinerea Nyl.
— — var. gyrizans (Nyl.) Nyl. ex
Brenner
— geophana Nyl.
— geophila Florke
— globifera Ach.
— globulificans Nyl.
— glomerans Nyl.
— griseoatra auct.
— grumosa Leight.
— hedinii H. Magn.
— humigena Taylor
— hydropica Korb.
— illita Nyl.
— impavida Th. Fr.
— incana (L.) Ach.
— inconcinna Nyl.
— incospicua (Elenkin) A. Zahlbr.
— insignis Nageli ex Hepp
— insita Stirt.
— insular is Nyl.
— insulatula Nyl.
— intrusa Th. Fr.
— intumescens (Florke in Flot.) Nyl.
— irrubata Ach.
— lapicida (Ach.) Ach.
— laricinella
— lepidotella Nyl.
— leptocline Flot.
— leucoxantha Spreng.
— limborina (Nyl.) Lamy
— limosa Ach.
— luridella Tuck.
— lygaeiza Vain.
— macrophylla Zahlbr.
— margaritella Hulting
— melancheima Tuck.
— melaphana Nyl.
— micro ter a Nyl.
— morbifera Vain.
— myriocarpa (DC.) Rohl.
— neglecta Nyl.
— nipponica Zahlbr.
— ochrorufa H. Magn.
— orphnaeilla Stirt.
— ostreata (Hoffm.) Schaer.
var. myrmecina (Ach.) Nyl.
— papillata Sommerf.
— parasema Ach.
var. disciform is Fr.
— parasemella Nyl.
— pelidna Ach.
— perpusilla Stizenb.
— petraea (Wulfen) Ach.
— — var. intumescens Florke in
Flot.
var. ocellata Flot.
— petrina Nyl.
— pezizoidea Ach.
var. coralloidea Nyl.
— piceicola Nyl.
— praecavenda Nyl. ex Cromb.
—praecox Vezda
— praeruptarum
— praestabilis Nyl.
— psephota Tuck.
— pulcherrima Vain.
— punctata Hoffm.
— quernea (Dicks.) Ach.
f. saxicola Nyl. in Brenner
— rhizocarpiza Zahlbr.
— rubidula Nyl.
— rubiformis (Ach.) Wahlenb.
— rubiginans (Nyl.) H. Magn.
— rugifera Vain.
— rufonigra (Tuck.) Nyl.
— rupestris (Scop.) Ach.
var. calva (Dicks.) Schaer.
var. incrustans (DC.) Schaer.
— sabuletorum Ach.
у fuscocinerea Schaer.
— sanguinaria Ach.
— sariolae Rasanen
— scalaris (Ach. ex Lilj.) Ach.
var. myrmecina Ach.
— serenior (Vain.) Vain. nom. illeg.
— siebenhaariana (Korb.) Th. Fr.
— silenii Vain.
— somphotera (Vain.) H. Olivier
f. epiiodiza (Nyl.) Vain.
— somphoterella Vain.
— sorediata Ach.
— sphacelata Th. Fr.
— spongiosula Nyl.
— spuria Schaer.
— squalens Nyl.
— stellulata Taylor
— subcinnabarina Tonsberg
— subconcava H. Magn.
— subdisciformis Leight.
— subflexuosa Nyl.
— subfurva Nyl.
— subgyratula Nyl.
— subrhagadiella Lynge
— symmictella Nyl.
— symmictella Nyl. var. albida
Vain.
— symplecta H. Magn.
— synothea Ach.
— tenebrosa Flot.
ssp. somphotera Vain.
var. serenior Vain.
var. lygaeoides Vain.
— terricola (Anzi) Th. Fr.
— terrigena Ach.
— testacea (Hoffm.) Ach.
— trichogena Norman
— triphragmia Nyl.
— trochodes (Taylor ex Leight.)
Cromb.
var. euopsoides Vain.
— umbonatula Nyl.
— umbriformis Nyl.
— umbrina Ach.
var. neotea Nyl.
— undulata H. Magn.
— xanthococca Sommerf
subsp. sorophora Vain.
LECIDEACEAE Chevall.
Lecidella Korb.
— borealis Korb.
— elabens Korb.
— eluta Korb.
— insularis (Nyl.) Korb.
— parasemella (Nyl.) Hertel
Lecideopsis (Almq.) Rehm
— intrusa (Th. Fr.) Zopf
Lecidoma Gotth. Schneid. & Hertel
in Hertel
— demissum (Rutstr.) Gotth.
Schneid. & Hertel in Hertel
Leciographa A. Massal.
— muscigenae (Anzi) Rehm
Lent aria
— mucida (Pers.) Corner
Lepidoma Gray
— demissa (Rutstr.) M. Choisy nom.
illegit.
Lepraria Ach.
— aeruginosa auct.
— alpina (de Lesd.) Tretiach & Ba¬
ruffo
— angardiana 0vstedal
— arctica (Lynge) Wetmore
— argena Ach.
— atlantica Orange
— atrotomentosa Orange & Wolse¬
ley
— bergensis Tonsberg
— borealis Lohtander & Tonsberg
— cacuminum (A. Massal.)
Lohtander
— caerulescens (Hue) Botnen &
Ovstedal
— caesioalba (de Lesd.) J. R. Laun¬
don
— candelaris (L.) Fr.
— celata Slavikova
— chlorina (Ach.) Ach. ex Sm.
— citrina (Schaer.) D. Dietr.
— crassissima (Hue) Lettau
— crassissima (Hue) Lettau
— crassissima auct.
— diffusa (J. R. Laundon) Kukwa
— — var. chrysodetoides (J. R.
Laundon) Kukwa
— eburnea J. R. Laundon
— elobata Tonsberg
—finkii (de Lesd.) R. C. Harris
—flava (Schreb.) Ach.
—frigida J. R. Laundon
— gelida Tonsberg & Zhurbenko
— glaucella (Florke) Nyl.
— humida Slavikova & Orange
— incana (L.) Ach.
— jackii Tonsberg
— lesdainii (Hue) R. C. Harris
— lobificans Nyl.
— membranacea (Dicks.) Vain.
— neglecta (Nyl.) Lettau
— nivalis J. R. Laundon
— nylanderiana Kummerl. & Leuck¬
ert
— obtusatica Tonsberg
— plumbeo-virescens Jatta
— rigidula (de Lesd.) Tonsberg
— sylvicola Orange
— toensbergiana Bayerova &
Kukwa
— umbricola Tonsberg
— vouauxii (Hue) R. C. Harris
— zonata Brodo
Leproloma Nyl. ex Crombie
— angardianum (0vstedal) J. R.
Laundon
— cacuminum (A. Massal.) J. R.
Laundon
— diffusum J. R. Laundon
— lobificans (Nyl.) Boistel
— membranacea (Dicks.) Vain.
— vouauxii (Hue) J. R. Laundon
Lichen L.
— agelaeus Ach.
— alboater Hoffm.
— caesius Hoffm.
— calicaris L.
— calvus Dicks.
— chlorinus Ach.
— corticola Ach.
— decipiens Hedw.
— distortus With.
— epigaeus Pers.
— epipolius Ach.
—farinaceous L.
—fastigiatus Pers.
—fraxineus L.
—fuscoiuteus Dicks.
— griseus Lam.
— ieucomeios L.
— membranaceus Dicks.
— pollinarius Westr.
— polymorphus Ach.
— querneus Dicks.
— runcinatus With.
— rupestris Scop.
— sanguinaria L.
— scalaris Ach. ex Lilj.
— scutelliferus With.
— speciosus Wulfen
Lichenomphalia Redhead, Lutzoni,
Moncalvo & Vilgalys
— alpina (Britzelm.) Redhead,
Lutzoni, Moncalvo &
Vilgalys
— hudsoniana (H. S. Jenn.)
Redhead, Lutzoni, Moncalvo
& Vilgalys
— umbellifera (L.: Fr.) Redhead,
Lutzoni, Moncalvo &
Vilgalys
— velutina (Quel.) Redhead,
Lutzoni, Moncalvo &
Vilgalys
Lilliputeana Serus.
— curvata Serus.
Lithographa Nyl.
Lobaria (Schreb.) Hoffm.
— scrobiculata (Scop.) DC.
Lopadium Korb.
— athaiioides Nyl.
— coralloidemum (Nyl.) Lynge
— disciforme (Flot.) Kullh.
—ferrugineum Mull.Arg.
—fuscoluteum (Dicks.)
— leucoxantha (Spreng.) Zahlbr.
— newtonii Samp.
— nipponicum M. Sato
— pezizoides (Ach.) Korb.
var. corailoideum (Nyl.) Th.
Fr.
var. discidorme (Flot.) Korb.
var. muscicola (Sommerf.) Th.
Fr.
— purpuratum Zahlbr.
Melanaspicilia Vain.
— aethalea (Ach.) Vain.
— ectolechioides Vain.
— microplaca Vain.
— tschuctschorum Vain.
Merismatium Zopf
— peregrinum (Flot.) Triebel
Micarea Fr.
— assimilata (Nyl.) Coppins
— chrysophthalma P. James
— intrusa (Th. Fr.) Coppins & H.
Kilias
— leprosa P. James
— peliocarpa (Anzi) Coppins & R.
Sant.
— prasina Fr.
Miriquidica Hertel & Rambold
— atrofulva (Sommerf.) A. J.
Schwab & Rambold in Ram¬
bold & A. J. Schwab
— lulensis (Hellb.) Hertel & Ram¬
bold
Mosigia A. Massal.
— gibbosa (Ach.) Fr. ex A. Massal.
— illita (Nyl.) R. Sant.
— intercedens (H. Magn.) R. Sant.
Multiclavula R. H. Petersen
— corynoides (Peck) R. H. Petersen
— mucida (Pers.) R. H. Petersen
— vernalis (Schwein.) R. H.
Petersen
Mycobilimbia Rehm
— lobulata (Sommerf.) Hafellner in
V. Wirth
MYCOBLASTACEAE Hafellner
Mycoblastus Norman
— affinis (Schaer.) T. Schauer
— alpinus (Fr.) Th. Fr. ex Hellb.
— caesius (Coppins & P. James)
Tonsberg
— fucatus (Stirt.) Zahlbr.
— melinus (Kremp. ex Nyl.) Hellb.
— sanguinarius (L.) Norman
— sanguinarius var. alpinus (Fr.)
Stein
— sterilis Coppins & P. James
Nesolechia A. Massal.
— geophana (Nyl.) Vouaux
Niebla Rundel &Bowler
Normandina Nyl.
— viridis (Ach.) Nyl.
Omphalina Quel.
— alpina (Britzelm.) Bresinsky &
Stangl
— ericetorum (Pers.: Fr.) M. Lange
— luteovitellina (Pilat & Nannf) M.
Lange
—flava (Sommerf) M. Lange
— hudsoniana (H. S. Jenn.) H. E.
Bigelow
— luteolilacina (J. Favre) D. M.
Hend.
— pseudoandrosacea (Bull.) М. M.
Moser
— pseudoandrosacea auct.
— umbellifera (L.: Fr.) Quel.
— velutina (Quel.) Quel.
Orceolina Hertel
Parmelia Ach.
— adglutinata (Florke) Florke
— albicans Pers.
— argena Spreng.
— aquila Ach.
var. stippaea Ach.
— bockii Fr.
— bouteillei Desm.
— corallophora Taylor
— desertorum Rupr.
— diademata Taylor
— elaeina Gray
—farrea Ach.
— granulifera Ach.
— minor Fee
— muscigena Ach.
— ostreata (Hoffm.) Fr.
— ostreata (Hoffm.) Wallr.
— podocarpa Bel.
— pulverulenta (Schreb.) Ach.
var. deters a Nyl.
var. leucoleiptes Tuck.
— speciosa Ach.
b. hypoleuca Ach.
— venusta Ach.
Patellaria Pers.
— in crus tans DC.
— ostreata (Hoffm.) Wallr.
Peltigera Willd.
— collina (Ach.) Schrad.
Peltiphylla M. Choisy
— albilabra auct.
Phaeophyscia Moberg
— adiastola (Essl.) Essl.
— cernohorskyi (Nadv.) Essl.
— chloantha (Ach.) Moberg
— ciliata (Hoffm.) Moberg
— strigosa (Poelt & G. Buschardt)
N. S. Golubk.
— confusa Moberg
— constipata (Norrl. & Nyl.)
Moberg
— decolor (Kashiw.) Essl.
— denigrata (Hue) Moberg
— dissecta G. Urban, I. Urban & T.
Otn.
— echinata (Frey) N. S. Golubk.
— endococcina (Korb.) Moberg
— — var. decolor (Kashiw.)
Clauzade & Cl. Roux
— — var. endococcinodes (Poelt)
Moberg
— endococcinodes (Poelt) Essl.
— endophoenicea (Harm.) Moberg
— erythrocardia (Tuck.) Essl.
— exornatula (Zahlbr.) Kashiw.
— fumosa Moberg
— hirsuta (Mereschk.) Essl.
— hirsuta (Mereschk.) Moberg
— hirtella Essl.
— hirtuosa (Kremp.) Essl.
— hirtuosa (Kremp.) N. S. Golubk
— hispidula (Ach.) Essl.
— hispidula (Ach.) D. D. Awasthi
— hispidula (Ach.) Moberg
— imbricata (Vain.) Essl.
var. hispidula
— — var. exornatula (Zahlbr.)
Moberg
— insignis (Mereschk.) Moberg
— kairamoi (Vain.) Moberg
— leana (Tuck.) Essl.
— limbata (Poelt) Kashiw.
— melanchra (Hue) Hale
— nadvornikii (Frey & Poelt) N. S.
Golubk.
— nigricans (Florke) Moberg
— orbicularis (Neck.) Moberg
— poeltii (Frey) Nimis
— poeltii (Frey) Clauzade & Cl.
Roux
— primaria (Poelt) Trass
— pusilloides (Zahlbr.) Essl.
— pyrrhophora (Poelt) D. D.
Awasthi & M. Joshi
— pyrrhophora (Poelt) Kashiw.
— rubropulchra (Degel.) Essl.
— rubropulchra (Degel.) Moberg
— saxatilis Kashiw.
— sciastra (Ach.) Moberg
— spinellosa Kashiw.
— squarrosa Kashiw.
— stiriaca (Poelt) Clauzade & Cl.
Roux
— trichophora (Hue) Essl.
PHLYCTIDACEAE Poelt & Vezda
ex J.C. David & D. Hawksw.
Phlyctis (Wallr.) Flot.
— agelaea (Ach.) Flot.
— argena (Ach.) Flot.
— erythrosora Erichsen
— speirea G. Merr.
— willeyi Tuck.
— schizospora Zahlbr.
Physcia (Schreb.) Michx.
— adiastola Essl.
— adglutinata (Florke) Nyl.
— adscendens H. Olivier
— aipolia (Humb.) Fiirnr.
f. alnophila Vain.
subsp. scopulorum Lambinon
& Vezda
var. alnophila (Vain.) Lynge
var. subincisa (Th. Fr.) Lynge
— albicans (Pers.) J. W. Thomson
— albinea (Ach.) Nyl.
— albonigra (Schleich.) Dalla Torre
& Sarnth.
— alnophila (Vain.) Lohtander & al.
— asiana Nyl.
— astroidea (Clem.) Nyl.
— biziana (A. Massal.) Zahlbr.
— caesia (Hoffm.) Furnr.
var. caesia
var. albinea (Ach.) Torss.
— — var. caesiella (de Lesd.)
Clauzade & Cl. Roux
var. melops auct. non (Dufour)
Vain.
var. rhaetica Frey
— caesiella (de Lesd.) Suza
— capitata Mereschk.
— caucasica J. Steiner
— cernohorskyi Nadv.
var. erosa Nadv.
— ciliata (Hoffm.) Du Rietz
— — var. crinalis Schleich. ex
Schaer.
var. nigrescens (Bory) Zahlbr.
— clementei (Turner) Maas Geest.
— concrustans Nyl.
— constipata Norrl. & Nyl.
— cycloselis (Ach.) Vain.
— decolor Kashiw.
— denigrata Hue
— desertorum (Rupr.) Savicz
— deter sella Nadv.
— dimidiata (Arnold) Nyl.
— dubia (Hoffm.) Lettau
— echinella (Poelt) R. Schubert &
Klem.
— elaeina (Sm.) A. L. Sm.
— elbursiana (Szatala) Szatala
— endoaurantiaca Barchal.
— endochroidea Nyl.
— endochrysoides Nyl.
— endococcina (Korb.) Th. Fr.
var. latiloba H. Magn.
— endococcinodes Poelt
var. megalospora Poelt
— endophoenicea (Harm.) Santha
pseudoplatani (Britz.) Nadv.
— enteroxantha Nyl.
— erumpens Moberg
— erythrocardia (Tuck.) Vain.
— grisea (Lam.) Zahlbr.
var. elbursiana Szatala
— hirsuta Mereschk.
var. echinella Poelt
— hirtuosa Kremp.
— hispidula (Ach.) Frey
— — subsp. exornatula (Zahlbr.)
Poelt
subsp. limbata Poelt
subsp. primaria Poelt
— hueana (Harm.) Rasanen
— imbricata Vain.
— insignis Mereschk.
— intermedia Vain.
— intricata (Desf.) Schaer.
—japonica (Hue) Vain.
var. denigrata Vain.
—jenisejensis H. Magn.
— kairamoi Vain.
— karakorina Poelt
— kansuensis H. Magn.
— labrata Mereschk.
— — var. endophoenicea (Harm.)
Mereschk.
var. intermedia Mereschk.
f. minor Mereschk.
— labrata sensu Frey
— leptalea (Ach.) DC.
— leptothallina (Vain.) Zahlbr.
— leucoleiptes (Tuck.) Lettau
— lithotea auct.
— lithotodes Nyl.
— luganensis Mereschk.
— lyngei Nadv.
— magnussonii Frey
— marina (A. Nyl.) Lynge
— melanchra Hue
— melops Dufour ex Nyl.
— mereschkowskii Tomin
— minor (Fee) Vain.
— muscigena (Ach.) Nyl.
— nadvornikii Frey & Poelt
f. echinata Frey
— nepalensis Poelt
— nigricans (Florke) Stizenb.
— nipponica Asahina
— norrlinii Vain.
— obscura auct.
— obscura ([Ehrh. ex] Humb.)
Fiirnr.
var. chloantha (Ach.) Rabenh.
var. endophoenicea Harm.
var. erythrocardia Tuck.
var. nigricans (Florke) Hazsl.
— obscurata Nyl.
— ocellata Erichsen
— orbicularis (Neck.) Poetsch
f. rubropulchra Degel.
var. pulvinata (Korb.) Santha
— orientalis Kashiw.
— oxneri Inasch.
— palmulata (Michx.) Nyl.
— parvula Vain.
— perisidiosa Erichsen
— phaea (Tuck.) J. W. Thomson
— poeltii Frey
— pragensis Nadv.
— pulverulenta Hamp.
— pulverulenta (Schreb.) Hamp.
var. tenuis (Korb.) Th. Fr.
var. venusta auct.
— pusilla Mereschk.
— pusilloides Zahlbr.
— pyrrhophora Poelt
— rubropulchra (Degel.) Moberg
— sciastra (Ach.) Du Rietz
— sciastrella (Nyl.) Harm.
— scopulorum (Lambinon & Vezda)
Poelt & Nimis
— semipinnata (J. F. Gmel.)
Moberg
— servitii Nadv.
— setosa (Ach.) Nyl.
var. albociliata de Lesd.
var. japonica (Hue) Zahlbr.
— speciosa (Wulf.) Nyl.
f. isidiophora Nyl.
— stellaris (L.) Nyl.
var. aipolia (Humb.) Hazsl.
var. subincisa Th. Fr.
— stiriaca Poelt
— strigosa Poelt & G. Buschardt
— subalbinea Nyl.
— subobscura Nyl.
— subpulverulenta Szatala
— suzai Nadv.
— syncolla Tuck, ex Nyl.
— tenella (Scop.) DC.
var. marina (A. Nyl.) Lynge
— teretiuscula (Ach.) Lynge
— ticinensis (Mereschk.) Frey
— tremulicola Nyl.
— tribacia (Ach.) Nyl.
— tribacella Nyl.
— tribacoides Nyl.
— trichophora Hue
var. cinereoalbida Vain.
— ulothrix (Ach.) Nyl.
— ulotrichoides Vain.
— undulata Moberg
— ventosa (Lynge) Santha
— virella (Ach.) Flagey
— vitii Nadv.
— wahlenbergii Lynge
— wainioi Rasanen
PHYSCIACEAE Zahlbr.
Physciella Essl.
— austrosibirica G. Urban.
— chloantha (Ach.) Essl.
— denigrata (Hue) Essl.
— melanchra (Hue) Essl.
— nepalensis (Poelt) Essl.
Physciopsis M. Choisy
— adglutinata (Florke) M. Choisy
— elaeina (Sm.) Poelt
— minor (Fee) B. J. Moore
— granulata Poelt
— syncolla (Tuck, ex Nyl.) Poelt
Physconia Poelt
— americana Essl.
— californica Essl.
— chinensis J. B. Chen & G. R. Hu
— detersa (Nyl.) Poelt
— distorta (With.) J. R. Laundon
— elegantula Essl.
— enteroxantha (Nyl.) Poelt
— fallax Essl.
—farrea auct.
— grisea (Lam.) Poelt
— grumosa Kashiw. & Poelt
— hokkaidensis Kashiw.
— isidiigera (Zahlbr.) Essl.
— isidiomuscigena Essl.
— jacutica G. Urban., Ahti &
Lohtander
— kurokawae Kashiw.
— leucoleiptes (Tuck.) Essl.
— lobulifera Kashiw.
— muscigena (Ach.) Poelt
var. petraea Poelt
— perisidiosa (Erichs.) Moberg
— petraea (Poelt) Vezda & Poelt
— pulverulenta (Schreb.) Poelt
— pulverulacea Moberg
— rossica G. Urban.
— servitii (Nadv.) Poelt
— subpallida Essl.
— subpulverulenta (Szatala) Poelt
— thomsonii Essl.
— venusta (Ach.) Poelt
Phytoconis Bory
— ericetorum (Pers.: Fr.) Redhead
& Kuyper
— luteovitellina (Pilat & Nannf.)
Redhead & Kuyper
— velutina (Quel.) Redhead &
Kuyper
— viridis (Ach.) Redhead & Kuyper
PILOCARPACEAE Zahlbr.
Placinthiella Elenkin
— uliginosa (Schrad.) Coppins & P.
James
Placodium (Pers.) Poelt
— ostreatum (Hoffm.) Link
— leucoleprosum Vain.
Placopsis (Nyl.) Linds.
Pleolecis Clem.
— geophana (Nyl.) Clem.
Protoanaptychia Poelt
Protoblastenia (Zahlbr.) J. Steiner
— areolata H. Magn.
— aurata Poelt & Vezda
— calva (Dicks.) Zahlbr.
— coniasis (A. Massal.) Poelt
— cinnabarina (Sommerf.) Rasanen
— cyclospora (Hepp ex Korb.) Poelt
— globulificans (Nyl.) Zahlbr.
— incrustans (DC.) J. Steiner
— — var. coniasis (A. Massal.)
Poelt
— rupestris (Scop.) J. Steiner
var. calva (Dicks.) J. Steiner
var. incrustans (DC.) Zahlbr.
var. irrubata (Ach.) Szatala
— — var. sanguinea (Arnold)
Zahlbr.
— siebenhaariana (Korb.) J. Steiner
var. alpina (Arnold) Clauzade
& Cl. Roux
var. terricola (Anzi) Hafellner
& Turk
— terricola (Anzi) Lynge
Protomicarea Hafellner in Hafellner
& Turk
— limosa (Ach.) Hafellner
Protoparmelia M. Choisy
Psathyrophlyctis Brusse
Pseudephebe M. Choisy
— minuscula (Nyl. ex Arnold) Bro¬
do & D. Hawksw.
Psilolechia A. Massal.
— lucida (Ach.) M. Choisy
Psora Hoffm.
— acutula (Nyl.) Walt. Watson
— albilabra auct.
— anthracophila (Nyl.) Arnold
— cinereorufa (Schaer.) Hellb.
— cladonioides (Fr.) Elenkin
var. albocervina Rasanen
var. castaneocinerea Rasanen
— concava de Lesd.
— conglomerata (Ach.) Korb.
— coroniformis Krmp.
— crenata (Taylor) Reinke
— decipiens (Hedw.) Hoffm.
— demissa (Rutstr.) Hepp
— elenkinii Rass.
—friesii (Ach.) Hellb.
— globifera (Ach.) A. Massal.
— himalayana (C. Bab.) Timdal
— incospicua Elenkin
— indigirkae (Zahlbr.) Timdal &
Zhurbenko
— luridella (Tuck.) Fink
— nipponica (Zahlbr.) Gotth.
Schneid.
— ostreata Hoffm.
— pulcherrima (Vain.) Elenkin
var. elegantula Elenkin
— rubiformis (Ach.) Hook.
— rugifera (Vain.) Dombr.
— saviczii (Tomin) Follmann & A.
Crespo
— scalaris (Ach. ex Lilj.) Hook.
— — var. myrmecina (Ach.)
Rasanen
— silenii (Vain.) Dombr.
— subconcava (H. Magn.) Bredkina
— subrubiformis (Vain.) Timdal
— tenuifolia Timdal
— testacea Rasanen
— tuckermanii Timdal
— vallesiaca (Schaer.) Timdal
PSORACEAE Zahlbr.
Psorinia Gotth. Schneid.
— conglomerata (Ach.) Gotth.
Schneid.
— lepidotella (Nyl.) Gotth. Schneid.
Psoroma Michx.
— palmulata Michx.
Psorula Gotth. Schneid.
— rufonigra (Tuck.) Gotth. Schneid.
Ptychographa Nyl.
Pycnora Hafellner
— leucococca (R. Sant.) R. Sant.
— praestabilis (Nyl.) Hafellner
— sorophora (Vain.) Hafellner
— xanthococca (Sommerf.)
Hafellner
Pyrenula A. Massal.
— gibbosa Ach.
Pyrrhospora Korb.
— cinnabarina (Sommerf.) M.
Choisy
— elabens (Fr.) Hafellner
— quernea (Dicks.) Korb.
— rubiginans (Nyl.) P. James &
Poelt in Poelt & Vezda
— subcinnabarina (Tonsberg)
Hafellner
Pyxine Fr.
— berteriana (Fee) Imshaug
— chrysantha Vain.
— chrysanthoides Vain.
— endochrysina Nyl.
— endochrysoides (Nyl.) Degel.
— limbulata Mull. Arg.
— meissneri Tuck.
— meissneriana Nyl.
— sibirica Tomin
— sorediata (Ach.) Mont.
— subcinerea Stirt.
Quercus L.
Ramalina Ach.
— almquistii Vain.
— angustissima (Anzi) Vain.
— asahinana Zahlbr.
— baltica Lettau
— calicaris (L.) Fr.
ssp. elegans (Bagl. & Carestia)
Stizenb.
var. crispa
— — var. evernioides (Anzi)
Motyka
var. japonica Hue
var. subfastigiata Nyl.
f. fibril osa Th. Fr.
— canaliculata Fr.
— canariensis J. Steiner
— capitata (Ach.) Nyl.
— carpatica Korber
— commixta Asahina
— conduplicans Vain.
— crinalis (Ach.) Gyeln.
— dilacerata (Hoffm.) Hoffm.
f. turgida Rasanen
— elegans (Bagl. & Carestia) Jatta
— exilis Asahina
f. папа Asahina
—fallax Motyka
— farinacea (L.) Ach.
var. angustissima Anzi
var. dendroides Mull. Arg.
var. rubescens Rasanen
var. squarossa Mull. Arg.
— fastigiata (Pers.) Ach.
— fraxinea (L.) Ach.
— geniculata Hook f. & Taylor
— hokkaidensis Kashiw.
— hypoprotocetrarica W. L. Culb.
& C. F. Culb.
— intermedia (Delise ex Nyl.) Nyl.
— intermediella Vain.
— kardakovae Oxner
— kazakhorum Oxner
— koreana Kashiw. & К. H. Moon
— kurokawae Kashiw.
— landroensis Zopf
— ligulata (Ach.) Brandt
— ligulata Asahina
— litoralis Asahina
— minuscula (Nyl.) Nyl.
— — subsp. intermedia Delise ex
Nyl.
var. obtusata Arnold
— montana Barkh.
— motykana Bystrek
— nervosa Rasanen
— obtusata (Arnold) Bitter
var. ventricosa (Eitner) Keissl.
ex Hillmann & Grummann
— obtusata auct.
— pacifica Asahina
— pertusa Kashiw.
— peruviana Ach.
— pollinaria (Westr.) Ach.
var. insularis Vain.
— pollinariella (Nyl.) Nyl.
— polymorpha (Lilj.) Ach.
var. capitata (Ach.) Clauzade
& Cl. Roux
— pontica Vezda
— populina (Ehrh.) Vain.
— pulvinata (Anzi) Nyl.
— pumila Mont.
— rjabuschinskii Savicz
— roesleri (Hochst. ex Schaer.) Hue
— scoparia Vain.
— scropulorum (Retz.) Ach.
— seawardii Aptroot & Sipman
— shinanoana Kashiw.
— siliquosa (Huds.) A. L. Sm.
— sinensis Jatta
— strepsilis (Ach.) Zahlbr.
— subbreviuscula Asahina
— subcomplanata (Nyl.) Kashiw.
— subfarinacea (Nyl. ex Cromb.)
Nyl.
— sublitoralis Asahina
— thrausta Nyl.
— throwerae Aptroot & Sipman
— tinctoria auct.
— vogulica Vain.
— yasudae Rasanen
RAMALINACEAE C. Agardh
Rhaphiospora A. Massal.
—flavovirescens A. Massal.
var. alpina Kremp.
Rhexophiale Th. Fr.
— rhexoblephara (Nyl.) Hellb.
Rhizocarpon Ramond ex DC.
— alboatrum (Hoffm.) Anzi
var. epipolium (Ach.) Anzi
— betulinum Zw.
— copelandii (Korb.) Th. Fr.
— efflorescens Th. Fr.
— ferax H. Magn.
— geminatum Korb.
— grande (Florke in Flot.) Arnold
— inarense (Vain.) Vain.
— intermediellum Rasanen
— pusillum Runemark
— superficial (Schaer.) Vain.
Rimularia Nyl.
— badioatra (Hepp ex Kremp.)
Hertel & Rambold
— caeca (Lowe) Rambold &
Printzen
— furvella (Nyl. ex Mudd) Hertel &
Rambold
— fuscosora Muhr & Tonsberg
— gibbosa (Ach.) Coppins, Hertel &
Rambold
— gyrizans (Nyl.) Hertel &
Rambold
— impavida (Th. Fr.) Hertel &
Rambold
— insularis (Nyl.) Rambold & Her¬
tel
— intercedens (H. Magn.) Coppins
in Purvis, Coppins & P.
James
— limborina Nyl.
— psephota (Tuck.) Hertel &
Rambold in Hertel
— sphacelata (Th. Fr.) Hertel &
Rambold
— subconcava (H. Magn) Timdal
RIMULARIACEAE Hafellner
Rinodina (Ach.) Gray
— albana (A. Massal.) A. Massal.
— altissima H. Magn.
— archaea (Ach.) Arnold
— archaeoides H. Magn.
— arctica H. Magn.
— arenaria (Hepp) Arnold
— arenaria auct.
— arnoldii H. Mayrhofer & Poelt
— aspersa (Borrer) J. R. Laundon
— atrocinerea (Hook.) Korb.
— atrocinerella (Nyl.) Boist.
— atropallidula (Nyl.) Arnold
— badiella (Nyl.) Th. Fr.
— balanina (Wahlenb.) Vain.
— biatorina Korb.
— bischoffii (Hepp) A. Massal.
— budensis (Nyl.) Zahlbr.
— cacuminum (Th. Fr.) Malme
— calcarea (Arnold) Arnold
— calcigena (Th. Fr.) Lynge
— canella Arnold
— capensis Hampe
— castanomela (Nyl.) Arnold
— cinereovirens (Vain.) Vain.
— chionea Th. Fr.
— cinnamomea (Th. Fr.) Rasanen
— collectica (Florke) Arnold
— colobina (Ach.) Th. Fr.
— colobinoides (Nyl.) Zahlbr.
— confragosa (Ach.) Korb.
— conradii Korb.
— consocians H. Magn.
— convexula H. Magn.
— crassescens (Nyl.) Arnold
— cretica H. Mayrhofer
— degeliana Coppins
— dem iss a auct.
— discolor (Hepp) Arnold
— disjuncta Sheard & Tonsberg
— dispersella (Vain.) Vain.
— dubyana (Hepp) J. Steiner
— efflorescens Malme
— endophragmia 1. M. Lamb
— epimilvina H. Mayrhofer
— excrescens Vain.
— exigua (Ach.) Gray
— exiguella (Vain.) H. Magn.
— fimbriata Korb.
— flavosoralifera Tonsberg
— gennarii Bagl.
— granulans Vain.
— griseosoralifera Coppins
— guzzinii Jatta
— hyperborea H. Magn.
— immersa (Korb.) Zahlbr.
— immersata (Nyl.) H. Olivier
— insularis (Arnold) Hafellner
— intermedia Bagl.
— interpolata (Stirt.) Sheard
— isidioides (Borrer) H. Olivier
— kansuensis H. Magn.
— kozukensis (Vain.) Zahlbr.
— laevigata (Ach.) Malme
— lecanorina (A. Massal.) A. Mas¬
sal.
— lecideina H. Mayrhofer & Poelt
— lecideoides (Nyl.) Kernst.
— lignaria H. Magn.
— luridata (Korb.) H. Mayrhofer,
Scheid. & Sheard
— luridescens (Anzi) Arnold
— malangica (Norman) Arnold.
— mayrhoferi A. Crespo
— melanocarpa Mull. Arg.
— metabolica auct.
— metaboliza Vain.
— milvina (Wahlenb.) Th. Fr.
— — var. subdeflectens (Vain.)
Zahlbr.
— mniaraea (Ach.) Korb.
var. cinnamomea Th. Fr.
— — var. mniaraeiza (Nyl.) H.
Magn.
— mniaraeiza (Nyl.) Arnold
— mniaraeoides (Nyl.) H. Olivier
— mucronatula H. Magn.
— muscicola H. Magn.
— obnascens (Nyl.) H. Olivier
— occidentalis Lynge
— occulta (Korb.) Sheard
— ocellata (Hoffm.) Arnold
— olivaceobrunnea C. W. Dodge &
G. E. Baker
— orbata (Ach.) Vain.
— orculata Poelt & M. Steiner
— oreina (Ach.) A. Massal.
— oxydata (A. Massal.) A. Massal.
— parasitica H. Mayrhofer & Poelt
— pinicola (Ach.) Zahlbr.
— pityrea Ropin & H. Mayrhofer
— plana H. Magn.
— polyspora Th. Fr.
— polysporoides Giralt & H.
Mayrhofer
— pruinella Bagl.
— pycnocarpa H. Magn.
— pyrina (Ach.) Arnold
— radiata (Tuck.) Tuck.
f. fimbriata (Tuck.) Zahlbr.
— rinodinoides (Anzi) H. Mayrhofer
& Scheid.
— roscida (Sommerf.) Arnold
— septentrionalis Malme
— sheardii Tonsberg
— sibirica H. Magn.
granulans (Vain.) H. Magn.
— sophodes (Ach.) A. Massal.
f. cacuminum Th. Fr.
— sophodioides H. Magn.
— sorediata H. Magn.
— straussii J. Steiner
— subfusca H. Magn.
— subnigra H. Magn.
— subobscura H. Magn.
— subrubescens (Vain.) Zahlbr.
— teichophila (Nyl.) Arnold
— tephraspis (Nyl.) Here
— terrestris Tomin
— tominii H. Mayrhofer
— trachytica (A. Massal.) Bagl. &
Carestia
— tunicata H. Mayrhofer & Poelt
— turfacea (Wahlenb.) Korb.
var. cinereovirens (Vain.) H.
Mayrhofer
— — var. ecrustacea (Vain.) H.
Olivier.
— umbrinofusca (Nyl.) H. Olivier
— vezdae H. Mayrhofer
— victoris (Harm.) H. Olivier
— violascens H. Magn.
— xylophila H. Magn.
— zwackhiana (Kremp.) Korb.
Rinodinella H. Mayrhofer
— controversa (A. Massal.) H.
Mayrhofer & Poelt
— dubyanoides (Hepp) H.
Mayrhofer & Poelt
SA CCOMORPHA CEA E
Sagiolechia A. Massal.
— protuberans (Ach.) A. Massal.
— rhexoblephara (Nyl.) Zahlbr.
Sarcogyne Flot.
— geophana (Nyl.) Boistel
Sarea Fr.
Scalidium Hellb.
— ophiosporum Hellb.
Schaereria Th. Fr.
— bullata Kantvilas
— cinereorufa (Schaer.) Th. Fr.
— corticola Muhr & Tonsberg in
Tonsberg
— endocyanea (Stirt.) Hertel &
Gotth. Schneider
— fabispora Hertel & Ziirn
— fuscocinerea (Nyl.) Clauzade &
Cl. Roux
var. sorediata (Houmeau & Cl.
Roux) Coppins
— lugubris Korb.
— ostreata (Hoffm.) Gyeln.
— parasemella (Nyl.) Lumbsch
— serenior (Vain.) Vitik.
— tenebrosa (Flot.) Hertel & Poelt
— xerophila Rambold & H. Mayrho¬
fer
SCHAERERIACEAE M. Choisy ex
Hafellner
Scoliciosporum A. Massal.
— chlorococcum (Graewe ex Stenh.)
Vezda
— comp actum Korb.
— corticolum (Anzi) Arnold.
— curvatum Serus.
— gallurae Vezda & Poelt
— holomelaenum (Florke) A.
Massal.
— intrusum (Th. Fr.) Hafellner
— ophiosporum (Hellb.) Hav.
— perpusillum J. Lahm ex Korb.
— pruinosum (P. James) Vezda
— sarothamni (Vain.) Vezda
— schadeanum (Erichsen) Vezda
— turgidum Korb.
— umbrinum (Ach.) Arnold
var. corticolum (Anzi) Bagl. &
Carestia
var. marinum (Degel.) Degel.
Solorina Ach.
Spilonema Bornet
— revertens Nyl.
Sporopodium Vain.
— caucasicum Elenkin & Woron.
Sporastatia A. Massal.
— testudinea (Ach.) A. Massal.
Squamaria Poelt
— biziana A. Massal.
Squamarina Poelt
— pamirica N. S. Golubk.
Steinia Korb. in Stein
— geophana (Nyl.) Stein
— luridescens Korb.
STEREOCAULACEAE Chevall.
Stereocaulon Hoffm.
Tephromela M. Choisy
— atra (Huds.) Hafellner in Kalb
Toninia A. Massal.
— caradocensis (Leight. ex Nyl.) J.
Lahm
— conglomerata (Ach.) Boist.
— loitlesbergeri Zahlbr.
Tornabea 0sth.
— atlantica (Ach.) 0sth.
var. intricata (Desf.) Clauzade
& Cl. Roux
— ephebea (Ach.) 0sth.
— scutellifera (With.) J. R. Laundon
Tornabenia Trevis.
— atlantica (Ach.) Kurok.
— ephebea (Ach.) Kurok.
— intricata (Defs.) Trevis.
— montagnei Trevis.
Tornabeniopsis Follmann
— atlantica (Ach.) Follmann
Trapelia M. Choisy
TRAPELIACEAE M. Choysy ex
Hertel
Trapeliopsis Hertel & Gotth.
Schneid.
TRICHOLOMATACEAE R. Heim
ex Pouzar
Umbilicaria Hoffm.
Variolaria Ach.
— griseovirens Turn. & Borr. ex
Sm.
Verrucaria Schrad.
— rupestris (Scop.) F. H. Wigg.
VEZDAEACEAE Poelt & Vezda ex
J. C. David & D. Hawksw.
Vezdaea Tscherm.-Woess & Poelt
— acicularis Coppins
— aestivalis (Ohlert) Tscherm.-
Woess
— leprosa (P. James) Vezda
— retigera Poelt & Dobbeler
— reocarpa Poelt & Dobbeler
— stipitata Poelt & Dobbeler
Xanthoria (Fr.) Th. Fr.
— adglutinata (Florke) Horw.
— intricata (Desf.) Horw.
Xylographa (Fr.) Fr.
— vitiligo (Ach.) J. R. Laundon
СОДЕРЖАНИЕ
Стр.
Перечень авторов 3.
Предисловие 5.
Глава I. Три века русской лихенологии. История
изучения лишайников России (Андреев М. П,
Ахти Т.) 7.
Глава II. Основные лихенологические гербарии Рос¬
сии и важнейшие зарубежные коллекции рос¬
сийских лишайников (Андреев М. П, Ахти Т.) 48.
Глава III. Таллом и репродуктивные структуры ли¬
шайников (Гимельбрант Д. Е, Кузнецова Е. С.) 63.
Таллом 65.
Морфология таллома 67.
Жизненные формы лишайников 67.
Накипные лишайники 85.
Листоватые лишайники 87.
Кустистые лишайники 88.
Неприкрепленные лишайники 90.
Форма таллома 92.
Характер ветвления талломов ку¬
стистых и листоватых ли¬
шайников 93.
Поверхностные структуры таллома 94.
Анатомия таллома 105.
Стратификация таллома лишайника 108.
Репродуктивные структуры 112.
Структуры вегетативного размножения
лишайников 113.
Структуры полового размножения микобио¬
нта
119.
Конидиальные спороношения микобионта 133.
Глава IV. Водоросли-фотобионты лишайников
(Войцехович А. А, Надеина О. В.) 143.
Разнообразие фотобионтов 143.
Корреляция между таксономической принад¬
лежностью мико- и фотобионта 147.
Источники фотобионтов. Основные, дополни¬
тельные и «потенциальные» фотобионты
154.
Изоляция и культивирование фотобионтов 162.
Глава V. Особенности вторичного метаболизма и
хемосистематика лишайников (Рандлане Т,
Сааг А.) 176
Лишайниковые вещества 176
Роль лишайниковых веществ в лишайнико¬
вых талломах 178
Хемотаксономия лишайников 194
Основные методики определения лишайни¬
ковых веществ 197
Цветные реакции («spot»-test) 198
Ультрафиолетовая флюоресценция 200
Микрокристаллизация 202
Тонкослойная хроматография (TLC) 202
Тонкослойная хроматография высокой
точности (HPTLC) 204
Жидкостная хроматография высокой
точности (HPLC) 204
Глава VI. Разнообразие и распространение лишай¬
ников (Урбанавичюс Г. П.) 209
Уровень богатства лихенофлоры России 209
Важнейшие таксономические группы ли¬
хенофлоры России 215
Экологические основы региональных и зо¬
нальных различий лихенофлор России 224
Характер распространения лишайников на
территории России. Эндемичные и эв-
ритопные роды 230
Глава VII. Экология лишайников (Макрый Т. В.) 242
Экологические факторы 242
Экологическая обусловленность жизненных форм
253
Экология и особенности распространения
лишайников 255
Лишайники и растительный покров 257
Роль лишайников в экосистемах 258
Экология и эволюция 259
Глава VIII. Сбор и определение лишайников, хра¬
нение лихенологических коллекций (Гагари¬
на Л. В, Степанчикова И. С.) 262
Методы полевой работы 263
Сбор материала и необходимые инструменты.
Хранение образцов 266
Определение лишайников 269
Регистрация данных об образце 272
Правила работы в гербарии 274.
Глава IX. Номенклатура и типификация лишайников
(Гагарина JI. В, Степанчикова И. С.) 282.
Наиболее часто употребляемые понятия и термины
285.
Аббревиатуры и условные обозначения, использу¬
емые при работе с гербарием 287.
Типификация 288.
Глава X. Молекулярно-генетические методы изучения
лишайников (Давыдов Е. А.) 933.
Основные вопросы систематики, решаемые при
помощи молекулярных методов 296.
Геном грибов и лишайников 297.
Выделение и клонирование ДНК 298.
Полимеразная цепная реакция 300.
Разделение фрагментов ДНК, имеющих раз¬
ную длину 301.
Основные молекулярные методы 302.
Молекулярные методы, базирующиеся на
анонимном полиморфизме ДНК 302.
Методы изучения последовательностей
фрагментов ДНК 307.
Методы филогенетического анализа 310.
Фенетические методы 311.
Кладистические методы 313.
Филогенетические сети 317.
Интерпретация результатов филогенетических по¬
строений 318.
ДНК-баркодинг 318.
Глава XI. Систематическая классификация таксонов ли¬
хенофлоры России (Урбанавичюс Г. П.) 323.
Глава XII. Ключи для определения родов лишайников
флоры России (Урбанавичюс Г. П, Макрый Т. В,
Кузнецова Е. С, Гагарина Л. В, Гимельбрант Д. Е,
Степанчикова И. С, Конорева JI. А, Андреев М.
П.) 353.
Морфологические группы родов лишайников фло¬
ры России 354.
Синопсис (вспомогательный ключ для определе¬
ния групп родов) 355.
Родовые ключи 356.
Стерильные лишайники 430.
Литература 435.
Приложения 477.
Приложение I. Деление территории Российской
Федерации на регионы, принятое в настоя¬
щем издании 477
Приложение II. Терминологический словарь 483
Приложение III. Принятые сокращения фамилий
авторов при таксонах 504
Приложение IV. Индекс латинских названий 509
Содержание 532
Contents 536